

















































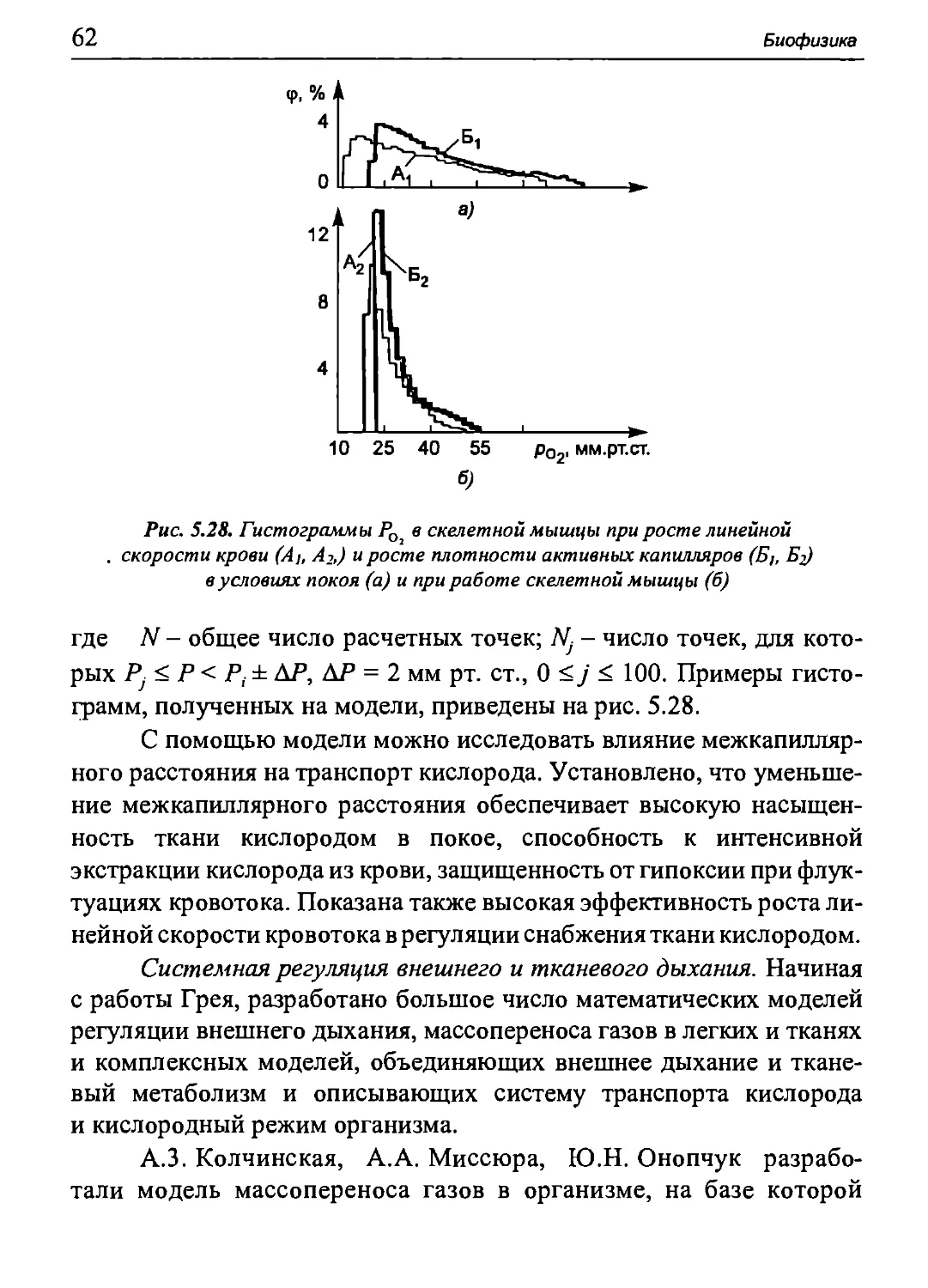
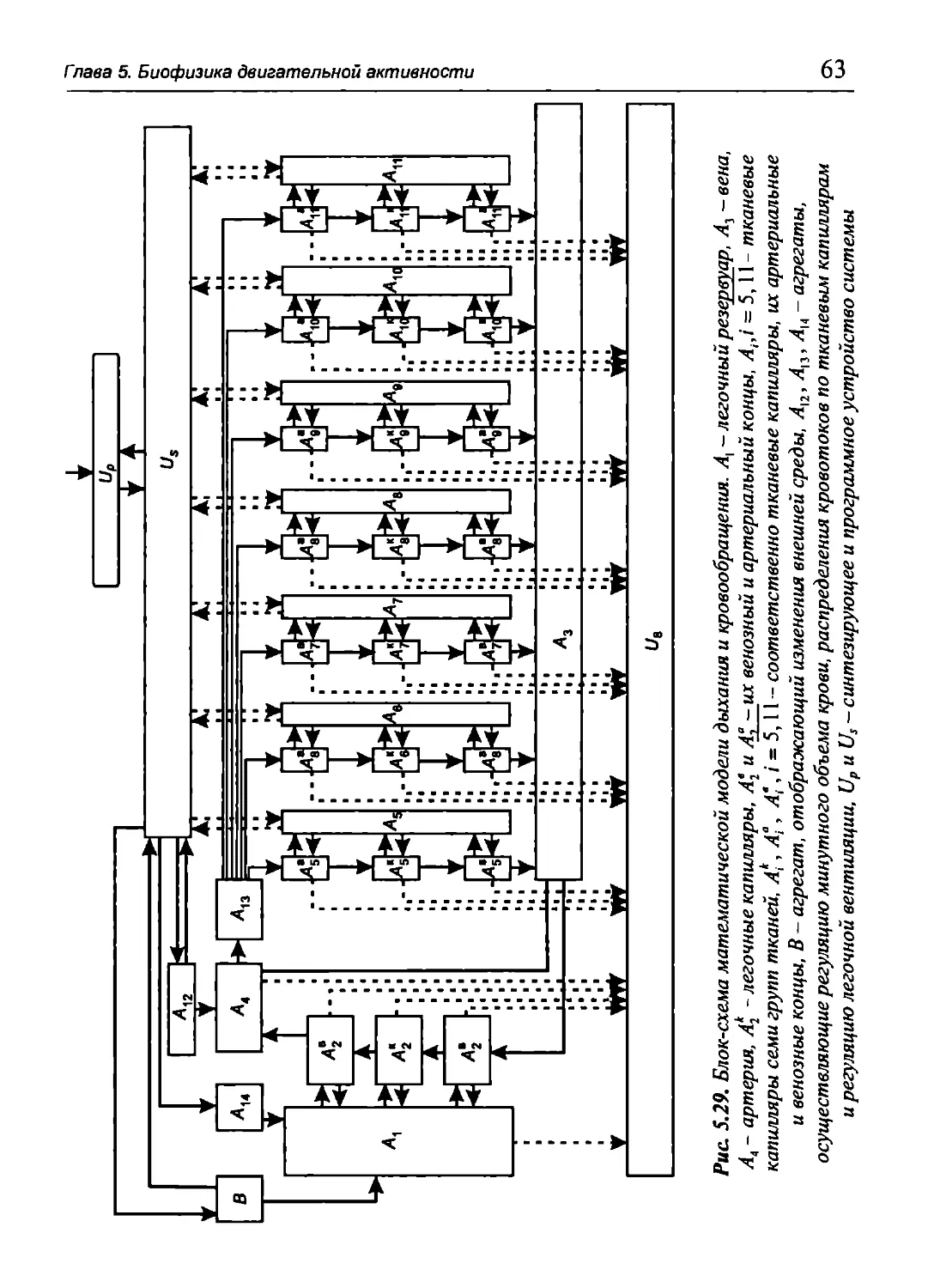





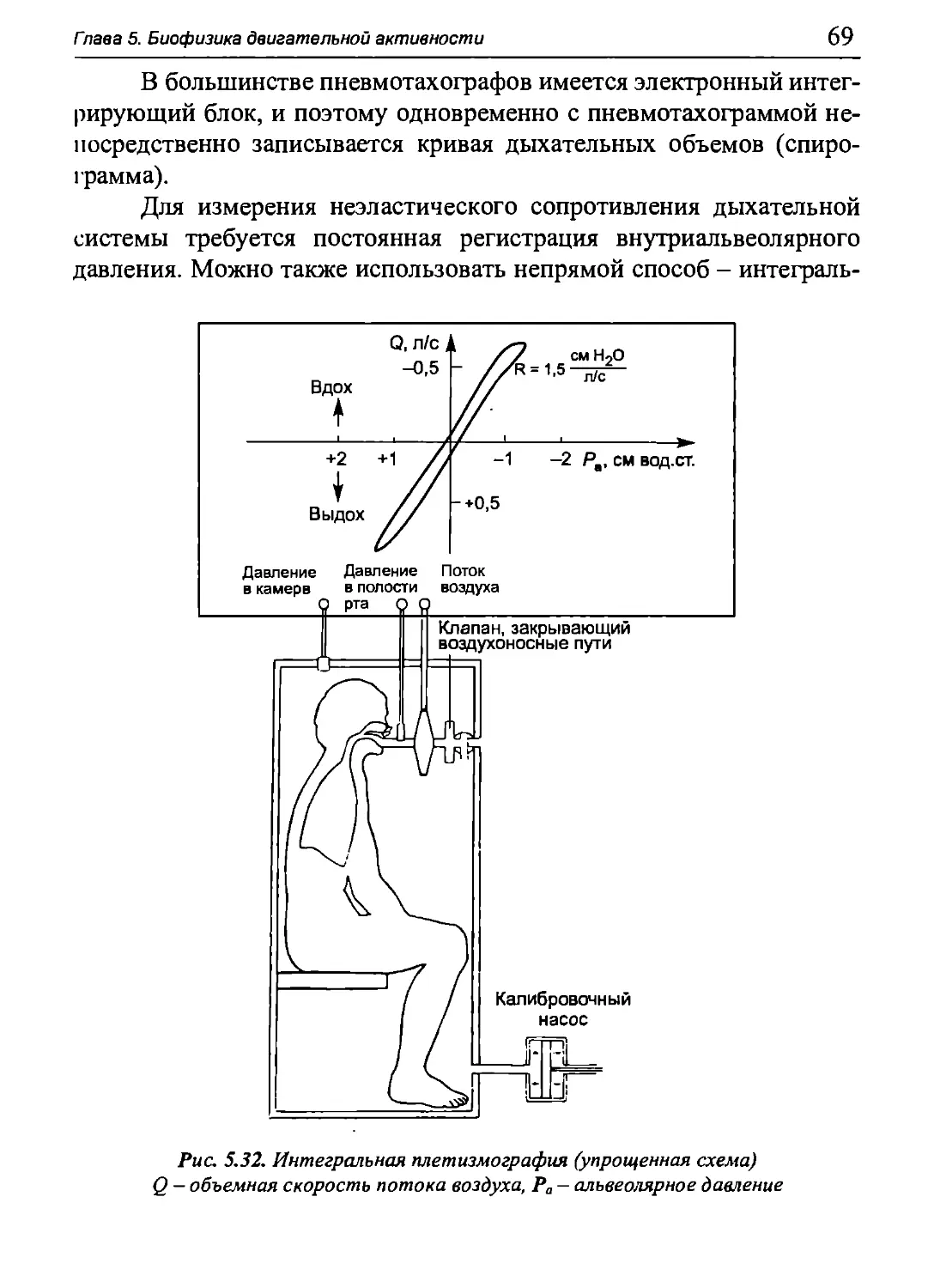









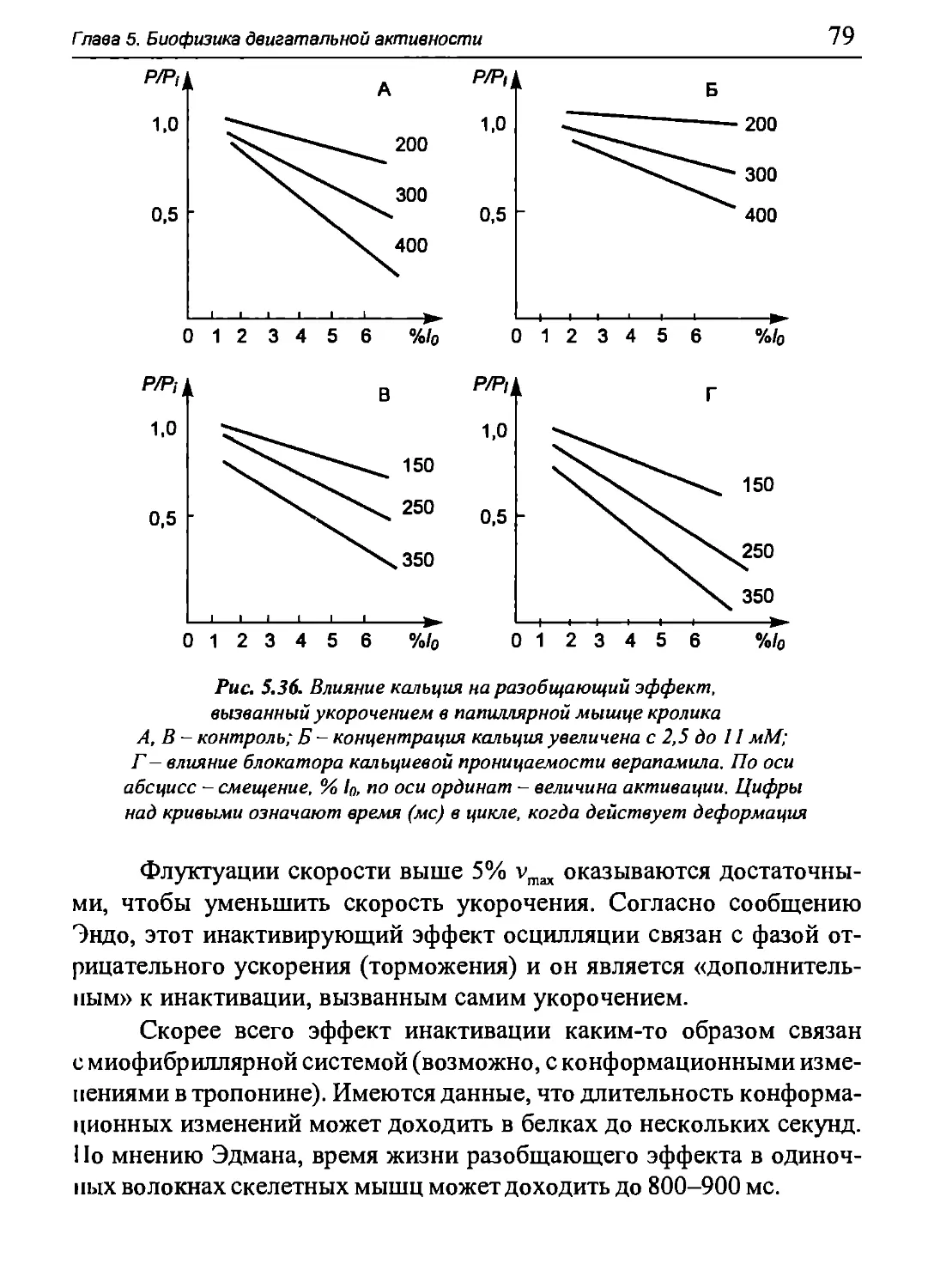




































































































































































































































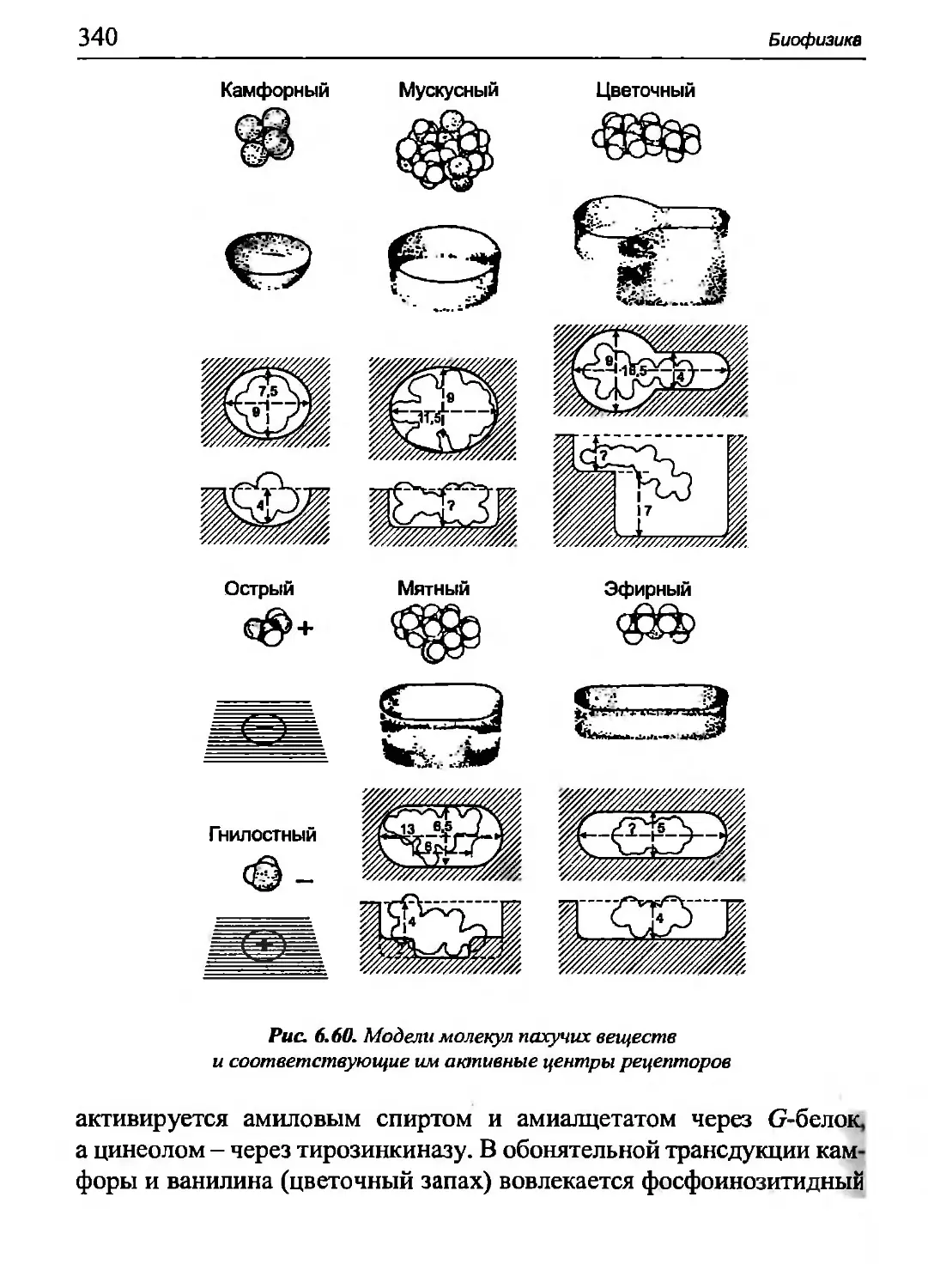
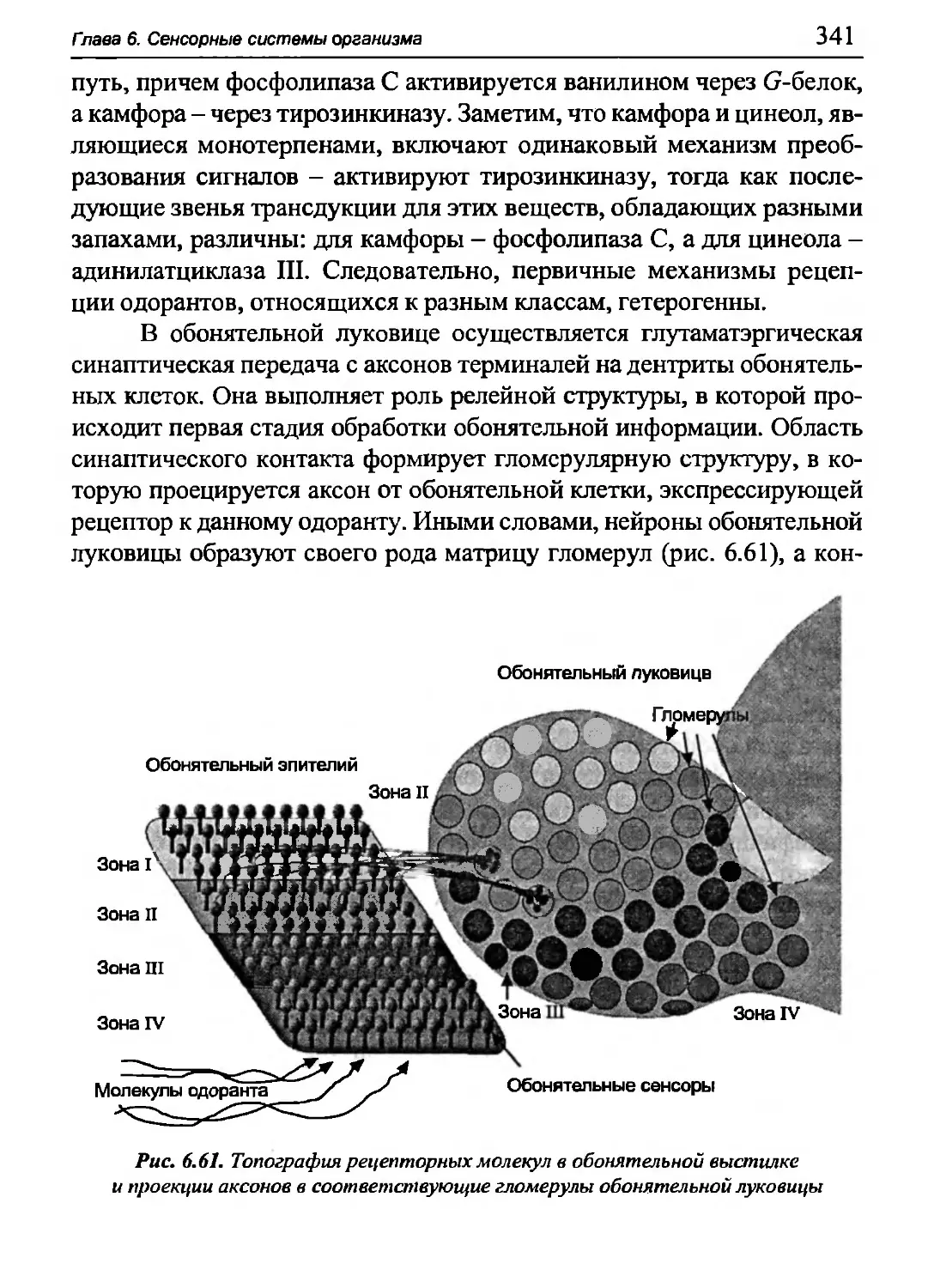













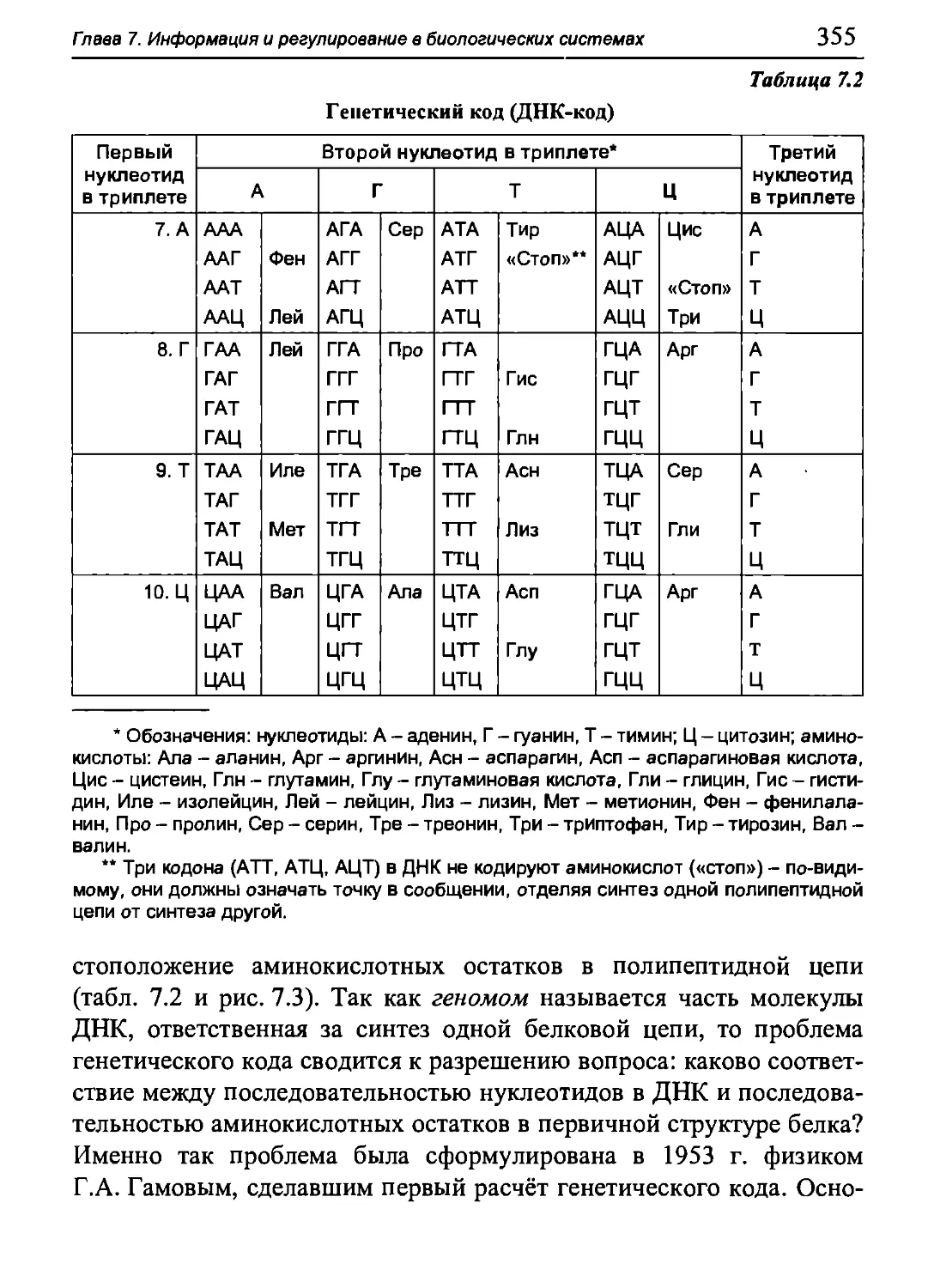






















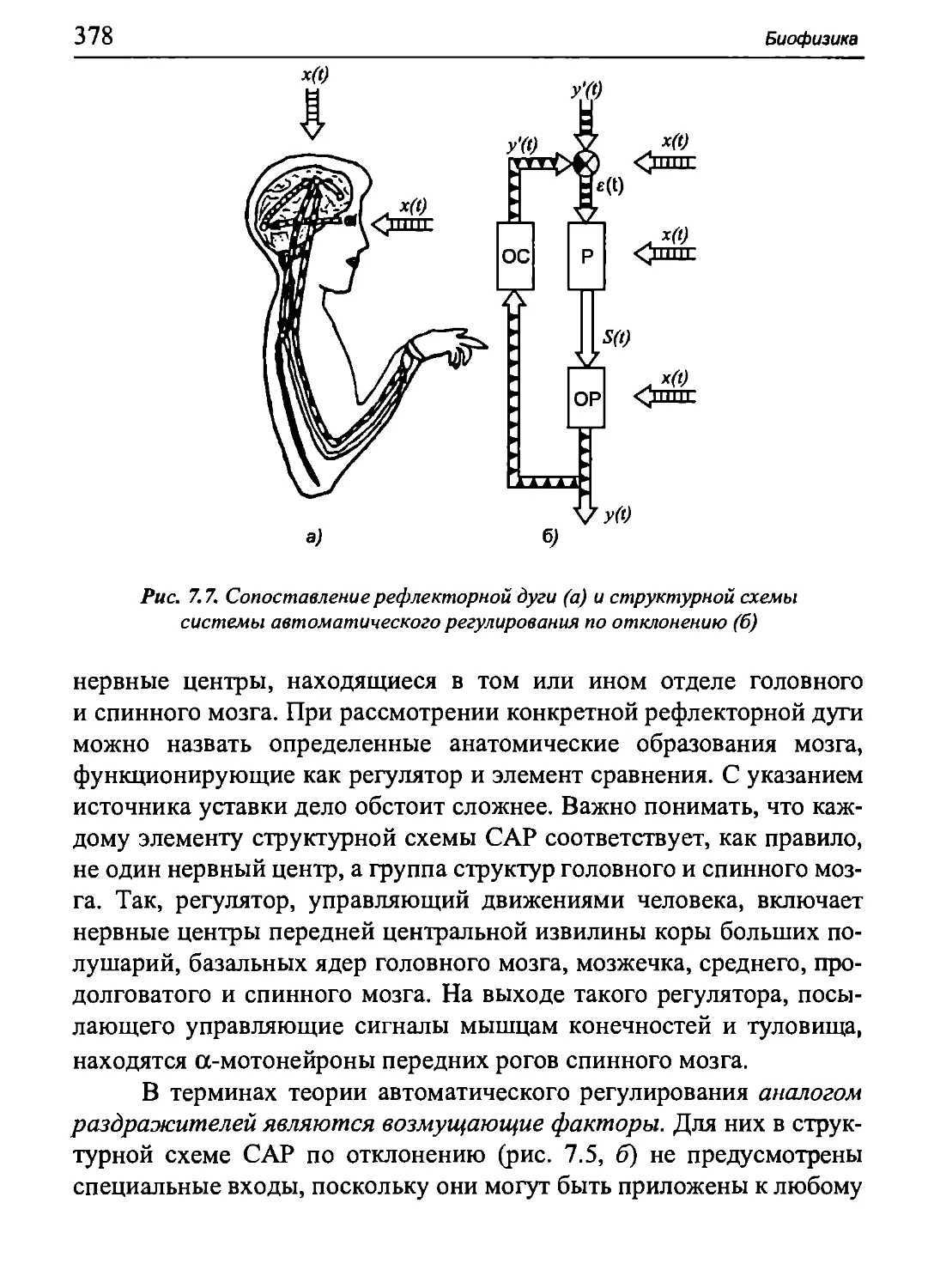


















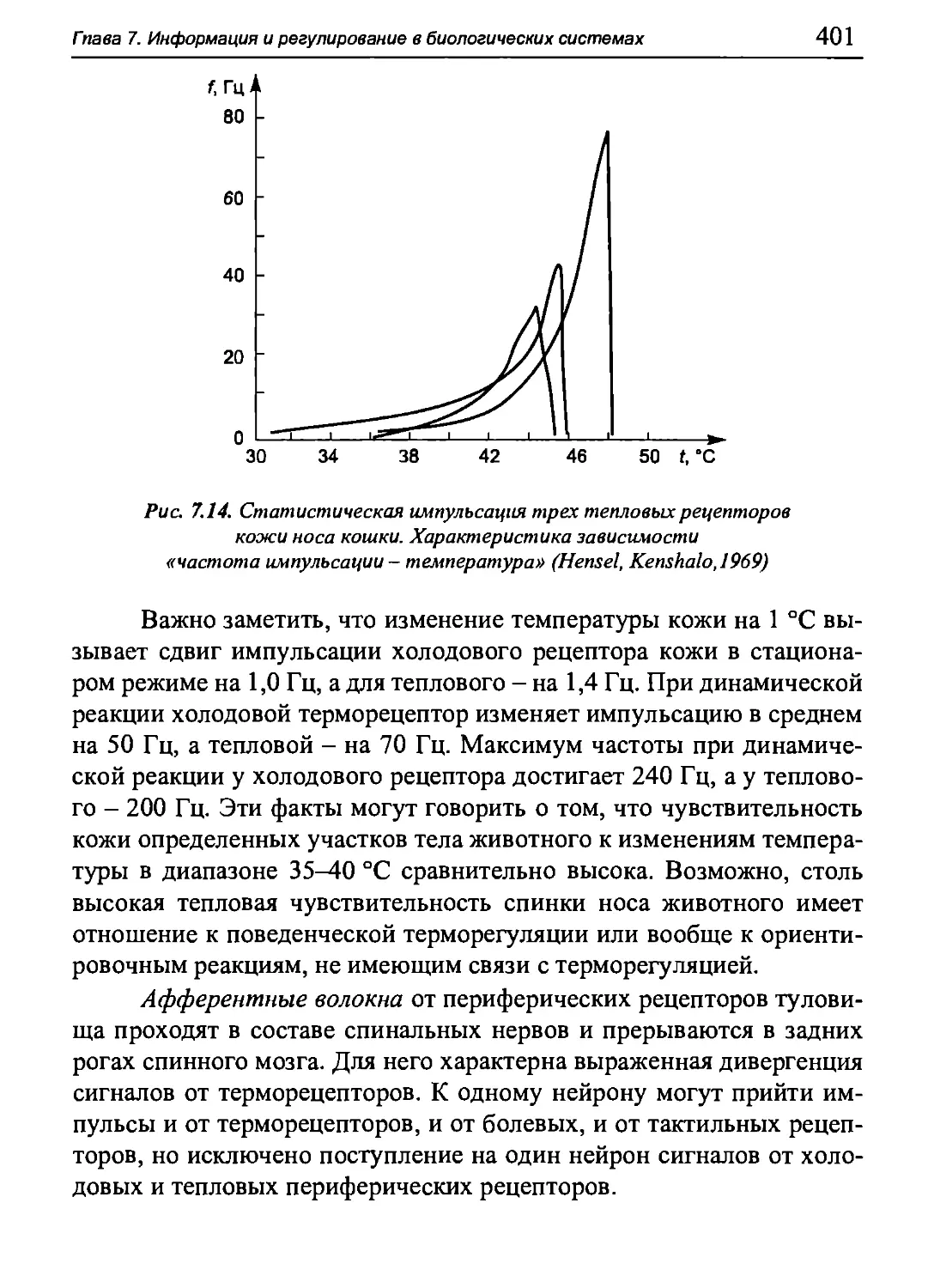







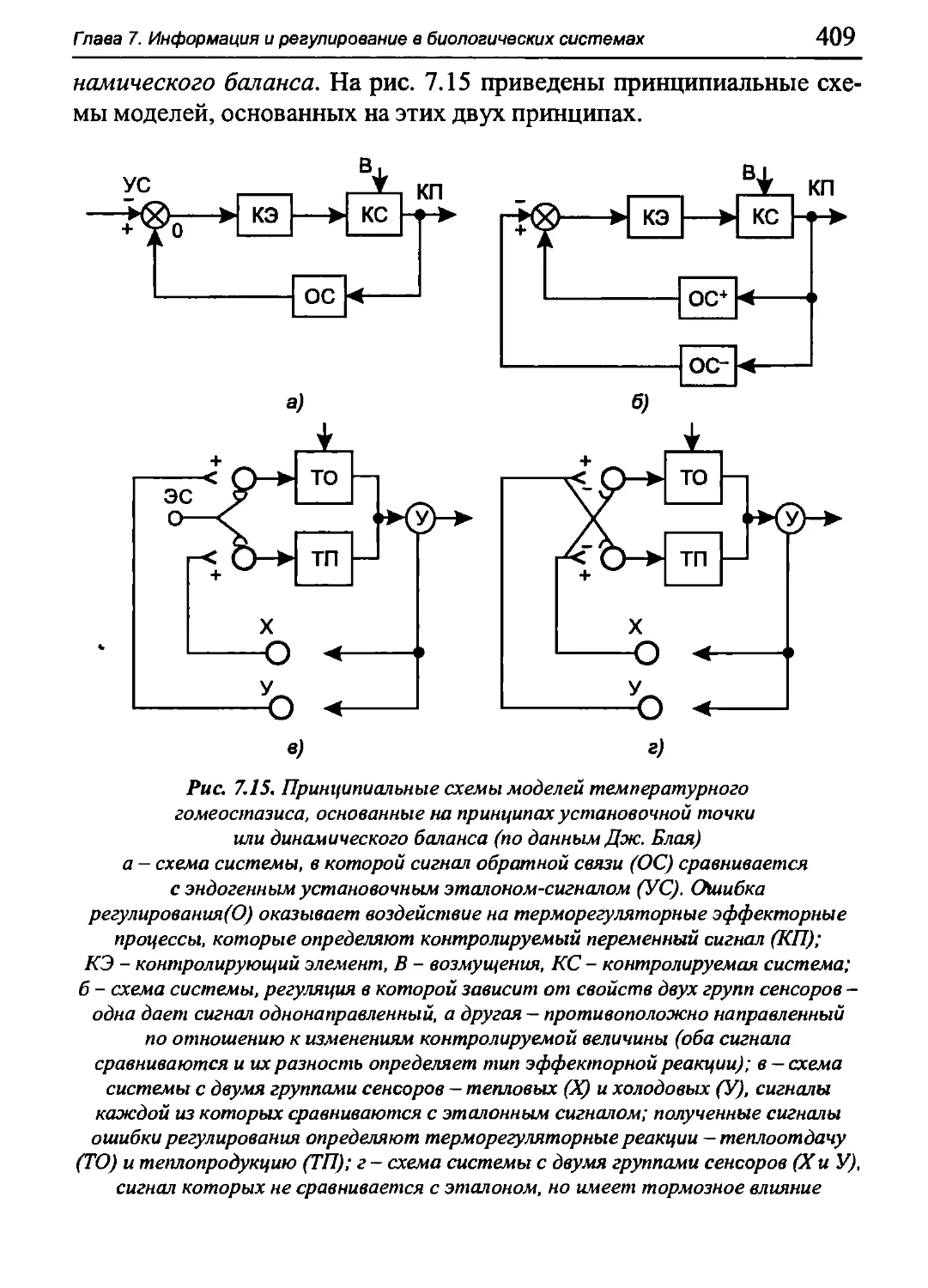


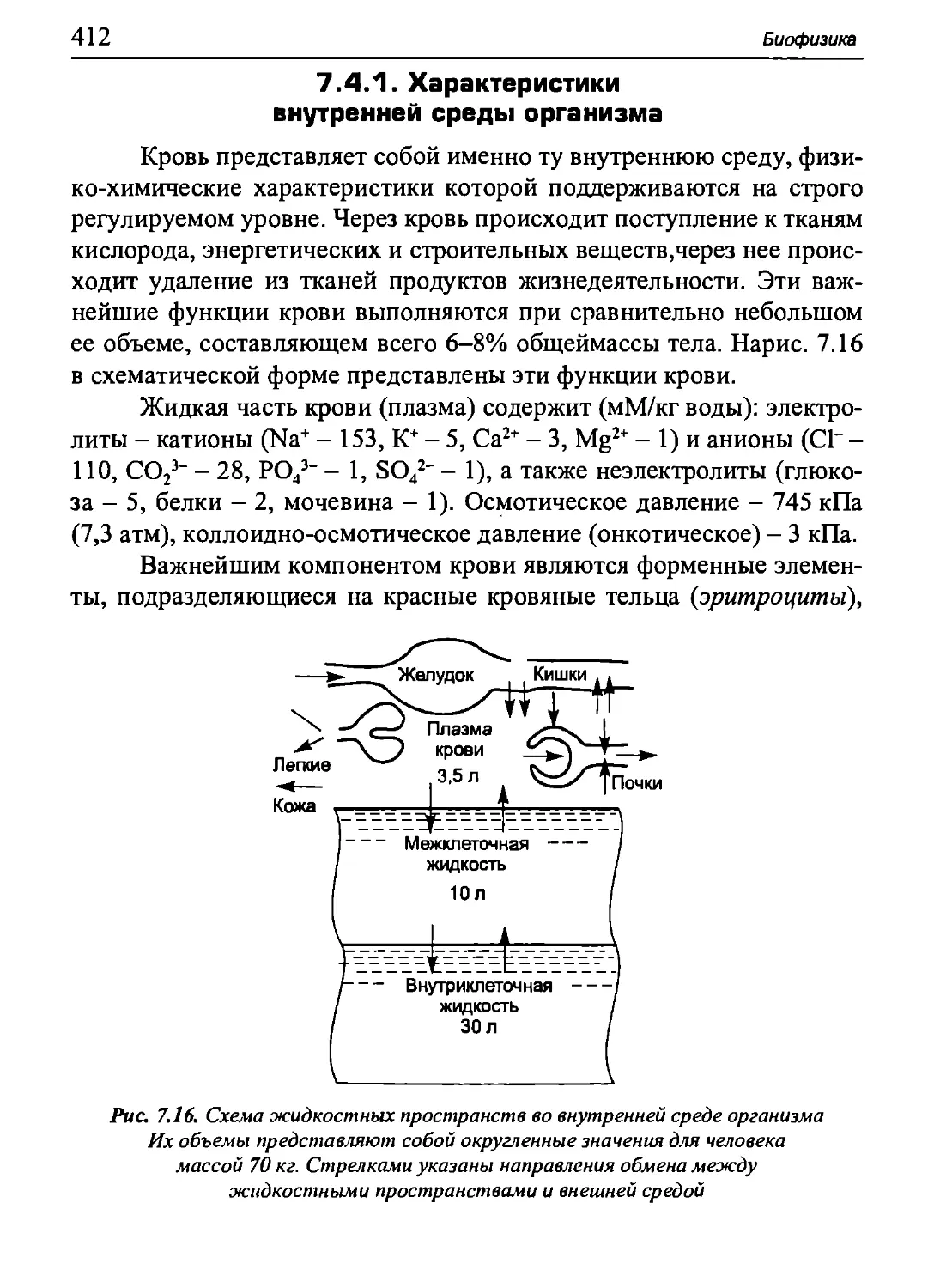

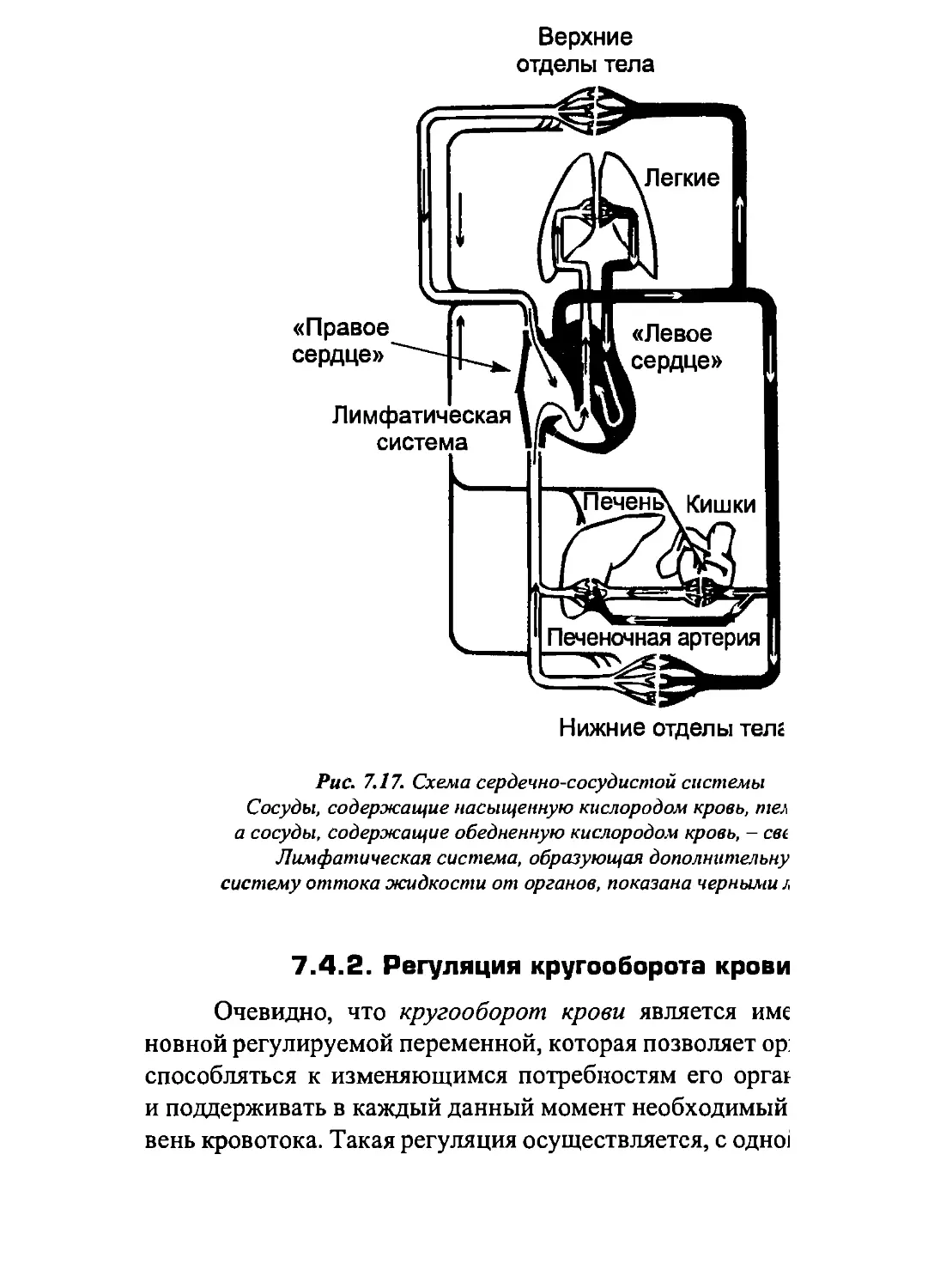








































Author: Бигдай Е.В. Вихров С.П. Гривеннаи П.В. Редькин В.М. Самийлив В.В. Чигирев Б.И.
Tags: науки о земле геологические науки общая биофизика инженерия биофизика биомеханика
ISBN: 978-5-9912-0049-3
Year: 2008




для ι
ι
ДЛЯ НЖЕНЕР В
— Ιιιι 2
ι Ι
μ
t
/τ
Ε. Β. Бигдай, смирив, П. В. Гривеннаи,
В. М. Редымн, В. В. Самкйлкв, Б. И. Чкгнрев
БИОФИЗИКА
ДЛЯ ИНЖЕНЕРОВ
Под редакцией
С. П. Вихрова и В. О. Самойлова
В 2 томах
1вм I Киомешико, информация
и регулирввввие в живых системах
Рекомендовано УМО по образованию
в области радиотехники, электроники, биомедицинской
техники и автоматизации для студентов
высших учебных заведений, обучающихся
по направлению подготовки дипломированных
специалистов 653900 - «Биомедицинская техника»
и направлению подготовки бакалавров
и магистров 553400 - «Биомедицинская инженерия»
Москва
Горячая линия - Телеком
2008
УДК 557.3
ББК 28.071
Б63
Рецензент: доктор биолог, наук, профессор Е. В. Чурносов
Авторы: Е. В. Бигдай, С. П. Вихров, Н. В. Гривенная, В. М. Редькин,
В. О. Самойлов, Б. И. Чигирев
Б63 Биофизика для инженеров: Учебное пособие. В 2 томах. Том 2. -
Биомеханика, информация и регулирование в живых системах /
Е. В. Бигдай, С. П. Вихров, Н. В. Гривенная и др. Под ред. С. П. Вихрова
и В. О. Самойлова- М.: Горячая линия-Телеком, 2008. -456 с: ил.
ISBN 978-5-9912-0049-3.
В учебном пособии систематизированы сведения о физических и физико-
химических процессах, лежащих в основе жизнедеятельности организмов
на всех уровнях их организации, необходимые для инженеров и
специалистов, занимающихся разработкой и обслуживанием биомедицинской техники.
В первом томе двухтомного пособия изложены основы биоэнергетики
и термодинамики биологических процессов, рассмотрены структура,
свойства и функции биологических мембран и биоэлектрогенез. Второй том
посвящен биофизическим основам двигательной активности человека,
информационных и регуляторных процессов в биологических системах, а также
общим принципам функционирования сенсорных систем.
Для студентов, обучающихся по направлению «Биомедицинская техника»,
будет полезна инженерам и специалистам, занимающимся
моделированием физиологических процессов и разработкой аналитической,
диагностической и лечебной аппаратуры.
ББК 28.071
Учебное издание
Бигдай Елена Владимировна, Вихров Сергей Павлович,
Гривенная Наталья Владимировна, Редькин Владимир Михайлович,
Самойлов Владимир Олегович, Чигирев Борис Иванович
БИОФИЗИКА ДЛЯ ИНЖЕНЕРОВ
Учебное пособие
Том 2. - Биомеханика, информация и регулирование в живых системах
Редактор И. М. Андреева
Компьютерная верстка О. В. Ушакова
Обложка художника В. Г. Ситникова
Подписано в печать 04.04.08. Формат 60 χ 90'/,». Усл. псч. л. 28,5. Изд. № 849. Тираж 1000 экз.
ООО «Научно-техническое издательство «Горячая линия Телеком». Заказ № 9844.
Отпсчатио с готовых диапозитивов в ОАО ордена «Знак Почета» «Смоленская областная
типография им. В. И. Смирнова». 214000, г. Смоленск, проспект мм. Ю. Гагарина, 2.
ISBN 978-5-9912-0049-3 (Т. 2) © Е. В. Бигдай, С. П. Вихров,
ISBN 978-5-9912-0050-9 Η. В. Гривенная и др., 2008
© Оформление издательства
«Горячая линия-Телеком», 2008
ВВЕДЕНИЕ
Учебное пособие состоит из двух томов. В первом томе
изложены основы биоэнергетики, рассмотренные с позиций
термодинамики и законов квантовой механики, а также вопросы
структурно-функциональной организации биомембран и биоэлектрогенеза.
Второй том учебного пособия состоит из трех глав (гл. 5-7),
которые посвящены основам биомеханики, двигательной активности
рида систем живых организмов (гл. 5), сенсорным биосистемам (гл.
(>), а также процессам регулирования в системах организма (гл. 7).
Биомеханика изучает кинематику, динамику движений биоло-
I ичсских систем, механику мягких и твердых биологических тканей,
механику кровообращения и дыхания, механику массопереноса на
различных уровнях организации биосистем.
Глава 6 ориентирована на читателей, которые знакомы с рядом
рачделов высшей математики и физики, основами биологии
человека, молекулярной биофизики, электробиологии и биофизики клетки.
11собходимо иметь в виду, что основные процессы жизнедеятельно-
ι,Ίΐι организмов протекают на уровне биологических мембран. Любая
живая клетка имеет мембранную разность электрических потенциа-
11011 (потенциал покоя), которая может изменяться дискретно
(потенциал действия) или относительно плавно (рецепторный потенциал).
Частота следования потенциалов действия по нервным волокнам
ограничена рефрактерным периодом и другими свойствами и
особенностями волокна. В данной главе рассмотены также и
нерешенные проблемы рецепторных систем, что может вызвать у читателя
интерес и желание поработать самостоятельно с научно-технической
литературой по этим проблемам.
Теория информации представляет собой совокупность
математических концепций, которые позволяют оценить количество
информации, содержащейся в том или ином сообщении, определить пропу-
4
Биофизика
скную способность каналов связи, количественно сопоставить
эффективность применяемых систем кодирования. Живой организм
является открытой системой. Это означает, что он обменивается
с окружающей средой веществом, энергией и информацией.
Биофизика, исследуя обмен информацией между организмом и
окружающей средой, проникает прежде всего в механизмы воздействия ее на
биологические системы, для чего изучает особенности
физико-химических процессов, происходящих в организме человека и животных
под влиянием информации, поступающей из внешней среды.
Обращение к рефлексам потребовалось нам для того, чтобы
понять, как в рефлекторной теории проявляю гея основные принципы
кибернетики. Среди них одним из важных является принцип
обратной связи. Изучение обратных связей как в рефлекторных реакциях,
так и в других формах регуляции функций в организме
(гуморальной, местной) привело к выводу о замкнутости контуров
биологического регулирования. При этом весьма полезным оказалось
применение принципов теории автоматического регулирования к изучению
разнообразных регуляторных механизмов в биологических системах.
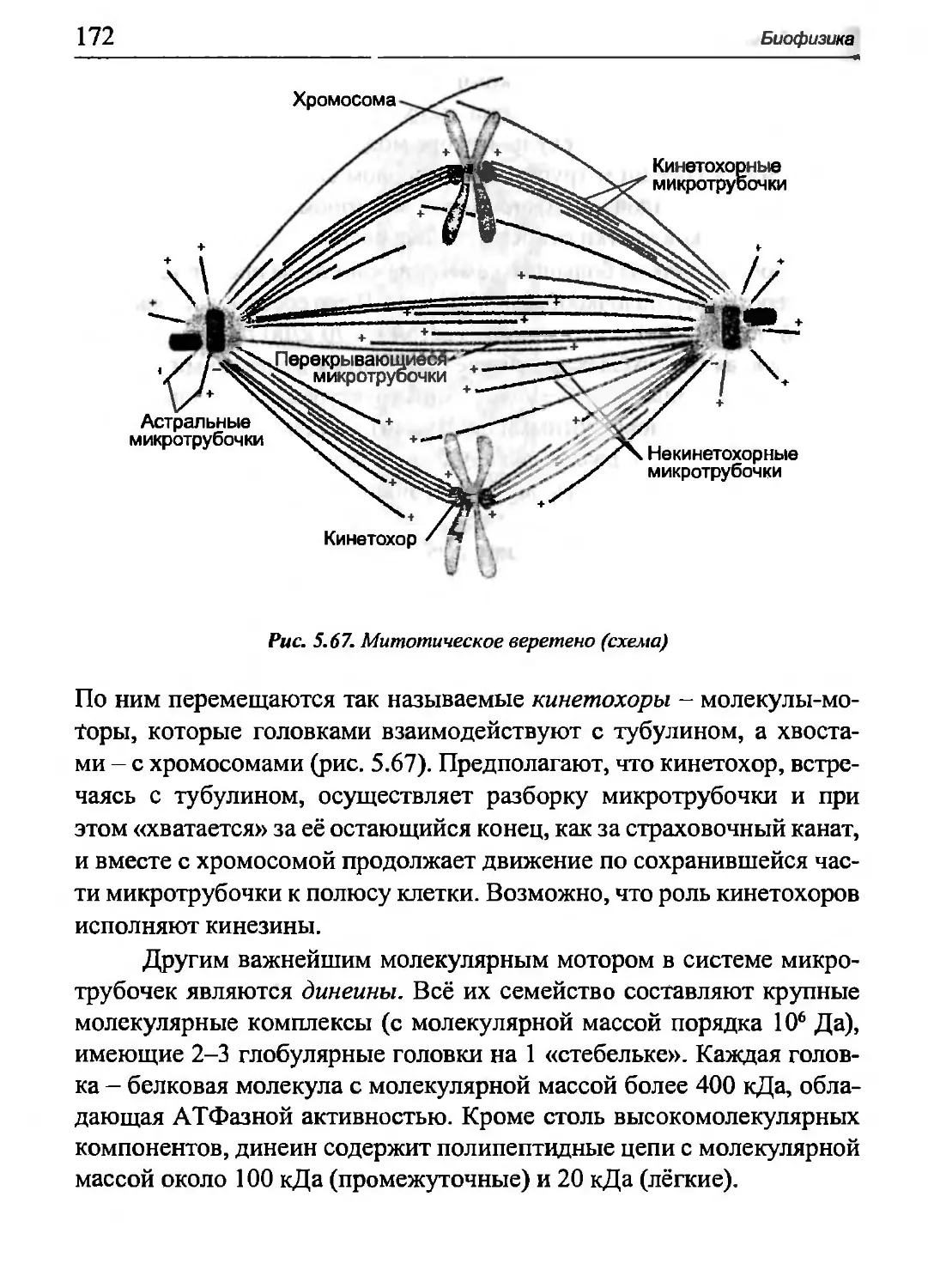
Глава 5. БИОФИЗИКА
ДВИГАТЕЛЬНОЙ АКТИВНОСТИ
Механические свойства живых тканей и органов, а также
механические процессы, обеспечивающие двигательную активность, изу-
Ίιΐι-ι биомеханика. Биомеханичкое моделирование исходит из того
иннпжения, что пассивные механические свойства биологических
niii.i'KTou отображаются в модели упругим и вязкостным элементами,
и иidi отображения способности живых тканей генерировать
механическую энергию вводится активный («сократительный») элемент.
Ιιΐκ,ΐΜ трехэлементная модель мышцы (рис. 5.1) была предложена
и I') И)-с годы выдающимся биофизиком, лауреатом Нобелевской
премии А.В. Хиллом.
1 2
Вязкий Упругий
элемент элемент
ι
vW/A-
Π
Активный
элемент
Рис. 5.1. Модель Хилла
Позднее модель Хилла была распространена на другие ткани
■ >р| пинзма, поскольку та или иная двигательная активность
присущи иссм животным и растительным клеткам. Теперь известны не
milι.κο актин-миозиновая, но и тубулин-динеиновая системы
подии жности, причём первая из них присуща не только мышечной
ткани, как думали прежде. Молекулярные компоненты систем
подвижности всех эукариотических организмов (от амёбы до человека)
кодируются генами, которые сохранились неизменными на
протяжении исей эволюции. Рассмотрим молекулярные механизмы подвиж-
ΚΙл ти, которые обеспечиваются актин-миозиновой и тубулин-ди-
непновой системами.
6
Биофизика
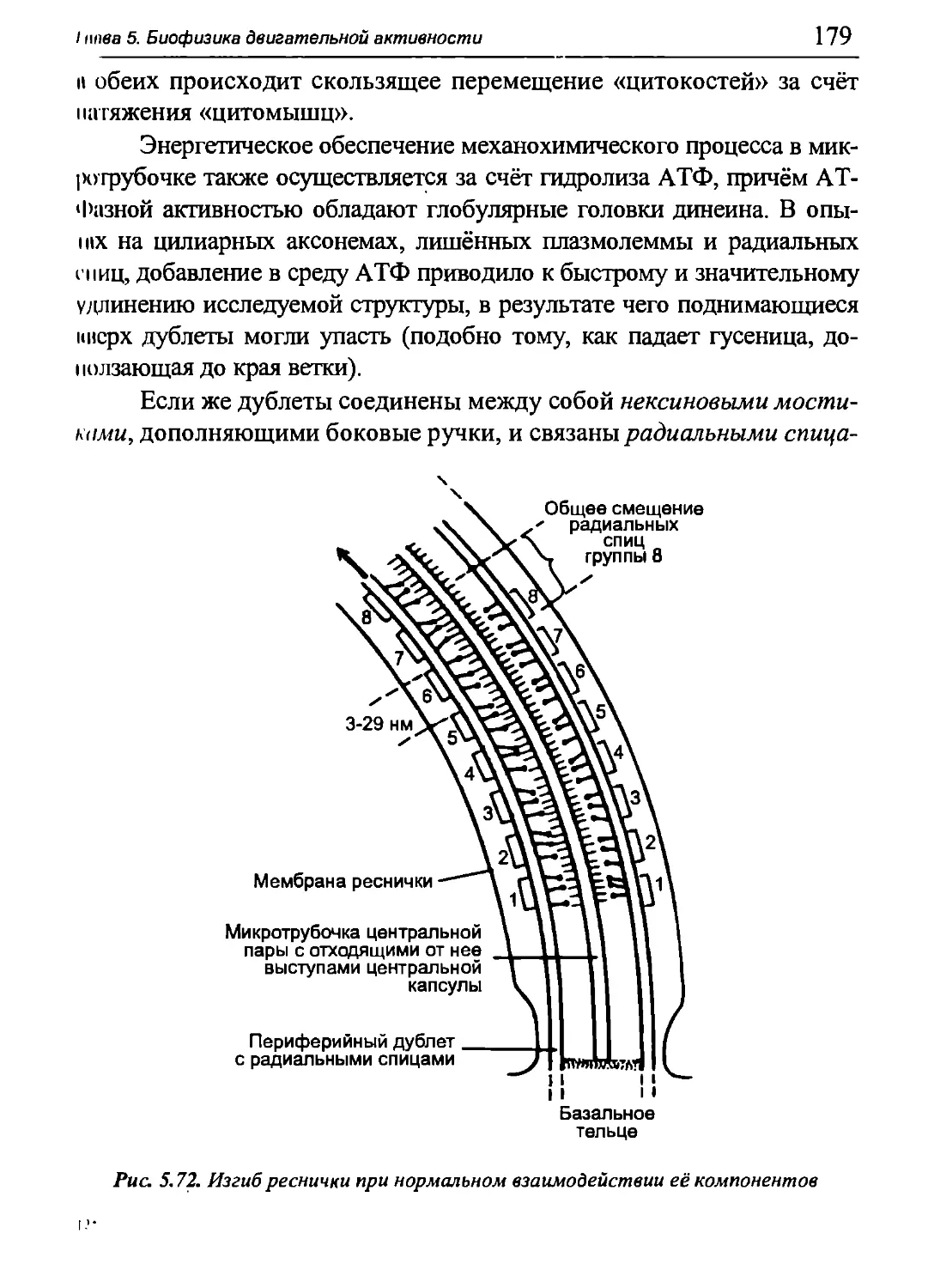
5.1. Биофизика мышечного сокращения
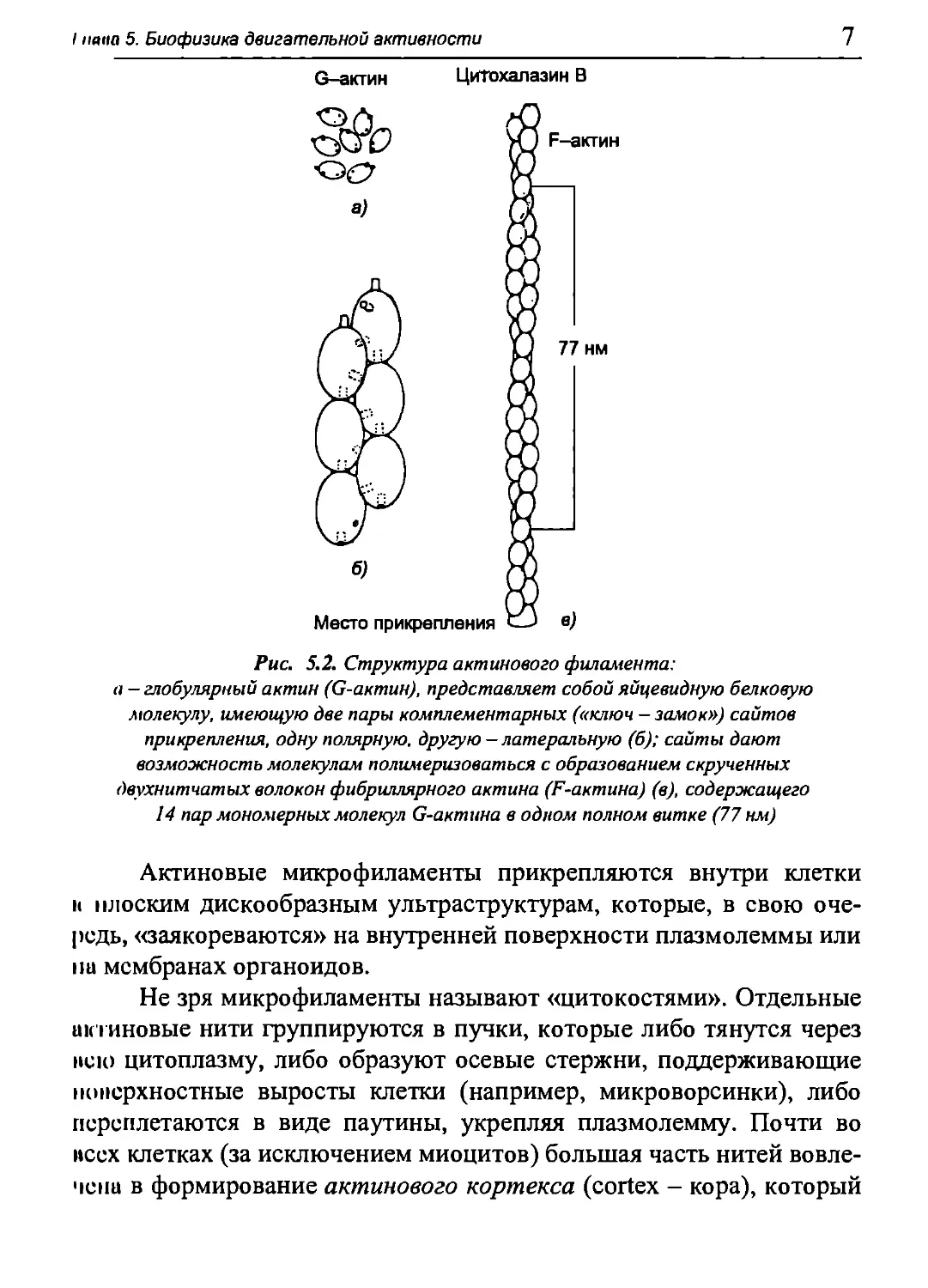
5.1.1. Актин-миозиновая система миоцита
поперечнополосатых мышц
Актин-миозионовая система образована иктиишшми и миози-
новыми нитями. Актиновые нити образуются при полимеризации
молекул глобулярного белка актина (G-актипа). Πιο полипептидная
цепь (первичная структура) включает 374 аминокислотных остатка.
Молекулярная масса 41,8 кДа. Третичная структура представляет
собой глобулу с бугристой поверхностью, изрезанной щелями, самая
глубокая из которых пролегает в середине молекулы и содержит
активные центры связывания АТФ и АДФ, а также Са2' и Mg2+. Кроме
того, на молекуле G-актина есть сайты связывания тропомиозина,
тропонина, головки миозина (об этих белках пойдет речь ниже),
а также соседних молекул G-актина.
Благодаря активным центрам связывания G-актина на нем
глобулы объединяются в фибриллы, образуя F-актин (рис. 5.2).
Контакты между соседними мономерами G-актина обеспечиваются
гидрофобными и электростатическими взаимодействиями. Фибриллы
F-актина объединяются попарно (наподобие пары перекрученных
ниток бус), и таким образом формируется актиповый микрофила-
мент длиной до 1 мкм и диаметром (толщиной) около 6-8 нм. Шаг
спирали (полный виток) F-актина составляет 77 нм и на его
протяжении содержится примерно 14 пар молекул G-актина.
Актиновый микрофиламент - весьма динамичная полярная
структура, причем на её положительном полюсе происходит
полимеризация (присоединение новых G-мономеров к F-актину), а на
противоположном (отрицательном полюсе) поляризованной молекулы -
деполимеризация. Поэтому актиновая нить непрерывно растет в
сторону своего положительного полюса и столь же непрерывно
укорачивается на отрицательном полюсе (противоположном конце).
Полимеризация актина регулируется несколькими белками и обеспечивается
энергией посредством гидролиза АТФ, причём образующийся в этой
реакции АДФ остается на F-актине. Он замещается молекулами АТФ
на отрицательном полюсе актиновой нити при её укорочении.
I nana 5. Биофизика двигательной активности
7
G-актин
а)
Цитохапазин В
F-актин
77 нм
Место прикрепления
в)
Рис 5.2. Структура актинового филамента:
а — глобулярный актин (G-актин), представляет собой яйцевидную белковую
молекулу, имеющую две пары комплементарных («ключ - замок») сайтов
прикрепления, одну полярную, другую — латеральную (б); сайты дают
возможность молекулам полимеризоваться с образованием скрученных
двухнитчатых волокон фибриллярного актина (F-актина) (в), содержащего
14 пар мономерных молекул G-актина в одном полном витке (77 нм)
Актиновые микрофиламенты прикрепляются внутри клетки
и плоским дискообразным ультраструктурам, которые, в свою
очередь, «заякореваются» на внутренней поверхности плазмолеммы или
пи мембранах органоидов.
Не зря микрофиламенты называют «цитокостями». Отдельные
икгиновые нити группируются в пучки, которые либо тянутся через
нею цитоплазму, либо образуют осевые стержни, поддерживающие
икиерхностные выросты клетки (например, микроворсинки), либо
переплетаются в виде паутины, укрепляя плазмолемму. Почти во
исех клетках (за исключением миоцитов) большая часть нитей
вовлечена в формирование актинового кортекса (cortex - кора), который
8
Биофизика
залегает под плазмолеммой наподобие паутинообразной сетки.
Благодаря тому, что в кортексе микрофиламенты сплетаются в сложные
сети, он при сжатии клетки деформируется во всех направлениях.
Средняя продолжительность жизни актиновой нити коротка -
например, в кортексе лейкоцита всего 5 с.
Актиновый кортекс испытывает существенные перестройки
при образовании клеткой псевдоподий (ложных ножек) - выростов
цитоплазмы в сторону интерстиция. На их возникновение уходит
несколько минут, а то и секунд. Внутри каждой псевдоподии находятся
кортикальные микрофиламенты, которые на протяжении сё жизни
беспрестанно полимеризуются и деполимеризуются. Кончик
псевдоподии способен временно прикрепляться к поверхностям, по
которым ползёт клетка (например, лейкоцит). В псевдоподии для
перемещения клетки должен находиться не только актин, но и миозин,
взаимодействующий с актиновыми нитями, выполняющими
функцию рельсов, по которым движется (скользит) миозин, относящийся
к так называемым белкам-моторам («цитомышцам»). Выделено
несколько типов миозинов. Одни из них обеспечичают
внутриклеточный транспорт, а другие - сокращение миоцитов.
Каждая из молекул миозинов, осуществляющих сокращение
миоцитов, образована четырьмя полипептидными цепями, две из
которых (тяжёлые) имеют молекулярную массу по 200 кДа, а две
лёгкие - 20 кДа и 17 кДа. Следовательно, молекулярная масса этого
гиганта достигает почти 500 кДа. Каждая из двух тяжёлых цепей
образует вторичную структуру типа α-спирали. Спирали обеих цепей
скручены в двухнитчатую (левозакрученную) суперспираль с шагом
в 7,5 нм (рис. 5.3, а). Она является стержнем миозиновой молекулы,
который по своей длине (135 нм) при толщине в 2 нм превосходит все
аналогичные молекулярные структуры, встречающиеся в природе.
На верхнем конце стержня две нити расходятся, чтобы
образовать подвижные «стебельки», на вершинах которых тяжёлые
полипептидные цепи сплетаются с дополнительными (лёгкими) цепями
и формируют глобулярные молекулярные структуры, которые как
поникшие головки выступают на поверхности. За счёт них общая длина
миозиновой молекулы достигает 155 нм. Молекула напоминает
двойной цветок на длинной ножке. Соединение между головкой и стерж-
/ пава 5. Биофизика двигвтельной активности
9
нем молекулы подобно шарниру, что позволяет «цветам» наклоняться
к той или иной степени. Важно заметить, что головки миозина
обладают АТФазной активностью, т.е. активируют гидролиз АТФ.
Миозиновые молекулы самопроизвольно собираются в пучок.
При таком объединении стержни нескольких сотен молекул,
располагаясь параллельно друг другу с некоторым сдвигом по длине
(относительно расположения головок), образуют своеобразный ствол
толщиной 11-14 нм, из которого в верхней его части выступают
головки (рис. 5.3, б). Важно знать, что эта надмолекулярная структура
биполярна, поскольку в её середине соединяются (конец в конец) два
■ч-
б)
е)
Рис. 5.3. Миозиновые нити.
a - двухнитчатая структура миозина II (белковая молекула с двумя
головками); б - пучки (стержни) толщиной 15-20 нм с многочисленными
головками, выступающими над поверхностью и образующими регулярные
спиральные ряды, в середине стержня - оголённый сегмент без головок;
в — молекулы миозина I (M), которые прикреплены короткими хвостами
к мембране транспортируемого органоида (О), а головками — к актиновой нити (А)
10
Биофизика
«ствола» при том, что «кроны» обращены в противоположные
стороны. Поэтому в середине толстой нити, в которой объединены сотни
молекул миозина, имеется участок длиной около 300 нм, свободный
от головок. Общая длина всего такого филамента составляет в
разных клетках от 2,5 до 15 мкм.
В миозинах, которые обеспечивают внутриклеточный
транспорт органоидов (митохондрий, лизосом и др.), «хвосты» короткие.
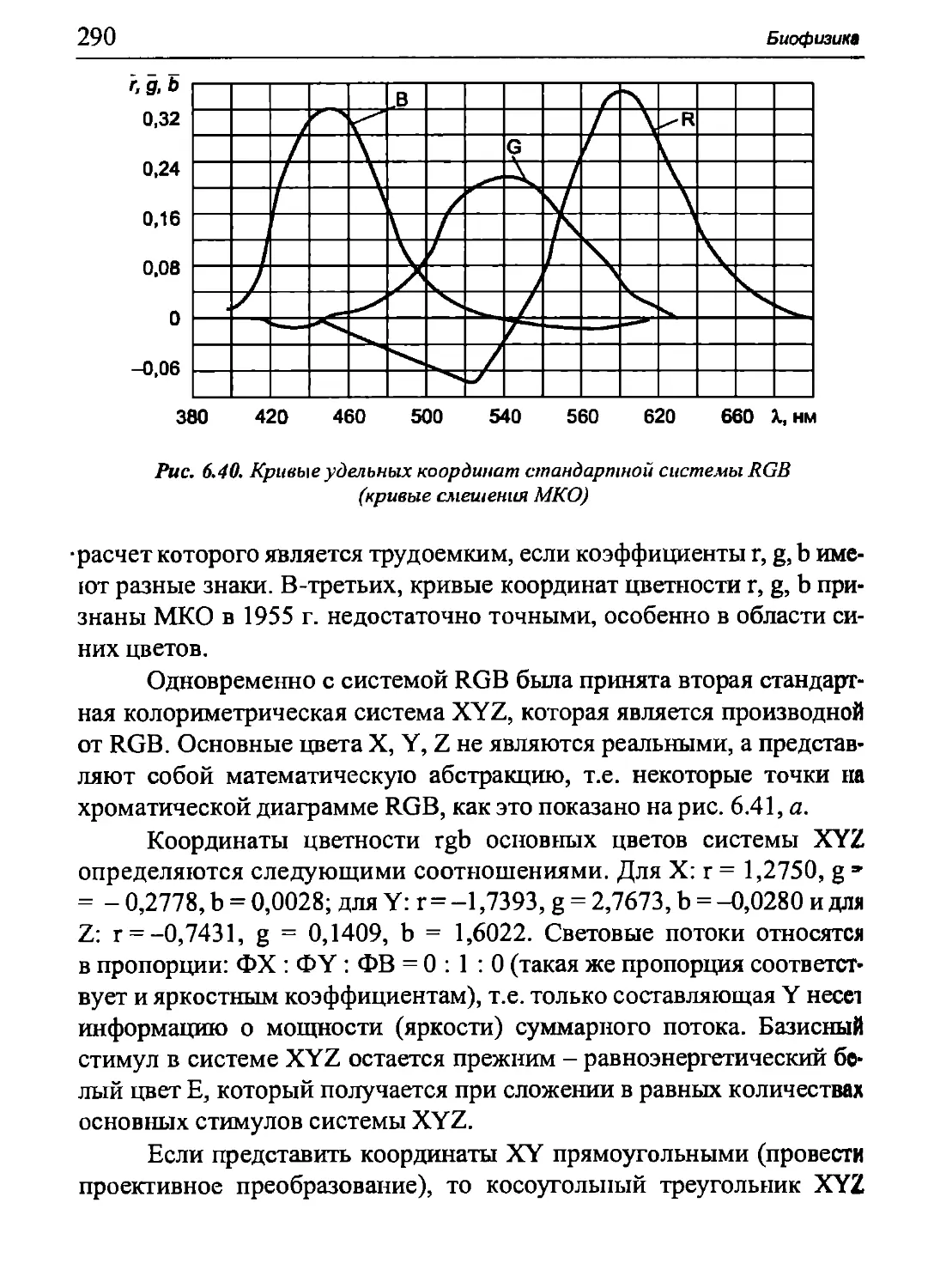
Ими миозиновые молекулы присоединяются к мембранам
органоидов и, скользя затем по актиновым нитям, перемещают их в
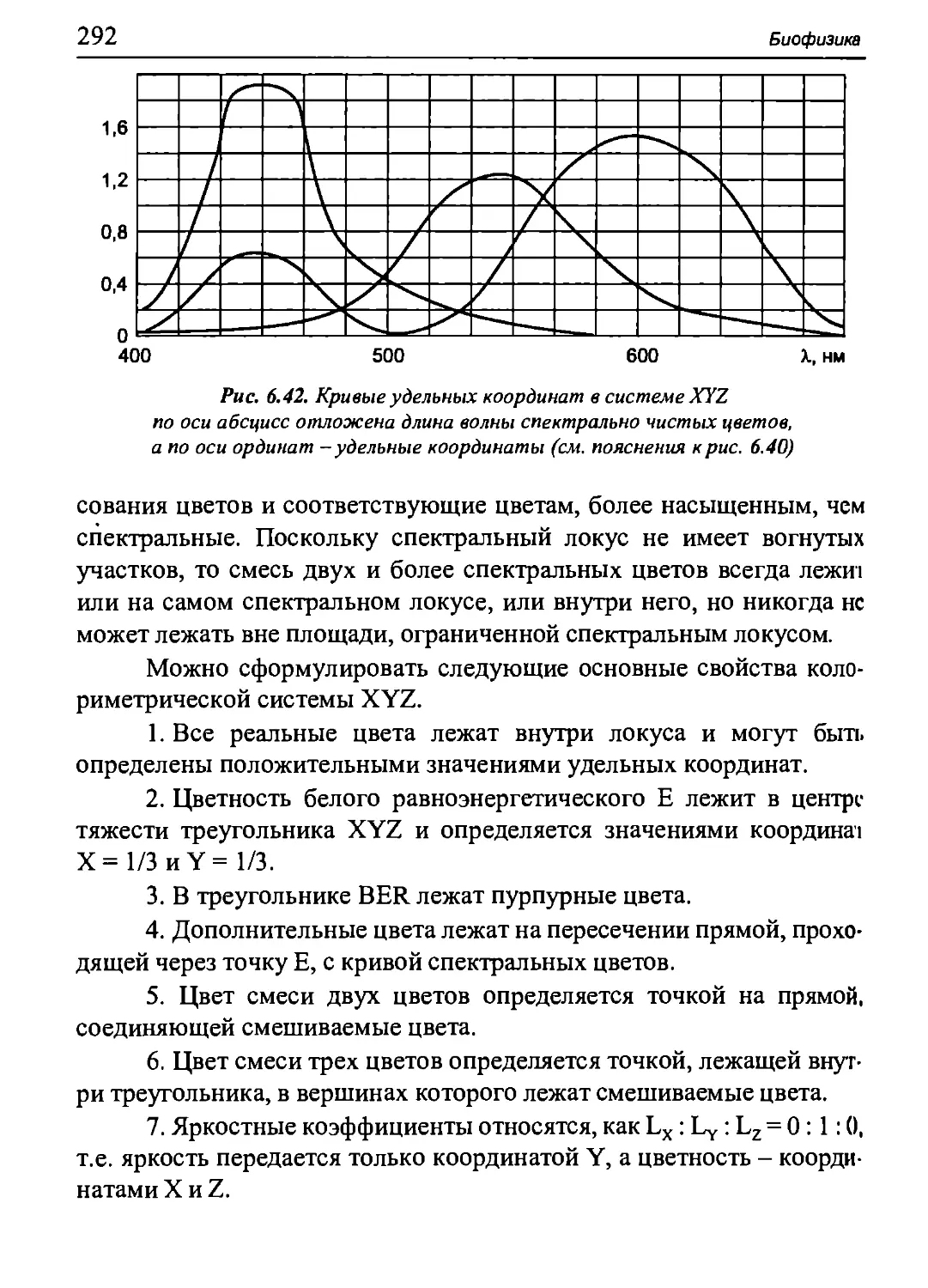
различные участки клетки (рис. 5.3, в). В таком варианте могут работать
даже отдельные молекулы миозина или тонкие пучки, образованные
их малым количеством.
Благодаря способности к беспрестанным процессам сборки
и разборки, протекающим очень быстро, нити F-актина могут
достигать любого участка клетки, а по ним, как по рельсам, туда
устремляется миозин, который подобно локомотиву тянет за собой различные
клеточные компоненты, прикрепляющиеся к нему. За счёт скольжения
миозина I типа по микрофибриллам движутся органоиды в цитоплазме
(например, митохондрии подтягиваются к тем местам, где есть нужда
в макроэргах), а при скольжении миозина II типа перемещаются
отдельные участки клетки друг относительно друга. Взаимодействие
миозина с актином обеспечивает животным и человеку преодоление
земного притяжения и перемещение в пространстве, кровообращение,
внешнее дыхание, двигательную активность желудочно-кишечного
тракта и других внутренних органов, родовую деятельность и другие
виды подвижности, которые обеспечиваются миоцитами разных типов.
Характерной особенностью актин-миозиновой системы мио-
цита (мышечного волокна) является строго параллельное
расположение актиновых и миозиновых нитей (рис. 5.4), благодаря чему их
скольжение происходит в одном направлении, и в клетке развивается
большое напряжение. Это свойственно всем трём типам миоцитов:
миоцитам скелетных мышц, кардиомиоцитам, гладкомышечным
клеткам. Рассмотрим прежде всего биомеханические процессы в мио-
ците скелетных мышц.
Его длина достигает в разных мышцах от 5 до 100 мм, а диаметр
значительно уступает длине, составляя от 10 до 100 мкм. Преоблада-
ι ινημ II биофизика двигательной активности
11
иш< дмпмы над толщиной в такой веретенообразной клетке позволяет
ни н.пмгп. сё волокном. Вместе с тем следует отметить, что мышечное
иишшпо образуется путем слияния множества отдельных клеток. По-
ниму и миоците (весьма крупной клетке) содержится несколько ядер.
Нпутри миоцита находится порядка 103 миофибрилл. Диаметр
MrtuinN из них составляет 1-3 мкм, а длина такая же, как у всей
клетки I la пом протяжении в миофибрилле помещается огромное коли-
ίργ ι но так называемых саркомеров (sarx - мясо, meros — часть), вы-
Z-линия
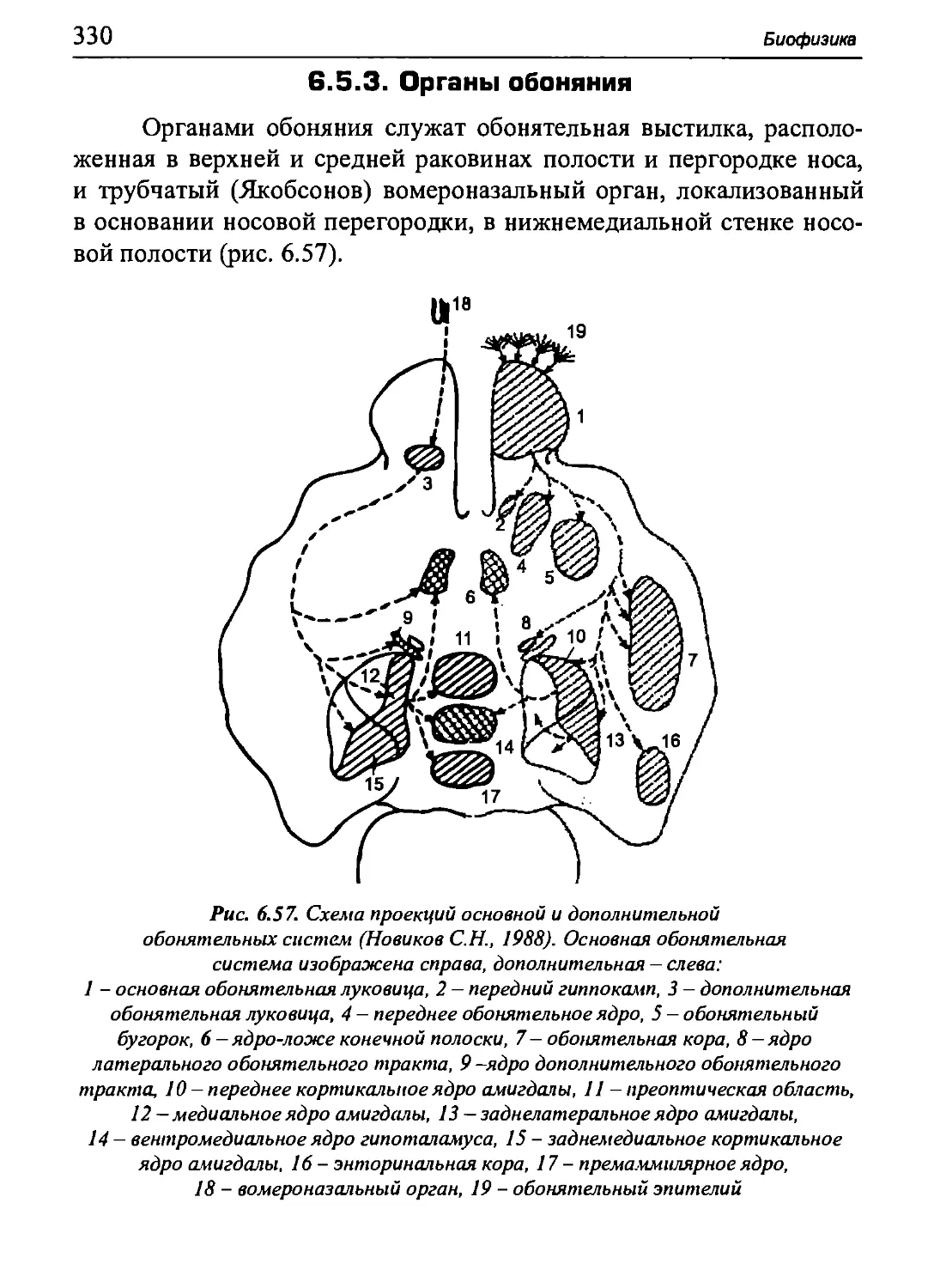
-ΎΊΤ
Рис. 5.4. Схематическое изображение миофибриллы
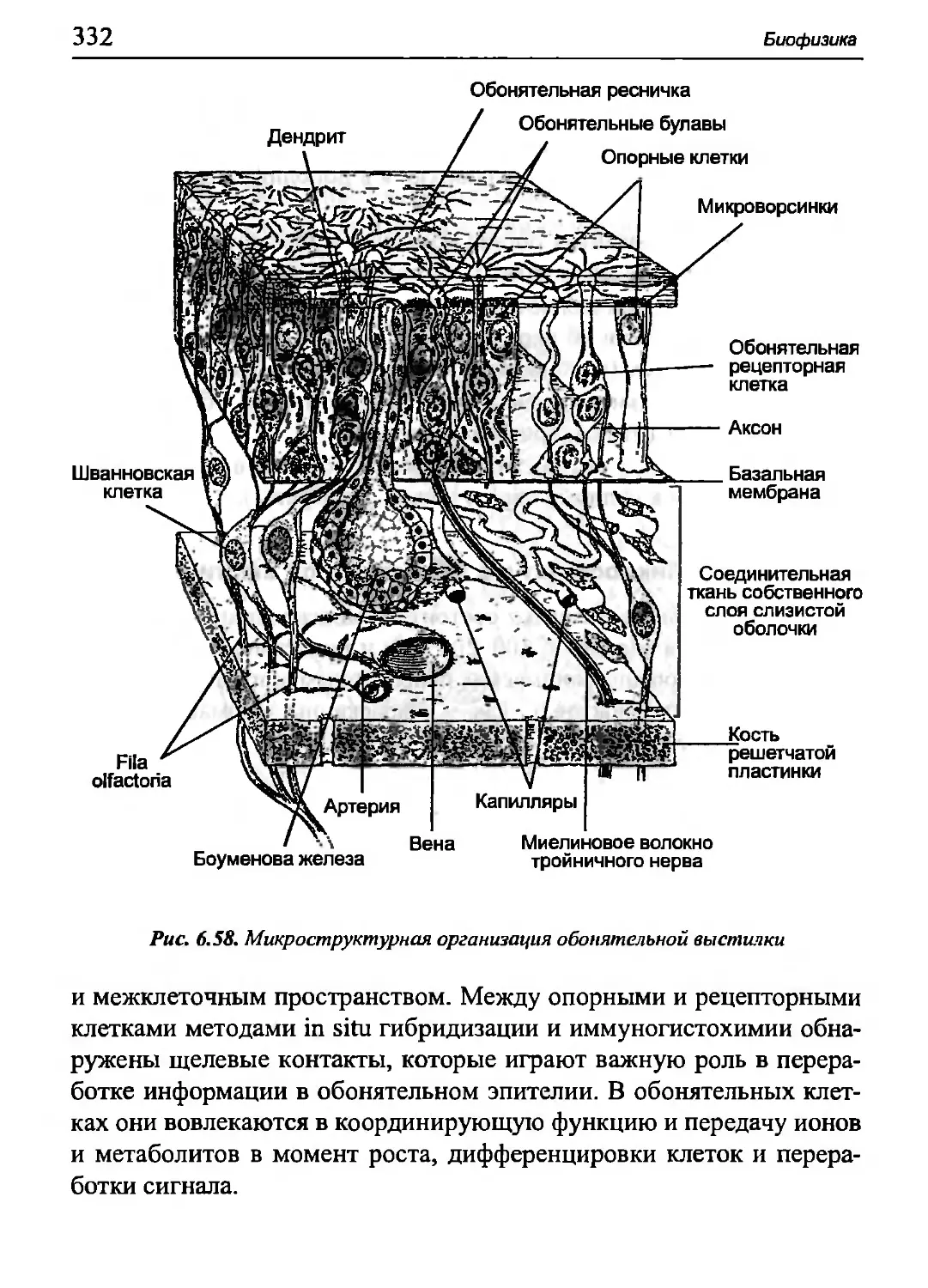
в миоците поперечнополосатых мышц
1—изотропный диск, А — анизотропный диск, Н-зона, Z-линия
12
Биофизика
Рис 5.5. Электронограмма миоцита поперечнополосатых мышц
Обозначения: А - анизотропный диск; 1— изотропный диск; Ζ - Z-линия (полоска)
строенных в цепочку благодаря последовательному соединению
друг с другом. Соседние саркомеры «склеиваются» друг с другом
белком десмином, который вместе с актиновыми нитями образует
актин-десминовую ячеистую сеть. В растянутом состоянии
мышечного волокна длина его саркомеров может достигать 3,5 мкм.
Разделив длину волокна (а, значит, и составляющих его миофибрилл) на
эту величину (длину саркомера при растяжении), находят число
саркомеров в миофибрилле. Саркомер является элементарной
надмолекулярной сократительной единицей мышечного волокна.
Благодаря упорядоченности молекулярной структуры миоцита
границы саркомеров в соседних миофибриллах совпадают,
следствием чего является характернейшая особенность поперечнополосатых
(скелетных) мышц, давшая им такое название, - регулярная исчер-
ченность за счёт чередования светлых (изотропных - 7) и тёмных
(анизотропных - А) полос, наблюдаемых под микроскопом (рис. 5.5).
Анизотропные диски обладают двойным лучепреломлением: в
обычном свете выглядят тёмными, а в поляризованном - прозрачными
(светлыми) в продольном и непрозрачными (тёмными) в поперечном
направлениях. Изотропным дискам не свойственно двойное
лучепреломление. Затемнение в центре /-диска (/-полосы) обусловлено
контактами между соседними саркомерами. Эта тёмная полоска
толщиной 80-160 нм называется Z-линией (или Z-диском). Она служит
границей между соседними саркомерами в миофибрилле. Под
электронным микроскопом в 1953 г. увидели, что каждый саркомер
состоит из тонких (актиновых) и толстых (миозиновых) филаментов. Они
называются протофибриллами, число которых в среднем составляет
2500 в одной миофибрилле. Протофибриллы расположены паралле-
ι мияи ft. Биофизика двигательной активности
13
■4* *Ч
z-диск
Μ
ίίλΤ
z-диск
Рис. 5.6. Схема строения саркомера при продольном (1)
и поперечном (2) срезах в зоне перекрывания актиновых (А)
и миозиновых (М) нитей
щ.но друг другу, причём /-диск соответствует тому участку саркоме-
|ш. где присутствуют только актиновые нити (см. рис. 5.4).
В Л-диске актиновые нити перекрываются миозиновыми.
II центре Л-диска наблюдается светлый участок (//-зона), в котором
н|)исугствуют только миозиновые нити. Следовательно, анизотропия
обусловлена совместным присутствием и миозиновых, и актиновых
фпламентов. На поперечном срезе саркомера нити образуют
регулярную гексагональную систему, в которой каждая миозиновая прото-
фибрилла взаимодействует с шестью актиновыми, а каждая актино-
iiidi с тремя миозиновыми (рис. 5.6).
Взаимодействие между тонкими и толстыми филаментами
осуществляется посредством так называемых поперечных мостиков, об-
|щ юнанных головками миозиновых молекул. Они расположены по
отношению к продольной оси миозиновой нити под углом 120°.
Расстояние между соседними миозиновыми мостиками, лежащими на
одной оси, составляет 43 нм (см. рис. 5.3, б). Поперечный мостик
работает как химиомеханический преобразователь.
Стоит ещё раз заметить, что в миоците актиновые и миозиновые
фпламенты лежат строго параллельно друг другу. Промежутки межу
ними составляют примерно 13 нм. В них-то и выступают головки мио-
шповых молекул — поперечные мостики, которые подобно миниа-
порпым веслам могут совершать гребковые движения, перемещая
юнкие нити относительно толстых. Для того чтобы это произошло,
14
Биофизика
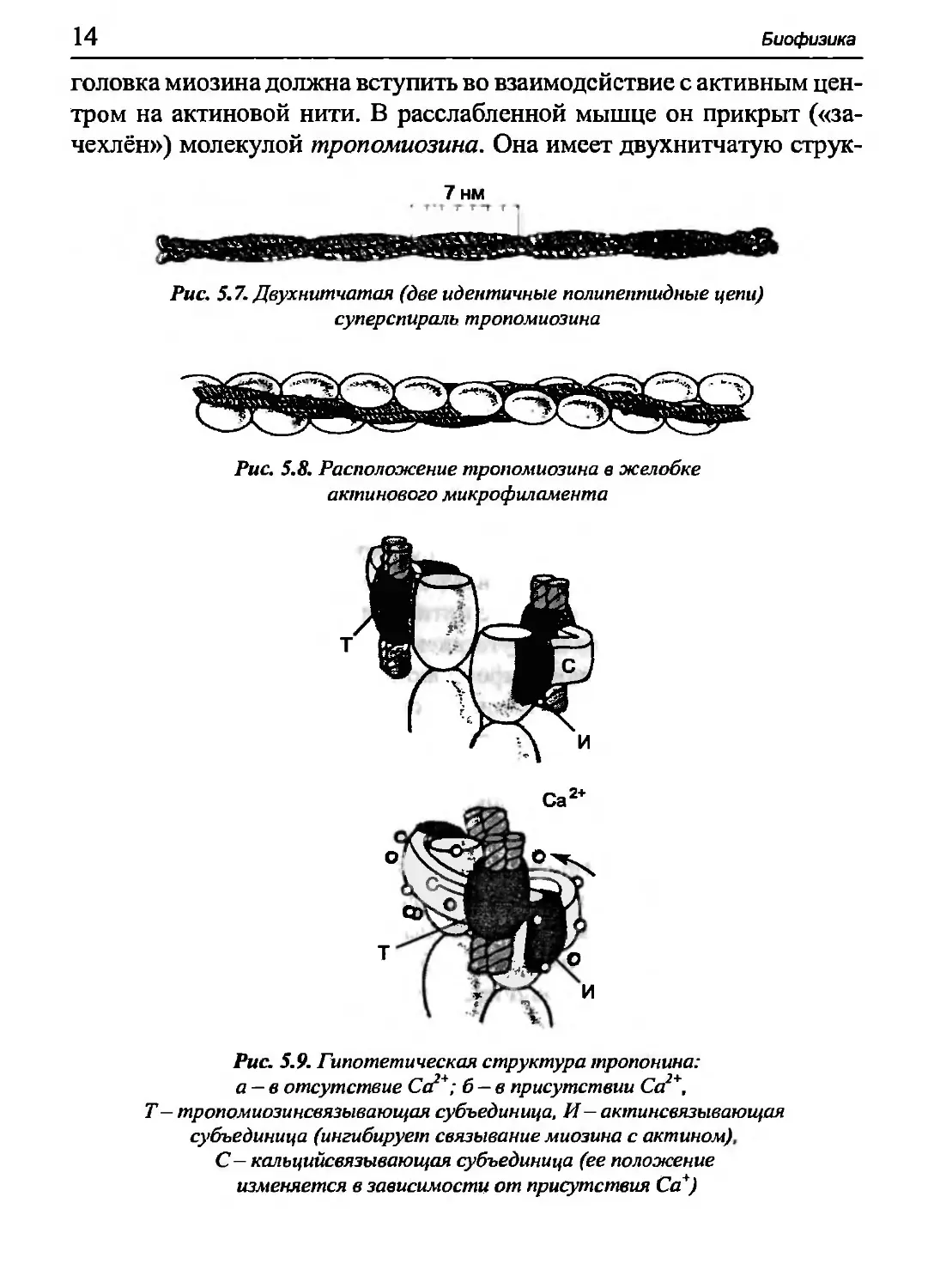
головка миозина должна вступить во взаимодействие с активным
центром на актиновой нити. В расслабленной мышце он прикрыт
(«зачехлён») молекулой тропомиозина. Она имеет двухнитчатую струк-
7нм
Рис. 5.7. Двухнитчатая (две идентичные полипептидные цепи)
суперспираль тропомиозина
Рис 5.8. Расположение тропомиозина в желобке
актинового микрофиламента
Са2+
О °"*\
CD
Τ О
И
Рис 5.9. Гипотетическая структура тропонина:
а —в отсутствие Са*; б —в присутствии Са *,
Τ — тропомиозинсвязывающая субъединица, И — актинсвязывающая
субъединица (ингибирует связывание миозина с актином)
С— калъцийсвязывающая субъединица (ее положение
изменяется в зависимости от присутствия Са*)
ι паяя fi, Биофизика двигательной активности
15
ι v|»v. образованную двумя субъединицами с молекулярными массами
Μ II .16 кДа. Молекула длиной 41 нм скручена с шагом 7 нм (рис. 5.7)
и ишимает спиральный желобок между двумя нитями ^-актина
||Ш1'. 5.8), где расположены активные центры, с которыми должны
hi ι у нить во взаимодействие миозиновые головки, чтобы произошло
ι ι·ΐιращение мышцы.
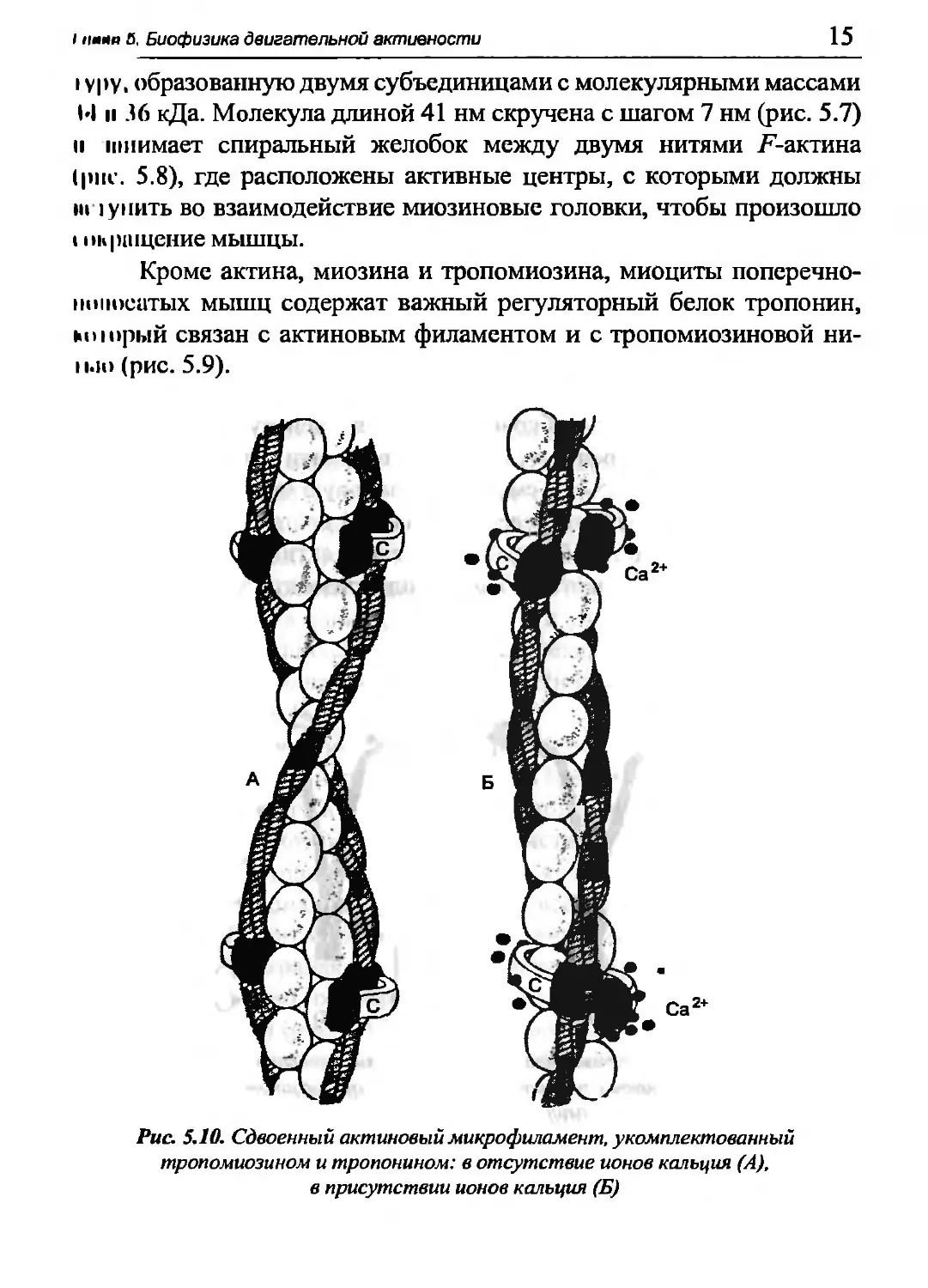
Кроме актина, миозина и тропомиозина, миоциты поперечно-
ншюситых мышц содержат важный регуляторный белок тропонин,
мчормй связан с актиновым филаментом и с тропомиозиновой ни-
ii.li) (рис. 5.9).
Рис. 5.10. Сдвоенный актиновый микрофиламент, укомплектованный
тропомиозином и тропонином: в отсутствие ионов кальция (А),
в присутствии ионов кальция (Б)
16
Биофизика
Три субъединицы тропанина имеют в сумме молекулярную
массу 76 кДа. Одна из субъединиц способна связываться с 4 ионами
Са2+. Когда такое взаимодействие происходит, тропонин
воздействует на тропомиозин, и тот освобождает на актиновом филаменте
активный центр для миозина (рис. 5.10). Тем самым устраняется
препятствие их взаимодействию.
5.1.2. Механизм мышечного сокращения
Благодаря снятию ионами кальция тропомиозиновой блокады
активных центров F-актина с ними взаимодействуют миозиновые
головки - они совершают гребковые движения к центру саркомера.
Головка временно прикрепляется к актиновой нити и наклоняется на
своем стебельке (рис. 5.И), смещая миозиновую нить вдоль
актиновой (как локомотив тянет железнодорожный состав по рельсам) на
один шаг (примерно на 10 нм). Вместе с тем актин активирует
способность миозина в качестве АТФазы гидролизовать АТФ. При взаи-
Рис. 5.11. Механизм скольжения — продольного перемещения
микрофилментов относительно центральной миозиновой нити:
а - положение головок миозина при низкой концентрации Са * в миоплазме,
б — положение головок миозина (сгибание в шарнирном соединении головок
с двунитчатым стеблем миозиновой молекулы) при повышении содержания Са'
в миоплазме. Стрелками указано направление перемещения аюпиновых нитей
ι пики Л йчофизика двигательной активности
17
мнщмстнии с актином каждая миозиновая молекула ежесекундно
ι наролпзует до 10 молекул АТФ. Следовательно, головки молекул
мно ппш обеспечивают как связывание толстых нитей с тонкими, так
к ι ид|)олиз АТФ. В конце гребка миозиновой головки по актиновой
ни in к молекуле миозина присоединяется новая молекула АТФ, что
нрниодит к отделению головки от актина. При последующем гидро-
шне АТФ происходит восстановление исходной конформации мио-
ииюиой молекулы, благодаря чему её головка может снова присое-
шинпься к актиновой нити, но уже ближе к центру саркомера, чем
при предыдущем взаимодействии с ней.
В каждом толстом филаменте содержится около 500 миозино-
ιιι.ιχ головок. Каждая из них при быстром сокращении мышцы
совершает примерно 5 циклических гребков (смыканий и размыканий
поперечных мостиков) в 1 с. Естественно, что не все мостики работают
одновременно: одни замыкаются на актиновых нитях, другие в этот
миг отделены от них. По расчетам М.В. Волькенштейна, каждый мио-
шпопый мостик должен создавать усилие порядка 5 · 10~12 Н. Такая
типа и пересчёте на единицу площади и с учетом всех миозиновых
мостиков, приходящихся на бицепс человека, даёт напряжение,
равное 2 · 105 Па, что близко совпадает с фактическим значением
напряжения этой мышцы (1,8 · 105Па).
Однократное прикрепление миозиновой головки к актиново-
му филаменту укорачивает саркомер на 1% его первоначальной
длины и развивает силу в 3-5 пН с КПД примерно 20%. При
многократном актин-миозиновом взаимодействии, что и происходит
и естественном мышечном сокращении, каждый саркомер укорачи-
нистся пропорционально укорочению всей мышцы. Сокращение
мышцы не приводит к изменению собственной длины ни толстых, ни
кхжих нитей. Сохраняя свои размеры и характер упаковки
образующих их субъединиц, они скользят друг относительно друга. Теория
скользящих нитей (рис. 5.12), сформулированная в 1954 г. А. Хаксли
н Г. Хаксли, подтверждена многими экспериментами. Иногда её
формулируют как теорию лазания или теорию гребков, но
принципиально существо современных представлений о механизме
мышечного сокращения не изменилось за истекшие полвека: максимальная
сила сокращения мышцы пропорциональна степени взаимного пере-
1 4X44
18
Биофизика
Актиновая нить /-диск Миоэиновая нить
Рис 5.12. Перемещение актиновых нитей
относительно окружённой ими миозиновой нити:
а -расслабление; б-сокращение
крытия толстых и тонких нитей. При уменьшении длины саркоме-
ра укорачивается только /-диск, тогда как Л-диск не изменяет своих
размеров.
Для максимального сокращения саркомера миозиновые
мостики должны совершить примерно 50 гребков. Дискретность процесса
скольжения обеспечивает дозирование степени сокращения. Оно
определяется количеством одновременно замкнутых поперечных мос-
тиков,тогда как его скорость зависит от частоты их замыкания (от
числа замыкаемых мостиков в единицу времени). Скорость
укорочения миоцитов скелетных мышц составляет 10-20 мкм · с-1, а кардио-
миоцитов - на порядок меньше.
Как уже говорилось, саркомер может быть растянут до 3,6-
3,8 мкм. Тогда мышечное волокно не развивает никакого
напряжения. Обычно при естественном расслаблении мышцы длина
саркомера составляет около 2,5 мкм. Мышца развивает максимальную силу,
когда саркомер укорачивается до 2,0-2,2 мкм. Однако его длина
может уменьшаться и до 1,5-1,6 мкм. В таком случае напряжение
мышцы ослабевает, поскольку концы миозиновых филаментов упираются
в Z-зону (актин-дисминовую сеть), что приводит к нарушению
правильной ориентации миозиновых мостиков относительно актиновых
филаментов и ослаблению тянущего усилия.
111.inn .') Биофизика двигательной активности
19
Только при наличии в миоплазме (рядом с миофибриллами)
Гц" и АТФ осуществляется правильная двигательная активность
М1.Ш1Ц, в которой чередуются процессы замыкания и размыкания
мпониювых мостиков на актиновых нитях. Если в миоплазме
недоспи <>к АТФ, то развивается окоченение мышц. Примером служит
ι рунное окоченение, которое при наступлении смерти приходит на
ι Mi'iiy первоначальному расслаблению мускулов, когда в них исчеза-
ΐΊ ΛΤΦ, поскольку он обеспечивает размыкание замкнутых
поперечных мостиков.
Источником энергии, необходимой для мышечной деятельно-
■ mi. служат реакции окисления субстратов клеточного дыхания, в ходе
мнорых образуются макроэрги (АТФ). Для мышечного сокращения
необходим гидролиз АТФ. КПД сократительной деятельности мышц
примерно 20%. Таким образом, в работающей мышце пятая часть
химической энергии преобразуется в механическую, а 80% выделяется
к и пде тепла. Энергетические процессы в мышцах неодинаковы при
оипомоментном (кратковременном) усилии и при длительной работе.
Кратковременное мышечное сокращение происходит за счёт запасов
ΛΤΦ в волокне, тогда как продолжительная мышечная деятельность
ipcGyer пополнения АТФ при помощи интенсификации окислитель-
пню фосфорилирования в процессе совершения этой работы. Суб-
ι ι раты клеточного дыхания и кислород поступают в мышечное
волокно из крови. Поэтому при тренировке выносливости (способности
ι ннершать длительную работу) необходимо развивать прежде всего
хорошее кровоснабжение мышц. При тренировке силы (способности
ι онершать значительную работу за короткое время, т.е. работать с
вышкой мощностью) требуется увеличение числа миозиновых мости-
пои, которые могут синхронно включаться в сократительный акт. Это
качество, необходимое, например, штангистам, достигается главным
образом путем значительного утолщения мышечных волокон за счёт
уисличения количества актиновых и миозиновых протофибрилл в во-
мокне. Сами миоциты дифференцированы и не способны к делению.
11ри нагрузке развивается их гипертрофия, а не гиперплазия.
