









































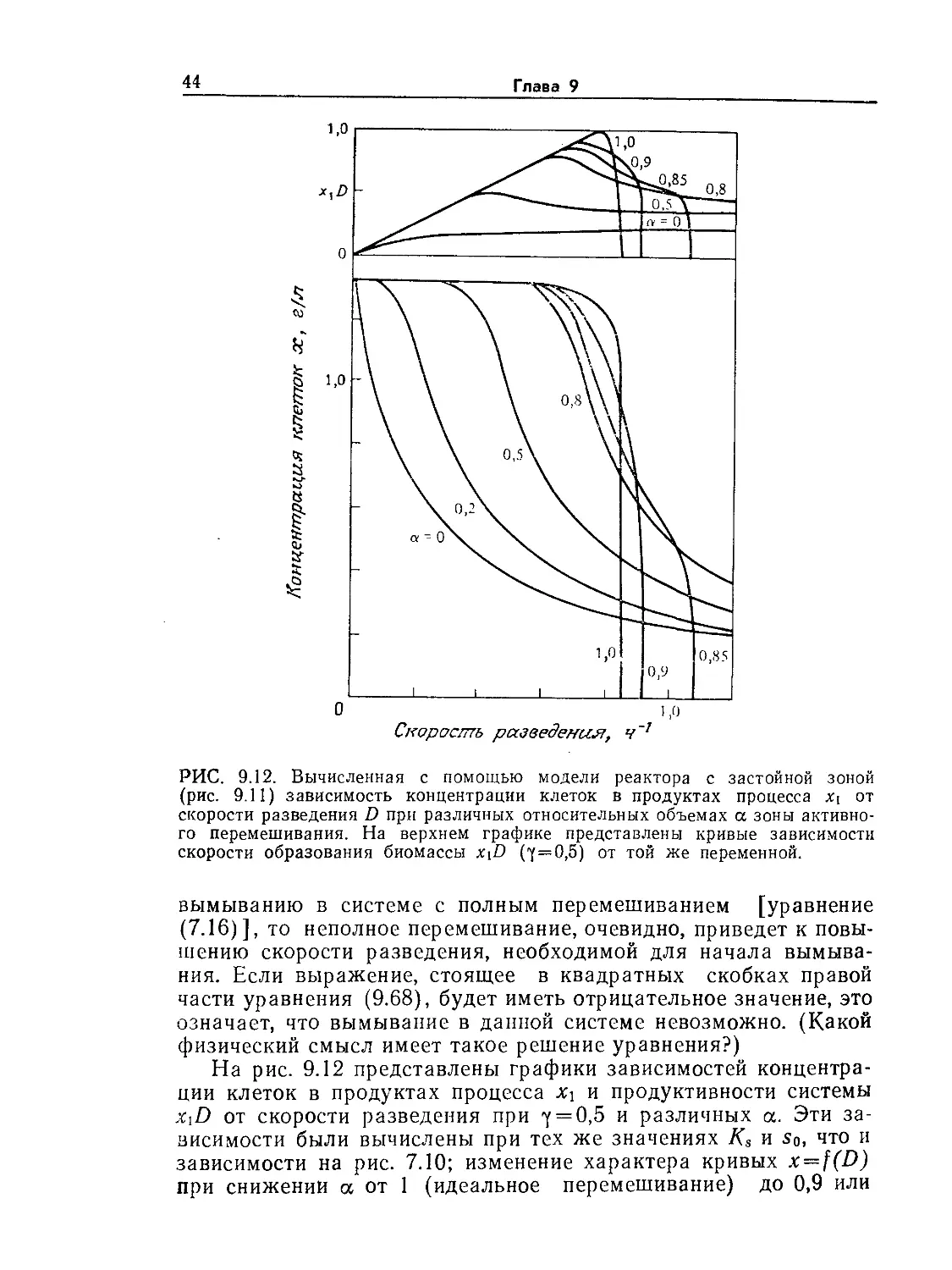




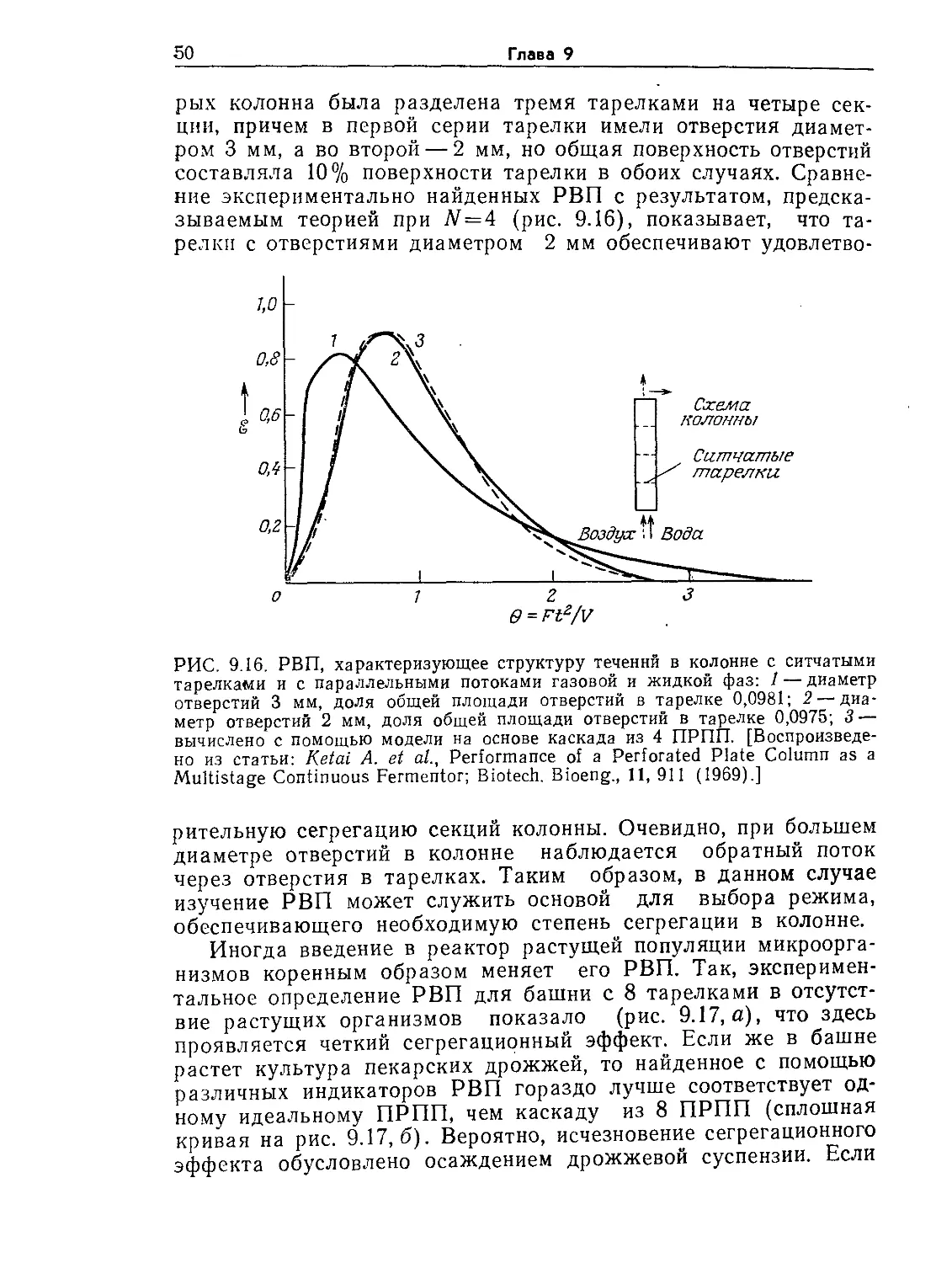

















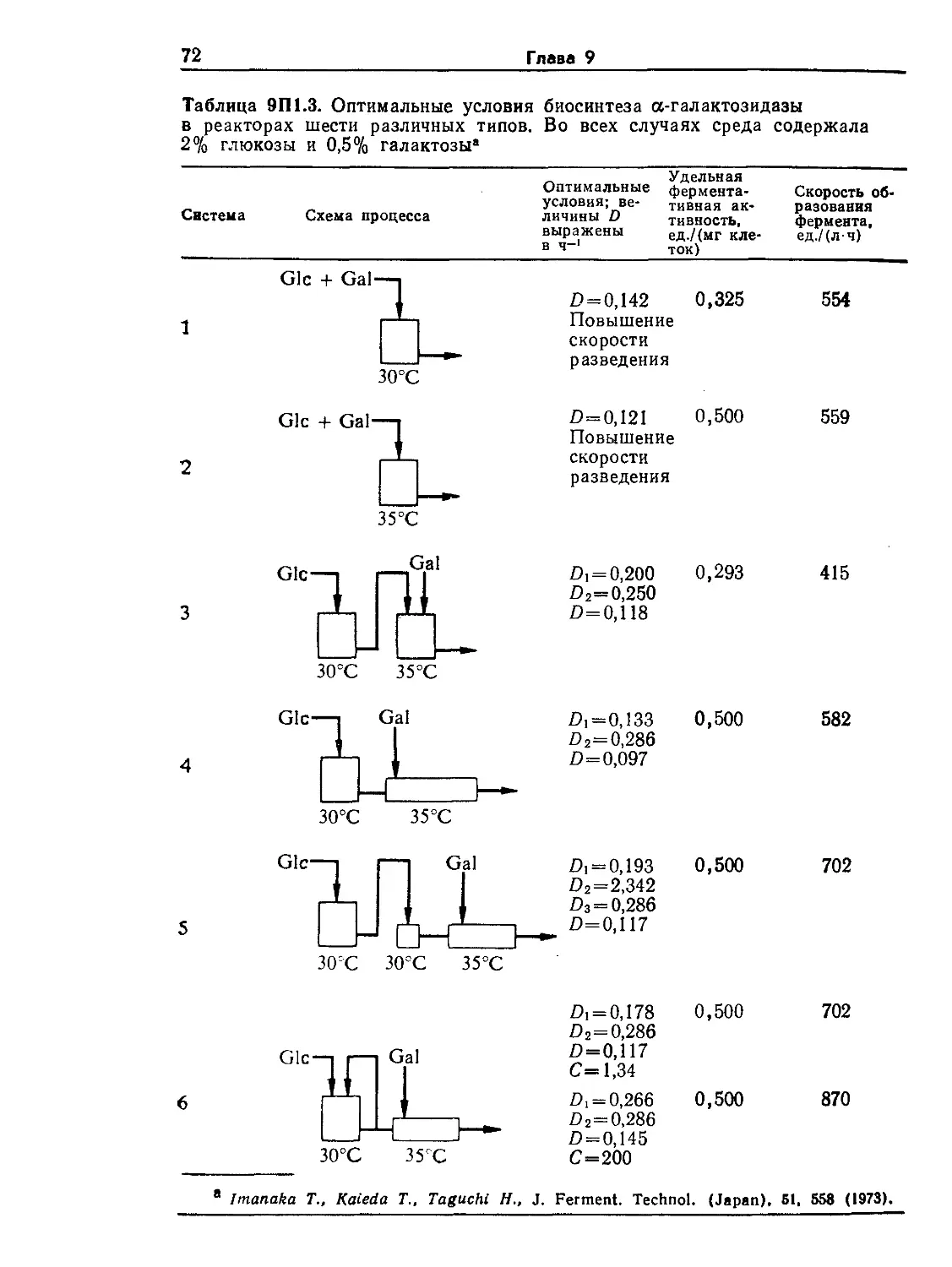
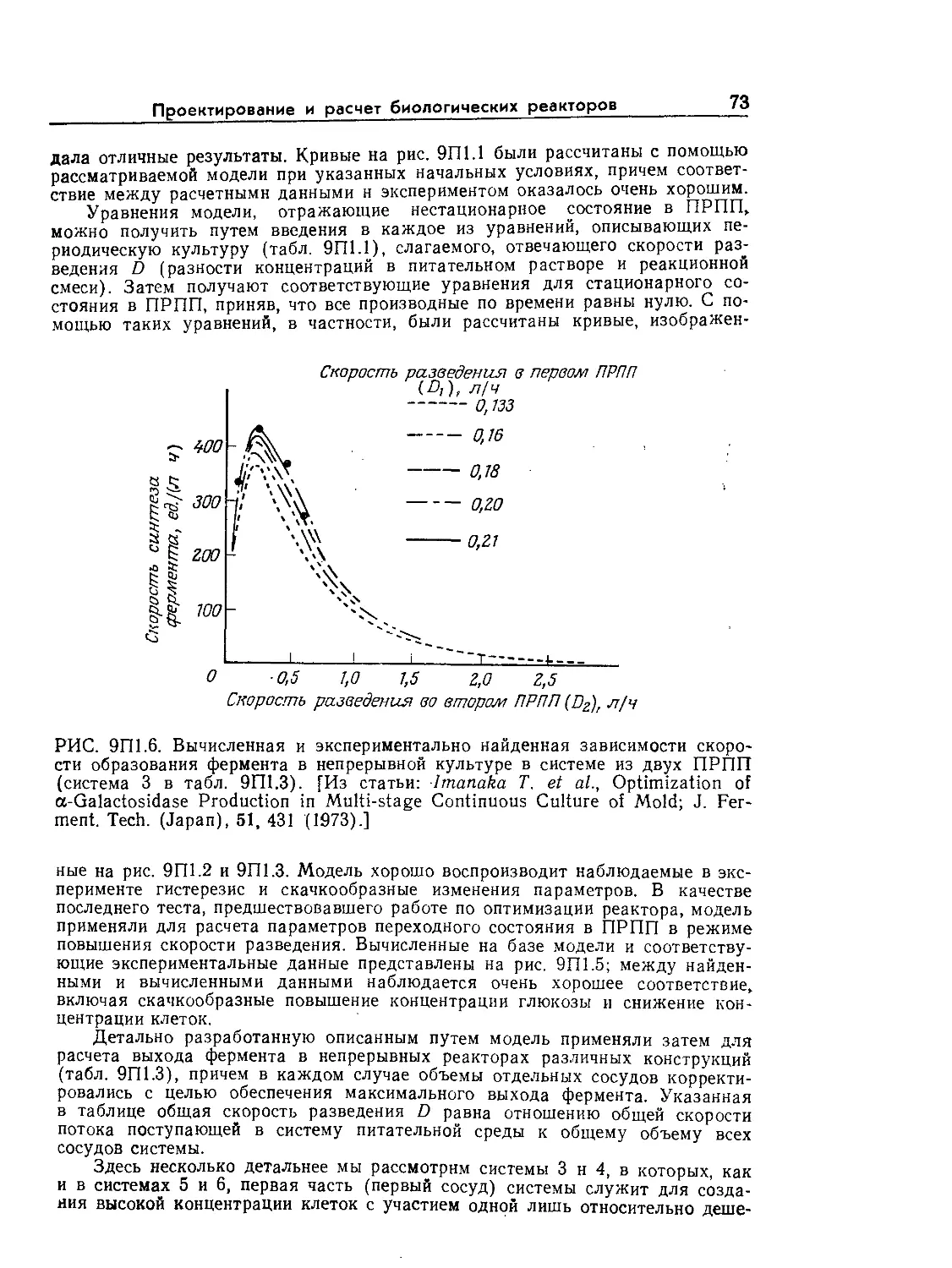






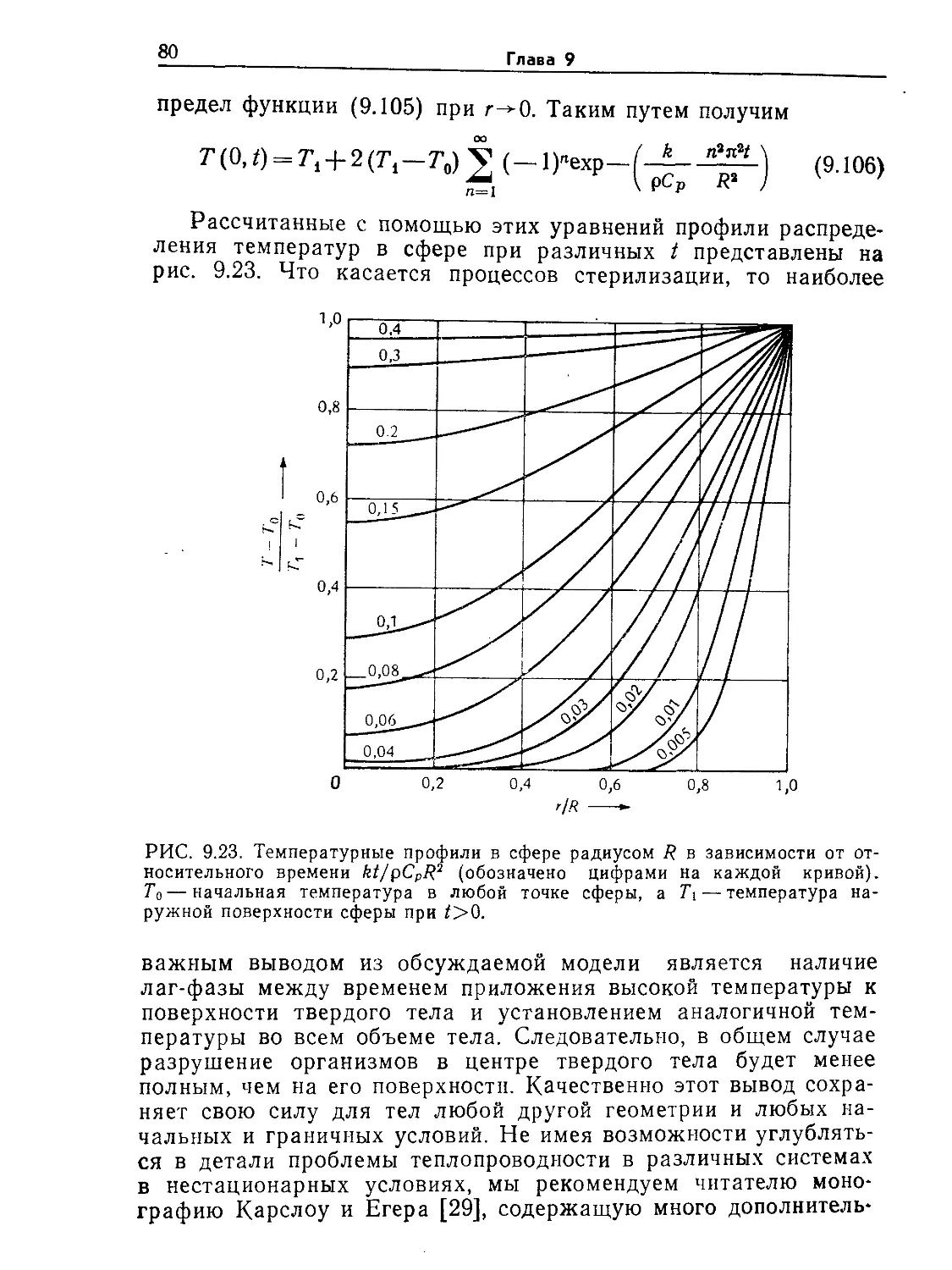


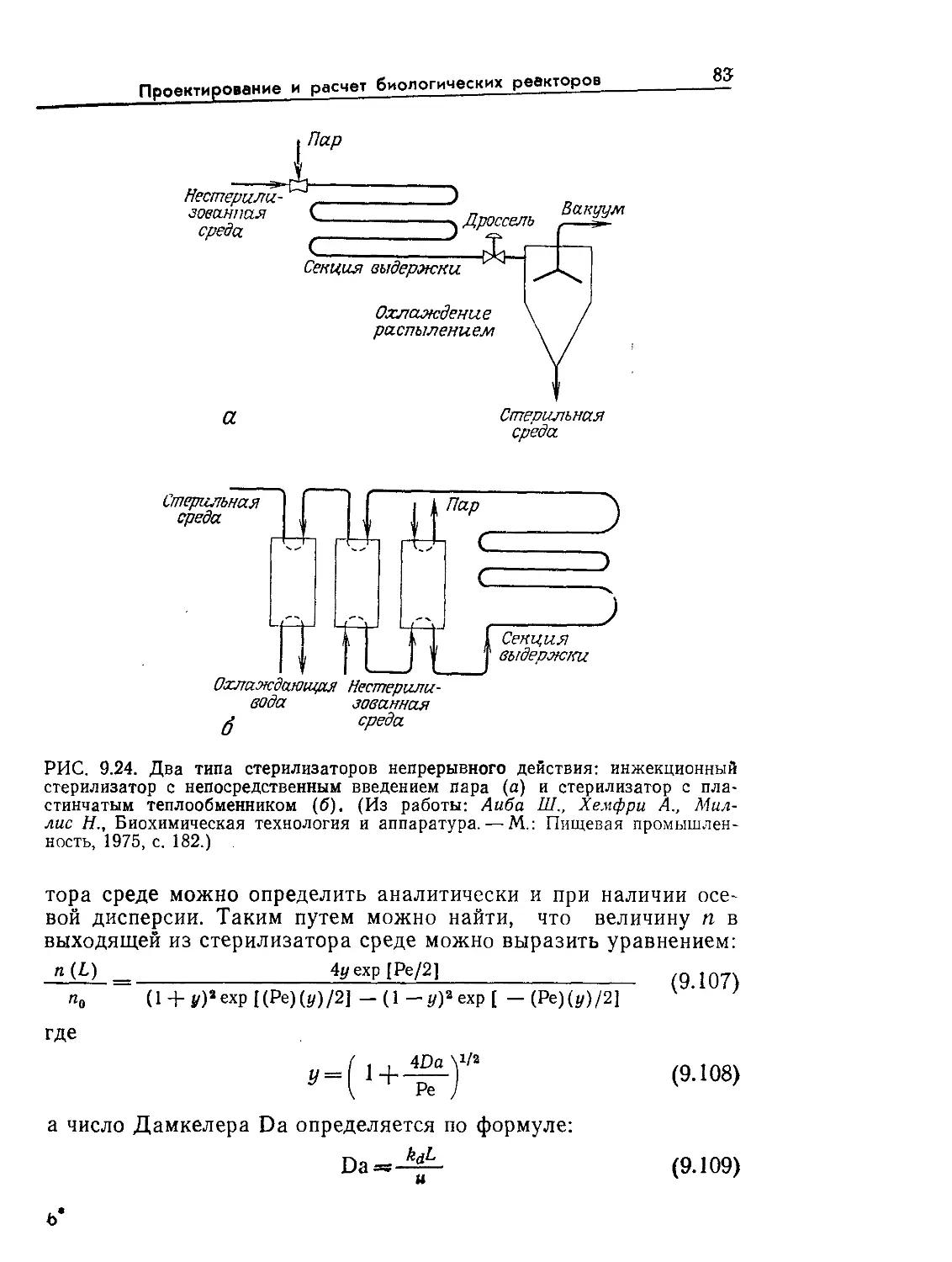






















































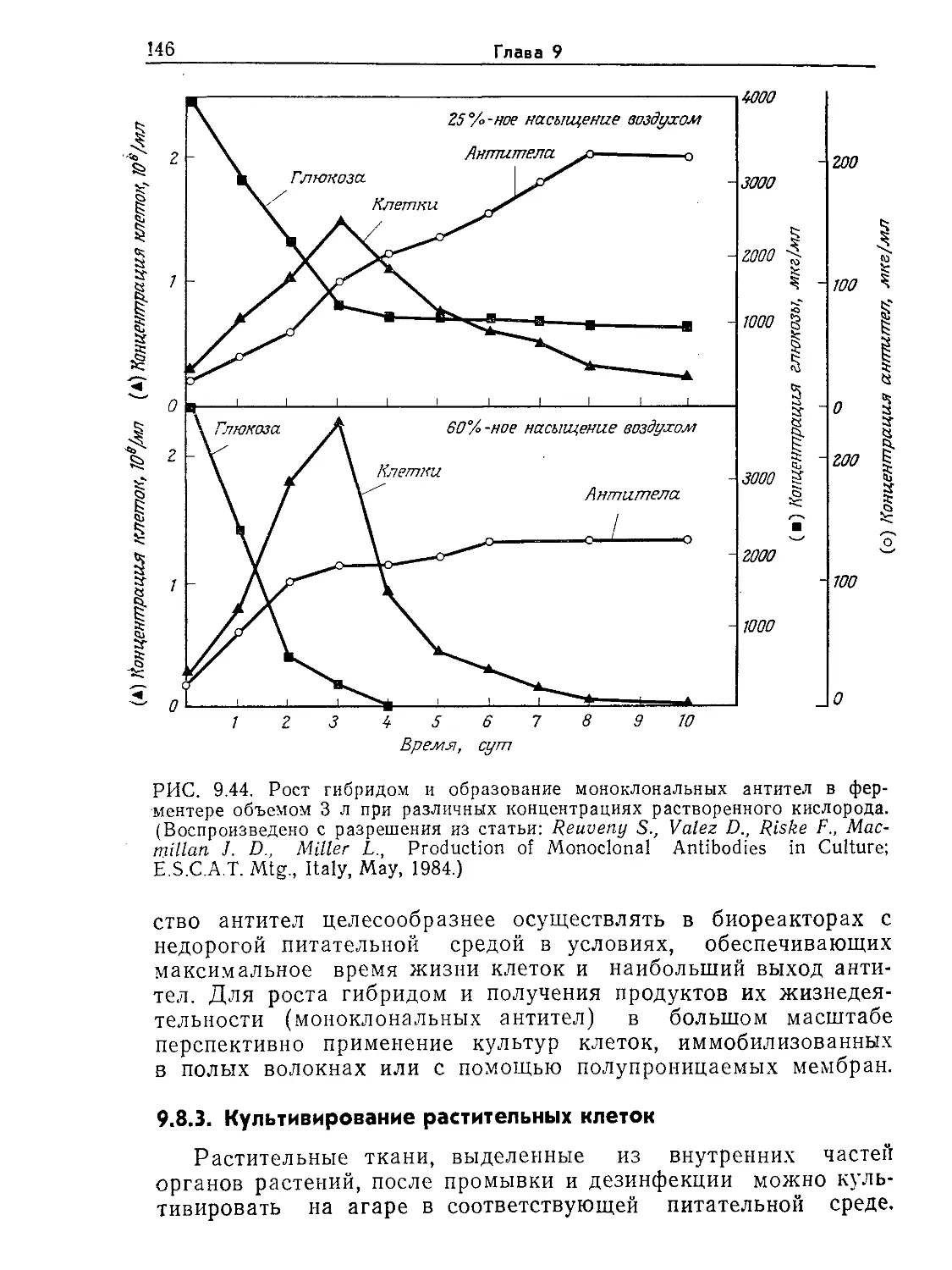
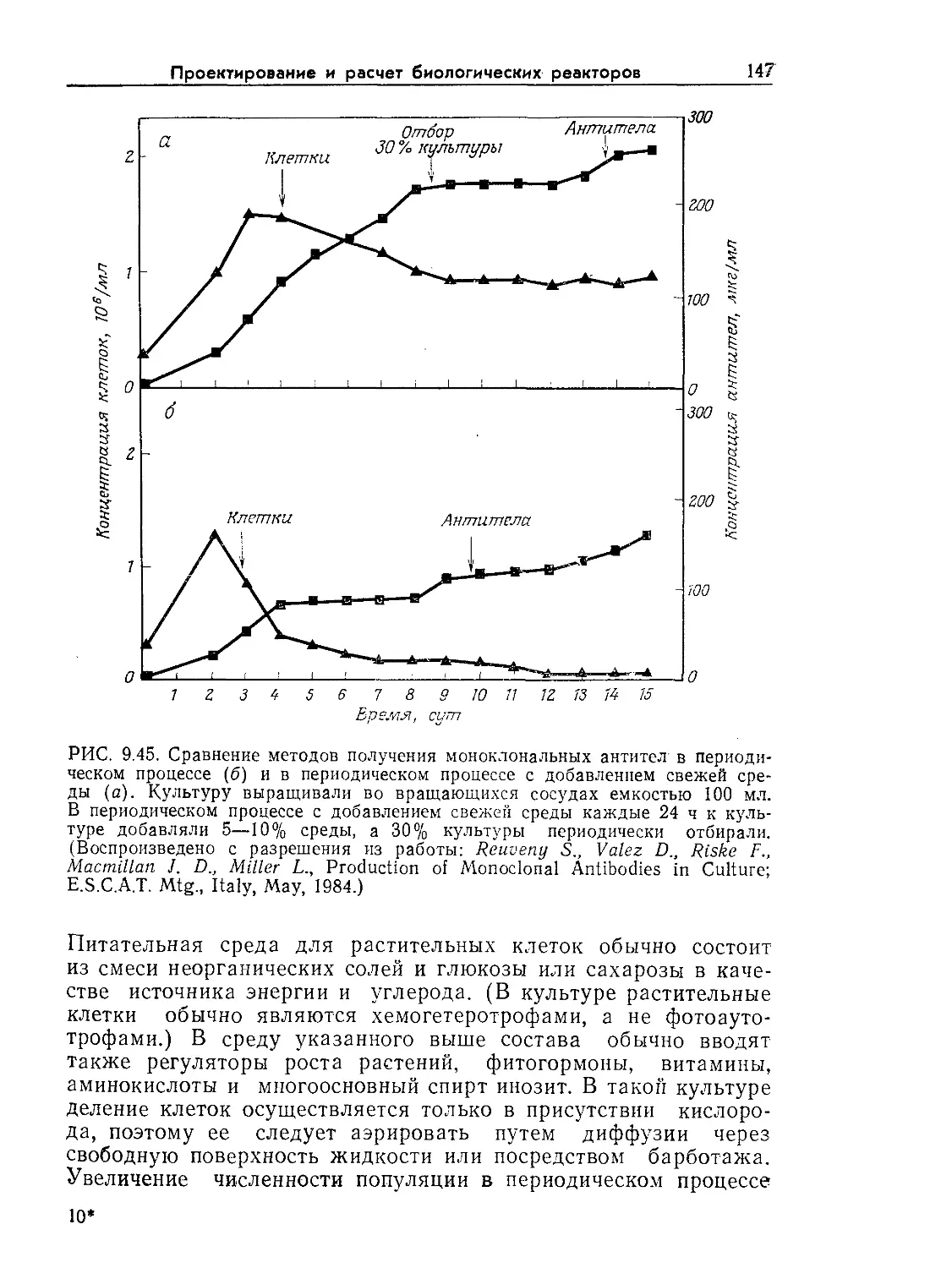








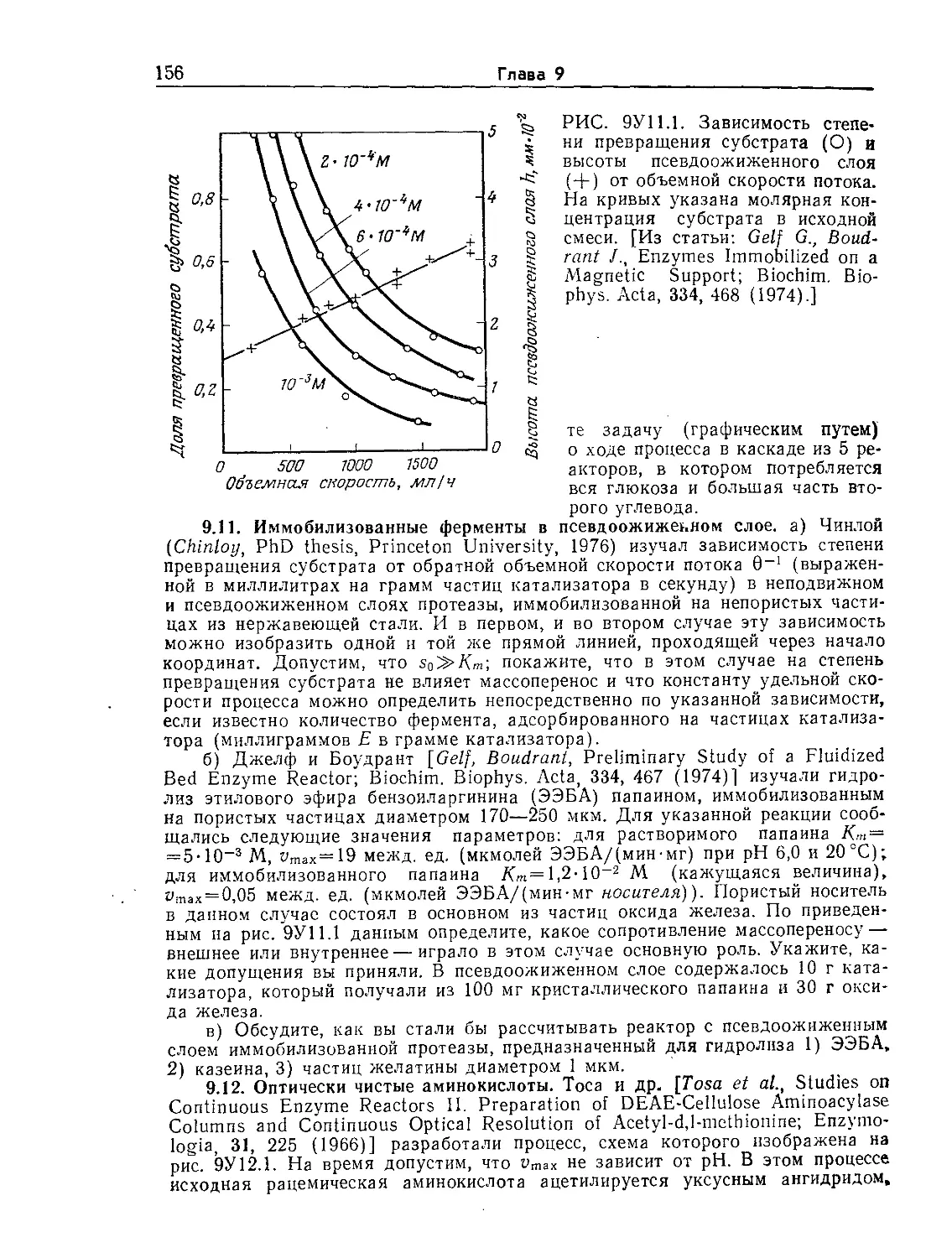


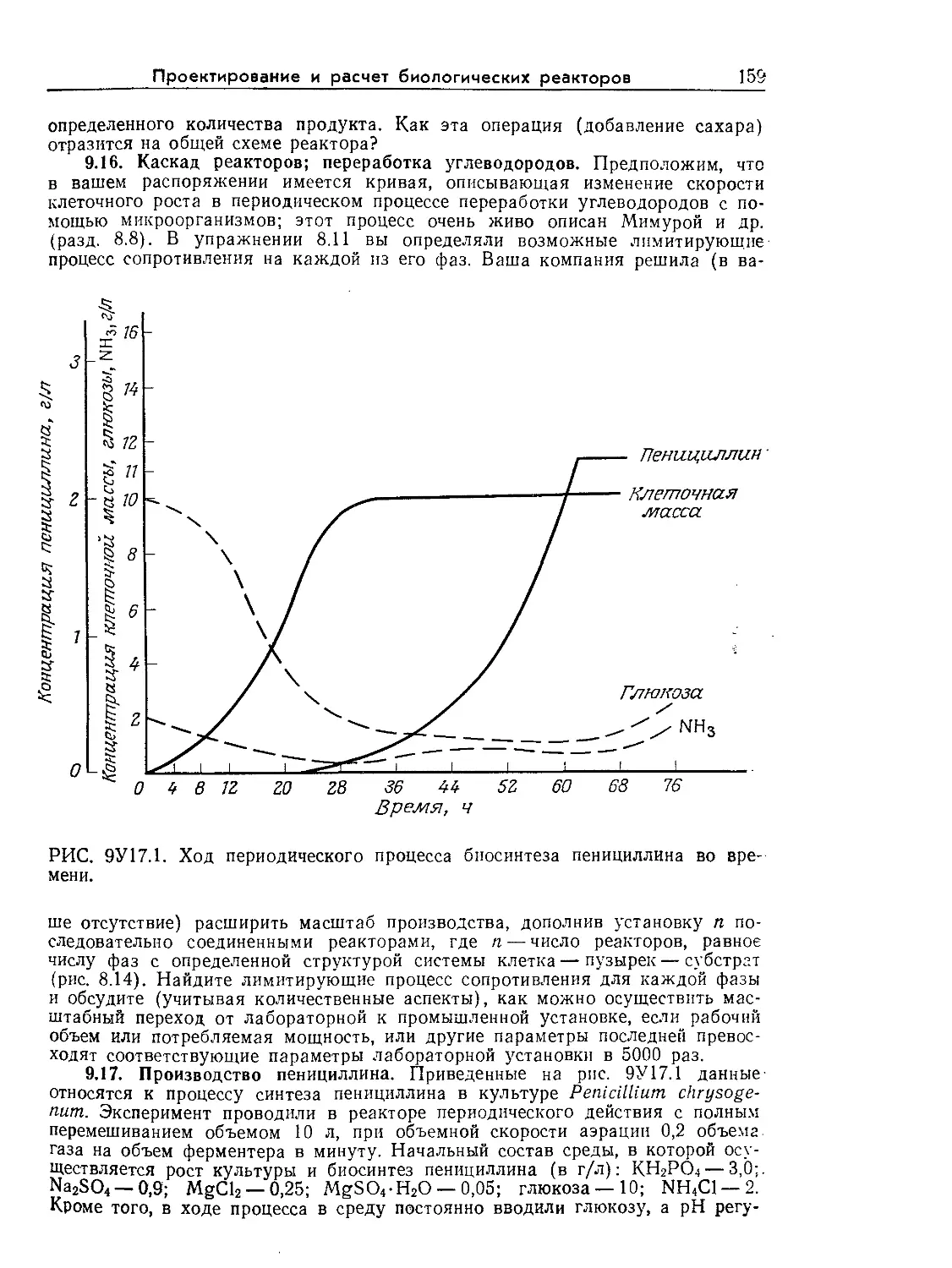










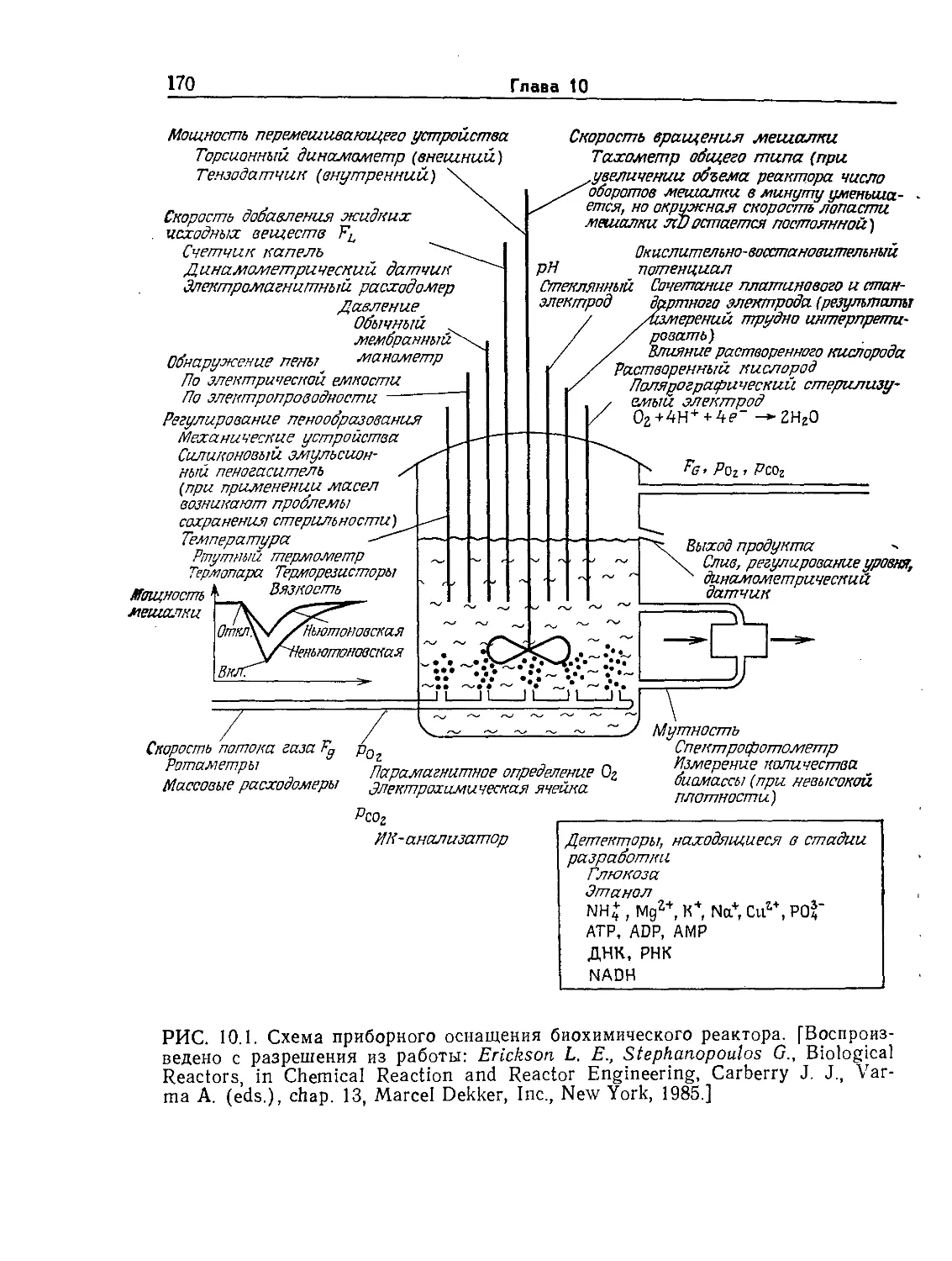






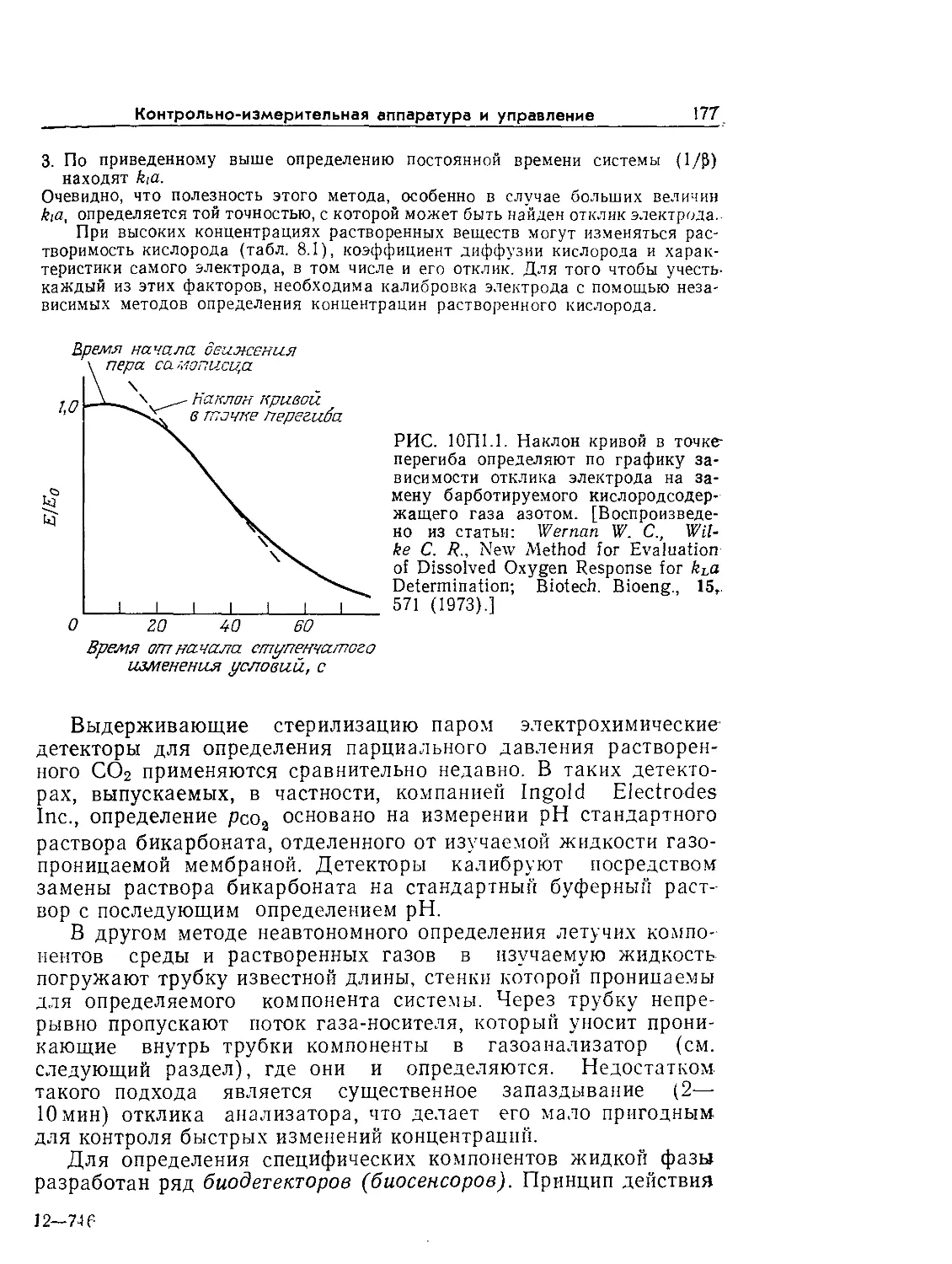















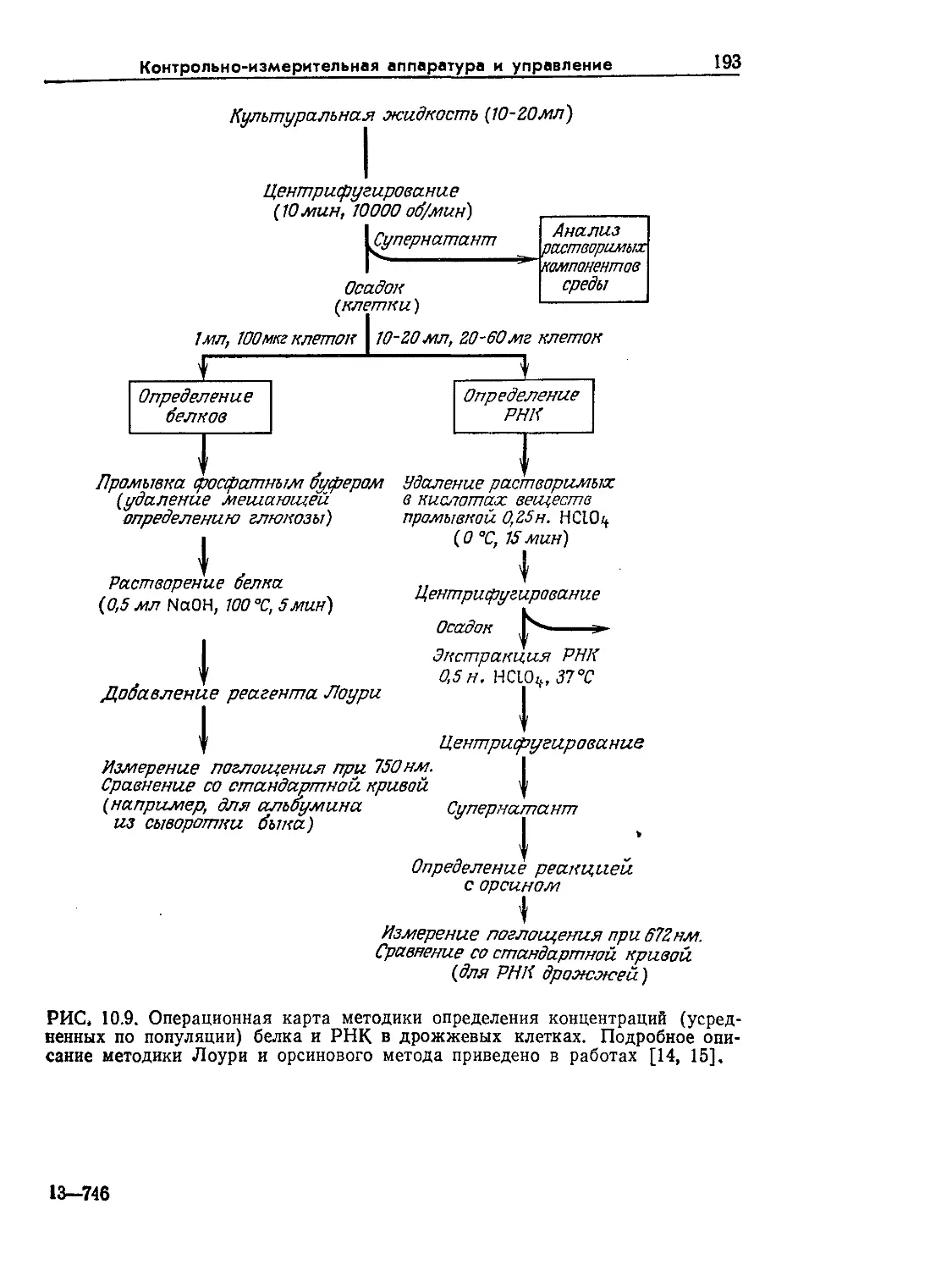






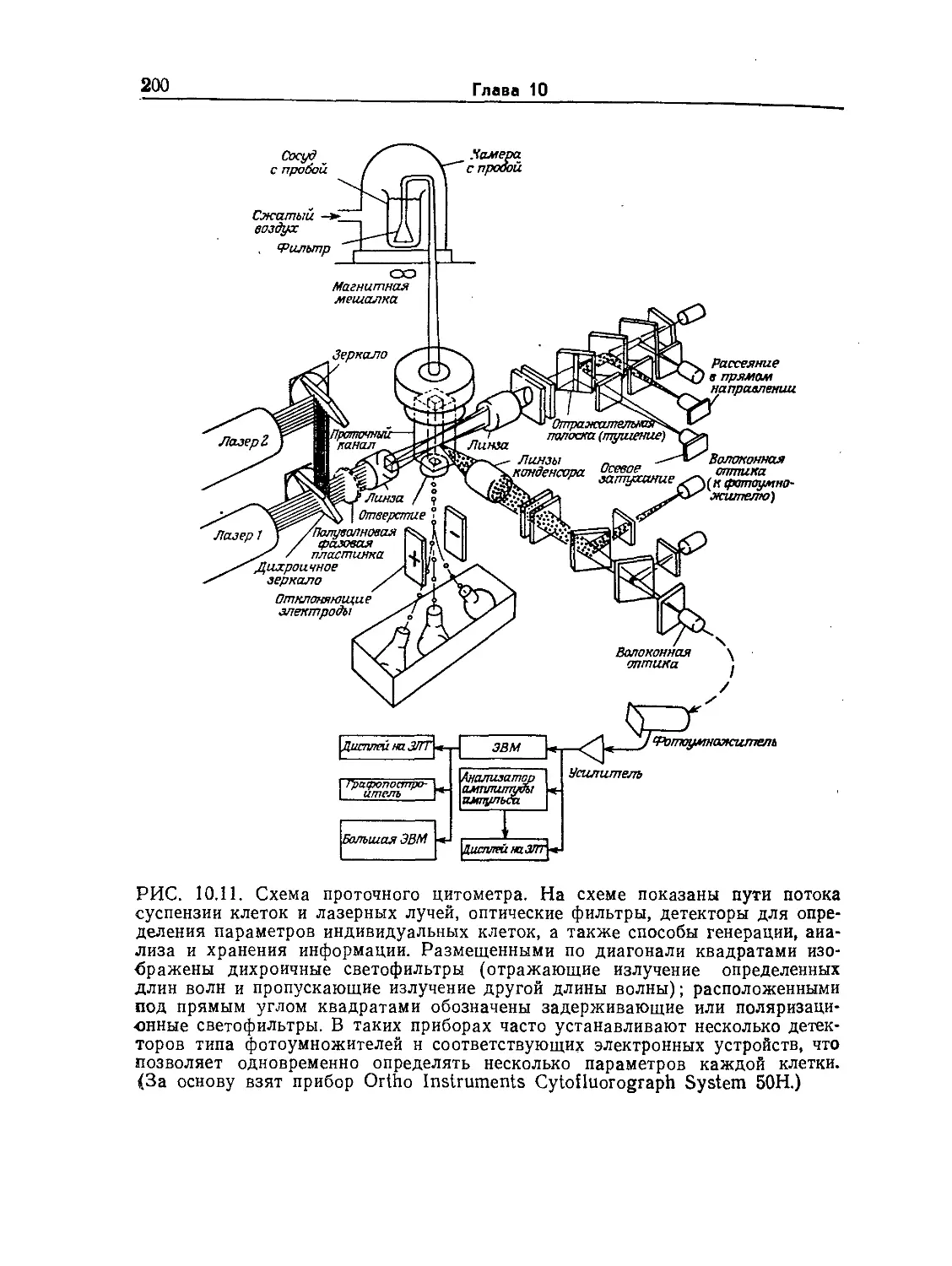


















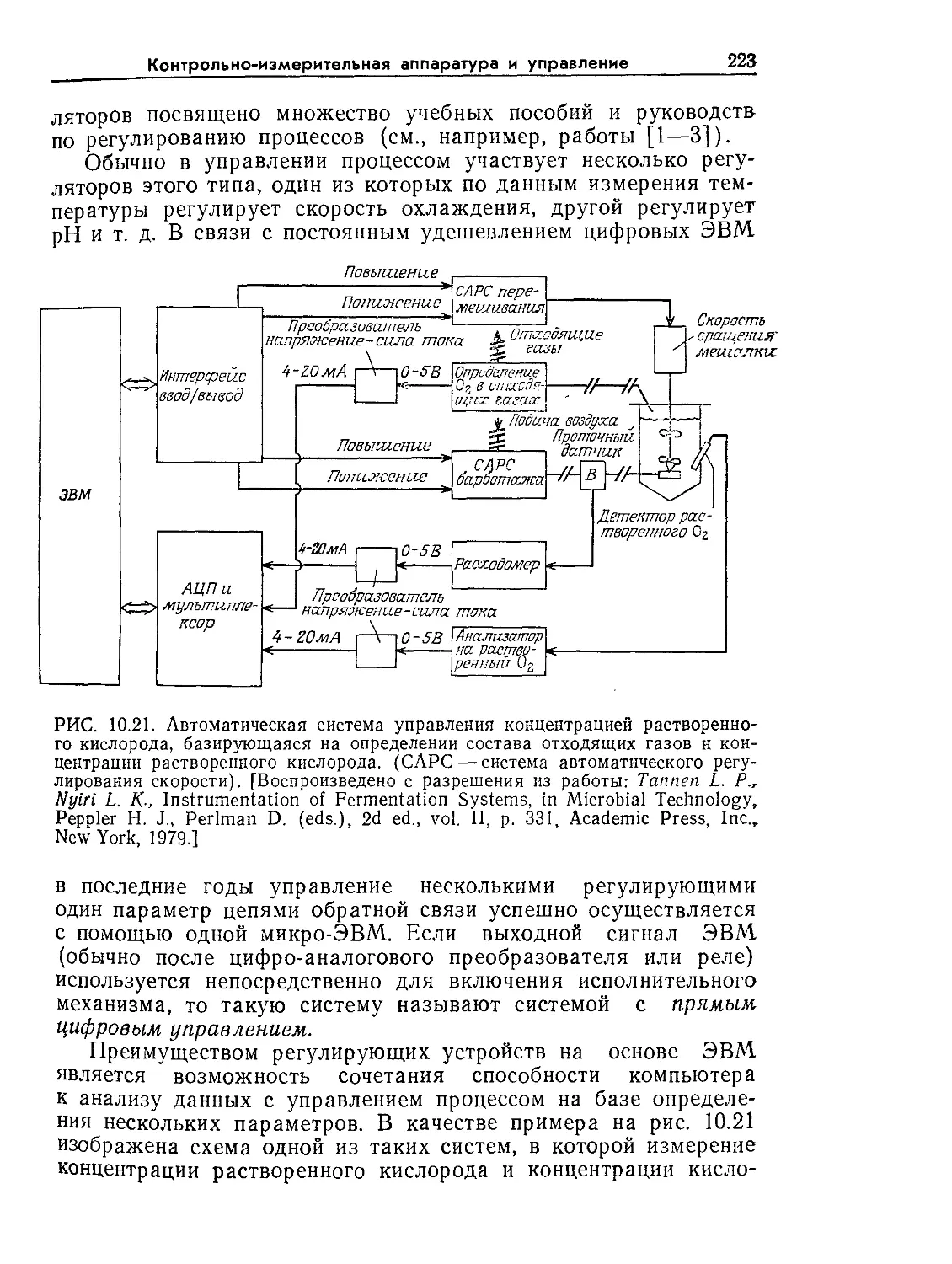

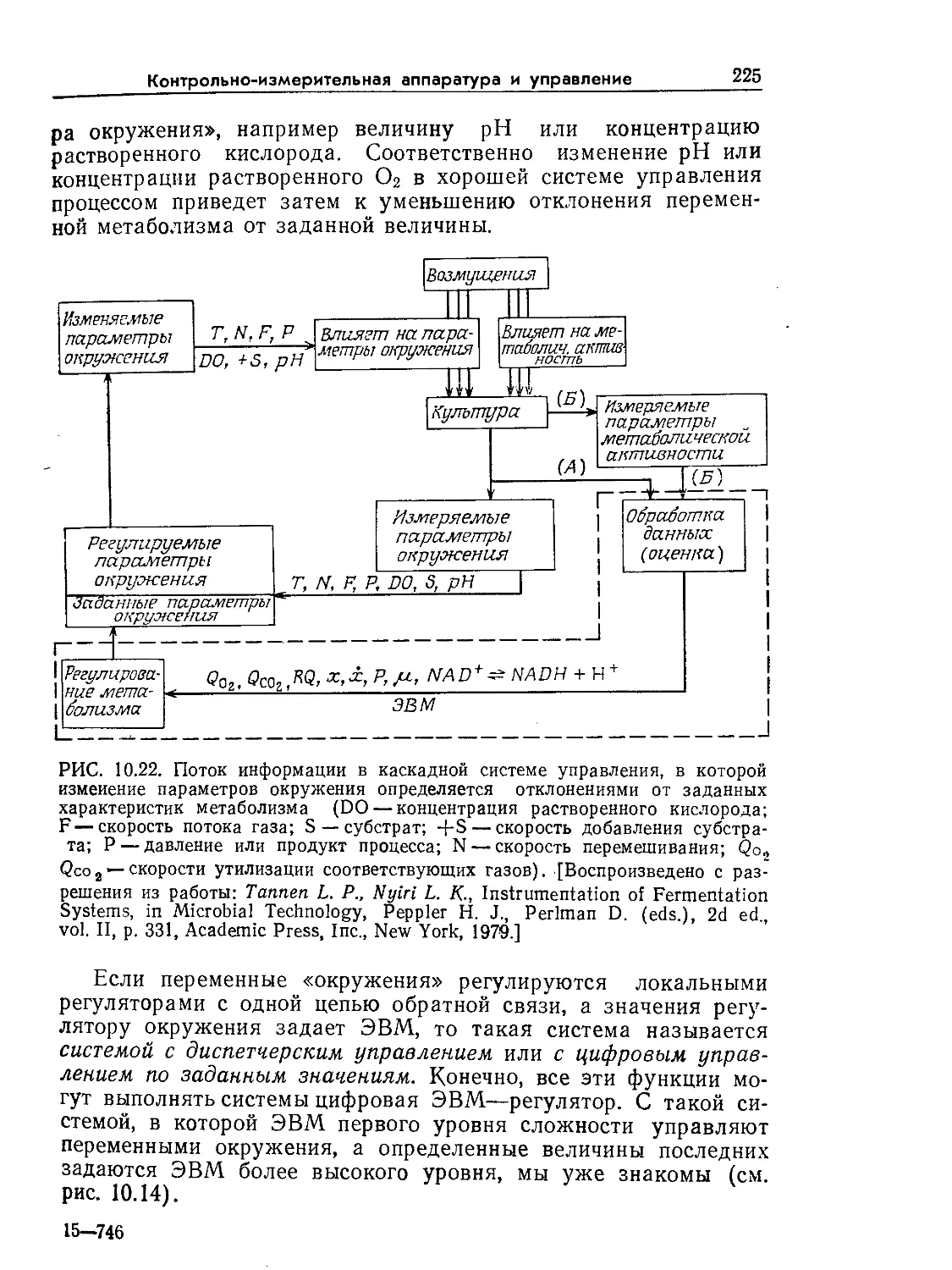








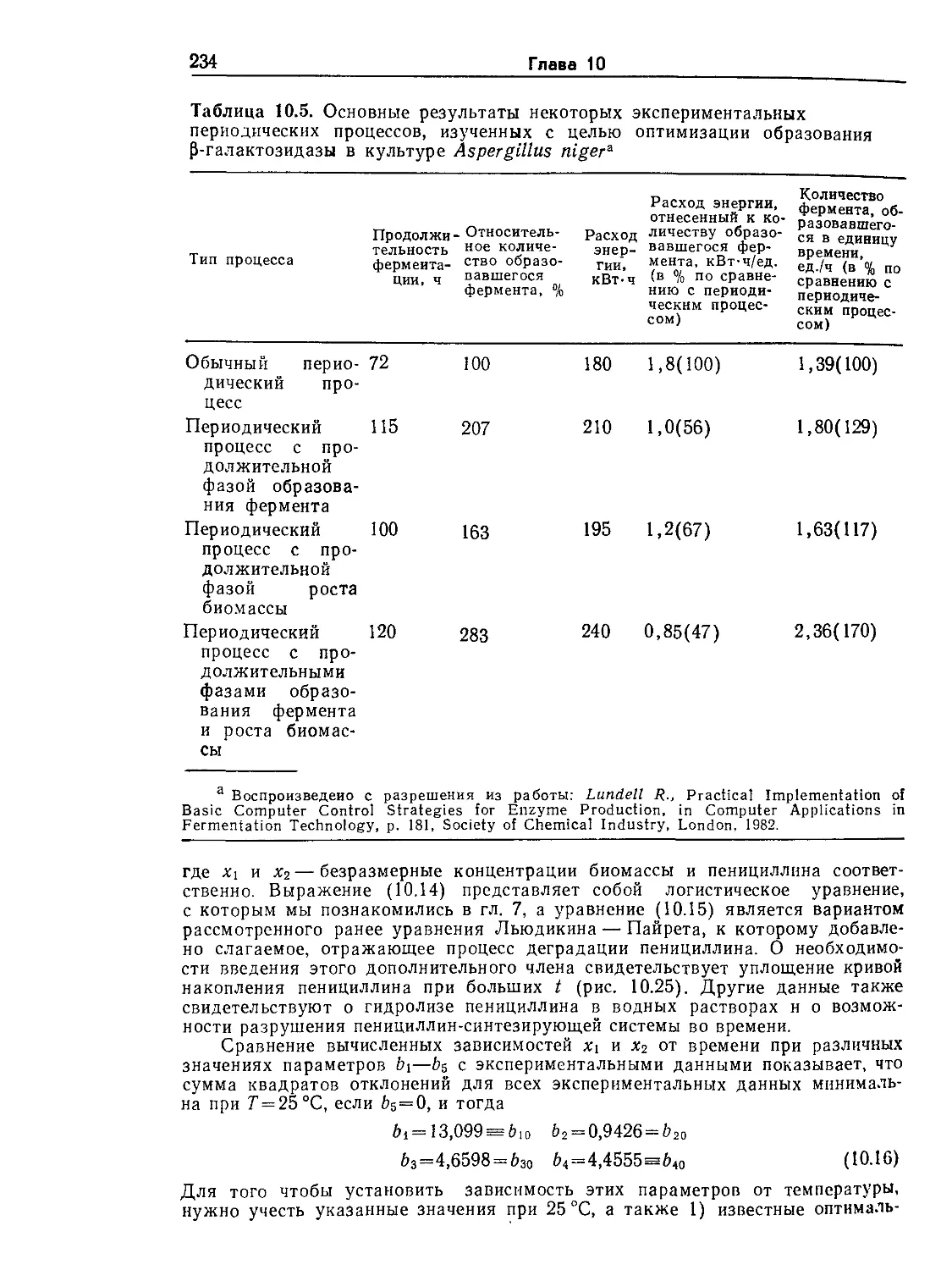























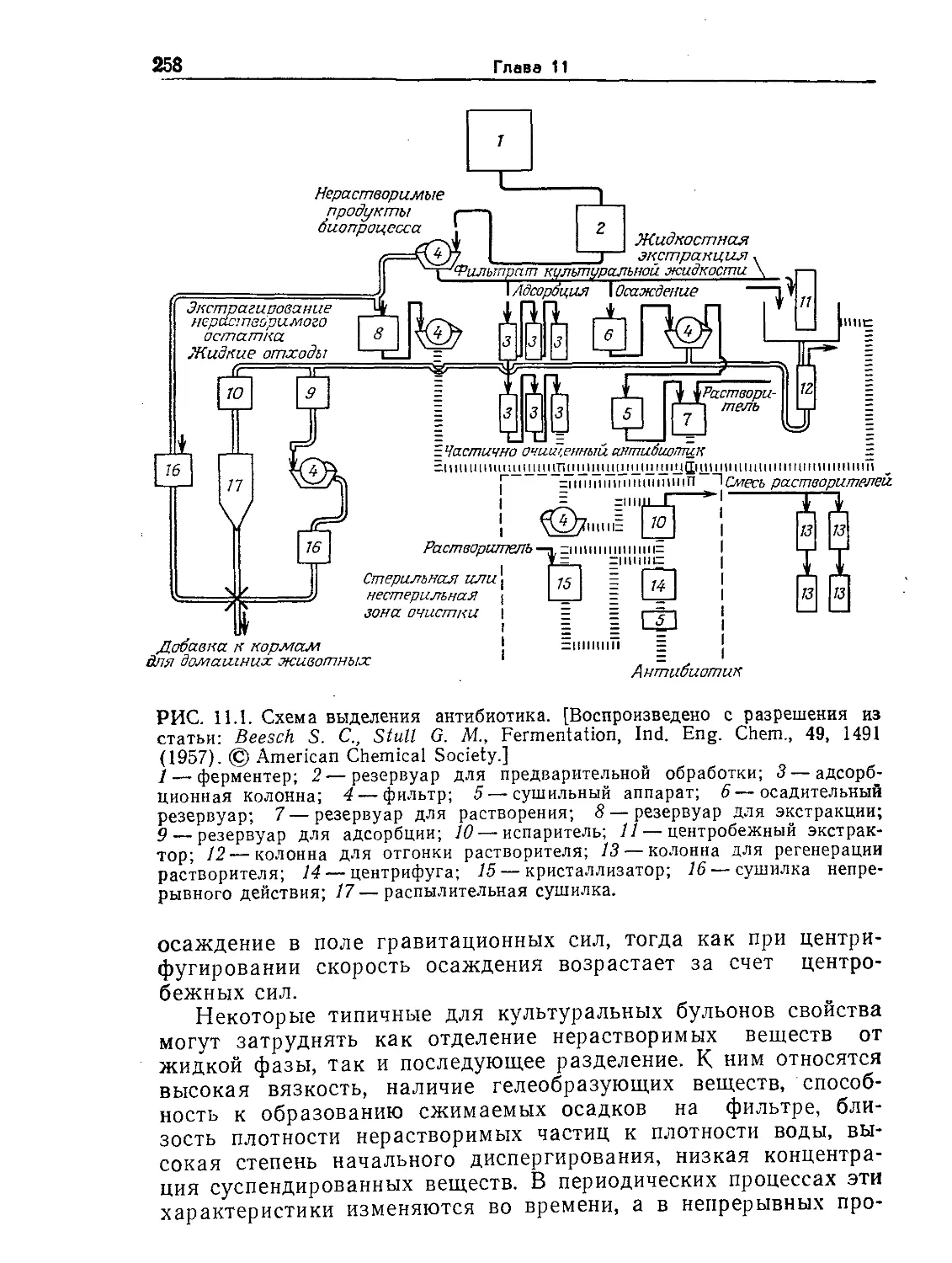



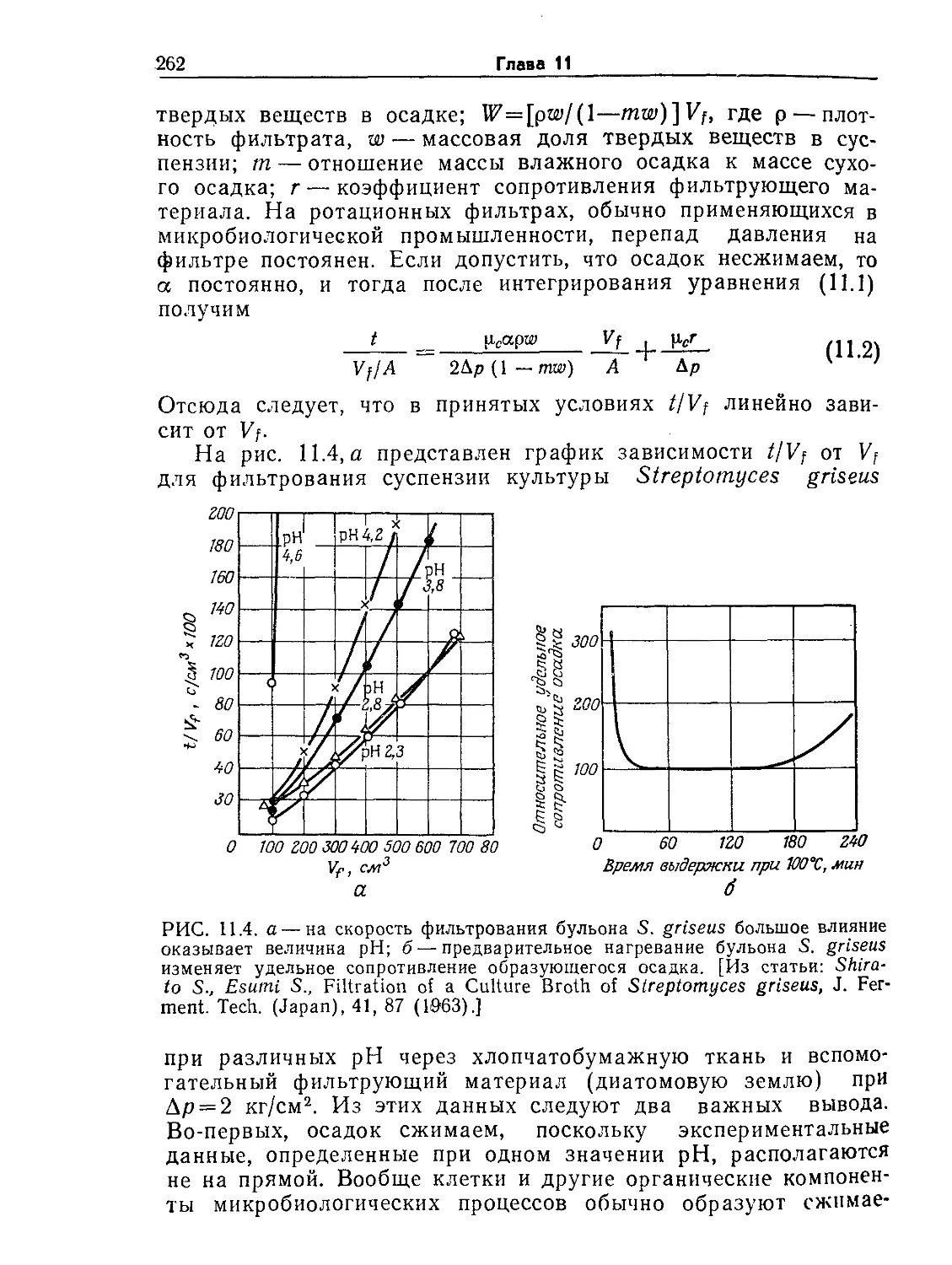


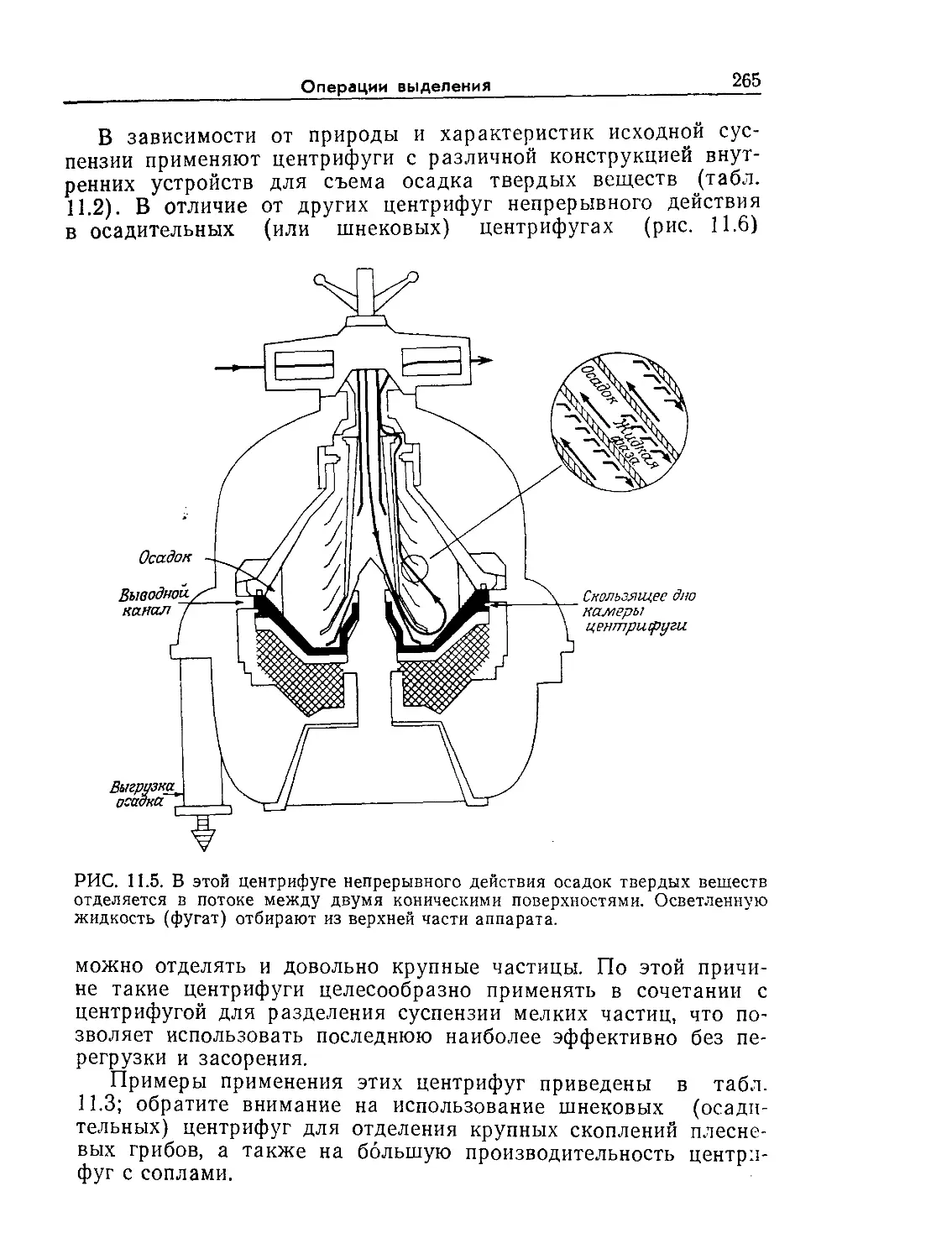
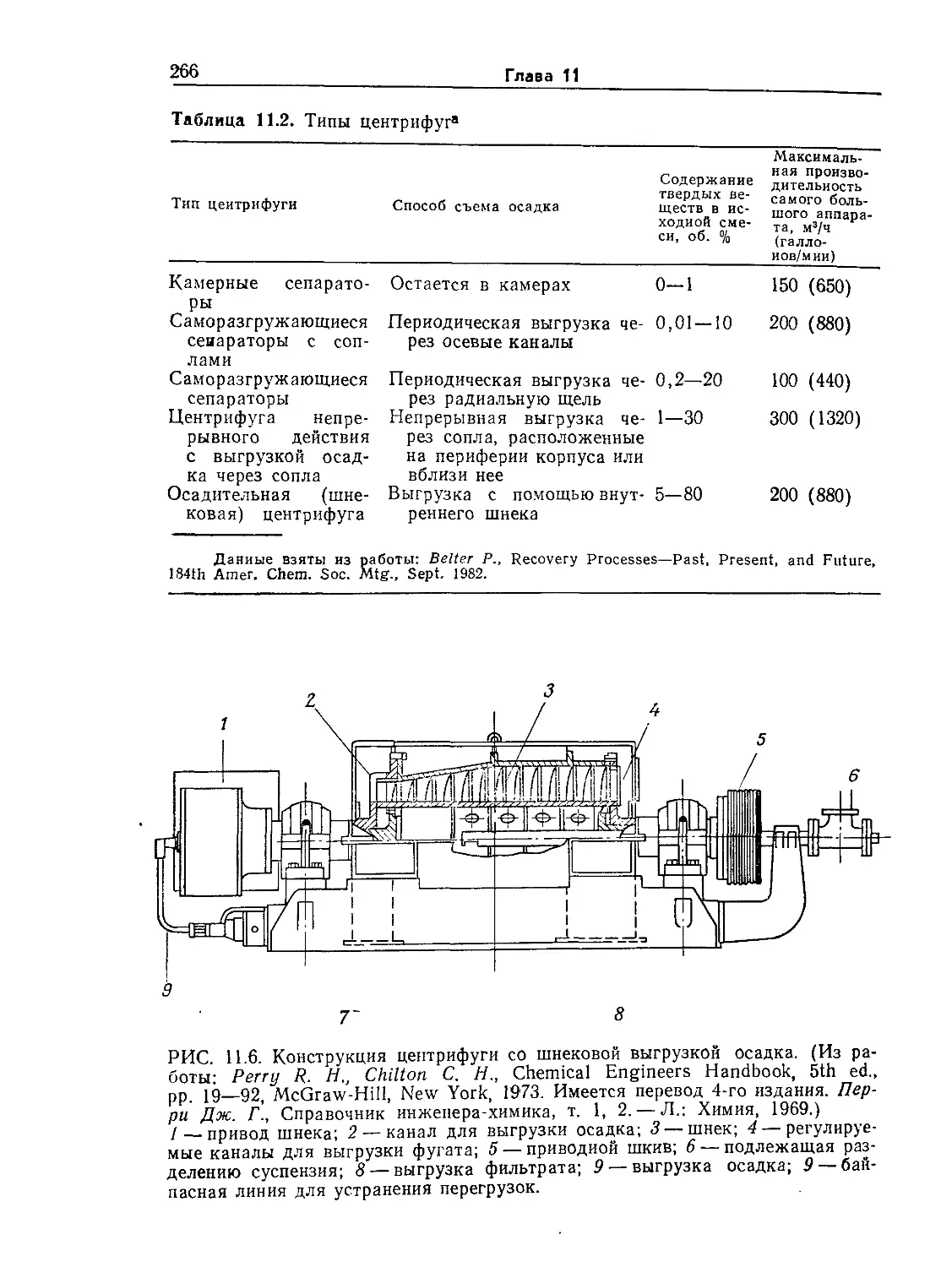

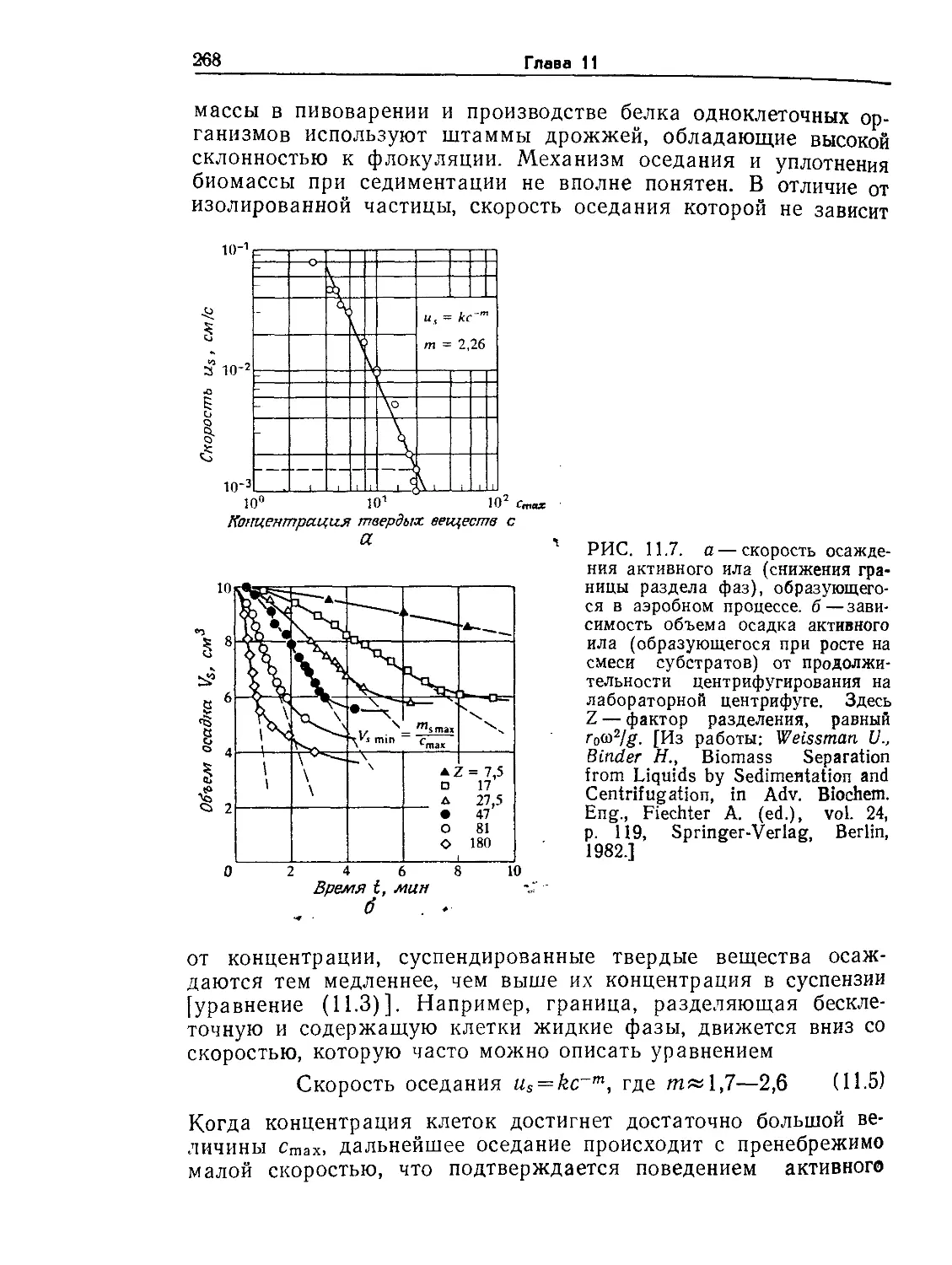












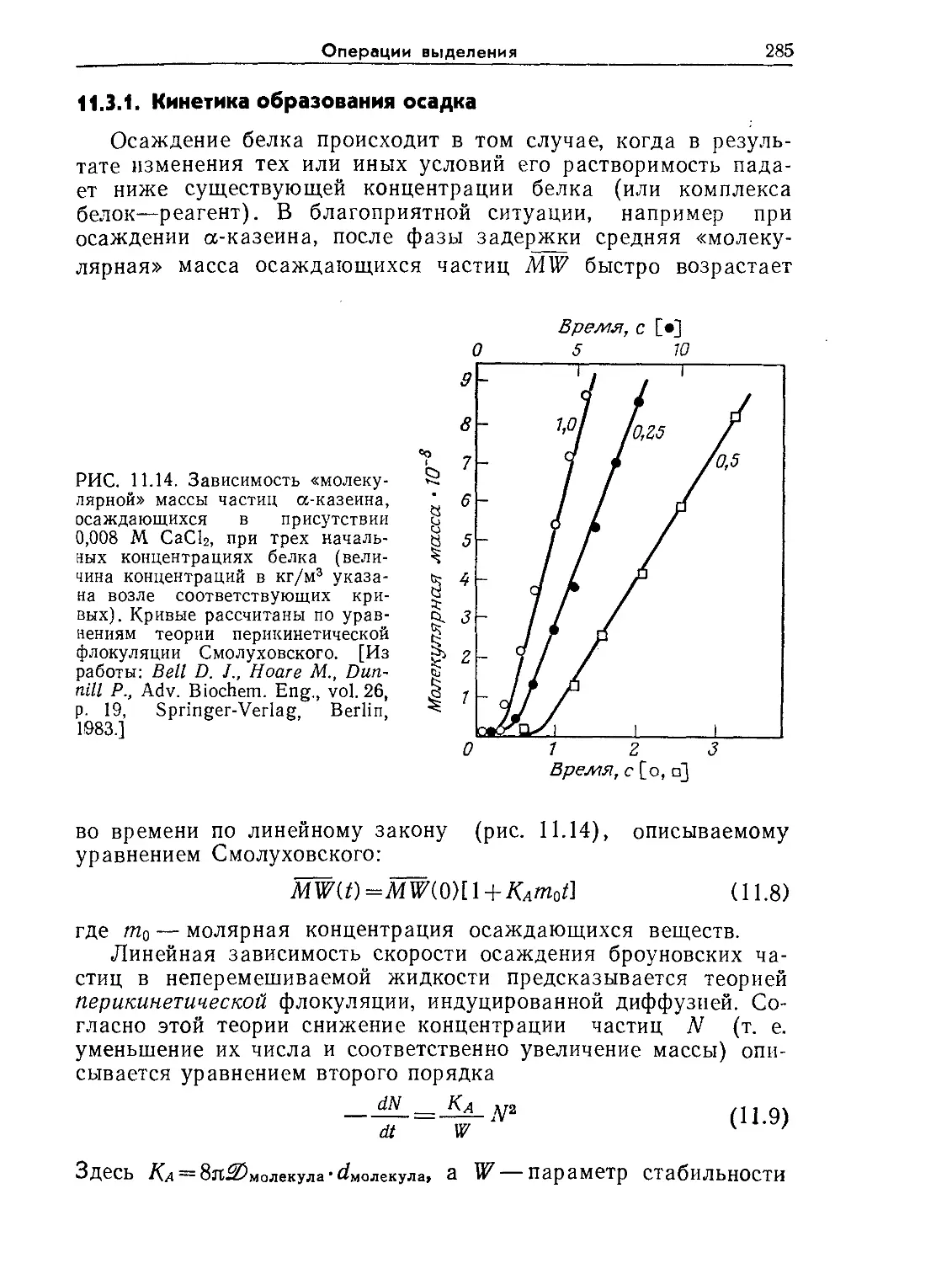

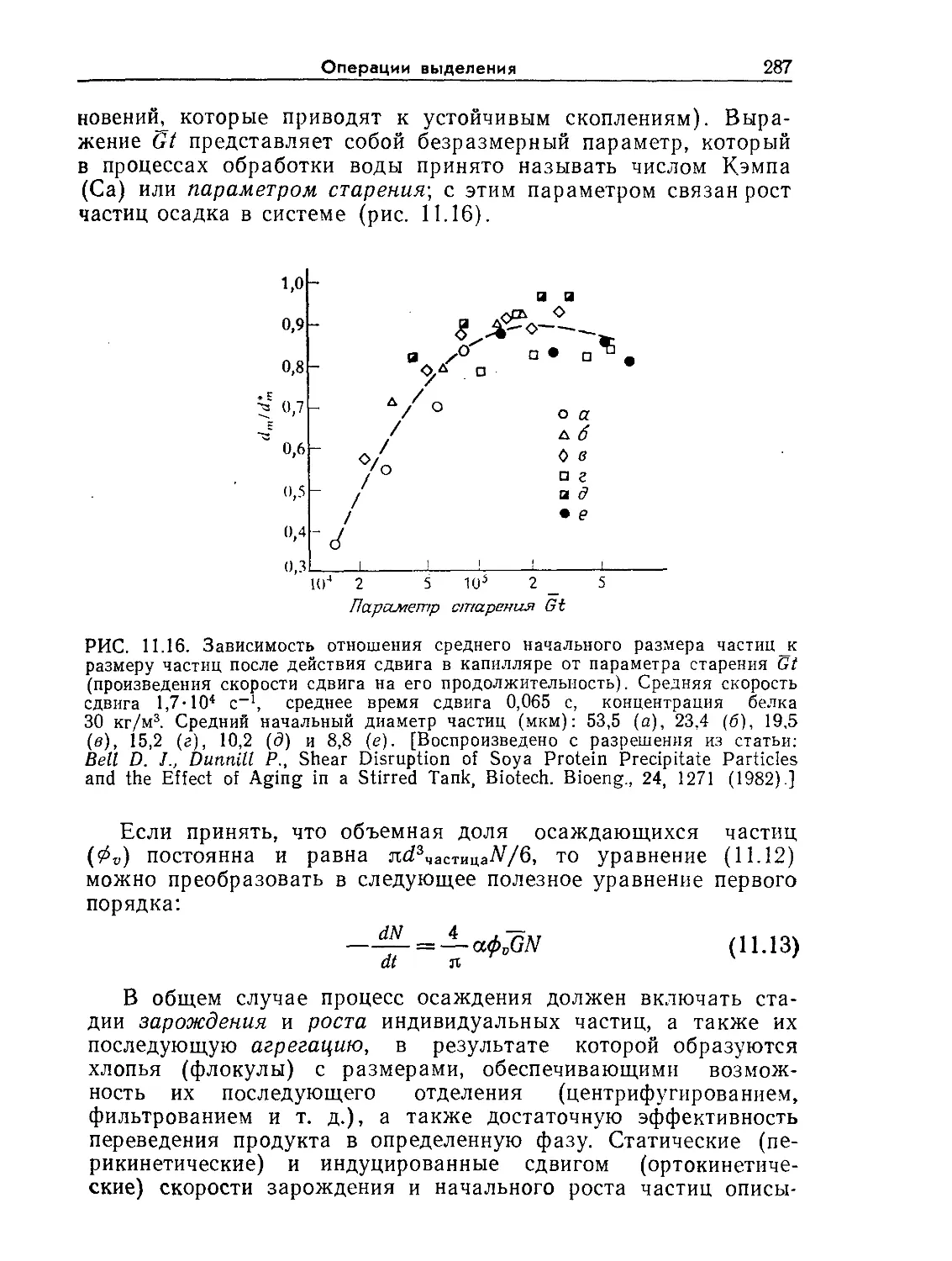



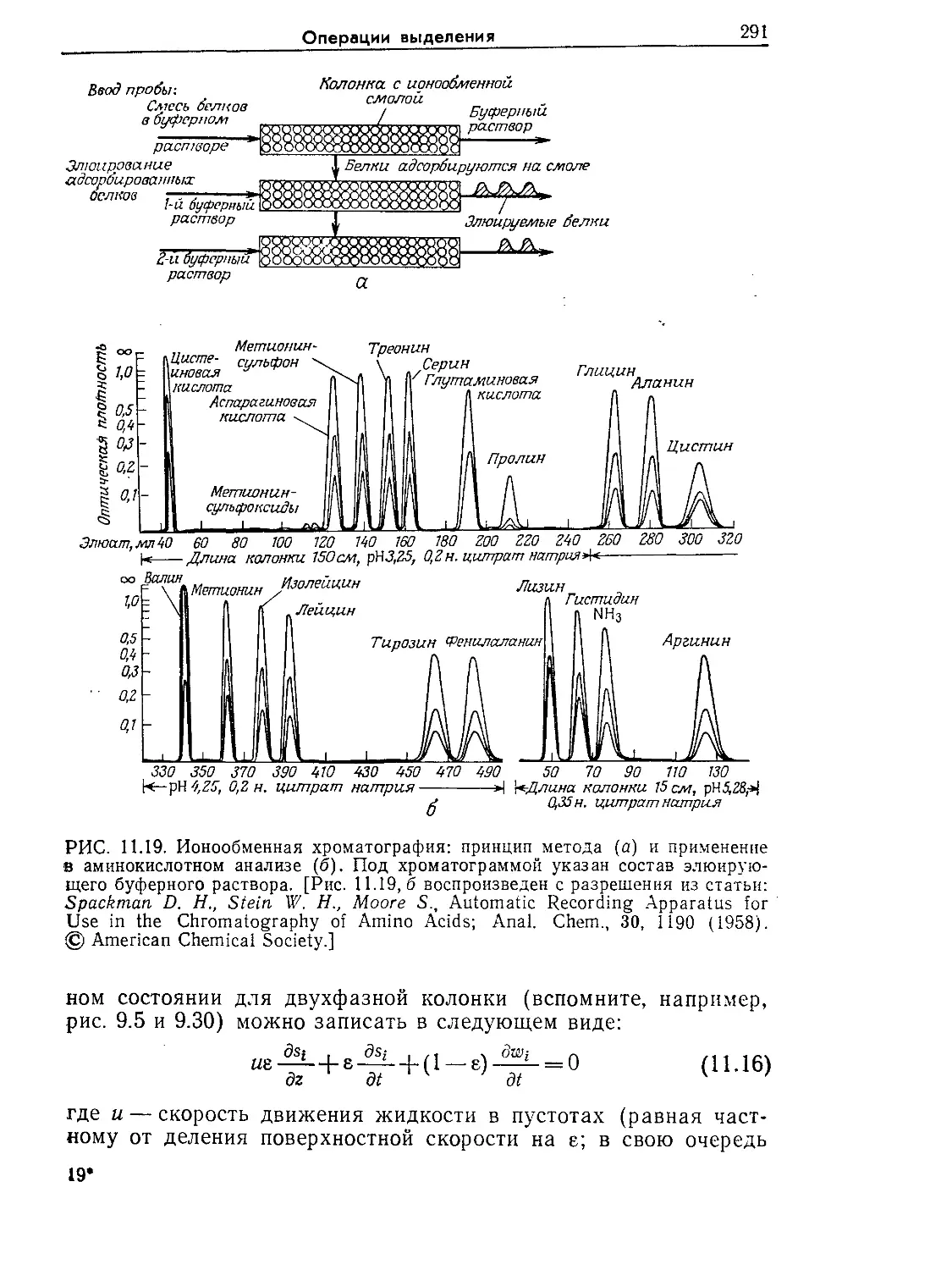



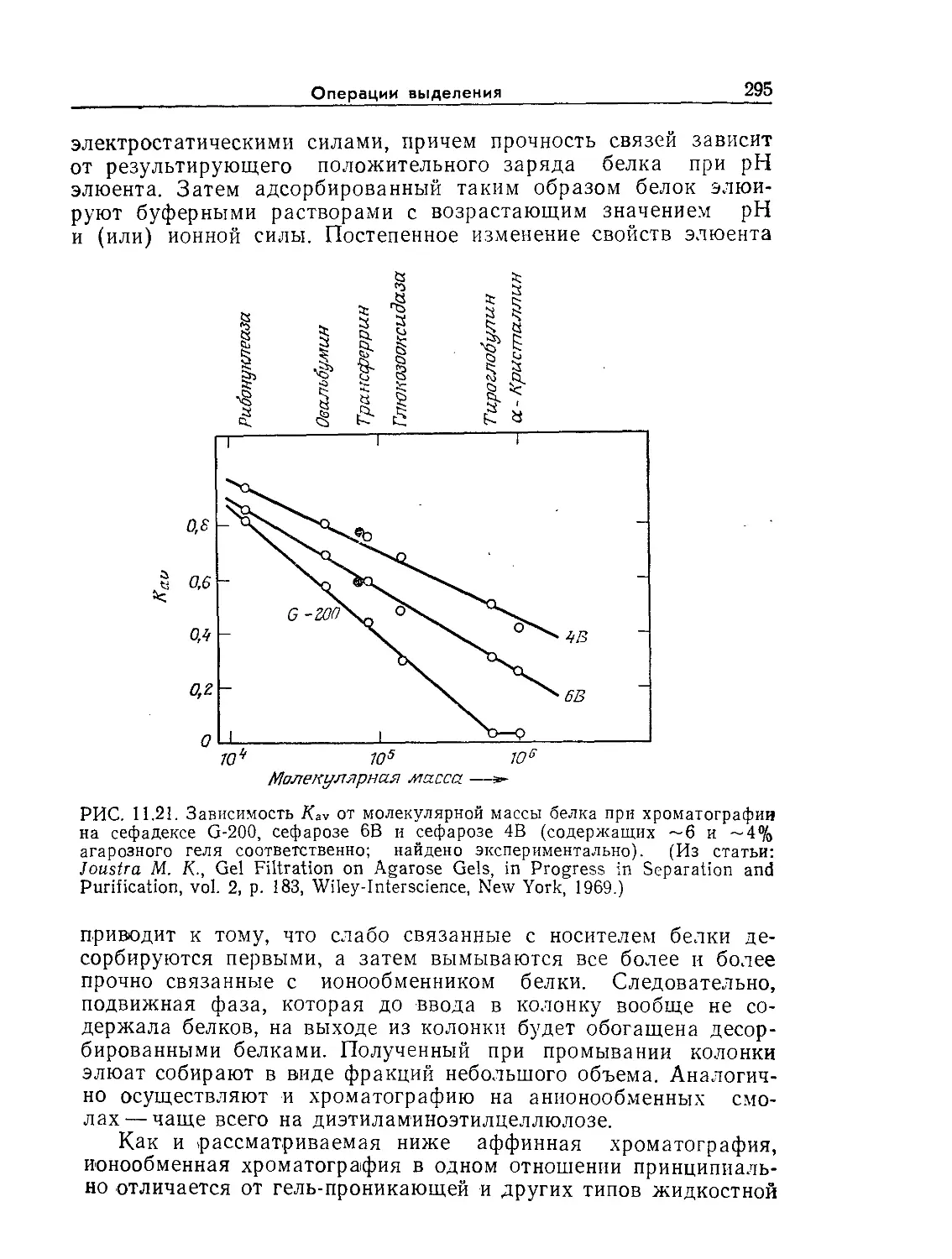
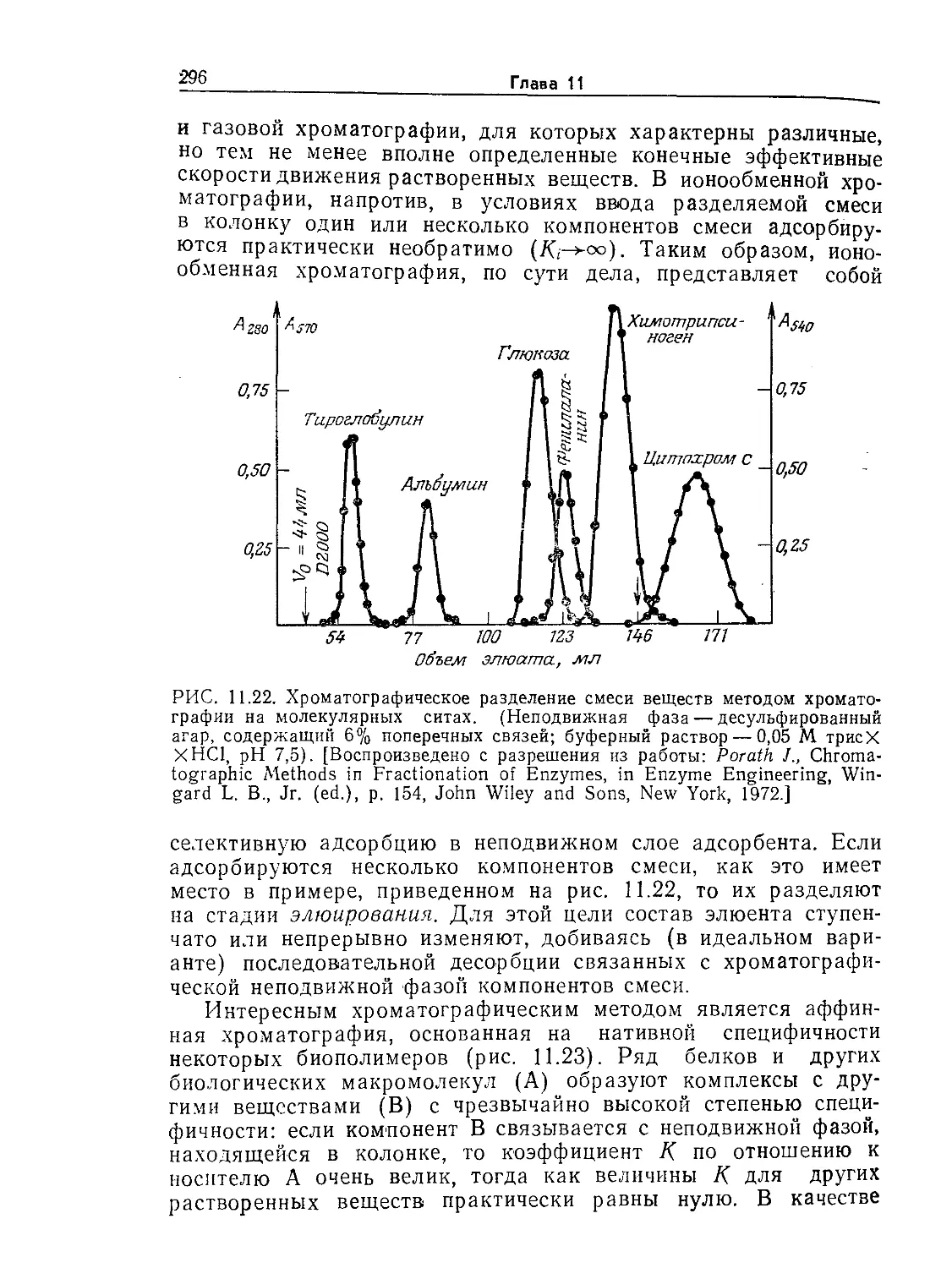
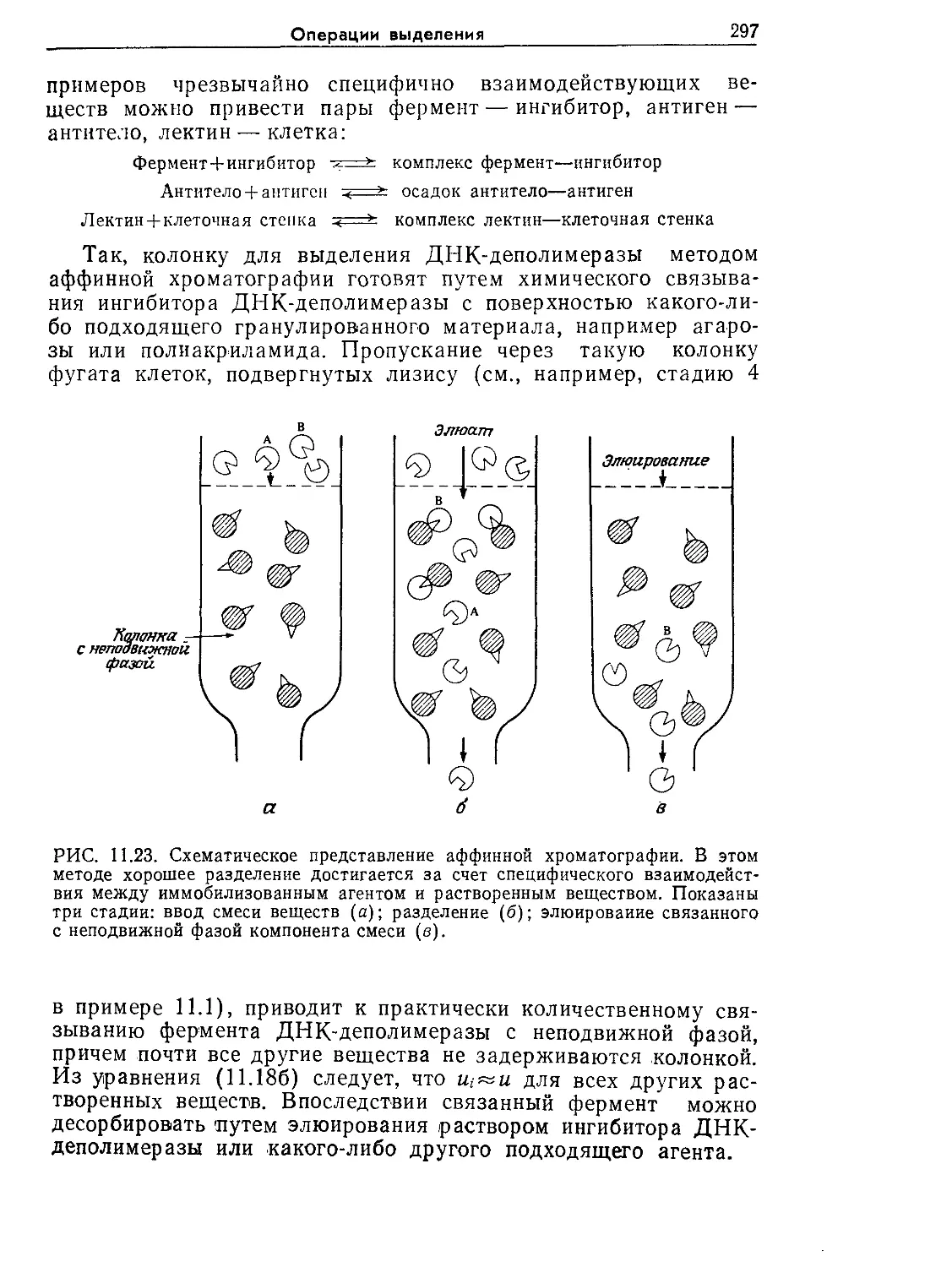


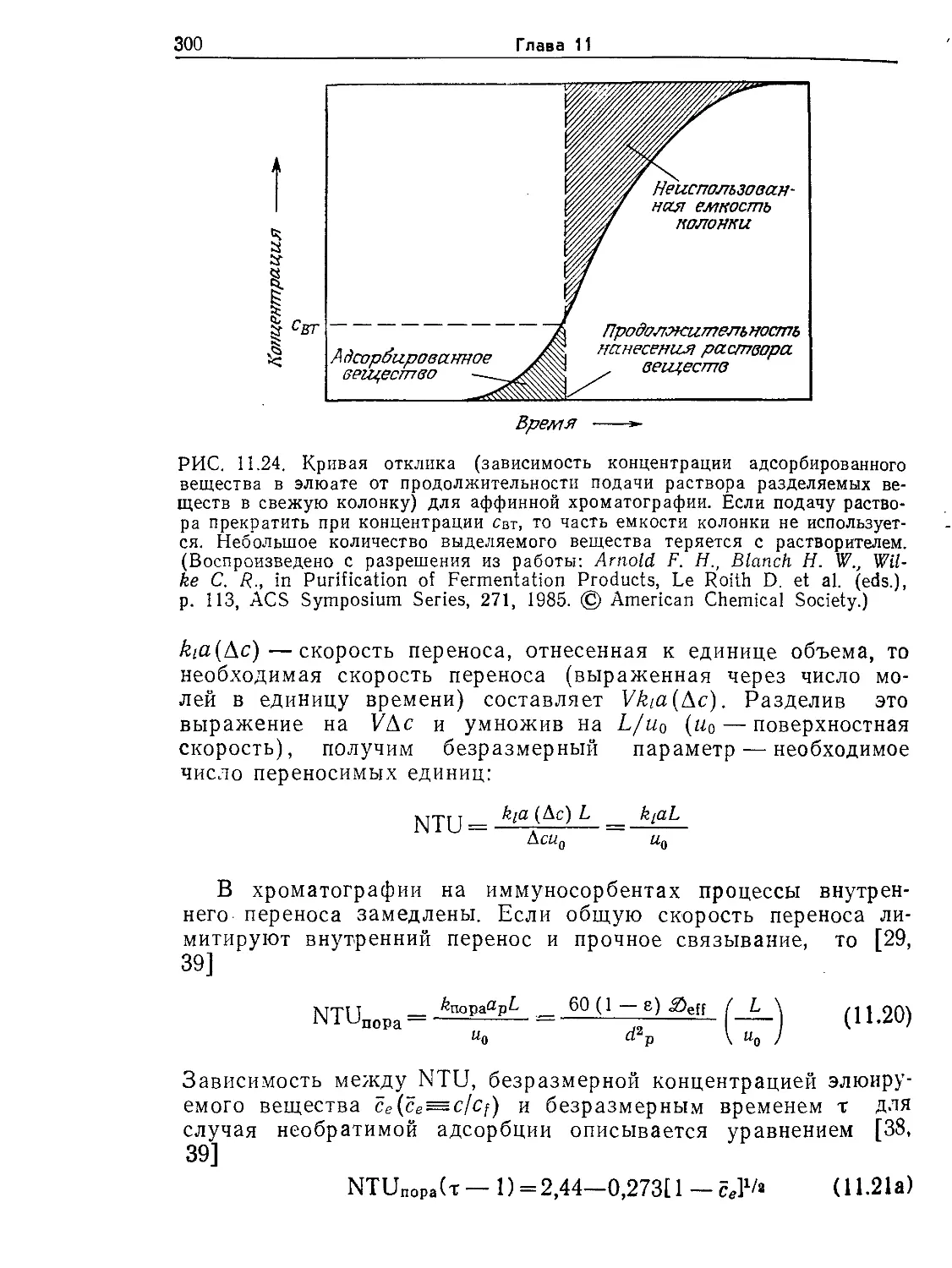
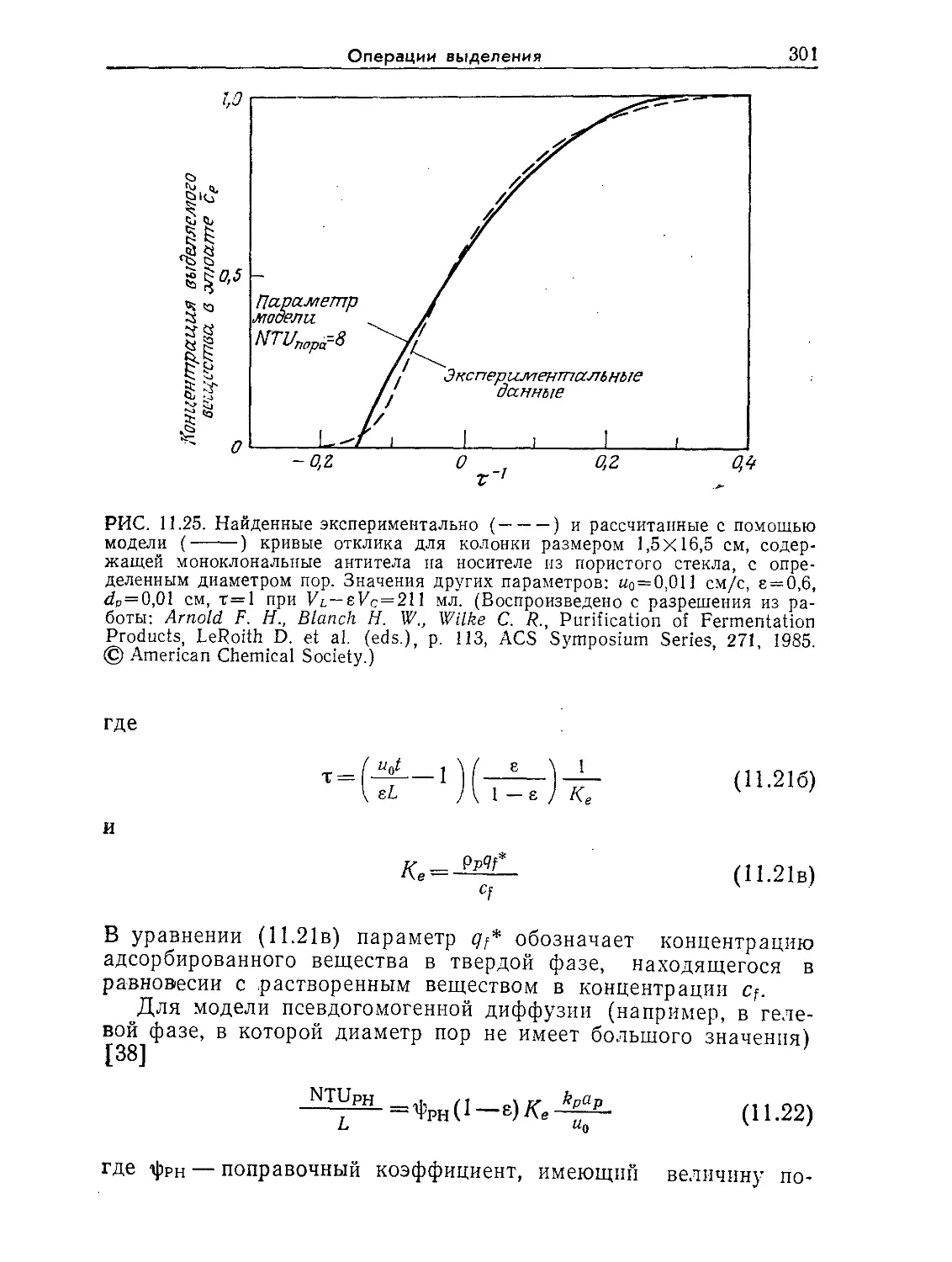






































































































































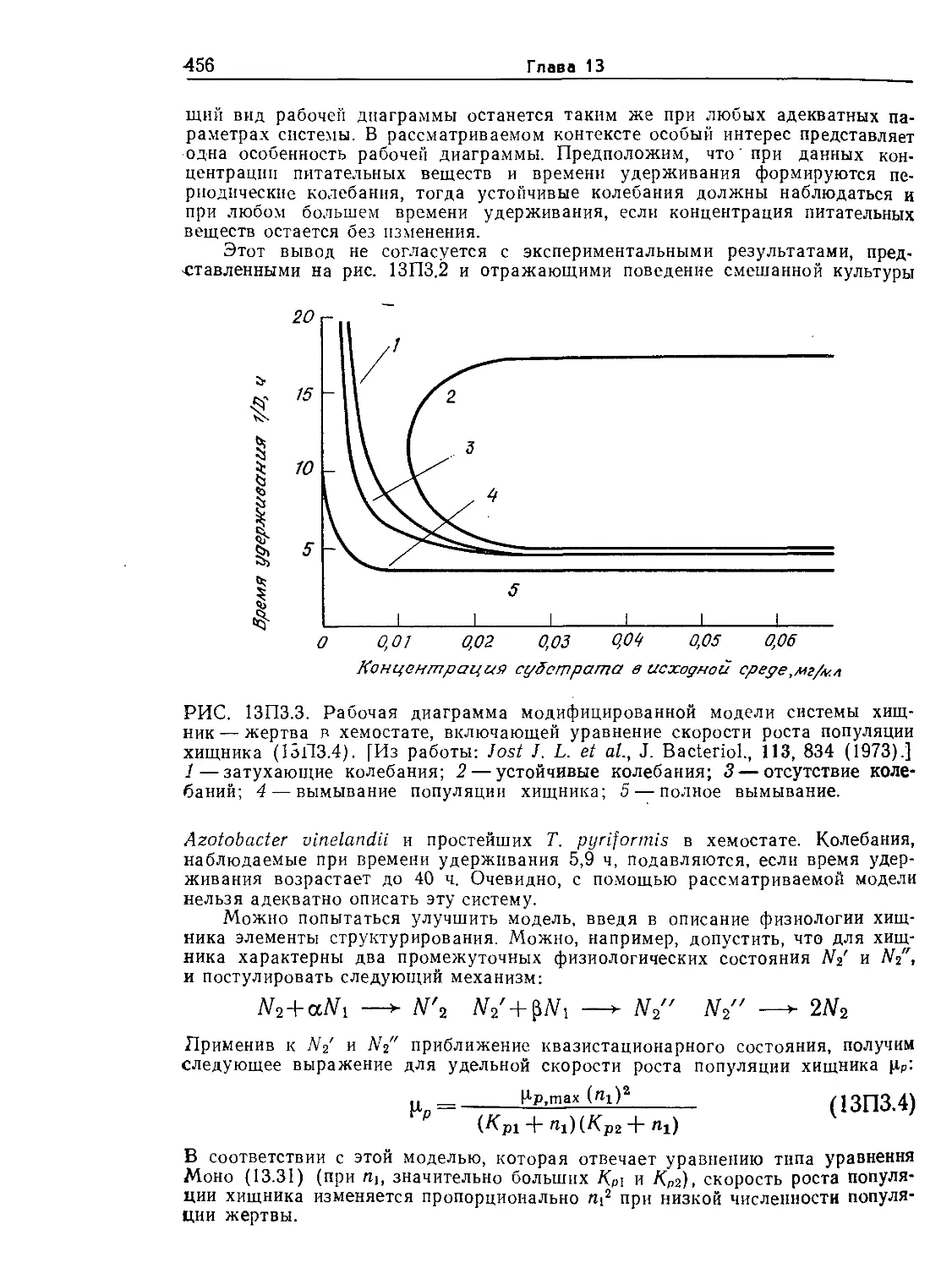












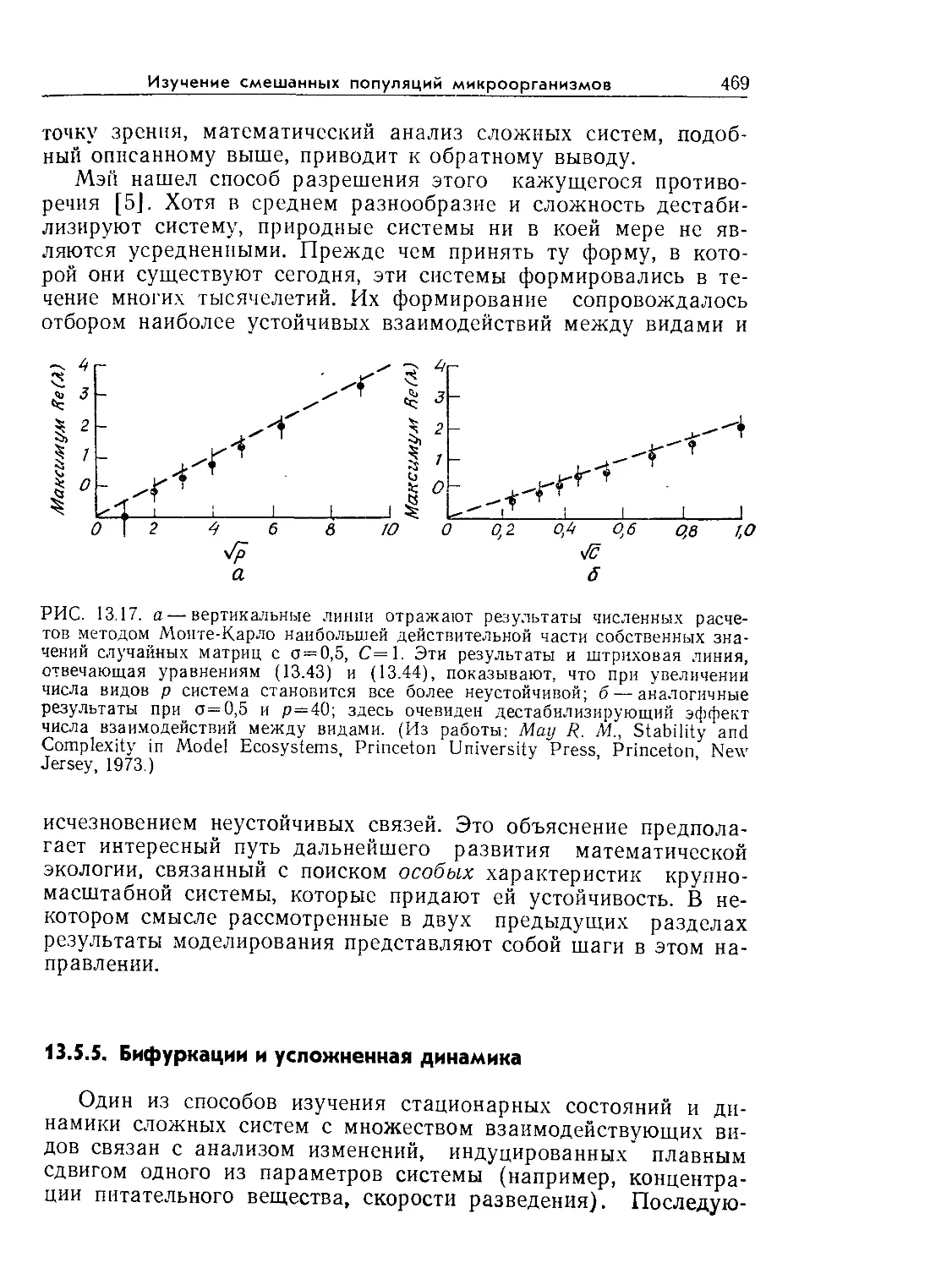





























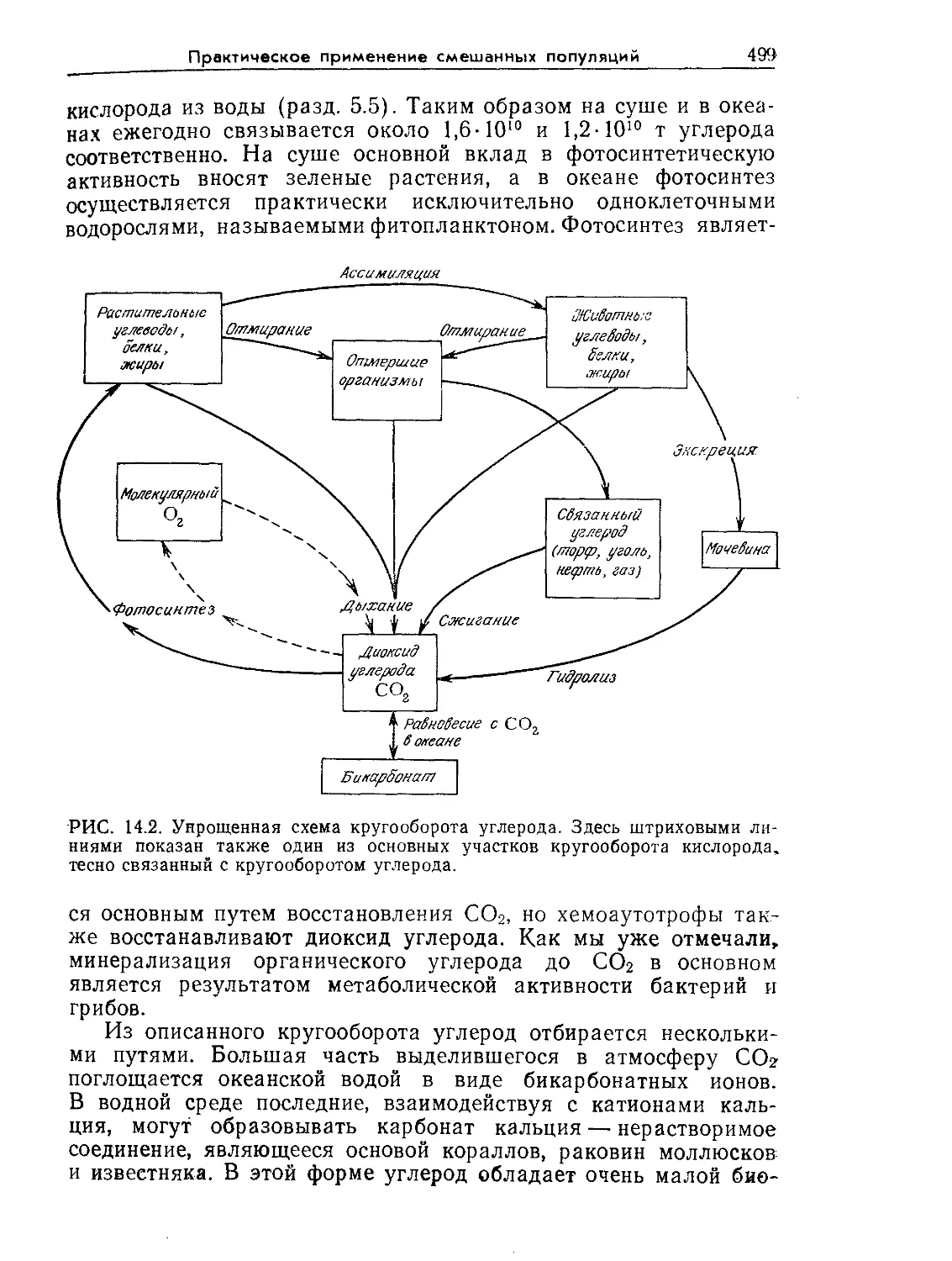




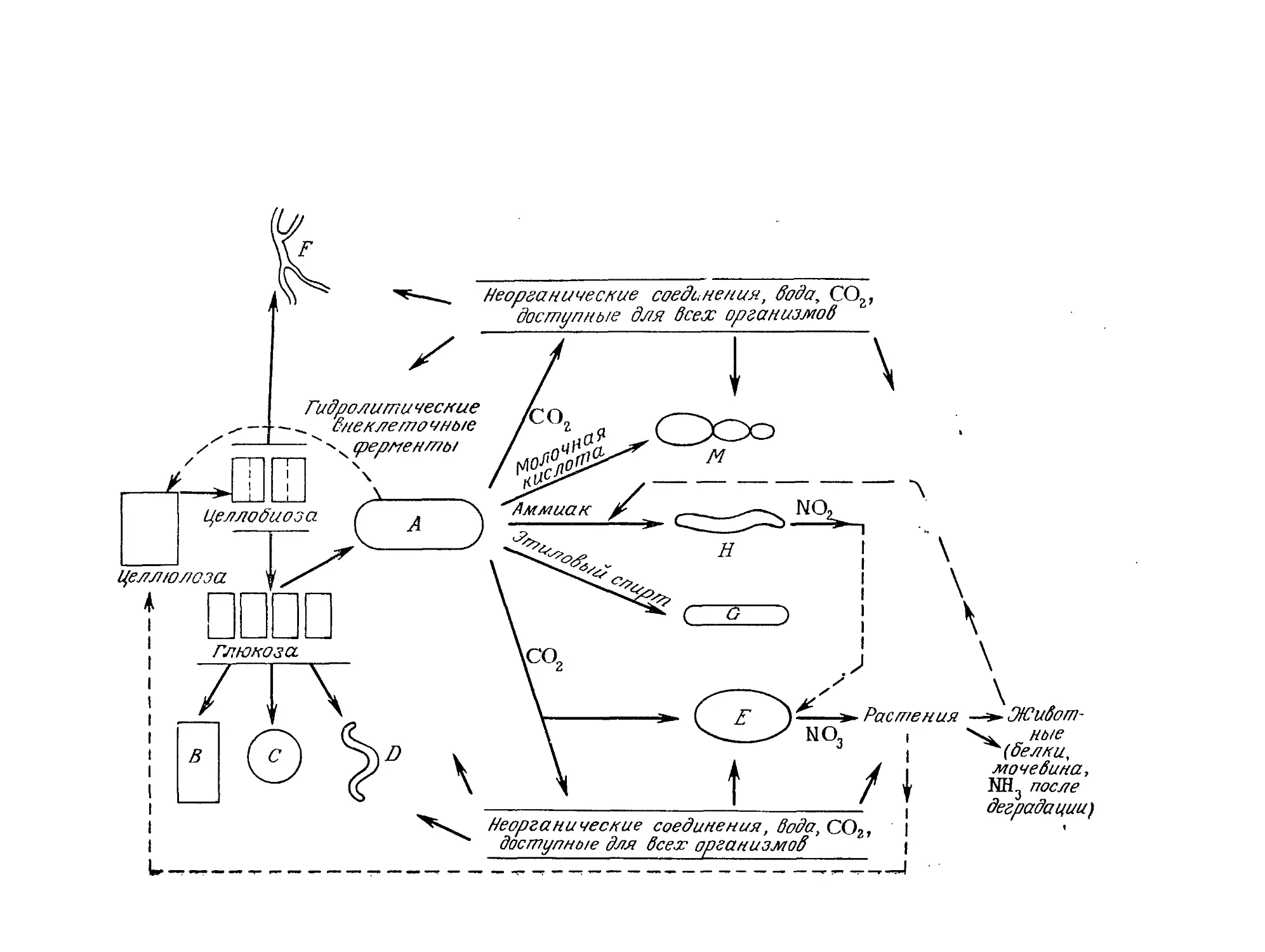



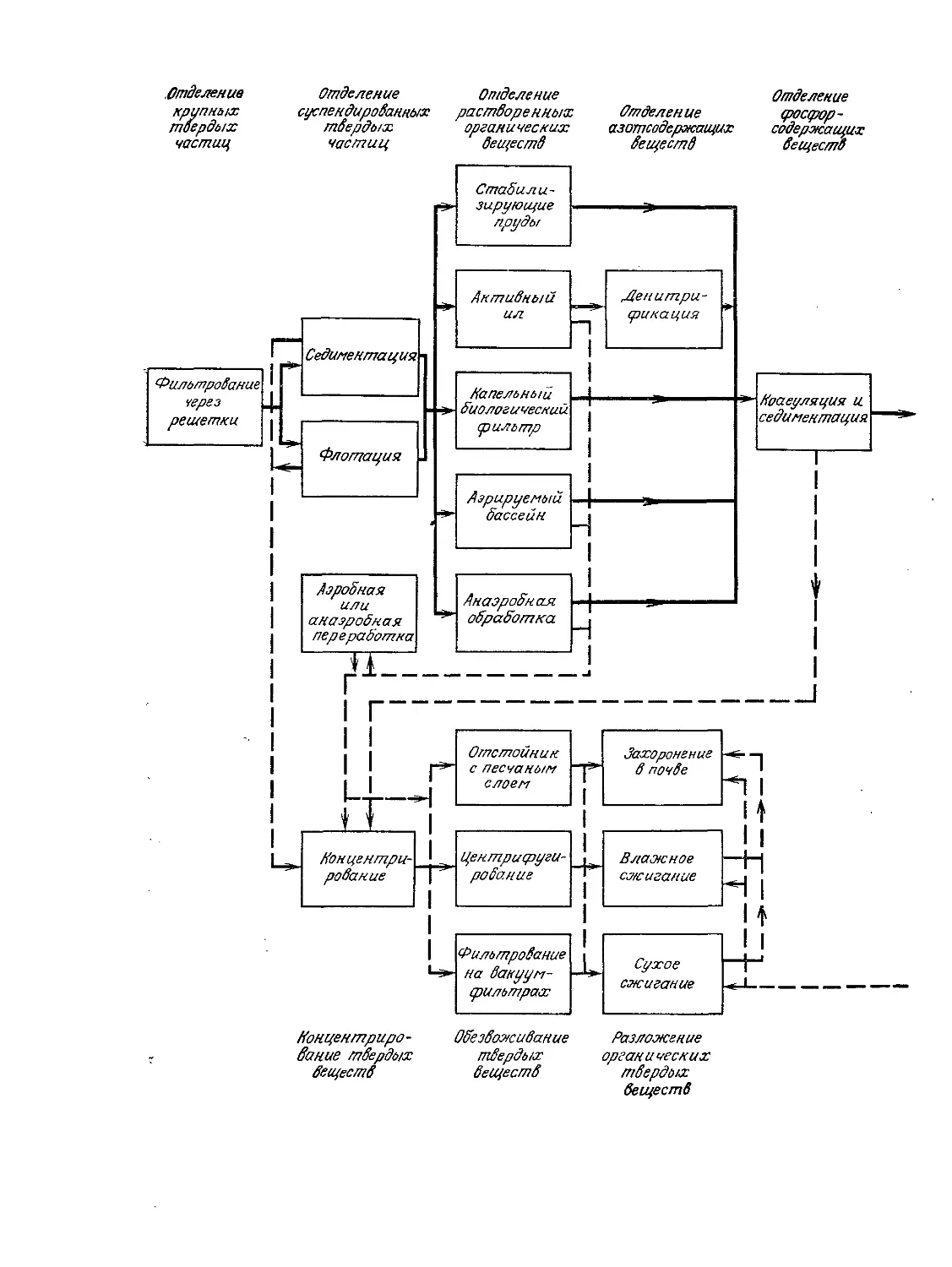







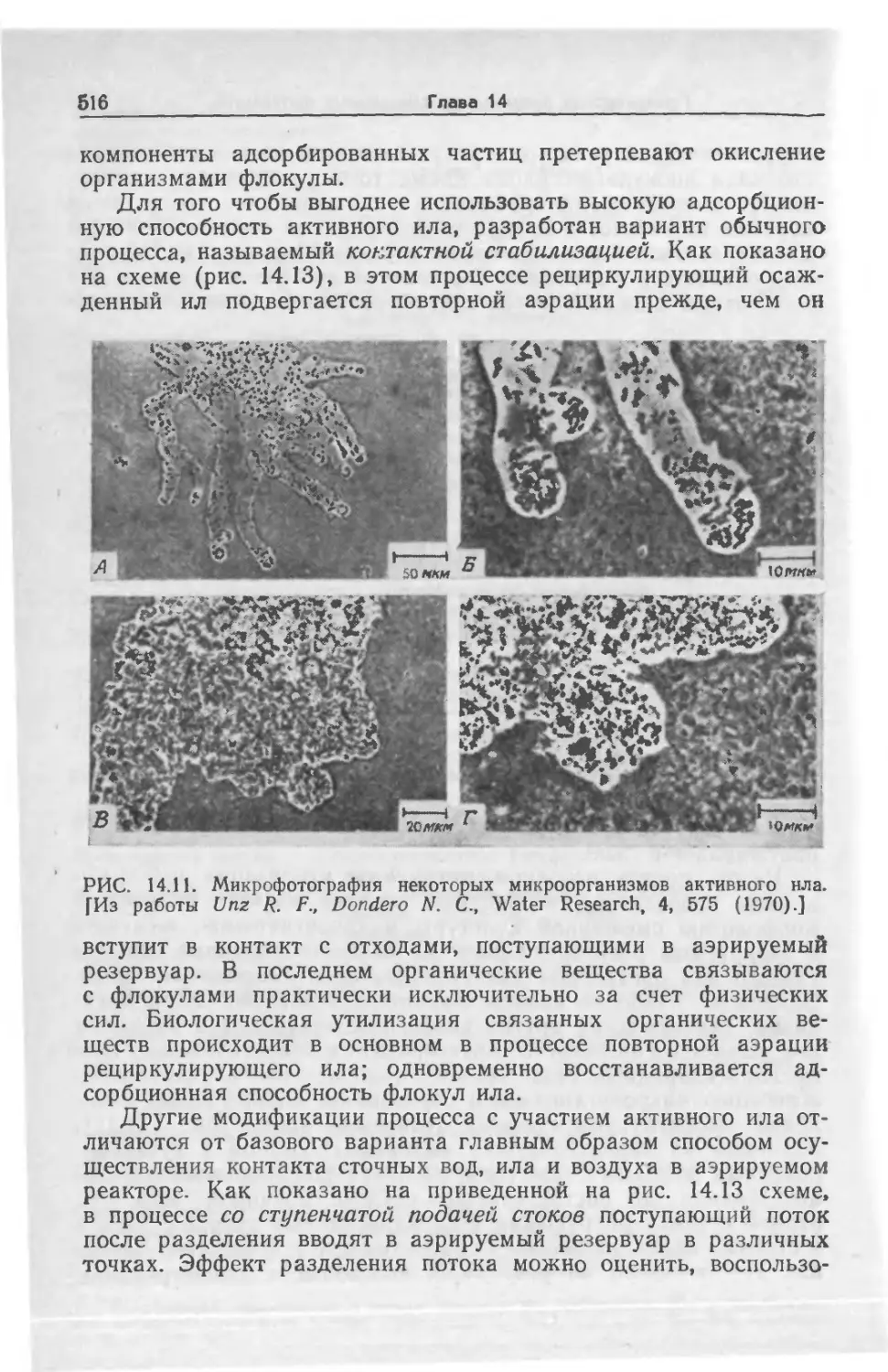
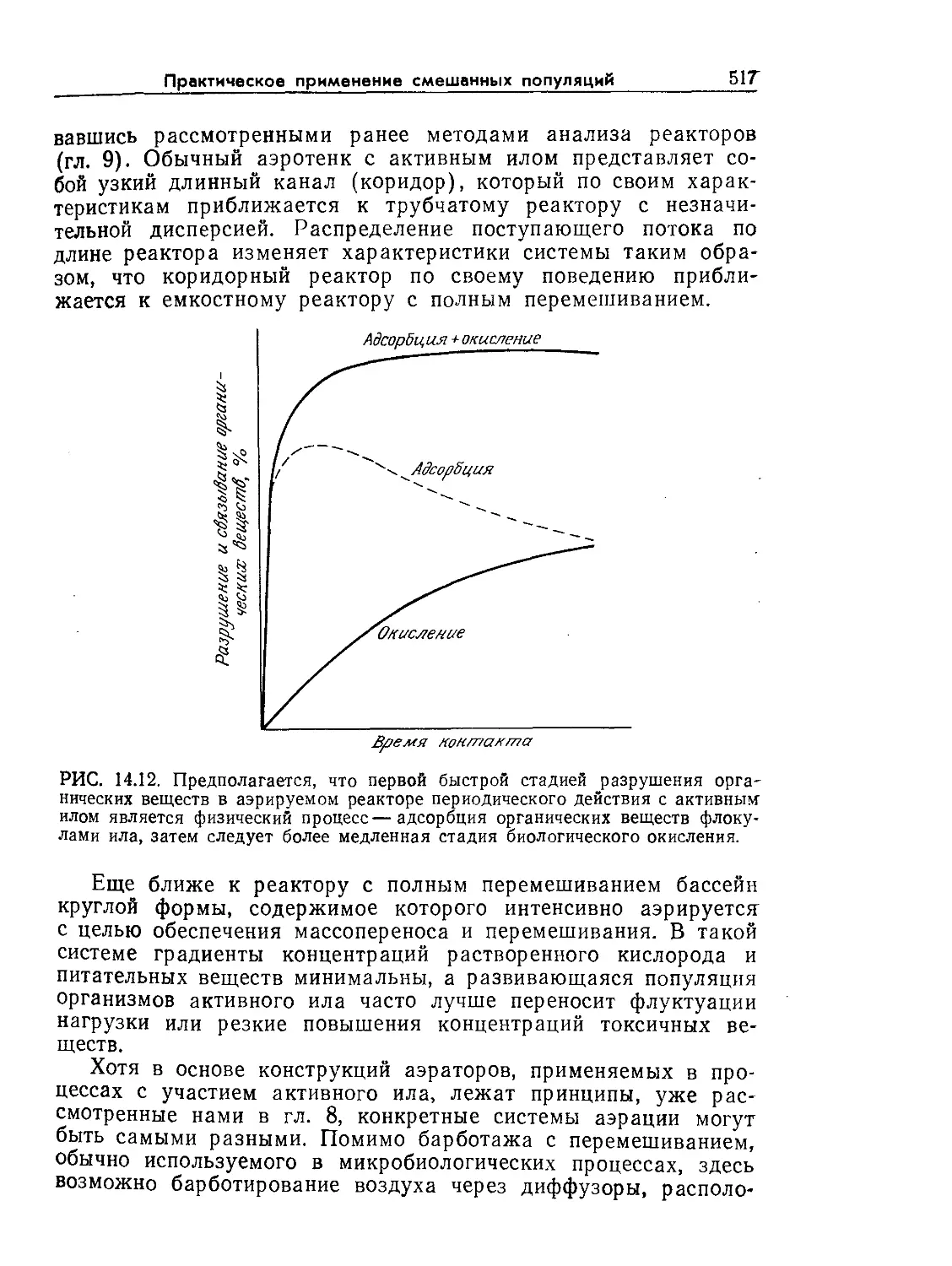






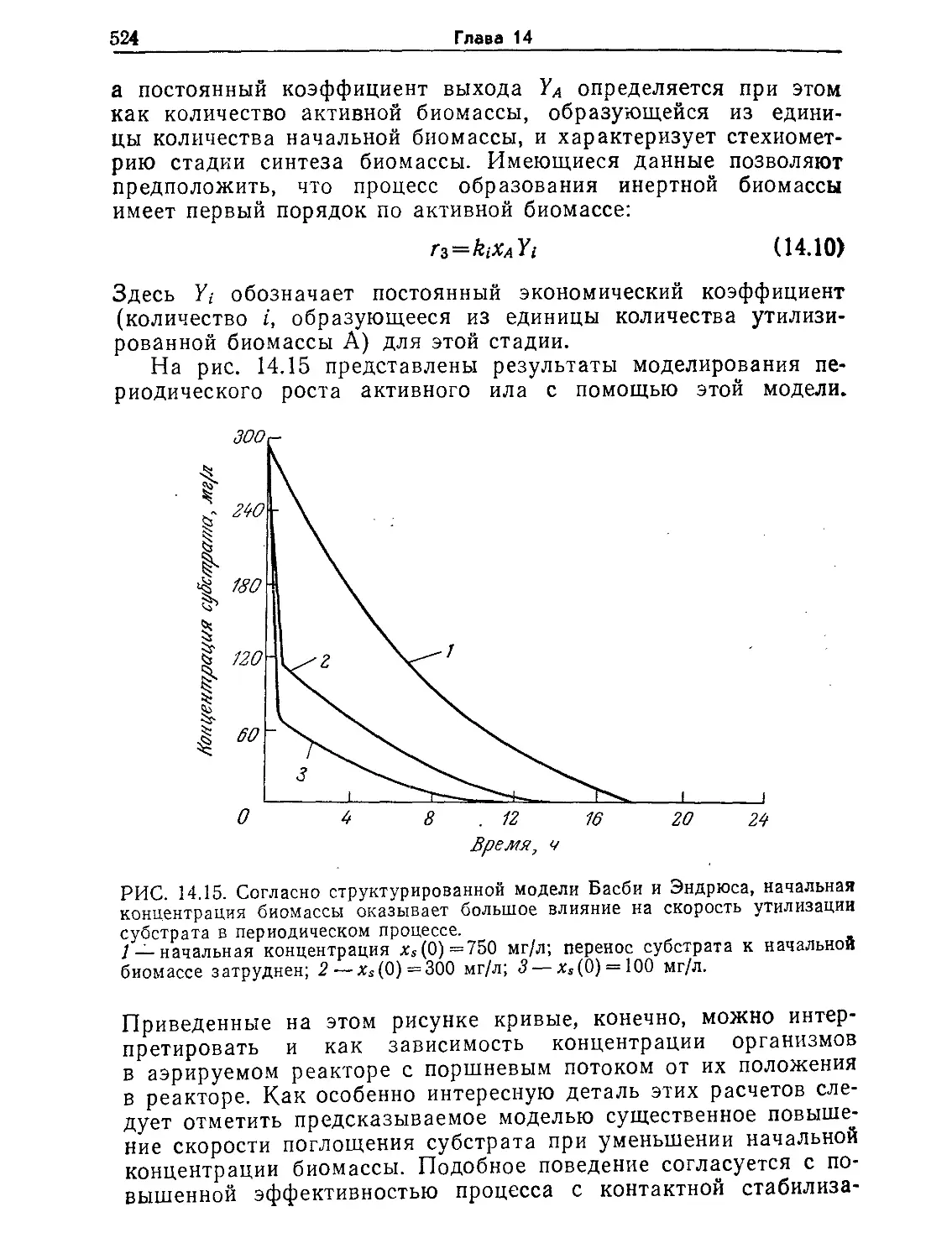
































































Tags: биологические науки в целом молекулярная биология общая биофизика, общая биохимия и общая физиология химия биология биохимия биофизика издательство мир биохимическая инженерия
ISBN: 5-03-001029-7
Year: 1989