Как следует из сказанного, для обеспечения молекулярного
механизма взаимодействия актиновых и миозиновых нитей необходи-
20
Биофизика
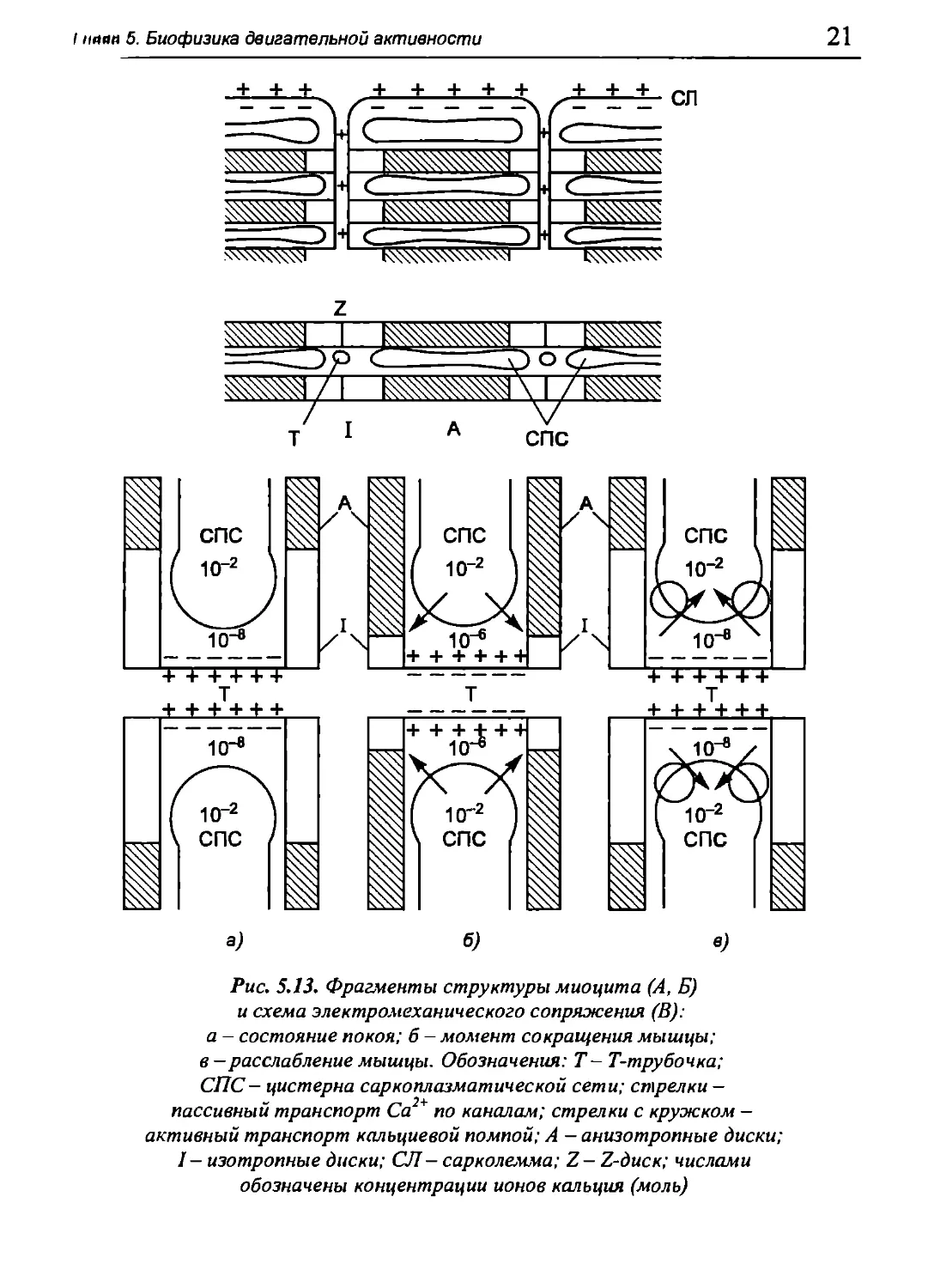
мы как макроэрги, так и ионы кальция. Именно Са2+ является
важнейшим компонентом электромеханического сопряжения (рис. 5.13).
Миофибриллы начинают сокращаться, когда вокруг них в ци-
тозоле концентрация свободных ионов кальция станет равна (0,4-
1,5) · Ю-6 моль. Максимальное сокращение мышечного волокна
происходит при её повышении до 5 · Ю-6 моль. Между тем, в покое
концентрация Са2+ в цитозоле мышечного волокна не превышает 10~7
моль. Для возникновения сокращения скелетной мышцы ионы
кальция должны поступить к миофибриллам из саркоплазматической
сети (СПС). Так называют систему пузырьков и цистерн, отделённых
мембранами от остальной саркоплазмы. СПС занимает примерно
10% объёма мышечного волокна, а суммарная площадь её мембран
в миоците приблизительно в 100 раз больше поверхности
сарколеммы. СПС служит кальциевым депо в мышечном волокне -
содержание в ней ионов кальция достигает 10~2 моль. Следовательно, на
мембране СПС поддерживается колоссальный градиент Са2+, но в покое
она совершенно непроницаема для этого иона.
СПС расположена поблизости от миофибрилл, причём в
электромеханическом сопряжении важнейшую роль играют её цистерны,
примыкающие к диску Ζ (рис. 5.13, а, б). Здесь же находятся впячи-
вания сарколеммы внутрь волокна, имеющие форму трубочек
диаметром около 50 нм. Они образуются за счёт того, что сарколемма во
многих местах «ныряет» вглубь и заканчивается в саркоплазме
слепыми концами наподобие пальцев вывернутой перчатки. Такой
мешочек (трубочка) идёт поперек волокна, достигая в длину более
10 мкм. В мышечном волокне диаметром 80 мкм, имеющем форму
цилиндра, мембраны этих трубочек образуют поверхность,
суммарная площадь которой в 2,5 раза превосходит поверхность плазмолем-
мы миоцита. В зоне Z-диска каждая трубочка вместе с двумя
соседними цистернами СПС образует так называемую Т-систему. Они
окружают каждую миофибриллу. Г-система с миофибриллой служат
основным звеном в электромеханическом сопряжении.
Сигналом к сокращению скелетной мышцы являются нервные
импульсы, поступающие из спинного или головного мозга к
концевой моторной пластинке (нервно-мышечному синапсу). Далее
следует синаптическая передача с участием ацетилхолина и холинорецеп-
I /iitoo 5. Биофизика двигательной активности
21
+ + +
—_) ^
шш
+ + + + +
г- ν
► С )
шшш
+ + +
f
г
ш
ι* с > , г ~-—
мш
ЕШ"
шш
>4 < > * г
*шт$
Ь^тЧ\ЧЧЧЧЧЧЧ
ктЧЧЧЧ^
ел
а;
б)
°)
Рис. 5.13. Фрагменты структуры миоцита (А, Б)
и схема электромеханического сопряжения (В):
а - состояние покоя; б - момент сокращения мышцы;
в - расслабление мышцы. Обозначения: Т- Т-трубочка;
СПС- цистерна саркоплазматической сети; стрелки —
пассивный транспорт Са по каналам; стрелки с кружком —
активный транспорт кальциевой помпой; А — анизотропные диски;
1 — изотропные диски; СП— сарколемма; Z—Z-диск; числами
обозначены концентрации ионов кальция (моль)
22
Биофизика
торов. Потенциалы действия, образовавшиеся на внесинаптических
участках сарколеммы под действием потенциала концевой пластин-
ки,распространяются бездекрементно по всей плазматической
мембране мышечного волокна, проникая и в Г-систему (по мембране
трубочки). С трубочки деполяризация распространяется на
мембрану СПС и открывает в ней потенциалзависимые кальциевые каналы.
Через открывшиеся каналы Са2+ пассивно (в сторону более низкого
электрохимического потенциала) устремляется из цистерн СПС
в саркоплазму и достигает миофибрилл. Около них создаётся
достаточная для замыкания миозиновых мостиков концентрация ионов
кальция.
Выход кальция из СПС прекращается сразу вслед за реполяри-
зациией сарколеммы, но миофибриллы пребывают в сокращённом
состоянии до тех пор, пока содержание кальция подле них не
снизится до 10"7 моль. Для этого Са24 должен возвратиться в СПС, но такой
транспорт приходится осуществлять вопреки действию огромного
концентрационного градиента. Следовательно, расслабление
миофибрилл в миоците скелетных мышц после их сокращения
невозможно без участия системы активного транспорта - кальциевой помпы. Её
работа - неотъемлемый элемент сократительного процесса в мышце.
Из мембраны СПС выделена Са-активируемая АТФаза, которая
служит основным компонентом кальциевого насоса. Таким образом,
электромеханическое сопряжение включает цепь последовательно
идущих процессов: распространение ПД по сарколемме —»
деполяризация мембран трубочек Г-системы —> открытие кальциевых каналов
в мембране СПС —» выход Са2+ из цистерн СПС в цитозоль
мышечного волокна —> замыкание миозиновых мостиков. Расслабление
мышцы связано с реполяризацией сарколеммы и последующей
активизацией работы кальциевого насоса, локализованного в мембране СПС.
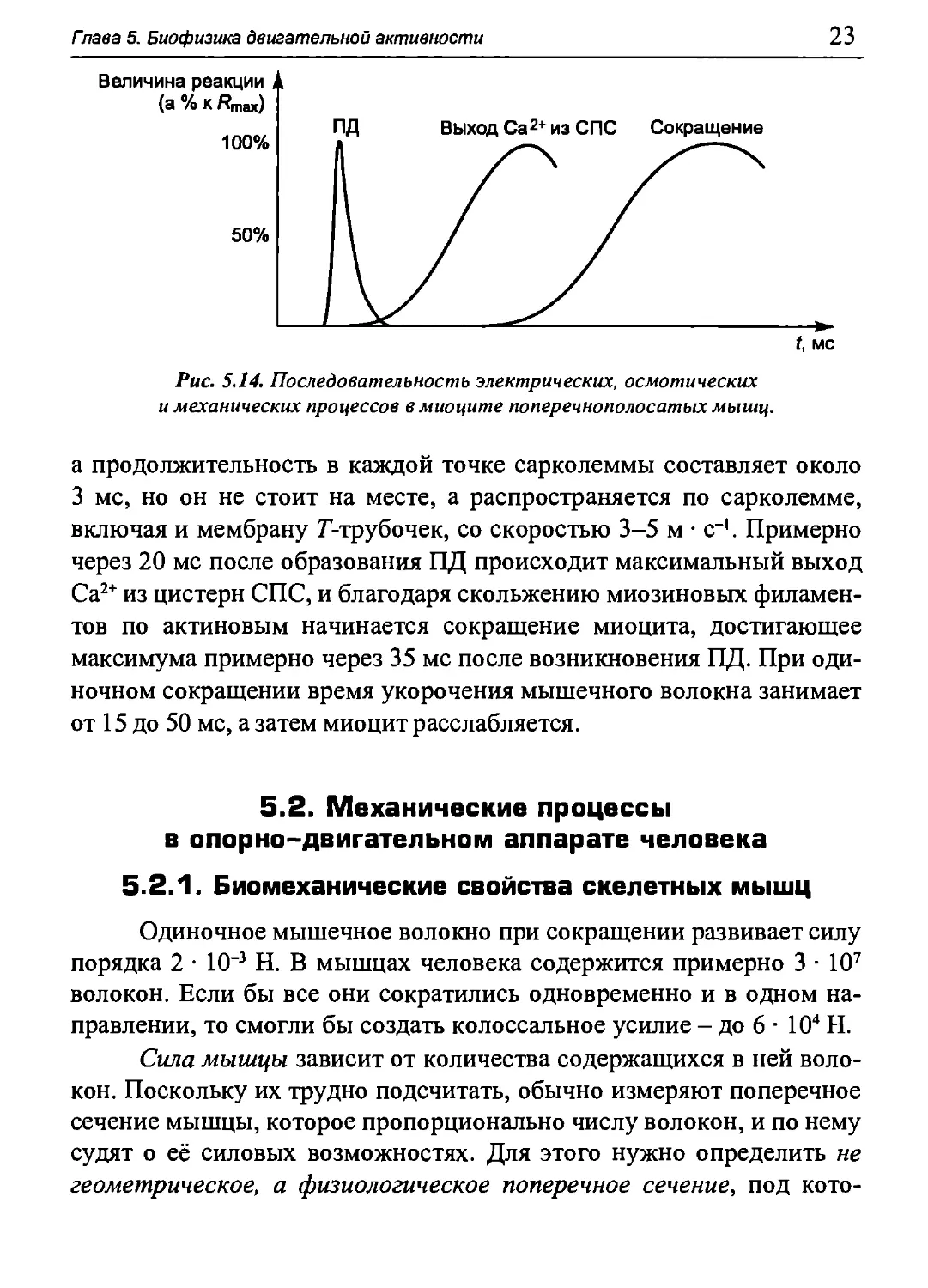
Последовательность электрических, осмотических и
механических процессов в миоците скелетных мышц представлена на рис. 5.14.
После синаптической передачи возбуждения с нервного волокна на
миоцит в его субсинаптической мембране возникает потенциал
концевой пластинки (ПКП), который инициирует генерацию потенциала
действия (ПД) на сарколемме. Его амплитуда достигает 120-130 мВ,
Глава 5. Биофизика двигательной активности
23
Величина реакции
(а % к Rmax)
100%
50%
f, мс
Рис. 5.14. Последовательность электрических, осмотических
и механических процессов в миоците поперечнополосатых мышц.
а продолжительность в каждой точке сарколеммы составляет около
3 мс, но он не стоит на месте, а распространяется по сарколемме,
включая и мембрану Г-трубочек, со скоростью 3-5 м · с-1. Примерно
через 20 мс после образования ПД происходит максимальный выход
Са2+ из цистерн СПС, и благодаря скольжению миозиновых филамен-
тов по актиновым начинается сокращение миоцита, достигающее
максимума примерно через 35 мс после возникновения ПД. При
одиночном сокращении время укорочения мышечного волокна занимает
от 15 до 50 мс, а затем миоцит расслабляется.
5.2. Механические процессы
в опорно-двигательном аппарате человека
5.2.1. Биомеханические свойства скелетных мышц
Одиночное мышечное волокно при сокращении развивает силу
порядка 2 · Ю-3 Н. В мышцах человека содержится примерно 3 ■ Ю7
волокон. Если бы все они сократились одновременно и в одном
направлении, то смогли бы создать колоссальное усилие - до 6 ■ ΙΟ4 Η.
Сила мышцы зависит от количества содержащихся в ней
волокон. Поскольку их трудно подсчитать, обычно измеряют поперечное
сечение мышцы, которое пропорционально числу волокон, и по нему
судят о её силовых возможностях. Для этого нужно определить не
геометрическое, а физиологическое поперечное сечение, под кото-
ПД Выход Са2+из СПС Сокращение
24
Биофизика
рым понимают сумму поперечных сечений всех волокон,
образующих данную мышцу. В мышце, все волокна которой идут
параллельно продольной оси и своими концами прикрепляются к костям
(непосредственно или через сухожилия), геометрическое и
физиологическое сечения совпадают. Такие мышцы при сокращении
испытывают значительное укорочение, но развивают малую силу (выигрыш
в расстоянии, но проигрыш в силе).
В мышцах другого типа волокна располагаются косо по
отношению к продольной оси и прикрепляются с обеих сторон к
сухожилию, которое тянется вдоль неё на всем протяжении мышцы. Волокна
таких (перистых) мышц намного короче самой мышцы.
Физиологическое поперечное сечение перистых мышц больше геометрического.
При мышечном сокращении сухожилие перемещается на малое
расстояние, но при этом развивается значительное тяговое усилие
(выигрыш в силе при проигрыше в расстоянии). Соотношение сил,
развиваемых мышцами с параллельным ходом волокон, и перистыми,
выражает уравнение:
Fx 2/ . г-2 г^—
—L= — since ·ν« -sin α,
F2 nb
где F, - сила сокращения перистой мышцы, F2 - сила сокращения
мышцы с параллельным расположением волокон, / - длина мышцы
в покое, η - доля этой длины при сокращении, Ъ - средняя толщина
мышцы, α - угол между сухожилием и волокнами перистой мышцы.
Лишь немногие скелетные мышцы могут сокращаться более
чем до 70% первоначальной длины, т.е. до η = 0,7. Если в перистой
мышце угол α был бы равен 45° (sin α = 0,7), то сила, направленная
вдоль сухожилия, обращалась бы в 0, поскольку сократившиеся
волокна располагались бы перпендикулярно сухожилию. Обычно в
перистой мышце α < 30°. Она сильнее мышцы с параллельным ходом
волокон при условии, что 0,9 / > Ъ. Перистые мышцы практически не
утолщаются.
Отношение максимального веса груза, который может поднять
мышца, к её физиологическому поперечному сечению называется
абсолютной мышечной силой и выражается в единицах напряжения
/ пава 5. Биофизика двигательной активности
25
(11 · м-2 = Па). Абсолютная мышечная сила большинства скелетных
мышц человека имеет порядок 106 Па, а гладких - 105 Па.
Сокращения скелетных мышц обеспечивают человеку, во-пер-
и ых, поддержание определённой позы, для чего необходимо
противодействовать прежде всего силе земного притяжения, во-вторых,
перемещение частей тела друг относительно друга, в-третьих,
передвижение человека в пространстве. Перечисленные процессы
осуществляются при разной степени укорочения скелетных мышц. По
этому признаку различают три типа мышечных сокращений:
тометрический, изотонический, ауксотонический {или анизотони-
ческий).
При изометрическом сокращении длина мышцы почти не
изменяется (теоретически — вообще не изменяется), и вся сила
затрачивается на развитие напряжения, т.е. на совершение статической
работы (в частности, на поддержание позы). Для исследования этого типа
сокращения мышцы её концы удерживают при возбуждении в том
положении, какое им было свойственно в покое (рис. 5.15, а). Тогда
в возбуждённой мышце развивается значительное напряжение,
которое затем уменьшается по экспоненциальному закону.
При изотоническом сокращении (рис. 5.15, б) мышечные
волокна укорачиваются в условиях постоянной (неизменной) внешней
нагрузки. Происходит быстрое укорочение мышцы, сменяющееся
расслаблением.
Между нагрузкой (Р) и скоростью укорочения (υ) мышцы при
сё изотоническом сокращении установлена следующая
зависимость -уравнение Хилла: (Р + а)х> = Ь(Р0 - Р), где а, Ъ и Р0 -
константы (рис. 5.15, в). Произведение α υ отображает теплопродукцию
мышцы за единицу времени (так называемая тепловая мощность);
произведение Ρ ■ υ выражает полезную мощность. Следовательно,
левая часть уравнения в целом характеризует полную мощность
изотонического мышечного сокращения. Величина Р0 соответствует
максимальной нагрузке, которую мышца ещё может удержать, но
уже не в состоянии поднять (передвинуть). Другими словами, Р0 -
максимальная сила, развиваемая мышцей в изотоническом режиме.
Когда нагрузка мало отличается от максимальной (Р0), мышца не мо-
26
Биофизика
0,10
0,30
0,50
0,70 F, Η
e)
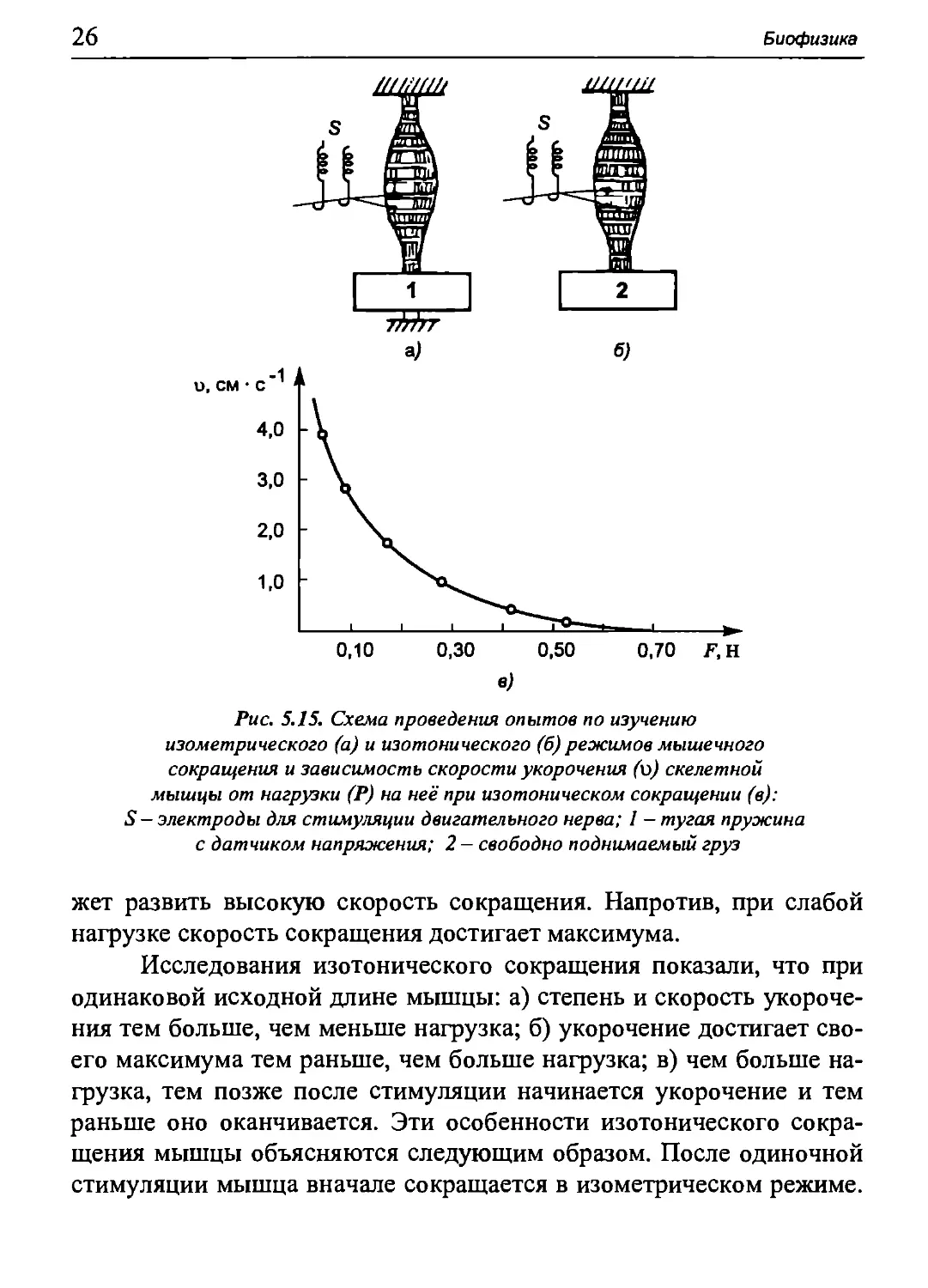
Рис. 5.15. Схема проведения опытов по изучению
изометрического (а) и изотонического (б) режимов мышечного
сокращения и зависимость скорости укорочения (ό) скелетной
мышцы от нагрузки (Р) на неё при изотоническом сокращении (в):
S - электроды для стимуляции двигательного нерва; I - тугая пружина
с датчиком напряжения; 2 - свободно поднимаемый груз
жет развить высокую скорость сокращения. Напротив, при слабой
нагрузке скорость сокращения достигает максимума.
Исследования изотонического сокращения показали, что при
одинаковой исходной длине мышцы: а) степень и скорость
укорочения тем больше, чем меньше нагрузка; б) укорочение достигает
своего максимума тем раньше, чем больше нагрузка; в) чем больше
нагрузка, тем позже после стимуляции начинается укорочение и тем
раньше оно оканчивается. Эти особенности изотонического
сокращения мышцы объясняются следующим образом. После одиночной
стимуляции мышца вначале сокращается в изометрическом режиме.
Глава 5. Биофизика двигательной активности
27
В это время сократительный элемент, укорачиваясь, растягивает
последовательный упругий элемент. После того как изометрическая
сила мышцы (активное напряжение) достигает массы груза, мышца
начинает укорачиваться, поднимая груз. Чем больше груз, тем
меньше изотоническое укорочение мышцы. И если величина груза будет
равна максимальному изометрическому напряжению, то никакого
укорочения мышцы не произойдет. Таким образом, подбором груза
можно при данной длине мышцы определить ее максимальное
изометрическое напряжение.
При изотоническом сокращении мышцы она, укорачиваясь,
поднимает груз и, таким образом, выполняет внешнюю работу.
Величина этой работы равна произведению степени укорочения
мышцы на величину поднимаемого груза.
Кривую «укорочение — нагрузка» можно получить двумя
путями. В первом случае мышца предварительно растягивается грузом и,
таким образом, исходная ее длина будет различной в зависимости от
величины груза. Это показано на рис. 5.16 кривой напряжения покоя
(I). Предположим, что при нагрузке 120 г на кривой 1 длина мышцы
соответствует точке А. При титаническом раздражении мышцы она
укорачивается до длины, показанной на рис. 5.16 точкой В.
Подвешивая к мышце различные грузы, получаем кривую 2 изотонических
максимумов.
Во втором случае исследуется укорочение мышцы при
различных нагрузках, но исходная длина мышцы сохраняется постоянной -
метод постнагрузочного сокращения. Пассивное растяжение мышцы
под влиянием груза предотвращается стопорным винтом, как это
показано в верхней части рисунка 5.16. Таким путем получаем кривую
3 постнагрузочных изотонических максимумов. При каждой
нагрузке в мышце в ответ на раздражение развивается сначала
изометрическое напряжение. Когда последнее достигает величины груза, мышца
начинает укорачиваться, производя работу. Как видно из рис. 5.16,
при нагрузке, например, 90 г выполненная мышцей работа будет
равна площади фигуры ОВВТ. При нагрузках ~ 160 и 30 г работа
мышцы будет равна площади фигур ОДЦ'Е и ОЖЖ'З соответственно.
Площадь ОВВТ больше, чем площадь ОДЦ'Е или ОЖЖ'З.
28
Биофизика
1,5 Г Ε 2,5
3,5 L, см
б)
Рис. 5.16. Изотоническое сокращение мышцы:
а -установка для регистрации
изотопического сокращения: I - мышца; 2 - стимулятор;
3 - показатель длины мышцы; 4 - груз; 5 - ось крепления рычага;
6 — винт, ограничивающий натяжение мышцы; б — зависимость
между нагрузкой Ρ и длиной L мышцы: I - кривая пассивного
напряжения (исходная длина мышцы 2,5 см); 2 — кривая
изотонических максимумов; 3 -кривая постнагрузочных
изотонических максимумов расслабления
Таким образом, максимальную работу мышца выполняет при
умеренной и средней нагрузках. Увеличение или уменьшение
нагрузки ведет к уменьшению работы, а при максимальной (равной
изометрическому напряжению) и нулевой нагрузках работа,
выполненная мышцей, будет стремиться к нулю.
Важной характеристикой изотонического сокращения
является скорость укорочения мышцы V. Оказалось, что при одинаковой
исходной длине мышцы скорость укорочения при изотоническом
сокращении тем больше, чем меньше нагрузка (рис. 5.17). При
нулевой нагрузке скорость максимальна, а при максимальной
нагрузке (когда она равна максимальному изометрическому напряжению
Р0) укорочение мышцы отсутствует, поэтому и скорость его равна
нулю.
Если откладывать по оси абсцисс нагрузку, а по оси ординат
скорость укорочения мышцы в изотоническом режиме сокращения,
получим кривую сила (нагрузка) - скорость, которая имеет форму
гиперболы (рис. 5.17, б).
/ /тез 5. Биофизика двигательной активности
29
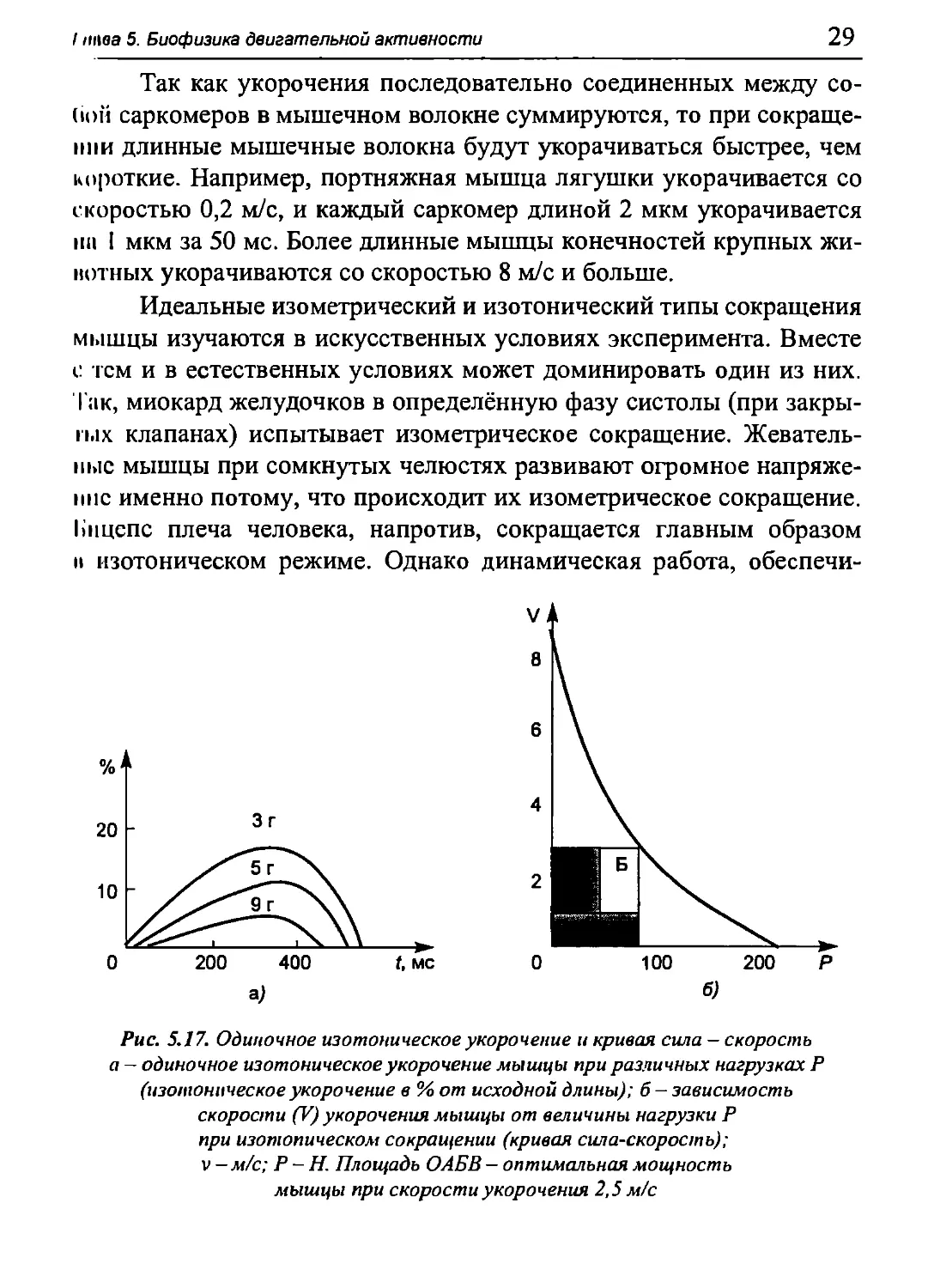
Так как укорочения последовательно соединенных между со-
(ioii саркомеров в мышечном волокне суммируются, то при
сокращении длинные мышечные волокна будут укорачиваться быстрее, чем
короткие. Например, портняжная мышца лягушки укорачивается со
скоростью 0,2 м/с, и каждый саркомер длиной 2 мкм укорачивается
пи I мкм за 50 мс. Более длинные мышцы конечностей крупных жи-
нотных укорачиваются со скоростью 8 м/с и больше.
Идеальные изометрический и изотонический типы сокращения
мышцы изучаются в искусственных условиях эксперимента. Вместе
ι: тем и в естественных условиях может доминировать один из них.
Гак, миокард желудочков в определённую фазу систолы (при
закрытых клапанах) испытывает изометрическое сокращение.
Жевательные мышцы при сомкнутых челюстях развивают огромное
напряжение именно потому, что происходит их изометрическое сокращение.
1)пцепс плеча человека, напротив, сокращается главным образом
it изотоническом режиме. Однако динамическая работа, обеспечи-
aj
t, мс
Рис. 5.17. Одиночное изотоническое укорочение и кривая сила — скорость
а — одиночное изотоническое укорочение мышцы при различных нагрузках Ρ
(изотоническое укорочение в%от исходной длины); б - зависимость
скорости (V) укорочения мышцы от величины нагрузки Ρ
при изотопическом сокращении (кривая сила-скорость);
v—м/с; Ρ -Η. Площадь ОАБВ — оптимальная мощность
мышцы при скорости укорочения 2,5 м/с
30
Биофизика
вающая перемещение тела и его частей в пространстве, совершается
преимущественно благодаря ауксотоническому типу мышечных
сокращений, когда мышцы в процессе сокращения и укорачиваются,
и напрягаются.
Различают два режима сократительной деятельности скелетных
мышц: одиночное и тетаническое сокращения. Первое из них (рис. 5.18)
возникает в ответ на однократную стимуляцию. В нём чётко
различают две стадии: 1) напряжение (укорочение) и 2) расслабление
(удлинение). В соотношении их продолжительности прослеживается так
называемая золотая пропорция, свойственная гармонии в природе:
в двухфазных процессах отношение продолжительности релакси-
рующей (восстанавливающей) части к длительности рабочей части
равно 1,618. Этому же числу равно отношение времени полного
цикла сокращения к длительности релаксирующей части. Заметим, что
в архитектуре и музыке золотая пропорция между составляющими
элементами воспринимается как гармония. Она характерна для
структуры и функционирования тела человека. При патологии
золотая пропорция нарушается,
Различают быстрые и медленные мышечные волокна и в
зависимости от преобладания тех или других - быстрые и медленные
мышцы. Среди первых лидером является мышцы глазного яблока -
у них стадия напряжения в одиночном сокращении занимает 7 -
10 мс. У одной из самых медленных мышц - камбаловидной
укорочение затягивается до 50-100 мс.
Режим одиночных сокращений устанавливается не только при
однократной, но и при редкой ритмическойстимуляции, причём её
максимальная частота, обеспечивающая одиночные сокращения,
неодинакова для быстрых и медленных мышц. Если у медленных она
ниже 10 Гц, то у быстрых достигает 50 Гц. В режиме одиночных
сокращений мышца способна работать долго, но их амплитуда всегда
меньше максимальных возможностей.
При тетанических сокращениях, возникающих в ответ на
ритмическую стимуляцию определенной частоты, мышца сокращается
в 2-4 раза сильнее, чем в режиме одиночных сокращений. Их
амплитуда увеличивается благодаря наложению (суперпозиции)
следующих друг за другом одиночных сокращений. Их суперпозиция осо-
Глава 5. Биофизика двигательной активности
31
0,5
0,25
0 0,5 1 f,"c
Рис. 5.18. Режимы сократительной деятельности скелетных мышц:
I - одиночное сокращение; 2 - зубчатый тетанус; 3 — гладкий тетанус
бснно эффективна, когда последующий стимул приходит в фазу
экзальтации предшествующего возбуждения.
По мере повышения частоты стимуляции сначала развивается
зубчатый тетанус, если последующий стимул поступает к мышце
па стадии её расслабления (рис. 5.18), когда же интервал между
стимулами меньше длительности стадии укорочения мышцы, возникает
гладкий тетанус (рис. 5.18). В быстрой мышце это происходит при
частоте стимуляции 150-200 Гц, а в медленной - уже при 30 Гц.
В патологии примером тетанических сокращений являются
судороги. Работа в тетаническом режиме быстро приводит мышцу к
утомлению, что проявляется вначале в уменьшении силы сокращений
и удлинении стадии расслабления, а затем — в прекращении
сократительной деятельности.
При оценке усилий, создаваемых теми или иными мышцами,
следует учитывать особенности их прикрепления к костям.
Сочлененные суставами, кости скелета вместе с мышцами, которые
прикрепляются к ним в определённых точках, составляют
опорно-двигательный аппарат. Сила тяги, возникающая при сокращении мышц,
приводит в движение кости, которые действуют как рычаги с точками
опоры в суставах.
Рычагом называется твёрдое тело, имеющее ось вращения. Он
характеризуется моментом сил, равным произведению силы на плечо
рычага, к которому она приложена. Различают рычаги двух типов
(родов). У рычага первого рода силы направлены в одну сторону
Рис. 5.19. Рычаги в опорно-двигательном аппарате человека
а-рычаг первого рода; б, в -рычаги второго рода
и точка опоры (ось вращения) находится между точками приложения
сил. Примером является сочленение черепа с первым позвонком
(рис. 5.19, а). Условием равновесия такого рычага служит равенство
моментов сил: Fa = Ρ ■ b, где Ρ - сила тяжести головы, Ъ - плечо
этой силы, F - сила тяги мышц и связок, прикрепляющихся к
затылочной кости, а - плечо этой силы. У рычага второго рода силы
направлены в противоположные стороны, а ось вращения расположена
по одну сторону от точек приложения сил (рис. 5.19, б, в).
Когда рассматривают движения в суставах, оценивают
кинетическую энергию (W) вращающегося тела по формуле: W =-/ω2, где
/ - массовый момент инерции тела относительно оси вращения, -
его угловая скорость. Момент инерции определяется произведением
массы вращающегося тела на квадрат радиуса окружности, которую
/ пава 5. Биофизика двигательной активности
33
она описывает в плоскости вращения: I = ^(тг2).Чем больше I, тем
большая кинетическая энергия требуется для обеспечения
вращательного движения. Понятно, что / возрастает по мере удаления
массы от оси вращения. Поэтому при движениях животных и человека
■шергия тратится более экономно, если масса мышечной ткани
сосредоточена преимущественно в проксимальных частях конечностей
(ближе к оси вращения, т.е. к суставу), а в дистальных частях - она
невелика. У млекопитающих, способных к быстрому бегу, мощные
бедра и сравнительно тонкие голени. Проводя отбор спринтеров,
педагоги детских спортивных школ учитывают конституцию детей,
прогнозируя лучшие скоростные качества при врождённом
преобладании бедренных мышц над мышцами голени. При прочих равных
условиях нижние конечности таких людей обладают лучшими
скоростными качествами.
5.2.2. Энергетика мышечного сокращения
При сокращении мышцы химическая энергия превращается
в механическую и тепловую. С помощью высокочувствительных
термопар А. Хилл впервые измерил тепловые потери в этом процессе.
При одиночном изометрическом сокращении мышцы выделяется
энергия, равная 12,54 Дж · кг-1. При изотоническом сокращении
общее изменение энергии в мышце можно разбить на несколько
компонент. Быстрое начальное выделение теплоты происходит на самой
ранней стадии активации сокращения, когда еще отсутствует
заметное укорочение мышцы. Это - теплота активации А. Она
выделяется в результате освобождения ионов Са2+ из саркоплазматического
рстикулума.
По мере укорочения мышцы выделяется основное количество
теплоты, которая называется теплотой укорочения Qy. Небольшая
часть теплоты выделяется или поглощается при расслаблении
мышцы ±Qp. С учетом выполняемой механической работы W общий
энергетический баланс [Е) для мышцы будет
E=A±Qy+W±Qp.
(5.1)
34
Биофизика
А. Хилл установил важный факт, что при изотоническом
сокращении теплота укорочения Qy пропорциональна величине
укорочения ΔΖ, мышцы:
Qy =oAL, (5.2)
где α - коэффициент теплоты укорочения. Так как W =PbL, (5.1)
можно записать в следующем виде:
E=A+aAL + PAL±Qp=A+(P + a)AL±Qp. (5.3)
Продифференцировав по времени (5.3), находим мощность
мышцы
N=—=(P + a)—=(P + a)V, (5.4)
dt dt
где К-скорость укорочения мышцы.
При одинаковом изменении длины мышцы скорость выхода на
новый изометрический уровень уменьшается с увеличением
нагрузки Р. При этом избыточная мощность по сравнению с
изометрической была пропорциональна Р0 - Р. На основании этого А. Хилл
получил выражение:
(P + a)V=b(P0-P), (5.5)
где Ъ - константа; Р0 - максимальноеизометрическоенапряжение
мышцы. Сравнивая (5.4) и (5.5), получим
PV+aV=bP0-bP. (5.6)
Таким образом, уравнение Хилла (5.6), полученное из
термодинамических экспериментов, устанавливает связь между
напряжением и скоростью укорочения мышцы в изотонических условиях.
Оно справедливо в стационарном режиме работы мышцы.
Преобразуем (5.6) следующим образом:
PV+aV=bP0-bP
(5.7)
/ пава 5. Биофизика двигательной активности
35
и перенесем переменные величины Ρ и V в левую часть уравнения.
11олучим
PV+aV+bP = bP0. (5.8)
Добавим слева и справа уравнения произведение постоянных
Хиллаяб:
PV +aV+bP+ab = bP0 +οώ.
После чего получим преобразованное уравнение Хилла в
следующем виде
Ρ + aXV + b) = b(P0+ a) = const. (5.9)
Эта гиперболическая зависимость хорошо подтверждена
экспериментально биомеханическими экспериментами. При нагрузке
I' = Р0 мышца вообще не сокращается и V= 0. Ненагруженная
мышца {Р = 0) сокращается с максимальной скоростью V0 и
b=±V0. (5.10)
•"о
ОС
Для портняжной мышцы лягушки — = 0,25. Такое же значение
"о
было получено для исчерченных мышц крысы, кошки, жабы и др.
В более точных экспериментах А. Хилл установил, что
параметр α не является константой, а зависит от Р:
α =0Д6Р0 +0Д8Р. (5.11)
Это значит, что мощность сокращения мышцы зависит от Ρ не-
нинейно.
5.2.3. Элементы теории мышечного сокращения
Теория мышечного сокращения, особенно для
поперечно-полосатых мышц, в настоящее время хорошо разработана и подтвер-
36
Биофизика
ждена экспериментально. Существенную роль в ней играет
кинетика ионов Са2\ Реакции, в которых участвует Са2+, изучаются в курсах
биохимии.
Рассмотрим реакции взаимодействия актина и миозина на
основе теории В.И. Дещеревского. Предположим, что в данном
положении миозиновой нити количество мостиков в разомкнутом,
тянущем и тормозящем состояниях соответственно равно: γ, η, т
(рис. 5.20). Константы скоростей перехода из свободного состояния
в тянущее (замыкание мостиков) кх, из тянущего состояния в
тормозящее (взаимное скольжение филаментов) к, из тормозящего в
свободное (размыкание мостиков) к^.
Второй закон Ньютона можно записать в виде:
dL , . у. _
— =(n-m)f+Pt,
at
(5.12)
где Ζ,, - импульс, даваемый одной миозиновой нитью на полусар-
комере, / - сила, развиваемая нитью на полусаркомере, Ру - часть
внешней нагрузки Р, приходящейся на эту нить в полусаркомере, t -
время.
Полагая, что соединение актина и миозина есть химическая
реакция первого порядка, запишем скорость изменения количества
мостиков в тянущем состоянии:
dn
dt
= к{у-кп.
(5.13)
Аналогично можно получить и уравнение реакции распада ак-
тин-миозинового комплекса:
dm
It
= кп -к2т.
(5.14)
Рис. 5.20. Кинетика перехода мостиков между состояниями
/ нова 5. Биофизика двигательной активности
37
В стационарном состоянии производные по времени в
уравнениях (5.12) - (5.14) равны нулю. Следовательно, имеем три
алгебраических уравнения:
(n-m)f-Px =0, (5.15)
kxy-kn=0, (5.16)
bi-k2m = 0. (5.17)
Величину γ можно выразить через тип, используя общее
количество мостиков на миозиновой нити полусаркомера θ =const:
у = В-п-т. (5.18)
Следовательно, уравнение (5.16) примет вид:
кх(Ъ-п-т)-кп=0. (5.19)
Решая систему (5.17), (5.19), находим:
kik2d
£,(£, +k) + kkx
(5.20)
т = Щ . (5.21)
Jfc2(Jfc, +k) + kk{
Подставляя (5.20) и (5.21) в (5.15), находим:
[р{ +/ *'θ \= к^ (Θ/-Ρ,). (5.22)
Переход мостиков из тянущего в тормозящее состояние
происходит в процессе развития ими силового эффекта. Константа
скорости этого процесса к - это величина, обратная времени процесса /:
*=-=?. (5-23)
/ δ
38
Биофизика
где и - скорость относительного скольжения актиновои и миози-
новой нитей, 5 - величина скольжения при однократной
перестыковке мостиков.
Подставляя (5.23) в (5.22) и умножая уравнение (5.22) на
количество полусаркомеров 2Ν по длине мышцы и число миозиновых
волокон в мышце Ν0, получим:
( к θ ^
2NNn\P+f !—
!ί=2ΝΝ0-^—(θ/-Ρ<). (5.24)
δ (к. +к2)
Учитывая, что скорость сокращения мышцы при
последовательном расположении саркомеров, V = 2Nu, а также сила,
развиваемая мышцей Ρ = Ρ,Λ'ο, получим:
(P + a)V=b(P-P0), (5.25)
где Р0 = N0Qf- сила, развиваемая мышцей при изометрическом
сокращении:
fl=/^oM.=_^s (526)
b=2N5-^-. (5.27)
kt+k2
Уравнение (5.25) представляет собой известное
экспериментально полученное уравнение Хилла для мышечного сокращения.
5.3. Биомеханика дыхания
5.3.1. Легочная вентиляция
Газообмен между альвеолярной газовой смесью и
атмосферным воздухом, обеспечивающий эффективную диффузию кислорода
и углекислого газа через альвеоло-капиллярную мембрану,
осуществляется благодаря работе аппарата вентиляции, который состоит из
/ пава 5. Биофизика двигательной активности
39
диух анатомофизиологических образований: грудной клетки с
дыхательными мышцами и лёгких с дыхательными путями.
Грудная клетка представляет собой жёсткий подвижный
футляр для лёгких, сердца и сосудов, обладающий упругостью. Грудная
клетка активно изменяет свой объём посредством сокращения
диафрагмы и других дыхательных мышц. При сокращении диафрагмы
се' купол уплощается и смещается в сторону брюшной полости, что
приводит к увеличению объёма грудной клетки, а вслед за нею
и легких. Согласно закону Бойля-Мариотта (р · V = const),
увеличение объёма лёгких неизбежно сопровождается понижением давления
ннутри них. Оно становится ниже атмосферного (примерно на 6 мм.
рг. ст., т.е. на 0,8 Па при форсированном вдохе) и воздух
засасывается внутрь лёгких (в альвеолы). Так совершается вдох.
При вдохе воздух поступает в трахею и затем по бронхам
достигает альвеол. Все дыхательные пути в совокупности составляют
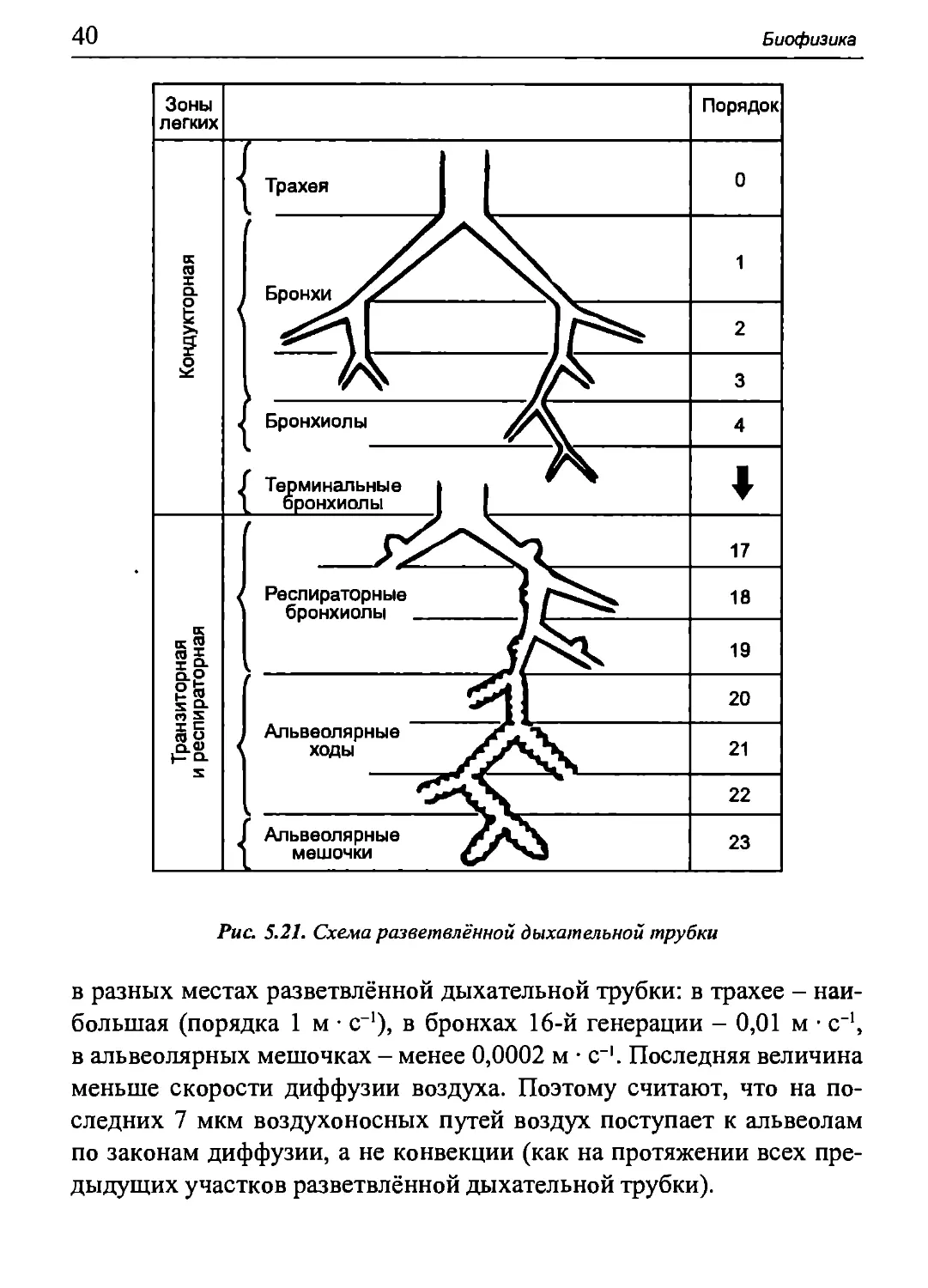
так называемую разветвлённую дыхательную трубку (рис. 5.21).
В ней рассчитывают суммарные поперечные сечения каждой
генерации бронхов. Наименьшей площадью поперечного сечения обладает
трахея (в среднем, 2,54 см2 у взрослого человека). Суммарное
сечение двух главных (левого и правого) бронхов больше этой величины.
По направлению к альвеолам суммарное сечение каждой
последующей генерации бронхов становится всё больше. Так, площадь
суммарного сечения всех бронхов 16-й генерации достигает 180 см2,
а всех альвеолярных мешочков - 11800 см2.
Для сплошного потока воздух по воздухоносным путям живот-
пых и человека в физиологических условиях выполняется условие
неразрывности струи: при сплошном течении несжимаемой среды
через любое сечение струи в единицу времени переносятсяодинаковые
объёмы перемещаемой среды (в данном случае, воздуха).
Применительно к разветвлённой дыхательной трубке это означает, что
величина Q =V 11 (объёмная скорость воздушного потока) одинакова во
псех её суммарных поперечных сечениях (на уровне каждой
генерации). Между объёмной (Q) и линейной (υ) скоростями воздушного
потока установлена следующая зависимость: Q = υ · S, где S -
площадь суммарного сечения данной генерации бронхов. Отсюда
следует вывод, что линейная скорость воздушного потока неодинакова
40
Биофизика
Рис. 5.21. Схема разветвлённой дыхательной трубки
в разных местах разветвлённой дыхательной трубки: в трахее -
наибольшая (порядка 1 м · с-1), в бронхах 16-й генерации - 0,01 м · с-1,
в альвеолярных мешочках - менее 0,0002 м · с-1. Последняя величина
меньше скорости диффузии воздуха. Поэтому считают, что на
последних 7 мкм воздухоносных путей воздух поступает к альвеолам
по законам диффузии, а не конвекции (как на протяжении всех
предыдущих участков разветвлённой дыхательной трубки).
/ пава 5. Биофизика двигательной активности
41
При спокойном дыхании глубина вдоха на 70-80% обеспечи-
мастся сокращением диафрагмы, а на 20-30% - сокращением наруэ/с-
иых межрёберных мышц. Усиление дыхания связано с включением
и работу вспомогательных дыхательных мышц (мышц шеи и плече-
иого пояса, прикрепляющихся к грудной клетке). Они могут способ-
стновать форсированию как вдоха, так и выдоха. В форсированный
иыдох наибольший вклад вносят внутренние межрёберные мышцы
и мышцы живота (брюшной пресс). При спокойном дыхании
сокращение дыхательных мышц обеспечивает только вдох, тогда как
выдох совершается пассивно - за счёт возникновения (при вдохе) силы
упругости как в лёгких, так и в тканях грудной клетки. Мощность
дыхательной мускулатуры при спокойном дыхании составляет
0,05 Вт, а при форсированном дыхании возрастает на порядок. В пер-
ном случае на энергообеспечение дыхательных мышц затрачивается
исего 2%, тогда как во втором - более 20% поглощённого
организмом кислорода.
Последовательность биофизических процессов,
обеспечивающих вентиляцию лёгких, можно представить в виде следующей схемы:
акт вдоха: поступление нервного импульса к дыхательным
мышцам —> синаптическая (нервно-мышечная) передача —>
сокращение дыхательных мышц —» увеличение объёма грудной полости —>
увеличение объёма лёгких —> снижение давления в лёгких (по закону
1>ойля-Мариотта) —> всасывание воздуха из атмосферы в лёгкие;
акт выдоха: расслабление дыхательных мышц (вслед за
сокращением при вдохе) —> уменьшение объёма грудной полости —>
уменьшение объёма лёгких —» повышение давления в лёгких (по
закону Бойля-Мариотта) —> выдавливание воздуха из лёгких в
атмосферу.
Основной вклад в эластические свойства грудной клетки
вносит упругость рёбер, особенно их хрящевых частей, и дыхательных
мышц. Эластическое сопротивление грудной клетки зависит от
степени их растяжения, а она, в свою очередь, повышается с
увеличением объёма лёгких (эта зависимость близка к параболической). При
•дополнении лёгких примерно на 55% их максимального объёма (VtmJ
шастические структуры грудной клетки полностью расслаблены.
42
Биофизика
Увеличение объёма лёгких (относительно 55% Vmax) приводит к
растяжению эластических (упругих) компонентов грудной клетки,
а уменьшение - к их сжатию. Как растяжение, так и сжатие
происходят только при сокращении дыхательных мышц (в первом случае -
мышц вдоха, во втором - мышц форсированного выдоха).
Сила упругости в лёгких, которая заставляет их спадать на
выдохе, называется эластической тягой лёгких - ЭТЛ. Она имеет два
основных компонента. Во-первых, тканям лёгких присущи упругие
свойства (они зависят не только от коэффициента упругости их
компонентов как таковых, но и от степени кровенаполнения лёгких, от
тонуса гладкомышечных волокон и т.д.).
Вторым компонентом ЭТЛ является сила поверхностного
натяжения, возникающая на границе между альвеолярной газовой
смесью и внутренней поверхностью альвеол, выстланной слоем
жидкости. Давление (р), создаваемое поверхностным натяжением,
вычисляют по формуле Лапласа: ρ - —, где г - радиус альвеолы (её
г
форма близка к усечённой сфере), - коэффициент поверхностного
натяжения.
Под действием этого давления газы, находящиеся в альвеоле
и сжимаемые им, стремятся покинуть её и выйти через дыхательные
пути наружу. Средний радиус альвеолы составляет 100-150 мкм, а
на вдохе около 0,05 Η · м~'. Следовательно, давление, обусловленное
поверхностным натяжением, достигает на вдохе 800 Па. Им
обеспечивается от 50 до 70% всей энергии выдоха. Вторая часть (30-50%)
приходится на силу упругости, развивающуюся при деформациях
лёгочной паренхимы, воздухоносных путей, тканей грудной клетки.
Зависимость силы упругости, возникающей при деформациях
легочной паренхимы и стенок воздухоносных путей, от величины
деформации описывается линейными функциями. Упругость
обусловлена преимущественно эластическими волокнами,
растягивающимися в лёгочной паренхиме при дыхании (на вдохе). При этом
коллагеновые волокна, уложенные волнообразно («со слабиной»),
только распрямляются, но не растягиваются. Их миссия состоит
в обеспечении прочности (при перерастяжении лёгочной ткани). Су-
Глава 5. Биофизика двигательной активности
43
хая масса лёгких состоит на 20% из коллагена и на 5-12% из
эластина. Минимальное разрушающее напряжение лёгочной паренхимы
составляет 6 · 103 Па, а предел текучести - 4 · 103 Па. Лёгкие ведут
себя как пластические тела.
Энергия сокращающихся дыхательных мышц затрачивается не
только на преодоление эластического сопротивления грудной клетки
и лёгких. Значительны энерготраты на преодоление сил
сопротивления движению воздуха по дыхательными путям. Они зависят от
характера воздушного потока. При ламинарном движении силы
сопротивления пропорциональны объёму перемещающегося воздуха
в единицу времени, а при турбулентном - почти квадрату этого
объёма. При спокойном дыхании в бронхах преобладает ламинарный
поздушный поток. Когда лёгочная вентиляция усиливается
(например, во время физической работы) или возникает спазм бронхов,
движение воздуха может стать турбулентным. Это ведет к усилению
•шерготрат, связанных с дыханием.
Энергия, затрачиваемая дыхательной мускулатурой на
осуществление спокойного дыхания, ежеминутно составляет 2-3,5 Дж,
причём 70% этой энергии расходуется на преодоление эластического
сопротивления грудной клетки и лёгочной ткани, а остальные 30%
идут на совершение работы по перемещению воздуха против сил
грсния, которые пропорциональны скорости перемещения
воздушной массы. Поэтому второй компонент энерготрат возрастает при
учащении дыхания. Благодаря работе дыхательных мышц
преодолевается так называемый лёгочный резистанс, т.е. сопротивление
воздухоносных путей колебаниям потока воздуха в них. Оно составляет
сравнительно небольшую величину - 0,2 Па · с · мл-1. Сравните её
с аналогичным показателем в гемодинамике. Величина, обратная
лёгочному резистансу, называется растяжимостью (комплансом).
У взрослого человека она составляет 200 мл -(смн 0)_|, а у детей
меньше. Сурфактант увеличивает растяжимость.
Сила сокращений дыхательной мускулатуры при вентиляции
легких направлена на преодоление упругих и вязких сопротивлений.
При очень медленном дыхании вязкие сопротивления весьма
невелики, и поэтому соотношение между объемом и эффективным давлени-
44
Биофизика
ем в дыхательной системе почти целиком определяется упругими
(эластическими) свойствами легких и грудной клетки. Для того
чтобы построить статические кривые объема-давления, необходимо
исключить влияния дыхательной мускулатуры. Лишь при этом условии
можно исследовать действие упругих сил в отдельности. Для этого
либо исследуемый должен научиться на короткий срок полностью
расслаблять дыхательные мышцы, либо следует применять миоре-
лаксанты в сочетании с искусственным дыханием.
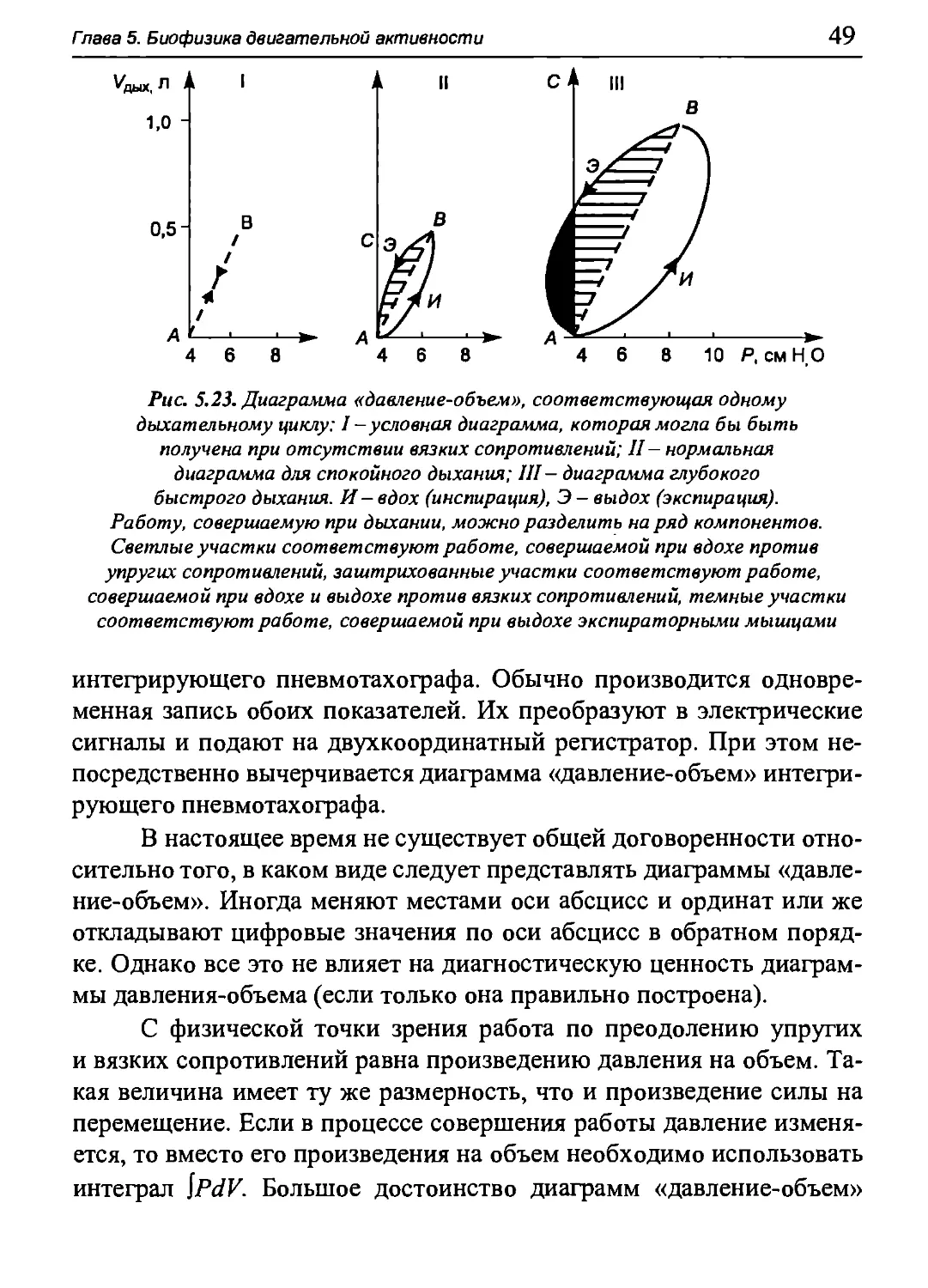
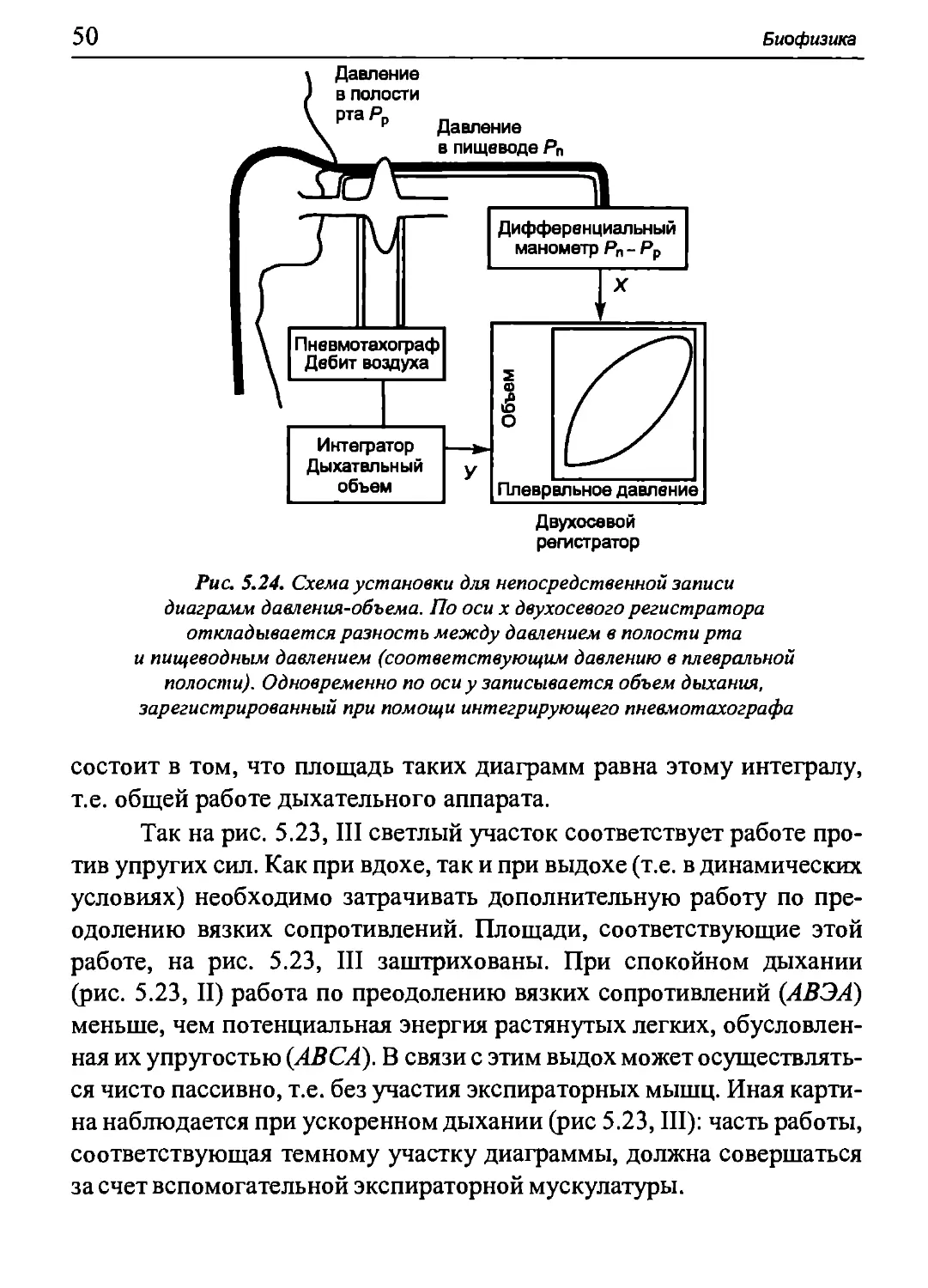
Статические кривые «объем-давление» для дыхательной
системы в целом, т.е. для легких и грудной клетки, можно построить
следующим образом. Пациент вдыхает из спирометра определенный
объем воздуха (при этом носовые пути должны быть перекрыты).
Затем соединение со спирометром перекрывается, и испытуемый как
можно глубже расслабляет дыхательные мышцы при открытой
голосовой щели. При этом посредством датчика, соединенного со ртом
пациента, можно измерить давление, оказываемое на альвеолы со
стороны сил упругости легких и грудной клетки (релаксационный
метод измерения давления). Разность давлений между альвеолярным
и атмосферным воздухом называется альвеолярным давлением Ра.
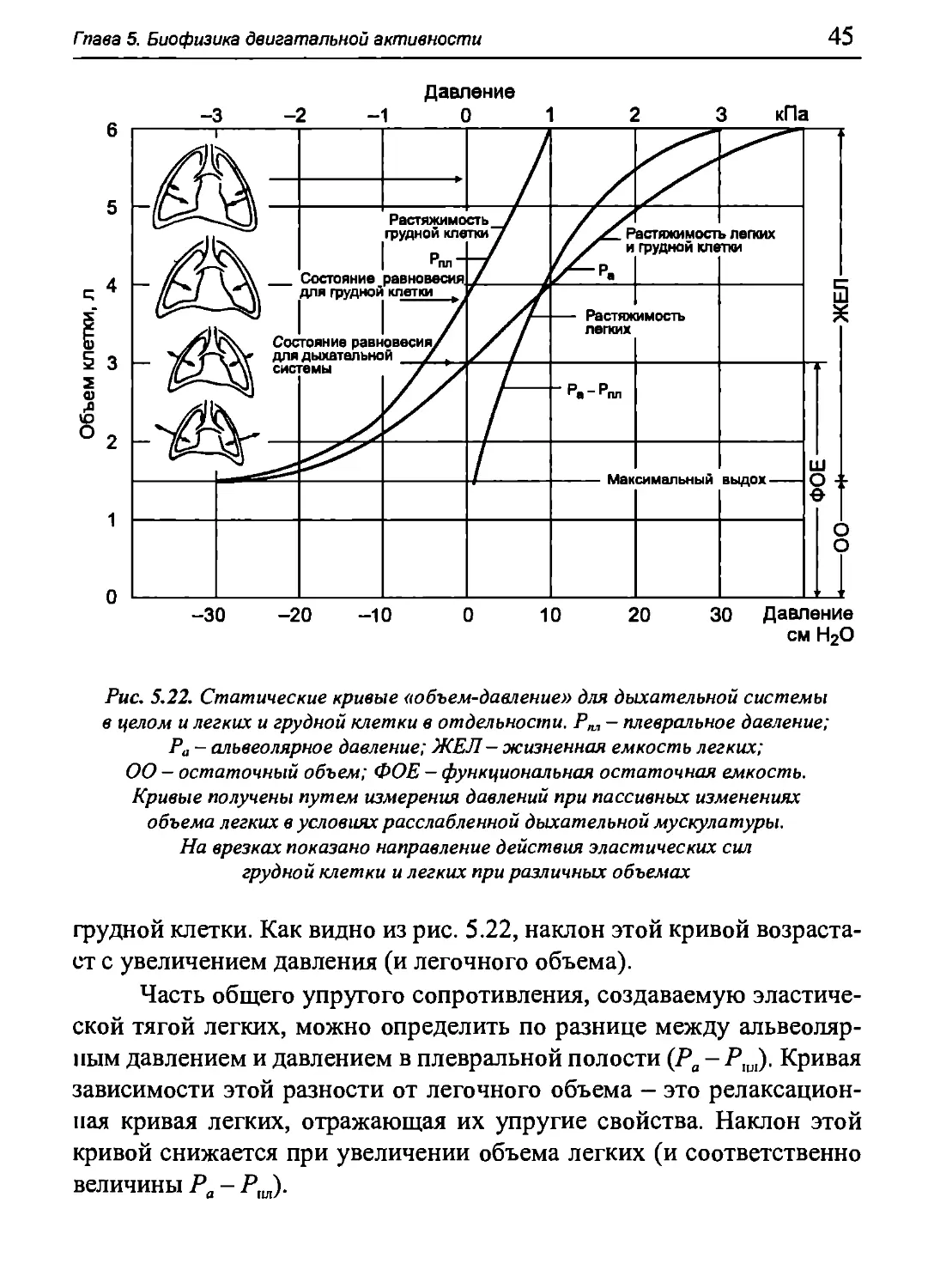
На рис. 5.22 изображена типичная кривая зависимости
альвеолярного давления от объема легких, полученная при помощи
описанного выше метода. Эта релаксационная кривая легких и грудной
клетки имеет ^-образную форму, причем ее участок,
приближающийся к линейному, соответствует большинству значений, в
пределах которых изменяются объем и давление в легких при нормальном
дыхании. Следовательно, упругое сопротивление дыхательной
системы при вдохе в этих пределах почти постоянно.
Часть общего упругого сопротивления, приходящуюся на
долю упругого сопротивления грудной клетки (в том числе опорного
аппарата, мышц и внутренностей брюшной полости), можно
определить по разнице между атмосферным и внутриплевральным
давлением. Как указывалось выше, эту разницу принято сокращенно
называть давлением в плевральной полости Рт. Если при проведении
измерений, о которых шла речь выше, одновременно регистрируется
давление в плевральной полости (или пищеводе) при различных
значениях объема легких, то можно построить релаксационную кривую
Гпава 5. Биофизика двигательной активности
45
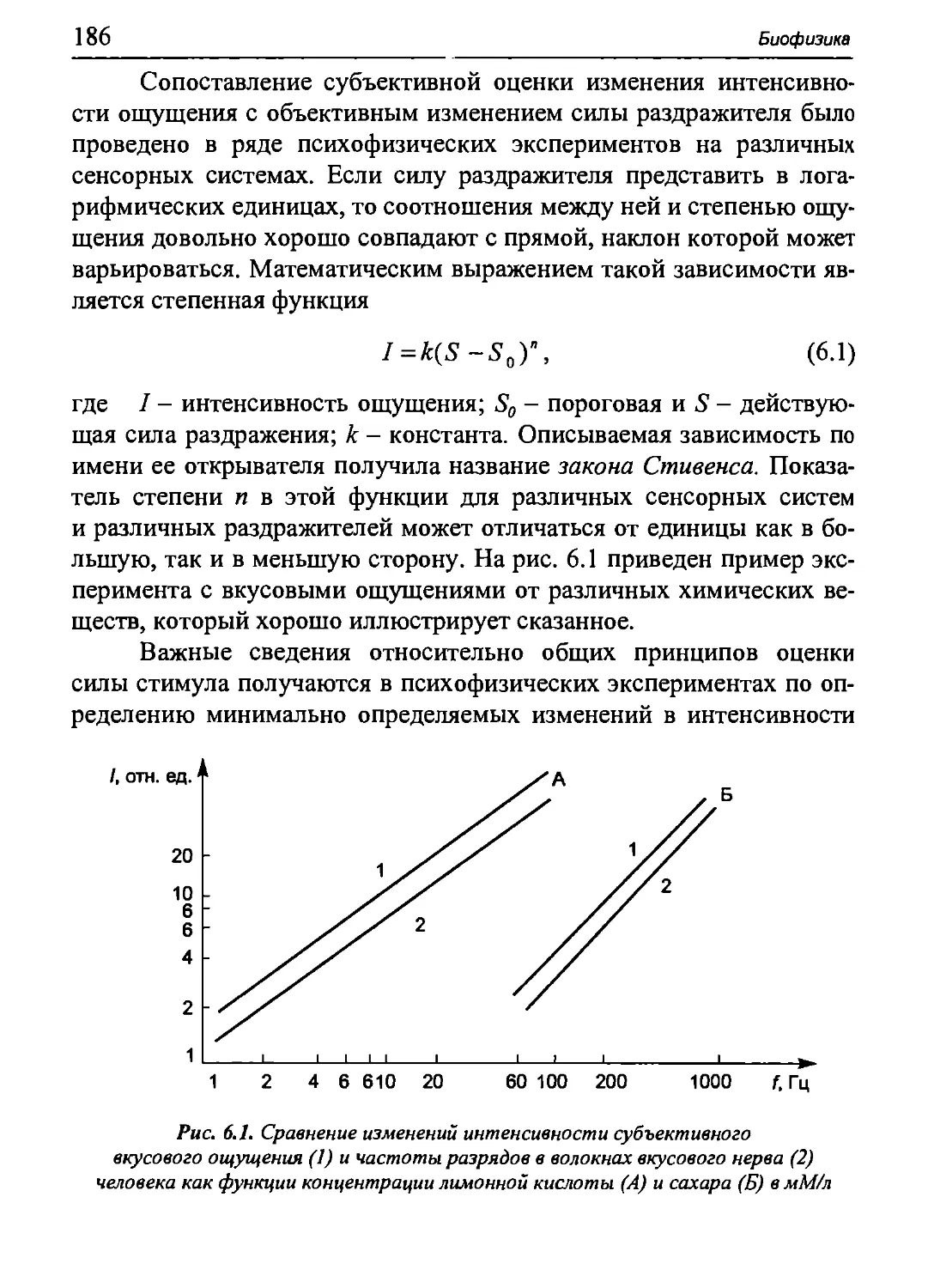
Давление
-3-2-1 0 1 2 3 кПа
-30 -20 -10 0 10 20 30 Давление
смН20
Рис. 5.22. Статические кривые «объем-давление» для дыхательной системы
в целом и легких и грудной клетки в отдельности. РЛ, - плевральное давление;
Ра - альвеолярное давление; ЖЕЛ - жизненная емкость легких;
00 - остаточный объем; ФОБ - функциональная остаточная емкость.
Кривые получены путем измерения давлений при пассивных изменениях
объема легких в условиях расслабленной дыхательной мускулатуры.
На врезках показано направление действия эластических сил
грудной клетки и легких при различных объемах
грудной клетки. Как видно из рис. 5.22, наклон этой кривой
возрастает с увеличением давления (и легочного объема).
Часть общего упругого сопротивления, создаваемую
эластической тягой легких, можно определить по разнице между
альвеолярным давлением и давлением в плевральной полости (Ра-Рш). Кривая
зависимости этой разности от легочного объема - это
релаксационная кривая легких, отражающая их упругие свойства. Наклон этой
кривой снижается при увеличении объема легких (и соответственно
величины Ра - Рт).
46
Биофизика
Сравнение всех трех кривых на рис. 5.22 позволяет оценить
влияние различных упругих сил при разной степени наполнения
легких. Состояние равновесия всех упругих сил для дыхательной
системы в целом (Ра = 0) наблюдается в конце выдоха, когда объем легких
соответствует функциональной остаточной емкости (ФОБ). В этом
случае силы, расправляющие грудную клетку, и эластические силы
легких, под действием которых они стремятся спасть,
уравновешивают друг друга. Когда при вдохе объем легких увеличивается, их
эластическая сила, направленная внутрь, возрастает, а сила упругости
грудной клетки, направленная наружу, снижается. При объеме
легких, равном около 55% жизненной емкости, наблюдается состояние
равновесия для грудной клетки (Р^ = 0). При дальнейшем
увеличении легочного объема направление силы упругости грудной клетки
меняется на противоположное.
Поскольку растяжимость есть величина, обратная упругому
сопротивлению растяжения, это сопротивление для дыхательной
системы в целом равно сумме упругих сопротивлений грудной клетки
(&.«.) и легких (Rn).
Как видно из рис. 5.22, наклон релаксационной кривой (а,
следовательно, и растяжимость) дыхательной системы (грудной клетки
и легких) больше в области значений, соответствующих спокойному
дыханию. У здорового взрослого человека растяжимость
дыхательной системы и ее компонентов при спокойном дыхании составляет:
С = 0,1 л/см вод. ст. = 1 л/кПа, Сгк = 0,2 л/см вод. ст. = 2 л/кПа, Сл =
= 0,2 л/см вод. ст. = 2 л/кПа, где С - растяжимость всего аппарата
внешнего дыхания, С^ - растяжимость грудной клетки, Сл -
растяжимость легких. Эти величины связаны соотношением:
С С С
Любое изменение этих величин, а особенно их снижение при
патологии, имеет диагностическое значение. Однако точно измерить
растяжимость трудно, так как при ее определении дыхательная
мускулатура должна быть полностью расслаблена. Поэтому часто
ограничиваются измерением растяжимости легких, используя более про-
Глава 5. Биофизика двигательной активности
47
стой способ, который заключается в том, что исследуемый вдыхает
определенный объем воздуха, а затем при помощи дыхательных
мышц фиксирует грудную клетку при открытой голосовой щели.
В этих условиях давление в альвеолах равно атмосферному (т.е. Ра -
- 0)и тогда
Сл=--^-. (5.29)
Таким образом, для определения растяжимости легких
достаточно измерить изменение внутриплеврального давления (или,
проще, внугрипищеводного давления) и подставить полученные цифры
в это уравнение. При этом вычисляют так называемую статическую
растяжимость легких, зависящую не только от их эластических
свойств, но также от их объема. При прочих равных условиях
изменение объема меньше при меньшем исходном объеме. У детей в
возрасте 9-12 лет растяжимость в 2-3 раза ниже, чем у взрослых. В
связи с этим при диагностическом определении растяжимости легких
необходимо учитывать их исходный объем, т.е., как правило,
функциональную остаточную емкость (ФОБ). Для этого используют так
называемую специфическую растяжимость легких:
1 AV
СЛ(сп) = —■ (5-3°)
Л(СП) ФОЕ ДРШ
Если бы при вдохе приходилось преодолевать только упругое
сопротивление, то изменения объема легких в любых пределах были
бы примерно пропорциональны изменениям внутриплеврального
давления; на диаграмме «давление-объем» подобные
взаимоотношения выглядят в виде восходящей прямой (рис. 5.23, I). При выдохе
значения отношений объема к давлению лежат на той же прямой, но
ц эту фазу дыхательного цикла она уже нисходящая.
Однако в связи с влиянием вязкого сопротивления кривая
«давление-объем» при вдохе становится выпуклой кверху (рис. 5.23, II).
Это означает, что при вдыхании одного и того же объема воздуха
виутриплевралыюе давление при наличии вязкого сопротивления
48
Биофизика
должно быть более отрицательным, чем в случае, если бы объем был
прямо пропорционален давлению. Только после того, как вдох
завершается (точка В), кривая вдоха совпадает с прямой АВ, ибо в этот
момент дыхательные движения отсутствуют и действуют только силы
упругости. Кривая «давление-объем» для выдоха под влиянием
вязких сопротивлений образует выпуклость книзу, возвращаясь к
исходной точке (точка А) лишь в конце выдоха. Таким образом,
динамическая диаграмма «давление-объем» имеет форму петли.
На рис. 5.23, II изображена диаграмма для спокойного
дыхания. При более глубоком и быстром дыхании форма петли этой
диаграммы несколько изменяется (рис. 5.23, III), поскольку дыхание
становится глубже и осуществляется быстрее (наклон инспираторной
и экспираторной кривых круче). Увеличение выпуклости этих
кривых обусловлено тем, что при быстрых изменениях внутриальвео-
лярного давления объемная скорость воздушного потока возрастает
недостаточно быстро. Таким образом, при большой скорости
дыхательных движений вязкое сопротивление воздухоносных путей
играет большую роль, чем при спокойном дыхании.
У человека со здоровыми легкими диаграмму
«давление-объем» можно использовать для вычисления их растяжимости.
Поскольку в поворотных точках А и В дыхательные движения отсутствуют,
параметры этих точек отражают действие статических упругих сил
легких. Следовательно, наклон прямой, соединяющей точки А и В,
т.е. AV/AP, соответствует растяжимости легких Сл. В случае,
представленном на рис. 5.23, Сл = 0,2 л/см вод. ст. Однако при
заболеваниях легких этот метод определения растяжимости не применяется,
так как из-за увеличенного сопротивления не все альвеолы успевают
равномерно заполниться воздухом за время вдоха. При этом в точках
А и В статическое состояние не достигается. В связи с этим у лиц
с легочной патологией растяжимость легких можно определить
только статическим методом, требующим задержки дыхания.
На рис. 5.24 показана схема установки для одновременного
измерения давления и объема. По оси абсцисс отложено давление
в плевральной полости, или в данном случае разница между
атмосферным и внутрипищеводным давлением, по оси ординат -
дыхательные объемы, измеряемые при помощи либо спирометра, либо
Глава 5. Биофизика двигательной активности
49
4 6 8 10 Р, см Η О
Рис 5.23. Диаграмма «давление-объем», соответствующая одному
дыхательному циклу: I -условная диаграмма, которая могла бы быть
получена при отсутствии вязких сопротивлений; II— нормальная
диаграмма для спокойного дыхания; III — диаграмма глубокого
быстрого дыхания. И—вдох (инспирация), Э - выдох (экспирация).
Работу, совершаемую при дыхании, можно разделить на ряд компонентов.
Светлые участки соответствуют работе, совершаемой при вдохе против
упругих сопротивлений, заштрихованные участки соответствуют работе,
совершаемой при вдохе и выдохе против вязких сопротивлений, темные участки
соответствуют работе, совершаемой при выдохе экспираторными мышцами
интегрирующего пневмотахографа. Обычно производится
одновременная запись обоих показателей. Их преобразуют в электрические
сигналы и подают на двухкоординатный регистратор. При этом
непосредственно вычерчивается диаграмма «давление-объем»
интегрирующего пневмотахографа.
В настоящее время не существует общей договоренности
относительно того, в каком виде следует представлять диаграммы
«давление-объем». Иногда меняют местами оси абсцисс и ординат или же
откладывают цифровые значения по оси абсцисс в обратном
порядке. Однако все это не влияет на диагностическую ценность
диаграммы давления-объема (если только она правильно построена).
С физической точки зрения работа по преодолению упругих
и вязких сопротивлений равна произведению давления на объем.
Такая величина имеет ту же размерность, что и произведение силы на
перемещение. Если в процессе совершения работы давление
изменяется, то вместо его произведения на объем необходимо использовать
интеграл \PdV. Большое достоинство диаграмм «давление-объем»
50
Биофизика
Давление
в полости
ртаРр
Давление
в пищеводе Рп
Ά
Дифференциальный
манометр Р„ - Рр
Интефатор
Дыхательный
объем
—*>
У
V
Плевральное давление
Двухосввой
регистратор
Рис. 5.24. Схема установки для непосредственной записи
диаграмм давления-объема. По оси χ двухосевого регистратора
откладывается разность между давлением в полости рта
и пищеводным давлением (соответствующим давлению в плевральной
полости). Одновременно по оси у записывается объем дыхания,
зарегистрированный при помощи интегрирующего пневмотахографа
состоит в том, что площадь таких диаграмм равна этому интегралу,
т.е. общей работе дыхательного аппарата.
Так на рис. 5.23, III светлый участок соответствует работе
против упругих сил. Как при вдохе, так и при выдохе (т.е. в динамических
условиях) необходимо затрачивать дополнительную работу по
преодолению вязких сопротивлений. Площади, соответствующие этой
работе, на рис. 5.23, III заштрихованы. При спокойном дыхании
(рис. 5.23, II) работа по преодолению вязких сопротивлений (АВЭА)
меньше, чем потенциальная энергия растянутых легких,
обусловленная их упругостью (АВСА). В связи с этим выдох может
осуществляться чисто пассивно, т.е. без участия экспираторных мышц. Иная
картина наблюдается при ускоренном дыхании (рис 5.23, III): часть работы,
соответствующая темному участку диаграммы, должна совершаться
за счет вспомогательной экспираторной мускулатуры.
Глава 5. Биофизика двигательной активности
51
При спокойном дыхании на работу дыхательных мышц
затрачивается примерно 2% поглощенного кислорода. Однако при
физической нагрузке энергетические потребности дыхательных мышц
возрастают в большей степени, чем минутный объем дыхания и
поглощение 02. В связи с этим при тяжелой физической работе на
деятельность дыхательной мускулатуры затрачивается до 20% общего
потребления кислорода.
5.3.2. Показатели легочной вентиляции
Энерготраты на преодоление эластического сопротивления
органов дыхания почти не зависят от скорости движения воздуха,
а значит, и от частоты дыхания. Они определяются объёмом
воздуха, поступающего в лёгкие при вдохе, т.е. глубиной дыхания.
Неодинаковая зависимость каждого из компонентов работы дыхательного
аппарата от частоты и глубины дыхания приводит к установлению
оптимального соотношения между ними, при котором работа
поддерживается на минимальном уровне при данном объёме
вентиляции. В условиях физического покоя глубина дыхания (дыхательный
объём) составляет 0,5 л, а частота - 12-16 мин-1 (у детей - 20-
30 мин-1, а у новорождённых - 40-50 мин-1). Произведение
глубины дыхания (дыхательного объёма) на его частоту (в расчете на 1
мин) определяет минутный объём дыхания (МОД). При спокойном
дыхании он составляет 6-8 л · мин-1, а при тяжёлой физической
нагрузке увеличивается более чем до 100 л · мин-1 благодаря
углублению и учащению дыхания. Между длительностями вдоха и выдоха
существует золотая пропорция (ίωοχ „ выдох: ?выдох = *выдох: /вдох =
1,618).
Глубина и частота дыхания широко варьируют в зависимости
от потребностей организма. В покое дыхательный объем мал по
сравнению с общим объемом воздуха в легких. Таким образом,
человек может как вдохнуть, так и выдохнуть большой дополнительный
объем. Однако даже при самом глубоком выдохе в альвеолах и
воздухоносных путях легких остается некоторое количество воздуха.
Для того чтобы количественно описать все эти взаимоотношения,
общую емкость легких разделили на несколько компонентов (объе-
52
Биофизика
мов и емкостей). При этом под термином емкость понимают
совокупность двух и более объемов.
Дыхательный объем - объем воздуха, который человек
вдыхает и выдыхает при спокойном дыхании.
Резервный объем вдоха - объем воздуха, который человек
может дополнительно вдохнуть после нормального вдоха.
Резервный объем выдоха - объем воздуха, который человек
может дополнительно выдохнуть после спокойного выдоха.
Остаточный объем - объем воздуха, остающийся в легких
после максимального выдоха.
Жизненная емкость легких - объем воздуха, который можно
выдохнуть после максимального вдоха (равна сумме первых трех).
Функциональная остаточная емкость - объем воздуха,
остающийся в легких после спокойного выдоха (равна сумме резервного
объема выдоха и остаточного объема).
Общая емкость легких - объем воздуха, содержащийся в
легких на' высоте максимального вдоха (равна сумме остаточного
объема и жизненной емкости легких).
Из параметров легочной вентиляции наибольшее практическое
значение имеют дыхательный объем, жизненная емкость легких
и функциональная остаточная емкость. Жизненная емкость легких
(ЖЕЛ) является показателем подвижности легких и грудной клетки.
Несмотря на ее название, она не имеет жизненно важного значения,
так как даже при самых высоких потребностях, предъявляемых
организмом к дыхательной системе, глубина дыхания никогда не
достигает максимально возможного значения.
С практической точки зрения нецелесообразно устанавливать
«единую» норму для ЖЕЛ, так как эта величина зависит от ряда
факторов, и в частности от возраста, пола, размеров и положения тела
и степени тренированности.
Как видно из рис. 5.25, жизненная емкость легких с возрастом
(особенно после 40 лет) уменьшается. Это связано со снижением
эластичности легких и подвижности грудной клетки. У женщин ЖЕЛ
в среднем на 25% меньше, чем у мужчин. Совершенно очевидно, что
ЖЕЛ зависит от роста, так как величина грудной клетки
пропорциональна остальным размерам тела. У молодых людей ЖЕЛ можно
Глава 5. Биофизика двигательной активности
53
вычислить, исходя из следующего эмпирического уравнения:
ЖЕЛ (л) = 2,5 · рост (м).
Таким образом, у мужчины ростом 180 см жизненная емкость
легких должна составлять примерно 4,5 л. ЖЕЛ зависит от
положения тела: в вертикальном положении она несколько больше, чем
в горизонтальном (это связано с тем, что в вертикальном положении
в легких содержится меньше крови). Наконец, жизненная емкость
легких зависит от степени тренированности: у лиц, занимающихся
такими видами спорта, где необходима выносливость, ЖЕЛ
значительно выше, чем у нетренированных людей. Она особенно велика
у пловцов и гребцов (до 8 л), так как у этих спортсменов сильно
развиты вспомогательные дыхательные мышцы (большие и малые
грудные). Определение жизненной емкости легких имеет значение
главным образом для диагностики.
Функциональная остаточная емкость. Физиологическое
значение функциональной остаточной емкости (ФОЕ) состоит в том,
что благодаря наличию этой емкости в альвеолярном воздухе
выравниваются колебания содержания 02 и С02, связанные с разной
концентрацией этих газов во вдыхаемом и выдыхаемом воздухе.
Если бы атмосферный воздух поступал непосредственно в
альвеолы, не смешиваясь с воздухом, уже содержащимся в легких, то
содержание 02 и С02 в альвеолах претерпевало бы колебания в
соответствии с фазами дыхательного цикла. Однако этого не происхо-
0 20 40 60 80 годы
Рис. 5.25. Кривые зависимости общей и жизненной емкости легких
и остаточного объема от возраста для людей среднего роста
54
Биофизика
дит - вдыхаемый воздух смешивается с воздухом, содержащимся
в легких, и, поскольку ФОЕ в несколько раз больше дыхательного
объема, изменения состава альвеолярной газовой смеси
относительно невелики.
ФОЕ равна сумме остаточного объема и резервного объема
выдоха, и величина ее зависит от ряда факторов. В среднем у молодых
людей она составляет 2,4 л, а в более пожилом возрасте - 3,4 л.
У женщин ФОЕ примерно на 25% меньше, чем у мужчин.
Анатомическим мертвым пространством называют объем
воздухоносных путей - трахеи, бронхов и бронхиол вплоть до их
перехода в альвеолы. Это пространство является «мертвым» лишь
в том смысле, что в нем не происходит газообмена. Однако
структуры, образующие мертвое пространство, выполняют важные
вспомогательные функции - они обеспечивают вентиляцию легких и
очищение, увлажнение и согревание вдыхаемого воздуха.
. Вдыхаемый воздух частично очищается уже при прохождении
через носовую полость, слизистая оболочка которой улавливает
мелкие частицы, пыль и бактерии. В связи с этим лица, постоянно
дышащие через рот, наиболее подвержены воспалительным заболеваниям
дыхательных путей. Важную роль в очищении воздуха играет
эпителий воздухоносных путей. В результате ритмических движений его
ресничек слизь и погруженные в нее мелкие частички удаляются по
направлению к глотке. Более крупные частицы, попавшие в
воздухоносные пути, раздражают их и вызывают кашель, при этом они
выбрасываются со струей воздуха. Выдыхаемый воздух на 70%
насыщен водяными парами. При вдохе воздух увлажняется уже в верхних
дыхательных путях, в значительной степени насыщаясь водяными
парами в носу и гортани. В нижних дыхательных путях насыщение
водяными парами достигает 100%.
Одна из функций дыхательных путей состоит в согревании
вдыхаемого воздуха. Длительность вдоха достаточно велика для
того, чтобы к моменту поступления в легкие воздух нагрелся почти
до температуры тела. В связи с этим температура выдыхаемого
воздуха обычно составляет 37° С.
Измерение объема мертвого пространства. Экспираторный
дыхательный объем (V3) состоит из двух компонентов: объема воз-
Глава 5. Биофизика двигательной активности
55
духа, поступающего из мертвого пространства (VM), и объема воздуха
из альвеолярного пространства (KJ:
V,= VM±Va. (5.31)
Для изучения функции легких важно измерить оба этих
компонента отдельно. При этом, как и для определения функциональной
остаточной емкости, используют непрямые методы. Они основаны
на том, что содержание дыхательных газов (02 и С02) в воздухе из
мертвого и альвеолярного пространства различно. Содержание газов
в воздухе мертвого пространства аналогично содержанию в воздухе,
поступившем при вдохе (инспирации) (FJ. Содержание же газов
в воздухе из альвеолярного пространства такое же, как и в самой
альвеолярной газовой смеси (FJ. Если выразить парциальный объем
газа в виде произведения общего объема газовой смеси V и
содержания этого газа F, то для любого дыхательного газа будет справедливо
равенство:
V^=VMFn+V^ (5.32)
где К, - экспираторный объем; VM - объем мертвого пространства;
Ка - выдыхаемый объем воздуха из альвеолярного пространства; F, -
содержание газа в выдыхаемом воздухе; Fa - содержание газа в
альвеолярном воздухе, Fu - то же при выдохе.
В соответствии с выражением (5.31) УЛ можно представить, как
к-к-
V3-F3=VMFH±(V3-VM)Fa, (5.33)
или после преобразования
V F -F
-ϊ- = -2 s.. (5.34)
V V -V
з и а
Это равенство, называемое уравнением Бора, справедливо для
любого дыхательного газа. Однако для С02 его можно упростить, так
как содержание этого газа во вдыхаемом воздухе FuC0 близко к 0:
56
Биофизика
V м _ "СО; эСОг (535)
V F
' э * аС02
Отношение объема мертвого пространства к экспираторному
объему можно вычислить, исходя из уравнения (5.34) или (5.35).
Значения объемных содержаний газов, представленные в правой части
уравнения, можно определить путем газового анализа. Пусть
газовый анализ дал следующие числа: FaCO = 0,056; F3CO = 0,04 мл С02
на 1 мл смеси. Отсюда VJV3 = 0,3, т.е. объем мертвого пространства
составляет 30% от экспираторного объема.
Минутный объем дыхания, т.е. объем воздуха, вдыхаемый (или
выдыхаемый) за 1 мин, равен по определению произведению
дыхательного объема на частоту дыхательных движений. Экспираторный
объем обычно несколько меньше ииспираторного, так как
поглощение С02 больше, чем выделение С02 (дыхательный коэффициент < 1).
Таким образом, для большей точности следует различать инспиратор-
ный и экспираторный минутный объем дыхания. При расчетах
вентиляции принято исходить из экспираторных объемов, помечаемых «э».
Экспираторный минутный объем дыхания V3 составляет
V,=V3f, (5.36)
где точка над символом означает, что речь идет об этой величине
за единицу времени; Уэ - экспираторный дыхательный объем;/-
частота дыхательных движений.
Как уже говорилось, частота дыхательных движений у
взрослого человека в покое в среднем равна 14 в 1 мин. Эта величина
может претерпевать значительные колебания (от 10 до 18 в 1 мин).
У детей она достигает 20-30 в 1 мин; у грудных - 30-40 в 1 мин,
а у новорожденных - 40-50 в 1 мин.
У взрослого человека при дыхательном объеме 0,5 л и частоте
дыхания 14 в 1 мин минутный объем дыхания равен 7 л. При
физической нагрузке в соответствии с увеличением потребности в кислороде
повышается и минутный объем дыхания, достигая в условиях
максимальной нагрузки 120 л в 1 мин. Основные показатели дыхания у
здорового молодого человека и их отклонения представлены в таблице 5.1.
Глава 5. Биофизика двигательной активности
57
Таблица 5.1
Показатели дыхания у здорового молодого человека
(площадь поверхности тела 1,7 м!) в покое. Отклонения от этих
показателей и факторы, влияющие на них, обсуждаются в тексте
Легочные объемы, л.
Общая емкость
Жизненная емкость
Функциональная остаточная емкость
Остаточный объем
Дыхательный объем
6,0
4,5
2,4
1,2
0,5
Параметры вентиляции
Частота дыхания
Минутный объем дыхания
Альвеолярная вентиляция
14 мин-1
7 л/мин
5 л/мин
Параметры механики дыхания
Внутриплевральное давление
В конце выдоха
В конце вдоха
Растяжимость легких
Растяжимость легких и грудной клетки
Сопротивление дыханию
-4 см вод. ст (-0,4 кПа)
-6,5 см вод. ст (-0,65 кПа)
0,2 л на 1 см вод.
0,1 л на 1 см вод.
ст. (2 л/ кПа)
ст. (1 л/ кПа)
0,15 см вод. ст. · с-1 · л-1 (0,15 кПа · с
-1-л-1)
5.3.3. Моделирование дыхания
Исследование закономерностей распределения Р0 в тканях
с помощью математических моделей были начаты еще в 1915 г. Эр-
лапгом и Крогом. Модель тканевого цилиндра, предложенная Крогом
и 1936 г., используется и в настоящее время. В результате
дальнейшего развития в этом направлении модели диффузионных процессов
существенно усложнены и приближены к реальным условиям
газообмена и кровообращения различных тканей организма. Математическую
модель распределения кислорода и углекислоты в головном мозге
разработал Ю.Я. Кисляков в содружестве с К.П. Ивановым.
При построении модели приняты следующие основные
допущения: а) распределение кислорода рассматривается в пространстве
ячейки, включающей капиллярную сеть, нервную клетку и
окружающую ее ткань; б) в капилляре перенос кислорода в осевом направле-
58
Биофизика
нии с током крови значительно превышает его диффузию; в)
зависимость между степенью насыщения крови кислородом и уровнем его
напряжения описывается кривой диссоциации оксигемоглобина; г)
распределение кислорода в радиальном сечении капилляра
равномерно; д) транспорт кислорода из капилляра в ткань осуществляется
за счет диффузии по его концентрационному градиенту, который
применительно к газам называют градиентом напряжений. В
модели учтено различие потребления кислорода нервными клетками и
окружающей тканью.
Распределение напряжения (U) кислорода в ткани мозга в
стационарных условиях описывается уравнением диффузии:
AU(x,y,z)=r*X'y>z)a, (5.37)
Do2 ·°4
где . т - потребление кислорода; DQ - коэффициент диффузии;
сс0 - растворимость кислорода в ткани мозга; х, у, ζ -
пространственные координаты; Δ - оператор Лапласа; Р0 - напряжение
кислорода в крови.
Распределение Р0 в каждом капилляре Рк0 ячейки,
содержащей N капилляров с суммарным потреблением кислорода, равным
М, описывается следующими уравнениями:
Ркп = In
xJHboi
(5.38)
I loo )
η =а,(НЬОг)2 +аг{НЬОг) + аъ, (5.39)
fl,= VW +сп '' = 1,2,3, (5.40)
НЬО,=— 100, (5.41)
2 КЕ
dC _С\_Р«>№.М\
dl U
(5.42)
Глава 5. Биофизика двигательной активности
59
vUr
АС= VUNr ^ . (5.43)
(νπΓ2)£ί7λΓλ
Здесь С - содержание кислорода в единице объема; АС - арте-
риально-венозная разность содержания кислорода; г - радиус
капилляра; ν - линейная скорость движения крови; КЕ - кислородная
емкость крови; Ъ„ с, - эмпирические коэффициенты аппроксимации
кривых диссоциации оксигемоглобина; U - суммарный градиент Р0
между капилляром и окружающей тканью в плоскости,
перпендикулярной оси капилляра:
U =)(P^(t)-U]{tj)dt, (5.44)
о
где L - длина капилляра; / - расстояние по оси капилляров от
артериального конца; U] - среднее значение Р0 в точках нервной ткани,
нблизи поверхности капилляра.
Граничные условия налагаются в предположении равенства
нулю градиентов PQ на границах ячейки s по направлению к нормали:
?р|,=а (5.45)
дп
Расчет распределения Р0 проведен на ПЭВМ для модели
ячейки капиллярной сети (рис. 5.26). С помощью этой модели можно
определить особенности, вносимые нервной клеткой в распределение
Р0 , исследовать влияние направления кровотока в капиллярах на
распределение Р0 и условия функционирования клеток различной
нсличины. При моделировании использованы экспериментальные
данные о расположении капилляров в направлении кровотока,
полученные с помощью методики прижизненного микроскопирования.
Исследования с помощью математической модели транспорта
кислорода в ткани скелетной мышцы (рис. 5.27) выполнены Е.Г. Ля-
бах. Уравнения транспорта кислорода имеют вид:
60
Биофизика
I^AO,
ΫΪΣΛ
Рао2
Рис 5.26. Модель ячейки капиллярной сети с шаровидной нервной клеткой
(трубками показаны капилляры, штрихами - окружающая ткань)
Ρ Ρ
*Λ<ννο2
- напряжения кислорода в артериальной и венозной крови;
АА, ВВ - вспомогательные линии
Щ7
В А
Рис. 5.27. Модель кислородного снабжения скелетной мышцы
А, В— артериальные и венозные концы капилляров
Глава 5. Биофизика двигательной активности
61
Ц+Ц+Щ./^ПР% (5.46)
дх2 ду2 dz2 OLD
mAVD(dPi ВР{
dCi _ viri у дх ду
ι-Λ v,r, Цдх ду )
(5.47)
дР
ψ=0, (5.48)
дп
С,(0) = Са(или C,{L) = СД (5.49)
у
ADV=-^IIJf(P)dxdydz, (5.50)
*£ χ у ζ
где Q - кровоток; Ρ - напряжение кислорода в крови Р0 ; С, -
содержание кислорода; v;, г, - линейная скорость крови и радиус
поперечного сечения i-ro капилляра из общего числа т капилляров; η -
нормаль к поверхности; f(F) — фактор подавления потребления
кислорода при гипоксии (0 <f(P) < 1).
Зависимость Р0 в крови от содержания 02 описывается
выражением
Р=(-ьд-Лс+цррв»
0,039
Множество численных значений N решений системы
уравнений на ПЭВМ можно анализировать по кривым частотного
распределения (гистограммам) Р0 - «р,(Р), которые рассчитываются по
формуле:
<Pj(P) = Nj(PjAP) ■ 100/NAP,
(5.52)
62
Биофизика
φ, % J
4
О
12
^k
aj
L
10 25 40 55
9
Po2, мм.рт.ст.
Рис. 5.2А Гистограммы Р0 в скелетной мышцы при росте линейной
. скорости крови (Αι, Αί,) и росте плотности активных капилляров (Б/, £^
в условиях покоя (а) и при работе скелетной мышцы (б)
где N - общее число расчетных точек; Л^ - число точек, для
которых Pj< Р< Р, ± АР, АР = 2 мм рт. ст., 0 <j < 100. Примеры
гистограмм, полученных на модели, приведены на рис. 5.28.
С помощью модели можно исследовать влияние
межкапиллярного расстояния на транспорт кислорода. Установлено, что
уменьшение межкапиллярного расстояния обеспечивает высокую
насыщенность ткани кислородом в покое, способность к интенсивной
экстракции кислорода из крови, защищенность от гипоксии при флук-
туациях кровотока. Показана также высокая эффективность роста
линейной скорости кровотока в регуляции снабжения ткани кислородом.
Системная регуляция внешнего и тканевого дыхания. Начиная
с работы Грея, разработано большое число математических моделей
регуляции внешнего дыхания, массопереноса газов в легких и тканях
и комплексных моделей, объединяющих внешнее дыхание и
тканевый метаболизм и описывающих систему транспорта кислорода
и кислородный режим организма.
А.З. Колчинская, А.А. Миссюра, Ю.Н. Онопчук
разработали модель массопереноса газов в организме, на базе которой
Глава 5. Биофизика двигательной активности
63
з з
* з
ε й-
§-,-
ST"1
δ "
^■~.
'3 ~"
3 ^
ϊ
ί-
Ο
Si
ч
I
я
Ji
il
a _
Si. a,
α S 3
§ a s
» 13 u
g 3 ε
α Η !
S 3 "
Ό ...
I,
Si
о
©\
«N
О
a
|4
3
i § s-
$ s =
5 3«
a I
■*■ δ
3" ε
- § 3
з- &· ν
So1»
1^ \
0 Ρ -
§ 3
в" Ε Ι
ι
Ι
1 3-§
* ii
ч1§
II
64
Биофизика
Ю.Н. Онопчук предложил модель регуляции дыхания и
кровообращения (рис. 5.29), основанная на принципах оптимального
управления доставкой кислорода. Модель описывает следующие основные
явления: газообмен между легкими и внешней средой за счет вдоха
и выдоха, газообмен между альвеолярным пространством и кровью
легочных капилляров, транспорт газов кровью, газообмен между
кровью и тканями через капиллярную мембрану. Управляемая
система включает легочный резервуар, кровь легочных и тканевых
капилляров, артериальную и смешанную венозную кровь, тканевые
резервуары. Выделено семь групп тканей: мозг, сердце, печень и
желудочно-кишечный тракт, почки, скелетные мышцы, кожа, жировая
и костная ткани. В капиллярной сети выделены собственно
капилляры и их артериальные и венозные концы.
Функциональным назначением синтезирующего устройства
является анализ состояний агрегатов, выбор режимов
функционирования, для них в зависимости от программы достижения заданной
цели, определяемой программным устройством. Управляющие
процессы в модели были организованы на принципе оптимального
управления доставкой кислорода в соответствии с запросами
организма. Модель реализована на ПЭВМ. С помощью этой модели
проводится анализ динамики кровообращения и кислородного режима
организма при имитации физической нагрузки, изменений состава
дыхательной смеси и др.
5.3.4. Методы и технические средства
для исследования легочной вентиляции
Объем вдыхаемого и выдыхаемого воздуха можно
непосредственно измерить при помощи спирометра или пневмотахометра. Что
же касается остаточного объема и функциональной остаточной
емкости, то их можно определить лишь косвенными методами.
Спирометрия. Спирометрами называются приборы,
измеряющие количество воздуха при постоянном давлении (рис. 5.30).
Измерение осуществляется за счет того, что камера изготавливается из
эластичного материала с гофрированным покрытием (сильфоны),
либо применяются камеры, состоящие из двух частей: внешней - не-
Глава 5. Биофизика двигательной активности
65
После
разведения
Рис 5.30. Схема определения функциональной
остаточной емкости по методу разведения гелия.
Вверху - аппаратура и дыхательная система исследуемого
в исходном состоянии; гелий (точки) находится только в спирометре,
где содержание его составляет 10 об.%. Внизу — полное и равномерное
распределение гелия между легкими (функциональная остаточная
подвижной и внутренней - подвижной. Для фиксации объема между
фазами входа и выхода и разделения фаз дыхания применяются
клапаны. В спирометрах с подвижной частью роль клапана выполняет
иода, налитая во внешний корпус. Поэтому такие спирометры назы-
иаются водяными или жидкостными.
Спирометры с эластичной камерой называются сухими. Управ-
иоиие газовыми потоками, проходящими через сильфоны,
производится механическими клапанами различной конструкции.
Наиболее распространен водный спирометр. Этот прибор пред-
ι ганляет собой цилиндр, помещенный дном кверху в резервуар
ι ΊΝ44
66
Биофизика
с водой. Воздух, попавший в этот цилиндр, не сообщается с внешней
средой. Сам цилиндр уравновешивается противовесом.
Воздухоносные пути испытуемого соединяются посредствомширокойтрубкисза-
губником с пространством внутри цилиндра. Во время выдоха объем
воздуха в цилиндре увеличивается, и он всплывает, а при вдохе
цилиндр погружается. Эти перемещения могут быть измерены при
помощи калиброванной шкалы или зарегистрированы посредством
самописца на барабане кимографа (в последнем случае получают так
называемую спирограмму). Премещение будет пропорционально
объему воздуха при выдохе.
В сухом спирометре в зависимости от объема выдыхаемого
воздуха изменяется длина сильфона.
Измерение функциональной остаточной емкости (ФОЕ).
Поскольку ФОЕ - это количество воздуха, остающееся в легких в конце
выдоха, ее можно измерить только непрямыми методами. Принцип
таких методов заключается в том, что в легкие либо вводят
инородный газ типа гелия (метод разведения), либо вымывают
содержащийся в альвеолярном воздухе азот, заставляя испытуемого дышать
чистым кислородом (метод вымывания). И в том, и в другом случае
искомый объем вычисляют, исходя из конечной концентрации газа.
На рис. 5.30 проиллюстрирован метод разведения гелия.
Спирометр закрытого типа заполняется газовой смесью. Пусть общий
объем смеси равен 3 л, а объемы 02 и Не - 2,7 и 0,3 л соответственно.
При этом исходное содержание гелия FHc составит 0,1 мл Не на 1 мл
смеси. После спокойного выдоха испытуемый начинает дышать из
спирометра, и в результате молекулы гелия равномерно
распределяются между объемом легких, равным ФОЕ, и объемом спирометра
VQU. Гелий очень медленно диффундирует через ткани, и для
практических целей переходом его из альвеол в кровь можно пренебречь.
Через несколько минут, когда содержание гелия в легких и
спирометре выравнивается, измеряют это содержание (FHe ) при помощи
специальных приборов. Предположим, что в нашем случае оно
составляет 0,05 мл Не на 1 мл смеси. При вычислении ФОЕ исходят из
того, что общее количество гелия, равное произведению объема Кна
содержание F, должно быть одинаковым как в исходном состоянии,
так и после перемешивания:
Vпава 5. Биофизика двигательной активности
67
^„^„е,=(^П±^ОЕ^Нсг)· (5-53)
Подставляя в это уравнение приведенные выше данные, можно
рассчитать ФОБ:
OQE=^(^-^,)^(0,l-0;05)=3ji
При использовании метода вымывания азота испытуемый
после спокойного выдоха в течение нескольких минут дышит чистым
кислородом. Выдыхаемый воздух поступает в спирометр. И вместе
с ним в спирометр переходят молекулы азота, содержащиеся в
легких. Зная объем выдыхаемого воздуха, начальное содержание N2
и легких и конечное содержание N2 в спирометре, можно, используя
уравнение, подобное (5.54), вычислить ФОБ.