Основы биохимической инженерии
BIOCHEMICAL
ENGINEERING
FUNDAMENTALS
Second Edition
James E. Bailey
California Institute of Technology
David F. Ollis
North Carolina State University
McGraw-Hill Book Company
New York St. Louis San Francisco Auckland Bogota
Hamburg Johannesburg London Madrid Mexico
Montreal New Delhi Panama Paris Sao Paulo
Singapore Sydney Tokyo Toronto
Дж.Бейли, Д.Оллис
ОСНОВЫ
биохимической
инженерии
В 2-х частях
2
Перевод с английского
А. А. Кирюшкина
Москва «Мир» 1989
ББК 28.07
Б 40
УДК 57.04
Бейли Дж., Оллис Д.
Б 40 Основы биохимической инженерии. Пер. с англ. В 2-х час-
тях. Ч. 2. — М.: Мир, 1989. — 590 с., ил.
ISBN 5-03-001029-7
Фундаментальный труд, написанный известными американскими специали-
стами в области проектирования, разработки и применения процессов, связанных
с переработкой биологических материалов и использованием биологических аген-
тов, в первую очередь ферментов и клеток. Ои написан с целью заполнить про-
бел, часто существующий между биологией и технологией на их пути к био-
технологии.
В русском переводе книга выходит в двух частях.
Вторая часть посвящена собственно вопросам биотехнологии — рассмотрению
особенностей биологических реакторов, систем управления и контроля, методов
выделения и очистки конечных продуктов, а также экономике биопроцессов.
Предназначена для студентов и преподавателей биологических и технологи-
ческих специальностей, научных работников, инженеров, работающих в микро-
биологической и медицинской промышленности.
„ 1901000000—370
Б 041 (01)—89 93—89
ББК 28.07
Редакция литературы по химии
ISBN 5-03-001029-7 (русск.) © 1986, 1987 by McGraw-Hill, Inc.
ISBN 5-03-001027-0
ISBN 0-07-003212-2 (англ.)
© перевод на русский язык, «Мир», 1989
Глава 9
Проектирование и расчет
биологических реакторов
Знание кинетики биологических превращений и процессов
массопередачи (этим проблемам были посвящены гл. 7 и 8)
необходимо для понимания основных принципов работы биоло-
гических реакторов. Чтобы создать полную картину работы
биологического реактора, эти два фундаментальных явления
необходимо связать с данными о перемешивании газовой и жид-
кой фаз и о контакте между фазами в реакторе. Для реакторов
с различными характеристиками течений и перемешивания
нужны различные способы проектирования, расчета и масштаб-
ного перехода. Из сказанного следует, что нашей основной за-
дачей в этой главе должна быть разработка общих подходов
к расчету биологических реакторов, в которых учитывалась бы
совокупность всех указанных явлений, факторов и характери-
стик.
При изучении кинетики клеточного роста в гл. 7 мы подчер-
кивали сложную, многофазную, согласованную природу про-
цессов с участием живых клеток, происходящих в биореакто-
рах. В этой связи мы рассмотрели различные приближения и
допущения, которые могли бы упростить описание кинетики ро-
ста популяций клеток и вывод математических выражений, при-
менимых в практических расчетах; в то же время мы стара-
лись свести к минимуму погрешности, обусловленные этими
приближениями и допущениями. С подобными проблемами мы
столкнемся и в ходе изучения методов проектирования, расчета
и анализа биологических реакторов. Здесь мы рассмотрим
многосторонние взаимосвязи между обсуждавшейся ранее слож-
ной кинетикой клеточного роста и сложными процессами тече-
ния жидкой фазы (или газожидкостной дисперсной системы),
перемешивания и теплопередачи. В этой главе мы изучим влия-
ние масштаба процесса или объема реактора на перемешива-
ние, структуру течений, явления тепло- и массообмена в реак-
торе, а также многостороннее взаимное влияние течений и яв-
лений переноса, с одной стороны, и кинетики биокаталитиче-
ских процессов, с другой. Основное внимание в настоящей гла-
ве будет уделено описаниям контакта взаимодействующих фаз
в реакторе и взаимосвязи между характером этого контакта и
6
Глава 9
биохимическими превращениями. Как и ранее при изучении
кинетики клеточного роста, для вывода полезных в практиче-
ской работе математических выражений нам придется восполь-
зоваться обоснованными приближениями и упрощениями.
Что касается рассматриваемых в настоящей главе методов
приближенного математического описания биореакторов, то в
качестве введения к этой теме чрезвычайно полезно рассмотреть
Растительные
и животные
клетки
Дрожжи
Бактерии Плесени.
f Н |
Продолжительность
клеточного цикла
<-------------
Репликация
хромосомы
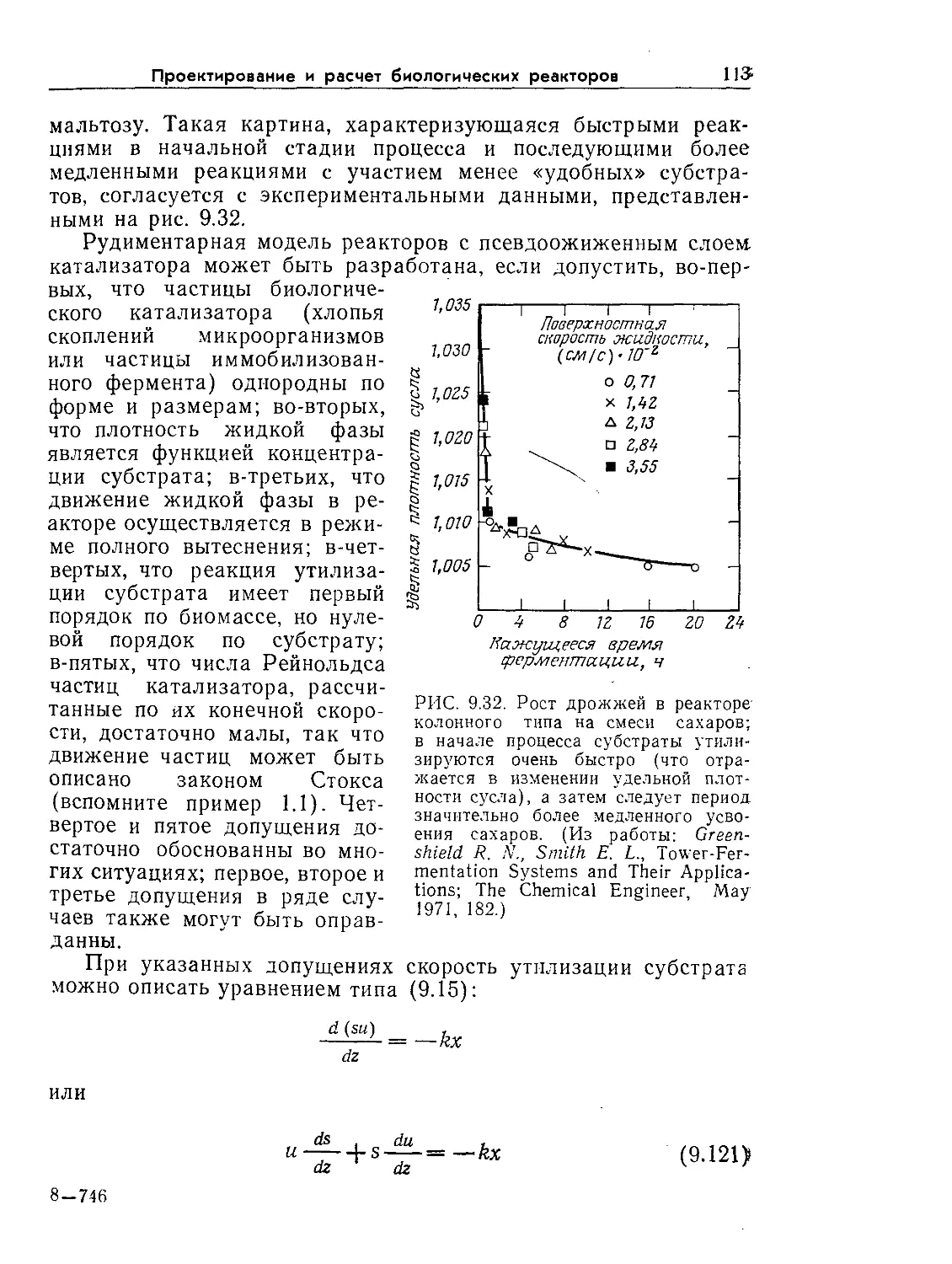
10'610'510'* 10'31О'г 1O'J 10' 10г 103 10* Ю5 Ю6
-Н+ +Н -I I I I I I I I I------peSSw, с
10°
--------> <---------------------->
Элементарные Регуляция
химические транскрипции
реакции
<--------------------
Аллостери ческая
регуляция белков
Возникновение
мутантов
<------->
Изменение
концентрации
ферментов
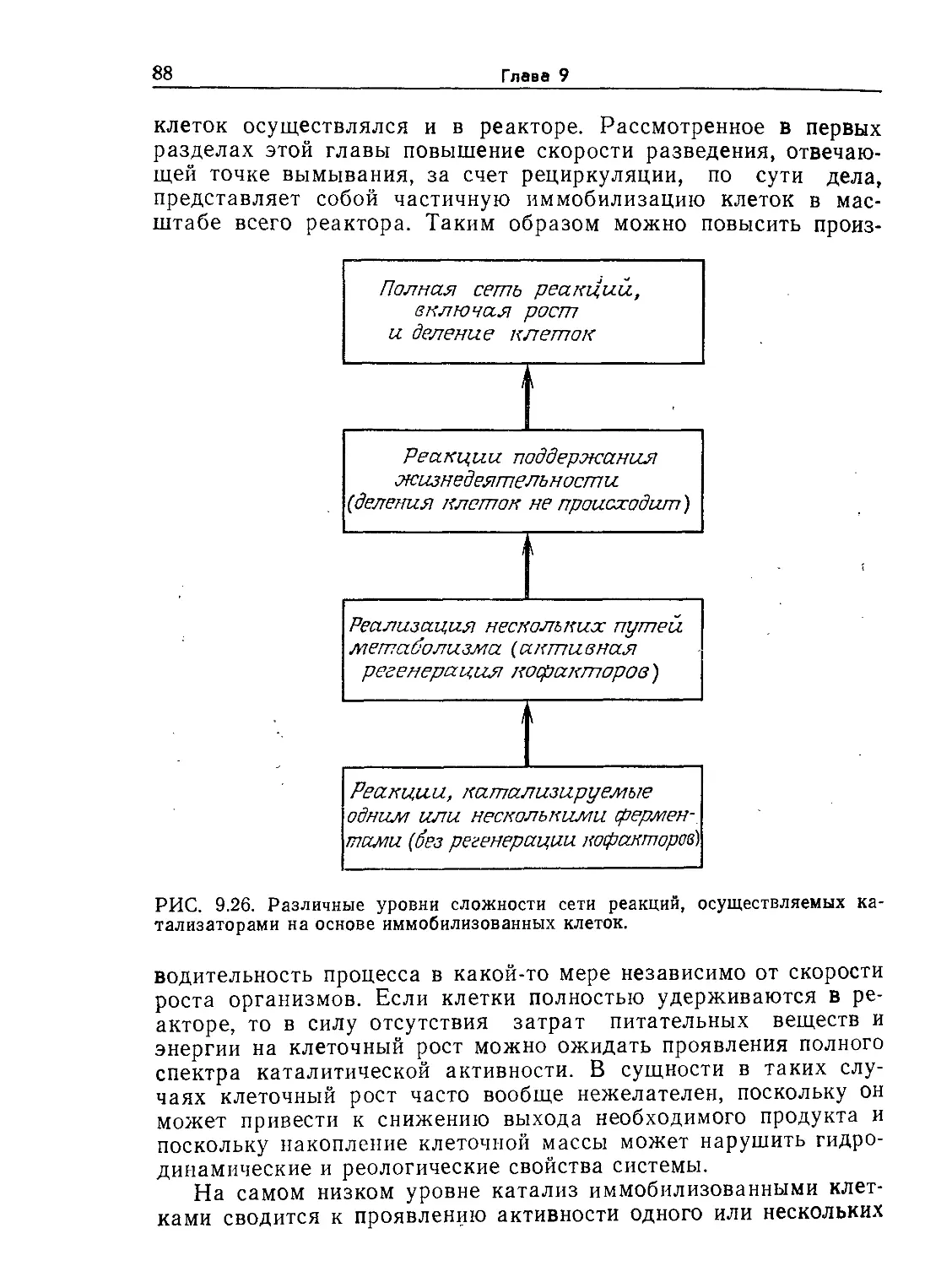
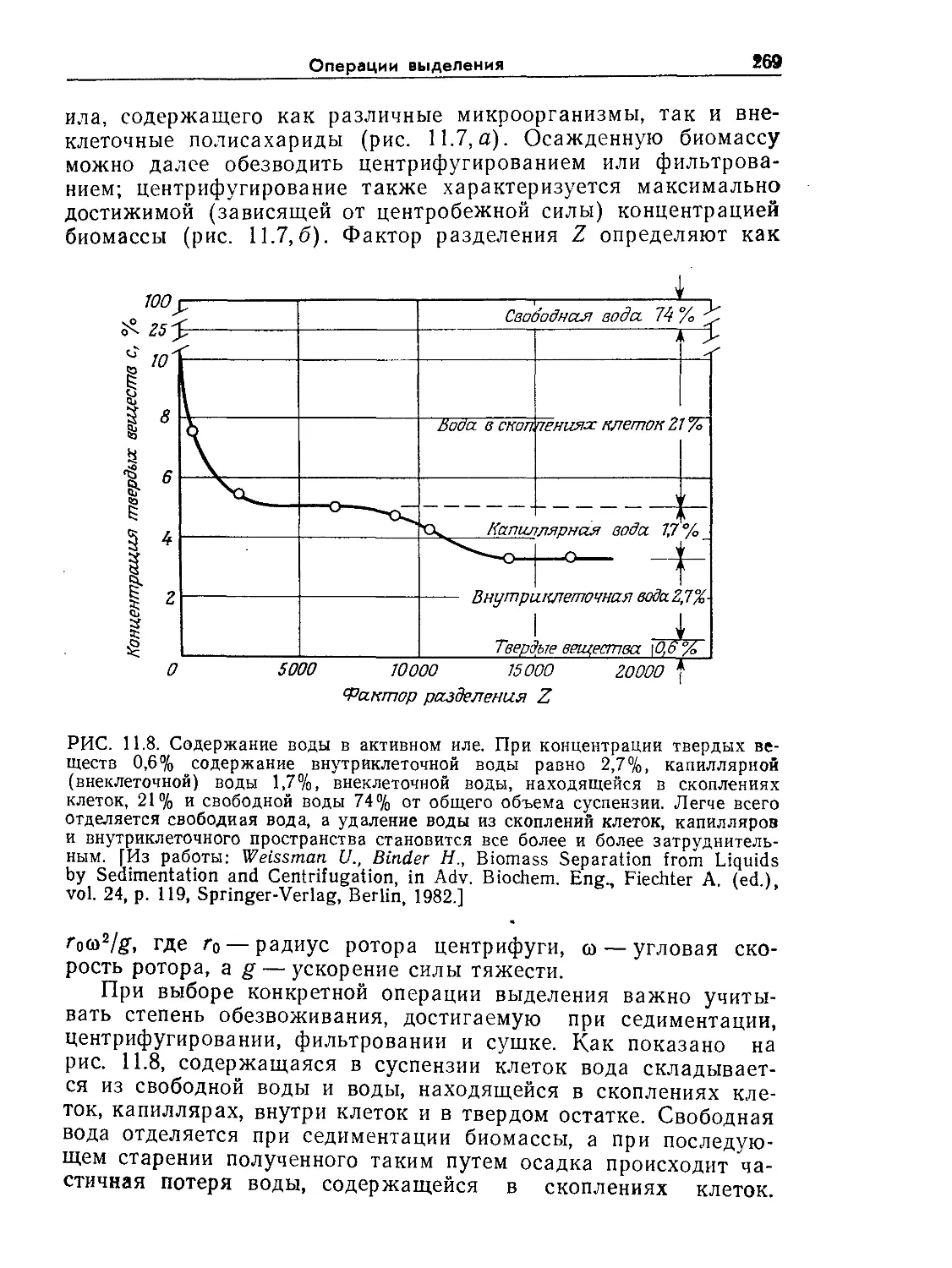
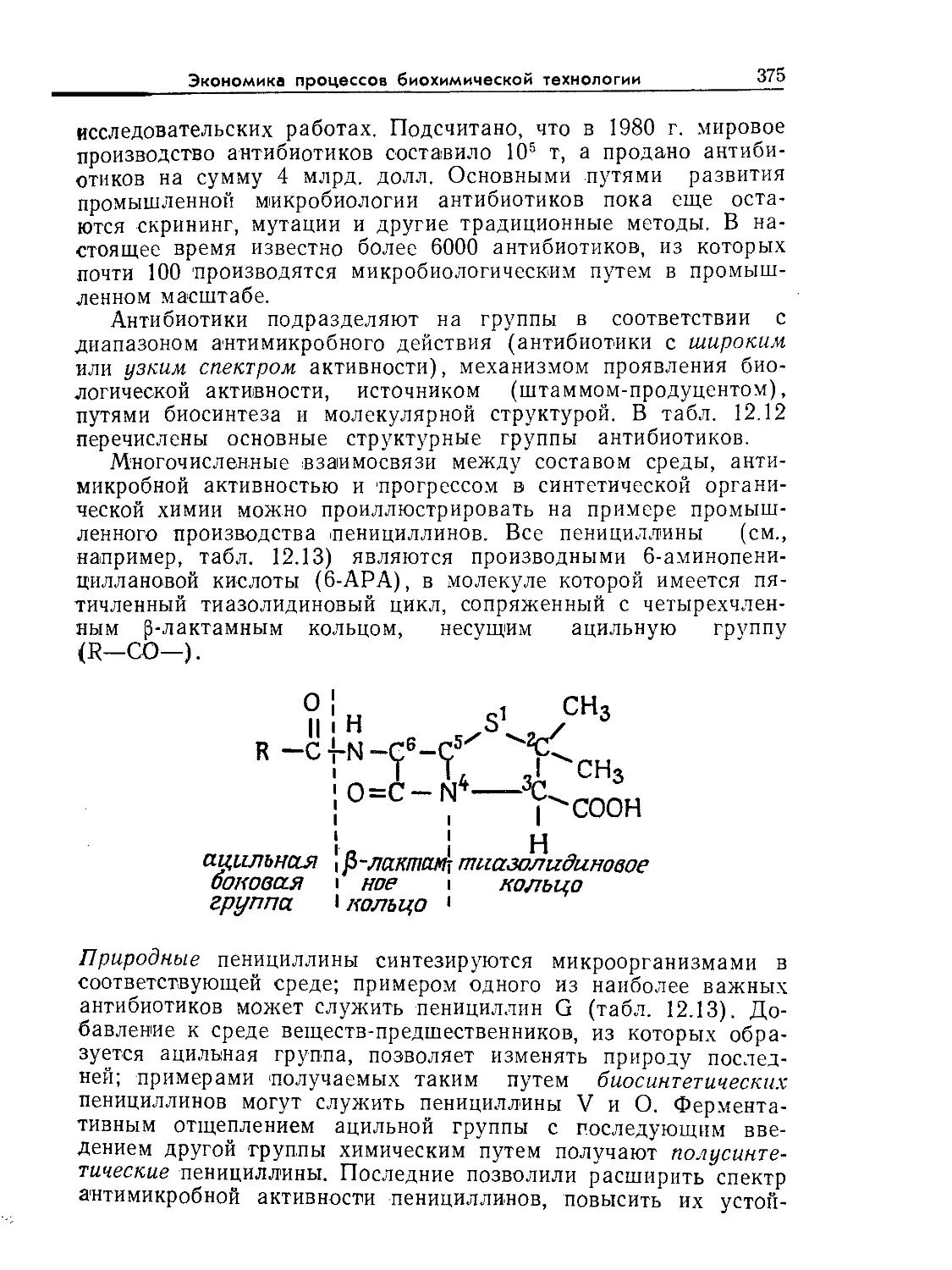
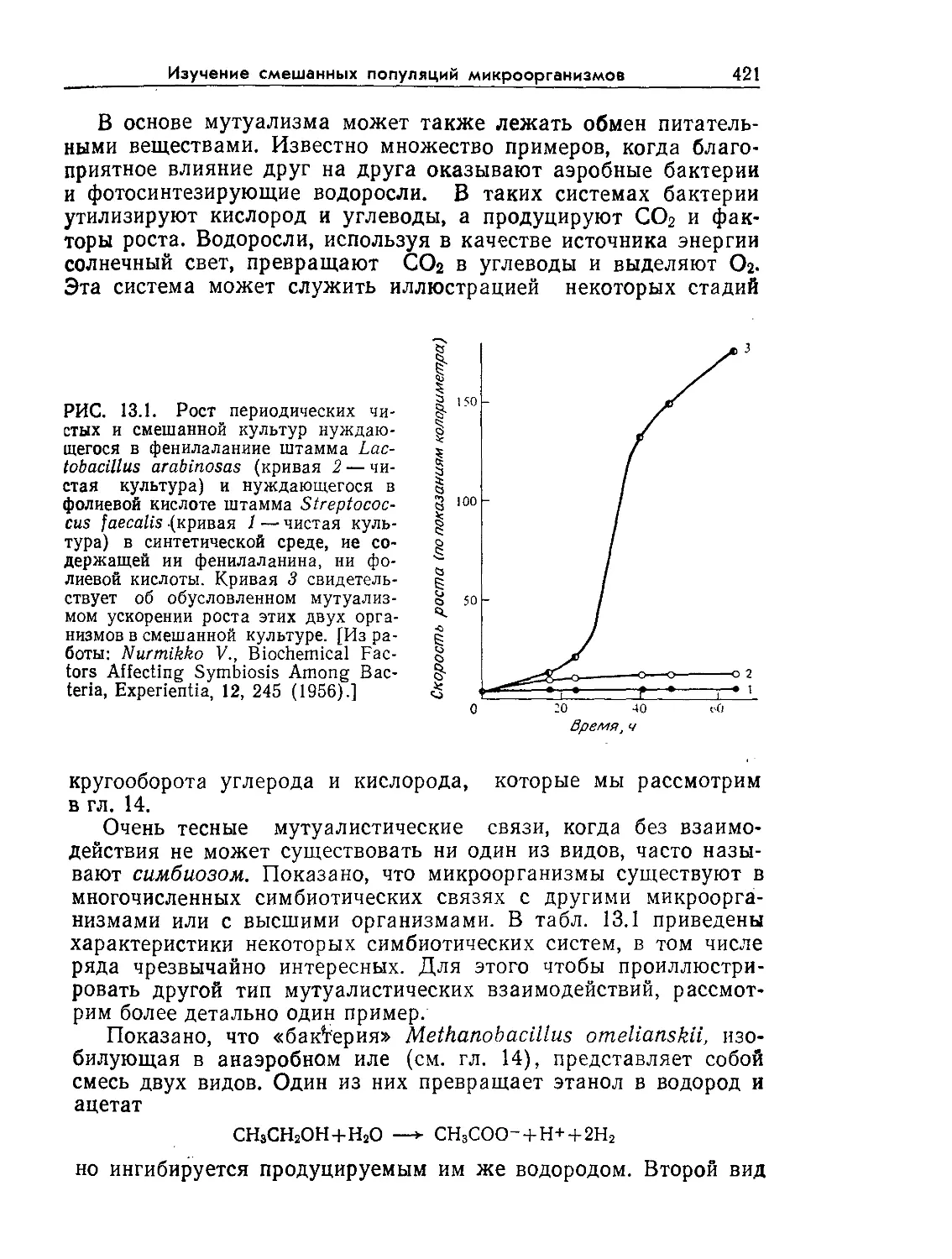
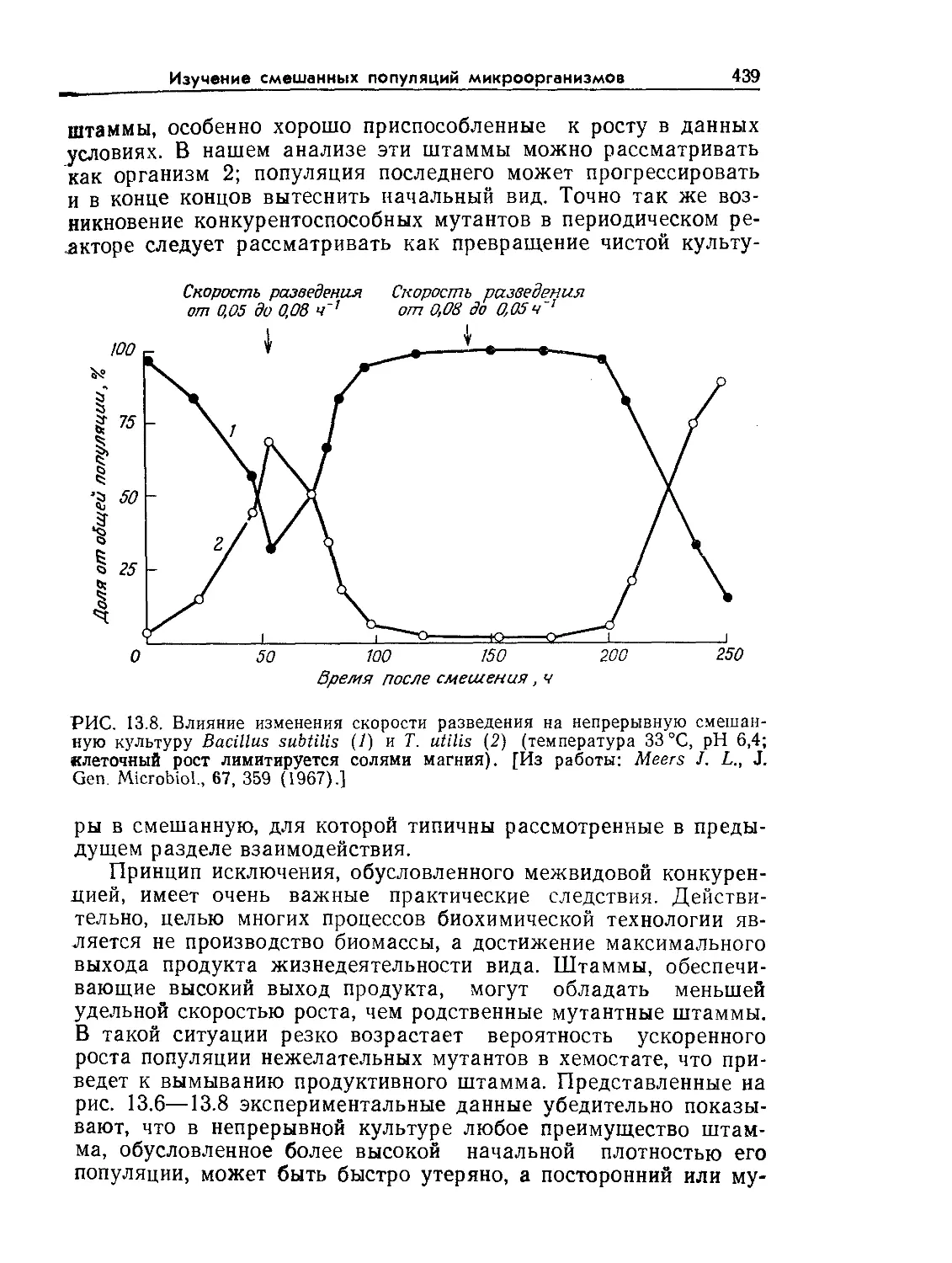
РИС. 9.1. Характерное время биологических явлений и процессов, играющих
большую роль в биохимической технологии (по данным Роуэлса [9]).
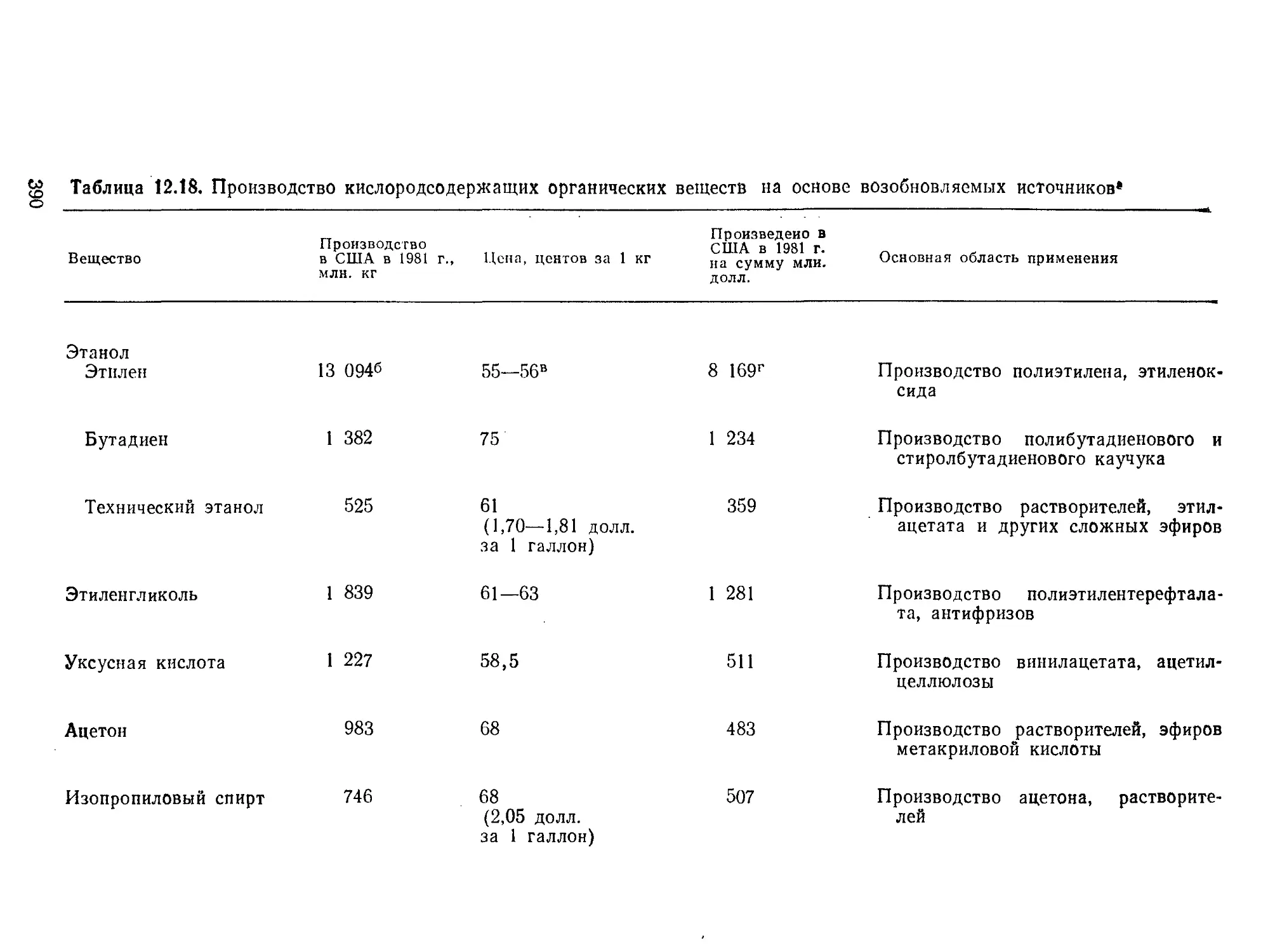
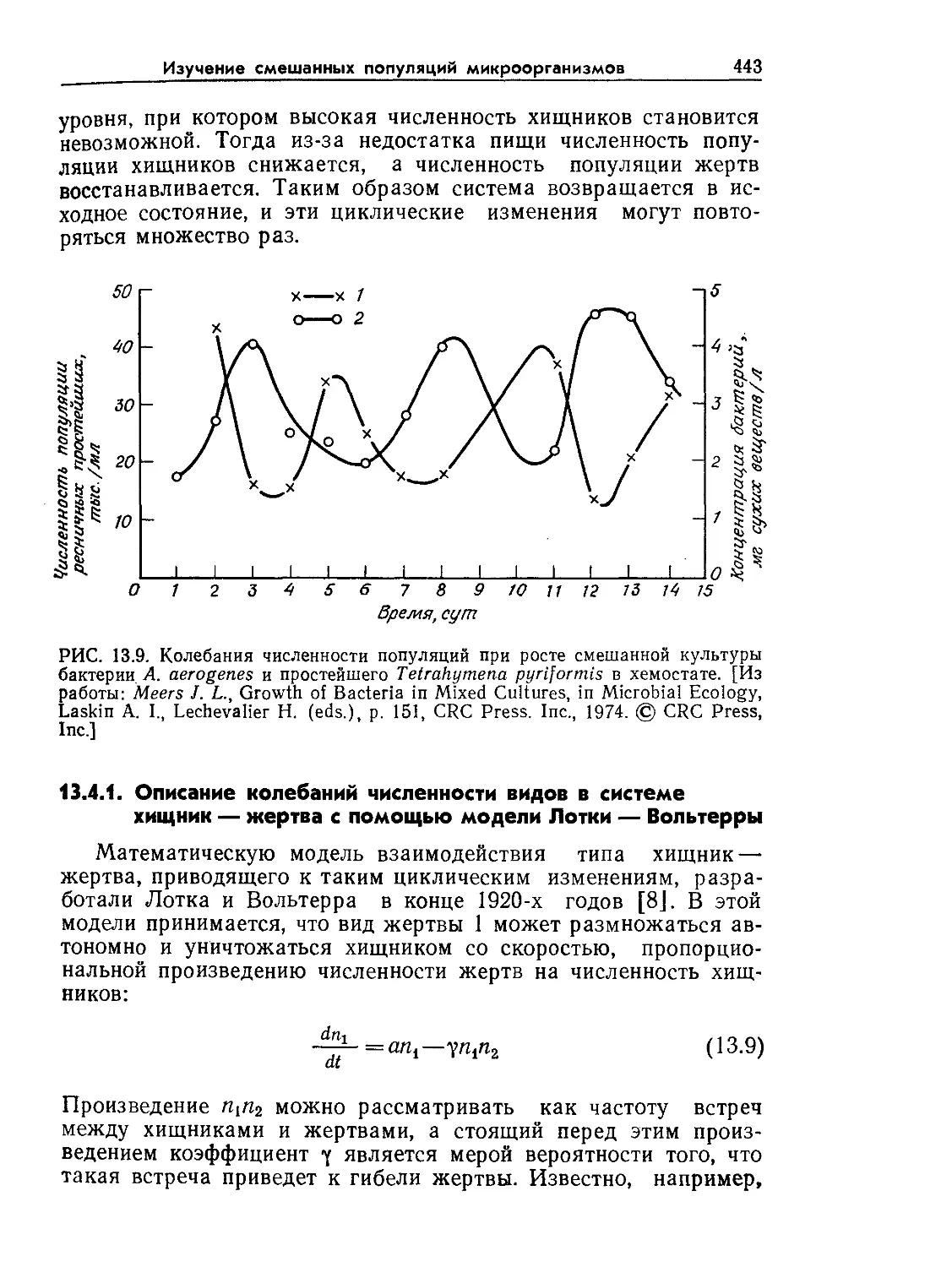
относительные масштабы времени и длины в интересующих
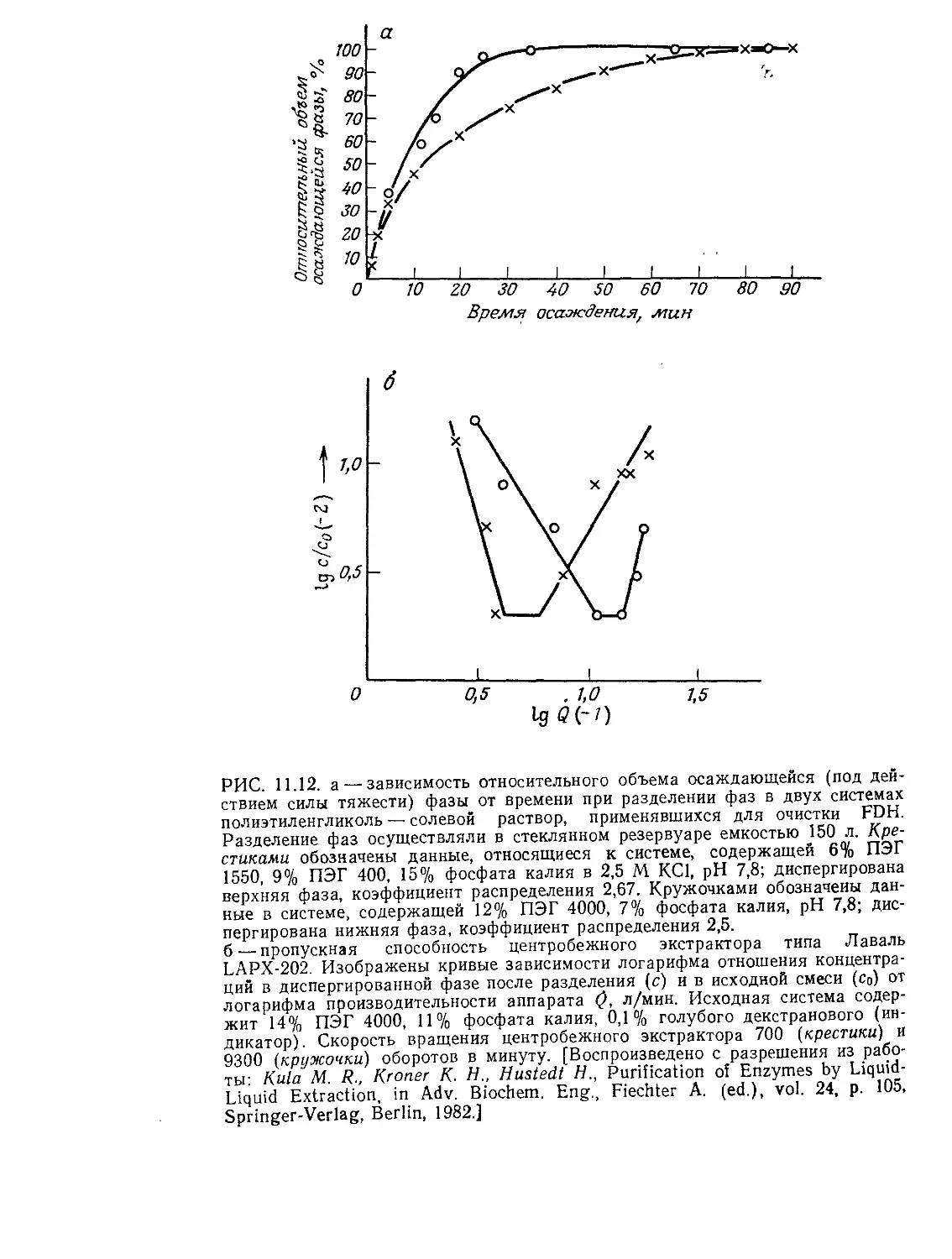
нас процессах. Как схематично показано на рис. 9.1 и 9.2, диа-
пазоны масштабов времени и длины, которые встречаются при
проектировании и расчете реакторов, чрезвычайно широки.
Ключ к анализу биореакторов — определение масштабов време-
ни и длины для явлений, представляющих наибольший интерес
в расчете конкретного реактора. После этого явления с мас-
штабами времени или длины, значительно большими или зна-
чительно меньшими по сравнению с соответствующими масшта-
бами, представляющими основной интерес в данном процессе,
часто удается анализировать с помощью относительно простых
приближений. С таким подходом мы уже сталкивались при об-
суждении приближения квазиравновесного состояния в гл. 3 и
Проектирование и расчет биологических реакторов
7
структурированных моделей клеточного роста в разд. 7.4.1.
Здесь же путем сравнения масштабов длины и времени попы-
таемся создать четкую картину процессов, наиболее важных
при описании биореактора с точки зрения определенного проек-
та или анализа.
При принятии решения о соответствующем описании реак-
тора следует учитывать, кроме того, степень доступности экспе-
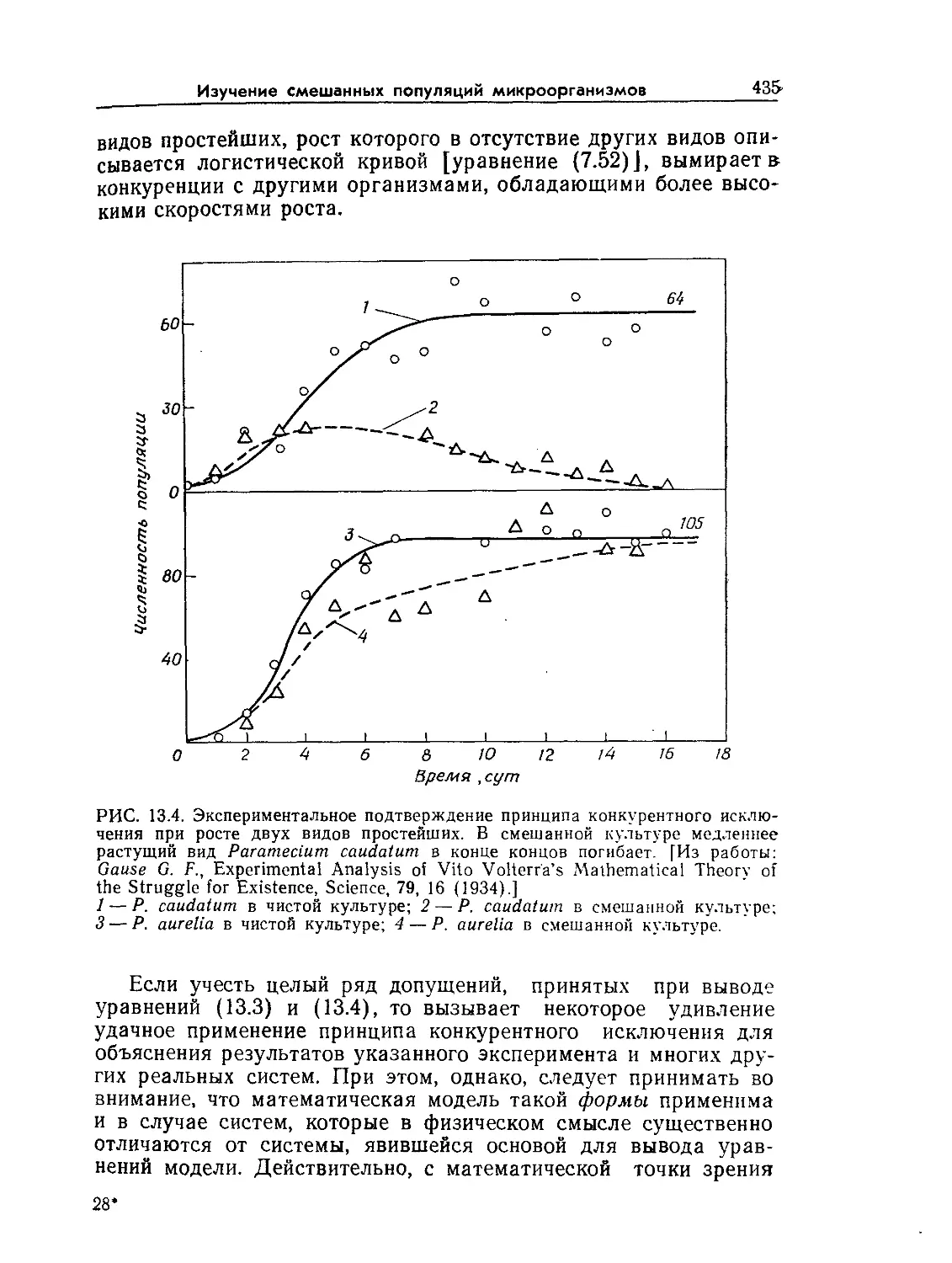
______Явления, связанные
Молекулярная с течением жидкостей.
^диффузия > Турбулентные вихри
Граничные
слои
-2 -7
+ 2 1g (метры)
-I ------->
"Животные
клетки
Биополимеры
Газовые пузырьки
Микроорга- Скопления
назмы микроорганизмов,
Органоиды растительные клетки
Биореакторы
Меоаалки
Гранулы
биокатализаторов
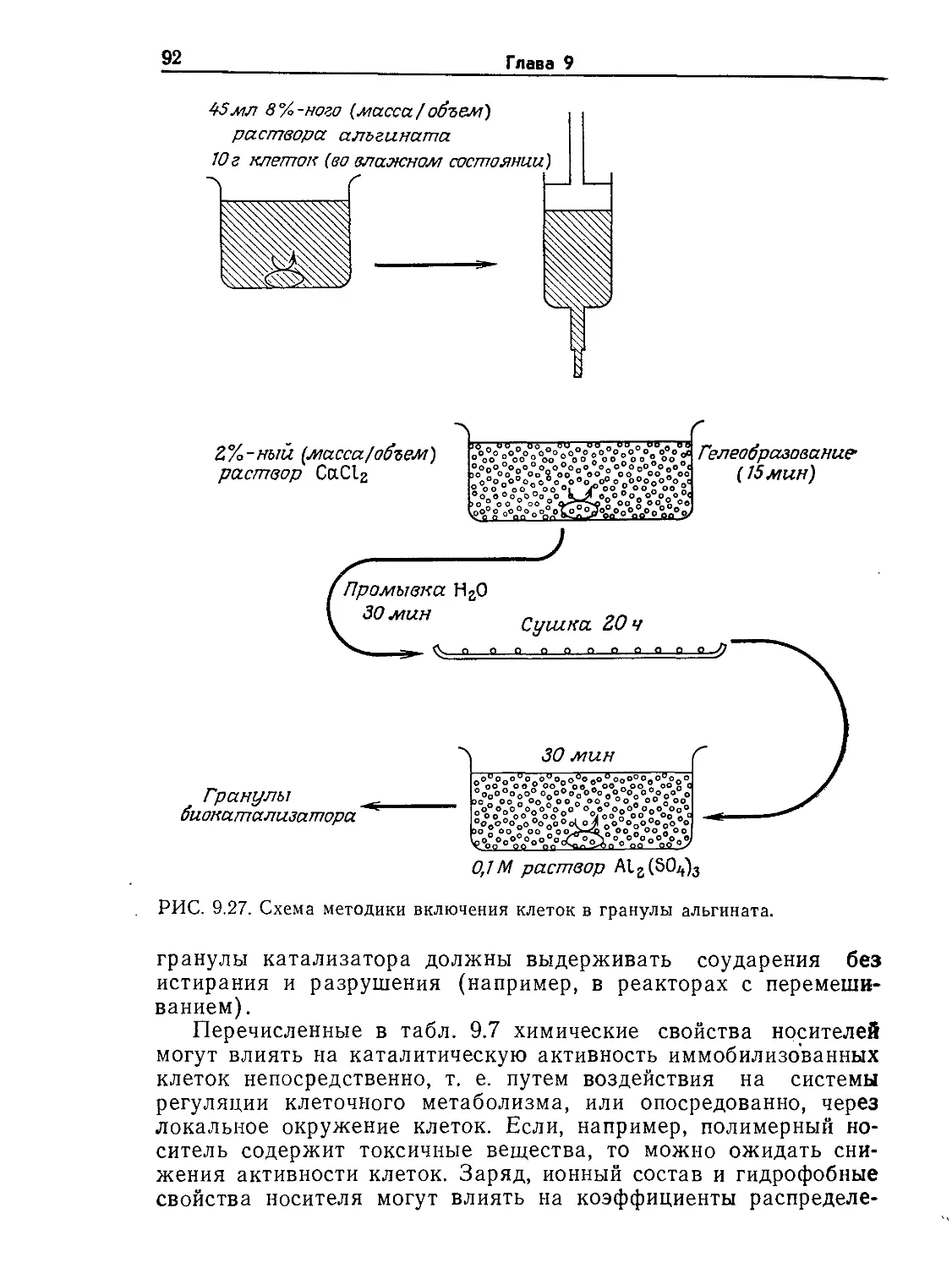
РИС. 9.2. Характерные диапазоны длин, часто встречающиеся при проектиро-
вании и расчете биореакторов (по данным Коссена [8]).
риментальных методов количественной оценки важных транс-
портных явлений и химических реакций, а также возможность
решения основанных на определенном описании реактора мате-
матических моделей с помощью имеющихся методов в числен-
ной форме и в приемлемое время.
В 9 главе мы увидим, что возможность создания надежных
математических моделей биореакторов наиболее реальна в отно-
сительно простых ситуациях, когда одна или несколько реакций
катализируются одним или несколькими ферментами. Напротив,
анализ процессов с участием культур клеток в сложной много-
фазной перемешиваемой системе из-за недостатка данных о ки-
нетике таких процессов и структуре течений жидкой фазы в
настоящее время затруднен. В этой связи становится понятной
насущная необходимость новых работ по изучению взаимосвя-
зей между биореакциями и течениями в многофазных системах.
Не обладая достаточно разработанной теорией этих сложных
8
Глава 9
процессов, мы всегда будем вынуждены в существенной степени
опираться на эмпирический подход и осуществлять масштабные
переходы только с помощью большого числа экспериментов по-
следовательно возрастающего масштаба, причем даже в этом
случае всегда существует опасность, что разработанный нами
режим будет весьма далек от оптимального.
Изучение проектирования, расчета и анализа биореакторов
мы начнем с идеальных реакторов периодического действия с
полным перемешиванием и проточных реакторов с полным пе-
ремешиванием (ПРПП), упоминавшихся ранее в связи с изу-
чением кинетики клеточного роста в гл. 7.
9.1. Идеальные биореакторы
В разд. 7.1 мы познакомились с идеальными биореакторами
с полным перемешиванием. Предполагается, что в таких реак-
торах перемешивание настолько эффективно, что концентрации
биокатализаторов и условия протекания реакций практически
одинаковы в любой точке объема реактора. Это приближение
будет справедливым только в том случае, если любые сущест-
вующие в реакторе градиенты достаточно малы, так что ло-
кальная скорость реакции для данной клетки или данной части-
цы биокатализатора в существенной степени не меняется при
перемещении частицы катализатора (или клетки) из одной зо-
ны реактора в другую. Возможна и альтернативная ситуация,
когда частицы биокатализатора очень быстро циркулируют
между различными зонами реактора, и тогда расчет реактора
на базе усредненных условий, считающихся одинаковыми во
всем объеме реактора, обычно дает удовлетворительные резуль-
таты. Эта ситуация может реализоваться (в зависимости от ти-
па изучаемого процесса) в лабораторных и даже в пилотных
промышленных установках небольшого объема. Напротив, при
росте культур филаментозных организмов или организмов,
продуцирующих внеклеточные полимерные соединения, и при
высокой плотности организмов культуральная жидкость при-
обретает в высшей степени неньютоновские свойства, поэтому
даже в случае небольших лабораторных реакторов система ни-
когда не приближается к режиму идеального перемешивания.
Изучение теории идеальных биореакторов с полным переме-
шиванием важно с нескольких точек зрения. Во-первых, такие
реакторы обеспечивают достаточно определенные условия, что
необходимо для изучения кинетики соответствующих процессов
в лабораторных условиях. Во-вторых, модели на основе идеаль-
ных реакторов часто могут сравнительно успешно применяться
даже в тех случаях, когда требуемые этими моделями условия
выполняются не полностью. Наконец, реакторы с полным пе-
Проектирование и расчет биологических реакторов
9
ремешиванием являются отправной точкой для изучения и оп-
ределения параметров реакторов с неполным перемешиванием
и со значительной объемной неоднородностью условий реакций.
Как мы увидим позднее, иногда в лаборатории удается рассчи-
тать и смоделировать крупномасштабные реакторы с неполным
перемешиванием, воспользовавшись модельными системами,
состоящими из каскада взаимосвязанных реакторов. Прежде все-
го мы рассмотрим идеальные реакторы периодического дейст-
вия и идеальные ПРПП, с которыми познакомились в разд. 7.1,
а затем перейдем к изучению идеальных реакторов полного вы-
теснения (с поршневым потоком), в которых обратное переме-
шивание (осевое смешение) пренебрежимо мало.
9.1.1. Реакторы периодического действия с добавлением
субстрата
Часто в ходе микробиологического процесса возникает не-
обходимость во введении в биореактор периодического действия
потоков жидких веществ, например растворов предшественни-
ков синтезируемых продуктов метаболизма, соединений с регу-
ляторной активностью (в частности, индукторов, потребность в
которых может возникнуть только в определенный момент про-
цесса), растворов веществ, способствующих поддержанию низ-
ких концентраций питательных веществ (в частности, для по-
давления катаболитной репрессии), или растворов питательных
веществ (что позволяет увеличить продолжительность стацио-
нарной фазы и тем самым повысить выход продукта). Когда
поток жидких питательных веществ поступает в реактор, есте-
ственно, изменяется и объем культуры; этот факт необходимо
учитывать и в уравнениях, описывающих работу реактора.
Пусть F(t)—объемная скорость поступающего в реактор рас-
твора питательных веществ во время t, a Cif(t)— концентра-
ция компонента i в этом растворе; тогда уравнение материаль-
ного баланса по компоненту i можно записать в такой форме
l + F(t)-cll (9.1)
Если принять, что и плотность поступающего в реактор раство-
ра, и плотность культуральной жидкости в реакторе равны р,
то уравнение полного материального баланса содержимого ре-
актора примет вид
(9.2)
(Как можно модифицировать это уравнение, чтобы в нем учи-
тывалось различие плотностей поступающего в реактор раство-
10
Глава 9
ра и содержимого реактора, обусловленное, например, аэраци-
ей культуральной жидкости?) При условии, что р практически
не меняется в ходе периодического процесса, уравнение (9.2)
упрощается
-^- = Г(0 (9.3)
Дифференцируя выражение, стоящее в левой части уравне-
ния (9.1) (с учетом того, что Vr теперь является функцией от
времени), и подставляя dVRldt из уравнения (9.3), после пре-
образований можно получить следующее удобное рабочее вы-
ражение для материального баланса по компоненту i:
d-ci _ F (0 r„ z,.i I f
---------— fo,-C,]+r,(
(9.4)
Уравнения (9.3) и (9.4) описывают материальный баланс по
всему содержимому реактора и одному из компонентов соот-
ветственно. Если известны математические выражения, описы-
вающие кинетику rfi, то эти уравнения можно использовать для
моделирования влияния различных режимов введения веществ
F(t) на работу реактора.
9.1.2. Реакции в ПРПП, катализируемые ферментами
В зависимости от способа обеспечения ферментативной ак-
тивности при осуществлении катализируемых ферментами ре-
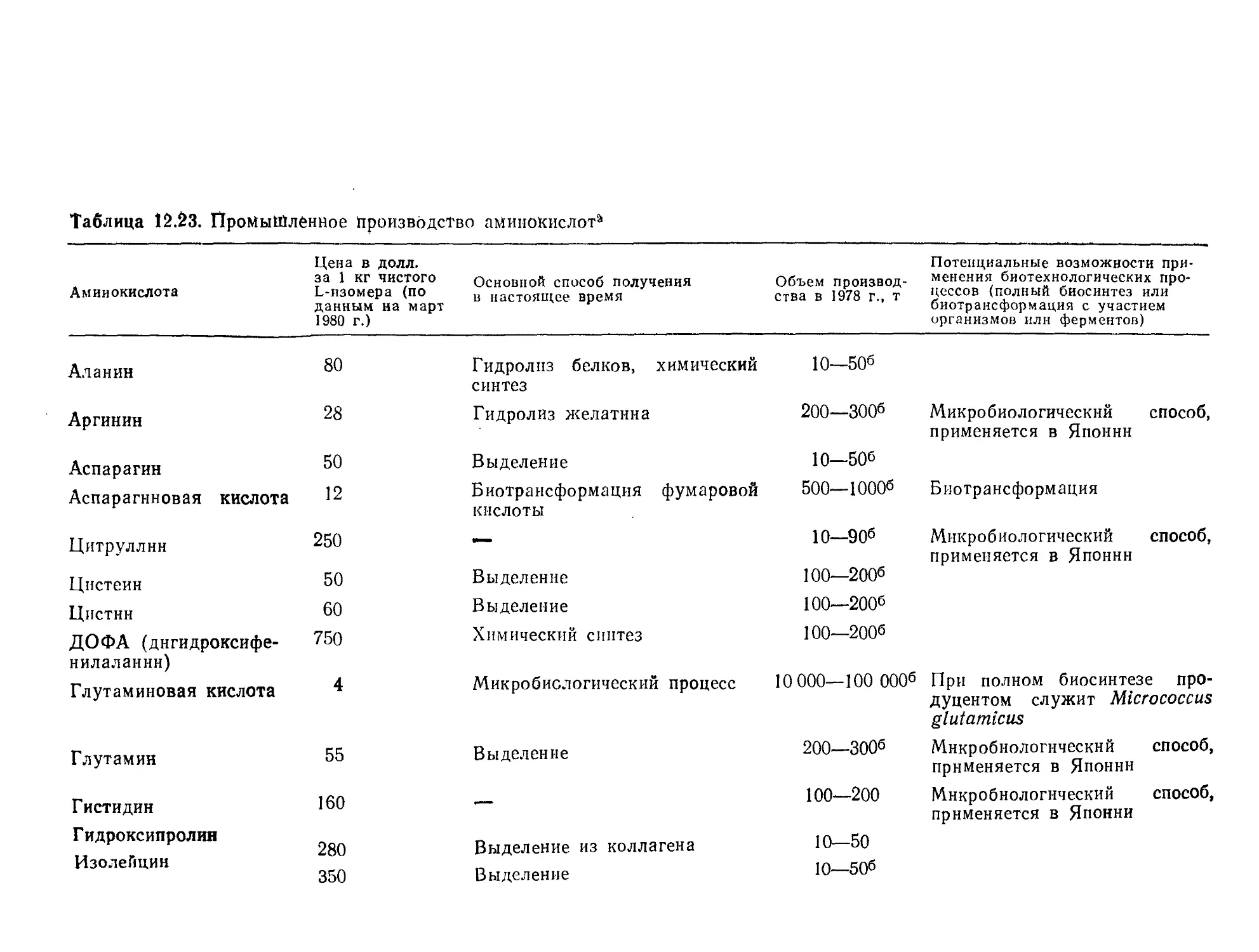
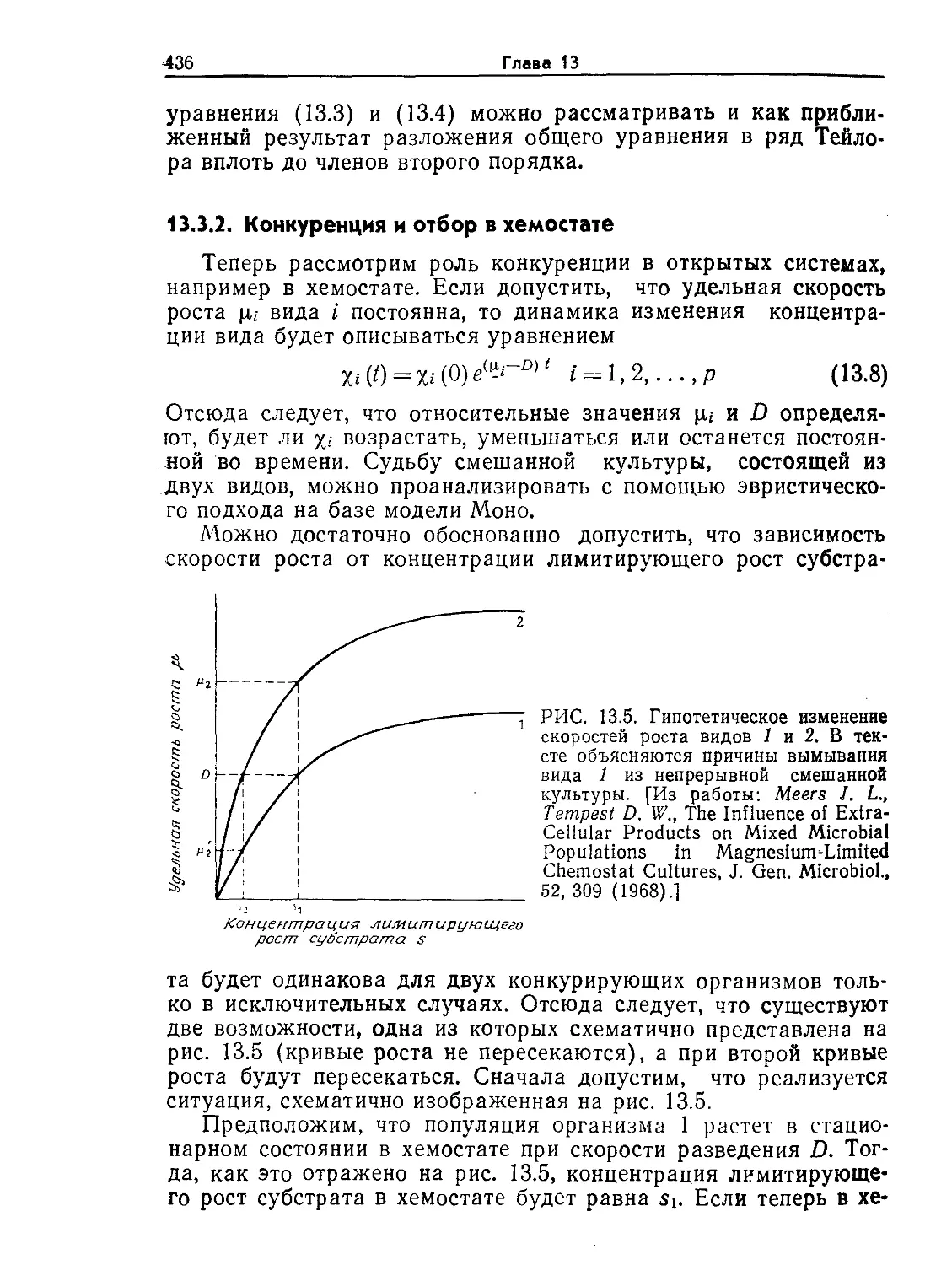
акций используются ПРПП различных конструкций (рис. 9.3).
В проточном реакторе простейшей конструкции ферменты по-
стоянно вводятся в реактор и выводятся из него по соответст-
вующим линиям (рис. 9.3, а). Очевидно, такая конструкция
оправдана только в том случае, если ферменты настолько де-
шевы, что их расходом можно пренебречь.
Более дорогостоящие ферменты необходимо удерживать в
реакторе или возвращать в него путем рециркуляции. Ознако-
мившись в гл. 4 с методами иммобилизации ферментов, мы уже
знаем, что эту цель можно достигнуть несколькими путями.
Во-первых, (рис. 9.3, б) на пути вытекающего из реактора пото-
ка можно установить ультрафильтрационную мембрану с пора-
ми, диаметр которых позволяет удерживать относительно боль-
шие молекулы ферментов в растворе. Если фермент иммобили-
зован на частицах нерастворимого носителя, суспендированного
в реакционной смеси, то для предотвращения потери фермента
с вытекающим из реактора потоком достаточно установки про-
стого сетчатого фильтра (рис. 9.3, в). Принцип другого подхода
к удерживанию иммобилизованных ферментов в реакторе схе-
матично изображен на рис 9.3, а; здесь фермент удерживается в
Проектирование и расчет биологических реакторов 11
сетчатых контейнерах, укрепленных на валу мешалки. Реакто-
ры такой конструкции, широко применявшиеся также для изу-
чения реакций в газовой фазе на металлических катализаторах
на носителях, позволяют свести к минимуму сопротивление мас-
сопередаче между жидкой фазой и гранулами иммобилизован-
ного фермента. Та же цель может быть достигнута и более
удобным путем — посредством циркуляции реакционной смеси
РИС. 9.3. Схематическое изображение ПРПП различного типа, применяемых
для проведения катализируемых ферментами реакций.
а — реактор с непрерывной подачей раствора ферментов; б — реактор с удер-
живанием ферментов в растворе с помощью полупроницаемой мембраны; в —
реактор с удерживанием гранул иммобилизованного фермента с помощью сет-
чатого фильтра; г — реактор с удерживанием гранул иммобилизованного
фермента в контейнерах, размещенных на валу мешалки; д—реактор с ре-
циркуляцией реакционной массы через короткую колонну, наполненную им-
мобилизованным ферментом.
в системе реактор с полным перемешиванием — короткая ко-
лонна с насадкой иммобилизованного фермента (рис. 9.3, д).
Если скорость рециркуляции достаточно высока и в каждом
цикле при однократном прохождении реакционной смеси через
колонну превращению подвергается небольшая часть субстрата
(менее 1%), то вся система становится эквивалентной проточ-
ному реактору с полным перемешиванием [4]. Отсюда следует,
что эта конструкция реактора особенно удобна для изучения
кинетики реакций в лабораторных исследованиях.
Рециркуляция фермента возможна только при условии, что
фермент легко выделить из вытекающего из реактора потока
продуктов реакции. Есть два перспективных подхода к реше-
нию этой задачи, один из которых связан с включением фер-
мента в мембрану из жидкого поверхностно-активного вещест-
12
Глава 9
ва или фосфолипида, а второй — с иммобилизацией ферментов
на магнитных носителях.
Независимо от типа и конструкции реактора основная зада-
ча всегда сводится к поддержанию заданной концентрации фер-
мента в ПРПП. Если эта задача решена, то мы можем скон-
центрировать наше внимание на расчете концентраций субстра-
та и продукта реакции в вытекающем из реактора потоке. Здесь
применимы рассмотренные выше общие принципы и уравнения
материального баланса, используемые для описания роста по-
пуляции микроорганизмов, а также дополнительные ограниче-
ния, налагаемые относительно простой стехиометрией изучае-
мых реакций.
Например, для простой реакции
S —Р
из каждого моля прореагировавшего вещества S образуется
один моль вещества Р, и, следовательно, концентрации этих ве-
ществ на входе в реактор (s0, р0) и на выходе из реактора
(s, р) связаны соотношением
s0 — s=p — ро (9.5)
Если условие уравнения (9.5) выполнено, то скорость реакций
v(s, р), которая является функцией как s, так и р, можно вы-
разить только через s, что упрощает соответствующие выраже-
ния. В этом случае уравнение материального баланса по суб-
страту принимает форму
E(s0 — s) — Vrv{s, po + so — s)=0 (9.6)
В табл. 9.1 даны решения этого уравнения для различных ти-
пов кинетики реакций. Приведенные в таблице уравнения про-
ще использовать непрямым путем; для этого нужно в правую
часть уравнения перенести необходимые параметры превраще-
ния субстрата и рассчитать искомое время пребывания и (или)
концентрацию фермента. Аналогичными способами можно по-
лучить уравнения для более сложных реакций и типов кинетики
превращений.
9.1.3. Проточные реакторы
с полным перемешиванием для культур клеток
и пристеночный рост клеток
Для повышения выхода биомассы и продуктов жизнедея-
тельности организмов в единице объема реактора в единицу
времени ПРПП можно снабдить сепаратором (гл. 11) и устрой-
ством для рециркуляции концентрированной суспензии клеток.
В соответствии с принятыми на рис. 9.4 обозначениями, симво-
Проектирование и расчет биологических реакторов
13
Таблица 9.1. Зависимости между степенью превращения субстрата
6= (s0—s)/s0, средним временем пребывания и концентрацией катализатора
в случае катализируемых ферментами реакций в ПРППа
Уравнение, описывающее
скорость реакции
Уравнение, описывающее Vvm3x/F в ПРПП
Уравнение Михаэлиса — Мен-
тен:
^maxs
Уравнение Михаэлиса — Мен-
тен для обратимой реакции:
ртах($—Р/К)
К-р
Конкурентное ингибирование
продуктом реакции:
_____^тах$____
а~1гКт(
Ингибирование субстратом:
^тах
о
АГ„ , л । ^т(Ро + so5)
О Кт + so — dso Н"
1 -d(l+ 1/К)
А Г v । х । 4~ Sq6)
О Лт > S0 os0 -р
1-6
а Messing R. A., Immobilized Enzymes for Industrial Reactors, p. 158, table 1, Acade-
mic Press, New York, 1975.
лами Fo и Fr мы будем обозначать объемные скорости потоков
поступающих в систему питательных веществ и рециркуляции
соответственно, а символами хь х0 и х— концентрации био-
массы в реакторе, рециркулирующем потоке и в выходящем из
реактора потоке соответствен-
но. Эти величины могут разли-
чаться в силу специфики рабо-
ты сепаратора (которым мо-
жет быть, например, отстой-
ник), расположенного в точке
разделения потока, вытекаю-
щего из реактора. При а —
= Fr/F0 и Ь=х0/х( в стацио-
нарном состоянии уравнение
материального баланса по
Гг,Л0
РИС. 9.4. Схема ПРПП с рецир-
куляцией.
14
Глава 9
биомассе для системы с рециркуляцией выглядит следующим
образом:
FrXo+цх!^— (Fo+Frjx^G (9.7)
Тогда общая (или внешняя) скорость разведения D (равная
Fo/Vr) составит:
Поскольку концентрация микроорганизмов в циркулирую-
щем потоке обычно выше концентрации на выходе из реактора,
то Ь>1. В этом случае, как показывает уравнение (9.8), ско-
рость разведения выше удельной скорости роста организмов.
Следовательно, рециркуляция при той же скорости роста орга-
низмов позволяет перерабатывать за единицу времени больше
питательных веществ в единице объема реактора. Эта особен-
ность проточных реакторов с рециркуляцией с большим успе-
хом используется в процессах биологической переработки от-
ходов, подробнее рассматриваемых в гл. 14. (Каков эффект
рециркуляции при 6=1? Объясните физический смысл получен-
ного вами результата.)
Другие важные преимущества системы с рециркуляцией об-
наруживаются после ряда преобразований уравнений материаль-
ного баланса по компонентам системы. Если принять, что эко-
номический коэффициент постоянен, то уравнение материально-
го баланса по субстрату можно записать в виде
О(5»-«)--^ = 0 (9-9)
Из уравнений (9.9) и (9.8) нетрудно найти, что скорость обра-
зования биомассы в единице объема реактора (pxi) равна:
|iK(s0 —s)_ (910)
г 1 1 -а (&- 1)
Эта величина больше скорости образования биомассы без ре-
циркуляции в [1—а(Ь—I)]-1 раз. Если принять, что ц подчи-
няется уравнению Моно, то можно показать, что и скорость
разведения в точке вымывания при рециркуляции возрастает
во столько же раз.
В экспериментах с растущими популяциями клеток в ПРПП
иногда и в отсутствие рециркуляции можно добиться более вы-
соких скоростей разведения без вымывания по сравнению с ве-
личиной, предсказываемой теорией идеальных ПРПП (вспом-
ните разд. 7.1.2). Это явление может быть обусловлено ростом
организмов на стенках реактора (так называемым пристеноч-
ным ростом). В общем случае в реакторе может быть несколь-
Проектирование и расчет биологических реакторов 15
ко центров образования твердых пленок организмов. Колонии
микроорганизмов могут возникать, например, над уровнем жид-
кой фазы там, где разбрызгиваемые капли культуры попадают
на стенки резервуара, или в трещинах и щелях в относительно
неперемешиваемых (застойных) зонах биореактора. Если допу-
стить, что концентрация клеток в пленках на стенках реактора
Xf постоянна во времени, то размножение организмов в этих
пленках должно сопровождаться переносом клеток со стенок в
жидкую перемешиваемую культуру. В этом случае уравнения
материального баланса для стационарного состояния в проточ-
ном реакторе примут следующую общую форму:
Dx = nx+nfXf (9.11)
D(s0—s) = -t-iix+-±- pfXf (9.12)
Здесь pf и Yf — удельная скорость клеточного роста и экономи-
ческий коэффициент в пленке соответственно. Эти параметры
по целому ряду причин отличаются от соответствующих пара-
метров основной массы культуры ц и У, в том числе из-за влия-
ния диффузии на скорость реакции.
Здесь необходимо отметить, что слагаемое р/Х/ в уравнении
(9.11) отражает второй источник поступления клеток в культу-
ральную жидкость, который практически не зависит от D, по-
этому пристеночный рост организмов предотвращает вымыва-
ние. В этой связи следует также подчеркнуть, что у лаборатор-
ных реакторов отношение поверхности к объему значительно
выше, чем у крупномасштабных промышленных реакторов, и,
следовательно, вклад пристеночного роста в лабораторных уста-
новках также в общем случае должен быть большим. Таким
образом, для систем, в которых наблюдается существенный
пристеночный рост организмов, последний необходимо учиты-
вать при масштабном переходе от данных по кинетике роста
микроорганизмов, полученных в лаборатории, к промышленным
реакторам большого объема.
9.1.4. Идеальный трубчатый реактор полного вытеснения (ТРПВ)
Если при течении жидкости по трубопроводу сравнительно
большого диаметра или по каналу число Рейнольдса достаточ-
но велико (например, больше 2100 для трубы), то течение в
такой системе приближается к режиму полного вытеснения
(поршневому потоку), при котором в поперечном сечении пото-
ка осевые скорости не изменяются. Если допустить, что режим
полного вытеснения приближенно описывает движение жидко-
сти через реактор, то, воспользовавшись понятием об элемен-
16
Глава 9
тарном объеме, можно вывести уравнения материального ба-
ланса по компонентам системы в трубчатом реакторе полного
вытеснения (ТРПВ). Как показано на рис. 9.5, при таком под-
ходе принимается, что в трубчатом реакторе стационарное со-
стояние сохраняется в элементарном объеме потока, перпенди-
кулярном оси реактора. Материальный баланс
произвольному компоненту С в элементарном
выразить следующим уравнением:
по некоторому
объеме можно
Аис
—Аис
г
+ AAzrfc
(9.13)
г+Дг
Здесь г/с — скорость образования
через его количество в единице
= 0
Z
компонента
объема за единицу времени.
z z + ZJz
С, выраженная
ис|2
и
ИС\г+Л
и
z= О
z = L
РИС. 9.5. Реактор полного вытеснения.
Преобразование и деление на ЛДг дает
lz+Дг ис 1г_______________________,
-------------
(9.U)
Az
Вспомнив определение производной, нетрудно найти, что в пре-
деле это уравнение преобразуется, давая в окончательном виде
(„<;) = Г (9.15)
dz
Если в результате реакции плотность жидкости не изменяется
(справедливо ли это допущение для микробиологического про-
цесса?) и если осевая скорость постоянна, то уравнение (9.15)
принимает вид:
de
dz
Величина z/w равна времени, которое затрачивает элементар-
ный объем жидкости на движение от входа в реактор до осе-
вого положения z. Если в качестве новой независимой перемен-
ной использовать время движения элементарного объема t,
равное
(9.16)
t== —
и
(9.17)
Проектирование и расчет биологических реакторов 17
то уравнение материального баланса по компоненту С примет
вид:
de
(9.18)
Последнее уравнение полностью совпадает с уравнением мате-
риального баланса по компоненту С в реакторе периодического
действия. Приведенное выше математическое доказательство
можно дополнить следующей аргументацией: в режиме поршне-
вого потока при постоянной скорости каждый элементарный
объем жидкой фазы движется вдоль трубчатого реактора без
какого-либо взаимодействия с соседними объемами. Следова-
тельно, система полностью сегрегирована, и каждый элементар-
ный объем ведет себя как реактор периодического действия.
Отсюда следует, что если исходная смесь в реакторе периоди-
ческого действия имеет тот же состав, что и смесь, подаваемая
в реактор полного вытеснения, и если среднее время пребыва-
ния Lfu в последнем равно времени реакции в реакторе перио-
дического действия, то и вытекающая из трубчатого реактора
масса будет идентична по своему составу продукту в реакторе
периодического действия. Этой модели отвечает граничное
условие
с ~cQ (9.19)
z=0
Здесь г = 0 отвечает точке ввода исходных веществ в реактор,
а с0 — концентрация компонента С в исходной смеси.
Допустим, например, что кинетика процесса в хемостате
Моно не отличается от кинетики процесса в ТРПВ (или в экви-
валентном реакторе периодического действия). В этом случае
уравнения материального баланса по клеточной массе и субст-
рату [уравнение (9.18)] преобразуются следующим образом:
dx PmaxXS
dt s -f- Ks
ds _ 1 PmaxXS
~dT~~ Y s+Ks
с начальными условиями
%(0)=Xo s(O)=so
Логическим путем или посредством преобразования уравнений
(9.20) и (9.21) нетрудно найти, что концентрации s и х связа-
ны стехиометрически
х+ Уз = %о+ (9.23)
(9.20)
(9.21)
(9.22)
2-746
18
Глава 9
Если при помощи уравнения (9.23) выразить х через s и под-
ставить полученное выражение в уравнение (9.21), то мы полу-
чим обычное дифференциальное уравнение:
__ Ртах 1хо Т~ Е (s0 s) ] S
dt ~ Y s + Ks '
Это уравнение при условии (9.22) можно интегрировать анали-
тическим путем; тогда получим:
+Y («,+К.) In _ К,у In 2- - (x0+1%)
Л0 d0
(9.25)
Концентрации субстрата на выходе из реактора отвечает зна-
чение 5 при t=Lju-, соответствующее значение х легко найти с
помощью уравнения (9.23). Если считать, что уравнение (9.20)
описывает кинетику периодического процесса, нетрудно видеть,
что эта математическая модель не учитывает лаг-фазу и фазу
отмирания клеток, но отражает стационарную фазу клеточного
роста.
В отличие от ПРПП стерильность исходных веществ в ТРПВ
автоматически предполагает и нулевую концентрацию биомас-
сы на выходе из реактора, поскольку поршневой характер по-
тока предотвращает инокуляцию движущегося по трубчатому ре-
актору элементарного объема жидкости. Этот недостаток мож-
но устранить, например, путем предварительной инокуляции по-
ступающего в реактор потока исходных веществ.
Не представляет затруднений интегрирование уравнений ма-
териального баланса по отдельным компонентам в реакторе
полного вытеснения; таким образом можно получить выраже-
ния, описывающие связь между концентрациями компонентов в
продуктах и общим временем пребывания в реакторе L/u для
некоторых обычных реакций, катализируемых ферментами. Ре-
зультаты таких расчетов приведены в табл. 9.2.
Сравнительные характеристики идеальных ПРПП и ТРПВ
зависят от типа осуществляемых в них реакций и их кинетики.
При протекании одностадийной реакции с обычной кинетикой
(когда скорость реакции снижается при увеличении превраще-
ния субстрата; примером могут служить реакции, описываемые
уравнением Михаэлиса — Ментен) ТРПВ обеспечивает более
высокую степень превращения субстрата и большую концентра-
цию продукта реакции на выходе из реактора по сравнению с
ПРПП равного объема. Для автокаталитических реакций (ког-
да скорость реакции возрастает при снижении концентрации
субстрата) справедливо обратное соотношение характеристик
ТРПВ и ПРПП. В микробиологических процессах ТРПВ обыч-
Проектирование и расчет биологических реакторов
19'
Таблица 9.2. Взаимосвязь между степенью превращения субстрата
6= (s0—s)/s0 и важнейшими параметрами ферментативных реакций в ТРПВа
Данные относятся к ферментам в растворе или к гранулам
иммобилизованных ферментов с пренебрежимо малым сопротивлением
массопередаче; для ферментов в растворе выражение (1—е)/е
равно единице
Уравнение, описывающее
скорость реакции и
Уравнение, описывающее [(1— е)/е] • [Lv;nax/u]
в ТРПВ
Уравнение Михаэлиса — Мен-
тен:
^тах5 s06—Amln(l -б)
Уравнение Михаэлиса — Мен-
тен для обратимой реакции:
flmax(s—Р/К) Кт + s + Ктр!Кр s° G \т — ~' \ Ар / L — ^Ат + $о + 1п(1 — &б) \ А р / 0 , К + 1 где b = К
Конкурентное ингибирование
продуктом реакции:
Атах$ H-Km(l + P/Ki) So6-^ [6-{-ln(I — б)]- \ Ai / / Am \ — 1 Am + So + „ Po I ln( 1 \ Aj J
Ингибирование субстратом:
gmar 1 4" Kmls + s/Ki ( 62 \ S06- Amln( 1 - 6) + -Мб- — A i \ /
а Воспроизведено из работы: Messing R. A., Immobilized Enzymes for Industrial Re-
actors, p. 158, Academic Press, Inc., New York, 1975.
но обеспечивает большую концентрацию продукта в вытекаю-
щем из реактора потоке. В то же время необходимость непре-
рывного внесения посевного материала и ряд затруднений тех-
нического характера при осуществлении газового обмена в
ТРПВ часто приводят к тому, что на практике выгоднее при-
менять аналогичные реакторы периодического действия, даже
если высокая концентрация продукта в вытекающем из реак-
тора потоке является важным фактором. В фазе экспоненциаль-
ного роста ПРПП эффективнее ТРПВ или реакторов периоди-
ческого действия. Сравнительное изучение идеальных ТРПВ и
2
20
Глава 9
ПРПП при их использовании для различных простых сочета-
ний реакций составляет основную тему учебных пособий по хи-
мической технологии, перечисленных в списке литературы в
конце этой главы. С другими примерами сравнительного изуче-
ния различных реакторов мы познакомимся в упражнениях.
9.2. Динамика процессов в реакторах
В этом разделе мы рассмотрим динамические характеристи-
ки биореакторов. Хотя основное внимание мы будем уделять
динамике процессов в ПРПП, многие из анализируемых здесь
принципов и концепций можно успешно перенести и на ре-
акторы других типов. Сначала выведем основные уравнения,
необходимые для описания работы реактора в нестационарном
состоянии, а затем рассмотрим возможность применения этих
уравнений для изучения процессов в биореакторах в неустано-
вившемся состоянии. Как мы увидим, успешному применению
изучаемых здесь подходов часто препятствует отсутствие кине-
тических моделей, достаточно точно описывающих особенности
переходного состояния.
9.2.1. Динамические модели
В основу изучения динамики процессов в ПРПП может быть
положено уравнение материального баланса (7.4), преобразо-
ванное в соответствующее уравнение для нестационарного со-
стояния:
---- (общее количество компонента в реакторе) =
dt
(скорость \ /скорость
поступления I — I выведения
в реактор) / \ из реактора
скорость
образования
в реакторе
Тогда для компонента i в реакторе с полным перемешиванием
получим:
^-(rR-ei) = K(ci/-ci)+Vf< (9.26)
dt
Если считать плотности потока исходных веществ и содержи-
мого реактора равными, то при одинаковых объемных скоро-
стях на входе в реактор и на выходе из него объем содержимо-
го реактора будет постоянным и уравнение (9.26) можно при-
вести к такому виду:
dc{
"dt
=D(cif—ct)+rfi
(9.27)
Проектирование и расчет биологических реакторов
21
Это уравнение материального баланса по компоненту i в не-
стационарном состоянии является отправным пунктом при изу-
чении динамики процессов в реакторе. Прежде чем приступить
к изучению некоторых общих математических приемов, успеш-
но используемых при анализе динамики процессов в реакторах,
следует привести ряд новых доводов, подтверждающих обосно-
ванность применения модели ПРПП для соответствующих рас-
четов. Дело в том, что здесь мы сосредоточим внимание только
на явлениях перемешивания, а соответствующую модель кине-
тики биологических превращений для анализа процессов в пе-
реходном состоянии рассмотрим позднее. Как мы уже упомина-
ли в гл. 8, одним из важных параметров перемешивания явля-
ется время циркуляции. Этот параметр приближенно характе-
ризует время, необходимое, чтобы элемент жидкости, циркули-
рующий в соответствии с существующей в реакторе структурой
течений, возвратился в тот же участок объема реактора. Этот
параметр применим к ПРПП только в том случае, если время
циркуляции мало по сравнению с масштабом времени других
процессов в ПРПП.
При изучении динамики процессов в ПРПП появляется не-
обходимость во введении нового параметра времени. Действи-
тельно, теперь мы сталкиваемся с возможностью изменения во
времени скорости ввода или концентрации питательных ве-
ществ. Тогда приближение идеального перемешивания будет
справедливым только в том случае, если время циркуляции бу-
дет намного меньше характерного масштаба времени флуктуа-
ций в составе поступающего в реактор потока исходных ве-
ществ или любых других веществ, например раствора основа-
ния, с помощью которого поддерживается необходимое значе-
ние pH. Все сказанное относится и к только что рассмотрен-
ным идеальным реакторам периодического действия с добавле-
нием субстрата.
Уравнение (9.27) применимо к любому компоненту, рас-
сматриваемому в модели биореактора. Следовательно, в самом
общем (и самом сложном) случае динамическая модель реак-
тора должна состоять из системы уравнений типа уравнения
(9.27), каждое из которых обычно связано с другими через па-
раметры скоростей образования . Таким образом, в общем
случае скорость образования компонента I зависит от концент-
раций всех других компонентов в реакторе. Чтобы не записы-
вать большое количество уравнений, здесь удобно воспользо-
ваться методами векторного исчисления и матричной алгебры.
В дальнейшем строчными жирными буквами (например, с) мы
будем обозначать векторы, а прописными жирными буквами
(например, А) — матрицы. Тогда систему уравнений, состоя-
22
Глава 9
щую из уравнений типа (9.27), можно записать в форме:
-^-=Г(с(О.Р) (9.28)
Здесь с — вектор концентраций m-го измерения (т— число
элементов или компонентов; m-вектор), которое равно числу
компонентов, рассматриваемых в модели; р — ^-вектор пара-
метров модели (в том числе концентраций питательных ве-
ществ, скорости разведения, кинетических параметров); i-й
компонент векторной функции f равен правой части уравне-
ния (9.27).
Поскольку описываемая уравнением (9.28) система в общем
случае нелинейна, то при ее анализе без тех или иных прибли-
жений обычно не удается продвинуться очень далеко. Часто
основной интерес представляют динамические свойства систе-
мы вблизи какого-либо конкретного стационарного состояния
cs. В системе уравнений (9.28) вектор концентраций в стацио-
нарном состоянии должен удовлетворять условию:
f(cs, р)=0 (9.29)
Можно попытаться определить поведение системы вблизи cs
путем разложения правой части уравнения (9.28) в ряд Тейло-
ра относительно cs, пренебрегая всеми членами второго и выс-
шего порядков в отклонениях Ci(t)—cis, поскольку они должны
быть малы. Тогда мы получим следующее приближенное ли-
нейное описание нашей системы:
-^- = Ах(0 (9.30)
dt
где x(Q—вектор отклонений от стационарного состояния с8:
х(/)=с(/)—cs (9.31)
Элемент ац в i-м столбце матрицы А определяется выражением:
ди== (9.32)
dcj
Следует подчеркнуть, что А отвечает какому-либо конкретному
стационарному состоянию. Для некоторых систем при данном р
характерно несколько стационарных состояний, каждому из ко-
торых отвечает своя матрица А.
Определение динамических свойств линеаризованной систе-
мы не представляет особых затруднений, поскольку все реше-
ния уравнения (9.30) обычно имеют форму:
х(0 = 2 <9-33)
Z=1
Проектирование и расчет биологических реакторов
23
Параметры & и X; представляют собой соответствующие пары
собственных векторов и собственных значений А. Таким обра-
зом, условию Х = Х»- удовлетворяет следующее характеристиче-
ское уравнение:
det (А — XI) =0 (9.34)
Здесь I — единичная матрица, а р,- удовлетворяют
(А—Хг1)0,- = О i-l,..., т (9.35)
Символом аг обозначены константы, выбранные таким образом,
чтобы они отвечали заданным начальным условиям; следова-
тельно, они удовлетворяют алгебраическим уравнениям
т
£афг = х(0) (9.36)
i=i
где х(0)—заданный вектор начальных отклонений.
Описанная линеаризованная динамическая модель реактора
представляет собой систематическую основу для оценки харак-
терных времен отклика системы. Что касается рассматривае-
мой здесь локальной динамики, то из уравнения (9.33) следует,
что масштаб времени затухания возмущений относительно
стандартного стационарного состояния cs характеризуется соб-
ственными значениями Xi матрицы А. Таким образом, для ло-
кального поведения системы свойствен спектр характерных вре-
мен, приближенно описываемых уравнением:
Ц = |Хг|-1 i = l,...,/n (9.37)
Этими значениями можно пользоваться, например, для сравне-
ния относительных масштабов времени изменений в реакторе и
в поступающих в реактор потоках.
Хотя уравнение (9.37) позволяет систематически и локаль-
но достаточно строго оценить масштабы времени, полученным
таким путем оценкам трудно приписать определенный физиче-
ский смысл. Собственные значения Xi в общем случае являются
функциями всех переменных, входящих в матрицу А, и как та-
ковые зависят от всего вектора стационарного состояния cs и
всего вектора параметров р. В такой ситуации трудно сравни-
вать масштабы времени перемешивания, реакции и других
взаимодействий в системе. Можно показать, что в случае ПРПП
собственные значения имеют форму —(значение, характер-
ное для данной сети реакций)*. Эта закономерность в каком-то
* Fjeld М., Asbjernsen. О. A., Astrotn К- Reaction Invariants and their
Importance in the Analysis of Eigenvectors, State Observability, and
Controllability of the Continuous Stirred Tank Reactor; Chem Ene Sci 29
1917 (1974). '
24
Глава 9
смысле может оказаться полезной, но тем не менее она не поз-
воляет выяснить сложные зависимости между другими парамет-
рами и Л,;. Поэтому для разработки достаточно обоснованных и
в то же время математически не вполне строгих подходов к оп-
ределению различных характерных масштабов длины и време-
ни необходимы прежде всего здравый смысл, большой опыт и
своего рода искусство. Впрочем, в химической технологии хоро-
шо известен систематический подход к решению таких задач,
основанный на переводе всех уравнений системы в безразмер-
ную форму с последующими преобразованиями, позволяющи-
ми определить безразмерные параметры, которые характери-
зуют поведение системы. Часто в качестве базразмерных пара-
метров используют отношения масштабов характерных длин
или времен.
9.2.2. Устойчивость
В этом разделе мы рассмотрим зависимость динамических
характеристик системы в реакторе от функции f и ряда задан-
ных значений параметров р. Для нас наибольший интерес здесь
будет представлять локальная устойчивость определенного ста-
ционарного состояния cs. Если стационарное состояние локаль-
но асимптотически устойчиво, то после небольшого возмущения,
вызывающего малые отклонения концентраций компонентов
системы от изучаемого стандартного стационарного состояния,
они вновь примут исходные значения стационарного состояния.
В неустойчивом стационарном состоянии некоторые небольшие
возмущения приведут к необратимому отклонению концентра-
ций в системе от значений, характерных для стационарного со-
стояния. Во избежание двусмысленного толкования полезно пе-
ревести эти положения на более строгий язык математики.
Мы будем называть стационарное состояние cs локально
асимптотически устойчивым, если lim^oo cs при условии,
что начальное состояние с0 достаточно близко к cs. [Здесь ма-
тематической мерой близости векторов является евклидова нор-
ма, определяемая как
(m X 1/2
V с<2
/=1 /
Тогда выражение «с0 и cs достаточно близки» означает, что
| Со—cs| представляет собой достаточно малое действительное
число.] Стационарное состояние cs называют глобально асимп-
тотически устойчивым, если lim^0Oc(Z) =cs при любом Со (за
исключением бессмысленных значений, например отрицатель-
ный концентраций). Если cs — неустойчивое стационарное со-
Проектирование и расчет биологических реакторов
25
стояние, то некоторые сколь угодно близкие к cs начальные
состояния Со будут приводить к зависимости с от t, причем с
не будет приближаться к cs и не будет располагаться сколь
угодно близко от cs. Таким образом, состояние неустойчивости
характеризуется тем, что при некоторых начальных отклонени-
ях величина отклонения х(/) стремится к росту по сравнению
с ее начальным значением.
Локальная устойчивость в большинстве случаев определяет-
ся собственными значениями Хг матрицы А [уравнение (9.32)].
Стационарное состояние cs локально асимптотически устойчи-
во, если действительная часть всех собственных значений А
отрицательна:
Re(A,;)<0 i=l, т (9.38)
Напротив, cs неустойчиво, если действительная часть любого
собственного значения А положительна:
Re(A,/)>0 при любом / (9.39)
Эти выводы легко понять, если обратиться к уравнению (9.33).
В самом деле, если наибольшая действительная часть собствен-
ных значений системы равна нулю, то мы имеем дело с крити-
ческим состоянием и для определения локальных динамических
характеристик системы необходим дальнейший анализ (см., на-
пример, работу [Ю]). К счастью, для проверки указанных вы-
ше неравенств нет необходимости рассчитывать все собствен-
ные значения. Можно, например, преобразовать определитель
уравнения (9.34) в алгебраическое уравнение m-го порядка:
+ ... +Bm-^ + Bm = 0 (9.40)
Теперь можно применить критерий Гурвица*, который устанав-
ливает, что все корни уравнения (9.40) имеют отрицательные
действительные части в том и только в том случае, если удов-
летворяются следующие условия (9.41):
В, > О
det
В3~
В2
О
В,
В3 В5
det 1
в2 В4 > О
О в. в3
* Walter С. F., Kinetic and Biological and Biochemical Control Mechanisms,
p. 335 in Biochemical Regulatory Mechanisms in Eucaryotic Cells, Kun E., Gri-
sola S. (eds.), John Wiley and Sons, Inc., New York, 1972.
26
Глава 9
Bi В3 Bs
1 в2 В,
О В1 в3
о
о
(9.41}
В качестве примера рассмотрим ситуацию, когда клеточный
рост лимитируется одним субстратом, и изучим динамический
вариант модели хемостата Моно. Применение как к биофазе,,
так и к субстрату общего уравнения материального баланса
для нестационарного состояния [уравнение (9.27)] и использо-
вание уравнения Моно [уравнение (7.10)] для удельной скоро-
сти роста клеток приводит к следующим выражениям:
-^- = D(x0-x) + -!^- (9.42а>
dt s -J- /\§
И
— = D (s0—s)------!----
dt Ух/s x + Ks
При стерильности исходных питательных веществ (хо=0) воз-
можны два стационарных состояния, одно из которых (нетри-
виальное) описывается уравнениями (7.14) и (7.15), а другое
отвечает состоянию вымывания (х = 0; s=so). Определив устой-
чивость каждого из этих стационарных состояний, можно опре-
делить, какое из них будет наблюдаться в проточной культуре.
Локальную устойчивость можно изучать с помощью линеаризо-
ванных уравнений типа (9.42). Результаты такого изучения ло-
кальной стабильности в хемостате Моно приведены ниже:
(9.42б>
„ Чтах«о ^тах5° Ks-t-$o
Нетривиальное стационарное состояние [уравнения (7.14) и Неустойчиво Устойчиво
(7.15)1 Стационарное состояние вы- Устойчиво Неустойчиво
мывания
Из этого анализа следует также, что концентрации компо-
нентов системы не могут достигать величин, соответствующих
стационарному состоянию, в режиме затухающих колебаний.
Проектирование и расчет биологических реакторов
27
Поскольку подобные колебательные изменения концентраций на-
блюдались экспериментально, то описываемая модель системы
субстрат — клетка, очевидно, не может предсказывать все дина-
мические особенности этой системы в некоторых специфических
условиях.
Известны и другие недостатки динамической модели хемо-
стата Моно. Так, она предсказывает мгновенный отклик удель-
ной скорости клеточного роста на изменение концентрации суб-
страта, а в эксперименте обнаружена лаг-фаза (см. упражне-
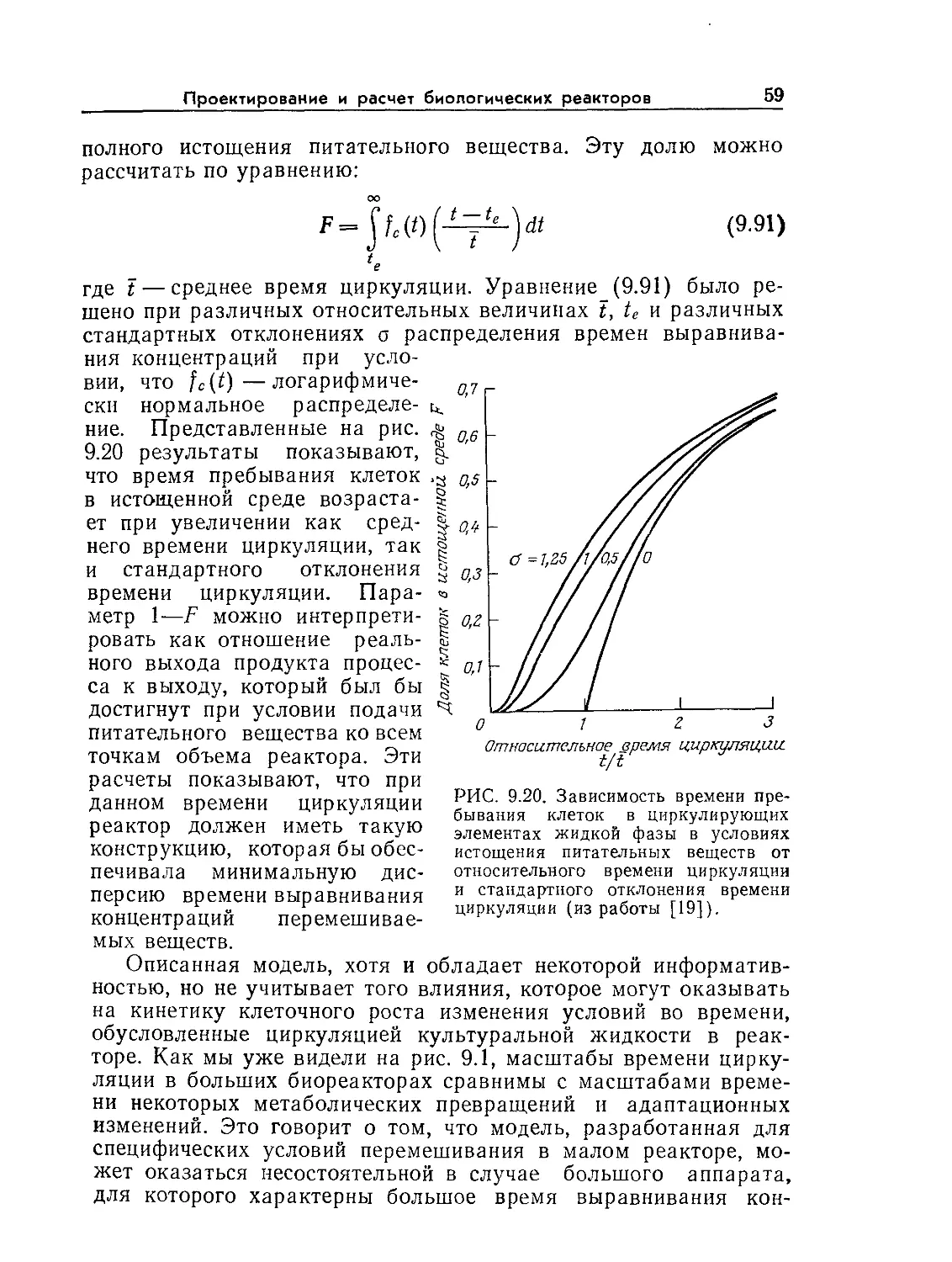
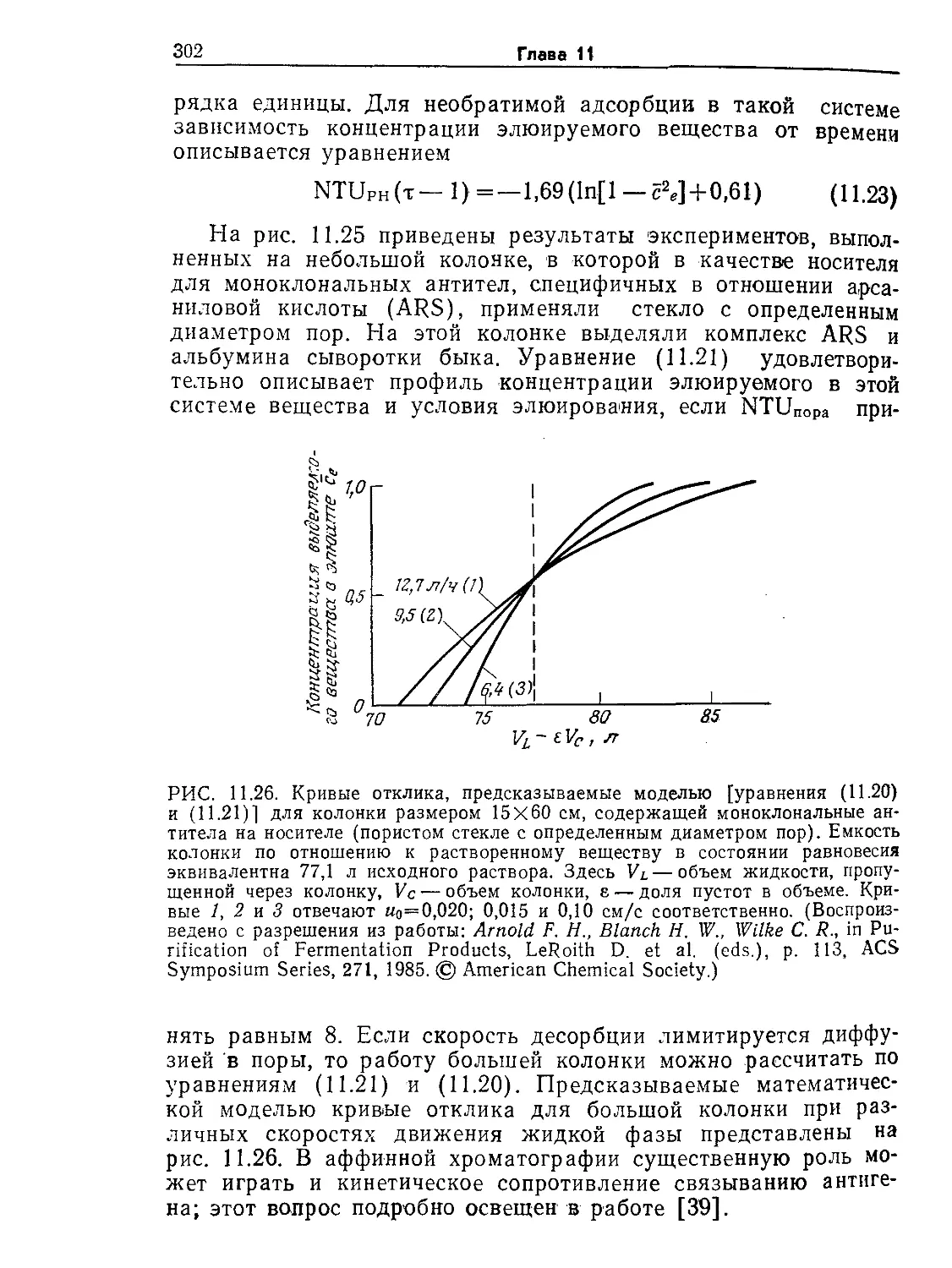
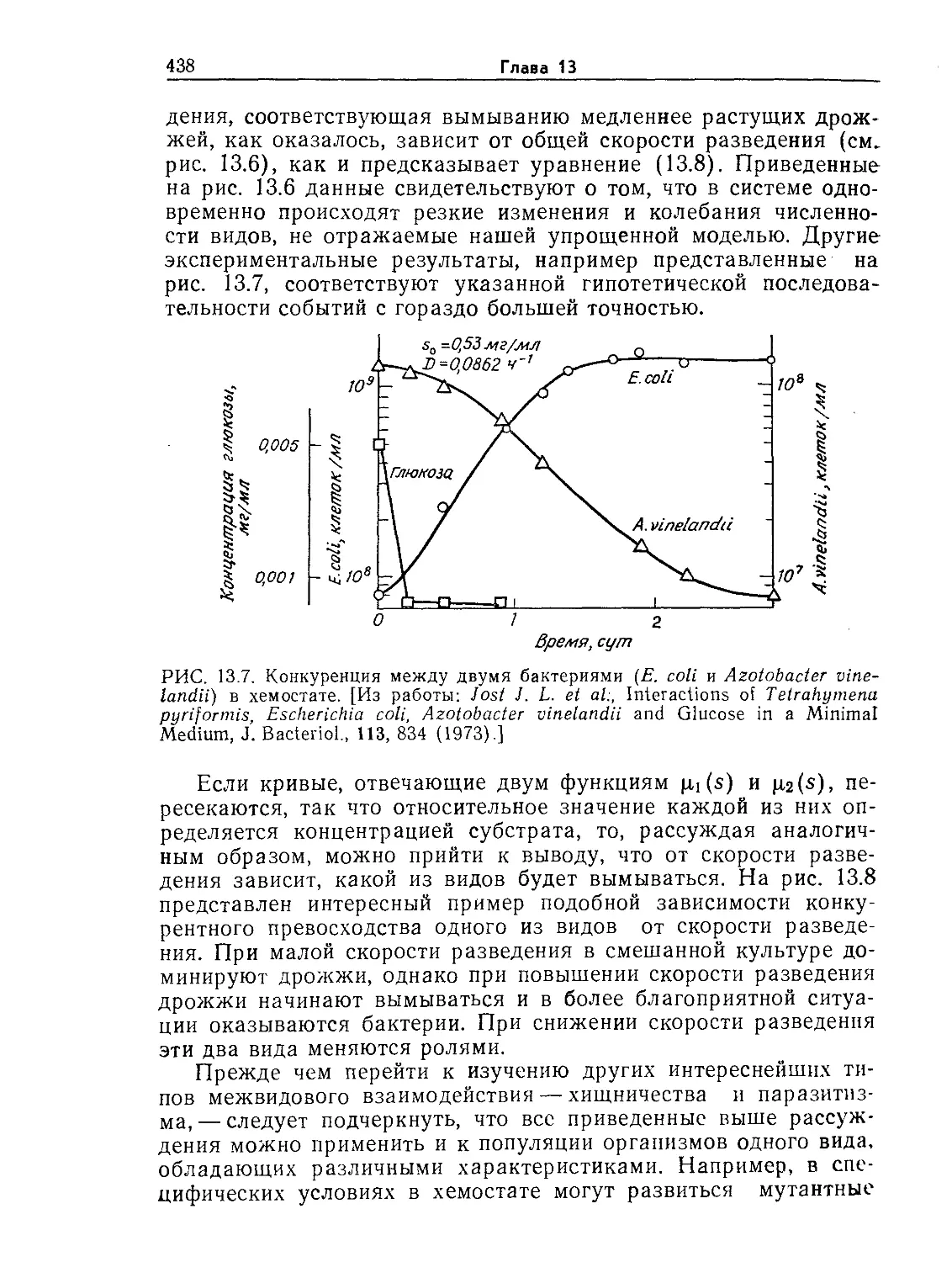
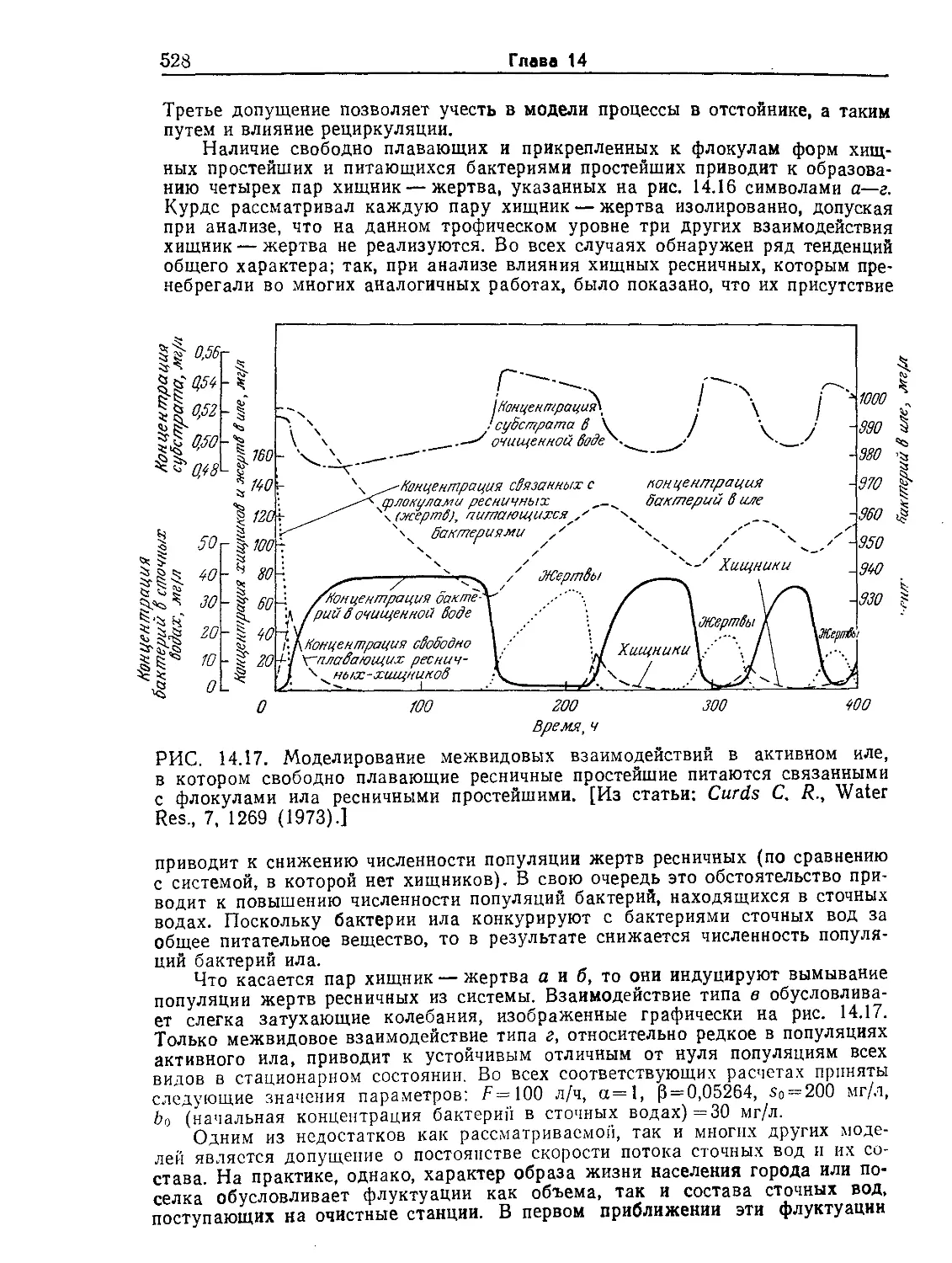
ние. 9.6. Устойчивые колебания концентрации пирувата (жирные точки)
в непрерывной культуре Е. coli. Обратите внимание на то, что концентрация
клеток (отмечена крестиками) остается приблизительно постоянной. [Воспро-
изведено из статьи: Sikyta В., Continuous Cultivation of Microorganisms; Suom.
Kemistil., 38, 180 (1965).]
ние 10.6). Кроме того, экспериментально наблюдались гистере-
зис скорости клеточного роста и нестабильность экономического
коэффициента, а в некоторых экспериментах обнаружены и
устойчивые колебания концентраций (рис. 9.6). Следовательно,
модель хемостата Моно, в ряде случаев вполне приемлемая для
анализа стационарных состояний, имеет многочисленные недо-
статки, когда речь идет об изучении динамики процессов.
Некоторые явления, которые нельзя объяснить при помощи
модели Моно, могут быть учтены путем введения в модель до-
полнительных переменных (посредством ее «структурирова-
ния»), В таких случаях необходимость структурированных мо-
делей обусловливается некоторыми соображениями общего по-
рядка, упоминавшимися ранее в гл. 7 и во введении к этой гла-
ве. В переходном состоянии приближение сбалансированного
роста не применимо, если масштаб времени изменений в среде
сравним с масштабом времени биологического отклика (напри-
мер, посредством индукции или репрессии синтеза ферментов).
В таких случаях кинетическую биологическую модель необхо-
димо расширить, включив в нее большее число компонентов
(или псевдокомпонентов; разд. 7.4) клеточной фазы.
В разд. 7.4.1 мы уже познакомились с двухкомпонентной
структурированной моделью Уильямса. В применении к динами-
ке роста проточных культур эта модель воспроизводит некоторые
наблюдаемые в эксперименте особенности систем, не отражае-
мые моделью Моно. Расширяя аналогию между уравнением
Моно для клеточного роста [уравнение (7.10)] и уравнениями
кинетики ферментативного катализа, Джеффресон и Смит
включили в свою динамическую модель хемостата промежуточ-
ные компоненты, в известной степени отвечающие фермент-суб-
стратным комплексам [11]. В модели Рамкришны, Фредриксо-
на и Цучии учтено ингибирование клеточного роста [12]. В ка-
ком-то смысле такой подход можно рассматривать как струк-
турирование фазы питательного вещества.
Совершенно другой подход применили Ли, Джекман и Шрё-
дер, изучавшие влияние флокуляции клеток на процесс их ро-
ста [13]. Мы уже указывали, что во многих микробных систе-
мах индивидуальные клетки образуют скопления (иногда назы-
ваемые флокулами или хлопьями). Считается, что в таких
скоплениях метаболические процессы могут отличаться от ана-
логичных процессов в отдельных диспергированных клетках.
Действительно, для того чтобы питательные вещества могли
войти в контакт с клетками, расположенными внутри скопле-
ний, они должны сначала диффундировать внутри этих скопле-
ний. Поэтому биофаза рассматривается как двухкомпонентная
система (скопления клеток и индивидуальные клетки), в ко-
торой каждый компонент характеризуется своей кинетикой и в
которой в то же время возможен обмен отдельными клетками
между двумя морфологически различными формами. Эта мо-
дель отражает флуктуации общего экономического коэффициен-
та, гистерезис скорости роста и характеризуется замедленным
откликом по сравнению с моделью Моно. Вообще такая модель
гораздо лучше согласуется с экспериментальными данными,
чем модель Моно.
Для модели Янга, Брюли и Бангэя характерен другой под-
ход [14]. Эти исследователи предположили, что в силу сопро-
тивления процессам массопередачи, обеспечивающим клетку
питательными веществами, концентрация субстрата внутри
клетки не равна концентрации питательного вещества в среде,
причем на скорость роста клетки непосредственно влияет имен-
но первая величина. Основанная на таком подходе модель от-
ражает часто наблюдаемую в эксперименте лаг-фазу после из-
менения условий среды.
Проектирование и расчет биологических реакторов 29
В завершение нашего обзора динамики процессов в хемо-
стате отметим еще один потенциально важный фактор, не учи-
тываемый моделью Моно. Если клеточный рост ингибируется
избытком питательного вещества, то для удельной скорости
роста клеток следует применять уравнение (7.32):
..____Hmaxs____
^s + s+sWp
Поведение системы в хемостате, в котором удельная скорость
клеточного роста определяется этим уравнением, может суще-
ственно отличаться от поведения системы в классическом хемо-
стате Моно: теперь в некоторых условиях могут наблюдаться
три стационарных состояния. Динамическое поведение такой си-
стемы может быть весьма сложным, а вклад нелинейных эффек-
тов, не учитываемых в анализе локальной устойчивости, очень
велик. Предполагалось, что эта модель с ее необычными харак-
теристиками окажется полезной в объяснении ряда трудностей,
возникающих обычно в процессах анаэробной переработки от-
ходов. Некоторые другие проблемы, связанные с эффектами
субстратного ингибирования в ПРПП, будут рассмотрены в
гл. 14.
Сложное динамическое поведение может быть характерным
и для смешанных культур, состоящих из клеток различных ви-
дов. Биологические превращения в подобных системах мы рас-
смотрим детальнее в гл. 13. Там же мы познакомимся с други-
ми общими математическими методами и результатами их при-
менения в анализе и в описании динамики процессов в реак-
торах.
9.3. Реакторы с неидеальным перемешиванием
Закончив изучение идеальных реакторов с полным переме-
шиванием или трубчатых реакторов полного вытеснения, кото-
рые можно воспроизвести в лабораторных мелкомасштабных
экспериментах, рассмотрим теперь более реальную ситуацию,
типичную для больших промышленных реакторов. Здесь основ-
ное внимание будет уделено методам изучения параметров пе-
ремешивания и структуры течений в резервуарах реакторов,
применению полученных данных в проектировании биореакто-
ров, а также исследованию некоторых взаимосвязей между био-
логическими или биокаталитическими реакциями и характером
перемешивания и структурой течений в реакторе. Сначала рас-
смотрим вопрос о времени выравнивания концентраций компо-
нентов в перемешиваемых резервуарах, что необходимо для
знакомства с масштабами времени; здесь мы познакомимся так-
30
Глава 9
Без отражательных
перегородок
ектрод
Введение
индикаторов
|— Электрод
Несколько отражательных
перегородок, большая
лопастной меш-алка.
Время
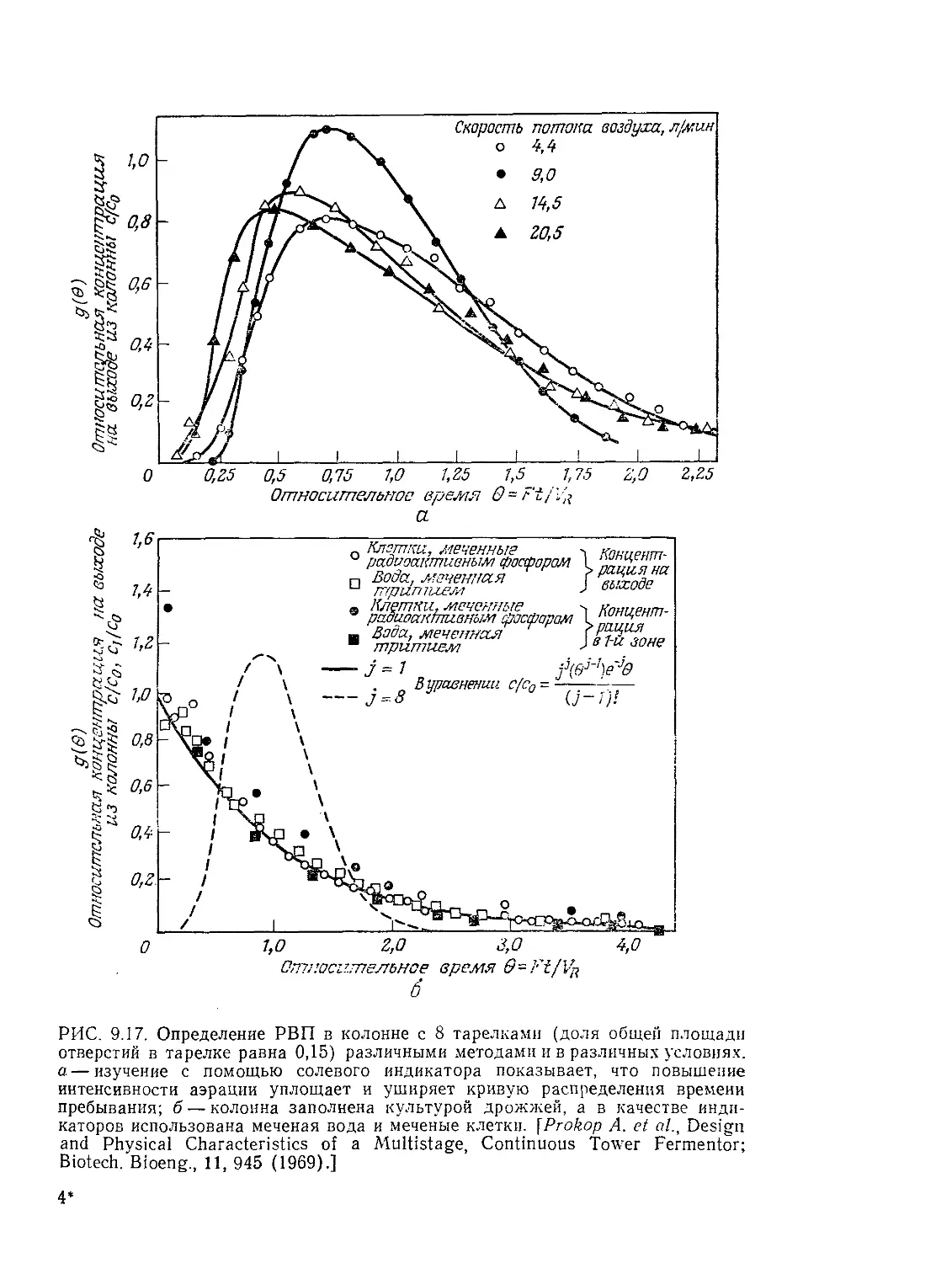
Проектирование и расчет биологических реакторов ЗВ
же с существованием крупномасштабных циркуляций в реакто-
рах и получим некоторое представление о характерном для био-
реакторов порядке величин времени циркуляции.
9.3.1. Время выравнивания концентраций в реакторах
с перемешиванием
Под временем выравнивания концентраций понимают время,,
необходимое для достижения определенного уровня гомогенно-
сти содержимого реактора после импульсного введения индика-
тора в определенную зону реактора. Функции индикатора мо-
жет выполнять раствор соли, кислоты или основания, нагретая
или охлажденная жидкость. Циркуляционные характеристики
реактора и время выравнивания концентраций можно опреде-
лить путем непрерывного мониторинга концентрации индикато-
ра в одной или нескольких точках реактора. Как схематично
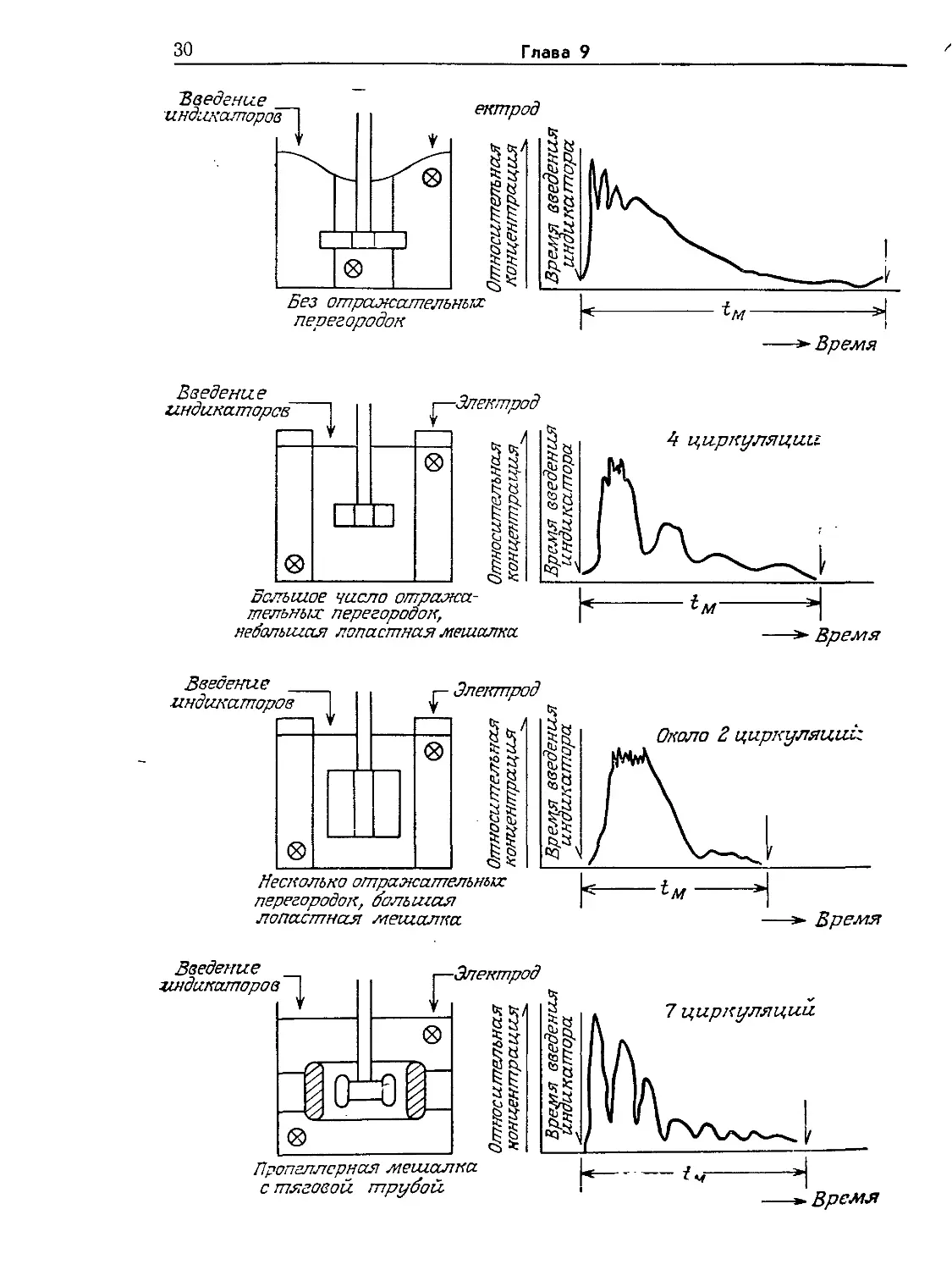
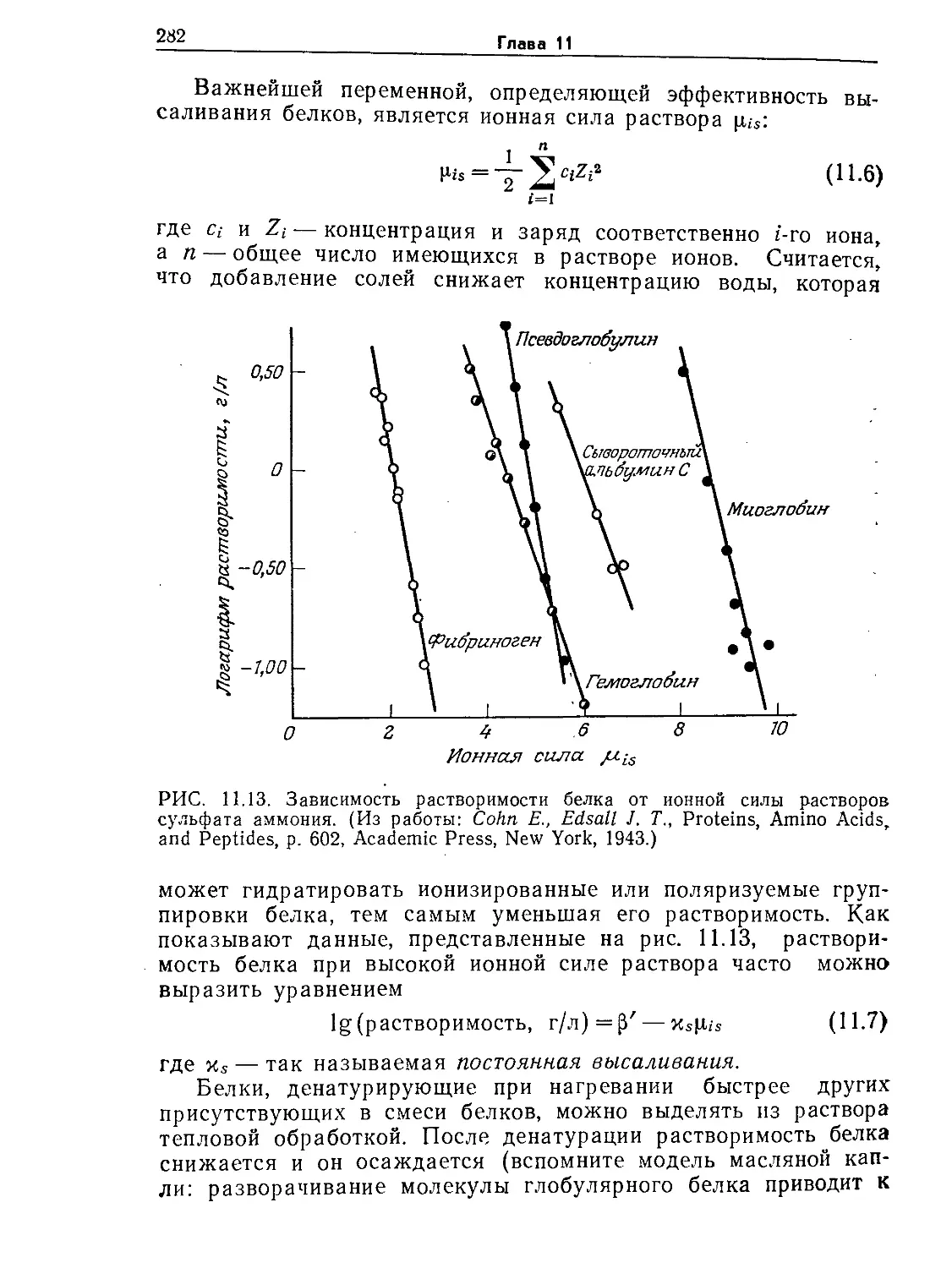
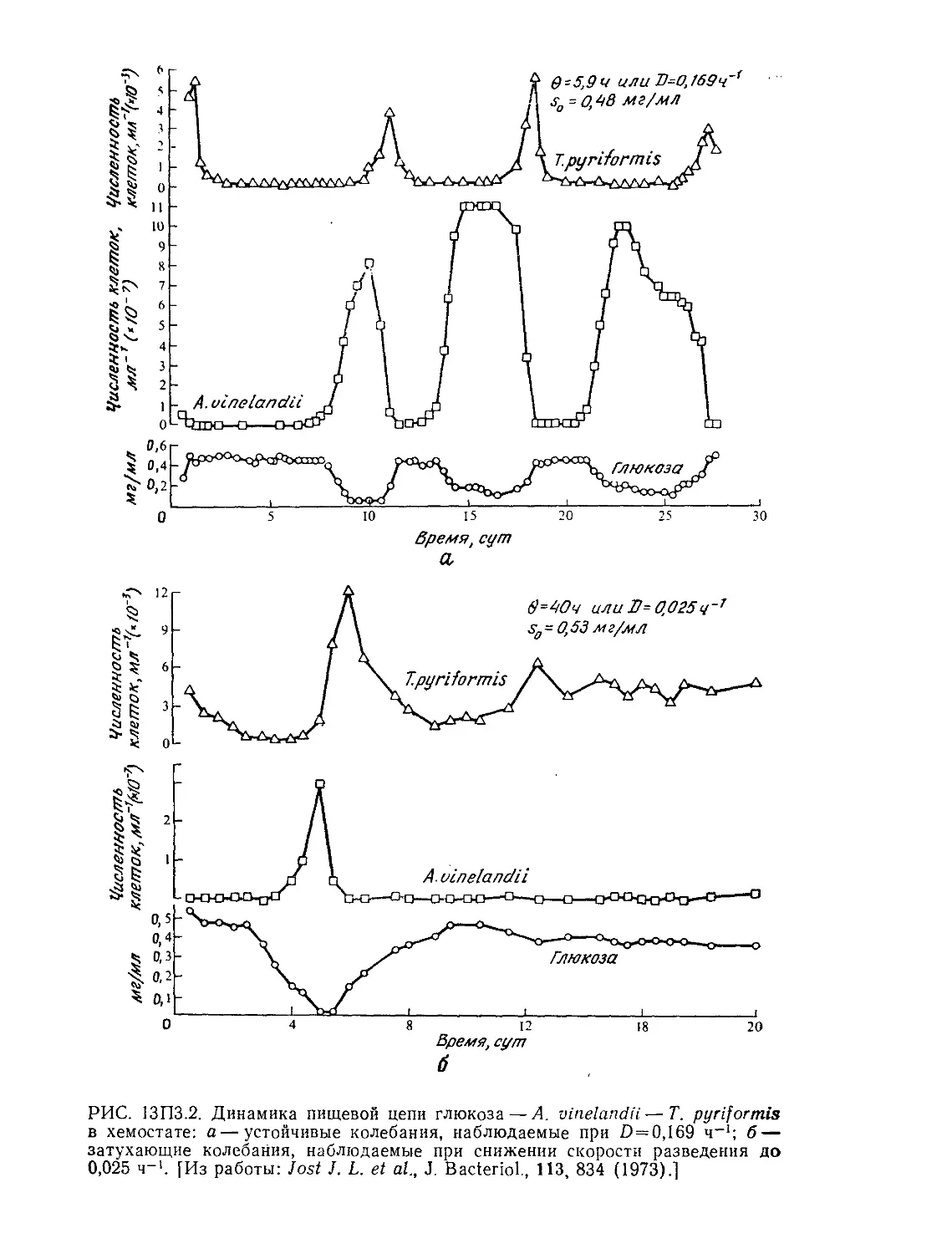
показано на рис. 9.7, характер циркуляции и время выравнива-
ния концентраций в общем случае зависят от конструкции ре-
актора (в особенности от наличия и конструкции внутренних,
приспособлений) и перемешивающих устройств. На представ-
ленных на том же рисунке графиках отражен периодический
характер изменения концентрации индикатора, указывающий
на то, что для выравнивания концентраций компонентов смеси
необходимо определенное число циркуляций всего содержимо-
го реактора. Время циркуляции является важным параметром
и с другой точки зрения: оно приближенно указывает харак-
терный временной интервал, в течение которого суспендирован-
ные в перемешиваемой жидкости клетки или частицы биоката-
лизатора циркулируют через различные зоны реактора, воз-
можно, сталкиваясь на этом пути с различными условиями
реакций. Затем, как упоминалось выше, необходимо опреде-
лить, могут ли эти флуктуации условий в силу своей интенсив-
ности и временного диапазона в существенной степени влиять
на локальное кинетическое поведение системы. К вопросу о
времени выравнивания концентраций вы вернемся в заключи-
тельной части этого раздела при анализе некоторых экспери-
ментальных работ, повященных изучению влияния перемеши-
вания на активность биокатализаторов.
С математическими выражениями, позволяющими опреде-
лить время выравнивания концентраций в ньютоновских жидко-
РИС. 9.7. Влияние различных перемешивающих и других внутренних
устройств в резервуарах с перемешиванием на соответствующие характерные
отклики системы, наблюдаемые после импульсного введения индикатора. От-
клик системы здесь представлен как отклонения от концентрации индикатора'
в конечном, полностью перемешанном состоянии. (По данным Нагаты [15]
32
Глава 9
стях и мицелиальных культурах, и с экспериментальными дан-
ными по оценке времени выравнивания концентраций в раство-
рах полисахаридов, продуцируемых микроорганизмами, можно
познакомиться в обзоре Чарльза [18]. Установлено, что в не-
которых ферментерах емкостью 2,5—160 м3 время выравнива-
ния концентраций составляет 29—104 с. С другой стороны, со-
общалось, что в ряде случаев время выравнивания концентра-
ций достигает нескольких минут. В реакторе объемом 25 м3 при
глубокой струйной аэрации время выравнивания концентрации
составило 80 с (в случае воды). Чарльз указывал, что время
выравнивания концентраций в 1%-ном растворе ксантана рав-
но приблизительно 6 мин при скорости вращения мешалки
300 об/мин в отсутствие аэрации и около 1 мин при скорости
вращения мешалки 500 об/мин и аэрации воздухом, когда объ-
ем последнего составляет 0,25% объема смеси. С другой сторо-
ны, упоминалось, что в небольших реакторах время выравнива-
ния концентраций составляет 2—3 с. Эти величины дают пред-
ставление о порядке величин времени выравнивания концентра-
ций в реакторах различной емкости в зависимости от природы
содержащейся в биореакторе реакционной массы. В данном
разделе мы будем изучать закономерности циркуляции в боль-
ших объемах жидкости и связанную с этим возможность неод-
нородности состава смеси и ее температуры. Анализ меньших
реакторов, где главную роль играют турбулентность и ее взаи-
мосвязь с массопередачей и ростом клеток, был дан в гл. 8.
Очевидно, что некоторое единое время циркуляции, напри-
мер в перемешиваемом резервуаре, является приближенным по-
нятием. Если мы проследим за движением элементов жидкости
из области, прилегающей к мешалке, то различные элементы
будут перемещаться в реакторе по различным траекториям и,
следовательно, будут иметь различное время циркуляции.
Брайант описал метод экспериментального определения распре-
деления времен циркуляции fc(t) с помощью небольшого сво-
бодно плавающего радиопередатчика и контролирующей его
положение антенны, размещенной в реакторе [19]. По опреде-
лению fc(t)dt — доля циркуляций с продолжительностью в ин-
тервале от t до t-\-dt. Брайант указывал, что для реакторов с
перемешиванием распределение времен циркуляции обычно
можно с хорошим приближением описать функцией логариф-
мически нормального распределения:
f ft) =___!__exp Г___(ltU~ fr-)2-l (9.43)
lc щ УГл Р L 2а^ J V
В этом выражении два параметра — среднелогарифмическое
время циркуляции h и стандартное отклонение среднелогариф-
мического времени циркуляции щ — связаны со средним време-
Проектирование и расчет биологических реакторов
33
нем циркуляции t и стандартным отклонением а времени цир-
куляции fc уравнениями
t = e{ii+ai2/2) (9.44а)
o = t^e°i2—l) (9.446)
Экспериментально можно определить t и о, а затем с помощью
уравнений (9.44) найти значения параметров для fc в уравне-
нии (9.43). Позднее в этой же главе мы познакомимся с ис-
пользованием понятия о времени циркуляции для расчета влия-
ния циркуляции жидкой фазы на общие характеристики био-
реактора.
9.3.2. Распределение времени пребывания
Попытаемся представить себе, что случится с небольшим
объемом жидкости после его введения в проточный биореактор
непрерывного действия. Благодаря перемешиванию этот малый
объем будет разбит на еще более мелкие объемы, которые бу-
дут отделены друг от друга и распределены во всем объеме
реактора. Таким образом, какая-то часть первоначально мало-
го объема жидкости достаточно быстро достигнет вытекающего
из реактора потока, а другие части будут в течение различного
времени перемещаться по реактору, пока не попадут в выхо-
дящую из реактора линию. Другими словами, вытекающий из
реактора поток можно представить как смесь элементов жид-
кой фазы с разным временем пребывания в реакторе. Распре-
деление времени пребывания элементов жидкой фазы в покида-
ющем реактор потоке характеризует режим перемешивания и
структуру течений в реакторе. Ниже мы познакомимся с мето-
дами оценки распределения времени пребывания.
Рассмотрим сначала произвольный сосуд с одной линией
подачи исходных веществ и одной линией выведения продуктов
процесса. Допустим на мгновение, что в этой системе совершен-
но отсутствует обратная диффузия в подающую линию из со-
суда и в сосуд из линии выведения продуктов процесса. Для
определения характеристик перемешивания в сосуде можно вы-
полнить эксперимент типа стимул — отклик с использованием
инертного индикатора. Для этого в нулевой момент времени
(/ = 0) введем в линию подачи исходных веществ индикатор в
концентрации с* и продолжим его подачу в течение времени
£>0, а затем проконтролируем отклик системы (в данном слу-
чае концентрацию индикатора на выходе из сосуда) на этот
специфический стимул. На рис. 9.8 схематично изображены ос-
новные узлы такого эксперимента, а также типичная кривая
3—746
3*
Глава 9
зависимости отклика (концентрации индикатора на выходе из
сосуда) от времени c(t).
В таких условиях отклик системы на однократный ввод ин-
дикатора c0(t)=H(t), где
"Н? ti0 (9'45)
определяется как отношение полученной в описанном выше экс-
перименте зависимости c(t) к использовавшейся в этом экспери-
РИС. 9.8. Экспериментальное определение отклика системы (^"-функции) на
импульсный ввод индикатора.
менте концентрации индикатора с* в линии подачи. Рассчитан-
ный таким путем однократный отклик сосуда с перемешивани-
ем называют ^“-функцией (рис. 9.8):
(Г) = —-— = отклик на однократный ввод индикатора
(9.46)
Во многих случаях удобнее применять не функцию
а функцию распределения времени пребывания (РВП)* <^(0>
которая определяется как
$(/)б# = доля выходящего из реактора потока жид- (9.47)
кости, время пребывания которой в реак-
торе составляет от t до i+dt
* Согласно принятой в статистике терминологии, S — функция плотно-
сти, а — соответствующее распределение. В литературе по химической тех-
нологии, однако, закрепилась употребляемая здесь терминология, которой во
избежание недоразумений мы и будем пользоваться в дальнейшем.
Проектирование и расчет биологических реакторов 35
Таким образом, доля выходящего из реактора потока, время
пребывания которого в реакторе меньше t, равна:
t
J $ (х) dx
о
Из определения (9.47) следует, что
Jg(x)dx=l (9.48)
о
Взаимосвязь между функциями и ST можно выяснить с
помощью несложного рассуждения. Возвращаясь к эксперимен-
ту типа стимул — отклик (рис. 9.8), представим, что содержи-
мое сосуда состоит из двух различных жидкостей, причем жид-
кость I содержит индикатор в концентрации с*, а жидкость II
вообще не содержит индикатора. Следовательно, все элементы
жидкости I должны были бы поступить в сосуд за некоторое
время t>Q и тогда любой элемент жидкости I на выходе из ре-
актора в момент времени t находился бы в системе в течение
времени, меньшего t. С другой стороны, жидкость II должна
была бы находиться в реакторе при t = 0, поскольку после это-
го в реактор поступала только жидкость I. Следовательно, вре-
мя пребывания всех элементов жидкости II, покидающих со-
суд в момент времени t, больше t. Допустим, что нам известна
^-функция, тогда мы можем выразить концентрацию индикато-
ра на выходе из реактора с(/) как сумму вкладов жидкостей
I и II:
t со
c(t) = c*- p(x)dx4-0. p(x)dx (9.49)
6 't
Из уравнений (9.49) и (9.46) следует искомое выражение:
t
f(t) = ^(x)dx (9.50)
о
которое путем дифференцирования по t можно преобразовать
в такую форму:
(9.51)
Прежде всего отметим, что, согласно уравнению (9.51), функ-
цию &> (/) можно определить путем дифференцирования функ-
ции ££"(/), найденной экспериментальным путем. Кроме того,
теория линейных систем утверждает, что производная одно-
3*
36
Глава 9
кратного отклика по времени есть одноимпульсный отклик, от-
куда следует, что <S (0 можно интерпретировать как отклик на-
ходящейся в реакторе реакционной смеси на однократное вве-
дение (импульс) индикатора в нулевое время. Хотя «импульс»
представляет собой абстрактное математическое понятие, в экс-
перименте ему приближенно соответствует введение определен-
ного количества концентрированного раствора индикатора в те-
чение очень короткого промежутка времени.
Функцию РВП можно определить не только описанным вы-
ше экспериментальным путем, но иногда и теоретически, если
известна математическая или логическая модель процесса пе-
ремешивания. Например, в случае идеального ПРПП уравне-
ние материального баланса по индикатору (не взаимодействую-
щему с компонентами системы) в нестационарном состоянии
можно записать в такой форме:
= (952)
Для определения результата ^“-эксперимента для этой системы
допустим, что
с(0) = 0 (9.53)
= (9.54)
Решение уравнения (9.52) при условиях (9.53) и (9.54) дает:
= (9.55)
с*
Из уравнений (9.51) и (9.55) следует, что функцию РВП для
ПРПП можно выразить уравнением:
$(/) = — e~Ft'v* (9.56)
VR
Что касается реакторов полного вытеснения, то функция
РВП вытекает уже из определения такого типа реакторов. Дей-
ствительно, если в поток исходных веществ введен импульс ин-
дикатора, то последний проходит через реактор, не смешиваясь
с соседними элементами жидкости, и выходит из реактора че-
рез время L/u. Следовательно, импульс индикатора на выходе
из реактора имеет точно такой же профиль, что и на входе в
реактор, но смещен во времени на однократное время удержи-
вания реактора. Отклонения от такого поведения свидетельст-
вуют о нарушении режима идеального вытеснения.
При работе с функциями распределения типа <§ (t) часто
полезно учитывать и моменты распределения; k-и момент $*(0
Проектирование и расчет биологических реакторов
37
определяется по формуле:
= 6 = 0,1,2... (9.57)
о
Поскольку для определения ^-функции в реактор вводят еди-
ницу количества индикатора и поскольку весь индикатор в кон-
це концов должен покинуть реактор, то
т0=1 (9.58)
Первый момент является средним РВП или средним време-
нем пребывания t. Можно показать, что при отсутствии обрат-
ной диффузии (это допущение мы приняли в начале настояще-
го раздела) для однофазной жидкости в произвольном сосу-
де [2]:
= (9.59)
Иными словами, среднее время пребывания равно номинально-
му времени удерживания реактора. Это соотношение неприме-
нимо ни к одной фазе многофазной системы, ни к одной фазе
системы с нерастворимым катализатором или адсорбентом.
Второй момент т2 часто используется для оценки отклонения
распределения а2 (а2 = т22—mi2); о представляет собой средне-
квадратичное отклонение от среднего времени пребывания.
Из других сходных функций, применяющихся в анализе па-
раметров перемешивания реакторов, следует упомянуть функ-
цию распределения времени пребывания в реакторе /(/); I(t)dt
представляет собой долю содержащейся в реакторе жидкости,
находящейся в реакторе в промежутке времени от t до
Уравнение материального баланса для этой доли жидкости
можно записать в виде:
/(f) = F-1[l—^(/)] (9.60)
Производная A(Q dt функции интенсивности Л(/) отражает ве-
роятность того, что элемент жидкости, находившийся в реакто-
ре в течение времени t, покинет реактор в следующий за t ко-
роткий интервал времени dt. Эта функция полезна при изуче-
нии отклонений от режима идеального перемешивания. С ранее
рассмотренными функциями A(Q связана соотношениями [16]:
1 — еЛ (t) at
Для идеального ПРПП функция Л(7) постоянна. На рис. 9.9
показано поведение функции Л(() при некоторых типах откло-
нений от идеальных режимов в реакторах с перемешиванием.
38
Глава 9
В общем случае наличие максимума, после которого функция
Л(0 убывает, свидетельствует о том, что в реакторе имеются
застойные зоны или зоны с быстрым и медленным течением на
пути от входа в реактор до выхода из него.
Не имея возможности обсуждать здесь все детали, мы все
же должны отметить, что РВП, как это в настоящее время на-
дежно установлено, не характеризует процесс перемешивания
РИС. 9.9. Функция интенсивности, характеризующая различные варианты
неполного смешения в емкостных реакторах с перемешиванием.
а — время между поступлением в реактор и выходом из него невелико (нор-
мальная ситуация); б — задержка в реакторе поступающего раствора обуслов-
лена неэффективным перемешиванием; в — поступающий в реактор раствор
выводится из него, не перемешиваясь с реакционной массой; г — то же, что
в пункте «в» и, кроме того, в силу неэффективного перемешивания в реакторе
имеются застойные зоны. (Из работы [16]).
со всех сторон (подробнее и детальнее эта проблема рассмотре-
на в приведенной в конце главы литературе). РВП содержит
данные о том, как долго различные элементы вытекающей из
реактора жидкой фазы находились в реакторе, но ничего не го-
ворит о том, когда произошло выравнивание концентраций
между элементами жидкости с различным временем пребыва-
ния в реакторе.
Эти выводы можно пояснить, рассмотрев два предельных
варианта. Сначала предположим, что все элементы жидкой
фазы независимо от их предыстории постоянно и эффективно
перемешиваются. Другими словами, поступающие в реактор
исходные вещества сразу же контактируют с другими элемен-
тами жидкой фазы независимо от времени их пребывания в ре-
акторе. Такая ситуация, называемая состоянием максимально-
Проектирование и расчет биологических реакторов
39
го смешения, преобладает в идеальном ПРПП. В другом край-
нем случае элементы жидкости с различным временем пребы-
вания в реакторе вообще не смешиваются и контактируют друг
с другом только в выходящем из реактора потоке. В этом слу-
чае, называемом состоянием полной сегрегации, реакции проте-
кают независимо в каждом элементе жидкости, причем на ход
реакции в одном элементе жидкости не влияют условия и ско-
рости реакций в соседних элементах. Между этими двумя пре-
дельными случаями располагаются различные промежуточные
случаи перемешивания в небольшой части объема реактора или
микросмешения.
РВП реактора совершенно не зависит от характеристик мик-
росмешения. Часто это обстоятельство не играет большой роли,
поскольку микросмешение обычно мало влияет на функциони-
рование реактора в целом. Впрочем, в особых случаях микро-
смешение может оказать существенный эффект на ход процес-
са. Предполагалось [16], что степень влияния микросмешения
на данный реактор можно достаточно надежно оценить путем
расчета работы реактора в особых случаях максимального сме-
шения и полной сегрегации. Если режимы работы в этих экст-
ремальных ситуациях существенно различаются, то проектируе-
мый реактор будет чувствителен к микросмешению, а соответ-
ствующий масштабный переход будет вызывать большие за-
труднения. В таких случаях надежность масштабного перехода
можно повысить за счет применения ТРПВ или реактора близ-
кого типа (см. следующий раздел), поскольку микросмешение
не влияет на параметры процессов в ТРПВ независимо от типа
реакций или их кинетики.
В реакторе с полной сегрегацией каждый независимый эле-
мент жидкости ведет себя подобно небольшому реактору перио-
дического действия. Продукт процесса представляет собой смесь
продуктов, образовавшихся в каждом из этих микрореакторов,
причем время пребывания каждого из продуктов в реакторе
различно. Переходя на язык математики, обозначим символом
Cib(t) концентрацию компонента i в реакторе периодического
действия спустя время t после начала процесса. Допустим, что
начальный состав реакционной смеси в реакторе периодическо-
го действия идентичен составу поступающей в проточный реак-
тор смеси исходных веществ. Если только реакция (или реак-
ции) не вызывает существенного повышения температуры или
заметного изменения объема, то в данном случае не играет ро-
ли, происходит ли здесь одна или несколько различных реак-
ций. Доля вытекающего из реактора потока содержит
элементы жидкости с временем пребывания в реакторе около t
и, следовательно, с концентрацией компонента i около <?,&(/).
Суммирование всех таких долей жидкости дает выражение для
40
Глава 9
концентрации Ct компонента i на выходе из реактора при усло-
вии полной сегрегации:
ci=Jcib(t)g (i)dt (9.62)
о
При выводе уравнения (9.62) принималось, что условия ре-
акции (температура, pH, концентрация растворенного кислоро-
да и т. д.) практически одинаковы как в «микрореакторе пе-
риодического действия», так и во всем реакторе с перемешива-
нием, обладающим определенным РВП. Говоря «практически
одинаковы», мы подразумеваем, что любые имеющиеся разли-
чия в условиях оказывают пренебрежимо малое или допустимо
малое влияние на изучаемые биологические (биохимические)
процессы. Это допущение может оказаться несостоятельным в
случае реакторов большого объема, для которых характерна
существенная объемная неоднородность условий реакции.
Функция РВП для данного реактора сама по себе не говорит
о том, как циркулирует в нем жидкость по зонам с различными
условиями реакций. Отсюда следует, что если реакционная
смесь в биореакторе заметно неоднородна, то уравнение (9.62)
применимо только для грубой оценки системы.
Понятно, что индивидуальные клетки или их скопления в
биохимических реакторах отделены друг от друга (сегрегиро-
ваны), поэтому применение уравнения (9.62), по-видимому,
оправдано в случае любого микробиологического процесса. Это
предположение, однако, не столь обосновано, как это может
показаться на первый взгляд. Не надо забывать, что живые
клетки содержат сложные контрольные системы, с помощью
которых они приспосабливаются к изменениям в среде и пере-
страивают свои функции в соответствии с этими изменениями.
Следовательно, происходящие в периодической культуре изме-
нения отражают взаимное влияние и взаимодействие жидкой
фазы (среды) и биологической фазы (клеток). Состав любой
одной из этих фаз в периодическом процессе изменяется только
в непосредственной связи с изменениями в другой фазе. По-
этому клетка или скопление клеток будут вести себя в проточ-
ной системе как небольшой реактор периодического действия
только в том случае, если окружающая клетку среда (жид-
кость) также будет сегрегирована. Если изменения в составе
среды не оказывают существенного влияния на биологические
реакции в периодическом процессе, то это условие необязатель-
но и применение уравнения (9.62) становится более обоснован-
ным. С примерами такого типа мы познакомимся позднее в этой
же главе.
Проектирование и расчет биологических реакторов
41
Возвращаясь к вопросу о влиянии микросмешения, рассмот-
рим простую необратимую реакцию порядка 0,5:
S —> Р r=ks'h (9.63)
Пусть эта реакция осуществляется в перемешиваемом реакто-
ре с полной сегрегацией, который имеет, однако, то же РВП,
что и ПРПП. Реакцию порядка 0,5 можно приближенно рас-
сматривать как реакцию, описываемую уравнением Михаэли-
са— Ментен в довольно узком интервале концентраций субст-
рата. Путем расчета зависимости s(t) для реакции (9.63) в ре-
акторе периодического действия с учетом уравнений (9.56) и
(9.62) получим:
( . kt 1 \ 1
s = s0 1------1—exp---------f-
l so L \ kt / jj
(9.64)
С другой стороны, если та же самая реакция происходит в иде-
альном ПРПП, в котором по определению достигается макси-
мальное смешение, то концентрация субстрата на выходе из ре-
актора будет равна:
s = s0 1
(ki)2
2s0
4Sq 11
(^)2 ]|
(9.65)
Общий метод расчета степени превращения субстрата в усло-
виях максимального смешения при произвольном времени пре-
бывания в реакторе описан в работе [20].
Часто микросмешение не оказывает большого влияния на
режим работы реактора; очевидно, что это обстоятельство об-
легчает проектирование соответствующих реакторов. Например,
при протекании в ПРПП необратимой реакции второго порядка
наибольшая разница между степенью превращения субстрата
при полной сегрегации и при максимальном смешении состав-
ляет менее 10%. Таким образом, даже если уравнение (9.62)
заведомо не отражает реальную ситуацию, с его помощью
можно иногда с хорошим приближением описать работу реак-
тора. [Путем ряда преобразований уравнений (9.64) и (9.65)
найдите выражение для разности |1—s (9.64)/s (9.65) |.J
Максимальное превращение субстрата во всех простых ре-
акциях с порядком меньше единицы достигается при макси-
мальном смешении. Напротив, полная сегрегация обеспечивает
наивысшую степень превращения для реакций с порядком боль-
ше единицы. Как можно показать с помощью принципа супер-
позиции для линейных систем, степень микросмешения не влия-
ет на реакции первого порядка. Следовательно, в случае одной
или нескольких реакций первого порядка уравнение (9.62) мож-
но использовать независимо от степени микросмешения.
42
Глава 9
Данные о РВП можно также применять для определения
параметров различных моделей неидеальных проточных реак-
торов. Ряд моделей такого типа мы рассмотрим в следующем
разделе.
9.3.3. Модели неидеальных реакторов
Очевидно, РВП содержит полезную информацию о структу-
ре течений и характере перемешивания содержимого реактора.
Для оценки степени отклонения поведения реактора от иде-
ального режима можно использовать, например, функции <S и
SF. Выше мы рассчитали функции <S и для идеального ПРПП;
если теперь эти функции сравнить с соответствующими функ-
циями для реального реактора, то можно получить представ-
РИС. 9.10. ST- и ^-функции для идеального ПРПП (а); ПРПП с обводной
(байпасной) линией (б); ПРПП с застойной зоной (в).
ление о том, насколько режим в реальном аппарате приближа-
ется к режиму полного перемешивания.
Типичные отклонения от идеального режима часто проявля-
ются в виде определенных зависимостей характеристик реак-
тора. Оценить эти зависимости можно, построив соответствую-
щие модели, которые отражают различные типичные отклоне-
ния от системы с идеальным перемешиванием. На рис. 9.10 при-
Проектирование и расчет биологических реакторов
43
ведены модели а) идеального ПРПП, б) реактора с обводной
(байпасной) линией потока исходных веществ и в) реактора с
застойной зоной (1—a) VR. При наличии байпасной линии влия-
ние индикатора сказывается на З^-функции немедленно, а при-
сутствие застойной зоны приводит к более быстрому (по срав-
нению с идеальным реактором) спаду кривой ^-функции.
Другая полезная и часто применяемая модель неидеального
проточного реактора с перемешиванием состоит из двух свя-
занных идеальных ПРПП (рис. 9.11). В этой модели содержи-
мое системы разделено на две меньшие полностью перемеши-
ваемые зоны. Потоки исходных веществ и продуктов процесса
проходят через зону 1, объ-
ем которой составляет а (в
долях единицы) от общего
объема реактора. В свою
очередь содержимое зоны 1
обменивается с застойной
зоной 2 с объемной скоро-,
стью F'. Если допустить, что
кинетика данного процесса
подчиняется уравнению Мо-
но с постоянным экономиче-
ским коэффициентом, то
РИС. 9.11. Модель реактора с неполным
перемешиванием и с застойной зоной.
стационарное состояние в
этой системе можно описать следующими уравнениями матери-
ального баланса:
М —^x/s(so—si) субстрат в зоне 1 (9.66а)
х2—M=;^x/s(si—5г) субстрат в зоне 2 (9.666)
х24-ауртах—;----М = (1 +?^)М клетки в зоне 1 (9.66в)
As si
М + (1—а) YРтах-7-7—х?~х2 клетки в зоне 2 (9.66г)
Здесь
V п— F
U= номинальная скорость разведения (9.67)
В этой модели скорость разведения, отвечающую вымыванию,
можно определить, подставив в уравнения (9.66)
D
вымывания
PmaxSp
+ So
J (! — «)2YPmaxS0
(1 a) Y|.iniaxso
(9.68)
Поскольку первое выражение в правой части этого уравнения
идентично выражению для скорости разведения, отвечающей
44
Глава 9
РИС. 9.12. Вычисленная с помощью модели реактора с застойной зоной
(рис. 9.11) зависимость концентрации клеток в продуктах процесса х1 от
скорости разведения D при различных относительных объемах а зоны активно-
го перемешивания. На верхнем графике представлены кривые зависимости
скорости образования биомассы xiD (7=0,5) от той же переменной.
вымыванию в системе с полным перемешиванием [уравнение
(7.16)], то неполное перемешивание, очевидно, приведет к повы-
шению скорости разведения, необходимой для начала вымыва-
ния. Если выражение, стоящее в квадратных скобках правой
части уравнения (9.68), будет иметь отрицательное значение, это
означает, что вымывание в данной системе невозможно. (Какой
физический смысл имеет такое решение уравнения?)
На рис. 9.12 представлены графики зависимостей концентра-
ции клеток в продуктах процесса хг и продуктивности системы
x^D от скорости разведения при у = 0,5 и различных а. Эти за-
висимости были вычислены при тех же значениях Ks и s0, что и
зависимости на рис. 7.10; изменение характера кривых x = f(D)
при снижении а от 1 (идеальное перемешивание) до 0,9 или
Проектирование и расчет биологических реакторов
45
0,85 (небольшая застойная зона) очень напоминает различия
между теоретическими и экспериментальными данными для
хемостата (рис. 7.10), причем значение а = 0,9 примерно соот-
ветствует находящейся над мешалкой доле объема эксперимен-
тального реактора, использовавшегося для получения представ-
ленных на рис. 7.10 опытных данных.
При изучении РВП до сих пор мы ограничивались реактора-
ми с непрерывными потоками среды и культуры клеток, забы-
вая о том, что в периодических процессах с аэрацией также
участвуют газовые потоки. Функция РВП применялась и для
оценки газосодержания и степени смешения газа в биореакто-
рах периодического действия. Результаты исследований пока-
зали, что в этом случае многое определяется конструкцией
применяемого смесителя и реологией реакционной смеси. На-
пример, РВП в системе ТРПВ — ПРПП очень близко к РВП
газа в реакторах с механическим перемешиванием, если эти
реакторы заполнены маловязкими жидкостями (водой, водными
растворами солей, суспензией Saccharomyces cerevisiae). Более
сложные функции РВП газа характерны для перемешиваемых
высоковязких растворов. На рис. 9.13 приведены результаты
изучения распределения времен пребывания газа в реакторе,
когда поток газа поступал через барботер в стандартный реак-
тор с перемешиванием, содержащий суспензию Penicilliutn
chrysogenum (10 г/л). Соответствие этих экспериментальных
данных расчетным было достигнуто только после введения в
математическую модель (рис. 9.11) выражения, соответствую-
щего элементу полного вытеснения.
Модели, построенные на основе идеальных реакторов разно-
го типа, обычно называют комбинированными, или смешанными
моделями; они представляют собой только один из множества
способов описания неполного перемешивания. Комбинирован-
ные модели имеют то преимущество, что они могут одновремен-
но являться полезным инструментом для теоретического изуче-
ния структуры течений и перемешивания в реакторе, служить
основой для расчета параметров неидеального сосуда в качест-
ве реактора, а также при конструировании лабораторных реак-
торов для изучения эффектов, вызываемых отклонениями от
идеального режима в аппаратах большого объема.
Если для описания данного неидеального биореактора мы
выбрали одну из указанных выше комбинированных моделей,
то ее можно применять и для расчета параметров этого реак-
тора. Иными словами, в рамках выбранной комбинированной
теории можно записать и решить уравнения, характеризующие
кинетику реакций и материальные балансы по субстратам, про-
дуктам процесса и эффекторам реакций. Примером может слу-
жить приведенная выше система уравнений (9.66), основанная
46
Глава 9
_£о
Fc
8FC,
РИС. 9.13. Результаты экспериментального определения распределения вре-
мени пребывания газа в сосуде с перемешиванием (Д). Сплошная кривая
отвечает расчетным данным для того же распределения, полученным с по-
мощью описанной в тексте комбинированной модели при указанных на рисун-
ке значениях параметров. (Воспроизведено с разрешения из работы: Popo-
vic М., Papalexiou A., Reufi М., Gas Residence Time Distribution in Stirred
Tank Reactors, VI International Fermentation Symposium, London, Canada,
1980.)
на модели типа изображенной на рис. 9.11 и уравнении Моно.
При этом, однако, необходимо помнить о свойственных такому
подходу ограничениях, поскольку даже если РВП комбиниро-
ванной модели согласуется с РВП реального реактора, то в лю-
бом случае характеристики микросмешения и циркуляции для
модели и реактора будут различными. Следовательно, комби-
нированная модель в некоторых случаях не может удовлетво-
Проектирование и расчет биологических реакторов 47
рительно описывать функционирование системы как реактора.
Например, комбинированные модели типа изображенных на
рис. 9.11 будут обеспечивать одно и то же РВП независимо от
того, располагается ли трубчатый элемент полного вытеснения
в начале или в конце системы, но в случае процесса с нелиней-
ной кинетикой расположение элемента полного вытеснения бу-
дет влиять и на параметры всей системы. Напротив, если все
реакции и физические явления (например, массообмен) имеют
первый порядок, то один и тот же результат будет получен не-
зависимо от положения элемента полного вытеснения.
Несмотря на указанные ограничения, комбинированные мо-
дели в некоторых ситуациях можно распространить и на такие
процессы, в которых условия реакции пространственно неодно-
родны. Часто чисто интуитивно можно найти соответствие меж-
ду различными идеальными компонентами комбинированной
модели и реальными зонами и участками объема биореактора.
Так, при обсуждении комбинированной модели, изображенной
на рис. 9.11, мы уже отмечали, что один компонент модели мо-
жет соответствовать зоне реактора, примыкающей к мешалке,
а другой — остальному объему реактора. Если поток воздуха
вводится в систему с помощью барботера вблизи мешалки, то
можно допустить, что концентрация растворенного кислорода
будет относительно высокой в первом элементе комбинирован-
ной модели, но сравнительно низкой во втором элементе и, со-
ответственно, во всех удаленных от мешалки зонах жидкой фа-
зы большого аппарата. Следовательно, в расчетах режимов ра-
боты реактора можно учитывать как результаты теоретической
оценки различных сочетаний связанных реакторов, так и экспе-
риментально найденные параметры условий процесса в различ-
ных зонах биореактора.
Для надежного моделирования процесса, осуществляющегося
в однородных условиях, одних только данных о кинетике био-
логических превращений может оказаться недостаточно. К этой
теме мы вернемся в следующем разделе. Возможность описа-
ния реальных крупномасштабных биореакторов с помощью мо-
дели с различными условиями реакций в различных идеаль-
ных элементах модели подсказывает один из подходов к реше-
нию проблемы обратного масштабного перехода. Под обрат-
ным масштабным переходом мы понимаем достаточно обосно-
ванный метод разработки лабораторных экспериментов с целью
моделирования и изучения функционирования больших неиде-
альных реакторов. Поскольку в лабораторных экспериментах с
помощью ограниченного числа реакторов, насосов и соедини-
тельных устройств удается с достаточно хорошим приближени-
ем воспроизвести идеальные системы, особенно ПРПП, то та-
кие эксперименты могут выполнять функции физической моде-
48
Глава 9
ли, отвечающей комбинированной математической модели.
В отдельных небольших реакторах этой установки можно со-
здать разные интенсивности перемешивания, степени аэрации,
значения pH и другие условия и тем самым попытаться моде-
лировать и изучить влияние изменения этих параметров на ход
биологических процессов (в том числе на их кинетику) в боль-
в
каскада последовательно
РИС. 9.14. Различные варианты модели на основе
соединенных реакторов: а — последовательные реакторы; б — последователь-
ные реакторы с рециркуляцией; в — последовательные реакторы с обратным
потоком.
ших аппаратах. Такой подход применим к изучению как реак-
торов периодического действия, так и проточных реакторов.
Для моделирования каскадов реакторов и колонных реакто-
ров, а также режимов перемешивания, приближающихся к ре-
жиму полного вытеснения, применяются различные типы ком-
бинированных моделей с последовательно связанными идеаль-
ными ПРПП (рис. 9.14). Функции РВП для этих моделей мож-
но определить с помощью уравнений материального баланса
по индикатору в каскаде реакторов с перемешиванием путем
определения распределения этого индикатора после его одно-
кратного импульсного введения. Если, как обычно, допустить,
что общий объем системы Vr поделен между N равными по
объему реакторами, т. е.
у у =. ,>==у (9.69)
12 А
Проектирование и расчет биологических реакторов
49
РИС. 9.15. Кривые функции РВП для
каскада из N идеальных ПРПП
(Ук — общий объем системы, Vr/N—
объем одного реактора).
то после несложного математического анализа системы, схема-
тично изображенной на рис. 9.14, а, получим:
g(0= (Z"1)| (тгГ‘/Л,~'ехр(—vr') (9-70’
Графики этой функции при различных N приведены на рис.
9.15. Нетрудно видеть, что при увеличении У функция & (t) из-
меняется от экспоненциально затухающей (М=1) до функции с
резким максимумом при t=VR!F (N — 20).
Отклонение РВП, описываемого уравнением (9.70), равно:
а= = А. (9.71)
Это уравнение полезно при разработке модели с последователь-
ными ПРПП для произвольного сосуда, РВП которого опреде-
лено экспериментально. Если
принять, что общий объем ка-
скада ПРПП и скорость пото-
ка F равны соответствующим
параметрам в реальном про-
цессе, то среднее время пре-
бывания для модели будет со-
ответствовать среднему вре-
мени пребывания в реальном
реакторе. Если, кроме того,
принять N равным экспери-
ментально найденному обрат-
ному отклонению РВП реаль-
ного сосуда, то моменты вто-
рого порядка РВП модели ка-
скада ПРПП и изучаемого со-
суда также будут одинаковы.
Модель с последовательными
ПРПП, вероятно, будет удов-
летворительно описывать
функционирование реального
реактора в том случае, когда
кривая функции РВП этого
реактора имеет ту же форму, что и кривые на рис. 9.15. (На-
помним, что идентичные РВП гарантируют идентичные пара-
метры работы реакторов только при полной сегрегации или в
случае реакций первого порядка.) Обратите внимание на то,
что при N->oo РВП этой модели приближается к РВП реакто-
ра полного вытеснения.
На рис. 9.16 приведены результаты экспериментального изу-
чения РВП в колонне с ситчатыми тарелками с параллельными
потоками. Были выполнены две серии экспериментов, в кото-
4—746
50
Глава 9
рых колонна была разделена тремя тарелками на четыре сек-
ции, причем в первой серии тарелки имели отверстия диамет-
ром 3 мм, а во второй — 2 мм, но общая поверхность отверстий
составляла 10% поверхности тарелки в обоих случаях. Сравне-
ние экспериментально найденных РВП с результатом, предска-
зываемым теорией при N = 4 (рис. 9.16), показывает, что та-
релки с отверстиями диаметром 2 мм обеспечивают удовлетво-
рив 9.16. РВП, характеризующее структуру течений в колонне с ситчатыми
тарелками и с параллельными потоками газовой и жидкой фаз: 1 — диаметр
отверстий 3 мм, доля общей площади отверстий в тарелке 0,0981; 2 — диа-
метр отверстий 2 мм, доля общей площади отверстий в тарелке 0,0975; 3 —
вычислено с помощью модели на основе каскада из 4 ПРПП. [Воспроизведе-
но из статьи: Ketal A. et al., Performance of a Perforated Plate Column as a
Multistage Continuous Fermentor; Biotech. Bioeng., 11, 911 (1969).]
рительную сегрегацию секций колонны. Очевидно, при большем
диаметре отверстий в колонне наблюдается обратный поток
через отверстия в тарелках. Таким образом, в данном случае
изучение РВП может служить основой для выбора режима,
обеспечивающего необходимую степень сегрегации в колонне.
Иногда введение в реактор растущей популяции микроорга-
низмов коренным образом меняет его РВП. Так, эксперимен-
тальное определение РВП для башни с 8 тарелками в отсутст-
вие растущих организмов показало (рис. 9.17, а), что здесь
проявляется четкий сегрегационный эффект. Если же в башне
растет культура пекарских дрожжей, то найденное с помощью
различных индикаторов РВП гораздо лучше соответствует од-
ному идеальному ПРПП, чем каскаду из 8 ПРПП (сплошная
кривая на рис. 9.17,6). Вероятно, исчезновение сегрегационного
эффекта обусловлено осаждением дрожжевой суспензии. Если
1,4
1,Z
1,0
0,6
1,6
Клетка, меченные > кониент-
u радиоактивным фоарором 1
и Вода, меченная \ Р%£пде
u ГГфиГПи&М > 6^X000
а s Кпнирит-
9 рядаоакггывньж фосфорам I ^Г7Г„
И вода, меченная^ Р А f/Жяе
тритием >а'и '}ине
В уравнении с/с0 ----
U-i)i
0,8-
0,4 -
о,г.
\
\
о
1,0 Z,0 3,0 4,0
Сггшосигпельное время 9-l't/Vn
б
РИС. 9.17. Определение РВП в колонне с 8 тарелками (доля общей площади
отверстий в тарелке равна 0,15) различными методами и в различных условиях.
а. — изучение с помощью солевого индикатора показывает, что повышение
интенсивности аэрации уплощает и уширяет кривую распределения времени
пребывания; б — колонна заполнена культурой дрожжей, а в качестве инди-
каторов использована меченая вода и меченые клетки. \Prokop A. et al., Design
and Physical Characteristics of a Multistage, Continuous Tower Fermentor;
Biotech. Bioeng., 11, 945 (1969).]
4
52
Глава 9
в четырех тарелках колонны площадь отверстий будет состав-
лять только 3%, то РВП колонны с растущей популяцией дрож-
жей будет практически таким же, что и в случае каскада из
4 ПРПП.
Теперь перейдем к изучению изображенных на рис. 9.14 кас-
кадов ПРПП как биореакторов. Уравнение материального ба-
ланса по произвольному компоненту с в /-м реакторе можно за-
писать в следующем виде:
Fci-x-^/+Р/Гм|реактор/ =0 (9.72)
Если, например, мы рассмотрим рост культуры микроорганиз-
мов в каскаде реакторов при условии нестерильности питатель-
ного вещества (х^О), то уравнения материального баланса по
биомассе для реакторов от 1- до N-ro будут выглядеть следую-
щим образом:
F(.Xf — Xi) +
F(Xj-i— Xj) + Vj[ijXj = Q j = 2, 3, ..., N (9.73)
Решение этих уравнений дает:
Fxf
Е-у1И1
(9-74)
и
Fi~1x1
Xj (F - Н2У2) (f - НзЕ3) . . . (F — pjVJ
j=2,...,N (9.75)
Если объемы реакторов равны, то уравнение (9.75) упрощается:
Xj (^1-Н2)(^1-Нз) . • • (^1-HJ)
(9.76)
Здесь Di — скорость разведения в каждом реакторе (Di =
= F/Vl).
Влияние сегрегационного эффекта и рециркуляции на ско-
рость разведения в точке вымывания Dmax можно оценить пу-
тем изучения системы, изображенной на рис. 9.14,6. Показано,
что если кинетика клеточного роста подчиняется уравнению
Моно с учетом метаболизма поддержания жизнедеятельности
клеток [уравнение (7.22) J, то при стерильной питательной сре-
де, равных объемах реакторов и постоянстве экономического
коэффициента DmZx удовлетворяет уравнению:
Hrnax N (1 + /<S/SO) {1 -[r/(l + r)]1/jV)
Pmax
1 — (^e/P'max)(l 4“ Ks/Sf)
(9.77)
Проектирование и расчет биологических реакторов
53
Здесь D, как обычно, выражено через параметры системы в
целом:
р F _ F _ О*
VR лгух — W
Подстановка в уравнение (9.77) г = 0 дает выражение для кри-
тической скорости разведения в каскаде реакторов равных объ-
емов без рециркуляции. Если, кроме того, принять ke = 0, т. е.
допустить, что кинетика клеточного роста описывается уравне-
нием Моно в его первичной
стится до уже известного
(7.16)J:
(9.78)
форме, то уравнение (9.77) упро-
нам выражения [ср. уравнение
D'
max
Pmaxso
Ks + so
(9.79)
Такой результат можно было бы предполагать интуитивно; дей-
ствительно, если вымывание происходит в первом из ряда по-
следовательно соединенных реакторов равного объема, то куль-
тура будет вымываться и из всех последующих реакторов.
(Что произойдет, если объемы реакторов не равны и если
F~>VjD*max При /=1, 2, ...,& — 1 И F< 14Е>*тах?)
Теоретической модели каскада ПРПП с обратным потоком
(рис. 9.14, в) близка дисперсионная модель, которая представ-
ляет собой модификацию модели идеального ТРПВ; в ней до-
полнительно учитывается процесс осевой диффузии, налагаю-
щийся на процесс конвективного потока через трубу. Для этого
модель элементарного объема ТРПВ, изображенную на рис. 9.5,
нужно дополнить членом, описывающим дисперсионный поток
A(Dzdcldz)z\ аналогичное слагаемое А(—Dzdc/dz)z+!iZ вводится
в левую часть уравнения (9.13). Произведя описанные выше
преобразования с учетом лимитирующих процессов, это уравне-
ние можно перевести в следующее уравнение материального ба-
ланса по компоненту с дисперсионной модели:
d (ис) _ d [п de ''i t
3 Г~ uz “3 “Г rfc
dz dz \ dz J
(9.80)
В случае жидкостей и и коэффициент эффективной осевой дис-
персии Dz обычно приблизительно постоянны и уравнение (9.80)
принимает вид:
de n d.zc .
и-------= D,-------------Urf(.
dz z dz^ i c
(9.81)
Поскольку (9.81) является дифференциальным уравнением
второго порядка, необходимы два граничных условия. Обычно
54
Глава 9
принимают следующие условия:
ucf = f ис—Dz-^-\
1 к dz Л=о
— =0
dz 2=1
(9.82)
(9.83)
Если, как это часто бывает, Dz не слишком велико, то сложное
условие (9.82) можно упростить:
с |2=о = cf
В этом случае отклонение РВП дисперсионной
равно:
о2 = ——— Г 1--— (1 — е~Ре)"
Ре |_ Ре _
(9.84)
модели будет
(9.85)
Здесь осевое число Пекле Ре определяется как
Ре = -^- (9.86)
С точки зрения физического смысла Ре можно рассматривать'
как отношение конвективной массопередачи (жис) к массопе-
редаче за счет дисперсии i^cDzc!L)- Анализируя пределы урав-
нения (9.85), нетрудно найти, что при Ре->0, как и в случае
отклонения РВП идеального ПРПП, о2-^1. При очень боль-
ших числах Пекле о2->0, что отвечает режиму идеального вы-
теснения. Как и в случае моделей с каскадами реакторов, урав-
нение (9.85) можно использовать для определения числа Ре дис-
персионной модели, если величина о2 найдена эксперименталь-
но. При условии, что кинетика клеточного роста описывается
уравнением Моно с учетом метаболизма поддержания, скорость
разведения в точке вымывания Z)max для дисперсионной модели
определяется уравнением:
Ртах 1 4~ Ks/Sf) (9 87)
^тах 1 (^e/Pinax) (1 4" Ks/Sf)
Когда влияние дисперсии очень мало (Ре—-0), то Dmax умень-
шается до нуля. Этот вывод согласуется с очевидным фактом:
в идеальном ТРПВ не поддерживается существование популя-
ции при условии стерильности исходных веществ.
Следует подчеркнуть, что коэффициент осевой дисперсии D:
обычно не равен коэффициенту молекулярной диффузии. Dz
представляет собой модельный параметр, отражающий разно-
сторонние эффекты ряда физических явлений. Из такого рода
явлений мы упомянем три — ламинарное течение в трубах, тур-
булентное течение в трубах и течение в слоях насадки.
Проектирование и расчет биологических реакторов 55
Если осевая и радиальная диффузия, характеризующаяся
коэффициентом диффузии S>, налагается на осевой конвектив-
ный перенос, обусловленный ламинарным течением, то можно
показать, что коэффициент эффективной осевой дисперсии бу-
дет определяться уравнением:
D‘=®+-^ (9-88)
Здесь й — средняя осевая скорость (й равно половине скоро-
сти в средней линии). Дисперсионная модель, Dz которой опре-
деляется уравнением (9.88), хорошо описывает РВП ламинар-
ного потока с учетом диффузии для труб достаточно большой
длины (Л»й;2/40.2)).
При турбулентном течении в трубах (Re>2100) перенос
масс, количества движения и энергии осуществляется главным
образом путем движения макроскопических вихрей жидкой фа-
зы. Влияние вихревого переноса напоминает влияние процессов
молекулярной диффузии, но турбулентные потоки обычно гораз-
до более интенсивны. Следовательно, в этом случае коэффици-
ент эффективной осевой дисперсии зависит главным обра-
зом от режима потока жидкой фазы. Число Пекле для турбу-
лентного потока обычно равно примерно 3, но уменьшается при
снижении числа Рейнольдса. Такое поведение турбулентных по-
токов, а также ряд данных, относящихся к ламинарным пото-
кам, представлены на рис. 9.18. Одним из наиболее известных
биологических трубчатых реакторов без насадки является, веро-
ятно, проточный стерилизатор жидкостей, подробно рассмат-
риваемый в следующем разделе.
В некоторых трубчатых реакторах применяется насадка из
твердых частиц иммобилизованного фермента или иного биока-
тализатора. Частицы катализатора составляют неподвижный
слой, и текущая по трубе жидкость омывает эти частицы. По-
скольку жидкость течет в промежутках между твердыми части-
цами, то любой элемент жидкой фазы при осевом движении со-
вершает также беспорядочные перемещения в радиальных на-
правлениях. Влияние такого прерываемого твердыми частицами
движения на РВП системы может быть описано с помощью
дисперсионной модели. Теория предсказывает, что осевое число
Пекле Рер, учитывающее диаметр частиц, т. е.
Рер=^ (9.89)
в данном случае равно приблизительно 2 (dp — диаметр час-
тиц). Это предположение в ряде случаев было подтверждено
экспериментально, но, как показано на рис. 9.19, в общем слу-
чае Рер зависит от Sc и Rep.
56
Глава 9
РИС. 9.18. Зависимость осевого числа Пекле (Ре) от чисел Рейнольдса (Re)
и Шмидта (Sc) для течения жидкости в трубах.
В заключение нашего анализа процессов, связанных с дис-
персией, следует отметить, что характер течения жидкостей
может осложняться в результате влияния других факторов, на-
пример изгибов трубопроводов, изменений профиля рек и дру-
гих водных потоков. В таких сложных случаях установить Dz
расчетным путем не представляется возможным, и для опреде-
ления этого параметра приходится опираться только на данные
о РВП, найденные экспериментальным путем.
Описанные выше модели с каскадом последовательно соеди-
ненных реакторов и дисперсионного типа содержат по одному
параметру (N и Dz соответственно) и описывают режимы не-
полного перемешивания. Каждая из этих моделей относится к
определенному диапазону состояний смешения и сегрегации,
Проектирование и расчет биологических реакторов
57
начиная от идеального ПРПП и кончая идеальным ТРВП. Та-
ким образом, в каждом конкретном случае мы должны выбрать
модель определенного типа. С точки зрения удобства расчета
и анализа модель каскада последовательно соединенных реак-
торов обычно намного выгоднее. Кроме того, применение дис-
персионной модели при большом обратном смешении (о2) обыч-
но связано с рядом трудностей. С другой стороны, как показа-
но на рис. 9.14, для описания режима, близкого к режиму пол-
РИС. 9.19. Зависимости осевого числа Пекле Ре„ от Рер и Sc для течения
в трубах с насадкой. Обратите внимание на то, что в выражения для крите-
риев подобия в данном случае входит диаметр частиц dp, а не диаметр тру-
бы dt.
кого вытеснения, необходимо очень большое число реакторов в
каскаде. По этой причине дисперсионная модель предпочтитель-
нее при небольших отклонениях от режима полного вытеснения
(например, при о2, равном или меньшем 0,05), а модель с кас-
кадом последовательно соединенных реакторов точнее описы-
вает режимы при существенном обратном смешении (о2 ^0,2).
Последняя ситуация типична для ферментеров и бассейнов для
биологической переработки отходов. Небольшие отклонения от
режима полного вытеснения встречаются при непрерывной сте-
рилизации жидкостей, которую мы рассмотрим позднее в этой
же главе.
9.3.4. Взаимосвязь между перемешиванием
и биологическими превращениями
Характер течений и явления переноса влияют на кинетику
клеточных процессов в различных масштабах. Так, эффекты,
проявляющиеся при определенных масштабах длины (вспомни-
58
Глава 9
те рис. 9.2), могут различными путями влиять на наблюдаемую
кинетику роста популяции клеток. Эти влияния необходимо
учитывать с тем, чтобы изучение кинетики и разработка моде-
лей осуществлялись в условиях, аналогичных в основном усло-
виям в большом реакторе. В настоящем разделе мы изучим не-
которые стороны взаимосвязи между кинетикой клеточного
роста и перемешиванием в биореакторах.
Изучение начнем с анализа наибольшего масштаба, вклю-
чающего циркуляцию всей жидкой фазы, в результате которой
клетки переносятся в различные зоны реактора, где, как мы
уже отмечали, в случае больших аппаратов обычно имеются
разные концентрации растворенного кислорода и различные
уровни турбулентности. Влияние циркуляции жидкой фазы на
общую кинетику клеточного роста можно понять, обратившись
к простому примеру. Предположим, что кислород подается ло-
кально в какую-то определенную зону реактора (как обычно и
бывает в случае реакторов с перемешиванием) и что утилизация
кислорода происходит в изолированных элементах жидкости,
циркулирующих в реакторе. Такая ситуация может возникнуть
в высоковязких микробиологических системах. Если начальная
концентрация кислорода в воде близка к уровню насыщения
(0,3 моль/м3) и если кислород поглощается с обычной скоро-
стью 10—100 моль/(ч-м3), то в отдельном элементе системы
кислород истощится через И—50 с. Эта величина имеет тот же
порядок, что и время циркуляции в большом реакторе.
Для того чтобы оценить влияние циркуляции жидкой фазы
с количественной стороны в более общем виде, а также чтобы
учесть статистическое распределение времен циркуляции, рас-
смотрим работу реактора периодического действия с распреде-
лением времени циркуляции Допустим далее, что в пол-
ностью сегрегированных элементах жидкой фазы осуществля-
ется реакция нулевого порядка. Распространение этой модели
на другие типы локальной кинетики реакций, в том числе и на
реакции, описываемые уравнением Михаэлиса — Ментен, не
представляет принципиальных затруднений, а только немного
усложняет соответствующие расчеты, но все выводы качествен-
ного характера при этом сохраняют свою силу. В реакции ну-
левого порядка, константа скорости которой равна k0, при
начальной концентрации реагента s0 его концентрация в эле-
менте жидкой фазы будет изменяться согласно уравнениям:
s(/) =
t ^==
t>te
(9.90)
Здесь мы впервые ввели параметр времени истощения питатель-
ного вещества te, равный &o/so. Далее символом F обозначим
долю времени, которую элемент жидкости находится в условиях
Проектирование и расчет биологических реакторов
59
полного истощения питательного вещества. Эту долю можно
рассчитать по уравнению:
t — te
1
(9.91)
где t — среднее время циркуляции. Уравнение (9.91) было ре-
шено при различных относительных величинах t, te и различных
стандартных отклонениях о распределения времен выравнива-
ния
вии,
ски
ние.
9.20
что
Л»
Ъ
Qj
Cj
а
о i г з
Относительное^рремя циркуляции
РИС. 9.20. Зависимость времени пре-
бывания клеток в циркулирующих
элементах жидкой фазы в условиях
истощения питательных веществ от
относительного времени циркуляции
и стандартного отклонения времени
циркуляции (из работы [19]).
I
§
i
концентраций при усло-
что fc(0—логарифмиче-
нормальное распределе-
Представленные на рис.
результаты показывают,
время пребывания клеток
в истощенной среде возраста-
ет при увеличении как сред-
него времени циркуляции, так
и стандартного отклонения
времени циркуляции. Пара-
метр 1—F можно интерпрети-
ровать как отношение реаль-
ного выхода продукта процес-
са к выходу, который был бы
достигнут при условии подачи
питательного вещества ко всем
точкам объема реактора. Эти
расчеты показывают, что при
данном времени циркуляции
реактор должен иметь такую
конструкцию, которая бы обес-
печивала минимальную дис-
персию времени выравнивания
концентраций перемешивае-
мых веществ.
Описанная модель, хотя и обладает некоторой информатив-
ностью, но не учитывает того влияния, которое могут оказывать
на кинетику клеточного роста изменения условий во времени,
обусловленные циркуляцией культуральной жидкости в реак-
торе. Как мы уже видели на рис. 9.1, масштабы времени цирку-
ляции в больших биореакторах сравнимы с масштабами време-
ни некоторых метаболических превращений и адаптационных
изменений. Это говорит о том, что модель, разработанная для
специфических условий перемешивания в малом реакторе, мо-
жет оказаться несостоятельной в случае большого аппарата,
для которого характерны большое время выравнивания кон-
60
Глава 9
центраций перемешиваемых компонентов и большие флуктуации
условий процесса.
Эксперименты, выполненные с помощью встроенного непо-
средственно в реактор флуориметрического устройства, описы-
ваемого подробнее в разд. 10.2, наглядно показали близость
времени выравнивания концентраций и биологического отклика
системы. Время выравнивания концентраций в аэрируемом
(0,5% по объему) 70-литровом реакторе с механическим пере-
мешиванием (рабочий объем 40 л) определяли путем импульс-
ного введения раствора хинина в 0,05 М. H2SO4 и последующего
Таблица 9.3. Сравнение времени выравнивания концентраций в аэрируемом
(0.5% по объему) реакторе объемом 70 л (рабочий объем 40 л) и времени
отклика процесса утилизации глюкозы дрожжами (в секундах)
при различных скоростях перемешивания3
Скорость вращения мешалки, об/мин
200 500 700
Время отклика флуоресценции внутриклеточной 8,5 6,8 5,9
Время выравнивания концентраций в жидкой фазе 4,2 2,5 1,5
Разность (время ческой системы) отклика биологи- 4,3 4,3 4,4
а Воспроизведено с разрешения из статьи: Einsele A., Risiroph D. L., Humphrey А. Е.г
Mixing Times and Glucose Uptake Measured with a Fluorometer; Biotech. Bioeng., 20.
1487 (1978).
контроля изменения флуоресценции культуры во времени. Эти
изменения позволили определить процессы циркуляции жидкой
фазы и смешивания в реакторе. Затем флуоресценцию измеря-
ли после импульсного введения глюкозы в культуру дрожжей.
В этом случае источником флуоресценции являлись внутрикле-
точные восстановленные нуклеозиды, концентрации которых в
большой степени зависят от скорости превращений глюкозы
в клетке. Таким образом, результаты второго эксперимента
дают ценную информацию о времени отклика полной последо-
вательности процессов переноса (смешение с жидкой фазой -*
-> диффузия к клеткам транспорт в клетки) и метаболических
превращений, связанных с утилизацией питательного вещества.
В табл. 9.3 приведены время выравнивания концентраций
и время отклика процесса утилизации субстрата, найденные для
этого реактора при различных скоростях перемешивания. Инте-
ресно отметить, что время отклика процесса утилизации глюко-
зы, как и можно было предположить, практически одно и то же
во всех трех случаях, а время выравнивания концентраций при
повышении скорости перемешивания существенно снижается.
Проектирование и расчет биологических реакторов
61
Эти данные интересны и с еще одной точки зрения — обращает
на себя внимание сходство порядка величин времени отклика
процесса локального поглощения и утилизации глюкозы и вре-
мени выравнивания концентраций в растворе. Отсюда следует,
что в ходе циркуляции клетки сталкиваются с различными
условиями в разных зонах реактора и, вероятно, постоянно
находятся в некотором переходном метаболическом состоянии,
поскольку время отклика метаболизма мало отличается от ха-
рактерного масштаба времени изменения условий в среде, обус-
ловленного циркуляцией культуры.
В некоторых экспериментах наблюдалось заметное влияние
неоднородности среды на метаболизм клеток. Так, в трубчатом
реакторе с линией циркуляции, в которой наблюдаются колеба-
ния концентрации растворенного кислорода, в культуре Candida
tropicalis повышается экономический коэффициент образования
биомассы относительно субстрата, снижается скорость потреб-
ления кислорода, расходуемого на образование биомассы, по-
нижается дыхательный коэффициент. При колебании концент-
рации растворенного кислорода в проточной культуре Pseudo-
monas methylotropha ASI, растущей на метаноле как источнике
углерода и энергии, наблюдались противоположные эффекты —
снижение скорости клеточного роста и экономического коэффи-
циента образования биомассы по субстрату. В другом экспери-
менте в культуре Penicillium chrysogenum Pl концентрацию
растворенного кислорода изменяли периодически (с периодом
2 мин), так что средняя концентрация О2 составляла 30% от
концентрации при насыщении воздухом. В данном случае удель-
ная скорость биосинтеза пенициллина существенно снижалась,
причем в общих чертах этот эффект напоминал влияние пони-
жения средней концентрации растворенного кислорода, что
свидетельствовало о нелинейном характере влияния изменений
условий на биореакцию.
Если перейти к анализу более мелкомасштабных флуктуаций
скорости турбулентного течения жидкой фазы, то логично ожи-
дать существенного влияния интенсивности турбулентных вих-
рей различных масштабов на морфологию (и, вероятно, на
характер метаболизма) некоторых организмов. Можно далее
предположить, что эта зависимость будет иметь наибольшее
значение в случае организмов, вырастающих до размеров, мас-
штаб которых сравним с ожидаемым масштабом турбулентных
вихрей. Последние в свою очередь изменяются от крупномас-
штабных (соответствующих масштабу высоты лопасти мешалки,
например 0,1 м) до самых мелкомасштабных, образующихся в
каскаде передачи энергии турбулентного потока (разд. 8.3).
В биореакторах с перемешиванием размер самых мелких вих-
рей равен примерно 20—100 мкм. Чтобы определить, на какие
62
Глава 9
биологические системы флуктуации скоростей в этом масштабе
будут влиять в наибольшей степени, можно обратиться к рис.
9.2. Нетрудно видеть, что размеры скоплений микроорганизмов
(особенно мицелиальных) соответствуют среднемасштабным
турбулентным вихрям, и, следовательно, интенсивность переме-
шивания и распределение зон турбулентности в реакторе будут
оказывать влияние в основном именно на эти образования.
РИС. 9.21. Зависимость концентрации лимонной кислоты, образовавшейся че-
рез 7 сут в культуре Aspergillus niger, от скорости вращения турбинной ме-
шалки (кривые 1, 2 и 3 отвечают штаммам S59, N233 и Е81 соответственно),
а также зависимость количества сухой биомассы, образующейся в культуре
штамма S59, от того же параметра (кривая 4). [Воспроизведено с разреше-
ния из статьи: Ujcovd Е., Fencl Z., Musi’lkovd М„ Siechert L„ Dependence of
Release of Nucleotides from Fungi on Fermentor Turbine Speed; Biotech.
Bioeng., 22, 237 (1980).]
В предыдущих главах мы уже обсуждали связанное с диф-
фузионными ограничениями влияние общего размера клеточных
скоплений на общие скорости процессов. Здесь же основное
внимание мы уделим более скрытым эффектам, включая изме-
нения морфологии и метаболического состояния организмов,
в существенной степени влияющие на микробиологические про-
цессы. Интересным примером такого рода эффектов является
влияние интенсивности перемешивания на рост, образование
продуктов жизнедеятельности и секрецию нуклеотидов в среду
в случае различных мутантов Aspergillus niger — эффективных
продуцентов лимонной кислоты. Как показано на рис. 9.21, при
повышении интенсивности перемешивания возрастает урожай
Проектирование и расчет биологических реакторов 63
биомассы, однако зависимость скорости биосинтеза лимонной
кислоты от интенсивности перемешивания носит более сложный
характер и в случае всех трех изученных мутантов характери-
зуется наличием четко выраженных максимумов. Параллельное
изучение зависимости процесса секреции нуклеотидов в среду
от скорости перемешивания показало, что характер этой зави-
симости определяется природой штамма. Для ряда штаммов
убыстрение вращения мешалки сопровождалось увеличением
концентрации нуклеотидов в среде, а для другого штамма со-
вершенно неожиданно была обнаружена обратная зависимость,
когда при более высоких скоростях вращения мешалки скорость
секреции нуклеотидов снижалась. Влияние скорости вращения
мешалки на морфологию мицелия этого необычного мутанта
представлено на рис. 9.22. На микрофотографиях видно, что при
низких скоростях перемешивания мицелий имеет длинные, тон-
кие и мало разветвленные филаменты с небольшим числом септ,
в то время как при высоких скоростях перемешивания преобла-
дают толстые, в высшей степени септированные, более разветв-
ленные и скрученные гифы. Этот пример показывает, что интен-
сивность перемешивания может оказывать на организмы боль-
шое влияние, в существенной степени определяющее кинетику
всего процесса.
В обсуждении материала этого раздела и в приведенных
примерах мы старались подчеркнуть потенциально сложную
реакцию растущих клеток на физические и химические пара-
метры окружения и невозможность разработки общей матема-
тической модели кинетики, применимой при любых обстоятель-
ствах. Поэтому необходимы тщательный учет совместного-
влияния химических и физических параметров среды в данном
биореакторе и применение соответствующего математического
описания кинетики процессов, необходимого именно в данной
ситуации. Очевидны трудности, связанные с масштабным пере-
ходом в случае систем, кинетика которых отличается высокой
чувствительностью к флуктуациям параметров окружения.
В этих случаях нужно попытаться найти такую конструкцию
реактора, которая обеспечивала бы определенные (но не обя-
зательно одинаковые во всем объеме) параметры окружения,
что может быть чрезвычайно трудной задачей, если речь идет
о крупномасштабных реакторах. Выбрав реактор иной конст-
рукции (см. разд. 9.7), иногда удается снизить разброс в зна-
чениях времени выравнивания концентраций и степени флук-
туаций параметров окружения и тем самым получить более
определенную картину смешения, для которой легче подыскать
соответствующее математическое выражение, описывающее ки-
нетику процесса. Однако в общем случае нельзя рассчитывать
на успех рационального, заранее спланированного масштабиро-
64
Глава 9
РИС. 9.22. На этих микрофотографиях отчетливо видно различие в морфоло-
гии и септации клеток A. niger S59, выращенных при 400 (а) и 1200 (б) обо-
ротах в минуту (увеличение Х400). [Воспроизведено с разрешения из статьи:
Ujcova Е.. Fencl Z., Musi'lkova М., Siechert L., Dependence of Release of
Nucleotides from Fungi on Fermentor Turbine Speed: Biotech Bioene 22 237
(1980).] ’
Проектирование и расчет биологических реакторов 65
вания биореакторов до тех пор, пока мы не будем лучше пони-
мать процессы переноса в различных масштабах и кинетику
биореакций на нескольких условиях сложности и пока не сможем
объединить эти данные с тем, чтобы достаточно надежно рас-
считать работу реактора на основе одних лишь теоретических
данных.
Пример 9.1. Моделирование и оптимизация процесса производства «-га-
лактозидазы с помощью плесневого гриба Monascus sp. Фермент а-галакто-
Продолжитпелъносгпъ культивирования, ч
РИС. 9П1.1. Результаты изучения роста периодической культуры плесневого
гриба Monascus sp. на глюкозно-галактозной среде [начальный состав среды;
1% (по массе) глюкозы, 0,3% галактозы, 0,5% NH4NO3, 0,5% КН2РО4, 0,1%
MgSO4-7H2O, 0,01% дрожжевого экстракта]. Посевной материал выращен на
глюкозной среде. При расчете исходили из следующих начальных условий:
х=5-10-4 г/мл, $1 = 1-10-2 г/мл, $2=3-10-3 г/мл; $2/=0 мкгДмг клеток),
г$2/=0,910 мкг/(мг клеток), е=0 единицДмг клеток). [Воспроизведено из
статьи: Imanaka Т. et al., Unsteady-State Analysis of a Kinetic Model for Cell
Growth and a-Galactosidase Production in Mold; J. Ferment. Tech. (Japan),
51, 423 (1973).]
зидаза применяется в производстве сахара из сахарной свеклы благодаря его
способности разрушать рафинозу — ингибитор кристаллизации сахарозы. В ря-
де интересных работ Иманака, Кайеда, Сато и Тагучи изучили процесс про-
изводства этого внутриклеточного фермента с помощью плесневого гриба, вы-
деленного ими из почвы*. Опубликованные этими исследователями данные
заслуживают тщательного анализа, мы же суммируем некоторые основные
результаты.
* Приведенные в этом примере данные заимствованы главным образом
из статей: Imanaka Т., Kaieda Т., Sato К., Taguchi Н., a-Galactosidase Produc-
tion in Batch and Continuous Culture and a Kinetic Model for Enzyme Produc-
tion, J. Ferment. Technol. (Japan), 50, 633 (1972); Imanaka T., Kaieda T., Ta-
guchi H., Optimization of a-Galactosidase Production by Mold II III- J Fer-
ment. Technol. (Japan), 51, 423, 431 (1973).
5—746
66
Глава 9
На первом этапе разработки высокоэффективного процесса биосинтеза
фермента были изучены различные условия культивирования плесени в пе-
риодическом и непрерывном процессах. К числу основных результатов изуче-
ния периодической культуры относятся следующие:
1. Из 20 изученных источников углерода (в том числе глюкозы, фруктозы»
маннита, крахмала) только четыре вещества углеводной природы доста-
РИС. 9П1.2. Концентрации клеток и субстрата, а также удельная фермента-
тивная активность в проточной культуре при 30 °C в стационарных состояниях
после ступенчатого повышения скорости разведения. Начальные концентраций
глюкозы и галактозы составляли 2 и 0,5% соответственно. [Воспроизведена
из статьи: Imanaka Т. et al., Optimization of a-Galactosidase Production by
Mold; J. Ferment. Tech. (Japan), 50, 633 (1972).]
точно эффективно индуцируют высокий уровень а-галактозидазной актив-
ности. Эффективными индукторами являются галактоза, мелибиоза, рафи-
ноза и стахиоза.
2. Что касается источников азота, то большие концентрации фермента проду-
цируются в присутствии нитрата аммония; мочевина, KNO3, (NH^SO*
и пептон в этом отношении менее эффективны. Оптимальная концентрация
NH4NO3 в среде составляет 0,3—0,5% (по массе).
3. При росте на галактозной среде урожай клеточной массы пропорционалев-
а-галактозидазной активности. Если в качестве источника углерода исполь-
зуется смесь глюкозы и галактозы, то наблюдается явление диаукси»
(рис. 9П1.1): до тех пор, пока не истощится почти вся глюкоза, галактоз»
практически не утилизируется.
er
Проектирование и расчет биологических реакторов
л Как покачано на пис 9П1.1, продуцирование а-галактозидазы начинается
* ™ьк"сктощи™ почти вся глюкоза. В отдельных аксперимен-
тах было показано, что глюкоза в концентрациях, превышающих 0,05 /0 (по
массе), подавляет синтез фермента. „„„„„
Изучению непрерывной культуры в стационарном состоянии были посвя-
щены две серии экспериментов, которые были выполнены в одном ПРПП.
В первой серии экспериментов скорость разведения вначале удерживали на
РИС. 9П1.3. Концентрации клеток и субстрата, а также удельная фермен-
тативная активность в стационарных состояниях после ступенчатого сниже-
ния скорости разведения проточной культуры при 30 °C. Начальные концентра-
ции глюкозы и галактозы составляли 2 и 0,5% соответственно. [Воспроизве-
дено из статьи: Imanaka Т. et al., Optimization of a-Galactosidase Production
by Mold; J. Ferment. Tech. (Japan), 50, 633 (1972).]
очень низком уровне, а затем ступенчато повышали, определяя параметры
системы в стационарном состоянии при каждом значении D. Полученные ре-
зультаты приведены на рис. 9Ш.2. Особенно интересно резкое изменение
свойств системы при £>=0,142 ч-1. При скоростях разведения ниже этой вели-
чины происходит утилизация галактозы и синтез а-галактозидазы. Если же D
превышает 0,142 ч-1, то оба процесса прекращаются и начинается утилизация
глюкозы как единственного источника углерода для плесени. Очевидно, это
скачкообразное изменение обусловлено влиянием глюкозы, уже упоминавшим-
ся при анализе поведения периодической культуры данного организма. При
относительно большой удельной скорости роста (больших величинах £>) пле-
сень утилизирует преимущественно глюкозу.
На рис. 9П1.3 представлены результаты аналогичной серии опытов
® ПРПП при высокой начальной скорости разведения, которую постепенно и
5*
68
Глава 9
также ступенчато уменьшали. В новой серии экспериментов снова обнаруже-
но скачкообразное изменение параметров, хотя и менее резкое, чем в первой
серии, причем на этот раз скачок наблюдался приблизительно при D=t
= 0,008 ч-1. Ниже этой критической скорости разведения ассимилируется га-
лактоза и продуцируется фермент; при £>>0,008 ч-1 а-галактозидазной актив-
ности не наблюдалось, что не согласуется с результатами первой серии экспе-
риментов, в которых фермент продуцировался вплоть до £> = 0,142 ч-1.
РИС. 9П1.4. Здесь еще раз воспроизведены некоторые кривые из рис. 9П1.2
и 9П1.3, отражающие наличие нескольких стационарных состояний и гистере-
зиса в проточной культуре. Два устойчивых стационарных состояния наблю-
даются в диапазоне скоростей разведения от 0,008 до 0,142 ч-1. Какое из
этих стационарных состояний реализуется, зависит от начальных условий.
[Воспроизведено из статьи: Itnanaka Т. et al., Optimization of a-Galactosidase
Production by Mold, J. Ferment. Tech. (Japan), 50, 633 (1972).]
На рис. 9П1.4 воспроизведены также некоторые данные из двух предыду-
щих рисунков, которые достаточно наглядно показывают, что в диапазоне
между £> = 0,008 и £> = 0,142 ч-1 система имеет несколько устойчивых ста-
ционарных состояний. В этом диапазоне скоростей разведения способность си-
стемы продуцировать а-галактозидазу зависит от начальных условий работы
реактора, причем синтез а-галактозидазы обеспечивается только при подходе
к этому диапазону со стороны более низких скоростей разведения.
На базе этих и ряда других экспериментальных данных была разработана
математическая модель, описывающая утилизацию субстрата, рост клеток и
синтез фермента. Приведенные в табл. 9П1.1 индивидуальные параметры мо-
дели в общих чертах нам уже знакомы: так, например, удельная скорость
клеточного роста Цг на галактозе описывается уравнением Моно, а уравнение
Проектирование и расчет биологических реакторов
69
Таблица 9П1.1. Математическая модель процесса образования
а-галактозидазыа’* 6
Утилизация субстрата
ds}
~dt
1
~y-V-ix
Yi
j=\ (глюкоза), 2 (галактоза)
(9П1.1)
Скорость роста биомассы dx — =(Н1 + М* (9П1.2) си
где в Ц. = ^ma*-1S1 (9П1.3) Si+Kid+s,/^) ' 1 P-max, 2S2 /пт H2=—(9П1.4) од “г Да
Оперонная модель биосинтеза
Внутриклеточная галактоза:
f ^2^2 \
— (S2J-X) = Ux\T------;--- — S2/ I — «1S2/* При Si<Sic
dt \Ams + s2 /
= — kiS2lX при S1>S1C
Репрессор:
d
~(r-x) = k2s-k3r-x —kir.sirx + k3(rs2l)x'}
Комплекс галактоза — репрессор:
d
~1("2/)X1 = V-S2/-X —*B(rSa/)X
мРНК галактозидазы:
d ( kg(rc—r)x — k,m-x при rc>r
~—(m-x) = 1
dt ( —kjtn-x при r ^rc
Фермент:
d
= kgtn-x
(9П1.5а)
(9П1.56)
(9П1.6)
(9П1.7)
(9П1.8а)
(9П1.86)
(9П1.9)
а Imanaka T., Kaieda T., Sato K„ Taauchi
(1972).
1
J. Ferment. Technol. (Japan), 50,633
б В уравнениях модели концентрации компонентов системы обозначены следующими
символами: биомасса — xt глюкоза — внеклеточная галактоза ~~ внутриклеточная
галактоза — s2p внутриклеточный репрессор — г, внутриклеточный комплекс индуктор —
репрессор — rs2p внутриклеточная мРНК — т, внутриклеточная а-галактозидаза — е. Дру-
гими символами обозначены параметры кинетики процессов, выходов и транспортных
явлений.
70
Глава 9
удельной скорости роста клеток на глюкозе Ц1 включает член, учитывающий
конкурентное ингибирование галактозой. Все константы, входящие в эти урав-
нения, были определены в экспериментах с непрерывной культурой в двух
различных средах. В табл. 9П1.2 параметры с индексом G отвечают глюкозной
среде (20 г глюкозы, 5 г NH4NO3, 5 г КН2РО4, 1 г MgSO4-7H2O, 0,1 г дрож-
жевого экстракта в 1000 мл водопроводной воды при pH 4,5), а индекс р
отвечает галактозной среде (5 г галактозы, 5 г NH4NO3, 5 г КН2РО4, 1 г
MgSO4-7H2O в 1000 мл водопроводной воды, pH 4,5), в которой преимущест-
венно и осуществляется синтез фермента.
Таблица 9П1.2. Параметры математической модели биосинтеза
а-галактозидазыа
Находящиеся в правом столбце параметры определены экспериментально;
другие параметры скорректированы таким образом, чтобы достигалось
соответствие результатам экспериментального изучения периодической и неп-
рерывной культур. Здесь индексами G и р обозначены параметры, относящие-
ся к глюкозной (30 °C) и галактозной (35 °C) средам соответственно
£1 = 40 ч-1
£2=1 мг/(мг клеток-ч)
£з=1 ч-1
£4 = 0,1 (мг клеток)/(мг-ч)
£5 = Н0-4 ч-1
£е=1 ч-1
£? = 8 ч-1
£sg = 3,2787 ед./(мг мРНК-ч)
£вр = 5,0442 ед./(мг мРНК-ч)
(7=100 ч-1
б2=1 мг/(мг клеток)
/Сп2=1-10_8 мг/(мг клеток)
sic = 2,25-10-4 г/мл
гс=0,934 мг/(мг клеток)
gmax.lG = 0,215 Ч-1
gmax,2G = 0,208 Ч~‘
Kig= 1,54-10-“ г/мл
K2g = 2,58 • 10-4 г/мл
|Хтах,1р = 0,190 Ч-1
Цтах,2р = 0,162 Ч *
К1р= 1,45-10"4 г/мл
/С2р=3,07- 10-4 г/мл
«1=1,39-10-4 г/мл
У1(?=0,530
y2G = 0,516
У1Р=0,377
У2Р=О,361
в Imanaka Т., Kaieda Т., Sato К.. Taguchi Н., J. Ferment. Technol. (Japan), Б0, 558
(1972).
Модель биосинтеза фермента основана на оперонной теории индукции
синтеза ферментов, рассмотренной нами в гл. 6. Считается, что удельная ско-
рость синтеза а-галактозидазы пропорциональна внутриклеточной концентра-
ции кодирующей этот фермент мРНК. Далее допускается, что эта мРНК
разрушается в реакции первого порядка, а продуцируется только при условии,
что внутриклеточная концентрация репрессора R меньше некоторого порого-
вого значения гс. Ниже этой пороговой величины удельная скорость синтеза
мРНК повышается при снижении концентрации г. Репрессор образуется с по-
стоянной удельной скоростью £2 и разрушается по реакции первого порядка
с удельной скоростью £3г. Концентрация репрессора снижается также за счет
его связывания с индуктором — внутриклеточной галактозой.
В уравнении материального баланса по внутриклеточной галактозе ско-
рость транспорта галактозы в клетку описывается членом с коэффициентом U.
Допускается, что при концентрации глюкозы si выше некоторой критической
величины Sic (принимается равной 2,25-10-4 г/мл) этот член равен нулю;
таким образом учитывается влияние глюкозы.
Прямое определение констант скоростей в модели оперона затруднено
из-за отсутствия соответствующих данных. Для оценки значений £3 и £7 еде-
Проектирование и расчет биологических реакторов
71
лано допущение, что периоды полураспада репрессора и мРНК составляют
40 и 5 мин соответственно. Приведенные в табл. 9П1.2 значения других пара-
метров определены методом проб и ошибок так, чтобы в конце концов дости-
галось приемлемое соответствие экспериментальным данным.
Эта модель обладает рядом преимуществ, обусловленных высокой сте-
пенью структурирования, базирующейся в основном на хорошо известных био-
логических особенностях системы. С другой стороны, известные возражения
может вызвать большое число корректируемых параметров. Для изучения
диапазона применимости этой модели можно использовать несколько тестов.
РИС. 9П1.5. Переходное состояние в ПРПП, инициируемое изменением ско-
рости разведения от £> = 0,140 до £> = 0,142 ч-1. Среда содержит два источника
углерода — глюкозу (2%) и галактозу (0,5%). В расчетах были приняты сле-
дующие начальные условия: х=1,30-10~2 г/мл, si=2,23-10“4 г/мл, s2=
=5,01-10“5 г/мл, s2/ = 2,5 мкг/(мг клеток), г=0,718 мкг/(мг клеток), rs2! —
= 1,28 мкг/(мг клеток), т= 1,04-10-2 мкг/(мг клеток), е = 0,297 ед./(мг кле-
ток). [Из статьи: Imanaka Т. et al., Unsteady-state Analysis of a Kinetic Model
for Cell Growth and a-Galactosidase Production by Mold; j. Ferment. Tech.,
(Japan), 51, 423 (1973).]
Один из них сводится к ответу на вопрос, могут ли другие модели, основанные
на иных допущениях и содержащие аналогичное число нуждающихся в кор-
ректировке параметров, так же хорошо соответствовать экспериментальным
данным? Если это так, то у нас нет оснований предпочитать именно эту мо-
дель. Иманака и сотрудники провели несколько подобных тестов, включая
варианты, когда внутриклеточная концентрация галактозы считается пропор-
циональной концентрации галактозы в среде, когда скорость образования
репрессора принимается пропорциональной внутриклеточной концентрации
глюкозы или когда скорость образования мРНК считается обратно пропор-
циональной концентрации репрессора. Любое из указанных изменений модели
приводило к серьезному расхождению между расчетными и эксперименталь-
ными данными.
Другие тесты, рассмотренные Иманакой и сотрудниками, включали ана-
лиз соответствия моделй экспериментальным данным в широком диапазоне
условий. Действительно, именно этот критерий определяет право на существо-
вание сложной высокоструктурированной кинетической модели, поскольку
только достаточно универсальная модель может использоваться для опреде-
ления оптимальных параметров процесса. Здесь описываемая модель также
72
Глава 9
Таблица 9П1.3. Оптимальные условия биосинтеза а-галактозидазы
в реакторах шести различных типов. Во всех случаях среда содержала
2% глюкозы и 0,5% галактозы*
Система Схема пр Glc + Gal- 1 Glc + Gal- 2 Glc—। 3 fl 30°C Glc—i 4 оце 3 30° 3 35° 3 Ga Оптимальные фермента- Скорость об- условил; ве- тивная ак- разования сса личины D тивность, фермента, выражены ед./(мг кле- ед./(лч) в 4 Ток) £> = 0,142 0,325 554 — । Повышение скорости — 1 * разведения С £> = 0,121 0,500 559 Повышение — скорости разведения С GiaI £>1 = 0,200 0,293 415 , 1 £>2=0,250 £> = 0,118 5 °C 1 £>1=0,133 0,500 582 £>2=0,286 £>=0,097
1 »
30°C Glc—। 5 _ 30=C Glc—I 6 30 G 35°С Gal £>1 = 0,193 0,500 702 £>2=2,342 I £>3 = 0,286 Lj j-1— । £>=0,117 °C 35°С £>1 = 0,178 0,500 702 £>2=0,286 я! £> = 0,117 С=1,34 г 01 = 0,266 0,500 870
| _ и 2 — U,zou 1 * л — л 145
30°C 35СС С=200
а Imanaka Т., Kaieda Т., Taguchi Н., J. Ferment. Technol. (Japan), 51, 558 (1973).
Проектирование и расчет биологических реакторов
73
дала отличные результаты. Кривые на рис. 9П1.1 были рассчитаны с помощью
рассматриваемой модели при указанных начальных условиях, причем соответ-
ствие между расчетными данными н экспериментом оказалось очень хорошим.
Уравнения модели, отражающие нестационарное состояние в ПРПП»
можно получить путем введения в каждое из уравнений, описывающих пе-
риодическую культуру (табл. 9П1.1), слагаемого, отвечающего скорости раз-
ведения D (разности концентраций в питательном растворе и реакционной
смеси). Затем получают соответствующие уравнения для стационарного со-
стояния в ПРПП, приняв, что все производные по времени равны нулю. С по-
мощью таких уравнений, в частности, были рассчитаны кривые, изображен-
Скоростъ разведется в первом ПРПП
W,n/4
-----0/133
Скорость разведения во втором ПРПП (Dz), л/ч
РИС. 9П1.6. Вычисленная и экспериментально найденная зависимости скоро-
сти образования фермента в непрерывной культуре в системе из двух ПРПП
(система 3 в табл. 9П1.3). [Из статьи: Imanaka Т. et al., Optimization of
a-Galactosidase Production in Multi-stage Continuous Culture of Mold; J. Fer-
ment. Tech. (Japan), 51, 431 (1973).]
ные на рис. 9П1.2 и 9П1.3. Модель хорошо воспроизводит наблюдаемые в экс-
перименте гистерезис и скачкообразные изменения параметров. В качестве
последнего теста, предшествовавшего работе по оптимизации реактора, модель
применяли для расчета параметров переходного состояния в ПРПП в режиме
повышения скорости разведения. Вычисленные на базе модели и соответству-
ющие экспериментальные данные представлены на рис. 9П1.5; между найден-
ными и вычисленными данными наблюдается очень хорошее соответствие,
включая скачкообразные повышение концентрации глюкозы и снижение кон-
центрации клеток.
Детально разработанную описанным путем модель применяли затем для
расчета выхода фермента в непрерывных реакторах различных конструкций
(табл. 9П1.3), причем в каждом случае объемы отдельных сосудов корректи-
ровались с целью обеспечения максимального выхода фермента. Указанная
в таблице общая скорость разведения D равна отношению общей скорости
потока поступающей в систему питательной среды к общему объему всех
сосудов системы.
Здесь несколько детальнее мы рассмотрим системы 3 н 4, в которых, как
и в системах 5 и 6, первая часть (первый сосуд) системы служит для созда-
ния высокой концентрации клеток с участием одной лишь относительно деше-
74
Глава 9
вой глюкозы; позднее в систему вводят галактозу, индуцирующую биосинтез
фермента. Поскольку процесс индукции происходит не мгновенно, а в течение
какого-то времени, то необходимо учитывать и этот период адаптации внут*
риклеточных систем регуляции. В математической модели стадия индукции
рассматривается изолированно, а ферментативная активность на выходе из
реактора рассчитывается по уравнению (9.62), которое для данного случая
можно записать в форме:
оо
ee=\eb{t)^{t)dt (9П1.10)
6
Здесь вь{{) —концентрация фермента в момент времени t, рассчитанная с по-
мощью математической модели периодической культуры. При расчете послед-
ней в качестве начальных условий принимаются значения концентраций ком-
понентов питательной смеси перед стадией индукции.
ь боо
500
5} С
^ЮО
£ В 300
О Ж
£ I 100
о loo гоо зоо боо
Время пребывания во Z-м реакторе,
мин
РИС. 9П1.7. Скорость образова-
ния фермента в системе из ПРПП
и трубчатого реактора (система 4
в табл. 9П1.3). [Из статьи: 1та-
naka Т. et al., Optimization of
a-Galactosidase Production in Mul-
ti-stage Continuous Culture of
Mold; J. Ferment Tech. (Japan), 51,
431 (1973).]
Описанным путем была рассчитана скорость образования фермента
(Deex) в двухстадийной системе 3; результаты расчетов представлены на
рис. 9П1.6. Расчеты показали, что скорость образования фермента должна
быть максимальной при 01 = 0,20 ч-1; результаты соответствующих экспери-
ментов практически полностью совпадали с расчетной зависимостью выхода
фермента от Di. В системе 4 на второй стадии применялся биореактор, при-
ближенно соответствующий реактору полного вытеснения. И в этом случае
(рис. 9П1.7) расчетные данные (сплошная линия) хорошо согласовались
с результатами эксперимента (точки).
Приведенные в табл. 9П1.3 данные показывают, что соответствующим
образом подобранная многостадийная система для культивирования организ-
мов обеспечивает существенное повышение выхода фермента по сравнению
с лучшим одностадийным процессом. Например, в системе 6 синтезируется на
55% больше фермента, чем в простом ПРПП. Наличие обоснованной кинети-
ческой модели и применение общих принципов расчета реакторов в этом
случае, очевидно, оказались важнейшими взаимодополняющими элементами
В создании и оптимизации высокоэкономичного непрерывного процесса.
9.4. Стерилизаторы
Жидкости, обычно водные растворы, можно стерилизовать
несколькими методами, в том числе облучением (ультрафиоле-
товым или рентгеновским излучением), воздействием ультра-
звука, фильтрованием, нагреванием и обработкой особыми
Проектирование и расчет биологических реакторов
75
химическими агентами. В крупномасштабном производстве широ-
ко применяются только два последних метода, хотя сравнитель-
но небольшие количества растворов неустойчивых витаминов
и других сложных соединений иногда стерилизуют путем про-
пускания через пористые мембраны. В этом разделе основное
внимание будет уделено процессам и аппаратам для стерилиза-
ции путем тепловой обработки.
Необходимые для разрушения жизнеспособных микроорга-
низмов и вирусов условия могут изменяться в очень широких
пределах в зависимости от природы стерилизуемого объекта и
сферы его последующего использования. Иногда, например при
квашении капусты или при биологической очистке сточных вод,
необходимые превращения обеспечивают уже присутствующие
в системе микроорганизмы. В процессах производства спирта,
винного уксуса и заготовки силоса быстро вырабатываются ин-
гибиторы, подавляющие рост нежелательных организмов, по-
этому здесь требования к стерилизации также невысоки. Пасте-
ризация молока сопровождается гибелью большинства, но да-
леко не всех активно растущих микроорганизмов. Более жесткая
обработка молока опасна из-за деструкции его полезных ком-
понентов. Вообще оптимальный баланс между разрушением
полезных компонентов и гибелью нежелательных микроорганиз-
мов играет основную роль в выборе метода стерилизации и
пастеризации, а также при расчете соответствующих аппаратов.
Процессы с участием чистых культур, работа с культурами
тканей и обработка некоторых пищевых продуктов требуют
гораздо более жесткой стерилизации. В таких случаях систему
необходимо практически полностью освободить от всех нежела-
тельных микроорганизмов, хотя «полнота» стерилизации также
зависит от конкретной системы. По экономическим соображе-
ниям, например, для периодического процесса допустима веро-
ятность заражения [(1—Ро) в уравнении (7.131)] порядка 10~2.
Это означает, что один процесс из каждых 100 может быть за-
бракован именно из-за недопустимо высокого заражения.
С таким ущербом можно примириться, если стоимость потерь
сравнима со стоимостью дополнительного стерилизующего обо-
рудования.
Значительно более жесткие правила приняты в консервной
промышленности. Здесь необходима действительно полная сте-
рилизация, поскольку единственная жизнеспособная спора
Clostridium botulinum может в конце концов привести к смер-
тельному отравлению. Обычно в такой ситуации основное тре-
бование, предъявляемое к процессу стерилизации, состоит в
снижении вероятности выживания спор (1— Ро) до величины
менее 10~12. Этот пример наглядно демонстрирует опасность
даже небольших отклонений от совершенно полного превраще-
76
Глава 9
ния субстрата (спор и вегетативных клеток) в продукты про-
цесса (инактивированные споры, погибшие клетки) в стерили-
зационных реакторах. Отсюда следует, что расчет таких реак-
торов необходимо выполнять чрезвычайно тщательно. Сначала
мы рассмотрим периодические, а затем непрерывные процессы
стерилизации.
9.4.1. Периодическая стерилизация
Изучение процессов стерилизации мы начнем с анализа за-
крытого сосуда с полным перемешиванием, содержащего суспен-
зию клеток или спор. Жидкость должна стерилизоваться при
нагревании, а затем охлаждаться до температуры, отвечающей
требованиям дальнейших этапов процесса. Концентрацию орга-
низмов, оставшихся жизнеспособными после такой обработки,
можно вычислить по уравнениям (7.125) и (7.126):
= —kdoe-Ed/RT (9.92)
dt
В это уравнение в явной форме включена зависимость темпера-
туры жидкости от времени. Путем разделения переменных в
уравнении (9.92) и интегрирования находим
*f
\n-^-^(e~Ed/RTii}dt (9.93)
nt J
о
Индекс f указывает, что данный параметр отражает конечные
условия.
Конструкция стерилизатора периодического действия обыч-
но включает одно или несколько нагревающих устройств, где
нагревание может осуществляться путем барботирования пара
(пропускания острого пара через стерилизуемую среду), с по-
мощью электрических нагревателей, посредством нагревания
или охлаждения в жидкостном теплообменнике. Дейндорфер и
Хамфри* предложили подразделять различные способы нагре-
вания или охлаждения в соответствии с формой кривой измене-
ния температуры во времени (температурным профилем); соот-
ветствующие функции приведены в табл. 9.4. Интеграл в правой
части уравнения (9.93) можно определить, разделив весь диа-
пазон интегрирования на три интервала — нагревание (повыше-
ние температуры), выдержку и охлаждение. В конечном счете
мы получим четыре интегральных уравнения, одно из которых
♦ Deindoerfer F. Н., Humphrey А. Е., Analytical Method for Calculating
Heat Sterilization Times; Appl. Microbiol., 7, 256 (1959).
Проектирование и расчет биологических реакторов
77
Таблица 9.4. Температурные профили периодических процессов стерилизации’
Тип теплообмена Температурный профиль Параметры
Барботаж пара
Электрические
нагреватели
гиперболический
7=7о(1,О+аО
линейный
Пар (в тепло-
обменниках)
Охлаждение
(с помощью
теплообмен-
ников)
7=7w(l-f-&e-°‘)
эксп оненци ал ьны й
7=7ео(1+&е-“‘)
экспоненциальный
hs $
--------- __ —
МТорСр М
_ я
МТцрСр
и А , T0-TN
= — ь ----------
Me Тк
= WC'- П _e-UA/wc\
МрСр ( >
__ Тр Тео
Обозначения: И — разность между энтальпией пара (при температуре барботажа)
и энтальпией среды; s — массовая скорость потока пара; М— начальнаи масса среды;
То— начальная температура среды; q — скорость теплопередачи, ккал в единицу вре-
мени; U — коэффициент общей теплопередачи, ккал/(м2-ч-град); А — площадь поверхно-
сти теплообмена, м2; TN— температура источника тепла; w — массовая скорость потока
охлаждающего агента; с'—удельная теплоемкость охлаждающего агента; Те0 — темпе-
-ратура охлаждающего агента на входе; р — плотность среды; Ср — средняя теплоемкость
среды.
а По данным статьи: Deindoerjer F. Н., Humphrey А. Е., Appl. Microbiol., 7, 256
(1959).
описывает интервал постоянной температуры, а три других —
интервалы изменения температуры по гиперболическому, линей-
ному или экспоненциальному законам.
Постоянная температура:
In = Г kd.e~EdIRTdi = kdotfe~EdfRT (9.94)
rt,f J
о
Изменение температуры по гиперболическому закону:
1П ”о . _ kdOa(Ed/RTo) c-(E,,IRTa)fbl(a+bH
nf (a-\-b)2
у P f ^d . ____1 ~b Mf________1_______p I Ed_____a
_ 21 RT0 L 1 + (a+b)tf a+ b ]j fl+Jj ^9'95^
Линейное повышение температуры:
Ini (p Ed^r,-----
nf RTpa \ 1 + atf RT0 /
78
Глава 9
Изменение температуры по экспоненциальному закону:
In »о ( Ед/РТн \__f Ed/RTfj \ ___
nf ~ a |_ 4 1 + & ) 41 + bef~at}_
kd0 (.-EJRT/A g ( —Ed b \ ( —Ed bef~at y
a L 4 RTh i + b J-r 4 RTh 1 + bef~at /
(9.97)
Здесь En — интеграл экспоненциальной функции:
oo
£n(z)=C^ (9.98)
J
Величины En(z) приведены в различных справочниках и вклю-
чены в стандартные программы компьютеров. Соответствующие
выражения для охлаждения получают, заменяя Тн на Тс0 и ис-
пользуя отвечающие охлаждению определения параметров а и
b (табл. 9.4).
В каждом случае в результате решения уравнения получа-
ется логарифм отношения конечной и начальной концентраций
1п(п^/по). Если, например, за электрическим нагреванием сле-
дует выдерживание при повышенной температуре, а затем
охлаждение с помощью жидкостного теплообменника, то отно-
шение конечной и начальной концентраций жизнеспособных
клеток можно выразить в виде уравнения:
nf nf (выдержка) nf (нагревание) "1 ggj
_ п0 (охлаждение) п0 (выдержка) п0 (нагревание) J
ln-2L=ln
n0
поскольку «о (охлаждение) =п} (выдержка) и п0 (выдержка) =
= nf (нагревание). Если уравнение (9.99) записать в другой
форме:
1пА = 1пДНЧ-+1п-Щ^- + 1пЛН£1 (9.100)
п0 По (с) «о (Л) по (е)
где с — охлаждение, h — выдержка и е — нагревание, то ста-
новится очевидным, что общий результат 1п(п^/п0) получается
путем сложения указанных выше трех индивидуальных реше-
ний, каждое из которых относится к определенному временному
интервалу, охватывающему определенный режим теплопередачи.
Здесь целесообразно изучить также математическую модель
другого процесса, а именно тепловой стерилизации твердых тел
или жидкостей в состоянии покоя. Одной из областей использо-
вания тепловой стерилизации является разрушение токсичных
организмов в герметично упакованных пищевых продуктах. Час-
то возникает необходимость и в максимальном удалении попу-
Проектирование и расчет биологических реакторов
79
ляций микроорганизмов, способных разлагать или каким-либо
другим образом вызывать порчу продуктов в закрытых контей-
нерах. В подлежащих стерилизации жидкостях часто содержат-
ся суспендированные твердые частицы или скопления микроор-
ганизмов. Если внутри этих частиц или скоплений имеются не-
желательные организмы, то они труднее поддаются тепловой
обработке, и поэтому для их стерилизации необходимо более
продолжительное нагревание или более высокая температура.
Математический анализ этих двух типов стерилизации, по
крайней мере для случая простого контейнера той или иной гео-
метрии, можно выполнить в упрощенном виде с помощью дву-
стадийной модели, аналогичной рассмотренной выше. Прежде
всего необходимо определить температуру в твердом теле как
функцию положения и времени, т. е. решить задачу о теплопро-
водности в нестационарном состоянии. Если допустить, что
удельная теплопроводность k постоянна, то эту задачу можно
сформулировать в виде уравнения:
f>C„-^ = ky‘T (9.101)
Здесь V2 — оператор Лапласа, а определения символов р и Ср
даны в табл. 9.4. Помимо частного дифференциального уравне-
ния (9.101), Т определяется также как функция положения
внутри твердого тела при / = 0; кроме того, известно, что тем-
пература наружной поверхности твердого тела при t>0 явля-
ется функцией времени.
Если, например, изучать сферическое твердое тело, то для
определения температуры внутри этого тела при £>0 необходи-
мо решить следующие уравнения:
дТ k 1 д / 2 дТ\
dt рСр г2 dr \ dr J
Т(г, 0) =f(r) ОсгсА?
T(R,t)=g(f) />0
(9.102)
(9.103)
(9.104)
Здесь г —расстояние от центра сферы, a f и g — соответствую-
щие функции. Если первоначально температура в любой точке
сферы равна То и если температуру ее поверхности T(t,R) при
/>0 поддерживать на одном и том же уровне 7\, то эту задачу
можно решить путем разделения переменных:
Т (G t) = 7\ + у J-1)” Sin 2ЕК- exp f-—
пг Й n R \ pCp
Для определения температуры в центре сферы (г=0)
R* J
(9.105)
найдем
80
Глава 9
предел функции (9.105) при г->0. Таким путем получим
Т (0, t) = Л 4- 2 (7\ - То) 2 (-1 )"ехр - М-
П= 1 ' Р^Р '
(9.106)
Рассчитанные с помощью этих уравнений профили распреде-
ления температур в сфере при различных t представлены на
рис. 9.23. Что касается процессов стерилизации, то наиболее
РИС. 9.23. Температурные профили в сфере радиусом R в зависимости от от-
носительного времени kt/pCpR2 (обозначено цифрами на каждой кривой).
То — начальная температура в любой точке сферы, а Т\— температура на-
ружной поверхности сферы при />0.
важным выводом из обсуждаемой модели является наличие
лаг-фазы между временем приложения высокой температуры к
поверхности твердого тела и установлением аналогичной тем-
пературы во всем объеме тела. Следовательно, в общем случае
разрушение организмов в центре твердого тела будет менее
полным, чем на его поверхности. Качественно этот вывод сохра-
няет свою силу для тел любой другой геометрии и любых на-
чальных и граничных условий. Не имея возможности углублять-
ся в детали проблемы теплопроводности в различных системах
в нестационарных условиях, мы рекомендуем читателю моно-
графию Карслоу и Егера [29], содержащую много дополнитель-
Проектирование и расчет биологических реакторов
81
ной информации, описывающей теории и аналитические ре-
шения.
Если зависимость температуры от времени и положения
внутри твердого тела определена, то затем можно рассчитать
и долю погибших микроорганизмов и спор в этом теле. Для
этой цели в пищевой промышленности использовались два под-
хода. В первом из них рассматривается концентрация организ-
мов в центре твердого тела. Поскольку центр нагревается в
последнюю очередь, то, если его нагреть до соответствующей
температуры, будет обеспечена достаточная степень стерилиза-
ции и всего твердого тела. Далее для расчета доли выживших
организмов в центре нам достаточно ввести в уравнение (9.93)
функцию зависимости температуры от времени в этой же точке
и вычислить соответствующий интеграл. Последнюю операцию
часто удобнее выполнять в численной или графической форме,
поскольку зависимость Т от t обычно выражается довольно
сложным уравнением, например (9.106).
В другом подходе описанные выше вычисления повторяют
много раз и таким путем определяют концентрацию выживших
организмов в каждой точке внутри твердого тела. Затем, инте-
грируя эти концентрации по всему объему твердого тела, можно
определить численность выжившей популяции или вероятность
выживания организмов. Хотя принцип такого подхода очевиден
и несложен, соответствующие расчеты довольно трудоемки.
Поэтому для решения конкретных задач пищевой промышлен-
ности были разработаны упрощенные методики расчета, осно-
ванные на том же принципе. Объяснение этих методик невоз-
можно без введения новых понятий и определений; интересую-
щийся этой проблемой читатель может ознакомиться с ней
детальнее в книге Чарма [30].
Преимущество периодической стерилизации состоит в отно-
сительной простоте процесса, но вместе с тем ей свойствен и
ряд недостатков. Один из них связан с продолжительным на-
греванием и охлаждением, другой вытекает из первого и за-
ключается в возможности разрушения полезных компонентов
стерилизуемой системы. Действительно, при нагревании разру-
шаются многие витамины, а белки подвергаются денатурации.
Необходимо подчеркнуть, что кинетика разрушения этих ком-
понентов системы часто описывается теми же уравнениями, что
и кинетика гибели организмов [уравнение (9.92)], а энергия
активации этих нежелательных побочных превращений обычно
намного ниже энергии «реакции» стерилизации. Приведенные
в табл. 9.5 величины, например, на 50—100 ккал/моль меньше
энергии, обычно необходимой для разрушения клеток и спор.
Поскольку энергия активации необходимого процесса выше
энергии активации побочной реакции, то повышение температу-
б-746
<82
Глава 9
ры благоприятно влияет на отношение скоростей необходимого
и побочного процессов. Отсюда следует, что для подавления
реакции Майяра или предотвращения разрушения лабильных
соединений процесс стерилизации нужно проводить при возмож-
но более высокой температуре в течение минимального времени,
обеспечивающего гибель нежелательных организмов (высоко-
температурная кратковременная обработка, или режим ВТКВ).
.В процессах периодической стерилизации при медленном нагре-
Таблица 9.5. Приближенные величины энергии активации
некоторых нежелательных побочных реакций, происходящих
при тепловой обработке
Реакция Энергия ак- тивации Е, ккал/моль
Реакция Майяра между белками и уг- 31,2
леводами
Разрушение витамина Bi 21,0
Разрушение рибофлавина (витамина В2) 23,6
Денатурация пероксидазы 23,6
вании и медленном охлаждении эти цели не достигаются. В рас-
сматриваемой ниже непрерывной стерилизации в потоке режим
ВТКВ обеспечивается значительно легче.
9.4.2. Непрерывная стерилизация
На рис. 9.24 схематично изображены два основных типа
непрерывных стерилизаторов. В первом из них (рис. 9.24, а)
нагревание осуществляется путем введения струи пара, затем
нагретая жидкость проходит через секцию с постоянной темпе-
ратурой и, наконец, охлаждается путем распыления. В схеме,
изображенной на рис. 9.24,6, стерилизуемая жидкость нагрева-
ется в пластинчатом теплообменнике. Предложено много вари-
антов стерилизаторов каждого типа; многие из них отличаются
особенно высоким отношением поверхности теплообмена к объ-
ему аппарата. Такие стерилизаторы обеспечивают относительно
быстрое нагревание и охлаждение, поэтому в них может быть
реализован режим ВТКВ.
Если допустить, что температура в каждой точке непрерыв-
ного стерилизатора приблизительно одинакова (это допущение
справедливо по крайней мере для секции выдержки температу-
ры), то для математического анализа и расчета стерилизатора
применимы два подхода. В первом подходе используется опи-
санная выше дисперсионная модель, в которой в уравнениях
(9.81) — (9.83) заменено на —kdn, а с — на п. Тогда кон-
.центрацию организмов в выходящей из трубчатого стерилиза-
Проектирование и расчет биологических реакторов
среда
РИС. 9.24. Два типа стерилизаторов непрерывного действия: инжекционный
стерилизатор с непосредственным введением пара (а) и стерилизатор с пла-
стинчатым теплообменником (б). (Из работы: Аиба Ш„ Хемфри А., Мил-
лис Н., Биохимическая технология и аппаратура. — М.: Пищевая промышлен-
ность, 1975, с. 182.)
тора среде можно определить аналитически и при наличии осе-
вой дисперсии. Таким путем можно найти, что величину п в
выходящей из стерилизатора среде можно выразить уравнением:
п (L) =__________________4t/exp [Ре/2]_______________ (9 107)
«о (1 + у)аехр[(Ре)(!/)/2]-(1-у)2ехр[ -(Pe)(t/)/2] 7
где
! 4Da\1/а
£/ = ( 1+-^-) (9.Ю8)
\ Ре /
а число Дамкелера Da определяется по формуле:
Da=-^- (9.109)
ь
-84
Глава 9
При небольших отклонениях от режима полного вытеснения
(при малых Ре-1) уравнение (9.107) упрощается:
п (L) f Т> I Da2 \ /л 1
—x_2_ = eXp^_£)a_|____j (9.110)
Эти уравнения удобно решать графически в виде зависимо-
сти доли выживших клеток n(L)/n0 от критерия подобия Da
для различных значений числа Пекле (рис. 9.25). Эти зависи-
мости показывают, что при Ре->-оо (и, следовательно, при при-
ближении к режиму полного вытеснения) необходимая степень
стерильности может быть достигнута в самом коротком труб-
чатом стерилизаторе. Отсюда далее следует, что конструкция
непрерывного стерилизатора должна обеспечивать минимальную
осевую дисперсию.
Если по какой-либо причине поток через изотермический
непрерывный стерилизатор нельзя удовлетворительно описать
дисперсионной моделью, то мы можем обратиться к общей тео-
рии РВП. Трудно представить себе лучший пример полностью
сегрегированной системы, чем суспензия клеток или спор, под-
вергающаяся тепловой обработке. Поэтому, если обратная диф-
фузия организмов в линию подачи в стерилизатор исходной
суспензии ничтожно мала, то концентрацию выживших орга-
низмов п на выходе из стерилизатора можно рассчитать с по-
мощью уравнения (9.62). Записав это уравнение в параметрах
концентраций организмов, получим
n = Jnb(tyt(t)dt (9.111)
о
Здесь <S(t)—функция РВП непрерывного стерилизатора. На-
помним, что п&(0—концентрация организмов в периодическом
стерилизаторе в момент времени t, причем Пь(О)=по. Из урав-
нений (7.125) и (9.111) следует, что
оо
-А-= f (9.112)
п0 J
о
Иногда вычисление правой части уравнения (9.112) облегчается
тем, что формально оно идентично преобразованию Лапласа
функции S, в котором обычный параметр преобразования Лап-
ласа 5 заменен на ka.
В заключение следует отметить некоторые другие (помимо
упоминавшейся возможности осуществления режима ВТКВ)
преимущества непрерывной стерилизации. Во-первых, непрерыв-
ная стерилизация по сравнению с периодическим процессом
обычно менее трудоемка. Во-вторых, аппараты для непрерывной
Проектирование и расчет биологических реакторов
85
РИС. 9.25. Влияние осевой дисперсии на разрушение организмов в непрерыв-
ном стерилизаторе. (Из работы: Аиба Ш., Хемфри А., Миллис Н., Биохимиче-
ская технология и аппаратура. — М.: Пищевая промышленность, 1975, с. 187.)
86
Глава 9
стерилизации обеспечивают большую однородность условий и
воспроизводимость результатов, что особенно важно в пищевой
промышленности, где даже небольшие отклонения от режима
обработки могут существенно влиять на вкусовые качества
продукта.
В микробиологической промышленности периодическая сте-
рилизация обычно осуществляется непосредственно в реакторе
(in situ). В таких случаях скорость нагревания и охлаждения
зависит от отношения площади поверхности реактора к его
объему, а эта величина в свою очередь изменяется при мас-
штабном переходе. В непрерывной стерилизации может быть
снижена зависимость влияния стерилизации на организмы и
компоненты среды от размеров аппарата, и, кроме того, при
проектировании реактора отпадает необходимость учитывать
требования, предъявляемые к эффективному периодическому
стерилизатору.
Конечно, стерилизация в потоке имеет и свои недостатки.
Так, стерилизация путем прямого ввода пара в среду может
привести к излишнему разведению системы водой, а использу-
ющиеся для нагревания или охлаждения поверхностные тепло-
обменники могут засоряться в результате осаждения суспенди-
рованных твердых частиц. При непрерывной стерилизации по-
вышается вероятность вспенивания культуральной жидкости.
Другие сведения о преимуществах и недостатках различных
стерилизаторов можно найти в работе [33], приведенной в гл. 7.
9.5. Иммобилизованные биокатализаторы
Работа биологического реактора в основном определяется
свойствами применяющихся биокатализаторов. Ранее мы об-
суждали различные методы и способы использования фермен-
тов, при помощи которых последние могли бы успешно выпол-
нять свои каталитические функции. Кроме того, нами уже
изучены методы генетической модификации клеток, позволяю-
щие повысить выход синтезируемых ими ценных соединений.
Прежде чем перейти к изучению реакторов других типов, позна-
комимся с еще одним типом биокатализаторов.
Этот раздел мы посвятим в основном изучению катализато-
ров на основе иммобилизованных клеток. В таких катализаторах
клетки связаны или включены в структуры, позволяющие удер-
живать их в определенной зоне реактора. Клетки, как и фер-
менты, могут быть иммобилизованы путем их включения в не-
большие частицы или посредством закрепления на поверхности
этих частиц, или с помощью того или иного макроскопического
барьера, препятствующего переносу клеток из одной зоны реак-
тора в другую.
Проектирование и расчет биологических реакторов 87
Прежде чем перейти к изучению методов иммобилизации
клеток и некоторых экспериментов, выполненных к настоящему
времени с участием такого рода катализаторов, мы должны
-ответить на вопрос, почему вообще возникает необходимость в
иммобилизации клеток. Одно из преимуществ иммобилизован-
ных клеток связано с возможностью достижения гораздо боль-
шей их плотности в культуре по сравнению с суспендированны-
ми клеточными системами. В биореакторах периодического
действия иммобилизованные клетки обладают и некоторыми
другими преимуществами. Так, некоторые клетки млекопитаю-
щих растут только в том случае, если они связаны с поверхно-
стью, и, следовательно, для них иммобилизация является обя-
зательным условием, а не одним из возможных вариантов.
Кроме того, иммобилизованные клетки могут использоваться
для создания специфических электродов, предназначенных для
определения концентраций питательных веществ, метаболитов,
лекарственных препаратов и токсичных веществ в биореакторах,
а также в других процессах и в клинической практике. Иммо-
билизация клеток может оказаться полезной как средство для
регуляции морфологии клеток и реологии бульонов. Действи-
тельно, рост микроорганизмов ограничен пустотами и поверх-
ностями частиц носителя, что позволяет достаточно точно регу-
лировать реологические свойства бульона и соответствующие
параметры массопереноса, которые уже не изменяются в ходе
процесса в такой степени, в какой это типично для периодиче-
ского роста отдельных культур. Иммобилизация клеток создает
возможность для осуществления непрерывного процесса без
вымывания организмов и изменения их генетической природы,
з в некоторых случаях позволяет непрерывно отделять продукт
и удалять ингибирующие вещества. Как и в случае других им-
мобилизованных катализаторов, стоимость иммобилизованных
клеток может быть довольно высока и не обязательно бу-
дет компенсироваться указанными преимуществами.
В случае биореакторов непрерывного действия применение
иммобилизованных клеток дает возможность расширить вре-
менной диапазон, в течение которого клетки проявляют свои
каталитические свойства при осуществлении необходимой хи-
мической реакции или последовательности реакций. В отличие
от иммобилизованных ферментных катализаторов иммобилизо-
ванные клетки выполняют целый спектр каталитических функ-
ций самой разнообразной сложности. Как схематично показано
на рис. 9.26, катализаторы на основе иммобилизованных клеток
могут быть нескольких типов в зависимости от того, какая часть
метаболического аппарата клетки сохранила свою активность
в данном процессе. При работе с суспензиями клеток для пред-
отвращения их частичного вымывания необходимо, чтобы рост
88
Глава 9
клеток осуществлялся и в реакторе. Рассмотренное в первых
разделах этой главы повышение скорости разведения, отвечаю-
щей точке вымывания, за счет рециркуляции, по сути дела,
представляет собой частичную иммобилизацию клеток в мас-
штабе всего реактора. Таким образом можно повысить произ-
РИС. 9.26. Различные уровни сложности сети реакций, осуществляемых ка-
тализаторами на основе иммобилизованных клеток.
водительность процесса в какой-то мере независимо от скорости
роста организмов. Если клетки полностью удерживаются в ре-
акторе, то в силу отсутствия затрат питательных веществ и
энергии на клеточный рост можно ожидать проявления полного
спектра каталитической активности. В сущности в таких слу-
чаях клеточный рост часто вообще нежелателен, поскольку он
может привести к снижению выхода необходимого продукта и
поскольку накопление клеточной массы может нарушить гидро-
динамические и реологические свойства системы.
На самом низком уровне катализ иммобилизованными клет-
ками сводится к проявлению активности одного или нескольких
Проектирование и расчет биологических реакторов
89
ферментов (без участия кофакторов). В этом случае иммобили-
зованные клетки выполняют ту же функцию, что и иммобили-
зованные ферментные катализаторы, и применение целых клеток
целесообразно только с точки зрения снижения трудоемкости
и стоимости процесса изготовления иммобилизованного катали-
затора. При использовании таких катализаторов следует, одна-
ко, учитывать трудности, связанные с дополнительным сопро-
тивлением клеточной оболочки массопереносу и с возможностью
расщепления необходимого фермента внутриклеточными проте-
азами. Впрочем, в других случаях нативное клеточное окруже-
ние может способствовать стабилизации некоторых ферментов.
Далее, нерастущие иммобилизованные клетки могут катали-
зировать многостадийные превращения, включая стадии утили-
зации и регенерации кофактора. Если в системе присутствуют
все кофакторы, ферменты, субстраты и регенерирующие вещест-
ва, то в общем случае связанные с этими каталитическими про-
цессами биосинтетические реакции и клеточный рост не нужны;
если к тому же в организме уже существует определенный путь
метаболизма с участием кофакторов, то иногда целесообразно
использовать этот естественный путь, а не пытаться создать
другой механизм утилизации и регенерации кофактора.
На следующем по степени сложности каталитической актив-
ности уровне осуществляются процессы биосинтеза и поддержа-
ния жизнедеятельности клеток, но их рост и деление несущест-
венны. Наконец, в самом сложном случае активна вся метабо-
лическая сеть, клетки растут и делятся. В упоминавшихся выше
реакторах периодического действия с участием культур иммо-
билизованных клеток, например, возникает каталитическая
активность именно этого наивысшего уровня сложности. Такой
же уровень каталитической активности иммобилизованных кле-
ток иногда целесообразен и в реакторах непрерывного действия
с регенерацией катализатора, поскольку некоторые организмы
плохо реагируют на среду, которая не способствует росту,
и в таких условиях быстро теряют активность. Как мы увидим
в приведенных ниже примерах, чередование условий процесса
с менее сложным уровнем каталитической активности клеток
до режима полного роста представляет собой обычный прием,
позволяющий продлить срок службы катализатора.
Катализатор, который выполняет необходимые каталитиче-
ские функции, но не инициирует большое число побочных реак-
ций, имеет два преимущества. Первое из них связано с увели-
чением выхода продукта. Если субстрат можно стехиометриче-
ски превратить в конкретный продукт определенного пути
метаболизма микроорганизмов, избежав при этом клеточный
рост или образование других конечных веществ, то выход необ-
ходимого продукта в таких условиях, очевидно, будет макси-
90
Глава 9
мальным. Другое преимущество обсуждаемых катализаторов*
заключается в более высокой общей скорости реакций. По мере
усложнения каталитического процесса от простейшего однофер-
ментного до наиболее сложного — полного клеточного роста
(рис. 9.26)—резко возрастает и характерное время пребывания,
в реакторе, необходимое для существенного превращения суб-
страта. На одноферментном уровне скорости реакций определя-
ются скоростями катализируемых ферментом отдельных эле-
ментарных стадий, которые обычно завершаются примерно за
одну минуту. Напротив, время удвоения клеток, характеризую-
щее время осуществления всей сети метаболических превраще-
ний, обычно составляет от нескольких часов до нескольких
суток во всех случаях за исключением наиболее быстро расту-
щих бактерий. Поэтому- по мере усложнения метаболических
превращений (см. рис. 9.26) в общем случае значительно воз-
растает и время пребывания реакционной смеси в реакторе,
необходимое для превращения значительной доли субстрата.
Отсюда следует, что выгоднее использовать только ферменты и
кофакторы, необходимые для данного превращения; если этот
путь удается реализовать, то тем самым удается существенно
сократить расчетное время пребывания компонентов в реакторе.
Закончив на этом изложение основ проблемы использования
иммобилизованных клеток, перейдем теперь к изучению катали-
заторов на основе иммобилизованных клеток, методов их полу-
чения и изучения, а затем рассмотрим ряд примеров, иллюстри-
рующих различные области применения этих катализаторов.
9.5.1. Типы биокатализаторов на основе иммобилизованных
клеток и их свойства
В этом разделе мы рассмотрим методы, применявшиеся для
иммобилизации клеток и изучения свойств полученных катали-
заторов, играющих важнейшую роль в процессах с их участием.
Иммобилизация клеток чаще всего осуществляется путем их
включения в полимерный носитель. В табл. 9.6 перечислены раз-
личные методы, применявшиеся для создания сшитых попереч-
ными связями полимерных носителей. Гораздо чаще других
применяется метод включения клеток в слой или в гранулу аль-
гината (природного полисахарида) путем сшивки последнего
ионными поперечными связями. Мягкие условия процедуры
сшивки (в отличие от методов химической полимеризации)
обеспечивают значительно более высокий уровень жизнеспособ-
ности иммобилизованных клеток. На рис. 9.27 представлена
схема одной из методик получения сферических гранул альги-
ната кальция, содержащих иммобилизованные клетки.
Проектирование и расчет биологических реакторов
91
Таблица 9.6. Полимерные носители, применяющиеся
для иммобилизации клеток
Способ получения носителя Тип поперечных связей Примеры
Осаждение Неспецифические Коллаген, полистирол, каррагинан
Гелеобразование, связан- Ионные ное с ионным обменом Поликонденсация Ковалентные Полимеризация Ковалентные Альгинаты алюминия Эпоксидные смолы Полимеры на основе акриловой и метакри- ловой кислот
При выборе методики и конкретных условий включения в
полимерный носитель необходимо учитывать и конечные свой-
ства получаемого иммобилизованного катализатора. В табл. 9.7
перечислены некоторые основные свойства носителей, представ-
ляющие наибольший интерес с точки зрения процесса, в кото-
ром будет использоваться катализатор. На проницаемость но-
сителя по отношению к субстратам, ингибиторам, продуктам
реакции и другим компонентам среды могут влиять как хими-
ческие, так и механические характеристики носителя. Так, на-
Таблица 9.7. Важнейшие свойства полимерных носителей,
применяющихся для иммобилизации клеток
Механические Химические
Проницаемость Геометрия частиц Токсичность Сжимаемость, механическая Гидрофобность, гидрофиль-
прочность частиц Чувствительность к сдвигу ность Ионный состав
пример, механические свойства носителя могут налагать опре-
деленные ограничения на допустимую геометрию частиц ката-
лизатора. Сжимаемость и другие механические свойства ката-
лизатора, не столь существенные в лабораторных эксперимен-
тах, могут оказаться чрезвычайно важными в крупномасштабных
процессах. Гранулированные полисахаридные носители, широко
использующиеся в лабораторных исследованиях благодаря сво-
ей высокой биосовместимости, мало подходят для крупномас-
штабного производства из-за большой сжимаемости и по этой
причине их можно применять только в невысоких реакторах
колонного типа. Точно так же неустойчивые к ударам иммоби-
лизованные катализаторы непригодны в тех случаях, когда
92
Глава 9
45 мл 8 %-нога (масса / объем}
раствора альгината
Юг клеток (во влажном состоянии)
В
Z7o-miii (масса/объем)
раствор CaClg
0,1 М раствор AlgCSO^
РИС. 9.27. Схема методики включения клеток в гранулы альгината.
гранулы катализатора должны выдерживать соударения без
истирания и разрушения (например, в реакторах с перемеши-
ванием).
Перечисленные в табл. 9.7 химические свойства носителей
могут влиять на каталитическую активность иммобилизованных
клеток непосредственно, т. е. путем воздействия на системы
регуляции клеточного метаболизма, или опосредованно, через
локальное окружение клеток. Если, например, полимерный но-
ситель содержит токсичные вещества, то можно ожидать сни-
жения активности клеток. Заряд, ионный состав и гидрофобные
свойства носителя могут влиять на коэффициенты распределе-
Проектирование и расчет биологических реакторов
95
ния компонентов реакционной смеси и тем самым вызывать из-
менение их концентрации в зонах непосредственного контакта
с клетками по сравнению с концентрациями в жидкой фазе,
не контактирующей с частицами катализатора.
Другой метод иммобилизации клеток состоит в их включе-
нии в полупроницаемые или пористые материалы. В качестве
носителей для растущих микроорганизмов использовали очень-
мелкие сита из нержавеющей стали. Кроме того, как посевной,
материал для периодических процессов применяли споры плес-
Таблица 9.8. Примеры иммобилизации клеток путем
связывания с поверхностью носителей
Клетки Носитель
Адсорбция
Е. coll, Cl. acetobutylicum Asp. orysae Streptomyces Lactobacilli, дрожжи S. carlsbergensis Pseudomonas sp. Клетки тканей Ионообменные смолы Модифицированная целлюлоза Модифицированный сефадекс Желатин Поливинилхлорид Антрацит Полисахариды
Ковалентное связывание
В. subtilis Е. coli Агароза и карбодиимид Оксид титана(IV)
невых грибов, включенные в автоклавированные гранулы порис-
того фильтрующего материала. Клетки также включали в мак-
ропористые зоны асимметричных полых волокон. Помимо ука-
занных методов включения клеток в носители полная или;
частичная иммобилизация клеток, как мы уже не раз упомина-
ли, может быть достигнута в масштабе всего реактора путем их
рециркуляции. В альтернативном подходе относительно низко-
молекулярные продукты выделяют из суспензии клеток методом
диализа или ультрафильтрации с помощью помещенного в сус-
пензию мембранного устройства; таким же путем можно вводить
субстрат в суспензию клеток.
Клетки многих типов иммобилизовали путем адсорбции или
ковалентного связывания с поверхностями твердых носителей.
Примеры иммобилизованных клеток (от бактерий до клеток
тканей животных) и соответствующих природных или синтети-
ческих носителей приведены в табл. 9.8. Другие сведения о
методах иммобилизации читатель может найти в приведенной
в конце главы литературе, а также в литературе, цитируемой-
в рассматриваемых ниже примерах.
94
Глава 9
9.5.2. Применение биокатализаторов
на основе иммобилизованных клеток
Рассматривая примеры, иллюстрирующие степень сложности
процессов катализа с участием иммобилизованных клеток, мы
начнем с самого простого случая и постепенно будем переходить
ко все более и более сложным случаям. Путем иммобилизации
целых клеток, предварительно обработанных с целью повышения
их проницаемости литическими ферментами, спиртами или ди-
метилсульфоксидом (ДМСО), мы, по сути дела, иммобилизуем
входящие в их состав индивидуальные ферменты, в той или иной
мере сохраняя их естественное окружение; такие катализаторы
в принципе пригодны для использования в непрерывных процес-
сах. К настоящему времени получено большое число катализа-
торов типа «иммобилизованные ферменты в иммобилизованных
РИС. 9.28. Инактивация аспартазы и фумаразы в суспендированных и иммо-
билизованных клетках. (Воспроизведено с разрешения из работы: Immobilized
Enzymes, Chibata I. (ed.), p. 140, Kodansha Ltd., Tokyo, 1978.)
клетках»; некоторые из этих катализаторов нашли применение
в промышленности. Такой подход к иммобилизации ферментов
проще, чем выделение ферментов из клеток с последующей им-
мобилизацией. С точки зрения практического использования,
по-видимому, важнее другое преимущество иммобилизованных
клеток, заключающееся в возможном повышении стабильности
ферментов.
На рис. 9.28 представлены результаты экспериментов по
изучению инактивации аспартазы и фумаразы в иммобилизован-
Проектирование и расчет биологических реакторов 95
ных клетках Е. coll и в суспендированных интактных клетках
того же организма. В иммобилизованных клетках инактивация
как первого, так и второго ферментов происходит значительно
медленнее. Возможно, что отчасти эта стабильность является
кажущейся и обусловлена ограничениями ферментативных про-
цессов за счет сопротивления массопереносу (см. разд. 4.4).
Действительно, если катализатор на основе иммобилизованных
клеток (или ферментов) функционирует в режиме, строго лими-
тируемом диффузией, то наблюдаемая скорость инактивации
будет значительно ниже действительной скорости этого процес-
са. Однако в дальнейших экспериментах Шибата и др. показа-
ли, что в данном конкретном случае имеет место истинная ста-
билизация фермента вследствие его включения в нативное
клеточное окружение. Интактные клетки Е. colt были обрабо-
таны ультразвуком, затем растворимая и нерастворимая фрак-
ции продуктов обработки были разделены и иммобилизованы.
В иммобилизованной растворимой фракции аспартазная актив-
ность снижалась значительно быстрее, чем в иммобилизованной
нерастворимой фракции. Кроме того, обработкой интактных
клеток агентами, солюбилизирующими связанные с мембранами
белки (дезоксихолатом или тритоном Х-100), также были по-
лучены растворимая и нерастворимая фракции, которые после
иммобилизации быстро теряли активность. Эти результаты сви-
детельствуют о возможности стабилизации аспартазы путем
связывания или ассоциации с клеточными мембранами или с их
компонентами. Следует отметить, однако, что в других случаях
иммобилизация ферментов в интактных клетках сопровождается
и обратным эффектом. Так, в проницаемых иммобилизованных
клетках некоторых организмов глюкозоизомераза теряет актив-
ность быстрее, чем тот же самый фермент, выделенный из этих
клеток. Этот эффект объясняли протеолизом фермента под дей-
ствием внутриклеточных ферментов.
Проблема внутриклеточной протеолитической активности
очень важна при любом применении иммобилизованных клеток.
Как мы упоминали в гл. 6, для нормальной жизнедеятельности
клеткам необходимы внутриклеточные гидролазы. Очевидно,
что потеря ферментативной активности в результате внутрикле-
точного гидролиза, а также разрушение клеточной мембраны
или утрата клеткой способности транспортировать вещества за
счет какого-либо другого процесса играют основную роль в оп-
ределении полезного времени жизни катализатора на основе
иммобилизованных клеток. Для оптимизации генетической при-
роды клеток и разработки более совершенных катализаторов на
основе иммобилизованных клеток, соответствующих конструк-
ций реакторов и оптимальных режимов их эксплуатации, оче-
видно, необходимо лучше понимать механизмы процессов внутри-
96
Глава 9
клеточного гидролиза, а также других процессов инактивации,
связанных с изменением конформаций белковых молекул или
с повреждением мембран вследствие химического взаимодейст-
вия с компонентами среды. Действительно, на разработку штам-
мов микроорганизмов, успешно применяющихся в настоящее
время в культурах суспензий клеток, были затрачены десятиле-
тия; в этой связи представляется вполне вероятным, что для
получения генетически модифицированных штаммов, специаль-
но предназначенных для их использования в иммобилизованной
форме, потребуются еще очень большие усилия.
Следующий по степени сложности уровень каталитической
активности включает многостадийные превращения, связанные
о использованием и регенерацией кофакторов. Из процессов
такого типа наиболее широко изучалось превращение глюкозы
в анаэробных условиях. Много работ посвящено изучению пре-
вращения глюкозы в этанол иммобилизованными дрожжами
(Saccharotnyces cerevisiae) или бактериями (в первую очередь
Zymomonas mobilis)-, по меньшей мере один процесс с участием
биокатализаторов на основе иммобилизованных клеток реали-
зован на пилотной установке [34].
Изучению превращения глюкозы в этанол при участии им-
мобилизованных клеток предшествовали работы по исследова-
нию непрерывных процессов пивоварения в присутствии склон-
ных к флокуляции штаммов дрожжей; по сути дела, флокуляция
обеспечивает достаточно эффективное удерживание (иммоби-
лизацию) клеток в реакторе с псевдоожиженным слоем ката-
лизатора (разд. 9.6.4). Позднее дрожжи иммобилизовали путем
адсорбции на желатиновой пленке и включения в х-каррагинан,
альгинат или полиакриламид. Показано, что для реактора ко-
лонного типа с клетками Saccharomyces carlsbergensis, иммо-
билизованными в х-каррагинане, характерен стартовый период
с неустановившимся режимом, продолжающийся около 7 сут,
после чего колонна функционирует в стационарном режиме с
постоянными концентрациями жизнеспособных клеток в геле,
а также глюкозы и этанола в продуктах процесса. После ряда
конструкционных доработок концентрацию этанола в продуктах
процесса удалось довести до 100 г/л и более.
Японская компания Kyowa Hakko Kogyo Company сообщила
о создании пилотной установки для производства этанола с по-
мощью иммобилизованных дрожжей в реакторе с псевдоожи-
женным слоем катализатора (разд. 9.6.4). Схема этой установ-
ки представлена на рис. 9.29. После трехмесячных испытаний
удалось найти режим, при котором разбавленная меласса са-
харного тростника (содержащая 14% глюкозы) при 30 °C пре-
вращалась в 8,5%-ный (по объему) водный этанол с объемной
скоростью [(объем раствора исходных веществ) • (объем ге-
Проектирование и расчет биологических реакторов
97
ля)-1^-1] от 0,4 до 0,5 ч-1. Степень превращения сахара в
этанол составляла 95%, а производительность установки в рас-
чете на единицу объема колонны 20 г/ (л-ч).
Более сложен процесс превращения глюкозы в ацетон, бута-
нол и этанол с помощью бактерии Clostridium acetobutylicum
и родственных организмов. Пути катаболизма в^этой бактерии
контролируются сложной регуляторной системой, которая до
Воздух
Охлаждающая Ферментер
вода
РИС. 9.29. Схема установки для производства этанола с помощью иммоби-
лизованных дрожжей [34].
настоящего времени детально не изучена, несмотря на большой
практический опыт по применению этого организма в традици-
онных ферментациях с участием суспендированных культур.
В зависимости от состава и условий среды и состояния клеток
в одних случаях основными конечными продуктами метаболиз-
ма глюкозы могут быть органические кислоты, а в других об-
разуются необходимые органические растворители. По сравне-
нию с катализируемым дрожжами превращением глюкозы в
этанол эта система значительно сложнее, поскольку здесь мы
должны уметь регулировать селективность процесса, направляя
его в сторону образования ценных веществ.
Возможность практической реализации непрерывного дли-
тельного процесса биопревращения глюкозы в органические
растворители была продемонстрирована Фёрбергом и др. [35].
7-746
98
Глава 9
В разработанном ими процессе катализатором служил организм
Cl. acetobutylicum, включенный в гель на основе альгината,
кальция, а глюкозная среда была лишена факторов, необходи-
мых для клеточного роста. Время от времени, сначала с интер-
валом 2—4 ч, а затем через каждые 8 ч в культуру вводили
полную питательную среду, что позволяло регенерировать час-
тично инактивированный катализатор. Сообщалось, что в таком
режиме, сочетающем не связанное с клеточным ростом много-
стадийное биологическое превращение с короткими периодами
Таблица 9.9. Выход бутанола в расчете на единицу клеточной массы,
массы субстрата или объема культуры с участием иммобилизованных3
и суспендированных6 клеток
Биокатализатор УР/Х, г бутанола из 1 г сухнх веществ клетки Tp/S, г бутанола из 1 г глюкозы /у г бутанола нз 1 л культуры в сутки
Растущие иммобилизован- ные клетки (полная сре- да) 1,3—1,9 0,11 16,8
Иммобилизованные клетки (чередование сред, обеспе- чивающих только клеточ- ный рост или только пре- вращение субстрата) 50 0,20
Периодическая суспензион- ная культура 3,6 0,19 6,9
а Forberg С., Enfors S.-О., Haggstrom, Eur. Appl. Microbiol. Biotech., 17, 143 (1983).
6 Moreira A. R., Ulmer D. C., Linden J. C., Biotech. Bioeng. Symp. no. 11, 567 (1981).
полного клеточного роста, система устойчиво функционировала
в течение 8 недель. В этом процессе существенно подавлялось
образование биомассы, а выход бутанола в расчете на единицу
массы утилизированной глюкозы был значительно выше, чем в
процессе, в котором иммобилизованные клетки контактировали
с полной средой. В табл. 9.9 сравниваются важнейшие парамет-
ры процессов с использованием иммобилизованных клеток в
полной среде, иммобилизованных клеток с чередованием сред
(обеспечивающих только клеточный рост или только превраще-
ние субстрата), а также обычного периодического процесса с
суспензией бактерий.
Очевидно, что для дальнейшего совершенствования катали-
заторов на основе иммобилизованных клеток необходимы фун-
даментальные исследования и проектные разработки. Вместе с
тем приведенный пример достаточно наглядно свидетельствует
о возможности изменения кинетики и стехиометрии процесса
путем подбора типа катализатора и соответствующего режима;
Проектирование и расчет биологических реакторов
99
этот путь создает дополнительные возможности для управления
биокаталитпческими процессами и их оптимизации.
В связи с рассмотренными примерами целесообразно изучить
и интересную проблему переноса газов в иммобилизованных
клеточных катализаторах и реакторах с такими катализатора-
ми. Здесь мы рассмотрим только частную проблему массопере-
носа в самом биокатализаторе. Допустим, что стехиометрию
данного биопревращения можно описать уравнением простой
химической реакции и что перенос субстрата S и продукта про-
цесса Р в частицы катализатора и из них характеризуется ко-
эффициентами эффективной диффузии Ds и Dp соответственно.
С помощью основных уравнений материальных балансов и гра-
ничных условий на частице катализатора нетрудно показать,
что концентрации продукта процесса ср и субстрата cs внутри
катализатора связаны уравнением
ср = сро+ n s (cso~cs) (9.113)
в котором а обозначает число молей продукта, образующегося
из 1 моля утилизированного субстрата, а подстрочный индекс О
указывает на условия на наружной границе (поверхности) час-
тицы катализатора. Подставив в уравнение (9.113) cs = 0 и ре-
шив полученное уравнение, можно оценить максимальную для
данного катализатора концентрацию продукта, основываясь
только на данных о стехиометрии и массопереносе. .
Если в системе происходит поглощение и образование газов,
это простое уравнение позволяет сделать важные выводы. Пред-
положим сначала, что субстрат представляет собой ограниченно
растворимый газ, например кислород. Тогда из уравнения
(9.113) следует, что в результате реакции с одной частицей ка-
тализатора произойдет лишь незначительное повышение кон-
центрации продукта. Если мы далее рассмотрим другой вариант,
когда в биокаталитическом процессе образуется умеренно рас-
творимый газ, например СО2, и когда используется хорошо
растворимый субстрат, например глюкоза, то, очевидно, может
наступить такой момент, когда концентрация СО2 в катализа-
торе превысит уровень насыщения, что приведет к зарождению
и росту газовых пузырьков внутри частицы катализатора. Со-
общалось, что такое явление вызывает серьезные затруднения
в работе реакторов с иммобилизованными клетками, так как
газовые пузырьки механически разрывают частицы носителей
с включенными клетками, продуцирующими СО2, или патроны
с дрожжевыми клетками, иммобилизованные в полых волокнах.
Итак, мы сталкиваемся с определенными трудностями как
при получении продукта из малорастворимых субстратов, так
и при выведении из катализатора малорастворимых продуктов.
7*
100
Глава 9
Очевидно, эти трудности можно преодолеть путем снижения
толщины частиц катализатора. Тогда отпадет необходимость
в использовании неэффективной внутренней зоны частиц в слу-
чае ограниченно растворимого газа-субстрата, а ограниченно
растворимые продукты не будут в избытке образовываться
внутри частицы катализатора. В свою очередь для снижения
толщины катализатора нужно или уменьшить размер частиц,,
или использовать катализаторы с тонким слоем клеток, иммо-
билизованных путем включения или связывания на наружной
поверхности большей частицы или иного объекта, которые бы
удерживали клетки в реакторе. Выполнение требований, предъ-
являемых к вводу и выводу ограниченно растворимых соедине-
ний, является одним из основных условий при проектировании
всего процесса и соответствующего реактора.
Интересно отметить, что, как было обнаружено эксперимен-
тально, процессы получения этанола из глюкозы с помощью
иммобилизованных или суспендированных дрожжей в сходных
условиях имеют различные общие скорости. В некоторых из этих
экспериментов, возможно, существенный вклад в общую ско-
рость процесса вносили диффузионные ограничения и связанные
с ними эффекты; вместе с тем в тщательно контролируемых
идентичных условиях и в отсутствие ограничений массопереносу
наблюдалось обусловленное иммобилизацией клеток повышение
удельной скорости образования этанола на 30—50%. Высказы-
валось предположение, что причиной этого явления может быть
или локальное изменение активности воды, вызванное матрицей-
носителем или ее поверхностью, или изменение химического ми-
кроокружения носителя, рассмотренное ранее.
В общем случае нас не должно удивлять существенное изме-
нение метаболической активности клеток, обусловленное их
необычным окружением в иммобилизованном состоянии. Свя-
зывание или сцепление клетки с твердой поверхностью может
узнаваться специфическими клеточными рецепторами, влияю-
щими на метаболические функции клетки. Точно так же нару-
шение нормального морфологического развития клетки вследст-
вие ее связывания в нескольких точках с твердой поверхностью,
влияния полых волокон, в которые включены клетки, или меж-
клеточных контактов может обусловливать заметное изменение
картины метаболизма клетки, которое не наблюдается в отно-
сительно неконцентрированных суспензиях. Кроме того, в плотных
скоплениях иммобилизованных клеток некоторые секрети-
руемые клетками продукты жизнедеятельности могут накапли-
ваться в значительно более высоких по сравнению с суспензия-
ми концентрациях. Наконец, можно достаточно обоснованно
ожидать, что каталитические свойства относительно независимо
развивающихся клеток и катализатора на основе иммобилизо-
Проектирование и расчет биологических реакторов
101
ванных клеток, который по клеточной плотности ближе к тка-
невым препаратам, будут заметно различаться. Следовательно,
применению катализаторов на основе иммобилизованных клеток
должно предшествовать изучение их собственных кинетических
характеристик, которые могут отличаться от кинетики анало-
гичных превращений в суспензии клеток, а поэтому результаты
изучения последних даже в идентичных условиях могут ока-
Таблица 9.10. Примеры процессов биосинтеза, осуществляемых
с помощью иммобилизованных клеток
Субстраты Иммобилизованные клетки Продукт процесса Выход продукта процесса
Глюкоза, неорга- Corynebacterium Глутаминовая 15 г/л за 144 ч
нические соли аммония, ионы металлов Пантотеновая кис- glutamicum Brevibacterium кислота Кофермент А 500 мкг/мл
лота, цистеин, ATP, MgSO4 Глюкозная среда ammoniagenes Penicillium Пенициллин 1,5 ед./(мл-ч)
Пептонная (1 %) chrysogenum Bacillus sp. Бацнтрацнн 16—19 ед./мл
среда Мясной экстракт (KY 4515)a Bacillus subtilis а-Амилаза 15 000 ед./мл
(1%), дрожже- вой экстракт (0,05%) Бульон LB FERM-P № 2040 Escherichia coli В-Лактамаза 1-ю-11
а Включенные в 1 & Включенные в i C600(pBR 322)6 толиакриламидный гель. полые полупроницаемые волокна. ед./(клетка-ч) 8 ед./(мл-ч)
заться неприменимыми к иммобилизованным клеткам. При рас-
смотрении более сложных биокаталитических функций мы впра-
ве ожидать еще более сложных зависимостей биохимической
активности клеток от морфологии.
Пока что в области применения иммобилизованных клеток
для биосинтеза продуктов метаболизма и биополимеров успехи
не очень велики. Это и не удивительно, если принять во внима-
ние указанные выше осложнения и тот факт, что разработка
методов осуществления многостадийных превращений, катали-
зируемых иммобилизованными клетками, по сути дела, пока еще
только начинается. В табл. 9.10 перечислены некоторые резуль-
таты экспериментального изучения методов биосинтеза ряда
веществ с участием иммобилизованных клеток. В некоторых из
приведенных примеров рост клеток был существенно ограничен,
102
Глава 9
в других для расширения периода активной жизни клеток при-
менялось чередование фаз полного клеточного роста и подавле-
ния роста, а в третьих рост клеток вообще не ограничивался.
Разумеется, при росте клеток с некоторой ограниченной скоро-
стью должны иметь место вымывание или лизис клеток, причем
скорость этих процессов должна уравновешиваться скоростью
клеточного роста; в противном случае не будет достигнуто ста-
ционарное состояние. При культивировании на твердых поверх-
ностях клеток тканей животных регуляторные механизмы оста-
навливают рост клеток примерно на стадии монослоя и затем
переключают метаболизм с роста на образование продуктов
метаболизма. Однако для достижения идеала — системы с им-
мобилизованными клетками, способной синтезировать сложные
вещества с высокими выходом и скоростью в проточных реак-
торах, — нам еще предстоит исследовать множество проблем.
9.6. Биореакторы с многофазными системами
В биореакторах реакционная масса почти всегда состоит из
нескольких фаз (клетки, малорастворимые субстраты или про-
дукты процесса, газовые пузырьки или частицы катализатора);
тем не менее во многих случаях целесообразно приближенно
описывать реакционную массу как практически гомогенную.
В других случаях, однако, при расчете или анализе работы ре-
актора важно принимать во внимание многофазную природу
содержимого реактора. В этом разделе мы сосредоточим вни-
мание именно на таких подходах к изучению реакторов и на
некоторых методах математического описания многофазных
систем.
9.6.1. Превращение нерастворимых субстратов
Превращение нерастворимых субстратов типично для про-
цессов утилизации крахмала и целлюлозы, модификации стерои-
дов, а также для роста микроорганизмов на парафиновых
углеводородах. В таких случаях мы прежде всего должны уста-
новить (или принять допущение), где происходит реакция: или
на границе раздела фаз, или в жидкой фазе с участием очень
небольшого количества растворенного субстрата. Иногда могут
реализоваться оба варианта одновременно. Так, например, час-
тицы целлюлозы при трансформации сначала взаимодействуют
с адсорбированными целлюлазными ферментами, а далее глюко-
за утилизируется микроорганизмами в растворе. В случае не-
растворимых субстратов важнейшим параметром системы явля-
ется площадь межфазной поверхности, отнесенная к единице
объема реактора; в свою очередь этот параметр иногда зависит
Проектирование и расчет биологических реакторов
103
от способа предварительной обработки субстрата, а чаще всего
непосредственно связан с режимом работы реактора. Приведен-
ный ниже пример роста микроорганизмов на нерастворимом
источнике углерода наглядно иллюстрирует различные факторы,
которые необходимо учитывать при анализе, расчете и эксплуа-
тации реакторов с такими системами.
Пример 9.2. Расчет ПРПП для микробиологической переработки жидких
углеводородов*. Некоторые микроорганизмы, например дрожжи Candida lypo-
lytica, могут расти на практически нерастворимых в воде додекане и других
парафиновых углеводородах. Для роста микроорганизмов на границе раздела
углеводородная — водная фаза предложены два альтернативных механизма.
Во-первых, если характерный диаметр клеток Dc значительно меньше диамет-
ра Dtl диспергированных капель углеводорода, то клетки могут концентриро-
ваться вблизи капель. С другой стороны, если Dh<PDc, то можно допустить,
что капельки углеводорода будут адсорбироваться на наружных поверхностях
относительно больших микроорганизмов.
В заключительной части разд. 8.4 мы указывали, что при микробиологиче-
ской трансформации углеводородов с аэрацией в ходе периодического процес-
са меняются соотношения между адсорбцией и флокуляцией в системе воз-
душные пузырьки — клетки — углеводородные капли. Можно ожидать,
однако, что в проточном процессе будет преобладать один определенный тип
клеточного роста и одна соответствующая структура системы пузырек — клет-
ка — углеводород; это облегчит количественное описание процесса.
Му-Янг и сотрудники изучили обе ситуации и предложили следующее
модифицированное уравнение Моно, учитывающее фактор доступности поверх-
ности раздела фаз:
Вариант I; Dc<zpDh
s/Dh
В — Вшах ! /п (9П2.1)
As “Г Ь/Чь.
Вариант II-, Dc~^>Dh
s/(Dft)2
кАА^ <9П2-2>
Здесь Ks и Кз' — модифицированные константы K.s. Как показано на
рис. 9П2.1, экспериментальные данные для системы С. lipolytica— додекан
(изображенные в координатах Лайнуивера — Бэрка) свидетельствуют в поль-
зу второго гипотетического механизма.
Уравнения, пригодные для расчета ПРПП, можно получить, связав мощ-
ность, расходуемую перемешивающим устройством, с размером диспергиро-
ванных капелек углеводорода (гл. 8). Если диаметр Dtl капель углеводород-
ного субстрата определяется напряжением сдвига в области, прилегающей
к концам лопасти мешалки, то уравнения, приведенные в примере 8.2, пока-
зывают, что
Л Г ( Р \~°’4
UhZ=(jAA) (9П2.3)
Найдено, что это уравнение удовлетворяет опытным данным описываемого
* Из статьи; Moo-Young М., Microbial Reactor Design for Synthetic Protein
Production; Can. J. Chem. Eng., 53, 113 (1975).
104
Глава 9
эксперимента при С=0,023. Подстановка этого выражения в уравнение
(9П2.2) дает
s (P/V)°,s
= P-max (9П2.4)
Если описываемую уравнением (9П2.4) удельную скорость клеточного роста
отнести к процессу в идеальном ПРПП, то при постоянстве экономического
коэффициента и стерильности питательных веществ получим
х = У
DKS" ( Р \~0’8'
Ртах D \, V /
(9П2.5)
Скорость разведения, отвечающая максимальной скорости роста биомассы,
будет определяться уравнением:
D = ----S( р ?-,8 • (9П2.6)
Ks " + so
На рис. 9П2.2 представлены зависимости скорости образования биомассы Dx
от скорости разведения D, рассчитанные по уравнению (9П2.5). Эти графики,
как и уравнения (9П2.5) и (9П2.6), наглядно демонстрируют важность учета
РИС. 9П2.1. Результаты экспери-
ментального определения скорости
роста дрожжей С. lipolytica на
додекане (в двойных обратных
координатах) согласуются с мо-
делью, в которой принимается, что
микрокапли углеводорода адсор-
бируются на поверхности клеток.
[Из статьи: Moo-Young М., Micro-
bial Reactor Design for Synthetic
Protein Production; Can. J. Chem.
Eng, 53, 113 (1975).]
взаимосвязи между биологическими реакциями и гидродинамическими харак-
теристиками при расчете и анализе микробиологических реакторов.
Хотя для периодического процесса типично изменение характера адсорб-
ционных явлений (см. разд. 8.4 и рис. 8.14), модель, в которой принимается,
что клетки С. lipolytica, растущие на углеводородных каплях, непрерывно ад-
сорбируются на их поверхности, образуя монослой, в некоторой степени
согласуется с результатами экспериментального изучения периодического про-
цесса. [См. работу: Erickson L. Е, Humphrey А. Е.. Prokop A, Growth Models
of Cultures with Two Liquid Phases. I. Substrate Dissolved in Dispersed Phase;
Biotech. Bioeng, 11, 449 (1969).]
Проектирование и расчет биологических реакторов
105
РИС. 9П2.2. Зависимость скоро-
сти образования биомассы в про-
цессе ферментации жидкого угле-
водорода от скорости разведения
и расхода мощности на перемеши-
вание, отнесенного к единице
объема (P/V), л. с./м3.
9.6.2. Реакторы с неподвижным слоем биокатализатора
Колонны с насадкой иммобилизованного катализатора в на-
стоящее время используются в нескольких промышленных про-
цессах, и есть все основания полагать, что в ближайшее время
область их применения существенно расширится. В таких реак-
торах, называемых реакторами с неподвижным слоем катализа-
тора, с помощью иммобилизованных ферментов осуществляют
изомеризацию глюкозы, частичный селективный гидролиз пени-
циллина, селективное расщепление смеси производных рацеми-
ческих аминокислот. В реакторах с неподвижным слоем изуча-
лись также процессы с участием иммобилизованных клеток.
В простейшем и часто довольно успешно применяющемся
математическом описании работы реактора с неподвижным
слоем катализатора в основу положена модель реактора полно-
го вытеснения, модифицированная с целью учета влияния ка-
талитической насадки на структуру течений и кинетику реакций.
Поверхностную скорость потока через реактор определяют как
объемную скорость потока исходных веществ, отнесенную к пло-
щади поперечного сечения пустот, которая представляет собой
произведение общей площади поперечного сечения колонны на
долю пустот е. Соответствующее уравнение скорости реакции,
пригодное для трубчатых реакторов, основано на учете коэф-
фициентов эффективности, описанных в гл. 4. Например, для
простой реакции S-*P, протекающей с собственной скоростью
v = v(s, р), скорость образования продукта в единице объема
гранулы иммобилизованного катализатора в какой-либо опре-
деленной точке реактора равна:
(общая в единице объема ~ nCWsMs8? Ps)
I гранулы катализатора
(9.114)
106
Глава 9
Здесь ss и ps — концентрации субстрата и продукта соответст-
венно на наружной поверхности частицы катализатора в данной
точке объема реактора. Как указано в уравнении (9.114),
в общем случае коэффициент эффективности -q, определяющий
скорость диффузии в частицу катализатора, и скорость реакции
v зависят как от ss, так и от ps.
Прежде чем перейти к анализу сопротивления массопере-
носу между жидкой фазой и поверхностью гранулы катализа-
тора, запишем уравнение материального баланса по субстрату
в сферической частице катализатора радиусом R в стационар-
ном состоянии:
Скорость диффузии субстрата из жидкой фазы =
=скорость трансформации субстрата внутри частицы
в результате реакции
или
4nR2ks(s — ss) =4/3n7?3T](Ss, ps)v(ss,ps) (9.115)
С помощью этого уравнения и учитывая стехиометрию реакции
величины ss и ps можно выразить через концентрацию субстра-
та s в жидкой фазе. Последующая подстановка полученных
выражений в уравнение (9.114) дает выражение, позволяющее
определить общую скорость утилизации субстрата, отнесенную
к единице объема частиц катализатора, если известна концент-
рация субстрата в жидкой фазе.
При выводе уравнения материального баланса по субстрату
в элементарном объеме реактора полного вытеснения с непод-
вижным слоем катализатора нельзя забывать о том, что жидкая
фаза и катализатор занимают е-ю и (1—е)-ю доли этого объ-
ема соответственно. Применяющийся в этом уравнении параметр
концентрации s отнесен к единице объема жидкой фазы, поэто-
му уравнение материального баланса по субстрату принимает
вид:
= — р-21— ps)v(ss, Ps) (9.116)
dz \ & J
Здесь, как мы отмечали выше, параметры правой части уравне-
ния можно выразить через s, что позволит проинтегрировать
уравнение (9.116) по заданным концентрациям субстрата и
продукта.
Ситуация существенно упростится, если сопротивление мас-
сопереносу внутри частицы и вне ее пренебрежимо мало, по-
скольку в таких условиях т]->1 и s.s->s соответственно. В такой
ситуации основные уравнения материального баланса можно
интегрировать аналитически и прийти к приведенным ранее в
табл. 9.2 уравнениям, описывающим процессы в реакторах пол-
ного вытеснения.
Проектирование и расчет биологических реакторов 107
Как мы уже упоминали выше в ходе обсуждения дисперси-
онной модели проточных реакторов, течение вокруг частиц, со-
ставляющих слой насадки, и особенности смешения жидкой
фазы в пустотах между частицами создают обратное (осевое)
смешение, которое может вызвать отклонения от режима пол-
ного вытеснения. В таких случаях можно применять дисперси-
онную модель (коэффициент дисперсии можно определить так,
как мы указывали выше) или модель на основе каскада реак-
торов. Влияние небольшой дисперсии на работу реактора в
сравнении с режимом идеального вытеснения мы уже обсуждали
при изучении стерилизаторов. Другие подходы к математиче-
скому моделированию реакторов с неподвижным слоем катали-
затора с различными степенями обратного смешения и с раз-
ными типами взаимодействий между неподвижной и подвижной
фазами можно найти в приведенной в конце главы литературе.
9.6.3. Биореакторы типа барботажных колонн
Под биореакторами типа барботажных колонн мы подразу-
меваем реакторы с большим отношением высоты к диаметру,
которые в отличие от реакторов с перемешиванием, обычно
имеющих менее вытянутую форму, по форме напоминают ко-
лонны или башни, а перемешивание в них осуществляется ис-
ключительно за счет восходящего потока газа, подаваемого в
реактор под давлением. Реакторы такого типа уже давно ши-
роко распространены в химической промышленности благодаря
относительно малым затратам на их изготовление и монтаж,
простоте конструкции и невысокому расходу энергии в процессе
эксплуатации. В промышленности, связанной с использованием
биологических процессов, башенные реакторы довольно редки;
в то же время такие биореакторы широко используются в про-
изводстве пива и винного уксуса. Кроме того, близкие по кон-
струкции башенные реакторы входят в состав весьма крупно-
масштабных установок, предназначенных для культивирования
микроорганизмов при производстве белка одноклеточных орга-
низмов (БОО) в качестве корма для скота.
В некоторых случаях применяют только одну колонну, кото-
рую иногда снабжают внутренними тарелками или даже пере-
мешивающими устройствами на отдельных или всех ступенях.
Колонные реакторы могут функционировать как в периодиче-
ском, так и в непрерывном режимах. В последнем случае воз-
можны два варианта, в первом из которых направления потоков
жидкой фазы и газа параллельны (т. е. совпадают), а во втором
варианте используется принцип противотока. В разработанных
сравнительно недавно так называемых эрлифтных реакторах с
помощью наружного устройства осуществляется рециркуляция
108
Глава 9
жидкой фазы. Рециркуляционные устройства обеспечивают вы-
сокоэффективный теплообмен, необходимый в крупномасштаб-
ных микробиологических процессах с участием парафиновых
или метанольных субстратов. Рециркуляционное устройство,
кроме того, способствует формированию устойчивой структуры
течений и определенных характеристик перемешивания в реак-
торе. Особенности барботажных колонн, в том числе с внешни-
ми рециркуляционными устройствами, с одной или несколькими
ступенями были детально изучены Шюгерлем и сотрудниками;
результаты этих работ опубликованы в ряде статей [36, 37].
Здесь мы ограничимся рассмотрением ряда основных концепций
и понятий, которые полезны для вывода уравнений при расчете
таких реакторов и анализе полученных с их помощью экспери-
ментальных данных.
В гл. 8 мы показали, что при достаточной плотности куль-
туры быстро растущих аэробных организмов общая скорость
клеточного роста обычно лимитируется скоростью переноса кис-
лорода из газовых пузырьков в жидкую фазу. Анализ переноса
кислорода, лимитирующего скорость всего процесса, требует
сведений о параметрах перемешивания газовой и жидкой фаз
в башне. Изучение системы воздух — вода в барботажной ко-
лонне без рециркуляционного устройства показало, что жидкая
и газовая фазы полностью перемешиваются, если скорость га-
зового потока намного выше скорости течения жидкой фазы и
если высота башни L близка ее диаметру dt. В случае более
обычных высоких колонн необходимую скорость переноса О2
можно определить по уравнению (8.115) при L = z.
В интегральной форме уравнение (8.115) справедливо при
практически постоянной величине удельной площади межфазной
поверхности а по всей высоте колонны. Это требование в свою
очередь выполняется только при сохранении пузырьковой струк-
туры газового потока. Эксперименты с системой воздух — вода
показали, что если объемная доля газа е превышает критиче-
скую ВеЛИЧИНу бтах, равную приблизительно 0,3, то поднимаю-
щиеся через слой жидкости газовые пузырьки коалесцируют
вплоть до образования воздушных пробок. Для того чтобы пе-
ревести на язык математики требование о необходимости под-
держания объемной доли газа ниже етах и связать его с диа-
метром колонны, отметим, что в любой точке башни
(9.117)
Здесь Fg и ug — объемная и линейная скорости потока газа
соответственно. Достаточно обоснованно можно принять, что
равна конечной скорости щ отдельного газового пузырька в не-
перемешиваемой жидкости и что FG приблизительно равна ско-
Проектирование и расчет биологических реакторов
109
рости поступающего в реактор газа FGf. Последнее допущение
основывается на том факте, что поглощающийся из пузырьков
Ог по меньшей мере частично замещается на СО2. При этих
допущениях из уравнения (9.П7) следует, что е меньше етах,
когда
¥/! (9.118)
\ uGsmax J
Это неравенство можно использовать для оценки геометриче-
ских размеров колонны. В нашем упрощенном анализе мы не
учитывали увеличение размеров воздушных пузырьков, обуслов-
ленное снижением гидростатического давления в верхней части
колонны. Этот фактор может привести к тому, что сначала воз-
растет величина Н/, а затем пузырьки примут грибообразную
форму.
Достаточно малый размер пузырьков по всей высоте колон-
ны обеспечивают ситчатые тарелки и (или) перемешивающие
или другие внутренние устройства, разрушающие все воздушные
пробки и таким образом способствующие сохранению высокой
величины площади контакта между газовой и жидкой фазами.
Сведения об иных механических приспособлениях, использую-
щихся в биореакторах, приведены в разд. 9.7.3.
На рис. 9.30 представлена схема, положенная в основу ма-
тематической модели башенного реактора с рециркуляционным
устройством и с параллельными потоками газовой и жидкой
фаз (биореактор эрлифтного типа). В собственно башне реак-
тора (на рисунке изображенной справа) в одном направлении
движутся потоки жидкой и газовой фаз. В верхней части башни
газ отделяется, а жидкая фаза через рециркуляционное устрой-
ство (изображенное слева) возвращается в нижнюю часть ре-
актора, где расположено барботирующее устройство. При мате-
матическом описании жидкость и газ рассматриваются как от-
дельные фазы, поднимающиеся снизу вверх по башне с
различными линейными скоростями uL и uG соответственно.
Занимаемые жидкой и газовой фазами доли объема обозначают
символами ед и 8g соответственно; эти величины можно опре-
делить экспериментально или расчетным путем с помощью
уравнений для оценки величины газосодержания. На движение
газожидкостной дисперсной системы в режиме полного вытес-
нения налагается осевая дисперсия как в газовой, так и в жид-
кой фазах, характеризующаяся различными коэффициентами.
В жидкой фазе в результате микробиологических превращений
происходит утилизация субстрата и кислорода, а газовая фаза
обедняется кислородом за счет его переноса в жидкость.
Переходя теперь к изучению элементарного объема башни
между положениями z и z + dz (см. рис. 9.30,6), можно вывести
РИС. 9.30. Схема, положенная в основу математической модели реактора колонного типа с барботажем и рециркуляци-
ей (а), и элементарный объем этого реактора (б); указаны термины и параметры, применяющиеся при выводе уравнений
материального баланса в газовой (g, G) и жидкой (I, L) фазах [37],
Проектирование и расчет биологических реакторов
111
указанные ниже уравнения материального баланса по концент-
рации кислорода в жидкой фазе и мольной доли кислорода в
газовой фазе. Для этого нужно воспользоваться теми же мето-
дами, которые мы применяли при анализе стационарного со-
стояния системы в реакторах полного вытеснения. В оконча-
тельной форме уравнения этих материальных балансов можно
записать следующим образом:
Жидкая фаза:
дС1 '* = О, (0 ----uL (0 S“ {г' ° -г, (х, S, с„ г, t) +
dt d9-z L dz r
+ (z, f) a\z, t) [сг* (г, t)—ct (z, /)] (9.119)
Газовая фаза:
p (z, t) dxs =Dg(t)—lp (z, t) —
' dt gy’dz[K dz J
---~(p(z,f) X (z, t) uG (z,rt) — .уел(Д -)—(z, t) a(z, t)[ct* (z,t)—
dz \ s - Mo2eG(t) J
-cdz,t)] (9.120)
В сочетании с аналогичными уравнениями для утилизации суб-
страта и клеточного роста и с такими же уравнениями матери-
альных балансов для жидкой фазы в рециркуляционном уст-
ройстве эти уравнения можно использовать для описания
экспериментов по изучению массопереноса в нестационарном
состоянии, работы биореактора в периодическом или непрерыв-
ном режиме как в стационарном, так и в переходном состояниях
с непрерывной подачей исходных веществ и непрерывным отбо-
ром продуктов. Результаты обширных экспериментальных ис-
следований и методы определения параметров рассматриваемой
модели описаны в статье [37].
9.6.4. Биореакторы с псевдоожиженным слоем катализатора
Процессы в псевдоожиженном слое катализатора обычно
осуществляют в реакторах колонного типа, рассмотренных в
предыдущем разделе, поэтому если такие процессы включают
подачу или отвод газа, то расчет газовых потоков и массопере-
носа должен выполняться так, как мы только что описали.
В то же время в реакторах с псевдоожиженным слоем катали-
затора появляется еще одна фаза.
В башенном реакторе с псевдоожиженным слоем катализа-
тора, например в изображенном на рис. 9.31, поток жидкости
направлен снизу вверх по высокому вертикальному цилиндру.
112
Глава 9
РИС. 9.31. Башенные ферментеры,
применяющиеся в непрерывных про-
цессах пивоварения. (Фотография
любезно предоставлена компанией
А. Р. V. Со., Ltd.)
Частицы нерастворимого био-
катализатора (скопления мик-
роорганизмов, частицы иммо-
билизованных ферментов или
клеток) суспендируются, увле-
каемые восходящим потоком
жидкости. Вовлеченные в этот
поток частицы катализатора в
верхней расширяющейся части
реактора прекращают подъем
и затем вновь поступают в баш-
ню. Если тщательно подо-
брать режим работы реактора
с учетом характеристик орга-
низма, то биокатализатор уда-
ется удерживать в реакторе,
несмотря на то что через ре-
актор непрерывно протекает
среда.
Биологические реакторы с
псевдоожиженным слоем ка-
тализатора намного сложнее
рассмотренных нами ранее
ПРПП и ТРПВ. Например, в
башенных ферментерах, ис-
пользующихся в непрерывных
процессах пивоварения, созда-
ется определенный градиент
концентрации дрожжевых кле-
ток по высоте башни, причем
вблизи от дна реактора кон-
центрация микроорганизмов
(определенная по отношению
массы влажного осадка, обра-
зующегося после центрифуги-
рования и отделения суперна-
тантной жидкости, к массе
бульона) может достигать 35%, а в верхней части башни
этот параметр снижается до 5—10%. Более того, в зависи-
мости от высоты в реакторе постепенно изменяются и ха-
рактеристики среды. Так, вблизи зоны поступления ис-
ходных питательных веществ превращениям подвергаются
прежде всего легко ферментируемые сахара (глюкоза, фрукто-
за, сахароза, частично мальтоза), что приводит к снижению
плотности среды. В средней и верхней зонах башни скопления
дрожжевых клеток трансформируют мальтотриозу и отчасти
Проектирование и расчет биологических реакторов
113
мальтозу. Такая картина, характеризующаяся быстрыми реак-
циями в начальной стадии процесса и последующими более
медленными реакциями с участием менее «удобных» субстра-
тов, согласуется с экспериментальными данными, представлен-
ными на рис. 9.32.
Рудиментарная модель реакторов с псевдоожиженным слоем
катализатора может быть разработана, если допустить, во-пер-
вых, что частицы биологиче-
ского катализатора (хлопья
скоплений микроорганизмов
или частицы иммобилизован-
ного фермента) однородны по
форме и размерам; во-вторых,
что плотность жидкой фазы
является функцией концентра-
ции субстрата; в-третьих, что
движение жидкой фазы в ре-
акторе осуществляется в режи-
ме полного вытеснения; в-чет-
вертых, что реакция утилиза-
ции субстрата имеет первый
порядок по биомассе, но нуле-
вой порядок по субстрату;
в-пятых, что числа Рейнольдса
Кажущееся время
ферментации., ч
РИС. 9.32. Рост дрожжей в реакторе'
колонного типа на смеси сахаров;
в начале процесса субстраты утили-
зируются очень быстро (что отра-
жается в изменении удельной плот-
ности сусла), а затем следует период
значительно более медленного усво-
ения сахаров. (Из работы: Green-
shield R. N., Smith Е. L., Tower-Fer-
mentation Systems and Their Applica-
tions; The Chemical Engineer, May
1971, 182.)
частиц катализатора, рассчи-
танные по их конечной скоро-
сти, достаточно малы, так что
движение частиц может быть
описано законом Стокса
(вспомните пример 1.1). Чет-
вертое и пятое допущения до-
статочно обоснованны во мно-
гих ситуациях; первое, второе и
третье допущения в ряде слу-
чаев также могут быть оправ-
данны.
При указанных допущениях скорость утилизации субстрата
можно описать уравнением типа (9.15):
d (su)
dz
ИЛИ
ds
и-----
dz
du
dz
kx
(9.121)
8-746
114
Глава 9
Если движение частиц (клеток) описывается законом Стокса,
то зависимость концентрации суспендированной биомассы от
скорости потока жидкости в псевдоожиженном слое должна
подчиняться уравнению:
Г , ( U XTI/4.65
х = р0 1-Мч (9.122)
\ / J
Здесь ро — плотность культуры микроорганизмов (масса сухого
клеточного вещества в единице объема), a ut— конечная ско-
рость сферы в стоксовом потоке [уравнение (8.34)]. (Какую
плотность биомассы — рассчитанную по сухой или влажной
массе клеток — следует учитывать в формуле для определения
конечной скорости?) Обратите внимание на то, что при анализе
процессов в реакторах с псевдоожиженным слоем катализатора
мы допускаем, что локальная концентрация биомассы опреде-
ляется только гидродинамическими факторами. Этим рассмат-
риваемые реакторы резко отличаются от реакторов других
типов, в которых определяющую роль играют биохимические
реакции и метаболизм организмов. Подстановка уравнения
(9.122) в уравнение (9.121) приводит к выражению с двумя
неизвестными 5 и и, величина которых зависит от положения z
в реакторе.
Построение модели завершается применением уравнения
(9.15) при условии, что /7 = 0:
— (pw) = 0 (9.123)
Раскрыв уравнение (9.123) и подставив p(s) вместо р, получим
, ч du . (
Р s) ~ + и
dz \
dp \ ds
d s J dz
(9.124)
0
Чтобы перевести это уравнение в форму, пригодную для инте-
грирования в численной форме, рассмотрим уравнения (9.121)
и (9.124) как систему алгебраических уравнений с неизвестными
dsjdz и du/dz. Решение этой системы уравнений, приводить
которое здесь полностью нет необходимости, дает выражения
для dsfdz и du[dz, определяемые через переменные s и и, т. е.
систему двух дифференциальных уравнений, которые следует
интегрировать при начальных условиях:
s(0) = s^ дг (0) = (9.125)
Здесь Af — площадь поперечного сечения нижней секции баш-
ни. Концентрация субстрата в продуктах процесса se зависит
от высоты реактора L: se = s (z = L).
Проектирование и расчет биологических реакторов
115-
Положение существенно упрощается, если допустить, что
любое изменение плотности жидкой фазы мало сказывается на
величине и. Если и не зависит от положения в реакторе, то
уравнение (9.121) можно проинтегрировать непосредственно
и таким путем получить
sc = sf—&р0 1—V/4,65 _L (9.126>
L \ Uf J и
Здесь L — высота башни; при выводе этого уравнения прини-
малось, что х определяется уравнением (9.122). Отражаемая
уравнением (9.126) линейная зависимость концентрации суб-
страта от среднего времени реакции L/u (если допустить, что
р также линейно зависит от s) действительно наблюдается по
меньшей мере на некоторых участках соответствующей кривой
(рис. 9.32).
Как показывают данные, представленные на рис. 9.32, основ-
ным недостатком этой модели является обезличивание субстра-
тов. Действительно, в обсуждаемой модели различные сахара,,
утилизируемые в ходе анаэробного спиртового брожения, сгруп-
пированы в некий гипотетический единый или средний субстрат.
При таком подходе мы лишены возможности учесть эффект
глюкозы, играющий очень важную роль в процессах пивоваре-
ния в башенных ферментерах непрерывного действия.
Что касается потока жидкой фазы через псевдоожиженный
слой, то обычно желательно поддерживать режим полного вы-
теснения. Нестабильная структура течений в слое в ряде случаев
может вызывать существенное обратное смешение, нарушающее
ход процесса и нормальную работу реактора. Вероятность об-
ратного смешения возрастает при уменьшении диаметра колон-
ны и снижении скорости потока жидкой фазы. В то же время
в биореакторах с псевдоожиженным слоем катализатора в силу
малых размеров его частиц и небольшого различия между
плотностями жидкой фазы и катализатора приходится ограни-
чиваться относительно невысокими линейными скоростями по-
тока жидкости. Кроме того, при понижении скорости потока
жидкой фазы повышается концентрация катализатора в реак-
торе. Показано, что введение в биореактор с псевдоожиженным
слоем катализатора статических элементов перемешивания мо-
жет значительно улучшить характеристики расширения слоя и.
снизить нежелательное обратное смешение [44].
Поскольку реакторы с неподвижным слоем катализатора в
общем случае ближе к реакторам полного вытеснения, может
возникнуть вопрос о целесообразности и преимуществах биоре-
акторов с псевдоожиженным слоем катализатора. Прежде всего
преимущества реакторов с псевдоожиженным слоем очень ярко
проявляются при необходимости контакта реакционной смеси с
8*
116
Глава 9
газами. В реакторах с неподвижным слоем катализатора до-
вольно трудно добиться эффективной аэрации (особенно при
большом объеме реактора), а если в ходе процесса образуются
газообразные продукты, например СО2, то нелегко и предупре-
дить избыточное накопление газа в верхней части реактора с
неподвижным слоем. Реактор с псевдоожиженным слоем ката-
лизатора обеспечивает режимы течений, в большей степени
способствующие межфазному контакту в системе газ — жид-
кость — твердое тело. Хороший контакт между газовой и жид-
кой фазами, с одной стороны, и биокатализатором, с другой,
обеспечивают также реакторы со струйным течением жидкости,
которые мы и рассмотрим в заключение этого раздела.
9.6.5. Реакторы с неподвижным слоем катализатора
и со струйным течением жидкости
Содержимое реакторов с неподвижным слоем катализатора
и струйным течением жидкости представляет собой трехфазную
систему, состоящую из неподвижного слоя нерастворимого ка-
тализатора, а также подвижных газовой и жидкой фаз. Посту-
пающие в реактор жидкая и газовая фазы содержат по одному
или несколько реагентов, поэтому скорость биохимической ре-
акции зависит от характеристик контакта между жидкостью,
в которую переносится ограниченно растворимый реагент из
газовой фазы, и поверхностью катализатора. Отсюда следует,
что на работу таких реакторов в существенной степени влияет
физическое состояние газожидкостного потока, проходящего
через неподвижный слой катализатора, и связанные с этим про-
цессы массопереноса.
К числу важных характеристик таких реакторов и содержа-
щихся в них систем относятся площадь поверхности катализа-
тора, эффективность смачивания катализатора подвижной жид-
кой фазой, структура течений газожидкостной смеси, массопе-
реноса ограниченно растворимых реагентов из газовой в жидкую
фазу, массопереноса реагентов к поверхности катализатора,
а в случае пористого или проницаемого катализатора—диффу-
зия реагентов к каталитическим центрам, находящимся внутри
частиц катализатора.
Одной из первых областей применения биореакторов с на-
садкой и струйным течением жидкости, сохраняющей свое зна-
чение и в настоящее время, является обработка сточных вод с
помощью биологических капельных фильтров, описанных де-
тальнее в гл. 14: вращающееся распределительное устройство
разбрызгивает поток жидких отходов по кольцевому слою гра-
вия, на котором находится пленка микроорганизмов. Жидкость
Проектирование и расчет биологических реакторов 117
стекает через неподвижный слой в почти ламинарном режиме,
а воздух поднимается через слой катализатора благодаря ес-
тественной конвекции за счет выделяющейся в микробиологи-
ческом процессе теплоты. Аналогичный принцип лежит в основе
традиционного способа производства винного уксуса (биологи-
ческого окисления этанола до уксусной кислоты), где применя-
ются прямоугольные колонны с насадкой из древесной щепы.
Для ламинарного течения жидкой фазы и упрощенной геомет-
рии слоя, например для плоского слоя, можно создать деталь-
ную математическую модель, описывающую характеристики
потоков и процессов переноса, и решить соответствующие урав-
нения. Детальнее ознакомиться с этой проблемой можно в мо-
нографии Аткинсона [5].
В промышленности встречаются и другие конструкции реак-
торов со струйным течением жидкости и неподвижным слоем
катализатора, в частности такие, в которых параллельные по-
токи газовой и жидкой фаз движутся: сверху вниз или снизу
вверх. Подобные реакторы уже давно применяются в нефтепе-
рерабатывающей и нефтехимической промышленности для гид-
рокрекинга, гидроочистки и других многофазных процессов. При
изучении режима работы таких реакторов и разработке соот-
ветствующих моделей необходимо помнить, что в зависимости
от относительных скоростей газовых и жидкостных потоков
(и в некоторой степени от других свойств газожидкостной сис-
темы) можно получить самые разные дисперсные системы, на-
чиная от непрерывной жидкой фазы с диспергированными в ней
газовыми пузырьками и кончая непрерывной газовой фазой с
диспергированными каплями жидкости (туманом) (рис. 9.33).
На этом рисунке выделена и зона нестабильности потока, когда
через реактор попеременно проходят газ и жидкость в виде
крупных газовых пузырей и жидких поршней соответственно.
Участки графика, обозначенные как «пилотная установка» и
«промышленная установка», заимствованы из опытных данных,
полученных при изучении процессов переработки нефти. В не-
которых режимах работы биореактора применяются низкие
скорости потока воздуха. Так, в процессах биологической обра-
ботки отходов на капельных фильтрах аэрация осуществляется
за счет естественной конвекции, обусловленной небольшой экзо-
термичностью происходящих реакций.
Конструкционно реакторы с неподвижным слоем катализа-
тора и со струйным течением жидкости напоминают реакторы,
рассматривавшиеся ранее в этом разделе. При математическом
моделировании систему обычно условно рассматривают как твер-
дую фазу, находящуюся в контакте с жидкой пленкой, которая в
свою очередь контактирует с газовой фазой. В сущности такой
подход к моделированию является расширенным вариантом уже
118
Глава 9
упоминавшейся двухфазной модели барботажной колонны.
Затем рассматриваются процессы переноса между фазами и в
каждой из фаз, а также ограничения, налагаемые на скорость
реакций диффузионными эффектами. Блестящим примером
инженерного подхода к анализу биологических реакторов со
Скорость жидкости, кс/(.мг>с)
РИС. 9.33. Различные состояния двухфазной дисперсной системы с параллель-
ными потоками газовой и жидкой фаз в неподвижном слое катализатора.
[Воспроизведено с разрешения из работы: van de Vusse J. G„ Wesselingh J. A.,
Multiphase Reactors, p. 561 in Chemical Reaction Engineering: Survey Papers
(4th International/6th European Symposium on Chemical Reaction Engineering),
DECHEMA, Frankfurt, 1976.]
струйным течением жидкости могут служить работы Бриффо
и Энгассера, посвященные изучению превращения глюкозы в
лимонную кислоту в биореакторе со струйным течением жидко-
сти и с неподвижной пленкой катализатора [46].
Как обычно, при выборе конкретной конструкции реактора
приходится учитывать самые различные характеристики проек-
тируемого процесса и эксплуатационные параметры. В табл.
9.11 суммированы преимущества и недостатки трех типов реак-
торов, применяемых для осуществления процессов в трехфазных
Проектирование и расчет биологических реакторов
119
Таблица 9.11. Сравнение основных конструкционных и эксплуатационных
характеристик различных реакторов для трехфазных систем
(_|_ положительные характеристики, — отрицательные характеристики)"
Реактор
Характеристики С неподвижным слоем катализа- тора и струй- ным течением жидкости С перемешивае- мой суспензией С псевдоожи- женным слоем катализатора
Разделение реактора на —h — +
ступени
Перепад давлений — + 4~
Максимальная скорость по- — + н—н
тока
Отвод теплоты (+) 4- 4-
Замена катализатора + 4-
Истирание катализатора (+) — (+)
Утилизация катализатора — + +
Простота конструкции 4- •— 4—h
Масштабирование (+) + ——
а Воспроизведено с разрешения из статьи: van de Vusse J. F., Wesselingh J. A., Mul-
tiphase Reactors, p. 561 in “Chemical Reaction Engineering: Survey Papers” (4th Internatio-
nal/6th European Symposium on Chemical Reaction Engineering), DECHEMA, Frankfurt,
1976.
системах: реакторов с неподвижным слоем катализатора и
струйным течением жидкости, реакторов с перемешиваемой
суспензией и реакторов с перемешиваемой суспензией и с бар-
ботажем (с псевдоожиженным слоем катализатора).
9.7. Технология микробиологических процессов
Для того чтобы получить некоторое представление о различ-
ных практических аспектах расчета и эксплуатации биореакто-
ров, а также об осуществляемых в них процессах, рассмотрим
ряд вопросов, связанных с промышленным применением микро-
биологических реакторов (которые по традиции часто называют
ферментерами). Основное внимание мы будем уделять типичным
материалам и методам, используемым в периодических процес-
сах. В завершающей части этого раздела мы изучим некоторые
альтернативные конструкции реакторов, разработанные для
лабораторных и пилотных установок и в некоторых случах ус-
пешно перенесенные на крупномасштабные промышленные
установки.
На рис. 9.34 представлена схема важнейших этапов типич-
ного микробиологического, процесса. Ранее мы уже упоминали
о подходах к выбору соответствующей среды (разд. 7.1.2), спо-
собах ее стерилизации (разд. 9.4) и различных газах, приме-
120
Глава 9
РИС. 9.34. Основные этапы и операции типичного аэробного микробиологиче-
ского процесса.
няющихся в таких процессах (гл. 8). В следующем разделе мы
дадим дополнительные сведения о подборе состава среды. Хотя
в гл. 7 мы уже обсуждали некоторые стороны влияния природы
посевного материала на ход процесса, ряд проблем микробио-
логического характера, связанных с введением инокулята в сре-
ду, целесообразно несколько детальнее рассмотреть и здесь.
Разд. 9.7.2 посвящен изучению конструкций собственно фермен-
теров. Контрольно-измерительная аппаратура и соответствую-
щие методы измерений рассматриваются в гл. 10, а операции
по выделению клеточной массы и других продуктов — в гл. 11.
9.7.1. Подбор состава среды
При подборе необходимого для определенного микробиоло-
гического процесса состава среды следует принимать во внима-
ние множество факторов. Один из них связан со стехиометрией
Проектирование и расчет биологических реакторов 121
клеточного роста и количеством биомассы, которое мы хотели
бы получить по завершении процесса. Основой любых расчетов
здесь является простой материальный баланс, связанный с пре-
вращением в процессе клеточного роста низкомолекулярных
органических и неорганических соединений, например глюкозы
и аммиака, в биомассу. Для синтеза заданного количества био-
массы (продукта процесса) в систему необходимо ввести доста-
точное количество питательных веществ (реагентов), взятых в
определенном соотношении. Очевидно, что для расчета требую-
Таблица 9.12. Элементный состав различных микроорганизмов3
Элемент Содержание, г в 100 г сухого клеточного вещества
Бактерии Грибы Дрожжи
Фосфор 2,0—3,0 0,4—4,5 0,8—2,6
Сера 0,2—1,0 0,1—0,5 0,01—0,24
Калий 1,0—4,5 0,2—2,5 1,0-4,0
Магний 0,1—0,5 0,1-0,3 0,1—0,5
Натрий 0,5—1,0 0,02—0,5 0,01—0,1
Кальций 0,01 — 1,1 0,1—1,4 0,1—0,3
Железо 0,02—0,2 0,1—0,2 0,01—0,5
Медь 0,01—0,02 0,002—0,01
Марганец 0,001—0,01 0,0005—0,007
Молибден 0,0001—0,0002
Всего золы 7—21 2—8 5—10
а Воспроизведено из работы: Aiba S., Humphrey А. Е., Millis N. F., Biochemical En-
gineering, 2d ed., p. 29, Academic Press, Inc., New York, 1973.
щегося количества различных субстратов необходимо знать эле-
ментный состав продукта (биомассы). Примеры элементного
состава ряда микроорганизмов приведены в табл. 5.10. В среде
должны присутствовать и необходимые неорганические вещест-
ва (табл. 9.12).
Если требования к элементному составу питательных ве-
ществ определены, то мы должны далее выбрать конкретные
соединения, содержащие необходимые для клеточного роста эле-
менты. Многие из применяемых в промышленности микроорга-
низмов являются хемогетеротрофами, потребности которых в
энергии и углероде удовлетворяются простыми сахарами. В про-
мышленности в качестве источников энергии и углерода часто
применяют не очищенные сахара, а те или иные полупродукты,
например свеклосахарную, тростниково-сахарную или кукуруз-
ную мелассу (содержащую от 50 до 70% ферментируемых са-
харов). В некоторых случаях дешевым, но в то же время вполне
удовлетворительным источником углерода для микроорганизмов
могут служить отходы, например сыворотка или отходы консерв-
ного производства. Так, один из видов пищевых дрожжей полу-
122
Глава 9
чают в промышленном масштабе путем роста культуры на
побочном продукте бумажного производства, сульфитно-спирто-
вой барде, содержащей всего лишь около 2% способных к фер-
ментации гексоз и пентоз.
К числу доступных источников азота относятся аммиак, мо-
чевина и нитрат. Вместе с тем микроорганизмы, продуцирующие
протеолитические ферменты, могут усваивать азот и из разно-
образных смесей, содержащих белки. Из потенциально ценных
источников такого рода следует упомянуть фильтрат барды,,
зерно хлебных злаков, пептоны, мясные отходы, соевую муку,,
казеин, дрожжевые экстракты, муку из жмыха семян хлопчат-
ника, арахисовый шрот, муку из жмыха льняного семени, куку-
рузный настой. В производстве пенициллина особенно важен
кукурузный настой, представляющий собой концентрированный
(50% твердых веществ) водный раствор, образующийся в ка-
честве отхода при замачивании кукурузы в процессах получения
крахмала, клейковины и других продуктов.
В предыдущих главах мы упоминали, что для нормального
роста и жизнедеятельности некоторых микроорганизмов необхо-
димо наличие в питательной среде определенных аминокислот
и факторов роста. Другие микроорганизмы, не столь требова-
тельные к составу среды, тем не менее растут значительно
быстрее в присутствии определенных (хотя и не обязательных)
факторов роста и источников азота и углерода. Что касается
промышленных процессов, то необходимые факторы роста обыч-
но обеспечивает какой-либо из полупродуктов, входящих в со-
став среды, например кукурузный настой или дрожжевой авто-
лизат. Кроме того, полупродукты часто являются источниками
различных минеральных веществ, необходимых для нормальной
жизнедеятельности клеток. Другие минеральные вещества до-
бавляют в среду по мере необходимости.
Если основной целью процесса является синтез продукта
жизнедеятельности микроорганизмов, то к среде для повышения
выхода или для улучшения качества продукта процесса можно
добавлять определенные вещества-предшественники. Обычно
молекула предшественника или его близкого производного
включается в молекулу продукта. Так, например, в производст-
ве новобиоцина, пенициллина G и витамина B[2 в качестве
предшественников используют бензойную кислоту, фенилуксус-
ную кислоту и 5,6-диметилбензимидазол соответственно. Входя-
щие в состав среды полупродукты также могут содержать
полезные вещества-предшественники.
Более подробные сведения о подборе состава среды можно
найти в приведенной в конце главы литературе. Теперь мы мо-
жем переключить наше внимание на другие аспекты технологии
микробиологических процессов.
Проектирование и расчет биологических реакторов
123
9.7.2. Проектирование типичного асептического
аэробного микробиологического процесса и его ведение
Большинство промышленных микробиологических процессов
имеют те или иные общие черты, однако на практике появляют-
ся существенно различающиеся проекты процессов и способы их
ведения, что часто обусловлено разными требованиями к зара-
жению системы вредными организмами. Если заражение вообще
нежелательно, то процесс необходимо вести в асептических ус-
ловиях при поддержании чистоты культуры. В некоторых случа-
ях, например при росте дрожжей в слабокислой среде или при
ферментации углеводородов тщательно подобранными штамма-
ми бактерий, требования к соблюдению асептических условий
могут быть несколько снижены, так как условия процесса сами
по себе не стимулируют рост многих потенциально вредных
микроорганизмов.
В этом разделе основное внимание будет уделено способам
поддержания асептического режима, поскольку именно здесь
очень многое зависит от умения и искусства биохимика-техно-
лога. В нашем обсуждении мы будем придерживаться последо-
вательности операций, принятой в периодическом процессе,
и начнем с получения посевного материала (инокулята) из чис-
той культуры.
Подготовка посевного материала сводится к пролиферации
небольшого числа клеток в строго контролируемых условиях
до плотной суспензии, объем которой составляет 1—20% объема
промышленного ферментера. Эта операция выполняется ступен-
чато путем постепенного увеличения масштаба. Исходным ма-
териалом служит чистая культура-—тщательно поддерживаемая
популяция данного штамма микроорганизма. Обычно нужный
штамм получают в результате многочисленных экспериментов,
включающих выведение множества мутантов, их скрининг и
селекцию. Поскольку полученный таким путем штамм микро-
организмов может обладать целым рядом преимуществ, то при
хранении необходимо обеспечить неизменность его генетической
природы. Обычно генетическая стабильность штамма микроорга-
низмов достигается за счет максимального снижения его мета-
болической активности при хранении. Микроорганизмы, как
правило, хранят в состоянии покоя в частично обезвоженном
виде; обезвоживание осуществляют путем лиофилизации (суб-
лимационной сушки) суспензии клеток в жидкости или путем
тщательного высушивания дисперсии спор или клеток в сте-
рильной почве или песке. Склонные к мутациям организмы
часто подвержены обратным мутациям или иным нежелатель-
ным генетическим изменениям. В таких случаях важно регуляр-
но контролировать чистоту культуры.
124
Глава 9
Если мы в качестве примера возьмем лиофилизованную куль-
туру, то следующей стадией процесса получения посевного ма-
териала будет приготовление суспензии клеток в стерильной
жидкости. Затем каплю этой суспензии переносят на скошенный
агар, который приготавливают желатинизацией стерильной пи-
тательной среды в наклоненной пробирке с помощью агара —
полисахарида, выделяемого из водорослей. После инкубации,
обеспечивающей достаточный рост, клетки опять суспендируют
в жидкости и наносят на агаровую поверхность большей площа-
ди в плоских сосудах Блэйка или Ру или переносят во встряхи-
ваемый сосуд. Эти сосуды затем устанавливают на качалках,
обеспечивающих перемешивание за счет вращательного или
вращательно-поступательного движения; таким путем достига-
ется рост погруженной культуры, а также адекватное поступле-
ние кислорода в культуру и выведение газов из нее. Обычно эту
операцию повторяют несколько раз с сосудами все больших
объемов и только затем переходят к следующей стадии. Все
описанные операции переноса культуры должны выполняться в
строго стерильных условиях. Для осуществления этих тонких
операций на предприятиях микробиологической промышленно-
сти обычно предусматривают специальные помещения со сте-
рильной атмосферой, асептическими условиями и регулирова-
нием температуры.
Дальнейшая пролиферация культуры осуществляется в одном
или нескольких сосудах для посевного материала — небольших
ферментерах, снабженных почти всеми контрольно-измеритель-
ными приборами, которые применяются в крупномасштабных
процессах. В этих реакторах подбираются такие условия, кото-
рые бы обеспечивали максимальную скорость роста культуры.
На этой стадии получения посевного материала мы перехо-
дим от лабораторных экспериментов к заводским установкам и
условиям. Здесь уместно ознакомиться с некоторыми основными
принципами и методами поддержания асептического режима,
а также с устройствами и операциями, применяющимися для
достижения этой цели. Прежде всего система должна иметь та-
кое устройство, чтобы можно было проводить независимую
стерилизацию всех входящих в ее состав узлов и компонентов.
В качестве примера чрезвычайных мер предосторожности, кото-
рые должны предусматриваться в проекте системы и соблюдать-
ся в процессе ее эксплуатации, рассмотрим частную проблему
переноса инокулята из сосуда для посевного материала в про-
мышленный реактор. На представленной на рис. 9.35 схеме
изображены основные узлы и устройства, а также последова-
тельность операций. Все клапаны системы должны быть доступ-
ны для проверки, чистки и стерилизации; по этой причине здесь
наиболее популярны шаровые клапаны. Рис. 9.35 дает представ-
Проектирование и расчет биологических реакторов
125
ление и о некоторых других общих принципах асептического
процесса. Особое внимание должно быть обращено на пароне-
проницаемость всех соединений системы; в системе должны быть
Резервуар
для посев-
РИС. 9.35. Система трубопроводов и запорно-регулирующей арматуры, ис-
пользуемая для асептической инокуляции крупномасштабного ферментера.
Последовательность операций: 1) установить участок трубопровода АВ\
2) стерилизовать этот участок паром при давлении 1,055 кг/см2 в течение
20 мин; клапаны D и J открыты, клапан С закрыт, а конденсат собирается
в ловушках, расположенных в узлах И и 1\ 3) клапаны С, G, Н, I, J закрыть;
клапаны D, Е, F открыть; охладить ферментер под давлением стерильного-
воздуха, при этом стерильная среда заполняет соединительный трубопровод;
4) в резервуаре для посевного материала повысить давление до 0,7 кг/см2,
а в ферментере снизить до 0,14 кг/см2; 5) перевести инокулят в ферментер,
открыв клапан С; 6) отключить резервуар для посевного материала и фермен-
тер от системы подачи пара, закрыв вентили F и С; открыть вентили G и J,
спустить пар и конденсат, частично открыв вентили D и Е.
полностью исключены любые контакты между стерильными и
нестерильными зонами. Поддержание в системе небольшого
избыточного давления способствует тому, что все случайные
утечки будут происходить только из системы, а не в обратном
направлении.
На рис. 9.36 приведен чертеж общего вида (с указанием
основных узлов и размеров) промышленного ферментера, вы-
пускаемого в серийном масштабе. Обычно такие ферментеры,
изготавливают из нержавеющей стали, что существенно снижает’
ООО? 1 ooos
Привод
электродвигателя
Редуктор
Пеногаситель
Отвод газов
Система
подшипников
Асептическое
уплотнение
Оэслаждаюьций
змеевик
Подача
стерильного
воздуха
4
_____Cs
РИС. 9.36, а. Ферментер объемом 100 000 л для получения пенициллина. [Из
работы: Аиба Ш., Хемфри А., Миллис Н., Биохимическая технология и аппа-
ратура.— М.: Пищевая промышленность, 1975.]
1700
wo-^
300j^
3100
ШО’
3000'
300
I
Отражательные
' перегородки
Мешалка
турбинного типа
с плоскими
лопастями
Барботер
Проектирование и расчет биологических реакторов
127
РИС. 9.36, б. Большой промышленный ферментер; вид сверху. [Из статьи'
Milller R., Kieslich К., Technology of the Microbiological Preparation to Orga-
nic Substances; Angew. Chem., Intern. Edit., 5, 653 (1966).]
возможность коррозии аппаратуры и сводит к минимуму опас-
ность загрязнения культуральной жидкости нежелательными
ионами металлов (вспомните приведенные в разд. 5.9.2 данные
о влиянии ионов железа на процесс микробиологического син-
теза лимонной кислоты). Как в самом ферментере, так и в лю-
бых других аппаратах и элементах системы не должно быть
застойных зон, щелей, трещин и других участков, в которых
могут накапливаться устойчивые к стерилизации твердые веще-
ства и в которых растущие микроорганизмы могут образовывать
плотные скопления и пленки. Решить эти проблемы помогает
применение цельносварных конструкций с тщательно зачищен-
ными и отполированными сварными швами.
Перемешивающее устройство должно обеспечивать переме-
шивание и аэрацию системы в той степени, в какой это необхо-
димо (гл. 8). Чтобы исключить возможность заражения культу-
ры, при проектировании и в процессе эксплуатации реакторов
особое внимание следует уделять асептическим уплотнениям.
В лабораторных ферментерах обычно достаточно одной лопаст-
ной мешалки, а в больших промышленных реакторах могут по-
требоваться несколько перемешивающих устройств.
Обычно только 70—80% объема реактора занимает жидкая
фаза, а верхнюю его часть заполняют газы. Часто перемешива-
128
Глава 9
ние и аэрация жидкости способствуют образованию пены на ее
поверхности, особенно если в среде в больших концентрациях
содержатся пептиды. Пена препятствует газообмену между
культуральной жидкостью и находящимся над ней пространст-
вом; она может выйти из ферментера и загрязнить систему,
если составляющие пену пузырьки коалесцируют и образующая-
ся жидкая фаза вновь поступает в ферментер. В изображенном
на рис. 9.36 ферментере для разрушения пены над уровнем
жидкой фазы устанавливают дополнительную мешалку. В слу-
чае особенно устойчивых пен к культуральной жидкости добав-
ляют особые вещества — пеногасители, дестабилизирующие пену
путем снижения поверхностного натяжения. Как мы уже отме-
чали в гл. 8, поверхностно-активные свойства пеногасителей
могут явиться причиной снижения скорости переноса кислорода.
Змеевик теплообменника в реакторе может выполнять не-
сколько функций. Если предполагается периодически стерили-
зовать среду в самом ферментере, то змеевик должен обеспечи-
вать необходимые скорости нагревания и охлаждения. Тепло-
обменное устройство должно также справляться с пиковыми
нагрузками, куда входят тепловые эффекты микробиологическо-
го процесса, а также вязкое рассеяние теплоты, выделяющейся
при перемешивании (разд. 8.10). Коэффициенты теплопередачи
для неинокулированной среды примерно равны коэффициенту
теплопередачи воды, в то время как плотная мицелиальная
культура по этим характеристикам приближается к пастам.
Процесс в ферментере может продолжаться от суток (или
даже менее) до 2 нед и более; чаще всего процесс биосинтеза
антибиотика длится от 4 до 5 сут. В течение этого времени
необходимо поддерживать строгий режим процесса, а в ряде
случаев те или иные параметры необходимо изменять в соответ-
ствии с определенной программой. Такие проблемы в эксплуа-
тации реакторов мы рассмотрим подробнее в гл. 10. Тем не
менее даже наш краткий обзор может дать представление о
сложности и трудоемкости асептических процессов ферментации,
а также о высокой стоимости этих работ. Очевидно, в такой
ситуации потеря продуктов даже одного периодического про-
цесса может быть связана с очень большими расходами, что
оправдывает отмечавшееся выше большое внимание, уделяемое
стерилизации.
9.7.3. Биореакторы других типов
В табл. 9.13 перечислен ряд факторов, стимулировавших
разработку новых типов и конструкций биореакторов. Многие
.из этих факторов сыграли свою роль при разработке компанией
JCI чрезвычайно крупномасштабного процесса производства
Проектирование и расчет биологических реакторов
129
белка одноклеточных организмов. Общий объем соответствую-
щего реактора составляет 2300 м3 (колонна диаметром 7 м и
высотой 60 м, рабочий объем реактора 1560 м3). Более того,
в этом реакторе микроорганизмы растут на метаноле, что при-
водит к выделению чрезвычайно большого количества тепла.
При таком объеме ферментеры обычной конструкции с механи-
ческим перемешиванием и барботажем совершенно непригодны,
что и послужило стимулом для разработки новых реакторов
эрлифтного типа, упоминавшихся в предыдущем разделе.
Предлагались и другие конструкции реакторов, которые про-
шли испытания в различных масштабах — от лабораторного
Таблица 9.13. Факторы, оказавшие влияние на разработку реакторов
новых типов [50]
Изготовление и монтаж реакторов очень большого объема
Проблемы, связанные с расчетом и проектированием
Высокий расход мощности (при постоянном P/V)
Большие затраты на энергию и охлаждающую воду
Проблемы, связанные с необходимостью отвода теплоты
Снижение удельных капиталовложений
Снижение удельного расхода энергии
Снижение степени механического повреждения клеток
Снижение потерь субстрата (обусловленных испарением и дыханием)
Повышение степени превращения субстрата * В
прибора и пилотной установки до крупномасштабного промыш-
ленного аппарата. Хорошая обзорная статья, посвященная срав-
нительному изучению реакторов различных конструкций и их
характеристик с точки зрения эффективности использования
энергии для диспергирования газа и перемешивания, опублико-
вана Шюгерлем [50]. В этом обзоре предложено подразделять
реакторы на три основных группы в соответствии со способом
потребления энергии.
В реакторах первой группы энергия расходуется на механи-
ческое перемещение внутренних устройств (рис. 9.37). В неко-
торых из реакторов этой группы с помощью внутренних каналов
устанавливается определенная структура течений, например
циркуляция жидкости. Реактор 1.6 представляет собой горизон-
тально расположенный аппарат с циркуляцией жидкой фазы
и с пеноотделителем, заполненным газожидкостной смесью.
В реакторе типа 1.9 перемешивание происходит за счет пульса-
ции жидкой фазы, а в реакторах конструкции 1.11 (представлен
вид сбоку) путем периодического погружения размещенных на
вращающемся валу дисков в жидкую реакционную смесь, нахо-
дящуюся в нижней части реактора.
На рис. 9.38 представлены конструкции реакторов другой
группы, в которых энергия, расходуемая на циркуляцию жидко-
9—746
Пульсация
1.9 ^0
Проектирование и расчет биологических реакторов 131
сти, подается с помощью насоса, расположенного вне реактора.
В реакторе 2.1 направленная сверху вниз струя жидкости стре-
мительно уходит в реакционную смесь, а в конструкции 2.7
имеются сопла, распыляющие жидкость и расположенные на
концах вращающегося стержня.
Схемы реакторов последней группы, в которых энергия рас-
ходуется на сжатие газа, поступающего в реактор, изображены
на рис. 9.39. В большинстве реакторов этой группы имеются
те или иные внутренние устройства, способствующие более тща-
тельному диспергированию газа, а также различные устройства
для циркуляции жидкой фазы. Дополнительные сведения о всех
упомянутых конструкциях реакторов и расширенный перечень
литературы читатель может найти в обзоре Шюгерля [50].
9.8. Особенности технологии процессов
с участием растительных и животных клеток
и соответствующих реакторов
В настоящее время культуры животных клеток использу-
ются для производства ряда ценных продуктов, в том числе вак-
цин, протеолитического фермента урокиназы, моноклональных
антител и интерферонов. С помощью культур животных клеток
можно также получать лимфокины (белки, оказывающие регу-
ляторный эффект на иммунную систему), многие ферменты,
факторы роста, факторы свертывания крови и гормоны. Техно-
логия рекомбинантных ДНК позволяет получать некоторые из
этих веществ и другими путями, и в то же время открывает
новые перспективы для синтеза различных веществ с помощью
культур животных клеток. С одной стороны, возможность эк-
спрессии чужеродных белков в микроорганизмах означает, что
для производства этих белков в значительных количествах
можно пользоваться микробиологическими процессами. В то же
время, как отмечалось в разд. 6.4.4, белок, синтезируемый в
животных клетках, часто подвержен различным посттрансля-
ционным модификациям; прокариоты не способны осуществлять
такие превращения. Отсюда следует, что для получения веществ,
проявляющих необходимую активность или устойчивость только
после посттрансляционных модификаций, нужны эукариотиче-
ские клетки-хозяева. Экспрессия некоторых эукариотических
РИС. 9.37. Биореакторы, в которых энергия расходуется на механическое дви-
жение внутренних устройств [50].
Обозначения-. Г —газ (воздух); К—внутренний канал; Д —двигатель; ОП—
отражательные перегородки; ПГ — пеногаситель; Ж —жидкая фаза; ВЦ —
вращающийся цилиндр,
9*
132
Глава 9
РИС. 9.38. Реакторы, в которых энергия расходуется внешним насосом, вклю-
ченным в систему рециркуляции [50].
Обозначения-. Г—газ; Ж — жидкая фаза; ИС — инжекционное сопло; П —
поплавок.
Проектирование и расчет биологических реакторов
133
РИС. 9.39. Реакторы, предназначенные для проведения процессов с участием
погруженных культур; расход энергии Связан со сжатием и подачей га-
за [50].
Обозначения-. Г — газ; Ж—жидкая фаза; 1 — газовый клапан; 2 — статиче-
ское перемешивающее устройство; клапан 1 периодически закрывают и откры-
вают.
белков в прокариотах-хозяевах затруднена также в силу про-
теолиза этих белков или невозможности их складывания в
определенную пространственную структуру. Наряду с совершен-
ствованием методов экспрессии чужеродных генов в животных
клетках расширяются перспективы промышленного применения
рекомбинантных систем с участием животных клеток.
Многие полезные и интересные вещества в принципе можно
синтезировать в культурах растительных клеток. Кроме того,
культивирование растительных клеток может облегчить реше-
ние задач генной инженерии растений и в конце концов приве-
134
Глава 9
сти к реализации возможности регенерации целого продуктив-
ного растения из клеток культуры тканей.
Культивированию растительных и животных клеток свойст-
венны некоторые общие черты, осложняющие проектирование
реакторов, предназначенных для роста культур таких клеток.
Многие типы растительных и животных клеток в естественной
сфере их обитания существуют в виде тканей — плотных струк-
тур, построенных из множества одинаковых клеток. В нативном
окружении ткани постоянно контактируют с жидкостями орга-
низма, которые имеют определенный состав и содержат множе-
ство регуляторных соединений, оказывающих существенное
влияние на функционирование клеток. Наконец, скорости роста
многих растительных и животных клеток в естественных усло-
виях очень низки. При росте погруженных культур таких
клеток возникают трудности в обеспечении окружения, способ-
ствующего росту клеток; многие типы растительных и живот-
ных клеток, помещенные в культуральную среду, вообще не
растут. Если эти трудности устранены, то наша следующая цель
будет заключаться в подыскании таких условий культивирова-
ния, которые обеспечивают достижение высокой плотности
клеток в культуре за минимальное время при сохранении мета-
болических механизмов клеток, ответственных за необходимые
реакции.
В этом разделе основное внимание будет уделено изучению
подходов к созданию сред и условий, поддерживающих жизне-
способность животных клеток; здесь мы дадим также краткий
обзор результатов некоторых работ с животными клетками.
Изучение культур растительных клеток пока только начинается,
и поэтому мы уделим ему лишь несколько строк в конце разде-
ла. Количественные параметры кинетики процессов с участием
культур животных клеток практически неизвестны из-за высо-
кой стоимости таких процессов, малой скорости роста клеток,
что приводит к существенному увеличению продолжительности
экспериментов, и отчасти из-за недостаточной изученности этих
объектов. Во многих случаях неизвестны даже лимитирующие
рост клеток питательные вещества, а описания зависимости
кинетики роста животных клеток и образования продуктов их
жизнедеятельности от концентрации питательных веществ и ин-
гибиторов, pH, температуры и других параметров окружения
вообще недоступны. Хотя при проектировании реакторов опре-
деляющим фактором считается влияние механических сил на
культуры растительных и животных клеток, до сих пор не
проводилось систематического изучения влияния механического
воздействия на жизнеспособность, рост и морфологию клеток
с тем, чтобы результаты этих исследований были бы положены
в основу инженерных расчетов. Вместо этого были предложены
Проектирование и расчет биологических реакторов
135
конструкции аппаратов и конкретные формы биокатализато-
ров, обеспечивающие повышение скорости роста клеток, дости-
жение более высокой плотности клеток в культуре и более
высокого выхода продукта процесса с участием животных и рас-
тительных клеток. Хотя на базе такого чисто эмпирического
подхода достигнуты значительные успехи, тем не менее оче-
видно, что выяснение закономерностей кинетики роста живот-
ных и растительных клеток и их технологических характеристик
позволит добиться еще более существенного прогресса.
9.8.1. Культивирование животных клеток;
требования к среде
По сравнению с микроорганизмами для культивирования
животных клеток требуются более сложные и дорогие среды.
Обычно для предотвращения заражения в среду вводят анти-
биотики. В состав большинства сред входит сыворотка живот-
ных в концентрации 5—20%, которая способствует репликации
клеток. Известны и другие (но далеко не все) функции, выпол-
няемые сывороткой в среде для роста животных клеток.
Сыворотка обеспечивает возможность применения данной среды
для роста животных клеток различных типов, которые требуют
различных питательных веществ и факторов роста. Показано,
что потребность в питательных веществах может зависеть от
условий культивирования; в некоторых случаях применение
очень богатой и высококонцентрированной среды в какой-то
степени компенсирует отсутствие точных данных о необходимо-
сти определенных компонентов. По не вполне понятным пока
причинам клетки, выросшие в среде с более высоким содержа-
нием сыворотки, часто оказываются более устойчивыми к ме-
ханическим повреждениям.
Вместе с тем использование сыворотки для культивирования
животных клеток не свободно от ряда проблем, важнейшей из
которых является высокая стоимость сыворотки; это особенно
сильно сказывается на общей стоимости крупномасштабных
процессов. Так, например, если содержание сыворотки в среде
составляет 10%, то затраты на сыворотку стоимостью до 300
долларов за литр составляют 80% общих материальных затрат.
Сыворотка, кроме того, представляет собой главный источник
заражения культуры вирусами, микоплазмой (паразитными
или патогенными грамотрицательными прокариотами) и бак-
териями. Она может содержать ингибиторы, затрудняющие
репликацию вирусов (в производстве вакцин) или биосинтез
ферментов. Как и сложные природные компоненты сред для
роста микроорганизмов, сыворотка представляет собой компо-
нент непостоянного и отчасти непредсказуемого состава. Нако-
136
Глава 9
нец, вместе с сывороткой в среду поступают пирогенные (вызы-
вающие лихорадку) вещества, затрудняющие выделение
продукта. Содержащийся в сыворотке фоновый белок альбумин
также затрудняет выделение белков, синтезируемых в неболь-
ших количествах; именно по этой причине для облегчения
РИС. 9.40. Культивирование клеток с применением в качестве одного из ком-
понентов среды профильтрованной лимфы крупного рогатого скота. (Воспро-
изведено с разрешения Bio-Response, Inc.)
выделения продукта (например, моноклональных антител) иног-
да прибегают к бессывороточным средам.
На рис. 9.40 представлена схема недавно предложенного
иного подхода к снижению стоимости сложных компонентов
среды. В этом методе цельную лимфу живой коровы фильтру-
ют и вводят в камеру клеточного роста, где лимфа через полу-
проницаемые полые волокна контактирует с клетками. На ри-
сунке указано также положение устройств для введения других
компонентов среды и газообмена.
Большие успехи достигнуты в создании бессывороточных
сред определенного гормонального состава. К потенциальным
Проектирование и расчет биологических реакторов
137
преимуществам таких сред относятся возможность регулирова-
ния состава среды в соответствии с особенностями клеток
данного типа и условий культивирования, снижение опасности
заражения, лучшая воспроизводимость результатов. С другой
стороны, подобные среды определенного состава имеют более
высокую стоимость и к тому же могут оказывать то или иное
воздействие на организмы, которое проявляется лишь спустя
некоторое время и которое трудно предвидеть заранее или
обнаружить в ходе сравнительно кратковременных эксперимен-
тов по подбору состава среды.
Животные клетки не имеют типичных для микроорганиз-
мов клеточных стенок, обеспечивающих механическую проч-
ность; кроме того, животные клетки обычно значительно крупнее
клеток микроорганизмов. Поэтому на любое механическое
воздействие на культуру животных клеток (перемешивание
суспензии клеток или частиц носителей, содержащих клетки на
поверхности или во всем объеме, с целью поддержания одно-
родности реакционной массы или ускорения переноса кислоро-
да к клеткам) должны налагаться определенные ограничения.
Для культур животных клеток концентрация кислорода долж-
на составлять приблизительно 25—40% от концентрации
кислорода в насыщенной воздухом системе, поэтому проекти-
рование систем подачи кислорода в большой объем культуры
животных клеток всегда представляет собой сложную инженер-
ную проблему. Жесткие требования, предъявляемые к насыще-
нию системы кислородом, могут быть несколько смягчены благо-
даря низким скоростям роста животных клеток и протеканию
метаболических процессов в клетках.
При типичной плотности культуры, составляющей 106 кле-
ток в 1 мл, потребность в кислороде составляет 0,05—0,6 ммоль
О2 на литр культуры в час. В небольших сосудах, обычных для
лабораторных экспериментов, потребность в кислороде может
быть удовлетворена за счет диффузии кислорода из газовой
смеси, находящейся над поверхностью жидкости, в жидкую
фазу. При переходе к реакторам большего объема такой меха-
низм переноса кислорода уже не может обеспечить потребность
культуры в кислороде. Иногда в таких случаях с успехом при-
меняется прямой барботаж газа в жидкую фазу. В других
ситуациях непосредственный барботаж повреждает клетки и
может вызвать интенсивное пенообразование, особенно если
в состав среды входит значительное количество сыворотки. Для
насыщения культуры животных клеток кислородом предлагался
и другой метод, заключающийся во введении в культуру сили?
коновых трубок, через стенки которых кислород диффунди-
рует в среду, не образуя пузырьков. Общий коэффициент
массопередачи кислорода через стенку силиконовой трубки
138
Глава 9
составляет около 0,35 ммоль Ог/(атм-см2-ч); по этой величине
можно рассчитать необходимое количество трубок. По мере
увеличения масштаба процесса возможность обеспечения куль-
туры кислородом с помощью силиконовых трубок становится
все более и более проблематичной.
В относительно хорошо отработанных технологических про-
цессах с участием культур животных клеток pH культуры пос-
тоянно контролируется и поддерживается на постоянном
уровне, обычно около 7,0. К настоящему времени накоплен
лишь очень небольшой опыт работы с культурами животных
клеток в реакторах, снабженных всеми необходимыми контроль-
но-измерительными приборами, обеспечивающими получение
наиболее полного набора аналитических данных и самые раз-
нообразные методы контроля. Как мы увидим в следующей
главе, многие контрольно-измерительные приборы и методы,
которые находят применение при работе микробиологических
реакторов, в будущем можно будет использовать и для оптими-
зации работы реакторов с культурами животных клеток.
Регулирование pH культуры клеток осуществляется несколь-
кими способами. Введение в состав среды компонентов с буфер-
ными свойствами может сглаживать флуктуации pH, часто
обусловленные продуцируемой клетками молочной кислотой.
Величиной pH среды можно также управлять путем изменения
концентрации СОг в газовой фазе, находящейся в контакте с
культурой. Другим методом регулирования pH является непос-
редственное добавление основания. Флуктуации pH в реакто-
рах могут быть сглажены и путем постоянной или периодиче-
ской замены среды в реакторе на свежую.
9.8.2. Промышленные реакторы для крупномасштабных
процессов с участием животных клеток
Все животные' клетки по способности к росту в суспензии
можно разделить на две группы. Так, клетки крови, лимфы,
опухолевой ткани и многие трансформированные клетки могут
расти в суспендированной культуре. Клетки других типов растут
только в том случае, если они связаны с совместимой твердой
поверхностью; в этом случае регуляторный механизм контакт-
ного ингибирования обычно останавливает клеточный рост, как
только на поверхности твердого носителя образуется монослой
клеток. В небольшом масштабе для перемешивания суспензи-
онных культур достаточно магнитной мешалки и колбы или
сосуда, частично заполненного средой, вращающегося в гори-
зонтальном положении со скоростью около 1 об/мин.
По своей конструкции многие реакторы, предназначенные
Проектирование и расчет биологических реакторов
139
для культивирования больших количеств животных клеток, на-
поминают микробиологические биореакторы. Обычные для
культур микроорганизмов турбинные мешалки с плоскими ло-
пастями в случае культур животных клеток часто заменяют на
пропеллерные. Предложена конструкция эффективного реакто-
ра с рабочим объемом около 100 л, в котором для перемеши-
вания суспензионной культуры применяется не лопастная
мешалка, а устройство с «парусами» из моноволокна. В отличие
от культур микроорганизмов для перемешивания культур жи-
вотных клеток в реакторах сравнительно часто применяется
устройство типа «вибромиксер», представляющее собой плоский
круглый диск с несколькими отверстиями, укрепленный в гори-
зонтальном положении на конце вала у дна реактора. Быстрое
перемещение вала вверх и вниз и соответствующее колеба-
тельное движение перфорированного диска вызывают циркуля-
цию перемешиваемой жидкости. Для культур животных клеток
применялись и эрлифтные системы, аналогичные рассмотрен-
ным выше.
В случае культур животных клеток понятие о крупномасш-
табном процессе приобретает несколько иной смысл. В силу
высокой стоимости продукта процесса и высоких эксплуатацион-
ных расходов реактор объемом 10 л уже может считаться круп-
номасштабным, а реактор объемом 100 л, безусловно, относится
к числу очень больших.
Преимущественное применение относительно небольших
реакторов отчасти обусловлено и довольно низкой скоростью
роста животных клеток. На рис. 9.41 представлены данные,
характеризующие рост периодической культуры клеток адено-
карциномы печени человека (линия SK-HEP-1). В табл. 9.14
суммированы экспериментально найденные удельные скорости
клеточного роста и скорости поглощения кислорода для не-
скольких линий клеток; в первых двух случаях клетки выра-
щивали в суспензии в ферментере эрлифтного типа, а клетки
двух других типов представляли собой линии, растущие только
на твердых поверхностях; особенности культивирования таких
клеток мы вкратце рассмотрим ниже.
Показано, что очень высокая локальная плотность клеток
может быть достигнута двумя различными методами иммоби-
лизации— с помощью ультрафильтрационных мембран или
путем микрокапсулирования. Хотя принципы иммобилизации
животных клеток и клеток микроорганизмов в общих чертах
одинаковы, к иммобилизации относительно больших по разме-
рам животных клеток предъявляются и свои специфические
требования. В одном очень перспективном способе микро-
капсулирования [60] клетки сначала включают в гранулы
но
Глава 9
РИС. 9.41. Изменения, происходя-
щие в периодической культуре
суспензии клеток SK-HEP-1 (аде-
нокарциномы печени человека),
во времени; объем культуры 100 л.
Штриховая линия на верхнем
графике отвечает экспоненциаль-
ному росту со временем удвоения
62 ч. Спустя трое суток начинают
барботировать чистый кислород со
скоростью 0,2 мл/мин. [Воспроиз-
ведено с разрешения из статьи:
Tolbert W. R., Schoenfled R. А.,
Lewis С., Feder J., Large-Scale
Mammalian Cell Culture: Design
and Use of an Economical Batch
Suspension System; Biotech, Bio-
eng., 24, 1671 (1982).]
альгината (см. рис. 9.27).
Затем на гранулы наносят
слой полилизина и послед-
ний сшивают поперечными
связями, после чего альги-
натный гель можно удалить
растворением. В этом случае
находящиеся внутри полили-
зиновой капсулы клетки по-
сле стадии роста могут за-
полнить весь объем внутри
капсулы, и их плотность
приблизится к плотности в
тканях. Подобные катализа-
торы перспективны с точки
зрения возможности дости-
жения более высокой произ-
водительности реакторов (в
расчете на единицу объема).
Специфические проблемы возникают при увеличении масш-
таба биореакторов с клетками, растущими на поверхностях.
Для вращающихся сосудов отношение площади поверхности к
объему мало, поэтому переход к аналогичным сосудам боль-
шего объема практически нецелесообразен. Предложено не-
сколько способов увеличения отношения площади поверхности
биореактора к его объему (S/V) и, следовательно, объемной
плотности растущих на поверхностях клеток. Ряд конструктив-
ных решений этой задачи схематично изображен на рис. 9.42;
здесь же приведены соответствующие величины S/V. Из раз-
Проектирование и расчет биологических реакторов
141
Таблица 9.14. Удельные скорости клеточного роста и поглощения
кислорода для клеток различных линий. Рост непрерывной культуры
осуществляли в реакторе колонного типа с барботажем11
Тип клеток Метод культиви- рования Удельная скорость по- глощения Оа [мкмоль О2/(10в кле- ток-ч)]10-’ Удельная скорость клеточного роста ц, ч-1 Давление С>2, мм рт. ст.
ВНК 21С13 (из почек новорож- денных хомя- ков) Клетки лимфобла- стов человека (Namalwa) Клетки мыши L929 Препуциум чело- века FS-4 Непрерывная культура Непрерывная культура На микроно- сителе На микроно- сителе На микроно- сителе 85 НО 54 75 20 65 50 85 0,018 0,033 0,012 0,020 Стационарная фа- за после форми- рования монослоя клеток Экспоненциальная фаза роста 0,010 0,025 60 60 50 50 60—80 ~60 ~60 ~60
а Воспроизведено с разрешения из статьи: Katinger Н. W. D., Scheirer W., Status and
Development of Animal Cell Technology using Suspension Culture Techniques; Acta Bio-
technologica, 2, 3 (1982).
личных принципиально возможных путей решения задачи в пос-
ледние годы наибольшее внимание уделяется микроносителям —
небольшим бусинам, на поверхности которых могут расти
клетки и которые можно суспендировать в культуральной жид-
кости. Основное преимущество микроносителей связано с
возможностью их использования в тех же реакторах и с теми
же перемешивающими устройствами, которые применяются для
процессов с участием животных клеток без носителей. Потен-
циальные возможности микроносителей впервые были проде-
монстрированы на заряженных декстрановых бусинах. Наблю-
давшееся в первых экспериментах ингибирование клеточного
роста удалось позднее подавить путем изменения состава бусин
и снижения их поверхностного заряда. Один из выпускаемых
в промышленном масштабе и поступающих в продажу мик-
роносителей состоит из бусин поперечно-связанного декстрана,
на которые нанесен слой денатурированного коллагена, кова-
142
Глава 9
материал
Отдельный капилляр
связанные
с наружной
поверхностью
капилляра
Неподвижный
слой
Реактор с
перемеш иванием
6
о ° о о
о О о оо
о о о о
° , о о
0 oo°g°
О о о °о
о о о °
Псевдоожиженный
слой
Питатель ные
вещества
Продукты
метаболизма
в
Отдельный
кидающийся сосуд
Фильтр Отходы (можно
стерилизовать
или сжигать}
Параллельные
пластины
V 00
Магнитная
мешалка
Стерильный Двига-
воздух
тель
Меилалка
Отдельный
сосуд
/Оо°оо\
/Ооо°оооо'
Г0о°ооо00
°Оо°оооооу
Го°оооооо/
\°ООООо/
Вращающийся
стенд
Проектирование и расчет биологических реакторов
143
лентно связанного с декстрановой основой. Сообщалось, что
плотность такого катализатора в реакторе для культур клеток
может составлять от 3 до 25 г/л.
Переход от исходной чистой культуры к культуре с высокой
плотностью клеток в большом реакторе обычно осуществляют
ступенчато с постепенным повышением масштаба. На рис. 9.43
схематично изображены основные стадии операции по повы-
шению масштаба для суспензионной и растущей на поверхно-
стях культур животных клеток. Обратите внимание на то, что
здесь объем инокулята, переносимого в следующий по масштабу
реактор, довольно велик по сравнению с системами на основе
культур микроорганизмов.
Культуры животных клеток выращивают в различных режи-
мах. Наряду с простым периодическим режимом применяют и
ступенчатый периодический режим, при котором по окончании
одного периодического процесса определенную часть культуры
используют как посевной материал для другого реактора, где
и происходит затем окончательное созревание клеток. Дру-
гим вариантом являются периодические процессы с добавлени-
ем субстрата, в которых по мере роста клеток в культуру
вводят питательные вещества по определенной программе или
в соответствии с данными анализов. В перфузионной системе
клетки удерживаются в реакторе, а среда частично замещается
на свежую; такая система представляет собой вариант рассмот-
ренных ранее методов иммобилизации клеток в масштабе
всего реактора. Наконец, использовались и непрерывные культу-
ры с постоянным обновлением как клеток, так и среды. Обнов-
ление среды позволяет снизить эффекты ингибирования, обус-
ловленные накоплением продуктов клеточного метаболизма.
Действительно, аммиак ингибирует клеточный рост в концент-
рациях выше 4 мМ.
Скорость образования продуктов метаболизма животных
клеток может варьировать в очень широких пределах в зави-
симости от типа процесса и пути образования данного продук-
та метаболизма. В некоторых случаях скорость образования
РИС. 9.42. Аппараты и устройства, обеспечивающие высокую площадь поверх-
ности контакта животных клеток со средой, а — пучок искусственных капил-
ляров (обычно полупроницаемых) (S/V=31 см-1); б — гранулированные но-
сители (S/V= 120 см-1 при плотности 20 г гранул в 1 л); в — свернутый лист
(с прокладками, обеспечивающими постоянное расстояние между'витками)
(S/V=4 см-1); г — культиватор с параллельными пластинами (и эрлифтным
устройством) (S/P=l,7 cm-i)j д — наклоненные вращающиеся сосуды (от-
дельный сосуд или вращающийся стенд, вмещающий до нескольких сотен со-
судов) (S/l/=0,2—0,7 см-1).
144
Глава 9
Культура на поверхности
| Культура в суспензии
О Чистая культура в атмосфере Ng
А (3-Ю5 клеток/смг) | \(5~ 10)-Ю5 клеток/мл\
М Культура в сосудах
АААА кУльтУРа
иииив сосудах
Культура, в сосудах \
со встряхиванием >
(7-Ю9)
(<? • Юв)
Промышленный масштаб
1 биореактор
(7,5 - Ю")
1 биореактор
(7,5- Юп)
Iсистема для
типового процесса
роста клеток
на поверхности
(1-1(11!) u
--------1 (.
1 биореактор
(3,7-Ю72)
т----1—
Культура на
микроносителях?
Технологи чест гй
предел достигнут?
РИС. 9.43. Увеличение масштаба культур животных клеток (на примере ВНК
21—клеток из почек новорожденного хомяка). В скобках указано общее ко-
личество клеток, образующихся на данной стадии. [Воспроизведено с разре-
шения из статьи: Katinger Н. W. D., Scheirer W., Status and Development of
Animal Cell Technology using Suspension Culture Techniques; Acta Biotechno-
logica, 2, 3 (1982).]
метаболита связана с клеточным ростом. В других ситуациях
после того, как клетки прошли определенную стадию роста, в
систему вводят те или иные вещества-предшественники, из
которых позднее и образуется продукт метаболизма. В произ-
водстве вирусов, в котором клетки, по сути дела, являются для
них питательной средой, может наблюдаться очень сложная
Проектирование и расчет биологических реакторов
145-
кинетика. В литературе, посвященной изучению применения
культур животных клеток для получения вирусов, клеточный
слой обычно называют «субстратом». Посев вируса часто
производят только после того, как культура клеток достигнет
высокой плотности. Затем вирусы развиваются в соответствии
со своим жизненным циклом (см. разд. 6.2.1), включающим
стадии проникновения вируса в клетку, синтеза его отдельных
компонентов и сборки частиц вируса из этих компонентов;
только после этого вирус можно обнаружить в клетках или
среде.
Разработка способов производства моноклональных анти-
тел может служить примером тех потенциальных возможностей,
которые заложены в оптимизации процессов с участием живот-
ных клеток. Гибридомы клеток мыши выращивали в периодиче-
ской, периодической с добавлением субстрата и в непрерывной
культурах с целью получения моноклональных антител для
поверхностного антигена клеток Rhizobium japonica NR-7*.
Изучение различных сред показало, что для данного случая
подходит недорогая среда типа Dulbecco’s Modified Eagle
Medium, к которой добавлено 0,25% приматона RL, 0,01%
детергента плюроник F-68 и совсем немного (1%) сыворотки
эмбрионов крупного рогатого скота. В этой среде концентра-
ция клеток в суспензии достигает 2-106 мл-1 при времени
удвоения 24 ч. Для этого процесса не было предложено деталь-
ной модели кинетики клеточного роста, но полученные резуль-
таты тем не менее представляют большой интерес (рис. 9.44
и 9.45). Снижение концентрации растворенного кислорода (с
60 до 25% относительно концентрации при насыщении возду-
хом) привело к уменьшению максимальной плотности клеток
в культуре, но способствовало увеличению времени жизни
культуры, так что окончательная концентрация моноклональ-
ных антител оказалась более высокой (рис. 9.44).
. Высказывалось предположение, что в любых условиях об-
разуется ингибитор клеточного роста. Если это предположение
справедливо, то периодический процесс с добавлением субстра-
та должен дать лучшие результаты в силу ступенчатого сниже-
ния концентрации ингибитора. С этой гипотезой согласуются
результаты сравнительного изучения периодических процессов
с добавлением и без добавления субстрата (рис. 9.45); действи-
тельно, и жизнеспособность клеток, и концентрация антител
выше в периодическом процессе с добавлением субстрата. Не-
большие количества моноклональных антител удобнее получать
в брюшной полости мыши, однако крупномасштабное производ-
* Reuveny S., Velez D., Riske F., Macmillan J. D., Miller L., Production of
Monoclonal Antibodies in Culture; E.S.C.A.T. Mtg., Italy, May, 1984.
10-746
146
Глава 9
(А) Концентрации клеток, 106[мл (л) Концентрация клеток, Ю^/лт
гоо
100
о
гоо
100
о
РИС. 9.44. Рост гибридом и образование моноклональных антител в фер-
ментере объемом 3 л при различных концентрациях растворенного кислорода.
(Воспроизведено с разрешения из статьи: Reuveny S., Valez D., Riske F„ Mac-
millan, J. D., Miller L., Production of Monoclonal Antibodies in Culture;
E.S.C.A.T. Mtg., Italy, May, 1984.)
ctbo антител целесообразнее осуществлять в биореакторах с
недорогой питательной средой в условиях, обеспечивающих
максимальное время жизни клеток и наибольший выход анти-
тел. Для роста гибридом и получения продуктов их жизнедея-
тельности (моноклональных антител) в большом масштабе
перспективно применение культур клеток, иммобилизованных
в полых волокнах или с помощью полупроницаемых мембран.
9.8.3. Культивирование растительных клеток
Растительные ткани, выделенные из внутренних частей
органов растений, после промывки и дезинфекции можно куль-
тивировать на агаре в соответствующей питательной среде.
Проектирование и расчет биологических реакторов
147
РИС. 9.45. Сравнение методов получения моноклональных антител в периоди-
ческом процессе (б) и в периодическом процессе с добавлением свежей сре-
ды (а). Культуру выращивали во вращающихся сосудах емкостью 100 мл.
В периодическом процессе с добавлением свежей среды каждые 24 ч к куль-
туре добавляли 5—10% среды, а 30% культуры периодически отбирали.
(Воспроизведено с разрешения из работы: Reuveny S., Valez D., Riske F.,
Macmillan J. D„ Miller L., Production of Monoclonal Antibodies in Culture;
E.S.C.A.T. Mtg., Italy, May, 1984.)
Питательная среда для растительных клеток обычно состоит
из смеси неорганических солей и глюкозы или сахарозы в каче-
стве источника энергии и углерода. (В культуре растительные
клетки обычно являются хемогетеротрофами, а не фотоауто-
трофами.) В среду указанного выше состава обычно вводят
также регуляторы роста растений, фитогормоны, витамины,
аминокислоты и многоосновный спирт инозит. В такой культуре
деление клеток осуществляется только в присутствии кислоро-
да, поэтому ее следует аэрировать путем диффузии через
свободную поверхность жидкости или посредством барботажа.
Увеличение численности популяции в периодическом процессе
10*
148
Глава 9
подчиняется обычным закономерностям клеточного роста, рас-
смотренным в гл. 7, но здесь обычно удается добиться только
трех- или четырехкратного удвоения популяции, что соответству-
ет увеличению клеточной массы в 10—15 раз по сравнению с
массой посевного материала.
Культуры растительных тканей могут быть полезными при
решении многих задач. Так, ферментные и метаболические
системы растений можно использовать для проведения как
сравнительно простых, так и очень сложных биохимических
превращений. Примером таких реакций может служить полу-
чение дигоксина (важного стимулятора сердечной деятельности)
из дигитоксина путем введения гидроксильной группы в 12^-по-
ложение последнего с помощью клеток наперстянки (Digitalis).
Растительные клетки можно также использовать в биосинтезе
сложных соединений из сравнительно простых предшественни-
ков; в частности, культуры растительных тканей способны
осуществлять полный синтез сложных веществ, обычно вторич-
ных метаболитов, из простых компонентов среды. К числу
вторичных растительных метаболитов, представляющих интерес
с точки зрения их промышленного производства, относятся
фармацевтические препараты (в настоящее время около одной
четверти всех разрешенных к применению в США лекарствен-
ных препаратов имеют растительное происхождение), красите-
ли (например, шиконин), камеди, природные инсектициды
группы пиретринов, вкусовые вещества, а также пряности и
душистые вещества типа продуцируемых ванилью и жасмином.
Культуры растительных тканей, кроме того, служат ценным
инструментом в генной инженерии растений: с их помощью
можно существенно повысить урожайность, а в будущем воз-
можно добиться выращивания целых растений хлебных злаков
из культуры клеток. По сравнению с обычным методом выра-
щивания растений в грунте и последующего сбора урожая
культуры растительных тканей могут обеспечить более высокую
продуктивность, надежность и предсказуемость урожая.
В суспензионной культуре растительные клетки находятся
в изолированном состоянии или в виде крупных скоплений.
Отмечалось, что в скоплениях растительные клетки в некото-
рой степени дифференцированы так, что клетки, находящиеся
на наружной поверхности и внутри скопления, морфологически
различны. В некоторых случаях необходимый продукт метабо-
лизма вэобще не синтезируется отдельными клетками, а только
их скоплениями. Отсюда следует, что при определенных обстоя-
тельствах некоторая степень дифференциации клеток способст-
вует реализации тех путей метаболизма, в которых синтезиру-
ется необходимое вещество. В других случаях предпочтительнее
недифференцированное состояние клеток в культуре. Дальней-
Проектирование и расчет биологических реакторов
149
шее практическое развитие этой области биотехнологии требует
более глубокого изучения процессов регуляции дифференциации
клеток, обусловленных дифференциацией эффектов, и возмож-
ности управления дифференциацией в культурах клеток путем
изменения условий роста.
О кинетике роста растительных клеток в культуре опублико-
вано мало данных. Одно из немногих исключений представля-
ют результаты работ Уилсона и сотрудников по изучению
культивирования растительных клеток в хемостате. В табл. 9.15
Таблица 9.15. Время удвоения и экономические коэффициенты
для культур растительных клеток3
Лимитирующее Экономический ко- Время удвое-
Культура клеток питательное ве- эффициент, НИЯ, ч
щество 10s клеток/мкмоль
Galium (подмаренник) Фосфат (РО4~) 3,95 40
Galium (подмаренник) Фосфат (РО4~) 2,47 25
Acer (белый клен) Фосфат (РО4_) 3,47 182
Acer (белый клен) Фосфат (РО4_) 1,28 36
Acer (белый клен) Нитрат (NO3-) 0,263 109
Acer (белый клен) Глюкоза 0,039 109
а Воспроизведено с разрешения из статьи: Wilson. G., Continuous Culture of Plant
Cells Using the Chemostat Principle, in Adv. Biochem. Eng., Fiechter A. (ed ), vol. 16,
p. 1. 1980.
приведены найденные этими исследователями экономические
коэффициенты и время удвоения для культур растительных
клеток двух типов при росте в средах с различными лимитирую-
щими питательными веществами. Обратите внимание на чрез-
вычайно большое время удвоения популяции, что является
одним из основных препятствий как в экспериментальном изу-
чении культур растительных клеток, так и в их промышленном
применении в качестве катализаторов. Оценка константы насы-
щения Ks (или константы Моно) показала, что в случае роста
культуры клеток Acer (клена белого) на лимитирующих субст-
ратах (нитрате, глюкозе и фосфате) величина Ks составляет
0,13, 0,5 и 0,032 мМ соответственно [66].
Рост культур растительных клеток осуществляли не только
во встряхиваемых сосудах, но и в реакторах с механическим
перемешиванием, а также в реакторах колонного типа с барбо-
тажем. Для того чтобы повысить объемную плотность клеток в
культуре, обеспечить возможность повторного их использования
и сохранить в определенной стадии морфологического развития
и дифференциации, а также для лучшего регулирования контак-
та между средой и клетками, несколько исследовательских
групп изучали реакторы с иммобилизованными растительными
150
Глава 9
клетками. В частности, исследовалось включение растительных
клеток в полупроницаемые полые волокна, гранулы альгинат-
ного геля и другие гранулированные носители [67, 68]. Многие
продукты метаболизма растений накапливаются только внутри
клеток, что может затруднять осуществление промышленных
процессов с участием иммобилизованных клеток. Для решения
этой проблемы предлагалось попеременно изменять проницае-
мость клеточных стенок путем изменения состава среды, чтобы
Реактор 1-й стадии.
{Z00л, 9 су гл}
Реактор 2~й стадии.
{750л, сут}
РИС. 9.46. Схема установки для производства вторичного метаболита шико-
нина с помощью культуры клеток растений {Lithospermum erthrorhizon). (Вос-
произведено с разрешения из работы; Curtin М. Е., Harvesting Profitable
Products from Plant Tissue Culture; Bio/Technology, October, 1983, p. 649.)
одна среда обеспечивала рост клеток, а другая образование и
секрецию продукта метаболизма [68].
Как мы уже отмечали при изучении способов производства
вторичных метаболитов с помощью периодических культур мик-
роорганизмов, в этом случае необходимо разделять во време-
ни или в объеме фазу роста клеток и фазу образования вторич-
ного метаболита. Такой подход был осуществлен и в первом
промышленном крупномасштабном процессе с участием куль-
туры растительных клеток, разработанном компанией Mitsui
Petrochemical Industries. В этом процессе, включающем три
последовательные операции общей продолжительностью более
трех недель, в культуре Lithospermum erythrorhizon получают
ценный краситель и фармацевтический препарат шиконии (рис.
9.46). Клетки сначала выращивают в аэрируемом реакторе, а
затем переносят в меньший реактор со средой типа М-9, которая
Проектирование и расчет биологических реакторов
151
стимулирует образование шиконина. Далее культуру переносят
в третий емкостный реактор, в котором в течение 14 сут шико-
нин накапливается в клетках. О выходе продукта компания,
производящая шиконин, не сообщала; согласно независимым
оценкам, производительность установки составляет около 5 кг
шиконина за один периодический процесс. Очевидно, трудоза-
траты, расходы на сооружение установки и приобретение доро-
гой среды должны окупаться высокой стоимостью продукта.
Действительно, продажная цепа шиконина в октябре 1983 г.
составляла 4000 долларов за килограмм!
9.9. Заключение
В этой главе мы рассмотрели основы расчета и проектирова-
ния реакторов и основные принципы соответствующих техноло-
гий, которые находят применение в настоящее время и, воз-
можно, будут использоваться в будущем для производства тех
или иных продуктов с помощью микроорганизмов или клеток
высших организмов. Мы попытались показать, как на основе
данных о кинетике биологических процессов и параметров,
характеризующих течения и перемешивание в реакторе, можно
составить математическое описание процесса, пригодное для его
расчета и оптимизации. Такой традиционный и успешно приме-
нявшийся в классической химической технологии подход к
решению проблем технологии химических реакций уходит сво-
ими корнями в химическую и нефтеперерабатывающую про-
мышленность. К сожалению, вследствие отсутствия достаточно-
го количества данных о кинетике биологических превращений,
применения сложных сред с неопределенным составом и непред-
сказуемыми эффектами, использования реакторов с нечетко
определенными структурой течений и параметрами перемеши-
вания, а также в силу того, что до сих пор не проводилось
комплексных работ по поиску более систематических универ-
сальных и надежных методов расчета биореакторов, разработан-
ные с помощью описанной в этой главе систематической методо-
логии режимы работы биореакторов и спецификации соответ-
ствующих проектов встречаются крайне редко. Масштабирова-
ние биопроцессов чаще всего осуществляют в нескольких
последовательных экспериментах с постепенным увеличением
масштаба (что, естественно, приводит к удорожанию проектных
работ), причем масштабы последовательных экспериментов
гораздо меньше отличаются один от другого, чем в случае
аппаратов и процессов химической технологии. При разработке
биологического реактора периодического действия в настоящее
время основная задача, скорее всего, будет заключаться в обес-
печении необходимой скорости массопереноса, а не в расчете
152
Глава 9
оптимального режима работы реактора на основе данных о ки-
нетике процесса.
Опыт работы отраслей промышленности, в которых широко,
применяются методы химической технологии., показывает, что
за счет оптимизации процессов можно добиться существенного
снижения капитальных вложени?! и эксплуатационных расхо-
дов, уменьшения количества побочных продуктов и повышения
выхода основного продукта. Кроме того, в нефтеперерабатываю-
щей промышленности благодаря хорошо разработанной теории
процессов и аппаратов масштаб процесса можно обычно увели-
чивать в 10 000 раз, что соответствует непосредственному пере-
ходу от лабораторной установки к крупномасштабному аппара-
ту. Поскольку в ближайшем будущем откроется возможность-
получения многих новых продуктов с помощью новых организ-
мов и новых процессов, мы вправе надеяться на развитие
технологии биохимических процессов, что позволит обеспечить
большую эффективность масштабирования, проектирования и
эксплуатации биореакторов. Это в свою очередь будет способ-
ствовать повышению их надежности и ускорению экономически
оправданного внедрения биопроцессов в промышленность.
Следующая глава будет посвящена знакомству с контроль-
но-измерительной аппаратурой для биореакторов. Отсутствие
данных о явлениях переноса основных компонентов системы
и химических процессах в реакторе ограничивают число мето-
дов контроля для установления оптимального режима работы
реактора. Хотя в следующей главе основное внимание будет
уделено контролю и регулированию процессов в емкостных
реакторах, большая часть рассматриваемых контрольно-измери-
тельных приборов может с тем же успехом применяться и для
аппаратов с восходящим или нисходящим потоком. Закончив
изучение контрольно-измерительной аппаратуры, мы перейдем
к рассмотрению основных процессов разделения, позволяющих
превратить выходящий из биореактора поток смеси веществ в
достаточно чистый целевой продукт.
Упражнения
9.1. Анализ ПРПП. а) Проверьте справедливость всех приведенных
в табл. 9.1. уравнений, описывающих процессы в ПРПП.
б) Как с помощью соответствующих графиков можно определить все па-
раметры, входящие в уравнения скоростей реакций в ПРПП (табл. 9.1)?
9.2. Анализ ТРПВ. а) Проверьте справедливость приведенных в табл. 9.2
выражений путем их интегрирования.
б) Какие переменные удобнее применять в каждом из уравнений для
ТРПВ? Начертите графики, по которым можно определить каждый кинетиче-
ский параметр в уравнении скорости реакции.
в) Параметры какой модели — ПРПП или ТРПВ — в общем случае легче
поддаются определению? Почему?
Проектирование и расчет биологических реакторов
153
9.3. Производство белка одноклеточных организмов. Экспериментально
определены следующие значения параметров кинетики роста Methylomonos
methanolica на метаноле при 30 °C и pH 6: р,тах = 0,53 ч-1; Ys (экономический
коэффициент по метанолу) =0,48 г/г; Уо2=0,53 г/г; эффективность утилизации
углерода (сбНомасса/сметаноЛ) =0,57 г/г; кислородный коэффициент — 0,90 молей
О2 на ’моль СН3ОН; дыхательный коэффициент (RQ)=0,52 моль СО2 на моль
О2; коэффициент поддержания т = 0,35 г СН3ОН/г; 7G = 2,0 мг/л \Dostolek М.,
Molin N., Studies of Biomass Production of Methanol Oxidizing Bacteria, p. 385
in Single Cell Protein II, Tannenbaum S. R., Wang D. I. C. (eds.), MIT Press,
Cambridge, Mass., 1975]. Приведенные выше экономические коэффициенты
отвечают скорости разведения 0,52 ч-1.
а) Запишите уравнения, описывающие зависимости скоростей образова-
ния клеточной массы, поглощения кислорода и образования СО2 от скорости
разведения в ПРПП.
б) Постройте графики зависимостей х, xD и s от D (ч-1) и определите
максимальные значения х и xD при концентрации метанола в питательной
среде s=7,96 г/л.
в) На том же чертеже постройте график зависимости скорости потребле-
ния кислорода от D. Определите расходуемую на перемешивание мощность
(в расчете на единицу объема), которая обеспечивает максимальную продук-
тивность системы (xD)max. Сформулируйте принятые допущения.
г) На том же чертеже постройте график зависимости расчетной скорости
выделения теплоты в единице объема от D.
д) Показано, что удельная скорость роста ц линейно зависит от концент-
рации субстрата и снижается от 0,53 ч-1 при s = 3 г/л до нуля при s= 13 г/л
(последняя величина найдена путем экстраполяции). Повторите решение за-
дачи 9.3. б с учетом указанных данных.
е) При каких условиях на выходе каскад реакторов выгоднее, чем один
реактор того же объема?
9.4. Стерилизация жидкостей. Составьте уравнение, позволяющее опреде-
лить вероятность достижения полной стерильности (и<1,0) в непрерывном
стерилизаторе, работающем в режиме полного вытеснения при следующих
параметрах: удельная скорость гибели клеток 10 мин-1; объем стерилизатора
10 л; средняя скорость потока 10 л/мин; температура среды 131 °C; время
сбора 5 мин; начальная концентрация клеток п0=Ю3 л-1. Как, по вашему мне-
нию, изменится вероятность достижения полной стерильности, если при тех
же указанных переменных стерилизатор будет короче и шире? Поясните ваш
ответ и приведите соответствующие уравнения.
9.5. Клеточный рост при непостоянном экономическом коэффициенте.
Выведите уравнения для зависимостей x(Z)) и s(D). аналогичные уравнениям
(7.14) и (7.15), если Y определяется уравнением (7.26).
9.6. Микробиологический процесс в реакторе с перемешиванием в присут-
ствии инертного носителя. Предположим, что питательный раствор содержит
суспендированные инертные частицы, а также субстрат для анаэробной фер-
ментации. Перемешивание обеспечивает полное диспергирование частиц. Рас-
пределение микроорганизмов (одного штамма) между частицами и жидкой
фазой описывается уравнением:
Л\(клеток/см2) = КХжидкая фаза (КЛвТОК/мл)
где К имеет размерность мл/см2. Адсорбция не изменяет максимальной удель-
ной скорости роста Цтах, но влияет на Ks (будем считать, что рост клеток
описывается уравнением Моно).
а) Приняв диаметр частиц равным 0,1 мм, а их объемную долю равной
1%, определите во всем доступном диапазоне скоростей разведения, во-пер-
вых, отношение скорости утилизации субстрата в присутствии инертных час-
тиц к скорости утилизации субстрата в их отсутствие и, во-вторых, влияние
154
Глава 9
суспендированных частиц на скорость разведения, отвечающую началу вымы-
вания.
б) Предложите конструкцию устройства для регенерации клеток на вы-
ходе из реактора, которое бы обеспечивало максимальный выход биомассы.
9.7. Метод быстрого определения Ks. Уилльямсон и Мак-Карти [William-
son, McCarty, A Rapid Measurement of Monod Half Velociity Coefficients for
Bacterial Kinetics; Biotech. Bioeng., 17, 915 (1975)] разработали метод относи-
тельно быстрого определения Ks — константы в уравнениях, описывающих ки-
нетику микробиологических процессов. В этом методе в микробиологический
реактор постепенно вводят концентрированный раствор питательных веществ
в таком количестве, чтобы общий объем смеси существенно не увеличивался
в течение нескольких часов. Если величина s такова, что скорость утилизации
субстрата клетки ниже максимально возможной (ЦшахК/У), то стационарное
состояние достигается менее чем за 1 ч.
а) Покажите, что с помощью графической зависимости типа графика
Лайнуивера — Бэрка (зависимости обратной скорости клеточного роста от
обратной концентрации субстрата в «стационарном состоянии») можно опре-
делить Ks.
б) В каком диапазоне времени отбора проб применим указанный метод?
9.8. Реактор периодического действия с добавлением субстрата, а) Взяв
за основу уравнение (9.4), выведите общее уравнение, описывающее зависи-
мость х от t, если при ц —Цтах клетки находятся в экспоненциальной фазе роста.
б) При каких условиях параметр х будет возрастать, уменьшаться или
оставаться постоянным?
в) Найдите выражения, описывающие х при t<ti, при Ь<1<12 и при
сколь угодно большом t, если концентрация питательных веществ F изменяет-
ся во времени следующим образом: F=FQ при F=0 при
F=F0 при t2<t<t3 и т. д.
г) Пусть удельная скорость роста штамма микроорганизмов, утилизирую-
щих метанол, описывается уравнением (7.32) (т. е. уравнением Моно плюс
выражение, учитывающее субстратное ингибирование). Объясните, почему
периодический процесс с добавлением субстрата дает лучшие результаты по
сравнению с периодическим процессом с введением всего метанола в начале
процесса.
9.9. Последовательная утилизация субстратов. Как упоминалось в тексте
настоящей главы, при наличии нескольких субстратов характер их утилизации
в каскаде реакторов или реакторе башенного типа может быть весьма слож-
ным. Рассмотрим очень простую модель диауксии, т. е. роста клеток на двух
субстратах, одним из которых является глюкоза (G), а вторым — другой уг-
левод (S), так что
rx = p(g,s)x =
PGg . HSS
_ Kg + g Ks + s
Кк + g Г
а) Если, как обычно, глюкоза играет роль репрессора, то следует ожи-
дать, что И Ks~Kg»Kr. Нарисуйте графики зависимостей g, s и 1пх
от времени в периодическом процессе; укажите взаимосвязь между измене-
нием концентрации биомассы и двух субстратов G и S. [Считайте, что
(go, So)»(Ks, Kg).]
б) Какой реактор лучше применять — реактор полного вытеснения, кас-
кад хемостатов или один хемостат, — если целью микробиологического про-
цесса с участием двух субстратов является их полная утилизация?
в) Поскольку Kr<Kg, Ks, то уравнение клеточного роста можно значи-
тельно упростить, получив в конечном итоге два выражения для g>K*
и КК>ё~0. Выведите упрощенное уравнение, описывающее процесс в потоке
полного вытеснения, и путем интегрирования найдите зависимости x(z), g(z)*
и s(z).
Проектирование и расчет биологических реакторов 155
г) Начертите графики зависимостей, найденных в предыдущем упражне-
нии, при следующих значениях параметров: |Хо=1 ч_, = 1,1 p,s, Kg = Ks =
= 10 ммоль, Kr = 0,1 ммоль, хо=О,1 г/л, Уо = Уз = 0,5 г/г, go=So=O,l ммоль,
а отношение Uo/L (отношение скорости потока к длине реактора) равно 1, 3
или 5 ч~*.
д) В вертикальных ферментерах башенного типа микроорганизмы могут
образовывать медленно осаждающиеся скопления. Как модифицировать про-
стое уравнение, найденное при решении упражнения 9.9, б, чтобы оно учиты-
вало и осаждение клеток со средней скоростью ис (ис>ио)7 (Результирующая
скорость осаждения клеток равна ис—и0.)
9.10. Расчет хемостата на основе данных для периодического процесса.
Уравнения материального баланса по компоненту С в реакторе периодическо-
го действия (или в реакторе полного вытеснения) и в хемостате (в ПРПП)
можно записать в следующей форме:
Периодический процесс (или реактор полного вытеснения) = f(c) (I)
dt
Процесс в хемостате (в стационарном состоянии) D(c;.—с) = —)(с) (II)
Поскольку зависимость f(c), согласно уравнению (I), можно найти диффе-
ренцированием, то график зависимости f от с можно построить по результа-
там изучения периодического процесса. С другой стороны, уравнение (II) по-
казывает, что график зависимости D(c—с0) от с будет пересекаться с кривой
зависимости [(c) от с при с*, отвечающем решению уравнения (II). [Этот ме-
тод, позволяющий найти с* для любого заданного D, применим только в том
случае, если процесс ферментации описывается одной переменной, например с;
он описан в статье: Luedeking R., Piret Е. L., Transient and Steady States in
Continuous Fermentation; Theory and Experiment; J. Biochem. Microbiol. Tech-
nol. Eng., 1, 431 (1959).] В этой статье приведены результаты сравнения рас-
четных и экспериментальных данных, иллюстрирующие потенциальные воз-
можности метода:
Найдено (единиц оптической плотности в 1 мл) Вычислено графическим методом по результатам изучения периодического процесса
5,37 5,35
2,4 2,3
8,0 7,15
а) Рассмотрим логистическое уравнение dx/dt=\ix(\ — x/xmax). Описанным
выше графическим методом определите х* при ц=1 ч-1, xmax=10 г/л и х0
(в исходных веществах) равном нулю, если скорость разведения D составляет
1,5, 0,75 или 0,25 ч-1. Проверьте правильность полученных вами результатов
прямым аналитическим решением.
б) В каскаде ферментеров продукты, полученные в одном реакторе, яв-
ляются исходными веществами для следующего реактора. Как графически
определить х3* (концентрацию биомассы в последнем реакторе каскада, со-
стоящего из трех ПРПП) при общей скорости разведения £> = 0,75 ч~‘?
в) Внимательно изучите данные, относящиеся к росту культуры на двух
субстратах (рис. 7.14). Путем дифференцирования найдите зависимость dx/dt
от х и начертите график этой зависимости. Чем этот график отличается от кри-
вой простого логистического уравнения? С помощью описанного метода реши-
156
Глава 9
РИС. 9У11.1. Зависимость степе-
ни превращения субстрата (О) и
высоты псевдоожиженного слоя
(+) от объемной скорости потока.
На кривых указана молярная кон-
центрация субстрата в исходной
смеси. [Из статьи: Gelf G., Boud-
rant Enzymes Immobilized on a
Magnetic Support; Biochim. Bio-
phys. Acta, 334, 468 (1974).]
те задачу (графическим путем)
о ходе процесса в каскаде из 5 ре-
акторов, в котором потребляется
вся глюкоза и большая часть вто-
рого углевода.
9.11. Иммобилизованные ферменты в псевдоожиженном слое, а) Чинлой
(Chinloy, PhD thesis, Princeton University, 1976) изучал зависимость степени
превращения субстрата от обратной объемной скорости потока 0-1 (выражен-
ной в миллилитрах на грамм частиц катализатора в секунду) в неподвижном
и псевдоожиженном слоях протеазы, иммобилизованной на непористых части-
цах из нержавеющей стали. И в первом, и во втором случае эту зависимость
можно изобразить одной и той же прямой линией, проходящей через начало
координат. Допустим, что s0»7(m; покажите, что в этом случае на степень
превращения субстрата не влияет массоперенос и что константу удельной ско-
рости процесса можно определить непосредственно по указанной зависимости,
если известно количество фермента, адсорбированного на частицах катализа-
тора (миллиграммов Е в грамме катализатора).
б) Джелф и Боудрант [Gelf, Boudrant, Preliminary Study of a Fluidized
Bed Enzyme Reactor; Biochim. Biophys. Acta, 334, 467 (1974)] изучали гидро-
лиз этилового эфира бензоиларгинина (ЭЭБА) папаином, иммобилизованным
на пористых частицах диаметром 170—250 мкм. Для указанной реакции сооб-
щались следующие значения параметров: для растворимого папаина К,п =
= 5-10-3М, щПах = 19 межд. ед. (мкмолей ЭЭБА/(мин-мг) при pH 6,0 и 20°C);
для иммобилизованного папаина /Ст=1,2-10—2 М (кажущаяся величина),
Umax = 0,05 межд. ед. (мкмолей ЭЭБА/(мин-мг носителя}}. Пористый носитель
в данном случае состоял в основном из частиц оксида железа. По приведен-
ным на рис. 9У11.1 данным определите, какое сопротивление массопереносу —
внешнее или внутреннее—играло в этом случае основную роль. Укажите, ка-
кие допущения вы приняли. В псевдоожиженном слое содержалось 10 г ката-
лизатора, который получали из 100 мг кристаллического папаина и 30 г окси-
да железа.
в) Обсудите, как вы стали бы рассчитывать реактор с псевдоожиженным
слоем иммобилизованной протеазы, предназначенный для гидролиза 1) ЭЭБА,
2) казеина, 3) частиц желатины диаметром 1 мкм.
9.12. Оптически чистые аминокислоты. Тоса и Др. [Tosa et al., Studies on
Continuous Enzyme Reactors II. Preparation of DEAE-Cellulose Aminoacylase
Columns and Continuous Optical Resolution of Acetyl-d,l-methionine; Enzymo-
logia, 31, 225 (1966)] разработали процесс, схема которого изображена на
рис. 9У12.1. На время допустим, что Umax не зависит от pH. В этом процессе
исходная рацемическая аминокислота ацетилируется уксусным ангидридом.
Проектирование и расчет биологических реакторов
157
а в колонне с L-аминоацилазой идет обратная реакция, в результате которой
образуется свободная L-аминокислота, очищаемая перекристаллизацией из
водного спирта.
а) Допустим, что начальная концентрация аминокислоты намного больше
Кт. Найдите уравнения, описывающие гидролиз ацетил-Ь-аминокислоты под
действием фермента в колонне в режиме полного вытеснения, а также при на-
личии осевой дисперсии.
б) Можно принять, что реакция рацемизации обратима и имеет первый
порядок, так что ее скорость пропорциональна cDa —c*Da, где c*Da— концен-
Мсходная
ИЬ-ациламиио-
кислота
РИС. 9У12.1. Каталитическое разделение рацемических аминокислот. [Из
статьи: Tosa Т., Mori Т., Fuse N., Shibata I., Enzymologia, 31, 225 (1966).]
трация ацетил-В-аминокислоты в растворе при равновесии. Какой объем дол-
жен иметь ПРПП, в котором осуществляется рацемизация, если заданная
степень приближения к равновесию равна 95% и если с продуктами из систе-
мы выводится 90% L-аминокислоты, 1% ацетил-D-аминокислоты и 10% вод-
ной фазы?
в) При ферментативном деацетилировании выделяется уксусная кислота.
Если процесс инактивации фермента имеет pKi = 5 и рК2=8, то при каком pH
превращение ацетил-DL-аминокислоты в 10-6, 10~4 и в 10-1 М растворах бу-
дет максимальным? (Примите, что процесс происходит в реакторе полного
вытеснения.) Четко сформулируйте принятые допущения.
9.13. Превращение нерастворимых субстратов. Для выращивания куль-
туры дрожжей на газетной бумаге в качестве нерастворимого субстрата была
предложена следующая последовательность операций: механическое измель-
чение субстрата, его кислотный гидролиз, нейтрализация среды, введение не-
большого количества других питательных веществ, необходимых для роста1
Дрожжей, рост дрожжей в аэробном процессе, фильтрование под вакуумом
для отделения жидкой фазы от биомассы.
а) Нарисуйте технологическую схему указанного процесса, на которой
стрелками укажите места введения компонентов, а кружками выделите от-
дельные типовые операции. При необходимости укажите также расположение-
транспортеров для подачи твердых веществ и насосов для подачи жидкостей.
б) Нарисуйте аналогичную схему процесса переваривания пищи в орга-
низме человека, взяв за основу любой учебник по физиологии человека.
158
Глава 9
в) Бионика — это наука об изучении природных систем с целью разработ-
ки искусственных аналогов. Обсудите сходство и различие между процессами,
описанными в упражнениях 9.13, а и 9.13,6. Предложите схему устройства'
для обработки твердых веществ (упражнение 9.13, а), воспользовавшись прин-
ципом подачи «транспортером» из упражнения 9.13, б.
9.14. Поддержание жизнедеятельности клеток; вымывание при небольших
S. Для достижения нетривиального стационарного состояния популяций неко-
торых микроорганизмов субстрат может потребоваться в минимальной кон-
центрации. Рассмотрим, например, систему, в которой кинетика клеточного
роста описывается уравнениями:
Цтах5^ , 1 Цтах5-^
гх — jz I о %еХ rs — v- jz ।
As + s Ks /Cs + S
.Допустим, что к этой системе применим аналитический подход, описанный
в упражнении 9.10.
а) Покажите, что при концентрации субстрата ниже keKs/famax—ke) един-
ственное стационарное состояние в ПРПП достигается при х=0.
б) Постройте график зависимости dxfdt от х (для периодического про-
цесса) при gmax = 0,5 ч-1, 7G = 0,2 г/л, ke=0,1 ч~’, /5=0,6 г клеток/г субстра-
та; аналитическим решением указанных уравнений покажите, что для соответ-
ствующего непрерывного процесса dx/dt={) при малых величинах s и при
D > Двымывания.
в) Определите устойчивость каждого стационарного состояния по отно-
шению к небольшим возмущениям.
9.15. Ферментация сыворотки. Показано, что процесс превращения содер-
жащейся в сыворотке лактозы в молочную кислоту в присутствии Lactobacil-
lum bulgaricus при 44 °C и pH 5,6 можно описать уравнением Льюдикина —
Пайрета [уравнение (7.93)], если его модифицировать следующим образом:
1. Максимальная скорость роста равна
,.О (,______Р__
I* max I 1
\ Ртах
2. g°max=0,48 Ч-1, Ртах = 5% При £<3,8%
g°max=l,l Ч-1, £тах = 4,3°/о ПрИ £>3,8%
3. Параметры непрерывного процесса: а=2,2; [3 = 0,2 ч-1; У=0,88 г про-
дукта/г субстрата; /<=50 мг/л.
а) Напишите уравнения, описывающие изменение s, х и р в непрерывном
процессе.
б) Покажите, что в стационарном состоянии при общем времени удержи-
вания 15 ч субстрат превращается полнее в каскаде из двух реакторов, однако
дальнейшее увеличение числа реакторов практически не влияет на степень
превращения субстрата. Относится ли этот вывод и к урожаю биомассы?
(Рассмотрите вариант, когда So=5,O%, хо=ро=О.)
в) Келлер и Герхардт [Keller, Gerhardt, Continuous Lactic Acid Fermenta-
tion of Whey to Produce a Ruminant Feed Supplement High in Grude Protein;
Biotech. Bioeng., 17, 997 (1975)] отмечали, что при s0 меньше 5% ингибирова-
ние продуктом процесса не слишком велико, и поэтому, по их мнению,
«...с практической точки зрения... сыворотку сыра чеддер (4,9% лактозы,
•0,2% молочной кислоты) можно с удовлетворительным результатом ферменти-
ровать в одностадийном ферментере, тогда как при получении сыра коттедж
(5,8% лактозы, 0,7% молочной кислоты) этот процесс выгоднее осуществлять
в каскаде из двух ферментеров». Оцените количественно преимущества систе-
мы из двух ферментеров для переработки сыворотки сыра коттедж, повторив
^решение упражнения 9.15,6 при s0=5,8%, ро=О,7%.
г) Эти исследователи отмечали также, что добавление сахара может при-
вести к снижению количества воды, которое нужно удалить для получения
Проектирование и расчет биологических реакторов
159
определенного количества продукта. Как эта операция (добавление сахара)
отразится на общей схеме реактора?
9.16. Каскад реакторов; переработка углеводородов. Предположим, что
в вашем распоряжении имеется кривая, описывающая изменение скорости
клеточного роста в периодическом процессе переработки углеводородов с по-
мощью микроорганизмов; этот процесс очень живо описан Мимурой и др.
(разд. 8.8). В упражнении 8.11 вы определяли возможные лимитирующие
процесс сопротивления на каждой из его фаз. Ваша компания решила (в ва-
РИС. 9У17.1. Ход периодического процесса биосинтеза пенициллина во вре-
мени.
ше отсутствие) расширить масштаб производства, дополнив установку п по-
следовательно соединенными реакторами, где п — число реакторов, равное
числу фаз с определенной структурой системы клетка — пузырек — субстрат
(рис. 8.14). Найдите лимитирующие процесс сопротивления для каждой фазы
и обсудите (учитывая количественные аспекты), как можно осуществить мас-
штабный переход от лабораторной к промышленной установке, если рабочий
объем или потребляемая мощность, или другие параметры последней превос-
ходят соответствующие параметры лабораторной установки в 5000 раз.
9.17. Производство пенициллина. Приведенные на рис. 9У17.1 данные
относятся к процессу синтеза пенициллина в культуре Penicillium chrysoge-
пит. Эксперимент проводили в реакторе периодического действия с полным
перемешиванием объемом 10 л, при объемной скорости аэрации 0,2 объема
газа на объем ферментера в минуту. Начальный состав среды, в которой осу-
ществляется рост культуры и биосинтез пенициллина (в г/л): КН2РО4— 3,0;.
Na2SO4 —0,9; MgCl2 —0,25; MgSO4-H2O — 0,05; глюкоза—10; NH4C1 —2.
Кроме того, в ходе процесса в среду постоянно вводили глюкозу, а pH регу-
160
Глава 9
.лнровали добавлением NH4OH. Бензилпенициллин имеет брутто-формулу
C16H18N2O4S.
а) Объясните характер изменения концентраций биомассы, пенициллина,
глюкозы и NH3 в этом периодическом процессе.
б) Совершенно неожиданно биосинтез пенициллина резко прекратился,
хотя организм был способен синтезировать еще в 5 раз больше пенициллина,
чем было накоплено в системе к моменту прекращения синтеза. Как главного
консультанта компании Antibiotics Unlimited, Ltd., вас попросили найти при-
чину прекращения синтеза пенициллина.
9.18. Полупериодические процессы. В общем случае поддерживать низкую,
строго контролируемую скорость поступления питательных веществ в реактор
труднее, чем периодически добавлять субстрат, одновременно отбирая равный
объем культуральной жидкости. Допустим, что рост микроорганизмов лими-
тируется одним субстратом в соответствии с уравнением типа уравнения Моно
и что скорость клеточного роста пропорциональна скорости изменения кон-
центрации субстрата. Пусть процесс начинается при f=0 (т. е. продолжитель-
ность лаг-фазы пренебрежимо мала). В момент времени t из ферментера
(с объемом жидкой фазы V) отбирают объем Vi и добавляют точно такой же
объем свежей питательной среды с концентрацией субстрата Sf.
а) Покажите, что в момент времени t (до отбора УД концентрация суб-
страта s описывается уравнением:
1 +
(х0/Г) 4~ s0 _
1 — s/sQ 1 _ Ks Г_5
*o/(>%) J (x0/K) -Ho L sc
б) В момент времени t отбирают объем Vi, добавляют свежую питатель-
ную среду, после чего процесс возобновляется. Выразите «о, х0 через s, xt.
в) Покажите также, что эффективность утилизации субстрата описывает-
ся выражением
P = (Sf — S1)/sf= 1 -—(W/Vi) sfg>/As
1 —<р
где <р= V\/V.
г) Покажите, что при ф->0 эффективность утилизации субстрата стано-
вится такой же, как и в случае непрерывной культуры:
sf [рт (V/Vx) t - 1]
где Vt/Vi — обратная скорость разведения.
д) Покажите графически, что при Sf//G=l,0 и конечном значении Ф (при
периодическом добавлении свежей среды) параметр р больше, чем для про-
точного реактора, если (|XmW/Ui) >4.
9.19. Культура тканевых клеток в псевдоожиженном слое. Непрерывные
культуры тканевых клеток предлагалось выращивать в псевдоожиженном слое
на непористом гранулированном носителе (рис. 9У19.1). При этом питательная
среда движется снизу вверх, поддерживая носитель в суспендированном со-
стоянии, выходит из реактора через ситчатый фильтр, задерживающий грану-
лы носителя, и затем поступает в насос. Если разность плотностей гранул
носителя и воды невелика, то процесс может идти и при невысокой объемной
скорости жидкой фазы, и тогда насос не повреждает содержащиеся в ней
макромолекулы. Допустим, что жидкая фаза движется в режиме полного
вытеснения и что гранулы носителя полностью диспергированы во всем объеме
реактора. Гранулы, проходящие через выходную воронку, осаждаются на ней
и постоянно отбираются; выходящая из реактора жидкая фаза после фильтро-
вания вновь поступает в реактор.
Проектирование и расчет биологических реакторов 161
а) Допустим, что приведенные на рис. 9У19.2 данные отражают кинетику
клеточного роста на каждой грануле, причем р,=0 при />160 ч [вследствие
завершения образования монослоя клеток (контактное ингибирование)]. Най-
дите выражение, описывающее концентрацию биомассы, отнесенную к одной
грануле, на выходе из реактора через объем пустот в псевдоожиженном слое
е, скорость введения в реактор инокулированных гранул носителя (Z ч-1),
объем реактора VR и соответствующее уравнение скорости клеточного роста.
Подача, инокулированных
гранул со скоростью Z-
Насос
РИС. 9У19.1. Биореактор с псевдоожиженным слоем микроносителя для про-
цессов с участием культур клеток ткаией.
Примите, что начальная концентрация клеток на гранулах равна Со (в расчете
на одну гранулу).
б) Повторите решение предыдущего упражнения для случая, когда кон-
центрация клеток на поступающих в реактор частицах носителя характери-
зуется распределением типа:
Вероятность нахождения у клеток на одной грануле= Ае~^у~ут)2
где А — нормализующий фактор.
в) Предложите конструкцию системы, обеспечивающей «непрерывную»
подачу в реактор свежих гранул носителя и непрерывную регенерацию жидкой
среды в стерильных условиях.
9.20. Ингибирование роста культур тканевых клеток носителем. Экспе-
риментально показано, что твердые носители в высоких концентрациях
(см2/см3) могут ингибировать клеточный рост в силу адсорбции на поверх-
ностях этих носителей факторов роста и сывороточных белков из среды, при-
чем адсорбированные белки не могут взаимодействовать с клетками
\Horng С. В., McLimens W., Biotech. Bioeng., 17, 713 (1975)]. Рассмотрим
11—746
162
Глава 9
реактор с полным перемешиванием, содержащий культуру клеток тканей на
микроносителе, частицы которого равномерно распределены в жидкой фазе.
а) Найдите выражение, описывающее изменение общего количества био-
массы в системе во времени, если рост клеток подчиняется логарифмическому
закону (все питательные вещества присутствуют в избытке) до тех пор, пока
на всей доступной поверхности частиц носителя не образуется монослой кле-
ток (см. рис. 9У19.2).
РИС. 9У19.2. Изменение количества клеток и титра вируса во времени, и —
зависимость с//со от времени (с/ и со — концентрация связанных с носителем
клеток в момент времени t и при /=0 соответственно; ------- 75 %-ное насы-
щение О2;-------5%-ное насыщение О2); б — зависимость TCID50 от времен»
(TCID50 — доза вирусов, приводящая к 50%-ному заражению культуры тка-
ни в 1 мл; ----- начальная концентрация клеток 13-105 в 1 мл;---------на-
чальная концентрация клеток 3,5" 105 в мл). (Из статьи: van Wezel A. L.,
Microcarrier Cultures of Animal Cells, in Tissue Culture: Methods and Appli-
cations, p. 372, Academic Press, Inc., New York, 1973.)
б) Теперь допустим, что скорость роста клеток завнснт от концентрация
фактора роста sg, не расходующегося в процессе роста, н что зависимость
скорости клеточного роста от sg описывается уравнением типа уравнения
Моно. Выведите аналогичное уравнение для зависимости количества биомассы
от времени, если фактор роста обратимо адсорбируется на твердой поверх-
ности (и при этом обратимо инактивируется) и если процесс адсорбции опи-
сывается линейным уравнением:
Sg (ty = Ksg
в жидкой на твердой
фазе поверхности
в) Покажите, что при конечном времени существования периодической
культуры (например, 6 сут) существует оптимальная величина начальной
загрузки носителя в посевной материал культуры клеток ткани, если в конце
концов рост клеток прекращается из-за отсутствия свободной поверхности
носителя, Выведите уравнение, с помощью которого можно определить эту
величину.
г) Поскольку частицы носителя в известном смысле выполняют роль ин-
гибитора клеточного роста, казалось бы, в этом случае целесообразно вместо
Проектирование и расчет биологических реакторов
163
одного реактора использовать каскад реакторов. С какими трудностями столк-
нется экспериментатор, если ои приступит к изучению системы с каскадом
реакторов?
9.21. Деацилирование пенициллина V; реакции, протекающие по несколь-
ким направлениям. Ферментативное деацилирование пенициллина V (фенокси-
метилпенициллина) до 6-аминопенициллановой кислоты (6-АРА; гл. 12), яв-
ляющейся исходным веществом для получения полусинтетических пеницилли-
нов, включает следующие последовательные и параллельные реакции:
Г1
А-(пенициллин V) —> Р-(б-АРА)+<ЭОН(феноксиуксусная кислота)
Г2
А- —> R2- (неактивное вещество)+Н+
гз
р- —>- HS- (неактивный продукт расщепления кольца)
Ki
QOH QO- + H+
Р-+Н+ HP (устойчивое соединение)
Существует некоторое оптимальное для этого процесса значение pH, посколь-
ку протонирование Р_ приводит к стабильному соединению HP, однако даль-
нейшее повышение кислотности снижает скорость первой реакции гь
а) Допустим, что скорость первой реакции г\ можно описать уравнением
Михаэлиса — Ментен с учетом неконкурентного ингибирования протонами,
а гг и гз можно выразить через простые уравнения скорости необратимой
реакции первого порядка. Запишите эти уравнения, а также уравнения, опи-
сывающие скорости двух последних (предположительно обратимых) про-
цессов.
б) Выведите уравнение для стационарного режима в ПРПП с полной
рециркуляцией фермента, описывающее концентрацию А- в исходной смеси,
при которой обеспечивается максимальная степень превращения А- в Р-.
в) Для обеспечения очень высокой степени превращения дорогого исход-
ного вещества представляется целесообразным применение каскада из не-
скольких ПРПП. Обсудите преимущества и недостатки такой системы; приве-
дите соответствующие расчеты. Считайте, что величины г2 и г3 составляют при-
мерно 2—5% от скорости fi в первом реакторе. [Karlsen L. G., Villadsen J.,
Optimization of a Reactor Assembly for the Production of 6-APA from Penicil-
lin V; Biotech. Bioeng., 26, 1485 (1984).]
9.22. Производительность реактора с полыми волокнами. Реакторы раз-
личных типов резко отличаются друг от друга по плотности микроорганизмов
и производительности. Так, например, по сравнению с реактором с суспензион-
ной культурой в реакторе с полыми волокнами достигается в 5 раз меньшая
удельная 0-лактамазная активность:
Реактор Производительность, ед. фермента в 1 ч, в расчете на 1 клетку
Сосуд с перемешиванием (встряхиванием) Реактор с полыми волок- нами 1 • 10"10 2-10-11
11
164
Глава 9
а) Вычислите производительность реактора каждого типа (в единицах
ферментативной активности в объеме реактора за 1 ч), если концентрация
биомассы в реакторе с полыми волокнами в 1000 раз выше, чем в реакторе
с суспендированной культурой. Примите, что концентрация биомассы х в со-
суде с перемешиванием (встряхиванием) равна 109 клеток/мл.
б) Считается, что выход р-лактамазы несколько повышается за счет лизи-
са клеток в полых волокнах и секреции белка. Объясните, как бы вы опреде-
лили параметры кинетики лизиса клеток, чтобы затем с помощью этих пара-
метров описать процесс образования р-лактамазы путем секреции и лизиса.
[Inloes D. S. et al., Hollow Fiber Membrane Bioreactors Using Immobilized
E. coll for Protein Synthesis; Biotech. Bioeng., 25, 2653 (1983).]
9.23. Образование не связанного с клеточным ростом продукта жизне-
деятельности в периодическом процессе. Периодический микробиологический
процесс обычно включает фазы роста биомассы и образования продукта жиз-
недеятельности клеток, причем субстрат утилизируется в каждой из фаз.
Предположим, что рост периодической культуры описывается логистическим
уравнением, а хтах соответствует решению такого уравнения при начальной
концентрации второго субстрата зг (не используемого для образования про-
дукта метаболизма). Тогда для процесса образования не связанного с клеточ-
ным ростом продукта жизнедеятельности микроорганизмов (например, L-глу-
тамата в культуре Micrococcus glutamicus) можно записать
dx/d t=jix (1 — х/хт ах)
dp/dt=nx
ds/dt=—(dp/dt)/Yp — (dx/dt)/Yx
а) Путем интегрирования каждого из указанных уравнений найдите вы-
ражения, описывающие зависимости x(t), p(t) и s(t).
б) Обсудите, как вы смогли бы найти хтах, если вам необходимо добиться
максимальной величины р(0) или р(0)/Дз(0), где 0 — известное время окон-
чания процесса ферментации.
в) При каких обстоятельствах вы бы стали искать максимум р(0) и при
каких — максимум р(0)/Дз(9)?
Литература
Довольно много сведений о проектировании и анализе реакторов содер-
жится в литературе, приведенной в гл. 7.
Дополнительные данные можно найти в следующих источниках:
1. Levenspiel О., Chemical Reaction Engineering, 2d ed., John Wiley and Sons,.
Inc., New York, 1972. Эта книга, возможно, является лучшей монографией
по проблемам перемешивания и распределения времени пребывания; в та
же время здесь рассматриваются и многие другие вопросы проектирова-
ния реакторов. Имеется перевод первого издания этой книги: Левен-
шпиль О., Инженерное оформление химических процессов. — М.: Химия,
1969.
2. Smith J. М., Chemical Engineering Kinetics, 2d ed., McGraw-Hill Book Com-
pany, New York, 1970. Одна из наиболее популярных книг в этой области;
ее достоинством является множество конкретных примеров, рассмотренных
в применении к реальным реакторам.
3. Carberry J. J., Chemical and Catalytic Reaction Engineering, McGraw-Hill
Book Company, New York, 1976. Книга очень информативна; особое вни-
мание в ней уделяется гетерогенному катализу и многофазным реакторам.
4. Hill С. С., Jr., An Introduction to Chemical Engineering Kinetics and Reac-
tor Design, John Wiley and Sons, Inc., New York, 1977. Здесь хорошо пред-
Проектирование и расчет биологических реакторов
165
ставлены многие темы, начиная от проблем кинетики и кончая технологией
реакторов.
В перечисленных ниже источниках широко трактуются многие вопросы проек-
тирования и анализа биореакторов:
5. Аткинсон Б., Биологические реакторы. — М.: Пищевая промышленность,
1979.
6. Erickson L. Е„ Stephanopoulos G., Biological Reactors, Chap. 13 in Chemical
Reaction and Reactor Engineering, Carberry J. J., Varma A. (eds.), Marcel
Dekker, Inc., New York, 1985.
Влияние пристеночного роста на работу биологических реакторов рассмотрено
в статье:
7. Howell J. A., Chi С. Т., Pawlowsky U., Effect of Wall Groth on Scale-Up
Problems and Dynamic Operating Characteristics of the Biological Aerator;
Biotech. Bioeng., 14, 253 (1972).
Хорошие обзоры по проблеме масштабов времени и длины и ее роли в проек-
тировании реакторов даны в статьях:
8. Kossen N. W. F., Models in Bioreactor Design, p. 23 in Computer Applica-
tions in Fermentation Technology, Society of Chemical Industry, London,
1982.
9. Roels J. A., Mathematical Models and the Design of Biochemical Reactors;
J. Chem. Tech. Biotechnol., 32, 59 (1982).
Элементарная теория устойчивости и динамика реакторов рассмотрены
в статьях:
10. Amundson N. К., Aris К., An Analysis of Chemical Reactor Stability and
Control, Parts I—III; Chem. Eng. Sci., 7, 121 (1958).
11. Jeffreson С. P., Smith J. M., Stationary and Nonstationary Models of Bacte-
rial Kinetics in Well-Mixed Flow Reactors; Chem. Eng. Sci., 28, 629 (1973).
12. Ramkrishna D., Fredrickson A. G., Tsuchiya H. M., Dynamics of Microbial
Propagation: Models Considering Inhibitors and Variable Cell Composition;
Biotech. Bioeng., 9, 129 (1967).
13. Lee S. S., Jackman A. P., Schroeder E. D., A Two-State Microbial Growth
Kinetic Model; Water Res., 9, 491 (1975).
14. Young T. B., Bruley D. F., Bungay H. R., Ill, A Dynamic Mathematical
Model of the Chemostat; Biotech. Bioeng., 12, 747 (1970).
Проблемы, связанные с неидеальным перемешиванием в реакторах, освещают-
ся в следующих источниках:
15. Nagata S., Mixing: Principles and Applications, Wiley, New York, 1975.
16. Shinnar R., Residence Time and Contact Time Distributions in Chemical
Reactor Design, in Chemical Reaction and Reactor Engineering, Carberry J. J.,
Varma A. (eds.), Marcel Dekker, Inc., New York, 1985.
17. Villermaux J., Mixing in Chemical Reactors, p. 135 in Chemical Reaction
Engineering —Plenary Lectures, Wei J., Georgakis C. (eds.), American Che-
mical Society, Washington, D.C., 1983.
18. Charles M., Technical Aspects of the Rheological Properties of Microbial
Cultures, p. 1 in Advances in Biochemical Engineering, vol. 8, Ghose T. K.,
Fiechter A., Blakebrough N. (eds.), Springer-Verlag, New York. 1978.
19. Bryant J., The Characterization of Mixing in Fermenters, p. 101 in Advances
in Biochemical Engineering, vol. 5, Ghose T. K., Fiechter A., Blakebrough N.
(eds.), Springer-Verlag, New York, 1977.
166
Глава 9
20. Zweitering Th. N., The Degree of Mixing in Continuous Flow Systems;
Chem. Eng. Sci., 11, 1 (1959).
21. Popovic M., Papalexiou A., Reuss M., Gas Residence Time Distribution in
Stirred Tank Bioreactors; Chem. Eng. Sci., 38, 2015 (1983).
Модели нендеальных реакторов:
22. Sinclair C. G., Brown D. E., The Effect of Incomplete Mixing on the Analy-
sis of the Static Behavior of Continuous Cultures; Biotech. Bioeng., 12, 1001
(1970).
23. Gschwend K., Fiechter A., Widmer F., Oxygen Transfer in a Loop Reactor
for Viscous Non-Newtonian Biosystems; J. Ferment. Technol., 61, 491
(1983).
Взаимосвязь между перемешиванием и биореакцней:
24. Vardar F., Problems of Mass and Momentum Transfer in Large Fermen-
ters; Process Biochem., 18, 21 (1983).
25. Ujcovd E., Fend Z., Musi’lkovd M., Seichert L., Dependence of Release of
Nucleotides from Fungi on Fermentor Turbine Speed; Biotech. Bioeng., 22,
237 (1980).
26. van Suihdam J. C., Metz B., Influence of Engineering Variables upon the
Morphology of Filamentous Molds; Biotech. Bioeng., 23, 111 (1981).
27. Wang D. I. C„ Fewkes R. C. J., Effect of Operating and Geometric Para-
meters on the Behavior of Non-Newtonian, Mycelial, Antibiotic Fermenta-
tions; Devel. Ind. Microbiol., 18, 39 (1977).
28. Vardar F., Lilly M. D., Effect of Cycling Dissolved Oxygen Concentrations
on Product Formation in Penicillin Fermentations; European J. Appl. Micro-
biol. Biotechnol., 14, 203 (1982).
Вопросы теплопроводности в неустановившемся состоянии в твердых телах
рассмотрены в известной монографии:
29. Карслоу Г. С., Егер Д., Теплопроводность твердых тел. — М.: Наука,
1964.
Процессы и аппараты для тепловой обработки в пищевой промышленности
рассматриваются в монографии:
30. Charm S. Е., The Fundamentals of Food Engineering, 2d ed., Avi Publishing
Co., Inc., Westport, Conn., 1971.
Проблемы получения, изучения и применения биокатализаторов на основе
иммобилизованных клеток изложены в ссылках [14, 15] гл. 4, а также в сле-
дующих источниках:
31. Immobilized Microbial Cells, Venkatsubramanian К. (ed.), ACS Symposium
Series 106, American Chemical Society, Washington, D.C., 1979.
32. Jack T. R., Zajic J. E.t The Immobilization of Whole Cells, p. 125 in Advan-
ces in Biochemical Engineering, vol. 5, Ghose T. K-, Fiechter A., Blake-
brough N. (eds.), Springer-Verlag, New York, 1977.
33. Gbewonyo K., Wang D. I. C., Confining Mycelial Growth to Porous Micro-
beads. A Novel Technique to Alter the Morphology of Non-Newtonian Myce-
lial Cultures; Biotech. Bioeng., 25, 967 (1983).
34. Nagashima M., Azuma M., Noguchi S., Technology Developments in Bio-
mass Alcohol Production in Japan: Continuous Alcohol Production with
Immobilized Microbial Cells; Ann. N.Y. Acad. Sci., 413, 457 (1983).
35, Forberg C., Enfors S.-О., Haggstrbm L., Control of Immobilized, Non-Gro-
wing Cells for Continuous Production of Metabolites; Eur. J. Appl. Microbiol.
Biotechnol., 17, 143 (1983).
Проектирование и расчет биологических реакторов 167
Башеииые и эрлифтные реакторы:
36. Schugerl К., LUcke J., Oels U., Bubble Column Bioreactors (Tower Bioreac-
tors Without Mechanical Agitation), p. 1 in Advances in Biochemical Engi-
neering, vol. 7, Ghose T. K., Fiechter A., Blakebrough N. (eds.), Springer-
Verlag, New York, 1977.
37. Schugerl K., Characterization and Performance of Single- and Multistage
Tower Reactors with Outer Loop for Cell Mass Production, p. 93 in Advan-
ces in Biochemical Engineering, 22, Fiechter A. (ed.), Springer-Verlag, New
York, 1982.
38. Kiiai A., Tone H„ Ozaki A., Performance of a Perforated Plate Column as
a Multistage Continuous Fermentor; Biotech. Bioeng., 11, 911 (1969).
39. Prokop A. et al., Design and Physical Characteristics of a Multistage Con-
tinuous Tower Fermentor; Biotech. Bioeng., 11, 945 (1969).
40. Falch E. A., Gaden E. L., Jr., A Continuous, Multistage Tower Fermentor,
I: Design and Performance Tests; Biotech. Bioeng., 11, 927 (1969); II: Analy-
sis of Reactor Performance; ibid., 12, 465 (1970).
41. Ault R. G. et al., Biological and Biochemical Aspects of Tower Fermenta-
tion; J. Inst. Brewing, 75, 260 (1960).
42. Greenshields R. N., Smith E. L., Tower-Fermentation Systems and Their
Applications, Chem. Eng. London, May 1971, p. 182.
Реакторы с псевдоожиженным слоем, насадкой и струйным течением
жидкости:
43. Scott С. D., Fluidized-Bed Bioreactors Using a Flocculating Strain of Zymo-
monas mobilis for Ethanol Production; Ann. N.Y. Acad. Sci., 413, 448
(1983).
44. Flaschel E., Fauquex P.-F., Renken A., Zum Verhalten von Fhissigkeits-
Wirbelschichten mit Einbauten; Chem.-Ing.-Tech., 54, 54 (1982).
45. van de Vusse J. G., Wesselingh J. A., Multiphase Reactors, p. 561 in 4th
International/6th European Symposium on Chemical Reaction Engineering,
vol. 2, DECHEMA, Deutsche Gesellschaft fur Chemisches Apparatewesen,
e. V., Frankfurt, 1976.
46. Briffaud J., Engasser J., Citric Acid Production from Glucose. II. Growth and
Excretion Kinetics in a Trickle-Flow Fermentor; Biotech. Bioeng., 21, 2093
(1979).
Технология микробиологических процессов:
47. Microbial Technology, Peppier H. J., Perlman D. (eds.), 2d ed., vols. I and
II, Academic Press, Inc., New York, 1979. Необычно обширный сборник
обзорных статей по технологии, продуктам и применению процессов мик-
робиологической технологии.
48. Casida L. Е., Jr., Industrial Microbiology, John Wiley and Sons, Inc.. New
York, 1968. Хороший обзор продуктов, процессов и методов микробиоло-
гической промышленности, в том числе проблем патентоведения и эконо-
мики. Многочисленные иллюстрации дают достаточно полное представле-
ние о практической стороне этой отрасли промышленности.
49. Уэбб Ф. Ч., Биохимическая технология и микробиологический синтез.—
М.: Медицина, 1969. В этой монографии обсуждены проблемы проектиро-
вания аппаратов для микробиологической промышленности, рассмотрены
получаемые в них продукты. Особенностью этой книги является анализ
ряда других важных тем, в том числе посвященных коллоидным систе-
мам, эмульсиям, редокс-потенциалам, химической дезинфекции, дегидра-
тации, радиации и производству вакцин.
50. Schhgerl К., New Bioreactors for Aerobic Processes; Int. Chem. Eng 22
591 (1982).
168
Глава 9
Культуры животных клеток:
51. Feder J., Tolbert W. R., The Large-Scale Production of Mammalian Cells;
Scientific American, 248, 36 (1983).
52. Tolbert №. R., Schoenfeld R. A, Lewis C., Feder J., Large-Scale Mammalian
Cell Culture: Design and Use of an Economical Batch Suspension System;
Biotech. Bioeng., 24, 1671 (1982).
53. Fiechter A., Batch and Continuous Culture of Microbial, Plant and Animal
Cells, p. 453 in Biotechnology, vol. 1, Rehm H.-J., Reed G. (eds.), Veriag
Chemie, Weinheim, 1982.
54. Katinger H. W. £>., Scheirer W., Status and Development of Animal Cell
Technology using Suspension Culture Techniques; Acta Biotechnologica, 2
3 (1982).
55. Glacken M. W., Fleischaker R. J., Sinskey A. J., Large-scale Production of
Mammalian Cell and Their Products: Engineering Principles and Barriers to
Scale-up; Ann. N.Y. Acad. Sci., 413, 355 (1983).
56. Kruse P. F., Jr., Patterson M. K., Jr., Tissue Culture: Methods and Appli-
cation, Academic Press, Inc., 1973.
57. Johnson I. S., Boder G. B., Metabolites from Animal and Plant Cell Culture;
Adv. Microbiol., 15, 215 (1973).
58. Cell Culture and its Application, Acton R., Lynn J. D. (eds.), Academic
Press, New York, 1977.
59. Acton R. T., Lynn J. D., Description and Operation of a Large-Scale, Мань
malian Cell, Suspension Culture Facility, in Advances in Biochemical Engi-
neering, vol. 7, Ghose T. K-, Fiechter A., Blakebrough N. (eds.), Springer-
Verlag, New York, 1977.
60. Lim F., Moss R. D., Microencapsulation of Living Cells and Tissues; J.
Pharm. Sci, 70, 351 (1981).
61. Ku K-, Kuo M. J., Delente J., Wildi B. S, Feder J., Development of a Hollow-
Fiber System for Large-Scale Culture of Mammalian Cells; Biotech. Bioeng,
23, 79 (1981).
62. Scattergood E. M., Schlabach A. J., McAleer W. J., Hilleman M. R., Scale-Up
of Chick Cell Growth on Microcarriers in Fermenters for Vaccine Production;
Ann. N. Y. Acad. Sci, 413, 332 (1983).
63. White R. J., Klein F., Chen J. A, Stroshane R. M., Large-Scale Production
of Human Interferons, in Annual Reports on Fermentation Processes, vol. 4,
Tsao G. T. (ed.), Academic Press, 1980.
Культуры растительных клеток:
64. Tissue Culture and Plant Science, Street H. E. (ed.), Academic Press, Lon-
don, 1974.
65. Advances in Biochemical Engineering, Fiechter A. (ed.), vols. 16, 18, Sprin-
ger-Verlag, Berlin, 1980.
66. Wilson G„ Continuous Culture of Plant Cells Using the Chemostat Principle,
p. 1 in Advances in Biochemical Engineering, vol. 16, Fiechter A. (ed.),
Springer-Verlag, New York, 1980.
67 Shuler M L., Sahai О. P„ Hallsby G. A, Entrapped Plant Cell Tissue Cultu-
res; Ann. N.Y. Acad. Sci, 413, 373 (1983).
68. Brodelius P., Production of Biochemicals with Immobilized Plant Cells:
Possibilities and Problems; Ann. N.Y. Acad. Sci, 413, 383 (1983).
69. Curtin M. E., Harvesting Profitable Products from Plant Tissue Culture;
Bio/Technology, 1, 649 (1983).
Глава 10
Контрольно-измерительная аппаратура
и управление процессами
биохимической технологии
В предыдущих главах мы уже неоднократно имели возмож-
ность убедиться в том, что активность и полезное время жизни
ферментного катализатора или популяции клеток непосредствен-
но зависят от их окружения. Следовательно, для проектирова-
ния, оптимизации и наиболее эффективной эксплуатации био-
реакторов необходимо сначала изучить влияние окружения на
свойства катализатора, а затем контролировать и регулировать
параметры окружения. Эту цель в свою очередь можно достичь
путем определения параметров среды, анализа полученных
данных и соответствующего управления параметрами. В настоя-
щей главе мы опишем имеющуюся в настоящее время контроль-
но-измерительную аппаратуру для биореакторов и ее работу,
а затем после беглого рассмотрения быстро развивающихся
методов регистрации и расчета данных ознакомимся с некото-
рыми способами их анализа и управления параметрами процес-
са. Хотя в этой главе основное внимание уделяется контроль-
но-измерительной аппаратуре и регулированию процессов,
протекающих в биореакторах, рассматриваемые в ней общие
принципы приложимы и к другим процессам, в том числе к
операциям получения исходных веществ.
10.1. Детекторы для определения физических
и химических параметров среды и газов
На рис. 10.1 представлена одна из последних схем прибор-
ного оснащения биохимического реактора. Она предназначена в
основном для определения химического состава и физических
свойств среды и газовой фазы и лишь очень немногих особен-
ностей популяции клеток. Следовательно, при анализе резуль-
татов определения параметров системы в биореакторе жела-
тельно (или даже необходимо) оценить особенности популяции
клеток по доступным данным физико-химического анализа
газовой фазы и среды. В этом разделе мы сконцентрируем
внимание на неавтономной аппаратуре, работающей в реальном
масштабе времени и предназначенной для контроля физических
и химических параметров системы в биореакторе.
170
Глава 10
Мощность перемешивающего устройства
Торсионный динамометр (внешний}
Тензодатчик (внутренний}
Скорость добавления жидких
. исходных веществ FL
Счетчик капель
Динамометрический датчик '
Электромагнитный расходомер
Давление
Обычный
мембранный''-^
Обнаружение пены манометр
По электрической емкости
По электропроводности ----
Регулирование пенообразования
Механические устройства
Силиконовый эмульсион-
ный пеногаситель
(при применении масел
возникают проблемы
сохранения стерильности}
Температура
Ртутный термометр
Термопара Терморезисторы
Мощность |
мешалки
Скорость вращения мешалки
Тахометр общего типа (при
увеличении объема реактора число
м'оборотов мешалка в минуту уменыиа- -
м ется, но окружная скорость лопасти
мешалки ли остается постоянной}
Окислительно-восстановительный
pH потенциал
Стеклянный Сочетание платинового и стан-
электрод дрртного электрода (результаты
ерений трудно интерпрети-
ровать )
Влияние растворенного кислорода
Растворенный кислород
Полярографический стерилизу-
емый электрод
02+4Н+ + 4р" ->ZHzO
Вязкость
Ет Рог, Рсог
У
Ньютоновская
ньютоновская
Выход продукта
Слив, регулирование уровня,
динамометрический
датчик
Скорость потока газа Fg pQz
Ротаметры Парамагнитное определение 02
Массовые расходомеры Электрохимическая ячейка
рсог
ИН- анализатор
Мутность
Спектрофотометр
Измерение количества
биомассы (при невысокой
плотности}
Детекторы, находящиеся в стадии
разработка
Глюкоза
Этанол
NHj, Mgz+, К+, Na*, Сиг+, РО^
ATP, ADP, АМР
ДНК, РНК
NADH
РИС. 10.1. Схема приборного оснащения биохимического реактора. [Воспроиз-
ведено с разрешения из работы: Erickson L. Е., Stephanopoulos G., Biological
Reactors, in Chemical Reaction and Reactor Engineering, Carberry J. J., Var-
ma A. (eds.), chap. 13, Marcel Dekker, Inc., New York, 1985.]
Контрольно-измерительная аппаратура и управление
171
10.1.1. Детекторы для определения физических свойств среды
и газов
Из параметров, влияющих на жизнедеятельность клеток и
экономичность биопроцесса, в ходе процесса можно непрерывно
определять температуру, давление, мощность, расходуемую на
перемешивание, скорость вращения мешалки, вязкость бульона,
объемные скорости газовой и жидкой фаз, пенообразование,
объем или массу содержимого реактора. На небольших лабора-
торных установках обычно измеряют только температуру и
объемную скорость воздуха. Определение и регулирование
давления более типично для больших ферментеров.
В качестве датчика температуры чаще всего применяют
терморезисторы — полупроводниковые устройства, электричес-
кое сопротивление которых зависит от температуры. Хотя зави-
симость сопротивления терморезистора от температуры носит
нелинейный характер, это обстоятельство не является серьез-
ным препятствием в узком температурном интервале (25—
45°C), в котором проходит подавляющее большинство микро-
биологических процессов. В качестве датчиков температуры
также используются платиновые термометры сопротивления,
ртутные термометры в стальном чехле и термопары.
Контроль давления особенно важен при стерилизации, когда
в реакторе необходимо поддерживать небольшое избыточное
давление (около 1,2 атм), чтобы предотвратить заражение
системы. В микробиологических реакторах давление обычно
контролируют с помощью мембранных манометров, дающих
пневматический сигнал, который может быть преобразован в
электрический.
Для измерения мощности, расходуемой реактором с механи-
ческим перемешиванием, применяются различные методы.