При практическом применении этих методов необходимо
вносить некоторые поправки. Кроме того, недостатком обоих методов
является то, что у больных с неравномерной вентиляцией некоторых
участков легких полное разведение или вымывание газов требует
очень длительного времени. В связи с этим в последнее время
получило широкое распространение измерение ФОБ при помощи
интегрального пневмотахографа и плетизмографа.
Пневмотахография. Для исследования дыхания в течение
длительного времени значительно удобнее так называемые спирометры
открытого типа. В этом случае регистрируют не сами дыхательные
объемы, а объемную скорость воздушной струи (рис. 5.31). Для этого
используют пневмотахографы - приборы, основной частью которых
служит широкая трубка с малым аэродинамическим
сопротивлением. Для определения скорости пользуются разными методами:
1) При прохождении воздуха через трубку между ее началом
и концом создается небольшая разность давлений, которую можно
чарегистрировать при помощи двух манометрических датчиков. Эта
разность давлений прямо пропорциональна объемной скорости
воздушной струи, т.е. количеству воздуха, проходящего через
поперечное сечение трубки в единицу времени. Кривая изменений этой
68
Биофизика
Пневмотахограммз
Спирограмма
Выдох
Рис. 5.31. Принцип действия пневмотахографа
Разность давлений между двумя концами трубки, обладающей
определенным аэродинамическим сопротивлением и соединенной
с загубником, пропорциональна объемной скорости потока
воздуха V. Кривая изменений этой скорости называется
пневмотахограммой, а кривая изменений интеграла этой скорости
во времени т.е. объема дыхания, представляет собой спирограмму
объемной скорости называется пневмотахограммой. Интегрируя
пневмотахограмму, представляющую собой запись dVldt, можно
получить искомый объем воздуха
-/(£>·
dt )
(5.55)
2) В трубке может быть установлена малоинерционная турбин-
ка, частота вращения которой зависит от скорости воздушного потока
в ней. Частота вращения измеряется бесконтактно с помощью опто-
электронного или емкостного датчика. Интегрируя частоты вращения
турбинки за промежутки времени можно определить расходы воздуха
при вдохе или выдохе.
С помощью турбинки можно идентифицировать циклы выдоха
и вдоха, так как при изменениях направления воздушного потока
изменяется направление ее вращения.
Гпава 5. Биофизика двигательной активности
69
В большинстве пневмотахографов имеется электронный
интегрирующий блок, и поэтому одновременно с пневмотахограммой
непосредственно записывается кривая дыхательных объемов (спиро-
грамма).
Для измерения неэластического сопротивления дыхательной
системы требуется постоянная регистрация внутриальвеолярного
давления. Можно также использовать непрямой способ - интеграль-
Давление Давление Поток
в камере в полости воздуха
g рта ς) Q
Клапан, закрывающий
воздухоносные пути
Калибровочный
насос
Рис. 5.32. Интегральная плетизмография (упрощенная схема)
Q — объемная скорость потока воздуха, Ра — альвеолярное давление
70
Биофизика
ную плетизмографию. Основным элементом интегрального
плетизмографа (рис. 5.32) служит герметичная камера, в которой человек
может удобно сидеть. При дыхательных движениях давление в легких
испытуемого изменяется, и обратно пропорционально ему меняется
давление в замкнутой плетизмографической камере. Откалибровав
измерительную систему, можно судить об изменениях внутриальвео-
лярного давления Ра по изменениям давления в камере.
Одновременно можно регистрировать расход воздуха Q при помощи пневмотахо-
графа. Соотношение этих двух величин представляет собой искомое
сопротивление. Обычно производят его непрерывную запись
посредством перьевого регистратора.
При помощи интегрального плетизмографа можно также
определить функциональную остаточную емкость (ФОБ). Для этого на
короткий срок перекрывают трубку, идущую от загубника, в
результате чего воздух в легких перестает общаться с атмосферным. После
этого исследуемый совершает попытку вдохнуть, при этом измеряют
изменения давления в полости рта и объема легких. ФОБ можно
вычислить, исходя из закона Бойля-Мариотта.
5.4. Биомеханика миокарда
Кровообращение происходит в сердечно-сосудистой системе.
С позиций механики, сердце является комбинированным
многокамерным насосом, нагнетающим кровь в большой и малый круги
кровообращения, образованные сосудами разных типов (артериями,
капиллярами, венами).
5.4.1. Сокращение миокарда
Механическая работа, совершаемая сердцем, развивается за
счёт сократительной деятельности миокарда. Вслед за
распространением возбуждения происходит сокращение миокардиальных волокон.
Механизмы электромеханического сопряжения и сократительного
процесса в кардиомиоцитах и миоцитах скелетных мышц аналогичны,
но не идентичны. Эти процессы в кардиомиоцитах имеют важные
особенности.
Глава 5. Биофизика двигательной активности
71
Функциональным элементом миокарда служит мышечное
волокно, образованное цепочкой кардиомиоцитов, соединённых «конец
в конец» и заключённых в общую сарколемму. В такие цепочки
объединяются от 2 до 15 кардиомиоцитов, каждый из которых имеет
длину от 100 до 200 мкм. Общая сарколемма имеет выраженный гли-
кокаликс толщиной примерно 50 нм. Мышечные волокна
собираются в пучки, отделённые друг от друга тонкими прослойками
соединительной ткани. Пучки группируются в тяжи, взаимодействующие
между собой посредством сложных нерегулярных контактов.
Сократительной способностью обладает 99% всех
кардиомиоцитов. Их называют сократительными (типичными, рабочими).
В этих клетках миофибриллы упорядочены (направлены вдоль их
длинной оси и параллельны друг другу), хорошо развиты саркоплаз-
матическая сеть (СПС) и система Г-трубочек, много митохондрий.
Толщина сократительных кардиомиоцитов у человека составляет 15—
25 мкм. Поверхность каждого из них примерно 8,5 · Ю-5 см2, а объём —
1,94 · 10~10 л, причём 34% объёма приходятся на митохондрии.
В проводящих (атипичных, узловых, специализированных) кар-
диомиоцитах, на долю которых приходится всего 1% миокардиаль-
ных клеток, сократительный аппарат развит слабо.
В дальнейшем пойдет речь только о сократительных кардио-
миоцитах. Именно их способность активно укорачиваться и
развивать напряжение обеспечивает механическую (насосную) функцию
сердца.
Возбуждающий сигнал (потенциал действия) поступает без
декремента к сократительным кардиомиоцитам от пейсмекера по
миокардиальному синцитию. Потенциал действия на мембране
Г-трубочек инициирует повышение концентрации Са2+ в миоплазме
подле миофибрилл. Са2+ поступает к миофибриллам миоцита
скелетной мускулатуры исключительно из цистерн СПС. В отличие от
этого, в кардиомиоцитах СПС развита слабее, и ключевым фактором
в электромеханическом сопряжении является вход Са2+ в миоплазму
из интерстиция через потенциалзависимые каналы в сарколемме.
Кальциевый (медленный входящий) ток (/„·) течёт через сарколемму
в течение существования «плато» на потенциале действия. Чем оно
продолжительнее, тем больше Са2+ входит в миоплазму. Входящий
72
Биофизика
Интерстиций
Сарколемма
Рис. 5.33. Схема электромеханического сопряжения в кардиомиоците
СПР — саркоплазматический ретикулум
кальциевый ток выполняет тригтерную функцию по отношению
к потенциалзависимым кальциевым каналам в мембране СПС. При
небольшом повышении концентрации Са2+ в миоплазме
деполяризация мембраны Г-трубочек открывает кальциевые каналы в мембране
СПС кардиомиоцитов млекопитающих (рис. 5.33).
Поток Са2+ из СПС в миоплазму на 2 порядка интенсивнее
потока через сарколемму, но кальциевый ток из СПС в саркоплазму не
пойдёт без /,,·. В этом состоит важное отличие электромеханического
сопряжения от аналогичного процесса в миоците скелетных мышц.
Этот обнаруженный в конце XX века факт ответил на вопрос,
поставленный С. Рингером более века назад: почему сокращение миокарда
требует внеклеточного кальция (в окружающем растворе), а для
сокращения скелетных мышц он не обязателен. В кардиомиоцитах
лягушек СПС развита очень слабо. Там входящий ток ионов кальция
через сарколемму обеспечивает электромеханическое сопряжение
полностью - без участия СПС.
Как и в миоцитах скелетных мышц, повышение концентрации
Са2+ от Ю-7-10"* Μ до (0,2-1,5) · ΙΟ"6 Μ устраняет тропомиозиновую
блокаду сокращения, «расчехляя» активные центры для миозина на
актиновых нитях. Однако в кардиомиоцитах нет тропонина, и
процесс их сокращения происходит без его участия. Максимальное со-
Глава 5. Биофизика двигательной активности
73
кращение кардиомиоцита наблюдается при достижении
концентрации Са2+ вблизи миофибрилл около 5 · ΙΟ-6 Μ. Однако в условиях
покоя напряжение миокарда не превышает 70% от максимально
возможного.
После закрытия кальциевых каналов в сарколемме (в этот
момент обрывается «плато» на потенциале действия) прекращается Isi,
а избыток Са2+ удаляется из миоплазмы. В кардиомиоцитах
выведение ионов кальция осуществляется двумя путями. Во-первых,
кальциевый насос, сосредоточенный в мембране СПС, обеспечивает их
активный транспорт в цистерны СПС - так же, как в миоцитах
скелетной мускулатуры. Во-вторых, кардиомиоцитам присущ
своеобразный механизм выведения кальция в интерстиций посредством
кальций-натриевого обменника, находящегося в сарколемме. Эта
система пассивного транспорта осуществляет во время диастолы: обмен
одного иона Са2+, который выходит из миоплазмы, на 2 иона Na+,
входящих в неё из интерстиция (рис. 5.34).
При нарушении электромеханического сопряжения в сердце
наблюдается явление электромеханической диссоциации -
разобщения электрической активности и сокращений миокарда вследствие
ослабления поступления Са2+ в миоплазму из интерстиция. Это
происходит при понижении концентрации ионов кальция в интерстиций
(например, вследствие недостатка их в пище и воде), при блокаде
кальциевых каналов в сарколемме кардиомиоцитов (солями Со2+, Cd2+,
Ni2+, La3+ и некоторых других элементов). В таких случаях бывает
внезапная остановка сердца.
Интерстиций
2Na+
Сарколемма
Са2+
Миоплазма кардиомиоцита
Рис. 5.34. Натрий-кальциевый обменник в сарколемме кардиомиоцита
74
Биофизика
Особенности сократимости миокарда
1. Зависимость силы сокращения миокарда от силы стимула
подчиняется закону «всё или ничего». Этот закон, характерный для
отдельных клеток, свойствен всему миокарду, поскольку он
представляет собой синцитий.
2. В зависимости силы сердечных сокращений от частоты
стимула наблюдается феномен «лестницы Боудича» — нарастание
силы сердечных сокращений по мере повышения частоты
стимуляции. Такой (хроноинотропный) эффект объясняется тем, что при
частоте стимуляции не весь Са2+, вошедший в миоплазму во время
систолы, удаляется в диастолу. Его концентрация вблизи миофибрилл
поддерживается на повышенном уровне, что и обусловливает
усиление сокращений.
3. Сила сокращений желудочков сердца прямо
пропорциональна исходной длине мышечных волокон - иными словами, чем больше
расслабление миокарда в диастолу, тем сильнее сокращение в
систолу. Эту зависимость называют основным законом сердца Франка-
Старлинга. Он был установлен О. Франком в 1895 г. в опытах на
лягушках и Е. Старлингом в 1912-1914 гг. в опытах на млекопитающих.
Закон гласит, что ударный объем прямо связан с изменениями конеч-
нодиастолического объема желудочка.
4. Зависимость скорость-сила: чем больше нагрузка на
сердце, тем оно медленнее сокращается. Такая зависимость следует из
уравнения А. Хилла, которому миокард подчиняется так же, как
скелетная мышца.
Эксперименты на одиночных волокнах скелетных мышц
показали, что развиваемая сила пропорциональна изменению мгновенной
длины саркомера и не зависит от пути, по которому саркомер
изменял свою длину. Отсюда вытекает важный вывод, что в
изотоническом режиме мышца должна укорачиваться до длины, при которой
сила равна весу поднимаемого груза. Действительно, в папиллярной
мышце кошки связь «длина-напряжение» в изометрическом режиме
совпадает с отношением «конечная длина-груз» при изотонии.
Однако в других опытах было обнаружено, что изометрическая сила
при данной длине мышцы больше, чем изотонически поднимаемый
груз при той же самой длине (рис. 5.35).
Гпава 5. Биофизика двигательной активности
75
Р.гм
5,4
/ Л
ж ^ш ι . . .
X 4 ..... .
Х- ■...■
7,4 /, мм
Рис 5.35. Функция «длина-напряжение» для изометрических
(непрерывная линия) и изотонических (пунктир) сокращений
При изометрии длина мышцы устанавливается до стимуляции.
При изотонии мышца поднимает различные грузы.
По оси абсцисс - длина мышцы; по оси ординат - сила
Начальная длина влияет не только на амплитуду, но и на
другие параметры изометрических сокращений. Зависимость скорости
изотонического укорочения от силы (нагрузки) является, как и связь
«длина-сила», фундаментальным свойством мышцы. Как уже
упоминалось, часто для описания функции «сила-скорость» используется
уравнение Хилла:
(p + a)v=b(p-p0),
которое в безразмерных величинах р/р0 и vmax имеет вид
(5.56)
_Р_
Ро
1-
1 + р.
или
. Ро;
a )v.
^_+_Р_
Ро
(5.57)
\Ро Poj
В миокарде эта функция лучше описывается отношением:
(p + a)v=b(p-Poy, (5.58)
где 0 < η < 1 .
76
Биофизика
Казалось бы, снимая зависимость «сила-скорость» в миокарде,
можно найти фундаментальные константы α и Ь, значения р0
(максимальной силы) и vMaKC (максимальной скорости ненагруженного
укорочения) и охарактеризовать ими сократительную деятельность
и энергетические процессы. Однако в сердечной мышце, в отличие
от скелетной, имеется ряд особенностей, которые затрудняют
интерпретацию функции «сила-скорость» и характеристик механохимиче-
ских процессов.
Поскольку сердечную мышцу необходимо предварительно
растянуть, т.е. задать определенную преднагрузку, максимальную
скорость укорочения можно обнаружить путем экстраполяции.
Заметим, что чем больше мышца предварительно растянута, тем менее
точно определяется vmax. Хотя ранее сообщалось, что vmM не зависит
от начальной длины (преднагрузки), ряд данных свидетельствует об
обратном.
В скелетной мышце при длине саркомера 2,0-2,2 мкм р0 не
зависит от длины, тогда как в миокарде максимальное изометрическое
напряжение сильно зависит от длины, в силу чего смысл констант a
и Ъ не вполне ясен. Кроме того, как упоминалось выше, при / близких
к /тах сами изменения длины влияют на ход активации.
Скорость укорочения даже при малых грузах устанавливается
не сразу, а требует времени, в течение которого происходит
укорочение контрактильного элемента. Таким образом, при определении
функции «сила-скорость» длина становится неконтролируемой
переменной. Поскольку податливость сердечной мышцы выше, чем
скелетной, внутреннее укорочение приводит к большему
уменьшению способности развивать силу. Если в скелетной мышце
измерение скорости укорочения производится после достижения плато
активного состояния, то в сердце это не так.
В миокарде влияние напряжения покоя не вполне ясно, а
оценка влияния зависит от типа выбранной для анализа реологической
модели. При изотоническом укорочении сила в параллельном
элементе падает и переносится на контрактильный элемент, а в покое
в последовательном элементе имеется определенное напряжение.
В изометрическом режиме напряжение падает и перераспределение
напряжений между ними зависит от преднагрузки. Так как напряже-
Глава 5. Биофизика двигательной активности
77
пие покоя является нелинейной функцией длины, оно должно
учитываться каждый раз, особенно при больших нагрузках.
Поскольку активное состояние в миокарде развивается
относительно медленно, то в ряде случаев поднятие груза происходит при
разных уровнях активации. Исследование функции «сила-скорость»
в начальных фазах сердечного цикла дает заниженные значения уже
в силу неполной активации контрактильного элемента. Медленное
нарастание активного состояния заставляет при анализе функции
«сила-скорость» в миокарде обязательно учитывать фактор времени.
Работая на скелетной мышце, Хилл установил, что ее
деформации вызывают уменьшение силы сокращений. Аналогичное явление
было обнаружено в миокарде: высвобождение или растяжение во
нремя изометрического сокращения приводило к падению
напряжения ниже контрольной величины. Этот эффект Бреди назвал
разобщением, или инактивацией сокращений. В скелетной мышце
изменения длины, помимо уменьшения величины напряжения, приводят
к укорочению длительности активного состояния и более раннему
чавершению расслабления.
Имеющиеся данные свидетельствовали, что деформации во
нремя сокращения приводят к уменьшению числа активно
включенных мостиков.
Как показал Бреди, влияние кратковременных деформаций не
чависит от направления деформации (укорочение или растяжение) -
и обоих случаях имеется уменьшение последующего напряжения.
Степень выраженности эффекта разобщения зависит от момента
сердечного цикла, в который действует механическое возмущение.
Установлено, что если смещение действует через 0-50 мс после
стимула, то разобщающий эффект отсутствует. Чем позже от начала
сокращения начинает действовать деформация, тем больше величина
инактивации. Деформация в 5-10%, попадающая в фазу
расслабления, вообще может уменьшить напряжение до нуля. Для данной
деформации инактивация (оцененная как процент уменьшения
напряжения по отношению к невозмущенному сокращению) линейно зависит
от времени после стимула, а для данного момента линейно связана
с величиной деформации при деформациях, не превосходящих 7%
длины. При больших деформациях, когда напряжение при разгрузке
78
Биофизика
падает ниже 20-30% р0, инактивация на пике изометрической силы
перестает зависеть от деформации.
В отношении влияния скорости деформации на величину
инактивации данные противоречивы. Хотя разобщающий эффект
увеличивается при уменьшении скорости рывка от 8 до 0,5 с-1, при
скоростях деформации, не превосходящих 1,2 с-1, влияние этого параметра
на инактивацию не установлено. Разобщающий эффект не зависел от
начальной длины и в сердечной и в скелетной мышце при / = 1,9-
2,7 мкм. Увеличение внеклеточной концентрации Са^ уменьшает
относительный эффект инактивации (рис. 5.36). Кофеин в присутствии
высоких концентраций кальция также уменьшает эффект
инактивации или даже обращает его. Подавление активации верапамилом или
гипокальциевой средой увеличивает инактивацию.
При исследованиях обнаружена линейная зависимость между
величиной инактивации и постоянной скорости расслабления (τ).
Вне зависимости от вида воздействующего фактора (температура,
изменение частоты стимуляции, адреналин, кальций) уменьшение
константы скорости расслабления мышцы приводит к уменьшению
эффекта инактивации. Однако корреляции между величиной
инактивации и максимально развиваемым напряжением не обнаружено.
С этим созвучны факты, что эффекты инактивации не
обнаруживаются при контрактурах вне зависимости от способов, которыми они
индуцируются (гиперкалиевые, гипонатриевые, бариевые
контрактуры), и в тетанусе, который в сердечной мышце может быть получен
в специфических условиях (кофеин + гиперкальциевая среда). На
скицированных препаратах миокарда, где сокращение запускалось
ионофоретической инъекцией кальция, инактивация деформацией
отсутствовала. Удлинение сокращений уменьшает деактивирующий
эффект или даже обращает его. Разобщающий эффект был недавно
получен на изолированном сердце.
Процесс инактивации весьма существен в методе быстрого
высвобождения. При этом вследствие инактивации величины скоростей
укорочения оказываются заниженными, а степень влияния быстрого
высвобождения тем больше, чем на большую величину мышца
отпускается.
Глава 5. Биофизика двигательной активности
79
0 12 3 4 5 6
°/Р^
1,0
0.5
• В
v^-^^
\^^150
\^\250
^N.350
I I I I I 1 w-
0 12 3 4 5 6
%/о
0 1 2 3 4 5 6
Рис 5.36. Влияние кальция на разобщающий эффект,
вызванный укорочением в папиллярной мышце кролика
А, В- контроль; Б - концентрация кальция увеличена с 2,5 до 11 мМ;
Г- влияние блокатора кальциевой проницаемости верапамила. По оси
абсцисс - смещение, % 10, по оси ординат - величина активации. Цифры
над кривыми означают время (мс) в цикле, когда действует деформация
Флуктуации скорости выше 5% vmax оказываются
достаточными, чтобы уменьшить скорость укорочения. Согласно сообщению
Эндо, этот инактивирующий эффект осцилляции связан с фазой
отрицательного ускорения (торможения) и он является
«дополнительным» к инактивации, вызванным самим укорочением.
Скорее всего эффект инактивации каким-то образом связан
с миофибриллярной системой (возможно, с конформационными
изменениями в тропонине). Имеются данные, что длительность конформа-
ционных изменений может доходить в белках до нескольких секунд.
Ι Ιο мнению Эдмана, время жизни разобщающего эффекта в
одиночных волокнах скелетных мышц может доходить до 800-900 мс.
80
Биофизика
Об этом, возможно, свидетельствует следующий факт.
Инактивация во время первого сокращения в зубчатом тетанусе
воздействует на силу сокращений в последующих циклах. Важно, что
пассивные деформации не влияют на амплитуду сокращений. Скорее всего
количество кальция, связанного с тропонином, контролируется
скольжением нитей, и в момент быстрых деформаций сродство тропо-
нииа к кальцию уменьшается и высвобождающийся кальций быстро
поглощается ретикулумом. Это должно означать, что сродство к
кальцию в изотонии должно быть меньше, чем в изометрическом
сокращении.
Эдманом была предложена математическая модель сопряжения
возбуждения с сокращением, учитывающая процессы поглощения
и высвобождения ионов кальция и их связывание с тропонином.
В случае низкой частоты стимуляции уравнение активации имеет вид:
έ + γ=/(Ο-φψ(<0, (5.59)
y=k_x[F(kc)-yl (5.60)
где γ - отклонение концентрации кальций-тропонинового
комплекса от его полной концентрации х0; с - концентрация свободного
кальция в саркоплазме, отнесенная Kx0;flt) - функция, описывающая
зависимость скорости высвобождения кальция; у(с) - зависимость
поглощения кальция ретикулумом от его концентрации с константой
φ„ к - безразмерная константа сродства реакции связывания кальция
с тропонином; &_, - константа скорости этой реакции; F -
равновесное состояние этой реакции.
Оценки показывают, что 1/ &_, намного меньше характерного
времени механических процессов, а к_х « 1. В рамках данной
модели имеющиеся гипотезы о природе инактивации сводятся к
предположению, что величины φ или к являются функцией модуля скорости
деформации, т.е.
φ=φ(|ε|) или к = к(\в\). (5.61)
Считая реакцию связывания кальция с тропонином
квазиравновесной, получим:
/ пава 5. Биофизика двигательной активности
81
C=^,C=J_-HZ). (5.62)
k kF'(y) к2
После преобразований имеем
γ=/(0-φψ F 1(γ)
(5.63)
Уравнение (5.63) показывает, что по данным механических
измерений в одиночном сокращении при одновременном
функционировании системы тропонина и оттока кальция невозможно
различить, что влияет на комплекс тропонин-кальций: деформация или
поглощение кальция ретикулумом.
Термин «сократимость» миокарда, «контрактильное
состояние», или «инотропное состояние», широко употребляется в
литературе и чаще всего в интуитивном смысле без точного определения.
Ι Ιο сути дела под изменением сократимости большинство
исследователей понимает именно те сдвиги в сократительной способности
сердца, которые определяются внутренними, а не внешними
обстоятельствами (пост- и преднагрузками). Это важно потому, что
необходимо отличать изменения производительности сердца, связанные
с изменениями давления в аорте, конечнодиастолического давления,
механическими дефектами клапанов или самого сердца, от тех,
которые связаны с подавлением сократимости. Даже нормальное сердце,
находящееся в хорошем функциональном состоянии, не может
работать при большой перегрузке давлением или объемом.
Серьезные трудности возникают при попытке измерить
сократимость с помощью одного параметра, даже такого, как ν^, ибо
оценка контрактильной деятельности должна по меньшей мере
нключать в себя способность к развитию силы и укорочения. Связь
между этими двумя способностями задается с помощью
соотношения сила-скорость, которое могло бы характеризовать данное
контрактильное состояние. Выше уже указывались реальные трудности,
нозиикающие при экспериментальном определении (р-у)-связи даже
па полосках миокарда. Ситуация в целом сердце осложняется еще
82
Биофизика
и теми обстоятельствами, что сила сокращения во время выброса не
является постоянной и прямая трансляция данных механики полоски
на целое сердце ограничена. Кроме того, в патологии необходимо
характеризовать и сегментарные нарушения сократительной
деятельности (асинхронность, акинезия).
В последнее время появились сомнения в принципиальной
возможности выделить показатель (индекс), который не зависел бы от
изменений длины. По мнению Брэди, это совсем неразрешимая
задача, поскольку в миокарде имеется несколько причин,
обусловливающих связь сила-скорость. Поэтому общеупотребляемые индексы не
позволяют различать влияния, обусловленные начальной длиной
и активацией, и концептуально и фактически.
5.4.2. Расслабление миокарда
Фаза релаксации сокращения, которую можно определить как
процесс, посредством которого мышца после сокращения
возвращается к первоначальной длине или напряжению, изучена значительно
хуже, чем фаза сокращения. Природа расслабления не менее сложна,
чем природа сокращения. Доказано также, что фаза расслабления
изменяется при увеличении частоты сердцебиений, под влиянием ка-
техоламинов, в гипернатриевых растворах. Кроме того, имеются
указания, что расслабление мышцы находится под контролем
мембранного потенциала. Иными словами, фаза расслабления регулируется
физиологическими факторами. Полагают, что скорость расслабления
отражает скорость удаления кальция из контрактильного материала,
а изменения в фазе расслабления объясняются изменениями
скорости поглощения кальция саркоплазматическим ретикулумом и (или)
другими мембранными структурами.
Работами последних лет установлено, что в процессе удаления
кальция из саркоплазмы саркоплазматическим ретикулумом можно
различить быстрое связывание и поглощение. Быстрое связывание
наблюдается в отсутствие преципитирующих анионов, и в
оптимальных условиях скорость удаления кальция составляет 250-
300 · 106 мл/с (для полной активации необходимо 50-100 · 108 мл/с),
что вполне достаточно для адекватного объяснения скорости рас-
Глава 5. Биофизика двигательной активности
83
слабления. В присутствии преципитирующих анионов кальций
поглощается значительно медленнее, но кальций поглощающая емкость
ретикулума возрастает в 50-100 раз. Найдено также, что поглощение
кальция из миоплазмы регулируется АМФ-зависимой киназой мик-
росом. Этот факт объясняет ускорение расслабления, которое
наблюдается в сердцах теплокровных под влиянием катехоламинов. Для
«линейного» расслабления максимальная скорость определяется в
точке начала экспоненциального участка кривой.
Найдено, что увеличение начальной длины от L0 до LmM в
предсердиях теплокровных и холоднокровных животных в
исследованном температурном диапазоне приводит к уменьшению
соответствующих констант расслабления. В предсердиях кролика к уменьшалась
и 1,5-2,0 раза. Перерастяжение приводило к дальнейшему падению
неличины к. В препаратах желудочков черепах и кроликов не
обнаружено существенного влияния начальной длины на константы
расслабления. При увеличении температуры от 10 до 35 °С возрастают
константы расслабления и заметно изменяется процесс
расслабления. Так, в миокарде при 35 °С почти всё расслабление идет по
экспоненциальному закону. При 10 °С почти все время расслабления
приходится на «квадратичное расслабление». В миокарде
предсердий кролика чувствительность констант расслабления к растяжению
растет с понижением температуры.
Если, варьируя концентрации кальция и растяжение, добиться
одной и той же амплитуды сокращений, то на константу
расслабления повышение концентрации кальция оказывает воздействие,
противоположное растяжению.
В миокарде человека (при экспоненциальном расслаблении) к
увеличивается от 4 до 7,7 с-1 в частотном диапазоне 0,05-0,2 Гц.
При «квадратичном» типе расслабления в препаратах
предсердия человека к возрастет от 6,6 до 10,0 с -|, когда частота
увеличивается от 0,07 до 2,0 Гц. Наконец, при смешанном типе расслабления
(например, в одном из препаратов предсердия человека) при частоте
0,07 Гц к = 7,1 с-1, а при частоте 0,2 Гц к = 9,5 с-1.
До сих пор не ясно, как объяснить различную чувствительность
к растяжению кардиомиоцитов желудочков и предсердий. Показано,
что клетки предсердий имеют особые секреторные гранулы, ответст-
84
Биофизика
венные за выделение физиологически активных веществ. Кроме того,
в предсердиях имеется много симпатических и парасимпатических
волокон. Ясно, что растяжение может влиять на подобные структуры
и регулировать тем самым выделение веществ, влияющих на
расслабление (например, путем воздействия на кальцийсвязывающие
структуры).
В основе модели изометрического расслабления лежит
предположение, что напряжение линейно связано с числом кальций-тропо-
ниновых комплексов. В свою очередь, концентрация кальций-тро-
понинового комплекса определяется кинетикой взаимодействия
кальция с тропонином и кинетикой поглощения кальция саркоплаз-
матическим ретикулумом или его функциональными эквивалентами.
В отношении «квадратичного расслабления» есть основания
полагать, что отток кальция может быть связан с деятельностью Са—
Na обменного механизма. Предполагают, что при деполяризации
этот механизм выключается, а после окончания деполяризации
происходит постепенное (линейное) восстановление от инактивации.
С постулируемой точки зрения, процесс расслабления в
сердечной мышце может регулироваться за счет трех механизмов:
1) скорости восстановления от инактивации (рис. 5.37, а); 2)
изменений констант скорости поглощения кальция при тех же
характеристиках инактивации (рис. 5.37, б); 3) изменения длительности
потенциалов действия (рис. 5.37, в). До сих пор не ясно, связан ли
смешанный тип расслабления с «сосуществованием» двух
механизмов удаления кальция или с наличием инактивации кальциевого
насоса в мембране саркоплазматического ретикулума.
Возможно, что расслабление может регулироваться за счет
константы распада кальций-тропонинового комплекса, которая,
в свою очередь, может быть функцией нагрузки или скорости
укорочения. Нельзя также исключить, что «константа» поглощения
кальция ретикулумом также является зависимой от деформации или
скорости движения. Вероятно, в одной и той же клетке функционируют
несколько систем удаления кальция: саркоплазматический ретику-
лум; натрийзависимое удаление; удаление за счет митохондрий.
По-видимому, вклад этих систем в расслабление зависит от
удельной плотности соответствующих органелл, от соотношения их
Гпвва 5. Биофизика двигательной активности
85
Π
t ' t t
а) б) в)
Рис. 5.37. Варианты изменения коэффициента оттока кальция
а — скорость расслабления изменяется за счет хода инактивации;
б - скорость расслабления изменяется за счет вариации
стационарного значения; в - скорость расслабления зависит
от длительности потенциала действия (изображено ступенькой)
поверхности к объему и активности кальцийпоглощающих систем
м каждой структуре.
Была изучена кинетика изометрического расслабления в пост-
нагрузочных сокращениях. В этом режиме расслабление состоит из
двух фаз: 1) изотонического растяжения, во время которого мышца
нозвращается к исходной длине; 2) изометрического падения силы,
когда в мышце восстанавливается первоначальное напряжение.
Найдено, что изометрическое расслабление после изотонического
сокращения имеет две фазы: фазу быстрого падения и фазу
экспоненциального расслабления. В обычных изометрических сокращениях
релаксация изометрического сокращения была не экспоненциальна.
Ι Ιο мере увеличения нагрузки, как и следовало ожидать,
изометрическое расслабление удлинялось, крутизна фазы начального быстрого
падения релаксации падала. Чем больше груз, тем более была
выражена экспоненциальная фаза.
В сердце млекопитающих экспоненциальный ход
изотонического расслабления имеется при нагрузках, меньших 0,8 Р0, а при
больших нагрузках оно не экспоненциально. В скелетной мышце и
миокарде наличие укорочения в изотонической фазе постнагрузочных
сокращений оказывает существенное влияние на последующую фазу
изометрической релаксации. Так, после изотонической фазы изомет-
кЛ
86
Биофизика
Цмм ι \
6
4
2
^
и
0,2
P. г/мм2 ι
9
5
3
0
L
/
/
/
/
0,2
/^
^^
0.4
0,4
,»·"""1^у
>
^
0,6
й
N
0,6
б)
\
Л
0.6
ν
^
^
0,6
1,0
Ss^
1,0
f.c
f.c
Рис. 5.38. Ход изометрического расслабления
после изотонической фазы в скелетной мышце лягушки:
а — изменения длины; б - напряжение
рическая фаза расслабления начинается раньше, чем в обычных
изометрических сокращениях (рис. 5.38).
В миокарде изометрическое расслабление после
изотонической фазы из неэкспоненциального становится экспоненциальным.
Если в скелетной мышце длительность изотонического расслабления
возрастает по мере уменьшения груза (рис. 5.38), то в миокарде по
мере уменьшения груза длительность изотонической фазы вначале
возрастает, однако при дальнейшем уменьшении груза время
изотонического расслабления уменьшается, вероятно, вследствие
разобщающего эффекта укорочения (рис. 5.39), выраженность которого
в сердце значительно больше, чем в скелетных мышцах.
Было также найдено, что постоянная времени
экспоненциального расслабления в изометрической фазе не меняется после
предварительного укорочения как в скелетной, так и в сердечной мышцах.
Эти факты означают, что укорочение приводит к сдвигу кривых
падения напряжения, тогда как крутизна спада напряжения в
полулогарифмических координатах не зависит от величины укорочения.
Каков бы ни был режим нагружения и характеристики укорочения,
Глава 5. Биофизика двигательной активности
87
Рис. 5.39. Ход расслабления в скелетной (1) и сердечной (11) мышцах лягушки
а - обычное расслабление, б - расслабление в условиях фиксации длины.
Стрелками показаны начало и конец фиксации; 1 - начальная линейная
фаза расслабления, 2 — конечная экспоненциачьная фаза (схема)
постоянная времени изометрического расслабления остается
постоянной.
Исследования функции «сила-скорость» для фазы
расслабления крайне малочисленны. Установлено, что если скелетную мышцу
иысвободить в разные фазы цикла, то активное укорочение
происходит вплоть до конца расслабления (рис. 5.40).
Для каждого момента времени высвобождения кривая
«сила-скорость» описывается гиперболической функцией. Аналогичный
процесс происходит в миокарде. Это свидетельствует, что активация
м поздние фазы цикла все еще не закончена.
Много новой дополнительной информации о влиянии
механических условий на ход расслабления получено с помощью метода
фиксации длины саркомеров в центральных участках полоски с
помощью специальных сервосистем. На одиночных волокнах скелетных
мышц показано, что в условиях фиксации длины саркомеров
напряжение затягивается за счет ранней фазы расслабления. Последующая
жепоненциальная фаза расслабления совпадает с началом удлинения
саркомеров. Интересно, что изменение напряжения при фиксации
длины в полоске внешне напоминает напряжения в целом сердце во
нремя выброса крови. Во время первой фазы спад механического
напряжения линеен. В обычном изометрическом сокращении во время
фазы экспоненциального падения напряжения всегда имеется
удлинение саркомеров. Сходство падения напряжения в изометрических со-
88
Биофизика
кращениях в скелетной и сердечной мышцах свидетельствует, что
поведение миофибрилл в обоих случаях качественно одинаково. Однако
в скелетной мышце постоянство длины саркомеров при расслаблении
можно поддерживать растяжением препарата, а в сердечной мышце -
L, мм
8
L, мм
9
200 мс
L, мм А
6 Г
Р, г/мм 2
4 г
2
0 L
/ t
/ \\
/ ^\
/ **г^\
-»-
//
t
Рис. 5.40. Изометрическое и изотоническое
расслабление в капиллярной мышце кролика
А - сравнение изометрического и изотонического расслабления: кривая 1 -
изотоническое расслабление, кривая 2 - изометрическое расслабление;
Б— семейство постнагрузочных изменений силы; В — семейство изотонических
сокращений, в которых на разное время движение останавливается
(изометрическая фаза); Г- изотония при разных нагрузках. I-изотония
при различных нагрузках, II - изометрия при различных фиксациях длины
Глава 5. Биофизика двигательной активности
89
ча счет отпускания. Кроме того, фиксация длины саркомеров в
миокарде не приводит к удлинению напряжения, как в скелетной мышце.
В сердечной мышце не найдено дисперсии длин саркомеров во
нрсмя фазы релаксации напряжения. Более того, обнаружено, что
сразу после сокращения дифракционные линии более четкие, чем к
началу следующего сокращения. При частотах стимуляции выше 0,4 Гц
дисперсия длин саркомеров исчезала. Таким образом, дисперсия длин
саркомеров имелась только во время длительных пауз, а наличие
сокращений восстанавливало однородность после каждого сокращения.
Вероятно, в миокарде должны быть системы, стабилизирующие
популяцию расслабляющихся саркомеров и препятствующие удлинению.
Такой структурой, по-видимому, является параллельный упругий
элемент и его составная часть - внеклеточная эластическая ткань. На
основании этих данных было сделано предположение, что причина
падения напряжения заложена в самих миофибриллах, ибо абсолютная
скорость падения напряжения не зависит от начальных условий и,
самое главное, от начальной длины.
Интересно, что кальций оказывает неодинаковые эффекты на
изометрическое и изотоническое расслабление, т.е. не изменяет
абсолютной скорости изометрического сокращения, но увеличивает
константу изотонического расслабления. Вероятно, это говорит о том,
что в изометрии и изотонии управление расслаблением
осуществляется разными механизмами. Отметим, что изотоническое
расслабление под малыми нагрузками заканчивается до начала второй фазы
изометрического расслабления. По-видимому, первая фаза
изометрического расслабления (так же как и изотоническое расслабление)
протекает на фоне определенной внутриклеточной концентрации
кальция и, следовательно, зависит от деятельности саркоплазматиче-
ского ретикулума. Рианодин, влияющий на скорость поглощения им
кальция, замедляет изотоническое расслабление. Вторая фаза
изометрического расслабления начинается, когда концентрация
внутриклеточного кальция близка к нулю, и поэтому нечувствительна к
воздействиям на саркоплазматический ретикулум.
Для объяснения отсутствия влияния начальной длины на абсо-
шотную скорость расслабления и влияния Са на относительную
скорость падения напряжения в первую фазу было выдвинуто предполо-
90
Биофизика
жение о кооперативное™, согласно которой сродство миофибрилл
к кальцию возрастает по мере увеличения числа мостиков. Поэтому,
когда напряжение высоко, относительная скорость падения
напряжения падает даже в условиях фиксации длины саркомеров, когда одна
из существенных переменных находится на постоянном уровне,
анализ природы расслабления представляет большие трудности.
Ситуация становится еще более сложной при «естественном» ходе
расслабления, при котором механические события через одну или
несколько систем обратной связи влияют на образование и распад
мостиков, а особенно в условиях наличия кальция в митоплазме.
В этом случае на ход расслабления влияют многие функции, такие,
как «сила-скорость», «длина-сила» контрактильного элемента,
«длина-сила» последовательного элемента и т.д. При постоянной длине
саркомеров и отсутствии дисперсии их длин некоторые
неопределенности устраняются.
5.4.3. Работа сердца
Вследствие ритмического характера автоматизма сердцу
свойственна циклическая сократительная деятельность. Каждый цикл
сердечных сокращений состоит из ряда механических процессов,
идущих в строгой последовательности. Прежде чем характеризовать
эти процессы, отметим, что левый желудочек нагнетает кровь в
большой, а правый - в малый круги кровообращения. Кроме того, левое
предсердие совершает работу по наполнению левого желудочка и тем
самым способствует эффективности его нагнетательной функции. То
же делает правое предсердие по отношению к правому желудочку.
Поэтому принято условно говорить о левом сердце и правом сердце,
как о двух насосах, соединённых друг с другом последовательно
(выход одного является входом другого). Сразу заметим, что оба они
перекачивают примерно одинаковые объёмы крови за каждый цикл.
Проанализируем цикл сердечной деятельности при средней
частоте пульса (75 мин-1). В этом случае полное время сердечного цикла
(рис. 5.41) составляет 0,8 с. Сначала сокращаются предсердия.
Благодаря атрио-вентрикулярной задержке их сокращение (систола
предсердий) при расслабленных желудочках продолжается 0,1 с. Затем на-
Глава 5. Биофизика двигательной активности
91
Рис. 5.41. Цикл сердечной деятельности
Обозначения: внутренний круг - временная шкала (каждый сектор — 0,1 с);
внутреннее кольцо - систола (заштрихована) и диастола предсердий; наружное
кольцо — систола (заштрихована) и диастола желудочков; числами обозначены
различные фазы систолы (1—4) и диастолы (5-9) желудочков (1 - фаза
асинхронного сокращения, 2 — фаза изометрического сокращения,
3 — фаза быстрого изгнания крови, 4 - фаза медленного изгнания крови,
5 — протодиастолический интервал, 6 - фаза изометрического расслабления,
7 - фаза быстрого наполнения, 8 - фаза медленного наполнения, 9 - пресистола);
вертикальная стрелка указывает начало сердечного цикла;
стрелка по кругу - последовательность смены фаз
чинают сокращаться желудочки. В желудочковой систоле выделяют
ряд стадий (их называют фазами). Первая из них - фаза асинхронного
сокращения длится 0,05 с. В течение её нет полной синфазности в
возбуждении миокардиальных волокон. Из-за неодновременности
сокращений отдельных из них желудочки развивают слабое усилие, но
к концу фазы асинхронного сокращения степень синфазности
нарастает, многие волокна вовлекаются в сокращение, вследствие чего
достигается достаточная сила, чтобы создать внутри желудочков большее
кровяное давление (КД), чем в предсердиях (около 5 мм. рт. ст., что
эквивалентно 0,66 кПа.). В этот момент КД в магистральных
артериальных стволах, куда кровь затем должна быть выброшена желудочками,
шачительно выше: в аорте - примерно 60 мм. рт. ст. (8 кПа), в
лёгочном артериальном стволе-15 мм. рт. ст. (2 кПа).
92
Биофизика
Каждый из желудочков отделен от своего предсердия
створчатым, а от соответствующего магистрального ствола - полулунными
клапанами. Клапаны открываются и закрываются в зависимости от
соотношения КД в полостях, которые ими разделяются. Створчатые
клапаны закрыты тогда, когда КД внутри желудочков выше, чем
в предсердиях. Полулунные клапаны, напротив, открываются при
высоком давлении в желудочках (при КД в левом желудочке выше,
чем в аорте, и при КД в правом желудочке выше, чем в лёгочном
артериальном стволе). Когда в конце фазы асинхронного сокращения
внутрижелудочковое КД станет выше, чем внутрипредсердное,
створчатые клапаны закрываются, после чего желудочки
продолжают сокращаться в условиях герметичности своих полостей,
заполненных кровью. Следовательно, первая фаза систолы желудочков
сменяется второй - фазой изометрического сокращения.
Изометрический режим обусловлен тем, что сердечная мышца сокращается
вокруг несжимаемого тела - крови, содержащейся в желудочках, из
которых она не может выйти, так как обе пары отверстий перекрыты
клапанами. Существование фазы изометрического сокращения
является важным условием нормальной сердечной деятельности. Всё
усилие, развиваемое миокардом, идёт на повышение его
потенциальной энергии и, следовательно, на создание напряжения. В результате
КД внутри желудочков стремительно нарастает: в левом желудочке -
от 5 до 60 мм. рт. ст., в правом - от 5 до 10 мм. рт. ст. Это происходит
примерно за 0,03 с - такова продолжительность фазы
изометрического сокращения.
Благодаря достижению внутри желудочков КД, большего, чем
в магистральных артериальных стволах, открываются полулунные
клапаны, и кровь устремляется из сердца в сосуды. Вначале КД в
желудочках ещё продолжает повышаться, несмотря на открытие
клапанов. Этот режим сокращения называют изотоническим, что не
совсем точно выражает его характер, поскольку при нём КД не
удерживается на постоянном уровне. Например, в левом желудочке
давление повышается примерно на такую же величину, как и в
течение фазы изометрического сокращения (около 60 мм. рт. ст.), но не
за 0,03, а за 0,12 с. За это время кровь быстро вытекает из
желудочков в сосудистое русло. Поэтому третья фаза сердечного цикла назы-
Гпава 5. Биофизика двигательной активности
93
нается фазой быстрого изгнания. Затем ещё 0,13 с кровь вытекает из
желудочков в фазу медленного изгнания, причём в течение четвёртой
фазы скорость истечения крови неуклонно понижается, поскольку
падает внутрижелудочковое давление. Так, КД в полости левого
желудочка, достигшее к началу фазы медленного изгнания 120-130 мм.
рт ст. (16,0-17,3 кПа), к концу её становится меньше диастолическо-
ΐΌ давления в аорте. После расслабления - диастолы, в которой
также выделяют несколько фаз (см. рис. 5.41), наступает новое
сокращение. Сердечный цикл повторяется.
Как уже говорилось, сила, возникающая в сердечной мышце
при её сокращении, зависит от исходной длины миокардиальных
волокон, поскольку по мере укорочения волокна увеличивается число
миозиновых мостиков, участвующих в сокращении. Это общее
свойство мышечной ткани. В миокарде оно находит выражение в законе
Франка-Старлинга: сила сердечного сокращения прямо
пропорциональна исходной длине миокардиальных волокон (перед началом
систолы), а исходная длина зависит от степени растяжения их кровью,
поступающей из предсердий. Следовательно, сила сокращения
миокарда желудочков тем значительнее, чем больше крови поступает
н них во время диастолы. От диастолического наполнения
желудочков зависит основной показатель (параметр) сократительной
функции сердца - систолический (ударный) объём (Vc). Так называют
объём крови, выбрасываемый каждым из желудочков при одном
сокращении. Систолические объёмы левого и правого желудочков
одинаковы. Для каждого их желудочков Vc составляет 70-100 мл в усло-
11иях покоя, причём изгоняется 60-70% крови, содержавшейся там
и период диастолы. При физической работе и эмоциях Vc
увеличивается в 2-3 раза. От систолического объёма зависит работа сердца, но
сё значение может быть разным при одной и той же величине Vc
Работа, совершаемая сердцем, затрачивается, во-первых, на
выталкивание крови в магистральные артериальные сосуды против
сил давления и, во-вторых, на придание крови кинетической энергии.
Первый компонент работы называется статическим
(потенциальным), а второй - кинетическим.
Статический компонент работы сердца вычисляется по
формуле Аст = рср ■ Vc, где рср - среднее давление крови в соответствую-
94
Биофизика
рт.ст.
U
ί»
А
j
/
Рл
к
у
^
'
£
'
^
h
I
ν
\
Ε
β
С
д
'г
t, 1CH
с
КД, мм
150
100
50
Рис. 5.42. Кривая динамики кровяного давления (КД) в артерии
На графике показан способ планиметрического определения среднего давления:
определив площадь под кривой кровяного давления за время сердечного цикла
(площадь фигуры АабвгД) при помощи планиметра, строим равновеликий
ей прямоугольник АВСД, основание которого равно основанию фигуры АабвгД
(т.е. АД = At); тогда высота этого прямоугольника (ЕН) численно равна
величине среднего давления. Ρ с — систолическое давление;
Рд- диастолическое давление; tc = АД-время сердечного цикла (At)
щем магистральном сосуде (аорте - для левого желудочка, лёгочном
артериальном стволе - для правого желудочка). Поскольку
изменение КД в артериях является сложной периодической функцией,
среднее давление равно не полусумме максимального (систолического)
и минимального (диастолического) давлений, а среднему из
бесконечно малых изменений давления от максимального до
минимального в течение одного сердечного цикла (рис. 5.42).
Следовательно, среднее давление определяется так:
1 '2
Рср -— [ p(t)dt„ где At = t2 -1\ - время сердечного цикла. В отличие
Ί
от систолического и диастолического давлений, изменяющихся в
довольно широких пределах, /?ср характеризуется постоянством. И.П.
Павлов относил его к гомеостатическим константам организма.
Величина ρψ в большом круге кровообращения составляет
приблизительно 100 мм. рт. ст. (13,3 кПа). В малом круге рср « 15 мм. рт. ст.
(2 кПа), т.е. примерно в 6 раз меньше, чем в большом. Поскольку Vc
обоих желудочков одинаков, а давление, против которого они
совершают работу, имеет шестикратное различие, то и статический компо-
/ пава 5. Биофизика двигательной активности
95
мент работы левого желудочка приблизительно в 6 раз больше:
■1ИлевоГОудочка = 13>3 ' Ю3 Па χ 70 ■ 10^ м3 « 0,9 Дж; ^етправогожш1удочка «
« 0,15 Дж. В этих расчетах Vc принят равным 70 мл.
Кинетический компонент работы сердца определяется по фор-
. тм2 V м ,
муле: А. =р ——, где ρ - плотность крови (примерно
2 2
Ι03 кг · м~3), υ - скорость кровотока в магистральном артериальном
стволе (в среднем 0,7 м · с-1).
Следовательно, А. =- · 103кг ■ м"3 · 70 · 1(г6 м3 · 0,49 м2 · с"2 =
2
0,02 Дж.
В целом работа левого желудочка за одно сокращение в усло-
ниях покоя составляет около 1 Дж, а правого - менее 0,2 Дж, причём
статический компонент доминирует, достигая 98% всей работы,
тогда как на долю кинетического компонента приходится всего 2%.
Средняя мощность миокарда поддерживается на уровне 1 Вт.
При физической работе и эмоциях повышается КД в
магистральных сосудах, увеличивается Vc, но в ещё большей степени
возрастает скорость кровотока. К тому же Ак зависиг от квадрата этой
скорости. Поэтому при физических и психических нагрузках вклад
кинетического компонента в работу сердца становится весомее (до
30% всей работы), чем в покое. Например, при выполнении тяжёлой
физической работы тренированным человеком его рср достигает
16 кПа, Vc - 200 мл, υ - 3 м · с-1. Тогда работа левого желудочка
достигает .4 = 16· 103 · 200· 10-6 + -· 103· 200· 10"6 · 9 = 3,2 + 0,9 =
2
" 4,1 Дж. Средняя мощность возрастает до 8,2 Вт. Приведённые
числа не являются предельными.
5.4.4. Методы исследования
механической активности сердца
Баллистокардиография (от греч. balk - бросаю, kardia - сердце
и grapho - пишу) - метод исследования механических проявлений
сердечной деятельности, выражающихся в смещениях тела человека.
96
Биофизика
Баллистокардиография состоит в регистрации этих смещений,
вызванных «отдачей» при сокращении сердца и выбросе крови в аорту
и лёгочную артерию и движением крови по сосудистому руслу.
Баллистокардиография получила распространение в медицинских
исследованиях начиная с 1939 г., после работ американского исследователя
А. Старра с сотрудниками. Для баллистокардиографии используют
специальные приборы - баллистокардиографы Различают два типа
баллистокардиографии: 1) непрямую, при которой регистрируются
перемещения подвижного стола с находящимся на нем пациентом,
и 2) прямую, когда регистрируются движения тела непосредственно.
В обоих случаях перемещение тела пациента обусловлены выбросом
крови из сердца в магистральные артерии.
Баллистокардиографы состоят из механической части,
воспринимающей движения исспытуемого, преобразователя,
превращающего эти механического движения в электрические сигналы, и
электронной части, обеспечивающей усиление и регистрацию этих
сигналов. При непрямой баллистокардиографии механической
частью является подвижный стол, на котором располагается пациент,
при этом датчик (механоэлектрический преобразователь)
помещается на подвижном столе. При прямой баллистокардиографии датчик
накладывается на любой участок тела (чаще на голень) и с его
помощью регистрируется смещение этого участка тела. В зависимости от
частотной характеристики различают ультранизкочастотные
(собственная частота 0,25 и 0,5 Гц), низкочастотные {\-А Гц) и
высокочастотные (15-30 Гц) баллистокардиографы.
Обычно регистрируют смещения тела вдоль продольной оси.
Можно также регистрировать скорость и ускорение перемещений
тела, возникающих в результате сердечной деятельности. Движения
записываются в виде кривой - баллистокардиограммы (БКГ). На этой
кривой различают волны (зубцы), обозначаемые буквами латинского
алфавита от Η до О. Волны Н, I, J, к возникают во время сокращения
(систолы) желудочков, L и следующие - во время расслабления
(диастолы). По изменению БКГ судят о состоянии сократительной
функции миокарда и функции сердечно-сосудистой системы в целом.
Изменения баллистокардиограммы указывают на нарушения сердечной
деятельности.
( пава 5. Биофизика двигательной активности
97
Динамокардиография - метод исследования механических про-
инлсний сердечной деятельности, основанный на регистрации
перемещений центра тяжести грудной клетки в результате сердечной
кинематики и движения крови в крупных сосудах. Динамокардиография
разработана в 1951 г. Е.Б. Бабским с сотрудниками. Динамокардио-
I раф состоит из тензометрического устройства, преобразующего
динамические усилия в электрические сигналы и
усилительно-регистрирующей аппаратуры. Тензометрическое устройство вмонтировано
и крышку стола, на котором лежит пациент, но на подвижной части
крышки располагают только его грудную клетку. Посредством дина-
мокардиографа регистрируют перемещения центра тяжести грудной
клетки вдоль продольной оси тела и перпендикулярно ей.
Обычно при регистрации синхронно записывается продольная
(ДКГ-1) и поперечная динамокардиограммы (ДКГ-2). Основные
зубцы кривой (рис. 5.43), соответствующие точкам перегиба,
обозначаются буквами А, В, С, D, E, F, G, а интервалы между ними - римскими
цифрами. В интервалах I, II, III, IV имеются расщепления (q, ζ, χ,у).
Каждый зубец и интервал соответствует определенной фазе
или периоду в работе сердца. АВ (I)- отражение систолы
предсердий; точка В - начало систолы желудочков; точка ζ - начало
изгнания крови из желудочков; zC - перемещение сердца в начальный
период изгнания крови из желудочков; CD (III) - перемещение массы
крови по аорте и легочной артерии; DEFG (IV-VI)-период
расслабления желудочков; GHA (VII) - наполнение желудочков кровью;
интервалы q-B и B-z отражают соответственно длительность фаз
асинхронного и изометрического сокращения; интервал z-y - период
изгнания; интервал Ε - протодиастола.
Динамокардиограмма обнаруживает характерные изменения
деятельности сердца при некоторых сердечных заболеваниях и в
сочетании с электрокардиограммой позволяет определять фазы
сердечного цикла.
Кинетокардиография (от греч. kinetos - движущийся,
подвижный, кардио... и...графия) - метод электрической регистрации
низкочастотных вибраций грудной стенки, обусловленных сокращениями
сердца. Кинетокардиография основана на преобразовании
механических колебаний в изменение какого-либо электрического параметра
98
Биофизика
ДКГ-1
ДКГ-2
Рис. 5.43. Продольная (ДКГ-1) и поперечная (ДКГ-2) динамокардиограммы
датчика, приложенного к грудной клетке обследуемого. Полоса
регистрируемых частот - в пределах 1-25 Гц. Чаще регистрируют
вибрации в двух точках грудной клетки, соответствующих проекции
левого и правого желудочков (на 2 см левее грудины, на уровне 5-го
ребра, и справа от грудины, у места прикрепления 4-5-го ребер).
Регистрируемая кривая состоит из серии зубцов, отражающих
различные фазы сердечного цикла: систолу предсердий, периоды
асинхронного и изометрического сокращения желудочков, быстрого
и медленного изгнания из них крови, быстрого и медленного их
наполнения. Кинетокардиография позволяет выявить изменения
сердечной деятельности при некоторых заболеваниях и оценить
эффективность лечения.
Фонокардиография (от греч. phone - звук и кардиография) -
метод графической регистрации звуков, рождаемых сердцем при его
( пава 5. Биофизика двигательной активности
99
механической активности: сердечных тонов и сердечных шумов. Она
применяется в дополнение к аускультации (выслушиванию),
позволяет объективно оценить интенсивность и продолжительность тонов
и шумов, их характер и происхождение, записать неслышимые при
аускультации 3-й и 4-й тоны.
Специальный аппарат для фонокардиографии - фонокардиог-
раф — состоит из микрофона, усилителя электрических колебаний,
системы частотных фильтров и регистрирующего устройства.
Микрофон прикладывают к разным точкам грудной клетки над областью
сердца. После усиления и фильтрации электрические колебания
поступают на различные каналы регистрации, что позволяет
избирательно фиксировать низкие, средние и высокие частоты. Запись ФКГ
производят в звукоизолированном помещении при задержке
дыхания на выдохе (при необходимости - на высоте вдоха) в положении
лёжа, после отдыха исследуемого в течение 5 мин (рис. 5.44).
На ФКГ прямая (изоакустическая) линия отражает
систолические и диастолические паузы. Нормальный 1-й тон состоит из 3
ipynn осцилляции: начальной (низкочастотной), обусловленной
сокращением мышц желудочков; центральной (большей амплитуды),
обусловленной закрытием митрального и трикуспидальнего
клапанов; конечной (малой амплитуды), связанной с открытием клапанов
аорты и лёгочной артерии и колебаниями стенок крупных сосудов.
2-й тон состоит из 2 групп осцилляции: первая (большая по
амплитуде) обусловлена закрытием аортальных клапанов, вторая связана
с закрытием клапанов лёгочной артерии. Нормальные 3-й (связан
с мышечными колебаниями при быстром наполнении желудочков)
и 4-й (встречается реже, обусловлен сокращением предсердий) тоны
ι π ι и
Рис. 5.44. Фонокардиограмма
I - нормальный тон, обусловленный сокращением мышц желудочков,
II-второй тон, обусловленный закрытием артериальных клапанов
г
100
Биофизика
определяются преимущественно у детей и у спортсменов.
Характерные изменения ФКГ (ослабление, усиление или расщепление 1-го
и 2-го тонов, появление патологических 3-го и 4-го тонов,
систолических и диастолических шумов) помогают распознавать пороки
сердца и некоторые др. заболевания.
Поликардиография, представляющая собой синхронную запись
фонокардиограммы (ФКГ), электрокардиограммы и кривой
центрального пульса, позволяет определить длительность фаз
сердечного цикла, т.е. получить косвенные данные о сократительной
способности миокарда.
5.5. Биофизические закономерности
движения крови по сосудам
Движение крови по кровеносным сосудам изучает
гемодинамика, являющаяся одним из разделов биомеханики. Принято выделять
две группы показателей, интегрально характеризующих
кровообращение. Их называют основными гемодинамическими показателями.
Первую группу составляет скорость кровотока, вторую - кровяное
давление (КД).
5.5.1. Основные показатели гемодинамики
Различают объёмную и линейную скорости кровотока.
Объёмной скоростью (Q) называют объём жидкости (V),
протекающей в единицу времени: Q = VI t. Применяются разные единицы
измерения этой величины: мл · с-1, л · мин-1 и др. Линейная скорость
(ν) представляет собой путь (/), проходимый частицами крови в
единицу времени: ν =//1 [м · с-1]. Объёмная и линейная скорости
связаны простым отношением Q = ν ■ S, где S - площадь поперечного
сечения потока жидкости.
Для сплошного течения несжимаемой жидкости выполняется
условие неразрывности струи: через любое сечение струи в единицу
времени протекают одинаковые объёмы жидкости: Q = υ · S = const.
Так формулируется в гидродинамике закон сохранения массы.
Поскольку для кровообращения характерно сплошное течение, условие
Глава 5. Биофизика двигательной активности
101
неразрывности струи выполняется и в гемодинамике, где принята его
следующая формулировка: в любом сечении сердечно-сосудистой
системы объёмная скорость кровотока одинакова: Q = const. В покое
она составляет примерно 5 л · мин-1. Словосочетание «любое сечение
сердечно-сосудистой системы» требует пояснения. Одной из моделей
круга кровообращения (как большого, так и малого) служит так
называемая разветвлённая сосудистая трубка (рис. 5.45, а). Каждое её
сечение представляет собой поперечный разрез всех кровеносных
сосудов одного уровня ветвления. Например, в большом круге
кровообращения первое сечение проходит через аорту, второе - через все
артерии, на которые разветвляется аорта непосредственно, третье -
через все ветви этих артерий и т.д. В одно сечение попадают все
капилляры большого круга кровообращения. Площадь последнего
сечения большого круга (перед правым предсердием) равна сумме
площадей поперечных разрезов верхней и нижней полых вен, которыми он
оканчивается.
Из рис. 5.45, а следует, что самым узким сечением в большом
круге кровообращения обладает аорта (S около 4 см2). Суммарная
площадь сечения обеих полых вен немного больше. Самое обширное
сечение сосудистой трубки приходится на уровень кровеносных ка-
а) б)
Рис. 5.45. Схема разветвления сосудов в большом круге кровообращения (а)
(модель разветвлённой сосудистой трубки) и соотношение между
суммарным поперечным сечением каждой генерации сосудов большого
круга кровообращения (S) на разных уровнях (сплошная линия) и линейной
скоростью кровотока (ν) в соответствующих сосудах (штриховая линия) (б)
Цифрами обозначены уровни сечения: I - аорта; 2 — капиллярное русло;
3 - полые вены (верхняя и нижняя)
102 Биофизика
пилляров (примерно 11 000 см2, из которых лишь через 3 000 см2
течёт кровь, а остальные капилляры в покое находятся в спавшем
состоянии). Следовательно, в большом круге кровообращения площадь
суммарного просвета капилляров, в которых есть кровоток, в 700-
800 раз больше поперечного сечения аорты. С учётом условия
неразрывности струи это означает, что линейная скорость кровотока в
капиллярной сети в 700-800 раз меньше, чем в аорте, и составляет
около 1 мм · с-1. В покое средняя ν в аорте лежит в пределах от 0,5 до
1 м · с-1, а при большой физической нагрузке может достигать
20 м · с-1. Ускорения в аорте также максимальны и колеблются от 1 g
до 8 g, где g - ускорение свободного падения (9,81 м · с-2). Средняя
линейная скорость кровотока в полых венах 0,10 - 0,16 м · с-1, но
может достигать 0,5 м · с-1. Время полного кругооборота крови равно 27
систолам сердца, т.е. при частоте сердечных сокращений около
75 мин-1 кругооборот крови занимает 20-23 с. Из них 4/5
затрачивается на движение в большом и 1/5 - в малом кругах кровообращения.
В разных точках сечения сосуда кровь движется с
неодинаковой скоростью. Она максимальна на продольной оси сосуда и
постепенно падает до нуля у стенки (пристеночный слой текущей
жидкости неподвижен). Между слоями движущейся по сосуду крови
существует градиент скорости. Его принято называть скоростью
сдвига: γ = — [с-1 ].Из формулы Ньютона следует, что сила внутрен-
dy
него трения (вязкость) текущей жидкости определяется скоростью
F
сдвига: F =—η· s-y. Коэффициент вязкости: η = —— [Па-с]. У цель-
s-y
ной крови η, измеренный на вискозиметре, составляет около
5 мПа ■ с (5 сП), что в 5 раз больше вязкости воды. При патологии η
крови колеблется от 1,7 до 22,9 мПа · с. При скоростях сдвига
меньше 100 с-1 наиболее соответствует опытным данным в случае
движения крови эмпирическое уравнение Кессона:
I пава 5. Биофизика двигательной активности
103
где ση - характерная константа (предел текучести): кровь приходит
|| движение лишь при ас > ση. Значение параметра кс для крови 0,05-
0,15 Па05 · с0·5, а ση составляет 0,001-0,005 Н/м2. При высоких
величинах напряжения сдвига (а значит, и при высоких градиентах
скорости) ас » ση, и уравнение (5.64) превращается в уравнение Ньютона
при η = к]. Все эти свойства крови обусловлены тем, что при низких
скоростях сдвига в ней имеются агрегаты эритроцитов в виде
монетных столбиков. Эти агрегаты распадаются по мере увеличения
скорости сдвига, и поэтому эффективная вязкость снижается и
приближается постепенно к определенному пределу.
Между вязкостью крови и воды обнаружена не только
количественная, но и качественная разница. В воде, как и других
однородных жидкостях, η не зависит от скорости сдвига, а значит, и от
линейной скорости. Такие жидкости называют ньютоновскими.
У крови же вязкость принимает разные значения при различных
скоростях сдвига. Так, η = 5 мПа ■ с при сравнительно большой υ крови,
присущей методике определения вязкости (вискозиметрии) и
движению крови в аорте. При малых скоростях сдвига, свойственных более
мелким сосудам, η оказывается заметно больше: в мелких артериях -
10 мПа · с, в капиллярах - до 800 мПа · с. Кровь вместе с другими
жидкостями, вязкость которых зависит от скорости сдвига,
относится к неньютоновским жидкостям. Они представляют собой
неоднородные жидкие среды, в основном, суспензии.
В крови зависимость η от скорости сдвига связана с наличием
форменных элементов (прежде всего, эритроцитов). При самых
низких скоростях сдвига (γ порядка 1 с-1) большая часть эритроцитов
объединяется в группы из нескольких клеток, разделённые плазмой.
')ти конгломераты образуют жёсткие структуры (рис. 5.46), что
придаёт крови некоторые свойства твёрдого тела. Когда же напряжение
сдвига превосходит предел текучести такого твёрдого тела (это
достигается при повышении υ), конгломераты изгибаются и
разрушаются, вследствие чего вязкость крови, движущейся достаточно
быстро, начинает уменьшаться, причём тем в большей степени, чем выше
скорость кровотока. Следовательно, различия вязкости крови при
разных скоростях (и, значит, в разных сосудах) обусловлены обрати-
104
Биофизика
а)
6)
в)
г)
Рис. 5.46. Образование конгломератов (а, б) и монетных
столбиков (в, г) из эритроцитов (Э) при медленном кровотоке
а, б — сканирующая электронная микроскопия; в — электронная
микроскопия; г — схема образования монетных столбиков
мой агрегацией эритроцитов. Поскольку эти агрегаты распадаются
с увеличением скорости сдвига, эффективная вязкость Э снижается
и постепенно приближается к определенному пределу (рис. 5.47).
Глава 5. Биофизике двигательной активности 105
При большой скорости сдвига кровь можно рассматривать
просто как суспензию и изучать ее механические свойства на двух
моделях: суспензиях эритроцитов в физиологическом растворе и
суспензиях других частиц. Это справедливо для крови, текущей в круп-
пых артериях. Течение крови по таким сосудам зависит от
концентрации и физических свойств эритроцитов.
Вязкость суспензии сферических твердых частиц и суспензии
эритроцитов сильно увеличивается при повышении концентрации
(рис. 5.48). Концентрационная зависимость вязкости суспензии
эритроцитов незначительно отличается от концентрационной зависимо-
стисуспензии сферических частиц, если оболочки эритроцитов
сделать жесткими. Последнее может быть достигнуто, например, путем
фиксации белков, выдерживанием эритроцитов в глутаровом
альдегиде. Однако кривая для нормальных, нефиксированных эритроцитов
тметно отличается от кривой для сферических частиц или «жестких»
эритроцитов (рис. 5.48). При этом вязкость суспензии эритроцитов
при значении гематокрита, соответствующем нормальной крови, т.е.
немного более 40%, почти вдвое ниже вязкости жестких эритроцитов
/)э, ПаАс
0,06
0,04
0,02
0
0,1 1 10 100 1000 dvldx.c-i
Рис 5.47. Эффективная вязкость крови человека (1)
и вязкость некоторой ньютоновской жидкости (2)
при разных скоростях деформации сдвига. Величины скорости
сдвига dv/dx представлены в логарифмическом масштабе.
Вязкость ньютоновской жидкости взята равной предельной вязкости крови
106
Биофизика
и тем более ниже вязкости жестких сферических частиц. Благодаря
дисковидной форме клеток и эластичности оболочки суспензия
эритроцитов обладает сравнительно невысокой вязкостью, что важно для
уменьшения нагрузки на сердце, которое прокачивает кровь по
кровеносным сосудам. Увеличение жесткости стенок эритроцитов при
патологических процессах приводит к возраста*шю вязкости крови
и к ухудшению кровообращения.
Вязкость медленно текущей крови (ηκ) можно приближенно
рассчитать по формуле
(1-*,С)
где η0 - вязкость кровяной плазмы (около 1,2 мПа · с); А", -
коэффициент > 2,5; С- объём форменных элементов крови в 1 см3 (около
ю
б
б
4
з
2
0
20 40 60 60 с. %
Рис. 5.48. Концентрационная зависимость вязкости
суспензии эритроцитов собаки в норме (1), жестких эритроцитов (2)
и суспензии твердых сферических частиц (3)
η/ηο - относительная вязкость в логарифмическом масштабе;
с - объемная концентрации (гематокрит). Градиент скорости 230 с"1
I лава 5. Биофизика двигательной активности
107
0,42-0,45 см3). Величина цк («кажущейся вязкости») зависит прежде
псего от свойств форменных элементов крови, 93% которых
составляют эритроциты. Поэтому вязкость крови определяется главным
образом свойствами эритроцитов, причём не только их агрегацией. При
повышении жёсткости эритроцитарной мембраны, например, при
атеросклерозе, вязкость крови также возрастает, что приводит к уве-
ничению нагрузки на сердце.
Вязкость крови неодинакова в широких и узких сосудах,
причём влияние диаметра кровеносного сосуда на вязкость начинает
сказываться при просвете менее 1 мм. В сосудах тоньше 0,5 мм η
уменьшается прямо пропорционально укорочению диаметра (эффект
Фареуса-Линдквиста), поскольку в них эригроциты выстраиваются
идоль центральной оси в цепочку наподобие змейки и окружены
слоем плазмы, изолирующей «змейку» от сосудистой стенки. В этой
(»ссклеточный краевой зоне вязкость невелика (около 2 мПа · с), что
несколько понижает величину кажущейся вязкости крови в артерио-
лах и капиллярах, но всё-таки она достигает 800 мПа · с.
От вязкости крови зависит характер её течения по сосудам,
которое может быть ламинарным или турбулентным. Если слои жидко-
ι· πι движутся параллельно друг другу, не смешиваясь между собой, то
такое течение называют ламинарным. Когда же ток жидкости сопро-
иождается перемешиванием слоев, обусловленным образованием
нихрей в ней, то говорят о турбулентном (вихревом) движении.
Переход одного вида течения в другой определяется числом Рейнольдса
... λ _ ρ · d d η
(Ke): Re =- ν = —ν, где ρ - плотность жидкости, ν = — - кинемати-
η ν ρ
ческая вязкость, d- диаметр сосуда, ν - линейная скорость движения
жидкости. Существует критическое значение числа Рейнольдса
(Кс ), которое служит граничным параметром перехода ламинарного
ючеиия в турбулентное. Если фактическое значение Re < Re^, то
жидкости свойственно ламинарное течение, тогда как при Re > Re^ в
потоки нозникают вихри, и движение приобретает турбулентный характер.
Для однородной жидкости Re^ = 2300, для крови Re^ = 970 ± 80, но
уже при Re > 400 возникают локальные завихрения в разветвлениях
иргерий и в области их крутых изгибов. Ламинарное течение крови
108
Биофизика
создаёт меньшую нагрузку на сердце, поскольку при нём работа
сердца (А) прямо пропорциональна объёмной скорости кровотока (Q),
тогда как при турбулентном движении крови между ними установлена
почти квадратичная зависимость (А ~ β1,8). В физиологических
условиях кровоток в магистральных сосудах имеет преимущественно
ламинарный характер. При тяжёлой физической нагрузке он может
стать турбулентным из-за повышения скорости. Переход к
турбулентному течению крови в сосудистом русле сопровождает ряд
заболеваний, отягощая их. Например, при анемии (малокровии) Re возрастает
за счёт понижения кинематической вязкости крови вследствие
уменьшения количества эритроцитов и нарушения их свойств.
А.Л. Чижевский предполагал, что в сосудах определётюго
диаметра (порядка 100-1000 мкм) эритроциты совершают не только
поступательное, но и вращательное движение. В конце XX века это удалось
увидеть при прижизненной микроскопии сосудистого русла in situ.
Вязкость крови оказывает существенное влияние и на второй
основной гемодинамический показатель - кровяное давление (КД).
Это сила, с которой движущаяся кровь воздействует (давит) на
единицу площади стенки кровеносного сосуда. Анализ факторов,
определяющих КД, целесообразно проводить исходя из уравнения Пуазейля:
8η/
где Q - объёмная скорость кровотока, г - радиус сосуда, / - его
длина, η - вязкость крови, Ар - разность давлений на концах сосуда.
Если кровь течёт по нескольким одинаковым сосудам, включенным
параллельно, то
8η/
где η - число сосудов данного типа (калибра).
Для анализа факторов, от которых зависит КД, решим
уравнение Пуазейля относительно Ар: Ар = η —— · Q и, обозначив сомножи-
/ лава 5. Биофизика двигательной активности
109
гель —— символом RT, запишем: Ар = Rr · Q. При такой записи урав-
мспие Пуазейля обнаруживает сходство с законом Ома: AU = R · I,
гели признать аналогию потенциальных (AUu Ар), а также потоковых
(/ и Q) величин. Заметим, что между ними существует не формальное,
и смысловое сходство. Тогда RT аналогично электрическому
сопротивлению^). И действительно, величина .К г = —— отображает сопро-
ι пиление сосудистого русла кровотоку, включая все факторы, от ко-
1п|)ых оно зависит. Поэтому RT называют гемодинамическим
сопротивлением (или общим периферическим сопротивлением
сосудов - ОПСС). Его единица измерения в системе СИ - [Н · м-5 · с =
lia · с · м-3]. В гемодинамике обычно используют другую единицу:
|11а · с ■ млг1]. В большом круге кровообращения ОПСС и
140 Па · с · мл-1, а в малом - на порядок меньше (около 11 Па · с · мл"').
И клад в ОПСС большого круга различных его частей характеризуют
ι'недующие величины: аорта и крупные артерии - 19%, артериолы -
М)%, капилляры - 25%, вены - 4%, артерио-венозные анастомозы
и еосуды-сфинктеры - 3%.
Аналогия, существующая между законом Пуазейля и законом
I )ма, позволяет моделировать кровообращение при помощи электри-
■ц'ских цепей. В электротехнике хорошо разработаны методы расчёта
м женериментального исследования сложных (разветвлённых) элек-
фических цепей. Применяя эти методы к изучению кровообращения
пи его электрических моделях, приходят к важным теоретическим
и практическим выводам о закономерностях гемодинамики.
Электрическое моделирование сердечно-сосудистой системы оказалось
тчч.ма плодотворным при создании аппаратов искусственного
кроит вращения, в протезировании сердца и сосудов, а также во многих
'||>угпх приложениях.
Анализ уравнения Пуазейля, разрешённого относительно Ар,
ι имдетельствует, что КД зависит от объёмной скорости кровотока и,
ι щ-донательно, от массы циркулирующей крови и сократительной
к'шелыюсти миокарда, определяющих эту скорость. Ещё более вы-
|||Ыичшое влияние на динамику КД оказывает гемодинамическое со-
по
Биофизика
противление (ОПСС), прежде всего такой его компонент, как радиус
сосуда, поскольку в формулу он входит в четвертой степени (г4).
Поэтому изменение радиуса на 20% (при Q = const) приводит к
повышению КД в сосуде в (1,2)4 раз, т.е. более чем вдвое. Даже небольшие
колебания просвета кровеносных сосудов сильно сказываются на
кровообращении. Не случайно регуляция уровня КД в организме
связана с нервными и гуморальными влияниями прежде всего на
гладкомышечную оболочку кровеносных сосудов в целях активного
изменения их просвета. Сюда же направлены основные
фармакологические средства нормализации КД.
Проанализируем изменения Rr в системе кровообращения на
модели разветвлённой сосудистой трубки (рис. 5.45, а). Все крупные
артерии имеют большой радиус, который к тому же мало изменяется
в обычных условиях. Поэтому их вклад вЛги его изменения
незначителен, хотя общая длина всех крупных артерий сравнительно
велика. По мере удаления от желудочков сердца число артерий,
включённых параллельно кровотоку, возрастает. Поскольку при
параллельном включении в кровообращение многих (и) сосудов Rr
падает в η раз, вклад этого звена артериального русла должен быть,
казалось бы, меньшим по сравнению с вкладом крупных артерий, тем
более, что по мере удаления от сердца после каждого разветвления
артерии становятся всё короче. Однако по мере ветвления артерий
уменьшается их просвет и, поскольку Rr зависит от г4, гемодинами-
ческое сопротивление становится тем больше, чем дальше от сердца
расположено данное артериальное звено. Особенно резкое
увеличение Rr наблюдается на уровне артериол (рис. 5.49).
Переход от артериол к капиллярам характеризуется
значительным увеличением количества параллельно включенных сосудов,
тогда как радиусы прекапилляра (сосуда, принадлежащего последнему
звену артериол) и капилляра примерно одинаковы. Поэтому общее
Rr капиллярной сети приблизительно в четыре раза меньше, чем
артериол. В венозном русле гемодинамическое сопротивление меньше,
чем в капиллярном. Отмеченные особенности Rr в различных
звеньях кровеносного русла определяют распределение КД в
сердечно-сосудистой системе человека (рис. 5.50).
I пава 5. Биофизика двигательной активности
111
Рис. 5.49. Гемодинамическое сопротивление (Rp)
разных отделов кровеносного русла
I - аорта; 2 —магистральные артерии; 3 - артериолы; 4 — капилляры; 5 - вены
Артериальные Венозные
сосуды сосуды
КД, мм рт. ст.'1
100
80
60
40
20
7060 5040 30 2010Ю20 3040 50 6070 d сосуда, мкм
б)
Рис. 5.50. Распределение кровяного давления в разных сосудах
большого круга кровообращения (а) и в микрососудах брыжейки (б)
I - аорта; 2 - крупные вены; 3 - мелкие артерии; 4 - артериолы;
5 - капилляры; б - венулы; 7 - вены; 8 - полые вены
112
Биофизика
В аорте и крупных артериях падение давления невелико (КД
в начале и конце таких сосудов почти одинаково). В артериолах
наблюдается максимальный перепад давления - свыше половины
общего падения КД в сосудистом русле. На рис. 5.50, α показано, что
в крупных и средних артериях КД неодинаково в систолу и диастолу.
Принято различать систолическое (максимальное) и диастолическое
(минимальное) КД, а также пульсовое давление крови, равное
разности их и составляющее в большом круге примерно 40 мм рт. ст.
(5,3 кПа), поскольку первое из них 110-130, а второе-60-80 мм рт ст.
В малом круге кровообращения эти величины составляют
соответственно 20-30 и около 10 мм, а пульсовое давление- 10-20 мм рт. ст.
Кроме того, измеряют среднее КД. Названные виды кровяного
давления составляют вторую группу основных гемодинамических
показателей - показателей давления крови. Понятно, что между
артериальным, капиллярным и венозным давлениями крови существует
большая разница (рис. 5.50, б). В капиллярах большого круга КД
падает (от 30 до 15 мм рт. ст. на протяжении всего 0,75 мм), а в малом
круге капиллярное давление около 7 мм рт. ст. В венулах большого
круга КД = 15-20 мм рт. ст., а в крупных венах (за пределами
грудной полости) падает до 5-6 мм рт. ст. В предсердиях КД имеет
отрицательное значение (на 2—4 мм рт. ст. ниже атмосферного).
Волна давления, как установил Η. Ε. Жуковский,
распространяется по жидкости со скоростью звука (порядка 103 м · с-1).
Следовательно, в самых удалённых от сердца артериях большого круга
кровообращения уровни систолического и диастолического давлений
достигаются мгновенно после того, как они устанавливаются в аорте.
То же происходит и в малом круге относительно лёгочного
артериального ствола. Нельзя путать огромную скорость распространения
давления с низкой линейной скоростью кровотока (1-10_3 м · с-1),
а также со скоростью распространения пульсовой волны по
сосудистой стенке - порядка Юме-1.
В начале говорилось, что в физиологии и биофизике принято
выделять 2 группы основных гемодинамических показателей.
Первый из них (скорость) служит потоковой характеристикой, а второй
(кровяное давление) - потенциальной. Они исчерпывающим образом
характеризуют линейные процессы массопереноса, поскольку коэф-
/ паев 5. Биофизике двигательной активности 113
фициент связи между ними - постоянная величина. В гемодинамике
W, является функцией скорости, что свидетельствует о нелинейном
характере движения крови по сосудам. Поэтому Rr следует считать
фстьим основным гемодинамическим показателем, так как он не
определяется однозначно из первых двух.
5.5.2. Линейная теория движения крови
по кровеносному руслу
Примем, что кровеносный сосуд имеет цилиндрическую
форму с радиусом просвета (поперечного сечения) г. Рассмотрим
движение цилиндрического элемента крови размером dx (рис. 5.51)
(величина dx - бесконечно малая). Движение этого элемента вдоль
ишшной оси χ сосуда слева направо вызывается силой, которая
обусловлена разностью давлений dxp между правой и левой границей,
и равна S ■ dxp; S = nr2- площадь просвета сосуда). В dxp нижний ин-
Ш'кс обозначает то, что дифференциал частный, т.е. находится при
тменении χ на dx при постоянных других переменных.
I , , , I
Рис. 5.51. Схема к выводу уравнений движения крови
по кровеносному сосуду (1) и схема электрической линии,
эквивалентной наполненному кровью сосуду (2)
г - внутренний радиус сосуда; h - толщина стенки сосуда;
х—расстояние вдоль продольной оси сосуда; R, L, С — активное
сопротивление, индуктивность и емкость (соответственно).
На вход электрической линии подается импульс, эквивалентный пульсу
< ЧК'Н
114
Биофизика
Эта сила создает ускорение массы указанного элемента крови
и преодолевает силу вязкостного трения, так что по второму закону
Ньютона без учета силы тяжести имеем выражение:
-Sdxp = dx^ + dxp, (5.65)
at
в котором ν - средняя (по площади просвета) скорость движения
частиц крови; — ускорение; d/n - масса элемента крови, равная pSdx
dt
(ρ - плотность крови; Sdx - объем элемента крови).
Будем считать, что вязкое движение крови по сосуду
подчиняется закону Пуазейля. Тогда
dxf=n^Qdx, (5.66)
где η - вязкость крови.
Величина Q = Sv представляет собой объемную скорость
кровотока. Подстановка выражения (5.66) в уравнение (5.65), деление
уравнения (5.65) на dx и S и замена отношений дифференциалов на
частные производные фр I дх вместо dj)ldx и т.д.) дают:
ψ. = ρψ. + *ψΩ. (5.67)
дх dt S2
Важно далее, что крупные кровеносные сосуды при изменении
внутрисосудистого давления в ходе сокращений сердца способны
к быстрой упругой деформации, в результате чего возникает
распространяющаяся по сосудам пульсовая волна. Это можно пояснить
следующим образом. Под действием сокращения сердца в отрезок
сосуда dx за небольшой период dt подается порция крови и давление
здесь возрастает. Из-за инерции крови это вызовет не ее движение
вдоль сосуда, а расширение сосуда и вход крови в образовавшееся
расширение. Затем упругая сила стенки сосуда начнет выталкивать
избыток крови в соседний участок, где все описанные события
повторяются: по сосуду распространяется импульс давления, скорости
' нова 5. Биофизика двигатепьной активности
115
кровотока и деформации сосудистой стенки. Скорость
распространения этого импульса (т.е. пульса) намного выше средней скорости
течения крови.
Рассмотрим теперь уравнение, описывающее количественно
и вменение во времени давления крови и скорости кровотока по ходу
сосуда в связи с его деформацией. Пусть деформация сосуда
происходит за время dt. Получаем связь между скоростью изменения дав-
мсния и скоростью деформации сосуда:
dlL=EaJLd2l
dt 2r2S dt
где Е - эффективный модуль упругости стенки сосуда в
рассматриваемый момент времени; нижний индекс t - частный дифференци-
iiji по времени, aQ -r-h.
Выразим άβ/dt через изменение объемной скорости кровотока.
Рассмотрим участок сосуда длиной dx. Объем этого участка за время
ill в результате расширения сосуда увеличивается на d,Sdx. Так как
кровь практически несжимаема и течет по сосуду неразрывной
пруей, указанное изменение объема равно разности между объемом
крови Q{dt, втекающей в участок dx, и объемом крови Q2dt, которая
имтекает (т.е. втекает в соседний участок):
dl=Sdx=Qidt-Q2dt = -dIQdt, (5.69)
где dxQ - изменение объемной скорости кровотока по длине
сосуда. Уравнение (5.69) можно переписать в виде
d'S~ d*Q (5.70)
dt dx
При подстановке (5.70) в (5.68) получаем
Ър _ Еа0 dQ
dt 2r2S dx '
(5.71)
Дифференциальные уравнения (5.67) и (5.71) образуют
систему, которая отражает взаимную зависимость давления и объемной
116
Биофизика
скорости кровотока и описывает их изменение по ходу сосуда и во
времени.
Строго говоря, величины параметров η, Ε и тем более S и г
в уравнениях (5.67) и (5.71) могут изменяться при сдвиге давления
в сосуде. Однако при высоких скоростях кровотока, характерных для
магистральных сосудов, вязкость крови можно считать постоянной
величиной. При быстрых изменениях давления, характерных для
пульса, величина эффективного модуля упругости стенки кровеносных
сосудов Ε достигает больших величин и, следовательно, S
изменяется незначительно. Наконец, сам модуль Ε можно считать постоянной
величиной при небольших изменениях S. Таким образом, можно
принять за постоянные величины все параметры сосуда и крови,
стоящие перед производными в выражениях (5.67) и (5.71). Введем
новые постоянные:
8πη 2rS_ p
S2 Eh S '
Теперь уравнения (5.67) и (5.71) приобретают простой вид:
дх dt
dp _ IdQ
я, RQ> (573)
οχ dt
(5.74)
dt С дх
Такой же вид имеют уравнения, известные в электротехнике
и описывающие изменение электрического потенциала вдоль элект-
рическои цепи
и во времени
удх) ν
dt
если эта электрическая цепь
составлена из элементов, имеющих в расчете на единицу длины
сопротивление R, емкость Си индуктивность L (см. рис. 5.51).
Для сосудов системы микроциркуляции С—»0 вследствие их
низкой способности к расширению при изменении давления, и
поэтому уравнение (5.74) теряет смысл. Для этих сосудов отношение
вязкостного сопротивления к индуктивному г I &1 = 8πη / copS
оказывается во много раз выше, чем для артерий, что обусловлено неболь-
I пава 5. Биофизика двигательной активности
117
Q
3
2
1
О
6 12 18 Ар, 103Па
Рис. 5.52. Зависимость объемного расхода крови
в системе микроциркуляции от давления
Q — объемный расход крови в условных единицах
Ар -разность давлений на входе и выходе системы
той величиной S отдельных артериол и капилляров по сравнению
ι артериями. Поэтому первым членом в правой части уравнения
(5.73) можно пренебречь, и в итоге кровоток в микрососуде
описывайся дифференциальным уравнением Пуазейля. Это уравнение при
интегрировании в пределах длины сосуда от *, до х2 и давления отрх
цо р2 переходит в выражение р{ — р2 =R(x2 -*,) или
Pl-p2=RTQ, (5.75)
которое аналогично закону Ома для участка электрической цепи
ι: проводником длиной (х2 - χ,) и сопротивлением R(x2 - ху) = Rt.
Для системы параллельно соединенных сосудов величина,
обратная общему вязкостному сопротивлению (проводимость), равна
1:умме проводимостей отдельных сосудов.
Описанное представление кровотока в системе
микроциркуляции называют чисто резистивной моделью кровообращения.
Выражение (5.75) часто именуют формулой периферического гемодинами-
ческого сопротивления.
Экспериментальная зависимость (рис. 5.52) Q от Δρ =ρ2 -pi для
иртериальной части системы микрососудов в целом имеет
нелинейный характер: кровоток растет по мере увеличения давления более
резко, чем это следует по уравнению (5.75). Это объясняется тем, что
118
Биофизика
Рис 5.53. Разложение импульса давления
в бедренной артерии собаки на гармонические составляющие
I, 2, 3 - гармонические колебания с частотой I, 2 и 3 Гц; 4 — сумма гармоник
(сплошная кривая) и экспериментальные значения давления (кружочки);
5 - постоянная составляющая; ρ - давление в относительных единицах; t — время
при повышении Δρ величина RT снижается. Падение сопротивления
при повышенных давлениях может быть обусловлено несколькими
причинами, например, расширением микрососудов или увеличением
количества действующих капилляров, подключающихся параллельно
к функционировавшим сосудам.
Распространение пульсовых волн. Сокращения сердца
сопровождаются выбросом крови в кровяное русло, что приводит к
периодическим изменениям давления. Эти изменения, как и всякий
периодический процесс, могут быть выражены в форме суммы гармонических
колебаний с частотой nf0, где η - натуральные числа, af0 - частота 1 Гц
(разложение сложного колебания на сумму гармонических колебаний
называется Фурье-анализом). На рис. 5.53 в качестве примера
показано, что пульс в бедренной артерии собаки хорошо аппроксимируется
рядом Фурье, состоящим всего из четырех слагаемых (гармоник).
Таким образом, целесообразно сначала рассматривать распространение
по сосуду простых гармонических волн, а затем просуммировать их
для описания распространения естественных (пульсовых) волн.
Итак, предположим, что давление в точке χ = О изменяется по
гармоническому закону с круговой частотой ω = 2π/ (f - частота
в герцах):
I пава 5. Биофизика двигательной активности
119
/7(0) = /70COSCUi. (5.76)
При выполнении этого условия решение системы уравнений
(^.73) и (5.74) приводит к уравнению затухающей волны:
p=A0exp(-xx)cos((ut-$x), (5.77)
причем коэффициенты χ и β связаны с величинами R, L и С в
уравнениях (5.73) и (5.74) соотношениями:
β2-χ2=ω2ΖΧ:; 2Рх=соЛС. (5.78)
Величина А0 в уравнении (5.77) представляет собой
максимальную амплитуду колебаний давления и равна ρ при cos(o>i-|ix) = l
и ν = 0. Коэффициент β показывает, как быстро затухает колебание
но ходу сосуда. В самом деле, амплитуда колебаний в точке χ равна
■'Ίι схр(~9С.х),т.е. уменьшается по ходу сосуда по экспоненте, показа-
1сль которой пропорционален х. Величина β* в уравнении (5.77)
показывает сдвиг фазы на расстоянии χ от начала отсчета, а
коэффициент β связан со скоростью распространения волны а и, следова-
I сльно, с длиной волны λ, соотношениями:
β ω β
Легко найти величины α и λ, решив систему уравнений (5.78)
и подставив величину β в (5.79). Однако выражения получаются
громоздкими. Если сопротивление току крови носит в основном вязко-
1П1ЫЙ характер ((uL«R), можно получить приближенное
выражение для скорости:
·'&=№■
В этом случае, как видим, фазовая скорость распространения
упругих волн растет с увеличением частоты. Если, наоборот, сопро-
120
Биофизика
тивление току крови обусловлено в основном ее инерцией (ωΖ, » К),
что характерно для крупных сосудов, приближенное выражение для
скорости будет иным:
Скорость распространения упругих волн уже не зависит от
частоты, т.е. все гармонические слагаемые сложного колебания
(пульса) будут распространяться по сосудам с одинаковой скоростью.
Изучение кровеносных сосудов и кровотока может
осуществляться на основании анализа распространения искусственных
гармонических волн давления в опытах in situ. Измеряя амплитуды
давления искусственной волны в месте расположения датчиков давления,
находят фазовую скорость распространения волны, a~x / At.
Эти опыты показали, что фазовая скорость практически не
зависит от частоты (рис. 5.54). Это говорит о справедливости
уравнения (5.81).
Экспериментально был подтвержден и другой теоретический
вывод об экспоненциальном характере затухания волны давления по
ходу сосуда. Из уравнения (5.77) следует, что при данном χ
амплитуда давления А=А0 е\р(-%х), или после логарифмирования
lnfj-V-χ*. (5.82)
Это и наблюдалось в опыте: в полулогарифмических
координатах амплитуда давления оказалась линейно зависимой от х.
Существует много других подтверждений правильности
линейной модели кровотока. Например, согласно уравнению (5.81),
скорость распространения волны давления по крупным сосудам
должна возрастать с увеличением модуля упругости сосудистой
стенки Е.
Выводы линейной теории кровотока по упругим сосудам тем
не менее могут использоваться для изучения ряда характеристик
системы кровообращения. Например, измеряя скорость распростране-
/ пива 5. Биофизика двигательной активности
121
пня пульса, можно определить модуль упругости сосудистой стенки
/■.' но уравнению (5.81). Измеряя давление в двух близко
расположенных участках сосуда и зная R и L, можно рассчитывать по уравнению
0,73) импульс объемного расхода крови.
Моделирование работы кровеносной системы проще всего
осуществить, объединив каждую совокупность сосудов данного типа
и один гидродинамический элемент. В модели О. Франка, созданной
|| 1899 г., система крупных сосудов артериальной части большого
круга кровообращения моделируется одной упругой камерой, а
οποί см а мелких сосудов с вязкостным сопротивлением - жесткой
трубной (рис. 5.55). Электрическим аналогом этой гидродинамической
РЬ
га
С
о
о
а, м/с
5 Г
4 -
поип00ио0°о-
40
80
120 f, кГц
Рис. 5.54. Схема определения фазовой скорости и коэффициента
затухания искусственных волн давления и зависимость фазовой
скорости распространения этих волн в аорте собаки от частоты
Ао, А- амплитуда давления в месте расположения датчика β,ι
и после прохождения расстояния 4 см (в месте расположения датчика Д2);
I - время прохождения максимума волны давления (фазы) от одного до другого
датчика; а — фазовая скорость распространения волны давления; f— частота
122
Биофизика^
Рис. 5.55. Гидродинамическая модель с сосредоточенными параметрами,
описывающая артериальную часть большого круга кровообращения
I -упругая камера (крупные артерии), представляющая собой цилиндр
с поршнем, который соединен супругой пружиной; 2 - жесткая трубка
(периферические сосуды); 3 -клапан; 4 - насос (левый желудочек сердца).
Стрелки С и Д показывают направление поступления крови в разные
части системы в период систолы и диастолы (соответственно)
модели служит цепь, состоящая из параллельно соединенных
конденсатора и активного сопротивления. Давление/?, создаваемое
сердцем при сокращении, воздействует сразу на все крупные артерии (на
единую гидродинамическую емкость С0). Теоретический анализ
кровотока в такой гемодинамической системе с сосредоточенными
параметрами приводит к формулам, на основе которых, измеряя
показатели кровяного давления, можно рассчитать ударный объем крови
в большом круге кровообращения.
Объем (Qdt) крови, выходящей из сердца со скоростью Q за
время dt {dt > 0), равен сумме объема (Qodt) крови, протекающей
с объемной скоростью Q0 через жесткую трубку, и изменения объема
(dV) крови в упругой камере:
Qdt = Q0dt + dV.
(5.83)
Очевидно, что dV = s ■ dl, где s - площадь поверхности поршня
упругой камеры; dl - изменение длины камеры (упругой пружины).
ι нова 5. Биофизика двигательной активности
123
11ыразив dl через модуль упругости Ε и длину / упругой камеры
(пружины) и учитывая при этом, что в данном случае do равна
изменению давления dp, имеем dV -sldp IΕ.В результате уравнение (5.83)
переходит в выражение:
Qdt=Q0dt + C0dp, (5.84)
it котором С0 =sl ΙΕ - гидродинамическая емкость упругой камеры
(совокупности крупных сосудов).
Объемная скорость кровотока через периферические сосуды
(жесткую трубку):
β0=^^ (5-85)
где ρ - давление в крупных сосудах (в упругой камере), рв -
венозное давление, Rr - общее вязкостное сопротивление периферических
сосудов
Из последних двух уравнений при рв = О следует
Qdt=^i + C0dp. (5.86)
Rr
Запишем определенный интеграл этого выражения в
промежутке времени, равном одному периоду пульса Тп, т.е. в промежутке
между моментами минимального диастолического давления pR
(рис. 5.56):
JQdt = j-\Pdt+C0jdp. (5.87)
Второй член правой части уравнения (5.87) равен нулю. Левая
часть этого уравнения и есть ударный объем крови (Vc), который, как
нидим, может быть найден как площадь под кривой p{t) за период
пульса (рис. 5.56), деленная на вязкостное сопротивление RT. Введя
параметр среднее давление рп за период пульса, получаем:
124
Биофизика
Рис. 5.56. Определение в модели с сосредоточенными параметрами
гидродинамических характеристик артериальной части системы
кровообращения по данным измерения пульсовой волны
1,2 — пульс сонной и бедренной артерий (соответственно)
(рд - диастолическое давление, рс - систолическое давление)
vc =
ΡπΤπ
(5.88)
Теперь предстоит найти Лг. В основе дальнейших рассуждений
лежит тот факт, что сердце в основном заканчивает выброс крови
к моменту достижения максимального систолического давления (рс
( ппва 5. Биофизика двигательной активности
125
ни рис. 5.56). Дальнейшее изменение давления до минимального диа-
сюлического связано с выходом крови из крупных сосудов (упругой
камеры). Таким образом, в период Тп (рис. 5.56) Q = О, и для этого
периода уравнение (5.86) приобретает вид C0dp--pdt I Rr.
Проинтегрировав это выражение в пределах давления отрс яора для левой
части и в промежутке времени от 0 до Гд для правой части и выразив
штем интеграл правой части через среднее давление, как это сделано
нмше для всего периода пульса, приходим к уравнению:
С0(Рс-Ра) = ^> (5-89)
КГ
еде ρ - среднее давление за период Гп, приближенно равное рп
и формуле (5.88).
Емкость С0 в этом уравнении равна С · 1а, где 1а - некая
эффективная длина крупных артерий, а С - их средняя емкость на единицу
длины. Величина С может быть найдена, если известна скорость
распространения пульсовых волн а:
С=-Ц-, (5.90)
где S0 - эффективная площадь поперечного сечения, которую
обычно принимают равной площади поперечного сечения
восходящей дуги аорты, и определяют по данным рентгенографии.
Из уравнений (5.88), (5.89) и (5.90) можно получить выражение
для ударного объема крови, которое при р — рп имеет следующий
иид:
kSJpr -ρπ)Τ„1
у = ov^c уд! и ^ (591)
Ра%
где k - эмпирический коэффициент, близкий к единице, введение
которого связано со сделанными упрощениями. Наибольшая
трудность при использовании уравнения (5.91) связана с тем, что
эффективная длина артерий 1а неизвестна. Существует несколько более или
126
Биофизика
менее произвольных методов ее оценки. О. Франк предположил,
например, что la равна половине длины волны основного колебания
пульса, т.е. величине a ■ Т/2, где Τ - период указанного колебания,
определяемого обычно по пульсу в бедренной артерии (рис. 5.56).
Другие исследователи делают иные допущения. Но если даже
параметры к и la неизвестны, уравнение (5.91) дает возможность
определять изменения ударного объема крови у данного испытуемого, так
как к, la и даже S0 и ρ при этом обычно меняются незначительно.
5.5.3. Нелинейная теория движения крови
по кровеносному руслу
При создании этой теории основное внимание уделено двум
вопросам:
- связи характера потока жидкости с изменяющейся под
действием этого потока геометрией эластичной трубки;
- роли нелинейности уравнений гидродинамики при течении
жидкости по эластичным трубкам.
В исследуемой гидродинамической модели сделаны
существенные упрощения. Во-первых, принято приближение идеальной
жидкости, во-вторых, используется безмоментпая теория для
эластичной трубки.
Течение жидкости по тонкостенным эластичным трубкам
можно условно разделить на три относительно самостоятельных
гидродинамических явлений: перенос объема жидкости по трубке,
распространение волны давления, скорость которой много выше скорости
жидкости, и возникновение высокочастотных колебаний вследствие
потери устойчивости по типу «поток-стенка» (флатгер).
Течение идеальной жидкости в трубках с эластичными
стенками. Для получения замкнутой системы уравнений гидродинамики
необходимо задать связь между деформацией эластичной трубки и
избыточным давлением внутри нее. В простейшем случае небольших
деформаций эта связь определяется законом Гука. Однако
формулировка закона Гука зависит от характера решаемой задачи. Она должна
быть различной для различных скоростей изменения площади
сечения трубки. В случае относительно небольших скоростей потока и из-
ι пяла 5. Биофизика двигательной активности
127
мщений площади течения трубки можно использовать формулу связи
м 1('>ыточного давления в трубке и площади ее сечения в виде:
Р,=С^-, (5.92)
ι цс AS = S - S0, S - площадь сечения трубки в данном месте, S0 -
пмощадь сечения трубки при нулевом избыточном давлении р„
Οι мистичность стенок трубки. Из теории тонких оболочек следует, что
(' _ Ehld, где Ε - эффективный модуль упругости стенок трубки, h -
юлщина ее стенки, d - средний диаметр.
В случае более значительных изменений площади сечения
ι рубки можно использовать закон Гука в виде:
Ар,=С — . (5.93)
Здесь сохраняется линейная связь изменения избыточного дав-
нсмия от относительной деформации площади сечения трубки, одна-
к1» зависимость площади от избыточного давления носит
нелинейный характер:
S=S0 exp
'Р.^
(5.94)
В первом порядке разложения в ряд формула (5.94) переходит
η (5.92).
Если скорость изменения площади сечения трубки достаточно
нслика, то в закон Гука должны входить параметры,
характеризующие движение жидкости по эластичной трубке. В этом случае закон
I 'ука можно применять в виде :
Р^С^П, (5.95)
dV
ι дс dV = SdX - элемент объема трубки, рис. 5.57. Если жидкость
неподвижна, то dV = Sdl, где dl - неизменяющаяся длина участка
128
Биофизика
dV=Sdx
Рис. 5.57. Деформация элемента объема эластичной трубки dV
при перемещении жидкости вдоль нее (V*t - V* - изменение
элемента объема трубки при перемещении жидкости)
трубки. Тогда формула (5.95) переходит в (5.93). Если же жидкость
движется, то преобразование носит более сложный характер
(рис. 5.57):
A(dV)_V{ -V
dV
dV
dV JF,
+ dV
dV
ldV =
(5.96)
_dV _d(uS)
~ dV ~ S ■ dx '
где ε - относительная деформация элемента объема трубки dV
в процессе смещения жидкости вдоль нее на dx, а V* - V* -
изменение элемента объема трубки при этом.
При S= const формула (5.96) переходит в обычное выражение
для относительной деформации в твердом теле ε = ди/ .
Относительно смещения и необходимо сделать следующее
замечание. Если в эластичной трубке возникли высокочастотные
автоколебания стенки и потока, то на осредненное движение жидкости со
скоростью ν0 накладывается колебательное движение со скоростью
ν*, так что суммарная скорость
ν = ν0+ν,.
(5.97)
I пава 5. Биофизика двигательной активности
129
Величина и - это смещение элемента объема жидкости в
процессе продольных колебаний относительно точки, движущейся со
скоростью v0 так, что vk =^% · Таким образом, из (5.95) следует,
ΊΙΌ
Р, =С9-^. (5.98)
S ■ αχ
Формулы (5.92)-(5.95) и (5.98) связывают избыточное
давление с геометрией трубки. Однако, если записывается уравнение
колебаний, то необходимо использовать связь между реакцией стенки
частичной трубки и ее геометрией. Реакция стенки трубки ρ - это
ι ила, действующая со стороны единицы площади внутренней по-
иерхности трубки на жидкость. Так как ρ = -рр то в формулах
(S.92) - (5.95) и (5.98) необходимо учесть минус.
Для возникновения флаттерного режима течения совсем не-
пЬизательно наличие волн давления. Он может появиться и при
неизменном расходе в эластичной трубке, т.е. когда перепад давления на
ι рубке постоянен. Течение жидкости по эластичной трубке принципи-
ιΐΐΜ.ικ) неустойчивый процесс. Любое случайное увеличение скорости
жидкости приводит в соответствии с «эффектом Бернулли» к
снижению статического давления в ней. Это ведет к уменьшению площади
ιч-чения трубки, что еще больше увеличивает скорость потока. Про-
нсес нарастает лавинообразно (положительная обратная связь), что
приводит к схлопыванию трубки. Необходимость прокачивания
определенного объема жидкости раскрывает трубку. Возникают автоколе-
инпия по типу неустойчивости «поток-стенка» или флаттер оболочеч-
ιιοίί моды. Процесс непосредственно не связан с наличием вязкости
жидкости, поэтому будем рассматривать возникновение автоколеба-
1ПП.НОГО (автоволнового) режима течения в идеальной жидкости,
и нлпяние вязкости отметим в дальнейшем.
Выведем уравнение импульса для рассматриваемого процесса.
11 ииснение площади поперечного сечения трубопровода на полуво-
iiie возникших колебаний состоит из двух частей: диффузорной
и копфузорной (рис. 5.58). Второй закон Ньютона в проекции на ось
\ для элемента объема жидкости dV можно записать в виде
130
Биофизика
Рис. 5.58. Схема продольных составляющих сил, действующих на элемент
объема dV со стороны диффузорной и конфузорной частей стенок трубки
dFr + N-(N-dN)-dF. = pdV- — ,
g dt
(5.99).
где dF% = dFk = PdS - продольные составляющие сил, действующих
на элемент объема dV со стороны диффузорной и конфузорной час^
тей стенок трубки. По 3-му закону Ньютона они равны между собой;
N = pS - продольная составляющая сил давления, обеспечивающая
течение жидкости, ρ - ее плотность, t - время.
Сила F, действующая на элемент объема dV по всей
внутренней поверхности диффузорной части, уравновешивается силой Fh
действующей со стороны конфузорной части. Если dFk = 0, т.е.
диффузор открытый, уравнение (5.99) преобразуется к виду:
dp dv
dx dt
(5.100)
Такой же вид имеет уравнение импульса при S = const. Однако
в случае существования диффузорной и конфузорной частей
симметричной геометрии уравнение импульса (5.100) преобразуется к виду:
d(PS)_ dv
S-дх dt'
(5.101)
Следовательно, в случае трубки неизменного сечения,
расширяющегося либо сужающегося сечений, можно пользоваться
уравнениями Эйлера (5.100). Если же расширение трубки сменяется
сужением, то необходимо использовать уравнение (5.101). Подобный вид
I пава 5. Биофизика двигательной активности
131
уравнения используется, например, при решении задачи Жуковского
и гидравлическом ударе в упругом «трубопроводе».
Подставляя в (5.101) закон Гука (5.95) с учетом знака, а также
используя (5.96) и учитывая, что dvjdt = 0, получаем:
С
d\uS)_(dvk dv
S-dx2
э. +ν· » ' (5102)
В случае колебаний небольшой амплитуды нелинейным кон-
мсктивным слагаемым в правой части (5.102) можно пренебречь.
('корость распространения автоволн по трубопроводу
a=J(C/p)=J(E-/pd). (5.103)
Используя vk =ди /dt,a также (5.103) получим:
дх2 dt2
Получено двухпараметрическое волновое уравнение. Еще одно
уравнение, связывающее неизвестные величины и и S, - это
уравнение неразрывности:
^+*^=о. (5.105)
dt дх
В уравнение неразрывности входит только колебательная
составляющая скорости, так как изменение площади и расхода Q = VS
происходит только за счет колебательной составляющей скорости vk.
Будем искать решение системы (5.104) и (5.105) методом
разделения переменных Фурье. Примем:
u = Fl{x)-Ol(t)i
S=F2(jc)-<D2(0; (5.106)
ιώ'=^(Λ)·Φ1(θ·^(*)·Φ2(0=^)-Φ(0·
132
Биофизика
Подставим (5.106) в (5.104) и сократив на Φ2(ι), разделим
переменные:
_^.^ФЛ0^^^М = ^=СОП51. (5.107)
Ф,(0 dt2 F(x) dx2
Получаем два уравнения
Л (5.108)
VaJ
d2F(x) (ω
dx2
Решение этих уравнений имеет вид:
Ф1(0=А0-зт(Ш + <р0), (5Л09)
F(x)=A^ -sin(roi+90), (5.110)
где ω - циклическая частота колебаний, А{ ц>у- постоянные
интегрирования. Функции Ф2(0> Fy(x), F2(x) найдем из уравнения
неразрывности (5.105). Учитывая, что скорость vk ~^xAt имеем:
F2(x)-^^l^2(t)d-^ldJM^. (5.111)
dt2 dt dx
Следовательно, уравнение неразрывности распадается на два
уравнения:
1 dF(x)_ 1 9Φ2(0_Θ (5Π2)
F2(x) dx Ф2(0 5Ф,(0 '
где θ - постоянная. Решения уравнений (5.112) с учетом (5.110)
имеют вид:
I пава 5. Биофизика двигательной активности
133
Ф2(1)=А2-ехр(-0Ф1({))
FAx).
А2 ω
-cos
ω
-+φ,
(5.113)
Функция F, находится из условия:
„ , ч F(x) θα t (ω , ^ ,....,
^i(*)=-=Vt =—-tgl-x+vA (5·114)
F,(x) ω \α )
Система функций (5.109), (5.113), (5.114) позволяет получить
решение уравнений (5.104), (5.105):
5=^Ос)Ф2(0-^^°
cos
ω
ехр(-0Ло sin(o>i + φ 0)),
—χ +φ,
V«
(5.115)
u=Fl(xyi>l(t)=^-^.
ω
tg
ω
-Jf+φ,
•sin(coi+(p0). (5.116)
При записи решений уравнений (5.104), (5.105) необходимо
учитывать, что эти уравнения обладают слабой нелинейностью. Они
линейны относительно каждого из параметров, но в них входят
произведения этих параметров. Указанное обстоятельство налагает
некоторые ограничения на результат решения системы (5.104), (5.105).
11лощадь эластичной трубки величина положительная, поэтому
необходимо использовать знак абсолютной величины. Абсолютную
величину в соответствии с уравнением неразрывности (5.105) нужно
использовать и в уравнении для перемещения (5.116).
Найдем скорость жидкости и давление в ней; для скорости
имеем:
ν=ν° +^7 = νο+α·θ'Λ
at
tg
(ω
-*+φ,
Va
cos(cof+(p0). (5.117)
Давление определим исходя из соотношения (5.98):
134
Биофизика
х=0
Рис. 5.59. Возникновение автоколебаний стенок эластичной трубки
(к схеме измерения артериального давления крови по методу Короткова;
манжета сфигмоманометра не показана)
Ру
Pi
=са(И5)=сэ(^-Ф)=свф
S · dx Р2Ф2дх
-CQA0 sin(coi +φ0).
(5.118)
Как и следовало ожидать, градиент давления (так как жидкость
идеальная) получился равным нулю. Таким образом, в эластичных
трубках возможно возникновение автоколебаний площади, скорости
и давления.
Такие автоколебания, или флаттер возникают, например, при
измерении артериального давления в кровеносных сосудах сфигмо-
мапометром - тоны Короткова.
Если принять максимальную отпосителыгую деформацию ε,,^
и учесть, что амплитудное значение колебаний давления Ρ^,,χ ~
= С ■ егаах, то из (5.118) следует егаах = θ · А0.