С помощью ваттметра, работающего на принципе эффекта
Холла, измеряется общая энергия на якоре электродвигателя,
потребляемая перемешивающим устройством. Для определения
мощности можно также использовать торсионный динамометр,
установленный на валу мешалки. Как первый, так и второй
метод измерения позволяют определять только суммарную
мощность, куда входят также потери на подшипниках и уплот-
нениях. Изучение перемешивания в 270-литровом ферментере
с рабочим объемом 200 л показало, например, что 30% энергии
двигателя расходуется на преодоление трения в соединитель-
ных и герметизирующих деталях между двигателем и частью
вала мешалки в реакторе. Показано также, что доля таких
потерь возрастает при увеличении скорости вращения мешалки.
Непосредственно определить мощность, передаваемую мешал-
172
Глава 10
кой реакционной массе, можно с помощью балансированных
тензодатчиков, установленных на валу мешалки в реакторе.
Устройства для непрерывного измерения (в процессе рабо-
ты) вязкости и других реологических характеристик культураль-
ного бульона пока еще не разработаны. Одно из возможных
решений заключается в измерении мощности, потребляемой при
нескольких скоростях вращения мешалки. Предлагался также
динамический метод, в котором потребляемую мешалкой мощ-
ность измеряют во время и после короткого (продолжительно-
стью менее 30 с) отключения питания электродвигателя, вра-
щающего вал мешалки. Как показано на рис. 10.1 (и подтверж-
дено экспериментально), ньютоновские и неньютоновские
жидкости в течение этого короткого нестационарного периода
характеризуются различным поведением.
Существует несколько типов приборов для измерения скоро-
стей газовых потоков (поступающего в реактор воздуха или
отходящих газов). На простейшем из них — расходомере с
переменной площадью проходного сечения (ротаметре) можно
считывать показания непосредственно или преобразовывать
их с помощью соответствующего устройства в электрический
сигнал. В последнее время все большую популярность приобре-
тают термические массовые расходомеры, особенно в лабора-
торных и пилотных установках. В этих приборах поток газа
проходит через нагретый участок трубки, разность температур
в сечении которого зависит от массовой скорости потока.
Точность этих приборов составляет около 1 % от измеряемого
диапазона; их целесообразно применять при скоростях потока
ниже 500 л/мин. Имеются также устройства для определения
скорости ламинарных потоков, которые измеряют перепады
давления на матричном устройстве, разделяющем общий поток
на множество параллельных капиллярных потоков. Важность
определения скоростей газовых потоков определяется тем об-
стоятельством, что эти параметры часто применяются при рас-
четах материальных балансов (разд. 10.5).
Скорость потока жидкостей можно контролировать электро-
магнитными расходомерами, но эти приборы не находят широко-
го применения из-за их высокой стоимости. Иногда, особенно
в лабораторных установках, необходимую скорость потока
жидкости устанавливают с помощью дозиметра или тщательно
откалиброванного насоса. В другом варианте жидкость вводят
в реактор порциями строго определенного объема или массы.
Длительный контроль скорости поступающего в реактор потока
можно осуществлять посредством непрерывного взвешивания
заполненного реактора с помощью тензометра (в случае реак-
торов объемом более 250 л) или весов (в случае реакторов
меньшего объема). Для контроля за количеством находящейся
Контрольно-измерительная аппаратура и управление
173
в реакторе жидкой фазы мож-
но также использовать датчик
уровня жидкости, измеряю-
щий электрическую емкость
на заданном уровне в реакто-
ре. Аналогичные зонды, дейст-
вие которых основано на из-
мерении электрической емкос-
ти или электропроводности,
можно применять и для конт-
роля пенообразования на по-
верхности находящейся в ре-
акторе жидкой фазы. Иногда
измерение осуществляют в
циркуляционной линии (см.
ниже), которая служит для
отделения продукта процесса,
рециркуляции клеток, для теп-
ло- или газообмена (см. гл. 9
и 14). В таких случаях опре-
деление скорости потока жид-
кой фазы могут затруднять
суспендированные нераствори-
мые вещества, а также не-
постоянство реологических
свойств бульона.
10.1.2. Детекторы для
определения
химического
состава среды
В настоящее время разра-
ботаны электроды для опреде-
ления pH, окислительно-вос-
становительного потенциала
(Ен), парциального давления
Электролит
солевого
мостика
Отверстие
для ввода
электролита
сравнения
---Навинчива-
ющийся
колпачок
Герметичные
[J-g пробки
Отверстие для
ввода электролита
солевого мостика
Электролит
сравнения
Элемент
сравнения
-Диафрагма
- Стартовый
элемент
Мембрана
растворенного кислорода и
СОг, которые можно подвер-
гать неоднократной стерили-
зации паром на месте уста-
новки. Чаще других применя-
ют pH-электроды, отличаю-
РИС. 10.2. Комбинированный элект-
род типа Ingold 465 для определения
pH, выдерживающий стерилизацию
паром в автоклаве. (С любезного
разрешения Ingold Electrodes Inc.)
щиеся высокой надежностью; обычно в качестве рН-электрода
применяют стеклянный мембранный электрод. На рис. 10.2
изображено устройство pH-электрода, предназначенного для
174
Глава 10
контроля процесса стерилизации в автоклаве. Электроды, сте-
рилизуемые на месте установки, должны быть заключены в че-
хол, защищающий их от повышения давления. Окислительно-
восстановительный потенциал среды можно измерять платино-
вым электродом в сочетании с электродом сравнения. Имеются
и комбинированные зонды для определения pH и окислитель-
но-восстановительного потенциала. Неоднократно показано, что
pH влияет на кинетику биохимических превращений, поэтому
необходимость измерения pH очевидна. Напротив, интерпретация
результатов определения окислительно-восстановительных по-
тенциалов и выяснение взаимосвязи между окислительно-вос-
становительным потенциалом и активностью клеток могут вы-
зывать затруднения. Одной из перспективных областей приме-
нения электродов для измерения окислительно-восстановитель-
ных потенциалов является определение низких концентраций
кислорода (<1 млн-1) в анаэробных микробиологических про-
цессах (—450 mB<£\<—150 мВ), когда скорость образования
продукта может быть очень чувствительной к изменению Eh.
Для определения концентрации растворенного кислорода
применяют гальванические (потенциометрические) или поляро-
графические (амперометрические или электрод Кларка) зонды.
По сути дела, эти электроды измеряют не концентрацию раство-
ренного кислорода, а его парциальное давление (или актив-
ность). И в первом, и во втором зонде собственно электрод
обычно отделен от среды мембраной (рис. 10.3), а на поверхно-
сти катода осуществляется восстановление кислорода
Катод:
Pt
‘ЛОг+НгО+ге- 2ОН" (10.1 >
Реакция на аноде в гальваническом электроде:
Анод (гальванический электрод):
РЬ РЬ2++2е- (Ю.2>
Катод и анод образуют гальванический элемент, напряжение
которого зависит от количества кислорода, достигающего по-
верхности электрода. В кислородном электроде полярографиче-
ского типа на катоде также происходит реакция (10.1), а на
аноде осуществляется другой процесс
Анод (полярографический):
Ag+CP —* AgCl + e- (Ю.3>
Между катодом и анодом устанавливают постоянное напряже-
ние и измеряют силу тока, которая зависит от притока кислоро-
да к катоду. Недостатком как первого, так и второго электрода
является дрейф сигнала (силы тока или напряжения), обуслов-
ленный накоплением гидроксильных ионов или ионов металла»
Контрольно-измерительная аппаратура и управление
175
отверстие
Анод
Изоляция
- Кольцо
Платиновая
проволока
Платиновый
катод
Раствор
-электролита
РИС. 10.3. Устройство и основные элементы электрохимического зонда для
определения парциального давления растворенного кислорода. (Воспроизве-
дено с разрешения из работы: Wang N. S., Stephanopoulos G., Computer Appli-
cation to Fermentation Processes, CRC Critical Reviews in Biotechnology, vol. 2,
p. 1. © CRC Press, Inc., 1974.)
Мембрана
или истощением Cl-. Дрейфу способствует также загрязнение
наружной поверхности мембраны.
В стационарном состоянии приток кислорода к катоду зави-
сит от ряда процессов переноса, включая перенос кислорода из
жидкой фазы к наружной поверхности мембраны, диффузию
через мембрану и, наконец, диффузию через раствор электро-
лита к поверхности катода, где реакция происходит практически
мгновенно. Если общая скорость переноса, а следовательно, и
концентрация кислорода вблизи катода лимитируется первой
стадией, то сигнал электрода будет зависеть от свойств жидкой
фазы (например, от ее вязкости) и локальных гидродинамиче-
ских условий вблизи электрода. По этой причине, в частности,
рекомендовалось, чтобы скорость движения жидкости вблизи
176
Глава 10
полярографического электрода была не менее 0,55 м/с. Чув-
ствительность сигнала электрода к переносу через наружный
граничный слой можно снизить за счет применения менее про-
ницаемой мембраны. (Почему?) Однако в этом случае увели-
чивается запаздывание отклика электрода на изменение пар-
циального давления кислорода, которое в случае мембранных
электродов и без того довольно велико (10—100 с). В то же
время, как показано в приведенном ниже примере, такие элект-
роды можно применять и для изучения процессов массопереноса
в биореакторах в переходном состоянии, если при анализе
данных учитывать диффузию через мембрану электрода.
Пример 10.1. Электрохимическое определение kia. В аэрируемом с посто-
янной скоростью реакторе периодического или непрерывного действия уста-
навливают кислородный электрод. После того, как отклик электрода достиг-
нет некоторого постоянного уровня, ток кислорода резко прерывают и сразу
же с такой же скоростью начинают пропускать азот, который постепенно вы-
тесняет кислород из среды. В этом случае изменение напряжения на электро-
де во времени можно описать уравнением [11]
Е (О = Ео
т1/2 ехр ( — р/)
sin т1/2
СО
П = 1
( — l)rt ехр ( — £)o„t / L?)
1 — п2л2/т
(10П1.1>
Здесь T = pL2/S5o2; E(t) —напряжение на электроде в момент времени Ео —
напряжение на электроде в нулевое время; Fs— скорость потока газа; V —
объем жидкости в реакторе; М — константа в уравнении закона Генри; L —
толщина мембраны электрода; —коэффициент диффузии О2 через мем-
брану электрода; 2 = Pm/L, где Рт— проницаемость мембраны.
1 1 VRT
— = постоянная времени системы (реактора) — —----+ ~ ~~ (10П1.2)
Р kia FgM
Если постоянная времени системы велика (т. е. kia мало), то величину р опре-
деляют по напряжению на электроде при времени, значительно превышающем
время отклика электрода; в этом случае вклад второго слагаемого в правой
части уравнения (10П1.1) пренебрежимо мал. Большие значения kia (50—
500 ч-1) Вернан и Уилке [11] предложили определять по наклону касатель-
ной к кривой зависимости отклика электрода от времени в точке ее перегиба
(где вторая производная Е по t равна нулю). Как было показано, этот метод
обеспечивает такую же точность результатов, как и подбор эмпирической
кривой с помощью вычислительных машин во всем диапазоне E(t). При
непостоянном времени релаксации электрода обратное время релаксации мож-
но определить по графику зависимости этого параметра от наклона касатель-
ной к кривой E(f) в точке перегиба.
Далее искомую величину kia находят в три этапа:
1. Определяют наклон касательной к кривой зависимости отклика Е от t
в точке перегиба (рис. 10П1.1).
2. По обобщенной графической зависимости, описываемой уравнением
(10П1.2), (рис. 10П1.2), определяют [} для данных величин наклона каса-
тельной к кривой и постоянной времени электрода.
Контрольно-измерительная аппаратура и управление 177
3. По приведенному выше определению постоянной времени системы (1/$)
находят kia.
Очевидно, что полезность этого метода, особенно в случае больших величин
kia, определяется той точностью, с которой может быть найден отклик электрода.-
При высоких концентрациях растворенных веществ могут изменяться рас-
творимость кислорода (табл. 8.1), коэффициент диффузии кислорода и харак-
теристики самого электрода, в том числе и его отклик. Для того чтобы учесть-
каждый из этих факторов, необходима калибровка электрода с помощью неза-
висимых методов определения концентрации растворенного кислорода.
Времfi от начала ступенчатого
изменения условий, с
РИС. 10П1.1. Наклон кривой в точке-
перегиба определяют по графику за-
висимости отклика электрода на за-
мену барботируемого кислородсодер-
жащего газа азотом. [Воспроизведе-
но из статьи: Wernan W. С., Wil-
ke С. R., New iMethod for Evaluation
of Dissolved Oxygen Response for kLa
Determination; Biotech. Bioeng., 15T
571 (1973).]
Выдерживающие стерилизацию паром электрохимические
детекторы для определения парциального давления растворен-
ного СО2 применяются сравнительно недавно. В таких детекто-
рах, выпускаемых, в частности, компанией Ingold Electrodes
Inc., определение рсо2 основано на измерении pH стандартного
раствора бикарбоната, отделенного от изучаемой жидкости газо-
проницаемой мембраной. Детекторы калибруют посредством
замены раствора бикарбоната на стандартный буферный раст-
вор с последующим определением pH.
В другом методе неавтономного определения летучих компо-
нентов среды и растворенных газов в изучаемую жидкость
погружают трубку известной длины, стенки которой проницаемы
для определяемого компонента системы. Через трубку непре-
рывно пропускают поток газа-носителя, который уносит прони-
кающие внутрь трубки компоненты в газоанализатор (см.
следующий раздел), где они и определяются. Недостатком
такого подхода является существенное запаздывание (2—
10мин) отклика анализатора, что делает его мало пригодным
для контроля быстрых изменений концентраций.
Для определения специфических компонентов жидкой фазы
разработан ряд биодетекторов (биосенсоров). Принцип действия
12—74 6
178
Глава 10
РИС. 10П1.2. Эти графики позволяют определить величину по наклону кри-
вой в точке перегиба и параметрам транспорта через мембрану. Затем по
уравнению (10П1.2) находят kia. [Из статьи: Wernan W. С., Wilke С. R., New
Method for Evaluation of Dissolved Oxygen Response for kLa Determination;
Biotech. Bioeng., 15, 571 (1973).]
биосенсоров заключается в определении с помощью того или
иного аналитического устройства конкретного продукта реакции,
катализируемой иммобилизованными ферментом или клетками.
Примеры сочетания катализируемых ферментами реакций с
электрохимическими детекторами мы уже рассматривали в
разд. 4.3.3. Широко изучаются также ферментные терморези-
сторы, в которых с помощью калориметра определяется
количество теплоты, выделяющейся в процессе катализируемой
ферментом реакции. В табл. 10.1 перечислены некоторые опре-
делявшиеся этим методом соединения и соответствующие
Контрольно-измерительная аппаратура и управление
17?
Таблица 10.1. Примеры применения ферментных терморезисторов
в аналитической химии3
Определяемое соединение Иммобилизованный фермент Диапазон опреде- ляемых концентра- ций или предел обнаружения, ммоль/л
Аскорбиновая кислота Оксидаза аскорбиновой кисло- ты 0,05—0,6
Альбумин (антиген) Иммобилизованные антитела и связанный с ферментом ан- тиген 10-ю
АТР Апираза или гексокиназа 1—8
Целлобиоза ^-Глюкозидаза, глюкозооксида- за и каталаза 0,05—5
Цефалоспорин Цефалоспориназа 0,005—10
Холестерин Холестерпиоксидаза 0,03—0,15
Сложные эфиры холе- Холестеринэстераза и холесте- 0,03—0,15
стерина риноксидаза
Креатинин Креатининиминогидролаза 0,01—10
Этанол Алкогольоксидаза 0,01—1
Галактоза Г алактозооксидаза 0,01—1
Гентамицин (антиген) Иммобилизованные антитела и связанный с ферментом антиген 0,1 мкг/мл
Глюкоза Глюкозооксидаза и каталаза 0,002—0,8
Ионы тяжелых металлов (например, РЬ2+) Уреаза 10"в
Инсектициды (например, паратион) Ацетилхолинэстераза 5-Ю-3
Инсулин (антиген) Иммобилизованные антитела и связанный с ферментом антиген 0,1—1,0 ед./мл
Лактат Лактат-2-монооксигеназа 0,01 — 1
Лактоза Лактаза, глюкозооксидаза и каталаза 0,05—10
Щавелевая кислота Оксалатоксидаза 0,005—0,5
Пенициллин G Пенициллиназа 0,01—500
Фенол (субстрат) Тирозиназа 0,01—1
Сахароза Инвертаза 0,05—100
Триглицериды Липаза, липопротеин Уреаза 0,1—5
Мочевина 0,01—500
Мочевая кислота Уриказа 0,5—4
а По данным, приведенным в работе: Danielsson В., Mosbach, К., The Prospects for
Enzyme-Coupled Probes in Fermentation, in Computer Application in Fermentation Techno-
logy. P- 137, Society of Chemical Industry, London, 1982.
ферменты. Иммобилизованные ферменты можно применять и в
биосенсорах других типов, например в таких, в которых продук-
ты реакции (водород) изменяют электронную проводимость
полупроводниковых устройств (кремниевых кристаллов, на ко-
торые нанесен слой SiO2 и пленка Pd).
12
180
Глава 10
Сферу применения биосенсоров можно существенно расши-
рить за счет использования различных биохимических превраще-
ний, в том числе многостадийных и разветвленных реакций,
в результате которых образуется детектируемое вещество. Во
многих случаях с помощью иммобилизованных клеток анализи-
руемое соединение можно превратить в более подходящее для
непосредственного определения соединение. В основу разработ-
ки и применения биосенсоров с иммобилизованными клетками
была положена дыхательная активность интактных иммобили-
Таблица 10.2. Микробиологические детекторы для определения
компонентов жидкой фазы
Определяе- мое соеди- нение Микроорганизм ,, Принцип из- Метод иммобилизации мерения Детектирую- щее устрой- ство
Уксусная Trichosporon Связывание с пори- Одновремен- Кислород-
кислота, brassicae стой ацетилцеллюло- ное погло- ный элект-
этанол зой щение О2 род
Аммиак Nitrosomonas Удерживание газо- Одновремен- Кислород-
sp., Nitrobac- проницаемой мембра- ное погло- ный элект-
ter sp. ной щение О2 род
Цефало- Citrobacter Включение в колла- Образова- рН-Элект-
спорин freundii геновую мембрану ние Н+ [род
Муравьи- Clostridium Включение в агар на Образова- Топливный
пая кислота butyricum ацетилцеллюлозном ние Н2 электрод
фильтрате
Глюкоза Pseudomonas Включение в колла- Одновре- Кислород-
fluorescens геновую мембрану менное пог- ный элект-
лощение О2 род
Глутамино- Escherichia coli Удерживание целло- Образова- СО2-элект-
вая кислота фановой мембраной ние СО2 [род
зованных клеток (определение с помощью кислородного элект-
рода), а также образование или поглощение интактными
клетками электроактивных метаболитов (определение с помо-
щью топливного электрода, pH- или СО2-электродов). В табл.
10.2 приведены свойства ряда биосенсоров на основе иммобили-
зованных клеток; подробнее подобные аналитические устройства
описаны в работе [12].
Шульц и сотрудники [13] разработали и описали другой
интересный и перспективный подход к определению индивиду-
альных метаболитов, основанный на применении специфичных
«аффинных сенсоров». Для реализации этого подхода необходи-
мы агент, специфично связывающий определяемое соединение;
меченое соединение, конкурентно связываемое тем же самым
агентом; метод выделения связывающего агента из анализируе-
мого раствора. Так, например, для определения глюкозы с по-
мощью иммобилизованного конкопавалина А (Con-А) [белка
Контрольно-измерительная аппаратура и управление 181
из канавалии мечевидной (растения семейства бобовых), селек-
тивно связывающего сахара] создан флуоресцентный детектор
на основе волоконной оптики, в котором Con-А иммобилизован
на внутренней поверхности аналитической ячейки. Последняя
отделена от анализируемого раствора диализационной мембра-
ной, проницаемой для глюкозы, и содержит также декстран,
меченный флуорохромом — флуоресцеинизотиоцианатом (ФИЦ).
Мембрана непроницаема для ФИЦ-декстрана, который конкури-
рует с глюкозой за связывание с Соп-А:
Глюкоза+Соп-А Соп-А-глюкоза
ФИЦ-декстран+Con-А Соп-А-ФИЦ-декстран
Таким образом, количество несвязанного ФИЦ-декстрана, а
следовательно, и интенсивность флуоресценции зависят от кон-
центрации глюкозы в растворе. Основной недостаток этого
метода обусловлен возможностью взаимодействия связываю-
щего агента с другими сахарами (например, с мальтозой, са-
харозой и фруктозой в случае Con-А), но принцип этого инте-
ресного подхода в силу его общности, безусловно, заслуживает
дальнейшего изучения.
При использовании любых сенсоров на основе клеток, фер-
ментов или других объектов биологического происхождения
необходимо принимать во внимание также возможность
инактивации биосенсора в ходе стерилизации реактора. Для
решения этой проблемы были сконструированы механические
устройства, позволяющие вводить биосенсор в реактор и выво-
дить из него в асептических условиях. Как и в случае других
датчиков, сигнал которых зависит от транспорта определяемого
соединения через мембрану, загрязнение мембраны биосенсоров
клетками или иными компонентами среды и увеличение сопро-
тивления внешнему массопереносу может вызвать дрейф сиг-
нала или изменение калибровки биосенсора.
10.1.3 . Газовый анализ
Концентрация СО2 в отходящих газах биореактора, содер-
жащего культуру клеток, связана с дыхательной или иной
ферментативной активностью клеток. Неудивительно, что этот
параметр широко и успешно применяется для контроля и регу-
лирования биореакторов. Концентрацию СОг в отходящих газах
биореактора чаще всего определяют с помощью инфракрасного
спектрофотометра. Для предотвращения повреждений стекол
ячейки анализируемый газ до поступления в спектрофотометр
необходимо высушить. Концентрацию СОг в потоке газа можно
также определять по теплопроводности, методом газовой хрома-
тографии или масс-спектрометрии.
182
Глава 10
Парциальное давление кислорода в газовом потоке обычно
измеряют с помощью парамагнитного анализатора. Приборы
этого типа также требуют предварительного высушивания и
тщательного регулирования скорости газового потока; только
в этом случае обеспечиваются минимальный дрейф и достаточ-
ная надежность результатов. Парамагнитные анализаторы,,
кроме того, очень чувствительны даже к небольшим колебани-
ям общего атмосферного давления, поэтому необходим одно-
временный контроль барометрического давления, чтобы внести,
соответствующие поправки в результаты анализа на кислород.
В силу дрейфа показаний парамагнитных анализаторов их
применение в микробиологических процессах часто требует
повторной калибровки в ходе процесса.
С помощью газовой хроматографии можно определять не-
сколько компонентов отходящих газов: Ог, СО2, СН4 (образу-
ющегося, например, в анаэробных процессах) и Н2 (выделяемо-
го, например, культурами Hydrogenomonas). Этим методом
можно также определять парциальное давление летучих ком-
понентов газовой фазы — этанола, ацетальдегида и карбоновых
кислот. Таким образом, газовая хроматография дает ценную
информацию о ходе микробиологического процесса в целом,
а также о концентрациях перечисленных выше веществ в жид-
кой фазе. Применению газовой хроматографии для контроля
процессов в нестационарном состоянии препятствует периодиче-
ский характер измерений, поскольку интервал между двумя
последовательными измерениями составляет примерно 15 мин.
Для контроля состава газового потока в последнее время все
чаще применяют масс-спектрометрический метод. Появление
сравнительно недорогих серийных масс-спектрометров позволи-
ло расширить область применения этих приборов в исследова-
тельских работах, а выпуск надежных, устойчивых к внешним
воздействиям промышленных масс-спектрометров обусловил
целесообразность их использования в промышленности. Масс-
спектрометры выгодно отличаются малым временем отклика
(менее 1 мин), высокой чувствительностью (предел обнаруже-
ния около 10-5 М), способностью определять несколько компо-
нентов одновременно, линейной зависимостью отклика от
концентрации в достаточно широком диапазоне и очень малым
дрейфом калибровки. Ввиду высокой стоимости масс-спектро-
метров целесообразно обслуживать одним прибором несколько
биореакторов с помощью управляемого ЭВМ переключающего
устройства, которое последовательно вводит в масс-спектрометр
потоки из различных реакторов (рис. 10.4). Как показано на
рис. 10.4, с помощью этой же ЭВМ можно управлять и процес-
сом (здесь показаны три реактора, но практически один масс-
спектрометр может обслуживать до 30 реакторов).
Контрольно-измерительная аппаратура и управление
183
РИС. 10.4. Схема управляемой ЭВМ системы отбора проб при использовании
масс-спектрометра для контроля процессов в нескольких реакторах. (Воспро-
изведено с разрешения из работы: Buckland R. С., Fastert Н.г Analysis of
Fermentation Exhaust Gas Using a Mass Spectrometer, in Computer Applications
in Fermentation Technology, p. 119, Society of Chemical Industry, London,
1982.)
Часто в анализах принимают, что поступающий в систему
воздух имеет стандартный состав (20,91% Ог, 0,03% СО2),
однако иногда в целях повышения надежности и достоверности
результатов измерений целесообразно определять состав и
поступающего в реактор воздуха, включив, как это показано на
рис. 10.4, его поток в распределяющее устройство. Конечно,
такое комплексное и многостороннее использование аналити-
ческих приборов с помощью распределяющих устройств выгод-
но не только в случае масс-спектрометров.
10.2. Детекторы для непрерывного контроля
характеристик популяции клеток
К сожалению, в настоящее время имеется очень ограничен-
ное число приборов, предназначенных для непрерывного конт-
роля за поведением популяции клеток в биореакторе. Чаще
184
Глава 10
всего возникает потребность в определении содержания или
концентрации биомассы или, что еще важнее, концентрации
активной биомассы. Существует несколько принципиально-
возможных подходов к определению этих параметров, однако
надежный и достаточно универсальный метод, применимый к
самым различным организмам и средам, пока что не разрабо-
тан.
Широко изучались оптические методы, основанные на изме-
рении поглощения света (спектрофотометрические методы) или
рассеяния света (нефелометрические методы, определение-
коэффициента отражения). Можно, например, непрерывно про-
Ввод _------/
пробы *-------
D d
Трубка, внутри,
кюветы
Вывод
пробы
Дистиллированная
вода
РИС. 10.5. Схема проточной кюветы с внутренним разведением, для чего внут-
ри кюветы имеется трубка, заполненная дистиллированной водой. При диа-
метре кюветы D=12 мм и диаметре трубки d=8 мм это устройство обеспечи-
вает линейную зависимость оптической плотности при 600 нм от концентрации1
клеток, даже если величина последней превышает 1,8 г/л. [Из статьи: Lee С.,
Lim Н., New Device of Continuously Monitoring the Optical Density of Con-
centrated Microbial Cultures; Biotech. Bioeng., 22, 636 (1980).]
пускать поток культуры через ячейку спектрофотометра, но в
этом случае возникают затруднения, связанные с нелинейным
характером зависимости оптической плотности от концентраций
биомассы, если последняя превышает 0,5 г/л или если оптиче-
ская плотность превышает 0,5. Следовательно, более концент-
рированные культуры можно изучать, разбавив изучаемую
пробу или уменьшив длину пути светового луча. На рис. 10.5'
приведена схема проточной кюветы, разработанной Лимом и
сотрудниками; такая конструкция кюветы хорошо зарекомендо-
вала себя в ряде лабораторий. Предлагались также принципи-
ально иные аналитические устройства, в которых оптическая
плотность культуры определяется с помощью зондов, погружае-
мых в изучаемую культуральную жидкость.
Единственным методом, пригодным для непрерывного кон-
троля биохимического или метаболического состояния популя-
ции клеток непосредственно в реакторе, является флуорометрия.
В этом методе культуральную жидкость облучают ультрафиоле-
товым светом с длиной волны 366 нм. Это излучение вызывает
флуоресценцию восстановленных пиридиновых нуклеотидов
(NADH и NADPH), причем максимальная интенсивность
Контрольно-измерительная аппаратура и управление 185
•флуоресценции наблюдается приблизительно при 460 нм.
Флуоресценцию культуры измеряют соответствующим детекто-
ром, например фотодиодом или фотоумножителем. Первоначаль-
но такие измерения проводили через кварцевые окна, вмонти-
рованные в стенки лабораторных ферментеров. Создание флуо-
ресцентных зондов, которые могут быть установлены в реакто-
рах так же, как и электроды, должно существенно расширить
•область применения флуорометрических методов в биотехноло-
гии (рис. 10.6).
Охлаждающее К самописцу Подвод
устройство и ЭВМ электроэнергии
РИС. 10.6. Принципиальная схема зонда для непосредственного изучения
флуоресценции культуральной жидкости. [Воспроизведено с разрешения из
работы: Beyeler W„ Einsele A., Fiechter A., On-Line Measurements of Culture
Fluorescence: Method and Application, European J. Appl. Microbiol. Biotechnol.,
13, 10 (1981).]
A — усилители; LA — источник УФ-излучения; D — фотоприемник; Fj —
фильтр, пропускающий коротковолновое УФ-излучение (336 нм); F2 —
фильтр, пропускающий излучение с длиной волны 450 нм; Li-6 — линзы; СС —
градуировочная кювета; М — дихроичное зеркало, пропускающее излучение с
длиной волны 460 нм и отражающее излучение с длиной волны 366 нм.
Интенсивность флуоресценции зависит от плотности популя-
ции клеток, среднего состояния метаболической активности и
эмиссии флуоресценции, а также от поглощения света средой.
Эксперименты в свободной от нерастворимых частиц среде
показали, что изучение флуоресценции культуры дает полезную
информацию о концентрации биомассы, переносе кислорода,
а также о времени выравнивания концентраций, истощении
субстрата и изменении характера метаболизма. Так, Айнзеле
й сотрудники изучали динамику перемешивания жидкости в
ферментере рабочим объемом 40 л с механическим перемеши-
186
Глава 10
ванием (при скорости вращения мешалки 200 об/мин) в срав-
нении с динамикой перемешивания в том же реакторе при.
условии, что в нем находится культура дрожжевых клеток в
глюкозной среде. В первом эксперименте в раствор, содержащий
0,05 М H2SO4, в импульсном режиме вводили порцию хинина
РИС. 10.7. Изменение флуоресценции системы после импульсного добавления
раствора хинина в 0,5 М H2SO4 (результаты характеризуют процесс выравни-
вания концентрации растворенного вещества) (а) и после добавления глюко-
зы к культуре дрожжей (результаты характеризуют как процесс выравнива-
ния концентраций, как и утилизацию глюкозы клетками) (б). [Воспроизведе-
но с разрешения из статьи: Einsele A., Ristroph D. L., Humphrey А. Е., Mixing
Times and Glucose Uptake Measured with a Fluorometer; Biotech. Bioeng., 20,
1487 (1978).]
и затем измеряли флуоресценцию (хинин имеет примерно такой
же спектр флуоресценции, что и NADH). Результаты этого
эксперимента представлены на рис. 10.7, а, где виден колеба-
тельный характер изменений концентрации индикатора (хини-
на), отражающий структуру циркуляции жидкости в реакторе
и показывающий, что новое стационарное состояние в реакторе
достигается только после довольно продолжительного переход-
ного периода. На рис 10.7, б представлены результаты изуче-
Контрольно-измерительная аппаратура и управление
187
ния флуоресценции восстановленных пиридиновых нуклеотидов
в культуре дрожжей после введения глюкозы (при t=0). Во
втором эксперименте время отклика системы больше, что
свидетельствует о существенном динамическом запаздывании
процесса утилизации глюкозы клетками. О необходимости учета
подобных эффектов при операциях масштабного перехода мы
уже упоминали в разд. 9.3.4.
о 4,00
^.3,50
S 3,00
с:
В 2,50
С
is 2,00
Ч
У 1,50
§
1.00
^4,00
3,50
м
ts 3,00
§
§ 2,50
Е
"> 2,00
Ч
§ 1,50
Б 1,00
? °’50
В 2,50
£ 2,00
§
S’ 1,50
i
1,00
g
0,50
§
s 0
e-e®
^-1,00
-1,50
14,00
* 13,00 Y
^12,00 -
§11,00
g 10,00
Is,00
is
8,00 ~
&
7,00 -
&
6,00'—
23 24 25 26 27 28
Время от начала процесса, ч
v 1; л 2; a 3 ; о 4
РИС. 10.8. Взаимосвязь между флуоресценцией культуры (4), концентрацией
этанола (2), дыхательным коэффициентом (3) и концентрацией ацетата (1)
в культуре С. utilis при периодическом добавлении субстрата (этанола). [Вос-
произведено с разрешения из статьи: Watteeuw С. М., Armiger W. В., Ris-
troph D. L., Humphrey A. E., Production of Single Cell Protein from Ethanol by
a Fed-Batch Process; Biotech. Bioeng., 21, 1221 (1979).]
Другой областью применения флуорометрического метода
в биотехнологии является контроль метаболического состоя-
ния клеток в культуре, отражающего ход ферментации. При
экспериментальном изучении роста культуры Candida utilis на
этаноле в периодическом процессе с добавлением субстрата с
помощью флуорометрии определяли время истощения субст-
рата (этанола) и контролировали добавление свежих порций
этанола к системе. На рис. 10.8 представлены зависимости флу-
оресценции культуры, дыхательного коэффициента и концентра-
ции этанола от времени в этом процессе. Кружками выделена
кривая флуоресценции культуры, найденная при непрерывном
контроле этого параметра. В представленной на этом рисунке
части эксперимента в систему импульсно вводили 1,85, 1,85 и
3 г/л этанола спустя 23, 24,5 и 26,15 ч после начала процесса.
Очевидно, что истощение субстрата (этанола) сопровождается
снижением интенсивности флуоресценции.
188
Глава 10
На результаты непосредственного измерения оптических
свойств культур клеток может влиять ряд факторов. Так, по-
верхности оптических приборов, находящиеся в контакте с
культуральной жидкостью, могут загрязняться клетками или
компонентами среды. Проведению измерений в многофазных
системах могут мешать газовые пузырьки или частицы нераст-
воримых веществ. Что касается самого флуорометрического
метода, то ряд компонентов среды или продуктов процесса мо-
жет флуоресцировать при длинах волн, используемых для
контроля состояния клеток, что, очевидно, затрудняет интерпре-
тацию результатов измерений. Тем не менее надо полагать, что
флуорометрические и другие оптические методы в будущем
найдут гораздо более широкое применение, поскольку концент-
рация биомассы и метаболическая активность клеток являются
ключевыми параметрами в контроле, регулировании и оптими-
зации микробиологических процессов.
В следующем разделе рассматриваются некоторые автоном-
ные, периодические методы изучения поведения клеток и свойств
среды, играющих важную роль в биотехнологических процес-
сах. Следует отметить, что между непрерывными методами
мониторинга (осуществляемыми обычно с помощью встроен-
ных в реактор или другой аппарат детекторов) и периодически-
ми, автономными методами измерения нет четкой границы.
Если время между последовательными анализами с помощью
автономного аналитического прибора меньше характерного
масштаба времени изменений определяемого параметра, то
периодический отбор проб с некоторым запаздыванием во вре-
мени и их последующий анализ дадут точно такую же инфор-
мацию, как и непрерывные измерения изучаемой системы
в процессе работы (в реальном масштабе времени). Поскольку
масштабы времени изменений свойств ферментов, культур кле-
ток и жидкой фазы могут варьировать от нескольких минут до
нескольких часов и даже дней, то некоторые автономные мето-
ды анализа с предварительным отбором проб дают ценную для
контроля и регулирования процесса информацию.
10.3. Автономные методы анализа
В этом разделе мы рассмотрим принципы работы некоторых
аналитических методов определения свойств культуральных
жидкостей, биокатализаторов и биосорбентов. Вообще говоря,
для этой цели можно использовать любые методы аналитиче-
ской химии, спектроскопии и биохимии; очевидно, что в настоя-
щем пособии не представляется возможным дать сколько-ни-
будь полное описание этой темы. Поэтому мы сосредоточим
внимание на некоторых новых методах определения тех свойств
Контрольно-измерительная аппаратура и управление 183^
клеток, которые наиболее важны для контроля микробиологиче-
ских процессов и управления ими, а также рассмотрим другие-
наиболее часто применяемые методы анализа.
10.3.1. Определение свойств среды
Первая стадия обработки пробы, отобранной из биореактора
или аппарата, где происходит разделение продуктов, обычно-
заключается в отделении твердой фазы (клеток или любых,
других нерастворимых компонентов) от жидкой путем фильтро-
вания или центрифугирования. Применение затем той или иной
конкретной аналитической методики зависит от природы изу-
чаемого материала и цели анализа. Действительно, методы,
анализа, пригодные для изучения синтетической среды, могут
оказаться неточными или вообще неприменимыми в случае-
сложной среды неопределенного состава, которая может содер-
жать мешающие ходу анализа вещества.
Из параметров находящейся в биореакторе системы обычно-
наибольший интерес представляют концентрации субстратов ц
других компонентов, влияющих на скорости реакций, а также
концентрации продуктов реакции и ингибиторов. В микробио-
логических процессах часто желательно также определять,
содержание источников углерода и азота, а иногда и ряда ио-
нов, например ионов магния или фосфатного иона. По свойствам:
и степени сложности химической структуры продукты жизнеде-
ятельности клеток могут варьировать в очень широких пределах,,
например, от этанола до гораздо более сложных соединений
типа пенициллина и биологических макромолекулярных ве-
ществ типа ферментов и других белков. Соответственно и-
методы анализа продуктов микробиологических процессов очень,
разнообразны.
Для количественного определения компонентов жидкой фа-
зы обычно применяют методы, основанные на измерении пока-
зателя преломления с помощью рефрактометрических детекто-
ров, поглощения света при определенной длине волны с помо-
щью спектрофотометра или флуоресценции (после возбуждения,
при одной длине волны и последующей эмиссии при больших:
длинах волн) с помощью спектрофлуорометра. Сахара, напри-
мер, практически не поглощают свет и не флуоресцируют, по-
влияют на показатель преломления раствора. С другой стороны,,
белки и нуклеиновые кислоты удобно определять спектрофото-
метрическим и спектрофлуорометрическим методами. Послед-
ний метод обычно более чувствителен и позволяет определять
более низкие концентрации веществ, однако спектрофлуоро-
метры намного дороже спектрофотометров, поэтому спектро-
фотометрия остается одним из самых распространенных и попу-
190
Глава 10
лярных методов. Чтобы устранить помехи со стороны других
растворенных веществ, часто приходится прибегать к операциям
концентрирования или отделения определяемого соединения от
других растворенных веществ. Иногда отделение удается осу-
ществить путем химической обработки, в результате которой
мешающие вещества разрушаются или осаждаются. Так, на-
пример, РНК экстрагируют из лизатов клеток перхлорной
кислотой НС1О4 при 37 °C и определяют с помощью орсина
(специфического реагента на рибозу). Мешающие определению
сахара отделяются в процессе экстракции.
В ходе изучения химического состава среды обычно приме-
няют и более тонкие методы разделения близких по химической
природе веществ, например хроматографические. Хроматогра-
фические методы разделения, подробнее рассматриваемые в
разд. 11.4, основаны на селективном удерживании некоторых
соединений неподвижной фазой, находящейся в колонке. Селек-
тивность удерживания обеспечивается предпочтительным срод-
ством по отношению к неподвижной фазе одних веществ по
сравнению с другими. Так, при анализе смесей сахаров (напри-
мер, мальтозы и глюкозы) применяют метод высокоэффективной
жидкостной хроматографии (ВЭЖХ), в котором разделение
достигается за счет различного сродства этих двух углеводов
к первичным аминогруппам на поверхности носителя (непод-
вижной фазы), содержащегося в обычной выпускаемой серийно
колонке. Различные сахара вымываются с колонки в разное
время; их можно обнаружить и определить количественно с по-
мощью рефрактометрического детектора. Метод ВЭЖХ приме-
няется и для определения многих других компонентов среды.
Помимо ВЭЖХ для разделения смесей близких веществ ис-
пользуют ионообменную и эксклюзионную хроматографию (в
отличие от ВЭЖХ в этих хроматографических методах разделе-
ние осуществляется при атмосферном или слегка повышенном
давлении) с последующим количественным определением ин-
дивидуальных компонентов среды.
Как мы уже отмечали в гл. 4, полезным методическим прие-
мом органического анализа является селективное превращение
определяемых соединений в производные, легко поддающиеся
непосредственному определению. На этом принципе основан
один из стандартных лабораторных методов определения глю-
козы, в котором ферменты глюкозооксидаза и пероксидаза
селективно превращают глюкозу в окрашенное соединение,
концентрацию которого определяют спектрофотометрически.
глюкозооксидаза
Глюкоза+2Н2О----------------- глюконовая кислота+2Н2О2
пероксидаза
Н2О2+о-дианизидин ----------* продукт окисления о-дианизидииа
(бесцветный) (коричневый)
Контрольно-измерительная аппаратура и управление 191
В последние годы для определения ряда биологически важ-
ных ионов широко применяют ионоселективные электроды. Так,
азотсодержащие клеточные вещества можно определять с по-
мощью КНз-электрода непосредственно (NH3, а после соответ-
ствующего изменения pH также 1NH4+) после восстановления
(NO2~, NO3“) или после обработки ферментами (аминокислоты,
мочевина). Разработаны ионоселективные электроды для опре-
деления многих других ионов, влияющих на биохимические
структуры и их функции, в том числе К+, .Na+, Са2+.
Иногда концентрацию того или иного вещества в растворе
определяют путем измерения его биологической активности.
Именно таким способом обычно определяют, например, кон-
центрацию пенициллина в культуральной жидкости при произ-
водстве этого антибиотика. Здесь мерой активности пеницилли-
на в растворе является размер зоны погибших бактерий,
окружающей пористый диск, пропитанный изучаемым раство-
ром. Содержание ферментов в подавляющем большинстве слу-
чаев определяют путем измерения их активности. Для этой цели
раствор пробы обрабатывают стандартным субстратом в стан-
дартных условиях с последующим измерением скорости
исчезновения субстрата или образования продукта реакции;
соответствующие измерения часто осуществляют спектрофото-
метрическим или спектрофлуорометрическим методами.
Другой подход к анализу биологически важных веществ осно-
ван на их переведении в газовую фазу и последующем опреде-
лении. Глюкозу, например, превращают в триметилсилильное
(ТМС) производное, которое затем переводят в газовую фазу
в системе ввода газового хроматографа, а потом определяют с
помощью пламенно-ионизационного детектора. Понятно, что
таким же путем без модификации изучаемых веществ можно
определять содержание в культуральной жидкости относительно
летучих компонентов, например этанола, ацетона и бутанола.
В аналитических лабораториях при пилотных и крупномас-
штабных промышленных ферментерах часто имеется один или
несколько автоанализаторов для проведения мокрого химиче-
ского анализа, где автоматически осуществляются операции
экстракции, разведения и определения нескольких веществ.
Время выполнения анализов составляет 10—30 мин, что во
многих случаях вполне достаточно для успешного контроля за
ходом процессов в биореакторе.
10.3.2. Анализ состава популяции клеток
Методы анализа популяций клеток можно классифициро-
вать примерно таким же образом, как и математические моде-
ли, описывающие кинетику роста культур клеток (вспомните
•192
Глава 10
тл. 7 и рис. 7.2). Большинство классических методов биохимии
дает усредненную по всей популяции и, следовательно, несегре-
тированную информацию. Такие методы можно распространить
на очень большое число компонентов клетки вплоть до отдель-
ных белков, молекул РНК и ДНК и даже последовательностей
-их фрагментов. Если же экспериментальные данные относятся
к одной клетке или могут дать информацию о состоянии отдель-
ных клеток, так что в конечном счет удается получить сведения
о распределении свойств отдельных клеток, то такие данные
можно назвать сегрегированными.
Прежде всего мы обсудим методы, дающие информацию не-
•оегрегированного типа. К наиболее грубым результатам таких
методов после данных по определению массовой или численной
плотности популяции относятся данные об элементном составе
клеток, включая содержание углерода, водорода и азота. Раз-
работаны автоматические анализаторы, предназначенные,
например, для определения общего азота в пробах. Известно,
что в биологических процессах важную роль играет ряд ионов,
поэтому в анализах на элементы обычно определяют также
общее содержание железа, магния, фосфора и кальция. Общее
содержание белков, РНК, ДНК и других компонентов клеток
большой молекулярной массы определяют с помощью хорошо
известных методов. В качестве примера на рис. 10.9 перечисле-
ны основные операции методики определения макромолекуляр-
ных компонентов дрожжей. Здесь мы не имеем возможности
детально описывать каждый из методов анализа различных
макромолекулярных соединений; подробные сведения можно
найти в работах [14, 15], а также в многочисленных статьях,
посвященных определению состава клеток в процессе их роста.
С помощью этих методов получен ряд ценных результатов, на-
пример данные, приведенные на рис. 7.17. Хотя любую конкрет-
ную методику определения какого-либо вещества, например бел-
ка или ДНК, можно применять для анализа различных организ-
мов, в зависимости от вида последних может потребоваться
та или иная модификация этой аналитической методики.
Усредненное содержание конкретных белков в популяции
клеток можно определять несколькими методами. Что касается
«ферментов, то для контроля за изменением их концентрации в
ходе процесса обычно измеряют соответствующую ферментатив-
ную активность. В табл. 10.3 перечислены характерные реакции
и принципы измерений, используемые для определения концент-
раций основных ферментов, участвующих в метаболическом
превращении глюкозы в органические растворители в микробио-
логическом процессе производства ацетона и бутанола с помо-
щью Clostridium acetobutylicum. На рис. 10.10 представлены
.зависимости активности ряда основных ферментов этого орга-
Контрольно-измерительная аппаратура и управление
193
Культуральная жидкость (10-20 мл)
Центрифугирование
(10 мин, 10000 об/мин)
Анализ
растворимых
' компонентов
среды
Осадок
(клетки)
1мл, 100мкг клеток 10-20 мл, 20-60мг клеток
Определение
белков
Определение
РНК
Промывка фосфатным буфером
(удаление мешающей
определению глюкозы)
Удаление растворимых
в кислотах веществ
промывкой 0,25 н. НСЮ 4
( О °C, 15 мин}
Растворение белка
(0,5 мл NaOH, 100 °C, 5мин}
Центрифугирование
Добавление реагента Лоури
Экстракция РНК
0,5 н, НС104, 37°C
| Центрифугирование
Измерение поглощения при 750нм. I
Сравнение со стандартной кривой |
(например, для альбумина Супернатант
из сыворотки быка) I
Определение реакцией
с арсином
Измерение поглощения при 672нм.
Сравнение со стандартной кривой
(для РНК дрожжей)
РИС, 10.9. Операционная карта методики определения концентраций (усред-
ненных по популяции) белка и РНК в дрожжевых клетках. Подробное опи-
сание методики Лоури и орсинового метода приведено в работах [14, 15].
13-746
194
Глава 10
низма от времени периодического процесса. Как показывают эти
данные, на последних фазах процесса активности ферментов,,
участвующих в биосинтезе кислот, снижаются, а активности
ферментов, связанных с образованием бутанола и ацетона, на-
против, возрастают. Подобная информация может быть полез-
Таблица 10.3. Типичные субстраты и методы определения активности
основных ферментов, участвующих в путях катаболизма
в Clostridium acetobutylicum
Фермент Субстрат Реакция Метод определения активности фермента
Ацетоаце- татдекарбо- ксилаза Фосфофрук- токиназа Дегидро- геназа Пируват— ферредок- син-оксидо- редуктаза Бутироки- наза Ацето- ацетат Фруктозо- 6-фосфат Метилвио- логен Ацетил-СоА Бутирофос- фат Ацетоацетат->ацетон+ +со2 Реакция осуществляется в присутствии альдола- зы, триозофосф атизоме- разы и «-глицерофосфа- та Дегидрогеназа+метил- виологен+2Н+->метил- виологен (окисленный)+ +н2 14СО2+ацетил-СоА+ +ферредоксин (восста- новленный)-^- [* 4С] пи- руват+СоА+ферредок- син (окисленный) Бутирофосфат+гидрок- силамин->бутирогидрок- самовая кислота-|-фос- фат Спектрофотометрическое определение непрореаги- ровавшего ацетоацетата (по поглощению при 270 нм) Флуорометрическое оп- ределение непрореагиро- вавшего NADH (воз- буждение при 340 нм» эмиссия при 460 нм) Окисление метилвиоло- гена, сопровождающееся образованием газооб- разного Н2, контролиру- ется манометрически Образование пирувата» меченного радиоактив- ным 14С, контролирует- ся сцинтилляционным счетчиком Спектрофотометрическое определение бутирогидр- оксамовой кислоты (па поглощению при 540
нм)
ной при выяснении зависимостей между изменением клеточного’
метаболизма, природой штамма микроорганизма и эксплуата-
ционными параметрами биореактора; эти данные в свою очередь
необходимы для оптимизации как организма, так и процесса.
Индивидуальные белки иногда можно определять хромато-
графическими (см. разд. 11.4) или электрофоретическим метода-
ми путем измерения интенсивностей отдельных полос. Для того
чтобы повысить разрешающую способность и чувствительность
электрофоретического метода при самых разных концентрациях
белков, можно применять двумерный электрофорез в геле; в
этой методике белки разделяют в одном направлении по вели-
Контрольно-измерительная аппаратура и управление
195
чине их молекул, а во втором направлении — по заряду. При
таком разделении индивидуальные белки концентрируются в
виде пятен на плоскости, что позволяет идентифицировать и
количественно определять большое число белков одновременно.
Эта методика применялась, например, для изучения скоростей
синтеза 140 различных белков в процессе роста бактерии Е. coll
[17].
Основой другого метода анализа индивидуальных оелков
является связывание антител с определенным участком молеку-
лы изучаемого белка [18]. Если в распоряжении исследователя
имеются антитела, специфичные по отношению к изучаемому
белку, то последний можно определить количественно путем
осаждения комплекса белок — антитело или посредством детек-
тирования антител, меченных радиоактивными изотопами, или
путем измерения активности фермента, связываемого с изучае-
мым белком (метод иммуносорбционного ферментативного ана-
лиза, ELISA). Аналогично можно определять и другие компо-
ненты системы, в том числе и высокомолекулярные соединения,
если только удается найти антитела, специфичные в отношении
этих компонентов. Меченые антитела часто применяют для
идентификации на поверхности клеток определенных соединений
или структур, а в некоторых случаях таким путем удается оп-
ределить и количество этих соединений на наружной поверхно-
сти клеток. Связывание антител с теми или иными соединениями
на поверхности клеток и последующие измерения могут быть
выполнены и без потери жизнеспособности организмов, что осо-
бенно важно при скрининге и селекции мутантных штаммов.
Этот метод пригоден и для обнаружения определенных штаммов
в смешанных культурах, поскольку каждый организм обычно
имеет на своей поверхности специфические соединения-марке-
ры, которые с помощью соответствующих антител можно селек-
тивно идентифицировать и определять количественно.
Важность плазмид как носителей генетической информации,
используемой для синтеза необходимых веществ в рекомбинант-
ных организмах, обусловливает потенциальную ценность мето-
дов определения содержания плазмид в клетках. Наиболее
надежный метод количественного определения плазмид в бакте-
риях заключается в выделении суммарной ДНК организма с
последующим разделением плазмидной и хромосомальной ДНК
ультрацентрифугированием в градиенте хлорида цезия. Отно-
сительные количества хромосомальной и плазмидной ДНК
можно определять несколькими методами. При использовании
меченных радиоактивными изотопами препаратов ДНК можно,
например, отделить осадок после центрифугирования и опреде-
лить его радиоактивность с помощью сцинтилляционного счет-
чика. Содержание плазмидной ДНК в клетках дрожжей или
13*
196
Глава 10
Время, ч
РИС. 10.10. Рост Clostridium acetobutylicum в периодическом процессе и из-
менение концентраций ферментов, участвующих в образовании продуктов ме-
таболизма.
а — параметры клеточного роста и образования продуктов метаболизма: 1 —
оптическая плотность; 2— значение pH; 3 — концентрация ацетата; 4— кон-
центрация бутирата; 5 — концентрация ацетона; 6 — концентрация бутанола.
б — концентрации ферментов, участвующих в биосинтезе ацетата и бутирата,
а также гидрогеназы: 1 — фосфотрансацетилаза; 2—ацетаткиназа; 3— фос-
фотрансбутирилаза; 4 — бутираткиназа; 5 — гидрогеназа.
в — концентрации ферментов, участвующих в образовании ацетона и бутано-
ла: 1 или 2 — ацетоацетил-СоА: СоА — трансфераза с ацетатом или бутиратом
Контрольно-измерительная аппаратура и управление
197
животных клетках можно оценить методом гиоридизации с по-
мощью меченого зонда, комплементарного специфической нук-
леотидной последовательности плазмиды. Другой! подход к оп-
ределению содержания плазмид в организмах основан на
включении в плазмиду в качестве маркера гена какого-либо
конкретного фермента, активность которого можно было бы
затем легко определить. Такой подход применялся для оценки
количества плазмид в рекомбинантных клетках бактерий, дрож-
жей и животных организмов.
Все рассмотренные выше методы анализа дают информацию
о составе клеток и в некоторой степени о характере их мета-
болизма, но не сообщают непосредственно никаких данных
об энергетическом состоянии организма в данный момент. Коли-
чество внутриклеточного АТР можно определить с помощью
анализатора типа Биометр. Этот прибор измеряет интенсивность
люминесценции, генерируемой в реакции с участием АТР,
которая катализируется ферментом люциферазой. Поскольку
концентрация АТР может очень быстро изменяться в зависимо-
сти от окружения клетки и ее метаболической активности,
необходимо до анализа этим (или иным) методом быстро обра-
ботать образцы популяции клеток фосфорной кислотой; в про-
тивном случае нельзя гарантировать сохранение концентрации
АТР в процессе анализа. Так как в нежизнеспособных клетках
АТР отсутствует, то путем определения содержания АТР иногда
можно оценить и долю метаболически активной биомассы в
популяции.
Для определения внутриклеточных концентраций АТР, ADP,
фосфатов сахаров, полифосфата и величины pH успешно при-
менялась спектроскопия ядерного магнитного резонанса (ЯМР)
на ядрах 31Р. Этим методом изучали различные микроорганиз-
мы, в том числе бактерии Е. coll [19] и Clostridium thermocellum
[20], дрожжи Saccharomyces cerevisiae, Candida utilis и Zygo-
saccharomyces bailii [21]. Для изучения путей внутриклеточного
метаболизма углерода и азота можно применять также спектро-
скопию ЯМР 13С и 15N, если изучаемый организм выращен на
субстратах, обогащенных изотопами 13С и (или) 15N [23, 24].
В некоторых процессах с участием мицелиальных микроорга-
низмов наблюдалась зависимость эффективности и кинетики
этого процесса от морфологического состояния плесени или ак-
в качестве акцептора соответственно; 3 — ацетоацетатдекарбокснлаза; 4 — бу-
тиральдегиддегидрогеназа; 5 — бутанолдегпдрогеназа. [Воспроизведено с раз-
решения из статьи: Andersch Bahl И., Gottschalk G., Level of Enzvmes In-
volved in Acetate, Butyrate, Acetone and Butanol Formation by Clostridium
acetobutylicum; Eur. J. Appl. Microbiol. Biotechnol., 18, 327 (1983)']
198
Глава 10
тиномицета. В примере 7.2 мы уже обсуждали некоторые
данные о связи между морфологией и образованием антибио-
тика цефалоспорина в культуре плесени Cephalosporiutn асгето-
тит. Непосредственное качественное визуальное изучение и
количественный контроль морфологии мицелия очень сложны и
трудоемки, поскольку они связаны с большим объемом работ
по микроскопии и с ручным или автоматическим анализом
полученных данных. Морфология мицелия, как было показано,
влияет на его поведение при фильтровании. Ванг и сотрудники
[25, 26] использовали эту зависимость для создания остроумно-
го метода определения морфологии мицелия на основе данных
по изучению фильтрования пробы культуры, который далее был
усовершенствован Лимом и сотрудниками [27]. В этом методе
периодически отбирают небольшую пробу культуральной сус-
пензии, фильтруют ее и затем определяют объем фильтрата и
толщину слоя осадка на фильтре. Полученные таким путем дан-
ные в совокупности с найденными ранее корреляциями между
параметрами процесса фильтрования и морфологическими
свойствами и плотностью изучаемой культуры плесени могут
служить основой для периодического непосредственного контро-
ля за ходом ферментации (см. упражнение 11.6).
Существует несколько методов изучения и определения рас-
пределения характеристик индивидуальных клеток в популяции
одноклеточных организмов. Такие распределения приближенно
можно оценить методом микроскопии, а в сочетании с методами
анализа изображений можно получить и информацию количе-
ственного характера, хотя сбор характеристик достаточно
большого числа клеток, необходимых для получения статисти-
чески достоверной информации, очень трудоемок. Для быстрого
определения свойств большого числа индивидуальных клеток
более удобны непрерывные методы измерения в потоке. Име-
ются два основных типа таких методов. В приборах типа счет-
чика Коултера объем отдельных клеток определяют путем
измерения сопротивления небольшого отверстия, через которое
протекает изучаемая проба (суспензия клеток), разбавленная
раствором электролита. Известна зависимость между объемом
сферической частицы и изменением удельного сопротивления
отверстия; таким путем можно определить размеры большого
числа быстро протекающих через отверстие суспендированных
частиц. Некоторые затруднения при интерпретации результатов
измерений может вызвать изменение морфологии клеток; тем
не менее этот метод позволяет получить ценную информацию о
распределении клеток популяции по размерам.
Более универсальны приборы, называемые проточными цито-
метрами. В этих приборах разбавленная суспензия клеток так-
же проходит через детектирующее устройство, однако здесь
Контрольно-измерительная аппаратура и управление 199
измеряются оптические свойства суспензии. Как показано на
схеме прибора (рис. 10.11), в проточном цитометре поток про-
бы облучают с помощью лазера или другого источника света, а
затем измеряют поглощение, рассеяние и (или) флуоресценцию
индивидуальных клеток. Результаты изучения рассеяния света
можно использовать для получения информации о распределе-
нии клеток по размерам. Поскольку интенсивность рассеяния
света под прямым углом зависит от наличия и концентрации
специфических внутриклеточных структур, этот метод успешно
применялся для контроля накопления в индивидуальных бакте-
риальных клетках частиц резервного углевода, полигидрокси-
бутирата, обладающих характерным показателем преломления
[28]. С помощью специфических флуоресцентных красителей,
выполняющих роль метки, в клетках микроорганизмов и живот-
ных клетках определяли распределение концентраций различ-
ных биополимеров, в том числе суммарного клеточного белка,
двухцепочечной ДНК и клеточной РНК [29, 30]. С помощью
проточных цитометров можно также следить за накоплением в
отдельных клетках внутриклеточного флуоресцирующего соеди-
нения, образующегося при действии конкретного фермента.
Таким путем можно устанавливать активность одного фермен-
та в индивидуальных клетках, изучать in vitro кинетику фер-
ментативных реакций, определять (путем клонирования гена,
отвечающего данному ферменту) содержание плазмид в клет-
ке [31]. Методом цитометрии в потоке, кроме того, можно
идентифицировать и определять содержание конкретных штам-
мов в смешанных культурах и обнаруживать посторонние штам-
мы в посевном материале [32, 33]. Поскольку метод цитометрии
в потоке дает информацию не только об усредненных парамет-
рах популяции, но и о распределении характеристик индивиду-
альных клеток, это позволяет получить более детальные данные
о состоянии популяции микроорганизмов.
Хотя в большинстве случаев методом цитометрии в потоке
измеряли лишь один параметр микробиологического процесса,
при помощи этого метода можно определять и несколько
параметров индивидуальных клеток одновременно, что позво-
ляет получать еще более детальную информацию о состоянии
популяции клеток. При одновременном определении двух пара-
метров результаты измерений изображают на плоскости в виде
зависимости частоты или относительного числа клеток от двух
изучаемых параметров, величины которых откладывают на осях
координат, ограничивающих указанную плоскость. В качестве
примера на рис. 10.12 приведены результаты одновременного
определения интенсивности рассеяния света (связанной с раз-
мером клеток) и внутриклеточной флуоресценции, эмиссия
которой происходит с участием фермента, кодируемого плаз-
200
Глава 10
камера
с правой
Зеркало
Магнитная
мешалка
Отклоняющие
электроды
Линза
канал
I
Волоконная
оптика
Рассеяние
в прямом
направлении.
Волоконная
оптика
>(« фотоумно-
жителю}
Отра
полоска (тушение}
Линзы
конденсора °^юние
Лазере
Лазер 1
Линза
Полуволновая
фазовая
пластинка
ЭВМ
Фотоумножитель
Усилитель
I Грофопостро- I
I ителъ п"
ампул
Большая ЭВМ ♦
РИС. 10.11. Схема проточного цитометра. На схеме показаны пути потока
суспензии клеток и лазерных лучей, оптические фильтры, детекторы для опре-
деления параметров индивидуальных клеток, а также способы генерации, ана-
лиза и хранения информации. Размещенными по диагонали квадратами изо-
бражены дихроичные светофильтры (отражающие излучение определенных
длин волн и пропускающие излучение другой длины волны); расположенными
под прямым углом квадратами обозначены задерживающие или поляризаци-
онные светофильтры. В таких приборах часто устанавливают несколько детек-
торов типа фотоумножителей н соответствующих электронных устройств, что
позволяет одновременно определять несколько параметров каждой клетки.
(За основу взят прибор Ortho Instruments Cytofluorograph System 50H.)
Контрольно-измерительная аппаратура и управление
201
РИС. 10.12. Изучение популяции рекомбинантных клеток Saccharomyces сеге-
visiae путем определения двух параметров методом цитометрии в потоке. На
осях координат базисной плоскости отложены интенсивность рассеяния света
индивидуальными клетками (связанная с размером клеток) и интенсивность
флуоресценции индивидуальных клеток (в данном случае связанная с числом:
плазмид в одной клетке). Флуорометрия позволяет легко различать клетки,
с плазмидами и клетки, не содержащие плазмид.
мидным геном. Таким способом можно найти корреляцию*
между количеством флуоресцирующих веществ, накопленных
клеткой, и наличием или даже числом плазмид в одной дрож-
жевой клетке. На рис. 10.12 видны два четких пика, отвечаю-
щие клеткам разных типов в изучаемой культуре. Клетки с низ-
кой интенсивностью флуоресценции не содержат плазмид, по-
скольку в условиях этого эксперимента для нативных клеток
типичен низкий уровень естественной флуоресценции. Часты
популяции клеток с более интенсивной флуоресценцией, заре-
гистрированных в виде второго четкого пика, содержит плазми-
ды, а следовательно, и фермент, ответственный за повышение
интенсивности флуоресценции. Этот метод позволяет быстро
оценить относительное содержание клеток с плазмидами и без
плазмид в культуре и таким образом получить ценную инфор-
мацию о репликации плазмид и процессах расщепления
рекомбинантного штамма.
202
Глава 10
10.4. ЭВМ и интерфейсы
Сочетание контрольно-измерительной аппаратуры с цифро-
выми ЭВМ. выгодно в нескольких отношениях. Во-первых, ЭВМ
может разносторонне усовершенствовать работу по сбору дан-
ных. Статистические методы и методы цифрового фильтрования
позволяют повысить надежность и точность результатов изме-
рений. ЭВМ могут сравнивать отклики нескольких параллель-
ных детекторов и анализировать результаты сравнения; в част-
ности, таким путем в ходе измерений можно проверять
правильность калибровки детекторов и находить детекторы, вы-
шедшие из строя. ЭВМ позволяют увеличить число аналитичес-
ких приборов и использовать более сложные аналитические
методы. Например, управляемая ЭВМ аналитическая система
может автоматически отбирать пробы, проводить хроматогра-
фический анализ, интерпретировать результаты анализа с по-
мощью хранящихся в ее памяти градуировочных шкал или
алгоритмов и выдавать выходной сигнал в удобной для опера-
тора форме. Хотя простейшие операции согласования и
коррекции сигналов (например, линеаризация) можно выпол-
нять также с помощью специальных электронных схем, ЭВМ
позволяют осуществить эти операции без всяких дополнитель-
ных приборов. Другое преимущество ЭВМ при сборе данных
связано с их способностью хранить в памяти большое число
результатов измерений в цифровой форме. При необходимости
эти данные можно вызывать из памяти машины в любое удоб-
ное время, подвергать анализу и отображать в необходимом
виде.
ЭВМ позволяют существенно улучшить качество анализа и
интерпретации данных. С их помощью можно, например, по
результатам нескольких измерений практически мгновенно вы-
числить такие важные параметры, как скорость утилизации
кислорода и дыхательный коэффициент. Дополнительную
информацию о ходе процесса можно получить по результатам
ограниченного числа измерений с помощью современных неав-
тономных методов расчета состояния и параметров системы.
Несколько детальнее проблема применения ЭВМ для анализа
данных освещена в следующем разделе; там же приведен ряд
конкретных примеров.
ЭВМ в очень большой степени расширяют возможности
оптимизации процессов и управления ими. Одна ЭВМ, заменяя
множество традиционных аналоговых регуляторов, может с
помощью стандартных алгоритмов обратной связи одновремен-
но регулировать несколько переменных, например pH и темпе-
Контрольно-измерительная аппаратура и управление
203
ратуру. ЭВМ облегчают внедрение сложных методов регули-
рования, в которых учитывается несколько параметров одно-
временно, например регулирование определяемого расчетным
путем дыхательного коэффициента. Их можно применять для
оценки и совершенствования математических моделей процес-
сов, которые затем в свою очередь могут быть использованы
для нахождения оптимальных условий и режима процесса. Бо-
лее того, блок памяти и расчетный блок ЭВМ обеспечивают
выполнение, например, программы скорости подачи питатель-
ных веществ или изменения pH в периодическом микробиологи-
ческом процессе.
Управление периодическим процессом, по сути дела, заклю-
чается в тщательно контролируемом и координируемом после-
довательном включении, выключении и переключении насосов,
вентилей и клапанов. Ранее все эти операции выполнялись
различными таймерами и реле, а теперь с ними успешно справ-
ляются ЭВМ. Использование ЭВМ для управления операциями
переключения целесообразно в тех случаях, когда возникает
необходимость в регулировании нескольких параллельных
периодических процессов (например, ферментеров), продукты
которых далее последовательно поступают на ту или иную
периодическую операцию обработки (например, осаждение,,
хроматографическое разделение и т. д.). Управление отдельны-
ми операциями, а также более сложные методы и цели управ-
ления мы рассмотрим в разд. 10.6 и 10.7 соответственно.
Прежде чем перейти к интереснейшей теме применения
ЭВМ, вкратце ознакомимся с некоторыми принципами устрой-
ства цифровых компьютеров и соответствующих интерфейсов.
Здесь мы рассмотрим только самые общие принципы и поня-
тия, а также ряд конкретных примеров сочетания ЭВМ с про-
цессами. Поскольку совершенствование конструкций и удешев-
ление ЭВМ и их программного обеспечения происходят в-
настоящее время очень быстрыми темпами, то описание какой-
либо конкретной ЭВМ, скорее всего, устареет к тому времени,
когда оно будет опубликовано (во всяком случае, в книге).
По этой же причине читатель должен отдавать себе отчет в
том, что в тот момент, когда он знакомится с теми или иными
примерами, приводящимися в этой главе, весьма вероятно, что
уже существуют более дешевые и более эффективные способы
решения той же или гораздо более сложной аналогичной проб-
лемы.
204
Глава 10
10.4.1. Основные элементы цифровых ЭВМ
Основные блоки цифровой ЭВМ представлены на рис. 10.13.
Центральный процессор принимает команды, передаваемые
блоком управления в соответствии с заданной программой, и
выполняет указываемые арифметические и логические опера-
ции в арифметическом и логическом блоке, используя внутрен-
ние регистры для кратковременного хранения информации.
Работа процессора регулируется и синхронизируется с помо-
щью встроенных кварцевых часов. Время цикла процессора,
которое может изменяться от 10 (и даже менее) до ПО4 нс
(1 нс=10-9 с), а также число байтов информации (один байт
содержит 8 битов — чисел в двоичной системе, 0 или 1), пере-
даваемых в одном цикле (длина слова), определяют скорость
обработки информации в процессоре. В выпускавшихся в
1980 г. микро-ЭВМ использовались слова из 8 бит, к 1982 г.
длина слова микро-ЭВМ возросла до 16 бит, а в 1984 г. неко-
торые фирмы уже выпускали компьютеры с длиной слова,
равной 32 бит.
Для хранения команд и данных используется несколько
типов памяти. В постоянном запоминающем устройстве (ПЗУ)
хранятся определенные наборы команд, например программы
компилятора и интерпретатора, а запоминающее устройство
с произвольным доступом (оперативное ЗУ) применяется для
кратковременного хранения программ, вводимых данных и ре-
зультатов расчета. В процессе работы ЭВМ центральный про-
цессор часто считывает информацию с оперативного ЗУ и за-
писывает на нем данные. В выпускавшихся в большом количе-
стве в середине 80-х годов микро-ЭВМ информационная емкость
оперативного ЗУ составляла от 64 000 до 512 000 байт и выше.
Дополнительной памятью обладают также одно или несколько
периферийных (внешних) устройств. В качестве последних в
случае микро-ЭВМ применяют кассеты с магнитными лентами
(память 80 000—300 000 байт; время обращения около 10 с),
гибкие диски (100 000—600 000 байт, время обращения около
0,5 с), жесткие диски (500 000—20 000 000 байт, время обраще-
ния около 3 с) (все данные относятся к 1984 г.).
Блок управления вводом данных позволяет соединять ЭВМ
с внешними периферийными устройствами. В самой ЭВМ шина
передачи информации соединяет центральный процессор, блок
памяти и блок управления вводом и выводом данных.
Перечисленные выше блоки и функциональные элементы
имеются у любых ЭВМ, но скорость переработки информации
и объем памяти различны для разных типов ЭВМ. По этим
параметрам ЭВМ часто подразделяют на супер-ЭВМ (самые
большие, быстродействующие и дорогие), большие ЭВМ, мини-
Контрольно-измерительная аппаратура и управление
205
РИС. 10.13. Основные функциональные элементы вычислительной машины
(ЦП — центральный процессор, АЛУ—арифметическое и логическое устрой-
ство, ЗУ — запоминающее устройство). Связь между различными элементами
осуществляется с помощью шины передачи информации с временным разде-
лением сигналов. [Из статьи: Hampel W. A., Application of Microcomputers in
the Study of Microbial Processes, in Advances in Biochemical Engineering,
vol. 13, Ghose T. K-, Fiechter A., Blakebrough N. (eds.), p. 1, Springer-Veriag
Berlin, 1979.]
ЭВМ и микро-ЭВМ (самые маленькие, медленнодействующие и
дешевые). Границы между этими типами ЭВМ постоянно сме-
щаются и сегодняшние микро-ЭВМ по своим возможностям
отвечают большим ЭВМ 70-х годов. Доступность дешевых и
высокоэффективных ЭВМ послужила толчком к их чрезвычайно
быстрому и широкому распространению в производстве това-
ров широкого потребления, в средствах связи, процессах
обработки информации, научном приборостроении и в биотех-
нологии. В 60-е годы управляемые ЭВМ ферментеры были
новшеством; в ближайшем же будущем, надо полагать, практи-
чески каждый биореактор, аналитический прибор или другой
аппарат в биотехнологии будут контролироваться и управлять-
ся цифровыми ЭВМ.
206
Глава 10
10.4.2. Интерфейсы и периферийные устройства ЭВМ
Запоминающие, вычислительные и логические возможности
ЭВМ останутся бесполезными, если она не соединена с каким-
либо другим устройством или аппаратом. Такие соединения-
осуществляются с помощью интерфейсов. В соответствии с
объектом, с которым связаны ЭВМ, интерфейсы можно подраз-
делить на несколько типов:
1. ЭВМ—>ЭВМ; межмашинная связь
2. Оператор—>ЭВМ; передача команд
3. ЭВМ—^-оператор; передача информации
4. Датчик—>ЭВМ; ввод данных
5. ЭВМ—^-исполнительный механизм; управление
Ниже мы рассмотрим каждый тип интерфейсов и ту ролщ
которую они играют в применении ЭВМ в биохимической тех-
нологии. Однако прежде чем обратиться к изучению конкрет-
ных интерфейсов, следует отметить, что как приборное обес-
печение интерфейсов, так и программное управление ими
не стандартизованы; поэтому различные ЭВМ, периферийные
устройства и датчики в общем случае нельзя соединять в про-
извольном порядке. Всегда следует предварительно убедиться
в том, что соединяемые блоки и устройства имеют совместимые
характеристики на входе и выходе.
Важность межмашинной связи типа ЭВМ—ЭВМ определя-
ется тем обстоятельством, что ЭВМ имеют разные стоимость и
возможности. Наибольшая эффективность при наименьших за-
тратах достигается путем использования ЭВМ с минимальными
возможностями, обеспечивающими решение основной задачи,
в то время как более мощные ЭВМ следует привлекать только'
в тех случаях, когда возникает потребность в решении более
сложных задач, в более быстром решении той же задачи или
в хранении большего количества информации. На рис. 10.14
представлен один из вариантов классификации уровней слож-
ности расчетов при проектировании, разработке и оптимизации
процессов биохимической технологии. Первый, самый низкий
уровень сложности (задачи такого уровня сложности обычна
решаются с помощью микро-ЭВМ) включает отображение на
дисплее и хранение в памяти машины данных о параметрах
биотехнологического процесса, а регулирование этих парамет-
ров на таком уровне осуществляется с помощью обычной систе-
мы обратной связи (см. разд. 10.6). На втором уровне, когда
необходим более детальный анализ данных и дальнейший диа-
лог оператора с ЭВМ, можно работать с мини-ЭВМ. На этом
же уровне можно выполнять расчеты, необходимые в случае
более сложных алгоритмов управления.
Контрольно-измерительная аппаратура и управление
207
. 2**
Автономная
обработка данных '
(анализа и.т.д.’) I
База данных г*~ Последующее изучение (моделирование^
Усложненные системы
управления (в соответствии
с программой или алгорат-
, МОМ)
Помощь оператору
Изучение в реальном масштабе
времени
Обработка, данных
в реальном мас-
штабе времени
'Запись данных в
аналоговой форме
Измерение,
отображение
и хранение
данных
Обычные системы управления
(П-И-Д, пропори, времени
,________ит. а.)
>а
Микробиологический процесс
С) Оператор
РИС. 10.14. Функции, выполняемые ЭВМ, в зависимости от их уровня слож-
ности. Уровни 1, 2 и 3 приблизительно соответствуют микро-ЭВМ, мини-ЭВМ
и большой ЭВМ соответственно. (Воспроизведено с разрешения из статьи-
Valentini L., Andreoni F„ Buvoli M., Pennella P., A Computer System for Fer-
mentation Research: Objectives, Fulfillment Criteria and Use, in Computer Appli-
cation in Fermentation Technology, p. 175, Society of Chemical Industry Lon-
don, 1982.) '
208
Глава 10
Функции первого и в очень большой степени второго уров-
ней выполняются в реальном масштабе времени, т. е. ввод
информации или команды в ЭВМ и подача сигнала (на основе
этой информации) на регулирующее устройство происходят
практически одновременно. На высшем (третьем) уровне для
решения сложных модельных уравнений, определения парамет-
ров модели на базе полученных данных, разработки более
совершенных методов управления и оптимизации может исполь-
зоваться большая ЭВМ. При этом информация поступает от
ЭВМ первого и второго уровней, но результаты не обязательно
будут получены в реальном масштабе времени. Целью этих
расчетов может быть планируемое в будущем совершенство-
вание режима процесса.
Связь между компьютером и оператором (диалог) может
осуществляться в различных формах. Для этой цели применя-
ются цифровые измерительные приборы, дисплеи на электрон-
но-лучевых трубках, печатающее устройство, графопостроитель,
а в последнее время искусственная речь. Информация на дис-
плее, печатающем устройстве или графопостроителе представ-
ляет собой текст или график, выполненный в одном или несколь-
ких цветах. Часто оператор вводит информацию в ЭВМ с
помощью клавиатуры, которая связана с ЭВМ непосредственно
(в случае терминала) или посредством клавишного перфорато-
ра. Другие способы ввода информации оператором включают
переключатели, сенсорные экраны и устройство ввода коорди-
нат («мышь»). Движение «мыши» по твердой поверхности
передает движение курсору на экране дисплея. Широко изучае-
мые в настоящее время устройства, узнающие речь человека,
уже сейчас позволяют передавать некоторые команды голосом.
Ввод предварительно записанных программ или данных осу-
ществляется с помощью одного из описанных выше периферий-
ных запоминающих устройств.
Удобство и гибкость связи между оператором и ЭВМ в
сочетании со способностью ЭВМ хранить и обрабатывать огром-
ное количество данных можно выгодно использовать в биотех-
нологии и других отраслях промышленности в нескольких
направлениях. Во-пёрвых, данные о ходе процесса, состоянии
системы и регулирующих устройств могут быть сообщены
оператору в четкой и эффективной форме, что в свою очередь
дает ему возможность гораздо внимательнее и детальнее сле-
дить (в соответствии с его командами) за теми или другими
конкретными характеристиками процесса. Во-вторых, сигналы
тревоги и контрольные сигналы ЭВМ могут обратить внимание
оператора на существующие и только что возникшие отклоне-
ния и неполадки. Наконец, оператор обладает очень большими
возможностями в подборе параметров регулирующих устройств.
Контрольно-измерительная аппаратура и управление
209
а при необходимости и в замене алгоритмов или даже конст-
рукции таких устройств.
Существующие нормы и правила обычно предписывают сбор
и хранение детальной информации о каждом периодическом
процессе, связанном с производством лекарственных препара-
тов. Применение ЭВМ значительно облегчает решение этой
задачи и работу по подготовке отчета о процессе. Если аппара-
ты, участвующие в каком-либо процессе, связаны с ЭВМ, то
это значительно ускоряет и облегчает сбор данных о расходе
питательных веществ и энергии, времени каждой операции,
выходах продуктов и других величинах и параметрах, необ-
ходимых для проведения исследовательских и конструкторских
работ, для руководства всем предприятием, а также для обес-
печения максимальной экономической эффективности производ-
ства. Одни только преимущества, связанные с хранением необ-
ходимой информации и составлением отчетов, послужили до-
статочным основанием, чтобы несколько фармацевтических
компаний сочли экономически целесообразным контролировать
ход процессов с помощью ЭВМ.
Тип применяемого интерфейса датчик — ЭВМ зависит от
определяемого параметра и природы сигнала датчика. Некото-
рые важные переменные процесса имеют только два состоя-
ния— «включено» и «выключено». Это относится, в частности,
к работе насосов, компрессоров, клапанов. Такие переменные
ЭВМ считывают непосредственно в цифровой форме. В цифро-
вой форме в ЭВМ могут передаваться также данные об импуль-
се, генерируемом тахометром или массовым расходомером при
данной скорости вращения вала или заданной массе. Соответ-
ствующие скорости затем определяют путем подсчета числа
импульсов в единицу времени.
С помощью многих детекторов контролируют параметры,
изменяющиеся непрерывно в определенном диапазоне величин.
В таких случаях помимо аналогового измерительного устрой-
ства устанавливают аппаратуру для измерения изменения си-
гнала детектора (силы тока или напряжения) во времени и
регистрации результатов измерений на диаграммной бумаге
с помощью самописца или для передачи сигнала электронному
регулятору. Аналоговые сигналы вводятся в вычислительную
машину с помощью аналого-цифрового преобразователя
(АЦП). Обычно аналоговый сигнал детектора аналитической
аппаратуры должен быть соответствующим образом согласо-
ван, т. е. преобразован, усилен или ослаблен, а иногда сигнал
силы тока преобразован в сигнал напряжения; только после
этого аналоговый сигнал подается в аналого-цифровой преобра-
зователь, принимающий его в заданном диапазоне величин.
Разрешение аналогового сигнала непосредственно связано с
14—746
210
Глава 10
РИС. 10.15. Схема газоанализатора на основе микро-ЭВМ. Здесь символами
КГ-1 и КГ-2 обозначены градуировочные газы (с повышением и снижением
концентрации соответственно), которые можно время от времени вводить
вместо анализируемого газа, отходящего из биореактора. Этим переключени-
ем может управлять ЭВМ с помощью реле с втяжным сердечником (Р-1 и
т. д.). [Воспроизведено с разрешения из статьи: Forrest Е. Н., Jansen N. В.,
Flickinger М. С., Tsao G. Т., A Simple Hobby Computer-Based Off-Gas Analysis
System; Biotech. Bioeng., 23, 455 (1981).]
количеством содержащейся в нем информации. Если цифровой
сигнал содержит п битов, то диапазон поступающего аналого-
вого сигнала будет разделен преобразователем на 2п дискрет-
ных поддиапазонов.
На рис. 10.15 представлена схема лабораторной системы
сбора данных на основе микро-ЭВМ. Как мы уже указывали,
выходные аналоговые сигналы Ог- и СОг-анализаторов адапти-
руют до поступления в аналого-цифровой преобразователь.
Интересно отметить, однако, что здесь сигнал (напряжение),
Контрольно-измерительная аппаратура и управление 21Г
генерируемый детектором па растворенный кислород, вводится
непосредственно в усилитель, минуя какое-либо промежуточ-
ное устройство или блок считывания показаний. Изменяя неко-
торые функции аналоговых и контрольно-измерительных при-
боров, можно реализовать все рассмотренные выше возмож-
ности микро-ЭВМ и с меньшими затратами получить те же
данные, что и с помощью традиционного набора аналоговых
приборов.
Для введения в ЭВМ нескольких аналоговых сигналов ис-
пользовались два подхода. В первом из них для ввода каждого
сигнала применяется свой аналого-цифровой преобразователь
(обычно в одном блоке АЦП имеется несколько таких прибо-
ров). В другом подходе может использоваться сканирующее
устройство с несколькими реле (релейный мультиплексор),
которое путем переключения выбирает определенный сигнал и
передает его единственному аналого-цифровому преобразовате-
лю. Первый подход за счет нескольких АЦП обеспечивает более
высокую скорость выборки и считывания каждого аналогового
сигнала. В последнее время аналитическую аппаратуру все
чаще снабжают цифровыми выходами, обычно в виде двоич-
ного цифрового сигнала, который может быть непосредственно
введен в ЭВМ.
Интерфейсы типа ЭВМ—исполнительный механизм анало-
гичны только что рассмотренным интерфейсам измерительная
аппаратура—ЭВМ. Цифровые выходные сигналы включают и
выключают реле и шаговые двигатели. Цифро-аналоговые пре-
образователи (ЦАП) направляют исполнительным механизмам
сигналы, соответствующие численным величинам выходных
сигналов ЭВМ. На рис. 10.16 приведена схема соединений и
интерфейсов в системе ЭВМ—пилотная установка. Хотя по
сложности и сфере применения обе системы существенно отли-
чаются друг от друга, основные блоки, потоки информации и
сигналов в системе ЭВМ—пилотная установка и микро-ЭВМ—
газовый анализатор (рис. 10.15) очень близки.
10.4.3. Системы программного обеспечения
Под программным обеспечением подразумевается набор
программ и команд, с помощью которых осуществляется управ-
ление работой ЭВМ, соответствующих интерфейсов и перифе-
рийных устройств. Программное обеспечение является одним
из основных компонентов системы ЭВМ—микробиологический
процесс. Именно программы определяют работу ЭВМ, а в ко-
нечном итоге и ход биопроцесса. От программного обеспече-
ния зависит, какие данные будут отображаться и храниться в
памяти ЭВМ и в каком формате, какие команды или вмеша-
14*
21'?
Глава 10
тельства оператора необходимы или допустимы, а в некоторых
случаях, как будет протекать процесс. Если вы имеете опыт
работы с системами типа ЭВМ—аппарат или ЭВМ—процесс, то
вы должны знать, что необходимым условием успешного функ-
ционирования системы являются хорошие программы и что
РИС. 10.16. Схема ЭВМ, управляющей пилотным биореактором (УПП — уст-
ройство построчной печати, ФС — фиксированные соединения, СП — система
переключения, Р—распределитель). Контактные сенсоры получают информа-
цию об аналитическом оборудовании и команды от оператора. [Воспроизве-
дено с разрешения из статьи: Meiners М., Rapmundt W., Some Practical Aspects
of Computer Applications in a Fermentor Hall; Biotech. Bioeng., 25, 809
(1983).]
разработка программ может быть самой трудной задачей и
самым дорогостоящим элементом при создании системы биопро-
цесс—ЭВМ. Такое соотношение затрат в последнее время усу-
губляется еще и постоянным снижением цен на ЭВМ. Поэтому
как исследователям, так и руководителям предприятий следует
тщательно изучать возможности имеющегося программного
обеспечения, имея в виду его вероятное использование в других
ЭВМ, а также сравнивать цены программного и аппаратного
обеспечения ЭВМ и выбирать оптимальный вариант.
Операционная система ЭВМ контролирует выполнение про-
грамм, хранение файлов, инвентаризацию и распределение
памяти и координирует эти функции. Имеется несколько типов
Контрольно-измерительная аппаратура и управление
213
программ для ЭВМ: сервисные программы, выполняющие вспо-
могательные функции (запуск системы, создание файлов);
языковые программы, позволяющие использовать языки более
высокого уровня (БЕЙСИК, ФОРТРАН, АПЛ и другие); при-
кладные программы, предназначенные для выполнения специ-
альных расчетов и других прикладных задач. Некоторые из за-
дач прикладных программ (сбор данных, передача информации
оператору, составление отчета) мы уже обсуждали, а другие
задачи таких программ (обработка данных, управление процес-
сом) мы рассмотрим в остальных разделах этой главы. Множе-
ство примеров конкретных алгоритмов и функций прикладных
программ приведено в литературе, перечисленной в конце главы.
При выборе той или иной конкретной системы аппаратного и
программного обеспечения важно учитывать ее способность
выполнять несколько задач одновременно и работать в режиме
с разделением времени. Если система обладает такой способ-
ностью, то она, по сути дела, может выполнять несколько про-
грамм одновременно, например определять параметры процес-
са, анализировать полученные данные и управлять несколькими
операциями. Такие системы, кроме того, позволяют записы-
вать новые и отлаживать или модифицировать старые програм-
мы, не прерывая управление операциями. Способность ЭВМ
решать несколько задач одновременно существенна на втором
и третьем уровнях (рис. 10.14), а специализированные системы
сбора и анализа данных на базе микро-ЭВМ (рис. 10.15) не
обязательно должны работать в режиме с разделением времени.
Ознакомившись с основными положениями, касающимися
систем процесс—ЭВМ, перейдем теперь к рассмотрению при-
менения ЭВМ и полученных с их помощью данных для изуче-
ния состояния биопроцесса и создания условий, обеспечиваю-
щих его максимальную производительность.
10.5. Анализ данных
Хотя в настоящее время удается измерить лишь ограничен-
ное число параметров системы в биореакторе, все же на осно-
вании этих параметров в сочетании с уравнениями общего ма-
териального и энергетического балансов и с уравнениями со-
ответствующих математических моделей можно рассчитать зна-
чения тех переменных и параметров биопроцесса, которые нель-
зя наблюдать и измерить непосредственно. Как сказал в 1971 г.
Хамфри, совокупность всех результатов измерений открывает
«доступ» к другим характеристикам, которые нельзя наблюдать
непосредственно. Мы уже говорили, например, что измерение
скоростей потоков газа на входе в реактор и выходе из него
в сочетании с определением состава газов позволяет вычис-
214
Глава 10
лить средний коэффициент объемной скорости массопередачи
kia в биореакторе (см. разд. 8.2.1). Кроме того, в разд. 5.10 мы
показали, что суммарные изменения концентраций компонентов
системы можно рассчитать с помощью уравнений общего ма-
териального баланса для клеточного роста, если известна об-
щая стехиометрия последнего. На рис. 10.17 представлена схе-
Регистрсщия
РИС. 10.17. Результаты непосредственных первичных измерений (показанных
в верхней части рисунка) могут служить основой для расчета различных ха-
рактеристик и параметров процесса. [Воспроизведено с разрешения из рабо-
ты: Таппеп L. Р., Nyiri L. К., Instrumentation of Fermentation Systems, in Mic-
robial Technology, Peppier H. J., Perlman D. (eds.), 2d ed., vol. II, p. 331, Aca-
demic Press, Inc., New York, N. Y., 1979.]
ма, на которой показано, как с помощью непосредственно из-
меряемых параметров можно рассчитать характеристики про-
цесса, связанные с перемешиванием и аэрацией, клеточным ро-
стом и метаболизмом.
Основной целью разработки методов анализа данных явля-
ется непрерывное определение концентрации биомассы и удель-
ной скорости клеточного роста в ходе микробиологического
процесса. В этом разделе на примере определения концентрации
биомассы мы продемонстрируем несколько подходов к анали-
зу данных о системе в биореакторе.
10 .5.1. Сглаживание и интерполяция данных
Часто на результаты измерений накладывается шум. Кро-
ме того, существенные флуктуации результатов измерений при-
водят к тому, что непосредственные показания прибора уже
Контрольно-измерительная аппаратура и управление
215
и щ — па-
частности,
двумя от-
(10.5)
недостаточно точно отражают мгновенное значение определяе-
мого параметра. В таких случаях необходимы обработка или
сглаживание сигнала.
В простейшем способе сглаживания данных применяют
фильтр первого порядка (аналогичный резистивно-емкостному
фильтру), который равномерно ослабляет флуктуации входно-
го сигнала, причем степень ослабления возрастает по мере по-
вышения частоты флуктуаций. Такой фильтр характеризуется
постоянной времени tf, входные сигналы, частота которых на-
много меньше tf-1, ослабляются до пренебрежимо малой вели-
чины на выходе, а флуктуации с частотами, намного превыша-
ющими tf-1, проходят через фильтр практически без измене-
ний. В дискретно-временной форме, удобной для ввода в циф-
ровую ЭВМ, действие фильтра описывается уравнением [2]
Wk=aouk+aiUk-i — bxWk-\ (10.4)
где и— сигнал на входе в фильтр (сигнал с шумами); w —
сигнал на выходе из фильтра; индексы k и k—1 обозначают
время анализируемого и предыдущего отсчета; До, Щ
раметры, определяющие характеристики фильтра. В
tf связано с временем отсчета Ts (временем между
счетами) и параметром bi уравнением
t -
Другой расчетный метод, называемый определением сколь-
зящего среднего, принципиально проще, но позволяет добить-
ся примерно такой же эффективности сглаживания. В этом ме-
тоде измерения проводят чаще и усредняют какое-либо задан-
ное число последовательных измерений (например, 10 измере-
ний); представительным результатом измерений для всего диа-
пазона времени измерений считают это среднее значение. Та-
кой подход представляется вполне оправданным в тех случа-
ях, когда значительное изменение определяемых параметров
происходит за значительно больший промежуток времени, чем
время, необходимое для однократного измерения.
Джефферис и сотрудники [35] предложили анализировать
данные о процессах ферментации с помощью более сложного
фильтра, обеспечивающего также полиномную интерполяцию.
В качестве примера рассмотрим применение этого фильтра для
оценки концентрации биомассы и скорости клеточного роста по
зашумленным результатам периодических турбидиметрических
измерений. Изменение концентрации биомассы во времени в
течение временного интервала т, предшествовавшего измере-
нию, можно описать полиномом второго порядка
х(/)=а1 + аг/+аз/2 (10.6)
216
Глава 10
Нам необходимо определить значения коэффициентов аь а2.
и аз с помощью измеренного значения х, x(tk), найденных ра-
нее значений х и, возможно, той или иной информации о ха-
рактере шумов. Для этого можно, например, методом наимень-
ших квадратов определить отклонения от величин, предсказы-
ваемых уравнением (10.6), в интервале времени от tk—х до
Такой рекурсивный метод обработки данных описан в работе
[35]. Другие методы обработки зашумленных данных рассмот-
рены в литературе, приведенной в конце главы.
10 .5.2. Оценка параметров и состояния системы
Если накоплением кислорода в реакторе пренебречь, то-
уравнение материального баланса по кислороду для периоди-
ческого процесса принимает форму
FfcO2,f ~FecO<i,e
=Qo2 “ ^о2/хх+
1_____dx
Ex/O2 dt
(10.7>
где индексами f и е обозначены параметры на входе в реактор
и на выходе из него соответственно. В этом уравнении учтен
расход кислорода как на клеточный рост, так и на метаболизм
поддержания жизнедеятельности клеток. Путем определения
состава газов, поступающих в реактор и выходящих из него,.
можно экспериментально определить скорость утилизации кис-
лорода Qo2 (в литературе этот параметр часто сокращенно?
обозначают OUR). Если зависимость Qo2 от времени опреде-
лена и если принять, что коэффициенты Л4о2/х и Ух/о2 постоян-
ны, то после интегрирования уравнения (10.7) получим
х
Ух/О2 ехр (Л1О2/хУх/о2т) QO2(t) dx
(10.8>
Теперь с помощью этого уравнения можно по найденной зави-
симости Qo„ (0 определить зависимость x(t), а затем по урав-
нению (10.7") найти скорость роста dx/dt и удельную скорость
клеточного роста.
Прежде чем применять описанный метод расчета к конкрет-
ному процессу, необходимо определить стехиометрические пара-
метры утилизации кислорода. Для этого проинтегрируем урав-
нение (10.7) по времени и после преобразований получим [29]
t
f Qo2 (т) dx
b_____________
t
j x (t) dx
о
= Мэ2/х + у ~
Ex/O2
r.v(/)-x(0)
(10.9)
Контрольно-измерительная аппаратура и управление 217
Если величины Qo2 и x(t) определены в независимом автоном-
ном эксперименте, то параметры Л4оа/х и Ух/оа можно найти по
графику зависимости левой части уравнения (10.9) от выраже-
ния, стоящего в квадратных скобках правой части того же
уравнения.
Забриски и Хамфри [36] описанным методом изучали пе-
риодические культуры нескольких видов микроорганизмов. На
рис. 10.18 представлены результаты изучения роста культур
РИС. 10.18. Экспериментально найденная (точки) и вычисленная (сплошная
кривая) зависимости изменения концентрации биомассы от времени в периоди-
ческих культурах Streptomyces sp. (а) и Saccharomyces cerevisiae (б). [Вос-
произведено с разрешения из статьи: Zabriskie D. W„ Humphrey А. Е., Real-
Time Estimation of Aerobic Batch Fermentation Biomass Concentration by
•Component Balancing; AIChE J., 24, 138 (1978).]
Streptomyces (а) и Saccharomyces cerevisiae (б), приведены
величины экономических коэффициентов и коэффициентов под-
держания для каждой культуры, а также коэффициент откло-
нения v экспериментальных данных (точки) от концентрации
биомассы, рассчитанной по уравнению (10.8) (непрерывная
кривая). Очевидно, что соответствие между экспериментальны-
ми и расчетными данными очень хорошее в случае культуры
Streptomyces. Что же касается 5. cerevisiae, то здесь наблюда-
ются заметные количественные расхождения между найденны-
ми и вычисленными величинами. Дело в том, что в последней
культуре экспериментально обнаружено явление диауксии, от-
ражающейся на общей скорости клеточного роста, но не учи-
тываемой в расчетах по уравнениям материального баланса.
Расхождения между найденными и вычисленными величинами
объясняли изменением Л1о2/х и Ух/о2, сопровождающим вариа-
218
Глава 10
ции в характере метаболизма утилизации глюкозы в ходе мик-
робиологического процесса [36].
Приведенный пример можно считать типичным для большо-
го числа методов анализа данных на основе уравнений общего
материального баланса. Эти методы успешно выполняют роль
связующего звена между непосредственно измеряемыми пара-
метрами и важными переменными процесса, определяемыми
путем расчета. Возможные трудности здесь могут быть связа-
ны с накоплением погрешностей.
Общие методы оценки состояния процесса, т. е. методы оп-
ределения переменных, входящих в дифференциальные уравне-
ния соответствующей математической модели, разработаны спе-
циалистами в области систем управления и прикладной мате-
матики. Как и рассмотренные выше методы на основе уравне-
ний материального баланса, эти более совершенные методы
учитывают известные взаимосвязи между переменными процес-
са. В то же время в общих методах оценки состояния процесса
учитываются и шумовые эффекты и размножение ошибок, т. е.
математический аппарат этих методов базируется на теории
стохастических процессов.
Изучение современных способов многомерного оценивания
переменных, например модифицированного фильтра Калмана,
не входит в нашу задачу. Мы ограничимся лишь тем, что отме-
тим некоторые особенности фильтра Калмана, имеющие осо-
бенно большое значение при его использовании для управле-
ния биореакторами, которые были выявлены Саном и Стефано-
пулосом в ходе теоретических и экспериментальных исследова-
ний [37, 38].
1. Оценка быстро и точно реагирует на изменения параметров
процесса.
2. Оценка не чувствительна к погрешностям начальных значе-
ний параметров состояния системы.
3. Метод оценки состояния можно непосредственно распростра-
нить и на оценку изменяющихся во времени параметров про-
цесса.
4. При работе с оценками такого типа не возникает проблем,
связанных с ростом и распространением погрешностей.
На рис. 10.19 представлены результаты проверки предло-
женного Саном и Стефанопулосом [31] метода оценки состоя-
ния системы с помощью модифицированного фильтра Калмана.
В этом тесте в математической модели хемостата ступенчато
повышали скорость разведения, причем переменные модели бы-
ли разрушены генерируемым ЭВМ шумом до их оценки. Обу-
словленное разведением изменение плотности и удельной ско-
рости роста клеточной массы прослеживается с достаточной
Контрольно-измерительная аппаратура и управление
219
точностью, даже если скорость разведения превысит цтах
и начнется процесс вымывания.
Данные, полученные путем непосредственного измерения,
переменные и параметры процесса, найденные расчетным путем
на основе этих данных, дают необходимую для управления
РИС. 10.19. Сравнение вычисленных на основе математической модели
(сплошная линия) и оцененных с помощью устройства оценки на основе мо-
дифицированного фильтра Калмана (отмечены крестиками) параметров куль-
туры микроорганизма в ПРПП при последовательном ступенчатом повышении
скорости разведения. [Воспроизведено с разрешения из статьи: San K.-Y.,
Stephanopoulos G., Studies of On-Line Bioreactor Identification II. Numerical
and Experimental Results; Biotech. Bioeng., 26, 1189 (1984).]
процессом информацию. В следующих разделах мы рассмот-
рим различные способы регулирования параметров находящей-
ся в реакторе системы, обеспечивающие оптимальное течение
процесса.
10.6. Управление процессами биохимической
технологии
Для успешного осуществления биотехнологического про-
цесса необходимо условия и параметры процесса поддер-
живать на уровне проектных величин. Для этого в свою оче-
220
Глава 10
редь необходимо регулировать указанные в проекте условия
и параметры, поскольку они могут непредсказуемым образом
меняться в силу неизбежных флуктуаций скоростей подачи ис-
ходных веществ, скорости перемешивания и других эксплуата-
ционных параметров, а также в силу изменения химического-
состава системы в ходе процесса. Иногда ход периодического-
процесса можно оптимизировать путем изменения значений
ряда параметров системы, например pH или температуры, в
соответствии с заданной программой. Для программированного
изменения параметров процесса также необходимы соответст-
вующие методы управления. В этом разделе мы сначала рас-
смотрим способы поддержания на заданном уровне непосред-
ственно измеряемых параметров, а затем перейдем к изучению
способов управления процессом на базе расчетных параметров.
10.6.1. Непосредственное управление процессами
Если процесс осуществляется, например, в биореакторе, то
часто возникает необходимость в регулировании pH, темпера-
туры, скорости аэрации и перемешивания, иногда также парци-
ального давления растворенного кислорода с тем, чтобы вели-
чины этих параметров соответствовали заданным значениям.
Поскольку все эти параметры можно определять в ходе процес-
са постоянно, в реальном масштабе времени, то для регулиро-
вания каждого из них можно использовать обычный регулятор
с обратной связью, основные элементы которого изображены
на рис. 10.20. Здесь измеряется регулируемый или выходной
параметр, например pH, и сигнал анализатора вводится в ре-
гулирующее устройство, в котором величина измеренного сиг-
нала сравнивается с заданной или необходимой величиной. Да-
лее по найденному отклонению между измеренной и заданной
величинами с помощью того или иного алгоритма регулируется
собственно параметр процесса.
Регулирование может выполнять и оператор, наблюдающий
за показаниями прибора и в зависимости от этих показаний
принимающий конкретное решение. Чаще, однако, регулирова-
ние осуществляется пневматическим или электронным устрой-
ством или цифровой ЭВМ. Простейшим типом управления яв-
ляется двухпозиционное регулирование (регулирование по прин-
ципу «включено — выключено»). В этом случае исполнитель-
ный механизм включается, когда разность между измеренной
и заданной величинами превышает установленный предел,
и выключается, когда она становится ниже другого установлен-
ного предела, или наоборот. Такой тип регулирования применя-
ют тогда, когда блок управления или регулирующее устройст-
во, непосредственно воздействующее на процесс с целью кор-
Возмущение
РИС. 10.20. Основные элементы системы управления с обратной связью,
222
Глава 10
ректирования того или иного параметра, также являются двух-
позиционными, как, например, в случае односкоростных насо-
сов. При регулировании pH по принципу двухпозиционного уп-
равления насос, подающий основание в биореактор, включает-
ся при снижении pH ниже заданного значения на определенную
величину (обычно 0,25 единицы pH). Если же pH становится
выше заданного значения на 0,25 единицы, то насос выключа-
ется. (В последнем случае можно подключить насос, подающий
в систему кислоту, но обычно в этом нет необходимости, по-
скольку в ходе большинства микробиологических процессов pH
среды понижается.) Таким же способом обычно регулируют
температуру (по крайней мере в небольших реакторах).
Если параметры регулирующего устройства, например уста-
новленного на валу мешалки электродвигателя с переменной
скоростью вращения или регулирующего клапана на линии по-
дачи воздуха, можно изменять непрерывно, то в таких случаях
обычно применяют пропорционально-интегрально-дифференци-
альное регулирование или один из вариантов регулирования
этого типа. В данном случае выходной сигнал регулирующего
устройства описывается уравнением
о(0=Оз + Кс е(04
J e(w) dw-\-xD
de (t)
dt~
(10.10)
Здесь Os — номинальный выходной сигнал регулирующего уст-
ройства в невозмущенном состоянии (в проектных условиях),
a e(t) —погрешность:
в (0 = (заданное значение — измеренное значение) в момент времени t
(10.11)
Из уравнения (10.10) следует, что регулирующее действие
пропорционально сумме трех слагаемых: погрешности, интег-
рала погрешности и производной погрешности. Относительная
важность этих трех типов регулирования определяется пара-
метрами т/ и то, называемыми интегральным и дифференци-
альным временем соответственно. Общая «сила» регулирующе-
го действия определяется величиной коэффициента усиления
Кс- Если дифференциальное время равно нулю (то = 0), то ре-
гулятор называют пропорционально-интегральным. Возможны
и другие варианты.
Хорошо настроенный регулятор этого типа часто обеспечи-
вает очень хорошее управление измеряемой переменной. На-
против, плохо настроенный регулятор может дестабилизировать
систему, вызывая нежелательные сильные флуктуации. На-
стройке пропорционально-дифференциально-интегральных регу-
Контрольно-измерительная аппаратура и управление
223
ляторов посвящено множество учебных пособий и руководств
по регулированию процессов (см., например, работы [1—3]).
Обычно в управлении процессом участвует несколько регу-
ляторов этого типа, один из которых по данным измерения тем-
пературы регулирует скорость охлаждения, другой регулирует
pH и т. д. В связи с постоянным удешевлением цифровых ЭВМ
РИС. 10.21. Автоматическая система управления концентрацией растворенно-
го кислорода, базирующаяся на определении состава отходящих газов н кон-
центрации растворенного кислорода. (САРС — система автоматического регу-
лирования скорости). [Воспроизведено с разрешения из работы: Таппеп L. Р.,
Nyiri L. К., Instrumentation of Fermentation Systems, in Microbial Technology,
Peppier H. J., Perlman D. (eds.), 2d ed., vol. II, p. 331, Academic Press, Inc.,
New York, 1979.]
в последние годы управление несколькими регулирующими
один параметр цепями обратной связи успешно осуществляется
с помощью одной микро-ЭВМ. Если выходной сигнал ЭВМ
(обычно после цифро-аналогового преобразователя или реле)
используется непосредственно для включения исполнительного
механизма, то такую систему называют системой с прямым
цифровым управлением.
Преимуществом регулирующих устройств на основе ЭВМ
является возможность сочетания способности компьютера
к анализу данных с управлением процессом на базе определе-
ния нескольких параметров. В качестве примера на рис. 10.21
изображена схема одной из таких систем, в которой измерение
концентрации растворенного кислорода и концентрации кисло-
.224
Глава 10
рода в отходящих газах позволяет определять в реальном
масштабе времени величину kia\ полученную таким путем ин-
формацию вместе с результатами определения концентрации
растворенного кислорода можно использовать для соответст-
вующего регулирования скорости вращения мешалки и (или)
скорости поступающего в реактор потока газа с тем, чтобы
концентрация О2 в растворе поддерживалась на необходимом
уровне. Эта система относится к типу систем с прямым цифро-
вым управлением. Обратите внимание на то, что здесь аналого-
цифровой преобразователь принимает сигнал в виде изменяю-
щейся силы тока, что вызывает необходимость преобразовывать
сигнал напряжения в сигнал силы тока. Чтобы избежать су-
щественных потерь в линии передачи сигнала, если только они
не слишком короткие (как это бывает, например, в лаборатор-
ных установках), электрические аналоговые сигналы лучше пе-
редавать в виде силы тока.
К проблеме использования найденных расчетным путем па-
раметров состояния системы для управления процессом мы
еще вернемся в следующем разделе.
10.6.2. Каскадное управление метаболизмом
Конечной задачей системы управления любым биореактором
с культурой клеток является обеспечение таких условий, ко-
торые в конце концов способствуют максимальному использо-
ванию систем регуляции метаболических путей клеток для эф-
фективного образования необходимых соединений. Поэтому
вместо того, чтобы все усилия направлять на поддержание за-
данных величин температуры и pH, иногда более целесообраз-
но путем регулирования системы в биореакторе создать опре-
деленную скорость клеточного роста или заданную величину
дыхательного коэффициента. В принципе это вполне возможно,
поскольку, как мы показали в разд. Ю.5, некоторые характе-
ристики метаболических процессов можно рассчитать непосред-
ственно по измеряемым параметрам. Затем, как показано на
приведенной на рис. 10.22 схеме, вычисленную величину харак-
теристики метаболизма можно сравнить с заданной величиной.
Расхождение между вычисленной и заданной величинами опре-
деляет степень «метаболического регулирования» (рис. 10.22),
осуществляемого, например, с помощью одного из только что
•описанных алгоритмов регулятора обратной связи.
Выходной сигнал регулятора метаболизма может использо-
ваться для непосредственного регулирования одного из пара-
метров процесса, например скорости подачи питательных ве-
ществ. В альтернативном варианте с помощью регулятора ме-
таболизма можно изменять установленное значение «регулято-
Контрольно-измерительная аппаратура и управление
225
ра окружения», например величину pH или концентрацию
растворенного кислорода. Соответственно изменение pH или
концентрации растворенного О2 в хорошей системе управления
процессом приведет затем к уменьшению отклонения перемен-
ной метаболизма от заданной величины.
РИС. 10.22. Поток информации в каскадной системе управления, в которой
изменение параметров окружения определяется отклонениями от заданных
характеристик метаболизма (DO — концентрация растворенного кислорода;
F—скорость потока газа; S — субстрат; +S — скорость добавления субстра-
та; Р — давление или продукт процесса; N — скорость перемешивания; Qo4
Qco2 — скорости утилизации соответствующих газов). [Воспроизведено с раз-
решения из работы: Таппеп L. Р„ Nyiri L. К., Instrumentation of Fermentation
Systems, in Microbial Technology, Peppier H. J., Perlman D. (eds.), 2d ed.,
vol. II, p. 331, Academic Press, Inc., New York, 1979.]
Если переменные «окружения» регулируются локальными
регуляторами с одной цепью обратной связи, а значения регу-
лятору окружения задает ЭВМ, то такая система называется
системой с диспетчерским управлением или с цифровым управ-
лением по заданным значениям. Конечно, все эти функции мо-
гут выполнять системы цифровая ЭВМ—регулятор. С такой си-
стемой, в которой ЭВМ первого уровня сложности управляют
переменными окружения, а определенные величины последних
задаются ЭВМ более высокого уровня, мы уже знакомы (см.
рис. 10.14).
15—746
226
Глава 10
Интересным примером регулирования метаболизма путем
соответствующего управления окружением популяции клеток
может служить контроль с помощью ЭВМ периодической куль-
туры пекарских дрожжей (Saccharomyces cerevisiae) с добав-
лением субстрата. Катаболизм глюкозы в этом организме ре-
гулируется очень сложным путем. При соответствующих кон-
центрациях глюкозы и растворенного кислорода глюкоза рас-
ходуется на дыхание, что обеспечивает максимальный выход
биомассы в расчете на единицу количества утилизированной
глюкозы (вспомните разд. 5.3). Если концентрация глюкозы
возрастает выше некоторого порогового значения, то метабо-
лизм дрожжевых клеток переключается на ферментацию даже
в присутствии кислорода. Этот процесс, называемый аэробной
ферментацией, обусловлен особой регуляцией метаболической
системы, называемой эффектом Крэбтри. При аэробной фермен-
тации выход биомассы (отнесенный к количеству утилизирован-
ной глюкозы) сокращается, а в качестве конечных продуктов
метаболизма образуются этанол и СОг. Как отмечалось выше
(разд. 7.2.3), этанол ингибирует рост дрожжевых клеток.
Поэтому если целью процесса является получение дрожже-
вой биомассы (как это часто и бывает в реальном процессе),
то аэробной ферментации следует избегать. Для этого можно,
например, постепенно добавлять глюкозу в ходе периодическо-
го процесса. Соответствующую программу введения глюкозы в
систему можно составить заранее по опыту работы с аналогич-
ными процессами. Поскольку, однако, характеристики как по-
севного материала, так и среды (которую в производственных
процессах готовят на основе мелассы, а не чистой глюкозы)
могут меняться от загрузки к загрузке, то заданная программа
введения глюкозы может не удовлетворять изменяющимся
в ходе процесса потребностям культуры в глюкозе, что приво-
дит к повышению концентрации сахара в культуре и как след-
ствие к развитию аэробной ферментации и снижению выхода
биомассы.
Для того чтобы глюкоза поступала в систему только в необ-
ходимом в данное время для данной загрузки количестве,
Ванг, Куни и Ванг [39, 40] контролировали ход процесса
ферментации путем непрерывного определения соотношений
различных компонентов в системе и соответственно регулиро-
вали скорость введения раствора глюкозы. Оказалось, что
удобным индикатором процесса утилизации глюкозы является
дыхательный коэффициент RQ [уравнение (5.51)]. Если вели-
чина RQ превышает 1,0, то это свидетельствует о преобладании
процесса образования этанола. Если величины RQ меньше 0,6
или равны 0,7—0,8 или 0,9—1,0, то это свидетельствует о пре-
Контрольно-измерительная аппаратура и управление
227
имущественной утилизации этанола, эндогенном метаболизме
или окислительном росте клеток соответственно.
Ванг и другие [39, 40] описывали процессы образования
этанола и биомассы пекарских дрожжей следующими стехио-
метрическими уравнениями:
аСбН^Об+й О2+С NH3 —>- СбНю.эМьозОз.об + е НгО+f СО2 (10.12)
С6Н12О6 —* 2C2HsOH + 2CO2 (10.13)
Эмпирическая формула биомассы [стехиометрический коэффи-
циент которой в уравнении (10.12) равен единице] найдена
путем определения элементного состава дрожжей на различ-
ных стадиях периодического процесса. Такое описание системы
включает семь неизвестных: пять стехиометрических коэффици-
ентов в уравнении (10.12) и относительные степени утилизации
глюкозы в процессах (10.12) и (10.13). Все семь неизвестных
можно определять в ходе процесса, исходя из уравнений мате-
риального баланса по С, Н, О и N в соответствии с уравнени-
ем (10.12), а также из результатов непосредственного измере-
ния скоростей утилизации О2, выделения СО2 и добавления
NH3.
Регулирование скорости введения глюкозы, чтобы величина
RQ была не более 1,0, привело к результатам, показанным на
рис. 10.23. В этом случае сочетание указанного метода оценки
состояния культуры и регулирования путем введения питатель-
ных веществ дало очень хорошие результаты. Концентрации
этанола и глюкозы в ходе всего процесса, за исключением
очень короткого периода при /«16 ч (когда наблюдалось не-
большое повышение концентрации С2Н5ОН), поддерживались на
низком уровне. Результаты экспериментального определения
плотности биомассы хорошо согласуются с соответствующими
данными, вычисленными на ЭВМ с помощью уравнений мате-
риального баланса.
В других экспериментальных периодических процессах
с добавлением субстрата, в которых наблюдалось образование
большего количества этанола, такого соответствия между вы-
численными и экспериментально найденными величинами полу-
чить не удалось. В таких ситуациях дрожжи расходовали эта-
мол и на клеточный рост, а этот процесс не учитывался в ме-
тодике оценки количества образующейся биомассы. Этот при-
мер указывает на возможные источники погрешностей в мето-
дах, ^базирующихся на непосредственном использовании урав-
нений материального баланса. Однажды допущенные при оцен-
15*
228
Глава 10
РИС. 10.23. Результаты управления периодическим процессом роста дрожжей
с добавлением субстрата (глюкозы); количество добавляемого субстрата ре-
гулировали с помощью ЭВМ по заданной величине дыхательного коэффициен-
та. Приведены измеренные и вычисленные концентрации биомассы. [Воспроиз-
ведено с разрешения из статьи: Wang Н. Y., Cooney С. L„ Wang D. I. С.,
Computer-Aided Baker’s Yeast Fermentations; Biotech. Bioeng., 19, 69 (1977).]
ке состояния системы ошибки впоследствии умножаются, а на-
личие путей метаболизма, неучтенных в принятой стехиометрии
процесса, может привести к совершенно неверным результатам.
10.7. Прогрессивные методы управления
биопроцессами
В завершение нашего краткого обзора методов управления
процессами биохимической технологии и соответствующей конт-
рольно-измерительной аппаратуры рассмотрим некоторые пути
обеспечения максимального выхода продукта в реакторах пе-
риодического действия и регулирования и стабилизации режи-
мов в реакторах непрерывного действия. Кроме того, вкратце
рассмотрим некоторые интересные проблемы, возникающие
при расчете и проектировании биопроцессов, которые состоят
из нескольких последовательных операций. Во всех указанных
случаях регулирование процесса и расчеты, связанные с разра-
боткой соответствующей схемы управления процессом, сущест-
венно упрощаются при использовании ЭВМ.
Контрольно-измерительная аппаратура и управление
229
10.7.1. Программированное управление процессами
в биореакторах периодического действия
Обеспечить максимальный выход продукта в периодическом
процессе с участием определенного штамма микроорганизмов
можно только в том случае, если заранее известны те среда и
условия, в которых этот организм проявляет наибольшую ак-
тивность или, говоря иными словами, в которых генетический
потенциал организмов используется в максимальной степени.
В зависимости от имеющихся контрольно-измерительной аппа-
ратуры и систем управления, проблему оптимизации условий
биопроцесса можно решить различными путями. В простейшем
варианте, например в случае колбы, установленной на качалке,
в ходе процесса обычно не измеряют и не регулируют ни одно-
го параметра, кроме температуры, а управление процессом сво-
дится к нахождению оптимального начального состава среды
и наилучшего температурного режима. Если непосредственно
регулируются только параметры окружения, то оптимизации
процесса в биореакторе может способствовать контроль pH, ин-
тенсивности перемешивания или других параметров окружения.
В таких случаях регулируемый параметр обычно поддержива-
ют на постоянном уровне в ходе всего периодического процес-
са, если только необходимое для данной системы значение па-
раметра вообще достижимо. (Если, например, потребность
культуры в кислороде не обеспечивается даже при максималь-
ной скорости переноса кислорода в биореакторе, то регулиро-
вать концентрацию растворенного кислорода, очевидно, не име-
ет смысла.)
Как известно, однако, во многих случаях постоянство усло-
вий и состава среды не обеспечивает оптимального течения
микробиологического процесса. Так, быстрый рост клеток не
способствует образованию вторичных метаболитов, но в то же
время на начальной стадии периодического процесса низкая
скорость клеточного роста нежелательна, если плотность ис-
ходной культуры невелика. Поэтому сначала условия процесса
должны обеспечивать максимальную скорость клеточного ро-
ста, а затем, когда будет достигнута достаточно высокая кон-
центрация биомассы, их необходимо изменить так, чтобы они
стимулировали образование максимального количества про-
дукта метаболизма. На этой стадии важно учитывать не толь-
ко скорость синтеза продукта метаболизма, но и скорость его
инактивации или разложения.
Аналогично при производстве белков с помощью культур
генетически видоизмененных клеток в начале периодического
процесса обычно выгоднее затормозить биосинтез белка, по-
скольку последний часто ингибирует клеточный рост и может
230
Глава 10
осложнить решение проблемы генетической нестабильности. По
достижении необходимой плотности культуры генетический ап-
парат клеток можно переключить на образование белка путем
введения индуктора (или посредством подавления ингибитора
экспрессии соответствующего гена). Таким образом, режимы
периодических процессов биосинтеза вторичных метаболитов
и синтеза белков с помощью рекомбинантных ДНК могут быть
сходными, несмотря на принципиальные различия в биохимии
и механизмах регуляции этих процессов.
В любой данный момент активность периодической культу-
ры зависит как от состава среды и условий окружения клеток,
так и от предыстории культуры и среды. Для того чтобы куль-
тура к определенному моменту достигала наибольшей продук-
тивности, могут потребоваться определенные профили измене-
ния pH, концентрации растворенного кислорода и других пере-
менных во времени. Часто необходимые зависимости перемен-
ных от времени находят эмпирическим путем. В то же время
достаточно глубоко структурированная математическая модель
процесса в принципе позволяет обеспечить максимальную про-
изводительность путем моделирования и оптимизации с по-
мощью ЭВМ. В последующих разделах мы рассмотрим ряд
примеров, иллюстрирующих преимущества программированного
регулирования периодических процессов, а также методы со-
ставления оптимальных программ для таких процессов.
Образование фермента ^-галактозидазы в периодической культуре Asper-
gillus niger подчиняется тем же закономерностям, что и биосинтез вторичных
метаболитов. Лунделл [41] предложил способ управления этим процессом
с помощью ЭВМ, который базируется на следующих принципах:
Фаза клеточного роста
1. Для повышения концентрации клеточной массы и увеличения продолжи-
тельности фазы роста необходимо добавлять в систему питательное веще-
ство — источник углерода.
2. Источник углерода необходимо добавлять периодически по мере его исто-
щения, что определяют по снижению скорости выделения СОг и дыхатель-
ного коэффициента до 20% от соответствующих максимальных величин.
3, Температуру и pH необходимо установить на уровне, обеспечивающем мак-
симальную скорость клеточного роста.
4. Во избежание излишнего расхода энергии скорость вращения мешалки
и скорость поступления воздуха должны быть минимальными, что опреде-
ляется скоростью переноса кислорода в системе, скоростью выделения СО2
и дыхательным коэффициентом.
Фаза биосинтеза фермента
1. Когда скорость роста клеток упадет (снизятся скорость выделения СОз
и дыхательный коэффициент), режимы процесса следует переключить на
условия, способствующие образованию фермента.
2. Температура и pH должны обеспечивать оптимальную скорость биосинтеза
фермента.
Контрольно-измерительная аппаратура и управление 231
Таблица 10.4. Основные характеристики некоторых периодических процессов,
изучавшихся в ходе работ по оптимизации биосинтеза ^-галактозидазы
штаммом Aspergillus niger. В каждом процессе общее давление составляло
13Э кПа, а концентрация растворенного кислорода 20% от уровня насыщения’
1. Обычный периодический процесс, использовавшийся для сравнения
температура 30 °C
pH 4,5
скорость вращения мешалки 200 об/мин
скорость подачи воздуха 30 м3/ч
2. Периодический процесс с продолжительной фазой образования фермента
(ПФОФ) за счет добавления избыточных количеств органического пита-
тельного вещества
температура 30 °C
pH 4,5
скорость вращения мешалки 175—200—150 об/мин
скорость подачи воздуха 15—30—20 м3/ч
добавление избытка органического питательного вещества
3. Периодический процесс с продолжительной фазой роста биомассы
(ПФРБ) за счет добавления избытка источника углерода в фазе кле-
точного роста
температура 35—30 °C
pH 4,8—4,5
скорость вращения мешалки 175—250—200 об/мин
скорость подачи воздуха 15—30—20 м3/ч
добавление избытка крахмала и (NH4)2HPO4
4. Периодический процесс типа ПФРБ/ПФОФ с продолжительными фазами
роста биомассы и образования фермента
температура 35—30 °C
pH 4,8—4,5
скорость вращения мешалки 175—250—200 об/мин
скорость подачи воздуха 15—30—20 м3/ч
добавление крахмала и (NH4)2HPO4 в фазе роста биомассы
добавление избытка органического питательного вещества в фазе образо-
вания фермента
а Воспроизведено с разрешения из работы: Lundell R., Practical Implementation of
Basic Computer Control Strategies for Enzyme Production, in Computer Application in Fer-
mentation Technology, p. 181. Society of Chemical Industry, London, 1982.
3. Скорость вращения мешалки и подачи воздуха следует снизить до необхо-
димых величин (фаза биосинтеза фермента характеризуется малой ско-
ростью потребления кислорода и пониженной вязкостью бульона).
4. При снижении скорости биосинтеза фермента следует добавить соответст-
вующий индуктор.
5. Для продолжения клеточного роста необходимо добавлять питательные ве-
щества.
6. После добавления поверхностно-активного вещества при резком снижении
скорости образования фермента процесс следует прекратить.
В табл. 10.4 суммированы основные характеристики изученных экспери-
ментально режимов ведения процесса. В периодическом процессе с продолжи-
тельной фазой образования фермента (ПФОФ) питательное вещество добав-
ляют в фазе биосинтеза фермента. В периодическом процессе с продолжитель-
ной фазой роста биомассы (ПФРБ) питательное вещество добавляют не толь-
ко в начале процесса, но и в фазе роста. В процессе ПФОФ/ПФРБ объедине-
Периодический, процесс
Периодический процесс с добавлением субстра'па
□
5,0р 40г-1300
4,0
3,0
2,0
1,0
0,0
д
в
Контрольно-измерительная аппаратура и управление
233
ны условия как первого, так и второго процессов. На рис. 10.24 представлены
профили изменения количества образовавшегося фермента в обычном перио-
дическом процессе и в ПФОФ. Эксплуатационные характеристики всех четырех
процессов перечислены в табл. 10.5. Тщательный подбор оптимальных условий
периодического процесса в сочетании с автоматическим анализом данных и ре-
гулированием позволил повысить выход фермента на 70% и одновременно
снизить расход энергии на 50%• Обратите внимание на то, что в основном
последовательные изменения режима программируются заранее, хотя конкрет-
ный момент переключения с одного режима на другой определяют по резуль-
татам непосредственных измерений и последующего расчета параметров про-
цесса. Последнее обстоятельство очень существенно с практической точки зре-
ния, особенно если принять во внимание, что состав как посевного материала,
так и среды непостоянен и может меняться от одной загрузки к другой.
Известно, что повышение температуры (30 °C) способствует ускоренному
росту плесневого гриба Penicillium, в то время как более низкие температуры
(около 20°C) в большей степени благоприятствуют ускоренному синтезу пе-
нициллина. Ранее промышленное получение пенициллина проводили преиму-
щественно при температуре, лежащей между этими двумя предельными зна-
чениями (от 24 до 25 °C), однако программированное изменение температуры
в ходе периодического процесса позволяет существенно повысить выход пени-
циллина по сравнению с процессом, проводимым при любой постоянной тем-
пературе. Для разработки оптимальной программы изменения температуры
применяли стандартные математические методы [42, 43]; ниже суммированы
наиболее интересные результаты этих работ.
Применение теории оптимального управления требует создания соответ-
ствующей математической модели. На рис. 10.25 крестиками отмечены экспе-
риментальные данные, полученные при проведении ферментации в промышлен-
ных аппаратах при постоянной температуре (25°C). Во избежание отображе-
ния характеристик, относящихся только к какому-либо конкретному процессу,
эти данные приведены в безразмерной форме. На графиках не отражена лаг-
фаза, так как первые данные относятся ко времени, когда после начала про-
цесса прошло 50 ч. Характер изменения измеренных параметров во времени
предполагает следующее общее математическое описание кинетики клеточного
роста и образования продукта метаболизма:
dxi / х, \
— =М1 (10.14)
\ C/g /
rfxg dxr
=b3x1 — bix2~\-Ьъ—^~ (10.15)
РИС. 10.24. Образование ^-галактозидазы в культуре Aspergillus niger в пе-
риодическом процессе (а) и в периодических процессах с добавлением субст-
рата (б и в). Цифрами на двух нижних рисунках обозначены управляемые
ЭВМ изменения режима периодического процесса: 1 — повышение скорости
вращения мешалки; 2 — снижение скорости вращения мешалки; 3 — постоян-
ная скорость вращения мешалки; 4, 5, 6 — добавление органического пита-
тельного вещества; 7 — добавление поверхностно-активного вещества. (Вос-
произведено с разрешения из работы: Lundell R., Practical Implementation of
Basic Computer Control Strategies for Enzyme Production, in Computer Appli-
cations in Fermentation Technology, p. 181, Society of Chemical Industry Lon-
don, 1982.)
а —температура, °C; 6— P-галактозидазная активность; в — концентрация
СО2 в отходящих газах, об.%; г — масса ферментера, кг; д — скорость враще-
ния мешалки, с-1; е — дыхательный коэффициент, кг/кг.
234
Глава 10
Таблица 10.5. Основные результаты некоторых экспериментальных
периодических процессов, изученных с целью оптимизации образования
0-галактозидазы в культуре Aspergillus niger3
Тип процесса Продолжи - Относитель- Расход энер- гии, кВТ’Ч Расход энергии, отнесенный к ко- личеству образо- вавшегося фер- мента, кВт-ч/ед. (в % по сравне- нию с периоди- ческим процес- сом) Количество фермента, об- разовавшего- ся в единицу времени, ед./ч (в % по сравнению с периодиче- ским процес- сом)
тельность фермента- ции, ч ное количе- ство образо- вавшегося фермента, %
Обычный перио- дический про- цесс 72 100 180 1,8(100) 1,39(100)
Периодический процесс с про- должительной фазой образова- ния фермента 115 207 210 1,0(56) 1,80(129)
Периодический процесс с про- должительной фазой роста биомассы 100 163 195 1,2(67) 1,63(117)
Периодический процесс с про- должительными фазами образо- вания фермента и роста биомас- сы 120 283 240 0,85(47) 2,36(170)
а Воспроизведено с разрешения из работы: Lundell R., Practical Implementation of
Basic Computer Control Strategies for Enzyme Production, in Computer Applications in
Fermentation Technology, p. 181, Society of Chemical Industry, London, 1982.
где Xi и x2— безразмерные концентрации биомассы и пенициллина соответ-
ственно. Выражение (10.14) представляет собой логистическое уравнение,
с которым мы познакомились в гл. 7, а уравнение (10.15) является вариантом
рассмотренного ранее уравнения Льюдикина — Пайрета, к которому добавле-
но слагаемое, отражающее процесс деградации пенициллина. О необходимо-
сти введения этого дополнительного члена свидетельствует уплощение кривой
накопления пенициллина при больших t (рис. 10.25). Другие данные также
свидетельствуют о гидролизе пенициллина в водных растворах н о возмож-
ности разрушения пенициллин-синтезирующей системы во времени.
Сравнение вычисленных зависимостей Xi и х2 от времени при различных
значениях параметров Ь\—Ь$ с экспериментальными данными показывает, что
сумма квадратов отклонений для всех экспериментальных данных минималь-
на при 7 = 25 °C, если (?5=0, и тогда
bi = 13,099 = b । о Ь2 = 0,9426 — Ь2$
6з=4,6598 = 6зо Ь< = 4,4555=^40 (10.16)
Для того чтобы установить зависимость этих параметров от температуры,
нужно учесть указанные значения при 25 °C, а также 1) известные оптнмаль-
Контрольно-измерительная аппаратура и управление
235
Относительное время
Относительное время
РИС. 10.25. Результаты экспериментального изучения промышленного пе-
риодического процесса производства пенициллина при 25 °C. [Воспроизведено
с разрешения из статьи: Constantinides A., Spencer J. L., Gaden Е. L.. Jr., Opti-
mization of Batch Fermentation Processes. I. Development of Mathematical Mo-
dels for Batch Penicillin Fermentations; Biotech. Bioeng., 12, 803 (1970).]
ные температуры для скорости роста клеток (30 °C) и скорости синтеза пе-
нициллина (20 °C) и 2) экспериментальные данные, показывающие, что кон-
станта скорости деградации пенициллина описывается уравнением Аррениуса
с энергией активации 12—15 ккал/моль. Перечисленные выше факты позво-
ляют предложить следующие уравнения:
bi(Q)=b!og(e) 1=1,2
g(9) = 1,143[1—0,005(30—9)2]
&з(9) = 1,143ЬЗО[1—0,005(9 — 20)2]
(10.17)
+ (9) = fe40 exp
( 1 1
— 6145 —--------------------
I 273,1 + 9 298
где 9 — температура в градусах Цельсия.
236
Глава 10
Теперь, объединив все указанные уравнения, можно записать математи-
ческую модель процесса в общем виде
dxi (f)
—(°’*2 (0.0(01 ^=1.2 (10.18)
с начальными значениями
х,(0) =0,0294 хй(О)=О (10.19)
После этого нашу основную цель — максимальный выход пенициллина — мож-
но выразить в строгой математической форме. Для этого необходимо найти 9
как функцию t в диапазоне от /=0 до окончания процесса (t=T) так, чтобы
хг(Т) было максимальным. Согласно принципу максимума [5] для оптималь-
ной программы изменения температуры, которую мы обозначим символом 9*,
функция гамильтониана Н
2
Н (9) = (0 h 1*1* (0 > Л2* (0 ’ 01 (10.20)
i=l
должна принимать максимальное значение при 9 = 0* (/) для всех t от 0 до Т.
Надстрочный знак звездочка (*) в уравнении (10.20) указывает, что соответ-
ствующие параметры определены с помощью 0*, а входящие в то же уравне-
ние переменные Л,- удовлетворяют дифференциальным уравнениям
2
А- = _ у Kj (0 dfi [X1 (0 ’ *2 - (10.21)
dt dxi
/=1
и условиям
X1(7')=0 X2(T) = 1 (10.22)
Эти необходимые условия и другие теоретические результаты предпола-
гают следующий итерационный алгоритм для расчета оптимальной темпера-
турной программы:
1. Пусть 0(л)(^) обозначает n-е приближение программы.
2. Решить уравнения (10.14) и (10.15) относительно Х1(л)(/), Х2(л)(/), приняв,
что 0 = 0(п).
3. Используя Xi(n), Х2(п), 0(п), определить зависящие от времени коэффициенты
в правой части уравнения (10.21), проинтегрировать эти уравнения в обрат-
ном направлении в численной форме от t—T до t=0 и таким образом опре-
делить A-i(n)(0, А-2(л)(0 (обратное интегрирование необходимо, чтобы из-
бежать численной нестабильности, встречающейся в тех случаях, когда ин-
тегрирование начинают при ^ = 0).
4. Вычислить
дЯМ (t} у %.(п) (t} dfj(X1W (t),x^ (Q, 0<») (Q) (10 23)
50 U 3 k 3 50
Определить (п-Н)-е приближение оптимальной программы при помощи
0(«+1)(/) = 0(п)(/)_|_8 (10.24)
50
где в — небольшое положительное число.
Контрольно-измерительная аппаратура и управление
237
Константинидес, Спенсер и Гэйден [42, 431 применили эту методику
(в несколько модифицированном виде), чтобы предотвратить снижение кон-
центрации биомассы. Результаты этих исследований приведены на рис. 10.26.
Обратите внимание на то, что вначале устанавливали более высокую темпе-
ратуру (чтобы обеспечить максимальную скорость клеточного роста), а затем
ее снижали (чтобы добиться оптимальной скорости образования пеницилли-
на). В конечном итоге программированное изменение температуры позволило
повысить выход пенициллина на 76,6% по сравнению с наиболее эффективным
процессом при постоянной температуре (25°C).
РИС. 10.26. Оптимальная программа изменения температуры и соответствую-
щие профили клеточного роста и образования пенициллина, вычисленные с
помощью математической модели. [Воспроизведено с разрешения из статьи:
Constantinides A., Spencer J. L., Gaden Е. L., Jr., Optimization of Batch Fermen-
tation Processes. II. Optimum Temperature Profiles for Batch Penicillin Fermen-
tations; Biotech. Bioeng., 12, 1081 (1970).]
В цитированных выше работах осуществлена оптимизация и других мо-
делей, основанных на различных базах данных. Таким путем программиро-
вание температурного режима ферментации позволило повысить выход пени-
циллина примерно на 15%. Найденные расчетными методами программы из-
менения температуры могут быть внедрены в крупномасштабное производство
с очень малыми затратами. Очевидно, что сочетание методов математическо-
го моделирования и теории оптимизации окажется плодотворным и при раз-
работке других процессов биохимической технологии.
10.7.2. Расчет и стратегия эксплуатации
промышленных периодических процессов
Промышленный процесс состоит из ряда периодических опе-
раций (предварительной обработки субстрата, стерилизации,
ферментации, выделения продукта, расфасовки). Разработка
такого процесса предполагает решение ряда задач — от рас-
чета отдельных операций до расчета всего процесса как комп-
лекса взаимосвязанных операций и расчетов, связанных с про-
238
Глава 10
изводством нескольких продуктов. Эти задачи можно сформу-
лировать следующим образом [44]:
1. Изучение и оптимизация отдельных операций и аппаратов.
2. Оптимизация процесса, состоящего из ряда последователь-
ных периодических операций, целью которых является по-
лучение одного продукта.
3. Спецификация оборудования, необходимого для производст-
ва одного или нескольких продуктов.
4. Выбор схемы соединений отдельных аппаратов, обеспечива-
ющей наиболее эффективное образование продукта.
5. Выбор режима эксплуатации, позволяющего производить не-
сколько различных продуктов в течение заданного времени.
Первые из этих задач мы уже рассматривали с нескольких
точек зрения. Чтобы понять, как изучение взаимосвязанных по-
следовательных операций может повлиять на процесс в целом,
рассмотрим простой пример. Пусть функция f(t) описывает
долю исходных питательных веществ, превращающихся за
время t в вещества, являющиеся исходными в следующей опе-
рации. Допустим также, что время очистки аппарата и его за-
грузки для проведения следующей операции равно tci. Рас-
сматриваемая здесь целевая функция F описывает количество
исходных веществ, превращаемых в единицу времени, следова-
тельно:
F(t) = (10.25)
Путем дифференцирования уравнения (10.25), приравнивания
результата дифференцирования нулю и преобразования этого
условия оптимального времени цикла (обозначенного символом
t*) получим
г а*—— <10-26>
Как показано на рис. 10.27, это условие имеет очень простую
графическую интерпретацию: оптимальное время цикла перио-
дической операции равно абсциссе точки, в которой прямая,
проходящая через начало координат, касается кривой, отвеча-
ющей рабочей функции этой операции f (t—tci). На рис. 10.27
представлена также графическая интерпретация условия (10.25)
для второй периодической операции с рабочей функцией g-
Теперь рассмотрим две последовательные периодические
операции, характеризующиеся функциями f и g соответственно.
Доля исходных веществ, которые за эти две операции превра-
щаются в необходимый продукт, равна f(t—tci)g(t—обо-
значим эту функцию h (t). Здесь t'ci — время, необходимое для
Контрольно-измерительная аппаратура и управление 239
чистки и зарядки аппаратов перед второй операцией. Для оп-
ределения максимальной степени превращения исходных ве-
ществ в единицу времени найдем максимум функции
F = (10.27)
Так же, как и в предыдущем случае, максимальная степень
превращения исходных веществ выражается графически как
абсцисса точки касания, однако кривая в данном случае отве-
чает сложной рабочей функции h (рис. 10.27, в). Представлен-
ные на рис. 10.27 графики позволяют сделать следующий очень
важный вывод: оптимальное время цикла для процесса, со-
стоящего из двух последовательных периодических операций,
может отличаться от оптимального времени цикла для любой
из этих периодических операций.
Если оптимальное время цикла отдельных последователь-
ных операций в периодическом процессе существенно различа-
ется, то возможно, что оптимальным вариантом для процесса
в целом будут различные времена циклов отдельных повторя-
ющихся операций. В простейшем случае для самой медленной
операции выбирают время цикла, равное произведению про-
должительности более быстрой операции на некоторое целое
число т; тогда непрерывность процесса в целом будет обеспе-
чена, если мы установим т аппаратов для проведения самой
медленной операции.
В производственных условиях максимальную гибкость в вы-
боре продолжительности повторяющихся периодических опера-
ций обеспечивают промежуточные хранилища для временного
хранения продукта одной периодической операции до тех пор,
пока он не потребуется в следующей операции. В периодиче-
ских процессах промежуточные хранилища могут выполнять и
другие полезные функции [45]. Подобно тому, как хранилища
или уравнительные резервуары в непрерывных процессах могут
сглаживать флуктуации, обусловленные неустановившимся ре-
жимом, промежуточные хранилища в периодических процессах
позволяют сгладить помехи, вызванные поломкой оборудова-
ния, колебаниями во времени начала отдельных операций,
а также довольно обычными в периодических микробиологиче-
ских процессах флуктуациями параметров от цикла к циклу.
Кроме того, промежуточные хранилища помогают устранить
некоторые неудобства, которые могут возникнуть при переводе
процесса на производство другого продукта.
При изучении вопроса о необходимости промежуточных хра-
нилищ следует иметь в виду несколько предъявляемых к ним
требований. Во-первых, на предприятии должно быть установ-
лено достаточное число хранилищ каждого типа с тем, чтобы
240
Глава 10
Г
Рис. 10.27. Кривые рабочих функций
и определение оптимального време-
ни цикла периодических операций
(а, б) и процесса, включающего две
последовательные периодические опе-
рации (б).
средняя пропускная способ-
ность всех операций процесса
была одинаковой. Во-вторых,
необходимо учитывать устой-
чивость исходных л промежу-
точных веществ при их хране-
нии. Понятно, что неустойчи-
вые соединения хранению не
подлежат. Наконец, требова-
ние постоянства состава одной
партии может перевесить лю-
бые экономические преимуще-
ства определенной технологи-
ческой схемы периодического
процесса. Постоянство соста-
ва партии требует, чтобы ма-
териалы, используемые в од-
ной загрузке, обрабатывались
отдельно от материалов дру-
гой загрузки и ни при каких
обстоятельствах не смешива-
лись друг с другом. Это тре-
бование, обычное в фармацев-
тической промышленности, га-
рантирует, ч.то какую-либо
часть продукта можно совер-
шенно однозначно отнести к
определенной партии. Однако,
даже если необходимо соблю-
дать постоянство состава пар-
тии, промежуточные хранили-
ща могут оказаться полезны-
ми для согласования различий
в продолжительности отдель-
ных операций и для выполне-
ния других амортизирующих
функций.
Множество интересных
проблем возникает при проек-
тировании, оптимизации и
эксплуатации процессов на
предприятиях, производящих несколько конечных продуктов;,
в таких случаях расширяются и возможности подбора оборудо-
вания. Результаты изучения таких и ряда родственных проблем-
периодических и полупериодических процессов рассмотрены в
литературе, приведенной в конце этой главы.
Контрольно-измерительная аппаратура и управление 241
10.7.3. Управление непрерывными процессами
При проведении непрерывных процессов возникают специ-
фические проблемы регулирования и особые возможности при-
менения прогрессивных методов управления. В непрерывном
процессе система обычно должна функционировать в том или
ином стационарном состоянии. Одной из основных задач управ-
ления непрерывными процессами является сокращение продол-
жительности стартового периода (или связанных со стартовым
периодом расходов). По сути дела, эта задача аналогична за-
даче оптимизации периодического процесса с тем отличием,
что здесь мы должны изменить состояние системы от стартово-
го до рабочего (стационарного) или близкого ему так, чтобы
некоторая целевая функция была минимальной. При наличии
хорошей модели эту проблему можно решить практически та-
кими же методами, которые мы рассмотрели в применении к
оптимизации температурного режима производства пеницилли-
на в периодическом режиме.
По завершении стартового периода целью управления про-
цессом обычно является сохранение заданного стационарного
состояния. Часто эту задачу удается решить с помощью не-
скольких регуляторов, каждый из которых управляет одним па-
раметром и работает по принципу обратной связи (см. разд.
10.6). В настоящее время такой подход к управлению непре-
рывными процессами биохимической технологии применяется
очень часто. В то же время опыт других работ по управлению
системами и процессами показал, что между различными це-
пями регулирования могут возникать нежелательные взаимо-
действия, т. е. регулирование одной переменной процесса с
целью уменьшения ее отклонения от заданного значения может
вызвать изменения других переменных. В общем случае причи-
на этого явления заключается в том, что многие переменные
процесса связаны друг с другом и с несколькими входными
данными процесса.
Для решения проблемы взаимодействия между отдельными
цепями регулирования предложено несколько подходов.
С большей частью из них читатель может познакомиться в
приведенной в конце главы литературе; здесь же вкратце будет
рассмотрен только метод оптимального многомерного регули-
рования.
Напомним прежде всего, что систему или процесс, описываемые уравне-
нием
dc(t)
-^ = f [c(/),d(/)] (10.28)
где с — вектор переменных состояния процесса, ad — вектор входных данных
16—746
242
Глава 10
процесса вблизи рабочей точки стационарного состояния (cs, ds), удовлетво-
ряющего условию
f(cs> <М=0 (10.29)
можно приближенно описать линеаризованным дифференциальным уравне-
нием
dy (/)
-^^AxW + BvW (10.30)
at
где i и v—отклонения состояния системы и входных данных соответственно
(см. разд. 9.2):
X(0=c(0-cs (10.31)
v(0=d(0— ds (10.32)
Схема многомерного управления базируется на локальном линеаризованном
приближенном уравнении (10.30).
Проблема оптимизации при условии, что с(/) или %(/) остается близким
к cs или к нулю соответственно, математически сводится к нахождению ми-
нимума скалярной целевой функции.
00
J = 7г f 7) Сх Ц) + vT (0 Rv (/) ] dt (10.33)
б
где С и R — положительные определенные матрицы. В подынтегральном выра-
жении уравнения (10.33) первое слагаемое имеет вполне определенный физи-
ческий смысл: в некотором роде оно является мерой отклонения состояния
от вектора рабочей точки cs, а интегрирование суммирует все мгновенные
отклонения от заданного режима. Способ выбора С зависит от решения, при-
нятого при проектировании системы управления. Исходя из технологических
требований процесса или экономических соображений, можно решить, что
важнее предотвратить отклонение одних параметров, а не других. В простей-
шем варианте С представляет собой единичную матрицу, и тогда первое сла-
гаемое в подынтегральном выражении является просто суммой квадратов всех
отклонений переменных состояния.
Слагаемое v7Rv целевой функции иногда называют ценой регулирования,
но в отношении к управлению химическими процессами такое определение
обычно ничего не говорит о физическом смысле. Это слагаемое выполняет
важную математическую и практическую функции: без него оптимальное
управление было бы неограниченным и поэтому физически невозможным. Це-
лесообразно рассматривать R как матрицу, определяющую масштаб регули-
рующего действия, причем большие значения нормы матрицы R отвечают
меньшим отклонениям входных данных процесса от номинальных значений,
принятых для расчетных условий стационарного состояния.
С помощью принципа максимума (разд. 10.7.1) можно показать, что регу-
лирование, связанное с минимизацией J в уравнении (10.33) при условии, что
взаимосвязи между регулированием и состоянием системы описываются урав-
нением (10.30) (которое часто называют линейно-квадратичной задачей опти-
мального управления) , можно свести к уравнению
v(0=— R-'BrMx(0 (10.34)
где постоянная во времени матрица М удовлетворяет следующему нелинейно-
му алгебраическому уравнению:
МА+АГМ+С— MBR->BrM = 0 (10.35)
Контрольно-измерительная аппаратура и управление 24$
Таким путем мы пришли к интересному результату; действительно, уравнение
(10.34) имеет форму уравнения многомерного управления с обратной связью.
Следовательно, регулируемые параметры здесь выражаются через имеющиеся
в данное время значения переменных состояния системы.
Если переменную состояния нельзя измерить непосредственно, то соответ-
ствующие значения можно найти с помощью одного из рассмотренных выше-
методов анализа данных. Здесь особенно удобны методы типа фильтра Кал-
мана, поскольку математический аппарат, используемый для расчета фильтра,
близок аппарату для расчета оптимального многомерного управления с обрат-
ной связью. В работе [46] описаны результаты сравнительного изучения
управления модельным микробиологическим процессом с помощью регулято-
ров с прямым измерением параметров состояния, а также с помощью расчет-
ных данных, полученных на основе фильтра Калмана.
Фэн, Эриксон и сотрудники [47] с помощью математических моделей изу-
чали отклик одного биологического ПРПП, кинетика роста клеток в котором
описывается уравнением Моно с учетом метаболизма поддержания жизне-
деятельности клеток. Для управления скоростью потока через реактор при-
меняли метод оптимального регулирования только что рассмотренного типа,
а в основу, расчетов были положены результаты измерения концентраций суб-
страта и биомассы в выходящем из реактора потоке. При этом принималось,
что матрицы управления Q и R равны
/0 \
Q= R=1 (скалярная матрица) (10.36)-
\ 0 ^22/
При постоянной R увеличение одного или двух отличных от нуля составляю-
щих Q соответствует усилению регулирующего действия.
На рис. 10.28 отображены изменения объемной скорости потока (регули-
руемый входной параметр или управляемая переменная) и соответствующие
изменения относительных концентраций клеток и субстрата в вытекающем из
реактора потоке при повышении концентрации питательных веществ на 12,5%
за период от относительного времени (=0 до относительного времени (=2.
Цифры на кривых отвечают различным величинам £/ц при постоянном q22=
= 1000. Обратите внимание на то, что при повышении qn регулируемая пере-
менная изменяется в большей степени, отклонения относительной концентра-
ции субстрата (yi) на выходе из реактора уменьшаются, а флуктуации отно-
сительной концентрации биомассы (у2) возрастают. Если qn оставить постоян-
ным, a q22 повышать, то усиление регулирующего действия в процессе возму-
щения приведет к снижению отклонений у2 и к повышению отклонений у\.
Отсюда следует, что отклик и эффекты регулятора можно варьировать в соот-
ветствии с требованиями процесса.
Другой возможной целью управления непрерывным процес-
сом является стабилизация стационарного состояния, неустой-
чивого в отсутствие регулирования. В этой связи можно приве-
сти два недавно описанных в литературе примера. Если рост
организмов на метаноле в ПРПП ингибируется субстратом, то
соответствующие модели допускают существование в некоторых
условиях трех стационарных состояний. ДиБиасио, Лим и Вей-
ганд [48, 49] показали теоретически и подтвердили эксперимен-
тально, что применение пропорционального управления позво-
ляет добиться устойчивой работы реактора в промежуточном
нестабильном стационарном состоянии. Измеряемым парамет-
ром в данном случае было помутнение культуры, а регулируе-
мой переменной — скорость введения субстрата.
16*
244
Глава 10
РИС. 10.28. Результаты моделирования управления микробиологическим
ПРПП методом оптимального линейно-квадратичного регулирования. На кри-
вых цифрами обозначены значения параметра q\\, а штриховые линии соответ-
ствуют величинам относительной концентрации субстрата (yi = s/«fs) и отно-
сительной концентрации биомассы (y2=x/Yx/sSjS) в стандартном стационар-
ном состоянии. Во всех случаях принято, что параметр равен 1000. [Вос-
произведено с разрешения из статьи: Fan L. Т., Shah Р. S., Pereira N. С.,
Erickson L. E.t Dynamic Analysis and Optimal Feedback Control Synthesis
Applied to Biological Waste Treatment; Water Research, 7, 1609 (1973).]
Хотя при эксплуатации проточных реакторов обычно стара-
ются добиться работы в определенном стационарном состоя-
нии, в ряде теоретических и экспериментальных работ было по-
казано, что в некоторых случаях лучшие эксплуатационные ха-
рактеристики можно получить в динамическом режиме, созда-
ваемом изменением скорости подачи питательных веществ или
условий процесса [50]. Из динамических режимов наиболее ши-
К о н т р о льно-измерительная аппаратура и управление
245
роко изучались колебательные режимы, включающие периоди-
ческое изменение определенных входных параметров и соответ-
ствующие периодические колебания переменных системы. В ос-
нове возможности улучшения эксплуатационных характеристик
процесса путем намеренного перехода к нестационарному со-
стоянию лежит очень простой принцип: в переходном состоя-
нии состав клеток и среды, а также скорости реакций могут
существенно отличаться от соответствующих характеристик лю-
бого стационарного состояния, когда состав смеси определяет-
ся стехиометрией процесса. Показано, например, что создавае-
мые искусственно периодические флуктуации параметров про-
цесса позволяют повысить выход цитохрома в культуре Candi-
da utilis, водорода в биофотолитическом процессе, катализируе-
мом Anabaena cylindrica, а также модифицировать макромоле-
кулярный состав Е. coli [51].
Как мы увидим далее в гл. 13, рост двух различных орга-
низмов, конкурирующих за общее лимитирующее рост пита-
тельное вещество, обычно приводит к вымыванию одного из
штаммов. Хэтч, Кэдмен и Уайлдер [52] путем моделирования
показали, однако, что с помощью пропорционального алго-
ритма управления, согласно которому скорость добавления
питательных веществ и общая скорость разведения регулиру-
ются в соответствии с найденными плотностями клеток двух
штаммов, может быть достигнуто устойчивое стационарное со-
стояние, в котором оба штамма сосуществуют. Далее эти ис-
следователи экспериментально показали возможность быстро-
го определения концентраций как Candida utilis, так и Согупе-
bacterium glutamicum методом цитометрии в потоке (путем
измерения рассеяния света).
10.8. Заключение
Для управления биотехнологическим процессом независимо
<от того, осуществляется ли это управление оператором или
ЭВМ, необходима информация о состоянии процесса. В свою
очередь такую информацию можно получить только при нали-
чии соответствующей аналитической аппаратуры и при тща-
тельном систематическом анализе и интерпретации полученных
данных. В ближайшие годы здесь должны произойти большие
сдвиги благодаря внедрению в промышленные процессы новых
мощных методов анализа, заимствованных из химии, биохимии
и биологии клетки. В основанных на моделях методах анали-
за ^данных найдут применение не только принципы элементар-
ной стехиометрии, но и дополнительные данные, базирующиеся
на оценке параметров в реальном масштабе времени, и более
детальные оценки метаболического состояния, в основу кото-
246
Глава 10
рых положены достаточно универсальные структурированные
модели.
Прежде чем перейти к теме следующей главы — к процес-
сам разделения, следует упомянуть несколько важных понятий
и тем, связанных с управлением процессами, но не рассматри-
вавшихся в этой главе. На низшем уровне задача технолога
сводится к проведению процесса в соответствии с некоторым
проектом или технологической инструкцией. Эти проект или
технологическая инструкция в свою очередь отвечают решению
проблемы оптимизации или регулирования на более высоком
уровне, на котором цель данного процесса сводится к дости-
жению того или иного максимального вклада (например, мак-
симальной прибыли) в результате деятельности всего пред-
приятия, затем соответствующей корпорации и, наконец, все-
го общества. Рассматриваемые инженером-технологом целевые
функции и ограничения представляют собой, таким образом,
естественное продолжение целей и требований на более высо-
ких уровнях управления.
Часто важными составными частями целевых функций или
эксплуатационных ограничений являются надежность и безо-
пасность. При управлении процессами часто намного важнее
обеспечить максимальную однородность продукта, свести к ми-
нимуму возможные потери и вероятность работы во вредных
условиях, чем повысить выход продукта на несколько процен-
тов или допустить небольшое отклонение от установленных зна-
чений параметров. Изучение этих вопросов в целом еще толь-
ко начинается, и мы вправе ожидать здесь новых достижений,
которые окажут влияние и на развитие проблемы управления
процессами биохимической технологии.
Упражнения
10.1. Аналитическая аппаратура. Дайте определение и кратко объясните
принцип действия следующих аналитических приборов:
а) терморезистора, термопары, мембранного манометра, ваттметра Хол-
ла, торсионного динамометра, тензометра, газового ротаметра, термического
массового расходомера, датчика емкости;
б) pH-электрода, гальванического и полярографического зондов для опре-
деления концентрации растворенного кислорода, электрохимического зонда
для определения давления растворенного СО2, электрода на основе иммобили-
зованного фермента;
в) масс-спектрометра, газового хроматографа, парамагнитного Оо-анали-
затора;
г) спектрометра, нефелометра, флуорометрии.
10.2. ЭВМ и функции управления. Дайте определения следующим поня-
тиям: а) аналого-цифровой преобразователь, мультиплексор, цифро-аналого-
вый преобразователь; б) аппаратное обеспечение, программное обеспечение;
в) процесс, датчик, детектор, монитор, регулятор, исполнительный элемент.
Контрольно-измерительная аппаратура и управление
247
10.3. Оценка состояния и расчет параметров системы, а) Объясните раз-
личие между непосредственно измеряемыми н оцениваемыми (рассчитываемы-
ми) переменными системы.
б) Какое аппаратное н (илн) программное обеспечение необходимо для
оценки состояния системы?
10.4. Управление процессом и преобразование Лапласа. С помощью пре-
образования Лапласа удобно проводить оценку линейных нлн линеаризован-
ных систем управления. По определению преобразованием любой функции
времени f(t) является F(s):
О©
F (s) == J f (t) e~stdt
о
Это тождество часто изображают сокращенно: F(s) = L{f (/)}.
а) Опишите свойства преобразования производной и интеграла
L{df/dt}=sF(s) если f(/=0) =0
t
J f W dt
о
L
= F (s)/s
б) Покажите, что преобразование Лапласа для отклонения выходного
сигнала регулятора o(t)—os [уравнение (10.10)] равно
( E(s) )
О (s) = Кс (s) + + xdE (s) |
и что таким образом передаточная функция нли отношение трансформации
выходного сигнала (коррекции) к трансформации входного сигнала (погреш-
ности) в s-области описывается уравнением
1
T;S
О (s)
Е (s)
= К0
+ == Gc (s)
10.5. Регулирование pH. Льюднкин н Пайрет описали кинетику образова-
ния молочной кислоты в периодическом микробиологическом процессе урав-
нением (7.93):
dp п dx
—— = fix + а----------
dt н dt
а) Предположим, что зависимость x(t) при приблизительно постоянном
pH описывается логистическим уравнением
dx/dt=p,x(\ — Х/Хщах)
Найдите соответствующую зависимость p(t), если р(0)=0.
б) В нерегулируемом процессе накопление молочной кислоты (pFa = 3,88)
приводит к изменению pH. Предположим, что мы решили установить pH око-
ло 6,5. Найдите выражения, описывающие изменение pH в нерегулируемом
процессе:
е(/)=рН(0 —6,5
Предложите программу добавления концентрированного раствора основания,
позволяющую поддерживать pH на таком уровне, чтобы отклонения (pH —
—рН0) не превышали 0,1.
248
Глава 10
в) С помощью уравнения (10.10) и выражения для зависимости e(f) в не-
регулируемом процессе найдите выражение, описывающее выходной сигнал
O(t) пропорционально-дифференциально-интегрального регулятора.
г) Рассмотрим пропорциональный регулятор [уравнение (10.10) при тс=
= 0 и Т/ = оо]. Наибольшее допустимое отклонение (pH — рНо = О,1) должно
привести к такой скорости добавления основания тв, которая соответствует
максимальной скорости образования продукта VR{dp/dt)m^. Какой нужен
коэффициент пропорциональности Кс, чтобы обеспечить удовлетворительное-
регулирование, если выходной сигнал O(t) равен скорости добавления осно-
вания и»?
д) Найдите выражение, описывающее зависимость рН(/) для регулируе-
мого процесса, упрощенная схема управления которым приведена на рис.
10.У5.1. Увеличением объема системы за счет добавляемого концентрирован-
ного основания тв можно пренебречь. Примите, что рН,п = рН, т. е. определе-
рН0=5,5
РИС. 10У5.1.
ние pH производится мгновенно и с достаточно высокой точностью. (Данные
о принятых способах изображения схем управления можно найти в любом
руководстве по управлению процессами, например в книге: Coughanowr D. R.,
Koppel L. В., Process Systems Analysis and Control, McGraw-Hill, New York,
1965.)
10.6. Время запаздывания в переходном состоянии. В случае нестационар-
ного состояния предлагалось вводить константу времени запаздывания (1/7),
учитывающую запаздывание отклика мгновенной удельной скорости клеточно-
го роста ц(/) на изменение концентрации субстрата \Young Т. В., Bruley D. F.,
Bungay Н. R., A Dynamic Mathematical Model of the Chemostat; Biotech. Bio-
eng., 12, 747, (1970)]. Постепенное приближение ц(/) к соответствующему
значению для стационарного состояния Цо описывается уравнениями
__ P-maxs tfp ____ Ио В
Ио “ Ks + s Л Y
е О r>/Vs (/') dt'
а) Покажите, что если s является функцией времени s(t), то решение
указанных уравнений приводит к выражению
t
о
s (t')
б) Предположим, что s(f) =s0(l+a cos wt), причем a<l и /Gi»s(f). Най-
дите явное решение приведенного выше уравнения для р,(/) при этих условиях,
постройте график зависимости ц(/)/|*тах от t при а=0,5, 7=1 и 0 = 0,17, 1,07
Конт рольно-измерительная аппаратура и управление 249
и 107. Объясните различие между полученными кривыми. Для каких переход-
ных состояний учет временной константы 7 обязателен?
в) Найдите уравнения, описывающие поведение системы в ПРПП, если
Sf=Sfo(l+cos wt) и если клеточный рост подчиняется уравнению rx=p,xs.
10.7. Контроль хемостата с аэрацией. При росте аэробных организмов
в хемостате через систему пропускают сжатый воздух со скоростью, обычно
равной одному объему воздуха на объем жидкости в минуту. Поступающий
в реактор воздух практически не содержит влаги, а выходящий из реактора
воздух насыщен парами воды. Следовательно, скорости потоков жидкостей,
поступающих в реактор и вытекающих из него, должны быть различными.
Как описанное выше явление скажется на интерпретации аналитических
данных, полученных при изучении вытекающего из реактора потока? Чему бу-
дет равна истинная концентрация продукта на выходе из реактора (с учетом
испарения), если объем реактора равен 10 л, скорость разведения составляет
0,1 ч-1, а другие параметры системы равны: so=5O г/л (глюкоза): Ks=10 мг
глюкозы/л; цшах=0,7 ч-1; Ух/з=0.5 г клеток/(глюкозы): т=0,02 г глюко-
зы/[(г клеток)-ч]; УР/5 = 0,5; дР = 0,2 г продукта/[(г клеток)-ч]; 7=60 °C. Дав-
ление паров воды при 60 °C равно 150 мм рт. ст. Примите, что скорость кле-
точного роста описывается уравнением Моно. Можно также принять, что кон-
центрация СОг в воздухе пренебрежимо мала, а дыхательный коэффициент
изучаемого процесса равен единице.
10.8. Автоматическое управление с прямой связью. Рост микроорганизмов
в периодическом процессе описывается уравнением dx/dt=[ix до тех пор, пока
при (х—хо)=Узо не истощится лимитирующее рост питательное вещество S.
В соответствии со стехиометрией аэробного процесса потребность в кислороде
на рост биомассы и образование продуктов метаболизма учитываются коэф-
фициентами Ух/о2 и Ур/о2.
а) Найдите выражения, описывающие мгновенную потребность в кисло-
роде Qo2 и удельную потребность в кислороде (отнесенную к концентрации
клеток) Qo2/x, если образование продукта не сопряжено с клеточным ростом,
так что dp/dt=\>x. При каких условиях эти выражения максимальны?
б) Выведите уравнение, позволяющее рассчитать величину Kia, при кото-
рой обеспечивается удовлетворение максимальной потребности системы в кис-
лороде.
в) По какой программе следует изменять Fg (вход), чтобы необходимая
скорость аэрации достигалась при минимальной общей утилизации газа, если
Ki приблизительно постоянно, а значение а изменяется пропорционально ско-
рости поступающего в реактор потока газа в степени 3/4. Найдите выражение,
описывающее отношение всего объема воздуха, введенного в систему в про-
цессе роста биомассы от х0 до Хтах, к объему воздуха, который был бы введен
в систему, если бы Fg поддерживали на максимальном уровне в течение всего
периода клеточного роста.
г) Управление с прямой связью часто легче осуществимо, чем управление
с обратной связью, но поскольку при первом способе управления отсутствует
непрерывный контроль процесса, то фактически управление проводится «всле-
пую». Обсудите, какие последствия будет иметь неправильная калибровка
расходомера для воздуха в ситуации, описанной в задаче 10.8, в, если пока-
зания расходомера на 10% ниже или на 50% выше запрограммированной
скорости подачи газа. Как бы вы модифицировали систему управления, чтобы
решить эту проблему?
10.9. Оценка состояния системы в реальном масштабе времени с помощью
уравнений материального баланса, а) Рассмотрим процесс быстрого роста кле-
ток, в ходе которого скорость образования СО2 (СОО) пропорциональна ско-
рости роста биомассы. Найдите выражения для определения х и р, а) если
СОО измеряют непрерывно и б) если СОО измеряют периодически с интерва-
лами Д/.
250
Глава 10
б) При низких скоростях клеточного роста, благоприятных для образова-
ния пенициллина, пропорциональная зависимость между СОО и скоростью
роста биомассы нарушается и возникает необходимость в определении более
полного баланса по углероду. Превращения в периодическом процессе с до-
бавлением субстрата описываются общим уравнением
Субстрат-(-предшественники пенициллина-(-посевной материал —>- непрореаги-
ровавший субстрат-)- непрореагировавший предшественник + образовавшаяся
биомасса-(-растворимые неидентифицированные продукты-)-пенициллин-)-СОг
Выразите концентрацию биомассы с помощью уравнения материального ба-
ланса по углероду через найденные концентрации субстрата, предшественни-
ка, пенициллина и СО2, если у,-— доля углерода в t-м компоненте и если угле-
род в заметных количествах входит только в состав биомассы, субстрата,
предшественника, СО2 и пенициллина. Изменением объема реакционной смеси
во времени можно пренебречь.
в) Предположим, что концентрацию пенициллина нельзя определить непо-
средственно, но в то же время пенициллин вносит существенный вклад в об-
щий баланс по углероду. Найдите выражение, с помощью которого можно
определить концентрацию пенициллина р, если известны концентрации суб-
страта, предшественника и СО2 и если кинетика накопления пенициллина опи-
сывается указанным выше уравнением:
dp dx
— пх-\- т — kp
dt-------------dt
[Пример использования уравнений материального баланса для управления
процессом производства пенициллина детально описан в работе: Мои D.-G.,
Cooney С. L., Biotech. Bioeng., 25, 225, 257 (1983).]
10.10. Автоматическое управление добавлением неорганических веществ
в периодическом процессе с добавлением субстрата. Определение in situ кон-
центраций NH3 или летучего углеродсодержащего субстрата (например, этано-
ла) позволяет осуществить управление с обратной связью скоростью добавле-
ния питательных веществ, содержащих азот и углерод. К сожалению, кон-
центрации многих неорганических веществ, необходимых для роста и жизне-
деятельности клеток, нельзя определять непрерывно в ходе процесса. С дру-
гой стороны, добавление всех неорганических компонентов при t = Q часто
ингибирует рост инокулята. В таких случаях выгодно добавлять неорганиче-
ские компоненты в ходе процесса по определенной программе, в основу кото-
рой положены стехиометрические соотношения между теми компонентами, кон-
центрации которых можно определить in situ, и другими компонентами си-
стемы.
Рассмотрим простое уравнение типа
a (NH3) +₽ (источник углерода) -И (неорганический компонент) —>-биомасса
а) Какое соотношение объемных скоростей растворов исходных веществ
будет соответствовать указанной стехиометрии образования биомассы, если
концентрации питательных веществ в исходных растворах равны по (источник
азота), с0 (источник углерода), т0 (источник неорганических веществ)? Сколь-
ко детекторов потребуется для управления потоками всех питательных ве-
ществ?
б) Выведите уравнение, описывающее адекватную скорость добавления
раствора неорганических веществ Fm(t) через регулируемые по принципу об-
ратной связи скорости добавления растворов источников азота и углерода
Fn(t) и Fc(t), если в результате процесса образуются как биомасса, так н про-
дукт (C7NO3H10). Сколько детекторов потребуется теперь? [См. статью: Suzu-
ki Т. et al., Automatic Supplementation of Minerals in Fed-Batch Culture te
High Biomass Concentrations; Biotech. Bioeng., 27, 192 (1985).]
Контрольно-измерительная аппаратура и управление
251
10.11. Ферментативный метод контроля клеточного роста. Изучение про-
цесса утилизации целлюлозы микроорганизмами затруднено из-за нераство-
римости субстрата в воде. Проконтролируйте процесс накопления фермента
глюкозооксидазы в культуре целлюлолитической бактерии, растущей на цел-
люлозе. Глюкозооксидаза катализирует реакцию
Глюкоза+ О2 —> б-глюконолактон + Н2О
Кинетику этой реакции можно изучать колориметрическим или манометриче-
ским (например, с помощью респирометра Варбурга) методами. Вам известно,
что изучаемый организм содержит глюкозооксидазу в количестве около 1 еди-
ницы активности на 1 грамм клеточного белка (1 единица активности равна
такому количеству фермента, которое трансформирует 1 мкмоль глюкозы
в минуту). В этой бактерии целлюлазная активность сконцентрирована в кле-
точной стенке, поэтому для роста необходим непосредственный контакт клет-
ки с субстратом.
а) Составьте план эксперимента для изучения кинетики клеточного роста
и образования глюкозооксидазы бактериями, растущими на целлюлозном по-
рошке (частицы диаметром 100 мкм). Этот эксперимент должен охватывать
как экспоненциальную, так и стационарную фазы. В частности, вы должны
описать необходимую для эксперимента аппаратуру, методы анализа, количе-
ство целлюлозы и источника азота (сульфат аммония), которое нужно доба-
вить к среде.
б) Как вы думаете, сколько времени займет этот эксперимент и как будут
выглядеть кривые клеточного роста и удельной ферментативной активности?
Можно принять следующие значения параметров: Yx/s = 0,5; Yx/n=10; Ух/о =
= 1; площадь поверхности одной бактерии около 5-Ю-8 см2, масса бактерии
около 10-12 г; желательная конечная концентрация клеток 10 г/л; рШзх~
= 0,2 ч~'; клетки на 60% состоят из белков.
Литература
Полезные обзоры по управлению процессами химической технологии даны
в следующих монографиях:
1. Stephanopoulos G., Chemical Process Control. An Introduction to Theory and
Practice, Prentice-Hall, Inc., Englewood Cliffs, N. J., 1984.
2. Luyben W. L., Process Modeling, Simulation, and Control for Chemical
Engineers, McGraw-Hill, N. Y., 1973.
3. Coughanowr D. R., Koppel L. B., Process Systems Analysis and Control,
McGraw-Hill, N.Y., 1965.
Фильтры Калмана и теория оптимального управления рассматриваются во
многих сборниках и монографиях, в том числе в следующих:
4. Jazwinski А. Н., Stochastic Processes and Filtering Theory, Academic Press,
New York, 1970.
5. Атанс M„ Фалб П. Л., Оптимальное управление. — M.: Машиностроение,
Проблемы контрольно-измерительной аппаратуры и управления биопроцесса-
ми изложены во многих опубликованных недавно обзорных статьях и моно-
графиях:
6. Wang N. S., Stephanopoulos G., Computer Application in Fermentation
Processes; CRC Critical Reviews in Biotechnology, 2, 1, (1984).
7. Computer Applications in Fermentation Technology, 3rd Int. Conf, on Com-
puter Applications in Fermentation Technology, Manchester, England, 1981;
Society of Chemical Industry, London, 1982.
252
Глава 10
8. Computer Applications to Fermentation Processes (Biotechnology and Bio-
engineering Symposium Series, No. 9), John Wiley, N.Y, 1979.
9. Tannen L. P., Nyiri L. K., Instrumentation of Fermentation Systems, in Mic-
robial Technology, 2d ed., vol. 2, Peppier H. J., Perlman D. (eds.), p. 331,
Academic Press, Inc., New York, N. Y., 1979.
10. Armiger IF. B., Humphrey A. E., Computer Application to Fermentation.
Technology, in Microbial Technology, 2d ed., vol. 2. Peppier H. J., Perk
man D. (eds.), Academic Press, Inc,, New York, N. Y., 1979.
Ниже приведена литература к отдельным темам, рассмотренным в этой главе:
Детекторы:
11. Wernan W. С., Wilke С. R., New Method for Evaluation of Dissolved Oxy-
gen Response for kLa Determination; Biotech. Bioeng., 15, 571 (1973).
12. Karube I., Suzuki S., Application of Biosensor in Fermentation Processes, in
Annual Reports on Fermentation Processes, vol. 6, Tsao G. T. (ed.), p. 203,
Academic Press, N. Y., 1983.
13. Schultz J. S., Sims G., Affinity Sensors for Individual Metabolites, in Biotech.
Bioeng. Symposium 9, Armiger W. B. (ed.), p. 65, Wiley, N. Y, 1979.
Автономные методы анализа:
14. Методы общей микробиологии, под ред. Герхарда Ф., т. 1—3. — М.; Мир,
1983.
15. Methods in Cell Biology, vol. XII, Yeast Cells, Prescott D. H. (ed.), Acade-
mic Press, N. Y, 1975.
16. Rodriguez R. L.t Tait R. C., Recombinant DNA Techniques: An Introduction,
Addison-Wesley Publishing Co., Reading, MA, 1983.
17. Pedersen S., Bloch P. L., Reeh S., Neidhardt F. C., Patterns of Protein
Synthesis in E. coli: A Catalog of the Amount of 140 Individual Proteins at
Different Growth Rates; Cell, 14, 179 (1978).
18 Methods in Immunodiagnosis, Rose N. R., Bijayzi P. E. (eds.), 2d ed., Wiley,
N.Y, 1980.
19. Ugurbil K., Rottenberg H., Glynn P., Shulman R. G., 31P Nuclear Magnetic
Resonance Studies of Bioenergetics and Glycolysis in Anaerobic Escherichia
coli Cells; Proc. Natl. Acad. Sci. USA, 75, 2244 (1978).
20. Herrero A. A.t Gomez R. F., Snedecor B., Tolman C. J., Roberts M. F., Growth
Inhibition of Clostridium thermocellum by Carboxylic Acids: A Mechanism
Based on Uncoupling by Week Acids; Appl. Microbiol. Biotechnol, 22, 53
(1985).
21. den Hollander J. A., Ugurbil K., Brown T. R., Shulman R. G., Phosphorus-31
Nuclear Magnetic Resonance Studies of the Effect of Oxygen upon Glycolysis
in Yeast; Biochemistry, 20, 5871 (1981).
22. Nicolay K., Sheffers W. A., Bruinenberg P. M„ Kaptein R., Phosphorus-31
Nuclear Magnetic Resonance Studies of Intracellular pH, Phosphate Com-
partmentation, and Phosphate Transport in Yeasts; Arch. Microbiol, 133,
83 (1982).
23. Ugurbil K., Brown T.R., den Hollander J. A., Glynn P., Shulman R. G.,
High-Resolution 13C Nuclear Magnetic Resonance Studies of Glucose Meta-
bolism in Escherichia coli-, Proc. Natl. Acad. Sci. USA, 75, 3742 (1978).
24. Legerton T. L., Ranamori K., Weiss R. L„ Roberts J. D., 15N NMR Studies
of Nitrogen Metabolism in Intact Mycelia of Neurospora crassa: Proc. Natl.
Acad. Sci. USA, 78, 1495 (1981).
25. Nestaas E., Wang D. I. C., Suzuke H., Evans L. B., A New Sensor — the
«Filtration Probe» — for Quantitative Characterization of the Penicillin Fer-
mentation. II. The Monitor of Mycelial Growth; Biotech. Bioeng, 23, 2815
(1981).
26. Nestaas E., Wang D. I. C., A New Sensor — the «Filtration Probe» —for
Контрольно-измерительная аппаратура и управление 253
Quantitative Characterization of the Penicillin Fermentation. III. An Auto-
matically Operating Probe; Biotech. Bioeng., 25, 1981 (1983).
27. Thomas D. C., Chittur V. K-, Cagney J. W„ Lim H. C., On-Line Estimation
of Mycelial Cell Mass Concentrations with a Computer-Interfaced Filtration
Probe; Biotech. Bioeng., 27, 729 (1985).
28. Srienc F„ Arnold B., Bailey J. E., Characterization of Intracellular Accumu-
lation of Poly-P-Hydroxybutyrate (PHB) in Individual Cells of Alcaligenes
eutrophus H16 by Flow Cytometry; Biotech. Bioeng., 26, 982 (1984).
29. Flow Cytometry and Sorting, Melamed M. R., Mullaney P. F., Mendel-
sohn M. L. (eds.), John Wiley and Sons, N. Y., 1979.
30. Bailey J. E., Single-Cell Metabolic Model Determination by Analysis of
Microbial Populations, in Foundations of Biochemical Engineering: Kinetics
and Thermodynamics in Biological Systems, Blanch H. W., Papoutsakis E. T.,
Stephanopoulos G. (eds.), American Chemical Society, Washington, D. C ,
1983.
31. Srienc F., Campbell J. L.t Bailey J. E., Detection of Bacterial ^-Galactosidase
Activity in Individual Saccharomyces cerevisiae Cells by Flow Cytometrv;
Biotech. Lett., 5, 43 (1983).
32. Hatch R. T., Wilder C., Cadman T. W., Analysis and Control of Mixed Cultu-
res, Biotech. Bioeng. Symp. No. 9, Armiger W. B. (ed.), p. 25, John Wiley
and Sons, 1979.
33. Hutter R.-J., Punessen U„ Eipel H. E., Flow Cytometric Determination of
Microbial Contaminants; Biotech. Lett., 1, 35 (1979).
Микро-ЭВМ:
34. Hampel W. A., Application of Microcomputers in the Study of Microbial
Processes, in Advances in Biochemical Engineering, vol. 13, Ghose T. K.,
Fiechter A., Blakebrough N. (eds.), p. 1, Springer-Verlag, Berlin, 1979.
Фильтрация и рекурсивная оценка:
35. Jefferis R. P., Winter H., Vogelmann H., Digital Filtering for Automatic-
Analysis of Cell Density and Productivity, in Workshop Computer Applica-
tions in Fermentation Technology, Jefferis III R. P. (ed.), Verlag Chemie.
N.Y., 1977.
Методы оценки материальных балансов:
36. Zabriskie D. W., Humphrey A. E., Real-Time Estimation of Aerobic Batch
Fermentation Biomass Concentration by Component Balancing; AIChE J,
24, 138 (1978).
Применение фильтра Калмана:
37. Stephanopoulos G., San K.-Y., Studies on On-Line Bioreactor Identification.
Part I. Theory; Biotech. Bioeng., 26, 1176 (1984).
38. San K.-Y., Stephanopoulos G., Studies on On-Line Bioreactor Identification.
Part II. Numerical and Experimental Results; Biotech. Bioeng. 26, 1189
(1984).
Управление биореакторами периодического действия с добавлением суб-
страта:
39. Cooney С. L., WangH. Y., Wang D. I. C., Computer-Aided Material Balan-
cing for Prediction of Fermentation Parameters; Biotech. Bioeng. 19, 55
(1977).
40. Wang H. Y., Cooney C. L., Wang D. I. C., Computer-Aided Baker’s Yeast
Fermentations; Biotech. Bioeng., 19, 69 (1977).
254
Глава 10
Управление биореакторами периодического действия:
41. Lundell R., Practical Implementation of Basic Computer Control Strategies
for Enzyme Production, in Computer Applications in Fermentation Techno-
logy. P- 181, Society of Chemical Industry, London, 1982.
42. Constantinides A., Spencer J. L., Gaden E. L., Jr., Optimization of Batch
Fermentation Processes. I. Development of Mathematical Models for Batch
Penicillin Fermentations; Biotech. Bioeng., 12, 803 (1970).
43. Constantinides A., Spencer J. L., Gaden E. L., Jr., Optimization of Batch
Fermentation Processes. II. Optimum Temperature Profiles for Batch Penicil-
lin Fermentations; Biotech. Bioeng., 12, 1081 (1970).
Проектирование и эксплуатация периодических процессов:
44. Rippin D. W. T., Simulation of Single- and Multiproduct Batch Chemical
Plants for Optimal Design and Operation; Computers and Chem. Eng., 7,
137 (1983).
45. Karimi I. A., Reklaitis G. V., Intermediate Storage in Non-continuous Proces-
sing, in Foundations of Computer Aided Process Design, Westerberg A. W.,
Chien H. H. (eds.), p. 425, CACHE Publications, Ann Arbor, Michigan,
1984.
Линейно-квадратичное оптимальное управление:
46. Fawzy A. S., Hinton O. R., Microprocessors Control of Fermentation Pro-
cess; J. Ferment. Technol., 58, 61 (1980).
47. Fan L. T., Shah P. S., Pereira N. C.t Erickson L. E., Dynamic Analysis and
Optimal Feedback Control Synthesis Applied to Biological Waste Treatment;
Water Research, 7, 1609 (1973).
Управление биореакторами при наличии нескольких стационарных состояний:
48. DiBiasio D., Lim Н. С., Weigand W. A., Tsao G. Т., Phase-Plane Analysis
of Feedback Control of Unstable Steadv States in a Biological Reactor;
AIChE J., 24, 686 (1978).
•49. DiBiasio D., Lim H. C., Weigand W. A., An Experimental Investigation of
Stability and Multiplicity of Steady States in a Biological Reactor; AIChE
J„ 27, 284 (1981).
Биореакторы вынужденно-периодического действия:
50. Bailey J. E., Periodic Phenomena, in Chemical Reactor Theory: A Review,
Lapidus L., Amundson N. R. (eds.), p. 758, Prentice-Hall, Englewood Cliffs,
N. J., 1977.
51. Pickett A. M., Bazin M. J., Topiwala H. H., Growth and Composition of
Escherichia coli Subjected to Square-Wave Perturbation in Nutrient Supply:
Effect of Varying Frequencies; Biotech. Bioeng., 21, 1043 (1979).
Управление ростом смешанных культур:
52. Wilder С. Т., Cadman Т. W., Hatch R. Т., Feedback Control of a Competitive
Mixed-Culture System; Biotech. Bioeng., 22, 89 (1980).
Глава 11<
Операции выделения в процессах
биохимической технологии
Культуральный бульон представляет собой сложную смесь
клеток (биомассы), растворимых внеклеточных продуктов ме-
таболизма, внутриклеточных продуктов жизнедеятельности ор-
ганизмов, а также не подвергшихся превращениям (или не
способных к превращениям) компонентов субстрата. Как пра-
вило, биопроцесс состоит из двух основных этапов — биотранс-
формации (осуществляемой в биореакторе) и последующего
выделения продукта. Применяемый в каждом конкретном био-
процессе метод выделения продукта определяется не только
тем, где он находится (внутри клетки или вне ее), молекуляр-
ной массой, зарядом и растворимостью, но и масштабом про-
цесса и даже стоимостью продукта. Например, в лабораторных
условиях для выделения и очистки ценных фармацевтических
препаратов или таких биологически активных веществ, как
гормоны, антитела и ферменты, с успехом применяются раз-
личные хроматографические методы, которые обычно невыгод-
ны в производственных масштабах из-за их высокой стоимо-
сти и ряда затруднений при масштабировании.
Эта глава, посвященная операциям разделения и очистки в
биопроцессах, построена следующим образом. Сначала будут
рассмотрены отдельные методы выделения продуктов, получен-
ных с участием микроорганизмов, растительных или животных
клеток. Затем мы покажем, как сочетать эти методы, чтобы
в конце концов добиться необходимой для данного продукта
степени чистоты, которая может варьировать в очень широких
пределах. Понятно, что требования, предъявляемые к методам
выделения, например, дрожжевых клеток (путем фильтрова-
ния), сырого экстракта фермента или свободного от пирогенов
инсулина, продуцируемого рекомбинантными организмами
Е. coli, будут существенно различаться. Мы также будем обра-
щать внимание на высокую стоимость многих операций разде-
ления и на необходимость учета этого фактора при проекти-
ровании процесса в целом. С этой точки зрения, например,
удобнее работать со штаммами микроорганизмов, секретирую-
щих продукт в среду, а не накапливающих его в клетках.
С другой стороны, применение более дешевых неочищенных
256
Глава 11
субстратов может быть экономически нецелесообразным из-
за усложнения и удорожания операций выделения продукта.
Принципы и основные операции выделения одинаковы для про-
цессов с участием ферментов, культур клеток или микроорга-
низмов, хотя в процессах последнего типа операции выделения
изучены наиболее тщательно.
В конечном итоге основная задача биотехнолога заключа-
ется в выборе наиболее выгодного для получения данного про-
дукта сочетания субстрата, фермента или организма, биореак-
тора и метода выделения. Основным критерием при выборе
оптимального варианта является экономическая сторона про-
ектируемого биотехнологического процесса; эту проблему мы
рассмотрим в следующей главе.
Необходимость применения конкретного метода или опера-
ции разделения зависит от начальных свойств культурального
бульона (вязкости, концентрации продукта, наличия примесей
и нежелательных нерастворимых веществ и т. д.), а также от
требуемой степени чистоты и конечной формы продукта (кри-
сталлическое вещество, его концентрированный раствор, высу-
шенный порошок и т. д.). Неочищенный продукт можно выде-
лить, например, путем простого упаривания культуральной
жидкости. Последовательность операций выделения и очистки
при получении высокоочищенного продукта биопроцесса вы-
глядит обычно следующим образом:
1. Отделение нерастворимых веществ. Для этой цели обычно
используют фильтрование, центрифугирование и (или) от-
стаивание, седиментацию и декантацию.
2. Первичное выделение. Здесь чаще всего применяют экстрак-
цию растворителями, сорбцию, осаждение и ультрафильтра-
цию. Последний метод позволяет разделять вещества в соот-
ветствии с их молекулярными массами. На стадии первич-
ного выделения значительно возрастает концентрация про-
дукта, причем продукт отделяется в первую очередь от ве-
ществ, существенно отличающихся от него по полярности.
3. Очистка. В ходе операции очистки обычно происходит отде-
ление примесей, а также дальнейшее концентрирование про-
дукта. Для этой цели используют фракционное осаждение,
различные хроматографические методы и адсорбцию.
-4. Окончательная очистка продукта. После этой операции (или
нескольких операций) продукт готов к смешению с другими
составными частями композиции или к непосредственной от-
правке потребителю. На этой стадии применяют центрифу-
гирование и последующее высушивание кристаллического
вещества, сушку распылением или в барабане, сушку лио-
филизацией (вымораживанием) или отгонку органического
растворителя.
Операции выделения
257
Типичная последовательность операций выделения и очист-
ки фармацевтического препарата (полученного методами био-
химической технологии) приведена в табл. 11.1, где указаны
как концентрация, так и относительная чистота продукта по-
сле каждой операции, в том числе после отделения нераство-
римых веществ, первичного выделения, предварительной и
окончательной очистки (кристаллизации).
На рис. 11.1 в качестве примера изображена схема всей
системы выделения и очистки. Эта система, в которой при-
менено 15 различных методов выделения и помимо этого
Таблица 11.1. Последовательность операций выделения
и очистки3
Операции Концентрация продукта, г/л Чистота про- дукта, %
Исходный бульон 0,1—5,0 0,1—1,0
Фильтрование 0,1—5,0 0,1—2,0
Первичное выделение 5,0—10,0 1,0—10,0
Очистка 50—200 50—80
Кристаллизация 90—100
а По данным из работы: Belter Р., General Procedures for Isolation
of Fermentation Products, in Microbial Technology, Peppier H. J., Perl-
man D. (eds.), 2d ed., vol. 2, p. 403, Academic Press, Inc., New York,
1979.
предусмотрена предварительная обработка культуральной жид-
кости, предназначена для производства антибиотика в виде не-
очищенного препарата (твердого вещества, полученного путем
распылительной или непрерывной сушки) или в виде высоко-
очищенного кристаллического вещества.
11.1. Отделение клеток и нерастворимых твердых
материалов
Нерастворимые компоненты системы (от клеток до индиви-
дуальных веществ) можно отделить от раствора, воспользовав-
шись особенностями их основных физико-химических свойств,
например размером частиц, плотностью, растворимостью или
диффузионной способностью. Используемые для этой цели ме-
тоды (в зависимости от размера отделяемых частиц) суммиро-
ваны на рис. 11.2, где, в частности, показано, что частицы, раз-
меры которых соответствуют размерам клеток (от 0,3 до
10 мкм), можно выделять фильтрованием, центрифугировани-
ем и (или) седиментацией, воспользовавшись их характерным
размером или плотностью. Седиментация представляет собой
17—746
258
Глава 11
1
Нерастворимые
продукты
биопроцесса
f=^4
70
Экстрагирование
нерастворимого
остатка
Жидкие отходы
Добавка к кормам
аля домашних животных
Жидкостная
\ АдсорбцияI Осаждение
1*
; г
7 _________ I экстракция.
Фильтрат культуральной жидкости \
Л-i ..у" \Раствори-
ш И 0 тель
3
3
г Частично очищенный антибиотик =
—Hill UlllllllUUIUimjJIHniUinUIUIUIU[£lll.llllllllllllllllll|IIIIIIIIIIHIH
gniiiiiiiiiiiiuiiiiiinn _~\смесь растворителей.
=1НШ_1
’iniiE 70
1 ft*
I
Растворитель —
Стерильная или j
нестерильная ।
зона очистки I
76
74
| Ш 1
llllllllll = [
Антибиотик
РИС. 11.1. Схема выделения антибиотика. [Воспроизведено с разрешения из
статьи: Beesch S. С., Stull G. М., Fermentation, Ind. Eng. Chem., 49, 1491
(1957). © American Chemical Society.]
1 — ферментер; 2—резервуар для предварительной обработки; 3—адсорб-
ционная колонна; 4 — фильтр; 5 — сушильный аппарат; 6—осадительный
резервуар; 7—резервуар для растворения; 8 — резервуар для экстракции;
9— резервуар для адсорбции; 10 — испаритель; 11 — центробежный экстрак-
тор; 12 — колонна для отгонки растворителя; 13— колонна для регенерации
растворителя; 14 — центрифуга; 15—кристаллизатор; 16 — сушилка непре-
рывного действия; 17 — распылительная сушилка.
осаждение в поле гравитационных сил, тогда как при центри-
фугировании скорость осаждения возрастает за счет центро-
бежных сил.
Некоторые типичные для культуральных бульонов свойства
могут затруднять как отделение нерастворимых веществ от
жидкой фазы, так и последующее разделение. К ним относятся
высокая вязкость, наличие гелеобразующих веществ, способ-
ность к образованию сжимаемых осадков на фильтре, бли-
зость плотности нерастворимых частиц к плотности воды, вы-
сокая степень начального диспергирования, низкая концентра-
ция суспендированных веществ. В периодических процессах эти
характеристики изменяются во времени, а в непрерывных про-
Свойство или параметр,
лежащие в основе
разделения
Размер
Диффузионная
способность
Ионный заряд
Давление паров
Растворимость
Поверхностная
активность
Плотность
Обратный
осмос
-<-------Диализ -
Электродиализ-
<---------------Ионный обмен
----------Микрофилыпры—
Ультрафильтрация--------
-------Гель-хроматография
Тканевые и волок-
нистые фильтры
«—Дистилляция, лиофилизациям
Экстракция
-* ~растворителями~
«--------Фракционирование в пене и пузырьках
-«— Ультра^нт^ифугиро---*-
-*-------Центрифугирование ---------->•
-Жидкостные циклонные
"* сепараторы
«-------- Седиментация--------
Ачгстремов, A 1 10 10 2 Ю3 10*
Нанометров, нм 10~' 1,0 10 ю2 10s
Макрометров, мкм 10 ~3 10'2 10'1 1,0
Миллиметров, мм io-7 1О'е 10 5 10'* 1О'С
10 5 106 10 7
10* 10s 10 6
10 10 2 103
10'2 Ю'1 1
Н Низкомолекулярные
вещества
Высокомолекулярные \ Частицы ______
вещества *1* размером 1мкм
_Мелкие
частицы
\ Крупные *
I частицы
РИС. 11.2. Принципы и методы выделения нерастворимых частиц в зависимости от их размера. (Из работы: Atkinson В.,
Maviiuna F„ Biochemical and Biotechnology Handbook, p. 935, Macmillan Publ., Ltd., Surrey, England, 1983.)
60
Глава 11
цессах свойства бульона зависят от скорости разведения и дру-
гих условий. В гл. 14 мы познакомимся с успешным исполь-
зованием непрерывного осаждения клеток в биологической
очистке сточных (бытовых) вод.
Предварительная обработка бульона для облегчения после-
дующего отделения биомассы может включать старение (в кон-
це стационарной фазы клетки обычно образуют скопления),
тепловую обработку, изменение pH и (или) добавление ве-
ществ, способствующих флокуляции клеток (полиэлектролитов,
хлорида кальция, коллоидной глины). Затраты на предвари-
тельную обработку окупаются повышением скорости седимен-
тации или фильтрования; обычно после предварительной обра-
ботки удается ускорить эти операции примерно в 10 раз, а в
отдельных случаях сообщалось о 100—200-кратном повышении
скорости осаждения (фильтрования).
Выбор конкретного отстойника, фильтра или центрифуги
зависит от свойств исходного бульона и заданного конечного
содержания воды в отделяемой биомассе:
Отстаивание применяют в крупномасштабных процессах пере-
работки отходов, а также в традиционных процессах броже-
ния (почему хорошее шампанское оставляют дозревать
в почти перевернутых бутылках?). Образующийся хлопье-
видный осадок обычно содержит несколько процентов твер-
дых веществ.
Центрифугирование позволяет получать концентрат биомассы,
часто называемой пастой, поскольку он содержит значитель-
ное количество твердых веществ (около 15% по массе) и
тем не менее сохраняет некоторые характерные для жидко-
стей свойства.
Фильтрованием получают более концентрированные (более
обезвоженные) осадки (20—35% твердых веществ по массе)
или твердую биомассу (более 40% твердых веществ по мас-
се). В некоторых случаях скапливающийся на фильтре оса-
док биомассы имеет хорошую проницаемость, что позволяет
отделять ее прямым фильтрованием. Гораздо чаще биомасса
оказывает большое сопротивление фильтрованию, и тогда
применяют фильтрующий слой на подкладке.
11.1.1. Фильтрование
Небольшие количества бульона, полученные в мелкомас-
штабных периодических процессах, можно фильтровать на
рамном фильтр-прессе, на котором постепенно накапливается
биомасса; затем фильтр открывают и снимают осадок био-
массы. В более крупномасштабном производстве применяют
Операции выделения 261
непрерывное фильтрование. Эту операцию выполняют, напри-
мер, на ротационных (барабанных) вакуум-фильтрах; иногда
такие фильтры лучше функционируют, если на фильтрующую
поверхность нанесен еще один фильтрующий слой. На
рис. 11.3 изображена схема ротационного вакуум-фильтра неп-
рерывного действия, где для снятия накапливающегося на вра-
щающемся барабане осадка (слоя твердых веществ) исполь-
%
Выводйой
канал
Валики
"Уровень разделяемой,
суспензии
РИС. 11.3. Ротационный вакуум-фильтр со шнуровым съемом осадка. (С лю-
безного разрешения компании Ametek, Inc.)
Выводной, канал
Неподвижное кольцо
автоматического распре-
делительного устройства
Затвердевший осадок
после фильтрования
Воздушный канал
Шнуры,
возврайрамсциеся
на барабан
зуются шнуры, которые удовлетворительно снимают осадок
мицелия PeniciUium. При снятии мицелия Streptomyces фильт-
рующую ткань необходимо покрывать вспомогательным фильт-
рующим материалом, например диатомовой землей, а слой ми-
целия с вращающегося барабана снимают с помощью ножа.
Характеристики фильтрования взвеси твердых частиц
в жидкости часто описывают с помощью элементарной теории
фильтрования. Приняв, что фильтрат течет через слой отфильт-
рованного материала в ламинарном режиме, мы можем за-
писать
1 dVf Вр
-------------- --------- Н 1 П
A dt--------------------'
где А — площадь фильтрующей поверхности; Vf — объем
фильтрата; t — продолжительность фильтрования; Др— пере-
пад давлений на фильтре; цс— вязкость фильтрата; а — сред-
нее удельное сопротивление осадка; W— масса накопленных
262
Глава 11
твердых веществ в осадке; IF=[pw/(l—mw)]Vf, где р — плот-
ность фильтрата, w — массовая доля твердых веществ в сус-
пензии; т — отношение массы влажного осадка к массе сухо-
го осадка; г — коэффициент сопротивления фильтрующего ма-
териала. На ротационных фильтрах, обычно применяющихся в
микробиологической промышленности, перепад давления на
фильтре постоянен. Если допустить, что осадок несжимаем, то
а постоянно, и тогда после интегрирования уравнения (11.1)
получим
t vf I № /ц 2)
Vf/A 2Др(1—mu>) А Др ’ '
Отсюда следует, что в принятых условиях t/Vf линейно зави-
сит от Vf.
На рис. 11.4, а представлен график зависимости t/Vf от Vf
для фильтрования суспензии культуры Streptomyces griseus
Уг, см3
а
РИС. 11.4. а — на скорость фильтрования бульона S. griseus большое влияние
оказывает величина pH; б — предварительное нагревание бульона S. griseus
изменяет удельное сопротивление образующегося осадка. [Из статьи: Shira-
to S., Esutni S., Filtration of a Culture Broth of Streptomyces griseus, J. Fer-
ment. Tech. (Japan), 41, 87 (1963).]
Время выдержки при WOX, мин
б
при различных pH через хлопчатобумажную ткань и вспомо-
гательный фильтрующий материал (диатомовую землю) при
Др = 2 кг/см2. Из этих данных следуют два важных вывода.
Во-первых, осадок сжимаем, поскольку экспериментальные
данные, определенные при одном значении pH, располагаются
не на прямой. Вообще клетки и другие органические компонен-
ты микробиологических процессов обычно образуют сжимае-
Операции выделения
263
мые осадки. Во-вторых, экспериментальные данные свидетель-
ствуют о том, что фильтрующие свойства сильно зависят от
предварительной обработки бульона и условий фильтрования.
Очень большое влияние на скорость фильтрования, очевидно,
оказывает pH. Приведенные на рис. 11.4,6 данные показыва-
ют, что предварительная тепловая обработка бульона может
значительно снизить удельное сопротивление осадка за счет
коагуляции мицелиальных белков.
Для повышения объемной производительности биореакто-
ров в последние годы все чаще применяют рециркуляцию
культур клеток (вспомните гл. 9, разд. 9.1.3). В крупномас-
штабных установках по аэробной переработке бытовых отходов
(гл. 14) флокулирующую биомассу (состоящую из различных
организмов) отделяют отстаиванием и образовавшуюся кон-
центрированную суспензию клеток подвергают рециркуляции.
В мелкомасштабных процессах с участием нефлокулирующих
организмов можно применять метод фильтрования, при кото-
ром параллельный поверхности фильтра непрерывный поток
жидкости уносит накапливающуюся клеточную массу, тем са-
мым создавая почти стационарный поток фильтрата [в отличие
от изменяющегося во времени поведения системы при периоди-
ческом фильтровании; см. уравнение (11.2)]. Скорость потока
фильтрата через фильтровальную перегородку зависит как от
перепада давления на перегородке Др, так и от установившего-
ся сопротивления слоя биомассы a(W7/4). Следовательно,
уравнение (11.1) применимо и для фильтрования с параллель-
ным фильтрующей поверхности потоком, однако теперь устано-
вившаяся масса осадка на фильтре W и удельное сопротивле-
ние а являются функциями скорости поперечного потока жид-
кости (или касательного потока).
Состав осадка при обычном фильтровании и фильтровании
с параллельным фильтрующей поверхности потоком может
быть различен, особенно если исходная смесь помимо клеток
содержит другие нерастворимые компоненты. Последний спо-
соб фильтрования позволяет повысить концентрацию биомас-
сы в 10 и более раз. Сообщалось, что при фильтровании с па-
раллельным фильтрующей поверхности потоком через мембра-
ну, задерживающую вещества с молекулярной массой более
100 000, наблюдается 15—20-кратное повышение концентрации
Е. coli, микоплазмы (при производстве ветеринарных вакцин)
и вируса гриппа (при производстве вакцин на основе интакт-
ных вирусов). Если фильтрованию с параллельным фильтрую-
щей поверхности потоком подвергается периодическая культу-
ра клеток, то в непрерывно рециркулирующем потоке культу-
ры концентрация клеток возрастает во времени; этот факт не-
обходимо учитывать при анализе таких систем.
264
Глава 11
Как и в случае центрифугирования при больших скоростях
вращения ротора, иногда перед фильтрованием с параллель-
ным фильтрующей поверхности потоком необходимо предвари-
тельное фильтрование на грубых фильтрах. Так, в производст-
ве антибиотиков неутилизированные соевую крупу и карбонат
кальция удобнее всего отделять с помощью сетчатого или дру-
гого фильтра с большим диаметром пор, а затем отделять био-
массу методом фильтрования с параллельным фильтрующей
поверхности потоком.
Если внеклеточный продукт метаболизма можно выделить
экстракцией несмешивающимся с водой растворителем непо-
средственно из культурального бульона, то операции отделения
биомассы и выделения продукта можно объединить. Такие
комбинированные операции мы рассмотрим в одном из следую-
щих разделов.
11.1.2. Центрифугирование
Из культурального бульона биомассу можно выделить цент-
рифугированием; таким методом иногда выделяют, например,
дрожжи. На рис. 11.5 приведено схематическое изображение
одного из типов центрифуг непрерывного действия. В разбав-
ленных суспензиях каждую клетку можно рассматривать как
индивидуальную частицу в непрерывной жидкой фазе; к такой
системе применимо математическое описание, данное в приме-
ре 1.1. Для концентрированных суспензий, в которых на дви-
жение каждой частицы влияют соседние частицы, этот подход
неприменим.
Выведены уравнения, связывающие скорость движения ча-
стицы в концентрированной суспензии Uh со скоростью движе-
ния индивидуальной частицы в отсутствие влияния соседних
частиц «о и с объемной долей частиц гр. Для общего урав-
нения
«д 1
«о 1 + Рер1/3
(Н.З)
эмпирическим путем найдены следующие корреляции между
Р и ер:
l + 3,05gp2’84 0,15<gp<0,5; частицы неправильной формы
1 + 2,29ер3’43 0,2<ер<0,5; сферические частицы
1—2 разбавленные суспензии (ер<0,15)
(11.4)
Операции выделения
265
В зависимости от природы и характеристик исходной сус-
пензии применяют центрифуги с различной конструкцией внут-
ренних устройств для съема осадка твердых веществ ^(табл.
11.2). В отличие от других центрифуг непрерывного действия
в осадительных (или шнековых) центрифугах (рис. 11.6)
РИС. 11.5. В этой центрифуге непрерывного действия осадок твердых веществ
отделяется в потоке между двумя коническими поверхностями. Осветленную
жидкость (фугат) отбирают из верхней части аппарата.
можно отделять и довольно крупные частицы. По этой причи-
не такие центрифуги целесообразно применять в сочетании с
центрифугой для разделения суспензии мелких частиц, что по-
зволяет использовать последнюю наиболее эффективно без пе-
регрузки и засорения.
Примеры применения этих центрифуг приведены в табл.
11.3; обратите внимание на использование шнековых (осади-
тельных) центрифуг для отделения крупных скоплений плесне-
вых грибов, а также на большую производительность центри-
фуг с соплами.
266
Глава 11
Таблица 11.2. Типы центрифуг’
Максималь-
Тип центрифуги Способ съема осадка Содержание твердых ве- ществ в ис- ходной сме- си, об. % ная произво- дительность самого боль- шого аппара- та, м3/ч (галло- иов/мии)
Камерные сепарато- ры Саморазгружающиеся сепараторы с соп- лами Остается в камерах 0-1 150 (650)
Периодическая выгрузка рез осевые каналы че- 0,01-10 200 (880)
Саморазгружающиеся сепараторы Периодическая выгрузка рез радиальную щель че- 0,2—20 100 (440)
Центрифуга непре- рывного действия с выгрузкой осад- ка через сопла Непрерывная выгрузка че- 1—30 рез сопла, расположенные на периферии корпуса или вблизи нее 300 (1320)
Осадительная (шне- ковая) центрифуга Выгрузка с помощью внут- 5—80 реннего шнека 200 (880)
Даниые взяты из работы: Belter Р., Recovery Processes—Past, Present, and Future,
184th Amer. Chem. Soc. Mtg., Sept. 1982.
РИС. 11.6. Конструкция центрифуги co шнековой выгрузкой осадка. (Из ра-
боты- Perry R. Н., Chilton С. Н„ Chemical Engineers Handbook, 5th ed.,
pp 19—92, McGraw-Hill, New York, 1973. Имеется перевод 4-го издания. Пер-
ри Дж. Г., Справочник инженера-химика, т. 1, 2. — Л.: Химия, 1969.)
/ — привод шнека; 2 —канал для выгрузки осадка; 3 — шнек; 4 — регулируе-
мые каналы для выгрузки фугата; 5 — приводной шкив; 6 подлежащая раз-
делению суспензия; 8 — выгрузка фильтрата; 9 выгрузка осадка, 9 бай-
пасная линия для устранения перегрузок.
Операции выделения
267
В продуктах биопроцессов часто встречаются внеклеточные
твердые вещества. Так, при производстве ферментов из расти-
тельной биомассы последнюю обычно сначала измельчают и
экстрагируют раствором с высокой ионной силой при охлаж-
дении, после чего большую часть твердых веществ необходимо
отделить. Точно так же при частичной утилизации твердых суб-
Таблица 11.3. Примеры использования центрифугирования
для выделения биомассы3
Продукт Микроорганизм Относитель- ная произ- водитель- ность цент- рифуги Тип центрифуги
Тип Размер, мкм
Пекарские дрож- жи Saccharomyces 7—10 100 С соплами
Пивные дрожжи Saccharomyces 5-8 70 С соплами, само- разгружающаяся
Винные дрожжи Saccharomyces 5-8 60 С соплами, само- разгружающаяся
Белок одноклеточ- ных организмов Антибиотики Candida Плесневые грибы 4—7 50 10—20 С соплами, осади- тельная Осадительная
Антибиотики Лимонная кисло- та Actinomyces Плесневые грибы 10—20 7 20—30 Саморазгружаю- щаяся Саморазгружаю- щаяся, осадитель- ная
Ферменты Bacillus 1—3 7 С соплами, само- разгружающаяся
Вакцины Clostridia 1—3 5 Камерная, само- разгружающаяся
а Данные взяты из работы: Belter Р., 184th Amer. Chem. Soc. Mtg., Sept. 1982. Recovery Processes—Past, Present, and Future,
стратов (например, целлюлозы) в бульоне могут остаться мак-
роскопические частицы субстрата. Если плотность последних
заметно отличается от плотности жидкой фазы, то для первич-
ного отделения твердых веществ применяют шнековые центри-
фуги (рис. 11.6).
11.1.3 . Седиментация
Если под влиянием многозарядных катионов или внекле-
точных полимеров клетки легко образуют коагулирующие
скопления или хлопья, то биомассу можно отделять седимен-
тацией. Этот очень дешевый способ разделения используется
для обеспечения рециркуляции биомассы в процессах биологи-
ческой переработки отходов методом активного ила (гл. 14).
Кроме того, именно из-за простоты и дешевизны отделения био-
268
Глава 11
массы в пивоварении и производстве белка одноклеточных ор-
ганизмов используют штаммы дрожжей, обладающие высокой
склонностью к флокуляции. Механизм оседания и уплотнения
биомассы при седиментации не вполне понятен. В отличие от
изолированной частицы, скорость оседания которой не зависит
Концентрация твердых веществ с
а
РИС. 11.7. а — скорость осажде-
ния активного ила (снижения гра-
ницы раздела фаз), образующего-
ся в аэробном процессе, б — зави-
симость объема осадка активного
ила (образующегося при росте на
смеси субстратов) от продолжи-
тельности центрифугирования на
лабораторной центрифуге. Здесь
Z — фактор разделения, равный
ro®2/g. [Из работы; Weissman U.,
Binder Н., Biomass Separation
from Liquids by Sedimentation and
Centrifugation, in Adv. Biochem.
Eng., Fiechter A. (ed.), vol. 24,
p. 119, Springer-Verlag, Berlin,
1982.]
от концентрации, суспендированные твердые вещества осаж-
даются тем медленнее, чем выше их концентрация в суспензии
[уравнение (11.3)]. Например, граница, разделяющая бескле-
точную и содержащую клетки жидкие фазы, движется вниз со
скоростью, которую часто можно описать уравнением
Скорость оседания us = kc~m, где т«1,7—2,6 (11.5)
Когда концентрация клеток достигнет достаточно большой ве-
личины Стах, дальнейшее оседание происходит с пренебрежимо
малой скоростью, что подтверждается поведением активного
Операции выделения
269
ила, содержащего как различные микроорганизмы, так и вне-
клеточные полисахариды (рис. 11.7, а). Осажденную биомассу
можно далее обезводить центрифугированием или фильтрова-
нием; центрифугирование также характеризуется максимально
достижимой (зависящей от центробежной силы) концентрацией
биомассы (рис. 11.7,6). Фактор разделения Z определяют как
РИС. 11.8. Содержание воды в активном иле. При концентрации твердых ве-
ществ 0,6% содержание внутриклеточной воды равно 2,7%, капиллярной
(внеклеточной) воды 1,7%, внеклеточной воды, находящейся в скоплениях
клеток, 21% и свободной воды 74% от общего объема суспензии. Легче всего
отделяется свободная вода, а удаление воды из скоплений клеток, капилляров
и внутриклеточного пространства становится все более и более затруднитель-
ным. [Из работы: Weissman U., Binder Н., Biomass Separation from Liquids
by Sedimentation and Centrifugation, in Adv. Biochem. Eng., Fiechter A. (ed.),
vol. 24, p. 119, Springer-Verlag, Berlin, 1982.]
roet2lg, где r0 — радиус ротора центрифуги, со — угловая ско-
рость ротора, a g — ускорение силы тяжести.
При выборе конкретной операции выделения важно учиты-
вать степень обезвоживания, достигаемую при седиментации,
центрифугировании, фильтровании и сушке. Как показано на
рис. 11.8, содержащаяся в суспензии клеток вода складывает-
ся из свободной воды и воды, находящейся в скоплениях кле-
ток, капиллярах, внутри клеток и в твердом остатке. Свободная
вода отделяется при седиментации биомассы, а при последую-
щем старении полученного таким путем осадка происходит ча-
стичная потеря воды, содержащейся в скоплениях клеток.
270
Глава 11
Центрифугирование при Z = 3000 позволяет удалить всю воду,
содержащуюся в скоплениях клеток, и приводит к образова-
нию осадка с 5% воды. Дальнейшее обезвоживание (удаление
капиллярной воды) возможно только с помощью небольших
лабораторных центрифуг, а внутриклеточную воду можно уда-
лить высушиванием только после лизиса клеток. Аппараты
с наклонными трубками или узкими каналами позволяют по-
высить скорость седиментации благодаря вторичному эффекту
(эффекту Бойкотта), способствующему быстрому осветлению
жидкой фазы в верхней части канала или трубки.
11.1.4 . Перспективные методы выделения биомассы
Твердую фазу можно удалить из водной суспензии с по-
мощью восходящего потока пузырьков воздуха, к которым при-
липают нерастворимые твердые частицы. Такой метод (флота-
ция) широко применяется для извлечения мелких частиц из
водных суспензий минералов. Флотация используется также
в пивоварении. В близком методе, пенной флотации, раство-
ренные белки отделяют в виде высококонцентрированной ус-
тойчивой белковой пены благодаря поверхностному натяжению
на границе раздела фаз воздух — вода (именно этот эффект
ответствен за образование устойчивой пены на поверхности ев-
ропейского или английского пива). Флотацию использовали
также для концентрирования клеток Acinetobacter cerificans
при производстве белка одноклеточных организмов.
В методе электрокинетического осаждения образованию
плотной биомассы, содержащей до 40% (по массе) клеток,
способствует разность потенциалов 10—50 В/см; затраты на
электроэнергию компенсируются на последующих операциях
обезвоживания или сушки. Эффективность метода снижается
из-за частичного расхода электроэнергии на электролиз бульо-
на, сопровождающий осаждение биомассы. Этот метод пыта-
лись использовать для выполнения обратной задачи — предот-
вращения осаждения твердых веществ на фильтрующей по-
верхности в процессе обычного фильтрования. Электрохимиче-
ское осаждение биомассы нашло применение в производстве
белка одноклеточных организмов с участием Methylomonas cla-
га (компания Hoechst).
11.1.5 . Заключение
Необходимость в выделении биомассы может возникнуть по
разным причинам. Во-первых, биомасса может представлять
собой конечный продукт; во-вторых, она может являться ис-
ходным материалом в последующих операциях лизиса и виде-
Операции выделения
271
ления внутриклеточного продукта; в-третьих, отделение био-
массы необходимо после экстракции дезинтегрированных лизи-
сом или механическим способом клеток; в-четвертых, оно
должно предшествовать выделению или регенерации следовых
количеств непрореагировавшего растворимого субстрата, а так-
же регенерации и повторному использованию клеток в биоре-
акторе; наконец, может возникнуть потребность и в отделении
биомассы от реакционной массы, из которой уже выделен (на-
пример, экстракцией) продукт. Выбор конкретного способа вы-
деления биомассы — фильтрования (периодического, непрерыв-
ного в вакууме, с поперечным потоком и др.), центрифугирова-
ния (с вертикальным ротором или горизонтальным шнеком)
или коагуляции и седиментации — зависит от состояния бульо-
на (температуры, pH, ионной силы), состава реакционной мас-
сы (наличия или отсутствия биомассы, полимеров, многозаряд-
ных ионов, других нерастворимых частиц) и от состояния,
в котором желательно получить продукт.
11.2. Первичное выделение
11.2.1. Экстракция
Для экстракции необходимо наличие двух жидких фаз. При
выделении антибиотиков применяют в основном экстракцию
органическими растворителями из водной фазы, а для выделе-
ния белков недавно были предложены новые двухфазные вод-
но-водные системы.
11.2.1.1. Экстракция органическими растворителями. Многие
антибиотики хорошо растворяются в органических раствори-
телях, практически не смешивающихся с водой. Многостадий-
ная экстракция — сначала из водной фазы в органическую, за-
тем из органической фазы и т. д. — обеспечивает как концент-
рирование, так и очистку выделяемых веществ.
В 1 л обычного пенициллинового бульона содержится 20—
35 г антибиотика. После отделения биомассы мицелия фильт-
рованием фильтрат охлаждают до температуры не выше 4°C
и затем иногда повторно фильтруют через полимерный фильт-
рующий слой. Величина рЛа пенициллинов лежит в диапазоне
2,5—3,1. Поэтому подкисление бульона до pH 2,0—3,0 приво-
дит к протонированию (нейтрализации) пенициллинов, после
чего их можно экстрагировать органическими растворителями.
Противоточное распределение в системе бульон—амилацетат
(или бутилацетат) (10 : 1) позволяет перевести в органиче-
скую фазу практически весь пенициллин. Для того чтобы по-
высить концентрацию пенициллина, проводят еще одну экст-
272
Глава 11
ракцию водным раствором фосфатного буфера (pH 5—7,5).
Антибиотик выделяют в виде натриевой соли осаждением из
водно-бутанольной смеси.
Непрерывную экстракцию осуществляют в противоточном
аппарате непрерывного действия — центробежном экстракторе
Подбильняка (рис. 11.9). Органическая (легкая) и водная
(тяжелая) фазы поступают в аппарат и выводятся из него через
вращающийся вал.
РИС. 11.9. В центробежном экстракторе Подбильняка две жидкие фазы по-
ступают через вращающийся вал вблизи средней линии вала. Более легкая
(органическая) фаза выводится первой. Быстрое вращение обеспечивает дви-
жение двух фаз в противоположных направлениях при постоянном межфаз-
ном контакте. [Из работы: Todd W. J., Podbielniak D. В., Centrifugal Extrac-
tion, Chem. Eng. Progress, 61 (5), 69 (1965).]
Быстрое вращение создает центробежное ускорение, благо-
даря которому две жидкие фазы движутся в противоположных
направлениях.
Аналогично экстракцией из водной фазы органическим рас-
творителем и повторным переведением в водную фазу выделя-
ют и очищают другие практически важные антибиотики. Так,
эритромицин выделяют экстракцией амилацетатом, а полипеп-
тидный антибиотик бацитрацин непосредственно из бульона
экстрагируют бутанолом, затем переводят в водный буферный
раствор, концентрируют упариванием и высушивают. Еще раз
отметим, что необходимость конкретных операций разделения
Операции выделения
273
РИС. 11.10. Распределение пенициллинов и других содержащихся в культу-
ральной жидкости веществ между органической и водной фазами. [Из работы:
Queener S., Swartz R., Secondary Products of Metabolism, in Economic Micro-
biology, vol. 3, Rose A. H. (ed.), Academic Press, London, 1979.]
определяется сферой применения продукта; так, бацитрацин,
выпускаемый в качестве добавки к кормам для домашних жи-
вотных, получают простым упариванием культурального бульо-
на (без отделения биомассы) и последующей сушкой получен-
ного осадка.
Не все антибиотики выделяют и очищают экстракцией. Так,
например, стрептомицин выделяют из культуральной жидкости
методом ионного обмена (см. разд. 11.2.2, 11.4 и 11.7.2).
На стадии первой экстракции культуральную жидкость
обычно подкисляют, чтобы перевести антибиотик в неионизп-
рованную форму, в которой он легко переходит в органическую
фазу. В то же время выбор соответствующего диапазона pH
может обеспечить не только экстракцию данного антибиотика,
но и его отделение от других экстрагируемых (при других зна-
чениях pH) веществ. Как показано на рис. 11.10 при pH ниже
18—746
274
Глава 11
2,0 обеспечивается максимальная селективность экстракции пе-
нициллина по сравнению с другими (менее кислыми) вещест-
вами с более высокими рКа. С другой стороны, увеличение ско-
рости гидролиза пенициллинов, сопровождающее повышение
кислотности среды, является причиной того, что на практике
выгоднее применять экстракцию при рН>2,0. Другое решение
этой дилеммы — проводить экстракцию в экстракторе Подбиль-
няка, что позволяет свести к минимуму продолжительность экст-
ракции, а следовательно, и степень гидролиза антибиотика.
Предлагалось проводить экстракцию непосредственно из
культуральной жидкости и при выделении других сложных
органических соединений. Соответствующие примеры мы рас-
смотрим позднее в разделе, посвященном изучению методов
переработки цельного бульона (разд. 11.7.2).
11.2.1.2. Экстракция в двухфазных водных системах. По-
требность в экстрагентах и методах экстракции, которые не
повреждают лабильных ферментов и других белков, легко пе-
реносятся на крупномасштабные операции и позволяют выде-
лять целые органоиды и другие надмолекулярные структуры,
вызвала необходимость в изучении и разработке систем жид-
костной экстракции, где каждая фаза является в основном вод-
ной. Две водные фазы образуются при растворении двух не-
совместимых полимеров, например полиэтиленгликоля (ПЭГ)
и декстрана (рис. 11.11,а). Образующиеся фазы содержат бо-
лее 75% воды, а также большие количества декстрана или
ПЭГ; на рис. 11.11, а показано, что ПЭГ-содержащая фаза мо-
жет быть практически свободной от декстрана. Предлагались
и другие аналогичные системы, в том числе ПЭГ — фосфат ка-
лия (рис. 11.11,6), декстран — фикол — ПЭГ, поливиниловый
спирт — декстран, декстран — метил целлюлоза; к настоящему
времени лучше других изучены системы, изображенные на
рис. 11.11.
В системе ПЭГ — декстран коэффициенты распределения
\К=е (ПЭГ-фаза)/е(декстрановая фаза)] восьми ферментов
лежат в диапазоне от 1,0 до 3,7. Для увеличения коэффициен-
та распределения предлагалось применять производные поли-
меров (чтобы вызвать ионный обмен в растворе), растворимые
низкоспецифичные аффинные адсорбенты (см. следующий раз-
дел) или добавлять соли. В системе ПЭГ — сульфат аммония
добавление сульфата аммония или фосфата калия до концентра-
ции 12% или 0,3 М соответственно приводит к повышению ко-
эффициента распределения пуллуланазы в 10 раз (см. работу
[16]).
Полимерсодержащие фазы можно разделять декантацией
(умеренная продолжительность отстаивания; рис. 11.12, а) или
кратковременным центрифугированием (рис. 11.12,6). Затем
Операции выделения
275
Массовая доля декстрана, %
РИС. 11.11. а — фазовая диаграмма системы декстран — полиэтиленгликоль
(D48 — ПЭГ 6000) при 20 °C (кружочки) и при 0°С (крестики). б — фазовая
диаграмма и фазовый состав системы фосфат калия — ПЭГ 6000 при 0°С.
(Воспроизведено с разрешения из работы: Albertsson P.-А., Partition of Cell
Particles and Macromolecules, 2d ed., p. 41, Wiley-Interscience, New York,
1971.)
18
___________I__________L_
0 0,5 , 1,0
1,5
РИС. 11.12. a — зависимость относительного объема осаждающейся (под дей-
ствием силы тяжести) фазы от времени при разделении фаз в двух системах
полиэтиленгликоль — солевой раствор, применявшихся для очистки FDH.
Разделение фаз осуществляли в стеклянном резервуаре емкостью 150 л. Кре-
стиками обозначены данные, относящиеся к системе, содержащей 6% ПЭГ
1550, 9% ПЭГ 400, 15% фосфата калия в 2,5 М. КО, pH 7,8; диспергирована
верхняя фаза, коэффициент распределения 2,67. Кружочками обозначены дан-
ные в системе, содержащей 12% ПЭГ 4000, 7% фосфата калия, pH 7,8; дис-
пергирована нижняя фаза, коэффициент распределения 2,5.
б — пропускная способность центробежного экстрактора типа Лаваль
LAPX-202. Изображены кривые зависимости логарифма отношения концентра-
ций в диспергированной фазе после разделения (с) и в исходной смеси (с0) от
логарифма производительности аппарата (?, л/мин. Исходная система содер-
жит 14% ПЭГ 4000, 11% фосфата калия, 0,1 % голубого декстранового (ин-
дикатор). Скорость вращения центробежного экстрактора 700 (крестики) и
9300 (кружочки) оборотов в минуту. [Воспроизведено с разрешения из рабо-
ты: Kula М. R., Kroner К. И., Hustedt Н., Purification of Enzymes by Liquid-
Liquid Extraction, in Adv. Biochem. Eng., Fiechter A. (ed.), vol. 24, p. 105,
Springer-Verlag, Berlin, 1982.]
Операции выделения
277
выделяют (напрймер, ультрафильтрацией) необходимый про-
дукт, одновременно регенерируя полимер (ПЭГ). Поскольку
все операции смешения и разделения легко и быстро выполня-
ются при комнатной температуре, то дополнительные затраты,
связанные со стоимостью полимера (расходуемого как в пер-
воначальной загрузке, так и на возмещение частичных потерь
в ходе экстракции), невелики по сравнению с затратами на
охлаждение в более продолжительных методах выделения бел-
ков обычной экстракцией или осаждением. Стоимость декстра-
на, даже неочищенного, вносит основной вклад в стоимость
всей системы, поэтому регенерация декстрана с экономической
точки зрения обязательна. В табл. 11.4 перечислены ферменты,
выделенные из культур микроорганизмов методом экстракции
в двухфазных водных системах.
11.2.2. Сорбция
Под сорбцией понимают распределение растворенного ве-
щества между жидкой и твердой (обычно пористым или обла-
дающим большой поверхностью материалом) фазами. Одним
из наиболее известных примеров сорбции является выделение
стрептомицина с помощью ионного обмена. Ионообменники
представляют собой диссоциирующие ионные пары, в которых
один из ионов иммобилизован, например
Катионообменник
(твердый носитель) _Н++(растворенное вещество)+ —.
(твердый носитель)_(растворенное вещество) + + Н+
Анионообменник
(твердый носитель)+С1~+(растворенное вещество)- ч—--
ч (твердый носитель) + (растворенное вещество)- + С1"
Стрептомицин можно адсорбировать на катионообменной смо-
ле, несущей группы СООН:
п(смола)-Н++стрептомицинп+ ч п(смола)~стрептомицинп++Н+
Смолу, содержащую связанный антибиотик, отделяют от куль-
туральной жидкости, а антибиотик выделяют элюированием
подкисленной водой; при этом происходит процесс, обратный
связыванию антибиотика, и одновременно регенерируется ионо-
обменная смола.
Роль ионообменников могут выполнять и растворимые по-
лимеры, образующие водные двухфазные системы (см. пре-
дыдущий раздел). Растворы смесей полиэтиленгликоля (ПЭГ)
и декстрана неустойчивы и разделяются на две фазы. Если
вместо ПЭГ взять его производное, обладающее катионо- или
‘'8 Глава 11
Таблица 11.4. Выделение ферментов из биомассы микроорганизмов
методом экстракции в двухфазной водной системе3
Фермент®
Организм
Кон-
пняТРа" ДвУхФазная
биомас- система
сы, %
ентмс-И’ оВ,ЫХОД’
ент рас о/ ент
пределеиия очисткв
Изолейцил— тРНК-синте- таза Фумараза Escherichia coli 20 25 ПЭГ — соле- вой раствор ПЭГ — соле- вой раствор 3,6 3,2 93 93 2,3 3,4
Аспартаза 25 ПЭГ — соле- вой раствор 5,7 96 6,6
Пенициллин- ацилаза 20 ПЭГ — соле- вой раствор 2,5 90 8,2
а-Глюкозидаза Saccharomyces 30 cerevisiae ПЭГ — соле- вой раствор 2,5 95 3,2
Глюкозо- 6-фосфатде- 30 ПЭГ — соле- вой раствор 4,1 91 1,8
гпдрогеназа
Алкогольде- гидрогеназа Гексокиназа 30 30 ПЭГ — соле- вой раствор ПЭГ — соле- вой раствор 8,2 96 92 2,5 1,6
Глюкозоизо- мераза Streptomyces 20 sp‘ ПЭГ — соле- вой раствор 3,0 86 2,5
Пуллуланаза Klebsiella pneu- 25 moniae ПЭГ — декст- ран 3,0 91 2
Фосфорилаза 16 ПЭГ — декст- ран 1,4 85 1
Лейциндегид- рогеназа Bacillus sphaeri- 20 cus ПЭГ — декст- ран (неочи- щенный) 9,5 98 2,4
Диацетатдегид- рогеназа Lactobacillus sp. 20 ПЭГ — соле- вой раствор 4,8 95 1,5
L-2-гидрокси- изокапроат- Lactobacillus 20 confusus ПЭГ — соле- вой раствор 10 94 16
дегидрогена- за
-2-гидрокси- изокапроат- Lactobacillus 20 casei ПЭГ — соле- вой раствор И 95 4,9
дегидрогена-
за
а Данные любезно предоставлены д-ром М.-Р. Кулой. См. также сборник: Partitioning
in Aqueous Two-Phase Systems: Theory. Methods, Uses and Applications to Biotechnology,
Walter H„ Brooks D. E., Fisher D. (eds.), Academic Press, New York, 1985.
За исключением пуллуланазы выделению всех ферментов предшествовала дезинте-
грация клеток гомогенизацией под высоким давлением или размалыванием во влажном
состоянии.
Операции выделения
279
анионообменными свойствами, то образуется двухфазная си-
стема, по диэлектрическим свойствам близкая воде, которая
поэтому способствует повышению стабильности белков. С по-
мощью таких двухфазных систем легко разделить ^-интерфе-
рон и сопутствующие белки [17].
Как и другие операции концентрирования продуктов мик-
робиологических процессов, ионный обмен применяли для не-
посредственной обработки цельного бульона, например при вы-
делении антибиотика новобиоцина (см. разд. 11.7.2).
11.3. Осаждение
Растворимость органических веществ зависит от темпера-
туры, pH, состава, ионной силы и диэлектрической проницаемо-
сти растворителя. Осаждение можно индуцировать различными
способами:
1. Добавлением осадителя, реагирующего с растворенным ве-
ществом и образующего с ним нерастворимое соединение,
чаще всего соль. В качестве примера можно привести сле-
дующие операции выделения антибиотиков:
Гидрохлорид прока-
прокаин
натрий
калий
ина
Буферный раствор
натриевых и калие-
+ пени-
циллин
пени-
циллин
вых солей
Органический растворитель-]- , сульфат стрептоми-
-]-стрептомицин-|-Н28О4 цина
Органический растворитель-]- , гидрат эритроми-
+Н2О-|-эритромицин цина
Биополимеры также можно выделить в виде солей. Так, на-
пример, ксантан представляет собой полианион и при до-
бавлении ионов двухвалентных металлов (например, каль-
ция) осаждается в виде геля. Альгинаты можно выделять
из биомассы водорослей путем отделения (фильтрованием)
биомассы с последующим осаждением биополимера хлори-
дом кальция. Вспомните, что именно такой способ создания
альгинатных гелей применяется обычно для иммобилизации
клеток (гл. 9).
2. Индуцированное растворителем осаждение применяется
в операциях выделения микробных полисахаридов, в том
числе декстрана и ксантана. Эти биополимеры обычно об-
разуются в аэробных процессах, и получающаяся культу-
280
Глава 11
ральная жидкость, как правило, имеет очень высокую вяз-
кость. При производстве ксантана клетки Xanthamonas по-
гибают в процессе стерилизации бульона. После добавления
хлорида калия (для уменьшения количества используемого
затем растворителя) полисахарид осаждают путем разведе-
ния бульона метанолом или изопропанолом. Осадок, содер-
жащий биополимер и отмершие клетки, обезвоживают, про-
Таблица 11.5. Изоэлектрические точки ряда ферментов
и других белкова
Белок р/
Пепсин Яичный альбумин Сывороточный альбумин Уреаза Р-Лактоглобулин Yi-Глобулин Гемоглобин Миоглобин Химотрипсиноген Цитохром с Лизоцим 1,0 4,6 4,9 5,0 5,2 6,0 6,8 7,0 9,5 10,65 11,0
а Из работы: Lehninger A. L., Biochemistry, 2d ed., table 7.1, Worth
Publishers, Inc., New York, 1975; есть перевод более раннего издания:
Ленинджер А., Биохимия.—М.: Мир, 1976.
мывают и сушат. Декстран выделяют из бульона осаждени-
ем спиртом или ацетоном. В производстве сравнительно не-
дорогих неочищенных полисахаридов после осаждения рас-
творитель должен быть регенерирован и использован пов-
торно; при производстве полисахаридов для нужд пищевой
или фармацевтической промышленности необходимо обеспе-
чить достаточно полное удаление растворителя.
3. Методы осаждения белков сводятся к таким изменениям ус-
ловий, которые нарушают устойчивость исходной термодина-
мически стабильной однофазной системы (раствора белка)
и тем самым приводят к осаждению белка. К числу методов
снижения растворимости белков относятся:
а) добавление концентрированных растворов солей (так на-
зываемое «высаливание»);
б) изменение pH до величины, соответствующей изоэлектри-
ческой точке (т. е. результирующему нулевому заряду
белковой молекулы; см. табл. 11.5), в которой белок
имеет наименьшую растворимость;
Операции выделения
281
в) снижение диэлектрической проницаемости среды, напри-
мер, путем добавления смешивающегося с водой органи-
ческого растворителя; таким путем достигается усиление
электростатических взаимодействий;
г) добавление неионных полимеров, снижающих степень
сольватации белка;
д) добавление полиэлектролитов, действующих, по-видимо-
му, как флокулирующие агенты;
е) добавление солей многозарядных ионов металлов, обра-
тимо осаждающих белки.
При выборе конкретного метода осаждения необходимо
учитывать не только требуемую степень обогащения и затраты
на осаждение, но и требуемую степень чистоты белка.
Понятно, что растворимость белков зависит от pH раство-
ра и обычно минимальна в изоэлектрической точке. Поскольку
при р/ результирующий заряд молекулы белка равен нулю,
а при иных значениях pH молекулы белка имеют тот или иной
заряд, то силы электростатического отталкивания между моле-
кулами растворенного вещества минимальны при р/. Такой ме-
ханизм предполагает возможность разделения белков с различ-
ными изоэлектрическими точками путем фракционированного
осаждения-, при данном pH будут осаждаться белки, р/ кото-
рых наиболее близко этому pH (если другие характеристики
белков, например молекулярная масса, близки). Путем изме-
нения pH сложную смесь белков разделяют на фракции, со-
держащие различные белки.
В то же время при слишком высоких или слишком низких
значениях pH многие белки могут денатурироваться. По этой
причине чаще применяют другой метод осаждения, называе-
мый высаливанием. На растворимость белков заметно влияет
концентрация солей в растворе. Еще в 1889 г. Гофмейстер от-
метил, что наиболее эффективно белки осаждаются в присут-
ствии солей с многозарядными анионами или катионами. Ря-
ды ионов Гофмейстера (или лиотропные ряды), в которых
ионы расположены примерно в порядке уменьшения высалива-
ющей способности, выглядят следующим образом:
Анионы: цитрат3-, тартрат2-, F-, Ю2-, Н2РО4-,
ацетат-, ВгОз”, С1~*, С10з-, Вт-, NO3-, СЮ4-, I-, CNS-
Катионы: Th4+, АР+, Н+, Ва2+, Sr2+, Са2+, Mg2+, Cs+, Rb+,
1NH4+, К+, Na+, Li+
Для осаждения белков чаще всего применяют хорошо раство-
римый в водных средах сульфат аммония, хотя ряды Гофмей-
стера показывают, что для этих же целей можно использовать
и многие другие соли.
282
Глава 11
Важнейшей переменной, определяющей эффективность вы-
саливания белков, является ионная сила раствора p£S:
1 п
(п-б)
i=l
где Ci и Zi—концентрация и заряд соответственно г-го иона,
а п — общее число имеющихся в растворе ионов. Считается,
что добавление солей снижает концентрацию воды, которая
РИС. 11.13. Зависимость растворимости белка от ионной силы растворов
сульфата аммония. (Из работы: Cohn Е., Edsall J. Т., Proteins, Amino Acids,
and Peptides, p. 602, Academic Press, New York, 1943.)
может гидратировать ионизированные или поляризуемые груп-
пировки белка, тем самым уменьшая его растворимость. Как
показывают данные, представленные на рис. 11.13, раствори-
мость белка при высокой ионной силе раствора часто можно
выразить уравнением
1g(растворимость, г/л)=£' — xs|iis (11.7)
где xs — так называемая постоянная высаливания.
Белки, денатурирующие при нагревании быстрее других
присутствующих в смеси белков, можно выделять из раствора
тепловой обработкой. После денатурации растворимость ^белка
снижается и он осаждается (вспомните модель масляной кап-
ли: разворачивание молекулы глобулярного белка приводит к
Операции выделения
283
контакту его гидрофобных групп с водной фазой). Этот ме-
тод с успехом применяли, например, для очистки термоста-
бильного белка, полученного экспрессией соответствующего
гена в мезофильной клетке-хозяине; термостабильный белок
остается в растворе, а сопутствующие белки клетки-хозяина
селективно осаждаются. При другом методе осаждения в ка-
честве осадителя применяют казеин, диатомовую землю (со-
стоящую из пористых скелетов диатомовых водорослей) или
желатин; белки осаждаются из раствора, адсорбируясь на
сравнительно крупных частицах этих агентов.
Как показано в примере 11.1, при выделении белков часто
сочетают несколько методов осаждения. Выделение дегидроге-
назы начинают с экстракции фермента буферным раствором.
За этим следует умеренное нагревание для осаждения белков
и других компонентов смеси, легче подвергающихся денатурации,
чем выделяемая дегидрогеназа. Большую часть фермента
(а также другие вещества с близкой растворимостью) осаж-
дают ацетоном; при этом отделяются многие низкомолекуляр-
ные соединения, экстрагирующиеся вместе с белком. Последую-
щее растворение и повторное осаждение сульфатом аммония
приводят к получению препарата с большей удельной фермен-
тативной активностью. Такой способ очистки, однако, сопро-
вождается потерей некоторого количества фермента на каждой
операции (см. таблицу в примере 11.1).
Дальнейшую очистку для отделения дегидрогеназы от дру-
гих белков с близкой растворимостью выполняют путем после-
довательного суспендирования осадка, полученного на ста-
дии 8 (пример 11.1), во все менее и менее концентрированных
растворах сульфата аммония. Поскольку растворимость белков
возрастает при снижении концентрации соли до величин, зна-
чительно уступающих концентрации насыщенного раствора, то
таким путем удается очистить фермент от все более и более
близких белков. Полученная в результате ряда таких операций
супернатантная жидкость имеет удельную активность (фермен-
тативную активность отнесенную к общему количеству раство-
ренных белков), которая примерно в 100 раз превышает удель-
ную активность исходного продукта экстракции буферным рас-
твором. Кроме того, в этих операциях отделяется большая
часть примесей небелковой природы.
Пример 11.1. Последовательность операций при выделении ферментов из
биомассы (алкогольдегидрогеназы из сухих пекарских дрожжей)*.
1. Измельчить дрожжи в шаровой мельнице.
2. Смешать 100 г мелкоизмельченных сухих дрожжей со 100 мл 0,066 М
* Из работы: Chaykin S., Biochemistry Laboratory Techniques, pp. 22—32,
John Wiley and Sons, Inc., New York, 1966.
284
Глава 11
динатрийфосфатного буфера, содержащего 0,001 М этилендиаминтетра-
уксусной кислоты (EDTA).
3. Добавить 200 мл буферного раствора, перемешивать 2 ч при 37 °C и оста-
вить на 3 ч при комнатной температуре.
4. Отделить дрожжевую биомассу центрифугированием (28 000 g, 20 мин)
(определить ферментативную активность).
5. Отделить термически неустойчивые белки нагреванием при —55°C в тече-
ние 15 мин (алкогольдегидрогеназа относительно устойчива) (определить
ферментативную активность).
6. К прошедшему тепловую обработку раствору добавить ацетон из расчета
50 мл ацетона на 100 мл раствора (смешивание производить при темпера-
туре от —2 до —4°C), осадок отбросить. Добавить еще 55 мл ацетона,
центрифугировать при 28 000 g и супернатант отбросить (определить фер-
ментативную активность).
7. Осадок суспендировать повторно в 50 мл 10-3 М фосфатного буфера
(pH 7,5), содержащего 10~3 М EDTA, дважды диализовать против 3 л
свежего фосфатного буфера в течение 1 ч.
8. При 0 °C постепенно добавить сульфат аммония (36 г на 100 мл диализо-
ванного раствора) и центрифугировать при этой же температуре (опреде-
лить ферментативную активность).
9. Полученный на предыдущей стадии осадок суспендировать в 30 мл раз-
бавленного вдвое насыщенного раствора (NH4)2SO4. Осадок отделить от
супернатанта и провести повторное осаждение последовательно раствора-
ми сульфата аммония, концентрация которых составляет 45, 40, 35 и 25%
от насыщенного раствора (определить ферментативную активность).
10. Объединить высокоактивные супернатанты и добавить насыщенный рас-
твор сульфата аммония до помутнения. Оставить на несколько суток при
0 °C для кристаллизации. Растворить осадок и перекристаллизовать из
10 мл 10~3 М EDTA (определить ферментативную активность).
Получены следующие результаты выделения:
Ферментативная активность, единиц
Общая (ХЮ-8) Удельная
Первый экстракт 25 4 000
После тепловой обработки 30 8 000
После осаждения ацетоном 20 45 000
После первого осаждения суль- фатом аммония 14 70 000
Концентрация (МНчЬЭОч, в % от насыщенного раствора Белок в растворе
Удельная тивность ак- Всего белков, %
50 180 000 14,9
45 200 000 12,5
40 650 000 12,0
35 440 000 6,6
25 430 000 19,5
Операции выделения
285
11.3.1. Кинетика образования осадка
Осаждение белка происходит в том случае, когда в резуль-
тате изменения тех или иных условий его растворимость пада-
ет ниже существующей концентрации белка (или комплекса
белок—реагент). В благоприятной ситуации, например при
осаждении а-казеина, после фазы задержки средняя «молеку-
лярная» масса осаждающихся частиц MW быстро возрастает
РИС. 11.14. Зависимость «молеку-
лярной» массы частиц а-казеина,
осаждающихся в присутствии
0,008 М СаСЬ, при трех началь-
ных концентрациях белка (вели-
чина концентраций в кг/м3 указа-
на возле соответствующих кри-
вых). Кривые рассчитаны по урав-
нениям теории перикинетической
флокуляции Смолуховского. [Из
работы: Bell D. J., Hoare М., Dun-
nill Р., Adv. Biochem. Eng., vol. 26,
p. 19, Springer-Verlag, Berlin,
11983.]
Время, c [•]
Время, c [o, □]
во времени по линейному закону (рис. 11.14), описываемому
уравнением Смолуховского:
MW(f) =W(0)[l + KAmotl (11.8)
где т0 — молярная концентрация осаждающихся веществ.
Линейная зависимость скорости осаждения броуновских ча-
стиц в неперемешиваемой жидкости предсказывается теорией
перикинетической флокуляции, индуцированной диффузией. Со-
гласно этой теории снижение концентрации частиц N (т. е.
уменьшение их числа и соответственно увеличение массы) опи-
сывается уравнением второго порядка
___^У_ = Акдг2 (Н.9)
dt W v '
Здесь ?Ci — 8л^)молекула'^/молекула» э W параметр стабильности
286
Глава 11
коллоидной системы, учитывающий барьер электрического по-
ля вокруг каждой из идентичных молекул.
В течение короткого периода времени зависимость
от t линейна, затем повышение «молекулярной» массы осажда-
ющихся частиц точнее описывается уравнением (11.10)
d(t) = <J(0)[l +а In (/)] (11.10)
где d — усредненный диаметр осаждающихся частиц. Приме-
ром может служить осаждение казеина сульфатом аммония
(рис. 11.15).
РИС. 11.15. Старение осадка; зави-
симость размера частиц от времени
при фракционированном осаждении
казеина 1,8 М раствором сульфата
аммония. Штриховая линия отвечает
расчетным данным, предсказываемым
перикинетической теорией. [Воспро-
изведено с разрешения из работы:
Hoare М., Protein Precipitation and
Precipitate Ageing. Part I. Salting Out
and Ageing of Case in Precipitates,
Trans. I., Chem. E., 60 (2), 79 (1982).]
Механическое воздействие может повысить скорость осаж-
дения, а также способствовать созданию более плотных хлопь-
ев. Образующиеся в результате механического воздействия
устойчивые частицы могут переносить центрифугирование без
разрушения. Увеличение частоты столкновений частиц друг с
другом приводит к возрастанию скорости их флокуляции. Так,
в турбулентном режиме, создаваемом перемешиванием при
расходе мощности на единицу объема P/V, средняя скорость
сдвига G определяется выражением
Goc(P/V)7‘ (п.и)
Поскольку частота столкновений частиц друг с другом пропор-
циональна G, то в случае индуцированной сдвигом или орто-
кинетической флокуляции уравнение (11.9) преобразуется в та-
кую форму:
—= — ad34ac™«a -G.№ (11.12)
at 3
где а — коэффициент эффективности столкновений (доля столк-
Операции выделения
287
новенищ которые приводят к устойчивым скоплениям). Выра-
жение Gt представляет собой безразмерный параметр, который
в процессах обработки воды принято называть числом Кэмпа
(Са) или параметром старения-, с этим параметром связан рост
частиц осадка в системе (рис. 11.16).
Параметр старения Gt
РИС. 11.16. Зависимость отношения среднего начального размера частиц_к
размеру частиц после действия сдвига в капилляре от параметра старения Gt
(произведения скорости сдвига на его продолжительность). Средняя скорость
сдвига 1,7-104 с-1, среднее время сдвига 0,065 с, концентрация белка
30 кг/м3. Средний начальный диаметр частиц (мкм): 53,5 (а), 23,4 (б), 19,5
(в), 15,2 (г), 10,2 (д) и 8,8 (е). [Воспроизведено с разрешения из статьи:
Bell D. J., Dunnill Р., Shear Disruption of Soya Protein Precipitate Particles
and the Effect of Aging in a Stirred Tank, Biotech. Bioeng., 24, 1271 (1982).]
а
б
в
г
д
е
Если принять, что объемная доля осаждающихся частиц
(0V) постоянна и равна ш/3Частица^/6, то уравнение (11.12)
можно преобразовать в следующее полезное уравнение первого
порядка:
__^. = Аа^сдг (ПЛЗ)
dt jr
В общем случае процесс осаждения должен включать ста-
дии зарождения и роста индивидуальных частиц, а также их
последующую агрегацию, в результате которой образуются
хлопья (флокулы) с размерами, обеспечивающими возмож-
ность их последующего отделения (центрифугированием,
фильтрованием и т. д.), а также достаточную эффективность
переведения продукта в определенную фазу. Статические (пе-
рикинетические) и индуцированные сдвигом (ортокинетиче-
ские) скорости зарождения и начального роста частиц описы-
288
Глава 11
ваются приведенными выше уравнениями (11.9) и (11.12),
а последующая более низкая скорость флокуляции осадка,
т. е. образования более крупных агрегатов из осажденных ча-
стиц (соответствующая стадия часто осуществляется в отдель-
ном сосуде с перемешиванием), может быть описана уравне-
нием первого порядка. Предложены уравнения, связывающие
время пребывания в сосуде с перемешиванием со снижением
концентрации частиц осадка, как для режима полного вытес-
нения, так и для каскада из т ПРПП.
Режим полного вытеснения:
In f———= — OC0DG/TpnB
\ N(t) J ji трпв
Каскад из т ПРПП:
1 _ п т 17 No
ПРПП Л , I a? t)
4 L\ N (/) /
(11.14)
(11.15)
Применимость этих простых уравнений для изучения процес-
сов осаждения белков зависит от точности, с которой извест-
ны величины а и на последние размер частиц не влияет.
Сдвиговые напряжения способствуют также формированию
более компактных осадков. Как показано на рис. П.16, наибо-
лее плотные частицы осаждаются при величинах G7(Ca), пре-
вышающих 105. Впрочем, чрезмерно большие сдвиговые напря-
жения могут разрушать непрочные хлопья; механизм этого эф-
фекта аналогичен механизму эффекта, проявляющегося при
критическом числе Вебера (см. гл. 8). Максимальный диаметр
устойчивых частиц почти обратно пропорционален средней ско-
рости сдвига, т. е. dmaxoeG-1, если dmax больше масштаба тур-
булентных вихрей; при значительно меньших величинах dmax
этот параметр при изменении скорости сдвига практически не
меняется.
Выделению осадков центрифугированием должно предшест-
вовать достаточно длительное время, необходимое для обра-
зования хлопьев и их стабилизации. Как показано на рис. 11.17,
продолжительность и скорость сдвига (перемешивания и т. д.)
должны удовлетворять следующим требованиям:
1. Центрифугированию может подвергаться осадок, диаметр
частиц которого не ниже некоторой минимальной величины.
2. Величина параметра Gt (произведения скорости сдвига на
время) должна обеспечивать образование прочных хлопьев.
3. Эффективное перемешивающее устройство должно обеспе-
чивать хорошее перемешивание жидкой фазы.
4. Продолжительность старения не должна быть слишком
большой.
Операции выделения
289
Продолжительность старения t, с
РИС, 11.17. Зависимость свойств осадка (размеров частиц) от времени старе-
ния t при осаждении в периодическом режиме. Заштрихованная область отве-
чает осадку, удовлетворяющему всем требованиям, предъявляемым к осадкам,
отделяемым центрифугированием. Границы этой области определяются: а —
минимальным размером частиц, необходимым для их отделения центрифуги-
рованием; б—оптимальной величиной параметра Gt (Gf=105); в — мини-
мальной скоростью перемешивания, обеспечивающей выравнивание концент-
раций компонентов; г — максимально допустимым временем старения.
[Воспроизведено с разрешения из работы: Bell D. J., Ноаге Л1., Dunnill P.t
The Formation of Protein Precipitates and their Centrifugal Recovery, in Adv.
Biochem. Eng., Fiechter A. (ed.), vol. 26, p. 65, Springer-Verlag, Berlin, 1983.]
Все эти требования и ограничения в графической форме
отображены на рис. 11.17, где заштрихованная область отве-
чает значениям параметров, удовлетворяющим всем перечис-
ленным требованиям.
11.4. Хроматография и адсорбция в неподвижном
слое; периодические операции
с селективной адсорбцией
При хроматографии сравнительно небольшую порцию рас-
твора смеси веществ (или продуктов мелкомасштабного про-
цесса) разделяют на хроматографической колонке на фракции,
содержащие растворы практически чистых компонентов смеси
(рис. 11.18). Вещества биологического происхождения, напрп-
19—746
290
Глава 11
мер белки, имеют различную тенденцию к связыванию с таки-
ми адсорбентами, как крахмал, диатомовая земля, полиакрил-
амидный гель. Поэтому если содержащий смесь белков рас-
твор пропустить через колонку, заполненную таким адсорбен-
том, то каждый белок будет двигаться по колонке с эффектив-
ной скоростью, зависящей от склонности этого белка к адсорб-
ции. Для объяснения основного принципа хроматографического
разделения часто прибегают к следующей аналогии. Предста-
вим себе, что группа водителей едет по дороге, вдоль которой
имеется множество баров. Очевидно, что самые большие лю-
Ввод
г пробы
Белок У \
Белок 2/
Злюирующуги.
буферный,
раствор
Колонка
с адсорбентом
Спустя
некоторое
время
Элюируюш,и.и.
буферный,
раствор
>qOqOqOqOqOqO рО о
РИС. 11.18. Схематическое представление хроматографического разделения
белков.
бители пива закончат путешествие последними, а те из води-
телей, кто вообще не пьет пива, уйдут далеко вперед! Если же
водитель отличается особенно большим пристрастием к спирт-
ным напиткам, то он застрянет в одном из баров до тех пор,
пока его не выпроводят оттуда и пока его не подберет случай-
ный проезжий. Последняя ситуация отвечает адсорбции в не-
подвижном слое и последующему элюированию (см., например,
рис. 11.19); такой способ разделения в биохимической и био-
технологической литературе также часто называют хромато-
графией.
Для количественного математического описания хромато-
графического процесса допустим, что жидкая фаза, содержа-
щая растворенные вещества Si, S2, ..., Sat, движется в режиме
полного вытеснения (в поршневом потоке) по колонке с насад-
кой, доля е общего объема которой занята жидкостью, нахо-
дящейся в пустотах. Предполагается, что концентрация рас-
творенных веществ в жидкой фазе St достаточно низка, поэто-
му обмен растворенным веществом с неподвижной фазой прак-
тически не изменяет общей концентрации жидкой фазы Ст
(а следовательно, и поверхностной скорости движения жидкой
фазы по колонке). Тогда дифференциальное уравнение мате-
риального баланса по растворенным веществам в нестационар-
Операции выделения
291
Колонка с ионообменной.
смолой
Ввод пробы:
Смесь белков
в буферном
растворе
Буферный,
раствор
раствор
7,0 =
0,5
0,4
Ой
0.Z
Метионин-
Цисте- сильфон
уновал 3
\ кислота
I Аспарагиновая
| кислота
0,1
Треонин
Серин Глииин
Глутаминовая Аланин
Метионин-
сульфоксиды
1 1 L zvJ
Элюат,мл40 60 80 100 1Z0 140 160 180 ZOO ZZO Z40 Z60 Z80 300 320
----Длина колонки 150см, pH3,Z5, O,Zh. цилтрат натрия*f*------------
РИС. 11.19. Ионообменная хроматография: принцип метода (а) и применение
в аминокислотном анализе (б). Под хроматограммой указан состав элюирую-
щего буферного раствора. [Рис. 11.19, б воспроизведен с разрешения из статьи:
Spackman D. Н., Stein IF. Н., Moore S., Automatic Recording Apparatus for
Use in the Chromatography of Amino Acids; Anal. Chem., 30, 1190 (1958).
© American Chemical Society.]
S
а
ном состоянии для двухфазной колонки (вспомните, например,
рис. 9.5 и 9.30) можно записать в следующем виде:
d$i , ds; . ., s OW; г, ., ,
,к1Г+еУГ+(1-Е)ЛГ = 0 (11ле)
где и — скорость движения жидкости в пустотах (равная част-
ному от деления поверхностной скорости на е; в свою очередь
19*
292
Главе 11
поверхностная скорость равна общей объемной скорости, де-
ленной на общую площадь поперечного сечения колонки). Пе-
ременная W; в уравнении (11.16) обозначает число молей ком-
понента i в единице объема неподвижной фазы.
Скорость адсорбции растворенного вещества i неподвижной
фазой dwi/dt в общем случае является функцией режима тече-
ния, скорости диффузии растворенного вещества в неподвиж-
ную фазу, кинетики адсорбции, емкости адсорбента и других
параметров. В простейшем случае адсорбированное неподвиж-
ной фазой вещество находится в локальном равновесии с рас-
творенным веществом в жидкой фазе, находящейся в пустотах
колонки, т. е.
= (11.17)
si
В общем случае Ki зависит от концентраций всех веществ, на-
ходящихся в неподвижной фазе (оц,..., wn). При малых кон-
центрациях, однако, можно принять, что /G постоянно. Тогда
подстановкой Wi из уравнения (11.17) в уравнение (11.16) и
последующим преобразованием получим
= ° (Ч.18а)
dz dt
где
щ=и( l-J-l^-^y1 (11.186)
По форме уравнение (11.18а) идентично уравнению мате-
риального баланса по растворенному веществу i, движущемуся
со скоростью щ по трубе без насадки. Следовательно, уравне-
ние (11.186) описывает эффективную скорость движения ком-
понента i в хроматографической колонке с насадкой. Как и в
аналогии с водителями, уравнение (11.186) указывает, что
компоненты с большим сродством к неподвижной фазе (с
большой величиной ТС) будут двигаться по колонке относитель-
но медленно, а компоненты, обладающие малой тенденцией к
связыванию с неподвижной фазой (с малой величиной Kt},
будут двигаться со скоростями, близкими к скорости раствори-
теля. В более общем виде эта теория учитывает также эффек-
ты осевой дисперсии, неравновесное связывание и диффузион-
ный перенос в неподвижную фазу (см. литературу). Анализ
явлений необратимой адсорбции или десорбции необходим при
изучении адсорбции в неподвижном слое с последующим селек-
тивным элюированием растворителем.
В другом методе разделения, называемом хроматографией
на молекулярных ситах (гель-проникающей хроматографией
Операции выделения
293
или гель-фильтрацией), разделяют вещества с различными мо-
лекулярными массами. В этом случае насадка колонки состо-
ит из частиц геля с определенным диаметром пор. Если размер
молекул больше диаметра пор, то они не могут диффундиро-
вать в гель и быстро проходят через колонку, тогда как моле-
кулы меньшего размера проникают в гель и поэтому движутся
более медленно (рис. 11.20). Следует иметь в виду, что меха-
низм разделения путем молекулярной эксклюзии в соответст-
вии с диаметром пор (или диаметром волокон, если гель рас-
• ••••••
ООО
о о
ООО
О о
РИС. 11.20. В хроматографии на молекулярных ситах молекулы большего
размера быстрее проходят через колонку, а молекулы меньшего размера за-
держиваются, проникая в частицы геля.
сматривать как множество переплетенных твердых волокон)
не является единственным. На скорость движения растворен-
ного вещества могут влиять также коэффициент эффективной
диффузии растворенного вещества в геле, способность раство-
ренного вещества адсорбироваться на внутренних поверхностях
адсорбента, а также осмотическое давление. В настоящее вре-
мя оценить с достаточной точностью вклад каждого из этих
факторов не представляется возможным. Тем не менее можно
считать, что в, большинстве случаев преобладает процесс моле-
кулярной эксклюзии. Тогда соответствующую константу равно-
весия Кау, отвечающую константе в уравнении (11.186), можно
вычислить по уравнению
Кач, i = exp [— лЛ(г^ + гг-)2] (11.19)
где Kav, i — доля общего внутреннего объема геля, доступная
для сферических молекул радиусом n; L — концентрация "воло-
кон геля, выраженная в единицах длины на единицу объема,
см/см3; гё — радиус волокон геля; п— радиус сферических мо-
лекул растворенного вещества i. В табл. 11.6 приведен ряд
величин rg и г/. Величину эффективного радиуса волокон геля
можно изменять путем увеличения или уменьшения числа попе-
294
Глава 11
речных связей между цепями в геле. Новый гелевый носитель с
неизвестными характеристиками можно охарактеризовать, по-
строив график зависимости —(lnAav)0,5 от г,. Из уравнения
(11.19) следует, что наклон этой кривой равен (л±)°>5, а на оси
координат она отсекает отрезок, равный (лЛ)0-5 rg. На рис.
11.21 приведены величины Кау для ряда гелей типа сефадекс и
сефароза (сефадекс-агароза). Если значение г. неизвестно, то
гель можно охарактеризовать по графику зависимости KZN от
молекулярной массы.
Таблица 11.6. Определение радиусов молекул ряда белков с помощью
коэффициентов диффузии8- 6
Белок * Молекуляр- ная .масса Коэффициент диффузии £>-107, см2/с О Гр А
Рибонуклеаза 13 683 11,9 18,0
Лизоцим 14 100 10,4 20,6
Химотрипсиноген 23 200 9,5 22,5
Сывороточный альбумин 65 000 5,94 36,1
Каталаза 250 000 4,1 52,2
Уреаза 480 000 3,46 61,9
а Характерный радиус волокон О в геле г_ (А) для сефадекса 7, для агарозы 25.
& По данным, приведенным в работе: Тенфорд Ч., физическая химия полимеров.—
М.: Химиия, 1965.
На рис. 11.22 приведены результаты разделения смеси бел-
ков методом хроматографии на молекулярных ситах. Обратите
внимание на то, что различные белки появляются на хромато-
грамме в виде довольно широких пиков. Уширению пиков спо-
собствуют многие факторы, в том числе молекулярная диффу-
зия, сопротивление массопереносу и непостоянство осевой ско-
рости потока в поперечном сечении колонки. Изображенные на
рис. 11.22 результаты разделения типичны для любых хрома-
тографических методов. Аналогично на неподвижной хромато-
графической фазе с различным сродством к разным компонен-
там смеси можно разделять белки с очень большой молеку-
лярной массой, вирусы, компоненты клеток и даже смеси раз-
личных клеток.
В ионообменной хроматографии раствор смеси белков про-
пускают через колонку с неподвижным слоем ионообменной
смолы (рис. 11.19). Одним из наиболее широко используемых
для очистки белков ионообменников является карбоксиметил-
целлюлоза — катионообменная смола, получаемая посредством
введения карбоксиметильных групп (несущих отрицательный
заряд) в целлюлозную матрицу. Белки в катионной форме
(несущие положительный заряд) связываются с этой смолой
Операции выделения
295
электростатическими силами, причем прочность связей зависит
от результирующего положительного заряда белка при pH
элюента. Затем адсорбированный таким образом белок элюи-
руют буферными растворами с возрастающим значением pH
и (или) ионной силы. Постепенное изменение свойств элюента
РИС. 11.21. Зависимость Kav от молекулярной массы белка при хроматографии
на сефадексе G-200, сефарозе 6В и сефарозе 4В (содержащих ~6 и —4%
агарозного геля соответственно; найдено экспериментально). (Из статьи:
Joustra М. К., Gel Filtration on Agarose Gels, in Progress in Separation and
Purification, vol. 2, p. 183, Wiley-Interscience, New York, 1969.)
приводит к тому, что слабо связанные с носителем белки де-
сорбируются первыми, а затем вымываются все более и более
прочно связанные с ионообменником белки. Следовательно,
подвижная фаза, которая до ввода в колонку вообще не со-
держала белков, на выходе из колонки будет обогащена десор-
бированными белками. Полученный при промывании колонки
элюат собирают в виде фракций небольшого объема. Аналогич-
но осуществляют и хроматографию на анионообменных смо-
лах— чаще всего на диэтиламиноэтилцеллюлозе.
Как и рассматриваемая ниже аффинная хроматография,
ионообменная хроматография в одном отношении принципиаль-
но отличается от гель-проникающей и других типов жидкостной
296
Глава 11
и газовой хроматографии, для которых характерны различные,
но тем не менее вполне определенные конечные эффективные
скорости движения растворенных веществ. В ионообменной хро-
матографии, напротив, в условиях ввода разделяемой смеси
в колонку один или несколько компонентов смеси адсорбиру-
ются практически необратимо (Кг+оо). Таким образом, ионо-
обменная хроматография, по сути дела, представляет собой
РИС. 11.22. Хроматографическое разделение смеси веществ методом хромато-
графии на молекулярных ситах. (Неподвижная фаза — десульфированный
агар, содержащий 6% поперечных связей; буферный раствор — 0,05 М трисХ
ХНС1, pH 7,5). [Воспроизведено с разрешения из работы: Porath J., Chroma-
tographic Methods in Fractionation of Enzymes, in Enzyme Engineering, Win-
gard L. B., Jr. (ed.), p. 154, John Wiley and Sons, New York, 1972.]
селективную адсорбцию в неподвижном слое адсорбента. Если
адсорбируются несколько компонентов смеси, как это имеет
место в примере, приведенном на рис. 11.22, то их разделяют
на стадии элюирования. Для этой цели состав элюента ступен-
чато или непрерывно изменяют, добиваясь (в идеальном вари-
анте) последовательной десорбции связанных с хроматографи-
ческой неподвижной фазой компонентов смеси.
Интересным хроматографическим методом является аффин-
ная хроматография, основанная на нативной специфичности
некоторых биополимеров (рис. 11.23). Ряд белков и других
биологических макромолекул (А) образуют комплексы с дру-
гими веществами (В) с чрезвычайно высокой степенью специ-
фичности: если компонент В связывается с неподвижной фазой,
находящейся в колонке, то коэффициент К по отношению к
носителю А очень велик, тогда как величины К для других
растворенных веществ практически равны нулю. В качестве
Операции выделения
297
примеров чрезвычайно специфично взаимодействующих ве-
ществ можно привести пары фермент—ингибитор, антиген —
антитело, лектин — клетка:
Фермент+ингибитор комплекс фермент—ингибитор
Антитело-)-антиген < > осадок антитело—антиген
Лектин-)-клеточная стенка < комплекс лектин—клеточная стенка
Так, колонку для выделения ДНК-деполимеразы методом
аффинной хроматографии готовят путем химического связыва-
ния ингибитора ДНК-деполимеразы с поверхностью какого-ли-
бо подходящего гранулированного материала, например агаро-
зы или полиакриламида. Пропускание через такую колонку
фугата клеток, подвергнутых лизису (см., например, стадию 4
РИС. 11.23. Схематическое представление аффинной хроматографии. В этом
методе хорошее разделение достигается за счет специфического взаимодейст-
вия между иммобилизованным агентом и растворенным веществом. Показаны
три стадии: ввод смеси веществ (а); разделение (б); элюирование связанного
с неподвижной фазой компонента смеси (е).
в примере 11.1), приводит к практически количественному свя-
зыванию фермента ДНК-деполимеразы с неподвижной фазой,
причем почти все другие вещества не задерживаются колонкой.
Из уравнения (11.186) следует, что щ^и для всех других рас-
творенных веществ. Впоследствии связанный фермент можно
десорбировать 'путем элюирования раствором ингибитора ДНК-
деполимеразы или какого-либо другого подходящего агента.
298
Глава 11
Широкому распространению аффинной хроматографии спо-
собствовало создание методов получения моноклональных ан-
тител, обладающих чрезвычайно высокой константой связыва-
ния с соответствующим антигеном (определенным белком или
другим веществом). Примером может служить рассматривае-
мый ниже метод очистки лейкоцитарного интерферона человека
из лизата рекомбинантной бактерии Е. coli на колонке с им-
мобилизованными моноклональными антителами. Такая разно-
видность аффинной хроматографии, которая называется хро-
матографией на иммуносорбентах, существенно отличается от
описанных выше традиционных типов хроматографии. Ниже
перечислены основ!ные особенности хроматографии на иммуно-
сорбентах:
1. Стоимость антител, используемых в качестве неподвижной
фазы в хроматографии на иммуносорбентах, может во много
раз превышать стоимость бульона, содержащего антигены.
2. Отсюда следует, что применение хроматографии на иммуно-
сорбентах экономически выгодно только в случае небольших
колонок многократного использования, обладающих боль-
шой емкостью.
3. Элюированию адсорбированного вещества должно предшест-
вовать разрушение комплекса антиген—антитело, что часто
сопровождается денатурацией белков. При этом антитела
обычно частично теряют связывающую активность, что при-
водит к постепенному снижению емкости колонки.
4. Наиболее удачным носителем для иммуносорбентов оказа-
лась агароза, применение которой позволило регулировать
поверхностную плотность антител. Недостатком как этого,
так и других полисахаридных носителей является отсутст-
вие механической прочности (высокая сжимаемость), что
ограничивает высоту колонки, которую можно использовать
без чрезмерного перепада давления (вспомните пример 4.2).
5. Очистка смеси на предколонке не является обязательной;
тем не менее в каждом конкретном случае следует предва-
рительно проверить возможность непосредственного разде-
ления смеси на иммуносорбенте (поскольку пока еще не
накоплен достаточный опыт работы с иммуносорбентами).
6. Первое разделение на новой колонке обычно дает худшие
результаты по сравнению с последующими разделениями,
по-видимому, из-за частичного необратимого связывания ан-
тигена.
7. С экономической точки зрения основными задачами при хро-
матографии на иммуносорбентах является определение оп-
тимальных объема и состава элюирующих буферных раство-
ров.
Операции выделения
299
Между аффинной хроматографией и другими более тради-
ционными методами адсорбционной или ионообменной хрома-
тографии существуют и другие различия. В традиционных хро-
матографических методах сначала адсорбируются все компо-
ненты смеси, а их разделение осуществляется на стадии де-
сорбции посредством, например, постепенного повышения pH
или концентрации солей в элюенте (рис. 11.19). Напротив, спе-
цифичность аффинной хроматографии проявляется в основном
на стадии адсорбции (рис. 11.23). Поэтому в аффинной хрома-
тографии через колонку целесообразно пропускать раствор
разделяемой смеси в течение достаточно длительного проме-
жутка времени, пока почти не будет достигнуто насыщения не-
подвижной фазы, поскольку на ней адсорбируется практически
только выделяемое соединение. Таким образом, проведение
разделения в аффинной хроматографии приближается к обыч-
ной сорбции в неподвижном слое вплоть до насыщения слоя
адсорбента и резко отличается от обычного разделения много-
компонентной смеси, вводимой в колонку однократно в виде
концентрированного раствора. Высокая стоимость сорбентов
для аффинной хроматографии частично окупается их повышен-
ной специфичностью, позволяющей перерабатывать значитель-
но большие объемы смеси за один цикл.
Благодаря высокой специфичности связывания четкий фронт
адсорбированного компонента движется через слой иммуносор-
бента практически без изменения профиля концентраций. По
этой причине, как уже отмечалось выше, разделяемую смесь
целесообразно пропускать через аффинную колонку до насы-
щения, которое обнаруживается по появлению выделяемого ве-
щества в элюате. Чрезмерная загрузка колонки вызывает по-
терю вещества с растворителем, а недостаточная загрузка при-
водит к тому, что нижняя часть этой обычно очень дорогой
колонки не используется (рис. 11.24).
Изучение сорбции при связывании антигена с антителом в
зависимости от молекулярной массы антитела показывает, что
пористые носители оказывают существенное сопротивление
массопереносу. В таких случаях простая равновесная модель,
например уравнение (11.18), неприменима. К тому же недавно
было экспериментально показано, что скорость образования
комплекса антиген—антитело также может заметно влиять на
общую скорость сорбции антигена.
При описании явлений массопереноса вместо длины колон-
ки удобно использовать другой параметр, который называется
количеством переносимых единиц (A'umber of Transfer t/nits,
NTU), необходимым для достижения заданной степени разде-
ления. Рассмотрим в качестве примера ситуацию, когда массо-
перенос лимитируется внешним сопротивлением. Если
300
Глава 11
§
йг
t
СВТ
Л ^сорбированное
вещество -—
Неиспользован-
ная емкость
колонки
Продолжительность
нанесения раствора
веществ
Время
РИС. 11.24. Кривая отклика (зависимость концентрации адсорбированного
вещества в элюате от продолжительности подачи раствора разделяемых ве-
ществ в свежую колонку) для аффинной хроматографии. Если подачу раство-
ра прекратить при концентрации свт, то часть емкости колонки не использует-
ся. Небольшое количество выделяемого вещества теряется с растворителем.
(Воспроизведено с разрешения из работы: Arnold F. Н., Blanch Н. W., Wil-
ke С. R., in Purification of Fermentation Products, Le Roith D. et al. (eds.),
p. 113, ACS Symposium Series, 271, 1985. © American Chemical Society.)
kia(Ac)—скорость переноса, отнесенная к единице объема, то
необходимая скорость переноса (выраженная через число мо-
лей в единицу времени) составляет УА/а(Дс). Разделив это
выражение на ЁДс и умножив на L/u0 (z/0— поверхностная
скорость), получим безразмерный параметр — необходимое
число переносимых единиц:
NTU =
kia (Ac) L kiaL
Acu0 u0
В хроматографии на иммуносорбентах процессы внутрен-
него переноса замедлены. Если общую скорость переноса ли-
митируют внутренний перенос и прочное связывание, то [29,
39]
NTUnopa —
kn<ypa.apL
«0
rf2p
60 (1 — е) £>eff
(11.20)
Зависимость между NTU, безразмерной концентрацией элюиру-
емого вещества се(се^с/С[) и безразмерным временем т для
случая необратимой адсорбции описывается уравнением [38,
39]
NTUnopaCx — 1) =2,44-0,27311 -cJVa (11.21а)
Операции выделения
301
где
РИС. 11.25. Найденные экспериментально (-----) и рассчитанные с помощью
модели (-----) кривые отклика для колонки размером 1,5X16,5 см, содер-
жащей моноклональные антитела на носителе из пористого стекла, с опре-
деленным диаметром пор. Значения других параметров: «о=О,О11 см/с, 8 = 0,6,
^р=0,01 см, т=1 при Vl~eVc=2ll мл. (Воспроизведено с разрешения из ра-
боты: Arnold F. Н., Blanch Н. W„ Wilke С. R., Purification of Fermentation
Products, LeRoith D. et al. (eds.), p. 113, ACS Symposium Series, 271, 1985.
© American Chemical Society.)
uot | W e \ 1
eL J \ 1 — 8 / Ke
И
cf
(11.216)
(11.21b)
В уравнении (11.21в) параметр qf* обозначает концентрацию
адсорбированного вещества в твердой фазе, находящегося в
равновесии с растворенным веществом в концентрации cf.
Для модели псевдогомогенной диффузии (например, в геле-
вой фазе, в которой диаметр пор не имеет большого значения)
[38]
L wo
(11.22)
где фРН — поправочный коэффициент, имеющий величину по-
302
Глава It
рядка единицы. Для необратимой адсорбции в такой системе
зависимость концентрации элюируемого вещества от времени
описывается уравнением
NTUph(t— 1) =—1,69 (1п-[1 — сМ+0,61) (Ц.23)
На рис. 11.25 приведены результаты экспериментов, выпол-
ненных на небольшой колонке, в которой в качестве носителя
для моноклональных антител, специфичных в отношении арса-
ниловой кислоты (ARS), применяли стекло с определенным
диаметром пор. На этой колонке выделяли комплекс ARS и
альбумина сыворотки быка. Уравнение (11.21) удовлетвори-
тельно описывает профиль концентрации элюируемого в этой
системе вещества и условия элюирования, если NTUnopa при-
РИС. 11.26. Кривые отклика, предсказываемые моделью [уравнения (11.20)
и (11.21)] для колонки размером 15X60 см, содержащей моноклональные ан-
титела на носителе (пористом стекле с определенным диаметром пор). Емкость
колонки по отношению к растворенному веществу в состоянии равновесия
эквивалентна 77,1 л исходного раствора. Здесь Vl — объем жидкости, пропу-
щенной через колонку, Vc — объем колонки, е — доля пустот в объеме. Кри-
вые 7, 2 и 3 отвечают Ио=О,О2О; 0,015 и 0,10 см/с соответственно. (Воспроиз-
ведено с разрешения из работы: Arnold F. Н., Blanch Н. W., Wilke С. R., in Pu-
rification of Fermentation Products, LeRoith D. et al. (eds.), p. 113, ACS
Symposium Series, 271, 1985. © American Chemical Society.)
нять равным 8. Если скорость десорбции лимитируется диффу-
зией в поры, то работу большей колонки можно рассчитать по
уравнениям (11.21) и (11.20). Предсказываемые математичес-
кой моделью кривые отклика для большой колонки при раз-
личных скоростях движения жидкой фазы представлены на
рис. 11.26. В аффинной хроматографии существенную роль мо-
жет играть и кинетическое сопротивление связыванию антиге-
на; этот вопрос подробно освещен в работе [39].
Операции выделения 303
11.5. Разделение с помощью мембран
Принципиальное преимущество методов разделения с по-
мощью мембран заключается в возможности их осуществле-
ния без фазового перехода или межфазового переноса, и по-
этому любое выделяемое вещество постоянно находится в вод-
ном окружении. Один из принципов разделения с помощью
мембран основан на различной проницаемости мембран для
молекул разных размеров. Существуют два основных метода
разделения с помощью мембран — обратный осмос и ультра-
фильтрация; возможность их применения определяется диамет-
ром молекул выделяемых веществ, который изменяется в сле-
дующих пределах:
Вещество
Диаметр, нм
Дрожжи и грибы 10s—104
Бактерии 300—103
Коллоидные частицы 100—1000
Макромолекулы (белки, полисахариды с молеку- 2—10
лярной массой 104—106)
Антибиотики (молекулярная масса 300—1000) 0,6—1,2
Моно-, дисахариды (молекулярная масса 200— 0,8—1,0
400)
Органические кислоты (молекулярная масса 0,4—0,8
100—500)
Неорганические ионы (молекулярная масса 10— 0,2—0,4
100)
Вода (молекулярная масса 18) 0,2
11.5.1. Обратный осмос
Если раствор какого-либо вещества и чистый растворитель
разделить мембраной, непроницаемой для растворенного веще-
ства, но проницаемой для растворителя, то растворитель будет
диффундировать через мембрану в раствор до тех пор, пока
химические потенциалы растворенного вещества в двух фазах
не уравновесятся. Это явление называют осмосом. Если к
раствору приложить избыточное давление Др, то, когда прило-
женное давление Др уравновесит осмотическое давление рас-
твора л, осмос прекратится; осмотическое давление раствора
л равно:
л = с/?П1 + В2[с]+5з[с]2+ ...] (11.24)
Здесь В2, В3—вириальные коэффициенты растворенных ве-
ществ.
304
Глава 11
Таким образом, при низких концентрациях растворенных
веществ и нулевом потоке растворителя через мембрану Др =
= zi^cRT. Если Др больше л, то растворитель переносится про-
тив градиента концентрации из разбавленного раствора в чис-
тый растворитель. Такое явление, приводящее к повышению
концентрации растворенного вещества, называют обратным,
осмосом.
В реальной ситуации мембраны не обладают абсолютной
селективностью по отношению к размерам молекул, поэтому
Таблица 11.7. Основные характеристики трех типов мембран8
Вещество Моле- куляр- ная масса Моле- куляр- ный ра- диус, о А Коэффициент отражения
Диализаци- онные труб- ки (целлю- лоза Visking) Целлофан (DuPont 450-РТ-62) Влажный вискозный гель (Sylva- nia 300)
d2o 20 1,9 0,002 — 0,001
Мочевина 60 2,7 0,024 0,006 0,004
Глюкоза 180 4,4 0,20 0,044 0,016
Сахароза 342 5,3 0,37 0,074 0,028
Рафиноза 595 6,1 0,44 0,089 0,035
Инулин 991 12 0,76 0,43 0,23
Альбумин сыворотки 66 000 37 1,02 1,03 0,73
быка
Вычисленный радиус 23 41 82
пор, А
Константа мембраны 1,7 6,5 25
Lp, 10_°-г/(см2-с-атм)
а Данные заимствованы из работы: Durbin R. Р., J. Gen. Physiol., 44, 315 (1960).
при анализе осмоса удобнее рассматривать совместно как
пассивную проницаемость растворенного вещества Р, так и ха-
рактеризующий перенос через мембрану коэффициент отраже-
ния о для каждого растворенного вещества. Последний пара-
метр представляет собой долю молекул растворенного вещест-
ва, не проникающих через мембрану; так, для случая полного
отражения о = 1,0, а если растворенное вещество вообще не за-
держивается мембраной, то а = 0. Коэффициенты отражения
характеризуют мембрану; кажущийся диаметр пор мембраны
может служить мерой селективности осмоса (табл. 11.7).
Если к раствору приложено давление, то скорости переноса
растворителя против градиента концентрации и растворенного
вещества описываются уравнениями (11.25) и (11.26) соответ-
ственно:
2Vi (растворитель) = Lp(&p — л) (11.25)
^(растворенное вещество) =сг(1 —с)М-|-РДс2 (Н-26)
Операции выделения
305
где Lp и Р — проницаемость мембраны для растворителя и
растворенного вещества соответственно, Дс2 — разность кон-
центраций растворенного веществ1а по разные стороны мембра-
ны, а С2 — средняя концентрация растворенного вещества в ра-
створе. Соответствующая концентрация растворенного веще-
ства в жидкости, проходящей через мембрану, равна
ЗД=Г;2(1-ОН
L
Если о приблизительно равно 1,0, Р мало и (или) Ni велико,
то эта величина будет значительно меньше N2.
Жидкости практически несжимаемы, поэтому само по себе
приложение даже большого избыточного давления не связано
с большим расходом энергии. Однако повышение скорости пе-
реноса растворителя против градиента концентрации через-
ssssssss
sssssssss
SSSSSSSSSS
s SSSSSSSSS S
SSSSSSSSSS
sssssssssssssss
РИС. 11.27. Концентрационная поляризация, вызванная повышением концент-
рации растворенного вещества (или веществ) (S) вблизи обращенной к по-
току поверхности мембраны.
мембрану приводит к образованию вблизи границы раздела
фаз зоны с повышенной концентрацией растворенного вещест-
ва (рис. 11.27), что в свою очередь обусловливает повышение
скорости переноса растворенного вещества через мембрану
[уравнение (11.26)], а также снижение скорости переноса рас-
творителя против градиента концентрации в силу локального
повышения осмотического давления [уравнение (11.25)]. Это
явление, называемое концентрационной поляризацией, уста-
навливает верхний предел скорости переноса растворителя че-
20—746
306
Глава 11
рез мембрану против градиента концентрации. Как показано
на рис. 11.2S, перемешивание слоя раствора, примыкающего
к обращенной к раствору поверхности мембраны, может спо-
собствовать снижению сопротивления переносу. Механизм
этого эффекта аналогичен снижению сопротивления осадка
0,10
<s>
I
£ 0,06
OM
I
|вЛ!
11
$ I 0,08
8 £
РИС. 11.28. Индуцированное концентрационной поляризацией отклонение от
линейной зависимости скорости переноса растворителя через мембрану от
давления; повышение давления в конце концов приводит к постоянной ско-
рости переноса растворителя против градиента концентрации. [Воспроизведено
с разрешения из работы: Porter М. С., in Enzyme Engineering, Wingard L. В.,
Jr. (ed.), p. 119, John Wiley and Sons, Inc., New York, 1972,]
вблизи фильтрующей поверхности при фильтровании с парал-
лельным фильтрующей поверхности потоком.
Общее осмотическое давление многокомпонентного раствора
(если пренебречь всеми возможными взаимодействиями между
растворенными веществами) равно сумме осмотических давле-
ний отдельных компонентов:
= 2 лi = Z CiRT [ 1 + B2i [с] i + B3i [c]2f +...] (11.27)
i i
В этом случае скорость переноса растворителя через мембрану
против градиента концентрации опять-таки пропорциональна
(Др—лц), так что перенос каждого растворенного вещества
зависит как от общего осмотического давления, так и от про-
Операции выделения
307
ницаемости и коэффициента отражения данного вещества; сле-
довательно, концентрационная поляризация любого растворен-
ного вещества снижает скорость переноса всех растворенных
веществ.
В отсутствие заметного переноса растворителя против гра-
диента концентрации при циркуляции и постоянном обновле-
нии растворителя происходит диализ — непрерывная диффузия
небольших молекул в циркулирующий растворитель и удержи-
вание больших молекул. На принципе диализа основана ра-
бота аппарата типа «искусственной почки», который удаляет
соли я низкомолекулярные вещества, являющиеся отходами
жизнедеятельности организма человека (например, мочевину;
см. табл. 11.7), и в то же время эффективно удерживает вы-
сокомолекулярные вещества, например белки и полисахариды.
Если удаление одного из низкомолекулярных веществ по ка-
ким-либо причинам нежелательно, то оно должно иметься в
достаточной концентрации в циркулирующей жидкости; так,
при очистке диализом фермента (пример 11.1, стадия 7) для
отделения низкомолекулярных веществ применяют 10~3 М фос-
фатный буфер.
11.5.2. Ультрафильтрация
Если средний диаметр пор мембраны превышает размер
пор в процессе обратного осмоса, то через мембрану проника-
ют все вещества с диаметром молекул 1 —10 А, а белки и дру-
гие высокомолекулярные соединения задерживаются. Следова-
тельно, вещества с молекулами диаметром от 10 до 500—
1000 А с помощью ультрафильтрации можно сконцентрировать
(т. е. частично удалить растворитель) и очистить от низкомоле-
кулярных примесей.
Если концентрационная поляризация обусловлена макро-
молекулами (т. е. значение MW велико), то обычно в соответ-
ствии с уравнением (11.24) осмотическое давление мало. В то
же время накопление поляризационного слоя высокомолекуляр-
ного растворенного вещества может оказывать сопротивление
массопереносу.
Для анализа процессов ультрафильтрации в зависимости
от конкретной ситуации применялось несколько подходов:
1. Для очень разбавленных растворов скорость переноса рас-
творителя через мембрану определяется собственным сопро-
тивлением мембраны в соответствии с упрощенными уравне-
ниями, описывающими обратный осмос при пренебрежимо
малом осмотическом давлении, т. е.
(растворитель) —Lp\p
(растворенное вещество) &С2(1—(f) LpAp-^~ РАс2
20*
.308
Глава 11
2. Накапливающийся поляризационный слой макромолекул
при благоприятных обстоятельствах может привести к обра-
зованию гелевой фазы, которая может оказывать заметное
пли даже решающее влияние на скорость процесса ультра-
фильтрации.
Коэффициент отражения амилазы К
Функция отражения амилазы (
РИС. 11.29. Зависимость скорости переноса растворителя через ультрафиль-
трационную мембрану типа HFA-300 от отражения амилазы (верхняя шкала),
pH и присутствия ^-лактоглобулина; данные относятся к раствору амилазы
при pH 6,0 (7); при pH 5,2 (2) и к раствору смеси амилазы с ^-лактоглобули-
ном при pH 6,0 (3) и pH 5,2 (4). Другие параметры системы: температура 30 °C;
скорость вращения мешалки 900 об/мин (Re=50 000); р = 0,35—1,41 кг/см2
(раствор амилазы), 1,055 кг/см2 (амилаза в присутствии ^-лактоглобулина);
0,01 М ацетатный буфер, содержащий 0,002 М СаС1г. [Из работы: Butter-
worth Т. A., Wang D. I. С., in Fermentation Technology Today, Terui G. (ed.),
p. 195, Soc. Ferm. Technol., Japan, 1972.]
Если мы введем параметр молекулярнго сита Ф = сР!с^,
а наблюдаемый коэффициент отражения растворенного вещест-
ва R примем равным (q—cp)!Cf, где cw, Cf и ср — концентрации
вещества в геле, исходной смеси и в прошедшем через мембра-
ну растворе соответственно, то
ЛГ1 = рмИп[(-^)(-4^-)] (11.28)
Д Ф ) \ К /.
где k — коэффициент массопереноса жидкой фазы, а рм — мо-
лярная плотность растворителя. Приведенные на рис. 11.29
экспериментальные данные, относящиеся к амилазе, подтверж-
дают линейную зависимость In [ (1—RjfR} от N\. Добавление
второго высокомолекулярного соединения, [3-лактоглобулина,
опять приводит к линейной зависимости In [ (1—R)!R\ от Ni,
Операции выделения
309
но величина (1—0)/0 в этом случае больше. Следовательно,
при данной скорости переноса растворителя коэффициент от-
ражения R амилазы при диализе смеси высокомолекулярных
веществ возрастает. Обратите внимание на то, что наклоны со-
ответствующих кривых также несколько различаются, что
обусловлено зависимостью сопротивления слоя геля от его со-
става.
Другое доказательство сопротивления геля массопереносу
представлено на рис. 11.30; приведенные данные показывают,
РИС. 11.30. Зависимость скорости переноса растворителя через мембрану от
разности концентраций ^-лактоглобулина Cf—ср (%). Параметры системы: ско-
рость вращения мешалки 900 об/мин, р= 1,055 кг/см2, 0,01 М. ацетатный буфер,
содержащий 0,002 М СаСЬ. [Из работы: Butterworth Т. A., Wang D. I. С., in
Fermentation Technology Today, Terui G. (ed.), p. 195, Soc. Ferm. Technol.,
Japan, 1972.]
что диализ при pH 5,2, равном p.K р-лактоглобулина, приводит
к максимальной концентрации геля. Интуитивно можно было
бы предположить, что более плотный гель скорее будет форми-
роваться из нейтральных (т. е. находящихся в изоэлектричес-
кой точке) молекул, чем из молекул, несущих определенный
результирующий заряд. Это предположение подтверждается
очень резким повышением коэффициента отражения сс-амила-
зы при увеличении концентрации ^-лактоглобулина, если pH
равен рД" последнего (рис. 11.31).
Эти результаты показывают, что для данного фильтрующе-
го материала скорость переноса растворителя и отражение
растворенного вещества могут быть описаны уравнением
(11.28). В то же время наклон кривой рм& и отсекаемый на
оси абсцисс отрезок pM^ln[(1—0)/0] зависят от концентрации,
а эффективность ультрафильтрации определяется не только
величиной молекулярной массы веществ. Действительно, в
310
Глава 11
U U,UQ U,1Z. U,1U U,X.V
Концентрация ft-лакто глобулина,
в исходной смеси, масс- °/о
РИС. 11.31. Зависимость коэффициента отражения амилазы R от концентра-
ции [}-лактоглобулина в исходной смеси. Параметры системы: скорость вра-
щения мешалки 900 об/мин; р= 1,055 кг/см2; 0,01 М ацетатный буфер, содер-
жащий 0,002 М СаС12. [Из работы: Butterworth Т. A., Wang D. I. С., in Fer-
mentation Technology Today, Terui G. (ed.), p. 195, Soc. Ferm. Technol., Japan,
1972.]
приведенном выше примере более высокомолекулярная а-ами-
лаза (молекулярная масса 48000) при низкой концентрации
проникает через мембрану быстрее, чем гелеобразующий £-
лактоглобулин (молекулярная масса 36000) при более высокой
концентрации. (Это только кажущееся противоречие; на самом
деле молекулярная масса геля представляет собой довольно
неопределенную величину.)
11.6. Электрофорез
Электрофорезом называют разделение веществ благодаря
различной скорости их перемещения в электрическом поле.
Постоянная скорость иЕ, достигаемая частицей с зарядом q в
жидкой среде под влиянием электрического поля с напряжен-
ностью Е, определяется балансом моментов
Сопротивление жидкой \ /Электппстати- \ г /1 i
среды, действующее = (,сскаяРсила )=№ (11.29)
на частицу / ' '
Для глобулярных белков можно использовать закон Стокса в
применении к сопротивлению сферы радиуса гр, движущейся в
ньютоновской жидкости с вязкостью рс:
/ Сопротивление жидкой среды, \ _ g^.^ г ц (11.30)
\ действующее на частицу ) № р Е \
Операции выделения
311
так что
и£ = —2ь- (11.31)
6пр.сГр
Поскольку в общем случае каждый белок имеет свойственный
только ему результирующий заряд q, то наложение электриче-
ского поля приводит к тому, что разные белки движутся с
разными скоростями. Таким путем смесь нескольких белков
можно разделить на индивидуальные компоненты (рис. 11.32).
Посредством изменения pH можно регулировать электрофоре-
тическую подвижность белка. Если р/ данного белка меньше
pH среды, то его заряд и скорость будут отрицательными. На-
против, белки с р/>рН будут двигаться в положительном на-
правлении. Этот принцип положен в основу одного из методов
определения р/ белков и других веществ; в градиенте pH р/
белка равна pH, при котором его электрофоретическая под-
вижность Ue равна нулю.
В методе электрофореза в потоке жидкая фаза движется
перпендикулярно направлению электрического поля, что позво-
ляет осуществлять непрерывное разделение. При электрофоре-
зе на бумаге или в геле на движение молекул веществ влияют
процессы адсорбции и десорбции, а также сопротивление
диффузии в пористый носитель или гель.
При разделении методом электрофореза больших количеств!
веществ основные затруднения связаны с омическим нагревом.
Растворы, стабилизирующие белки, обычно обладают высокой
проводимостью, и поэтому при электрофорезе в таких раство-
рах возникают высокие плотности тока, а адекватные эффек-
тивные теплообменные устройства для таких крупномасштаб-
ных систем пока еще не разработаны. Напротив, электрофоре-
тическая аппаратура для непрерывного разделения небольших
количеств веществ, выпускается серийно.
11.7. Сочетание нескольких операций разделения
Если в одном резервуаре проводить несколько операций,
то таким путем можно сократить как время процесса (т. е.
эксплуатационные расходы), так и число аппаратов (т. е. ка-
питаловложения). Эти возможности, однако, редко удается
реализовать, поскольку совмещение различных операций воз-
можно только в случае одновременного выполнения требова-
ний, предъявляемых к каждой из операций. В этом разделе
мы приведем несколько примеров, иллюстрирующих как пре-
имущества, так и недостатки некоторых способов сочетания
различных операций разделения.
312
Глава 11
Нисходящий,
патак
Поверхность
раздела I
Раствор смеси
белков (компоненты А, Б,С]'
Буферный
раствор
Восходящий,
патак
Поверхность
раздела А
а
РИС. 11.32. Локализация белковых компонентов в электрофоретической ячей-
ке до (а) и после (б) электрофореза. (С любезного разрешения компании
Beckman Instruments, Inc.)
Операции выделения
313
11.7.1. Иммобилизованные клетки
Иммобилизованные клетки (гл. 9) одновременно выполня-
ют операции биотрансформации (или биосинтеза) и отделения
биокатализатора от культуральной жидкости. Они могут про-
являть каталитическую активность в псевдоожиженном или в
неподвижном слое. Преимуществом процессов с участием им-
мобилизованных клеток является отсутствие необходимости в
отдельной операции отделения биомассы, а к числу их недос-
татков относится высокая стоимость носителя клеток и их им-
мобилизации. Кроме того, иммобилизованные клетки могут
расти и в конце концов частично просачиваться в среду, что,
естественно, сводит на нет только что отмеченные преимущест-
ва таких биокатализаторов. Этот недостаток можно устранить
путем предотвращения клеточного роста в среде, практически
лишив их того или иного незаменимого питательного вещества
(например, источника азота); в таких случаях время от време-
ни, когда каталитическая активность клеток заметно снижает-
ся, катализатор регенерируют.
11.7.2. Операции переработки цельного бульона
При переработке цельного бульона продукт выделяют без
предварительного отделения биомассы или иных твердых ве-
ществ. В таких операциях применяют методы экстракции, ион-
ного обмена, диализа, адсорбции (на активированном угле),
разделения с помощью мембран и (или) фильтрования.
В качестве примера на рис. 11.33 представлены результаты
изучения процесса биосинтеза антибиотика циклогексимида в
присутствии гидрофобной ионообменной смолы и в ее отсутст-
вие. На смоле адсорбируется более 98% циклогексимида; в
промышленных процессах смолу необходимо регенерировать.
Выход антибиотика после его десорбции со смолы бутилацета-
том практически такой же, как и после прямой экстракции
растворителем из культуральной жидкости, но в первом методе
выделения чистота продукта значительно выше.
Летучий продукт метаболизма, обладающий ингибиторными
свойствами (например, этанол), можно непрерывно удалять
отгонкой в вакууме; при низкой температуре в вакууме дрож-
жевые клетки (вместе с культуральной жидкостью) могут без
потери жизнеспособности рециркулировать с одновременной от-
гонкой этанола.
Непосредственную экстракцию растворителем применяют
при выделении пенициллинов и стероидов. Как было показано,
для этих целей можно использовать и экстракцию в системе
из двух водных фаз с участием полиэтиленгликоля, хотя этот
314
Глава 11
3 ‘3
§ §
? $
<Ь Qj
О •
8,0 -
6,0
Добавление смолы
4, (XAD-4) (6°/о по массе)
tu
£
2400 А
V
2000
1600
12(Х)
\ А
Xх/'
о а ~
А 6
• в
A Z
KJ4
‘О
50 I
40 В
I
§
зо Йк
а
у
800-
400-
I
/
/
• /
/
/
О 24 48 72 96 120 144 158 192 216
Продолжительность процесса, ч
РИС. 11.33. Зависимость концентрации циклогексимида от времени в обычном
процессе (а, б) и в присутствии 6% (по массе) смолы XAD-4 (в, г), добавлен-
ной через 48 ч после начала процесса. [Из статьи: Wang Н., Ann. N. Y. Acad.
Sci., 413, 313 (1983).]
метод, по-видимому, намного дороже из-за высокой стоимости
полиэтиленгликоля.
Непосредственное выделение продукта из бульона путем
адсорбции на ионообменных смолах применялось в производстве
антибиотиков стрептомицина и новобиоцина на пилотных и
крупномасштабных промышленных установках [37]. В этом
случае через ряд последовательно соединенных адсорберов с
неподвижным слоем ионообменной смолы непрерывно пропус-
кают поток цельного бульона (рис. 11.34). Такую систему мож-
но описать с помощью простой модели переходного состояния
типа
vc~^-=Fcn_t-Fcn~vR-^ (1L32)
где Vc-—объем жидкой фазы в n-й колонне, л; сп-ъ сп—кон-
Операции выделения
315
центрация антибиотика на входе и выходе из n-й колонны со-
ответственно; F — объемная скорость потока, л/ч; Vr— объем
смолы в п-й колонне, л; wn — концентрация антибиотика в
смоле, г/(л смолы); t— время, ч. Смола очень прочно связы-
вает антибиотик, поэтому в каждой частице ионообменника
антибиотик распределяется неравномерно, а связывание анти-
биотика осуществляется не с постоянной скоростью, а таким
образом, что сопротивление его переносу внутрь частицы воз-
растает во времени. Для описания такой ситуации удобно ис-
пользовать зависимость кажущегося коэффициента массопере-
РИС. 11.34. Каскад адсорберов для выделения антибиотика. [Воспроизведено
с разрешения из статьи: Belter Р. A., Cunningham F. С., Chen J. W., Develop-
ment of a Recovery Process for Novobiocin, Biotech. Bioeng., 15, 533 (1973).]
носа от степени адсорбции. Следовательно, скорость адсорбции
антибиотика можно выразить уравнениехМ
^L = £(Cn —с*) (11.33)
dt
где кажущийся коэффициент массопереноса /г, определяемый
как k = f (ау/a'max), равен
k = Де-B(te’/C£’niax) De~ E(w^wmax) ( ] 1.34)
Наконец, с* описывается изотермой Фрейндлиха
c*=b{wn)a (11.35)
Когда концентрация антибиотика на выходе из последнего
адсорбера достигает заданной величины, первый адсорбер пе-
реключают на выделение антибиотика, а в конце системы уста-
навливают еще один адсорбер со свежей ионообменной смолой.
Полуэмпирическая модель [уравнения (11.33) — (11.35)] по-
лезна в том отношении, что она позволяет достаточно точно
описать процесс выделения антибиотика из цельного бульона
путем полупериодической адсорбции на ионообменной смоле.
Такой процесс выделения был разработан, чтобы избежать
дорогостоящей операции отделения мицелия фильтрованием,
сопровождающейся потерей существенного количества антибио-
тика вместе с отфильтрованным мицелием.
316
Глава 11
При экстракции цельного бульона возникают проблемы,
связанные с адсорбцией других растворенных веществ (сопут-
ствующих белков, непрореагировавшего субстрата и т. д.), что
может уменьшить емкость сорбента и привести к снижению
степени чистоты элюируемого продукта; с загрязнением мемб-
ран, если их регулярно не очищать; с образованием в системе
типа растворитель — бульон трудноразделяющихся эмульсий,
способствующих осаждению твердых веществ в аппаратуре,
а также потере растворителя с отбрасываемой водной фазой.
В общем случае удачная система выделения должна быть
достаточно селективной, нетоксичной по отношению к расту-
щей культуре, способной функционировать в асептических ус-
ловиях, а в случае ее высокой стоимости должна регенериро-
ваться и использоваться многократно.
11.7.3. Рециркуляция и операции выделения
В системах с рециркуляцией совмещены операции выделе-
ния каких-либо компонентов из содержимого биореактора или
обрабатываемого бульона и повторного добавления последнего
в биореактор как дополнительного исходного материала. При
этом возможны три основных варианта, когда рециркуляции
подвергается газовая фаза, водная фаза или биомасса (био-
массу можно также удерживать в реакторе в процессе рецир-
куляции). В первом варианте, рассмотренном ранее в гл. 9,
используют только такие биореакторы, конструкция которых
позволяет вновь вводить в реактор отходящие газы после их
разделения. Рециркуляция биомассы применяется в основном
в процессах аэробной переработки отходов (гл. 14), а соответ-
ствующие теоретические проблемы были рассмотрены в гл. 9.
Различные операции с применением мембран (фильтрование с
параллельным фильтрующей поверхности потоком, ультрафиль-
трация) обеспечивают удерживание клеток в биореакторе и
приблизительно такие же скорости процессов, что и рециркуля-
ция клеток.
Перспективы эксплуатации крупномасштабных биореакто-
ров, предназначенных, например, для производства белка од-
ноклеточных организмов или этанола, требуют специальной
аппаратуры для рециркуляции воды. Так, установка произво-
дительностью 50 000—70 000 т сухого белка одноклеточных ор-
ганизмов в год расходует за тот же период 2—2,8 млн. т воды.
Сброс такого количества воды привел бы к потере неполностью
утилизированных питательных веществ и полезных (не выде-
ленных) органических побочных продуктов, а также к повыше-
нию стоимости процесса обработки сточных вод, если их
Операции выделения
317
предполагается использовать, например, в системах оборотного
водоснабжения. В регионах с засушливым климатом рецирку-
ляция воды может стать решающим фактором, определяющим
целесообразность биопроцесса. Так, указанное выше количест-
во воды, необходимое для крупномасштабного биопроцесса,
составляет одну четверть годового потребления пресной и оп-
ресненной воды в основном промышленном районе Кувейта.
С рециркуляцией связано несколько факторов., которые мо-
гут существенно влиять на работу биореактора:
1. Накопление секретируемых в среду метаболитов или про-
дуктов лизиса клеток, обладающих ингибиторными свойст-
вами. (Эти вещества могут образовываться как в биореакто-
ре, так и при выделении продукта.)
2. Накопление нежелательных или не используемых компонен-
тов питательных веществ.
3. В случае смешанных культур селективная рециркуляция
биомассы может привести к изменению состава культуры.
Аналитическое изучение работы реакторов с помощью ма-
тематических моделей типа рассмотренных в гл. 9 показывает,
что в системах с ингибированием
1. Рециркуляция воды всегда приводит к снижению степени
превращения по сравнению с системой без рециркуляции
при той же скорости разведения.
2. При постоянной скорости подачи питательных веществ (от-
несенной к единице объема) рециркуляция также приводит
к снижению степени превращения субстрата.
3. Для каждой системы с рециркуляцией существует опреде-
ленное критическое отношение рециркуляции, ниже которого
влияние рециркуляции на степень превращения относительно
невелико, а выше которого степень превращения субстрата
существенно снижается.
Выше мы отмечали, что рециркуляция в биологическом реакто-
ре приводит к обратному смешению; как и в других областях
химической технологии, на превращения положительного по-
рядка (например, описываемые уравнением Моно, в котором
скорость превращения субстрата пропорциональна его концент-
рации в дробной степени) накладывается обратное смешение.
Отсюда следует, что рециркуляция воды во всех случаях при-
водит к необходимости использовать больший (и более доро-
гой) реактор; связанные с этим дополнительные расходы долж-
ны перекрываться экономией, достигаемой за счет сокращения»
количества необходимой воды.
318
Глава 11
11.8. Последовательность операций выделения
продуктов процессов
биохимической технологии
В этом разделе мы изучим последовательность операций
выделения и очистки продуктов биопроцессов и приведем не-
сколько примеров таких типичных для биохимической техноло-
гии последовательностей, включающих отделение твердых ве-
ществ, первичное концентрирование, очистку и окончательную
обработку.
Прежде чем перейти к изучению деталей технологии выде-
ления и очистки, полезно оценить взаимосвязи между биореак-
цией и операциями выделения. Поступающие в реактор пита-
тельные вещества в предельных случаях могут быть чрезвы-
чайно сложной смесью или вполне определенным химическим
соединением (или смесью нескольких соединений). Сложная
среда часто обеспечивает более высокий выход продукта или
биомассы, но образующийся в таком случае бульон более
неоднороден, и поэтому выделение продукта может вызвать
большие затруднения. Применение более дорогих сред с опре-
деленным химическим составом может быть оправдано упро-
щением операций выделения при производстве дорогостоящих
препаратов. Аналогично обеспечение высокой производитель-
ности биореактора несовместимо с одновременной полной ути-
лизацией субстрата. Стремление добиться наиболее полного
использования питательных веществ, с одной стороны, приво-
дит к снижению удельной скорости осуществляющихся в био-
реакторе процессов, а с другой — к тому, что культуральная
жидкость практически не содержит ряда мешающих растворен-
ных веществ (например, сахаров), которые могут затруднить
операции выделения продукта или снизить устойчивость гото-
вого сухого продукта.
Судя по богатому опыту, накопленному в фармацевтической
промышленности, при проведении биопроцессов необходимо
придерживаться следующих принципов:
1. Количество вводимых в систему пеногасителей должно быть
минимальным, поскольку поверхностно-активные вещества
могут стабилизировать эмульсии в последующих операциях
экстракции.
2. Для регулирования pH лучше вводить кислоты и основания,
а не буферные растворы.
3. Для стерилизации лучше использовать тепловую обработку,
а не химические агенты.
4. Выделять продукт легче в тех случаях, когда он образуется
только в одной фазе (внутри клеток или в среде); напротив.
Операции выделения
31»
выделение затруднено, если продукт распределен между био-
фазой и культуральной жидкостью.
5. Количество непрореагировавшего субстрата должно быть
минимальным. Чтобы свести к минимуму требования, предъ-
являемые к последовательным операциям выделения про-
дукта, следует избегать присутствия непрореагировавшего
субстрата, непревращенных питательных веществ и любых
других нерастворимых твердых веществ.
11.8.1. Выделение ферментов в промышленных процессах
Ферменты выделяют в виде неочищенных сухих препаратов,
разбавленных или концентрированных растворов или высоко-
очищенных (иногда даже перекристаллизованных) твердых ве-
ществ. На рис. 11.35 представлена общая схема выделения
фермента из поверхностной или погруженной культуры расти-
тельных или животных клеток. Для изображенных в верхней
части рисунка источников требуется предварительная обработ-
ка и прямая экстракция фермента буферным раствором с по-
следующими операциями отделения твердых веществ (если не-
обходим раствор фермента или очищенные препараты). На
рис. 11.36 в качестве примера приведена более детальная схе-
ма выделения растительного фермента, где показано, что для
обеспечения максимальной эффективности первой экстракции
необходимы хорошее перемешивание (в змеевиках) и охлаж-
дение (для стабилизации внутриклеточных продуктов). Нерас-
творимые компоненты отделяют последовательно на двух цент-
рифугах, причем сначала более крупные частицы отделяют на
шнековой центрифуге, чтобы более дорогостоящая высоко-
скоростная камерная центрифуга не забивалась крупными час-
тицами. Затем большую часть растворимых белков осаждают
подкислением; при этом подкисленный раствор должен нахо-
диться в охлаждаемой секции выдержки (охлаждаемом зме-
евике) достаточно долго, чтобы сформировался осадок, кото-
рый можно выделить центрифугированием (вспомните
рис. 11.17). Образовавшийся осадок сопутствующих белков от-
деляют на дисковой центрифуге; повторные подкисление и вы-
держка в охлаждаемом змеевике приводят к осаждению выде-
ляемого белка, который отделяют в виде влажного осадка на
второй дисковой центрифуге. Этот пример показывает, как с
помощью двух последовательных идентичных или близких
операций (центрифугирование в первом случае и осаждение
подкислением—во втором) можно фракционировать сложные
смеси.
В связи с тем что по мере повышения степени чистоты
фермента масштабы операций выделения (объемы обрабаты-
Источники Экстракция Фильтрование Концентрирова-
фермента кие
Вспомогательный
i фильтрующий
материал
Железы
. животных
Раститель-
ные ткани
Вода
Осаждение Сушка Стандартизация,
стабилизация
Ацетон, спирт
(NH4)2SOIf
Растительный
экстракт
Филыпр-
пресс
Вакуумный
концентратор
Рост микроорганизмов
Поверхности
культура ~] ♦________
___________ — Экстрактор —
Вращающ. Авспомогат.
оараоан фильтрующий I
материал |^~J|
Емкостный.
биореактор
Инертные
ингредиенты
Ротационный
вакуум-фильтр
Смеситель
^уишлка\
Сухой нео-
чащенный.
фермент
ХолодитА
ник
_ Обратный
осмос
Центрифуга
Распылитель-
ная сушилка
Лотковая
сушилка
Шаровая мельница или
молотковая дробилка
Стабилизирующие
соли, инертные
ингредиенты
Стабилизаторы,
консерванты
Электрофорез,
хроматография
Смеситель
Разбавленный
раствор
очищенного
фермента
,жЯШтЯ Фракционированный Стандартизо-
очищенного Юепмент (для трип- яанний нитирип
фермента
фермент (для специ- ванный фермент
алъных целей) (сухой)
РИС. 11.35. Выделение ферментов в промышленном масштабе,
(сухой)
Операции выделения
321
ваемых растворов) уменьшаются, на более поздних стадиях
очистки, как показано на рис. 11.37 для протеазы, от непре-
рывной переработки переходят к работе в периодическом режи-
ме. Обратите внимание на следующие дополнительные опера-
ции, введенные с целью повышения выхода фермента: 1) мно-
гократная промывка биомассы; 2) двустадийная ультрафильт-
РИС. 11.36. Непрерывный процесс выделения фермента пролил-тРНК-синтета-
зы из маша (фасоли золотистой). [Из работы: Lilly М. D„ Dunnill Р., Science
J., 5, 59 (1969).] 1 — охлаждаемый змеевиковый экстрактор; 2 — центрифуга
со шнековой выгрузкой; 3 — центрифуга; 4 — рН-метр; 5 — секция выдержки
при охлаждении; 6 — дисковая центрифуга.
рация, позволяющая ступенчато уменьшить объем смеси в
5 раз; 3) переход от непрерывной к периодической ультра-
фильтрации, позволяющей уменьшить объем смеси еще в
40 раз.
Важной отраслью микробиологической промышленности яв-
ляется производство протеаз (и других гидролаз) в качестве
добавок к синтетическим моющим средствам. В 1978—1979 го-
дах во всем мире производилось около 500 т (в расчете на чис-
тый фермент) бактериальных протеаз из Bacillus, стоимость
которых составляла 40% стоимости всех выпущенных мировой
промышленностью ферментов. Вначале их выпускали в виде
сухого порошка, в состав которого входили частицы диаметром
менее 10 мкм. Образуемые протеазами аэрозоли вызывали у
21—746
1 Бульон (ZOOm3) (активность Z000 ед./мл
биомасса 3000кг, вода и растворенные
вещества 198000 кг) р
Охлаждение (5V,
Бульон (90м3/ч)
Непрерывная
переработки
Суспензия
фильтрующего
материала
Флокулянт
(20м3/ч)
Осветленная
кцлътиральная
жидкость (189 л
Вода для
промывки (50м3)
Концентрирование
, (99м3)
Концентрирование
. (Юм3)
Суспензия отрабо-
танного фильтрую- г---—
' щего материала
Фильтр для „ ,
окончательной ->
очистки (150м2)
J Концентрирование (10м3)
[Осадитель
—^.или—
Безводный
(1,5м3)
Промёжуточь
емкость (10м3)'
Ротационный г„ _э
фильтр(1мг) -^Фильтрат* Отходы
Переработка
в периодическом
режиме
сульфат натрия
(WOO не)
РИС.11.37. Выделение внеклеточного фермента (протеазы). Исходную смесь
для выделения получали в периодическом биореакторе рабочим объемом
200 м3 в течение 10 ч с последующей чисткой реактора в течение 2 ч. (Воспро-
изведено с разрешения из работы: Atkinson В., Mavituna F., Biochemical Engi-
neering and Biotechnology Handbook, p. 918, Macmillan Publishers Ltd., Surrey,
England, 1983.)
Операции выделения
323
Соль фермента
Добавки Вода.
г—I——.
|/\/VWl Перемеишвание
псевдоожиженном
слое
1 Сито
Парафин
Добавки
а
—I Лппарат
__I для парафи-
нирования
] Охлаждение
Фермент
Парафин
Готовая частица
препарата
Сито
Расфасовка
РИС. 11.38. Процесс производства бактериальной протеазы, в котором не об-
разуются аэрозоли; на частицы фермента наносят парафиновое покрытие.
[Воспроизведено с разрешения из работы: Austrup К.., Andersen О., Falch Е. А.,
Nielsen Т. К-, in Microbial Technology, Peppier H. J., Perlman D. (eds.), 2d ed.,
vol. I, p. 281, Academic Press, Inc., N.Y., 1979.]
рабочих, занятых в их производстве, аллергическую реакцию,
что явилось причиной временного запрета на продажу детер-
гентов с добавками ферментов в США. Последующая разра-
ботка технологического процесса получения гранулированных,
не образующих аэрозоли ферментов (рис. 11.38) позволила от-
менить этот запрет. В настоящее время содержащие протеазы
гранулированные препараты выпускают в виде покрытых пара-
фином сфер диаметром около 500 мкм (рис. 11.38) или в виде
порошка в парафиновой матрице.
21
ABC
РИС. 11.39. Основные операции очистки грех белков человека, синтезирован-
ных в культуре рекомбинантных Е. coli. [Воспроизведено с разрешения из
статьи: McGregor С., Large-Scale Isolation and Purification of Proteins
from Recombinant E. coli, Ann. N. Y. Acad. Sci., 413, 231 (1983).]
Операции выделения
325
11.8.2. Выделение внутриклеточных чужеродных белков
из культур рекомбинантных Е. colt
Если белок синтезируется в генетически модифицированном
организме и предназначается для инъекций животным, то его
необходимо тщательнейшим образом очистить. Так, пирогены
из Е. coli, в том числе липополисахариды (LPS) внешней обо-
лочки бактерии, необходимо отделить или инактивировать.
(В организме человека LPS вызывают лихорадку в дозах, пре-
вышающих 0,5 нг/кг массы тела.) Кроме того, продукт должен
быть защищен от действия протеаз Е. coll. Для этой цели при
выделении и очистке продукта к буферному раствору часто
добавляют ингибитор сериновых протеаз—метилсульфанил-
фторид.
На рис. 11.39 приведены последовательности операций при
очистке инсулина человека, гормона роста человека и лейкоци-
тарного интерферона человека, синтезированных в культурах
рекомбинантных Е. coll. В каждом процессе для достижения
необходимой степени чистоты использовали ряд операций осаж-
дения и хроматографического разделения [22].
Если белок синтезируется в рекомбинантной Е. colt в боль-
ших количествах, то он часто накапливается в виде внутрикле-
точных структур, обладающих специфической преломляющей
способностью (см. рис. 6.27). Эти упорядоченные образования
легко отделяются от дезинтегрированных путем лизиса клеток
центрифугированием. В таких образованиях, однако, молекулы
белков связаны множеством поперечных связей, поэтому для
выделения белка в нативном виде осадок после центрифугиро-
вания необходимо растворить и ренатурировать. На рис. 11.40
приведена методика растворения этих структур методом окис-
лительного сульфитолиза с последующей ренатурацией раство-
римого денатурированного белка за счет его складывания в
соответствующую третичную структуру и образования дисуль-
фидных связей. Эти методы были разработаны исследователь-
ской группой компании Genencor, Inc. в связи с работами по
клонированию и экспрессии гена реннина в Е. coll.
11.8.3. Выделение полисахаридов
Основным методом выделения полисахаридов и их произ-
водных является осаждение спиртом. Декстраны представляют
собой чрезвычайно ценные полисахариды, широко используе-
мые, в частности, в пищевой промышленности и медицине, по-
этому процесс выделения и очистки декстранов (рис. 11.41)
включает несколько операций осаждения спиртом и промывки
свободной от пирогенных веществ водой. Образующийся при
326
Глава 11
первом осаждении осадок подвергают частичному гидролизу
затем отделяют (негидролизующиеся) нерастворимые твердые
вещества. Продукт очищают двукратной фракционной кристал-
лизацией, а оставшиеся примеси удаляют с помощью ионооб-
менных смол. При получении ксантана бульон стерилизуют /в
Окислительный сулъфигполиз
SH
S— белок — S
|s°r
SH
'S— белок— sso;
ф
белок —sso; ----------—г----------->
J 5O5
SSO32
'O3SS— белок— SSOj
д
so,
А
I
(О)
S'
~o3ss— белок -sso3
Образование дисульфидных связей
GS'
\ р Мочевина Мочевина
।—S—SO3 GSH/GSSG I—S—SG GSH/GSSG
Ls—SO3 * i— s—so3 >
Мочевина
S—SG gsh/gssg I—7
------> ф P
S—SG I— S--SG
Мочевина
gsh/gssg r~S
> L_s
РИС. 11.40. Превращение внутриклеточных образований в растворимый актив-
ный белок путем окислительного сульфитолиза и образования дисульфидных
связей. (GSH и GSSG — восстановленная и окисленная форма глутатиона
соответственно). (С любезного разрешения Б. Лоулиса н К. Хайенги, Genencor,
Inc.)
результате чего погибают все клетки), осаждают изопропано-
лом, затем высушивают распыливанием и измельчают; полу-
ченный таким путем ксантан можно использовать в пищевой
промышленности.
11.8.4. Выделение антибиотиков
Антибиотики выделяют или в виде сравнительно неочищен-
ных препаратов (примером может служить натриевая соль пе-
нициллина; см. рис. 11.42) или в виде высокоочищенных ве-
Декстран (25 кг)
РИС. 11.41. Выделение декстрана из бульона с низким содержанием нераство-
римых веществ. Установка для производства декстрана состоит из 4 фермен-
теров периодического действия объемом по 4 м3; рабочий цикл ферментера
длится 1 сутки. (Из работы: Atkinson. В., Mavituna F., Biochemical Engineering
and Biotechnology Handbook, p. 911, Macmillan Publishers Ltd. Surrey, Eng-
land, 1983.)
Penicillium
chrysogenum
Биореактор
Ротационный
фильтр
Ферментер
для роста
культуръ
Отработанный
Растворитель растворитель
Колонна для
очистки
Первый
экстрактор
I-------
Второй и
г-япретий
Испаритель экстракто-
ры
'I
Промывка
кристаллов
Керамический. рилътр
Испаритель
Y Отработанный
растворитель
Ленточный
фильтр
Растворитель
И
Фильтр
Раствор
гидрохлорида
прокаина
Ленточный
фильм/.
v
Кристаллическая
калиевая соль
пенициллина
► Раство-
ритель
Резервуар
для
смешения
л
Центрифуга
н
\ Сетчатый
\ фильтр
Вакуум
Лиофильная
сушка
Прокаиновая соль
пенициллина
РИС. 11.42. Процесс производства пенициллина. (Из работы: Atkinson В.,
Mavituna F., Biochemical Engineering and Biotechnology Handbook, p. 990,
Macmillan Publishers Ltd., Surrey, England, 1983.)
Операции выделения
329
ществ (например, в виде прокаиновой соли пенициллина),
предназначенных для клинического применения. Обратите вни-
мание на то, что здесь для отделения от растворимых и нерас-
творимых примесей используются адсорбционные колонны и
ленточные фильтры соответственно. Окончательную распыли-
тельную сушку осадка прокаиновой соли пенициллина осущест-
вляют при пониженной температуре методом лиофилизации.
Биомассу можно выделить практически количественно и ис-
пользовать как добавку к кормам для домашних животных.
11.8.5. Выделение органических кислот
Лимонную, глюконовую и итаконовую кислоты выделяют
(в виде цитрата кальция, глюконата натрия или кальция, сво-
бодной итаконовой кислоты) путем осаждения после предвари-
тельного отделения биомассы фильтрованием. В производстве
лимонной кислоты цитрат кальция обрабатывают серной кисло-
той (рис. 11.43), осадок сульфата кальция отбрасывают, а ли-
монную кислоту или ее натриевую соль осаждают в кристал-
лическом виде и выделяют центрифугированием. Последую-
щая операция сушки зависит от требований, предъявляемых
к продукту (безводная кислота, моногидрат, натриевая соль)
При получении чистой итаконовой кислоты ее раствор обраба
тывают активированным углем; эта операция не является обя-
зательной в процессе производства технической кислоты.
11.8.6. Выделение этанола
Этанол образует азеотропную смесь с водой, содержащую
95,7 масс.% (89 мол.%) спирта. Выделение этанола дистилля-
цией осуществляют в два этапа. Сначала на установке из трех
обычных ректификационных колонн получают продукт с высо-
ким содержанием спирта. В производстве абсолютного спирта
(применяемого, например, в качестве добавки к моторному
топливу) полученный спирт смешивают с бензолом и вновь
подвергают ректификации; вода отгоняется в первую очередь
в виде тройной азеотропной смеси, что позволяет получить
100%-ный этанол (рис. 11.44). Расход энергии на выделение
этанола зависит от качества получаемого продукта; в табл. 11.8
приведены основные характеристики различных способов выде-
ления этанола из культуральных жидкостей при его производ-
стве для промышленных целей и в качестве добавки к мотор-
ному топливу.
330
Глава 11
Промывные
воды (29м3)
Ротационный
фильтр
Бульон (100 м3), содержащий
лимонную кислоту [Ю%(масса/объем),
10000 кг] и биомассу [2°/о(масса/объем)]
, Блажная
веществ),
кислота
биомасса (ZOOO кг сухих
вода (1Z м3), лимонная
(100 кг)
Раствор (11Z м3)
лимонной кислоты {9900кг)
Известь (WOOкг),
содержащая
3800 кг Са(ОН)2
Резервуар с пере-
мешиванием
Отходы
Суспензия
Ротационный
фильтр
Серная кислота_
вода или маточный
раствор (59 м3)
Влажная паста: цитрат
кальция (10700кг), другие
твердые вещества (800кг),
вода (18 м3)
Сточные воды
(105м3)
I
Отходы
Суспензия: твердые вещества
(25%, 17600 кг), вода (70 м3),
растворенная лимонная
кислота (9800 кг)
Промывка
водой (26м3)
Кристаллизация
Рециркуляция
маточного
раствора
Центрифугирова-
ние и промывка
Сушка
Лимонная кислота (9Z00 кг)
РИС. 11.43. Выделение органических кислот. Приведена схема производства
лимонной кислоты в периодическом режиме; объем ферментера 100 м8, произ-
водительность 9200 кг лимонной кислоты за один цикл. (Из работы: Atkin-
son В., Mavituna F., Biochemical Engineering and Biotechnology Handbook,
p. 910, Macmillan Publishers Ltd., Surrey, England, 1983.)
11.8.7. Выделение белка одноклеточных организмов
При выделении биомассы, являющейся главным, а не по-
бочным (как, например, при производстве глутаминовой кис-
лоты или антибиотиков) продуктом производства, применяют
очень простые операции. В двух микробиологических процес-
сах, разработанных недавно компаниями ICI и Hoechst, выра-
щенную на метаноле биомассу отделяют с помощью простого
Операции выделения
331
концентрирования. Для этой цели в процессе ICI применяют
коагуляцию и декантацию, после чего получают продукт, со-
держащий 25 г биомассы (сухих веществ) в литре, а в процес-
се Hoechst биомассу выделяют электрокоагуляцией. Получен-
ную биомассу, обладающую достаточно высокой плотностью,
можно далее обезводить центрифугированием, минуя промежу-
точную стадию фильтрования. Процесс завершается распыли-
Рециркуляция азеотропной слоеси. 3 В
Исходная
смесь
89°АЭ
11"/» В
Этанол
Азеотропная
Вистилляирся
Регенерация
бензола
Отгонка,
воды
РИС. 11.44. Производство чистого этанола методом азеотропной дистилляции
(Э — этанол, В — вода, Б — бензол). (Из работы: Maiorella В., Wilke С. R.,
Blanch Н. W., Alcohol Production and Recovery, Adv. Biochem. Eng., Fiechter A.
(ed.), vol. 20, p. 43, Springer-Verlag, Berlin, 1981.)
тельной сушкой. Разработка способа выделения биомассы без
фильтрования была обусловлена, во-первых, необходимостью
непрерывной переработки больших объемов, что исключало
применение рамных фильтр-прессов; во-вторых, постоянным за-
сорением ротационных вакуум-фильтров мелкими микроорга-
низмами; в-третьих, невозможностью применения вспомога-
тельных фильтрующих материалов, которые загрязнили бы
продукт процесса (биомассу). В этих разработанных в послед-
ние годы процессах применяют не дрожжи, а бактерии, обла-
дающие, как правило, более высокой удельной скоростью
роста.
Биомассу сравнительно медленно растущих дрожжей выде-
лять легче, чем биомассу бактерий, что обусловлено большим
размером дрожжевых клеток (обычно tZ=5—8 мкм) по сравне-
нию с бактериальными клетками (d=\—2 мкм). Так, пекарские
332
Глава 11
Таблица 11.8. Расход энергии на операции выделения этанола®
Процесс Расход пара, кг на 1 л этанола Примечания
Простая дистилляция на 1 или 2 колоннах; исходная смесь — 6,25% этанола (по объему); продукт — 96% этанола (по объему) 2,4 Ректификационные колонны с 20 и 30 тарелками
Регенерация 96%-ного (по массе) технического спирта высшего качества из вод- ного раствора (6,25% по объему) на установке из трех колонн 2,4 Ректификационная колонна (20 тарелок), колонна для отделения альдегидов (45 тарелок), ректификаци- онная колонна (54 тарел- ки)
Установка Барбе для про- изводства 96%-ного (по объему) технического спир- та из 6,25%-ного (по объ- ему) водного раствора 4,1 Ректификационная колонна (20 тарелок), колонна для очистки (30 тарелок), рек- тификационная колонна (54 тарелки)
Обезвоживание 96%-ного спирта до 99,9%-ного (по объему) с помощью бен- зола 1,0 Колонна для обезвожива- ния (50 тарелок), обогре- ваемая паром колонна (30 тарелок), дополни- тельная ректификационная колонна (45 тарелок)
Комбинированная установ- ка для регенерации этано- ла из 6,25%-ного раствора (3 колонны) и для обез- воживания до 99,9% с по- мощью эфира под давле- нием 2,5 Ректификационная колонна (20 тарелок), колонна для отделения альдегидов (45 тарелок), ректификацион- ная колонна (54 тарелки), колонна для обезвожива- ния (30 тарелок), стрип- пинг-колонна (20 тарелок)
Абсолютирование 96%-ного этанола до 99,9%-ного (по объему) с помощью пента- на 0,9 Колонна обезвоживания (18 тарелок), стриппинг- колонна (14 тарелок); не- обходима также ректифика- ционная колонна
Получение 95%-ного (по массе) этанола из 13%- ного ректификацией в ва- кууме 1,1 Ректификационная колонна (40 тарелок) (при отсутст- вии альдегидов и сивуш- ных масел)
Получение этанола (99,9 1,1 (без учета Простая колонна (20 та-
мол. %) из 5%-ного (по энергии, расходуе- редок) (при отсутствии
массе) раствора путем эк- мой на регенера- альдегидов и сивушных
страктивной дистилляции с цию солей, кото- масел); процессы регене-
помощью солей рая зависит от из- бранного процесса регенерации) рации солей, не требую- щие больших затрат энер- гии, пока не изучены
Операции выделения
333
Продолжение табл. 11.8
Процесс Расход пара, кг „ на 1 л этанола Примечания
Получение 99,9%-ного спирта из 85%-ного путем адсорбции воды из паро- вой фазы Экстракция Расход энергии на Процесс осуществлен толь- регенерацию ад- ко в масштабе небольшой сорбента не уста- лабораторной установки новлен Лабораторный процесс; нужда- ется в дальней- шем изучении
Разделение с помощью мембран Разделение с помощью мо- лекулярных сит Лабораторный процесс Изучен только принцип процесса; необходимо даль- нейшее изучение
а По данным из работы: Maiorella В., Wilke С. R., Blanch Н. W., Alcohol Production
and Recovery, in Adv. Biochem. Eng., Fiechter A. (ed.) vol. 20, p. 43, Springer-Verlag, Ber-
lin, 1981.
дрожжи Saccharomyces cerevisiae (p= 1,133 г/см3) можно вы-
делять из 3—5%-ного бульона с помощью вертикальных цент-
рифуг непрерывного действия или центрифуг с соплами. Один
цикл такой обработки обеспечивает трехкратное концентриро-
вание, а после нескольких операций центрифугирования полу-
чают дрожжи в виде пастообразной массы, поддающейся пере-
качке насосами и содержащей 18—20% сухих веществ дрож-
жевых клеток.
Плесневые и высшие грибы можно выделять с помощью
корзиночных (отжимных) центрифуг, ротационных вакуум-
фильтров или сетчатых фильтров; таким путем получают био-
массу, содержащую 22—45% твердых веществ, которую затем
сушат. Экономия, достигаемая за счет более простого способа
выделения, может, однако, перекрываться дополнительными
расходами, связанными с более медленной скоростью роста
этих организмов; напомним, что время удвоения бактерий и
дрожжей равно 20—30 мин и 2—3 ч соответственно, а время
удвоения плесней, водорослей и высших грибов составляет не-
сколько часов.
На завершающей стадии обработки биомассы перед ее рас-
фасовкой и упаковкой отделяют нежелательные непрореагиро-
вавшие субстраты. В случае углеводов не возникает никаких
проблем. Метан всегда регенерируют и повторно используют в
биопроцессе, а следовые количества метанола легко удаляются
334
Глава 11
в процессе сушки. Если же при производстве белка одноклеточ-
ных организмов в качестве субстрата используется газойль,то
отделение непрореагировавшего субстрата от биомассы всегда
вызывает трудности, несмотря на сложную систему очистки,
включающую операции декантации, разделения фаз с помо-
щью растворителей, промывки растворами поверхностно-актив-
ных веществ, экстракции растворителями. При получении бел-
ка одноклеточных организмов на чистом алкане для полного
отделения непрореагировавшего субстрата достаточно промыв-
ки раствором поверхностно-активных веществ после первого
центрифугирования. Завершающая операция обработки белка
одноклеточных организмов должна обеспечивать практически
полную гибель как основного, так и всех других типов орга-
низмов, но в то же время, чтобы не снизилось качество про-
дукта, она не должна быть слишком жесткой, как, например,
при сушке на барабане, которая приводит к чрезмерному по-
темнению биомассы.
Если биомассу предполагается использовать в пищевой
промышленности, то необходимо предусмотреть способы сни-
жения содержания ДНК. Изучались два подхода к решению
этой задачи, в одном из которых ДНК, находящуюся в клет-
ках, разрушают путем теплового удара или экстракции щелоча-
ми, а в другом клетки дезинтегрируют и выделяют белковую
фракцию. Недостатком второго подхода является обычно низ-
кий выход белков.
11.9. Заключение
Основной задачей процессов выделения (независимо от то-
го, состоит ли этот процесс из одной операции или ряда после-
довательных операций) является получение продукта, облада-
ющего заданной степенью чистоты, необходимой концентрацией
и устойчивостью. Эта глава представляет собой введение в
изучение принципов, лежащих в основе отдельных операций
выделения; в ней также приведено несколько конкретных при-
меров выделения продуктов биохимической технологии.
Как показано на рис. 11.45, расходы на выделение продукта
непосредственно зависят от его концентрации в бульоне, посту-
пающем bi систему выделения и очистки. Прослеживаемая на
этом рисунке четкая зависимость стоимости вещества от его
концентрации в исходной смеси напоминает о том, что в на-
стоящее время совершенствование процессов биохимической
технологии должно быть направлено в первую очередь на мо-
дернизацию начальных операций в биореакторе с тем, чтобы
повысить концентрацию продукта в реакционной смеси и таким
образом снизить расходы на последующие операции выделения,
Операции выделения
335
очистки и регенерации. В конечном итоге сравнивать все ре-
сурсы, необходимые для каждой операции (хранения и обра-
ботки исходных материалов, биореакции, разделения и очист-
ки), можно только тогда, когда все затраты на каждую опера-
цию представлены в денежном выражении. Основам экономи-
10 3 •- Вода
10"
101
10°
10'1
1О'2
иг3
10^
Этанол
Аминокис-
~ лоты
Антибиоти
Бактериальные
протеазы
Амилазы
Лимонная кислота, моноглутамат натрия
Пенициллин
Треонин
Цефалоспорин
Генталгацин
—Гиббереллиновая кислота
Неочищенные ферменты
Глюкозооксидаза
Ферменты для исследова-
~ твльоких работ и диагности-
ческих целей
Моноклинальные
антитела
Ренни
Люцифераза
!
10~5
10~6
Фактор VIJ.
Урокиназа
Ферменты для терапии
ю-7\______I I______I____I____I____I____I____!____I___
1СГ2 КГ1 10° 107 102- 103 10^ 1G5 10е 107 10s
Продажная цена, долл/кг
РИС. 11.45. Зависимость продажной цены продуктов биопроцесса от их на-
чальной концентрации в бульоне или среде, поступающих в систему выделе-
ния и очистки. [Воспроизведено с разрешения из статьи: Dwyer J. L., Scaling
Up Bioproduct Separation with High Performance Liquid Chromatography, Bio/
Technology, 2, 957 (1984).]
ки процессов биохимической технологии посвящена первая
часть следующей главы, а во второй ее части будет приведен
ряд конкретных примеров.
Упражнения
11.1. Осаждение биомассы, а) Вычислите кажущийся размер изолирован-
ных частиц, если известны их плотности (р) н скорости осаждения (us):
первичный осадок в сточных водах (гл. 14) (р= 1,001 г/см3, us =
=0,042 см/с);
частицы активного ила (гл. 14) (р= 1,005 г/см3, «s=0,2 см/с);
дрожжи (р= 1,001, «s=0,0003 см/с). (Данные из работы: Physicochemical
336
Глава 11
Processes for Water Quality Control, Weber W. J. (ed.), p. 128, Wiley Inter-
science, 1972.)
б) Целесообразно ли выделять бактерии (р= 1,001, эффективный диаметр
1,0 мкм) отстаиванием, если продолжительность отстаивания составляет ме-
нее 1 ч? Почему (нет)?
11.2. Фильтрование биомассы; периодическая операция. Необходимо от-
фильтровать бульон, находящийся в перемешиваемом (встряхиванием) сосу-
де, при следующих условиях:
А = 10 см2; цс=2 сП; Др=0,3 атм; р = 1,1 г/см3; а; = 0,001 и т=2,5.
а) Рассчитайте удельное сопротивление осадка а и коэффициент сопротив-
ления фильтра г, если в разное время найдено, что [f(s), У(мл)] = (30,
38) (60,52).
б) Масштабирование. Фильтр какой площади Af потребуется для перера-
ботки 105 л такого бульона в течение 10 мин?
11.3. Ротационный вакуум-фильтр; непрерывное фильтрование. Вращаю-
щийся вакуум-фильтр частично погружен в культуральную жидкость. Процесс
фильтрования и накопления осадка начинается, как только фильтр погрузит-
ся в культуральную жидкость.
а) Найдите выражение, описывающее зависимость локальной плотности
накапливающегося осадка (W/Л) от времени (/) при вращении фильтра в
бульоне. Как упростится это выражение, если скорость фильтрования опреде-
ляется 1) только сопротивлением фильтра; 2) только сопротивлением осадка?
Набросайте графики всех трех зависимостей.
б) Предположим, что на фильтре в конечном итоге накапливается осадок
с плотностью у. Найдите выражение, описывающее абсолютную скорость
фильтрования бульона, если радиус фильтра равен R, а скорость его враще-
ния составляет <а рад/мин. Примите во внимание необходимость интегрирова-
ния локальных скоростей по всей погруженной фильтрующей поверхности.
11.4. Центрифугирование. Пусть суспензия сферических частиц радиусом R
и плотностью рр помещена в центрифужную пробирку, содержащую жидкую
среду с плотностью pf и вязкостью p,f. Если пробирка вращается с угловой
скоростью со рад/с, то движение изолированной частицы в разбавленной сус-
пензии описывается уравнением (1П1.4).
а) Разбавленную суспензию дрожжей (R=5 мкм, р= 1,001) подвергают
центрифугированию со скоростью 10 000 об/мин, причем высота слоя суспен-
зии в пробирке равна 4 см. При вращении ротора пробирки располагаются
перпендикулярно оси вращения, а расстояние от дна пробирки до оси враще-
ния составляет 8 см. Сколько времени потребуется для полного разделения
суспензии? Сформулируйте все принятые допущения.
б) В условиях, описанных в предыдущей задаче, центрифугируют кон-
центрированную суспензию сферических частиц (объемная доля частиц в сус-
пензии равна ер). Найдите выражение, с помощью которого можно опреде-
лить время, необходимое для разделения суспензии, если ер постоянно. Спра-
ведливо ли, по вашему мнению, допущение ер=const? Почему (иет)?
11.5. Фильтрование мицелия. Удельное сопротивление осадка при фильтро-
вании мицелиальных организмов (например, при производстве пенициллина)
можно оценить по параметрам сопротивления потоку через неподвижный слой,
состоящий из цилиндров. Удельное сопротивление осадка г описывается урав-
нением
r=K"s2p(l — е)2/е3
где /("— коэффициент сферичности Козени, sP— удельная площадь поверх-
ности частиц осадка, е — объем пустот в осадке. Параметр sp зависит от диа-
метра гифов (клеток) dh согласно выражению
sp=nd/,/(. ncPh/4) = 4/c?h
Операции выделения
337
а объем пустот е определяют по уравнению
е = 1—l/(vpn)
где рл—плотность гифов, a v — удельный объем осадка.
а) Покажите, что из приведенных выше уравнений и основного уравне-
ния фильтрования (11.2) следует, что
- _ ip ______ Г 1 . 1L ' /_______р/;У_____\
VFVC ~ ЛДр L А Ф Vc \ (рЛ^- j
где L — среднее сопротивление фильтра.
б) Мицелий образует сжимаемые осадки, поэтому при изменении пара-
метра фильтрования т меняется и V. В данных условиях фильтрования это
изменение описывается уравнениями
v = a — Ь(1/т) или v=a — 6(1/т) + с(1/т)2
Найдите уравнения, описывающие зависимость удельного сопротивления осад-
ка на фильтре г от модифицированного времени фильтрования т, для каждого
типа зависимости м(т).
в) В уравнениях предыдущей задачи используются переменные, имеющие
определенную размерность. Для практических целей удобнее уравнения, выра-
женные с помощью безразмерных переменных. Переведите уравнения, описы-
вающие т и г, в безразмерную форму.
Такие уравнения можно применить при проектировании фильтров, а так-
же при расчете детекторов для определения концентрации биомассы in situ
(см. следующее упражнение). (Nestaas Е., PhD thesis, MIT, 1980.)
11.6. Фильтрующий детектор для определения концентрации биомассы.
В упражнении 11.5 приведено модифицированное уравнение (11.2), описы-
вающее процесс фильтрования сжимаемого мицелия. Разработан детектор для
неавтономного определения концентрации биомассы in situ-, принцип действия
детектора заключается в измерении времени, необходимого для заполнения
небольшого сосуда определенного объема, расположенного непосредственно
над фильтром. Момент заполнения сосуда регистрируется, когда световой
луч прерывает граница раздела фаз осадок — среда; таким путем определяют
tp и Vр(/р).
а) Покажите, что концентрация биомассы в среде определяется уравне-
нием I000Pc/[v( Vc+Ef)].
б) Объясните, как можно определить х, если с помощью описанного детек-
тора измерено одно значение tp и одно значение Vf^f) для данного фиксиро-
ванного объема пробы.
При эксплуатации биореактора х определяют путем однократного изме-
рения tF, Vf, Vc. Затем фильтр промывают обратным потоком, смещая осадок,
после чего детектор готов к следующему измерению (извлекать осадок не
обязательно). [Из работы: Thomas D. С., Chitter V. К., Cagney J. W., Lim Н. С.
Biotech. Bioeng., 27, 729 (1985).]
11.7. Гомогенизатор высокого давления. Даннилл и Лилли (Dunnill, Lilly,
Protein Extraction from Microbial Cells, in Single Cell Protein II, Tannen-
baum S., Wang D. I. C. (eds.), MIT Press, Cambridge, Mass., 1975) показали,
что выход белков при пропускании суспензии клеток пекарских дрожжей че-
рез небольшой объем (гомогенизатор) при большом перепаде давления описы-
вается уравнением
1п
Rm
Rm ~~ R
22—746
338
Глава 11
где К — константа скорости процесса первого порядка; А— число пропуска-
ний через гомогенизатор; Rm — максимальный выход белков; Р— выход бел-
ков после N пропусканий через гомогенизатор; К=хр“, где х и а зависят от
штамма микроорганизма, ар — рабочее давление.
а) Покажите, что уравнение скорости процесса первого порядка приводит
к указанному выражению, если N рассматривать как непрерывную пере-
менную.
б) Покажите, что на кривой зависимости Q (выход белка в процентах,
отнесенный к киловатту расходуемой мощности) от давления имеется макси-
мум, если расход мощности на одно пропускание W пропорционален рабоче-
му давлению р.
в) Вычислите х и W/p и рассчитайте профили зависимости q от р при
ЛГ=1, 2 для пекарских дрожжей, если а=2,9, a Q достигает максимума (7,0)
при р=570 кг/см2 и А=1.
11.8. Экстракция с химической реакцией. Экстракцию неустойчивого со-
единения можно ускорить, если к растворителю добавить агент, обратимо
связывающий экстрагируемое вещество. Так, для связывания пенициллина
можно использовать растворимые в органических растворителях амины
Амин (органическая фаза) + пенициллин- (водная фаза) +
Ч-Н+ (водная фаза) амин-пенициллин( + Н)(органическая фаза)
а) Обсудите принципиальное сходство этого процесса с процессом сопря-
женного транспорта (гл. 5), например с транспортом кислорода в красные
кровяные тельца путем образования комплекса с гемоглобином.
Полный анализ массопереноса в сочетании с химической реакцией может
быть очень сложен. Экстракция в сочетании с химической реакцией может
способствовать ускорению реакции двумя путями: во-первых, за счет повы-
шения скорости экстракции и, во-вторых, за счет изменения коэффициента
распределения в сторону большей равновесной концентрации в органической
фазе. Анализ каждого из этих факторов в отдельности как ограничивающих
скорость экстракции не представляет затруднений.
б) Рассмотрим следующие элементарные стадии, приводящие к повыше-
нию скорости экстракции пенициллина (Р):
&d
Р —>- R (продукт деградации)
fei
Р Ps (массоперенос к границе раздела фаз)
ko
Ps —► Ро (перенос в органическую фазу)
ОДА]
Ps —>- АНР (экстракция в органическую фазу в сочетании
с химической реакцией)
Запишите уравнения для dp/dt и dps/dt. Приняв приближение псевдоста-
ционарного состояния по отношению к промежуточному соединению Ps (р« —
концентрация Ps на границе раздела фаз), покажите, что частное от деления
результирующей скорости переноса Р к границе раздела фаз на скорость
деградации Р определяется уравнением
Результирующая скорость переноса______/ _______kd_______\
Скорость деградации ~ kd \ ~~ + k0 + й2[А] /
Начертите график зависимости этой функции от концентрации амина [А].
Чему будет равно отношение скоростей переноса и деградации при обычной
экстракции ([А]=0) и в тех случаях, когда преобладает экстракция в соче-
тании с химической реакцией ([А] велико)?
Операции выделения
339
в) При равновесном повышении эффективности экстракции достаточно
рассмотреть элементарные стадии
Ко
Р =₽*= Ро
А+Ро ч* АРо
Покажите, что при данных объемах водной и органической фаз (Va и Vo
соответственно) отношение общей концентрации пенициллина в водной фазе
к общей концентрации пенициллина в органической фазе в состоянии равно-
весия описывается выражением (Уа/Уо)/Ка(1+Кн[А]). Начертите график за-
висимости этой функции от [А].
Параметры скоростей деградации и равновесные константы распределения
пенициллина в присутствии различных реагентов приведены в работе: Resch-
ke М., Schugerl К., Chem. Eng. J., 28, Bl, Bl 1 (1984).
11.9. Выделение внутриклеточного и внеклеточного продуктов. Для нера-
стущих клеток изменение концентраций внутриклеточного и внеклеточного
продуктов во времени можно описать такими уравнениями
Pl(O“Pimax(l — <Г,Л1
p.(0=aU-<„(I-e-'P-)]
а) Начертите графики этих функций.
б) Объясните приведенные выше уравнения, исходя из известных функ-
ций микроорганизмов.
в) При каких обстоятельствах вы стали бы проектировать процесс выде-
ления внутриклеточного продукта, а при каких — внеклеточного?
г) Какие пути совершенствования биокатализаторов вы могли бы реко-
мендовать для получения внутриклеточного и внеклеточного продуктов?
д) Набросайте схемы процессов получения внутриклеточного и внеклеточ-
ного продуктов; укажите основные операции получения выделяемого экстрак-
цией кристаллического продукта и (в качестве побочного продукта) сухой
биомассы.
11.10. Выделение антибиотика из цельного бульона. Высокая эффектив-
ность адсорбции антибиотика (и, возможно, других растворенных веществ)
приводит к следующему выражению для коэффициента скорости зависящего
от адсорбции массопереноса k [общая форма дана в уравнении (11.34)]:
k = 130е~-|- 46б—З^^гпах) ч-1
а) Рассмотрим смеситель, состоящий из одного резервуара, в который
непрерывно добавляют и из которого непрерывно отбирают содержащий анти-
биотик бульон и смолу. В такой ситуации движущей силой процесса является
постоянная разность концентраций с—с*, а в уравнении (11.33) время следует
заменить на время удерживания смолы т. Для случая одного резервуара
аналитическим путем, т. е. интегрированием уравнения (11.33), найдите зави-
симость T=fcn(q). (Для этой цели проще всего воспользоваться соотноше-
нием dq/dx= (dx/dq)-\ т. е. принять, что т является зависимой переменной.
Тогда интегрирование методом разложения на элементарные дроби не пред-
ставит затруднений.)
б) В периодическом режиме ионообменную смолу и бульон вводят в ре-
зервуар одновременно, так что операция смешения описывается уравнением
dq ( de \
Vrir-Vi[~S') <»
а зависимость с* от времени выражается уравнением c* = bqa [уравнение
22*
340
Глава 11
(11.35)]. Повторив решение предыдущей задачи, исходя из уравнений (1)
и (11.35), найдите необходимое время t путем интегрирования функции ад-
сорбции q.
в) Линейные уравнения, описывающие простые переходные состояния,
часто легко решить с помощью преобразования Лапласа. Можно ли решить
таким путем систему уравнений (11.32) — (11.35) для многостадийного про-
тивоточного процесса? Почему (нет)? [См. статью: Belter Р. A., Cunnin-
gham F. L., Chen J. 117., Development of a Recovery Process for Novobiocin;
Biotech. Bioeng., 15, 533 (1973).]
11.11. Разделение в капиллярах. В бумажной и тонкослойной хроматогра-
фии скорость подвижной фазы изменяется во времени. Перепад давления,
обусловленный движущейся в капиллярах бумаги границей раздела подвиж-
ной и неподвижной фаз, описывается уравнением Ap=4ocos (0)/dc, где о —
поверхностное натяжение, dc — диаметр капилляра, 0 — угол контакта жид-
кой и твердой фаз. Вязкое сопротивление скорости потока т пропорционально
цг (произведению вязкости на длину капилляра, заполненного жидкой фа-
зой) и обратно пропорционально поперечному сечению капилляра («=dc2).
а) Покажите, что z2 (z— положение фронта растворителя) пропорцио-
нально t, если т пропорционально dz/dt.
б) Покажите, что та же зависимость z от t характерна и для расстояния
между двумя пиками растворенных веществ.
в) Чтобы более реально описать хроматографический процесс, предполо-
жим, что частота распределения диаметров капилляров d определяется урав-
нением N(d) —А ехр[— (d—dm)2], где dm — диаметр, встречающийся чаще дру-
гих. Найдите выражение, описывающее зависимость относительного содержа-
ния воды в капиллярном носителе от времени и расстояния.
г) Найдите выражение, описывающее кажущееся расстояние между пика-
ми двух растворенных веществ; другими дисперсионными эффектами можно
пренебречь.
11.12. Хроматография. В хроматографии каждое растворенное вещество
характеризуется равновесным коэффициентом распределения Ki=Si/ci, где
$,• и с,- — концентрации вещества i в неподвижной и подвижной фазах соот-
ветственно. Покажите, что если проба (смесь веществ) в момент нанесения
занимает объем колонки высотой d, причем проба должна быть разделена иа
индивидуальные вещества так, чтобы расстояние между пиками было не ме-
нее d, то
а) Среднее время удерживания любого одного компонента i определяется
уравнением
L
где и — скорость подвижной фазы и L — длина колонки.
б) Необходимая максимальная длина колонки определяется максимумом
функции
d (Kj - Kj)
{l + Kj^l + Ki)
где Kf>Ki
в) В жидкостной хроматографии коэффициент диффузии сравнительно
высокомолекулярных веществ имеет величину порядка 10-6 см2/с (табл. 11.5).
С помощью уравнения Эйнштейна для среднеквадратичного расстояния, ко-
торое проходит молекула за время /, <22>,/2= (Z)/)l/2 покажите, что при ско-
рости подвижной фазы, превышающей 10-4 см/с, потребуется колонка боль-
шей длины, чем в задаче 11.126, если эта колонка наполнена частицами диа-
метром 10-2 см.
Операции выделения
341
г) Чарм и Вонг [Charm, Wong, An Immunosorbent Process for Removing
Hepatitis Antigen from Blood and Plasma; Biotech. Bioeng., 16, 539 (1974)]
с помощью аффинной хроматографии выделили антиген, специфичный по от-
ношению к вирусам эпидемического гепатита, из большого объема плазмы
крови. При таком высокоспецифичном разделении (г — антиген),
Пусть колонка для аффинной хроматографии заполнена непористыми
частицами радиусом R, а концентрация антигена составляет п частиц в 1 л;
покажите, что в этих условиях максимальный объем плазмы крови, который
можно очистить от антигена в единице объема колонки (без ее регенерации),
определяется выражением
3(1 -е)
nnr2R
где е— объем пустот в колонке, аг — радиус антигена.
11.13. Хроматография. По данным, приведенным на рис. 11.21 и в табл.
11.5, найдите значения L и rs для трех хроматографических носителей, пере-
численных на рис. 11.21.
11.14. Осаждение белков «высаливанием», а) Определите параметры {3
и Ks в уравнении растворимости [уравнение (11.7)] для каждого из белков,
перечисленных на рис. 11.13.
б) Предложите методику фракционного осаждения, позволяющую раз-
делить смесь представленных на этом рисунке белков на практически чистые
белки.
в) Ионы, неэффективные при высаливании белков, эффективны в обрат-
ной операции — растворении полимеров. Так, растворы LiCNS способны рас-
творять шелковые чулки! Прочтите в учебнике по физической химии главу,
посвященную растворению полимеров в присутствии солей, и кратко обсуди-
те явления, участвующие в «высаливании» и растворении белков.
11.15. Эффективность выделения ферментов. Предположим, что выход
фермента на 4 стадии в примере 11.1 составляет 50%. Вычислите долю на-
чального количества фермента, выделяемого на последней стадии.
11.16. Емкость иммуносорбентов. Сообщалось, что связывающая способ-
ность С некоторых колонок с иммуносорбентом снижается по мере увеличения
числа циклов адсорбция — элюирование (п) в соответствии с уравнением
С(п)=С(0)е-ал (п>0).
а) Покажите, что общее количество белка, которое можно выделить за N
циклов, составляет
N—1
С(Т)= ^С(п)
п=0
б) Если а мало, то указанную выше зависимость можно приближенно
описать выражением
(N-1)
С(Т) = J C(0)e~andn
о
Покажите, что общее количество белка, которое можно выделить на колонке,
равноС(0)/а.
в) Как вы определите максимальное число циклов, которые с экономиче-
ской точки зрения целесообразно осуществлять на данной колонке?
11.17. Рециркуляция компонента питательных веществ с ингибиторными
свойствами. Рециркуляция воды или биомассы сопровождается рециркуляцией
растворенных веществ; некоторые из них могут обладать ингибиторными
342
Глава 11
свойствами. Предположим, что ингибитор I влияет на удельную скорость кле-
точного роста согласно уравнению
14 “Wh
а) Рассмотрим простои реактор с рециркуляцией (рис. 9.4), в котором
концентрация растворенного ингибитора Покажите, что в этом случае
рециркуляция не влияет на концентрацию I в реакторе.
б) Теперь предположим, что основной продукт (например, этанол) и во-
да непрерывно отгоняются и что ингибитор I нелетуч. Какое максимальное
отношение рециркуляции Fr/F0 можно допустить, чтобы |A(i) /р. (i=0) не было
меньше 0,95? [Пример. В производстве этанола некоторые растворенные в
тростниково-сахарной мелассе соли являются ингибиторами по отношению
к Saccharomyces cerevisiae; см. работу Maiorella В. L., Blanch H. W., Wil-
ke C. R., Biotech. Bioeng., 26, 1155 (1984).]
в) Пусть ингибитор I продуцируется биомассой co скоростью nx. Найди-
те выражение, позволяющее определить (Fr/Fo)max через п, а (отношение обо-
гащения клеток в сепараторе) н минимально допустимое значение ц ((i)/р. (1=
= 0)).
11.18. Кинетика реакции, катализируемой иммобилизованным субтилнзи-
ном, в двухфазной системе с регенерацией. Оптически активные D-арилглицн-
ны, применяемые в качестве ацилирующих агентов в производстве полусин-
тетическнх пенициллинов н цефалоспоринов, можно получать из соответству-
ющих рацемических соединений с помощью иммобилизованной протеазы
(субтилнзина). В условиях реакции рацемические вещества-предшественники
растворяют в органической фазе, которую затем смешивают с приготовленной
заранее суспензией иммобилизованного фермента в воде. Ограниченно рас-
творимое исходное вещество легко переходит в водную фазу н там превра-
щается в соответствующую кислоту. Скорость реакции легко контролировать
методом титрования основанием.
Условия: 143,3 ммоль метилового эфира DL-2-ацетамидофенилуксусной кис-
лоты растворены в 250 мл метилизобутнлкетона. Добавлена суспен-
зия иммобилизованного катализатора в 250 мл воды.
Титрование основанием:
1 н. NaOH, мл 0,0 33 45 53 58 66 69 71 73
Время, ч 0,0 0,5 1,0 1,5 2,0 3,0 4,0 5,0 9,0
а) Какому уравнению скорости реакции, по вашему мнению, соответству-
ют эти данные?
б) Прн использовании дорогостоящего исходного вещества очень важен
выход оптически активного соединения. Так, весьма существенно определить
время, необходимое для 90 и 95 %-кого превращения субстрата. Можно ли
получить такую информацию на базе приведенных выше данных?
в) Набросайте схему непрерывного технологического процесса, включаю-