Зададим следующие граничные условия исходя из модели
измерения артериального давления по методу Короткова (рис. 5.59):
при χ = 0 величины S - S0; и = и0 = 0; ν = ν0, где S0, u0, v0 - площадь
сечения, перемещение и скорость жидкости вне манжетки
сфигмоманометра. Из (5.117) следует φ1 = 0. Принимая начальные условия:
при t = 0 величина и = щ = 0, S = S0, из (5.116) получаем φ0 = 0, а из
(5.115) А[ А2 ω/(θ · а) = S0. Следовательно, окончательно решение
системы уравнений (5.104), (5.105) при данных начальных и
граничных условиях имеет вид:
I пава 5. Биофизика двигательной активности
135
V, SL
(со/а)х
Рис. 5.60. Расчетные кривые изменений площади сечения
эластичной трубки и скорости жидкости по ее длине
Сплошные линии соответствуют идеальной жидкости, штриховые — вязкой
tg\ -х
а
v=v0 +a-emax
tg
S=S,
cos
ω
sin(coi),
cos(coi),
exp(-s m sin(coi)),
—χ
Ka )
(5.119)
На рис. 5.60 показаны графики изменения площади сечения
пластичной трубки и скорости жидкости в ней в зависимости от ее
длины в момент времени, соответствующий sin(cot) = 0 и cos(coi) = 1,
т.е. t = 2k7t/co, где к = 0, 1, 2,..., построенные в соответствии с
формулами (5.119). Для удобства построения по оси абсцисс взята
величина сох/а.
Из рис. 5.60 видно, что в моменты схлопывания трубки, там
где S~ 0, скорость жидкости возрастает до бесконечности. Это свя-
чапо с тем, что формулы для скорости и площади относятся к
невязкой жидкости. Такой же эффект имеет место, например, при рас-
136
Биофизика
смотрении вынужденных колебаний, когда амплитуда колебаний
в резонансе стремится к бесконечности при отсутствии
сопротивления среды. Пунктиром показан реальный ход кривых с учетом
вязкости жидкости.
Определим возможные частоты автоколебаний при измерении
артериального давления по методу Короткова. Пусть длина
манжетки сфигмоманометра равна 1. Тогда дополнительное граничное
условие будет: при χ = 1, ν = ν0 в любой момент времени. Из (5.119)
следует ωΐ/α = kn, где к = О, 1, 2,..., т.е. целое число. Следовательно, частота
автоколебаний будет равна:
7, . „ 7, I Z7 . 7.
(5.120)
Полученное выражение дает возможность определить
эффективный модуль упругости сосудистой стенки в живой системе путем
измерения частоты тонов Короткова.
Вязкий peoicuu течения в эластичных трубках. До сих пор
рассматривали течение идеальной жидкости по эластичной трубке.
Реально кровь обладает довольно большой вязкостью. Вязкость крови
примерно в 4,2 раза больше вязкости воды.
При рассмотрении уравнения импульса для вязкой крови
примем, что амплитуда волн в эластичной трубке мала по сравнению
с длиной волны. В этом случае можно не учитывать нелинейные
конвективные члены в уравнении импульса. Если считать скорость
распространения пульсовой волны α и 6 м/с, а длину волны λ
вычислить исходя из характеристик тонов Короткова, частота которых/=
= 60... 150 Гц, то, исходя из этого, получим: λ = aJf= 10.. .4 см.
Амплитуда колебаний не может быть больше радиуса артерии,
т.е. менее 0,5 см. Следовательно, условие, при котором можно
пренебречь нелинейными членами в уравнении импульса, выполнимо.
Учитывая формулу члена с давлением (5.98), запишем
уравнение импульса в виде
δν _ d(pS) 1 δ
г
+ -
dt Sdx r дг \ дг) дх
δνλ δ
^—1 +
Г я,Л
δν
дх
(5.121)
I пава 5. Биофизика двигательной активности
137
где г - радиальная координата, η - динамическая вязкость.
Усредним уравнение (5.121) в соответствии с профилем скорости Пуазейля
г =2ν (l-λ·2 / R2), где vcp - средняя по сечению скорость в трубке,
R - внутренний радиус сосуда.
В результате уравнение импульса принимает вид:
δν _ d(pS) 8πην δ
Ρδ7~ Sdx S ~д~Х
\^- L (5.122)
V
дх
В (5.122) у скорости ν опущен индекс ср., так как в дальнейшем
будем иметь дело только со средней скоростью кровотока по
сечению.
Для анализа влияния вязкости на поток крови рассмотрим
течение очень вязкой жидкости, т.е. примем в (5.122):
dv 8πην д ( δν
р_«_Г^11+_ η_ (5.123)
3t S дх{ дх)
Такое течение аналогично пуазейлевскому, но с учетом
возможности изменения площади сосуда за счет его эластичности. Пер-
иопачально пренебрежем последним диффузионным членом в
уравнении (5.122) и предположим отсутствие пульсового движения
жидкости, т.е. используем приближение пограничного слоя. Кроме
того, левую часть вследствие малости не учитываем. Тогда
уравнение импульса можно записать в виде
djpS) _ 8πην (5 124)
Sdx S
Используя (5.98) с учетом знака, получаем
cd2(uS) _8πην ди f5 125Ϊ
Sdx2 S 3t'
Так как при принятых условиях течение не имеет
колебательного характера, разделять скорость на среднюю и колебательную со-
138
Биофизика
ставляющие нет необходимости. Обозначим γ —С 18πη, тогда
уравнение (5.125) будет иметь вид
d\uS)^du
дх2 dt
Это двухпараметрическое уравнение типа уравнения
теплопроводности. Используя (5.96), будем искать решение уравнения
(5.126) методом разделения переменных Фурье:
1 </Ф, _ 1 d2F
Φγ dt Fl dx:
=0, =const. (5.127)
Для нахождения функций Φ, ΦγΦ2 используем (5.127) и
уравнение (5.112), являющееся следствием уравнения неразрывности:
дА + д±^1^. (5.128)
dt дх
Для функции Ф„ используя (5.113) и (5.127), получаем урав-
θ, ·γ·Φ = θ, ·γ·Φ, ·Λ2·βχρ(-θ2·Φ,). (5129)
нение:
άΦ
ι -ι
dt
В уравнении (5.129) постоянная интегрирования, обозначенная
ранее в (5.112) как Θ, теперь обозначена θ2. Решение уравнения
(5.129) имеет вид
Φ, = ±ΕΓι(λ2ί), (5.130)
θ2
где обозначено -λ2 = θ, уА2, £Т' (λ21) - модифицированная обратная
интегральная показательная функция, так что
' пава 5. Биофизика двигательной активности
139
При этом учтено, что при t = О функция Ф,(0) = 0, так как
перемещение и = 0. Функция Ф2 получается из уравнения (5.112):
Φ2=Λ2εχρ[-£Γ'(λζ0]· (5.131)
Для нахождения зависимостей переменных от х, умножив
(V127) на (5.122), получим уравнение для F:
d2F dF
—- = 0,·02·ί=θ·ί. (5.132)
dx2 dx
Решение уравнения (5.132) через элементарные функции не
иыражается. Функции Fy(x) и F2(x) можно выразить через F, исполь-
■уя (5.127) и (5.112):
1 d2F 1 dF
, F,(x)=
θ, άχ2 θ2 dx
F'w=;rT-r' F^-~-^-· (5·133)
Используя (5.130), (5.131), (5.133), находим решение системы
(5.126) и (5.128):
u=F1(x) — Er1(k2t). (5.134)
θ2
При t —> оо имеем: ΕΓι(λ2ί) —> 0, поэтому для λ2 > 0 имеем:
.V -» 0. Таким образом, решение (5.134) при λ2 > 0 указывает на зажа-
I и с трубки со временем.
Если Л[ = uS, Λ2 = и - решения уравнения (5.134), а Л3 = S и Л4 =
-^решения уравнения (5.126), то
Λ,* =^B(X)uSdX, R\ =JB(X)udX
также являются решениями (5.124), а
140
Биофизика·
R] = $D(X)SdX, R'4 = |Ζ)(λ)ν&/λ
-решения уравнения (5.126).
Следовательно, учитывая, что функции Fy и FMoryr зависеть от
λ, получим
1 d2F
u = \B(X)-^-^Erl(X2t)dX,
J ft ^7-v -
0
θ dx
S = \D(X) — — ехр[-ЕГ1(Х2()Щ, (5.135)
ν =ЁИ= \Β(λ)—^-ζ-ΕΓι (Х2()ехр[-ЕГ1 (X2t)]dX.
dt { θ dx
Функции В(Х) и D(X) определяются начальными и граничными
условиями.
Рассмотрим стационарный режим при вязком течении.
Формулы (5.135) описывают переходный процесс. Однако опыт
показывает, что при течении очень вязкой жидкости через короткое
время форма эластичной трубы достигает стационарного состояния
и в дальнейшем не меняется, т.е. S = S(x) при t -» оо. Уравнение
(5.126) принципиально не может описать этот режим течения.
Действительно, из (5.128) следует vS = Q Φ fix). Следовательно,
SdU/dt =d^US)dt * f(*}'T-e· uS*f№ и левая часть УРавнения (5.126)
равна нулю. Таким образом, получаем ν = 0, т.е. стационарный
режим возникает при полной остановке потока, что тривиально.
Для получения формы трубы при наличии скорости учтем
последний член в уравнении (5.123). Тогда уравнение импульса
приобретает вид
*^ = ^. (5.136)
S дх2
/ пива 5. Биофизика двигательной активности
141
Учитывая ν = Q/S и φ = 1/5, получим
8πφ2=^Φ. (5.137)
дх2
Интегрируя уравнение (5.137) методом понижения порядка
производной, находим
Используя условие, что — =0 при φ = 1/5гаах, находим Аъ =
дх
-16/7t(35max), где 511ШХ в данном случае - максимальная площадь
сечения эластичной трубки, возникающая в некотором месте.
Умножим обе части уравнения (5.138) на 5^х и, перейдя к
безразмерному аргументу х* ~x.yj[4n / 35 „,^], получим:
d(4>Smax)
dx
2
= ^™)3-4. (5.139)
Решением уравнения (5.139) является эллиптическая функция
Нсйерштрасса: φ5^, -Β(χ' +ΑΑ).Таким образом,
5= — 5« (5.140)
Постоянная АЛ находится из условия: при χ = 0 имеем 5 = 50,
г.е. В(АЛ) = SUJS0. Графически (5тах/50)1/2 как функция от
Z=x I +А4 представлена на рис. 5.61.
\Р^тах
142
Биофизика
(S/Smax)s|
Рис 5.61. Изменение площади сечения эластичной трубки
при устойчиво вязком течении жидкости
Переход от автоколебательного к вязкому режиму течения.
Анализируя возможные ламинарные непульсовые режимы течения
жидкости в эластичной трубке, можно сделать вывод, что в случае
малой вязкости жидкости, или когда скорость ее достаточно велика,
наблюдается автоколебательное течение, описываемое
дифференциальными уравнениями (5.104) и (5.105). При высокой вязкости или
малой скорости течения жидкости поток описывается
дифференциальными уравнениями (5.126) и (5.128).
Рассмотрим роль параметра γ в уравнении (5.126). При увели-
:%^=раХ^'гдеа-скорость
чении вязкости, т.е. уменьшении γ =
пульсовой волны, течение жидкости все лучше описывается
уравнением (5.126), а при уменьшении вязкости или увеличении γ -
уравнением (5.104).
Найдем критерий подобия для уравнения (5.123). С учетом
(5.98) в приближении пограничного слоя его можно записать в виде:
Эу_сЭ2(^)
Э/ Sdx2
8πην
(5.141)
/ imna 5. Биофизика двигательной активности
143
Примем масштабы переменных в уравнении (5.141) для
смещения Мц, для скорости Mv, для времени Mt, для площади сосуда Ms,
чин продольной координаты Мх. Тогда в безразмерном виде
уравнение (5.141) запишется
Mvdv' _cd2(uS')MuMs 8πην' Μν
Μ, *' S'dx'2 Κ S' Μ/
|дс безразмерные переменные Μ' '* "*
Μν Μ, Ms
и
II = , Χ
Ми Μχ
Учитывая, что ν =°uAt, имеем Μν = MJMr
Найдем безразмерную форму уравнения неразрывности
(S.128):
MsdS' [Μ,Μ53(ν,·^,)_0
M,dt' M\ Ъх
Следовательно, масштабы времени, скорости и продольной
координаты связаны соотношением:
^- = 1 (5.143)
Мх
С учетом (5.143) после несложных преобразований уравнение
(5.142) можно записать в виде
М>· _b\u'S') %τπ\ΜνΜχν'
a2dt' S'dx'2 pa2 MSS' '
где α = Jc/p.Обозначим N=(alMv)2,K=— —— ·
у 8ττη MSM
144
Биофизика
Величина К в рассматриваемой задаче играет роль,
аналогичную числу Рейнольдса Re. При больших γ, а следовательно, и К
течение автоколебательное, при малых К- вязкое.
Примем в качестве масштаба площади площадь сечения сосуда
с невозмущенным диаметром Μ s ~π А, масштаб продольной
координаты Af„ = d, где d - диаметр кровеносного сосуда при равенстве
внешнего и внутреннего давлений/70 =р. В качестве масштаба
скорости примем среднюю скорость потока по сосуду Μν = ν .
Следовательно,
K=-i-=-^- = ^-t (5.145)
4Уср 32η% 4τ
nd
о
«ην,
где τ0 = - напряжение сдвига на стенке сосуда, если про-
d
филь скорости удовлетворяет закону Пуазейля.
Характеристикой перехода к турбулентному течению является
число Рейнольдса:
p\'„d Spv*d 8pv„n
Re =L-*-=J-2-= -!-*-. (5.146)
ημ 8ηνορ τ0
Так как число Рейнольдса Re является общим критерием
подобия уравнения (5.121), то значения Re и К можно сравнивать
непосредственно. Очевидно, что автоколебательный режим ламинарного
течения может наблюдаться до тех пор, пока поток не станет
турбулентным, т.е. должен существовать диапазон, где
Re >К, ν >- a ' С
* ΚΌ ' CD
V32 V 32р'
При скорости распространения пульсовой волны a ~ 5 м/с
автоколебания могут наблюдаться, если скорость потока крови
ν > 0,88 м/с. Таким образом, если скорость потока выше 0,88 м/с
/ imea 5. Биофизика двигательной активности
145
м не произошел переход к турбулентности, может возникнуть
неустойчивость типа «поток-стенка», т.е. автоколебания жидкости
м стенки сосуда.
Оценим величину критерия перехода К от вязкого к
автоколебательному режиму течения. Уравнение (5.144) можно представить
и ьиде:
1 dv' _d2(u'S') 1 ν'
N dt' S'-dX'2 KS''
Автоколебания могут наблюдаться, если член в левой части
(.S. 147) и последний в правой части одного порядка. Это возможно
при Ν ~ Ккр. Обычные скорости крови в артериях vcp » 0,3...0,5 м/с.
С'корость распространения пульсовой волны α к 5...6 м/с,
следовательно, KKp~N= (a/vcp)2 = 200.. .400.
Необходимо отметить, что отождествлять величину
Ν = (α/ν )2 с критерием перехода к автоколебательному режиму
течения нельзя. Она скорее играет противоположную роль, поэтому
при увеличении ν величина N падает, а соответственно левая часть
и (5.147) растет.
Опыт показывает, что в приведенном диапазоне скоростей при
нарушении механизмов антифлаттерной стабилизации потока крови
могут возникнуть автоколебания, но может сохраниться и устойчи-
иый режим течения. Большое значение здесь имеет наличие
пульсового потока крови. Поэтому в диапазоне К = 200.. .400 и должно лежать
критическое число перехода К^ от вязкого к автоколебательному
режиму течения, причем для пульсового потока, который является про-
иоцирутощим фактором, справедлива нижняя граница диапазона.
С точки зрения флаттера поток в аорте и крупных артериях имеет
переходные характеристики от вязкого к автоколебательному режиму
течения.
Если принять, что критическое число перехода к
турбулентности ReKp и 2300, то практически всегда в аорте и крупных артериях
существует диапазон чисел К, в пределах которого возможно
развитие автоколебательного режима течения.
146
Биофизика
5.5.4. Биофизические особенности
кровеносных сосудов разного типа
В биофизике принято различать четыре типа кровеносных
сосудов: артерии эластичного типа, артерии мышечного типа, капилляры,
вены. Их функциональное предназначение в системе кровообращения
неодинаково. Артерии эластического типа (или амортизирующие
кровеносные сосуды) поддерживают кровоток во время диастолы
сердца и тем самым обеспечивают непрерывность движения крови
в сосудистой системе. Артерии мышечного типа (или резистивные
кровеносные сосуды) создают переменное сопротивление кровотоку
и, следовательно, регулируют уровень КД в системе кровообращения,
а также объёмную скорость кровотока в каждом из органов.
Капилляры
В капиллярах (обменных кровеносных сосудах)
осуществляется обмен веществ между кровью и тканями, ради чего существует вся
сердечно-сосудистая система. Поэтому капилляры зачастую
называют центральным (главным) её элементом. Их характерной
особенностью является то, что сосудистая стенка образована одним слоем
эндотелия, содержит тонкую базальную мембрану при отсутствии
мышечной и адвенитициальной оболочек.
Диаметр капилляра от 5-7 до 10 мкм, а длина от 0,5 до 1,1 мм,
но если все капилляры человека приложить друг к другу конец в
конец, то общая длина составит 100000 км. Ими можно опоясать Землю
по экватору 2,5 раза. Общая площадь обмена через стенки
капиллярного русла превышает 1000 м2. В 1 мм3 тканей человека содержится
в среднем 600 капилляров, но в разных тканях этот показатель
неодинаков: в миокарде, головном мозге, почках и печени он достигает
3000, в скелетных мышцах - 300-400 (в тонических даже 100), а в
костной и жировой тканях и того меньше.
Линейная скорость капиллярного кровотока порядка 1 мм · с-1
(от 0,05 до 1,93 мм · с~') и, следовательно, каждая частица крови
проходит капилляр от начала до конца примерно за 1 с. Эритроциты
могут двигаться в нём как поодиночке, так и объединёнными в группы,
содержащие от 3-4 до 8-10 штук. Прослойки плазмы внутри такой
группы имеют размер вдоль оси капилляра от 0,5 до 1 мкм, а между
I пава 5. Биофизика двигательной активности
147
группами - по 10-15 мкм. Упакованные в одну группу эритроциты
имеете с находящейся между ними плазмой составляют единое целое
и кинетическом отношении, т.е. не изменяют своих параметров при
прохождении через капилляры.
Основная масса эритроцитов подвергается в капиллярах
продольной деформации. Деформированный эритроцит имеет форму ко-
иокола, обращенного выпуклым концом по направлению кровотока.
11ри колокообразной деформации эритроцита площадь его
поверхности увеличивается примерно на 10%. Эритроциты могут протиснуться
через капилляры и меньшего диаметра, чем 7 мкм. При исследовании
угого процесса на поликарбональных фильтрах было установлено, что
•ритроциты проникают через поры, диаметр которых 2,4 мкм. Если
скорость капиллярного кровотока падает до 0,1-0,2 мм · с-1, то
эритроциты образуют агрегаты по 20-50 клеток в виде монетных столбиков.
И них эритроциты сохраняют форму двояковогнутых дисков, но
утрачивают способность к пластической деформации, вследствие чего
могут закупорить микрососуды. Их локальную закупорку нередко
создают лейкоциты, даже не объединённые в агрегаты, поскольку они
деформируются в гораздо меньшей степени, чем эритроциты, под
действием как продольных, так и радиальных сил.
В зависимости от ультраструктуры организации стенки
различают 3 типа капилляров: 1) с непрерывной стенкой, причём эндо-
тслиальные клетки с диаметром пор в плазмолемме 4-5 нм
уложены наподобие черепицы - проницаемость стенки таких капилляров
наименьшая по сравнению с другими (капилляры с непрерывной
стенкой присущи мышцам, лёгким, жировой ткани); 2) с фенестри-
рованной (окончатой) стенкой - диаметр «окон» в ней достигает
100 нм, что обеспечивает их лучшую проницаемость, например,
и почечных капиллярных клубочках и в слизистой оболочке кишки;
3) с прерывистой стенкой - за счёт крупных пространств (с
линейными размерами более 10 мкм), например, между эндотелиальными
клетками в капиллярах костного мозга, печёночных и селезёночных
синусах, где проницаемость капиллярной стенки наибольшая.
Через эндотелиальную стенку капилляра может «протиснуться» даже
лейкоцит.
148
Биофизика
Вены
Вены являются ёмкостными кровеносными сосудами.
Благодаря выраженной пластичности они могут сильно деформироваться
(растягиваться) и вмещать много крови без развития напряжения
в сосудистой стенке. При повышении венозного КД на 10 мм рт. ст.
вместимость венозного русла большого круга кровообращения
увеличивается в 6 раз. Поэтому вены называют резервуаром крови с
переменной (регулируемой) ёмкостью. Ими образованы кровяные депо
организма, которые в покое содержат 40-50% всего объёма крови
(примерно 1,5 л в венозных синусах селезёнки, до 1 л в венозных
сплетениях кожи и т.д.). Во всех венах большого круга
сосредоточено до 80% крови, протекающей по нему.
Кровь течёт по венам под действием двух сил: «vis a tergo»
(действующая сзади) и «vis a fronte» (действующая спереди). Первым
термином прежде всего обозначают силу, которая обусловлена
механической энергией, сообщенной крови сердцем во время систолы
и сохранившейся после кровотока по артериям и капиллярам.
Остаточная энергия в венулах большого круга кровообращения
составляет в покое не более 13% работы левого желудочка (13% от 0,92 Дж за
1 систолу). Это основной компонент vis a tergo. Дополнительными
служат массирующее действие пульсирующих рядом с венами
артерий и, в большей степени, сокращения скелетных мышц,
сжимающие венозные сосуды и проталкивающие кровь. Такое
проталкивание действует в одном направлении (к предсердиям) благодаря
наличию в венах клапанов. Движению крови в венах конечностей
способствует то обстоятельство, что они образуют с артериями
сообщающиеся сосуды.
Вторая сила обусловлена присасывающим действием на кровь
грудной клетки («дыхательного насоса» и отчасти смещения в
сторону желудочков предсердно-желудочковой перегородки в
определённую фазу сердечного цикла). КД в правом предсердии
называется центральным венозным давлением (ЦВД). На выдохе оно
повышается, а на вдохе понижается. При понижении ЦВД от 0 до -
4 мм рт. ст. приток венозной крови в правое предсердие возрастает
на 20-30%. Однако дальнейшее снижение ЦВД ухудшает кровоток,
поскольку это приводит к спадению вен. Пределы изменений ЦВД,
/ пива 5. Биофизика двигательной активности
149
мри которых сердце работает устойчиво, лежат в диапазоне от 0,01-
0,05 кПа до 0,1-0,12 кПа. Средняя величина ЦВД у здорового чело-
иска в покое от 0,04 до 1,12 кПа.
Аорта
Под действием крови, выбрасываемой в систолу левым
желудочком, происходит растяжение аортальной стенки, обладающей
упругими свойствами. Согласно закону Гука, в ней развивается сила
упругости: Fynp = к ■ χ. Оценим значения χ и к для аортальной стенки.
( уществует связь между Fynp, давлением (р), под действием которого
госуд растягивается, и параметрами этого сосуда: Fynp = ρ · 2rl, где
/ радиус, / - длина сосуда. При колебаниях давления крови в сосуде
тменяется главным образом его просвет, а длина остается
практически неизменной. Следовательно, χ в уравнении Гука для аорты - ни
■по иное как изменение её диаметра. При рентгенографии аорты об-
ииружили, что в систолу её диаметр увеличивается примерно на 10%
относительно своего диастолического значения. Коэффициент
упругости (к) определяется преимущественно эластическими
волокнами, хотя в аортальной стенке присутствуют наряду с ними коллаге-
иовые волокна.
На гистологических препаратах аорты коллагеновые волокна
имеют волнистую (гофрированную) форму, обусловленную их
свободной (рыхлой) укладкой среди других структур, пребывающих
it недеформированном состоянии. Под действием повышения КД
и физиологических пределах коллагеновые волокна только
распрямляются, но не растягиваются. Деформация сети коллагеновых
волокон создаёт так называемые термокинетические упругие силы, но их
иеличина незначительна вследствие малого значения коэффициента
упругости в этом процессе. Высокая упругость коллагеновых
волокон, выражением чего является значительный модуль Юнга (108-
I О9 Па), свойственна им при растяжении, а не при распрямлении, при
котором возникают термокинетические упругие силы. Иными
словами, значительная упругость коллагена проявляется только под
действием больших усилий, выходящих за пределы физиологических
чначений КД. Благодаря коллагеновым волокнам стенки артерий
здорового человека не разрушаются даже при 5-10-кратном повышении
150
Биофизика
КД. Следовательно, коллагеновые волокна обеспечивают
артериальной стенке не упругость, а жёсткость и прочность.
Напротив, эластические волокна аортальной стенки
растягиваются при обычных колебаниях КД во время систолы сердца. В
эластических волокнах возникает сила упругости в соответствии с законом
Гука. Коэффициентом пропорциональности между F и величиной
растяжения стенки аорты при повышении КД служит модуль Юнга
эластических волокон, равный (0,4-1,0)· 106 Па. Эластическим
волокнам аорты в физиологических условиях свойственна
экспоненциальная зависимость силы упругости от степени растяжения. При
более сильном растяжении устанавливается линейная зависимость,
а чрезмерно растянутые эластические волокна разрываются.
Сопоставление вклада эластическихи коллагеновых волокон
в упругие свойства артериальной стенки подтверждает важный
принцип биомеханики: анализ механических свойств тканей организма
необходимо проводить с учётом реальных сил, действующих на них.
Так, если судить по модулю Юнга, коллагеновые волокна обладают
более выраженными упругими свойствами по сравнению с
эластическими. Однако упругость коллагеновых волокон не может
проявиться при естественных колебания КД в артериях, поскольку в таких
условиях, как говорилось выше, эти волокна не растягиваются, и сила
упругости за их счёт не развивается (в уравнении Гука χ = 0).
Внутреннее напряжение (σ,„,) аортальной стенки определяется главным
образом упругостью эластических волокон, хотя их основной белок
(эластин) является менее упругим материалом, чем коллаген. Не
случайно в аорте содержится много эластина, причём он сосредоточен
преимущественно в грудном отделе. Тамэластин составляет 30%
сухого остатка (в 1,5 раза больше, чем коллаген). В брюшном отделе
аорты содержание эластина ниже - 10%, а в бедренной артерии -
только 4-5%, хотя она наряду с аортой относится к артериям
эластического типа. Отношение эластина к коллагену в стенках
периферических артерий равно не 1,5 (как в грудном отделе аорты), а 0,5.
Поэтому модуль Юнга в артериях большого круга кровообращения
увеличивается к периферии. Периферические артерии обладают
большей жёсткостью.
ι imtin 5. Биофизика двигательной активности
151
Среди артерий эластического типа важнейшую роль играет
1>\<)ной отдел аорты. С ним главным образом связана основная
функция сосудов этого типа - обеспечение непрерывности
кровотоки Источником энергии служит миокард. Его работа имеет два ком-
щщсита: статический и динамический, причём на долю первого из
них (А^) приходится 98% всей энергии сократившегося сердца. Ап
ипрачивается на растяжение стенок артерий (прежде всего, грудно-
II) отдела аорты). Накопив энергию во время систолы, аортальная
ι ι гика отдает её крови, когда сердце пребывает в расслабленном со-
• шяиии и не поставляет кровь в сосудистое русло. Благодаря упру-
Iin:ι и артерий эластического типа кровь не останавливается и в диа-
| юлу сердца, когда потенциальная энергия растянутой стенки
|||»1срии эластического типа преобразуется в кинетическую энергию
кропи.
Механизм функционирования грудного отдела аорты был
изучен в прошлом веке физиологом Э. Вебером, в трудах которого он
ничывался феноменом «Windkessel». В отечественной литературе
tuti немецкий термин переводится как «воздушный колпак». Встре-
I ни такой термин, не стоит думать, будто в аорте содержится воздух.
Ili'fiep воспользовался технической терминологией, заподозрив ана-
П01 ию между артериями эластического типа и пожарной помпой,
и которой непрерывность струи при ритмичном качании двух руко-
•иок обеспечивается созданием воздушной подушки в стальном ре-
к-рвуаре насоса. Воздух там находится под давлением, создаваемым
циклической работой двух людей, попеременно пригибающих руко-
и ι ки. Этим давлением в пожарной помпе определяется потенциаль-
ипя энергия сжатого воздуха, которая преобразуется в кинетическую
шсргию водяной струи. В стенке аорты потенциальная энергия свя-
шпа не со сжатием воздуха, а с упругими усилиями, возникающими
и растянутых эластических волокнах. Поэтому эффект накопления
потенциальной энергии аортальной стенкой во время систолы лучше
инчывать феноменом не «воздушного колпака», а «компрессионной
кпмеры» (этот термин также используется в научной литературе).
Упругость аортальной стенки обуславливает ещё одно важное
пиление - возникновение и распространение пульсовой волны по
стенке артерий. Дело в том, что F , развивающаяся при растяже-
152
Биофизика
Ν.
Рис. 5.62. Разложение силы упругости (Fy^,), возникающей в стенке аорты
при её растяжении выбрасываемой сердцем кровью, на нормальную (Fh)
и тангенциальную (Ft) составляющие. Цифрами обозначены положения
аортальной стенки в диастолу (1) и систолу (2)
нии аорты, направлена не строго перпендикулярно к оси сосуда
(рис. 5.62) и может быть разложена на нормальную и
тангенциальную составляющие. Непрерывность кровотока обеспечивается
первой из них, тогда как вторая является источником артериального
импульса, под которым понимают упругие колебания артериальной
стенки. Заметим, что FH гораздо больше преобладает над FT, чем
показано на рис. 5.62. На создание пульса затрачивается
незначительная часть силы упругости растянутой аорты.
Пульсовая волна распространяется от места своего
возникновения до капилляров, где затухает. Скорость её распространения (а^)
можно рассчитать по формуле
\Е-Ъ
где Ε - модуль Юнга сосудистой стенки, Ъ - её толщина, г - радиус
сосуда, ρ - плотность тканей сосудистой стенки.
Общую характеристику пульсовой волны врач получает при
пальпации артерии, но более полные сведения даст регистрация
кривой артериального пульса, которая называется сфигмограммой
(рис. 5.63). Записав сфигмограммы в двух точках артериальной
магистрали и измерив сдвиг фазы между ними, можно определить ско-
I imea 5. Биофизика двигательной активности
153
|юсть пульсовой волны в стенках исследуемых артерий и по ней су-
цмть об их модуле Юнга. Из формулы, приведённой выше, следует,
'но при поддержании р, Ь, г неизменными яп однозначно
определяется модулем Юнга. Следовательно, яп можно принять в качестве ко-
ипчсственного показателя упругих свойств артерий эластического
пит - тех свойств, благодаря которым они выполняют свою
основную функцию. Скорость пульсовой волны в аорте составляет 4-6,
и н лучевой артерии - 8-12 м · с-1. При склеротических изменениях
мргерий повышается их жёсткость, что проявляется в нарастании яп.
(' иозрастом скорость пульсовой волны увеличивается. Чем выше
упругость артериальной стенки, тем больше амплитуда колебаний кро-
иипого давления в аорте и крупных артериях. Высокоамплитудные
колебания КД создают дополнительную нагрузку на сердце и усили-
иают деформацию сосудистых стенок.
Волна артериального пульса - сложное периодическое
колебание, которое по теореме Фурье раскладывается на сумму гармоник,
чьи частоты кратны основной, равной частоте сердечных сокраще-
ι
I
I
Рис 5.63. Сфигмограммы, записанные двумя датчиками
пульсовых колебаний, расположенными в разных точках
артериального русла. Под сфигмограммами — отметки времени,
Ц— время запаздывания сфигмограммы.зарегистрированной дистальнее
154
Биофизика}
ний. Достаточно 4-5 гармоник для хорошего аппроксимирования
сфигмограммы.
Более сложным является венный пульс - колебания стенок
венозных сосудов. Он возникает в венах, впадающих в предсердия,
и распространяется по направлению к капиллярам. Амплитуда
венного пульса ниже, чем артериального, что обусловлено прежде всего
меньшей упругостью оболочек венозных сосудов. Кривая венного
пульса называется флебограммой. Следовательно, от сердца к
капиллярному руслу движутся навстречу друг другу две пульсовые волны
(артериальная и венная), и обе угасают в капиллярах.
Артериолы большого круга кровообращения
Из артерий эластического типа кровь поступает в резистив-
ные сосуды, обладающие гладкомышечными клетками (ГМК),
которые способны, сокращаясь, активно изменять просвет сосудов.
Тем самым регулируется гемодинамическое сопротивление, от
которого, в свою очередь, зависят объёмная скорость кровотока в
органах, васкуляризируемых этими сосудами, и кровяное давление.
Следовательно, резистивные сосуды - важнейшие регуляторы
гемодинамики.
Типичными представителями артерий мышечного типа
являются артериолы большого круга кровообращения. Такое название
имеют мелкие артерии диаметром от десятка до сотни микронов
с общим структурным признаком - наличием выраженной гладкомы-
шечной оболочки, на долю которой приходится значительная часть
общего диаметра сосуда (табл. 5.2).
Мощной мышечной оболочкой обусловлено основное
функциональное свойство артериол - активный сосудистый тонус, под
которым понимают регулируемое тоническое сокращение гладкомы-
шечных клеток сосудистой стенки.
Они имеют веретенообразную форму со значительным
преобладанием длины (от 50 до 400 мкм; в среднем, 200 мкм) над
толщиной (диаметр у экватора - от 2 до 10 мкм; в среднем, 4,7 мкм).
Расположены циркулярно, т.е. ориентированы по окружности сосуда -
поэтому при их сокращении длина окружности, а значит, и просвет
сосуда уменьшаются.
I /швв 5. Биофизика двигательной активности
155
Таблица 5.2
Приблизительный тканевой состав микрососудов
(К. Каро, Т. Педли, Р. Шротер, У. Сид, 1981)
Типы
кровеносных
сосудов
Лртериола
Квпилляр
Ненула
Тканевой состав, %
эндотелий
(во внутренней
оболочке -
tunicae intimae)
10
100
20
мышечная ткань
(в средней
оболочке -
tunicae mediae)
60
0
20
соединительная
ткань (в наружной
оболочке -
tunicae adventitiae)
30
0
60
Между ГМК имеются щелевые контакты, обеспечивающие
•лектрическое взаимодействие посредством нексусов. Нервные во-
иокна, иннервирующие ГМК, не образуют концевых пластинок
υ нервно-мышечных синапсов на каждой клетке. Медиаторы
выделяются по ходу аксонов, особенно в их утолщениях (варикозах), и
воздействуют сразу на многие ГМК, усиливая или ослабляя их
сокращение. При электрической стимуляции аксонов, иннервирующих
гладкие мышцы, одиночными импульсами ГМК не сокращаются.
Для этого нужна серия импульсов частотой не менее 1 Гц.
Различают 2 типа сосудистых ГМК: 1) спонтанно активные
(например, в стенке воротной вены) и 2) спонтанно неактивные.
11 артериолах доминируют ГМК второго типа. Их сарколемма
обладает довольно низкой проницаемостью для К+, но всё же потенциал
покоя составляет от -50 до -70 мВ (из них около 8 мВ генерируется
благодаря стехиометрии K-Na-АТФазы). При возбуждении ГМК
нозникают потенциалы действия, в образовании которых потенциал-
швисимые натриевые каналы не участвуют. Считается, что их нет
и сарколемме ГМК. Зато там присутствуют потенциалзависимые
кальциевые каналы. Поэтому входящий ток исключительно кальцие-
иый - Isi. За счёт него деполяризация ГМК развивается медленно
с овершутом не более 5 мВ. Скорость распространения таких
потенциалов по сарколемме 0,02-0,1 м-с-1. Реполяризация очень вялая,
и общая продолжительность потенциала действия может достигать
нескольких секунд.
156
Биофизика
В течение всего потенциала действия ГМК сокращается,
поскольку в неё входит Са2+ из интерстиция через кальциевые каналы
в сарколемме, блокада которых расслабляет гладкие мышцы. Именно
потоку ионов кальция через сарколемму принадлежит ведущая роль
в электромеханическом сопряжении, происходящем в ГМК . Их сар-
коплазматическая сеть развита слабо. Для сокращения ГМК
необходимо повысить концентрацию Са2+ подле актиновых и миозиновых
филаментов от 10~7 до 10~5 М. При [Са2+],· около 2 · Ю-6 м наблюдается
сокращение в половину максимального, а при [Са2+], примерно Ю-4
Μ - максимальное.
Са2+ соединяется не с тропонином, которого в ГМК нет, а с лей-
отонином или кальмодулином, благодаря чему устраняется тропо-
миозиновая блокада активных центров на актиновых нитях для
головок миозина. Характерной особенностью ГМК является значительное
преобладание в содержании актина над миозином. Их соотношение
здесь в 10 раз больше, чем в миоцитах скелетной мускулатуры. В
актине ГМК иная последовательность аминокислотных остатков, чем
в актине кардиомиоцитов и миоцитов скелетных мышц мышц.
Другое важное отличие ГМК от них состоит в том, что с актином может
взаимодействовать только фосфорилированный миозин.
Катализатором такого фосфорилирования служит киназа лёгкой цепи миозина,
а её активирует комплекс Са2+ с кальмодулином и цАМФ.
Следовательно, взаимодействие с кальмодулином входящих в клетку ионов
кальция не только устраняет тропомиозиновую блокаду активных
центров для миозина на актиновой нити, но и позволяет ему проявить
свою АТФазную активность, которая на порядок ниже, чем у миозина
скелетных мышц.
Актиновые и миозиновые филаменты в ГМК расположены не
столь упорядоченно, как в других миоцитах, и по-видимому, не
образуют миофибрилл. Гладким мышцам не свойственна поперечная ис-
черченность. К особенностям сократительного процесса в ГМК
следует отнести весьма низкие скорости их укорочения и гидролиза
АТФ - они на 2-3 порядка ниже, чем в других мышечных клетках
млекопитающих. Вместе с тем по силе сокращения ГМК значительно
превосходят другие миоциты.
/ ппва 5. Биофизика двигательной активности
157
Сочетание низкой скорости гидролиза АТФ с большой силой
пжращения свидетельствует о невысоком уровне так называемой
стоимости силы. Не случайно запас макроэргов в ГМК сосудов
it I0-15 раз, а расход АТФ в 5-10 раз меньше, чем в миоцитах
скелетной мускулатуры. Общая масса всех сосудов тела человека на
порядок больше массы миокарда, но на выполнение своей работы они
расходуют вдвое меньше энергии, чем сердце. Соответственно, ГМК
сосудов потребляют кислорода примерно в 2 раза меньше, чем
сердце. Благодаря этим свойствам ГМК сосудов приспособлены к
длительному тоническому сокращению с небольшими энерготратами и без
развития утомления. Гладкие мышцы артериол находятся в состоя-
11ии непрерывной активности, что служит важным фактором
поддержания регулируемого сосудистого тонуса.
Сосудистый тонус создаётся и поддерживается несколькими
механизмами. Первым из них является возникновение силы
упругости при растяжении соединительнотканной наружной оболочки ар-
гсриолы - tunicae adventitiae (её называют «сумкой» артериального
сосуда, удерживающей его в определённом состоянии и пассивно
реагирующей на растяжение кровью созданием силы упругости в соот-
нетствии с законом Гука). Второй механизм связан с уникальным
свойством ГМК - способностью отвечать на растяжение их кровью
генерацией потенциалов действия, инициирующих активное
сокращение этих клеток. В ГМК, обладающих автоматией (в спонтанно
активных ГМК), третьим механизмом являются их автоматические
(спонтанные) сокращения. Три перечисленных процесса создают так
называемый базальный сосудистый тонус, для которого характерно
относительное постоянство.
Вместе с тем сосудистый тонус даже в покое обусловлен не
только базальным тонусом, но и сокращениями ГМК под влиянием
нервных импульсов, поступающих к ним по вегетативным (главным
образом, симпатическим) нервным волокнам. В совокупности с
базальным тонусом тонические сокращения ГМК под влиянием
управляющих сигналов формируют сосудистый тонус покоя. В
сократившейся ГМК модуль Юнга на порядок больше, чем в расслабленной,
а увеличение её жёсткости пропорционально повышению
внутреннего напряжения при сокращении.
158
Биофизика
Управление сосудистого тонуса сигналами, идущими к ГМК
по нервным волокнам, и физиологически активными веществами
(в частности гормонами), к которым чувствительны ГМК,
приобретает особое значение не в стационарном, а в переходном режиме,
когда возникает потребность в перераспределении крови между
сосудами, питающими его разные органы, если изменяется их
функциональная активность.
Изменения сосудистого тонуса приводят к сужению или
расширению просвета артериол и, следовательно, оказывают
выраженное влияние на гемодинамическое сопротивление, которое, согласно
уравнению Пуазейля, пропорционально четвёртой степени радиуса
кровеносных сосудов. За счёт активного сосудистого тонуса артери-
олы наиболее эффективно выполняют основные функции артерий
мышечного типа: 1) поддержание определённого уровня КД в
системе кровообращения; 2) перераспределение крови между органами
в зависимости от потребности в ней каждого из них.
Для эффективного обмена веществ через стенку капилляра
достаточно поддерживать КД на уровне 10-15 мм рт. ст. Однако в
сердечно-сосудистой системе создаётся примерно десятикратное
превышение этого уровня. На первый взгляд, оно невыгодно как
в энергетическом отношении, так и в связи с разрушающим
воздействием на сосуды. Чтобы понять необходимость избыточного КД
в артериальном русле, обратимся к роли водонапорной башни в
водопроводной сети. При наличии большого количества потребителей
воды (например, людей, пользующихся одной водопроводной сетью)
и случайном распределении во времени потребностей каждого из
них в воде необходим «подпор» в водопроводе, т.е. избыточное
давление, которое может обеспечить ток воды при открытии как одного,
так и всех кранов водопроводной сети. Можно- удовлетворить
большие потребности в воде иначе - за счёт подачи в водопровод
значительного запаса резервной воды. В водопроводных системах эти
способы комбинируются.
В организме ограничены возможности создания запаса крови
в большом объёме. Правда, существуют кровяные депо, которые в
состоянии покоя содержат до 2/3 всей крови. При нагрузках этот запас
уменьшается. И все же основным средством достаточной поставки
(imim 5. Биофизика двигательной активности
159
кропи всем органам, которые в данный момент испытывают в ней
noil (сбность, служит поддержание избыточного давления в системе кро-
иообращения. Это позволяет существенно изменять объёмную
скорость кровотока, обходясь сравнительно небольшим количеством
крови (масса крови у человека составляет 1/13 часть массы тела).
Таким образом, артерии мышечного типа предназначены для
поддержании в системе кровообращения такого избыточного давления, которое
обеспечивает надежное перераспределение крови между органами
м любых условиях. Поэтому артериолы по их функциональному
назначению могут быть уподоблены водонапорной башне - они создают
«подпор» кровяного давления в артериальном русле. Нарушение этой
функции артериол приводит к коллапсу (тяжёлому обмороку с потерей
пгшания при остро развивающейся сосудистой недостаточности, вы-
рижением которой является прежде всего падение КД). Таким образом,
именно ГМК артериол большого круга предопределяют работу,
которую должно совершить сердце (миокард левого желудочка), чтобы
преодолеть общее периферическое сопротивление сосудов (ОПСС).
Вторая функция артерий мышечного типа - перераспределение
крови между органами в зависимости от потребности в ней -
обеспечивается работой артериол в качестве сосудистых кранов. Это меткое
образное определение принадлежит И. М. Сеченову. Из уравнения
11уазейля, разрешенного относительно Q, прямо следует, что в
зависимости от просвета артериол меняется объёмная скорость кровотока
и органе, которому они принадлежат, причем её изменения
пропорциональны изменениям радиуса этого просвета в четвёртой степени.
За счёт изменения тонуса артериол, находящихся в скелетных
мышцах, объёмная скорость кровотока в них увеличивается при
физической работе в несколько десятков раз. Кроме того, с регуляцией
сосудистого тонуса непосредственно связано перераспределение крови
между внутренними органами и кожей при том или ином уровне теп-
пообмена организма с внешней средой. Даже в пределах одного органа
происходит непрерывная «игра сосудов»: одни артериолы
суживаются, другие - расширяются. В результате масса крови в разных участках
органа беспрестанно перераспределяется, чем достигается наилучшее
кровоснабжение при минимальном уровне работы сердечной мышцы.
160
Биофизика
Артериолам принадлежит большая роль в развитии ряда
патологических процессов. Гипертоническая болезнь связана с их
стойким сужением (спазмом). При гипотонических состояниях
сосудистый тонус ослаблен.
5.5.5. Математические модели кровообращения
Сердечно-сосудистая система. Первые модели
кровообращения описывали деятельность сердца в замкнутой гемодинамической
цепи. При этом контуры рефлекторной регуляции предполагались
разомкнутыми, а показатели насосной функции сердца и резистив-
но-емкостные свойства сосудистого русла могли изменяться по воле
экспериментатора. Эти свойства рассматривались как входы модели,
а выходами служили основные показатели гемодинамики (минутный
объем крови и кровяное давление) в различных частях системы,
прежде всего в участках, прилегающих к сердцу.
Во всех моделях сосудистое русло представлено в виде сети из
участков с сосредоточенными параметрами. Описания
гемодинамики в /-μ участке системы практически одинаковы в разных моделях
и имеют вид
Q,^P'~Pm\ (5.148)
R.
ι
ГС,·-",)
V>u.
Pi={ С,. (5.149)
0, Vt<u,
I
Г,-=v0,+/(?,_, -?,-)*, (5-150)
о
где gj, gj_, - объемная скорость на выходе и входе /-го участка; Vi -
объем крови в г'-м участке; /?, - давление; Ri - сопротивление между
ί-м и (г + \)-м участками, С, - растяжимость; щ - ненапряженный
объем; V0i - начальное наполнение участка. В некоторых моделях
учитывается также инерционность кровотока и вязкие свойства сосу-
(пива 5. Биофизика двигательной активности
161
Г """" ^-ч- ^ "" ^\ /^ ^^~ -'" ~"\
1 «1 /
\ +7
\ ι2
Ч
=1 J? у Ι* ^ Яз1 „, «1 ft « „. IPS ^ ft
ι—ι ил #<
о*" /
Е1=1/С/ Т
/ /4
г / -
У
/5
= ι =
ч μ-ι ι—ι '
■ 1 1 |^1 ,<
\ Π iS I
сз\ 1^ /по
г \ τ / 1
ч /
\ _ У
1__|
ί11
Ύ
Рис. 5.64. Электрический аналог
математической модели сердечно-сосудистой системы:
ρ - давление крови, i - кровоток, R - сопротивление
сосудов кровотоку, С - растяжимость сосудов,
Ε - объемная жесткость желудочков сердца
цистой стенки в участках аорты (R'). В этом случае уравнения
(.Ч. 148), (5.149) имеют соответственно вид:
dt L.
Ρι=-
dt K ■
",)
ι дс Z,, - инерционность кровотока, т е. Li = —, где р - плотность
u .
I
крови, S - площадь сечения сосуда.
Приведенные выше уравнения аналогичны описанию элек-
фических RLC-цепей, поэтому модели сердечно-сосудистой
системы нередко иллюстрируют соответствующими электрическими
схемами (рис. 5.64). Создаются такие модели для исследования ха-
рмктеристик сердечно-сосудистой системы как объекта управления.
Поскольку в замкнутую гемодинамическую цепь включен саморе-
I улирующийся насос (сердце), модели используются также для
определения общих саморегуляторных свойств сердечно-сосудистой
системы. Несмотря на то, что большинство моделей неуправляемой
системы кровообращения динамические, на практике исследования
с их помощью проводятся в установившихся режимах. При этом
162
Биофизика
изучаются статические зависимости таких важнейших переменных
гемодинамики, как минутный объем крови Q, средние величины
системного Ра и легочного артериального Рар, центрального Pv и
легочного венозного Pvp давлений от параметров, характеризующих
состояние сердца и сосудов. В качестве последних рассматривались
общее периферическое сопротивление R, растяжимость С и
ненапряженный объем и сосудистого русла или его частей, общий
объем крови V, частота сердечных сокращений, показатель
сократимости (инотропного состояния)сердца.
Целью модельного анализа было прежде всего определение
чувствительности переменных к изменению параметров и
относительной роли сердечных и сосудистых факторов в регуляции
гемодинамики. Статические модели сердечно-сосудистой системы
позволяют получить и исследовать аналитические выражения,
связывающие переменные кровообращения с параметрами,
характеризующими насосную функцию сердца и резистивно-емкостные
свойства сосудистого русла. Выведена формула для минутного объема
крови:
V„JP--P*C-+C-\ (5.151)
cvRv+ca(Rv+Ra)
где pms - среднее системное давление; индексами α и ν обозначены
величины, соответствующие артериальному и венозному участкам
системы кровообращения. Проведен анализ зависимости минутного
объема крови от условий притока крови к сердцу (венозного
возврата). Показано, что изменения венозного сопротивления оказывают
значительно большее влияние, чем артериального.
Модель статики сердечно-сосудистой системы, состоящей из
правого и левого желудочков сердца, двух системных
(артериального и венозного) и двух легочных участков, имеет вид:
(Ρ -Ρ Ϊ (Р -Р )
Q = aPv+b=aPvp+$ = K ' Г*>=К-ЛР vl. (5.152)
R Rp
I пива 5. Биофизика двигательной активности
163
Здесь
ρ _(г„-0 р _(^-Q
С ' ν С '
a v
(V -и ) (V -и )
ρ _ V яр др / ρ _УГ νρ "урУ ,- , --,4
^ор „ > Γψ > ij.ijj;
^ op ^ vp
V +V +V +V =V.
a v r ap vp
Индексом р обозначены величины, относящиеся к сосудам
легких. Коэффициенты а, Ъ и α, β характеризуют насосную способность
иевого и правого желудочков сердца в соответствии с описанными
иыше данными о статических характеристиках сердца и зависят от
чистоты сокращений н инотропного состояния сердца. Эта
зависимость может быть раскрыта, например, с помощью приведенной
иыше модели статики изолированного желудочка следующим обра-
юм (величины с индексами R и L соответствуют правому и левому
/келудочкам сердца):
а_ kCR(\-AR) ш b_(kuR-u0)(l-AR)_
(\-(\-к)Ая)Г {\-{\-k)AR)T '
AR=J^-\a- K^-V ■
RRCRj {}-{}-k)AL)T (5154)
(\-(\-k)AL)T yRRcR)
xd=a,T + b,{\-k); T = ^.
F
где к — показатель инотропного состояния сердца,
экспериментальным аналогом которого является изгоняемая фракция желудочка, F —
частота сердечных сокращений (ударов/мин.)
Решение приведенной системы уравнений дает формулы для
основных гемодинамических переменных:
164
Биофизика
α α
RCn+RpCnp+^ +
α
Λ=β(Λ+α-')--,
ΛΡ=β(Λ+α"')--:
α
ρ _Q-b
ρ _β-β
4>
α
У
\-J-i-J-J)
(5.156)
(5.157)
(5.158)
(5.159)
α
Здесь С - С + С - суммарная растяжимость малого, аС,=
= Ca + Cv - большого круга кровообращения; и - суммарный
ненапряженный объем сосудистого русла.
Формулы (5.155) - (5.159) позволили провести анализ
чувствительности Q и Р„ к изменению регулируемых параметров k, F, Dv =
= \ICV G=\IR, К по следующей методике. По литературным данным
были выбраны два исходных уровня параметров кровообращения,
соответствующие двум состояниям организма человека: а) в случае
покоя; б) в случае интенсивной физической нагрузки.
Для обоих состояний определялись функции чувствителыю-
dQ ЗРа г - ft 17
сти —=-, —— (где а, - варьируемый параметр из множества {к, F,
да, δα,.
Dr, G, V)), при условии постоянства (на одном из двух принятых
уровней) остальных параметров. Показано, что чувствительность Q
к изменению какого-либо параметра связана обратной зависимо-
/ пава 5. Биофизика двигательной активности
165
пью с этим параметром и прямой - с остальными. Аналогичный
Ш.1ВОД можно сделать для чувствительности Ра, если во множестве
параметров рассматривать не G (общую проводимость сосудистого
русла), a R.
Рассмотренные выше модели неуправляемой
сердечно-сосудистой системы представляются на первый взгляд крайне
упрощенными и в силу отсутствия регуляторных связей далекими от практики.
()диако именно они нашли в настоящее время наибольшее
применение в экспериментальных исследованиях и клинике, поскольку в ус-
кшовившихся режимах соотношения между переменными и пара-
мсфами действительно определяются формулами (5.155)—(5.159)
или близкими к ним. Следовательно, если известны истинные
значения парамегров, характеризующих сердце и сосуды (k, F, Rf, Д, V), то
по формулам (5.155) -(5.159) могут быть найдены истинные
значения переменных (Q, Pa, Pv и др.). Наоборот, если известны для
данного состояния организма значения перемегашх кровообращения, то
подставляя их в формулы (5.155)-(5.159), можно решить
получившуюся систему уравнений относительно ее параметров, определив
гаким образом состояние насосной функции сердца и сосудистого
русла.
Для изучения регионарного кровообращения использованы
математические модели, учитывающие особенности геометрии
сосудистого русла, биофизические свойства сосудистой стенки и
гетерогенность крови.
В принципе все известные математические модели
регионарного кровообращения можно разбить на модели кровотока в
единичном сосуде, модели кровотока в системе сосудов без учета регуля-
гариых и управляющих механизмов, модели управления сосудистой
сетью, учитывающие определенные регуляторные и управляющие
ноздействия. В первых двух типах используется биофизический
подход, а модели третьего типа представляют собой формализованные
системы, исследование которых основано на применении методов
теории автоматического управления. В задачу анализа последних
иходит определение зависимостей между гемодинамическими харак-
166
Биофизика
дЛ=
дх
-lV'-rS,.*·-^*·
' dt ' ' Эх ' Э/
теристиками сети, геометрическими параметрами, реологическими
свойствами стенки и крови, а также параметрами регулирования
кровотока в различных режимах работы органа.
Математическое описание регионарной и системной
гемодинамики базируется на методах механики сплошных сред, т.е.
уравнениях Навье - Стокса или на реологическом описании движения крови
с учетом ее неньютоновских свойств. Распространенным, хотя
и весьма приближенным способом описания гидродинамики,
является применение уравнения Пуазейля.
Гемодинамику в каждом из участков сосудистого русла можно
адекватно описать с помощью упрощенных линеаризованных
уравнений типа Навье-Стокса и уравнения неразрывности:
■Gf,. (5.160)
Здесь Р, - давление крови; g, - объемная скорость кровотока; Л„ С„
G„ Ζ,, - соответственно эквивалентные значения сопротивления,
податливости стенки, утечки и инерционности потока
рассматриваемого участка сети; / - номер участка; χ - фиктивная пространственная
координата, связанная с номером (порядком ветвления) участка
сосудистой сети. Коэффициенты уравнений (5.160) являются
переменными величинами, зависящими от геометрических и биофизических
свойств сосудов, и определяются из соотношений:
ϋ,-UL, L,--^, С,-Ц£. (5.16.)
тт.г. тт.г. Е,Ь.
II II I I
где т|( - динамическая вязкость крови; и, - количество сосудов
в данном участке; г, - средний радиус сосудов /-го участка; ρ -
плотность крови; Е-, - модуль упругости Юнга для сосудов данного
участка; δ, - толщина стенки сосуда.
Система уравнений микрогемодинамики решается вместе с
уравнениями, описывающими влияние упруговязких свойств стенки:
I пава 5. Биофизика двигательной активности
167
dt цс[ьк a) J г dt 3J
= — \-±(Р-Р0)-Е \ + a2^-aJ. (5.162)
ii уравнением, учитывающим сократительную активность гладкой
мускулатуры стенки под влиянием импульсации водителей ритма:
diA1r) = al(P-Pn)+ d^_
dt Ρ 2 dt iJ
В уравнениях (5.162) и (5.163) г0 - радиус сосуда, а 5 толщина
сосудистой стенки при нулевом трансмуральном давлении Ρ = Ре;
1\ - давление окружающей сосуды среды; η,. - модуль вязкости
материала стенки; А,г, А2г - изменение радиуса сосудов под действием
соответственно упруговязкой деформации и электрической
активности соответственно;/- эквивалентная частота пейсмекеров,
характеризующая влияние электрической активности водителей ритмов на
сокращение гладкомыщечных волокон стенки; Ркм - осмотическое
давление в ткани; РП - пороговое значение давления крови, при
превышении которого активируются гладкая мускулатура сосудистой
стенки, а,, аъ аъ - константы.
В модели принято допущение об аддитивном вкладе
различных регуляторных воздействий в изменение просвета сосудов. В
качестве внешнего воздействия на сосудистую сеть рассматривается
изменение давления крови на артериальном конце каждого
исследуемого участка. Гемодинамические характеристики рассчитывают при
фиксированных значениях объемной скорости кровотока на выходе
сети, соответствующих определенным функциональным состояниям
органов. Значения геометрических и биофизических параметров
сосудистой сети выбирают в соответствии с имеющимися
экспериментальными и клиническими данными. В частности, для нормы
диапазоны значений параметров для резистивного отдела сосудистой сети
выбраны в следующих пределах: Ε = 10s—107 дин/см2, Q = 2-
12 мл/мин на 100 г, η0 = Ι0Μ06 дин · с/см2, % = 1,5^ сП, η = 900-
5 · 103 мкм, г0 = 10-100 мкм, 5 = 3-60 мкм.
168
Биофизика
5.6. Тубулин-динеиновая и тубулин-кинезиновая
молекулярные системы подвижности
5.6.1. Системы микротрубочек
Микротрубочки (микротубулы) представляют собой полые
цилиндры, стенка которых образована молекулами глобулярного белка-
тубулина. Его не очень крупные (диаметром около 4 нм и
молекулярной массой примерно 115 кДа) молекулы обладают полярностью и по-
лимеризуются на стороне положительного полюса (подобно G-акти-
ну). Поэтому сборка микротрубочек посредством полимеризации
тубулина происходит путём «наступления» положительных полюсов
губулиновых глобул от центров к периферии. Наличие комплементар-
иой оси типа «замок-ключ» позволяет осуществлять безграничное
линейное объединение молекул тубулина.
Различают 2 модификации тубулина (а и β), причём соединения
этих близкородственных белков между собой более прочные, чем
между тубулинами одноимённой модификации. Поэтому распад
микротрубочек при их перестройках происходит не до элементарных
молекул тубулина (а или β), а до димеров - α-β-соединений (рис. 5.65, а),
вследствие чего аналогом (эквивалентом) G-актина, из которого
строится F-актин, считают не мономеры тубулина, а его α-β-гетеродимер.
Из таких димеров тубулина посредством их линейного объединения
за счёт образования β-α-связей в клетках собираются тубулиновые
протофиламенты, т.е. первичные нитеобразные ультраструктуры.
В них субъединицы в силу присущих им асимметрии и полярности
расположены вдоль оси полимера по принципу «голова к хвосту».
Поэтому каждый протофиламент напоминает длинную стрелу,
состоящую из коротких стрелок, направленных в одну и ту же сторону
Каждая молекула а- и β-тубулина имеет и вторую систему
«ключ-замок», т.е. обладает способностью образовать боковые связи
с тубулинами соседних протофиламентов (рис. 5.65, б). Эта
латеральная сборка происходит ступенчато, в результате чего каждая а-
β-субъединица одного протофиламента взаимодействует с α-β-субъ-
единицами соседних протофиламентов, сдвинутых примерно на
четверть её длины.
ι /шва 5. Биофизика двигательной активности
169
Формирующийся таким образом полимолекулярный пласт из
11 протофиламентов приобретает изогнутую поверхность и выстраи-
илстся в полый цилиндр, наружный диаметр которого составляет
,'К им, а внутренний (ограничивающий полость) - 14 нм (рис. 5.66).
(' ι упенчатый характер бокового соединения протофиламентов
приводит не только к возникновению цилиндрической структуры, но
и к спиральной периодичности идентичных субъединиц.
Энергетическое обеспечение сборки микротрубочек осуществляется посредст-
иом гидролиза гуанозинтрифосфата (ГТФ), а не АТФ.
Подобно микрофиламентам, микротубулы беспрестанно
распадаются и созидаются. Ими образованы многие внутриклеточные
ультраструктуры. Во всех клетках они играют роль внутриклеточно-
1 о каркаса с прекрасными механическими свойствами, причём пре-
β-тубулин
_- Гетеродимер
* ^
λ
\
Протофиламент
Латеральная сборка „
о)
Рис 5.65. Молекулы тубулина и гетеродимер
а - образован а- и ^-молекулами тубулина; б- образующийся протофиламент
а-тубулин
170
Биофизика
Рис. 5.66. Микротрубочка (цилиндр), образованная посредством
латерального ступенчатого соединения протофиламентов
имущества трубчатых опорных элементов биологические системы
«открыли» более миллиарда лет назад.
Микротрубочками укреплены отростки нейронов. В аксонах
по каналам в микротубулах перемещаются от тела нейрона на
периферию разнообразные вещества со скоростью до 8 мкм · с-1 (это
называется аксональным током).
Вместе с тем микротубулы, как и микрофиламенты, служат
«рельсами», по которым не внутри, как аксональный ток, а по
наружной поверхности перемещаются (скользят) белки-моторы {динеины
ι илп/ι 5. Биофизика двигательной активности
171
ι/ ышезины). Они одним своим концом контактируют с наружной по-
нгрхностью микротрубочки и при гидролизе АТФ могут двигаться
ми ней. Другой конец молекулы-мотора может прикрепляться к
органоидам или к соседним трубочкам. В первом случае обеспечивается
фипепорт органоидов по цитоплазме, а во втором - перемещение от-
щ'льных участков клетки относительно соседних.
Любой белок из большого семейства кинезинов представляет со-
iinii гетеротример палочкообразной формы. В его составе две тяжёлые
(миссой по 120 кДа) и две лёгкие (массой по 70 кДа) полипептидные
Ш'ии. Каждая из тяжёлых цепей увенчана двумя маленькими «голов-
инми» (глобулярными молекулярными структурами, образованными
примерно 340 аминокислотными остатками). Остальная часть тяжёлой
цепи напоминает стержень, заканчивающийся перообразным «хво-
I гом». С ним соединены две лёгкие полипептидные цепи. Этим «хво-
I и)м» кинезин прикрепляется к тем веществам и ультраструктурам,
которые необходимо транспортировать внутри клетки по поверхности
микротубул.
С тубулярной поверхностью взаимодействуют головки кинези-
|щ. На них есть активные центры (связывающие домены) для тубули-
ιιιι и АТФ. Механизм взаимодействия кинезина с микротрубочкой
ниалогичен скольжению миозина по актиновым микрофиламентам.
11ри каждом шаге, а он составляет 8 нм, развивается сила в 5 пиконь-
ιιπόηοβ. КПД такого перемещения достигает 50%.
Кинезины обеспечивают активные деформации клеток,
участвуют в работе эндоплазматической сети по синтезу макромолекул,
подтягивая мономеры по микротубуле к положительному полюсу со-
шдаемых полимеров. Кинезины переносят секреторные гранулы
и накуоли к апикальному концу секреторной клетки, перемещают уд-
иоившиеся хромосомы к противоположным полюсам родительской
клетки в процессе их митотического деления.
Ещё в XIX веке было установлено, что хромосомы,
расходящиеся в анафазе митоза к противоположным полюсам родительской
клетки, движутся по определённым траекториям вдоль системы стро-
ю упорядоченных фибрилл, которую за её конфигурацию назвали
митотическим веретеном. Позднее выяснилось, что его основным
компонентом являются микротрубочки, построенные из тубулина.
172
Биофизика
Рис. 5.67. Митотическое веретено (схема)
По ним перемещаются так называемые кинетохоры -
молекулы-моторы, которые головками взаимодействуют с тубулином, а
хвостами - с хромосомами (рис. 5.67). Предполагают, что кинетохор,
встречаясь с тубулином, осуществляет разборку микротрубочки и при
этом «хватается» за её остающийся конец, как за страховочный канат,
и вместе с хромосомой продолжает движение по сохранившейся
части микротрубочки к полюсу клетки. Возможно, что роль кинетохоров
исполняют кинезины.
Другим важнейшим молекулярным мотором в системе
микротрубочек являются динеины. Всё их семейство составляют крупные
молекулярные комплексы (с молекулярной массой порядка 106 Да),
имеющие 2-3 глобулярные головки на 1 «стебельке». Каждая
головка - белковая молекула с молекулярной массой более 400 кДа,
обладающая АТФазной активностью. Кроме столь высокомолекулярных
компонентов, динеин содержит полипептидные цепи с молекулярной
массой около 100 кДа (промежуточные) и 20 кДа (лёгкие).
I /шва 5. Биофизика двигательной активности
173
Уникальное свойство динеина - способность скользить по по-
исрхности микротрубочки к её отрицательному полюсу. Динеин
икшмодействует с тубулином подобно тому, как миозин с актином,
и и результате механического процесса перемещаются 2 дублета
микротрубочек, соединённых так называемыми боковыми ручками, обра-
юванными динеином, который к тому же катализирует гидролиз
ΛΤΦ, необходимый для энергетического обеспечения скольжения
микротрубочек. Поэтому боковые ручки микротрубочек называют
«цитомышцами», а сами микротрубочки - «цитоскелетом». Эти
термины адресуют также соответственно миозиновым и актиновым фи-
иаментам.
Все динеины устремляются в клетках к отрицательному, а бо-
ш.шинство кинезинов - к положительному полюсу микротрубочки.
11оэтому органоиды могут «ездить»по одним и тем же «рельсам»
(микротрубочкам) к центру и периферии клетки, пользуясь разными
«локомотивами» (динеинами или кинезинами). Благодаря таким
транспортным системам один и тот же органоид, меняя их
неоднократно, может изменять направление своего движения по
цитоплазме несколько раз в минуту.
Предполагают, что на одном и том же органоиде «сидят»
разные молекулы-моторы (миозины, динеины, кинезины), которые
нключаются и переключаются специальными регуляторами
(например, ферментами, присоединяющими к ним ортофосфат,
освобождающийся при гидролизе АТФ). По-видимому, микротрубочки служат
«рельсами кинезинам и динеинам для дальних дорог» в пространстве
клетки, а микрофиламенты обеспечивают транспорт органоидов по
актиновым нитям при помощи миозинов на меньшие расстояния.
5.6.2. Биомеханические процессы
в жгутиках и ресничках
Реснички и жгутики представляют собой важнейшие
двигательные органоиды немышечных клеток. Микроорганизмам и
простейшим (класса жгутиконосцев), а также гаметам морских водорослей
и сперматозоидам животных жгутики обеспечивают перемещение
в пространстве. Например, у кишечной палочки, как правило, 6 жгу-
174
Биофизика
тиков, каждый из которых в 2-3 раза длиннее тела клетки. При
синхронном движении против часовой стрелки все жгутики сплетаются
в единый пучок, который вращается подобно пропеллеру, в
результате чего бактерия, длина которой меньше 2 мкм, преодолевает за
1 с расстояние в 20 мкм, что эквивалентно бегу лошади со скоростью
180 км · ч-1. Немудрено, что жгутиковые бактерии в своём
большинстве обладают лучшей конкурентоспособностью и живучестью по
сравнению с микроорганизмами без жгутиков. Следует, правда,
заметить, что у бактерий (кроме спирохет) подвижность жгутиков
обеспечивается не тубулином, а флагеллином. Однако
биофизический механизм движения любых жгутиков, а также ресничек в
основных чертах сходен, хотя реснички у животных служат главным
образом для перемещения не клеток, которым они принадлежат, а среды
вокруг них. Уместно заметить, что и жгутики у некоторых
многоклеточных организмов создают циркуляцию внутренней среды, а
реснички инфузорий класса Ciliata, ресничных червей (турбеллярий)
и гребневиков (из типа морских позвоночных) перемещают их тела
в пространстве.
Жгутики и реснички представляют собой тонкие, но довольно
длинные выросты клетки, одетые у эукариот плазматической
мембраной. Толщина этих структур (0,20-0,25 мкм) находится почти на
пределе разрешающей способности световой микроскопии, а длина
составляет 5-10 мкм у реснички и до 150 мкм у жгутика. Ресничек
у клетки очень много (порядка 100), а жгутики располагаются
поодиночке или по несколько штук на хвостовом конце свободно
плавающей клетки.
Траектории движений жгутиков и ресничек также различны.
Жгутики движутся волнообразно или воронкообразно, а реснички -
маятникообразно. В маятникообразном движении реснички
различают 2 фазы: замаха и удара (рис. 5.68). По данным В. В. Шабалина,
соотношение их продолжительностей в норме соответствует золотой
пропорции: tnoJIHOro цнкла: tMMaxa = ^„„,: 1^,= 1,62 ± 0,24. При патологии
нарушается как это гармоническое соотношение, так и сама
траектория. Если при нормальной работе реснички замах и удар происходят
во взаимно перпендикулярных плоскостях, то при развитии
патологии они совершаются в одной плоскости, что приводит к колебаниям
I шша 5. Биофизика двигательной активности 175
Рис. 5.68. Полный гребной удар реснички
при последовательном микрофотографировании
Цифрами обозначена последовательность движения:
1-6 - стадия эффективного удара (а), 6-12- стадия замаха (б)
окружающей среды (вперёд и назад) относительно стартовой линии
(положения равновесия), а не к поступательному однонаправленному
диижению, обеспечивающему эвакуацию компонентов этой среды.
Дело в том, что биения ресничек на поверхности многореснич-
пых клеток координированы. Расположенные параллельными
рядами, они при движении создают над клетками волны, бегущие в
определённом направлении. Например, эти волны на поверхности клеток
респираторного (реснитчатого) эпителия «бегут» к глотке: сверху
«низ - в верхних и снизу вверх - в нижних дыхательных путях чело-
нека. Направления движения ресничек генетически
детерминированы. Поэтому в участке слизистой оболочки трахеи или бронхов,
вырезанном и затем пришитом «задом наперёд», реснички будут
совершать удар в сторону альвеол, а не глотки, т.е. сохранять
прежнюю траекторию движения (рис. 5.69). Поверхность реснитчатого
■щителия, над каждой клеткой которого колеблются 150-200
ресничек, напоминает волнующееся море или подвластное ветру
пшеничное поле, по которым бегут волны с определённой частотой (12-
176
Биофизика
Рис. 5.69. Поверхность реснитчатого эпителия
(сканирующая электронная микроскопия)
16 Гц - у человека). При патологии (например, при хроническом
бронхите) частота биений ресничек падает, и эвакуация содержимого
бронхов ухудшается.
Колебания ресничек, расположенных на поверхности
бронхиальных эпителиоцитов, формирует поток жидкости над ними. У
здорового человека за сутки в среднем секретируется в нижних
дыхательных путях 0,75 мл слизи на 1 кг массы тела. Благодаря
непрерывной работе секреторных клеток (бокаловидных и клеток
Клара) слизистая оболочка дыхательных путей человека и животных
выстлана слоем слизи. Толщина слизистого покрытия около 7 мкм.
Оно неоднородно в своём профиле: нижние 5 мкм - золь, а верхние
2 мкм - гель. Золь обладает гораздо меньшей вязкостью (около
1 мПа · с), чем гель (до 10 мПа ■ с), вследствие чего движения
ресничек в золе совершаются за счёт сравнительно небольших усилий.
Толщина этого слоя почти совпадает с длиной реснички, что
позволяет ей экономно расходовать энергию. Только верхними кончиками
реснички касаются геля, в котором обычно находятся частицы,
подлежащие эвакуации из дыхательных путей (рис. 5.70). Слой геля
в них удерживает на своей поверхности частицы массой до 12 мг.
Они вместе с гелем скользят по поверхности золя. Возможно, кончи-
/ nana 5. Биофизика двигательной активности
177
Рис. 5.70. Профиль просвета бронха (в пределах 10 мкм от поверхности)
I —реснички; 2 - слой золя; 3 - слой геля
mi ресничек контактируют с неровностями на нижней поверхности
ι сия, что способствует более эффективной эвакуации слизи.
Скорость её перемещения составляет в мелких бронхах около 1 мм ■ мин-
', а в трахее 10-15 мм мин-1.
Взаимодействие бьющихся ресничек со слизью в
дыхательных путях обеспечивает так называемый мукоцилиарный клиренс
(I'lcarance - очищение, mucus - слизь, cilia - ресничка), т.е.
очищение дыхательных путей посредством перемещения слизи к глотке,
происходящего при колебаниях ресничек. Заметим, что этот
процесс, называемый мукоцилиарным транспортом (МЦТ), не единст-
нспный механизм мукоцилиарного клиренса. В его осуществлении
пижная роль принадлежит также альвеолярным макрофагам
(особенно, в самых мелких бронхах и альвеолах). Очищению крупных
Оронхов и трахеи способствует кашель, а верхних дыхательных
путей - чихание и насморк.
Ради понимания биомеханических процессов в ресничке
рассмотрим её ультраструктурную организацию. Под плазмолеммой
находится аксонема - цилиндрическое образование, построенное из 9 пар
микротрубочек, связанных между собой динеиновыми ручками. В
центре аксонемы находятся ещё две микротрубочки, заключённые в
оболочку (центральную капсулу). Поэтому говорят, что реснички (равно
как и жгутики) имеют структурную модель: 9 + 2 (рис. 5.71). Девять
периферических дублетов соединяются с центральной парой
радиальными «спицами». В основании лежат базальные тельца: в ресничке одно,
к жгутиках - два, расположенные взаимно перпендикулярно.
Каждый из 9 периферических дублетов построен из полной
микротрубочки (А), образованной 13 тубулиновыми протофиламен-
тами и неполной (В), составленной из 10 тубулиновых протофила-
I.1 -9844
178
Биофизика
Субфибрилла В
Субфибрилла А
Нексин
Внутренняя
динсиновая
ручка
Наружная двойная
микротрубочка
Плазматическая мамбрана
Радиальный мостик
Центральная капсула
Наружная диней новая
ручка
Центральная одинарная
микротрубочка
Рис. 5.71. Схема ультраструктурной организации реснички
ментов. Полная и неполная микротрубочки соединяются между собой
таким образом, что 3 «лишние» протофиламента трубочки а
становятся для них общими (объединяющими обе трубочки).
Кроме того, полная микротрубочка (А) взаимодействует с
неполной микротубулой (В) прилежащего (соседнего) дублета
посредством пары тангенциально ориентированных боковых ручек. Они по
форме напоминают гантели и образованы динеином. В связи ручек
с трубочками участвует белок пектин. Как и другие динеиновые
(а также кинезиновые и миозиновые) надмолекулярные структуры,
ручки работают как «цитомышцы» на «цитоскелете». С ними
связано и энергетическое обеспечение движения реснички. Механохими-
ческий процесс взаимодействия динеина с тубулином приводит к
перемещению друг относительно друга двух дублетов микротрубочек,
соединённых боковыми ручками. Механизм этого перемещения
напоминает скольжение миозиновых нитей вдоль актиновых. Модель
взаимного скольжения компонентов реснички предложил П. Сатирп-
римерно через 10 лет после появления гипотезы скользящих нитей
в миоците (А. Хаксли и X. Хаксли). Обнаружено поразительное
сходство в работе двух основных клеточных систем подвижности:
' нпвв 5. Биофизика двигательной активности
179
it обеих происходит скользящее перемещение «цитокостей» за счёт
натяжения «цитомышц».
Энергетическое обеспечение механохимического процесса в
микротрубочке также осуществляется за счёт гидролиза АТФ, причём
ΑΤΙ >азной активностью обладают глобулярные головки линейна. В опы-
IHX на цилиарных аксонемах, лишённых плазмолеммы и радиальных
1'ииц, добавление в среду АТФ приводило к быстрому и значительному
удлинению исследуемой структуры, в результате чего поднимающиеся
инсрх дублеты могли упасть (подобно тому, как падает гусеница,
доползающая до края ветки).
Если же дублеты соединены между собой нексиновыми
мостиками, дополняющими боковые ручки, и связаны радиальными спица-
Общее смещение
радиальных
спиц
группы 8
Мембрана реснички
Микротрубочка центральной
пары с отходящими от нее
выступами центральной
капсулы
Периферийный дублет
с радиальными спицами
И ι;
Μ 11
Базальное
тельце
Рис. 5.72. Изгиб реснички при нормальном взаимодействии её компонентов
I."
180
Биофизика
ми, то при скольжении они не разделяются, а изгибаются (рис. 5.72).
Боковые ручки, расположенные в плоскости сгибания, которая
перпендикулярна плоскости центральной пары микротрубочек,
обеспечивают ресничке возвратно-поступательное движение. Для этого
ручки, расположенные на стороне изгиба, максимально
сокращаются, а противоположные - расслабляются. При замахе первые
расслабляются, а вторые - испытывают максимальное сокращение.
Ручки, лежащие в плоскости, перпендикулярной плоскости сгибания,
остаются пассивными. На самом деле сложная траектория
формируется разной степенью сокращения и расслабления многих динеино-
вых ручек, расположенных между взаимно перпендикулярными
плоскостями, т.е. между пассивными и наиболее активными динеи-
новыми ручками.
Изложенная схема весьма упрощает механизм биения
ресничек. В нём участвуют более сотни различных белков, входящих в
состав аксонемы. Однако ведущая роль принадлежит тубулину и дине-
"ину. Поражает отсутствие эволюции основных молекулярных
механизмов двигательной активности от спирохет до человека.
Мукоцилиарная система органов дыхания нарушается при
многих патологических состояниях, включая профпатологию. Работа
в атмосфере с высоким содержанием агрессивных веществ уже через
1-2 года приводит к угнетению мукоцилиарного транспорта в
верхних дыхательных путях. Позже подобные изменения происходят
в бронхах, а они, в свою очередь, опережают клинические
проявления пневмокониозов и бронхитов. Любые формы дисфункции
ресничек в дыхательных путях служат противопоказаниями к работе
в условиях запылённости и загазованности рабочих помещений.
С другой стороны, при хронических бронхитах (хронической об-
структивной болезни лёгких) содержание реснитчатых клеток в
слизистой оболочке бронхов заметно уменьшается. Если в норме они
составляют 80-90% эпителия дыхательных путей, то при бронхите -
около 50-60%. Потеря «шевелюры» слизистой оболочкой приводит
к нарушению мукоцилиарного клиренса, усугубляющему
патологический процесс. Наркоз угнетает двигательную активность ресничек,
что является одним из существенных факторов развития
послеоперационных пневмоний.
/ паев 5. Биофизика двигательной активности
181
Вместе с тем есть такие формы патологии, этиология которых
непосредственно связана с дефектами развития тубулин-динеиновой
ι нстемы. Яркий пример тому - синдром Картагенера (первичная ци-
инарная дискинезия). Это наследственная аномалия развития,
обусловленная генетическим дефектом тубулин-динеиновой системы,
приводящим к отсутствию в ресничках динеиновых ручек и транспо-
шции радиальных спиц. Происходит транслокация микротрубочек,
естественно, что двигательная активность таких ресничек, а следова-
I ельно, и мукоцилиарный транспорт резко нарушены. Однако прояв-
нения синдрома Картагенера не ограничиваются дыхательной
системой. Страдают и пищеварение, поскольку реснички на поверхности
шителиальных клеток желудочно-кишечного тракта обеспечивают
•ффективное перемешивание химуса в зоне пристеночного
пищеварения. Наконец, люди с синдромом Картагенера бесплодны, поско-
иьку жгутики сперматозоидов не способны быть для них
пропеллерами, и мужские половые клетки обречены на неподвижность
и невозможность встречи с яйцеклетками.
Глава 6. СЕНСОРНЫЕ СИСТЕМЫ
ОРГАНИЗМА
6.1. Анализ сенсорных систем
Живые организмы функционируют в условиях постоянно
изменяющейся внешней среды, и основой их выживания является
наличие у них способности реагировать на изменения среды
соответствующей перестройкой своей жизнедеятельности, позволяющей
определить и захватить пищевые вещества, избежать повреждения,
осуществить воспроизводство себе подобных организмов и т.д.
Начальным Звеном во всех этих реакциях, обеспечиваемых у
многоклеточных организмов в первую очередь деятельностью нервной
системы, является превращение внешнего воздействия (обозначаемого
обычно как раздражитель, или стимул) в определенный
физиологический процесс, который содержит в себе информацию об этом
воздействии.
Живые организмы для своего существования должны получать
информацию о внешней и внутренней средах (о пище, опасности,
температуре, химическом и других составах окружающей среды,
о возможности продолжения рода и т.д.). Передача информации
происходит между внешней средой и организмом, между различными
отделами организма, между клетками, органоидами и другими
составляющими клетки, т.е. на всех уровнях организации биообъектов,
причем внешняя среда может простираться от космического
окружения (космосферы) до субатомного уровня.
Стимуляция может заключаться в действии на организм самых
различных видов энергии (механической, тепловой, химической,
гравитационной, электрической, электромагнитной). В организме же
сведения об этом действии передаются посредством сигнальной
деятельности нервных клеток - потенциалами действия (нервными им-
ι пива 6. Сенсорные системы организма
183
пульсами). Трансформация внешней энергии в эту деятельность
обеспечивается особыми структурами, обозначаемыми как
рецепторы органов чувств (сенсорных систем).
Это слово образовалось от латинского recepto - брать,
принимать. Рецепторы реагируют на раздражители, которыми называют
факторы внешней среды, влияющие на изменения свойств или
состояния клеток, ткани, органа и целого организма. Раздражители делятся
ιι;ι адекватные (соответственные) и неадекватные для определенного
рецептора. Адекватность раздражителя для рецептора проявляется
II том, что пороговая интенсивность воздействия адекватного
раздражителя намного меньше, чем неадекватного. Рецепторам
свойственны чрезвычайно малые пороги восприятия адекватных стимулов,
например фоторецепторы глаза возбуждаются единицами квантов
енста, орган слуха реагирует на механические смещения на уровне
теплового движения молекул, обонятельные рецепторы способны
воспринимать появление единичных молекул пахучих веществ.
Классификация биосенсоров (рецепторов) проводится по раз-
пичным признакам. Например, по модальности адекватного
раздражителя рецепторные структуры подразделяются на фоторецепторы,
механорецепторы, хеморецепторы, терморецепторы, осморецепто-
ры, электрорецепторы и др. Фоторецепторы обеспечивают
восприятие света. Хеморецепторы воспринимают химический состав
внешней и внутренней среды организма. Механорецепторы реагируют на
механические стимулы и обеспечивают слух, восприятие гравитации
и угловых ускорений, осязание, механические процессы в
сердечно-сосудистой системе, желудочно-кишечном тракте и других
органах. Терморецепторы воспринимают изменения температуры. Осмо-
рецепторы реагируют на сдвиги осмотического давления в основном
но внутренних органах и тканях организма. Электрорецепторы
реагируют на появление или изменение электрического поля (надпоро-
говых напряженностей). У человека такие рецепторы не выявлены.
Псе рецепторы снабжают центральную нервную систему
информацией о внешних и внутренних факторах среды.
Другой принцип классификации рецепторов основан на их
локализации в организме: интерорецепторы реагируют на изменения
его внутренней, а экстероцепторы - внешней среды.
184
Биофизика
Очень важен структурный принцип классификации
рецепторов, который сейчас по праву называется
структурно-функциональным принципом, поскольку структурные особенности рецепторов
предопределяют биофизические механизмы преобразования
сигналов в них. Различают свободные нервные окончания,
инкапсулированные нервные тельца и рецепторы со специальными клетками.
Рецепторы со специальными клетками, в свою очередь, подразделяются
на первично-чувствующие и вторично-чувствующие рецепторы.
Рецепторы обладают свойством не только отмечать сам факт
действия раздражителя, но и отражать его определенные параметры,
осуществляя таким образом функцию перекодирования их в
определенные характеристики сигнальной деятельности нервных клеток.
Этот процесс кодирования, передачи и обработки информации
о внешних раздражениях обозначается как сенсорная (афферентная)
деятельность организма, а структуры, его осуществляющие, - как
чувствительные (сенсорные, афферентные) системы. Он является,
очевидно, тем биофизическим процессом, который должен быть
рассмотрен в первую очередь при жизнедеятельности организмов как
сложных биофизических систем.
6.1.1. Общие принципы
функционирования сенсорных систем
С деятельностью сенсорных систем связана вся наша
сознательная жизнь, поскольку конечным ее результатом у находящегося
в сознании человека является возникновение ощущений -
субъективных образов внешних раздражений. Сопоставление характеристик
этих ощущений с объективными свойствами вызвавших их
раздражений ранее всего было использовано для определения общих
принципов функционирования сенсорных систем. Такое сопоставление
объективных и субъективных параметров сенсорных раздражителей
обозначается как психофизическое направление биофизики
сенсорных систем.
При оценке сенсорных ощущений в первую очередь обращает
на себя внимание субъективно переживаемое их сходство в случае
активации одной чувствительной системы и качественное отличие -
/ ппва 6. Сенсорные системы организма
185
и случае активации различных. Эта особенность обозначается как
модальность ощущений, отражающая, очевидно, качественное раз-
инчие соответствующих внешних раздражителей. «Классическими»
модальностями являются зрение, слух, осязание, вкус, обоняние,
отражающие действие на рецепторы соответственно световых лучей,
туковых колебаний, механических воздействий, химических
веществ. К сенсорным модальностям необходимо отнести ощущения
ι сила и холода, вибрации, положения тела в пространстве и боли,
и также «внутренние» ощущения, отражающие изменения во
внутренней среде организма, являющиеся раздражителями для
рецепторов внутренних органов.
Очевидно, что качественная особенность ощущений каждой
модальности - явление субъективное, переживаемое каждым инди-
иидуумом и непередаваемое другому индивидууму. Правильное
отражение в ней объективных различий внешних раздражителей
промеряется всей ответной деятельностью организма, и возможные
нарушения соответствия между качеством раздражителя и его
субъективным отражением, возникающие, например, при некоторых
заболеваниях, обычно быстро определяются и корректируются в
поведении индивидуума.
Следующим основным параметром ощущения является его
интенсивность, отражающая количественные показатели раздражите-
н». В общем, чем больше сила стимула, тем выше интенсивность
ощущения. Однако эта зависимость не является линейной и также
отражает субъективный характер нашего отражательного процесса. Во
исех сенсорных системах ощущение возникает только после
достижения раздражителем определенной критической величины,
называемой абсолютным порогом ощущения. Затем интенсивность ощущения
увеличивается с усилением силы стимула, однако до определенных
пределов. При слишком сильных стимулах повышение интенсивности
ощущения прекращается (верхний порог ощущения). В наших
ощущениях мы можем оценить изменения их интенсивности, сопоставив
с объективными изменениями силы раздражителя. Однако в связи
с наличием порога ощущения для измерения его интенсивности
приходится проводить отсчет от порогового уровня раздражителя и
определять эту интенсивность как кратное порогового ощущения.
186
Биофизика
Сопоставление субъективной оценки изменения
интенсивности ощущения с объективным изменением силы раздражителя было
проведено в ряде психофизических экспериментов на различных
сенсорных системах. Если силу раздражителя представить в
логарифмических единицах, то соотношения между ней и степенью
ощущения довольно хорошо совпадают с прямой, наклон которой может
варьироваться. Математическим выражением такой зависимости
является степенная функция
I=k(S-S0Y,
(6.1)
где / - интенсивность ощущения; S0 - пороговая и S -
действующая сила раздражения; к - константа. Описываемая зависимость по
имени ее открывателя получила название закона Стивенса.
Показатель степени η в этой функции для различных сенсорных систем
и различных раздражителей может отличаться от единицы как в
большую, так и в меньшую сторону. На рис. 6.1 приведен пример
эксперимента с вкусовыми ощущениями от различных химических
веществ, который хорошо иллюстрирует сказанное.
Важные сведения относительно общих принципов оценки
силы стимула получаются в психофизических экспериментах по
определению минимально определяемых изменений в интенсивности
/, отн. ед.'1
6 610
60 100
Л Гц
Рис. 6.1. Сравнение изменений интенсивности субъективного
вкусового ощущения (1) и частоты разрядов в волокнах вкусового нерва (2)
человека как функции концентрации лимонной кислоты (А) и сахара (Б) вмМ/л
( пава 6. Сенсорные системы организма
187
ощущения при усилении раздражения. Как это было впервые
отмечено Э. Вебером еще в 1834 г., такое минимально определяемое
изменение (дифференциальный порог) находится, как правило, в
постоянном отношении к исходной силе раздражения (закон Вебера):
AS „
= const. (6.2)
S
Впоследствии Г. Фехнер по новому выразил эту
закономерность, проинтегрировав уравнение Вебера. В такой форме
интенсивность ощущения пропорциональна логарифму силы стимула (закон
Иебера-Фехнера):
I = k-\gS. (6.3)
Однако закономерности, обнаруженные Э. Вебером в
эксперименте, оказались справедливыми в сравнительно узких пределах,
и для некоторых сенсорных систем вообще не подтвердились. В этом
отношении установленная С. Стивенсом закономерность имеет
значительно более широкую область применения и может быть с
большим успехом перенесена на нейрофизиологические процессы,
лежащие в основе кодирования и передачи информации в сенсорных
системах. Итогом работ Э. Вебера, Г. Фехнера, С. Стивенса, В.О.
Самойлова и других исследователей стал вывод о том, ч,то на
сенсорных входах в центральную нервную систему происходит нелинейное
преобразование информации об интенсивности раздражителей. Это
обстоятельство предопределяет многие закономерности
функционирования мозга человека и животных. Нелинейное преобразование
сигналов на входе в систему позволяет ей заменять математические
операции второго порядка (умножение и деление) операциями
первого порядка (сложение и вычитание). Поэтому нейрон работает как
интегратор, совершая алгебраическое сложение многочисленных по-
стсинаптических потенциалов, образующихся на его дендритах
и соме.
Наряду с качественными и количественными
характеристиками, ощущения имеют также пространственные и временные
характеристики. Пространственные особенности ощущения могут быть из-
188
Биофизика
мерены дифференциальным пространственным порогом
наименьшим интервалом между раздражителями, при котором они
воспринимаются как раздельные. Вместе с тем площадь стимула
оказывает влияние на характер ощущения и в том случае, если оно
является слитным: ощущаемая интенсивность возрастает при увеличении
площади органа чувств (А), на которую действует раздражитель.
Очевидно, в общем смысле это явление можно рассматривать как
пространственную суммацию эффектов элементарных раздражений
в процессе формирования ощущения. В идеальном случае
произведение силы стимула на его площадь должно было быть постоянным
при инициации ощущения:
Is ■ А = const. (6.4J
где /, - интенсивность силы раздражителя.
Однако и для этой характеристики в эксперименте
наблюдаются существенные вариации, и указанная зависимость подтверждается
в определенных пределах изменения площади воздействия стимула.
Положение осложняется наличием противоположных эффектов -
ослаблением интенсивности ощущения в случае одновременной
стимуляции соседнего участка воспринимающей поверхности. Такое
явление получило название контраста. Оно хорошо знакомо по
зрительным ощущениям, но имеет место и в других сенсорных системах
и способствует пространственному различению раздражителей.
По аналогии с эффектом увеличения площади раздражителя
увеличение его длительности также влияет на интенсивность
ощущения. Здесь встречаемся с временной суммацией актов стимуляции
в процессе формирования ощущений. В некоторых случаях можно
показать постоянство произведения силы раздражителя на время его
действия в определении интенсивности ощущения:
ls ■ t = const. (6.5)
Это положение справедливо только для определенных
длительностей стимула. Когда же последняя превосходит некоторую
критическую величину, интенсивность ощущения перестает зависеть от
нее. Более того, при дальнейшем увеличении длительности стимула
( пава 6. Сенсорные системы организма
189
S, отн. ед.^
10
0
ι, отн. ед. f
40
20
Длинный
стимул
Короткие
стимулы
12
18
·', с
ί. с
Рис. 6.2. Адаптация к запаху
Вверху - предъявления стимула (сероводорода)
в концентрации 6,5 ■ КГ6 по объему. Внизу - изменения интенсивности
субъективного ощущения /, оцениваемые четырьмя различными испытуемыми
наступает противоположное явление - интенсивность ощущения
начинает ослабевать. Такое ослабление получило название адаптации.
На рис. 6.2 приведен пример адаптационных изменений ощущения
шпаха, полученный в психофизическом опыте. Как видно, после
прекращения действия раздражителя адаптационное снижение
интенсивности восстанавливается также постепенно.
Ход восстановления можно определить кратковременными
предъявлениями раздражителя той же интенсивности. Явления
адаптации чрезвычайно сильно проявляются в деятельности всех сенсор-
пых систем и играют важную роль в восприятии организмом
внешних стимулов, существенно расширяя диапазон воспринимаемых
интенсивностей последних. Так, в зрительной системе в ходе
адаптации пороговая сила света, вызывающая минимальное ощущение,
может измениться в десятки тысяч раз.
Хотя приведенные ранее общие характеристики функциониро-
иания сенсорных систем получены на основании отчетов испытуемых
о своих субъективных ощущениях, они имеют объективный характер,
поскольку подтверждаются на всех испытуемых. На протяжении
длительного времени такие психофизические данные были основой
вашего понимания принципов восприятия организмом действующих на
него внешних раздражителей. Поскольку невозможно получить
отчетов о субъективных ощущениях, возникающих у животных, то о функ-
190 Биофизика
ционировании сенсорных систем у последних можно было делать
заключения лишь по аналогии.
Эта принципиальная трудность была в значительной мере
преодолена путем использования условнорефлекторного метода И.П.
Павлова, открывшего возможности объективно регистрировать
способность животного отличать один раздражитель от другого на основе
дифференцировки выработанных на них условных рефлексов.
Использование условнорефлекторного метода показало, что
распространение установленных в результате психофизических измерений
общих принципов функционирования сенсорных систем человека на
аналогичные системы животных вполне правомочно.
Прямое исследование процессов, происходящих в различных
звеньях той или иной сенсорной системы, стало возможным с
введением в практику регистрации и анализа электрических феноменов,
наблюдаемых в этих звеньях и отражающих течение нервных
процессов, обеспечивающих сенсорную информацию в организме. В ходе
таких исследований удалось в большинстве случаев точно
определить те процессы, которые лежат в основе описанных общих
характеристик функционирования сенсорных систем.
6.1.2. Трансформация раздражителей
в рецепторах
Первым звеном в деятельности любой сенсорной системы
является трансформация энергии раздражителя (стимула) в
распространяющийся сигнал - нервный импульс. Рецепторные структуры,
в которых происходит эта трансформация, оказываются
приспособленными к реагированию на раздражителей определенного рода,
которые обозначаются как адекватные для данных рецепторов. Однако
эта приспособленность не является абсолютной, и в некоторой
степени рецепторы могут активироваться и неадекватными стимулами.
Специализация рецепторов определяется наличием у них особых
механизмов, позволяющих реагировать на очень слабые адекватные
раздражители. Чувствительность при этом может достигать в некоторых
случаях практически предельно возможной по физическим законам
величины («физического предела» - по выражению СИ. Вавилова).
/ пава 6. Сенсорные системы организма
191
Как уже говорилось, по модальности адекватного раздражителя
Пиосенсоры могут быть разделены на механо-, термо-, хемо-,
фоторецепторы. У некоторых водных животных имеются также
электрорецепторы, для которых адекватным раздражителем является
изменение электрического поля в проводящей внешней среде. Внутри
каждой из этих групп имеется ряд подразделений, рецепторы в
которых приспособлены для реагирования на те или иные более узкоогра-
пиченные параметры соответствующих раздражителей. Несмотря на
такую специализацию, у всех рецепторных структур есть общий
механизм, наличие которого определяется обшей конечной задачей
каждого рецептора - вызвать распространяющийся нервный импульс,
единственным путем инициации этого импульса является
деполяризация плазматической мембраны нервного окончания чувствительной
нервной клетки или специальной клетки во вторично-чувствующих
рецепторах. Поэтому во всех рецепторах внешняя энергия должна
быть трансформирована в аналоговый электрический процесс,
который обозначается как рецепторный потенциал, а он, в свою очередь,
нызывает возникновение дискретных сигналов - потенциалов
действия в афферентных волокнах.
В некоторых рецепторах сравнительно нетрудно
регистрировать и анализировать рецепторный потенциал. На их примере можно
ясно представить себе основные закономерности трансформации
внешнего раздражителя в нервные импульсы. К ним относится
рецептор растяжения у ракообразных, структура которого показана на
рис. 6.3.
При растяжении мышечных волокон (рис. 6.3, б. 1) происходит
деформация контактирующих с ними дендритных отростков
чувствительной нервной клетки, и в них возникает деполяризация
мембраны - рецепторный потенциал (рис. 6.3, б. 2), который электротониче-
ски распространяется на сому клетки, вызывая в ее плазмолемме
активацию потенциалзависимых ионных каналов и генерацию
потенциалов действия. Параметры всех звеньев этого электрического
процесса могут быть точно измерены путем регистрации самого
растяжения мышцы и отведения электрических реакций клетки с помощью
внутриклеточного микроэлектрода. Как видно на том же рисунке,
рецепторный потенциал представляет собой аналоговое отражение раз-
192
Биофизики
а) б)
Рис. 6.3. Схема трансформации внешнего раздражителя
в нервный процесс в рецепторе растяжения рака
а - схема рецептора: 1 -мышечные волокна, 2 - дендриты,
3 — нервная клетка, 4 — аксон; б — последовательные этапы
трансформации: J - внешний стимул
дражения, повторяющее амплитудные и временные характеристики
последнего (рис. 6.3, б. 3).
Такое отражение, которое часто обозначается как трансдукция
внешнего раздражителя, имеет ряд особенностей. Амплитуда и
длительность стимула преобразуются в амплитуду совершенно иного
физического процесса - изменения мембранного потенциала.
Соотношения между этими изменениями являются линейными, однако
лишь в определенных пределах. Эти соотношения претерпевают
изменение во времени, постепенно уменьшаясь (адаптация). Наконец,
после прекращения стимуляции появляется преходящее изменение
мембранного потенциала в противоположную сторону
(гиперполяризация). Как видно, в отмеченных особенностях рецепторного
потенциала приходится сталкиваться с чертами, отмеченными ранее при
характеристике субъективных ощущений (например, с адаптацией).
Появление рецепторного потенциала, в свою очередь, дает
начало следующему процессу -ритмическому разряду потенциалов
действия, которые далее распространяются в соответствующие нервные
центры и в которых в закодированном виде переносится информация
о предыдущих процессах. При этом интенсивность раздражителя и
соответственно амплитуда рецепторного потенциала
трансформируются в частоту разряда импульсов, а его длительность - в длительность
такого разряда. Функциональная зависимость силы стимула от часто-
I Dikia 6. Сенсорные системы организма
193
11.1 афферентой импульсации имеет, как правило, нелинейный харак-
ιι·ρ. Поскольку амплитуда рецепторного потенциала вследствие адап-
И1ЦИИ снижается со временем, несмотря на постоянство стимуляции,
чистота потенциалов действия также становится реже.
Преобразование рецепторного потенциала в разряд нервных
импульсов вносит свои ограничения в процесс трансформации.
Возникновение потенциала действия требует пороговой деполяризации
мембраны. В связи с дискретностью импульсации возникают
ограничения и в передаче изменений интенсивности раздражителя. При
и'юнь коротких сигналах такая передача вообще невозможна, так как
мри этом возникает одиночный импульс, подчиняющийся закону
«псе или ничего». По мере удлинения сигнала возможный объем
передаваемой информации возрастет. Подобную закономерность в
передаче сенсорных сигналов уже отмечали при анализе характеристик
интенсивности сенсорных ощущений.
Описанные принципы трансформации внешнего стимула
II нервные процессы являются общими для всех сенсорных систем
организма, и различия между ними связаны лишь с различиями кле-
I очных механизмов, определяющих процесс транедукции и
обеспечивающих максимальное повышение чувствительности рецептора
к инешнему воздействию. Далее познакомимся с особенностями это-
I о процесса в различных сенсорных системах.
6.1.3. Передача и переработка информации
в нервных центрах
После трансформации внешней энергии раздражителя в нер-
иные процессы в каждой сенсорной системе происходит сложная
последовательность превращений этих процессов, захватывающая ряд
надстроенных друг над другом нервных центров и заканчивающаяся
и самом высшем из них - коре больших полушарий головного мозга,
и которой на основе пространственно-временного распределения ак-
иншости нервных клеток возникает качественно новое явление -
субъективный образ раздражителя. Каждая сенсорная система имеет
спои промежуточные и высшие нервные центры, отличающиеся по
расположению и организации. Изучение их функционирования пред-
I I 1)8<М
194
Биофизика
ставляет собой предмет нейрофизиологии, и сведения о нем можно
найти в соответствующих руководствах. Здесь будут рассмотрены
только общие принципы передачи и переработки информации в
сенсорных системах.
Сенсорные пути, передающие сигналы от рецепторных клеток
до коры полушарий большого мозга, всегда состоят из нескольких
последовательно соединенных нейронов. При этом нейроны каждого
последующего звена собраны в структуру, которая имеет аналогию
с экраном: поступающие от различных рецепторных клеток
афферентные волокна образуют синаптические соединения с
определенными клетками этой структуры, так что пространственная
организация рецепторов оказывается как бы спроецированной на нейроны
следующего звена. Количество таких экранных структур
варьируется у различных сенсорных систем. Особенно велико оно у
зрительной системы, поскольку два последовательных переключения имеют
место еще в сетчатке.
Благодаря такой организации сенсорных систем, в них
возможна передача информации по принципу пространственного
кодирования, необходимая для отражения целого ряда характеристик
внешних раздражителей. Однако в каждом переключении в действие
включаются дополнительные механизмы, которые существенно
влияют на передачу через них сенсорных сигналов.
Каждое поступающее в переключающее звено афферентное
волокно заканчивается синаптическими окончаниями не на одном
нейроне, а охватывает ряд клеток (дивергенция возбуждения). В
результате участок возбуждения после переключения оказывается
расширенным, «размытым».
В свою очередь, в результате дивергенции к каждому нейрону
поступают сигналы не по одному афферентному волокну, а сразу по
нескольким. Это называется конвергенцией возбуждения. За счет нее
даже слабая стимуляция отдельных входов получает возможность
суммироваться на теле нейрона - возникает пространственная сум-
мация передаваемых сигналов. Конвергенция и дивергенция
представлены схематически на рис. 6.4.
Очевидно, при отсутствии какого-то дополнительного контроля
явлений дивергенции и конвергенции на нескольких последователь-
( ппва 6. Сенсорные системы организма
195
хххххх
хххххх
Рис. 6.4. Схема конвергенции и дивергенции афферентных волокон
в последовательных синоптических переключениях афферентных путей
I -рецепторы; 2 - первое переключение; 3 - второе переключение
пых звеньях переключения возможность пространственного кодиро-
иания информации свелась бы к нулю. Однако этому
противодействуют ограничивающие механизмы, которые также присутствуют во всех
переключающих структурах. Ограничение распространения
возбуждения осуществляется тормозными нейронами, которые
активируются поступающими афферентными сигналами и, в свою очередь,
затормаживают активность переключающих нейронов. Такое торможение
может осуществляться несколькими способами.
1. Опережающее торможение (рис. 6.5, а). В этом случае
тормозящие нейроны активируются прямо коллатералями афферентных
полокон и торможение в переключающем звене возникает
одновременно с возбуждением передающих клеток. Оно не зависит от
уровня их активности, а отражает активность рецепторных клеток или
предыдущего переключающего звена (feed-forward inhibition).
2. Возвратное постсинаптическое торможение (рис. 6.5, б),
β этом случае тормозные клетки активируются коллатералями
аксонов, отходящих от переключающих нейронов, и создают в этих же
нейронах тормозной постсинаптический потенциал (ТПСП). Чем
сильнее возбуждаются эти нейтроны, тем сильнее будет и возвратное
их торможение. Иными словами, осуществляется отрицательная
обратная связь.
I I*
196
Биофизик*
'{ψ
а) б) в)
Рис. 6.5. Различные виды торможения
а - опережающее торможение; б — возвратное
постсинаптическое торможение; в — возвратное
пресинаптическое торможение; I - входящие афферентные
волокна; 2 — тормозящие нейроны; 3 - выходящие нейроны
3. Возвратное пресинаптическое торможение. В этом случае
тормозные клетки активируются коллатералями аксонов
переключающих нейронов, но их действие направлено не на сами нейроны,
а на терминали контактирующих с ними афферентных волокон
(рис. 6.5, в). Деполяризуя их посредством аксо-аксональных
синапсов, такие клетки снижают эффективность синаптической передачи
возбуждения через переключающие нейроны.
Поскольку степени активации каждого из этих механизмов,
как и степени охвата соответствующим торможением
переключающих нейронов, могут быть самыми различными, то создается
исключительно динамичная система регуляции передаточных функций
соответствующего переключающего звена. Количественное описание
такой регуляции пока не представляется возможным из-за
отсутствия точных характеристик организации и активности каждой
системы торможения нервной деятельности. Качественно наиболее
определенно роль торможения выявляется в форме так называемого
латерального торможения, которое может создаваться за счет как
пост-, так и пресинаптических возвратных тормозных влияний.
Схематически эффект такого торможения представлен на
рис. 6.6. Поскольку тормозные нейроны обладают обычно широко
ветвящимися отростками и устанавливают синаптические контакты
со многими переключающими нейронами, то при поступлении по ка-
' tmtia 6. Сенсорные системы организма
197
Рис. 6.6. Эффект латерального торможения на пространственную
передачу сигналов через синоптические переключения
Справа показана схема возбуждающих синоптических связей между
рецепторами и вставочными нейронами. Черные - тормозящие вставочные
нейроны. Светлые - вставочные нейроны. Частоты импульсаций f
в возбужденном участке вокруг места стимуляции на уровне рецепторных
клеток (1), после первого переключения (2) и после второго переключения (3)
кому-либо афферентному входу волны возбуждения, наряду с акти-
кацией соответствующих нейронов, возникает более широко распро-
г ι раненное торможение, которое может ограничивать «размывание»
нозбуждения после переключения, связанное с описанной выше ди-
нергенцией возбуждающих влияний. Явления контраста, описанные
ранее при изложении данных психофизических экспериментов,
являются, по видимому, следствием латерального торможения.
Таким образом, при передаче информации через каждое
последующее переключающее звено в сенсорной системе происходит
взаимодействие двух противоположных тенденций - потери точности
пространственного кодирования в связи с дивергенцией возбуждения
и восстановления этой точности за счет латерального торможения.
Гикая система передачи является весьма динамичной и допускает,
м зависимости от интенсивности поступающего сигнала и уровня
активности тормозящих нейронов, широкие изменения в передаточных
функциях каждого переключающего звена. Более того, если предста-
нить себе, что пространственная организация тормозящих связей
198
Биофизик»
в переключающем звене может быть не случайной, а ориентирован
ной, то в результате в нем возникнут возможности для преимущест
венной передачи сигналов, имеющих определенные пространствен
ные характеристики.
Весьма четко эти особенности передачи сигналов через переклю
чающие звенья можно проследить в зрительной системе, в которой лег
ко наносить на рецептивное поле (сетчатку) стимулы любой
пространственной ориентации. При этом в соответствующих переключающих
звеньях при помощи микроэлектродного отведения можно точно
определить характеристики возбуждения нейрона в ответ на приложенные
стимулы. Наряду с клетками, имеющими простые рецептивные поля,
обнаруживаются клетки, отвечающие импульсным разрядом либо ни
определенную ориентацию стимулирующего засвета, либо на движе
ние стимула по сетчатке в определенном направлении.
Таким образом, в переключающих звеньях образуются
нейронные структуры, являющиеся детекторами соответствующих
сложных характеристик внешнего стимула. Они позволяют проводить
дополнительный анализ сигналов, поступающих от рецепторных
клеток, и выделять определенные их характеристики на основании
перекодирования пространственной и временной структуры
передаваемой импульсации.
Активный характер переработки передаваемых от рецепторон
сигналов в переключающих звеньях сенсорных систем проявляется
и в том, что эти звенья находятся под контролем влияний,
поступающих к ним из высших центров головного мозга. В составе сенсорных
систем имеются не только центростремительные (афферентные)
волокна, но и центробежные (эфферентные), устанавливающие синап-
тические связи с различными нейронами переключающих звеньев.
В результате нисходящих влияний может происходить ряд
изменений в передаточных функциях переключающего звена. Если
в результате активации тормозящих нейронов произойдет общее
повышение порога синаптической передачи, то это будет равноценно
ослаблению внешнего стимула, действующего на рецепторы. Такой
эффект может быть одним из механизмов широко известного
психологического явления внимания, когда при сосредоточении усилия на
восприятии какого-то одного типа раздражителей другие их типы пс·
(1 inea 6. Сансорныа системы организма
199
рсстают замечаться. Но изменения могут быть и более
специфическими, заключаясь, например, в сужении или расширении рецептивного
ноля переключающего нейрона или даже изменении модальности
нейрона, на котором конвергируют различные типы афферентных во-
нокон.
В высшем отделе головного мозга (коре больших полушарий
ι оловного мозга) различные сенсорные системы представлены в
пространственно разграниченных областях. Так, у человека первичная
соматосенсорная чувствительность локализована в основном в
ποτ ι центральной извилине, зрительная - в затылочной области коры,
слуховая - в верхней части височных долей, вкусовая - в постцентра-
ш.пой извилине и некоторых других участках коры, обонятельная -
преимущественно на внутренней поверхности полушарий (пирифор-
миая область). Кроме этих первичных сенсорных областей, для
большинства сенсорных систем характерно наличие дополнительных
(нгоричных и третичных) областей, пространственно отдаленных от
первичных.
В сенсорных областях коры сохраняется пространственная ор-
I янизация представительства соответствующей рецепторной
поверхности, характерная для переключающих звеньев. Наряду с клетками,
отличающимися простыми рецептивными полями, здесь имеются
нейроны со сложными и сверхсложными рецептивными полями,
к активности которых отражаются самые различные характеристики
действующих на рецепторы раздражителей.
Каким образом в коре происходит объединение активности
различных типов нейронов, дающее в результате цельное восприятие
ниешнего воздействия, пока неизвестно. Ясно, что предпосылкой для
>гого является исключительно сложная структурная организация
коры головного мозга, представляющая собой слоистую структуру,
it которой строго ориентированные клетки связаны между собой
системой тангенциальных и вертикальных связей, допускающих
практически беспредельные возможности межнейронных контактов.
О том, что в основе высшего анализа сенсорных сигналов
лежит активация сложных объединений корковых нейронов, говорит,
и частности, то обстоятельство, что появление сенсорных ощущений
можно вызвать и путем прямой стимуляции сенсорных областей
200
Биофизика
коры головного мозга электрическим током у больных при
нейрохирургических операциях, проводимых под местной анестезией.
Вместе с тем для возникновения субъективного ощущения
необходим, очевидно, определенный уровень такой активации,
поскольку отдельные сенсорные сигналы могут оставаться неощущаемыми,
несмотря на то, что, судя по электрофизиологическим показателям,
они несомненно поступают в корковые структуры. Существенную
роль играет и общий уровень активности коры большого мозга,
поддерживаемый неспецифическими влияниями из подкорковых
структур, в первую очередь из ретикулярной формации мозгового
ствола. При снижении этого уровня способность к сознанию
обратимо исчезает.
6.1.4. Биосенсоры
Сенсорные системы кожи. Рецепторы кожи представляют
собой весьма сложный комплекс и расположены на относительно
большой поверхности (примерно от 1,5 до 2,5 м2), в самых различных
участках кожного покрова. Толщина кожи (без подкожной клетчатки)
колеблется от долей миллиметра (веки, наружный слуховой проход)
до 4 мм (ладони и подошвы). Чтобы представить функционирование
сенсорных систем кожи, необходимо рассмотреть структуру кожного
покрова, так как различные рецепторы кожи расположены в разных
ее слоях, что обеспечивает их совместное специфическое
реагирование на внешние стимулы. Например, к кожным ощущениям
традиционно относят следующие: чувство легкого прикосновения и осязания,
давления, укола и боли, тепла и холода, вибрации, зуда, щекотки.