щего операции эффективного гидролиза рацемического соединения, выделения
оптически активного продукта кристаллизацией из воды, рециркуляцию орга-
нического растворителя н его регенерацию отгонкой нз раствора негидроли-
зованного исходного вещества (которое затем необходимо подвергнуть раце-
мизации). Какое основное оборудование требуется для осуществления собст-
венно реакции и операций выделения? [См. работу: Schutt Н. et al., Biotech.
Bioeng., 27, 420 (1985).]
11.19. Аффинная хроматография клеток. Костный мозг содержит стволо-
вые кроветворные клетки и зрелые Т-лимфоциты. Первые можно успешно
Операции выделения
343
трансплантировать пациенту с нарушенной функцией костного мозга с тем,
чтобы резко повысить число здоровых клеток и восстановить функции мозга.
Напротив, Т-лимфоциты могут принять клетки организма-хозяина за «чуже-
родные» и инициировать опасную реакцию против тканей человека, которому
был пересажен костный мозг. Поскольку на поверхности лейкоцитов имеются
специфические связывающие лектин рецепторы, то для отделения Т-лимфо-
цитов от популяции клеток костного мозга можно использовать колонки с им-
мобилизованным лектином.
а) Оказалось, что адсорбция клеток зависит от скорости потока, которая
определяет частоту соударений клеток с частицами адсорбента vp и среднее
время пребывания клетки тр (в отсутствие связывания). Пусть vp пропор-
ционально и“, а тр пропорционально (а, ?>0). Покажите, что в такой си-
туации концентрация клеток 6С в адсорбенте в стационарном состоянии про-
порциональна и“~₽.
б) Предполагается, что адсорбция клеток зависит от критического числа
связей типа клетка — неподвижная фаза, образующихся за время (т) пребы-
вания клеток на поверхности частиц неподвижной фазы. Предположим, что
связи В образуются за счет лектиновых центров L и клеточных центров S
согласно уравнению
db
= kfNLNs — kr
где L+S В, а на поверхности контакта клетка — неподвижная фаза имеет-
ся Nt.o и y^so независимых диффундирующих центров каждого типа.
I) Найдите выражение, описывающее число связей Ь* в состоянии равно-
весия.
II) Интегрированием уравнения скорости реакции db/dt (методом разло-
жения на элементарные дроби) найдите зависимость b(t) от t.
Ill) Пусть &crlt«b*) —число связей, необходимых для необратимого свя-
зывания клетки Т-лимфоцита с поверхностью частиц неподвижной фазы.. Най-
дите выражение, которое устанавливает зависимость между &Crit и максималь-
ной скоростью подвижной фазы, приводящей к связыванию клеток. [См. ра-
боту: Hertz С. М. et al., Biotech. Bioeng., 27, 603 (1985).]
11.20. Мембраны и искусственные органы. Для создания искусственных
имплантируемых органов предлагалось использовать микрокапсулирование
клеток млекопитающих или их включение в полые волокна. Поры мембраны
должны пропускать в окружение микрокапсул или волокон необходимые поли-
пептиды или белки и в то же время задерживать другие белки, способные
вызвать реакцию отторжения организма-хозяина.
а) Оцените необходимый средний диаметр пор мембраны, если мембрана
должна пропускать в окружение инсулин или гормон роста человека.
б) Включенные в капсулу или в полые волокна клетки снабжаются пи-
тательными веществами за счет организма-хозяина. Эти питательные вещества
должны диффундировать к иммобилизованным клеткам в таком количестве,
чтобы в идеальном варианте обеспечивать и метаболизм поддержания, и об-
разование продукта. Приняв, что коэффициент молекулярной диффузии кис-
лорода ®о2 равен приблизительно 10~б см2/с, оцените радиус частицы с им-
мобилизованными клетками, при котором модуль Тила процесса нулевого
порядка будет равен единице для случая: а) иммобилизованных растущих
дрожжей (tD=2 ч); б) иммобилизованных дрожжей в состоянии покоя (по-
требность в О2 на метаболизм поддержания составляет 3% потребности рас-
тущих клеток); в) иммобилизованных растущих животных клеток (/о=40 ч);
г) иммобилизованных животных клеток в состоянии покоя (потребность в О2
на метаболизм поддержания составляет 10% потребности растущих клеток).
Можно ли иммобилизовать покоящиеся клетки млекопитающих диаметром
10 мкм в микросферах диаметром 0,1, 0,5 и 2 мм?
344
Глава 11
Литература
Последовательные операции выделения продуктов:
1. Atkinson В., Mavituna F., Biochemical Engineering and Biotechnology Hand-
book, Macmillan Publishers Ltd., Surrey, England, 1983. (В главах 12—14
приведено множество интересных примеров.)
2. Belter Р., General Procedures for Isolation of Fermentation Products, in
Microbial Technology, Peppier H. J., Perlman D. (eds.), 2d ed., vol. 2,
p. 403, Academic Press, New York, 1979.
3. Edwards V., Recovery and Purification of Biologicals; Adv. Appl. Microbiol
11, 159 (1969).
Выделение биомассы и нерастворимых материалов:
4. Aiba S., Nagatani M., Separation of Cells from Culture Media; Adv. Biochem.
Eng., 1, 3 (1971).
5. Belter P., Recovery Processes: Past, Present, Future, 184th Amer. Chem. Soc.
Mtg., September, 1982.
6. Fermentation and Biochemical Engineering Handbook, Vogel H. C. (ed.),
Noyes Publications, Park Ridge, N. J., 1983. (Главы 5 и 10 посвящены
фильтрованию и центрифугированию соответственно.)
7. Weissman U., Binder EL, Biomass Separation from Liquids by Sedimentation
and Centrifugation, in Adv. Biochem. Eng., Fiechter A. (ed.), vol. 24, p. 119,
Springer-Verlag, Berlin, 1982. (Седиментация.)
8. Weber W. J., Jr., Physicochemical Processes for Water Quality Control,
Wiley-Interscience, N. Y., 1972. (Имеются главы, посвященные детальному
анализу коагуляции, седиментации и фильтрования.)
9. Shirato S., Esumi S., Filtration of a Culture Broth of Streptomyces griseus;
J. Ferment. Technol. (Japan), 41, 87 (1963).
10. Gabler R.. Ryan M., Processing Cell Lysate with Tangential Flow Filtration,
in Purification of Fermentation Products, LeRoith D. et al. (eds.), Amer.
Chem. Soc., Symposium Ser., 271, 1 (1985).
11. Zahka J., Leahy T. J., Practical Aspects of Tangential Flow Filtration in
Cell Separations, Purification of Fermentation Products, LeRoith D. et al.
(eds.), Amer. Chem. Soc. Symposium Ser., 271, 51 (1985).
12. Todd D. B., Podbielniak W. J., Centrifugal Extraction; Chem. Eng. Progress,
61, (5), 69 (1965).
13. Queener S.t Swartz R., Secondary Products of Metabolism, in Economic Mic-
robiology, Rose A. H., (ed.), vol. 3, p. 35, Academic Press, London, 1979.
(Экстракция пенициллина растворителями.)
14. Shreve R. N., Brink J., The Chemical Process Industries, McGraw-Hill, New
York, 1977. (Имеются главы, посвященные процессам фармацевтической
промышленности, в том числе выделению антибиотиков адсорбцией и экс-
тракцией.)
15. Альбертсон П.-О., Разделение клеточных частиц и макромолекул. — М.:
Мир, 1974. (Здесь впервые подробно описан метод экстракции в двухфаз-
ных водных системах.)
Выделение и очистка:
16. Kula М. R., Kroner К. И., Hustedt Н., Purification of Enzymes by Liquid-
Liquid Extraction; Adv. Biochem. Eng., 24, 103 (1982). (Выделение белков
экстракцией в двухфазной водной системе.)
17. Menge U., Kula М. R., Purification Techniques for Human Interferons; Enz.
Microb. Technol., 6, 101 (1984). (Экстракция в двухфазной водной системе
и высокоэффективная жидкостная хроматография.)
18. Darbi/shire JLarge Scale Enzyme Extraction and Recovery, Topics in
Операции выделения
345
Enzyme and Fermentation Biotechnology, Wiseman A. (ed.), vol. 5, p. 147,
Halsted Press/J. Wiley, N. Y., 1981.
19. Grabender G. C., Glatz С. E., Protein Precipitation: Analysis of Particle Size
Distribution and Kinetics; Chem. Eng. Commun., 12, 203 (1981).
20. Bell D. J., Hoare M., Dunnill P., Formation of Protein Precipitates and
Their Centrifugal Recovery, in Adv. Biochem. Eng., Fiechter A. (ed.), vol. 26,
p. 1, Springer-Verlag, Berlin/N. Y., 1983.
21. Flaschel E„ Wandrey Ch., Kula M. R., Ultrafiltration for the Separation of
Biocatalysts, in Adv. Biochem. Eng., Fiechter A. (ed.). vol. 26, p. 73, Sprin-
ger-Verlag, Berlin/N. Y., 1983.
22 McGregor IV C., Large Scale Isolation and Purification of Proteins from
E. coli\ Ann. N.Y. Acad. Sci., 413, 231 (1983).
23. Schoner R. G. et al., Isolation and Purification of Protein Granules from
E. coll Cells Overproducing Bovine Growth Hormone; Bio/Technology. 3,
151 (1985).
24. Joustra M. K., Gel Filtration on Agarose Gels, in Prog, in Separation and
Purification, vol. 2, p. 183, Wiley-Interscience, N. Y., 1969.
25. Dubin P. L., Aqueous Size Exclusion Chromatography, in Separation and
Purification Methods, 10(2), 287 (1981).
26. Graves D. J., Wu Y. T., The Rational Design of Affinity Chromatography
Separation Processes, in Adv. Biochem. Eng., Fiechter A. (ed.), vol. 12,
p. 219, Springer-Verlag, Berlin/N. Y., 1979.
27. Eveleigh J. W., Practical Considerations in the Use of Immunosorbents
' and Associated Instrumentation, in Affinity Chromatography and Related
Techniques, Gribneau T. C. J. et al. (eds.), Elsevier, Amsterdam, 1982.
28. Chase H. A., Affinity Separations Using Immobilized Monoclonal Antibodies;
Chem. End. Sci., 39, 1099 (1984).
29. Arnold F. H., Chalmers J. J., Saunders M. S., Croughan M. S., Blanch H. W„
Wilke C. R., A Rational Approach to Scale-Up of Affinity Chromatography,
in Purification of Fermentation Products, LeRoith D. et al. (eds), Amer.
Chem. Soc. Symp. Ser., 271, 113 (1985).
30. Safer G., Chromatographic Removal of Pyrogenic Material; Bio/Technology,
2, 1035 (1984).
31. Janson J. C., Large Scale Affinity Purification — State of the Art and Future
Prospects; Trends in Biotechnology, 2(2), 31 (1984).
32. Maiorella B„ Wilke C. R„ Blanch H., Alcohol Production and Recovery, Adv.
Biochem. Eng., vol. 20, p. 43, 1981. (Процессы и экономика выделения
этанола.)
33. Whitesides G. М. et al.. Magnetic Separations in Biotechnology; Trends in
Biotechnology, 1(5), 144 (1983).
34. Beaton N. C., Steadly H., Industrial Ultrafiltration, in Recent Developments
in Separation Science, vol. VII, p. 1, 1982.
35. Smith I. H., Pace G. H., Recovery of Microbial Polysaccharides; J Chem
Tech. Biotechnol., 32, 119 (1982).
36. Fish N. M., Lilly M. D., The Interactions between Fermentation and Protein
Recovery; Bio/Technology, 2, 623 (1984).
37. Belter P. A., Cunningham F. C., Chen J. W., Development of a Recovery
Process for Novobiocin; Biotech. Bioeng., 15, 533 (1973).
38. Vermeulen T„ Adsorption and Ion Exchange, in Chemical Engineers’ Hand-
book, Perry R. H„ Chilton С. H. (eds.), 5th ed., Section 16, McGraw-Hill,
New York, 1973. (Есть перевод 4-го издания: Перри Дж. Г., Справочник
инженера-химика, т. I, гл. VIII. —Л.: Химия, 1969.)
39. Arnold F. Н., Blanch Н. W., Wilke С. R., Analysis of Affinity Separations;
The Chem. Eng. J., 30, B9, B25 (1985).
Глава 12
Экономика процессов
биохимической технологии
В этой главе мы рассмотрим роль экономических факторов
в изучении, внедрении и эксплуатации процессов биохимичес-
кой технологии. В первом разделе описаны основные этапы
разработки процессов, а затем в качестве иллюстрации доста-
точно детально рассмотрен пример экономического расчета
биотехнологического процесса, включающего анализ общей
технологической схемы процесса, оценку необходимого обору-
дования, потребности в материалах, водо-, тепло-, газо- и энер-
госнабжении, оценку стоимости всего предприятия и расходов
на его эксплуатацию, расчет рентабельности (прибыли на ин-
вестированный капитал). После этого мы обсудим особенности
экономики конкретных биотехнологических процессов, в том
числе получения продуктов тонкой биотехнологии и химичес-
ких продуктов массового производства, а также белка одно-
клеточных организмов. В этих примерах мы сравним расходы
на приобретение субстратов и питательных веществ, оборудо-
вания, обеспечение водо-, тепло-, газо- и энергоснабжения, на
осуществление биотрансформации и выделение продукта, по-
скольку обнаружение конкретной наиболее дорогостоящей опе-
рации может указать самое слабое место в экономике всего
процесса, а следовательно, и наиболее целесообразные пути
его оптимизации, например, за счет технологических усовер-
шенствований или посредством поиска нового штамма.
Мы отметим также особенности каждого типа продуктов
процессов биохимической технологии. Так, если речь идет о
разработке процесса производства нового лекарственного пре-
парата, то в таких случаях следует учитывать расходы на
клинические испытания и на получение разрешения от Управ-
ления по контролю за качеством пищевых продуктов, медика-
ментов и косметических средств (подчиненного министерству
здравоохранения, просвещения и социального обеспечения
США). С другой стороны, в процессах получения химических
продуктов массового производства с помощью культур микро-
организмов в качестве побочного продукта образуется боль-
шое количество биомассы, для которой желательно найти ры-
нок сбыта; в противном случае необходимо учитывать расходы
Экономика процессов биохимической технологии 347
на ликвидацию отходов. При производстве этанола из биомас-
сы помимо ферментации и выделения может понадобиться так-
же требующая значительных затрат предварительная обработ-
ка субстрата, например гидролиз и солюбилизация компонен-
тов биомассы. Кроме того, во многих странах (и в США в
том числе) для спиртовых заводов введены в различных фор-
мах пониженные налоги или кредиты, которые, очевидно, так-
же влияют на экономику производства. Биологическая обра-
ботка отходов является основным типом процессов, цель ко-
торых не в производстве продукта или биомассы, а в превра-
щении субстрата; в таких случаях основные эксплуатационные
расходы связаны с ликвидацией биомассы (активного ила).
12.1. Экономика технологических процессов
Экономические факторы играют важную роль практически
на каждом этапе разработки проекта предприятия (табл. 12.1).
Для того чтобы проект в конечном итоге был реализован в
промышленном масштабе, этапы разработки проекта должны
Таблица 12.1. Основные этапы разработки проекта предприятия
и его создания
1. Идея создания процесса (предприятия)
2. Предварительная оценка экономики и рынка сбыта продукции3
3. Подбор и обработка данных, необходимых для создания окончательного
проекта3
4. Окончательный экономический анализ
5. Детальный технический проект
6. Материальное обеспечение
7. Строительно-монтажные работы
8. Пусконаладочные работы и пробный пуск
9. Эксплуатация (выпуск продукции).
Если для данной продукции необходимо разрешение Управления по контролю за
качеством пищевых продуктов, медикаментов и косметических средств (или министерства
сельского хозяйства США), то эти этапы включают также соответствующие испытания
(в том числе клинические), оформление запроса и получение разрешения от соответст-
вующих правительственных организаций (см. разд. 12.2).
включать идею создания процесса или выпуска определенного
продукта; предварительную оценку экономики и рынков сбы-
та; разработку всех дополнительных данных, необходимых для
•создания окончательного варианта детального проекта; окон-
чательную экономическую оценку' проектируемого предприятия
в свете всех полученных данных; создание детального техни-
ческого проекта; обеспечение предприятия территорией и обо-
рудованием; строительство зданий и сооружений и монтаж обо-
рудования; пусковые работы, пробный пуск и, наконец, плано-
мерную эксплуатацию предприятия.
348
Глава 12
Идея создания процесса может возникнуть под воздействи-
ем любого фактора — предложений торговых организаций, по-
явления конкурентоспособного продукта, требований покупате-
лей, новых предложений исследовательских центров и групп.
Мысль о создании нового процесса, родившаяся внутри сущест-
вующей компании может быть запатентована компанией-изго-
товителем; в альтернативном варианте идея нового процесса
может быть заимствована из других патентов и реализована в
соответствии с лицензионными соглашениями.
Предполагается, что в конечном итоге каждый проект дол-
жен окупить все затраты и принести соответствующую при-
быль. Следовательно, при возникновении идеи о разработке
нового процесса или выпуске нового продукта необходимо про-
вести предварительную оценку процесса (или продукта) с
точки зрения экономики и потенциального рынка сбыта. При
оценке рынка сбыта учитывают потенциальный спрос при раз-
личных предположительных ценах на продукт. Рентабельность
(или ее отсутствие) определяют путем изучения экономики,
сравнивая расходы, необходимые для производства продукта
в количестве, удовлетворяющем запросы рынка, и соответству-
ющую себестоимость.
Если предварительная оценка показала, что данный про-
цесс будет рентабельным, то обычно приступают к разработке
данных, необходимых для создания окончательного проекта.
При этом уточняют потребность рынка и себестоимость про-
дукта, проводят тщательное изучение рынка, причем потенци-
альным покупателям часто направляют образцы продукта,
чтобы убедиться, что он отвечает их требованиям и что пред-
варительные оценки объема спроса на этот продукт реальны.
Для определения себестоимости продукта разрабатывают пол-
ную технологическую карту процесса, определяют соответст-
вующий объем капитальных затрат и эксплуатационных расхо-
дов, а также ежегодную прибыль на предполагаемые период
выпуска продукта или на период выплаты субсидий и займов.
При оценке прибыли проводят также элементарный анализ
чувствительности, для того чтобы найти предполагаемые основ-
ные составные части себестоимости и эксплуатационных рас-
ходов, оказывающие наибольшее влияние на расчетную рента-
бельность (поскольку до выпуска продукта цены могут значи-
тельно измениться).
Прежде чем приступить к разработке детального проекта,
в обзоре для руководства дается окончательный экономичес-
кий анализ, в котором рассматриваются основные сравнитель-
ные характеристики проекта в целом. Экономический анализ
должен дать ответы на ряд вопросов, например, как выглядит
рентабельность изучаемого проекта по сравнению с рентабель-
Экономика процессов биохимической технологии 349
ностью других возможных проектов, требующих примерно та-
ких же капитальных затрат; можно ли осуществить планируе-
мый проект силами имеющегося в компании технического пер-
сонала и персонала системы сбыта или же планируемый про-
ект принципиально отличается от эксплуатируемых предприя-
тий и поэтому потребует привлечения нового персонала; како-
ва вероятность того, что на базе планируемого проекта можно
будет реализовать несколько сходных процессов, или же сфера
применения проекта ограничена одним процессом; обладает ли
разрабатываемый процесс достаточной гибкостью, чтобы его
можно было использовать в других близких проектах (изучае-
мых или проектируемых), или же его (и обслуживающий пер-
сонал) можно использовать только в применении к данному
проекту; каковы ожидаемые направления развития компании
в целом и способствует ли планируемый проект укреплению
положения компании и использованию накопленного ею техни-
ческого опыта в ожидаемых направлениях развития, или же
новый проект приведет к чрезмерному распылению сил компа-
нии на не связанных друг с другом направлениях. Если выводы
анализа положительны, то в этом случае отпускаются средства
на реализацию нескольких следующих этапов.
В детальном техническом проекте содержится вся информа-
ция, необходимая для последующего строительства предприя-
тия, в том числе спецификация и номенклатура основного обо-
рудования, контрольно-измерительной аппаратуры, оборудова-
ния вспомогательных служб, схемы трубопроводов и последние
курсы цен. В детальный проект должна быть включена вся
строительная документация, планировка расположения обору-
дования, а также любая другая информация, необходимая при
строительстве предприятия.
Материальное обеспечение включает приобретение необхо-
димого земельного участка и недостающего оборудования, по-
лучение разрешений на все виды планируемых работ — строи-
тельные, эксплуатационные, сооружение и подключение систем
водо-, тепло-, газо- и энергоснабжения, сброса отработанных
воздуха и воды, сооружения системы сточных трубопроводов
и т. д. Обратите внимание на то,1 что для выполнения этих
задач потребуется немало времени, особенно для приобретения
специального или выпускаемого по заказу оборудования.
На этапе строительно-монтажных работ полностью соору-
жается новое предприятие, после чего начинают пусковые ра-
боты и пробный пуск. На этом этапе осуществляют пробную
эксплуатацию и опробование всего предприятия и отдельных
его звеньев, обучают операторов, разрабатывают инструкции
по пуску, эксплуатации, остановке процесса, а также необходи-
мые мероприятия, обеспечивающие безопасность работы. Пос-
350
Глава 12
ле этого приступают к завершающему этапу — эксплуатации
предприятия (выпуску продукта).
На всех этапах неоднократно проверяют и уточняют исход-
ные данные для экономического анализа с целью учета любых
изменений в потребности рынка, доступности сырья, намерени-
ях компании, ходе оформления разрешений от Управления по
контролю за качеством пищевых продуктов, медикаментов и
косметических средств или от министерства сельского хозяйст-
ва США или в, других факторах, которые могут повлиять на
прогнозируемую рентабельность проекта. Поскольку соответст-
вующие полные расчеты трудоемки и сложны, то в таких слу-
чаях обычно используют стандартные методы объективного
анализа, например критерий оценки прибыли на инвестирован-
ный капитал или другой близкий критерий. Такой метод по-
зволяет получить наиболее объективную картину. Напротив,
пессимистичная или чрезмерно предвзятая оценка может при-
вести к отклонению проекта, который на самом деле может
оказаться решающим для дальнейшего развития всей компа-
нии, а чрезмерно оптимистичный подход может привести к та-
ким расходам, которые по завершении строительства нового
предприятия вызовут финансовую катастрофу.
12.2. Контроль за качеством продукции
биохимической технологии
Вмешательство правительственных организаций в контроль
за качеством продукции биотехнологии обусловлено установ-
ленной законодательными актами ответственностью этих орга-
низаций за здоровье и безопасность рабочих, безопасность по-
требителей, решение связанных с биотехнологией проблем ох-
раны окружающей среды, а также за разработку специальных
правил, касающихся новых аспектов биотехнологии (особенно
связанных с применением рекомбинантных ДНК). Экономичес-
кий анализ должен включать расходы, связанные с получением
разрешения от правительственных организаций на выпуск со-
ответствующей продукции (в том числе при необходимости и
на клинические испытания).
В США надзор за производством биотехнологической про-
дукции в основном осуществляется тремя правительственными
организациями — Управлением по контролю за качеством пи-
щевых продуктов, медикаментов и косметических средств
(Food and Drug Administration, сокращенно FDA), министерст-
вом сельского хозяйства США (U. S. Department of Agricul-
ture, сокращенно USDA) и Агентством по охране окружающей
среды (Environmental Protection Agency, сокращенно ЕРА).
Экономика процессов биохимической технологии 351
Надзор за производством медикаментов, биологических пре-
паратов, пищевых продуктов и добавок к ним, диагностических
средств является сферой деятельности FDA. Проверка новых
лекарственных препаратов и выдача разрешений на их выпуск
(включая испытания на животных и человеке) может потребо-
вать нескольких лет. Здесь важно уточнить понятие «новый
лекарственный препарат», поскольку, согласно принятой в
1984 г. интерпретации, «новым» считается каждый лекарствен-
ный препарат, выпускаемый с помощью технологии рекомби-
нантных ДНК, даже если этот препарат по химической приро-
де идентичен уже имеющемуся. Производству каждого из та-
ких медикаментов должна предшествовать проверка его
эффективности и безопасности. Под биологически активным по
отношению к человеку препаратом понимают «вакцины, тера-
певтические сыворотки, токсины, антитоксины или аналогич-
ные продукты, предназначенные для профилактики заболева-
ний, ухода за больными или лечения болезней или ран». Как
сам продукт, так и предприятие, выпускающее этот продукт,
должны быть зарегистрированы в FDA, причем разрешение
может выдаваться на выпуск каждой партии продукта. Анало-
гичные ограничения налагаются и на выпуск другой биотехно-
логической продукции. Краткие сведения о функциях и сфере
деятельности соответствующих контролирующих правительст-
венных организаций США приведены в табл. 12.2.
Таблица 12.2. Правительственные организации США, осуществляющие
надзор за выпуском биотехнологической продукции3
1. FDA
а) Отдел оценки новых лекарственных препаратов (все новые лекарст-
венные препараты должны быть испытаны на животных и человеке;
любой препарат, полученный с помощью рекомбинантных ДНК, счи-
тается «новым»)
б) Отдел биопрепаратов контролирует клинические испытания и затем
выдает разрешения как на продукт, так и на работу выпускающего
этот продукт предприятия
в) Национальный центр медицинской аппаратуры и радиологической за-
щиты; центр контролирует медицинскую аппаратуру и диагностиче-
ские средства (включая диагностические наборы на основе монокло-
нальных антител, если только диагностика не связана с работами по
созданию банка крови). В настоящее время системы для диагностики
на основе моноклональных антител должны проверяться не столько
на безопасность и эффективность, сколько на адекватность выпускав-
шимся ранее системам.
2. USDA
Осуществляет первичный контроль биологически активных веществ, пред-
назначенных для животных; только после этого к оценке биопрепаратов
подключается FDA. Что касается моноклональных антител и продуктов,
полученных с помощью рекомбинантных ДНК, то функции этих двух
организаций разграничены нечетко (так, USDA регулирует внешнетор-
352
Глава 12
Продолжение табл. 12.2
говые связи, a FDA может контролировать правила, принимаемые на
территории США),
3. ЕРА
Эта организация обязана «обнаруживать потенциально опасные вещества,
оценивать степень их опасности и затем контролировать производство,
применение, распределение и уничтожение этих веществ, включав хими-
каты, гербициды и пестициды».
4. Управление профессиональной безопасности и здравоохранения (Occupa-
tional Safety and Health Administration, сокращенно OSHA).
Эта организация отвечает за охрану труда и технику безопасности. До
1984 г. включительно организацией не издано ни одного стандарта по
бпопроцессам; не ясна и ее позиция по отношению к контролю биотехно-
логических процессов.
а Commercial Biotechnology: An International Analysis, Office of Technology Assess-
ment, January 1984, pp. 359—365.
12.3. Общий экономический анализ процессов
биохимической технологии
Несмотря на то что между отдельными биотехнологически-
ми процессами с участием культур микроорганизмов имеются
существенные различия, любой проект сначала удобнее всего
рассматривать с помощью общей схемы микробиологического
технологического процесса, приведенной на рис. 12.1. На этой
упрощенной схеме показано, что весь процесс можно разбить
в основном на две части — операции ферментации (или био-
реакции) и операции выделения продукта. На рисунке в левом
верхнем углу указаны операции приемки и хранения исходных
материалов, а в правой части — завершающие операции полу-
чения продукта (композиции), его упаковки и отправки потреби-
телям, Для выполнения любой из операций могут потребоваться
системы водо-, газо- и (или) энергоснабжения; в полном проек-
те процесса должно быть отражено и решение проблемы охраны
окружающей среды, в том числе операции размещения, хране-
ния и ликвидации стоков.
Разработка любого биопроцесса должна быть направлена
на реализацию той или иной конкретной идеи — обычно это
получение определенного продукта. Если цель определена,
можно приступать к разработке более детальной схемы тех-
нологического процесса. На этом этапе необходимы сведения
(которые используются только для качественной оценки проек-
та) об основном субстрате, других исходных материалах, типе
реакции (биотрансформации), основных и побочных продуктах,
Экономика процессов биохимической технологии
353
используемом организме, необходимой интенсивности переме-
шивания и аэрации, специальных операциях, необходимых или
желательных для функционирования биореактора или выделе-
ния продукта, а также о требованиях, предъявляемых к управ-
лению процессом.
В предварительном экономическом анализе удобно и це-
лесообразно заимствовать сведения о ценах из относительно
РИС. 12.1. Обобщенная технологическая схема биопроцесса. [Воспроизведено
с разрешения из работы: Bartholomew W. Н., Reisman Н. В., Economics of Fer-
mentation Processes, in Microbial Technology, Peppier H. J., Perlman D. (eds.),
2d ed., vol. 2, p. 481, Academic Press, Inc., New York, N.Y., 1979.]
стандартных и доступных источников; при последующем более
детальном анализе цены всех конкретных материалов и обо-
рудования могут быть уточнены. При анализе прогнозируемых
затрат общие затраты обычно разделяют на три составляю-
щие— капитальные затраты (здания и сооружения, оборудо-
вание для проведения биореакции и операций выделения), из-
держки производства (заработная плата рабочих, техников,
инженеров, служащих, налоги, амортизационные отчисления и
т. д.), стоимость материалов и вспомогательных систем (пита-
тельных веществ, присадок, упаковочных материалов и средств,
электроэнергии, газа, пара, воды).
Приблизительная стоимость оборудования для микробиоло-
гической промышленности приведена в табл. 12.3. Указанные
23—-746
354
Глава 12
в таблице данные получены в 1977 г. от компаний, выпускаю-
щих такое оборудование в США; со временем они могут из-
меняться. Соответствующие изменения цен (для предваритель-
ной оценки затрат) удобно определять с помощью одного из
двух прейскурантов: Engineering and News Record construction
cost index и Marshall and Swift Equipment cost index (MSE).
Эти прейскуранты дают возможность экстраполировать цены,
Таблица 12.3. Стоимость типового оборудования в ценах 1977 года
(в долларах США)а
Ферментеры (нержавеющая сталь 304; со змеевиками и отражательными
перегородками; рабочее давление до 2,75 атм)
15 м3 35 000
113 м3 95 000
225 м3 165 000
Резервуар (нержавеющая сталь 304)
190 м3 37 000
В установленном виде, 60 000
с контрольно-измеритель-
ной аппаратурой
Распылительные сушилки (нержавеющая сталь 304)
Номинальная скорость Стоимость установленного
обезвоживания', кг/ч оборудования
1800 600 000
3200 1 250 000
4500 1 600 000
(В комплекте с вентиляторами, трубопроводами, электрооборудованием,
подогревателем пара, газовой горелкой, сушильной камерой, системой сбора
влаги, системой упаковки в мешки, постройками.)
а Bartholomew ТГ. Н., Reisman. Н. В., Economics of Fermentation Processes, in Mi-
crobial Technology, Peppier H. J., Perlman D. (eds.), 2d ed., vol. 2, p. 463, Academic Press,
Inc., New York, N. Y., 1979.
установленные в конкретном году, на последующие годы. Если
для планируемого процесса выбраны типы и размеры необхо-
димого основного оборудования, то цены 1977 г. можно опреде-
лить по табл. 12.3, а затем с помощью прейскуранта MSE
найти поправки на цены текущего года. Впрочем, лучше всегда
за основу расчетов брать действующие цены, сообщаемые ком-
паниями, производящими соответствующее оборудование. Ни-
же вкратце рассмотрен предварительный экономический анализ
конкретного процесса.
Издержки производства, если их составляющие выражены
как доли от других затрат, обычно относительно мало зависят
от конкретных деталей процесса. В табл. 12.4 перечислены ос-
новные составляющие издержек производства п общие коэф-
фициенты затрат; эти данные могут служить основой при
оценке предстоящих расходов. Для того чтобы по данным, при-
Экономика процессов биохимической технологии
355
Таблица 12.4. Ориентировочная оценка издержек производства3
Издержки производства
Амортизационные отчисления: 6,7—10% от капитальных затрат (при неп-
рерывной эксплуатации в течение 10—15 лет)
Налоги и страхование: 2—6% от капитальных затрат (конкретная сумма
зависит от расположения предприятия)
Сумма заработной платы служащих за исключением персонала, занятого
техническим обслуживанием: 70—100% от оплаты труда производствен-
ных рабочих (администрация, контрольные службы, конторы, техниче-
ские службы, инженерный состав)
Заработная плата рабочих, не занятых непосредственно на производстве:
30—50% от оплаты труда производственных рабочих (включая работ-
ников лаборатории контроля и проверки качества, склада, занятых на
погрузочно-разгрузочных работах, уборке и т. д.)
Стоимость вспомогательной рабочей силы: 20—40% от фонда заработной
платы
Оплата труда работников материально-технического снабжения: 20—30%
от оплаты труда производственных рабочих
Другие накладные расходы: 50—70% от оплаты труда производственных
рабочих (телефон, внешние службы, арендная плата, оплата инструмен-
та, командировочные расходы, перевозка грузов, различные сборы, юри-
дическая служба, медицинское обслуживание, профессиональные служ-
бы, плата за сброс сточных вод и т. д.)
Техническое обслуживание: 4—8% от капитальных затрат (суммарная
заработная плата соответствующей категории рабочих, инженерного
персонала и служащих составляет 50% от этой суммы)
Общие коэффициенты затрат
Содержание служб сбыта, администрации, рас-
ходы на исследовательские работы и техни-
ческие усовершенствования
Резерв для уплаты налогов
Расходы на пусковые и наладочные работы
10—15% от общего объ-
ема товарооборота
50% от прибыли
5—10% от капитальных
Оборотные средства
затрат
20—30% от общего объ-
ема товарооборота
а Воспроизведено с разрешения из работы: Bartholomew W. Н.. Heisman Н. В. Eco-
nomics of Fermentation Processes, in Microbial Technology, Peppier H. J„ Perlman D.
(eds.), 2d ed., vol. 2, p. 486, Academic Press, Inc., New York, N. Y„ 1979.
веденным в табл. 12.4, определить издержки производства для
конкретного процесса, нужно знать соответствующие этому
процессу капитальные затраты и затраты на оплату труда про-
изводственных рабочих.
Определив объем капитальных затрат, расходы на оплату
труда производственных рабочих, издержки производства,
затраты на материалы, водо-, газо-, тепло- и энергоснабжение,
можно оценить ожидаемую прибыль на инвестированный капи-
тал, т. е. выполнить анализ рентабельности. В следующем раз-
деле мы детально рассмотрим пример полного экономического
анализа конкретного процесса. Последующие разделы посвя-
щены изучению специфических экономических проблем, типич-
23*
356
Глава 12
ных для процессов производства биопродуктов основных четы-
рех классов: химических продуктов тонкой биотехнологии, хи-
мических продуктов массового производства, белка одноклеточ-
ных организмов и метана (как продукта микробиологических
процессов).
12.4. Экономический анализ биопроцесса
В качестве примера рассмотрим среднемасштабный микро-
биологический процесс производства гипотетического вещества
(антимикробного агента, предназначенного для использования
в сельском хозяйстве) [1]. Предварительный анализ потенци-
ального объема производства показал:
1. Согласно предварительному изучению рынка, продажная
цена 2,5 долл, за килограмм продукта позволит завоевать
15—25% рынков сбыта.
2. Если допустить, что подлежащие обработке этим препара-
том культуры возделываются на площади 4,5—7,5 млн. гек-
таров, то потенциальная потребность рынка в ^продукте со-
ставит 13—22 млн. кг/год.
Теперь можно оценить потенциальный масштаб процесса:
Примем, что большой, но удобный для работы ферментер
имеет общий объем 225 м3. Объем жидкой фазы в ферментере
(рабочий объем) должен быть равен 180 м3. Если ферментер
будет непрерывно работать в течение 97% всего времени
(включая профилактические и ремонтные работы), тогда, со-
гласно результатам лабораторных исследований, за один цикл
в однохМ ферментере можно получить 20 000 кг продукта (при
стандартизированной активности в единице массы).
При объеме производства 17 млн. кг в год и производитель-
ности за один рабочий цикл 20 тыс. кг (по данным лаборатор-
ных исследований) потребуется (17-106/20-103) =850 рабочих
циклов в год. Если продолжительность одного рабочего цикла
составляет 40 ч (1,67 сут, включая 4 ч на профилактические
работы и ремонт), то в одном ферментере можно будет осу-
ществить 365/(5/3) =216 рабочих циклов в год. Следовательно,
для заданного объема 'Производства потребуется 850/216«4
ферментера объемом 225 м3.
Определив таким образом базисные 'показатели проектируе-
мого процесса, можно перейти к оценке необходимого количе-
ства материалов и их стоимости. Для этой цели можно вос-
пользоваться масштабированием результатов лабораторных
испытаний, а также анализом полученных ранее или опубли-
кованных данных, характеризующих исходные материалы и
Экономика процессов биохимической технологии
357
составные части рецептуры. Полученные результаты приведены
в табл. 12.5. Обратите внимание на то, что указанные в таб-
лице расходы определены путем простых стехиометрических
расчетов на основе лабораторных данных о выходе продукта
из конкретного количества питательных веществ и об оконча-
тельном составе продукта [активное начало (продукт микроби-
ологического процесса), разбавитель, стабилизатор, связующее
вещество, агент, предохраняющий от слеживания и т. д.]; дру-
гими словами, эти расходы не зависят от характера операций,
выбранных для осуществления биореакции и выделения про-
дукта.
В табл. 12.5 перечислено основное оборудование и матери-
алы, которые потребуются для реализации процесса в 4 фер-
ментерах при установленной производительности 17 млн. кг
продукта в год. С точки зрения экономики наибольший интерес
представляют три этапа разработки проекта этого процесса:
1. Определение конкретной последовательности технологичес-
ких операций и на этой основе необходимого масштаба все-
го предприятия. Последовательность операций и данные,
приведенные в табл. 12.3, позволяют рассчитать капиталь-
ные затраты на оборудование. Далее по литературным дан-
ным и опыту эксплуатации близких производств оценивают
людские ресурсы, необходимые для обслуживания основного
оборудования, что позволяет рассчитать стоимость труда
производственных рабочих.
2. По операционной карте процесса, на которой указаны точ-
ная спецификация оборудования и число единиц оборудова-
ния каждого типа, определяют потребности каждой опера-
ции в паре, технологической воде, воде для промывки и ох-
лаждения, стерилизованном воздухе, электроэнергии, при-
родном газе или другом топливе. Суммируя эти данные по
всем операциям, рассчитывают расходы на пар, воду, топли-
во и электроэнергию для всего процесса. (На основе этой
же подробной операционной карты составляют детальную
схему потоков исходных веществ, продукта, побочных про-
дуктов и отходов; в окончательном варианте на схеме
должны быть указаны расходы, связанные с переработкой
и (или) ликвидацией всех отходов процесса.)
3. На основе рассчитанных описанным путем стоимости труда
производственных рабочих, общих капитальных затрат и
других данных (стоимости оборудования цеха ферментации,
систем водо-, тепло- и энергоснабжения, материалов, расхо-
дов, связанных с хранением продукта, затрат на проектные
и строительные работы, резерва на непредвиденные расхо-
ды), а также с учетом данных, приведенных в табл. 12.4,
находят общие издержки производства.
358
Глава 12
Таблица 12.5. Потребность в материалах и их стоимость®
Исходные данные Объем ферментера Рабочий объем ферментера Число ферментеров Непрерывная работа ферментера Продолжительность рабочего цикла (вклю- чая 4 ч на профилактику и ремонт) Производительность за 1 рабочий цикл Число рабочих циклов в год Производительность предприятия за год 225 м3 180 м3 4 97% 40 ч 20 тыс. кг (в расчете на стандартную актив- ность продукта) 850 17 млн. кг
Потребность на Материалы 1 рабочий цикл, кг Цена долл. за 1 кг, США Стоимость тыс. долл.
Ферментация Меласса 11475 Вспомогательные мате- 3 668 0,07 0,30 683 935
риалы Различные соли и т. п. 922 0,70 (в среднем) 549
Другие ингредиенты компо- зиции Разбавитель 9100 Стабилизаторы Ю0 Связующее 400 Агент, предохраняющий 400 от слеживания 0,30 2,00 0,50 0,24 Итого 2167 2 320 170 170 82
Итого 2742
Упаковка 25 кг/’мешок 0,40 (1 мешок) 272
Всего 5181
а Воспроизведено с разрешения из работы: Bartholomew IF. II., Reisman. Н. В., Eco-
nomics oi Fermentation Processes, in Microbial Technology, Peppier H. J., Perlman D.
(eds.), 2d ed., vol. 2, p. 463. Academic Press, Inc., New York, N. Y., 1979.
Имея в своем распоряжении данные о стоимости труда про-
изводственных рабочих, оборудования, материалов, систем во-
до-, тепло- и энергоснабжения, а также об общих издержках
производства, на следующем этапе проводят анализ рентабель-
ности для рассматриваемого основного (базисного) варианта
и ряда близких вариантов, с помощью которых определяют
чувствительность результата расчетов рентабельности к изме-
нениям основных переменных (продажной цены продукта,
объема рынка сбыта, капитальных затрат, объема производ-
ства).
Теперь рассмотрим пример операционной карты конкретно-
го процесса и рассчитаем необходимые капитальные затраты,
Экономика процессов биохимической технологии 359
Лабораторный
посевной
материал
Первичный
посевной
материал
Приготовление
среды
Технологическая вода
Технологическая вода
Электроэнергия
•Пар
Охлаждающая вода
Стерильный воздух
Другие
составляющие
рецептуры
3 резервуара
для приготовле-
ния посевного
материала
Пар Технологичес- Охлаждающая
кал вода вода
Отходящие газы
Технологическая вода
Электроэнергия
Пар
Охлаждающая вода
Стерильный воздух
Испаритель
(с регенерацией
воды)
Сборник
для культуры
Пар ---*-
Вода для____
промывки
Водяное -*
охлаждение
2 сборника
для концентра-
та продукта
Сборник для
промывных вод
Отходы ______ {
Воздух
Пар ______» Распылительная
Газ -----* сушилка
Электроэнергия—-у _____________
Вода, для праМГйвки
Бункер
Смеситель
Расфасовка
2 бункера
Упаковка
и отправка
РИС. 12.2. Пример операционной карты процесса. [Воспроизведено с разре-
шения из работы: Bartholomew W. Н., Reisman Н. В., Economics of Fermenta-
tion Processes, in Microbial Technology, Peppier H. J., Perlman D. (eds.), 2d
ed., vol. 2, p. 488, Academic Press, Inc., New York, N.Y., 1979.]
стоимость труда производственных рабочих, систем водо-,
тепло- и энергоснабжения, а также заводскую себестоимость и
прибыль на инвестированный капитал.
Операционная карта процесса (рис. 12.2) в рассматривае-
мом примере представляет собой, по сути дела, всего лишь
соответствующим образом детализированный вариант общей
схемы процесса (рис. 12.1). В операционной карте указывают
последовательность операций, число идентичных аппаратов и
необходимость подачи воды, тепла (пара), газа и электроэнер-
гии для каждой операции.
360
Глава 12
Скорость переработки исходных материалов тесно связана
со скоростью потребления пара, воздуха, воды, образования
отходящих газов (СО2, Н2О) и твердых отходов (биомассы),
поэтому потребности в указанных вспомогательных средствах
и в мощностях для ликвидации отходов можно достаточно точ-
но отразить на детальной операционной карте расходования
материалов (рис. 12.3). На этой карте для каждой отдельной
операции указывают состав и объем всех потоков, в том числе
исходных материалов и компонентов окончательной компози-
ции, воды, пара, воздуха, газа, продукта и отходов. Для оп-
ределения потребности в электроэнергии суммируют расход
энергии каждым аппаратом; такие данные обычно сообщает
компания, изготовляющая эти аппараты.
В табл. 12.6 представлены результаты определения капи-
тальных затрат на процесс, схематично изображенный на
рис. 12.3. Затраты на контрольно-измерительную аппаратуру,
трубопроводы, здания и сооружения для цехов ферментации
и выделения продукта оценивают как определенную долю от
капитальных затрат на основное оборудование. В конечном
итоге все затраты на оборудование выражают с учетом расхо-
дов на монтаж и установку. Для процесса, изображенного на
рис. 12.3, общую стоимость основного оборудования можно
представить в виде соответствующей доли от суммарного ос-
новного капитала: цех ферментации (61%), цех выделения
продукта и приготовления композиции (26%), системы водо-,
газо-, тепло- и энергоснабжения, помещения для хранения ма-
териалов (13%). Приблизительные расходы на проектирование
и строительные работы, а также непредвиденные расходы в
каждом случае приняты равными 20% от основного капитала
(который включает стоимость основного оборудования, конт-
рольно-измерительной и регулирующей аппаратуры, технологи-
ческих трубопроводов, зданий и сооружений).
Для определения общего расхода воды, тепла, газа и элек-
троэнергии, необходимых для осуществления планируемого
процесса, суммируют соответствующие расходы на отдельные
операции, указанные на рис. 12.3; результаты такого расчета
приведены в табл. 12.7. Цех выделения продукта и составления
рецептуры потребляет 84% пара, 73% электроэнергии и 100%
природного газа, а в цехе ферментации расходуется 100%
технологической воды и 84% охлаждающей воды. Очевидно,
что наиболее вероятным источником экономии электроэнергии
на протяжении всего процесса в первую очередь может быть
цех выделения продукта. Поскольку в этом цехе почти все
энергетические затраты связаны с удалением воды (концент-
рирование, упаривание), то становится понятным стремление
к разработке новых штаммов микроорганизмов, позволяющих
РИС. 12.3. Карта потоков материалов в биопроцессе. [Воспроизведено с раз-
решения из работы: Bartholomew W. Н., Heisman Н. В., Economics of Fermen-
tation Processes, in Microbial Technology, Peppier H. J., Perlman D. (eds.), 2d.
ed., vol. 2, p. 489, Academic Press, Inc., New York, N. Y., 1979.)
362
Глава 12
Таблица 12.6 Расчет капиталовложений (в тысячах долларов)3
Оборудование цеха фер- Число еди- меитации ииц. обору- дования Основные ха- рактеристики Стоимость единицы оборудовании Общая стои- мость
Резервуары для состав- 2 38 м3 15 30
ления рецептуры Смеситель 2 3,75 кВт 4 8
Непрерывный стерилиза- 1 2,25 м3/мин 100 100
Компрессоры 2 85 м3/мин 225 450
Система пеногашения 1 100 100
Резервуары для хране- 3 15 м3 35 105
ния посевного мате-
риала Смеситель 3 19 кВт 14 42
Ферментеры 4 225 м3 165 660
Смеситель 4 150 кВт 70 280
Сборники 2 190 м3 40 80
Насосы 150
Сепараторы Специальное оборудова- 100
ние
Общая стоимость основного оборудования 2105
Стоимость основного оборудования с учетом монтажных 5263
работ (250% от стоимости основного оборудования)
Контрольно-измерительная и регулирующая аппаратура 526
(10% от стоимости основного оборудования)
Технологические трубопроводы (20% от стоимости основного 1053
оборудования) Здания и сооружения (40% от стоимости основного оборудо- 2105
вания)
Общие капиталовложения в цех ферментации 8947
Оборудова- Число еди- Основные ха- Стоимость^ Общая стой- Стоимость установлен-
иие цеха ииц оборудо- рактерпстики единицы обо- мость ного обору-
обработки вания рудованпя дования
Испаритель 1 18 000 кг/ч 400 400 600
Сборники 3 75 м3 24 72 180
Бункеры 3 75 м3 24 72 180
Смеситель 1 15 м3 75
Виброгрохот 1 4 м2, 50
3 слоя
Общая стоимость основного оборудования 1085
Контрольно-измерительная и регулирующая аппаратура 109
(10% от стоимости основного оборудования)
Технологические трубопроводы (20% от стоимости основно- 217
го оборудования)
Здания и внутренние сооружения (75% от стоимости основ- 814
ного оборудования)
Итого 2225
Экономика процессов биохимической технологии
363
'Продолжение табл. 12.6
Распылитель- 1 4500 кг/ч 1070 1070 1600
ная сушилка
Общие капиталовложения в цех обработки 382э
Системы водо-, газо-, паро- и энергоснабжения после мон- 1590
тажа (20% от стоимости основного оборудования)
Резервуары для хранения меласы, 2X2800 м3 270
Итого 1860
Общая сумма капиталовложений 14 632
Инженерные и строительные работы (20% от основного 2926
капитала)
Непредвиденные расходы: в среднем 20% от основного ка- 2926
питала (их величина может существенно меняться в зави-
симости от конкретного процесса)
Итого капиталовложения (без стоимости земельного участка) 20 484
Примечание: относительная стоимость контрольно-измерительной и регули-
рующей аппаратуры, трубопроводов, зданий и сооружений, систем водо-,
газо-, паро- и энергоснабжения, а также расходов на инженерные и строи-
тельные работы дана в соответствии с данными Чилтона (1949 г.) [Engi-
neering and contingency: Chilton (1949)].
a Воспроизведено с разрешения из работы: Bartholomew W. H., Reisman H. B., Eco-
nomics of Fermentation Processes, in Microbial Technology, Peppier H. J„ Perlman D.
(eds.), 2d ed., vol. 2, p. 463, Academic Press, Inc., New York, N. Y., 1979.
повысить концентрацию продукта в бульоне. О влиянии подоб-
ных факторов на экономику- процесса в целом мы уже упоми-
нали bi предыдущей главе. Обращают на себя внимание боль-
шие непредвиденные расходы (33%). В табл. 12.7 суммиро-
ваны потребности в воде, паре, газе и электроэнергии на каж-
дую операцию, а стоимость установленных систем снабжения
соответствующей производительности указана в нижней части
таблицы. Обратите внимание также на то, что принятая в ко-
нечном итоге производительность таких систем превосходит
предварительные оценки (включая непредвиденные расходы)
на 14% (для охлаждающей воды) и даже примерно на 50%
(для пара и электроэнергии). Причина этого заключается в
необходимости обеспечить достаточную гибкость процесса и
(или) в номинальной производительности имеющихся в про-
даже систем энерго- и пароснабжения.
Согласно расчетам, общие капиталовложения в предприя-
тие с общим объемом ферментеров 4x230 = 920 м3 (9,2-105 л)
составляют 20 484 тыс. долл, (без стоимости земельного участ-
ка). Если учесть затраты на системы водо-, тепло-, газо- и
энергоснабжения, составляющие 1550 тыс. долл., то общие рас-
ходы, отнесенные к 1 л емкости установленных ферментеров^
364
Глава 12
составят 24 долл. Для сравнения можно указать, что, согласно
опубликованным данным, затраты на 1 л емкости установлен-
ных ферментеров при приблизительно такой же производи-
тельности предприятия в ценах 1977 г. составляли от 20 до
50 долл. Если принять, что ежегодный средний рост цен на-
чиная с 1977 г. равен 10%, то в ценах 1986 г. расходы на 1 л
Таблица 12.7. Затраты на водо-, паро-, газо- и энергоснабжение3
Операция или аппарат Пар, кг Технологи- Охлаждаю- Электро- энергия, кВт-ч Газ, м3
ческая во- да, м3 щая вода, м3
Приготовление посевного 7 000 (на материала 1 рабочий цикл) 9 300 300
Ферментер 25 000 Концентрирование 53 000 Сушка 120 000 Компрессор — Смеситель — 147 6 000 1 200 5 700 650 5 400 9 250 900 1 800
Итого 205 000 156 7 500 22 200 1800
Другие цели и непредви- 68 000 денные расходы (33%) 52 2 500 7 325 600
Всего на 1 рабочий цикл 273 000 208 10 000 29 525 2 400
Всего за сутки 636 000 С учетом производительно- 1000 000 сти соответствующих си- стем 484 23 300 27 000 68 800 100 000 5 600
Затраты (оценочные) тыс. 600 долл. 100 450 373 25
а Воспроизведено с разрешения из работы: Bartholomew W. Н., Reisman. Н. В.,
Economics of Fermentation Processes, in Microbial Technology, Peppier H. J.. Perlman D.
(eds.), 2d ed., vol. 2, p. 463, Academic Press, Inc., New York, N. Y., 1979.
емкости установленных ферментеров будут на 135% выше,
т. е. 47—118 долларов.
Расходы на оплату рабочих, занятых в основном производ-
стве, определяют по опыту работы аналогичных предприятий
(табл. 12.8). Для конкретного процесса, схема которого приве-
дена на рис. 12.3 и 12.4, расчет заводской себестоимости про-
дукции представлен в табл. 12.9. Себестоимость продукции
складывается из стоимости сырья и материалов (37,0%), зара-
ботной платы (15%), снабженческих расходов и издержек
(5,3%), расходов на водо-, газо-, паро- и энергоснабжение
(22,3%) и других расходов (20,3%); таким образом находят,
что общая себестоимость продукции равна 0,82 долл, за 1 кг.
а ожидаемая продажная цена (для покупателя) составляет
2,5 долл./кг.
Экономика процессов биохимической технологии Зоб
Для завершения экономического анализа планируемого про-
цесса необходимо выполнить простой расчет прибыли на инвес-
тированный капитал (или любого другого эквивалентного эко-
номического показателя). Приведенная в табл. 12.10 схема
расчета (включающего 14 строк) показывает, что частное от
деления средней чистой прибыли за 10 лет после выплаты на-
логов (среднее из величин, приведенных в строках 10 и 11) на
сумму оборотного капитала (строка 12) и 'первоначального
Таблица 12.8. Оплата труда производственных рабочих3
Аппарат или операция
Число рабочих в смену
Ферментеры
Испаритель
Сушилка
Смеситель
Упаковка и отправка
Всего в 1 смену
Всего за 4 смены6
Заработная плата 1 оператора
Оплата труда производственных
рабочих, всего
3
1
1
1
2______
8
32
16 000 долл, в год
512 000 долл.
а Воспроизведено с разрешения из работы: Bartholomew W. H., Reisman. Н. В., Eco-
nomics of Fermentation Processes, in Microbial Technology, Peppier H. J., Perlman D.
(eds.), 2d ed., vol. 2, p. 463, Academic Press, Inc.. New York, N. Y., 1979.
б Продолжительность рабочей недели каждого оператора (согласно принятым в
1978 г. правилам) составляет 42 ч (168 ч: 4=42 ч).
инвестированного основного капитала (строка 13) дает при-
быль на инвестированный капитал в размере 21,2%. Далее
оценивают чувствительность расчетной величины прибыли на
инвестированный капитал к 20%-ным флуктуациям принятой
в расчетах продажной цены на продукцию (понижение), капи-
тальных затрат (перерасход; повышение), объема производст-
ва (снижение). Если определенную величину прибыли на ин-
вестированный капитал рассматривать как границу, разделяю-
щую допустимый и экономически неоправданный варианты, то
становится понятной важность результатов анализа прибыли
на инвестированный капитал по базовому варианту (табл. 12.10,
графа А) и другим вариантам, соответствующим изучению
чувствительности (графы Б—Д).
Конечным результатом всех типовых расчетов являются 8
простых таблиц и 2 схемы, которых достаточно для разработки
окончательного проекта (третий этап, табл. 12.1). После это-
го все результаты расчетов обычно направляют руководству
366
Глава 12
Таблица 12.9. Определение заводской себестоимости продукта3
Цели затрат Затраты, тыс. долл.
Исходные материалы для ферментации обработки продукта упаковки 2167 2742 272
Итого 5181
Оплата труда Производственные рабочие Вспомогательные рабочие (40% от оплаты труда производственных рабочих) Служащие и инженерно-технический персонал (85% от оплаты труда производственных рабочих) Сопутствующие расходы заработной платы (30% от фонда заработной платы) Заработная плата рабочих и служащих, занятых в сфере технического обслуживания (3% от капи- тальных затрат) 512 205 435 346 615
Итого Материально-техническое обеспечение и издержки Техническое обслуживание (3% от капитальных за- трат) Снабжение (25% от оплаты труда производственных рабочих) 2113 615 128
Итого Водо-, паро-, газо- и энергоснабжение Пар: 8,5 долл, за 100 кг (Х273 Х850) Электроэнергия: 0,035 долл, за кВт-ч (Х30000Х850) Вода: 0,008 долл, за 1 м3 (Х10000Х850) Газ: 0,09 долл, за 1 м3 (Х2400 Х850) 743 1942 893 68 184
Итого 3117
Другие затраты Амортизационные отчисления (12 лет) Налоги и страхование (4% от капитальных затрат) Другие заводские накладные расходы (60% от оп- латы труда производственных рабочих) 1708 820 307
Итого 2835
Всего 13 989 или 0,82 долл, на 1 кг продукции
а Воспроизведено с разрешения из работы: Bartholomew IT. //., Reisman Н. В.,
Economics of Fermentation Processes, in Microbial Technology, Peppier H. J., Perlman D
(eds.), 2d ed., vol. 2, p. 463, Academic Press, Inc., New York, N. Y., 1979.
Экономика процессов биохимической технологии
367
компании для окончательного экономического анализа (четвер-
тый этап, табл. 12.1).
Здесь целесообразно сделать небольшое отступление, каса-
ющееся взаимосвязи между организационной деятельностью и
проектированием процессов. Основные принципы планирования
организационной деятельности всей компании (и вообще любо-
РИС. 12.4. Поток информации в модели развития промышленных предприя-
тий. [Воспроизведено с разрешения из статьи: Rudd D. F., Modelling the Deve-
lopment of the Intermediate Chemical Industry; Chem. Eng. J., 9, 11 (1975).]
го планирования на ближайший период или более длительные
сроки) иллюстрируются представленной на рис. 12.4 схемой.
Рассмотренные в этой главе материалы по анализу экономики
и технологии проектируемого процесса являются лишь частью
той информации, которую должно рассмотреть и учесть руко-
водство компании. Следует обратить внимание на две цепи" об-
ратной связи в схеме планирования организационной деятель-
ности компании (рис. 12.4). В первой (короткой) цепи
доминируют количественные расчеты и анализы типа представ-
ленных в табл. 12.4—12.10 и на рис. 12.1—12.3. Во второй це-
пи главную роль играют проблемы стратегического и такти-
ческого характера, упоминавшиеся в начале этой главы. В час-
Таблица 12.10. Расчет прибыли на инвестированный капитал (при непрерывной эксплуатации предприятия в течение
**> 10 лет)а
Сумма, тыс. долл.
А Базовый вари- Основные статьи расходов н доходов ант (объем про- изводства 17 млн. кг в год, цена 2,5 долл, за 1 кг) Б Снижение про- дажной цены на 20% до 2 долл, за 1 кг в Снижение объ- ема производ- ства на 20% до 13,6 млн. кг в год г Повышение ка- питальных за- трат на 20% Д Планируемый объем произ- водства 13,6 млн. кг в год
1. Продажа продукции фермерам 42 500 34 000 34 000 42 500 34 000
2. Скидка посредникам, доставка, распре- (12 750) деление (30%) (10 200) (10 200) (12 750) (10 200)
3. Общий доход от продажи продук- 29 750 цип 23 800 23 800 29 750 23 800
4. Себестоимость проданной продукции (13 989) (13 989) (12 210)6 (14743)6 (11 818)6
5. Валовая прибыль 15761 9811 11 590 15 007 11982
6. Администрация, служба сбыта, иссле- (3 719) довательские работы, технические усо- вершенствования (12,5% от общего до- хода) (2 975) (2 975) (3 719) (2 975)
7. Доход от эксплуатации предприятия 12042 (в течение второго — десятого годов эксплуатации) 6 836 8 615 11 288 9 007
24-746
8. Расходы на пробный пуск и наладку в первый год эксплуатации (10% от капитальных затрат) (2 050) (2 050) (2 050) (2 460) (1770)
9. Доход от эксплуатации в первый год 9992 4 786 6 565 8 828 7 237
10. Чистый доход после уплаты налогов (50%) в первый год 4 996 2 393 3 283 4414 3 619
11. Чистый доход после уплаты налогов во второй — десятый годы эксплуата- ции 6 021 3418 4 308 5 644 4 504
12. Оборотный капитал (25% от общего дохода) 7 438 5950 5 950 7 438 5950
13. Начальный инвестированный капитал 20 500 20 500 20 500 24 600 17 000в
14. Прибыль на инвестированный капитал 21,2 12,5 15,9 17,2 18,7
(°/о)г
а Воспроизведено с разрешения из работы: Bartholomew W. Н., Heisman Н. В., Economics of Fermentation Processes, in Microbial Techno-
logy, Peppier H. J., Perlman D. (eds.), 2d ed., vol. 2, p. 463, Academic Press, Inc., New York, N. Y., 1979.
$ Стоимость материалов, систем водо-, газо-, паро- и энергоснабжения учтена с коэффициентом 0,8. Потребность в производственных ра-
бочих сокращена с 32 до 30 человек (т. с. все величины, связанные с трудом производственных рабочих, учтены с коэффициентом 0,94). За-
траты, связанные с объемом капиталовложений, рассчитаны с помощью обычных коэффициентов.
в Коэффициент для перерасчета капитальных затрат найден но формуле (17/13,6)0,67. В свою очередь коэффициент 0,67 является сред-
ней величиной между 0,75 (коэффициентом масштабирования для процессов ферментации) и 0,6 (часто применяемым прн масштабировании про-
цессов переработки).
г . (строка 10+9Хстрока 111/10
Прибыль на инвестированный капитал^ -—I-------------1-----------
строка 13+строка 12
-370
Глава 12
тности, здесь выясняется, возможно ли расширение (или
сужение) сферы применения планируемого процесса, имеет ли
компания опыт эксплуатации аналогичных процессов и соот-
ветствующие выходы на рынок, целесообразно ли производить
продукт на предприятиях компании или выгоднее продать ли-
цензию другой компании, насколько надежны источники ис-
ходных материалов и энергии, какова стоимость строительных
работ. Несмотря на то что на многие из этих вопросов нельзя
дать вполне определенные ответы, руководство компании
должно принять конкретное адекватное решение относительно
дальнейшей судьбы планируемого процесса и строительства
предприятия с учетом всех существующих и прогнозируемых
условий.
Располагая примером полного экономического анализа про-
цесса, можно перейти к рассмотрению конкретных примеров
существующих (эксплуатируемых), а также разрабатываемых
или перспективных промышленных процессов.
12.5. Химические продукты
тонких биотехнологических процессов
К химическим продуктам тонкой биотехнологии относят до-
вольно обширную группу ценных продуктов биохимической
технологии (витаминов, гормонов, ферментов, антибиотиков,
моноклональных антител и т. д.), а также ряд еще более цен-
ных продуктов последующей биотрансформации некоторых из
этих веществ (антибиотиков;, стероидов и т. д.). Основные ха-
рактеристики тонких биотехнологических процессов могут из-
меняться в очень широких пределах в зависимости от конкрет-
ного продукта. Некоторые продукты содержатся в культураль-
ной жидкости в следовых количествах, тогда как концентрация
пенициллина в готовом бульоне может превышать 50 г/л. Если
продукт предназначен для медицинских целей или в качестве
добавки к пищевым продуктам, то одной из основных задач (ча-
сто связанной с большими затратами) является его очистка.
Примером может служить отделение пирогенных веществ от
соединений, продуцируемых Е. coli, или тщательная очистка,
необходимая при получении высокоочищенных кристаллических
веществ, в том числе пенициллина.
12.5.1. Ферменты
Как мы уже указывали в гл. 4, промышленность выпускает
множество ферментов для пищевых, медицинских, диагности-
ческих и фармацевтических целей. В 1977—1978 годах промыш-
Экономика процессов биохимической технологии
371
Таблица 12.11. Мировое производство ферментов в 1977—1978 годах3
Фермент
Производство за год, Относительная про-
тонн чистого фер- дажная стоимость,
мента %
Протеаза из Bacillus 500 40
Глюкоамилаза 300 14
Амилаза из Bacillus 300 12
Глюкозоизомер аза 50 12
Бактериальный реинин 10 7
Грибковая амилаза 10 3
Пектиназы 10 10
Грибковая протеаза 10 1
Другие ферменты 1
а Воспроизведено с разрешения издательства из работы: Aunstrup К.., Andre-
sen О., Falch Е. A., Nielsen Т. К., Production of Microbial Enzymes, in Microbial
Technology, Peppier H. J., Perlman D., (eds.), 2d ed., vol. 2, p. 283, Academic Press,
Inc., New’York, N. Y., 1979.
ленность США производила ферменты на общую сумму 50—
55 млн. долл. В табл. 12.11 отражены результаты одной из по-
следних оценок объема продажи ферментов на мировом рынке.
Обратите внимание на то, что в 1977—1978 годах 88% суммар-
ной продажной стоимости ферментов приходилось на бактери-
альные протеазы’ (применяющиеся в качестве добавок к мою-
щим средствам), две амилазы, пектиназу и глюкозоизомеразу.
12.5.2. Производство белков
с помощью рекомбинантных ДНК
Технология рекомбинантных ДНК в организмах Е. coli по-
зволила разработать методы промышленного производства ин-
сулина (1979 г.), гормона роста (1981 г.) и лейкоцитарного ин-
терферона (1981 г.) человека в количествах, достаточных для
их широкого применения в медицинской практике. В 1984 г. в
продажу поступили инсулин человека и вакцина против вызы-
ваемого Е. coll поноса у новорожденных поросят; технология
получения многих других продуктов находится на различных
стадиях разработки и промышленного внедрения. Экономичес-
кая оценка этих разнообразных продуктов должна учитывать:
1. Стоимость исследовательских и проектных работ, предше-
ствовавших внедрению промышленного процесса. Получение
веществ с помощью рекомбинантных ДНК является принци-
пиально новым методом биохимической технологии, и стои-
мость таких продуктов должна окупать значительные рас-
ходы на предшествующие промышленному производству ис-
следования, в том числе на лабораторное оборудование, со-
держание персонала и создание опытных установок.
24
2>П
Глава 12
2. Стоимость обычных операций биореакции и выделения про-
дукта в крупномасштабном производственном процессе.
3. Расходы на клинические и другие испытания, требуемые
Управлением по контролю за качеством пищевых продуктов,
медикаментов и косметических средств, Министерством сель-
ского хозяйства США и другими правительственными орга-
низациями.
Биореакция при использовании рекомбинантных ДНК
представляет собой обычный микробиологический процесс, хо-
тя здесь может возникнуть ряд специфических проблем, свя-
занных с предотвращением заражения культуры и стабиль-
ностью рекомбинантных плазмид (гл. 6). В основу технологии
рекомбинантных ДНК положен принцип вектор—организм-хо-
зяин. В настоящее время в промышленном производстве при-
меняется (или считается перспективным) очень ограниченное
число организмов-хозяев (Е. coli, Saccharomyces cerevisiae,
несколько линий животных клеток, возможно также Bacillus
subtilis и некоторые штаммы Pseudomonas). Конструкцию при-
меняемого биореактора и схему процесса выделения определя-
ет в основном природа организма-хозяина. Для бактерии Е. co-
li, например, характерна простая генетика, отсутствие проблем
при идентификации и высокая скорость роста. В то же время
в тех случаях, когда организмом-хозяином является Е. coli,
при проведении биореакций и операций выделения возникают
следующие затруднения:
1. Е. coli содержит по меньшей мере восемь растворимых про-
теаз, которые, возможно, принимают участие в гидролизе
экспрессированных белков соматостатина и гормона роста
человека. Частично протеазы можно блокировать в ходе вы-
деления продукта с помощью соответствующего ингибитора
фенилметилсульфонилфторида, эффективного в отношении
сериновых протеаз, но не действующего на металлофермен-
ты. Снижение концентрации ионов железа понижает протео-
литическую активность металлоферментов, но одновременно
индуцирует связывающие металл агенты, которые могут за-
труднять очистку продукта.
2. Содержащиеся в оболочке клетки липополисахариды явля-
ются очень мощными пирогенами, вызывающими лихорадку
даже в дозах 0,5 нг/кг (0,5 трлн-1). В более старом про-
цессе получения ь-аспарагиназы с помощью Е. coli продукт,
как оказалось, был загрязнен такими пирогенами.
Последовательность операций выделения первых промыш-
ленных продуктов, полученных методами технологии рекомби-
нантных ДНК, была приведена на рис. 11.39. Поскольку боль-
Экономика процессов биохимической технологии
373
шие количества экспрессируемых в Е. coll белков накаплива-
ются в виде особых изолированных образований, в которых
белки находятся в частично денатурированном состоянии, то
при их выделении приходится вводить дополнительные опера-
ции солюбилизации и обработки, способствующей созданию на-
тивных вторичной и третичной структур. Последующую очист-
Стоимость,
долл. США
Фармацевтические препараты
{более 10000 долл, за 7г)
Ветеринарные препараты
(более 100 долл, за 1г)
Несколько
генов, другие
Техническая
сложность
Крупномас-
штабные
реакторы
Ферменты организмы-
(1-100 долл. хозяева
за 1 грунт)
Химические препараты
специального назначения
(Т50 долл, за 1 фунт)
Химические продукты
Один ген, широкого потреоления
Е- С°Ч- (5долл, за 1 фунт)
Начало 1980-х годов Середина и коней, 1990-е годы
1980-х годов
РИС. 12.5. Прогнозируемое развитие производства продуктов с помощью тех-
нологии рекомбинантных ДНК. (По данным В. Хустедтера, Genentech, Inc.,
1985. Воспроизведено с разрешения.)
ку производят с помощью хроматографических методов, хоро-
шо разработанных по меньшей мере в лабораторных
масштабах. Новшеством здесь является разделение на имму-
носорбентах; этот метод, вкратце рассмотренный в гл. 11, при-
менялся для очистки интерферона и других белков.
Метод рекомбинантных ДНК оказал большое влияние на
экономику процессов производства многих продуктов!, пред-
назначенных для использования в фармакологии, сельском хо-
зяйстве и т. п. Как показано на рис. 12.5, темпы развития в
каждой отрасли связаны с продажной ценой продукта и тех-
нической сложностью процесса с участием рекомбинантных
ДНК. Примером чрезвычайно ценного фармацевтического пре-
374
Глава 12
парата может служить инсулин, рынок сбыта которого к 1985 г.
(начиная с 1981 г.) может удвоиться, достигнув (в США и стра-
нах ЕЭС) двух третей миллиарда долларов. Компании Eli Lilly,
имеющей большой опыт в производстве и продаже инсулина и
в получении разрешений на производство фармацевтических пре-
паратов, удалось добиться разрешения на выпуск инсулина че-
ловека на базе разработанного компанией Genentech процесса
с участием рекомбинантных ДНК в течение четырех лет. Пре-
имущества инсулина человека перед инсулинами быка и свиньи
пока еще только изучаются.
Технология рекомбинантных ДНК лежит в основе разраба-
тываемых или внедряемых в промышленное производство про-
цессов получения других чрезвычайно ценных продуктов: регу-
ляторных белков (интерферонов, гормонов роста человека и
животных, нейроактивных пептидов и лимфокинов), факторов
крови (альбумина сыворотки человека, антигемофилического
фактора, тромболитического и фибринолитического ферментов),
вакцин (вирусных и бактериальных) и антибиотиков. Следует
подчеркнуть, что с экономической точки зрения будущее техно-
логии рекомбинантных ДНК несколько неопределенно даже
в применении к очень дорогим продуктам; так, в США объем
рынка сбыта вдкцины для животных в стоимостном выражении
обычно составляет 5—10 млн. долл, в год, что может оказать-
ся недостаточным для поддержания компании, поэтому может
возникнуть необходимость в одновременном выпуске несколь-
ких вакцин.
Общее потенциальное влияние технологии рекомбинантных
ДНК, затрагивающей практически все сферы биотехнологии,
очень велико. В начале 80-х годов эта отрасль биотехнологии
развивалась чрезвычайно быстрыми темпами; достаточно пол-
ный обзор состояния технологии рекомбинантных ДНК в на-
стоящее время и перспектив ее развития, а также оценка име-
ющихся и потенциальных рынков сбыта продукции, получаемой
по этой технологии, приведены в докладе Бюро технической
оценки США за 1984 г. — Commercial Biotechnology: An Inter-
national Analysis [2].
12.5.3. Антибиотики
Продукты вторичного метаболизма микроорганизмов, инги-
бирующие рост других микроорганизмов даже в низких кон-
центрациях, называются антибиотиками. Антибиотики применя-
ют в качестве антимикробных агентов (при лечении заболева-
ний челов,ека), противоопухолевых средств, фунгицидов и
пестицидов (для защиты растений), в качестве средств для
лечения и ускорения роста домашних животных, а также в
Экономика процессов биохимической технологии
375
исследовательских работах. Подсчитано, что в 1980 г. мировое
производство антибиотиков составило 105 т, а продано антиби-
отиков на сумму 4 млрд. долл. Основными путями развития
промышленной микробиологии антибиотиков пока еще оста-
ются скрининг, мутации и другие традиционные методы. В на-
стоящее время известно более 6000 антибиотиков,, из которых
почти 100 производятся микробиологическим путем в промыш-
ленном масштабе.
Антибиотики подразделяют на группы в соответствии с
диапазоном антимикробного действия (антибиотики с широким
или узким спектром активности), механизмом проявления био-
логической активности, источником (штаммом-продуцентом),
путями биосинтеза и молекулярной структурой. В табл. 12.12
перечислены основные структурные группы антибиотиков.
Многочисленные взаимосвязи между составом среды, анти-
микробной активностью и прогрессом в, синтетической органи-
ческой химии можно проиллюстрировать на примере промыш-
ленного производства пенициллинов. Все пенициллины (см.,
например, табл. 12.13) являются производными 6-аминопени-
циллановой кислоты (6-АРА), в молекуле которой имеется пя-
тичленный тиазолидиновый цикл, сопряженный с четырехчлен-
ным ^-лактамным к<
(R-CO-).
О
II
R —С
несущим ацильную группу
1 1 н
। fi-лактамх таазолидиновое
। ное । кольцо
* кольцо 1
ацильная
боковая
группа
Природные пенициллины синтезируются микроорганизмами в
соответствующей среде; примером одного из наиболее важных
антибиотиков может служить пенициллин G (табл. 12.13). До-
бавление к среде веществ-предшественников,, из которых обра-
зуется ацильная группа, позволяет изменять природу послед-
ней; примерами получаемых таким путем биосинтетических
пенициллинов могут служить пенициллины V и О. Фермента-
тивным отщеплением ацильной группы с последующим вве-
дением другой группы химическим путем получают полусинте-
тические пенициллины. Последние позволили расширить спектр
антимикробной активности пенициллинов, повысить их устой-
376
Глава 12
Таблица 12.12. Классификация антибиотиков в соответствии
с их химическим строением. Примеры конкретных антибиотиков
приведены в скобках3
1. Антибиотики углеводной природы
Сахара
Аминогликозиды
Ортозомицины
N-Гликозиды
С-Гликозиды
Гликолипиды
2. Макроциклические лактоны
Макролидные антибиотики
Полиеновые антибиотики
Ансамицины
Макротетролиды
3. Хиноны и родственные антибиотики
Тетрациклины
Антрациклины
Нафтохиноны
Бензохиноны
4. Аминокислотные и пептидные анти-
биотики
Производные аминокислот
P-Лактамные антибиотики
Пептидные антибиотики
Хромопептиды
Депсипептиды
Хелатообразующие пептиды
5. Гетероциклические азотсодержащие
антибиотики
Нуклеозидные антибиотики
6. Гетероциклические кислородсодержа-
щие антибиотики
Полиэфирные антибиотики
7. Алициклические антибиотики
Производные циклоалканов
Стероидные антибиотики
8. Ароматические антибиотики
Производные бензола
Производные конденсированных аро-
матических соединений
Ароматические эфиры
9. Алифатические антибиотики
Фосфорсодержащие соединения
(Нойиримицин)
(Стрептомицин)
(Эверниномицин)
(Стрептотрицин)
(Ванкомицин)
(Моеномицин)
(Эритромицин)
(Кандицидин).
(Рифамицин)
(Тетранактин)
(Тетрациклин)
(Адриамицин)
(Актинородин)
(Митомицин)
(Циклосерин)
(Пенициллин)
(Бацитрацин)
(Актиномицин)
(Валиномицин)
(Блеомицин)
(Полиоксины)
(Моненсин)
(Циклогексимид)
(Фузидиевая кислота)
(Хлорамфеникол)
(Гризеофульфин)
(Новобиоцин)
(Фосфомицины)
воспроизведено с разрешения из работы: Crueger W., Crtieger Л.. Bio-
technology: A Textbook of Industrial Microbiology (English edition), p. 198, Sci-
ence Tech., Inc., Madison WI, 1984. [Данные заимствованы из статьи: Berdy J.,
Adv. Appl. Microbiol., 18,'309 (1979).]
чивость к разрушению нативными р-лактамазами и действию
кислот.
Антибиотики цефалоспориновой группы также содержат
р-лактамное кольцо, а кроме того, дигидротиаз!иновый цикл и
три заместителя Rb R2 и R3. К преимуществам цефалоспоринов
Таблица 12.13. Химическое строение и свойства трех типов пенициллинов3
Активность в отношении
Тривиаль- ное назва- ние Всасывание
Группа R Химическое название при перо- ральном ВВС’ денин Staphylo- coccus Staphylococ- cus, продуди- Других рующих пе- кокков Грамотрица- тельных бак- терий Ограничения
ннциллнназу
С6Н6СНг- 6- (Фенилацетамидо) - пенициллин Пени- циллин G Плохое Высокая Отсутствует Высо- Отсутствует кая Переменное всасывание
с6н5—с - С— 1) II N С \о/\сн3 6- (5-Метил-З-фенил- 4-нзоксазолкарбокс- амидо)пенициллин Окса- циллин Хорошее Высокая Высокая Раз- Отсутствует лич- ная Переменное всасывание, уз- кий спектр
CeH6CH(NH2) — 6- [ D(—)-а-Амино- фенилацетамидо] пе- нициллин Ампи- циллин Хорошее Высокая Отсутствует Высо- Умеренная кая Грамотрица- тельные бак- терии могут приобрести устойчивость во время ле- чения
а Воспроизведено из работы: Stewart G. Т., The Penicillin Group of Drugs, Elsevier Publishing Company, Amsterdam, 1965.
378
Глава 12
относятся широкий спектр антимикробной активности, низкая
токсичность и устойчивость к действию нативных ферментов
пенициллиназ.
соон
структура цефалоспоринов
Полусинтетические цефалоспорины получают двумя путями:
во-первых, отщеплением одной из боковых цепей, приводящим
к образованию 7-аминоцефалоспорановой кислоты (7-АСА),
у которой Ri = NH2, Кг^СНзОСО, Rs = H, и последующим аци-
лированием, и, во-вторых, расширением цикла пенициллинов
(в данном случае являющихся предшественниками цефалоспо-
ринов) до 7-АСА с последующей модификацией.
Существуют и антибиотики, получаемые только синтетичес-
ким путем. Так, хлорамфеникол и пирролнитрин впервые были
; NHCOCHClz
\-кн-сн—СН2ОН
• он
п -нитрофенол алюнопроггандоол
структура, хлорамфеникола.
выделены из продуктов жизнедеятельности микроорганизмов,
но в настоящее время производятся только синтетическим пу-
тем. Следует ожидать, что число синтетических антибиотиков
будет все время возрастать; так, исследователи из Пенсиль-
ванского университета недавно закончили полный синтез эф-
ротомицина, молекула которого содержит 21 асимметрический
центр. Следовательно, на экономику производства антибиоти-
ков могут оказывать влияние чисто биологические, смешанные
и чисто химические процессы.
Методы производства антибиотиков в основном были рас-
смотрены в гл. 11, поскольку в соответствующих процессах
часто используются сложные многоступенчатые комбинации
Экономика процессов биохимической технологии 379
различных операций выделения и очистки. В силу генетической
нестабильности штаммов-продуцентов обычно антибиотики по-
лучают в периодических режимах, хотя при получении пени-
циллина успешно без снижения выхода применялся и отъемно-
доливной процесс. В настоящее время концентрация пеницил-
лина в бульоне достигает десятков граммов в литре; соответст-
вующая низкая цена и возможность дальнейших превращений
пенициллинов (в полусинтетические пенициллины, цефалоспо-
рины и т. д.) являются причиной того, что их можно считать
скорее продуктом промежуточного типа, а не химическим про-
дуктом тонкой биотехнологии.
12.5.4. Витамины, алкалоиды, нуклеозиды, стероиды
Микроорганизмы продуцируют не только белки (ферменты)
и антибиотики, но и многие другие сложные метаболиты. Про-
мышленный интерес представляют несколько процессов такого
типа, успешно конкурирующих с соответствующими химичес-
кими процессами полного синтеза или даже значительно пре-
восходящих последние.
Из витаминов, синтезированных микробиологически [вита-
мин В12, рибофлавин (В2), тиамин, фолиевая кислота, пантоте-
новая кислота, пиридоксаль], в промышленности таким путем
получают только BJ2 и В2, причем последний все чаще произво-
дят чисто химическим способом. Витамин Bi2, называемый так-
же цианокобаламином, в природе синтезируется только микро-
организмами, несмотря на то что это соединение необходимо
всем животным. Микробная флора толстой кишки может проду-
цировать В12, но организм человека не ассимилирует его, и поэто-
му человек должен получать Bi2 с пищей. В начале 1980-х го-
дов объем мирового производства BJ2 составлял примерно 10 т,
из которых около двух третей потребляла фармацевтическая
промышленность (в виде цианокобаламина, гидроксокобаламп-
на, кофермента BJ2 и метилкобаламина), а одна треть исполь-
зовалась в качестве добавок к растительным кормам для до-
машних животных, обедненным Bi2.
При производстве витамина Bi2 в периодическом режиме
используются главным образом организмы Propionibacteria
(freudenreichii и shermanii) (табл. 12.14), а весь процесс разде-
ляется на две стадии — анаэробную (2—4 сут) и аэробную (3—
4 сут). Продукт анаэробной стадии (5'-дезоксиаденозилкобин-
амидгуанозинфосфат) конденсируется с продуктом аэробной
стадии (5,6-диметилбензимидазолом), в результате чего в кон-
це концов образуется витамин Bi2. Операции выделения вклю-
чают нагревание (для высвобождения связанного с клетками
витамина Bi2) и химическое превращение в устойчивый циано-
380
Глава 12
Таблица 12.14. Характеристики микробиологических получения витамина В12а процессов
Микроорганизм Основные компоненты среды Выход В12, мг/л Примечания
Bacillus megateriutn Свекольная меласса, фосфат аммония, соли кобальта, не- органические соли 0,45 Аэробный процесс продолжитель- ностью 18 ч
Propionibacterium freudenreichii Кукурузный настой, кобальто- вое производное глюкозы; pH поддерживают около 7,0 до- бавлением NH4OH 19 Аэробная (3 сут) и анаэробная (3 сут) стадии
P. freudenreichii Кукурузный настой (или ав- толизат мицелия пеницилли- на), глюкоза, соли кобальта; pH поддерживают около 7,0 добавлением NH4OH 8 Непрерывный дву- стадийный про- цесс продолжи- тельностью 33 ч
Propionibacterium shermanii Кукурузный настой, глюкоза, соли кобальта; pH поддержи- вают около 7,0 добавлением NH4OH 23 Периодический процесс продолжи- тельностью 7 сут, (анаэробная ста- дия 3 сут, аэроб- ная стадия 4 сут)
Streptomyces olivaceus Глюкоза, соевая мука, раство- римые отходы спиртового про- изводства, кобальтовые соли, неорганические соли 3,3 Продолжитель- ность ферментаций с аэрацией 6 сут
Streptomyces sp. Соевая мука, глюкоза, ко- бальтовая соль, КгНРО4 5,7 Продолжитель- ность ферментаций с аэрацией 6 сут
a Из работы: Microbial Technology, Peppier H. J. (ed Corporation, New York, 1967. .), р. 286, Reinhold Publishing
кобаламин. Разработан и непрерывный двустадийный процесс.
Микроорганизм Pseudomonas denitr if leans может продуциро-
вать Bi2 в одну стадию, если к среде в качестве предшествен-
ников добавлены соли кобальта и 5,6-диметилбензимидазол.
За десять лет работы по отысканию более продуктивных штам-
мов выход витамина Bi2 удалось повысить с 0,6 до 60 мг/л.
В качестве питательного вещества используется меласса сахар-
ной свеклы, являющаяся дешевым источником бетаина. Образу-
ющийся при обработке отходов активный ил также содержит
В12 (4—10 мг/л), но его выделение затруднено из-за одновре-
менного образования множества близких Bi2 веществ.
Фармакологически активные эргоалкалоиды продуцируются
грибами Claviceps. Микробиологический способ позволяет до-
биться концентрации эргоалкалоидов в культуральной жидко-
сти более 5 г/л и этим выгодно отличается от чрезвычайно
Экономика процессов биохимической технологии
381
сложного полного химического синтеза и способа биосинтеза
путем заражения ржи. Обычной проблемой в микробиологиче-
ских способах производства эргоалкалоидов является деграда-
ция штамма. Масштабирование процессов биосинтеза эргоал-
калоидов затруднено из-за высокой потребности в кислороде и
чувствительности к скорости перемешивания.
Нуклеозиды и нуклеотиды применяют в качестве вкусовых
добавок к пищевым продуктам. В этом отношении наиболее эф-
фективны пурин-5'-монофосфаты— гуаниловая кислота
(5'-GMP), инозиновая кислота (5'-1МР) и ксантиловая кисло-
та (5'-ХМР), причем их эффективность возрастает еще боль-
ше в присутствии глутамата натрия (см. органические кислоты,
разд. 12.6.3). В Японии ежегодно производится примерно 3000 т
5'-1МР (66%) и 5'-GMP (34%). Поскольку биосинтез нуклеози-
дов обычно регулируется по принципу обратной связи, то для
их «перепроизводства» микроорганизмами необходимы либо
ауксотрофные мутанты и соответствующие лимитирующие их
рост конечные продукты метаболизма, либо особые аналоги пу-
риновых оснований, обеспечивающие подавление контроля био-
синтеза. В промышленном масштабе нуклеозиды получают, как
указано выше, микробиологическим путем или посредством гид-
ролиза дрожжевой РНК in vitro с помощью микробных фермен-
тов или in vivo с помощью эндогенных рибонуклеаз с последу-
ющей экскрецией мононуклеотидов в среду.
Осуществляемые с помощью микроорганизмов или фермен-
тов биохимические превращения обычно называют биотрансфор-
мацией или биопревращением. Биотрансформации выгодно от-
личаются от небиологических превращений, во-первых, высокой
специфичностью по отношению к субстрату, во-вторых, регио-
специфичностью (селективностью по отношению к определен-
ным группам молекулы), в-третьих, стереоспецифичностью
(в частности, возможностью разделения рацемических смесей;
вспомните процесс с участием иммобилизованной ь-аминоаци-
лазы, рис. 4.12) и, в-четвертых, мягкими условиями превраще-
ний. К недостаткам биотрансформаций (при наличии конкурен-
тоспособных химических методов) относятся необходимость
поддержания популяции жизнеспособных клеток в ходе био-
382
Глава 12
трансформации и высокая стоимость извлечения продукта из
сложной смеси. Биотрансформации стероидов обычно пред-
ставляют собой очень селективное частичное окисление одного
положения стероидного скелета (например, прогестерона). Так,
например, разработка и внедрение метода селективного микро-
биологического 1 la-гидроксилирования прогестерона позволили
в конце концов снизить цену кортизона с 200 долл, за 1 г
(в 1949 г.) до 1 долл, за 1 г (в 1979 г.).
11 а-гидроксилирование
Прогестерон •---------------* кортизол ----> кортизон
В табл. 12.15 приведены состав сред и условия роста культур,
используемые в таких биотрансформациях. Обращает на себя
внимание определенное время добавления вещества-предшест-
венника и большая продолжительность трансформаций. При-
родные стероиды и стеролы, а также их производные применя-
ются в терапии (эстрогены, прогестерон и андрогены), а также
в качестве противозачаточных средств (производные эстрогена
и прогестерона), транквилизаторов, противоопухолевых препара-
тов, ветеринарных препаратов, противовоспалительных средств
при лечении кожных заболеваний и артрита (кортизон), средств
для регуляции натрий-калиевого обмена. В последних исследо-
ваниях, включавших тысячи различных превращений стероидов
и стеролов, применялись иммобилизованные клетки и фермен-
ты, а также сочетания биологических превращений и химиче-
ских реакций.
12.5.5. Моноклональные антитела
Когда чужеродное вещество (антиген) попадает в организм
животного, например мыши, то часто иммунная система узнает
его и вырабатывает специфические антитела, которые селектив-
но связывают антиген и тем самым инактивируют и (или) ме-
тят его, а другие системы организма узнают комплекс, нейтра-
лизуют и выводят из организма. Обычно антитела продуциру-
ются специализированными клетками, называемыми лимфоци-
тами В (в селезенке, крови, лимфе); типичная иммунная реак-
ция приводит к образованию ряда (т. е. смеси) антител, специ-
фичных по отношению к данному антигену. Каждый лимфоцит
В продуцирует только один тип антител. Эти клетки можно вы-
делить (например, из ткани селезенки) и вырастить в культуре
вне организма мыши, чтобы в конечном итоге получить антите-
ла. Такая культура множества клеток имеет два недостатка:
во-первых, клетки продуцируют различные антитела, посколь-
Экономика процессов биохимической технологии
383-
ку культура поликлональна (т. е. получена из различных кле-
ток В селезенки); во-вторых, для клеток характерна тенденция
к дедифференцировке или обратному развитию и к потере спо-
собности продуцировать специфические антитела. Эти две про-
блемы решаются методом слияния клеток (разд. 6.2.3), а имен-
но путем слияния продуктивного лимфоцита с быстро растущей
клеткой миеломы (опухоли), в результате чего образуется гиб-
ридная клетка, или гибридома. Гибридомы могут быстро расти
в культуре, что позволяет клонировать их и получать индиви-
дуальные культуры, развившиеся из одного лимфоцита. В ре-
зультате непрерывного роста культуры таких «бессмертных»
клеток образуется только один тип антител, называемых моно-
клональными антителами.
В 1980-х годах моноклональные антитела быстро нашли
практическое применение в первую очередь благодаря тому об-
стоятельству, что первые препараты поступили в продажу в со-
ставе наборов для диагностики in vitro и оказались намного
удобнее, чем традиционные методы диагностики in vivo. Дру-
гая потенциально важная сфера применения моноклональных
антител охватывает диагностику in vivo, профилактику и лече-
ние заболеваний человека, но для этого моноклональные анти-
тела должны предварительно пройти такую же строгую провер-
ку, как и все лекарственные препараты и другие биологически
активные вещества (разд. 12.2).
Диагностика с применением моноклональных антител имеет
несколько преимуществ. Во-первых, моноклональные антитела
позволяют существенно сократить время диагноза. Действи-
тельно, традиционные методы диагностики, связанные с pocTOiM
культур, отнимают много времени, тогда как наборы с моно-
клональными антителами позволяют быстро произвести такие
же анализы. Моноклональные антитела применяются для диаг-
ностики венерических и других заболеваний человека (гонор-
реи, урогенитальных хламидиоз, вирусного герпеса) и ряда бак-
териальных инфекций, распространенных среди госпитализиро-
ванных больных (в том числе вызванных Pseudomonas aerugi-
nosa и стрептококками группы В). В будущем моноклональные
антитела, возможно, найдут применение и в диагностике рако-
вых заболеваний, если удастся найти специфичный по отноше-
нию к опухоли маркер; примером такого маркера может слу-
жить кислая фосфатаза, выделяемая опухолью предстательной
железы. Во-вторых, моноклональные антитела существенно по-
вышают чувствительность; сообщалось, что обнаружение гепа-
тита В с помощью системы на основе моноклональных антител
в 100 раз чувствительнее, чем определение с помощью поли-
клональных антител. Высокая чувствительность особенно важ-
Таблица 12.18. Условия биотрансфорМации некоторых стероидов и стеролов1
Условия
Субстрат
Продукт
Выход, % Микроорганизм Среда
Прогестерон 1-Дегидротесто- лактон 50 Cylindrocarpon radicicola 3 г (ё расчете на сухие вещества) Ку- 75 курузного настоя, 3 г NH4H2PO4, 2,5 г СаСОз, 2,2 г соевого масла, 0,5 г про- гестерона, дистиллированная вода до 1 л, pH 7,0 ч, 25 °C
Прогестерон Андростадиён-М- дион-3,17 85 Fusarium solatii 15 г пептона, 6 мл кукурузного настоя, 96 50 г глюкозы, дистиллированная вода до 1 л, pH 6,0; 0,25 г прогестерона (добавляют через 48 ч после начйлй Процесса) ч, 25 °C
Прогестерон 15а-Гидрокси- прегнен-4-ди- он-3,20 11 Streptomyces aureus 2,2 г соевого масла, 15 г соевой муки, 72 10 г глюкозы, 2,5 г СаСОз, 0,25 г про- гестерона, дистиллированная вода до 1 л ч, 25 °C
Андростен-4-ди- он-3,17 11а-Гидрокси- андростен-4-ди- он-3,17 25 Rhizopus arrhizus 20 г пептона, 5 Мл кукурузного настоя, 96 50 г глюкозы, водопроводная вода до 1 л, pH 5,5—5,9, 0,25 г андростендио- на (добавляют через 27 ч после начала процесса) ч, 28 °C
Прогестерон 11а-Гидрокси- прогестерон 90 Aspergillus ochraceus Эдамин, глюкоза, кукурузный настой, 72 20 г тонкоизмельченного прогестерона, суспендированного в 0,01 %-ном раство- ре твина 80 (добавляют через 18—24 ч после начала процесса) ч, 28 °C
jn Гидрокортизон
o>
Холестерин
(J-Ситостерин, хо-
лестерин, стигма-
стерин, кампесте-
рол
Преднизолон 93 Arthrobacter simplex 5 г пептона, 5 г кукурузного настоя, 120 ч, 28 ®С; 5 г глюкозы, дистиллированная вода псевдокристалли» до 1 л, pH 7,0; 1—50% тонкоизмель- ческая фермента- ченного гидрокортизона, суспендиро- ция ванного в этаноле (добавляют через 24 ч после начала процесса)
Андростадиен- 1,4-дион-3,17 90 Arthrobacter simplex 10 г кукурузного настоя, 2 г мясного 44 ч, 30 ®С; дО- экстракта, 5 г глюкозы, 0,5 г К2НРО4, бавляют хелато- дистиллированная вода до 1 л, pH 7,0; образующие аген- суспендированный в воде холестерин ты (1 г) добавляют через 20 ч, а 0,8 мМ этанольный раствор а,а'-дипиридила (связывающего ионы железа)—через 26 ч после начала процесса
9a-Г идроксианд- ростен-4-дион-3,17 Нет дан- ных Mycobacterium fortuitum 10 г глицерина, 8,4 г Na2HPO4, 4,5 г 336 ч, 30 °C; при- КН2РО4, 2 г NH4CI, следовые элемен- меняют мутант с ты, дистиллированная вода до 1 л; блокированной добавляют 1 г соевой муки и 10 г си- способностью к тостерина расщеплению сте- роидного скелета
О. K.. Perlman b., Microbial Transformation of Steroids and
p. 483. Academic Press, Inc., New York, N. Y., 1979,
Sterols, in Microbial Techno-
аВоспроизведено с разрешения из работы: Sebek
logy. Peppier Н. J., Perlman D. (eds.), 2d. ed., vol. 1,
386
Глава 12
на при обязательной регулярной проверке банков крови на ге-
патит.
Как уже отмечалось в этой главе, производство принципи-
ально новых продуктов, требующее проведения исследователь-
ских и проектных работ, связано со значительно большими за-
тратами, чем последующее производство аналогичных продук-
Таблица 12.16. Прогнозируемый объем производства моноклональных
антител в США (в миллионах долларов в ценах 1981 г.)а
Объем производства
Область применения
1982 1990
Диагностика
1. Наборы для диагностики in vitro 5—6 300—500 (40)5
2. Иммуногистохимические наборы биопсии, мазков и т. д.) (для 0 25
3. Диагностика in vivo (в первую интроскопия) Терапевтические препараты (включая очередь 0 реаген- до 100в
ты, меченные радиоактивными изотопами и 0 токсинами) Другие Исследовательские работы Небольшой Очистка веществ Небольшой 500—1000г 10 10
а Воспроизведено из работы: Commercial Biotechnology, An International Analysis,
Office of Technology Assessment, Washington, D. C., 20510, 1984.
б Приведена стоимость наборов, а в скобках указана стоимость собственно антител.
Истинная стоимость наборов будет зависеть от процесса их производства.
в За основу взята стоимость набора в 1981 г. Эта величина может изменяться в ту
или иную сторону в зависимости от затрат на получение разрешения на выпуск препа-
рата.
гСтоимость может изменяться в зависимости от процесса производства и затрат на
получение разрешения на выпуск препарата.
тов на базе уже освоенной технологии. Подсчитано, что в 1983 г.
разработка нового типа моноклональных антител (поставляе-
мого компаниям, выпускающим наборы для диагностики in
vitro) продолжалась более 3 лет и обошлась в 3,5 млн. долл.;
затраты на промышленный выпуск наборов были в 5—10 раз
выше.
Помимо диагностики гепатита В, опухоли предстательной,
железы, венерических заболеваний и больничных бактериаль-
ных инфекций, выпускавшиеся в начале 1980-х годов монокло-
нальные антитела предназначались для диагностики беремен-
ности (обнаружения хорионического гонадотропина человека),
а также краснухи, бешенства и других заболеваний.
Экономика процессов биохимической технологии 387
Прогнозируемый рост производства моноклональных анти-
тел для медицинских и ветеринарных целей представлен в табл.
12.16. Еще раз обращаем внимание на то, что моноклональные
антитела применялись для анализов in vitro уже в 1982 г., а в
конце этого десятилетия, как ожидается, моноклональные ан-
титела будут широко применяться и in vivo (для транспорта ле-
карственных препаратов, интроскопии опухолей и других
целей).
12.6. Кислородсодержащие химические продукты
массового производства
Для крупномасштабного производства кислородсодержащих
химических продуктов применяются как анаэробные, так и аэроб-
ные процессы. Сначала рассмотрим состояние экономики
традиционного производства алкогольных напитков, а затем
перейдем к экономическому анализу промышленности простых
кислородсодержащих веществ (спиртов, кетонов, кислот) и ами-
нокислот. В табл. 12.17 перечислены соединения, получаемые
микробиологическим путем, а также соответствующие штаммы
микроорганизмов и субстраты. Приведенные в табл. 12.18 дан-
ные о производстве простых кислородсодержащих веществ в
США, их стоимости и сферах применения свидетельствуют о
наличии колоссального потенциального рынка сбыта для этих
получаемых микробиологическим путем веществ. Самым ста-
рейшим из них является этанол, образующийся при любых тра-
диционных методах производства спиртных напитков; экономи-
ку производства этанола мы обсудим в следующем разделе.
Затем будут рассмотрены проблемы производства этанола в ка-
честве моторного топлива, а также микробиологические процес-
сы производства ацетона, бутанола, органических кислот и ами-
нокислот.
12.6.1. Пивоварение и виноделие
Производство пива может служить хорошей иллюстрацией
ряда рассмотренных выше общих технологических принципов
ферментативных процессов. Сначала ячмень инкубируют при
определенных температуре и влажности в течение 2—6 сут до
прорастания зерен. На этой стадии, называемой соложением,
индуцируется образование активных а- и [J-амилаз, а также
протеолитических ферментов. Затем зерна тщательно высуши-
вают.
Питательную среду для роста дрожжей, называемую пив-
ным суслом, приготавливают путем затирания солода — тща-
25*
Таблица 12.17. Микробиологические способы производства химических продуктов
массового производства (М— микробиологические процессы; X — химические процессы)*
Продукт
Процесс
Микроорганизм
Этанол С6Н12О6 м ->С2Н5ОН Saccharomyces cerevlsiae
Этилен Бутадиен-1,3 Этиленгликоль м —>сн2=сна х >сн2=снсн=сн2 > НОСН2СН2ОН Zymomonas mobiUs
Уксусная кислота Ацетон Бутанол С6Н12О6 С2Н5ОН С6Н12О6 м м м ->СН3СООН ->СНзСООН ->СНзСОСНз -»-СНз(СН2)зОН Clostridium thermoaceticum Acetobacter aceticum Clostridium acetobutylicum
Изопропиловый спирт С6Н12О6 м -> (СН3)2СНОН ->СНз(СН2)3ОН Clostridium aurianticum
Адипиновая кислота Акриловая кислота СНз(СН2)4СН9 >НООС(СН2)4СООН мх м8/х с6н12о6 >сн3сн(он)соон —► >сн2=^ снсоон Pseudomonas sp. Mi — Lactobacillus bulgaricus M2/X — Clostridium propionium
M/X
Метилэтилкетон СН2ОНСН(ОН)СН2ОН >сн2=снсоон М X Klebsiella pneumoniae (Aerobacter aerogenes) Klebsiella pneumoniae
С6Н12О6 >СН3СН(ОН)СН(ОН)СН3 -> ->СНзСОСН2СНз
Пропиленгликоль Глицерин X СН2ОНСН (ОН) СН2ОН > СН3СН (ОН) СН2ОН м С6Н12О6 >СН2ОНСН(ОН)СН2ОН Saccharomyces cerevislae
Лимонная кислота Н2О—|~СО2 С6Н12О6 м > СН2ОНСН (ОН) сн2он м >НООССН2С(ОН) (СООН)СН2СООН Dunaliella sp. Aspergillus niger
Воспроизведено с разрешения из работы: Ng Т. К., Busche R. М., McDonald С. С., Production of Feedstock Chemicals, Science, 219, 736
(1983). © American Association for the Advancement of Science.
Таблица 12.18. Производство кислородсодержащих органических веществ на основе возобновляемых источников*
Вещество Производство в США в 1981 г„ млн. кг Цена, центов за 1 кг Произведено в США в 1981 г. на сумму мли, долл. Основная область применения
Этанол Этилен 13 0946 55—56в 8 169г Производство полиэтилена, этиленок- сида
Бутадиен 1 382 75 1 234 Производство полибутадиенового и стиролбутадиенового каучука
Технический этанол 525 61 (1,70—1,81 долл, за 1 галлон) 359 Производство растворителей, этил- ацетата и других сложных эфиров
Этиленгликоль 1 839 61—63 1 281 Производство полиэтилентерефтала- та, антифризов
Уксусная кислота 1 227 58,5 511 Производство винилацетата, ацетил- целлюлозы
Ацетон 983 68 483 Производство растворителей, эфиров метакриловой кислоты
Изопропиловый спирт 746 68 507 Производство ацетона, растворите-
(2,05 долл. лей
за 1 галлон)
Адипиновая кислота 549 126 653 Производство найлона 66
Бутанол 375 74 251 Производство растворителей, бутил- акрилата
Акриловая кислота 313 128 276 Производство полимеров
Метилэтилкстон 284 82 260 Производство растворителей
Пропиленгликоль 218 97 208 Производство ненасыщенных поли- эфирных смол
Глицерин 168 178 259 Производство лекарственных и кос- метических препаратов
Лимонная кислота 107 157—171 192 Производство лекарственных препа-
ратов, пищевая промышленность
а Воспроизведено с разрешения из работы: Ng Т. К., Busche R. М., McDonald С. С., Production of Feedstock Chemicals, Science, 219, 737
(1983). © American Association for the Advancement of Science.
^Приведенные в этой графе данные заимствованы из Chemical Marketing Economics Handbook и материалов Комиссии по внешней торговле
США.
в По данным Chemical Marketing Reporter (20 сентября 1982 г.); реальная цепа зависит от качества, количества и места продажи про-
дукции.
г По данным Chemical Marketing Economics Handbook, U. S. Bureau of Census н Комиссии no внешней торговле США. Приведенные
в этой графе данные не отражают реальные суммы торговых операций или действующие цены.
392
Глава 12
тельно контролируемого нагревания смеси солода, крахмала и
воды. Успех операции затирания зависит от множества пре-
вращений в сложной смеси субстратов и ферментов. В смеси
солод — крахмал белки и углеводы должны быть гидролизова-
ны, поскольку участвующие в стадии ферментации дрожжи спо-
собны утилизировать в качестве питательных веществ только
простые сахара и аминокислоты. С другой стороны, полный гид-
ролиз нежелателен, поскольку декстрины, пептиды и пептоны
придают пиву специфический вкус и консистенцию. Ситуация
еще более осложняется из-за участия в образовании сусла раз-
Таблица 12.19. Три гидролитических фермента, участвующих в образовании
солода; ферменты обладают различными субстратной специфичностью
и кинетическими характеристиками [23]
Фермент Субстрат (или гидролизуе- мая связь) Продукты гидролиза Температу- ра макси- мальной ак- тивности, °C
а-Амилаза Крахмал (все связи) Высокомолекулярные 70—75
ф-Амилаза Амилоза (^-1,4-связи) декстрины, амилоза Мальтоза 57—65
Протеазы Белки Аминокислоты, неболь- шие пептоны Большие пептиды, 50 50
пептоны
личных гидролитических ферментов. Для оптимизации этого
сложного процесса используются тщательно отработанные ме-
тодики, включающие изменение температуры от 40 до 100 °C по
определенной программе (табл. 12.19). Полученный таким пу-
тем продукт содержит смесь питательных веществ для дрожжей
и вкусовых веществ в удовлетворительном соотношении.
На следующей стадии пивное сусло осветляют и затем кипя-
тят в течение примерно 2 ч, периодически добавляя хмель (спе-
лые, высушенные шишки хмеля), который придает пиву спе-
цифический вкус, аромат и окраску и одновременно оказывает
антибактериальное действие. Стерилизованное таким путем сус-
ло охлаждают и инокулируют соответствующим штаммом пив-
ных дрожжей (Saccharomyces cerevisiae). Дрожжи культивиру-
ют сначала в аэробных, а затем в анаэробных условиях, обес-
печивающих образование этанола и СО2. Наконец, пиво освет-
ляют, пастеризуют, выдерживают и расфасовывают.
Масштабы пивоваренной промышленности впечатляют.
В 1975 г. промышленность США продала 160 млн. баррелей
(25 438 млн. литров) пива. Затраты на производство этого ко-
личества пива распределялись приблизительно таким образом
Экономик а процессов биохимической технологии
393
(в млн. долл.): стоимость солода 610, других зерновых 188,
хмеля 49, оплата труда производственных рабочих 860, упаков-
ка и расфасовка 2500, топливо, электроэнергия и вода 80,
транспортные расходы 360, оборудование и модернизация 600,
реклама 230.
На рис. 12.6 приведена операционная карта обычного вино-
дельческого процесса. Сырьем для производства вин могут быть
практически любые спелые фрукты или растительные экстрак-
ты, содержащие от 12 до 30% сахара (в соке). Ферментацию
проводят или с участием дикого штамма дрожжей, имеющегося
в кожуре винограда или фруктов, или путем внесения опреде-
ленной культуры дрожжей в пастеризованные фруктовые соки.
Во многих анаэробных процессах брожения культуру дрож-
жей сначала выращивают в соке до необходимой массы в аэроб-
ных условиях, причем наряду с биомассой образуется диок-
сид углерода. Последующая ферментация, приводящая к полу-
чению 7—15 %-ного раствора этанола и выделению дополни-
тельного количества СО2, проводится в анаэробном режиме.
Некоторые органические кислоты, образующиеся в процессе
брожения, впоследствии разрушаются бактериями, что придает
винам характерный вкус и аромат («букет»). Тип полученного-
вина зависит от сырья (различные сорта винограда, абрикосы,
персики и т. д.), штаммов дрожжей и бактерий, участвующих в
брожении, послеферментационной обработки (например, креп-
ления, т. е. добавления спирта), а также от ряда операций,
предшествующих разливу, в том числе от механических воздей-
ствий, нагревания, охлаждения и старения (выдержки).
Мировое производство вин в основном сосредоточено в ев-
ропейских странах (почти 80%), а на американском континенте
производится только 15% вин. Вина производятся самыми раз-
личными предприятиями — от крупномасштабных до очень мел-
ких («семейного» типа). Оценка качества вин весьма сложна и.
включает дегустацию, т. е. оценку вкусовых качеств (кислотно-
сти, сахаристости, крепости и терпкости, обусловленной нали-
чием таннина), внешнего вида (цвета, прозрачности, отсутствия
осадка), аромата и букета, а также послевкусия (сложный по-
казатель, включающий остающийся привкус).
12.6.2. Производство спирта в качестве топлива
Производство этанола в качестве топлива привлекло широ-
кое внимание в последние десятилетия; впрочем, в Европе в го-
ды, предшествовавшие второй мировой войне, в сравнительно
широких масштабах этанол получали из сельскохозяйственных
культур. Последовавшее после 1945 г. снижение цен на нефть,
а также разработка новых нефтехимических процессов послу-
Современное винодельческое предприятие
Виноград I
виноградная фобилкаЛ
грсунеотделитель [ у1-?
С02 в
атмосферу
Емкость
для хране-
ния дрорс-
жеи
S02
Ферментер
для столового Столовое вино
вина
Плодоножки.
Охлаждающая
Дезинтегратор вода
| Промывные
\ х—<1 воды выжамкРы..
Н20
Ферментер
''
Столовое
вино
Ферментер
Э
Автоматический
сетчатый фильтр
В цех по производству
спирта ZS
Автоматический
Ю сетчатый
фильтр
Сырье
для
перегонки
Выжимки
Сырье -»
для пере-
гонки
Емкость для
хранения
сырья для
перегонки
Тартраты
виноградные выжимки (.корм)
Осадок
Диатомовая
земля
Сушилка для
выжимок
Магнитный сепаратор
циклон
Воздуходувка
м
. Из цеха по произ-
| водству спирта
Емкость для
хранения
(бренди '
Отстой
ник
Десертное
вино
Осадок f
i’"* Емкость
для
\оздух У™™*
Отстой -
ник
< Ферментер
Емкость для
хранения
р осадка
Осадок „
Ьентонитовая
суспензия । 1 f
Тартра-
ты
о \Дезинтпегратор
Аммиак
1 Холодильная
установка
Компоненты купажа
! I
Резервуар
для купажи-
рования
Охлаждаемая
емкость для
хранения
Фильтр
Пар
Емкость
для приго-
товления
. лиерри
Диатомовая
земля
Емкость
для
очистки
\ Оса док
Ем наст для
разливало
бутылкам
Яш, аки для
хранения
и транспор-
тировки
Разливочная
машина
Стерилизатор
Емкость для
выдержки
(старения)
Диатомовая
земля
‘Фильтр
Емкость для
хранения
вина
Отправка
в цистернах
Отправка
в бочках
Экономика процессов биохимической технологии 395
жили причиной того, что нефтехимическая промышленность ста-
ла основным поставщиком многих веществ, производившихся
ранее путем брожения, в том числе этанола и практически всех
простых кислородсодержащих органических соединений (ацето-
на, бутанола, уксусной кислоты и т. п.). Резкое повышение цен
на нефть в последнее десятилетие заставило пересмотреть отно-
шение к биотехнологическим способам получения этих веществ.
Потенциальная ценность этанола в качестве моторного топлива
обусловлена четырьмя обстоятельствами. Во-первых, этанол
представляет собой легко транспортируемую жидкость. Во-вто-
рых, его теплотворная способность довольно высока и состав-
ляет около двух третей от теплотворной способности бензина.
В-третьих, к этанолу можно добавлять до 10% бензина без из-
менения регулировки двигателя и теплотворной способности.
В-четвертых, этанол повышает октановое число бензина, не со-
держащего тетраэтилсвинец. Следует учитывать и тот факт,
что во многих странах этанол в качестве топлива может частич-
но заменить импортируемые нефть и нефтяные продукты.
Сырьем для биохимических методов производства этанола
служат сахара (сок сахарного тростника, маниок, мелассы),,
крахмал (зерновые культуры), древесина и отходы сельскохо-
зяйственных производств (солома зерновых культур), а также
промышленные и бытовые отходы, в том числе отходы лесной
промышленности (газетная бумага, сульфитно-спиртовая бар-
да, молочная сыворотка, отходы предприятий по переработке
фруктов и овощей и т. д.). При выборе конкретного сырья для
производства этанола следует искать компромиссное решение
между легче ферментируемым более чистым (но более доро-
гим) субстратом и более дешевым, но и значительно более ге-
терогенным исходным веществом. В последнем случае значи-
тельно возрастают расходы на предварительную обработку суб-
страта. Так, согласно одной из оценок экономических проблем
производства спирта из кукурузной соломы, стоимость оборудо-
вания для предварительной обработки субстрата (см. операции
предварительной обработки на рис. 12.7) составляет более 50%
всех капитальных затрат предприятия. Для крупномасштабного
завода по производству этанола из целлюлозы капитальные
затраты распределяются примерно следующим образом: подго-
товка сырья 5%, гидролиз 19%, регенерация кислоты 29,4%,
брожение и очистка 19,2%, водо-, паро- и энергоснабжение,
а также внезаводские системы 27,4%.
РИС. 12.6. Основные операции современного винодельческого процесса. (Вос-
произведено из статьи: Hull W. Q., Kite IV7. Е., Auerbach R. С., Modern Wine-
making; Ind. Eng. Chem., 43, 2182 (1951). © American Chemical Society.]
396
Глава 12
Солома
Предварительный
гидролиз
—.------(
f
] Реииркуляция
1 кислоты
Возврат
кислоты
Осадок
Промыв-
ка
Раствор
ксилозы
Регенерация
наслать?
-----------------------1
| Гидролиз
Раствор
глюкозы
Жидкость
Возврат
кислоты
РИС. 12.7. Схема процесса предварительной обработки кукурузной соломы на
предприятии по производству этанола в качестве топлива. [Воспроизведено из
статьи: Clausey Е. С., Gaddy J. L., Production of Ethanol from Biomass; Ann.
N.Y. Acad. Sci, 413, 435 (1983).]
Обычная промышленная операция представляет собой пе-
риодический процесс брожения продолжительностью около 50 ч
при начальном pH 4,5 и температуре 20—30 °C; выход этанола
составляет около 90% от теоретического, рассчитанного по со-
держанию сахаров. Конечная концентрация этанола в культу-
ральной жидкости составляет 10—16% (по объему); далее эта-
нол выделяют из культуральной жидкости описанными в пре-
дыдущей главе способами.
Ниже приведены основные характеристики традиционного
периодического процесса брожения.
Экономика процессов биохимической технологии
397
1. Стоимость сырья составляет основную долю общей себестои-
мости спирта.
2. Невысокая скорость реакции, обусловливающая необходи-
мость использования ферментеров больших объемов, стои-
мость которых составляет около 10 центов в расчете на
1 галлон (4,8 л) полученного спирта (по данным 1981 г.).
3. Необходимость выделения ценных побочных продуктов про-
изводства; так, доход от продажи дрожжей (используемых в
качестве добавки к кормам для домашних животных) мо-
жет снизить себестоимость спирта на 22 цента за 1 галлон.
4. Дистилляция представляет собой энергоемкий, но достаточ-
но эффективный процесс; типичный «старый» завод в Пео-
рии (шт. Иллинойс, США) расходует на дистилляцию менее
30% теплотворной способности получаемого этанола.
Возрождение интереса к этанолу как к топливу послужило
причиной разработки многих новых процессов и новых штам-
мов микроорганизмов, что в конечном итоге значительно улуч-
шило и экономические показатели процесса. В настоящее вре-
мя в нескольких штатах США в качестве моторного топлива
применяют спиртосодержащие смеси.
В последнее десятилетие введены следующие важные усо-
вершенствования:
1. Операции при пониженном давлении, что позволяет отгонять
спирт в ходе брожения. Таким путем достигается снижение
степени ингибирования процесса спиртом, повышение про-
должительности ферментации и повышение степени утилиза-
ции сахара.
2. Непрерывные процессы с использованием рециркуляции био-
массы и пониженного давления, что позволяет повысить про-
изводительность установки в 12 раз (по сравнению с перио-
дическим процессом). При давлении 50 мм рт. ст. и концент-
рации глюкозы 33% (масса/объем) достигнута производи-
тельность 40 г/(л-ч).
3. Процессы с участием бактерий (Zymotnonas mobilis), кото-
рые по сравнению с дрожжами обеспечивают более высокие
удельные скорости утилизации глюкозы и образования
спирта.
4. Применение иммобилизованных клеток, позволяющих зна-
чительно повысить объемную производительность процесса
(в расчете на этанол) по сравнению с обычной культурой в
суспензии.
5. Применение высоких башенных ферментеров с высокой сте-
пенью флокуляции дрожжей; в этих ферментерах в стацио-
нарном состоянии в нижней части башни обеспечивается
плотность биомассы до 50—80 г/л. Благодаря расширению в
398
Глава 12
верхней части башни в ней удерживается (путем седимента-
ции, без каких-бы то ни было специальных механических
устройств) практически вся биомасса. После стартового пе-
риода продолжительностью несколько недель такие фермен-
теры устойчиво работают в течение года.
6. Модифицированные операции выделения этанола, позволяю-
щие снизить высокие затраты на этой стадии. К ним отно-
сятся повышение давления паров, рекуперация теплоты, вы-
деляющейся при обработке сточных вод (на заводах по про-
изводству спирта из зерна), обезвоживание водно-этаноль-
ных смесей с помощью различных синтетических или при-
родных сорбентов.
Многие из перечисленных усовершенствований уже провере-
ны, а другие пока еще нуждаются в дополнительном изучении.
Только после этого можно будет оценить экономическую целе-
сообразность их использования в промышленных процессах.
В табл. 12.20 приведены сравнительные характеристики перс-
пективных процессов брожения.
Ввиду того, что в будущем предприятия по производству
спирта будут существенно отличаться от работающих в настоя-
щее время заводов, полезно сравнить экономические характе-
ристики четырех наиболее хорошо изученных процессов. Как
показывают приведенные в табл. 12.21 данные, для предприя-
тий, производящих 295 м3 95%-ного этанола в сутки из 50 %-но-
го раствора тростниково-сахарной мелассы, доля эксплуатаци-
онных расходов уменьшается в ряду: периодический процесс>
> непрерывный процесс> непрерывный процесс с рециркуляци-
ей культуры> вакуумный процесс с рециркуляцией культуры..
Отсюда следует, что повышение объемной производительности
установки намного перекрывает дополнительные расходы, свя-
занные с использованием более сложного оборудования. Для
всех трех непрерывных процессов наблюдается сокращение рас-
ходов на оплату труда производственных рабочих, а в случае
непрерывного вакуумного процесса с рециркуляцией культуры
неожиданно большая экономия достигнута за счет сокращения
расхода пара.
Политика правительственных органов (как в США, так и в
других странах) по отношению к производству этанола в каче-
стве добавки к моторному топливу зависит от целого ряда не-
предсказуемых факторов, в том числе от объема добычи нефти
(и производства бензина) и цен на эти товары, целесообразно-
сти (или нецелесообразности) использования основного пище-
вого продукта — зерна в качестве сырья для производства топ-
лива, от наличия (или отсутствия) твердых гарантированных
правительством цен и (или) субсидий. После окончания второй
Экономика процессов биохимической технологии
399
мировой войны никакая другая продукция биохимической тех-
нологии не рассматривалась в столь тесной связи с вопросами
национальных ресурсов и национальной безопасности многих
стран.
Микробиологические способы производства ацетона и бута-
нола изучались в свете возможного применения бутанола в ка-
честве добавки к дизельному топливу (дизооль). Расчеты, вы-
полненные для предприятия производительностью 45 млн. кг
растворителей в год, показали, что процесс, в котором исходным
сырьем является меласса, неэкономичен, поскольку стоимость
этого сырья составляет около 60% от всех издержек производ-
ства в течение года. Гораздо более благоприятная с экономиче-
ской точки зрения ситуация складывается при использовании в
качестве сырья жидкой молочной сыворотки (табл. 12.22), если
считать, что она является бесплатным отходом соответствую-
щих производств, и учитывать только транспортные расходы на
ее доставку. В последнем случае проект предприятия, перво-
начально предназначенного для переработки мелассы, пришлось
модернизировать, включив операции ультрафильтрации (для
выделения ценного побочного продукта — сывороточного белка)
и анаэробной переработки отходов (источник метана; см.
гл. 14).
В этих условиях расчетная величина чистой прибыли на ин-
вестированный капитал (с учетом различных продуктов) соста-
вила 30,8%. Расчетная валовая прибыль предприятия состави-
ла 15 млн. долл, в год; на такую сумму предприятие должно
производить только некоторые побочные продукты (сухие кор-
ма, водород, углекислый газ, этанол). Важными характеристи-
ками процесса являются низкий выход бутанола (1,9% по дан-
ным на 1980 г.), строго анаэробный режим, тенденция к зара-
жению фагами и молочнокислыми бактериями.
12.6.3 . Производство органических кислот и аминокислот
Производство органических кислот может служить показа-
тельным примером того влияния, которое оказывает выход про-
цесса на его экономику. Капитальные затраты на цех фермен-
тации при получении лимонной кислоты в погруженной культу-
ре производительностью 70 кг лимонной кислоты из 100 кг са-
хара на 15—25% ниже, чем в процессе с поверхностной фер-
ментацией, но в первом случае эксплуатационные расходы на
16—26% выше. Если же включить сюда стоимость операций
выделения и очистки, то эти величины уменьшатся примерно в
два раза. При производстве лимонной кислоты операции выде-
ления довольно просты, не требуют больших затрат и приводят
Таблица 12.20. Некоторые промышленные процессы производства этанола1
Процесс
Производительность
Характеристики процесса
Периодический процесс брожения Очень низкая (1,8— 2,5 г/(л-ч))
Один ферментер типа ПРПП Низкая [6 г/(л-ч)]
Каскад из нескольких ПРПП В 2—3 раза выше, чем в одном ПРПП
С перфорированными тарелками Нет данных
ПРПП с центробежной рециркуля- Высокая [30—40
цией культуры г/(л-ч)]
ПРПП с другой системой рецирку- Высокая
ляции Башенные ферментеры Высокая
Наклонные трубчатые ферментеры
Башенные ферментеры с неподвиж
ным слоем
Диализационный ферментер
Высокая
Высокая
Нет данных
Диализационный ферментер, рабо-
тающий под давлением
Роторный ферментер
Нет данных, но потен-
циально высокая
Высокая [36 г/(л-ч)]
ферментер с полыми волокнами
Нет данных, но потен-
циально высокая
Очень высокие капитальные затраты и эксплуатацион-
ные расходы
Конструктивно простое оборудование, простое обслужи-
вание непрерывного процесса
Простое обслуживание непрерывного процесса
Конструктивно сложное оборудование
Повышенный расход энергии; наблюдение за работой
системы рециркуляции требует дополнительного внима-
ния оператора
Отстойники слишком велики для промышленного при-
менения; вихревые сепараторы могут оказаться удобнее
Конструктивно простое оборудование, обслуживание
также простое, но продолжительность стартового перио-
да слишком велика
Конструктивно сложное оборудование
Основные проблемы связаны с закупоркой и движени-
ем параллельных потоков
Скорость реакции лимитируется диффузией субстрата
через мембрану. Загрязнение мембраны может вы-
звать частые остановки процесса
Основная трудность связана с загрязнением мембраны
Проблема загрязнения мембраны решена, но конструк-
ция аппарата очень сложна. Возможно также механи-
ческое повреждение мембраны
Дорогие аппараты. Производительность может лими-
тироваться сопротивлением массопереносу
26—746
Ферментер с поршневым потоком
Ферментер с включением дрожже-
вых клеток в гель
Ферментер с химически связанными
дрожжевыми клетками
Высокая (в 72 раза вы-
ше, чем в периодичес-
ком процессе)
Выход 90% из 10% -ко-
го раствора глюкозы в
течение 10 ч
Нет данных
Прямая экстрактивная ферментация
Мембранная экстрактивная фермен-
тация
Селективная мембранная фермента-
ция
Вакуумная ферментация
Потенциально очень вы-
сокая
Нет данных, но потен-
циально очень высокая
Нет данных, но потен-
циально очень высокая
Очень высокая
[80 г/(л-ч)1
Импульсная ферментация
Очень высокая
Необходимы частые остановки процесса для обновле-
ния дрожжей в аэробной среде. Необходимы высоко-
производительные питательные насосы
Очень простая система, не требующая перемешивания
или рециркуляции. Для повышения эффективности
процесса необходимо найти пути увеличения времени
жизни иммобилизованных клеток
После ковалентного связывания жизнеспособность
клеток остается высокой. Если при этом удастся со-
хранить и высокую производительность культуры, то
система в целом будет очень простой и одновременно
высокопроизводительной
Пока не найдено соответствующего экстрагента
Возможно загрязнение мембраны, но в остальном про-
стой процесс
Пока еще не найдено мембран с достаточно высокой
пропускной способностью
Механически сложное оборудование, требующее по-
стоянного контроля. Дополнительный небольшой рас-
ход энергии связан с сжатием паров. Возможно за-
грязнение вакуумированного резервуара. Для барбо-
тирования необходим чистый кислород
Механически и конструктивно очень сложное обору-
дование. Расход энергии не намного выше, чем в
обычных процессах. Проблема загрязнения намного
менее серьезна, чем в вакуумном процессе; нет необ-
ходимости в чистом кислороде
’Воспроизведено с разрешения из работы: Maiorella В., Wilke С. R., Blanch II. W., Alcohol Production and Recovery, in Adv. Biochem.
Eng., vol. 20, p. 72, 1981.
402
Глава 12
Таблица 12.21. Сравнительная экономическая оценка различных
процессов производства этанола3' 6
Статья расходов Расходы, млн. долл.
Периодиче- ский процесс Непрерывный процесс Непрерывный процесс с ре- циркуляцией Вакуумный процесс с ре- циркуляцией
Расходы, связанные с капитальными затра- тами 27,21 12,94 10,57 9,25
Оплата труда производ- ственных рабочих 8,45 2,38 1,32 1,06
Оплата работников конт- рольных и конторских служб Вспомогательные систе- мы: 0,53 0,26 0,26 0,26
Вода 1,58 1,58 1,58 1,06
Электроэнергия 3,17 1,58 2,38 1,58
Пар 26,68 25,10 25,10 17,96
Кислород — — — 1,32
Лабораторные расходы 0,26 0,26 0,26 0,26
Заводские накладные расходы 4,75 1,58 1,06 0,79
Всего 72,63 45,68 42,53 33,54
а Воспроизведено с разрешения из работы: Cysewski G. Ji., Wilke С. R., Process De-
sign and Economic Studies of Alternative Fermentation Methods for the Production of Etha-
nol; Biotech. Bioeng., 20, 1421 (1978).
б Производительность предприятия составляла 295 м3 95%-ного этанола в сутки из
тростниково-сахариой мелассы, содержащей 50% сахаров.
Таблица 12.22. Доходы предприятия, производящего 45-106 кг
растворителей в год (на основе ацетоно-бутанольного брожения)3
Продукт Объем произ- водства, 10е кг в год Стоимость, долл, за 1 кг Доходы, 10е долл, в год
w-Бутанол 26,4 0,53 13,9
Ацетон 13,2 0,44 5,8
Этанол 4,5 0,40 1,8
Сухие корма 64,3 0,094 4,7
Водород 1,8 0,29 0,52
Диоксид углерода 72,0 0,11 7,9
Сывороточный белок 10 0,46 9,3
Метан 0,46
Всего 44,4
3 По данным, приведенным в работе: Lenz Т., Moreira A., Economic Evaluation of
the Acetone-Butanol Fermentation; Ind. Eng. Chem. Prod. Res. Dev., 19, 478 (1980),
Экономика процессов биохимической технологии
403
к чистому продукту, поэтому любые модернизации процесса
должны затрагивать главным образом стадию ферментации.
Более умеренный выход итаконовой кислоты в непрерывном
процессе (40 г/л) явился причиной изучения различных методов
выделения этого продукта — дистилляции, ионного обмена и
электродиализа. Подсчитано, что доля затрат на строительно-
монтажные работы и оборудование для этих трех непрерывных
операций выделения в общей стоимости строительства составля-
ет 92% (дистилляция), 96% (ионный обмен) и 56,7% (элект-
родиализ). При использовании наименее дорогостоящей систе-
мы выделения (электродиализа) расчетные величины затрат
на сырье, а также водо-, паро- и энергоснабжение составляют
соответственно 49 и 13% эксплуатационных расходов. По-
скольку относительные преимущества различных операций вы-
деления зависят от концентрации продукта в культуральной
жидкости, было бы интересно определить, как изменятся ре-
зультаты этих расчетов, если путем совершенствования фер-
ментационного процесса удастся поднять выход итаконовой
кислоты до 60, 80 и 100 г/л.
Теперь рассмотрим экономические аспекты производства
аминокислот. Из двадцати белковых аминокислот в 1980 г.
только две (лизин и глутаминовая кислота) производились-
главным образом микробиологическим путем (в Японии), хотя
разработаны микробиологические методы получения и других
аминокислот (аргинина, глутамина, гистидина, лейцина, орни-
тина, фенилаланина, пролина, треонина и валина) (табл. 12.23).
Интересно, что при получении t-лизина в промышленном мас-
штабе применяются как химический, так и микробиологический
пути синтеза (табл. 12.23), причем в 1980 г. в Японии и Юж-
ной Корее 80% лизина производилось микробиологическим пу-
тем, а в США лизин получали только химическим синтезом.
Сравнение затрат при производстве лизина микробиологическим
и химическим способами приведено в табл. 12.24.
Новым синтетическим заменителем сахара является аспар-
там — метиловый эфир ь-аспартил-г.-фенилаланина. Недавно вы-
данное разрешение на применение этого препарата в качестве
добавок к пищевым продуктам и в особенности к безалкоголь-
ным напиткам привело к резкому расширению рынка t-фенил-
аланина — от 50 т в 1981 г. до 3000 т в 1984 г. и ориентировоч-
но до 8000 т в 1990 г. Первоначальная цена ь-фенилаланина,
составлявшая 50 долл./кг, упала до 35 долл./кг в 1984 г. и в
ближайшем будущем ожидается дальнейшее снижение цены до
20 долл./кг. Принятые в настоящее время способы получения
фенилаланина включают микробиологическую трансформацию
сахаров и ферментативное превращение коричной кислоты или.
гидантоина.
26*
Таблица 12.23. Промышленное Производство аминокислот*
Аминокислота Цена в долл, за 1 кг чистого L-изомера (по данным на март 1980 г.) Основной способ получения в настоящее время Объем производ- ства в 1978 г., т Потенциальные возможности при- менения биотехнологических про- цессов (полный биосинтез или биотрансформация с участием организмов или ферментов)
Аланин 80 Гидролиз белков, химический 10—50б
синтез
Аргинин 28 Гидролиз желатина 200—3006 Микробиологический способ, применяется в Японии
Аспарагин 50 Выделение 10—506
Аспарагиновая кислота 12 Биотрансформация фумаровой 500—10006 Биотрансформация
кислоты
Цитруллин 250 — 10—906 Микробиологический способ, применяется в Японии
Цистеин 50 Выделение 100—2006
Цистин 60 Выделение 100—2006
ДОФА (днгидроксифе- нилаланнн) 750 Химический синтез 100—2006 10 000—100 0006
Глутаминовая кислота 4 Микробиологический процесс При полном биосинтезе про- дуцентом служит Micrococcus
glutamicus
Глутамин 55 Выделение 200—3006 Микробиологический способ, применяется в Японии
Г истидин 160 .— 100—200 Микробиологический способ, применяется в Японии
Гидроксипролин 280 Выделение из коллагена 10—50
Изолейцин 350 Выделение 10—506
Лейций 55 4МЬ 50— ЮОб Микробиологический способ, применяется в Японии
Лизин 350 Микробиологический процесс (80%); химический синтез (20%) 10 0006 80% лизнна производится пол- ностью микробиологическим пу- тем с участием Corynebacterium glutamicum и Brevibacterium flavum
Метионин 265 Химический синтез нз акро- леина 17 000 (DL)B 20 000 (DL)°
Орнитин 60 — 10—50° Микробиологический процесс, применяется в Японии
Фенилаланин 55 Химический синтез из бенз- альдегида 50—100° Микробиологический процесс, применяется в Японии
Пролнн 125 Гидролиз желатина 10— 50° Микробиологический процесс, применяется в Японии
Сернн 320 — 10—50° Процесс с биотрансформаци- ей, применяется в Японии
Треоннн 150 — 50—10° Микробиологический процесс, применяется в Японии
Триптофан НО Химический синтез нз индола 55°
Тнрознн 13 Выделение 50—100°
Валнн 60 — 50—100° Микробиологический процесс,
применяется в Японии
а Воспроизведено из работы: Impacts of Applied Genetics: Microorganisms, Plants and Animals, p. 88. Office of Techology Assessment, Wa-
shington, D. C., 1981. Данные об объеме производства относятся главным образом к Японии ввиду малого объема производства в США.
б в
В Японии. D- и L-формы.
Таблица 12.24. Сравнение расходов на производство моногидрохлорида
L-лизина в США микробиологическим н химическим методами3
Расходы при производстве мюногидрохлорида L-лизина (содержащего 98% Ь-изом:ера)
Микробиологический
б Химический метод® метод **
Статья расхода
Оценка рас- Оценка рас-
Потребность ХОДОВ п° Дан' Потребность ходов по даи- (единиц иа нь‘м иа'^.6 г- (единиц на ным на 1976 г..
единицу пД^-пнн3, единицу на единицу
продукции) продукции, продукции) продукции, придукци / центов за к J центов за
1 КГ 1 КГ
Оплата труда, всего1. 18 20
Материалы
Меласса 44 16
Соевая мука, гидро- 0,462 9
лизованная
Циклогексанол 0,595 37
Аммиак безводный 0,645 14
Другие химнкатыд 15 10
Растворители оо 8
Материалы для упа- 22 21
ковки, обслуживания,
ремонта
Всего материалов 62 до
Водо-, паро-, газо- и 12 16
энергоснабжение®
Прямые эксплуатаци- 92 126
онные расходы, всего
Накладные расходы, на- 21 21
логи, страхование
Итого 113 147
Амортизационные отчие- 35 28
ленияж
Процент на оборотный 3 3
капитал
Заводская 151 178
себестоимость
аВоспроизведено из работы: Impacts of Applied Genetics: Microorganisms; Plants and
Animals, p. 89, Office of Technology Assessment, Washington, D. C., 1981. Источник: Stan-
ford Research Institute. Chemical Economic Handbook, 583, 3401, May 1979.
® В расчете на выход 23% из мелассы.
вВ расчете иа выход 65% из циклогексанола.
гВключая эксплуатацию, техническое обслуживание, контрольные и лабораторные-
пспытаиня.
ДВ расчетах принято, что и в микробиологическом, и химическом процессах приме-
няют соляную кислоту (36%) и аммиак (29%). В микробиологическом процессе используют
также дифосфат калия, мочевину, сульфат аммония, карбонат кальция и сульфат маг-
ния, а в химическом синтезе — нитрозилхлорид и серную кислоту; в последнем случае
учтены доходы от реализации побочного продукта (сульфата аммюиия).
еДля того и другого процесса учтены расходы на воду для охлаждения, пар для
технологических нужд, электроэнергию. В химическом процессе учтена также стоимость
природного газа.
ж 10% в год от основного капитала для нового предприятия производительностью
20 мли. фунтов в год, построенного в США в 1975 г., при расчетных капитальных за-
тратах 38,6-10е долл, для микробиологического процесса и 32,5-10® долл, для хнмиче-
ского процесса (без учета стоимости земельного участка).________________ .
406
Экономика процессов биохимической технологии
407
12.7. Белок одноклеточных организмов
Белок одноклеточных организмов (БОО)—это содержащие
белок материалы, представляющие собой высушенные клетки
микроорганизмов. В качестве добавок к пищевым продуктам
или кормам для домашних животных применяется сухая био-
масса водорослей, актиномицетов, бактерий, дрожжей, плесе-
ней и высших грибов. Человек употребляет в пищу белки мик-
роорганизмов с незапамятных времен, в том числе дрожжи в
дрожжевом хлебе, молочнокислые бактерии в кисломолочных
продуктах (молоке, сыре, некоторых сортах колбас), а также во-
доросли (Spirulina), растущие в прудах и озерах.
Таблица 12.25. Основные характеристики некоторых процессов
производства белка одноклеточных организмов3
Характери- стики процесса Процесс с участием
водорослей Spi- rulina maxima бактерий Metliy- lophilus methy- loirophus (суб- страт — метанол) дрожжей Candi- da utilis (суб- страт — этанол) плесневых гри- бов Paecilomyces varioti (суб- страт — суль- фитно-спиртовая барда)
Тип про- цесса Периодический или полунеп- рерывный Непрерывный Непрерывный или периодиче- ский Непрерывный
Стериль- ность Неасептиче- ский Асептический Асептический Неасептиче- ский
Ферментер Бассейновый Эрлифтного типа С перемеши- ванием С перемешива- нием
Утилизация субстрата Частичная или полная Полная Полная Частичная
Темпера- тура Комнатная 35—42 °C 30—40 °C 38—39 °C
pH 9—11 6,0—7,0 4,6 4,5—4,7
Метод выделения продукта Фильтрование Флокуляция II центрифугиро- вание Центрифуги- рование Фильтрование
аИз работы: Litchfield J., Single Cell Protein; Science, 219, 740 (1983). ф 1983 bv the
American Association for the Advancement of Science.
Развитие биохимической технологии позволило выращивать
культуры микроорганизмов в аэрируемых ферментерах на та-
ких субстратах, как газойль, парафины, природный газ, мелас-
са и целлюлозные материалы. Наиболее важные эксплуатаци-
онные характеристики некоторых процессов получения БОО
представлены в табл. 12.25. В этих процессах используются
асептические и неасептические режимы, открытые и закрытые
ферментеры, полная и частичная утилизация субстрата, различ-
408
Глава 12
Таблица 12.26. Некоторые материалы, использующиеся в качестве источников углерода и энергии в производстве белка одноклеточных организмов3
Исходные мате- риалы Тип и масштаб про- цесса 6 Организм Разработчик проекта или компания-произво- дитель
со2 Тростниковый сироп, мелас- са (сахаро- за) СО2 или NaHCO3, Na2CO3 Метанол Водоросли, 2 т в сут- ки® Фотосинтезирующий Нефотосинтезирую- щий Chlorella sp. Taiwan Chlorella Ma- nufacture Co., Ltd.,. Taipei
Водоросли, 320 т в годг, фотосинте- зирующий Бактерии 70 000 т в год Spir ulina maxi- ma Methylophilus methylotro- phus Methylomonas clara Candida utilis (Torula) Candida sp. Candida utilis Paecilomyces varioti Fusarium graminearum Kluyveromyces fragilis Penicillium cyc- lo pium Sosa Texcoco, S. A., Mexico City Imperial Chemical In- dustries, Billingham
Этаиол н-Алканы, гид- ролизат дре- весины Сульфитно- спиртовая барда Глюкоза Сыворотка (от- ходы произ- водства сы- ра) 1000 т в год Дрожжи, 6800 т в год Дрожжи (на некото- рых предприяти- ях), 20 000—40 000 т в год Дрожжи, 10,9 т в сутки Плесени, 10 000 т в год Плесени, 50—100 т в год Дрожжи, 4536 т в год Плесени, 300 т в год Hoechst-Uhde, Frank- furt, West Germany Pure Culture Products, Hutchinson, Minne- sota Всесоюзный научно- исследовательский институт биосинте- за белка, СССР Rhinelander Paper Corp., Rhinelander, Wisconsin Pekilo Process, Fin- nish Pulp and Paper Re- search Institute, Jamsankoski, Fin- land Rank Hovis MacDou- gall Research Limi- ted, High Wycombe, U. K. Amber Laboratories-, Juneau, Wisconsin Heurty, S. A., France
а Из работы: Litchfield J., Single Cell Protein; Science, 219, 740 (1983). © 1983 by the
American Association for the Advancement of Science.
® Указана производительность предприятия в тоииа.х в единицу времени.
в Общая площадь бассейна 83 400 м2.
г Площадь бассейна 900 гектар.
Экономика процессов биохимической технологии
409
ные оптимальные значения pH (от кислого до основного), раз-
ные способы выделения продукта (фильтрование, центрифуги-
рование). Не удивительно, что производство БОО лимитируется
в первую очередь доступностью соответствующих субстратов,
которыми могут быть углеводороды, а также сточные воды бу-
мажных, сельскохозяйственных, молочных и промышленных
предприятий (табл. 12.26). Практически все заводы по произ-
водству БОО продают свою продукцию в виде добавки к кор-
Отработанный воздух + СО г.
Инокулят_________
Стерилизация
Метанол
Аммиак________
Фосфорная кислота
Неорганические
соли _
Воздух ' Ц
Компрессор
Ферментер
эрлифтного типа
РИС. 12.8. Схема одного из предложенных недавно процессов производства
белка одноклеточных организмов. [Воспроизведено с разрешения из статьи:
Litchfield J., Single Cell Protein; Science, 219, 740 (1983). © by the American
Association for the Advancement of Science.]
мам для домашних животных. Для нужд пищевой промышлен-
ности БОО производится в значительно меньших масштабах,
причем здесь основную долю и сегодня составляют пекарские
дрожжи. Удельная скорость роста организмов, участвующих в
производстве БОО, изменяется в пределах от 0,11 до 0,7 ч-1.
Производство БОО развито во многих странах, а крупнейшим
производителем его является Советский Союз.
Недавно разработанный процесс производства БОО с уча-
стием бактерий, утилизирующих метанол (Methylophiltis met-
hylotrophus), (рис. 12.8) можно описать общим стехиометриче-
ским уравнением:
1,72СН3ОН+0,23ЫНз+1,5Ю2 1,ОСН,,68Оо,збН0,22+0,72С02+2,94Н20
Процесс характеризуется быстрым ростом клеток (ц = 0,5 ч-1),
густотой конечного бульона (вследствие агломерации в биоре-
акторе), который можно сразу подавать на центрифугирование,
образованием в качестве побочного продукта СО2 и экономиче-
ским коэффициентом более 0,5 г клеток/(г субстрата).
410
Глава 12
Совершенствование процесса производства БОО заключа-
лось в получении штаммов с улучшенными характеристиками
(в частности, в увеличении размеров клеток 3. cerevisiae, что
облегчает последующие операции выделения), генетической мо-
дификации организмов, утилизирующих метанол (в результате
чего повышается содержание белка в клетках), модификации
Таблица 12.27. Сравнение продажной цены белковых продуктов,
получаемых из некоторых одноклеточных организмов, растительных
и животных источников3
Продукт, субстрат п качество продукта Содержание белка в про- дукте, % Цена (долл. США) за 1 кг (в ценах 1979 г.)
Белок одноклеточных организмов
Candida utilis, этанол (пищевой продукт) 52 1,32—1,35
Kluyveromyces fragilis, молочная сыворотка (пищевой продукт) Saccharomyces cerevisiae: 54 1,32
Отходы пивоваренного производства, ли- шенные горьких примесей (пищевой про- дукт) 52 1,00—1,20
Кормовой продукт Растительные белки 52 0,39—0,50
Люцерна обезвоженная 17 0,12—0,13
Соевая мука 49 0,20—0,22
Концентрат соевого белка 70—72 0,90—1,14
Чистый соевый белок Животные белки 90—92 1,96—2,20
Рыбная мука (перуанская) 65 0,41—0,45
Костная мука и мука из мясных отходов 50 0,24—0,25
Сухое обезжиренное молоко 37 0,88—1,00
а Воспроизведено из работы Impacts of Applied Genetics: Microorganisms, Plants and
Animals, Office of Technology Assessment, Washington, D. C., 1981. Источник: Office of
Technology Assessment.
Methylophilus methylotrophus (обеспечивающей повышение эф-
фективности утилизации NH3), разработке штамма Bacillus
brevis, секретирующего белки в среду, а также в улучшении
операций выделения биомассы путем флокуляции и электроко-
агуляции и, наконец, автоматизации процесса.
С экономической точки зрения БОО должен успешно кон-
курировать с другими источниками белка (табл. 12.27). Осо-
бенно богатым источником дешевого белка являются соевые бо-
бы. Поскольку в 1980-х годах ожидается расширение производ-
ства этого вида сельскохозяйственной продукции, то вероятно»
что в большинстве стран и БОО также сохранит свои позиции
как основной побочный продукт ее производства. Перспективы
расширения производства БОО зависят от рынков сбыта. Так.
Экономика процессов биохимической технологии
411
Б00, используемый в качестве корма для домашних животных,
успешно конкурирует с традиционными кормами и в ряде случа-
ев обеспечивает такой же привес животных (например, цыплят,
индеек, свиней). Если БОО предназначен в качестве пищевого
продукта, то он должен обладать определенными качествами,
в первую очередь вкусовыми, а кроме того текстурой и пита-
тельной ценностью; для этих целей обычно применяют дрожжи
Torula. Производство БОО для пищевой промышленности свя-
зано с гораздо большими капитальными затратами; согласно
некоторым оценкам, в 1975—1976 годах производство пищевого
белка было экономически целесообразным при среднегодо-
вой продажной цене 660—1000 долл, за тонну.
При проведении крупномасштабных аэробных процессов ос-
новные затраты связаны с закупкой сырья (субстрата), расхо-
дом электроэнергии (на перемешивание, аэрацию и процессы
массопереноса) и охлаждением (теплообменом). При использо-
вании целлюлозных материалов в качестве сырья для произ-
водства БОО необходимо учитывать расход электроэнергии, во-
ды, пара и топлива на глубокую предварительную обработку
целлюлозы, чтобы превратить ее в поддающиеся микробиологи-
ческому воздействию соединения.
Сравнительная оценка различных способов производства
белка одноклеточных организмов показывает (табл. 12.28), что
основные расходы здесь связаны со стоимостью сырья, состав-
ляющей 44—77% всех затрат.
12.8. Анаэробные процессы производства метана
Широко изучались анаэробные процессы превращения суб-
стратов углеводно-целлюлозной природы (особенно отходов
сельскохозяйственных производств) в биогаз (метан), который
обычно содержит 50—65% СН4, 35—50% СО2, 30—160 г/м3 Н2О и
1,5—12,5г/м3Н25. В табл. 12.29 приведены сравнительные затра-
ты на производство биогаза (вариант А) и метана (биогаза после
поглощения СО2) (вариант Б). На самых крупных предприяти-
ях по производству биогаза себестоимость последнего составля-
ла 2,61—2,74 долл, за 1 гигаджоуль (ГДж) теплотворной спо-
собности; для сравнения отметим, что себестоимость природно-
го газа в 1980 году была равна 3,00 доллара за 1 ГДж тепло-
творной способности. Если побочные продукты этого процесса
использовать в качестве корма для домашних животных (по це-
не 60 долл, за тонну), то даже совсем небольшое предприятие
по производству биогаза будет вполне конкурентоспособным.
Таблица 12.28. Сравнение затрат при некоторых способах производства биомассы®
Затраты, %
Статьи затрат Дрожжи, пара- фины (Италия) Бактерии, мета- нол (Англия) Дрожжи, этанол (Чехословакия) Грибы, сульфит- но-спиртовая барда (Финлян- дия) Бактерии, багас- са (США)
Накладные расходы 9,3 5,8 5,8 9,1 11,5
Исходные материалы 58,5 73,8 77,1 55,1 43,6
в том числе:
Субстрат 29,4 47,4 63,9 17,0 25,7
Фосфорная кислота 11,1 11,8 3,2 16,2 5,7
Аммиак 9,9 12,0 [4,8 13,3 3,6(?)
Неорганические соли 2,9 2,6 1,9 4,2 4,2
Другие 5,2 3,3 4,4 4,4
Водо-, тепло-, газо- и энергоснабжение 23,8 14,2 12,0 24,8 36,6
Оплата труда и другие расходы 8,4 6,2 5,1 11,0 8,3
Итого: 100 100 100 100 100
аПо данным, приведенным в работе: Moo-Young М., Economics of SCP Production. Process Biochem., 12(4), 6—10 (1977).
Примечание: в расчетах принято, что цена парафинов в 1,3 раза больше цены метанола, а цена этанола в 2,6 раза выше цены ме-
танола.
Экономика процессов биохимической технологии 41?
Таблица 12.29. Расходы на производство биогаза (вариант А)а
и метана (вариант Б)6 в процессах анаэробной ферментации навоза
крупного рогатого скота при 55°СВ
Статьи расходов Относительная производительность предприятия 1 10 100 1000
Капитальные затраты,
тыс. долл. А 70,7 354 2185 13490
Б 89,6 449 2769 17090
Основной капитал, тыс. долл, в год А 17,0 85,0 525 3240
Б 21,5 107,7 665 4100
Расходы на водо-, теп- ло- и энергоснабже- ние, тыс. долл, в год А 2,24 21,9 213 2126
Б 2,86 28,2 276 2760
Оплата труда, тыс. долл. 11,8 22,9 44,3 85,9
в год Всего расходов, тыс. долл, в год А 31,0 130,1 782 5450
Б 36,2 158,8 985 6942
Себестоимость, долл, за 1 гигаджоуль тепло- творной способности А 13,94 5,72 3,41 2,61
Б 16,28 6,98 4,29 2,74
а Содержимое реактора и поступающую суспензию перемешивали в течение 12 ч;
биогаз сжат до 860 кПа. ®Содержимое реактора и поступающую суспензию перемешивали в течение 12 ч;
СО2 отделяли в скруббере; метан сжат до 860 кПа. в Chen Y.-R., Varel V. Н., Hashimoto A. G. Methane Production from Agricultural Re-
sidues; I & ЕС Product Res. & Dev., vol. 19, p. 476, 1980.
12.9. Заключение
Успех эпсплуатации микробиологического процесса зависит
как от технических, так и экономических факторов. Приведен-
ный в этой главе подробный анализ экономики одного из про-
цессов может служить моделью для изучения экономических
аспектов любого другого процесса. Экономика микробиологи-
ческого процесса определяется его типом; так, например, суще-
ственные различия наблюдаются в экономике тонких биотех-
нологических процессов, процессов получения химических про-
414
Глава 12
дуктов массового производства, спиртных напитков, белка одно-
клеточных организмов и анаэробных процессов производства
метана. Другие примеры будут приведены в упражнениях.
Упражнения
12.1. Этапы реализации процесса. Дайте определение (по памяти)
следующим этапам реализации процесса: идея его создания, предвари-
тельная оценка, разработка окончательных проектных данных, окончательный
экономический анализ, детальный технологический процесс, материально-тех-
ническое обеспечение, строительно-монтажные работы, пусконаладочные рабо-
ты, пробный пуск, промышленная эксплуатация.
12.2. Анализ чувствительности. Приведенные в табл. 12.10 данные убеди-
тельно показывают, что в случае существенных отклонений (20% в неблаго-
приятную сторону) переменных от принятого базового варианта чувствитель-
ность уменьшается в ряду: продажная цена, объем реализации продукта,
перерасход капитальных затрат. Лучше оценивать чувствительность или про-
центное изменение прибыли на инвестированный капитал относительно изме-
нения каждой переменной на ±5 и ±10%. Выполните такие расчеты для ука-
занных трех переменных и постройте графики зависимости прибыли на инве-
стированный капитал (в процентах) от отклонений (также в процентах) от
базовых величин продажной цены, объема реализации продукции и перерас-
хода капитальных затрат. Одинаков ли порядок чувствительности к различ-
ным переменным при небольших отклонениях (несколько процентов) и при
20%-ных отклонениях, рассмотренных в табл. 12.10? (Рассмотрите возмож-
ность применения разработочных таблиц в таких расчетах.)
12.3. Чувствительность экономического коэффициента. В графе Б табл.
12.10 приведены данные, отражающие результаты снижения продажной цены
продукта на 20%, однако причина снижения цены не указана.
а) Допустим, что продажная цена непосредственно связана с процентным
содержанием активного ингредиента в продукте и что этот фактор в свою
очередь непосредственно связан с обратным экономическим коэффициентом
(Ур/s)-1- Начертите график зависимости прибыли на инвестированный капи-
тал от отклонения Ур/s ( в процентах), соответствующего изменению продаж-
ной цены продукта на 0, ±5, ±10, ±20%.
б) Просмотрите в журнале Biotechnology and Bioengineering несколько
статей, в которых приведены графики, позволяющие определить экономиче-
ский коэффициент продукта. По приведенным там же исходным данным
подтвердите расчетным путем значение коэффициента и вычислите стандарт-
ное отклонение о2= (ц—ц)2/л. Какую долю (в процентах) составляет о от
Ур/s? С какой степенью достоверности можно предсказать величину Ур/s за
один цикл? Как этот результат повлияет на достоверность вашего расчета
прибыли на инвестированный капитал?
в) Предположим, что изменение Ур/s от цикла к циклу в «идентичных»
условиях можно описать распределением типа
#(yP/s) = e~{YP/S~Yp/s
Начертите график этой нормализованной функции распределения и оцените до-
полнительные расходы, связанные с тем, что приходится хранить продукцию
10 последовательных циклов, для того чтобы при смешении с другими ингре-
диентами средний состав смеси отклонялся от Ур/s на 5% или меньше.
12.4. Альтернативные процессы производства этанола. Недавно опубли-
кован обзор, посвященный сравнительному изучению различных процессов по-
лучения и выделения этанола \Maiorella В. L., Blanch Н. W., Wilke С. R., Bio-
tech. Bioeng., 26, 1003 (1984)]. Прочтите этот обзор, выберите какой-либо про-
Экономика процессов биохимической технологии
415
цесс и подготовьте соответствующие варианты таблиц 12.2—12.9. Суммируйте
все таблицы и составьте сводную таблицу объемом не более двух страниц
(частные таблицы представьте в виде приложения).
12.5. Конкурирующие процессы. Ваша компания открыла новый микробио-
логический продукт с заметной фармакологической активностью, который дол-
жен производиться в большом масштабе, с тем чтобы удалось завоевать 20%
потенциального рынка. Директор исследовательского центра компании, яв-
ляющийся специалистом в области органической химии, поставил задачу пе-
ред химиками-органиками и микробиологами за 3 месяца разработать проек-
ты «идеальных» полностью химического и микробиологического процессов
производства этого продукта. Экономический анализ представленных через-
3 месяца проектов показал, что с точки зрения расходов два пути эквивалент-
ны. Что бы вы рекомендовали руководству компании и почему?
12.6. Производство аминокислот. В справочном издании Kirk-Othmer
Encyclopedia of Chemical Technology прочтите раздел, посвященный произ-
водству аминокислот. Объясните, почему (как это указано в последнем столб-
це табл. 12.23) во всем мире только одна страна серьезно заинтересована
в развитии микробиологических способов производства аминокислот?
12.7. Экономика процесса производства L-лизина, а) Рассчитайте чув-
ствительность суммарных расходов (табл. 12.24) в процессе производства
L-лизина по отношению к каждой переменной (статье расхода). [Выразите
чувствительность как изменение суммарных расходов (в процентах) при из-
менении каждой переменной на 5%.]
б) Предположим, что принято решение о сооружении предприятия,
строительство и ввод в эксплуатацию которого потребует 5 лет. Взяв за ос-
нову данные табл. 12.24, найдите оптимальное решение, какое предприятие-
для производства L-лизина следует строить —• микробиологическое или хими-
ческое, если ожидается, что цена на мелассу будет возрастать на 10% в год.
(по сравнению с другими ценами), а цена на циклогексанол на 0, 10, 20
или 30% в год?
12.8. Совершенствование процесса; растворимые и иммобилизованные фер-
менты. Рассмотрим ферментативный процесс, включающий Nb рабочих циклов
в год, в каждом из которых расходуется количество фермента, равное Еь.
Сравним этот процесс с другим, в котором участвует иммобилизованный фер-
ментный катализатор и который также включает Nb рабочих циклов в год.
Иммобилизованный ферментный катализатор характеризуется массой носи-
теля Мс, удельной загрузкой катализатора С (отношение массы фермента Е
к массе носителя), коэффициентом активности иммобилизованного фермен-
та R (отношение массы иммобилизованного фермента к массе используемого
фермента) и временем жизни иммобилизованного фермента, равному ЕЕ рабо-
чих циклов.
а) Пусть Хе и Хс — стоимость единицы массы фермента и носителя соот-
ветственно, Еь — количество фермента, теряющегося после каждого цикла
с участием растворимого фермента, а Ес и Мс — количество фермента и но-
сителя, теряющихся после Nt циклов с участием иммобилизованного фер-
мента (без регенерации). Покажите, что разность между затратами на рабо-
чий цикл с участием растворимого фермента и такой же цикл с участием им-
мобилизованного фермента описывается выражением
XeEbNb — МсС (1 - Я) XeNi — McCRXeNi — McXcNi
б) С помощью приведенного в предыдущей задаче выражения покажите,
что процессы с участием иммобилизованного и растворимого ферментов эко-
номически равноценны, если степень превращения и стоимость субстрата рав-
ны в обоих процессах и если выполняется следующее условие:
(Nt\ 1
k E’b J
C
416
Глава 12
.где я — используемая доля общей активности иммобилизованного фермента
(фактор эффективности, гл. 4). Таким образом, иммобилизованный фермент
экономически выгоднее только в том случае, если время жизни иммобилизо-
ванного катализатора удовлетворяет условию
^•>^(1+Хг/С)/^
.Аналогичное сравнительное изучение периодического процесса с участием рас-
творимого фермента и непрерывного процесса с участием иммобилизованного
катализатора в зависимости от скорости инактивации катализатора и стоимо-
сти субстрата проведено в работе; Domach М. М., Chem. Eng. J., 29, Bl
(1984); здесь же приведены необходимые величины фактора эффективности т],
коэффициента активности иммобилизованного фермента R для ^-амилазы,
глюкозоизомеразы и лактазы.
12.9. Производство биологически опасных соединений. Рассмотренные в
этой главе проблемы экономики строительства и приборного оснащения, а так-
же оценки накладных расходов биохимических предприятий применимы к от-
носительно безопасным процессам и материалам. Производство потенциально
биологически опасных веществ, например цитотоксичных противоопухолевых
соединений или препаратов на основе вирусов человека, связано с дополни-
тельными расходами. Согласно рекомендациям Национального института по
.изучению рака, оценка этих расходов должна учитывать следующие факторы:
1. Контроль доступа к оборудованию (оборудование должно быть полностью
закрытым, огражденным).
:2. Поддержание отрицательного атмосферного давления (нельзя допускать
утечки воздуха через двери, окна или не снабженные фильтрами системы
вентиляции).
'3. Выброс воздуха должен осуществляться только через фильтры типа НЕРА,
обеспечивающие полную задержку пылевых частиц.
4. Полная местная изоляция всего оборудования, которое может быть источ-
ником образования аэрозолей (ферментеры, центрифуги, фильтры и т. д.).
5. Защита обслуживающего персонала: обеспечение одеждой, масками,
обувью, перчатками (расходы на закупку и ежедневную чистку или стирку)..
6. Регулярный медицинский осмотр обслуживающего персонала,
7. Постоянный контроль за загрязнением окружающей среды.
Предположим, что микробиологическое предприятие, производительность ко-
торого в четыре раза ниже производительности предприятия, детально рас-
смотренного в этой главе, предназначено для выпуска цитотоксичных соеди-
нений (в различных биологических процессах одинаковой продолжитель-
ности). С помощью приведенной в тексте схемы цеха ферментации составьте
все таблицы, необходимые для сравнения расходов на обычном предприятии
и предприятии, производящем цитотоксичные вещества. (Учтите стоимость
-оборудования, материалов, водо-, тепло-, газо- и энергоснабжения, эксплуа-
тационные расходы, прибыль на инвестированный капитал не рассчитывайте).
[Flickinger М. С., Sansome Е. В., Biotech. Bioeng., 26, 860 (1984).]
12.10. Экономика процесса с участием активного ила. Детальный анализ
аэрируемого реактора с рециркуляцией обогащенного биомассой потока, вы-
делением биомассы (отходы производства) центрифугированием и хлорирова-
нием обработанной воды дан в работе: Paterson R. В., Denn М. М., Chem.
Eng. J., 27, В13 (1983). Авторы этой работы показали, что общие расходы
предприятия практически нечувствительны в довольно широкохМ диапазоне
к двум основным переменным — концентрации биомассы в реакторе (повы-
шается при уменьшении объема реактора) и среднему времени пребывания
биомассы (в обогащенных биомассой системах с рециркуляцией эта перемен-
ная отличается от времени пребывания жидкости).
а) Ознакомьтесь с разд. 14.4, включая уравнение (14.5).
б) Чтобы понять кинетику процесса, решите упражнение 14.7.
Экономика процессов биохимической технологии
417
в) Прочтите указанную статью и обсудите, почему были получены такие
любопытные результаты. Обратите внимание на то, что типичные для всех
проектов эксплуатационные критерии основаны на одной и той же (99,25%)
степени восстановления поступающего в систему углеродсодержащего суб-
страта.
Литература
Общие вопросы
См. работу [5] в гл. 5, а также:
1. Bartholomew W. Н„ Reisman Н. В., Economics of Fermentation Processes,
Microbial Technology, 2d ed., Peppier H. J., Perlman D. (eds.), vol. 2.
pp. 463—496, Academic Press, Inc., New York, N. Y., 1979. (Здесь приведен
пример полного описания процесса, начиная с идеи создания и кончая
расчетом прибыли на инвестированный капитал.)
2. Commercial Biotechnology. An International Analysis, Off.ice of Technology
Assessment, Washington, D. C., January, 1984.
3. Stowell J. D., Bateson J. B., Economic Aspects of Industrial Fermentation,
in Bioactive Microbial Products 2: Development and Production, Nisbet L.J.,
Winstanley D. J. (eds.), p. 117, Academic Press, New York, 1983.
4. Rose A. H„ Economic Microbiology, vols. 1—5, Academic Press, New York,
1977—1981.
5. Peters M. S., Timmerhaus K. D., Plant Design and Economics for Chemical
Engineers, 3d ed., McGraw-Hill, N. Y., 1980. (Очень подробная монография,
в которой рассмотрены вопросы изучения начального проекта, оценки
стоимости, накладных расходов, налогов, прибыли на инвестированный ка-
питал, рентабельности, оптимального проекта, выбора материалов, обору-
дования для обработки материалов, теплообмена, массообмена и реак-
торов.)
6. Ulrich G., A Guide to Chemical Engineering Process Design and Economics,
John Wiley and Sons, New York, 1984. [Эта монография, состоящая из
двух разделов, построена аналогично работе [1]; в ней рассмотрены проб-
лемы проектирования процесса (идея создания; подготовка операционной
карты; спецификация и проектирование оборудования), экономического
анализа (расчет затрат на сооружение предприятия и эксплуатацию, опти-
мизация процесса, анализ рентабельности), подготовки технической доку-
ментации.]
7. Crueger №.. Crueger A., A Textbook of Industrial Microbiology (English
ed.), Science Tech., Inc., Madison, WI, 1984. (Многие главы, посвященные
различным типам биохимических трансформаций, содержат и полезную
информацию об экономике биопроцессов.)
8. Impact of Applied Genetics: Microorganisms, Plants, and Animals, Office
of Technology Assessment, Washington, D. C., 1981. (Обзор потенциальных
последствий и экономических проблем «новых» биотехнологических про-
цессов.)
9. Microbial Technology, 2d ed., vol. 1: Microbial Processes; vol. II: Fermenta-
tion Technology, Peppier H. J., Perlman D. (eds.), Academic Press, New
York. 1979. (Книга содержит множество данных о технической стороне
и экономике биопроцессов).
Частные проблемы:
10. Swartz R. W., The Use of Economic Analysis of Penicillin G Manufacturing
Cost in Establishing Priorities for Fermentation Process Improvement, Ann.
Reports on Fermentation Processes, 3, 75 (1979).
11. Avgerinos G. C., Wang D. I. C., Direct Microbiological Conversion of Cellu-
losics to Ethanol, Ann. Reports on Fermentation Processes, 4, 165 (1979).
27-746
418
Глава 12
(На с. 180—188 рассмотрена экономика процессов производства этанола.)
12. Ratledge С., Fermentation Substrates, Annual Reports on Fermentation
Processes, 1, 49 (1977). (Рассмотрены вопросы стоимости субстрата.)
13. Roberts R. S. et al.. Process Optimization for Saccharification of Cellulose by
Acid Hydrolysis, Biotech. Bioeng. Symp., 10, 125 (1980). (Приведен пример
получения гексоз из целлюлозы при минимальных затратах.)
14. Ackerson М. et al., Two Stage Acid Hydrolysis of Biomass, Biotech. Bioeng.
Symp., 10, 103 (1980). (Приведен пример превращения кукурузной соло-
мы в сбраживаемые сахара.)
15. Constantinides A., Application of Rigorous Optimization Methods to the
Control and Operation of Fermentation Processes, Ann. N.Y. Acad. Sci.,
326, 193 (1979).
16. Clausen E. C„ Gaddy J. L., Production of Ethanol from Biomass, Ann. N.Y.
Acad. Sci., 413, 435 (1983).
17. Litchfield J., Single Cell Protein, Science, 219, 740 (1983).
18. Aunstrup K., Andresen O., Falch E. A., Nielsen T. K., Production of Micro-
bial Enzymes, in Microbial Technology, 2d ed., Peppier H. J., Perlman D.,
(eds.), vol. 1, p. 283, Academic Press, Inc., New York, N.Y., 1979.
19. Ng T. K., Busche R. M., McDonald С. C., Production of Feedstock Chemicals,
Science, 219, 737 (1983). (Основное внимание уделено возобновляемым
ресурсам).
20. Maiorella В. L„ Wilke С. R., Blanch Н. W., Alcohol Production and Recovery,
Adv. Biochem. Eng., 20, 72 (1981). (Сравнительный анализ различных про-
цессов, включая их экономику.)
21. Maiorella В. L., Blanch Н. W., Wilke С. R., Economic Evaluation of Alter-
native Ethanol Fermentation Processes, Biotech. Bioeng., 26, 1003 (1984).
22. Lenz T., Moreira A., Economic Evaluation of the Acetone-Butanol Fermenta-
tion, Ind. Eng. Chem. Proc. Des. Dev., 19, 478 (1980).
23. Moo-Young M., Economics of SCP Production, Process Biochemistry, 12,
6 (1977).
24. Klausner A., Building for Success in Phenylalanine, Biotechnology, 3
(Anril), 301 (1985).
25. Cusida L. E., Industrial Microbiology, John Wiley and Sons, Inc., New York,
1968.
Глава 13
Изучение взаимодействий
в смешанных популяциях
микроорганизмов
Во всех предыдущих главах основное внимание мы уделили
системам, в которых доминирует один тип микроорганизмов,
и практически не касались чрезвычайно разнообразных и очень
широко распространенных систем, в которых примерно одинако-
во важны различные штаммы микроорганизмов. В качестве
примеров промышленных процессов с участием смешанных
культур микроорганизмов можно указать на биологическую очи-
стку сточных вод и сыроварение. Более того, в естественных си-
стемах смешанные популяции микроорганизмов являются ско-
рее правилом, чем исключением. Естественные кругообороты
углерода, азота, кислорода и многих других элементов требу-
ют активного участия многих типов микроорганизмов. Пробле-
мы практического использования смешанных культур будут де-
тальнее рассмотрены в следующей главе.
Здесь же мы сконцентрируем внимание на анализе различ-
ных взаимодействий микроорганизмов. При этом сначала попы-
таемся охарактеризовать взаимодействия в популяции, состоя-
щей из двух штаммов микроорганизмов, а затем распространим
наш анализ на более сложные системы. В следующем разделе
будут рассмотрены четыре из шести основных типов взаимодей-
ствий микроорганизмов. Позднее будет дан детальный матема-
тический анализ двух оставшихся типов взаимодействий. В за-
ключительных разделах главы мы рассмотрим более сложные и
большие системы, а также формирование пространственных
структур в смешанных популяциях.
13.1. Нейтрализм, мутуализм, комменсализм
и аменсализм
В случае смешанных культур, состоящих из двух штаммов
микроорганизмов, нейтрализм и мутуализм представляют со-
бой предельные варианты взаимодействий. Под нейтрализмом
понимают такое состояние системы, когда ни один из организ-
мов не оказывает влияния на скорость роста другого микроор-
ганизма. В такой ситуации, очевидно, не наблюдается никакого
27*
420
Глава 13
влияния одного штамма на другой —во всяком случае до тех
пор, пока принимается во внимание только скорость клеточного-
роста. Другим крайним типом взаимодействия является мутуа-
лизм, когда оба штамма растут быстрее в смешанной культуре,
чем в соответствующих чистых культурах. Наконец, при конку-
ренции, которую мы рассмотрим в разд. 13.3, каждый вид ока-
зывает отрицательное воздействие на скорость роста другого'
вида.
Известно лишь несколько примеров нейтрализма. Одним из
них является рост штаммов Streptococcus и Lactobacillus (вхо-
дящих в состав закваски для йогурта) в хемостате. При скоро-
сти разведения 0,4 ч-1 эти два вида микроорганизмов растут
практически с такой же скоростью, что и в чистых культурах.
Теоретически трудно представить, что может существовать мно-
жество ситуаций, когда поглощение питательных веществ и вы-
деление продуктов метаболизма одним видом не оказывает ни-
какого влияния на другой вид. Вероятно, нейтрализм может
наблюдаться только в особых системах, в которых каждый вид
утилизирует специфические питательные вещества, а конечные
продукты жизнедеятельности каждого вида эффективно нейт-
рализуются или разбавляются.
Говоря другими словами, нейтрализм — это такое состояние
системы, когда поведение организмов как первого, так и второ-
го видов не отличается от их поведения в соответствующих чи-
стых культурах. Тот факт, что нейтрализм встречается крайне
редко, заставляет усомниться в возможности описания смешан-
ных популяций с помощью данных, полученных при изучении
чистых культур. В общем случае предсказание поведения сме-
шанной культуры путем изучения чистых культур возможно
только тогда, когда используемые данные достаточно детально
характеризуют взаимосвязи между микроорганизмом и его окру-
жением. Суммирование такой информации по каждому виду
может дать сведения, полезные для описания смешанной куль-
туры.
По сравнению с нейтрализмом мутуализм распространен
гораздо шире и включает несколько различных механизмов.
Один из них заключается в обмене факторами роста. Показа-
тельным примером таких взаимодействий может служить рост
нуждающегося в фенилаланине штамма Lactobacillus и нужда-
ющегося в фолиевой кислоте штамма Streptococcus в смешан-
ной периодической культуре. На рис. 13.1 представлены ре-
зультаты изучения роста смешанной культуры на синтетиче-
ской среде, не содержащей ни фенилаланина, ни фолиевой кис-
лоты. Смешанная культура хорошо растет на этой среде, тогда
как соответствующие чистые культуры практически не растут
на ней.
Изучение смешанных популяций микроорганизмов 421
В основе мутуализма может также лежать обмен питатель-
ными веществами. Известно множество примеров, когда благо-
приятное влияние друг на друга оказывают аэробные бактерии
и фотосинтезирующие водоросли. В таких системах бактерии
утилизируют кислород и углеводы, а продуцируют СО2 и фак-
торы роста. Водоросли, используя в качестве источника энергии
солнечный свет, превращают СО2 в углеводы и выделяют О2.
Эта система может служить иллюстрацией некоторых стадий
РИС. 13.1. Рост периодических чи-
стых и смешанной культур нуждаю-
щегося в фенилаланине штамма Lac-
tobacillus arabinosas (кривая 2 — чи-
стая культура) и нуждающегося в
фолиевой кислоте штамма Streptococ-
cus faecalis (кривая 1 — чистая куль-
тура) в синтетической среде, ие со-
держащей ии фенилаланина, ни фо-
лиевой кислоты. Кривая 3 свидетель-
ствует об обусловленном мутуализ-
мом ускорении роста этих двух орга-
низмов в смешанной культуре. [Из ра-
боты: Nurmikko V., Biochemical Fac-
tors Affecting Symbiosis Among Bac-
teria, Experientia, 12, 245 (1956).]
кругооборота углерода и кислорода, которые мы рассмотрим
в гл. 14.
Очень тесные мутуалистические связи, когда без взаимо-
действия не может существовать ни один из видов, часто назы-
вают симбиозом. Показано, что микроорганизмы существуют в
многочисленных симбиотических связях с другими микроорга-
низмами или с высшими организмами. В табл. 13.1 приведены
характеристики некоторых симбиотических систем, в том числе
ряда чрезвычайно интересных. Для этого чтобы проиллюстри-
ровать другой тип мутуалистических взаимодействий, рассмот-
рим более детально один пример.
Показано, что «бактерия» Methanobacillus omelianskii, изо-
билующая в анаэробном иле (см. гл. 14), представляет собой
смесь двух видов. Один из них превращает этанол в водород и
ацетат
СН3СН2ОН+Н2О —* СН3СОО- + Н+ + 2Н2
но ингибируется продуцируемым им же водородом. Второй вид
Таблица 13.1. Примеры симбиотических взаимодействий с участием
по меньшей мере одного вида микроорганизма
Микроорганизм Другой организм или орган Примечания
Жгутиковые простейшие Термиты Простейшие гидролизуют целлюлозу для термитов, которые ее доставля- ют, но сами не могут переваривать. Простейшие являются хозяевами по отношению к бактериям, продуциру- ющим целлюлазные ферменты
Люминесцирую- Кальмары, неко- В железах кальмара обитают люми-
щие бактерии торые виды рыб несцирующие бактерии, которые обеспечивают кальмаров высокоспе- цифичным механизмом узнавания; взамен бактерии получают защиту и питательные вещества
Микроорганизмы Овцы, козы, круп- Первый отдел желудка коровы (ру-
рубца ный рогатый скот бец) содержит множество видов бактерий, которые в обмен на пита- тельные вещества помогают корове переваривать растительные материа- лы— целлюлозу, крахмал и липиды
Обычная мик- Кожа, гортань, Обычная флора, как было показа-
робная флора полость рта, ки- шечник но в опытах со стерильными живот- ными, играет важную, хотя и не вполне выясненную роль в предот- вращении многих заболеваний
Бактерии Простейшие Бактерии, живущие внутри орга- низма простейших (эндосимбиоз), вырабатывают питательные вещест- ва; по меньшей мере в одном слу- чае они обеспечивают простейших необходимыми аминокислотами и факторами роста
Водоросли Простейшие Каждое простейшее содержит от 50 до нескольких сотен водорослей; водоросли на свету связывают СО2 и выделяют О2, который использу- ется простейшими для окисления питательных веществ и регенерации СО2
Водоросли Грибы Эти организмы образуют тесное со- общество, называемое лишайником, которое выгодно главным образом в очень влажном или очень сухом окружении при ограниченном коли- честве питательных веществ; водо- росли обеспечивают грибы питатель- ными веществами; роль грибов не выяснена
Бактерии Бобовые рас- Бактерии живут в клубеньках, об-
Rhizobium тения разующихся на корнях растений, и питаются веществами, вырабатывае- мыми растениями; бактерии связы- вают атмосферный азот, делая его доступным для растений
422
Изучение смешанных популяций микроорганизмов 423
«бактерии» не способен расти на этаноле, но утилизирует во-
дород, превращая его в метан:
4Н2+СО2 —> СН4 + 2Н2О
Таким образом, здесь мы встречаемся с ситуацией, когда
второй вид разрушает токсин первого вида, являющийся одно-
временно питательным веществом для второго вида. Такой му-
туализм, связанный с нейтрализацией токсичного вещества,
возможен также тогда, когда аэробный организм защищает об-
лигатный анаэроб от избытка свободного кислорода. Другим
вариантом мутуализма является поддержание оптимального
значения pH в культуре организмов двух видов, один из кото-
рых повышает pH среды, а другой обеспечивает обратный эф-
фект.
Два последних из рассматриваемых в этом разделе типов
взаимодействий оказывают заметное влияние только на один
вид микроорганизмов. При комменсализме определенные пре-
имущества в смешанной культуре получает второй вид. Проти-
воположный эффект наблюдается при аменсализме, когда в ре-
зультате взаимодействия с первым видом второй вид оказыва-
ется в менее выгодной ситуации.
Иногда механизм комменсализма аналогичен последнему ме-
ханизму только что рассмотренного мутуализма. В этом случае
первый вид связывает вещество, являющееся токсичным для
второго вида, но в отличие от мутуализма второй вид не созда-
ет никаких благоприятных условий для первого. Как показыва-
ют данные, приведенные в табл. 13.2, такой тип комменсализ-
ма довольно обычен.
Еще более широко распространен другой вариант комменса-
лизма, когда один вид продуцирует соединения, ускоряющие
рост организмов другого вида. Конечные продукты жизнедея-
тельности одного вида, обеспечивающие эффект комменсализ-
ма, довольно многочисленны (см. табл. 13.3). В зависимости от
конкретной системы продуцируемое соединение может служить
источником энергии и (или) углерода для другого вида. По-
добные комменсальные взаимодействия часто объединяются в
цепи, отдельные звенья которых образуют комменсальные пары.
В периодической культуре, например, дрожжи могут превра-
щать глюкозу в спирт, являющийся питательным веществом для
Acetobacter. Этот организм продуцирует затем уксусную кисло-
ту, которая поглощается другими микроорганизмами и т. д.
В табл. 13.3 приведены также несколько примеров передачи
витамина или какого-либо другого фактора роста от одного ви-
да к другому. Это происходит особенно легко, если одним из
видов является ауксотрофный мутант. В таком случае коммен-
сализм будет обеспечен, если второй вид продуцирует метабо-
424
Глава 13
Таблица 13.2. Комменсализм; нейтрализация токсичных веществ®
Токсичное соединение
Механизм комменсализма
Концентрированные растворы сахаров Осмофильные дрожжи метаболизируют сахара и тем самым снижают осмотическое давление, позволяя расти видам, чувствительным к вы-
Кислород сокому осмотическому давлению Аэробные организмы могут снижать концентра- цию кислорода, позволяя таким образом ра-
Сероводород сти анаэробам Токсичный H2S окисляется фотосинтезирующими серными бактериями, что обеспечивает воз-
Пищевые консерванты можность роста других бактерий Ингибиторы роста бензоат и диоксид серы раз-
Молочная кислота рушаются биологическим путем Гриб Geotrichum candidum метаболизирует мо- лочную кислоту, продуцируемую Streptococcus lactis', в противном случае кислота накаплива-
Ртутные гермициды ется и ингибирует рост бактерий Организмы Desulfovibrio sp. превращают суль- фат в H2S; образующийся сульфид инактиви- рует ртутные гермициды и позволяет бактери-
Антибиотики ям расти Ферменты, продуцируемые некоторыми видами бактерий, разрушают антибиотики; таким об- разом могут расти чувствительные к антибио-
Фенолы тикам виды Некоторые бактерии способны окислять фенолы; благодаря этому обстоятельству другие виды могут расти в присутствии фенолов
Трихлорфенол Некоторые грамотрицательные бактерии могут адсорбировать трихлорфенол липидами клеточ- ных стенок и тем самым защищать Staphylo- coccus aureus от его действия
а Из работы: Meers J. L., Growth of Bacteria in Mixed Cultures, in Microbial Ecology,
Laskin A. J., Lechevalier H., (eds), p. 158, CRC Press, Cleveland, 1974.
лит, являющийся незаменимым веществом для роста первого
вида.
Большинство примеров комменсализма обнаружено при изу-
чении периодических культур. Действительно, строго коммен-
сальные взаимодействия в непрерывной культуре или в другой
открытой системе затруднены; в этом случае проблема связана
с возможностью появления конкурентных взаимодействий. Хо-
тя один организм может помогать другому, оба этих организма
могут конкурировать за одно питательное вещество, которое в
конце концов становится лимитирующим рост питательным ве-
ществом. Следовательно, в открытых системах комменсализм
можно ожидать только в тех случаях, когда взаимодействующие
Изучение смешанных популяций микроорганизмов
425
виды существенно различаются по своим потребностям в пита-
тельных веществах.
Аменсализм противоположен комменсализму; при аменса-
лизме рост одного вида подавляется в присутствии другого ви-
да. Ингибирующий эффект в таких случаях обычно обусловлен
или синтезом токсичных веществ, или поглощением незамени-
Таблица 13.3. Комменсализм; обеспечение незаменимыми питательными
веществами3
Соединение Организм, продуцирующий соединение Организм, нуждающийся в соединении
Пурин Bacillus subtilis Ауксотрофы В. subtilis
Органические кислоты Aerobacter cloacae Бактерия неназван- ного вида
Изобутират Corynebacterium diphthe- riae Тreponema microdentium
Никотиновая кислота Saccharomyces cerevisiae Proteus vulgaris
Витамин К Staphylococcus aureus Bacteroides melaninogenicus
Нитритный ион Nitrosomonas Nitrobacter
Сероводород Desulfovibrio Серные бактерии
Вода Bacillus mesentericus Clostridium botulinum
Полисахариды Водоросли Бактерии
Водород Бактерии рубца Methanobacterium rumina- tium
Метан Анаэробные метановые бактерии Окисляющие метан бактерии
Аммониевый ион Многие гетеротрофы Nitrosomonas
Нитрат Nitrobacter Денитрифицирующие бактерии
Ацетилфосфат Corynebacterium diphthe- riae Boreilia Vincenti
Фруктоза Acetobacter suboxydans Saccharomyces carlsbergen- sis
аИз работы: Meers J. L., Growth of Bacteria in Mixed Cultures, in Microbial Ecology,
Baskin A. J„ Lechevalier H. (eds.), p. 156, CRC Press, Cleveland, 1974.
мых питательных веществ. В большей части известных приме-
ров аменсализм обусловлен первым механизмом; в таких слу-
чаях второй организм создает окружение, в котором первый
вид с трудом поддерживает жизнеспособность или вообще по-
гибает.
Мы уже видели, что некоторые антибиотики продуцируются
микроорганизмами. Один из обычных лабораторных тестов на
биосинтез антибиотиков, по сути дела, представляет собой
тест на аменсализм; в этом тесте синтезирующий антибиотик и
чувствительный к антибиотику виды выращивают совместно на
426
Глава 13
агаровой пластинке. Наличие чистых зон вокруг продуцирую-
щих антибиотик колоний является свидетельством в пользу
антибиотической активности (рис. 13.2). (В частности, именно
таким методом был открыт пенициллин.) Чистые зоны образу-
ются в результате диффузии антибиотика из синтезирующей его
колонии, что приводит к ингибированию роста чувствительного
штамма.
О синтезе антибиотиков плесневыми грибами и актиномице-
тами упоминалось выше. Чтобы оценить возможную роль ан-
тибиотиков в экологии микроорганизмов, следует учесть, что БО-
РИС. 13.2. Аменсализм: диффузия антибиотика из колоний грибов, продуци-
рующих антибиотик, ингибирует рост организмов другого вида на агаровой
среде. (Фотография любезно предоставлена компанией Charles Pfizer and Со.,
Inc.)
доросли и другие бактерии также могут продуцировать анти-
биотики. Большая часть таких антибиотиков по химической
природе относится к полипептидам. Подобно пенициллину мно-
гие из них ингибируют синтез компонентов клеточной стенки,
а другие разрушают барьер проницаемости, обеспечиваемый
клеточной мембраной.
Близкий тип аменсализма обусловлен секрецией некоторыми
микроорганизмами ферментов, разрушающих полимеры кле-
точной стенки. Деполимеризуя с помощью литических фермен-
тов клеточные стенки организмов других видов, подобные орга-
низмы получают два преимущества. Во-первых, таким путем
один вид уничтожает другой потенциально конкурентноспособ-
ный вид и, во-вторых, разрушенные клетки являются источни-
ком питательных веществ, которые могут утилизироваться мик-
Изучение смешанных популяций микроорганизмов 427
роорганизмами, продуцирующими ферменты. Действительно,
если образующиеся таким путем питательные вещества вносят
существенный вклад в питание продуцирующих ферменты ви-
дов, то последние, очевидно, выигрывают в таком окружении.
Тогда аменсалитические взаимодействия трансформируются в
паразитические (см. разд. 13.4).
Синтез микроорганизмами органических кислот также мо-
жет быть причиной ингибирования, но не сголь эффективного,
как в предыдущем случае. Кислоты часто снижают pH и замед-
ляют рост других организмов. Возможны и более специфиче-
ские ингибирующие эффекты органических кислот. Так, муравьи-
ная и уксусная кислоты ингибируют рост Shigella flexneri, а про-
пионат и ацетат способны ингибировать рост бактерии Propioni-
bacteriutn shermanii. К другому типу аменсализма относится
ингибирование хемоаутотрофов небольшими количествами ам-
миака, продуцируемого другими видами, особенно использую-
щими в качестве источника энергии аминокислоты.
13.2. Классификация взаимодействий между двумя
видами
Рассматриваемая здесь классификация взаимодействий
между двумя различными видами базируется на изучении ди-
намических свойств смешанной культуры. Такие динамические
свойства можно описать с помощью линеаризованной модели
типа рассмотренной в разд. 9.2.1. Пусть п — вектор с элемента-
ми щ, каждый из которых равен числу (или численной кон-
центрации, т. е. числу клеток в единице объема) организмов
вида i в системе, тогда уравнения балансов по численностям
видов для системы с полным перемешиванием можно записать
в виде
-^Р-t (П (0, ш) (13.1)
at
где m — вектор параметров, характеризующих окружение орга-
низмов. В контексте экологии смешанных культур матрица ко-
эффициентов А соответствующей линеаризованной модели с
элементами
_ dff(ns,m)
Clij —
dnj
называется матрицей сообщества. Как уже упоминалось в гл. 9,
ns в уравнении (13.2) обозначает постоянный во времени век-
тор п, соответствующий уравнению (13.1) в применении к ста-
ционарному состоянию.
428
Глава 13
Переменные, используемые для описания концентраций
(плотностей) отдельных видов в системах на основе смешанных
культур (и в ряде других), заслуживают особого внимания.
Изучение роста чистых культур в переходном состоянии (напри-
мер. периодической культуры) показывает, что средняя масса
одной клетки изменяется во времени (вспомните разд. 7.3.1); от-
сюда следует, что массовая и численная концентрации клеток
изменяются во времени неодинаково. Говоря другими словами,
отношение х^щ не постоянно во времени (здесь Xi и nt- — мас-
совая и численная концентрации вида i соответственно). Изу-
чение моделей кинетики показывает, что описание кинетики ро-
ста существенно упрощается и может быть сведено к сравни-
тельно простым функциональным зависимостям, рассмотренным
в гл. 7, если оно базируется на массовой, а не на численной
концентрации организмов. Например, основанная на известных
особенностях клеточного цикла Е. coli кинетика синтеза био-
массы первого порядка подразумевает процесс повышения чис-
ленной концентрации первого порядка с лаг-фазой, что обуслов-
ливает значительно более сложное динамическое поведение и
существенно усложняет анализ переходных состояний (вспом-
ните разд. 9.2.2). Следовательно, в общем случае предпочти-
тельно опираться на общую концентрацию клеточной массы.
К сожалению, экспериментальные методы определения плот-
ностей различных видов в смешанных культурах обычно сво-
дятся к подсчету колоний, растущих на пластинках. Получен-
ные таким путем данные учитывают число клеток, но не их мас-
совую концентрацию. Одновременное измерение среднего раз-
мера клеток с помощью микроскопа или другим методом (по
рассеянию света или с помощью счетчика Коултера) в принци-
пе позволяет перевести полученные результаты в величины мас-
совой концентрации, но для смешанных культур такие данные
чрезвычайно скудны. Поскольку критерием справедливости лю-
бой теории служит сравнение с экспериментальными результа-
тами, то рассматриваемые в этой главе данные будут выраже-
ны в основном в величинах численной концентрации клеток.
Следует подчеркнуть, что некоторые затруднения, возникающие
при описании с помощью кинетических моделей динамики чис-
ленности клеток, могут быть обусловлены неудачными попыт-
ками использовать переменные, выраженные в единицах массы
клеток, и одновременно учесть переходные состояния в разме-
рах клеток.
Взаимодействия между двумя видами вблизи стационарного
состояния характеризуются двумя элементами в матрице сооб-
щества А. Если мы рассмотрим виды i и /, то нетрудно заме-
тить, что небольшие увеличения популяции / по сравнению с
соответствующим значением для стационарного состояния
Изучение смешанных популяций микроорганизмов
429
Тх,(0)>0] приводят к увеличению начальной производной
на выражение (Во избежание путаницы с концентра-
циями клеточной массы отклонение [tii(t)—tiis] мы будем
обозначать %/(0.) Таким образом, если численность популяций
всех других видов первоначально соответствует стационарному
состоянию [т. е. %/;(0) =0, k=£j], то знак будет зави-
сеть от знака а,ц Можно сказать, что если величина поло-
жительна, то организмы вида / оказывают положительное влия-
ние на рост организмов вида i, а если отрицательно, то на-
лицо ингибирующий эффект. Отсутствию взаимодействия меж-
Таблица 13.4. Классификация парных взаимодействий,
основанная на знаке элементов ац и ац матрицы сообщества A (b^j)*
Влияние вида / на вид i (знак а(-,)
— 0 4“
Влияние - - — 0 —ь
вида i Конкуренция Аменсализм Хищничество
на вид Д 0 0 — 00 0 +
(знак ац) Аменсализм Нейтрализм Комменсализм
+ 4— + 0 + +
Хищничество Комменсализм Мутуализм
* По данным, приведенным в работе: May R. М., Stability and Complexity in Model
'Ecosystems, p. 25, Princeton University Press, Princeton, N. J., 1973.
ду видами отвечает aij = 0. Те же самые доводы приложимы и
к изучению влияния вида i на вид /: соответствующий эффект
носит стимулирующий характер при положительном ац, ингиби-
рующий— при отрицательном ац, нейтральный — при нуле-
вом ац.
Определяемые таким путем все возможные сочетания взаи-
модействий между видами представлены в табл. 13.4. Нетрудно
видеть, что все четыре типа парных взаимодействий, рассмот-
ренных в предыдущем разделе, учитываются этой схемой. Кро-
ме того, в таблице появились два новых типа взаимодействий —
конкуренция и хищничество. Прежде чем в деталях рассматри-
вать эти типы взаимодействий, мы изучим некоторые стороны
•влияния типа взаимодействия на динамику смешанной популя-
ции вблизи стационарного состояния. Пока что ограничимся
изучением смешанных культур, состоящих из двух взаимодей-
ствующих видов.
Пример 13.1. Динамика системы, состоящей из двух видов, вблизи ста-
щионарного состояния. Если взаимодействуют только два вида, то, как следует
430
Глава 13
из соответств}-’ющего варианта уравнения (9.30) щества имеет форму 2x2: , в этом случае матрица сооб-
d(n,—nls) dY,
.. = — «11X1-г «12X2 dt dt
[(13П1.1)
d(n2 — n2s) dv2
J. — JI —^21X1+^22X2 at at
или
d Г Xi I Г«п «12 V ч
(13П1.2>
df L X2 J L«21 «22. L Хг J
Характерное уравнение (9.34) можно легко преобразовать в уравнение второй
степени
№ — (flii + 022) А+ (fli 1^22 — 0120ц) = X2 — (tr А) А + det А=0
(13П1.3)
где trA и detA обозначают след и детерминант матрицы А соответственно.
Применив к уравнению (13П1.3) критерий Гурвица—[уравнение (9.41)],
нетрудно найти, что линеаризованная система асимптотически устойчива
в том и только в том случае, если
detA>0 (13П1.4)
trA<0 (13П1.5)
Другие детали динамики системы можно найти, решив уравнение
(13П1.3):
tr А 1 _______________
А1>2 = —— ± — V(trA)2-4det А (13П1.6>
При trA = O (и detA>0) или при detA, равном нулю, по меньшей мере одно-
собственное значение будет иметь нулевую действительную часть. Исключим
этот критический вариант из нашего дальнейшего анализа и перейдем к ана-
лизу других возможностей. (Типичные для критических ситуаций бифуркации
рассматриваются в разд. 13.5.5.) Если
( tr А \2
------ >detA>0 (13П1.7)
\ 2 /
то оба собственных значения действительны и имеют один и тот же знак; со-
ответствующее стационарное состояние ns называется узловой точкой. Если
detA<0, то одно собственное значение положительно, а другое отрицательно;
такое стационарное состояние называют седловой точкой. Комплексные соб-
ственные значения наблюдаются в том случае, если 4 det А больше, чем квад-
рат trA. В этом случае стационарное состояние называют фокальной точкой.
Причину появления таких специфических терминов, заимствованных из
математического аппарата механики, легче понять, если обратиться к анали-
зу траекторий численности популяций в фазовой плоскости. По определению
координатами фазовой плоскости являются %i и %2. Определенные начальные
условия (Xi(0), %г(0)) можно представить точкой на этой плоскости. Анало-
гично если мы нанесем на фазовую плоскость все точки, относящиеся ко вре-
менам t (xi(0> Хг(О), то получим непрерывную кривую, называемую траекто*
рией. Следовательно, изображение траекторий в фазовой плоскости является
Изучение смешанных популяций микроорганизмов
431
удобным способом представления динамического поведения двух переменных
на одном графике.
На рис. 13П1.1 представлены траектории для трех обычных ситуаций.
Каждая из траекторий соответствует определенным начальным условиям,
а стрелки указывают направление увеличения времени. Для того чтобы отра-
зить поведение неустойчивых узловых и фокальных точек в фазовой плоско-
сти, нужно изменить направление стрелок на траекториях, изображенных на
рис. 13П1.1, а и б, на обратное. (Почему?)
Возвращаясь к классификации, представленной в табл. 13.4, нетрудно
видеть, что она включает только недиагональные элементы матрицы А. Сле-
РИС. 13П1.1. Некоторые характерные типы траекторий в фазовой плоскости,
отображающие линеаризованную динамику экосистем, состоящих из двух ви-
дов: а — устойчивая узловая точка; б — устойчивая фокальная точка; в — сед-
ловая точка.
довательно. без дополнительных данных мы ничего не можем сказать о trA
и очень мало о det А.
Теперь рассмотрим достаточно обоснованные допущения относительно ав-
тономных удельных скоростей роста йц и а22 в случае некоторых из уже ука-
занных типов взаимодействий. Мутуализм может привести к большей числен-
ности популяции в стационарном состоянии (по сравнению со стационарными
состояниями без взаимного благоприятного влияния видов друг на друга),
так что ац<0, а22<0. Очевидно, что выполняется также условие (13П1.5), но
условие (13П1.4) будет выполнено только в том случае, если
ttlltt22>ttl2®21 (13П1.8)
Если это неравенство не удовлетворяется, то такое мутуалистическое ста-
ционарное состояние неустойчиво.
В системе, состоящей из двух видов, как в случае комменсализма, так
и в случае аменсализма характеристики устойчивости полностью зависят от
автономных скоростей роста, поскольку detA = aIIa22 и tr А = аи+а22. Отсюда
следует, что асимптотическая устойчивость возможна только в том случае, ес-
ли и ап, и й22 отрицательны.
Как мы увидим в разд. 13.5, некоторые из полученных в этом примере вы-
водов можно распространить и на более сложные популяции.
432
Глава 13
13.3. Конкуренция: выживание наиболее приспособ-
ленного вида
В работах Дарвина о естественном отборе подчеркивалась
важность конкуренции между различными видами. В том смыс-
ле, в каком бы будем употреблять его здесь, термин конкурен-
ция означает зависимость двух видов от одного фактора, на-
пример от источника питательного вещества, света, наличия
пространства или от какого-либо другого лимитирующего рост
вида фактора. Утилизация этого общего ресурса одним видом
уменьшает его доступность для другого вида, что оказывает
отрицательное влияние на скорости роста обоих организмов.
При анализе конкуренции нас прежде всего должно интере-
совать, не имеет ли один из видов какого-либо естественного
преимущества перед другим видом. Чисто умозрительно можно
предположить, что вид с более высокой скоростью роста дол-
жен быть лучше приспособленным, поскольку именно благода-
ря быстрому росту он способен утилизировать больше общего
лимитирующего рост ресурса, чем медленнее растущий орга-
низм. Как мы увидим ниже, экспериментальные данные и ре-
зультаты математического анализа с помощью соответствующих
моделей подтверждают это предположение.
Рассмотрим пример, представленный на рис. 13.3. Растущие
в чистых культурах (незачерненные символы) Е. coli и Staphy-
lococcus aureus достигают примерно одинаковой максимальной
численности популяции, хотя Е. coll растут быстрее. Если же
эти два вида бактерий культивировать в смешанной периодиче-
ской культуре, то картина роста Е. coll практически не будет
отличаться от роста этого организма в чистой культуре. Напро-
тив, максимальная численность популяции S. aureus в смешан-
ной культуре намного ниже, чем в чистой культуре. Поскольку
Е. coli не продуцируют веществ, ингибирующих рост Staphy-
lococcus aureus, то этот эффект можно объяснить преимущест-
венным поглощением питательных веществ бактериями Е. colL
(При каком допущении справедлив этот вывод?)
13.3.1. Анализ межвидовой конкуренции по Вольтерра
Известный итальянский математик Вито Вольтерра, выяс-
нивший многие принципы математической экологии, изучал
рост двух конкурирующих организмов в изолированной системе.
В нелинейной модели Вольтерры предполагается, что автоном-
ные удельные скорости роста pi и р2 положительны. Далее при-
нимается, что первый вид утилизирует лимитирующее рост пи-
тательное вещество со скоростью h{nb а второй — со скоростью
Л2п2, причем и hlf и h2 также имеют положительное значение.
Изучение смешанных популяций микроорганизмов
433
РИС 13.3. Снижение скорости роста S. aureus в смешанной культуре вслед-
ствие конкуренции этого организма с Е. coll. [Из работы: Oberhofer Т. R.,
Frazier W. С., Competition of Staphylococcus aureus with Other Organisms, J.
Milk Food Techno!., 24, 172 (1961).]
1 — E. coli в чистой культуре; 2— E. coli в смешанной культуре; 3 — S. aureus
в чистой культуре; 4 — S. aureus в смешанной культуре.
Утилизация одного питательного вещества двумя видами при-
водит к снижению соответствующих удельных скоростей роста,
и поэтому
= Т1(Мг+Мг)М1 (13.3)
= [Нг~~?2 (^i^i^2^2)] п2 (13-4)
В этих уравнениях константы Yi>0 и у2>0 отражают различ-
ное влияние истощения питательного вещества на виды 1 и 2
соответственно. Вспомнив разд. 7.3.2, нетрудно заметить, что
эта модель является расширенным вариантом рассмотренной
28—746
434
Глава 13
нами ранее модели Ферлхурста и Перла, в котором учтена сме-
шанная природа культуры.
Окончательный результат конкуренции можно предсказать,
умножив уравнение (13.3) на y2Mi и уравнение (13.4) на 71/П2,
а затем сложив полученные выражения; таким путем получим
d In п, d In n2 -
V2 - — — Vi —= T2M1 — T1H2 (13.5)
at at
Интегрирование этого уравнения дает
—— = Cexp(T2L(1—y^t (13.6)
«Г
:где С — константа интегрирования, зависящая от /21(0) и /г2(0).
Теперь предположим, что —W2 имеет отрицательное
значение. (Теряется ли при этом общность модели?) Тогда ле-
вая часть уравнения (13.6) при увеличении времени приближа-
ется к нулю. Понятно, что причиной этого не может быть без-
граничный рост п2. Возвращаясь к уравнению (13.4), можно
видеть, что dn2/dt отрицательно, если п2~> р2/^2h2\ отсюда сле-
дует, что величина п2 ограничена. Тогда единственной причиной
обсуждаемого эффекта, предсказываемого уравнением (13.6),
может быть приближение П\ во времени к нулю.
Результат рассмотренного анализа известен под названием
принципа конкурентного исключения. Когда два вида конкури-
руют в общем окружении и yi/p 1 >72/Ц2, то численность попу-
ляции второго вида достигает предельного значения 112/72^2,
а первый вид в конце концов вообще вымирает. Путем анало-
гичных рассуждений нетрудно найти, что этот принцип может
быть распространен и на смешанные культуры, состоящие из
многих видов:
= (Pi— У1?(щ, п2, . . . , 77,,)] Пг 7=1,2,... , р (13.7)
dt
Опять-таки конечным результатом, согласно этому уравнению,
будет гибель всех видов за исключением одного. Обратите вни-
мание на то, что, поскольку величины ц, считаются независимы-
ми от плотности популяции, это предполагает, что источник пи-
тательных веществ неисчерпаем или питательные вещества по-
стоянно подаются в систему. Строго говоря, принцип конку-
рентного исключения применим только к открытым экосисте-
мам.
На рис. 13.4 приведены экспериментальные данные, нагляд-
но иллюстрирующие принцип конкурентного исключения. Здесь
представлены результаты изучения периодической системы,
в которую постоянно подаются питательные вещества; один из
Изучение смешанных популяций микроорганизмов
43S
видов простейших, рост которого в отсутствие других видов опи-
сывается логистической кривой [уравнение (7.52)], вымирает»
конкуренции с другими организмами, обладающими более высо-
кими скоростями роста.
РИС. 13.4. Экспериментальное подтверждение принципа конкурентного исклю-
чения при росте двух видов простейших. В смешанной культуре медленнее
растущий вид Paramecium caudatum в конце концов погибает. [Из работы:
Gause G. F., Experimental Analysis of Vito Volterra’s Mathematical Theory of
the Struggle for Existence, Science, 79, 16 (1934).]
1 — P. caudatum в чистой культуре; 2 — P. caudatum в смешанной культуре;
3 — Р. aurelia в чистой культуре; 4 — Р. aurelia в смешанной культуре.
Если учесть целый ряд допущений, принятых при выводе
уравнений (13.3) и (13.4), то вызывает некоторое удивление
удачное применение принципа конкурентного исключения для
объяснения результатов указанного эксперимента и многих дру-
гих реальных систем. При этом, однако, следует принимать во
внимание, что математическая модель такой формы применима
и в случае систем, которые в физическом смысле существенно
отличаются от системы, явившейся основой для вывода урав-
нений модели. Действительно, с математической точки зрения
28
436
Глава 13
уравнения (13.3) и (13.4) можно рассматривать и как прибли-
женный результат разложения общего уравнения в ряд Тейло-
ра вплоть до членов второго порядка.
13.3.2. Конкуренция и отбор в хемостате
Теперь рассмотрим роль конкуренции в открытых системах,
например в хемостате. Если допустить, что удельная скорость
роста рг- вида i постоянна, то динамика изменения концентра-
ции вида будет описываться уравнением
Х> (0 = »(«)«'- <' = 1.2..Р (13.8)
Отсюда следует, что относительные значения р; и D определя-
ют, будет ли %г- возрастать, уменьшаться или останется постоян-
ной во времени. Судьбу смешанной культуры, состоящей из
двух видов, можно проанализировать с помощью эвристическо-
го подхода на базе модели Моно.
Можно достаточно обоснованно допустить, что зависимость
скорости роста от концентрации лимитирующего рост субстра-
Концентрация лимитирующего
рост субстрата s
РИС. 13.5. Гипотетическое изменение
скоростей роста видов 1 и 2. В тек-
сте объясняются причины вымывания
вида 1 из непрерывной смешанной
культуры. [Из работы: Meers J. L.,
Tempest D. W., The Influence of Extra-
Cellular Products on Mixed Microbial
Populations in Magnesium-Limited
Chemostat Cultures, J. Gen. Microbiol.,
52, 309 (1968).]
та будет одинакова для двух конкурирующих организмов толь-
ко в исключительных случаях. Отсюда следует, что существуют
две возможности, одна из которых схематично представлена на
рис. 13.5 (кривые роста не пересекаются), а при второй кривые
роста будут пересекаться. Сначала допустим, что реализуется
ситуация, схематично изображенная на рис. 13.5.
Предположим, что популяция организма 1 растет в стацио-
нарном состоянии в хемостате при скорости разведения D. Тог-
да, как это отражено на рис. 13.5, концентрация лимитирующе-
го рост субстрата в хемостате будет равна Если теперь в хе-
Изучение смешанных популяций микроорганизмов
437
РИС. 13.6. Рост конкурирующих популяций A. aerogenes (□) и Т. utilis {As)
в хемостате при pH 6,4 и температуре 33 °C в среде, в которой рост организ-
мов лимитируется солями калия: а — В=0,05 ч-1; б — В=0,3 ч”1. Штриховые
линии отвечают вычисленным кривым вымывания нерастущих организмов.
[Из работы: Meers J. L., Growth of Bacteria in Mixed Cultures, in Microbial
Ecology, Laskin A. I., Lechevalier H. (eds.), p. 142, CRC Press, Inc., 1974.
© CRC Press, Inc.]
мостат ввести популяцию организма 2, то сначала эти организ-
мы будут расти со скоростью ц2- Поскольку то числен-
ность популяции 2 будет возрастать во времени, вызывая сни-
жение концентрации лимитирующего рост питательного веще-
ства. При s<$i удельная скорость роста вида 1 будет меньше D
и популяция организмов 1 начнет вымываться. Такая ситуация
сохранится до тех пор, пока концентрация субстрата не достиг-
нет величины $2, при которой популяция вида 2 достигает ста-
ционарного состояния, а вид 1 будет все так же вымываться.
Подобная картина неоднократно наблюдалась эксперимен-
тально. Так, при одновременном росте бактерий Aerobacter
aerogenes и дрожжей Torula utilis в хемостате скорость разве-
438
Глава 13
дения, соответствующая вымыванию медленнее растущих дрож-
жей, как оказалось, зависит от общей скорости разведения (см.
рис. 13.6), как и предсказывает уравнение (13.8). Приведенные
на рис. 13.6 данные свидетельствуют о том, что в системе одно-
временно происходят резкие изменения и колебания численно-
сти видов, не отражаемые нашей упрощенной моделью. Другие
экспериментальные результаты, например представленные на
рис. 13.7, соответствуют указанной гипотетической последова-
тельности событий с гораздо большей точностью.
РИС. 13.7. Конкуренция между двумя бактериями (Е. coli и Azotobacter vine-
landii) в хемостате. [Из работы: Jost J. L. et al:, Interactions of Tetrahymena
pyriformis, Escherichia coli, Azotobacter vinelandii and Glucose in a Minimal
Medium, J. Bacteriol., 113, 834 (1973).]
Если кривые, отвечающие двум функциям gi (s) и Ц2($), пе-
ресекаются, так что относительное значение каждой из них оп-
ределяется концентрацией субстрата, то, рассуждая аналогич-
ным образом, можно прийти к выводу, что от скорости разве-
дения зависит, какой из видов будет вымываться. На рис. 13.8
представлен интересный пример подобной зависимости конку-
рентного превосходства одного из видов от скорости разведе-
ния. При малой скорости разведения в смешанной культуре до-
минируют дрожжи, однако при повышении скорости разведения
дрожжи начинают вымываться и в более благоприятной ситуа-
ции оказываются бактерии. При снижении скорости разведения
эти два вида меняются ролями.
Прежде чем перейти к изучению других интереснейших ти-
пов межвидового взаимодействия — хищничества и паразитиз-
ма,— следует подчеркнуть, что все приведенные выше рассуж-
дения можно применить и к популяции организмов одного вида,
обладающих различными характеристиками. Например, в спе-
цифических условиях в хемостате могут развиться мутантные
Изучение смешанных популяций микроорганизмов 439
штаммы, особенно хорошо приспособленные к росту в данных
условиях. В нашем анализе эти штаммы можно рассматривать
как организм 2; популяция последнего может прогрессировать
и в конце концов вытеснить начальный вид. Точно так же воз-
никновение конкурентоспособных мутантов в периодическом ре-
акторе следует рассматривать как превращение чистой культу-
РИС. 13.8. Влияние изменения скорости разведения на непрерывную смешан-
ную культуру Bacillus subtilis (/) и Т. utilis (2) (температура 33 °C, pH 6,4;
клеточный рост лимитируется солями магния). [Из работы: Meers J. L., J.
Gen. Microbiol., 67, 359 (1967).]
ры в смешанную, для которой типичны рассмотренные в преды-
дущем разделе взаимодействия.
Принцип исключения, обусловленного межвидовой конкурен-
цией, имеет очень важные практические следствия. Действи-
тельно, целью многих процессов биохимической технологии яв-
ляется не производство биомассы, а достижение максимального
выхода продукта жизнедеятельности вида. Штаммы, обеспечи-
вающие высокий выход продукта, могут обладать меньшей
удельной скоростью роста, чем родственные мутантные штаммы.
В такой ситуации резко возрастает вероятность ускоренного
роста популяции нежелательных мутантов в хемостате, что при-
ведет к вымыванию продуктивного штамма. Представленные на
рис. 13.6—13.8 экспериментальные данные убедительно показы-
вают, что в непрерывной культуре любое преимущество штам-
ма, обусловленное более высокой начальной плотностью его
популяции, может быть быстро утеряно, а посторонний или му-
440
Глава 13
тантный штамм с большей по сравнению с исходным штаммом
величиной ц в конечном счете станет доминирующим. До сих
пор мутанты с нужными свойствами применялись в основном в
реакторах периодического действия; в будущем, очевидно, воз*
никнет необходимость получения продуктивных штаммов, спо-
собных успешно конкурировать с бесполезными мутантами и в
реакторах непрерывного действия.
В контексте межвидовой конкуренции в открытых системах
следует также упомянуть и об эффекте пристеночного роста»
рассмотренном в гл. 9. Мы уже говорили (разд. 9.1.3) о том»
что связанная с поверхностью популяция может существовать
в хемостате даже в том случае, если D превосходит максималь-
ную удельную скорость роста. Аналогично если медленно расту-
щий организм находится в удобной экологической нише, на-
пример в пленке, то как показано экспериментально, он может
успешно конкурировать с видами, обладающими более высокой
скоростью роста.
Пример 13.2. Конкуренция при росте культур неустойчивых рекомбинант-
ных организмов. При росте содержащих плазмиды клеток, которые недоста-
точно стабильно реплицируются или расщепляются, обнаруживаются многие
различные «виды» клеток, различающиеся числом имеющихся в них плазмид.
Здесь мы условно будем считать, что анализируемая система состоит из кле-
ток двух типов — содержащих плазмиды (их мы обозначим символом Х+)
и не имеющих плазмид (X-). Описывая рост этих двух «видов» в самом об-
щем виде, можно записать
Х+—> (2 — р)Х++рХ~ Скорость |х+х+ (13П2.1)
X- —> 2Х~ Скорость |х~х~ (13П2.2)
Параметр р, определяющий число свободных от плазмид клеток, которые
образуются в поколении клеток, содержащих плазмиды, является мерой сте-
пени расщепления при передаче плазмид. Предполагается, что два «вида»
характеризуются различными удельными скоростями роста, поскольку в несе-
лективной среде плазмидсодержащие клетки обычно растут медленнее, чем
клетки, не имеющие плазмид. (Почему?) Обратная ситуация типична для се-
лективных сред.
Рассматривая периодическую культуру нестабильного штамма с постоян-
ными удельными скоростями роста, легко показать, что через N поколений
плазмидсодержащих клеток доля (F) клеток с плазмидами составит
1_а'^<а4-.)р (13П2.3)
где а— отношение удельных скоростей роста
а=ц-/Ц+ (13П2.4)
Размножение чистой культуры до популяции, необходимой для большого био-
реактора, обычно требует не менее 25 удвоений. На рис. 13П2.1 представлены
зависимости доли плазмидсодержащих клеток после 25 удвоений популяции
(Раз) от параметров аир. Эти расчеты показывают, что опасность потери
существенной доли продуктивной плазмидсодержащей популяции достаточно
реальна (такие случаи иногда наблюдались и на практике).
Изучение смешанных популяций микроорганизмов
44!
РИС. 13П2.1. Доля плазмидсодержащих клеток в периодической культуре
после 25 поколений. (По данным, приведенным в работе [12].)
Такой тип генетической нестабильности существенно влияет и на работу
реакторов непрерывного действия. Если допустить, что в ПРПП ц+ = ц_ = ц, то
изменения масс двух популяций во времени можно описать дифференциаль-
ными уравнениями
//у+
——= (1 — р) р,х+ — Dx+ (13П2.5)
at
dx~
-----= р,х~ + рц.х+ — Dx (13П2.6)
dt
Обозначив символом х общую концентрацию биомассы (х=х++х~) и сложив
два последних уравнения, легко показать, что х имеет отличное от нуля зна-
чение для стационарного состояния только при £>=ц. В этих условиях, одна-
ко, решение уравнения (13ГТ2.5) дает
x+(/)==x+(0) —> О (13П2.7)
Следовательно, концентрация плазмидсодержащих клеток убывает экспонен-
циально, приближаясь в стационарном состоянии к нулю. Скорость вымыва-
ния плазмидсодержащих клеток определяется параметром неустойчивости р,
который, по сути дела, показывает, как долго может работать проточный реак-
тор (если р достаточно мало).
Опубликованы работы, в которых в модифицированных вариантах рас-
смотренных моделей учтены скорости утилизации субстрата и образования
продукта, а также зависимость ц от концентрации субстрата, описываемая
уравнением Моно [13, 14]. В других работах на основе уравнений материаль-
ных балансов по массам популяций были выявлены погрешности, обусловлен-
442
Глава 13
ные принятыми при выводе соответствующих уравнений приближениями
(обычно эти погрешности невелики), а также неучтенными параметрами цикла
роста индивидуальных клеток, репликации плазмид и расщепления плазмид-
содержащих клеток [15, 16]. Следует отметить, что рассмотренные модели
с тем же успехом могут применяться и для изучения популяций нерекомби-
нантных организмов, для которых характерны повторные мутации. Кроме то-
го, некоторые из математических методов анализа неустойчивых рекомбинант-
ных популяций являются вариантами или частными случаями методов теории
популяционной генетики [17, 18].
13.4. Хищничество и паразитизм
В случае хищничества и паразитизма один вид получает оп-
ределенные преимущества за счет другого вида. Различия меж-
ду этими двумя типами межвидовых взаимодействий заключа-
ются в относительных размерах взаимодействующих организ-
мов и в механизме уничтожения одного вида другим. Под хищ-
ничеством понимают поглощение одного вида (жертвы) другим
(хищником). Этот тип взаимодействия довольно обычен среди
микроорганизмов; типичным примером может служить поглоще-
ние бактерий простейшими. При паразитизме организм-хозяин
(обычно больший организм) повреждается паразитом, который
получает питательные вещества от хозяина. В мире микроорга-
низмов наиболее наглядным примером паразитизма может слу-
жить нападение фагов на микроорганизм, уже рассмотренное в
некоторых аспектах в гл. 6.
Хотя механизмы межвидовых взаимодействий при парази-
тизме и хищничестве различаются, концептуальное и матема-
тическое описания этих двух типов взаимодействия имеют мно-
го общего. Поэтому в целях упрощения дальнейшего обсужде-
ния мы будем говорить о хищнике и жертве, имея в виду орга-
низмы, которые выигрывают и проигрывают в этих типах взаи-
модействия соответственно.
Часто при взаимодействии типа хищник — жертва числен-
ности популяций хищника и жертвы не достигают постоянных
значений, а колеблются, как это показано на рис. 13.9 для си-
стемы бактерии—простейшие. Другими примерами колебаний
численности популяций в системах хищник — жертва (помимо
рассматриваемого в настоящей главе мира микроорганизмов)
могут служить пары рысь — заяц, сова — лемминг, а также си-
стема моль — хвойное дерево (см. с. 188 в работе [1J). Подоб-
ные устойчивые колебания качественно можно объяснить сле-
дующим образом. Если начальная численность популяции жерт-
вы высока, а хищника—низка, то хищники активно уничтожа-
ют жертв и в результате численность хищников возрастает,
а численность жертв падает. Такая тенденция продолжается до
тех пор, пока численность популяции жертв не снизится до
Изучение смешанных популяций микроорганизмов
443
уровня, при котором высокая численность хищников становится
невозможной. Тогда из-за недостатка пищи численность попу-
ляции хищников снижается, а численность популяции жертв
восстанавливается. Таким образом система возвращается в ис-
ходное состояние, и эти циклические изменения могут повто-
ряться множество раз.
РИС. 13.9. Колебания численности популяций при росте смешанной культуры
бактерии A aerogenes и простейшего Tetrahymena pyriformis в хемостате. [Из
работы: Meers J. L., Growth of Bacteria in Mixed Cultures, in Microbial Ecology,
Laskin A. I., Lechevalier H. (eds.), p. 151, CRC Press. Inc., 1974. © CRC Press,
Inc.]
13.4.1. Описание колебаний численности видов в системе
хищник — жертва с помощью модели Лотки — Вольтерры
Математическую модель взаимодействия типа хищник —
жертва, приводящего к таким циклическим изменениям, разра-
ботали Лотка и Вольтерра в конце 1920-х годов [8]. В этой
модели принимается, что вид жертвы 1 может размножаться ав-
тономно и уничтожаться хищником со скоростью, пропорцио-
нальной произведению численности жертв на численность хищ-
ников:
(13.9)
at
Произведение можно рассматривать как частоту встреч
между хищниками и жертвами, а стоящий перед этим произ-
ведением коэффициент -у является мерой вероятности того, что
такая встреча приведет к гибели жертвы. Известно, например,
444
Глава 13
что некоторые бактерии более устойчивы к атаке со стороны
простейших, чем другие.
Вид хищников 2 получает определенные преимущества при
встрече с жертвой; коэффициент пропорциональности 8 отража-
ет степень увеличения численности популяции хищников за счет
жертв. Поскольку предполагается, что жертвы являются един-
ственным источником пищи для хищников, то в отсутствие
жертв численность популяции хищников снижается. Таким об-
разом, уравнение баланса численности популяции хищников
принимает форму
=—Ьпг-^&уп^Пг (13.10)
dt
Уравнения (13.9) и (13.10) составляют основу модели Лот-
ки— Вольтерры. Допускаемое моделью нетривиальное стацио-
нарное состояние возможно при
nis= — n2S = — (13.11)
еу У
Для упрощения дальнейшего анализа можно несколько преоб-
разовать указанные выражения, введя новые переменные у\ и
у 2, которые связаны со стационарными состояниями, описывае-
мыми уравнениями (13.11):
У,= Т- = ~ (13-12)
'4s n2S
Выраженные в этих переменных уравнения модели Лотки —
Вольтерры принимают вид
^- = а(1-уг)у, (13.13)
-^- = -Ь(1-у,)уг (13.14)
at
Если из уравнений (13.13) и (13.14) исключить параметр
времени, то мы получим простое дифференциальное уравнение,
связывающее у у и У2 и описывающее поведение системы в фа-
зовой плоскости. Разделив уравнение (13.14) на уравнение
(13.13), получим
dy2/dt = — byz + Ьугу2 (is j 5)
dyi/dt ау! — ауху2
Последующее умножение на а(1—Уъ) (dy\ldt)ly2 приводит к
уравнению
д dy2 & dy2 . b dyx dyt __ g (13.16)
у2 dt dt yi dt dt
Изучение смешанных популяций микроорганизмов
445
(13.17)
(13.18)-
Интегрированием этого уравнения получим
а In z/г — ay2+b In У\ — Ьу{ = С
или
/ У1 \ь ( Уг \
( еУ1 М еУ2 J
где С — постоянная интегрирования, зависящая от начальной
численности популяций.
Теперь, приняв определенные значения для ух и вычислив
соответствующие значения у2> можно рассчитать траектории в
фазовой плоскости. При этом следует иметь в виду, что при
данном у\ это трансцендентное уравнение может иметь одно
или два положительных решения или вообще не иметь никаких
решений. На рис. 13.10 приведены примеры рассчитанных та-
ким путем траекторий; очевидно, что замкнутые кривые харак-
njs
РИС. 13.10. Траектории в фазовой плоскости, соответствующие модели Лот-
ки— Вольтерры. Точка Q отвечает стационарному состоянию. Колебательные
траектории, отмеченные символами X, Т, А и Ф, отвечают различным на-
чальным численностям популяций.
терны для любых начальных состояний системы за исключени-
ем стационарного. (Какой вывод можно сделать на основании
этих данных об асимптотической устойчивости стационарного
состояния?) Если выразить полученные таким путем данные на
графике зависимости численности популяций от времени, то
решив уравнения (13.13) и (13.14), мы найдем, что численно-
сти популяций хищника и жертвы колеблются во времени (см.
рис. 13.11); эти данные очень близки экспериментальным ре-
зультатам, полученным при изучении многочисленных систем
хищник — жертва, в том числе и системы, поведение которой бы-
ло представлено ранее на рис. 13.9.
446
Глава 13
Прежде чем перейти к изучению и критике расширенных и
модифицированных вариантов модели Лотки—Вольтерры, рас-
смотрим несколько детальнее некоторые из следствий модели.
Интегрируя уравнение (13.13) по периоду колебаний Т, полу-
чим
1/1(0)
т
= аТ—a j*y2(t)dt
о
(13.19)
Поскольку z/i изменяется периодически с периодом Т, то
г/1(0) = г/1(Г) (13.20)
и поэтому левая часть уравнения (13.19) равна нулю. После-
дующее преобразование дает
т
-Ьрг(0<й=1 (13.21)
о
Переходя к начальным переменным, получим
т
л, (/)<« = «,. (13.22)
о
Отсюда следует, что, хотя периоды популяционных циклов,
представленных на рис. 13.10, а также положение максимумов
(минимумов) численности популяции на шкале времени зави-
РИС. 13.11. Периодические колебания численности популяций жертвы П[ и
хищника «г, предсказываемые моделью Лотки — Вольтерры.
сят от начальных условий, средняя численность популяции
жертвы, определяемая уравнением (13.22), не зависит от на-
чальных условий и равна численности популяции в стационар-
ном состоянии. Исходя из уравнения (13.14) и повторив весь
ход рассуждений, нетрудно показать, что аналогичным уравне-
нием описывается и средняя численность популяции хищника:
т
J п2 (0 dt = n2s (13.23)
о
Изучение смешанных популяций микроорганизмов 447"
Из модели Лотки — Вольтерры вытекают некоторые интерес-
ные следствия, имеющие непосредственное отношение к регули-
рованию популяций микроорганизмов (или иных организмов),
для которых типичны межвидовые взаимодействия типа хищ-
ник— жертва. Предположим, что к такой системе добавили
агент, уничтожающий как хищников, так и их жертв пропор-
ционально их численности. Математически это соответствует
добавлению выражений —6i«i и —б2«2 (<5i >0 и б2>0) к урав-
нениям (13.9) и (13.10) соответственно. В этих условиях чис-
ленность популяций жертв и хищников в стационарном состоя-
нии будет описываться уравнениями
п —
ву
n2s
(13.24>
Следовательно, результатом такого регулирования численности
популяции будет повышение средней численности популяции
жертв и снижение средней численности популяции хищников.
13.4.2. Применение модели Лотки — Вольтерры к системам,
состоящим из многих видов
Экологов интересует изучение взаимосвязей между степенью
сложности системы и ее динамическим поведением. В частности,
очень большой интерес представляют ответы на вопрос, будет
ли система более устойчивой или менее устойчивой и будут ли
для нее характерны колебания численности популяций при воз-
растании числа видов или числа и интенсивности межвидовых
взаимодействий. Подобные проблемы мы рассмотрим детальнее
в разд. 13.5; здесь же ограничимся анализом предложенной
Мэем [5] обобщенной модификации модели Лотки — Воль-
терры.
Для системы с N видами жертв и N видами хищников по
аналогии с уравнениями (13.9) и (13.10) можно записать
(13.25)-
где Ль..., nN — численности популяций N видов жертв,,
а ть ..., mN— численности популяций N видов хищников.
448
Глава 13
Пусть а — матрица с элементами ац, р— матрица с элемента-
ми fiij, а — вектор столбца с элементами at-, а b — вектор столб-
ца с элементами Ьг. Тогда в системе, описываемой уравнениями
(13.25), численность популяций в стационарном состоянии
.., nN&, mls, ..mNg должна удовлетворять условию
pns=b ocms = a (13.26)
где ns и ms — векторы с элементами nis и mis соответственно.
Если попытаться оценить матрицу сообщества 2Ny.2N для
этой системы, то результат можно будет представить следую-
щим образом:
О : -а*
₽* V ”о
(13.27)
где
a*ij = niSaij = (13.28)
а элементы 0 в (13.27) обозначают нулевые матрицы ЛГХМ
Интересные сведения о стабильности этой системы можно по-
лучить на базе следующих общих формул матричной алгебры.
Для произвольной матрицы С (qXq)
q
trС= 2 ог (13.29)
i=i
где Oi — собственные значения С. Поскольку след матрицы Ав
выражении (11.40) равен нулю, то из уравнения (13.29) сле-
дует
2W
(1з.зо)
i=i
Что касается собственных значений то отсюда вытекают
две возможности. Во-первых, они могут быть равны нулю или
парам сопряженных мнимых чисел. Хотя этот вывод справедлив
для первоначальной модели Лотки — Вольтерры типа один хищ-
ник— одна жертва, он маловероятен при большом N. Другая
возможность заключается в том, что по меньшей мере часть соб-
ственных значений А существует в форме c + id, —с—id при
c=£.Q. В этом случае по меньшей мере одно собственное значе-
ние имеет положительную действительную часть и стационар-
ное состояние неустойчиво. Следовательно, в общем случае, со-
гласно результатам этого анализа, усложнение системы хищ-
ник— жертва должно приводить к ее дестабилизации.
Изучение смешанных популяций микроорганизмов
449
13.4.3. Другие модели системы
один вид хищников — один вид жертв
Работа Лотки и Вольтерры послужила стимулом для целого
ряда исследований, в результате которых были предложены
усовершенствованные варианты модели. Один из недостатков
модели Лотки — Вольтерры нетрудно заметить при изучении
уравнения (13.9); согласно последнему, в отсутствие хищников
популяция жертв гъ будет возрастать неограниченно по экспо-
Обратная плотность популяции Е. coli., л/мг
РИС. 13.12. Зависимость обратной скорости роста популяции хищника (про-
стейшее Colpoda steinii) от обратной плотности популяции жертвы (бактерия
Е. coli). Приблизительно линейная зависимость, представленная на этом ри-
сунке, дает возможность предположить, что скорость роста популяции описы-
вается уравнением типа уравнения Моно. [Из статьи: Proper G., Garver J. С.,
Mass Culture of the Protozoa Colpoda steinii, Biotech. Bioeng., 8, 287 (1966).]
ненциальному закону. Вероятность такой, очевидно, нереальной
ситуации можно отбросить, если учесть, что жертва утилизиру-
ет субстрат, ограничивающий рост ее популяции.
Кроме того, в некоторых экспериментальных работах было
показано, что удельные скорости роста популяций хищников из-
меняются не пропорционально численности популяции жертвы,
как это принималось в первоначальном варианте модели Лот-
ки— Вольтерры. Как показано на рис. 13.12, где приведены
представленные в координатах Лайнуивера — Бэрка данные
изучения роста популяций в системе простейшие—бактерии,
часто удельную скорость роста популяции хищника лучше опи-
сывать уравнением типа уравнения Моно:
= Нр,тахП1 (13.31)
Кр+«1
Если учесть указанные уточнения и ввести экономические
коэффициенты У5 и Ур, отражающие эффективность роста по-
29—746
450
Глава 13
пуляци соответ уравне! жертвы и жертвы на субстрате и популяции хищника на жертве ственно, то для системы в хемостате получим следующие яия, описывающие численности популяций хищника и i: JL=D(Sq—s) !—Fts’maxS-^- (13.32) dt v 0 J Ys Ks + s ' ' dtli Ps,maxs/ll 1 Цр.тахЩ^г (13 33V dt 1 Ks-p s Yp = —Dn,+ (J 3.34) dt Kp + th y '
Цучия вает бс НЫХ ХЭ Dictyos лив кив четным Модель и другие [19] показали, что эта модель хорошо описы- )льшинство найденных экспериментальным путем основ- рактеристик смешанной популяции типа хищник (амеба telium discoideum) — жертва (бактерия Е. coll). Опреде- [етические константы этой системы (табл. 13.5), они рас- путем нашли кривые, представленные на рис. 13.13. не только предсказывает колебания численности попу-
Таблица [уравнен: 13.5. Значения параметров в уравнениях модели хищник — жертва ня (13.32) —(13.34) ]а Максималь-
Организм ная удель- ~ . пяя гкппогть ТГ жж v Экономический КОЭффИЦИ- ная скорость Коэффициент насыщения К у роста цтах, ент г 4-1
D. disco- idetun Е. coll 0,24 4-Ю8 бактерий в 1 мл 1,4-103 бактерия/амеба 0,25 5-Ю-4 мг глюкозы/мл 3,3-10-10 мг глюкозы/ /бактерия
а Из работы: Tsuchiya Н. М. et al., J. Bacteriol., 110, 1151 (1972).
ляций в системе при заданных условиях (£> = 0,0625 ч-1, s0=
= 0,5 мг/мл), но также хорошо отражает найденные экспери-
ментально период и амплитуду всех трех переменных.
Теперь рассмотрим другой недостаток модели Лотки — Воль-
терры. Мы уже знаем, что предсказываемые моделью Лотки —
Вольтерры колебания зависят от начальных условий. Если на
этот факт взглянуть с другой точки зрения (см. рис. 13.14, а),
отсюда следует, что в соответствии с теорией после возмуще-
ния должны измениться и период, и амплитуда колебаний.
С физической точки зрения такое явление, называемое иногда
математиками мягкими колебаниями, по меньшей мере мало-
вероятно. Оно означает, что небольшое возмущение системы
Изучение смешанных популяций микроорганизмов
451
может изменить ее динамические характеристики на все время
дальнейшего существования этой системы.
Регулярные устойчивые колебания численности популяций в
естественных условиях, где часто происходят небольшие изме-
нения в ту или иную сторону, свидетельствуют о том, что есте-
ственные колебания популяций жертвы и хищника более устой-
РИС. 13.13. Расчетные (-----) и экспериментально найденные (О) данные
для системы хищник — жертва (D. discoideum — Е. coli) в непрерывной куль-
туре (25 °C, 72=0,0625 ч-1). [Из статьи: Tsuchiya Н. М. et al., J. Bacteriol,
110, 1147 (1972).]
чивы, чем предсказывает модель Лотки — Вольтерры. Устойчи-
вость колебаний, однако, предполагается моделью, которая
описывается уравнениями (13.32) — (13.34). Приведенные на
рис. 13.14,6 данные показывают, что предсказываемые этой
моделью колебания не зависят от начальных условий. Такие
колебания, называемые жесткими колебаниями, или предель-
ным циклом, больше отвечают реальному поведению системы
хищник — жертва, чем мягкие колебания, отвечающие решени-
ям уравнений Лотки—Вольтерры.
Пример 13.3. Дискриминация и совершенствование моделей путем анализа
устойчивости. В продолжение рассмотренных выше работ Цучия, Фредриксон
и сотрудники [19—21] попытались дискриминировать математические модели,
29*
452
Глава 13
Время, сут
б
РИС. 13.14. Сравнение характеристик моделей, описывающих колебания чис-
ленности популяций в системе хищник — жертва: а — модель Лотки — Воль-
терры, предполагающая мягкие колебания, характер которых зависит от на-
чальных условий; б — модель на основе уравнений (13.32) — (13.34), предпо-
лагающая жесткие колебания, характер которых не зависит от начальных
условий. В этих расчетах в точках А' численность популяций хищника н жерт-
вы принимает новые значения. [Из статьи: Tsuchiya Н. М. et al., J. Bacteriol.,
ПО, 1147 (1972).]
Изучение смешанных популяций микроорганизмов 453
описывающие системы хищник — жертва, путем анализа устойчивости. Эти ис-
следования взяли за основу работы Кэнала*, который показал, что система
различных стационарных состояния:
Полное вымывание:
721s —^2s”^) Ss^^Sp
Вымывание популяции хищника:
Mis>0 722s — 0 O<C^Ss'CSo
Выживание популяций жертвы и хищника:
721s>0 n2s>0 0<Ss<S0
Кинетические параметры системы определяют, может ли в данных условиях
(при определенных значениях D и s0) существовать более одного из этих
стационарных состояний.
Аналогично от всех этих параметров системы зависит и ее устойчивость.
Прежде чем углубиться в обсуждение анализа устойчивости, полезно сокра-
тить одну переменную в уравнениях (13.32) — (13.34), воспользовавшись неко-
торыми их особенностями. Разделив уравнение (13.34) на Yp, умножив урав-
нение (13.32) на и затем сложив полученные два выражения н уравнение
(13.33), получим
d
dt
Ys(s— «о) + «i +
Y p -
(13П3.1)
Здесь к левой части уравнения добавлено выражение d(—Yss0)/dt, равное
нулю; таким путем достигается идентичность стоящих в скобках выражений.
Далее интегрированием уравнения (13П3.1) получим
«»(/) (
W) - SO] + nrf) + = rs[s(0) - s0] + пг(0) +
Yp '
(13П3.2)
Следовательно, если с начала функционирования системы прошло время,
в 3—5 раз превышающее время удерживания хемостата, то с хорошим при-
ближением можно принять
yjs(O-so] + ni(O +^7Г-~0 (13ПЗ.З)
у р
С помощью уравнения (13ПЗ.З) можно избавиться от одной из зависимых
переменных, например s(7), в уравнениях (13.33) и (13.34). Попытаемся най-
ти решение ^нашей задачи в двумерном пространстве с привлечением уравне-
Одним из способов отображения общих динамических особенностей си-
стемы в хемостате является рабочая диаграмма, подобная представленной иа
рис. 13П3.1. Здесь для данных значений кинетических констант в плоскости So,
\/D отмечены области с различными характеристиками. Так, если точка, отве-
* Canale R. РAn Analysis of Models Describing Predator-Prey Interac-
tion, Biotech. Bioeng., 12, 353 (1970).
454
Глава 13
чающая данным рабочим условиям (so и D), располагается, например, в об-
ласти а, то существует только первое стационарное состояние Кэнала, отли-
чающееся асимптотической устойчивостью и вымыванием популяций как хищ-
ника, так и жертвы.
С помощью уже рассмотренных нами методов анализа локальной устой-
чивости можно показать, что при наличии стационарного состояния в другие
стационарные состояния а и б всегда неустойчивы. Три верхних области на
рис. 13П3.1 различаются собственными значениями, характеризующими ста-
Концвнтрация субстрата в исходной
среде, ллг/мл
РИС. 13П3.1. Качественные динамические характеристики модели системы
хищник — жертва [уравнения (13.32) — (13.34)] в применении к условиям
в хемостате. Из статьи: Tsuchiya Н. М. et al., J. Bacteriol., ПО, 1147 (1972).]
ционарное состояние в. В области el оба собственных значения действительны
и отрицательны; здесь стационарное состояние в представляет собой устойчи-
вую узловую точку (вспомните рис. 13.1). В условиях, отвечающих области в2,
стационарное состояние в представляет собой устойчивую фокальную точку.
Область еЗ отвечает неустойчивому стационарному состоянию в.
В последней области, очевидно, ни одно из стационарных состояний не
обладает устойчивостью. Эта дилемма может быть решена с помощью теоре-
мы Пуанкаре — Бендикссона, которая утверждает, что в данном случае
должны наблюдаться колебания типа предельного цикла. Указанную теорему,
однако, можно применять только в тех случаях, когда обе независимые пе-
ременные имеют нижнюю и верхнюю границы (в рассматриваемом примере
это условие выполняется) и когда модель имеет только две независимые пе-
ременные. В части работ, посвященных колебаниям в химических и биологи-
ческих системах, это обстоятельство часто упускалось из вида (детальнее дан-
ный вопрос рассмотрено работе [10]).
Хотя в зависимости от выбранных значений кинетических параметров по-
ложение граничных кривых на рис. 13П3.1 будет несколько изменяться, об-
время, сут
б
РИС. 13П3.2. Динамика пищевой цепи глюкоза — A. vinelandii—Т. pyriformis
в хемостате: а — устойчивые колебания, наблюдаемые при £>=0,169 ч-1; б —
затухающие колебания, наблюдаемые при снижении скорости разведения до
0,025 ч-1. [Из работы: Jost J. L. et al., J. Bacteriol., 113, 834 (1973).]
456
Глава 13
щий вид рабочей диаграммы останется таким же при любых адекватных па-
раметрах системы. В рассматриваемом контексте особый интерес представляет
одна особенность рабочей диаграммы. Предположим, что 'при данных кон-
центрации питательных веществ и времени удерживания формируются пе-
риодические колебания, тогда устойчивые колебания должны наблюдаться и
при любом большем времени удерживания, если концентрация питательных
веществ остается без изменения.
Этот вывод не согласуется с экспериментальными результатами, пред-
ставленными на рис. 13П3.2 и отражающими поведение смешанной культуры
РИС. 13ПЗ.З. Рабочая диаграмма модифицированной модели системы хищ-
ник— жертва в хемостате, включающей уравнение скорости роста популяции
хищника (13113.4). [Из работы: Jost J. L. et al., J. Bacterio!., 113, 834 (1973).]
1— затухающие колебания; 2 — устойчивые колебания; 3—отсутствие коле-
баний; 4 — вымывание популяции хищника; 5 — полное вымывание.
Azotobacter vinelandii и простейших Т. pyriformis в хемостате. Колебания,
наблюдаемые при времени удерживания 5,9 ч, подавляются, если время удер-
живания возрастает до 40 ч. Очевидно, с помощью рассматриваемой модели
нельзя адекватно описать эту систему.
Можно попытаться улучшить модель, введя в описание физиологии хищ-
ника элементы структурирования. Можно, например, допустить, что для хищ-
ника характерны два промежуточных физиологических состояния Nz' и УгЛ
и постулировать следующий механизм:
—> N'2 —*- N2" Л^2,х —*" 2Л^2
Применив к jV2z и Nf приближение квазистационарного состояния, получим
следующее выражение для удельной скорости роста популяции хищника ц₽:
„ ________Hp,max (их)2____
Р (^pi + ni) (^рг + «1)
В соответствии с этой моделью, которая отвечает уравнению типа уравнения
Моно (13.31) (при П1, значительно больших КР\ и Кр2), скорость роста популя-
ции хищника изменяется пропорционально ni2 при низкой численности популя-
ции жертвы.
Изучение смешанных популяций микроорганизмов
457
Точно таким же путем можно выполнить анализ стабильности модифици-
рованной модели системы в хемостате, включающей уравнение (13П3.4). На
базе кинетических параметров, определенных путем изучения периодической
смешанной культуры A. vinelandii и Е. coli, была получена рабочая диаграм-
ма, представленная на рис. 13ПЗ.З. В соответствии с экспериментальными
данными эти расчеты показывают, что при уменьшении D устойчивые коле-
бания подавляются. Таким образом, результаты экспериментальных исследо-
ваний в сочетании с теоретическим анализом устойчивости помогают выявить
недостатки одной математической модели и проверить адекватность другой,,
более совершенной модели.
13.5. Влияние количества видов
и сети их взаимодействий
В этом разделе мы перейдем от конкретных систем к срав-
нительно абстрактному изучению поведения систем, состоящих
из популяций более двух видов. Прежде чем приступить к этой,
иногда довольно сложной, проблеме, необходимо познакомить-
ся со специфической терминологией, применяемой в экологии и
смежных областях.
13.5.1. Трофические уровни, пищевые цепи и пищевые сети;
определения и пример
В естественных системах часто существует та или иная ие-
рархия взаимосвязей между потребителями пищи. Рассматри-
вавшиеся в предыдущем разделе бактерии, например, были по-
требителями глюкозы и одновременно сами являлись пищей для
простейших; последние в свою очередь поглощались более слож-
ными организмами. Подобные взаимосвязи называют пищевы-
ми цепями, а отдельные последовательные звенья цепи — тро-
фическими уровнями. Обычно пищевые цепи не изолированы од-
на от другой, а связываются и переплетаются, образуя пище-
вые сети. На рис. 13.15 представлен пример простой пищевой
сети, изучавшейся в лабораторных условиях; в этом при-
мере две пищевые цепи (субстрат—>-жертва 1—>-хищник и суб-
страт—^жертва 2—^хищник) совпадают на первом и третьем
трофическом уровнях.
Прежде чем перейти к обсуждению результатов эксперимен-
тального изучения пищевой сети, изображенной на рис. 13.15,.
мы должны вспомнить обсуждавшиеся выше результаты анали-
за отдельных компонентов этой системы. Так, если рассматри-
вать только первый и второй трофические уровни, нетрудно за-
метить, что они образуют систему, к которой применим прин-
цип конкурентного исключения. Действительно, данные, приве-
денные на рис. 13.7, показывают, что в этой системе может дей-
ствовать принцип конкурентного исключения, если иметь в ви-
458
Глава 13
ду только два первых трофических уровня. Экспериментально
изучалась также система глюкоза—>-жертва 2—>-хищник (рис.
13.15); показано, что для нее в некоторых условиях характер-
ны устойчивые колебания (см. рис. 13П3.2). Интересно просле-
дить за динамическим поведением сложной системы, в которой
Хищник
ТеtrahутепарупТо rm is Третий
тросу и чес кий
уровень
Жертва Т
Escherichia coli
Жертва 2
Azotobacter vinelandii
Второй
тросрическии
уровень
Лимитирующий Первый
рост субстрат (глюкоза) тросрическии
уровень
РИС. 13.15, Схема простой пищевой сети, состоящей из трех трофических уров-
ней. На втором уровне находятся два вида бактерий, потребляющих пита-
тельное вещество (первый уровень); на высшем (третьем) уровне оба вида
бактерий служат пищей для хищника (простейшего). [Из работы: Jost J. L.
et al., J. Bacteriol., 113, 834, (1973).]
указанная цепь переплетается с другой, образуя изображенную
на рис. 13.15 пищевую сеть.
На рис. 13.16 представлены результаты экспериментального
изучения этой пищевой сети в хемостате при £>=0,1 ч-1. Наибо-
лее интересен факт сосуществования двух конкурирующих ви-
дов бактерий при условии, что они имеют общего хищника. Та-
ким образом, введение в систему популяции хищника оказывает
стабилизирующее влияние на популяции конкурирующих видов
жертв, а вымывание популяции одного из видов жертв пред-
отвращается.
Хотя численность популяций всех трех видов подвержена
колебаниям, они гораздо менее интенсивны и более сглажены
по сравнению с колебаниями в изолированной пищевой цепи
Изучение смешанных популяций микроорганизмов
459
хищник — жертва. К сожалению, полученные эксперименталь-
ные данные охватывают только небольшой временной интервал
(8 сут), поэтому нельзя исключить возможность последующего
затухания колебаний. Эксперимент был прекращен в силу бы-
строго и интенсивного роста культур на стенках хемостата.
РИС. 13.16. Экспериментально обнаруженные колебания численности популя-
ций видов, участвующих в пищевой сети, изображенной на рис. 13.15. [Иэ
работы: Jost J. L. et al., J. Bacteriol., 113, 834, (1973).]
13.5.2. Изучение динамики популяций с помощью моделей
в форме закона действующих масс
В оставшейся части раздела 13.5 мы рассмотрим некоторые
теоретические методы анализа и изучения сложных взаимодей-
ствующих популяций. Следует подчеркнуть, что большинство
рассматриваемых здесь методов и результатов применимо к
изучению сложных систем биологических реакций. Действи-
тельно, исходным пунктом для создания экологических моделей
часто является допущение о существовании ряда микробиологи-
ческих «реакций» типа учитываемых в уравнении (13П3.4).
Изложенный в этом разделе методический подход имеет в ос-
новном химическую ориентацию; ниже мы рассмотрим пример
применения такого подхода к моделированию роста микроорга-
низмов.
460
Глава 13
Предположим, что в изотермической гомогенной системе по-
стоянного объема, к которой добавляют компонент В в таком
количестве, чтобы его концентрация оставалась постоянной,
происходят следующие реакции:
Г1 Г2 г3
Ai + B —»- 2Ai Ai + Аг —>- 2Аг Аг —>- С (13.35)
Допустим, кроме того, что каждая из этих реакций элементар-
на, т. е. кинетика реакции непосредственно связана с ее стехио-
метрией. Следовательно, для системы реакций (13.35) можно
записать
rx=kxaxb r2=k2axa2 (13.36)
Поскольку мы условились считать концентрацию В постоянной
во времени, то в нашем анализе соответствующий параметр
можно не учитывать. В связи с этим введем новый параметр
k* = kxb (13.37)
При этом условии Г1 равно:
rx=k*ax (13.38)
Теперь уравнения материальных балансов по Ах и А2 можно за-
писать в форме
(13.39)
^=^2-^2 (13.40)
Сравнение с уравнениями (13.9) и (13.10) показывает, что урав-
нения (13.39) и (13.40) по форме соответствуют модели Лот-
ки — Вольтерры.
Теперь перейдем к рассмотрению некоторых в высшей сте-
пени общих и очень полезных теорем, применимых к изучению
кинетики систем, формально описываемых законом действующих
масс. Результаты такого изучения часто позволяют на основе
только лишь алгебраической структуры, сети реакций сделать
вывод, что данная система имеет одно стационарное состояние,
характеризующееся глобальной асимптотической устойчиво-
стью. Здесь термин «алгебраическая структура» относится
только к путям превращения видов и способам их взаимодей-
ствия в сети реакций; полученные таким путем результаты
можно применять независимо от констант скоростей реакций и
других параметров.
Изучение соответствующей теории, разработанной Хорном,
Фейнбергом и Джексоном [9], невозможно без знакомства с
некоторыми новыми понятиями и определениями. Для этой цели
Изучение смешанных популяций микроорганизмов
461
удобнее обратиться к ряду примеров. В табл. 13.6 приведены
некоторые гипотетические механизмы, два из которых пред-
ставляют интерес для биологических исследований. Нетрудно
видеть, что механизм 3 эквивалентен модели Лотки — Вольтер-
ры, а механизм 6 соответствует одной из моделей гликолиза.
(Можете ли вы отнести компоненты Ai—Д7 к определенным со-
единениям, участвующим в метаболическом пути гликолиза?)
Таблица 13.6. Примеры механизмов реакций, иллюстрирующих принципы
и определения, которые используются в моделях в форме закона
действующих масс3
а Из работы: Feinberg М., Horn. F. J. М., Chem. Eng. Sci., 29, 775 (1974).
Нулевые символы в этой таблице имеют следующий смысл:
О—^Aj означает, что компонент Aj добавляют к системе с по-
стоянной скоростью. Такая ситуация может возникнуть, напри-
мер, при добавлении субстрата в потоке исходных веществ к
системе, находящейся в хемостате. Символ Ak—>-0 означает,
что Ak отбирают из системы со скоростью, пропорциональной
его концентрации; такая ситуация типична, например, для ча-
стичного вымывания вида из хемостата или для процессов от-
мирания вида. Таким образом, символ 0, используемый для
обозначения взаимосвязей с окружением системы, позволяет
применять модели в форме закона действующих масс к откры-
тым системам.
Теперь мы должны дать определения трем параметрам (вы-
ражаемым целыми числами) механизмов реакций. К этим па-
раметрам относятся: пс — число комплексов в механизме; щ —
число типов связей в механизме; ns — размерность механизма.
Для каждого из этих параметров необходимо дать соответст-
462
Глава 13
вующее определение. Во-первых, комплексом называют элемент»
стоящий в начале или в конце стрелки, соответствующей дан-
ной реакции. Например, механизмы (1) и (2) в табл. 13.6 име-
ют три комплекса-—2Аь А2 и А3-|-А4, так что в этом случае
пс = 3. Механизм модели Лотки — Вольтерры (3) имеет пс = 6
(Аь 2Ai, Ai+A2, 2А2, А2 и 0), а в механизмах 4 и 5 имеется
пять комплексов (пс = 5; Аь А2, Ai-J-Аз, А4, А2+А5).
Переходя к типам связей, не следует обращать внимание на
направление стрелок; здесь необходимо рассматривать только
комплексы, связанные стрелкой соответствующей реакции. На-
пример, рассматривая механизм 1, нетрудно заметить, что та-
ким образом непосредственно связаны комплексы 2А1 и А2. Кро-
ме того, хотя комплекс А2 непосредственно связан с комплексом
Аз+А4, можно сказать, что комплексы 2Ai и А3-|-А4 связаны
косвенно, поскольку от одного комплекса к другому можно пе-
рейти только через несколько непосредственно связанных ста-
дий. Тип связи представляет собой ряд комплексов, связанных
друг с другом или непосредственно, или косвенно, так что ни
один комплекс в ряду не связан ни с каким другим комплексом»
не входящим в этот ряд.
Из сказанного следует, что механизм 1 имеет один тип связи
{2АЬ А2, Аз+А4}; следовательно, для этого механизма щ=1.
Тот же самый тип связи характерен и для механизма 2. Меха-
низмы 4 и 5 (щ = 2) имеют два типа связей — {Ai, А2} и {Ai-f-
-}-Аз, А4, Аг+Аб}, а механизм 3 (п/ = 3)—три типа, а именно
{Аь 2AJ, {Ai+Аг, 2А2}, {А2, 0}.
С понятием о типах связей связана концепция слабой обра-
тимости. Чтобы дать определение этому понятию, рассмотрим
направление стрелок реакции и зададимся следующим вопро-
сом: если есть указываемый стрелкой путь, ведущий от одного
комплекса к другому, то существует ли и указываемый стрел-
ками путь, ведущий от второго комплекса к первому? Если от-
вет на этот вопрос положителен для любой пары комплексов
механизма, то такой механизм называют слабообратимым.
Примером слабообратимого механизма может служить ме-
ханизм 2 в табл. 13.6. От комплекса А3-|-А4 к комплексу 2Ai
можно перейти в одну стадию, а возвратиться от 2At к А3~|-А4
можно в две стадии через А2. Более того, аналогичные аргу-
менты применимы к каждой паре комплексов механизма. Рас-
суждая аналогичным образом, можно показать, что механизмы
1, 3, 4, 6 не относятся к числу слабообратимых, тогда как ме-
ханизм 5 является таковым.
Прежде чем перейти к основным теоремам, необходимо рас-
смотреть еще один вопрос. Для каждой реакции (т. е. для
каждой стрелки) данного механизма определим вектор реак-
ции. Если в механизме участвует М различных химических ком-
Изучение смешанных популяций микроорганизмов
463
понентов Аь А2, А3, ..Ам (для механизмов 1—5Л4 = 4, 4, 2, 5,
5 соответственно), то каждый вектор реакции будет иметь М
элементов. Эти элементы определяются следующим образом:
i-й элемент равен нулю, если Aj не появляется в реакции. Ес-
ли же в реакции участвует Аг-, то i-й элемент является стехио-
метрическим коэффициентом А,. Этому коэффициенту приписы-
вают отрицательный знак, если Аг- находится в начале стрелки,
и положительный, если стрелка реакции указывает на Аг-.
Например, механизм 1 в табл. 13.6 имеет три стрелки и че-
тыре компонента. Ниже каждая из этих реакций записана в
ином виде вместе с соответствующими векторами реакций
2Ai —А2 vi=(—2, 1, о, 0)
А2 -->- А3 + А4 V2= (0, —1, 1, 1)
А3 + А4 —>- А2 V3= (0, 1, —1, 1.)
Следующий шаг заключается в определении на базе найденных
таким путем векторов реакции максимального числа линейно
независимых векторов. Это число ns называют размерностью
механизма. Для механизма 1, например, ns = 2, поскольку ука-
занные Vi и v2 линейно независимы, но v3 зависит от двух пер-
вых векторов (v3+v2 = 0).
Дав таким образом необходимые определения, мы можем
перейти к формулировке теоремы нулевого ограничения [9].
Предположим, что для данного механизма
пс— гу — ns = Q (13.41)
Тогда:
1. При любой кинетике, в том числе формально отвечающей за-
кону действующих масс, невозможно ни одно стационарное
состояние с положительными концентрациями всех элемен-
тов М, ни периодические колебания концентраций, если ме-
ханизм не является слабообратимым.
2. Если выполнено условие (13.41), механизм слабообратим и
кинетика процесса формально соответствует закону действу-
ющих масс, то при всех стехиометрически эквивалентных по-
ложительных начальных составах существует одно глобально
асимптотически устойчивое стационарное состояние. Этот вы-
вод справедлив независимо от значений констант скоростей.
В последнем предложении два термина нуждаются в пояснени-
ях. Термин «положительный» означает, что каждый компонент
имеет положительную концентрацию. Под стехиометрически эк-
вивалентными составами понимают такие составы, которые мо-
гут быть превращены один в другой путем изменения направле-
ния одной или нескольких стрелок в механизме. Так, для реак-
ции А1=^2А2 стехиометрически эквивалентны следующие на-
чальные концентрации (а10) а20): (2, 4), (3, 2), (1, 6) и так да-
464
Глава 13
лее до тех пор, пока выполняется условие 2а10-{-а2о=8. Для
механизма I стехиометрически эквивалентны следующие поло-
жительные начальные составы (йю, а2о, азо, <з4о): (2, 1, 1, l)t
(3, V2) 1, 1), (2, «/2, 172, 17г), (2, 172, 72, V2) И т. д.
Возможности этой теоремы в конечном счете зависят от то-
го, сколько механизмов реакций удовлетворяют условию (13.41).
Этот вопрос изучался Хорном, который показал, что подавляю-
щее большинство возможных механизмов согласуется с услови-
ем (13.41). Следовательно, приведенные в табл. 13.6 примеры в
известной мере нетипичны, поскольку только четыре из них
(1, 2, 3 и 5) удовлетворяют условию (13.41). Теперь перейдем
к изучению возможной сферы применения этой теоремы.
Предположим, что перед нами поставлена задача моделиро-
вания смешанной популяции с колебаниями численности видов
путем постулирования механизма «реакции» между популя-
циями, включающего несколько элементарных стадий с кинети-
кой, соответствующей закону действующих масс. Согласно тео-
реме, для этого механизм должен либо нарушать условие
(13.41), либо не обладать слабой обратимостью. Такая ситуа-
ция, действительно, типична для механизмов 3 и 6, которые
связаны с устойчивыми колебаниями. Поскольку подавляющее
большинство механизмов не будет нарушать ни одного условия,
то мы можем использовать выводы теоремы для существенного
ограничения сферы наших поисков. С другой стороны, согласно
второй части теоремы для слабообратимых механизмов, фор-
мально соответствующих закону действующих масс и удовлет-
воряющих условию (13.41), асимптотическая устойчивость в
большинстве случаев обеспечивается независимо от типа един-
ственного стационарного состояния или величин скоростей ре-
акций. Следовательно, в таких случаях мы можем обойтись без
анализа локальной устойчивости и без поиска путей доказа-
тельства устойчивости в глобальном масштабе.
Первая часть теоремы имеет, кроме того, важные экологи-
ческие следствия. Согласно теореме, если условие (13.41) удов-
летворяется, а условие слабой обратимости не выполняется, то
в стационарном состоянии концентрации некоторых компонен-
тов будут равны нулю. Таким образом в рамках популяционной
динамики эта теория позволяет открыть механизмы, обеспечи-
вающие вымирание по меньшей мере одного вида.
Пример 13.4. Применение моделей в форме закона действующих масс*.
Для иллюстрации возможностей теоремы нулевого ограничения применим ее
* Этот пример заимствован из работы Фейнберга и Хорна [9] и предназ-
начен только для иллюстративных целей. Он не обязательно соответствует
какой-либо реальной микробиологической или биохимической системе.
Изучение смешанных популяций микроорганизмов
465
для изучения динамики системы, описываемой следующим механизмом:
ka ЗА! — А] + 2А2 (а>
kb за2 2А14-А2 (б)
А)+2 А2 kc ЗА2 (в>
2А)+А2 kd ЗА! (г)
Будем считать, что кинетика этих «реакций» формально подчиняется закону
действующих масс. Комплексами данного механизма являются 3Ai, At-}-2A2,.
ЗА2 и 2Aj-}“A2, поэтому Нс = 4. Обратим внимание на то, что этот механизм
слабообратимый. Действительно, следуя по стрелкам реакций от «а» до «г»,
легко обнаружить один путь, соединяющий все четыре комплекса. Таким об-
разом, в данном случае существует только один тип связи и ni — 1.
Для определения размерности механизма сформируем четыре вектора
реакций, каждый нз которых отвечает одной стрелке в механизме. Каждый
вектор имеет два компонента, поскольку в этой системе участвуют только два
вида, Ai и А2. Перечислим векторы реакций: va=( —2, 2), v6=(2, —2), vB=
= (— 1, 1), vr=(l, —1). Поскольку —2vB=2vr= —V6=va, то существует только
одна линейно независимая реакция и ns=l.
Проверив условие (13.41) для нашего механизма, получим нс —щ —щ =
= 4 —1 — 1 = 2. Таким образом, это условие не выполняется. Следовательно, для
данного механизма нельзя исключить возможность колебательных процессов
или нескольких стационарных состояний. В самом деле, Фейнберг и Хорн по-
казали, что в этой системе несколько стационарных состояний будут наблю-
даться при kc=kd=l и ka = kb<1/6.
13.5.3. Качественная устойчивость
В предыдущем разделе мы рассмотрели некоторые из са-
мых эффективных теоретических методов анализа нелинейных
моделей процессов. В конце изучения влияния масштаба экоси-
стемы и степени ее сложности рассмотрим одну из линеаризо-
ванных моделей. Сначала посмотрим, что можно сказать об
устойчивости системы, основываясь только на знаках элемен-
тов в матрице сообщества.
Если независимо от величин ненулевых элементов все соб-
ственные значения А имеют отрицательные действительные ча-
сти, то матрицу А называют качественно устойчивой. Необходи-
мыми и достаточными условиями для качественной устойчиво-
сти А являются:
1. О для всех /
2. <2гг<0 по меньшей мере для одного i
3. для всех
4. Для любой последовательности из трех или более различных
индексов /, &,..., s, t
^•ijajk ... = 0
5. det Ат^О
30—746
466
Глава 13
Как показал Мэй [5J, каждое из этих условий имеет непо-
средственную физическую интерпретацию. Условие 5 обеспечи-
вает существование единственного стационарного состояния ли-
нейной системы (9.30) при % = 0. Условие 1 требует, чтобы
удельная скорость автономного роста каждого вида была не-
положительной, а условие 2 ужесточает это требование, уста-
навливая, что по меньшей мере у одного вида удельная ско-
рость автономного роста должна быть отрицательной. Усло-
вие 4 устанавливает, что в системе не должно быть замкнутых
цепей взаимодействий, в которых участвуют более 2 видов.
В свете классификации типов взаимодействий (см. табл.
13.4) особенно интересно условие 3. С экологической точки зре-
ния оно показывает, что не может быть качественно устойчивой
-смешанной популяции при наличии парных мутуалистических
(положительные a2-j и aji) или конкурентных (отрицательные ац
и aji) взаимодействий. Напротив, условию 3 удовлетворяют все
три оставшихся нетривиальных типа взаимодействий, а именно
комменсализм, аменсализм и хищничество.
«Это математически строгое утверждение можно свести к бо-
лее широкому (но и менее строго обоснованному) утвержде-
нию о том, что конкуренция или мутуализм между двумя ви-
дами менее выгодны для общей устойчивости сети, чем от-
ношения типа хищник — жертва. Представляется интересным
проанализировать влияние факторов устойчивости на сооб-
щества, в которых сильные связи типа хищник — жертва бо-
лее распространены, чем симбиотические. Результат такого
анализа трудно предсказать заранее; тем не менее такое рас-
пределение взаимодействий между видами характерно для
многих реально существующих экосистем...» (см. работу [5J,
с. 73).
Пример 13.5. Качественная устойчивость простой пищевой сети*. Рассмот-
рим простую пищевую сеть, схематично изображенную на рис. 13П5.1а.
•Стрелка, направленная от вида 1 к виду 2, показывает, что вид 1 является
пищей для вида 2, так что й21>0 и й12<0. Аналогично интерпретируются и все
другие стрелки. Таким путем по рисунку можно определить знаки элементов
соответствующей этой сети матрицы сообщества Аа:
Аа= + - — (13П5.1)
-+ + --
Здесь принято, что все виды имеют отрицательные удельные скорости авто-
номного роста. Проверив соответствие этой системы знаков условиям качест-
венной устойчивости, можно, например, найти, что
С12й2зйз1>0 (13П5.2)
* Этот пример заимствован из работы Мэя [5].
Изучение смешанных популяций микроорганизмов
467
Следовательно, условие 4 не выполняется. Таким образом, при некоторых
значениях ац система неустойчива, поэтому далее необходимо проверить дей-
ствительные части собственных значений матрицы или эквивалентное условие,
например, выражаемое уравнением (9.41), с помощью определенных числен-
ных значений ац.
В случаях (3 и у (рис. 13П5.1), однако, структура знаков элементов в со-
ответствующих матрицах сообщества принимает вид
(13П5.3)
Очевидно, что условия 1—3 выполняются для каждой из этих матриц. Про-
веряя условие 4, нетрудно найти, что detAp<0 и detAT<0, так что соответст-
вующие матрицы не сингулярны. Наконец, физическая интерпретация усло-
РИС. 13П5.1. Три простых пищевых
сети, каждая из которых имеет три
трофических уровня.
вия 4 и анализ рис. 13П5.1 показывают, что в сетях ₽ и у нет замкнутых це-
пей взаимодействий, включающих более двух видов. Этот вывод можно про-
верить, применив условие 4 непосредственно к выражениям (13П5.3).
Следовательно, пищевые сети ₽ и у, образующиеся при удалении одной тро-
фической связи из сети а, устойчивы независимо от численных значений ац.
13.5.4. Устойчивость сложных неупорядоченных пищевых сетей
В этом разделе будут рассмотрены некоторые интересней-
шие работы Гарднера и Эшби*, Мэя [5] и Макмертри**, по-
священные изучению взаимосвязей между устойчивостью, раз-
мером и сложностью экосистемы. Предположим, что система
содержит р различных видов, которые в отсутствие других ви-
дов обладают отрицательными удельными скоростями автоном-
ного роста близкой величины. Подобрав соответствующий мас-
штаб времени, можно принять далее, что каждый вид вносит
элемент —1 в главную диагональ матрицы сообщества.
Влияние взаимодействий между видами можно представить
матрицей В так, что матрица сообщества А примет форму
А = В —I (13.42)
* Gardner М. R., Ashby W. R., Connectance of Large Dynamical (Cyberne-
tic) Systems: Critical Values for Stability, Nature, 228, 784 (1970).
** Работа, подготовленная к печати н цитируемая в монографии [5].
30*
468
Глава 13
где I — единичная матрица рХр. Наличие взаимодействий и их
эффективность будет определяться двумя параметрами — связ-
ностью С (0<С< 1) и силой о2 соответственно.
Теперь произвольно выберем матрицы В (для подбора каж-
дого элемента Ьц можно использовать генератор случайных чи-
сел) при следующих ограничениях:
Из р- элементов bij матрицы доля С будет не равна нулю. Точ-
нее, вероятность того, что любой &2-7- равен нулю, составляет
1—С.
Ненулевые элементы равновероятно могут быть положительны-
ми или отрицательными. Таким образом, среднее значение
этих bij равно нулю.
Среднеквадратичное значение ненулевых bij равно о2.
Для каждой выбранной таким образом матрицы В с помо-
щью условия (13.42) можно рассчитать А и затем выполнить
стандартные тесты на устойчивость. При этом наибольший ин-
терес представляет оценка вероятности того, что случайно вы-
бранная данная матрица В отвечает устойчивой модели. Из при-
веденных выше определений следует, что вероятность этого со-
бытия будет зависеть от трех параметров — р, С и о2. Мэй по-
казал, что при р>-1 вероятность того, что система будет устой-
чивой, приближается к единице, если
о/рС<1 (13.43)
Напротив, если 1 и
о/рС>1 (13.44)
то система почти наверное будет неустойчивой. На рис. 13.17
приведены результаты моделирования по методу Монте-Карло,
хорошо согласующиеся с уравнениями (13.43) и (13.44).
Из приведенного выше анализа следует, что чем больше чис-
ло присутствующих в системе видов (т. е. чем больше р), чем
больше число взаимодействующих видов (т. е. чем больше С)
и чем больше влияние взаимодействий на рост видов (т. е. чем
больше о), тем более вероятно, что данная экосистема неустой-
чива. Этот вывод хорошо согласуется с результатами анализа
стабильности многих динамических систем других типов; вме-
сте с тем следует подчеркнуть, что многие экологи придержи-
ваются противоположного мнения. Поскольку высокоустойчивые
естественные экологические системы обычно включают множе-
ство видов, связанных очень сложной сетью взаимодействий,
часто считают, что разнообразие и сложность экологической си-
стемы обусловливают ее повышенную устойчивость. Хотя эм-
пирические наблюдения естественных систем подтверждают эту
Изучение смешанных популяций микроорганизмов
469
точку зрения, математический анализ сложных систем, подоб-
ный описанному выше, приводит к обратному выводу.
Мэй нашел способ разрешения этого кажущегося противо-
речия [5J. Хотя в среднем разнообразие и сложность дестаби-
лизируют систему, природные системы ни в коей мере не яв-
ляются усредненными. Прежде чем принять ту форму, в кото-
рой они существуют сегодня, эти системы формировались в те-
чение многих тысячелетий. Их формирование сопровождалось
отбором наиболее устойчивых взаимодействий между видами и
РИС. 13.17. а — вертикальные линии отражают результаты численных расче-
тов методом Монте-Карло наибольшей действительной части собственных зна-
чений случайных матриц с о = 0,5, С=1. Эти результаты и штриховая линия,
отвечающая уравнениям (13.43) и (13.44), показывают, что при увеличении
числа видов р система становится все более неустойчивой; б — аналогичные
результаты при о = 0,5 и р=40; здесь очевиден дестабилизирующий эффект
числа взаимодействий между видами. (Из работы: May М., Stability and
Complexity in Model Ecosystems, Princeton University Press, Princeton, New
Jersey, 1973.)
исчезновением неустойчивых связей. Это объяснение предпола-
гает интересный путь дальнейшего развития математической
экологии, связанный с поиском особых характеристик крупно-
масштабной системы, которые придают ей устойчивость. В не-
котором смысле рассмотренные в двух предыдущих разделах
результаты моделирования представляют собой шаги в этом на-
правлении.
13.5.5. Бифуркации и усложненная динамика
Один из способов изучения стационарных состояний и ди-
намики сложных систем с множеством взаимодействующих ви-
дов связан с анализом изменений, индуцированных плавным
сдвигом одного из параметров системы (например, концентра-
ции питательного вещества, скорости разведения). Последую-
470
Глава 13
щие изменения часто имеют качественный характер и включа-
ют, в частности, появление новых стационарных состояний или.
предельных циклов. Кроме того, устойчивые предельные цик-
лы могут стать неустойчивыми, что приводит к усложненному,,
часто непериодическому поведению системы в переходном со-
стоянии. В предельном варианте соответствующие кривые из-
меняются почти случайно; в этом случае их называют хаотиче-
скими.
Математический анализ этих качественных изменений в ди-
намике систем, обусловленных вариациями одного или не-
скольких параметров системы, выполняется с помощью мето-
дов теории бифуркаций. Важность применения этой теории,
обусловлена тем обстоятельством, что она позволяет во многих
случаях проследить генезис нелинейных характеристик на базе-
относительно простых линеаризованных расчетов. Здесь мы смо-
жем привести только некоторые концептуальные начала этой,
теории, а с более подробными описаниями читатель может озна-
комиться в литературе, приведенной в конце главы.
В примере 13.1 мы рассматривали метод анализа локальной
устойчивости для системы, состоящей из двух видов. Напомним
(см. разд. 9.2.2), что в случае системы, состоящей из многих
видов, стационарное состояние ns является локально асимпто-
тически устойчивым в том и только в том случае, если все соб-
ственные значения матрицы сообщества [уравнение (13.2)}
имеют отрицательные действительные части. Любое собствен-
ное значение с положительной действительной частью обуслов-
ливает неустойчивость стационарного состояния ns.
Теперь предположим, что сначала система имеет единствен-
ное локально асимптотически устойчивое стационарное состоя-
ние; затем условия (т. е. параметры модели) изменяются и со-
ответственно изменяются собственные значения А. Если стацио-
нарное состояние теряет устойчивость, то может возникнуть ряд
нелинейных явлений; это означает, что ряд собственных значе-
ний матрицы переместится из левой части комплексной плоско-
сти в правую, пересекая при этом воображаемую ось.
Если эту ось пересекает одно собственное значение с нуле-
вой мнимой частью, то наблюдается бифуркация, т. е. появле-
ние дополнительных решений системы уравнений, описывающих
стационарные состояния. В этом случае колебания маловероят-
ны. С другой стороны, если сопряженная пара собственных зна-
чений пересекает воображаемую ось при ±iw, то возникают
колебания. Такое явление называют бифуркацией Хопфа. Ста-
ционарные состояния или колебания, возникающие при бифур-
кации, могут быть устойчивыми или неустойчивыми; для реше-
ния этой дилеммы необходим дальнейший анализ. Более слож-
ные бифуркации, в которых воображаемую ось пересекают дру-
Изучение смешанных популяций микроорганизмов
471
гие сочетания собственных значений, а также бифуркации,
связанные с потерей устойчивости предельных циклов, рассмот-
рены в литературе.
13.6. Расположение популяций в пространстве
До сих пор мы принимали, что все популяции распределены
в изучаемом объеме (пространстве) системы равномерно или
такое распределение обеспечивается эффективным перемешива-
нием. Допущение о пространственной однородности систем
очень удобно при моделировании и анализе, однако оно не обя-
зательно соответствует реальной структуре системы. Многие
системы обладают внутренними градиентами тех или иных па-
раметров, что обусловливает изменение численности популяций
в зависимости от положения в пространстве.
Еще больший интерес представляют довольно многочислен-
ные системы, в которых пространственно неоднородное распре-
деление одной или нескольких популяций возникает спонтанно
в отсутствие градиентов каких-либо параметров. Хотя для выяс-
нения .деталей механизмов, управляющих возникновением и под-
держанием пространственно неоднородного распределения попу-
ляций, необходимы широкие дополнительные исследования,
можно достаточно обоснованно допустить, что этот эффект
обусловлен взаимодействием между отдельными организмами
системы. Эти организмы могут принадлежать к двум различ-
ным видам (или клеткам) или к одному виду на разных ста-
диях его развития.
Показательный пример такого эффекта представлен на рис.
13.18, на котором зафиксирован процесс спорообразования в по-
пуляции гриба Nectria cinnabarincr, споры располагаются кон-
центрическими кругами с интервалами в 1 мм. Здесь очевидна
пространственная периодичность: по мере удаления от центра
колонии периодически появляются полосы спор гриба. Возмож-
но, что в основе пространственной периодичности и периодично-
сти во времени лежит один и тот же физико-химический меха-
низм. Некоторые результаты математического анализа указы-
вают на справедливость такой гипотезы, хотя ее ни в коей ме-
ре нельзя считать доказанной в общем случае. Косвенно эту
гипотезу подтверждает также существование реальной анало-
гичной химической системы. Действительно, в реакции Белоусо-
ва— Жаботйнского в отсутствие перемешивания проявляются
пространственные неоднородности, также изменяющие свой ха-
рактер во времени. При эффективном перемешивании в этой си-
стеме наблюдаются периодические флуктуации во времени.
Этой интересной проблеме посвящено много работ, но здесь
мы можем остановиться только на одной простой модели про-
472
Глава 13
РИС. 13.18. Фотография колонии (диаметр колонии 6 см) мицелиального ор-
ганизма N. cinnabarina, отражающая регулярное чередование в пространстве
зон спорообразования, при котором зоны с высокой концентрацией спор от-
стоят одна от другой приблизительно на 1 мм. [Из статьи: Winfree А. Т., J.
Theor. Biol., 38, 363 (1973).!
странственно неоднородного распределения. С другими работа-
ми в этой области можно ознакомиться из литературы, приве-
денной в конце главы.
Рассмотрим результаты работ Гэртина [11]. Допустим, что
два вида (1 и 2) существуют в одномерной полосе от 2 = 0 до
z = L. Это условие означает, что
= дПг - = 0 при 2 = 0 и z = L (13.45)
dz dz
Допустим также, что ни один из видов не размножается и не
погибает. Как мы вскоре покажем, из этого условия в сочета-
нии с указанным допущением следует, что общее число орга-
низмов «1 и п2 в системе постоянно.
Изучение смешанных популяций микроорганизмов
473
В силу ряда аргументов можно предположить, что потоки
видов (число организмов, пересекающих данное положение z
в единицу времени) /Дг) и /2(г) можно описать уравнением
Л (z) = —Dtl -дМ.г) — Вг2 ~ 2 i = l,2 (13.46)
дг дг
Обращает на себя внимание тот факт, что уравнение (13.46)
отвечает общему уравнению закона Фика для движения попу-
ляций. Далее в этом анализе будем принимать, что поток вида
1 более чувствителен к градиенту вида 2, чем к собственному
градиенту, и что вид 1 будет двигаться в направлении уменьше-
ния плотности вида 2. Это соответствует допущению
Вп=0 D12 = Dp>0 (13.47)
Как можно будет убедиться по завершении этого анализа, ука-
занное допущение справедливо во многих случаях. Аналогичное
допущение для вида 2 выражается в виде
D21=Dp>0 D22 = 0 (13.48)
В этих условиях изменение численности популяций видов 1
и 2 в нестационарном состоянии можно выразить уравнениями
^i_=DP^2. (13.49)
dt dz* 4
dn2 __jj
~дГ ~~>p dz*
(13.50)
Соответствующие граничные условия уже указаны в уравнении
(13.45), а начальными условиями являются начальные распре-
деления rzio(z), П20 (z) популяций двух видов:
пДг, 0) = nl0(z) i=l,2 (13.51)
Сложив уравнения (13.49) и (13.50), получим
где п — суммарная популяция в точке
n(z,t)=nl(z,t) + n2(z,t) (13.53)
Принимая во внимание условия (13.45) и (13.51), легко пока-
зать, что
-~- = Q при 2 = 0 и z = L (13.54)
и
n(z, O)=nio(z) + n2o(z) (13.55)
Эта задача о граничных значениях п стандартна и может быть
474
Глава 13
решена путем разделения переменных. Не останавливаясь на
промежуточных расчетах, отметим только, что решение прибли-
жается к постоянной величине Е при возрастании времени
lim n{z, t) =Е (13.56)
/—>оо
Постоянную Е можно найти, интегрируя уравнение (13.52)
от z = 0 до z = L:
— С п (г, /) dz = D„ С dz = D Г ------------------1
dt .) р] дг2 р]_ дг дг I
о о
(13.57)
Условие (13.54) показывает, что правая часть уравнения (13.57)
равна нулю, откуда следует, что
L
J п (г, t) dz = const (13.58)
о
Далее, применяя уравнение (13.58) при больших временах
[уравнение (13.56) ] и начальном времени t = Q, найдем
L L
J Edz = [/г10 (г) + /г20 (z)] dz (13.59)
о о
Левая часть уравнения (13.59) есть не что иное, как EL\ отсю-
да можно найти Е и преобразовать уравнение (13.56) следую-
щим образом:
L
limrz(z,f)=— ( [п10(?) + п20(z)] dz (13.60)
t^oo L J
о
Таким образом мы показали, что по мере увеличения времени
общая плотность популяции приближается к постоянной вели-
чине, равной средней плотности популяции.
Более интересные результаты можно получить, рассматривая
разность w между плотностями двух популяций
W = tl{— П2 (13.61)
Объединив описанным выше путем уравнения и условия ДЛЯ П\
и /г2, получим дм п д2м dt ~ р дг2 (13.62)
дм __ дг - 0 при z = 0 и z = L (13.63)
и
w(0, z) = w0(z) “Лю(г) — n2o(z)
(13.64)
Изучение с мешанных популяций микроорганизмов 475
На этом, однако, сходство с задачей определения п кончается.
Те, кто знаком с параболическими частными дифференциальны-
ми уравнениями, легко увидят в уравнениях (13.62) — (13.64)
не корректно поставленную задачу.
В любой точке z, где w0(z) не равно нулю, величина w бу-
дет возрастать во времени бесконечно. Этот общий вывод мож-
но проиллюстрировать на конкретном примере. Если мы при-
мем, что
w0 (z) = е cos (13.65)
то решением уравнений (13.62) — (13.65) будет выражение
w (z, t) = & cos —— exp —— t (13.66)
Очевидно, что при t—>-oo w(z, t) приближается к плюс или
минус бесконечности за исключением точек, где w0(z) равно
нулю. Конечно, этот результат не имеет физического смысла,
если П1 или пг приближается к нулю; остается признать, что
в таких ситуациях описываемая модель неприменима.
В совокупности с уравнением (13.60) этот результат показы-
вает, что если в точке z не выполняется условие Пю (?) =П2о (z),
то в этой точке со временем один из видов исчезнет, а числен-
ность популяции другого вида достигнет постоянной величины,
причем в данной области пространства выживет тот вид, чис-
ленность популяции которого сначала была более высокой.
Таким образом, конечным результатом будет полная изоляция
двух взаимодействующих популяций.
Этот пример, поддающийся полному математическому ана-
лизу, нетипичен в том смысле, что он не охватывает реально
существующих пространственно неоднородно распределенных
смешанных популяций. В другой важной ситуации основную
роль играет баланс между градиентами состава среды и хемо-
таксисом клетки (движение к питательным веществам или от
ингибиторов; см. например, работу [29] и упражнение 13.15).
Кроме того, нелинейные взаимодействия в пространственно
однородной системе могут приводить к неустойчивым состоя-
ниям, ведущим к «нарушению симметрии» и «образованию пат-
терна». Этой области исследований посвящается все большее
число работ. Интересно отметить, что системы, проявляющие
флуктуации в реакторах с полным перемешиванием, часто об-
разуют пространственные паттерны в отсутствие перемешива-
ния при наличии диффузионного переноса растворенных ве-
ществ (а возможно, и клеток). Флуктуирующие во времени
пространственные паттерны наблюдались, например, в случае
колоний слизистых грибов. В работе Писмена [30] дан вели-
476
Глава 13
колепный обзор математических аспектов явлений пространст-
венного и временного упорядочения, обусловленных неустойчи-
востью кинетики химических систем.
В этой главе мы рассмотрели некоторые биологические и ма-
тематические принципы, лежащие в основе изучения поведения
смешанных популяций с взаимодействующими видами.
Мы узнали, что в тщательно контролируемых условиях возмож-
но экспериментальное изучение таких систем и можно созда-
вать математические модели, согласующиеся в общих чертах
с экспериментальными результатами. В следующей главе мы
расстанемся с методами лабораторного исследования, матема-
тического моделирования и расчетов и рассмотрим некоторые
чрезвычайно сложные смешанные популяции, существующие
в естественных условиях и используемые в промышленности.
В гл. 14 мы узнаем, что приобретенные нами фундаментальные
знания чрезвычайно полезны при изучении и эксплуатации
таких систем, несмотря на то что имеющаяся теоретическая
основа все еще недостаточна для аналитического прогнозирова-
ния поведения сложных экосистем.
Упражнения
13.1. Конкуренция в периодической смешанной культуре. Два конкурирую-
щих вида А и В помещены в реактор периодического действия, содержащий
субстрат в концентрации s02>Aa, Ав, причем
Hmax,zs
Kz + s
i = А, В
Покажите, что при истощении субстрата
(rtAf)^max’А («Bf)|J'max’ в = const
где «аг, tisf — концентрации соответствующих популяций при $=0. (Этот ре-
зультат получен Толлингом при изучении смешанных культур диатомовых
водорослей [31].)
13.2. Простая модель мутуализма. Мейер, Цучия и Фредериксон [32] изу-
чали устойчивость популяций двух видов, находящихся в мутуалистических
взаимосвязях, и описали систему уравнениями
dtii
= — Dnt 4- i = 1,2
at
cis*
—Dsi +aipjnj— Ptwii i,/=l,2; i /
dt
При выводе этих уравнений допускалось, что в среде содержатся все необхо-
димые питательные вещества за исключением Si (продуцируемого Пг) и
(продуцируемого щ).
а) Покажите, что для каждого из уравнений роста ш = р.тах.<£. и |Ц-
= Hmax.iSi/(Ai+$i) существуют только два решения, соответствующих ста-
ционарным состояниям, и что решение, не отвечающее вымыванию, возможно
Изучение смешанных популяций микроорганизмов 477
только при <Х|а2>[1ф2; если же рост описывается уравнением Моно, то не
отвечающее вымыванию решение возможно и при D, меньшем некоторой ми-
нимальной величины gmax.f 2) .
б) Путем разложения в ряд Тейлора решения предыдущей задачи, соот-
ветствующего стационарному состоянию без вымывания, покажите, что оно
неустойчиво и, следовательно, чисто мутуалистические взаимодействия типа
рассмотренных в условии упражнения не могут существовать.
13.3. Сочетание мутуализма с конкуренцией. Мейер и другие [32] пред-
положили, что один из способов решения упражнения 13.2 заключается в уче-
те зависимости роста двух видов от общего субстрата S3; эта зависимость
может быть отражена в виде отдельного множителя в каждом выражении ти-
па уравнения Моно, например в уравнении (7.34).
а) Запишите пять уравнений, необходимых теперь для описания пере-
ходных состояний системы в хемостате.
б) Графическим или расчетным путем найдите решения при следующих
значениях параметров: Цтах.1 = 0,3 ч-1, цтах,2=0,2 ч-1, 7<1 = 10_3 г/л, К2=
= 0,002 г/л, /С1з=0,02 г/л, а! = 4,0, ^=2,0, а2=10,0, ^2=10,0; экономические
коэффициенты для s3 У!3=2,5 и У23=5.
в) Еще раз разложите начальную систему уравнений в ряд Тейлора и опре-
делите устойчивость каждой переменной после небольших возмущений.
13.4. Мутуализм и субстратное ингибирование. Второе решение упражне-
ния 13.2, соответствующее устойчивому состоянию системы в хемостате без
вымывания (сосуществованию видов), возможно в том случае, если допустить,
что рост видов подчиняется уравнению Эндрюса (7.32):
_______Hmax,is;____
14 ~ Ki + st + s^/Ki'
а) Запишите уравнения, описывающие поведение системы в хемостате
во времени.
б) Покажите, что можно найти четыре решения, отвечающие сосущество-
ванию видов в стационарном состоянии (в отсутствие вымывания), если
«1СС2>Р1^2 и D меньше максимальных абсолютных значений, которые могут
иметь указанные Ц; (i= 1, 2).
в) Методом линеаризованного анализа устойчивости покажите, что ста-
ционарное состояние будет устойчиво, если удовлетворяются следующие ус-
ловия:
PiAi+P2A2>0 и и
(или все неравенства изменены на обратные).
д _ Иглах,(KjKi' si2) . ________ %
1~ ке Ki + si + sW/]2 ’
(Черточка над обозначением переменной указывает, что значение этой пере-
менной относится к стационарному состоянию.)
г) Для стационарного состояния приведенные выше неравенства указы-
вают, что устойчивость возможна только в том случае, когда скорость роста
одного вида лимитируется субстратом, а скорость роста другого вида ингиби-
руется субстратом. С помощью соответствующих графиков покажите, почему
такая система устойчива к возмущениям si и s2 и почему одновременное огра-
ничение роста или ингибирование субстратом двух видов приводит к неустой-
чивому стационарному состоянию.
13.5. Methanobacillus omelianskii. В тексте этой главы при обсуждении,
мутуализма был упомянут пример совместного анаэробного роста двух видов,
478
Глава 13
которые, как теперь известно, составляют «вид» М. omelianskii (разд. 13.1):
вид 1
СН3СН2ОН+Н2О —>- СНзСОО-+Н++2Н2
вид 2
4Н2+СО2 —* СН4+2Н2О
Таким образом, первый вид продуцирует субстрат для второго вида, но инги-
бируется продуктом своей жизнедеятельности — водородом.
а) Почему такие взаимодействия относят к мутуализму, а не к коммен-
сализму'?
б) Запишите систему соответствующих уравнений для переходного состоя-
ния, учитывающих ингибирование водородом.
в) Обсудите устойчивость стационарных состояний системы с помощью
разложения соответствующих уравнений в ряды Тейлора.
13.6. Частицы с аэробными и анаэробными зонами. Облигатные анаэробы
могут существовать в центральной зоне влажной частицы почвы; возможность
продвижения их популяции к периферии частицы определяется процессами
восстановления кислорода, снижающими его концентрацию до субкритическо-
го уровня:
Воздух Аэробные условия { Анаэробные j
1 атм (NO 1 условия (N2) |
> 1
Поверхность Центр
частицы частицы
а) Пусть питательное вещество (источник углерода Si) постоянно прони-
кает через наружную поверхность частицы почвы. Найдите уравнения, описы-
вающие поведение системы в следующих случаях:
1. Вид Nj потребляет как питательное вещество Si, так и кислород, а вид N2
потребляет продукт жизнедеятельности вида N|, но взамен не дает ему
ничего.
2. Виды Ni и N2 конкурируют за питательное вещество Si; других типов
взаимодействий в системе нет.
б) Каким образом неоднородное распределение кислорода, обусловливаю-
щее существование аэробной и анаэробной экологических ниш в частице, из-
меняет устойчивость системы по сравнению с условиями, указанными в упраж-
нениях 13.2—13.4?
13.7. Экологические ниши, появление которых обусловлено неэффективным
перемешиванием. На рис. 13.8 приведены данные, показывающие, что в систе-
ме, которая состоит из двух поглощающих один субстрат видов, скорость раз-
ведения определяет, какой вид будет доминировать (если графики зависимо-
стей Ц1 и ц2 от концентрации субстрата пересекаются). На рис. 9.11 и в соот-
ветствующем разделе текста приведен анализ системы в реакторе с непол-
ным перемешиванием, что обусловливает разделение содержимого реактора
на две системы с полным перемешиванием, каждая из которых характеризует-
ся собственной скоростью разведения.
а) Найдите уравнения, описывающие поведение смешанной популяции
(состоящей из двух видов) в такой системе с неполным перемешиванием.
б) Определите, в каких условиях популяция одного вида будет домини-
ровать в одном сосуде, а другого во втором, т. е. при какой скорости разведе-
ния и других параметрах пространственное разделение стабилизирует неустой-
чивое в других условиях сосуществование двух видов (ср. рис. 13.8).
в) Почему студенты и преподаватели живут в раздельных помещениях?
13.8. Теория Фейнберга — Хорна — Джексона. Для условий упражнений
13.2—13.5
Изучение смешанных популяций микроорганизмов
479>
а) Опишите систему популяций микроорганизмов в рамках теории Фейн-
берга — Хорна — Джексона (табл. 13.6).
б) Определите каждый комплекс, тип связи и наличие или отсутствие
слабой обратимости.
в) С помощью теоремы нулевого ограничения (13.41) обсудите каждый
нз полученных результатов с точки зрения нетривиальностн состояния и ста-
бильности системы.
г) Как использовать эту теорию для анализа системы, находящейся в кас-
каде реакторов?
13.9. Устойчивость сети внутриклеточных превращений. Теория Фейнбер-
га— Хорна — Джексона применима к любой системе, кинетика которой может
быть описана законом действующих масс, а следовательно, и к сети внутри-
клеточных реакций.
а) Запишите в соответствующих символах элементарные реакции, участ-
вующие в биосинтезе нуклеотидов (рис. 6.20).
б) С помощью теоремы нулевого ограничения определите, какая нз сетей
реакций — нормальная (рис. 6.20,а) или мутантная (рис. 6.20,6) — устойчива,.,
а какая подвержена колебаниям.
13.10. Уравнения Лесли. Для описания системы хищник — жертва Лес-
ли [29] предложил следующую математическую модель:
dx , „ dy fy*
= ах — bx2 —- сху —— = еу —
dt-----------------------------------------dt-х
Последнее слагаемое второго уравнения учитывает отношение численностей
популяций хищника и жертвы.
а) Постройте графики зависимости у от х при dx/dt и dy[dt = ^. Выберите
некоторые простые положительные значения для а, b, с, е, f и затем путем
определения ряда значений dy/dt постройте график поведения системы во вре-
мени, когда начальные условия отличны от равновесных.
б) Справедливо ли уравнение для кажущейся удельной скорости роста у,-
еслн отношение у/х постоянно, а х уменьшается до очень малой величины?
Почему?
в) Докажите, что для этой системы характерны затухающие колебания.
13.11. Статистическая механика популяций. По мере того как при изуче-
нии предмета мы переходим к анализу математических моделей все более и
более сложных систем, часто возникает стремление провести аналогию с ка-
жущимися аналогичными системами. Напишите краткий обзор, посвященный,
обсуждению сходств и различий между кажущимся поведением «популяции
молекул» и популяции клеток. В обсуждение включите сравнение открытых
и изолированных систем, равновесных и стационарных состояний, анализ урав-
нений движения и соответствующих постоянных.
13.12. Кинетика изменения численности видов, связанных пищевой сетью.
Рассмотрим пищевую цепь 1^-2-^-3-^-4, в которой виды 2, 3 и 4 являются-
хищниками по отношению к видам 1, 2 и 3 соответственно.
а) Приняв, что скорость роста каждого вида описывается логистическим
уравнением, и включив в уравнения члены, учитывающие взаимодействия
хищник — жертва, найдите уравнения, описывающие стационарные состояния
в этой системе. В частности, покажите, что численность популяции вида 1 за-
висит от всех кинетических констант, а численность популяции вида 4 опре-
деляется только логистическим уравнением для вида 4 [35].
б) В естественных условиях углерод, азот и другие элементы включены
в соответствующие кругообороты. Дополните систему еще одним взаимодей-
ствием, при котором вид 4 является жертвой по отношению к виду 1; теперь
пищевая цепь^замкнута. Покажите, что в этом случае решения для стационар-
ных состояний имеют одну и ту же форму для всех видов, и поэтому числен-
ность популяции каждого вида зависит от всех кинетических констант сети,-
480
Глава 13
Оцените количественно сравнительную устойчивость этих двух систем — ли-
нейной и замкнутой пищевых цепей.
13.13. Скопления в слизистых грибах [36—381. Когда растущая смешан-
ная популяция слизистых грибов и амеб истощает свой запас питательных
веществ, наступает период относительного покоя, в котором первоначально
однородно распределенные в культуре амебы группируются в скопления, от^
стоящие одно от другого приблизительно иа 0,1 мм. В конце концов в этих
скоплениях возникают споры. Образование скоплений, как оказалось, обуслов-
лено реакцией амебы на изменение концентрации циклического АМР, выде-
ляемого голодающими амебами. Пусть а — численность популяции амебы
в единице объема, а р — концентрация аттрактанта (циклического АМР).
.Можно разработать математическую модель, которая при описанных условиях
приводит к следующему уравнению:
да д f да др \ др „ д2р
"л7" = Iх = fa—kp+D ——
dt дх \ дх дх ) dt дх2
а) Путем линеаризации этих уравнений покажите, что решения в форме
(sin qx)eQt для аир предсказывают устойчивое состояние, если %ао/<ц(^+
+Dq2~), q=£-0.
б) Возмущения какой частоты обусловливают скорейшее возникновение
неустойчивости?
в) Высказывалось мнение, что система вначале устойчива, но истощение
питательных веществ изменяет параметры системы и при а=const приводит
к неустойчивому состоянию. Определите физический смысл каждого пара-
метра модели. Изменение какого из этих параметров может привести к
неустойчивости системы?
г) Почему образование скоплений необходимо для спорообразования?
13.14. Конкуренция в непрерывной культуре. Эксперименты с непрерыв-
ной культурой, для которой лимитирующим рост питательным веществом яв-
ляется лактат, показали, что Ks для псевдомонад равно приблизительно
6 мг/л, для спирилл около 12 мг/л. Обсудите влияние концентрации лактата
в смешанной культуре, содержащей оба вида бактерий, если максимальная
скорость роста псевдомонад равна 0,6 ч-1, а спирилл 0,8 ч-1.
13.15. Хемотаксис и конкурентное сосуществование видов. Рассмотренный
в этой главе принцип конкурентного исключения не всегда реализуется, если
для системы характерны пространственные градиенты питательных веществ
или ингибиторов или если условия окружения изменяются во времени. Моде-
лирование и анализ систем при наличии градиентов питательных веществ или
ингибиторов описаны Лауффенбергером (Lauffenberger D. A., Effects of Cell
Motility Properties on Cell Populations in Ecosystems, in Foundations of Bioche-
mical Engineering: Kinetics and Thermodynamics in Biological Systems,
Blanch H. W., Papoutsakis E. T„ Stephanopoulos G. N. (eds.), p. 265, American
Chemical Society, Washington, D.C., 1983]. Градиенты концентраций питатель-
ных веществ обусловливают необходимость учета хемотактического движения
организмов. В такой ситуации поток клеток Jx в направлении A~z можно опи-
сать уравнением
dx ds
где х и s — концентрации клеток и лимитирующего рост питательного веще-
ства соответственно. Параметры 0 и % являются коэффициентами неупорядо-
ченной подвижности и хемотаксиса соответственно. Приняв, что поток суб-
страта описывается законом Фика и что удельную скорость роста клеток
Изучение смешанных популяций микроорганизмов 481
можно приближенно описать ступенчатой функцией
(u,0 — ke при s > %
и = <
( — ke при s sc
а) Определите распределение клеток одного вида и питательного вещества
в стационарном состоянии в ограниченной области между z = 0 и z = L. Клетки
не могут выходить за эти границы, а субстрат не диффундирует через грани-
цу z=0. Концентрация субстрата при z = L равна s0.
б) Рассматривая важные безразмерные группы и явления, учитываемые
при анализе описанной в предыдущей задаче системы, подумайте о возмож-
ных последствиях конкуренции двух видов в такой ограниченной области.
Могут ли два вида сосуществовать в такой системе, и, если могут, то при ка-
ких условиях? Проверьте правильность ваших догадок, ознакомившись с ра-
ботой Лауффенбергера. Сравните результаты моделирования с вашими соб-
ственными наблюдениями конкуренции в человеческом обществе.
Литература
Некоторые сведения о смешанных популяциях даны в руководствах по общей
микробиологии, перечисленных в гл. 1. В работе [4] имеются три раздела,
посвященных симбиозу. См. также:
1. Одум Ю., Основы экологии. — М.: Мир, 1975. Подробный учебник, напи-
санный популярным языком; в нем анализируются все проблемы эко-
логии.
2. Gause G. F., The Struggle for Existence, Williams and Wilkins Company,
Baltimore, 1934. Эта работа послужила исходной точкой для большинства
последующих экспериментальных и теоретических исследований смешан-
ных культур микроорганизмов. Основы многих из изложенных в настоя-
щей главе концепций, определений и представлений были заложены в этой
классической монографии.
3. Microbial Ecology, Laskin A. I., Lechevalier H. (eds.), CRC Press, Cleveland,
1974. Здесь в первых двух главах рассматривается влияние пестицидов на
популяции микроорганизмов, а третья глава посвящена экологии почвен-
ных микроорганизмов. Особенно рекомендуем ознакомиться с великолеп-
ной заключительной главой (Meers J. L., Growth of Bacteria in Mixed
Cultures).
4. Hattori T., Microbial Life in the Soil: An Introduction, Marcel Dekker, Inc.,
New York, 1973. Хороший обзор почвенных экосистем, отличающихся очень
большой сложностью; приведено множество интересных примеров меж-
видовых отношений в мире микроорганизмов.
5. May R. М., Stability and Complexity in Model Ecosystems, Princeton Univer-
sity Press, Princeton, N. J., 1973. Хорошая монография, посвященная ма-
тематическому анализу взаимодействий между популяциями; эта книга
послужила основой для большей части материала, изложенного в разд. 13.4
и 13.5. Здесь убедительно обсужден принцип нестабильности систем, обус-
ловленной их сложностью.
6, Bungay Н. R„ Bungay М. L., Microbial Interactions in Continuous Culture,
Adv. Appl. Microbiol., 10, 269 (1968). Несмотря на то что эта работа не-
сколько устарела, она по-прежнему является полезным дополнительным
материалом и источником различных идей.
7, Rescigno A., Richardson I. W., The Deterministic Theory of Population Dyna-
mics, in Foundations of Mathematical Biology, R. Rosen (ed.), vol. 3, Super-
cellular Systems, p. 283, Academic Press, Inc., New York, 1973. В этой рабо-
те дан хороший обзор работ Вольтерры по межвидовой конкуренции и
31—746
482
Глава 13
взаимодействиям типа хищник — жертва, а также некоторых более позд-
них работ по популяционной динамике, особенно работ Костицина.
8. Lotka A. J., Undamped Oscillations Derived from the Law of Mass Action,
J. Am. Chem. Soc., 42, 1595 (1920). В этой работе заложены основы извест-
ной модели Лотки — Вольтерры для взаимосвязей типа хищник — жертва.
9. Feinberg М., Horn F. J. М., Dynamics of Open Chemical Systems and the
Algebraic Structure of the Underlying Reaction Network, Chem. Eng. Sci.,
29, 775 (1974). Обзорная статья по теории закона действующих масс, раз-
работанной авторами и Р. Джексоном и рассмотренной в разд. 13.5.2.
(Более поздний обзор опубликован М. Фейнбергом; см. работу [10], с. 1.)
10. Bailey J. Е., Periodic Phenomena, in The R. H. Wilhelm Memorial Volume
on Chemical Reactor Theory, Lapidus L., Amundson N. R. (eds.), p. 758,
Prentice-Hall, Inc., Englewood Cliffs, N. J., 1977. В этой работе дано вве-
дение в математическую теорию спонтанных и вынужденных колебаний
в контексте химических реакций.
11. Gurtin М. Е., Some Mathematical Models for Population Dynamics That
Lead to Segregation, Q. Appl. Math., 32, 1 (1974). Здесь детально рассмот-
рен пример, приведенный в разд. 13.6, а также родственные проблемы.
Рост неустойчивых популяций рекомбинантных организмов и родственные
системы:
12. Imanaka Т., Aiba S., A Perspective on the Application of Genetic Engi-
neering: Stability of Recombinant Plasmids, Ann. N.Y. Acad. Sci., 369, 1
(1981).
13. Ollis D. F„ Chang H.-Т., Batch Fermentation Kinetics with (Unstable)
Recombinant Cultures, Biotech. Bioeng., 24, 2583 (1982).
14. Ollis D. F., Industrial Fermentations with (Unstable) Recombinant Cultures,
Phil. Trans. Roy. Soc. London, B297, 617 (1982).
15. Hjortso M. A., Bailey J. E., Plasmid Stability in Budding Yeast Populations:
Steady-State Growth with Selection Pressure, Biotech. Bioeng., 26, 528
(1984).
16. Seo J.-H., Bailey J. E., A Segregated Model for Plasmid Content and Pro-
duct Synthesis in Unstable Binary Fission Recombinant Organisms, Biotech.
Bioeng., 27, 156 (1985).
17. Kimura M., Ohta T., Theoretical Aspects of Population Genetics, Princeton
University Press, Princeton, N. J., 1971.
18. Stewart F. M., Levin B. R., The Population Biology of Bacterial Plasmids:
A priori Conditions for the Existence of Conjugationally Transmitted Factors,
Genetics, 87, 209 (1977).
Работы, посвященные изучению взаимодействий типа хищник — жертва:
19. Tsuchiya Н. М. et al., Predator-Prey Interactions of Dictyostelium discoideum
and Escherichia coli in Continuous Culture, J. Bacteriol., 110, 1147 (1972).
20. Jost J. L. et al., Interactions of Tetrahymena pyriformis, Escherichia coli, and
Azotobacter vinelandii and Glucose in a Minimal Medium, J. Bacteriol., 113,
834 (1973).
21. Jost J. L. et al., Microbial Food Chains and Food Webs, J. Theoret. Biol.. 41,
461 (1973).
Другие сведения о расположении популяций в пространстве:
22. Turing А. М., The Chemical Basis of Morphogenesis, Trans. Roy. Soc.
London, B237, 337 (1952).
23. Гленсдорф П., Пригожин И., Термодинамическая теория структуры, устой-
чивости и флуктуаций. — М.: Мир, 1973.
24. Othmer Н. G., Scriven L. Е., Instability and Dynamic Pattern in Cellular
Networks, J. Theoret. Biol., 32, 507 (1971).
Изучение смешанных популяций микроорганизмов
4С3
25 Feld R. J., A Reaction Periodic in Time and Space, J. Chem. Educ., 49, 308
(1972).
26. Winfree A. T., Rotating Chemical Reactions, Sci. Am., Jane, 1974, p. 82.
Явление бифуркации:
См. работу [10], а также:
27. Smith С. В., Kuszta В., Lyberatos G., Bailey J. Е., Period Doubling and
Complex Dynamics in an Isothermal Reaction System, Chem. Eng. Sci., 38,
425 (1983).
28. Lyberatos G., Kuszta B., Bailey J. E_, Versal Matrix Families, Normal Forms,
and Higher Order Bifurcations in Dynamic Chemical Systems, Chem. Eng.
Sci., 40, 1177 (1985).
Расположение популяций в пространстве:
29. Lauffenberger D. A., Effects of Cell Motility Properties on Cell Populations
in Ecosystems. ACS Symp. Ser., 207, 265 (1983).
30. Pismen L. M., Kinetic Instabilities in Man-Made and Natural Reactors,
Chem. Eng. Sci., 35, 1950 (1980).
Упражнения:
31. Talling J. F., The Growth of Two Plankton Diatoms in Mixed Culture,
Physiol. Plant., 10, 215 (1957).
32. Meyer J. M., Tsuchiya H. M., Frederickson A. G., Dynamics of Mixed Popu-
lations Having Complementary Metabolism, Biotech. Bioeng., 17, 1065
(1975).
33. Leslie P. FL, Some Further Notes on the Use of Matrices in Population
Mathematics, Biometrika, 35, 213 (1948).
34. Смит Дж. M., Модели в экологии. — М.: Мир, 1976.
35. Rapport D. J., Turner J., Predator-Prey Interactions in Natural Communities
J. Theoret. Biol., 51, 169 (1975).
'36. Bonner J. T., The Cellular Slime Molds, Princeton University Press, Prince-
ton, N. J., 1967.
37. Feller E. F., Segel L. A., Initiation of Slime Mold Aggregation Viewed as an
Instability, J. Theoret. Biol., 26, 399 (1970).
38. Lin С. C., Segel L. A., Mathematics Applied to Deterministic Problems in
the Natural Sciences, pp. 22—31, The Macmillan Company, New York, 1974.
31*
Глава 14
Смешанные популяции микроорганизмов
в естественных системах
и промышленных процессах
Смешанные популяции микроорганизмов играли важную
роль уже в период возникновения жизни на земле. Позднее
различные виды микроорганизмов выполняли важные функции
в биосфере и ее эволюции. Так, когда-то, не позднее 500 мил-
лионов лет и не ранее 2 миллиардов лет тому назад, развитие
примитивных водорослей, способных осуществлять фотосинтети-
ческие реакции, достигло такого уровня, что в атмосфере в зна-
чительном количестве (примерно 1% от концентрации в настоя-
щее время) появился кислород. До этого периода из микроор-
ганизмов существовали только облигатные анаэробы, появление
кислорода в атмосфере обусловило возникновение аэробных
организмов и факультативных анаэробов. Таким образом, воз-
никновение кислородной атмосферы и последующее появление
аэробных форм жизни стало возможным благодаря жизнедея-
тельности примитивных фотосинтезирующих микроорганизмов.
В разд. 14.3 мы вкратце рассмотрим роль мира микроорганиз-
мов в современной глобальной экосистеме.
Смешанные популяции микроорганизмов широко распро-
странены в природе — они встречаются в воздухе, почве, водо-
емах, растут на наружных поверхностях и внутри высших орга-
низмов. Одним из самых интересных примеров может служить
симбиоз микроорганизмов со жвачными, например крупным ро-
гатым скотом, овцами и козами (см. табл. 13.1). В рубце, пер-
вом отделе желудка жвачных, существует плотная смешанная
культура бактерий (около 1010 клеток в миллилитре) и простей-
ших. В этой очень сложной и чрезвычайно разнообразной попу-
ляции наблюдались мутуалистические, конкурентные, аменсали-
тические взаимодействия, а также межвидовые отношения типа
хищник — жертва.
В результате деятельности микроорганизмов в рубце проис-
ходит разложение растительных материалов, в том числе целлю-
лозы и других сложных углеводов, на более простые вещества,
которые поглощаются кровотоком животного. В организме
жвачных пищеварительные ферменты секретируются только
в отделах желудка, расположенных после рубца. Таким путем
Практическое применение смешанных популяций 435
обеспечивается, с одной стороны, высокая активность микроор-
ганизмов, а с другой — лизис и переваривание микроорганизмов
в последующих отделах желудка. Такая система пищеварения
позволяет жвачным усваивать и переваривать значительно более
простые (по сравнению с человеком и другими млекопитающи-
ми) азотсодержащие питательные вещества. Так, например, ам-
миак и мочевина под действием микроорганизмов рубца вклю-
чаются в такие сложные органические структуры, как белки, и
именно последние утилизируются организмом жвачного в после-
дующих отделах желудка.
Другим широко известным примером смешанных популяций
микроорганизмов является естественная флора, населяющая
организм человека. Для эффективного переваривания и усвое-
ния пищи человеку необходима помощь со стороны многих ви-
дов бактерий, обитающих в пищеварительном тракте. Среди
этих организмов наблюдались аменсализм и комменсализм,
а существенная часть популяции простейших питается бакте-
риями, находящимися в кишечнике. В зубных бляшках обитает
несколько видов микроорганизмов, из которых наиболее типич-
ны факультативные анаэробы. (Почему именно эти организмы
хорошо переносят обычную гигиеническую чистку зубов?)
На кожном покрове человеческого организма также обитает
множество видов протистов; так, бактерии изобилуют в волося-
ных мешочках, а дрожжи и грибы населяют наиболее влажные
участки. Изучение вопроса о том, как организм человека защи-
щается от многочисленных соседей, а часто извлекает выгоду
из этого сожительства, представляет очень большой интерес,
однако мы не будем углубляться в его изучение, поскольку это
увело бы нас слишком далеко от нашей основной темы. Вместо
этого мы перейдем к проблеме использования смешанных попу-
ляций микроорганизмов в промышленности.
История использования человеком смешанных популяций
микроорганизмов насчитывает несколько тысячелетий. Резуль-
таты археологических раскопок показывают, что человек полу-
чал вино и пиво путем сбраживания фруктов и зерновых куль-
тур задолго до 2000 года до новой эры. Эти процессы, во вся-
ком случае в том варианте, в котором их осуществляли в то
время, могут служить прекрасными примерами применения
естественных, смешанных культур. В процессах с участием есте-
ственных смешанных культур обычно нет необходимости во вне-
сении посевного материала, содержащего определенные орга-
низмы. В таких процессах все необходимые превращения осу-
ществляются организмами, уже имеющимися в исходном сырье.
Как мы подробнее покажем ниже, естественные смешанные
культуры особенно эффективны при утилизации смесей суб-
стратов.
486
Глава 14
Указанные особенности естественных смешанных популяций
предполагают, что одновременный рост нескольких видов обес-
печивает какие-то особые выгоды и преимущества, недостижи-
мые в случае чистых культур. В следующем разделе вкратце
будет рассмотрена технология процессов с участием смешанных
культур определенного состава. Последний раздел этой главы
будет посвящен использованию естественных смешанных куль-
тур в основном в процессах биологической очистки сточных вод
(разд. 14.4).
14.1. Применение смешанных культур
определенного состава
Самым наглядным примером использования смешанных
культур определенного видового состава является сыроделие.
Гастрономическая сторона этих процессов известна каждому,
но далеко не все знакомы с экономической важностью молочной
и сыродельческой промышленности. Согласно одной из оценок,
доля молочной промышленности составляет около 4% в вало-
вом национальном продукте США; из них около 15% приходит-
ся на производство сыра.
Ранее в сыроделии применяли местные дикие смешанные
культуры; теперь же для обеспечения строго стандартизирован-
ного качества сыров для закваски используют особые культуры.
Различные сорта сыров производят путем засева пастеризован-
ного свежего молока соответствующей культурой организма,
продуцирующего молочную кислоту. В кислой среде образуется
богатая белками творожистая масса; ее отделяют от сыворотки
и ставят на созревание в присутствии бактерий или плесеней.
При производстве твердых сортов сыра к инокулированному
молоку после того, как создалась слегка кислая среда, добавля-
ют смесь сычужных ферментов (содержащую протеолитический
фермент реннин). В конечном счете образуется резиноподобная
сырная масса (сгусток), которую разрезают на мелкие кусочки,
подвергают тепловой обработке, подсушивают, размалывают и
прессуют для отделения остатков сыворотки (процесс чеддари-
зации). Наконец, после второй серии операций высаливания,
подсушивания и прессования получают сыр, готовый к созрева-
нию. В процессе созревания в результате медленных фермента-
тивных (анаэробных) реакций частично расщепляются липиды
и белки, образуя частично окисленные продукты, в том числе
молочную, масляную и уксусную кислоты, которые по мере ста-
рения сыра придают ему специфический острый вкус.
Начальные стадии производства многих других сортов нату-
ральных сыров аналогичны описанным выше; консистенция и
Практическое применение смешанных популяций 487
вкус готового сыра определяются в основном на более поздних
стадиях; в частности, природой применяемого на этих стадиях
организма, концентрацией соли и влаги и температурой созре-
вания. В табл. 14.1 приведен перечень наиболее распространен-
ных сортов сыра и соответствующих организмов, используемых
при их получении. Многие из этих организмов синтезируют ви-
тамины, которые повышают питательную ценность сыров в ходе
i созревания.
Молочнокислые бактерии являются составными частями и
других смешанных культур определенного состава, использую-
щихся в пищевой промышленности. Так, при производстве вис-
ки добавление Lactobacillus к дрожжам способствует снижению
pH, а также уменьшению вероятности заражения посторонними
организмами, приданию необходимого вкуса и аромата. Другой
пример применения смешанных культур дрожжей и молочно-
кислых бактерий — производство имбирного пива. Наконец,
Д'
® Таблица 14.1. Микроорганизмы, применяющиеся в сыроделии’
1! __________________________________________________________________________
И а) Микроорганизмы, применяющиеся в сыроделии
I. Бактерии
А. Бактерии, использующиеся в основном в качестве продуцентов мо-
лочной кислоты
Е 1. Streptococcus lactis
К 2. Streptococcus cremoris
(js 3. Streptococcus thermophilus
4. Streptococcus durans (модифицированный министерством сельского
хозяйства США процесс производства сыра чеддер)
в 5. Lactobacillus bulgaricus
К Б. бактерии, использующиеся в основном для придания сыру вкуса и
м аромата; эти бактерии могут оказывать также влияние на текстуру
L, и консистенцию сыра
® 6. Brevibacterium linens
® 7. Propionibacterium shermanii
jg 8. Leuconostoc sp. (придающие специфический аромат или продуци-
рующие лимонную кислоту)
. 9. Streptococcus diacetilactis (придающие специфический аромат; ис-
“ пользуются также для специальных целей)
II. Дрожжи
Uf А. Применяющиеся для поддержания и ускорения роста бактерий в
культуре и сырах (не обязательные)
" 10. Candida krusei
11. Му coderma sp.
III. Плесени (применяются для усиления вкусовых качеств, аромата, при-
дания соответствующих консистенции, текстуры, состава и внешнего
вида)
А. Растущие на наружных поверхностях
р 12. Penicillium camemberti
13. Различные виды
Б. Растущие внутри сыра
, 14. Penicillium roqueforti и родственные виды
Г ----------------—----------------------------------------------------------
488
Глава 14
Продолжение табл. 14.1
б) Зависимость сорта сыра от типа заквасочной культуры
Сорт сыра Тип применяющейся культуры Способ прв* ,, т Классы II менения Класс I и ш
Брик А. 1—3 Б.6,8,9 П.А х, у, Z6
Камамбер А. 1,2 Б.8,9 III.A.12 X, Z
Чеддер А.1,2,4 Б.8,9 х, у, Z
Коттедж А. 1,2 Б.8,9 X, Z
Сливочный А. 1,2 Б.8,9 X, Z
Эдамский А. 1,2 Б.8,9 X, Z
Гауда А. 1,2 Б.8,9 X, Z
Лимбургер А. К-3 Б.6,8,9 П.А х, у, z
Невшатель А. 1,2 Б.8.9 X, Z
Пармезан А. 1—3,5 Б.8.9 х, у, Z
Проволоне А.1—3,5 Б.8.9 х, у, Z
Романе А. 1—3,5 Б.8.9 х, у, Z
Рокфор А. 1,2 Б.8.9 III.Б.14 X, Z
Швейцарский А.1—3,5 Б.7—9 X, у, Z
Трапистский А.1—3 Б.6,8,9 П.А х, у, z
аДанные любезно предоставлены Дж. У. Рейнболдом (Iowa State University, Ames,
Iowa).
б Обозначения: x— чистая культура; у — смешанная культура; z — комбинированная
смешанная культура (культура двойного назначения и (илн) чистая или смешанная куль»
тура).
применение двух видов Lactobacillus способствует повышению
выхода молочной кислоты.
Последовательную трансформацию питательного вещества
в необходимый конечный продукт иногда можно успешно осу-
ществлять с помощью особого методического приема, называе-
мого двойной ферментацией. Примером может служить процесс
получения ь-лизина из глицерина, в котором в качестве проме-
жуточного вещества образуется а,е-диаминопимелиновая кисло-
та (DAP). В одной из операций с участием ауксотрофного му-
танта Е. coli накапливают это промежуточное соединение.
В другой отдельной операции выращивают культуру Aerobacter
aerogenes. Этот организм синтезирует фермент DAP-декар-
боксилазу, которая превращает DAP в ь-лизин. Последующее
смешение двух культур и добавление толуола приводит к выде-
лению в среду как DAP, так и DAP-декарбоксилазы, что обес-
печивает образование ь-лизина.
Известно несколько других примеров промышленных процес-
сов с применением смешанных культур определенного состава.
Одним из таких процессов является производство ^-каротина.
В этом процессе различные скрещивающиеся типы одного и
Практическое применение смешанных популяций
489
того же организма обеспечивают повышение выхода каротина
приблизительно в 20 раз по сравнению с процессом, катализи-
руемым любым одним типом организмов. В приведенном ниже
примере рассмотрено повышение степени усвоения метана
в процессах производства белка одноклеточных организмов бла-
годаря применению смешанных культур.
Пример 14.1. Обусловленное мутуализмом ускорение роста утилизирую-
щих метан бактерий Pseudomonas sp. в хемостате*. В разд. 12.7 и приме-
ре 9.2 мы рассматривали некоторые аспекты проблемы производства белка
одноклеточных организмов. В соответствующих процессах в последнее время
большой интерес вызывает изучение возможности применения газообразных
углеводородных субстратов, в особенности метана. В некоторых эксперимен-
тальных работах было показано, что часто смешанные культуры растут на
метане быстрее, чем соответствующие чистые культуры организмов, утилизи-
рующих метан.
Этот факт объясняли, в частности, мутуалистическим взаимодействием
типа изображенного схематично на рис. 14П1.1. В данной смешанной культу-
ре, изучавшейся экспериментально Уилкинсоном, Топивалой и Хэймером, име-
ются в основном два вида бактерий — утилизирующие метан Pseudomonas sp.
и утилизирующие метанол Иyphomicrobium sp. Поскольку метанол является
конечным продуктом метаболизма Pseudomonas sp. и ингибирует рост этих
организмов, то псевдомонады существенно выигрывают от присутствия второ-
го вида, связывающего метанол. Очевидно, что Нyphomicrobium sp. в смешан-
ной культуре также получают определенные преимущества, поскольку этот
вид таким путем обеспечивается источником углерода. В смешанной культуре
присутствуют в относительно невысоких концентрациях два других вида бак-
терий, которые, как полагают, выполняют полезные функции, утилизируя
другие конечные продукты метаболизма Pseudomonas sp.
С детальным обсуждением предложенного механизма можно ознакомить-
ся в оригинальной работе; мы же ограничимся рассмотрением одного экспе-
римента, в ходе которого был выявлен ряд эффектов, представленных на
рис. 14П1.2. Смешанную культуру выращивали в хемостате; в нулевой момент
времени (/=0) в ферментер и резервуар со средой добавляли метанол до
концентрации 1,6 г/л. Как показано на рис. 14П1.2, добавление метанола вы-
зывает немедленное и очень резкое снижение скорости утилизации метана
и кислорода и несколько меньшее снижение общей массы сухих веществ мик-
роорганизмов, хотя относительное количество Нyphomicrobium sp. в фермен-
тере возрастает. Приблизительно через 17 ч концентрация метанола в фер-
ментере падает почти до нуля, а указанные выше тенденции изменения кон-
центраций компонентов системы меняются на обратные. Считается, что в этот
момент активность Pseudomonas sp. в существенной степени восстанавли-
вается.
Все составляющие математической модели этой системы должны быть нам
знакомы по материалу предыдущих разделов и глав, посвященных изучению
массопереноса и кинетики процессов в биологических системах. Обозначив
символами Xi и х% концентрации Pseudomonas и Нyphomicrobium sp. соответ-
ственно, можно записать соответствующие уравнения для изменения этих кон-
центраций в переходном состоянии в хемостате:
dxt
-Dxf\-rfi г =1,2 (14П1.1)
* Этот пример заимствован из работы: Wilkinson Т. G., Topiwala Н. Н
Hamer G.„ Interactions in a Mixed Population Growing on Methane in Conti-
nuous Culture; Biotech. Bioeng., 16, 41 (1974).
32—746
490
Глава 14
РИС. 14П1.1. Возможные межвидовые взаимодействия в смешанной культуре,
растущей на метане.
Выражения для скорости роста Pseudomonas и Hyphomicrobium выбраны та-
ким образом, чтобы они отражали ситуацию, схематично изображенную на
рис. 14П1.1. В частности, для лимитируемого кислородом роста бактерий
Pseudomonas принимается, что
Hl,maxcOa 1
1 = + Соа ~l + s/Ki Xl
где Со2 и s — концентрации растворенного кислорода и метанола соответст-
венно. Используемые в этом примере функции ингибирования аналогичны
рассмотренным в гл. 3 (неконкурентное ингибирование катализируемых фер*
(14Ш.2)
8
О 5 10 15 20 25 30 35
Время, ч
РИС. 14П1.2. Поведение утилизирующей метан смешанной культуры в нестационарном состоянии в хемостате после до-
бавления метанола до концентрации 1,6 г/л в ферментере и питательной смеси при /=0; D=0,08 ч”1. [Из работы: Wilkin-
son Т. G. et al., Interactions in a Mixed Bacterial Population Growing on Methane in Continuous Culture; Biotech. Bioeng.,
16, 14 (1974).]
1 — общая масса сухих веществ бактерий; 2 — концентрация кислорода в отходящих газах; 3— парциальное давление рас-
творенного кислорода; 4 — скорость образования СОг; 5 — скорость утилизации метана; 6 — относительная численность по-
пуляции Hyphomicrobium sp., %.
492
Глава 14
Таблица 14П1.1. Параметры математической модели, описывающей поведение смешанной культуры, утилизирующей метана
Параметр Физический смысл параметра Значение параметра
Р» I, m а х Максимальная удельная скорость роста штамма, утилизирующего метан 0,185 ч-1
J12,max Максимальная удельная скорость роста штамма, поглощающего ме- танол 0,185 ч-1
Константа Михаэлиса для погло- щения кислорода культурой Xi 1 • 10~5 г/л
Кг Константа Михаэлиса для поглоще- ния метанола культурой х2 5-10-6 г/л
Ki Константа ингибирования метано- лом Xi 1 • 10~4 г/л
Xi Стехиометрический экономический коэффициент поглощения метано- ла культурой х1 5,0 г бактерий/ /(г метанола)
У2 Стехиометрический экономический коэффициент поглощения метано- ла культурой х2 0,3 г бактерий/ /(г метанола)
Уз Стехиометрический экономический коэффициент поглощения кислоро- да Культурой Xi 0,2 г бактерий/ /(г кислорода)
kia Коэффициент массопереноса кисло- рода (произведение площади раз- дела фаз в единице объема систе- мы на коэффициент массоотдачи) 42,0 ч-1
Уровень насыщения растворенным кислородом 0,128 атм (0,008 г/л)
а Wilkinson. Т. G., Topiwala Н. II., Hamer G., Biotech. Bioeng., 16, 56 (1974).
ментами реакций) и в гл. 7 (ингибирование роста дрожжей этанолом). По-
скольку при низкой концентрации растворенного кислорода Hyphomicrobium
без труда могут использовать нитрат в качестве акцептора электронов, то мы
принимаем, что не зависит от Со2 и что зависимость г,2 от s описывается
уравнением Моно
Г =..H2,maxS- (14П1.3)
/2 K2 + s 2
Допустив далее, что экономические коэффициенты для обоих видов по
отношению к метанолу постоянны и что большая часть кислорода поглощает-
ся Pseudomonas, получим следующие уравнения материальных балансов по s
и Со2:
7Г = _Ds+J_ (14П1.4)
-d^2 = kta(COts ~ co2) - rfi - Dco2 (14П1.5)
ul I 3
Практическое применение смешанных популяций 493
С помощью экспериментов с периодическими культурами Уилкинсон, Топива-
ла и Хэймер определили значения всех параметров этой модели; скорость
поглощения кислорода определяли непосредственно. Предложенные ими зна-
чения параметров перечислены в табл. 14П1.1.
На рис. 14П1.3 приведены характеристики стационарных состояний, вы-
численные с помощью указанной модели, при различных скоростях разведе-
РИС. 14П1.3. Сплошные кривые — расчетные данные; точки — эксперименталь-
но найденные значения концентрации сухих веществ. [Из работы: Wilkin-
son Т. G. et al., Biotech. Bioeng., 16, 41 (1974).]
ния, а также некоторые результаты экспериментального изучения этой
системы. Соответствие между экспериментальными и расчетными данны-
ми хотя и не идеальное, но вполне удовлетворительное. Моделирование пере-
ходных состояний, возникающих при влиянии метанола на смешанную куль-
туру, также привело к результатам, качественно согласующимся с экспери-
ментальными данными (см. рис. 14П1.2). В то же время динамика реальной
системы значительно более замедленна, чем динамика модели. Очевидно, для
адекватного отражения происходящих в смешанной культуре взаимодействий,
в нестационарном состоянии необходима более структурированная модель.
494
Глава 14
14.2. Естественные смешанные популяции
микроорганизмов и их роль в производстве
и порче продуктов
Теперь перейдем к изучению процессов, в которых посевной
материал поступает из естественных источников. При этом
условии природа доминирующих видов определяется в основном
составом и количеством поступающих в систему питательных
веществ и другими параметрами окружения. Наиболее типич-
ным примером таких процессов является биологическая очистка
сточных вод. Для того чтобы понять превращения, происходя-
щие в таких и других близких системах с естественными сме-
шанными популяциями микроорганизмов, необходимо учесть
тот факт, что микроорганизмы чрезвычайно разнообразны и
очень широко распространены. Поэтому в качестве правила
можно принять следующее положение: если условия благоприят-
ствуют росту данного вида микроорганизмов, то он растет или
будет расти в этих условиях. Так, в смешанных популяциях,
возникающих в аэрируемом реакторе для обработки сточных
вод, в результате естественного отбора содержатся в основном
виды, хорошо приспособленные к росту в этих условиях.
По этой же причине смешанные популяции микроорганизмов
особенно эффективно утилизируют смеси субстратов. В процес-
сах очистки сточных вод эта способность смешанных популяций
чрезвычайно выгодна; напротив, последствия могут быть весьма
неприятными, если смешанная популяция микроорганизмов
атакует такие «субстраты», как пищевые продукты и древесину.
Ниже мы рассмотрим такие нежелательные стороны активности
микроорганизмов, а обсуждению проблемы биологической
очистки сточных вод будет посвящен разд. 14.4.
Порча продуктов или материалов обычно заключается в раз-
ложении органических веществ, в том числе полимеров, напри-
мер белков и полисахаридов. Иногда, например при древесной
гнили, порча обусловлена разрушением одного из основных
компонентов исходного продукта или материала. С другой сто-
роны, подавляющее большинство нежелательных эффектов пор-
чи пищевых продуктов обусловлено специфическими свойствами
продуктов метаболизма атакующих микроорганизмов. И в пер-
вом, и во втором случае начальной стадией являются реакции,
катализируемые внеклеточными ферментами, продуцируемыми
микроорганизмами.
Что касается древесной гнили, то в этом случае ферментная
система, выделяемая грибом, разрушает один или несколько из
трех основных высокомолекулярных компонентов древесины.
Наряду с бактериями грибы участвуют в разрушении пектина
Практическое применение смешанных популяций
495
в пищевых продуктах. Именно такой механизм ответствен за
порчу консервированных фруктов, размягчение маринованных
огурцов и гниение овощей. В то же время, как мы уже видели
ранее в разд. 4.1.4, пектические ферменты находят и очень удач-
ные сферы применения.
Разрушение белков, с которым связана другая проблема
в пищевой промышленности, можно рассматривать как двуста-
дийный процесс. На первой стадии, называемой протеолизом,
белки гидролизуются до пептидов и аминокислот. Примером
внешних эффектов, сопровождающих эту стадию, может слу-
жить разжижение желатина. Затем (обычно в анаэробных усло-
виях) аминокислоты метаболизируются с образованием ряда
веществ с отвратительным запахом, например путресцина:
-со2
H2N(CH2)3CH(NH2)COOH ----------> H2N(CH2)4NH2
орнитинде- путресцин
карбоксилаза
Именно этот процесс (гниение) ответствен за отвратительный
запах сгнивших мясных продуктов.
Самый известный пример порчи пищевых продуктов — это,
по-видимому, скисание молока. Хотя в большинстве случаев мо-
локо перед расфасовкой пастеризуют, таким путем достигается
только дезинфекция продукта, но ни в коей мере не стерилиза-
ция. Многие спорообразующие бактерии легко переносят пасте-
ризацию и спустя некоторое время вызывают скисание или порчу
молока. Особенно интересны последовательные микробиологиче-
ские превращения в сыром молоке при комнатной температуре.
Сначала стрептококки, бациллы и другие бактерии транс-
формируют лактозу (молочный сахар). В результате значе-
ние pH молока снижается (рис. 14.1), что в свою очередь инги-
бирует рост начальной смешанной популяции и создает благо-
приятные условия для роста кислотоустойчивых бактерий, в том
числе Lactobacilli. Дальнейшее повышение кислотности (ниже
pH 4,7) приводит к образованию и осаждению творожистой
массы (резиноподобного материала, состоящего главным обра-
зом из белка казеина).
На следующем этапе размножаются дрожжи и плесени, ко-
торые могут использовать молочную кислоту в качестве основ-
ного питательного вещества; в результате pH повышается. Рост
популяций этих организмов затем создает условия, благоприят-
ные для размножения других грибов и бактерий, утилизирую-
щих в качестве питательных веществ жир и казеин. В конце
концов запасы кислорода истощаются, и под влиянием анаэроб-
ных бактерий начинается процесс гниения.
В естественных смешанных популяциях часто наблюдается
подобная последовательная смена доминирующих видов микро-
496
Глава 14
организмов, каждый из которых превалирует в особенно благо-
приятных для него условиях. С другими аналогичными приме-
рами мы познакомимся позднее при изучении микробиологии
почвы и биологических капельных фильтров. Еще раз обратим
внимание на большую практическую важность контроля началь-
ных стадий заражения продукта микроорганизмами; так, как
мы уже знаем из материала предыдущего раздела, получение
сырной массы из молока под действием бактерий является пер-
вой стадией производства сыра.
Streptococcus
lactls
Lactobacilli
Спороодразию -
Дрожжевые щие Pseuctomo
пленки и др. паз и др.
'Рост гни-
/лостнь/л
бактерий
Образование молоч-
\ной кислоты t
Время
РИС. 14.1. Последовательное проявление активности различными видами
микроорганизмов в сыром молоке при комнатной температуре.
Известны и другие процессы пищевой промышленности,
в которых используется естественная ферментация, инициируе-
мая бактериями — продуцентами молочной кислоты. Так, ма-
ринованные огурцы приготовляют путем молочнокислого бро-
жения с участием смешанных популяций микроорганизмов.
В производстве сосисок и других подобных мясных продуктов
Lactobacillus sp. и другие микроорганизмы не только продуци-
руют молочную кислоту, но и восстанавливают нитраты до нит-
ритов; последний процесс обусловливает появление специфиче-
ской окраски и придание острого вкуса. Опару для дрожжевого
хлеба ферментируют в течение 1—2 сут; за это время образуют-
ся этанол и органические кислоты. Другим пищевым продук-
том, получаемым брожением при участии смешанных культур,
является квашеная капуста.
Хотя процессы виноделия, пивоварения и производства уксу-
са мы уже рассматривали ранее, в некоторых аспектах они
затрагивают и тему настоящей главы. Во многих случаях ми-
кробиологическая активность, обусловливающая необходимые
качества продукта, обеспечивается естественными смешанными
культурами. В настоящее время, однако, наметилась тенденция
Практическое применение смешанных популяций 497
к проектированию и эксплуатации более воспроизводимых про-
цессов с участием микробиологических систем определенного
состава. В связи с этим обычным становится использование по-
севного материала, состоящего из тщательно отобранных и со-
храняемых видов.
14.3. Участие микроорганизмов
в естественных кругооборотах веществ
Большая часть рассмотренных нами выше примеров приме-
нения микроорганизмов так или иначе была связана с процес-
сами, осуществляемыми и контролируемыми человеком. В связи
с возрастающей важностью проблемы охраны окружающей сре-
ды, однако, необходимо понять и основные проявления микро-
биологической активности в биосфере. Обладая этими данными,
мы сможем создать модели таких естественных процессов как
эвтрофикация озер, биодеградация загрязнений и очистка воды
в почве, динамика экосистем в естественных водных потоках и
эстуариях. Кроме того, искусственная биологическая очистка
сточных вод, представляющая собой один из наиболее важных
процессов, которые выполняет человек с помощью микроорга-
низмов, в очень большой степени воспроизводит элементы есте-
ственных экологических циклов. Изучение активности микроор-
ганизмов в природных условиях помогает понять, каким обра-
зом поддерживается жизнь на земле, и может служить ценной
основой для практически важных применений смешанных куль-
тур микроорганизмов.
Адекватное функционирование глобальной экосистемы пла-
неты невозможно без кругооборота необходимых для жизни хи-
мических элементов (так называемых биогеохимических цик-
лов). Так, органические вещества, образующиеся в качестве от-
ходов жизнедеятельности организмов или входящие в состав
отмерших организмов, должны разлагаться и превращаться
в неорганические вещества. Основные стадии этого процесса,,
называемого минерализацией, особенно эффективно осущест-
вляются с участием микроорганизмов. В этом отношении осо-
бенно ценны такие качества микроорганизмов, как метаболиче-
ская универсальность, высокие скорости осуществляемых
превращений и широкое распространение в природе. Микроор-
ганизмы в общем случае могут разлагать любое природное
органическое соединение.
Понятно, что для предотвращения накопления чрезмерных
количеств любых органических отходов необходимо, чтобы
реакции деградации протекали с соответствующими абсолютны-
ми скоростями (т. е. чтобы произведение удельной скорости на
498
Глава 14
плотность популяции было достаточно велико). Мы уже неод-
нократно отмечали, что микроорганизмы растут гораздо быст-
рее, чем любые высшие организмы. Соответственно, удельные
скорости утилизации субстрата микроорганизмами также очень
высоки, поэтому и химические трансформации, необходимые для
минерализации, обычно осуществляются с высокими скоростя-
ми. Важность микроорганизмов в природе обусловлена и еще
одним фактором — их чрезвычайно широкой распространенно-
стью и высокой плотностью популяций. Чрезвычайно много
микроорганизмов обитает в поверхностных слоях водоемов и
верхнем слое почвы; бактерии были обнаружены даже в образ-
цах почвы из Антарктиды. Показано, что в слое плодородной
почвы площадью 1 акр и глубиной до 6 дюймов содержится
около 2 тонн бактерий и грибов. Учитывая высокие скорости
метаболизма, нетрудно подсчитать, что находящиеся в этом
почвенном слое микроорганизмы обладают большей метаболи-
ческой активностью, чем 10 000 человек. Из поступающего
в биосферу углекислого газа около 90% вносится за счет дыха-
ния бактерий и грибов.
В кругооборотах большинства химических элементов микро-
организмы не только доминируют в процессах минерализации,
но и вносят существенный вклад в другие трансформации.
В следующих двух разделах мы ознакомимся с конкретными
примерами участия протистов в биогеохимических циклах; при
этом сначала будут рассмотрены общие характеристики круго-
оборота химических элементов в глобальном аспекте, а затем
мы перейдем к изучению некоторых сравнительно хорошо иссле-
дованных экосистем.
14.3.1. Кругооборот необходимых
для жизни химических элементов
Кругооборот биологически важных химических элементов
часто сопровождается циклическими изменениями степени их
окисления. Это обстоятельство немаловажно, поскольку мы уже
знаем, что биологическая роль элемента часто непосредственно
связана с его химическим состоянием (см. гл. 5). Сопряженные
кругообороты углерода и кислорода (как и рассматриваемые
ниже кругообороты всех других элементов) наглядно иллюстри-
руют общие принципы циклических превращений веществ.
Основные характеристики кругооборотов углерода и кислоро-
да представлены на рис. 14.2. Главной движущей силой этих
(как и ряда других) кругооборотов является фотосинтез, обес-
печивающий усвоение солнечной энергии и таким путем восста-
новление СОг, бикарбоната и карбоната (окисленных состояний
углерода), а также одновременное образование молекулярного
Практическое применение смешанных популяций
499
кислорода из воды (разд. 5.5). Таким образом на суше и в океа-
нах ежегодно связывается около 1,6-1010 и 1,2-1010 т углерода
соответственно. На суше основной вклад в фотосинтетическую
активность вносят зеленые растения, а в океане фотосинтез
осуществляется практически исключительно одноклеточными
водорослями, называемыми фитопланктоном. Фотосинтез являет-
Ассимиляция
РИС. 14.2. Упрощенная схема кругооборота углерода. Здесь штриховыми ли-
ниями показан также один из основных участков кругооборота кислорода,
тесно связанный с кругооборотом углерода.
ся основным путем восстановления СО2, но хемоаутотрофы так-
же восстанавливают диоксид углерода. Как мы уже отмечали,
минерализация органического углерода до СО2 в основном
является результатом метаболической активности бактерий и
грибов.
Из описанного кругооборота углерод отбирается нескольки-
ми путями. Большая часть выделившегося в атмосферу СО?
поглощается океанской водой в виде бикарбонатных ионов.
В водной среде последние, взаимодействуя с катионами каль-
ция, могут образовывать карбонат кальция — нерастворимое
соединение, являющееся основой кораллов, раковин моллюсков
и известняка. В этой форме углерод обладает очень малой био-
500
Глава 14
доступностью, но в конце концов в результате эрозии или дей-
ствия кислот карбонат кальция разрушается, а образующийся
СО2 вновь включается в кругооборот углерода. Микроорганиз-
мы также участвуют в разрушении карбоната кальция, синте-
зируя угольную, серную, азотную и другие кислоты.
Все мы знакомы со связанным углеродом в органической
форме. Так, важной составной частью плодородной почвы
является гумус—органический остаток, образующийся из
устойчивых к действию микроорганизмов компонентов растений.
Когда условия благоприятствуют накоплению большого количе-
ства гумуса, с течением времени образуются залежи торфа, ко-
торые в ходе целых геологических эпох могут постепенно пре-
вращаться в каменный уголь. Другими обычными формами свя-
занного органического углерода являются нефть и природный
газ. Содержащийся в угле, нефти и газе углерод в конечном
счете возвращается в биосферу благодаря постоянно возрастаю-
щей потребности человека в топливе.
Теперь перейдем к изучению кругооборота азота в биосфере
и, в частности, к изменению его химического состояния в ходе
соответствующих превращений. При этом полезно не забывать
несколько основных положений. Во-первых, находящийся в ат-
мосфере молекулярный азот практически совершенно инертен,
т. е. для подавляющего большинства организмов в таком виде
он не может служить питательным веществом. Во-вторых, в жи-
вых организмах азот в основном содержится в белках в восста-
новленной форме.
Общая схема кругооборота азота приведена на рис. 14.3.
Органический азот превращается в аммиак при действии микро-
организмов. Небольшая часть аммиака поступает в атмосферу
и далее непосредственно утилизируется растениями и микроор-
ганизмами; большая же часть аммиака окисляется до нитрата
(МОз-). Соответствующий процесс, осуществляющийся в две
стадии, называют нитрификацией. Каждая из стадий процесса
нитрификации катализируется своей группой бактерий, в пер-
вую очередь Nitrosomonas и Nitrobacter. И первые, и вторые
являются хемоаутотрофами и облигатными аэробами.
Образующиеся таким путем нитраты являются одним из
наилучших источников азота для растений, поэтому именно
через нитраты большая часть азота возвращается в пул вос-
становленного органического азота. Водоросли и растения
включают азот в органические соединения, и в этой форме
азот может далее усваиваться и животными. В то же время
микроорганизмы могут утилизировать нитрат и по другому
пути. При недостатке кислорода, например в микроокружении
бактерий, где в результате дыхания последних запасы кислоро-
да истощились, многие бактерии способны перестроить свой
Практическое применение смешанных популяций
501
РИС. 14.3. Упрощенная схема кругооборота азота.
метаболизм и использовать нитрат как акцептор водорода.
Результатом такой активности является денитрификация, т. е.
образование молекулярного азота.
К счастью, некоторые микроорганизмы способны утилизи-
ровать молекулярный азот и таким образом возвращать его
в биосферу в связанном виде. Известны два основных пути фик-
сации азота микроорганизмами. В первом из них азот связы-
вается бактериями Rhizobium в симбиозе с бобовыми растениями.
В другом пути азот связывается в несимбиотическом про-
цессе сине-зелеными водорослями, а также некоторыми аэроб-
502
Глава 14
ними (Azotobacter) и анаэробными {Clostridium pasteurianum}
бактериями.
В связи с описанными путями фиксации азота перед генети-
ческой инженерией микроорганизмов и (или) растений возни-
кают две важные и интересные задачи:
а) введение генов Nif, ответственных за фиксацию азота, в ге-
номы бактерий, населяющих корневые системы других (не
бобовых) растений;
б) создание растений, содержащих гены Nif непосредственно
в геноме растения.
Решение любой из этих задач привело бы к созданию сортов
растений, не нуждающихся в азотных удобрениях (во всяком
Утилизация сульфатов
(растениями, микроорга-
низмами)
Восстановленная органическая сера
в живых организмах (например,
SH- группы цистеина)
Растения---^-Животные-----— Микроорганизмы
| sb; |-----------
т
Окисление серы (бесцветными
и фотосинтезирующими
серными бактериями)
Daulfovi.brio
Деградации органических:
веществ (микроорганиз-
мами)
_L
Окисление Нг5 (бесцветными
и фотосинтезирующими
серными бактериями; возможно
и неферментативное окисление)
S
РИС. 14.4. Основные стадии кругооборота серы.
случае в таком количестве, в каком их применяют в настоящее
время).
В своем кругообороте сера также претерпевает стадии окис-
ления, восстановления, включения в органические вещества и
высвобождения из них (рис. 14.4). Сульфатредуцирующие и
окисляющие серу бактерии играют важную роль в процессах
микробиологической коррозии. Дополнительные сведения о кру-
гообороте серы и участвующих в нем организмах читатель мо-
жет найти в приведенной в конце главы литературе. В следую-
щем разделе мы рассмотрим различные пути реализации пол-
ного или частичного кругооборота элементов в специфическом
окружении.
14.3.2. Взаимосвязи микроорганизмов в почве
и некоторых других естественных экосистемах
Почва представляет собой сложную систему переменного со-
става, являющуюся отличной сферой обитания для микроорга-
низмов. Она состоит из тонкоизмельченных минералов (в основ-
Практическое применение смешанных популяций 503
ном алюмосиликатов), разлагающихся органических остатков
и смешанной популяции живых микроорганизмов. Кроме того,
в почве часто присутствует вода, а также газовая фаза, в со-
став которой могут входить N2, О2, СО2, H2S, NH3 и другие
газы. Большая удельная площадь поверхности мелких твердых
частиц почвы обусловливает концентрирование (за счет адсорб-
ции) некоторых питательных веществ и внеклеточных гидроли-
тических ферментов. Растворенные минералы и разлагающиеся
органические материалы являются источниками ионов, углево-
дов, азотсодержащих соединений и витаминов. Таким образом
создается богатая среда, способная поддерживать рост микро-
организмов.
Синтрофизмом называют тип межвидовых взаимосвязей, при
котором один организм продуцирует пищу для другого. Синтро-
физм очень широко распространен среди популяций организмов
в почве и играет важную роль в кругообороте химических эле-
ментов. Ряд примеров синтрофизма схематично представлен на
рис. 14.5. Обратите внимание на то, что один организм (обозна-
ченный символом А), продуцирующий целлюлолитические фер-
менты для самообеспечения питательными веществами, одно-
временно снабжает простыми сахарами и другие виды (В, С
и D). Конечные продукты метаболизма организмов А исполь-
зуются другими микроорганизмами, обладающими иными по-
требностями в питательных веществах.
Синтрофизм можно объяснить и с другой точки зрения,
проследив за последовательностью событий, которые наступают
вслед за вспашкой земли, засеянной травами или клевером (эти
события в какой-то степени напоминают рассмотренные в пре-
дыдущем разделе процессы, сопровождающие порчу молока).
Вспашка не только обеспечивает почву различными питатель-
ными веществами из растений, но и аэрирует ее. Создающиеся
таким путем условия способствуют быстрому росту гетеротроф-
ных организмов и факультативных аутотрофов. Метаболическая
активность этих организмов в свою очередь способствует повы-
шению температуры и кислотности почвы.
После истощения имеющихся запасов кислорода появляются
облигатные анаэробы, что приводит к еще большему повыше-
нию кислотности. В конце концов этот быстрый рост микроор-
ганизмов прекращается в результате истощения питательных
веществ или образования токсинов (вспомните разд. 7.3.1), и
множество клеток погибает, выделяя соединения, способствую-
щие росту растений. Остаточная относительно малочисленная
популяция микроорганизмов в основном состоит из видов, спо-
собных разрушать устойчивые вещества, например, из числа
содержащихся в гумусе.
По сути дела, здесь мы только коснулись проблемы изуче-
Неорганические соединения, вода, С0)ъ,
доступные для всех организмов
Практическое применение смешанных популяций
505-
ния микробиологической активности в почве. Так, мы не ука-
зали, что в анаэробных условиях могут реализоваться и реа-
лизуются на самом деле полные кругообороты всех основных
химических элементов. Необходимые трансформации могут осу-
ществляться даже в очень небольшой обедненной кислородом
экологической нише, поскольку все они осуществляются с уча-
стием микроорганизмов. На рис. 14.6 отражены тесные связи,
Нг5
Сулыратредуцирующие
бактерии
н.
сог
Фотосинтез в пурпурных
и зеленых бактериях
NH.
Бактерии,
продуцирующие
мешан
Фотосинтез в пурпурных
несерных бактериях
Ацетат,
простые
органические
соединения
Деградация бактериями
Клеточные комлонен- Clostridia и другими
ты в сротосинтези- ~ ~
рцющих бактериях сбраживающими
> ' организмами
РИС. 14.6. Упрощенная схема кругооборота элементов в анаэробных усло-
виях.
которые могут устанавливаться между фотосинтезирующими,
сбраживающими, сульфатредуцирующими и другими бактерия-
ми в анаэробном окружении. Дополнительные сведения по мик-
робиологии почвы можно найти в приведенной в конце главы
литературе. В заключение обсуждения следует отметить, что
подавляющее большинство промышленно важных микроорга-
низмов было выделено из почвы.
РИС. 14.5. Некоторые возможные синтрофические связи между микроорганиз-
мами, растениями и животными в почве. Конечные продукты метаболизма
микроорганизмов А являются питательными веществами для организмов М,
Н, G и Е, а секретируемые микроорганизмами А целлюлолитические фермен-
ты гидролизуют целлюлозу. Образующиеся простые сахара являются пита-
тельными веществами для микроорганизмов В, С и D.
33—746
506
Глава 14
Микроорганизмы являются основными продуцентами орга-
нических веществ в пресноводных водоемах и океане. Как схе-
матично показано на рис. 14.7, зеленые растения и крупные
многоклеточные водоросли могут осуществлять фотосинтез
только на небольшой глубине вблизи берегов. В открытом океа-
не только свободно плавающие одноклеточные водоросли усваи-
вают солнечную энергию в поверхностных слоях. По мере уве-
личения глубины фотосинтезирующая активность снижается и
РИС. 14.7: Звенья пищевой цепи в водной экосистеме, в которой основным
источником энергии является солнечная радиация. Римскими цифрами I, II,
III и IV обозначены абиотические компоненты системы, организмы-продуцен-
ты, организмы-потребители и микроорганизмы, разлагающие органические ве-
щества, соответственно.
в конце концов вообще исчезает в силу поглощения света во-
дой и суспендированными твердыми частицами.
Образующийся в поверхностных слоях океана фитопланк-
тон является питательным веществом для большой цепи орга-
низмов-потребителей, роль первого звена в которой выполняет
зоопланктон, а завершающими звеньями являются рыбы, киты
и другие морские животные. Органические вещества, образую-
щиеся в результате метаболической активности и гибели орга-
низмов, составляющих звенья этой цепи, в свою очередь раз-
лагаются микроорганизмами. Возникающие при этом относи-
тельно несложные вещества могут утилизироваться фитопланк-
тоном; этот этап замыкает пищевой цикл.
В опасном процессе, называемом эвтрофикацией, избыток
питательных веществ вызывает чрезвычайно интенсивный рост
водорослей. Некоторые из них продуцируют токсины; бурное
размножение водорослей часто сопровождается появлением не-
приятного запаха. В конечном счете большая часть водорослей
отмирает, высвобождая питательные вещества для гетеротроф-
ных организмов. На этом этапе общая скорость дыхания пре-
Практическое применение смешанных популяций 507
вышает скорость фотосинтеза, поэтому перенос растворенного
кислорода в водную фазу резко уменьшается. Это в свою оче-
редь вызывает массовую гибель рыб и многих других аэробных
организмов и таким образом нарушает локальные равновесия,
необходимые для существования экосистемы озера или пруда.
14.4. Биологическая очистка сточных вод
В сточных водах содержится сложная смесь твердых и рас-
творенных веществ, причем последние обычно присутствуют
в очень малых концентрациях. На очистных станциях концент-
рации всех этих веществ снижают до приемлемого уровня или
же химически трансформируют вредные вещества в безопасные
соединения. Конкретная схема очистной станции зависит от
степени загрязненности и количества обрабатываемых стоков,
а также от экономических и экологических соображений. Боль-
шая часть водоочистных станций, однако, имеет много общего,
что позволяет изобразить общую схему системы водоочистки,
как на рис. 14.8. Показанные здесь различные способы перера-
ботки ила и удаления загрязняющих веществ отражают разные
возможные пути достижения одной и той же цели. В типичной
станции по биологической очистке сточных вод используются
лишь некоторые из множества возможных вариантов.
Теперь перейдем к изучению общих задач и целей каждой
из основных операций (или последовательностей нескольких
операций). В операциях первичной обработки удаляют наибо-
лее легко отделяющиеся загрязнения, например крупные, легко
осаждающиеся частицы (см. рис. 14.9), масляные пленки и
другие «легкие» компоненты. Суспендированные твердые части-
цы и растворимые компоненты отделяют в процессе вторичной
обработки. Во многих случаях загрязняющие вещества имеют
органическую природу; в таких случаях обычно используют
биологическое окисление, которое мы рассмотрим детальнее
чуть позже. Цель третичной обработки заключается в полном
или частичном отделении всех оставшихся примесей. На этой
стадии используются такие методы, как электродиализ, обрат-
ный осмос, фильтрование через толстый слой и адсорбция.
В процессе первичной обработки отделяют влажные кон-
центрированные твердые вещества, называемые илом-, при вто-
ричной обработке образуется активный клеточный ил. Мы не-
однократно упоминали о взаимосвязи между утилизацией суб-
страта и образованием биомассы. Хотя процессы вторичной
биологической обработки с участием множества видов микроор-
ганизмов очень эффективны при деградации разбавленных сме-
сей органических отходов, не следует забывать, что при этом
образуется и биомасса. Таким путем очень мелкие нераствори-
«3*
.Отделение
крупных
твердых
частиц
Отделение
суспендированных растворенных
твердых
частиц
Отделение
органических
веществ
Отделение
азотсодержащих
веществ
Отделение
фосфор-
содержащих
веществ
Концентриро-
вание твердых
веществ
Обезвоживание
твердых
веществ
Разложение
органических
твердых
веществ
Отделение
тонкоизмелб-
ченнь/х сус-
пендированных
твердо/х
веществ
Стерилизация
Отделение сле-
довых количеств
органических
веществ
Отделение
неорганических
солей
РИС. 14.8. Типовые операции, применяющиеся для первичной, вторичной и
третичной очистки сточных вод. (Из работы: Eckenfelder W. W. Jr., Industrial
Water Pollution Control, pp. 6—7, McGraw-Hill Book Company, Inc., New York,
510
Глава 14
мые частицы и растворенные компоненты жидких отходов час-
тично превращаются в ил, который легче поддается отделению,
чем исходные загрязняющие вещества. Установки для перера-
ботки ила являются важной составной частью станций по очист-
ке сточных вод. Для уменьшения объема ила, образующегося
pL Размер Примеры
От 1м Валуны
1 - - 10см Камни
2 - - 1см Галька
3 - - 1мм Хлопьевидные частицы
4-- 100 мкм
5 - - Юмкм
Суспендированные
частицы
Надколлоидные частицы
G-- 1мкм Коллоидные частицы
7-- 0,1 мкм
Субколлоидные частицы
8-- 10нм Растворенные веьцества
9 - - 1 нм
10-*- 1А
РИС. 14.9. Отделение твердых частиц различной величины на различных ста-
диях очистки сточных вод. [Из работы: Helfgott Т. et al., J. Sanit. Eng. Div.
ASCE, 96, 79 (1970).]
при очистке воды, широко применяется операция анаэробной
переработки, при которой органические вещества подвергаются
биологической деградации в анаэробном окружении.
Не следует думать, что во всех случаях используются все
три уровня очистки сточных вод и переработки ила. Иногда
сточные воды спускают в природные водоемы (ручьи, реки,
пруды, озера и океан) без какой бы то ни было обработки.
В других случаях применяют только первичную обработку.
В то же время для большинства городских систем водоочистки
в США та или иная форма вторичной обработки является обя-
Практическое применение смешанных популяций 511
зательной, а третичная обработка в настоящее время применя-
ется лишь изредка.
Далее в соответствии с основной темой этой главы основное
внимание будет уделено биологическим методам очистки сточ-
ных вод. Рассмотрев в разд. 14.4.1 основные характеристики
сточных вод, мы перейдем далее к изучению активного ила
и его применению в различных методах вторичной биологиче-
ской очистки. Краткий обзор этих важных сфер применения
биохимической технологии мы завершим анализом операций
анаэробной переработки ила. Дополнительную информацию об
этих операциях и процессах, а также о физических и химиче-
ских способах очистки сточных вод читатель найдет в литерату-
ре, перечисленной в конце главы.
14.4.1. Основные характеристики сточных вод
Понятно, что природа и концентрация загрязняющих ве-
ществ в сточных водах зависят от их источника. Существуют
два основных вида сточных вод—промышленные и бытовые.
Последние загрязнены главным образом уличным мусором,
моющими средствами и экскрементами.
Бытовые сточные воды обычно содержат более 99% воды,
около 300 млн-1 (мг/л) суспендированных твердых веществ,
а также около 500 мг/л летучих веществ. Большая часть сус-
пендированных твердых веществ имеет целлюлозную природу,
а другие загрязняющие органические вещества включают (в по-
рядке убывания концентрации) жирные кислоты, углеводы и
белки. Как мы уже упоминали при обсуждении процессов порчи
пищевых продуктов, неприятный запах бытовых сточных вод
обусловлен разложением белков в анаэробных условиях.
Если учесть происхождение бытовых сточных вод, то не
должен вызывать удивления тот факт, что в них содержатся
различные виды почвенных и кишечных микроорганизмов, в том
числе аэробные организмы, облигатные и факультативные