Восприятие каждого раздражителя характеризуется не только
большим числом интенсивностей, но и значительным количеством
характерных оттенков. Например, различают до 250 разновидностей
болевых ощущений (резкая, свербящая, ноющая, колющая, острая,
жгучая, тупая и др.). Следует сразу отметить, что нет полного
соответствия между морфологическим строением концевых нервных
аппаратов кожи и характеристикой ощущения от воздействия внешнего
стимула. Кожа состоит из трех основных слоев: эпидермиса, дермы
и гиподермы (рис. 6.7).
/ лова 6. Сенсорные системы организма
201
Слои кожи связаны достаточно плотно, и четко выраженной
ι раницы между ними нет, что обеспечивает не только механическую
прочность кожи, но и ее другие функциональные особенности. В
самом верхнем или наружном слое кожного покрова (эпидермисе) вы-
и
чА
Л
г
41
\
^Ч
10 12 345
б)
Рис 6.7. Строение кожи
a — схематическое изображение основных компонентов кожи:
I -роговой слой (с изменяющимися по глубине
кератиноцитами), 2 — волос, 3 - волосяной фолликул, 4 — сальная
железа, 5 — потовая железа, б - дерма, 7 - кровеносные сосуды;
б - структура кожного покрова: 1 - эпидермис, II - дерма,
Ш - гиподерма; слои эпидермиса и дермы: I -роговой, 2 - стекловидный,
3 - зернистый, 4 - шиловидный,5 — базалъный, б - сосочковый; 7 —сетчатый;
202
Биофизика
деляют до 5 различных слоев. В зависимости от локализации
кожного покрова число слоев и их толщина могут существенно меняться.
Например, в области век толщина эпидермиса варьирует от 60 до
90 мкм, а в области ладоней и подошв от 0,5 до 0,8 мм. В эпидермисе
по мере удаления от поверхности в глубь кожи расположены
роговой, блестящий (стекловидный), зернистый, шиловидный и базаль-
ный (основной) слои.
Базальный слой - самый глубокий и граничит непосредственно
с дермой. Он состоит из одного ряда призматических
(цилиндрических) клеток, которые отделены друг от друга узкими канальцами.
Кроме этих клеток, в базальном слое находится небольшое число
ветвистых клеток с многочисленными отростками, которые
оплетают призматические клетки, контактируя таким образом с клетками
вышележащих слоев. Клетки базального слоя являются основными
рбстковыми элементами эпидермиса, из которых образуются клетки
всех вышележащих слоев. Кроме того, в цитоплазме клеток
базального слоя находится пигмент - меланин, располагающийся в виде
коричневых зерен различной величины. Меланин защищает кожу от
повреждающего действия солнечного света благодаря сильному
поглощению ультрафиолетовой части спектра.
Шиловидный слой, расположенный над базальным, состоит из
5-10 рядов клеток, которые в глубине имеют кубическую форму,
а по мере приближения к зернистому слою становятся более
плоскими. В шиловидном слое находится и ряд других клеток, имеющих
многочисленные отростки. Клетки шиловидного слоя без резких
границ переходят в зернистый слой, который содержит от 1 до 2-4
(ладони, подошвы) рядов клеток, вытянутых параллельно поверхности
эпидермиса. Блестящий (стекловидный) слой расположен выше
зернистого и состоит из клеток вытянутой формы, содержащих
белковое вещество (элеидин), сильно преломляющее свет. Предполагают,
что этот слой выполняет барьерную функцию кожи для воды и
электролитов. Верхним слоем эпидермиса является роговой слой,
который состоит из тонких безъядерных ороговевших клеток, вытянутых
в длину. Эти клетки прочно связаны между собой и заполнены
кератином. В наружной части роговой слой менее компактен, отдельные
/ пава 6. Сенсорные системы организма
203
пластинки отстают от общей массы, т.е. наблюдается процесс
физиологического шелушения.
В эпидермисе находится большое число нервных окончаний,
и кровеносные сосуды в нем отсутствуют, и питание клеток осущест-
нляется интерстициальной жидкостью и лимфой. Граница между
тидермисом и дермой волнистая вследствие так называемых
кожных сосочков, которые представляют собой выросты из поверхности
дермы. Промежутки между сосочками заполнены эпителиальными
отростками.
Дерма, или собственно кожа, находится между эпидермисом
и подкожной жировой клетчаткой и состоит из двух слоев: сосочко-
ного (подэпителиального) и сетчатого. Сосочковым слоем
считается часть дермы от эпидермиса до поверхностной сети кровеносных
сосудов. Волнообразная граница между эпидермисом и дермой
обеспечивает более плотное соединение этих слоев и улучшает
условия питания эпидермиса в результате увеличения поверхности
соприкосновения, не содержащей кровеносных сосудов
(эпидермис), и дермы, в которой находятся капилляры. Высота сосочков
связана с толщиной эпидермиса, в области ладоней и подошв сосочки
имеют наибольшую высоту. Структура дермы образована
волокнистой субстанцией, в которую входят коллагеновые, эластические
и другие волокна, а также бесструктурным аморфным
промежуточным веществом, расположенным между волокнами соединительной
ткани. В сосочковом слое расположены тонкие пучки коллагеновых
волокон и значительное количество тонких эластических и аргиро-
фильных (преколлагеновых) волокон. Сетчатый слой состоит из более
компактных толстых коллагеновых пучков, которые переплетаются
в густую петлистую сеть. Между этими волокнами располагаются
также более толстые эластические волокна, которые тоже образуют
густое сплетение. Структура сетчатого слоя в основном определяет
прочность кожи, неодинаковую на разных участках кожного
покрова. Сетчатый слой дермы без резких границ переходит в подкожную
жировую клетчатку.
Подкожная клетчатка, или гиподерма, состоит из толстых
пучков коллагеновых и эластических волокон, которые идут из
сетчатого слоя дермы. Они образуют широкопетлистую сеть, в петлях кото-
204
Биофизика
рой находятся скопления крупных жировых клеток - дольки
жировой ткани. В гиподерму входит и кожная фасция, состоящая из
плотной соединительнотканной пластинки. От верхней поверхности
фасции по направлению к дерме идут пучки соединительнотканных
волокон, которые образуют решетчатую структуру. От плотности
и длины коллагеновых и эластических пучков зависит подвижность
кожи над подлежащими тканями и органами. Толщина гиподермы
значительно колеблется как у разных лиц, так и на различных
участках тела одного и того же человека. Обычно в области черепа ее
толщина около 2 мм, в области спины 5... 10 мм, в области живота и
ягодиц - несколько сантиметров. Гиподерма отсутствует на веках, под
ногтевыми пластинками, на крайней плоти, малых половых губах,
мошонке и слабо выражена в области носа, ушных раковин, на
красной кайме губ. Подкожная жировая клетчатка защищает организм от
механических повреждений и холода. В ее толще расположены
сосуды и нервы, а в верхнюю часть могут проникать луковицы
некоторых волос и клубочки потовых желез.
Кровеносная система кожи состоит из нескольких сетей
кровеносных сосудов. Идущие от фасции крупные артериальные сосуды
проходят через гиподерму, по пути давая веточки для жировых
долек. На границе дермы и гиподермы эти артерии делятся на ветви,
которые идут в продольном направлении и соединяются между
собой анастомозами (шунтами). Так образуется глубокое кожное
артериальное сплетение, из которого отходящие веточки питают
клубочки потовых желез, волосяные фолликулы и жировые дольки. Кроме
того, от глубокого артериального сплетения отходят по направлению
к дерме еще довольно крупные артерии, достигающие подсосочково-
го слоя и образующие здесь поверхностное - подсосочковое -
артериальное сплетение.
Мелкие артериальные веточки, отходящие от этого сплетения,
питают мышцы, сальные и потовые железы, волосяные фолликулы.
От артерий отходят капилляры, которые достигают сосочков,
образуют там петли и возвращаются назад уже в виде венозных
капилляров, более широких, чем артериальные. Венозные капилляры,
собираясь от сосочков, сальных желез, выводных протоков потовых
желез волосяных фолликулов и мышц, образуют первое поверхност-
Гпава 6. Сансорныа системы организма
205
нос венозное сплетение. Всего до границы с гиподермой
располагается 4 венозных сплетения. Выходящие из четвертого сплетения
пены проходят через гиподерму и вливаются в подкожные вены.
Наиболее мощная сеть кровеносных сосудов кожи расположена на
лице, в области ладоней, подошв, в коже красной каймы губ, поло-
пых органов и вокруг анального отверстия.
Лимфатическая система в коже образует поверхностную и
глубокую сети. Поверхностная лимфатическая сеть начинается в сосоч-
ковом слое слепыми закругленными расширенными капиллярами,
между которыми имеются многочисленные анастомозы. Вторая сеть
лимфатических сосудов расположена в нижней части дермы. Это
широкопетлистая капиллярная сеть, образующая лимфатическое
сплетение, которое глубже переходит в лимфатические стволы.
В коже также находятся сальные и потовые железы, волосы
и разветвленная сеть нервных волокон. Сальные железы
распространены во всей коже, их нет только на ладонях и подошвах. В
большинстве случаев они связаны с волосяными фолликулами, около
каждого из которых имеется от 1 до 8 желез. Размер сальных желез
чначительно варьирует. Наиболее крупные расположены в коже
носа, щек, в области грудины, над и между лопатками. Потовые
железы разделяются на два вида: эккриновые (мерокриновые) и
апокриновые. В железах первого вида клетки при секреции сохраняются, а в
железах второго вида верхняя часть клеток разрушается и переходит
в секрет.
Количество потовых желез в коже человека превышает 2 млн
и выделяют они в сутки до 0,8 л пота, а при перегревании
организма-до 1,5 л и даже более. Потовые эккриновые железы
расположены на всем кожном покрове, кроме красной каймы губ и некоторых
участков половых органов. Особенно много их в коже ладоней и
подошв, а также на лбу, груди, животе, предплечьях. Тело железы
располагается в глубоких слоях дермы, иногда в верхних слоях
гиподермы и связано с поверхностью кожи длинным выводным протоком.
Апокриновые потовые железы находятся преимущественно в
подмышечных впадинах, вокруг заднего прохода и грудных сосков, в
наружных половых органах женщин, в паху, на лобке и вокруг пупка.
По размеру апокриновые железы больше эккриновых, их выводные
206
Биофизика
протоки открываются в волосяной фолликул, тело железы
расположено глубоко в подкожной жировой клетчатке. Функции
апокриновых и половых желез связаны, поэтому до полового созревания они
недоразвиты, а в преклонном возрасте их функции ослабевают.
В коже расположено большое число нервных волокон,
специальных концевых нервных аппаратов (нервных окончаний), которые
образуют обширное рецептивное поле кожи, выполняющее функцию
сенсорной системы. В глубоких отделах гиподермы заложено
основное нервное сплетение, от которого берут начало нервы кожи. Затем
они проходят через толщу собственно кожи, образуя по пути
многочисленные веточки, идущие к волосяным фолликулам, сальным и
потовым железам, мышечным волокнам и сосудам кожи. В подсосоч-
ковом слое находится сплетение, состоящее из густо расположенных
нервных волокон. От этого сплетения отдельные нервные веточки
проникают в сосочки и эпидермис. Подходя к эпидермису, тонкие
нервные волокна теряют свою миелиновую оболочку и проникают
в межклеточные канальцы основного и шиловидного слоев.
Выделяют две группы рецепторов кожи - свободные нервные
окончания и инкапсулированные нервные тельца. Свободные
нервные окончания имеют либо древовидную форму, либо форму
клубочков, иногда с утолщением воронкообразной или другой формы на
конце нервного волокна. Примеры этих рецепторов показаны на
рис. 6.8. Корни волос окружены кольцевой сеточкой нервных
элементов (рис. 6.8, а), которые раздражаются при сгибании волоса.
Корзинчатые окончания, оплетающие волосяные фолликулы,
отсутствуют на частях тела, лишенных волос (около 5% поверхности
кожного покрова). Их относят к рецепторам давления, а тактильные
точки на коже, покрытой волосами, обнаруживаются у корней волос.
Разветвленные свободные нервные окончания (рис. 6.8, б)
расположены по всему кожному покрову. Все области кожи иннервируются
большим числом нервных волокон, причем каждое из них ветвится
вновь и вновь так, что отростки одного волокна могут
распространяться на площади от долей до единиц квадратных сантиметров.
Результатом этого является сложная система перекрестной иннервации
кожи многочисленными нервными волокнами.
( пива 6. Сансорныа системы организма
207
а) б)
Рис. 6.8. Свободные нервные окончания в коже
а - нервное окончание вокруг волосяного фолликула,
б - ветвящиеся нервные окончания
Инкапсулированные нервные тельца имеют вокруг тонких нер-
Ш1ЫХ окончаний (терминален) нерва соединительнотканную
оболочку (капсулу). В зависимости от особенностей строения концевых раз-
нствлений нерва (внутренняя колба) и наружной капсулы различают
несколько разновидностей этих рецепторов, основные из них
представлены на рис. 6.9. В коже человека различают:
Пластинчатые тельца имеют капсулу из
соединительнотканных пластинок, весьма тонких, между которыми находится тканевая
жидкость. В центре капсулы располагается полость (внутренняя
колба), в которую входит нервное волокно, лишенное миелиновой
оболочки (рис. 6.9, а). Это волокно тянется вдоль всей внутренней
колбы, давая тонкие боковые веточки, и заканчивается утолщением
булавовидной формы. Тельца Почини являются самыми крупными
инкапсулированными рецепторами, достигающими в длину 3 мм
и более. Расположены они обычно в подкожной жировой клетчатке,
причем особенно много их в области ладоней, подошв и половых
органов. Этим рецепторам приписывают восприятие чувства глубокого
а) б) в)
Рис. 6.9. Инкапсулированные нервные тельца
а - пластинчатое тельце (Фатера-Пачини),
б - тельце Мейснера, в - колба Краузе
208
Биофизика
давления и проприорецептивное ощущение (мышечное чувство).
Выделяют еще видоизмененные пластинчатые тельца (тельца Голь-
джи-Маццони), имеющие сходную структуру, но существенно
меньших размеров, и расположенные не в гиподерме, а в дерме, почти
непосредственно под сосочками и в самих сосочках.
Тельца Мейснера, или осязательные тельца, имеют
удлиненно-овальную форму (рис. 6.9, б) и располагаются прямо в сосочках.
Наружная соединительнотканная капсула их тонкая, внутри нее
перпендикулярно к длинной оси расположены так называемые
«осязательные клетки». Миелинизированное (мякотное) нервное волокно,
подходя к краю капсулы, теряет свои оболочки и в виде цилиндра без
миелиновой оболочки проходит в капсулу, где образует утолщение
в форме менисков, прилегающих к «осязательным клеткам». Из
названия рецепторов понятно, что им приписывают чувство осязания
(легкого прикосновения). У человека, для которого осязание имеет
важное значение, эти рецепторы представлены весьма широко.
Больше всего осязательных телец Мейснера расположено в кончиках
пальцев, в губах и слизистой оболочке языка.
О природе процессов, происходящих в клетках капсулы и
непосредственно в терминали инкапсулированного нервного тельца,
пока нет точных сведений. Несущественные отличия в
характеристиках этих процессов в различных типах механорецепторов указывают
особенности той импульсной активности, которая распространяется
от них по нервному волокну. В одних случаях эта активность длится
на протяжении всего времени действия механического
раздражителя, лишь постепенно становясь более редкой в связи с адаптацией.
Такие рецепторы, очевидно, передают информацию о силе, а также
длительности стимула и могут быть обозначены как
пропорциональные. Сюда относится осязательный мениск (тельце Меркеля).
В других случаях разряд импульсов возникает лишь в начале
стимула. Такие рецепторы адаптируются очень быстро, и импульса-
ция быстро прекращается, несмотря на продолжающееся действие
стимула. Но она оказывается очень зависимой от скорости
нарастания силы стимуляции: при увеличении последней число
импульсов в начальном разряде возрастает. Иными словами, такой
рецептор передает информацию не столько об интенсивности стимула,
I пава 6. Сенсорные системы организма
209
сколько о скорости ее изменения. Поэтому его называют
дифференциальным рецептором (т.е. реагирующим на производную от
стимула). Некоторые механорецепторы занимают по этим
характеристикам промежуточное положение и соответственно могут быть
названы пропорционально-дифференциальными. На рис. 6.10
представлены примеры регистрации активности различных типов
кожных механорецепторов.
Наконец, в коже присутствуют механорецепторы,
приспособленные к оптимальной передаче сигналов о синусоидальных
механических колебаниях тканей (т.е. о вибрации). Сюда относятся
пластинчатые тельца (тельца Пачини), располагающиеся в подкожной
жировой клетчатке. Эти рецепторы относятся к быстроадаптирую-
щимся, поскольку при непрерывном механическом раздражении они
нсегда отвечают на него лишь одним начальным импульсом. Однако
при синусоидальной стимуляции такой импульс появляется в ответ
0,6 Н/СМ2
0,1 Н/сМ2 0ДН£м2 | 1
дщ и циннии JiilL
inn шипи
б)
110 Гц 55 Гц
тшт ΊΜ/wb
шипим
θ;
Рис. 6.10. Импульсная активность различных механорецепторов кожи кошки
а-медленно адаптирующийся (пропорциональный) рецептор. Вверху указана
сила давления на кожу; б - быстро адаптирующийся (дифференциальный)
рецептор, реагирующий на скорость надавливания на кожу; в - вибрационный
рецептор (рецептор ускорения). Вверху указана частота вибраций
210
Биофизика
на каждое колебание, причем сила ощущений зависит от частоты
колебаний. По мере повышения частоты (в определенных пределах)
порог снижается пропорционально квадратичной степени
увеличения частоты. По существу рецептор оказывается реагирующим на
ускорение прогибания расположенной над ним кожной поверхности
и поэтому может быть обозначен как рецептор ускорения.
Колбы Краузе тоже имеют тонкую соединительнотканную
капсулу овальной формы. Нервное волокно без миелиновой оболочки
входит в капсулу, внутри которой образует густой клубок (см,
рис. 6.9, в). Расположены колбы Краузе в сосочках и под ними, эти
рецепторы ответственны за восприятие чувства холода, т.е. являются
Холодовыми рецепторами. Тельца Руффини по своему строению
аналогичны колбам Краузе, но располагаются значительно глубже -
в глубоких отделах дермы и в верхних отделах гиподермы. Их
относят к тепловым рецепорам. Следовательно, колбы Краузе и тельца
Руффини являются терморецепторами.
Приведенный анализ характеристик активности различных
рецепторов кожи иллюстрирует второй важнейший принцип, на
котором строятся в организме кодирование, передача и последующий
анализ информации о раздражителях. Различные стороны одного и того
же по своей физической природе стимула оказываются
воспринимаемыми различными рецепторными структурами, и информация о них
передается дальше по различным проводникам. Эти рецепторы и
связанные с ними проводники могут быть обозначены как детекторы
соответствующих свойств раздражителя, сведения о которых
поступают и кодируются пространственным перераспределением активности
в разных сенсорных путях. Этот принцип пространственного
кодирования, наряду с упомянутым выше частотным кодированием,
является основой всей сенсорной деятельности.
Вестибулярная сенсорная система. Вестибулярный
анализатор участвует в регуляции положения и движения тела в
пространстве, в поддержании равновесия и регуляции мышечного тонуса.
Периферический отдел анализатора представлен рецепторами,
расположенными в преддверии улитки, находящейся внутри
височной кости. На рис. 6.11 изображено взаимное расположение улитки
и ее преддверия.
/ пава 6. Сенсорные системы организма
211
Рис. 6.11. Улитка и ее преддверие
1 - височная кость; 2 - вертикальные полукружные каналы;
3 - горизонтальный полукружный канал; 4 — купула;
5 -макулы вестибулярного аппарата; 6-улитка; 7 - полость среднего уха
Вестибулярные рецепторы возбуждаются при изменении
скорости вращательного движения, прямолинейном ускорении,
изменении направления силы тяжести, вибрации. Нервные импульсы,
возникшие в этих рецепторах, поступают по вестибулярному нерву
II ствол мозга, а оттуда по проводящим путям - к центральному отделу
нпализатора, расположенному в передних отделах височной доли
коры головного мозга. В результате возбуждения нейронов этого
отдела коры возникают ощущения, дающие представления о положении
тела и отдельных его частей в пространстве, способствующие
сохранению равновесия и поддержанию определенной позы тела в покое
и при движении. При этом действуют особые механизмы
трансформации внешней силы в механическое воздействие на так называемые
волоски рецепторной клетки, выступающие из ее апикального полюса
и эндолимфу. Поэтому рецепторные клетки называются волосковыми.
Будучи расположенными в полукружных каналах, они
реагируют на угловые перемещения эндолимфы, а значит, и всего тела чело-
иска и животного. В преддверии улитки, кроме полукружных каналов,
находятся еще две структуры: маточка и мешочек. Их волосковые
клетки реагируют на прямолинейные перемещения тела.
Рецепторным органом маточки и мешочка (см. рис. 6.12, а)
являются так называемые пятна. Волоски клеток, образующих пятна,
212
Биофизика
а) б) в)
Рис. 6.12. Упрощенная модель вестибулярного аппарата
а - аппарат, воспринимающий линейные ускорения (1 — отолиты, 2 - отолитовая
мембрана, 3 - волосковые клетки); б, в- аппарат, воспринимающий угловые
ускорения (1 - ампула, 2 - купула, 3 - полукружный канал с перилимфой,
4 - волосковая клетка, 5 - гребешок); б- в покое, в - при угловом перемещении
направлены внутрь заполненной эндолимфой полости, где они
погружены в желеобразную массу, заключающую в себе кристаллики
кальциевых солей (камешки - отолиты). Вследствие своеобразия
этих кристалликов пятна нередко называют отолитовым прибором.
При наклоне головы под действием силы земного притяжения
происходит смещение отолитов, что ведет к сгибанию волосков.
Сгибание волосков, как у других механорецепторных клеток, передает
сигнал на контактирующие с рецепторной клеткой нервные
окончания. Регистрация активности последних показала, что ритмическая
импульсная активность генерируется рецептором и в том случае,
когда волоски выпрямлены. При сгибании их в одну сторону она
прекращается, а при сгибании в другую - учащается. Так как макулы
мешочка и маточки расположены в разных плоскостях - одна примерно
в горизонтальной, а другая - в вертикальной, то при любом положении
головы в пространстве будет иметь место особое сочетание
импульсной активности в иннервирующих их афферентных нервных
волокнах, передающих в нервные центры информацию об этом положении.
Три полукружных канала, расположенные во взаимно
перпендикулярных плоскостях, образуют декартову систему координат. Их
механорецепторными органами являются так называемые гребешки,
образованные волосками рецепторных клеток и желеобразной
массой (рис. 6.12, б, в).
Гребешки сосредоточены в расширениях (ампулах) каждого из
трех каналов. Погруженные в желеобразную массу, гребешки пере-
I пава 6. Сенсорные системы организма
213
гораживают канал подвижной желеобразной перегородкой (купу-
лой). Как показали прямые наблюдения, смещение купулы и, следо-
иательно, сгибание или разгибание волосков происходит при
перемещениях эндолимфы в канале. В свою очередь, такие
перемещения возникают в случае ускорения или замедления вращения
головы в плоскости, в которой расположен соответствующий канал
вследствие инерции эндолимфы). При переходе вращения в
равномерное купула медленно возвращается в свое исходное положение.
Таким образом, эта рецепторная система оказывается сенсором
вращательного ускорения. Она сигнализирует о нем разрядом
импульсов, частота которого уменьшается и приобретает стационарный
характер при переходе к равномерному вращению или к покою.
И физиологических условиях ускорения вращения являются
постоянно встречающимися раздражителями, возникающими при каждом
движении головы. Равномерное же вращение - состояние, как
правило, искусственное, возникающее в связи с развитием техники,
поэтому оно и не является адекватным раздражителем для сенсорных
клеток полукружных каналов.
Двигательный анализатор. Положение тела или его отдельных
частей в пространстве определяется функционированием не только
нсстибулярного, но и так называемого двигательного анализатора.
Рецепторы двигательного анализатора расположены в скелет-
пых мышцах, связках, сухожилиях, суставных капсулах. Так, в
соединительнотканной капсуле сустава расположены ветвящиеся нер-
пные окончания диаметром 7-10 мкм. Расположение и свойства этих
рецепторов позволяют им информировать центральную нервную
систему об устойчивом положении суставов, направлении, скорости
и амплитуде движения суставных поверхностей. Чувствительность
II быстродействие их высоки, а адаптация развивается медленно.
Аналогичные рецепторы находятся вблизи связок. Их рецепторы,
расположенные в соединительной ткани, окружающей сустав,
быстро адаптируются, и от них отходят крупные афферентные волокна
нервов, иннервирующих сустав.
Рецепторы суставов и мышц обеспечивает формирование
сигналов о том, как совершаются движения, и о степени напряжения,
которое добавляется при сопротивлении движению, например при
214
Биофизика
1 .Л3 4 5
а) б)
Рис. 6.13. Мышечные веретена
а —мышца напряжена, кольца спирали нервного окончания сжаты;
б — мышца расслаблена, витки спирали растянуты (1 —мышечное волокно,
2 - витки спирали нервного окончания без миелиновой оболочки,
3 - миелиновая оболочка, 4 — перехват Ранвье, 5 - аксон)
подъеме тяжелого груза или нарушении кровообращения.
Восприятие движения конечностей зависит от стимуляции рецепторов
суставов, а не мышц или кожи. Доказано это было путем воздействия
кокаином на последние (кокаин обладает местным анестезирующим
действием), при этом оценка положения и движения сустава
сохранялась. Пропускание же электрического тока через сустав
существенно нарушало эту оценку. Измерение пороговой чувствительности
смещения суставов (при скорости смещения 0,3%) показало, что
у человека самым чувствительным является плечевой сустав (0,2-
0,4°), а наименее чувствительным - голеностопный (1,2-1,3°).
Рецепторы мышц (так называемые мышечные веретена)
представляют собой закрученные в виде спирали вокруг мышечного
волокна нервные окончания, лишенные миелиновой оболочки (рис. 6.\Ъ).
Когда мышца расслаблена (рис. 6.13, а), мышечное веретено не
генерирует рецепторный потенциал. При растяжении мышцы
(рис. 6.13, б) витки спирали отходят друг от друга, и в них
развивается деполяризация, т.е. рецепторный потенциал, который обусловлен
открыванием натриевых каналов, чувствительных к механической
деформации плазмолеммы нервного окончания, входящего в состав
мышечного веретена. Этот потенциал распространяется электрото-
нически (с декрементом) со скоростью электромагнитной волны до
ближайшего перехвата Ранвье того же нервного волокна, что
приводит к генерации потенциалов действия, следующих без декремента
в центральную нервную систему. После ряда синаптических передач
сигнал от проприоцепторов достигает коры головного мозга.
Корковая часть двигательного анализатора состредоточена преимущест-
Глава 6. Сенсорные системы организма
215
пенно в передней центральной извилине. Там осуществляются
высший анализ и синтез двигательных реакций. Наличие двусторонней
связи между передней центральной извилиной и мышцами создает
условия для информирования коры головного мозга о текущем
состоянии двигательного аппарата, что позволяет точно и тонко
управлять мышечными движениями.
Кроме мышечных веретен, проприорецепторами являются
луковицеобразные тельца (Гольджи), расположенные в сухожилиях,
пластинчатые тельца Паччини, обнаруженные в фасциях,
покрывающих мышцы, в сухожилиях, связках и надкостнице. Изменение
активности различных проприорецепторов происходит в момент
сокращения или расслабления мышц. Так, возбуждение мышечных веретён
отмечается при удлинении (в момент расслабления или растяжения)
мышечных волокон. Луковицеобразные тельца возбуждаются при
сокращении мышечных волокон. При повышении активности
мышечных веретен импульсы, поступающие от них в центральную нервную
систему, облегчают сокращения данной мышцы и тормозят
сокращение мышцы-антагониста. Импульсы, приходящие в центральную
нервную систему от луковицеобразных телец, вызывают
противоположную реакцию, т.е. тормозят сокращение данной мышцы и
способствуют сокращению мышцы-антагониста.
В обеспечении движения принимают участие не только
двигательный и вестибулярный анализаторы, но также кожная,
зрительная и слуховая сенсорные системы. Следовательно, любой
двигательный акт является итогом совместной деятельности различных
анализаторов.
Интерорецептивный анализатор. Этот анализатор
обеспечивает поддержание постоянства внутренней среды организма (гоме-
остазиса).
Интерорецептивный анализатор, как и другие сенсорные
системы, состоит из трех отделов. Однако каждый из них имеет свои
особенности. Периферический отдел образован разнообразными интеро-
рсцепторами, диффузно расположенными во внутренних органах.
Проводниковый отдел включает несколько различных по
функциональному значению нервов, которые иннервируют внутренние
органы, а также проводящие пути головного и спинного мозга. Централь-
216
Биофизика
ный отдел располагается в моторной и премоторной областях коры
головного мозга. Функционирование интерорецептивного
анализатора у здорового человека обычно не сопровождается ощущениями.
Хемосенсорные системы. Способность реагировать на
различные химические воздействия - важнейшая функция организма,
осуществляемая с помощью специализированных хеморецепторных клеток,
С одной стороны, она позволяет определить источник химического
воздействия на расстоянии путем восприятия летучих веществ из
воздуха (обоняние). Такая химическая чувствительность является не
только способом поиска пищи, но и широко используется в
коммуникации организмов, например для привлечения особей
противоположного пола при половом поведении или при отпугивании врагов. С
другой стороны, она служит для оценки химических веществ при
непосредственном контакте с ними (вкус). Наконец, химическая
чувствительность широко распространена во внутренних органах
(например, в стенках сосудов), где она воспринимает изменения химического
состава внутренней среды организма.
В связи с такими различиями в принципах функционирования
клеточные механизмы хеморецепции также оказываются очень
различными. Особенности интерорецептивной хеморецепции, а также
вкусовой и обонятельной сенсорных систем изложены далее.
Термосенсорная система. Восприятие температуры внешней
среды и ее изменений - также весьма важный фактор, определяющий
реакции организма, в особенности теплокровного. Как показывают
психофизические наблюдения, при этом речь идет о двух различных
рецепторных системах - рецепторах тепла и рецепторах холода. При
локальной тепловой стимуляции кожи можно легко показать
различное пространственное расположение этих рецепторов - так
называемые тепловые и холодовые точки.
Расположение терморецепторов на поверхности тела
обеспечивает включение регуляторных механизмов до того, как
температурные изменения достигнут ядра тела. В теории автоматического
регулирования такие системы называются системами с управлением
по возмущению (в отличие от систем с регулированием по
отклонению). Важно отметить, что терморецепторы кожи реагируют не
только на знак и величину изменения внешней температуры, но и на ско-
( /шоа б. Сенсорные системы организма
217
рость изменения, т.е. на производную по времени. Благодаря этому
при быстрых температурных возмущениях возможна особенно ин-
кпсивная регуляция.
Терморецепторы расположены не только в коже, но и в
промежуточном мозге, о чем пойдет речь при рассмотрении системы
терморегуляции организма.
Сенсорная система боли. Болевые рецепторы представляют
1'обой особую группу рецепторных структур, характеризующуюся
отсутствием специфической чувствительности к раздражителю
определенной модальности. Основным условием активации таких
рецепторов является большая интенсивность внешнего воздействия,
которая может повредить организм. Функциональная роль болевой
рецепции очевидна - она сигнализирует об опасности воздействия
II вызывает комплекс реакций, направленных на устранение такой
опасности. Много болевых рецепторов на внешних покровах тела -
коже и слизистых оболочках. Кроме того, они многочисленны и в
ряде внутренних органов - в основном в их оболочках (плевре,
перикарде, твердой мозговой оболочке и др.).
Вопрос о структуре болевых рецепторов до сих пор является
дискуссионным. До последнего времени преобладало мнение,
согласно которому ноцицептивные сигналы возникают не в специализиро-
нанных, а в любых рецепторах, когда на них- действуют сверхсильные
раздражители. Сигналом о повреждающем воздействии является
особый характер нервной импульсации, который каким-то образом
оценивается в центральных нервных структурах как болевой.
Однако в настоящее время появляются данные о наличии
особых (специализированных) ноцицепторов. Так, в коже при точечной
стимуляции острым предметом четко выделяются болевые точки,
ноздействие на которые не вызывает никаких иных ощущений,
кроме боли. С наибольшей вероятностью к ноцицепторам можно
отнести свободные нервные окончания. Особенно четко принадлежность
свободных нервных окончаний к ноцицепторам видна в роговице -
поверхности, обладающей весьма высокой болевой
чувствительностью при отсутствии в ней каких бы то ни было рецепторов, кроме
свободных нервных окончаний. Вероятно, среди ноцицепторов есть
все же некоторая специализация: определенные болевые точки эф-
218
Биофизика
фективнее раздражаются сильными механическими, а другие —
сильными термическими воздействиями, некоторые в равной мере
чувствительны к тем и другим.
В связи с ничтожными размерами ноцицепторов не
представляется пока возможным определить биофизические механизмы
трансформации в них энергии раздражителей в электрический
сигнал. Возможно, что в некоторых случаях этот процесс включает
промежуточный механизм, например сдвиги химического статуса
(изменение рН, выделение особых биологически активных веществ)
тканей, окружающих ноцицепторы.
Характерной особенностью ноцицепторов является отсутствие
в них выраженной адаптации. Более того, в ряде случаев отмечается
противоположное явление - повышение эффективности раздражения
при длительном его действии (сенситизация). Эта особенность
соответствует функциональному назначению болевой
чувствительности - предупреждению об опасности, которая ни в коей мере не
ослабевает при повторении повреждающего стимула.
Электрорецепция. Электрорецепторы были открыты и
исследованы сравнительно недавно - в середине XX в. Реакция рыб на
слабые электрические поля была известна ученым давно, но считалось,
что при прохождении тока через воду в ней изменяется
распределение ионов и это влияет на вкус воды. Однако экспериментального
подтверждения данное предположение не имело. Электрорецепторы
обнаружены у многих морских и пресноводных рыб (акул, скатов,
сомов и других), а также у миног. Позднее такие рецепторы были
открыты у амфибий (саламандр и аксолотля) и у млекопитающих
(утконосов).
У разных животных электрорецепторы имеют разное
расположение: на голове, вдоль тела, на плавниках и по своему строению
они отличаются, образуя иногда специализированные органы.
Рассмотрим один из таких органов - ампулу Лоренцини как наиболее
чувствительный электрорецепторный орган. Итальянский ученый
Лоренцини описал этот орган в 1678 г. и предположил, что ампула
вырабатывает слизь рыбы. Этот рецепторный орган представляет
собой подкожный канал, один конец которого открыт в наружную
среду, образуя пору, а другой оканчивается глухим расширением (ампу-
( пава 6. Сансорныа системы организма
219
ной Лоренцини). Просвет канала заполнен желеобразной массой,
и шектрорецепторные клетки выстилают в один ряд дно ампулы. Все
клетки эпителия, выстилающего стенки канала, прочно соединены
между собой плотными контактами, что обеспечивает высокое уде-
ш.ное электрическое сопротивление эпителия - около 6 МОм · см2
(поскольку рассматривается поверхность, а не объем, то удельное
сопротивление измеряется в Ом · м2). Канал, покрытый такой хорошей
и'юляцией, проходит под кожей и может иметь длину в несколько
десятков сантиметров. Напротив, желе, заполняющее канал ампулы
Лоренцини, имеет весьма низкое удельное сопротивление (около
Ю Ом · см), что обеспечивается высокой концентрацией ионов
калия, которая заметно больше, чем в морской воде или плазме крови
рыбы. Ионы закачиваются в просвет канала за счет их активного
транспорта. Таким образом, канал электрического органа
представляет собой отрезок хорошего кабеля с высоким сопротивлением
изоляции и хорошо проводящей центральной частью.
Дно ампулы выстилают в один ряд несколько десятков тысяч
шектрорецепторных клеток, которые тоже плотно соединены между
собой. Таким образом, рецепторная клетка одним концом обращена
ннутрь канала, а на другом конце образует синапс, где выделяется
нозбуждающий медиатор, действующий на постсинаптическую
мембрану подходящего к клетке окончания нервного волокна. К каждой
импуле подходит 10-20 афферентных волокон, каждое из которых
образует много веточек (терминалей), идущих к рецепторным
клеткам. В результате на каждое нервное волокно приходится примерно
2000 рецепторных клеток. Сами рецепторные клетки имеют разное
сопротивление: синаптическая мембрана, где выделяется медиатор,
имеет большое сопротивление, а противоположный участок
мембраны, обращенный внутрь ампулы, - малое.
Если любую клетку поместить в электрическое поле, то в
одной части мембраны знак потенциала покоя совпадет со знаком
напряженности поля, а в другой - окажется противоположным. Значит,
па одной половине клетки мембранный потенциал возрастет
(мембрана гиперполяризуется), а на другой, наоборот, снизится (мембрана
деполяризуется). Таким образом, механизм работы электрорецептор-
пых клеток достаточно прост: при соответствующем знаке внешнего
220
Биофизика
поля деполяризуется синаптическая мембрана клетки, и этот сдвиг
потенциала управляет выделением медиатора.
Естественные электрические поля чрезвычайно слабы, поэтому
электрорецепторные органы должны иметь очень высокую
чувствительность. И это действительно так: например, акулы, скаты и
некоторые другие рыбы реагируют на электрические поля
напряженностью 0,1 мкВ/см. Таким образом, все особенности функционирования
электрорецепторных органов направлены на то, чтобы, во-первых,
получить на синаптической мембране возможно большую разность
потенциалов, во-вторых, обеспечить высокую чувствительность
механизма выделения медиатора к изменению мембранного
потенциала, и, в-третьих, выделить полезный сигнал на фоне больших помех.
Обеспечению такой высокой чувствительности
способствует устройство ампул Лоренцини. Если напряженность поля Е~
=^ 0,1 мкВ/см, а длина канала ампулы 7=10 см, то на всю ампулу
придется разность потенциалов U = 1 мкВ. Практически все это
напряжение будет падать на слое рецепторов, так как его сопротивление
гораздо выше, чем сопротивление канала и окружающей его среды.
Следовательно, чем длиннее канал ампулы и чем меньше его
сопротивление, тем больше будет падение напряжения на
электрорецепторных клетках. Учитывая, что сопротивление внутренней
мембраны самой рецепторной клетки существенно больше сопротивления
синаптической мембраны, все падение напряжения приходится
именно на нее.
Для увеличения чувствительности электрорецепторных клеток
выгодно иметь возможно более высокое сопротивление мембраны,
чтобы на ней падала большая часть напряжения. И действительно,
сопротивление мембраны, которая выделяет медиатор, у электрорс-
цепторной клетки очень велико, порядка десятков ГОм. Однако
тепловой шум 1/ш такого сопротивления тоже велик. Его можно
определить по известному выражению: (£/ш)2 = 4kTRf, где к - постоянная
Больцмана, Г- температура, R - сопротивление,/- полоса частот,
в которой рассматривается шум.
С учетом приведенного сопротивления мембраны напряжение
шума составляет около 30 мкВ, т.е. в 30 раз больше, чем
минимальный воспринимаемый сдвиг мембранного потенциала, возникающий
I пава 6. Сенсорные системы организма
221
под действием внешнего поля. Поскольку одно нервное волокно
соединяется через синапсы с 2000 рецепторных клеток, то суммарное
напряжение шума всех N клеток определяется как корень
квадратный из N, т.е. из 2000. В результате получаем, что суммарный сигнал
шума (при UIU = 30 мкВ), поступающий на нервное волокно,
гостами г 1350 неких единиц (например, молекул медиатора). В то же
иремя полезный сигнал от всех 2000 клеток, формируемый
синхронно, составит 2000 тех же единиц. Значит, отношение сигнал/шум
составляет 1,5, что вполне достаточно для выделения сигнала на
уровне шума.
Большим числом экспериментальных исследований показано,
что рыбы используют электрорецепцию для ориентации в мутной
поде, для поиска добычи (акулы и скаты). Например, акула способна
обнаружить камбалу, зарывшуюся в ил, по электрическим полям,
генерируемым ее мышцами при дыхательных движениях. Скаты могут
обнаруживать крабов по их биопотенциалам, а сомы обнаруживают
даже электрические поля, генерируемые червями, закопавшимися
к землю.
6.2. Биофизика слуха
6.2.1.Слуховая рецепция
Значение слухового анализатора состоит в восприятии и
анализе звуковых воли. Вследствие этого возможно определение силы,
иысоты и тембра звука, его направления, а также степени
удаленности источника звука.
Рассмотрим некоторые понятия, используемые в биофизике.
Интенсивностью звука называют энергию, приходящуюся на
единицу площади поверхности перпендикулярно к направлению
распространения звуковой волны в единицу времени:
/=/-. (6-6)
5"·/
где Е- энергия, S- площадь, t- время, /- также называют плотно-
С1ью потока энергии звуковых колебаний (вектор Умова).
222
Биофизика
При решении практических задач проще измерить не энергию
колебаний, а звуковое давление (р), регистрируемое при помощи
микрофона и выражаемое в паскалях. Звуковое давление линейно
связано с амплитудой звуковых колебаний, и поэтому его квадрат
пропорционален энергии звука при любых двух интенсивностях:
77
Е_
р1
(6.7)
Порог слышимости чистого тона при частоте 1 кГц принято
считать равным 2 · 10-5 Па, хотя можно наблюдать многократные
индивидуальные колебания порога слышимости (в пределах 30 дБ),
Кроме того, порог слышимости значительно меняется с частотой
(рис. 6.14). Может также сильно различаться и наивысшая
воспринимаемая частота (от 8 до 25 кГц).
По закону Вебера-Фехнера ощущение громкости связано с
создающим его физическим раздражением логарифмической
зависимостью. Это стимулировало введение единиц уровня интенсивности
звука, выражаемых в децибелах:
Z, = 10Ig—ξ 20In— (в децибелах),
7о Ро
(6.8)
/.дБ А
120
80
40
Болевой порог
0,01 0,1
10
Порог нормального слуха
Порог поврежденного слуха
f. кГц
Рис 6.14. Характеристики слухового анализатора
' пава 6. Сенсорные системы организма
223
где р0 — пороговое значение звукового давления, равное 2 · 10~5 Па
при частоте звукового тона 1 кГц, ρ - регистрируемое значение
звукового давления, /0 - пороговая интенсивность звука. Таким образом,
нулевая точка шкалы громкости соответствует 0 дБ при 1 кГц. При
других частотах порог слуха отличается от 0 дБ (рис. 6.21).
Строение наружного и среднего уха. Наружное ухо включает
ушную раковину и слуховой проход длиной 2,7 см. С наружним
ухом связано свойство направленности слухового восприятия, под
которым понимают лучшее восприятие звуков, идущих в
определенном направлении. Это обеспечивает локализацию источника звука
и пространстве [ртотолику), кроме этого, слуховой проход служит
резонатором. Его резонансная частота у человека лежит в области
4 кГц у мужчин и 6 кГц у женщин, но резонансная кривая имеет
уплощенную форму, поскольку добротность слухового прохода, как
колебательной системы, невелика. Разница в коэффициенте передачи
туков резонансной и не резонансной частот не более одного
порядка. Границей между наружным и средним ухом служит барабанная
перепонка (БП). Наружное ухо защищает БП от механических
повреждений и, поддерживая определенные микроклиматические
условия (температуру, влажность и пр.), стабилизирует ее механические
свойства. Таким образом, наружное ухо выполняет следующие
функции: локализация звука в пространстве, усиление звукового сигнала
(резонансное), механическая защита БП, обеспечение микроклимата,
что необходимо для нормального функционирования БП.
Наружное ухо от среднего отделяется барабанной перепонкой.
С внутренней стороны барабанная перепонка соединена с рукояткой
молоточка. Колебания барабанной перепонки происходят тогда,
когда на нее падают звуковые колебания, улавливаемые наружным
ухом. Натяжение барабанной перепонки в различных частях
неодинаковое. Это приводит к тому, что она не имеет своего собственного
периода колебаний и колеблется при любом звуке соответственно
длине его волны.
Среднее ухо предназначено для трансформации звуковых
колебаний воздуха в звуковые колебания более инерционной жидкой
среды внутреннего уха. Это прежде всего импеданссогласующая
функция. Кроме того, среднее ухо обладает усиливающей функцией
224
Биофизика
Рис. 6.15. Схема строения уха
I - ушная раковина; 2 - наружный слуховой проход; 3 - барабанная перепонка;
4 - полость среднего уха; 5 - слуховая труба; 6 - молоточек; 7 - наковальня;
8 - стремечко; 9 - полукружный канал; 10-улитка; II - вестибулярный аппарат
как за счет разности площадей барабанной перепонки и овального
окна улитки, так и за счет системы рычагов слуховых косточек. При
отсутствии среднего уха огромное различие в акустических импедан-
сах воздуха и среды внутреннего уха не позволило бы проникнуть
звуковым колебаниям во внутреннее ухо.
В состав среднего уха входит система слуховых косточек -
молоточек, наковальня, стремечко, слуховая (евстахиева) труба
(рис. 6.15). Они работают как рычаг, обеспечивающий выигрыш в силе
в 1,3 раза. Одна из косточек - молоточек - встроена своей рукояткой
в барабанную перепонку, а его противоположная часть сочленена с
наковальней.
Наковальня соединена со стремечком, которое прилегает
к мембране окна преддверия (овального окна) внутренней стенки
среднего уха. Поскольку площадь барабанной перепонки человека
(66-69,5 мм2) значительно больше площади овального окна
внутреннего уха (3 мм2), общий коэффициент усиления по давлению
стремечка на овальное окно больше двадцати.
Слуховые косточки участвуют в передаче колебаний
барабанной перепонки, вызванных звуковыми волнами, окну преддверия,
( пава 6. Сенсорные системы организма
225
и чатем перилимфе и эндолимфе улитки внутреннего уха. Окно
преддверия расположено на стенке, отделяющей среднее ухо от
внутреннего. Там же имеется круглое окно. Колебания жидкости улитки,
начавшиеся у овального окна, распространяются по ходам улитки, не
штухая, до круглого окна.
При помощи особого канала - евстахиевой трубы полость
ереднего уха соединяется с носоглоткой, благодаря чему в полости
ереднего уха поддерживается давление, равное атмосферному. Если
давление в полости среднего уха отличается от атмосферного, то это
приводит к неприятным ощущуениям («закладывает уши») и
понижению слуха, так как нарушаются нормальные колебания
барабанной перепонки.
Другой важной функцией среднего уха является защита
внутреннего уха от чрезмерных механических нагрузок при воздействии
очень громких звуков. Это достигается изменением характера
колебаний стремечка в зависимости от силы звука. При пороговых
значениях звукового давления основание стремечка входит в овальное
окно и выходит из него, как педаль автомобиля. Абсолютные
значения смещения очень малы и повторяют движения барабанной
перепонки. Для барабанной перепонки величину смещения легко
рассчитать. Толщина барабанной перепонки (0,1 мм) очень мала по
сравнению с длиной звуковых волн, и скорость ее перемещения
совпадает со скоростью частиц в плоской волне в воздухе. Средняя
скорость смещения частиц в волне (ν) связана со значением звукового
давления (р), скоростью распространения звуковой волны (с) и
плотностью воздуха (р) выражением:
ν=^-. (6.9)
рс
Произведение рс называеся акустическим импедансом - гш.
В свою очередь, скорость смещения связана с амплитудой
смещения выражением:
IS — 9844
226
Биофизик»
где /- частота звуковой волны.
Подставив в (6.9) численные значения ρ = 1,3 кг/м\ с = 340 м/о
и порогового значения звукового давления/^ = 2 ■ 10~5 Па, получим
скорость смещения перепонки ν = 5 · Ю-8 м/с. Отсюда смещение
перепонки при пороговом звуковом давлении равно 10-1' м.
Если при низких значениях звукового давления смещения
основания стремечка практически повторяют движения барабанной
перепонки, то при возрастании силы звука характер колебаний
стремечка меняется. Причиной этого изменения является рефлекторное
сокращение тонких мышц внутри барабанной полости, вследствие
чего нарушается движение слуховых косточек в соединяющих их
суставах. При средней силе звука стремечко начинает совершать
колебательные движения вокруг вертикальной оси у одного конца
овального окна, а при очень громком звуке - вращательные движения
вокруг горизонтальной оси, т.е. когда один конец стремечка
вдавливается в овальное окно, другой движется в противоположную
сторону. Таким образом предотвращаются избыточно сильные движения
жидкости во внутреннем ухе.
Строение внутреннего уха. В состав внутреннего уха входят
преддверие и улитка, в которой расположены рецепторы,
реагирующие на звуковые волны. Преддверие не относится к органу слуха.
Вестибулярный аппарат, сосредоточенный в нем, участвует в
регуляции положения тела в пространстве и сохранении равновесия, о чем
говорилось ранее.
Улитка - это костный постепенно расширяющийся спиральный
канал, образующий у человека 2,5 витка. Длина развернутой улитки
около 35 мм, а объем - около 100 мкл. Костный канал на всем своем
протяжении разделен двумя перепонками: более тонкой, называемой
вестибулярной мембраной, или мембраной Рейспера, и более
плотной и упругой, получившей название основной (базилярной)
мембраны (рис. 6.16). Эти мембраны на вершине улитки соединяются.
Костный канал улитки за счет вестибулярной и основной мембран
разделяется на три параллельных наполненных жидкостью канала.
Вестибулярный и барабанный каналы отделены друг от друга
улитковым каналом (протоком). Улитковый канал заполнен эндолимфой,
тогда как в двух других каналах находится перилимфа, которая мо-
β 6 Сенсорные системы организма
227
ст перетекать из одного канала в другой через геликотрему - отвер-
шс в вершине (крючке) улитки. На базилярпой мембране находится
Кортиев орган, содержащий рецепторные и опорные клетки, а также
рвные окончания.
Рецепторные (волосковые) клетки ориентированы перпендику-
1ярно к базилярпой мембране. В каждой из двух улиток уха человека
смержится по 15500 волосковых клеток, из которых 3500 образуют
инутренний ряд и 12000 образуют 3-5 наружных рядов. По форме во-
юсковые клетки внутреннего ряда подобны эллипсоиду, вертикаль-
гая ось которого составляет в среднем примерно 40 мкм, а горизон-
Рис 6.16. Поперечный разрез витка улитки (по Расмуссену)
А — общий вид: 1 —лестница преддверия, 2 — барабанная лестница,
3 - вестибулярная мембрана. 4 - базилярная (основная) мембрана,
5 —улитковый канал, б — покровная мембрана, 7 -Кортиеа орган,
8 — секреторный эпителий, 9 — спиральная связка, 10 - спиральный
ганглий; Б - увеличенный в несколько раз участок спирального
органа: 11 — наружные волосковые клетки, 12 - внутренние волосковые
клетки, 13 - нервные волокна, подходящие к волосковым клеткам
228
Биофизика
тальная примерно 10 мкм. Наружная волосковая клетка имеет
цилиндрическую форму.
В связи с изменением по ходу улитки ее поперечного сечения
меняется и ширина натянутой поперек нее базилярной мембраны. Как
это впервые предположил Гельмгольц, последняя может
представлять собой структуру, резонирующую на поступающие во внутреннее
ухо звуковые колебания. При этом место максимальных резонансных
колебаний должно смещаться в зависимости от частоты звука.
Колебания мембраны будут передаваться на различные волосковые
клетки, и таким образом частота звука будет отражаться в активации
рецепторов, расположенных в разных местах базилярной мембраны:
около овального отверстия (у входа в улитку) - звуками высоких
частот, а на вершине улитки (в крючке) - звуками низких частот.
Поступающие во внутреннее ухо звуки вызывают ответные
колебания базилярной мембраны, причем они имеют характер бегущей
волны. Ее амплитудный максимум перемещается по мере изменения
частоты звука - при высоких частотах он находится у входа в улитку,
а при низких - у ее вершины. Рецепторные клетки раздражаются
сильнее всего там, где смещение базилярной мембраны максимально.
Поэтому под действием звуков разных частот возбуждаются
различные рецепторы.
Таким образом, в слуховой рецепции имеет место
пространственно-временное кодирование характеристик параметров внешнего
раздражителя. При изменениях амплитуды и частоты звуковых
колебаний изменяются как пространственная структура активности
рецепторов, так и частота разрядов в каждом из них. Извлечение
информации об этих параметрах звука осуществляется соответствующими
нервными центрами за счет сложного взаимодействия в них
процессов возбуждения и торможения. Среди различных свойств звукового
раздражителя мозг способен анализировать и направление звука, что
обеспечивается бинауральным слухом. Благодаря ему нейроны
центральной нервной системы способны оценивать разность фаз между
сигналами от правого и левого ушей.
При воздушной проводимости звука человек может
воспринимать звуки в очень широком диапазоне - от 16 до 20 000
колебаний в 1 с.
Глава 6. Сенсорные системы организма
229
Первичный частотный анализ сложной звуковой волны
происходит с помощью базилярной мембраны улитки внутреннего уха.
Максимальные амплитуды колебаний соответствуют тем областям
мембраны, которые отвечают воспринимаемому частотному
спектру звука. Следовательно, базилярная мембрана является
первичным частотно-амплитудным преобразователем. Более точная
селекция звукового спектра связана с работой ЦНС. На базилярной
мембране происходит сложение двух стоячих волн: волны,
возникающей в пространстве улиткового протока, и волны, возникающей
II пространстве барабанной лестницы, рис. 6.17.
Уравнение стоячей волны, возникающей в пространстве
улиткового протока, имеет вид:
S, =2^cos sinatf, (6.11)
ν
где S, - смещение в стоячей волне, А - амплитуда колебаний в
волне, ω - круговая частота волны, ν - скорость звуковой волны в эндо-
лимфе, X - координата, отсчитываемая от овального окна (4), t -
нремя.
Уравнение стоячей волны, возникающей в пространстве
барабанной лестницы (2-3), имеет вид:
Рис. 6.17. Схема внутреннего уха человека (улитка условно развернута)
I - улитковый проток; 2 - вестибулярная лестница; 3 - барабанная лестница;
4 - овальное окно; 5 - круглое окно; 6 - геликотрема; 7 - базилярная мембрана
230
Биофизика
с . . a(l-X) ■ ,
S2 = Z4C0S —: -SHICO/,
(6.12)
где S2 - смещение в стоячей волне, / - общая длина вестибулярной
и барабанной лестниц, которая примерно в два раза больше длины
улиткового протока, (1-Х)- координата, отсчитываемая от круглого
окна (5), ν, - скорость звуковой волны в перилимфе.
Эти две автоволны отличаются, во-первых, скоростными
характеристиками, так как плотности перилимфы и эндолимфы
различны, и, во-вторых, длиной каналов, в которых возникают автоволны.
Сложение стоячих волн происходит на базилярной мембране
(7), и в соответствии с принципом суперпозиции суммарное
смещение S равно:
= 5, +S2 =2Α\οο*^+οο*ω(1-Χ) kin см.
Ι ν ν. )
(6.13)
Как показывают расчеты по формуле (6.13), при сложении
стоячих волн возникает уединенный максимум, положение которого по
длине базилярной мембраны и острота зависят от частоты
воспринимаемого звука. На рис. 6.18 показаны графики зависимости
смещения S от координаты X, построенные по формуле
B(X)=2Acos(^+cos°i(1-X)
\ ν ν,
(6.14)
s A
АА
2,5/»
112
1=0.07 Х.м
Рис. 6.18. Смещение максимума колебаний базилярной мембраны по длине
базилярной мембраны: (1 - для частоты/ = 1& Гц; 2 - для частоты/ = 3000 Гц)
I пава 6. Сансорныа системы организма 231
/Гц
20000
10000
0 ΙΙΛ //2 = 0,035 Х,м
Рис. 6.19. Зависимость положения максимума колебаний
на данной частота от длины базилярной мембраны
для двух значений круговой частоты: кривая 2 - при ω = 18840 с-1 (/=
3 000 Гц) и кривая 1 - при ω = 62800 с"1 (/= 10 000Гц). Скорости
звука в эндолимфе ν = 1 400 м/с, в перилимфе ν, = 1 700 м/с. Длина
вестибулярной и барабанной лестниц / = 0,07 м.
Как видно из графика рис. 6.18, с уменьшением частоты
максимум смещения сдвинулся к геликотреме (6).
На рис. 6.19 показана зависимость положения максимума
смещения на данной частоте/= ω /2π от длины улитки. Расчет
проводился с использованием условия возникновения максимума:
—sin sin—: - = 0. (6.15)
/ ν /ι ν,
В соответствии с (6.15) строилась зависимость/ =βΧ).
Из рис. 6.19 видно, что наблюдается движение максимума при
уменьшении частоты в сторону геликотремы, что соответствует из-
пестным экспериментам.
Более точное количественное совпадение с экспериментами
требует учета расширения базилярной мембраны и, как следствие,
учета ее упругости, что ведет к большим расчетным затруднениям.
Однако даже такой приближенный анализ указывает, например, что
разрушение преддверной мембраны (мембраны Рейснера) приводит
к исчезновению восприятия тембра звука. Действительно, в этом
232
Биофизика
случае эндолимфа и перилимфа перемешиваются, скорости звука
в них выравниваются ν = ν, .
Благодаря зависимости местоположения максимальной
амплитуды бегущей волны на базилярной мембране от частоты,
вызывающей ее звукового тона, происходит распределение этих частот между
различными участками Кортиевого органа. Его волосковые клетки
возбуждаются, главным образом, в местах максимального смещения
базилярной мембраны. Следовательно, звуковой тон каждой частоты
возбуждает соответствующие слуховые рецепторы. Так, в улитке
осуществляется первый этап частотного анализа звука, который
основывается на пространственном разграничении участков
базилярной мембраны, колеблющихся с неодинаковой амплитудой под
действием звукового тона определенной частоты.
Звуки пороговой интенсивности вызывают бегущие волны,
амплитуда которых в Кортиевом органе не превышает 10~9 м. При
усилении звуков амплитуда колебаний базилярной мембраны больше
этой величины всего на 1-2 порядка, значит, слуховой рецептор
реагирует на ничтожные механические перемещения. Начало этой
реакции связано с движением волосков (цилий) на апикальном полюсе
волосковой клетки. Эти смещения обусловлены распределением
бегущей волны по базилярной мембране. Среди цилий в каждой
волосковой клетке различают один длинный волосок (киноцилия) и
множество (примерно 100-120) коротких (стереоцилий). Рецепция звука
обусловлена воздействием механических усилий на стереоцилий.
Они прикасаются к покровной мембране, один край которой
закреплен на костной спиральной пластинке, а другой, свободный, нависает
над волосковыми клетками. Модель изгиба стереоцилий под
действием звуковых тонов схематически показана на рис. 6.20.
При совместных колебаниях базилярной и покровной мембран
происходит изгиб стереоцилий. Небольшое смещение их свободных
концов приводит к значительной конформационной перестройке
молекул плазмолеммы волосковой клетки в тех местах, где залегают
«корни» изогнувшихся волосков, поскольку стереоцилиям присущи
свойства микроочагов. Конформации мембранных макромолекул
обусловливают изменение проницаемостей для ионов и, следовате-
/ пава 6. Сенсорные системы организма
233
С СУ
ксп
В состоянии покоя
Смещение а сторону УП
Смещение в сторону БЛ
Рис. 6.20. Модель изгиба стереоцилий под действием звука
КСП - костная спиральная пластинка; БМ- базилярная мембрана;
ПМ- покровная мембрана; СУ- стенка улитки, к которой прикрепляется
базилярная мембрана; С - стереоцилий волосковой клетки, УП-улитковый
проток, БЛ — барабанная лестница (канал под базилярной мембраной)
льно, приводят к возникновению ионного тока через мембрану
волосковой клетки.
При отклонении пучка стереоцилий в сторону киноцилии
возникает деполяризация волосковой клетки, а в обратную сторону -
гиперполяризация. Например, у ящериц и черепах расстояние, на
которое нужно отклонить волоски, чтобы получить ответную реакцию,
составляет 0,34 нм, а для волосковых клеток млекопитающих еще
меньше - около 0,1 нм. Исследования улитки лягушки показали, что
мембранный потенциал при смещении волосков в сторону киноци-
234
Биофизика
лии изменяется от -60 до -40 мВ. При отклонении стереоцилий
в противоположную сторону возникает обратная реакция -
гиперполяризация (от -60 до -65 мВ). Смещения стереоцилий в поперечном
направлении по отношению к киноцилии реакций рецептора не
вызывают. Характерно, что, когда пучок волосков надолго перемещали
в другое положение, скажем, на 1 мкм, значительного постоянного
возбуждения рецептора не наблюдалось. Через десятые доли
секунды рецепторная клетка вновь становилась чувствительной к
микроперемещениям волосков.
Большой интерес исследователей вызывает расшифровка
молекулярных механизмов рецепции, происходящих в волосковой клетке,
способной реагировать на стимулы с пикометровой амплитудой
и частотой до 20 (даже 100) кГц. Одна из гипотез предполагает
наличие каналов для ионов, которые открываются и закрываются вследст-
. вие неупорядоченного теплового движения лежащих рядом молекул.
Доля каналов, открытых в каждый момент времени, зависит от
разницы в энергетических уровнях открытых и закрытых каналов. Когда
эта разница превышает среднюю кинетическую энергию
столкновений с окружающими молекулами, каналы будут открываться редко,
и наоборот, если исходный энергетический уровень открытых
каналов в апикальной мембране волосковой клетки высок, то большая
часть каналов закрыта. При движении пучка волосков каким-то
образом изменяется энергетическое соотношение между каналами и,
например, число открытых ионных каналов быстро увеличивается под
влиянием теплового движения соседних молекул.
Уровень потенциала покоя волосковой клетки относительно
межклеточной среды (перелимфы), находится в пределах от -50 до -
70 мВ. Предполагают, что при перемещении пучка стереоцилий под
действием звука происходит открывание мембранных ионных
каналов на апикальном полюсе волосковой клетки. Это доказывается тем,
что в эксперименте зарегистрированы максимальные токи в среде
около концов движущегося пучка. Входящий в волосковую клетку
катионный ток вызывает деполяризацию ее мембраны, что и является
рецепторным потенциалом (РП). Значение рецепторного потенциала
зависит от интенсивности звука, и его амплитуда составляет
примерно 10 мВ. Максимальная величина РП, зарегистрированного с помо-
Глава 6. Сенсорные системы организма
235
щью микроэлектронной техники, составляет примерно 24 мВ. Со
скоростью электромагнитной волны РП электротонически (с
декрементом) распределяется по плазмолемме волосковой клетки от ее
апикального полюса к базальному. Там деполяризация приводит
к высвобождению медиатора (определенное химическое вещество),
поступающего далее через синаптическую щель на
субсиноптическую мембрану, которая принадлежит чувствительному нервному
окончанию, контактирующему с данной волосковой клеткой. Под
действием медиатора на субсинаптической мембране возникает
генераторный потенциал (ГП). Он, как и РП, электротонически
распространяется на внесинаптические участки афферентных волокон, где
вызывает образование ПД. В таком виде нервная импульсация
направляется в ЦНС по слуховому нерву. Следовательно, по механизму
преобразования адекватного стимула (звука) в нервные импульсы
слуховой рецептор является типичным представителем
вторично-чувствующих рецепторов.
Каждая волосковая клетка соединена с нервными
окончаниями, при этом каждое волокно слухового нерва начинается от узкого
ограниченного участка Кортиевого органа, в отдельных случаях, от
одной волосковой клетки. Поскольку слуховые рецепторы,
расположенные в том или ином месте базилярной мембраны, возбуждаются
звуками определенных частот, каждая небольшая группа нервных
волокон слухового нерва проводит импульсы в ответ на звуки
преимущественно одной частоты. Эта частота называется характерической
частотой волокна. Если звук представляет собой сложные
колебания, то в слуховом нерве активизируются все волокна, характериче-
ские частоты которых соответствуют гармоническому спектру
сложного звука. Следовательно, на уровне слуховых рецепторов звуки
разлагаются в гармонический спектр. Длительность звукового
сигнала кодируется временем активации афферентных волокон, которые
входят в состав слухового нерва.
Костная проводимость звука осуществляется через кости
черепа. Если поставить ножку звучащего камертона на темя или
сосцевидный отросток, то звук будет слышен даже при закрытом
слуховом проходе. Это связано с тем, что звуковые колебания хорошо
проводятся костями черепа, передаются сразу на перилимфу верхне-
236
Биофизика
го и нижнего ходов улитки внутреннего уха, а затем - па эндолимфу
слухового протока. Происходит колебание основной мембраны с во-
лосковыми клетками, в результате чего они возбуждаются, и
возникшие нервные импульсы в дальнейшем передаются к нейронам
головного мозга.
При исследованиях органов чувств стремятся установить
степень отклонения получаемых при эксперименте данных от
статистически установленных норм. Одним из основных физиологических
понятий, используемых при подобных исследованиях, является
порог. Под ним обычно понимают ту минимальную величину
раздражителя, которая вызывает его ощущение как сигнала на фоне помех,
Различают абсолютный и дифференциальный пороги
чувствительности. Абсолютный порог чувствительности измеряется в абсолютных
величинах раздражителя, который испытуемый ощущает на фоне
внутренних шумов и помех. Минимальная разница между двумя
раздражителями, действующими последовательно или параллельно,
которая воспринимается испытуемым, носит название
дифференциального порога. При оценке состояния звукового анализатора
исследуются как абсолютный, так и дифференциальный пороги.
В зависимости от характера и степени повреждения слуха
кривая порога слышимости (рис. 6.21) обычно отклоняется вверх, а
кривая боли или не меняется, или иногда отклоняется вниз. Если кривая
порога поврежденного слуха расположена выше диапазона речи, то
человек перестанет понимать то, что ему говорят.
Метод исследования абсолютных порогов восприятия
пациентом звуковых тонов разных частот называется аудиометрией.
Приборы, предназначенные для проведения таких исследований,
называются аудиометрами.
Поскольку различие в порогах восприятия разных звуковых
частот является важным показателем состояния звукового
анализатора, при аудиометрии изучают пороги для ряда «чистых тонов», т.е.
восприятие звуков определенной частоты, создаваемых генератором
чистых синусоидальных колебаний. В медицине этот метод
называют моноуральной аудиометрией по чистым тонам. Её обычно
проводят в диапазоне частот от 100 до 8000 Гц. Исследуют пороги слуха
для частот 125, 250, 500, 1000, 2000, 4000, 8000 Гц.
Глава 6. Сенсорные системы организма
237
Р. Па I. Вт
2-Ю2,- Ю2
1.Д6
Е, фон
2-101
2
2-Ю"1
2-Ю-2
2-10-3
2-10^ - 10
2-10"
2-10-8 L 10
1
ΙΟ"2
10^·
ю-8
ю-*
Ί-10
10
-12
-14
20 63 250 1000 4000 16000
31,5 125 500 2000 6000
Рис. 6.21. Контуры равных уровней громкости
(кривые равных громкостей, изофоны)
по оси абсцисс — частота звуковых тонов (f), no оси ординат —
слева приведены три шкалы с эквивалентными значениями -
уровня интенсивности (L), интенсивности (I) и звукового давления (р),
справа -шкала уровня громкости (Е) и эта же шкала на уровне/ = 1000 Гц
Используются также аудиометры, в которых исследование
слуха проводится не «чистыми» тонами, а акустическими
колебаниями «шума», спектр которого находится в узкой полосе частот.
6.2.2. Методы и средства проведения
аудиометрических исследований
В физиологической акустике принято сопоставлять
объективные (физические) и субъективные (психофизические)
характеристики звука (табл. 6.1).
Звук при распространении в среде представляет собой волны
сгущения и разрежения ее частиц. В участках сгущения давление
больше, чем в участках разрежения. Амплитуда периодических
колебаний давления в среде, происходящих под действием
распространяющего звука, называют звуковым давлениемр. Единицы его измере-
238
Биофизика
ния [Па] или [Н/м2]. Звуковым давлением определяется интенсивность
звука, которая находится по формуле:
ι=-Ε-, (6.16)
2ср
где ρ - давление; с - скорость распространения звука в данной
среде; ρ - плотность среды [кг/м3].
Таблица 6.1
Сопоставление физических и психологических характеристик звука
Физические характеристики
звука
1. Гармонический спектр (характеризуется
суммой всех гармоник Σ^):
а) основная гармоника f0, Гц;
б) высшие гармоники fb Гц
2. Интенсивность (плотность потока звуковой
энергии) / [Вт/м2]
Психофизические
характеристики
звука
1. Акустический спектр:
а) основной тон (высота);
измеряется в мелах и барках;
б) обертоны (определяют тембр)
2. Громкость, измеряется в сонах
Кроме того, интенсивность (вектор Умова) можно рассчитать
по формуле: I = w · V, если известна w - объемная плотность энергии
звука, т.е. энергия в единице объема среды (V), в которой звук
распространяется. Величина Ζ = 2ср есть акустический импеданс среды.
Для установления взаимосвязи между интенсивностью и
громкостью вводятся еще две физические величины, которые
характеризуют звук: уровень интенсивности (L) и уровень громкости (Е).
Уровень интенсивности, или уровень звукового давления,
определяется по формуле (8):
£ = *lg —, (6.17)
где 10 - эталонная интенсивность; / - интенсивность исследуемого
звука, 10 принимают равной Ю-12 Вт/м2. Такая интенсивность чистого
Глава 6. Сенсорные системы организма
239
юна частотой в 1 кГц установлена как средний порог слуховой
чувствительности для людей. Эта интенсивность соответствует
звуковому давлению, равному 2 · 10~5 Па. При к = 1 единицей уровня
интенсивности L служит бел [Б]. При А: = 10 единицей уровня
интенсивности L служит децибел [дБ]. В медицине принято исполь-
ювать децибельную шкалу. Если 1=\0'5 Вт/м2, то L = 0.
Следовательно, на частоте 1 кГц три величины ρ = 2 · 10~5 Па, и / = 10~12 Вт/м2 и
Λ = 0 дБ - характеризуют один и тот же звук. При I > 10 величина L
принимает положительно значение, а при К /0 имеем L < 0. Чем
меньше величина L для порога слышимости, тем выше слуховая чувст-
нительность. Человек лучше всего слышит звуковые тоны частотой
от 2 до 4 кГц.
Уровень громкости (Е) — это выражение различий в восприятии
человеком звуков разной частоты. Уровнем громкости звука данной
частоты называют уровень интенсивности звукового тона частотой
1 кГц, громкость которого при сравнении на слух эквивалентна
громкости исследуемого звука. Единицы измерения Ε - фон. Для звуков
частотой 1 кГц численные значения Ε и L совпадают: так, если первая
из них L составляет 0 дБ, то Ε = 0 фонов. Средний порог слухового
носприятия у людей составляет примерно 4 фона. Болевые ощущения
ι» ухе возникают под действием звуков, уровень громкости которых
достигает примерно 130 фонов. Таким образом, все частоты звуковых
тонов, которые слышит человек, ограничивается диапазоном от 16 Гц
до 20 кГц. В этом частотном диапазоне уровень громкости всех
слышимых звуков лежит в пределах от 4 до 130 фонов.
Громкость звука является психофизической (субъективной)
характеристикой. Она оценивается по шкале сонов. Для выражения
громкости того или иного звука в сонах за эталон принимается
звуковой тон частотой 1 кГц при уровне интенсивности L = 40 дБ. Дав
испытуемому послушать эталонный тон, исследователь предъявляет
ему тестируемый звук. При этом он просит определить, во сколько
раз тестируемый звук громче или тише эталонного. Так, например,
ломкость тестируемого звука, если испытуемый слышит в 2 раза
громче эталонного тона, будет равен 2 сонам, а если он слышит
тестируемый звук в 2 раза тише эталонного, то громкость тестируемого
звука составляет 0,5 сона.
240
Биофизика
Задача исследования слуха сводится не только к установлению
степени и причины его повреждения, но и к определению того, какой
слуховой аппарат пригоден в конкретном случае.
При использовании метода аудиометрии «чистого звука»
сигнал от электронного генератора электрических колебаний подается
в наушники. Порог слышимости определяется так: врач увеличивает
интенсивность звука на определенной частоте до тех пор, пока
пациент через наушники не уловит его и не просигнализирует об этом.
Воспринятая интенсивность звука будет являться порогом
возбуждения слухового анализатора на данной частоте. При использовании
наушников звуковые колебания через воздушную подушку между
барабанной перепонкой и мембраной наушника непосредственно
попадают во внутреннее ухо. Для исключения влияния внешних
акустических помех больного целесообразно помещать в
звукоизолированную камеру (сурдокамеру).
Используется также метод исследования слуха, когда звуковые
колебания с определенной частотой и интенсивностью подаются на
сосцевидный отросток (кость позади уха). При этом звук передается
черепной костью. Наличие акустической связи между ушами,
осуществляемой через черепную кость, приводит к тому, что при
обследовании характеристик одного уха звуковые колебания попадают
и в другое. Для уменьшения влияния такой перекрестной связи ухо,
которое в данный момент не исследуется, «маскируется». В него
подаются звуковые колебания определенной силы и хаотично
изменяющейся частоты («белый шум»). Для этой цели в состав аудиометра
вводится специальный электронный шумовой генератор, который
может подключаться к одному из двух наушников. В некоторых
случаях порог чувствительности определяют при одновременном
действии сигнала и шума. В этом случае на ухо больного (исследуемое или
противоположное - в зависимости от стоящей задачи) подается
широкополосный шум с одинаковой интенсивностью на всех частотах
и проводится определение «порогов чистых тонов». Большое
значение имеет время «предъявления» звука. Оно обычно порядка 1-2 с.
Оценка дифференциального порога выполняется чаще всего
с помощью методики Люшера. При ее использовании ритмически
с частотой два раза в секунду производится изменение интенсивно-
( пава 6. Сансорныа системы организма
241
гти звука. Для этого используется соответствующий модулятор
интенсивности колебаний. Устанавливается минимальное значение
амплитуды модуляции, которое испытуемый различает и отмечает как
колебание постоянного уровня звука.
Для большей объективизации результатов исследований
слухового анализатора иногда используется то обстоятельство, что
•лсктроэнцефалограмма обследуемого пациента меняется, если он
подвергается звуковому возбуждению. Если пациент глухой, то
•лектроэнцефалограмма ЭЭГ не меняется при воздействии на него
туковыми импульсами. По изменению ЭЭГ можно сделать выводы
о степени повреждения слуха. С помощью этого метода выявляются
повреждения слуха у младенцев.
Для диагностики ряда заболеваний исследуется явление
выравнивания громкости. При этом оцениваются субъективные ощущения,
нозникающие при воздействии звуковых колебаний разных интен-
сивностей. Так, например, считается установленным факт
уменьшения дифференциальных порогов при поражении звуковоспринимаю-
щего аппарата и их увеличение - при поражении центральных
структур.
Субъективность большинства результатов, получаемых в том
числе с помощью различных технических средств, приводит к
затруднениям с определением нормы и патологии. Поэтому точно
принятой полосы нормы для аудиограммы нет. Обычно считается
нормальным показатель отклонения от типовых принятых значений
π ±(3-5) дБ.
Используется также речевая аудиометрия. Целью ее является
изучение восприятия речи, т.е. идентификация не изолированных
сигналов, а потока раздражителей, в котором необходимо
идентифицировать и звуки, и слова, и фразы. При речевой аудиометрии
выясняются закономерности восприятия слов при изменениях уровня
речи.
Речевая аудиометрия осуществляется следующим способом.
На магнитофон записывается словесная или слоговая таблица.
Особенностями записи является то, что диктор отчетливо выговаривает
слова и звуки и старается поддерживать постоянный уровень
громкости. Промежутки между читаемыми словами или слогами делаются
%
100
80
60
40
20
20 40 60 80 100 дБ
Рис 6.22. Изменения разборчивости слов
в зависимости от уровня громкости речи
такими, чтобы испытуемый успевал их спокойно записывать. Вместо
диктора могут использоваться звукосинтезирующие устройства. При
обследованиях обычно необходимо иметь записанными 3-4 таблицы
(по 50 слов или слогов в каждой). Интенсивность звуковых сигналов
устанавливают такой, что испытуемый начинает слышать
неопределенные звуки.
Этот уровень берется за порог обнаружения. При увеличении
уровня звукового сигнала испытуемый начинает правильно
опознавать отдельные слова. Чем больше уровень звукового сигнала, тем
выше процент правильно опознанных слов.
По результатам повторных измерений при разных интенсивно-
стях звукового сигнала строится кривая изменения разборчивости
слов (рис. 6.22).
При отсутствии патологии 20-% разборчивость достигается
при превышенииуровня интенсивности над порогом обнаружения на
17-20 дБ, 50-% разборчивость получается при превышении над
порогом обнаружения на 24-26 дБ, 80% - на 30-34 дБ, 100% - на 37-
38 дБ.
Кривая речевой аудиограммы многое говорит врачу о природе
заболевания и возможностях его лечения. С ее помощьюможно
оценивать эффективность подбора слухового аппарата.
Существует достаточно большое количество различных
аудиометров. Среди них можно выделить:
( пава 6. Сенсорные системы организма
243
— клинические аудиометры - приборы, обеспечивающие
высокую точность измерений, генерирующие проверочные сигналы в
широкой полосе частот, в широком динамическом диапазоне со
многими градациями интенсивности;
-проверочные аудиометры - простые, переносные, дешевые
приборы с более узкими функциональными возможностями.
В клинических аудиометрах обычно генерируются звуковые
колебания фиксированных частот: 125, 250, 500, 1000, 2000, 4000,
К000 Гц. Переключение частот, как правило, ступенчатое. В простых
проверочных аудиометрах обычно генерируются 3-4 частоты
(например, 500, 1 000, 2 000, 4 000 Гц). В клинических аудиометрах
интенсивность звука в наушниках может быть изменена в пределах
110-120 дБ и 60 дБ при подаче сигнала на кости черепа. В
проверочных аудиометрах диапазон изменений интенсивности звука 0-60 дБ.
6.2.3. Акустические поля
Акустическое поле организма - динамическая система
акустических полей, генерируемых его органами и физиологическими
системами в процессе функционирования, а также при взаимодействии их
с физико-химическими факторами внешней среды. Диапазон собст-
нснного акустического излучения ограничен со стороны длинных волн
механическими колебаниями поверхности тела человека (0,01 Гц), а со
стороны коротких волн ультразвуковым излучением. Низкочастотное
излучение создается физиологическими процессами,
сопровождающимися колебаниями поверхности человеческого тела, и может быть
зарегистрировано контактными или бесконтактными методами. Однако
его практически невозможно зафиксировать с помощью микрофонов
(акустические волны от источников в глубине тела полностью
отражаются от границы раздела «воздух-тело человека»).
В организме существуют следующие источники акустических
иолн (в основном звукового диапазона).
1. Сердце и сердечно-сосудистая система. Акустическое
давление, развиваемое сердцем на поверхности тела, составляет 0,1 Η · м-2
η низкочастотной области звукового диапазона и 10~'-10~2Н · \г2
и высокочастотной.
244
Биофизика
2. Система органов дыхания.
3. Пищеварительный тракт.
4. Суставы опорно-двигательного аппарата.
5. Сердечно-сосудистая система плода.
6. Излучение из уха человека - кохлеарная акустическая
эмиссия {\ОъТи).
7. Неоднородности структуры, на которых возникают
комбинационные звуки, - результат взаимоналожения акустических волн,
генерируемых указанными ранее источниками.
Из уха животных и человека могут излучаться звуки - это
явление называют кохлеарной акустической эмиссией (КАЭ),
поскольку их источник локализован в улитке органа слуха. Эти звуки можно
зарегистрировать микрофоном, расположенном в ушном канале.
Обнаружен ряд видов КАЭ, среди которых выделяют спонтанную
эмиссию и акустическое эхо.
Спонтанная эмиссия - это самопроизвольное непрерывное
излучение звука из уха человека. Уровень интенсивности звукового
давления достигает 20 дБ, что в 10 раз выше порогового значения
2 · Ю-5 Па, которое способно воспринимать ухо человека на частоте
1 кГц. Частоты эмиссии у различных лиц отличаются и лежат в
диапазоне 0,5-5 кГц.
Кохлеарная акустическая эмиссия связана с деятельностью во-
лосковых клеток, расположенных в кортиевом органе улитки. В
ответ на приходящую звуковую волну они изменяют свои размеры
и вызывают во внутреннем ухе механические колебания, которые
способны выходить наружу через среднее ухо. Биофизический
механизм изменения геометрии пока не ясен, его быстродействие в 100
раз выше, чем у мышц. Из всех видов КАЭ применение в медицине
нашло явление акустического эха - излучения звуков из уха спустя
некоторое время после подачи в ухо короткого звукового сигнала.
Оно используется для диагностики слуха новорожденных в первые
несколько дней жизни, когда невозможно использовать обычные
методы аудиометрии. Отсутствие эха является тревожным симптомом
не только глухоты, но и зачастую сопряженных с ней поражений
других отделов нервной системы. Ранняя диагностика позволяет уже
/ пава 6. Сенсорные системы организма
245
с первых дней принять меры и в значительной степени ослабить
неблагоприятные последствия этого недуга.
Акустические поля сердечно-сосудистой системы. Во время
работы сердца в результате ритмических движений клапанного
аппарата, сокращения сердечных мышц и гидравлических факторов внут-
рисердечной динамики возникают колебательные движения,
которые распространяются через окружающие ткани к поверхности
грудной клетки.
В процессе распространения через негомогенную среду с
разными акустическими свойствами существенно меняются
характеристики и параметры исходного звука. Законы распространения звуко-
ных волн от места их генерации до органа восприятия зависят от
свойств источника и окружающей его внешней среды. Одним из
существенных источников акустических колебаний является
турбулентное движение кровяной струи. При этом параметры звука зависят
как от скорости кровотока, так и от неровностей поверхности и
выраженных изменений размеров «камер», через которые струя
протекает. Последние два фактора считают более важными.
Все звуковые явления, связанные с сердечной деятельностью,
с физической точки зрения являются шумами, которые возникают
и определенные моменты времени и различаются
продолжительностью. Звуки, характеризующие работу сердца, в норме представляют
собой короткие по длительности шумы. Патологические шумы
длятся значительно дольше (табл. 6.2).
Так, на нормальной фонокардиограмме различают четыре
гона, которые обозначают I, II, III и IV (рис. 6.23). I и II тоны возни-
IV тон
I тон II тон III тон
Рис. 6.23. Нормальная фонокардиограмма
246
Биофизике
кают в фазе систолы сердца. Они обычно хорошо воспринимаются
при аускультации сердца и называются нормальными
систолическими тонами, III и IV тоны возникают в фазе диастолы и называются
нормальными диастолическими тонами.
Таблица 6.2
Частотный спектр топов различного гепеза
Механизмы генерации тонов сердца
Сокращение и расслабление миокарда желудочков
Вибрация крови при быстром диастолическом наполнении
желудочков
Открытие и закрытие полулунных клапанов
Открытие и закрытие створчатых клапанов
Вибрации сухожильных нитей при их натяжении
f-Гц
М1п
12,5
31,5
80
125
200
Мах
20
63
100
160
400
Тон I возникает в начале систолы желудочков и кончается
к моменту начала изгнания крови. Его условно разделяют на 3 (1, 2,
3) или 5 участков. Первая часть (1) состоит из колебаний низкой
частоты, характеризующих напряжения в миокарде, в стенках
желудочков и створках клапанов. Эти колебания длятся 0,02-0,04 с. Вторая
часть (2) представляет собой высокочастотные колебания и длится
0,05 с. Предполагается, что эти звуки имеют клапанное
происхождение. Третья часть (3) состоит из низкочастотных колебаний
длительностью 0,02-0,04 с. Она имеет сосудистое происхождение.
Интенсивность I тона характеризует сократительные функции миокарда
левого желудочка.
II тон возникает к концу систолы желудочков и длится в
течение их изометрического расслабления. Начальная низкочастотная
часть (1) предшествует закрытию клапанов. Центральная
высокочастотная часть (2) отражает закрытие клапанов аорты и легочной
артерии. Конечная низкочастотная часть (3) отражает открытие
трехстворчатого и митрального клапанов.
III и IV тоны плохо слышны, являются низкочастотными (10-
50) Гц и могут быть зарегистрированы не всегда.
/ iiaea 6. Сенсорные системы организма
247
III тон возникает в начале быстрого пассивного наполнения
желудочков кровью. Он содержит одно или два небольших коле-
Пания.
IV тон возникает в фазе быстрого активного наполнения
желудочков. По параметрам он аналогичен звуку III тона.
Акустическая эмиссия клеточных и молекулярных структур.
Па клеточном уровне обнаружено существование упругих волн
и нервном волокне. Процесс распространения нервного импульса
сопровождается пульсацией поверхности нервного волокна до 30 нм
по амплитуде при частоте следования импульсов 4 Гц. При
прохождении нервного импульса по аксону краба наблюдаются поперечные
упругие волны. Скорость проведения нервных импульсов (1-
100 м · с-1) одного порядка по величине со скоростью упругих
поперечных волн (1-20 м · с1) частотой 2 кГц, распространяющихся
и агаровых стержнях.
Существование клеточных и молекулярных источников
упругих волн предполагается, но еще не доказано экспериментально. Счи-
N, отн. ед.
5
4
з
2
1
5
4
3
2
1
5 10 f, с
б)
Рис. 6.24. Акустическая эмиссия N в диапазоне 30-800 кГц при реакции замещения
FeCU + CuSOj CuCU + FeSO^ (а) и при окислительно-восстановительной реакции
AI + CaS(?4 Ca + AI2(S04)3 (б) (началореакции по истечении 1 с)
_/А^\^
^-
248
Биофизш
6.3. Биофизика зрения
6.3.1. Светопреломляющая система глаза
Органом зрения принято называть глаз. Однако в зрительную
сенсорную систему (зрительный анализатор) вместе с
фоторецепторами (палочками и колбочками) и рядом нейронов, которые
находятся в сетчатой оболочке глаза, входят определенные отделы ЦНС.
Восприятие света есть один из наиболее важных для жизни
факторов, очень нужных для получения информации об
окружающем мире как для защиты, так и для продолжения рода, питания
и других процессов жизнедеятельности. Для анализа окружающего
распределения светового поля почти для всех живых организмои
в ходе эволюции был выработан весьма совершенный вид
рецепции - фоторецепция. Свет является непревзойденным носителем
информации, которая может быть заключена в интенсивности, частоте,
поляризации, фазе, спектральном составе, во временной и
пространственной структурах светового потока. Процесс зрительного
восприятия состоит из 3 этапов:
- образование оптического изображения на сетчатке глаза;
- преобразование оптического изображения в нервные импульсы;
- передача и обработка сигналов зрительными нейронами
центральной части зрительного анализатора.
Глаз включает не только фоторецепторы и нейроны, но также
разнообразные вспомогательные элементы. Они сами не воспринима-
/ ипеа 6. Сенсорные системы организма
249
Рис. 6.25. Схема горизонтального разреза глаза человека
1 -роговица; 2—хрусталик, 3 - стекловидное тело, камерная влага;
4 - сетчатка; 5 - центральная ямка желтого пятна; 6 — зрительный
нерв; 7 - сосудистая оболочка; 8 - склера; ОО'—главная оптическая
ось; КК' - зрительная ось; у и 5° —угол между ОО' и КК'
км свет и не участвуют в переработке (передаче) сигналов от фоторе-
цситоров, но способствуют восприятию света. Важнейшим вспомога-
ιι-льным аппаратом фоторецепции является светопреломляющая
циоптрическая система глаза. Упрощенная схема глаза человека в
разрезе показана на рис. 6.25.
Глазное яблоко представляет собой шарообразное тело
диаметром около 24 мм с довольно прочной внешней оболочкой -
склерой (см. рис. 6.25). Передняя часть склеры (роговица) прозрачна, ее
радиус кривизны около 7,7 мм, показатель преломления света η =
1,376, под роговицей находится передняя камера, которая заполне-
П11 прозрачной жидкостью сл= 1,336. Радужная оболочка
расположена перед хрусталиком и имеет диафрагму (зрачок), диаметр
которой может меняться от 2 до 8 мм. Хрусталик с я = 1,386 представляет
гпбой линзу, радиус кривизны которой может изменяться большим
или меньшим напряжением цилиарной мышцы. Стекловидное тело
или задняя камера имеет η = 1,336. Сопоставление показателей пре-
иомления элементов оптической системы глаза позволяет говорить
к достаточно хорошем их согласовании
250
Биофизик»
Светопреломляющая система глаза фокусирует изображение
окружающего мира на сетчатой оболочке (сетчатке), в которой
расположены фоторецепторы. Диоптрический аппарат глаза подобен
сильной собирающей линзе, и при рассмотрении далеких предметов
сводит в точку на сетчатке входящие в глаз параллельные лучи, т.е,
действует наподобие объектива зрительной трубы Фокусированные
изображения осуществляются за счет преломляющей системы
глаза, к которой относятся: роговица, хрусталик, камерная влага,
стекловидное тело. Преломляя световые лучи, они обеспечивают
формирование на сетчатке действительного уменьшенного и обратного
(перевернутого) изображения рассматриваемого предмета. Отрезок
прямой, проходящий через геометрические центры роговицы и
хрусталика, называют главной оптической осью глаза (00'). Она
объединяет весь его диоптрический аппарат в центрированную
светопреломляющую систему. Центрированной называют такую систему
светопреломляющих поверхностей, геометрические центры которых
лежат на одной прямой.
Глаз представляет собой саморегулирующуюся систему, ее
оптические недостатки в значительной мере компенсируются регуля-
торными механизмами, оптимизирующими работу глаза.
Важнейшими механизмами регулирования являются система фокусировки
изображения на сетчатке и механизм регуляции количества света,
попадающего на сетчатку. Фокусировка осуществляется путем
автоматического изменения радиуса кривизны хрусталика. Этот эффект
называется аккомодацией. Система аккомодации представляет собой
следящую систему, которая удерживает в фокусе изображение
удаляющегося или приближающегося объекта. Управление
осуществляется цилиарной мышцей. Количество света регулируется
изменением размеров оптического отверстия - зрачка, причем освещенность
сетчатки изменяется пропорционально квадрату диаметра зрачка.
Диаметр зрачка регулируется двумя мышцами-антагонистами:
кольцевая (сфинктер) сужает зрачок, а радиальная (дилататор) его
расширяет. Системы работают по принципу получения сигнала ошибки
между формируемым сигналом и некоторым эталонным (требуемым
или оптимальным). Система регуляции состоит из регулятора и
объекта управления и содержит контур отрицательной обратной связи.
/ пава 6. Сенсорные, системы организма
251
Моделью светопреломляющего аппарата органа зрения служит
гак называемый приведенный редуцированный глаз. Редукция
означает упрощение, т.е. сведение сложного к простому, более доступному
для анализа. Физическим аналогом приведенного редуцированного
глаза является стеклянная линза, которая одной поверхностью
контактирует с воздухом, а другой - с жидкостью, обладающей η =
1,336. Первая поверхность линзы направлена в пространство
предметов, а другая - к пространству изображений.
Диоптрический аппарат глаза не поддается точному
математическому описанию. Это происходит из-за того, что глаза разных
людей сильно различаются, а также из-за подвижности хрусталика и по
ряду других причин. Так, например, главная оптическая ось глаза
человека проводится довольно приблизительно. Она не совпадает со
зрительной осью КК', которая проходит через геометрические
центры роговицы и желтого пятна сетчатки. В направлении зрительной
оси глаз имеет наилучшую разрешающую способность. Угол между
главной оптической осью 00' и зрительной осью КК', как правило,
не превышает 5 градусов. Он обозначается γ и обычно учитывается
и офтальмологии при назначении очков.
Итак, человеческий глаз можно рассматривать в качестве
центрированной преломляющей системы весьма условно. На рис. 6.26
0,25
Рис. 6.26. Глаз как центрированная светопреломляющая система
Fi и F2- передний и задний фокусы соответственно, Ht и Н2- передняя
и задняя главные точки, NtuN2- передняя и задняя узловые точки,
/, - расстояние от передней главной точки Н, до переднего фокуса F/,
/2 - расстояние от задней главной точки Н2 до заднего фокуса F2
252
Биофизика
приведена количественная характеристика одной из наиболее
адекватных физических моделей человеческого глаза.
Из рис. 6.26 видно, что расстояние между //, и Н2 составляет
примерно 0,25 мм. Столь близкое расстояние допускает замену
двух главных плоскостей (//, и Н2) на одну главную плоскость.
Точки N{ и N2 практически также совпадают в единую узловую
точку глаза, ее называют оптическим центром глаза (между JV,
и N2). В замене каждой из названных пар кардинальных точек на
единые угловую и узловую точки и состоит смысл редукции
центрированной диоптрической системы реального глаза. В схеме
Гульстранда единая плоскость Η и единая узловая точка N
находятся на расстоянии 1,6 мм и 7,5 мм соответственно от передней
поверхности роговицы. Все расстояния, кроме фокусных (fi,f2), от-
считываются от передней поверхности роговицы. По ходу лучей
внутрь глаза эти расстояния имеют знак «+». В противоположном
(наружном) направлении расстояния имеют знак «-». Числа,
характеризующие расстояние единых главной и узловой точек глаза,
необходимы для расчета диоптрического аппарата человеческого
глаза. За эту работу Альвар Гульстранд был удостоен Нобелевской
премии в 1911 г.
6.3.2. Биофизический механизм
восприятия света рецепторами
Свет, попавший в глаз, фокусируется при помощи хрусталика
на светочувствительных клетках сетчатки - палочках и колбочках.
У человека в сетчатке имеется около 6-7 млн колбочек и 11
ΟΙ 25 млн палочек. Палочки и колбочки распределены в сетчатке
неравномерно. Центральная ямка сетчатки содержит только колбочки
(до 140 тыс. на 1 мм2). По направлению к периферии сетчатки
количество колбочек уменьшается, а палочек соответственно возрастает.
Периферическая часть сетчатки содержит только палочки. Участок
сетчатки глаза, где сосредоточены только колбочки, называется
желтым пятном. Место выхода зрительного нерва из глазного яблока -
диск зрительного нерва - совсем не содержит фоторецепторов. Это
так называемое слепое пятно (рис. 6.27).
/ 1ща 6. Сенсорные системы организма
253
полностью транс-ретинапь
Рис. 6.27. Структура органа зрения
1 -разрез глаза; 2 - колбочки; 3 - палочки (М - скопление митохондрий);
4 - диск наружного сегмента палочки; 5 - фрагмент мембраны диска
со встроенной в нее молекулой родопсина; 6 - хромофорная группировка
родопсина -ретиналъ: а - 11-цис-ретиналь. б- полностью транс-ретиналь
Колбочки обеспечивают дневное зрение и восприятие цвета,
палочки - сумеречное, ночное зрение.
Строение сетчатки человека и позвоночных животных может
показаться парадоксальным, так как светочувствительные клетки
находятся в заднем слое сетчатки. Прежде чем свет попадет на них,
он должен пройти через несколько слоев различных клеток
сетчатки. Кроме того, сами палочки и колбочки ориентированы к свету
своим внутренним сегментом, не содержащим зрительного
пигмента (рис. 6.28). Однако такая организация не снижает существенно
чувствительности глаза к свету, так как клетки прозрачны для
видимого света.
254
Биофизика
Абсолютный порог чувствительности зрения человека (т.е.
минимальная еще обнаруживаемая величина яркости светового
пятна при отсутствии светового фона и в условиях полной темновой
адаптации) равен 2,1-5,7 · 10~17 Дж на поверхности роговицы глаза,
что соответствует 58-148 квантам сине-зеленого света. Из них
только 5-15 квантов поглощаются молекулами зрительного пигмента
в палочках сетчатки, а остальные «теряются». По данным некоторых
исследователей, например Пинегина, для того чтобы вызвать
ощущение света у человека, достаточно, чтобы фоторецепторов достигли 2
фотона. Отдельная палочка возбуждается всего одним квантом
света - в связи с этим СИ. Вавилов утверждал, что глаз в своей
чувствительности к свету «достиг физического предела».
Диаметр палочки составляет примерно 2 мкм, а длина -
примерно 50-60 мкм. Длина наружного сегмента (НС) лежит в пределах
примерно 20-30 мкм. Вся цитоплазма НС заполнена своеобразными
органеллами, которые называют дисками. Промежутки между ними
не превышают 15 нм. В НС каждой палочки содержится от 700 до
1000 дисков, которые напоминают собой сплющенные воздушные
шары, сложенные в стопку. Диаметр каждого диска почти
соответствует диаметру наружного слоя (~ 2 мкм), а толщина диска составляет
15-18 нм. Его внутреннее пространство представляет собой узкую
щель шириной около 1 нм. Эта щель расширяется до нескольких
нанометров у краев диска. Таким образом, на внутридисковое
содержимое вместе с цитозолем приходится не более половины объема НС,
а другую половину занимают мембраны дисков. Следовательно,
наружный сегмент буквально нафарширован мембранами, и именно
в них осуществляется первичный процесс восприятия света. Поэтому
мембрану дисков называют фоторецепторной мембраной.
Молекулярная организация фоторецепторной мембраны у
позвоночных и беспозвоночных животных практически одинакова.
Подобно всем биомембранам, она состоит из липидов и белков
(протеинов). Вместе с тем ей присущи две кардинальные особенности.
Во-первых, ее основным белковым компонентом является
зрительный пигмент - хромогликопротеид родопсин. Его концентрация
достигает 90% общего содержания протеинов в фоторецепторной
мембране. Во-вторых, более 40% жиро кислотного состава фосфолипидов,
I пава 6. Сенсорные системы организма
255
Падающий свет
Рис. 6.28. Строение сетчатки
образующих её, приходится на полиненасыщенную докозагексаено-
вую кислоту, в составе которой 22 атома углерода. Среди связей
между ними 6 двойных. Кроме нее, в состав мембранных фосфолипидов
нходят и другие полиненасыщенные длинноцепочечные жирные
кислоты, содержащие от 1 до 4 двойных связей. Вместе они составляют
более половины всех жирнокислотных остатков. Высокая степень
ненасыщенное™ жирнокислотных «хвостов» фосфолипидных молекул
придает гидрофобному бимолекулярному каркасу фоторецепторной
мембраны необычайно высокую подвижность, вследствие чего
данная мембрана обладает малой вязкостью по сравнению с другими
мембранами.
256
Биофизик!
Первичный механизм возбуждения палочки светом связан со
сложными превращениями родопсина в фоторецепторной мембране.
Родопсин - это высокомолекулярное соединение, состоящее из двух
основных компонентов: альдегида витамина а (ретиналя) и липопро·
теина под названием опсин (белок глаза). Спектр поглощения
родопсина показан на рис. 6.29.
Спектр поглощения родопсина хорошо приспособлен к
восприятию света в достаточно широкой области солнечного излучения, но
лучше всего этот зрительный пигмент поглощает видимый свет в
зеленой части этого спектра (500 нм). Красные и синие лучи он
отражает, причем в отраженном свете преобладает красный компонент, хотя
присутствует и синий. Поэтому пигмент имеет пурпурный цвет,
определивший и его название. Родопсин относится к наиболее интенсивно
окрашенным соединениям из всех известных в настоящее время в
органической химиии. Степень поглощения фотонов родопсином
близка к теоретически возможному значению.
В темноте ретиналь пребывает в 11-цис-форме, для которой
характерна некоторая скрученность молекулы. Скрученность
объясняется тем, что в планарной форме между близко расположенными
метильной группой ν углерода (С 13) и водородом у углерода (СЮ)
возникает сильное отталкивание. Сила отталкивания не допускает
плоского расположения π-связи. Скрученность в молекуле 11-цис-ретиналя
обеспечивает наибольшую энергию взаимодействия с опсином,
вследствие чего комплекс ретиналя и опсина весьма устойчив. Для
D, отн. ед.
10
8
6
4
2
0
250 350 450 550 λ, нм
Рис 6.29. Спектр поглощения родопсина
По оси абсцисс - длина волны (к); по оси ординат - оптическая плотность (D)
/ пава 6. Сенсорные системы организма
257
по разрушения необходимо преобразовать 11-цис-ретиналь в
другой изомер: в так называемый полностью транс-ретиналь. Такое
преобразование из цис- в транс-ретиналь происходит под дейст-
II нем света. Этот процесс называется фотоизомеризацией
(рис. 6.27, а, б).
При поглощении фотона (Αν) молекула родопсина претерпева-
ν ι электронный переход «π -» π*», при этом молекула родопсина
оказывается в синглетном возбужденном состоянии. Необходимо
отмстить, что в органических молекулах π-орбитали, которые свойст-
испмы основному энергетическому состоянию, являются
связывающими, т.е. они стремятся к перекрытию и образованию двойной
пики. Напротив, π'-орбитали, характерные для возбужденного
состояния молекулы, относятся к разрыхляющим орбиталям. Их
перекрытие энергетически невыгодно. В силу этого части молекулы,
соединенные двойной связью с участием π'-орбитали, разворачиваются
па 90°, и двойная связь превращается в ординарную. Поэтому в
молекуле ретиналя, возбужденной светом, происходит разворот атомов
иокруг двойной связи. Такое изменепние пространственного распо-
ножения атомов, входящих в состав ретиналя, требует затрат
энергии. Эти энергозатраты характеризуются преодолением
потенциального барьера. Для разворота атома вокруг двойной связи в 11-
цис-рстипале достаточно энергии одного фотона. Для фотоизомери-
|ации достаточно и времени, в течение которого молекула родопсина
пребывает в сигпетном возбужденном состоянии (»10~9 с), поскольку
цис-тран-изомеризация ретиналя происходит всего за 0,1-0,3 пс,
киантовый выход фотоизомеризации 11-цис-региналя в родопсине
достигает 0,7.
Фотоизомеризация - первая и единственная фотохимическая
реакция в зрительном акте. Она протекает внутри отдельной
молекулы родопсина и, в отличие от фотосинтеза, не сопровождается
никакими химическими реакциями. Зрение отличается от фотосинтеза,
который происходит в растениях, еще и тем, что между молекулами
родопсина невозможен перенос энергии, так как в фоторецепторной
мембране молекулы родопсина удалены друг от друга на расстояние
около 7 нм. Непосредственным следствием фотоизомеризации, при-
258
Биофизика
водящей к появлению у человека ощущения света, служат конформа-
ционные перестройки молекулы родопсина. Дело в том, что
пространственно 11-цис-ретиналь точно соответствует конфигурации
активного центра на опсине. При образовании транс-ретиналя это
соответствие (комплементарность) нарушается, в результате чего рети-
наль отщепляется от опсина и родопсин обесцвечивается, т.е.
претерпевает фотолиз.
Распавшийся родопсин восстанавливается, когда на него
прекращает действовать свет. В ходе восстановления транс-ретиналь
преобразуется в цис-форму. Такой переход, обратный фотоизомеризации,
называется реизомеризацией. У позвоночных животных
транс-ретиналь, отщепившись от опсина, поступает в пигментный эпителий
сетчатки, где подвергается ферментативным превращениям в 11-
цис-ретиналь, который возвращается из пигментного эпителия к
фоторецепторам. Каждая палочка ежесуточно синтезирует (8-9) · 105
молекул родопсина из поступающего в нее 11-цис-ретиналя, который
соединяется с опсином. Суточный синтез этого зрительного пигмента
в целом оценивается величиной примерно ~ 2 · 10й молекул.
Конформационная перестройка родопсина, называющаяся
фотоизомеризацией ретиналя, довольно существенна. Естественно, что
перестройку сопровождают значительные изменения
физико-химических свойств фоторецепторной мембраны, в которую встроен
родопсин. Одним из важных последствий этих изменений служит сдвиг
мембранного потенциала фоторецепторной клетки.
6.3.3. Рецепторные потенциалы
Сдвиг мембранного потенциала фоторецепторной клетки,
который принято называть рецепторным потенциалом, происходит на
ее плазмолемме, тогда как фотоизомеризация родопсина
совершается преимущественно во внутриклеточных мембранах, т.е. в
мембранных дисках, находящихся в наружном сегменте.
Свойства фоторецепторных мембран изменяются
одновременно с конформационной перестройкой в них опсина. Через 1,5-2 мс
после поглощения кванта света наблюдается гиперполяризация плаз-
молеммы фоторецепторной клетки. Следовательно, в палочке (и кол-
Глава 6. Сенсорные системы организма
259
Почке) существует механизм передачи сигнала с фоторецепторных
мембран дисков на плазмолемму. В передаче участвует так
называемый внутриклеточный посредник. Первоначальная гипотеза
относительно природы этого посредника отводила его роль ионам Са.
Согласно «кальциевой гипотезе» в ответ на действие света ионы Са
ныбрасываются изнутри фоторецепторных дисков в цитоплазму
наружного сегмента, диффундируют к плазматической мембране
и блокируют в ней Ка+-каналы. В темноте же Са2+ активно
«закачивается» из цитоплазмы снова внутрь дисков. Однако опыты Фесенко
(1985 г.) показали, что проводимость наружной мембраны палочки
не изменяется под действием Са2+, который, следовательно, не
является клеточным медиатором в процессе зрительной рецепции. В то
же время оказалось, что циклический нуклеотид цГМФ (3', 5' -
циклический гуанозин-монофосфат) способен непосредственно откры-
нать натриевые каналы мембраны палочек. В темно-адаптированном
рецепторе концентрация цГМФ в цитоплазме наружного сегмента
иесьма велика, поэтому очевидно, что циклический нуклеотид цГМФ
и темноте поддерживает Ка+-каналы в открытом состоянии. Если
происходит гидролиз цГМФ (под действием фермента фосфодиэсте-
разы ФДЕ), то концентрация цГМФ резко падает, а натриевые
каналы переходят в закрытое состояние.
Именно на этом основана роль цГМФ как клеточного
медиатора. Изменение его концентрации в наружном сегменте регулирует
еостояние Ыа+-каналов внешней мембраны и, как следствие,
величину трансмембранного электрического потенциала палочки. Между
еветовым возбуждением родопсина и гидролизом цГМФ фосфодиэ-
егеразой существует сложная многоступенчатая функциональная
евязь, природа которой установлена в конце 30-х годов благодаря
работам целого ряда лабораторий. Было показано, что на свету в
наружном сегменте действительно резко уменьшается уровень цГМФ
при одновременной активации нескольких сот молекул фермента
ФДЕ. Для этого требуется присутствие также ГТФ (гуанозинтрифос-
фат), как источника энергии. Фактором, связывающим ГТФ в
клетке, является особый G-белок транедуцин Т, ответственный за акти-
нацию ФДЕ. Молекула транедуцина состоит из трех субъединиц а,
|1, γ (Та, β, γ).
260
Биофизика
Трансдуцин может также связывать ГДФ (гуанозиндифосфат),
но в этом состоянии он не способен активировать ФДЕ. Переход
трансдуцина в активное состояние из неактивного происходит при
обмене ГДФ на ГТФ, а именно
Т-ГДФ + ГТФ -> Т-ГТФ + ГДФ.
В образованном комплексе Τα, β, γ -ГТФ отщепляется
активная α - субъединица трансдуцина Та и связанная с ней молекула
ГТФ. Отделенный от Τα, β, γ комплекс Т-ГТФ в свою очередь
активирует фермент ФДЕ. Активация ФДЕ также основана на разной
функциональной роли субъединиц α, β, γ, входящих в состав
молекулы фермента, из которых γ-субъединица подавляет активность ФДЕ.
Именно отщепление γ-субъединицы, ингибирующей фермент,
приводит его в активное состояние.
Сложная последовательность реакций запускается при
фотоизомеризации ретиналя в родопсине и переводе родопсина R в
активированное состояние R*. Активированный родопсин мигрирует ла-
терально в фоторецепторной мембране, обладающей крайне низкой
вязкостью, и образует комплекс с трансдуцином
R* + Т-ГДФ -> R* - Т-ГДФ,
в которой затем ГДФ замещается на ГТФ
R* - та> р, т-гдф + гтф -> r* - та, Ρι υ-γτφ + гдф,
с последующей его диссоциацией
R* - Τα, ρ, γ-ΓΤΦ -> R* + Тр> γ + Τα-ΓΤΦ.
Фрагмент Т-ГТФ активирует непосредственно ФДЕ,
«отбирая» у нее γ-субъединицу
Та-ГДФ + ФДЕ(а, β, γ) -> ФДЕ(а, β) + Та-ГДФ-ФДЕу.
Последовательность этих событий представлена на рис. 6.30.
I пава 6. Сенсорные системы организма
261
Рис. 6.30. Каскад реакций, лежащих в основе зрительной рецепции
I - поглощение кванта света активирует родопсин R*, который затем реагирует
с белком-трансдуцином Т; II - происходит замена ГДФ на ГТФ в а-субъединице
Та трансдуцина; III - отщепленная Та-ГТФ активирует фосфодиэстеразу ФДЕ,
отщепляя ее у-субъединицу; IV- активированная ФДЕ гидролизует тысячи
молекул цГМФ. Через некоторое время ГТФ в а-субъединице Та расщепляется
и превращается в ГДФ, Та-ГДФ присоединяется к Γβγ. Трансдуцин и ФДЕ
восстанавливаются. Родопсин R* инактивируется и регенерирует в исходное состояние
Центральное место в этом каскаде реакций занимает стадия IV
(рис. 6.30), где возникает эффект многократного усиления действия
кванта света за счет того, что одна активная молекула ФДЕ (α, β)
гидролизует тысячи молекул цГМФ.
Регенерация активированного родопсина R* осуществляется
и ходе последовательных реакций его фосфорилирования родоп-
син-киназой РК с образованием R-πΦ, где π - число фосфатных
ipynn (п и 9). Я*-пФ связывается с белком S, образуя комплекс R*-
иФ · S. В этом состоянии уже не происходит дальнейшая активация
("/-белка трансдуцина родопсином, который релаксирует из активи-
262
Биофизика
рованного состояния R*-nOS —> R°-nOS. Окончательная репарация
родопсина осуществляется через ~ 30 минут после его дефосфорили-
рования (фосфатазой) и присоединения новой молекулы 11-цис-ре-
тиналя взамен покинувшего родопсин транс-ретиналя.
Вследствие конформационной перестройки молекул
родопсина, в фоторецепторной мембране изменяется содержание
внутриклеточного посредника в цитозоле наружного сегмента
фоторецепторной клетки. Это приводит к закрытию 100-300 натриевых каналов
в плазмолемме наружного сегмента, которые в отсутствие светового
раздражителя пребывали в открытом состоянии. Они обладают
свойствами потениалнезависимых ионных каналов и, будучи
управляемыми светом, получили название фотозависимых натриевых
каналов. Когда закрываются 100-300 каналов, электропроводность
плазмолеммы наружного сегмента падает на 1-3%. Падение
мембранной электропроводности приводит к гиперполяризации
фоторецепторной клетки. Принято считать, что потенциал покоя
фоторецепторной клетки генерируется в темноте на плазмолемме внутреннего
сегмента. В темноте потенциал покоя палочки (колбочки) составляет
примерно от -20 до -40 мВ, хотя по расчету он по модулю должен
быть примерно вдвое больше.
Причина расхождений расчетной и измеряемой величин
заключается в том, что в системе генерации рецепторного потенциала
плазмолемма наружного сегмента выполняет роль шунта с
переменной электропроводностью. Электропроводность регулируется
(управляется) светом при помощи фотозависимых натриевых
каналов. Когда все натриевые каналы плазматической мембраны
наружного сегмента одной палочки оказываются заблокированными
светом, удельное поверхностное сопротивление этой мембраны
достигает величины =106Ом/см2. В темноте ионный ток, входящий
через каналы в цитоплазму, довольно значителен и составляет
примерно 40 пА, а при освещении сетчатки он блокируется, при этом
степень его ослабления зависит от интенсивности света. Чем слабее
натриевый ток через плазматическую мембрану наружного сегмента,
тем больше выражена гиперполяризация в фоторецепторной клетке.
Изменение потенциала палочки или колбочки (U) во времени
показано на рис. 6.31.
Глава б. Сенсорные системы организма
263
U.mB
/7V
\
3,1 0.2Л, 0,3 0,4
0,5 0,6
Рис 6.31. Рецепторные потенциалы одиночной палочки,
возникающие в ответ на короткие (10 мс) вспышки света при трех
разных интенсивностях света: I,, h, h - интенсивность света;
чем больше I, тем больше гиперполяризация, I, >12 >1з
При поглощении одного фотона (Αν) гиперполяризация
фоторецепторов разных представителей животного мира неодинакова.
Так, в палочках сетчатки позвоночных животных гиперполяризация
составляет десятки и сотни мкВ, тогда как в фоторецепторных
клетках членистоногих она достигает 10 мВ. Приведенные числа
подтверждают положение об усилении сигнала в рецепторных
аппаратах. Действительно, квант света, несущий энергию = 4 · 10~19 Дж,
вызывает фотолиз только одной молекулы родопсина, что, в свою
очередь, приводит к закрытию в плазмолемме фоторецепторной
клетки позвоночных животных 100-300 натриевых каналов. Это
приводит к образованию рецепторного потенциала, энергия которого на
3 порядка превосходит энергию вызвавшего его фотона.
Амплитуда рецепторного потенциала (РП) возрастает при
повышении интенсивности света, падающего на сетчатку, причем
в пределах 1 000-кратного изменения интенсивности эта зависимость
имеет логарифмический характер:
MJ
РП
= *-lg-
(6.18)
264
Биофизика
где к - коэффициент пропорциональности; /„ - интенсивность
последующего воздействия света; /„_, - интенсивность предыдущего
воздействия света.
При ярком освещении амплитуда рецепторного потенциала
может достигать 25 мВ. Величина РП зависит также и от длины
волны света. Фоторецепторные клетки в ответ па зеленый свет (λ -
= 500 нм) генерируют наибольший РП. Среди колбочек выделено 3
типа: 1) дает максимальный ответ РП на синий цвет; 2) дает
максимальный ответ РП на зеленый цвет; 3) дает максимальный ответ РП
на красный цвет.
Использование микроэлектронной техники для изучения
реакции фоторецепторов па облучение светом позволило обнаружить
деполяризацию фоторецепторов, при этом деполяризация предшествует
гиперполяризации. Амплитуда деполяризации очень мала и
составляет примерно 0,1 мкВ. Учитывая временные соотношения де- и
гиперполяризации, первую из них стали называть ранним, а вторую -
поздним РП и обозначили соответственно, РРП и ПРП. Однако позднее
установили, что РРП обусловлен перемещениями электрических
диполей в мембранах дисков наружного сегмента. Эти перемещения
диполей связаны с фотолизом молекул родопсина. Другими словами,
РРП обусловлен током смещения в фоторецепторных мембранах и он
не служит промежуточным звеном передачи сигналов к синаптиче-
ской зоне фоторецептора при действии света. В отличие от других ре-
цепторных аппаратов, где таким звеном служат деполяризационные
сдвиги мембранного потенциала, в фоторецепторах эту функцию
выполняет гиперполяризация. Таким образом, только поздний РП
является истинным рецепторным потенциалом (в полном значении этого
термина).
РП, возникнув в наружном сегменте, распространяется к синап-
тической зоне палочки или колбочки по закону электротонической
передачи (с декрементом). При этом постоянная длины плазмолеммы
фоторецепторной клетки, по которой «движется» с затуханием волна
гиперполяризации, достигает 10 мкм. Достигнув синаптической зоны,
гиперполяризация изменяет свойства ее мембраны и инициирует ме-
диатор1гую систему, обеспечивая передачу сигнала к биполярному
нейрону. Химическая природа медиатора, выделяемого фоторецеп-
/ пава 6. Сенсорные системы организма
265
горной клеткой, пока еще неизвестна точно, однако наиболее
вероятными кандидатами на эту роль признаются глутамат и аспартат. Обе
аминокислоты обладают деполяризующим действием на постсинап-
шческую мембрану, но при облучении сетчатки светом биполярные
нейроны испытывают гипреполяризацгао. Принято считать, что фото-
рсцепторной клетке в темноте свойствен определенный темп
выделения деполяризующего медиатора (глутамата или аспартата), а гипер-
иоляризация синаптической зоны при освещении замедляет темп
иыброса медиатора, вследствие чего постсинаптические мембраны,
принадлежащие биполярным нейронам, не деполяризуются, а гипер-
I юляризуются. Гиперполяризационный сдвиг мембранного
потенциала биполярного нейрона электротонически распространяется по его
илазмолемме к синапсу на теле третьего нейрона, который также
находится в сетчатке. Медиатор, поступающий от биполярного нейрона
к ганглиозной (мультиполярной) клетке, вызывает ее деполяризацию.
Деполяризационный сдвиг потенциала распределяется
электротонически с тела по аксонам ганглиозной клетки. Плазматическая
мембрана аксона ганглиозной клетки (аксолемма) обладает возбудимостью,
благодаря чему там формируется потенициал действия, когда
деполяризация участка аксолеммы, в котором есть потениалзависимые
ионные каналы, достигает порогового уровня. Потенциалы действия,
неся информацию от фоторецепторов, поступают в центральную
нервную систему по волокнам зрительного нерва. Эти волокна
представляют собой аксоны ганглиозных клеток сетчатки. Схематически
передача поздних рецепторных потенциалов по сетчатке показана на
рис. 6.32.
Зрительный нерв включает примерно 106 волокон. По этим
волокнам в мозг направляются сигналы от 1,3*108 фоторецепторов.
Следовательно, в среднем один аксон ганглиозной клетки собирает
информацию от множества рецепторных клеток. Все
фоторецепторы, посылающие сигналы к одной ганглиозной клетке, составляют ее
рецептивное поле. Рецептивное поле, образованное палочками,
содержит гораздо больше фоторецепторов, чем колбочковое рецептор-
пое поле. Каждая колбочка, находящаяся в центральной ямке
желтого пятна сетчатки, связана только с одной ганглиозной клеткой.
Другими словами, рецептивное поле ганглиозной (мультиполярной)
266
Биофизика
/S\
Рецептор Щ
(полочка)
_РП
(гиперполяризация)
Электротоническое
распространение
Синаптические
потенциалы
(гиперполяризация)
Биполярный
нейрон
Электротоническое
распространение
Синаптические
потенциалы
(деполяризация)
Рис. 6.32. Схема передачи фоторецепторного сигнала
по афферентному пути в пределах сетчатки
РП—рецепторный потенциал; ПД—потенциал действия
клетки включает только одну колбочку. С этим связана наилучшая
разрешающая способность центрального колбочкового зрения.
Другие ганглиозные клетки получают сигналы больше, чем от одного
фоторецептора. Существуют такие ганглиозные клетки, каждая из
которых собирает сигналы от сотен и даже тысяч палочек.
Следовательно, рецептивные поля сетчатки весьма разнообразны.
6.3.4. Чувствительность и адаптация глаза
Чувствительностью (Е) глаза называют величину, обратно
пропорциональную пороговому стимулу:
Я =!//„,
(6.19)
Глава 6. Сансорныа системы организма
267
где /„ - пороговая сила света.
Чтобы пороговая сила света была минимальной, т.е. соответст-
новала своему определению, используют мелькающий свет. После
длительного пребывания в темноте (темновой адаптации) глаз
человека способен реагировать на единицы квантов света, что говорит об
очень высокой чувствительности. Чувствительность уменьшается
при увеличении освещенности. Минимальную чувствительность, на
которую реагирует глаз, можно рассмотреть на примере наблюдения
звезд. Звезды относятся к так называемым точечным источникам
света, которые видны под очень малыми углами (малые доли
секунды). Когда наблюдатель смотрит на звезду, то площадь сетчатки,
освещаемая падающим пучком света, определяется не угловым
размером источника, а в основном дифракцией света на зрачке и оказы-
нается того же порядка, что и площадь, занимаемая одним
фоторецептором. Световой поток, который падает на эту площадь,
определяется освещенностью, создаваемой звездой на зрачке
наблюдателя. Эту освещенность принято называть блеском звезды.
Минимальная освещенность на зрачке (пороговый блеск), которую может
тметить наблюдатель, зависит от ряда факторов, включая условия
наблюдения, например яркость фона, а также от того, известно или
ист местоположение источника света (в последнем случае пороговый
блеск выше). В случае полной темноты пороговый блеск составляет
примерно 1 нлк. Такой слабый источник света может быть замечен
только периферическим зрением, т.е. палочковым аппаратом.
Ночное небо не является совершенно черным фоном, поскольку в ясную
безлунную ночь суммарная освещенность горизонтальной
поверхности от звезд и неба составляет около 0,3 млк. Самая неяркая звезда,
которую глаз замечает на этом фоне (звезда шестой величины),
создает блеск около 10 нлк, а яркие звезды (первой величины) - 1 млк.
При освещенности до 3 лк работает палочковый аппарат, затем
до 30 лк работают и палочковый, и колбочковый аппараты, а свет
и 30 лк и более воспринимают колбочки. Учитывая, что при высоком
стоянии и безоблачном небе солнечные лучи создают освещенность
на плоскости, перпендикулярной к их падению, около 100 клк,
можно оценить реальный динамический диапазон воспринимаемых гла-
юм освещенностей. Он составляет 10V10-9 = 1014. Это весьма сущест-
268
Биофизика
венная величина, которая еще не достигнута в лучших технических
устройствах.
Воспринимать такой диапазон входных сигналов (освещенно-
стей) можно только с помощью системы адаптации - работы
соответствующих регуляторных механизмов подстройки физиологических,
биохимических, психологических и других факторов. Процессы тем-
новой и световой адаптации можно объяснить, рассматривая
фотохимические реакции родопсина. Покажем, что при темновой адаптации
чувствительность глаза возрастает. Предположим, что глаз
переходит от света к темноте, например, из освещенной комнаты в темную,
и скорость изменения концентрации родопсина пропорциональна
разности между его максимально возможной и текущей
концентрациями: dCldt - a(C0 - С), где С - концентрация родопсина в сетчатке
глаза в текущий момент времени t; C0 - максимально возможная кон-
.центрация родопсина; a - коэффициент пропорциональности.
Учитывая, что при t —> оо, т.е. при длительном нахождении
в темноте, С = С0, а при t = 0 концентрация родопсина С,
определяется освещенностью, т.е. С= С,, решение дифференциального
уравнения находим в виде:
С = С0{ 1 - [(С0 - С,)/С0]ехр(-а0}· (6.20)
Скорость разложения родопсина, которую можно определить
как производную по времени от количества продуктов распада
dCp/dt, будет пропорциональна силе света /, падающего на сетчатку
глаза, и коэффициенту поглощения света α (коэффициент
поглощения света определяется как отношение поглощенного светового
потока к падающему). Это соотношение будет справедливо для весьма
коротких интервалов времени, сразу после попадания света па
сетчатку, а затем начинается процесс восстановления родопсина,
поэтому можно написать:
dCpldt = ki alC -k2Cp, (6.21)
где k{ и 1^ - соответственно константы скорости реакции
разложения родопсина и его восстановления из продуктов распада.
Глава 6. Сансорныа системы организма
269
Условием стационарности будет равенство нулю выражения
(6.21); откуда найдем Ср = k^aJCIk^.
Последнее соотношение применимо и для порогового стимула.
Если обозначить через Сп концентрацию продуктов распада
родопсина, соответствующую пороговой стимуляции, т.е. при / = /п, то
можно написать, что Сп = k^oJJZIk^.
Учитывая выражения (6.19), (6.20) и последнюю формулу,
а также введя обозначение (С0 - Су)/С0 = А, найдем: Ε = С0(1 -А)
схр (-at) kyaljkj.
Максимальная чувствительность глаза Е0 будет при t -> оо, т.е.
при длительной адаптации, поэтому из последнего выражения
получим:
Ео = С0 kioJn/k2, (6.22)
или Ε = £0(1 - A)e\p(-at).
Световую адаптацию (когда глаз переходит от темноты к
свету) тоже можно рассмотреть на основе фотохимических процессов.
Принимая во внимание, что текущая концентрация родопсина (С)
определяется разностью между его максимально возможной
концентрацией (С0) и концентрацией продуктов распада (Ср), т.е. С = С0-
- Ср, и учитывая выражение (6.21), можно написать dCJdt = &,a/(C0 -
- Ср) - kjCp откуда dCJdt + (kpJ + к2)Ср - £,a/ C0 = 0.
С учетом того, что при Ср = 0 t = 0, после интегрирования
получим:
Ср = [A,a/Co/(Aia/ + к2)] {1 - ехр[- (jfc,a/ + k2)t]}. (6.23)
Изменение чувствительности от времени в процессе световой
плантации можно найти, если воспользоваться определением (6.19),
соотношением С = С0 - Ср и выражением (6.22), с учетом того, что
i\pnE = E01= 0:
βχρΚ*,α/ + Α2)]. (6.24)
д. *■*' +
{kxal + k2)
E0kxal
(λ, a/ + k2)
270
Биофизика
Рис 6.33. Изменение чувствительности глаза при адаптации:
слева - темповой, справа - световой
Обе полученные зависимости увеличения или уменьшения
чувствительности глаза соответственно при темновой и световой адапта-
циях показаны на рис. 6.33. Кривые имеют экспоненциальный
характер и приведены для двух значений силы света: кривая 1 соответствует
меньшей силе света по сравнению с кривой 2. Следует отметить, что
темновая адаптация протекает существенно медленнее световой.
Спектральная чувствительность колбочек разных типов
позволяет человеку ощущать цвет. Глаз по-разному воспринимает
излучения разных длин волн: коротковолновые излучения соответствуют
фиолетовым цветам, а длинноволновые - красным. Плавные
переходы и примерные спектральные границы представлены в табл. 6.3.
Таблица 6.3
Длины волн монохроматического излучения
Цвет
Фиолетовый
Синий
Сине-зеленый
Зеленый
Длины волн
монохроматического
излучения, нм
370-455
455-485
485-505
505-550
Цвет
Желто-зеленый
Желтый
Оранжевый
Красный
Длины волн
монохроматического
излучения, нм
550-575
575-587
587-610
610-780
Излучения, равные по мощности, но с разной длиной волны,
совершенно различны по своему световому действию. Можно устано-
Глава 6. Сенсорные системы организма 271
N, отн. ед.)
ν,(λ)
0,6
0,6
0,4
0,2
400 500 600 λ, нм
Рис. 6.34. Кривые видности стандартного наблюдателя МКО:
I - фотопического зрения, 2 - скотопического зрения
вить некоторую среднюю кривую, которая характеризует
чувствительность глаза человека. Международная комиссия по освещению
(МКО) в 1924 г. приняла стандартную кривую относительной
видности (эта кривая еще называется кривой спектральной
чувствительности стандартного наблюдателя МКО), которая показана на рис. 6.34.
Под V(k) понимается зависимость от длины волны λ ощущаемой
яркости монохроматического света при постоянной энергии излучения,
или, что то же самое, V(k) есть величина, обратная энергии,
воздействующей на глаз на разных λ при ощущении постоянной яркости
светового потока.
Максимум кривой спектральной чувствительности зрения,
адаптированного к дневной яркости, или, как принято называть,
фотопического зрения (кривая 1 на рис. 6.34), соответствует 554 нм; 0,5
ширины спектра лежит в пределах от 510 до 610 нм.
Таким образом, ширина фильтра спектральной
чувствительности составляет 100 нм. Для палочкового аппарата, который реагирует
на сумеречное (или скотопическое) зрение (кривая 2 на рис. 6.34),
максимум спектральной чувствительности соответствует 507 нм,
а 0,5 спектральной характеристики лежит в диапазоне от 455 до 550
нм. Максимумы по абсолютному значению разные и отличаются
приблизительно в 100 раз, а спектральная характеристика сдвинута
в сторону коротковолновой области на 50 нм. Этот сдвиг
объясняется специфическим явлением - эффектом Пуркине, который заклю-
272
Биофизика
чается в том, что при заходе или восходе солнца, когда
одновременно действуют колбочковый и палочковый аппараты, происходит
сдвиг максимума чувствительности. Например, красная и синяя
поверхности, которые днем кажутся примерно одинаковыми по
яркости (красный мак и василек), ночью становятся разными: синий
предмет светлее красного, красный - существенно темнее (красный
нам кажется черным).
Эффект цветового зрения возникает за счет того, что в сетчатке
глаза имеются колбочки с тремя видами спектральной
чувствительности. Это подтверждается, например, следующим экспериментом.
К сетчаткам кошек и лягушек прикладывали микроэлектроды,
позволяющие отводить потенциалы действия от отдельных волокон
зрительного нерва или от небольших групп таких волокон. В результате
получено 4 вида кривых, одна из которых охватывает почти весь
спектр видимого излучения, а три остальные воспринимают крас-
■ ный, зеленый и синий диапазоны. Существование в сетчатке
колбочек с различной спектральной избирательностью подтверждено для
птиц, черепах, ящериц, насекомых. Некоторые авторы считают, что
существует не 3, а до 7 спектрально-чувствительных видов колбочек.
У человека и приматов в результате исследования спектров
поглощения зрительных пигментов найдено 3 вида колбочек, которые
характеризуются различными пигментами. Их спектральные
характеристики поглощения приведены на рис. 6.35. Кривые
соответствуют фиолетово-синей - 1, зеленой - 2 и желто-красной - 3 областям
спектра. Их максимумы, соответственно, расположены на длинах
волн около 449, 500 и 562 нм. Хромофорной группой всех колбочек
служит ретиналь, а отличаются они опсином, который и придает
различия в спектральных свойствах. Таким образом, три вида колбочек
отвечают за цветовое зрение. Перекрытие спектральных
характеристик дает возможность ощущать все плавные переходы цвета.
Контрастная чувствительность глаза позволяет определять
различия в яркости предметов. Самый простой пример контраста можно
представить, когда на сером фоне с яркостью В^ расположен такой
же серый объект, например круг, полоска или область другой
конфигурации, но с яркостью В0. Для этого объекта контраст определяется
соотношениями: К] = (5ф - В0)/В0 или К2 = (бф - В0)/(В± + В0). Как пра-
Глава 6. Сенсорные системы организма
273
A,i
Ο.θ.
1
0,5
ι
1
ί\/
β>
2
W/N
<s
3
ν
\
\.
—►
400
500
600
700 λ, ημ
Рис. 6.35. Спектральные характеристики поглощения света
фоторецепторами сетчатки глаза: I, 2, 3 —различные виды колбочек
вило, контраст не должен иметь отрицательных значений, поэтому
числитель берется по абсолютной величине.
Наименьший контраст, который наблюдатель способен
заметить, называется порогом контрастной чувствительности глаза.
При наиболее благоприятных условиях, когда света достаточно,
наблюдаемый объект виден под углом, большим 0,5°, и его
местоположение хорошо известно, порог контрастной чувствительности
составляет приблизительно 0,01-0,02.
6.3.5. Разрешающая способность глаза
Разрешающая способность характеризуется тем
минимальным углом (в угловых минутах), под которым глаз видит две
расположенные рядом детали, как раздельные. Остротой зрения
называется величина, обратная тому углу зрения, при котором еще можно
раздельно видеть две близкие точки предмета, т.е. это величина,
обратная разрешающей способности. Для нормального глаза этот угол
составляет 1-1,5'. Разрешающую способность, равную Г, принимают
ча 100%. Разрешающая способность зрительного анализатора
определяется тремя факторами: дифракцией света, структурой сетчатки
и аберрацией.
Дифракция света в зрительном анализаторе происходит за счет
того, что перед хрусталиком расположена радужная оболочка, в
центре которой находится зрачок, представляющий собой диафрагму.
Когда наблюдатель смотрит на далекую светящуюся точку (напри-
274
Биофизика
мер, звезду), пучок лучей, проходящий внутрь глаза, не собирается
в одну точку, а освещает на сетчатке маленький кружок. Размер этого
кружка как раз и определяется в нормальном случае сферической
и хроматической аберрациями оптической системы глаза и
дифракцией света на краю зрачка. При прохождении света через диафрагму
диаметром D происходит его дифракция, или явление отклонения
света от прямолинейного распространения, когда свет, огибая
препятствие, заходит в область геометрической тени. В результате
прохождения света через зрачок на сетчатке образуется кружок рассеяния.
Предположим, что на сетчатку проецируются две точки.
Можно считать, что одна точка воспринимается отдельно от другой, если
центральная зона дифракционного изображения первой точки
совпадает с минимумом первого порядка (радиуса г) второй
дифракционной картины. В диапазоне между максимумами яркость определяется
суммарной кривой. Если «провал» яркости в центре диапазона
составляет 27%, то считается, что это соответствует наилучшей
разрешающей способности глаза. При дневном освещении, приняв λ = 600
нм, D = 3 мм и фокусное расстояние/глаза около 17 мм, по известной
формуле для круглого отверстия г = 1,22λ/7Ζ) получим г = 4 мкм.
Структура сетчатки тоже влияет на разрешающую способность,
Поскольку фоточувствительные рецепторы представляют собой
отдельные клетки, то структура сетчатки является дискретной, причем
в центральной части (желтое пятно) почти каждая колбочка
соединена с отдельным нервным окончанием. Чтобы получить изображение
двух точек, необходимо возбудить два фоторецептора, но между ними
должен находиться хотя бы один невозбужденный. Следовательно,
два дифракционных изображения должны возбудить две колбочки,
а минимум в этой двойной дифракционной картине должен попасть
на третью - между ними (чтобы была достаточная неравномерность
освещения). Если принять, как и ранее,/ = 17 мм, то при угловом
размере между двумя видимыми точками в Г линейное расстояние
между центрами их изображений будет 4 мкм. Гистологические
исследования показали, что диаметр колбочек соответствует примерно 2 мкм
и, если колбочки располагаются вплотную, минимальное расстояние
на сетчатке, способное сформировать ощущение изображения двух
точек, составляет примерно 4 мкм. Таким образом, разрешающая спо-
Глава 6. Сенсорные системы организма
275
собность глаза определяется хорошо согласующимися размерами
светочувствительных элементов сетчатки и структурой дифракционного
изображения светящихся точек.
Третий фактор, влияющий на разрешающую способность, -
аберрация света. Наиболее значимые для разрешающей
способности глаза аберрации - хроматическая и сферическая. Хроматическая
аберрация состоит в том, что излучения различных длин волн,
входящих в состав падающего потока света, преломляются по-разному
и образуют не одно, а множество изображений предмета. Накладыва-
ясь друг на друга, эти изображения создают нерезкую картину,
которая не позволяет достичь предельного различения деталей. Этот вид
аберрации сказывается, если фотоприемник (глаз) чувствителен
η широком диапазоне длин волн. Поскольку диапазон
воспринимаемого глазом излучения относительно мал (учитывая еще и
спектральные свойства колбочек), то влияние хроматической аберрации на
разрешающую способность незначительно. Сферическая аберрация
заключается в том, что параллельные лучи, идущие около
оптической оси, имеют большее фокусное расстояние, чем лучи, удаленные
от этой оси. Кома - сферическая аберрация, возникающая при косом
попадании пучка света. Дисторсия - искривление прямых линий
объекта. Недостаточность кривизны преломляющих поверхностей
глаза (астигматизм роговицы) компенсируется равным по значению
и обратным по направлению астигматизмом хрусталика.
Дифракция и аберрация света не зависят от освещенности, а
разрешающая способность глаза - зависит. Следовательно, этот параметр
связан с явлениями, происходящими в сетчатке. Освещенность может
быть настолько малой, что энергии, получаемой колбочкой,
недостаточно для возбуждения нервного волокна, но зрение не пропадает, т.е.
происходит пространственное суммирование на уровне синаптиче-
ских слоев. Следует отметить, что глазное яблоко находится в
непрерывном (автоматическом) движении, в результате которого
изображения внешних предметов все время скользят по поверхности сетчатки.
Такое перемещение сглаживает зернистую структуру, приближая
действие сетчатки к действию, которое обеспечивает идеальный
светочувствительный слой. Известно, что оптическое изображение намато-
ιχ·
276
Биофизика
вом стекле становится заметно более четким, если стекло приводится
в быстрое движение (вращается).
6.3.6. Кодирование информации в органе зрения
При изучении кодирования информации рассматривают
кодирование длины волны света, интенсивности света, размеров и формы
предметов, расстояния до предметов. Длина волны кодируется тем,
что в сетчатке присутствуют три вида колбочек с разной
спектральной чувствительностью. Интенсивность света кодируется частотой
потенциалов действия в волокнах зрительного нерва, которая
определяется параметрами рецепторных потенциалов фоторецепторов.
Кодирование информации о расстоянии до предметов
происходит при монокулярном (работает один глаз) или бинокулярном
(работают два глаза) зрении. При монокулярном зрении основное значение
в оценке расстояния имеет аккомодация. Кривизна хрусталика
изменяется за счет изменения степени напряжения ресничной мышцы,
которая воспринимается мышечным веретеном (проприорецептором),
передающим информацию о кривизне хрусталика. При
бинокулярном зрении основное значение имеет диспарация или расхождение
изображений предметов на сетчатках обоих глаз. Когда глаза
наведены на предмет, изображения всех его точек попадают на идентичные
участки сетчаток обоих глаз и в представлении человека два
изображения сливаются в одно. В то же время изображения предметов,
удаленных и приближенных, попадают уже на разные (диспаратные)
участки двух сетчаток и представляются раздвоенными. Возникаег
ощущение большей или меньшей удаленности, т.е. глубина рельефа
кодируется значением диспарации. При бинокулярном зрении
точность оценки расстояния значительно выше, чем при монокулярном.
Линейные размеры предметов кодируются количеством
возбуждаемых фоторецепторов. В зависимости от величины предмета
на сетчатке будет формироваться изображение большей или
меньшей площади и будет возбуждаться разное число рецепторов.
Величина изображения предмета на сетчатке зависит от расстояния до
предмета, т.е. от углового размера, поэтому оценка величины будет
зависеть и от расстояния.
Гпава 6. Сенсорные системы организма
277
Как уже отмечалось, важную роль в кодировании информации
в органе зрения имеют процессы обработки сигналов на уровне
сетчатки, особенно латеральное торможение, за счет которого
повышается контраст, в том числе и цветовой. У некоторых животных
(например, лягушек) уже в сетчатке происходит выделение информации
о движущихся объектах.
6.3.7. Цветовое зрение
Трехкомпонентная теория цвета. Всю гамму спектральных
цветов Ньютон в 1669 г. представил графически, расположив их на
окружности, в центре которой находится белый цвет. Поверхность,
ограниченная окружностью, называется цветовым кругом Ньютона,
он изображен на рис. 6.36, а. В 1756 г. М.В. Ломоносов первым
к мире сформулировал трехкомпонентную теорию цветового зрения,
Томас Юнг в 1802 г. доказал в эксперименте, что смешением трех
Красный
Фиолетовый
Синий
Оранжевый
L* Б /\ Желтый
\Л\ ^Зеленый
й ^и—
Голубой
а;
Желтый
Зеленый Ι КРасныи
Белый
Голубой ^у Пурпурный
Синий
б)
Зеленый
530 _
Желтый
580
Оранжевый
610
630
Красный
Синий
Рис. 6.36. Аддитивное смешение цветов
а - цветовой круг Ньютона, б - смешение цветов по Юнгу, в - цветовой
треугольник Максвелла. Ц- выбранный цвет, Щ - дополнительный к цвету Ц цвет
278
Биофизика
основных цветов можно получить все другие цвета, Д. К. Максвелл
построил один из первых колориметров, а Г. Гельмгольц в 1852 г.
его усовершенствовал.
Классический эксперимент, поставленный Юнгом, заключался
в том, что на экран проектировались с перекрытием световые потоки
трех проекционных фонарей, предварительно пропущенные через
зеленый, красный и синий светофильтры. В середине изображения, как
показано на рис. 6.36, б, получился белый, перекрытие синего и
зеленого давало голубой и т.д.
Максвелл в 1861 г. фотографировал окрашенный объект на три
черно-белые фотопластинки: на первую - через красный
светофильтр, на вторую - через зеленый, а на третью - через синий. В
результате получались три цветоделенных негатива. Полученные
затем три позитива проектировались на один экран, каждый через
.свой светофильтр, причем от источника белого света, как и при
фотографировании. В результате совмещения трех изображений на
экране воспроизводилась картина объекта в натуральных цветах. В
качестве основных цветов Максвелл выбрал красный, зеленый и синий
спектральные цвета с длинами волн соответственно λκ = 630, λ3 = 528
и λ,. = 457 нм. Расположив основные цвета в углах равностороннего
треугольника, он графически показал смешение цветов. Этот график
(рис. 6.36, в) называется цветовым треугольником Максвелла. В нем
спектрально чистые цвета лежат на сплошной линии, расположенной
за пределами треугольника. Цвета, которые могут быть получены
смесью основных, расположены внутри треугольника или на его
сторонах. В центре треугольника (как и в центре цветового круга
Ньютона) расположен белый цвет (Б). Цифры обозначают длину волны
(в нанометрах) монохроматического света.
Многие экспериментально установленные факты, включая
приведенные выше, легли в основу современной трехкомпонентной
теории цветового зрения. В 1959 г. Лэнд (США) попытался
опровергнуть трехкомпонентную теорию, используя в экспериментах два
светофильтра (зеленый и красный) при фотографировании цветного
объекта. После воспроизведения исходной картины вся гамма
цветовых оттенков получалась только для ограниченного сочетания
площадей цветных объектов, у остальных частей наблюдались искаже-
Гпава 6. Сенсорные системы организма
279
пия цвета, малая гамма, низкая насыщенность. Считается, что этот
отрицательный результат экспериментального исследования служит
нсским подтверждением теоретической основы трехкомпонентной
теории цветового зрения, т.е. того, что автор хотел опровергнуть. Так
Пыла заложена основа современных колориметрических систем,
относящихся к низшей метрике цвета, которая широко используется
и различных прикладных областях техники, дизайна, искусства и др.
В цветовом круге Ньютона и треугольнике Максвелла каждая
точка на прямой, соединяющей точку Ц и центр (точка белого Б),
имеют тот же цветовой тон (оттенок), что и цвет Ц (см. рис. 6.35, а,
(Т), но насыщенность цвета при приближении к центру уменьшается.
Г.сли эту прямую продлить, то на противоположной окружности
круга (стороне треугольника) она пройдет через точку Цд,
обозначающую цвет, дополнительный цвету Ц. Каждому цвету соответст-
нует дополнительный, обладающий тем свойством, что при
смешении в определенной пропорции световых потоков данного цвета
и дополнительного получается белый цвет - это один из законов
Грассмана, сформулированных в 1853 г. Например, для желтого
цвета дополнительным является синий, для оранжевого - голубой,
для зеленого - пурпурный (смесь красного и синего). Связь длин
ноли основного и дополнительного спектрально чистых цветов при-
нсдена на рис. 6.37.
Ощущения одинакового цвета могут создаваться при различных
спектральных составах поступающего в глаз светового потока. Это
явление называется метамеризмом. Два световых потока, различные по
λ, нм
620
600
580
560
430 450 470 490 λ, нм
Рис 6.37. Примерное соотношение длин волн
спектрально чистых и дополнительных цветов
280
Биофизика
составу спектров, но создающие ощущения одинакового цвета,
называются метамерной парой. Однако световой поток определенного
спектрального состава при одинаковых условиях всегда создает
ощущение одного и того же присущего ему цвета. Это следующий закон
Грассмана, из которого получаются весьма важные следствия,
рассмотренные далее.
Определим субъективные и физические параметры цвета. Цвег
характеризуется тремя параметрами - яркостью, цветовым тоном
и насыщенностью, поэтому он является величиной трехмерной.
Яркость цвета определяется световым потоком, излучаемым в данном
направлении единицей площади поверхности (излучающей или
отражающей) в пределах единичного телесного угла. Яркость цвета
принято определять по эквивалентности зрительного восприятия
интенсивности свечения окрашенной поверхности и поверхности
с соответствующей ступенью серой шкалы. Для оценки
интенсивности цвета наряду с яркостью иногда применяется понятие светлоты
цвета - это относительная величина, определяемая путем сравнения
яркости цвета с яркостью белой поверхности в тех же условиях
освещения и совпадающая с коэффициентом отражения (пропускания,
если объект прозрачен). Цветовой тон цвета позволяет оценивать его
как красный, зеленый, синий и т.д.
Насыщенность цвета есть степень его свободы от примеси
белого цвета. Ее еще можно определить как число световых порогов
(едва заметных переходов), которые отделяют данный цвет от
белого, равного с ним по яркости. Спектральные цвета имеют
максимальную насыщенность, а для белого насыщенность нулевая. Цветовой
юн, яркость и насыщенность не являются полностью независимыми.
Например, цветовой тон слегка изменяется при изменении яркости
и насыщенности. Цветовой тон и насыщенность характеризуют
цветность цвета независимо от его яркости. Таким образом, цвет может
быть охарактеризован яркостью и цветностью.
Яркость является физической величиной, а цветовой тон и
насыщенность - субъективными. Последним соответствуют
физические величины, называемые преобладающей длиной волны цвета λη
и чистотой цвета рк. Под преобладающей длиной волны цвета
понимают длину волны монохроматического света (излучения с очень
Гпава 6. Сенсорные системы организма
281
узким спектральным составом), цветовой тон которого такой же, как
у данного цвета. Для пурпурных цветов, которые не могут быть
воспроизведены только монохроматическим излучением, под
преобладающей длиной волны λ]^ понимают длину волны дополнительных
к ним монохроматических световых потоков.
Под чистотой цвета р,. (колориметрической) понимают
относительное содержание в нем спектрального цвета
(монохроматического светового потока) ФЦ по отношению к сумме
полихроматического излучения ФБ и монохроматического ФЦ, т.е. рк = ФЦ /(ФЦ +
ι ФБ). Для спектральных цветов рк = 1, так как ФБ = 0, а для белого
циета/7к = 0, поскольку ФЦ = 0.
Способы смешения цветов и цветовое уравнение. Смесь
нескольких цветовых потоков называется аддитивной (от латинского -
сложение). Существуют следующие способы аддитивного смешения
циетов: одновременное смешение, рассмотренное ранее;
поочередное смешение, когда частота предъявления изображения больше
критической частоты мельканий; пространственное смешение, когда по-
исрхпость покрыта мелкими разноцветными пятнами, например
текстильная ткань или мозаика, экраны цветных телевизоров или
циетпых мониторов компьютеров; бинокулярное смешение - когда
каждый глаз в отдельности воспринимает разные цвета (стереоцвет-
ное кино и специальное цветное телевидение). s
Кроме аддитивного, используется субтрактивное (вычитание)
смешение. Его суть состоит в том, что при прохождении света через
различные цветные (относительно прозрачные) среды в каждой из
них поглощается некоторая часть исходного спектрального состава
)гого цвета. В результате на выходе остается спектральный состав
сиета за вычетом поглощенных составляющих. Такой способ чаще
исего используется, например, в живописи и полиграфии, когда свет,
проходя через частицы красителей, избирательно поглощается и в
оставшемся (отраженном от несущей поверхности) потоке сохраняются
пепоглощенные цвета.
Рассмотрим согласование цвета. Предположим, что в
простейшем колориметре, упрощенная схема которого приведена на рис. 6.38,
есть призма Π (например, выполненная из гипса или другого
материала с нанесенным на него отражающим покрытием - окида магния или
282
Биофизика
др.). На одну грань направляют изучаемый световой поток Фц от
лампы Л, прошедший через объект Об, цвет (Ц) которого необходимо
согласовать. На другую грань подаются световые потоки трех основных
цветов, например красного, зеленого и синего, формируемые
лампами Л, - Л3 и соответствующими светофильтрами К, 3, С. Совмещение
световых потоков производится с помощью полупрозрачных зеркал
3,, 32 и обычных зеркал 33,34. Наблюдатель Η через отверстие в экране
Э видит два поля, показанных на рис. 6.38.
Зеркала 33 и 34 должны по возможности полностью отражать
падающий на них свет, а зеркала 3, и 32 - частично пропускать и
частично отражать свет. Если, например, 3, пропускает 50% света, а 3;
пропускает 66%, то до призмы дойдет около трети каждого светового
потока (без учета потерь при пропускании и отражении). Регулируя
освещенность полей 1 и 2, которые видит наблюдатель (например,
изменяя расстояния между каждым из источников и призмой, или
другим способом), можно добиться исчезновения грани между полями
сравнения (рис. 6.38), т.е. выполнить условие согласования по ярко
сти и цветности. Процесс согласования цвета происходит при
различной мощности излучений, но смесь двух или трех основных цветои
дает цвет менее насыщенный, чем спектральные цвета.
Согласование по спектральным цветам невозможно, поэтому
расширим понятие согласования. Добавим к цвету Ц один или дви
основных цвета. Это можно осуществить, например, перемещением
зеркал на пути от излучателей Л, - Л3. Тогда цвет (Ц) будет менее
насыщенным и согласование становится возможным. Можно считать,
что смеси цветов содержат основные цвета К, 3, с справа со знаком
«плюс», а слева - «минус». Три основных цвета позволяют получить
пЩ^ ^wf ^->г- -тЧ®*
,vh—Ч® л2
э3
Рис. 6.38. Схема простейшего колориметра и поле сравнения
Гпава 6. Сенсорные системы организма
283
согласование с любым цветом. Согласование сохраняется в широком
диапазоне яркостей.
Из упомянутого ранее закона Грассмана о том, что результат
смешения нескольких излучений определяется цветами этих
излучений, а не их спектральными составами, хотя изменение спектра
излучения может вызвать изменение его цвета, вытекают три важных
следствия.
1. Если цвет Ц[ согласован с цветом Ц2 и цвет Ц3 согласован
с ГЦ, то Ц, + Цз согласуется с цветом смеси Ц2 + 1Ц, т.е. Ц, + Цз =
Ц2 + Ц4·
2. Это правило справедливо и при вычитании цветов: Ц, - Цз =
Ц2-Ц4·
3. Если цвет Ц[ согласован с цветом Ц2, то увеличение или
уменьшение яркости обоих цветов в одинаковое число раз не
нарушает согласования.
Приведенные следствия называют законами линейности при
установлении цветовых равенств. Первые два указывают на
сохранение аддитивности при согласовании цветов, а последнее - на
сохранение пропорциональности.
Если основшле цвета обозначить через К, 3, С и принять за
единичные количества этих цветов значения [К], [3],JC], то получим,
что число этих единичных количеств для согласования с цветом Ц
нужно взять соответственно К, 3, С, т.е. можно написать уравнение
Ц = К[К] + 3[3] + С [С], (6.25)
где К, 3, С - координаты цвета или абсолютные трехцветные
коэффициенты, т.е. число единичных количеств основных цветов.
Справедливо будет и уравнение
Ц, + Ц2 = (К, + К2)[К] + (3, + 32)[3] + (С, + С2)[С]. (6.26)
Обозначив через [Ц] единичное количество цвета Ц в системе
основных цветов К, 3, С, получим, что для цвета Ц необходимо m
них единичных количеств, или Ц = т[Ц]. Пользуясь последним
равенством и выражением (6.25), напишем
284
Биофизик»
т[Ц] = К[К] + 3[3] + С [С], (6.27)
где т = К + 3 + С-цветовой модуль цветаЦ.
Уравнение (6.27) определяет трихроматическую систему
количественной оценки цвета. Единичное количество цвета [Щ,
соответствующее основным цветам, называется трихроматическои
единицей Τ цвета и является относительной величиной. Абсолютное
значение Τ определяется абсолютным количеством единичных
основных цветов (например, их световым потоком, яркостью и др.),
Цвет, для которого К = 3 = С, называется равностимульным ЦР, а его
модуль - тР. Если К = 3 = с = IT, то тР = ЗТ.
Разделив обе части уравнения (6.27) на т, получим
[Ц] = (К/т)[К] + (3/т)[3] + (С/т) [С] = К' [К] + 3' [3] + С [С], (6.28)
где К', 3', С - относительные трехцветные (трихроматические)
коэффициенты или координаты цветности.
Учитывая, что m = К + 3 + С, получаем
К' + 3' + С'=1. (6.29)
Таким образом, достаточно знать два коэффициента, поскольку
третий можно вычислить. Уравнение (6.29) называется единичным
или унифицированным цветовым уравнением и определяет цветность
цвета независимо от яркости. Так как цветность определяется двумя
параметрами (цветовым тоном и насыщенностью или
преобладающей длиной волны и чистотой), а не тремя, то название
коэффициентов К', 3', С координатами цветности вполне оправданно.
Обозначив яркости единичных количеств основных цветов
ЦК], L[3] и L[C], можно, по аналогии с выражениями (6.25) и (6.28),
определить яркость Щ цвета Ц и яркость ЦЦ] его единичного
количества:
Щ = К ЦК] + ЗЦЗ] + СЦС], (6.30)
ЦЦ] = Щ /т = К'ЦК] + З'ЦЗ] + СЦС], (6.31)
Глава 6. Сенсорные системы организма
285
где m = Ц/[Ц] = LIJ/ЦЦ] - модуль цвета, как и в выражении (6.27).
Величины ЦК], L[3], L[C] в формулах (6.30 и 6.31) могут быть
выражены как в абсолютных единицах (кд/м2), так и в
относительных, например долях яркости равностимульного цвета.
Поскольку цвет является трехмерной величиной
(характеризуется тремя параметрами - яркостью, цветовым тоном и
насыщенностью), его можно представить в виде точки в трехмерном
пространстве. Положение точки задается тремя координатными осями основных
цветов, образующими цветовое пространство, в котором расположен
вектор Ц. Его начало помещается в начале координат, а конец - в
данной точке пространства, отображающей цвет Ц. Длина вектора
характеризует яркость цвета, направление определяет цветность (цветовой
гон и насыщенность). Пространство, в котором находятся цветовые
векторы, называется цветовым пространством (цветовым телом).
Единичные цветовые векторы (т.е., векторы, представляющие
единичное количество цвета, равное IT), как правило, имеют различную
геометрическую длину. Это обусловлено тем, что единичные
количества цветов могут иметь различную яркость.
Длина цветового вектора, представляющего любой цвет,
определяется его яркостью, а не трихроматическим количеством цвета.
Модуль цветового вектора есть его длина, выраженная в трихромати-
ческих единицах. Единичные количества основных цветов могут
быть представлены единичными векторами (ортами) [К], [3], [С],
например в традиционной декартовой системе координат. Состав
спектра излучения или длин волн может быть разным, но сумма количеств
единичных цветов всегда принимается равной одной трихроматиче-
ской единице. Точка 0 представляет собой «черное», так как нулевая
длина соответствует нулевой яркости. Поскольку при аддитивном
смешении цветов яркость результирующего цвета не может
уменьшаться, то не может быть и цветовых векторов отрицательных
направлений. Совокупность всех реальных цветовых векторов занимает
в пространстве телесный угол, меньший 2π, поэтому векторы должны
быть некомпланарными. На ортах в цветовом пространстве
расположен единичный параллелепипед, диагональ которого, проходящая
через начало координат, представляет собой вектор равностимульного
цвета Ц с координатами 1 Т.
286
Биофизике
На концах единичных векторов основных цветов можно
определить плоскость, которая называется единичной плоскостью в
данном цветовом пространстве. На этой плоскости определим
треугольник, опирающийся своими углами на концы ортов основных цветов,
и называемый единичным цветовым треугольником. Отрезок вектора
цвета от начала координат до точки его пересечения с единичной
плоскостью определяет единицу измерения модуля единичного
вектора цвета. Модуль единичного вектора определяет ту единицу,
которой измеряют модуль вектора соответствующего направления
(цвета). Поскольку яркость пропорциональна модулю, то единичный
вектор не несет информации о яркости цвета. Поскольку выражение
(6.29) позволяет однозначно определить цветность только двумя
координатами, то ее (цветность) можно найти по проекции единичной
плоскости на одну из координатных плоскостей. Однако для
формирования базовой (или опорной) точки, относительно которой следует
определять координаты цветности, необходимо выполнить
конкретные условия.
В качестве основного (базового) цвета берут равноэнергетиче-
ский белый цвет источника Е, т.е. такой белый, у которого все
спектральные составляющие имеют одинаковую энергию. Приняты 6
стандартных источников белого цвета. Четыре введены в 1931 г.
МКО, это источники А, В, С и Е, в дальнейшем ввели D. Они
отличаются цветовыми координатами или цветовой температурой ТЦ,
которой называется цветность излучения абсолютно черного тела
в пределах видимого спектра, нагретого до указанной температуры
по шкале Кельвина. Источник типа а соответствует излучению
вольфрамовой лампы накаливания (средний искусственный свет, ТЦ =
= 2856°К); В - рассеянному полуденному солнечному свету
(искусственный дневной свет, ТЦ = 4874°К); С - прямому солнечному свету
(смешанный цвет излучения солнца и неба при малооблачном небе),
он близок к среднему дневному свету северного неба (искусственный
дневной свет, ТЦ = 6774°К). Следует учесть, что спектральный состав
излучения различных ламп «дневного света» (уличных и комнатных)
основан на люминесцентных эффектах покрытий внутренней
поверхности колбы. Источником белого Ε утвержден тот, у которого спектр
излучения равноэнергетический.
I пава 6. Сенсорные системы организма
287
Источники белого D65 и D75 соответствуют цвету голубого
неба и содержат в своем спектре ультрафиолетовую область (начиная
с 300 им). Они были введены позднее, поскольку при разработке
и применении люминесцентных красителей и покрытий возникла
необходимость учитывать стандартизованное соотношение в источнике
освещения спектральных составов излучения ультрафиолетовой час-
I и спектра. Например, применяют так называемые оптические
отбеливатели, т.е. люминофоры, которые, поглощая ультрафиолетовое
ичлучение, люминесцируют голубым светом и делают желтоватую
поверхность (ткани или бумаги) значительно белее. Добавление
к красному красителю люминофора, который, поглощая
коротковолновое излучение, тоже светится красным, повышает яркость
поверхности так, что она невольно бросается в глаза. Ни один из первых
четырех источников не создает этих эффектов, поэтому их применение
для измерения цвета или цветности образцов, в окраске которых
содержится люминофор, дает заведомо неправильные результаты. Два
источника D отличаются цветовой температурой: D65 имеет ТЦ =
- 6500°К, а у D75 ТЦ = 7500°К.
Зависимость координат цветности спектральных цветов от
длины волны изображается графически кривыми смешения.
Спектральные цвета, которые изображаются на единичной плоскости
векторами, образуют кривую, называемую спектральным локусом. Рисунок,
представляющий собой изображение цветностей на единичной
плоскости или ее проекциях, называется хроматической диаграммой, или
диаграммой цветности. Если основные цвета существуют реально,
то кривые смешения имеют для отдельных участков спектра
отрицательные значения, поскольку сумма основных цветов дает цвет менее
насыщенный, чем спектральные цвета. Можно в качестве основных
выбрать условные, удобные для расчетов цвета, причем так, чтобы
кривые смешения не имели отрицательных значений. Тогда они на-
чываются кривыми сложения основных возбуждений.
Стандартные колориметрические системы МКО. В 1931 г.
МКО на 8-й сессии утвердила две стандартные системы: RGB и XYZ.
В системе RGB приняты за основные цвета красный (R) с λR = 700 нм,
челеный (G) с XG = 546,1 нм и синий (В) с λΒ = 435,8 нм. Единичное
цветовое уравнение в системе RGB выглядит так:
288
Биофизика
Ц = rR + gG + ЬВ,
где г, g, b - координаты цветности, a R, G и В - единичные
количества основных цветов.
В качестве единичных количеств основных цветов выбраны
такие их количества, смесь которых согласована с равноэнергетичс-
ским белым Е:
Е= 1,0 R+ 1,0 G+1,0 В,
т.е. R = G = B= 1Тит = ЗТ. Здесь, как и ранее, m - модуль, а Т - три-
хроматическая единица цвета. Световые потоки OR, OG, ФВ
единичных основных цветов относятся, как OR : OG : ФВ = 1 : 4,5907 :
0,0601, т.е., если яркости единичных количеств цветов (условно):
L R = 1 кд/м2; L G = 4,5907 кд/м2; L В = 0,0601 кд/м2, тогда яркость рав-
ностимульного белого Ε будет: LE = LR + LG + LB = 1+ 4,5907 +
+ 0,0601 = 5,6508 кд/м2. Коэффициенты L R, L G и L В являются яр-
костными коэффициентами.
Расположив в прямоугольной системе координат основные
цвета RGB, получим равносторонний цветовой треугольник (в
проекции на одну из координатных плоскостей). Вид диаграммы цветности
на плоскости rg показан на рис. 6.39. Здесь координаты г и g
изображаются отрезками одинаковой длины, так что шкалы значений г и g
равномерны. Для точек, лежащих на стороне RG цветового
треугольника, справедливо b = 0 и г ± g = 1). На этой стороне находятся
насыщенные красный, оранжевый, желтый, зеленый цвета. На стороне BR
расположены насыщенные пурпурные цвета.
Спектральный локус выходит за пределы треугольника
основных цветов. Это связано с тем, что любая цветность с меньшей
насыщенностью, чем у основных цветов, может быть согласована смесью
положительных количеств этих цветов и изображается внутри
цветового треугольника RGB. Цветность с насыщенностью, большей
насыщенности основных цветов, согласуется смесью трех основных, из
которых один или два берутся с отрицательным знаком, т.е.
добавляются к данному цвету. Поэтому такая цветность изображается
точкой, лежащей вне треугольника.
Глава 6. Сенсорные системы организма
289
Рис. 6.39. Диаграмма цветности rg
Стрелкой показано увеличение длины волны спектрально чистых цветов,
в центре треугольника RGB находится равноэнергетический белый цвет Е,
в вершинах треугольника находятся основные цвета R, G и В
Рассмотрим согласование цвета в системе RGB. Предположим,
что вместо источника Л (см. рис. 6.38) стоит монохроматор, который
последовательно выделяет узкие спектральные диапазоны излучения
и направляет их на левую грань призмы. Мощность излучения в
каждом диапазоне всегда поддерживается постоянной, например 1 Вт.
При каждом предъявлении спектрального диапазона выполняется
согласование цветов. В результате получены таблицы и кривые,
которые показывают, сколько единиц соответствующего основного цвета
(R, G или В) нужно взять, чтобы получить цвет излучения с
соответствующей длиной волны и мощностью 1 Вт. Эти зависимости назы-
иаются кривыми удельных координат стандартной системы RGB
(рис. 6.40) и обладают такими свойствами, что площади под кривыми
одинаковы и сумма световых потоков основных цветов,
необходимых для согласования со спектральным цветом мощностью 1 Вт,
соответствует кривой стандартной относительной видности (см.
рис. 6.40).
Несмотря на наглядность и другие достоинства системы RGB,
она имеет и три существенных недостатка. Во-первых,
отрицательные значения координат цветности г, g, b и удельных координат
затрудняют расчеты для сложных излучений. Во-вторых, если цвет
задан значением яркости (или светового потока) и координатами
цветности г, g, b, то для вычисления координат цвета R, G, В по
формулам, аналогичным (6.28), необходимо знать цветовой модуль т,
290
Биофизика
—■
-^
.в
ν
V
λ
/
I
ι
^
**·
•^
^ /
^*
G
\,
4
/
V
л
/
\
s
V
^R
s
Л ft Ь
0,32
0,24
0,16
0.0В
О
-0,06
380 420 460 500 540 560 620 660 λ, нм
Рис. 6.40. Кривые удельных координат стандартной системы RGB
(кривые смешения МКО)
•расчет которого является трудоемким, если коэффициенты г, g, b
имеют разные знаки. В-третьих, кривые координат цветности г, g, b
признаны МКО в 1955 г. недостаточно точными, особенно в области
синих цветов.
Одновременно с системой RGB была принята вторая
стандартная колориметрическая система XYZ, которая является производной
от RGB. Основные цвета Χ, Υ, Ζ не являются реальными, а
представляют собой математическую абстракцию, т.е. некоторые точки на
хроматической диаграмме RGB, как это показано на рис. 6.41, а.
Координаты цветности rgb основных цветов системы XYZ
определяются следующими соотношениями. Для X: г = 1,2750, g »
= - 0,2778, b = 0,0028; для Υ: г = -1,7393, g = 2,7673, b = -0,0280 и для
Ζ: г = -0,7431, g = 0,1409, b = 1,6022. Световые потоки относятся
в пропорции: ФХ : ΦΥ : ФВ = 0:1:0 (такая же пропорция
соответствует и яркостным коэффициентам), т.е. только составляющая Υ несе!
информацию о мощности (яркости) суммарного потока. Базисный
стимул в системе XYZ остается прежним - равноэнергетический
белый цвет Е, который получается при сложении в равных количествах
основных стимулов системы XYZ.
Если представить координаты XY прямоугольными (провести
проективное преобразование), то косоугольный треугольник XYZ
Глава 6. Сенсорные системы организма
291
Рис. 6.41. Хроматическая диаграмма XYZ
а — изображение координат цветности в системе rg; б - спектральный локус
(рис. 6.41, а) превратится в прямоугольный. Спектральный локус
расположен внутри треугольника XYZ, поэтому кривые сложения
основных стимулов в системе XYZ, показанные на рис. 6.42, не
содержат отрицательных участков.
На хроматической диаграмме XYZ (рис. 6.41, б) область
зеленых и голубых цветов помещается на криволинейном участке от 510
до 480 нм, а область синих и фиолетовых расположена на коротком
криволинейном участке от 480 нм и менее. В области от 510 до 550 нм
кривизна спектрального локуса наибольшая, поэтому насыщенные
цвета в этой области получаются только при смешении двух близких
по длине волны спектральных цветов. Высоконасыщенные желтые,
оранжевые и красные цвета могут получаться при смеси более
широких по спектральному диапазону световых потоков, так как
спектральный локус в этой области почти прямолинеен.
Пурпурные цвета с максимальной насыщенностью (в спектре
солнца пурпурных цветов нет) размещены на прямой, соединяющей
концы спектрального локуса (т.е. точки, соответствующие красному
и фиолетовому цветам). Точки, лежащие вне области, ограниченной
спектральным локусом и линией пурпурных цветов, следует
рассматривать не как реальные цвета, а как точки, отвечающие закону согла-
I1)·
292
Биофизика
1,6
1,2
0,8
0,4
Л
У
/
/
/
/
/
s~
(
'
"Nl
>
*ч
——
\
\
\
N
s„
\
—*
ч
S
У
^
/
У
/
^
—-
/
'
*>^<
,>с
/
/
/
—.
400
500
600
Рис. 6.42. Кривые удельных координат в системе XYZ
по оси абсцисс отложена длина волны спектрально чистых цветов,
а по оси ординат -удельные координаты (см. пояснения к рис. 6.40)
сования цветов и соответствующие цветам, более насыщенным, чем
спектральные. Поскольку спектральный локус не имеет вогнутых
участков, то смесь двух и более спектральных цветов всегда лежи'1
или на самом спектральном локусе, или внутри него, но никогда не
может лежать вне площади, ограниченной спектральным локусом.
Можно сформулировать следующие основные свойства
колориметрической системы XYZ.
1. Все реальные цвета лежат внутри локуса и могут быть
определены положительными значениями удельных координат.
2. Цветность белого равноэнергетического Ε лежит в центре·
тяжести треугольника ΧΥΖ и определяется значениями координа'1
Х=1/ЗиУ=1/3.
3. В треугольнике BER лежат пурпурные цвета.
4. Дополнительные цвета лежат на пересечении прямой,
проходящей через точку Е, с кривой спектральных цветов.
5. Цвет смеси двух цветов определяется точкой на прямой,
соединяющей смешиваемые цвета.
6. Цвет смеси трех цветов определяется точкой, лежащей
внутри треугольника, в вершинах которого лежат смешиваемые цвета.
7. Яркостные коэффициенты относятся, как Lx: Ly: Lz = 0: 1:0,
т.е. яркость передается только координатой Y, а цветность -
координатами X и Z.
Глава 6. Сенсорные системы организма
293
Последнее свойство является важным преимуществом системы
XYZ, поскольку выражения, аналогичные (6.25)—(6.31), в этой
системе существенно упрощаются. Для примера на хроматической
диаграмме XYZ (рис. 6.41, а) показаны линии постоянной яркости (от
L = 0 до L = 6), которые параллельны стороне ZX.
Законы колориметрии, относящиеся к смешению цветов и
соответствующие свойствам глаза при согласовании (уравнивании)
цветов, относятся к низшей метрике цвета. Под высшей метрикой
цвета понимается часть колориметрии, относящаяся к изучению
отличий между цветами. Для этого вводятся понятия цветового порога
и меры различия между цветами. Порогом изменения ощущения
называется наименьшее изменение ощущения, испытываемое
наблюдателем. Если известен такой порог, то можно определить число
порогов между ощущениями от двух различных раздражителей. Это
число считается мерой различия таких ощущений.
По имеющимся данным о чувствительности глаза к
изменениям цветности Джадд (США) и Мак-Адам построили зоны, соответст-
нующие 10 порогам цветоразличения. Оказалось, что диаграмма XY,
показанная на рис. 6.43, обладает тем свойством, что в различных ее
областях пороги цветоразличения занимают разные зоны. В области
синих цветов число различимых цветов, приходящееся на единицу
Рис. 6.43. Эллипсы равного числа цветовых порогов
294
Биофизика
площади диаграммы, примерно в 400 раз больше, чем в области
зеленых цветов. Это связано с тем, что глаз воспринимает изменения
координат цветности XY не по линейному закону. Таким образом, хотя
на хроматической диаграмме расстояние между двумя точками
всегда является критерием различия цветностей, по величине этого
расстояния нельзя точно судить о количестве содержащихся в нем
порогов цветоразличения.
Нужно создать такую хроматическую диаграмму, в которой
пороги цветоразличения (при неизменной яркости) во всех
направлениях выражаются отрезками прямой постоянной длины, независимо от
места на диаграмме. Такая диаграмма называется равноконтрастной
диаграммой цветности. Она может быть получена путем
проективного преобразования плоскости XYZ на плоскость UVW,
расположенную так, чтобы все эллипсы преобразовались бы в окружности
и область зеленых цветов сократилась бы, а синих - расширилась,
Нужно поменять основные цвета на U, V, W. В 1960 г. диаграмма
Мак-Адама была рекомендована МКО в качестве весьма точной
равноконтрастной диаграммы.
Равноконтрастная диаграмма Мак-Адама была построена,
исходя из уравнений: U = 2Х/3; V = Y; W = 1,5Y-0,5X + 0,5Z,
откуда модуль цвета в системе UVW
m = U + V + W = (X + 15Υ + 3Ζ)/6
Координаты цветности связаны дробно-линейными зависимостями:
u = U/m = 2r/(6Y - X + 1,5);
v = V/m = 3Y/(6Y- X+l,5);w=l- (u + v).
Если эллипсы, показанные на рис. 6.41, построить на
равноконтрастной диаграмме, то равновеликих кругов получить не удастся,
что подтверждает приближенный характер этой диаграммы. Однако
отношение наиболее длинного отрезка между определенным числом
порогов цветоразличения к наиболее короткому на разных участках
диаграммы составляет 2:1, тогда как на диаграмме XY это
отношение 20: 1.
Глава 6. Сансорныа системы организма
295
Измерение цвета и субъективные эффекты цветовосприятия.
Способы измерения цвета делятся на субъективные и объективные.
Первые основаны на сравнении цвета объекта с цветом эталона
(образца) или использовании визуальных колориметров. Вторые - на
применении вместо глаза человека (как фоторегистратора)
фотоэлектрических преобразователей, как правило, совмещенных с
автоматизированными компьютерными средствами.
Среди субъективных способов в мировой практике приобрели
известность атласы образцов цвета Манселла. Атласы Манселла
содержат около 1500 образцов цвета, в зависимости от издания (полный
атлас содержит 3500 образцов). Например, в одном из изданий размер
каждой из карт цветового тона составляет 25,4 χ 33 см2, а размер
отдельного образца - 1,8 х 2,1 см2. Цвета расположены так, что
различия в их яркости, насыщенности и цветовом тоне везде
воспринимаются как примерно одинаковые. Яркость называют интенсивностью
цвета, а насыщенность - хромой. Координаты Χ, Υ для цветов
образцов Манселла известны. Цветовые тона расположены по окружности,
а образцы цвета помещены рядом друг с другом в том же порядке, что
и в спектре, так что взаимодополнительные цвета находятся на
концах этой окружности. Между красным и синим введены образцы
пурпурного цвета. В центре расположен серый. Круг разделен на 10
равных главных секторов, представленных в таблице.
Каждый главный сектор разделен на 10 равных малых
секторов. Насыщенность (хрома) показана расстоянием от центра по
радиусу, который делится на 16 одинаковых частей (16-е деление
имеют не все образцы). По яркости круги Манселла располагаются
по высоте цилиндра. Для образцов яркости шкала квадратичная
и содержит 11 кругов, но используются обычно от 1 до 9. Например,
цвет 6GY4/5 означает, что цвет с интенсивностью 4 помещается
в малом секторе 6 главного сектора GY и имеет хрому 5.
В 1956 г. в СССР был издан атлас Е. Б. Рабкина,
представляющий собой таблицы, в которых содержатся образцы цветов, каждый
в виде кружка диаметром 12 мм. Каждая таблица (их 12)
характеризуется цветовым тоном, в ней 55 кружков, расположенных
треугольником, так образуется конус, ось которого соответствует ахромата-
296
Биофизика
ческим цветам. Верхний кружок соответствует наиболее чистому
цвету.
Название главного
сектора
Красный
Желто-красный
(оранжевый)
Желтый
Желто-зеленый
Зеленый
Обозначение
R
YR
Υ
GY
G
Название главного
сектора
Сине-зеленый (голубой)
Синий
Синевато-пурпурный
Пурпурный
Красновато-пурпурный
Обозначение
BG
В
РВ
Ρ
RP
Кроме этих атласов и таблиц, существуют, например, атлас
Тейлора, система Оствальда, атлас архитектурных цветов и др.
Объективные измерения цвета производятся с помощью
колориметров, которые традиционно делятся на субъективные (пример
показан на рис. 6.44, а) и объективные. В последних цвет измеряется
с помощью электронного прибора, в котором используется один
фотоприемник (с соответствующей спектральной энергетической
характеристикой фототока) с тремя сменными светофильтрами
(последовательный колориметр) или тремя фотоприемниками, каждый со
своим светофильтром (параллельный колориметр).
Обобщенные структуры параллельного и последовательного
приборов для измерения цвета (спектральных свойств объектов)
показаны на рис. 6.44. В первом приборе (рис. 6.45, а) световой поток
от источника излучения ИИ направляется и формируется обычно при
помощи конденсоров или других наборов оптических элементов (от
световодов до лазеров). Формирование потока может производиться
по спектральному составу, направленности, мощности излучения,
распределению во времени и пространстве (для сканирующих
приборов). В зависимости от конкретного назначения устройства часть из
перечисленных функций оптической системы ОС1 может
отсутствовать. Сформированный поток излучения поступает на объект
исследования Об, оптические свойства которого (цвет) могут
определяться в проходящем, рассеянном или отраженном излучении. После
Глава 6. Сансорныа системы организма
297
взаимодействия с объектом исследования преобразованный световой
поток собирается и формируется второй оптической системой ОС2
II направляется в канал, содержащий полупрозрачные зеркала П31
и П32 (рис. 6.44, а). Полупрозрачные зеркала делят световой поток
на три части. На пути каждой из частей разделенного светового
потока находятся светофильтры СФ1, СФ2 и СФЗ, которые выделяют
части световых потоков, соответствующие основным цветам. Эти
составляющие поступают на три фотоприемника ФП1-ФПЗ, которые
преобразуют лучистую энергию в электрические сигналы. Затем
сигналы фотоприемников, прошедшие через электронное устройство
согласования с фотоприемниками ЭУС, поступают на вход устройст-
на математической обработки электрических сигналов УМОС,
которое, как правило, включает электронную вычислительную технику
(от специального, чаще серийного программируемого
микропроцессора до универсальной ЭВМ, в том числе персональной).
Для достаточно точного измерения координат цветности нужно
поставить в соответствие оптические свойства Об и выходные
сигналы фотоприемников. Поэтому необходимо учитывать спектральное
распределение мощности излучения ИИ, спектральные
коэффициенты пропускания оптических систем, полупрозрачных зеркал и
светофильтров, спектральные чувствительности фотоприемников и другие
оптические свойства измерительного тракта. Коррекцию
электрических сигналов уже на этом этапе проще выполнить с помощью
вычислительной техники. На выходе прибора находится устройство
регистрации УР, которое предназначено для формирования электронной
копии результатов измерения и записи, например на бумажном,носи-
тсле, или отображения на другом визуально воспринимаемом
источнике информации (цифровой или буквенной, чаще смешанной).
В последовательном колориметре (рис. 6.44, б) часть тракта от
ИИ до ОС2 включительно выполняет те же функции, что и в
параллельном. После ОС2 расположен вращающийся диск Д, на котором
находятся три светофильтра СФ1-СФЗ, функции которых такие же,
как и в параллельном устройстве. Вращением диска управляет
устройство управления УУ. При вращении в измерительный тракт
последовательно помещается один из светофильтров и выделяется свето-
ной поток, соответствующий одному из основных цветов. Поскольку
298
Биофизика
потоки формируются последовательно, то необходимы только один
фотоприемник ФП и устройство согласования ЭУС с одним входом.
Назначение остальных частей прибора соответствует рис. 6.44, а.
Связь между блоками УУ и УМОС обеспечивает передачу
информации о номере светофильтра, находящегося в оптическом канале.
Таким образом, в обоих колориметрах светофильтрами
выделяются световые потоки, соответствующие основным цветам.
Последующий расчет координат цветности представляется достаточно
сложной процедурой, связанной с математической обработкой
электрических сигналов. Например, на рис. 6.41, б, показан треугольник,
в вершинах которого находятся основные цвета, регистрируемые
фотоприемниками Rn, Gn, Вп. Преобразование измеренных координат
цветности объекта в координаты стандартных колориметрических
систем можно выполнить различными способами, например,
графически, как представлено на рис. 6.41, б. Для этого необходимо
провести прямые через точку Ε из точек Gn, Вп и определить отрезки GnP2,
RnP, и ВПР,, RnP,. Затем по отношениям длин отрезков определяют
относительные яркости основных цветов и цвета Ε относительно
яркости цвета Rn. По полученным соотношениям уже можно
определить координаты цветности измеряемого цвета. Эти преобразования
выполняются с помощью компьютерных систем, поэтому такие
колориметры часто называют компьютерными.
Рассмотрим субъективные эффекты при цветовых
ощущениях. Цветовая адаптация состоит в том, что источник яркого цвета
снижает чувствительность глаза к этому цвету и повышает
чувствительность тех рецепторов, которые возбуждаются слабо. В результате
изменяется восприятие ряда цветов и увеличивается цветовой
контраст. Например, яркий синий цвет снижает чувствительность
«синего» рецептора и делает голубой цвет зеленоватым. Если оба цвета
имеют одинаковый цветовой тон, но отличаются по насыщенности,
восприятие цвета также изменяется - цветовой тон менее
насыщенного цвета изменяется в сторону дополнительного цвета.
Цветовая адаптация приводит к явлению константности цвета -
восприятие цвета известных предметов не изменяется при изменении
в некоторых пределах спектрального состава источника света.
Например, красные воздушный шарик, свитер, кофта (но именно крас-
Глава 6. Сансорныа системы организма
299
ИИ
ОС1
N
Об
N
ОС2
П31
I
I
Π 32
^к-
СФЗ
СФ1
"Г
СФ2
ФП1
ФП2
ФПЗ
УР
- УМОС (ЭВМ) (-<
ЭУС
ИИ
ОС1
л
"7
Об
"7
ОС2
Щ-
УУ
УР
УМОС (ЭВМ)
-а
СФ2·
СФЗ·
СФ1
д
ФП
ЭУС
б)
Рис. 6.44. Структуры параллельного (а)
и последовательного (б) колориметров
(пояснения в тексте)
ные) всегда будут красными и при дневном, и при искусственном или
вечернем освещении (последнее существенно отличается по спектру
от дневного в связи с более длинным путем прохождения солнечных
лучей через атмосферу). Так, цвет привычного предмета кажется
одинаковым при разном освещении, это и есть константность воспри-
300
Биофизика
ятия окраски. Способность вносить коррективу непосредственно
в воспринимаемый цвет, поправку на особенности данного
освещения подтверждается многими экспериментами. Существует она
у многих животных, не только у позвоночных, но и у насекомых
(например, у пчел). Константность восприятия окраски биологически
чрезвычайно полезна, поскольку этот механизм позволяет
автоматически пересчитывать воспринимаемый цвет с учетом спектрального
состава каждого конкретного освещения и приводить его к какому-то
одному, привычному для организма. Сложность этого процесса
доказывается тем, что константность восприятия окраски далеко не
абсолютна, она до известной степени приближает наблюдателя к
определению истинной окраски предмета, а иногда вообще отсутствует.
Последовательный и одновременный контрасты цветов тоже
основаны на цветовой адаптации и заключаются в следующем. Если
после напряженного наблюдения небольшой зеленой полоски
перевести взгляд на большую белую поверхность, то в течение
некоторого времени будет видна полоска, окрашенная в цвет, дополнительный
к зеленому (пурпурный) - этот эффект называется
последовательным образом. Последовательный образ тем лучше заметен и тем
дольше сохраняется, чем ярче предмет, на который первоначально
был обращен взгляд. Особенно хорошо он проявляется после
прямого взгляда на источник света.
Если яркий предмет имел ясно выраженную окраску и
определенный цветовой тон, то последовательный образ виден в цвете,
близком к дополнительному к цвету яркого предмета. Это явление
называется последовательным цветовым контрастом. Проверить
его можно, взяв квадратную пластинку, разделенную на четыре
части, каждая из которых окрашена соответственно в зеленый, синий,
желтый и красный цвета. Если в течение 15-20 с фиксировать взгляд
на середине пластинки, а потом быстро заменить пластинку листом
белой бумаги, то на нем будут видны четыре цветных квадрата,
окрашенных в цвета, близкие к дополнительным цветам четырех
квадратов пластинки: красно-пурпурный, оранжевый, пурпурновато-синий,
сине-зеленый. Цвета последовательного контраста связаны с
локальной адаптацией колбочек сетчатки: направляя взгляд на другую точку
Глава 6. Сенсорные системы оргенизме
301
бумаги, можно ясно видеть, что вся картина перемещается по бумаге
вместе с перемещением глаз.
Известно другое явление: если положить синюю полоску на
большом красном фоне, а вторую такую же полоску на большом
зеленом фоне, то эти полоски будут восприниматься как поверхности
разного цвета, причем цвет каждой из них сдвинется в сторону цвета,
дополнительного к цвету фона. Это явление называется одновременным
цветовым контрастом. Из него вытекает важное следствие: любой
цвет в окружении дополнительного к нему кажется более
насыщенным. На фоне теплых цветов этот эффект проявляется сильнее. Если
цветовой круг (рис. 6.36, а) разделить на части радиусами,
соединяющими красный и зеленые цвета, то в одной части круга будут так
называемые теплые цвета (красно-оранжевые, желтые, желто-зеленые),
а в другой - холодные (пурпурно-синие, голубые,
голубовато-зеленые). В некоторых случаях одновременный цветовой контраст якобы
уничтожает цвет окрашенной поверхности, делает ее серой.
Рассмотрим чувствительность глаза к изменениям цветового
тона, насыщенности и особенности восприятия мелких по размеру
предметов. Чувствительность глаза к изменениям цветового тона
«первые была исследована в 1867-1872 гг. (Е. Мандельштам и В.
Добровольский). Определено, что для поля зрения 2° зависимость
различаемой пороговой разности длин волн Δλ от длины волны λ выглядит
и виде графика, показанного на рис. 6.45, а. На длине волны 500-
600 нм (голубовато-зелено-оранжевый) глаз различает изменения
преобладающей длины волны λ = 1 нм. При увеличении
насыщенности число различаемых цветовых тонов возрастает. Этот же эффект
проявляется при низкой и большой яркостях (например, при яркостях
дневного света, когда освещенность поверхности, перпендикулярной
к солнечным лучам, составляет около 133 000 лк).
Колебания чувствительности к изменениям насыщенности
цвета, показанные на рис. 6.45, б, проявляются для сильно- и
малонасыщенных цветов. Для средней насыщенности глаз мало чувствителен.
Носприятие изменения насыщенности для желтого цвета наименьшее
и составляет около 4 порогов, а чувствительность к изменению
насыщенности синего и красного - около 25 порогов.
302
Биофизика
Δλ,
0
420 460 500 540 580 620 660 λ, нм
20
10
^
■-^ ζ
_5 г-
π
V
400 500 600 700 λ, нм
б)
Рис. 6.45. Зависимость пороговой чувствительности глаза
к изменениям длины волны от длины волны спектрально-чистых
цветов (а) и график чувствительности к изменению насыщенности
цвета (б) (по оси ординат отложено число порогов η цветовосприятия)
Изучение порогового блеска (освещенности на зрачке от
точечного источника света) показало, что наблюдатель, по мере
приближения к источнику света, вначале замечает светлую точку и только
потом может опознать его цвет. На рис. 6.46 приведена зависимость
порогового блеска Е, позволяющего распознать цвет огня в
зависимости от длины волны монохроматического света, а пунктиром показан
ахроматический порог для красного цвета при большом угловом
зрении. В области желто-зеленого цвета порог существенно больше, чем
в области красного цвета. Этот эффект определяет повсеместный
выбор красного как запрещающего сигнала или цвета опасности. Такой
выбор обусловлен еще и тем, что коротковолновое излучение сильнее
Е, лк
10
10
10
"ν:
-π
-β
400
500
600
λ, нм
Рис. 6.46. Хроматический порог: график зависимости
порога чувствительности от длины волны точечного источника света
Гпава 6. Сенсорные системы оргенизме
303
Рис. 6.47. Изменение цвета предметов по мере уменьшения их размеров
На диаграмме XY показан спектральный локус с указанием увеличения
длин волн спектральных цветов и направления изменения цвета
предметов по мере уменьшения их размеров (стрелками)
длинноволнового поглощается атмосферой, поэтому свет, проходя
через атмосферу, в той или иной мере краснеет. Например, если
заметен огонь (но его цвет еще не различается) с расстояния 1 км, то цвет
красного будет различим с расстояния 830 м, зеленого - с 650 м,
а желтого - с 550 м.
Изучение восприятия цвета мелких предметов показало, что
все спектральные цвета могут быть согласованы только двумя (а не
тремя) основными цветами, если поле зрения составляет 10'—25'. Эти
основные цвета являются спектральными красным (650 нм) и синим
(460 нм). Для рассматриваемых условий были получены две кривые
смешения г и b вместо обычных трех. На длинах волн фиолетового
и желтого диапазонов глаз не различает цвета, а видит предмет серым
(нейтральным) с температурой цвета Тц, которая близка к 4800°К.
Лучше всего цветовой тон различается в области оранжевых и
близких к ним красных цветов, где цветовой порог составляет около 7 нм,
тогда как в других областях он доходит до 48 нм.
Было обнаружено, что по мере уменьшения размеров предметов
их цвета воспринимаются как смесь оранжевого и голубого цветов,
что соответствует оси I на рис. 6.47. Например, цвета трех крупных
304
Биофизика
предметов Ц1, Ц2 и ЦЗ, показанные на рис. 6.47, по мере уменьшения
размеров вначале изменяются вдоль оси Q, приближаясь к оси I, а
затем стремятся вдоль оси I, приближаясь к ахроматическому Е.
Количество воспринимаемых глазом цветов зависит от многих
факторов (условий наблюдения, состояния наблюдателя и т.д.).
Некоторые исследователи считают, что глаз человека различает до 17 000
отдельных цветностей, другие - 7 300, из них в области пурпурных,
красных и синих - 5 500, причем спектральных цветов - 150, и 30
насыщенных пурпурных. По данным Мак-Адама, который является
одним из наиболее известных исследователей цветового зрения,
человек воспринимает 250 спектральных и 140 насыщенных пурпурных
цветов. Если учесть различие цветов по яркостям, то глаз человека
может воспринимать примерно 20 000. различных цветов,
отличающихся по яркости, цветовому тону и насыщенности.
Таблица 6.4
Сопоставление энергетических и светотехнических параметров света
N
1
2
3
4
Энергетические
параметры света
Поток излучения (лучистый поток)
<b=Wlt, Вт
Энергетическая сила света
1э = Фд/ш, Вт/ср - сила излучения
Энергетическая яркость
Вэ = l^S, Вт/ср ■ м2
Энергетическая освещенность
(облученность) Еэ = Фэ/S, Вт/м2
N
1
2
3
4
Светотехнические
параметры света
Световой поток Φ = / ■ со, лм,
лм = кд · ср,
ω - телесный угол, в который
распространяется свет,
лм - люмен; ср - стерадиан
Сила света /, кд
Яркость В = I/S, кд/м2
Освещенность Ε = Φ/S, лк, лк = лм/м2,
лк - люкс
Сопоставление энергетических и светотехнических
параметров света (табл. 6.4). Основным энергетическим параметром света
служит поток излучения - это энергия квантов света, излучаемая
в единицу времени. Энергетическая сила света представляет собой
поток излучения, приходящийся на единицу телесного угла в данном
направлении. Под энергетической яркостью понимают энергетиче-
Глава 6. Сенсорные системы организма
305
скую силу света, отнесенную к единице площади проекции
поверхности излучающего тела на направление, перпендикулярное к
распределению света. Энергетическая освещенность - это поток излучения,
падающий на единицу площади облучаемой поверхности.
Следовательно, энергетическая яркость является характеристикой источника
излучения, а энергетическая освещенность характеризует
облучаемую поверхность.
За основную светотехническую величину принята сила света.
Ее размерность - кандела [кд]. Значение канделы принимают таким,
что яркость полного излучения при температуре затвердевания Pt
(2042°К или 1769°С) равна 60 кд/см2. Применявшаяся ранее
международная свеча составляет 1,005 кд. Производными параметрами от
силы света являются яркость и световой поток. Единицей яркости
служит кд/м2, т.е., 1 кд/м2 характеризует яркость источника каждого
м2 излучающей поверхности, которая имеет в данном направлении
силу света, равную 1 кд. В медицине широко используются и
внесистемные единицы яркости света: нит, стильб (сб), апостильб (асб),
ламберт (Лб):
1 нит = 1, кд/м2; 1 сб = 1, кд/см2;
1 асб = 3,18· 10"5 сб = 0,318 ныт = 10^ Лб
Яркость снега в солнечный день достигает примерно
2,5 · 104 кд/м2, а яркость земляного грунта беззвездной ночью -
примерно 2 ■ 10~5 кд/м2.
Световой поток определяется произведением силы света на
телесный угол, в который он распределяется. Производной величиной
от светового потока является освещенность, представляющая собой
световой поток, приходящийся на единицу облучаемой поверхности.
Б.4. Биофизика хемосенсорных систем
К хемосенсорным системам человека относятся обонятельная,
вкусовая, интерорецептивная.
В первой из них присутствуют первичночувствующие, во
второй - вторичночувствующие, а в интерорецептивной - свободные
306
Биофизика
нервные окончания и вторичночувствующие рецепторы.
Естественно, что механизмы преобразования химических стимулов в нервную
импульсацию в разных хемосенсорных системах неодинаковы.
Вместе с тем неоднородность хеморецепторных механизмов проявляется
не только в наличии (во вторичночувствующих рецепторах) или
отсутствии (в первичночувствующих рецепторах и свободных нервных
окончаниях) синаптической передачи от той структуры, где
происходит взаимодействие химического агента с рецептором, к месту
генерации потенциала действия. Неоднородны и сами процессы
взаимодействия стимулов с рецепторами. Они зависят от химической
природы адекватных раздражителей хеморецепторов. Такое
представление развивается сторонниками теории гетерогенности хемо-
рецепции (В.О.Самойлов, 1983).Она формулировалась как
своеобразная альтернатива теориям, каждая из которых постулировала
единообразный механизм первичного взаимодействия любого
химического агента с хеморецепторами. Среди них наибольшее
распространение получили адсорбционная (стереохимическая) и
метаболическая (ферментативная) теории.
Согласно первой из них химический агент взаимодействует
с рецепторными молекулярными комплексами (преимущественно
гликопротеидами) в мембране специальной клетки или нервного
окончания. Результатом такого взаимодействия является изменение
конформации мембранного рецептора, обеспечивающее его реакцию
с G-белками, которая приводит к открыванию в плазмолемме
рецептора ионных каналов. Влияние конформационных перестроек
мембранных рецепторов на изменения проницаемости ионных каналов
осуществляется либо при участии различных вторичных мессендже-
ров (для одних стимулов), либо без них (для других стимулов).
В свою очередь, потоки ионов через каналы, открывающиеся под
действием раздражителей, создают рецепторные потенциалы, а они
обеспечивают дальнейшее преобразование сигналов в потенциалы
действия.
Основные положения адсорбционной теории вкуса
сформулировал Л. Бейдлер (1961), а стереохимической теории обоняния -
Д. Эймур (1963). Согласно предположению первого из них адсорбция
Глава 6. Сенсорные системы организма
307
химического агента на вкусовых рецепторах определяется
следующим выражением:
К= - , (6.32)
C(N-Z)
где К - константа равновесия, Ζ - количество активных центров на
плазмолемме вкусовой клетки, занятых химическим веществом, N -
общее количество активных центров на этой мембране, С -
концентрация химического вещества (вкусового стимула).
Рецепторный ответ вкусовой клетки (R) прямо
пропорционален количеству активных центров на её плазмолемме, занятых
вкусовым стимулом (Ζ), причем R достигает максимума, когда Ζ = Ν, т.е.
при заполнении им всех активных центров.
Результатом исследований Л. Бейдлера стало уравнение вкуса,
отображающее зависимость величины реакции вкусового рецептора
от концентрации химического стимула:
С С 1
- = -^+—!—. (6.33)
R Л ЯИ
max max
Заметим, однако, что уравнение Бейдлера подтвердилось
в опытах далеко не для всех вкусовых агентов и в довольно узком
диапазоне их концентраций. Не меньше недоразумений возникает
при использовании аналогичных уравнений для обонятельной
рецепции.
Что же касается интерорецептивных хемосенсорных систем,
то применительно к ним адсорбционная теория даже не
рассматривается. Многие исследователи (СВ. Аничков и М.Л. Беленький,
1962) объясняют механизмы трансдукции в интерорецепторах
(например, в клетках каротидного тельца) с помощтю метаболической
теории. Согласно ей под действием химических стимулов в рецеп-
торных клетках происходят сдвиги метаболизма, что и служит
механизмом преобразования раздражителя в электрические реакции
рецептора. Существуют аналогичные взгляды и на механизмы транс-
дукцииво вкусовых рецепторах (ферментативная теория А. Баради
и Г. Бурна, 1951).
308
Биофизика
6.4.1. Хеморецепция внутренней среды организма
Обратимся вначале к интерорецептивным хемосенсорным
системам. Они реагируют на изменения химического состава
внутренней среды организма.
Рассмотрим, к примеру, комплекс рефлекторных реакций,
возникающих с интерорецепторов при нарушении энергетического
баланса организма, когда расход свободной энергии преобладает над ее
восполнением. Изменения химического состава внутренней среды,
являющиеся следствием этого состояния, воспринимаются хеморе-
цепторами, с которых возникают безусловные рефлексы,
направленные на восстановление в тканях нормального баланса энергии. Среди
них в первую очередь следует назвать дыхательные рефлексы,
проявляющиеся в усилении внешнего дыхания, поскольку для компенсации
затрат свободной энергии клетки нуждаются в усиленной доставке им
кислорода. Вследствие усиления внешнего дыхания и легочного
кровообращения повышаются темпы поступления кислорода в кровь.
Наряду с дыхательными и сосудодвигательными рефлексами,
происходят рефлекторные сокращения селезенки, приводящие к
увеличению количества эритроцитов в циркулирующей крови и тем
самым способствующие лучшему транспорту кислорода от легких
к тканям. Безусловно-рефлекторные связи между интерорецепторами
и селезенкой осуществляются при участии мозгового вещества
надпочечников, создающего под действием интерорецептивных сигналов
гиперадреналемию (повышенное содержание в крови адреналина).
Вместе с влиянием на селезенку адреналин усиливает распад
гликогена в печени, что приводит к гипергликемии. Эффективному
извлечению тканями глюкозы из крови способствует повышенная
продукция инсулина, которая также регулируется рефлекторно при
возбуждении интерорецепторов.
При нарушении энергетического баланса организма возникают
и другие интерорецептивные безусловные рефлексы, а
перечисленные реакции были экспериментально изучены выдающимся
советским фармакологом СВ. Аничковым и его учениками.
Установленные ими факты были квалифицированы как одно из первых открытий,
официально зарегистрированных в Советском Союзе. СВ. Аничков
Глвва 6. Сенсорные системы оргенизме
309
Рис 6.48. Каротидный клубочек кошки. Микрофото.
Съемка на МФН—5,10-кратное увеличение
стал лауреатом Ленинской премии. В течение многих лет он
исследовал функцию каротидного тельца — органа, в котором сосредоточены
интерорецепторы со специальными (гломусными) клетками Именно
в каротидном тельце лучше, чем в других органах изучены механизмы
химического анализа внутренней среды организма интерорецептора-
ми. Рассмотрим их в качестве примера.
Каротидное тельце млекопитающих расположено в развилке
общей сонной артерии (рис. 6.48).
Хеморецепторы, сосредоточенные в этом органе,
предназначены для проточного химического анализа артериальной крови,
которая течет по его капиллярам. В пространстве между капиллярами
находятся специальные (гломусные) клетки, образующие синапсы
с чувствительными нервными окончаниями. Следовательно, каро-
тидные хеморецепторы относятся к вторичночувствующим, и
нетрудно подметить аналогию их с вкусовыми рецепторами. На эту
аналогию первым обратил внимание Б.И. Лаврентьев (1944),
который образно назвал каротидное тельце «органом вкуса крови».
На рис. 6.49 схематически представлен механизм
преобразования информации о химическом составе крови в каротидном хеморе-
цепторе.
310
Биофизика
Генерация РП под действием поваренной соли происходит при
неизменяющейся ионной проницаемости плазмолеммы гломусной
клетки. Эта деполяризация осуществляется за счет сдвига
концентрационного градиента ионов натрия на мембране при возрастании
концентрации поваренной соли в плазме крови. Такая возможность
прямо следует из уравнения Гольдмана - Ходжкина - Катца. В ответах
на поваренную соль выявлена логарифмическая зависимость
амплитуды РП от концентрации стимула. Только под действием NaCl
генерируется РП с крутым передним фронтом, что является необходимым
условием надежного декрементного распространения потенциала по
невозбудимой мембране.
Создается впечатление, будто деполяризационный сдвиг
мембранного потенциала гломусной клетки выполняет функцию
кодирования сигнала (химического раздражителя) только в ответах на
поваренную соль. Иными словами, только под действием этого стимула
на плазмолемме гломусной клетки возникает рецепторный
потенциал в настоящем значении (подлинном смысле) этого термина.
При воздействии кислот и алкалоидов деполяризация
гломусной клетки развивается вяло, вследствие чего так называемый РП
Рис 6.49. Схема преобразования химического стимула
в потенциалы действия каротидными хеморецепторами
I- действие стимула, II — рецепторный потенциал (РП), III — генераторный
потенциал, IV- потенциалы действия^ - стимул (химический агент),
U — разность потенциалов, t — время, I — кровеносный капилляр, 2 — гломусная
клетка, 3 — чувствительное нервное окончание, 4 — афферентное волокно
Глава 6. Сенсорные системы организма
311
достигает амплитудного значения позже, чем возникают потенциалы
действия в афферентных нервных волокнах, связанных синапсами
с гломусными клетками.
В 1970-е годы установили, что кислоты и алкалоиды, в отличие
от поваренной соли и Сахаров, вызывают в гломусных клетках очень
быстрые сдвиги метаболизма. Их латентный период оказался короче,
чем у генерации РП, и тем более, потенциалов действия.
Интенсивность метаболических реакций гломусных клеток имеет
логарифмическую зависимость от концентрации Н+ - ионов и исследованных
алкалоидов.
По-видимому, кислоты проникают из крови в гломусные
клетки и вмешиваются в их метаболизм. Особая роль при этом
принадлежит буферным системам крови и карбоангидразе гломусных клеток.
При закислении крови бикарбонатный буфер нейтрализует Н+-ионы
с образованием С02. Это вещество в силу почти абсолютной
неполярности легко проникает через клеточные мембраны. В течение
нескольких миллисекунд С02 оказывается в цитозоле гломусных
клеток. Благодаря присутствию там карбоангидразы происходит
быстрая гидратация углекислого газа и образуется угольная кислота,
диссоциирующая на Н+ и НСО~3.
Так, водородные ионы проникают в гломусные клетки,
«переодевшись» в С02, с тем чтобы в их цитоплазме сбросить с себя эти
«одежды» и вступить во внутриклеточные реакции, итогом которых
будут усиление биологического окисления, мобилизация Са2+ из
клеточных депо и другие метаболические сдвиги. Накопление Са2+ в ба-
зальном полюсе гломусной клетки поблизости от чувствительного
нервного окончания обеспечивает синаптическую передачу сигнала
с рецепторной клетки на афферентное волокно, которое проводит
информацию о химической стимуляции каротидного рецептора
в центральную нервную систему.
Алкалоиды в силу липофильности легко проникают через
плазмолемму гломусной клетки и в ее цитоплазме ингибируют фос-
фодиэстеразу - один из ключевых ферментов системы циклического
аденозин-монофосфата (цАМФ). При угнетении фосфодиэстеразы
содержание цАМФ в гломусной клетке возрастает, что приводит
к усилению в ней гликолиза, липолиза и других метаболических про-
312
Биофизика
цессов. Гломусную клетку, образно говоря, охватывает
«метаболическая буря», которая затихает столь же быстро, как и возникает.
В ходе этих метаболических процессов происходит кодирование
информации о действии на рецепторные клетки алкалоидов. В синапти-
ческой передаче сигнала с гломусной клетки на афферентное
волокно участвуют ионы кальция, освобождающиеся из внутриклеточных
депо в области пресинапса при «метаболической буре». Са2+
способствует выходу медиатора в синаптическую щель.
Следствием «метаболической бури» является изменение
мембранной проницаемости для ионов натрия (посредством повышения
протеинкиназной активности под действием цАМФ), приводящее
к деполяризации гломусной клетки. Деполяризационный сдвиг
регистрируется при ее микроэлектродном исследовании, причем его
принято называть рецепторным потенциалом (РП). По-нашему мнению,
он в таком случае, в отличие от реакции на NaCl, не выполняет
функции кодирования информации о химической стимуляции и не
участвует в передаче сигнала по гломусной клетке.
Таким образом, первичные механизмы восприятия гломусны-
ми клетками разнообразных химических веществ имеют не
универсальную, а различную физико-химическую природу. Поваренная
соль сдвигает мембранный потенциал рецепторной клетки за счет
изменения концентрационного градиента ионов натрия (в соответствии
с уравнением Гольдмана-Ходжкина-Катца). Ионы водорода
проникают в гломусные клетки и на короткое время нарушают их
метаболизм. Алкалоиды делают то же самое, но более сложным путем. Для
ацетилхолина имеются специфические белки-рецепторы в плазмо-
лемме не гломусных клеток, а нервных окончаний, сосредоточенных
в каротидном теле. Так можно вкратце сформулировать теорию
гетерогенности хеморецепции (В.О. Самойлов, 1983). Аналогичные
механизмы рецепции перечисленных выше классов химических
веществ установлены во всех хемосенсорных системах организма.
Рецепторы со специальными клетками, в которых происходят
описанные выше события, присутствуют не только в
специализированных рецепторных органах (тельцах). Морфологи обнаружили их
почти во всех тканях, причем эти клетки оказались весьма
полиморфными (рис. 6.50). Многие из них обеспечивают не хеморецепцию,
Глава 6. Сенсорные системы организме
313
Рис 6.50. Интерорецепторы языка кошки. Цитохимическое выявление
карбоангидразы по методике Курата в модификации Хаузлера.
Микрофото. Съемка на фотомикроскопе «Оптон-3».
Нервные волокна погружены в «кисет», образованный специальными
клетками, содержащими карбоангидразу — фермент, выполняющий
важную функцию в рецепции кислотного стимула. Со специальных клеток
осуществляется синоптическая передача сигнала на нервную терминаль
а другие виды чувствительности, но всегда и всюду с ним связана
высокая избирательная восприимчивость определенных стимулов.
За счет существования как свободных нервных окончаний, так
и рецепторов с более сложной организацией любая интерорецептив-
ная зона обладает разнопороговыми афферентными системами.
Предполагают, что при возбуждении высокопороговых интерорецеп-
торов возникают болевые ощущения, а также генерализованные
вегетативные реакции. Низкопороговые афферентные системы служат
формированию более ограниченных и специализированных
рефлекторных реакций (например, усиление гемопоэза в костном мозге при
повышении активности мононуклеарных фагоцитов).
Подобно другим рецепторным приборам, интерорецепторы
являются нелинейными преобразователями информации об
интенсивности раздражителя. Для многих интерорецептивных зон
установлена логарифмическая зависимость величины афферентных
ответов и рефлекторных реакций от концентрации химических стимулов.
314
Биофизика
Например, так кодируют интенсивность стимуляции каротидные хе-
морецепторы.
Сейчас пересматривается бытовавшее прежде мнение о низкой
чувствительности интерорецептивного анализатора. Полагали, будто
самой чувствительной хемосенсорной системой является обоняние,
второе место отдавали вкусу, думая, что интерорецепция им сильно
уступает. Сравнивать разные хеморецепторы довольно сложно хотя бы
потому, что им свойственны разные адекватные_раздражители.
Например, на кислоты обонятельные рецепторы реагируют слабо, вкусовые -
лучше, но самой высокой чувствительностью к кислотному стимулу
обладают каротидные, тканевые и медуллярные хеморецепторы. Так,
и каротидные хеморецепторы, и тканевые рецепторы кошки
возбуждаются, когда рН крови снижается на 0,1. Никакие сенсорные системы не
могут сравниться с каротидной по чувствительности к снижению
напряжения кислорода. Перечень подобныхпримеров уже сейчас
довольно велик. Словом, при внимательном рассмотрении этого вопроса
выяснилось, что интерорецептивный анализатор отнюдь не такой
низкочувствительный, как считали прежде.
Нередко роль интерорецептивной сигнализации ограничивают
только участием в регуляции вегетативных функций. В связи с этим
уместно вспомнить, что еще в 1864 г. в докторской диссертации
«Хорея, ревматизм и эндокардит» и позднее в монографии «Учение
о tabes dorsalis» (1867) И.Ф. Цион писал о существовании
рефлекторных влиянийрецепторов внутренних органов на скелетные мышцы.
В этом он усматривал механизм локомоторных расстройств при ряде
внутренних болезней, например подергиваний скелетных мышц при
глистных инвазиях. Кстати говоря, именно И.Ф. Цион в лаборатории
К. Людвига открыл знаменитый депрессорный нерв и первым
высказал мысль о саморегуляции кровяного давления при помощи
рефлексов с механорецепторов дуги аорты. И.М. Сеченов утверждал, что
«смутное валовое чувство общего благополучия» зависит прежде
всего от сигналов, поступающих в мозг от внутренностей и мышц.
Учение об интерорецепции составляет в сенсорнойфизиологии
особую главу, написанную преимущественно отечественными
естествоиспытателями. Причина приоритета заключена, по-видимому,
в том, что отечественная физиология развивалась в Военно-медицин-
Глава 6. Сенсорные системы организма
315
ской академии и российских университетах прежде всего в связи
с потребностями клинической медицины, которая с давних пор
ставила перед учеными проблему чувствительности внутренних органов.
СП. Боткин и И.М. Сеченов без обиняков провозгласили тезис о
родстве органов чувств и рецепторных приборов всех органов и тканей.
Эти парадоксальные для XX века идеи, когда деятельность органов
чувств считалась неотделимой от ощущений, развил И.П. Павлов.
В 1894 г. он писал: «Все ткани богато снабжены рецепторами.... Эти
окончания необходимо представлять как крайне разнообразные,
специфические, подобно окончаниям нервов органов чувств,
приспособленное каждое к своему раздражителю механического, физического
или химического характера образования».
Объединение закономерностей реагирования органов чувств
с особенностями работы тканевых рецепторов явилось одной из
важных предпосылок создания И.П. Павловым учения об анализаторах,
в котором было преодолено мнение, будто обязательным атрибутом
функционирования органов чувств служат специфические
ощущения. Результатом анализаторной деятельности могут быть также
разнообразные рефлекторные акты, многие из которых не осознаются
человеком, и «неосознаваемые эмотивные состояния» (М.И. Аства-
цатуров, 1936). Открытие интерорецептивных условных рефлексов
укрепило позиции сторонников такого мнения. В конце 1<930-х годов
Г.В. Гершуни доказал, что и работа органов чувств (в частности,
слуха) может не осознаваться человеком, особенно при воздействии
стимулов, подпороговых относительно ощущений, но пороговых и над-
пороговых относительно возбуждения рецепторов. В таком случае
рецепторы являются источником безусловных рефлексов и
подсознательных «эмотивных состояний».
Учение об интерорецептивном анализаторе в том виде, как его
разработали отечественные физиологи, не получило за рубежом
всеобщего признания. Основные разногласия касаются вопроса о
распространенности интерорецепторов. Многие зарубежные ученые
поддерживают мнение К. Гейманса (К. Heymans), будто интероре-
цепторы сосредоточены только в отдельных участках тела человека
(например, в каротидном синусе, в дуге аорты), а не во всех органах
и тканях. Такому взгляду противоречат прежде всего данные морфо-
316
Биофизика
логического изучения интерорецепторов, обнаруженных
повсеместно. При проведении же физиологических изысканий вначале
казалось, будто тканевые хеморецепторы уступают в чувствительности
рецепторам сосудистых хемосенсорных зон, в частности каротид-
ным. Позднее было установлено, что сказанное справедливо только
для некоторых раздражителей, например для снижения напряжения
кислорода в крови. Для большинства стимулов чувствительность
тканевых артериальных хеморецепторов примерно одинакова и, что
важнее всего, есть раздражители, адекватные только для
интерорецепторов определенных органов, что связано с особенностями их
метаболизма. Этот путь исследования интерорецептивного
анализатора - изучение специфичности интерорецепторов разных органов
и тканей - был указан И.П. Павловым, но исследователи встали на
него совсем недавно. Успехи на указанном пути важны не только для
развития физиологии, но и для прогресса практической медицины.
6.4.2. Первичные вкусы и их пороги
Многообразие вкусовых ощущений принято сводить к
четырём основным (первичным, элементарным): соленому, кислому,
горькому и сладкому. Из их сочетаний складываются более сложные
вкусовые оттенки. Это отображает модель формирования сложных
вкусов - пирамида вкусов Хеннинга (рис. 6.51). В пространстве
между вершинами пирамиды, соответствующими четырём основным
вкусам, находятся сложные вкусовые оттенки.
Пороговые концентрации веществ, вызывающих у человека
типичные вкусовые ощущения, неодинаковы. Самым низким порогом
обладают агенты, имеющие вкус горечи. Он характеризуется
концентрацией в 8 мкмоль · л-1 для сернокислого хинина и в 16 мкмоль ■ л"1
для никотина. Для возникновения кислого вкуса концентрация
кислот в ротовой полости человека должна достигать более высоких (на
2-3 порядка) величин: для НС1 - 0,9 ммоль · л-1, а для лимонной
кислоты - 2,3 ммоль · л-1. Следует заметить, что приведенные
концентрации являются пороговыми при наличии в ротовой полости слюны.
Если же во рту нет слюны, то пороги кислого вкуса на 1-2 порядка
выше и становятся примерно такими, какие характерны для сладких
Глава 6. Сенсорные системы организма
317
Соленое
/ \\ Кислое
Сладкое Г°Рьков
Рис. 6.51. Пирамида вирусов Хеннинга
и соленых веществ (для NaCl - 10 ммоль · л-1, для сахарозы -
10 ммоль · л-1, для глюкозы - 80 ммоль ■ л-1). Правда, пороговая
концентрация сахарина (23 мкмоль · л"') по сравнению с глюкозой более
чем на 3 порядка ниже.
При стимуляции вкусовых рецепторов химическими агентами
возникают не только ощущения, но также рефлекторные реакции
(слюноотделительные, сосудодвигательные, дыхательные и др.),
подсознательные «эмотивные состояния», вкусовые предпочтения при
пищевом поведении. Наиболее низким порогом безусловных
рефлексов, вызываемых воздействием химических агентов на вкусовые
рецепторы, обладают слюноотделительные рефлекторные реакции. Для
них пороговые концентрации всех типичных вкусовых агентов
примерно такие же, как и для ощущений. Для неосознаваемых вкусовых
предпочтений пороги ниже.
6.4.3. Вкусовые почки
Вкусовые рецепторы входят в состав периферического органа
вкуса, который имеет несколько названий: вкусовая почка, вкусовая
рюмка, вкусовая луковица. Анатомическая номенклатура
рекомендует последнее из них, но в данном учебном пособии будет
применяться первое.
Форма вкусовой почки различна у разных представителей
животного мира: у земноводных - блюдцеобразная (рис. 6.52 и 6.53),
а у млекопитающих она имеет форму эллипсоида и похожа на апель-
318
Биофизика
Рис. 6.52. Схема клеточной организации вкусовой почки лягушки
tc — вкусовые клетки, ptc — предвкусовые клетки, s - опорные клетки,
gc — секретирующие опорные клетки, ее - цилиарные клетки,
тс - клетки Меркеля, be - базапьные клетки, gbc - бокаловидные клетки,
of— афферентные волокна, рп — периневрий, ср - кровеносный капилляр
стрелками указаны синапсы на вкусовых клетках
Рис 6.53. Грибовидный сосочек языка лягушки. Прижизненное окрашивани
метиленовым синим, 5 · 10~ моль, ионофорез. Витальный препарат
Между контуром наружной поверхности грибовидного сосочка и базальнои
мембраной находится вкусовая почка, поперек которой расположены
вкусовые клетки. Под базальнои мембраной — петля кровеносного капилляра
Глава 6. Сенсорные системы оргвнизма
319
Рис. 6.54. Схема вкусовой почки млекопитающих (Taste роге)
Вкусовые клетки (темные) лежат между опорными (светлыми). Базолатеральные
участки вкусовых клеток контактируют с чувствительными нервными волокнами
син (рис. 6.54), среди «долек» которого присутствуют рецепторные
и опорные клетки.
В свою очередь, вкусовые почки лежат в эпителии
грибовидных, желобоватых и листовидных сосочков, представляющих собой
выпячивания слизистой оболочки ротовой полости (на дорсальной
и боковых поверхностях языка, мягкого нёба), надгортанника,
передних отделов глотки и гортани. У человека половина всех вкусовых
почек сосредоточена в желобоватых сосочках. Они расположены на
корне языка в V-образной бороздке. Каждый такой сосочек окружен
ровиком, на дне которого находятся отверстия выводных протоков
Эбнеровских желез, секретирующих жидкий сок, в котором
растворяются вкусовые агенты для лучшего взаимодействия с
рецепторами. В языке человека примерно 10 (от 7 до 12) желобоватых
сосочков, а в каждом их них около 250 вкусовых почек.
Грибовидные сосочки расположены в передних 2/3 языка, они
мельче желобоватых (диаметр не более 1 мм и высота до 2 мм).
320
Биофизика
У человека их количество не превосходит 200. В 60% грибовидных
сосочков нет вкусовых почек, а в тех, где они присутствуют, их
число неодинаково (максимум - 30, как и у лягушки). Листовидные
сосочки расположены на боковых поверхностях языка, ближе к его
корню. Их много у кролика, а у человека - очень мало, причем
у взрослых людей они встречаются крайне редко. Заметим, что по
мере старения человек утрачивает часть вкусовых почек.
С топографией вкусовых сосочков отчасти коррелируют зоны
преимущественной вкусовой чувствительности: кончик языка
наиболее чувствителен к сладким и соленым веществам, боковые
поверхности - к кислым, корень - к горечам.
У рыб вкусовые почки рассредоточены по всей поверхности
тела.
У млекопитающих вкусовая почка расположена таким образом,
что ее длинная ось, равная 50-80 мкм, перпендикулярна к
поверхности-языка. Вкусовая почка простирается на всю толщу эпителия от
базальной мембраны до свободной поверхности, где находится
вкусовая пора. Она заполнена гомогенной массой, образованной глико-
заминогликанами и белками.
Ниже вкусовой поры рецепторные и опорные клетки
соединены между собой плотными контактами и десмосомами, благодаря
чему химические агенты не проникают в глубь вкусовой почки. Они
взаимодействуют с апикальным отростком рецепторной клетки,
который проходит через вкусовую пору, и доступны химическим
веществам, растворенным в слюне.
6.4.4. Вкусовые клетки
Рецепторные (вкусовые) клетки составляют 10% клеточного
состава вкусовой почки. Еще меньше базальных клеток, а львиная
доля приходится на опорные клетки. Предполагают, что многие из
них представляют собой «несозревшие» рецепторные, т.е.
промежуточную форму развития вкусовых клеток, срок жизни которых
составляет 250 ± 50 ч. Происходит непрерывное обновление рецеп-
торных клеток, которое управляется центральной нервной
системой. При денервации языка новые рецепторные клетки не образу-
Глава 6. Сенсорные системы организма
321
ются, и по мере отмирания старых животное лишается вкусовой
рецепции.
Длина вкусовой клетки около 50 мкм, а толщина - всего 5-
6 мкм. На ее базальном полюсе сосредоточены синапсы. Их большая
часть обеспечивает эфферентный контроль, так называемые
трофические влияния центральной нервной системы на развитие,
метаболизм и чувствительность рецепторных клеток.
Вместе с тем базальный полюс вкусовой клетки содержит
типичные элементы пресинаптической ультраструктуры - гранулы
и везикулы диаметром от 40 до 60 нм. На одной рецепторной клетке
находится от 1 до 6 таких пресинаптических участков, каждый из
которых контактирует с чувствительными нервными волокнами.
Предполагается, что медиатором в таких (афферентных) синапсах
вкусовой клетки служит либо серотонин, либо норадреналин. Именно эти
синапсы обеспечивают передачу информации о свойствах
химических (вкусовых) веществ из ротовой полости в ЦНС. Следовательно,
вкусовые рецепторы по структуре и механизму транедукции
относятся ко вторичночувствующим.
6.4.5. Афферентные пути
От хеморецепторов, расположенных в передних 2/3 языка,
сигналы поступают в ЦНС по афферентным волокнам, входящим в
состав язычного нерва. Большую часть его волокон составляют дендри-
ты нейронов Гассерова узла, т.е. принадлежат системе тройничного
нерва. Они обеспечивают тактильную, температурную, болевую, но
не вкусовую чувствительность языка.
Сигналы от хеморецепторов проводят волокна так называемой
барабанной струны, отходящие от языка в составе язычного нерва,
а хемосенсорные волокна представляют собой дендриты нейронов не
Гассерова, а коленчатого ганглия. Их центральные отростки
образуют Врисбергов нерв (его называют XIII парой черепно-мозговых
нервов) и следуют к нейронам ядра одиночного пучка, где происходит
синаптическая передача на следующий нейрон (нейрон II порядка)
вкусовой сенсорной системы. Это место считается первичным
переключающим реле для афферентной импульсации от вкусовых рецеп-
322
Биофизика
торов. К нему поступают сигналы и от вкусовых рецепторов задней
трети и корня языка. Их проводят афферентные волокна языкогло-
точного и блуждающего нервов. Они представляют собой дендриты
и аксоны псевдоуниполярных нейронов нижнего (каменистого) узла.
6.4.6. Механизмы вкусовой трансдукции
Как уже говорилось, механизмы вкусовой трансдукции
пытаются объяснить с позиций унитарной адсорбционной теории вкуса
(Л. Бейдлер, 1961). Однако во вкусовых клетках до сих пор выявлены
мембранные рецепторы только для сладких веществ. Методом
молекулярного клонирования получен специфический для вкусовых
клеток G-белок, одна из субъединиц которого (сс-гастдуцин) оказалось
сходной с сс-трансдуцином, выявленным в фоторецепторах.
Механизм трансдукции сладкого стимула представляется
следующим образом (рис. 6.55). Вкусовой агент связывается со сладко-
чувствительным мембранным рецептором в апикальном отростке
вкусовой клетки. Следствием адсорбционного взаимодействия является
изменение конформации мембранного рецептора (сладкочувстви-
тельного белка), что обеспечивает его реакцию с G-белком (сс-гастду-
цином), вследствие чего активируется аденилатциклаза,
катализирующая превращение АТФ в цАМФ, который, в свою очередь, активирует
протеинкиназу А, а она блокирует калиевые каналы в плазмолемме
вкусовой клетки, за счет чего мембрана деполяризуется. Деполяриза-
ционный сдвиг мембранного потенциала на апикальном полюсе
вкусовой клетки является рецепторным потенциалом. Он
распространяется электротонически (с декрементом) к базальному полюсу
рецепторной клетки, где инициирует синаптическую передачу на
чувствительные волокна барабанной струны. В них генерируются
потенциалы действия, в паттернах которых закодирована информация
о свойствах вкусового агента, которая поступает в мозг.
По аналогии с рецепцией сладких веществ пытаются
рассматривать трансдукцию других типичных вкусовых агентов. Однако ни для
одного из них не обнаружен специфический молекулярный рецептор
в мембране вкусовой клетки. Согласно теории гетерогенности хеморе-
цепции (В.О. Самойлов, 1983), трансдукция химических стимулов,
Глвва б. Сенсорные системы оргенизма
323
2
/
β У
С ι . ·
Рис. 6.55. Механизм трансдукции веществ, обладающих сладким вкусом
1 - сладкий рецептор; 2 - аденилатциклаза; 3 - протеинкиназа А
вызывающих основные (типичные) вкусовые ощущения,
осуществляется посредством различных (неоднородных) физико-химических
процессов. Сладкие вещества рецептируются так, как утверждает
адсорбционная теория вкуса (см. выше).
При исследовании рецепции поваренной соли было
установлено, что если ее концентрации на поверхности вкусовых клеток
достигают пороговой величины (10 ммоль · л-1), то происходит их
деполяризация в соответствии с уравнением Гольдмана-Ходжкина-Катца
за счет увеличения концентрационного градиента. При этом
проницаемость плазмолеммы не изменяется (В.О. Самойлов, А.Н. Лотарев,
1986). Следует отметить, что передний фронт деполяризационного
сдвига мембранного потенциала под действием NaCl довольно
крутой - величина Vmax гораздо больше, чем в ответах на другие
вкусовые агенты.