анаэробы, бактерии, дрожжи, плесени и грибы. Поскольку в бы-
товых сточных водах часто присутствуют также патогенные
организмы и различные вирусы, чрезвычайно важна полная
изоляция, источников и трубопроводов для подачи питьевой во-
ды от загрязнения сточными водами. Популяции микроорганиз-
мов в сточных водах служат постоянным смешанным посевным
материалом для процессов биологической очистки и, кроме
того, источником метаболической активности в стандартных
методах определения степени загрязнения сточных вод.
Наиболее распространенным критерием концентрации за-
грязняющих веществ в бытовых сточных водах является пока-
затель биохимической потребности в кислороде (БПК), равный
512
Глава 14
количеству растворенного кислорода, поглощаемого единицей
объема сточных вод за определенное время при 20 °C. Продол-
жительность периода инкубации обычно указывают в виде под-
строчного индекса; так, если БПК определяют по результатам
инкубирования в течение 5 сут (один из принятых периодов),
то соответствующий показатель обозначают символом БПК5.
Количество растворенного кислорода, поглощаемого в ходе ин-
кубации вплоть до полного прекращения биологического окис-
ления органических веществ, называют предельней (или пол-
ной) БПК (БПКп). Этот тест, разработанный еще в 1898 г.
Британской Королевской комиссией по ликвидации отходов,
должен был моделировать условия в водных потоках и обеспе-
чивать относительно прямое определение одного из наиболее
вредных и опасных последствий сброса сточных вод — истоще-
ния растворенного кислорода в водных бассейнах, куда сбрасы-
ваются отходы. Снижение концентрации растворенного кисло-
рода быстро приводит к гибели множества аэробных организмов,
а также животных; конечным результатом истощения рас-
творенного кислорода будет грязная, неприятно пахнущая река,
зараженная патогенными микроорганизмами.
Другим критерием потенциального снижения общей концент-
рации растворенного кислорода в водоемах, в которые посту-
пают сточные воды, служит химическая потребность в кислоро-
де (ХПК), равная числу миллиграммов кислорода, поглощае-
мого 1 литром пробы (сточных вод) из горячего подкисленного
раствора бихромата калия. В общем случае химическому окис-
лению подвергается больше веществ, чем биологической дегра-
дации, и, следовательно, величина ХПК должна быть больше
величины БПК Для одного и того же образца. Измерение ХПК
связано с возможной степенью загрязнения естественных водое-
мов сточными водами не столь непосредственно, как определе-
ние БПК; с другой стороны, ХПК можно определить с помощью
доступной простой аппаратуры за 2 ч, а с помощью сложных
приборов — за несколько минут. БПК и ХПК являются общими
и самыми грубыми индикаторами состава сточных вод. Тем не
менее они дают полезную информацию о степени опасности,
которую представляют сточные воды для окружающей среды.
Другим преимуществом показателей БПК и ХПК является
возможность их определения с минимальным количеством не-
сложной аппаратуры, причем выполнение соответствующих ана-
лизов требует лишь кратковременного обучения персонала.
Чтобы охарактеризовать качество воды, часто применяют и
другие параметры, в том числе концентрации фосфорсодержа-
щих веществ (общего фосфора), азотсодержащих веществ (об-
щего азота) и суспендированных нерастворимых веществ.
В табл. 14.2 приведены характерные параметры потоков, посту-
Практическое применение смешанных популяций 513
пающих на станцию очистки бытовых сточных вод, и очищенной
воды. В этой таблице не указаны некоторые важные загрязняю-
щие вещества, в первую очередь соли тяжелых металлов и ток-
сичные органические вещества, например пестициды, которые
часто присутствуют в очень малых, но способных принести
большой вред концентрациях.
Состав промышленных сточных вод определяется их проис-
хождением. Как показывают данные, приведенные в табл. 14.3,
стоки промышленных предприятий часто загрязнены в гораздо
большей степени, чем бытовые сточные воды. В стоках про-
мышленных предприятий, связанных с переработкой материа-
Таблица 14.2. Некоторые параметры, характеризующие качество воды
(содержание азота и фосфора в воде после очистки проверяется не всегда)
_ „ Требования к воде после
Параметр Сточные воды очистки
БПК, мг/л 100—250 5-15
ХПК, мг/л 200—700 15-75
Общий фосфор, мг/л 6—10 0,2—0,6
Азот, мг/л 20—30 2—5
Суспендированные нерастворимые 100—400 10—25
вещества, мг/л
лов углеводородной природы, часто содержатся и ядовитые ве-
щества, например формальдегид, аммиак или цианиды. Здесь
возникают две взаимосвязанные проблемы: во-первых, эти сто-
ки чрезвычайно опасны для живых организмов в водоемах,
куда они сбрасываются, во-вторых, они могут убивать микроор-
ганизмы, участвующие в аэробных и анаэробных процессах пе-
реработки отходов. Эффективные и достаточно экономичные
методы обезвреживания подобных токсичных веществ пока еще
не разработаны.
В остальных разделах этой главы основное внимание будет
уделено процессам обработки бытовых сточных вод и их ре-
жимам. Такие же процессы часто применяются и для очистки
промьГшленных стоков. Более того, часто промышленные стоки
обрабатывают совместно с бытовыми сточными водами.
14.4.2. Процессы с участием активного ила
В процессах с участием активного ила основным типом обо-
рудования является проточный аэрируемый биологический реак-
тор. Как показано на рис. 14.10, этот аэробный реактор (аэро-
тенк) связан с отстойником, в котором вода осветляется. Часть
ила, собирающегося в отстойнике, обычно вновь поступает
Таблица 14.3. Сравнительная степень загрязненности стоков
промышленных предприятий3
Предприятие или процесс Основные загрязняющие вещества БПК 5 хпк
Скотобойня Суспендированные нераствори- мые вещества, белки 2600 4150
Производство свеклович- ного сахара Суспендированные нераствори- мые вещества, углеводы 850 1150
Лесопильный завод Пивоваренный завод Суспендированные нераствори- мые вещества, углеводы Углеводы, белки 430 550 1400
(мытье бутылок)
Консервный завод (мя- со) Суспендированные нераствори- мые вещества, жиры, белки 8000 17940
Химические предприятия Суспендированные нераствори- мые вещества, чрезвычайно высокая щелочность или кислотность, органические химикаты 500 980
Коксование угля
Коксовые печи Фенолы, цианиды 780 1670
Коксовый газ Бездымное топливо Спирто-водочный завод Производство молочных Тиоцианаты, тиосульфаты Аммиак Суспендированные твердые ве- щества, углеводы, белки Углеводы, жиры, белки 6500 20 000 7000 600 16 400 10 000
продуктов
Бытовые сточные воды Суспендированные твердые ве- щества, масла, углеводы, белки 350 300
Промывка зерна Суспендированные твердые ве- щества, углеводы 1500 1800
Отбелка тканей Суспендированные твердые ве- щества, углеводы, лигнин 1600 3600
Прачечные Суспендированные твердые ве- щества, углеводы, мыла 1600 2700
Производство солода Суспендированные твердые ве- щества, углеводы Суспендированные твердые ве- щества, углеводы, лигнин, сульфаты 1240 1480
Целлюлозный завод 25 000 76000
Микробиологическая промышленность
Микробиологические 4560 4120
цехи
Цехи химического 960 1580
синтеза
Цехи по приготов- 145 217
лению композиций и упаковке
Неф теп ерер а б атыв а ющий завод Фенолы, углеводороды, соеди- нения серы 840 1500
Производство смол Фенолы, формальдегид, моче- вина 7400 12 900
Снижение концентрации крахмала в муке Суспендированные твердые ве- щества, углеводы, белки 12 000 17 150
Кожевенный завод Суспендированные твердые ве- щества, белки, сульфиды 2300 5100
а По данным, приведенным в работе: Abson J. W., Todhunter К. Н-. in Biochemical
and Biological Engineering Science, Biakebrough N. (ed.), vol. 1, pp. 318—319, Academic
Press, London, 1967.
Практическое применение смешанных популяций 515
в биологический реактор, в результате чего обеспечивается по-
стоянная инокуляция илом. Кроме того, рециркуляция увели-
чивает среднее время пребывания ила в системе, давая таким
образом возможность присутствующим в нем микроорганизмам
адаптироваться к имеющимся питательным веществам. Ил дол-
жен оставаться в аэробном биореакторе достаточно долго и для
того, чтобы окислились все адсорбированные органические ве-
Вторичный
отстойник Осветленная
Сточные s0
боды после
первичного
отстойника
<xF
Суспендированные в жидкости
микроорганизмы
Подача воздуха для перемешива-
ния и обеспечения кислородом
биореакций
(a+p)F
Хг, 5_
F
Рециркуляция ила
«с--------
__________
Избыточный ил
РИС. 14.10. Схема процесса очистки воды с участием активного ила.
щества. Другие преимущества реакторов с рециркуляцией уже
обсуждались в гл. 9.
Чтобы понять основные механизмы утилизации субстрата
в этом биореакторе, необходимо сначала изучить природу и
морфологию смешанной культуры микроорганизмов, живущих
в аэрируемом реакторе. Одним из наиболее типичных для ак-
тивного ила организмов является бактерия Zoogloea ramigera.
Возможно, наиболее важной характеристикой как этого орга-
низма, так и многих других видов, существующих в активном
иле, является способность синтезировать и секретировать в сре-
ду полисахаридный гель. Именно наличие геля обусловливает
агрегацию микроорганизмов и образование хлопьевидных скоп-
лений (флокул), называемых активным илом (рис. 14.11).
Активный ил характеризуется высоким сродством к суспенди-
рованным твердым веществам, включая коллоидальные части-
цы. Именно это обстоятельство служит причиной того, что
первой стадией разрушения суспендированных твердых частиц
в сточных водах является их присоединение к флокулам. Затем,
как это показано на рис. 14.12, способные к биодеградации
516
Глава 14
компоненты адсорбированных частиц претерпевают окисление
организмами флокулы.
Для того чтобы выгоднее использовать высокую адсорбцион-
ную способность активного ила, разработан вариант обычного
процесса, называемый контактной стабилизацией. Как показано
на схеме (рис. 14.13), в этом процессе рециркулирующий осаж-
денный ил подвергается повторной аэрации прежде, чем он
РИС. 14.11. Микрофотография некоторых микроорганизмов активного нла.
[Из работы Unz R. F., Dondero N. С., Water Research, 4, 575 (1970).]
вступит в контакт с отходами, поступающими в аэрируемый
резервуар. В последнем органические вещества связываются
с флокулами практически исключительно за счет физических
сил. Биологическая утилизация связанных органических ве-
ществ происходит в основном в процессе повторной аэрации
рециркулирующего ила; одновременно восстанавливается ад-
сорбционная способность флокул ила.
Другие модификации процесса с участием активного ила от-
личаются от базового варианта главным образом способом осу-
ществления контакта сточных вод, ила и воздуха в аэрируемом
реакторе. Как показано на приведенной на рис. 14.13 схеме,
в процессе со ступенчатой подачей стоков поступающий поток
после разделения вводят в аэрируемый резервуар в различных
точках. Эффект разделения потока можно оценить, воспользо-
Практическое применение смешанных популяций
517
вавшись рассмотренными ранее методами анализа реакторов
(гл. 9). Обычный аэротенк с активным илом представляет со-
бой узкий длинный канал (коридор), который по своим харак-
теристикам приближается к трубчатому реактору с незначи-
тельной дисперсией. Распределение поступающего потока по
длине реактора изменяет характеристики системы таким обра-
зом, что коридорный реактор по своему поведению прибли-
жается к емкостному реактору с полным перемешиванием.
РИС. 14.12. Предполагается, что первой быстрой стадией разрушения орга-
нических веществ в аэрируемом реакторе периодического действия с активным’
илом является физический процесс—адсорбция органических веществ флоку-
лами ила, затем следует более медленная стадия биологического окисления.
Еще ближе к реактору с полным перемешиванием бассейн
круглой формы, содержимое которого интенсивно аэрируется
с целью обеспечения массопереноса и перемешивания. В такой
системе градиенты концентраций растворенного кислорода и
питательных веществ минимальны, а развивающаяся популяция
организмов активного ила часто лучше переносит флуктуации
нагрузки или резкие повышения концентраций токсичных ве-
ществ.
Хотя в основе конструкций аэраторов, применяемых в про-
цессах с участием активного ила, лежат принципы, уже рас-
смотренные нами в гл. 8, конкретные системы аэрации могут
быть самыми разными. Помимо барботажа с перемешиванием,
обычно используемого в микробиологических процессах, здесь
возможно барботирование воздуха через диффузоры, располо-
518
Глава 14
женные на дне или в стенках резервуара. В другом варианте
на поверхности бассейна вращается мешалка с лопастями, соз-
дающая турбулентные течения и способствующая поглощению
газа. Третий вариант предусматривает перемешивание и аэра-
цию с помощью конуса, который забирает жидкость со дна бас-
сейна и разбрызгивает ее на стенки резервуара. Во всех слу-
чаях основной задачей системы аэрации и перемешивания явля-
ется снабжение кислородом микроорганизмов, суспендирование
Сточные
а) Схема процесса со ступенчатой подачей
стоков
Сточные
5) Схема процесса с контактной стабилизацией
(реаэрацией ила)
РИС. 14.13. Схемы двух процессов биологического окисления. (Схема стан-
дартного процесса с участием активного ила представлена на рис. 14.10.)
и перемешивание ила и других нерастворимых компонентов
системы, а также удаление летучих продуктов метаболизма ор-
ганизмов ила, например СО2.
Помимо высокой адсорбционной и метаболической актив-
ности хороший ил должен также быстро оседать. Например,
в цилиндре через 30 мин объем осевшего активного ила дол-
жен быть примерно в 40 раз больше объема суспендированных
твердых компонентов. Если этот показатель намного выше и
объем осевшего ила превышает объем суспендированных твер-
дых частиц, например, в 200 раз, то такой ил не удовлетворяет
предъявляемым к нему требованиям, поскольку он будет выте-
кать из отстойника вместе с очищенными сточными водами.
Такое состояние называют объемной перегрузкой', в этом случае
обработанные сточные воды не будут отвечать соответствую-
щим стандартам.
Хотя причины, вызывающие объемную перегрузку и меха-
низм этого явления пока еще не выяснены, изучение неудовлет-
Практическое применение смешанных популяций
51»
верительного ила часто показывает, что в нем содержатся фи-
ламентозные бактерии и жгутиковые простейшие. Напротив,
хороший ил обычно не содержит сколько-нибудь многочислен-
ные популяции филаментозных организмов, а из простейших
в нем присутствуют главным образом стебельчатые ресничные
виды. В процессе очистки воды эти простейшие выполняют по-
лезную функцию, захватывая свободные, т. е. не включенные
в флокулы, бактерии и таким образом осветляя обработанные
сточные воды.
В нормальных условиях эксплуатации очистных станций
филаментозные бактерии и грибы не могут конкурировать с гете-
ротрофными бактериями, присутствующими в хорошем иле. Рез-
кие изменения концентраций загрязняющих веществ в посту-
пающих сточных водах или грубое нарушение режима эксплуа-
тации системы водоочистки могут, однако, привести к условиям,
неблагоприятным для роста полезных популяций, что в свою
очередь позволит другим видам микроорганизмов занять доми-
нирующее положение в системе. Отсюда следует, что результа-
ты как объемной перегрузки, так и нормального режима работы
системы водоочистки представляют собой проявления принци-
пов конкуренции видов в смешанных популяциях.
14.4.3. Проектирование и моделирование процессов
с участием активного ила
Хотя стоимость водоочистных станций большого города пре-
вышает 100 млн. долл., входящие в состав этих станций биоло-
гические реакторы обычно проектируют с помощью чрезвычай-
но упрощенных и идеализированных моделей. Обычно аэрируе-
мый резервуар рассматривают как реактор с полным перемеши-
ванием, а ил считают одним условным видом организмов, ско-
рость роста которых описывается уравнением Моно (с учетом
затухающего эндогенного метаболизма). Как мы упоминали
в разд. 14.4.2, концентрацию субстрата или лимитирующего
рост питательного вещества обычно выражают в единицах
БПК.
Приняв условные обозначения, приведенные на рис. 14.10
(они соответствуют символам, использовавшимся в гл. 7—9),
можно записать уравнение материального баланса по активным
твердым компонентам в стационарном состоянии в следующем
виде:
(l-P)FXe+₽Fxr = Vxef-i^--^ (14.1>
\ Sa + Ks J
-520
Глава 14
После несложных алгебраических преобразований этого выра-
жения получим
(14.2)
1 __ д
ТГ=’"'" ’
где 0S — среднее время пребывания активного ила (отношение
времени удерживания активных твердых компонентов к скоро-
сти их вытекания), иногда называемое также возрастом ила:
0S =---------------- (14.3)
На большинстве водоочистных станций возраст ила составляет
от 6 до 15 сут.
С помощью уравнения (14.2) можно рассчитать время пре-
бывания активного ила 0$, необходимое для достижения задан-
ного значения БПК (sa) для данных сточных вод, если известны
кинетические константы соответствующей системы. Типич-
ные значения таких параметров приведены в табл. 14.4. Не сле-
дует забывать, однако, что, как уже указывалось ранее
(разд. 3.4.2. и 9.4.1), результатом искусственного объединения
Таблица 14.4. Параметры модели Моно для процессов утилизации
различных субстратов аэробной смешанной культурой3
Органический суб- страт Y ^тах’ сут_1 Ks, мг/л &e> сут-1 Основа для расчета Темпера- тура, °C
Бытовые отходы 0,5 0,055 бпк5
0,67 0,048 бпк5 20
0,67 3,7 22 0,07 хпк
Обезжиренное 0,48 2,4 100 0,045 бпк5
молоко
Глюкоза 0,42 1,2 355 0,087 бпк5
Пептон 0,43 6,2 65 БПК5 30
а Lawrence А. W., McCarthy Р. L., Unified Basis for Biological Treatment Design and
Operation; J. Sanit. Eng. Div. ASCE, 96, 768 (1970).
всех организмов в один условный «вид» является зависимость
кинетических «констант» таких упрощенных моделей от соста-
ва сточных вод и условий эксплуатации очистной станции. От-
сюда следует, что соответствующие значения pmax, Ks, ke и Y
необходимо определять экспериментально для каждого типа
сточных вод.
Если принять, что экономический коэффициент постоянен, то
теперь можно сформулировать уравнение материального балан-
са по субстрату в аэрируемом биореакторе в стационарном со-
стоянии. Допустив, что субстрат не отделяется в процессе освет-
ления воды (Sa = se = sr), и вспомнив, что первое слагаемое
Практическое применение смешанных популяций
521
в правой
получим
части уравнения (14.2) отражает скорость роста ила,
Vxa =
VFQs (so — sa)
1 + keQs
(14.4)
Это уравнение выражает массу активных твердых компонен-
тов Vxa в аэрируемом биореакторе через рассмотренные выше
переменные. На следующем этапе для определения хг необхо-
димо экспериментально найти характеристики оседания ила,
после чего необходимый объем бассейна (биореактора) V при
данном отношении рециркуляции а можно рассчитать по фор-
муле
V = JF0sfl4-a—(14.5)
\ ха )
Это выражение получено на основе уравнения материального
баланса по активным твердым веществам в биореакторе с уче-
том уравнения (14.2).
Чрезвычайная простота описанной методики моделирования
настолько резко контрастирует со сложностью физических и
биологических процессов, осуществляющихся в активном иле,
что возникает необходимость привести некоторые эксперимен-
тальные данные, подтверждающие обоснованность применения
этой модели. На рис. 14.14 представлены результаты экспери-
ментального изучения смешанной культуры микроорганизмов
сточных вод в хемостате. Здесь же сплошными линиями изо-
бражены расчетные данные, вычисленные с помощью модели
роста типа уравнения (14.1) и приведенных на рисунке пара-
метров; нетрудно видеть, что расчетные данные хорошо согла-
суются с экспериментальными. Обратите внимание на то, что
использовавшиеся в этом случае значения параметров сущест-
венно отличаются от приведенных в табл. 14.4, хотя порядок
величин сохраняется. Этот факт еще раз подтверждает только
что упоминавшуюся особенность рассматриваемой модели, па-
раметры которой должны определяться для каждого потока
сточных вод, поскольку они зависят от характеристик по-
следних.
Как мы уже знаем из материала гл. 7 и 9, зависимость па-
раметров модели от состава исходной смеси и конструкции
системы можно снизить путем структурирования модели/Хотя
в этой области предстоит сделать еще очень многое, необходимо
обратить внимание на блестящие работы по созданию структу-
рированных моделей для процессов с участием активного ила
и других близких процессов биологической обработки, выпол-
ненные Эндрюсом и сотрудниками [5—7]. Основанием для этих
работ послужили указанные выше причины, а также необходи-
522
Глава 14
мость разработки методов управления водоочистными станция-
ми. Большинство таких станций страдает от ежедневных
существенных флуктуаций нагрузки и степени загрязнения очи-
щаемых сточных вод, что требует соответствующего регулиро-
вания режима работы станции. Недостаточно структурирован-
ная модель не может точно отразить влияние флуктуаций про-
РИС. 14.14. Сравнение расчетных данных, полученных на основе уравнения
Моно, и результатов экспериментального изучения непрерывной смешанной
культуры. [Из статьи: Chiu S. У. et al., Biotech. Bioeng., 14, 207 (1972).]
цесса, и поэтому с ее помощью невозможно изучать динамику
очистной станции и систем управления ею. К этой проблеме мы
возвратимся в примере 14.3; сейчас же рассмотрим структури-
рованную модель Эндрюса для системы с активным илом и ее
основные характеристики.
В этой модели биомассу условно разделяют на три компо-
нента, образующиеся с участием субстрата и взаимопревра-
щающиеся в соответствии со схемой:
дыхание
дыхани е
адсорбция
Субстрат ----------
- х, •
начальная
биомасса
синтез
остаток
-
активная
биомасса
X/
инертная
биомасса
6)
Практическое применение смешанных популяций
523
Скорость стадии адсорбции субстрата принята равной
= /jg (х? Xg'j
\ S + Kg . J
(14.7)
где ks — коэффициент массопереноса, a xs — начальная кон-
центрация биомассы в фазе флокулы. Общую концентрацию
биомассы в жидкой фазе и суспендированных твердых частицах
обозначают символом хт‘, в свою очередь хт равно сумме кон-
центраций начальной биомассы xs, активной биомассы Ха и
инертной биомассы хг.
Xt=Xs~{' ХаЧ~ Xi (14.8)
Наконец, fs—максимальная доля общей концентрации биомас-
сы в жидкой и твердой фазах, которую может составить началь-
Таблица 14.5. Значения параметров, предложенные Басби и Эндрюсом
для структурированной кинетической модели процессов с участием
активного илаа
Параметр
Значение
параметра
Определение параметра
ks 3,0 Коэффициент массопереноса субстрата [время-1]
f. 0,45 Максимальная доля общей концентрации био- массы в жидкой и твердой фазах, которую мо- жет составить начальная биомасса
Ks 150 Коэффициент сорбции [мг/л3]
1U 0,06 Максимальная удельная скорость превращения начальной биомассы в активную биомассу [время-1]
Ka 80,0 Константа насыщения [мг/л3]
Ya 0,66 Количество активной биомассы, образующейся из единицы количества начальной биомассы
Yt 0,25 Количество инертной биомассы, образующейся из единицы количества активной биомассы
ki 0,03 Удельная скорость разрушения активной био- массы [время-1]
(1975^ раб°ты: I-
В., Andrews J. F., J. Water Pollut. Control Assoc., 47, 1067
ная биомасса, s — концентрация субстрата, a Ks — константа
насыщения. На базе ряда литературных данных и собственных
работ по моделированию Эндрюс и сотрудники предложили
значения параметров, приведенные в табл. 14.5.
Считается, что удельная скорость синтеза активной биомас-
сы г2 описывается уравнением Моно:
<2= (14.9)
А А + xs
524
Глава 14
а постоянный коэффициент выхода Ул определяется при этом
как количество активной биомассы, образующейся из едини-
цы количества начальной биомассы, и характеризует стехиомет-
рию стадии синтеза биомассы. Имеющиеся данные позволяют
предположить, что процесс образования инертной биомассы
имеет первый порядок по активной биомассе:
Гз=^хлУ/
(14.10)
Здесь У, обозначает постоянный экономический коэффициент
(количество /, образующееся из единицы количества утилизи-
рованной биомассы А) для этой стадии.
На рис. 14.15 представлены результаты моделирования пе-
риодического роста активного ила с помощью этой модели.
РИС. 14.15. Согласно структурированной модели Басби и Эндрюса, начальная
концентрация биомассы оказывает большое влияние на скорость утилизации
субстрата в периодическом процессе.
/ — начальная концентрация xs(0)=750 мг/л; перенос субстрата к начальной
биомассе затруднен; 2 — xs(0)=300 мг/л; 3 — xs(0) = 100 мг/л.
Приведенные на этом рисунке кривые, конечно, можно интер-
претировать и как зависимость концентрации организмов
в аэрируемом реакторе с поршневым потоком от их положения
в реакторе. Как особенно интересную деталь этих расчетов сле-
дует отметить предсказываемое моделью существенное повыше-
ние скорости поглощения субстрата при уменьшении начальной
концентрации биомассы. Подобное поведение согласуется с по-
вышенной эффективностью процесса с контактной стабилиза-
Практическое применение смешанных популяций 525
цией, в котором начальная биомасса превращается в другие
формы биомассы главным образом в аэротенке, куда вода по*
ступает после осветления.
С помощью рассматриваемой кинетической модели Эндрюс
и сотрудники успешно описали большинство вариантов процес-
сов с участием активного ила в стационарном состоянии. В со-
ответствующих расчетах перемешивание в аэротенке моделиро-
вали с помощью каскада из 4 ПРПП. Первичный и вторичный
отстойники описывали моделями, которые подробно рассмотре-
ны в указанной в конце главы литературе. В реальном диапа-
Таблица 14.6. Основные эксплуатационные характеристики некоторых
процессов биологического окисления (во всех случаях скорость поступления
сточных вод принята равной 1000 м3/ч)а
Тип процесса Снижение БКПП, % Интенсив- ность на- грузки, кг/(кг-сут) Активная биомасса, % Общий объ- ем аэротен- ка, м3 Снижение БПКП. кг/(ед. объ- ема сут)
Обычный 88 0,5 34 4500 1,22
С длительной аэ- 96 0,15 16 16 000 0,392
рацией
Ускоренный 80 0,75 40 2200 2,21
Контактный 88 0,29 28 3000 1.8
Со ступенчатой 91 0,27 28 4500 1,3
подачей
а Из рвботы: Busby J. F., Andrews J. F., J. Water Pollut. Control Assoc., 47, 1055
(1975).
зоне нагрузок и масштабов процесса вычисленные величины
эффективности, скорости снижения ВПК и другие эксплуата-
ционные характеристики хорошо согласовывались с экспери-
ментальными данными для соответствующих процессов. Неко-
торые из полученных результатов приведены в табл. 14.6.
Поскольку структурированная модель успешно описывала
множество альтернативных процессов с участием активного ила
в стационарном состоянии, она оказалась незаменимой при
анализе динамики систем водоочистки с участием активного
ила. Подтверждением могут служить данные, представленные
на рис. 14.15, на котором показано, что динамика изменения
концентрации субстрата чрезвычайно чувствительна к измене-
нию общей концентрации биомассы в определенном диапазоне
последней. Динамику и регулирование таких процессов мы рас-
смотрим позднее при изучении методов анаэробной переработки
ила. Читатели, особенно интересующиеся управлением и регу-
лированием систем с активным илом, могут ознакомиться с этой
проблемой в обзорных статьях Эндрюса [5—7].
526
Глава 14
Начаты работы по созданию еще более детальных и глубоко структури-
рованных моделей, исчерпывающе описывающих взаимодействия между по-
пуляциями бактерий и простейших в активном иле. В ряде работ, посвящен-
ных моделированию динамики активного ила, Курде изучал пищевую цепь,
изображенную на рис. 14.16. Как показано на этом рисунке, в систему посту-
пают только те субстраты и бактерии, которые находятся в потоке сточных
вод. В то же время, еще раз обратившись к рис. 14.10, нетрудно заметить, что
поступающий в аэротенк поток представляет собой смесь сточных вод и ре-
циркулирующего ила, поэтому весьма важно учитывать и судьбу организмов,
обитающих в фазе флокул и затем участвующих в рециркуляции ила. В моде-
РИС. 14.16. Пищевая цепь, положенная в основу изучения межвидовых взаи-
модействий микроорганизмов в активном иле в работах Курдса.
ли Курдса к этим видам (на рис. 14.16 они подчеркнуты) относятся бактерии
ила, связанные и свободно плавающие простейшие, питающиеся суспендиро-
ванными, но не включенными во флокулы бактериями, и, наконец, связанные
с флокулой хищные простейшие, питающиеся как свободными, так и связан-
ными с флокулой простейшими.
Поскольку теперь мы уже знакомы с методикой вывода уравнений ма-
териального баланса по отдельным компонентам, то приведем только основ-
ные допущения, необходимые для вывода уравнений Курдса:
1. Аэротенк представляет собой идеальный ПРПП.
2. Зависимость удельной скорости роста каждого из представленных на
рис. 14.16 видов от концентрации питательных веществ описывается урав-
нением Моно. Применявшиеся при моделировании кинетические константы
перечислены в табл. 14.7; считается, что экономический коэффициент иа
всех стадиях равен 0,5.
3. Концентрация флокулирующих организмов при рецпркуляппи ила прямо
пропорциональна их относительной концентрации в вытекающем из аэро-
тенка потоке. Так, например,
Xif==2biXia (14.11)
где bi — коэффициент концентрирования в отстойнике для вида I. Эти кон-
станты также перечислены в табл. 14.7. Принимается, что для субстрата
6-1.
Таблица 14.7. Параметры популяций различных микроорганизмов в модели активного ила по Курдсу!
Организм
Диапазон изме-
Концентрация нения концент- Максималь-
в потоке стоя- раций в сточных ная удельная Константа
ных вод посто- водах перемен- скорость насыщения,
янного состава, кого состава, роста, ч—1 мг/л
мг/л мг/л
Коэффици-
Основной источник питания ент концеит-
организмов рироваиия в
отстойнике
on Бактерии сточных вод 15—45 0,5 10,0 Субстрат
Бактерии ила 0,3 15,0 Субстрат 1,90
Жгутиковые Питающиеся бактериями ресничные 0,4 12,0 Субстрат
Свободно плавающие 0,35 12,0 Бактерии сточных вод
Передвигающиеся по флокуле 0,35 12,0 Бактерии сточных вод 1,27
Связанные с флокулой Хищные ресничные 0,35 12,0 Бактерии сточных вод 1,90
Свободно плавающие 0,35 12,0 Ресничные, питающиеся бактериями
Связанные с флокулой 0,35 12,0 Ресничные, питающиеся 1,90 бактериями
а Из работы: Curds С. R., Water Res., 7, 1269 (1973).
528
Глава 14
Третье допущение позволяет учесть в модели процессы в отстойнике, а таким
путем и влияние рециркуляции.
Наличие свободно плавающих и прикрепленных к флокулам форм хищ-
ных простейших и питающихся бактериями простейших приводит к образова-
нию четырех пар хищник — жертва, указанных на рис. 14.16 символами а—г.
Курде рассматривал каждую пару хищник — жертва изолированно, допуская
при анализе, что на данном трофическом уровне три других взаимодействия
хищник — жертва не реализуются. Во всех случаях обнаружен ряд тенденций
общего характера; так, при анализе влияния хищных ресничных, которым пре-
небрегали во многих аналогичных работах, было показано, что их присутствие
Время, ч
РИС. 14.17. Моделирование межвидовых взаимодействий в активном иле,
в котором свободно плавающие ресничные простейшие питаются связанными
с флокулами ила ресничными простейшими. [Из статьи: Curds С. Water
Res., 7, 1269 (1973).]
бактерий в иле, мг/л
приводит к снижению численности популяции жертв ресничных (по сравнению
с системой, в которой нет хищников). В свою очередь это обстоятельство при-
водит к повышению численности популяций бактерий, находящихся в сточных
водах. Поскольку бактерии ила конкурируют с бактериями сточных вод за
общее питательное вещество, то в результате снижается численность популя-
ций бактерий ила.
Что касается пар хищник — жертва а и б, то они индуцируют вымывание
популяции жертв ресничных из системы. Взаимодействие типа в обусловлива-
ет слегка затухающие колебания, изображенные графически на рис. 14.17.
Только межвидовое взаимодействие типа г, относительно редкое в популяциях
активного ила, приводит к устойчивым отличным от нуля популяциям всех
видов в стационарном состоянии. Во всех соответствующих расчетах приняты
следующие значения параметров: /7=100 л/ч, а=1, [3 = 0,05264, So = 2OO мг/л,
Ьо (начальная концентрация бактерий в сточных водах) =30 мг/л.
Одним из недостатков как рассматриваемой, так и многих других моде-
лей является допущение о постоянстве скорости потока сточных вод я их со-
става. На практике, однако, характер образа жизни населения города или по-
селка обусловливает флуктуации как объема, так и состава сточных вод,
поступающих на очистные станции. В первом приближении эти флуктуации
Практическое применение смешанных популяций 529
можно считать периодическими с периодом 24 ч, а их амплитуда зависит в
первую очередь от конструкции системы трубопроводов, по которым сточные
воды поступают на очистные станции. Так, если очистная станция обслужива-
ет крупный город, то трубопроводы, соединяющие источники сточных вод со
станцией, могут иметь самую разную длину, поэтому сама сеть трубопрово-
дов обеспечивает существенное сглаживание флуктуаций. Большие флук-
туации возможны на небольших очистных станциях. При флуктуациях отно-
шение максимального значения какого-либо параметра к минимальному обыч-
но равно примерно 3, хотя в отдельных случаях величина этого отношения
может достигать более 10.
В одной из первых попыток изучения влияния периодических флуктуаций
количества и состава сточных вод Курде рассматривал поведение описанной
выше системы при следующих периодических изменениях параметров посту-
пающего в систему водоочистки потока:
F = 100 + 50 sin 2л/ л/ч
s0 = 200 + 60 sin 2 л/ мг/л (14.12)
Ьо — 30 4- 15 sin 2 л/ мг/л
где / — время, сут; популяции хищников здесь не рассматриваются. Результа-
ты моделирования, представленные на рис. 14.18, свидетельствуют о последо-
вательной смене доминирующих популяций, в этом отношении изучаемая си-
стема напоминает процессы порчи пищевых продуктов и периодическую смену
преобладающих видов в частицах почвы. Другим интересным результатом
моделирования является вывод о выживании связанных с флокулами реснич-
ных, в то время как свободно плавающие и передвигающиеся по поверхностям
ресничные должны вымываться. На практике в хорошем иле свободно пла-
вающие ресничные не обнаружены, однако передвигающиеся по поверхностям
организмы присутствуют в нем. Следовательно, описываемая модель нуждает-
ся в дальнейшей доработке. В заключение следует отметить, что между коле-
баниями численности популяций, представленными на рис. 14.17 и 14.1'8, име-
ются принципиальные различия. В последнем случае колебания являются
вынужденными, поскольку они обусловлены периодическими флуктуациями
количества и состава поступающих в систему стоков. Напротив, колебания
первого типа, наблюдающиеся при постоянстве количества и состава сточных
вод, являются автономными и отражают специфические нелинейные характе-
ристики системы. Дальнейшее обсуждение этой проблемы читатель можег
найти в обзоре Бейли (ссылка [10] в гл. 11).
Хотя разработанные в последние годы Эндрюсом, Курдсом
и другими структурированные модели весьма перспективны
в отношении как совершенствования методов проектирования
процессов, так и создания систем управления этими процесса-
ми, они пока еще не нашли широкого применения. Можно на-
деяться, однако, что после дальнейшей аналитической и экспе-
риментальной проверки модели этого типа будут использовать-
ся значительно шире.
14.4.4. Аэробная обработка ила
Активный ил с большим содержанием биопродуктов, обра-
зующийся в рассмотренных выше процессах, часто подвергают
еще одной операции аэробной обработки; фактически она по-
вторяет описанную в предыдущих разделах в отсутствие по-
34—746
530
Глава 14
РИС. 14.18. Моделирование динамики системы с активным илом, в которой
скорость потока поступающих сточных вод, концентрация субстрата и бакте-
рий изменяются синусоидально с периодом 1 сут. [Из статьи: Curds С.
Water Res., 7, 1269 (1973).]
Концентрация оактериа б иле, мг/л Концентрация сдязаннггл с /рлокулоц
ресничных, мг/л
ступления свежих сточных вод. В таких условиях биомасса
в результате эндогенного дыхания утилизирует свои же источ-
ники углерода, так что в конечном счете содержание твердых
компонентов уменьшается обычно на 50%. В этой операции ре-
циркуляцию биомассы не применяют, а время пребывания по-
следней в реакторе составляет от 15 до 25 сут. Основной целью
этой операции является уменьшение общей массы ила, подле-
жащего перевозке (сухопутным или речным транспортом) и
уничтожению.
Практическое применение смешанных популяций
531
14.4.5. Нитрификация
В обычных процессах обработки отходов с аэрацией в числе
подвергающихся биологическому окислению субстратов имеют-
ся и азотсодержащие органические вещества. Из последних при
биологическом окислении обычно сначала образуется аммиак,
который затем необходимо окислить до нитрита и, наконец,
до нитрата; только в этом случае очищенная вода будет обла-
дать достаточно низкой БПК- Как было показано на рис. 14.3,
эти превращения осуществляются при участии двух видов мик-
роорганизмов — Nitrosomonas и Nitrobacter. Соответствующие
превращения описываются следующими схемами реакций:
Nitrosomonas
NH3+CO2+O2 ----------->- биомасса+ NO2-
Nitrobacter
NO2-4-CO2+O2 ---------> биомасса + МОз-
Удельные скорости роста каждого из этих видов описывают-
ся обычным уравнением Моно; согласно данным Лоуренса и
Маккарти*, соответствующие параметры имеют следующие зна-
чения:
TN> г клеток/(г азота) лтах- сут 1 Ks, мг/л
Nitrosomonas 0,05 0,33 1,0
Nitrobacter 0,02 0,14 2,1
Для оценки концентраций аммиака и нитрита в сточных во-
дах, прошедших обработку в системе водоочистки с активным
илом (эффективность работы которой оценивают по снижению
величины БПК до заданного уровня), за основу можно взять
уравнения материальных балансов по популяциям Nitrosomonas
и Nitrobacter. Ниже приведен пример такого расчета. Если вре-
мя пребывания биомассы в системе с активным илом слишком
мало, то для завершения процесса нитрификации можно исполь-
зовать второй аэрируемый биореактор.
Пример 14.2. Расчет процесса нитрификации. Шредер [10] рассматривал
следующие условия очистки сточных вод:
Максимально допустимые величины БПК и концентрации азота аммиака
в очищенной воде составляют 10 и 0,5 мг/л соответственно. Необходимо опре-
делить степень нитрификации в биореакторе с активным илом и установить,
необходим ли еще один реактор для нитрификации.
* Lawrence A. W., McCarty Р. L., Unified Basis for Biological Treatment
Design and Operation, J. Sanitary Eng. Div., Proc. ASCE, 96, 757 (1970).
34*
532
Глава 14
Обработка промышленных сточных вод
Характеристика сточ- 400 мг/л (растворенные
ных вод: БПКп
Органический азот вещества) 60 мг/л
Фосфат (РО43~) 30 мг/л
pH 7
Температура 20 °C
Стехиометрия и кине- тика процесса: Утилизация 1 V/ субстрата | vm к /набл 2,0 сут-1 5,0 мг/л 0,4/(1,0+0,06 9с)
Скорость оседания ин
ила: 2,7 — 0,00026% см/мин
Если рост микроорганизмов лимитируется только источником углерода
(но не азота или фосфора), то из определения ВПК следует
_ F (10 —400) х хц
Скорость снижения ВПК =--------Z— = ~~ “--------------------------
VR КнаблОс Унабл
(14П2.1)
С другой стороны, из уравнения материального баланса по биомассе следует
% УнаблЩ5-*
Скорость образования биомассы — —— = ——— ---------- (14П2.2)
Ос А/п + s
Минимальное среднее время пребывания ила, необходимое для достиже-
ния заданной величины ВПК, рассчитывают на базе s= 10 мг/л. Если УНабл=
= 0,4/(1+0,060с), то
п х {Кт +s) 5+10
Ос (минимальное) =------------~——--------=---------------------------в
2 0,4p,gS —0,06 (Km + s) 0,4-2-10 — 0,06(5+10)
=- 2,1 сут
Следовательно, при 9С>2,1 сут достигается заданный уровень ВПК.
Начальная концентрация азота в системе с активным илом равна 60 мг/л;
14
концентрация ассимилированного азота составляет у|з~(Унабл) (s(—s). Таким
образом, концентрация ассимилированного азота составит 16,6 и 14,3 мг/л при
9с равном 3 и 6 сут соответственно.
Согласно уравнению материального баланса по биомассе Nitrosomonas’.
1____Prnax.NH^NHg (14П2 3)
6; ~ ^NH3 + SNH3
Расчет удельной скорости роста при приведенных выше параметрах и задан-
ной концентрации аммиака после обработки дает
1 0,33-0,5
V = T+ET = 0’llcyT'1
Следовательно, заданная степень превращения аммиака обеспечивается при
Практическое применение смешанных популяций
533
0С>9 сут. Приняв 0с равным 10 сут, с помощью уравнения такой же формы
с параметрами превращения нитрита можно рассчитать соответствующую кон-
центрацию нитрита
__J_____Hmax, NO2SNO2 (14П2.4 )
% ~ ^NO2 + sNO2
илн
Kno2
Qcprnax.NOg 1
Подстановкой указанных выше для Nitrobacter параметров получим Sno2=
= 5,25 мг/л.
Выполненные расчеты показывают, что в этом случае снижение величины
БПК осуществляется быстрее, чем нитрификация. Следовательно, для дости-
жения заданной концентрации азота в очищенной воде необходимо или уве-
личить объем биореактора с активным илом, или установить после него аппа-
рат для нитрификации.
В результате нитрификации общая потребность в кислороде снижается
в соответствии со следующей стехиометрией реакций:
NH3+3/2O2 —> no2-+h2o+h+
NO2- + V2O2 —*- NO3-
В более точных расчетах должны быть учтены процессы нитрификации в от-
стойнике, расположенном непосредственно после биореактора с активным
илом.
14.4.6. Вторичная очистка сточных вод
с помощью капельных биологических фильтров
В довольно распространенном варианте очистки сточных вод
с участием активного ила применяют так называемые капель-
ные, или перколяционные биологические фильтры. В биологиче-
ском фильтре популяции микроорганизмов существуют в виде
пленки или слизистого слоя на поверхности твердой насадки,
неплотно заполняющей резервуар (доля пустот составляет око-
ло 0,5); в таких условиях воздух легко поступает в нижние
слои насадки. Типичная конструкция биологического фильтра
представлена на рис. 14.19.
Использование термина «фильтр» для описания этой систе-
мы водоочистки во многих отношениях неудачно, поскольку ме-
ханизм обезвреживания примесей здесь связан не с их механи-
ческим удерживанием, а с теми же самыми последовательными
процессами связывания и биологического окисления, которые
реализуются в системах с активным илом. Прежде чем присту-
пить к изучению популяций микроорганизмов, участвующих
в очистке сточных вод в биологических фильтрах, необходимо
остановиться на некоторых конструктивных и эксплуатацион-
ных особенностях соответствующих' аппаратов.
534
Глава 14
Подлежащие очистке сточные воды контактируют прежде
всего с верхней частью неподвижного слоя, толщина которого
составляет обычно от 1 до 3 м; сточные воды подают непрерыв-
но через расположенные над неподвижным слоем насадки сопла
или периодически с помощью вращающегося разбрызгивателя,
подобного изображенному на рис. 14.19. И в том и в другом
случае скорость потока сточных вод должна быть достаточно
низкой, чтобы слой насадки не оказался под водой. Для обеспе-
чения нужной скорости переноса кислорода поступающие в си-
РИС. 14.19. Биологический капельный фильтр. [Из работы: Abson. J. W.,
Todhunter К. Н., in Biochemical and Biological Engineering Science, Blake-
brough N. (ed.), vol. 1, p. 326, Academic Press, London, 1967.1
/ — вращающийся разбрызгиватель сточных вод; 2 — насадка; 3 — трубопро-
вод для подачи сточных вод; 4—-дренаж; 5—бетонная ограждающая стена;
6— отверстия для поступления воздуха.
стему сточные воды должны обтекать покрытую слизью насад-
ку достаточно тонким слоем, не препятствующим дыханию
аэробных организмов, находящихся на наружной поверхности
пленки микроорганизмов. В отличие от процессов с участием
активного ила, обычно требующих принудительной аэрации, че-
рез биологический фильтр воздух циркулирует благодаря есте-
ственной конвекции. Движущей силой конвекции является раз-
ность температур, создающаяся в фильтре за счет биологиче-
ского окисления загрязняющих веществ, присутствующих
в сточных водах; отверстия для поступления воздуха и связан-
ные с ними вентиляционные трубопроводы (расположенные
внутри фильтра) обеспечивают поступление воздуха в нижние
и промежуточные слои насадки.
К биологическим фильтрам применимы основные принципы
взаимосвязей между скоростью диффузии субстрата и ско-
ростью реакции, рассмотренные в разд. 4.4.2. В то же время
сложность систем со смешанными популяциями микроорганиз-
мов и смесями субстратов препятствует разработке ана-
Практическое применение смешанных популяций
535
литических методов изучения биологических фильтров, для
которых типичны как локальные (внутри пленки микро-
организмов), так и общие (по толщине слоя насадки)
градиенты концентраций субстратов и плотности популя-
ций. Тем не менее некоторые выводы качественного ха-
рактера, сделанные на основе анализа диффузии и реакции
в пленке (разд. 4.4.2), могут оказаться полезными. Так, можно
Таблица 14.8. Характеристики высоконагружаемых и низконагружаемых
биологических фильтров3
Характеристика Низконагружаемые биологические фильтры Высоконагружаемые биологические фильтры
Гидравлическая нагрузка, 1000—4000 8000—40000
л/(сут-м2)
Органическая нагрузка, кг БПК5/(1000 м3-сут) Высота неподвижного слоя насадки, м 80—400 400—4800
При одноступенчатом 1,5—2,4 0,9—1,8
фильтре
При многоступенчатом 0,8—1,2 0,5—1,2
фильтре
Режим подачи сточных вод Периодический Непрерывный
Рециркуляция Обычно без рецирку- ляции Всегда с рециркуля- цией
Очищенная вода Высоконитрифициро- ванная; БПК5= = 20 мг/л Неполностью нитри- фицированная; БПК5=30 мг/л или больше
а Из работы: Rich L. G., Environmental Systems Engineering, p. 370. .McGraw-Hill
Book Company, New York, 1973.
ожидать, что возникновение и развитие анаэробных областей
в толще пленки микроорганизмов приведут к формированию
газовых пузырьков, которые в свою очередь вызовут частичное
отделение пленки от носителя. Образовавшиеся таким путем и
унесенные из биологического фильтра потоком воды организмы
часто называют гумусом; последний необходимо отделять в от-
стойнике, установленном непосредственно после биологического
фильтра. С другой стороны, в результате этого процесса регу-
лируется толщина пленки микроорганизмов, среднее значение
которой зависит от множества факторов. В правильно эксплуа-
тируемом биологическом фильтре толщина пленки микроорга-
низмов обычно составляет около 0,35 мм.
В табл. 14.8. приведены диапазоны производительности и
эффективности биологических фильтров. В типичном процессе
гидравлическая нагрузка такова, что время пребывания стоков
в фильтре составляет 20—60 мин. В табл. 14.8 приведены так-
536
Глава 14
же характеристики высоконагружаемого биологического филь-
тра (иногда его называют импульсным капельным фильтром)-,
в этом случае высокие скорости потока сточных вод ограничи-
вают толщину формирующейся слизистой пленки микроорга-
низмов. Недостатком высоконагружаемых биологических
фильтров является вымывание большого количества гумуса,
который необходимо отделять в отстойнике.
Для того чтобы понять принцип работы биологического
фильтра, полезно проследить за происходящими в фильтре пре-
вращениями в пространстве и времени. Предположим, что мы
перемещаемся внутри фильтра сверху вниз вместе с каплей
жидкости. По мере движения через неподвижный слой насадки
состав жидкости изменяется во времени, что обусловлено погло-
щением разных компонентов различными микроорганизмами.
Теоретически эти изменения во многих отношениях близки со-
бытиям, происходящим в скисающем молоке (см. рис. 14.1) и
вспаханной почве. По мере изменения состава жидкой среды
в ней поочередно развиваются преимущественно определенные
виды микроорганизмов, что в свою очередь приводит к изме-
нению ее состава и затем к замене одной доминирующей попу-
ляции другой.
Теперь перенесем наблюдения в фиксированную в простран-
стве систему координат. То, что раньше представлялось нам
как изменения в капле во времени, теперь будет иметь харак-
тер распределения в рабочем пространстве фильтра, эксплуати-
руемого в стационарном состоянии. Организмы, наиболее при-
способленные к утилизации питательных веществ сточных вод,
доминируют в верхней части слоя насадки; здесь же изобилуют
прочно связанные с насадкой грибы и свободно плавающие рес-
ничные. В нижней части фильтра преобладают стебельчатые
ресничные и нитрифицирующие бактерии. Среди обитателей
биологических фильтров можно обнаружить и высших живот-
ных, из которых наиболее многочисленны популяции червей и
личинок насекомых. Эти животные питаются организмами сли-
зистого слоя, растущими на насадке фильтра; регулирование
численности их популяций является важным фактором при
управлении работой фильтра.
Разделение организмов в пространстве биологического филь-
тра позволяет каждому виду полностью адаптироваться к соот-
ветствующему окружению. По этой причине, в частности, низко-
нагружаемые биологические фильтры обычно обеспечивают
большую прозрачность и большую степень нитрификации очи-
щенной воды, чем системы с активным илом. Кроме того, опыт
эксплуатации водоочистных станций показал, что по сравнению
с системами с активным илом биологические фильтры менее
чувствительны к пиковым нагрузкам токсичных веществ. В то
Практическое применение смешанных популяций
537
же время, как показано в табл. 14.9, в некоторых отношениях
системы с активным илом превосходят биологические фильтры.
Предпочтение той или иной системе водоочистки можно отдать
только после тщательного изучения характеристик сточных вод,
стоимости оборудования и требований к качеству очищенной
воды. В некоторых случаях оптимальный вариант проекта
включает использование как первого, так и второго методов.
Таблица 14.9. Сравнение методов очистки сточных вод с помощью
биологических фильтров и активного илаа
Характеристики
Ьиологические фильтры Системы с активным илом
Капитальные затраты
Эксплуатационные рас-
ходы
Площадь, занимаемая
биореакторами
Регулирование аэрации
Регулирование темпера-
туры
Чувствительность к ко-
лебаниям концентра-
ций загрязняющих ве-
ществ в сточных во-
дах
Прозрачность очищенной
воды
Неприятный запах
Высокие
Низкие
Большая
Частичное (за исключе-
нием систем с прину-
дительной аэрацией)
Затруднено в силу боль-
ших потерь тепла
Низкая; восстановление
чувствительности про-
исходит медленно
Хорошая
Присутствует
Низкие
Высокие
Небольшая
Полное
Полное; потери тепла
невелики
Высокая, но восстанов-
ление чувствительнос-
ти происходит быстро
Не очень хорошая
Отсутствует
Из работы: Abson J. W., Todhunter К. Н., in Biochemical and Biological Engineering
Science, Blakebrough N. (ed.), vol. 1, p. 337, Academic Press, London, 1967.
В окончательном варианте проекта определяют также конкрет-
ные типы отстойников и устройств для рециркуляции; более
подробно с этим вопросом читатель может ознакомиться в ли-
тературе.
Основой другого метода очистки сточных вод являются так
называемые биологические пруды-, этот метод очистки намного
проще, чем водоочистка с помощью активного ила или биологи-
ческих фильтров. В биологических окислительных прудах,
напоминающих естественные водные экосистемы, в процессе фото-
синтеза водоросли выделяют кислород; тем самым поддержи-
вается аэробный режим, который необходим для бактерий, ути-
лизирующих органические загрязняющие вещества. Для пред-
отвращения образования анаэробных зон окислительные пруды
обычно делают неглубокими, от 0,6 до 1,2 м глубиной. Напро-
538
Глава 14
тив, в стабилизирующих прудах для обработки сточных вод,
содержащих осаждающиеся примеси, поддерживается анаэроб-
ный режим или чередование во времени аэробного и анаэробно-
го режимов. Дополнительные сведения о таких процессах водо-
очистки приведены в работе Рича [3].
14.4.7. Анаэробная переработка отходов*
Отходы, содержащие значительные количества ферментируе-
мых органических соединений, можно подвергать биологической
обработке в анаэробных условиях. Хотя анаэробная обработка
применяется во многих процессах, основной сферой использова-
ния этого метода является переработка избыточного активного
ила (рис. 14.10 и 14.13), образующегося при биологической
очистке сточных вод. Как мы уже знаем из материала предыду-
щих разделов, концентрированный ил образуется на нескольких
стадиях, в том числе при отделении твердых частиц на решет-
ках и в первичном отстойнике, а также при росте микроорганиз-
мов в ходе биологического окисления (при вторичной очистке
сточных вод). Ил далее концентрируют или сгущают часто пу-
тем простой седиментации; ликвидации ила обычно предшест-
вует операция анаэробной биологической переработки, являю-
щаяся одним из этапов водоочистки.
Механизм анаэробной переработки отходов, в котором
участвует множество видов микроорганизмов, в самом общем
и упрощенном виде можно описать следующей схемой:
Нерастворимые солюбилизация внекле- Растворенные __>
органические точными гндролнтнче- органические
вещества скимн Фермами вещества
кислотообразующие Бактериальная биомасса
----ба ин-------> Другие продукты
Летучие органические кислоты+ СО2+Н2
газификация
метанобразующими
,, бактериями
СН4 + СО2 + бактериальная биомасса (14.13)
На первой стадии твердые частицы ила солюбилизируются или
диспергируются внеклеточными ферментами, синтезируемыми
самыми различными бактериями. В системах для анаэробной
обработки ила обнаружены протеолитические, липолитические
и некоторые целлюлолитические ферменты. Поскольку в био-
реакторах для анаэробной переработки ила твердые вещества
* В основу этого раздела положено обсуждение проблемы математическо-
го моделирования процессов анаэробной обработки отходов, приведенное в ра-
боте: Graef S. Р., Andrews J. F., СЕР Symp. Ser., [136) 70, 101 —127 (1974).
Практическое применение смешанных популяций 539
не накапливаются, то, очевидно, реакции солюбилизации осу-
ществляются достаточно быстро и эта стадия не лимитирует
скорость всей последовательности превращений (14.13).
Экспериментальное изучение следующей стадии анаэробной
переработки ила, а именно микробиологического синтеза низко-
молекулярных жирных и летучих кислот из растворенных орга-
нических веществ, показало, что скорость осуществляющихся
на этой стадии реакций также довольно высока. По вполне по-
нятной причине ответственные за эти превращения организмы
называют кислотообразующими бактериями-, они являются фа-
культативными анаэробными гетеротрофами и лучше всего
функционируют в диапазоне pH от 4,0 до 6,5. Главным продук-
том этой стадии является уксусная кислота, хотя в некоторых
количествах образуются также пропионовая и масляная кис-
лоты.
Важнейшим субстратом для последней стадии процесса
является уксусная кислота; показано, что около 70% всего ме-
тана образуется именно из этого субстрата. Стадия газифика-
ции осуществляется с участием метанобразующих бактерий,
являющихся облигатными анаэробами. Эти организмы проявля-
ют наибольшую активность в гораздо более узком диапазоне
pH от 7,0 до 7,8; их сложно выделить в виде соответствующих
чистых культур, но в адекватно эксплуатируемом биореакторе
(метантенке) смешанная культура этих бактерий находит очень
хорошие условия для своей жизнедеятельности. Имеющиеся
данные свидетельствуют о том, что превращение летучих кислот
в СН4 и СО2 лимитирует скорость всей последовательности пре-
вращений, описываемой уравнением (14.13).
На рис. 14.20 представлена схема аппарата для анаэробной
переработки ила (метантенка). Для предотвращения чрезмер-
ного повышения локальных концентраций кислот содержимое
метантенка перемешивают. Создание условий, удовлетворитель-
ных как для кислотообразующих, так и для метанобразующих
бактерий, обеспечивается поддержанием pH около 7. На рис. 14.20
указан также выносной теплообменник для поддержания по-
вышенной температуры в резервуаре метантенка. В настоящее
время в большинстве случаев температуру содержимого метан-
тенка поддерживают на уровне мезофильного диапазона (около
32—38 °C), который обеспечивает максимальную скорость пере-
работки ила. Имеются указания на то, что скорость процесса
можно повысить в еще большей степени, если осуществлять его
в термофильном диапазоне (около 55°C). Впрочем, такой тем-
пературный режим применяют сравнительно редко; одной из
причин предпочтения, отдаваемого мезофильному диапазону
температур, является меньший расход энергии на нагревание
метантенка. При эффективном перемешивании и средней тем-
540
Глава 14
РИС. 14.20. Схема установки для анаэробной переработки ила. (Из работы:
Аткинсон Б., Биохимические реакторы. — М.: Пищевая промышленность,
1979.)
1 — смотровые окна; 2 — труба для выхода газа; 3— предохранительный кла-
пан для регулирования давления (вакуума); 4— пламягаситель; 5 — трубо-
провод для отвода газа; 6— возвратная вода; 7 — возвратная циркулирующая
вода и расширительная камера; 8 — регулируемый слив суспензии ила; 9—
регулятор уровня; 10 — вывод из камеры с илом; 11 — возврат воды в нагре-
ватель; 12 — выпуск переработанного ила; 13— дренажные трубы; 14 — пода-
ча сырого ила; 15 — газ; 16 — подача циркулирующей воды; 17 — выносной
теплообменник; 18— возврат циркулирующей воды; 19 — верхний уровень ила.
пературе (32—35 °C) необходимое для полной переработки ила
время его пребывания составляет от 10 до 30 сут.
К счастью, при анаэробной переработке ила образуется топ-
ливо, которое можно использовать для снижения эксплуатацион-
ных расходов водоочистных станций. Иногда образующийся при
анаэробной переработке ила метан используют вне водоочист-
ной станции для выработки тепла и электроэнергии. Газовая
Практическое применение смешанных популяций 541
смесь, образующаяся при анаэробной переработке ила и на-
капливающаяся, как это показано на рис. 14.20, в верхней части
метантенка, состоит в основном из метана (65—70%) и угле-
кислого газа. В небольших концентрациях в этой смеси содер-
жатся также сероводород (продуцируемый сульфатредуцирую-
щими бактериями), Н2 и СО. Газовая смесь обладает тепло-
творной способностью от 5800 до 6700 ккал/м3 и образуется
с выходом 0,75—1,12 м3 из 1 кг переработанных органических
веществ. Поскольку по своей теплотворной способности биогаз
Таблица 14.10. Изменение состава ила бытовых сточных вод
после анаэробной переработки2
Относительное содержа-
ние
Компонент
В исходном иле в иле после анаэробной переработки
Эфирорастворимые вещества 34,4 8,2
Водорастворимые вещества 9,5 5,5
Спирторастворимые вещества 2,5 1,6
Гемицеллюлоза 3,2 1,6
Целлюлоза 3,8 0,6
Лигнин 5,8 8,4
Белки 27,1 19,7
Зола 24,1 56,0
Из работы: Abson J. W., Todhunter К. Н., in Biochemical and Biological Engineering
Science, Blakebrough N. (ed.), vol. 1, p. 339, Academic Press, London, 1967.
значительно уступает природному газу (около 8900 ккал/м3), то
при наличии достаточных запасов последнего биогаз не пред-
ставляется удобным или привлекательным топливом. В связи
с повышением цен на топливо, однако, процессам анаэробной
переработки ила как потенциальному источнику топлива (после
обязательного удаления H2S) уделяется все большее внимание.
В результате анаэробной переработки ил легче поддается
последующим операциям. Во-первых, содержание органических
веществ в иле снижается на 50—60%. Во-вторых, существенные
изменения претерпевают и концентрации других компонентов
ила (см. табл. 14.10). После анаэробной переработки ил в го-
раздо меньшей степени подвержен гниению и легче обезвожи-
вается. После обезвоживания (эту операцию часто осуществля-
ют с помощью ротационного вакуум-фильтра) ил высушивают
и затем используют в качестве удобрения, складируют или сжи-
гают. На рис. 14.8 упомянуты некоторые другие методы перера-
ботки) ила; сведения о них можно найти в литературе.
542
Глава 14
14.4.8. Математическое моделирование динамики процесса
анаэробной переработки ила
Несмотря на очевидные выгоды, связанные с образованием
газообразного топлива и ценного удобрения (твердых отходов),
метантенки заслужили плохую репутацию в силу ряда проблем,
возникающих при их эксплуатации. Опубликованы данные
о многочисленных нарушениях режима работы метантенков;
чаще всего неполадки связаны с гидравлической, органической
или токсической перегрузками. В первом случае (при гидрав-
лической перегрузке) скорость разведения превышает скорость
роста микроорганизмов, и в результате последние вымываются
из системы. Чрезмерно высокие концентрации органического
субстрата, с другой стороны, вызывают накопление летучих
кислот, что приводит к ингибированию роста метанобразующих
бактерий и таким образом к нарушению нормального режима
работы метантенка по мере падения pH. К тому же результату
ведет и повышенная концентрация веществ, токсичных по отно-
шению к метанобразующим бактериям; в этом случае они также
вымываются из системы.
Поскольку работа в более стабильных эксплуатационных
режимах способствовала бы более широкому распространению
и более успешному использованию процессов анаэробной пере-
работки ила, очевидна необходимость изучения динамики соот-
ветствующих систем с целью разработки адекватных методов
управления. В этом разделе мы обсудим очень интересную ма-
тематическую модель процесса анаэробной переработки ила,
разработанную Граефом и Эндрюсом. Кроме того, в примере 14.3
мы рассмотрим некоторые принципы управления такими процес-
сами. Эта модель является как бы кульминационным пунктом
всего обширного материала, изложенного в настоящей книге,
поскольку в ней учтены теснейшие взаимосвязи между физиче-
скими, химическими и биологическими факторами; она пред-
ставляет собой синтез классического инженерного искусства и
основных результатов изучения биологии системы и успешно
решает конкретную биохимическую инженерную задачу.
Выше уже упоминалось, что в последовательности биологи-
ческих реакций (14.13) стадией, определяющей скорость всей
последовательности, является трансформация летучих кислот
метанобразующими бактериями. Допустив, что все летучие кис-
лоты можно представить в виде уксусной кислоты и что состав
метанобразующих бактерий приближенно выражается форму-
лой C5H7NO2, Граеф и Эндрюс [7] предложили следующую сте-
хиометрию «реакции» газификации:
CH3COOH+0,032NH3 —> 0,032C5H7N02+0>92CH4 +
+ 0,92С02+0,096Н20 (14.14>
Практическое применение смешанных популяций 543
Считается, что лимитирующим субстратом для этой «реак-
ции» являются неионизированные летучие кислоты. В общем
случае в силу ионизации концентрация неионизированного суб-
страта (HS) отличается от его общей концентрации s:
HS S-+H+ Ка (14.15)
Здесь символом S~ обозначен ионизированный субстрат. По-
скольку —1g /<а = р/Са = 4,5, а метантенк функционирует при pH
выше 6, то практически вся кислота должна находиться в иони-
зированной форме, так что
s~^s (14.16)
и
(14.17)
Ка
Для того чтобы в модели было учтено известное ингибирующее
действие высоких концентраций субстрата, Граеф и Эндрюс
[7] модифицировали уравнение Моно для удельной скорости
роста микроорганизмов следующим образом:
Н=ФшахГ" , „ ... \ (14.18)
L 1 4-Ks/(/?s) + (/is)//^
<3 такой формой уравнения Моно мы уже познакомились при
изучении кинетики клеточного роста в гл. 7. В уравнении бак-
териального роста отражена также скорость гибели микроорга-
низмов; считается, что этот процесс имеет первый порядок по
концентрации токсина [fox]*:
rD = —Zej-ltox] (14.19)
На базе ранее опубликованных данных и собственных оценок
Граеф и Эндрюс предложили следующие значения для парамет-
ров, входящих в состав указанных уравнений: цтах = 0,4 сут-1,
Ks = 0,0333 ммоль/л, /(, = 0,667 ммоль/л, /?7- = 2,0 сут-1.
Если далее принять, что экономические коэффициенты обра-
зования биомассы по лимитирующему рост субстрату, СО2 и
СН4 (Ух/s, Усо2/х и Усн4/х соответственно) постоянны, то урав-
нения материальных балансов по субстрату и биомассе в изу-
чаемой системе примут форму, представленную в нижней части
рис. 14.21. Параметры /?в и Qch4 обозначают скорости образо-
вания СО2 и метана в ходе биологической газификации. В усло-
виях, типичных для процессов анаэробной переработки ила,
* Во избежание недоразумений здесь концентрации некоторых компонен-
тов обозначены соответствующими символами в квадратных скобках; кон-
центрации других компонентов, как обычно, обозначены строчными буквами.
V, 1ц
Pl-P,
Газовая (раза
dPco, _ V Рсо2 п
л, ~ ?т-----т7 *С----(Г~ Q
« Р9 Уа Уа
л V
Qco2 — TG; Q = Qcot + 2сн, + 2н,о
Ря
Q
Рсо2
Рсо2
2сн4
Жидкая (раза
.. , s(h + )
(hs) = ~7Г
К.[СО2]Р
[НСО3-]
dz F
[НСО3-]=--5 _ = _(2о_г)
[нсо;]^
[СО:]Ро
Пох]о
F. V
TG = kL a([CO2JS - [CO2]D>
[CO2]P — КнРсо2
</[toxl F
—— = -([tox]0 - [tox]>
dt V
</[CO,]b F
= у ([CO2]Po - [CO2]P) + TG + Rg + Rc
[HCO;]
[CO2]P
(V)
Ка-К I-
--------->
ki.a
F ds dz
rc = ~([нсо3-]0 - [hco;]) + - - -
V dt dt
(/is)
[tox]
Kb
Qch4
РИС. 14.21, Схема математической модели анаэробной переработки ила,
Стрелками указаны потоки информации между подсистемами и взаимодей-
ствия с внешним окружением. [Из работы; Graef S. Р., Andrews J. F., Mathe-
matical Modeling and Control of Anaerobic Digestion, in Water-1973*
Bennet G. F, (ed), CEP Symp. Ser. No. 136, 76, 101 (1974).]
Практическое применение смешанных популяций
545
плотность газа ро равна 0,0389 моль/л, а Усн4/х и УСо2/х равны
28,8 моль/моль. Как показано на рис. 14.21, растворимость ме-
тана в среде очень низка и весь образующийся метан перехо-
дит в газовую фазу.
Напротив, СО2 существует как в жидкой фазе (в двух фор-
мах), так и в газовой фазе. Скорость переноса СО2 из газовой
фазы в жидкую описывается известным уравнением (гл. 8)
T0=kLa([CC)2]D*- [СО2]с) (14.20)
где [С02]о—концентрация растворенного СО2, а [С02]о*—
концентрация растворенного СО2 в состоянии равновесия.
Из закона Генри следует
[СО2]с*=Кнрсо2 (14.21)
где рсо2 — парциальное давление СО2 в газовой фазе. Для па-
раметров kLa и Кн Граеф и Эндрюс предложили значения
100 сут-1 и 3,25-10-5 моль/(л/мм рт. ст.) соответственно.
Другой способ существования СО2 в жидкой фазе обуслов-
лен образованием бикарбонатного иона согласно уравнению
НСОз_+Н+ ч=ь Н2О+СО2 константа равновесия= 1/Ki (14.22}
Если скорость этой реакции обозначить символом Rc, то урав-
нение материального баланса по бикарбонатному иону в жид-
кой фазе можно записать в следующем виде:
V Л 1Н^.°3 1 = F ([НСО8-]0 - [НСО3-]) - VRC (14.23)
at
Независимое выражение для [НСО3~]/<Д можно найти, исходя
из требования электронейтральности:
[Н+] +с= [НСОз-] + [S-] + [ОН-] +а+2[СОз2-] (14.24)
где с — общая концентрация катионов, включая катионы каль-
ция, натрия, магния и аммония, и а — общая концентрация
анионов (хлорида, фосфата, сульфида и т. п.). Если система
функционирует в нормальном диапазоне pH (от 6 до 8), то
концентрации [Н+], [ОН-] и [СО32-] пренебрежимо малы и
уравнение (14.24) преобразуется в выражение
z=[HCO3-]+s (14.25)
где результирующая концентрация катионов z определяется как
?=с-а (14.26)
а вместо s- в соответствии с уравнением (14.16) использует-
ся s. Если концентрация сульфидов не слишком высока, то z
приближенно соответствует концентрации аммониевого иона.
35—746
£>46
Глава 14
Предположим, что уравнение материального баланса по Z мож-
но записать в виде
d (z) F
— (14.27)
Далее, дифференцируя уравнение (14.25) по времени, получим
d [НСО3-] _ d (z) d (s)
dt dt dt
Исключение с?[НСОз~]/Д из уравнений (14.28) и (14.23) дает
F , d (s) d (z)
Rg = ([HCO3-]0 - [HCO3-]) + —(14.29)
V ilk Hk
Как показано на рис. 14.21, эта скорость учтена в уравнении
материального баланса по СОг в жидкой фазе.
Вывод уравнений материальных балансов по компонентам
газовой фазы не представляет затруднений (см. рис. 14.21).
Обратите внимание на то, что общая скорость газового потока
определяется на базе различных скоростей газообразования для
отдельных компонентов, рассчитываемых с помощью тех или
иных уравнений модели.
Теперь перейдем к рассмотрению некоторых результатов,
полученных с помощью описываемой модели. В табл. 14.11 пе-
речислены значения параметров модели и стандартные условия
(для стационарного состояния), принятые в соответствующих
расчетах. Моделирование периодического процесса анаэробной
переработки ила (соответствующие уравнения легко получить,
подставив F = 0 в уравнения для системы непрерывного дейст-
вия) показывает, что повышение начальной плотности популя-
ции организмов или начального значения pH, или снижение на-
чальной концентрации субстрата должно приводить к уменьше-
нию продолжительности периодического процесса. Действитель-
но, такая закономерность наблюдалась в эксплуатируемых
системах.
Граеф и Эндрюс изучали также особенности стартового пе-
риода работы биореактора для анаэробной переработки ила.
Они показали, что модель прогнозирует, во-первых, уменьшение
продолжительности стартового периода при повышении началь-
ного значения pH или концентрации ила в поступающем в си-
стему потоке, во-вторых, нарушение нормального режима био-
реактора, если начальное значение pH или концентрация ила
слишком низки, в-третьих, уменьшение вероятности нарушения
режима биореактора при медленном увеличении нагрузки
(в стартовом периоде) до установленной величины. И в этом
случае результаты расчетов хорошо согласовывались с особен-
££ Таблица 14.11. Условия стандартного стационарного состояния и значения параметров,
* использовавшиеся при моделировании процесса анаэробной переработки илаа
Параметры поступающего в систему ила Условия стационарного состояния Другие параметры и константы
Параметр Значение Параметр Значение Параметр Значение
So 167 мМ (в виде СНзСООН) 10 г/л (в виде СНзСООН) S 2,0 мМ (в виде СНзСООН) 120 мг/л (в виде СНзСООН) V Vg 10 Л 2 Л
Zo F 50 мэкв/л 1,0 л/сут (М 0,0112 мМ (в виде СНзСООН) 0,672 мг/л (в виде СНзСООН) Цшах Рг 0,4 сут-1 760 мм рт. ст.
[СО2] Do 0 мМ [НСОз-] 48 мМ D 25,7 л/моль
хп 0 мМ 24 000 мг/л (в виде СаСО3) I 0.1 моль/л
[HC03-]o 0 мМ [tox]0 0 мМ аИз работы: Graef S. Р., Andrews J. F., X Z [СО2] D /'СО2 U рн Qcyxoft Qch4 СЕР Symp. 5.28 мМ (в виде C5H7NO2) 597 мг/л 50 мэкв/л 9,0 мМ 273 мм рт. ст. 0,1 сут-1 6,91 6,35 л/сут 3,91 л/сут Ser., No. 136, 70, 130 (1974). Ка Ks Kt К> Ух/$ Усо2/Х Тсп4/х 100 сут-1 0,0333 мМ/л 0,667 мМ/л 6,5-10~7 (при 38 °C, 7=0,1) 0,032 моль организмов/ /моль субстрата 28,8 моль СО2/моль образующихся организмов 28,8 моль СН4/моль образующихся организмов
548
Глава 14
костями эксплуатируемых биореакторов для анаэробной пере-
работки ила.
Рассматриваемая модель отражает три типа нарушения ре-
жима, упомянутые в начале этого раздела. Результаты модели-
рования органической и гидравлической перегрузок представле-
ны на рис. 14.22 и 14.23. В соответствующих расчетах принято,
что ступенчатое изменение количества или состава поступаю-
щего в биореактор ила происходит при t=l сут. Если величина
этого изменения достаточно мала, например менее 35,7 г/л для
концентрации субстрата и менее 2,5 л/сут для скорости потока
суспензии ила (при номинальной скорости потока 1 л/сут), то
система вскоре переходит в новое устойчивое стационарное со-
стояние, параметры которого близки параметрам начального
стационарного состояния. Более глубокие вариации состава или
количества поступающего в систему ила вызывают серьезные
нарушения хода процесса, связанные с резким падением значе-
ния pH и скорости образования метана, а также с быстрым
повышением концентрации летучих кислот в продукте процесса.
Как показывают расчеты, резкое повышение концентрации ток-
сичного агента также приводит к нарушению режима процесса,
если концентрация этого агента превышает некоторое пороговое
значение.
Итак, рассматриваемая модель качественно вполне удовлет-
ворительно описывает некоторые стороны динамики процесса
анаэробной переработки ила. Дальнейшее изучение и совершен-
ствование этой модели будет способствовать, с одной стороны,
развитию наших представлений о таком сложном процессе,
а с другой — улучшению методов проектирования соответствую-
щих биореакторов и управления ими. Действительно, в работах
Эндрюса и других исследователей содержится ряд конкретных
предложений, направленных на разработку принципов управле-
ния процессом анаэробной переработки ила и методов обнару-
жения возможных отклонений от режима [5—7]. В приведен-
ном ниже примере рассмотрена одна из наиболее интересных
схем управления, предложенных этими исследователями. Сле-
дует подчеркнуть, что необходимой составной частью разработ-
ки систем управления процессом является математическая мо-
дель, отражающая известные важнейшие детали динамики
реального процесса.
Пример 14.3. Моделирование методов управления процессами анаэробной
переработки ила. Граеф и Эндрюс рассматривали следующие четыре метода
управления процессами анаэробной переработки ила: 1) частичное поглоще-
ние и рециркуляция газа; 2) добавление основания; 3) рециркуляция био-
массы; 4) снижение скорости потока суспензии ила. Поскольку первый метод
управления наименее типичен и поскольку в нем используется часть модели,
описывающая ионное равновесие, то здесь мы сосредоточим внимание именно
на методе управления путем частичного поглощения и рециркуляции газа.
Время, сути
РИС. 14.22. Характер отклика системы для анаэробной переработки ила зависит от масштаба изменения концентрации суб-
страта So в поступающем в систему потоке. Если изменение величины s0 превышает 36,3 г/л, то работа системы иарушает-
вЯ’ if Ра«°™:, 9r™L Р ' Abrews J. F„ Mathematical Modeling and Control of Anaerobic Digestion, in Water-1973,
Bennett G. F., (ed.) CEP Symp. Ser. No. 136, 76, 101 (1974).]
Парциальное давление COj,
мм pm. cm.
РИС. 14.23. Вычисленный с помощью математической модели отклик системы для анаэробной переработки ила на ступен-
чатое изменение гидравлической нагрузки F (Е1 = 1 л/сут). [Из работы: Graef S. Р., Andrews J. F., Mathematical Modeling
«nd Control of Anaerobic Digestion, in Water-1973, Bennett Q. F. (ed.), CEP Symp. Ser. No- 136, 76, 1QI (1974).]
Парциальное давление CQZ>
мм pm. cm.
Практическое применение смешанных популяций
551
(Третий и четвертый методы управления рассмотрены в упражнении 14.9.)
На рис. 14143.1 представлена схема, иллюстрирующая принцип этого ме-
тода управления. Как показано на рисунке, часть газа, образующегося в про-
цессе анаэробной переработки ила, отделяют, пропускают через скруббер для
поглощения СО2 и затем вновь возвращают в биореактор. Значение pH в био-
реакторе определяет, какая доля СО2 будет поглощаться при прохождении
через эту рециркуляционную линию. Если значение pH в биореакторе падает
ниже определенной пороговой величины, то скорость газового потока через
РИС. 14П3.1. Система селективного поглощения и рециркуляции отходящих
газов. [Из работы: Graef S. Р., Andrews J. F., Mathematical Modeling and
Control of Anaerobic Digestion, in Water-1973, Bennett G. F. (ed.), CEP Symp.
Ser. No. 136, 76, 101 (1974).]
скруббер повышают. Удаление CO2 из отходящих газов приводит к снижению
концентрации углекислоты и в жидкой фазе, что вызывает повышение pH.
Такой довольно своеобразный подход к регулированию pH имеет несколь-
ко потенциальных преимуществ перед обычными методами, основанными на
добавлении основания. Добавление концентрированных растворов щелочей
может приводить к образованию (по крайней мере временному) зон с чрез-
вычайно высоким значением pH, в то время как pH всей системы при этом
практически останется на прежнем уровне. Более того, содержащиеся в щело-
чи катионы металлов могут оказаться токсичными по отношению к популя-
циям микроорганизмов в биореакторе. При другом способе подщелачивания —
добавлеиии извести — возникает ряд трудностей, связанных с образованием
нерастворимого карбоната кальция.
После проверки предложенной ими системы управления Граеф и Эндрюс
пришли к выводу, что лучшие результаты достигаются при одновременном
регулировании на нескольких уровнях. В предложенной ими схеме имеются
два частично перекрывающихся диапазона pH, в которых производится вклю-
чение или выключение потока рециркулирующего газа (скорость потока рав-
на Qr):
РИС. 14П3.2. Эффективность управления биореактором для анаэробной переработки ила при перегрузке органическими
отходами путем регулирования селективного поглощения отходящих газов. [Из работы: Graef S. Р., Andrews J. F., Mathe-
matical Modeling and Control of Anaerobic Digestion in Water-1973, Bennett G. F., (ed.) CEP Symp, Ser. No. 136, 76, 101
(1974).]
6 8
Время, сут
Практическое применение смешанных популяций
553
Первая ступень Qru
Qri = 0 рН<6,75
Qri = Qi 6,75<pH<7,00
QRi = 0 pH>7,00
Вторая ступень Qr2:
Qr2=0 pH<6,65
Qr2=Q2 6,65<pH<7,00
Qr2=0 pH>7,00
причем
Qr = Qr1 + Qr2
На рис. 14П3.2 представлены результаты изучения отклика биореактора
для анаэробной переработки ила (с регулированием скорости потока через
скруббер по принципу обратной связи) в зависимости от органической пере-
грузки. Обратите внимание на то, что система с управлением без последствий
переносит резкое повышение концентрации субстрата (питательных веществ)
до 40 г/л; при отсутствии управления такой перегрузки более чем достаточно,
чтобы вывести биореактор из строя. К сожалению, этот способ управления не
может предотвратить нарушение режима работы биореактора при гидравли-
ческой перегрузке или при резком повышении концентрации токсичных ве-
ществ. Преимущества и недостатки этих и других методов управления био-
реакторами для анаэробной переработки ила суммированы в табл. 14П3.1,
Таблица 14П3.1. Методы управления процессами анаэробной
переработки ила
В каждом случае регулирование переменной осуществляли способом
«включено — выключено»3
Измеряемая переменная Эффективность регулирования с целью * предотвращения выхода из строя биореактора в случае Регулируемая перемен- 2 “
ная органической перегрузки токсической перегрузки гидравличе- ской пере- грузки
Величина pH содержимого биореактора Скорость обра- зования ме- тана Скорость потока газа -{- через рециркуляци- онную линию с поглощением СО2 Скорость добавления раствора щелочи Скорость потока Неизвестна жидкости, поступа- ющей в реактор и выходящей из не- го Скорость рециркуля ции ила, отбирае- мого на выходе из реактора Неизвестна + 1 1 + 1
а П° данным, приведенным в работе: Graef S. Р., Andrews /. F., СЕР Svmo Ser No. 136, 70, 101 (1974). у ’
554
Глава 14
Из указанных здесь результатов моделирования следует, что для устойчивой
работы биореактора для анаэробной переработки ила при наличии перегрузок
всех трех типов необходима схема управления с несколькими измеряемыми
переменными, например pH и скоростью образования метана. В соответствии
с результатами этих измерений далее следует регулировать другие параметры
системы (например, скорость рециркулирующего газа и время пребывания;
газа и ила).
14.4.9. Анаэробная денитрификация
В анаэробных условиях многие бактерии, которые могут
утилизировать органические вещества и использовать нитрат и
нитрит в качестве акцепторов электронов, восстанавливают
азотсодержащие соединения. В конечном счете при этом реали-
зуются два независимых механизма восстановления:
1. При ассимилирующем восстановлении нитрата часть азотсо-
держащих соединений превращается в аммиак, который
включается в клеточную биомассу.
2. При диссимилирующем восстановлении конечным продуктом
является молекулярный азот.
Поскольку не все бактерии могут осуществлять оба превра-
щения, можно записать две независимые реакции:
NO3_ + органические вещества —> биомасса + МОг-Ч-СОг
КО2- + органические вещества —>• биомасса+N2+CO2
Наблюдаемые на практике концентрации нитрита очень низки,
поэтому часто достаточно и одного суммарного уравнения, пре-
дусматривающего образование биомассы и iN2.
Для достижения необходимой степени превращения нитрата
можно добавлять к системе органические вещества. Ранее в ка-
честве источника углерода и энергии применяли метанол, обес-
печивающий относительно низкий выход биомассы, но вследст-
вие повышения его стоимости использование метанола стало
невыгодным.
Изучение процесса денитрификации в лабораторных усло-
виях в неподвижном слое показало, что концентрации NO3- и
NO2~ в растворе изменяются в зависимости от расстояния (прой-
денного раствором в неподвижном слое) в соответствии с про-
стой схемой последовательных реакций, в результате которых
обеспечивается эффективная денитрификация в режиме потока
с полным вытеснением. Рост биомассы в конце концов приводит
к засорению реактора; следовательно, для успешного использо-
вания этого нового метода денитрификации необходимы даль-
нейшие проектные и конструкторские разработки.
Практическое применение смешанных популяций
555
14.4.10. Отделение фосфорсодержащих соединений
В необработанных сточных водах фосфор обычно содержится
в концентрации около 10 мг/л в виде ортофосфата, дегидра-
тированного ортофосфата (полифосфата) и органических фос-
форсодержащих веществ. Процессы биологической очистки сточ-
ных вод приводят к превращению большинства фосфорсодержа-
щих соединений в ортофосфат (Н2РО4~, НРО42~, РО43~). Эти
ионы можно отделить осаждением, если соответствующие инст-
рукции требуют снижения содержания фосфора в очищенной
воде. Для осаждения ортофосфата можно применять соли каль-
ция или алюминия:
ЗНРО42- + 5Са2+ + 4ОН- —* Са5(ОН) (РО4)3+ЗН2О
НРО42~ + А13+ —А1РО4+Н+
Если в качестве источника Са2+ применяют известь, то осаж-
дение ортофосфата обычно осуществляют после биологической
очистки. Если же осаждающим агентом являются соли алюми-
ния (или железа), то эта операция может быть совмещена
с операцией образования активного ила или же осуществлена
в первичном отстойнике (рис. 14.8).
Упражнения
14.1. Проектирование процессов нитрификации, а) Какова величина ВПК
очищенной воды, если возраст ила определяли по заданной концентрации
аммиака после очистки, т. е. если 0С = 9 сут (см. пример 14.2)? б) В одной из
форм уравнения Моно учтена скорость утилизации углеродсодержащего суб-
страта: rs—nss/(Km+s). С помощью уравнения (7.25) покажите, что УнаблХ
Х|Ла = |Лт и что У7 [уравнение (7.25)] и Унабл представляют собой одну и ту же
переменную, в) Интерпретируйте поведение системы, если У'=а/(1,О+&0С)
(У'= Унабл) (см. пример 14.2).
14.2. Сравнение промышленных биореакторов и биореакторов для очистки
сточных вод. В табл. 14У2.1 суммированы сравнительные характеристики про-
мышленных микробиологических реакторов и биореакторов для очистки сточ-
ных вод.
а) Обсудите характерные сходства и различия двух типов биореакторов;
рассмотрите такие ситуации, когда определяющие для проектирования реак-
торов критерии будут, по всей вероятности, близкими или различными.
б) Показано, что kia изменяется пропорционально (P/V)n, где п=1,33
(сточные воды), 0,72 (дрожжевой бульон), 0,5 (культуральная жидкость, со-
держащая эндомицеты или мицелиальные организмы), 0,4 (культуральная
жидкость при ферментации углеводородов). Попытайтесь объяснить измене-
ние величины п. Каким образом величина п влияет на решение предыдущей
задачи?
14.3. Бифункциональность в синергизме бактерий. Если два различных
вида осуществляют совершенно различные химические трансформации, то та-
кое явление можно назвать бифункциональностью (по аналогии с бифунк-
циональным катализом, включающим два каталитических эффекта, например
556
Глава 14
Таблица 14У2.1а
Характеристика или параметр
Биореактор для очист- Промышленный био-
ки сточных вод реактор
Температура, °C
Колебания температуры, °C
Реология
10—30
±5
Ньютоновская
Диапазон изменения вязкости, 1—10
сП
Концентрация субстрата, г/л 0,1—5
Органическая нагрузка (БПК), 100—5000
млн-1
Объем реактора, м3 190—1500
Мощность, расходуемая на 0,013—0,13
единицу объема, л. с./м3
Скорость роста, ч-1 0,05—0,1
Нагрузка, г БПК/м3 160—16000
20—50
±0,5
Ньютоновская или
неньютоновская
1—1000
5—40
5000—40 000
0,1—150
0,27—5,3
0,1—1,0
аCooney С. L., Wang D. I. С., Biotech. Bioeng. Symp., 2, 63 (1971).
дегидрирование и изомеризацию). В отношении к микроорганизмам примером
бифункциональности может служить система
S. faecalis
Аргинин ------------>
—СО2 Е. coli
орнитин
—СО2 Е. coll
агматин путресцин
а) Допустив, что каждое из этих трех превращений описывается уравне-
нием Моно, найдите уравнения для всей системы в хемостате.
б) Набросайте график отклика системы (на выходе из хемостата) на
непрерывное повышение скорости разведения при низкой концентрации ар-
гинина.
в) При каком отношении объемов (V1/V2) будет обеспечен максимальный
выход путресцина в каскаде из двух ПРПП (при постоянном общем объеме),
если цтах для Е. coli имеет одно и то же значение для обеих стадий, превы-
шающее gmax для S. faecalis. Повторите решение задачи при p,max(£. coli)<
<Hmax(S. faecalis).
г) При каких условиях концентрация путресцина будет минимальной
(если исходные питательные вещества нестерильны)? Какие условия, найден-
ные в задаче «в» или «г», будут наиболее благоприятными для второго вида
микроорганизмов?
14.4. Стабильность взаимодействий в активном иле. Рассчитайте результат
парных межвидовых взаимодействий в активном иле типа а, б, в и г (см.
рис. 14.16) методом Фейнберга — Хорна — Джексона. Согласуются ли эти ре-
зультаты с результатами расчетов с помощью уравнений Курдса?
14.5. Биореактор с активным илом. Поступающий в биореактор с актив-
ным илом поток сточных вод имеет БПКз 220 мг/л, а на выходе из реактора
этот параметр не должен превышать 15 мг/л (табл. 14.2). Завершите описа-
ние реактора с активным илом при следующих дополнительных условиях: кон-
центрация активных твердых компонентов 3000 мг/л; отношение рециркуля-
Практическое применение смешанных популяций
557
ции 0,46; F= 17,03-106 л/сут; У, gmax, Ks, kd — нз табл. 14.4. За исключением
БПК все питательные вещества имеются в избытке.
а) Вычислите: 1) возраст ила; 2) необходимый объем биореактора;
3) концентрацию активных твердых компонентов в линии рециркуляции;
4) скорость аэрации, если степень утилизации кислорода равна 7,5%. Согласно
уравнению (14.5), минимальная величина V достигается при повышении хг/ха.
Очевидно, что повышение хг возможно только при увеличении объема второго
отстойника Vs. Хотя осаждение ила представляет собой сложный многоста-
дийный процесс (см., например, работу [16] и гл. 11), концентрацию активных
твердых компонентов в линии рециркуляции можно приближенно описать
уравнением
Xr=xr, тах(1 — е-₽')+^е-₽' хг, тж=хг при /=оо
где t ((e=Vs/F)—среднее время оседания ила, a [J— характерная констан-
та (Р>0).
б) Оцените величину хг (оптимальное), если все эксплуатационные расхо-
ды (отнесенные к единице объема биореактора и отстойника) распределяются
между биореактором и отстойником в отношении у (1,0<'{<10,0) (расходами,
на строительно-монтажные работы можно пренебречь).
14.6. Структурирование субстрата и биомассы при проектировании про-
цессов с участием активного ила. Уравнения (14.1) — (14.4) представляют со-
бой модель процесса с участием активного ила, в которой все субстраты и вся
биомасса описываются одним уравнением Моно с учетом эндогенного метабо-
лизма. Предлагалась и другая модель [Paterson R. В., Denn М. М., Computer-
Aided Design and Control of an Activated Sludge Process; The Chem. Eng. J.,
27, В 13 (1983)], в которой субстрат подразделен на углеродсодержащий,,
содержащий органический азот и нитрит компоненты, трансформируемые со-
ответствующими компонентами биомассы хс, хо и xn. Основные рабочие
параметры системы: F=2,0-104 м3/сут, а=0,15, [1=0,0015, $са=16,8 мг/л
(БПК5) , $со=1811 мг/л, $оо=250 мг/л, sno=0,5 мг/л. Параметры уравнений
Моно (с учетом эндогенного метаболизма): 1Хтах=5,0, 0,33, 0,80 сут-1;
£е=0,055, 0,05, 0,05 сут-1; А=100, 1,0, 2,1 мг/л; У = 0,5, 0,05, 002 г/г для ХсТ
хо и xn соответственно.
а) Вычислите возраст ила 0S для углеродсодержащей биомассы Хс-
б) Пусть вторичный отстойник одинаково эффективен по отношению ко
всем компонентам биомассы и, следовательно, 0S имеет одно и то же значение
для всех превращений. Вычислите концентрации органического азота ($оа)
и нитритного азота ($Na) в очищенной воде.
в) На базе данных для углеродсодержащей биомассы вычислите Vx и V.
Если аналогичные расчеты осуществить на базе данных для других компонен-
тов биомассы, будет ли получено такое же значение V? Почему? Для расчетов
необходимо знать характеристики вторичного отстойника. Начертите график
зависимости V (вычислено) от (хг/ха) в диапазоне 100< (хг/ха) < 1000.
14.7. Возможность управления системой обработки сточных вод методом
активного ила. Возможность управления системой очистки сточных вод зави-
сит от целей управления, динамики управления и системы, а также от взаимо-
связей между различными целями управления, которые в общем случае могут
быть несовместимыми. Так, целью управления системой может быть, во-пер-
вых, минимальное отклонение общей концентрации биомассы в жидкой и твер-
дой фазах [уравнение (14.8)] в реакторе и, во-вторых, поддержание относи-
тельно постоянной высоты «подушки» ила во вторичном отстойнике
(рис. 14.10).
а) Ознакомьтесь с моделью Патерсона и Денна (см. предыдущее упраж-
нение), сформулируйте испытанные авторами методы управления и выводы
авторов. Почему указанные две цели управления являются несовместимыми
в данном случае?
б) Первой целью управления является поддержание примерно постоянно-
'558
Глава 14
го значения общей концентрации биомассы в жидкой и твердой фазах; тем
не менее экономический анализ, приведенный в той же работе и основанный
на изучении работы системы в стационарном состоянии, показал, что колеба-
ния общей концентрации биомассы в очень широком диапазоне не сказывают-
ся на экономической стороне процесса. Какой важный для себя вывод может
извлечь инженер из этих результатов?
14.8. Анаэробный биореактор с двумя резервуарами. Поланд и Гхош [13]
обсуждали взаимодействие между кислотообразующими и метанобразующи-
ми бактериями в анаэробных биореакторах; теоретически эта проблема близ-
ка проблеме, рассмотренной в упражнении 13.5. Авторы предложили следую-
щие стехиометрические уравнения:
Образование кислот:
4C3H7O2NS + 8H2O 4CH3COOH-|-4CO2+4NH3+4H2S + 8H+-|-8e-
Образование метана:
8Н+ + 8е- + ЗСН3СООН + СО2 —>- 4СН4+ЗСО2+2Н2О
Поланд и Гхош предположили, что управление процессом образования метана
(путем регулирования параметров окружения) облегчается при использовании
биореактора, состоящего из двух последовательно соединенных резервуаров.
Параметры уравнений роста микроорганизмов на глюкозе (2 г/л) в первом
резервуаре для кислотообразующих бактерий: gmax(глюкоза) = 1,25 ч-1, Kg==
= 22,5 мг глюкозы/л, Yg=0,2; для метанобразующих бактерий: р.тах(кисло-
та)=0,14 ч-1, Кя = 600 мг уксусной кислоты/л, Уа = 0,05. Другие параметры си-
стемы; экономический коэффициент превращения глюкозы в уксусную кисло-
ту 0,8, константа ингибирования кислотообразующих бактерий 100 мг уксус-
ной кислоты/л; эффектами эндогенного метаболизма можно пренебречь.
а) Запишите уравнения, описывающие поведение системы в двух резер-
вуарах в стационарном состоянии, если кинетика роста двух видов описывает-
ся уравнениями, приведенными в упражнении 13.56.
б) Покажите, что метанобразующие бактерии вымываются, если скорость
разведения в первом резервуаре превышает 0,14 ч-1. Какой вывод можно сде-
лать относительно целесообразности использования реактора с двумя резер-
вуарами для организмов, не связанных (большей частью) пищевой цепью?
в) Предположим, что скорость потока превышает 0,14 ч_|. Определите
состав жидкости на выходе из первого и второго резервуаров, если V2=Vi
или если V2=5Vi; начертите график зависимости концентрации непревращен-
ного субстрата от D. Какие недостатки и преимущества имеет разделение ви-
.дов путем повышения Д?
г) В реальных процессах очистки сточных вод осевшая биомасса и непе-
реработанные твердые отходы будут циркулировать между выходом из каж-
дого резервуара реактора и отстойником. Запишите уравнения материальных
балансов по субстрату и биомассе для системы с двумя резервуарами.
14.9. Управление анаэробным биореактором для переработки ила. Найди-
те уравнения, описывающие динамику системы в биореакторе для анаэробной
переработки ила, если биореактор управляется путем а) рециркуляции био-
массы; б) изменения (повышения) скорости потока. Включите сюда уравнение,
описывающее изменение соответствующей регулируемой переменной; объясни-
те форму этого уравнения.
14.10. Кинетика межвидового взаимодействия типа хищник — жертва.
Обобщенная модель системы хищник — жертва, предложенная Розенвейгом
и Макартуром [17], сводится к уравнениям
dx du
— =Кх)—уФ(х) — = —еу + куф(х)
dt dt
где х и у — плотности популяций хищника и жертвы соответственно.
П ра кт и ч еское применение смешанных популяций
559
а) Сравните эти уравнения с уравнениями модели Лотки — Вольтерры
[уравнения (13.9) и (13.10)] и перечислите максимально возможное число фи-
зических и биологических ситуаций, когда Ф (х) и f(x) отличаются от соот-
ветствующих зависимостей, предсказываемых моделью Лотки — Вольтерры.
Опишите эти ситуации возможно точнее. Каков физический смысл Ф (х) и f(x)r
б) Полученные в 1973 г. Лакинбиллом результаты экспериментального-
изучения системы хищник — жертва (Paramecium и Didinium) описаны Сми-
том (с. 33 в работе [34], гл. 13) (ни в одном случае найденная плотность по-
пуляции жертвы не приближалась к плотности, достигаемой в чистой куль-
туре) :
1. «...сначала наблюдалось быстрое повышение численности жертв, позднее
начался рост популяции хищников, которые уничтожили всех жертв и за-
тем вынуждены были голодать ...»
2. «Продолжительность периода сосуществования двух видов можно увели-
чить, добавив к среде метилцеллюлозу; в результате среда становится более-
вязкой, и скорость передвижения организмов первого и второго видов за-
медляется. Тем не менее и в этом случае наблюдались колебания числен-
ности с повышенной амплитудой, приводящие в конце концов к гибели по-
пуляции хищника».
3. «Устойчивое сосуществование двух видов было достигнуто путем добавле-
ния метилцеллюлозы и одновременного снижения в два раза концентрации:
питательных веществ для популяции жертв».
Объясните эти экспериментальные наблюдения. Какие дополнительные экспе-
рименты вы могли бы предложить для подтверждения или опровержения лю-
бого из принятых вами допущений?
14.11. Нитрификация в почве. Макларен [14] предположил, что в первом
приближении нитрификацию под воздействием почвенных микроорганизмов,
можно представить в виде последовательных реакций
kl k%
NH4+ —> no2- no3-
B лабораторных экспериментах с обогащенной почвой первую и вторую реак-
ции можно отнести к видам Nitrosomonas и Nitrobacter соответственно.
а) Предположим, что в стационарном состоянии эти питательные вещест-
ва утилизируются для поддержания клеток в реакциях нулевого порядка по-
NH4+ и NO2~ с указанными константами скоростей реакций /г, и /г2, которые
пропорциональны локальным концентрациям микроорганизмов. Пусть соответ-
ствующие концентрации биомассы Xi и х2 не зависят от глубины г; опреде-
лите вертикальные профили концентраций NH4+, NO2~ и NO3~ (нормализо-
ванные по отношению к концентрации NH4+ при z=0), если скорость пере-
носа жидкости в глубину почвы равна и и если ионным обменом между
компонентами почвы и питательными веществами можно пренебречь. Вырази-
те результаты в графической форме для £1//г2=0,1, 1,0 и 10,0, используя без-
размерный параметр расстояния zk\/u. Рассмотрите две ситуации: af когда
можно пренебречь утилизацией азота для образования новой биомассы;
б) когда каждый вид для поддержания жизнедеятельности использует свою
реакцию утилизации NH4+ нулевого порядка с константами скорости [3xi
и 13x2 соответственно.
б) Предположим, что ион NH4+ действительно является лимитирующим
питательным веществом для популяции Nitrosomonas' с помощью решения
предыдущей задачи найдите безразмерную глубину, на которой допущение-
Xi=const не выполняется ни при каких условиях. В одном случае эксперимен-
тально найдено, что профиль концентрации организмов, окисляющих NH4+
изменяется следующим образом: 2-Ю5 на 1 см3 (поверхностные водык
2-Ю5 на 1 г почвы (на глубине от 0 до 1 см), 2-103 на 1 г почвы (на глубине
•560
Глава 14
от 1 до 3 см), 2-102 на 1 г почвы (на глубине от 3 до 5 см) (см. работу [1]
и приведенную в ней литературу).
в) Понятно, что допущение о постоянной плотности биомассы является
упрощением. Покажите, что указанная модель не позволяет определить зави-
симость X] И Х-2 от глубины.
г) Обсудите, как можно разработать модель, четко описывающую профи-
ли изменения плотности популяций Nitrosomonas и Nitrobacter, если учесть
материал предыдущей главы и следующую цитату из работы 1923 г.:
«Показано, что в почве обычно обитают в высшей степени смешанные
популяции организмов размером от мельчайших бактерий, нематод и дру-
гих организмов, которые едва можно различить невооруженным глазом,
до гораздо более крупных организмов и, наконец, земляных червей, кото-
рых совсем нетрудно рассмотреть и потрогать руками. Все они живут в
почве и поэтому должны находить условия, необходимые для их
роста [15]».
14.12. Аэробные реакторы в трубопроводах. «Во многих городах состав-
ной частью систем водоочистки являются трубопроводы, работающие как при
повышенном, так и при атмосферном давлении. Предлагалось использовать
эти трубопроводы в качестве аэробных биореакторов с тем, чтобы снизить
биохимическую потребность в кислороде (ВПК) на соответствующих водо-
очистных станциях». \Koch С. М., Zandt I., Use of Pipelines as Aerobic Biologi-
cal Reactors; J. Water Pollut. Control Fed., 45, 2537 (1973).]
а) Рассматривая только движущуюся жидкую фазу, Пауэлл и Лоу пред-
ложили следующее выражение для трубчатого реактора с полным вытесне-
нием;
, 1 + у , о — у
rmax у cr0 — у
In
О
где Со — безразмерная концентрация субстрата при t = Q (на входе), о — без-
размерная концентрация субстрата в момент времени t (на выходе), у=Оо+
4~х0, Хо—безразмерная концентрация биомассы на входе, цтах — максималь-
ная удельная скорость клеточного роста, t — время пребывания. Найдите при-
нятое для клеточного роста уравнение и выразите каждую переменную этого
уравнения через безразмерные переменные.
б) Кох и Занди предположили, что аэробный биореактор в трубопроводах
можно описать как двухфазную проточную систему с начальными объемными
скоростями потоков воздушной и жидкой (суспензии) фаз Qa и Qi соответст-
венно. Найдите два уравнения, описывающие зависимость концентрации кис-
лорода от расстояния, если допустить, что, во-первых, газовая и жидкая фазы
движутся в режиме полного вытеснения с одинаковыми скоростями и, во-вто-
рых, что с/(общее давление)/dz=7 = const, где z—расстояние в трубопроводе.
[Допустите также, что скорость поглощения кислорода суспензией линейно
зависит от плотности биомассы, т. е. что концентрация растворенного кислоро-
да со 2 всегда больше ссг (табл. 8.2).] Четко укажите другие допущения.
в) Интегрированием этих уравнений найдите длину реактора в трубопро-
водах, при которой с0=Ссг, т. е. ту точку, в которой газовую фазу нужно
•обновлять для поддержания активности аэробных организмов. Примите сле-
дующие значения параметров: Qa = 0,142 м3/с, Q/=14,2 м3/с, х0=Ю мг/л, внут-
ренний диаметр трубопровода 305 мм, начальная Со2 = 8 мг/л, ссг = 0,5 мг/л,
$о=начальная БПК=150 мг О2/л, 7=300 К, Ks = 100 мг О2/л, цтах = 0,3 ч_|,
Ys = 0,4 г биомассы/г БПК, скорость дыхания 0,375 г О2/(ч-г биомассы), kta=
= 0,4 мин-1, /7 = 4-104 атм/мольная доля, 7 = 0,005 м вод. ст./м трубопровода.
14.13. Флуктуации скорости потока сточных вод. На водоочистных станци-
ях скорость поступающего потока может изменяться с периодом от нескольких
часов до нескольких суток. С помощью уравнения Моно для одного лимити-
рующего субстрата и допущения, что S7 = const, £> = £>0 (1-]-ct sin со/), а>0:
Практическое применение смешанных
популяций
561
а) Покажите, что если х и соответствующие производные изменяются во
времени периодически, то
2л/<о
С ( ^axS(^) _ D{t,} 1 = 0
J I +s (t') )
о
Решение x(f)=0 при любом t’ отвечает состоянию вымывания. Укажите, как
должно измениться выражение, стоящее в фигурных скобках, в тех случаях,
когда существует нетривиальное решение, т. е. х(/')>0 при 0<У<2л/<а.
б) Если и а достаточно мало, то вблизи области вымывания s(t')
всегда близко S;. Покажите, что в этих условиях нетривиальное решение воз-
можно при £>о=Цтах и что изменение х в зависимости от времени в интервале
0^/<2л/<» описывается уравнением
' Ра
а
х = х(0) ехр
(1 — cos at)
в) Укажите, какие уравнения нужно решить (с помощью ЭВМ) для того,
чтобы найти x(t) для нетривиального случая, если
14.14. Биодеградация детергентов. Полное разложение устойчивых суб-
стратов, какими являются, например, некоторые поверхностно-активные веще-
ства, наблюдается иногда при ограниченной концентрации субстрата, т. е.
когда скорость клеточного роста линейно зависит от концентрации субстрата,
а скорость гибели клеток составляет заметную величину. Уэйман [Way-
тап С. Н., Prog. Ind. Microbiol., 10, 219 (1971)] предложил уравнение, опи-
сывающее зависимость между концентрациями биомассы х и субстрата-детер-
гента s в таких условиях:
dx , „ ,, ds
— + /?х = |imxs = — V —
at at
где R— скорость дыхания в отсутствие роста, т. е. s (в отсутствие роста) =
а) С помощью второго равенства найдите точное решение уравнения $=
= f(x).
б) Приближенно можно считать, что ] xdt^xt'/2^ если t' — время, при
о
котором х достигает максимального значения,- Покажите, что при этом усло-
вии плотность биомассы х описывается уравнением Бернулли:
dx
—— + ф хх = ф 2хп
dt
в) Путем замены переменной ф=1/х найдите решение для х, справед-
ливое в течение периода времени, непосредственно следующего после t'. [При
большом t решение становится менее точным, поскольку ошибка приближения
возрастает; это ограничение, однако, не представляется слишком существен-
ным, так как наибольший интерес представляет время (или место) вблизи
точки полного истощения субстрата.]
г) Начертите примерный график изменения s для Г«1, если 01/5>2=О,1,
1,0 или 10,0. Насколько важна кинетика процесса поддержания жизнедеятель-
ности, если речь идет о медленно метаболизирующихся субстратах?
14.15. Ультрафиолетовые стерилизаторы. Если на домашние нужды рас-
ходуется много воды, то степень контаминации воды микроорганизмами мож-
но снизить с помощью нескольких УФ-ламп, расположенных в виде шести-
угольника параллельно стеклянному участку водопроводной трубы, Предпо-
36—746
562
Глава 14
ложим, что микроорганизмы погибают в процессе первого порядка, причем
«константа» скорости этого процесса пропорциональна локальной интенсив-
ности ультрафиолетового излучения /. Допустим, что ультрафиолетовое излу-
чение радиально-симметрично и что его интенсивность изменяется радиально
в соответствии с законом Бера /(г) =I(R)e~a<-R~r\ где R— внутренний диа-
метр стеклянной трубы.
а) Для ламинарного потока «=«шах[1 — (r/R)1 2 3 4 5]. Определите долю остав-
шихся жизнеспособными организмов, усредненную по сечению трубы на выхо-
де, если длина трубы равна L. Какая средняя величина представляет наиболь-
ший интерес?
б) Повторите решение задачи 14.15а для случая турбулентного потока
в той же трубе (допустите, что
в) Согласно предъявляемым к питьевой воде требованиям, концентрация
живых организмов не должна превышать с организмов в литре. Каков верх-
ний предел концентрации организмов в поступающей в стерилизатор воде
в случае ламинарного и турбулентного потоков?
г) Определите Re для трубы диаметром 12,7 мм при обычной для вашего'
дома объемной скорости.
Литература
Довольно подробные данные о смешанных популяциях, а также некоторые
дополнительные сведения о естественных кругооборотах веществ приведены
в работах [1, 3], гл. 1. В этом отношении полезна также большая часть лите-
ратуры, перечисленной в гл. 13. Другая литература по применению смешанных
культур микроорганизмов;
1. Hattori Т., Microbial Life in the Soil: An Introduction, Marcel Dekker, Inc.,
New York, 1973. Расширенный вводный курс в микробиологию почвы, в ко-
тором везде, где это только возможно, дается количественная трактовка
явлений. Помимо глав, посвященных почвенным микроорганизмам (в том
числе их физиологии, межвидовым взаимодействиям и роли в росте расте-
ний и геохимических процессах), в книге приведены также довольно
подробные данные о составе почвы.
2. Mitchell R., Introduction to Environmental Microbiology, Prentice-Hall, Inc.,
Englewood Cliffs, N.J., 1974. Подробный обзор, посвященный изучению
жизни микроорганизмов в биосфере. Среди многих других тем здесь рас-
смотрены циклы питательных веществ, эвтрофикация, экология сообществ
и проблема переработки отходов.
3. Rich L. G.t Environmental Systems Engineering, McGraw-Hill Book Company,
New York, 1973. В этой монографии, посвященной проблемам охраны окру-
жающей среды, подчеркивается важность применения современных мето-
дов моделирования и системного анализа для изучения различных про-
цессов, в том числе процессов переработки отходов.
4. Abson J. W., Todhunter К.. Н., Effluent Disposal, in Biochemical and Biolo-
gical Engineering Science, Blakebrough N. (ed.), vol. 1, chap. 9, Academic
Press, London, 1967. Хороший краткий обзор методов очистки промышлен-
ных сточных вод. Материал гл. 9 хорошо дополняет гл. 10, написанная
Уиллсом и посвященная седиментации и флокуляции.
5. Andrews J. F., Review Paper: Dynamic Models and Control Strategies for
Wastewater Treatment Processes, Water Res., 8, 261—289 (1974). Велико-
лепный обзор проблем, связанных с описанием и регулированием динами-
ки водоочистных систем, а также подходов к решению этих проблем.
Практическое применение смешанных популяций
563
Детальная информация по специальным и частным вопросам дана в работах:
6. Busby I. В., Andrews J. F.. A Dynamic Model and Control Strategies for
the Activated Sludge Process, J. Water Pollut. Control Fed., 47, 1055 (1975).
7. Graef S. P., Andrews J. W., Mathematical Modeling and Control of Anaero-
bic Digestion, CEP Symp, Ser. [1361 70, 101—127 (1974).
3. Curds C. R., A Theoretical Study of Factors Influencing the Microbial Popu-
lation Dynamics of the Activated-Sludge Process, I: The Effects of Diurnal
Variations of Sewage and Carnivorous Ciliated Protozoa, Water Res., 7,
1269—1284 (1973).
9. Chiu S. Erickson L. E., Fan L. T., Kao I. C., Kinetic Model Identification
in Mixed Populations Using Continuous Culture Data, Biotech. Bioeng., 14,
207—231 (1972).
10. Schroeder E. R., Water and Waste Water Treatment, McGraw-Hill, New
York, 1977.
Упражнения:
11. Calam С. T„ Russell E. W., Microbial Aspects of Fermentation Process Deve-
lopment, J. Appl. Chem. Biotech., 23, 225 (1973).
12. Blanch H. W., Dunn I. J., Modelling and Simulation in Biochemical Engi-
neering, Adv. Biochem. Eng., 3, 159—162 (1974).
13. Pohland F. G., Ghosh S., Developments in Anaerobic Treatment Processes,
in Biological Waste Treatment, Biotech. Bioeng. Symp., 2, 85 (1971).
14. McLaren A. D„ Soil Sci. Soc. Am. Proc.. 33, 55 (1969).
15. Russel E. J., Microorganisms of the Soil, Longmans Greeen and Co., Lon-
don, 1923.
16. Canale R. P., Borchardt J. A., in Weber W. J., Jr. (ed.), Physicochemical
Processes for Water Quality Control, pn. 120—121, Wiley-Interscience New
York, 1972.
3 7. Rozenweig M. L., MacArthur R. H., Graphical Representation and Stability
Conditions of Predator-Prey Interactions. Am. Nat., 97, 209 (1963); 103,
81 (1969).
36*
Предметный указатель
Авицель 1, 220
в очистке сточных вод 2, 513 и сл.
Агароза как носитель для иммобили
зованных ферментов 1, 240
Агентство по охране окружающей
среды США, функции 2, 350 и сл.
Аденилатдеаминаза, применение
211
Аденин 1, 65 и сл.
Аденозин 1, 66 и сл.
Аденозин-5'-дифосфат 1, 66 и сл.
Аденозин-5'-монофосфат 1, 66 и сл.
Аденозинмонофосфатдеаминаза иммо«
билизованная, применение 1, 258
Аденозинтрифосфат 1, 66 и сл., 300,
307 и сл.
определение 2, 179
— в клетках 2, 197
Аденозинтрифосфатаза 1, 123
инактивация 1, 186
кинетика реакций 1, 145
Аденокарцинома печени человека,
рост в культуре 2, 139 и сл.
Адипиновая кислота, производство 2,
388, 391
Азот в микробиологических процессах
2, 122
Акриловая кислота, производство 2,
388, 391
Активированный уголь как носитель
для иммобилизованных ферментов
1, 240
Активный ил 1, 382; 2, 513 и сл.
анаэробная переработка 2, 538 и сл.
математические модели роста 2, 519
и сл.
обезвоживание 2, 269
осаждение 2, 268
1, Активный транспорт 1, 344 и сл.
Актиномицеты 1, 35 и сл.
Р* Аланин 1, 82
биосинтез 1, 333
Алиэстеразы 1, 230
Алкалоиды Claviceps purpurea, моде-
ли биосинтеза 1, 552 и сл.
Алкогольдегидрогеназа 1, 80, 98
выделение 2, 278, 283 и сл.
кинетика реакций 1, 152
Аллостерическая регуляция 1, 165
и сл.
Аллостерические ферменты см. Фер-
менты
Альбумин 1, 98
денатурация 1, 572
кинетика реакций 1, 152
определение 2, 179
Альгинаты, применение для иммоби-
лизации клеток 2, 90 и сл., 96
Альдолаза 1, 98
Амебы 1, 36
Аменсализм 2, 423 и сл.
а-Амилаза 1, 123; 2, 392
иммобилизованная 1, 258
получение с помощью иммобилизо-
ванных В. subtilis 2, 101
стабилизация 1, 195
-----математические модели 2, 542
и сл.
аэробная переработка 2, 529 и сл.
возраст 2, 520
[3-Амилаза 1, 215 и сл.; 2, 392
Амилазы 1, 215 и сл.
выделение 2, 308
грибковые, производство 2, 371
Предметный указатель
565
из Bacillus, производство 2, 371
коэффициенты отражения 2, 310
применение 1, 208
Амилоглюкозидаза 1, 216 и сл.
кинетика реакций 1, 158
применение 1, 208
Амилоза 1, 57 и сл.
Амилопектин 1, 58
Аминоацилаза 1, 237
иммобилизованная 1, 254 и сл., 258
Аминокислотная последовательность
белков, определение 1, 87 и сл.
Аминокислотный остаток 1, 84
Аминокислотный состав Е. coll 1, 86
Аминокислоты 1, 82 и сл.
разделение с помощью иммобилизо-
ванных ферментов 2, 156
экономика производства 2, 403 и сл.
Аммиак, определение с помощью мик-
робиологических детекторов 2, 180
Ампициллин 2, 377
Амплификация плазмид см. Плазмиды
Аналого-цифровые преобразователи 2,
209 и сл.
Анаэробная переработка отходов 2,
510
модели управления 2, 548 и сл.
Анаэробное дыхание 1, 320
Анаэробные процессы 1, 27
Анаэробный метаболизм, классифика-
ция типов см. Метаболизм
Анаэробы 1, 298
Андростадиен-1,4-дион-3,17, получение
2, 384, 385
Анионообменные смолы как носители
для иммобилизованных ферментов
1, 240
Антибиотики 1, 359, 558 и сл.
выделение 2, 325 и сл., 339
— в каскаде адсорберов 2, 315
— общая схема 2, 258
— центрифугированием 2, 267
классификация 2, 375 и сл.
производство 2, 374 и сл.
Антиген к вирусу гепатита, выделение
2, 341
Антигены 2, 382
Антикодон 1, 400
Антиметаболиты 1, 437 и сл.
Антитела 1, 80, 104 и сл.; 2, 328
моноклональные 1, 432
Апоферменты 1, 122
Аргиназа 1, 123
D-Арилглицины, получение с помощью
иммобилизованного субтилизина 2,
342
Арифметический и логический блок
ЭВМ 2, 204
Асептические условия 2, 123 и сл.
Аскорбиновая кислота, определение с
помощью ферментных терморези-
сторов 2, 179
Аскоспоры 1, 30 и сл.
Аспарагиназа 1, 195
применение 1, 234 и сл.
Аспартаза
выделение 2, 278
иммобилизованная 1, 258
инактивация 1, 134; 2, 94
применение 1, 237
Аспартам 2, 403
Аспартат, биосинтез 1, 333
Аспартаттранскарбамилаза 1, 98, 165
и сл.
Na+—К+-АТРаза 1, 345
Ауксотрофные мутанты см. Мутанты
Аутотрофы 1, 301
Аффинная хроматография см. Хрома-
тография
Аффинные сенсоры 2, 180 и сл.
Ацетаткиназа 2, 196
Ацетат, концентрация в культуре
С. acetobutylicum 2, 196
N-Ацетилацетамид, влияние на синтез
амидазы 1, 434
N-Ацетилглюкозамин 1, 101 и сл.
N-Ацетилмурамовая кислота 1, 101
и сл.
Ацетилхолинэстераза, кинетика реак-
ций 1, 152
Ацетоацетатдекарбоксилаза 2, 194,
196
566
Предметный указатель
Ацетоацетил-СоА: СоА-трансфераза
2, 196
Ацетон
концентрация в культуре С. aceto-
butylicum 2, 196
производство 2, 388, 390
— с помощью иммобилизованных
бактерий 2, 97 и сл.
Аэробное дыхание 1, 320
Аэробные процессы 1, 27
Аэробы
облигатные 1, 300
факультативные 1, 300
Аэротенк 2, 513 и сл.
Бактериальные инфекции, диагности-
ка с помощью моноклональных ан-
тител 2, 386
Бактерии 1, 26 и сл.
Бактериофаги 1, 76, 423 и сл., 448
Барботажная колонна 2, 107 и сл.
определение газосодержания 1, 623,
679
— коэффициента массопередачи 1,
632
Бацитрацин
выделение экстракцией 2, 272
получение с помощью иммобилизо-
ванных Bacillus sp. 2, 101
Белки 1, 78 и сл., 85
биосинтез 1, 338 и сл.
высаливание 2, 280, 281
глобулярные 1, 78 и сл.
посттрансляционная модификация
1, 408
простые 1, 84
растворимость 2, 282
структура вторичная 1, 87
— первичная 1, 87
— спиральная 1, 90
— третичная 1, 87 и сл.
— четвертичная 1, 87 и сл., 97 и сл.
^-структура 1, 90 и сл.
Белок-активатор катаболизма 1, 80
Белок одноклеточных организмов
выделение 2, 267, 330 и сл.
расчет процесса 2, 103 и сл.
экономика производства 2, 407 и сл.
Бензоиларгинина этиловый эфир, гид-
ролиз иммобилизованным папаином
2, 150
Бензойная кислота как предшествен-
ник новобиоцина 2, 122
Бимолекулярный слой 1, 49
Биогеохимические циклы элементов
2, 497 и сл.
Биогомополимеры 1, 44
Биодетекторы 2, 177 и сл.
Биологические пруды 2, 537
окислительные 2, 537
стабилизирующие 2, 538
Биомасса
влияние скорости перемешивания на
скорость образования 2, 62
определение концентрации 2, 337
Биополимеры 1, 44 и сл.
Биопроцессы
динамические модели для ПРПП 2,
20 и сл.
обобщенная технологическая схема
2, 353
экономический анализ 2, 356 и сл.
Биореакторы 2, 5 и сл.
дисперсионная модель 2, 53
для иммобилизованных клеток 2, 86
и сл.
— культур растительных и живот-
ных клеток 2, 131 и сл.
идеальные 2, 8 и сл.
— проточный с полным перемеши-
ванием (ПРПП) 1, 492 и сл.; 2,
10 и сл.
— периодического действия 1, 491
и сл.; 2, 8
классификация 2, 128 и сл.
колонного типа 2, 107 и сл.
комбинированные модели 2, 45 и сл.
неидеальные 2, 45 и сл.
периодического действия 2, 8 и сл.
-----с добавлением субстрата 2, 9
и сл.
Предметный указатель
567
полного вытеснения 2, 15 и сл.
смешанные модели 2, 45 и сл.
с многофазными системами 2, 102
и сл.
— неподвижным слоем катализато-
ра 2, 105 н сл.
— неполным перемешиванием 2, 29
и сл.
— псевдоожиженным слоем 2, 111
и сл.
— струйным течением жидкости 2,
116 и сл.
тепловой баланс 1, 666 и сл.
трубчатые, элементарный объем 2,
16
эрлифтные 2, 107, 109, 128
Биосенсоры 2, 177 и сл.
Биосинтез 1, 330 и сл.
Биосинтетическая активность Е. coll
1, 349
Биотин 1, 51
Биотрансформации 2, 381 и сл.
Бисдиазобензидин как бифункцио-
нальный реагент 1, 244
Бислой 1, 49
Бифункциональные реагенты 1, 195
Бифуркация Хопфа 2, 470
Брожение
ацетоно-бутиловое 1, 356
бутандиольное 1, 356
кислотное 1, 356
молочио-кислое 1, 356
пропионовокислое 1, 356
спиртовое 1, 355
Бромелаин 1, 124
применение 1, 209, 229
Бутанолдегидрогеназа 2, 196
Бутанол, производство 2, 97 и сл., 196,
388, 391, 399
Бутиральдегиддегидрогеназа 2, 196
Бутират 2, 196
Бутираткиназа 2, 196
Вакуоли 1, 21
Варидаза, применение 1, 209
Ваттметр, применение для измерения
мощности мешалки 2, 171
Вегетативная форма клеток 1, 27
Векторная алгебра в расчетах биоре-
акторов 2, 21 и сл.
Векторы 1, 440 и сл., 446 и сл.
Вероятность выживания организмов
при стерилизации см. Стерилизация
Вибромиксер 2, 139
Вид организма 1, 25 и сл.
Винный уксус 1, 359; 2, 117
Винодельческий процесс 2, 393 и сл.
Вирусы 1, 423 и сл.
гриппа 1, 424
карликовости томата 1, 424
полиомиелита 1, 424
табачной мозаики 1, 424
тутового шелкопряда 1, 425
Вискозный гель как материал для
мембран 2, 304
Витамины 1, 51 и сл.
незаменимые 1, 51
производство 2, 379 и сл.
Водород, повышение выхода в биофо-
толизе 2, 245
Водоросли 1, 36 и сл.
Восстанавливающие эквиваленты 1,
311
Время
выравнивания концентраций 2, 31
и сл., 60
децимальной редукции 1, 573
истощения питательного вещества
2, 58
пребывания в реакторе, среднее 2,
37
процесса оптимальное, определение
2, 241 и сл.
релаксации 1, 531
циркуляции в реакторе 2, 31
Выживаемость популяции 1, 480
Вымывание 1, 501; 2, 43 и сл.
Высаливание белков см. Белки
Высокоэнергетические соединения 1,
307 и сл.
Вязкость среды, определение в реаль-
ном масштабе времени 2, 172
568
Предметный указатель
Газлифтная колонна, расчет газосо-
держания 1, 624
Газовая хроматография, применение
для анализа отходящих газов см.
Хроматография
Газовый анализ, применение в био-
процессах 2, 181 и сл.
Газосодержание 1, 623
Галактоза 1, 54
определение с помощью фермент-
ных терморезисторов 2, 179
а-Галактозидаза, моделирование про-
цесса производства 2, 65 и сл.
|3-Галактозидаза 1, 98
биосинтез, управление процессом 2,
230 и сл.
регуляция биосинтеза 1, 411 и сл.
стабилизация 1, 195
Галактозооксидаза 1, 123
применение 1, 237
Гаплоидные клетки Г, 30, 417
Гексаметилендиизоцианат как би-
функциональный реагент 1, 244
Гексозомонофосфатный путь метабо-
лизма см. Пути метаболизма
Гексокиназа 1, 98
выделение 2, 278
Гель-фильтрация 2, 293 и сл.
Гемицеллюлоза 1, 60 и сл.
Гемоглобин 1, 79, 80, 86, 98, 114, 152
Гемэритрин 1, 98
Ген 1, 76, 398
Генетическая инженерия 1, 440 и сл.
Генетическая карта 1, 430
Генетический код 1, 398, 400, 402
Генотип 1, 411
Гентамицин, определение 2, 179
Гепатит В, Диагностика с помощью
моноклональных антител 2, 386
Гетеротрофы 1, 301
Гиалуронидаза, применение 1, 210,
234 и сл.
Гибридизация ДНК 1, 75
Гибридомы 1, 432; 2, 145, 383
Гидрирование 1, 310 и сл.
Гидрогеназа 2, 196
А^Гидрогеназа иммобилизованная 1,
258 и сл.
11 а-Гидроксиандростен-4-дион-3,17,
получение 2, 384, 385
D-L-2-Гидроксиизокапроатдегидро-
геназа, выделение 2, 278
1 lp-Гидроксилаза иммобилизованная
1, 259
15аТидроксипрегнен-4-дион-3,20, по-
лучение 2, 384
11а-Гидроксипрогестерон, получение
2, 384
Гидролитические ферменты, примене-
ние 1, 213 и сл. см. Ферменты
Гистоны 1, 75
Гифы 1, 32, 34, 561 и сл.
Глазок водорослей 1, 36
Гликоген 1, 44, 58 и сл.
биосинтез 1, 340
Гликогенфосфорилаза, стабилизация
1, 195
а-1,4-Гликозидная связь 1, 56
Гликолиз 1, 314 и сл.
Гликолипиды 1, 99
Гликопротеины 1, 80, 99, 104 и сл.
Глина как носитель для иммобилизо-
ванных ферментов 1, 240
Глицеральдегидфосфатдегидроге-
наза 1, 98
Глицериновый альдегид 1, 54
Глицерин, производство 2, 389, 391
Глицилглициндипептидаза, активация
1, 228
Глобин, кинетика реакций 1, 152
у-Глобулин 1, 98
кинетика реакций 1, 152
Глобулярные белки см. Белки
Глутамат, роль в биосинтезе амино-
кислот 1, 333
Глутамат — аспартат-трансаминаза,
кинетика реакций 1, 152
Глутаматдегидрогеназа 1, 98
Глутаминовая кислота
биосинтез 1, 331 и сл., 434
определение 2, 180
производство 2, 101
Предметный указатель
569
Глутаровый альдегид как бифункцио-
нальный реагент 1, 244
Глюкан 1, 103
1,3-Глюканаза 1, 103
Глюкоамилаза 1, 133
иммобилизованная 1, 258
стабилизация 1, 195
Глюкоза 1, 54, 55
биосинтез 1, 335 и сл.
катаболизм 1, 312 и сл.
определение 1, 261; 2, 179, 180
а-Глюкозидаза, выделение 2, 278
Глюкозоизомераза 1, 80, 123
выделение 2, 278
иммобилизованная 1, 249 и сл., 258
инактивация 2, 95
применение 1, 210, 237
производство 2, 371
Глюкозооксидаза 1, 104, 219, 220
применение 1, 210, 212, 234 и сл.
а-Глюкозо-6-фосфатдегидрогеиаза,
выделение 2, 278
Глюконеогенез 1, 335 и сл.
Глюконовая кислота, выделение 2, 329
Глюкуроновая кислота 1, 62
Голоферменты 1, 122
Гормон роста 1, 80
человека, получение с помощью ре-
комбинантных ДНК в Е. coll 2,
324 и сл., 371
Гормоны 1, 52 и сл.
Грама реакция 1, 26, 28 и сл.
Грамотрицательные клетки 1, 26
Грамположительные клетки 1, 26
Граничный слой 1, 267
Грибы 1, 30 и сл.
Гуаниловая кислота, производство 2,
381
Гуанин 1, 65 и сл.
Гумус 2, 500
Датчик уровня жидкости 2, 173
Двоичный цифровой сигнал 2, 211
Дегидрирование 1, 310 и сл.
Дегидрогеназа 2, 194
1-Дегидротестолактон, получение 2,
384
Дезоксирибонуклеаза 1, 123
Дезоксирибонуклеиновая кислота
(ДНК) 1, 63 и сл.
биосинтез 1, 339
двойная спираль 1, 69 и сл.
Декстраназа, применение 1, 233
Декстраисукраза иммобилизованная
1, 258
Декстраны
выделение 2, 280, 325 и сл.
как носители для иммобилизован-
ных ферментов 1, 240
Декстрины 1, 58
Деления 1, 417
Денатурация 1, 95 и сл.
белков 1, 181 и сл., 191 и сл.
Денитрификация 2, 501
анаэробная 2, 554
Деполимераза Р. lemoignei 1, 201
Десатураза, применение 1, 237
Диализ 2, 307
Диаметр (средний) газовых пузырь-
ков по Заутеру 1, 623
Диастаза, применение 1, 208
Диатомовые водоросли 1, 36
Диауксия 1, 514; 2, 65 и сл.
использование 1, 434
Диапетатдегидрогеназа, выделение 2,
278
Дигидрокортизон, получение 1, 391
Дигидроксиацетои 1, 54
Днгоксии, получение из дигитоксина
2, 148
Дизооль 2, 399
Дилатантные системы 1, 647
5,6-Диметилбензимидазол как пред-
шественник витамина В]2 2, 122
Динамометр торсионный, применение
для определения мощности 2, 171
Динеин 1, 80
Диоксид углерода, определение в га-
зах 2, 181 и сл.
Диплоидные клетки 1, 30, 417
Дисахариды 1, 56 и сл.
570
Предметный указатель
Дисульфидная связь 1, 93
Диффузионный слой Нернста 1, 267
Диэтиламиноэтилцеллюлоза 2, 295
ДНК-деполимераза, выделение 2, 297
ДНК-лигаза 1, 444, 481
ДНК-полимераза 1, 445
Додекан, утилизация дрожжами 2,
103 и сл.
Дрожжи 1, 30 и сл.
выделение 2, 267
применение 1, 32
рост на бумаге 2, 157
----и-гексадекане 1, 655
---- сахарах 2, 113
содержание белков и РНК 2, 193
управление метаболизмом 2, 226
Дыхание 1, 319 и сл.
Дыхательная цепь 1, 311, 323 и сл.
Дыхательный коэффициент 1, 370
в регулировании роста дрожжей 2,
226
Евклидова норма 2, 24
Енолаза нз дрожжей 1, 98
Естественный отбор 1, 480
Жгутики одноклеточных организмов
1, 79
Жгутиковые простейшие в системах
очистки сточных вод 2, 526 и сл.
Жесткие колебания 2, 451
Животные клетки см. Клетки
Жизнь, определение понятия 1, 392
Жирные кислоты 1, 46
биосинтез 1, 333 и сл.
Жиры 1, 48 и сл.
Закон
Бера — Ламберта 1, ПО
Генри 1, 598; 2, 545
Мальтуса 1, 523
Стокса 1, 23; 2, 310
Чика 1, 579
Зеин 1, 80
Зрелая мРНК 1» 407
Идеальный проточный реактор с пол-
ным перемешиванием (ПРПП) см.
Биореакторы
Идеальный реактор периодического
действия см. Биореакторы
Издержки производства 2, 353, 354
и сл., 357
Изоамнлаза, применение 1, 233
Изознмы 1, 97
Изолейцнл-тРНК — синтетаза, выде-
ление 2, 278
Изолейцин, биосинтез 1, 162, 334
Изопропиловый спирт, производство
2, 388, 390
Изопропнл-р-Э-тногалактозид 1, 434
Изотерма Фрейндлиха 2, 315
Изоферменты 1, 97, 354
Изоэлектрическая точка
аминокислот 1, 82 и сл.
белков 2, 280
Ил в очистке сточных вод см. Актив-
ный ил
Иммобилизация
клеток 2, 90 и сл., 139 и сл.
ферментов 1, 248 и сл.
— физические методы 1, 238 и сл.
— химические методы 1, 238 и сл.
Иммобилизованные ферменты 1, 237
и сл.
кинетика реакций 1, 265 и сл.
применение 1, 257 и сл.
скорость инактивации 1, 289 и сл.,
292
удельная активность 1, 275
Иммуноглобулины 1, 104 и сл.
Иммуносорбцнонный ферментативный
анализ 2, 195
Инактивация
клеток и спор 1, 570 и сл., 575
ферментов 1, 178, 186 и сл.
— энтальпия 1, 178
Инвертаза 1, 211, 217
иммобилизованная 1, 258
Ингибирование ферментов 1, 161 и сл.
См. также Субстратное ингибирова-
ние
Предметный указатель
571
конкуретное 1, 163 и сл., 171 и сл.
— математические модели 1, 169
— частичное 1, 163 и сл.
неконкурентное 1, 163 и сл., 171
и сл.
необратимое 1, 161 и сл.
обратимое 1, 161 и сл.
по принципу обратной связи 1, 162
смешанное 1, 163 и сл., 171 и сл.
согласованное 1, 437
субстратное 1, 153 и сл.
Ингибиторы конкурентные 1, 164
и сл.
Индукция 1, 409 и сл.
Индуцированная синхронизация 1,
466
Инозинмонофосфат 1, 437
производство 2, 381
Инсектициды, определение с помощью
ферментных терморезисторов 2,
179
Инсерционная инактивация 1, 448
Инсерция 1, 417
Инсулин 1, 80, 98
быка, строение 1, 94
выделение из культуры рекомби-
нантных Е. coli 2, 324 и сл.
определение 2, 179
производство с помощью рекомби-
нантных ДНК 2, 371
Интерфейсы 2, 206 н сл.
Интерфероны 1, 80
выделение 2, 279, 324 и сл.
очистка 2, 298
производство с помощью рекомби-
нантных ДНК 2, 371
установление строения 1, 442
Интроны 1, 406 и сл.
Информационная РНК 1, 76 и сл., 398
Ионная сила, влияние на kia 1, 643
Ионоселективные электроды, примене-
ние для анализа среды 2, 191
Искусственная почка 2, 307
Итаконовая кислота
выделение 2, 329
производство 2, 403
а-Казеин 1, 80
осаждение CaCh 2, 285
Кальциферол 1, 51
Каменный уголь 2, 500
Капельные биологические фильтры 2,
116
в очистке сточных вод 2, 533 и сл.
Капсид 1, 423
Капсула микроорганизмов 1, 103
Карбаматкиназа иммобилизованная
1, 258
Карбоксиметилцеллюлоза 2, 294
как носитель для иммобилизован-
ных ферментов 1, 240
Карбоксипептидаза 1, 98, 123, 156
иммобилизованная 1, 258
кинетика реакций 1, 145
Карбонилангидраза 1, 124
|3-Каротин, синтез в смешанной куль-
туре 2, 488
Каррагинан, применение для иммоби-
лизации дрожжей 2, 96
Карциномы 1, 38
Каскады реакторов как модели 2, 48
и сл.
Катаболизм 1, 312 и сл.
лактозы в Е. coli 1, 318
рибозы 1, 319
Катаболитная репрессия 1, 414
Каталаза 1, 123
денатурация 1, 191 и сл.
кинетика реакций 1, 152
применение 1, 210, 236
скорость разложения 1, 178
Катализаторы иммобилизованные 1,
273 и сл.
Катализ
ферментативный 1, 119 и сл.
кислотно-основной 1, 129
Катионообменные смолы
как носители для иммобилизован-
ных ферментов 1, 240
применение в операциях разделения
2, 291 и сл.
Качество продуктов биохимической
технологии, контроль 2, 350 и сл.
572
Предметный указатель
Кератиназа, применение 1, 210, 233
Кислород
определение в газах 2, 182
— — растворах 2, 174
-------электрохимическим мето-
дом 2, 174 и сл.
перенос в биосистемах 1, 592 и сл.
потребность биохимическая 2, 511
и сл.
— культур клеток 1, 595 и сл.
— химическая 1, 382; 2, 512
Кислотно-основной катализ 1, 129
Классы организмов 1, 26
Клен белый, рост клеток в культуре
2, 149
Клетки 1, 13 и сл. См. также Клеточ-
ный рост, Клеточный цикл
возраст 1, 464
вторичные линии 1, 37
животные 1, 37 и сл.
иммобилизованные 2, 86 и сл., 313
компетентные 1, 428
первичные линии 1, 37
перманентные линии 1, 38
половое размножение 1, 30 и сл.
прокариотические 1, 14 и сл.
растительные 1, 37 и сл.; 2, 146
и сл.
рост пристеночный 2, 14 и сл.
— сбалансированный 1, 488, 495
и сл.
установившиеся линии 1, 36
элементный состав 1, 45 и сл.
эукариотические 1, 17 и сл.
Клеточная оболочка Е. colt, строение
1, 248
Клеточная теория 1, 13 и сл., 21
Клеточные мембраны см. Мембраны
Клеточный рост
компа ртментальные модели см. Мо-
дели
лимитирующие скорость вещества
1, 365 и сл.
логистическое уравнение 1, 580; 2,
234
метаболические модели см. Модели
стехиометрия 1, 360 и сл.
удельная скорость 1, 496
Клеточный цикл 1, 463 и сл., 467 и сл.
Клонирование 1, 440 и сл.
Коалесценция газовых пузырьков 1,
615 и сл.; 2, 108 и сл.
Кодон 1, 400
Кодон-инициатор 1, 404 и сл.
Колибактерии 1, 28
Коллаген 1, 80
применение для иммобилизации
клеток 2, 91
Коллагеназа 1, 123
Комменсализм 1, 52; 2, 423 и сл.
Компартментализация 1, 21
Комплекс Гольджи 1, 21
Комплементарные ДНК 1» 454 и сл.
Комплементарные пары оснований 1,
72 и сл.
Конканавалин-сефароза, носитель для
иммобилизованных ферментов 1,
240
Конкурентное ингибирование 1, 163
и сл. См. также Ингибирование
ферментов
математические модели 1, 169
частичное 1, 163 и сл.
Конкуренция межвидовая 2, 420, 429,
432 и сл.
в открытых системах 2, 436 и сл.
рекомбинантных организмов 2, 440
и сл.
Константа
мембраны 2, 304
Михаэлиса 1, 137
скорости элементарных фермента-
тивных реакций 1, 151 и сл.
Контактная стабилизация 2, 516
Концентрационная поляризация 2, 305
и сл.
Координаты
Лайнуивера — Бэрка 1, 143, 168 и
сл., 201
Эди — Хофсти 1, 143
Кортизон 1, 53
получение 2, 382
Предметный указатель
573
Космиды 1, 448
Кофакторы 1, 122 и сл.
регенерация в непрерывных процес-
сах 1, 262 и сл.
Кофермент
А, 1, 68; 2, 101
Q 1, 323
Коферменты 1, 68, 122
Коэффициент
диффузии субстрата 1, 273 и сл.,
283, 642 и сл.
извилистости пор иммобилизованно-
го катализатора 1, 273 и сл.
Козени 2, 336
массоотдачи 1, 598
массопередачи 1, 599, 628 и сл.
осевой дисперсии 2, 53
отражения 2, 304
теплопередачи 1, 664
эффективности 1, 269
— иммобилизованного фермента 1,
276
экономический 1, 366 и сл.
Крафт-процесс 1, 219
Крахмал 1, 54, 58
как носитель для иммобилизован-
ных ферментов 1, 240
Креатинин, определение 2, 179
Критерий
Гурвица 2, 25, 430
оценки влияния эффектов массопе-
редачи 1, 279
Критическая величина объемной доли
газа в колонне 2, 108
Критический диаметр газовых пу-
зырьков 1, 618
Критическое состояние 2, 25
Кроссинговер 1, 426
Кругооборот
азота 2, 500 и сл.
в анаэробных условиях 2, 505
кислорода 2, 498 и сл.
серы 2, 502
углерода 2, 498 и сл.
Ксантан
время выравнивания концентраций
в растворах 2, 32
выделение 2, 280, 325
перенос Ог в растворах 1, 651
Ксантинмонофосфат 1, 437
производство 2, 381
Кукурузная солома как сырье для по-
лучения этанола 2, 396
Культуральный бульон, предваритель-
ная обработка 2, 260
Культуры клеток
животных 1, 37 и сл.; 2, 131 и сл.,
141
растений 1, 37 и сл.; 2, 141
Лаг-фаза 1, 511
Лактаза, применение 1, 217, 233
а-Лактальбумин, денатурация 1, 182
р-Лактамаза, получение с помощью
иммобилизованных Е. coll 2, 101
Лактат, определение 1, 26Г, 2, 179
Лактатдегидрогеназа 1, 98
стабилизация 1, 195
р-Лактоглобулин, диализ 2, 309
Лактоза 1, 57
определение 2, 179
Лактозопермеаза 1, 80, 345
Латентная фаза 1, 511
Лаурилсульфат натрия, влияние на
перенос кислорода 1, 645
Лейкемия 1, 38
Лейцинаминопептидаза
активация 1, 228
иммобилизованная 1, 258
Лейциндегидрогеназа, выделение 2,
278
Лигнин 1, 61 и сл.
Лигноцеллюлоза 1, 61 и сл.
Лизин 1, 82
биосинтез 1, 334
производство 2, 403, 405, 436, 488
экономика производства 2, 406
Лизис клеток 1, 101
Лизогения 1, 423
Лизосомы 1, 21
Лизоцим 1, 98, 101 и сл., 426
гидролиз клеточных стенок 1, 157
574
Предметный указатель
применение 1, 234 и сл.
структура 1, 87 и сл., 127
Лимонная кислота 1, 32, 358 и сл.
выделение 2, 267, 329, 330
получение из глюкозы 2, 118
синтез в культуре A. niger 2, 62
Лимфа, в культурах животных клеток
2, 136
Т-Лимфоциты, отделение с помощью
иммобилизованного лектина 2, 343
Линейно-квадратичная задача опти-
мального регулирования 2, 242
Лиотропные ряды 2, 281
Липазы 1, 123, 230 и сл.
гидролиз метилбутирата 1, 197 и сл.
применение 1, 210
Липидная мицелла 1, 47
Липиды 1, 46 и сл.
Липополисахариды 1, 99 и сл.
Липопротеины 1, 99 и сл.
Литические фаги 1, 423
Литический цикл 1, 423
Литотрофы 1, 300, 320
Логарифмическая фаза 1, 512
Логистическая кривая 1, 524 и сл.
Люцифераза, зависимость инактива-
ции от температуры 1, 190
Макроэргические связи 1, 308
Малатдегидрогеназа, кинетика реак-
ций 1, 152
Малеинового ангидрида сополимеры
как носители для иммобилизован-
ных ферментов 1, 240
Мальтоза 1, 56
Маниан 1, 103
Манноза 1, 54
Манометры мембранные, применение
в биореакторах 2, 171
Маркер селекции, генетический 1, 446
и сл.
Маслянокислое брожение 1, 356
Массообмен 1, 590 и сл.
Массоперенос 1, 590 и сл.
в неньютоновских жидкостях 1, 647
и сл.
— присутствии поверхностно-актив-
ных веществ 1, 644 и сл.
— суспензиях 1, 648 и сл.
между газовой и жидкой фазами 1,.
593 и сл.
кислорода 1, 603 и сл.
при принудительной конвекции
625 и сл.
через свободные поверхности 1, 638
и сл.
Масс-спектрометрия, применение для
анализа газов 2, 182
Масштабный переход 1, 656 и сл., 678
обратный 2, 47
Масштабы
времени 2, 6
длины 2, 7
Матрица сообщества 2, 427, 430, 448
качественно устойчивая 2, 465
Матричная РНК 1, 76 и сл., 398
Мезофильные организмы 1, 509
Межвидовые взаимодействия, класси-
фикация 2, 427 и сл.
Меласса как источник энергии и угле-
рода 2, 121
Мембраны
искусственные 1, 113
— применение для разделения ве-
ществ 2, 303 и сл.
клеточные 1, 16 и сл., 99 и сл.
Метаболизм 1, 299 и сл. См. также
Пути метаболизма
анаэробный, классификация типов
1, 374
Е. coli 1, 299
максимальный выход продуктов 1г
137
микроорганизмов, влияние среды 2,
61
поддержания 1, 368 и сл., 506
регулирование в биопроцессах 2,
224 и сл.
Метаболиты 1, 298 и сл.
вторичные 1, 355, 359 и сл.
промежуточные 1, 331 и сл.
частично окисленные 1, 358, 360
Предметный указатель
575
Метаболическая инженерия 1, 459
Метаболические модели клеточного
роста см. Модели
Металлоферменты 1, 194
Метан
как субстрат для производства бел-
ка 2, 489 и сл.
образование в анаэробных процес-
сах 2, 411 и сл., 538 и сл.
Метантенк 2, 539 и сл.
Метилэтилкетон, производство 2, 389,
391
Метионин 1, 82
биосинтез 1, 334
Метициллин 1, 434
Метод
блоттинга 1, 450
изучения кинетики предстационар-
иого состояния 1, 149
определения нуклеотидной последо-
вательности Максима — Г илберта
1, 450 и сл.
— скользящего среднего 2, 215
Методы релаксационные 1, 149 и сл.,
209
Микробиологические процессы
классификация по Дейндоерферу 1,
550
основные операции 2, 120
Микробиология 1, 13
Микрокапсулирование
животных клеток 2, 139
ферментов 1, 246, 294
Микроорганизмы, элементный состав
1, 45, 364; 2, 121
Микросмешение 2, 39
Микрофибриллы целлюлозы 1, 60
Минерализация 2, 497
Миоглобин 1, 79, 80, 86, 98
структура 1, 92
Миозин 1, 98
зависимость гидролиза АТР от тем-
пературы 1, 177
кинетика реакций 1, 152
Миссенс-мутации 1, 417
Митоз 1, 471
Митохондрии 1, 20
Мицелий 1, 32, 33
отделение фильтрованием 2, 261,
336 и сл.
Модели См. также Система хищ-
ник— жертва, модели
автолиза химотрипсина 1, 190
влияния степени кристалличности
целлюлозы на ее гидролиз 1, 223
и сл.
Вольтерры для периодического рос-
та 1, 525
-----межвидовой конкуренции 2,
432 и сл.
динамики популяций в форме зако-
на действующих масс 2, 459 и сл.
«замка и ключа» для ферментов 1,
94, 126
инактивации ферментов 1, 185 и сл.
кинетики ферментативных гетеро-
генных реакций 1, 199 и сл.
клеточного роста 1, 495 и сл.
-----в лаг-фазе 1, 585
----- кибернетические 1, 541 и сл.
-----компартментальные 1, 530 и сл.
----- метаболические 1, 535 и сл.
-----несегрегированные 1, 487 и сл.
------ неструктурированные 1, 487,
523 и сл.
-----сегрегированные 1, 566 и сл.
----- структурированные 1, 529
и сл., 555 и сл.
------ — образования продуктов 1,
529
— Уильямса 1, 531 и сл., 586; 2,
28
-----Хардера и Роуэлса 1, 534
образования продуктов метаболиз-
ма 1, 544 и сл.
— — — генетически структуриро-
ванные 1, 555 и сл.
обратимой термической инактива-
ции ферментов 1, 177 и сл.
отравления ферментов ядами 1, 191
периодической культуры в фазе от-
мирания 1, 520
576
Предметный указатель
процессов в ПРПП 2, 20 и сл.
псевдогомогенной диффузии 2, 301
роста периодической культуры в
присутствии токсина 1, 518
согласованных переходных состоя-
ний 1, 114
сферической гранулы иммобилизо-
ванного фермента 1, 274
ферментативного катализа 1, 131
и сл.
хемостата 1, 492
— Джеффресона и Смита 2, 28
— Ли, Джекмана и Шредера 2, 28
— Моно 1, 500 и сл.
— Рамкришны, Фредриксона и Цу-
чии 2, 28
— Шулера для роста Е. coli 1, 539
и сл.
— Янга, Брюли и Бангея 2, 28
Модуль Тила 1, 277
клетки 1, 581
Молекулярная генетика 1, 397 и сл.
Молоко, превращения при скисании
2, 495 и сл.
Молочнокислые бактерии 1, 29
применение 2, 486 и сл., 496
C-Моль клеток 1, 370
Моноклональные антитела 1, 432
получение 2, 145 и сл„ 298, 382 и сл.
Моносахариды 1, 55
Монослой 1, 47 и сл.
Морфология
микроорганизмов 1, 25
мицелия, зависимость от скорости
перемешивания 2, 63 и сл.
— методы изучения 2, 198
Мочевая кислота, определение 2, 179
Мочевина, определение 2, 179
Мощность, расходуемая на перемеши-
вание, определение 1, 635 и сл.; 2,
171
Моющие средства 1, 47 и сл.
Мультиплексоры 2, 211
Муравьиная кислота, определение 2,
180
Мутагены 1, 418 и сл.
Мутанты 1, 34 и сл., 417 и сл.
Мутации 1, 417 и сл. См. также Мис-
сенс-мутации, Нонсенс-мутации
ауксотрофные 1, 435 и сл.
бессмысленные 1, 418
конститутивные 1, 439
Мутуализм 2, 420 и сл., 489
Мыла 1, 48
Мягкие колебания 2, 450
Нарингиназа, применение 1, 211
Нейтрализм 2, 419 и сл.
Неньютоновские жидкости
математические модели 1, 647 и сл.
массоперенос 1, 646 и сл.
Непрерывные процессы, управление 2,
241 и сл.
Нефть 2, 500
Ниацин 1, 51
Никотинамидадениндинуклеотид 1, 6&
310 и сл.
Никотиновая кислота 1, 51
Нитрификация 2, 500
сточных вод 2, 531
Новобиоцин, выделение 2, 314
Нонсенс-мутации 1, 418
Нуклеаза 1, 123
Нуклеиновые кислоты, структура I,
69 и сл.
Нуклеозиды 1, 65 и сл.
Нуклеоид 1, 17
Нуклеотиды 1, 65 и сл.
биосинтез 1, 335
секреция A. niger 2, 63
Оборудование типовое, стоимость 2,
354
Обратный осмос, применение для раз-
деления веществ 2, 303 и сл.
Овальбумин 1, 80
Ограниченная диффузия 1, 273 и сл.
Окислительно-восстановительные ре-
акции 1, 306
Окислительное фосфорилирование 1»
323
Окружная скорость мешалки 1, 623
Предметный указатель
577
Оксациллин 2, 377
Оксид алюминия
влияние на перенос кислорода 1,
653
как носитель для иммобилизован-
ных ферментов 1, 240
Оксидаза L-аминокислот иммобилизо-
ванная, применение 1, 258
D-Оксинитрилаза иммобилизованная,
применение 1, 258
Олеиновая кислота 1, 47
Оперативное ЗУ 2, 204
Оператор 1, 412
Операционная карта процесса 2, 359
Операционная система ЭВМ 2, 212
Оперон 1, 339, 414
Оперонная модель биосинтеза а-га-
лактозидазы 2, 69 и сл.
Оптическая активность 1, 81
Организмы-хозяева в технологии ре-
комбинантных ДНК 2, 372
Органоиды 1, 20 и сл.
Органотрофы 1, 300
Орсин, применение для определения
РНК 2, 190
Осаждение 2, 279 и сл.
Осахаривающий фермент 1, 216
Осевая дисперсия 2, 84 и сл.
Осмос 2, 303 и сл.
Отжиг ДНК 1, 74, 444
Отношение Р/О 1, 326
Отстаивание 2, 260
Отъем но-доливные культуры 2, 379
Очистка продуктов 2, 256
Панкреатическая протеаза, примене-
ние 1, 209
Пантотеновая кислота 1, 51
Папаин 1, 98, 124
иммобилизованный, применение 1,
258
применение 1, 209, 229
стабилизация 1, 195
Паразитизм 2, 427, 442 и сл.
Парамагнитные анализаторы 2, 182
37-746
Параметр
молекулярного сита 2, 308
насыщения 1, 277
старения 2, 287
Пассивный транспорт 1, 341 н сл.
Пектиназы
применение 1, 211, 232 и сл.
производство 2, 371
Пектины 1, 232
Пенициллиназа, применение 1, 210,,
234 и сл.
Пенициллинамидаза иммобилизован-
ная, применение 1, 258
Пенициллинацилаза
выделение 2, 278
применение 1, 233
Пенициллины
биосинтез 1, 35
выделение 2, 313, 338
определение 2, 179
производство 2, 101, 233 и сл.
схема процесса получения 2, 328
типы 2, 375, 377
управление процессом биосинтеза 2Г
250
ферментативное деацилирование 2Г
163
Пеногасители 2, 128
Пентозофосфатный путь (цикл) мета-
болизма см. Пути метаболизма
Пепсин
активация 1, 291
кинетика реакций 1, 145
применение 1, 208
Пепсиноген 1, 228
Пептидогликаны 1, 99, 100 и сл.
Пептидная связь 1, 84, 90
Периплазматическое пространство 1„
100 и сл.
Периферийные устройства ЭВМ 2, 205
и сл.
Перколяционные биологические филь-
тры 2, 533
Пермеазы 1, 79, 344
Пероксидаза 1, 123
кинетика реакций 1, 152
578
Предметный указатель
Пивоварение 2, 387 и сл.
Пиранозы 1, 54
Пиридоксальфосфат 1, 333
Пиримидин 1, 65
Пирролнитрин 2, 378
Пируват 1, 310 и сл., 315, 357; 2, 27
Пируват •— ферредоксин-оксидоредук-
таза 2, 194
Питательные вещества 1, 12
Пищевые сети 2, 457 и сл.
Пищевые цепи 2, 455, 457 и сл.
Плазматическая мембрана 1, 17, 18
и сл.
Плазмиды 1, 75, 428 и сл., 446 и сл.
амплификация 1, 447
определение содержания в бактери-
ях 2, 195
Плазминогена тканевого активатор,
применение 1, 234 и сл.
Пластинчатая разводка 1, 42
Пластичные жидкости Бингама 1, 647
Плесневые грибы 1, 32 и сл.
Поверхностно-активные вещества,
влияние на перенос кислорода 1,
644 и сл.
Поверхностное натяжение, влияние на
денатурацию белков 1, 193
Подмаренник, рост клеток в культуре
2, 149
Полиакриламид как носитель для им-
мобилизованных ферментов 1, 240,
245 и сл.
Полиакрилаты как носители для им-
мобилизованных ферментов 2, 91,
96
Полиаминополистирол как носитель
для иммобилизованных ферментов
1, 240
Поли-(3-гидроксимасляная кислота 1,
53
Полинуклеотидфосфорилаза иммоби-
лизованная 1, 258
Полипептиды 1, 84
Полисахариды 1, 53 и сл.
внеклеточные 1, 103
выделение 2, 325
Полистирол как носитель для иммо-
билизованных ферментов 2, 91
Популяция клеток 1, 21
численность 1, 573
Постоянная высаливания 2, 282
Постоянное запоминающее устройство
2, 204
Почва как среда для микроорганиз-
мов 2, 502 и сл.
Почкование 1, 30, 472 и сл.
Праймер 1, 445, 481
Пребелки 1, 408
Предельный цикл 2, 451
Преднизолон, получение 1, 259; 2, 385
Предшественники биосинтетические 2,
122
Преобразование Лапласа, использо-
вание в управлении процессами 2,
247
Прибыль на инвестированный капи-
тал, расчет 2, 365, 368 и сл.
Прикладные программы 2, 213
Принцип конкурентного исключения
2, 434 и сл., 457
Прогестерон 1, 53
Программное обеспечение 2, 211 и сл.
Проинсулин 1, 409, 410
Прокариоты 1, 15 и сл.
Пролил-тРНК-синтетаза, выделение
2, 321
Пролин 1, 82 и сл., 333
Промотор 1, 399
Проницаемость мембран 1, 341
Пропиленгликоль, производство 2,
389, 391
Пропорционально-интегрально-диф-
ференциальное управление 2, 222
Простейшие 1, 36 и сл.
Простетические группы 1, 86
Протеазы
бактериальные 1, 209, 228, 434; 2,
321 и сл., 371
грибковые 1, 209; 2, 371
из актиномицетов 1, 209
Протеогликаны 1, 99
Противоточное распределение 2, 271
Предметный указатель
57»
Протисты 1, 24 и сл.
Протопласты I, 101 и сл., 430 и сл.
Прототрофы 1, 20, 435
Профаг 1, 423
Псевдопластичные жидкости 1, 647
Психрофильиые организмы 1, 508
Пуллуланаза 1, 217
выделение 2, 278
Пурин 1, 65
Пути метаболизма
амфиболические 1, 314
гексозомоносфатный 1, 316
Кребса 1, 321 и сл.
лимонной кислоты 1, 321 и сл.
пентозофосфатный 1, 316 и сл.
трикарбоновых кислот 1, 321 и сл.
фосфокетолазный 1, 318 и сл.
Эмбдена — Мейергофа — Парнаса
1, 302, 312 и сл., 356
Энтнера — Дудорова 1, 317
Путресцин 2, 495
Рабочая функция 2, 238 и сл.
Радиусы молекул белков, определение
2, 294
Распределение
времени пребывания 2, 33 и сл.
— циркуляции 2, 32 и сл.
С3-Растения 1, 386 и сл.
С4-Растения 1, 386 и сл.
Растительные клетки 1, 37 и сл.; 2,
146 и сл.
Расход мощности
на аэрацию 1, 657
— перемешивание 1, 630 и сл.
Расходомеры 2, 172
Расходы на оплату труда производ-
ственных рабочих 2, 357, 365
Реакторы см. Биореакторы
Реакция
Белоусова — Жаботинского 2, 471
Грама 1, 115
Майяра 1, 236; 2, 82
Сэнджера 1, 116
37*
Регулятор с обратной связью 2, 220
и сл.
Режим
высокотемпературной кратковре-
менной стерилизации 2, 82
полного вытеснения 2, 15
Резервные гранулы 1, 17
Рекомбинантные ДНК 1, 441 и сл.
Рекомбинантные организмы, струк-
турная неустойчивость 1, 457
Ренатурация 1, 95
Реннин 1, 208, 229; 2, 486
бактериальный 1, 210; 2, 371
Репликационная вилка 1, 468, 481
Репликация ДНК 1, 73, 415
Репрессия 1, 409 и сл.
/ас-Репрессор 1, 80
Рестриктазы 1, 442 и сл.
Рестрикционные карты 1, 450
Рефрактометрия 2, 189 и сл.
Рибоза 1, 54
Рибонуклеаза 1, 98, 124
иммобилизованная 1, 258
применение 1, 233
Рибосомальная РНК 1, 77
Рибосомы 1, 17, 19, 77, 398
Рибофлавин 1, 51
Рибулоза 1, 54
Рибулозо-1,5-дифосфат 1, 337, 386
Рибулозодифосфаткарбоксилаза 1, 98»
Рицин 1, 189
РНК 1, 65 и сл.
биосинтез 1, 339
РНК-полимераза 1, 80, 398
Род организма 1, 25 и сл.
Ротаметр 2, 172
Ротационные вакуум-фильтры 2, 26Г;
и сл.\
Ряды ионов Гофмейстера 2, 281
Сайт
связывающий рибосому 1, 400, 404-
узнавания 1, 443
Сахара 1, 53 и сл.
Сахароза 1, 57
определение 2, 179
580
Предметный указатель
Световые реакции фотосинтеза 1,
337
Свинец, определение с помощью фер-
ментных терморезисторов 2, 179
Себестоимость продукции 2, 364, 36b
Сегрегационная неустойчивость 1, 457
Седиментация 2, 267 и сл.
Седловая точка 2, 430
Секвенаторы 1, 87
Семейства организмов 1, 26
Сервисные программы 2, 213
Серповидно-клеточная анемия 1, 418
Сигнальные последовательности бел-
ков 1, 408
Силикагель как носитель для иммоби-
лизованных ферментов 1, 240
Силиконовые трубки, применение для
введения Ог в культуры 2, 137
Симбиоз 2, 421 и сл., 484
Сине-зеленые водоросли 1, 30, 36
Синтетические среды 1, 497
Синтрофизм 2, 503
Синхронные культуры 1, 465 и сл.
Система хищник — жертва, модели
Лесли 2, 479
Лотки — Вольтерры 2, 443 и сл.
Розенвейга и Макартура 2, 558
Цучии 2, 450 и сл.
Скорость
переноса кислорода, определение 1,
606 и сл.
разведения 1, 494
реакции 1, 119
Скошенный агар 2, 124
Скрининг клонов 1, 452
Сложные среды 1, 497
Солнечники 1, 36
Соложение 2, 387
Сопряженный транспорт 1, 343 и сл.
Сорбитдегидрогеназа, применение 1,
237
Сорбция 2, 277 и сл.
Сосуд
Блэйка 2, 124
Ру 2, 124
'Спектрофотометрия, применение для
контроля культур 1, ПО; 2, 184
Сплайсинг 1, 407 и сл.
Среды
для роста животных клеток 2, 135
и сл.
-----растительных клеток 2, 146
и сл.
— ферментации, основные требова-
ния 2, 120
методы анализа 2, 189 и сл.
синтетические 1, 497 и сл.
сложные 1, 497 и сл.
Стабильная линия клеток 1, 38
Старение осадков 2, 286
Стационарная фаза 1, 512
Стационарные состояния, устойчивые
и неустойчивые 2, 24 и сл.
Стеариновая кислота 1, 47
Стенка клетки 1, Гб
Степенной закон 1, 647, 681
Степень восстановленности 1, 372
Стерилизаторы 2, 74 и сл.
Стерилизация 1, 186; 2, 74 и сл.
вероятность выживания микроорга-
низмов 2, 75
кинетика 1, 570 и сл.
непрерывная 2, 82 и сл.
периодическая 2, 76 и сл.
— температурные профили 2, 76
и сл.
твердых тел 2, 78
тепловая 1, 30; 2, 74 и сл.
фильтрованием 1, 674 и сл.
хлорированием 1, 579 и сл.
Стероиды 1, 52 и сл.
выделение 2, 313
Стехиометрия
образования продуктов 1, 374 и сл.
роста хемогетеротрофов 1, 371 и сл.
энергетического обмена 1, 378 и сл.
Сточные воды 2, 507 и сл.
биологическая очистка 1, 37; 2, 507
и сл.
вторичная обработка 2, 507 и сл.
объемная перегрузка при очистке
2, 518
Предметный указатель
581
первичная обработка 2, 507 и сл.
промышленных предприятий 2, 514
и сл.
----- характеристики 2, 511 н сл.
третичная обработка 2, 507 и сл.
Стоячая пленка 1, 267
Структура течений при перемешива-
нии 1, 633
Структурированные модели см. Мо-
дели
Стрептодорназа, применение 1, 234
Стрептокиназа, применение 1, 209, 234
и сл.
Стрептомицин, выделение 2, 314
Субстратное ннгибированне 1, 155
и сл.
Субстратные аналоги 1, 164
Субстраты 1, 94
Сукцинилдегндрогеназа 1, 164
Сульфаниламид 1, 164
Сульфитно-спиртовая барда как суб-
страт для дрожжей 2, 122
Сусло пивное 2, 387
Сферопласты 1, 101
Счетчик Коултера 2, 198, 428
Сыворотка как составная часть сред
2, 135
Сыроделие 2, 486 и сл.
Сычужные ферменты 2, 486
Такаамилаза 1, 98
Такадиастаза, применение 1, 208
Таксономия 1, 25
Таксоны 1, 25
Танназа, применение 1, 233
Таннинаминогексилцеллюлоза как но-
ситель для иммобилизованных фер-
ментов 1, 240
Таутомерные формы оснований ДНК
1, 418
Темновые реакции фотосинтеза 1, 337
Температура
плавления ДНК 1, 74
среды, влияние на рост клеток 1,
508 и сл.
Тензодатчики, применение для опре-
деления мощности 2, 172
Теорема
Бэкингема 1, 681
нулевого ограничения 2, 463
Пуанкаре — Бендикссона 2, 454
Теория
бифуркаций 2, 470 и сл.
мишеней 1, 478 и сл.
ортокинетической флокуляции 2,
286
перикинетической флокуляции 2,
285 и сл.
турбулентности, статистическая 1,
619
Теплообмен, уравнения 1, 668 и сл.,
672 и сл.
Теплообменные аппараты, типы 1, 663
Терминатор 1, 399
Терминирующий кодон 1, 404 и сл.
Термометры сопротивления 2, 171
Термопары 2, 171
Терморезисторы 2, 171
ферментные 2, 178 и сл.
Термофильные организмы 1, 509
Тиамин 1, 51
Тимин 1, 65 н сл.
Р-Тнрозиназа иммобилизованная 1,
258
Тирозин — фенол-лиаза 1, 237
Токсины бактерий 1, 80
Торф 2, 500
Траектории численности популяций 2,
430, 445
Транзиция 1, 417
Трансаминаза 1, 237
Трансверсия 1, 417
Трансдукция 1, 426 и сл.
Трансдуцирующие фагн 1, 426
Транскриптаза обратная 1, 445
Транскрипция 1, 398 и сл.
Транслокация групп 1, 345
Трансляция 1, 398 и сл.
Трансмембранные белки 1, 345
Транспорт через мембраны 1, 51, 341
и сл.
582
Предметный указатель
активный 1, 344 и сл.
пассивный 1, 341 и сл.
сопряженный 1, 343 и сл.
Транспортные РНК 1, 77, 398, 400
и сл.
Трансфекция 1, 460
Трансфераза концевая 1, 445
Трансформация 1, 428
Треонин 1, 82, 334
Треониндезаминаза 1, 98
Триглицериды, определение 2, 179
Триозиназа 1, 123
Трипаносомиаз 1, 36
Трипаносомы 1, 36
Трипсин 1, 80, 98, 124
гидролиз лизоцима 1, 199
— тиогеля 1, 199 и сл.
иммобилизованный 1, 271
кинетика реакций 1, 145
применение 1, 208, 234
стабилизация 1, 195
Трипсиноген 1, 228
Триптофаназа 1, 98
иммобилизованная 1, 258
Триптофансинтетаза 1, 98
Тромбин 1, 80
Трофические уровни 2, 457
Углеводородные субстраты для дрож-
жей 2, 103 и сл.
Углеводы 1, 53 и сл.
Узловая точка 2, 430, 454
Уксусная кислота
определение 2, 180
производство 2, 388, 390
Уксуснокислые бактерии 1, 28
Ультрафильтрация 2, 307 и сл.
применение для иммобилизации
ферментов 1, 247
Умеренные фаги 1, 423
Уравнение
Анри 1, 135
Аррениуса 1, 176
Бриггса — Холдейна 1, 138 и сл.
Джонсона, Эйринга и Полиссара 1,
578
Кассона 1, 647, 649
Конту а 1, 507
Ленгмюра — Хиншелвуда 1, 142
Льюдикина — Пайрета 1, 548, 586;
2, 234
Михаэлиса — Ментен 1, 135 и сл.; 2,
13, 19
Мичела и Миллера 1, 638, 653
Мозера 1, 507
Моно 1, 497 и сл.; 2, 519 и сл.
О’Коннора — Даббинса 1, 642
Оствальда — Вейля 1, 647
Риккати 1, 524
Сведберга 1,111
Смолуховского 2, 285
Стокса — Эйнштейна 1, 676
Стритера — Фелпса 1, 680
Теиссье 1, 507
Уилки— Чанга 1, 643
Хигби 1, 641
Хилла 1, 205
Эйнштейна 2, 340
Эндрюса 1, 508
Урацил 1, 65 и сл.
Уреаза
денатурация 1, 191 и сл.
иммобилизованная, применение 1,
259
кинетика реакций 1, 145, 152
Урокиназа, применение 1, 234 и сл.
Участок начала репликации 1, 446
и сл., 468
Фаги 1, 76, 423 и сл.
Фазы клеточного роста 1, 511 и сл.
Фактор разделения 2, 269
Фактор F 1, 428
R-Факторы 1, 428
Факторы денатурации белков 1, 183
и сл.
Фенилуксусная кислота как предшест-
венник пенициллина 2, 122
Фенол, определение 2, 179
Фенотип 1, 411
Предметный указатель
583
Ферментативная активность, единицы
1, 134 и сл.
•Ферментативные реакции 1, 118 и сл.
гистерезис 1, 292
кинетика 1, 132 и сл.
максимальная скорость 1, 137
модуляторы 1, 161
Ферментативный катализ 1, 118 и сл.
Ферментация
аэробная 2, 226
двойная 2, 488
Ферментеры, см. Биореакторы
Фермент-субстратные комплексы 1,
126 и с л.
Ферменты 1, 45 и сл., 78 См. также
Иммобилизованные ферменты,
Ингибирование ферментов
активация 1, 153 и сл., 227 и сл.
активный центр 1, 122, 126 и сл.
аллостерические 1, 165 и сл.
внеклеточные 1, 212
внутриклеточные 1, 212
выделение 2, 267, 319 и сл.
гидролитические, применение 1, 213
и сл.
инактивация 1, 178, 186 и сл.
индуцибельные 1, 411
конститутивные 1, 411
номенклатура 1, 119 и сл.
промышленное производство 2, 370
и сл.
протеолитические 1, 129, 227 и сл.
репрессибельные 1, 412
стабилизация 1, 194 и сл.
Фибриллы целлюлозы 1, 60
Фибриллярные белки 1, 78 и сл.
Фибриноген 1, 98
Фиксация азота 2, 501 и сл.
Филаментозные организмы, модели
роста 1, 527 и сл.
Фильтр
Калмана 2, 218; 243
первого порядка 2, 215
Фильтрование 2, 260 и сл.
Фимбрии бактерий 1, 80
Фицин
иммобилизованный 1, 270
применение 1, 209
Флавинадениннуклеотид 1, 68, 323
Флавопротеииоксидаза иммобилизо-
ванная, применение 1, 258
Флавопротеины 1, 323
Флотация 2, 270
Флуорометрия, применение для изуче-
ния состояния культур 2, 184 и сл.
Фокальная точка 2, 430, 454
Фолиевая кислота 1, 51
Формилтетрагидрофолатсинтетаза 1,
98
Фосфатаза щелочная 1, 98, 123
5'-Фосфодиэстераза, применение 1,
211
Фосфодиэфирные связи 1, 70
Фосфоглицериды 1, 49
Фосфокетолазный путь метаболизма
1, 319
Фосфорилаза 1, 98
выделение 2, 278
иммобилизованная, применение 1,
258
Фосфорилирование на уровне суб-
страта 1, 314
Фосфотрансацетилаза 2, 196
Фосфотрансбутирилаза 2, 196
Фосфофруктокиназа 2, 194
Фотоаутотрофы 1, 302
Фотогетеротрофы 1, 302
Фотодыхание 1, 386 и сл.
Фотосинтез 1, 17, 327 и сл.; 2, 498
стехиометрия 1, 384 и сл.
эффективность 1, 386
Фотосистемы I и II 1, 327 и сл.
Фототрофы 1, 300
Фотофосфорилирование 1, 328 и сл.
циклическое 1, 329
Фракционированное осаждение бел-
ков 2, 281
Фруктоза 1, 54
Фумараза 1, 98
выделение 2, 278
инактивация в иммобилизованных
клетках 2, 94
584
Предметный указатель
кинетика реакций 1, 152
применение 1, 237
Фумаратгидратаза 1, 124
^“-Функция 2, 34
pH-Функция Михаэлиса 1, 175
Функция
интенсивности 2, 37
распределения времени пребывания
газа в вязких жидкостях 2, 45
------- в емкостных реакторах 2,
37
----------каскаде ПРПП 2, 49
---------- колонне 2, 50
----------непрерывном стерили-
заторе 2, 84
Хемиосмотическая теория 1, 345
Хемоаутотрофы 1, 302
Хемогетеротрофы 1, 302
Хемостат 1, 492
рабочая диаграмма системы 2, 453
Хемотаксис 2, 480
Хемотрофы 1, 300
Химерные белки 1, 456
Химотрипсин 1, 98, 129, 140
иммобилизованный 1, 243
инактивация 1, 190, 205
кинетика реакций 1, 130, 243
механизм действия 1, 130
применение 1, 208
стабилизация 1, 195
Химотрипсиноген 1, 228
Хитин 1, 103 и сл.
Хищничество 2, 429, 442 и сл.
Хлорамфеникол 2, 378
Хлоропласты 1, 20, 327
Хлорофилл 1, 327 и сл.
Холестерин 1, 53
определение 2, 179
Холин 1, 51
Холинэстеразы 1, 161
Хориогонический гонадотропин, опре-
деление 2, 386
Хроматин 1, 75
Хроматографические методы разделе-
ния 2, 289 и сл.
аминокислот 1, 112
Хроматография
афинная 2, 296 и сл.
высокоэффективная жидкостная 2,
190
газовая, применение для анализа
отходящих газов 2, 182
гель-проникающая 2, 292 и сл.
ионообменная 2, 291, 294 и сл.
на иммуносорбентах 2, 298, 373
— молекулярных ситах 2, 292 и сл.
Хромосомы 1, 75, 416
Целевая функция 2, 238 и сл.
Целлобиаза 1, 220
Целлобиоза, определение 2, 179
Целлофан как материал для мембран
2, 304
Целлюлаза 1, 222 и сл.
иммобилизованная 1, 258
применение 1, 211
Целлюлазная система Trichoderma 1,
218 и сл.
Целлюлоза 1, 54, 58 и сл.
аморфная 1, 60
делигнификация 1, 220 и сл.
как материал для мембран 2, 304
— носитель для иммобилизованных
ферментов 1, 240
кристалличность 1, 220
сульфатная варка 1, 219
сульфитная варка 1, 219
ферментативный гидролиз 1, 215,
291
Целлюлозная пульпа, перенос кисло-
рода в ней 1, 655
Цельный бульон, методы переработки
2, 313 и сл.
Центральный процессор ЭВМ 2, 204
Центрифуги
осадительные 2, 265
типы 2, 266
шнековые 2, 266
Центрифугирование 1, 22; 2, 264 и сл.
Предметный указатель
585
дифференциальное 1, 23
применение для выделения биомас-
сы 2, 267
с угловым ротором 1, 42
Цефалоспорины 2, 378
биосинтез 1, 561 и сл.
определение 2, 179, 180
Цианобактерии 1, 30
Цианурхлорид, как бифункциональ-
ный реагент 1, 244
Цикл
Калвина 1, 337
Кребса 1, 321 и сл.
лимонной кислоты 1, 321 и сл.
трикарбоновых кислот 1, 321 и сл.
Циклический АМР 1, 67
Циклогексимид, выделение 2, 313
Цитозин 1, 65 и сл.
Цитометры 2, 198 и сл.
Цитоплазма 1, 17
Цитоплазматическая мембрана 1, 100
и сл.
Цитохромы 1, 123, 323
повышение выхода 2, 245
применение 1, 210
Цифро-аналоговые преобразователи
2, 211
Частотная функция возраста клеток
1, 566 и сл.
Чеддеризация 2, 486
Челночные векторы 1, 460
Число
Био 1, 286
Бонда 1, 623
Бринкмана 1, 670
Вебера 1, 618, 620
Галилея 1, 623
Грасгофа 1, 611, 671
Дамкелера 1, 268; 2, 83
Кэмпа 2, 287
мощности 1, 635
Нуссельта 1, 670
оборотов катализатора 1, 124
Пекле 1, 612; 2, 83
— зависимость от Sc и Re 2, 56, 57
— осевое 2, 54 и сл.
переносимых единиц 2, 299 и сл.
Прандтля 1, 670
Рейнольдса 1, 13, 41, 612, 617, 627,
656, 670
— мешалки 1, 652
Рэлея 1, 613
Фруда 1, 617, 623, 627, 670
Шервуда 1, 611, 627, 640
Шмидта 1, 611, 627
Чистые культуры 2, 123
Шиконин, получение с помощью куль-
туры растительных клеток 2, 150
и сл.
Шунт (путь метаболизма) 1, 316
Щавелевая кислота
образование в культуре 1, 32
определение 2, 179
Щелочная фосфатаза 1, 434
Эвгленовые водоросли 1, 36
ЭВМ, использование в управлении
биопроцессами 2, 202 и сл.
Эвтрофикация 2, 506
Экзо-[3-1,4-глюканглюкогидролаза 1,
219
Экзоны 1, 406 и сл.
Экзопептидазы 1, 227
Экзо-0-1,4-целлобиогидролаза 1, 219,
220
Экзэргонические реакции 1, 307
Экономический коэффициент клеточ-
ного роста 1, 366 и сл., 582
зависимость от субстрата 1, 381
Эксклюзия молекулярная 2, 293
Экспоненциальный рост организмов 1,
512
Экспрессия
генов 1, 398 и сл.
эукариотических белков в прока-
586
Предметный указатель
риотах 1, 412 и сл.
Экстрактор Подбильняка 2, 272, 274
Экстракция 2, 271 и сл.
в двухфазных водных системах 2,
274 и сл.
органическими растворителями 2,
271 и сл.
с химической реакцией 2, 338
Эластин 1, 80
Электроды
для определения pH 2, 173 и сл.
— — окислительно-восстанови-
тельного потенциала 2, 174
Кларка 2, 174
Электрокинетическое осаждение 2,
270
Электрокоагуляция 2, 331
Электрофорез 2, 310 и сл.
Эндо-Р-1,4-глюканаза 1, 219, 220
Эндогенный метаболизм 1, 504 и сл.
Эндопептидазы 1, 227
Эндоплазматическая сеть 1, 19
Эндоплазматический ретикулум 1, 19
Эндоспоры 1, 27, 32
Эндотоксины 1, 101
Эндэргонические реакции 1, 307
Энергетический заряд клетки 1, 352
Энергия поддержания 1, 368 и сл.
Энтеробактерии 1, 28
Эписомы 1, 428
Эпоксидные смолы, применение для
иммобилизации клеток 2, 91
Эргоалкалоиды, производство 2, 380
и сл.
Эргостерол 1, 51
Эритромицин, выделение 2, 272
Эрлифтные реакторы 2, 107, 109 и сл.
Эстеразы 1, 141
применение 1, 230 и сл.
Этанол
выделение 2, 329 и сл., 331 и сл.
как топливо 2, 393 и сл.
определение 2, 179, 180
производство 2, 388, 390
— с помощью иммобилизованных
дрожжей и бактерий 2, 96 и сл.
Этилен, производство 2, 388, 390
Этиленгликоль, производство 2, 388,
390
Эукариоты 1, 17 и сл.
Эфротомицин, полный синтез 2, 378
Эффект
Бойкотта 2, 270
Крэбтри 2, 226
ориентации 1, 128
сближения 1, 128
Эффекторы ферментативных реакций
1, 161
Ядерный магнитный резонанс, опре-
деление ATP, ADP и фосфатов 2,
197
Ядро клетки 1, 17, 19, 20 и сл.
Яды
нервнопаралитические 1, 161
ферментные 1, 161
Оглавление
Глава 9. ПРОЕКТИРОВАНИЕ И РАСЧЕТ БИОЛОГИЧЕСКИХ РЕАК-
ТОРОВ ....................................................... 5
9.1. Идеальные биореакторы......................................... 8
9.1.1. Реакторы периодического действия с добавлением субстрата 9
9.1.2. Реакции в ПРПП, катализируемые ферментами ... 10
9.1.3. Проточные реакторы с полным перемешиванием для куль-
тур клеток и пристеночный рост клеток.................... 12
9.1.4. Идеальный трубчатый реактор полного вытеснения (ТРПВ) 15
9.2. Динамика процессов в реакторах................................20
9.2.1. Динамические модели.....................................20
9.2.2. Устойчивость............................................24
9.3. Реакторы с неидеальным перемешиванием.........................29
9.3.1. Время выравнивания концентраций в реакторах с переме-
шиванием .....................................................31
9.3.2. Распределение времени пребывания........................33
9.3.3. Модели неидеальных реакторов............................42
9.3.4. Взаимосвязь между перемешиванием и биологическими пре-
вращениями ...................................................57
9.4. Стерилизаторы . 74
9.4.1. Периодическая стерилизация..............................76
9.4.2. Непрерывная стерилизация................................82
9.5. Иммобилизованные биокатализаторы..............................86
9.5.1. Типы биокатализаторов на основе иммобилизованных кле-
ток и их свойства.............................................90
9.5.2. Применение биокатализаторов на основе иммобилизованных
клеток....................................................94
9.6. Биореакторы с многофазными системами....................102
9.6.1. Превращение нерастворимых субстратов..............102
9.6.2. Реакторы с неподвижным слоем биокатализатора . . . 105
9.6.3. Биореакторы типа барботажных колонн...............107
9.6.4. Биореакторы с псевдоожиженным слоем катализатора 111
9.6.5. Реакторы с неподвижным слоем катализатора и со струйным
течением жидкости........................................116
9.7. Технология микробиологических процессов.................119
9.7.1. Подбор состава среды...................................120
9.7.2. Проектирование типичного асептического аэробного микро-
биологического процесса и его ведение.......................123
9.7.3. Биореакторы других типов...............................128
9.8. Особенности технологии процессов с участием растительных и
животных клеток и соответствующих реакторов..................131
9.8.1. Культивирование животных клеток; требования к среде 135
9.8.2. Промышленные реакторы для крупномасштабных процессов
с участием животных клеток.............................. 138
9.8.3. Культивирование растительных клеток....................146
588
Оглавление
9.9. Заключение.................................................151
Упражнения......................................................152
Литература.................................................164
Глава 10. КОНТРОЛЬНО-ИЗМЕРИТЕЛЬНАЯ АППАРАТУРА И УП-
РАВЛЕНИЕ ПРОЦЕССАМИ БИОХИМИЧЕСКОЙ ТЕХНО-
ЛОГИИ .....................................................169
10.1. Детекторы для определения физических и химических парамет-
ров среды и газов..............................................169
10.1.1. Детекторы для определения физических свойств среды и
газов..................................................171
10.1.2. Детекторы для определения химического состава среды 17$
10.1.3. Газовый анализ..................................181
10.2. Детекторы для непрерывного контроля характеристик популя-
ции клеток.................................................183
10.3. Автономные методы анализа.............................188
10.3.1. Определение свойств среды.......................189
10.3.2. Анализ состава популяции клеток.................191
10.4. ЭВМ и интерфейсы......................................202
10.4.1. Основные элементы цифровых ЭВМ..................204
10.4.2. Интерфейсы и периферийные устройства ЭВМ . . . 206
10.4.3. Системы программного обеспечения................211
10.5. Анализ данных.........................................213
10.5.1. Сглаживание и интерполяция данных...............214
10.5.2. Оценка параметров и состояния системы...........216
10.6. Управление процессами биохимической технологии . . . . 219
10.6.1. Непосредственное управление процессами .... 220
10.6.2. Каскадное управление метаболизмом...............224
10.7. Прогрессивные методы управления биопроцессами .... 228
10.7.1. Программированное управление процессами в биореакто-
рах периодического действия ......................... 229
10.7.2. Расчет и стратегия эксплуатации промышленных периоди-
ческих процессов..........................................237
10.7.3. Управление непрерывными процессами..................241
10.8. Заключение................................................245
Упражнения......................................................246
Литература..................................................251
Глава 11. ОПЕРАЦИИ ВЫДЕЛЕНИЯ В ПРОЦЕССАХ БИОХИМИЧЕ-
СКОЙ ТЕХНОЛОГИИ............................................255
11.1. Отделение клеток и нерастворимых твердых материалов . 257
11.1.1. Фильтрование........................................260
11.1.2. Центрифугирование...................................264
11.1.3. Седиментация........................................267
11.1.4. Перспективные методы выделения биомассы .... 270
11.1.5. Заключение..........................................270
11.2. Первичное выделение ......................................271
11.2.1. Экстракция..........................................271
11.2.2. Сорбция.............................................277
11.3. Осаждение.................................................279
11.3.1. Кинетика образования осадка.........................285
11.4. Хроматография и адсорбция в неподвижном слое; периодиче-
ские операции с селективной адсорбцией ....................... 289
11.5. Разделение с помощью мембран..............................303
Оглавление 589*
11.5.1. Обратный осмос.......................................303
11.5.2. Ультрафильтрация.....................................307
11.6. Электрофорез...............................................310
11.7. Сочетание нескольких операций разделения...................311
11.7.1. Иммобилизованные клетки .............................313
11.7.2. Операции переработки цельного бульона .... 313
11.7.3. Рециркуляция и операции выделения....................316
11.8. Последовательность операций выделения продуктов процессов
биохимической технологии ........................................ 318
11.8.1. Выделение ферментов в промышленных процессах . . . 319"
fГ.8.2. Выделение внутриклеточных чужеродных белков из куль-
тур рекомбинантных Е. coli.............................325
11.8.3. Выделение полисахаридов..............................325
11.8.4. Выделение антибиотиков...............................326
11.8.5. Выделение органических кислот........................329
11.8.6. Выделение этанола....................................329
11.8.7. Выделение белка одноклеточных организмов . . . 330
11.9. Заключение.................................................334
Упражнения.......................................................335
Литература...................................................344
Глава 12. ЭКОНОМИКА ПРОЦЕССОВ БИОХИМИЧЕСКОЙ ТЕХНО-
ЛОГИИ ..........................................................346
12.1. Экономика технологических процессов.................347
12.2. Контроль за качеством продукции биохимической технологии 350
12.3. Общий экономический анализ процессов биохимической техно-
логии .........................................................352
12.4. Экономический анализ биопроцесса....................356
12.5. Химические продукты тонких биотехнологических процессов 370
12.5.1. Ферменты......................................370
12.5.2. Производство белков с помощью рекомбинантных ДНК 371
12.5.3. Антибиотики...................................374
12.5.4. Витамины, алкалоиды, нуклеозиды, стероиды . . . 379
12.5.5. Моноклональные антитела........................ . 382
12.6. Кислородсодержащие химические продукты массового производ-
ства ..........................................................387
12.6.1. Пивоварение и виноделие............................387
12.6.2. Производство спирта в качестве топлива.............393
12.6.3. Производство органических кислот и аминокислот . . 399
12.7. Белок одноклеточных организмов ......................... 407
12.8. Анаэробные процессы производства метана..................411
12.9. Заключение...............................................413
Упражнения.....................................................414
Литература.................................................417
Глава 13. ИЗУЧЕНИЕ ВЗАИМОДЕЙСТВИЙ В СМЕШАННЫХ ПО-
ПУЛЯЦИЯХ МИКРООРГАНИЗМОВ...............................419
13.1. Нейтрализм, мутуализм, комменсализм и аменсализм . . . 419
13.2. Классификация взаимодействий между двумя видами . . . 427
13.3. Конкуренция: выживание наиболее приспособленного вида 432
13.3.1. Анализ межвидовой конкуренции по Вольтерра . . . 432
13.3.2. Конкуренция и отбор в хемостате..................436
13.4. Хищничество и паразитизм...............................442
590
Оглавление
13.4.1. Описание колебаний численности видов в системе хищ-
ник — жертва с помощью модели Лотки — Вольтерры 443
13.4.2. Применение модели Лотки — Вольтерры к системам, со-
стоящим из многих видов...................................447
13.4.3. Другие модели системы один вид хищников — один вид
жертв.....................................................449
13.5. Влияние количества видов и сети их взаимодействий . . . 457
13.5.1. Трофические уровни, пищевые цепи и пищевые сети; оп-
ределения и пример........................................457
13.5.2. Изучение динамики популяций с помощью моделей в фор-
ме закона действующих масс................................459
13.5.3. Качественная устойчивость..........................465
13.5.4. Устойчивость сложных неупорядоченных пищевых сетей 467
13.5.5. Бифуркации и усложненная динамика...............469
13.6. Расположение популяций в пространстве....................471
Упражнения.....................................................476
Литература.................................................481
Глава 14. СМЕШАННЫЕ ПОПУЛЯЦИИ МИКРООРГАНИЗМОВ
В ЕСТЕСТВЕННЫХ СИСТЕМАХ И ПРОМЫШЛЕННЫХ
ПРОЦЕССАХ.........................................................484
14.1. Применение смешанных культур определенного состава . . 486
14.2. Естественные смешанные популяции микроорганизмов и их роль
в производстве и порче продуктов ............................. 494
14.3. Участие микроорганизмов в естественных кругооборотах веществ 497
14.3.1. Кругооборот необходимых для жизни химических эле-
ментов ................................................. 498
14.3.2. Взаимосвязи микроорганизмов в почве и некоторых дру-
гих естественных экосистемах..............................502
14.4. Биологическая очистка сточных вод........................507
14.4.1. Основные характеристики сточных вод...............511
14.4.2. Процессы с участием активного ила.................513
14.4.3. Проектирование и моделирование процессов с участием
активного ила.............................................519
14.4.4. Аэробная переработка ила...........................529
14.4.5. Нитрификация.......................................531
14.4.6. Вторичная очистка сточных вод с помощью капельных
биологических фильтров .................................. 533
14.4.7. Анаэробная переработка отходов....................538
14.4.8. Математическое моделирование динамики процесса ана-
эробной переработки ила...................................542
14.4.9. Анаэробная денитрификация..........................554
14.4.10. Отделение фосфорсодержащих соединений .... 555
Упражнения................................................... 555
Литература.................................................562
Указатель........................................................564
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформ-
лении, качестве перевода и другие просим присы-
лать по адресу:
129820, Москва, И-110, ГСП, 1-й Рижский пер.г
д. 2, издательство «Мир».
Учебное издание
Джеймс Э. Бейли, Дэвид Ф. Оллис
ОСНОВЫ БИОХИМИЧЕСКОЙ ИНЖЕНЕРИИ
ч. 2
Заведующий редакцией академик О. А. Реутов
Зам. зав. редакцией |з. Ф. Ходецка?.
Научный редактор Б. М. Комарова
Мл. научный редактор И. И. Землячева
Художник А. Д. Смеляков
Художественный редактор М. Н. Кузьмина
Технический редактор Е. В. Алехина
Корректор Н. А. Мистрюкова
ИБ Xs 6715
Сдано в набор 22.12.88. Подписано к печати 24.05.89.
Формат 60X90'/ie. Бумага книжно-журнальная. Печать
высокая. Гарнитура Литературная. Объем 18,5 бум. л.
Усл. печ. л. 37,00. Усл. кр.-отт. 37,00. Уч.-изд. л. 39,37.
Изд. Xs 3/5891. Тираж 7500 экз. Зак. 746.
Цена 4 р. 70 к.
Издательство «Мир» В/О «Совэкспорткнига» Государст-
венного комитета СССР по делам издательств, полигра-
фии и книжной торговли. 129820, ГСП, Москва, И-110,
1-й Рижский пер., 2.
Московская типография Xs 11 Союзполиграфпрома при
Государственном комитете СССР по делам издательств,
полиграфии и книжной торговли. 113105, Москва, Нага-
тинская ул., д. 1.
Фотографировал Семенюченко Владимир
chem vova@mail. univ. kiev. ua; vova2002@mail. ru
Дж.Бейли, Д.Оллис
основы
биохимической
инженерии
Издательство „Мир‘