Достаточно выраженная крутизна переднего фронта рецептор-
ного потенциала необходима для его надежного декрементного
распространения по невозбудимой мембране вкусовой клетки. При
малой величине Fmax электрический сигнал затухает настолько, что не
может инициировать синаптическую передачу. По нашим данным,
деполяризация вкусовой клетки при действии на нее адекватных кон-
21-
324
Биофизика
центраций кислых и горьких веществ, в отличие от NaCl, развивается
слишком вяло, чтобы ее можно было считать рецепторным
потенциалом, а в рецепции поваренной соли деполяризационный сдвиг
мембранного потенциала может выполнять миссию рецепторного
потенциала (в функциональном смысле этого термина). Его роль
заключается в том, что деполяризация пресинаптической мембраны
открывает в ней кальциевые каналы. Са2+ входит из интерстиция
и обеспечивает пиноцитоз медиатора.
Кислоты в высоких концентрациях могут сдвигать
мембранный потенциал вкусовой клетки по тому же механизму, что и NaCl.
Однако при более высоком рН он не может обеспечить рецепцию
кислых веществ. С давних пор было известно, что присутствие в
ротовой полости слюны понижает порог реагирования вкусовых
рецепторов на кислоту. Оказалось, что аналогичный эффект обусловливает
раствор бикарбоната во рту. Эффект повышения вкусовой
чувствительности к кислым агентам в присутствии слюны и раствора
бикарбоната устраняется после аппликации на язык ингибиторов карбоан-
гидразы - фермента, катализирующего реакцию гидратации С02.
Гистохимическое изучение вкусовых клеток земноводных и
млекопитающих выявило в них высокое содержание карбоангидразы.
Совокупность приведенных фактов привела к выводу, что
кислые агенты, попав в ротовую полость, вступают в реакцию с
бикарбонатами, входящими в состав слюны. Продуктом реакции является
углекислый газ, который легко проникает через плазмолемму
вкусовой клетки, где гидратируется. Благодаря карбоангидразе реакция
образования Н2С03 протекает в цитозоле вкусовой клетки менее чем
за 10 мс. Синтезированная угольная кислота диссоциирует на Н4
и НСО"3. Ион водорода вмешивается в метаболизм рецепторной
клетки, создавая эффект кратковременной гипоксии за счет избытка
субстрата биологического окисления. Следствием является усиление
клеточного дыхания, что было обнаружено посредством
исследования собственной флуоресценции восстановленного никотинамидаде-
ниндинуклеотида НАД · Η и окисленных флавопротеидов в рецеп-
торном эпителии языка при действии на него кислот (В.О. Самойлов,
В.Н. Соловьев, 1978).
Глава 6. Сенсорные системы организма
325
Инициация синаптической передачи с вкусовой клетки на
чувствительное волокно при рецепции кислот может быть обеспечена не
рецепторным потенциалом, который открывает каналы для
поступления Са2+ из интерстиция, а выделением ионов кальция из
внутриклеточных депо (эндоплазматической сети и митохондрий) за счет
ослабления кальцийаккумулирующей способности внутриклеточных
мембран. Именно такая реакция вкусовых клеток на кислые агенты
была обнаружена при помощи исследования вторичной
флуоресценции комплекса: Са2+ - хлортетрациклин-биомембрана (В.О.
Самойлов, Н.Г. Новикова, 1999).
Под действием горсчей во вкусовых клетках происходят
качественно такие же изменения клеточного дыхания и
кальцийаккумулирующей способности внутриклеточных мембран, как и при
рецепции кислых агентов. К таким выводам пришли в результате
прижизненной флуориметрии вкусовых клеток (В.О. Самойлов,
1983). Вместе с тем механизм воздействия горечей на клеточное
дыхание и метаболизм кальция оказался иной, нежели у кислот.
Вещества, обладающие горьким вкусом, являются ингибиторами фосфо-
диэстеразы - фермента, уменьшающего содержание в клетке цАМФ
за счёт превращения его в нециклический АМФ. Ингибирование
фосфодиэстеразы даёт такой же результат, как и активация аденилат-
циклазы, - повышение содержания цАМФ.
В свою очередь, цАМФ активирует протеинкиназы,
обеспечивающие перенос фосфатных групп на различные вещества, которые
благодаря фосфорилированию приобретают высокую реакционную
способность. В клетке резко усиливаются катаболические процессы,
приводящие к повышению содержания субстратов биологического
окисления. Клеточное дыхание кратковременно, но резко
усиливается, что и обнаруживается в ходе прижизненной флуориметрии
НАДН и окисленных флавопротеидов в рецепторном эпителии
языка. Повышение содержания Са2+ в пресинаптической области
вкусовой клетки может обеспечиваться не деполяризацией мембраны за
счет декрементного распространения рецепторного потенциала,
а выходом ионов кальция из внутриклеточных депо благодаря
ослаблению кальцийаккумулирующей способности мембран, что также
удалось установить посредством флуориметрического анализа.
326
Биофизика
А А
Б 1—
ΓΠ-η-π-Γ
"ϊ-1 »
;Γ -п-гПт-П.
80
40
0
100
40
20
О
100
40 I- \—l
20
О
100
40
20
О
123456789 10
Рис. 6.56. Относительные величины рецепторных потенциалов
10 вкусовых клеток лягушки в ответ на типичные вкусовые раздражители
(по данным: Sato, 1969): А - хлорид натрия (500 ммоль), Б-уксусная кислота
(16ммоль), В - сахароза (250 ммоль), Г-хинин (4 ммоль). По оси абсцисс -
номера исследованных клеток; по оси ординат - сдвиги мембранных
потенциалов вкусовых клеток (в % к исходному уровню мембранного потенциала)
Фосфодиэстераза содержится в цитозоле. Для воздействия на
неё горечам необходимо проникать внутрь вкусовой клетки. Для
подавляющего большинства горьких агентов это не является проблемой
в силу их липофильности. Однако есть несколько гидрофильных горе-
чей. Их рецепцию может обеспечивать небольшая мембранная
фракция фосфодиэстеразы, обнаруженная во вкусовых клетках Р.А. Певз-
нер, сотрудницей лаборатории Я.А. Винникова. Наличие мембранной
фракции фосфодиэстеразы является характерной особенностью
вкусовых рецепторов.
Таким образом, рецепция качественно различных вкусовых
агентов (соленых, кислых, горьких, сладких) осуществляется
посредством разнообразных физико-химических процессов. Вместе с тем
механизмы восприятия одних и тех же веществ разными хеморецеп-
торами (вкусовыми, каротидными, медуллярными, тканевыми) оди-
Глава 6. Сенсорные системы организма
327
наковы, за исключением рецепции веществ, обладающих сладким
вкусом. Адсорбционный механизм присущ вкусовым клеткам и
отсутствует в интерорецепторах. Они реагируют на глюкозу как осмо-
рецепторы.
По-видимому, разнообразные механизмы хеморецепции
формируются в развивающейся вкусовой клетке не одновременно.
Косвенным аргументом в пользу такого предположения служат данные
об электрических реакциях вкусовых клеток на NaCl, кислоту,
горечь и сахар (рис. 6.56). На поваренную соль реагируют все вкусовые
клетки, на кислоту - 90%, на горький и сладкий агенты - 60-70%.
Можно полагать, что весьма примитивным механизмом рецепции
NaCl вкусовая клетка обладает, едва зародившись, тогда как
активность карбоангидразы достигает необходимого уровня чуть позже,
а система цАМФ и рецепторные белки и сопряженные с ними G-бел-
ки и вторичные мессенджеры нуждаются в еще большем времени
для своего развития в течение тех 250 ± 50 часов, которые живёт
вкусовая клетка
Гетерогенность хеморецепторных механизмов присуща и
обонятельной рецепции.
6.5. Обоняние
Обоняние представляет собой древний вид сенсорной
рецепции. Не даром существует удивительное сходство в организации
обонятельной системы от насекомых до позвоночных. Обонятельная
система выполняет не только сенсорную функцию, но и
осуществляет модуляторно-регулирующее влияние на деятельность
разнообразных систем мозга, поскольку через обонятельную луковицу
осуществляется связь с центральными структурами лимбической системы.
6.5.1. Запахи и их классификация
Стимулами в обонянии являются химические вещества,
обладающие запахами. Однако качественная их оценка представляет
собой довольно трудную задачу, так как представление о качестве
запаха у человека предметное: предъявляемый стимул характеризуется
328
Биофизика
как запах конкретного цветка (роза, фиалка), фруктов (яблоко,
груша, банан), вещества (камфорный, синильный), а также по тому
ощущению, которое это пахучее вещество вызывает (приятный или
отталкивающий).
Попытки классифицировать известные человеку запахи,
группируя их по сходству вызываемых ощущений, предпринимаются
с XVIII века. К. Линней (1756 г.) предложил все запахи объединить
в 7 классов. В 1895 г. Г. Цваардемакер свел их к 9 типам: эфирный,
ароматический, цветочный (бальзамный), амбро-мускусный,
чесночный, пригорелый, каприловый, отталкивающий, тошнотворный.
Однако ощущение запаха вещества человеком зависит от
многих обстоятельств. В частности, некоторые одоранты пахнут
по-разному в зависимости от их концентрации, а также от
физиологического состояния человека. Пахучесть вещества определяется его
летучестью и способностью проникать в обонятельный орган.
Поэтому в предложенной классификации существует некоторая
произвольность, субъективность в отнесении веществ к тому или иному
классу.
6.5.2. Теории обоняния
В настоящее время существует несколько теорий, пытающихся
объяснить механизм восприятия запаха.
1. Колебательная теория. Исходной посылкой Р.Х. Райта,
автора этой теории, было предположение, что молекулам всех пахучих
веществ свойственны внутримолекулярные колебания, в результате
которых они испускают электромагнитные излучения в
инфракрасной области спектра. Веществам с определенными запахами
присущи специфические спектры излучения. Эти излучения по механизму
резонанса взаимодействуют с молекулами обонятельной выстилки.
В результате этого первичного процесса запускается механизм
генерации потенциала действия. Однако эта теория не нашла
экспериментального подтверждения.
2. Ферментатаивная теория. Согласно мнению Д.В. Ланцета,
в обонятельном эпителии присутствуют два фермента, один из
которых устраняет старые запахи, а второй подготавливает обонятельную
Глава 6. Сенсорные системы организма
329
клетку к восприятию новых. У разных людей активность этих
ферментов различна, что и обусловливает различия в чувствительности.
3. Теории, основанные на взаимодействии молекул пахучего
стимула с каротиноидами или липидами обонятельных клеток. В
соответствии с этими теориями протеинсвязываюшие каротиноиды
обонятельного эпителия являются рецепторами пахучих веществ,
проникающих в носовую полость. Эта теория в дальнейшем
встретила серьезные возражения, так как в обонятельном эпителии многих
позвоночных животных каротиноиды не обнаружены.
4. В настоящее время сохранила актуальность только стерео-
химическая теория, предложенная Д. Эймуром. Она основана на
предположении о существовании семи основных «первичных»
запахов: камфорного, мускусного, цветочного, мятного , эфирного,
острого, гнилостного. Исследовав 600 пахучих веществ, Д. Эймур
предложил теорию, в которой каждому из семи типов запахов соответствует
семь типов стереоспецифических «активных мест» на молекулярном
рецепторе. Запах определяется формой и размером молекулы одоран-
та, которая имеет стереоструктуру, сходную с активным центром
рецептора. Например, молекулы с камфороподобным запахом имеют
форму, близкую к сферической, с диаметром около 0,7 мкм. Такой
молекуле должен соответствовать участок на рецепторе
полусферической формы с таким же диаметром.
Эксперименты подтвердили справедливость стереохимической
теории для пяти из семи первичных запахов. Ей не соответствуют
острые и гнилостные запахи. Для их восприятия определенное
значение имеет электрический заряд: у молекул веществ с острым запахом
он положителен, а у веществ с гнилостным запахом - отрицателен.
Вместе с тем зачастую не существует сколько-нибудь
закономерной связи запаха со структурой молекулы данного вещества.
Пахучие молекулы с различной структурой могут обладать сходными
запахами. Например, розетон, фенилэтанол, гераниол и пеларгол
имеют розоподобный аромат. Около 300 различных веществ пахнут
камфорой, около 5 различных типов молекул - мускусом. И
наоборот, имея сходную молекулярную структуру, одоранты могут
пахнуть по-разному.
330
Биофизика
6.5.3. Органы обоняния
Органами обоняния служат обонятельная выстилка,
расположенная в верхней и средней раковинах полости и пергородке носа,
и трубчатый (Якобсонов) вомероназальный орган, локализованный
в основании носовой перегородки, в нижнемедиальной стенке
носовой полости (рис. 6.57).
Рис. 6.S7. Схема проекций основной и дополнительной
обонятельных систем (Новиков С.Н., 1988). Основная обонятельная
система изображена справа, дополнительная - слева:
1 - основная обонятельная луковица, 2 — передний гиппокамп, 3 - дополнительная
обонятельная луковица, 4 — переднее обонятельное ядро, 5 — обонятельный
бугорок, 6 — ядро-ложе конечной полоски, 7— обонятельная кора, 8-ядро
латерального обонятельного тракта, 9 -ядро дополнительного обонятельного
тракта, 10 - переднее кортикальное ядро амигдалы, 11 - преоптическая область,
12 — медиальное ядро амигдалы, 13 — заднелатеральное ядро амигдалы,
14 — вентромедиальное ядро гипоталамуса, 15 - заднемедиальное кортикальное
ядро амигдалы, 16- энторинальная кора, 17 - премаммилярное ядро,
18 - вомероназальный орган, 19 - обонятельный эпителий
Глава 6. Сенсорные системы организма
331
В состав вомероназального органа входят микровиллярные
рецепторные клетки, которые специализируются на восприятии
феромонов. Сигналы из вомероназального органа, в отличие от
обонятельного эпителия, идут не в основную, а в дополнительную
обонятельную луковицу.
По степени развития органа обоняния и способности
воспринимать запахи все виды животных делят на три категории:
- макросматики, для которых обоняние является основным
источником информации об окружающей среде; они обладают сильно
развитым обонянием (пресмыкающиеся, млекопитающие);
- микросматики, которым свойственна низкая обонятельная
чувствительность (птицы, приматы, человек);
- аносматики, у которых отсутствует обоняние в связи,
например, с переходом к водному образу жизни (дельфины).
6.5.4. Микроструктура обонятельной выстилки
Обонятельная выстилка состоит из многорядного
цилиндрического эпителия толщиной 100-200 мкм и подлежащей
соединительной ткани. Площадь, занимаемая рецепторным эпителием,
различна у разных типов животных. Так, у человека она занимает по 2 см2
в каждой ноздре, а у собаки и свиньи - до 100 см2.
В состав обонятельной выстилки входят три типа клеток:
рецепторные, опорные и базальные (рис. 6.58).
Базальные клетки имеют неправильную кубовидную или
конусовидную форму, не достигают поверхности обонятельного
эпителия, а распластываются над базальной мембраной, которая отделяет
обонятельный эпителий от подлежащей соединительной ткани.
Делящиеся недифференцированные базальные клетки заменяют'отми-
рающие старые рецепторные, срок жизни которых составляет от 5 до
8 недель.
Над базальными клетками располагаются рецепторные и
опорные. Последние имеют форму удлиненной рюмки. Между
мембранами соседних опорных клеток и вершиной каждого периферического
отростка обонятельной клетки имеются плотные контакты.
Благодаря этому создается диффузионный барьер между наружной средой
332
Биофизика
Дендрит
Обонятельная ресничка
Обонятельные булавы
Опорные клетки
Шванновская
клетка
Fila
olfactoria
Микроворсинки
Обонятельная
рецепторная
клетка
Аксон
Базальная
мембрана
Соединительная
ткань собственного
слоя слизистой
оболочки
Кость
решетчатой
пластинки
Боуменова железа
Вена
Миелиновое волокно
тройничного нерва
Рис. 6.58. Микроструктурная организация обонятельной выстилки
и межклеточным пространством. Между опорными и рецепторными
клетками методами in situ гибридизации и иммуногистохимии
обнаружены щелевые контакты, которые играют важную роль в
переработке информации в обонятельном эпителии. В обонятельных
клетках они вовлекаются в координирующую функцию и передачу ионов
и метаболитов в момент роста, дифференцировки клеток и
переработки сигнала.
Глава 6. Сенсорные системы организма
333
Щелевые контакты между опорными клетками участвуют
в поддержании ионного состава интерстициальной жидкости,
которая омывает их и рецепторные клетки, а также обеспечивают
удаление избыточных внеклеточных ионов калия в пероид высокой
активности обонятельных клеток.
Опорные клетки обонятельного эпителия не имеют жгутиков
и снабжены микроворсинками длиной от 1,5 до 3 мкм. В состав
этих клеток входят крупные секреторные гранулы, которые
участвуют в образовании обонятельной слизи. Опорные клетки являются
не только поддерживающим аппаратом, но и выполняют
трофическую и секреторную функции, а также участвуют в удалении из
обонятельного эпителия дегенерировавших обонятельных клеток.
Таким образом, опорные клетки обладают полифункциональными
свойствами.
У всех наземных позвоночных в соединительнотканном слое
располагаются Боуменовы железы, выводные протоки которых
пронизывают ольфакторный эпителий. Благодаря им на поверхность
обонятельной выстилки выделяется белково-полисахаридный секрет,
участвующий в формировании обонятельной слизи. Роль секрета этих
желез важна для функционирования обонятельных клеток, поскольку
в нем растворяются и диффундируют молекулы пахучих веществ,
связываются и удаляются прореагировавшие молекулы. Благодаря
наличию этого секрета на поверхности обонятельного эпителия
поддерживается определенный ионный состав, в частности в секрете Бо-
уменовых желез содержится много натрия.
Слизь ведет себя как структурированный матрикс и играет роль
фактора, определяющего интенсивность, крутизну нарастания и в
некоторой степени качество запаха. Она представляет собой сложный
многокомпонентный раствор биополимеров, обладающий высокой
степенью упорядоченности своей структуры. Это полиферментная
система, обогащенная одновалентными ионами, в состав которой входят
белки, гидролитические ферменты, кислые и нейтральные мукополиса-
хариды и гликопротеиды.
Важным компонентом слизи являются одорантсвязывающие
белки (ОСБ). У всех млекопитающих все известные на сегодняшний
день ОСБ относятся к большому семейству липокалинов. Они явля-
334
Биофизика
ются водорастворимыми, а не мембраносвязанными белками,
которые обратимо связывают несколько типов одорантов и выполняют
функцию неспецифических переносчиков пахучих молекул. В
обонятельной слизи обнаружено несколько различных одорантсвязываю-
щих белков. У млекопитающих они связываются с одорантами
селективно и могут служить фильтрами до того, как пахучий стимул
достигнет рецепторов. ОСБ образуют комплексы с одорантами,
которые могут взаимодействовать с молекулярными рецепторами в
молекуле пахучего вещества. Считают, что ОСБ инактивируют одоранты,
чтобы предотвратить десенситизацию и обеспечить восприятие
нового стимула. Одорантсвязывающим белкам приписывают
различные механизмы инактивации одорантов, включая удаление избытка
пахучих молекул из обонятельной слизи.
В обонятельной выстилке выявлены и локализованы как ре-
цепторные, так и нерецепторные белки. К ним относятся
обонятельный маркерный белок, фермент карнозинсинтетаза, карнозин, пира-
зинсвязываюший белок, анизолсвязывающий белок и другие. Таким
образом, слизевой секрет играет активную роль в периферических
процессах обонятельной рецепции.
В соединительнотканном слое слизистой носа проходят
веточки безмякотного обонятельного нерва, а также мякотные и безмя-
котные волокна тройничного нерва. В составе этого нерва в
обонятельную выстилку носа проникают волокна вегетативной нервной
системы, оканчивающиеся на кровеносных сосудах, Боуменовых
железах и в основном веществе соединительной ткани.
6.5.5. Строение обонятельных клеток
Клетками, непосредственно взаимодействующими с пахучим
стимулом, являются рецепторные (обонятельные) клетки. Они
являются первично чувствующими рецепторами и в среднем живут от 5 до
8 недель, после чего замещаются новыми. Тела их имеют
веретенообразную форму с поперечным диаметром 5-8 мкм. От верхнего полюса
биполярной клетки отходит периферический отросток (дендрит)
диаметром 1-3 мкм. Он простирается до апикальной поверхности оль-
факторного эпителия и имеет длину от 20 до 90 мкм у различных жи-
Глава 6. Сенсорные системы организма
335
вотных. От нижнего полюса начинается тонкий аксон толщиной 0,1-
0,3 мкм, который не разветвляясь, тянется в обонятельную луковицу
переднего мозга. Аксоны обонятельных клеток образуют примерно
20 тонких веточек, которые в совокупности составляют обонятельный
нерв, непосредственно связывающий рецепторные клетки с
митральными клетками обонятельной луковицы.
Количество обонятельных клеток в ольфакторном эпителии
различно у разных животных. Например, у собаки и свиньи их по
225 млн, тогда как у человека - только 10 млн.
Периферический отросток на всем протяжении содержит
микротрубочки, митохондрии, аппарат Гольджи и вакуоли. Часть
обонятельных клеток на своем апикальном полюсе покрыта
микроворсинками. Таких клеток в обонятельном эпителии свиньи насчитывается
до 250 000 на 1 мм2 эпителия, а у человека - около 30 000. У
большинства обонятельных клеток на вершине, обращенной к апикальной
поверхности эпителия, образуется утолщение - булава, увенчанная
жгутиками. Поэтому рецепторные клетки как низших, так и высших
позвоночных относятся к рецепторам жгутикового типа. Булава
особенно богата митохондриями. Она представляет собой один из
важнейших метаболических центров обонятельного рецептора.
Различают три разновидности жгутиков: 1) короткие (20-
30 мкм в длину) и тонкие; 2) средние - такой же длины, но более
толстые, поскольку их общая плазмолемма объединяет от 2 до 8
жгутиков; 3) длинные (50-200 мкм), утончающиеся к вершине.
Количество обонятельных жгутиков на одной рецепторной клетке различно
и может достигать 150 (например, у собак).
Все три разновидности жгутиков содержат набор из 9x2 + 2
микротрубочек, которые начинаются в цитоплазме булавы от базаль-
ного тельца, снабженного ножкой, спицеобразными выростами и
поперечно-исчерченным корешком. Эти ультраструктуры характерны
для всех локомоторных ресничек и жгутиков и обеспечивают их
движения. Тубулярный цитоскелет расположен в проксимальных
отделах жгутиков, тогда как их дистальные участки лишены тубулина
и динеина, вследствие чего пассивно следуют за проксимальным
отделом. В отсутствие одорантов обонятельные жгутики совершают
неупорядоченные асинхронные, изгибательные, волнообразные и ворон-
336
Биофизика
кообразные движения. Двигательная активность их обеспечивается
энергией АТФ. Генератор движений сосредоточен в проксимальном
отделе жгутика, а дистальный участок бьется наподобие хлыста в руке
погонщика.
С цилиарной подвижностью связывают способность
обонятельных клеток воспринимать пахучий стимул. По мнению А.А.
Бронштейна (1966), цилиарные аппараты обонятельных клеток являются
специализированными антеннами, обеспечивающими активный
поиск и обнаружение одорантов, растворенных в слизи. Обнаружив одо-
рант, жгутик начинает совершать упорядоченные движения навстречу
одоранту. Для этого в дистальных участках обонятельных жгутиков
под действием одорантов, как полагают, инициируется
полимеризация актина, т.е. построение актиновых нитей (F-актина) из G-актина,
в результате чего они приобретают самостоятельное упорядоченное
движение. По актиновым нитям («цитокостям») перемещаются
молекулы миозина, что и обеспечивает упорядоченные движения жгутика
в направлении одоранта.
Обонятельным жгутикам принадлежит основная роль в
рецепции одорантов. При их удалении исчезает электроольфактограмма,
которая восстанавливается после регенерации. Именно в их
дистальных участках сосредоточены рецепторные молекулы и другие
компоненты механизма трансдукции обонятельного стимула.
6.5.6. Механизм обонятельной трансдукции
Восприятие пахучего стимула начинается с взаимодействия
его молекулы с мембранным рецептором обонятельной клетки.
Методами биохимического анализа показано, что этот рецептор
локализован в жгутиковой мембране и принадлежит большому семейству
белков, сопряженных с G-белками, содержащих домены, семь раз
пронизывающих плазмолемму.
В обонятельном эпителии методом клонирования выделена
специфическая для него изоформа аденилатциклазы III. Ее
идентифицировали с помощью моноклональных антител и показали, что
она принадлежит семейству G-белокчувствительных аденилатцик-
лаз. В ходе эмбриогенеза аденилатциклаза III никогда не распростра-
Глава 6. Сенсорные системы организма
337
няется ниже слоя обонятельных жгутиков, будучи сосредоточенной
преимущественно в их дистальных отделах.
Вторым ключевым ферментом, участвующим в обонятельной
трансдукции, является специфическая фосфолипаза С, которая
локализуется в жгутиках и вовлекается в фосфоинозитидный путь
передачи информации в клетке.
В обонятельном эпителии обнаружен и специфический для
него ГТФ-связывающий белок (Goi/). По своим свойствам он сходен
со стимуляторными G-белками других тканей, но является особой их
разновидностью, характерной для обонятельного эпителия. Его
свойства обусловливаются уникальной для ольфакторного G-протеина
конфигурацией ГТФ-связывающего сайта.
Таким образом, в обонятельной трансдукции возбуждение ре-
цепторной молекулы передается на аденилатциклазу III или фос-
фолипазу С посредством чувствительного к холерному токсину сти-
муляторного специфического ольфакторного ГТФ-связывающего
протеина (Golj). Раньше полагали, будто внутриклеточной сигнальной
системой, обеспечивающей трансдукцию сигналов об одорантах,
служит система цАМФ (аденилатциклаза III), тогда как фосфоинози-
тидная система, включающая фосфолипазу С, осуществляет
трансдукцию феромонов. Недавно доказано (Е.В. Бигдай, 2004), что не
только обе названные сигнальные системы, но также и тирозинкиназ-
ная внутриклеточная сигнальная система участвует в трансдукции
одорантов, но для разных пахучих веществ комбинации систем,
осуществляющих обонятельную трансдукцию, различны (рис. 6.59).
Внутриклеточные сигнальные системы передают сигнал об
одорантах на кальциевые каналы, которые находятся в плазмолемме
обонятельных жгутиков и открываются под действием цАМФ и 1, 4,
5-трифосфоинозитидов. По этим каналам из межклеточной среды
в цитозоль обонятельной клетки могут поступать как Са2+, так и Na"1".
Таких каналов нет в мембранах микровилл опорных клеток и
ресничек мерцательного эпителия. Лучше изучены циклонуклеотидзави-
симые ионные каналы. Они идентифицированы и охарактеризованы.
Плотность этих каналов в мембране жгутиков обонятельных клеток
тритона составляет 920 каналов на 1 мм2, а у жабы - 2400 каналов на
1 мм2, тогда как в дендрите и соме выявлено всего 2 канала на 1 мм2
338
Биофизика
Рис 6.59. Механизм обонятельной трансдукции с участием
аденилатцикпазного (а) и фосфоинузитидного (б) путей
у тритона и 6 каналов на 1 мм2 у жабы. В отсутствие одорантов
подавляющее большинство цАМФ-регулируемых каналов закрыто
и поток катионов в рецепторные клетки невелик. Им обеспечивается
стабилизация мембранного потенциала на обонятельных жгутиках.
Одорант вступает во взаимодействие с рецепторными
молекулами мембраны жгутиков. Сопряженные с рецепторными
молекулами G-белки (Go//) активируют внутриклеточные сигнальные системы,
что приводит к открытию цАМФ- или 1Р3 - регулируемых кальциевых
каналов. Входящие катионные токи деполяризуют мембрану
обонятельной клетки. Деполяризационный сдвиг мембранного потенциала
является рецепторным потенциалом, который электротонически
(с декрементом) распространяется со скоростью электромагнитной
волны по обонятельной клетке - от жгутика до аксона, где
инициирует возникновение потенциалов действия (нервных импульсов),
следующих по веточкам обонятельного нерва к обонятельной луковице.
Обонятельные клетки существенно различаются по степени их
специфичности к стимулу. Данные последних лет показывают, что
Глава 6. Сенсорные системы организма
339
в одной обонятельной клетке экспрессируется один обонятельный
рецептор для данного одоранта, поскольку каждая рецепторная
клетка содержит ген только одного обонятельного рецептора. Р. Аксель
и Л. Бак, лауреаты Нобелевской премии 2004 г., открыли семейство
генов (более 1000), ответственных за обоняние. Оказалось, что
каждый ген из ста в организме млекопитающих участвует в синтезе
белков, осуществляющих обоняние. Все обонятельные рецепторы
принадлежат к классу так называемых семидоменных рецепторов (как
и родопсин), т.е. белковый комплекс рецепторной молекулы семь раз
пронизывает плазмолемму жгутика.
Имеется определенная топографическая организация
обонятельной выстилки в ее реагировании на разные классы одорантов.
У млекопитающих гены рецептора к одоранту экспрессируются в
одной из четырех основных зон в обонятельном эпителии, и экспрессия
одиночного гена рассеивается в пределах определенной зоны
случайно. Таким образом, на поверхности обонятельного эпителия рецеп-
торные клетки образуют «пространственные карты запахов». В
каждой из зон ольфакторный рецептор может взаимодействовать со
своим специфическим лигандом, а клетки объединяются в группы,
имеющие рецепторы со сходной лигандной специфичностью.
Обонятельным рецепторам свойственна неоднородность
механизма трансдукции сигналов об одорантах разных классов. Как уже
говорилось, для веществ, обладающих острым (аммиак) и
гнилостным (меркаптаны) запахами, специфических обонятельных
рецепторов не обнаружено. В их трансдукцию не вовлекаются и сигнальные
системы обонятельных клеток. Эти одоранты проникают через
плазмолемму в цитоплазму рецепторных клеток и действуют
непосредственно на митохондрии. Для одорантов, относящихся к пяти другим
классам, выявлены мембранные рецепторы, которые конгруэнтны по
своей стереоструктуре соответствующему одоранту, т.е. пахучие
вещества подходят к ним, как ключ к замку (рис. 6.60).
Вместе с тем для этих пяти классов одорантов,
воспринимаемых молекулярными рецепторами, присущи разные
внутриклеточные сигнальные системы. Трансдукция амилового спирта
(прогорклый запах), цинеола (эвкалиптовый запах) и амилацетата (фруктовый
запах) осуществляется системой цАМФ, причем аденилатциклаза III
22*
340
Биофизике
Камфорный Мускусный
Цветочный
Острый Мятный
> +
Эфирный
шшшш ш
Гнилостный
Рис. 6.60. Модели молекул пахучих веществ
и соответствующие им активные центры рецепторов
активируется амиловым спиртом и амиалцетатом через G-белок
а цинеолом - через тирозинкиназу. В обонятельной трансдукции кам
форы и ванилина (цветочный запах) вовлекается фосфоинозитидный
Глава 6. Сенсорные системы организма
341
путь, причем фосфолипаза С активируется ванилином через G-белок,
а камфора - через тирозинкиназу. Заметим, что камфора и цинеол,
являющиеся монотерпенами, включают одинаковый механизм
преобразования сигналов - активируют тирозинкиназу, тогда как
последующие звенья трансдукции для этих веществ, обладающих разными
запахами, различны: для камфоры - фосфолипаза С, а для цинеола -
адинилатциклаза III. Следовательно, первичные механизмы
рецепции одорантов, относящихся к разным классам, гетерогенны.
В обонятельной луковице осуществляется глутаматэргическая
синаптическая передача с аксонов терминалей на дентриты
обонятельных клеток. Она выполняет роль релейной структуры, в которой
происходит первая стадия обработки обонятельной информации. Область
синаптического контакта формирует гломсрулярную структуру, в
которую проецируется аксон от обонятельной клетки, экспрессирующей
рецептор к данному одоранту. Иными словами, нейроны обонятельной
луковицы образуют своего рода матрицу гломерул (рис. 6.61), а кон-
Обонятельный луковице
омеру
Обонятельный эпителий
Зона Π
Зона II
Зона III
Зона IV
Зона Зона IV
Молекулы одоранта _у jS Обонятельные сенсоры
Рис. 6.61. Топография рецепторных молекул в обонятельной выстилке
и проекции аксонов в соответствующие гломерулы обонятельной луковицы
342
Биофизика
кретный запах воспринимается мозгом как возбуждение определенной
комбинации нейронов в обонятельной луковице. В Нобелевской речи
Р. Аксель утверждал: «Мозг как будто говорит: «Я наблюдаю
активность в нейронах NN 1, 15, 54 - это соответствует возбуждению
рецепторов №№ 1,15, 54, а значит, пахнет жасмином».
Таким образом, в гломерулах сосредоточена информация
о свойствах отдельных молекул пахучих веществ, которая
локализуется в определенном месте обонятельной луковицы. Это
классический пример кодирования качества раздражителя по принципу
меченой линии.
6.5.7. Чувствительность органов обоняния
Высокая чувствительность к запахам - это важное свойство
органов обоняния, которое характеризуется абсолютным порогом. Он
определяется минимальной концентрацией пахучего вещества,
вызывающей обонятельное ощущение.
Многие рыбы ощущают пахучее вещество при его
концентрации 10"" мг/мл. Примерами пахучих веществ с низкими пороговыми
концентрациями для человека могут служить скатол (4 · 10~7 мг/см3)
и этилмеркаптан (4,4 · 10~8 мг/см3). Это означает, что для
возбуждения одной обонятельной клетки этим гнилостным запахом требуется
от одной до восьми молекул в одном кубическом сантиметре воздуха.
При очень малых концентрациях обонятельные ощущения
неспецифичны: человек не определяет качество одоранта. Поэтому
различают порог обнаружения и разпознавания запаха. Пороги
распознавания запахов лежат выше порогов обнаружения.
6.5.8. Техническое моделирование обоняния
Технические устройства, предназначенные для химического
анализа, значительно уступают по чувствительности обонятельной
сенсорной системе человека и животных. Однако в последнее время
наметился путь к созданию высокочувствительного химического
анализатора, который получил название электронного носа. Он
сконструирован на базе растрового атомно-силового микроскопа и пред-
Глава 6. Сенсорные системы организма
343
ставляет собой одно из выдающихся достижений современной нано-
гехнологии.
Конструктивным элементом электронного носа служат канти-
леверы - упругие балочки толщиной в десятые - сотые доли
микрона. Один конец такой балочки закреплен, а второй может свободно
колебаться, поэтому конструкция напоминает расческу. На каждый
кантилевер наносится тончайший слой полимера, способного
селективно адсорбировать на себе молекулы только одного вещества,
например, определенного одоранта.
При оседании одоранта балочка слегка изгибается, и эти
ничтожные колебания исследуют посредством растрового атомно-си-
лового микроскопа. Тот или иной запах дает свою специфическую
спектральную картину. В «расческе» нет двух кантилеверов,
покрытых одним и тем же полимером. Поэтому набор кантилеверов в
данной «расческе» реагирует как на несколько простых одорантов, так
и на сложные запахи.
К сожалению, прибор узнает и различает только те запахи,
которые ему уже знакомы. В этом видит главный недостаток электронного
носа основатель направления сенсорной техники в IBM Кристофер
Гербер. Еще более пессимистично оценил способности электронного
носа Вольфганг Вальстер, руководитель германского
исследовательского центра по созданию искусственного интеллекта. По его мнению
машины не могут достичь способности человека, так как компьютер
бессилен осуществить сложный анализ нетипичных процессов;
преимущество человека и животных перед машиной состоит в том, что
они при анализе сигналов внешней среды использут все органы
чувств одновременно.
До сих пор не найден механизм автоматического распознавания
сложных разномодальных сигналов, поступающих в мозг по разным
сенсорным входам. Проблему взаимодействия сенсорных систем
(анализаторов) сформулировал и начал разрабатывать в 1930-е годы
выдающийся отечественный физиолог Л.А. Орбели. На многие
вопросы, поставленные им, ответил его ученик А.С. Мозжухин со
своими сотрудниками. Однако физиологические и биофизические
механизмы взаимодействия сенсорных систем еще выяснены.
Глава 7. ИНФОРМАЦИЯ И РЕГУЛИРОВАНИЕ
В БИОЛОГИЧЕСКИХ СИСТЕМАХ
7.1. Информация и живой организм
7.1.1. Элементы теории информации
Теория информации представляет собой совокупность
математических концепций, которые позволяют оценить количество
информации, содержащейся в том или ином сообщении, определить
пропускную способность каналов связи, используемых для передачи
сигналов, количественно сопоставить эффективность применяемых
систем кодирования для выбора лучшей из них. Эта теория возникла
в ходе решения важнейших практических задач электросвязи
математическими методами, и первый фундаментальный труд К. Шеннона,
основоположника теории информации, увидел свет под названием
«Математическая теория связи» (1948). В статье предлагался принцип
пиально новый способ обобщения человеческого опыта в передаче
сообщений. Теория информации - математическая концепция, но ей
основные идеи можно выразить без использования сложного математ
тического аппарата.
В обиходе широко употребляется слово «информация», причём
в разных значениях. Зачастую его адресуют непосредственно сообч
щению, порой даже отождествляют сообщение и информацию. В на*
учном же определении информации должна содержаться характерна
стика не сообщения как такового, а процесса взаимодействия между.
сообщением и его получателем. Поэтому информативность сообще»
ния нельзя оценивать количеством единиц носителя информации (на*
пример, числом слов в докладе или букв в книге). Информативна
только такое сообщение, которое приносит что-то неизвестное ранееу
в чем нуждается тот, кому оно адресовано. С учётом этого К. Шенно^
Глава 7. Информация и регулирование в биологических системах
345
предложил определение информации, на котором построена теория
информации.
Информация, содержащаяся в сообщении, есть мера того
количества неопределённости, которое ликвидируется после
получения данного сообщения.
Из определения следует, что количество информации,
полученной человеком, следует оценивать степенью уменьшения
неопределённости в его сведениях об информируемом событии. В свою
очередь, неопределенность того или иного результата разрешения
неопределённой ситуации тем больше, чем меньше его вероятность.
Заметим, что в теории информации принято различать априорную (/?,)
и апостериорную (р2) вероятности. Априорной называют такую
вероятность сообщаемого события, которой оно характеризовалось до
получения сообщения. Вероятность события, установившаяся после
получения сообщения о нем, называется апостериорной вероятностью.
Если содержание получаемого сообщения было заведомо точно
известно его адресату, то априорная вероятность события,
информацию о котором несло сообщение, равнялась единице (pj = I), и такое
сообщение абсолютно не информативно (J = 0). Нулевой считается
избыточная и повторная информация, а также всё, что содержится
в сообщениях, не имеющих отношения к достижению цели, ради
которой адресат стремился получить информацию. Ценность
информации принимает отрицательное значение, когда она имеет характер
дезинформации.
Прир, < 1 сообщение содержит информацию, но только при
условии, что после его получения повысится вероятность события,
о котором информируют, т.е. когда апостериорная вероятность
станет больше априорной (р2 > pt). Например, до получения сообщения
нероятность события, о котором нас информируют, равнялась 1/8
(р{ = 1/8), а после приёма сообщения значение вероятности достигло
\12{р2 = 1/2). В таком случае сообщение повысило вероятность
грядущего события и уменьшило неопределённость в наших знаниях
о нём. Следовательно, сообщение оказалось информативным.
Информация максимальна, если при данной априорной вероятности,
меньшей единицы, апостериорная вероятность (после получения
сообщения) достигает единицы.
346
Биофизика
Понятия максимума и минимума информации определяются не
столько абсолютными значениями р1 ир2, сколько их соотношением,
которое само по себе может служить мерой информативности
сообщения. Однако для количественной оценки информационных
процессов удобнее пользоваться логарифмом отношения Ρη/ρν
Основание логарифмов выбирается в соответствии с выбором единицы
измерения информации и зависит от структуры линии связи
(информационного канала). Нередко человек приобретает информацию
путем совершения последовательных шагов двоичного выбора, т.е.
выбирает одно из двух возможных решений, давая ответы «да» или
«нет». Аналогичный путь реализуется в технике при помощи двухпо-
зиционных переключателей (реле или мультивибраторов): в одном
положении они пропускают сигнал («да»), в другом - блокируют его
передачу («нет»). Такому переключателю можно уподобить нейрон,
.когда он проводит нервные импульсы. Если в результате
суммирования всех постсинаптических потенциалов устанавливается пороговая
или надпороговая деполяризация мембраны в начальном участке
аксона (аксонном холмике), возникает импульсация, направляющаяся
к другому нейрону или к исполнительному органу, неся сигнал для их
возбуждения («да»). Когда же интегрирование постсинаптических
потенциалов приводит к гиперполяризации или подпороговой
деполяризации мембраны аксонного холмика, передача импульсации бло1
кируется («нет»).
Рассмотренные системы передачи сообщений называются дво*
ичными. В них информация оценивается двоичным логарифмом
отношения Ρη/ρ{. J = log2 —-. Если сообщение исчерпывает всю неоп-
Р] и:
ределённость ситуации, τορ2 = 1. Именно такие сообщения наиболее\
информативны (при данному,). Для них упрощается расчёт количе-f
1 "
ства принесенной информации: J - log2 — = - log2 ρ,.
Ρχ
При использовании логарифмов с основанием, равным двум|.|
единицей измерения информации служит бит. Этот термин прем
изошёл от английского словосочетания binary digit - двоичная единил
ца (от первого слова взяты две первые буквы, от второго - одна поя
Глава 7. Информация и регулирование в биологических системах
347
следняя). 1 бит количественно характеризует такую информацию,
которая полностью ликвидирует неопределённость выбора между
двумя равновероятными возможностями, т.е. в условиях, когда /?, =
= 1/2,а/?2 = 1.
Для уяснения смысла введенных понятий обратимся к
примерам. Предположим, что идем по дороге к определённой цели. В
каком-то месте дорога делится на две, расходящиеся в разные стороны,
причём мы не знаем, какая из них ведет к цели. Очевидно, априорная
вероятность правильного выбора составляет 1/2 (рг = 1/2). Если
повстречается знающий человек и точно укажет дальнейший путь
к цели нашего пугешествия, то апостериорная вероятность
достижения цели станет равной единице (р2 = 1). Информативность такого
сообщения J = -log2 p^ =-log,- = 1бит.
Может случиться так, что повстречавшийся человек на вопрос
о пути к цели нашего путешествия даст такой ответ: «Не знаю, но
этот путь не приведет вас к цели» - и укажет на одну из двух
расходящихся дорог. Информативность подобного сообщения также
оценивается в 1 бит, поскольку и оно при априорной вероятности,
равной 1/2, приводит кр2 = 1. Следовательно, независимо от конкретной
формы ответа, любое сообщение, полностью устраняющее
неопределённость выбора из двух равновероятных возможностей, содержит
информацию в 1 бит.
Предположим теперь, что дорога, по которой мы шли,
разделилась не на 2, а на 8 путей, уходящих в разные стороны. В таком
случае априорная вероятность правильного выбора пути составляет 1/8
(р] = 1/8) и, следовательно, сообщение, определённо указывающее
путь к цели и тем самым позволяющее сделать правильный выбор
одной из восьми дорог, приносит нам больше информации по
сравнению с теми же словами в первом примере. Выразим эту
информацию в битах: J =-log2 - =3 бит. Таким образом, одна и та же фраза
о
в разных ситуациях (при неодинаковых значениях /?,), заключает
в себе разное количество информации. Если в обоих случаях р2 = 1,
то эта фраза во второй ситуации оказывается в 3 раза
информативнее, чем в первой.
348
Биофизика
Какова же во второй ситуации информативность другого
ответа, который отвергает одну из дорог, не приводящих нас к цели (когда
неопределённость выбора только уменьшается, но не устраняется)?
Обратимся к величинам априорной и апостериорной вероятностей:
ρ, = 7/8,р2 = 6/7. Отсюда J = log, = log, — = 0,03 бит.
7-7 49
Итак, в первой ситуации (при выборе одной из двух дорог)
разные по содержанию сообщения одинаково информативны - каждое
приносит информацию в 1 бит. При выборе одного из восьми путей
информативность двух разных сообщений существенно различна:
указание на путь, приводящий к цели, содержит информацию
в 3 бита, тогда как устранение одной из возможных ошибок - только
0,03 бита. Следовательно, разница в содержании информации
достигает двух порядков.
Важным понятием теории информации является сообщение.
Так называется любая функция от времени a(i), описывающая
характер сигналов, приходящих к получателю информации. В системах
электросвязи сообщение заключается в определённой форме
электрического тока (или напряжения) на выходе кодирующего
устройства. В нервных каналах связи сообщениями служат
последовательности нервных импульсов, в которых отображены сигналы о среде
или состоянии организма.
Для оценки эффективности работы канала связи принято
рассчитывать среднее количество информации, приходящееся на одно
сообщение, передаваемое каналом за определённое время. Среднее
количество информации, приходящееся на одно сообщение, названо
К. Шенноном информационной энтропией (Н). Для расчёта
информационной энтропии предложена следующая формула:
H=-]p{x)-\og2 p(x)dx=-^p. · log2 pt,
-ее '=1
где М- количество сообщений, переданных данным каналом связи,
Ρι - априорные вероятности каждого из Μ сообщений (в данном случае
апостериорная вероятность принимается равной единице).
Глава 7. Информация и регулирование в биологических системах
349
Информационная энтропия возрастает по мере того, как
увеличивается количество сообщений (Л/), из которых получатель
информации делает выбор. Наивысшее из всех возможных значений
энтропии сообщения, переданного по каналу связи, называется пропускной
способностью этого канала. Зная величину информационной
энтропии источника сообщений, можно найти минимальное значение
среднего количества двоичных чисел на каждое сообщение,
достаточного для надёжного кодирования сигналов, создаваемых данным
источником.
7.1.2. Примеры применения теории информации
к анализу процессов передачи информации
в нервных каналах связи
Определим, как зависит среднее количество информации на
символ (сигнальный признак в нервной импульсации) от того, какие
сигнальные признаки различает нейрон, принимающий сообщения
в форме нервных импульсов.
Допустим, к нейрону поступают с равной частотой 8
сообщений - 8 определённых последовательностей нервных импульсов
(рис. 7.1). Заметим, что в импульсации содержатся «пачки»
импульсов двух типов - короткие (по 3 импульса) и длинные (по 5
импульсов), причём длинные по-разному распределены среди коротких:
в первом сообщении (1) нет длинных пачек, во втором, третьем,
четвёртом (2, 3,4)- одна длинная «пачка» приходится на две
коротких (во втором сообщении она предшествует коротким, в третьем -
находится между ними, в четвертом - следует после них), в пятом,
шестом, седьмом (5, 6, 7) сообщениях содержатся по две длинных
и одной короткой «пачке», а в восьмом (8) - присутствуют только
длинные «пачки».
Если нейрон, принимающий эту импульсацию, способен
различать «пачки» по их длительности, то он может по-разному
реагировать на каждую из восьми последовательностей нервных
импульсов. Вероятность поступления к нему каждого из таких сообщений
равна 1/8 Q?, = 1/8). Для упрощения расчёта примем апостериорную
вероятность равной единице (р2 = 1). Тогда приход к нейрону любой
350
Биофизика
Рис. 7.1. Последовательности нервных импульсов
(иллюстрация к примеру расчёта информационной энтропии)
/= и ι
ставляет 3 бита на сообщение: Я = -]Г--log,- = -8
из последовательностей импульсов, изображенных на рис. 7.1, даёт
ему 3 бита информации.
Понятно, что в таком случае информационная энтропия со-
= 3 бит · (сообщение)-1.
Если же нейрон потеряет способность реагировать на короткие
«пачки», то для него информативными будут только длинные. Такой
нейрон станет работать как счетчик числа длинных «пачек». В тех же
последовательностях нервных импульсов он сможет различить не 8,
а 4 сообщения: 1 - отсутствие длинных «пачек»; 2 - наличие одной
(длинной) «пачки» (для нейрона станут одинаковыми вторая, третья
и четвёртая последовательности импульсов); 3 - наличие двух
(длинных) «пачек» (нейрон не сможет различить пятую, шестую и
седьмую последовательности импульсов); 4 - наличие трёх (длинных)
«пачек» (восьмая последовательность импульсов). Следовательно,
Глава 7. Информация и регулирование в биологических системах
351
в таком случае Μ = 4, априорные вероятности первого и четвёртого
сообщений равны 1/8, а второго и третьего - 3/8. Рассчитаем
информационную энтропию:
гт Πι 13, 3 3, 3 1, П
Н = log2- + -log2- + -log2- + -log2- =
U 28 8 8 8 28 8 28j
= /llog 1 + ^log2 ^ = -(-025·3-0,75· 1,42) =
V. 4 8 8 oj
= 1,81 бит(сообщение)"1.
Подобные расчёты позволяют не только сделать вывод, что
в нервных каналах связи эффективнее работают нейроны,
различающие «пачки» импульсной активности по их продолжительности, но
и оценить количество информации, анализируемой нейронами
разных типов. Так, у нейронов первого типа информационная энтропия
составляет 3 бита на сообщение, тогда как у нейронов второго типа
Η = 1,81 бит · (сообщение)-1. Когда такие числа характеризуют один
и тот же нейрон при изменении его функционального состояния,
получается количественная оценка потерь информации в этом
процессе. Например, при нарушении возбудимости нейрон способен
реагировать на более длительные стимулы (при прежней их амплитуде),
утрачивая тем самым реакции на короткие раздражители. Поэтому
даже частичная потеря возбудимости может сопровождаться
потерями в передаче информации. Их удается оценить количественно
путем расчёта изменения информационной энтропии.
Однако далеко не всякий патологический процесс в нервной
системе приводит к понижению информационной энтропии,
поскольку организм обладает рядом защитных механизмов,
предупреждающих потери информации даже в условиях патологии. Наиболее
эффективным способом предупреждения потерь информации во всех
сферах человеческой деятельности служит создание
информационной избыточности (как и сама информация, её избыточность
измеряется в битах). Избыточность информации свойственна, например,
любому тексту. Так, в письменных текстах на немецком языке
содержится, в среднем, по 1,3 бита на букву, тогда как средняя избыточ-
352
Биофизика
ность достигает 3,4 бита на букву. Следовательно, в таких текстах
избыточность значительно больше содержания информации. Эта
закономерность характерна для всех языков. Благодаря избыточности
подобная форма передачи информации весьма надежна.
Один из эффективных конкретных способов реализации
принципа избыточности для повышения надёжности передачи
информации состоит в использовании нескольких параллельных каналов
связи. Он присущ нервной системе. Так, количество рецепторов гораздо
больше числа нейронов, к которым от них поступают сигналы.
Пропускная способность любых рецепторных аппаратов значительно
выше возможностей осознания сенсорной информации.
Человеческое сознание пользуется очень малой долей информации,
поставляемой в мозг периферическими отделами анализаторов -
сенсорными входами (табл. 7.1). Например, максимальный поток информации
в процессе зрительного восприятия, осознаваемого человеком,
составляет всего 40 бит · с-1, тогда как информационная ёмкость потока
нервных импульсов в зрительных нервах достигает 107 бит ■ с-1.
Сопоставление этих чисел свидетельствует об огромной избыточности
и, следовательно, высокой надёжности информационных процессов
в зрительной сенсорной системе. То же свойственно другим
анализаторам (табл. 7.1) и всем функциональным аппаратам мозга.
Таблица 7.1
Сравнение потоков информации (ёмкости каналов) на уровне афферентных
волокон и сознательного восприятия (М. Циммерманн, 1984)
Сенсорная
система
Зрительная
Слуховая
Тактильная
Обонятельная
Вкусовая
Число
рецепторов
2 108
3-104
ю7
7-107
3-107
Число
афферентных
волокон
2 106
2.104
106
105
103
Общая
ёмкость
каналов,
бит · с-1
107
ю5
106
ю5
103
Психофизическая
оценка емкости каналов (по
осознанному
восприятию), бит · с-1
40
30
5
1(?)
1(?)
Применение теории информации к сенсорным системам
позволяет сравнивать эффективность различных способов кодирования
Глава 7. Информация и регулирование в биологических системах
353
информации о внешней среде и информационные возможности
каждого анализатора, сопоставлять органы чувств с техническими
системами передачи информации. Язык теории информации пытаются
употреблять для количественной оценки даже тех информационных
систем организма человека, которые осуществляют психическую
деятельность. На этом пути совершается много ошибок. Среди них
следует назвать попытки применения понятий и математического
аппарата теории информации к процессам, обеспечивающим
осмысление передаваемой информации.
Для определения допустимых границ грамотного приложения
теории информации нужно усвоить, что в ней нет положений,
которые позволили бы количественно оценить смысл сообщения.
«Теория информации, - предупреждал К. Шеннон, - пригодна лишь для
систем, в которых нет никакой семантики». Она позволяет строго
изучать только такие каналы связи в организме, которые подобно
машине передают всю поступающую к ним информацию без
проникновения в её содержание и без выражения субъективного отношения
к ней. Например, телеграфист может рассматриваться в качестве
элемента системы связи, анализируемой с позиции теории информации,
только в том случае, если при передаче сигналов он не вдумывается
в их смысл. Результаты изучения сложных психических актов на
основе теории информации пока не удовлетворяют исследователей.
Однако существует мнение, что её дальнейшее развитие приведёт
к созданию математического аппарата, который окажется
пригодным для проникновения в сложнейшие механизмы человеческой
психики. Сегодня эта теория применима к относительно простым
проявлениям деятельности мозга.
7.1.3. Информация, заключенная
в генетическом коде
Важное медицинское и общебиологическое значение имеет
количественная оценка информационных свойств генетического кода.
Его кодирующей единицей служит триплетный кодон, образованный
тремя из четырёх нуклеотидов, из которых «собраны» молекулы ДНК
и РНК. Иными словами, «тексты» нуклеиновых кислот «написаны»
354
Биофизика
Про
- fwT^y -
Про Сер
-Ц-Ц-У-У-Ц-У-
Про во всех случаях
- ц-
-ц-
- ц-
-ц-
-ц.
■ ц-
■ц-
■ц-
•ц-
■ц-
■ц-
■ц-
■ У -
■ А -
- г -
Рис 7.2. Три характерные особенности генетического кода:
1) кодирующей единицей (специфической комбинацией нуклеотидов,
определяющей включение аминокислоты в молекулу белка) является
триплетный кодон; 2) универсальность кода для всех биологических систем;
3) вырожденность кода (одна и та же аминокислота кодируется более
чем одним триплетом, но первые два нукпеотида для данной аминокислоты
всегда одни и те же). Обозначения нуклеотидов: А - аденин; Г - гуанин;
У - урацил, который в РНК занимает место тимина, содержащегося в ДНК;
Ц- цитозин. Обозначения аминокислот: Про - пролин; Сер - серин.
четырёхбуквенным алфавитом: в каждой из них четыре нуклеотида
(аденин - А, гуанин - Г, тимин - Т, цитозин - Ц) складываются в
различные комбинации по 3, и с каждым таким триплетом (строго
фиксированной последовательностью из 3 нуклеотидов: например, АГТ,
ΑΓΑ, ЦТГ и т.д.) может взаимодействовать определённая
аминокислота (рис. 7.2 и табл. 7.2). Общее число разных кодонов равно 43 = 64.
Аминокислоты разнообразнее нуклеотидов. Природные белки
состоят из 20 разных аминокислот, которые, образуя различные
сочетания, формируют белковые молекулы, включающие сотни и тысячи
аминокислотных остатков. Место каждой аминокислоты в первичной
структуре белка определяется триплетным кодоном; а именно,
последовательность аминокислот в белковой молекуле, за синтез которой
ответствен данный участок ДНК, предопределена
последовательностью нуклеотидных триплетов в нем. Информация, «записанная»
4-символьным нуклеотидным кодом, перекодируется в 20-символ1.
ный аминокислотный код в тех участках ДНК, которые кодируют мс
Глава 7. Информация и регулирование в биологических системах
355
Таблица 7.2
Генетический код (ДНК-код)
Первый
нуклеотид
в триплете
7. А
8. Г
9. Τ
10. Ц
Второй нуклеотид в триплете*
А
AAA
ААГ
ААТ
ААЦ
ГАА
ГАГ
ГАТ
ГАЦ
ТАА
ТАГ
ТАТ
ТАЦ
ЦАА
ЦАГ
ЦАТ
ЦАЦ
Фен
Лей
Лей
Иле
Мет
Вал
Г
ΑΓΑ
АГГ
АГТ
АГЦ
ГГА
ГГГ
ГГТ
ггц
ТГА
ТГГ
ТГТ
тгц
ЦГА
ЦГГ
ЦГТ
ЦГЦ
Сер
Про
Тре
Ала
Τ
ΑΤΑ
ΑΤΓ
ATT
АТЦ
ГТА
ГТГ
ГГТ
ГТЦ
ТТА
ТТГ
ТГГ
ТГЦ
ЦТА
ЦТГ
ЦТТ
цтц
Тир
«Стоп»**
Гис
Глн
Асн
Лиз
Асп
Глу
ц
АЦА
АЦГ
АЦТ
АЦЦ
ГЦА
ГЦГ
ГЦТ
ГЦЦ
ТЦА
ТЦГ
ТЦТ
ТЦЦ
ГЦА
ГЦГ
ГЦТ
ГЦЦ
Цис
«Стоп»
Три
Apr
Сер
Гли
Apr
Третий
нуклеотид
в триплете
А
Г
Τ
Ц
А
Г
Τ
Ц
А
Г
Τ
Ц
А
Г
Τ
Ц
* Обозначения: нуклеотиды: А - аденин, Г - гуанин, Τ - тимин; Ц - цитозин;
аминокислоты: Ала - аланин, Apr - аргинин, Асн - аспарагин, Асп - аспарагиновая кислота,
Цис - цистеин, Глн - глутамин, Глу - глутаминовая кислота, Гли - глицин, Гис - гисти-
дин, Иле - изолейцин, Лей - лейцин, Лиз - лизин, Мет - метионин, Фен - фенилала-
нин, Про - пролин, Сер - серии, Тре - треонин, Три - триптофан, Тир - тирозин, Вал -
валин.
** Три кодона (АТТ, АТЦ, АЦТ) в ДНК не кодируют аминокислот («стоп») -
по-видимому, они должны означать точку в сообщении, отделяя синтез одной полипептидной
цепи от синтеза другой.
стоположение аминокислотных остатков в полипептидной цепи
(табл. 7.2 и рис. 7.3). Так как геномом называется часть молекулы
ДНК, ответственная за синтез одной белковой цепи, то проблема
генетического кода сводится к разрешению вопроса: каково
соответствие между последовательностью нуклеотидов в ДНК и
последовательностью аминокислотных остатков в первичной структуре белка?
Именно так проблема была сформулирована в 1953 г. физиком
Г.А. Гамовым, сделавшим первый расчёт генетического кода. Осно-
356
Биофизика
вополагающие исследования по его расшифровке провели М.У. Ни-
ренберг и Р.У. Холли, а окончательно расшифровал генетический код
и синтезировал ген тРНК Х.Г. Корана в 1966 г. Эти учёные
удостоены Нобелевской премии (1968 г.).
Каждый участок ДНК, с которым связан синтез белковой
молекулы, включающей N аминокислотных остатков, образован 3N нук-
леотидами. Например, генетический аппарат, обеспечивающий
синтез белковой молекулы из 200 аминокислот (N = 200), состоит из 600
нуклеотидов. Количество информации (/,), заключённой в участке
ДНК, ответственном за синтез макромолекулы из N компонентов,
рассчитывается по формуле: Iг =log2 и'" , где пн - число символов
в нуклеотидном коде (ин = 4). Поскольку ячейкой, содержащей 2 бита
информации, в молекуле ДНК служит каждая пара комплементарных
нуклеотидов (аденин - тимин, гуанин - цитозин), то в
рассмотренном примере 1г = log24600 = 2 · 600 = 1200 бит.
Для количественной оценки информации (1б), содержащейся
в уже синтезированной белковой молекуле, воспользуемся аналогич-
Двойная спираль ДНК
Υ////////////////////////////////////////Λ
Рис. 7.3. Схема синтеза белка
а - код ДНК; б - матричная РНК (мРНК), образованная
соответственно той цепью ДНК, которая изображена непрерывной линией;
в — образование полипептида в рибосоме. Обозначения аминокислот.
Мет — метионин; Гли - глицин; Лей—лейцин; Асн - аспарагин; Г - гуанин;
Τ - тимин; У - урацил; Ц - цитозин; тРНК - транспортная РНК.
Глеве 7. Информеция и регупировение в биологических системех
357
ной формулой 16 = log2 η " , где па - число символов в
аминокислотном коде (па = 20), N - число аминокислот, места которых строго
фиксированы в первичной структуре белка. Обратимся к той же
белковой молекуле, включающей 200 аминокислотных остатков. Если
бы при её синтезе каждой из аминокислот следовало занять строго
определённое ей место, то 1б = log220200« 4 · 200 « 800 бит1. Это явно
завышенная оценка, поскольку любой белок состоит не более чем из
20 разных аминокислот, причём определённая аминокислота
(скажем, лизин), для которой отведено несколько строго фиксированных
мест в первичной структуре белка, может занять любое из них.
Можно рассчитать 16, исходя из предположения, что для выполнения
белком присущей ему функции (например, ферментативного катализа)
достаточно строго зафиксировать в его первичной структуре
положение всего 2-3 аминокислотных остатков, образующих активный
центр. Тогда получится значение 1б, в котором не учтена
информация, необходимая для формирования вторичной, третичной и
четвертичной структур белковой молекулы. Так, число водородных связей,
стабилизирующих белковую глобулу, имеет, порядок 102. К
наиболее вероятному значению количества информации, заключённой
в белковой молекуле 1б , приводит предположение, что хотя бы
половина этих водородных связей строго фиксирована. Следовательно,
для расчёта Iб нужно принять число строго фиксированных
положений аминокислотных остатков в белковой молекуле
приблизительно равным 50. Тогда 1б =log2 2050 «4· 50«200 бит.
Из сопоставления значений 1г и 1б следует вывод об
огромной избыточности генетической информации. Если принять
1б =1б =200 бит, то количество информации, содержащейся в
генетическом аппарате, которым обеспечивается синтез такой
белковой молекулы, в 6 раз (на 1000 бит) превосходит информацию,
заключённую в ней. Следовательно, большая часть информации,
которой обладает ген для синтеза белка, оказывается избыточной.
Избыточность генетического кода повышает надёжность
биосинтеза, а надёжность синтеза белковых молекул жизненно необходима,
' В расчёте log220 взят равным 4 (с точностью до 1), поскольку 4 = log216,
а 5 = log232.
358
Биофизика
поскольку ошибки в нём приводят к продукции чужеродных
организму белков и, вследствие этого, к тяжелейшим заболеваниям.
Другим выводом из подобных расчётов является
сомнительность взгляда о возможности зарождения жизни на Земле благодаря
случайному образованию на ней белковых тел. Вероятность
случайного синтеза молекулы определяется выражением ρ = 2~', где / -
информация, заключённая в молекуле. Для рассмотренного выше
случая 16 = 200 бит и, следовательно, ρ = 2~ш ~ Ю-60. Вероятность
любого события на Земле, меньшая Ю-30, признается абсурдно
малой. Таким образом, случайный характер (без определённой
программы) синтеза белка теоретически невероятен.
7.2. Регулирование биологических процессов
Информация и регулирование неотделимы друг от друга. Они
рассматриваются как компоненты единого процесса управления в
технике, биологии и социологии. Изучение биологических форм
управления составляет содержание биофизики сложных систем. В
биологических системах приём, передача, хранение и переработка информации
обеспечивают управление функциями организма. Поэтому проблемы
связи исследуются не только в рамках теории информации, но и с
позиций более общей науки - кибернетики.
7.2.1. Содержание кибернетики и бионики
Термин «кибернетика» появился в современной научной
литературе после того как основоположники этой науки пришли к
заключению «о принципиальном единстве ряда задач, в центре которых
находились вопросы связи и управления, причём как в машине, так
и в биологических объектах» (Винер Н., 1948). Чтобы подчеркнуть
отмеченное единство и устранить разрозненность существовавшей
тогда терминологии для обозначения принципов и понятий теории
связи и управления, Норберту Винеру пришлось, как он пишет,
«изобрести» неологизм - «кибернетика». Автор произвёл его от
греческого слова χιιβερνητηξ, которым в Древней Греции называли моряка,
управляющего кораблем (кормчего). Нужно заметить, что задолго до
Глава 7. Информация и регулирование в биологических системах
359
Винера это слово употреблялось и в переносном смысле. Так,
древнегреческий философ Платон пользовался им для характеристики
человека, способного управлять людьми. Тот же корень делает
созвучными названия правительств в английском, французском и испанском
языках. Слова «губернатор» и «гувернантка» проникли в русский
язык после сложной эволюции этого греческого слова. А. Ампер
в своей единой системе классификации знаний (1843 г.) назвал
кибернетикой науку об управлении государством, а Д.К. Максвелл в 1868 г.
обозначил близким по звучанию термином (governors)
автоматические регуляторы в технике.
Однако только в 1948 г. с появлением книги Н. Винера
«Кибернетика» наука обогатилась новой огромной областью знаний.
Толчком к развитию кибернетики послужило изучение
математиками и инженерами не столько технических, сколько биологических
систем. Во время Второй мировой войны Н. Винер и его коллеги
разработали принципиально новые саморегулирующиеся комплексы
противовоздушной обороны. Вскоре после войны Винер был
приглашен к решению ряда задач практической медицины и неожиданно
для себя понял, что изобретённые им принципы управления
системами противовоздушной обороны давно реализованы природой в
организме животных и человека. Эти принципы управления автор
обобщил в «Кибернетике», включавшей восемь глав, четыре из которых
были посвящены нервной системе. Поэтому книга вышла с таким
подзаголовком: «Управление и связь в животном и машине».
Уже в 1948 г. Н. Винер определил основной теоретический
аппарат современной кибернетики: 1) теория информации, 2) теория
сглаживания и фильтрации «помех» (сигналов, мешающих системам
управления принимать, перерабатывать, хранить и передавать
информацию), 3) теория обнаружения и прогнозирования поведения
управляющего сигнала, 4) теория обратной связи, 5) теория
автоматов и сложных машин.
На основе перечисленных теорий стали создаваться новые
технические системы управления и глубоко анализироваться регуляторные
процессы в организме человека и животных. Позднее область
применения кибернетических принципов охватила социологию и экономику,
поскольку «основное содержание этих наук заключается в изучении
360
Биофизика
связей» (Винер Н., 1969). Следовательно, кибернетика - безграничная
наука, так как охватывает многие области человеческого знания, каждая
из которых сама по себе весьма обширна. Поэтому определение
кибернетики после первых работ, посвященных этой науке, не только
расширялось, но и уточнялось. Основоположник кибернетики в нашей стране
академик А.И. Берг определил кибернетику, как науку об управлении,
об управляющих системах, а также об организации как управляющих
систем, так и самого процесса управления.
Таким образом, кибернетика - это наука об управлении, о
связях, при помощи которых оно осуществляется, об организации как
управляющих систем, так и самого процесса управления.
Основные кибернетические законы универсальны, но их
конкретные проявления в системах различной природы довольно
специфичны, чем предопределяется выделение в кибернетике трёх областей
исследований: технической кибернетики, биологической кибернетики,
социологической кибернетики. Приведённая классификация весьма
условна, поскольку названные области взаимосвязаны. Внутри каждой
из них есть направления, стоящие ближе к целям и задачам смежных
областей, чем к проблемам своей области. Так, автоматизированные
системы искусственного кровообращения развиваются на основе
представлений биологической кибернетики и достижений технической
кибернетики.
Важным направлением социологической кибернетики является
разработка автоматизированных систем управления (АСУ)
производством и другими формами общественной деятельности. Принципы
социологической кибернетики положены в основу проектирования
АСУ здравоохранения, хотя при этом учитываются закономерности
биологической и технической кибернетики. Отчасти поэтому нельзя
считать медицинскую кибернетику ветвью биологической
кибернетики. Медицинская кибернетика, с одной стороны, уже, так как
включает те принципы биологической кибернетики, которые могут быть
использованы для сохранения и восстановления здоровья человека,
но, с другой стороны, шире, поскольку составными частями
медицинской кибернетики служат некоторые направления технической
и социологической кибернетики.
Глава 7. Информация и регулирование в биологических системах
361
В рамках медицинской кибернетики биофизика
ограничивается в основном разработкой некоторых проблем биологической
кибернетики. Биофизические исследования в области кибернетики
вскрывают физические и физико-химические основы
регулирования физиологических процессов в организме. Поэтому в курсе
биофизики рассматриваются далеко не все стороны медицинской
и биологической кибернетики.
Физические и физико-химические основы процессов
регулирования биологических систем актуальны как для биофизики, так
и для бионики. Термин «бионика» принят в 1960 г. на симпозиуме,
проходившем под девизом: «Живые прототипы - ключ к новой
технике». Так была чётко сформулирована цель бионики - создание
новой техники по аналогии с биологическими объектами, в
частности путём использования принципов управления биологическими
системами. В отличие от биофизиков, специалисты в области
бионики изучают организацию биологических объектов не ради того,
чтобы разобраться в физических и физико-химических основах
жизнедеятельности, а с целью применения этих знаний в интересах
развития техники. Бионика не ограничивается разработкой
кибернетических проблем, поскольку при построении технических
аналогов биообъектов используются знания об особенностях
структуры и функции не только управляющих систем организма. Создано
немало технических устройств, в которых реализованы сугубо
анатомические аналогии. Например, есть машины с подвижными
узлами, устроенными так же, как конечности некоторых животных (в
частности членистоногих). С бионикой связаны многие достижения
робототехники.
Кибернетические идеи весьма полезны биологии и медицине.
Они позволили по-новому рассматривать традиционные
физиологические проблемы регуляции функций.
7.2.2. Стратегия управления функциями организма
Физиологи изучают организм в единстве с окружающей
средой. При её изменениях возникают разнообразные реакции
организма. Стратегия управления функциями биологической системы состо-
362
Биофизика
ит в том, чтобы при любых изменениях внешней среды обеспечить
сохранение её жизнеспособности. В этом заключается смысл
физиологического представления о приспособлении организма к внешней
среде.
В докладе на XV Международном медицинском конгрессе
(1903 г.) И.П. Павлов говорил: «... что, собственно, есть в факте
приспособления? Ничего..., кроме точной связи элементов сложной
(биологической) системы между собою и всего их комплекса с
окружающей обстановкой.»
Позднее, в 1918 г., в цикле лекций «Об уме вообще, русском
в частности» он утверждал: «Что такое жизнь? Если стоять на
фактической стороне дела, то придётся выразиться до некоторой степени
забавно. Жизнь есть сохранение жизни.»
В начале XIX века французский естествоиспытатель К. Биша
создал учение о сферах проявления жизнедеятельности человека,
которое можно положить в основу классификации механизмов
приспособления организма к внешней среде. По мысли автора,
жизнедеятельность человека проявляется в вегетативных, соматических
и психических процессах.
Суть вегетатики составляет обмен веществ и энергии между
организмом и окружающей средой. Благодаря обмену, точнее его
изменениям в разных условиях, биологическая система может
приспособиться к изменяющейся среде. Характер и уровень обмена регулируются
у человека и животных посредством управления так называемыми
вегетативными функциями: дыханием, кровообращением, пищеварением,
выделением и др. Возможности такой формы приспособления
довольно ограничены. При значительных отклонениях параметров среды от
нормальных для жизни условий изменения вегетатики не могут
обеспечить поддержание жизнеспособности. По мнению К. Биша (1802),
вегетативные процессы, как форма приспособления организма к среде,
присущи растению, животному и человеку.
Принципиальным отличием животных от растений является
способность передвигаться и за счёт этого избегать действия
неблагоприятной среды (движение позволяет уходить из «плохой» среды
в «хорошую»). К. Биша говорил о появлении у животного новой
сферы проявления жизнедеятельности - соматики (двигательной актив-
Глава 7. Информация и регулирование в биологических системах
363
ности). Зачатки двигательной активности, позволяющей в какой-то
степени избегать действия неблагоприятной среды, есть и у растений
(примером служат «ползающие» или «лазающие» растения), однако
у животных двигательная функция качественно иная. Её
приспособительные возможности несравненно выше.
В свою очередь, у животных намечаются зачатки новой формы
приспособления, которая обеспечивается психической сферой.
Однако у человека высшая нервная деятельность приобрела качественно
новые свойства. Она даёт возможность людям не только
приспосабливаться к существующей среде путем более тонкой регуляции веге-
татики, не только уходить из одной среды в другую и тем самым
избегать воздействия неблагоприятных факторов на организм, но
и активно влиять на среду, устраняя из неё неблагоприятные для
организма факторы. Разумное преобразование внешней среды
способствует сохранению жизнеспособности человеческого организма.
Следовательно, развитие психики позволило человеку
приспосабливаться к окружающей среде за счёт изменения её в интересах
поддержания жизни. Психическая деятельность считается высшей формой
приспособления организма к среде.
Исполнителем всех регуляторных функций в организме
является клетка, её генетический аппарат и метаболические системы.
Последние обеспечивают быстрые приспособительные реакции
организма на изменения окружающей среды.
Как правило, в эти реакции включаются внутриклеточные
сигнальные системы (системы вторичных мессенджеров, т.е.
посредников). Концепция о внутриклеточных сигнальных системах начала
развиваться после открытия в 1958 г. Ф. Сазерлендом роли
циклического аденозинмонофосфата (цАМФ) в гликогенолизе. Позднее
установили участие системы цАМФ в регуляции секреции, подвижности,
пролиферации, мембранного транспорта и других физиологических
процессов. Параллельно накапливались научные факты и о других
внутриклеточных сигнальных системах (циклического гуанозинмо-
нофосфата - цГМФ, тирозинкиназы, фосфоинозитидов, арахидоно-
вой кислоты). Развитие представлений о вторичных мессенджерах
объединило доселе разрозненные усилия физиологов, биофизиков
и биохимиков в изучении регуляторных процессов, свойственных
364
Биофизика
биологическим системам. Принципы функционирования
внутриклеточных систем частично рассмотрены в учебнике при изложении
таких вопросов, как фосфорилирование кальциевых каналов и
механизм фоторецепции.
Внутриклеточные сигнальные системы являются ключевым
звеном всех форм регуляции физиологических процессов: местной,
гуморальной и нервной.
7.2.3. Понятие о местной регуляции
физиологических процессов
Местной регуляцией называют информационные
взаимодействия между соседними клетками. Они могут взаимодействовать при
помощи сигналов самой разнообразной природы. Однако роль
механических и тепловых сигналов в межклеточном взаимодействии,
по-видимому, незначительна. С 20-х годов XX века
предпринимаются попытки доказать предположение А.Г. Гурвича о межклеточном
обмене информацией в форме так называемых митогенетических
лучей, отождествляемых сейчас со сверхслабой биолюминесценцией.
И все же доводы в пользу существования такого информационного
канала пока недостаточно убедительны. Основными формами
передачи информации от клетки к клетке считаются сигналы химической
и электрической природы.
Каждая клетка способна передать сообщение о своем
состоянии при помощи определённых химических веществ, выделяемых ею
в интерстиций. У соседей по клеточному сообществу есть два
основных способа восприятия таких сигналов. Со многими химическими
агентами они взаимодействуют при помощи мембранных
рецепторов, присутствующих в их плазмолемме. В результате возникают
разнообразные внутриклеточные (преимущественно
метаболические) реакции. Связь мембранных рецепторов с метаболическими
системами осуществляют внутриклеточные посредники (вторичные
мессенджеры).
Другие химические агенты проникают через плазмолемму
клетки и образуют комплексы с компонентами цитозоля и
содержимого органоидов. Такие комплексы оказывают воздействие на раз-
Глава 7. Информация и регулирование в биологических системах
365
личные внутриклеточные процессы. Например, в клеточном ядре они
взаимодействуют с хроматином, вызывая депрессию или индукцию
определённых генов.
Обнаружены и такие формы межклеточного взаимодействия,
которые осуществляются без выхода химических веществ в интер-
стиций. Речь идёт прежде всего о щелевых контактах. Через сквозные
каналы из клетки в клетку могут перемещаться химические
соединения, молекулярная масса которых не превышает 1,8 кДа (сахара,
аминокислоты, низкомолекулярные пептиды, нуклеотиды и т.п.). Таким
образом, щелевые контакты обеспечивают непрерывные и точно
направленные межклеточные химические коммуникации,
изолированные от интерстиция.
Щелевым контактам принадлежит важная роль и в
электрическом взаимодействии между клетками, поскольку
электропроводность плазмолеммы в электрических синапсах на 5-6 порядков выше,
чем в других участках плазмолеммы. Вместе с тем электрические
взаимодействия между клетками не ограничиваются щелевыми
контактами. Изменения мембранного потенциала отдельной клетки,
очевидно, оказывают некоторое влияние на её окружение.
К местной регуляции сейчас привлечено более пристальное
внимание исследователей, чем это было прежде. Показано, что на
ранних стадиях развития патологического процесса возможна
нормализация свойств дефектных клеток за счёт обмена информацией
с окружающими их нормально функционирующими клеточными
структурами. Клеточные сообщества существуют только благодаря
передаче информации от клетки к клетке. Её нарушение разобщает
клетки, и каждая из них начинает жить сама по себе. В таком
угнетении информационных процессов на уровне местной регуляции
некоторые исследователи видят причину безудержного клеточного роста
при развитии злокачественной опухоли. В экспериментах
установлено, что клетки злокачественных опухолей утрачивают способность
обмениваться информацией. Дальнейшее изучение нарушений
информационных процессов при межклеточных взаимодействиях
рассматривается как один из перспективных путей разрешения
кардинальных проблем онкологии.
366
Биофизика
7.2.4. Понятие о гуморальной регуляции
физиологических процессов
Наряду с местной существует гуморальная регуляция
физиологических процессов (её название происходит от латинского слова
humor - жидкость). Под гуморальной информационной системой
подразумевают перенос физиологически активных веществ
жидкостями, перемещающимися в организме. Гуморальная регуляция
осуществляется прежде всего благодаря кровообращению, а также
движению лимфы и ликвора. Некоторое значение имеют перемещения
межклеточной жидкости, но такую гуморальную регуляцию трудно
отличить от местной.
Физиологически активные агенты, доставляемые в органы
и ткани кровью и лимфой, оказывают регуляторное влияние на
клетки посредством одного из двух механизмов, охарактеризованных
в предыдущем параграфе. Химические соединения могут, во-первых,
взаимодействовать с мембранными рецепторами и, во-вторых, -
проникать в цитоплазму.
Большинство гормонов, обладающих пептидной структурой
(адреналин, инсулин, паратгормон, глюкагон, кальциотонин и т.п.),
а также энкефалины и эндорфины не проникают в клетки, а
связываются специализированными рецепторами их плазматических
мембран. Все мембранные рецепторы, известные сегодня, относятся
к гликопротеинам и гликолипидам (ганглиозидам). В связывании
гормонов основную роль играют полисахаридные цепи этих
соединений. Специфичность взаимодействия физиологически активных
агентов с мембранными рецепторами чрезвычайно высока.
Некоторые из пептидных гормонов действуют на клетки в концентрации 10"
1' моль ■ л-1. Каждая молекула гормона, провзаимодействовав с
мембранным рецептором, запускает сложные внутриклеточные
химические процессы, в которых участвуют миллионы других молекул.
Посредником таких реакций обычно служат системы вторичных
мессенджеров (внутриклеточные сигнальные системы).
Стероидные гормоны действуют на клетки иначе. В плазмо-
лемме присутствуют рецепторы стероидных гормонов, но их
функция состоит не в запуске внутриклеточных процессов, а в обеспече-
Глава 7. Информация и регулирование в биологических системах
367
нии трансмембранного переноса гормона, который распознан
рецептором. Связавшись с мембранным рецептором, стероидный
гормон сохраняет неизменной свою химическую структуру, в таком
виде проникает в цитозоль и далее - в клеточное ядро. Уже через 30-
60 мин он накапливается в ядре в максимальной концентрации.
Стероидные гормоны оказывают регуляторное влияние на генетический
аппарат клетки. После воздействия на хроматин они покидают ядро
и выводятся из клетки в интерстиций, откуда доставляются кровью
в органы выделения и удаляются из организма.
К изучению гуморальной регуляции также применяется
аппарат теории информации. Рассчитывается информативность
различных гуморальных агентов и таким образом сопоставляются их
информационные возможности. Подобная оценка циклического аде-
нозинмонофосфата (цАМФ) подтвердила справедливость для
организма основной идеи К. Шеннона: то, что передается часто, должно
кодироваться короткими сигналами. Использование вездесущего
цАМФ -пример краткости кодирования информации в клетке.
Приведенные примеры свидетельствуют, что изучение
информационных процессов в организме вышло за рамки нервных
каналов связи. Эта тенденция имеет важное значение как для
теоретической биологии, так и для практической медицины, поскольку
разумное вмешательство врача в информационные системы
больного человека служит эффективным средством лечения
разнообразных болезней.
7.2.5. Нервная регуляция
физиологических процессов
Гуморальная и местная формы регуляции свойственны
вегетативным процессам в организме человека. Вместе с тем в
управлении вегетатикой важная роль принадлежит нервной системе. Вся
соматическая активность обеспечивается нервной регуляцией.
Психика связана с функционированием высших отделов центральной
нервной системы. Универсальным механизмом нервной регуляции
вегетатики и соматики, а также психической деятельности служит
рефлекс.
368
Биофизика
Рис. 7.4. Схема рефлекторной дуги
А — афферентное звено; Ц- центральное звено; Э — эфферентное звено;
1 - рецепторный аппарат; 2 - афферентные волокна; 3 - нейроны
центральной нервной системы; 4 - эфферентные волокна; 5 - эффектор
В физиологии и биофизике рефлексом называют реакцию
организма на изменения внешней среды, протекающую при участии
центральной нервной системы. Участие ЦНС в рефлексе
обеспечивает наряду с приёмом информации её переработку и хранение,
а также формирование сигналов к исполнительным органам.
Структурной основой рефлекса является рефлекторная дуга^
состоящая из 3 основных звеньев: афферентного, центрального,
эфферентного (рис. 7.4). Афферентное {центростремительное) звено
включает рецепторные аппараты, воспринимающие адекватные
раздражители, и афферентные волокна (отростки чувствительных
нейронов), проводящие нервные импульсы от рецепторов в ЦНС.
Следовательно, афферентное звено рефлекторной дуги обеспечивает приём
информации и передачу её туда, где она обрабатывается. Приём
информации сопряжен с её первичной обработкой, выражением чего
является кодирование. Однако основная обработка информации
осуществляется в центральном звене рефлекторной дуги.
Глава 7. Информация и регулирование в биологических системах
369
В центральное звено входят все нейроны ЦНС, участвующие
в осуществлении данного рефлекса. Важным элементом
центрального звена являются синапсы между нейронами. С их функцией связаны
и приём сигналов, и интегративная деятельность нейронов, и
хранение информации, и формирование команды исполнительному
органу. Командный сигнал поступает в третье - эфферентное звено
рефлекторной дуги, включающее эфферентные (центробежные) нервные
волокна и исполнительный орган - эффектор (мышца, железа и т.д.).
Развитие рефлекторной теории связано с именами Ивана
Михайловича Сеченова и Ивана Петровича Павлова. До работ Сеченова
физиологи считали, что рефлекс служит механизмом нервной регуляции
вегетативных и соматических процессов. Психика изучалась отдельно,
причём большинство учёных не относило психические явления к
предмету физиологических исследований. И.М. Сеченов высказал
предположение, что механизмом психической деятельности является
рефлекс. Эта гипотеза, сыграв значительную роль в формировании
материалистического мировоззрения современников, долго оставалась
«гениальным взмахом мысли», не подкрепленным экспериментальными
доказательствами. Идею И.М. Сеченова доказал И.П. Павлов,
разработавший метод физиологического (объективного) изучения
психических явлений. Речь идёт о методе условных рефлексов.
И.П. Павлов установил, что те рефлексы, на которых строится
психическая деятельность, отличаются от врожденных форм
рефлекторных реакций. Поэтому он подразделил все рефлексы на две
большие группы: условные и безусловные. Первые из них обеспечивают
психические процессы. Различия между условными и безусловными
рефлексами изучаются подробно в курсе нормальной физиологии.
Здесь уместно остановиться лишь на главном различии двух групп
рефлекторных реакций. Они имеют принципиально разное
биологическое значение в регуляции функций организма.
Примером безусловного рефлекса может служить отдергивание
руки при уколе кожи. В этом случае биологическое значение реакции
очевидно - она направлена на то, чтобы избежать действия
вредоносного фактора, причём он сам является непосредственным
источником информации о своем действии. В качестве примера условного
рефлекса, изучаемого в лаборатории, рассмотрим слюноотделение,
370
Биофизика
возникающее в ответ на звонок или вспышку света, в
слюноотделительной реакции на которые организм не нуждается. Такая реакция
появляется только в том случае, если включение звонка или вспышка
света «подкрепляются» воздействием на организм ещё одного
стимула, на который организм реагирует слюноотделением по механизму
безусловного рефлекса (например, вливание в рот растворов
раздражающих веществ). После нескольких сочетаний индифферентного
(звук или свет) и биологически значимого (безусловного) по
отношению к слюноотделению раздражителей слюна начинает выделяться
и на отдельное предъявление индифферентного стимула (он теперь
становится условным раздражителем). Биологический смысл
условного раздражителя состоит в том, что он выполняет роль сигнала
о будущих событиях, к реагированию на которые организму
необходимо подготовиться соответствующим образом.
Условный раздражитель — это сигнал о грядущем событии,
побудительный мотив к предстоящей деятельности, а условнореф-
лекторные реакции обеспечивают организму предуготовленность
к ним. В момент действия условного раздражителя в среде ещё не
произошли изменения, к которым организм должен
приспосабливаться (избегать опасности, питаться, размножаться и т.д.), но
условный раздражитель служит для организма сигналом,
предупреждающим о том, что через определённое время произойдёт биологически
значимое изменение среды и к нему следует подготовиться.
7.2.6. Обратные связи в рефлекторных актах
Обращение к рефлексам потребовалось нам для того, чтобы
понять, как в рефлекторной теории проявляются основные принципы
кибернетики. Среди них одним из важнейших является принцип
обратной связи.
Развитие кибернетики привело к тому, что физиологи и
биофизики вспомнили одну из интереснейших идей выдающегося анатома
и физиолога начала XIX в. Чарльза Белла, который внёс весомый
вклад в развитие рефлекторной теории. Он утверждал, что
рефлекторная деятельность не заканчивается совершением работы исполни»
тельным органом. В нём есть собственные рецепторы, для которых
Глава 7. Информация и регулирование в биологических системах
371
результаты этой работы служат раздражителями. Рецепторы
исполнительного органа (эффектора) посылают в ЦНС сигналы,
содержащие информацию о том, как выполнена команда, и являющиеся
источником новых рефлексов, которые корректируют деятельность
эффектора в целях наиточнейшего выполнения команды.
Например, в скелетных мышцах присутствуют механорецепто-
ры, возбуждаемые перемещениями мышечных волокон. Эти так
называемые проприоцепторы кодируют информацию о сокращении и,
следовательно, о том, как мышца при своем сокращении выполнила
команду, посланную ей мозгом. Центральная нервная система
анализирует сигналы от проприоцепторов, что и служит основой для
коррекции дальнейшей работы исполнительного органа.
Таким образом, однократное стимулирование рецепторного
аппарата, приводящее к рефлекторному акту, в реальных условиях не
заканчивается однократным возбуждением исполнительного органа.
Работа эффектора является источником новых рефлекторных реакций,
протекающих до тех пор, пока не будет достигнуто точное достижение
цели определённого рефлекса. Информация циркулирует в
своеобразном «рефлекторном кольце»: рецептор - мозг - эффектор - рецепторы
самого эффектора - мозг - эффектор и т.д. В ЦНС сигналы от
рецепторов исполнительного органа взаимодействуют с новыми сигналами от
рецепторных аппаратов, служивших источником первоначальной
реакции. Кроме того, благодаря информации, поступающей от
рецепторов исполнительного органа, мозг может сравнивать результат
исполнения своей команды с моделью (эталоном) идеального результата,
формируемой в ЦНС на основании как сиюминутных сведений о
среде, так и предшествующего опыта, запёчатлённого в памяти.
Часть «рефлекторного кольца», по которой сигналы
передаются от эффектора в мозг (навстречу командам, поступающим от мозга
к эффектору), называется обратной связью. Если она обеспечивается
рецепторами самого исполнительного органа, говорят о внутреннем
контуре обратной связи. Вместе с тем контроль за деятельностью
эффекторов осуществляется не только их собственными рецепторными
аппаратами. Например, при письме движения руки контролируются
как рецепторами её мышц и сухожилий, так и другими сенсорными
системами - в первую очередь зрительным анализатором. В необхо-
372
Биофизика
димости такой обратной связи легко убедиться, попытавшись писать
с закрытыми глазами. Следовательно, наряду с внутренним
существуют и внешние контуры обратных связей. В данном случае слово
«внешний» подразумевает участие таких рецепторных аппаратов,
которые не принадлежат эффектору первичной рефлекторной дуги.
Обратные связи в рефлекторных актах весьма разнообразны, но все они
имеют одно предназначение - коррекцию рефлекторной
деятельности исполнительных органов. Без обратных связей все реакции были
бы менее четкими, не столь целесообразными и целенаправленными,
какими они являются в организме человека и животных.
7.2.7. Приложения теории
автоматического регулирования
к анализу рефлекторной деятельности
Изучение обратных связей как в рефлекторных реакциях, так
и в других формах регуляции функций в организме (гуморальной,
местной), привело к выводу о замкнутости контуров биологического
регулирования. При этом весьма полезным оказалось применение
принципов теории автоматического регулирования к изучению
разнообразных регуляторных механизмов в биологических системах.
Одним из основных понятий, которыми оперирует теория
автоматического регулирования (управления), является понятие системы.
В кибернетике системой принято называть любую совокупность
взаимодействующих устройств и процессов, выполняющих в целом
определённую функцию. Каждый из элементов системы может
выполнять одну или группу подчинённых функций (подфункций), но
только в совокупности и взаимодействии элементы, образующие систему,
способны осуществить полную функцию, изучаемую
исследователем. Например, система кровообращения в целом выполняет
транспортную функцию. Составные элементы системы кровообращения -
сердце и кровеносные сосуды - по-разному участвуют в
осуществлении транспортной функции. Сердце нагнетает в сосуды кровь и тем
самым служит насосом (это подфункция относительно всей
транспортной функции). Кровеносные сосуды - магистрали для движения
крови. Вместе с тем артерии эластического и мышечного типов, ка-
Глава 7. Информация и регулирование в биологических системах
373
Рис. 7.5. Структурные схемы систем автоматического регулирования
(а - по возмущению, б - по отклонению, в - комбинированной)
ЭС - элемент сравнения; Ρ - регулятор; ОР - объект
регулирования; ОС - обратная связь; И- измеритель
возмущающего фактора; КУ— корректирующее устройство
пилляры и вены выполняют свои дополнительные функции.
Феномены «воздушного колпака» в аорте, «сосудистых кранов» - артериол,
обмена веществ через капиллярную стенку, ёмкостных свойств вен -
все это подфункции общей транспортной функции
сердечно-сосудистой системы.
Систему автоматического регулирования (САР) изображают
обычно в виде структурной схемы, представляющей собой набор
элементов, каждый из которых выполняет определённую подфункцию,
описан математическим выражением и соединён с другими
элементами линиями со стрелками, указывающими на однонаправленные
причинно-следственные связи, существующие в системе (рис. 7.5).
Различают системы автоматического регулирования по
возмущению (рис. 7.5, а) и отклонению (рис 7.5, б). Важными блоками
обеих систем являются объект регулирования (ОР) и регулятор (Р).
Объект регулирования - это элемент САР, в котором протекает процесс,
подлежащий регулированию. На выходе ОР можно зарегистрировать
так называемую выходную характеристику системы -y(t). Она
представляет собой зависимость от времени регулируемого параметра
(иными словами, его изменения во времени). Для управления
объектом регулирования система содержит регулятор. В нём формируется
управляющий сигнал (команда), который также можно представить
374
Биофизика
в виде функциональной зависимости сигнального признака команды
от времени - S(t).
В системах автоматического регулирования по возмущению
непосредственным сигналом к изменению режима работы
регулятора служит реакция на возмуи(ающие факторы -x(t). К ним относятся
любые изменения внешней среды, способные нарушить исходные
режимы работы объекта регулирования или регулятора. Для
предупреждения таких нарушений в САР по возмущению присутствуют
измеритель возмущающего фактора (И) и корректирующее
устройство (КУ). Под влиянием x(t) в и возникает сигнал, включающий КУ,
а его выходные сигналы - ε'(ί) корректируют работу регулятора,
вследствие чего управляющая функция - S(t) изменяется таким
образом, чтобы обеспечить функционирование объекта регулирования
в оптимальном режиме, несмотря на существование возмущающих
факторов. Можно сказать, что ОР приспосабливается к их
присутствию в среде.
В системах автоматического регулирования по отклонению
непосредственным сигналом к изменению режима работы
регулятора служит изменение выходной характеристики —y(t). Как только она
изменится под влиянием возмущающих факторов - x(t) или
собственных «шумов» системы, произойдёт коррекция сигналов в цепи
ЭС-Р-ОР. Для осуществления такой коррекции САР по отклонению
имеет цепь обратной связи (ОС). Преобразованная выходная
характеристика -y(t) подается по цепи ОС па элемент сравнения (ЭС),
обладающий двумя входами. Один из них служит для приёма сигнала
обратной связи -y'(t), а другой - эталонной функции -g(t), с которой
в ЭС сравнивается выходная характеристика. Эталонная функция
является образцом для выходной характеристики, и вся работа САР по
отклонению заключается в том, чтобы y(t) не отличалась от своего
образца. Эталонную функцию принято называть задающим
воздействием или уставкой. В большинстве технических САР уставка
имеет электрическую природу, представляя собой аналоговую или
дискретную функцию тока или напряжения от времени. В сложных
системах уставка формируется электронными вычислительными
машинами. В ней учтены как задачи, так и возможности данной САР,
Глава 7. Информация и регулирование в биологических системах 375
а также текущие изменения условий её работы, которые могут
возникнуть под действием возмущающих факторов.
В отличие от уставки, выходной характеристике свойственна
зачастую неэлектрическая природа. Например, у такого ОР, как
электромотор, выходной характеристикой является механическая сила,
изменяющаяся во времени. В подобных случаях цепь ОС
осуществляет не только передачу части выходного сигнала на вход системы (на
ЭС), но и преобразование разномодальпых сигналов в электрические.
Только в таком виде выходная характеристика может сравниваться
с уставкой. Поэтому цепь ОС включает, как правило, измерительный
преобразователь, обеспечивающий кодирование разномодальпых
выходных характеристик - y(t) электрическими сигналами (функциями
тока или напряжения от времени). В ЭС поступает выходная
характеристика, преобразованная в электрические сигналы, -y'(t). He зря на
рис. 7.5, б в цепи обратной связи показаны две функции: до
измерительного преобразователя (ОС) -y(t), а после него -У (О·
Элемент сравнения выполняет операцию алгебраического
сложения, вычитая в каждый момент времени из g[t) соответствующие
значения y\t). В результате на выходе ЭС формируется сигнал
рассогласования (иначе его называют ошибкой регулирования) - ε(ί) = g[i) -
y\t). Значение ε(/) может быть равным нулю. Отсутствие ошибки
регулирования означает полное соответствие преобразованной выходной
характеристики уставке. В этом случае на регулятор и далее на объект
регулирования корректирующие сигналы не поступают. Режим работы
ОР не изменяется. Как только ε(ί) станет отличной от нуля, регулятор
изменит свой выходной сигнал - S(t), вследствие чего ОР будет
работать по-другому, подгоняя выходную характеристику к уставке. Таким
образом, ε(/) обеспечивает коррекцию режима работы объекта
регулирования и, следовательно, играет роль корректирующего сигнала.
В САР по отклонению нет нужды измерять и преобразовывать
возмущающие факторы -x(t), как это делает- САР по возмущению. Для
работы в среде с большим количеством разнообразных x(t)
практически невозможно использовать САР по возмущению, поскольку каждый
возмущающий фактор должен восприниматься отдельным
измерителем. Такая система не способна отреагировать и на появление в среде
376
Биофизика
новых x(t). В этом отношении САР по отклонению обладает
несомненным преимуществом, так как обеспечивает регулирование за счёт
оценки величины отклонения выходной характеристики от уставки
независимо от причины рассогласованияy'(t) и g{t). Однако простые САР
по отклонению начинают корректировать работу ОР только после
того, как в системе уже совершится такое рассогласование. Можно
сказать, что регулирование по отклонению - это регулирование вслед
за ошибкой. Когда же и кратковременное рассогласование g(t) viy'(t)
недопустимо, приходится усложнять структуру САР. Этому служат
комбинированные системы, сочетающие оба основных принципа
автоматического регулирования: по отклонению и по возмущению
(рис. 7.5, в). Ошибку регулирования можно уменьшить за счёт
усложнения устройств, формирующих уставку. Применение ЭВМ позволяет
формировать уставку с прогнозированием дальнейших условий работы
САР. Регулирование с предвосхищением грядущих событий на
основании их прогнозирования называется форпостным регулированием.
В зависимости от характера функции g{t) разнообразные САР
подразделяются на три основные группы. Если уставка - постоянная
величина, то это система стабилизации (рис. 7.6, а). В организме
человека и животных к системам стабилизации можно отнести
механизмы поддержания гомеостазиса. Если g(t) подчиняется
определённой программе, то САР называется системой программного
управления. Простой программой можно считать периодический
закон, которому следуют изменения уставки в системах регуляции
биоритмов (рис. 7.6, б). В следящих системах уставка изменяется
случайным образом. Многие процессы в организме имеют
вероятностный характер. Они регулируются следящими системами.
Сопоставим рефлекторную дугу и структурную схему САР по
отклонению (рис. 7.7). В рефлекторной дуге объектом регулирования
является исполнительный орган {эффектор). Деятельность любого
эффектора можно описать математически в виде функциональной
зависимости его основного параметра от времени. Такая функция
служит выходной характеристикой системы регулирования
(рефлекторной дуги).
Реакция исполнительного органа оценивается как его
собственными рецепторами, так и рецепторными аппаратами, принадле-
Глава 7. Информация и регулирование в биологических системах
377
\лаалааа>Кал/
у%г<
а)
6)
Рис. 7.6. Графики уставки (1) и выходной характеристики (2)
в системах стабилизации (а) и программного управления (б)
жащими другим анализаторам. Рецепторы, воспринимающие те или
иные проявления реакции эффектора на сигналы, поступившие из
центральной нервной системы (от регулятора), входят в цепь
обратной связи. Здесь рецепторные аппараты выполняют свою обычную
функцию - преобразуют разномодальные раздражители (те или иные
следствия рефлекторного акта, например мышечные усилия) в
нервные импульсы. Только в форме нервной импульсации мозг способен
получить информацию о выходной характеристике и сравнить её
с уставкой. Следовательно, в рефлекторных дугах элементами
сравнения являются нейроны центрального звена. Ошибочно мнение,
будто роль элемента сравнения принадлежит рецепторам. Эта
ошибка приводит к другой, не менее существенной. Глубоким
заблуждением является утверждение, что уставка - это раздражители,
воздействующие на рецепторы органов чувств.
Уставка формируется в центральной нервной системе. Она
представляет собой идеальную модель результата рефлекторного
акта. При её создании мозг синтезирует информацию о среде в
данный момент, учитывает состояние организма и на основе прежнего
опыта подобных реакций (памяти) формирует сигналы - g(t), с
которыми сравнивается преобразованная выходная характеристика -у'(().
Обе функции закодированы нервными импульсами.
Таким образом, элемент сравнения, источник уставки и
регулятор, показанные на рис. 7.7, локализованы в центральном звене
рефлекторной дуги (рис.7.4). В разных рефлекторных дугах миссию
каждого из перечисленных компонентов САР выполняют различные
378
Биофизика
а)
Щ
VX»
Рис. 7.7. Сопоставление рефлекторной дуги (а) и структурной схемы
системы автоматического регулирования по отклонению (б)
нервные центры, находящиеся в том или ином отделе головного
и спинного мозга. При рассмотрении конкретной рефлекторной дуги
можно назвать определенные анатомические образования мозга,
функционирующие как регулятор и элемент сравнения. С указанием
источника уставки дело обстоит сложнее. Важно понимать, что
каждому элементу структурной схемы САР соответствует, как правило,
не один нервный центр, а группа структур головного и спинного
мозга. Так, регулятор, управляющий движениями человека, включает
нервные центры передней центральной извилины коры больших
полушарий, базальных ядер головного мозга, мозжечка, среднего,
продолговатого и спинного мозга. На выходе такого регулятора,
посылающего управляющие сигналы мышцам конечностей и туловища,
находятся сс-мотонейроны передних рогов спинного мозга.
В терминах теории автоматического регулирования аналогом
раздражителей являются возмущающие факторы. Для них в
структурной схеме САР по отклонению (рис. 7.5, б) не предусмотрены
специальные входы, поскольку они могут быть приложены к любому
Глава 7. Информация и регулирование в биологических системах
379
элементу системы регулирования. В реальных условиях действие
большинства возмущающих факторов на мозг опосредуется рецеп-
торными аппаратами. Конечно, яды и механические перегрузки,
жара и холод способны нарушить нервную деятельность и за счет
непосредственного действия на мозговые структуры. Однако любые
изменения среды улавливаются прежде всего рецепторами, и ЦНС
информируется об этом сообщениями в форме афферентной импуль-
сации. Следовательно, подавляющее большинство возмущающих
факторов - x(t) внедряется в САР физиологических процессов через
рецепторные аппараты сенсорных систем организма. Иными
словами, входами для x(t) служат афферентные звенья рефлекторных дуг.
Сигналы от рецепторов вызывают изменения выходной
характеристики и, следовательно, отклонения её от уставки, за счёт чего в
элементе сравнения возникает сигнал рассогласования - (/),
корректирующий режим работы регулятора. В результате такой коррекции
исполнительный орган (объект регулирования) начинает работать
иначе, обеспечивая уменьшение возникшего рассогласования между
уставкой и выходной характеристикой.
На первый взгляд, возмущающие факторы мешают
автоматическому регулированию. Без них гораздо меньше рассогласований
между g(t) vLy\t) и, следовательно, ОР нуждается в меньшей
коррекции. Однако от такого мнения придётся отказаться, вспомнив, что
задача регулирования биологической системы состоит в её
приспособлении к среде. Ведь именно в x(t) содержится информация о тех
изменениях среды, которые требуют перевода системм на новый
уровень функционирования, обеспечивающий поддержание
жизнедеятельности в изменившихся условиях.
Рассмотрим, например, приспособление организма к
недостатку кислорода в окружающей среде. Понижение содержания 02 во
вдыхаемом воздухе может нарушить гомеостазис. Однако при
умеренном недостатке кислорода организм сохраняет гомеостазис за
счёт изменения режима регулирования вегетативных функций:
усиливается внешнее дыхание, большую мощность развивает сердце,
увеличивая объёмную скорость кровотока, расширяется капиллярная
сеть и т.д. Все эти реакции возникают вследствие рассогласования
380
Биофизика
между параметрами гомеостазиса и соответствующих уставок (в
физиологии их называют должными параметрами функции).
В сложных системах регулирования (например, при
управлении движениями) изменения среды приводят к перестройке и самой
уставки. Однако перестройка g(t) происходит также под влиянием
сигнализации от рецепторных аппаратов сенсорных систем
организма. В этом случае рассогласование уставки и выходной
характеристики происходит значительно сложнее.
Таким образом, возмущающие факторы заставляют САР
изменять регуляторные режимы для перевода объектов регулирования на
новые уровни функционирования. Так достигается приспособление
организма к меняющейся среде. Чем сильнее изменяется внешняя
среда, тем глубже и сложнее перестройка процессов регулирования
в биологических системах.
Важнейшим параметром ответа САР на возмущающие
факторы является время. Речь идет и о латентном (скрытом) периоде
реакции, и о скорости перехода на новый режим регулирования. Только
при достаточно быстром реагировании система точно следует за
непрерывно меняющимися условиями функционирования.
Беспрестанная динамика свойственна среде обитания живых организмов,
причём х(() имеет преимущественно случайный (вероятностный)
характер. Поэтому для биологических систем особое значение
приобрела реализация принципа форпостного регулирования.
7.2.8. Форпостное регулирование
функций организма
Форпостное (предвосхищающее грядущие события)
регулирование является более совершенной формой регуляции функций
организма по сравнению с механизмами управления, включающимися
вслед за изменением выходной характеристики. В свете
кибернетических представлений условный рефлекс можно рассматривать в
качестве механизма форпостного регулирования.
В условном рефлексе проявляется основной принцип
психической деятельности - образование временных связей между
организмом и окружающей средой. Тот или иной фактор среды после ряда
Глава 7. Информация и регулирование в биологических системах
381
сочетаний с биологически значимым изменением её параметров
приобретает роль предупреждающего сигнала. Такое предупреждение
предшествует действию безусловного раздражителя, в силу чего
организм оказывается заранее подготовленным к нему. Если это
неблагоприятный фактор, то на основе психической деятельности в
организме формируются реакции, устраняющие пагубные влияния еще до
того, как они начали действовать. Благодаря реализации принципа
форпостного регулирования образование временных связей между
организмом и внешней средой относится к высшим формам
приспособления биологических систем. Эти реакции составляют высшую
нервную деятельность, которая у человека приобрела качественно
новые свойства по сравнению с подобными процессами у животных
и получила название психической деятельности.
Реакции, предвосхищающие изменения внешней и внутренней
сред организма, сформировались в ходе эволюции уже на уровне
безусловных рефлексов. Однако механизмы форпостного регулирования
гораздо совершеннее и разнообразнее при образовании временных
связей. Психическая деятельность дала человеку неисчерпаемые
возможности реализации принципа форпостного регулирования в
приспособлении к внешней среде, изменяющейся стохастически.
Современную философию обогащают и кибернетические идеи,
одним из источников которых явилось изучение управления и связи
в биологических системах. Применение к ним кибернетических
принципов ярко продемонстрировало внутреннюю диалектику жизни.
Кибернетический подход способствует объединению разнообразных
и зачастую разрозненных фактов естествознания, помогая
естествоиспытателям продвигаться в направлении глубоких философских
обобщений явлений жизни на атомном, молекулярном, клеточном,
органном и организменном уровнях.
7.3. Регуляция температуры тела
гомойотермного организма
Постоянство температуры «ядра» тела у гомойотермного
животного поддерживается благодаря слаженному взаимодействию
различных регуляторных процессов. Их рассмотрение на конкретном
382
Биофизике
примере способствует глубокому пониманию принципов
регулирования биологических процессов.
7.3.1. Эффекторы теплопродукции
Изменение уровня теплопродукции в метаболических
процессах (т.е. химическая терморегуляция) осуществляется прежде всего
за счет усиления или ослабления двигательной активности человека
и животного. Для выработки организмом дополнительного тепла
посредством сократительного термогенеза особенно эффективна
активная произвольная мышечная деятельность (физическая работа).
Даже при слабой физической работе теплопродукция возрастает на
50-80%, а при тяжелой в 4—5 раз.
Сократительный термогенез имеет место и при
непроизвольных тонических и ритмических (фазических) сокращениях скелетной
мускулатуры. Тоническая мышечная активность на холоде
называется терморегуляционным тонусом скелетной мускулатуры. У
замерзающего человека вначале тонически сокращаются мышцы
подбородка, потом шеи, верхнего плечевого пояса и, наконец, туловища.
Такая сократительная активность мышц развивается подсознательно.
Вследствие непроизвольного сокращения сгибателей конечностей
животное на холоде сворачивается в клубок («калачиком»). Нечто
подобное наблюдается и у человека, особенно во сне.
Терморегуляционный тонус приводит к усилению теплообразования на 20-45%
от исходного уровня.
При более сильном охлаждении организма
терморегуляционный тонус сменяется мышечной дрожью. Она начинается с
жевательной мускулатуры, что проявляется у людей в стуке зубами, а
потом охватывает нижележащие мышцы туловища и конечностей,
вплоть до бедер.
Повышение уровня теплопродукции при замерзании
посредством непроизвольных ритмических сокращений поверхностно
расположенных мышц называют дрожательным термогенезом. Дрожь
приводит к пятикратному возрастанию скорости метаболических
процессов в скелетных мышцах, что значительно (в 2-3 раза)
усиливает химическую терморегуляцию. Считается, что эффективность
Глава 7. Информация и регулирование в биологических системах
383
дрожательного термогенеза в 2,5 раза выше произвольной
активности локомоторного аппарата, но это не очень экономный механизм,
поскольку сопровождается расширением кровеносных сосудов и
усилением теплоотдачи.
В физиологической литературе зачастую встречается термин
недрожательный термогенез. Строго говоря, к нему можно отнести
терморегуляционный тонус, но под недрожательным термогенезом
понимают теплопродукцию за счет ускорения метаболических
процессов, не связанных с какими бы то ни было мышечными
сокращениями. Поэтому лучше называть этот механизм теплопродукцией
несократительным термогенезом. Вместе с тем к недрожательному
(несократительному) термогенезу не принято относить возрастание
на 10-30% продукции тепла при приёме пищи, хотя её
специфическое динамическое действие вносит вклад в химическую
терморегуляцию.
К недрожательному {несократительному) термогенезу
относят усиление теплопродукции вследствие активации гликолиза,
гликогенолиза и липолиза, особенно при разобщении процессов
окисления и фосфорилирования в дыхательной цепи митохондрий.
Недрожательный термогенез особенно эффективен в печени, почках
и буром жире. Кроме того, ткани организма способны окислять сук-
цинат. В этом случае транспорт пары электронов по дыхательной
цепи обеспечивает синтез не трёх, а двух молекул АТФ, что,
естественно, снижает коэффициент полезного действия клеточного
дыхания и приводит к более сильной диссипации свободной энергии
и образованию избыточного тепла. К усилению теплопродукции
посредством перехода на окисление сукцината организм прибегает
только в экстремальных условиях, когда испытывает сильное и
продолжительное переохлаждение.
Разобщение окисления и фосфорилирования в дыхательной
цепи митохондрий на холоде служит более распространённым
механизмом химической терморегуляции. Он присущ так называемому
бурому жиру - своеобразной разновидности жировой ткани, которая
в сравнительно большом объеме имеется у животных, впадающих
в зимнюю спячку, и у новорожденных детей. Она располагается
в подмышечных впадинах, затылочной области, средостении и на
384
Биофизика
спине между лопатками. У взрослого человека бурый жир
присутствует в ничтожно малом количестве в окружении внутренних органов
и аорты, поскольку подвергается почти полной редукции при
половом созревании, но при длительном проживании в условиях
стабильно низкой температуры окружающей среды может вновь появиться.
Восстановление бурого жира в таких условиях было доказано в
экспериментах на кроликах. Бурая жировая ткань содержит в своих
клетках довольно крупные капли жира, окружённые большим
количеством митохондрий. В буром жире присутствует специфический
полипептид с молекулярной массой 32 кДа, обладающий
способностью разобщать окисление и фосфорилирование в митохондриях.
Полипептид выделяется при охлаждении. При этом уменьшается
синтез АТФ, и большая, чем обычно, доля свободной энергии
возбуждённых Солнцем электронов преобразуется в тепло. За счет
бурого жира у животных, впадающих в зимнюю спячку,
теплопродукция может усиливаться в 500 раз, тогда как у взрослого человека не
более чем на 10-15%.
Все перечисленные механизмы изменения теплопродукции
относятся к химической терморегуляции. Они служат выработке
дополнительного тепла при охлаждении организма. В жару химическая
терморегуляция ослабляет термогенез.
7.3.2. Эффекторы теплообмена (теплоотдачи)
Физическая терморегуляция обеспечивается регулированием
теплоизолирующих свойств тканей организма для внутреннего и,
главным образом, для наружного потоков тепловой энергии.
Динамика уровней теплопроводности, конвекции и теплоизлучения
определяется прежде всего объёмной скоростью кровотока в коже и слизистых
оболочках (для наружного потока тепловой энергии), а
кровообращение, в свою очередь, зависит от сосудистого тонуса и регулируется
изменениями просвета кровеносных сосудов.
От кровотока в поверхностных тканях зависит также неощу-
щаемое испарение (perspiratio insensibilis), однако количество
жидкости, выделяющейся на кожные покровы и подлежащей там
испарению для охлаждения организма в жару и при усилении химической
Глава 7. Информация и регулирование в биологических системах
385
терморегуляции, определяется у человека главным образом
секреторной активностью потовых желёз.
Следовательно, в процессах физической терморегуляции
основными эффекторами (исполнительными органами) регуляторных
влияний служат гладкомышечные клетки кровеносных сосудов
(преимущественно артериол) и потовые железы.
7.3.3. Физиологические
механизмы терморегуляции
Скелетным мышцам, которые служат основным эффектором
в химической терморегуляции, присуща только нервная регуляция.
В функционировании гладкомышечных клеток сосудов важную
роль играют и нервная, и гуморальная, и местная регуляция. Однако
местная регуляция сосудистого тонуса, имеющая важное значение
в поддержании необходимых уровней давления в системе
кровообращения и объёмного кровотока в том или ином органе, играет
сравнительно малую роль в терморегуляции. Правда, гладкомышечные
клетки кровеносных сосудов способны сами реагировать на понижение
температуры расслаблением, что приводит к вазодилятации и
усилению кровообращения в коже. Такая нерациональная реакция обычно
запаздывает относительно рефлекторной констрикции сосудов кожи
в ответ на охлаждение. Принято считать, что рефлекторная вазоконст-
рикция в таком случае способствует удержанию тепла в организме,
а сменяющая ее вазодилятация способствует улучшению
кровообращения в коже для восстановления ее метаболизма после ее ослабления
вследствие холодовой рефлекторной ишемии. Гуморальная регуляция
обеспечивает в химической и физической терморегуляции не столько
быстрые механизмы, сколько долгосрочные.
7.3.3.1. Нервная регуляция
постоянства температуры «ядра» тела
Универсальным механизмом нервной регуляции служит
рефлекс. Как и другие функции организма, терморегуляция
осуществляется посредством безусловных и условных рефлексов.
386
Биофизика
Условнорефлекторная регуляция
Вполне очевидна условнорефлекторная регуляция
произвольных движений, обеспечивающих терморегуляцию. Всякое усиление
двигательной активности приводит к повышению теплопродукции
(химической терморегуляции), но такие, к примеру, движения, как
обмахивания веером, способствуют лучшему теплообмену с
окружающей средой (физической терморегуляции) и обычно приводят
к некоторому охлаждению организма.
Были выявлены и другие терморегуляционные условные
рефлексы, причем эффекторами условнорефлекторных влияний
являются как скелетные мышцы, так и другие органы (печень, почки),
которым принадлежит важная роль в химической терморегуляции,
а также гладкие мышцы стенок сосудов и потовые железы,
влияющие на физическую терморегуляцию.
Условные рефлексы обеспечивают адекватное поведение
человека и животных при изменениях температурного режима. Речь идет
о выборе жилища, одежды и обуви человеком, укрытия от холода
и жары животными. В высших отделах центральной нервной
системы формируется ощущение замерзания и перегревания, теплового
комфорта. Условнорефлекторная деятельность служит важнейшим
механизмом форпостного (упреждающего) регулирования
теплового баланса. Например, у человека, который только еще собирается
выйти из теплого помещения в холодную атмосферу,
заблаговременно усиливается теплопродукция. Такая реакция не является
врожденной, а вырабатывается по мере приобретения жизненного опыта.
Безусловнорефлекторная регуляция
Рассмотрим более подробно рефлекторные дуги безусловных
терморегуляционных рефлексов (рис. 7.8).
Эфферентное звено. Начнём с эфферентного звена. Оно
включает исполнительные органы-эффекторы: скелетные мышцы,
печень, бурую жировую ткань, гладкие мышцы стенок сосудов,
потовые железы, а также иннервирующие их эфферентные нервные
волокна. Для скелетной мускулатуры ими служат аксоны (X -
мотонейронов) спинного мозга, сигнал с которых на миоциты обеспечивает
холинэргическая передача. Естественно, что миорелаксанты ослаб-
Глава 7. Информация и регулирование в биологических системах
387
Терморецепторы
гипоталамуса
Термо·
рецепторы
кожи
Температура
Другие
периферические
терморецепторы
Корв больших
полушарий
головного мозга
Гипотвлвмус /^\J:
,^]^памус у ' |
) Средний мозг ί
Продолговатый мозг
Центральное звено
Спинной
мозг
Афферентное
звено
Скелетные
мышцы
Эфферентное
звено
Рис 7.8. Схема безусловнорефлекторнойрегуляции
температуры тела человека и животных
ляют теплообразование, блокируя как произвольную, так и
непроизвольную термогенезную активность локомоторного аппарата
(терморегуляционный тонус и дрожь).
Недрожательный термогенез происходит преимущественно
в печени и бурой жировой ткани, клетки которых иннервируются
симпатическими нервными волокнами, причём в плазмолемме
клеток бурого жира и гепатоцитов присутствуют р-адренорецепторы.
При их активации в гепатоцитах усиливаются гликогенолиз и
гликолиз, а в буром жире - липолиз.
Гладкомышечные клетки кровеносных сосудов (в частности,
аритериол и аритерио-венозных анастомозов кожи) содержат в сар-
25·
388
Биофизика
колемме α-адренорецепторы, и эфферентные сигналы к ним
поступают по постузловым симпатическим волокнам.
В плазмолемме клеток потовых желез сосредоточены М-холи-
норецепторы, а иннервирующие их симпатические волокна
обеспечивают холинэргическую передачу, что свойственно предузловым
волокнам вегетативной нервной системы. Поэтому потоотделение
блокируется атропином и стимулируется парасимпатомиметиками
(ацетилхолином, пилокарпином и другими). К потогонным
средствам относятся аспирин и другие салицилаты, соцветия липы, ягоды
малины, листья мать-мачехи.
Заметим, что в термонейтральных условиях (при тепловом
комфорте) тепловой баланс гомойотермного организма поддерживается
преимущественно посредством сосудодвигательных реакций в
кожных покровах тела: при повышении средней температуры тела тонус
симпатических нервных волокон понижается, что приводит к
расширению сосудов и усилению теплообмена с окружающей средой; при
понижении средней температуры тела тонус симпатических нервных
волокон повышается, подкожные сосуды суживаются и теплообмен
с окружающей средой ослабляется.
По мере нарушения теплового комфорта к сосудодвигатель-
ным рефлексам в той или иной степени присоединяются
рефлекторные реакции других эффекторов терморегуляции.
Центральное звено дуг терморегуляционных безусловных
рефлексов включает совокупность нейронов на всех «этажах»
центральной нервной системы, участвующих в регуляции теплового
баланса организма. В спинном мозге к ним относятся вставочные
нейроны задних рогов, α-мотонейроны передних рогов и
симпатические нейроны боковых рогов. На эффекторных нейронах спинного
мозга образуют синапсы аксоны вставочных нейронов спинного
мозга, а также вышележащих отделов центра терморегуляции. К ним
принадлежат кора больших полушарий головного мозга, лимбиче-
ская система (амигдалярый комплекс и гиппокамп), гипоталамус,
мозжечок, средний и продолговатый мозг. Каждый отдел играет
свою роль в терморегуляции - свою подфункцию в терморегулятор-
ной функции всего центра.
Глава 7. Информация и регулирование в биологических системах
389
Спинной мозг обеспечивает не только непосредственный выход
сигналов из центральной нервной системы к эффекторам
терморегуляционной системы. У млекопитающих и птиц в нем найдены два типа
нейронов, реагирующих на понижение температуры самого спинного
мозга, что бывает в том случае, когда к нему по венозным сосудам
притекает кровь, охлажденная в коже спины в области лопаток.
Нередко в литературе спинной мозг называют вторым по
значимости отделом терморегуляционного центра, отдавая пальму
первенства гипоталамусу. При его разрушении гомойотермное животное
превращается в пойкилотермное. Участие гипоталамуса в
терморегуляции установил К. Бернар в середине XIX века - при уколе иглой
гипоталамуса повышалась температура тела кролика, чего не было при
механической стимуляции других отделов мозга. Такое воздействие
на гипоталамус автор открытия назвал «тепловым уколом». При
стимуляции электрическим током преоптической области гипоталамуса
(пара-вентрикулярного, супраоптического, супрахиазмального ядер)
расширяются сосуды кожи, возникают потоотделение и тепловая
отдышка, тогда как разрушение этих ядер делает животных
неспособными переносить жару. Следовательно, с передним гипоталамусом
преимущественно связана регуляция теплоотдачи (физическая
терморегуляция).
Электростимуляция заднего гипоталамуса (вентромедиального
и дорсоме-диального ядер) приводит к гипертермии, усилению липо-
лиза, гликолиза и гликогенолиза, возникновению дрожи. При
разрушении ядер заднего гипоталамуса животные плохо переносят холод.
По-видимому, задний гипоталамус обеспечивает химическую
терморегуляцию - его считают центром теплопродукции.
Согласно нейронной модели функционирования
терморегуляционного центра (Bligh, 1972), сигналы, поступившие в гипоталамус,
обрабатываются строго специфичными вставочными нейронами: от
Холодовых рецепторов - только «Холодовыми» нейронами, от
тепловых рецепторов - только «тепловыми» нейронами. От вставочных
нейронов информация передается так называемым интегрирующим
нейронам, которые тоже строго специфичны, причем «тепловые»
интегрирующие нейроны возбуждаются сигналами от «тепловых»
и тормозятся сигналами от «Холодовых» вставочных нейронов. Для
390
Биофизика
«Холодовых» интегрирующих нейронов характерны
противоположные влияния. Значит, между «тепловыми» и «Холодовыми»
интегрирующими нейронами существуют реципрокные отношения.
С интегрирующих нейронов сигналы поступают непоспредственно
на эффекторные нейроны гипоталамуса, сосредоточенные в его
задней (каудалъной) части. Они тоже специфичны по отношению
к информации о нагревании или охлаждении. Следовательно, в
системе терморегуляции кодирование качества стимула (холод или
тепло) осуществляется по принципу «меченой линии».
Аксоны эффекторных нейронов гипоталамуса следуют к ядрам
ретикулярной формации среднего и продолговатого мозга в
нисходящем медиальном пучке. Далее сигналы направляются к ядрам
передних и боковых рогов спинного мозга, регулируя активность
источников соматической (анимальной) и симпатической иннервации
эффекторов терморегуляции. Регулируются механизмы
теплопродукции и теплообмена, т.е. управляется как химическая, так и физическая
терморегуляция.
В медиальной преоптической области переднего гипоталамуса
имеются нервные клетки, формирующие так называемую уставку.
В кибернетике уставкой называют уровень регулируемой функции,
которой должна обеспечить система регуляции по отклонению.
В системе терморегуляции уставка является функцией
пороговых температур в конкретных условиях жизнедеятельности. С
уставкой сравнивается выходная характеристика системы, т.е. реальный
результат регулирования. Уставка и выходная характеристика
поступают в элемент сравнения. Если между ними нет тождества, то
элемент сравнения выдаёт сигнал на регулятор, который подгоняет
выходную характеристику под уставку.
Исследователи терморегуляции предполагают, что в переднем
гипоталамусе присутствуют как нейроны, формирующие уставку,
так и нейроны, играющие роль элементов сравнения. По-видимому,
эту функцию выполняют интегрирующие нейроны. С последних
сигнал об ошибке регулирования (расхождении между уставкой и
выходной характеристикой) поступает в задний гипоталамус, который,
в свою очередь, управляет нижележащими отделами центра
терморегуляции. Забегая вперёд, следует заметить, что переднему гипотала-
Глава 7. Информация и регулирование в биологических системах
391
мусу свойственна и терморецепция, обеспечивающая анализ
температуры крови в сосудах мозга.
Кроме того, гипоталамус осуществляет регуляторные влияния
на эффекторы терморегуляции через эндокринную систему. Так, ги-
поталамические нейроны продуцируют трипептид под названием ти-
реотропин - рилизинг-гормон, который приносится кровью в
переднюю долю гипофиза и стимулирует там выработку тиреотропного
гормона, а он, в свою очередь, в щитовидной железе усиливает синтез
тироксина и трийодтиронина. Эти гормоны весьма эффективно
регулируют теплообразование и тем самым участвуют в поддержании
определенной температуры тела. Аналогичные, опосредованные
другими гормонами, влияния на метаболизм, а значит, и на химическую
терморегуляцию, оказывают гипоталамические кортико-тропин-ри-
лизинг-гормон, гонадотропин-рилизинг-гормон, рилизинг-фактор
гормона роста. Однако их роль в регуляции температуры тела,
по-видимому, меньше, чем у тиреотропин-рилизинг-гормона.
В физиологии существует представление о «гипоталамическом
термостате». К нему относят весь терморегуляционный комплекс
гипоталамуса, в котором интегрируется информация от
терморецепторов кожи, мышц, внутренних органов и переднего гипоталамуса для
создания адекватной (взвешенной) выходной характеристики
терморегуляционного процесса, которая сравнивается с выработанной
в этом же «термостате» уставкой, и формируются сигналы к
эффекторам системы терморегуляции.
В этой системе уставка - не постоянная величина. Она
изменяется под влиянием внутренних и внешних факторов, из которых
в нормальных условиях главным является температура окружающей
среды. При лихорадке такими факторами служат эндогенные пироге-
ны. На холоде уровень уставки повышается, т.е. устанавливается
такой уровень регулирования, который обеспечивает большее
напряжение терморегуляционного комплекса для поддержания нормальной
температуры «ядра». Поэтому у всех гомойотермных животных во
время острого холодового воздействия температура «ядра»
увеличена на несколько десятых долей градуса.
Высшие отделы центра терморегуляции сосредоточены в сома-
тосенсорной коре. Она даёт возможность организму регулировать
392
Биофизика
температуру в форпостном режиме, т.е. не подгонять под уставку
выходную характеристику после её отклонения от должного уровня,
а предупреждать эти отклонения, когда условия существования
организма могут к ним привести (при изменении температуры
окружающей среды, физической работе и т.п.). Упреждающее (форпостное)
регулирование осуществляется системами автоматического
управления по возмущению. Важнейшим элементом таких систем является
кора головного мозга высших животных и человека.
Как уже говорилось, в системах регулирования по отклонению
на элемент сравнения подают уставку и информацию о выходной
характеристике. В системе терморегуляции значительная доля этой
информации содержится в сигналах, поступающих от терморецепторов
по афферентным путям.
Афферентное звено безусловных терморегуляционных
рефлексов включает афферентные волокна (дендриты чувствительных
нейронов) и разнообразные терморецепторы. Среди них принято
выделять центральные и периферические терморецепторы.
К первым относятся главным образом терморецепторы
переднего гипоталамуса. Их открыли в 1961 г. японские физиологи (На-
каяма с соавторами). Они представляют собой модифицированные
нейроны, которые приобрели способность реагировать на
повышение температуры протекающей мимо них крови. Их
чувствительность характеризуется пороговым сдвигом этой температуры,
который составляет всего 0,011 °С. Когда температура крови находится
в пределах 37,2-37,5 °С, в нервных волокнах, отходящих от термо-
рецепоров переднего гипоталамуса, регистрируется импульсация
частотой около 7 имп · с-1. При возрастании температуры до 38-
40 °С частота повышается примерно вдвое. Подобную ситуацию
моделируют локальным нагреванием переднего гипоталамуса. В
результате расширяются сосуды кожи, повышаются кровяное
давление в артериях, частота сердечных сокращений и дыхательных
движений, возрастает минутный объем дыхания, усиливается
испарение воды кожей.
На рис. 7.9 представлены данные о том, что у 8 различных
термочувствительных нейронов из гипоталамуса обнаруживаются рез-
Глава 7. Информация и регулирование в биологических системах
393
f. ГЦ η
20 -
Рис. 7.9. Реакции разных нейронов гипоталамуса на температуру
(Cabanacetai, 1968)
кие различия в ходе кривых, отражающих зависимость «частота -
температура».
Функциональная индивидуальность различных
термочувствительных нейронов гипоталамуса проявляется не только в частоте и
характере зависимости «частота - температура». Термочувствительные
нейроны имеют еще индивидуальный паттерн временной
последовательности импульсов и индивидуальные особенности изменений
этого паттерна при температурных воздействиях. На рис. 7.10 приведены
образцы такого рода паттерна для 7 отдельных термочувствительных
нервных клеток.
Такие изменения паттерна импульсации могут происходить и без
существенных сдвигов общей частоты импульсации. Следовательно,
можно предполагать, что наряду с частотным кодом, существует еще
и особый «код» температурного восприятия, который заключается
в специфических изменениях характера (временных
последовательностей, паттернов) импульсации.
Изучение изменений частоты их импульсации в диапазоне
физиологических температур у животных (от 34 до 40 °С) показало, что
температурная характеристика типичных нейронов имеет колоколо-
образный вид. Каждый термочувствительный нейрон имеет
некоторый температурный максимум частоты. По обе стороны от
максимума в типичных случаях частота уменьшается.
394
Биофизика
1-Н Ы—I 1 И
2 м—■ ιιιιι ι—ι m
3 II II II II I 111 II II I II И III!
4 HI I I) I
5 щипниниишшцмшимmwui
6 ι и m μ 11 ι ι μ
7 \ l Ml i II H—h-M—H-
Рис. 7.7ft Примеры импульсации семи различных
термочувствительных нейронов гипоталамуса
К периферическим терморецепторам относят терморецепторы
кожи, слизистых оболочек дыхательных путей и
желудочно-кишечного тракта, кровеносных сосудов, мышц, внутренних органов. Это
огромное терморецепторное поле. Морфологи описали ряд
инкапсулированных нервных телец (например, колбы Краузе, цилиндрические
окончания Руффини), а также свободные нервные окончания,
способные отвечать на изменения температуры окружающих их тканей.
Наибольшая плотность терморецепторов находится в коже лица и шеи.
В отличие от центральных терморецепторов, весьма чувствительных
к нагреванию и потому являющихся преимущественно тепловыми,
среди периферических доминируют холодовые, но есть и тепловые.
Один нервный ствол может давать несколько терминалей,
дихотомически разветвляясь несколько раз. Терминали имеют диаметр
всего 2-4 мкм и содержат по нескольку митохондрий. Известно,
однако, что для одного нервного волокна, отходящего от
терморецепторов, площадь рецептивного поля составляет около 1 мм2 (Duclaux,
Kenshalo, 1980).
Большой интерес представляет глубина залегания
терморецепторов кожи, так как от этого могут зависеть их физиологические «за-
Глава 7. Информация и регулирование в биологических системах
395
дачи». До недавнего времени считали, что они залегают сразу под
эпидермисом, не глубже 400 - 600 мкм от поверхности. Однако с
помощью особых, достаточно точных биофизических методов
исследования удалось установить, что терморецепторы расположены во всей
толще кожи, вплоть до границы кожи с подкожной клетчаткой.
Однако наибольшая плотность расположения этих рецепторов
выявлена на глубине 0,17 мм от кожной поверхности.
Если температуру кожи повышать ступенчато, т.е. давать
время терморецептору для установления постоянной импульсации, то
терморецепторы обнаруживают характерную колоколообразную
характеристику для отношений «температура - частота импульсации».
Иначе говоря, они повышают частоту импульсации от какой-то
исходной величины, достигают максимума частоты и затем снова
снижают ее до нуля.
Вторая важнейшая черта работы терморецептора - фазная
реакция. При скорости изменений температуры кожи примерно от
0,5 °С/с и выше они дают резкое увеличение (или уменьшение)
частоты импульсации, которая затем снижается (или повышается) до
уровня, свойственного данному рецептору при данной (постоянной)
температуре кожи.
Третья важная черта - значительная индивидуальность свойств
по порогам и характеристикам «температура-частота импульсации».
Частота импульсации в афферентных волокнах, проводящих
в центральную нервную систему сигналы от Холодовых рецепторов,
повышается при охлаждении. Так, понижение температуры кожи от
33°-34 °С до 27 °С приводит к максимальному учащению
афферентной импульсации. В коже человека находится примерно 250 тысяч
Холодовых рецепторов.
Как следует из рис. 7.11, холодовые рецепторы кожи имеют
различные частоты и различные углы наклона кривых повышения
частоты при понижении температуры кожи от 35-^40 до 20-25 °С.
Видно, что при температуре кожи 20-25 °С частота импульсации
остается еще достаточно высокой по сравнению с величиной
экстремума.
На рис. 7.12 представлены два Холодовых рецептора кожи
спинки носа кролика с классической колоколообразной формой кри-
396
Биофизика
f. Гц м
Рис. 7.11. Индивидуальные характеристики зависимости
«частота импульсации - температура»
10 Холодовых рецепторов кожи (Kenshalo et al.1976)
По оси ординат - частота импульсации Гц,
по оси абсцисс - температура кожи °С
вой зависимости «температура - частота» и два рецептора с резко
асимметричным распределением. Вообще говоря, экстремум у
Холодовых рецепторов может располагаться в очень широкой зоне
температур от 10-15 °С (и даже ниже) и до 30-35 °С. В последнем случае
холодовые рецепторы приближаются по свойствам к тепловым.
Индивидуальность кожных терморецепторов проявляется
также в характере временной последовательности импульсной
активности или в паттерне импульсации.Большая и далеко не решенная
проблема - «кодирование» температуры терморецепторами. Дело в том,
что при колоколообразной кривой зависимости «температура -
частота» по обе стороны от экстремума частота импульсации будет
одинаковой при различных температурах кожи. Это происходит потому,
что частота падает как при понижении, так и при повышении
температуры кожи. Так, например, если экстремум в «классическом»
случае достигается при 30 °С, то при снижении температуры на 5 °С
(т.е. до 25 °С) и при повышении температуры кожи на 5 °С (т.е. до
35 °С) частота импульсации окажется равной. Возникает вопрос: как
же «различают» нервные центры две эти разные температуры, если
Глава 7. Информация и регулирование в биологических системах
397
40 i, °C
Рис. 7.12. Различные характеристики зависимости «частота импульсации -
температура» Холодовых рецепторов кожи спинки носа у кролика при
непрерывном изменении температуры кожи со скоростью 0,2-0,4 "С/мин.
А - два Холодовых рецептора с классическим распределением частот;
Б - два других Холодовых рецептора с асимметричным распределением частот
частота импульсации, достигающая нервных центров
терморегуляции, оказывается одинаковой?
Интересно, что хотя специфическая импульсация кожных
терморецепторов изучается более 50 лет, такой вопрос возник
сравнительно недавно. Тщательное изучение особенностей временной
последовательности (паттерна) импульсации терморецепторов дает
ответ на этот вопрос. Дело в том, что хотя частота импульсации
действительно может падать довольно равномерно по обе стороны от
экстремума, паттерн временной последовательности импульсов на обоих
«склонах колокола» оказывается различным.
На рис. 7.13 приводятся данные, из анализа которых видно, что
постепенное и непрерывное понижение температуры кожи
сопровождается изменением не только частоты, но и характерного паттерна
импульсации.
У Холодовых рецепторов кожи спинки носа кошки, конечностей
обезьян и человека частота колеблется в зависимости от температуры
кожи в стационарном режиме от нуля до 10 Гц и выше. Имеется в виду
диапазон температур кожи от 10 до 40 °С. При медленном
непрерывном изменении температуры кожи, что имело место в опытах,
изменение частоты Холодовых терморецепторов было более значительным
(рис. 7.12).
398
Биофизика
А
Г=23,2°С 13 Гц
7=21,5'С 18 Гц
7 = 15,5 °С 25 Гц
1 с
7=14°С
7=9,8°С
Б Т=27°С
7=19,5°С
7=14°С
7= 11,5 "С
7=10,5°С
18 Гц
3 Гц
5 Гц
11 Гц
9 Гц
4 Гц
4-5 Гц
1 с
Рис. 7.13. Два рецептора (А и Б) кожи морды кролика с разным
характером импульсации и разной температурой частного экстремума
Воздействие - непрерывное изменение температуры кожи со скоростью
0,2—0,4 "С/мин Отметка времени —1 с. Над каждой нейрогралшой
обозначены температура кожи (Τ) υ средняя частота импульсации
за несколько секунд. Затем импульсации восстанавливается
и приобретает частоту, свойственную установившейся температуре кожи
Важным свойством терморецепторов является способность
резко изменять частоту импульсации при очень быстрых изменениях
Глава 7. Информация и регулирование в биологических системах
399
температуры кожи. Так, например, понижение температуры кожи со
скоростью 0,4-2 °С/с вызывает в Холодовых рецепторах кожи спинки
носа кошки быстрый подскок частоты до 20-80 Гц и более. Затем
частота устанавливается на уровне, свойственном стационарной им-
пульсации при данной температуре.
Аналогичные реакции по форме и величине были найдены для
Холодовых рецепторов кожи обезьян и человека. Надо заметить, что
величина динамического ответа зависит от быстроты и величины
(диапазона) понижения температуры кожи. В общем реакция тем
больше, чем больше скорость и диапазон перепада.
В коже наряду с Холодовыми присутствуют и тепловые
рецепторы (к ним относятся в частности тельца Руффини), но в меньшем
количестве (около 30 тысяч), чем холодовые. Сигналы от них
достигают максимальной частоты при повышении температуры кожи от
33-34 °С до примерно 45 °С. Тепловые рецепторы лежат глубже
Холодовых. Наибольшая плотность тепловых рецепторов отмечена на
глубине около 0,3 мм. Это совпадает с расстоянием, на котором
фокусируется инфракрасное излучение Солнца. Применяемые в
радиолокации СВЧ-лучи проникают на большую глубину и нагревают
ткани преимущественно вне зоны расположения тепловых рецепторов,
в связи с чем говорят о «коварстве СВЧ-излучения», поскольку на их
тепловой эффект организм реагирует неадекватно.
Специфические терморецепторы внутренних органов. Вопрос
о наличии во внутренних органах или в глубоких тканях тела
специфических терморецепторов поставлен очень давно. Однако в отличие от
совершенно ясных и четких фактов наличия специфических
термосенсоров в гипоталамусе, среднем, продолговатом, спинном мозгу, а
также на огромной кожной поверхности организма, данные о
существовании специфических терморецепторов во внутренних органах пока
неубедительны.
Нельзя отрицать существование температурной
чувствительности во внутренних органах и кровеносных сосудах. Однако не имеется
достаточно веских аргументов, что эта температурная
чувствительность имеет какое-либо значение для регуляции и поддержания
постоянства температуры у гомойотермных животных. Отметим, что в
тщательных исследованиях Кранстона и соавторов (Cranston et al., 1977,
400
Биофизика
1978) не было найдено специфических терморецепторов в правом
сердце, легких, задней полой вене, печени, печеночных венах.
Имеются сведения о высокой температурной чувствительности каротидных
телец (Gallego et al., 1979), а также мышечных веретен (Meuse, 1978).
В итоге можно сказать, что существуют отдельные факты
о температурной чувствительности глубоких тканей и органов.
Однако участие соответствующих структур в терморегуляции не
доказано. Поэтому в настоящее время нет веских оснований включать
в схему терморегуляции гомойотермного организма гипотетические
терморецепторы внутренних органов.
Периферические терморецепторы классифицируются по
температурному порогу (DT) на высокочувствительные (DT<0,1 °C),
среднечувствительные (DT около 1 °С) и низкочувствительные (DT
порядка 10 °С).
В передаче информации от периферических терморецепторов
участвуют тонкие миелиновые волокна типа Аб и безмякотные
волокна типа С. Каждое волокно имеет свой довольно узкий (примерно 5°)
оптимум температурной чувствительности. Изменение температуры
на 1° в пределах этих 5° вызывает повышение частоты афферентной
импульсации в отдельном волокне на 2 имп-с-1 (на 20%
максимальной частоты). Различия в диапазонах оптимальной чувствительности
у разных терморецепторов в пределах одного рецепторного поля даёт
возможность организму с достаточной точностью «измерять»
изменения температуры тела.
У человека частота импульсации в экстремуме достигает
ΙΟΙ 3 Гц. Экстремум отмечается при температуре кожи 40-47 °С. При
повышении температуры кожи от 32 до 37 °С со скоростью от 0,5 до
1,5 °С в секунду наблюдается динамический эффект, который тем
больше, чем с большей скоростью происходят изменения
температуры. При скорости 1,5 °С/с частота импульсации составляет 20-22 Гц.
При повышении температуры кожи со скоростью 1,5 °С/с от 37 до
42 °С частота импульсации достигает даже 40 Гц (рис. 7.14).
Теперь коснемся некоторых общих проблем терморецепции.
Отметим, что вообще в коже животных и человека тепловых
рецепторов меньше, чем Холодовых. Этому соответствует и число
Холодовых и тепловых точек на коже человека.
Гпава 7. Информация и регулирование в биологических системах 401
80
60
40
20
0
30 34 38 42 46 50 f, °С
Рис. 7.14. Статистическая импульсация трех тепловых рецепторов
кожи носа кошки. Характеристика зависимости
«частота импульсации — температура» (Hensel, Kenshalo,1969)
Важно заметить, что изменение температуры кожи на 1 °С
вызывает сдвиг импульсации холодового рецептора кожи в
стационаром режиме на 1,0 Гц, а для теплового - на 1,4 Гц. При динамической
реакции холодовой терморецептор изменяет импульсацию в среднем
на 50 Гц, а тепловой - на 70 Гц. Максимум частоты при
динамической реакции у холодового рецептора достигает 240 Гц, а у
теплового - 200 Гц. Эти факты могут говорить о том, что чувствительность
кожи определенных участков тела животного к изменениям
температуры в диапазоне 35-40 °С сравнительно высока. Возможно, столь
высокая тепловая чувствительность спинки носа животного имеет
отношение к поведенческой терморегуляции или вообще к
ориентировочным реакциям, не имеющим связи с терморегуляцией.
Афферентные волокна от периферических рецепторов
туловища проходят в составе спинальных нервов и прерываются в задних
рогах спинного мозга. Для него характерна выраженная дивергенция
сигналов от терморецепторов. К одному нейрону могут прийти
импульсы и от терморецепторов, и от болевых, и от тактильных
рецепторов, но исключено поступление на один нейрон сигналов от
Холодовых и тепловых периферических рецепторов.
402
Биофизика
Скорости проведения импульсов от Холодовых рецепторов
существенно зависят от места расположения этих рецепторов. Так,
например, от кожи ладони скорость проведения импульсов в среднем
составляет 14,5 ± 4,6 м/с, а от волосистой части кожи верхней и
нижней конечностей - 0,6-0,7 м/с. Для импульсов от тепловых
рецепторов скорость проведения (от 0,4 до 1,2 м/с) в среднем меньше, чем
для Холодовых. Можно полагать, что импульсы от тепловых
рецепторов проводятся немиелинизированными волокнами, тогда как от
Холодовых рецепторов в проведении участвуют как бсзмякотные, так
и миелиновые волокна.
Проводящие пути холодовой и тепловой чувствительности
в стволе мозга функционируют также независимо друг от друга. От
нейронов задних рогов нервные импульсы распространяются контр-
латерально по спиноталамическому тракту к вентро-базальным ядрам
таламуса и по спиноретикулярному тракту к нейронам ретикулярной
формации. В таламус стекается информация от терморецепторов кожи
головы и слизистых оболочек носа и ротовой полости, афферентную
импульсацию с которых проводят волокна тройничного нерва. После
синаптической передачи в таламусе сигналы от терморецепторов
могут достигать нейронов соматосенсорной коры больших полушарий
головного мозга и гипоталамуса. В коре формируются теплоощуще-
ния, причём ощущение теплового комфорта создаётся на основе
сигналов от периферических терморецепторов.
7.3.3.2. Гуморальная регуляция
Среди гуморальных регуляторов температуры тела необходимо
назвать такие вещества, как адреналин, ацетилхолин, гормоны
щитовидной железы, глюкокортикоиды, глюкагон, мелатонин (гормон
эпифиза), ненасыщенные жирные кислоты, липопротеиды низкой
плотности. Адреналин влияет на физическую терморегуляцию,
суживая подкожные сосуды и тем самым ослабляя теплообмен с
окружающей средой. Вместе с тем он, наряду с глюко-кортикоидами и глюка-
гоном, оказывает воздействие на химическую терморегуляцию,
усиливая теплопродукцию за счет активации гликогенолиза и
гликолиза в печени, а также липолиза в жировой ткани (включая и бурый
Глава 7. Информация и регулирование в биологических системах
403
жир) и окисления жирных кислот. Гормоны щитовидной железы
резко усиливают метаболизм, преимущественно процессы катаболизма,
коэффициент полезного действия которых составляет 20-25%.
Остальная энергия (80-75%) идет на теплообразование в организме.
Тироксин и трийодтиронин, повышая валовый уровень энергетических
процессов, увеличивают термогенез при неизменном коэффициенте
полезного действия. Поэтому при тиреотоксикозе (базедовой
болезни) больные испытывают чувство жара. Содержание гормонов
щитовидной железы регулируется тиреотропным гормоном гипофиза,
продукция которого находится под контролем тиреотропин-рили-
зинг-гормона, синтезируемого нейронами гипоталамуса.
Участие мелатонина в терморегуляции было выявлено при
вступлении животных в зимнюю спячку и выходе из нее. Гормон
оказывает прямое действие на гипоталамус, смещая уровень уставки
для обеспечения охлаждения организма. Смещение уставки
происходит постепенно и довольно медленно, благодаря чему температура
«ядра» плавно понижается до 5-15 °С. Угнетаются все функции
мозга, половая активность, гормонообразование в щитовидной железе
и надпочечниках, тогда как скорость секреции (инкреции) инсулина
возрастает. Во время спячки энергообеспечение организма
осуществляется за счет окисления жира, а гликоген не расходуется. Частота
сердечных сокращений и дыхательных движений составляет всего
10% от уровня бодрствования.
В отличие от погружения в спячку, пробуждение происходит не
медленно, а быстро (за 60-80 минут). Уровень уставки повышается за
считанные минуты, что резко активирует центр теплопродукции
гипоталамуса, а он, в свою очередь, быстро стимулирует окисление бурого
жира. Температура «ядра» повышается с 5-15° до 38 °С за 1-1,5 часа.
7.3.4. Адаптация организма
к низкой и высокой температуре
Для лучшего понимания взаимодействия разных механизмов
терморегуляции рассмотрим в качестве примера адаптацию
организма к охлаждению. На понижение температуры окружающей среды
прежде всего реагируют периферические холодовые рецепторы. Сиг-
404
Биофизика
налы с них поступают к нейронам задних рогов спинного мозга
преимущественно по спинальным нервам. Синаптическая передача
обеспечивает контрлатеральное распространение нервных импульсов по
восходящему спиноталамическому тракту. Из ядер таламуса
информация поступает как в соматосенсорную область коры головного мозга
и лимбическую систему, так и в гипоталамус. В результате «гипота-
ламический термостат» формирует систему реакций,
обеспечивающих и физическую, и химическую терморегуляцию. Они направлены
на поддержание постоянства температуры «ядра» гомойотермного
животного даже на холоде.
Исполнительные органы системы терморегуляции
активируются в определенной последовательности. Прежде всего происходит
рефлекторное сужение артериол кожи посредством возбуждения
сигналами, формирующимися в заднем гипоталамусе, симпатических
нейронов в боковых рогах спинного мозга. Повышение частоты им-
пульсации в симпатических волокнах приводит к констрикции глад-
комышечных клеток кожных артериол и уменьшению теплоотдачи.
Нередко спазм подкожных кровеносных сосудов сменяется их
расширением, что обусловлено чувствительностью самих гладкомы-
шечных клеток к холоду. Такая реакция предупреждает ишемию
кожи, но усиливает теплоотдачу, что не способствует сохранению
тепла в организме и не препятствует переохлаждению. Целесообразно
предупреждать этот неблагоприятный эффект, но заботиться
одновременно о состоянии кожных покровов.
При усилении холода рефлекторно открываются артерио-ве-
нозные анастомозы в коже, благодаря чему уменьшается объем крови,
поступающей в капилляры, а следовательно, ограничивается
теплоотдача, и тепло сохраняется в организме. Параллельно с ослаблением
кровообращения в подкожных сосудах усиливается кровоток во
внутренних органах (особенно в брюшной полости), благодаря чему
сберегается тепло в «ядре». Животные обычно свертываются
«калачиком», чем уменьшается (примерно на 35%) поверхность теплообмена
с окружающей средой. Благодаря повышению активности
симпатических волокон, иннервирующих гладкие мышцы, поднимающие
волосы, у них возникает пилоэрекция, улучшающая теплоизоляцию за
счет улучшения воздушной прослойки.
Глава 7. Информация и регулирование в биологических системах
405
Если температура «ядра» станет меньше 35° С, то наступает
так называемая гипотермия. При ней к рефлекторному ограничению
теплоотдачи (физической терморегуляции) присоединяется усиление
теплопродукции (химическая терморегуляция) за счет
непроизвольного сокращения скелетных мышц (терморегуляционного тонуса
и дрожи) и механизмов несократительного термогенеза. Вместе с тем
в таком состоянии человек и животные прибегают к усилению
произвольной мышечной деятельности (ходьбе, бегу, прыжкам и т.п.),
а также к приему пищи (лучше горячей). К нервной регуляции
присоединяется гуморальная: активируется продукция гормонов
щитовидной железы глюкокортикоидов, катехоламинов, глюкагона,
которые стимулируют преимущественно несократительный термогенез.
На жару реагируют периферические тепловые рецепторы, но
если высокая температура окружающей среды приводит к
повышению температуры крови, притекающей к гипоталамусу, хотя бы на
сотую долю градуса, то возбуждаются центральные терморецепторы.
Сигналы от тепловых рецепторов активируют «гипоталамический
термостат», который включает в определенной последовательности
систему реакций, обеспечивающих тепловой баланс гомойотермного
организма.
При высокой температуре окружающей среды возрастает
теплоотдача и прежде всего за счет испарения. Для его интенсификации
усиливается потоотделение. Эту рефлекторную реакцию обеспечивают
холинэргические симпатические волокна. Вслед за гипергидрозом
расширяются кровеносные сосуды благодаря урежению импульсации
в адренэргических симпатических волокнах, воздействующих на а-ад-
ренорецепторы, в результате усиливается кровоток как в коже, так
и в сосудах, отводящих тепло от внутренних органов, что неизбежно
приводит к увеличению внешнего и внутреннего тепловых потоков
и повышению теплоотдачи организма. Этому способствует
повышение объемной скорости кровообращения благодаря рефлекторному
выбросу крови из депо и дегидратации тканей (с переходом воды из
них в кровеносные сосуды). При дальнейшем нагревании понижается
метаболическая активность, а следовательно, угнетаются механизмы
теплообразования, чему способствуют также ограничение
двигательной активности, создание конвективных потоков вокруг тела и другие
406
Биофизика
произвольные способы охлаждения. При повторяющихся
воздействиях высокая температура окружающей среды снижает секрецию
гормона роста, тироксина и трийодтиронина, глюко-кортикоидов, половых
гормонов, а все это также ослабляет метаболическую активность и,
следовательно, теплопродукцию.
Не зря системы, регулирующие температуру тела у гомой-
термных животных, формируются в ранней стадии онтогенеза.
Теоретически жизнь возможна в узком диапазоне температур
окружающей организм среды, поскольку белки денатурируют при 45 °С,
а кристаллизуются, как и вода, при 0 °С. Однако жизнь существует
и в тропиках (при температуре в тени до 65 °С), и на полюсах Земли,
где температура опускается ниже —65 °С.
Совершенство системы терморегуляции, рассмотренной выше,
позволяет преодолеть теоретический запрет на жизнь в
неблагоприятных климатических условиях. Мало того, животные в процессе
эволюции приобрели способность управлять своим тепловым состоянием
для борьбы с другими живыми организмами. Речь идет о лихорадке.
7.3.5. Лихорадка
По определению Международного общества физиологических
наук (IVPS), лихорадка принципиально отличается от
гипертермического состояния организма, поскольку при гипертермии
терморегуляция направлена на понижение высокой температуры, а при
лихорадке повышенная температура тела поддерживается на высоком
уровне своеобразной активностью центральных механизмов
терморегуляции.
Суть различий состоит в том, что при гипертермии не
изменяется уровень уставки. Когда же организм человека и животных
включает свой термогенез в борьбу с микробами, вирусами, грибками и
другими агентами, он повышает уровень уставки («терморегуляционной
установочной точки») в гипоталамусе. При этом организм, имея
нормальную температуру, испытывает состояние гипотермии, и его «ги-
поталамический термостат» включает реакции, направленные на
устранение не гипертермии, а гипотермии, поскольку нормальная
температура воспринимается как пониженная относительно нового
Глава 7. Информация и регулирование в биологических системах
407
уровня уставки. И на гипотермию организм реагирует вазоконстрик-
цией в коже и пилоэрекцией (для ограничения теплоотдачи), а также
мышечной дрожью (для усиления теплопродукции). Человек при
переходе на новый уровень уставки испытывает ощущение озноба. Его
перестает знобить, когда достигается новый уровень терморегуляции.
Благодаря активации терморегуляторных механизмов
температура «ядра» повышается до возросшего уровня уставки. Наступает
состояние якобы нормотермии (относительно нового уровня уставки),
но на самом деле (относительно прежнего уровня уставки)
температура тела при лихорадке выше обычной нормы. Когда врачи убедились
в благотворном эффекте лихорадки в борьбе человека с инфекцией,
началась разработка технологий пирогенотерапии. Были изучены
экзогенные пирогены (от микробов, вирусов, грибков и их токсинов до
синтетических низкомолекулярных соединений, примером которых
является мурамилди-пептид).
Под влиянием таких агентов в организме образуются
эндогенные пирогены, представляющие собой протеины с молекулярной
массой от 10 до 40 кДа. Их синтезируют различные лейкоциты,
альвеолярные макрофаги, клетки печени и селезёнки. Посредством
изменения темпа и объёма их продукции регулируется высота лихорадки.
Именно эндогенные пирогены действуют на гипоталамус и
повышают уровень уставки.
Предполагают, что пирогены стимулируют в гипоталамусе
образование простагландина Ε из арахидоновой кислоты, поскольку его
содержание там возрастает под действием пирогенов. Аргументы
в пользу участия простагландина Ε в повышении уровня уставки
и развитии лихорадки сводятся главным образом к двум фактам.
Во-первых, при его инъекции в гипоталамус развивается типичная
лихорадка. Во-вторых, аспирин препятствует развитию лихорадки, а его
действие обусловлено подавлением активности циклооксигеназы -
фермента, катализирующего образование простагландина Ε из
арахидоновой кислоты. Поэтому уже более века аспирин служит
эффективным жаропонижающим лекарством. Учитывая благотворность
лихорадки в определенных пределах, врач и фармацевт должны понимать,
когда жаропонижающие препараты полезны, а когда их применять не
следует.
408
Биофизика
Что же касается лихорадки, то изучение её патогенеза
подтвердило современную физиологическую концепцию механизмов
терморегуляции человека.
7.3.6. Математическое
моделирование терморегуляции
Исследование процессов терморегуляции человека с помощью
математических моделей ведутся более 30 лет. Первой работой такого
типа была модель процессов теплообмена в руке человека. В
определенной степени приведенное в этой работе математическое описание
явилось исходным для последующих моделей, так как в нем
учитывались основные тепловые процессы: теплообразование в результате
обменных процессов, теплообмен кондукцией, перенос тепла кровью,
теплообмен с внешней средой. В последующих работах большинство
исследователей рассматривали систему терморегуляции как систему
автоматического управления, выделяя в ней объект управления и
регулятор.
Модели объекта (пассивная система по Столвику и Харди,
физическая модель по Аткинсу и Вайндхему, управляемая система по
Столвику) отражали тепловые процессы в теле человека с учетом его
основных теплофизических свойств и физиологических процессов,
участвующих в теплообмене. В этом плане модели различных
авторов в принципе близки между собой и отличаются степенью
детальности описания геометрической формы и послойной температурной
топографии тела.
Регулятор формирует величины терморегуляторных реакций -
сосудодвигательные реакции кожи, потоотделение, сократительный
и несократительный термогенез в зависимости от отклонения
регулируемой величины и информации с температурных рецепторов кожи.
В основу алгоритма управления закладывается та или иная гипотеза
о цели регулирования в системе, в первую очередь о том, какой
показатель теплового режима организма может быть выбран
регулируемой переменной. В физиологии терморегуляции и соответственно
в работах по математическому моделированию существует два
основных подхода к этому вопросу: принцип установочной точки и ди-
Глава 7. Информация и регулирование в биологических системах
409
комического баланса. На рис. 7.15 приведены принципиальные
схемы моделей, основанных на этих двух принципах.
УС
ivc>\
ϊψΊΓ
i
к ■
w
КЭ
w
ОС
ψ
КС
I
кп
_ы<
*
КЭ
—►
ос+
КС
<—
ос--«—
К
—1
.*<у)-^
<*<у>-*-
в)
Рис. 7.15. Принципиальные схемы моделей температурного
гомеостазиса, основанные на принципах установочной точки
или динамического баланса (по данным Дж. Блая)
а - схема системы, в которой сигнал обратной связи (ОС) сравнивается
с эндогенным установочным эталоном-сигналом (УС). Ошибка
регулирования(О) оказывает воздействие на терморегуляторные эффекторные
процессы, которые определяют контролируемый переменный сигнал (КП);
КЭ - контролирующий элемент, В - возмущения, КС - контролируемая система;
б - схема системы, регуляция в которой зависит от свойств двух групп сенсоров -
одна дает сигнал однонаправленный, а другая - противоположно направленный
по отношению к изменениям контролируемой величины (оба сигнала
сравниваются и их разность определяет тип эффекторной реакции); в —схема
системы с двумя группами сенсоров — тепловых (X) и Холодовых (У), сигналы
каждой из которых сравниваются с эталонным сигналом; полученные сигналы
ошибки регулирования определяют терморегуляторные реакции — теплоотдачу
(ТО) и теплопродукцию (ТП); г - схема системы с двумя группами сенсоров (Хи У),
сигнал которых не сравнивается с эталоном, но имеет тормозное влияние
410
Биофизика
Рассмотрим модель системы терморегуляции человека,
входящую как подсистема в комплексную модель внутренней сферы
организма человека. При построении модели объекта (системы
теплообмена) приняты общие для всех моделей терморегуляции следующие
допущения: реальная система теплообмена представлена в виде сети
из участков с сосредоточенными параметрами, каждый участок
системы имеет форму цилиндра, ткань любого участка гомогенна и
обладает изотропными свойствами, не учитывается теплопередача кон-
дукцией между торцевыми поверхностями цилиндра, не учитывается
теплообмен между поверхностными венами и близлежащими
артериями, а также сосудами с разнонаправленными кровотоками.
Математическое описание представляет собой систему
дифференциальных уравнений, включающую уравнение теплового
баланса, которое для i-ro участка имеет вид:
dT
C'mi~dt = Mi +Р*С**лЛЛ -pBcBKnFnT, +
+ Κ,,ι.ισί-Τι.ι)+Κυ+ι(Τι-ΤΜ)+ (7.1)
+HKAI(TI-TA)+HCAI(TI-TA)-Ei,
и уравнение смешивания
dT "
cB?BV<:-^ = YJ?BcBFJi-pBcBQTB -E„ (7.2)
at ,·=ι
где Tt - температура г'-ro участка; Тв - температура смешанной
венозной крови; Μ - скорость образования тепла; с,, св - удельная
теплоемкость крови и ткани; m - масса участка; FA, Fv - объемная
скорость артериальной и венозной крови; Ε - скорость теплоотдачи
испарением; А - площадь поверхности участка; ТА - температура
окружающей среды; рв - плотность крови; Vc - центральный объем
крови; Q - суммарный кровоток в системе кровообращения; Ет -
отдача тепла испарением с верхних дыхательных путей; KAJCV -
коэффициенты использования тепла, переносимого артериальной и
венозной кровью; Ки_„ Ки+1 - коэффициенты коммутации; л - число
участков.
Глава 7. Информация и регулирование в биологических системах
411
Основной регулируемой переменной в модели принята
температура мозга. Отклонение ее от исходного значения приводит к
изменениям сопротивления сосудов кожи и скорости потоотделения и
соответственно компенсаторным изменениям процессов теплоотдачи.
Изменение процессов теплообразования происходит в модели
пропорционально разности температур кожи и окружающей среды.
Особенностью моделирующего алгоритма при реализации
модели на ПЭВМ является то, что он позволяет путем ввода
соответствующих исходных данных (число участков л и коэффициенты
коммутации Ки_„ Kii+I) реализовать модели различной сложности,
в принципе сколь угодно точно отражающие теплофизическую
структуру тела человека.
7.4. Постоянство внутренней среды
организма и его регуляция
Важнейшим свойством живых организмов является, наряду
с непрерывным реагированием на внешние воздействия, способность
поддерживать свое внутреннее постоянство (гомеостаз). Это
постоянство выражается в первую очередь в поддержании строго
сохраняемой активной реакции внутренних сред, их температуры,
содержания в них кислорода и углекислого газа, основных энергетических
и строительных веществ (углеводов, белков, жирных кислот).
Нормальное функционирование всех клетоиных структур
организма является в высшей степени зависимым от указанных
факторов, и сдвиг их показателей даже в небольшой степени может
привести к глубоким нарушениям жизненных функций. Вместе
с тем клеточные процессы характеризуются образованием ряда
побочных продуктов, могущих привести к нарушениям
жизнедеятельности, и их удаление из организма также является жизненно
необходимым.
Поэтому в основе способности высших организмов к
длительному существованию лежит мощная система регулирования своего
внутреннего постоянства, основой которой является система
кровообращения и связанная с ней система лимфообращения.
412
Биофизика
7.4.1.Характеристики
внутренней среды организма
Кровь представляет собой именно ту внутреннюю среду,
физико-химические характеристики которой поддерживаются на строго
регулируемом уровне. Через кровь происходит поступление к тканям
кислорода, энергетических и строительных веществ.через нее
происходит удаление из тканей продуктов жизнедеятельности. Эти
важнейшие функции крови выполняются при сравнительно небольшом
ее объеме, составляющем всего 6-8% общеймассы тела. Нарис. 7.16
в схематической форме представлены эти функции крови.
Жидкая часть крови (плазма) содержит (мМ/кг воды):
электролиты - катионы (Na+ - 153, К+ - 5, Са2+ - 3, Mg2+ - 1) и анионы (О- -
110, С023" - 28, Р043" - 1, S042_ - 1), а также неэлектролиты
(глюкоза - 5, белки - 2, мочевина - 1). Осмотическое давление - 745 кПа
(7,3 атм), коллоидно-осмотическое давление (онкотическое) - 3 кПа.
Важнейшим компонентом крови являются форменные
элементы, подразделяющиеся на красные кровяные тельца (эритроциты),
Рис 7.16. Схема жидкостных пространств во внутренней среде организма
Их объемы представляют собой округленные значения для человека
массой 70 кг. Стрелками указаны направления обмена между
жидкостными пространствами и внешней средой
Глава 7. Информация и регулирование в биологических системах
413
белые кровяные тельца (лейкоциты), в свою очередь
подразделяющиеся на ряд форм, и кровяные бляшки (тромбоциты). У человека
и высших животных эритроциты представляют собой безъядерные
сравнительно короткоживущие (100-120 суток) клеточные
структуры, около 90% сухого веса которых составляет гемоглобин и
которые служат основным посредником в переносе кровью кислорода.
Ядерные клетки (лейкоциты) являются основой защитной функции
крови, позволяющей организму бороться с микробными и
вирусными инвазиями. Наконец, тромбоциты - также безъядерные элементы
благодаря наличию в них определенный ферментов существенна
важны для свертывания крови при повреждениях сосудистого русла.
Если газы, неорганические и органические вещества могу ι
в большей или меньшей степени свободно проходить через стснк>
сосудов и обмениваться с таковыми в тканевой жидкости, в полостях
пищеварительного и выделительного каналов и воздушной среде
легочных альвеол, то форменные элементы - постоянные компонент!,
крови и в свою очередь обмениваются своими ингредиентами с ei
плазмой (хотя лейкоциты могут покидать сосудистое русло за cici
амебидных движений).
Гомеостатическая функция крови возможна лишь благодаря α
непрерывному движению (циркуляции) в сосудах, поддерживаемо!
ритмическими сокращениями сердечной мышцы (миокарда). У вые
ших организмов (млекопитающих и птиц) циркуляция кровипроис
ходит через полностью разделенные большой («левое сердце») и ма
лый («правое сердце») круги кровообращения и четырехкамершх
сердце с двумя желудочками (рис. 7.17). Оттекающая от тканей не
нозная кровь поступает в правое предсердие и оттуда в правый жслу
дочек. При его сокращении она выбрасывается в легочную артерию
Протекая через капилляры легких, оплетающие концевые структурь
воздушных путей - альвеолы, кровь отдает углекислый газ и наем
щается кислородом. Обогащенная кислородом кровь из легких по ле
точным венам поступает в левое предсердие и из него в левый желу
дочек. При сокращении последнего она нагнетается в аорту и затеи
распределяется по артериям, артериолам и капиллярам всех opraiioi
и тканей с тем, чтобы затем вновь собраться в венозную систему.
Верхние
отделы тела
«Правое
сердце»
Лимфатическая
система
Печеночная артерия ||
&—>
Нижние отделы тел г
Рис. 7.17. Схема сердечно-сосудистой системы
Сосуды, содержащие насыщенную кислородом кровь, тел
а сосуды, содержащие обедненную кислородом кровь, - cet
Лимфатическая система, образующая дополнительну
систему оттока жидкости от органов, показана черными /,
7.4.2. Регуляция кругооборота крови
Очевидно, что кругооборот крови является име
новной регулируемой переменной, которая позволяет ор:
способляться к изменяющимся потребностям его oprat
и поддерживать в каждый данный момент необходимый
вень кровотока. Такая регуляция осуществляется, с одно]
Глава 7. Информация и регулирование в биологических системах
415
счет изменения насосной деятельности сердца, и с другой - за счет
регуляции периферического кровотока. Управляющими же
параметрами являются те изменения физических и химических характеристик
внутренней среды, которые возникают в ходе жизнедеятельности.
Внутрисердечные механизмы регуляции кровотока
обеспечивают в первую очередь изменение интенсивности деятельности
сердечной мышцы (миокарда). Основной из них известен как закон
сердца Франка - Стерлинга. Суть его заключается в зависимости силы
сокращения миокарда от количества притекающей к сердцу крови.
Основой такой зависимости служит связь силы сокращения
сердечных мышечных волокон и их исходной длины, т.е. степени их
растяжения во время фазы расслабления (диастолы). Более сильное
растяжение сердца при диастоле соответствует усиленному притоку крови
к сердцу. При этом соответственно возрастает сила его сокращения
(систола). В результате сердце каждый раз перекачивает в
артериальную систему то количество крови, которое притекает к нему из вен.
В последнее время получены данные, что наряду с этим
мышечным регуляторным механизмом в сердце функционирует также
нервный механизм, замыкающийся через внутрисердечные (интрамураль-
ные) нервные узлы. Эта система включает афферентные нейроны,
дендриты которых образуют рецепторы растяжения на мышечных
волокнах сердечной стенки, а также вставочные и эфферентные
нейроны, аксоны которых иннервируют миокард и неисчерченные
(гладкие) мышечные волокна венечных (коронарных) сосудов. Наличие
таких внутрисердечных нервных связей может быть установлено на
функционирующем пересаженном сердце, у которого все экстракар-
диальные нервные элементы дегенерируют. За счет внутрисердечных
рефлекторных влияний при увеличении растяжения стенки
предсердия также происходит усиление сокращения желудочков. Она
затрагивает не только тот отдел сердца, который непосредственно
растягивается притекающей кровью, но и другие его отделы.
Оба внутрисердечных механизма, таким образом,
обеспечивают стабильность кровенаполнения артериальной системы и создают
тем самым важнейшее условие для успешного ее функционирования.
Внесердечные нервные (рефлекторные)регуляторные
механизмы кровотока осуществляются в отличие от скелетной мускулатуры за
416
Биофизика
счет двух систем обратной связи. Одна из них образована
симпатической, другая - парасимпатической нервной системой. Это разделение
первоначально сложилось на основе анатомических критериев,
однако затем было подтверждено также функциональными и
фармакологическими различиями. Центральные структуры симпатической
нервной системы располагаются в грудных и верхних поясничных
сегментах спинного мозга, а ее эфферентные волокна разветвляются
фактически по всему телу, проходя в основном вдоль кровеносных
сосудов. Центральные структуры парасимпатической нервной системы
локализованы в стволе головного мозга и крестцовом отделе спинного
мозга. Наиболее значительным парасимпатическим эфферентным
путем является блуждающий нерв, спускающийся от ствола мозга
в грудтгую и брюппгую полости и ишюрвирующий там большинство
внутренних органов. Парасимпатические волокна имеют более
ограниченное распространение; так, в конечностях они отсутствуют.
Функциональное разделение заключается, как правило, в
противоположных (антагонистических) влиянияхиаэффекторпые органы, при
которых одна система приводит к усилению их функции, а другая -
к ее ослаблению. Однако в ряде случаев обе системы могут вызывать
сходные эффекты, отличающиеся лишь некоторыми качественными
особенностями.
Для упрощения анализа механизма рефлекторной регуляции
кровотока можно не учитывать легочное кровообращение (малый
круг), которое регулируется лишь в небольшой степени, а также
принять постоянным венозное давление, которое всегда остается
относительно стабильным. В этом случае:
кровоток — артериальное давление / общее периферическое сопротивление
Это соотношение аналогично закону Ома, связывающему
электрический ток, напряжение и сопротивление вэлектрической цепи.
Основными переменными в системе регуляции артериального
давления оказываются выброс крови левым желудочком {сердечный
выброс) и сопротивление трения, создаваемого сосудистым руслом
большого круга кровообращения. Сердечный выброс является
произведением частоты сокращений сердца на ударный объем. Как ука-
Глава 7. Информация и регулирование а биологических системах
417
зывалось выше, ударный объём регулируется внутри сердечными
механизмами. Вместе с тем он подвержен мощной внесердечной
регуляции, увеличиваясь под влиянием симпатической импульсации
и уменьшаясь под влиянием парасимпатической (поступающей
к сердцу по волокнам блуждающего нерва).
Регулируемой переменной является и второй фактор,
определяющий артериальное давление - сопротивление сосудистого русла
большого круга кровообращения. Оно зависит в основном от диаметра
прекапиллярных сосудов - мелких артерий и артериол. Они имеют
в своей оболочке слой неисчерченных мышц, клетки которого иннер-
вируются симпатическими нервными волокнами. Сокращение этих
клеток приводит к сужению сосудов и соответственно к повышению
периферического сопротивления. Однако в отличие от регуляции
сердечного выброса, в этом случае отсутствуют антагонистические
парасимпатические влияния, поскольку большинство сосудов не имеет
парасимпатической иннервации.
Расширение сосудов становится возможным благодаря тому,
что и в условиях покоя по симпатическим волокнам к сосудам
поступает периодическая импульсация, поддерживающая их мышечные
клетки в состоянии некоторого постоянного (тонического)
напряжения. При таких условиях расслабление сосудистой мускулатуры
и расширение сосуда можно вызвать посредством уменьшения
частоты фоновой импульсации, поступающей из мозга.
Таким образом, меняя сердечный выброс и периферическое
сопротивление, организм может в широких пределах варьировать
давление в кровеносной системе. Для того чтобы гарантированно
поддерживать его в заданных пределах, необходима, очевидно, тщательная
координация тех рефлекторных влияний, которые действуют на
сердце и сосудистую систему. Обязательным условием для осуществления
такой координации является наличие постоянной информации об
уровне давления в сосудах.
Эта информация обеспечивается системой рецепторов
давления (барорецепторов), расположенных в различных участках
сосудистого русла (в стенке дуги аорты, месте разветвления общей сонной
артерии - сонном (каротидном) синусе и др.). Барорецепторы
представляют собой вторичные механорецепторы, которые по афферент-
418
Биофизика
Артериальное давление
_1Ш i
Синокаротидный нерв
Рис 7.18. Импульсная активность в афферентных волокнах
синокаротидного нерва, отражающая активацию
сосудистых барорецепторов: 1 — при пониженном;
2 - при нормальном; 3 - при повышенном артериальном давлении
ным волокнам посылают в ствол мозга импульсацию, частота
которой отражает уровень артериального давления (рис. 7.18).
В продолговатом мозге существует несколько нейронных
структур («центров»), которые на основе информации, поступающей
от сосудистых барорецепторов, а также из ряда других источников
формируют команды к эффекторным элементам (сердцу и сосудам).
Одна из них обозначается как сосудодвигателъный {вазомоторный)
центр, ответственный, как следует из названия, в основном за
регуляцию сократительной деятельности мышц сосудистой стенки
посредством активаций симпатической нервной системы. Проведенные
к настоящему времени микроэлектродные исследования импульсной
активности отдельных нейронов, входящих в эту структуру,
показали, что они четко разделяются на две группы. Одна из них дает
разряд импульсов при повышении артериального давления, другая,
наоборот, при его снижении.
Эти результаты дают основание полагать, что
сосудодвигателъный центр состоит из двух частей - прессорной и депрессорной,
которые находятся между собой в реципрокных взаимоотношениях.
Нейроны, входящие в прессорную группу, через нисходящие пути
активируют симпатические нейроны спинного мозга и таким образом
Глава 7. Информация и регулирование а биологических системах
419
индуцируют сокращение сосудистых мышц и повышение
артериального давления. Наоборот, нейроны депрессорной группы тормозят
активность симпатических клеток, и тем самым приводят к
расслаблению сосудистых мышц и падению давления. В зависимости от
характера информации, поступающей к сосудодвигательному центру от
барорецепторов, происходит перераспределение активности между
нейронами прессорной и депрессорной групп, что и является основой
формирования соответствующей регулирующей команды.
Наряду с сосудистыми, реакциями при активации сосудодвига-
тельного центра происходит также изменение в симпатической
регуляции деятельности сердечной мускулатуры. При возбуждении де-
прессорных нейронов симпатическая стимуляция сердца ослабевает,
и сердечный выброс соответственно уменьшается. При возбуждении
прессорных, наоборот, усиленная активность симпатической
нервной системы стимулирует сердечную деятельность и увеличивает
сердечный выброс, что вносит свой вклад в повышение
артериального давления.
Особой нейронной структурой продолговатого мозга, также
принимающей непосредственное участие в формировании указанной
команды, является относящееся к парасимпатической нервной
системе ядро блуждающего нерва. Активация, нейронов этого ядра
угнетает сократительную функцию сердца и уменьшает сердечный
выброс (тормозящий центр сердца).
Все указанные структуры и взаимодействия между ними в
процессе стабилизации артериального давления схематически
представлены на рис. 7.19. Снижение давления в сосудах уменьшает частоту
разряда барорецепторов, тем самым активируя прессорный центр
путем снятия его фонового торможения. Одновременно ослабление
возбуждающих влияний от барорецепторов на депрессорную часть
сосудодвигательного центра и тормозящий центр сердца угнетает
активность последних. В результате происходят увеличение
сердечного выброса и повышение периферического сосудистого
сопротивления, приводящие к восстановлению сниженного давления. Наоборот,
повышение давления тормозит прессорный центр, но возбуждает де-
прессорный и сердечно-тормозящий; сердечный выброс и перифери-
420
Биофизике
Артериальное
давление
Барорецепторы
сонного синуса
и другие аорты
Уменьшение
Сердечный
выброс
Увеличение
Активация
Буферные нервы / ^"^
(Тоническая активность) Торможение
Повышение
Общее
периферическое
сопротивление
Понижение
Активация
Центры
продолговатого
мозга
Тормозящий
центр
сердца
Прессорная
часть
Депрессорная
часть
2
ζ
*£
9*
1
Рис. 7.19. Общая схема прямых и обратных связей,
участвующих в регуляции артериального давления
ческое сопротивление падают, и артериальное давление снижается
до исходного уровня.
Такая генеральная схема регуляции кровотока не исключает,
естественно, и участия в последней региональных механизмов,
перераспределяющих в определенных пределах общий кровоток между
различными органами и системами в зависимости от потребности
последних при данной деятельности организма. Локальными
факторами, которые могут вызывать расширение сосудов, являются
снижение рН и парциального давления кислорода, повышение
парциального давления С02, выделение ряда продуктов метаболизма.
Гуморальные механизмы регуляции кровотока. Описанный ау-
торегуляторный гомеостатический механизм, использующий
рефлекторные обратные связи, долгое время рассматривался как
основной механизм регуляции кровотока. Однако в результате работ
последних лет стало ясно, что он представляет собой лишь первое
самое быстрое звено этого механизма. Кроме него, в поддержании сер-
дечнососудиртого гомеостаза участвуют и другие механизмы дли-
Глава 7. Информация и регулирование в биологических системах
421
тельного действия, строящиеся на использовании не рефлекторного,
а гуморального пути передачи сигналов (т.е. на использовании
физиологически активных соединений, которые выделяются в кровь
при изменении параметров кровотока, разносятся кровью по
организму и достигают таким образом тех органов-мишеней, которые
обладают к ним специфической чувствительностью. Одним из них
является ренинангиотензиновый механизм, сущность которого в общих
чертах заключается в следующем.
Ключевым компонентом этого механизма является фермент
ренин. Ренин вырабатывается в юкстазломерулярных клетках,
расположенных в приносящих артериолах почечных клубочков {почечным
клубочком называется шаровидный начальный отдел основной
функциональной единицы почки - нефрона). В покое он хранится в этих
клетках, однако при уменьшении артериального давления в
почечных артериях (и при некоторых других обстоятельствах) выделяется
из них в кровоток. В крови ренин расщепляет вырабатывающийся
в печени полипептид ангиотензиноген. В результате образуется
меньший полипептид ангиотензин I, который далее превращается в ангио-
тензин П. Этот небольшой полипептид обладает мощным
сосудосуживающим действием. Кроме того, он возбуждает центральные
и периферические симпатические структуры. В результате
периферическое сопротивление и артериальное давление повышаются. Эффект
этот развивается медленно (десятки минут) и длится долго без
значительного ослабления.
Совсем недавно обнаружен еще один гуморальный фактор,
принимающий участие в функционировании этой системы.
Оказалось, что мышечные клетки предсердий сердца также обладают
гуморальной активностью, синтезируя и накапливая полипептид,
получивший название предсердного натрийуретического фактора (ПНФ).
Этот пептид выделяется в кровоток при растяжении мышечных
волокон, в связи с переполнением предсердий кровью. Попав в кровь, он
переносится по артерям к различным органам-мишеням, в том числе
к почкам. Поступая к последним, этот гормон каким-то образом тор-
мрзит выработку ренина, т.е. противодействует появлению связанной
с последним сосудосуживающей реакции. Кроме того, ПНФ
оказывает прямое сосудорасширяющее действие путем расслабления гладких
422
Биофизика
мышц крупных артерий. В результате обеспечивается ускорение
оттока крови от сердца и предотвращается развитие в нем застойных
явлений.
Таким образом, гуморальный механизм регуляции кровотока
оказывается, в свою очередь, состоящим из контролирующих друг
друга подсистем, что и позволяет организму эффективно
регулировать его на протяжении длительного функционирования организма
и поддерживать его даже при таких патологических ситуациях, как
значительная кровопотеря. Гуморальные факторы, участвующие в
регуляции кровообращения, одновременно эффективно влияют на
экскреторную функцию почек - выделение ими воды и солей. Тем самым
регуляторные механизмы длительного действия могут изменить еще
одну переменную в кровообращении, которую пока рассматривали
как постоянную, а именно: объем циркулирующей крови.
7.4.3. Регуляция газового состава крови
Как указывалось выше, важнейшей функцией кровотока
является снабжение тканей кислородом и избавление их от С02.
Потребности тканей в кислороде сильно меняются в ходе
жизнедеятельности, так же как и количество образующегося С02. Поэтому
поддержание постоянства парциальных давлений этих газов является
важнейшим компонентом гомеостатической функции
кровообращения. Поскольку поступление кислорода в кровь и выведение из нее
С02 происходит через конечные разветвления воздухоносных путей
легких - альвеолы, то основным механизмом в поддержании такого
постоянства оказывается именно поддержание постоянства состава
альвеолярного воздуха.
Альвеолы вентилируются притоком и оттоком воздуха при
ритмическом расширении и сжатии легких в результате увеличения
и уменьшения объема грудной клетки за счет сокращения диафрагмы
и межреберных мышц. В отличие от ритмической деятельности
сердечной мышцы, которая основана на внутренней способности к авто-
матии и регулируется реципрокными влияниями со стороны
симпатических и парасимпатических эфферентных волокон, ритм дыхания
имеет центральное происхождение и порождается командами из осо-
Глава 7. Информация и регулирование в биологических системах
423
бой системы нейронов продолговатого мозга (дыхательного центра).
Эти команды передаются по соматическим эфферентным путям,
аналогичным путям активации других типов скелетных мышц.
Частота дыхания и глубина каждого вдоха (дыхательный
объем) зависят от активности этого центра, которая, таким образом,
определяет состав альвеолярного воздуха и, следовательно, парциальное
давление 02 и С02 в артериальной крови. При этом обеспечивается
огромный диапазон регулирования такого постоянства. Во время
максимальной физической нагрузки площадь альвеол может обеспечивать
перенос кислорода со скоростью до 5000 мл/мин, а в покое - всего
около 250 мл/мин.
При регуляции артериального давления задающий входной
сигнал формируется в специальных барорецепторных структурах
артериального русла. В случае дыхания ситуация сложнее, так как речь
может идти по меньшей мере о двух кандидатах на роль
регулируемой переменной (Р0, и Рсо в крови), регистрация изменений которых
может происходить различными рецепторными структурами как на
периферии, так и в центральной нервной системе. Кроме того, роль
каждой из этих переменных может существенно изменяться в
зависимости от условий функционирования организма.
В условиях покоя и нормального атмосферного давления из
указанных двух переменных основную роль, как показывают
эксперименты, играет вторая - парциальное давление в крови С02. Это
хорошо видно из приведенных на рис. 7.20 кривых, на которых
представлены изменения дыхания (его минутного объема) при снижении
во вдыхаемом воздухе концентрации 02 или повышении
концентрации С02. Как видно, минутный объем дыхания не изменяется
практически до тех пор, пока содержание 02 не сократится до половины
нормального уровня. Таким образом, гипоксия (снижение Р02 в
артериальной крови) оказывается в этих условиях крайне
малоэффективным стимулом для усиления дыхания. Наоборот, повышение
содержания С02 во вдыхаемом воздухе (гиперкапния) стимулирует
дыхание сразу же, как только оно превысит нормальный уровень.
На первый взгляд такая ситуация может показаться
парадоксальной, так как именно недостаток кислорода является тем
основным фактором, который может быстро привести к нарушениям жиз-
424
Биофизике
Минутный объвм дыхания, л/мин
180
120
80
40
0
Расог40 50 60 70 кПа
Ра со 90 70 50 30 кПа
Рис 7.20. Изменения вентиляции лёгких у человека
при снижении концентрации Οι или повышении
концентрации СО2 во вдыхаемом воздухе
Заштрихованная полоса вверху - максимальная
произвольная вентиляция, на которую был способен испытуемый
ненных процессов. Сам по себе избыток С02 к таким нарушениям не
приводит. Однако дело в том, что в естественных условиях
функционирования организма оба параметра не являются независимыми, и их
изменения всегда тесно связаны - недостаточная вентиляция легких
приводит как к накоплению углекислоты, так и к нехватке
кислорода. По-видимому, с точки зрения биофизических механизмов
использование датчков изменения уровня углекислоты является более
надежным, чем датчиков изменения уровня кислорода, и именно они
в ходе эволюции закрепились как основные источники информации
в системе регуляции дыхания.
При подъеме на большую высоту положение меняется. Такой
подъем сопровождается нарастающей гипоксией и одновременно ги-
покапнией (снижением уровня углекислоты в крови) за счет снижения
Максимальная вентиляция
Глава 7. Информация и регулирование в биологических системах
425
общего барометрического давления. В этом случае стимулирующее
влияние гипоксии на дыхание приобретает первостепенное значение,
и поддержание уровня кислорода во вдыхаемом воздухе приобретает
ведущую роль в поддержании газового гомеостаза крови.
Что же представляют собой соответствующие датчики?
Наибольшее значение имеют рецепторы, расположенные прямо над
вентральной поверхностью продолговатого мозга и реагирующие на
изменения Рсо2 в спинномозговой жидкости (в свою очередь точно
отражающие изменения Рс02 в артериальной крови). Эти рецепторы
могут быть отнесены к хеморецепторам. При этом Рсо2 и рН
артериальной крови оказывают на их активность неодинаковый эффект
(первое более эффективно). Это не значит, что на поверхности мозга
имеются два различных типа хеморецепторных образований,
чувствительных либо к С02, либо к Н+. Возможно, такие хеморецепторы
являются именно рецепторами протонов, а действие С02 связано
с образованием последних в ходе диссоциации угольной кислоты.
В этом случае разница в эффекте Рс02 и рН может быть связана с
различной скоростью их транспорта в тканях. Известно, что С02 очень
быстро диффундирует из крови через клеточные мембраны, тогда
как для переноса ионов Н+ необходимы особые мембранные
структуры - ионные каналы или переносчики.
Рецепторы изменений газового состава крови имеются и
непосредственно в сосудистой стенке, в частности в сонном (каротидном)
синусе. Эти хеморецепторы реагируют именно на снижение
артериального давления и в меньшей степени на повышение РС02 или
снижение рН. Поэтому реакция дыхания на повышение Рсо2 оказывается
почти целиком зависящей от рецепторов самого мозга, более слабая
же реакция на снижение Рсо2, - от периферических хеморецепторов.
Комплекс нейронов, идентифицируемый как дыхательный
центр, имеет в своей функциональной организации много общего
с рассмотренным выше сосудодвигательным центром - он состоит из
двух связанных нейронных сетей, одна из которых приходит в
активность во время вдоха, а другая - во время выдоха. Эти нейронные
системы при помощи нисходящих путей связаны с мотонейронами
соответствующих дыхательных мышц, и возникающие в них команды
приводят к периодическому сокращению последних. Две системы
426
Биофизика
нейронов в дыхательном центре находятся между собой в реципрок-
ных тормозящих отношениях, когда одна из них приходит в
состояние активности, другая тормозится. Затем каким-то образом
активность первой системы ограничивается и торможение ослабевает, что
создает возможность для появления активности в другой системе.
Эта способность к периодическому переключению активности
из одной системы в другую сохраняется даже в изолированном
продолговатом мозге. Однако для стабилизации ритмической деятельности
и ее согласования с газовым составом крови необходимо поступление
к ней информации от описанных выше центральных и периферических
хеморецепторов, приводящей либо к усилению и учащению
дыхательных движений и соответственно к повышению минутного объема
дыхания, либо, наоборот, к его снижению.
Кроме описанных систем обратной связи, функционирующей
на основе сигналов о химическом составе артериальной крови,
важную роль в регуляции дыхания играют и обратные связи,
использующие сигналы от механорецепторов, расположенных непосредственно
в легких. Если слишком раздуть легкие при вдохе, последний рефлек-
торно тормозится, и начинается выдох. Напротив, если существенно
уменьшить объем легких, то это стимулирует глубокий вдох
(рефлексы Геринга-Брейера). Основой указанных рефлексов являются меха-
норецепторы, расположенные непосредственно в легочной
паренхиме, а также в трахее, бронхах и бронхиолах. Некоторые из этих
рецепторов являются позиционными и реагируют на степень
растяжения легочной ткани пачками импульсов, другие же являются
скорее дифференциальными и возбуждаются лишь при уменьшении или
увеличении растяжения.
Таким образом, нервная система получает постоянную
информацию как об уровне растяжения лёгких, так и об изменениях этого
растяжения. Афферентные волокна от таких рецепторов поступают
в спинной мозг и оказывают свое влияние на деятельность нейронных
систем дыхательного центра. При пересечении этих афферентных
путей дыхание становится редким и глубоким. Следовательно, в
физиологических условиях их влияния ограничивают дыхательные
экскурсии и делают работу дыхательной системы более экономной.
Глава 7. Информация и регулирование в биологических системах
427
Чрезмерное
сжатие
Артериальная
кровь
^v
^~— ^^
Венозная
кровь
|рсо2
Урсо^^
Усиление
венти-
Чрезмерное Л(ШИИ
расширение
Центральная
хеморецепция
Сосудистые
хеморецвпторы
Дыхательные
мышцы
\Торможение
Активация
/ Тор
Экспираторная
часть
Инспира-
торная
часть
о.
t
I
β
Цыхат
иожение
Рис. 7.21. Общая схема прямых
и обратных связей, участвующих в регуляции дыхания
На рис. 7.21 суммированы описанные выше главные системы
обратных связей, ответственных за поддержание газового гомеостаза
крови.
7.4.4. Регуляция уровня
энергетических веществ в крови
Все живые клетки нуждаются в непрерывном снабжении
веществами, которые могут быть для них источником метаболической
энергии. Эта потребность, как и потребность в кислороде, меняется
в зависимости от функциональной активности клетки. У
млекопитающих энергия поступает из крови в клетку в основном в виде глюкозы,
и поэтому поддержание ее гомеостаза в крови является необходимым
условием поддержания жизни. Регулирование этого гомеостаза
обеспечивается высокосовершенной системой, в которой управляющие
сигналы передаются гуморальным путем - через выделяемые в ток
крови физиологически активные соединения.
Рассмотрим принцип поддержания в крови необходимого
уровня глюкозы в самой общей форме. Этот уровень определяется,
во-первых, притоком глюкозы периодически из кишок после каждого прие-
28*
428
Биофизика
ма пищи в результате расщепления в них углеводов и всасывания
образовавшихся моносахаридов, и более постоянного - в результате
расщеплений печеночного гликогена (представляющего собой
внутреннее депо углеводов). Во-вторых, он зависит от постоянного
потребления глюкозы тканями для покрытия их энергетических трат.
Несмотря на эти процессы, в норме уровень глюкозы в крови
оказывается весьма постоянным (около 5 мМ/кг воды плазмы). Лишь сразу
после приема пищи во время всасывания он может возрастать до
6,Ιό,? мМ/кг, а во время умеренного голодания снижаться до 3,5 мМ/кг.
При пищевом повышении уровня глюкозы активируется
синтез гликогена в печени (гликогенез) и уровень ее в крови вскоре
стабилизируется за счет пополнения этого депо углеводов. Наоборот,
при снижении уровня глюкозы ниже нормы активируется
противоположный процесс распада гликогена (гликогенолиз), пополняющий
такой дефицит.
Направление перемещения глюкозы и является той
регулируемой переменной, которая управляется выделением в кровь
физиологически активных соединений (гормонов). Поступление глюкозы из
крови в клетки большинства тканей (в том числе и в клетки печени)
определяется гормоном инсулином, вырабатываемым В-клетками
панкреатических островков (островков Лангерганса) поджелудочной
железы. Если уровень глюкозы в крови ниже 3,3 мМ/кг, то инсулин
вовсе не выделяется. По мере превышения этого порога он выделяется
во все больших количествах за счет прямой стимуляции глюкозой
секреторной функции В-клеток. Вследствие этого во время дефицита
глюкозы последняя не может попасть в клетки инсулинчувствитель-
ных тканей, однако она сберегается для таких тканей, в которых
утилизация глюкозы не зависит от инсулина (нервная ткань). Инсулин-
чувствительные ткани при этом могут покрывать свои энергетические
потребности только за счет других источников (жиров или белков).
Инсулин, помимо влияния на поступление глюкозы в клетки,
стимулирует также гликогенез (синтез гликогена) и угнетает гликогенолиз
(его распад).
Таким образом, именно выделение инсулина оказывается тем
механизмом гуморальной обратной связи, который обеспечивает
снижение уровня глюкозы при появлении ее избытка в крови (гипог-
Глава 7. Информация и регулирование в биологических системах
429
ликемизирующее действие). Лишь при очень значительном
повышении этого уровня глюкоза начинает выводиться почками в мочу (глю-
козурия). Инсулин является единственным гормоном в организме,
обладающим таким действием. Наоборот, существует ряд гормонов,
которые стимулируют расщепление гликогена и тем самым
способствуют повышению уровня глюкозы (гипергликемизирующее
действие). К ним относятся глюкагон (вырабатываемый D-клетками
поджелудочной железы), адреналин, глюкокортикоиды (гормоны коры
надпочечников), тироксин (гормон щитовидной железы), гипофизар-
ный гормон роста. Выброс этих гормонов в кровь стимулируется
недостатком в ней глюкозы (гипогликемией), и они представляют собой
функционально антагонистическую систему обратных связей,
которая в сочетании с инсулиновой системой обеспечивает эффективный
гомеостаз глюкозы в организме.
Аналогичные регуляторные системы участвуют в гомеостазе
других возможных источников энергии - продуктов расщепления
жиров и белков, которые через специальные биохимические механизмы
могут превращаться в глюкозу. Жиры составляют самый обширный
резерв метаболической энергии и могут включаться в циркуляцию
в виде триглецеридов и свободных жирных кислот. Для
энергетических целей в крайнем случае могут расходоваться и аминокислоты.
Как и в случае мобилизации и запасания глюкозы, расходование
жировых и белковых запасов также регулируется системой
гормональных обратных связей, действие которых состоит в регуляции функции
специфических ферментов. В частности, инсулин способствует
образованию жировых запасов и подавляет липолиз. Он способствует
также синтезу белков. Наоборот, глюкагон, адреналин и гормон роста
стимулируют липолиз.
Все сказанное можно наглядно представить себе на схеме, на
которой уровни соответствующих энергетических веществ
представлены в виде уровней жидкости в баках, обладающих системами
притока и оттока (рис. 7.22).
Ясно, что в ходе жизнедеятельности происходит постепенное
исчерпание имеющихся в организме энергетических веществ, и
нормальное ее продолжение возможно только в случае постоянного
поступления их из внешней среды в виде пищи. Этот процесс также эф-
430
Биофизика
Рис 7.22. Общая схема регуляции уровней
углеводов, жиров и белков в организме
Гормоны, стимулирующие отдельные превращения, обозначены
знаком (+), угнетающие - знаком (-),СЖК— свободные жирные кислоты
фективно регулируется за счет активации высших мозговых центров,
ответственных за поведенческие реакции поиска и поедания пищи.
Датчиками сигналов, включающих такое поведение, являются те же
уровни энергетических веществ (в первую очередь глюкозы) в крови,
поступающей к соответствующим центрам. Снижение их вызывает
чувство голода и тем самым побуждает организм к пополнению
запасов энергетических веществ.
7.4.5. Регуляция осмотического давления крови
В заключение рассмотрения вопроса о механизмах
поддержания постоянства физических характеристик внутренней среды
организма коротко остановимся на принципах поддержания ее
осмотического гомеостаза. Эта сторона гомеостатической функции отличается
особенной сложностью и включает в себя целый ряд нервных и
гуморальных механизмов.
Глава 7. Информация и регулирование в биологических системах
431
В основе поддержания постоянства осмотического давления
крови лежат два тесно связанных регуляторных механизма -
поддержание постоянства объема крови (и межклеточной жидкости)
и поддержание постоянства ее электролитного состава. Обе
величины сохраняют при нормальной жизнедеятельности поразительное
постоянство, несмотря на воздействие внешних и внутренних
факторов, стремящихся его нарушить.
Объем крови зависит от поступления воды через
пищеварительный канал при питье и ее потери с мочой, потом и калом. Обе
переменные, в свою очередь, связаны между собой, поскольку
избыточное поступление или потеря воды будут изменять концентрацию
электролитов крови, а нарушение электролитного состава повлечет
осмотические изменения соотношения между объемами крови и
тканевой жидкости. Между всеми этими процессами поступления и
потери должно поддерживаться строгое равновесие, регулируемое, как
и в рассмотренных выше случаях, сигналами от соответствующих
внутренних датчиков.
Как и в случае регуляции ряда других характеристик крови,
основными датчиками нарушения электролитного баланса крови (ос-
морецепторами) являются центральные структуры (определенные
группы нейронов гипоталамуса), расположенные в области его суп-
раоптического ядра. Кроме того, осморецепторы имеются также на
вентральной поверхности головного мозга возле третьего желудочка.
Осморецепторы гипоталамической области реагируют на изменения
осмолярности крови, поступающей в эту область, а осморецепторы
мозговой поверхности - на аналогичные изменения спинномозговой
жидкости. Регуляторные сигналы могут исходить также от уже
упоминавшихся барорецепторов сосудистого русла, а также внутрисер-
дечных барорецепторов, которые реагируют на растяжение стенки
крупных артериальных сосудов и сердца, сопровождающее
увеличение объема циркулирующей крови. Напротив, при уменьшении
объема крови активность этих барорецепторов и соответственно
интенсивность поступающих от них в центральную нервную систему
сигналов снижаются.
Регуляция потерь воды осуществляется главным образом
почками. Другие перечисленные механизмы играют менее существен-
432
Биофизика
ную роль и могут здесь не рассматриваться. Особенностью функции
почки является двухстадийный процесс образования мочи. Сначала
практически вся вода плазмы крови, проходящей через сосуды
почечного клубочка, фильтруется в его просвете, образуя первичную
мочу. Однако затем, по мере прохождения последней по извилистым
канальцам, она частично всасывается в кровь. Этот процесс
обратного всасывания регулируется особым антидиуретическим гормоном
(АДГ), вырабатываемым задней долей гипофиза.
При увеличении содержания в крови этого гормона процесс
реабсорбции воды усиливается, вплоть до почти полного
прекращения образования мочи. Наоборот, при недостатке гормона реабсорб-
ция подавляется, и организм теряет воду в виде избыточного
мочеотделения. Очевидно, что основная роль сигналов, возникающих
в осморецепторах (а также сосудистых барорецепторах) при
изменениях осмолярности крови или ее объема, заключается в подавлении
или стимуляции секреции АДГ. Речь идет, таким образом, о
смешанном нейрогормональном эффекторном механизме.
Наряду с регуляцией потери воды особый механизм
регулирует и потерю электролитов (натрия), также происходящую в основном
через почки. Фильтрующийся в первичную мочу натрий также реаб-
сорбируется в почечных канальцах, однако этот процесс
регулируется раздельно, в основном посредством гормона коры надпочечников
альдостерона. Секреция альдостерона, в свою очередь, подвержена
гормональной регуляции через посредство уже упоминавшейся
выше ренинангиотензиновой системы, которая включается при
снижении артериального давления и уменьшении объема крови
протекающего через почки, а также при недостатке натрия в крови. В
результате усиливается выброс в кровь альдостерона и уменьшается
потеря натрия в связи с усилением его реабсорбции.
Что касается регуляции противоположного механизма
поступления воды и электролитов в организм, то датчиками для него
являются те же осморецепторы (и, возможно, барорецепторы), сигналы
от которых используются для регуляции потерь воды и
электролитов. Как и при регуляции потребления энергетических веществ, при
этом возникают ощущение жажды и активация соответствующих
нервных центров, ответственных за поведенческие реакции поиска
Глава 7. Информация и регулирование в биологических системах
433
воды. Вероятно, по крайней мере у ряда животных следует
специально выделять и «натриевый голод» - комплекс реакций,
возникающих при недостатке в пище соли и направленных на ее поиск.
Человек же, как правило, потребляет избыточное количество соли по
сравнению с физиологической потребностью, и у него такие реакции
отсутствуют.
ЗАКЛЮЧЕНИЕ
В последние годы медицинская техника занимает центральное
место в диагностических, терапевтических и аналитических
медицинских технологиях. Подготовка инженерных специалистов для
разработки и обслуживания биотехнического, медицинского и
экологического оборудования является важной задачей. Авторы
настоящего учебного пособия надеются, что материалы, изложенные в нем,
помогут студентам, аспирантам и инженерам, специализирующимся
в этом направлении, разобраться в процессах, протекающих в живых
организмах. Несмотря на большой объем пособия, авторы не
претендуют на полноту излагаемых вопросов. По ряду вопросов существует
литература, более подробно рассматривающая конкретные
проблемы. Заинтересованный читатель по вопросам теоретической
биофизики и биофизики клеточных процессов может обратиться к
учебнику А.Б. Рубина «Биофизика» (Т. I. М., 1999 г. и Т. П. М., 2000 г.),
а медицинские работники - к учебнику В.О. Самойлова
«Медицинская биофизика» (СПб., 2004 г.).
Дальнейшее развитие биофизики, биохимии и методов
исследования живых объектов приведет к углублению знаний в этих
областях, что станет базой для разработки нового медицинского
оборудования и технологий на их основе.
список
ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Антонов В.Ф., Черныш A.M., Пасечник В.К, Вознесенский
С.А., Козлова Е.К. Биофизика. - М.: Гуманит. изд. центр ВЛАДОС,
1999.-288 с.
2. Артюхов В.Г., Ковалева Т.А., Шмелев В.П. Биофизика. -
Воронеж: Изд-во ВГУ, 1994. - 336 с.
3. Бегун П.И., Шукейло Ю.А. Биомеханика. - СПб.: изд-во
«Политехника», 2000. - 463 с.
4. Биомеханика сердечной мышцы. [В.Я. Изаков, Г.П. Иткин,
B.C. Мархасян и др.; отв. ред.Г.Р. Иваницкий]. - М.: Наука, 1981. -
325 с.
5. Биофизика/ Под. общ. ред. акад. АН СССР ПГ.Костюка. -
Киев:Выща шк., 1988. - 504 с.
6. Биофизика сенсорных систем / Под ред. В.О. Самойлова. -
СПб.: Изд-во СПбГПУ, 2004.
7. Бэгшоу К. Мышечное сокращение: Пер. с англ. - М.: Мир,
1985.-128 с.
8. Вестерхофф Х.В., К ван Дам. Термодинамика и регуляция
превращений свободной энергии в биосистемах. - М.: Мир, 1992.
9. Винер Н. Кибернетика (Управление и связь в животном и
машине). - М.: Мир. 1958.
10. Владимиров Ю.А., Рощупкин Д.И., Потапенко А.Я., Де-
ев А.И. Биофизика. - М.: Медицина, 1983. - 272 с.
11. Волобуев А.Н. Биофизика. - Самара: «Самар. Дом печати»,
1999. -168 с.
12. Гордон Д.Г. Мышечная механика. - М.: 1985.
13. Гусев В.Г. Методы и технические средства для
медико-биологических исследований: учебное пособие. 4.2. - Уфа: УГАТУ,
2001.-119 с.
436
Биофизике
14. Дещеревский В. И. Математические модели мышечного
сокращения. - М.: Наука, 1977. - 160 с.
15. Инженерная физиология и моделирование систем
организма I Под ред. В.М. Ахутина. -Новосибирск: Наука, 1987.
16. Математические модели и компьютерное моделирование
в биомеханике. / Под ред. А.В.Зинковского и В.А.Палъмова. - СПб.:
Изд-во Политехи, ун-та, 2004. - 516 с.
17. Плонси Р., Барр Р. Биоэлектричество: Количественный
подход: Пер. с англ. - М.: Мир, 1991. - 366 с.
18. Рубин А.Б. Биофизика. - М., 2000. - 468 с.
19. Самойлов В.О. Медицинская биофизика. -СПб.: «Спец-
Лит», 2004. - 495 с.
20. Физиологическая и медицинская информатика/ Сб. научи.
Трудов. Под ред. П.М. Амосова. - Киев, 1990.
21. Черныш A.M. Биомеханика сердечной мышцы. - М.: Наука,
1993.
22. Чигирев Б.И. Биофизика органов чувств. - СПб.: Изд-во
СПбГЭТУ «ЛЭТИ», 2001. - 80 с.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Ζ
Z-диск 12
Z-линия 12
А
Аберрация света 276
Абсолютная мышечная сила 24
Абсолютный порог чувствительности зрения 255
Абсолютный порог ощущения 186
Адаптация 190
Адаптация организма к охлаждению 404
Адсорбционная (стереохимическая) 307
Аккомодация 251
Аксональный ток 172
Акт вдоха 41
Акт выдоха 41
Актиновый кортекс 7
Актиновый микрофиламент 6
Акустическая эмиссия клеточных и молекулярных структур 248
Акустическое поле сердечно-сосудистой системы 246
Акустический импеданс 226, 239
Акустическое излучение 249
Альвеолярное давление 44
Ампула Лоренцини 219
438
Биофизике
Анатомическое мертвое пространство 54
Апостильб 306
Артериальный импульс 154
Артериолы 155
Атлас архитектурных цветов 297
Атлас Тейлора 297
Атлас Манселла 296
Аудиометры 237
Аудиометрия 237
Ауксотонический (или анизотонический) 25
Афферентное волокно 402
Б .
Базальный сосудистый тонус 158
Баллистокардиограмма 96
Баллистокардиография 95
Баллистокардиограф 96
Барабанная струна 322
Барорецептор 418
Безусловный рефлекс 3 70
Безусловный терморегуляционный рефлекс 387
Бинауральный слух 229
Биологическая кибернетика 361
Биомеханика 5
Бионика 362
Биосенсор 184
Биофизика сложных систем 359
Блуждающий нерв 417
Боковая ручка 174, 180
Бурый жир 384
Предметный указатель
439
В
Вегетативная функция 363
Вектор Умова 222, 239
Венный пульс 155
Вентиляция 38
Вентиляция лёгких 41
Верхний порог ощущения 186
Вестибулярная сенсорная система 211
Вкусовая луковица 318
Вкусовая почка 318
Вкусовая рюмка 318
Внимание 199
Внутриклеточный посредник 260
Водонапорная башня 160
Возвратное постсинаптическое торможение 196
Воздушная проводимость 229
Волна давления 112
Волосковый 212
Временная суммация 188
Выходная характеристика 374
Г
Гемодинамика 100
Гемодинамическое сопротивление 109
Геном 356
Гипоталамус 407, 390
Гипотермия 406
Гладкий тетанус 31
Глубина дыхания 51
Градиентое напряжение 58
Грудной отдел аорты 152
440
Биофизике
Гуморальная регуляция 367
Гуморальный механизм 421
д
Давление в плевральной полости 44
Двигательный анализатор 214
Детектор 199
Диаграмма цветности 288
Диастолы 416
Диафрагма 41
Дивергенция 195
Дилататор 251
Динамический баланс 409
Динамокардиография 97
Динеины 172, 174, 180
Диск зрительного нерва 253
Диск 255
Дисторсия 276
Дифракция 275
Дифференциальный порог 188, 237
Дифференциальный пространственный порог 189
Дрожательный термогенез 383
Дыхательный центр 424
Дыхательный объем 52
Ε
Единичная плоскость в данном цветовом пространстве 287
Единичный цветовой треугольник 287
Ж
Жгутик 175
Желтое пятно 253
Жизненная емкость легких 52
Предметный указатель
441
3
Закон Вебера 188
Закон Гука 127
Закон сохранения массы 100
Закон Стивенса 187
Законы линейности при установлении цветовых равенств 284
Закон Франка-Старлинга 93
Закон Грассмана 280
Закон Вебера-Фехнера 223
Запах 329
Звено 369
Звуковое давление 223, 238
Золотая пропорция 30
И
Измерение объема мертвого пространства 54
Измерение цвета и субъективные эффекты цветовосприятия 296
Инактивация сокращений 77
Интенсивность 186
Интенсивность звука 222
Интенсивность цвета 296
Интерорецептивный анализатор 216
Интерорецепторы 184
Информационный канал 347
Информационная избыточность 352
Информация 346
К
Кальциевая помпа 22
Кальциевые каналы 22
Кандела 306
Капилляр 147
Кашель 180
Кибернетика 359
Кинезин 173
Кинетокардиография 97
Кинетохоры 173
Кислоты 325
Колбочки 253
Колбы Краузе 211
Колебательная теория 329
Колориметров 279
Комбинированные системы 377
Константность восприятия окраски 299
Контраст 189
Координаты цветности 285
Костная проводимость 236
Кохлеарная акустическая эмиссия 245
Кривые смешения 288
Кривые удельных координат стандартной системы 290
Кровяное давление 100
Кругооборот крови 415
Л
Ламберт 306
Латеральное торможение 197
Лёгочный резистанс 43
Лейкоциты 414
Лестницы Боудича 74
Линейная скорость 100
Люминофоры 288
Предметный указатель
443
Μ
Медицинская кибернетика 361
Медленные мышцы 30
Мембранные рецепторы 365
Местная регуляция 365
Метаболическая (ферментативная) 307
Метамеризм 280
Метамерная пара 281
Метод вымывания 66
Метод разведения 66
Метод условных рефлексов 370
Методика Люшера 241
Механизм восприятия запаха 329
Микротрубочки 170
Минутный объём дыхания 51
Миозин 8
Митотическое веретено 173
Молекулярные механизмы подвижности 5
Моноуральная аудиометрия 237
Мукоцилиарный клиренс 179
Мукоцилиарный транспорт 179
Мышечное волокно 71
Η
Наружные межрёберные мышцы 41
Насморк 180
Насыщенность цвета 281
Недрожательный термогенез 384
Нейрофизиология 195
Нексиновые мостики 181
Нексус 156
444
Биофизика
Нектин 180
Неньютоновские жидкости 103
Нервный импульс 191
Нефрон 422
Hum 306
Нормальные диастолические тоны 247
Нормальные систолические тоны 247
Ноцицепторы 218
Ньютоновские жидкости 103
О
Обеспечение непрерывности кровотока 152
Обоняние 328
Обонятельная выстилка 332
Обратная связь 372
Общая емкость легких 52
Объёмная скорость 100
Одновременный цветовой контраст 302
Озноб 408
Опережающее торможение 196
Опорно-двигательный аппарат 31
Оптические отбеливатели 288
Оптический центр глаза 253
Осморецепторы 184
Основной закон сердца 74
Основные гемодинамические показатели 100
Остаточный объем 52
Островки Лангерганса 429
Острота зрения 274
Относительная видность 272
Ототолик 224
Предметный укезетель
445
Ошибка регулирования 376
Π
Палочки 253
Парасимпатическая нервная система 417
Периферический терморецептор 395
Пирамида вкусов Хеннинга 317
Плетизмограф 67
Плетизмография 70
Пневмотахограф 67
Пневмотахография 67
Поликардиография 100
Поперечные мостики 13
Порог 237
Порог слышимости 223
Пороги восприятия 184
Пороговый блеск 303
Порог изменения ощущения 294
Порог контрастной чувствительности 274
Последовательный образ 301
Последовательный цветовой контраст 301
Постоянство электролитного состава 432
Постоянство объема крови 432
Постоянство осмотического давления крови 432
Почечный клубочек 422
Предсердный натрийуретический фактор 422
Преобладающая длина волны цвета 281
Приведенный редуцированный глаз 252
Принцип установочной точки 409
Проприоцепторы 372
Пространственная суммация 195
446
Биофизика
Протофибрилл 12
Псевдоподие 8
Психическая деятельность 382
Психическая сфера 364
Пульс 115
Пучки 71
Ρ
Равноконтрастная диаграмма цветности 295
Радиальные спицы 181
Разветвлённая сосудистая трубка 101
Разветвлённая дыхательная трубка 39
Раздражитель 183
Разобщение 77
Разрешающая способность 274
Растяжимость 43
Регионарное кровообращение 166
Регуляция артериального давления 417
Резервный объем вдоха 52
Резервный объем выдоха 52
Реизомеризация 259
Релаксационный метод измерения давления 44
Ренинангиотензиновый механизм 422
Реснички 175
Рефлекс 368, 370,386
Рефлекторная дуга 369
Рецепторный потенциал 192, 259
Рецепторы 184
Речевая аудиометрия 242
Родопсин 255
Рычаг 31
Предметный указатель
447
С
Саркомер 11
Саркоплазматическая сеть 20
Светлота цвета 281
Световой поток 306
Свободные нервные окончания и инкапсулированные нервные тельца 207
Семидоменные рецепторы 340
Сенсорная система боли 218
Сенсорные системы кожи 201
Сердечно-сосудистая система 161
Сердечный выброс 417
Сетчатка 253
Сигнал рассогласования 380
Сила мышцы 23
Сила поверхностного натяжения 42
Сила света 306
Сила сокращений 74
Сильфоны 64
Синдром Картагенера 182
Система Оствальда 297
Система стабилизации 377
Система программного управления 377
Система автоматического регулирования 3 74
Систола 416
Систолический (ударный) объём 93
Скорость кровотока 100
Скоростью сдвига 102
Скотопическое зрение 272
Сладкий стимул 323
Следящие системы 377
Слепое пятно 253
448
Биофизика
Согласование цвета 282
Сократительный термогенез 383
Сокращение миоцитов 8
Соматика 363
Соны 239
Сопротивление сосудистого русла большого круга кровообращения 418
Сосудистый тонус покоя 158
Сосудистые краны 160
Социологическая кибернетика 361
Спектр излучения равноэнергетический 287
Спектральный локус 288
Специфическая растяжимость легких 47
Спинной мозг 390
Спирограмма 66, 68
Спирометры 64
Спирометрия 64
Спирометры открытого типа 67
Спонтанная эмиссия 245
Стандартные колориметрические системы МКО 288
Стандартные источники белого цвета 287
Статическая растяжимость легких 47
Стереоцветное кино 282
Стильб 306
Стимул 183
Стоимость силы 158
Строение внутреннего уха 22 7
Строение наружного и среднего уха 224
Субъективные эффекты 299
Суммация 189
Сфигмограмма 154
Сфинктер 251
Предметный указатель
449
Τ
Таламус 403
Тельца Гольджи—Маццони 209
Тельца Мейснера 446
Тельца Пачини 208, 210
Тельца Руффини 211
Тельце Меркеля 209
Теория вкуса 307
Теория гетерогенности хеморецепции 307
Теория обоняния 307
Теория информации 345
Тепловая мощность 25
Тепловые рецепоры 211
Теплопродукция несократительным термогенезом 384
Теплота активации 33
Теплота укорочения 33
Термокинетические упругие силы 150
Терморегуляционный тонус 383
Терморецепторы 184, 211
Термосенсорная система 217
Тетанические сокращения 30
Техническая кибернетика 361
Тоны Короткова 134
Трансдукция 193
Трехкомпонентная теория цвета 278
Триплетный кодон 354
Трихроматическая единица 285
Тромбоциты 414
Тропомиозин 14
Тубулин 170
Тубулиновые протофиламенты 180
450
Биофизика
Тяжи 71
У
Унифицированное цветовое уравнение 285
Упругие свойства 42
Уравнение вкуса 308
Уравнение неразрывности 131
Уравнение Хилла 25
Уравнением Бора 55
Уравнения Пуазейля 108
Уравнениями Эйлера 131
Уровень громкости 239
Уровень интенсивности 239
Ускорения 211
Условие неразрывности струи 39, 100
Условнорефлекторная регуляция 387
Уставка 375
Φ
Фаза асинхронного сокращения 91
Фазная реакция 396
Фаза быстрого изгнания 93
Фаза изометрического сокращения 92
Фаза медленного изгнания 93
Физическая терморегуляция 385
Флаттерный режим 129
Флебограмма 155
Фон 240
Фонокардиограф 99
Фонокардиография 98
Формула периферического гемодинамического сопротивления 117
Предметный укезетель
451
Форпостное регулирование 377
Фотоизомеризация 258
Фотолиз 259
Фотопическое зрение 272
Фоторецепторная мембрана 255
Фоторецепторы 184
Фоторецепция 249
Функциональная остаточная емкость 52, 53
Функциональный элемент 71
Функция организма 359
Фурье-анализ 118
X
Характерическая частота волокна 236
Хеморецепторы 184
Хемосенсорные системы 217
Химическая терморегуляция 385
Холодовые точки 217
Хроматическая диаграмма 288
ц
Цветное телевидение 282
Цветовая температура 287
Цветовой круг Ньютона 278
Цветовое пространство 286
Цветовой тон 281
Цветовой треугольник Максвелла 279
Центр 419
Центральное звено 370, 389
Центральные и периферические терморецепторы 393
Центральное венозное давление 149
452
Биофизика
ч
Частота стимула 74
Чистота цвета 281, 282
э
Экстероцепторы 184
Эластические волокна 42
Эластическая тяга лёгких 42
Электромеханическое сопряжение 20
Электронный нос 343
Электрорецепторы 184
Электрорецепция 219
Энергетическая сила света 305
Эритроциты 413
Эффект Пуркине 272
Эффектор 370,372,377
Эфферентное звено 387
Я
Яркость 28! 306
ОГЛАВЛЕНИЕ
Том II. БИОМЕХАНИКА, ИНФОРМАЦИЯ
И РЕГУЛИРОВАНИЕ В ЖИВЫХ СИСТЕМАХ
Введение 3
ГЛАВА 5. Биофизика двигательной активности 5
5.1. Биофизика мышечного сокращения 6
5.1.1. Актин-миозиновая система миоцита
поперечнополосатых мышц 6
5.1.2. Механизм мышечного сокращения 16
5.2. Механические процессы
в опорно-двигательном аппарате человека 23
5.2.1. Биомеханические свойства скелетных мышц 23
5.2.2. Энергетика мышечного сокращения 33
5.2.3. Элементы теории мышечного сокращения 35
5.3. Биомеханика дыхания 38
5.3.1. Легочная вентиляция 38
5.3.2. Показатели легочной вентиляции 51
5.3.3. Моделирование дыхания 57
5.3.4. Методы и технические средства
для исследования легочной вентиляции 64
5.4. Биомеханика миокарда 70
5.4.1. Сокращение миокарда 70
Особенности сократимости миокарда: 74
5.4.2. Расслабление миокарда 82
5.4.3. Работа сердца 90
5.4.4. Методы исследования механической активности сердца 95
5.5. Биофизические закономерности движения крови по сосудам .... 100
5.5.1. Основные показатели гемодинамики 100
454
Биофизике
5.5.2. Линейная теория движения крови
по кровеносному руслу 113
5.5.3. Нелинейная теория движения крови
по кровеносному руслу 126
5.5.4. Биофизические особенности
кровеносных сосудов разного типа 146
Капилляры 146
Вены 148
Аорта 149
Артериолы большого круга кровообращения 154
5.5.5. Математические модели кровообращения 160
5.6. Тубулин-динеиновая и тубулин-кинезиновая
молекулярные системы подвижности 168
5.6.1. Системы микротрубочек 168
5.6:2. Биомеханические процессы в жгутиках и ресничках 173
ГЛАВА 6. Сенсорные системы организма 182
6.1. Анализ сенсорных систем 182
6.1.1. Общие принципы
функционирования сенсорных систем 184
6.1.2. Трансформация раздражителей в рецепторах 190
6.1.3. Передача и переработка информации
в нервных центрах 193
6.1.4. Биосенсоры 200
6.2. Биофизика слуха 221
6.2.1.Слуховая рецепция 221
6.2.2. Методы и средства проведения
аудиометрических исследований 237
6.2.3. Акустические поля 243
6.3. Биофизика зрения 248
6.3.1. Светопреломляющая система глаза 248
6.3.2. Биофизический механизм
восприятия света рецепторами 252
6.3.3. Рецепторные потенциалы 258
Огпевление 455
6.3.4. Чувствительность и адаптация глаза 266
6.3.5. Разрешающая способность глаза 273
6.3.6. Кодирование информации в органе зрения 276
6.3.7. Цветовое зрение 277
6.4. Биофизика хемосенсорных систем 305
6.4.1. Хеморецепция внутренней среды организма 308
6.4.2. Первичные вкусы и их пороги 316
6.4.3. Вкусовые почки 317
6.4.4. Вкусовые клетки 320
6.4.5. Афферентные пути 321
6.4.6. Механизмы вкусовой трансдукции 322
6.5. Обоняние 327
6.5.1. Запахи и их классификация 327
6.5.2. Теории обоняния 328
6.5.3. Органы обоняния 330
6.5.4. Микроструктура обонятельной выстилки 331
6.5.5. Строение обонятельных клеток 334
6.5.6. Механизм обонятельной трансдукции 336
6.5.7. Чувствительность органов обоняния 342
6.5.8. Техническое моделирование обоняния 342
ГЛАВА 7. Информация и регулирование
в биологических системах 344
7.1. Информация и живой организм 344
7.1.1. Элементы теории информации 344
7.1.2. Примеры применения теории информации к анализу
процессов передачи информации в нервных каналах связи. . . . 349
7.1.3. Информация, заключенная в генетическом коде 353
7.2. Регулирование биологических процессов 358
7.2.1. Содержание кибернетики и бионики 358
7.2.2. Стратегия управления функциями организма 361
7.2.3. Понятие о местной регуляции физиологических процессов. . . . 364
456 Биофизика
7.2.4. Понятие о гуморальной регуляции
физиологических процессов 366
7.2.5. Нервная регуляция физиологических процессов 367
7.2.6. Обратные связи в рефлекторных актах 370
7.2.7. Приложения теории автоматического регулирования
к анализу рефлекторной деятельности 372
7.2.8. Форпостное регулирование функций организма 380
7.3. Регуляция температуры тела
гомойотермного организма 381
7.3.1. Эффекторы теплопродукции 382
7.3.2. Эффекторы теплообмена (теплоотдачи) 384
7.3.3. Физиологические механизмы терморегуляции 385
7.3.3.1. Нервная регуляция
постоянства температуры «ядра» тела 385
Условно-рефлекторная регуляция 386
Безусловнорефлекторная регуляция 386
7.3.3.2. Гуморальная регуляция 402
7.3.4. Адаптация организма к низкой и высокой температуре 403
7.3.5. Лихорадка 406
7.3.6. Математическое моделирование терморегуляции 408
7.4. Постоянство внутренней среды организма и его регуляция 411
7.4.1. Характеристики внутренней среды организма 412
7.4.2. Регуляция кругооборота крови 414
7.4.3. Регуляция газового состава крови 422
7.4.4. Регуляция уровня энергетических веществ в крови 427
7.4.5. Регуляция осмотического давления крови 430
Заключение 435
Список использованной литературы 436
Предметный указатель 437
можно заказать через почтовое агентств DESSY: 107113, г.Москва, а/я 10,
а также интернет-магазин: www.dessy.ru
Е. В. Ьигяай, С. Π Вихров, П. В. Гривеииав,
В. М. Рсдькии. В. 0. Самвнлвн. Ь. И. Чшшрев
БИОФИЗ
ДЛЯ ИНЖЕНЕРОВ
1вм 2 Биимехаша, информации и регулирование в ним системах
В учебном пособии систематизированы сведения о физических
и физико-химических процессах, лежащих в основе
жизнедеятельности организмов на всех уровнях их организации, необходимые для
инженеров и специалистов, занимающихся разработкой и
обслуживанием биомедицинской техники
В первом томе двухтомного пособия изложены основы
биоэнергетики и термодинамики биологических процессов, рассмотрены
структура, свойства и функции биологических мембран и биоэлектро-
гвнвз Второй том посвящен биофизическим основам двигательной
активности человека, информационных и регуляторных процессов
в биологических системах, а также общим принципам
функционирования сенсорных систем
Для студентов, обучающихся по направлению «Биомедицинская
техника», будет полезна инженерам и специалистам, занимающимся
моделированием физиологических процессов и разработкой
аналитической, диагностической и лечебной аппаратуры
ISBN 978-5-9912-0049-3
9"785991"200493
Сайт издательства: