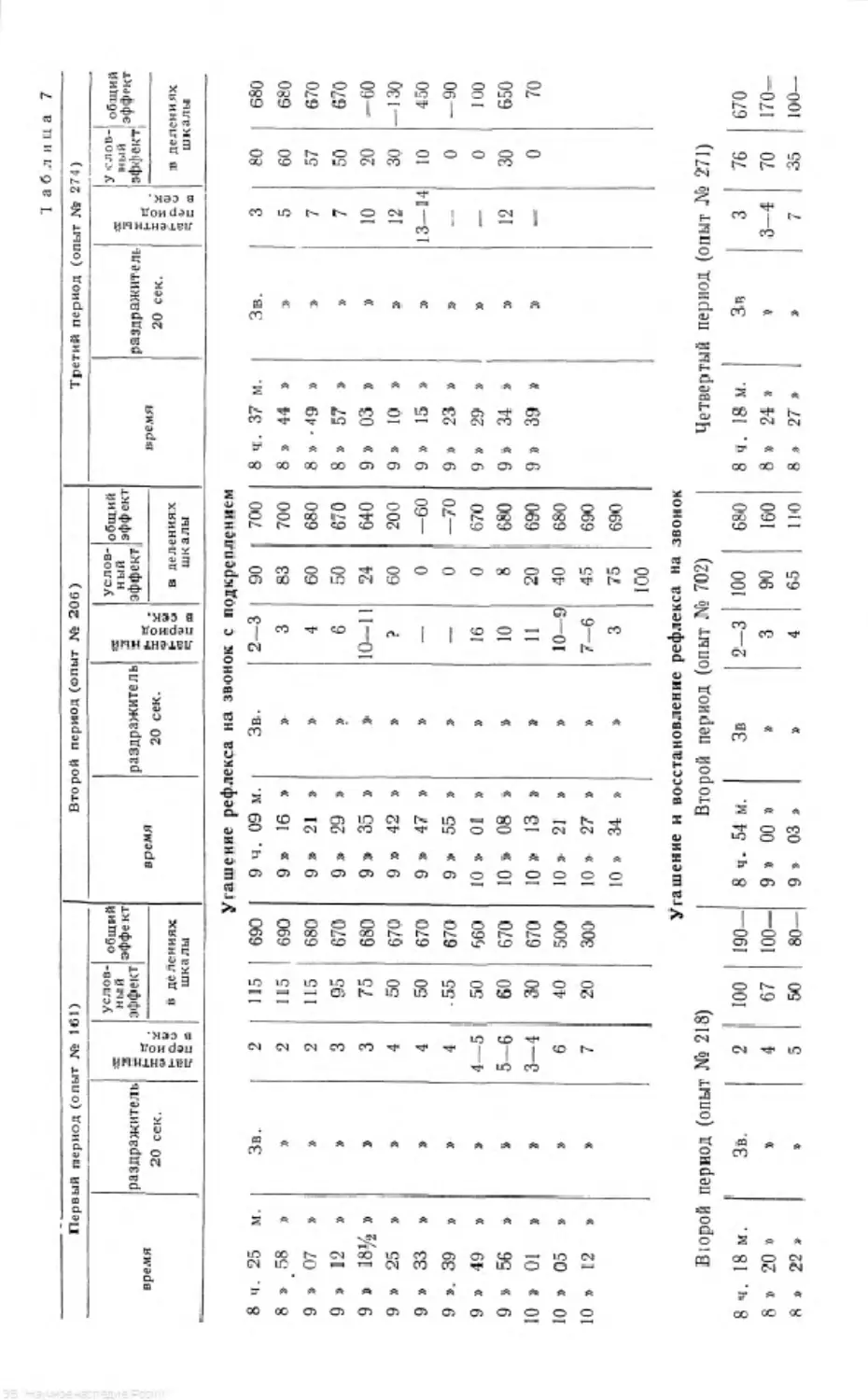
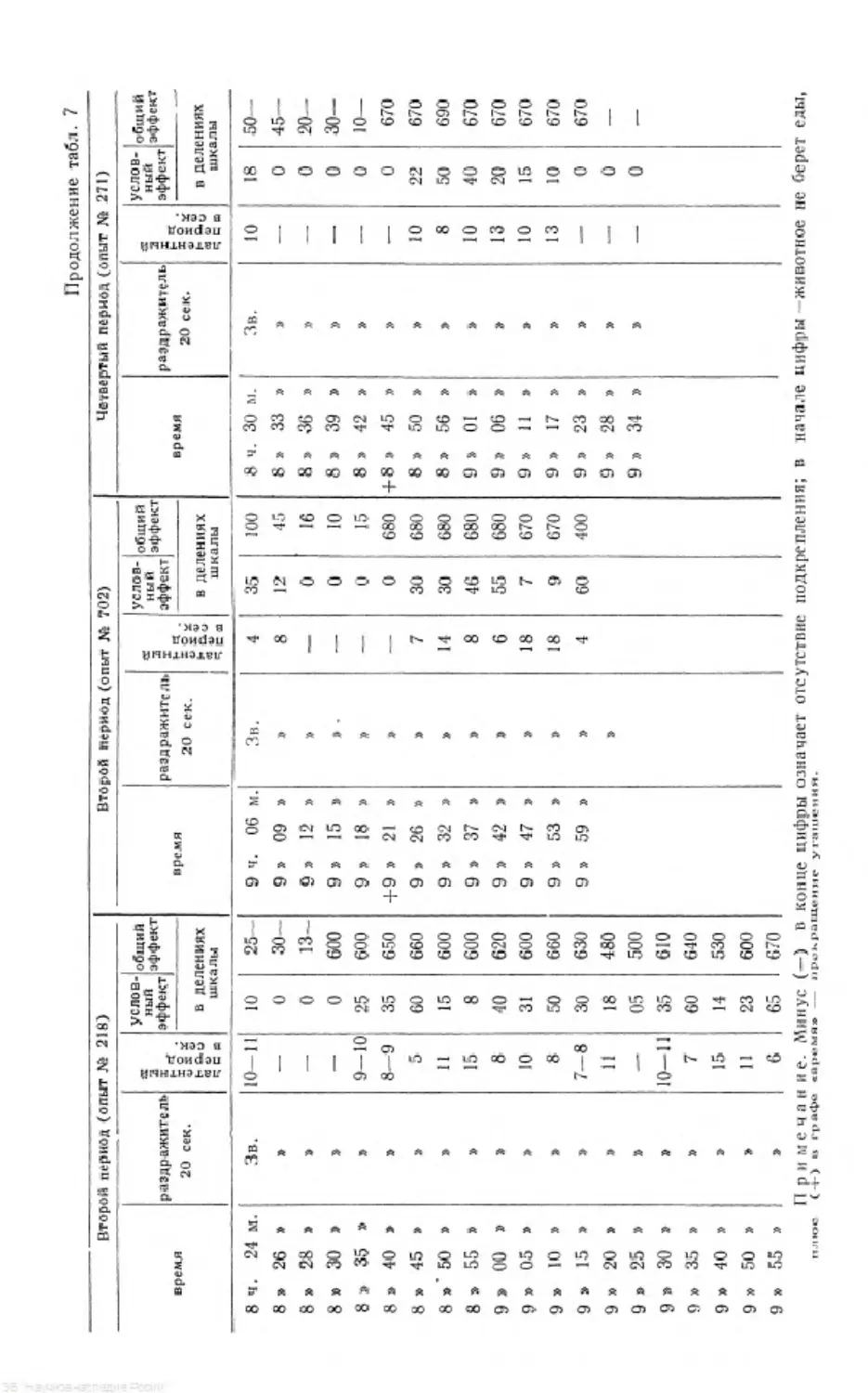
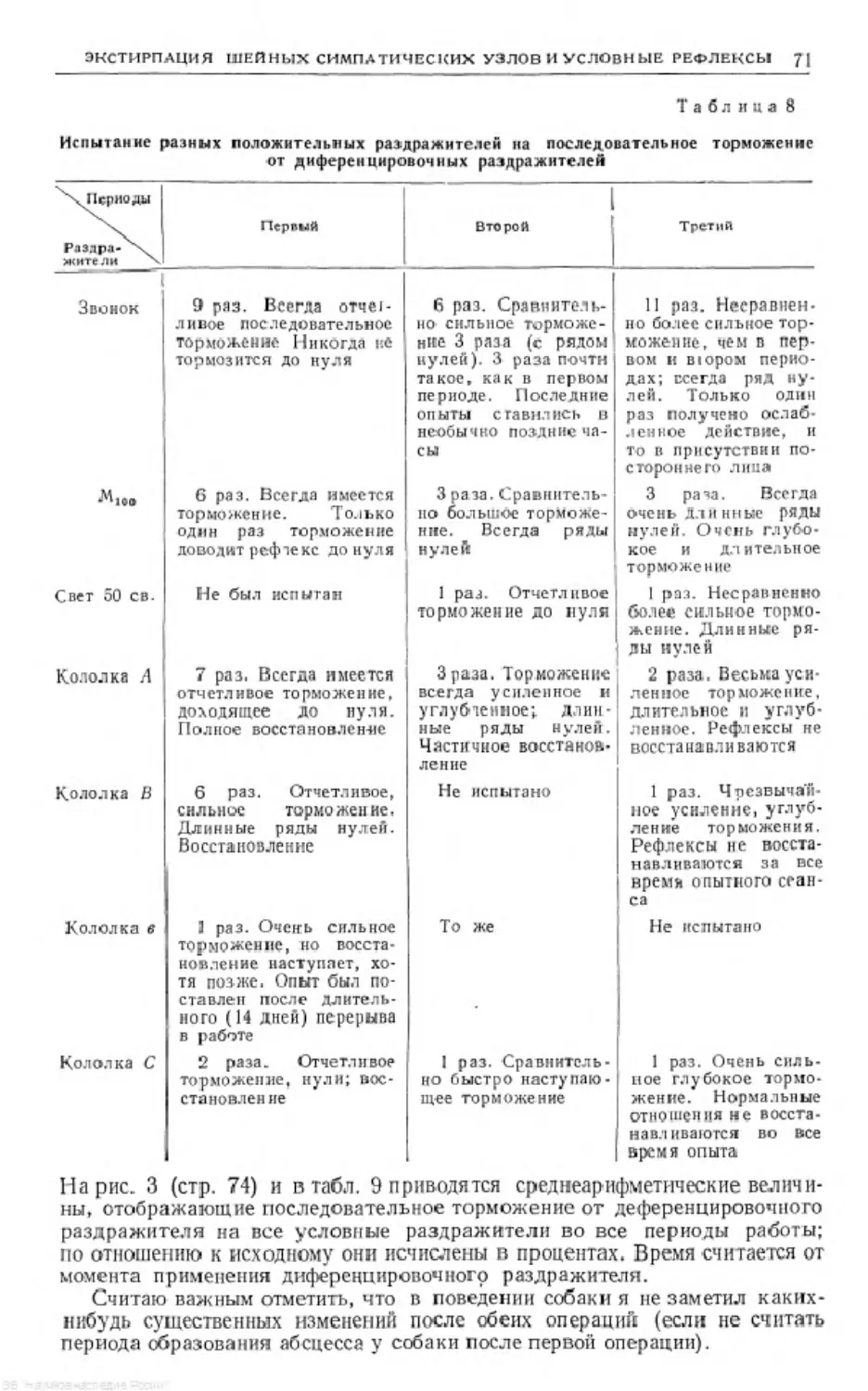
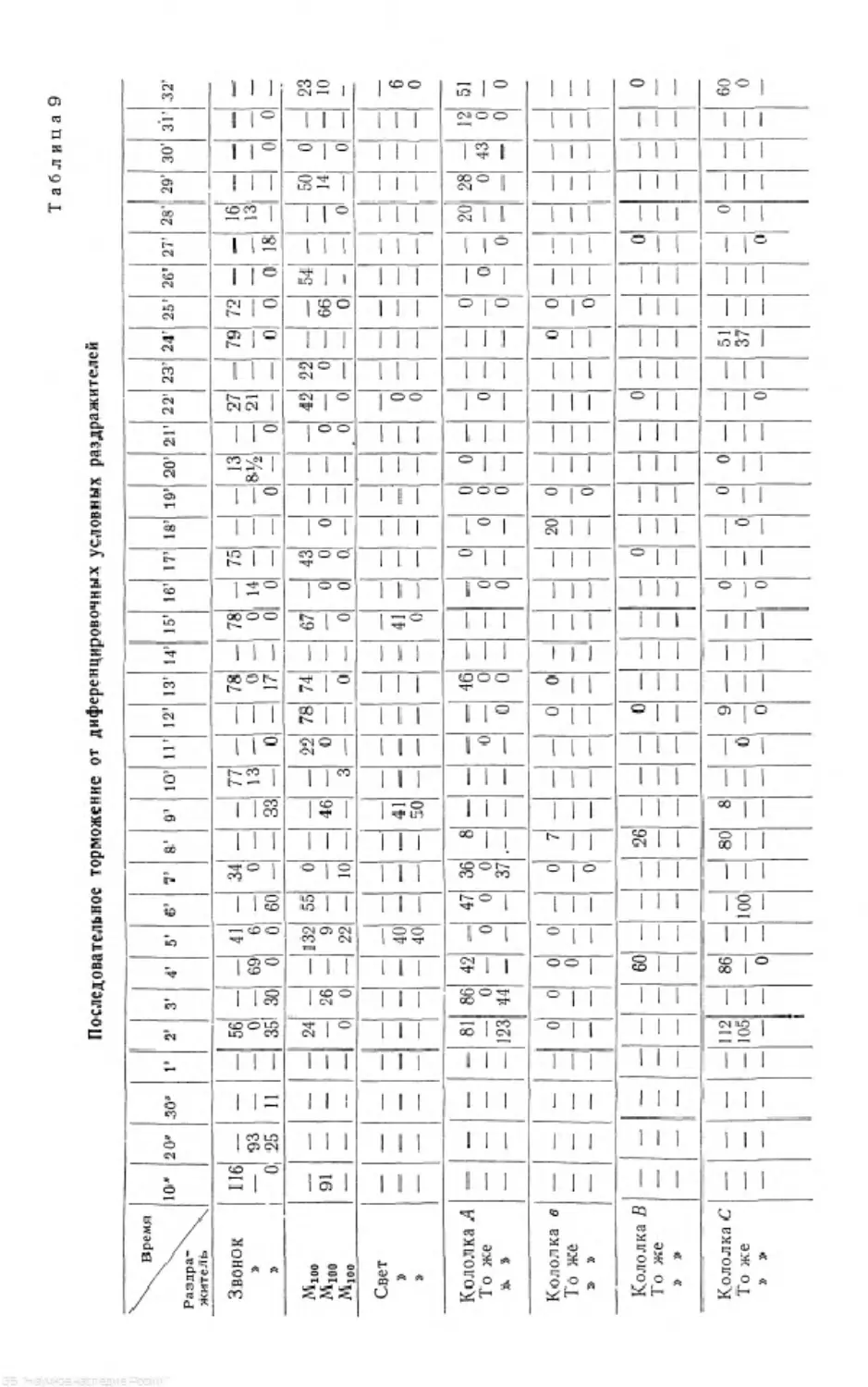
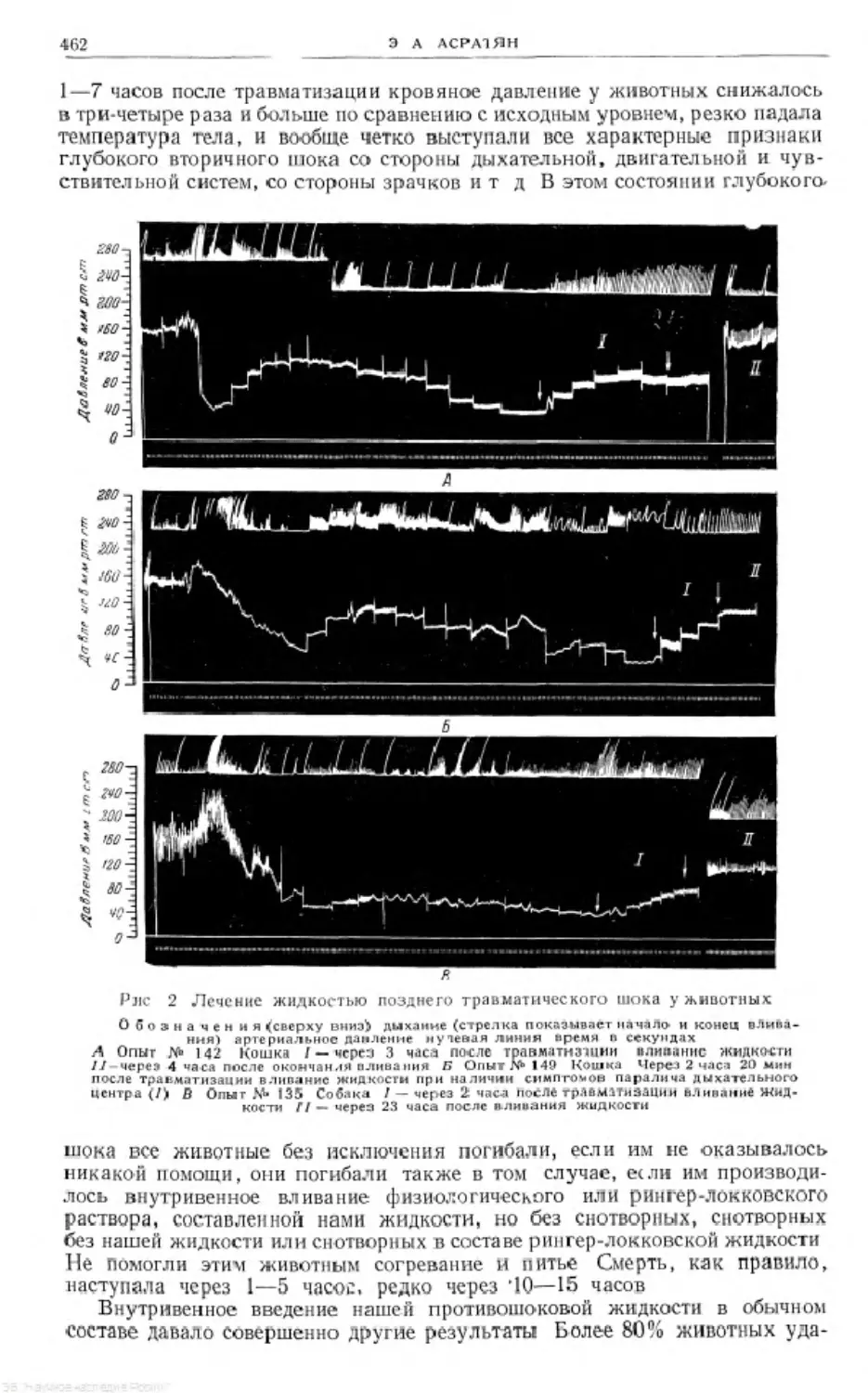
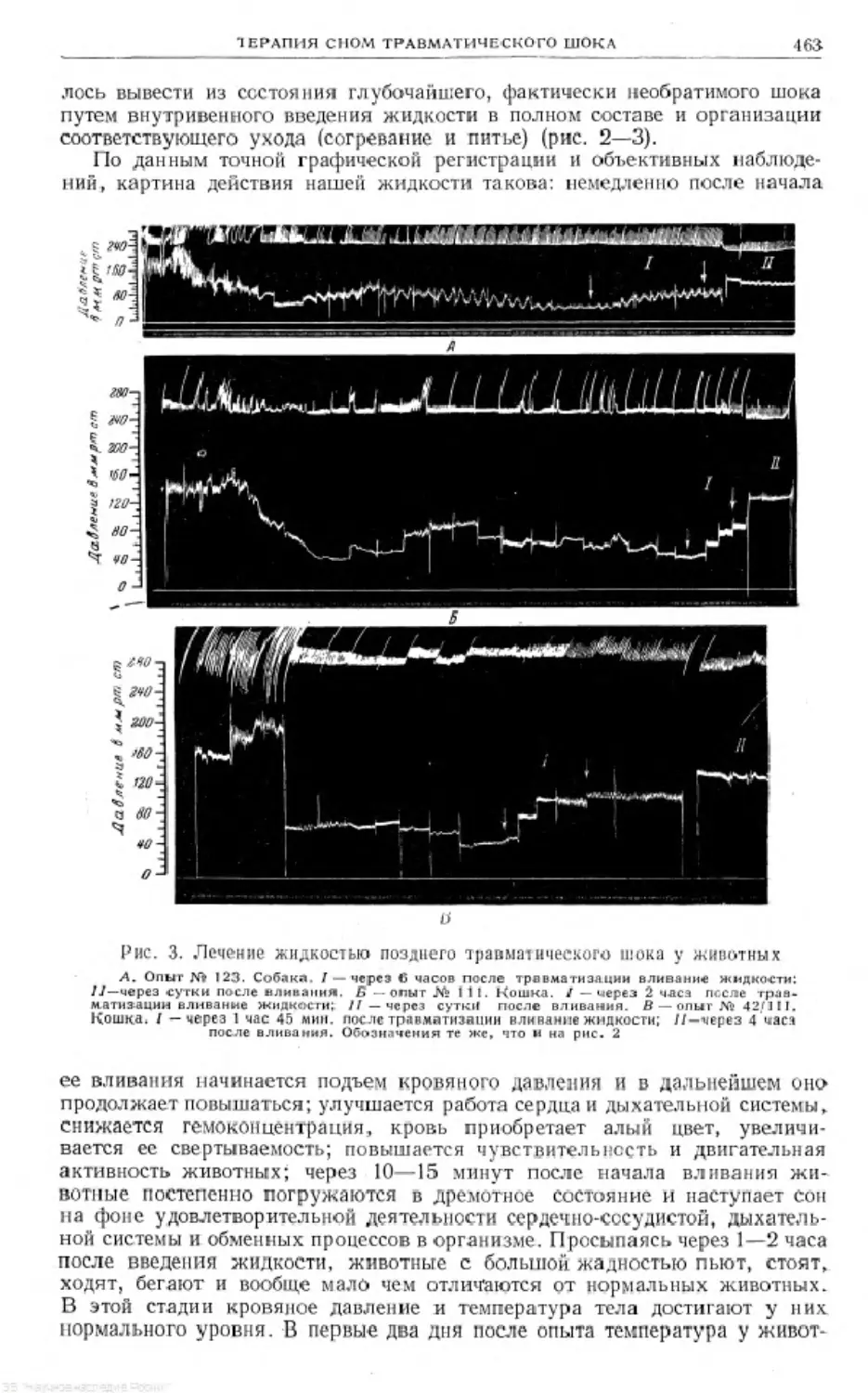
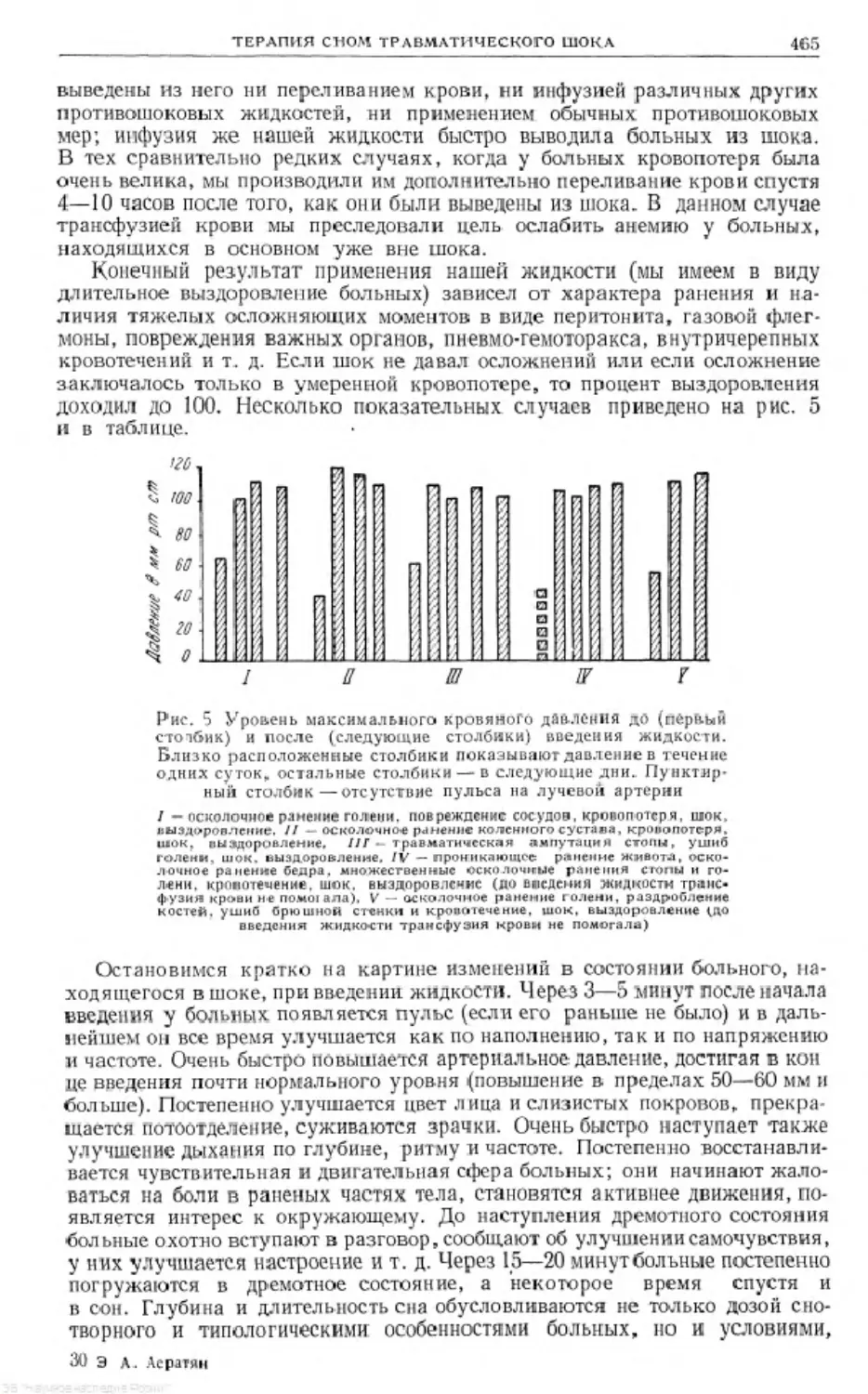
/

































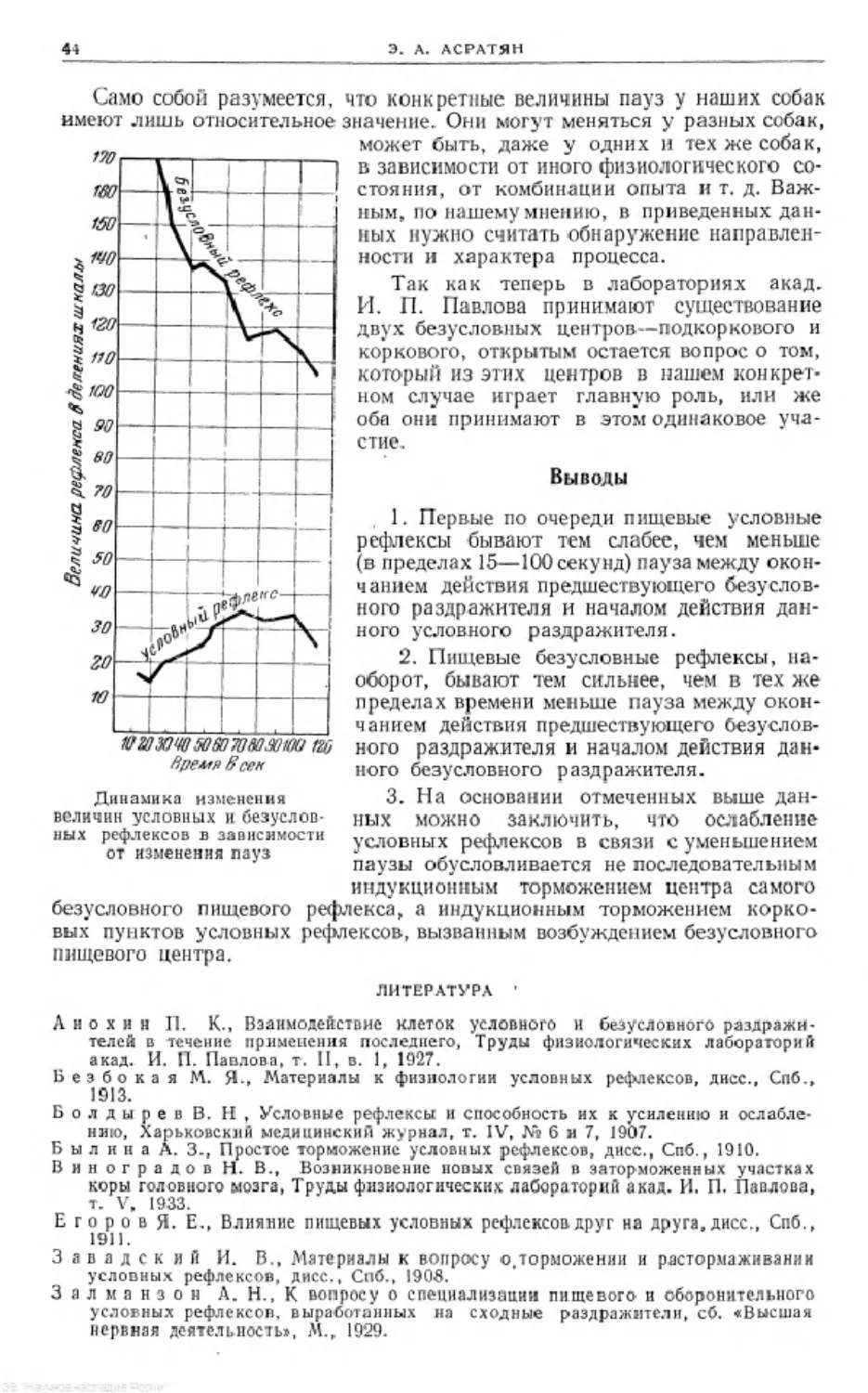









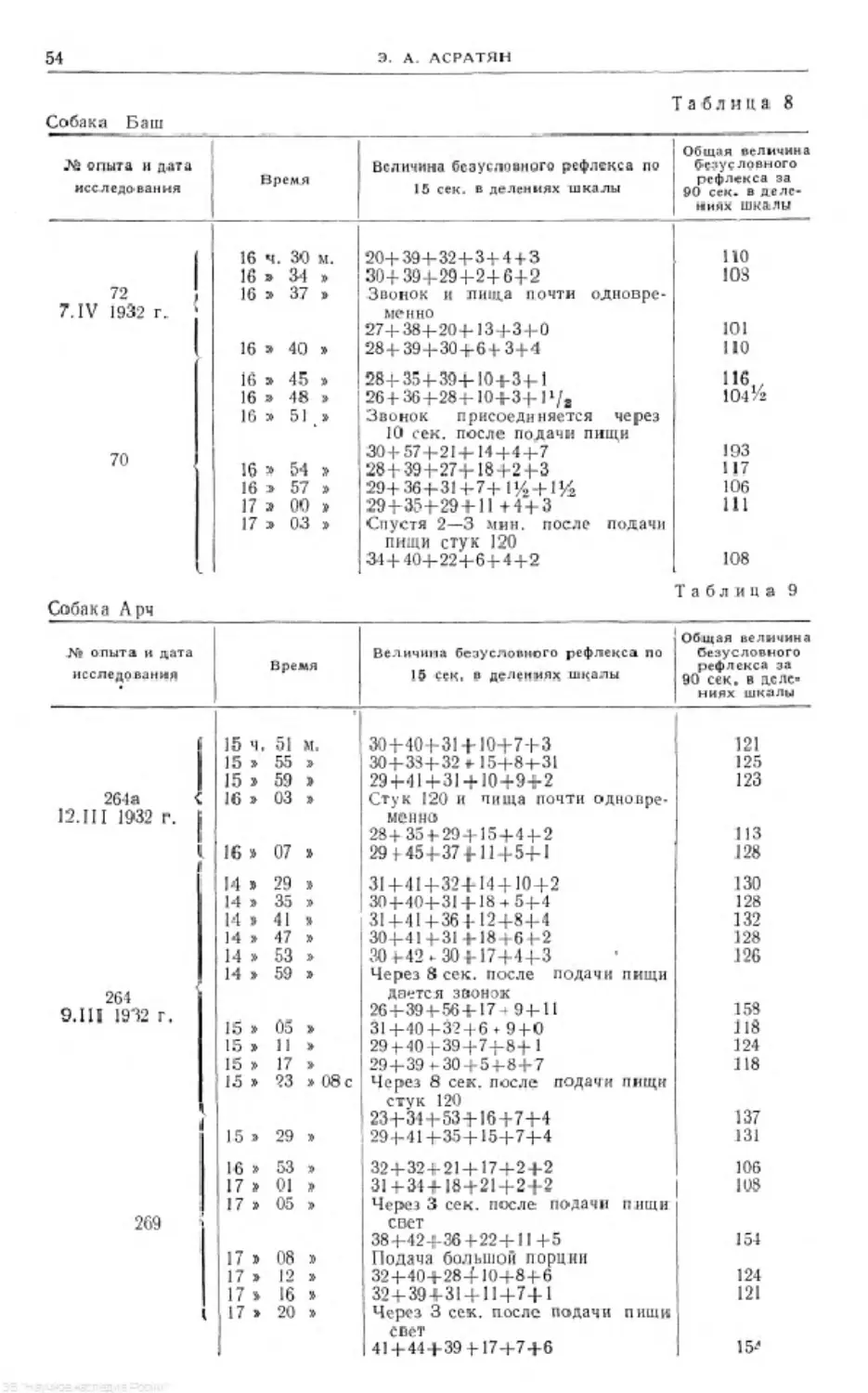

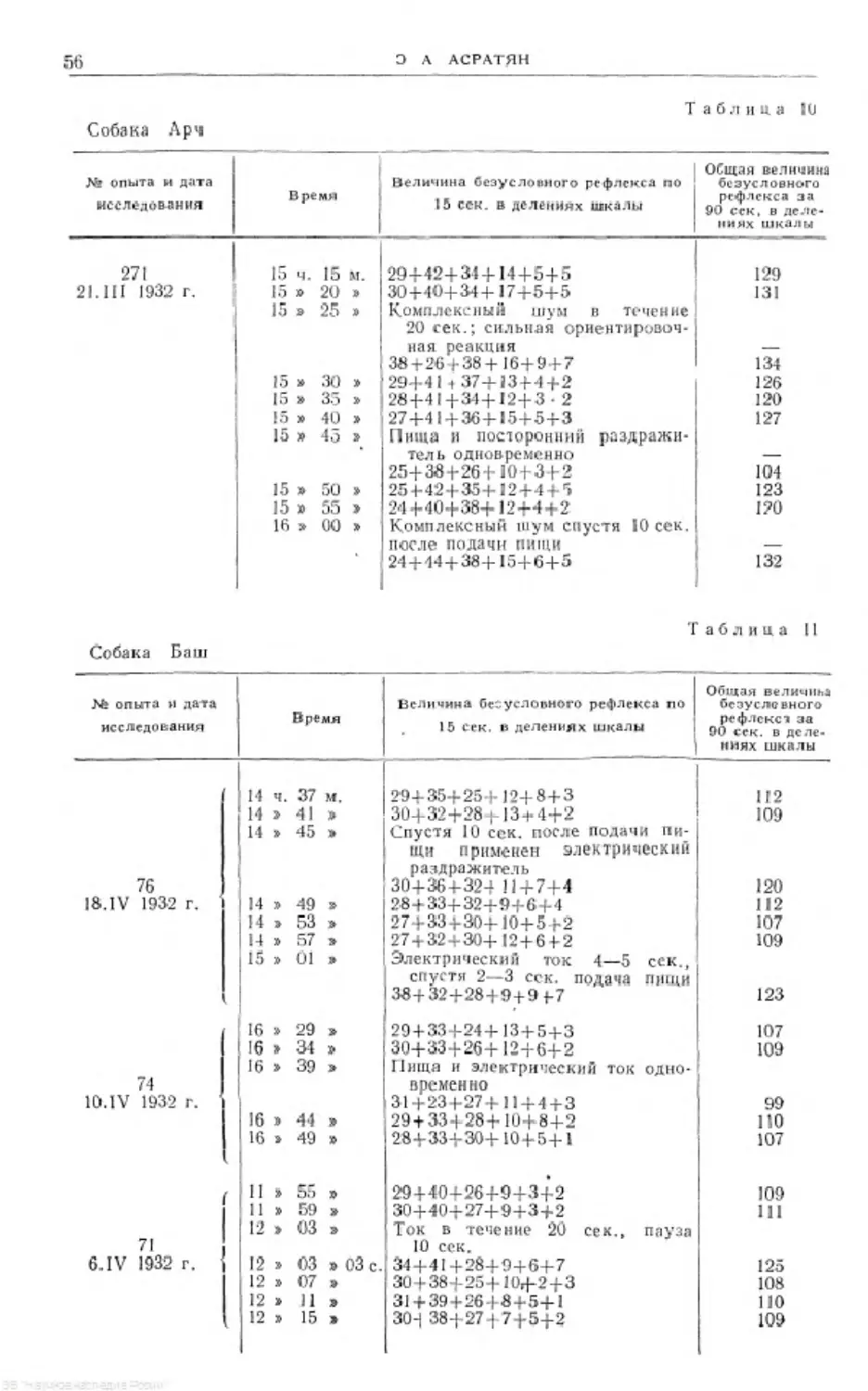



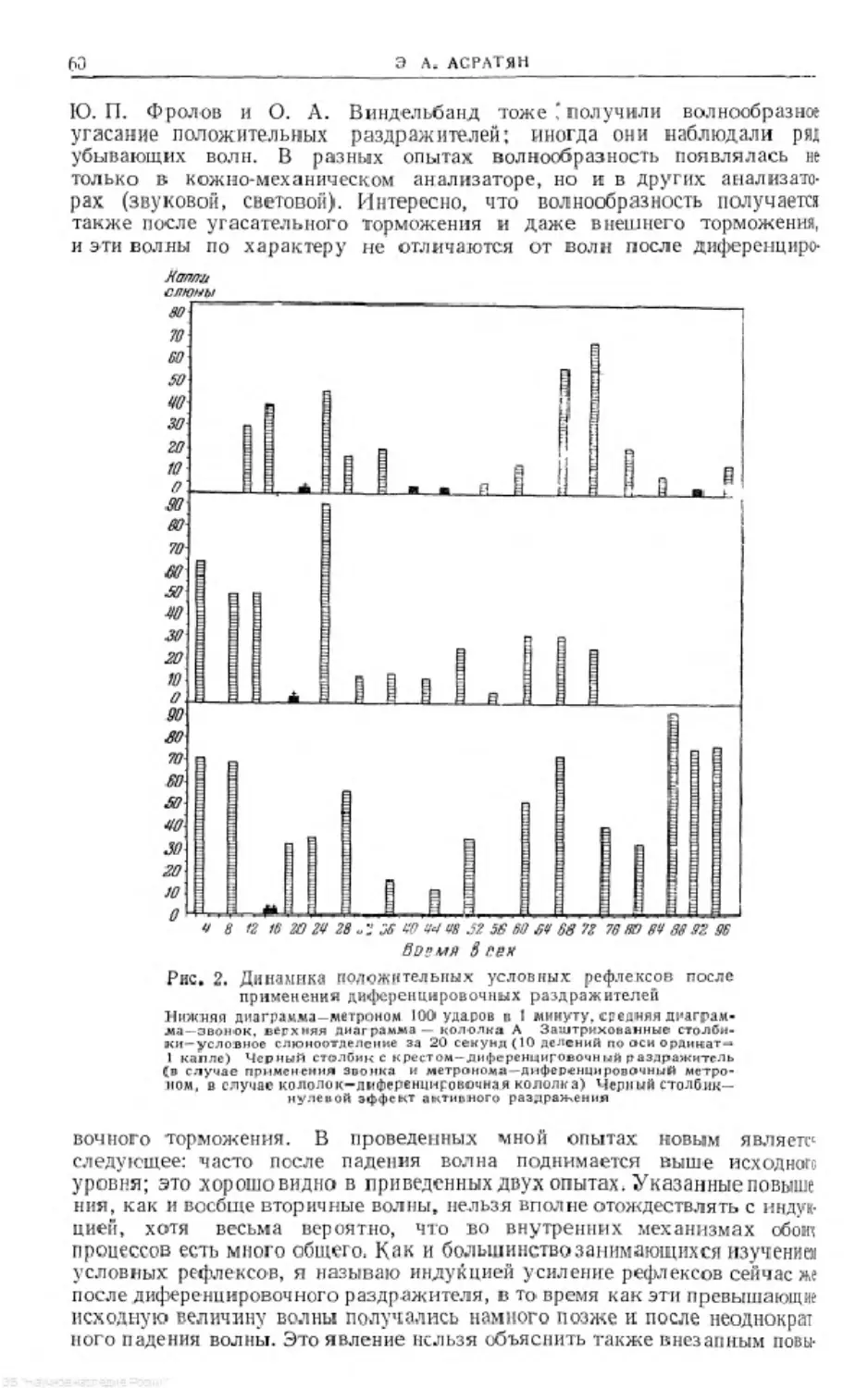
























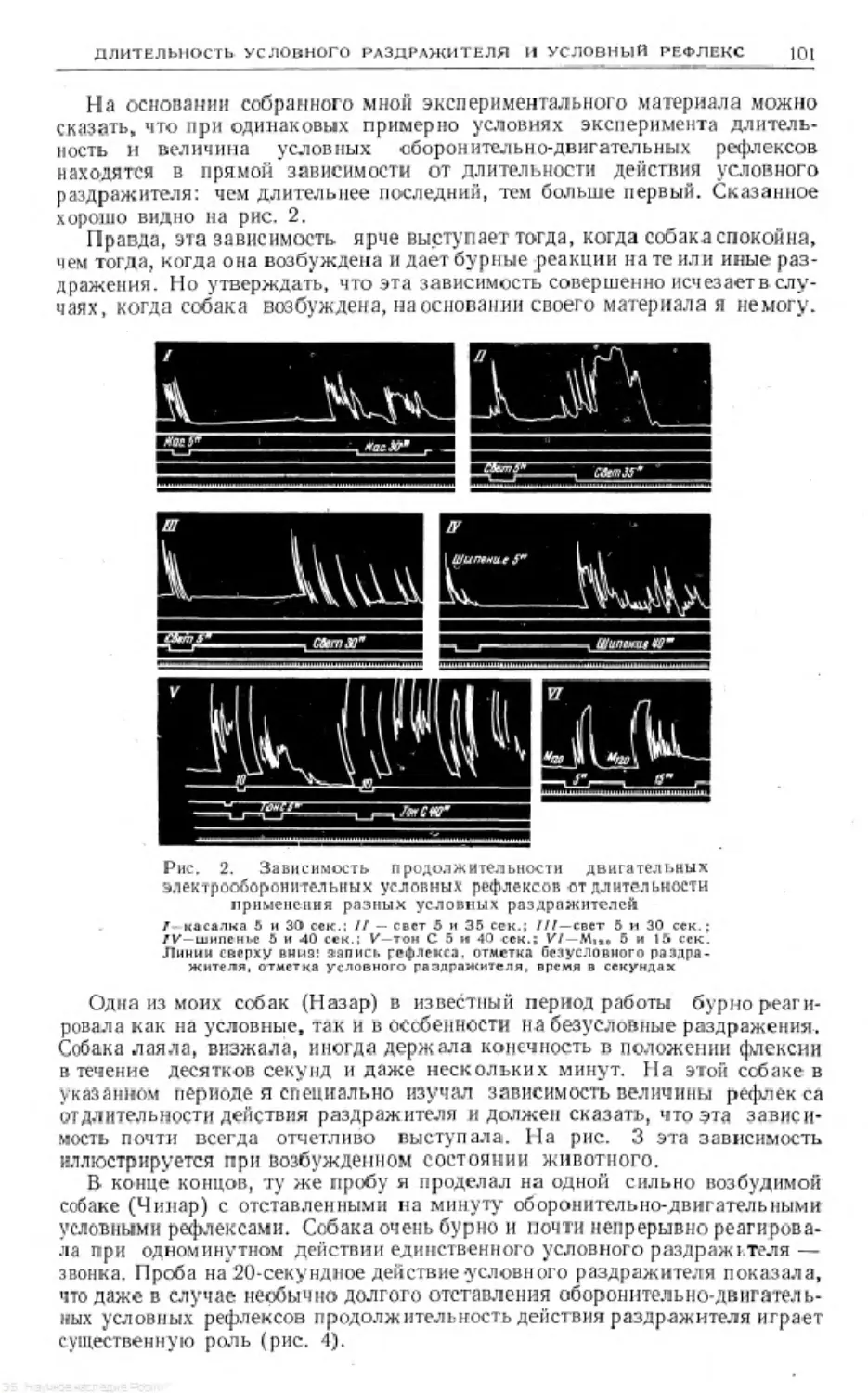













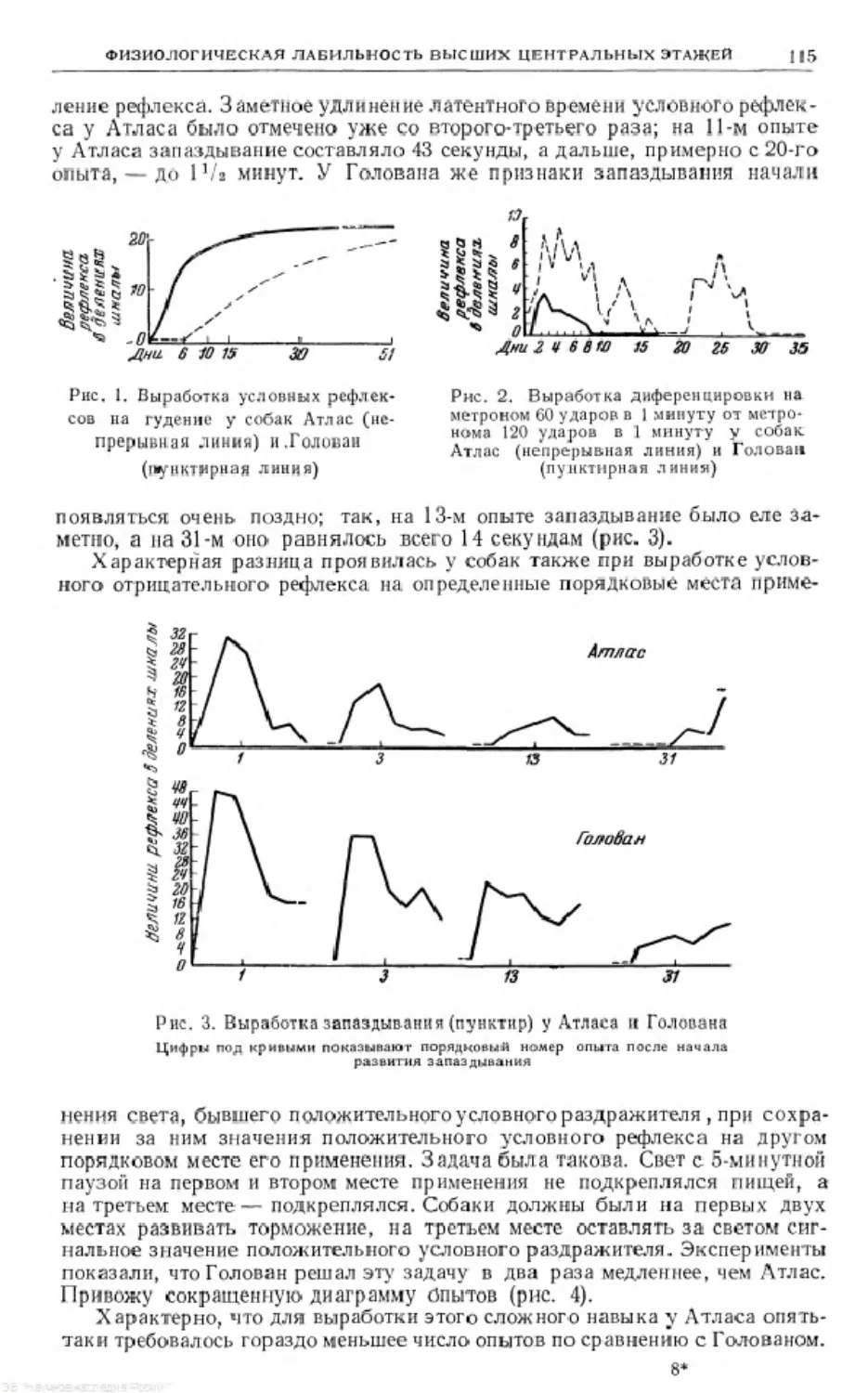
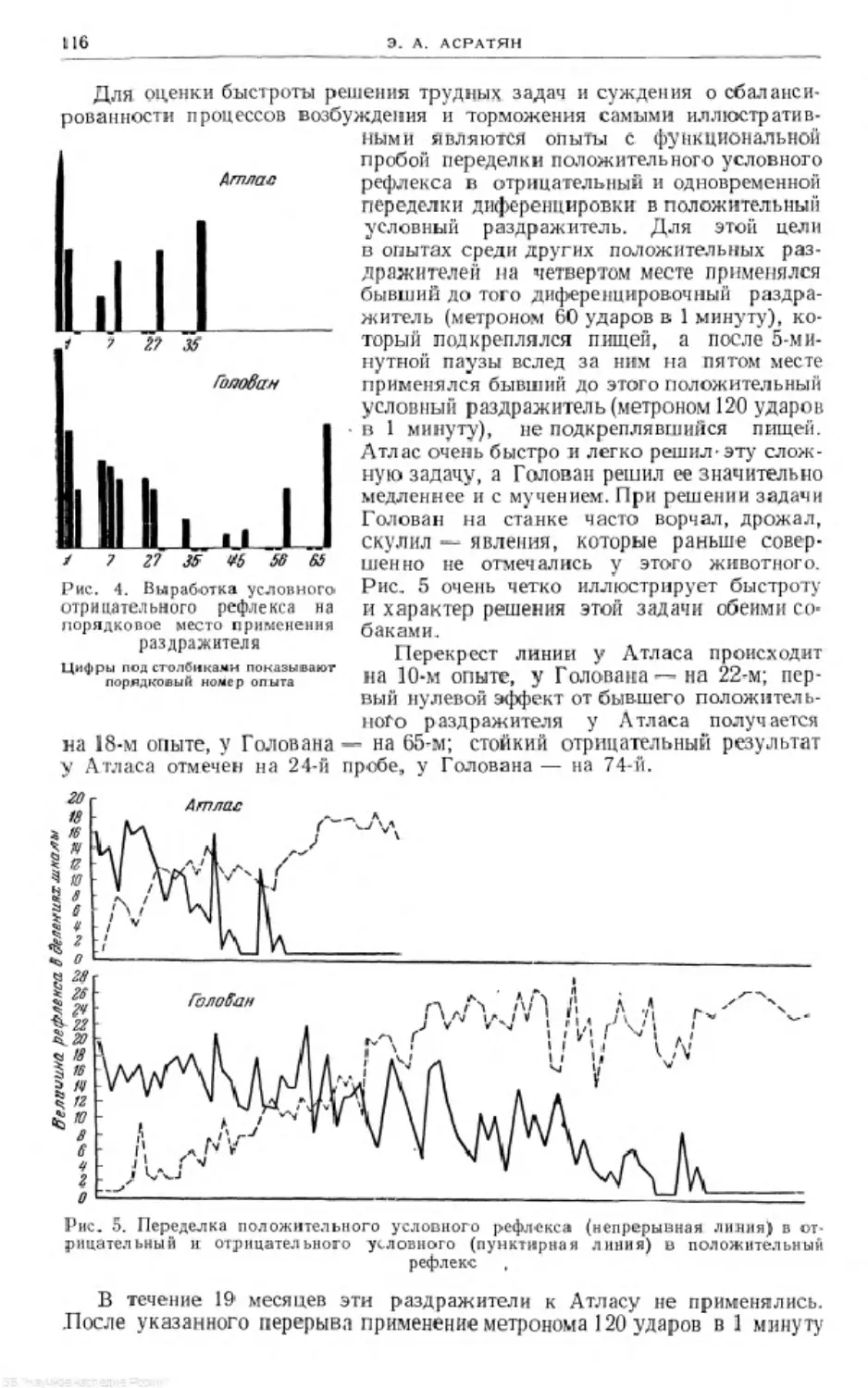

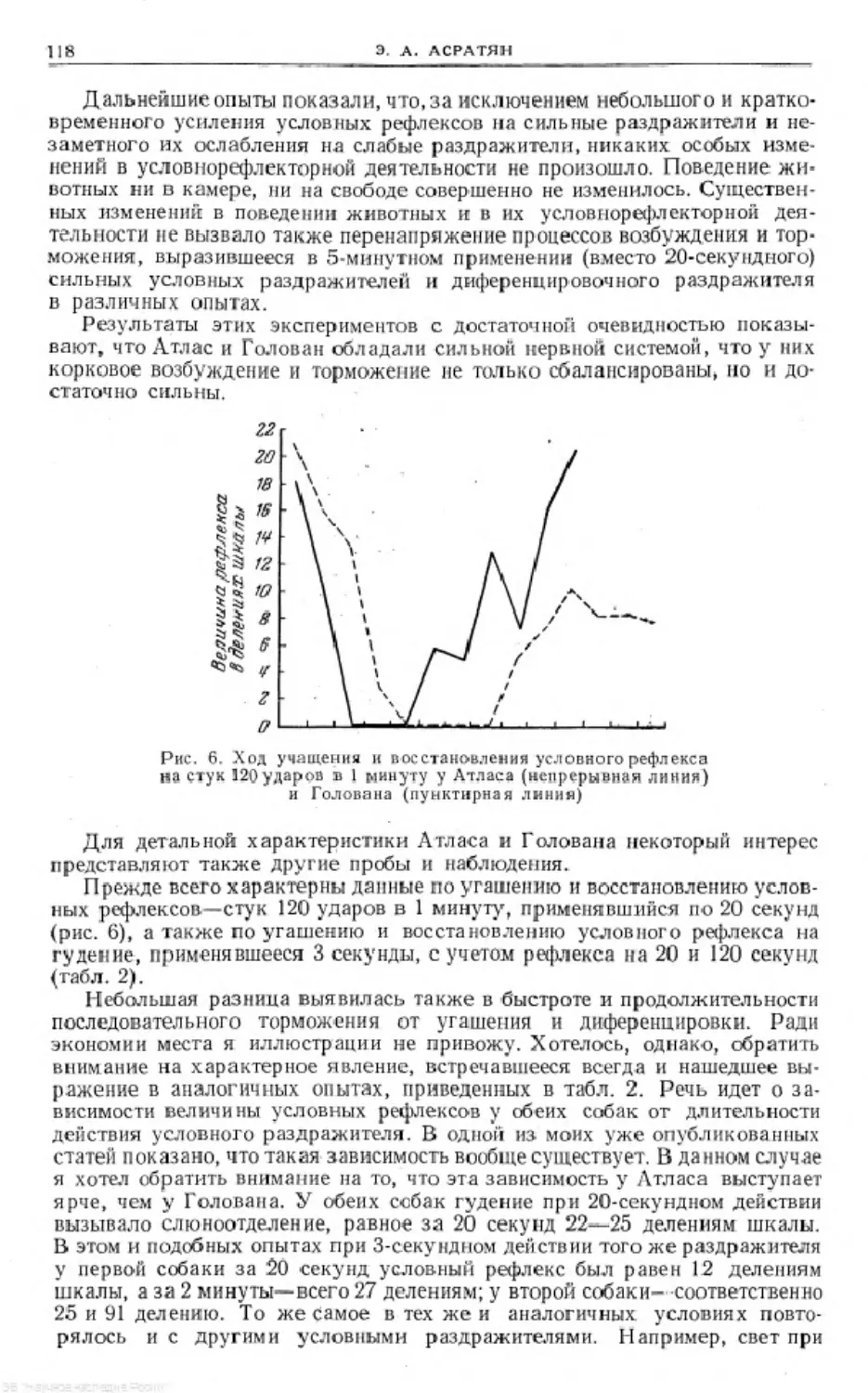














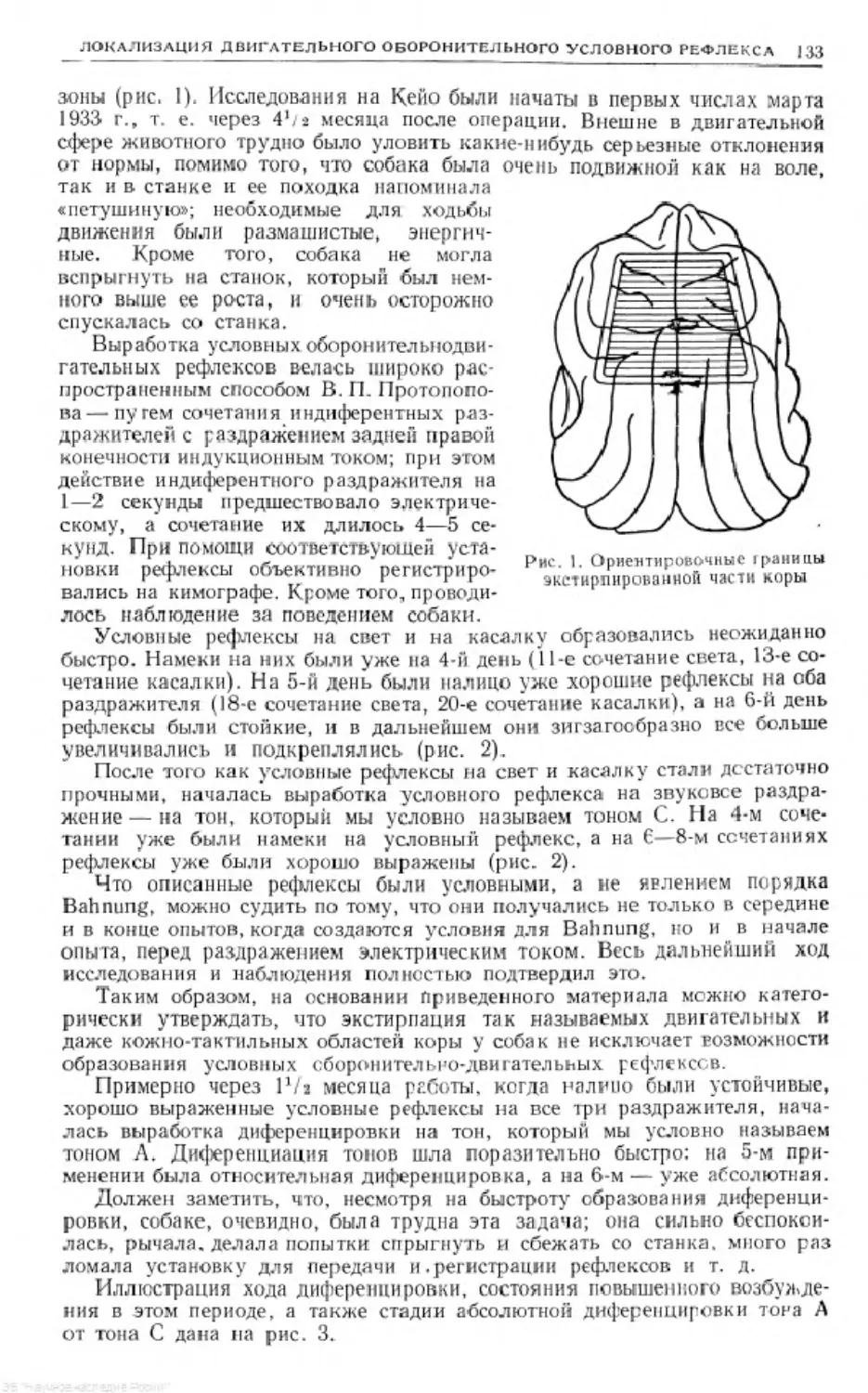



































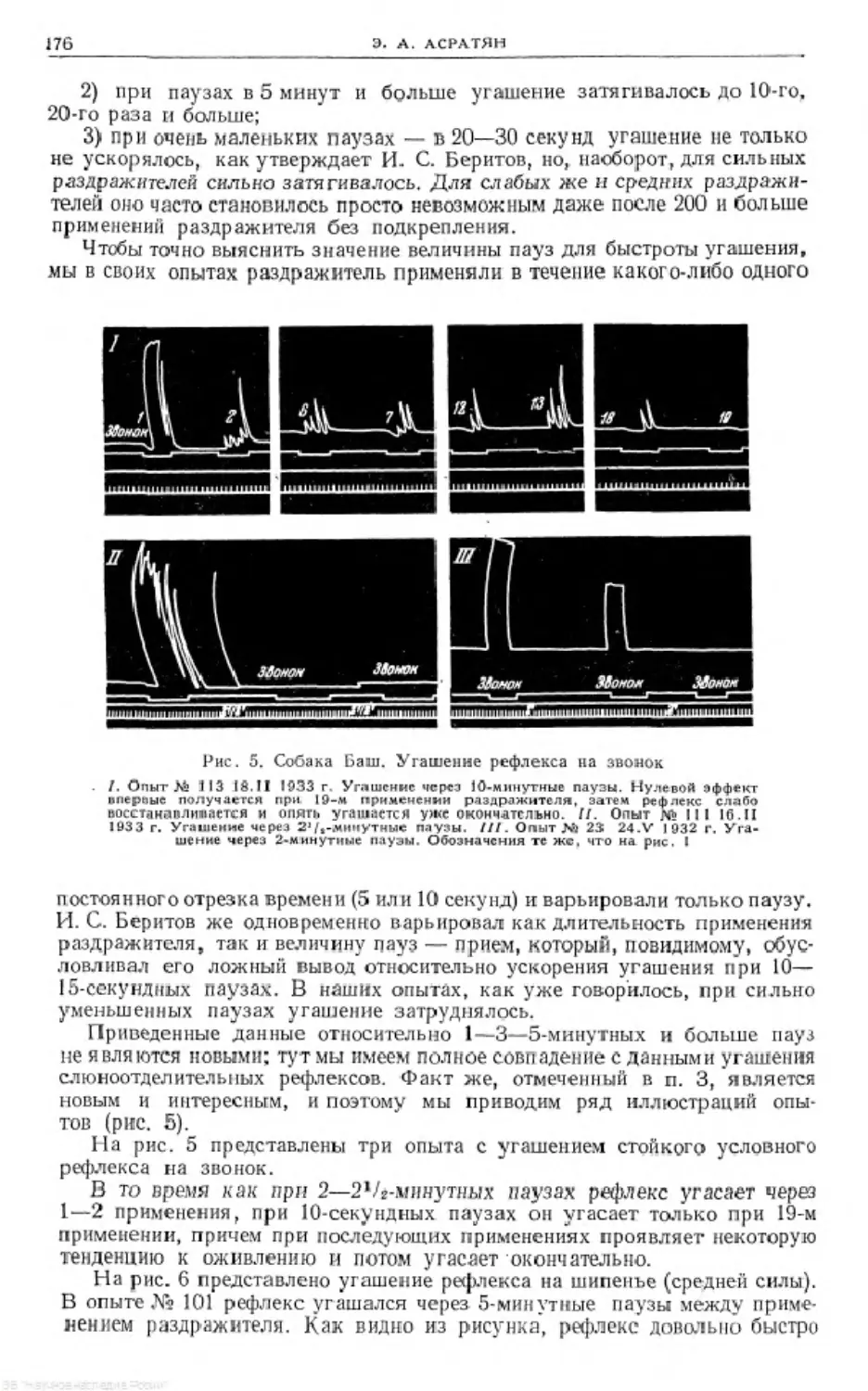
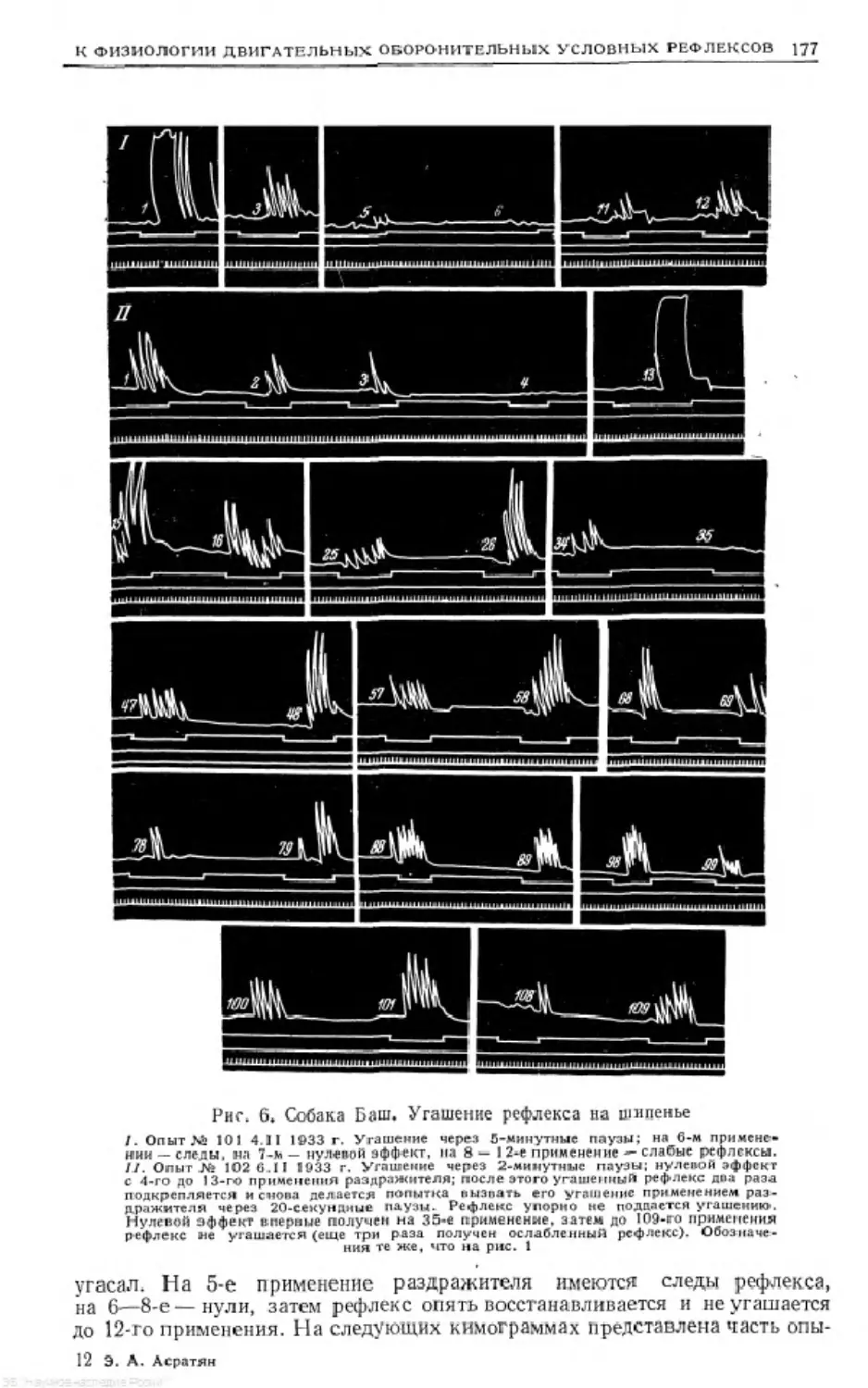






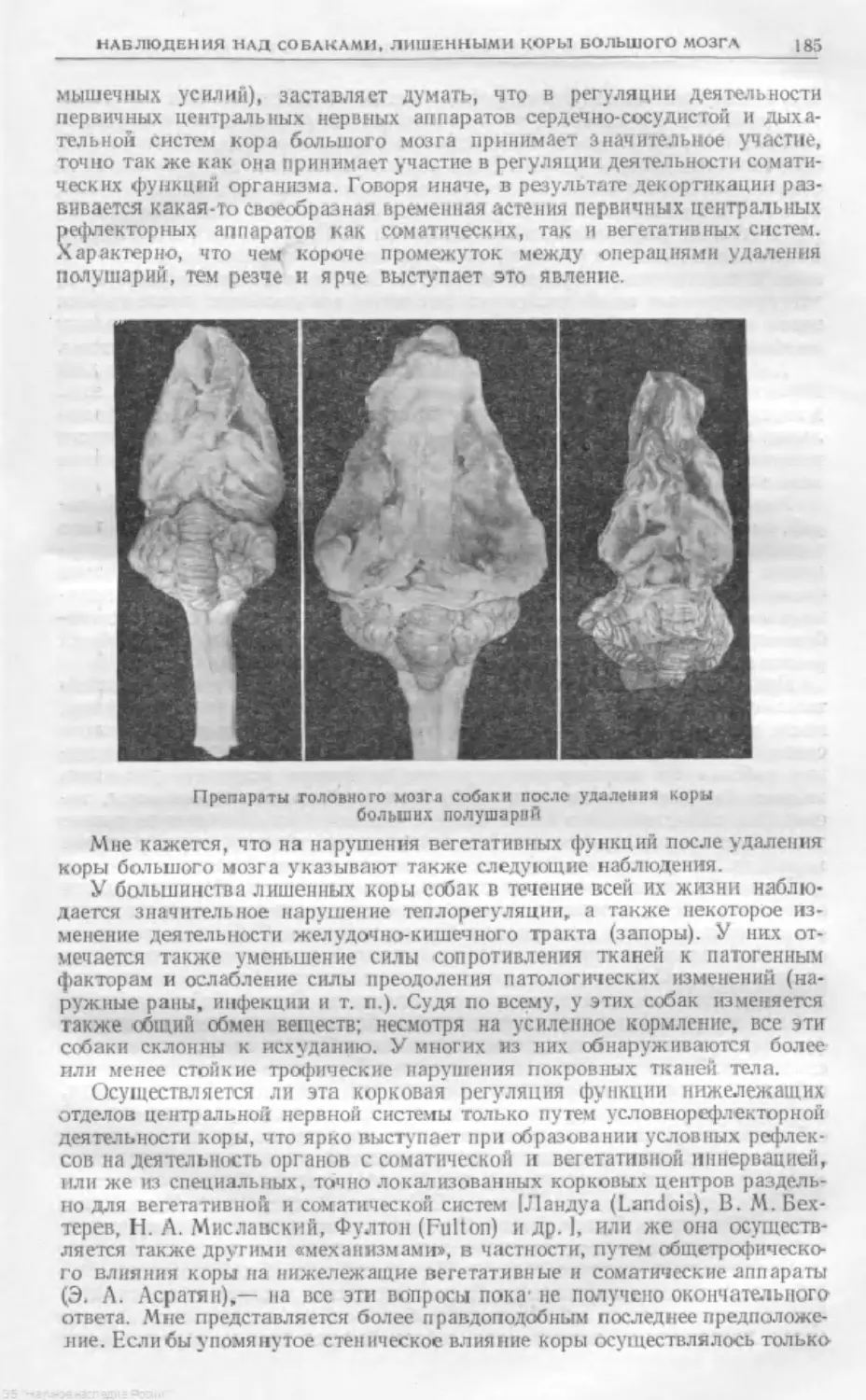


















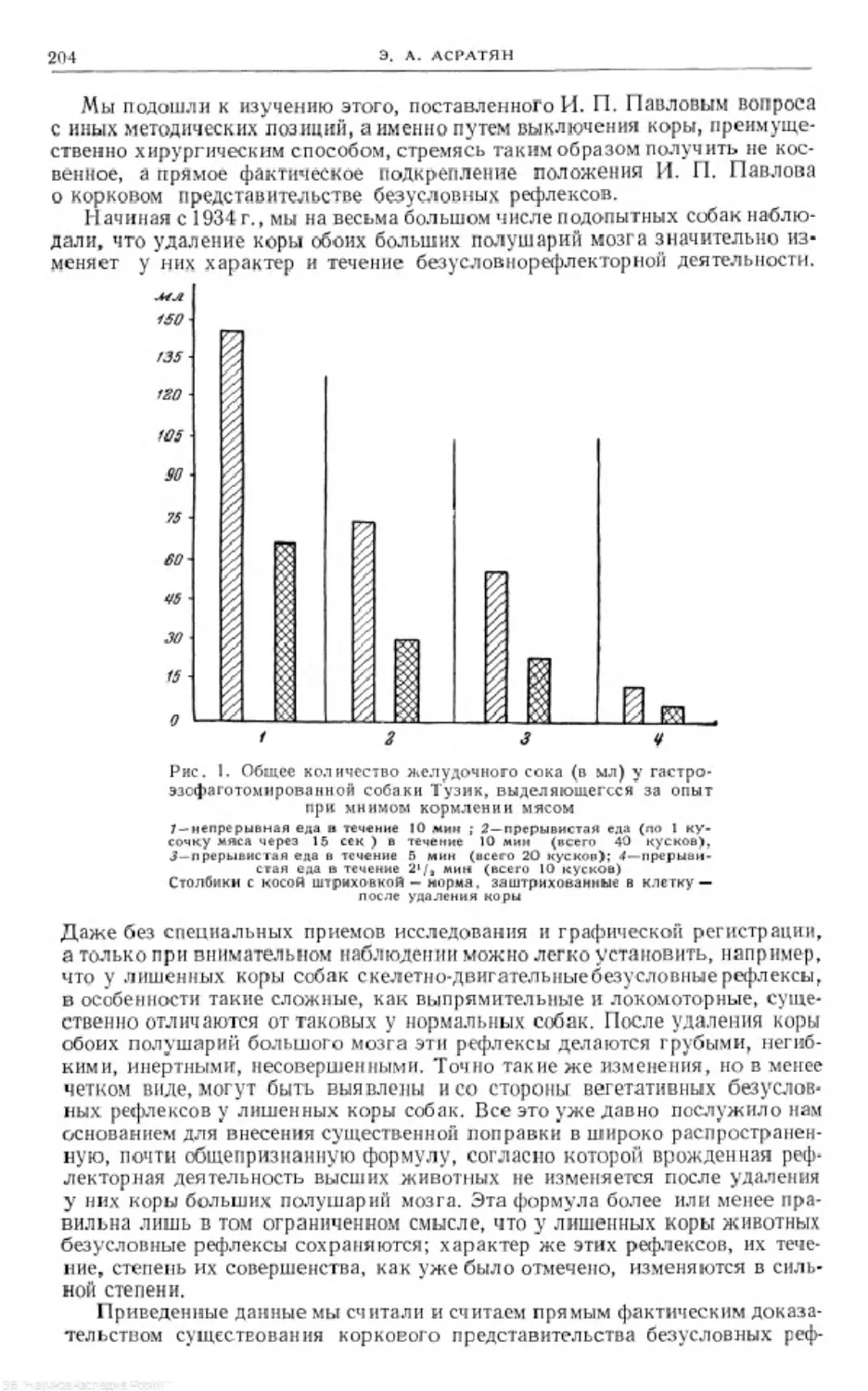
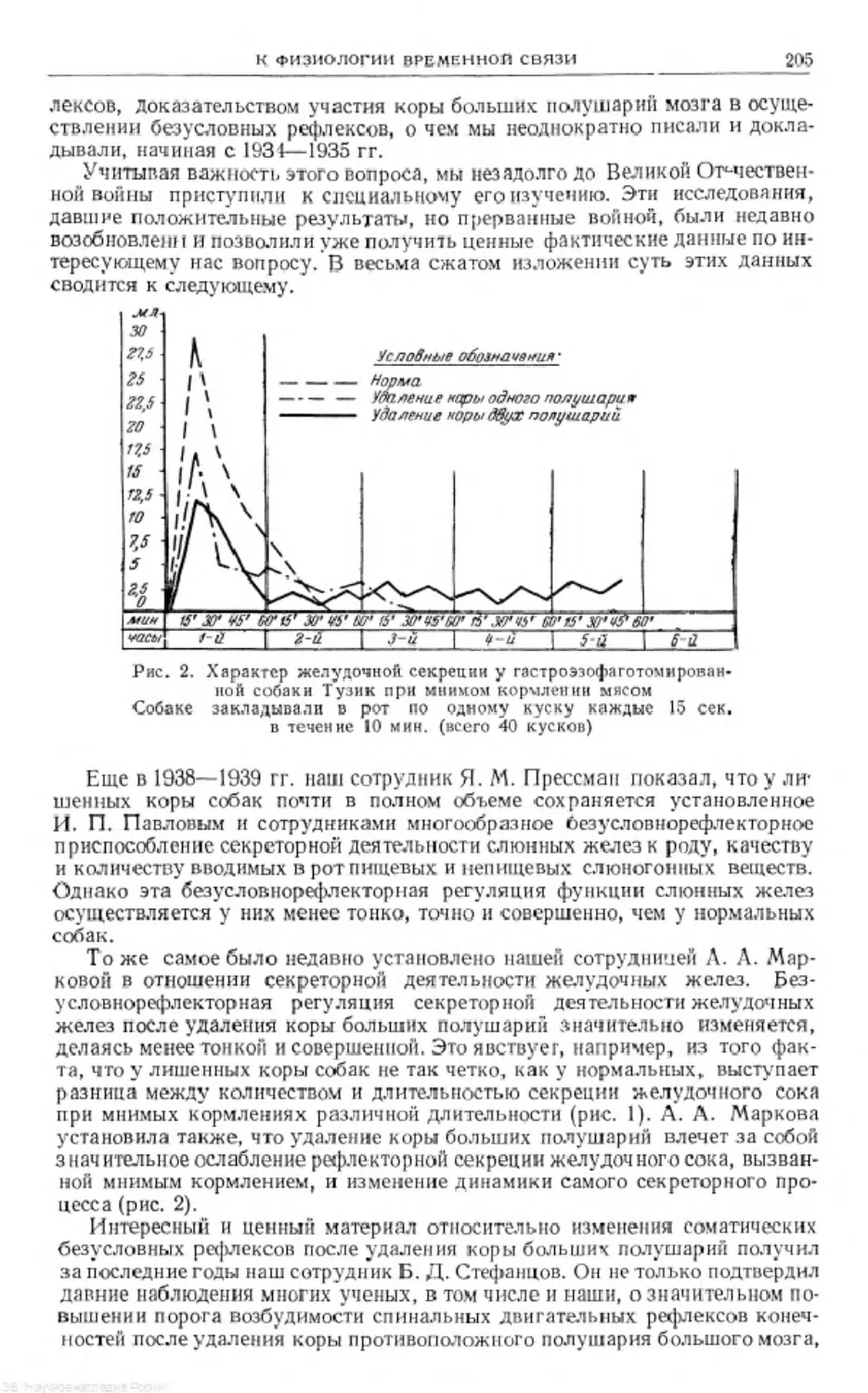
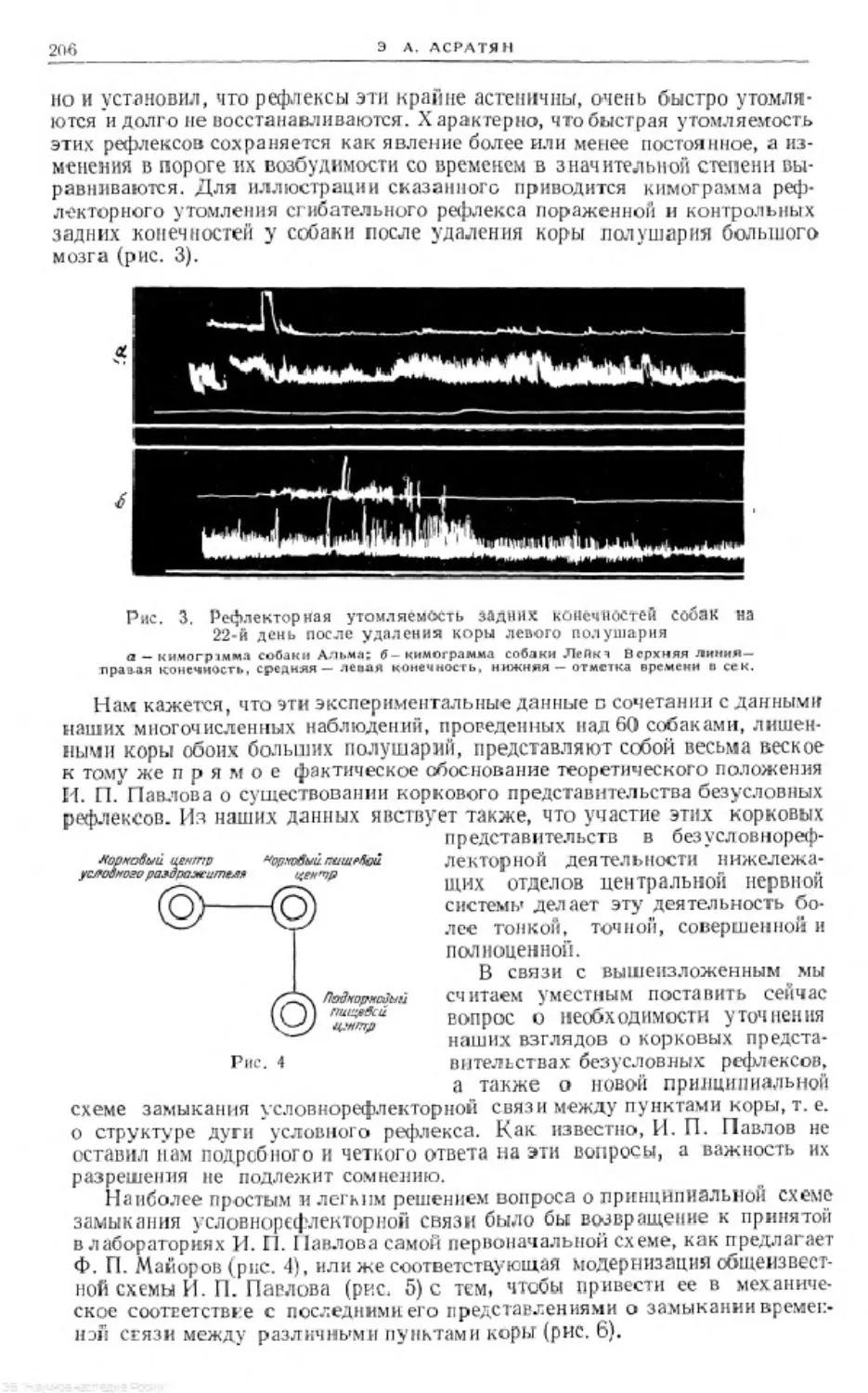


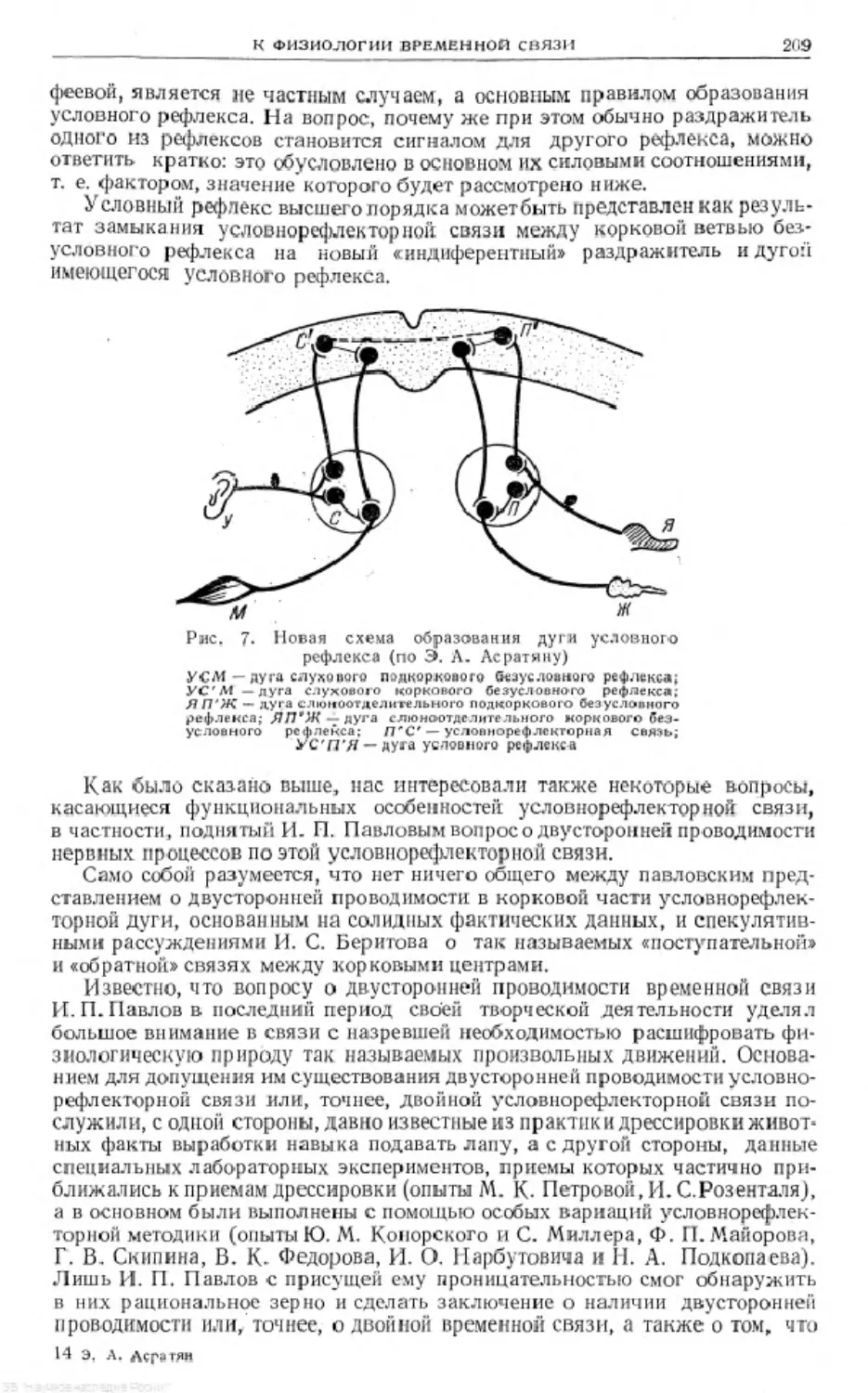









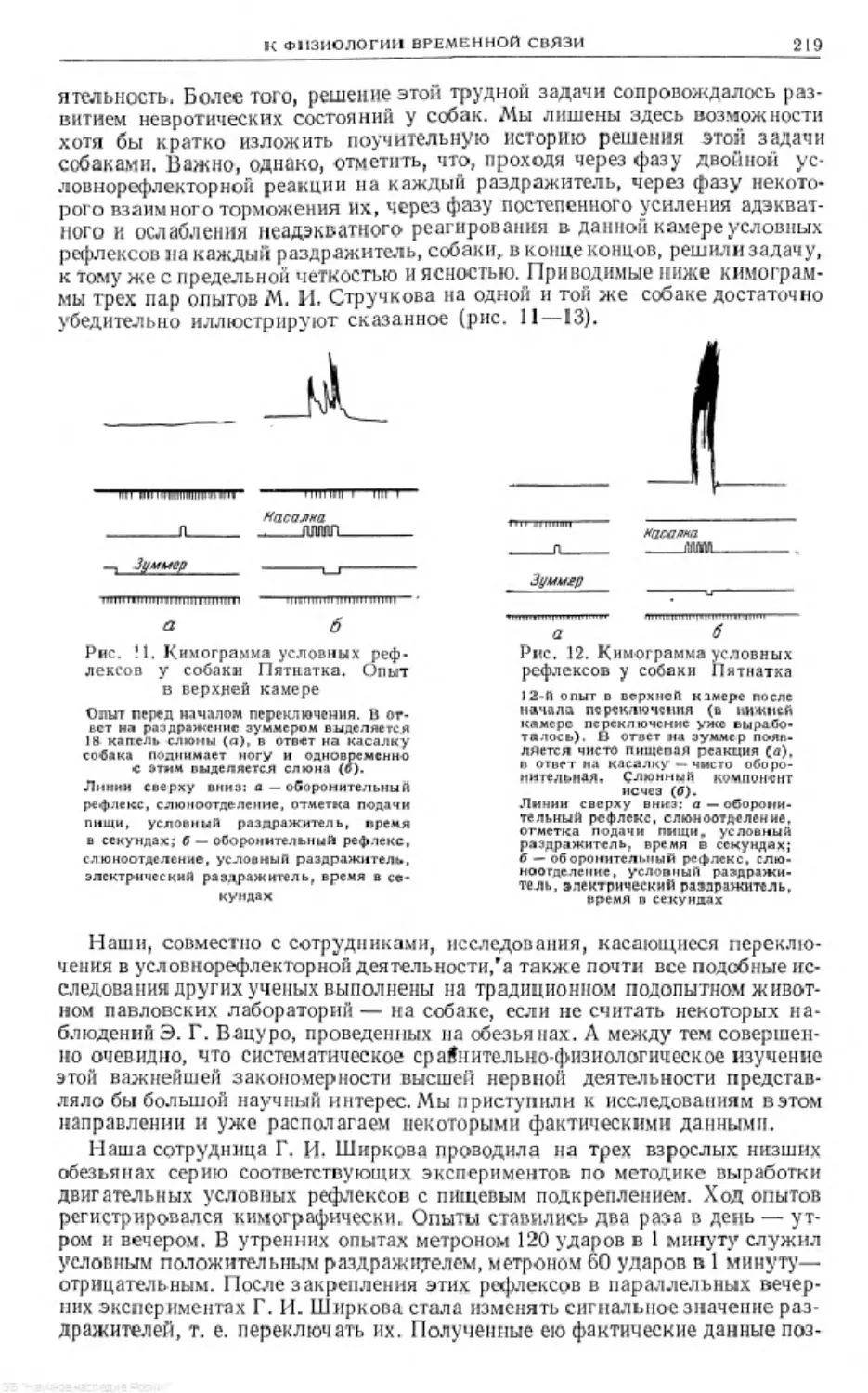
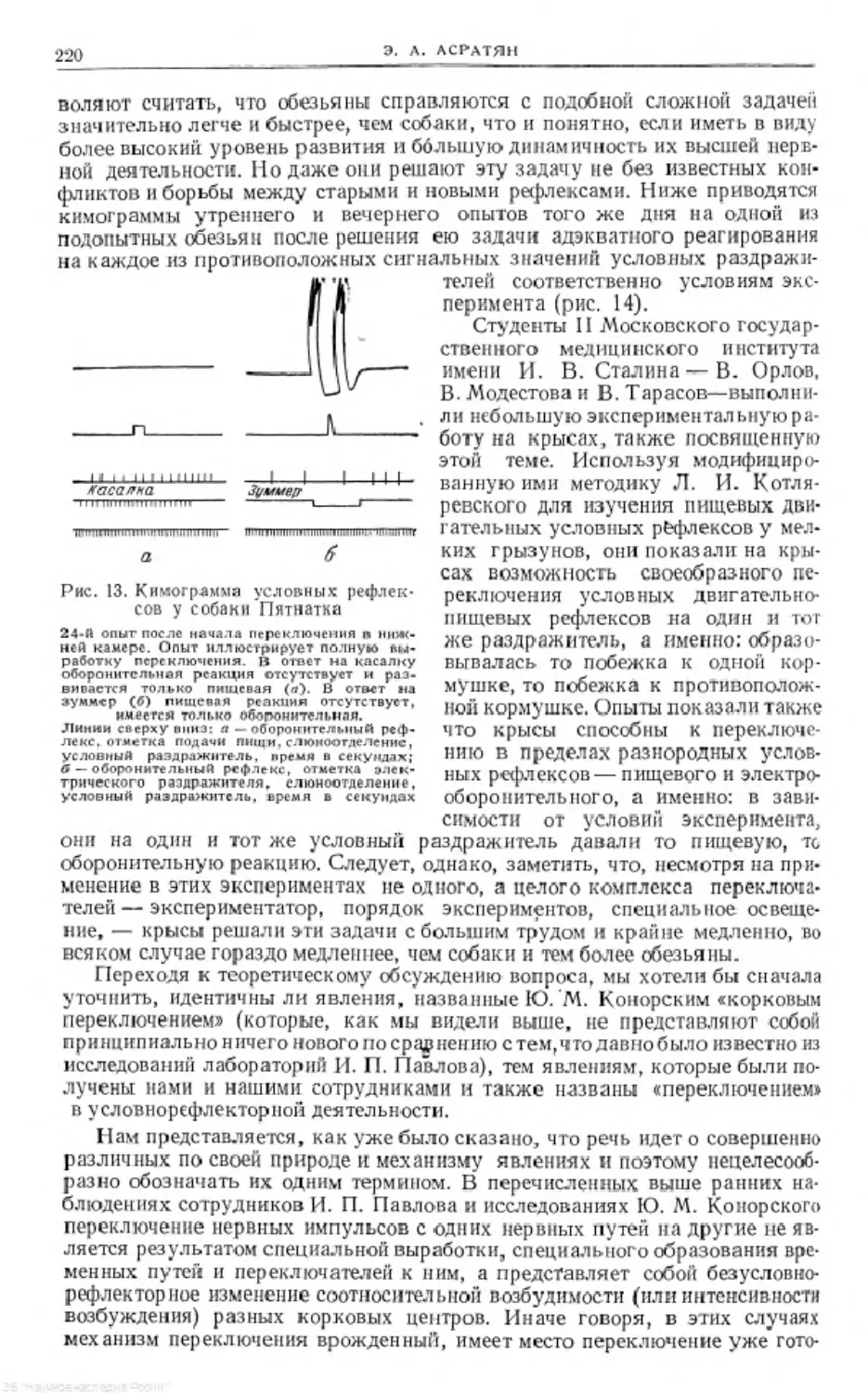



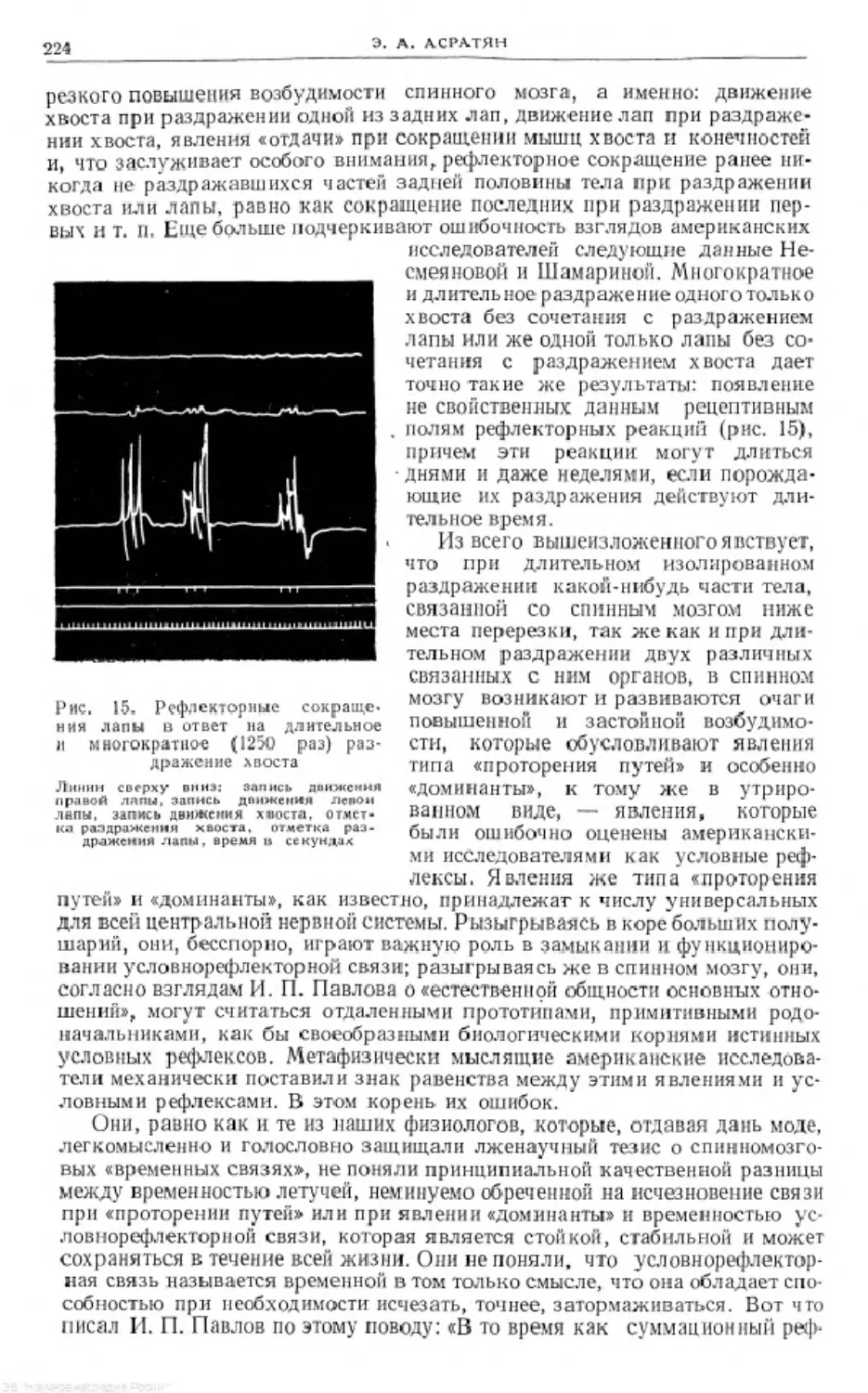




































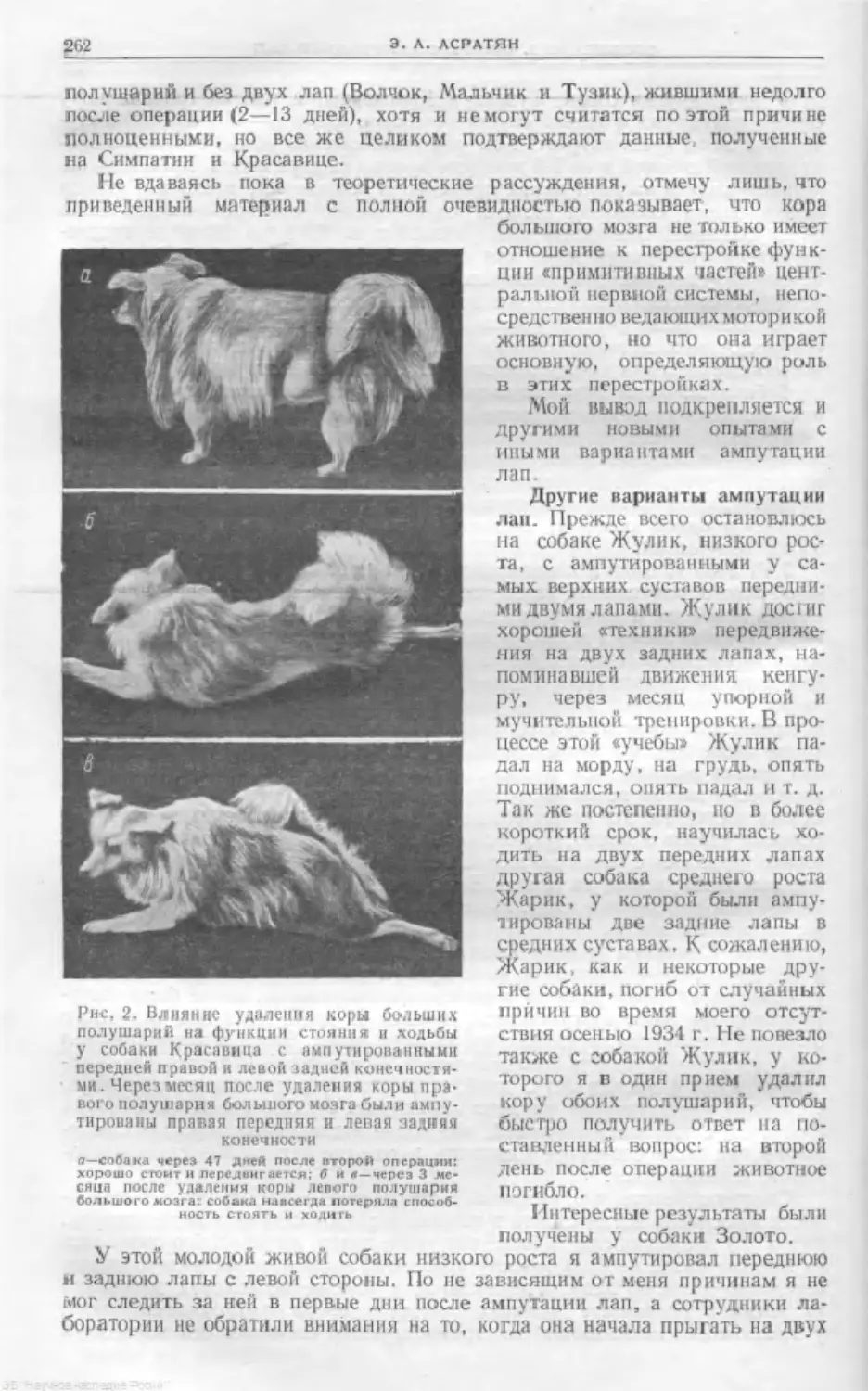










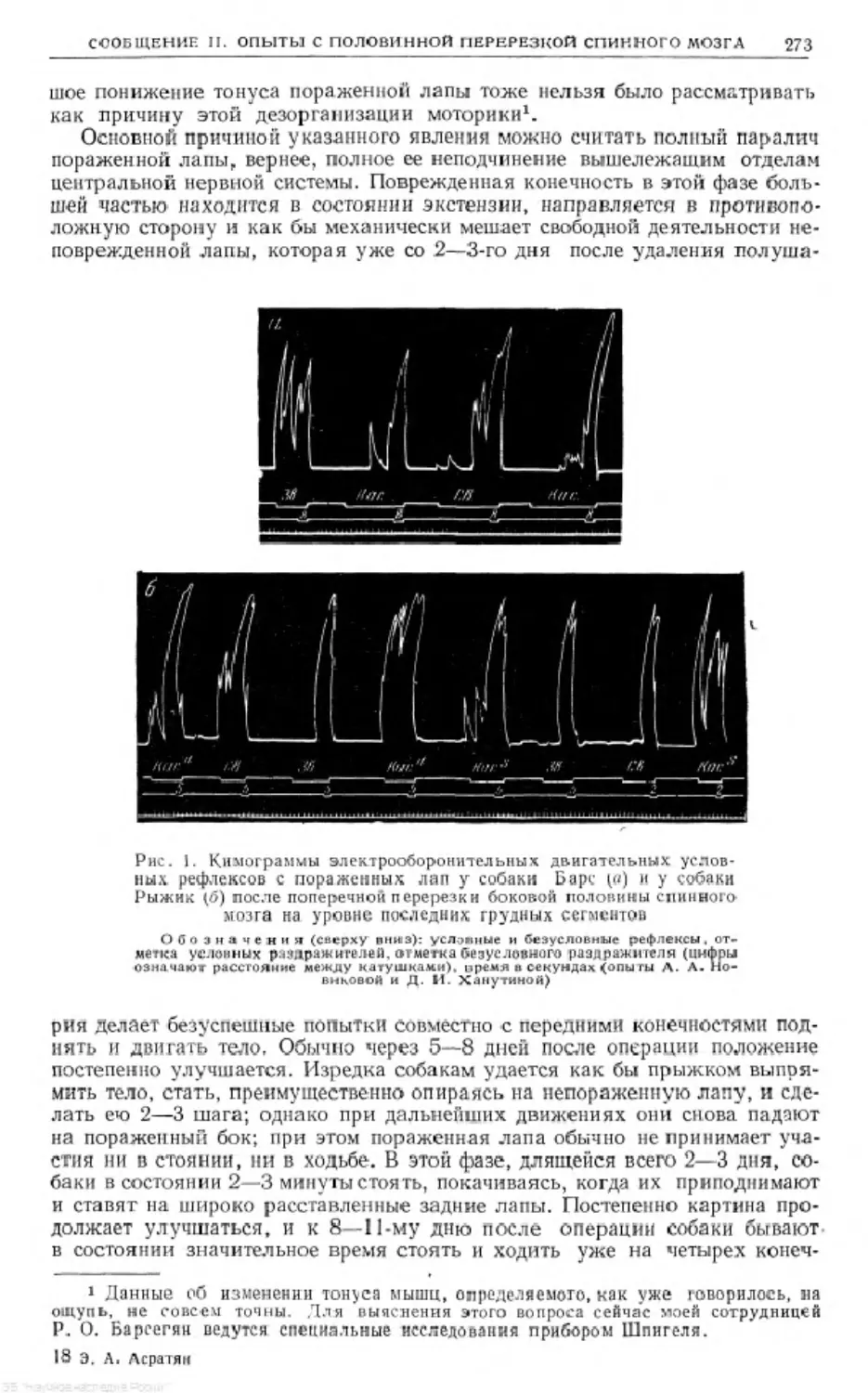






















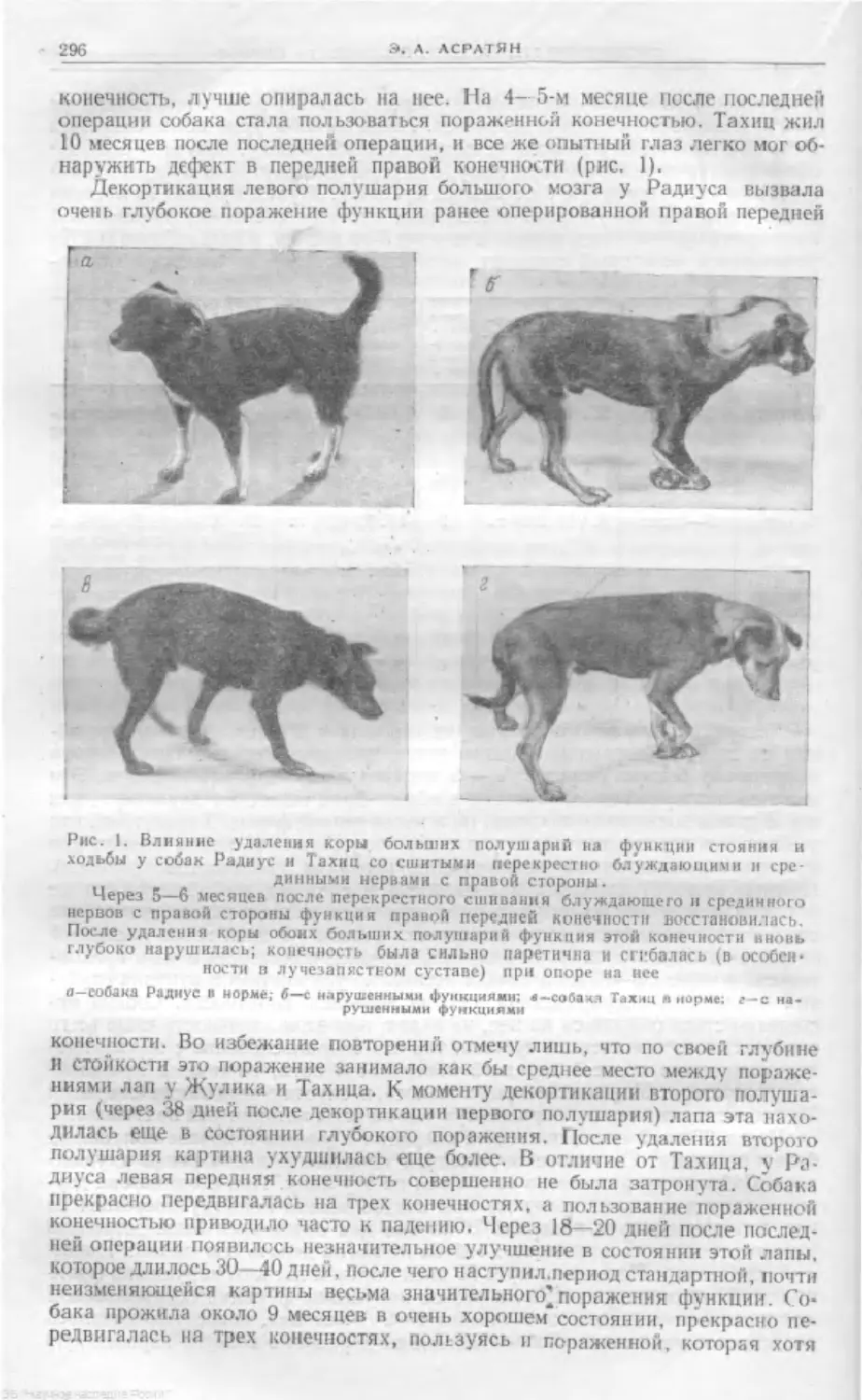


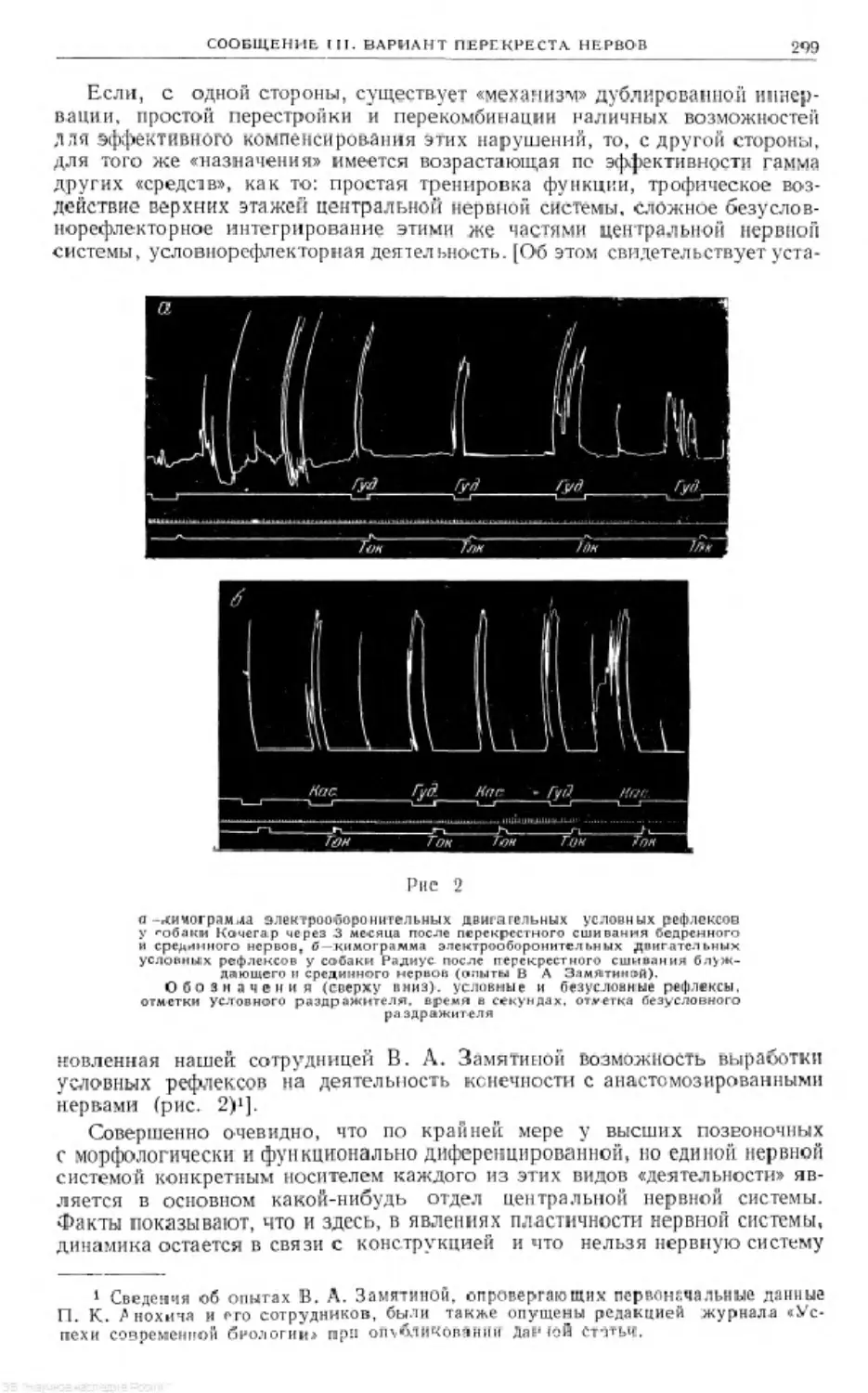




































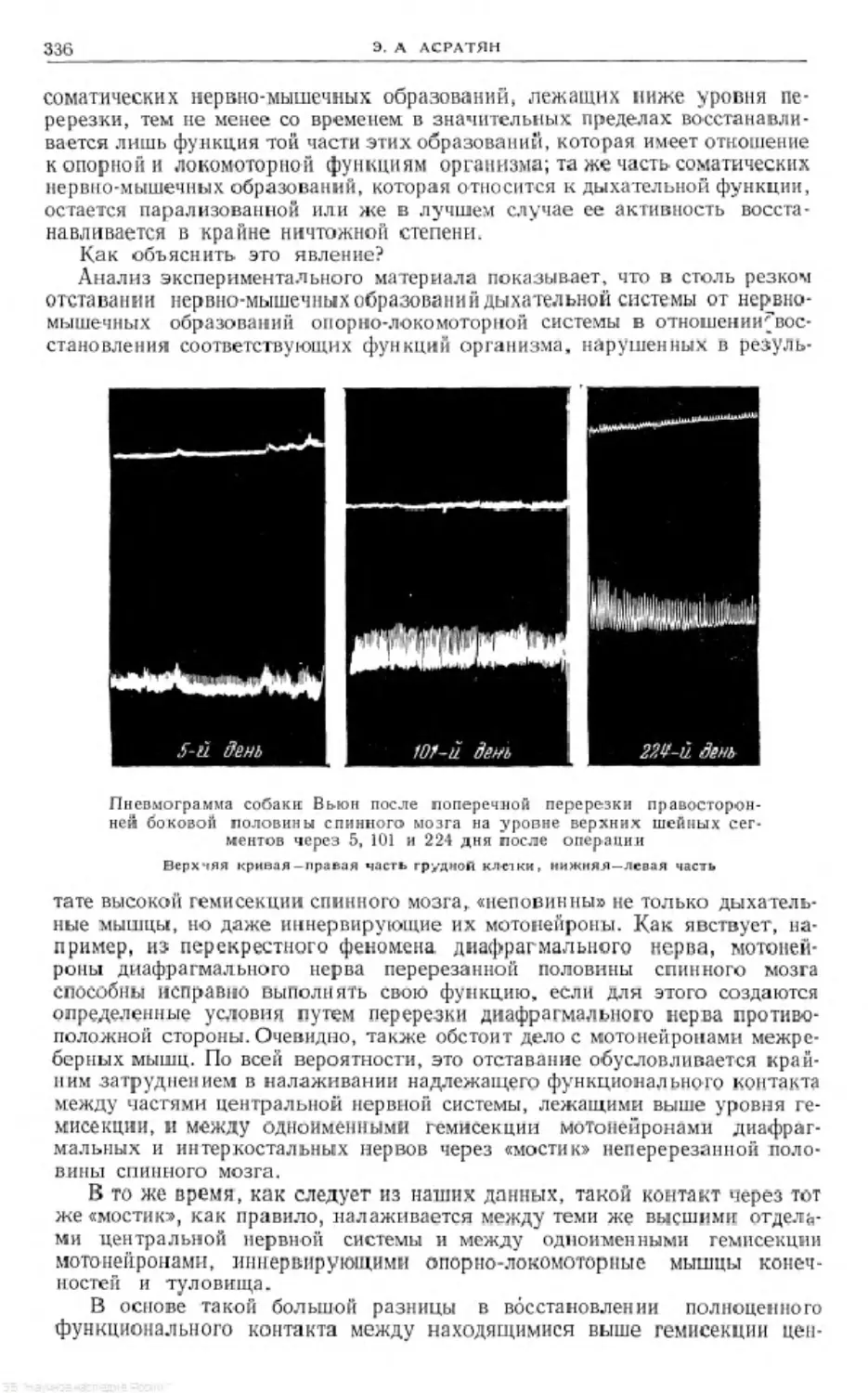










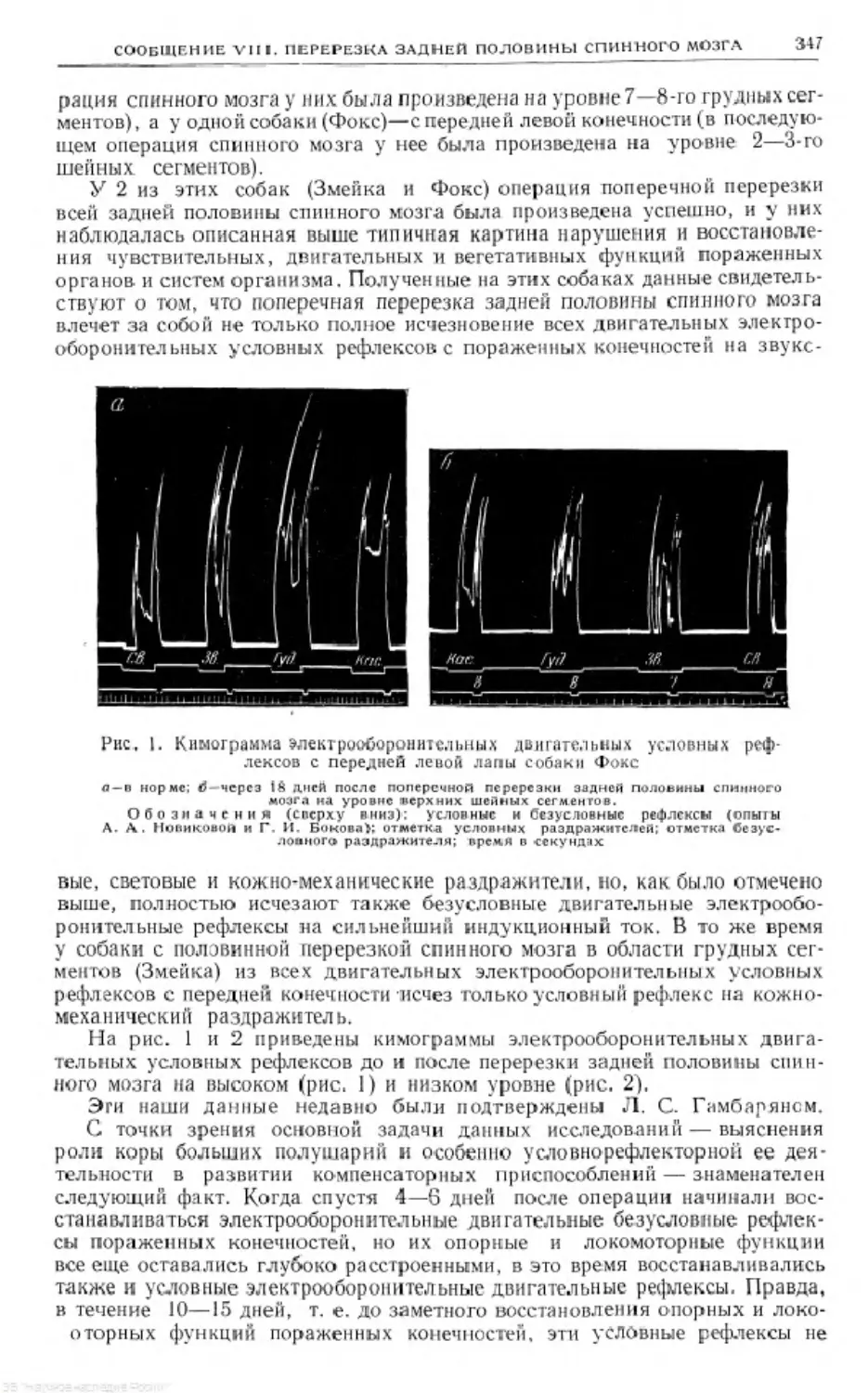



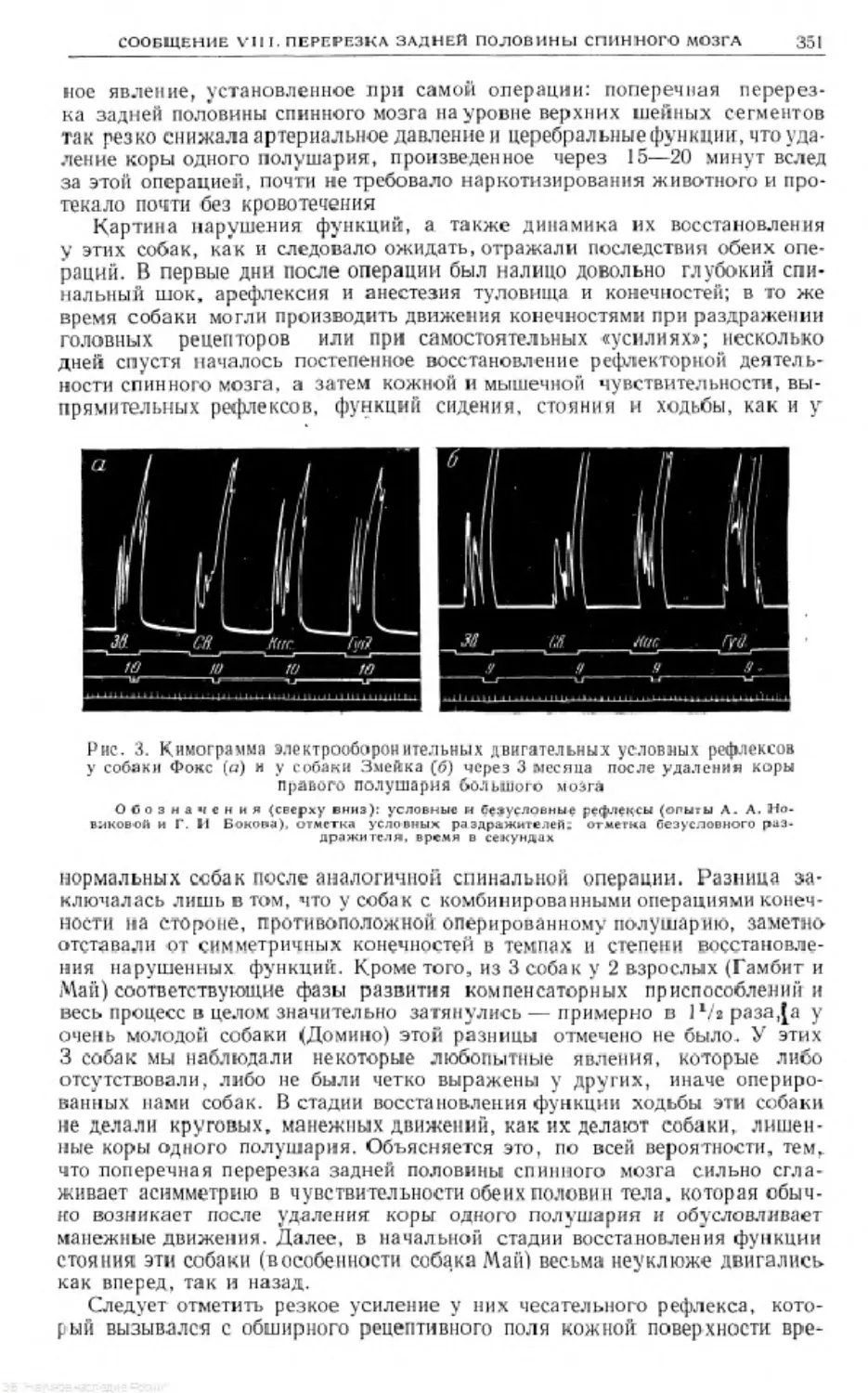

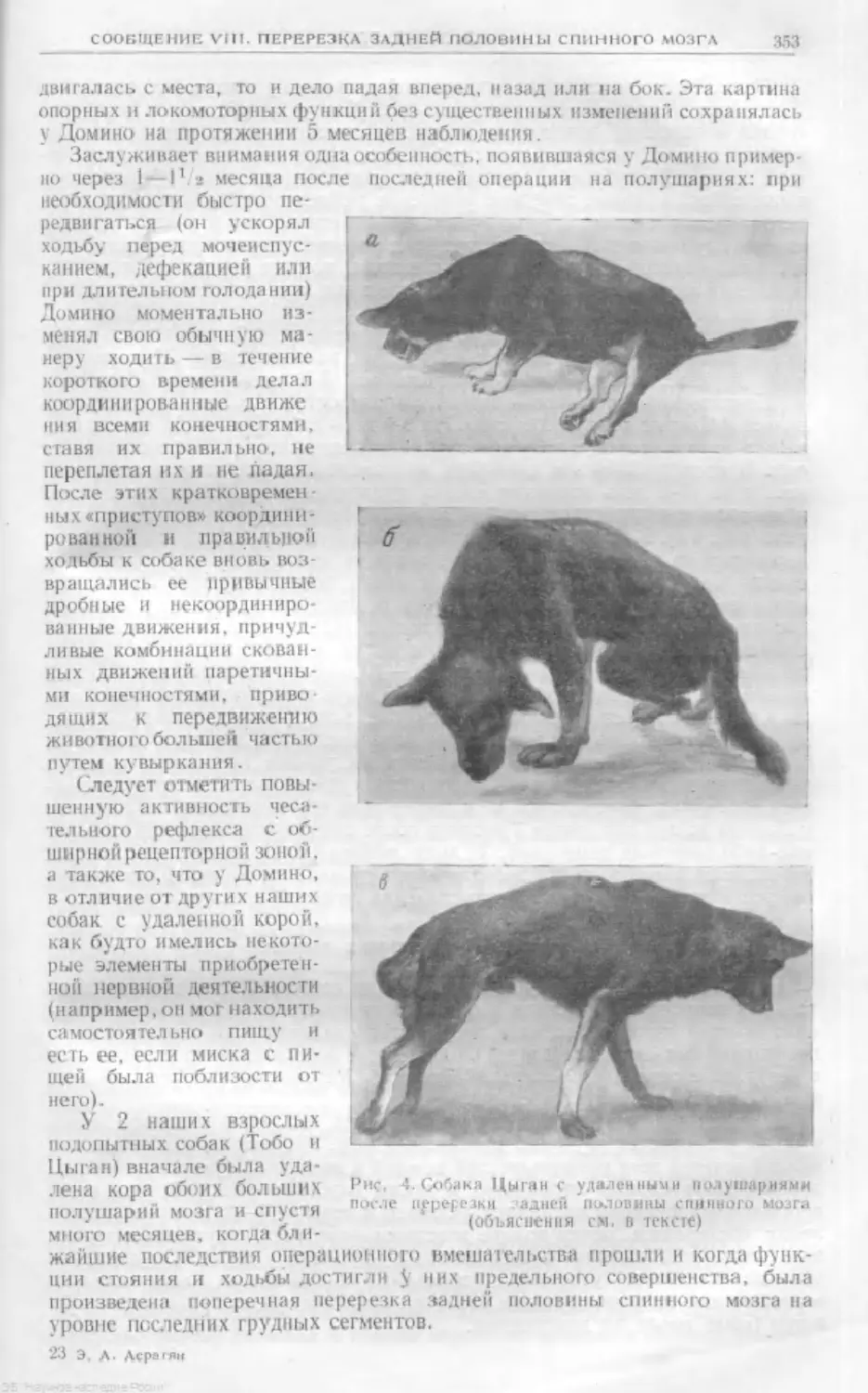


































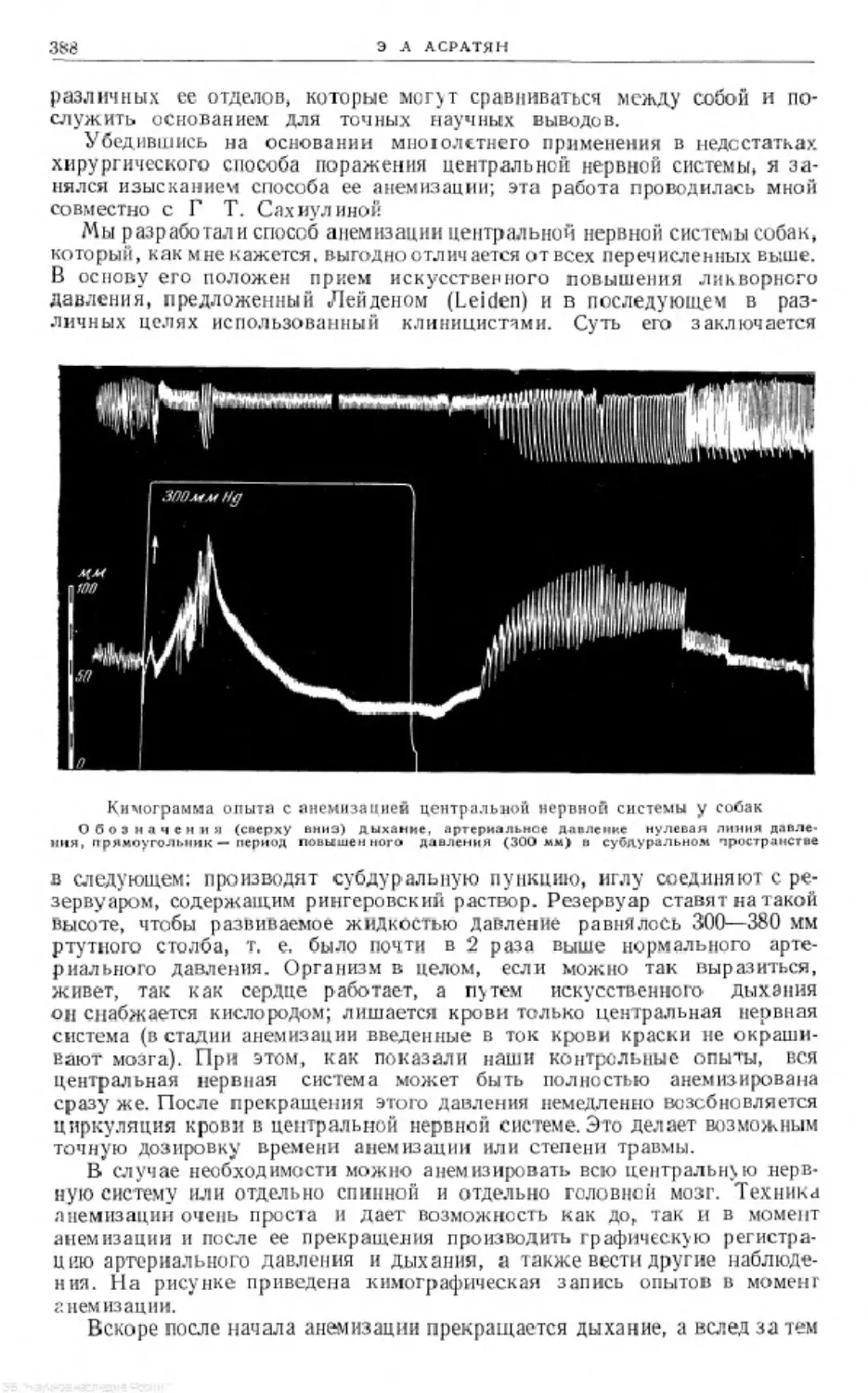



























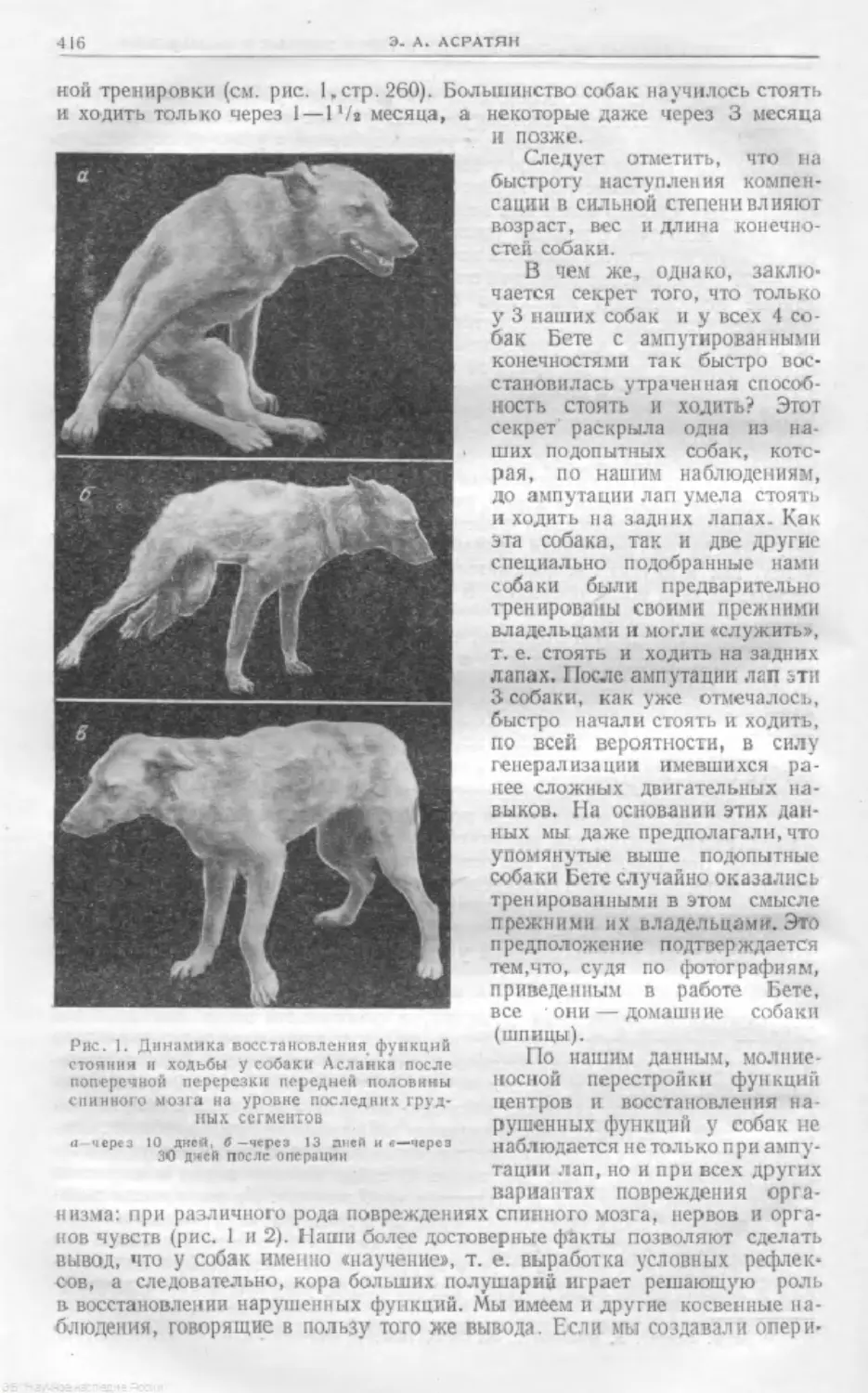

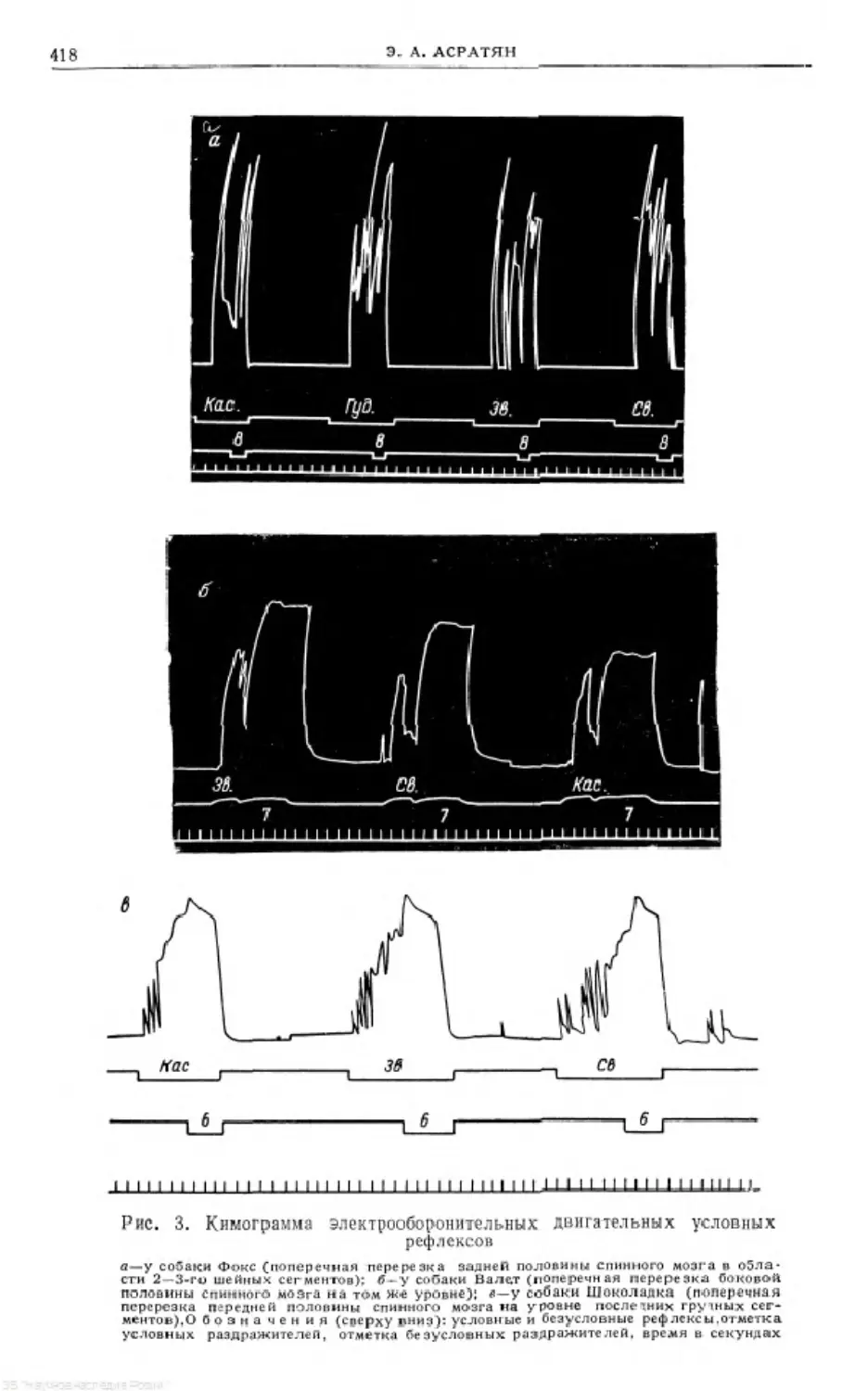


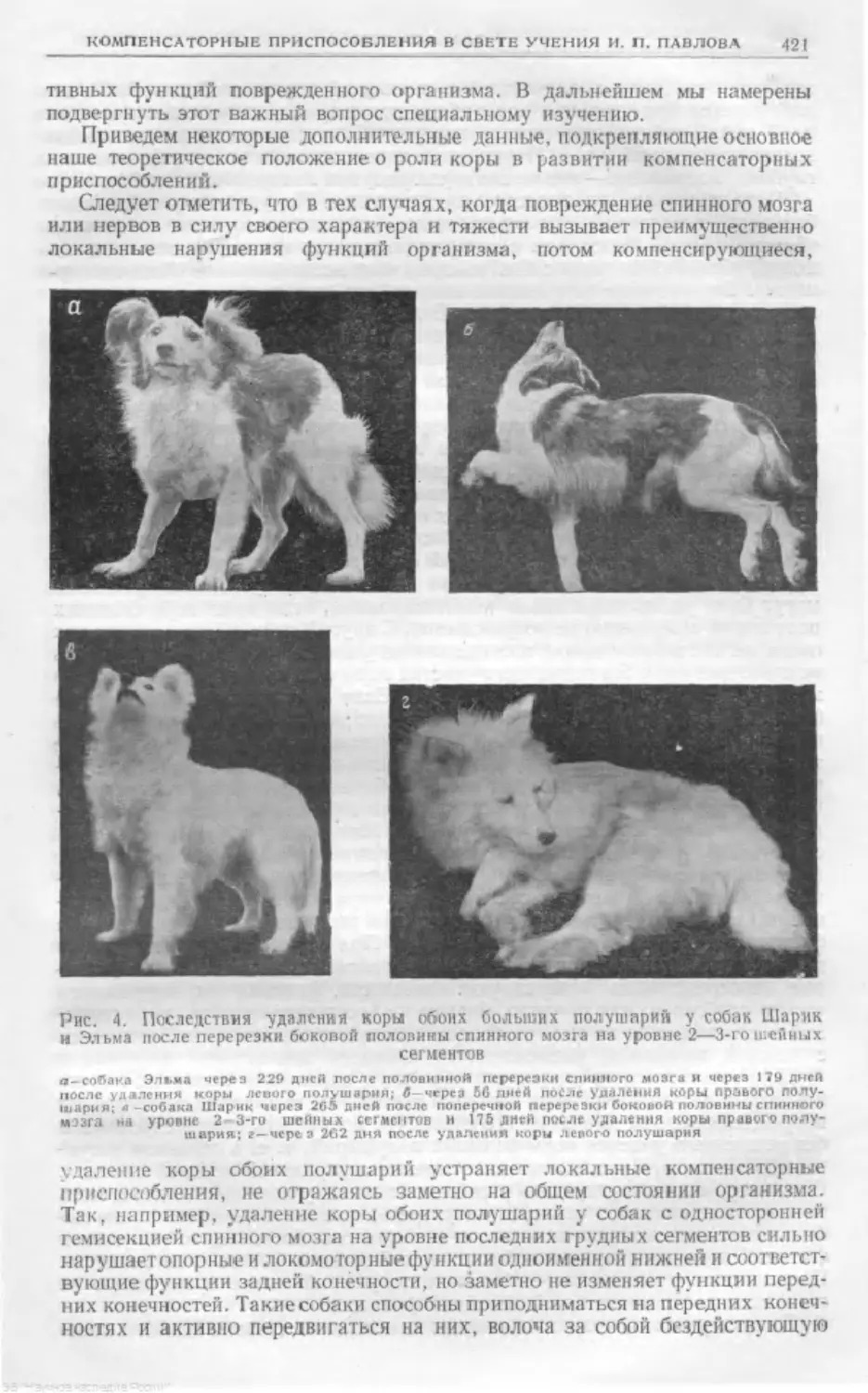


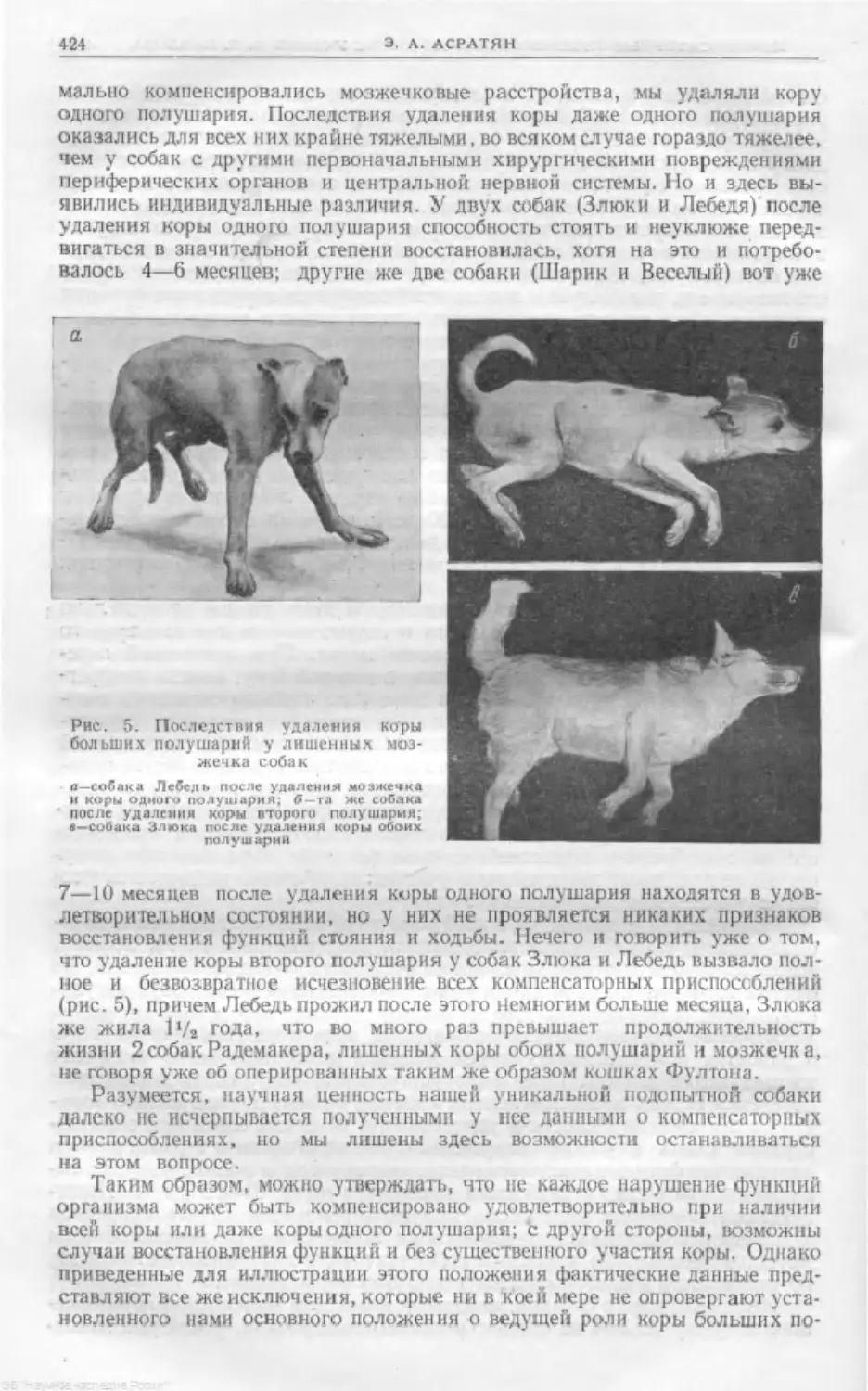
















































































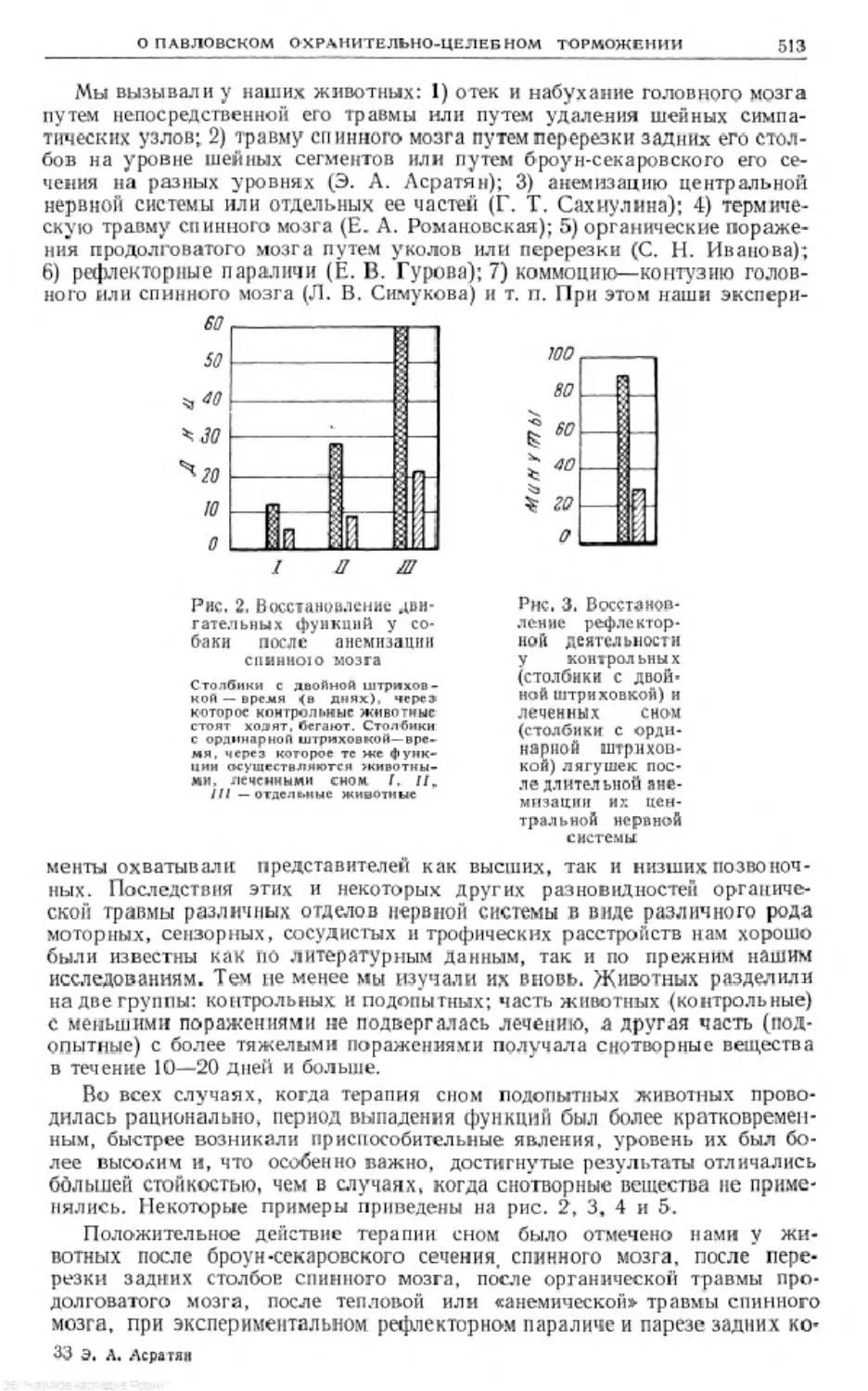
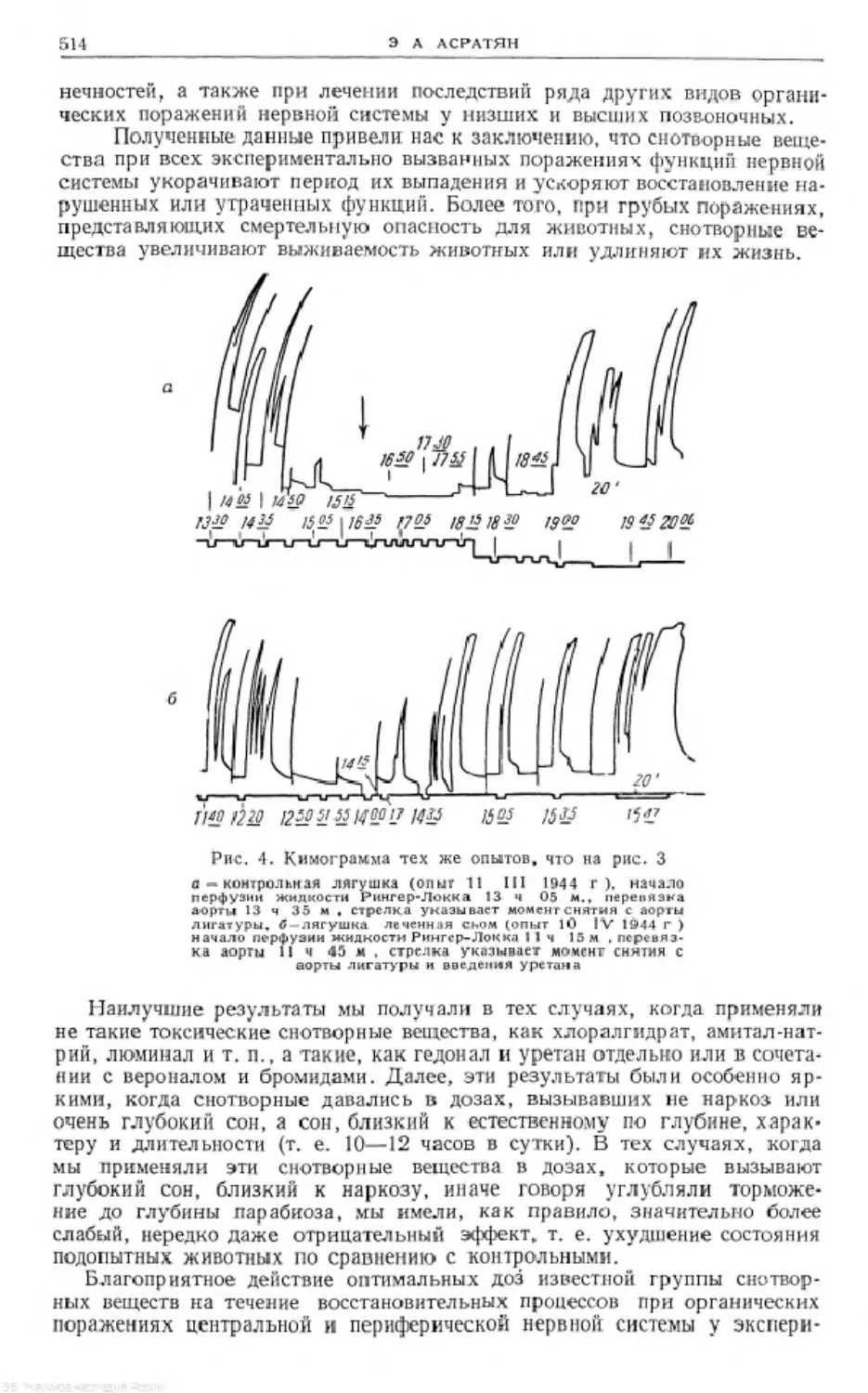




















































Text
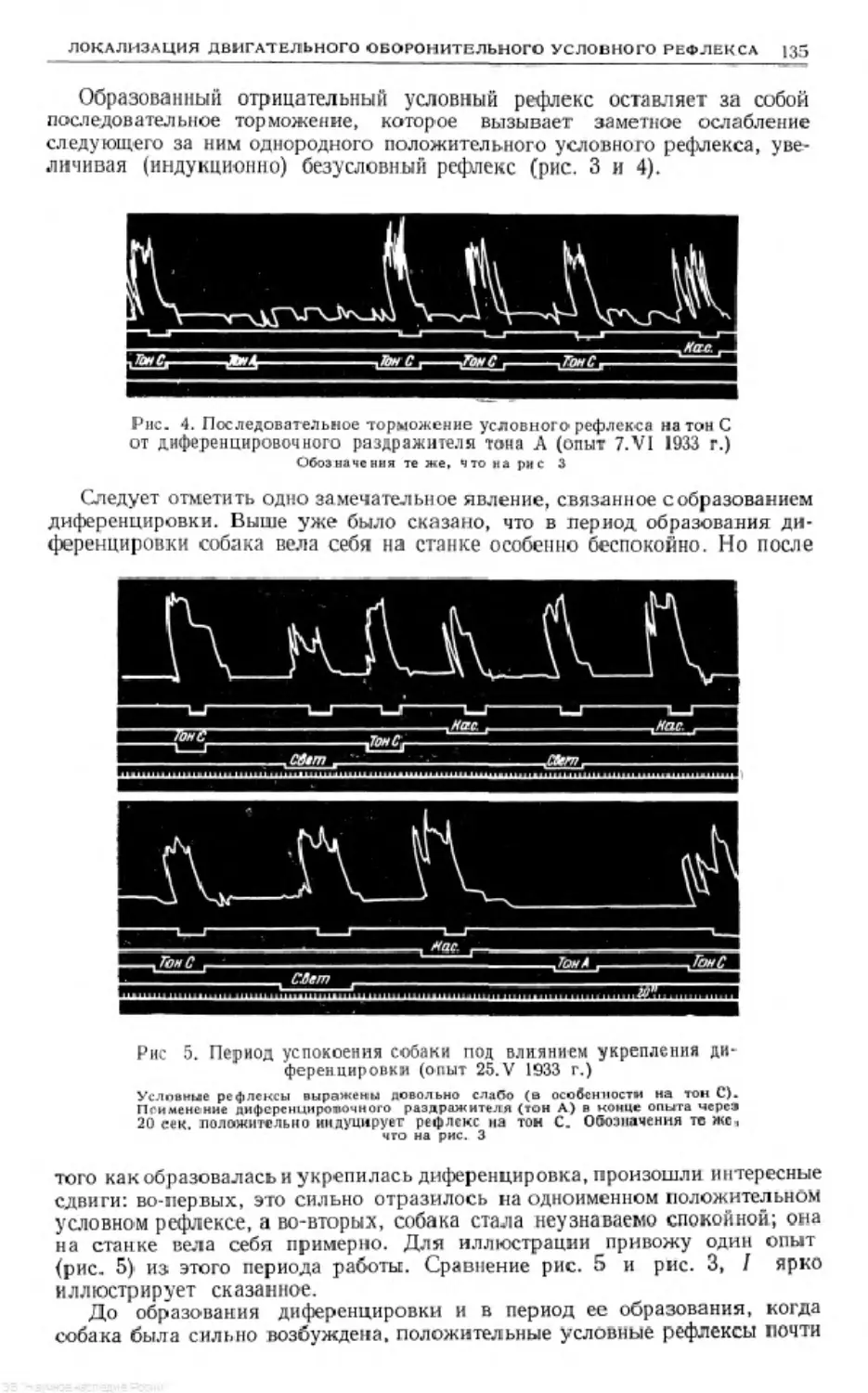
G'';
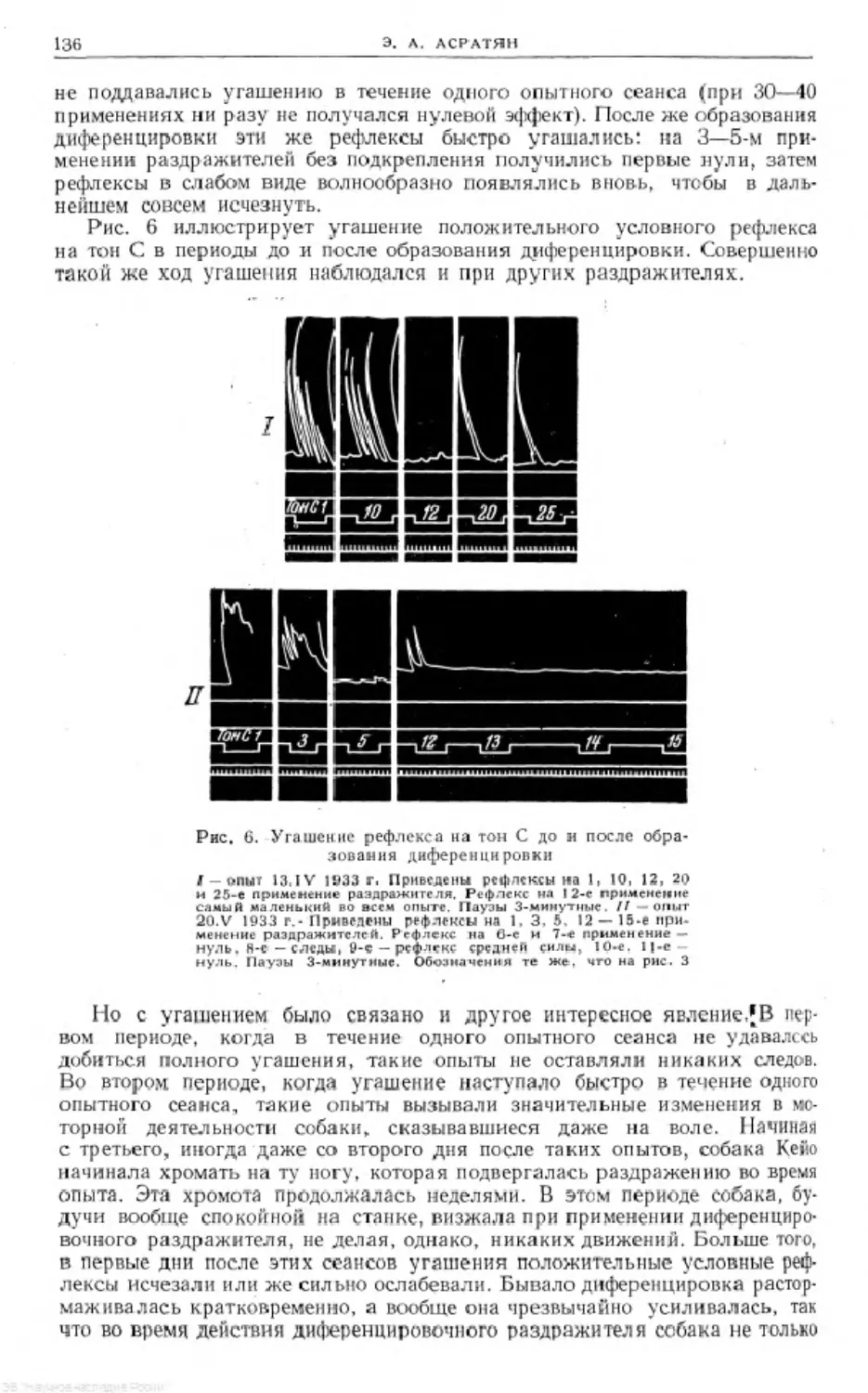
ФИЗИОЛОГИЯ
ЦЕНТРАЛЬНОЙ
НЕРВНОЙ
СИСТЕМЫ
и
АКАДЕМИЯ МЕДИЦИНСКИХ НАУК СССР
Э. А. АСРАТЯН
ФИЗИОЛОГИЯ
ЦЕНТРАЛЬНОЙ НЕРВНОЙ
СИСТЕМЫ
( Научные работы J
ъдательств о
АКАДЕМИИ МЕДИЦИНСКИХ НАУК СССР
-W о с k в а
* 9 5 3
Посвящаю
дорогой памяти
незабвенного учителя
великого
ИВАНА ПЕТРОВИЧА
ПАВЛОВА
Автор
ОТ АВТОРА
сторическая объединенная сессия Академии наук СССР и Ака-
демии медицинских наук СССР поставила перед всеми работни-
ками отечественной медицины, биологии и других родственных
отраслей знания важную и ответственную задачу: всемерно
развивать научное наследие И. П. Павлова, внедрять его идеи в
различные отрасли знания и практической деятельности, в. пер-
вую очередь—и теорию и практику здравоохранения, смелее перестраивать
медицину и ряд биологических и педагогических дисциплин на основах
павловской физиологии. Решения объединенной сессии двух академий нашли
широкий отклик со стороны не только заинтересованных специалистов, но и
всей советской общественности. Небывалый интерес широких кругов науч-
ных работников в области биологии и медицины к творчеству И. П. Павлова
находит выражение, в частности, в том, что они стремятся в целях изучения
тех или иных частных проблем: и вопросов, разработанных великим
физиологом, руководствоваться не только его классическими трудами,
но и использовать работы его учеников. Это и побудило меня переиздать
некоторые мои работы, посвященные трем органически связанным меж-
ду собой актуальным проблемам павловской физиологии: собственно
физиологии высшей нервной деятельности, физиология компенсаторных
приспособлений и охранительно-целебной роли процесса торможения.
Хотя все работы по физ иологии компенсаторных приспособлений и ох-
ранительно-целебной роли торможения, а также подавляющее большинст-
во работ по физиологии высшей нервной деятельности выполнены мной
самостоятельно, тем не менее все они овеяны гением любимого учителя,
под могучим влиянием которого формировались не только мои мысли и
идеи, мои взгляды на коренные вопросы физиологии, но и мой научный
«вкус», стиль и тактика ведения мною научно-исследовательской работы,
более того — самое желание стать физиологом. То было в 1925 г., когда
я, будучи еще студентом молодого Ереванского университета, ознако-
мился с «Двадцатилетним опытом», с большим трудом одолевая его
страницы, обращаясь то и дело к русско-армянскому словарю. Неот-
разимая тяга к великому физиологу привела меня в 1926 г. в далекий
Ленинград, к встрече с И. П. Павловым и мне вскоре посчастливилось
стать его учеником и постоянным сотрудником. Первоначальная искра
любви к его гениальному научному творчеству, подкрепленная обая-
нием его личности, стремительно разгоралась во мне, чтобы вскоре стать
направляющей силой в моей жизни, основным содержанием моего ин-
теллектуального мира, компасом моего поведения в мире науки, источ-
ником силы и вдохновения для моей научно-творческом деятельности.
Работы, включенные в раздел физиологии высшей нервной деятельности,
относятся преимущественно к начальному периоду моей научной лея гель-
нести. Мне хотелось обратить внимание читателей на пять из этих работ,
имеющих ближайшее отношение к коренным вопросам физиологии высшей
нервной деятельности. Три из них были выполнены еще при жизни
И. П. Павлова и получили с его стороны весьма положительную оценку.
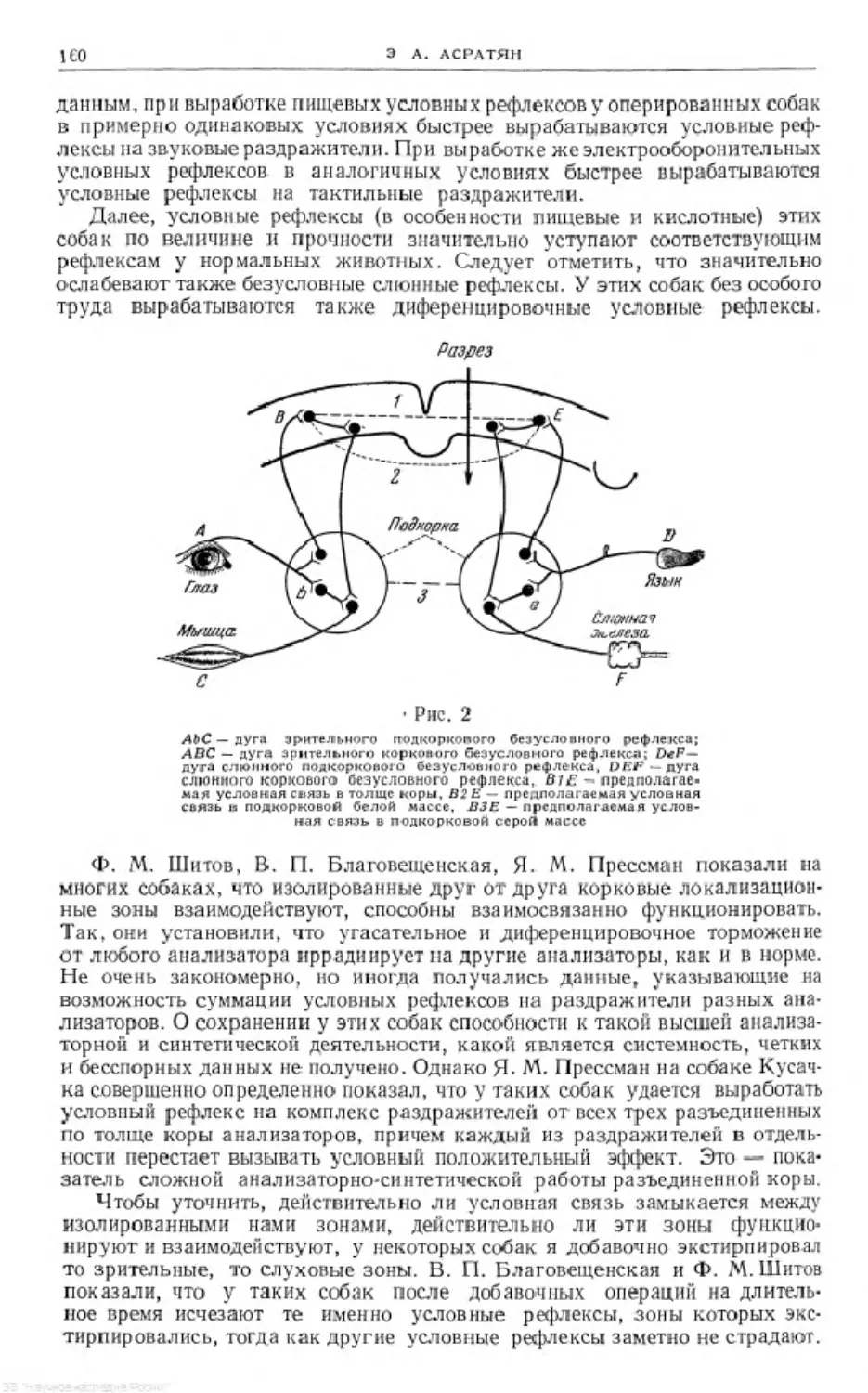
6
Э. А. АСРАТЯН
Я имею ввиду, во-первых, выполненную в 1930—1931 гг. работу «Си-
стемность работы больших полушарий головного мозга», касающуюся той
в.ысшей и сложной формы синтетической деятельности коры больших полу-
шарий, которую И. П. Павлов предпочитал называть «динамическим стерео-
типом» и которой он в последние годы своей жизни уделял особенное вни-
мание. Не могу без волнения вспомнить, что мой доклад на эту' тему в Ле-
нинградском обществе физиологов в январе 1933 г. был рекомендован
И. П. Павловым в специальном письме в правление общества.
К числу указанных работ я отношу также работу' «К физиологии двига-
тельных оборонительных условных рефлексов»,выполненную в 1932—1933 гг.
и направленную как в своем экспериментальной, так и теоретической части
против аитипавловских взглядов и недостаточно точных и проверенных
фактических данных И. С. Беритова, составляющих основное содержание
выпущенной нм в 1932 г. монографии «Индивидуально приобретенная дея-
тельность центральной нервной системы». В конце 1933 г. эта работа, с одоб-
рения И. 11. Павлова, была защищена мной в Институте физиологии
Акдемии наук СССР в качестве диссертации на соискание звания ученого
специалиста. Это было чуть ли не первым публичным выступлением
ученика И. II. Павлова против аитипавловских концепций Беритова,
к тому же выступлением не голословным, а основанным на точных экспе-
риментальных данных, доказавших ошибочность экспериментальных фак-
тов Беритова и несостоятельность сделанных из них выводов. Приходится
сожалеть,* что не было застенографировано заключительное выступление
председательствовавшего на заседании И. П. Павлова по поводу
защиты Диссертаций. Он не Только дал Положительную оценку моей рабо-
ты, но с присущей ему силой, выразительностью и меткостью охаракте-
ризовал всю антинаучную сущность исследований Беритова по физиологии
высшей нервной деятельности.
К этим работам относится также исследование «К учению о физиологи-
ческой лабильности высших центральных этажей», выполненное в 1932—
1933 гг. В этой работе впервые была сделана попытка применить глубокую
и оригинальную идею выдающегося отечественного физиолога Н. Е. Вве-
денского о лабильности к разрешению некоторых сложных вопросов, ка-
сающихся физиологических основ типов нервной системы и сильно инте-
ресовавших в те годы Павлова. Мои экспериментальные данные и теоре-
тические положения были полностью одобрены И. П. Павловым. По его
рекомендации работа эта была доложена мной как па IV Всесоюзном съезде
физиологов (1934), так и на XV Международном конгрессе физиологов (193-5).
Мне хочется обратить внимание читателей также па две близкие, а по
своему фактическому содержанию даже идентичные работы относительно
правила переключения в высшей нервной деятельности, выполненные после
кончины И. П. Павлова. В этих работах приводится сжатое изложение
установленного нами весьма важного правила высшей нервной деятель-
ности животных и краткий обзор относящихся сюда фактических данных,
полученных моими сотрудниками в 1936—1937 гг. в руководимом тогда
мной отделе физиологии центральной нервной системы Государственного
института мозга им. В. М. Бехтерева в Ленинграде. Фактическое содержа-
ние и теоретические положения этих работ представляют, по моему мнению,
новое доказательство правильности идей И. П. Павлова об изменчивости
сигнального значения условных раздражителей, а также его идей
о существовании общих генетических корней в закономерностях деятель-
ности низших и высших отделов центральной нервной системы. Они
являются дальнейшим развитием этих идей учителя применительно к важ-
ному правилу рефлекторной деятельности нервной системы — правилу
переключения, установленному впервые И. М. Сеченовым в отношении
спинного мозга, а мной и сотрудниками руководимой мной лаборатории —
ОТ АВТОРА
7
в отношении коры больших полушарий мозга, причем не в виде меха-
нического подобия, а в виде закономерности с присущими корковой дея-
тельности специфическими особенностями. Некоторые новые фактические
данные относительно этого правила условнорефлекторной деятельности, по-
лученные сотрудниками руководимой мной лаборатории за последи не годы,
а также дальнейшее развитие моих взглядов о его физиологических основах
нашли свое отражение и в последней из упомянутых выше пяти работ,
посвященной экспериментальному и теоретическому анализу ряда вопросов
физиологии временной связи,— актуальной и кардинальной проблеме мате-
риалистического учения великого Павлова о высшей нервной деятельности.
Среди работ по физиологии высшей нервной деятельности, выполнен-
ных мной в ранние годы научной деятельности, имеются и такие,
которые на современном уровне знаний меня не удовлетворяют во многих
отношениях и в первую очередь теоретическим освещением изложенных в
них фактических данных. Если в некоторых работах проводился не-
достаточно глубоким Теоретический анализ и осмысливание фактиче-
ского материала, то в отдельных работах давалось ошибочное теоретиче-
ское освещение приведенных фактических данных. Я имею в видудвеработы
о влиянии удаления верхних шейных симпатических узлов или перерезки
шейных симпатических нервов на условнорсфлекторную деятельность,
выполненные под руководством акад. Л. А. Орбели и под явным влиянием
его идей (первая относится к 1927—1928 гг., а вторая к 1932—1933 гг.,
т. е, к периоду, когда я одновременно работал и Bi руководимом И. П, Пав-
ловым институте, и в руководимой Л. А. Орбели лаборатории). Допущенная
мной в этих работах ошибка заключается в том, что я некритично вос-
принял теоретические положения Л. А. Орбели, односторонне и упрощенно
понимал роль симпатической нервной системы в регуляции различных си-
стем и функций организма, в частности, функций разных отделов централь-
ной нервной системы. Я не сумел тогда оттенить как факт взаимодействия
между большими полушариями и симпатической нервной системой, так
и ведущую роль больших полушарий в этом взаимодействии, что соответ-
ствует фактическому положению вещей и павловскому пониманию веду-
щей роли коры больших полушарий в регуляции функций организма.
Должен озметить, что на основании результатов другой обширной груп-
пы систематических исследований, проводившихся первоначально мной
(1932—1934), а позднее мной совместное сотрудниками (1935—1941), я в боль-
шой степени исправил эту ошибку*. В этих исследованиях было установлено,
в частности, что кора больших полушарий путем мощного трофического воз-
действия на симпатическую нервную систему способна в значительных пре-
делах изменятьеефункциоиальное состояние.Результаты этих исследований
были суммированы в двух моих работах: «К физиологии кортикальных и
мозжечковых центров автономной нервной системы»1 2 и «Влияние мозжечка
и коры большого мозга на вегетативные функции организма»3. Но должен
сознаться, что в полном объеме ошибку в понимании взаимоотношений сим-
патической нервной системы и коры больших полушарий мозга я осознал
значительно позже. Я решил включить эти ранние работы в настоящий сбор-
ник, во-первых, по тон причине, что, как мне кажется, содержащийся в них
фактический материал является оригинальным и представляет определен-
ный научный интерес, а также для того, чтобы показать, что пройденный
мной научный путь небыл свободен от известных ошибок. Оставляя первона-
чальный текст некоторых таких работ в основном без изменений, я снабдил
1 9т и исследования, начатые по моей инициативе в лаборатории Л. А. Орбели,
приводили к серьезным разногласиям между мной и им в понимании характера
зависимости симпатической нервной системы от высших центральных образований,
в частности, от мозжечка, что и заставила меня покинуть лабораторию весной 1933 г.
8 Труды V Кавказского съезда физиологов, Эриаань, 1935.
’ Невропатология и психиатрия, т. 10, № 3, 1941.
8
Э. А АСРАТЯН
их подстрочными примечаниями, уточняющими современное мое понимание
сущности обсуждаемых в них фактов и явлений.
В сборник включены также основные мои работы по проблеме пластич-
ности или компенсаторных приспособлений нервной системы, а также
основные работы по проблеме охранительно-целебной роли торможения.
Двадцатилетняя целеустремленная исследовательская моя работа в
области физиологии компенсаторных приспособлений—этой важной и
актуальной проблемы современной медицины и биологии — проводилась
в ожесточенной И непримиримой борьбе с антипйвловским направлением
группы зарубежных и некоторых отечественных исследователей. Основ-
ная руководящая идея всей моей многолетней экспериментальной и теоре-
тической работы в этой области — доказательство ведущей роли коры
больших полушарий мозга в возникновении компенсаторных приспособ-
лений организма при различного рода повреждениях. Эти работы являлись
творческим развитием гениальных идей И. П. Павлова о функциях коры
больших полушарий мозга применительно к особому классу приспособи-
тельных явлений, возникающих в поврежденном организме. Этим, несом-
ненно, обусловливался живой и постоянный интерес И. П. Павлова к моим
работам в указанном направлении, которые проводились в те годы в руко-
водимом мной отделе физиологии центральной нервной системы Инсти-
тута мозга им. В. М. Бехтерева.
В дни Великой Отечественной войны, когда каждый советский гражданин
считал своим долгом принимать посильное участие во всенародном деле по-
мощи фронту, я направил свою научно-исследовательскую работу на изуче-
ние актуальных вопросов, связанных с восстановлением здоровья героиче-
ских защитников нашей родины. Я изучал с позиций павловской физиоло-
гии происхождение и природу некоторых болезненных состояний организма,
возникающих в результате боевых травм, и на этой основе разрабо-
тал новые способы и средства профилактики и терапии этих болезненных
состояний. В поисках путей для решения этой ответственной и сложной за-
дачи я вновь обратился к неиссякаемому источнику животворных идей ве-
ликого Павлова и на сей раз — к его оригинальной и глубокой концепции
об охранительном и целебной роли торможения. В третий раздел включены
работы, в которых излагаются результаты проведенных мной и моими со-
трудниками систематических экспериментальных и теоретических исследо-
ваний в указанном направлении. Мы сделали попытку значительно расши-
рить границы теоретического и практического применения концепций учи-
теля об охранительной и целебной роли торможения и распространить Их на
всю нервную систему и на обширную область органических ее поврежде-
ний. Не мне судить, насколько это удалось осуществить.
Помещаемые в каждом разделе работы приведены в основном в хро-
нологической последовательности, в порядке их выполнения и публи-
кации. Не внося в них существенных изменений, мы сочли, однако, необ-
ходимым произвести в отдельных работах некоторые сокращения, а также
внести стилистические исправления.
Пользуюсь случаем принести сердечную благодарность своим дорогим
сотрудникам: Р. О. Барсегян, Г. И. Бокову, М. Е. Варга, И. В. Введенской,
Ф. М. Вишневецкой, В. Д. Дмитриеву, В. Н. Дроздовой, В. А. Замятиной,
С. Н. Ивановой, Е. М Лихиной, А. И. Карамяну, А. А. Марковой,
А. А. Новиковой,?. П. Несмеяновой, Е. А. Романовской, К. М. Ротарь,
Г. Т. Сахнулиной, Г. А. Степаискому, Б. Д. Стефанцову, М. И. Стручкову,
Я. М. Прессману, Д. И. Ханутиной, Н. И.-Шамариной, Ф. М. Шитову,
В. В. Яковлевой и др., принимавшим активное участие в творческой разра-
ботке некоторых актуальных вопросов гениального учения И. П. Павлова.
Раздел
УСЛОВНЫЕ
РЕФЛЕКСЫ
НОВАЯ МЕТОДИКА ИЗУЧЕНИЯ УСЛОВНЫХ РЕФЛЕКСОВ
У ЧЕРЕПАХ ‘
а последние Годы в физиологии изучению условных рефлексов
у низших животных уделяется особое внимание. Это объяс-
няется, с одной стороны, бурным развитием учения об условны х
рефлексах, а с другой стороны, тем растущим интересом, кото-
рый за последнее время проявляется к области сравнительной
физиологии.
В 1923 г. 10. П. Фролов впервые разработал методику изучения условных
рефлексов у рыб. Ценность этой методики заключается в том, что она удов-
летворяет всем тем требованиям, которые ставит современная физиология
перед всякой методикой, применяемой для научных исследований, а именно:
объективная и точная регистрация условного и безуслов.ного рефлексов и
одноименных раздражителей. С помощью этой методики Фролову удалось
установить ряд интересных явлений из физиологии высшей нервной деятель-
ности рыб.
В 1926 г. на II физиологическом съездеП. М. Никифоровский и Н. Л. Го-
лубев (из лаборатории И. С. Цитовича) предложили свои методики исследо-
вания условных рефлексов у черепах. Особенности этих методик заключа-
ются в следующем.
1. Объективной регистрации условных и безусловных рефлексов и одно-
именных раздражителей они не производят. Рефлексы исследуются путем
наблюдений. Если не говорить о некотором субъективизме, который может
иметь место при исследованиях этими методиками, то следует отметить, что
даже при точнейшем наблюдении не исключены неточности в исследовании
величины, времени и течения условных и безусловных двигательных реф-
лексов у черепах. Мы не говорим уже о том, что щит черепах и их особое
свойство реагировать на все окружающие раздражения осложняют и без
того сложную работу экспериментатора.
2. Исследователи (П. М. Никифоровский) несколько ограничены в
своих действиях, так как, принимая в качестве безусловного рефлекса
втягивание головы черепахи, экспериментаторы вынуждены ждать неопре-
деленное время, пока черепаха высунет голову; тот же, кто имел дело
с черепахами, знает, насколько непостоянным является это движение го-
ловы у черепах.
3. Н. А. Голубев в качестве безусловного рефлекса использовал дви-
жение черепахи к пище. Однако и эта методика страдала рядом недостат-
ков (невозможность точного количественного учета,непостоянствобезуслов-
ного рефлекса и т. д.).
Следует указать, что, несмотря на несовершенство приведенных методик,
с помощью их получены определенные данные относительно образования
условных рефлексов у черепах.
1 Совместно с Араратом Алексаняном и Ракель Барсегян. Доложено на
IV Всесоюзном съезде физиологов в Харькове в 1930 г. Физиологический журнал
СССР, т. XVI, № 3, 1933.
1?
Э. А. АСРАТЯН
В 1928 г. К. М. Леутский в Одессе предложил методику изучения услов-
ных рефлексов у лягушек. В качестве безусловного рефлекса был использо-
ван прыжок лягушки, вызванный электрическим раздражением, т. е. такая
сложная реакция, которая и без того вызывается под влия нием многочислен-
ных внешних н внутренних импульсов и не совсем удобна для этой цели. Дан-
ная методика, хотя и отличается в деталях от трех описанных, пострадает
теми же основными недостатками.
Насколько нам известно, этим исчерпываются методики изучения выс-
шей нервной деятельности у низших животных, которые за последнее время
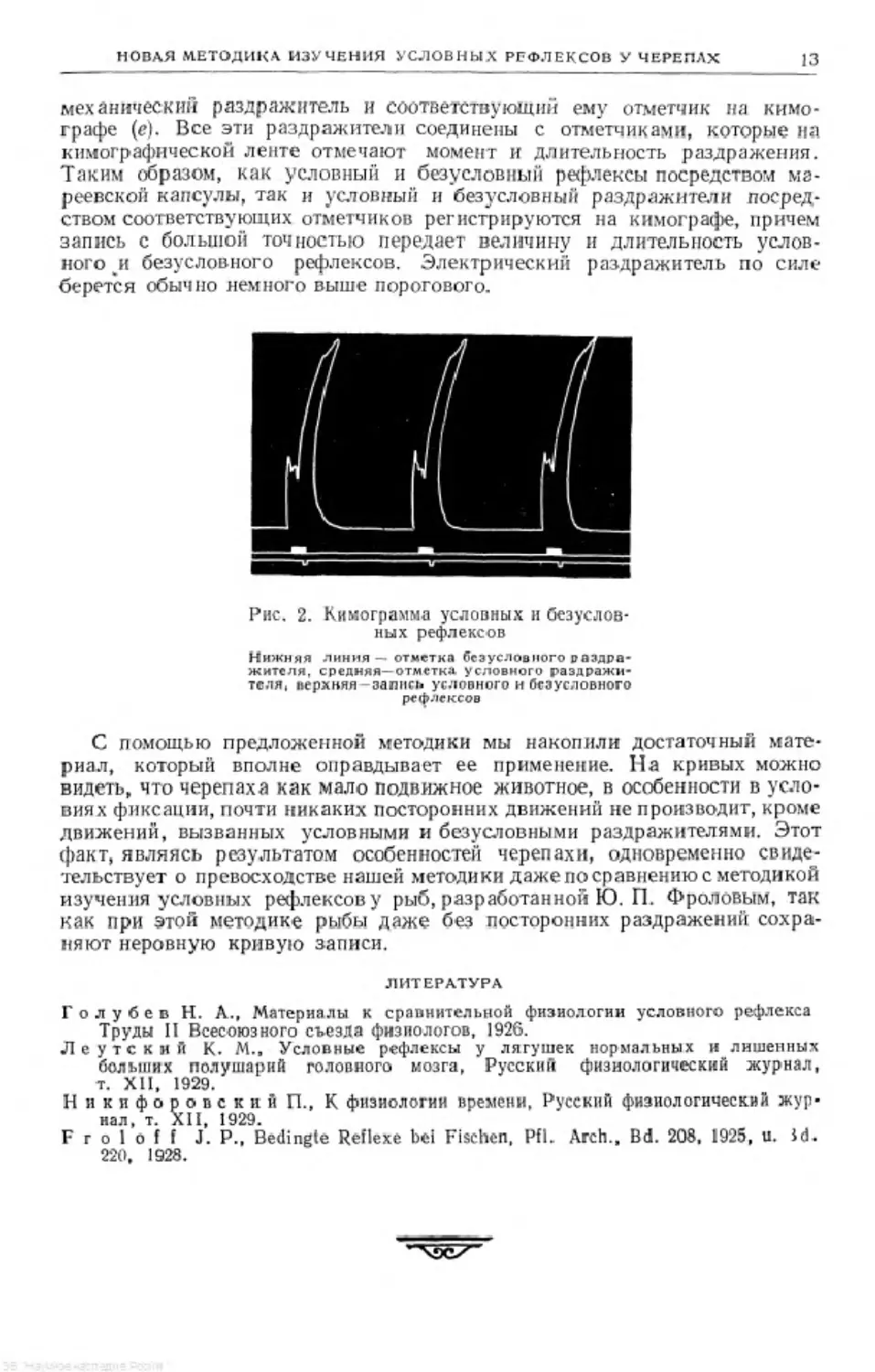
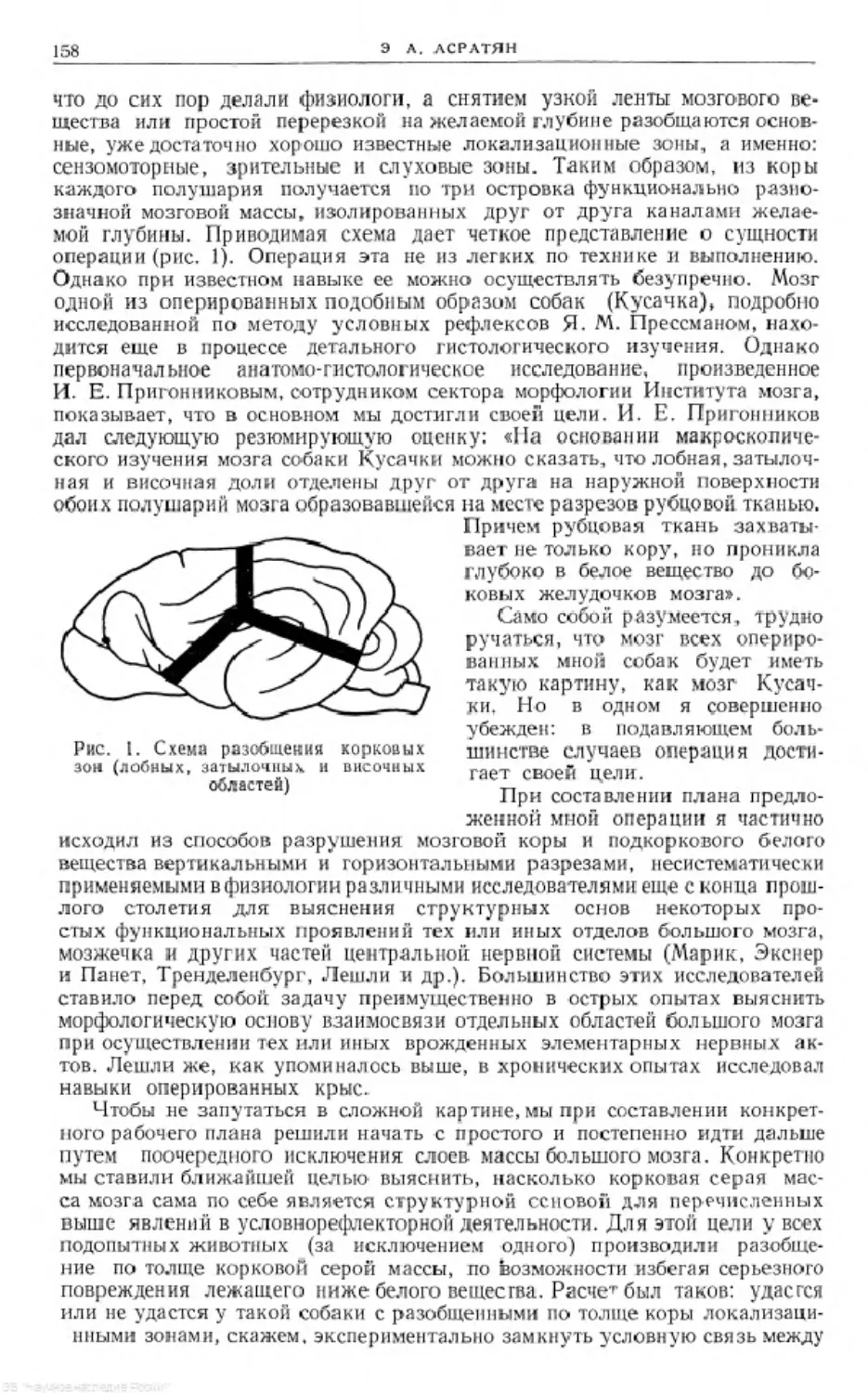
Рис. 1. Схема установки для изучения электрооборонительных
двигательных условных рефлексов у черепах
я станок; Ь и с— установка воздушной передачи для регистрации реф-
лексов; d- электроды; АКК—аккумулятор
предложены разными учеными. Для птиц и других позвоночных также
предложено несколько методик изучения условных рефлексов, но на них
МЫ останавливаться не будем,
Начиная с 1929 г., мы начали заниматься изучением условных рефлексов
у черепах в физиологической лаборатории Государственного университета
Армении и разработали методику, сущность которой заключается в сле-
дующем. Методика основана на объективной регистрации условного и безус-
ловного рефлексов и соответствующих мм раздражителей в условиях воз-
можно большей изоляции от раздражителей внешнего мира. Как безуслов-
ный нами взят оборонительный двигательный рефлекс черепахи, вызывае-
мый электрическим раздражителем. Это обстоятельство освобождает нас
от«капризов» черепахи.
В специальной комнате (в худшем случае в хорошо изолированном
ящике; см. рис. 1) черепаху укрепляют на станке (а); перед передней конеч-
ностью фиксируют барабан с натянутой резиновой пленкой (Ь), который
посредством резиновой трубки соединяется с мареевской капсулой (с).
На одной из задних конечностей черепахи укрепляютэлектроды (d), соеди-
ненные с источни ком электричества и сигнализатором Депре, отмечающим
момент и длительность раздражения. Вокруг черепахи расстанавливают
раздражители: кожно-механические, щито-механические, термические,
световые, звуковые и др. В качестве примера на схеме нарисован щито-
НОВАЯ МЕТОДИКА ИЗУЧЕНИЯ УСЛОВНЫХ РЕФЛЕКСОВ У ЧЕРЕПАХ
13
механический раздражитель и соответствующий ему отметчик на кимо-
графе (е). Все эти раздражители соединены с отметчиками, которые на
кимографнческой ленте отмечают момент и длительность раздражения.
Таким образом, как условный и безусловный рефлексы посредством мв-
реевской капсулы, так и условный и безусловный раздражители посред-
ством соответствующих отметчиков регистрируются на кимографе, причем
запись с большой точностью передает величину и длительность услов-
ного и безусловного рефлексов. Электрический раздражитель по силе
берется обычно немного выше порогового.
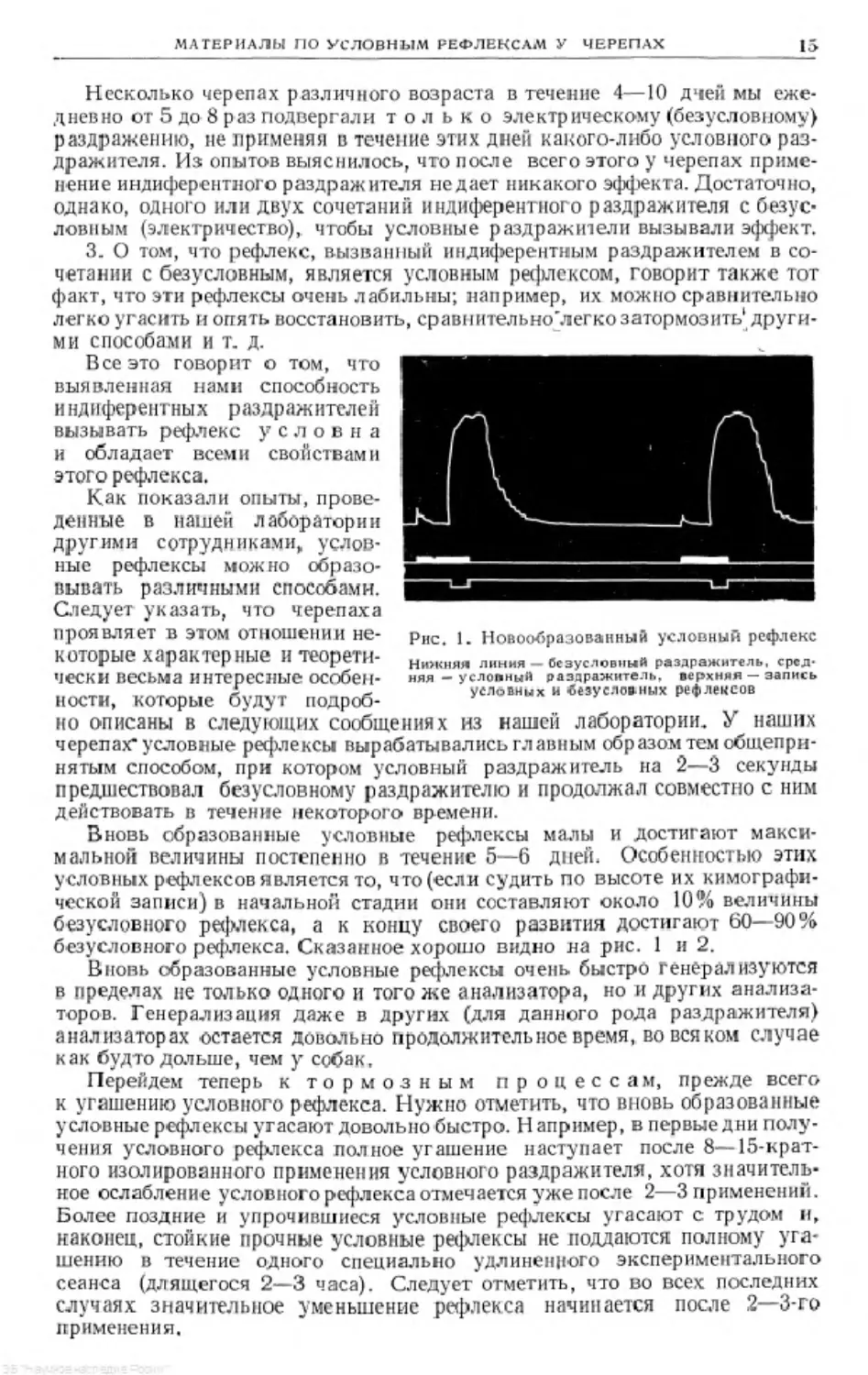
Рис. 2. Кимограмма условных и безуслов-
ных рефлексов
Нижняя линия — отметка безусловного раздра-
жителя, средняя—отметка условного раздражи-
теля, верхняя записи условного и безусловного
рефлексов
С помощью предложенной методики мы накопили достаточный мате-
риал, который вполне оправдывает ее применение. На кривых можно
видеть, что черепаха как мало подвижное животное, в особенности в усло-
виях фиксации, почти никаких посторонних движений не производит, кроме
движений, вызванных условными и безусловными раздражителями. Этот
факт, являясь результатом особенностей черепахи, одновременно свиде-
тельствует о превосходстве нашей методики даже по сравнению с методикой
изучения условных рефлексов у рыб, разработанной Ю. П. Фроловым, так
как при этой методике рыбы даже без посторонних раздражений сохра-
няют неровную кривую записи.
ЛИТ ЕРАТУРА
Голубев Н. А., Материалы к сравнительной физиологии условного рефлекса
Труды II Всесоюзного съезда физиологов, 1926.
Леутский К. М., Условные рефлексы у лягушек нормальных и лишенных
больших полушарий головного мозга, Русский физиологический журнал,
т. XII, 1929.
Н и к и ф о р о в с к и й П., К физиологии времени, Русский физиологический жур-
нал, т. XII, 1929.
F г о 1 о f I J. Р., Bedingte Reflexe bei Fischon, Pfl. Arch., Bd. 208, 1925, u. id.
220, 1928.
МАТЕРИАЛЫ ПО УСЛОВНЫМ РЕФЛЕКСАМ У ЧЕРЕПАХ '
о разработанной совместно с Р. Барсегян методике нами изу-
чены условные рефлексы на кож но-механические и щито-меха-
нические раздражители у черепах. Условные рефлексы обра-
зуются очень быстро как при так называемом короткоотстав-
ленном способе, так и при одновременном сочетании условных
и безусловных раздражителен, а именно: на кожномеханиче*
ский раздражитель после 4—20 сочетаний, на щито-механический—после
10—20 сочетаний.
Возникшее вначале сомнение в том, действительно ли полученное нами
явление относится к условным рефлексам , быстро рассеялось в процессе ра-
боты.
В условном характере рефлекса можно было убедиться на основе того,
что впервые примененный индиферентный раздражитель только после соче-
тания с безусловным раздражителем (электричество) давал положительный
эффект. Однако, чтобы окончательно рассеять сомнение, нами было про-
ведено следующее исследование.
1. Каждый индиферентный раздражитель, прежде чем превратить его
в условный, в течение нескольких дней применялся до 30—40 раз без
подкрепления безусловным раздражителем. Более 95% испытанных нами
черепах на кожно-мехэпические раздражители давали не положительный
эффект (экстензия), а отрицательный оборонительный эффект (флексия).
Под словами положительный и отрицательный эффект мы усло-вно подра-
зумеваем их аналогию или отличие от двигательного оборонительного реф-
лекса на электрическое раздражение. 5% черепах, дававших при контроль-
ных испытаниях едва заметные следы положительного эффекта на кожно-
мсханичсские раздражители, судя по внешности и величине, были особо
возбудимыми молодыми черепахами. Такие черепахи не применялись в ка-
честве объектов для дальнейших исследований. Следует, однако, упомянуть
что при проверочно-контрольных опытах не у всех молодых черепах полу-
чен подобный эффюкт.
Щито-механические раздражители при контрольном применении никогда
не вызывали даже в слабой степени того характерного экстензорного эф-
фекта, который они давали после того, как становились условными раздра-
жителями.
2. Можно было подозревать, что многократное раздражение электри-
ческим током (при сочетаниях условного и безусловного раздражителей)
повышает возбудимость черепахи, и поэтому индиферентный раздражитель,
н сдававший до этого никакого эффекта, теперь, после раздражения электри-
чеством, в силу понижения порога возбудимости начинает вызывать без-
условную двигательную оборонительную реакцию. Это подозрение рас-
сеялось после следующих опытов,
’ Совместно с Араратом Алексаняном. Доложено на IV Всесоюзном съезде физио-
логов в Харькове в 1930 г. Физиологический журнал СССР, т. XVI, № 3. 19 33.
МАТЕРИАЛЫ ПО УСЛОВНЫМ РЕФЛЕКСАМ У ЧЕРЕПАХ
15
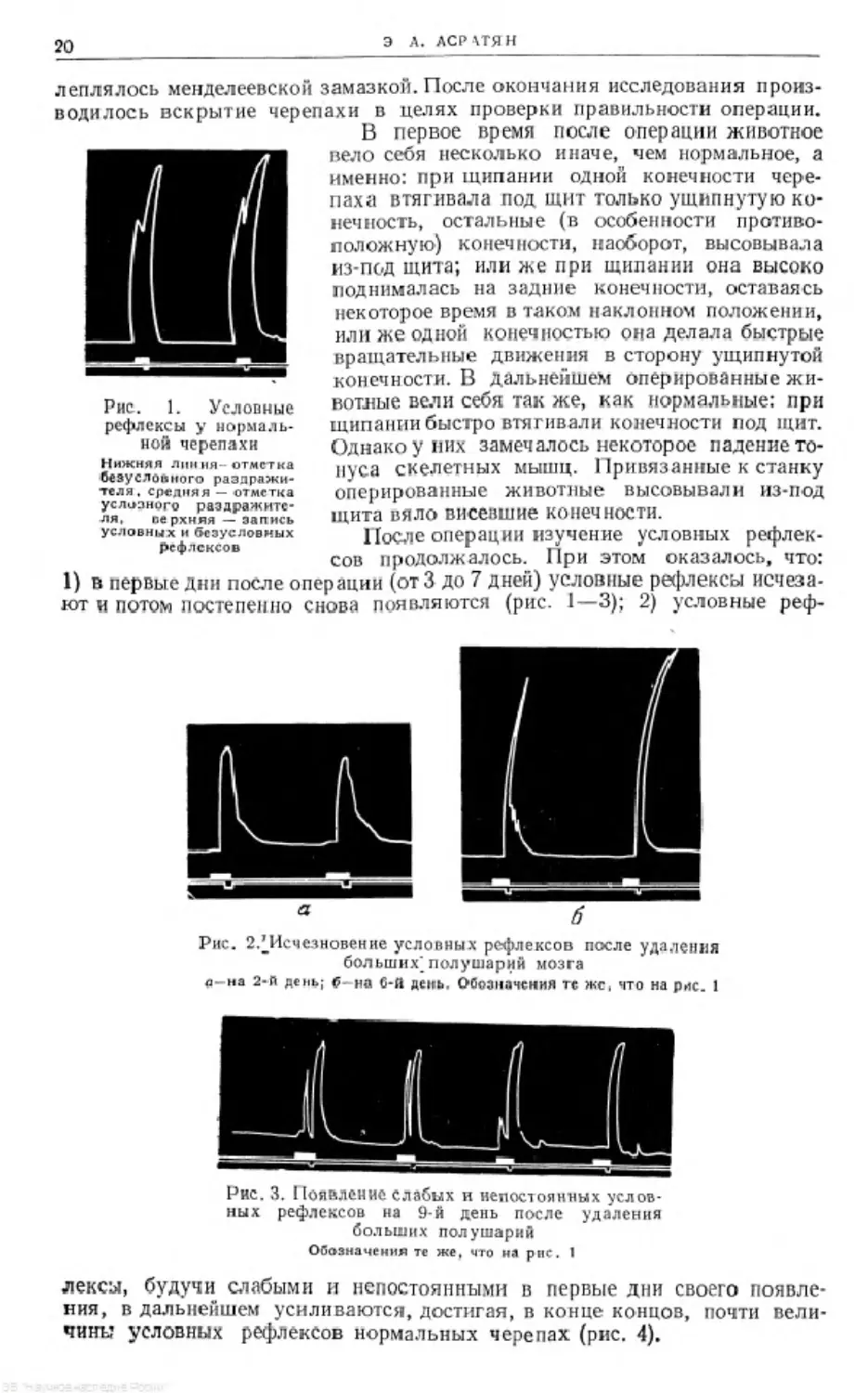
Рис. 1. Новообразованный условный рефлекс
Несколько черепах различного возраста в течение 4—10 дчей мы еже-
дневно от 5 до 8 раз подвергали только электрическому (безусловному}
раздражению, не применяя в течение этих дней какого-либо условного раз-
дражителя. Из опытов выяснилось, что после всего этого у черепах приме-
нение индиферентиого раздражителя не дает никакого эффекта. Достаточно,
однако, одного или двух сочетаний индиферентиого раздражителя с безус-
ловным (электричество), чтобы условные раздражители вызывали эффект.
3. О том, что рефлекс, вызванный индиферентным раздражителем в со-
четании с безусловным, является условным рефлексом, говорит также тот
факт, что эти рефлексы очень лабильны; например, их можно сравнительно
легко угасить и опять восстановить, сравнительно'легко затормозить* други-
ми способами и т. д.
Все это говорит о том, что
выявленная нами способность
и ндиферентных раздражителей
вызывать рефлекс условна
и обладает всеми свойствами
этого рефлекса.
Как показали опыты, прове-
денные в нашей лаборатории
другими сотрудниками, услов-
ные рефлексы можно образо-
вывать различными способами.
Следует указать, что черепаха
проявляет в этом отношении не-
КОТОрЫе характерные И теорети- Нижняя линия — безусловный раздражитель, сред-
чески весьма интересные особен- ияя - условный раздражитель, верхняя — запись
. „ 2 „ ytrwiHux и ««условных рефлексов
пости, которые будут подроб-
но описаны в следующих сообщениях из нашей лаборатории, У наших
черепах" условные рефлексы вырабатывались главным образом тем общепри-
нятым способом, при котором условный раздражитель на 2—3 секунды
предшествовал безусловному раздражителю и продолжал совместно с ним
действовать в течение некоторого времени.
Вновь образованные условные рефлексы малы и достигают макси-
мальной величины постепенно в течение 5—6 дней. Особенностью этих
условных рефлексов является то, что (если судить по высоте их кимографи-
ческой записи) в начальной стадии они составляют около 10% величины
безусловного рефлекса, а к концу своего развития достигают 60—90%
безусловного рефлекса. Сказанное хорошо видно на рис. 1 и 2.
Вновь образованные условные рефлексы очень быстро Генерализуются
в пределах не только одного и того же анализатора, но и других анализа-
торов. Генерализация даже в других (для данного рода раздражителя)
анализаторах остается довольно продолжительное время, во вся ком случае
как будто дольше, чем у собак.
Перейдем теперь к тормозным процессам, прежде всего
к угашению условного рефлекса. Нужно отметить, что вновь образованные
условные рефлексы угасают довольно быстро. Н апример, в первые дни полу-
чения условного рефлекса полное угашснис наступает после 8—15-крат-
ного изолированного применения условного раздражителя, хотя значитель-
ное ослабление условного рефлекса отмечается уже после 2—3 применений.
Более поздние и упрочившиеся условные рефлексы угасают с трудом и,
наконец, стойкие прочные условные рефлексы не поддаются полному уга-
шенню в течение одного специально удлиненного экспериментального
сеанса (длящегося 2—3 часа). Следует отметить, что во всех последних
случаях значительное уменьшение рефлекса начинается после 2—3-го
применения.
Ifi
Э. А. АСРАТЯН
Рис. 2. Укрепившийся услов-
ный рефлекс
Обозначения те же. что на рис. 1
Особенностью угашения условных рефлексов является также то, что оно
носит волнообразный характер, У некоторых черепах расстояние волн друг
от Друга имело как будто закономерный характер. Далее, чем короче
паузы, тем скорее наступает угашение.
У черепах, как и у собак и других высших животных, восстановле-
ние угашенного условного рефлекса происходит очень быстро, даже
после первого подкрепления (рис. 3
и 4).
Второй вид торможения — внеш-
ние тормозы (удар по стазу, по
щиту и др.). Эти посторонние раздра-
жители оказывают не очень сильное дей-
ствие; они тормозят в большинстве слу-
чаев только следующий за ним услов-
ный рефлекс; иногда тормозящее влия-
ние распространяется на два последую-
щих условных рефлекса (рис. 5).
Мы испытывали на черепахах также
диференци ровоч ное тор м_о-
ж е и и е.
Диференцировались друг от друга два
щито-механических условных рефлекса
при применении раздражителей почти
одинаковой силы, но расположенных в
различных местах на щите.
щита над задней лапкой положительный
условный рефлекс на 20 ударов в течение 20 секунд, мы начали применять
диференцировку подобного же раздражителя на расстоянии 5 см от первого
места раздражения в сторону передней лапки. Выяснилось, что, по закону
генерализации, со второго места
пачучались почти такой же
величины условные рефлексы,
как и с первого места, где услов-
ный раздражитель подкреплял-
ся. Продолжая диференциров-
ку обычным способом, т. е. пол*
крепляя раздражение первого
пункта и не подкрепляя второго
пункта, нам, хотя и с трудом,
все же удалось отдиференциро-
вать эти два раздражителя. Ди-
ференциация, которая является
показателем анализаторной спо-
собности животного, у черепах
оказалась несовершенной, хотя днференцировочный раздражитель при-
менялся длительное время — около 100 раз.
Последовательное торможение полученной нами несовершенной Диферен-
цировки на положительные условные рефлексы также незначительно, мож-
но даже сказать, что оно отсутствует.
На этом мы кончаем краткое изложение собранного нами материала.
Следует отметить, что полученный нами материал целиком укладывается
в понятия, установленные современной сравнительной физиологией цент-
ральной нервной системы низших позвоночных.
Сделаем некоторые сопоставления.
Во-первых, нескатько слов относительно скорости образования услов-
ных рефлексов у низших позвоночных вообще и у черепах в частности.
ав на левой
Рис. 3. Начало угашеиия рефлекса
Обозначения те же. что на рис. I
МАТЕРИАЛЫ ПО УСЛОВНЫМ РЕФЛЕКСАМ 5 ЧЕРЕПАХ
17
Ю. П. Фралов с помощью своей достаточно точной методики показал,
что условные рефлексы у рыб образуются довольно быстро, почти так же
быстро, как они получены нами у черепах, т. с. через 5—20 сочетаний
условного и безусловного раздражителей. Результаты наших исследова-
ний относительно скорости образова-
ния условных рефлексов у черепах на-
ходятся в соответствии с данными
Ю. П. Фролова и лаборатории, руко-
водимо» И. С. Беритовым, которые были
получены ими у рыб.
Данные, полученные II. М. Никифо-
рове к нм, как будто не согласуются с на-
шими данными. В лаборатории П. М. Ни-
кифоровского условные рефлексы появ-
лялись и упрочивались после 300—
600 сочетаний.
На наш взгляд, причиной такого рас-
хождения вскорости образования услов-
ных рефлексов у одних и тех же живот-
ных является существенная разница в
силе безусловного рефлек-
са, а также условных раз-
дражителей. Как известно, в ка-
Рис. 4.’Восстанечлсние угашен-
ного условного рефлекса
Середина (нижний ряд} и конец (верх-
ний ряд) угашен ия. Обозначения те
ЖС, ЧТО ИЛ рис. I
честве безусловного рефлекса мы избрали оборонительный рефлекс чере-
пахи на электрический раздражитель, П. М. Никифоровский же поль-
зовался сравнительно слабым, хотя по существу тоже оборонительным, но
весьма неслецифическим рефлексом, — рефлексом втягивания головы под
щит, вызываемым в ответ на
удар по щиту. Разница между
силой этих двух оборонитель-
ных рефлексов очевидна. Что же
касается условных раздражите-
лей, то очевидно, что тактиль-
ные раздражители, которыми мы
преимущественно пользовались,
являются для черепах биологи-
чески более сильными, чем зву-
ковые и обоия тельные раздраж и-
тели. Мы полагаем, что силу
раздражителей всегда нужно
учитывать при сравнительной
оценке скорости образования
условных рефлексов. Может
РИС. 5. Внешние торможения
1 — контрольный условный и безусловный реф-
лексы; 2 — торможение условного рефлекса
после внешнего тормоза — а (удар по щиту);
j и 4 — последующие рефлексы. Обозначения
тс же, что на рис. 1
быть, некоторую роль в этих расхождениях сыграла также сравнительно
грубая и недостаточно точная методика, которой пользовался П. М. Ники-
форовский прн изучении условных рефлексов у черепах.
Другим более важным моментом в нашей работе является констатиро-
вание слабых тормозных процессов у черепах.
Эти наши данные находят подтверждение в аналогичных данных
Ю. II. Фролова, И. С. Беритова, Б. М. Завадовского, М. Л. Рохлиной
и др. относительно слабости тормозов у низших позвоночных и птиц..
В этом аспекте представляют некоторый интерес исследования
Ф. П. Майорова и Ш. Топурия, выявившие относительную слабость тор-
мозных процессов у щенков, и общеизвестные многочисленные данные
•о слабости этих же процессов у детей ^(Н. И. Красногорский.
Л. Г. Иванов-Смоленский и др.).
2 Э. А. Асратян
!8
Э. А. АСРАТЯН
На основании приведенных данных нам кажется, что нельзя проводить
грубый параллелизм между скоростью образования условных рефлексов и
степенью развития resp. функционального удельного веса коры в централь-
ной нервной системе, как это делают многие. Нам представляется более ве-
роятным скорость образования условных рефлексов в указанных выше слу-
чаях ставить в связь со сравнительной слабостью тормозных процессов.
Выводы
Сравнивая данные об условных рефлексах, полученные у черепах,
с результатами подобных исследований, проведенных на собаках и дру-
гих высших животных, мы можем отметить следующее.
1. Положительный условный рефлекс у черепах образуется так же
быстро, как у многих высших животных.
2. Положительные условные рефлексы у черепах немного более стойки,
чем у вышеупомянутых животных.
3. Процессы торможения у черепах выражены несравненно слабее, чем
у высших животных.
4. По многим особенностям условные рефлексы у черепах занимают как
бы среднее место между условными и безусловными рефлексами у высших
позвоночных животных.
ЛИТЕРАТУРА
Асратян Э. А.,’Б а рсегянР. О., Алексанян А. М., Новая методика Из-
учения условных рефлексов у черепах, Физиологический журнал СССР, т. XVI,
№ 3, 1933.
БерптовИ. С., Индивидуально приобретенная деятельность центральной нервной
системы, Тифлис, 1932.
Майоров Ф. П., Условные рефлексы у щенят различного возраста, Архив биоло-
гических наук, т. 29, 1929.
3 ав адовски й Б, М, и Р о х Л И И 9 М- Л., Условные рефлексы у нормальных
и гнпертиреондизированных кур, Медико-биологический журнал, в. 6, 1927.
Осипова В. Н., Скорость образования сочетательных рефлексов у детей школь-
ного возраста, Новое в рефлексологии и физиологии нервной, системы, в. 2, 1926.
Топ у р ня Ш.» Образование и диферекциация индивидуальных оборонительных
рефлексов на щенках, Известия Тифлисского университета, т. X, 1929.
УСЛОВНОРЕФЛЕКТОРНЛЯ ДЕЯТЕЛЬНОСТЬ У ЧЕРЕПАХ
БЕЗ БОЛЬШИХ ПОЛУШАРИЙ И БЕЗ ПРОМЕЖУТОЧНОГО МОЗГА >
а последние годы все чаще и настойчивее ставится вопрос о воз-
можности образования у различных животных условных реф-
лексов вне коры больших полушарий головного мозга. С этим
же связан вопрос о роли больших полушарий вообще в образо-
вании условных связей. Что касается высших животных (собак),
то, как известно, в лабораториях И. П. Павлова принято счи-
тать, что местом образования условных рефлексов у собак являются боль-
шие полушария и именно кора больших полушарий.
Г. П. Зеленый и сотрудники на основании своих опытов пришли к выводу,
что у собак возможно образование условных рефлексов вне коры головного
мозга. Этого взгляда придерживается и ряд других авторов. В нашу задачу
не входит подробный разбор экспериментального материала этих авторов,
но нам кажется,что выводы их все еще не являются бесспорно доказанным и,
хотя мы не считаем, что такая возможность у собак исключена.
Из других позвоночных подобные опыты были поставлены на лягушках
(К. М. Леутский, А. Брегадзе и И. С. Беритов), на голубях (И. С. Беритов),
на рыбах [Ю. П. Фролов, Буль (Bull), Штейнер (Steiner)].
На основании экспериментальных данных большинством авторов было
высказано мнение, что образование условных связей у этих животных воз-
можно и без наличия больших полушарий.
Исходя из этих данных и некоторых соображений, высказанных нами
в предыдущих работах, мы поставили опыты на черепахах с полной
экстирпацией сперва больших полушарий, а затем (у тех же черепах) и
среднего мозга.
Результаты наших исследований полностью согласуются с выводами
вышеперечисленных авторов относительно условнорефлекторной деятель-
ности у низших животных без наличия больших полушарий.
Опыты были поставлены на б черепахах. У 3 из них по нашей методике
сначала были выработаны условные рефлексы на щито-механическое раз-
дражение. После того как достигалась обычная величина условного реф-
лекса, т. е. когда условный рефлекс становился достаточно прочным,
у этих черепах описанным ниже способом удалялись большие полуша-
рия. Операция производилась без наркоза. Вскрыв череп и обнажив
подлежащую удалению часть мозга, препаровальной иглой мы удаляли
большой мозг, а в последующих операциях — средний мозг. Образо-
вавшаяся полость заливалась парафином, что служило также для оста-
новки кровотечения, а сверху оперированное пространство черепа за-
1 Совместно'с Араратом Алексаняном. Доложено на IV Всесоюзном съезде физио-
логов в Харькове в 1930 г., Физиологический журнал СССР, т. лVI, № 6, 1933.
3*
20
Э А. АСРАТЯН
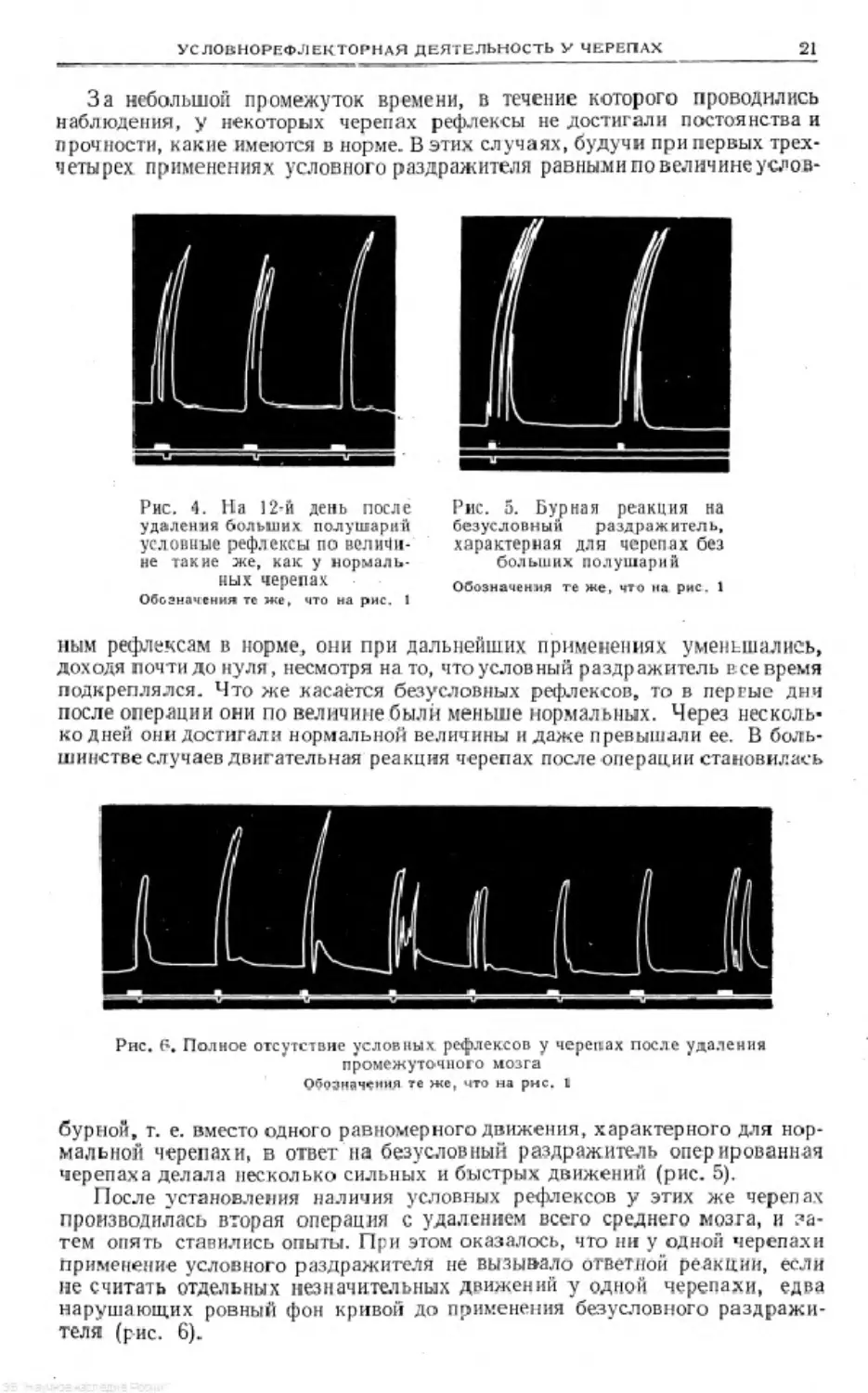
Рис. 1. Условные
рефлексы у нормаль-
ной черепахи
Нижняя лип ня-отметка
безусловного раздражи*
теля , средняя — отметка
услозного раздражите-
ля, верхняя — запись
условных и безусловных
рефлексов
леплялось менделеевской замазкой. Пелле окончания исследования произ-
водилось вскрытие черепахи в целях проверки правильности операции.
В первое время после операции животное
вело себя несколько иначе, чем нормальное, а
именно: при щипании одной конечности чере-
паха втягивала под щит только ущипнутую ко-
нечность, остальные (в особенности противо-
положную) конечности, наоборот, высовывала
из-под щита; или же при щипании она высоко
поднималась на задние конечности, оставаясь
некоторое время в таком наклонном положении,
или же одной конечностью она делала быстрые
вращательные движения в сторону ущипнутой
конечности. В Дальнейшем оперированные жи-
вотные вели себя так же, как нормальные; при
щипании быстро втягивали конечности под щит.
Однако у них замечалось некоторое падение то-
нуса скелетных мышц. 11ривязанные к станку
оперированные животные высовывали из-под
щита вяло висевшие конечности.
После операции изучение условных рефлек-
сов продолжалось. При этом оказалось, что:
1) й первые Дин после операции (отЗ до 7 дней) условные рефлексы исчеза-
ют н потом постепенно снова появляются (рис. 1—3); 2) условные реф-
а
Рис. 2? Исчезновение условных рефлексов после удаления
больших" полушарий мозга
п- на 2-й день; f- „а 0-Я день. Обозначения те же, что на рис. 1
Рис. 3. Появление Слабых и непостоянных услов-
ных рефлексов на 9-й день после удаления
больших полушарий
Обозначения те же, что на рнс. I
б
лексы, будучи слабыми и непостоянными в первые дни своего появле-
ния, в дальнейшем усиливаются, достигая, в конце концов, почти вели-
чина? условных рефлексов нормальных черепах (рис. 4).
УСЛОВНОРЕФЛЕКТОРНАЯ ДЕЯТЕЛЬНОСТЬ У ЧЕРЕПАХ
21
За небольшой промежуток времени, в течение которого проводились
наблюдения, у некоторых черепах рефлексы не достигали постоянства и
прочности, какие имеются в норме. В этих случаях, будучи при первых трех-
четырех применениях условного раздражителя равными по величине услов-
Рис. 4. На 12-й день после
удаления больших полушарий
условные рефлексы по величи-
не такие же, как у нормаль-
ных черепах
Обозначения те же, что на рис. 1
Рис. 5. Бурная реакция на
безусловный раздражитель,
характерная для черепах без
больших полушарий
Обозначения те же, что на рис. 1
ним рефлексам в норме, они при дальнейших применениях уменьшались,
доходя почти до нуля, несмотря на то, что условный раздражитель все время
подкреплялся. Что же касается безусловных рефлексов, то в пергыс днч
после операции они по величине были меньше нормальных. Через несколь-
ко дней они достигали нормальной величины и даже превышали ее. В боль-
шинстве случаев двигательная реакция черепах после операции становилась
Рис. 6. Полное отсутствие условных рефлексов у черепах после удаления
промежуточного мозга
Обозначения те же, что на рнс. 1
бурной, т. е. вместо одного равномерного движения, характерного для нор-
мальной черепахи, в ответ на безусловный раздражитель оперированная
черепаха делала несколько сильных и быстрых движений (рис. 5).
После установления наличия условных рефлексов у этих же черепах
производилась вторая операция с удалением всего среднего мозга, и за-
тем опять ставились опыты. При этом оказалось, что ни у одной черепахи
Применение условного раздражителя не вызывало ответной реакции, если
не считать отдельных незначительных движений у одной черепахи, едва
нарушающих ровный фон кривой до применения безусловного раздражи-
теля (рис. 6).
22
Э Л ЛСР \ гя н
Во второй серии опытов (3 черепахи) вначале удалялись большие полу-
шария таким же образом, как ив первой серии. Условные рефлексы выра-
батывались только после удаления больших полушарий. Нам удалось уста-
новить, что у этих черепах, так же как у нормальных, образовавшиеся
условные рефлексы не отличались от рефлексов у черепах первой серии.
При удалении всего среднего мозга эти условные рефлексы исчезали.
Таким образом, нами было установлено следующее:
1) условные рефлексы, выработанные до экстирпации больших полуша-
рий, сохраняются также после их экстирпации;
2) условные рефлексы у черепах образуются без наличия больших по-
лушарий и
3) при удалении среднего мозга выработанные условные рефлексы исчс-
з ают.
Заканчивая этим описание экспериментальной части, мы считаем необ-
ходимым остановиться на следующем вопросе: являются ли вышеописан-
ные рефлексы у лишенных полушарий черепах условными рефлексами
или они представляют собой безусловные рефлексы, вызванные раздра-
жителями в результате наступившего вследствие удаления больших полу-
шарий повышения возбудимости оставшихся частей центральной нервной
системы, как пытаются объяснить аналогичные опыты на голубях неко-
торые ученые.
Мы считаем более правильным первое предположение (для наших слу-
чаев) на основании следующих данных.
1. Для получения условных рефлексов после экстирпации больших полу-
шарий (следовательно, при наличии повышенной возбудимости) все же нужно
их вырабатывать. Черепахи реагируют на индиферентный раздражитель не
ср згу, а лишь после некоторого сочетания его с безусловным раздражителем.
2. После удаления среднего мозга (тут тоже сохраняется повышенная
возбудимость) условные рефлексы исчезают.
Наши данные и выводы, что большие полушария не являются единствен-
ным местом образования условных связей у черепах, вполне согласуются
с выводами других авторов, работавших с низшими позвоночными
(Ю. П. Фролов, К. М. Леутский и др.).
Результаты наших исследований, как и материалы, имеющиеся в лите-
ратуре, позволяют присоединиться к тому взгляду, что у животных, нахо-
дящихся на низкой ступени зоологической лестницы, возможно образован не
условных рефлексов, не только в больших полушариях, но и в- других от-
делах головного мозга.
В дальнейшем, при развитии больших полушарий, условнорефлекторная
деятельность становится их функцией. Таким образом, условнорефлектор-
ная деятельность является функцией высших отделов нервной системы дан-
ного животного. В одних случаях таким высшим отделом у животного
может быть нервный ганглий, как, например, у асцидий (Е. М. Крепе), у
других животных, у которых кора еще не развита (рыбы),—древние нерв-
ные образования больших полушарий и, наконец, кора больших полу-
шарий у высших животных. При этом, чем ниже стоит животное на эво-
люционной лестнице, чем менее диференцнрована нервная масса, тем менее
разнятся Друг от друга функции разных отделов нервной системы, Други-
ми словами, чем менее развито животное, тем больше шансов за то, что
функции высших отделов в той ИЛИ ИИой форме мы встретим и в низших
-отделах. Этот факт мы наблюдали на черепахах.
Можно полагать, что сравнительно-физиологическое изучение, выяв-
ление качественных различий, своеобразие условнорсфлек тор ной деятель-
КССТИ у различных представителей зоологической лестницы, а также из-
учение этой деятельности при удалении разных отделов нервной системы
дадут нам возможность выяснить генезис условнорефлекториой функции.
УСЛОВНОРЕФЛЕКТОРНАЯ ДЕЯТЕЛЬНОСТЬ У ЧЕРЕПАХ
23
ЛИТЕРАТУРА
Асратян Э. А. и Алекса яя и А. М., Новая методика изучения условных реф*
лексов у черепах. Физиологический журнал СССР, т. XVi, в. 3, 1933.
Бер и т о в И. С., Индивидуально приобретенная деятельность центральной нервной
системы, 1932.
Крепе Е. М., К физиология и биологии асцидий, Работы Мурманской биологиче-
ской станнин, т. 1, 1925.
ЛеутскнйК. М., Условные рефлексы у лягушек нормальных н лишенных боль,
ших полушарий головного мозга, Русский физиологический журнал, 12, 1929.
Павлов И. П. Дваднатилетний опыт объективного изучения высшей нервной
деятельности (поведения), животных, 1932.
Павлов И. П., Лекции о работе больших полушарий, 1927.
Алексееве. А. и П о л т ы р е в С. С., Cber die Moglichkeit der Bildung be-
dingter Reflexe bei Hunden von der extirpierter Hemisphare, Труды Общества рус-
ских физиологов им. Сеченова, в. 3, 1929.
Ба ш м урин А. Е. и Мюльберг Б. М., Zur Frage uber des Moglichkeit der
Bildung von bedingten Reflexe auf die der Hemisphare mit extirpierten Hirnrinde
gegen uberliegenden Extremitaten, там же.
Зеленый Г. П. и Пол ты ре а С. С., Groshirnrinde und Assoiiationsfunc lien,
Ztschr. f. Biol., Bd. 190, 1930.
Фролов Ю. П., Bedingte Reflexe bei Fischen, 11. Mitteilung. Pfhiger's Arch.,
Bd. 220, 1928.
Bull N. 0., Journ. of the Marine Biol. Assoc., v. 15, 1928.
Schrader M., Zur physiologic des Froschgehirns, Pfluger1's Arch., 44, 1889.
S t e i ne r I., Die Function des zentr. Nervensysten’ u. ihre Phylogenese, 1885.
СИСТЕМНОСТЬ РАБОТЫ БОЛЬШИХ ПОЛУШАРИЙ
ГОЛОВНОГО МОЗГ/Х 1
те в самом начальном периоде развития павловского учения
об условных рефлексах некоторые сотрудники И. П. Павлова
описали явление так называемого периодического слюноотде-
ления как результат раздражения ротовой полости слюногон-
ными веществами через определенные промежутки времени
(Г. П. Зеленый и др.). Очевидно, что в этих опытах имелись
первые определенные намеки на условные рефлексы на время.
В 1908 г. К. Н. Кржишковский методикой условных слюноотделитель-
ных рефлексов обнаружил очень интересное явление в физиологии коры
больших полушарий, явление, тоже родственное условным рефлексам на
время. Работая с кислотными оборонительными слюноотделительными
условными рефлексами, Кржишковский попутно проводил следующие
опыты: независимо от порядкового места условных раздражителей в опыте
он условный тормоз применял спустя 19—20 минут после вливания кислоты
в рот собаки, т. е. после подкрепления предшествующего положительного
условного рефлекса. Когда после двухнедельного применения такого
сочетания Кржишковский через 19—20 минут после вливания кислоты
в. рот собаки применил только условный положительный раздражитель без
комбинации с условным тормозом, он получил почти наполовину умень-
шенный эффект. Когда он тоже самое делал через более длинные или более
короткие паузы, условный положительный раздражитель вызывал полный
эффект. На основании этих весьма интересных опытов Кржишковский при-
шел к следующему выводу: «Таким образом, было ясно, что мы имеем
дело действительно с особым состоянием центральной нервной системы, раз-
вившимся, очевидно, под влиянием длительного применения условного тор-
моза, приуроченного к определенному промежутку времени»3.
С целью анализа этого явления Кржишковский ставил еще очень инте-
ресные опыты следующего характера. После того как совершенно исчезли
следы старых взаимоотношений положительных и отрицательных условных
раздражителей, он оставил условный тормоз и стал применять только поло-
жительные условные раздражители — то через 19—20 минут, то через
32—33 минуты, причем в первом случае он их не подкреплял, а во втором
Случае подкреплял. После определенного Числа таких опытов ему удалось
достигнуть того же результата, что и раньше, т.е. на один и тот же раздражи-
тель после паузы в 19—20 минут он получил сильно уменьшенный услов-
ный рефлекс, а через 32—33 минуты — полный эффект. Кржишковский
же указал на непрочный характер полученного нм феномена.
’ Доложено на 171-м заседании Ленинградского общества физиологов им.
И. М. Сеченова в январе 1933 г. Труды физиологических лабораторий акад. И. П Пав-
лоза, т. VIII, 1938. Предварительное сообщение напечатано в «Докладах Академии
«вук СССР», № 8, 1934, по представлению акад. И. П. Павлова 20 I! 1934 г.
8 Труды: Общества русских врачей в С.-Петербурге, т. 76, 1909, с гр. 325.
СИСТЕМНОСТЬ РАБОТЫ БОЛЬШИХ ПОЛУШАРИИ ГОЛОВНОГО МОЗГ \
25-
Совершенно очевидно, что в опытах Кржшиковского мы имеем дело уже
с такими отрицательными условными рефлексами, в которых решающую
роль играет время; порядковое место раздражителен, как видно из его про-
токолов и описаний опытов, никакой роли в описываемом явлении не играло.
Позже понятие «условные рефлексы на время» получило
права гражданства в учении об условных рефлексах; этому актуальному
вопросу учения о высшей нервной деятельности были посвящены даже
специальные диссертации и труды (Ю. П. Феокритова, М. М. Стукова,
В С. Дерябин, Ю. П. Фролов и др.). Следует отметить, что время как реаль-
ный положительный и отрицательный условный раздражитель играет реша-
ющую роль и в так называемых следовых условных рефлек-
сах (П. П. Пименов, Ф. С. Гроссман, И. В. Завадский, В. М. Доброволь-
ский, В. И. Павлова, М. Ф. Белиц, К). П. Фролов и др.).
С другой стороны, И. Я. Перельцвейг еще в 1907 г. наблюдал, что
порядок условных раздражителен оставляет некоторый след. Долгое время
он в своих опытах раньше всего применял к а с а л к у; затем, когда он
стал применять на первом месте холод, последний условный раздра-
житель на этом порядковом месте вначале вовсе не вызывал условного
рефлекса, тогда как, будучи применен в середине опыта, он всегда вызы-
вал значительное слюноотделение. Совершенно то же самое, но на более
убедительном и обширном материале получил П. С. Купалов. Он показал,
что применяемые на одном и том же месте в течение длительного времени
условные раздражители оставляют след в коре больших полушарий. На
большое значение порядка условных раздражителей указывает также
Д. И. Соловейчик,
Во избежание этих последствий и образования условного рефлекса на
время в лабораториях акад. И. П. Павлова в течение длительного периода
рекомендовали при работе с условными рефлексами возможно больше разно-
образить порядок условных раздражителей и изменять паузы между ними.
Следует отметить, что в первые же месяцы нашей работы по условным реф-
лексам, в 1927 г., мы на практике убедились в правильности этих положе-
ний и всячески избегали одинаковых пауз между применением условных
раздражителен. Больше того, чтобы не вызвать осложнений, мы в своих
опытах избегали пользоваться условными раздражителями в. определенной
последовател ь н ости или ж:е при измененной последовательности
их старались не применять раздражителя па каком-нибудь определенном
месте. Поэтому поводу в нашей работе сказано: «В разных опытах место
диферепцировочного раздражителя в ряду раздражений менялось, чтобы
не образовался условный рефлекс на место и на порядок
раздражен и й»1.
В силу того, что за последние годы в лабораториях акад. И. П. Павлова
для специальных целей широко использовался такой прием работы по услов-
ным рефлексам, при котором условные раздражители применялись каждо-
дневно в известном фиксированном порядке, по определенному шаблону,
естественно, возник интерес к более подробному изучению тех последствий
в коре большого мозга, которые обусловливаются фиксированным порядком
раздражителей. Такую задачу мы перед собой и поставили.
I
Наши первые подопытные собаки Арч иМасисбыли молодыми животны-
ми и, как потом выяснилось, отличались сильным уравновешенным ти-
пом высшей нервной деятельности.
У обеих собак были образованы новые условные положительные рефлек-
1 Архив биологических наук, т. 30, в. 2, 1930, стр 245.
2п
Э. А. АСРАТЯН
сы на звонок, шипенье, свет, стук 120 ударов в 1 минуту и затем был отди-
ференцирован от последнего стук 60 ударов в 1 минуту.
В общем как положительные условные рефлексы, так и днференцировка
на частоту стука у сбеих собак образовывались довольно быстро и без осо-
бых отклонений от нормы.
У Арча, например, условные рефлексы появились в. 4-м опыте, достигли
почти нормальной величины в 20—21-м опыте; днференцировка впервые
дала нулевой эффект на 12-м применении. То же самое с некоторым опозда-
нием имело место и у Маснса.
После того как уже были выработаны положительные условные рефлек-
сы, мы начали ежедневно применять шаблон раздражителей с 5-минутными
паузами между ними. Диференцировочный раздражитель, будучи еще не
абсолютным, занимал место в середине системы. Когда после 8-дневного
применения шаблона раздражителей испытывался какой-нибудь один
условный положительный раздражитель подряд в.течение всего опыта,
получалось интересное явление, отчасти ожидавшееся, отчасти неожидан-
ное. Можно было ожидать, что на месте диференцировочного раздражителя
положительный условный раздражитель сильно уменьшит свой условный
эффект; неожиданное заключалось в том, что примененный раздражитель
вызвал почти полное повторение всей картины опыта с системой раздражите-
лей, воспроизвел опыт с шаблоном в целом. Следует отметить, что все
положительные условные раздражители, входящие в систему, воспроизво-
дили это интересное явление вболылей или меныпей степей и. При этом заслу-
живает особого внимания двигательная условная реакция собак. При
многократном повторном применении какого-нибудь условного положи-
тельного раздражителя с целью воспроизведения системности собаки на
месте воздействия положительных раздражителей в стереотипе проявляли
явную положительную двигательную реакцию к кормушке; на месте же
диференцировочного раздражителя, т. е. на месте основного показателя
системности, они давали чрезвычайно сильную отрицательную двигатель-
ную реакцию, точно такую, как при применении самих диференцировоч-
ных раздражителей. Больше того, одна из собак (Масис) на этом месте
иногда даже отказывалась от поданной пищи, пока раздражитель продол-
жал действовать, и медленно подходила к кормушке лишь после прекра-
щения действия раздражителя.
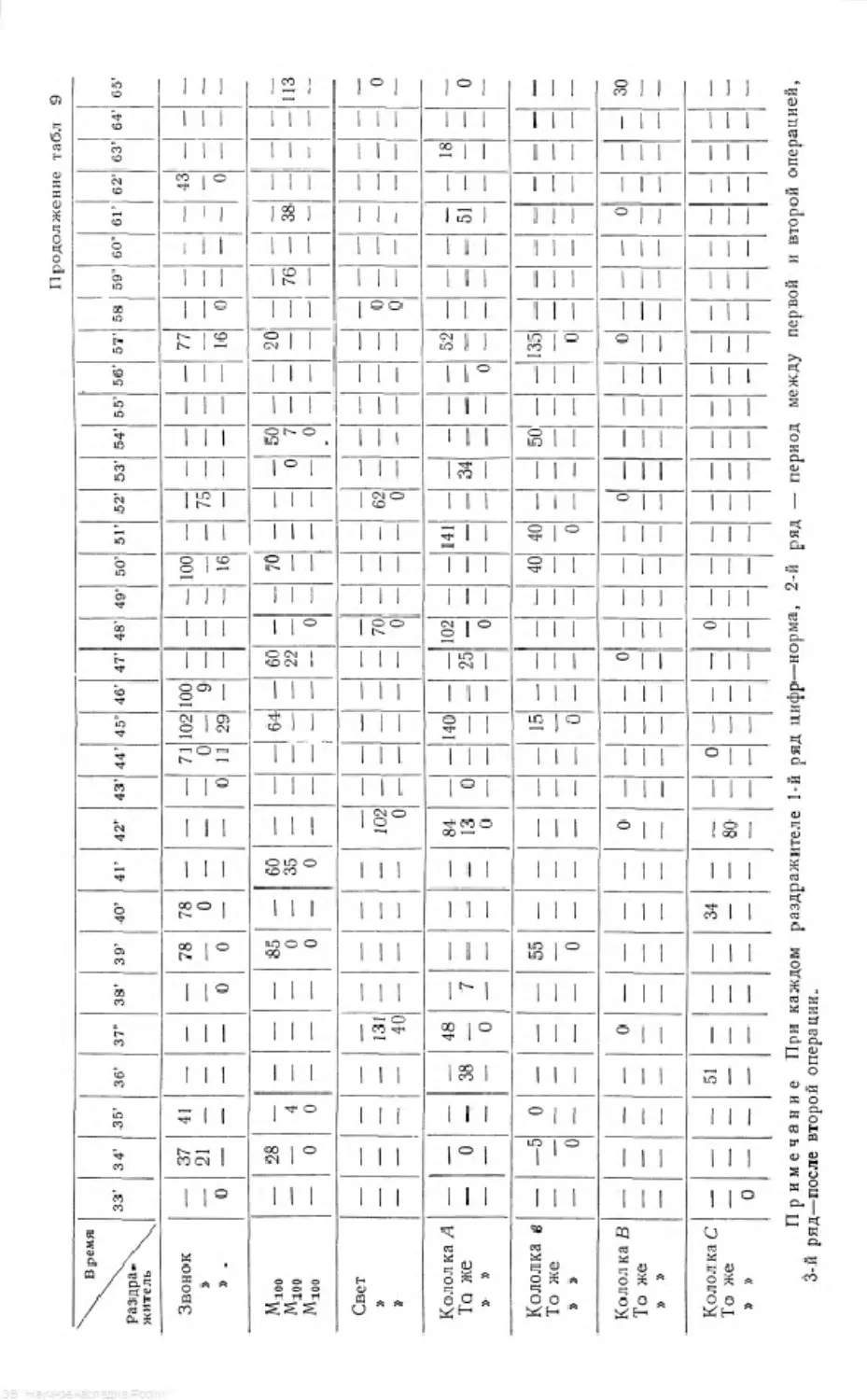
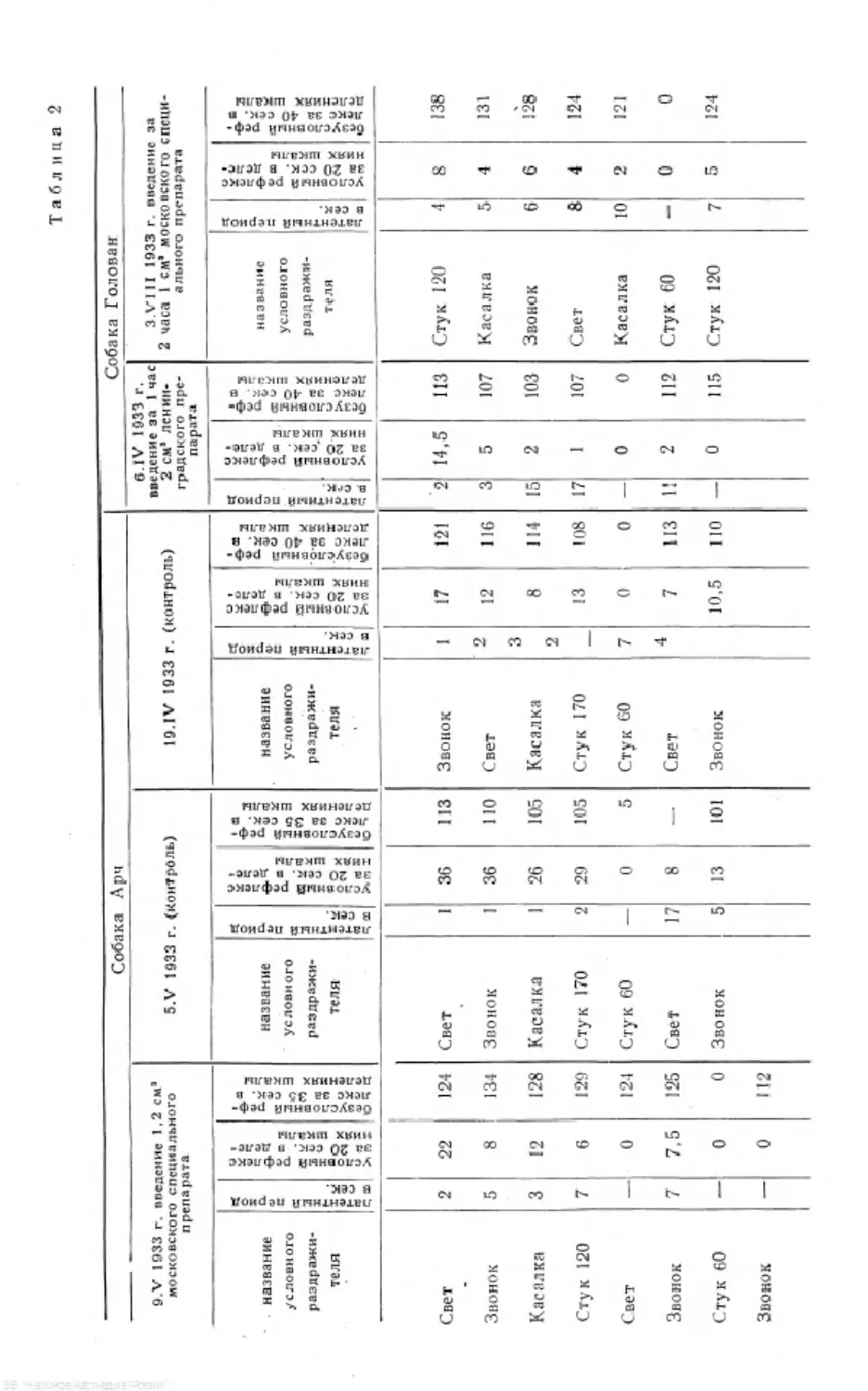
Для иллюстрации сказанного приводим табл. 1 (паузы между раздражи-
телями всюду 5-минутные).
Тоже явление можно- видеть и на приводимых диаграммах (рис. 1).
Так как в лабораториях акад. И. П. Павлова для характеристики ком-
плекса раздражителей, применяемых в определенном фиксированном по-
рядке, употребляются выражения «система», «шаблон» и «стереотип», мы
назвали систем ностью свойство коры больших полушарий
•объединять отдельные условные рефлексы в определенной последователь-
ности в единое целое.
Ввиду закономерного характера полученного явления и большого инте-
реса, проявленного к нему со стороны акад. И. П. Павлова, мы решили
продолжить работу в этой области. Полученный материал в течение годичной
работы на трех собаках кратко излагается в данной статье.
Минимальное время, необходимое для образования у наших собак систе-
мности в более или менее уловимой форме, находилось в пределах 3—4
дней, а 5—6-дневное применение системы раздражителей вызывало уже
отчетливую реакцию. Выраженность и стойкость нового феномена явно за-
висят от длительности применения системы раздражителен; повидимому,
тип нервной системы собак тоже оказывает известное влияние.
Наиболее отчетливо и длительно воспроизводят системность те раздра-
жители, которые близки к диференцировочному. У собак Арч и Масис
СИСТЕМНОСТЬ РАБОТЫ БОЛЬШИХ ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА
27
Таблица 1
Слюноотделение в делениях шкалы за 20 сек.
Условный раздражитель Опит № 32 8. I 1031 г. Опыт № 36 14 1 1931 г. Применяется ЗВОНОК Опыт № 37 1 В 1 1931 г. Применяется свет Примечание
Собака Арч
Звонок 28 27 26 В опытах № 36 и 37 у
Свет 24 27 22 Арча всегда имеется по-
Шипенье 18 22 19 ложительная двигатель-
Стук 120- в 1 мин. 20 19 18 ная реакция, кроме того
Стук 60 в 1 мин. 4 11 4Уг места применения ус-
Звонок 24 19 13 ловного положительного
Свет 16 «4 15 раздражителя, которое
Шипенье 10 8 114 совпадает с местом ди-
Стук 120 в 1 мин. 13 18 14’4 фереецировки в системе.
На этом месте имеется
ясная отрицательная
двигательная реакция
Опыт № 119 Опыт № 120. Опыт Хв 121.
Применяет- Применяет-
ся стук 120 ся звонок
в 1 мин.
Собака Масис
Стук 120 в 1 мин. 13 11 пу2
Шипенье 8 7’4 9
3-ROHOK 94 8% 9
Стук 60 в 1 мин. 0 1” 2*/г* • Отказ от еды на
Свет 5’4 6 в 25-й секунде
Стук 120 в 1 мни. 6’4 7 7 •• Отказ от еды на
Шипенье 1 3 4% 10-й секунде
Звонок 534 6 64
эту роль играл стук метронома 120 ударов в 1 минуту’. Что же касается дру-
гих раздражителей, то можно сказать, что слабые раздражители ее воспро-
изводят лучше сильных. Поразительно, — и это получилось в первых же
опытах, — что слабые раздражители при воспроизведении системности на
месте сильных раздражителей вызывают гораздо больший условный эффект,
чем обычно, почти близкий к эффекту сильных раздражителей. Для
иллюстрации приводится контрольный опыт, показывающий силовые со-
отношения условных раздражителей в норме (опыт № 174), одни из
опытов с системой (опыт№ 203), а также опыты, иллюстрирующие степень
воспроизведения системности каждым отдельным условным раздражителем
(опыты № 205—211; рис. 1).
Возник чрезвычайно важный вопрос, воспроизводится ли системность
только пунктами коры, соответствующими входящим в систему раз-
дражителям, или же она воспроизводится и другими участками коры, а то
и всеми ее пунктами.
Вопрос этот был решен следующим образом. Когда имевшаяся си-
стемность была полностью разрушена (о разрушении системности
будет специально сказано ниже), был образован новый пищевой услов-
ный положительный рефлекс из области нового анализатора — рефлекс
на касалку. После того как новый условный рефлекс был выработан и укреп-
лен, мы приступили к образованию повой системности в коре, причем си-
стема раздражителен была составлена из старых условных раздражителей,
а новый условный рефлекс на касалку не только не был включен в систему,
но его применение вовсе прекращалось во все время действия системы. После
нескольких опытов со стереотипом раздражителей впервые в течение всего
опыта была применена касалка с целью воспроизведения образованной си-
28
Э. А. АСРАТЯН
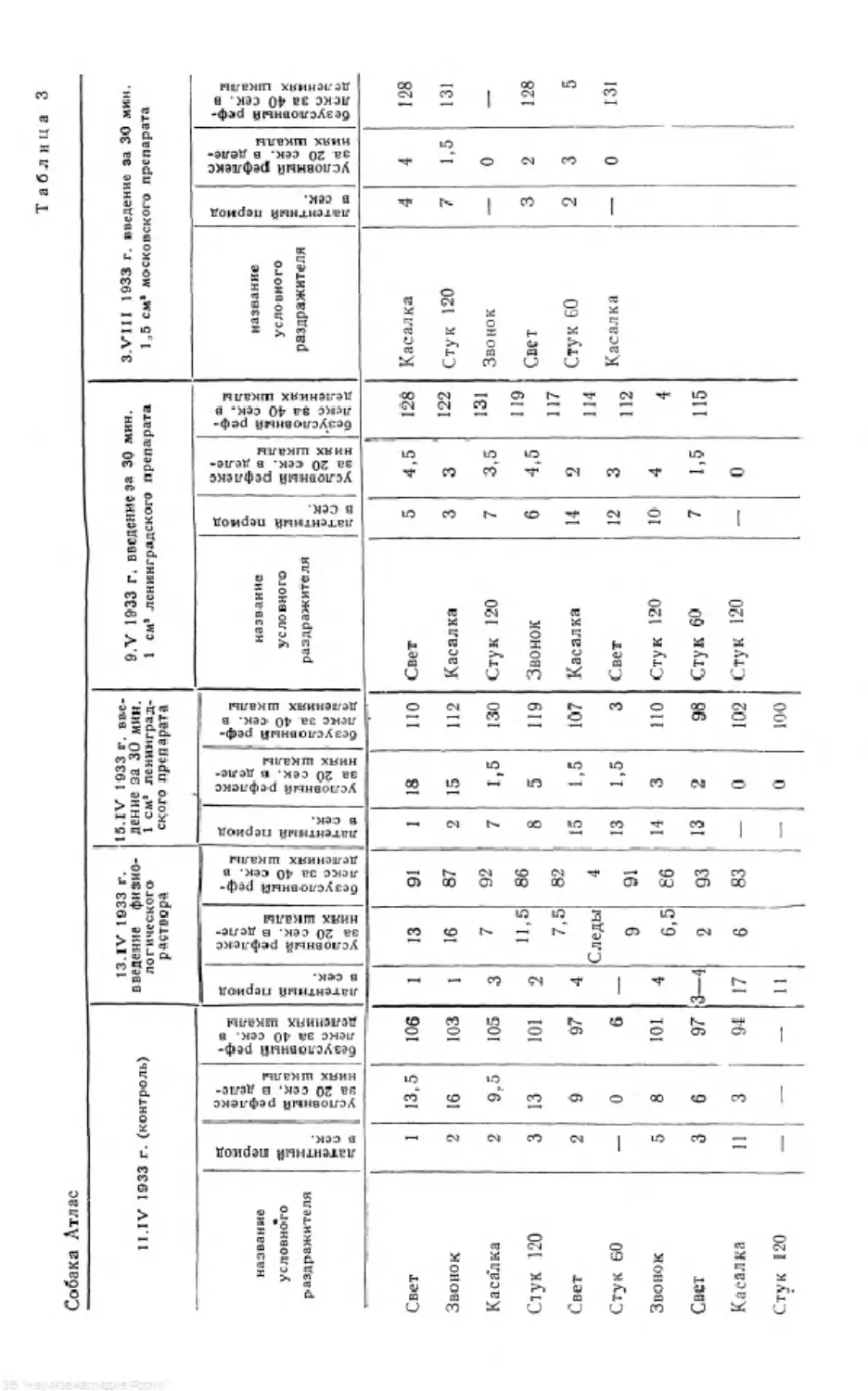
стоимости. И, как видно из табл. 2, касалка прекрасно воспроизвела систем-
ность в коре, хотя в образовании этой системности она не принимала уча-
стия. В таблице приводится контрольный опыт, показывающий картину
условных рефлексов на непрерывное применение касалки до образования
Ц i'A/
Условные обозначения'.
Q Свет | Касалка Звонок
| Стук 12О уда роб 6 Тмин. g С тун 60ударов в Тмин.
Рис. 1. Воспроизведение системности
системности (№ 175), после ее образования (опыт Мг 187) и, наконец, опыт
с самой системой (опыт№ 180). Паузы всюду 5-минутные.
Таблица 2
Слюноотделение у собаки Арч за 20 сек. в делениях шкалы
Условный раздражитель Опыт № 175 17.X 1931 Г, .Многократно применяется касалка Опыт Aft 18-7 I.XI 1931 г. Мши ок рати о применяется касалка Опыт № 180 23 X 1931 г.
Шипенье 25 29 32
» 23 2ft 29
Стук 120 в 1 мин. 21 >; 25 28
» 60 » 1 » 22 9 0
Свет 21 20 17
Звонок 20 17% 26
На основании приведенного материала можно сделать вывод, что систем-
ность воспроизводится и соседними участками коры, которые не принимают
СИСТЕМНОСТЬ РАБОТЫ БОЛЬШИХ ПОЛУШАРИИ ГОЛОВНОГО МОЗГА
29
непосредственного участия в се образовании, и. возможно, что эта способ-
ность генерализуется во всей коре—в одних участках в большей степени,
в других в меньшей.
Разрушается системность лучше и быстрее всего при каждодневном при-
менении отдельных условных раздражителей в течение всего опыта. Хотя
мы этого не испытывали, но можно полагать, что они будут также быстро
разрушаться при применении ряда раздражителей в течение опыта, но
в ином порядке, чем в системе. При применении вышеназванного приема
удается разрушить системность приблизительно в течение стольких же дней,
во сколько она была образована; маленькие отклонения, конечно, всегда
могут быть. Невидимому, играет роль и тип собаки. По в основном мож-
но сказать, что системность разрушается тем быстрее и легче, чем моложе
собаки, чем меньше опытов поставлено для ее образования.
Имеются интересные моменты и в разрушении системности.
Во-первых, если системность разрушается каждодневным применением
какого-нибудь одного раздражителя, то она оказывается уже разрушенной
и для всех остальных раздражителен, не принимавших участия в ее разру-
шении. В табл. 3 представлены опыты из раннего периода нашей работы,
когда пищевые условные положительные рефлексы были несколько слабо-
ваты и световой диференцировочный раздражитель еще вызывал сильное
последовательное торможение их. После 11-кратного применения системы
мы приступили к ее разрушению каждодневным включением одного только
положительного раздражителя — стука 120 в 1 минуту.
Таблица 3
Слюноотделенье у собаки Арч за 20 сек. & делениях шкалы
Условный раздражитель Опыт № 87 Опыт № 88. Применяется стук 120 » 1 МИН, Пер- вый день после пре- к ращения системы Опнт № 101. Применяется стук 120 н 1 МИН. 14-й лень после прекращения системы Опыт № 105. Приме- няется г«ет после разруше- ния сис- темы стуком Опыт № 1045. 11риме- няется Э BOW&R после раз- рушения системы стуко м Опыт № 107. Приме- няется ШИПИМ после раз- рушения системы стуком
Стук 120 в 1 мин. 25 22 32 14 34 31
Шипенье 24 21 27 10 26ft 26
Стук 60 в 1 мин. 0 6 14 Ц 16 17
Звонок 17 13 9 б'/2 13 12
Свет 6 6 5 4 6 1УЯ
Стук 120 в 1 мин. 2% 3 0 3 б’/2 5
Шипенье 8% 3 0 0 6/2 5
Этим приемом системность была целиком разрушена на 13—15-й день.
Следует упомянуть, что такое многократное применение только одного
раздражителя в течение всего опыта являлось довольно трудной зада-
чей для собаки; условные рефлексы, особенно к концу опыта, значи-
тельно падали в силу развившегося тормозного состояния. Состояние
торможения оставалось и некоторое время после этого приема. Когда после
разрушения системности стуком 120 в 1 минуту были применены другие
раздражители, принимавшие участие в се образовании, но не участвовав-
шие в разрушении ее, оказалось, что системность больше не воспроизводит-
ся ни одним из раздражителей. Это показывает, что не только при образова-
нии системности, но и при ее разрушении кора функционирует как целое.
Обращает на себя внимание то,что при многократном применении раздра-
жители не только не вызывают характерного хода рефлексов системы, но
каждый из них обусловл ивает эффект, примерно соответствующий по вели-
чине нормальному. Сильное ослабление рефлексов в течение опыта, очеаид
30
Э. А. АСРАТЯН
но, обусловливается, как уже был о сказано, длительным каждодневным при-
менением стука.
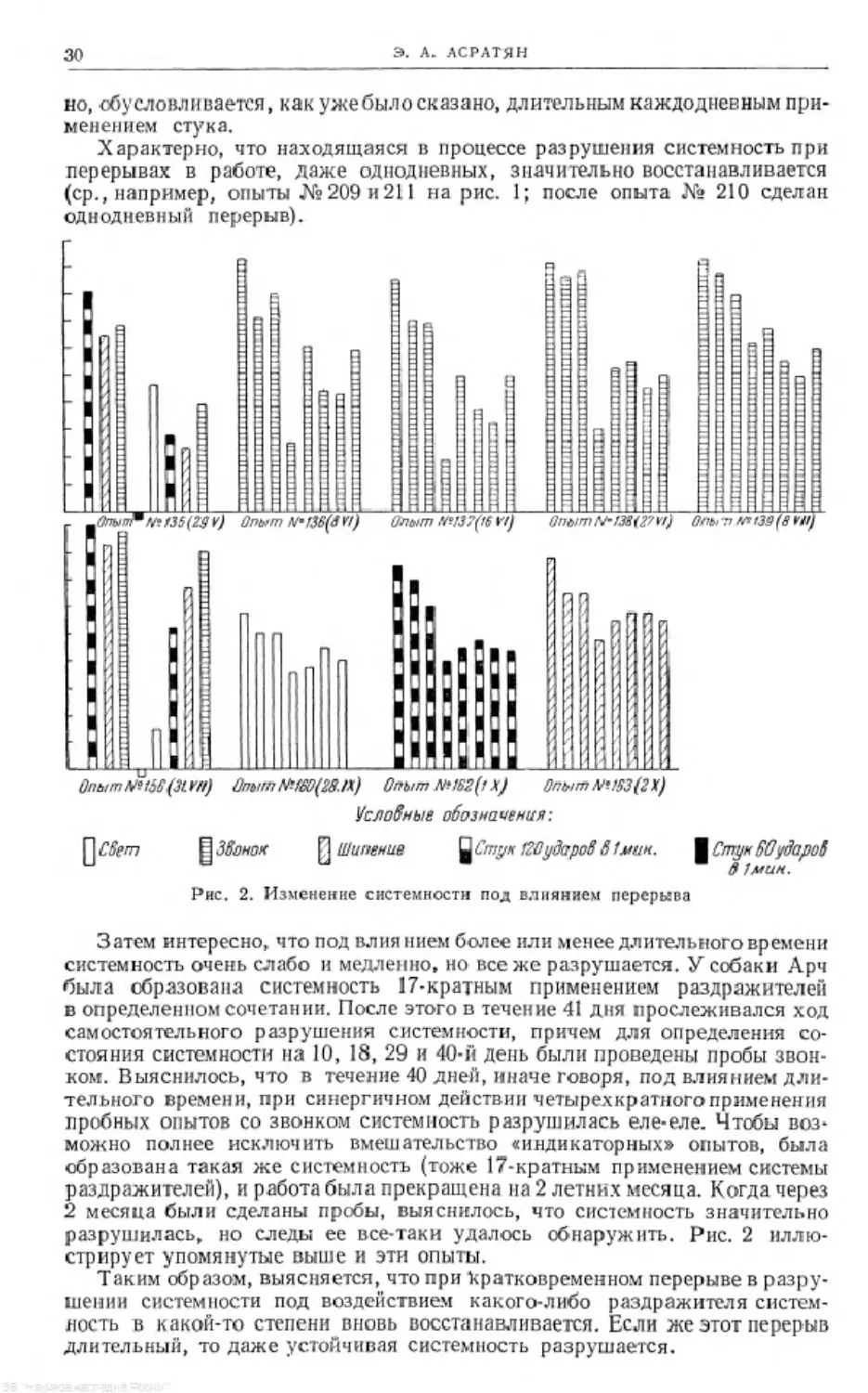
Характерно, что находящаяся в процессе разрушения системность при
перерывах в работе, даже однодневных, значительно восстанавливается
(ср., например, опыты №209 и211 на рис. 1; после опыта № 210 сделан
однодневный перерыв).
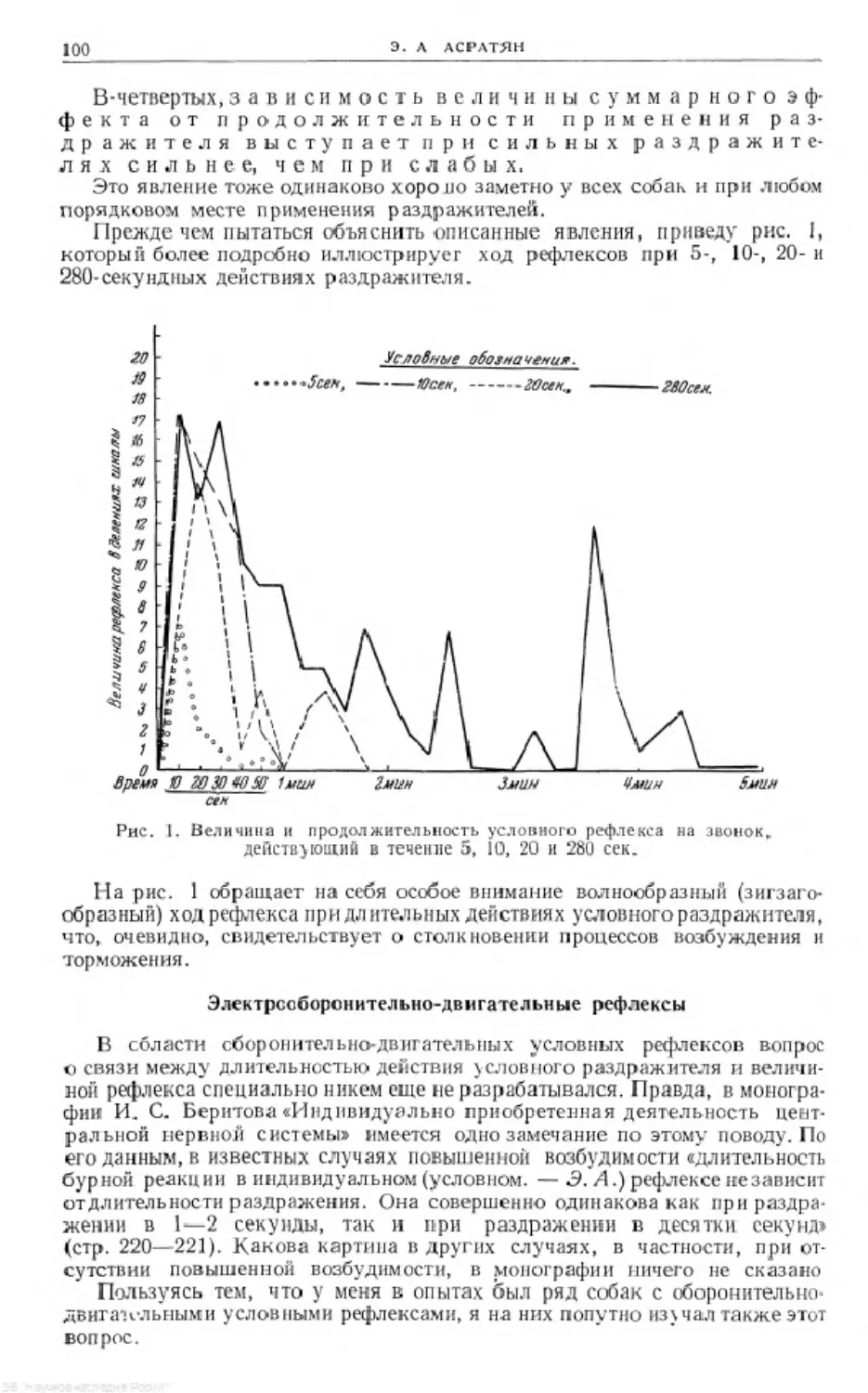
П.Ж'даг П Шипение Qtmy*Яйударов^Стук 50удароб
в в 1мин.
Рис. 2. Изменение системности под влиянием перерыва
Затем интересно, что под влиянием более или менее длительного времени
системность очень слабо и медленно, но все же разрушается. У собаки Арч
была образована системность 17-кратным применением раздражителей
в определенном сочетании. После этого в течение 41 дня прослеживался ход
самостоятельного разрушения системности, причем для определения со-
стояния системности на 10, 18, 29 и 40-й день были проведены пробы звон-
ком. Выяснилось, что в течение 40 дней, иначе говоря, под влиянием дли-
тельного времени, при синергичном действии четырехкратного применения
пробных опытов со звонком системность разрушилась еле-еле. Чтобы воз-
можно полнее исключить вмешательство «индикаторных» опытов, была
образована такая же системность (тоже 17-кратным применением системы
раздражителей), и работа была прекращена на 2 летних месяца. Когда через
2 месяца были сделаны пробы, выяснилось, что системность значительно
разрушилась, но следы ее все-таки удалось обнаружить. Рис. 2 иллю-
стрирует упомянутые выше и эти опыты.
Таким образом, выясняется, что при кратковременном перерыве в разру-
шении системности под воздействием какого-либо раздражителя систем-
ность в какой-то степени вновь восстанавливается. Если же этот перерыв
длительный, то даже устойчивая системность разрушается.
СИСТЕМНОСТЬ РАБОТЫ БОЛЬШИХ ПОЛУШАРИИ ГОЛОВНОГО МОЗГА
31
II
Мы занялись и кардинальным для работы вопросом, какова интимная
сторона возникновения системности; что лежит в основе ее образования:
время или порядковые места раздражителей.
С этой целью на собаке Арч, главном объекте настоящей работы, была
образована новая системность 11-кратным применением системы раздраж и-
телей с 5-минутными паузами между ними. Пробы с отдельными индикатор-
ными раздражителями для выявления системности работы коры ставились
следующим образом: индикаторные раздражители в разных опытах приме-
нялись не через 5-минутные паузы, а через 3-, За/г 7V1- и 10-минутные
паузы.
Эти опыты с большой убедительностью показали, что системность вос-
производится не по временному признаку, а по признаку порядковых мест
раздражителей в системе. Во всех опытах без исключения основной пока-
затель системности—след дифёренцировочного раздражителя—воспроизвел-
ся на своем порядковом месте в системе раздражителей независимо от пауз.
Для иллюстрации приводится диаграмма опытов (рис. 3).
Имея в виду исключительную важность вопроса, мы решили продол-
жить свои исследования на свежей собаке Баш.
Условные рефлексы у Баша мы образовали, избегая применения равных
пауз и какой-нибудь фиксированной последовательности раздражителей.
После того как был налицо ряд положительных условны?; пищевых раздра-
жителей и одна диференцировка, мы приступили к выработке системности
с помощью такого сочетания раздражителен, в котором порядок раздражи-
телей фиксировался, но паузы между ними постоянно различным образом
менялись. После 17-кратпого применения системы были начаты пробы с вос-
произведением предполагаемой системности, причем индикаторы-раздражи-
тели в одних местах применялись через равные паузы, в других через раз-
ные и т. д.
Результаты оказались очень интересными; выяснилось, что системность
может образоваться и без применения равных пауз, только на основании
фиксированной последовательности раздражителей. Сказанное хорошо
видно на рис. 4.
На основании этих показательных и убедительных опытов можно ска-
зать, что ведущим условием образования системности в коре больших полу-
шарий является порядковое место раздражителей в с и-
с т е м е; что касается времени, то, невидимому, его роль настолько второ-
степенна, ничтожна, что ее не удается даже обнаружить более или менее
заметным образом. Не исключена возможность, что имеется какая-то связь
между системностью и условным рефлексом на время, — эго покажут даль-
нейшие исследования в этой интересной области; однако сейчас уже можно
категорически утверждать, что это совершенно разные явления, совершенно
разные виды работы коры больших полушарий. Грубо говоря, можно счи-
тать, что условный рефлекс на время основывается на способности коры
«учитывать» время, реагировать на время как па реальный раздражитель,
системность же как более сложное явление, как явление качественно
иного, более высокого порядка основано на способности коры «учитывать»
порядок и место раздражителей. Ясно, что в первом случае тоже играют
роль порядок и место раздражителей, тем более во втором случае, в случае
системности, раздражители находятся и в определенной временной по-
следовательности. Итак, в первом случае определяющим, ведущим
является время, а остальные моменты относятся к второстепенным,
во втором же случае ведущим, основным является порядковое место,
а остальные моменты, в том числе и время, играют роль второстепен-
ных.
32
Э. А. АСРАТЯН
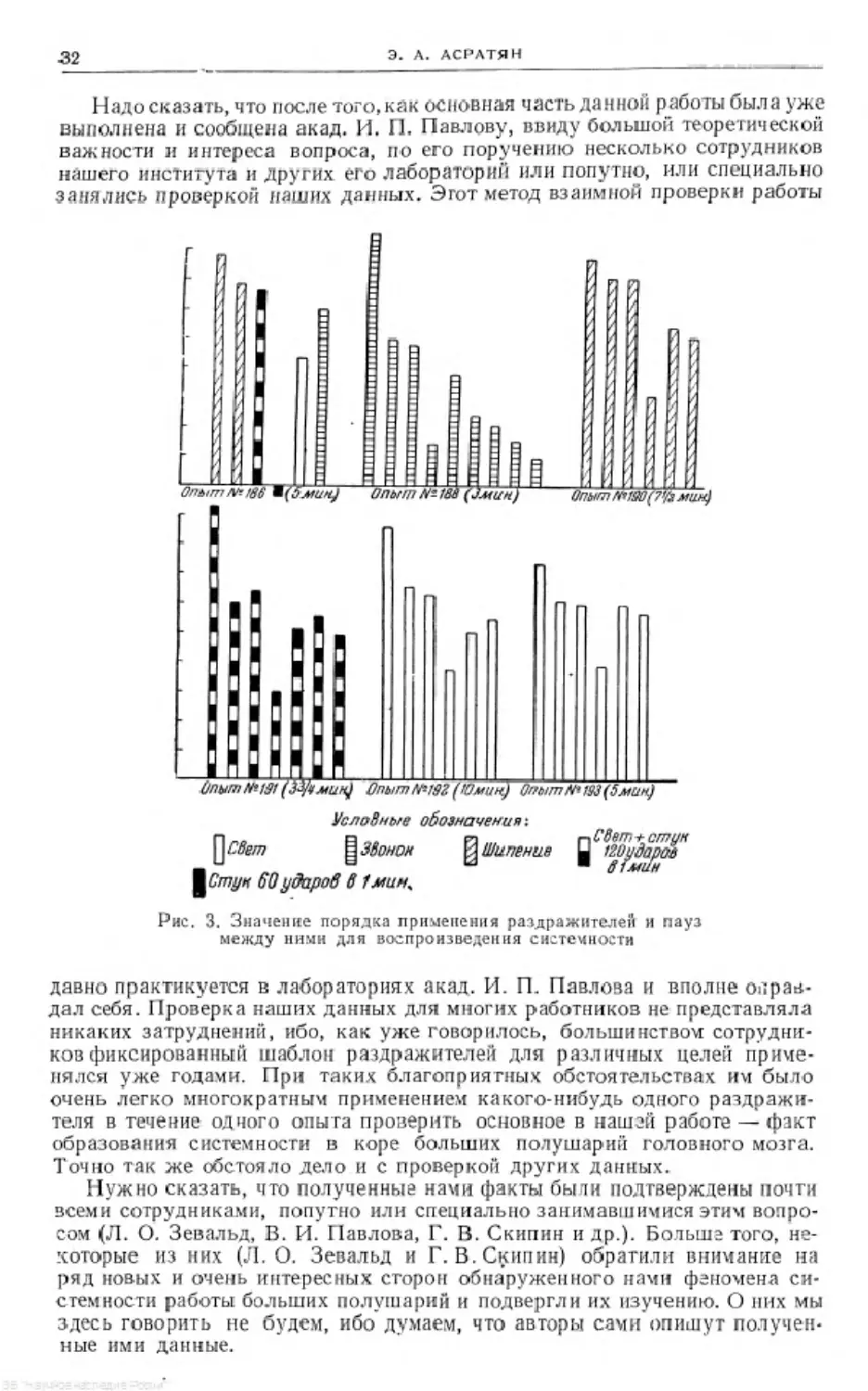
Надо сказать, что после того, как основная часть данной работы была уже
выполнена и сообщена акад. И. 11. Павлову, ввиду большой теоретической
важности и интереса вопроса, по его поручению несколько сотрудников
нашего института и Других его лабораторий или попутно, или специально
занялись Проверкой наших данных. Этот метод взаимной проверки работы
УслоВные обозначения:
flzBern ^УВонол Ши пение izuydapw
60 уборов в Гмин*
Рис. 3. Значение порядка применения раздражителей и пауз
между ними для воспроизведения системности
давно практикуется в лабораториях акад. И. FL Павлова и вполне оправ-
дал себя. Проверка наших данных для многих работников не представляла
никаких затруднений, ибо, как уже говорилось, большинством сотрудни-
ков фиксированный шаблон раздражителей для различных целей приме-
нялся уже годами. При таких благоприятных обстоятельствах им было
очень легко многократным применением какого-нибудь одного раздражи-
теля в течение одного опыта проверить основное в нашей работе —факт
образования системности в коре больших полушарий головного мозга.
Точно так же обстояло дело и с проверкой других данных.
Нужно сказать, что полученные нами факты были подтверждены почти
всеми сотрудниками, попутно или специально занимавшимися этим вопро-
сом (Л. О. Зевальд, В. И. Павлова, Г. В. Скипин и др.). Больше того, не-
которые из них (Л. О. Зевальд и Г. В. Скипин) обратили внимание на
ряд новых и очень интересных сторон обнаруженного нами феномена си-
стемности работы больших полушарий и подвергли их изучению. О них мы
здесь говорить не будем, ибо думаем, что авторы сами опишут получен-
ные ими данные.
СИСТЕМНОСТЬ РАБОТЫ БОЛЬШИХ ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА
33
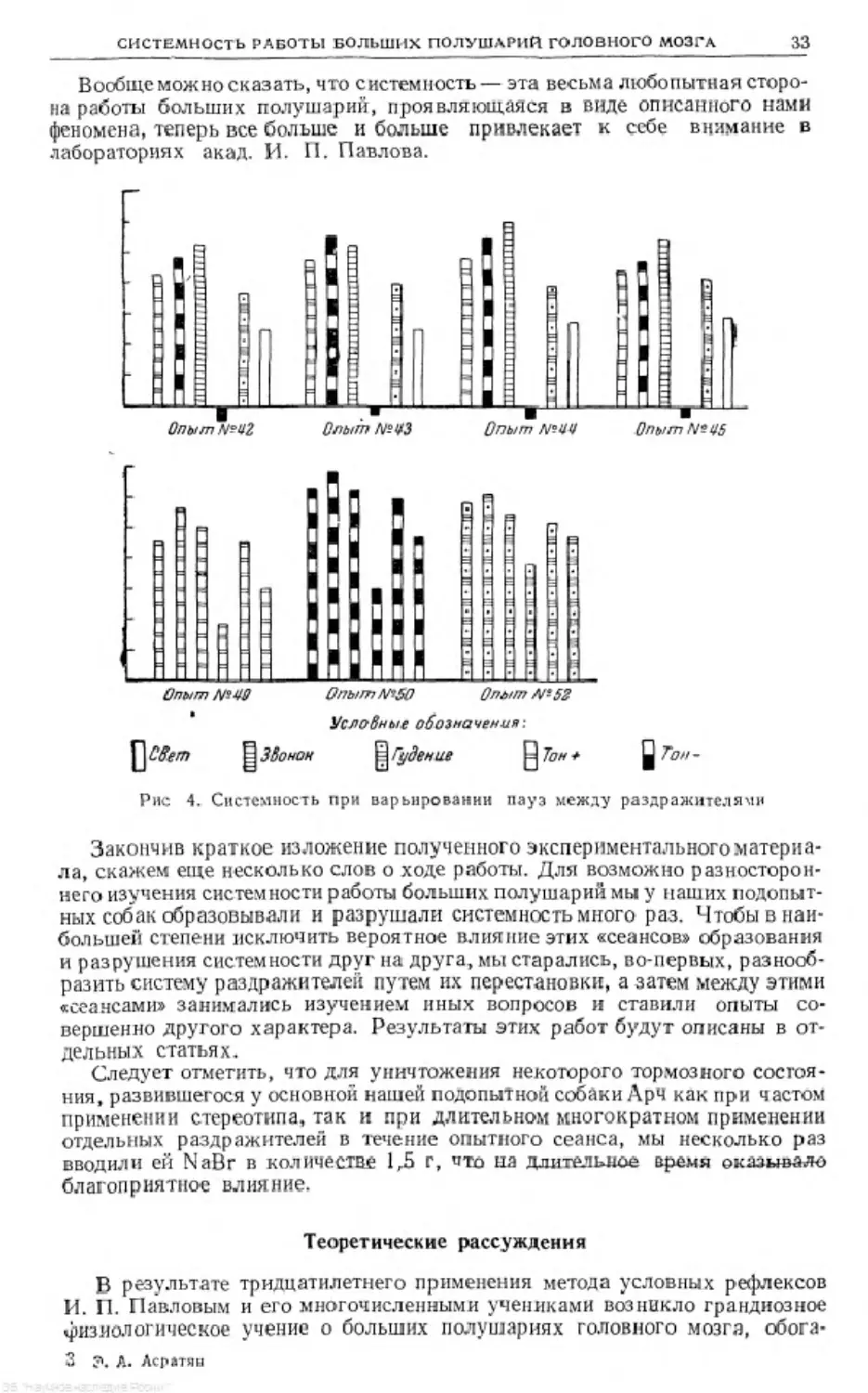
Вообще можно сказать, что системность— эта весьма любопытная сторо-
на работы больших полушарий, проявляющаяся в виде описанного нами
феномена, теперь все больше и больше привлекает к себе внимание в
лабораториях акад. И. II. Павлова.
Условные обозначения-.
Ц 3Во нон || Гудение Q Тон
Рис 4. Системность при варьировании пауз между раздражителями
Закончив краткое изложение полученного экспериментального материа-
ла, скажем еще несколько слов о ходе работы. Для возможно разносторон-
него изучения систем пости работы больших полушарий мы у наших подопыт-
ных собак образовывали и разрушали системность много раз. Чтобы в наи-
большей степени исключить вероятное влияние этих «сеансов» образования
и разрушения системности друг на друга, мы старались, во-первых, разнооб-
разить систему раздражителен путем их перестановки, а затем между этими
«сеансами» занимались изучением иных вопросов л ставили опыты со-
вершенно другого характера. Результаты этих работ будут описаны в от-
дельных статьях.
Следует отметить, что для уничтожения некоторого тормозного состоя-
ния, развившегося у основной нашей подопытной собаки ЛрЧ как при частом
применении стереотипа, так и при длительном многократном применении
отдельных раздражителей в течение опытного сеанса, мы несколько раз
вводили ей NaBr в количестве 1Д г, что па длительное время оказывало
благоприятное влияние.
Теоретические рассуждения
В результате тридцатилетнего применения метода условных рефлексов
И. II. Павловым и его многочисленными учениками возникло грандиозное
физиологическое учение о больших полушариях головного мозгя, обога-
3 У. Л. Лсратяв
34
Э. А. АСРАТЯН
тившее все разделы физиологии и массу сопредельных физиологии и от-
даленных от нее дисциплин.
Будучи построено на принципах материализма1, павловское учение об
условных рефлексах по своему фактическому 'научному материалу и по
своему существу диалектично. Это учение, величайшее достижение со-
временной физиологической науки, адэкватпо отражает диалектику при-
роды, объективным научным методом вскрывает физиологические
процессы в высокоорганизованной материи—в больших полушариях
головного мозга высших животных.
В маленькой статье, посвященной специальному вопросу, невозможно
дать более или менее удовлетворительное изложение основных законо-
мерностей учения об условных рефлексах, да это и не входит в нашу
задачу; мы остановимся только на тех из них, которые имеют близкое
отношение к изучаемым нами вопросам.
По методу условных рефлексов весьма подробно изучается анализатор-
ная деятельность больших полушарий головного мозга, Акад, И, П. Павлов
говорил: «...величайшая деятельность нервной системы, деятельность
больших полушарий — анализаторная деятельность. Эго главная задача
больших полушарий»’.
Синтетической работой больших полушарий с помощью метода условных
рефлексов начали заниматься сравнительно позже; однако за небольшой
период работы в этой области накоплен ценнейший материал. Несмотря на
все это, говоря словами акад. И. П. Павлова, «механизм и локализация
с и я т е т и ч еской нервной деятельности, в противоположность аналити-
ческой, остаются до сих пор мало известными... конечно, сейчас первая
задача—накоплять факты, относящиеся к синтетической деятельности»8.
Нам кажется, что в этом отношении описанная нами систем ность
работы больших полушарий является новым экспериментальным материа-
лом о высшей синтетической работе больших полушарий.
Далее, многочисленные работы павловской школы об анализаторной
деятельности больших полушарий полностью опровергли старые мета-
физические представления об узкой локализации, о строгой изолирован-
ности центров в коре, об их независимости друг от друга. Эти классические
исследования одновременно показали, что даже при осуществлении какой-
нибудь специфической анализаторной работы кора функционирует как
целое при ведущей роли в этом анализе каких-нибудь отдельных участ-
ков ее. Гипотетически можно было сказать, что это относится и к синтетиче-
ской работе коры, ибо известно, что в деятельности центральной нервной
системы анализ и синтез неотделимо связаны друг с другом, протекают
как неразрывное единство определенного рода нервной деятельности.
Описанная системность работы коры является, как нам кажет-
ся, экспериментальным доказательством этого положения, доказатель-
ством того, что высшая анализаторная и синтетическая работа совер-
шается корой в целом. Кроме того, приведенный здесь эксперименталь-
ный материал, полученный методом условных рефлексов, показывает,
что системность не является простой арифметической суммацией анали-
тической и синтетической работы отдельных участков коры больших
полушарий, скажем, слуховой, звуковой, кожно-механической и других
корковых анализаторов. Она является результатом монолитной высшей
анализаторной и синтетической работы всей коры как целого; и работа от-
1 В первоначальном тексте статьи явно ошибочно написано: «Будучи в основ-
ном построенным на принципах механического материализма...».
* И. П. П а в л о в, Двадцатилетний опыт объективного изучения высшей нервной
деятельности (поведения) животных, 1925, стр. 177.
’ И. П. П а в л о в. Лекции о работе больших полушарий головного мозга, 1927.
стр 124—125.
СИСТЕМНОСТЬ РАБОТЫ БОЛЬШИХ ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА
15
дельных ее частей, различных, более или менее сйособленных ее участков
проявляется в общей высшей анализаторно-синтетической деятельности—
систем пости.
В этой связи нам кажется, что описанное нами физиологическое явле-
ние— системность—имеет немалое значение для психологии.
Как себе представить ближайшую, интимную сторону образования и
разрушения системности в коре? Мы не претендуем на исчерпывающее объ-
яснение этого явления, тем более при теперешнем, пока что- недостаточном
уровне наших знаний о предмете. Однако считаем нелишним привести наше
далеко еще не достаточное понимание этого важнейшего вопроса.
Нам кажется, что системность имеет тесное отношение главным
образом к трем основным закономерностям физиологии нервной системы
вообще и физиологии больших полушарий в частности; это: 1) взаимодейст-
вие различных частей функционального целого, 2) свойство сохранения сле-
дов от испытываемых воздействий, 3) свойство фиксации, закрепления этих
следов.
1. Современная физиология нервной системы, основывающаяся на ра-
ботах И. М. Сеченова, И. П. Павлова, II. Е. Введенского, Магнуса, Лучи а-
ни и др., принимает как одно из основных положений взаимную функцио-
нальную св.язь и взаимодействие как между различными системами организ-
ма, осуществляемые в основном посредством нервной системы, между
отдельными частями самой нервной системы как целого, так и между отдель-
ными частями одного и того же морфологически и функционально более или
менее сбоссбленного отдела центральной нервной системы (больших полу-
шарии, мозжечка, спинного мозга и т. д.). Что же касается непосредст-
венно больших полушарий, то здесь мы имеем огромное количество новых и
старых работ, вышедших главным образом из лаборатории акад. И. П. Пав-
лова, которые отчетливо демонстрируют существующие взаимодействие и
взаимосвязь между различными частями коры больших полушарий, меж-
ду корой и подкорковыми образованиями и т. д. (И. Я. Перельцвейг,
С. Б. Хазен, Л. Л. Савич, Я. Е. Егоров, Д. С. Фурсиков, Г. П. Конради
и др.). Словами И. П. Павлова можно сказать, чтоввсякое новое местное воз-
действие на эту систему дает себя знать более или менее во всей системе»1.
2, По раннему еще замечанию акад, И. П, Павлова, от всякого раздраже-
ния в нервной системе остается некоторое время след. Действительно, спо-
собность временного сохранения следов испытываемого возбуждения и
торможения свойственна всем нервным центрам и вообще всем нервным
элементам. Возможно даже, что зачатки этого свойства присущи и осталь-
ному живому веществу. Что касается нервных центров, то ради примера
укажем на такие общие для всех них явления, как суммация, протарение
путей и доминанта, основанные на свойстве кратковременного сохране-
ния следов изменений, испытываемых нервными центрами.
Что же касается больших полушарий, то еще в самых ранних работах
по условным реф>лексам, вышедших из лабораторий акад. И. П. Павлова,
было установлено, что корко-вые процессы возбуждения и торможения,
возникшие под действием положительных и отрицательных условных
раздражителей на соответствующие рецепторы животного, не прекра-
щаются сразу же после прекращения действия этих раздражителей; в виде
о т к рытых или скрытых следов нервные процессы на некоторое
время остаются в возникших пунктах и даже временно распространяются
на остальные, главным образом соседние отделы коры, а то и на под-
корку (иррадиация). В виде следов кратковременно сохраняются также
возникшие индукционные взаимоотношения.’ Больше того, почти целиком
1 И, П. Павлов, Лекции о работе больших полушарий головного иоэга, 1927,
стр. 202.
з*
36
3. А. АСРАТЯН
на этом свойстве сохранения следов испытываемого возбуждения базирует-
ся образование специальных видов условных рефлексов — следовых услов-
ных рефлексов (П. И. Пименов, Ф. С. Гроссман, И. В. Завадский и др.).
В конце концов, о свойстве сохранения следов испытываемого клетками
коры больших полушарии возбуждения и торможения говорят давнишние
и новейшие многочисленные работы с электрическим раздражением так
называемой двигательной зоны коры и других ее отделов (Фрич
(Fritsch), Гитциг (Hitzig), Н. Е. Введенский, Шеррингтон (Sherrington),
А. А. Ухтомский и др. ].
3. Однако методом условных рефлексов доказано, что в коре больших
полушарий головного мозга свойство сохранения следов нс только развито
до высокой, качественно новой ступени (опыты Я. Е. Егорова и др.), но,
наряду с этим, появляется новое качество, качество фиксации на более
или менее длительное время как следов локальных возбуждений и торможе-
ний, так и следов различных стереотипных комбинаций качественно новых
видов нервной деятельности— положительных и отрицательных условных
рефлексов.
На сохранение и фиксирование следов деятельности в виде стереотип-
ной работы коры больших полушарий указывают еще старые наблюдения
И. Я. Перельцвейга и др. о глубоком следе длительно применявшегося
стереотипа раздражителей, а также опыты Ю. П. Фролова с превращением
следовых условных рефлексов в наличные условные рефлексы.
Интимную сторону образования системности, невидимому, можно пред-
ставить себе следующим образом. Отдельные специфические условные
рефлексы, входящие в систему и занимающие в ней определенные фиксиро-
ванные места, несомненно, находятся в определенном взаимодействии. При
повторном применении системы изо дня в день случайный порядок и
взаимодействие раздражителей фиксируются, синтезируются вединое целое.
В результате образуется единая стройная цепь рефлексов, отдельные
звенья которой находятся в сложной, но уже определенной фиксированной
взаимной связи и взаимодействии, примерно так же, как в цепных безуслов-
ных рефлексах. Этим путем определенные действующие элементы коры в
определенной последовательности во времени и по отношению друг к другу
фиксируют эффекты следов условных раздражителей в виде следов цепи.
В коре создается цепь заряженных скрытых очагов, которые способны про-
являться в определенном фиксированном порядке под влиянием какого-
нибудь'стимула, причем, благодаря сложному взаимодействию, каждое
функционирующее звено, в отличие от звена в цепном безусловном реф
лексе, не вызывает активного действия следующего за ним звена, а лишь
только подготавливает его к действию.’
По нашим представлениям, при воспроизведении образованной в коре
системности индикаторный условный раздражитель действует главным об-
разом как стимулятор в определенной последовательности и вызывает услов-
ные рефлексы, примерно характерные для каждого из этих очагов, т. е
на местах сильных раздражителен—сильный эффект, па местах слабых —
слабый эффект и т. д. Следует отметить, однако, что на месте диференци-
ровочного раздражителя индикаторные условные раздражители вместо
нулевого эффекта вызывают некоторый положительный эффект или силь-
ные индикаторы-раздражители на местах слабых положительных условных
раздражителей дают положительные эффекты немного больше свойствен-
ных последним. Слабые индикаторные раздражители на местах сильных
раздражителей хотя и вызывают эффекты больше свойственных им, но в-се же
немного меньше эффектов этих раздражителей. Одним словом, на осно-
вании того, что индикаторы-раздражители только приблизительно вос-
производят эффекты каждого из раздражителей, входящих в систему
СИСТЕМНОСТЬ РАБОТЫ БОЛЬШИХ ПОЛУШАРИИ ГОЛОВНОГО МОЗГА
27
можно думать, что роль индикаторного условного раздражителя не сводится
целиком к роли стимулятора заряженных центров.
Надо полагать, что индикаторные условные раздражители, помимо сти-
муляции заряженных центров, помимо посредственного услов-
ного эффекта, в слабом виде вызывают л непосредственные,
прямые условные эффекты, которые, суммируясь с первым, дают нам кар-
тину получаемых в действительности фактов. Больше того, надо полагать,
что в процессе разрушения системности, по мере ее ослабления, когда заря-
женные центры в силу отсутствия подкрепления или других моментов по-
степенно разряжаются, индикаторы-раздражители все больше и больше
уменьшают посредственные условные эффекты и все больше в
больше увеличивают непосредствен н ые условные эффекты, при-
чем в конце полного разрушения системности первые сводятся к нулю и
остаются только вторые.
В конце хотелось бы еще раз напомнить, что наше толкование этого но-
вого интересного и сложнейшего вида работы иысшего отдела центральной
нервной системы — больших полушарий — не претендует на исчерпываю-
щую полноту. Нам кажется, однако, что при теперешнем уровне знаний
предмета в целом оно может считаться одним из самых вероятных объясне-
ний и более или менее соответствует фактическому положению вещей.
ЛИТЕРАТУРА
Асратян Э. А., Влияние экстирпации верхних шейных симпатических узлов на
пищевые условные рефлексы, Архив биологических наук, т. 30, в. 2, 1930.
Бели ц М. Ф.» О следовых условных рефлексах, дисс., Птр., 1917.
ГроссмапФ С., Материалы к физиологии следовых условных рефлексов, дисс.,
Птр., 1909.
Добровольский В. М., О пищевых следовых рефлексах, дисс., Птр., 1911.
Д е р я б и и D. С., Дальнейшие материалы к физиологии времеин как условного воз-
будителя слюнных желез, дисс., Птр., 1916.
Е г о р о в Я- Е., Влияние пищевых условных рефлексов друг на друга, дисс., Птр.,
1911.
Завадский И. В., Материалы к вопросу о торможении и рас торможении условных
рефлексов, дисс., Птр., 1908.
К у п а л о в П. С., О состоянии коры больших полушарий в интервалах между при-
менением условных раздражителей. Архив биологических наук, т. 31, в. 4. 1931.
Кржишковский К. Н.. К физиологии условного тормоза, Труды Общества
русских врачей в С.-Петербурге, т. 76, 1909.
Павлов И П., Двадиатилетний опыт объективного изучения высшей нервной
деятельности (поведения) животных, 1925.
П а в л о в И. П., Лекции о работе больших полушарий головного мозга, 1927.
Павлова В. И., О следовых условных рефлексах, Труды Общества русских врачей,
т. 81, 1913 — 1914.
11 и м е и о в П. П., Особая группа условных рефлексов, дисс., Птр., 1907.
Перед ьцвей г И. Я., Материалы к учению об условных рефлексах, дисс., Птр.,
1907.
Соловейчик Д. И-, Нарушение нормальной деятельности больших полушарий
при изменении выработанного порядка исследования условного раздражения,
Труды И Всесоюзного съезда физиологов, 1926.
СтуковаМ. М., Дальнейшие материалы к физиологии времени как условного воз-
будителя слюнных желез, дисс., Птр., 1914.
Фролов Ю. П., К физиологии так называемого «чувства времени». Доклады на
11 съезде по психоневрологии, 1924.
Фролов Ю. П., О переводе следовых условных раздражителей неделовых условных
тормозов в различные условные раздражители, Труды физиологических лабора-
торий акад. И. П. Павлова, т. 1, в. 2—3, 1926.
Фёокритова Ю. П., Время как условный возбудитель слюнной железы, дисс.,
Птр., 1912.
ВЛИЯНИЕ ПИЩЕВОГО БЕЗУСЛОВНОГО РЕФЛЕКСА
НА СООТВЕТСТВУЮЩИЕ УСЛОВНЫЕ РЕФЛЕКСЫ1
опрос о взаимодействии и взаимной связи различных нервных
центров является интереснейшим вопросом физиологии цент-
ральной нервной системы вообще и физиологии больших
полушарий головного мозга в частности.
В лабораториях акад. II. П. Павлова и его учеников в про-
цессе 30-летней работы по физиологии больших полушарий
многие сотрудники неизбежно сталкивались с фактами взаимодействия раз-
личных частей высших отделов центральной нервной системы. И одним из
наилучших в наше время методов изучения физиологии больших полушарий
головного мозга—методом условных рефлексов множество и разнохарактер-
ность этих взаимоотношений и взаимодействий подверглись исследованию
с различных точек зрения. Мы вкратце напомним о некоторых работал,
представляющих интерес с точки зрения разбираемого нами вопроса.
Еще в самом начальном периоде развития учения сб условных рефлексах многими
исследователями было установлено, что образованные на почве различных безусловных
рефлексов условные рефлексы находятся в сложном взаимодействии, в большинства
случаев взаимно друг друга тормозят или, наоборот, растормаживают, если они былг
заторможены. Тогда же было найдено, что и безусловные рефлексы действуют тормо
зяте на условные рефлексы, образованные на базе других безусловных рефлексов
(И. Я. Перельцвейг, Е. А. Нейц, С. Б. Хазен, А. 3 Былина, Я. Е. Егоров, А. А. Са-
вич и мн. др.). О сложном взаимодействии корковых клеток говорят также инте-
ресные работы Г. ГГ. Зеленого, И Я. Пере.чьцвейга, А. В. Палладина и др. с ус-
ловными рефлексами на комплексные раздражители. Далее идут работы М. Я- Без
бокой, И. В. Завадского, Б. А. Когана, Д. С. Фурсикова и др. относительно суммации
растормаживания, ясно выявленной взаимной индукции и других взаимоотношений
корковых клеток.
Из новейших работ в этсйдбласти следует отметить интересные работы Д. С. Фур-
снкова, А. Н. Зальманзона, Н. И. Михельсон и М. Н. Юрман, а также работу
Г. П. Конради относительно днференцировки и взаимодействия положительных ус-
ловных рефлексов, образованных на базе различных безусловных рефлексов. Из но-
вых детально разработанных данных этих исследователей последовательно «ытекаег
то, что в основных чертах било отмечено старыми авторами. Они показали наличие
сложного взаимодействия, борьбы между центрами безусловных рефлексов различного
рода и характера, а также между различными пунктами коры, связанными с первыми
(безусловными центрами). Они же показали отрицательное индукционное влияние
одних безусловных рефлексов иа условные, основанные на других безусловных реф-
лексах.
Среди накопленного богатейшего материала по условным рефлексам существуют
также работы, которые имеют косвенное или прямое отношение к интересующему нас
конкретному вопросу, т. е. влиянию безусловного рефлекса на условные. В качестве
одного вида влияния безусловного рефлекса на условный можно упомянуть давно из-
вестные в лабораторной практике и в последнее время специально исследованные фак-
ты (С. В. Клещоъ и В. X. Гент) относительно тесной зависимости величины условного
рефлекса от величины безусловного. В этом отношении интересна также работа
Ф. II. Майорова об изменении величины условного рефлекса в зависимости от продол-
жительности совпадения условного раздражителя с безусловным. Это является пока-
1 Физиологический журнал СССР, т. XVIII. № 5, 1934. Предварительное сообще-
ние напечатано в Докладах АН СССР, т. 11, №1,1934, по представлению акал
И. П Павлова 20. 11 1934 г. г д
ВЛИЯНИЕ ПИЩЕВОГО БЕЗУСЛОВНОГО РЕФЛЕКСА НА УСЛОВНЫЕ РЕФЛЕКСЫ 39
зателем положительного влияния безусловного рефлекса на условный. Однако су-
ществует и отрицательное влияние. Работа А. Н. Крестовникова о невозможности
образования условного рефлекса при предшествовании безусловного раздражителя
условному показывает, что деятельность центра безусловного рефлекса тормозит ус-
ловные рефлексы. Из новых работ Д. И. Соловейчика, Р. Я - Райт, П. К. Анохина и
Н. А. Подкопаева с так называемым «покрытием* условных и индифферентных раздра-
жителей с полной очевидностью вытекает то же заключение. Работа Н. В. Виногра-
дова, внося точность в детальные формулировки вопроса, исследованного А. Н. Крес-
товниковым, в основном подкрепляет это заключение. Однако упомянутые работы
с «покрытием» лишь косвенно указывают на торможение условных рефлексов цент-
ром безусловного рефлекса, причем фактический материал не выясняет характера
этого торможения. Некоторые указания в этом отношении имеются и более старых
работах Е. А. Нейца; при недавней проверке В. В. Рикмана и пашей они оказались
вполне правильными. Дело в том, что в. опытах Нейца укорочение паузы между при-
менением условных раздражителей приводило к уменьшению условных рефлексов.
Хотя автор, обнаруживший этот факт, не да.т ему никаких объяснений, но его пред-
положительно можно было толковать двояко: во-первых, по принципу последователь-
ной индукции на месте—центр безусловного рефлекса после возбуждения на некоторый
отрезок времени попадает в тормозное состояние, и условные раздражители, приме-
ненные после малых пауз, вызывают меньшие условные рефлексы в силу пониженной
возбудимости центра безусловного рефлекса; во-вторых, по принципу индукции на
расстоянии—корковые пункты1 условных рефлексов кратковременно тормозятся при
возбуждении безусловного центра, и поэтому действующий в этот отрезок времени ус-
ловный раздражитель вызывает слабый условный рефлекс. Материал этих исследова-
телей не давал прямого и ясного ответа на вопрос, какое из этих двух вероятных
предположений является правильным. На основании определенного фактического
материала по данному вопросу, попутно собранного нами при выполнении других
задач, мы считали правдоподобным скорее второе предположение. С целью получе-
ния более ясного ответа на поставленный вопрос мы специально изучали его на наших
подопытных животных Арче к Масисе в феврале —октябре 1931 г. Полученный нами
материал излагается в данной статье.
В начале работы мы занялись уточнением и детализацией данных
относительно влияния пауз на величину условных рефлексов. Чтобы из-
бежать искажающего влияния фазовых изменений, влияния насыщения и
других моментов на состояние коркового пункта условного рефлекса и
центра безусловного рефлекса и чтобы по возможности отчетливее выявить
влияние безусловного рефлекса на условные, мы для наших проб дейст-
вующим безусловным агентом избрали первую порцию
мясо-сухарного порошка, которая без сочетания с каким-нибудь раздра-
жителем давалась собаке после ее подготовки к опыту и непосредственно
перед нашим выходом из камеры. В качестве же и н д и к а т о р а со-
стояния корковых пунктов условных рефлек-
сов в подавляющем большинстве опытов мы пользовались по очереди
условным рефлексом на тот или другой давний И хорошо Известный ус-
ловный раздражитель. Следует упомянуть, что как многие другие работ-
ники по условным рефлексам, так и мы раньше перед началом опыта и не-
посредственно перед нашим выходом из камеры давали собаке чашку пищи
С целью, как говорится, «зарядить» ее большие полушария к работе. Ис-
пользование уже известного приема облегчило разрешение поставленной
задачи. Для выявления характера и хода влияния безусловного рефшекса
на условные мы всячески изменяли длительность промежутка между дей-
ствием безусловного и условного раздражителей. С целью точного учета
времени мы, в отличие от предшествующих исследователей, учитывали
не паузы между началом применения двух раздражителей, а отрезок
времени между концом безусловного раздражения (практически поднятие
морды собаки от чашки) и началом применения условного раздражителя.
Паузы эти мы варьировали от 15 секунд до 3*/г минут.
У обеих собак для первого условного рефлекса закономерно получа-
1 Применявшийся в первоначальном тексте статьи термин «центр» условных
рефлексов мы сочлк необходимым заменить более правильным термином «корковые
пункты» условных рефлексов
40
Э. А. АСРАТЯН
лось следующее: в пределах около P/s минут чем меньше была пау-
за между окончанием действия безусловного раздра-
жителя и началом следующего за ним условного
раздражителя, тем меньше была величина услов-
ного рефлекса, вызванного последним. В случае же,
когда названные паузы были больше Р/s минут, условные рефлексы соот-
ветственно делались слабыми. Сказанное иллюстрируется данными, полу-
ченными у собаки Арч (табл. 1). Совершенно аналогичные данные получены
и у собаки Маске.
Таблица 1
2 В о £ Величина паузы в сек. Величина условного рефлекса за 20 Сек. в делениях WK9 ЛЫ "Условный раздражи- тель Nt опыта Величина паузы » сек. Вслнчиин условного рефлекса за 20 сек в деле ни>1х Ш^алы Условный раздражи- тель
102 94 88 95 96 98 101 101 119 122 89 120 126 103 93 100 15 25 40 50 50 55 60 60 60 60 60 67 85 90 90 90 1Г’| 22 24 25 28 32 32 30 31 30 32 30 32 30 33 Стук 120 в 1 мин. 119 121 124 126 116 118 116 178 177 117 115 158 106 115 117 117 105 103 77 105 120 150 210 20 30 60 75 75 25 22 40 75 90 105 120 35 40 60 34 26 22 17 12 20 29 34 36 18 20 (4) 27 3-4 32 27 23 | 14 1 20 25 ) Сгук 120 в 1 мин. Шипенье- Звонок Свет
Весьма существенным для понимания характера взаимоотношений
условного и безусловного рефлексов является следующий наблюдаемый в
лаборатории факт. Как после еды, так и после съедания первой порции
сильное последовательное слюноотделение продолжается еще до 30 секунд.
И вот часто, когда первый условный раздражитель применялся вовремя
еще продолжающегося последовательного слюноотделения, т. е. через 15,
20 и 25 секунд после конца непосредственного безусловного раздражения,
через 1—2 Секунды последовательное слюноотделение резко обрывалось;
затем, спустя 2—3 секунды, вновь начиналось слюноотделение, повиднмому,
чисто условнорефлекторное, которое и продолжалось в течение всего
времени действия условного раздражителя. Ради точности мы в таблице в
скобках приводим объем последовательного слюноотделения до его оста-
новки, а перед скобками—остальную часть.
Приведенный факт свидетельствует о влиянии условного рефлекса на
безусловный. На этом вопросе мы здесь подробно не останавливаемся, ибо
намереваемся опубликовать собранный нами по этому вопросу материал
в другой статье. Отметим лишь, что между условными и безусловны-
ми рефлексами существует взаимодействие в настоящем смысле
этого слова, что это действие носит двусторонний характер, конечно,
при доминирующем значении безусловного рефлекса.
После того как в основных чертах были выяснены характер и ход влия-
ния безусловного рефлекса на величину условного рефлекса, мы приступили
к выяснению внутренней, интимной стороны этого действия, т. е. к разреше-
ВЛИЯНИЕ ПИЩЕВОГО БЕЗУСЛОВНОГО РЕФЛЕКСА ИА УСЛОВНЫЕ РЕФЛЕКСЫ
41
нию основного вопроса. Конкретно нужно было выяснить причину умень-
шения условных рефлексов после маленьких пауз: было ли это обуслов-
лено последовательным индукционным торможением самого пищевого
безусловного центра или же индукционным торможением условных рефлек-
сов вследствие возбуждения центра безусловного рефлекса.
С этой целью мы поставили следующие опыты. В ряде контрольных опы-
тов ТОЧНО установили величину рефлекса на безусловный раздражитель стан-
дартной величины, состава и физического состояния (для Арча — 12 г
мясо-сухарного порошка 4-10 см3 воды, а для Масиса — 35 г мясо-сухар-
ного порошка +30 см3 воды). Безусловный стандартный раздражитель при-
менялся без сопровождения искусственными условными раздражителями.
В течение одного опытного сеанса через равные паузы вызывалось и измеря-
лось несколько таких безусловных рефлексов на эти стандартные порции.
Измерение рефлекса у Арча производилось каждые 15 секунд и не только
в процессе еды, которая длилась примерно 30—40 секунд, но в течение 90
секунд, т. е. до полного’прекращения безусловного слюноотделительного
рефлекса; у Масиса рефлекс измерялся каждые 3 секунды в течение 3 минут.
Эти опыты по-казлли, что при равных паузах достаточной величины (4—
5 минут) между этими безусловными рефлексами при возможно одинаковых
условиях (время года, время дня, сорт порошка и т. д.) стандартные без-
условные раздражители как в течение одного опытного сеанса, так и иногда
в опытах ряда дней вызывали одинаковые по величине безусловные реф-
лексы. Безусловные рефлексы на такие стандартные раздражители были
равны по величине и в тех опытах, в которых эти порции чередовались с
обыкновенными большими порциями мясо-сухарного порошка и также по-
давались через равные паузы. Как в первом, так и во втором случае ко-
лебания были в пределах от 3 до 8 делений шкалы при общей величине бе-
зусловного рефлекса около 110—130 делений в разных опытах. Сказанное
иллюстрирует табл. 2.
Таблица 2
Время подачи по-рцин Величина безусловного рефлекса на стандартный раздражитель за каждые 15 сек. в делениях шкалы Суммарная величина рефлекса на стан- дартный раздражи- тель эа 90 сек. е делениях шкалы
1 Ч. 17 М. 4 » 22 » 4 » 27 » 4 » 32 » 4 » 37 > 4 » 42 » 4 * 47 » •1 > 52 » 4 » 57 * Опыт № 215 от 14.XII 1931 г. Дача большой порции 30 + 42 + 32+14 +84-12 32+40 +31 +12+6+14 Дача большой порции 28+43+30+11+7+17 Дача большой порции 29 + 43 + 314-12+14+10 30 + 40 + 32+ 9 + 9+16 31 +40+334-12+ 4+17 138 136 136 139 136 137
С целью разрешения вышеупомянутой задачи были поставлены спе-
циальные опыты. Стандартные порции чередовались с обыкновенными
большими порциями пищи, но подавались собакам не через равные, а че-
рез разные паузы после окончания еды нестандартных порций (большие
порции). Паузы изменялись от 15 секунд до 6 минут. Как нетрудно за-
метить, в этих опытах для выявления состояния возбудимости центра
безусловного рефлекса, вернее, для выявления характера и хода изме-
нений, претерпеваемых центром безусловного рефлекса под влиянием собст-
венного возбуждения, применялся почти такой же прием, как в прежних
опытах для обнаружения состояния возбудимости корковых пунктов услов-
42
Э. Л. ЛСРЛТЯН
ных рефлексов в связи с работой центра безусловного рефлекса. Ряд опытов
этого характера, проведенных на двух собаках, с полной очевидностью по-
казал, что в пределах от 15 секунд до 21/а минут чем меньше была
пауза между окончанием действия предшествую'
щ, е г о нестандартного безусловного раздражи-
теля и началом действия стандартного безуслов-
ного раздражителя, тем сильнее был безусловный
рефлекс на последний. Надо сказать, что вьнпеприв.еденную
характеристику, которая охватывает почти все случаи с паузами от 15
секунд до 21/i минут, можно было бы распространить и на случаи с больши-
ми паузами, если бы не было одного исключения. Исключением являлось
то, что при паузах, лежащих в пределах 3 минут, безусловный рефлекс
на стандартную порцию был сильнее, чем при немного меньших паузах.
Однако при дальнейшем увеличении пауз это исключение исчезало. Думаем,
что оно связано было с условным рефлексом на время дачи пищи, ибо точно
совпадало с. паузой, которая в течение года с лишним применялась в нашей
работе. На это указывала характерная в таких случаях двигательная реак-
ция собак до подачи пищи, а также то, что в этих случаях после окончания
еды собаки примерно па 5—10 секунд дольше обычного продолжали облизы-
вать пустую чашку. Для иллюстрации сказанного о величине безусловных
рефлексов на стандартные порции в зависимости от величины пред-
шествующих пауз приводятся подробные протоколы типичных опытов
(табл. 3 и табл. 4). Безусловные рефлексы на стандартные порции ре-
гистрировались каждые 15 секунд (цифры перемежаются знаком +). Про-
межуточные большие порции, которые применялись в качестве сильных
возбудителей центра безусловного рефлекса, подавались в сопровождении
какого-нибудь условного раздражителя, предшествовавшего безусловному
на 2—3 секунды.
Таблица 3
Время дачи больших порций Паузы между окончанием еды больших порций и дачей стан- да ртных порций в сек. Безусловный рефлекс на стандартные порции на каждые 16 сек. в делениях шкалы Общая величина безусловного рефлекса за 90 сек. в деле- ниях шкалы
Опыт № 114 от 30.IV 1931 г.
3 ч. 10 м. 110 46+36+34+7+14-0 124
3 » 15 > 20 Дача промежуточной порции 52+44+35-21 +8+8 168
3 ч. 20 м. — Дача большой порции
— 165 37 4- 33 -f' 40 10 -|-4 4- 3 127
3 ч. 27 м. — Промежуточная большая порция ——
— 50 42 + 38 + 39 1-23+ 7+6 155
3 ч. 33 м. — Промежуточная большая порция —
— 210 37+34+35 + 16 + 4+3 129
3 ч. 40*£ м. —» Промежуточная большая порция —
— 165 40+35+26+ 17+ 2«/г + ‘/2 121
3. ч. М м. — Промежуточная большая порция —
— 30 44 —|- 44 -1-42 1 (> 4" 5 4" 5 156
В табл. 3 не приведены данные относительно величин безусловных реф-
лексов на большие порции, так как вследствие малого размера шкалы не-
возможно было точно измерить величину этих •рефлексов; имеющиеся же
данные об их величине за маленький отрезок времени в данном случае не
представляют никакого интереса и не приводятся, дабы не осложнять кар-
тины.
ВЛИЯНИЕ ПИЩЕВОГО БЕЗУСЛОВНОГО РЕФЛЕКСА НА УСЛОВНЫЕ РЕФЛЕКСЫ 43
В табл. 4 протоколы опытов немного упрощены. Приводятся только
паузы, предшествовавшие стандартным порциям, и суммарные величины
безусловных рефлексов. _. , ,
J it Таг» ли п я 4
Опыты на собаке АРЧ Опыты на собаке Масле
А? опыта и дата Величина пред- шествую- щих пауз н сек» Величина безуслов- ного рс<>- лекса за 90 сек. № опыта и дата 1 Величина п редшест- вующих пауз к сек. Величина безуслов- ного реф- лекса за 90 сек. № опыта И дата Величина предшест- вующ»^ пауз в сек. Величина безус лон- ного реф- лекса за во сек.
112 28.1 V 1931 г. 120 ео 30 120 00 30 112 128 148 116 5.V 1931 г. 25 90 180 139 152 96 22. IV 15 143
108 126 144 • 240 60 116 153 97 23. IV 40 134
117 6 V 40 270 45 135 133 115 139 117
123 29.IV 30 165 45 210 60 2Ю 172 112 146 161 137 136 98 24 IV 1 8 122
118 7.V 40 210 270 40 131 136 107 139 99 25. IV S 1 143
115 4. IV 145 138 117 115 103 134
25 45 75 105 120 45 119 8.V 40 270 135 111 100 27, IV 75 20 129 150
210 360 45 128 112 137 101 , 28.1V 120 35 121 142
При сопоставлении данных об изменении величин безусловных
рефлексов с данными об изменении величин условных рефлексов, описан-
ных нами выше, нетрудно заметить, что изменения эти, будучи связаны с од-
ними и теми же моментами, имеют диаметрально противоположное направ-
ление. В то время как с нарастанием пауз в некоторых пределах нарастают
и условные рефлексы, безусловные рефлексы, наоборот, в тех же Пределах
с нарастанием пауз уменьшаются (см. кривые на схеме стр. 44).
Нам думается, что в условиях нашего эксперимента величины безуслов-
ных рефлексов на стандартные раздражители можно считать показателями
состояния возбудимости центра безусловного рефлекса. В таком случае
с уверенностью можно ответить на поставленный перед нами вопрос,
во-первых, что уменьшение условны* рефлексов в связи с меньшими па-
узами (15—40 секунд) зависит от заторможенного состояния корковых
пунктов условных рефлексов, состояния, которое вызвано по принципу
индукции на расстоянии возбуждением центра безусловного рефлекса; во-
вторых, что в пределах отмеченного времени последовательная отрицатель-
ная индукция не имеет места в самом центре безусловного рефлекса, и по-
тому этот момент нс мог даже косвенно служить причиной описанного
выше уменьшения величины условных рефлексов.
Во избежание недоразумения оговоримся, что наши данные вовсе не
отрицают возможности последовательной отрицательной индукции в центре
безусловного пищевого рефлекса в иных временных отношениях. Вопрос
этот остается открытым и требует дальнейших исследований.
44
Э. А. АСРАТЯН
Само собой разумеется, что конкретные величины пауз у наших собак
имеют лишь относительное значение. Они могут меняться у разных собак,
Динамика изменения
величин условных и безуслов-
ных рефлексов в зависимости
от изменения пауз
может быть, даже у одних и тех же собак,
в зависимости от иного физиологического со-
стояния, от комбинации опыта и т. д. Важ-
ным, по нашему мнению, в приведенных дан-
ных нужно считать обнаружение направлен-
ности и характера процесса.
Так как теперь в лабораториях акад,
И. П. Павлова принимают существование
двух безусловных центров—подкоркового и
коркового, открытым остается вопрос о том,
который из этих центров в нашем конкрет-
ном случае играет главную роль, или же
оба они принимают в этом одинаковое уча-
стие.
Выводы
1. Первые по очереди пищевые условные
рефлексы бывают тем слабее, чем меньше
(в пределах 15—100 секунд) пауза между окон-
чанием действия предшествующего безуслов-
ного раздражителя и началом действия дан-
ного условного раздражителя.
2. Пищевые безусловные рефлексы, на-
оборот, бывают тем сильнее, чем в тех же
пределах времени меньше пауза между окон-
чанием действия предшествующего безуслов-
ного раздражителя и началом действия дан-
ного безусловного раздражителя.
3. На основании отмеченных выше дан-
ных можно заключить, что ослабление
условных рефлексов в связи с уменьшением
паузы обусловливается не последовательным
безусловного пищевого
вых пунктов условных
пищевого центра.
индукционным торможением центра самого
рефлекса, а индукционным торможением корко-
рефлексов, вызванным возбуждением безусловного
ЛИТЕРАТУРА
Л и о х и и П. К., Взаимодействие клеток условного и безусловного раздражи-
телей в течение применения последнего, Труды физиологических лабораторий
акад. И. 11. Павлова, т. II, в. 1, 1927.
Безбокая М. Я., Материалы к физиологии условных рефлексов, дисс., Спб.»
1513.
Болдырев В. Н , Условные рефлексы и способность их к усилению и ослабле-
нию, Харьковский медицинский журнал, т. IV, №6и 7, 1907.
Б ы л и и а А. 3-, Простое торможение условных рефлексов, дисс., Спб., 1910.
Виноградов Н. В., Возникновение повых связей в заторможенных участках
коры головного мозга, Труды физиологических лабораторий акад. И. П. Павлова,
т. V. 1933.
Е г о р о в Я. Е., Влияние пищевых условных рефлексов друг нй друга, дисс., Спб.,
1911.
Завадский И. В., Материалы к вопросу «.торможении и растормаживании
условных рефлексов, дисс., Спб., 1908.
Залманзон А. Н., К вопросу о специализации пищевого и оборонительного
условных рефлексов, выработанных на сходные раздражители, сб. «Высшая
нервизя деятельность», М., 1929.
ВЛИЯНИЕ ПИЩЕВОГО БЕЗУСЛОВНОГО РЕФЛЕКСА НА УСЛОВНЫЕ РЕФЛЕКСЫ 45
Зелепий Г. П.. Особый вид условных рефлексов, Архив биологических наук,
т. XIV, 1909.
Крестовников А. Н., Существенное условие при выработке условных
рефлексов, Труды Общества русских врачей в С.-Петербурге, 1913.
Коган Б. А., Об иррадиации и концентрации угасательного торможения, днсс.,
Спб., 1914.
Конради Г. П., Дифе рении ров ка и взаимодействие активных условных реф-
лексов, связанных с различными безусловными рефлексами, Труды физиологи-
ческих лабораторий акад. И. П. Павлова, т. IV, 1932.
М и л е .1 ь с о л Н. И, и Ю р ы а и М, Н-, Изучение условные рефлексов, вы-
работанных на основе различных безусловных. Иррадиация угасательиого тор-
можения. Сб. «Высшая нервная деятельность», М., 1929.
Майоров Ф. П. О влиянии продолжительности совпадения условного рефлекса
с безусловным на величину условного рефлекса. Труды физиологических ла-
бораторий акад. И. П. Павлова, т. Ill, в. 1, 19'28.
Ней « Е. Л„ К вопросу о влиянии условных рефлексов друг на друга, Известия
Военно-медицинской академии, 1908.
Пал ладин А. В., Образование искусственных условных рефлексов от суммы раз-
дражений, Труды Общества русских врачей, Спб., 1906.
Перельцвейг И. Я., Материалы к учению об условных рефлексах, дисс.,
Спб.. 1907.
Подкопаев И. А., Дальнейшие материалы к вопросу о восстановлении угашен-
ного условного рефлекса своим безусловным, Труды физиологических лабораторий
акад. И. П. Павлова, т. II, в. I, 1927.
Р а й т Р, Я-, Влияние безусловного рефлекса на условный рефлекс, Труды физиоло-
гических лабораторий акад. И. П. Павлова, т. II, в. 2, 1928.
С а в и ч А. А., Дальнейшие материалы к вопросу о влиянии пищевых рефлексов друг
на друга, дисс., Спб., 1913.
Сол ов ей чи к Д. И., Состояние возбудимости корковых клеток во время действия
безусловного раздражителя, Труды физиологических лабораторий акад.
И. П. Павлова, т. 11, п. 2. 1928.
Фу рс и ко в Д. С., Статическая иррадиация торможения. Архив биологических
наук, т. XXIII. 1923.
Ф-урсиковД. С., О соотношении процессов возбуждения и торможения, Русский
физиологический журИаЛ, Т-. IV, 1922.
X а з е н С. Б., О соотношении размеров безусловного и условного слюноотделитель-
ных рефлексов, дисс., Спб, 1908.
ВЛИЯНИЕ\ПОСТОРОННИХ И УСЛОВНЫХ РАЗДРАЖИТЕЛЕЙ
НА ВЕЛИЧИНУ БЕЗУСЛОВНОГО РЕФЛЕКСА1
нашей работе о влиянии безусловного рефлекса на условные
мы подчеркнули важность вопроса о взаимоотношении и взаи-
модействии нервных центров друг на друга, а также дали
краткий перечень важнейших работ в области условных реф-
лексов, имеющих непосредственное отношение к этому воп-
росу. В настоящей статье мы намереваемся изложить полу-
ченный нами» новый материал с иной стороны. Здесь речь будет идти о
влиянии посторонних и пищевых условных раздражителей на величину
соответствующего безусловного рефлекса.
В протокольном материале старой литературы по условным рефлексам
иногда можно встретить данные, указывающие, что условные рефлексы
могут ВЛИЯТЬ на величину безусловного рефлекса. Эти данные в большин-
стве случаев оставались незамеченными. В новой литературе уже име-
ются специальные исследования по этому вопросу.
В этой связи нельзя не привести очень интересных данных Ю. П. Фро-
лова о сильном и длительном влиянии условных рефлексов на безусловный
рефлекс, которое проявляется в процессе переделывания следовых услов-
ных рефлексов в наличные. Это влияние было настолько сильным, что ла-
тентный период действия безусловного раздражителя измерялся десятками
секунд. Совсем недавно этим вопросом занялась В. Ф. Шмидт в московской
лаборатории, руководимой проф. Д. С. Фурснковым. На основании своей
работы Шмидт пришла к заключению, что ни продолжительность изоли-
рованного действия метронома (метроном был условным положительным
раздражителем.—Э. А.), ни величина условного слюноотделения, ни уга-
шение искусственного и натурального условных рефлексов, Е1И запазды-
вание по сравнению с началом действия безусловного и полное отсутствие
условного рефлекса, ни сильный [внешний ряздражитель не оказывают
почти никакого влияния на количество безусловного слюноотделения. Из-
ложение нашего экспериментального материала покажет, что последний
нс согласуется с выводами и данными Шмидт. Здесь же, однако, обратим
внимание на важные моменты в работе автора,, которые бросаются в глаза
при чтении ее статьи. Речь идет о значительных колебаниях в величине
первых безусловных рефлексов у ее подопытных собак даже в тех опытах,
которые должны были служить в качестве контрольных, так как в НИХ не
оказывалось никакого воздействия на безусловные рефлексы. Пример:
на одно и то же количество пищи безусловные рефлексы выразились
в единицах 50,8, затем 37,7; или же в другом опыте соответственно
1 Труды физиологических лабораторий акад. И. П. Павлова т. X, 1941.’Пред-
варительное сообщение напечатано в «Докладах Академии налк СССР» за 1934 г.
по представлению акад. И. П. Павлова, 20. II 1934 г.
ВЛИЯНИЕ УСЛОВНЫХ РАЗДРАЖИТЕЛЕЙ НА БЕЗУСЛОВНЫЙ РЕФЛЕКС
47
44,4, 32,7 й 30,5. Чем обусловлено такое положение: тем ли, ЧТО ка-
мера, как свидетельствует автор статьи, была звукопроницаема для
внешнего раздражения; тем ли, что она применяла достаточно большие па-
узы, или чем-нибудь другим, — нам сказать трудно. Однако нельзя не
отметить, что данные Шмидт не совсем согласуются С установленными еще со
времени С. Б. Хазена правилами, что в приблизительно одинаковых усло-
виях равные порции нищи вызывают более или менее одинаковые рефлек-
сы в течение коротких опытов. Во всяком случае такого рода контрольные
опыты крайне затрудняют какие-либо выводы относительно влияния различ-
ных агентов на величину безусловных рефлексов.
Последняя, самая новая работа по этому вопросу вышла после того,
как мы уже закончили экспериментальную часть наших исследований. Эта
работа исходит из лаборатории проф. Г. В. Фольборта. А. М. Воробьев и
Л. Л. Линдберг исследовали изменение величины безусловного рефлекса
в процессе образования.условных рефлексов. Они нашли, что новые агенты
как до, так и после их превращения в условные раздражители по-разному
влияют на величину безусловного рефлекса у различных собак: у одних
безусловные рефлексы тормозились, у других они не изменялись, а у не-
которых даже увеличивались. Авторы все три случая объясняют разницей
в степени возбудимости безусловного пищевого центра у различных собак.
По нашему мнению, помимо причины, отмеченной авторами статьи,
возникновение такой пестрой картины зависело также от другого момента.
Судя по приведенным протоколам, не совсем одинаковы были условия
экспериментирования с различными собаками. Так, например, для изучения
влияния посторонних раздражителей на величину безусловного рефлекса
у разных собак эти раздражители применялись авторами то 2, то 5, го 15,
то 30 секунд. Затем одни собаки давали условный эффект даже при первом
применении постороннего раздражителя, другие нет. Как мы в дальней-
шем убедимся, оба эти момента являются далеко не безразличными для ве-
личины безусловного рефлекса.
Экспериментальные данные, полученные нами попутно при выпол-
нении наших прежних работ по условным рефлексам, бесспорно, гово-
рили, что условные раздражители оказывают заметное влияние на ве-
личину безусловного рефлекса, на состояние возбудимости центра безу-
словного рефлекса. Было небезынтересно подробно н систематически
исследовать этот важный вопрос физиологии головного мозга, тем более
что имеющиеся данные были противоречивы. Мы на наших подопытных
собаках Арч и Баш в звуконепроницаемых камерах разрабатывали этот
вопрос экспериментально. Обе собаки были уравновешенными, с силь-
ным типом нервной системы, причем Арч был сангвиником, но имел
черты флегматичности, а Баш также сангвиник и немножко трусоват.
С целью точного учета величины и характера проявления безусловного
рефлекса мы во все время работы применяли безусловный пищевой раздра-
житель постоянного состава, величины и физического состояния (для обеих
собак по 10 г мясо-сухарного порошка, смоченного 10 см3 воды). Такую пор-
цию пищи собаки съедали полностью, причем Арч—в течение 40 секунд,
а Баш — в течение 1 минуты. Безусловный ; рефлекс регистрировался за
15 секунд в течение Р/з минут.
Вначале мы ставили ряд контрольных опытов, причем убедились, что
эти стандартные безусловные раздражители при возможно одинаковых усло-
виях (время года, время дня, сорт порошка и т. д.) вызывают совершенно
равные безусловные рефлексы как в течение одного опытного сеанса, так
и в течение опытов, проводившихся ряд дней. Так, например, при общей
величине рефлекса в среднем НО—130 делений шкалы в разных опытах ко-
лебания эти находились в пределах 5—8 делений (табл. 1).
48
Э. А. АСРАТЯН
Собака Арч Таблица I
Время Величина безусловного рефлекса за 15 сек. в делениях шкалы Общая величина безусловного реф- лекса за 00 сек. в делениях шкалы
Опыт № 216 от 16.x н 1931 г.
15 ч. 57 ы. Подача большой порции
16 » 01 > 29+ 70 + 99+1164-122+127 127
16 » 05 з 28+40+30+ 19+4+8 129
16 » 09 » 30+40-1-29+ 16 + 5+8 128
16 » 13 » 28+39+28+19+7+4 125
16 » 17 » 29+40+28+19+4+6 126
16 » 21 в 26+42+27+22+5+0 121
16 » 27 > 27+43+28+16+7+6 127
16 » 31 » 27+41 + 18+4+7 128
В дальнейшем опыты ставились по следующему плану: в начале опыта
подачей первых двух или трех стандартных порции пищи устанавливалась
контрольная величина безусловного рефлекса в данный опытный сеанс,
затем в каком-нибудь виде испытывалось влияние одного из трех названных
агентов — постороннего раздражителя, положительного или отрицатель-
ного условного раздражителя. Чтобы проследить последовательное дейст-
вие примененного агента на безусловные рефлексы, несколько раз подряд
через разные паузы измерялись величины безусловных рефлексов на стан-
дартный раздражитель.
Для более детальной разработки вопроса изменялась также форма
испытания влияний названных факторов на величину и характер безуслов-
ного рефлекса; факторы эти применялись до подачи безусловного раздражи-
теля (причем иногда после некоторой паузы), одновременно с ним или же
некоторое время спустя после начала действия безусловного раздражителя
и до конца его действия. Полученные результаты в определенном порядке
кратко излагаются ниже.
Влияние отрицательных условных раздражителей на величину
безусловного рефлекса
В качестве отрицательного условного раздражителя мы применяли
диференцировочное торможение как концентрированный и остро дейст-
вующий вид торможения. У собаки Лрч дифереицирована была частота стука
метронома 60 ударов в 1 минуту от частоты 120 ударов в 1 минуту, а у Баша
был диференцирован музыкальный тон той же октавы. Получены следую-
щие результаты.
1. Когда днференцировочный условный раздражитель непосредственно
предшествует безусловному раздражителю или когда между концом дей-
ствия дифереицировочногораздражителя и началом действия условного
раздражителя делается небольшой перерыв — 10—30 секунд, то без-
условный рефлекс по сравнению с нормой значительно увеличивается
(табл. 2)1.
Как видно из приведенного материала, применение дифереицировочного
раздражителя вызывает не только усиление последующего безусловного
рефлекса, но это усиление распространяется и на следующие за послед*
ним безусловные рефлексы. Это явление наблюдалось во всех аналогич-
ных опытах.
* Ради экономии места мы как в табл. 2, так и в следующих, за редкими иск-
лючениями, будем приводить не полные протоколы опито»,'а характерные отрывки
из них.
ВЛИЯНИЕ УСЛОВНЫХ РАЗДРАЖИТЕЛЕЙ НА БЕЗУСЛОВНЫЙ РЕФЛЕКС
49
Таблица 2
Собака Арч
№ опыта и дата исследования Время Величина безусловного рефлекса по 15 сек. в делениях шкалы Общая величина безус до вного рефлекса за -90 сек. в делениях шкалы
/ 15 ч. 39 ы. 29 +34 + 27f 134-8 1-4 115
15 » 43 » 30 4~ 33 4- 24 lf> 4* 3 V 8 116
220 15 » 47 » Стук 60 и течение 20 сек , пауза
23..Х II 1913 г. 10 сек.
15 » 47 » 30 с. 38 +32 + 28+18+9+6 131
15 > 51 » 30 + 34 + 27 + 17+12+4 124
14 » 31 » 24 +36 + 27+ 16+1 +1 105
2">6 14 > 35 » Стук 60 в течение 20 сек., затем
1.1 1932 г. 14 * 36 » пауза 20 сек. 28+33+34 + 2Ц-3+3 122
I 14 » 39 » 24+37+33 + 15+5+1 115
14 » 17 » 28+39+32+15+5+2 121
14 » 21 » 27-1-38 +32+19+ 2+ уг 118У2
283 14 » 25 » Стук 60 в. течение 20 сек., пауза 20 сек.
6. IV 1932 г. 14 » 25 » 30 с. 29 +-41 +39+17+5+5 135
14 » 29 » 26 + 37 + 33 + 24 + 12+5«/2 137/j
1. 14 » 33 » 21+35 +30 |-17+16+1 123
2. В случаях, когда пауза между концом действия дмференцирово-чного
раздражителя и началом действия безусловного раздражителя имеет величи-
ну около 1 минуты и больше, безусловный рефлекс не только не усиливает-
ся, но даже ослабевает от действия диференцировочного раздражителя.
3. Когда же эта пауза увеличивается до 2 минут и больше, то безуслов-
ный рефлекс не претерпевает заметного изменения от предшествующего дей-
ствия диференцировочного раздражителя.
4. Если же диференцировочный раздражитель применяется подряд
два раза и больше, то следующий за ним безусловный рефлекс всегда умень-
шается в значительной степени как при маленьких паузах, так и после боль-
ших пауз. Характерно также, что в таких случаях уменьшается не только
безусловный рефлекс, следующий непосредственно за применением лифе-
ренцировочного раздражителя, но что это уменьшение распространяется
также на последующие безусловные рефлексы (табл. 3).
5. Применяемый одновременно с безусловным раздражителем диферен-
цировочный раздражитель вызывает ослабление безусловного рефлекса;
если же он присоединяется к безусловному раздражителю некоторое время
спустя после начала его действия (от 7 до 10 секунд), диференцировочный
раздражитель немного усиливает безусловный рефлекс, но зато оставляет
сильное и длительное последействие на следующих по очереди безусловных
рефлексах (табл. 4).
Должен сказать, что в своих последних опытах по выяснению влияния
угасателыюго торможения на величину безусловного рефлекса сотруд-
ник нашей лаборатории В. В. Рикман тоже получил данные, нс идентич-
ные нашим, но по общему смыслу близкие описанным в первом пункте.
Рикман наблюдал, что угашение условных рефлексов в начальной стадии
заметно усиливает безусловный рефлекс.
Интимную физиологическую сторону описанных фактов, пожалуй, мож-
но себе представить в таком виде. В первом случае— случай предшествова-
ния диференцировочного раздражителя безусловному — вызванное в коре
торможение положительно индуцирует центр безусловного рефлекса, а по-
тому происходит усиление безусловного рефлекса. Во втором случае, когда
увеличивается пауза между применением диференцнровки и безусловного
4 Э. А. Асратян
50
Э. Л. АСРАТЯН
Собака Арч
Таблипа 3
.S& опыта и дата исследования Время Величина безусловного рефлекса по 15 сек в делениях шкалы Общая величин! безусловного рефлекса за 90 сек. в деле* ниях шкалы
227 2.1 1932 г. 16 ч. 16 » 16 » 16 » 16 > 02 м. 06 > 10 » 11 » 15 > Нс. 28 + 34 + 34+ 15+1+0 27 +36 + 34+ 16+ 3 +2 Стук 60 в течение 20 сек., пауза 50сек. 26+32+29+11 + 2+0 28+11+33+14+3+4 113 118 102 116
231 17.1 1931 г. 14. » 17 » 15 » 15 » 15 » 15 » 15 » 15 » 15 » 15 » 15 » 56 . 59 » 03 » 07 > 11 9 13 > 17 » 20 » 23 » 25 • 27 » 45 с. 30 > 15» 45 с. 30» 27+39+29+ 14+ 10+3% 27+41 + 32+ 12+ 11-» 2 264-40+36+ 15+8+2 27+40+32+ 15+7+2 Стук 60 в течение 20^сек., пауаа 100 сек. 22+44+34+16+4+4 23+41+35+18+8+3 24+41 + 33+ [4+5+4 Стук 60 н течение 20 сек. Стук 60 в течение 20 сек., пауза 100 сек. 20+ 42+ 27+ 7+2+0 И2% 125 127 123 124 128 121 97
233 1 15 > 16 » 1S » 16 » 16 » | 16 » 16 » 16 » 56 » (К) » 04 » 05 » 05 » 09 » 13 » 17 » 40 с. 27 + 38+35+6+3+3 28+42+31 + 6+4+2 Стук 60 в течение 20 сек. Стук 60 в течение 20 сек., пауза 20 сек. 19+31 + 201-3+1 + Ц 22+ 39+26+7+3+1 %+0 25+40+28+ 8+2+1 + % 25+ 42 + 30+ 6+ 1 + 1 + 1 III 113 74% 98% 104% 108 '
Собака Арч 1 Таблица 4
№ опыта и дата исследования Время Величина безусловного рефлекса по 15 сек. -в делениях шкалы Общая пеличкм безус лои1сого рефлекса за 90 сек. в ле.^- ННЯХ шкалы
4 Отрывок из опыта 238 । 16 16 16 16 16 ч. 21 а 1 1 » 29 » 33 » 37 м. » > > 25+41+29+8+ 1 + 1+1 25+41 + 28+2+3+1+% 25+ 424-29+5+ 3+ 1 % +1 % Стук 60 и пиша применяются одновременно 22+ 40+23%+1 %+!%+% + 0 24 + 41+29+ 4 + 8 + 2+0 106 104% 107 94 108
1 1 288 14.1V 16 16 16 17 17 • » 48 » 53 » 57 » 01 » 05 о 09 » 13 » » » » » » 26+ 38+27+13+ 3+1 + 1% 25+37+28+12+2+1 + 1% 25 + 36 4-27+15+2+2+0 Спустя 10 сек. после подачи пищи присоединяется ст. к 60 20+ 32 л 33 + 28 + 4 +0 19+25+18 + 5 + 3+2 + 0 22+ 30+ 15 +3-1-2+2+2% 23+ 33 +22+11 + 1 + 2+2 109% 106% 107 117 72 76% 94
ВЛИЯНИЕ УСЛОВНЫХ РАЗДРАЖИТЕЛЕЙ НА БЕЗУСЛОВНЫЙ РЕФЛЕКС 5|
раздражителя, очевидно, тормозной процесс от коркового пункта приме-
ненной диференцировки уже успевает иррадиировать к центру безуслов-
ного рефлекса и вызывает ослабление последнего. То же объяснение от-
носится и к четвертому случаю, когда диференцировочпый раздражитель
применяется несколько раз подряд. Случай же третий, невидимому, пред-
ставляет фазу концентрации торможения и освобождения от него центра
безусловного рефлекса. Что же касается пятого случая, то мы затрудняемся
объяснить, почему диференцировха уменьшает безусловный рефлекс при
одновременном применении С безусловным раздражителем. Некоторое же
увеличение безусловного рефлекса при позднем присоединении диферен-
цировочного раздражителя к безусловному (случай 6) можно представить
себе так, что возбужденный пищевой центр притягивает к себе появившийся
позже стимул в коре и, суммируясь с ним, дает увеличенный эффект. Что
же касается волнообразного последействия, то, повидимому, оно является
или результатом борьбы процессов возбуждения и торможения в самом
центре безусловного рефлекса, или же, может быть, отражением борьбы
тех же процессов в корковых пунктах условных рефлексов.
Влияние положительных условных раздражителей на величину
безусловного рефлекса
1. Прежде всего мы хотели бы привести сделанное нами раньше
интересное наблюдение, что очень часто, когда условный раздражитель при-
меняется во время продолжающегося еще последовательного слюноотделе-
ния от предшествующего безусловного раздражителя, он почти сразу же на
несколько секунд останавливает это слюноотделение; затем, спустя некото-
рое время, снова начинается слюноотделение уже условного характера.
2. Условные положительные раздражители вызывают значительнее ослаб-
ление безусловного рефлекса, когда непосредственно предшествуют действию
безусловного раздражителя, т. е. подаче пищи. Это находится в согласии с
некоторыми недавно опубликованными данными Л. М. Воробьева
и А. Л. Линдберга, а также с еще не опубликованными данными
Л. М. Павловой, хотя мы должны сказать, что постановка наших опытов
существенно отличалась от постановки опытов этих исследователей. Раз-
ница заключалась в. том, что мы приостанавливали применение условного
раздражителя и затем только подавали пищу, а упомянутые исследователи
продолжали, как обычно, условные раздражите.’!и применять и во время
действия безусловного раздражителя. Этот момент, как увидим дальше,
далеко не может считаться безразличным.
В соответствии с нашей общей схемой мы и здесь ставили такие опыты,
в которых делалась некоторая пауза между окончанием применения ус-
ловного раздражителя и началом включения безусловного раздражителя.
Этиновые опыты показали, что тормозящее влияние предшествующих полс-
жигельных условных раздражителей па соответствующие безусловные реф-
лексы как будто даже усиливается, когда пауза составляет 10—30 секунд
(табл. 5 и 6).
Следует сказать, что на безусловные рефлексы очень сильное тормозя-
щее влияние оказывают также «условные» движения и действия экспери-
ментатора во время подготовки собак к опыту: приклеивание баллончика,
наполнение проводящей системы водой, движения, связанные с подачей пер-
вой чашки пищи в камеру, и т. д. Несомненно, с этим надо связать парадок-
сальный факт, что первый по очереди безусловный слюнный рефлекс на
стандартный раздражитель, как правило, всегда бывает слабее, чем следую-
щие за ним безусловные рефлексы, несмотря на то, что собаки всегда едят
их более жадно, чем последующие порции.
4*
52
Э. А. АСРАТЯН
Собака Арч
Таблица 5
№ опыта и дата исследования ! Величина безусловного рефлекса по 15 сек. в делениях шкалы Общая величина, безусловного рефлекса за 90 сек- в деле- ниях шкалы
244 10.П 1932 г. 258 2.111 1932 г. Собака Баш 14 ч. 45 м. И » 49 » 14 » 53 » 14 » 53 >20 с. 14 » 57 > | 15 » 01 » 15 » 05 » 1 15 > 09 > 15 » 23 » 15 » 29 » 15 » 29 »40 с. 15 » 35 » 15 » 41 » 15 » 47 » 1 26 4- 39 4-27 + 154-11+0 27 + 40+ 25+18+8+IJ-2 Стук 120 в течение 20 сек., ус- ловный рефлекс 27 делений 28+35+16+5+9+2 25+34+27+164-1+1 25+37+3! +16 + 7+0 34+39+30+23+13+2 23+ 39 +29 + 14 +2+1 30 + 42 +30+10+6 + 1 Звонок в течение 20 сек., услов- ный рефлекс 34 деления, пауза 10 сек. 27+36+12+6 +2+2 31 + 42+ 33 +11 + 11+6 28+40+40+13+7+6 26+40+39+19 + 17+4 118 ИЭ’/г 95 104 116 131 108 119 85 134 34 145 Таблипа 6
№ опыта и дата исследования Время Величина безусловного рефлекса по 15 сек. в делениях шкалы Общая величина безусловного рефлекса за 90 сек. в деле- ниях шкалы
70 5. IV 1932 г. 17 ч. 05 м. 17 » 09 » 17 » 12 » 17 » 15 » 17 » 15 > 30 с. 27+35+2-9+11 +2 +1 26 4 38+26 +10+2+ ‘/s 26 + 35 + 30 + 10 +2+Г Стук 120 в течение 20 сек., условный рефлекс 25 делений, пауза 10 сек, 29 + 27 + 26 +3+ 0+1 105 1041/s 104 86
В ряде опытов, например, величина первого по порядку безусловного
рефлекса равнялась 105, 106, 130 делениям шкалы, величина же следую-
щих безусловных рефлексов соответственно выражалась числами 128, 130,
139 и т. д.
Как только это было выяснено, мы на первом, иногда и на втором месте
стали давать большие порции пищи, как бы для сглаживания следов и шеро-
ховатостей, оставленных «условными» действиями экспериментатора в про-
цессе подготовки собак к опытам. В приведенных опытах об этих первых
больших порциях не упоминается, чтобы не осложнять картины.
3. Когда же отмеченная пауза, отделяющая условный раздражитель
от безусловного, увеличивается, уменьшающий эффект воздействия услов-
ного раздражителя на безусловный рефлекс постепенно ослабевает и при
паузах в IV»— 2 минуты почти целиком исчезает.
Характерно, что в случае, когда условные раздражители почему-либо
временно утрачивают свое действие и почти не вызывают условного эффекта,
предшествующее применение этих раздражителей не оказывает почти ника-
кого влияния на безусловные рефлексы. Это говорит о том, что важным яв-
ляется не простой факт предшествования условного раздражителя, а
действительное возбуждение корковых пунктов условных рефлексов
(табл. 7).
ВЛИЯНИЕ УСЛОВНЫХ РАЗД°АЖИТЕЛЕЙ НА БЕЗУСЛОВНЫЙ РЕФЛЕКС
53
Собака Арч
Таблица 7
№ ОПЫТ* и дата исследования Бремя Величина бсауслоан^го рефдекед пр 15 сек. а делениях шкалы Общая величина безусловного рефлекса за 90 сск в деле- ниях шкалы
14 ч. 40 м. 28440431+11-+841 110
14 » 44 » 14 » 48 » 30+40 + 36 +1448+1 Стук 120 в течение 30 сек., ус- ловный рефлекс 36 делений, пауза 2 мин 119
14 » 50 > .30+41 + 33+14 + 6+1 125
254 26.11 1932 г. 14 » 54 » 28+ 41 + 37+14 + 8+1 129
14 » 58 » 274-40+37+15+7+1 127
15 1» 02 » 15» 06 » 26+ 42+ 36+16 + 6+2 Звонок в течение 30 сск , услов- ный рефлекс 32 деления, пауза 1 мин. 128
15 j 07 з 30 с. 24 + 32 + 32+17 + 3+1 109
15 > 12 » 26+40+84+15+6 + 3 124
16 » 44 » 24 + 31 + 28+19 + 3 +3 108
16 » 47 » 16 » 50 » 25 + 304 29+19 + 3+3 Стук 120 в течение 20 сек , ус- ловный рефлекс 2 деления 109
16 » 50 » 2Юс. 26 + 29+ 29+19 + 3+4 НО
24G 13 11 1932 г. 16» 53 » Звонок в течение 20 сек., \слов- ный рефлекс 2J.4 деления, пауза 10 сек.
16 » 53 » 30с. 25+31+27+17 + 4+2 1Q6
16 » 56 » 24 +29 + 31 +18 + 2+0 101
16 » 59 » 23+33+ 26 + 16 + 341 102
4. Когда положительные условные раздражители и безусловный раз-
дражитель применяются почти одновременно и продолжают действовать
совместно до конца, то величина безусловного рефлекса немного умень-
шается.
5. Когда же положительный условный раздражитель присоединяется
к безусловному спустя некоторое время после начала действия последнего
(5—10 секунд) и продолжает действовать совместно с ним до конца, то
безусловный рефлекс сильно увеличивается как в слюнном своем компонен-
те, так и в двигательном. При этом собаки совершенно отчетливо удли-
няют акт еды (табл. 8 и 9).
Очень интересными являются факты, постоянно имевшие место также
при испытании влияний положительных условных рефлексов на безуслов-
ные, а именно: последействие, последовательное влияние, которое в виде
волн дает себя чувствовать достаточно длительное время от момента при-
менения условного раздражителя.
Интимно-физиологическую сторону описанных явлений, на наш взгляд,
можно себе представить следующим образом. Первый случай — случай
остановки последовательного безусловного слюноотделения в связи с приме-
нением условного раздражителя — указывает на отрицательную индукцию,
которая в центре безусловного рефлекса вызывается возбуждением корковых
пунктов условных рефлексов. Во втором и третьем случаях, когда безуслов-
ный рефлекс ослабевает от предшествующего применения условных раздра-
жителей, мы, повидимому, имеем дело с тЪй же формой воздействия, т. е.
с отрицательной индукцией центра безусловного рефлекса с возбужденных
корковых пунктов условных рефлексов. При этом, повидимому, неболь-
шая пауза между применением условных и безусловных раздражителей
54
Э. А АСРАТЯН
Таблица 8
Собака Баш
-Nj опыта и дата исследования Время Величина безусловного рефлекса по 15 сек. в делениях шкалы Общая величина безусловного рефлекса за 90 сек. в деле- ниях шкалы
16 ч. 30 м. 204-394-324-3 + 4 + 3 по
16 > 3-1 » 30+39+29+2+6+2 108
72 ! 16 » 37 » Звонок и пища почти одновре-
7. IV 1932 г. мон но
27+ 384-20 4-13 +3 +0 101
16 » 40 » 28 + 394-30 + 6 + 3+4 110
16 » 45 » 58+35+394-10+3+1 116,,
16 » 48 » 26 + 36 +28-ь Ю+3+ 1*/а 104!’
16 » 51» Звонок присоединяется через
10 сек. после подачи пищи
70 30+57+21+14+4+7 193
16 » 54 » 28 + 39 + 27+18 + 2 +3 117
16 > 57 » 29+ 36+ 31+7+П/г + 1^ 106
17 э 00 » 29+35+29 + 11 +4 + 3 111
17 э 03 » Спустя 2—3 мин. после подачи
пищи стук 120
А4+40+22+6+4+2 108
Таблица 9
Собака А рч
N? опыта и дата
исследовании
Время
Величина безусловного рефлекса по
16 сек, в делениях шкалы
15 ч, 51 ’1 30+40+31 +10+7+3
15 » 15 > 55 59 > > 30+33+32 г 15+8+31 29+41+31 + 10+9+2
264а < 12.111 1932 г. 264 9.111 1932 Г. 269 16 > 16 » И > 14 > 14 » 14 » 14 » 14 » 15 » 15 > 15 а 15 » 15 » 16 » 17 » 17 » 17 » 17 > 17 * 03 07 29 35 41 47 53 59 05 11 17 23 29 53 01 05 08 12 16 » » » » > » » > > > 08с » » » » Стук 120 и пища почти одновре- 28 + 35 + 29+15 + 4 +2 29 f 45 + 37 +11 + 5+1 31+4I+32+14+1O+2 30+40+31 + 18 + 5+4 31+41+36 + 12+8+4 30+41+31 +18 + 6+2 30 <-42-30J-17+4+3 Через 8 сек. после подачи пищи дается звонах 26 +39 + 56+17 - 9 + 11 31+40+32 + 6 «• 9+0 29 + 40 + 39+7+8+1 29+ 39 <-30+5+8+7 Через 8 сек. после подачи пищи стук 120 23 +31 + 53 + 16 + 7 +4 29 + 41 +35 +15+ / + 4 32+32+21 + 17+2+2 31 +34+18+21 + 2+2 Через 3 сек. после подачи пищи свет 38 +42+ 36 +22+11 +5 Подача большой порции 32 +40+ 28410+8+6 32 + 39 + 31 + 11+7+1
17 » 20 » Через 3 сек. после подачи свет 41+44 + 39 + 17+7+6 ПИЩ К
Общая величина
везусловкого
рефлекса за
80 сек. в мле-
ниях шкалы
121
125
123
113
128
130
128
132
128
126
158
118
124
118
137
131
106
108
154
124
121
15?
ВЛИЯНИЕ УСЛОВНЫХ РАЗДРАЖИТЕЛЕЙ НА БЕЗУСЛОВНЫЙ РЕФЛЕКС
55
благоприятно действует потому, что индукция достигает своего наиболь-
шего выражения некоторое время спустя после применения условных раз-
дражителей. В этом отношении характерно то, что при исчезновении услов-
ного рефлекса на условный раздражитель применение последних не вызы-
вает никаких изменений в величине безусловного рефлекса. Очевидно, при-
чиной указанного явления служит отсутствие должного возбуждения кор-
ковых пунктов условных рефлексов и связанной с этим отрицательной
индукции в центре безусловного рефлекса.
Случаи пятый с увеличением безусловного рефлекса вследствие запо-
здалого присоединения условного раздражителя к безусловному можно
себе представить в виде суммации возбуждения, возникшего в корковом
пункте условного рефлекса, с возбуждением уже доминирующего центра
безусловного рефлекса. На это указывают также ход и характер увели-
чения безусловного рефлекса.
Влияние посторонних раздражителей на величину
безусловного рефлекса
Испытанные нами некоторые посторонние раздражители (гудение, звук,
бульканье) далеко небыли индиферентными для наших собак. Оказалось,
что они вследствие генерализации вызывали некоторый условный положи-
тельный эффект и соответственно этому уменьшали безусловный, непос-
редственно следующий за ним, правда, в меньшем объеме, чем это было
при применении настоящих положительных условных раздражителей.
Но эти посторонние раздражители отличались от последних в другом от-
ношении. Они при запоздалом присоединении и при одновременном приме-
нении с безусловными раздражителями всегда уменьшали величину без-
условного рефлекса, как будто обладали некоторыми свойствами отрица-
тельных раздражителей. Что же касается тех посторонних раздражителей,
которые не вызывали услов-ного рефлекса, а обусловливали сильную
ориентировочную реакцию, то они оказывали своеобразное воздействие
на величину безусловного рефлекса.
Мы испытывали влияние слабого электрического тока, ударов и разных
комплексных шумов, вызываемых вне камеры (дверь в таких случаях
оставалась полуоткрытой).
1. Когда эти посторонние раздражители с паузой или без паузы пред-
шествовали безусловному раздражителю, безусловный рефлекс заметно
усиливался.
2. Когда же посторонний раздражитель начинал Действовать одновре-
менно с безусловным, безусловный рефлекс немного ослабевал.
3. Когда посторонний раздражитель присоединялся к безусловному
раздражителю немного позже после начала действия последнего (5—10 се-
кунд), безусловный рефлекс чаще всего усиливался, иногда же он не пре-
терпевал заметного изменения или немного ослабевал.
Приведенные опыты (табл. 10 и 11) можно объяснить так: посторонний
раздражитель, по всей вероятности, вызывает сильную ориентировочную
реакцию, отрицательно индуцирует центр безусловного рефлекса, т. е.
действует точно так же, как и отрицательный условный раздражитель.
Сходство между воздействием этих раздражителей на центр безусловного
рефлекса и воздействием отрицательных условных раздражителей на тот
же центр имеется также в случае одновременного их применения, а также
в случаях запоздалого присоединения этих раздражителей к безусловным.
Это значительное совпадение в характере воздействия обоих вндов раздра-
жителей на велич ину безусловного рефлекса может считаться лишним пока-
зателем того, что в основе внешнего и внутреннего торможения имеется
много общего.
56
Э А АСРАТЯН
Таблица 10
Собака Арч
№ опыта и дата исследования В ре мя Величина безуслонного рефлекса по 15 сек. в делениях шкалы Общая величина безусловного рефлекса за 90 сек, в деле- ниях шкалы
271 15 ч. 15 м. 29+42+ 34+14+5+5 129
21.111 1932 г. 15 » 20 » 30 +40+ 34+17+5+5 131
15 » 25 > Комплексный шум в течение
20 сек.; сильная ориентировоч-
пая реакция —
38 + 26 1-38+ 16+9+7 134
15 » 30 » 29+4 1 + 37+13+4+2 126
15 > 35 » 28+41+34+12+3-2 120
15 » 40 » 27+41 + 36+15+5+3 127
15 я 4о » Пиша и посторонний раздражи-
тель одновременно —
25+ 38 + 26+10 + 3 I 2 104
15 » 50 » 25 + 42+35+12+4 + 5 123
15 в 55 > 24 +40+38+ 12+4 + 2 1?0
16 » 00 » Комплексный шум спустя 10 сек.
после подачи пищи —
2 4 4" 4 4 4" 3815-1-6Ч-5 132
Таблица II
Собака Баш
Afe опыта и дата 1 исследовании Время Величина безусловного рефлекса ио 15 сек. и делениях шкалы Общая величина безусловного рефлекс! за 90 сек. в деле- ниях шкалы
14 ч. 37 м. 29+ 35+ 25-1-12+ 8+3 1Г2
14 > 41 » 30+32+28 1-13+4+2 109
14 » 45 » Спустя 10 сек. после подачи пн-
щи применен электрический
раздражитель
76 30+ 36 + 324 11+7+4 120
18.1 V 1932 г. 14 » 49 » 28 + 33+ 32+9+6 +4 112
14 » 53 » 27+33 + 30+10+5 г2 107
14 » 57 » 27+32 + 30+ 12+6+2 109
15 » 01 » Электрический ток 4—5 сек.,
спустя 2—3 сек. прдзча пищи
36+32+ 28 + 9 + 9 +7 123
16 » 29 » 29 + 33+24+13+5+3 107
16 » 34 » 30+33 + 26+12+6+2 109
16 » 39 » Пища и электрический ток одно-
74 времен но
10. IV 1932 г. 31+23+27+11 + 4 + 3 99
16 » 44 » 29 + 33 + 28+10+8+2 НО
16 » 49 » 28+33+30+ 10 + 5+1 107
1 11 > 55 » 29+40+26+9+3+2 109
11 » 59 » 30+ 40+ 27+ 9+3+2 ill
1 12 > 03 » Ток в течение 20 сек., пауза
71 10 сек.
6. IV 1932 г. 12 » 03 » 03 с. 34+41+28+9+6+7 125
12 » 07 > 30+38+25+Юг+2+3 108
12 » 11 > 31+39 + 26+8+5+1 110
. 12 » 15 » ЗОН 38+ 27 +7 + 5+2 109
ВЛИЯНИЕ УСЛОВНЫХ РАЗДРАЖИТЕЛЕЙ на БЕЗУСЛОВНЫЙ РЕФЛЕКС 57
г!тсбы представить себе картину описанных явлений, ниже приводим
схематическое сводное описание всех упомянутых случаев (табл. 12).
Таблица 12
Изменение условного рефлекса в зависимоеih от разных комбинаций условных
н безусловных раздражителей
Раздражится и Непосредст- пенное пред- шествование Величина паузы и сек. между окончаньем применения разд- ражителя м появлением безус- ловкого рефлекса Одновремен- ное действие Общих раз- дражителей Позднее при- соединение раз- дражителя к безус лонному рефлексу
раздражителя 10-30 со 90—120
Отрицатель- ные условные Усиление Усиление Ослабле- ние Изме не- ний нет Ослабление Усиление
Положитель- ные условные Ослабление Ослабле- ние Малое ослабле- ние То же Почти не изменяется »
Посторонние Усиление Усиление Ослабление Усиление; иногда не из- меняется
В нашей работе дана лишь грубо ориентировочная схема сложной
картины влияния условных и посторонних раздражителей на без-
условные рефлексы. Несомненно, что разнообразные результаты наших
вариаций и комбинаций раздражителей получены в итоге борьбы, взаимной
индукции, суммации или же, может быть, и других форм взаимодействия
между центрами этих раздражителей. При этом, невидимому, в каждом от-
дельном случае ведущая форма взаимодействия обусловливается степенью
возбудимости взаимодействующих центров. В этом отношении было бы ин-
тересно у собак различных типов и характера экспериментально изменять
силовые соотношения названных центров путем повышения возбудимости
пищевого центра, путем вариаций сил условных и посторонних раздражите-
лей и т. д. В начале работы мы избегали большой детализации и старались
временно ограничиться установлением основных, наиболее общих законо-
мерностей в этой области. Больше того, мы в данной статье не упомянули
о некоторых интересных данных, попутно выявленных в ходе работы, что-
бы не осложнять изложения. Систематическое и подробное изучение дета-
лей— задача будущих исследований.
ЛИТЕРАТУРА
Асратян Э. А, К вопросу о локализации .центральной части рефлекторной дуги
оборонительно-двигательного рефлекса, Физиологический журнал СССР,
т. XVII, в. 5, 1934.
Воробьев А. М. и Л и и д б е р г А. А., Об изменениях величины безусловного
рефлекса при присоединении к нему раздражителей при выработке условных реф-
лексов, сб. «Условные рефлексы», Харьков, 1932.
Фролов IO. П.» О переводе следовых условных раздражителей и следовых услошых
тормозов в наличные условные раздражители. Труды физиологических лабора-
торий акад. И. П. Павлова, т. I, в. 2—3, 1926.
Ха з е н С. Б., О соотношении размеров безусловного И условного слюноотДеЛНтеЛЬ-
ных рефлексов, дисс., 1908.
Шмидт В. Ф., Влияние условных пищевых рефлексов на величину безусловной
слюноотделительной реакции, сб. «Высшая нервная деятельность», 1929.
Посвящается И, П. Павлову
ко дню его 80-летнсго юбилея
ВЛИЯНИЕ ЭКСТИРПАЦИИ ВЕРХНИХ ШЕЙНЫХ
СИМПАТИЧЕСКИХ УЗЛОВ НА ПИЩЕВЫЕ УСЛОВНЫЕ
РЕФЛЕКСЫ СОБАКИ »
чение об условных рефлексах, созданное И. П. Павловым
и многочисленными его сотрудниками, достигло такого уровня
развития, что теперь оно составляет отдельную очень интерес-
ную и важную главу физиологии. Это учение охватывает и со-
судодвигателъные рефлексы, и деятельность почки и т. д. Уче-
ние это приобрело общебнологическое значение.
В последнее десятилетие рядом работ учеников и сотрудников
проф. Л. А. Орбели установлена адаптационная роль симпатической нерв-
ной системы для поперечнополосатой мускулатуры, центральной нервной
системы и периферических рецепторов. Осенью 1927 г. проф. Л. А. Орбели
предложил мне заняться вопросом о влиянии симпатической нервной си-
стемы на условные рефлексы (r.esp. набольшие полушария головного мозга).
Работа велась над собакой Бек (некровный сеттер-гордон), у которой мной с
помощью М. П. Бресткина был выведен по Павлову проток gl. parotis. В течение
лета я занимался, но не систематически, выработкой у собаки отдельных условных
рефлексов и упражнялся сам в технике этой работы. Условные рефлексы выраба-
тывались плохо (так как за неимением мясного порошка я старался образовать услов-
ный рефлекс на черный хлеб, а собака в собачнике получала гораздо более вкусную
пищу). В дальнейшем я стал подкреплять имеющиеся раздражител и мясо-сухарным по-
рошком и, кроме того, за редкими исключениями, начал ставить опыты ежедневно
в определенные часы дня. Подкрепления делались в течение 20 секунд; собака ела
в этот период сколько могла, после чего чашка бесшумно убиралась. Со времени
применения мясо-сухарного порошка я начал нумерацию опытов, считая этот момент
началом работы. Собака, невидимому, уже привыкла к обстановке и вскоре (на 12-м
опыте) у нее появились условные рефлексы на звонок, метроном (100 ударов в 1 ми-
нуту) и свет (лампочка в 50 свечей). Довольно быстро (после 11 применений) образо-
валась и постепенно укрепилась диференцнровка на метроном (60 ударов в I минуту).
С опыта № 25 я присоединил к имеющимся раздражителям две колодки с 17 уколами
в течение 20 секунд (на уровне задней правой лапы при расстоянии 10 см друг юг
друга). Применение одной из них (а) подкреплялось СДОЙ. Довольно быстро образо-
вался рефлекс на первую (о) и на вторую (нижнюю) кололку (в); так же быстро удалось
отдиференцировать верхнюю кололку (0), которая таким образом была сделана тормо-
зным агентом. Нужно отметить, что колол к и довольно сильно поколебали и величину
других уже имеющихся условных рефлексов. С опыта № 69 я прибавил еще две ко-
лодки, а с опыта№ 75 уже имелось 6 кололок, распределенных, как указано на рис. 1.
В силу генерализации новые кололхи уже давали эффект почти со второго примене-
ния, и выравнинанис эффектов всех кололок не потребовало много временя. Считаю
нужным отмстить, что в период, когда собака еще не вполне пришла в норму после
применения 6 кололок, произошло следующее осложнение; случайно в станок моей
собаки была поставлена сука с течкой, о чем я мог судить по следам свежей крови
на станке и по характерному поведению моей собаки. Эго сильно повлияло иа работу
Бека и в-ывелоее из равновесия на срок больше месяца. Затем только началась более
или менее постоянная, непрерывающаяся работа по укреплению условных пищевьц
рефлексов; это продолжалось вплоть до экстирпации верхних шейных симпатических
Ганглиев.
* Архив биологических наук. т. 30. в. II, 1930.
ЭКСТИРПАЦИЯ ШЕЙНЫХ СИМПАТИЧЕСКИХ УЗЛОВ И УСЛОВНЫЕ РЕФЛЕКСЫ 59
Рис. I. Собака со схематическим обозна-
чением расположения кололок на кожной
поверхности 1
Характеризуя собаку, нужно сказать, что она обладала сильной нервной системой
и, насколько можно судить по общему поведению и результатам однократной «сшибки»,
принадлежала к сангвиническому типу (по классификации И. П. Павлова): выработан-
ные днференцировкн устойчиво сохранялись; растормаживание дифсренцирово-к на-
С'и<1л.-1.юе ь и случаях, когда опыты почему-либо ставились из 4—7 часов П0ЭЯ№ ОбЫЧ-
него времени; восстановление происходило очень быстро. Опыты длились 1—I1/,
часа с большим числом подкреплений (8—15, иногда и больше).
Опыты собака перекосила довольно
легко. Во избежание разных недоразу-
мений собаку вне станка кормили только
черным хлебом, который давали ей в оп-
ределенном количестве. Хлеб она полу-
чала 20—30 минут спустя после оконча-
ния опыта, т. е. утром. Этим, может
быть, объясняется, что ничтожное ото-
двигание времени опыта сильно повы-
шало пищевую возбудимость собаки.
В станке собака стояла хорошо и спо-
койно. На внешние раздражения'(проезд
автомобилей, звуки со двора, коло-
кольный звон, гудки) собака, особенно
вначале, реагировала быстро, но они
только немного уменьшали величину ус-
ловного рефлекса и почти никогда не
доводили его до нуля. Шумы же из кори-
дора лаборатории (которые в начале ра-
боты слышались довольно часто, а затем.
после принятия ряда мер. почти прекратились) вызывали у собаки более сильное
уменьшение условного эффекта.
Главное мое внимание было обращено на соотношение положительных и отрица
тельных условных рефлексов. Практически это соотношение я пытался выявить
испытанием на последовательное торможение от диферениировочных раздражителей
и от угасательного торможения, на индукцию и г. д. Опыты пелись так: применялся
какой-нибудь положительный раздражитель один, два, три раза, затем следовал дифе-
реннировочный, после чего опять прежний положительный до конца опыта. Такне
опыты чередовались с опытами, в которых все раздражители были коротко отстав-
лены. Кроме того, в разных опытах место диференцировочного раздражителя в ряду
раздражений менялось, чтобы не образовался условный рефлекс на место и на порядок
раздражений. Для примера привожу опыты, показывающие характер и силу ирра-
диации и концентрации торможения от днференцировкн. Все раздражители действо-
вали изолированно 20 секунд (рис. 2).
В приведенных опытах последовательное торможение ясно выражено;
отчетливо видна также волнообразность движения торможения. Явление
волнообразности торможения отмечалось еще в исследованиях сотрудни-
ков лаборатории И. II. Павлова (Б. А. Коган), а в работах последних лет
оно выявляется все чаще и чаще (О. Л. Виндсльбанд, Н. А. Подкопаев,
Л. А. Андреев, Е. М. Крепе, И. С. Розенталь и др.). И. II. Павлов считает
это явление «результатом встречи, уравновешивания двух противополож-
ных нервных процессов; раздражения и торможения»1 2.
У вышеприведенных авторов в большинстве случаев, получалась одна
волна или же волнообразность выступала при сопоставлении данных нес-
кольких опытов. У меня, как видно из рис. 2, получается больше одной
волны (иногда три), притом в течение одного и того же опыта. Сопостав-
ляя опыты, можно заметить, что продолжительность волны, по кранной
мере в пределах одного раздражителя, почти одинакова (для звонка, на-
пример, 20—21 минута первая волна, 45—50 минут—вторая и т. д.; почти
то же самое получено и с другими раздражителями). Характерно, что
волны постепенно увеличиваются, по амплитуде и по длительности напо-
миная геринговские волны возбуждения сетчатки глаза. Упомяну, что
1 Колодка на плече обозначена в тексте буквой В, кололка на заднем голеностоп-
ном суставе — буквой в.
2 И. П. Павло в, Лекции о работе больших полушарий головного мозга, 1927
•стр. 182.
60
Э А. АСРАТЯН
Ю. П. Фролов и О. А. Виндельбанд тоже * получили волнообразное
угасание положительных раздражителей: иногда они наблюдали рял
убывающих волн. В разных опытах волнообразность появлялась не
только в кожно-механическом анализаторе, но и в других анализато-
рах (звуковой, световой). Интересно, что волнообразность получается
также после угасагельного торможения и даже внешнего торможения,
и эти волны по характеру не отличаются от волн после диференциро
Капля
слюны
6d« мй 3 сел
Рис, 2. Динамика положительных условных рефлексов после
применения диференцнровочных раздражителей
Нижняя диаграмма—метроном 100 ударов n I мииуту, средняя диаграм-
ма—звонок, верхняя диаграмма — колодка А Заштрихованные- столби-
ки-условное слюноотделение за 20 секунд (10 делений по осн ординат—
I капле) Черный столбик с крестом—лиференциговочный раздражитель
(в случае применения звонка н метронома-дифервнцировочнмй метро-
ном, в случае кололок-лкференцнровочна я кололи а) Черный столбик—
нулевой эффект активного раздражения
вочного торможения. В проведенных мной опытах новым являетг
следующее: часто после падения волна поднимается выше исходного
уровня; это хорошовидно в приведенных двух опытах. Указанные повыше
ння, как и воебще вторичные волны, нельзя вполне отождествлять с индук-
цией, хотя весьма вероятно, что во внутренних механизмах обоих
процессов есть много общего. Как и большинство занимающихся изучения
условных рефлексов, я называю индуйцией усиление рефлексов сейчас же
после диференцировочного раздражителя, в то время как эти превышающие
исходную величину волны получались намного позже и после неоднократ
ного падения волны. Это явление нельзя объяснить также внезапным повы-
ЭКСТИРПАЦИЯ ШЕЙНЫХ СИМПАТИЧЕСКИХ УЗЛОВ И УСЛОВНЫЕ РЕФЛЕКСЫ 61
шенпем пищевой Возбудимости, так как общеизвестно, что к концу опыта,
особенно длительного, общая пищевая возбудимость надает. А вместе с тем
повышение пищевой возбудимости наблюдается довольно регулярно и не
можетбыть объяснено случайностью. По аналогии с другими случаями по-
вышения возбудимости, известными в физиологии нервной системы под
названием последовательной экзальтации, я буду называть эту фазу фазой
экзальтации, не предрешая вопроса о внутреннем ее механизме. Данные,
аналогичные моим, можно иногда встретить в протоколах даже ранних работ
по условным рефлексам, но они оставались незамеченными исследова-
телями. Второе явление, которое обратило мое внимание при установле-
нии нормы условных рефлексов на собаке Бек, касается распростране-
ния тормозного процесса по кожному анализатору
Па основании работ ряда сотрудников лаборатории И. П. Павлова воз-
никло представление, что торможение от исходного пункта кожного ана-
лизатора распространяется по всем направлениям с одинаковой быстротой.
У моей собаки получились иные отношения. Как видно из рис. 1, дифэрен-
цнровочная кололка у собаки находилась на бедре, а вперед по туловищу
и вниз по передней конечности, как и вниз по задней конечности, были рас-
положены положительные колодки. В отличие от большинства опытоз дру-
гих авторов, помещав.ших диференцировочную кололку в конце ряда,
положительных колодок, у моей собаки она находилась посредине этого
ряда. Кололки А небыли расположены на равном расстоянии от колол-
кн О (диференцировочной), именно на расстоянии 28 см. При неоднократных
пробах последовательного торможения на эти колодки после применения
кололки О я констатировал, что кололка в тормозится в два-три раза бы-
стрее, чем кололка Л , т. е. что торможение по корковой проекции ко-
нечности продвигается быстрее, чем по проекции вдоль туловища. Должен
сказать, что скорость распространения тормозного процесса у исследован-
ной мной собаки совпадает с данными Б. А. Когана, а именно: 1) скорость
исчислялась минутами (2—12 минут и больше) и 2) во всех случаях
раньше тормозились близкие, а затем более удаленные пункты. То же
сходство наблюдалось и в концентрации торможения, ио и обратном по-
рядке (табл. 1).
Приведенные из протоколов опытов выдержки № 1, 2, 3 относятся
к раннему периоду, а № 4 и 5—к позднему периоду применения днференци-
ровки (кололка О), так что в них последовательное торможение выражено
слабовато (а кололка А даже по механизму положительной индукции уве-
личивает эффект через такой промежуток времени, при котором раньше
не давала индукции,—2*А> минуты). В норме, т. е. в доопсрационном пери-
оде, опытов такого рода с испытанием последовательного торможения от
кололки О на кололки в и А и на кололки В и С было поставлено всего 27.
Это явление везде выступает с той или иной степенью ясности. Обратного
явления ( т. е. чтобы А тормозилось быстрее, чем в) никогда не отмечалось.
Из приведенного примера видно также, что освобождение от торможения
совершается в пункте А быстрее, чем в пункте в. Все это говорит о том
будто кололка А находится от кололки О на гораздо большем расстоянии,
чем кололка а. В пользу моего толкования полученных фактов о разной бы-
строте распространения торможения по кожному анализатору, в зависи-
мости от исходного пункта торможения и от направления движения, гово-
рит и факт другого порядка: кололки В и С тормозятся от кололки О
почти через одинаковый промежуток времени, а именно: В через 8—10 ми-
нут, С через 9—11 минут, несмотря на то, что первая почти в два раза
дальше от кололки О, чем вторая (44 и 74 см).
Таким образом, определенное расположение кололок на кожной по-
верхности (днференцировка в середине положительных и в «углу») и мед-
ленное распространение тормозных процессов у исследованной собаки дали
Таблица 1
Последовательное торможение от диферениировочной колодки О на колодки 4, В
Опыт № 134 Опыт № 157 Отрывки нз опытов
время раздражитель 20 сек. С е- Е а услов- „ ный °®“»нй эффект эффект время раздражите ль 20 сек. 3 • SSg RCE услов- ный эффект общий эффек время раздражитель 20 сек. « 2 sh SaS rj V RCD услов- ный эффект общий эффект
о делениях шкалы в делениях шкалы в делениях шкалы
8 ч. 14 м. 8 » 19 » 8 » 23 р Я » 31 к 8 » 35 » 8 » 41 » 8 » 43%э 8 » 47%» 8 1 53 а 9 » 00 .. » 9 » 08 » 9 » 12 » 9 » 17 » 9 » 22 » 9 > 28%» 9 » 32%» 9 » 36%. 9 > 39 > 9 » 43 » 9 » 47 » Колодка А » в М10О Колодка Л То же Колодка О » А То же > » » » » * » » » » » » » » » » Колодка О » А То же > » 3 3 4 3 3 5-6 5-6 9 6 5 4 5 5 4 6 5 60 40 45 38 39 0 17 18 0 0 8 20 25 33 40 45 0 15 25 0 680 660 600 670 600 0 640 600 600 550 620 600 610 630 650 660 0 650 660 600 8 ч. 32 м. 8 > 42 » 8 » 46% > 8 » 52 » 8 » 55 » 8 ч. 59 м. 9 » 05 » 9 » II > 9 » 20 » 9 » 26 » 9 ч. 32 у. 9 » 38 » 9 » 44 » 9 » 50 » 9 ч. 47 м. 9 » 54 » 9 > 57 » 9 » 10 » Колодка г» » л » в . » . 0 > в То же » » » » » » в » » » » > » » > » № 4 Колодка в . О » в То же 1 3—4 17 8 11 2-3 7 3-4 11 19 0 50 45 0 0 0 0 0 3 25 0 17 60 30 50 0 9 2(?> 670 670 620 0 38* 640 520 620 660 660 540 600 690 600 670 0 670 670 8 ч. 00 и. 8 » 09 » 8 » 1’3 » 8 » 17 » 8 ч. 54 м. 8 > 59 » 9 » 03 » 9 » 04 %» 8 Ч. 54 М. 8 » 59%» 9 » 02%» 9 » Об1/.» 9ч. 43 м. 9 » 47 » 9 » 49%» 9 » 54 %» 9 > 59%» 10 » СО » № 1 Кололка А > 0 » А То же К? 2 Колслка в » О » в » А № 3 Кололка А » О » А То же № 5 Кололка А » О » А То же » > » » 4 7 4 4 11 4 4—5 16 5 — 4 7 б 40 0 17 3 31 0 0 8 36 0 22 7 40 0 47 18 36 0 660 0 630 &J0 570 20»’ 570 600 40* 400 670 670 670 680 670
Раздражение длилось 3 секунды
20 секунд.'Рефлекс
не подкреплялся. **Рефлекс не подкреплялся.
вместо
ЭКСТИРПАЦИЯ ШЕЙНЫХ СИМПАТИЧЕСКИХ УЗЛОВ И УСЛОВНЫЕ. РЕФЛЕКСЫ 63
возможность констатировать, что это движение по длине конечности совер-
шается быстрее, чем по длине туловища. Близкие явления наблюдались
также в опытах Е. А. Шевалева из лаборатории, руководимой В. М. Бех-
теревым. Я думаю, что это можно объяснить характером корковой проекции
кожного анализатора. И на самом деле, пункты в и О находятся почти в
одном сегменте спинного мозга, а пункт А отделяется от О рядом таких
сегментов. Кроме того, в коре по общему принципу проекции пункт в дач-
жен быть гораздо ближе к пункту'О, чем .4, На основе этого принципа
ясный объяснимы такие же случаи с колодками 13 и С.
Данные о возникновении последовательной экзальтации при выработке
условных рефлексов, как и данные о распространении торможения по кож-
ному' анализатору, разумеется, нуждаются в многократной проверке; ио
они у меня получались с такой регулярностью, что я считаю себя вправе
о них говорить. * * *
После того как был накоплен достаточный материал, характеризующий
соотношение тормозных и раздражительных процессов, у собаки Бек, а так-
же средние величины условных эффектов от разных раздражителей (опыт
№ 171), 30. VI 1928 г. под эфирно-хлороформным наркозом проф. Л, А. Ор-
бели был удален у собаки левый gangl. cervicale superius. Через 3 дня
после операции состояние собаки было хорошее, в соотношении и величи-
нах условных рефлексов значительных изменений заметно не было. Но
со следующего дня как общее состояние собаки, так и условные рефлексы
начали резко изменяться вследствие образования гнойника в опера-
ционной ране. Все условные рефлексы исчезли. Постепенно, с 3—4-го
дня после появления абсцесса, рефлексы начали понемногу восстанав-
ливаться. Позже других восстановились рефлексы на колодки. Еще
на 11 -и и 12-й день они были уменьшены. Но в общем к этому времени уста-
новилось прежнее соотношение между отдельными положительными и от-
рицательными агентами. Папученный материал, охватывающий период
от 15.VII до 12.IX (54 опыта), характеризуется так: не удается уловить
изменении в функции слюнной железы, наблюдается некоторое уменьше-
ние абсолютных величии условных рефлексов — усиление последователь-
ного торможения от диференцировочных раздражителен и вообще склон-
ность к быстрому торможению. Постепенно картина приблизилась к нор-
мальной. Конечно, ценность полученного материала пескапько снижается
наличием послеоперационного осложнения. Однако приписать эти поздние
явления быстро протекшему воспалительному процессу едва ли возможно.
12.Х 1928 г. был удален правый верхний шейный узел. Послеоперацион-
ных осложнений на этот раз не было. Уже на следующий день после опера-
ции собака была вполне бодра, операционная рана скоро зажила.
Но зато после этой операции изменения в условных рефлексах были выра-
жены очень резко. Материал, собиравшийся почти в течение двух месяцев
после последней операции (14. IX—7. XI; 48 опытов), позволяет характери-
зовать состояние собаки следующим образом.
1. Уменьшение абсолютных величин условных рефлексов от всех раздра-
жителей. Привожу средние арифметические для трех периодов работы: пер-
вый — до первой операции, второй — после первой операции, третий —
после второй операции; средние арифметические вычислены на основании
тех эффектов, которые получились до применения диференцировочного
условного раздражителя или какого-нибудь экстренного внешнего раз-
дражителя (табл. 2).
Противоречие в отношении света только кажущееся: оно объясняется
тем, что после операции световое раздражение почти всегда приходилось
применять на первом или втором месте, поэтому для него получается увели-
чение Эффекта. Делалось же это потому, что в середине опытного Дня свет
64
Э. А. АСРАТЯН
почти терял свой эффект и применение его угнетало и без того склонное
к быстрому утомлению животное. На самом деле рефлекс на свет пострадал
больше других раздражителен.
Таблица 2
Величина условных рефлексов до н после операции
Пс риоды 3 ВОНОК Метром ом 100 в 1 мин. Свет 50 св. Кололка (среднее ари фметиче скос данных всех КОЛОЛОК)
Первый 73 50 37 42
Второй 67 47 за 36
Третий 50 40 40 30
2. Быстрая остановка слюноотделения после прекращения акта еды.
3. Чрезвычайная реактивность собаки по отношению ко всякого рода
внешним раздражителям и быстрое возникновение и распространение тор-
можения под влиянием как этих раздражителей, так и длинных пауз.
4. Чрезмерное усиление последовательного торможения от диференци-
ровочных раздражителей. Это выражалось: 1) в быстром наступлении тор-
можения (вначале торможение наступало почти мгновенно, лаже отсутство-
вал период положительной индукции, которая хорошо была выражена как
в норме, так и после первой операции); 2) в углублении его-, судя по тому, что
собака часто небрала еды, и 3) в продолжительности его. Сказать, что собака
спала, нельзя, но двигательные реакции доходили до минимума. Собака
иногда в течение десятков минут или часа стояла неподвижно, лишь слегка
реагируя на подачу и взятие кормушки; слюна вытекала только после
взятия корма. Часто этому предшествовала кратковременная сильная дви-
гательная реакция со слюноотделением, будто собака с чем-то боролась.
Хотя я не наблюдал гипнотической фазы у других собак, но на основании
описания ряда авторов думаю, что моя собака часто после диференциро-
вочного агента впадала в такую фазу даже при очень высоких пищевых воз-
буждениях. Намека на это явление ранее совсем не бывало. Для иллюстра-
ции сказанного приведу сравнительные опыты из дооперациоиного периода,
после удаления одного ганглия и после удаления обоих узлов (табл. 3,
4 И 5).
Как видно из приведенных примеров, разница а продолжительности
последовательного торможения от Дифереицировочного раздражителя Ко-
лоссальная. Нужно сказать, что пищевая возбудимость в двух последних
периодах при прочих равных условиях вообщебыла выше; это можно объяс-
нить, во-первых, наступлением осенних холодов и,во-вторых, искусственным
повышением пищевой возбудимости путем некоторого уменьшения порции
хлеба внестанка. Чтобы собака не похудела, я удлинил время подкрепления
от 20 до 25 секунд. Но это повышение пищевой возбудимости но спасло по-
ложения—днференцировка всегда оказывала сильное тормозящее действие.
Волнообразный характер рефлексов сохранился во втором и третьем
периодах, нов слабой! степени (особенно в третьем периоде). Если условные
рефлексы дают сплошь нули, волнообразность можно проследить на ряде
цифр безусловных рефлексов (см. табл. 3, 4 и 5). Ради экономии места я не
буду приводить дальнейшие примеры, но весь имеющийся материал (в част-
ности, приведенные в качестве примера данные) показывает, что
длина волн после операции не изменилась; как будто явления волнооб-
разности возникают с такой же правильностью, как и в норме, но только
при более низком уровне возбудимости. Экзальтация получалась и во вто-
ром периоде, но в третьем ее никогда не приходилось наблюдать в течение
всего опыта.
Таблица 3
Последовательное торможение от диференцировочного метронома до и после операции
Первый периол (опыт № 1 23) Второй период (дпыТ № 209) Третий период (опыт № 247)
время раздражитель 20 сек. латентный Период в сек. ус лов-1 . „ ный °1“ий эффекТ|ЭФФскт в делениях шкалы время раздражитель 20 сек. латентный период в сек. УСЛОВ- НЫЙ Эффект в дел шк общий эффект еннях алы время раздражитель 20 сек. । латентный период в сек. УСЛОВ- НЫЙ дффек к деле общий эффект ниях шкалы
8 ч. 4 м. 8 » 15 » 8 » 22 » 8 » 29% » 8 » 32 > 8 » 36% » 8 > 43 > 8 » 47 » 8 » 53 » 9 » 0 » 9 » 5 » 9 » 9% » 9 » 16 » 9 » 39% » Зв. *"1е» Nte» м.« •М 14» Мюо ‘'Чое Мц* Мщо М100 Ml 00 М100 М1м Ml 00 3-4 3-4 4 11 7 7 10 12 7—8 5 6 5-6 60 50 48 0 12 0 37 30 25 25 28 35 32 35 6.30 620 630 0 590 660 620 610 440 500 610 620 600 8 ч. 52 и. 8 » 59 » 9 * 0В > 9 > 13 » 9 » 20 » 9 » 25 » 9 » 30 э 9 в 33 » 9 » 39 э 9 » 46 > 9 » 51 в 9 » 59 » 10 » 05 » 10 » 17 » 10 » 25 » 10 » 35 в 10 в 45 в 10 в 49% в 10 в 54 э Кололка в Зв. Св. Кололка С То же i и Мцо Мюо М1 оо М1О0 Мют М1.0 М1О0 ‘''1QQ Mjoo Ml он М1.0 Mi об 2 2 6 3 3 3-4 10 6 15 7 4 8 5 3 7 80 115 15 60 50 57 0 15 26 0 0 0 5 28 65 30 50 72 45 690 680 680 680 670 680 0 660 -НО — 40 — <Ю 350 60D 670 620 670 670 670 600 Эм. 05 м-. 9 з 57 » 10 » % » 10 > 07 » 10 » 11 » 10 » 13% » 10 в 18 » 10 3 24% » 10 » 28% » 10 » 33% » 10 , 41% » 10 , 45% » 10 з 50 » 10 » 56% » И » 05 » П » 05% » 11 » 11 > И з 15 » И > 21 » 11 » 28 > Св. Мюв Мюо Мц)» М,о Mio» Мюо M|M -Mino- -Ml 64 М.е. iMI40 М10б •М140 M1Q0 MIOO 2-3 3 6 6-7 18 I 6 60 56 40 40 0 0 4 0 0 0 0 0 0 0 0 0 70 34 0 0 700 700 700 700 0 —5 —46 -И -35 -20 -13 -26 — 6 - 3 —45 18 Я топаю но- гами н громко кричу: »ЕеК| Бек1» 670 670 70 20
Примечание. В графе «общи й эффект» знак минус перед цифрами в этой и других таблицах показывает, что собака еды не берет.
Таблица 4
Последовательное торможение от диференцировочното Метронома до и после операции
Первый период (опыт № 127) Второй период (опыт № IR8) Третий период (опыт № 1 81>
УСЛОВ- общий услов- общий услов- общий
раздражитель 2 ный эффект эффект время раздражитель 3 S а ный эффект эффект время раздражитель 3 ный эффект эффект
20 сек. - 5 х 20 сек. X $ X 20 сек, = S х
ы е. Ь и делениях к- с. * в делениях »- Ь у в делениях
5 V 5 Б « шкалы ~ с « 111 к л ды Е Е s шкал и
8 ч. 19 м. Кололка в * 9ч. 47 ы. Кололка е J — ♦ 9 ч. 05 м. Ml 00 —
8 > 28 » То же 2-3 30 410 9 в 54 > » с - — 9 » 11 » Св. — — —
8 » 31 » Зв. 2 -3 со 520 9 ж 57 > Зв. — — — 9 » 15 » Зв. 6 55 680
8 » 36 » В 2-3 60 580 10 » 02 » Кололка С — 0 50 9 > 19 » — — —
8 » 42 » М.0 — 0 0 10 » 05 » Зв. 3 85 680 9 » 25 » М1«и> 8 22 670
8 » 44% « Зв. 4 40 560 10 » 13 > в 4 80 680 9 » 32 » Зв. 7 47 670
8 » 49 » > 18 6 460 10 » 17 »30с. » 5-4 77 630 9 » 37 » В 7 45 670
8 » 55% э > 2-3 54 570 10 » 21 > > ш— 0 0 9 » 41 » М« — 0 0
8 > 59% » в 2-3 45 520 10 » 21 »20с. М.. 8 55 620 9» 41 >30 с. Зв. 18 5 560
9 » 04% » » 10 16 408 10 » 26 » Зв. 12 10 400 9 » 46 » В — 0 570
У > 10% » в II 11 450 10 » 31 » В — 0 600 9 > 54 » в — 0 13
9 > 17% » » 6-7 2о 500 10 » 37 > в — 0 -20 10 » 00 > в — 0 -20
9 » 21% » » 3 45 520 10 > 43 » — 9 570 10 » 05 » в — 0 • -40
9 » 28 » 2-3 60 520 10 » 49 » » — 0 650 10 > 12 » в — 0 500
У » 31% » в 2-3 60 520 10 » 55 » в 10 18 680 10 > 20 » в — 0 670
II » 01 » в — О 670 10.» 26 » » — 0 670
11 » 07 > 10 7 670 10 » 31 » в 16 13 670
II > 13 > в 3 58 400
* Прочеркнутые места (
) указывают па применение короткою отставления.
Таблица 5
Последовательное торможение от диференцировочной касалкн
Первый период (опыт № 138) Второй период (опыт № 2IO) Третий период (-опыт № 238)
время раздражитель 20 сек. 3 х 2 х ¥ — и <4 U ° Е Е со услов- ный эффект общий эффект время раздражитель 20 сек. в 3 S g . ? с. ЛЗ 4?'”' Е Е а У®лп“‘ [общий **“й 1 эффект эффект[ т г время раздражитель 20 сек. S 3 ® Ч х ® х - с> Е Е В услов- ный эффект общий эффект
в делениях шкалы в делениях шка лы в делениях шкалы
8 ч. 26 М. 8 » 33 > 8 > 39 » 8 » 44 » 8 » 48 » 8 » 52 э 8 » 56 э 9 » 00 » 9 » 04 » 9 » 08 э 9 » 12 » 9 » 16 а 9 » 20 » 9 » 24 » 9 » 28 » 9 » 32 » Кололка С То же > > > э » > > > > > » » » » > > 9 » 9 > » » Ю > » » » > 2-3 4 4 4 4-5 18-19 9 7-8 7—8 7—8 50 35 35 0 30 28 2 0 0 10 0 20 18 12 0 0 750 700 700 0 670 660 620 660 550 580 640 620 620 600 470 540 9 ч. 20 N 9 » 25 » 9 » 29% э 9 » 36 » 9 > 41 » 9 » 46 » 9 » 48% » 9 » 53 » 9 » 58 » 10 » 10 » 10 > 11 > 10 » 19 » 10 » 29 » Св. Кололо С Св. Колодка С То же Колодка 0 » С Го же » » » » » » » » » » 2 2 3-4 4 3 4 4 4 10 6-7 90 75 53 42 40 0 42 40 0 0 15 0 32 700 690 680 670 670 0 670 630 660 650 680 500 670 8 ч. 12 м. 8*19» б » 24 » 8 » 28 » 8 » 36 » 8 » 40 » 8 » 46 » 8 » 51 » 8 » 57 » 9 » 04 » Mie. Ко.’олка С » 0 » С То же » » » » » » » * > > 3 6 66 35 0 0 0 0 0 0 0 32 680 670 0 -40 -10 - 0 - 0 300 540 Вошли в коридор 600
63
Э. А. АСРАТЯН
Укрепление и усиление торможения от диференцировочных раздра-
жителей проявлялось еще в следующем: в норме, когда опыт ставился на
1—2 часа позже обычного или когда собака один день голодала (получая
мало хлеба или совсем не получая его), диференциров.кн, которые были
очень прочны, почти всегда растормаживались. Во втором периоде они
только ослабевали и не давали отчетливого последовательного торможения.
В третьем же периоде диференцировкн сохранялись полностью (иногда
только после прекращения раздражения наблюдалось незначительное слю-
ноотделение), а последовательное торможение выступало отчетливо, хотя
и менее сильно, чем свойственно данному периоду. Во всех трех перио-
дах работы присутствие посторонних лиц на опыте нарушало его при-
вычный ход. В указанных случаях обычно наступало ослабление диферел-
цировок, последовательного торможения от него, а также положительных
условных рефлексов, часто до полного их исчезновения. Это прямо
указывает на наличие внешнего, постороннего раздражителя, хотя, не-
сомненно, при опозданиях' время такл^е играло известную роль. Такне
факты не новы в практике изучения условных рефлексов. И. II. Павлов и
его многочисленные сотрудники часто упоминают об этом, особенно в ран-
них работах, проведенных еще в примитивных условиях эксперимента;
у меня же обстановка работы, особенно изоляция экспериментального жи-
вотного, была именно примитивной. Не могла не сказаться при этом
и суетливость собаки.
5. Быстрое торможение от ничтожных, иногда неуловимых внешних
причин, от длинных пауз и т. д.
6. Удлинение латентного периода слюноотделения.
7. Ускорение угашения положительных раздражителей и усиление по-
следовательного торможения от них (угашение как с подкреплением, так
и без подкрепления). К сожалению, в норме я специально не угашал поло-
жительных раздражителей без подкрепления, но угашение расторможен-
ных диференцировок иногда вызывалось с трудом (послеб—7-кратного при-
менения с одноминутной паузой). Сравнительные опыты из второго и треть-
его периодов дают четкое представление об этих изменениях (табл. 6),
Условный раздражитель не подкрепляется до 10-го применения, после
чего он подкрепляется.
Таблица 6
Упипение и восстановление рефлекса на М1ЛП
Время в мин. 0 2‘/. 5 10 12*/. 15 П7> Wh •• 37 43 49 55 61 67
Опыт № IS9 Опы т № 270 45 65 25 16 13 0 о 60 0 0 0 0 0 0 0 со 0 0 50 7 15 о 60 0 63 50 33 65 25 35 65 50
На угашение с подкреплением имеются опыта как из двух последних
периодов (метроном), так и из всех трех периодов (звонок). Приводимые
ниже данные представляют характерные примеры (табл. 7).
Каких-нибудь резких изменений в описанной картине третьего периода
с течением времени не наблюдалось. Некоторое усиление условных эффек-
тов в последнее время совпадает с применением мной нового приема для ис-
кусственного повышения возбудимости; удлинение срока еды до 25 секунд
вместо 20. Является ли это единственной причиной или произошли еще
какие-то изменения в условных рефлексах, не выяснено. Во всяком случае
сильное последовательное торможение диференцировкн и угашения, каки
некоторые другие признаки, почти с прежней силой наблюдались до пос-
ледних дней опыта. Как будет дальше, сказать трудно.
Для дополнения картины изменения условнорефлекторной деятель-
ности у собаки после удаления симпатических узлов привожу кривые
(рнс. 3) и сводные табл. 8 и 9.
Таблица 7
Пер в в ре.и я 8 ч. 25 и. 8 » .58 » 9 » 07 » 9 » 12 » 9 » 18% » 9 » 25 » 9 » 33 » 9 ». 39 » 9 » 49 » 9 * 56 » 10 > 01 > 10 » 05 » 10 > 12 > Вюрой 8 ч. 18 м. 8 » 20 » 8 » 22 * ый период (on раздражитель ат № И « 3 н Ч . 61) услов- ный эффект общий эффект Втор время 'гашение рефл 9 ч. 09 и. 9 » 16 » 9 > 21 > 9 > 29 > 9 > 35 » 9 » 42 » 9 » 47 » 9 > 55 » 10 » 01 » 10 » 08 > 10 » 13 » 10 > 21 » 10 » 27 » 10 > 34 » гашение н вое Втор 8 ч. 54 м. 9 » 00 » 9 » 03 » ой период (on раздражитель нт № 2< « * = И *2 и v ок с п 2-3 3 4 6 10-1! ? 16 10 11 10—9 7-6 3 рефлеи ишт .N 2-3 3 4 )б) услов- ный эффект общий эфф ект Тре время 8 ч. 37 м. 8 > 44 » 8 » - 49 > 8 » 57 » 9 » 03 > 9 > 10 > 9 » 15 » 9 » 23 » 9 » 29 » 9 * 34 > 9 » 39 » Четверт 8 ч. 18 м. 8 » 24 » 8 » 27 » тий период (опыт № раздражит елг £ 7П !у «ЛО1Г- | мый эффект общий эффект
20 сек. Зв- » > » > > > » > > > > период (опыт Зв. > * = ° X Й&5 5ES 2 2 2 3 3 4 4 4 4-5 5-6 3-4 6 7 № 21 2 4 5 п дел шк I 115 1 115 115 95 75 50 50 • 55 50 SO 30 40 20 8) 100 67 50 сниях алы 690 690 680 670 680 670 670 670 660 670 670 500 300 У 190— 100- 80- 20 сек. екса на звон За. » » » » » » > » » > » » » становление ОЙ период (( Зв » » в дел шк од креп. 90 83 60 50 24 60 0 0 0 8 20 40 45 75 100 са на » 702) 100 90 65 ениях алы пением 700 700 680 6*0 640 200 -60 -70 670 680 690 680 690 690 звонок 6ВД 160 ПО 20 сек. 3: > » > э э ый период (о Эй т > 1 2 х г Ч с ® : 7 7 10 12 13-14 12 пыт .№ 3 3-4 7 В дел шк 80 60 57 50 20 30 10 0 0 30 0 271) 76 70 35 СМИ ях алы 680 680 670 670 -60 -130 450 -90 100 650 70 670 170- 100—
Продолжение табл. 7
Второ» период (опыт № 218) Второй Период (опыт К; 702) MeyaepThin период Сопит № 271)
времл раздражитель 20 сек. латентный период в сек. услов- ный эффект в де; IUK общий эффект 1енмях алы время раздражитель 20 сек. латентный период в сек. услов- ный эффект в дел ШК общий эффект ениях алы время раздражитель 20 сек. латентный период в сек. углов- общий ный | эффект эффект! п делениях шкалы
8 ч. 24 м. Зв. 10-11 10 25 - 9 ч. 06 м. Зв. 4 35 100 8 ч. 30 м. Зв. 10 18 50 -
8 » 26 » » — 0 30 9 » 09 » 9 8 12 45 8 » 33 » > — 0 45-
8 » 28 » > — 0 13- 5» 12 > » — 0 16 8 » 36 » » — 0 20-
8 » 30 » > — 0 б(О 9 » 15 » > • — 0 10 8 > 39 » » — 0 30-
8 > 35 > э 9—10 25 60Q 9 1» 18 » — 6 15 8 » 42 > > — 0 10—
8 » 40 > » 8-9 35 650 +9 » 21 » > — 0 680 4-8 » 45 » » — 0 670
8 » 45 » > 5 60 660 9 » 26 » > 7 30 680 8 » 50 » » 10 22 670
8 »' 50 » > 11 15 600 9 » 32 » » 14 30 680 8 » 56 » > 8 50 690
8 » 55 » » 15 8 600 9 » 37 » > 8 46 G80 9 * 01 » > 10 40 670
9 > 00 > » 8 40 620 9 » 42 » 6 55 680 9 » 06 » 9 13 20 670
9 » 05 » » 10 31 600 9 » 47 > » 18 7 670 9 » 11 » > 10 15 670
9 » 10 » > 8 50 660 9 » 53 » » 18 9 670 9 » 17 » » 13 10 670
9 9 9 9 9 9 9 9 » 15 » » 20 * . 25 » в 30 » » 35 » » 40 » » 50 » » 55 » > > » » » » > 7-8 11 10-11 7 15 11 & 30 18 05 35 60 14 23 65 630 480 500 610 640 530 600 670 9 » 59 » » » 4 60 400 9 > 23 » 9 » 28 » 9 » 34 » » » > — 0 0 0 670
П римечанне. Минус (—) в койне цифры означает отсутствие подкрепления; в начале цифры животное не берет еды,
п-яюс » Грефа <ор*мя» — црокрашннис ymiuriiHH.
ЭКСТИРПАЦИЯ ШЕЙНЫХ СИМПАТИЧЕСКИХ УЗЛОВ И УСЛОВН ЫЕ РЕФЛЕКСЫ 7|
Таблица 8
Испытание разных положительных раздражителей на последовательное торможение
от диференцнровочных раздражителей
\ Периоды Разлра- 'х жите ли х Первый Второй Третий
1 Звонок 9 раз. Всегда отчеь ливое последовательное торможение Никогда иб тормозится до нуля 6 раз. Сравнитель- но сильное торможе- ние 3 раза (с рядом нулей). 3 раза почти такое, как в первом периоде. Последние опыты ставились в необычно поздние ча- сы 11 рая. Несравнен- но более сильное тор- можение, чем в пер- вом и в юром перио- дах; ссегда ряд ну- лей. Только один раз получено ослаб- ленное действие, и то в присутствии по- стороннего лица
М1»|> С раз. Всегда имеется торможение. Только один раз торможение доводит рефлекс до нуля 3 ра за. Сравнитель- но большое торможе- ние. Всегда ряды нулей 3 раза. Всегда очень Дли иные ряды нулей. Очень глубо- кое и длительное торможение
Свет 50 св. Не был испытан 1 раз. Отчетливое торможение до пуля I раз. Несравненно более сильное тормо- жение. Длинные ря- ды нулей
Колодка Л 7 раз. Всегда имеется отчетливое торможение, доходящее до нуля. Полное восстановлен-ие Зраза. Торможение всегда усиленное и углубленное; длин- ные ряды нулей. Частичное восстанов- ление 2 раза. Весьма уси- ленное торможение, длительное и углуб- ленное. Рефлексы ие восстанавливаются
Кололка В 6 раз. Отчетливое, сильное торможение, Длинные ряды нулей. Восстановление Не испытано 1 раз. Чрезвычай- ное усиление, углуб- ление торможения. Рефлексы не восста- навливаются за все время опытного сеан- са
Кололка в 1 раз. Очень сильное торможение, но восста- новление наступает, хо- тя позже. Опыт был по- ставлен после длитель- ного (14 дней) перерыва в работе То же Не испытано
Кололка С 2 раза. Отчетливое торможение, нули; вос- становлен ие 1 раз. Сравнитель- но быстро наступаю- щее торможение 1 раз. Очень силь- ное глубокое тормо- жение. Нормальные отношения не восста- навливаются во все время опыта
На рис. 3 (стр. 74) и в табл. 9 приводятся среднеарифметические величи-
ны, отображающие последовательное торможение от деференцировочиого
раздражителя на все условные раздражители во все периоды работы;
ио отношению к исходному они исчислены в процентах. Время считается от
момента применения диференцировочного раздражителя.
Считаю важным отмстить, что в поведении собаки я не заметил каких-
нибудь существенных изменений после обеих операций (если не считать
периода образования абсцесса у собаки после первой операции).
Т а б л и u а 9
Последовательное торможение от днференцировочны* условных раздражителей
\ Время Разлра- ЖИТС.'1Ь 10’ 20’ 20’ V 2’ 3' 4' 5' в' т 8.' В’ 10‘ 11' 12' 13' 14' 15’ 16’ 17’ 18’ 19’ 20’ 21' 22' 23' 24’ 25' 26* 27' »• 30' зг 32’
Звонок 116 56 41 34 77 —— —— 78 78 . 75 — 13 27 79 72 16 — м
> — 93 — — 0 -— 69 6 — 0 - - 13 — 0 — 0 14 — — — 8‘/г — 21 —— — —— —- 13 — — — —
» 0 25 11 — 35' 30 0 0 60 — — 33 —- 0 — 17 — 0 0 — — 0 0 — — 0 0 а 18 — — 0 0 —
Мио — . 24 132 55 0 22 78 74 67 43 __ —- 42 22 IW — 54 .— — 50 0 — 23
М1м 91 — — — — 26 9 — — — 46 -— 0 — — — — 0 0 0 — — 0 —- 0 — 66 — •— — 14 —1 — 10
м„, — — — — 0 0 — 22 — 10 — — 3 -— — 0 0 0 0 — — г 0 0 — — 0 — — 0 — 0 — —
Свет
» 40 41 41 0 6
> 40 50 0 0 0
Кололка Л ал __ 81 86 42 47 36 8 —— — 46 а— — ш 0 0 0 — -— — 0 — — 2’1 28 — 12 51
То же — — — — 0 —— 0 0 0 — —— 0 — 0 — 0 — 0 0 — — 0 — — 0 — — 0 43 0 —
ж » — — — — 123 44 — — — 37 .— — — — 0 0 — — 0 — — 0 — — — — 0 0 —а* — с 0
Кололка в — 0 и 0 0 0 7 .— - —— 0 0 — — —— — 20 0 — — — 0 0 — — .— — —
Го же 0 °
э » 0 0
Кололка В То же 60 — 26 1 0 — 0 0 0 0
» »
Кололка С . ___ 112 86 — 80 8 —— 9 0 — 0 0 — — 51 0 — — 60
То же 105 100 0 0 37 0
» » 0 0 0 0 0
Продолжение табл 9
В ремя раздра» х житель 33' 34' 35’ 36' 37" 39' ЗВ' 40’ 4Г 42’ 43' 44' 45' 46' 47' 48' 49' 50' 5Г •52' 53' 54' 55' 56' 57' 58 59' 61' 62' 63’ 65'
Звонок 37 41 __ 78 78 71 102 100 — 100 —- 77 43 __ —
> — 21 0 0 9 75
> . 0 — — — — и 0 —- — — с 11 29 — — — — 16 — — — — — — 16 0 — •— 0 —- —
М 100 — 28 — — — — 85 — 60 64 60 — 70 — Б0 — — 20 — — — — " —
м100 — — 4 — — — 0 —- 35 — — —— -— 22 — — — — — 0 7 — — — — 76 — 38 — — 113
М100 — 0 0 — ’— — 0 0 — — — 1 — — 0 — — •— — — ° — — — — — — —' — —- --
Свет
> — — — — 131 — — 102 — —— — — — 70 — — — 62 — — — — — 0 — — — — — — 0
> — — —* — 40 — — — — 0 •— г— — — — 0 — — — 0 — — — — — 0 — — — — — — —
Колол ка А — — 48 - -- - 81 140 102 141 .— 52 — .— —- 18 —
Та же — 0 —— Зв 7 м-1 13 0 25 — — — — - 34 ш— •W— •— — 51 — — — 0
» » 0 0 0 0
Кололка в — —5 0 __ . 55 15 40 40 50 135 — —- I
То же
> > — 0 0 0 0 0
Колол ка Н То же 0 0 0 0 0 0 30
> »
Колол ка С То же 51 34 80 0 0
» » 0
Примечание При каждом раздражителе 1-й ряд цифр—норма, 2-й ряд — период между первой и второй операцией,
3-й ряд—после второй операции.
74
Э. А. АСРАТЯН
Но однажды технический сотрудник лаборатории Мещеряков, ухажи-
вающий за собаками, сообщил, что Бек после второй операции стал
Рис. 3. Последовательное торможение от диференцнровочных раз-
дражителей до н после операции
I—ка кололку С от кололки О; 11—на свет от метронома 60; I И—на метро-
ном 100 от метронома СО; lY-на э»омок от м^тррнрмч 00; V—на кололку Д
от кололки О.
----- норма,---------после первой операции, — после второл
операции; по осн ординат отложена величина условного рефлекса в про-
центах к исходной величине, по оси абсцисс — время
очень спокойным, долго спит, почти все время лежит в ящике. И на станке
Бек стоял как будто спокойнее. Q состояниях, напоминающих гипнотиче-
скую фазу, я уже упоминал. Интересно, что сильные посторонние раздра-
жители моментально выводили Собаку из такого состояния; собака
ЭКСТИРПАЦИЯ ШЕЙНЫХ СИМПАТИЧЕСКИХ УЗЛОВ И УСЛОВНЫЕ РЕФЛЕКСЫ 7$
становилась бодрее, у нее появились хорошие условные рефлексы, она
охотно ела; новее это длилось очень короткое время, и затем собака опять
впадала в прежнее состояние.
Здоровье собаки нарушалось в течение всего периода исследования
три раза: до операции однажды наблюдалось припухание слизистой рта
и покраснение правого глаза; после первой операции развилось осложне-
ние (нагноение); после второй операции, в последние дни, возникла легкая
форма диспепсии, длившаяся один день. Все эти отклонения, кроме нагное-
ния, были кратковременными и особенно не отражались на ее условие-
рефлектором деятельности. Вес собаки колебался между 18,В и 20 кг.
В период диспепсии вес упал до 17,6 кг, но это, пожалуй, было вызвано
тем, что собаке ничего не давали есть. Все же после второй операции вес
приближался к низким цифрам (18,8 кг).
Заканчивая статью, считаю нужным отметить следующее: вследствие
моей малоопытности, примитивной обстановки работы, природной суетли-
вости собаки (на все реагирующей) и ряда других неблагоприятных моментов
технического характера, фон моих опытов получился довольно пестрым. Но
описанные выше изменения в условных рефлексах под влиянием экстирпа-
ции обоих верхних шейных симпатических узлов настолько значительны,
что даже на таком фоне они видны ярко и отчетливо. В дальнейших экс-
периментах надлежит выяснить долю влияния самой операции и наркоза
на эти изменения. Все это и главное чрезвычайная важность н сложность
вопроса заставляют меня воздерживаться от каких бы то ни было объясне-
ний механизма действия и от преждевременных выводов. Однако можно
утверждать, что основное изменение (усиление последовательного торможе-
ния) идет вразрез с общеизвестной динамикой развития этого явления
у нормальных собак. В норме последовательное торможение от дифе-
ренцировки и угашения со временем постепенно уменьшается, а иногда
совсем исчезает, что ясно наблюдалось и у моей собаки до операции (см.
табл. 1). Как мы видели, после операции процессы протекали необычно—
в направлении усиления последовательного торможения, хотя после обеих
операций (особенно после первой) вслед за скачкообразным усилением
наблюдалось некоторое постепенное ослабление и здесь.
Выводы
1. Волнообразный характер рефлексов, представляя собой результат
столкновения процессов возбуждения и торможения, является правилом
для течения этих процессов в коре больших полушарии. Часто эти волны
имеют одинаковую длительность.
2. На подъемах волн, особенно последних, можно уловить фазу воз-
будимости, превосходящей исходную. Я предлагаю назвать ее фазой эк-
зальтации.
3. У собак с очень медленным течением тормозного процесса можно
установить, что торможение распространяется по кожному анализатору
от исходного пункта с неравномерной быстротой: по корковой проекции
длины туловища медленнее, чем по корковой проекции длины конечно-
стей. Для концентрации торможения наблюдаются обратные отношения.
4. Экстирпация обоих верхних шейных симпатических узлов у собаки
сангвинического типа повлекла за собой резкое усиление тормозных про-
цессов, что проявилось в усилении последовательного торможения от
диферепцировок и угашения, уменьшении положительных условных реф-
лексов, склонности собаки вообще быстро переходить в состояние глубокого
торможения в-плоть до отказа от еды и до развития гипнотической фазы.
5. После экстирпации верхних шейных симпатических узлов явление
волнообразности течения мозговых процессов осталось неизменным; эк-
зальтацию же удалось наблюдать только после удаления одного узла.
76
3 А АСРАТЯН
ЛИТЕРАТУРА
ВиндельбанлО А и Фролов Ю П, Особый вид угасания искусственного
условного рефлекса, Архив биологических наук, т XXV, в 4—5, 1925
Коган Б А , Об иррадиации и концентрации угасательного тооможения, дисс ,
Спб, 1911
О рбел и Л А , Вегетативная нервная система, Большая советская энциклопедия,
Т 4, 1928
П а в л о в И П , Двадцатилетний опыт объективного изучения высшей нервной
деятельности (поведения) животных, 1925
П а в л о в И П , Лекции о работе больших полушарий головного мозга, 1927
Шевелен Е А,О процессе диференцирования сочетательно двигательного рефлек-
са на тактильное раздражение к о сегментарном типе распространения рефлектор-
ной возбудимости, дисс , Спб . 1913
ВЛИЯНИЕ ОДНОВРЕМЕННОЙ ПЕРЕРЕЗКИ ОБОИХ ШЕЙНЫХ
СИМПАТИЧЕСКИХ НЕРВОВ НА ПИЩЕВЫЕ УСЛОВНЫЕ
РЕФЛЕКСЫ У СОБАКИ1
исследованиях акад. Л. А. Орбели. посвященных вегетативной нерв-
ной системе, имеется раздел об адаптационном влиянии симпатиче-
ской нервной системы на деятельность центральной нервной системы.
Еще н старой литературе имелись отдельные указания на суще-
ствование сосудодвигательных нервов симпатического происхожде-
ния, иннервирую них сосуды оболочек центральной нервной системы.
Так, Валентин (Valentine) показал, что раздражение задних кореш-
ков вызывает сужение сосудов твердой мозговой оболочки спинного мозга. Гаскелл
(Gaskell), прослеживая внедряющиеся в спинной канал серые ветви, нашел, что они
заканчиваются только В оболочках спинного мозга, не проникая в самый мозг. Со-
здается впечатление, что Гаскелл не приписывает этим волокнам никакой другой
функции, кроме вазомоторной.
Совершенно аналогичные взгляды высказывались о роли тех симпатических во-
локон, которые в сопровождении мозговых сосудов проникают в черепную коробку
В литературе существуют указания, что эти вазомоторные нервы в некоторых отно-
шениях отличаются от других вазомоторных нервов. По данным Гертнера и Вагнера
(<j<irtner и Wagner), Бидля и Рингера (Biedt н Ringer), Виховского (WiechoaraHi)
и др., вазомоторы головы не обладают сосудотонизирующим влиянием. После их пе-
ререзки сосуды головы не расширяются; раздражение же этих нервов вызывает су-
жение сосудистого русла головного мозга. Имеются, однако, данные, говорящие
о том, что часто раздражение шейного симпатического нерва не вызывает каких-
либо вазомоторных изменений в сосудах головного мозга. Вебер (Weber), Хилл (Hill),
Рой и Шеррингтон (Roy и Sherrinclon) на основании своих экспериментов пришли
к выводу, что головной мозг вообще не имеет вазомоторов.
В литературе по рассматривав му вопросу не встречается, однако, даже намеков
па какие-нибудь другие функции симпатических волокон, внедряющихся в головной
мозг.
Необходимо отметить, что экстирпацией шейных симпатических волокон и пере-
резкой шейных симпатических нервов занимались еще со времени Пурфур-дю-Г1ти
(PouHour du Petit, 1727) многие исследователи. Я в доступной литературе обнаружил
имеющие отношение к изучаемому мною вопросу данные, или не подчеркивавшиеся
специально авторами, или являвшиеся недооцененными в протокольном материале.
Все указания, которые мне удалось найти, сводятся к следующему.
1. Экстирпация обоих верхних шейных симпатических узлов у высших животных
вызывает общую худобу, сопровождающуюся инфильтрацией членов, появлением
особого рода коросты, которая под конец поражает всю поверхность кожи [Дюпюи
(Dupuy)].
2. Экстирпация этих ганглиев часто вызывает смерть животного [Клод Бернар
(Cl. Bernard)].
3. Экстирпация: этих ганглиев у кошек и кроликов вызывает гиперестезию кожи
соответствующей стороны лица и уха.
4. Сюда относятся, наконец, опыты Клода Бернара, в которых повышение тем-
пературы самого мозгового вещества наступало после экстирпации симпатических
ганглиев или перереэкк симпатических нервов на шее. Следует отметить, что сам автор
объяснял увеличенную теплопродукцию, наступающую как в наружных органах
головы, так и внутри черепной коробки, не только вазомоторными изменениями в
этих органах; он даже не приписывал им главенствующей роли.
Работами А. В. Тонких, К. И. Кунстыан, А. Н. Крестовин кова и В. В. Савича было
1 Физиологический журнал СССР, т. XVIII. № 5, 1935. Предварительное сооб-
щение напечатано в «Докладах Академии наук СССР», № 9. 1931, по представлению
акад. И. П. Павлова 20. И 1934 г.
78 9. А. АСРАТЯН
установлено, что симпатическая нервная система оказывает адаптационное влияние
на спинальные и бульбарные центры, что экстирпация или раздражение симпатиче-
ских нервных образований существенно изменяет (функциональное состояние и работу
названных частей центральной нервной системы.
В 1927 г., йо предложению акад. Л. А. Орбелн и под его непосредственным руковод-
ством, я выполнил работу, которая выявила, что симпатическая нервная система ока-
зывает значительное влияние на условнорефлекторную деятельность собаки. В этой
же лаборатории один из учеников Л. А. Орбели — В. С. Стрельцов показал, что сим-
патическая нервная система оказывает влияние даже на thalamus, т. е. иа область,
которая многими считается высшим центром самой симпатической системы.
В 1928—'1930 гг. в Эривани я и ряд товарищей занялись вопросом о влиянии верх-
них шейных симпатических узлов на функцию больших полушарий головного мозга.
По моей просьбе, А. М. Алексанян и Д. Г. Гзгзян повторили на двух собаках выпол-
ненную мной в Ленинграде работу по изучению влияния экстирпации верхних
шейных узлов иа условнорефлекторную деятельность. Единственная вариация, вве-
денная нами в эту работу, заключалась в том, что перед иссечением ганглиев соба-
ки подвергались контрольной операции, что дало возможность выяснить влияние
самой операции. Эксперименты выявили, что контрольная операция не оказывает
какого-либо длительного существенного влияния на условнорефлекторную Деятель-
ность собак, тогда как двухыоментнаЯ экстирпация сначала одного, затем, после
некоторого перерыва, другого верхнего шейного симпатического узла вызвала серь-
езные и длительные изменения этой деятельности: у собак наступило значительное
ослабление условнорефлекторной деятельности, причем положительные условные реф-
лексы пострадали сильнее отрицательных, что дало явный перевес тормозным процес-
сам в работе больших полушарий головного мозга.
В Эриваии в те же годы я ’занялся изучением последствий экстирпации верхних
шейных симпатических ганглиев у щенят, интересуясь, в частности, поведением по-
следних. Полученные предварительные данные на 8 щенятах 2—3-месячного возраста
позволяют с некоторой осторожностью сказать, что; 1) одновременная экстирпация обо-
их ганглиев связана с риском для жизни щенят; 2) если экстирпация второго ганглия
делается через небольшой промежуток после экстирпации первого ганглия, то это тоже
связано с опасностью для жизни; 3) если же время между двумя операциями увеличи-
вается до 1—P/s месяцев, то щенки большей мастью выживают. Сравнение этих щен ят
с кооперированными или же со щенятами из того же помета, подвергшимися контроль-
ным операциям, показало с большой очевидностью, что первые являются вялыми,
мало подвижными, менее игривыми, больше лежат (спят) и как будто даже немного
отстают в росте.
Эти данные нуждаются в точной проверке, чем мы и займемся в ближайшее время;
однако О них Следует хотя бы ВСКОЛЬЗЬ упомянуть.
Все это не позволяет сомневаться в изменении работы головного мозга
под влиянием экстирпации шейных симпатических ганглиев. Но возникает
вопрос, как понять интимную сторону действия экстирпации ганглиев на
условнорефлекторну ю деятельность.
Для ответа на поставленный вопрос расчленим его условно на две части:
1) каково значение экстирпации самих ганглиев и перерыва путей для цен-
тральных симпатических импульсов к голове—перерыва, который неизбеж-
но обусловливается самой экстирпацией;. 2) каково ближайшее влияние
этой экстирпации (или перерезки): обусловливаются ли изменения сосудо-
двигательными эффектами или же они являются результатом наруше-
ния тонкого баланса сложной инкреторной системы, или, наконец, они обу-
словливаются прямым адаптационным влиянием симпатической нервной
системы на головной мозг и на рецепторы, или какой-нибудь другой причи-
ной? ЕСЛИ даже сделать весьма вероятное гипотетическое допущение, что
эти изменения являются результатом комплексного влияния всех пере-
численных моментов, все же имеет немаловажное значение хотя бы при-
близительно выяснить значение и место каждого ИЗ этих моментов.
Я в первую очередь занялся вопросами первой группы: выяснением
значения экстирпации ганглиев И Перерыва путей.
Составленный мной план работы был следующий. У возможно всесторон-
не изученной собаки в первую о-чередь пе’ререзать шейные симпатические
пути до вступления нх в верхние узлы, выяснить влияние этой операции,
а затем только во вторую очередь экстирпировать узлы и выяснить последст-
вия этой экстирпации. Одновременная перерезка обоих шейных симпатиче-
ПЕРЕРЕЗКА СИМПАТИЧЕСКИХ НЕРВОВ И ПИЩЕВЫЕ УСЛОВНЫЕ РЕФЛЕКСЫ 79
сник нервов тотчас же после (каудальнее) нижних симпатических узлов была
произведена Л. А. Орбели 15.IV 1932 г. у собаки Арч, которая почти два
года до того использовалась в опытах с условными слюноотделительными
рефлексами.
Прежде чем перейти к описанию полученных результатов, я позволю себе
привести некоторые данные о собаке и о проведенной до операции работе.
Арч, дворняжка, была прекрасным объектом для работы по условным рефлексам;
это собака среднего возраста, с сильной нервной системой, уравновешенная, санг-
виник. Как положительные, так и отрицательные рефлексы у нее хорошо выявлялись
и были стойкими. Совершенно бесперебойно она работала у меня в течение двух лет.
Несмотря на имевшиеся данные, незадолго до операции был проведен ряд специаль-
ных опытов-проб, чтобы можно было более обстоятельно судить об изменениях после
операции. Угашались и восстанавливались различные положительные пищевые услов-
ные рефлексы, испытывалось последовательное Торможение на положительные услов-
ные рефлексы от угасательного и днференнировочнога торможения, влияние сильного
экстрараздражителя (на собаку во время опыта набрасывалась кошка),перенапряжение
торможения (продолжительное действие дифереицировочного раздражителя — в те-
чение 5 минут), «сшибка» между положительными и отрицательными рефлексами.
Специально исследовались индукционные взаимоотношения между центрами услов-
ных и безусловных рефлексов, была установлена величина безусловного рефлекса
на стандартный по составу и величине безусловный раздражитель (мясо-сухарный по-
рошок). Кроме того, было исследовано влияние питуитрина.
Ради экономии места я об этих опытах подробно говорить не буду; они
будут разобраны вместе с аналогичными опытами, производившимися после
соответствующих операций.
Результаты перерезки шейных симпатических нервов
Изменения уСловнорефлекторНой деятельности Собаки после одновре-
менной перерезки обоих шейных симпатических нервов были глубокими
и длительными; они во всех отношениях были сильнее, чем у находившихся
в опытах ленинградских и эриваискнх собак после двух моментной экстир-
пации верхних шейных симпатических узлов.
Начиная со 2-го дня после операции и в течение 15 следующих дней
у собаки были рвота и кашель; собака ходила вяло, ела не особенно охотно,
не было характерного сильного стремления к камере; она почти не лаяла,
нейтрала, была апатичной, малоподвижной. Частота сердцебиений немного
увеличилась (100—130 в 1 минуту), а дыхание в первые дни было за-
медлено (10—12 в 1 минуту). Эти явления были особенна сильно выражены
в первые дни после операции, далее они постепенно проходили. Через
15 —20 дней рвота почти Совершенно исчезла, а кашель заметно ослабел. Со-
бака становилась все бодрее и живее, иногда лаяла и играла, стала охотно
входить в камеру и жадно ела, постепенно прибавляла в весе и почти до-
стигла дооперационного веса. В дальнейшем картина быстро улучшалась,
внешне собака хорошо поправилась. Однако, несмотря на это, через
40 дней после операции собака все-таки не достигла еще присущей ей до
операции живости, игривости и изредка кашляла.
Что касается условнорефлекторной деятельностей, то здесь изменения
оказались очень глубокими. Положительные условные рефлексы почти пол-
ностью отсутствовали в течение первых 15 дней. Правда, на 11—12-й день
после операции они появились в слабом виде, но затем быстро исчезли,
причем на 11-й день рефлекс получился только на первом месте, на 12-й
день — только на первом, втором и третьем месте. Несмотря на заметное
улучшение состояния собаки, наличие хорошего аппетита и сильного пище-
вого безусловного рефлекса, условные положительные рефлексы не появи-
лись и в течение последующих 5 дней.
80
Э. А. АСРАТЯН
После этого срока они стали постепенно появляться только „а первых
местах; на следующих местах их опять не было. Постепенно условные реф-
лексы появились также на втором, третьем, иногда даже на четвертом месте,
причем все они были очень слабо выражены. Даже эта слабая условнореф-
лекторная деятельность была нерегулярной — она то появлялась, тс
исчезала. Это явление наблюдалось в течение 40 дней после операции,
когда общее состояние собаки было вполне удовлетворительным. Данные,
приведенные в табл. 1, иллюстрируют сказанное.
В одном из опытов (5. V 1932 г.) при привычном 20-секу одном действии
условных раздражителей условные рефлексы совершенно исчезли, при
40-секундном действии условных раздражителен условные рефлексы в
слабом виде появились после больших латентных периодов.
Ясно отмечено, что условные рефлексы на слабые раздражители постра-
дали меньше, чем рефлексы на сильные раздражители; иногда получалась
парадоксальная фаза. Следует заметить, что наблюдавшееся у Арча раньше
промежуточное слюноотделение (особенно и начале опыта) после операции
совершенно прекратилось. Это относится к слюноотделительному ком-
поненту положительных условных ре<]>лексов. Двигательный же компонент
этих рефлексов поя вился сравнительно быстро — на 8—10-й день, быстро
усилился и укрепился в дальнейшей работе. В этом периоде опыта диссо-
циация между обоими компонентами условных рефлексов была очень ясна.
В упомянутом периоде отрицательные условные рефлексы на диферен-
цнровочный раздражитель (стук 60) оставались стойкими и совершенно не
растормаживались ни в слюноотделительном, ни в двигательном компонен-
тах. Судя по движению, они как будто даже усилились.
Отдых и дача небольших доз брома (1,5 г в день) не оказали сущест-
венного влияния, хотя создалось впечатление небольшого сдвига в сторону
улучшения. Следует заметить, что в период дачи брома собака стала очень
возбужденной, чуть не кусалась при работе, усиленно двигалась во время
опыта. Возбужденное состояние наблюдалось и вне камеры. Повышение
пищевой возбудимости (опыт па 2—3 часа позже обычного времени или даже
2—3-дневное голодание) тоже не дало значительного улучшения. Далее,
когда паузы между применением раздражителей были удлинены (7-минут-
ные вместо обычных 5-минутных, табл. 1), условиорефлекторная деятель-
ность почти совсем не улучшилась.
Для того чтобы составить более или менее полно-е представление о со-
стоянии условнорёфлекториой деятельности собаки в этом периоде, я испы-
тал влияние сильного экстрараздражителя на фоне отсутств.ия положитель-
ных условных рефлексов в одном опыте. В середине опыта один из товарищей
зашел в камеру с кошкой в руках, закрыл за собой дверь камеры и несколь-
ко р аз бросил кошку на собаку, ci оявщуво у стенки. Собака при этом прищла
в сильное возбуждение. После того как товарищ с кошкой вышел из камеры,
опыт продолжался. Оказалось, что это экстрараздражение кратковремен-
но вернуло исчезнувшие условные положительные рефлексы.
Привожу протокол этого опыта (табл. 2, стр. 82).
Перерезка шейных симпатических нервов оказала существенное влияние
также на безусловный слюноотделительный реф
л е к с. В первые 5—6 дней, когда состояние собаки было еще неудовлетво-
рительным, безусловные рефлексы значительно ослабели по сравнению
с дооперационным периодом; затем, однако, они быстро увеличились и в
скором времени превысили границы дооперационного уровня. Так, если
в дооперацнонном периоде величина безусловного рефлекса в среднем ко-
лебалась в пределах 100—120 делений шкалы за 35 секунд, то в первые 5—
6 дней после операции она колебалась в пределах 90—110 делений, а
потом получены цифры 120—150.
Таблица 1
<я g * 10 вного деле ни;- a S § а 3 i * gg ей О О =* слон- в де- за ►мод лскса 1 X i о * а и О ct gen Название V ng о X О X 1 ED Ы О ct i й о “ с 3 £ *
Назпаиие услов- Л Название услов- «т и ® ”53 Название уелав- ° о 3 >«з условного й *"* Й -*• Я я г> и у
кого раздражи- Чь 4» й 5 «в3 но го раздражи- u о. та 3 и ц О х я с ¥ ного раздражи- S л ? я □ «х е I s раздражителя - о. п ° X ё 5 =
теля и продол- з2 я сз 2J теля и продол- з2 «*« теля и продол- 3 2 Я - 2 Z »• 3 и продолжи- з 2 «S3 ?«•*
жительность его Р Z X - аз 5S.X3 жмтельмость его t s Величи! него ре< лениях 35 сек. жктельность его ► ? • Величи го рефл лениях тсльность его 5 5 • -Л. * т Чз
действия Ллтен уело® в сек, s п а е; Ль Л* * ей ь. к 5 х С * О оО a s ап действия Латеи услов в сек хини Jd oj ‘HL'» fl действия латы 1 УСЛО1 р сек “ 0 V10 3:“r. действия х а Вели го ре лени Вели кого деле» 35 се
Контрольный опыт Опыт 25. IV 1932 г. Опыт 5. V 1932 г. Опыт 22. V 1932 г
25. 1! 1932 г. (10-Й день после операции) (21-й день после операции) (38-и день после операции)
Звонок 20 сек. 1 33 112 Стук по 20 сек. 0 134 Свет 20 се к. 12 13 . 145 Касалка 20сек. 2 30 120
Стук по 20 » 2 28% ПО Звонок 20 » — 0 129 Стук по 20 » 17 4 148 Звонок 20 » 18 4 132
Свет 20 » 3 24 108 Касалка 20 » — 0 127» Звонок 20 » — 0 129 Свет 20 » 15 8 130
Шипенье20 » 3 26 105 Свет 20 » — 0 129 Касалка 20 » — 0 133 Стук по 20 » — 0 132
Звонок 20 » 3 28 102 Стук по 20 » — 0 — Свет 20 » — 0 — » » 20 » 0 134
Стук по 20 » — 0 .— Звонок 20 » — 0 134** Звонок 40 » 25 12 122 Свет 20 » 16 5 128
Свет 20 » 3 19 100 Стук по 20 » Свет 20 » — 0 126 Свет 40 » 30 5 — Звонок 20 » — 0 124
— 0 121 Стук по 40 » 27 8 — Свет 20 » — 0 120
• Паулы везде 5-минутные
Опыт 11. V 1632 г. Опыт 13. V 1032 г. Опыт 20. V 1932 г. Опыт 23. V 1932 г.
(на 3 часа позже обычного времени) (после однодневного отдыха) Бром в течение (в день 1,5 □ дней Г) (7-минутные паузы)
Стук по 20 сек. 1—2 28 120 Касалка 20сек. 1-2 26 130 Стук по 20 сек, 1-2 33 112 Свет 20 сек- 1-2 30 118
Свет 20 » 10 11 130 Стук по 20 » 2 27% 130 Свет 20 » 9 17 117 Звонок 20 » 17 3 129
Кйсал ка 20 » 17 2 332 Звонок 20 » 15 7% 124 Звонок 20 » 14 9 120 Стук по 20 » -~• 0 118
Звонок 20 » 0 137 Свет 20 » 14 6 131 Касалка 20 » 19 2 128-“ Касалка 20 » 0 IO»***
Стук по 20 » 0 Стук по 20 » — 0 124 Свет 40 » 19 17% 130 Звонок 20 » 0 107
Звонок 40 » 23 17 130 Касалка 20 » 0 120 Стук по 40 » 28 10 124 Сту к по 20 » — 0 106
» 40 » 0 122 Звонок 40 » — 0 120 Свет 20 » — 0 102
Стук по 40 » — 0 120 Свет 40 » 0 — Касалка20 » — 0 94
Свет 40 » — 0 116
* Не подходит сразу к пище.
•• Берет и жадно ест после специального приглашения и в моем присутствии.
••• После касалки даются три порции еды без предварительного действия условных раздражителей.
• ••• Появляется и постепенно увеличивается своеобразный «латентный период» безусловного рефлекса (2—5 сек.).
82
Э. А. АСРАТЯН
Таблица 2
Название условного раздражите- ля и продолжительность его действия и сек. Латентный пери- од условного рефлекса в сек. Величина условного рефлекса и делениях шкалы Величина безуслол- ного рефлекса за 35 еек. в деле- ниях шкалы
Стук ЗОсск. 23 17 133
Свет 30 » 0 135
Звонок 30 » — 0 136
Касалка 30 » —— О 128
Звонок 30 * * 15 14V. 113
Свет 30 » — 0 118
» 27 » 3 3 116
• Заходит сотрудник лаборатории с кошкой в руках и несколько раз бросает
ее на собаку; собака сильно возбуждена.
В приводимых ниже опытах сказанное иллюстрируется с достаточ-
ной ясностью. Было отмечено заметное усиление безуслов-
ного рефлекса на безусловный раздражитель одинаковой величины
и состава. Но ввиду недостаточных размеров шкалы в этих случаях учи-
тывался не весь безусловный рефлекс. С целью более точного измерения
величины безусловного рефлекса как в дооперацнонном, так и в после-
операционном периоде определялась суммарная величина безусловного
рефлекса на меньшее количество мясо-сухарного порошка стандартного веса
н состава (10 г порошка +10 см3 воды). Сравнение аналогичных проб по-
казывает, что, действительно, безусловные слюноотделительные рефлексы
усилились в противовес условным рефлексам. Соответствующие данные
приведены в табл. 3 (слюна измерялась за каждые 10 секунд, пауза
между кормлениями — 4 минуты)..
Таблица 3
Xs опыта Слюноотделения ?а каждые 10 сек. Р делениях шкалы Слюно- отделе- ние 3d 70 сек.
Часть опыта 13. IV 1932 г. (контроль)
№ 1 26 38 27 13 3 1 14 1094
А» 2 25 37 28 1 2 0 3 14 Ю64
№ 3 25 36 27 15 2 2 0 ‘ 107
№ 4 23 33 22 1 2 2 2 9И
Часть опыта 30.IV 1932 г. (16-й День после операции)
№ 1 36 I 62 I 31 I 30 I ' 0 1 0 I 0 152
А» 2 32 | 66 | 37 | 30 | 0 | 0 | 0 155
Звонок 20 сек.—берет сразу же еду
Ат 3 30 | 50 39 | 29 | 0 1 0 | 0 148
№ 4 Не берет нищи; вхожу, даю с руки 156
№ 5 » » » » > » » 151
№ 6 Берет сразу еду I 140
№ 7 35 | 50 1 25 1 7)510)0 122
Часть опыта 25.V 1932 г. (условные рефлексы значительно усилились,
см. этот же опыт в табл. 1)
№ 5 [33 50 36 12 0 0 — 131
№ 6 31 48 29 13 -0 1 - 122
№ 7 32 45 27 13 0 0 — 117
Характерно, что с появлением условных рефлексов безусловные рефлек-
сы постепенно уменьшались в величине. Затем установленное на этой со-
баке закономерное уменьшение безусловного рефлекса после предшествую-
щего действия усл овного раздражителя (отрицательная индукция на центр
ПЕРЕРЕЗКА СИМПАТИЧЕСКИХ НЕРВОВ И ПИЩЕВЫЕ УСЛОВНЫЕ РЕФЛЕКСЫ
S3
безусловного рефлекса) почти совершенно исчезло; условные раздражи-
тели не вызывали рефлекса и заметно не уменьшали непосредственно сле-
дующего за ним безусловного рефлекса (опыт 30.IV).
С безусловным рефлексом был связан еще ряд интересных моментов.
Первое, что обращало на себя особое внимание, —это быстрое прекращение
слюноотделения после окончания еды. В дооперационном периоде последо-
вательное безусловное слюноотделение длилось около 1 минуты, а после
операции — всего 20—25 секунд, несмотря на относительно большую вели-
чину рефлекса. Это хорошо видно на приведенных в табл. 3 специальных
пробах, но особенно ярко выступало при подкреплении условных рефлексов
большими порциями еды.
Другая особенность заключалась в появлении «латентных периодов»
безусловного рефлекса, чего раньше никогда не наблюдалось. В сере-
дине опытов и в особенности в конце, когда условные рефлексы совершенно
отсутствовали, безусловное слюноотделение начиналось спустя 5—7, иног-
да и больше секунд после того, как собака уже начинала есть поданную
пищу. Следует отметить, что этот «латентный период» особенно увеличился
после диференцировочного раздражителя. Иногда диференцировочный раз-
дражитель вызывал заметное последовательное торможение безусловных
рефлексов,что, очевидно, говорит об усилении тормозного процесса после
операции. И, наконец, еще одной особенностью, связанной с безусловным
рефлексом, являлось то, что собака при наличии хорошего аппетита (на
что указывали жадность и величина безусловного рефпекса) от 8-го до
15-го дня после операции часто отказывалась от еды во время опыта.
Характерно, что тут известную роль играло действие условного раздра-
жителя. В начале опыта собака брала еду даже тогда, когда перед пода-
чей пиши действовал условный раздражитель. Затем она не брала еды при
этой комбинации, но охотно брала пищу и жадно ела, если пища давалась
без предварительного действия условных раздражителей. В дальнейшем
она отказывалась брать пищу даже при таких обстоятельствах, но охотно и
жадно ела, если я входил в камеру и предлагал есть. В этой связи хотелось
отметить одно совпадение, может быть, случайного характера. Описанное
явление особспноярко было выражено в дни первого появления и исчезно-
вения первых условных рефтексов (11-й и 12-й день после операции).
В дальнейшем это явление исчезло.
Такова была картина, когда я сделал летний перерыв, который в силу
некоторых обстоятельств длился дольше, чем обычно, — почти 4 месяца.
За время этого 4-месячного перерыва собака хорошо поправилась.
Иногда опа все-таки кашляла (кстати говоря, данные о длительном каш-
ле у Арча совпадают с наблюдениями Клода Бернара, у которого собаки
после экстирпации шейных симпатических узлов кашляли в течение ряда
месяцев). Собака выглядел а гораздо бодрее, почти совершенно исчезли апа-
тия, вялость, малоподвижность. Она «рвалась» в камеру так же, как и в
дооперационный период. Судя по внешнему поведению, можно было смело
сказать, что почти все последствия операции исчезли. Я вления со стороны
обоих глаз (сужение зрачков, паралич третьего века, западение глазного
яблока, отвисание нижнего века) немного сгладились, но все-таки были
сильно выражены.
Условнорефлекторная деятельность тоже улучшилась,но до нормы было
еще далеко. В опытах на первый раздражитель были получены условные
рефлексы значительной величины, на второй раздражитель отмечалось уже
резкое падение, в дальнейшем рефлексы были еще меньше, а в конце опыта
(на 7—8-м месяце) получен уже нулевой эффект. При этом иногда наблюда-
лось парадоксальное отношение между рефлексами на слабые и сильные
раздражители. Диференцировочный рефлекс оставался попрежнему стой-
ким. Характерно, что по сравнению с дооперационным периодом немного
6*
М
Э. А. АСРАТЯН
ослабели также безусловные рефлексы; во всяком случае они очень быстро
уменьшались в течение одного и того же опыта. С целыобыстрого улучшения
состояния условных рефлексов мы перешли на некоторое время к опытам
с совпадающими условными рефлексами. Это существенно помогло укрепле-
нию условных рефлексов. Улучшение шло волнообразно, постепенно зна-
чительной величины условные рефлексы появились на втором, третьем и по-
следующих местах, а нулевой эффект даже в конце опытов стал исчезать.
Привожу примеры опытов (табл. 4).
Таблица 4
Название условного раздражителя Латентный период услов- ного рефлекса л сек. Величина условного рефлекса в делениях шкалы Величина без- условного реф- лекса за 35 сек. в делениях шкалы
Опыт 1.Х 1932 г.
Звонок 1—2 32 100
Стук 120 11 И 96
Шипенье 10 12 95
Свет 5 15 92
Звонок 14 6 90
Шипенье 12 9 93
Звонок 0 87
Свег — 0 83
Опыт 11.XI 1932 г.
Стук 1 30 92
Свет 2 20 96
Касалка 6 13 92
Звонок 4 19 87
Стук 0 —
Звонок 8 9 82
» 5 11 80
Стук 6 7 75
Касалка 9 3 72
Опит 24.XI 1932 г.
Шипенье 1 35 НО
Стук 1 30 116
Касалка 4 20 112
Звонок 3 27 114
Свет 4 1G 112
Стук •— 0
Звонок 6 10 109
Свет 2 19>/а 111
Касалка 6 14 108
Стук 7 юз
Как видно из табл. 4, параллельно увеличились и безусловные рефлексы.
Следует сказать, что с улучшением состояния условнорефлектороной дея-
тельности появилось также промежуточное слюноотделение в начале опыта.
Затем значительно удлинился период последовательного безусловно-
рефлекторного слюноотделения.
В период резкого ослабления условнорефлекторнон деятельности я не
ставил никаких специальных функциональных проб, во-первых, потому,
что и без них картина была ясна, во-вторых, потому, что нарочно избегал
осложнения и затруднения и без того ослабленной условнорефлекторной
деятельности собаки. Но когда, спустя 6—7 месяцев после операции, насту-
пил период улучшения, я начал ставить специальные функциональные
пробы.
ПЕРЕРЕЗКА СИМПАТИЧЕСКИХ НЕРВОВ И ПШЦЕВЫЕ УСЛОВНЫЕ РЕФЛЕКСЫ
85
Влияние перерыва
В норме недельные и даже месячные перерывы совершенно не ослабляли
условных рефлексов, скорее даже наоборот. В этом же периоде сравнитель-
ного улучшения условнорефлекторной деятельности, как ни странно, даже
3—4-дневный перерыв сильно ослаблял условные рефлексы. При этом услов-
ные рефлексы достигли прежней величины через 3—4 дня работы. С тече-
нием времени это ослабляющее действие перерыва в работе постепенно
уменьшалось, и спустя почти год после операции оно совершенно исчезло:
даже 23-дневный перерыв не оказал ослабляющего влияния (табл. 5).
Таблица 5
Название условного
раздражителя
Латентный
период услов-
ного рефлекса
в ск.
Величина
условного
ре фтекса в
делен и-вх
шкалы
Величина без-
условного реф-
лекса за 35 сек.
в делениях
шкаль*
Опыт 27.X1I 1932 г.
(после 4-дневного перерыва)
Звонок 3 18 72
Стук 120 4 11 68
Свет 6 7 64
Шипенье 5 9 63
Звонок 4 11 65
Стук 60 —. 0
Звонок — 0 56
Свет 9 64 60
Касалка 11 3 58
Стук 120 — 0 61
Опыт 14.1 1933 г.
Звонок 1 39 107
Стук 120 2 29 ПО
Свет 4 20 J10
Шипенье 2 25 108
Звонок 3 25 105
Стук 60 — 0 —
Звонок 9 114 103
Свет 5 19 105
Касалка 7 12 101
Стук 120 10 14 105
Опыт 23. L 1933 г.
Касалка 1 34 112
Звонок 1 35 112
Шипенье 2 23 116
Сгук 120 2 28 114
Касалка 3 17 И6
Свет 4 19 114
Звонок 3 25 109
Опыт 28.11 1933 г.
(после 3-дневного перерыва)
Касалка 2 20 97
Звонок 3 154 95
Шипенье 3 15 86
Стук 120 3 14 80
Касалка 4 94 81
Свет 3 11 80
Звонок 3 13 77
86
Э А. АСРАТЯН
Системность
В дооперационном периоде 5—6-дневного применения определенного
сочетания раздражителен было достаточно, чтобы у собаки образовалась
системность, которая как показатель высшей анализаторной и синтетиче-
ской деятельности проявлялась в том, что применение одного раздражителя
в течение всего опыта приблизительно повторяло ход условных рефлексов
с системой раздражителей. Даже в этом периоде сравнительного улучше-
ния условнорефлекторной деятельности через 7 месяцев после операции
была еще слабо выражена способность собаки образовывать систем-
ность. Через 11 месяцев после операции эта способность значительно уси-
лилась (табл, б).
Таблица 6
Название условного раздражителя Латентный период услов- ного рефлекса в сек. Величина условного рефлекса в делениях шкалы Величина безу- словного рефлек- са за 35 сек в делениях шкалы
Одни из 8 опытов Опыт 6.1 1933 г. с системой (через 7 мес. после операции)
Звонок 1 37 105
Стук 1 31 102
Свет 4 18 100
Шипенье 7 16 98
Звонок 3 24 98
Стук а 0 _1
Звонок 9 12 93
Касалка 4 11% 90
Стук 4 21 88
Опыт 9.1 1933 г.
Воспроизведение системности
Стук 120 1 36 109
То же 5 18 108
» > 6 17 —
» > 7 16 юз
» > 8 16 109
> > 7 17 112
» » 6 20 108
» » 9 16% 105
» > 9 14J? 102
Опыт 23,1V 1933 г.
Один из опытов с системой (через 11 мес. после операции),
уменьшена порция еды
Свет 1 36 115
Касалка 2 32 112
Звонок 2 31 109
Стук 2 30 112
» — 0 ——
Звонок 4 >4% 118
Свег 3 21 107
Касалка 6 10 102
Опыт 28.1V 1933 г.
Стук 120 То же 1 2 38 2854 108 105
» > 2 30 101
6 26 100
> > 15 8 105
> э 7 18 101
» > 6 22 103
» » 7 14 100
ПЕРЕРЕЗКА СИМПАТИЧЕСКИХ НЕРВОВ И ПИЩЕВЫЕ УСЛОВНЫЕ РЕФЛЕКСЫ 87
Влияние экстрараздражителей
Выше говорилось о влиянии экстрараздражителей (кошка) на условные
рефлексы в период сильного угнетения усповнорефлекторной деятельности
собаки. Через 9 месяцев после операции было еще раз испытано влияние
этого экстрараздражителя. Выяснилось, что условные рефлексы страдают
от этого воздействия больше, чем в дооперационном периоде. Для сравне-
ния привожу опыты (табл. 7).
Таблица 7
Називине условного
раздражителя
Латентный период
условного рефлекса
в сек.
Величина условного
рефлекса а делениях'
шкалы
Величина безуслов-
ного рефлекса
за 35 сек в делени-
ях шкалы
Опыт 5.IV 1932 Г.
Звонок 1 36 120
Свет 2 27 124
Звоне к 10 |<т 126
» 2 23 —
» 2 26’/, 1 18
Свет 4 114
Стук 3 22 115
» 3 24 113
» 5 12 123
» 6 18 98
Звонок 3 23 103
Опыт 22.11 1933 г.
Звонок 1 35 ПО
Свет 2 25 116
Звоно к 12 8 101
10 10 116
» 8 16/» 113
Свет 8 7 НО
Стук 9 10 1 13
» 10 8 110
» 12 4 86
» 13 зу» 98
Звонок 10 5 101
* Бросание кошки на собаку во время опыта.
Совершенно аналогичные данные имеются также о влиянии более сла-
бого экстрараздражителя — подкожного впрыскивания нескольких куби-
ческих сантиметров физиологического раствора перед началом опытов
(болевое раздражение от укола). В то время как в дооперационном периоде
эта процедура почти не оказывала влияния, после операции в период
улучшения, она вызвала заметное ослабление положительных условных
рефлексов и даже немного растормаживала диференцировочный рефлекс.
Влияние питуитрина
Влияние питуитрина на условнорефлекторную деятельность собаки
в данном случае представляет интерес лишь как проба; на этом вопросе я
здесь подробно останавливаться не буду. Питуитрин Т Московского
института экспериментальной эндокринологии), изготовленный специально
для научно-исследовательских работ, в средних дозах (1 — 1,5 см3) вызы-
вал сильное ослабление условных слюноотделительных рефлексов у Арча
вдооперационном периоде. Это влияние питуитрина, однако, на следую-
щий день исчезало. В описываемом периоде улучшения, спустя год после
88
Э Л АСРАТЯН
операции, тот же препарат в тех же дозах оказал совершенно аналогичное
влияние; однако оно не только не исчезло на следующий день, а даже как
будто усилилось; полностью исчезло оно только через день (табл. 8).
Таблица 8
Название условного
раздражителя
Латентный.
период ус-
ловного реф-
лекса п «к.
Величина
условного
рефлекса п
делениях
шкалы
! Величина безу-
словного рефлек-
са за 35 сек п
делениях шкалы
Опыт 11 IV 1933 г
Введен питуитрин за 2 часа до опыта
(опыт до симпатэктомии)
Звонок 7 9 117
Стук 120 9 6 124
Свет 12 5 176
Звонок 17 1 120
Свет гт 0 1 18
Стук 120 — 0 116
» 120 —4 0
Звонок — 0 109
Опыт 9.V 1932 г.
Введен питуитрин за 2 часа до опыта (опыт после
симпатэктомии)
Свет 2 22 124
Звонок 5 8 134
Касалка 3 12 128
Стук 120 7 6 129
Свет — 0 124
Звонок 7 7«/, 125
С1ук 120 — 0 —
Звонок — 0 112
Опыт 10 V 1933 г.
Свет 20 5 16 102
Стук 120 20 7 10 86*
Звонок 20 — Следы 76*
Касалка 20 12 2 71*
Свет 20 — 0 ——
Звонок 40 — 0 —
Свет 40
О п ы т 11.V 1933 г.
Касалкя 20 1 32 116
Звонок 20 2 25 118
Свет 20 4 18 120
Стук 120 20* 3 22/, 114
» 120 20
Свет 20 0
Звонок 20 0 106
• Не сразу берет пищу-
Угашение и восстановление рефлексов
Сравнение опытов с yi ашением положительных условных рефлексов
до операции и спустя год после нее показывает/что во втором случае услов-
ные рефлексы гораздо быстрее угасали и труднее восстанавливались, чем
в первом. Это относилось ко всем раздражителям.
ПЕРЕРЕЗКА СИМПАТИЧЕСКИХ НЕРВОВ И ПИЩЕВЫЕ УСЛОВНЫЕ РЕФЛЕКСЫ 89
Для рефлекса на метроном 120 ударов в 1 минуту соответствующие
цифровые величины условных рефлексов по делениям шкалы составляли
(через3-минутные паузы): 9.IV 1932г. (до операции) —32, 18, II, 9, 6*/а,
0,0,0: после подкрепления—3. 10, 16, 6, 18, 12; 15. II 1933 г. (через 8 ме-
сяцев после операции) —39, 20, О, О, О, О, О, 0; после подкрепления — О,
9, 10, 12, 9.
Для звонка соответствующие цифры составляли: 28. III 1932г.—29, 25,
12Vt, 6, 21/», 2, 0, 0; после подкрепления — 3, 9, 4, 13, 17, 19, 22; 13. II
1933 г.—34, 243/а, 12, 5, О, 0, 0, 0; после подкрепления—5, 8, 101/», 7, 11,8.
Последовательное торможение от дифсренцировочного раздражителя
было почти одинаково в норме и в послеоперационном периоде; может быть,
в последнем случае оно было даже немного сильнее, по эта разница не имела
существенного значения. Не было особой разницы также в результате
5-минутного действия диференцировочного раздражителя.
Этим кончается цикл тех специальных функциональных проб, которые
были проведены на Лрче до и после операций.
Удаление левого верхнего шейного симпатического узла
Спустя почти год после первой операции, 4. IV 1933 г., Арч подвергся
контрольной операции. Под эфирно-хлороформным наркозом у него был
сделан надрез кожи на бедре, и рана была зашита. Уже на 3-й день после
контрольной операции собака была совсем бодра, и рефлексы, хотя и в сла-
бом виде, были налицо. На 7-й день после операции рефлексы достигли нор-
мальной величины.
Спустя 2’/а месяца после контрольной операции, 17. VI 1933 г., у Арач
был удален левый верхний шейный симпатический узел. Общее состояние
ссбаки после этой операции изменилось больше, чем после контрольной
операции. Собака стала апатичнее; появился частый кашель, иногда была
рвота. Однако состояние собаки быстро улучшалось и через 10—15 дней
достигло почти прежнего уровня. Следует сказать, что уже на2-й день после
операции левый глаз отличался отправого: нижнее веко опускалось больше,
зрачок еще больше сузился, третье веко сильнее было выдвинуто’ вперед;
при надавливании на глазное яблоко сопротивление с левой стороны было
меньше; в величине глазной щели не было большой разницы. Однако еще
через 5—6 дней все эти явления сохранялись, лишь левый зрачок стал
шире правого. Эта разница была отчетливо выражена и сохранялась в те-
чение почти месяца.
Со стороны условнорсфлекторной деятельности изменения были выраже-
ны сильнее, чем посте контрольной операции, однако во много раз слабее,
чем посте первой операции» когда были перерезаны оба шейных симпати-
ческих нерва.
На 6-й день после операции появились первые рефлексы, на 9-й день
рефлексы на первых порядковых местах применения были значитель-
ными, а на последующих исчезли. В следующие дни они довольно быстро
увеличились, и через 20 Дней почти восстановилась дооперацнонпая картина.
Другие функциональные пробы не выявили каких-либо различий между
периодом до и после контрольной операции. Экстирпация левого шейного
симпатического узла, как и контрольная операция, по своим последствиям
значительно отличалась от операции перерезки шейных симпатических
нервов. По существу говоря, последние операции не вызывали серьезных
и длительных изменений в условнорефлекторной деятельности собаки.
При этом состоянии ссбаки был сделан летний перерыв на 53 дня. Ха-
рактерно, что указанный перерыв совершенно не ослабил условнорефлектор-
кой деятельности собаки.
После летнего перерыва у Арча почти исчезла разница между явлениями
со стороны обоих глаз, состояние его было хорошее, он только немного по-
90
Э А АСРАТЯН
худел. Условнорефлекторная деятельность была даже лучше, чем до послед-
ней операции. Снова испробован ряд функциональных проб (угашение,
последовательное торможение от угасательного и дифереицировочного тор-
можения и т. д.). Создалось впечатление, что состояние условнорефлектор-
ной деятельности достигло уровня дооперационного периода в отношении
баланса между процессами торможения и возбуждения (в смысле их под-
вижности и силы).
Удаление правого верхнего шейного симпатического узла
14. X 1933 г. был экстирпнрован второй, т. е. правый верхний шейный
симпатический узел. Со второго же ДНЯ появились сильная рвота и ка-
шель. Собака сильно исхудала, дыхание и сердцебиение у нее были
учащены. Ела неохотно, после еды сразу же Возникала рвота. Со стороны
правого глаза на 2-й день было отмечено слабое сужение зрачка, продвиже-
ние третьего века, западение глазного яблока и сужение глазной щели.
При надавливании на глазное яблоко правый глаз оказывал меньшее
сопротивление.
Характерны дальнейшие изменения со стороны глаза. На 3-й день после
операции эти явления усилились, с 4-годня и да смерти всеявления продол-
жали развиваться в том же направлении, а зрачок начал расширяться и
стал гораздо шире (раза в три) левого зрачка.
Постепенно состояние собаки ухудшалось, и 19.Х она погибла.
Вскрытие показало, что оба верхних шейных симпатических узла экс-
тирпированы полностью и что имеется воспалительный процесс в легких
(гепатизацня обоих легких, экссудат). Очевидно, это явилось одной из
глав-ных (если не самой главной) причин смерти.
Заключение
Суммируя полученный фактический материал, можно скатать, что од-
новременная перерезка обоих шейных симпатических нервов вызывает глу-
бокие и длительные нарушения в условнорсфлекторной деятельности ко-
ры головного мозга собаки. Характерной чертой этих нарушений являет-
ся ослабление условнорсфлекторной деятельности, что в наших опытах
выражалось в исчезновении условных рефлексов, в медленном их восста-
новлении, в их ослаблении после небольших перерывов, сравнительно
сильном ослаблении под влиянием экстрараздражителей, а также под влия-
нием питуитрина и ряда других моментов. Очевидно, этим же ослаблением
мож но объяснить нарушение равновесия между процессами возбуждения
и торможения в сторону усиления последнего и сравнительную инертность
процессов (быстрое угашение и медленное восстанавление положительных
условных рефлексов, усиленное последовательное торможение от угасатель-
нога и дифереицировочноготорможения ит. д.). Затем отмечается ослабле-
ииесложной тонкой синтетической деятельности (системность). Безуслов-
ные рефлексы в период полного исчезновения условных рефлексов и в пе-
риод их резкого ослабления, наоборот, значительно усилились, затем был
период, когда они, так же как и условные рефлексы, были слабы, а спустя
G—7 месяцев после операции они параллельно с условными рефлексами
постепенно возросли. Экстирпация левого верхнего шейного симпатичес-
кого узла спустя почти год после первой операции оказала сравнительно
слабое влияние на условнорефлекторную деятельность коры головного
мозга собаки.
Какие же можно сделать заключения и выводы из приведенного в статье
фактического материала?
Во-первых, как попять явления рвоты и кашля, которыми всегда сопро-
вождались операции на шейных симпатических нервах и узлах? Являлись
ли они результатом травматизацин волокон n. vagi или же результатом из-
ПЕРЕРЕЗКА СИМПАТИЧЕСКИХ НЕРВОВ И ПИЩЕВЫЕ УСЛОВНЫЕ РЕФЛЕКСЫ
91
менения возбудимости в соответствующих нервных элементах продолгова-
того мозга и вышележащих частей центральной нервной системы вслед-
ствие перерыва симпатических путей или экстирпации узлов? Хотя у
меня нет специального материала для ответа на поставленный вопрос, но
думается, что здесь играли роль оба момента. Исследования В. В. Савича
и А. Н. Крестовникова, а также эксперименты В. С. Стрельцова дают
основание полагать, что и второй момент МОГ участвовать в этом сим-
птомокомплексе. Не предрешая вопроса о первичном или вторичном
происхождении несомненного и значительного усиления безусловного реф-
лекса после перерезки шейных симпатических нервов, можно все же этот
факт привести в пользу указанного допущения.
Возникает также вопрос, можно ли описанные нарушения в условно-
рефлекторной деятельности собаки считать только вторично возникшими
в связи с происшедшими в организме общими изменениями, проявле-
нием которых отчасти являются рвота, кашель и т. д., или же эти отклоне-
ния являются результатом прямого или косвенного изменения функциональ-
ного состояния рецепторов и нервных механизмов (сосудодв ига тельные из-
менения, гуморальный баланс, адаптационные изменения)? Не исключая
совершенно возможности влияния изменения общего состояния организма
на условнорефлекторную деятельность собаки, я все-таки склонен думать,
что в данном случае оно не являлось единственной и даж:е основной причи-
ной. Общее состояние собаки после первой операции сравнительно быстро
улучшилось, а нарушения в условнорефлекторной деятельности продол-
жали существовать еще почти год после этой операции.
Очевидно, что основной причиной этих глубоких и длительных измене-
ний были сдвиги именно в функциональном состоянии соответствующих
нервных звеньев вследствие выключения притока тонических или часто
повторяющихся фазных симпатических нервных импульсов центрального
и ганглиозного происхождения. Весьма вероятно, однако, что изменение
функционального состояния рецепторов головы тоже играет некоторую
роль в этих нарушениях. Что симпатическая нервная система может изме-
нять функциональное состояние рецепторов, об этом свидетельствуют
наблюдения Клода Бернара и данные сотрудников лаборатории Л. А. Ор-
бели и др. Данные Клода Бернара говорят о том, что чувствительность
кожи головы и ушей посте экстирпации верхних шейных симпатических
узлов повышается. Имеются указания и относительно изменения функцио-
нального состояния глазного анализатора под влиянием симпатической
нервной системы [Гесс и Леман (Hess и Lehman)]. Возможны аналогич-
ные изменения также со стороны вкусового и обонятельного рецепторов.
На основании сказанного не приходится сомневаться в том, что у моей
подопытной собаки также имелись сдвиги в функциональном состоянии ре*
цепгоров головы. Что эти изменения оказали влияние на всю описанную
картину ее условнорефлекторной деятельности, тоже не подлежит сомнению;
почто и они не были основной причиной этих пертурбаций, об этом говорит
тот факт, что условный рефлекс на кожно-механическое раздражение бедра
(область, симпатическая иннервация которой не была затронута) в общем
изменялся одинаково со всеми остальными рефлексами на другие раздражи-
тели.
Что касается вопроса, чему придавать главное значение — перерыву
путей центральных импульсов или экстирпации ганглиев, то, повидимо-
му, основной причиной изменений следует считать перерыв путей,
по которым проходят центральные импульсы, хотя, как показывают опыта
с экстирпацией левого верхнего шейного узла, ганглии тоже играют из-
вестную роль. Возможно, что это влияние всюбщее сильнее, чем то, ко-
торое выявилось в случае с собакой Арч, где имела место компенсация
92
Э. А. АСРАТЯН
со стороны второго ганглия, но этот вопрос можно будет решить только
после удачной экстирпации обоих узлов.
Существует много различных точек зрения на физиологическую роль
вегетативных нервных узлов. Так, Ленгли (Lang-ley) приписывал им
функцию вспомогательной силы;Шульц (Schultz) полагал, что они усили-
вают импульсы преганглионарных волокон; имеется предположение, что
они расширяют область действия импульсов преганглионарных волокон;
раньше даже существовало мнение, что они являются своеобразными
центрами [Клод Бернар, Гарман (Hoffmann) и др. [.
Ряд старых исследований, в особенности наблюдения Клода Бернара
о том, что после экстирпации верхних шейных симпатических узлов тепло-
продукция в наружных головных органах сильнее, чем при простой пере-
резке шейных симпатических нервов, а также новейшие данные Эдриана
(Adrian) и сотрудников относительно увеличения волн тонических импуль-
сов в. постганглионарных волокнах шейного симпатического нерва с оче-
видностью говорят О том, что предположения Ленгли и Шульца близки к
истине. Я полагаю, что в пользу этого взгляда говорят как мои данные о
риске для жизни животных одновременной экстирпации обоих шейных
симпатических узлов и безопасности одновременной перерезки обоих шей-
ных симпатических нервов, так и данные о том, что шейные узлы после
перерезки преганглионарных волокон способны, хоть в слабой степени, к
самостоятельной деятельности (явления со стороны глаз, слабое измене-
ние условнорефлекторной деятельности).
Факт расхождения между условными и безусловными рефлексами в из-
вестный период работы (резкое ослабление первых и значительное усиле-
ние вторых) можно истолковать различно, допустив, что здесь имеет место
своеобразное «растормаживание» нервных механизмов безусловного рефлек-
са в силу отсутствия условных рефлексов или сильная отрицательная индук-
ция на нервные механизмы условных рефлексов в силу повышенной воз-
будимости центров безусловного рефлекса, илижечтооба явления обуслов-
лены какой-то третьей причиной. Какое из этих предположений соответ-
ствует истине, сейчас сказать трудно; решение вопроса — задача будущего.
Выводы
С помощью метода условных рефлексов удается вызвать и подробно ис-
следовать у собак ряд глубоких изменений в высшей нервной деятельности,
возникших в результате оперативного воздействия на шейные симпатиче-
ские нервы и узлы. В данной статье приводятся результаты дальнейшей
разработки ряда вопросов, которые были поставлены и частично разрешены
в прежней нашей работе.
У одной уравновешенной собаки с сильной нервной системой в течение
двух лет подробно исследовались пищевые условные рефлексы (взаимоот-
ношения корковых процессов возбуждения и торможения, влияние эксгра-
раздражителей, перерыва в работе, гормонов и т. д.). После одновременной
перерезки шейных симпатических нервов ниже (каудальнее) одноименных
нижних шейных узлов в течение 1 года и 4 месяцев были исследованы из-
менения в условнорефлекторной деятельности собаки.
Основные изменения следующие.
1. После одновременной перерезки обоих симпатических нервов поло-
жительные условные рефлексы оказались в течение 5—б месяцев сильно
ослабленными, причем в первые 15—20 дней они почти совершенно отсутст-
вовали.
2. В противовес условным положительным рефлексам безусловный реф-
лекс существенно усилился в первый месяц, затем с появлением и усилением
условных рефлексов он тоже ослабел.
ПЕРЕРЕЗКА СИМПАТИЧЕСКИХ НЕРВОВ И ПИЩЕВЫЕ УСЛОВНЫЕ РЕФЛЕКСЫ 93
3. В период наибольшего угнетения условнорефлекторной деятельности
кратковременный отдых и введение препаратов брома существенно не
улучшали ее. Сильные экстрараздражигели растормаживали хронически
заторможенные условные положительные рефлексы.
4. Несмотря на рано начавшееся и быстро прогрессирующее улучше-
ние общего состояния собаки, положительные условные рефлексы достигли
относительно хорошего состояния лишь через 6—7 месяцев после операции
(в смысле величины, стойкости, регулярности рефлексов). Однако специаль-
ные функциональные пробы показали, что в этом периоде относительного
улучшения условнорефлекторная деятельность собаки еще существенно
отличается от таковой дооперационного периода:
а) 3—4-дневный перерыв в работе вызывал силь ное ослабление условных
положительных рефлексов (в норме месячные перерывы не оказывали ослаб-
ляющего влияния на условные рефлексы);
б) сильно ослабела способность собаки образовывать системность в услов-
норсфлекторной деятельности;
в) экстрараздражители оказали более сильное задерживающее влияние
на условные рефлексы;
г) питуитрин оказывал более длительное ослабляющее действие на
условные рефлексы;
д) положительные рефлексы угасали быстрее и восстанавливались мед-
леннее, чем в норме.
5. В течение года после операции состояние условнорефлекторной дея-
тельности значительно улучшилось и, судя по новым показателям перечис-
ленных проб, это состояние по истечении года незначительно отличалось
от состояния до операции.
В этом периоде контрольная операция (наркоз, разрез кожи бедра, шов)
оказала незначительное и кратковременное (5—10 дней) ослабляющее влия-
ние на условнорефлекторную деятельность.
6. Последовательно произведенная экстирпация одного верхнего шей-
ного симпатического узла ослабила условпорефлекторную деятельность
значительно сильнее и длительнее, чем контрольная операция, по несрав-
ненно меньше, чем предшествовавшая одновременная перерезка обоих шей-
ных симпатических нервов. После экстирпации второго верхнего шейного
симпатического узла собака погибла (очевидно, от воспаления легких).
7. Анализ полученного экспериментального материала позволяет
прежде всего подтвердить основные данные моей первой работы в этой
области. Затем можно определенно сказать, что происшедшие изменения
в условпорефлектор пой деятельности нельзя приписывать изменениям в
общем состоянии собаки после операции, ибо улучшение общего состояния
началось очень скоро и протекало быстро, а условные рефлексы оставались
неудовлетворительными в течение многих месяцев.
Далее, не исключая возможности влияния сосудодвигательных изме-
нений в гуморальном балансе (gl. thyreoidea, hypophysis cerebri, pl. cho-
rioidei, gl. pinealis), а также изменений рецепторов на возникновение описан-
ной картины, я все же прихожу к выводу, что основной причиной всего опи-
санного являются адаптационно-трофические изменения больших полуша-
рий как следствие перерезки симпатических путей (или экстирпации узлов).
8. Сопоставляя эти данные с прежними моими данными и учитывая факт
сравнительно слабого изменения условнорефлекторной дея тельности собаки
после экстирпации верхнего шейного симпатического узла (вторично),
я прихожу к выводу, что изменения, которые происходят в высшей нервной
деятельности собаки после первичной экстирпации верхних шейных симпа-
тических узлов, обусловливаются главным образом не самой •> к ст и р-
нацией этих узлов, а перерывом путей цент-
ральных импульсов.
54
Э. А. АСРАТЯН
ЛИТЕРАТУРА
Асратян Э. А., Влияние экстирпации верхних шейных симпатических узлов
на пищевые условные рефлексы у собаки, Архив биологических наук. т. XXX
1930.
Клод Бернар, Лекции по физиологии нервной системы, 1866.
Крестовников А. Н. и Савич В. В., Влияние шейного симпатического
нерва на вазомоторный центр. Медико-биологический журнал, т. I, № 3,
Кунстман К. И., Влияние односторонней симпатэктомии на рефлексы с кожи
и сухожилий у собак, Известия Научного института им. Лесгафта, т. IX, 1924.
О р б е л и Л. А., Вегетативная нервная система, БСЭ, т. IV, 1928.
Тонких А. В., Влияние симпатической нервной системы на спинномозговые реф-
лексы лягушки, Русский физиологический журнал, т. VIII, 1925.
Ленгли Дж., Автономная нервная система. 1925.
A d г i a n Е., Journ. physiol., v. 74, 1932.
В г t’i с k е, Ergebnissc der Physiologic, Bd. 34, 1932.
Gaskell, The involuntary nervous system, 1916.
Langley, Ergebnisse der Physiologic, Bd. 2, 1903.
Sc hi If, Das autonome Nervensystem, 1926.
Schultz, Arch. Anat. u. Physiol., 1894.
Spiegel, Bcthe’s Handbuch d, norm. u. pathol. Physiol-, Bd. 10, 1927.
О ВЛИЯНИИ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
НА УСЛОВНОРЕФЛЕКТОРНУЮ ДЕЯТЕЛЬНОСТЬ СОБАКИ i
одной из наших собак (Кейо) несколько лет назад были экстнр-
пнрованы лобные доли коры большого мозга. У этой собаки
были выработаны прочные условные электрообороннтельные
рефлексы на зрительные, тактильные и звуковые раздражения,
очевидно, за счет так называемых вторичных, рассеянных кор-
ковых нервных клеток скелетно-мышечного и кожного ана-
лизаторов. Перерезка обоих шейных симпатических нервов резко нару-
шила условнорефлекторную оборонительную деятельность собаки: элект-
рооборониТельные рефлексы стали непрочными, слабыми, большей частью
нулевыми. Усиление безусловного раздражителя, введение брома, стрих-
нина, отдых и другие функциональные воздействия не привели к упро-
чению и усилению условных рефлексов.
У той же собаки были выработаны условные пищевые рефлексы, которые
обладали несравненно большей прочностью и силой, чем электрооборони-
тельные.
Материал, полученный у Кейо, дает некоторое основание предполагать,
что резкое ослабление электрообороннтельных условных рефлексов по срав-
нению с пищевыми после перерезки шейных симпатических нервов является
результатом того, что после снятия адаптационно-трофического влияния
симпатической нервной системы вторичные рассеянные нервные элементы
коры большого мозга страдают больше, чем «ядерпые» первичные нервные
элементы.
1 Сборник тезисов докладов первого совещания биологической группы Академии
наук СССР по физиологическим проблемам, 1937.
СВЯЗЬ МЕЖДУ ДЛИТЕЛЬНОСТЬЮ ДЕЙСТВИЯ УСЛОВНОГО
РАЗДРАЖИТЕЛЯ И ВЕЛИЧИНОЙ УСЛОВНОГО РЕФЛЕКСА1
Я
литературе по условным пищевым рефлексам встречаются
отдельные замечания о том, будто не наблюдается заметной
разницы в величине условных рефлексов, вызванных одним
и тем же раздражителем при его применении в течение 5—
10 и 20—60 секунд (Л. А. Орбели, В. М. Добровольский).
Эти замечания не привлекали к себе особого внимания в
течение продолжительного времени. В последнее время этим вопросом занял-
ся П. С. Купалов. Так как его исследования имеют ближайшее отноше-
ние к моей работе, я на них остановлюсь немного подробнее.
П. С. Купалов и Б. Н. Луков получили следующие данные. В различ-
ных однотипных опытах на одной и той же собаке они применяли один и тот
же раздражитель то в течение 1 секунды, то в течение 3—10 и 30 секунд
и следили за слюнным компонентом рефлекса в течение 30 секунд. Сопостав-
ление полученных результатов показало, что как в величине условных слюн-
ных рефлексов, так и в характере их течения во всех случаях не было
заметно особой разницы. Больше того, иногда они до мельчайших подроб-
ностей походили друг на друга.
В других опытах П. С. Купалова и Б. П. Лукова над теми же собаками
были измерены величины условных секреторных рефлексов на 3, 10 И 90-Й
секунде применения раздражителей, причем рефлекс всегда измерялся в те-
чение Р/а минут.
У одной собаки (возбудимой) получались совершенно одинаковые дан-
ные для 3- и 90-сскундного действия раздражителя, а у другой собаки (санг-
виника) условный рефлекс был значительно меньше при 3-секундном дейст-
вии условного раздражителя.
В отдельных опытах П. С. Купалов получил почти одинаковые по вели-
чине данные при 3-секундном и 3-минутном применении одного и того же
раздражителя, измеряя величину рефлекса в течение 3 минут.
Па основании этих данных П. С. Купалов приходит к выводу, «что для
условной секреторной реакции решающее значение имеет первый момент
действия условного раздражителя. Можно думать, что основное влияние
условного раздражителя состоит в том, что он дает короткий толчок, под
влиянием которого начинается сложная цепь нервных процессов, обеспечи-
вающая последующую длительную секреторную реакцию»2.
Позже Н. Н. Никитин тоже получил результаты, аналогичные данным
* Архив биологических наук, т. 37, в. f, 1935. Предварительное сообщение на-
печатано в «Докладах Академии наук СССР», т. IV, № 8—9, 1934, по представлению
акад. И. П. Павлова от 24. XI 1934 г.
’ Архив биологических наук, № 5—6, 1933, стр. 679.
ДЛИТЕЛЬНОСТЬ УСЛОВНОГО РАЗДРАЖИТЕЛЯ И УСЛОВНЫЙ рефлекс
97
Л. А. Орбели, В. М. Добровольского, П. С. Купалова и Б. Н. Лукова.
В объяснении фактов Н. Н. Никитин не выходит за пределы взглядов
П. С. Купалова. Подобные явления наблюдал также И. С. Беритов
в отношении электрооборонительных двигательных условных рефлексов.
Случайно замеченный мной факт аналогичного порядка приковал мое
внимание, а знакомство с ранними работами и с данными И. С. Бернтова
и II. С. Купалова усилило интерес к обсуждаемому вопросу как к одному
из кардинальнейших вопросов физиологии нервной системы вообще и фи-
зиологии больших полушарий в частности. Я приступил к изучению этого
вопроса как на электрооборонительно-двигательиых, так и на пищевых
слюнных рефлексах. С целью возможно полного изучения вопроса приме-
нялось гораздо большее число вариаций опытов, чем в опытах всех назван-
ных выше авторов. Эго дало возможность выявить некоторые новые сущест-
венные моменты, а также дать иное объяснение наблюдавшимся ранее
фактам.
Пищевые условные рефлексы
Опыты проводились над тремя собаками: Арчем, Атласом и Голованом.
Первая находится у меня а работе уже 3 года. Собака с сильной нервной
системой, уравновешенная и, судя по всему материалу, собранному мной
заЗ года работы с ней, — сангвиник, т. е. с лабильными, подвижными нерв-
ными процессами. Вторая собака в работе почти год, но когда с ней были
начаты специальные опыты в данной области, она находилась в работе около
3 месяцев. Это ссбака с сильной нервной системой, достаточно уравновешен-
ная и тоже сангвиник. Внешнее поведение собаки позволяет думать, что
в ней немного преобладает возбудимость, хотя у нее вполне хорошо выра-
жен и тормозной процесс. Т ротья собака была в работе еще меньше времени,
когда с ней начались опыта,—всего 2 месяца. Это спокойная собака с очень
сильной нервной системой, уравновешенная, и есть основание думать, что
флегматик, т. е. собака, у которой калейдоскопически разнообразная и
сложная игра основных нервных процессов совершается медленно, инерт-
но, не так лабильно, как у сангвиника.
На трех собаках поставлено всего около 60 опытов.
Большинство опытов проводилось примерно в. одни и те же часы дня.
Для тс го чтобы можно было сравнить данные, полученные для какого-
нибудь одного раздражителя, ставились однотипные опыты (последователь-
ность раздражителей, паузы) и испытуемый раздражитель применялся на
одном и том же порядковом месте то более короткий, то более длительный
период времени.
Испытывались все имеющиеся налицо раздражители — слабые и силь-
ные. Продолжительность применения этих раздражителен варьировала от
3 секунд до 5 минут.
Слюнный рефлекс регистрировался через каждые 10 секунд до полного
затухания (примерно от 1 до 5 минут). Эти специальные опыты чередовались
с обыкновенными опытами, чтобы предотвратить возможное угасание испы-
туемых раздражителей и образование системности — моменты, которые мог-
ли существенно затемнить картину.
Несмотря на разницу в лабораторном «стаже» трех собак и на различную
типовую их принадлежность, у них получены в основном одинаковые
данные, поэтому мы рассматриваем их вместе.
Основные закономерности, наблюдавшиеся нами, следующие.
Величина условного слюнного рефлекса тем
больше, чем больше д л и те л ь н ос ть д е й ст в и я раз-
дражителя.
Раздражители применялись главным образом 3, 5, 10, 20 и 280 секунд.
Указанная закономерность очень четко выявлялась в каждый из этих
7 Э. Л. Асратян
98
Э. А. АСРАТЯН
отрезков времени, хотя, само собой разумеется, строго математической про-
порциональности между ними н полученными цифровыми выражениями
условных рефлексов ие было (табл. 1).
Таблица 1
Дата опыта Название раздражи- теля Порядки- в-ое MeciHj раздра- жителя д ласте --П.- иисть приме* нения раздра ; жителя 1' сек Величина уологнлга рефлекса в делениях шкалы Примечания
1-я ми- нута 2-я ми- нута 3-я .ми- нута 4-я ми- нута 5-я ми- нута всего
Собака Лрч
f 6.Х II 1932г. II.II 1933г. м„. м„. Третье 2» ° 55 10% 2 — _— 61 10 Vi
7.V М,„ •> 10 36 — — 36
13. V ми. » 285 75 22 9 22 128
27.VII 280 66 18 12 зи 101U В камере
4.VIII ж 20 38 5 З’/а — 46' .. жарко, на-
20.Х 11 1932г. Касалка Первое 20 72 43 46 35 214 блюдается
6.II » э 5 51 9 18 60 слаба я
9.II 1933г. 9 > 10 69 24 5 — — 98 (>ДЫи ка и
29. XI 1932 г. Звонок Второе 20 82 25 12 — — 119 про межу
15.XII 3 32 2 — — 31 точное слю-
3.11 1933г. » э 10 30 — — — — 30 ноотделение
13.11 > > 20 71 4% I’- 77
10.Х > » 285 81 31 ll 7 2% 132'
12. XI 1932г. Свет » 3 65 14 1 —> 80
2 XI » » 20 85 20 6 м- —- ill
24.11 1933 г. > 3 38 —- — — — 38
2.III » > 20 58 10 П4 — — 69%
Собака Атлас
3.1 II 1933г. Зво» ок Первое 20 36 II — — — 46
5.111 » » 5 19 1 —— — — £0
16.111 > > 20 41 10 — — 51
22.111 > » 210 44 н 8 — — 66 После
27.111 Касалка 50 64 а— —- .— 64 50 сек. or-
9.III » 20 42 10 8 2 — 62 ставлен ия
8-IV » » 5 26 4 4 2 — 36 подкреп-
7.V » » 230 61 23 10 1 —— 95 ляется
26.1П Свет Второе 20 43 9 12* 2 — 66 * Облизы-
2.IV » 5 11 4 2 —— — 17 вается
17. V » 230 44 21 9 6 — 80
Собака Головян
3.III 1933г. Звонок Первое 5 25 и 2 38
13.III » » 20 39 ' 18 6% — 63%
29 IV D Второе 20 30 8 — 38
14.V » 5 14 2 - - — 16
Нужно отметить, что условные рефлексы были особенно длительными,
суммарные их величины особенно большими на первом и затем втором по-
рядковом месте применения условных раздражителей; на последующих
местах они постепенно уменьшались. Очевидно, что это явление зависит от
повышенной пищевой возбудимости в начале опыта и от постепенного по-
нижения этой возбудимости в. середине и в конце опыта. Естественно, что
при сравнении данных, относящихся к отдельным раздражителям, это
обстоятельство нужно учесть.
Следует сказать, однако, что основную закономерность— зависимость
эффекта от Продолжительности действия раздражителя — удается выявить
и ври другом характере опытов. На всех трех собаках в разное время ста-
вились следующие опыты: в середине опыта применялся какой-нибудь.
ДЛИТЕЛЬНОСТЬ УСЛОГ НО-ГО РАЗДРАЖИТЕЛЯ И УСЛОВНЫЙ РЕФЛЕКС
99
один раздражитель короткое время (3—5 секунд), измерялась врднчиет
рефлекса в течение 1 — Р/г минут и он подкреплялся безусловным раздра-
жителем. Затем этот же раздражитель 3—4 раза подкреплялся безусл» в-
ным раздражителем и применялся снова, но уже более длительный период
(20--30 секунд). Ставились опыты и в обратной последовательности — сна-
чала 20—30 секунд, затем 3—5 секунд. Результаты таких комбинирован-
ных опытов с полной очевидностью показали наличие зависимости между
величиной рефлекса и продолжительностью действия раздражителя (табл. 2).
При внимательном рассмотрении приведенных цифр (табл. 1 и 2) нельзя
не заметить ряд интересных явлений.
Во-первых, бросается в глаза, что суммарные величины
условных рефлексов для всех отрезков времени особенно
велики при первых пробах раздражителей; при
последующих же пробах на тех же порядковых местах,несмотря на предпри-
нятые меры (см. табл. 1), эти суммарные величины заметно уменьшаются.
Звонок, например, у Арча, на втором месте первый раз вызывает условный
рефлекс, равный 119 делениям шкалы за 20 секунд действия, а в следующие
разы — 77 и 69 делениям. Свету той же собаки на втором месте обусловли-
вает первый раз слюноотделение, равное 80 делениям шкалы за 3 секунды
действия, а в следующие разы — 39 и 30 делениям и т. д.
Из этого следует, что при сравнении данных лучше брать опыты, по вре-
мени более или менее близкие друг к другу.
Во-вторых, в начале опыта, в особенности на первом по-
рядковом месте, зависимость величины рефлекса
от продолжительности действия раздражителя
как будто проявляется слабее, чем в середине
нв конце опытов. Это довольно ясно видно при сравнении данных
табл. 1 с данными табл. 2.
• Таблица 2
Дата опыта Название раз1ражитсля Порядко- вое место раздражи- теля Длитель- ность при- менен ИЯ ра:д др;) жи- теля и «сек Величина условного рефлекса в делениях шкалы При мечаиис
1-я минута 2-я минута в-сего
2.11! 1933 г. 17. XII 6.II 11.11 5 III 14. V Касалка » Звонок » Шипенье Mis# Mu<, Звонок л Седьмое Двенад- цатое Третье Седьмое Шестое Девятое Т ретье Шестое Второе Четвер- то? Второе Седьмое 5 20 20 5 20. 5 10 5 20 5 2'3 29 14 13 44 10'/, 31% 19 34 И 22 е.-т | ] | | | J-?’ СЧ сч 31 48 13 14 15 39 ю% 31% 20 38% 16 24 Первые 4 от- рывка взяты из опытов на Ар- че, 5- й—нз опы- тов на Атласе, 6-й—из опытов на Головане
В-третьих, выявляется еще 'одна существенная подробность: создается
впечатление, будто даже при приблизительно одинаковых
прочих условиях слабые раздражители (свет,
касалка) вызывают более продолжительный и зна-
чительный суммарный у с л о в н ы и э ф ф е к т, чем силь-
ные (звонок, стук).
Это как бы парадоксальное явление выступает как в начале, так и в се-
редине опытов. Оно одинаково хорошо проявляется у всех трех собак.
7»
100
Э. Л АСРАТЯН
В-четвертых,зависимость величины суммарного эф-
фекта от продолжительности применения раз-
дражителя выступает при сильных раздражите-
лях сильнее, чем при слабых.
Это явление тоже одинаково хоропо заметно у всех собак и при любом
порядковом месте применения раздражителей.
Прежде чем пытаться объяснить описанные явления, приведу рис. 1,
который более подробно иллюстрирует ход рефлексов при 5-, 10-, 20- и
280-секундных действиях раздражителя.
Рис. 1. Величина и продолжительность условного рефлекса на звонок»
действующий в течение 5, 10, 20 и 280 сек.
На рис. 1 обращает на себя особое внимание волнообразный (зигзаго-
образный) ход рефлекса при длительных действиях условного раздражителя,
что, очевидно, свидетельствует о столкновении процессов возбуждения и
торможения.
Элсктрссборонительно-двигательные рефлексы
В области оборонительно-двигательных условных рефлексов вопрос
о связи между длительностью действия условного раздражителя и величи-
ной рефлекса специально ником еще не разрабатывался. Правда, в моногра-
фии И. С. Беритова «Индивидуально приобретенная деятельность цент-
ральной нервной системы» имеется одно замечание по этому поводу. По
его данным, в известных случаях повышенной возбудимости «длительность
бурной реакции в индивидуальном (условном. — Э. Л.) рефлексе ire зависит
от длительности раздражения. Она совершенно одинакова как при раздра-
жении в 1—2 секунды, так и при раздражении в десятки секунд»
(стр. 220—221). Какова картина в других случаях, в частности, при от-
сутствии повышенной возбудимости, в монографии ничего не сказано
Пользуясь тем, что у меня в опытах был ряд собак с оборонительно*
двигательными условными рефлексами, я на них попутно изучал также этот
вопрос.
ДЛИТЕЛЬНОСТЬ УСЛОВНОГО РАЗДРАЖИТЕЛЯ И УСЛОВНЫЙ РЕФЛЕКС Ю1
На основании собранного мной экспериментального материала можно
сказать, что при одинаковых примерно условиях эксперимента длитель-
ность и величина условных оборонительно-двигательных рефлексов
находятся в прямой зависимости от длительности действия условного
раздражителя: чем длительнее последний, тем больше первый. Сказанное
хорошо видно на рис. 2.
Правда, эта зависимость ярче выступает тогда, когда собака спокойна,
чем тогда, когда она возбуждена и дает бурные реакции на те или иные раз-
дражения. Но утверждать, что эта зависимость совершенно исчезает в слу-
чаях, когда собака возбуждена, на основании своего материала я немоту.
Рис. 2. Зависимость продолжительности двигательных
электрообороиительны.х условных рефлексов от длительности
применения разных условных раздражителей
г касалка 5 и 30 сек.; Ч — свет 5 и 35 сек.; ///—свет 5 и 30 сек.;
/V—шипенм; 5 и 40 сек.; V—тон С 5 и 40 сек.; VI—М,,. 5 н 15 сек.
Линин сверху вниз; вались рефлекса, отметка безусловного раздра-
жителя. отметка условного раздражителя, время в секундах
Одна из моих собак (Назар) в известный период работы бурно реаги-
ровала как на условные, так и в оссбенности на безусловные раздражения.
Собака лаяла, визжала, иногда держала конечность в положении флексии
в течение десятков секунд и даже нескольких минут. На этой собаке в
указанном периоде я специально изучал зависимость величины рефлек са
от длительности действия раздражителя и должен сказать, что эта зависи-
мость почти всегда отчетливо выступала. На рис. 3 эта зависимость
иллюстрируется при возбужденном состоянии животного.
В конце концов, ту же пробу я проделал на одной сильно возбудимой
собаке (Чинар) с отставленными на минуту оборонительно-двигательными
условными рефлексами. Собака очень бурно и почти непрерывно реагирова-
ла при одноминутном действии единственного условного раздражителя —
звонка. Проба на 20-секундное действие условного раздражителя показала,
что даже в случае необычно долгого отставления оборонительно-двигатель-
ных условных рефлексов продолжительность действия раздражителя играет
существенную роль (рис. 4).
102 Э. А. АСРАТЯН
Сравнивая полученные данные с секреторными и оборонительно-двига-
тельными рефлексами, можно сказать: в силу того, что обычно УСЛОВНЫЙ
(а также безусловный) оборонительный рефлекс вызывается главным обра-
зом во время действия раздражителя, зависимость между длитсль-
Рис. 3. Собака, Назар. Зарисммссть продолжительности дишаюльного
эдектрооборонительпого условного рефлекса на тон С от длительности
его применения (3 и 40 сек )
Линии сверху вниз: запись рефлекса, безусловный раздражитель, услсв пай
раздражитель <тон С), время в секундах
костью действия условного раздражителя и величиной условного реф-
лекса здесь выступает ярче, чем в случае выработки слюнных пищевых
рефлексов.
Рис. 4. Собака Чинар. Зависимость продолжительности двигатель-
ного электрооборонительноСО условного рефлекса на звонок От
длительности его применения (60 и 20 сек.)
Линии сверху вниз: запись рефлекса, безусловный раздражитель (uwifpu
расстояние между катушками в сантиметрах), условный раздражитель,
время в секундах
Теоретические рассуждения
Как понять полученные факты с точки зрения физиологии нервной си-
стемы?
Во-первых, мне думается, что полученный экспериментальный материал
находится в полном соответствии с основными фактами и теоретическими
положениями современной физиологии нервной системы. Не говоря уже
о простом нервном волокне, в особенности соматическом, длительность воз-
буждения которого в известных пределах находится в тесной зависимости от
длительности его раздражения, напомню о достоверных данных относитель-
но спинального рефлекторного аппарата. Из работ Шеррингтона (Sherring-
ton) известно, что длительность и сила спинальных рефлексов находятся
в тесной зависимости не только от силы стимулятора, нои от продолжи-
тельности его действия. То' же можно сказать и о бульбарных рефлексах,
не говоря уже о тонических. Больше того, в опытах Шеррингтона даже дли-
ДЛИТЕЛЬНОСТЬ УСЛОВНОГО РАЗДРАЖИТЕЛЯ И УСЛОВНЫЙ РЕФЛЕКС
103
тельность так называемого after-discharge в сгибательных, чесательных и
других рефлексах явно зависела от длительности применения стимулятора.
В приведенных же выше данных о слюнных рефлексах при длительности
раздражения 3—5 и 30—60секунд, как нетрудно заметить, разница в сум-
марных условнорефлекторных эффектах зависела не отвремени применения
раздражителя (в течение 30—GO секунд), а главным образом от последей-
ствия, иначе говоря, от того же after-discharge. Разумеется, положительное
последействие может проявляться только в известных пределах действия
раздражителя. Когда раздражитель действует очень долго, на сцену вы-
ступают другие закономерности (ив первую очередь торможение, адапта-
ция и т. д.), которые снимают вопрос о положительном последействии раз-
дражителя.
Что касается интимной стороны вопроса, то трудно сказать что-либо
определенное: пока мы еще мало знаем об истинной природе нервного воз-
буждения. На основании работ ряда физиологов л главным образом работ
Н. Е. Введенского, А. Ф. Самойлова и Эдриана (Adrian) можно допустить,
что в известных пределах длительное действ-ие раздражителя обусловливает
более продолжительную генерацию нервных импульсов в соответствующих
звеньях сложной рефлекторной дуги, чем кратковременное его действие,
и что, следовательно, для возникновения импульсов имеют значение не
только начален интенсивность раздражения, но и его д л и тел ь-
н о с т ь.
Однако приведенные выше данные показывают также, что не все зависит
от стимулятора и что для деятельности больших полушарий, так же как и
для функции нервов и низших отделов центральной нервной системы,сущест-
венное значение имеет их исходное функциональное состояние. Например,
повышенная возбудимость нервной системы в начале опыта обусловливает
более длительный и мощный эффект на один и тот же раздражитель; у собак
различных типов получаются различные вариации и т. п.
Очевидно, повышенной возбудимостью нервной системы следует объяс-
нить тотфакт, что па первых порядковых местах в ряду условных раздражи-
телей связь между длительностью действия раздражителя и суммарной
величиной слюнного условного рефлекса проявляется менее четко, чем на
срединных и последних местах этого ряда, когда возбудимость заметно
падает.
Труднее объяснить различия, наблюдаемые при применении слабых и
сильных раздражителей. Почему длительность и суммарная величина услов-
ных слюнных рефлексов при слабых раздражителях больше, чем при силь-
ных? Почему связь .между длительностью действия раздражителя н величи-
ной условного рефлекса в первом случае выступает менее ярко, чем во
втором? Я склонен объяснить это следующим образом: при действии силь-
ного раздражителя, наряду в процессом возбуждения, в звеньях рефлектор-
ной слюнной цепи возникает процесс торможения, который сильно ограни-
чивает, задерживает дальнейший рост и ход условного рефлекса. При дей-
ствии же слабого раздражителя возникшее торможение сравнительно слабее
возбуждения и поэтому не может его быстро и заметно затормозить.
В результате условный рефлекс действует Сравнительно долго и суммарно
получается больше. Это объяснение делает понятным также, почему связь
между величиной эффекта и длительностью действия стимулятора высту-
пает для сильных раздражителен ярче, чем для слабых.
Мне думается, что подобное объяснение имеет во всяком случае некото-
рое основание, поскольку эти данные аналогичны таким известным в физио-
логии данным, как феномен Введенского (pessimum) для нервного волокна,
явление торможения или извращения спинальных рефлексов при сверх-
сильных раздражениях, изученное II. М. Сеченовым и позже Шеррингтоном,
и, наконец, явление запредельного торможения, которое в лабораториях
104
Э Л. АСРАТЯН
акад. И. П. Павлова за последние годы приобретает характер закономерного
явления для физиологии больших полушарий. Можно даже сказать, что
в описанных нами конкретных случаях мы имеем дело со своеобразным
проявлением запредельного торможения.
Что касается согласования моих данных с фактическими данными
П. С. Купалова и Б. Н. Лукова, то, по моему мнению, противоречия между
ними нет: они друг друга не отрицают, а лишь дополняют. П. С. Купал»
свои данные получил в основном на возбудимой ссбакеи имел дело с непол-
ными, незаконченными рефлексами, с начальной их частью, я же изучал
полноценные,закопченные рефлексы.
С основными теоретическими соображениями и выводами Я. С. Ку-
палова и Б. Н. Лукова я никак не могу согласиться. Я полагаю, что на осно-
вании частных данных они делали довольно широкие обобщения о ряде важ-
нейших явлений условнорефтекторной деятельности—обобщения, подобные
цитированным выше, или такого рода, что «количество нервных импульсов,
идущих от эфферентной части дуги условного рефлекса, одинаково как
в случае короткого,так и в случае длительного раздражения», Мне думается,
что их фактический материал недостаточен для этого, так же как и для при-
веденных выше выводов относительно значения начала и длитель-
ности раздражения. Ведь в их опытах в случае кратковременного
применения раздражителей проявлялись эффекты как наличного действия,
так и последействия (27, 87 секунд и т. д.), а в случае длительного приме-
нения раздражителей — только эффекты наличного действия, эффект же
последействия совершенно не принимался в расчет! Это важное упущение.
Против этих выводов говорят все мои данные. Больше того, против этих
выводов и в особенности против их обобщений говорят некоторые данные
самих авторов — данные, которые ими небыли приняты в расчет при по-
строении ими теоретических положений. Речь идет о сохранении зависимо-
сти между длительностью действия условного раздражителя и велич иной
условного рефлекса при весьма кратковременных воздействиях раздражи-
телем (о чем авторы почему-то сообщают только в выводах) и о существо-
вании аналогичных соотношешгй у одной собаки (сангвиник) при приме
нении раздражителя в течение 10 и 90 секунд.
Выводы
1. При приблизительно одинаковых прочих условиях эксперимента ве-
личина пищевого условного рефлекса тел» больше, чем длительнее (в. из-
вестных пределах) применение условного раздражителя.
2, Связь между величиной рефлекса и длительностью действия раздра-
жителя при сильных и средних раздражителях выражена больше, чем при
слабых раздражителях.
3. Связь эта выступает в середине и конце опытов более четко, чем в на-
чале их, когда пищевая возбудимость сильно повышена.
4. Часто суммарный условный рефлекс под воздействием слабых
раздражителен больше и длительнее, чем при применении сильных раздра-
жителей.
5. Прямая зависимость между длительностью действия услов-ного раз-
дражителя и величиной условного рефлекса еще ярче выступает на электро-
оборонительно-двигательных условных рефлексах, — вероятно, в силу
сравнительно слабой выраженности последействия на этих рефлексах.
Эти данные говорят о том, что для величины условного рефлекса имеют
значение не только интенсивность и начало действия раздражи-
теля, но, в известных пределах, и продолжительность его дей-
ствия, точно так же, как это имеет место при раздражении нервного волокна,
при спинальных, бульбарных и тонических рефлексах. Факты, приведен-
ДЛИТЕЛЬНОСТЬ УСЛОВНОГО РАЗДРАЖИТЕЛЯ И УСЛОВНЫЙ РЕФЛЕКС
105
нЫё в л. 2 и 3, можно понимать как результат более быстрого воз-
никновения и более резкого вмешательства торможения при действии
сильных раздражителей, чем при действии слабых раздражителей.
«Pcssimum» II. Е. Введенского, установленный впервые И. М. Сечено-
вым факт торможения спинальных рефлексов И «запредельное тормо-
жение» условных рефлексов, установленное акад. И. П. Павловым,
дают основание ДЛЯ такого предложения.
ЛИТЕРАТУРА
Керитов И. С., Индивидуально приобретенная деятельность центральной нерв-
ной системы, 1932.
Введенский Н. Е., О соотношениях между возбуждением и торможением, 1896.
Добровольский В. М., О пищевых следовых рефлексах, дисс., 1911.
Купалов П. С. и Л у к о в Б. Н., Действие короткого применения условного.
раздражителя, Архив биологических наук, № 5—6, 1933,
Купа л о в П. С., Угашение условного рефлекса при длинном и коротком приме-
нении условного рздражитЬля, Архив биологических наук, № 5—6, 1933.
Н и к и т и н Н. Н., О анзиологнческом понимании природы инстинкта. Архив био-
логических наук, № 5—6, 1933.
Орбели Л. А., Условные рефлексы с глаза у собак, дисс., 1908.
Adrian Е., The mechanism of nervous action, 1932.
Magnus R.. KorpeTstllung, Berlin, 1994.
Sherrington Ch., RTh integrative action f the nervous system, 1911.
S h e г г i n g t о n Ch., Refelex activity of the spinal cord, 1932.
К ФИЗИОЛОГИИ ИРРАДИАЦИИ И
В КОРЕ БОЛЬШИХ
КОНЦЕНТРАЦИИ ПРОЦЕССОВ
ПОЛУШАРИЙ 1
сследованием иррадиации и концентрации процессов воз-
буждения и торможения в коре больших полушарий по
методу условных рефлексов занимаются в лабораториях
акад. И. П. Павлова примерно с 1910—1911 гг. Работами мно-
гочисленных учеников Н.П. Павловабыл изучен и выяснен ряд
существенных сторон иррадиации и концентрации этих про-
цессов, как то: последовательность, градация, начало, характер и быстрота
иррадиации и концентрации, значение силы процесса, типа собаки и т. д.
(Н. II. Красногорский, М. Я. Безбокая, Б. А. Коган, М. К. Петрова,
Д. С. Фурсиков, Г. В. Анреп, К- М. Быков, II. А. Подкопаев и др ). Что
же касается интимной стороны, «механизма» иррадиации и концентрации
процессов в коре, то этим вопросом почти никто специально не занимался
ни в наших лабораториях, ни в других. Однако па основании накопленного
огромного экспериментального материала было создано представление, что
при иррадиации какого-нибудь процесса из одного очага мы имеем радиар-
ное, постепенное распространение этого процесса на периферию, а при
концентрации — обратный ход, круговую концентрацию рассеянного про-
цесса в исходный очаг. Грубо говоря, при таком понимании вопроса все
энергетические затраты как бы приписываются центральному очагу, кото-
рый рассеивает, тратит энергию (процесс) и ослабевает, затем эта энергия
(процесс) вновь собирается в исходный пункт и вызывает его усиление.
С целью уточнения наших знаний об интимной стороне некоторых слу-
чаев иррадиации и концентрации процессов в коре больших полушарий я
занялся непосредственной экспериментальной проверкой правильности
приведенного выше Взгляда на Иррадиацию И концентрацию угасательного,
диференцировочного и условного торможения. Полученный эксперимен-
тальный материал, как мне кажется, не подтверждает этого взгляда и дает
основание взамен его выдвинуть другое, как будто более подходящее объяс-
нение «механизма» иррадиации и концентрации процессов в коре в назван-
ных случаях.
Опыты ставились иа трех собаках: Арче, Атласе и Головане. Две первые
собаки — сангвиники, третья — флегматик. В ряде специальных опытов
изучалась иррадиация и концентрация угасательного торможения. Такие
опыты ставились изредка по ходу выполнения других работ на тех же со-
баках в течение 1933—1934 гг. Число специальных опытов составляет око-
ло 30. Собранный фактический материал сводится к следующему.
Если взять два примерно одинаковых по величине условных положи-
тельных рефлекса на различные раздражители и в течение одного опытного
сеанса, не давая подкрепления, быстро угашать- их и затем путем под-
* Доклады Академии наук СССР, 1934. Представлено акад. И. П. Павловым
24.VIII 1934 г.
ИРРАДИАЦИЯ И КОНЦЕНТРАЦИЯ ПРОЦЕССОВ В КОРЕ БОЛЬШИХ ПОЛУШАРИЙ Ю7
крепления через 2—3-минутные паузы также быстро восстанавливать один
из рефлексов, то часто можно наблюдать явление такого порядка: в то
время как первично угашенный рефлекс успевает значительно восстано-
виться, второй рефлекс, охваченный торможением лишь вторичным, кос*
венным путем —путем иррадиации к нему торможения с первично угашен-
ного рефлекса, остается еще значительно заторможенным. Подобные дан-
ные более или менее четко получились у всех трех собак как с рефлексами
с различных анализаторов (большей частью на свет и на касалку), так и
с рефлексами из одного анализатора (на звонок и метроном).
Для иллюстрации привожу опыт на собаке Атлас (см. таблицу).
Время 11 мин. Название услов- ных раздражи- телей Время изоли- рованного действ-и я УС- ЛОВНЫХ раз- дражителей и мин. Латент- ный пери- од рефлек- са в мин. Величина ус- ловного пгф- лекса за 20 МИН. Ы де 1 - нмях шкалы Примечания
20 Касалка • 20 23 Подкрепление рефлекс
25 Звонок 20 1 25 » »
30 Свет 20 2 16 > »
35 Касалка ' 20 2 17 » »•
40 Спет 20 2 15*4
45 Касалка 20 2 16 Отсутнвнеподкреп пенни
50 » 20 5 6 э »
52«Л » 20 7 Следы
55 > 20 0 » »
Б7% » 20 — 0 » >
60 » 20 — 0 Подкрепление рефлекса
«2% » 2 — 0 » »
65 > 2 —— 0 » »
67 *» 2 — 0 » »
72% 20 3 12 > »
77*4 Свет 20 8 5 » 9»
82% Звонок 20 4 13 а к
87% Касалка 20 5
В некоторых опытах после угашения первого рефлекса и непосредст-
венно перед его восстановлением я, чтобы удостовериться в глубине за-
торможенности второго рефлекса, однократно применял второй раздражи-
тель, подкрепляя его. В самом начале экспериментирования, когда про-
цессы иррадиировали и концентрировались сравнительно медленно, даже
такой прием не помешал проявлению указанного феномена. Впоследствии,
очевидно, в силу тренировки и ускорения угашения., восстановления, ир-
радиации и концентрации, при таком приеме экспериментирования интере-
сующее нас явление выражалось менее четко. Прием же, иллюстрированный
в опыте, всегда вызывал это явление.
Если рассматривать явления иррадиации и концентрации торможения
как распространение этого активного процесса из первичного очага по
сторонам и возвращение того же процесса к исходному очагу, полученные
мной данные, как мне кажется, необъяснимы. При последовательном Про-
ведении этого принципа не должны наблюдаться случаи, когда первичный
•очаг торможения освобождается от последнего раньше, чем вторично за-
торможенные очаги на периферии. В проводимых мной опытах торможение
возникает в известном очаге, распространяется по сторонам и захватывает
периферические очаги; затем, благодаря особому приему экспериментиро-
вания, оно быстрее исчезает из первичного очага, чем из периферических.
На основании этих данных я считаю возможным предложить следующее
•объяснение иррадиации и концентрации торможения, объяснение, которое
108
Э А. АСРАТЯН
основано изданных современной физиологии центральной нервной си-
стемы.
С развиваемой нами точки зрения, иррадиация торможения не есть его
рассеивание из какого-либо очага периферии; это — явление, при котором
активное по своей природе фазное торможение одного очага коры обус-
ловливает возникновение особого состоя ния торможения в данном оча-
ге, непосредственно передающегося на прилегающие участки коры;
последние в свою очередь «заражают» соседние, и так чем дальше, тем сла-
бее и слабее. Это не пассивное проведение, а активная передача. Нерв-
ные элементы — не пассивные проводники активного процесса, а активные
передатчики своеобразного тонуса, вернее, функционального состояния,
подобного центральному состоянию торможения. При концентрации тормо-
жения происходит не возвращение этого активного процесса в исходный
очаг, а постепенное исчезновение этого состояния из всей охваченной им
области. И так как от центра к периферии мы имеем постепенно ослабеваю-
щие по силе «состояния», естественно, что при прочих равных условиях это
состояв не быстрее исчезнет на далекой периферии, затем постепенно из более
близких очагов и позднее всего из центрального очага. Если же применять
особые приемы с целью быстрой ликвидации этого состояния из централь-
ного очага, как это делалось мной в опытах, можно добиться того, что со-
стояние торможения быстрее исчезнет из центра и позже на периферии.
Таким образом, согласно этому представлению, «концентрация» тор-
можения является не истинной концентрацией активного процесса в исход-
ном очаге, а как бы последовательным и концентрическим исчезновением
состояния торможения из известной области,
Совершенно так же можно объяснить иррадиацию и концентрацию
торможения в случаях последовательного торможения от диференцировоч-
ного раздражения и условного тормоза.
В опытах с собаками Атлас и Голован я получил экспериментальные
данные другого порядка, которые представляют самостоятельный интерес
и одновременно подкрепляют высказанный взгляд об иррадиации и кон-
центрации торможения.
Если изо дня в день в опытах с условными рефлексами применять раз-
дражители в такой фиксированной последовательности (системе), когда
отрицательный условный раздражитель занимает срединное место, то
несколько дней спустя, по крайней мере у некоторых собак, возникает
следующее фазовое явление: ослабевают не только тс положительные услов-
ные рефлексы, которые следуют за отрицательным рефлексом, но становится
значительно слабее также тот положительный условный рефлекс, который
вызывается непосредственно перед применением тормозного условного
раздражителя. Иногда ослабевает даже следующий за ним рефлекс. Говоря
иначе, торможение во времени иррадиирует не только после, ноидосвоего
активного возникновения. Условно я второй вид торможения называю
ретроградным.
Как понять это парадоксальное явление?
Если исходить из старого представления об иррадиации и концентра-
ции, то ретроградное торможение не только необъяснимо, но кажется даже
абсурдом. Однако этот факт, кстати сказать, был подтвержден работами
некоторых других сотрудников наших лабораторий.
Мне это явление представляется следующим образом. Как было пока-
зано водной моей работе и исследованиях ряда других учеников И. П. Пав-
лова, при применении системы раздражителей изо дня в день рефлексы,
вызываемые этими раздражителями, синтезируются, образуя системность.
Возникает своеобразная цепь условных рефлексов, звенья которой на-
ходятся в тесном взаимодействии. Каждое действующее звено подготовляет,
адаптирует к действию следующее звено, а можетбыть, и следующие звенья.
ИРРАДИАЦИЯ И КОНЦЕНТРАЦИЯ ПРОЦЕССОВ В КОРЕ БОЛЬШИХ ПОЛУШАРИИ Ц)9
Если эту последовательную подготовку представить себе тоже в виде изме-
нения состояния (а другого представления не может быть), то можно до-
пустить, что действие звеньев друг на друга будет осуществляться (хотя бы
в ограниченных пределах) также в обратной последовательности. Легко
понять и механизм описанного случая ретроградного торможения во вре-
мени, когда стоящий на шестом порядковом месте тормозной рефлекс вы-
зывает ослабление положительных рефлексов, стоящих на пятом и даже на
четвертом месте: осуществление рефлексов, находящихся на втором или
третьем месте, и обусловливает уже соответствующее состояниев очаге тор-
мозного рефлекса; это состояние d свою очередь по известному нам способу
распространяется на очаги рефлексов, выработанных раньше его.
Впервые эти данные были получены мной с условным тормозом в на-
чале 1933 г.; затем они подтвердились и надиференцировочиоч торможении.
Фазовое явление держится у двух моих собак 8—12 дней, затем исче-
зает. Появление и исчезновение осуществляются постепенно.
Па основании своих опытов Г. В. Анреп пришел к выводу, что торможе-
ние от условного тормоза иррадиирует и концентрируется каким-то особым
образом, отличным от иррадиации и концентрации диференцировочного
и угасательного торможении. В первом случае будто бы наблюдается ие
постепенный захват торможением вторичных рефлексов и освобождение от
него, а имеют место одновременно захват и освобождение.
Мои данные, полученные на Атласе и Головане, позволяют утверждать,
что Анреп имел дело с совершенно особым, частным случаем. В моих опытах
торможение от условного тормоза иррадиировало и концентрировалось
в основном так же, как и другие виды торможения.
Кстати, хотя я не располагаю экспериментальным материалом по ирра-
диации и концентрации процесса возбуждения по коре, я думаю, что, исходя
из нового понимания, можно лучше объяснить по крайней мере некоторые
стучаи иррадиации и концентрации возбуждения. Действительно, если при-
держиваться взгляда, что при иррадиации возбуждения по коре мы имеем
распространение активного процесса из очага А на соседние и отдаленные
очаги В, С и D, то становится непонятным, почему вторично возбужденные
пункты В, С и D ничем не дают о себе знать. Законно было бы ожидать,
что если они охвачены активным процессом возбуждения, иначе говоря,
если они возбуждены, то они должны были это спонтанно проявлять по-
средством рефлекса. А между тем в действительности мы этого не видим.
О наличии иррадиации возбуждения в пункты В, С и D мы узнаем лищь
тогда, когда применяем соответствующие им раздражители (в случае ге-
нерализации, в известных опытах М. К. Петровой и т. д.). Если же стоять
на точке зрения, что в названных случаях при иррадиации возбуждения
по коре мы имеем постепенную .передачу известного как бы тонического
состояния возбудимости, а при концентрации—закономерное, постепенное
исчезновение этого состояния, то все становится ясным.
Что же касается сущности этих состояний торможения и возбуждения,
то трудно сказать, обусловливаются ли они какими-то гуморальными суб-
станциями, электротоническими изменениями или какими-нибудь другими
моментами.
Разумеется, что при известной степени интенсивности описанные спе-
цифические, как бы тонические состояния корковых очагов могут перейти
в явно деятельное состояние, как это допускается для соответствующих
состояний возбуждения («excitatory state») в спинном мозгу. Затем хотелось
бы оговориться, что для других случаев корковой деятельности, как, на-
пример, для Bahnunj,- и суммаций фазных нервных процессов, следует до-
пустить в основном такой вид Иррадиации, как это принято в учении об
условных рефлексах, т. е. распространение возбуждения по корковой массе
в виде нервных импульсов..
К УЧЕНИЮ О ФИЗИОЛОГИЧЕСКОЙ ЛАБИЛЬНОСТИ ВЫСШИХ
ЦЕНТРАЛЬНЫХ ЭТАЖЕЙ 1
лнжайшее знакомство с основными фактами, лежащими в
основе учения И. П. Павлова об условных рефлексах, а
также с фактами современной физиологии нервной системы
в целом делает очевидным, что и работа большого мозга выс-
ших животных, этого доминирующего во всей центральной
нервной системе высшего анализирующего и синтезирующего
органа, основана в значительной степени на закономерностях, общих для
всей нервной деятельности. Больше того, более интимный физиологиче-
ский анализ фактов, специфических для коры большого мозга высших жи-
вотных, как, например, замыкания новых временных связей, показывает,
что и эти особые виды кортикальной деятельности по павловскому прин-
ципу «естественной общности Основных Отношений» Имеют своих Отда-
ленных аналогов в деятельности более низких сегментов центральной
нервной системы. Речь идет не о внешних аналогиях, носплошь и рядом о
настоящем генетическом сродстве, о внутренней связи.
В качестве иллюстрации можно привести ряд хорошо известных при-
меров, как то: связь и родство между явлениями замыкания условной
связи и общими для всей центральной нервной системы явлениями про-
торения и доминанты (А. А. Ухтомский); почти тождество или во всяком
случае ближайшее родство установленных для условных рефлексов силовых
соотношений между стимулом и эффектом с такими же соотношениями, при-
сущими всем рефлекторным актам и даже деятельности еще более простых
нервных приборов (Н. Е. Введенский, Шеррингтон, Магнус и др.); анало-
гия и родство между явлениями суммации специфических корковых процес-
сов и процессов в других центральных и периферических образованиях;
родство или даже более тесная связь между корковыми явлениями индук-
ции и иррадиации нервных процессов, с одной стороны, и явлениями,
обозначаемыми теми же названиями в низших центральных этажах, — с
другой, не говоря уже о прототипе этих явлений в нервном проводнике в
виде электротоническнх, периэлектротонических явлений; аналогия после-
довательного протекания пар абиотических стадий или гипнотических фаз
в корей других более простых нервных образованиях; наконец, родство
и аналогия между многообразными проявлениями борьбы и взаимодей-
ствия процессов возбуждения и торможения в коре и других нервных
образованиях
Указывая на эти общеизвестные факты связи, родства и аналогии между
нервными процессами и явлениями в коре и нижележащих отделах централь-
ной нервной системы, я далек от стремления ставить знак равенства между
1 Ученые записки ЛГУ, № 41, 1939. Предварительное сообщение опубликовано
в сб. «Материалы к V Всесоюзному съезду физиологов, биохимиков и фармакологов»,
1934, и в сборнике тезисов докладов XV Международного физиологического конгресса,
1935, по представлению акад. И. П. Павлова.
ФИЗИОЛОГИЧЕСКАЯ ЛАБИЛЬНОСТЬ ВЫСШИХ ЦЕНЗ РЛЛЬНЫХ ЭТАЖЕЙ
1 II
ними или от механического нивелирования их. Очевидно, что не только кора
большого мозга, но каждая достаточно морфологически и функционально
отдиференцированная часть центральной нервной системы в процессе эво-
люции и специализации функции вносит свой собственный корректив, свой
качественно специфический характер в общие закономерности нервной
деятельности.
История развития теории и практики павловского учения об условных
рефлексах показывает, как полезно при исследовательской работе изучать
частное, специфическое для данного органа, не забывать об общем с дру-
гими родственными образованиями и о генетической, эволюционной связи
этого специфического и частного с тем общим, что было до его диференци-
ровкн и специализации. Рассматривая историю возникновения учения
об условных рефлексах, нельзя не заметит!, что попытки внедрения
в. него общих закономерностей и принципов физиологии нервной си-
стемы сыграли большую роль в его дальнейшем развитии. Сближение
закономерностей условных рефлексов с общими закономерностями физио-
логии нервной системы, проводившееся как самим И. П. Павловым, так
и некоторыми его учениками (Д. С. Фурсиков, И. П. Разенков), каждый
раз являлось новым стимулом для развития теории и практики условных
рефлексов. Трудно спорить против этого!
Во избежание возможных недоразумений повторяю еще раз, что все
это отнюдь не означает, будто кора большого мозга не имеет своих особых,
именно ей присущих законов действия. Речь идет лишь о той пользе, ко-
торую принесло учению об условных рефлексах, этому величайшему до-
стижению современного естествознания, критическое сопоставление с дан-
ными других отделов физиологии нервной системы. Это так же естественно,
как естественно и обратное влияние учения об условных рефлексах на по-
нимание явлений в других разделах нервной системы или даже в других
областях естествознания. Основной причиной, побудившей меня остано-
виться на этих элементарных явлениях, было желание на этот раз уже в пе-
чати отметить опасность тенденции, которая имеется у многих работни-
ков по условным рефлексам и которая заключается в забвении и даже
игнорировании практической и теоретической связи, существующей между
изучением условных рефлексов и изучением физиологии прочих отделов,
нервной системы или других систем организма. Эта опасная тенденция заб-
вения органической связи между отдельными частями нервной системы и
прочими физиологическими системами и игнорирование зависимостей,
установленных в трудах классиков общей: физиологии нервной системы
(И. М. Сеченов, И. П. Павлов, Н. Е. Введенский), тенденция везде и
всюду видеть только «специфшческое» для условных рефлексов льет воду
на мельницу тех немногочисленных зарубежных и отечественных ученых,
которые стали придерживаться ошибочного мнения, будто условные реф-
лексы — это не физиология.
Будучи горячим и убежденным сторонником укрепления связей между
физиологией условных рефлексов и общей физиологией нервной системы,
я всемерно старался выяснить эти связи для конкретных случаев. По ходу
моей работы я убедился, что при беспристрастном отношении многие из
кажущихся преград и противоречий между учением И. П. Павлова и уче-
ниями других передовых отечественных нейрофизиологов, в первую очередь.
Н. Е. Введенского, исчезают, и вместо этого обнаруживается глубокая
идейная близость. На собственном опыте я учился тому, чему учит вся исто-
рия развития нашей науки: взаимопонимание и контакт при носят огромную-
пользу дорогому для всех нас делу И. П. Павлова и других классиков ней-
рофизиологи.
В связи с этим я не могу нс отметить ту неоценимую пользу для моей
работы в области условных рефлексов, которую я постоянно извлекал из
1 12
Э. А. АСРАТЯН
богатого научного наследия выдающегося русского ученого Н. Е. Введен-
ского, по мере того как углублялось мое знакомство с его замечательным
научным творчеством. Вопрос об использовании для развития физиология
условных рефлексов принципа «относительной функциональной подвиж-
ности (лабильности)» Н. Е. Введенского, ныне так успешно развиваемого
его школой во главе с А. Л. Ухтомским, возник передо мной в 1932/33 г.
в связи с проблемой физиологической характеристики'тнпов собак. Стиму-
лирующим фактором было более углубленное, чем в 1927 г., знакомство с
научным наследием Н. Е. Введенского и с текущей научной работой лабо-
ратории Л. А. Ухтомского.
Известную роль сыграл и тот факт, что в физиологической лаборатории
Научного института им. Лесгафта, где я тогда работал, под моим наблюде-
нием были две собаки (Атлас и Голован), одна из которых была типичным
сангвиником, а другая типичным флегматиком, судя по их внешнему поведе-
нию. В процессе подготовки животных для намеченной темы я применял
одинаковые приемы работы с ними как при выработке положительных и
отрицательных условных рефлексов, так и при ряде функциональных проб.
При этом были получены очень интересные данные. Кроме того, работая
в Физиологическом институте им. И. П. Павлова, я не только теоретически,
как раньше, но и практически мог видеть, с каким трудом поддается теоре-
тической и экспериментальной диференциации физиологическая приро-
рода флегматика и сангвиника. Согласно учению И. II. Павлова, оба эти
типа обладают сильной нервной системой и у обоих имеется почти одина-
ковая степень сбалансированности процессов возбуждения и торможения,
и при всем том они друг от друга явно отличаются.
Это был тот период работы, когда зигзаги в физиологической характе-
ристике типов нервной системы для Ивана Петровича в основном были прой-
денным этапом, а по данному частному вопросу И. П. Павлов выдвигал
и отклонял ряд разнообразных предположительных объяснений.
Подробно и всесторонне разбирая вопрос в одной из бесед с И. П. Пав-
ловым, происходившей в узком кругу его сотрудников весной 1933 г., я вы-
сказал ему свои соображения о «функциональной подвижности (лабиль-
ности)», которую рассматривал как основной физиологический фактор дн-
ференцкации флегматиков и сангвиников, и вообще о желательности вне-
дрения этого принципа в учение об условных рефлексах. Иван Петрович
вначале отнесся к этому предложению с большой осторожностью и во время
этой беседы отклонил его. После двухнедельного колебания он, однако,
целиком принял мое предложение, о чем публично заявил на двух очеред-
ных и некоторых позднейших «средах».
После этого Иван Петрович со свойственным ему увлечением, горяч-
ностью и настойчивостью стал пользоваться этим новым принципом при
толковании многих до того не поддававшихся удовлетворительному объяс-
нению «запутанных» фактов нормальной и даже патологической деятель-
ности большого мозга у собак. Позже этот принцип был перенесен им в кли-
нику. В дальнейшем принцип «функциональной подвижности (лабиль-
ности)» был узаконен, стал быстро внедряться в практику работы лабора-
торий и клиник И. II. Павлова, причем применялся нс только в процессе
текущих работ, но и ретроспективно к ранее выполненным работам, сдан-
ным уже в печать *.
1 Как пример можно указать на работу В. В. Яковлевой, выполненную с собакой
Золотистая, и на ряд работ М. К. Петровой, проведенных с различными собаками.
По отношению к этим собакам И. П. Павлов давал и отклонял много различных объяс-
нений. После того как им был принят .введенный нарт в учение об условных рефлек-
сах принцип лабильности П. Е. Введенского, он «ретроспективно» по-новому объяс-
нял многое из сложного поведения этих собак и заставил сотрудников — хозяев этих
собак — коренным образом переделывать давно сданные в печать описания этих собак
ФИЗИОЛОГИЧЕСКАЯ ЛАБИЛЬНОСТЬ ВЫСШИХ ЦЕНТРАЛЬНЫХ ЭТАЖЕЙ
113
II. Е- Введенский под «функциональной подвижностью (лабильностью)»
подразумевал «ббльшую или меньшую скорость тех элементарных реак-
ций, которыми сопровождается физиологическая деятельность данного
аппарата».
Наиболее подходящим методом измерения лабильности он считал «то
наибольшее число электрических осцилляций, которое данный физиологи-
ческий аппарат может воспроизвести в 1 секунду, оставаясь в точном со-
ответствии с ритмом максимальных раздражений».
Простое сопоставление определения лабильности Н. Е. Введенским с
описанной еще Гиппократом картиной поведения флегматика и сангвиника
навело меня на соблазнительную мысль объяснить различие между этими ти-
пами различной степенью лабильности их нервных процессов. Но сложность
нервных процессов «аппарата высшей нервной деятельности» по сравнению
с деятельностью тех довольно простых «аппаратов», которые с непревзой-
денной глубиной и точностью были изучены Н. Е. Введенским, а также
ряд более деликатных соображений, которые будут высказаны в разделе
теоретических рассуждений этой статьи, заставили меня быть осторожным
и отказаться от категоричности в своих предположениях. Несмотря, одна-
ко, на это, принять данное предположение как руководящий рабочий прин-
цип, попытаться экспериментально проверить его я уже тогда считал не
только уместным, своевременным и полезным, но и возможным, учитывая
развитие современной физиологической техники (хронаксиметрия, опре-
деление рефрактерных фаз, характеристика тетанической кривой, электро-
грамма коры и т. д.).
Согласовав вопрос с И. П. Павловым, я перевел своих собак из Научного
института им. Лесгафта в Физиологический институт Академии наук и про-
должал накапливать экспериментальный материал по методу условных реф-
лексов, решив не удовлетворяться только показателями внешнего поведения
животных, как делалось в наших лабораториях до тех пор, а собрать точ-
ные данные о диференциальных функциональных особенностях этих двух
типов.
Для выполнения этой задачи я, естественно, воспользовался различными
функциональными показателями и пробами, разрабатывавшимися в ла-
бораториях И. П. Павлова в течение десятилетий и применявшимися раз-
л ичным и его сотрудн икам и для разнообраз пых целей: 1) быстрота вы раб отк и,
характер развития и прочность положительных и отрицательных условных
рефлексов; 2) испытание силы и прочности условных рефлексов; 3) быстрота
и характер угашения и восстановления условных положительных рефлек-
сов; 4) характер последовательного торможения на положительные услов-
ные рефлексы от дифереицировочного и. угасателыюго торможения; 5) пе-
ределка положительных и отрицательных условных рефлексов; 6) выработ-
ка и характер запаздывающих условных рефлексов; 7) выработка и характер
отрицательных условных рефлексов на определенное место применения од-
ного и того же условного положительного раздражителя; 8) выработка си-
стемности и ее прочность, а также ряд других мелких функциональных
проб.
При применении этих проб, разумеется, прочно моменты по возмож-
ности были выравнены.
с новой точки Зрения. И если в БЫШСДПШЙ НеДЗвно работе В. В. Яковлевой «Исследова-
ние высшей нервной деятельности собаки типа флегматика» (Труды павловских ла-
бораторий, t.VI, 193G) нет указаний на Н. Е. Введенского н на «виновника» перенесения
принципа лабильности в учение'об условных'рефлексах и даже отсутствуют ссылки
на мою работу по этому вопросу, являющуюся первой работой в данной области и опуб-
ликованную еще в 1934—1935 гг., то это вызывает не только удивление и недоумение,
но к сожаление.
<8 Э. А, Асратян
114
Э А. АСРАТЯН
Экспериментальная часть
Подопытные СобйкИ — Атлас и Голован — поступили в мое распоряже-
ние в 1932 г. «свежими», т. е. до этого времени для экспериментальной рабо-
ты не использовались. Обе— здоровые собаки-дворняжки выше среднего
роста. На вид Лтлас на Р/г —2 года моложе Голована. В процессе наблю-
дения за их поведением вне камеры, в свободной обстановке, а также в пе-
риод их адаптации к камере и к обстановке работы выявлено следующее.
Голован очень спокойный, мало подвижный, неповоротливый, нсреактнв-
иый. Ко мне, к камере и станку привык быстро, с первогоже раза вел
себя спокойно в станке и брал пищу из кормушки. Атлас, напротив, был
очень живой, вертлявый, реактивный, игривый, постоянно находился в
движении. Ко мне, к камере и к станку привыкал с некоторой осторож-
ностью, медленнее Голована, но все же довольно быстро. Пищу из кормуш-
ки Лтлас брал только на 3-й лень адаптирования к камере и к обстановке
работы. Подчеркнутая осторожность и реактивность иа новизну продлили
период адаптирования собаки к рабочей обстановке примерно на 8—10 дней.
Условные рефлексы на свет, на метроном 120, на звонок и на касалку
у обоих животных выработались довольно быстро.
Если формально подойти к показателям быстроты выработки положи-
тельных условных рефлексов, То трудно уловить существенную разницу
между обеими собаками. Все же у Атласа они выработались немного бы-
стрее. Учитывая, однако, сравнительно медленное адаптирование Атласа
к камере, станку, кормушке и раздражителям, а также небольшую разницу
в быстроте выработки рефлексов, нельзя было этим данным придавать боль-
шое значение. По этой причине позже, в 1934 г., когда собаки уже два года
были в работе, в строго одинаковых условиях я приступил к выработке
нового условного рефлекса — на гудение репродуктора, включенного в го-
родскую электрическую сеть. После пятикратного совпадающего сочетания
гудения и кормления у Атласа на тестой раз при отставлении раздражителя
на 20 секунд слюноотделение равнялось 14 делениям шкалы. В тех же усло-
виях у Голована условный рефлекс отсутствовал. После этого я продол-
жал у обеих собак совпадающее сочетание — гудения и пищи, а изредка
производил отставлепие раздражителя. На 10-е применение гудения у Ат-
ласа и Голована слюноотделение составляло 17 и 3 деления шкалы, на
15-е применение— 20 и 7 делений шкалы, на 30-е— 23 и 15 делений, на
51-е— 22 и 20 делений (рис. 1).
Вообще говоря, величины условных положительных рефлексов на один
и те же раздражители были почти одинаковы; кроме того, у обеих собак
одинаково отображался закон силовых соотношений условных рефлексов.
Достигнув своей предельной величины, положительные условные рефлексы
у обеих собак оставались стойкими.
У Атласа и Голована выявилась известная разница также при выработке
отрицательных условных рефлексов, условного тормоза и диференцировки.
Лтлас в этом отношении тоже быстрее Голована справился с задачами.
Особенно показательна в этом отношении выработка диференцировки на
метроном 60 и 120 ударов в 1 минуту (рис. 2).
Следует сказать,’что выработанные тормозные рефлексы у обеих собак
были одинаково стойкими и не колебались. Для характеристики быстроты
и легкости выработки условных рефлексов, а также ДЛЯ оценки сбаланси-
рованности корковых процессов возбуждения и торможения известный
интерес представляют данные по выработке запаздывающих условных реф-
лексов. С этой целью я взял одни из старых и устойчивых условных рефлек-
сов, именно рефлекс на звонок, получивший у каждой собаки около 409
подкреплений. Я стал отставлять звонок на 3 минуты, вместо обычного
отставлсния на 20 секунд. В течение опыта делались две пробы на отстав-
ФИЗИОЛОГИЧЕСКАЯ ЛАБИЛЬНОСТЬ ВЫСШИХ ЦЕНТРАЛЬНЫХ ЭТАЖЕЙ
115
ление рефлекса. Заметное удлинение .патентного Времени условного рефлек-
са у Атласа было отмечено уже со второго-третьего раза; на 11-м опыте
у Атласа запаздывание составляло 43 секунды, а дальше, примерно с 20-го
опыта, — до Р/з минут. У Голована же признаки запаздывания начали
л * si jS
ь S k 5
Рис. 1. Выработка условных рефлек-
сов иа гудение у собак Атлас (не-
прерывная линия) и.Голован
(i^-цктирная линия)
,!\ЛЛ
го г&
0
8
6
V
г
о
.Дни 2 1 6 8 гл
Рис. 2. Выработка днференцировкн на
метроном 60 ударов в 1 минуту от метро-
нома 120 ударов в 1 минуту у собак
Атлас (непрерывная линия) и Голован
(пунктирная линия)
появляться очень поздно; так, на 13-м опыте запаздывание было еле За-
метно, а па 31-м оно равнялось всего 14 секундам (рис. 3).
Характерная разница проявилась у собак также при выработке услов-
ного отрицательного рефлекса иа определенные порядковые места приме-
Рис. 3. Выработка запаздывания (пунктир) у Атласа и Голована
Цифры по,д кривыми показывают порядковый номер опыта после начала
развития запаздывания
нения света, бывшего положительного условно-го раздражителя , при сохра-
нении за ним значения положительного условного рефлекса на другом
порядковом месте его применения. Задача была такова. Свет с 5-минутной
паузой на первом и втором месте применения не подкреплялся пищей, а
на третьем месте—подкреплялся. Собаки должны были на первых двух
местах развивать торможение, на третьем место оставлять за светом сиг-
нальное значение положительного условного раздражителя. Эксперименты
показали, что Голован решал эту задачу в два раза медленнее, чем Атлас.
Привожу сокращенную диаграмму Опытов (рнс. 4).
Характерно, что для выработки этого сложного навыка у Атласа опять-
таки требовалось гораздо меньшее число опытов по сравнению с Голованом.
8*
116
Э. А. АСРАТЯН
Для оценки быстроты решения трудных задач и суждения о сбаланси
рованностн процессов возбуждения и торможения самыми иллюстратив-
ными являются опыты с функциональной
Рис. 4. Выработка условного
отрицательного рефлекса на
порядковое место применения
раздражителя
Цифры под столбиками показывают
порядковый номер опыта
пробой переделки положительного условного
рефлекса в отрицательный и одновременной
переделки дифоренцмровки в положительный
условный раздражитель. Для этой цели
в опытах среди других положительных раз-
дражителей на четвертом месте применялся
бывший до того диференцировочный раздра-
житель (метроном 60 ударов в 1 минуту'), ко-
торый подкреплялся пищей, а после 5-ми-
нутной паузы вслед за ним на пятом месте
применялся бывший до этого положительный
условный раздражитель (метроном 120 ударов
в 1 минуту), не подкреплявшийся пищей.
Атлас очень быстро и легко решил эту слож-
ную задачу, а Голован решил ее значительно
медленнее и с мучением. При решении задачи
Голован на станке часто ворчал, дрожал,
скулил — явления, которые раньше совер-
шенно не отмечались у этого животного.
Рис. 5 очень четко иллюстрирует быстроту
и характер решения этой задачи обеими со-
баками.
Перекрест линии у Атласа происходит
на 10-м опыте, у Голована на 22-м; пер-
вый нулевой эффект от бывшего положитель-
ного раздражителя у Атласа получается
на 18-м опыте, у Голована — на 65-м; стойкий отрицательный результат
у Атласа отмечен на 24-й пробе, у Голована — на 74-й.
В течение 19 месяцев эти раздражители к Атласу ие применялись.
После указанного перерыва применение метронома 120 ударов в 1 минуту
ФИЗИОЛОГИЧЕСКАЯ ЛАБИЛЬНОСТЬ ВЫСШИХ ЦЕНТРАЛЬНЫХ ЭТАЖЕЙ
117
вызывало слюноотделение, равное 20 делениям шкалы, а метронома 60 уда-
ров в 1 минуту—слюноотделение в 21 деление шкалы. У Голована же после
17-месячного перерыва при включении метронома 120 ударов в 1 минуту
величина рефлекса исчислялась 8Ч2 делениями шкалы, а метронома 60 уда-
ров в 1 минуту — 13 делениями шкалы.
Кроме перечисленных выше функциональных проб, указывающих глав-
ным образом на быстроту' выработки положительных и отрицательных ус-
ловных рефлексов и на степень сбалансированности процессов возбуждения
и торможения, поскольку об этом можно судить на основании выработанных
положительных и отрицательных рефлексов у обеих собак, я ставил спе-
циальные опыты для выяснения прочности этих рефлексов и для характе-
ристики силы нервной системы у собак. По существу говоря, проведенные
выше специальные опыты на животных, свидетельствующие о весьма отчет-
ливой регулярной условнорефлекторной деятельности в течение ЗЧ-г лет,
давали достаточное основание считать, что Лтлас и Голован обладали силь-
ной нервной системой й что рефлексы у них отличались достаточной проч-
ностью. Все же нами были поставлены специальные функциональные пробы.
Для определения силы нервной системы у собак я пользовался приня-
той в павловских лабораториях пробои на сверхсильный раздражи-
тель— трещотку.
Проба была применена в конце 1933 г., когда собаки находились в ра-
боте немного больше года. При действии трещотки в течение 26 секунд
Лтлас дал бурную двигательную реакцию, прижался к стенке, пристально
смотрел в сторону «страшного» раздражителя. Через5 минут после прекра-
щения действия трещотки была подана пища, которую Атлас взял лишь
через 2*/г минуты. В дальнейшем эта собака работала почти так же, как
обычно. Голован тоже сильно реагиров-ал на применение трещотки, при-
жался к стенке, смотрел в сторону раздражителя, но реакция егобыла сла-
бее, предложенную пищу он взял сразу. После опыта он тоже работал,
хак обычно. Для иллюстрации приведу два предыдущих контрольных
опыта, два опыта с применением трещотки и два последующих контроль-
ных опыта (табл. 1).
Таблица 1
Влияние сверхсильного экстрараздражителя на условные рефлексы
Раздражитель Длите ЯВ- НОСТЬ действия в сек. Атлас Голован
латентный период и рефлекс в делениях шкалы латентный период, и рефлекс в делениях шкалы
27-XII 1933 28 XII 1933 29.Х 1! 1933 27 XII 1 933 28. XII 1933 29 XII 1933
Свет 20 1сек.—19 1 сек.—20 1 сек.-20 1 сек.—17 I сек.-19 1 сек.—18
Звонок 20 1 » -21 1 » -22 1 » —25 2 »— 19% 2 » -21 1 » -22%
Касалка 20 2 » -17 2»- 17J4 2 » —15 3» - 14 2»—15% 2» -18
Стук 4- Г рещотк а Трещотка 1» -21
20 3 » —19 9 сек.— 7 2 » -18 2» — 17 3 сек.—13
Стук— 20 — 0 0 0 0 — 0 — 0
Свет 20 осек. —6 11 сек — 3 — 0 7 се к. — 9 18 сек.— 2 18 сек,—1%
Касалка 20 з»-юу2 4 » — 8 Зсек,- 9 6 » - 9 •1» -9% 2 » —14
Свет 20 2» -13 з» - ю% Зэ — 8 3»—13% 5» - 7 3 » —11
Стук-}- 20 4 > -12 2 » -13 31 —16 4» -11 3» —12% 3 » —12
Звонок 20 2 > -16 3 » -18 4 > —14 3» —15 4» - 9 2 » —Ц
118
Э. А. АСРАТЯН
Дальнейшие опыты показали, что, за исключением небольшого и кратко-
временного усиления условных рефлексов на сильные раздражители и не-
заметного их ослабления на слабые раздражители, никаких особых изме-
нений в условнорефлекторной деятельности не произошло. Поведение жи-
вотных ни в камере, ни на свободе совершенно не изменилось. Существен-
ных изменений: в поведении животных и в их условнорефлекторной дея-
тельности не вызвало также перенапряжение процессов возбуждения и тор-
можения, выразившееся в 5-минутном применении (вместо 20-секундного)
сильных условных раздражителей и диференцировочного раздражителя
в различных опытах.
Результаты этих экспериментов с достаточной очевидностью показы-
вают, что Атлас и Голован обладали сильной нервной системой, что у них
корковое возбуждение и торможение не только сбалансированы, по н до-
статочна сильны.
Рис. 6. Ход учащения и восстановления условного рефлекса
на стук 520ударов в 1 минуту у Атласа (непрерывная линия)
и Голована (пунктирная линия)
Для детальной характеристики Атласа и Голована некоторый интерес
представляют также другие пробы и наблюдения.
Прежде всего характерны данные по угашенмю и восстановлению услов-
ных рефлексов.—стук 120 ударов в 1 минуту, применявшийся по 20 секунд
(рнс. 6), а также по угашению и восстановлению условного рефлекса на
гудение, применявшееся 3 секунды, с учетом рефлекса на 20 и 120 секунд
(табл. 2).
Небольшая разница выявилась также в быстроте и продолжительности
последовательного торможения от угашения и днференцировкн. Ради
экономии места я иллюстрации не привожу. Хотелось, однако, обратить
внимание на характерное явление, встречавшееся всегда и нашедшее вы-
ражение в аналогичных опытах, приведенных в табл. 2. Речь идет о за-
висимости величины условных рефлексов у обеих собак от длительности
действия условного раздражителя. В одной из моих уже опубликованных
статей показано, что такая зависимость вообще существует. В данном случае
я хотел обратить внимание на то, что эта зависимость у Атласа выступает
ярче, чем у Голована. У обеих собак гудение при 20-секундном действии
вызывало слюноотделение, равное за 20 секунд 22—25 делениям шкалы.
В этом и подобных опытах при 3-секундном действии того же раздражителя
у первой собаки за 20 секунд условный рефлекс был равен 12 делениям
шкалы, а за 2 минуты—всего 27 делениям; у второй собаки- соответственно
25 и 91 делению. То же самое в тех же и аналогичных условиях повто-
рялось и с другими условными раздражителями. Например, свет при
ФИЗИОЛОГИЧЕСКАЯ ЛАБИЛЬНОСТЬ ВЫСШИХ ЦЕНТРАЛЬНЫХ ЭТАЖЕЙ
119
Таблица 2
Угашение и восстановление рефлексов
Атлас; опыт I 3. III 1 935 г. Гллоааи; опыт 18. XI 1935 г.
ПрбДОЛ- условные условные Пр<>доЛ- условные условные
рефлексы рефлексы за раздра- рефлексы рефлексы за
раздражи- житель- за 20 сек. |2О сек. житель- за 20 сек I 20 сек.
кость нскть
Тель действия жите л». действия
п сек. в делениях шкалы в сек. в делениях шкалы
Свег 20 24 Подкреп- Свет 1 20 21 Подкреп-
л ял с я ЛЯ Л с я
Гудение 3 12 27 нс под- Гудение 3 25 91 нс под-
креплялся креплялся
» 3 11 19 » 3 21 57
к> 3 Я 13 3 19 52
м 3 7 9 » 3 16 41
э 3 7 11 » 3 18 37
9 3 6’/i 10 » 3 14 33
* 3 4 8 3 11 25
3 7 9/, » 3 9 27
3 4 8 » 3 7 18/4
> 3 3 11 » 3 12 30/г
9 3 2 5 » 3 8 21
:♦ 3 ’V, 4 3 5 13/2
3 0 3 » 3 3 11
у, 3 1 6 э- 3 7 16
•л 3 0 2 » 3 4 11
> 3 0 3 » 3 2 7
» 30 ю/, Подкреп- » 3 0 13
л я лея
20 10 То же > 3 6 16
э —• 16 > > » 3 3 8
> — 19 > > > 3 0 9/,
» 3 2 7
• 3 0 10
» 3 0 8
* 3 0 6
30 8 Подкреп-
лился
я 20 15 То же
20 13 » »
* 20 Н/2 > в
20-секундном действии у Атласа вызывал за 5 минут слюноотделение,
соответствующее всего 65 делениям шкалы, а у Голована — 209 делениям,
причем у первого слюноотделение прекратилось еще в начале 2-й минуты,
а у второго продолжалось до середины 5-й минуты. Характерно, что после
прекращения еды безусловное слюноотделение у Атласа прекратилось
на 40—50 секунд раньше, чем у Голована.
Интересный материал дали опыты с удлинением и укорочением про-
межутков между применением раздражителен. Обе собаки в течение всех
опытов почти неизменно работали на 5-минутных паузах. В ряде опытов
я применял 10-, 9-, 8-, 61/#-МИНутные паузы, а в ряде других опытов —
3- н 2-минутные паузы. Оказалось, что у Атласа при увеличении пауз
в течение первых 2—3 дней незначительно увеличивалась величина услов-
ных рефлексов, а затем картина рефлексов Становилась прежней. У Голо-
вана же при этом начиналась какая-то хаотическая деятельность с общей
тенденцией к уменьшению рефлексов. При этом у Атласа в слабом виде
проявлялся условный рефлекс на время (5-минутная пауза) и затем быстро
исчезал, а у Голована этот рефлекс был выражен особенно сильно и упорно
не исчезал (табл. 3).
120
Э. А. АСРАТЯН
Таблица 3
Влияние увеличения пауз- на величину условных рефлексов
Атлас Голопан Примечание
дата опыта пауза между рефлекса* МИ D мин. суммар- ная вели- чина ше- сти реф- лексов и делени- ях шкалы дата опыта пауза между рефлекса- ми В МММ. суммарная величина пяти реф- лексов п делениях шкалы
23.11 1935 г. 25.11 27.11 1.111 4.Ill 6-1II 7-111 8 III 9.111 II.Ill 12.111 5 5 5 5 8 9 7 5 5 5 5 119 114 117 117 141 121 115 74 98 117 121 II.V 1935 г. 13. V 14. V 15. V 16. V 17. V 18. V 20. V 21. V 22. V 23. V 24. V 27. V 28. V 5 5 5 5 8 9 8 10 6*4 5 5 5 5 5 87 95% 97 92% 72% 94%* 5в»* 61 76 % 74 71 87 104% ИЮ • Сильный услов- ный рефлекс на время, который суммируется с рефлексом иа раздражитель * * Условный реф- лекс на время
При укорочении паузы до 3 минут у Атласа вначале резко снижался
уровень условных рефлексов и даже развивалось какое-то невротическое
состояние; при дальнейшей работе состояние животного постепенно улуч-
шалось, и величина условных рефлексов доходила до уровня, наблюдав-
шегося при 5-минутных паузах. На дальнейшее укорочение пауз до 2 ми-
нут Атлас реагировал так же, как и при первом укорочении, но менее
болезненно и меньшим снижением уровня рефлексов.
Голован же на укорочение пауз доЗ минут почти никак не реагировал,
если не считать ничтожного снижения уровня условных рефлексов. Однако
у него почему-то участились припадки (об этом подробно ниже). На даль-
нейшее укорочение паузы до 2 минут Голован в первые дни реагировал уже
заметным уменьшением условных рефлексов, а затем они возвращались
к прежнему уровню.
Характерно, что после того как обе собаки некоторое время (около
3 месяцев) работали при укороченных паузах, на возвращение к 5-минутным
паузам Атлас почти никак не реагировал (ничтожное повышение рефлек-
сов), а у Голована опять началась хаотическая деятельность с тенденцией
к ослаблению рефлексов.
В табл. 4 приводятся данные всех описанных выше вариаций рефлектор-
ной деятельности в зависимости от изменения пауз.
В процессе работы я несколько раз проводил также пробу на систем-
ность, причем каждый раз получал почти одинаковые результаты: у
Атласа лучше, чем у Голована, образовывалась системность, по она бы-
стрее и разрушалась. Трудно даже утверждать, что у Голована вообще
образовывалась системность в. четком виде.
Кромсэтих условнорсфлекторных проб и наблюдений, я производил так-
же измерение хронаксин у собак. Весной 1935 г. мы произвели несколько
измерений мышечной хронаксии у Атласа и Голована. Результаты оказа-
лись исключительно интересными. В 1936 г. я возобновил хронаксимстрию,
но, к сожалению, только у Голована, ибо другие мои собаки, в том числе
и Атлас, за время летнего отпуска погибли по неизвестным причинам. Для
расширения экспериментальной базы и с целью сравнения я за 1936/37 г.
производил параллельно измерения хронаксии больше чем у 20 собак нашего
института.
ФИЗИОЛОГИЧЕСКАЯ ЛАБИЛЬНОСТЬ ВЫСШИХ ЦЕНТРАЛЬНЫХ ЭТАЖЕЙ
121
Таблица 1
Влияние величины пауз на величину условных рефлексов
Атлас Голован
дата опыта пауза между рефлекса ми в мим. суммарная вели инна, семи условных рефлексов в делениях шкалы дата опыта пауза между рефлексами в мин. суммарная величина, ше- сти условных рефлексов в делениях шкалы Примечание
22.Х I 1955 г. 23.Х 1 26 . XI 28 .Х I 29 .Х I 30-XI 2.XII 4.XII 5.Х II 7.Х 11 9.Х11 11.Х II 13.Х 11 14.Х 11 16.XII 18.Х 11 19.Х II ЗО.ХП 2.1 1936 г 3.1 6.1 7.1 9.1 11.1 14.1 15.1 16.1 18.1 5 5 3 3 3 3 3 3 3 3 3 3 3 3 3 3 3 3 2 2 2 2 2 2 2 2 2 2 117 152 аок 72*2 56%* 6" 5154 • 61 51% 26%* 96 130 152 162 101 103 10454 98 104 63 109% 93 75 71% 23. XI 1935 г. 26.Х I 28 .Х I 29 .Х I 30 . XI 2.Х П 3.XII 4X11 5.Х 11 9.XII 11.X11 13 .Х 11 14 .Х II 16 .Х II 18 .XII 19 .XII 6.1 1936 г. 7.1 9.1 11.1 14,1 15.1 16.1 18.1 20.1 23.1 25.1 27.1 5 5 5 3 3 3 3 3 3 3 3 3 3 i 3 3 3 2 2 2 2 2 2 2 2 2 2 124% 119% 121 123 124 108 93% 102 ИЗ 110 82 115 122% 140 124 120% 123 128% 100% 88% 93 105% 95% 95% 112% 108% 120% 113% • Собака в депрес- сивном СО- СТОЯНИИ
Среди этих собак флегматиков не было, однако по ходу работы я на
моей установке измерял хронаксию у двух флегматиков из других лабо*
раторнй (Белогрудки из Колтушей и Крошки из руководимой мной
лаборатории в Институте мозга им. Бехтерева). Результаты хронаксимет-
рии в сопоставлении с данными типовой характеристики изученных собак
показывают, чтохронаксиметрия с удивительной точностью выявляет основ-
ную типологическую особенность этих собак—инертность нервных про-
цессов. У всех трех флегматиков хронаксия оказалась в два и в три раза
больше, чем у остальных исследованных до сих пор собак-нефлегматиков *.
В этой связи нельзя не упомянуть единственные из известных мне слу-
чаи резкой разницы в хронаксии у исследованных собак, описанные Бур-
гиньоном (1923). Хронаксию Бургиньон определял в острых опытах на
обнаженных седалищных нервах биполярной методикой. Две уличные
собаки дали в среднем хронаксию порядка 0,40s, две собаки породы фок-
стерьер — 0,10а. Разбирая свои данные, Бургиньон высказывает предполо-
жение, что, вероятнее всего, эта разность обусловливается неиндивидуаль-
ными особенностями, а особенностями различных пород. Он пишет по
этому поводу: «Мне кажется более правдоподобным думать, что различие
хронакСИЙ, наблюдаемое между двумя группами собак, происходит от раз-
личных пород, а не от различия индивидуальности».
Подробное описание этих данных будет приведено в специальном сообщении.
122
Э А. АСРАТЯН
На основании накопленного мной более разнообразного и тщательнее
разработанного экспериментального материала я не могу согласиться с вы-
сказанным Бургиньоном предположением. Мне кажется, что возрастной
фактор может иметь большее значение,, чем порода, хотя я не имею основа-
ний совершенно исключать влияние последнего фактора на хронаксию.
Но главное, мой материал позволяет утверждать, что индивидуальные —
типологические особенности оказывают в этом отношении большее
влияние, чем упомянутые выше два фактора.
Было бы очень интересно проверить пригодность хр онаксиметрии для
выявления типовых особенностей человека. В случае положительного ре-
зультата этой проверки мы получили бы простой и гмеете с тем дей-
ственный материал для изучения этой сложной и интересной проблемы
психологии и клиники.
Этим я заканчиваю краткое описание фактического материала, накоп-
ленного мной для физиологической .характеристики И определения инди-
видуальных особенностей у сангвиника Атласа и флегматика Голована.
Еще одно замечание. Ряд наблюдений как будто говорил о том, что у
Атласа тонус симпатической системы выше, чем у Голована. На это указы-
вала большая выпуклость глазных яблок, большая ширина зрачков у
Атласа, типичная мутная «симпатическая» слюна у Атласа в отличие от
типичной «хордовой» прозрачной слюны у Голована. Некоторая разница
получена также при пробе на окуло-кардиальный рефлекс: у Голована
ослабление сердечной деятельности как будто выявилось более четко. Ха-
рактерно, что у большинства наших собак-сангвиников слюна походила на
слюну Атласа, а слюна другого флегматика из числа собак лаборатории
И. П. Павлова — Золотистой была так же прозрачна, как у Голована
Теоретические сопоставления
Основная идея «относительной функциональной подвижности (лабиль-
ности)» возбудимых тканей высказывалась Н. Е. Введенским еще в самых
ранних его произведениях, из которых укажем на замечательную диссер-
тационную работу 1886 г. Значение точного физиологического параметра
лабильность приобрела в работе Н. Е. Введенского 1892 г.г напечатанной
на французском языке. Со свойственным автору высоким мастерством в этой
работе дается четкая формулировка закона «относительной функциональ-
ной подвижности (лабильности)» и открывается новый путь научного пред-
видения судьбы последовательных нервных импульсов по мере их проведе-
ния по нервному субстрату и по другим возбудимым субстратам. В дальней-
ших своих произведениях Н. Е. Введенский — искусный мастер тончай-
шего эксперимента и слова — применяет открытый им закон к новым и но-
вым областям, делает его краеугольным камнем созданного в дальнейшем
монолитного учения о высшей нервной деятельности. Это «основное поло-
жение», «основная теорема» Н. Е. Введенского продолжает оставаться в
центре внимания его школы и после смерти ее основоположника. Напря-
женные и плодотворные экспериментальные и теоретические исследования
учеников И. Е. Введенского во главе с А. А. Ухтомским, а в последнее
время также ряд работ других физиологов выдвинули учение об «отно-
сительной функциональной подвижности (лабильности)» в качестве одного
из руководящих принципов современной физиологии нервной системы.
Во вступлении я привел формулу этого закона, данную Н. Е. Введен-
ским. Кардинальное значение этого закона заключается в том, что он
«управляет всеми явлениями, вызванными прерывистым электрическим
1 Прозрачной оказалась также слюна у двух других флегматиков из других ла-
бораторий (Белогрудки и Крошки), которые были исследованы мной позже.
ФИЗИОЛОГИЧЕСКАЯ ЛАБИЛЬНОСТЬ ВЫСШИХ ЦЕНТРАЛЬНЫХ ЭТАЖЕЙ 123
раздражением, он относится как к функциональной деятельности, так и к
задержке этой деятельности, «состоянию бездействия (задержки)». Сам
Н. Е. Введенский придавал приведенному закону универсальное значение
для всех возбудимых тканей, в частности, для всех нервных образований,
что впоследствии получило подкрепление в многочисленных эксперимен-
тальных работах его учеников и внуков (укажем хотя бы на работы
Н. В. Голикова о лабильности нервных центров, на работы М. В. Кирзона
и И. А. Аршавского о лабильности вегетативных нервов и т. д.).
Акад. А. А. Ухтомский в недавно опубликованной статье совершенно
справедливо относит лабильность к важнейшим физиологическим факторам,
регулирующим интрацентральные взаимоотношения, определяющим обра-
зование доминант в центрах и организацию текущих рабочих ансамблей
в центральной нервной системе. А. А. Ухтомский обращает особенное вни-
мание как на кардинальный момент учения И. Е. Введенского о физиологи-
ческой лабильности на вариабильность величины лабильности в субстрате
по мере развития в нем физиологической реакции. Лабильность той или иной
функциональной единицы, например, нервного проводника, имеет стати-
стически относительно устойчивый уровень, который и определяет общую
дееспособность данного прибора в данный момент. По под действием теку-
щих рабочих импульсов и гуморальных влияний заданная в приборе ла-
бильность претерпевает сдвиги, вследствие которых существенно изменяется
н конечная реакция. Эти сдвиги по ходу реакции отнюдь не обязательно при-
обретают характер износа работоспособности субстрата в порядке утом-
ления от предыдущей деятельности. Сплошь и рядом они характеризуют
подъем лабильности и более выгодную установку субстрата к текущей
работе по ходу самой работы. Этим вариациям лабильности походу раз-
вития реакции и принадлежит главенствующее значение в определении
ответа ткани на текущие импульсы.
По характеристике А. А. Ухтомского, «лабильность есть коэфициент,
указывающий скорость, с которой данный физиологический субстрат успе-
вает переходить от состояния покоя к состоянию возбуждения и обратно
возвратиться от состояния возбуждения к состоянию физиологического
покоя с готовностью к новой реакции. Или иначе, лабильность есть ско-
рость, с которой ткань успевает закончить полный период отдельного воз-
буждения во времени. Или, наконец, лабильность есть показатель того,
сколько отдельных законченных периодов возбуждения способен субстрат
вместить в единицу времени» х.
Какое же отношение имеет лабильность к хронаксии? Этот вопрос имеет
для меня тем большее практическое значение, что я в своих экспериментах
пользовался для суждения о лабильности измерением хронаксии у собак.
Из всего сказанного вытекает, что закон относительной лабильности
основывается на существовании известкой, определенной во времени дли-
тельности каждого отдельного приступа возбуждения, возникающего в том
или ином звене сложного физиологического аппарата. Это и обусловливает
естественную внутреннюю близость между лабильностью Н. Е. Введен-
ского и хронаксией Лапика. Ведь хронаксии является как бы пороговым
временем действия раздражителя, характеризующим минимальное время,
необходимое для возникновения отдельного возбуждения. Вот почему
А. А. Ухтомский на XV конгрессе физиологов заявил: «Это основное пато-
жен ие нашей школы, развитое в 1892 г. его автором под названием закона
«относительной функциональной подвижности (лабильности)» физиологи-
ческих субстратов, очевидно, сближает нас счастливым образом с нынеш-
ними исканиями прекрасной физиологической школы Сорбонны» 4.
1 У х т о м с к и й А., Труды Физиологического научно-исследовательского ин-
ститута ЛГУ. № 14, 1934.
^Ухтомский А., Вестник Академии наук С-ССР, № 10, 1935.
124
Э. Л АСРАТЯН
Со своей стороны, как известно, Лапик сделал далеко идущие в этом на-
правлении признания; особенно ценно, что за последнее время школа
Лапика допускает вариабнльность хронаксии, что еще больше сближает ее
со школой Н. Е. Введенского.
Н. Е. Введенский закон относительной лабильности рассматривал
в аспекте эволюционной физиологии. Он отмечал, что ткани теплокровных
обладают большей лабильностью, чем ткани холоднокровных. Говоря о не-
обходимости раздражения блуждающего нерва более частыми токами для
остановки сердца у теплокровных, чем это нужно для остановки сердца у
холоднокровных, Н. Е. Введенский писал: «Смотря надело сточки зрения
большей лабильности раздражительных образований у теплокровных,
это становится понятно само собою, мало того, это даже требуется такой
точкой зрения»
Исходя из этого, многочисленные работы, выполненные сотрудниками
Лапика и другими исследователями по хронаксии различных физиологи-
ческих субстратов у разных животных, можно считать до известной степени
разработкой и развитием этого учения Н. Е. Введенского на сравнительно
широком экспериментальном материале по линии сравнительно-фнзиологи-
ческого и возрастно-физиологического подбора.
Учение Н. Е. Введенского находит подтверждение и получает дальней-
шее развитие в огромном экспериментальном материале, накопленном на
тех же субстратах путем применения других измерений с учетом латент-
ных периодов возбуждения тканей, характера реакции, количественной
характеристики биоэлектрических токов и т. д. Меня в данном случае больше
интересует центральная нервная система.
На основании ряда прямых и косвенных данных Н. Е. Введенский счи-
тал, что нервные центры обладают меньшей лабильностью, чем нервные
волокна, и даже меньшей, чем концевая пластинка скелетных мышц.
Это положение Н. Е. Введенского тоже находит прямое и косвенное
экспериментальное подтверждение в обширных данных, полученных други-
ми методиками исследования. Мне кажется, что в учении о лабильности есть
ряд вопросов, которые до сих пор не только не разрешены, но даже еще не
поставлены с достаточной остротой. К числу таких вопросов можно отнес-
ти, во первых, изучение сравнительной лабильности различных отделов
центральной нервной системы в аспекте сравнительной и возрастной физио-
логии; во-вторых, изучение вариаций лабильности в морфологически и
функционально достаточно обособленных частях центральной нервной си-
стемы в аспекте индивидуальной физиологии и индивидуальных вариаций.
Эти вариации — фактор, который на каждом шагу дает о себе знать экс-
периментатору па высших ступенях развития животного мира.
Свою работу' я рассматриваю не только как первую попытку использо-
вания кардинального закона Н. Е. Введенского об «относительной функ-
циональной подвижности» в области учения об условных рефлексах, но и
как первую попытку разработки вопроса о лабильности с точки зрения
новообразования временных связей.
Прежде всего возникает вопрос первостепенной важности: насколько
я прав, принимая за лабильность, в понимании Н. Е. Введенского, ту от-
носительную быстроту развития реакции, которая открывается методом
условных рефлексов у животных различных типов. Само собой понятно,
что даже отрицательный ответ на поставленный вопрос нисколько не ума-
лит огромного значения, которое может иметь в дальнейшем учение о ла-
бильности в понимании Н. Е. Введенского для разработки теории и
практики условных рефлексов в самых-широких пределах.
Что касается прямого ответа на поставленный выше вопрос, то мне
В Веденский И. Е., Возбуждение, торможение и наркоз, 1901.
ФИЗИОЛОГИЧЕСКАЯ ЛАБИЛЬНОСТЬ ВЫСШИХ ЦЕНТРАЛЬНЫХ ЭТАЖЕЙ
125
кажется, что сейчас, при имеющемся экспериментальном материале, на него
нельзя еще дать окончательный ответ. Пока я могу говорить лишь о большой
вероятности сделанного мной предположения на следующих основаниях.
Поскольку Н. Е. Введенский общей характеристикой лабильности считал
«большую или меньшую скорость тех элементарных реакций, которыми
сопровождается физиологическая деятельность данного аппарата», я вправе
сделать упомянутое выше предположение, ибо основной диференцирую-
щей особенностью условно- и безусловнорефлекторной детальности у иссле-
дованных мной собак как раз и была «большая или меныная скорость».
Несомненно, что здесь вопрос осложняется тем, что в актах условно-
даже спинальнорефлекторной деятельности мы имеем производное слож-
ного характера. Ведь целый комплекс различных органов принимает
участие в этой работе! Чему приписать у животных типологическую ди-
ференцирующую лабильность? Следует сделать допущение, что более
или менее устойчивая разница в лабильности у животных различных типов
определяется относительным постоянством статистических средних уров-
ней лабильности в тканях, от которых зависят текущие сдвиги, возникаю-
щие под действием эксцитаторных и гуморальных факторов. Статист- '
ческиесредние уровни лабильности у сангвиника и флегматика различны.
В случае условнорефлекторной деятельности мы сталкиваемся по
преимуществу с лабильностью большого мозга. Дело затрудняется еще
тем, что в моем распоряжении нет пока прямого и классического опреде-
лителя текущей лабильности тканей у собак — электрографического при-
бора. Болес тонкое наблюдение за текущей лабильностью я мог вести толь-
ко по показаниям хронаксиметра, и здесь мы стоим перед вопросом, на-
сколько вариации хронаксии могут точно свидетельствовать о вариациях
лабильности. Когда речь идет о больших разностях, как это наблюдается
в моих случаях, мне кажется, имеются все основания считать, что постав-
ленный вопрос должен быть решен положительно.
С этой точки зрения данные х ропак сим стр ии нервов у исследованных
нами собак также являются показателем правильности сделанного мной
предположения. Более решительно можно будет высказаться тогда, когда
измерение лабильности большого мозга мы сможем производить в соот-
ветствии с предложенным Н. Е. Введенским показателем — по способности
воспроизводить максимальные частоты осцилляции самого органа или
хотя бы нервов н скелетных мышц. Но это дело будущего.
Следует, однако, остановиться на другой стороне вопроса.
Согласно закону относительной лабильности Н. Е. Введенского, чем
лабильнее ткань, тем позже в ней возбуждение переходит в торможение,
тем меньше она склонна к торможению. При поверхностном подходе сле-
довало ожидать более быстрого перехода возбуждения в торможение, бдль-
шую склонность к торможению у флегматика, чем у сангвиника. Па самом
деле имеет место совершенно обратная картина: как и все остальные явле-
ния, переход возбуждения в торможение у сангвиника происходил быстрее,
раньше и легче, чем у флегматика. Мне думается, однако, что при более
глубоком рассмотрении вопроса это противоречие придется отнести к ка-
жущимся. В этом вопросе, как мне представляется, поможет разобраться
концепция Н. Е. Введенского, развиваемая А. А. Ухтомским, о том, что
лабильность ткани при нормальной работе последней сама изменчива,
что она изменяется под влиянием как текущих рабочих импульсов, так и
гуморальных, нервнотрофических и других факторов. На этом именно
основании я дальше развиваю свое предположение о различной лабиль-
ности моих собак в том смысле, что у флегматика и сангвиника лабиль-
ность отличается нс только разными исходными уровнями, но и различной
степенью реактивности, различными диапазонами вариации под действием
эксцитаторных, трофических и гуморальных факторов. С этой точки
|25 Э. А. АСРАТЯН
зрения, быстрое наступление торможения у сангвиника, животного более
лабильного типа, можно было бы попять как наклонность действующих
субстратов более быстро и в более широких, пределах варьировать исход-
ную лабильность. А. А. Ухтомский подчеркивал, что именно степень кон-
траста в двух соседних звеньях системы при прочих равных условиях
сказывается фактором торможения, поэтому быстрое возрастание лабиль-
ности в. предыдущем звене бывает столь же действенным условием для
блока импульсов в переходе на последующее звено, как и снижение ла-
бильности в последующем звене при неизменном исходном уровне ла-
бильности в предыдущем.
Диапазон, в пределах которого варьирует лабильность, и степень кон-
трастности ее в соседних звеньях системы могут зависеть от организации
нервных элементов сангвиника н флегматика, от гуморального баланса
у того и другого, от различной эффективности нервнотрофических «меха-
низмов», в частности, влияния симпатической нервной системы на нервные
центры. Что симпатическая система оказывает влияние на деятельность
нервных центров, это доказано работами сотрудников акад. Л. А. Ор-
белн. Это допущение по отношению к моим основным подопытным собакам
тем более уместно, что, судя по некоторым показателям, у сангвиника
тонус симпатической системы был выше, чем у флегматика. При наличии
высокого тонуса симпатическая система может, конечно, оказывать не
только прямое влияние на скорость реакций в центрах, но и влиять через
посредство тех разнообразных гуморальных факторов, которые этот тонус
за собой влечет1.
Я считаю, что проведенная экспериментальная работа является первым,
хотя бы пока и слабым, подкреплением моей попытки перенести учение
Н. Е. Введенского о лабильности в область учения об условных рефлексах.
Я ие сомневаюсь, что дальнейшие более детальные исследования в. этой
новой области физиологии еще больше расширят и углубят связь между
двумя великими школами пашей отечественной физиологии. Больше того,
не приходится сомневаться в том, что это «взаимопонимание» и контакт
окажут стимулирующее влияние как на дальнейшее развитие бессмерт-
ного учения И. П. Павлова, так и на развитие глубоких идей Н. Е. Вве-
денского.
Пользуясь случаем, я хочу выразить сердечную благодарность и при-
знательность глубокоуважаемому А. А. Ухтомскому не только за ценные
указания при литературном оформлении статьи и за постоянную под-
держку в изучении великого наследия Н. Е. Введенского в процессе моего
роста и формирования как физиолога, по и за неизменно теплое, чуткое
и внимательное отношение ко мне.
ЛИТЕРАТУРА
Аршавский И. А.. Учение о парабиозе и автономная нервная система, Труды,
Ленинградского общества естествоиспытателей, т. 62, в. 1—2, 1933.
Введенский Н. Е., Возбуждение, торможение и наркоз, 1901.
Введенский Н. Е., О соотношениях между возбуждением и торможением,
дисс., 1886.
Голиков Н. В., Изменения хронаксии и рефрактерной фазы нерва под влиянием
текущих импульсов. Сборник работ физиологической лаборатории
ЛГУ, посвященный 30-летию научной деятельности А. А. Ухтомского, 1930.
1 В этом моем положении, бесспорно, сказывается одностороннее и неправиль-
ное понимание взанмоотношений симпатической нервной системы и высших отделов,
центральной нервной системы. Я тогда упустил из виду, что симпатическая нервная
система, как и вся вегетативная нервная система в целом, подчинена коре больших
полушарий мозга и что эта система является одним из тех нервных образований,
посредством которых кора, этот верховный «распорядительи распределитель», осу-
ществляет тонкую, совершенную регуляиию.функции организма и его взаимоотно-
шений с окружающей средой.
ФИЗИОЛОГИЧЕСКАЯ ЛАБИЛЬНОСТЬ ВЫСШИХ ЦЕНТРАЛЬНЫХ ЭТАЖЕП
127
К и р з о и A. A., Optima и pcssima электрического раздражения в связи с вопросом
о лабильности последнего, Труды физиологической лаборатории Научно-иссле-
довательского института ЛГУ, № 14. 1934.
Лапик Л., Эволюция современных учений о нервной системе, Физиологический
журнал СССР, т. XIX, в. 1. 1935; т. XXI, в. 5—6, 1936.
Орбели Л, А-, Лекции по физиологии нервной системы, 1935-
Павлов И. И., Лекции о работе больших полушарий головного мозга, 1932.
Разенков И. П.. Изменение раздражительного процесса коры полушарий го-
ловного мозга собаки при трудных условиях. Труды физиологических лабора-
торий И. П. Павлова, т 1. в. 1, 1924.
Ухтомский А. А., Лабильность как условие срочности и координирования нерв-
ных актов, Труды физиологической лаборатории Научно-исследовательского
института ЛГУ, № 17, 1936.
Ухтомский А. А.. О показателе лабильности (функциональной подвижности)
физиологических приборов, Труды физиологической лаборатории Научно-иссле-
дов-ательского Института ЛГУ, № 14, 1934.
Ухтомский А. А-, Физиологическая лабильность и акт торможения. Вестник
Академии наук СССР, № 10, 1935.
Фурс и ков Д. С-, Явление взаимной индукции в коре головного мозга, Архив
биологических наук, -т. XXIII, в. 1—3, 1923.
Bourguignon G., La chronaxie cher 1'home, 1923.
Ex пег S., Pfluger's Archiv, Bd. 25, 1882.
Magnus R., Die Korperstellung. 1924.
Pfluger E., Die Sensorische Functionen <tes Ruckenmarks, 1853.
Sherrington Ch., The integrative action of nervous system, 1906.
ВЛИЯНИЕ УСЛОВНОГО ОБОРОНИТЕЛЬНО-ДВИГАТЕЛЬНОГО
РЕФЛЕКСА НА БЕЗУСЛОВНУЮ БОЛЕВУЮ РЕАКЦИЮ СОБАКИ'
1о ходу моей работы в области физиологии условных рефлек-
сов мне удалось наблюдать и подвергнуть специальному и
। подробному исследованию ряд моментов взаимно-индукцион-
ных отношений между условными и безусловными пищевыми
J рефлексами. Краткие сообщения о результатах были напеча-
таны в «Докладах» (т. II, № 1 и 2, 1934). Аналогичные взаимо-
отношения мной наблюдались между электрооборонительно-двигательными
условными и безусловными рефлексами. В частности, на ряде собак с
пищевыми и оборонительно-двигательными рефлексами было установлено,
что в известных временных пределах положительные условные рефлексы
вызывают значительное ослабление следующих за ними однородных безу-
словных рефлексов, в наоборот. В дальнейшей моей работе по оборони-
тсльно-двигательным условным рефлексам я подверг специальному изуче-
нию одно замечательное явление, которое наблюдалось мной раньше.
В 1932—1933 гг. по ходу выработки и изучения оборонительно-двига-
тельных условных рефлексов на собаках Баш, Кейо и в особенности на
собаке Назар я иногда замечал, что, когда действие условного раз
дражителя вызывало бурную реакцию собаки (усиленная общая двига-
тельная реакция, сильный лай, визг и т. д.), применение индукционного
электрического раздражения на этом фоне не только не усиливало эту
реакцию, но даже в значительной степени ослабляло ее, в особенности
лай и визг. В работе с этими собаками условные раздражители изолиро-
ванно действовали лишь в течение 5 секунд, и это явление, как говорилось,
наблюдалось нерегулярно.
В 1933—1934 гг. у собаки Чинар я вырабатывал запаздывающий оборо-
нительно-двигательный условный рефлекс на единственный раздражи-
тель—звонок. В течение 1 минуты действовал только звонок, затем к нему
присоединялось раздражение левой задней лапы индукционным током;
это комбинированное действие двух раздражителей длилось 5—6 секунд.
Время действия раздражителей и рефлексы регистрировались на кнмогра-
фнческой ленте. Параллельно велось наблюдение. В силу того, что звуко-
непроницаемая камера, в которой находилась подопытная собака, была
радиофицирована, я имел возможность по ходу опыта следить также за
голосовой реакцией собаки (лай, визг, скуление).
Чинар, молодая и живая собака, отличался более возбудимой нервной
системой по сравнению с остальными моими собаками. Условный оборони-
тельный рефлекс на звонок вырабатывался быстро, но запаздывание, т. е.
более или менее значительное удлинение скрытого периода появления услов-
ного рефлекса, образовывалось медленно и цоздно. Явление, о котором будет
1 Доклады Академии наук СССР, т, I, Xs 5, 1935. Представлено акад.
И. П Павловым 24.Х 1934 г.
УСЛОВНЫЙ ОБОРОНИТЕЛЬНО-ДВИГАТЕЛЬНЫЙ РЕФЛЕКС И БОЛЕВАЯ РЕАКЦИЯ 129
Схематическое изображение
взаимодействия рефлексов
идти речь, с достаточным постоянством наблюдалось почти с момента укреп-
ления условного рефлекса на звонок и в течение всего 4—5-месячного
периода работы с собакой.
При минутном изолированном действии звонка Чинар давал страшно
бурную реакцию: рвался всем телом, сгрыз цепочку, лямки, станок, все
что попало в поле вращения головы. Во время своих сильных и хаотиче-
ских движений собака не один раз ломала прикрепленную к ее лапе уста-
новку для регистрации рефлексов. При этом она изо всех сил лаяла, ску-
лила, визжала. Эта картина бурных явлений волнообразно, с небольшими
перерынами, вначале длилась во все время действия звонка; позже, по
ходу работы, параллельно с образованием запаздывания она развивалась
только в последние 40— 35 секунд; при этом, чем ближе к моменту раздра-
жения электрическим током, тем сильнее становилась реакция. И вот, когда
на фоне этих вулканических явлений производились раздражения электри-
ческим током, происходило резкое ослабление, иногда полное прекращение
голосовых реакций, собака успокаивалась,
исчезало насильственное перенапряжение
мускулатуры, прекращались движения
нераздражаемых лап, и только раздра-
жаемая левая задняя лапа продолжала
свое флексорное движение. Изолирован-
ное раздражение электрическим током той
же силы, без предшествующего действия
звонка, вызывало бурную реакцию. Затем,
если звонок почему-либо ие вызывал бур-
ной условной реакции, электрическое
раздражение не только не ослабляло, но
резко усиливало болевую реакцию со-
баки. Если же звонок вызывал бурную
условную реакцию и я подкреплял реф-
лекс электрическим раздражением не
обычной силы, а значительно сильнее
обычного, опять-так и реакция собаки не
ослабевала, а иногда даже усиливалась.
Эти явления неоднократно демонстрировались акад. И. П. Павлову,
а также многим работникам лаборатории (А. А. Линдбергу, И. В. Мал-
киману, Н. А. Подкопаеву, А. М. Павловой, В. Л. Васютину и Др.). Мои
товарищи по лаборатории, в го время сидевшие рядом со мной, через гла-
зок могли наблюдать собаку и по радио слушать ее голосовые реакции.
Демонстрации почти всегда были удачными. Ахад. И. П. Павлов по обык-
новению во время опыта сидел в камере рядом с собакой. Первые два раза
эти явления при начальных пробах вовсе не получились, а в конце на-
блюдались в очень слабом виде. Очевидно, непривычная обстановка опыта
для реактивной собаки была причиной смазывания описанных выше явле-
ний. Затем они в очень демонстративном виде получались даже в условиях
пребывания акад. И. П. Павлова в камере.
Физиологическую сторону явления я представляю в следующем пока
еще схематическом виде (для ясности изложения я буду пользоваться
упрощенной схемой рефлекторных дут, изображенной на рисунке).
1. При действии условного раздражителя — звонка — происходит воз-
буждение всех звеньев, всех рефлекторных дуг условнорефлекторной ком-
плексной оборонительной реакции, включая эфферентные части дуг соот-
ветствующих безусловных рефлексов. Этим обусловливаются лай, визг,
экскреторные и инкреторные изменения в организме, сложная двигатель-
ная оборонительная реакция и т. д. В частности, возбуждается дуга спе-
9 3. А. Асратян
130
Э А. АСРАТЯН
циально регистрируемого условнорефлекторного движения задней левой
лапы F -* Е -*~е-*С-+ d.
2. В результате этой отрицательной индукции понижается возбуди-
мость афферентной (и конвекторной?) части рефлекторных дуг сложной
безусловной реакции. В случае нашего регистрируемого рефлекса воз-
буждение F е отрицательно индуцирует Д-»- В-*- Ь'.
На это, по-моему, указывает тот факт, что после сильных условных
реакций порог раздражения безусловного рефлекса на некоторое время
повышается и рефлексы на индукционный ток средней силы ослабевают
(мои прежние данные, подтвержденные на этой собаке).
3. На фоне пониженной возбудимости афферентной части рефлектор-
ной дуги действие безусловного раздражителя (электрический ток) такой
силы, которая раньше сама по себе вызывала бурную оборонительную
реакцию собаки, теперь вызывает более слабую реакцию.
4. Однако это полностью не объясняет всей сложности явлений. По-
чему, например, при сочетанном действии обоих раздражителей прекра-
щается бурная картина и собака успокаивается^1 Здесь возможны два
объяснения: безусловное раздражение, недостаточное для вызывания
общей бурной реакции по корковой дуге (Л-* В-* £)-> С-*- d),
достаточно для того, чтобы вызвать слабую изолированную реакцию по
спинальной дуге (Л ^В d); возбуждение же спинальной дуги в свою
очередь отрицательно индуцирует корковый условный аппарат (Е ->е)
и прекращает обусловленную им бурную реакцию. Это наиболее вероятное
объяснение. Другое, менее вероятное объяснение заключается в следующем
Безусловное раздражение возбуждает афферентную часть безусловно-
рефлекторной дуги; это возбуждение дальше распространяется по двум
руслам: по спинномозговой и по корковой дуге (схематически это можно
представить так: Д-> 6-* d и ЛЬ -* D-»- С->- d). При этом
по первому руслу возбуждение протекает беспрепятственно, а по второму
руслу в звене Ь оно встречается с потоком возбуждения, идущим от услов-
ного раздражения (F -+Е- ё). Очевидно, здесь происходит стык, явление,
подобное тому, что некоторые физиологи описывали как интерференцию
волн возбуждения; в результате возникает торможение в разветвлен-
ном звене D и С, которое обусловливает резкое ослабление и порой пол-
ное прекращение бурной болевой условной реакции (успокоение, прекра-
щение лая и т. д,).
5. В пользу торможения в силу индукции или интерференции (не в фи-
зическом смысле слова) волн возбуждения примерно одинаковой силы,
а не торможения от суммирования возбуждений (запредельное торможение
или pessimum) говорят следующие факты: 1) увеличение силы раздражаю-
щего электрического тока не только не вызывает ослабления, но даже уси-
ливает бурную картину реакции; 2) если условный раздражитель почему-
либо вызывает не бурную, а умеренную реакцию, то электрическое раз-
дражение не только не ослабляет эту реакцию, ио даже усиливает ее. Го-
воря иначе, для того чтобы происходило описанное явление ослабления
условной бурной реакции под влиянием электрического раздражения,
требуется некоторое относительное равенство сил условного и безусловного
возбуждений, что, невидимому, необходимо для взаимной индукции (или
для интерференции), но вовсе не является необходимым для суммации.
К ВОПРОСУ О ЛОКАЛИЗАЦИИ ЦЕНТРАЛЬНОЙ ЧАСТИ
РЕФЛЕКТОРНОЙ ДУГИ ДВИГАТЕЛЬНОГО ОБОРОНИТЕЛЬНОГО
УСЛОВНОГО РЕФЛЕКСА >
ворец материалистического учения о высшей нервной деятель-
ности обогатил науку тончайшей и точной методикой услов-
ных рефлексов, благодаря которой современная физиология
постепенно освобождается от существовавших раньше метафи-
зических представлений о локализации функций в коре больших
полушарий головного мозга. Теперь редко кто из физиологов
говорит об узколокализованлых, строго отграниченных друг от друга об-
ластях в этом органе. Следует, однако, отметить, что многие неврологи,
в том числе и многие советские неврологи, сше не достаточно хорошо озна-
комились с этой интересной и важной частью учения акал. И. П. Павлова.
Досадной иллюстрацией этого является предисловие И, Сапнра к кните Лешли
(Lashley) «Мозг и интеллект», вышедшей недавно в русском переводе.( Согласно
мнению Сапнра. основной удар по традиционной механистической теории корковой
локализации функции нанесен работами Лешли. О работах павловских лабораторий,
выполненных больше чем два десятка лет назад, опубликованных на русском языке
и послуживших темой специальных докладов н статей акад. И. П- Павлова, автор пре-
дисловия ничего не говорит, несмотря ка то, что именно этим работам мы прежде всего
и больше всего обязаны своими теперешними правильными представлениями о дина-
мичности и пластичности корковой локализации функций.
На основании ряда классических работ, вышедших из лабораторий акад.И.II. Пав-
лова, была построена строгая 'теория динамической, пластической локализации
в коре большого мозга, основные положения которой примерно сводятся к следующему.
Для каждого качественно своеобразного анализатора в коре большого мозга имеются
более или менее ограниченные области с густым скоплением нервных элементов для
высшего и тончайшего анализа и синтеза адэкватных стимулов. По соседству с этими
специальными областями, на значительно большем пространстве, а то и по всей коре,
разбросаны нервные элементы для более грубого анализа и синтеза соответствующих
стимулов. Иначе говоря, в коре для каждого анализатора имеются «ядра» скопления
первичных нервных элементов и рассеянные вторичные элементы. Основанием и одно-
временно иллюстрацией эгнх положений для звукового анализатора послужили рабо-
ты И. И. Крыжановского, А. Н. Кудрина, М. И. Эльясона и др., для светового анали-
затора — работа А. Н. Кудрина, для кожно-механнчсского анализатора — работы
Н. П. Тихомирова, Л. А. Орбели, Н. И. Красногорского, И. П. Разенкова и др. Однако
до последнего времени как будто немного особняком стоял вопрос о локализации так на-
зываемой двигательной области коры. Попутно отметим, что на основании интересной
работы 14. И. Красногорского, в которой была показана возможность образования усло-
вных секреторно-пищевых рефлексов на проприоцептивные импульсы с двигательного
аппарата и серьезное нарушение этих рефлексов после экстирпации так называемой
двигательной зоны у собак, И. П. Павлов высказал гипотезу, что «двигательная область
коры есть такой же анализатор скеЛетОДвИгательнон энергии организма, как другие
области ее суть анализаторы разных видов внешней энергии, действующей на орга-
низм»2.
‘"Физиологический журнал СССР. т. XVII, № 6, 1935. Предварительное сообщение
напечатано в «Докладах АН СССР», т. I, № 2—3, 1935, по представлению акад.
И. II. Павлова 24 VIII 1934 г.
г И. П. Павлов, Лекции о работе больших полушарий, 1927, стр. 311
9“
132
Э. А. АСРАТЯН
Если Н. И. Красногорский ставил перед собой задачу доказать, что двигательная
область, с точки зрения функциональной конструкции, принципиально не отличается
от других областей коры, и для осуществления своей цели пользовался мышечным
аппаратом как рецептором, то В. П. Протопопов пользовался мышечным ап-
паратом как эффектором и, наряду с другими вопросами, пытался изучить также воп-
рос о характере и локализации двигательной области коры. В. П. Протопопов показал,
будто условные оборонительно-двигательные рефлексы у собак исчезают после экстир-
пации двигательной области (gg . sigmoideus и cruciatus) коры и что после этой операции
становится невозможным образование новых двигательно-оборонительных рефлексов.
Сравнительно недавно эти данные были подтверждены Д. С. Фурсиковым на обезьянах,
а в самое последнее Время В. П. Протопопов и его ученики вновь вернулись к этому
вопросу и на большом экспериментальном и клиническом материале снова подтвердили
прежние данные и выводы. Если даже оставить в стороне расхождение мнений о харак-
тере функциональной структуры двигательной области коры, о том, является ли эта
область рецепторным полем или эффекторным, то все же не устраняется противоре-
чие между павловским пониманием корковой локализации областей и теми выводами,
которые вытекали из работ В. П. Протопопова и других.
fi одной из лабораторий акад. И. Г1, Павлова в последнее время этим вопросом за-
нялся И. С. Розенталь в сотрудничестве с С. В. Сперанской и К. С. Абуладзе. Предметом
их изучения; был, однако, двигательный рефлекс не оборонительного порядка, а реф-
лекс подачи лапы на зов экспериментатора-хозяина. Данные, полученные у разных
собак с различными экстирпациями в соответствующих областях коры, привели
И.С. Розенталя к выводам, которые- целиком подтвердили взгляды И.П. 11анлова на этот
вопрос. И. С. Розенталь и сотрудники показали на многих собаках, что не только после
экстирпации двигательной области, но даже после экстирпации более значительной
массы вокруг этой области удается образовывать навыки подачи лэпы. Стедуст ска-
зать, что этим простым приемом исследования функции коры пользовались етде
Гольц (Goltz) я В. М. Бехтерев. При этом Гольц после экстирпации двигательной зоны
у собак не мог больше получить реакции подачи лапы, а В. М. Бехтерев указал на
возможность воспитания этого навыка даже после названной экстирпации. Впослед-
ствии В. М. Бехтерев перестал упоминать в своих трудах об этих данных, явно не
гармонирующих с поздними его взглядами на характер локализации функции
в коре больших полушарий мозга.
Немного более сложным приемом для разрешения этой задачи пользовали Гит-
циг (Hitzig) И Франц (Franz). Оши применяли методику, похожую на теперешнюю
методику многих бихевиористов, именно методику решения известных задач. Гитциг
отрицал возможность сохранения двигательных навыков после экстирпации двигатель-
ной зоны, а Франц придерживался противоположного взгляда. Впоследствии Франц и
Лешли на крысах подтвердили выводы Франца, а Лешли в своих позднейших ра-
ботах довел эти концепции до огульного отрицания всякой локализации в коре.
Нужно отметить, что эти методические приемы (подача лапы, решение
задач) слишком примитивны для окончательного разрешения таких тонких
задач, хотя известную пользу они могут принести. Если даже пренебречь
этой примитивностью, совершенно естественно могла возникнуть мысль
что причиной расхождения между данными экспериментов упомянутых ис-
следователей и данными В. М. Протопопова — Д. С. Фурснкова являются
разные объекты (рефлексы) исследования. Исходя из таких соображений
я с согласия И. П. Павлова занялся изучением этого вопроса, избрав
в качестве объекта экспериментирования оборонительно-двигательный ре-
флекс конечности. Позже на ту же тему начал работать и другой сотруд-
ник его лаборатории — К. С. Абуладзе.
Работа была начата мной в начале 1932 г. Но собака, у которой я успел
уже выработать положительные и отрицательные условные рефлексы, сбе-
жала из другой лаборатории, куда она была переведена для операции.
Чтобы больше не задерживать хода работы, я нс стал подготовлять новую
собаку для этой цели и, воспользовавшись любезным предложением И. С. Ро-
зенталя, взял оперированную им для других целей собаку Кейо и про-
должал исследования на ней.
Кейо была оперирована 14.X 1932 г. У нее одновременно с обеих сторон
из передней половины коры больших полушарий были экстирпириэваны
довольно большие участки, которые включали почти всю двигательную
область, область кожной чувствительности и даже захватывали немного
больше: gg. sygmoideus, coronarius, ectosylvius, cruciatus и прилегающие
ЛОКАЛИЗАЦИЯ ДВИГАТЕЛЬНОГО ОБОРОНИТЕЛЬНОГО УСЛОВНОГО РЕФЛЕКСА | 33
Рис. 1. Ориентировочные границы
экстирпированной части коры
зоны (рис. 1). Исследования на Кейо были начаты в первых числах марта
1933 г., т. е. через 4*/а месяца после операции. Внешне в двигательной
сфере животного трудно было уловить какие-нибудь серьезные отклонения
от нормы, помимо того, что собака была очень подвижной как на воле,
так и в станке и ее походка напоминала
«петушиную»; необходимые для ходьбы
движения были размашистые, энергич-
ные. Кроме того, собака не могла
вспрыгнуть на станок, который был нем-
ного выше ее роста, и очень осторожно
спускалась со станка.
Выработка условных оборонительнодви-
гатсльных рефлексов велась широко рас-
пространенным способом В. П. Протопопо-
ва— путем сочетания индиферентных раз-
дражителей с раздражением задней правой
конечности индукционным током; при этом
действие индиферентиого раздражителя на
1—2 секунды предшествовало электриче-
скому, а сочетание их длилось 4—5 се-
кунд. При помощи соответствующей уста-
новки рефлексы объективно регистриро-
вались на кимографе. Кроме того, проводи-
лось наблюдение за поведением собаки.
Условные рефлексы на свет и на касалку образовались неожиданно
быстро. Намеки на них были уже на 4-й день (11-е сочетание света, 13-е со-
четание касалки). На 5-й день были налицо уже хорошие рефлексы на оба
раздражителя (18-е сочетание света, 20-е сочетание касалки), а на 6-й день
рефлексы были стойкие, и в дальнейшем они зигзагообразно все больше
увеличивались и подкреплялись (рис. 2).
После того как условные рефлексы на свет и касалку стали достаточно
прочными, началась выработка условного рефлекса на звуковсе раздра-
жение— па той, который мы условно называем тоном С. На 4-м соче-
тании уже были намеки на условный рефлекс, а на С—8-м сочетаниях
рефлексы уже были хорошо выражены (рис. 2).
Что описанные рефлексы были условными, а не явлением порядка
Bahnung, можно судить по тому, что они получались не только в середине
и в конце опытов, когда создаются условия для Bahnung, но и в начале
опыта, перед раздражением электрическим током. Весь дальнейший ход
исследования и наблюдения полностью подтвердил это.
Таким образом, на основании приведенного материала можно катего-
рически утверждать, что экстирпация так называемых двигательных и
даже кожно-тактильных областей коры у собак не исключает возможности
образования условных сборонителы'о-двигателььых рефлексов.
Примерно через Р/з месяца работы, когда налнио были устойчивые,
хорошо выраженные условные рефтексы на все три раздражителя, нача-
лась выработка днференцировкн на тон, который мы условно называем
тоном А. Диференциация тонов шла поразительно быстро; на 5-м при-
менении была относительная днференцировка, а на 6-м — уже абсолютная.
Должен заметить, что, несмотря на быстроту образования днференци-
ровки, собаке, очевидно, была трудна эта задача; она сильно беспокои-
лась, рычала, делала попытки спрыгнуть и сбежать со станка, много раз
ломала установку для передачи и .регистрации рефлексов и т. д.
Иллюстрация хода днференцировкн, состояния повышенного возбужде-
ния в этом периоде, а также стадии абсолютной днференцировкн тона А
от тона С дана на рис. 3..
134
Э. А. АСРАТЯН
Рис. 2. Ход образования условных рефлексов у Кейо
Линии сверху вниз: запись рефлекса, безусловный раздражитель. так-
тильные условные раздражители, звуковые условные раздражители
и световые условные разражигели (цифры указывают расстояние между
катушками), время в секундах. I — свет 17-й раз; II — свет 19-й раз;
/// — касалка 20-й раз; /V —свет 24-й раз; / — касалка 37-Я раз;
свет SO-fl раз (начала опыта); VI — тон С 8-й раз
Рис. 3. Образование диферениировки
I и II (опыт от 4.V 1933 г.) — тон А на 5-е применение еще дает рефлексы,
на б-с рефлекс уже исчез. Собакя сильно беспокоится, ш (опыт от
13.V) — устойчивая диференцнровка оставляет за собой последовательное
торможение
Линии сверху вниз: запись рефлекса, отметка безусловного раздражителя,
отметка условных раздражителей, время в секундах
ЛОКАЛИЗАЦИЯ ДВИГАТЕЛЬНОГО ОБОРОНИТЕЛЬНОГО УСЛОВНОГО РЕФЛЕКСА
Образованный отрицательный условный рефлекс оставляет за собой
последовательное торможение, которое вызывает заметное оставление
следующего за ним однородного положительного условного рефлекса, уве-
личивая (нндукционно) безусловный рефлекс (рис. 3 и 4).
Рис. 4. Последовательное торможение условного рефлекса на тон С
от диференцировочного раздражителя тона А (опыт 7.VI 1933 г.)
Обозначения те же, что на рис 3
Следует отметить одно замеча тельное явление, связанное с образованием
днференцировкн. Выше уже было сказано, что в период образования ди-
ференцировки собака вела себя на станке особенно беспокойно. Но после
Рис 5. Период успокоения собаки под влиянием укрепления ди-
ференцировкм (опыт 25.V 1933 г.)
У сливные рефлексы выражены довольно слабо (в особенности на тон С).
Псимененне диференцировочного раздражителя (тон А) в конце опыта через
20 сек. положительно индуцирует рефлекс на той С. ОВоапачения те же,
что на рнс. 3
того как образовалась и укрепилась днференцировка, произошли интересные
сдвиги: во-первых, это сильно отразилось на одноименном положительном
условном рефлексе, а во-вторых, собака стала неузнаваемо спокойной; она
на станке вела себя примерно. Для иллюстрации привожу один опыт
(рис. 5) из этого периода работы. Сравнение рис. 5 и рис. 3, / ярко
иллюстрирует сказанное.
До образования днференцировкн и в период ее образования, когда
собака была сильно возбуждена, положительные условные рефлексы ПОЧТИ
136
Э. А. АСРАТЯН
не поддавались утешению в течение одного опытного сеанса (при 30—40
применениях ни разу не получался нулевой эффект). После же образования
диференцировки эти же рефлексы быстро угашались: на 3—5-м при-
менении раздражителей без подкрепления получились первые нули, затем
рефлексы в слабом виде волнообразно появлялись вновь, чтобы в даль-
нейшем совсем исчезнуть.
Рис. 6 иллюстрирует угашение положительного условного рефлекса
на тон С в периоды до и после образования диференцировки. Совершенно
такой же ход угашения наблюдался и при других раздражителях.
Рис. 6. Угашение рефлекса на тон С до и после обра-
зования диференцировки
Г — опыт 13.1V 1S33 г. Приведены рефлексы на 1, 10, 12, 80
и 25-е применение раздражителя. Рефлекс на 12-е применение
самый маленький во всем опыте. Паузы З-мииутные. It — опыт
20.V 1933 г,- Приведены рефлексы на 1.3. 5. 12 — 15-е при-
менение раздражителей. Рефлекс на в-е и 7-« применение —
нуль. Н-с — следы, 9-е — рефлекс средней силы, io-е. 1|-е
нуль. Паузы 3-мииутиые. Обозначения те же, что на рис. 3
Но с утешением было связано и другое интересное явление.JB пер-
вом периоде, когда в течение одного опытного сеанса не удавалссь
добиться полного угашения, такие опыты не оставляли никаких следов.
Во втором периоде, когда угашение наступало быстро в течение одного
опытного сеанса, такие опыты вызывали значительные изменения в мо-
торной деятельности собаки, сказывавшиеся даже на воле. Начиная
с третьего, иногда даже со второго дня после таких опытов, собака Кейо
начинала хромать на ту ногу, которая подвергалась раздражению во время
опыта. Эта хромота продолжалась неделями. В этом периоде собака, бу-
дучи вообще спокойной на станке, РИЗЖала при применении диференциро-
вочного раздражителя, не делая, однако, никаких движений. Больше того,
в первые дни поело этих сеансов угашения положительные условные реф-
лексы исчезали или же сильно ослабевали. Бывало днференцировка растор-
маживалась кратковременно, а вообще она чрезвычайно усиливалась, так
что во время действия диференцировочвого раздражи тел я’ссбака не только
ЛОКАЛИЗАЦИЯ ДВИГАТЕЛЬНОГО ОБОРОНИТЕЛЬНОГО УСЛОВНОГО РЕФЛЕКСА I37
стояла без движения, не только не сгибала конечности, а даже делала
слабые попытки вытянуть ее, что, очевидно, является показателем усиле-
ния торможения. Кроме того, последовательное торможение от действия
дифереицировочного раздражителя усиливалось к иногда даже распре?
странялось на положительные условные рефлексы, связанные с другими
анализаторами. Характерно, что применение дифереицировочного раздра-
жителя часто вызывало положительную индукцию на ослабленный рефлекс
на тон С (рис. 5).
Все сказанное о влиянии дифереицировочного раздражителя говорит
о том, что торможение, достигая известной степени интенсивности, как бы
вводит в определенные рамки повышенную возбудимость собаки, упорядо-
чивая ее хаотические движения. Но когда в дальнейшем торможение
усиливается еще угашением положительных условных рефлексов, это
перенапряжение торможения вызывает нечто вроде функционального нев-
роза в адэкватных элементах нервной системы.
В заключение следует отмстить некоторые другие показатели существо-
вания условных двигательных рефлексов у Кейо. Радемакер (Radeniaker)
в своей монографии «Das Stehen» описывает ряд двигательных реакций
условного порядка. Эти реакции суть: «Stehbereitschaft» [способность
животного упираться в подставку задними или передними конечностями
и вытягивать их, когда при висячем, положении в воздухе какая-нибудь
часть его тела (хвост или морда) приводится в соприкосновение с этой
подставкой или же когда оно видит эту’подставку ]; «Magnetreakt ion» (спо-
собность животного разгибать согнутую конечность, когда к ней прика-
саются пальцем или каким-нибудь предметом, и вытянутой конечностью
следовать за движением прикасающегося предмета); и, наконец, коррек-
ция ненормальных положений конечностей. У нормальных собак и даже
у собак, лишенных мозжечка, все эти реакции имеются в более или менее
выраженном виде, а у собак без больших полушарий они исчезают. На Кейо
были Испытаны все Эти реакции, и все они были налицо. К сожалению, реак-
ции не были сфотографированы, и мы не можем иллюстрировать сказанное.
Следует отметить, что мои данные и данные К. С. Абуладзе об образо-
вании условных рефлексов полностью совпадают.
Заключение
Как себе представить интимную сторону образования условных электро-
оборонительно-двнгательных рефлексов у собаки Кейо, а также существо-
вание «Stehhereitschaft», «Magnetreaktion» и способности корригировать
ненормальное положение конечностей?
Здесь возможны два объяснения: первое — условная связь образуется
между соответствующими элементами коры и подкорковыми двигательными
нервными элементами; второе — указанная связь возникает между’ ре-
цепторными элементами коры и теми рассеянными по их корковой массе
вторичными элементами коры, которые имеют отношение к двигательной
деятельности животного. Из этих двух допущений более вероятным нам
кажется второе. В пользу него, помимо всяких доводов гипотетического
характера, говорят и новые факты о том, что условная связь, вообще
говоря, образуется между элементами коры. Естественно возникает вопрос
каков же характер тех рассеянных вторичных нервных элементов, с кото-
рыми связываются условной связью различные рецепторные элементы коры
нашей собаки. Я склонен думать, что они не только афферентного, но и
эфферентного характера. Это, с одной стороны, говорит о том, что павлов-
ская концепция о первичных—ядерных и о разбросанных во коре вторичных
рецепторных элементах целиком распространяется и на эфферентные нерв-
ные элементы коры, по которым корковые импульсы передаются ниже-
138
Э Л. АСРАТЯН
лежащим отделам центральной нервной системы; с другой стороны,
делает понятным факт образования двигательного условного рефлекса
после удаления так называемых двигательных зон коры.
Как же понять то, что В. М. Протопопов и Д. С. Фурсиков па осно-
вании экспериментов отрицали возможность сохранения старых и обра-
зования новых оборонительно-двигательных условных рефлексов?
По всей вероятности, это объясняется следующим. После всякой серьез-
ной мозговой операции, как это хорошо показано в многочисленных
работах, вышедших из лаборатории акад. И. П. Павлова, условно-
рефлекторная деятельность ослабевает, а искусственные рефлексы со-
вершенно исчезают иногда на месяцы. Эти изменения выражены резче,
когда повреждаются те области коры, которые имеют близкое отношение
к изучаемым рефлексам. Чтобы вторичные эфферентные (и афферентные)
нервные элементы коры могли активизироваться после такого грубого
вмешательства, конечно, требуется время, причем для разных собак раз-
личное. Повидимому, упомянутые исследователи имели дело с такими
животными, которые при операции еще недостаточно поправились для
решения поставленной перед ними задачи.
Выводы
1. После экстирпации двигательных и кожно-тактильных зон коры
у собак возможно образование условных оборонитслызо-двигательных
положительных и отрицательных рефлексов.
2. Образование днференцировки значительно регулирует хаотическую
двигательную деятельность животного с экстирпацией указанных обла-
стей коры.
3. Перенапряжение торможения путем простого угашения условных
положительных рефлексов вызывает локальный функциональный невроз
раздражаемой конечности, что выражается в хромоте, ослаблении и даже
исчезновении условных положительных рефлексов, а иногда растормажи-
вании днференцировкн.
4. У собаки без названных областей коры сохраняется способность
корригировать ненормальные положения конечностей, «Stehbereitshaft*
и «Magnetreaction».
5. У собак без двигательных областей условная связь, видимо, обра-
зуется между соответствующими рецепторными нервными элементами
коры и рассеянными по коре вторичными нервными элементами афферент-
ного и эфферентного характера.
литература
Бехтерев В. М., Основы учения о функциях мозга, т, V, 1906,
Красногорский Н. И., О процессе задерживания и о локализации конечного
и двигательного анализатора в коре больших полушарий собаки, дисс., 1911,
Т1 а в л о в И. П., .Пекпии о работе больших полушарий головного мозга. 1927.
Протопопов В. П., О сочетательной двигательной реакции на звуковые раз-
дражения. днсс., Спб., 1909.
Протопопов В. II., Центральная часть рефлекторной дуги оборонительного
условного- рефлекса. Современная психоневрология, 1. 1931.
Розенталь И. С., Абуладзе К С.. Условнорефлекторная деятельность соба-
ки после частичного разрушения корковых зон, Труды физиологических лабо-
раторий И П Павлова, т XII1, 1948.
Фурсиков Д. С.. Изучение поведения и локализация функций коры мозга
у обезьян го методу условного рефлекса, Труды 11 Всесоюзного съезда
физиологов, 1926.
Н i t г i g. Arch. f. Psychiatr.. B-d. XV, 18841.
Franz, Amer, journ. of physiol., 1902.
Franz u. Lashley, Psychobiologie. 1920.
Lashley, Речь на заседании ИМ- Гарвея, 1930.
Rademaker, Das Stehen, Berlin, 1931.
ВЛИЯНИЕ ПИТУИТРИНА НА УСЛОВНЫЕ ПИЩЕВЫЕ
СЛЮНООТДЕЛИТЕЛЬНЫЕ РЕФЛЕКСЫ 1
Предварительное сообщение
опрос о взаимодействии нервной и эндокринной систем яв-
ляется одним из кардинальных вопросов современной фи-
зиологии. Современная клиническая и физиологическая ли-
тература насыщена данными по этому вопросу. Что же
касается частного вопроса — влияния эндокринных желез
на высшую нервную деятельность, — то, помимо обширного
клинического материала, в последнее время накапливается большой и
интересный экспериментальный материал. Углубленные работы В этом на-
правлении начались в лабораториях акад. И. П. Павлова. В то время как
путем простого наблюдения не удается заметить существенных изменений
в сфере высшей нервной деятельности у собак с различными отклонения-
ми и дефектами инкреторной системы ]Бндль (Biedl) и др.], могучий метод
условных рефлексов выявил у таких животных ряд серьезных изменений
в этой сфере. Работы К. Н. Кржишковского, Е. М. Крепса, Д. С. Фур-
сикова, И. С. Розенталя, А. В. Валькова показали, что течка и беремен-
ность существенно изменяют высшую нервную деятельность собаки.
Ф. П. Майоров такие же изменения отметил у самца в период полового
возбуждения. А. В. Вальков исследовал влияние экстирпации щитовидных
желёз на условнорефлекторную деятельность щенков и показал, что при
этом наступают очень глубокие изменения в сфере высшей нервной дея-
дельности.
Начиная с 1924 г., Б. М. Завадовский и его сотрудники, пользуясь
методом условных рефлексов, довольно обстоятельно изучили, с одной
стороны, влияние экстирпации gl. thyreoides, с другой стороны — влияние
дачи препаратов этой железы на условнорефлекторную деятельность собак
и кур. Примерно в 1927 г. в лаборатории акад. И. П. Павлова его сотруд-
никами Д. И. Соловейчиком, М. К. Петровой и А. М. Павловой было изу-
чено влияние кастрации и омолаживания на условнорефлекгорную дея-
тельность собаки. В самое последнее время, после того как началась из-
лагаемая работа, появились также данные относительно гмпсфиза.
В. Я. Кряжев исследовал влияние экстирпаций этой железы на условпо-
оборонительно-двигательную деятельность собаки. Основные результаты
В. Я. Кряжева сводятся к следующему. У собак сильно ослабевают как
безусловнорефлекторная, так и в особенности условнорефлекторная дви-
гательная электрооборонительная деятельность, провесы возбуждения и
торможения становятся очень инертными — «дискретными».
С целью анализа интересующего меня вопроса о влиянии симпатиче-
ской нервной системы на условнорефлекторную деятельность собаки я изу-
чал влияние питуитрина на условные слюноотделительные рефлексы у трех
1 Архив биологических наук,. 37, в. 1, 1935.
140
Э. А. АСРАТЯН
подопытных собак, у одной из которых были перерезаны оба шейных сим-
патических нерва. Так как полученные в этом направлении данные пред-
ставляют самостоятельный интерес, я их вкратце опишу ниже.
Я пользовался тремя препаратами питуитрина: московским «pitui-
trin Т», изготовленным специально для научно-исследовательских работ,
затем ленинградским н московским продажными препаратами. Однако
больше всего я пользовался первым. В основном действие всех пре-
паратов было одинаковым. Ленинградский препарат действовал слабее
других.
Чтобы исключить влияние самого акта подкожной инъекции, были
поставлены контрольные опыты с инъекцией физиологического раствора.
Во избежание возможных других осложнений такие опьиы чередовались
с опытами обычного порядка. Всего было проведено около 20 опытов над
тремя собаками.
Основные данные экспериментов сводятся к следующему. Через 40—
90 минут после подкожной инъекции первого препарата в дозах 1—1,5смя
условные слюноотделительные рефлексы сильно
уменьшаются, а иногда и вовсе исчезают. Если же взять увели-
ченную дозу — в 2—2,5 см3, то через такой же период условные положи-
тельные рефлексы совершенно исчезают. При дозах менее
1 см3 условные рефлексы только ослабевают. Во всех случаях безусловные
рефлексы заметно не изменяются, иногда создается даже впечатление,
что они увеличиваются. Действие дифереицировочного раздражителя осо-
бенно не изменяется, если не считать слабого его растормаживания у одной
собаки (Атлас). В табл. 1—3 приведены результаты обычных опытов, кон-
трольных опытов, опытов с инъекцией физиологического раствора и опытов
после инъекции.
В случаях, когда питуитрин вводился в еще меньших дозах или когда
опыт ставился не через 1 часа после подкожной инъекции питуитрина,
а через более короткие сроки, иногда удавалось улавливать периоды,
напоминающие парадоксальную фазу из цикла гипнотических фаз. В это
время безусловные рефлексы определенно увеличиваются. Затем скоро
эта ((таза уступает место фазе исчезновения всех рефлексов (см. опыт на
Атласе).
Из приведенных пока предварительных данных явствует, что иикрег
гипофиза оказывает сильное влияние на условнорефлекторную деятель-
ность собаки. Обстоятельные исследования в этой области еще впереди.
К сожалению, в моем распоряжении не было достаточного количества пре-
парата, изготовленного специально для научно-исследовательских работ, и
потому я был лишен возможности варьировать опыты, ставить контрольные
ОПыТы с инъекцией среды, в которой были Экстрагированы и консерви-
рованы ин крепя. Хотя продажный московский препарат сказал почти
такое же действие, я не мог руководствоваться полученными результатами,
поскольку неизвестно, все ли препараты одинаковой крепости, давности
изготовления и т. д.
Несмотря на все это, мне думается, что в грубых чертах установлен
основной факт— ослабление условнорефчекторной деятельности под влия-
нием значительных доз инкрета гипофиза.
Хотя мы не можем без оговорок сопоставлять эти предварительные
данные с данными В. Я. Кряжева, все же провести некоторую параллель
между ними возможно, поскольку в обоих случаях речь идет об условно-
рефлекторной деятельности.
Во-первых, у гипофнзэктомнрованиых’собак В. Я. Кряжева были силь-
но ослаблены как пищевые, так и электрооборонительные безусловные
рефлексы. Весьма вероятно, что это и является основной причиной осла-
бления условцорсфлекторной деятельности у этих животных.
Собака Арч
25.11 1933 г. (контроль) 0.IV 1932 г. подкожное введение 2,5 см* физиологического раствора
О а X V •“ж о X о 4> «“ 3
название а. IS &»S v Я название D. Ч — a jCj
условного с « О X ЯюЗ условного Е -а X и 2 2 ю 3
раздражи- 3 X С х 53 ?SI я rt X о 4 « раздражи- 3 X ь . « X “ в ЛОВ»- за 3 1ИЯХ
теля X X t° « Ш . X XU«f теля 5J “ о
ли в с Е х ш й I а Srt = з <>«» «=£«=(
Звонок 1 33 112 Стук 120 1—2 26 112
Стук 120 2 28,5 НО Звонок 1—2 28,5 108
Свдт 3 24 108 . Свет 1-2 22 113
Шипение 3—4 26 105 Звонок 7 26 ПО
Звонок 4 28 102 Свет 3 18 103
Стук 60 — 0 0 Звонок 3 24,5 104
Свет 2-3 18 100 Стук 120 3 22 98
Стук 60 4 1 12
Зво нок 4 19 103
Свет 4—5 14 97
- Стук 5 О 95
Таблица 1
9.1V 1932 г. по-дкожное введение 2,5 см* питуитрина 14.IV 1932 г. 1,1 см’ подкожное введение питуитрина
ех О W 1 г? •fa et V • а: о «“ 2
н азвание а. * название о 4-gS
условного с ье3 S U g Зщ 3 условного Е 3 >ый 5 с UIK
раздражи- теля 3 Z ь • z X ювный 2о сек lx шка «" X ?? х V Z 4» раздражи- теля 3 S •- . Т * h ° юпный 20 сек lx шка *<’> к 2 2 ж Ч Л — , О X >, и V
ь и п х е । 2 * 5
Z • 8« Г. £ ; с ч ч
Стук 120 1-2 Следы 106 Звонок 7 8 1 107
Звонок 2 1 104 Стук 120 9 5 104
Свет — 0 100 Свет 12 4 106
Звонок — 0 98 Звонок 18 1 100
Стук 60 — 0 0 Свет — 0 98
Касалка — 0 103 Стук 120 — 0 96
Стук 120 — 0 97 Стук 120 — 0 0
Звонок — 0 95 Звонок — 0 92
Свет — 0 93
Собака Арч
9.V 1933 г. введение 1,2 см’ московского специального препарата 5.V 1933 г. (Контроль)
название условного раздражи- теля латентный период в сек. условный рефлекс за 20 сек. п деле- ниях шкалы безусловный реф- лекс за 35 сек. в делениях шкалы название условного раздражи- теля латентный период 8 сек. условный рефлекс за 20 сек. в деле- ниях шкалы безусловный реф- лекс за 35 сек. в делениях шкалы
Свет 2 22 124 Свет 1 36 ИЗ
Звонок 5 8 134 Звонок 1 36 но
Касалка 3 12 128 Касалка 1 26 105
Стук 120 7 6 129 Стук 170 2 29 105
Свет — 0 124 Стук 60 — 0 5
Звонок 7 7,5 125 СвеТ 17 8 —
Стук 60 — 0 0 Зво нок 5 13 101
Звонок — 0 Н2
Таблица 2
__1_ Собака Голован
I5.IV 1033 г. (контроль) a.iV 1033 г. введение за 1 час 2 см1 пении- градского пре- парата 3.V1II 1933 г. введение за 2 часа 1 см* московского специ- ального препарата
название условного раздражи- теля Звонок Свет Касалка Стук 170 Стук G0 Свет Звонок tx о S а. о Б « 3 х h "• о я " а 1 2 3 2 7 4 _ _ _ условный рефлекс - •*-> О ciS оо го за 20 сек п делс- сл миях. шкалы безусловный реф- — ~ леке за 4и сек. в о да О да — О — делениях, шкалы латентный период 1 “ I СП CJ to 1 вегк. | — условный рефлекс о но О — ьэ сл . за 20 сек. в деле- сл ннях шкалы безусловный рсф» — — — — — •— леке за 40 сек. в сл to о -ч со ° “ делениях шкалы название условного раздражи- теля Стук 120 Касалка Звонок Свет Касалка Стук СО Стук 120 латентный период *sj о qo tn ел 4- в сек. условный рефлекс сл о ьэ «и о и* ос за 20 сек. в деле- ниях шкалы безусловный реф- “ Гз Г= леке за 40 сек. и m _ Yi гЙ — & делениях шкалы
Собака ?\тлас
II.IV 1933 Г. (контроль) 13.IV 1033 г. введение фи эно- логического раствора 15.IV 1933 г. вве- дение за 30 мии. 1 см‘ ленинград- ского препарата
название условного раздражителя латентный период п сек. условный рефлекс аа 20 сек. е деле- ниях шкалы безусловный реф- лекс за 40 сек. и делениях шкалы латентный период 1 в сек. условный рефлекс за 20 сек. в деле- ниях шкалы безусловный реф- лекс за 40 сек. в делениях шкалы латентный период в сек. условный рефлекс за 20 сек. о деле- ниях шкалы безусловный реф- I леке за 40 сек. в делениях шкалы
Свет Звонок Касалка Стук 120 Свет Стук 60 Звонок Свет Касалка Стук 120 1 2 2 3 2 5 3 11 13,5 16 9,5 13 9 0 8 6 3 106 103 105 101 97 6 101 97 94 1 1 3 2 4 4 3—4 17 11 13 16 7 11,5 7,5 Следы 9 6,5 2 6 91 87 92 86 82 4 91 £6 93 83 1 1 2 7 8 15 13 14 13 18 15 1,5 5 1,5 1,5 3 2 0 0 по 112 130 119 107 3 ПО 98 102 100
Таблица 3
9.V 19-33 г. введения» 3Q мин. 3.VIII 1933 г. введение за 30 мкн. 1,5 см* московского препарата
1 см’ ленннгрддскаг о препарата
название условного раздражителя атентмый период ССК. слоеный рефлекс 1 20 сек. в деле- иях шкалы езусловный реф- снс за 40 сек, в е.,ениях шкалы название уело иного раздражителя г( О X О. Е 3 X 1- . S 2 « словмый рефлекс а 20 сек. в деле- нях шкалы езусловный рсф- скс эй 40 сек. в делениях шкалы
Ч а >* Л Z о =; « н в п X Ю R Ч
Свет 5 4,5 126 Касалка 4 4 128
Касалка 3 3 122 Стук 120 7 1,5 131
Стук 120 7 3,5 13] Звонок — 0 —
Звонок 6 4,5 119 Свет 3 2 128
117
Касалка 14 2 114 Стук 60 2 3 5
Свет 12 3 112 Касалка — 0 131
Стук 120 10 4 4
Стук 69 7 1,5 115
Стук 120 — 0
144
Э А АСРАТЯН
У наших собак иод влиянием питуитрина условные рефлексы исчезли,
а безусловные рефлексы не только не ослабевали, но часто даже усили-
вались Возможно, что некоторое усиление безусловных рефлексов было
вызвано как раз исчезновением положительных условных рефлексов,
не исключена возможность и обратного влияния Выяснение этого —
дело будущего
Во всяком случае наличие парадоксального сходства фактов, а именно
значительное ослабление условнорефлекторной деятельности при отсут
ствим гипофиза (В Я Кряжев) и сильное ослабление этой деятельности
при даче значительных доз препаратов питуитрина (мои данные), пови-
димому, свидетельствует о различном механизме обусловливающего их
действия Физиология знает много таких примеров В этом кратком
предварительном сообщении я ограничусь лишь указанием на аналогичное
явление, полученное со щитовидной железой Работы А В Валькова,
Б Ц Завадовского и сотрудников, по существу говоря, тоже привели
к совершенно аналогичному парадоксальному явлению к сходным изме
нениям условнорефлекторнон деятельности как при экстирпации щиго
видной железы, так и при даче здоровым собакам препаратов этой
железы
ЛИТЕРАТУРА
Вальков А В, Опыт изучения высшей нервной деятельности у тнреоидэктоми
рованных щенков. Юбилейный сборник И П Павлова, 1925
Завидовский Б М , 3 а к А Л , К вопросу о влиянии щитовидок железы на
высшую нервную деятельность у собаки, Медико биологический журнал, в 2
1928, стр 15 ’
Завадовский Б М , Захаров В Р, Азимов Г., Lieber den Einfluss
der schilddruse auf die hoheren Nervenfunktionen d Hunde, Pllug Arch , Bd 223
Ъ 535, 1929.
Завадовский Б M и 3 л о т о в С М , О влнянии щитовидной железы на высшую
нервную деятельность у собак, Медико биологический журнал, в 4—5, 1929
Завадовский Б М . 3 л о то в С М, Захаров В Р,0 алия нии щита
видной железы на высшую нервную деятельность у собаки, Медико биологи
Мелкий журнал, в 3, 1929
Завадовский Б М , Р о х л и и а М Л , Условные рефлексы у нормальных и
гипертиреоидизированных кур. Медико биологический журнал, в 3, 1927
Крепе F м , К копросу о влиянии течки на РЫСШую нервную деятельность, Рус
ский физиологический журнал, т VI, в 4—6, 1924
КржишковскийК Н.' Die Verandcrtingen in der Funktion oberen Abschmtte
des Nervensystems bei ber Hunden wahrend der Brunst, Zbl f Physiol Bd 24,1909
Кряжев В Я, Характер высшей нервной деятельности у типофгзэктомирован
ных животных, Советская невропатология, т 9—10 1932.
Майоров Ф II . К вопросу о влияния полового возбуждения на выьшую нервную
деятельность у собак. Архив биологических наук, т , 38, в 2, 1935.
Розенталь И С, Влияние беременности м лактации на условные рефчексы.
Русский физиологический журнал, т V, 1922
Соловейчик Д И . Влияние так называемых операций омоложения на BbiCulViO
нервную деятельность Экспериментальное исследования на собаке по методу
условных рефтексов, Труды лаборатории акад И П Павлова, т 4 . в 1—2
1932
Фурсиков Д С . Влияние внешнего торможения на образование днференциров
ки и условного тормоза. Архив биологических наук, 21, 1922
О ПРОЧНОСТИ УСЛОВНЫХ РЕФЛЕКСОВ1
течение последних дет я вновь подверг тщательной про-
верке старое лабораторное наблюдение о том, что условный
пищевой рефлекс на кручение кормушки с пустыми чашками
является одним из наиболее постоянных.
При постановке экспериментов я руководствовался теоре-
тическими соображениями, выдвинутыми мной для объяс-
нения названного выше факта. Объяснение это, полностью поддержанное
в свое время Иваном Петровичем, заключалось в следующем: условный
рефлекс на кормушку является наиболее постоянным в силу того, что:
1) верчение кормушки подкрепляется большее число раз, чем любой
другой раздражитель;
2) действие кормушки никогда не отставляется, а наоборот,, постоянно
сразу же подкрепляется;
3) кормушка в силу своей близости к пище является как бы близким
к натуральным ее сигналам, во всяком случае ближе, чем другие искус-
ственные сигналы.
Поставленные на трех собаках эксперименты целиком подтвердили
правильность высказанного соображения.
1. У двух подопытных собак, давно находившихся у меня в экспери-
менте вновь были выработаны условные рефлексы на два примерно одина-
ковой силы раздражителя одного анализатора. Затем один из рефлек-
сов был оставлен остросовпадающим, другой — отставлен на 20 секунд.
Количество сочетаний было одинаково. Рефлексы подвергались испытанию
с помощью различных функциональных проб, совершенно идентичных
для обоих рефлексов 2. Оказалось, что:
а) при пробах отставленный рефлекс угашался намного быстрее, вос-
станавливался значительно позднее, чем рефлексы с остросовпадающим
подкреплением;
б) предварительная кормежка собаки перед опытом (с целью ослабле-
ния пищевой возбудимости) уменьшала отставленный рефлекс намного
сильнее, чем рефлекс с остросовпадающим сочетанием;
в) экстрараздражитель сильнее и длительнее действовал на отставлен-
ный рефлекс, чем на рефлекс, подкрепленный остросовпадающим со-
четанием.
2. Сравнение результатов проб при рефлексах, выработанных в раз-
личные сроки, показывает, что чем позже выработан рефлекс, тем легче
он поддается угашению, торможению под влиянием экстрараздражителя
и предварительного кормления животного. Однако позже известного срока
значение этих моментов ослабевает.
3. У одно»! вновь введенной в опыт собаки был выработан единственный
рефлекс на кручение кормушки.
1 Бюллетень ВИЭМ, № 3 — 4, 1936.
а При «робах остросов падающие условные рефлексы отставлялись вта 20 секунд.
10 Э А. Асратян
146
Э. А. АСРАТЯН
После 100-кратпого сочетания безусловного раздражителя с пустой
кормушкой были поставлены функциональные пробы.
Затем у этой собаки были выработаны два рефлекса таким же образом,
как у описанных выше собак. После 100-кратного сочетания они подвер-
гались испытанию с помощью тех же функциональных проб.
При сравнении данных функциональных проб двух этих рефлексов
были получены те же результаты, что и у описанных собак.
Сравнение показателей рефлекса на кормушку и на остросовпадающнй
раздражитель (гудение) показало, что рефлексы на кормушку несравненно
более стойки, чем на последний раздражитель.
На основании полученного материала и некоторых теоретических со-
ображений следует совершенно четко поставить вопрос о понятии «проч-
ности», «крепости» условных рефлексов наряду С понятием «силы». Это
понятие не ново в учении об условных рефлексах, но оно не определено
достаточно четко.
Что же касается сравнительного значения каждого из перечисленных
моментов, обусловливающих прочность условных рефлексов, то следует
сказать, что момент количества сочетаний с течением вре-
мени утрачивает свое решающее значение.
Момент о тс та влепи я, несомненно, является наиболее важным
и сохраняет свою силу более или менее постоянно.
Ближайший «механизм» явления представляется в следующем виде.
При изолированном действии отставленного условного раздражителя
процесс возбуждения сопровождается процессом торможения, почему
и рефлекс делается менее стойким. При условных рефлексах с остросов-
падающими сочетаниями возбуждение или совсем не сопровождается тор-
можением, или торможение сравнительно слабое, почему этот рефлекс
и остается более стойким.
«Вся органическая природа является
одним сплошным доказательством тождест-
ва или неразрывности формы и содержания.
Морфологические и физиологические явле-
ния, форма и функции обусловливают вза-
имно друг друга».
Ф. Энгельс, Диалектика природы
«После констатирования, возможного
анализа и систематизирования наших яв-
лений следующая фаза работы — и мы уже
вступили в нее — это систематическое дро-
бление и нарушение центральной нервной
системы, чтобы видеть, как будут при этом
изменяться уста новленные выше отношения.
Таким образом, произойдет анатоми-
ческий анализ механизма этих отношений».
И. П. II а в л о в, Двадцатилетний опыт
К ВОПРОСУ ОБ АНАТОМО-ГИСТОЛОГИЧЕСКОЙ ОСНОВЕ
УСЛОВНОРЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ ВЫСШИХ
ЖИВОТНЫХ 1
дна из основных биологических проблем соотношения фор-
мы и функции интересовала и интересует физиологию
не только и не столько в аспекте грубой коррелятивной свя-
зи между формой и функцией отдельных органов и систем,
но главным образом под углом зрения более топкой связи
между внутренней, деликатной архитектурой тканей и орга-
нов и между их функциональными проявлениями. Объединенным трудом
многочисленных физиологов, гистологов, клиницистов и эмбриологов за
последнее полустолетие в науке накоплен огромный материал, показываю-
щий неразрывную связь между тонкими структурными особенностями ма-
териальной организации и между интимными функциональными про-
явлениями тканей и органов, в том числе и в такой высокодиференциро-
ванной и сложной системе животного организма, какой является нервная
система. Этот богатейший и разнообразный материал, представляющий
собой новейшее достижение современного естествознания, вновь блестяще
доказывает и для более деликатных образований правильность приведен-
ного выше положения Энгельса о том, что «морфологические и физиоло-
гические явления, форма и функция обусловливают взаимно друг друга»,
а также положения И. П. Павлова о «приурочении динамики к струк-
туре».
Стремление к разрешению частного, но весьма важного для науки во-
проса из этой большой проблемы, именно попытки ученых найти морфо-
логические тонкоструктурные корреляты для разнообразных и много-
численных функциональных проявлений нервной системы, привело к за-
мечательным достижениям. Значительно расширились наши знания о ходе
афферентных и эфферентных волокон в нервах, о структурных особен-
ностях так называемых вегстатив.ных нервов, о ходе различных интрацен-
тральных Путей, соединяющих разные отделы центральной нервной си-
стемы между собой, о локализации функций в различных этажах цен-
тральной нервной системы, а также внутри каждого этажа в отдельности,
о характере этой локализации и т. и. Больше того, за последние десятиле-
тия, благодаря значительным достижениям гистологии и в особенности
молодой еще науки — гистофизиологин, выясняется история развития
1 «Природа», № 12, 1937. Первоначально статья пазыналась «Анатомо-гнстоло-
гнчсская основа условнорефлекторной деятельности высших животных.
10’
148 Э А АСРАТЯН
нервных путей и разнообразных контактов в нервной системе как по линии
филогенетического, так и по линии онтогенетического развития; выясняются
роль и значение отдельных микроструктурных частей нервных клеток
и нервных волокон, материальные сдвиги в них при покое, в деятельном
состоянии и т. д.
Какими же методами, какими способами в физиологии выясняли и
стараются выяснить этот важный вопрос, если можно так выразиться,
о макро- и микролокализации функций в нервной системе?
С этой целью физиологи пользовались и пользуются двумя излюблен-
ными методами исследования: методом раздражения и методом разрушения.
За последнее время успешно применяют также электрографическую ме-
тодику.
Казалось бы, действительно, что может быть легче того, как всевоз-
можными способами раздражать точно известные, зачастую легко доступ-
ные части тон или иной группы клеточной массы или проводящих путей
центральной нервной системы, наблюдать за соответствующими функцио-
нальными проявлениями в тех или иных частях организма и, сопоставляя
эти данные, делать соответствующие выводы? Метод раздражения, вообще
обогативший физиологию ценнейшими открытиями, немало способствовал
разрешению некоторых вопросов в интересующем нас аспекте. Стоит хотя
бы напомнить работы Фрича и Гитцига, Бубнова, В. Я. Данилевского,
Эвальда, Карплюса и Крейдла и многих других, которые, пользуясь этим
методом, обнаружили локализацию ряда функций в коре и подкорковых
образованиях. Однако, как правильно отметили еще Шифф, Гольц и в
особенности И. П. Павлов, метод раздражения в общем не является бе-
зупречным методом для разрешения таких вопросов, как нахождение
структурных коррелятов в центральной нервной системе для отдельных
функциональных проявлений в различных ее частях.
На первый взгляд безупречным казался второй метод — метод разру-
шения: хирургическим ножом или другими средствами производятся кон-
структивные нарушения в тех или иных отделах центральной нервной
системы, затем они сопоставляются с картиной функциональных наруше-
ний и делаются прямые выводы. Или же в клинической практике, каза-
лось, очень легко разрешить вопрос и придти к четким выводам путем
сопоставления картины болезненных нарушений в деятельности централь-
ной нервной системы у человека с картиной патологических изменений
в отдельных ее частях.
Этим методом пользовались и пользуются в широких пределах. Однако
история и современная практика клинической медицины и эксперименталь-
ной физиологии показывают, что и здесь дело обстоит далеко не так просто
При разрешении поставленной проблемы методом разрушений также
возникают многочисленные и многообразные препятствия, чрезвычайно
запутывающие и без того не совсем ясную картину.
Прежде всего пои этих экспериментальных и в особенности при болезнен-
ных нарушениях структуры в различных отделах центральной нервной си-
стемы трудно точно указать характер и границы этих разрушений и наруше-
ний. Бывают и такие случаи, когда при наличии ясного и даже глубокого
локального нарушения тех или иных функций нервной системы трудно найти
вообще какой-нибудь структурный коррелят этих нарушений в централь-
ной нервной системе. Даже в самых благоприятных случаях, когда удйеТСЯ
более или менее точно установить характер и локализацию происшедших
или экспериментально произведенных нарушений в тех или иных частях
центральной нервной системы, опять-таки нельзя сделать безоговорочные
выводы о функции этих именно частей на основании сопо-
ставления материально-структурных изменений с функциональными изме-
нениями. Благодаря большой реактивности нервной системы, благодаря
АНАТОМО-ГИСТОЛОГИЧЕСКАЯ ОСНОВА УСЛОВНОРЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 149
тесном}' взаимодействию всех частей центральной нервной системы и це-
лостного характера ее деятельности, всякие более или менее значительные
локальные изменения сказываются на всей системе или на большом коли-
честве звеньев этой системы, часто сильно нарушая, а порой функцио-
нально совсем выключая их из строя. К каким парадоксальным практи-
ческим результатам может привести этот момент, иллюстрирует отмечен-
ный Гольцем факт: «собака, лишенная головного мозга, может во многих
направлениях выявлять меньше расстройств, чем собака, у которой недо-
стает лишь одной четверти головного мозга. Ни один разумный исследо-
ватель не станет утверждать, однако, что одна четверть какого-либо органа
имеет больше функций, чем весь орган» *.
Даже в тех чрезвычайно благоприятных и вместе с тем исключительно
редких случаях, когда локальному нарушению в центральной нервной
системе сопутствуют более или менее локальные функциональные наруше-
ния, все-таки нельзя сделать прямые выводы о функции данного очага
центральной нервной системы.
Изучение этих явлений осложняется еще тем, что картина функциональ-
ных нарушений весьма изменчива. Для завершения описания препятствий
этого характера из многочисленных высказываний классиков я позволю
себе привести одно весьма яркое высказывание И. П. Павлова: «Чем
больше экстирпируешь большие полушария, тем больше удивляешься,
что этим приемом так много было получено прежними исследованиями.
Благодаря экстирпации, мы почти никогда не имеем постоянного, а всегда
только текучее, изменяющееся положение вещей. Вы наложили на мозг
свои руки, грубые руки, вы ранили мозг, удалив известные части. Это
ранение раздражает мозг и действие раневого раздражения длится не-
определенное время, и неизвестно, на какое расстояние оно распростра-
няется.,. Наконец, приходит желанный момент, раневое раздражение
проходит, рана заживает. Но тогда на сцену является новое раздраже-
ние — рубец» а.
Сказанное с полным правом можно отнести также к другим частям
центральной нервной системы.
Этим, однако, не исчерпывается перечень препятствий; имеются ослож-
нения совершенно другого рода. Нервная система, как и всякая живая
система, обладает значительной пластичностью, свойством, которое осо-
бенно ясно проявляется в случае значительных материально-структурных
изменений в нервной системе. Благодаря этому замечательному свойству,
этому природному «механическому иммунитету» против всевозможных
неожиданных повреждений, нервная система нередко так хорошо и полно
замещает выбывшие из строя части проводящих путей или клеточных
масс, что вызывает восхищение экспериментатора. «Такова специальная
особенность, таково своеобразное могущество живого вещества» * 3, — писал
И. П Павлов по этому поводу. Насколько указанное свойство препятствует
разрешению затронутой проблемы, явствует из того, что на почве многооб-
разных проявлений этого «своеобразного могущества» нервной системы
некоторые современные исследователи впали в такую крайность, что почти
отрицают, как это было и сто лет назад, всякую локализацию и специфич-
ность центров и путей в центральной нервной системе (Лешли, Бете, Гольд-
штейн и др.).
Сложность объекта и наличие разнообразных препятствий не могли,
однако, остановить прогресс наших знаний в этой области как в теорети-
ческом, так и в практическом отношении. Многие крупные исследователи
1 Pfluger's Archiv Physiologic, Bd. 51, 1892, S. 604.
3 И. П. Павлов, Двадцатилетний опыт, 1932, стр. 194,
а И. П. Павлов, Лекции о работе больших полушарий, 1927, стр. 280.
150
Э. А. АСРАТЯН
приложили свой труд к этому делу. Пользуясь пока что наиболее при-
годным для разрешения этой проблемы методом, именно методом разру-
шений, часто успешно сочетая метод экстирпации с методом искусствен-
ного раздражения и т. д., указанные исследователи значительно обогати-
ли наши знания поэтому вопросу. Практическая же медицина во многих
случаях локальных нарушений в центральной нервной системе не только
ставит точный диагноз, но часто своим вмешательством обеспечивает из-
лечение больного.
Не будем здесь вдаваться в подробности. В соответствии с названием
статьи остановлюсь несколько подробнее на самом высшем отделе цен-
тральной нервной системы — на большом мозге, органе условнорефлектор-
ной деятельности высших животных.
Произведенные с целью выяснения структурных особенностей большого
мозга хирургические вмешательства до последнего времени заключались
главным образом в полной или частичной экстирпации частей большого
мозга, в перерезке его- ножек, т. е. проводящих путей, в- перерезке гак
называемого мозолистого тела, г. е. большого пучка-проводящих путей
между двумя полушариями, в разрушении подкоркового белого вещества
и в линейных разрезах по коре большого мозга.
Большое значение для развития физиологии центральной нервной
системы имели работы Флуранса с частичной и полной экстирпацией боль-
шого мозга у птиц и последующим изучением состояния оперированных
животных и их поведения, а также не совсем удачные его попытки
экстирпации частей коры большого мозга у млекопитающих животных,
выполненные более 100 лет назад. Прежде всего они дали ценный экспе-
риментальный материал, а самое главное — эта смелая атака на высший
орган, на эту неприступную крепость сложного организма, еще на заре
экспериментальной физиологии открыла широкие перспективы перед
экспериментаторами, стимулировала их, послужила базой для дальней-
ших завоеваний. И, действительно, немного позже, после первых опытов
Флуранса, его современнику Булло (1830) удалось сохранить жизнь собаки
с частично удаленными полушариями большого мозга.
После некоторого затишья, примерно со второй половины прошлого
столетия, многие исследователи-физиологи (Гитцнг, Мунк, Гольц, Лучиани
и др.) уже начали систематически применять хирургическое вмешатель-
ство как метод исследования функций большого мозга высших млеко-
питающих. Среди этих имен я хочу особо подчеркнуть имя Гольца. Немец-
кий ученый Ф. Гольц последние почти 30 лет своей деятельности посвятил
систематическому изучению физиологии большого мозга. Еще около 50 лет
назад ему удалось удалить у трех собак кору большого мозга, у одной
собаки все полушарие целиком, а у.обезьяны переднюю половину коры
одного полушария и длительно сохранить их жизнь. Чтобы дать себе отчет
в громадном значении этих достижений даже с чисто технической стороны,
укажу на то, что, несмотря на колоссальный рост общей хирургической
техники за полстолетие, отделяющее нас от указанных опытов Гольца,
для многих современных физиологов простое повторение этих опытов
остается трудно выполнимой задачей.
Своих собак с частичной экстирпацией коры большого мозга Гольц
исследовал разносторонне: путем простого наблюдения, с помощью все-
возможных функциональных проб и тестов. Он разъяснил многочисленные
и разнообразные полученные им и его современниками факты, обобщил их
и высказал ряд теоретических положений о строении большого мозга,
если можно так выразиться, о <функциональной анатомии» этого органа.
Чтобы хорошо разобраться в запутанной картине послеоперационных
функциональных нарушений, Гольц предлагал эти нарушения делить
на две группы: на группу побочных явлений (Nebcnerschcimingen) и на
АНАТОМО-ГИСТОЛОГИЧЕСКАЯ ОСНОВА УСЛОВНОРЕФЛЕКТОРНОП ДЕЯТЕЛЬНОСТИ 15]
группу действительных явлений выпадения (Ausfallerscheinungen). Первая
группа явлений, выражающаяся главным образом в торможении (Нет-
mungerschei nungen) не затронутых операцией частей коры большого мозга
и нижележащих отделов центральной нервной системы в результате
раневого раздражения, как указывает их общее название, представляется
второстепенной и постепенно проходит. Более стойкими и вместе с тем
самыми важными оказываются явления выпадения, которые, по пред-
ставлению Гольца, могут частично сглаживаться путем тренировки ранее
существующих и функционирующих центров, освобожденных от раневого
торможения.
Значение результатов работ Гольца для «функциональной анатомии»
центральной нервной системы огромно. Прежде всего Гольц опроверг
упрочившуюся было в физиологии на основе работ Гитцнга, Мунка и др.
метафизическую теорию строгой и узкой локализации функций в коре
большого мозга высших животных, теорию, которая находила и находит
поддержку со стороны неврологов-клиницистов под впечатлением меха-
нистических концепций Джексона, Брока и др. Противники Гольца неспра-
ведливо обвиняли его в том, что он якобы совсем отрицает локализацию
функций в коре большого мозга, как это делал Флуранс. Гольц со всей
решительностью и свойственной ему резкостью отклонял эти и другие
необоснованные обвинения. «Про меня упорно говорят, что я утверждал,
будто мозговое вещество всюду равноценно. По это совершенно ошибочно»1,—
писал Гольц в одном из своих произведений. В другом он говорил: «Я ни-
когда не отрицал возможности локализации функции большого мозга».
И, действительно, он только не соглашался с искусственной мертвой схе-
мой локализационных зон, согласно которой кора представлялась в виде
политической карты стран с обособленными узкими участками и точными
границами. Сейчас это следует особенно подчеркнуть, потому что пред?
ставители современных идеалистических и мистических воззрений, отрицаю-
щие локализацию в коре большого мозга вообще, тоже ошибочно приписы-
вают Гольцу неверные взгляды и при обосновании своих положений
часто основываются на его авторитете в физиологии (см., например, книгу
Лешли «Мозг и интеллект»).
Частично на основании упомянутых выше экспериментов, частично на
основании экспериментов на собаках без коры одного полушария и без
цельного полушария Гольц сделал ряд других ценных выводов о функции
большого мозга. Прежде всего следует указать на его положение о связи
каждого полушария большого мозга с обеими половинами тела. Гольц
писал: «Для меня несомненно, что каждое полушарие посредством прово-
дящих путей состоит в связи со всеми мускулами и органами чувств всего
тела» 2. Далее он указал на различное влияние переднего и заднего мозга
собаки на ее пов-едение и на общую трофику.
Гольц дал нам правильное в основном представление об общей роли
коры большого мозга для организма. Он точнее, чем кто-либо до него,
представил картину связи между отдельными функциональными проявле-
ниями и анатомически более или менее обособленными частями большого
мозга. В этом отношении данные Гольца являются капитальным вкладом
в сокровищницу мировой науки, что неоднократно отмечалось
И. М. Сеченовым и И. П. Павловым.
Однако нн Гольц, ни Лучианн, ни Франц, ни другие дальше этих общих
констатаций не шли. Метод простого наблюдения и некоторых остроумных
функциональных проб и тестов, которыми они пользовались, не давали
им возможности вскрывать основные закономерности работы большого
1 Verhandlungen d. Congresses for innere Medizine. 1884, S. 261.
* Pfliiger’s Archiv Physiologic, Bd. 42, 1888, S. 436.
152
Э. Л, АСРАТЯН
мозга. А вопрос о значении более тонких структурных элементов полу-
шарии даже не был поднят. Вот почему не кто иной, как Гольц, в разгар
своих работ по физиологии большого мозга вынужден был констатировать,
что «каждый, кто основательно занимался физиологией головного мозга,
согласится со мной, что неоспоримое знание о процессах, протекающих
в этом важнейшем органе, немногим больше наших сведений о природе
планеты Марс» Ч
С тех пор как были произнесены эти слова, прошло 52 года, и сегодня
мы можем сказать словами нашего учителя И. П. Павлова, что в области
знаний о функциях мозга «теперь положение дела радикально изменилось».
Гениальный И. П. Павлов во главе огромной армии сотрудников, поль-
зуясь непревзойденным по совершенству и глубине методом условных реф-
лексов, одну за другой вскрывал, постоянно детализировал и уточнял
многие основные, внутренние закономерности работы большого мозга и в
результате более тридцатилетией неутомимой борьбы на этом фронте создал
свое цельное, монолитное, бессмертное учение об условных рефлексах.
По понятной причине я здесь не стану разбирать все стороны этого
обширного учения, а коснусь только тех его элементов, которые имеют
непосредственное отношение к интересующему нас здесь вопросу.
Девизом для боевого материалистического учения И. П. Павлова,
девизом, на основе которого выковывалось это учение, было строгое тре-
бование: как при изучении других систем организма, так и при изучении
функций сложнейшего его органа — большого мозга оставаться тем же
объективно наблюдающим и строго мыслящим натуралистом, все объяснять
«чисто физиологически, чисто материально, чисто пространственно»1 2, стре-
миться к тому, чтобы возможно было «пальцем указать», где именно про-
исходит тот или иной процесс.
Прежде всего в серии специальных работ большого числа учеников
И. II. Павлова, посвященных изучению нарушений условнорсфлектсрной
деятельности у собак с различными повреждениями тех или иных областей
коры большого мозга, школа И. П. Павлова непревзойденно тонким и
точным методом условных рефлексов более детально, обстоятельно и си-
стематически, чем это могли делать Мунк, Гитцнг, Ферриер, Гольц, Лу-
чиани и др., изучала проблему локализации функций в коре большого
мозга. Не вдаваясь в подробности, скажу только, что работы школы
И. П Павлова не только «языком точных цифр» подтвердили некоторые поло-
жения теории локализации Гольца — Лучиани, но и обнаружили ряд
весьма существенных фактов, имеющих отношение как к структуре коры
большого мозга, так и в особенности к физиологии органов чувств. Снова
И. П. Павлова о том, что «наши факты мыслятся в виде пространства и вре-
мени», полностью относятся к этой группе фактов, которые послужили
основанием для создания его передового учения о динамической лока-
лизации функций в коре больших полушарий мозга.
Каковы же наши представления о структурной основе условной
связи?
Еще в самых ранних своих исследованиях И. П. Павлов выдвинул
на первый план и заложил в фундамент будущего величественного здания
своего учения простой факт образования нового рефлекса, условного ре-
флекса как результат замыкания новой связи между двумя
активно действующими очагами головного мозга.
В 1908 г. впервые давая четкую схему выработки условных связен
в головном мозгу и сравнивая дугу безусловного рефлекса с дугой услов-
ного, он говорил: «Во втором случае, следовательно, мы имели дело с вре-
менным замыканием связи как основным свойством деятель-
1 Verhandlungen d. Congresses fur innere Medizirte, 1&84, S. 262.
2 И. П. Павлов, Двадиатилеткий опыт, 1932, стр. 293.
АНАТОМО-ГИСТОЛОГИЧЕСКАЯ ОСНОВА УСЛОВНО РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ | 53
пости высших отделов центральной нервной системы, как с первым капи-
тальным пунктом их механики»1 (разрядка моя. — Э. Л.).
Позже он постоянно возвращается к этой основе своего учения, все бо-
лее уточняя и развивая его. В своих выступлениях и статьях И. П. Павлов
неоднократно характеризует закон замыкания условной связи как «основ-
ной закон высшего отдела системы». Для деятельности больших полушарий
«основное явление — образование временных связей». Больше того, он
иногда вместо слов «условные рефлексы» употребляет слова «замыкатель-
ные рефлексы».
Закон замыкания условной связи сохранил свое кардинальное, пер-
венствующее значение в течение всего хода развитая учения об условных
рефлексах. Это явствует из последних блестящих итоговых выступлений
И. П. Павлова. Нет надобности приводить цитаты для обоснования ска-
занного; хотелось лишь подчеркнуть, что И. П. Павлов в этом замыкании
видел только частный, качественно новый момент более общего принципа
структурности. В прекрасной статье, направленной против мистических
воззрений некоторых современных психологов, воинствующий материалист-
физиолог И. П. Павлов выдвигает как один из общих принципов нервной
деятельности вообще материальную конструкцию, «принцип структур-
ности, т. е. расположения действий силы в пространстве, приурочение
динамики к структуре»®.
В одном из своих ранних докладов И. П. Павлов говорит, что «нервная
система не есть, как обыкновенно думают, только проводниковый прибор,
но и замыкательный.
И действительно, свойство замыкания, присущее в своем наиболее
завершенном виде и в своем качественном своеобразии только коре боль-
шого мозга высших животных, в задаточной, примитивной форме про-
является также в деятельности нижележащих отделов центральной нерв-
ной системы. С одной стороны, еще И. М. Сеченов, Гольц, позже Магнус
показали, что один и Те же раздражения одних и тех же рецепторов
организма могут вызывать различные, часто диаметрально противополож-
ные эффекты в зависимости от ситуации, установки, от Schaltung в цен-
тральной нервной системе, в зависимости от того, какие пути открыты
и какие закрыты для нервного импульса в данным момент. В этих случаях
своеобразного простого и кратковременного «замыкания» мы имеем дело
с «открытием» и «закрытием» уже готовых, законченных нервных путей.
Другое проявление элементарного, летучего «замыкания» мы имеем в
так называемом Bahnung, когда при известных ситуациях в центральной
нервной системе на очень короткое время проторяются новые пути, по
которым проводятся возникшие в различных ее частях нервные им-
пульсы, отклонившиеся от своих обычных, трафаретных путей, к из-
вестным центрам «большого возбуждения».
Еще более ярким выражением} такого рода «замыкания» в цент-
ральной нервной системе следует считать детально разработанное
А. А. Ухтомским явление доминанты. В центральной нервной системе очаг
доминирующего возбуждения притягивает к себе потоки нервных импуль-
сов из отдаленных источников, сопряженно затормозив те центры, в ко-
торых, благодаря более тесной структурной и функциональной связи, эта
импульсы протекают в обычных условиях, причем стойкость возбуждения
и своеобразная инерция средн моментов, характеризующих «доминанту»,
придают этому своеобразному виду временного «замыкания» намного более
стойкий характер и тем самым особенный интерес.
Как уже было сказано, во всех перечисленных случаях мы имеем дело 1 2
1 И, П- П а вл о в, Двадцатилетний опыт, 1932, стр. 77,
2 Там же, стр. 470.
154
Э. А. АСРАТЯН
с различными проявлениями кратковременного, неустойчивого, как бы
летучего «замыкания», нс оставляющего после себя следов в центральной
нервной системе.
Замыкание условной СВЯЗИ — ЭТО уже действительное, более или менее
стойкое замыкание связи между известными действующими очагами боль-
шого мозга. В сплошной цепи развития нервных процессов мы Имеем
скачок, переход количества в качество, возникновение качественно нового,
более высокого типа нервной деятельности.
Физиологические эксперименты не оставляют никакого сомнения в на-
личии замыкания новой связи, В каких-то материальных сдвигах и струк-
турных изменениях, обусловливающих это замыкание. Этот точно устано-
вленный гениальным И. П. Павловым факт является фактом огромнейшего
значения для всего современного естествознания.
В результате более тридцати летней непрерывной работы школы
И. П. Павлова достаточно точно выяснены и изучены условия, при ко-
торых происходит замыкание и укрепление этой связи, условия ее раз-
рушения и т. д. Нам известны отношения основных нервных процессов —
возбуждения и торможения к этой связи; мы кое-что знаем о некоторых
свойствах ее; мы с достаточной точностью знаем также, что у высших
животных кора большого мозга является главным, если не единственным
органом замыкания этой связи.
Кроме перечисленных и ряда других сведений об этой связи, мы имеем
ряд высказываний основоположника гениального учения об условных
рефлексах, касающихся более топких деталей условной связи. Так, не-
сколько лет назад И. П. Павлов отказался от первоначального предста-
вления о возникновении условной связи между корковыми рецепторными
клетками и подкорковыми нервными элементами, связанными с рабочими
аппаратами. На основании некоторых фактов он сделал косвенный вывод,
«что образование новой нервной связи, замыкательный процесс цел и-
к о м происходит в больших полушариях» *, т. е. между клеточными эле-
ментами коры. Точно так же на основании некоторых фактов И. П. Павлов
сделал гипотетическое предположение, что эта связь двойная: «Когда два
нервных пункта связаны, объединены, нервные процессы двигаются, идут
между ними в обоих направлениях. Если признать абсолютную законность
одностороннего проведения нервных процессов во всех пунктах централь-
ной нервной системы, то в данном случае придется принять добавочную
обратного направления связь между этими пунктами» ®. Больше того,
И. П. Павлов неоднократно высказывал предположение о микроструктуре,
о сущности этой связи: «Вероятно, место этой замыкательной способности
есть пункты сцепления нейронов (и специально коры больших полуша-
рий)»3. Он допускает также, что структурной основой такой связи
могут быть и тончайшие разветвления аксонов или «раздражительные
мемб раны»-син апсы.
Если подойти к последним предположениям с той объективностью и
строгостью, которой нас учил И. П. Павлов, то необходимо будет от-
метить, что они нуждаются в обстоятельной экспериментальной про-
верке и разработке, на что неоднократно указывал сам гениальный
создатель учения о высшей нервной деятельности. Затем нельзя не за-
метить, что, хотя многие из этих предположений относятся к структуре
условной связи, к «конструкции больших полушарий», все же они не осно-
ваны на фактах, полученных нгтиболес достоверным для разрешения таких
вопросов методом, именно методом хирургического вмешательства с по-
1 И. П. П а в л о в. Двадцатилетний опыт, 1932, стр. 406.
‘Там же, стр. 489.
•Там же, стр. 313.
АНАТОМО-ГИСТОЛОГИЧЕСКАЯ ОСНОВА УС ЛОВ НОРЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 155
следующим изучением у животных условно-рефлекторной деятельности.
Ведь не кто иной, как II. П. Павлов, говоря о том разделе своего уче-
ния, который касается подробного изучения структуры больших полуша-
рий, писал: «Перед нами для этой цели стоял пока единственный метод,
метод частичного разрушения» х.
Так обстояло дело непосредственно в лабораториях И. П. Павлова. Но
изучением этой проблемы занимались также его ученики и последова-
тели в других лабораториях.
Следует сказать, что за последние годы ряд исследователей Советского
Союза и зарубежных стран пользовался методом фармакологического,
термического и хирургического выключения тех или иных звеньев дуги
безусловного рефлекса и этим путем пытался выяснить их значение для
выработки, сохранения и проявления условных рефлексов.
В. А. Крылов, а вслед за ним Коллинс и Татум, Крислер и др. пока-
зали, что можно выработать условный рефлекс «автоматически, внутрен-
ними агентами, составом и свойствами крови раздражаемого центра*8.
Они показали, что можно условный рефлекс выработать на морфинное
отравление животного. Позже Лукс сообщил, что ему удалось выра-
ботать условный рефлекс на прямое и непосредственное раздражение
коры больших полушарий в условиях исключения афферентной части
дуги безусловного рефлекса. Однако совсем недавно из той же лабора-
тории сделал сообщение Гент, берущий под сомнение правильность перво-
начальных данных Лукса. К значению афферентной часта дуги безуслов-
ного рефлекса относится отчасти также сообщение А. А. Ющенко и
сотрудников о том, что холодовая «перерезка» задних столбов спинного
мозга, т. е. афферентных путей, не исключает наличия условного элек-
трообороннтельного рефлекса собаки на заднюю лапу.
Перечисленные разнородные данные говорят о том, что иногда аффе-
рентная часть дуги безусловного рефлекса нс является абсолютно необ-
ходимой как для выработки условного рефлекса на его основе, так и для
существования и проявления этого условного рефлекса.
С другой стороны, Крислер, Лайт, Гент и др. показали, что работа
последнего звена дуги, именно эффекторного органа, во время выработки
условного рефлекса не является необходимой для достижения поставленной
цели. Так, первому удалось выработать слюнный условный рефлекс на
фоне паралича секреторной деятельности слюнной железы, вызванного
атропином. А опыты Лайта и Гента были еще демонстративнее. Одну зад-
нюю конечность собаки они делали неподвижной путем перерезки соот-
ветствующих моторных корешков спинальных нервов. В период паралича
конечности некоторое время производили выработку условного электро-
оборонительного рефлекса. Задолго до восстановления моторной функции
парализованной конечности авторы прекратили эти манипуляции; однако
после восстановления ее деятельности оказалось все-таки, что условный
рефлекс налицо. Против этих данных О значении работы эффекторного ор-
гана говорят исследования Харлоу и Стагнера, показавшие, что манипуля-
ции по выработке условного электрообороннтелыюго рефлекса на курари-
зованных животных не приводят к цели, в чем авторы удостоверились
пробами после снятия влияния кураре физостигмином. У тех же живот-
ных, однако, условный рефлекс со стороны зрачка был налицо.
Среди работ этой категории особое место занимают работы В. П. Про-
топопова и сотрудников. На основании исследований больных с различ-
ного рода повреждениями центральной нервной системы, а также экспе-
риментов на животных с различного рода хирургически воспроизведен-
ными нарушениями различных частей коры и спинного мозга Протопопов
3 И. П. П а в л о в, Лекции о работе больших полушарий, 1927, стр. 279.
1 Там же, стр. 43.
156
Э А. АСРАТЯН
приходит к заключению, что для выработки, сохранения и проявления
моторного условного рефлекса необходимо наличие не только моторной
зоны коры, но и пирамидальных и экстрапирамидальных эфферентых
путей, т. е. всей эфферентной части дуги безусловного рефлекса. Далее,
он считает, что для той же цели необходимо наличие как кожной, так и
глубокой чувствительности, т. е. всей афферентной части дуги безусловного
рефлекса. Трудно согласиться с категорическими установками В. П. Прото-
попова, тем более что я и К. С. Абуладзе (из лаборатории И. П. Павлова)
не могли подтвердить важные фактические данные, приводимые В. П. Прото-
поповым. Не так легко, однако, согласиться и с установками Лешли, кото-
рый на основании своих экспериментов, проведенных сравнительно прими-
тивным методом исследования — методом выработки лабиринтного навыка
у крыс, пришел к выводу, что ни экстирпация моторной зоны, ни повреж-
дение сензорной зоны, ни повреждение пирамидных путей, а тем более ли-
нейные разрезы коры по любым направлениям не оказывают существенного
влияния на выработку, сохранение и проявление даже сложных навыков.
Противоречивость перечисленных выше данных и высказываний о зна-
чении тех или иных элементов дуги безусловного рефлекса для выработки,
сохранения и проявления условного рефлекса указывает, помимо всего
прочего, на чрезмерные трудности, связанные с изучением поставленной
проблемы. И хотя большинство перечисленных исследователей поставило
вопрос о выяснении необходимых наследственных структурных предпо-
сылок и условий для выработки условного рефлекса, все-таки общепри-
нятых, бесспорных данных по этому вопросу мы не имеем до сих пор. Бо-
лее же существенный вопрос о материальных основах самого замыкания
связи, о структурных основах самой связи авторами даже не ставился.
Из сказанного видно, что нам до сих пор не только ничего точно не-
известно об интимном характере материальных сдвигов при замыкании
условной связи или о структурных «микроэлементах», участвующих
в этом замыкании (в этом отношении не лучше обстоит дело даже в более
примитивных отделах центральной нервной системы), но мы также не знаем
точно места замыкания связи, не знаем тех анатомических и гистологи-
ческих образований, по которым прокладываются эти пути.
Совершенно недостаточны наши сведения и о тонких микроструктур-
ных, материальных, анатомо-гнстологических основах тех взаимодействий и
взаимной связи между отдельными частями большого мозга, которые
известны под названием иррадиации, концентрации, взаимной индукции
и суммации нервных процессов, хотя и эти виды мозговой деятельности,
их возникновение, развитие и другие свойства достаточно хорошо и полно
изучены в лабораториях И. П. Павлова. Какое значение придавал
И. П. Павлов выяснению материальных основ этих деятельностей, явст-
вует из его неоднократных высказываний по этому поводу. Вот одно из
них: «Центр тяжести в научном изучении нервной деятельности больших
полушарий лежит в определении путей, по которым нервный процесс
разливается и сосредоточивается, — задача исключительно пространствен-
ного мышления» !.
Справедливость требует отметить, что далеко не полны наши знания
аиатомо-гнсгологическнх основ нервной деятельности и более примитивных
отделов центральной нервной системы. Это, однако, не может служить пово-
дом к тому, чтобы мы в этом направлении ничего не предпринимали, пока в
низших отделах центральной нервной системы положение коренным образом
не улучшится и мы не будем в состоянии воспользоваться готовой схемой.
Мы не должны забывать ценнейшие указания классиков марксизма,
в частности, Энгельса, о преимущественном во многих случаях значении
1 И. П. Павлов, Двадватилетний опыт, 1932, стр. 213.
Л HAT ОМО-ГИС ТО ЛОГИЧЕСКАЯ ОСНОВА УСЛОВНОРЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 157
изучения сложного для познавания простого. О том же неоднократно гово-
рил и И. П. Павлов. «Если низший отдел сложен, то как же бесконечно
сложен должен быть высший! Несмотря на это отрицательное, неблагопри-
ятное обстоятельство, в высшем, отделе центральной нервной системы в
отношении исследования есть и свои преимущества» \—писал И. II. Павлов
еще 25 лет назад. Далее он обосновывает высказанное положение, говоря,
что в противоположность спинному мозгу, где рефлекторная деятель-
ность исследуется готовой, в высшем отделе, «мы видим самый про-
цесс образования этого отраженного акта». Я думаю, что при изучении
как некоторых других вопросов, так и анатомо-морфологических основ реф-
лекторной деятельности физиологи, исследующие большой мозг, должны
идти впереди своих коллег, изучающих деятельность низших отделов цен-
тральной нервной системы.
Гениальный И. П. Павлов на славном пути своей научной деятельности не
только вскрыл много глубоких тайн природы, но и поставил на разрешение
множество новых задач, стимулирующих исследовательский ум. Одна из
этих задач, являющаяся предметом данной статьи, кратко и четко им сфор-
мулирована таким образом: «Функция связи как внутренних, так и внеш-
них соотношений в организме осуществляется в нервной системе, пред-
ставляющей видимый аппарат. На этом, конечно, аппарате разыгрываются
динамические явления, которые и должны быть приурочены к тончайшим
деталям конструкции аппарата»2.
Бесспорно, что выяснение анатомо-гистологических основ замыкания
условной связи, этого базиса огромного разнообразия и богатства всей
условнорефлекторпой деятельности большого мозга, выяснение анатомо-
гистологических и другого рода материальных сдвигов при основных видах
работы большого мозга — задача исключительной важности, чрезвычай-
ной трудности, но вместе с тем и благодарная задача. Обдумывая поставлен-
ную проблему и руководствуясь указаниями Энгельса, что «в теорети-
ческом естествознании нельзя конструировать связи и вносить их в факты,
а надо извлекать их из последних и, найдя, доказывать и, поскольку это
возможно, опытным путем»3, я еще несколько лет назад эскизно наметил пути
решения некоторых из поставленных задач. К практическому осуществле-
нию этих предварительных проектов я приступил, однако, всего два года
назад, когда получил те минимальные возможности, которые были необхо-
димы для самостоятельной постановки и разрешения таких проблем.
Описание избранного пути для достижения поставленной пели, а также
описание пока еще первоначальных данных, полученных врезультае более
чем двухгодичной разведывательной и исследовательской работы совместно
с группой сотрудников, будут предметом дальнейшего изложения.
В качестве конкретных задач для экспериментального решения были
избраны следующие: действительно ли условная связь замыкается между
двумя корковыми пунктами или же между корковым и подкорковым пунк-
том; какую анатомо-морфологическую основу имеют замыкание условных
связей, иррадиация, взаимная индукция л суммация нервных процессов в
большом мозгу, а также целостная работа большого мозга, которая обуслов-
лена тесным взаимодействием и взаимосвязью между отдельными его частями.
Для решения этих задач был использован путь оперативного воздей-
ствия на большой мозг с последующим изучением его условнорефлектор-
ной деятельности. С этой целью два с лишним года назад я реализовал
придуманный раньше новый способ операции на коре больших полушарий
мозга— разобщение локализационных зон. Суть операции заключается в
том, что та или иная локализационная -область коры не экстирпируется,
* И. П. Павло в, Дваддатилетний опыт, 1932, стр. 168.
’Там же, стр. 471.
’ Ф. Энгельс, Диалектика природы, 1930, стр. 91.
158
Э Л. ЛСРАТЯН
что до сих пор делали физиологи, а снятием узкой ленты мозгового ве-
щества или простой перерезкой на желаемой глубине разобщаются основ-
ные, уже достаточно хорошо известные локализационные зоны, а именно:
сензомоторные, зрительные и слуховые зоны. Таким образом, из коры
каждого полушария получается по три островка функционально разно-
значной мозговой массы, изолированных друг от друга каналами желае-
мой глубины. Приводимая схема дает четкое представление о сущности
операции (рис. 1). Операция эта не из легких по технике и выполнению.
Однако при известном навыке ее можно осуществлять безупречно. Мозг
одной из оперированных подобным образом собак (Кусачка), подробно
исследованной по методу условных рефлексов Я. М. Прессманом, нахо-
дится еще в процессе детального гистологического изучения. Однако
первоначальное ана гомо-гистологическое исследование, произведенное
И. Е. Пригонниковым, сотрудником сектора морфологии Института мозга,
показывает, что в основном мы достигли своей цели. И. Е. Пригонников
дал следующую резюмирующую оценку: «Па основании макроскопиче-
ского изучения мозга собаки Кусачки можно сказать, что лобная, затылоч-
ная и височная доли отделены друг от друга на наружной поверхности
обоих полушарий мозга образовавшейся на месте разрезов рубцовой тканью.
Причем рубцовая ткань захваты-
вает не только кору, но проникла
.s' ’ "---С\ глубоко в белое вещество до бо-
Z/X. S I JL ковых желудочков мозга».
Л ( Л / //\\ Само собой разумеется, трудно
fxk 'у ) ручаться, что мозг всех опернро-
I ванных мной собак будет иметь
X, такую картину, как мозг Кусач-
ки. Но в одном я совершенно
убежден: в подавляющем боль-
Рис. 1. Схема разобщении корковых шннстве случаев операция дости-
зои (лобных, затылочных и височных гает своед "цели,
облйстсй) •
' При составлении плана предло-
женной мной операции я частично
исходил из способов разрушения мозговой коры и подкоркового белого
вещества вертикальными и горизонтальными разрезами, несистематически
применяемыми в физиологии различными исследователями еще с конца прош-
лого столетия для выяснения структурных основ некоторых про-
стых функциональных проявлений тех млн иных отделов большого мозга,
мозжечка к других частей центральной нервной системы (Марик, Экснер
и Панет, Тренделенбург, Лешли и др.). Большинство этих исследователей
ставило перед собой задачу преимущественно в острых опытах выяснить
морфологическую основу взаимосвязи отдельных областей большого мозга
при осуществлении тех или иных врожденных элементарных нервных ак-
тов. Лешли же, как упоминалось выше, в хронических опытах исследовал
навыки оперированных крыс.
Чтобы не запутаться в сложной картине, мы при составлении конкрет-
ного рабочего плана решили начать с простого и постепенно идти дальше
путем поочередного исключения слоев, массы большого мозга. Конкретно
мы ставили ближайшей целью выяснить, насколько корковая серая мас-
са мозга сама по себе является структурной основой для перечисленных
выше явлений в условнорефлекторной’деятельности. Для этой цели у всех
подопытных животных (за исключением одного) производили разобще-
ние по толще корковой серой массы, по возможности избегая серьезного
повреждения лежащего ниже белого вещества. Расчет был таков: удастся
или не удастся у такой собаки с разобщенными по толще коры локализаци-
нными зонами, скажем, экспериментально замкнуть условную связь между
АНАТОМО-ГИСТОЛОГИЧЕСКАЯ ОСНОВА УСЛОВНОРЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 15<>
зрительной и бензомоторной зоной; удастся или не удастся установить
между ними взаимодействие в виде иррадиации, взаимной индукции и
суммации процессов, в виде комплексных условных рефлексов от разоб-
щенных друг от друга анализаторов, и соответственно с этим сделать
выводы о роли корковой клеточной массы в этих явлениях.
За два с лишним года я произвел разобщение локализационных зон по
толще коры большого мозга примерно у 30 собак, из которых около 20
после благополучно перенесенной операции подверглись детальному наблю-
дению и исследованию по методу условных рефлексов и различными функ-
циональными пробами сотрудниками руководимой вами лаборатории:
Ф. М. Шитовым, В. П. Благовещенской, В. В. Яковлевой и Я. М. Пресс-
маном. В процессе работы изменялась последовательность проведения
операций и опытов по изучению условнорефлекторной деятельности у со-
бак. У некоторых собак вначале была произведена операция, затем нача-
лась выработка и изучение условных рефлексов на них; па других соба-
ках работа велась в обратном порядке — сначала выработка условных
рефлексов, изучение особенностей условнорефлекторной деятельности у
них, затем операция разобщения, а после этого вновь исследование услов-
норефлекторной деятельности. Были и такие вариации: то операция на
обоих полушариях производилась одномоментно, то сначала разобщались
зоны одного полушария, а некоторое время спустя (месяц, дв-а и больше)
разобщались зоны второго полушария или же кора второго полушария
полностью экстирпировалась.
Условнорефлекторная деятельность изучалась с помощью методики как
пищевых условных рефлексов. (Ф. М. Шитов), так и кислотных условных
рефлексов (В. В. Яковлева), а также методики электрооборонительных
условных рефлексов (В. П. Благовещенская, Я. М. Прсссман). Не вдаваясь
в подробности полученных первоначальных экспериментальных данных,
приведу основные результаты наблюдений и исследований около 20 собак
с разобщенными по толще коры локализационными зонами.
Даже простое наблюдение обнаруживает, что влияние названной опера-
ции на внешнее поведение и общее состояние животных неодинаково. На
молодых и крепких собаках это влияние сказалось в общем слабее, чем на
собаках со слабым здоровьем и собаках среднего возраста и старше. Как
более или менее типичный результат описанной операции можно отметить
уменьшение подвижности и активности животных, уменьшение интереса
к окружающему, некоторую неловкость движений как при стоянии и ходьбе,
так и при еде и чесании. Заметны нарушения зрительной функции. У многих
собак наблюдалось также некоторое похудание, как бы хорошо они ни
ели. В зависимости от указанных выше причин, а также от неизбежных
и незаметных индивидуальных вариаций при выполнении этой деликатной
корковой операции, от описанной картины имеются значительные откло-
нения в ту и другую сторону — от почти нормального поведения до глубо-
кой инвалидности. Можно отметить, однако, что в первые месяцы после
операции эти грубые нарушения сглаживаются, а затем устанавливается
более или менее стойкая картина, длящаяся год, полтора и больше.
Исследования показали, что у всех собак с разобщенными по толще
коры локализационными зонами удается выработать как пищевые, так
и кислотные и элсктрооборонительные условные рефлексы со всех трех
анализаторов, т. е. на звуковые, зрительные и тактильные раздражения
(Ф. М. Шитов, В. П. Благовещенская, Я. М. Пресеман, В. В. Яковлева)
С другой стороны, имеются некоторые данные, подвергающиеся сейчас до-
полнительной проверке, о том, что проявлявшиеся раньше пищевые услов-
ные рефлексы не претерпевают существенных изменений от производимой
нами операции (Ф. М. Шитов), тогда как электрооборонительные условные
рефлексы значительно сслабевают (Я. М. Пресеман).Согласно полученным
ICO
Э А. АСРАТЯН
данным, при выработке пищевых условных рефлексов у оперированных собак
в примерно одинаковых условиях быстрее вырабатываются условные реф-
лексы на звуковые раздражители. При выработке же электрооборонительных
условных рефлексов в аналогичных условиях быстрее вырабатываются
условные рефлексы на тактильные раздражители.
Далее, условные рефлексы (в особенности пищевые и кислотные) этих
собак по величине и прочности значительно уступают соответствующим
рефлексам у нормальных животных. Следует отметить, что значительно
ослабевают также безусловные слюнные рефлексы. У этих собак без особого
труда вырабатываются также диференцировочные условные рефлексы.
АЬС — луга зрительного подкоркового безусловного рефлекса;
А ПС — дуга зрительного коркового безусловного рефлекса; De/7—
дута слюнного подкорковою безусловного рефлекса, DEF -дуга
слюнного коркового безусловного рефлекса, В/£- предполагае-
мая условная связь в толще норы, В2 Е — предполагаемая условная
связь is подкорковой белой массе, ВЗЕ — предполагаемая услов-
ная связь в подкорковой ссроЯ массе
Ф. М. Шитов, В. П. Благовещенская, Я. М. Прессман показали на
многих собаках, что изолированные друг от друга корковые локализацион-
ные зоны взаимодействуют, способны взаимосвязанно функционировать.
Так, они установили, что угасательное и диференцировочное торможение
от любого анализатора иррадиирует иа другие анализаторы, как и в норме.
Не очень закономерно, но иногда получались данные, указывающие на
возможность суммации условных рефлексов на раздражители разных ана-
лизаторов. О сохранении у этих собак способности к такой высшей анализа-
торной и синтетической деятельности, какой является системность, четких
и бесспорных данных не получено. Однако Я. М. Прессман на собаке Кусач-
ки совершенно определенно показал, что у таких собак удается выработать
условный рефлекс на комплекс раздражителей от всех трех разъединенных
по толще коры анализаторов, причем каждый из раздражителей в отдель-
ности перестает вызывать условный положительный эффект. Это — пока-
затель сложной анализаторно-синтетической работы разъединенной коры.
Чтобы уточнить, действительно ли условная связь замыкается между
изолированными нами зонами, действительно ли эти зоны функцио-
нируют и взаимодействуют, у некоторых собак я добавочно экстирпировал
то зрительные, то слуховые зоны. В. П. Благовещенская и Ф. М. Шитов
показали, что у таких собак после добавочных операций на длитель-
ное время исчезают те именно условные рефлексы, зоны которых экс-
тирпировались, тогда как другие условные рефлексы заметно не страдают.
Л HATOMO-ГИСТОЛОГИЧЕСКАЯ ОСНОВА УСЛОВНОРЕ ФЛЕКТ0РНОЯ ДЕЯТЕ ЛЬНОСТИ 1QJ
Лишь через два-три месяца постепенно вновь появляются исчезнувшие
условные рефлексы. Даже при поверхностном наблюдении выявляется
резкое нарушение функций тех анализаторов., корковые изолированные
зоны которых экстирпировались.
Само собой разумеется, для того чтобы сделать окончательные выводы
по разрешаемой серьезной проблеме — установления тонких анатомо-ги-
стологических, или структурных, основ различных элементов условно-
рефлекторной деятельности сложнейшего органа, понадобится еще не один,
не два года и даже не одно десятилетие.
Кроме всего, первенствующее значение для нашей работы имеет ана-
томо-гистологическое детальное исследование оперированных мозгов,
к чему сектор морфологии нашего института уже приступает.
Но полученный на достаточном количестве подопытных животных ма-
териал дает возможность уже сейчас сделать двоякого рода выводы.
Во-первых, мы можем определенно сказать, что условная положитель-
ная и отрицательная связь, необходимая для осуществления простых и
даже комплексных рефлексов, а также процесс иррадиации торможения
могут осуществляться в условиях хирургического разобщения отдельных
зон коры большого мозга, по всей вероятности, через белое подкорковое
вещество или, еще вероятнее, через подкорковые узлы, а может быть, через
то и другое вместе (рис. 2). Это еще не говорит, конечно, о том, что через
кору невозможно замыкание условной связи, а также взаимодействие и нр-
радиация процессов. Вопрос этот будет решаться при другом варианте
операции. Затем мы можем определенно сказать, что разобщенные зоны
функционируют и выполняют анализ и синтез соответствующих им раз-
дражений, ибо, как и у нормальных собак, экстирпация каждой из этих
зон нарушает соответствующую им условнорефлекторную деятельность.
В заключение я хотел еще раз повторить, что часть статьи, посвященная
нашей работе, имеет целью не столько демонстрировать наши, пока что
предварительные данные и весьма осторожные выводы по затронутой проб-
леме структурной основы условнорефлекторной деятельности, сколько
поставить эту проблему и вынести на суд научного мира то средство, тот
способ и путь, которые нами избраны как наиболее подходящие на данном
этапе для се решения.
ЛИТЕРАТУРА
Крылов В. А., О возможности образования условного рефлекса на раздражитель
через кровь (автоматический раздражитель). Сборник, посвященный 75-летню
Павлова, 1925.
Лешли К., Мозг и интеллект, 1933.
Павлов И. П., Двадцатилетии!! опыт, объективного изучения высшей нервной
деятельности (поведения) животных, 1932.
Павлов И. П., Лекции о работе больших полушарий головного мозга, 1927.
Протопопов В. П., Центральная часть рефлекторной дуги оборонительного
условного рефлекса. Современная психоневрология, 1, 1931.
Раева Н. В. и Раппопорт Е. Я., Чувствительные пути в моторике, Физиоло-
гический журнал СССР, т. 27, № 3. 1934.
Ухтомский'А. А., Парабиоз и доминанта, 1927.
Bet he G., Hdb.d. norm. u. pathol. Physiol., Bd. 15. 2. Ilalftc, 1931.
Cri sler G„ Amer, journ. physiol., v. 94. p. 553, 1930..
Gantt W., Amer, journ. physiol., v. 1 19, p. 313, 1937.
E w a 1 d, Dtsch. med. Wschr., Vereinbeilage, 25, 1898.
Exner S., Pfluger's Arch. Physiol.. Bd. 28, 487. 1882.
Goltz F., Ueber die Verrichtungen des Grosshirns, 1882.
Goltz F., Pfluger's Arch. Physiol.. Bd. 51, 1891.
Harlow H. a. Stegner R., Journ. exp. physiol., v. 16, p. 283, 1933.
К ar p I us u. Kreldl, Pfluger's Arch. Physiol., Bd. 129, S. 133, 1909.
Magnus K., Korperstellung, 1924.
Light 1. u. Gantt W.. Journ. comp, psychol., v. 21, p. 19, 1936.
Louks R..Journ. psychol., v. 1. 5, 1935.
Lucia ni L., Human phyisiology, v. Ill, 1915.
11 Э А. Асратян
ПРИНЦИП «ПЕРЕКЛЮЧЕНИЯ» В УСЛОВНОРЕФЛЕКТОРНОЙ
ДЕЯТЕЛЬНОСТИ 1
ногочисленными работами по физиологии центральной нерв-
ной системы можно считать установленным положениеоб общ-
ности и родстве между основными закономерностями деятель-
ности низших и высших отделов централь нон нервной системы,
в частности, коры большого мозга. Одновременно с этим в со-
временной нейрофизиологии можно считать бесспорным и то
положение, что в процессе эволюции и каждой достаточно днференцирован-
ной части центральной нервной системы и особенно ярко в коре большого
мозга эти закономерности приобретают специфический характер, зачастую
даже качественно новую природу благодаря «скачку» в развитии. Оба эти
положения делают понятной плодотворность результатов исследований та-
ких корифеев нейрофизиологии, как И. М. Сеченов, II. П. Павлов, Н. Е. Вве-
денский, Магнус и др., которые при изучении специфических особенно-
стей какого-нибудь звена нервной системы постоянно учитывали общие
свойства всей цепи и прослеживали эволюцию изучаемых специфических
особенностей.
Нет нужды широко распространяться об особой полезности и необхо-
димости такого именно подхода к экспериментальной и теоретической раз-
работке богатей шего наследия И. П. Павлова об условных рефлексах. Исто-
рическим оправданием этого положения является не только вся научно-
творческая жизнь нашего учителя, но и деятельность группы его лучших
учеников. Будучи также убежденным сторонником этого принципа работы,
я в своих исследованиях постоянно стремился усилить связь между физио-
логией условных рефлексов и общей физиологией нервной системы и при
изучении отдельных специфических вопросов корковой физиологии не-
пременно исходил из предпосылки об органической связи различных частей
нервной системы, а Значит и их роДСтВе, общности их закономерностей.
Под этим же углом зрения я и группа моих сотрудников (Ф. М. Шитов,
В. В. Яковлева, Я. М. Прессман, В. Л. Замятина) изучали вопрос о прин-
ципе «переключения* (Schaltung) в деятельности коры большого мозга.
Средн многочисленных естественных факторов, обусловливающих вариа-
бильность, динамичность рефлекторной деятельности низших отделов
центральной нервной системы, важное место занимает исходное положение
конечности и вообще исходное состояние частей тела. Так, еще в середине
прошлого столетня И. М. Сеченов, за ним Сандерс-Эзн (Sanders-Ezn)
и Гольц (Colt?) экспериментально показали, что характер рефлекторного
ответа конечности лягушки на какое-нибудь определенное раздражение
зависит от исходного положения этой конечности. Например, в ответ на
1 Доложено на Зы физиологическом совещании Академии наук СССР в марте
1938 г.. Физиологический журнал СССР, т. XXX, в. I, 1941.
ПРИНЦИП «ПЕРЕКЛЮЧЕНИЯ» В УСЛОВНОРЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ ЮЗ
однотипное и одинаковое по силе раздражение какой-нибудь определенной
части одной из задних конечностей можно получить разгибание конечности,
если исходным было согнутое состоя вше конечности и, наоборот, сгибание,
если в момент раздражения конечность находилась в разогнутом положении.
Такое изменение характера рефлексов они объясняли влиянием на спин-
ной мозг импульсов из мышц и кожи конечностей. Позже аналогич-
ное явление было обнаружено Икскюлем (Ucxkkull) и Иорданом (Jordan)
на беспозвоночных; подвергая физиологическому анализу указанное яв-
ление, эти исследователи пришли к заключению, что состояние возбуди-
мости нервных центров обусловливается исходным состоянием соответ»
ствующих мышц на периферии. Они же показали, что чувствительные нерв-
ные пути будто не участвуют в этом процессе. На этом основании Икскюль
сформулировал свой общеизвестный закон «растянутой мышцы». Далее,
в процессе своих исследований в области физиологии спинного мозга тепло
кровных животных с подобным явлением встретился также Шеррингтон
(Sherrington). Детальному физиологическому анализу, однако, подверг эти
явления Магнус (Magnus), который и дал ему название Schaltung — «пе-
реключение». Па хронически оперированных спинальных собаках и кошках
Магнус показал, что исходное состояние задних конечностей и х поста пред-
определяет характер рефлекторного их ответа на какое-нибудь раздражение.
Он экспериментально показал такж:е, что это яркое выражение воздей-
ствия периферии на центральный нервный аппарат осуществляется через
чувствительные нервные пути, в основном через проприоцептивные пути.
Магнус установил также, что своеобразное влияние периферии на центр
носит тонический характер. Указывая на важное значение этого нового
принципа для различного рода деятельностей спинного мозга, Магнус сфор-
мулировал исключительно важное для физиологии нервной системы по-
ложение; «Спинной мозг ежеминутно изменяется, в каждый момент в
нем отображаются положение и установка различных частей тела и
всего тела в целом. Каждому положению тела соответствует определен-
ное распределение возбудимостей и наиболее легко доступных путей
в центральной нервной системе. Само тело устанавливает правильно
(in der richtigen Weise) свой центральный орган»1.
Отклоняясь немного в сторону, нельзя не указать на близость между
этим положением Магнуса и такими общими для физиологии централь-
ной нервной системы принципами, как «проторение путей» Экснера (Ехпег)
и доминанта А. А. Ухтомского. Не только проприоцептивные, но и интра-
цептивные и различного рода экстрацептивные импульсы, а также сдвиги
в гуморальном «хозяйстве» организма и ряд других факторов способны
путем изменения функционалыюго состояния нервных элементов создавать
различные функциональные «карты распределения» открытых и закрытых
путей, дееспособных и недееспособных в данный отрезок времени центров
и этим самым предопределять характер деятельности центрального аппа-
рата в целом или отдельных его звеньев.
Если в вариабильностн спинальной рефлекторной деятельности принцип
«переключения» играет такую важную роль, возникает вопрос, имеет ли
место «переключение» также в корковой деятельности, где, как известно,
изменчивость, динамичность выражены наиболее ярко по сравнению с
другими отделами центральной нервной системы и составляют важную
качественную характеристику условнорефлекторной деятельности.
Можно было с большой степенью вероятности ответить на этот вопрос
положительно на основании не только теоретических предпосылок,
но и отдельных, по преимуществу косвенных, отдаленных эксперимен-
тальных данных по физиологии условных рефлексов, полученных в лабо-
1 R Magnus, Korperstellung, Id24, S. 35.
II»
164
Э. А. АСРАТЯН
раториях И. П. Павлова. Прежде всего я имел в виду данные по
переделке условных рефлексов, когда один и тот же условный раздражи-
тель, в зависимости от рода безусловного подкрепления, приобретает то
одно, то другое сигнальное значение, порой, как это было в опытах
С. С. Фридемана, сохраняя как бы оба сигнальных значения одновре-
менно. Несомненно, к этому вопросу имеют отношение также много-
численные данные, согласно которым всякое изменение условий экспери»
мента так или иначе отражается на характере условнорефлекторной дея-
тельности собак и других животных. В этой связи нельзя не указать на
интересные данные Г. П. Зеленого, полученные па мышах; по этим данным,
изменение условий опыта коренным образом меняет характер высшей
нервной деятельности мышей *.
Тем по менее вопрос о принципе «переключения» в условнорефлекторной
деятельности специально не изучался, несмотря на его большой теоре-
тический интерес. Приступив к экспериментальной разработке этого
вопроса, мы исходили из того, что если в условиях экспериментов Магнуса
(изолированный спинной мозг) «переключателем» были проприоцептивные
импульсы, то в наших экспериментах количественный и качественный со-
став «переключателей» неизмеримо расширится: «переключение» приобре-
тет новое, более высокое физиологическое содержание.
Наши предположения полностью оправдались полученным экспери-
ментальным материалом, к краткому изложению которого я л перехожу.
На одной и той же собаке, в одной и той же камере, пользуясь одними
и теми же раздражителями и оборудованием, два моих сотрудника
Ф. М.. Шитов и В. В. Яковлева еще в 1936 г. изо дня в день проводили подва
опыта следующего характера.
Ф. М. Шитов в утреннем опыте сочетал действие метронома 120 ударов
в 1 минуту (М12о) с пищевым безусловным рефлексом, а действие метронома
60 ударов в I минуту (Mg0) не сочетал с пищевым безусловным рефлексом.
Посте того как были выработаны соответствующие пищевые условные
рефлексы на Mi20 н М^, уже в середине дня стала ставить опыты
В. В. Яковлева, сочетая Mi20 с электрооборонительиым рефлексом с лапы.
Через несколько дней собака выработала электрооборонительный условный
рефлекс на М12о, сохранив условный пищевой рефлекс на тот же раздра-
житель в утренних опытах с другим экспериментатором. Характерно,
что после выработки электрооборонигельного условного рефлекса на М12о
диференцировка (М60) к этому рефлексу с перв-ого же применения была на-
лицо. Далее, тем же путем и на той же собаке раздражители гудок и касалка
Ф. М. Шитовым были превращены в условные пищевые сигналы, а
В. В. Яковлевой —в электрооборонительные сигналы.
Таким образом, Ф. М. Шитов и В. В. Яковлева впервые эксперименталь-
ным путем получили интересные данные о том, что одни и те же раздражи-
тели одновременно могут иметь два совершенно различных условных зна-
чения, причем, в зависимости от условий опыта, раздражитель вызывает
то один, то другой вид деятельности. Экспериментально был получен чи-
стый случай «переключения» в условнорефлекторной деятельности. В даль-
нейших экспериментах Ф. М. Ш итов и В. В. Яковлева выяснили, что основ-
ным переключателем в этом варианте экспериментов были сами эксперимен-
таторы. Так, например, полное выравнивание других моментов опыта (при-
клеивание баллончика к щеке в вечерних опытах и прикрепление к лапе
электродов и регистрирующих приборов, в утренних опытах и т. д.), а
также перемена времени опыта (Ф. М. Шитов — днем, В. В. Яковлева —
* Более обстоятельный обзор истори и вопроса приводится в нашей статье
«К физиологии временной связи» напечатанной в данном сборнике.
ПРИНЦИП «ПЕРЕКЛЮЧЕНИЯ» В УСЛОВНОРЕФЛЕКТОРНОЙ. ДЕЯТЕЛЬНОСТИ [65
утром) не оказывали заметного влияния на характер описанной выше
условнорефлекторной деятельности. Когда же при опыте присутствовали
одновременно оба экспериментатора, собака выявляла хаотическую условно-
рефлекторную деятельность, давая на раздражитель то одну реакцию,
то другую, то обе реакции вместе в ослабленном виде.
Если в описанных выше опытах основным «переключателем» двух типов
рефлекторной деятельности был экспериментатор, то другие наши опыты
с той же и с другими собаками, но при другой постановке экспериментов
показали, что такими ж:с «переключателями» двух различных рефлексов
могут быть время дня, последовательность опытов в течение дня и даже
отдельные компоненты экспериментальной установки. Ради экономии вре-
мени я на всех этих вариантах останавливаться не буду. Скажу несколько
слов об интересном варианте экспериментов, проведенном моим сотруд-
ником Я. М. Прессманом. У собаки с левой задней лапы были выработаны
условные электрооборонительные рефлексы на свет и касалку. После упро-
чения этих рефлексов с левой лапы регистрирующие приборы и электроды
были перенесены на правую заднюю лапу. Выяснилось, что эта перемена
в условиях эксперимента достаточна, чтобы в ответ на условные раздра-
жители собака стала отвечать не левой, как раньше, а правой лапой. Реф-
лексы «переключались» на другую сторону. На другой собаке Я. М. Пресс-
мап получил ту же картину с той лишь разницей, что у этой собаки рефлек-
сы были старые и «переключение» осуществлялось не так гладко. Исключи-
тельный интерес представляют дальнейшие эксперименты Я. М. Прессмана.
К. обеим лапам Собак были прикреплены электроды и регистрирующие при-
боры. В этих условиях на первый условный раздражитель собака нс реаги-
ровала четким локальным ответом ни одной лапы. Первый электрический
удар в какую-нибудь лапу определял всю дальнейшую направленность
условнорефлекторной деятельности: раздражители вызывали условные
рсфлеюы с той лгпы, которая получила первый электрический удар. Со-
I ершгнно очевидно, что если в первой серин опытов Я. М. Прессмана «пе-
реключателем» было прикрепление приборов к лапе, то во второй серии
таким «переключателем» стал первый электрический удар*.
Дальнейшие наши искания были направлены на выяснение вопроса
о том, возможны ли «переключения» условного раздражителя с положи-
тельного на отрицательный и наоборот при помощи создания соответствую-
щей экспериментальной обстановки. Речь шла о возможности одновремен-
ной выработки условных рефлексов положительного и отрицательного
знака на один и тот же раздражитель и на базе одного и того же безуслов-
ного рефлекса.
Эксперименты Ф. М. Шитова и В. А. Замятиной ответили на этот
вопрос положительно. Выяснилось, что в. утренних, например, опытах
ссбаки без большого труда образуют Положительный условный пищевой
рефлекс на какой-нибудь раздражитель, а в дневных опытах с тем же эк-
спериментатором, в той же экспериментальной обстановке, на тот же самый
раздражитель образуют отрицательный пищевой условный рефлекс. Далее,
Ф. М. Ш итов показал, что собаки способны решать и более трудные задачи,
правда, уже не с такой легкостью. В своих утренних опытах Ф. М. Шитов
выработал положительный условный рефлекс на сильное гудение и отри-
цательный — на слабое гудение. В дневных же опытах на этой собаке
в той же камере он сильное гудение нс подкреплял пищей, а слабое гуде-
ние подкреплял, т. е. пытался придать этим раздражителям сигнальные
значения, обратные тем, которые они имели в утренних опытах. Выясни-
лось, что собаки, в конце концов, решают и эту нелегкую задачу.
* Опыты Я. XV Прессмана в нет оторой своей части представляют собой воспро-
изведение экспериментов Ю. М. Копорского. выполненных незадолго до этого.
166
Э А. АСРАТЯН
В описанных выше опытах по одновременной выработке однородных
условных рефлексов различного функционального знака на одни и те же
раздражители «переключателем» было время дня и последовательность
экспериментов; совершенно очевидно, однако, что в других вариациях опы-
тов при решении той же задачи «переключателями» могут стать также экс-
периментатор, камера, различные компоненты камеры и т. и. Предметом
дальнейшей работы моей и моих сотрудников является экспериментальная
проверка этих предположении, а также продолжение исследований ряда
других важных сторон этой интересной проблемы условнорефлекторной
деятельности высших животных.
Следует отметить, что за последнее время появились две работы из дру-
гих лабораторий, которые полностью соответствуют нашим данным. Я имею
в виду данные И. И. Лаптева, которому удалось выработать пищевые и
оборонительные условные речрлексы на один и тот же раздражитель, варьи-
руя только время, и данные работавшей раньше у меня по этой проблеме
В. В. Яковлевой и В. К- Федорова, которые получили подобные же ре-
зультаты, ставя опыты в различных камерах.
Прежде чем подытожить изложенный в кратких чертах эксперименталь-
ный материал, полученный в руководимой мной лаборатории, и сделать
некоторые теоретические заключения, во избежание недоразумений скажу
несколько слов о разнице между изученным нами «переключением» в услов-
норефлекторной деятельности и давно известными условными рефлексами
сложного типа, носящими название комплексных условных рефлексов.
Нет никакого сомнения, что это два совершенно различных типа деятель-
ности коры большого мозга. При комплексных условных рефлексах новый
компонент (раздражитель, порядок и т. д.) является частью условного
раздражителя, т. е. является условным пусковым фактором и как таковой
действует фазно, вызывает непосредственный эффект, тогда как в слу-
чае «переключения» этот компонент не является частью условного раз-
дражителя, не является фазнодействующим пусковым фактором, а путем
ряда функцио-нальных сдвшгов создает определенный фон тонического
характера, известные предпосылки к выявлению типа условного реф-
лекса.
Итак, можно считать бесспорно установленным, что закон «переключе-
ния» интрацентральных путей сохраняет свою силу также для деятельности
коры большого мозга. Этот закон относится, следовательно, к числу основ-
ных универсальных законов деятельности центральной нервной системы.
От простого закона «растянутой мышцы» через стадию более сложного спи-
нального «переключения» (возможно, также через некоторые другие, еще
не изученные стадии развития) закон «переключения» интрацентральных
путей достигаете деятельности коры наиболее совершенной формы. Прежде
всего неизмеримо расширяется количественный и качественный состав фак-
торов-переключателей. Кроме механического фактора, влияющего через
кожные и проприорецепторы, действенными делаются все раздражители,
влияющие через все без исключения рецептсры организма. Кроме того,
природа самого «переключения» в коре даже под влиянием одного какого-
либо фактора значительно изменяется: «переключение» перестает быть
шаблонным, однообразным, как в спинном мозгу, и становится значительно
вариабильнее, динамичнее, разнообразнее. Вес это делает понятным гро-
мадное значение свойства «переключения» именно для корковой деятель-
ности, одной из главных особенностей которой как раз н является вариа-
бнльность, гибкость, динамичность, временность. Нет сомнения, что, благо-
даря возможности «переключения», эти специфические особенности корко-
вой деятельности становятся физиологически более совершенными, значи-
тельно увеличивают способность организма к точному и тонкому приспо-
соблению к вечно изменяющимся условиям существования.
ПРИНЦИП «ПЕРЕКЛЮЧЕНИЯ» В УСЛОВНОРЕФЛЕКТОРНОП ДЕЯТЕЛЬНОСТИ 167
ЛИТЕРАТУРА
Павлов И. П., Двадцатилетний опыт объективного изучения высшей нервной
деятельности (поведения) животных, 1932.
Пресеман Я. М.. Функциональное значение ненрекратенных зрительных,
слуховых и моторных проводящих путей, тезисы диссертации, 1941.
Сеченов И. М.. Собрание сочинений, т. II. 1908.
Сеченов И. М., Физиология нервной системы, 1866.
Ухтомский Л. А.. Парабиоз и доминанта, еб. «Парабиоз», 1927.
Фридеман С. С., Дальнейшие материалы к физиологии диференцирования внеш-
них раздражений, дисс. 1912.
Шитов Ф. М., Фактор времени в принципах переключения условных связей,
Бюллетень экспериментальной биологий И медицины, VIII, 1939.
Шитов Ф. М. и Я к о в л е в. а В. В.. Одновременная выработка разнородных услов-
ных рефлексов, на один и тот же раздражитель, Бюллетень экспериментальной
биологии и медицины, IV, 1937.
Goltz F., Beitr. zur Lehre von den Function der Nervenzentren des Frosches, 1869.
Jordan, Pllug. Arch., 110, 1915.
Magnus. Korperstellung. 1924.
Sanders-Ezn, цит. rto Trcndelenburg «Method! k der Physiol, d. Zcntralnerv.»,
1923.
Sherr ingto'n, Proc. Roy. soc.> London., 66, 1899; Journ. physiol., 40, 1910.
Uexkttll. Ergebni d. Physiol., 1903; zbL Physiol., 1909.
Х5С/"
К ФИЗИОЛОГИИ ДВИГАТЕЛЬНЫХ ОБОРОНИТЕЛЬНЫХ
УСЛОВНЫХ РЕФЛЕКСОВ 1
етод условных рефлексов является наиболее совершенным и
плодотворным орудием современной физиологии при изучении
сложной функции больших полушарий головного мозга. Всем
известно, что основные физиологические закономерности выс-
шей нервной деятельности у высших животных установлены
в лабораториях основоположника и творца грандиозного уче-
ния о высшей нервной деятельности акад. И. П. Павлова главным
образом на пищевых и оборонительных слюноотделительных рефлексах
у собак с учетом сопровождающих их двигательных реакций.
Спустя некоторое время после возникновения и оформления учения об
условных рефлексах, под его влиянием как у нас, так и в других странах
ряд исследователей тоже начал проводить работу в этом направлении. Эти
исследования велись главным образом в аспекте расширения диапазона
методик экспериментов. В качестве основного безусловного рефлекса были
Избраны другие рефлексы (рефлексы на сердце — В. М. Бехтерев, А. Г. Ива-
нов-Смоленский; дыхательно-двигательные и оборонительно-двигательные
рефлексы — школа В. М. Бехтерева, И. С. Беритов, Д. С. Фурсиков и др.;
глотательные и слюноотделительные рефлексы—Н. И. Красногорский;
хватательный рефлекс—А. Г. Иванов-Смоленский; экскреторная дея-
тельность почек и секреторная деятельность ряда пищеварительных
желез—Л. Л. Орбели, К. М. Быков и corp.; зрачковые рефлексы —
Кэсон (Cason) в Америке и др.; так называемые «Magnetreaction» и «Stehbe-
reitschaft» — Радемакер (Rademaker) в Голландии; кожно-гальванический
феномен — Генкен, Фримен (Freemen) и др. и даже на такие реакции, как
рвота,—В. А. Крылов, лейкоцитоз—С. И. Метальников, Н. Л. Подкопаев
др. ]. Исследования были расширены также в области сравнительной и воз-
растной физиологии, причем изучались различные предстааите-лн зоологи-
ческой лестницы снизу доверху: от человека до асцидии, от младенческого
и до старческого возраста.
В этой статье мы нс будем вдаваться в подробности и нс будем разбирать
проблему в целом. Остановимся только на отдельных вопросах, тесно свя-
занных с полученными мной экспериментальнымн данными.
Средн всех указанных выше методических вариаций самой распростра-
ненной является методика оборонительно-двигательных рефлексов у собак
на электрическое раздражение. Этим путем получено много интересного
и ценного мате риала, который хороню согласуется сданными, выявленными
с помощью условных слюноотделительных рефлексов при учете своеобразия
1 Труды физиологических лабораторий акад. И. П Павлова, т. X, 1941. Данная
работа была защищена в декабре 1933 г. на звание ученого специалиста Академии наук
СССР.
К ФИЗИОЛОГИИ ДВИГАТЕЛЬНЫХ ОБОРОНИТЕЛЬНЫХ УСЛОВНЫХ РЕФЛЕКСОВ 169
физиологии двигательного аппарата (работы школы В. М. Бехтерева, в
особенности работа В, П. Протопопова, Н. Г, Изранльсоиа, Е, A. JJJ евелева,
А. Л. Шнирмана и др.; затем работы Д. С. Фурсикова, И. С. Беритова,
Ф. П. Майорова, Г. П. Конради, Л. И. Федорова, Ю. М. Коморского
и С. М. Миллера, В. Я- Кряжева и др.).
Совершенно естественно, что основные закономерности условнорефлек-
торной деятельности должны отражаться на индикаторах слюноотделитель-
ного и двигательного рефлекторных аппаратов организма с присущими им
некоторыми особенностями, свойственными этим сложным физиологическим
системам. Это совершенно законное физиологическое явление. Ведь доста-
точно указать на то, что названные рабочие аппараты имеют совершенно
различную первичную функциональную эфферентную иннервацию: один
вегетативную, другой — соматическую. Это важное обстоятельство учиты-
вается не всеми исследователями. Между тем не подлежит сомнению, что
коре больших полушарий вюсбше не с одинаковой легкостью н быстротой
удается овладевать той или другой системой, осуществлять свою функ-
цию через тот или другой эффекторный прибор.
Несмотря, однако, на глубокие морфологические и физиологические
различия между слюнными и двигательными индикаторами, все же следует
думать, что основные закономерности условнорефлекторной деятельности,
полученные отдельно посредством слюноотделительных и двигательных
рефлекторных аппаратов, нс могут по существу отличаться друг от друга.
В обоих случаях мы имеем дело с единой физиологией коры больш их полу-
шарий. Задача физиологии — диференцировать, расчленить то существен-
ное, что принадлежит коре, и то второстепенное, что тот или иной эффектор-
ный аппарат добавляет к этому существенному при его преломлении через
себя.
Среди новых исследований по условным оборонительно’двигательным
рефлексам следует остановиться на работе И. С. Беритова.
И. С. Беритов повторил и в основном подтвердил ряд работ из лабора-
тории В. М. Бехтерева, а также добавил новый экспериментальный мате-
риал. Экспериментальные данные И. С. Беритова в известной мере со-
гласуются сданными и основными принципиальными положениями павлов-
ского учения об условных рефлексах. Вместе с тем И. С. Беритов полу-
чил небольшое число отдельных фактов, которые находятся в явном про-
тиворечии с фактами, выявленными в лабораториях И. П Павлова.
Расхождения между этими данными И. С. Беритова и школы И. П. Пав-
лова таковы, что их: нельзя объяснить только специфическими особен-
ностями слюноотделительного и двигательного индикаторных аппаратов.
Разбирая эти противоречия и по-своему истолковывая гармонирующие
факты, И. С. Беритов развивает свою концепцию на предмет в целом,
которая им противопоставляется павловскому- учению об условных реф-
лексах. Он заявляет, что И. П. Павловым «устанавливаются такие новые
законы, которые в корне противоречат общеизвестным законам физиологии
центральной нервной системы»’. Эта мысль в других выражениях повто-
ряется на страницах его книги неоднократно-
Мы лишены возможности здесь подробно разбирать эти теорети-
ческие построения И. С. Беритова и будем касаться только отдельных
моментов его работы и частных его высказываний, которые имеют отно-
шение к изученным нами вопросам взаимной индукции, иррадиации про-
цессов и угашения элсктрооборонительных двигательных условных реф-
лексов. Скажем только, что при построении св-оих гипотетических положе-
ний И. С. Беритов в основном пользуется (правда, несколько своеоб-
1 И. С. Беритов, Индивидуально приобретенная деятельность нервной си-
стемы, 1932, стр. з.
170
Э. Л. АСРАТЯН
разно) экспериментальным материалом из богатейшего арсенала павлов-
ских лабораторий.
Изучением оборонительно-двигательных рефлексов у собак мы начали
заниматься с марта 1932 г. Под опытами были три собаки: среднего возраста
Баш— в высшей степени спокойная, уравновешенная собака, с сильной
нервной системой, но трусоватая; Кейо — очень подвижная собака, с не
очень сильной нервной системой (у Кейо 14.Л 1932 г. были экстирпированы
двигательные зоны коры с обеих сторон); Назар — спокойная, но трусливая
собака.
Методика работы была такова. Собаки стояли на станке в лямках.
Зашитые в кожаный лоскут электроды привязывались к плюсне их левой
задней конечности. В качестве безусловного раздражителя применялся
электрический прерывистый индукционный ток от санного аппарата Дюбуа-
Реймона. Сила тока бралась немного выше пороговой. Передача движения
ноги на кимограф производилась специальным рычагом, который привязы-
вался к раздражаемой конечности посредством манжета. Устройство ры-
чага не мешало собаке делать движения конечностью; на кимографической
ленте регистрировались только сгибание и разгибание конечности. Дви-
жения рычага передавались пишущему на кимографе пищику мареевского
полиграфа через воздушную передачу. Электромагнитными отметчиками
фиксировались условные и безусловные раздражения и время.
Полученные экспериментальные данные у всех трех собак в основном
совпадают. Материал, полученный у Кейо, опубликован в специальной
статье, и на нем мы здесь останавливаться не будем в силу того, что у Кейо
были удалены двигательные зоны коры большого мозга. Результаты ис-
следования двух других собак приводятся вместе.
Явления взаимной индукции и последовательного торможения
в условных оборонительно-двигательных рефлексах
Явления взаимной индукции и последовательного торможения достаточ-
но хорошо изучены в области слюноотделительных рефлексов многочислен-
ными сотрудниками павловских лабораторий (М. Н. Ерофеева, Б. А. Коган,
Д. С. Фурсиков, Н. А. Нарбутович, Н. А. Подкопаев и др.). Что же касается
условных оборонительно-двигательных рефлексов у собак, то эти явления
до сих пор остаются недостаточно изученными, если не считать отдельных
замечаний в работах бехтеревской школы. И. С. Беритов же совершенно
необоснованно отрицает существование индукции как закономерного явле-
ния в коре. В отношении положительной индукции он прямо говорит:
«В моих опытах с оборонительными или пищеводвигательными рефлексами
мне не приходилось наблюдать что-либо подобное положительной индук-
ции» ‘. Объясняет это И. С. Беритов, между прочим, тем, что он зани-
мался совпадающими рефлексами.
В наших опытах, в особенности в период образования и упрочения ди-
ференцировки, мы почти постоянно сталкивались с явлениями положи-
тельной последовательной индукции, хотя у наших собак рефлексы были
тоже совпадающими. Приводим иллюстрацию опытов (рис. 1).
Положительная индукция настолько ясно проявлялась, что даже в слу-
чаях, когда положительный рефлекс почему-либо ослабевал и исчезал,
достаточно было однократного применения дифереицировочного раздра-
жителя, чтобы этот исчезнувший рефлекс восстановился (рис. 1, опыт№7О).
Очевидно, этифакты правильнее рассматривать как проявление сукцессив-
ной положительной индукции от тормозного рефлекса на,положительные.
Как и в случае слюноотделительных рефлексов, здесь индукция лучше
’ И. С. Беритов, Индивидуально приобретенная деятельность нервной сис-
темы, 1932, стр. 85.
К ФИЗИОЛОГИИ ДВИГАТЕЛЬНЫХ ОБОРОНИТЕЛЬНЫХ УСЛОВНЫХ РЕФЛЕКСОВ 17 |
всего проявилась через 10 —30 секунд по окончании действия диференцн-
ровочного раздражителя; после этого времени обыкновенно начинались
явления иррадиации торможения оттормозного рефлекса на положительные.
Явления последовательного торможения в опытах И. С. Бсритова тоже не
получались в таком виде, в каком они получались в павловских лаборато-
риях. Диференцировочный раздражитель, а также посторонний раздражи-
тель (по его мнению, они вообще действуют совершенно одинаково) остав-
ляют после себя последовательное торможение только в течение нескольких
секу нд. Мы не говорим уже о том, что И. С. Бернтов это явление понимает
вовсе не как последовательную иррадиацию торможения, а совершенно
своеобразно, как иррадиацию возбуждения по обратным путям—из цент-
ров положительных рефлексов к сильно возбужденным центрам диферен-
цнровочного и постороннего раздражителей.
Рис. I. Собака Паш. Положительная индукаия дифсренцировочного
раздражителя на положительный условный рефлекс, затем последо-
вательное торможение на'положительные рефлексы
Опыт № 109 Cl-й и 2-й ряд) 14.11 1933 г. Через 20 сек. после применения
диференцмропочного раздражителя (стук 60) условный рефлекс заметно усили-
вается Далее отчетливо видна иррадиация торможения и концентрация его.
Опыт № 70(3-й ряд) 17X11 1932 г Р< флекс на стук 120 отсутствует. Действие
днфереицнровочного раздражителя через 20 сек . положительно индуцируя,
восстанавливает рефлекс на стук 120
Линии сверху вниз: условный двигательный оборонительный рефлекс, отметка
условных раздражителей, отметка безус лонного раздражителя (цифры указывают
расстояние между катушками), время о секундах (цифры указывают величину
пауз)
Оставляя в стороне его понимание явления, обратимся к фактам. Дол-
жен сказать, что у моих собак, в особенности у Баша, главным образом
в период, когда диференцировка была еще свежа, явление последователь-
ного торможения длилось минутами, иногда десятками минут. При этом
в полном соответствии со слюноотделительными условными рефлексами
и здесь волна торможения раньше, длительнее н сильнее всего захваты-
вала рефлексы из того же анализатора, что и диференцировка.
Обратный процесс—концентрация торможения, — как и в условных
слюноотделительных рефлексах, протекал в совершенно обратном порядке.
В конце концов, и здесь наблюдался волнообразный ход иррадиации и кон-
центрации торможения. Сказанное хорошо видно как в приведенном выше
опыте (рис. 1, опыт № 109), так и в опытах, приводимых на рис. 2.
Все эти опыты ставились на собаке Баш, Цо совершенно сходные дан-
ные как в отношении индукции, так и последовательного торможения были
получены и на ссбаке Кейо.
Затем следует сказать, что аналогичное последовательное торможение
и его концентрация наблюдались также от угасательного торможения.
172
Э. А. АСРАТЯН
У наших собак получались также данные, говорящие об индукционных
взаимоотношениях между условными и безусловными рефлексами. Явление
это раньше мимоходом наблюдалось некоторыми сотрудниками И. П. Пав-
лова (М. Н. Ерофеева и др.). Совсем недавно оно специально изучалось
многими его сотрудниками (Р. Я. Райт, Н. Л. Подкопаев, Д. И. Соловейчик,
П. К. Анохин, Н. В. Виноградов, А.М. Павлова, В. В. Рикман, Э. А. Лера-
тян). Если отрицательная и положительная последовательная индукция
между условными рефлексами в пределах одного анализатора соответствует
общепризнанной спинномозговой последовательной индукции, то этот
новый вид се, как и индукция между отдельными частями коры, очевидно,
Рис. 2. Собака Баш. Иррадиация торможения от диференцировочного
(стук 60) рефлекса на другие положительные рефлексы
/ - опыт № ТО* // — опыт № Н2; ill — опыт № Н Во всех трех опытах хорошо
видно. что торможение быстрее, сильнее и продолжительнее захватывает реф-
лексы от звукового анализатора: видна также волна концентрации торможения.
Обозначения те же, что на рнс. I
соответствует тому явлению в физиологии низших отделов центральной
нервной системы, которое описано Гольцем, А. А. Ухтомским и др. в виде
торможения от борьбы между отдельными рефлексами. Правда, ни Гольц,
ни кто другой данное явление не называет индукцией, но, во-первых,
закономерность этого явления всегда подчеркивается ими, а во-вторых,
сходство между этими явлениями и тем, что в павловской школе назы-
вается отрицательной индукцией на расстоянии, не может вызывать со-
мнения. Важно, конечно, не терминологическое сходство, а сходство по
существу. Это явление в коре уж:е давно было описано Бубновым и
Гейдеигайном (Bubnoff u. Heidenhein). Упомянутые исследователи в сво-
их острых опытах с корон собаки показали, что сильное раздражение
одних пунктов коры понижает возбудимость других ее частей. Все это
свидетельствует о том, что высказывания И. С. Беритова об ошибочности
павловских концепций по этому вопросу и о том, что в физиологии ниж-
них отделов центральной нервной системы подобные явления будто
неизвестны, совершенно не обоснованы.
В нашей работе по оборонительно-двигательным рефлексам, как уже го-
ворилось, наблюдались индукционные взаимоотношения между условными
и безусловными рефлексами: послеболее ил именем сильных положительных
условных оборонительно-двигательных рефлексов безусловный рефлекс
значительно ослабевает. Это ослабление особенно сильно заметно, когда
пауза между рефлексами не меньше 5 и не больше 20—30 секунд.
К ФИЗИОЛОГИИ ДВИГАТЕЛЬНЫХ ОБОРОНИТЕЛЬНЫХ УСЛОВНЫХ РЕФЛЕКСОВ 173
Что уменьшение рефлексов нельзя объяснить утомлением мускулатуры
или безусловнорсфлскторной дуги, на это указывает, во-первых, то, что
оно слабее проявляется при коротких паузах (1—5 секунд) и больше
выражено при 10—30-секундных паузах, а во-вторых, специальные опыты,
в которых безусловные рефлексы вызывались десятки раз подряд через
2—10 секунд и при этом не отмечалось никаких признаков уменьшения
рефлекса (рис. 3, опыты № ПО и 88). В некоторых опытах безусловные
рефлексы вызывались слабыми токами, и тогда условное раздражение на
целые минуты задерживало следующие за ним безусловные рефлексы
(рис. 3. опыт № 117).
Рис“ 3. Собака Баш. Отрицательно-индукционное влияние условных
положительных рефлексов на безусловные
I. Опыт № Юб 10.11 1933 г. Условный рефлекс на звонок через 15 сек. от на-
чала его действия вызывает заметное ослабление безусловного рефлекса;
явление быстро проходит. //.Олыт№!10 15-11 1933 г. Безусловный рефлекс
через паузы а 10 и затем 20 сек. вызывается 137 раз (величина всегда одина-
ковая); условный рефлекс на шипенье через 20 сек. приводит к «ильному ослаб-
лению безусловного рефлекса на несколько минут. III. Опыт № НН 17.1 1933 г.
Двойные раздражения электрическим током (2 и 5 сек. подряд) вызывают со-
вершенно одинаковый безусловный рефлекс; условный рефлекс на звонок При-
водит к ослаблению непосредственно следующего за ним безусловного рефлекса.
IV Опыт № 117 23 1 1 IS33 г. Слабым мектричегким током дна рала подряд
вызывается безусловный рефлекс; условный рефлекс на стук 120 целиком
снимает рефлекс на эту силу тока. Обозначения те же, что на рис. 1
Интересно, что если условные рефлексы на тот или иной раздражитель
почему-либо исчезали или если условный рефлекс специально угашался,
то часто в этих случаях безусловный рефлекс заметно усиливался. Это,
например, хорошо видно в опыте № 85, приведенном на рис. 2.
Как видно, в конце опыта в силу последовательного торможения от ди-
ференцировочного раздражителя (стука 50) стук 120 ударов в 1 минуту и
звонок утрачивали свое положительное действие, а безусловный рефлекс
после них заметно усиливался. Иногда безусловные рефлексы усиливались
также после применения дифереицировочного раздражителя, правда, по-
чему-то слабее, чем в указанных выше случаях. Совершенно очевидно, что в
описанных случаях мы имеем дело с индукционным влиянием условных
рефлексов на безусловные. Следует указать, что это целиком совпадает
с данными A. М. Павловой и нашими данными, касающимися слюноотде-
лительных условных рефлексов.
Но безусловный рефлекс со своей стороны также оказывает индукцион-
ное влияние на условные рефлексы. Если спустя несколько секунд (5—10)
после п рекращения безусловного рефлекса попытаться вызвать условный по-
ложительный рефлекс, то оказывается, что условные рефлексы значительно
174
Э А АСРАТЯН
ослаблены, а то и совсем исчезают. Сказанное иллюстрируется рис. 4 (опы-
ты № 90, 87, 117).
Должно сказать, что И. С. Беритов получил такие же данные, но он
объясняет их утомлением некоторых звеньев рефлекторных дуг. Трудно
согласиться с этим объяснением, ибо, как указывалось выше, безусловные
рефлексы значительной силы могли нами вызываться сотни раз через 10—
20-секундные паузы, 5 раз и больше — через 1-секундную паузу, причем
не наблюдалось никаких признаков уменьшения рефлексов, их утомления.
Следует еще раз указать на полное совпадение этих фактов с данными
исследования пищевых слюноотделительных рефлексов, полученными в ра-
ботах В. В. Рикмана и в специальной нашей работе на эту тему.
Рис. 4. Собака Баш. Отрицательно-индукционное влияние без-
условного рефлекса на положительные условные рефлексы
/. Часть опыта № 00. Через трехийнутиые паузы свет вызывает два
хороших условных рефлекса; сильный безусловный рефлекс через 10 сек.
приводит к значительному ослаблению рефлекса на свет, через 3 мин. зто
влияние прокалит. II. Опыт Гй 87. Безусловный рефлекс через 1—2 сек.
вызывает сильное ослабление условного рефлекса на тон С. f II Опыт № 117-
То же явление через 5 и 2 сек. выступает очень ярко, черви 1 мин. уже
не наблюдается. Обозначения те же, что на рис 1
Фактический материал, который был описан выше, ясно показывает,
что павловская концепция об индукционных взаимоотношениях между
положительными и отрицательными условными рефлексами, а также между
условными и безусловными рефлексами в полной мере сохраняет свое зна-
чение и ДЛЯ оборонительно-двигательных условных рефлексов. Взаимная
индукция является одной из основных закономерностей высшей нервной
деятельности и смеет Своего аналога в деятельности низших отделов цент-
ральной нервной системы.
Угашение условных сборе иительно-двигательиых рефлексов
Давно известный и хорошо изученный факт угашения условных слюно-
отделительных рефлексов рассматривался И. П. Павловым как один из
видов образования и проявления внутреннего торможения. Им же уга-
сатсльное торможение было отграничено от утомления и истощения нерв-
ных клеток и довольно подробно были описаны взаимоотношения между
этими основными явлениями в физиологии коры больших полушарий.
Но И. С. Беритов не совсем точно передает воззрения И. П. Павлова на
торможение вообще и на угасание торможения в частности. И, С. Беритов
пишет, что будто И. П. Павлову «торможение не представляется актив-
ным процессом», что будто «Павлов рассматривает внутреннее торможение
как явление истощения нервных элементов»1 и т. д.
1 И. С. Беритов, Индивидуально приобретенная деятельность нервной
системы, 1932, стр 78.
К ФИЗИОЛОГИИ ДВИГАТЕЛЬНЫХ ОБОРОНИТЕЛЬНЫХ УСЛОВНЫХ РЕФЛЕКСОВ |75
Еще в первые годы изучения условных рефлексов сотрудники И. П. Пав-
лова выявили особенности угашения пищевых слюноотделительных рефлек-
сов: быстрое угашенис при небольших (1—2 минуты) паузах и сравнительно
медленное угашёние при более длительных паузах. О. Л. Виндельбанд
и Ю. П. Фролов установили волнообразный ход угашения. Что же касается
оборонительно-двигательных условных рефлексов, то еще В. П. Прото-
попов в 1909 г. показал, что эти рефлексы, особенно свежевыработанные,
довольно быстро угашаются в течение одного опытного сеанса. На сле-
дующий день угашенные условные рефлексы были опять налицо. Для
окончательного угашения рефлексов потребовалось несколько Дней. Ха-
рактерно, что В. П. Протопопов аналогичного явления для соответствую-
щих безусловных рефлексов не наблюдал.
Эти данные впоследствии были подтверждены многими сотрудниками
лаборатории В. М. Бехтерева. И. С. Беритов. тоже занялся этим вопросом и
по существу подтвердил данные В. П. Протопопова и других относительно
«молодых» условных рефлексов. Что же касается крепких условных оборо-
нительно-двигательных рефлексов, то И. С. Беритов утверждает, что ему не
удавалось угашать эти рефлексы в течение одного опытного сеанса, что
для этого требовалось несколько дней: «Если рефлекс достиг боль-
шой прочности, тогда нужно применять индивидуальное раздражение сотни
раз в течение многих дней без единого сочетания с основным, чтобы полу-
чить полное угасание рефлекса»!. Очевидно, таков ход угашения при 1—
2-минутных паузах. Когда же И. С. Беритов хотел ускорить угашение,
то он удлинял время действия раздражителя и укорачивал паузы до 10—
15 секунд. В этих случаях он мог угашать через 16—20 применений. Если
при этом удвоить паузу — сделать ее более 20 секунд, то рефлекс снова
начнет получаться с большей интенсивностью. Больше того, И. С. Беритов,
считая ошибочным павловское понимание угашения, сам почти отождест-
вляет у гашение с утомлением; он находит, что нет никакой разницы между
утомлением безусловного рефлекса и этим утомлением, т. е. угашением.
Из его данных создается впечатление, что безусловный рефлекс угашается
даже быстрее условного. Явление вторичного угашения, которое школой
И. П. Павлова рассматривается как результат иррадиации торможения от
первичного очага на эти новые очаги, по И. С. Беритову, «обусловливается
утомлением общего для них двигательного участка в коре больших полу-
шарий» 4.
Следует сказать, что собранный нами фактический материал по этому
вопросу не согласуется ни с данными, ни с выводами И. С. Беритова.
Угашение условных оборонительно-двигательных стойких рефлексов,
хоти и имеет различный характер и течение у наших собак, в основном со-
ответствует угашению пищевых слюноотделительных рефлексов. У Баша,
например, при 1—3-минутных паузах рефлексы угасали через 2—10 при-
менений условного раздражителя без подкрепления, причем на сильные
раздражители через 2—3 применения, а на слабые через 6—10 примене-
ний. У Кейо то же получалось примерно через 7—15 применений, причем
угашение имело волнообразный ход и здесь. Что же касается зависимости
быстроты угашения от величины пауз, то здесь, помимо того, что подтвер-
дились закономерности, отмеченные в области пищевых условных рефлек-
сов, наблюдались и новые очень интересные закономерности,
В общем полученные данные можно обобщить так:
I) при паузах в 1—3 минуты между раздражителями рефлекс угасал
довольно быстро —примерно через 2—15 применений раздражителя без
подкрепления; t
1 И. С. Беритов, Индивидуально приобретенная деятельность нервной
системы, 1932, стр, 50,
! Т а м же, стр. 248.
176
3. А. АСРАТЯН
2) при паузах в 5 минут и больше угашение затягивалось до 10-го,
20-го раза и больше;
3) при очень маленьких паузах — в 20—30 секунд угашение не только
не ускорялось, как утверждает И. С. Беритов, но, наоборот, для сильных
раздражителей сильно затягивалось. Для слабых же н средних раздражи-
телей оно часто становилось просто невозможным даже после 200 и больше
применений раздражителя без подкрепления.
Чтобы точно выяснить значение величины пауз для быстроты угашения,
мы в своих опытах раздражитель применяли в течение какого-либо одного
Рис. 5. Собака Баш. Угашение рефлекса на звонок
/. Опыт .№ 113 18.11 1933 г. Угашение через 10-минутные паузы. Нулевой аффект
пперпые получается при 19-м применении раздражителя, затем рефлекс слабо
восстанаплииается и опять угашается уже окончательно. II. Опыт № III 16.11
1933 г. Угашение через 2’/,-минутные паузы. ///. Опыт Лй 23 24.V 1932 г. Уга-
шение через 2-минутиые паузы. Обозначения те же. что на рис. I
постоянного отрезка времени (5 или 10 секунд) и варьировали только паузу.
И. С. Беритов же одновременно варьировал как длительность применения
раздражителя, так и величину пауз — прием, который, невидимому, обус-
ловливал его ложный вывод относительно ускорения угашения при 10—
15-сскундных паузах. В наших опытах, как уже говорилось, при сильно
уменьшенных паузах угашение затруднялось.
Приведенные данные относительно 1—3—5-минутных и больше пауз
не являются новыми; тут мы имеем полное совпадение с данными угашения
слюноотделительных рефлексов. Факт же, отмеченный в п. 3, является
новым и интересным, и поэтому мы приводим ряд иллюстраций опы-
тов (рис. 5).
Па рис. 5 представлены три опыта с угашен ием стойкого условного
рефлекса на звонок.
В то время как при 2—2,/»-мияутнш паузах рефлекс угасает через
1—2 применения, при 10-секундных паузах он угасает только при 19-м
применении, причем при последующих применениях проявляет некоторую
тенденцию к оживлению и потом угасает окончательно.
На рис. 6 представлено угашение рефлекса на шипенье (средней силы).
В опыте № 101 рефлекс угашался через 5-минутные паузы между приме-
нением раздражителя. Как видно из рисунка, рефлекс довольно быстро
К ФИЗИОЛОГИИ ДВИГАТЕЛЬНЫХ ОБОРОНИТЕЛЬНЫХ УСЛОВНЫХ РЕФЛЕКСОВ 177
Рис, б. Собака Баш. Угашение рефлекса на шипенье
I. ОпытХ 101 4.II 1833 г. Угашение через 5-минутные паузы; на 6-м прнменс»
инн — следы, на 7-м — нулевой эффект, па 8—12-е применение — слабые рефлексы.
IJ. Опыт № 1O2 6.II 1033 г. Угашение через 2-мимутные паузы; нулевой эффект
с 4-го до 13-го применения раздражителя; после этого угашенный реф-лекс дна раза
подкрепляется и снова делается попытка вызнать его угашение применением раз-
дражмтеля через 20-секундиые паузы- Рефлекс упорно не поддается /гашению.
Нулевой эффект впервые получен на 35-е применение, затем до 109-го применения
рефлекс не угашается (еще три раза получен ослабленныП рефлекс). Обозначе-
ния те же, что па рис. 1
угасал. На 5-е применение раздражителя имеются следы рефлекса,
на &—8-е—нули, затем рефлекс опять восстанавливается и не угашается
до 12-го применения. На следующих кимограммах представлена часть опы-
12 9. А. Асратян
178
Э. А. АСРАТЯН
та (№ 102). В начале опыта рефлекс угашается через 2-минутные паузы.
Вскоре он угашается безвозвратно. Затем троекратным подкреплением
рефлекс снова восстанавливается и снова угашается, на этот раз уже через
20-секундные паузы. В течение 110 применений не удается погасить рефлекс,
хотя, правда, по ходу угашения два раза получался почти нулевой эффект
и два раза очень слабые рефлексы.
Особый интерес представляет кимограмма опыта № 74 (рис. 7). Раздра-
житель применялся 235 раз, и рефлекс на него не угашался (несколько раз
ослабевал и раза три почти исчезал). Характерно, что когда паузы удлиня*
лнсь от 20 до 50 секунд, рефлекс довольно быстро угашался.
В этом опыте обращает на себя внимание один момент, а именно что
частое применение раздражителя, через 20-секундные паузы, как будто
образовывало условный рефлекс на данный ритм; на это указывает двига-
тельная реакция на месте пропущенного 241-го применения.
Для более полного представления приводим сводную таблицу.
Название раздражителя Длитель- ность дей- ствия раз- дражителя а сек. Пауза между действием, раздражи- те ля в сск. На какое применение раздражи- теля получен первый нуле- вой эффект h.j каков применение раздражи- теля насту- пило окон- чательное угашение Дата
Звонок 10 240 2 2 24.V 1932 Г.
> 10 120 2 2 24.V
10 60 2 2 25.V
> 10 150 1 1 20.Х
» 5 300 5 19 13.11 1933 г.
> 5 60 1 1 16.11
» 5 60 11 11 17.11
в 5 1Й 20 31 1Я.П
М1И 10 60 2 2 25. V 1932 г.
М.,0 10 2-6 4 9 10.XI
м„. 10 120 3 5 14.Х 1
10 300 6 12 2.11 1933 г.
Шипенье 10 120 2 2 4.XI 1932 г.
» 10 150 3 3 22. XI
» 10 300 6 13 4. II
> 10 120 4 4 6.II 1932 г.
> 10 20 55 112 6.II 1933 г.
» 3-4 120 9 9 26.Х I
5 —— 1 ___
Свет 10 60 5 5 16.x 11 1933 г.
10 300 3 3 21.1 1934 г.
э 10 120 3 4 25.1
9 10 ?0 4 12 30.1
5 15 8 8 9.II
Тон С 10 20 39 232 29.Х II 1932 г.
» » 10 180 5 11 2.1 1933 г
> » 10 300 9 11 12.1
> » 10 300 12 16.1
э » 10 120 6 15 19.1
» » 5 300 3 3 5.1
» » 5 120 6 й 7.1
» > 5 10 —• 57 9.II
Что касается уменьшения рефлекса через 1—3-, а также 5-минутные и
большие паузы, то мы на них останавливаться не будем. Неугашаемость же
этих рефлексов через очень короткие паузы — явление, неизвестное для пи-
щевых условных рефлексов1. Нам думается, эти факты не согласуются со
1 Этим вопросом мы начали заниматься и на слюноотделительных рефлексах.
В слабый степени это явление как будто замечается й там.
К ФИЗИОЛОГИИ ДВИГАТЕЛЬНЫХ ОБОРОНИТЕЛЬНЫХ УСЛОВНЫХ РЕФЛЕКСОВ 179
взглядами И. С. Беритова, объяснявшего угашение условных рефлексов как
результат истощения, утомления нервных механизмов рефлекса. Что
утомление или истощение этого аппарата может способствовать угашению
и вообще развитию торможения в них, это очень вероятное предположение,
которое подтверждается большим фактическим материалом павловских
лабораторий, а также данными физиологии низших отделов центральной
нервной системы. Факт сравнительно трудного угашения слюноотделитель-
ных и оборонительно-двигательных рефлексов при больших паузах но
сравнению с таковым при меньших паузах, очевидно, частично является
Рис. 7. Собака Баш. Угашение рефлекса на тон С
Опыт № 74 29 XII 1932 г. Через короткие пауз*ы (20 сек.) рефлекс
упорно не угашается до 232-го применения раздражителя (заметно осла-
бев лет И ДДбГ нёСКЛЛькФ раз нулевой эффект , но снова восстанавливается
и продолжается с той же силой). На 211-м применении раздражитель
не вводится. Когда паузы увенчиваются до би сск., рефлекс довольно
быстро исчезает. Обозначения тс же. что на рис. 1
иллюстрацией этого положения. Но если бы все сводилось к утомлению,
как полагает И. С. Беритов, почему же при дальнейшем уменьшении
паузы угашение становилось особенно затруднительны!^ почти невозмож-
ным? Ведь этим создаются особенно хорошие условия для быстрого на-
ступления угашения.
Думается, установленное нами явление служит лишним доказательством
правильности павловской интерпретации угашения условных рефлексов
как проявления торможения.
Почему же все-таки оно так затрудняется в этих случаях? Мы пред-
ставляем себе это следующим образом: каждое действие условного раз-
дражителя вызывает в соответствующих звеньях условнорефлекторной
дуги волну возбуждения, которая исчезает не сразу после прекращения
действия раздражителя, а в виде следов некоторое время продолжает оста-
ваться в Этих Звеньях. На некоторое время в этих элементах создается
как бы состояние центрального возбуждения. Если следующий залп воз-
буждения попадает на свежие следы предшествующего залпа, то он сумми-
руется с ними и обусловливает то, что мы видим в действительности. Мож-
12*
180
Э Л АСРАТЯН
но допустить и другие предположения, но это предположение кажется
нам самым вероятным. Длительность сохранения этого состояния может,
видимо, колебаться под влиянием различных условий, а также индиви-
дуальности собак.
Вслед за сотрудниками лаборатории В. М. Бехтерева И. С. Беритов
утверждает, что при угашении рефлекса через паузы известной длитель-
ности рефлекс этот можно восстановить простым удвоением паузы. На осно-
вании главным образом этого факта он и отождествляет угашение с утом-
лением. Мы Должны сказать, что в действительности не всегда бывает так.
Если рефлекс угашен основательно, если нулевые эффекты получаются не-
сколько раз, то удвоение и даже утроение паузы нс приводит к восста-
новлению рефлекса. Сам И. С. Беритов получал такие же данные, но игно-
рирует их. Кроме того, удлинение паузы " в качестве нового фактора
может оказать такое же влияние на угашенный рефлекс, как любой
новый раздражитель, т. е. может вызвать ориентировочный рефлекс и рас-
тормаживать заторможенный угашением рефлекс. На рис. 7 (опыт №74)
Рис. 8. Собака Баш. После длительного многократного применения безусловного
раздражителя появляются затяжные безусловные рефлексы
хорошо’ видно', как при длительном применении раздражителя собака при-
выкает к определенному ритму: на «пропущенном месте» собака точно отве-
чает ориентировочной двигательной реакцией. В конце концов, то, что утом-
ление оказывает известное благоприятное влияние на ход угашения, еще не
может служить основанием для такой путаницы этих различных процессов.
Мы не можем согласиться с И. С. Беритовым, что безусловные рефлексы
так же быстро угашаются (утомляются, по Беритову), как и условные. Раз-
ница в угашении соответствующих слюноотделительных рефлексов столь
очевшдна, что о ней не стоит говорить. Что же касается оборонительно-
двигательных рефлексов, то здесь, по нашим данным, эта разница тоже от-
четлива. Угашение условного рефлексами видели. Безусловный же рефлекс
при известной оптимальной силе раздражителя нам не удавалось угасить
ни при больших, ни при малых, ни при очень малых паузах. Сотни раз мы
вызывали безусловный рефлекс, и почти все они как по величине, так и по
форме до поразительных подробностей были похожи друг на друга (рис. 3,
опыты № 106 и ПО). Иногда бывало, что несколько рефлексов получаются
необыкновенно длительными; скажем, при 5-секундном применении раз-
дражителей рефлекс длился 30—56 секунд (рис. 8). Это, очевидно, явилось
результатом именно истощения, вследствие чего процесс возбуждения вре-
менно делался особенно инертным в звеньях сложной цепи рефлекса.
Если же безусловный раздражитель брался очень слабый—немного выше
порога с паузой 10—20 секунд, то через 7—8 раз кратковременного при-
менения рефлекс внезапно уменьшался в течение 4—5 применений, затем
опять поднимался до прежнего уровня и оставался таковым долгое время-
до конца длительных опытов. Очевидно, быстрое уменьшение рефлекса
является результатом выключения коркового компонента безусловного
рефлекса, после чего субкортикальный механизм продолжает оказывать
длительное действие. Это вопрос особого порядка; поэтому подробно на нем
останавливаться мы не будем.
К ФИЗИОЛОГИИ ДВИГАТЕЛЬНЫХ ОБОРОНИТЕЛЬНЫХ УСЛОВНЫХ РЕФЛЕКСОВ 181
Заключение
На основании имеющегося в литературе материала, а также накоплен-
ных нами данных мы попытались показать, что обнаруженные и изученные
методом слюноотделительных рефлексов такие кардинальные закономер-
ности физиологии коры, как взаимная индукция (намосте и на расстоянии),
иррадиация и концентрация процессов, угашение, его особенности и сущ-
ность, в полной мере сохраняют силу также для оборони тельно-двигатель-
ных условных рефлексов. Если в опытах И. С. Беритова этого часто не по-
лучалось, то, вероятно, причиной являются некоторые особые приемы его
экспериментирования, возможно, даже индивидуальные особенности^ тех
немногочисленных собак, которые были у него в работе.
Выводы
1г Диференцировочный раздражитель повышает величину’быстро сле-
дующих за ним положительных оборонительно-двигательных рефлексов.
Затем начинаются иррадиация и концентрация торможения, которые иногда
могут длиться десятками минут.
2. Оборонительно-двигательные положительные условные рефлексы
отрицательно отражаются на безусловных оборонительных рефлексах; от-
мечено также обратное явление. Это явление, очевидно, правильнее всего
рассматривать как взаимную индукцию на месте и па расстоянии,
3. Оборонительно-двигательные условные рефлексы при больших пау-
зах (5 минут и больше) угашаются медленнее, чем при малых паузах (1—2
минуты). Но если паузу уменьшить до 10—20 секунд, то угашение рефлек-
сов сильно затрудняется и крайне затягивается.
4. При известной, силе раздражения условный оборонительный рефлекс
нс угашается ни при одной, из названных пауз.
ЛИТЕРАТУРА
Анохин П. К., Взаимодействие клеток условного и безусловного раздражителей
в течение применения последнего, Труды физиологических лабораторий акад.
И. П. Павлова, т. II. 1927.
Асратян Э. А., К вопросу о локализации центральной части рефлекторной дуги
оборонительно-двигательного условного рефлекса. Физиологический журнал
СССР. т. XVII, № 6, 193-4.
Асратян Э. А., Влияние нишевого безусловного рефлекса на соответствующие ус-
ловные рефлексы. Физиологический журнал СССР, т. XVIII, 1934.
Беритов И. С., Индивидуально приобретенная деятельность центральной нерв-
ной системы, Госиздат, Тифлис, 1932.
Бехтерев В. М.. Общие основы рефлексологии, 1926.
Болдырев В. Н.. Условные рефлексы и способность их к усилению и ослаблению,
Харьковский медицинский журнал, т. IV, 1907.
Быков К. М. и др.. Образование условных рефлексов на мочеотделение, Труды
физиологических лаборатории акад. И. П. Павлова, т. IV, 1932.
Виноградов Н. В.. Замаскирование закона силы раздражителей под влиянием
индукции, Труды физиологических лабораторий акад. И. П. Павлова, т. IV, 1932.
В и н д е л ь б а и д О. А., Ф р о л о в Ю. П., Особый вид угасания искусственного
условного рефлекса. Архив биологических наук, т. XXV. 1925.
Г е и к е н И., Обозрение психиатрии, № 2, 1909, стр. 123.
Ерофеева М. А., Электрическое раздражение кожи собак как условный возбу-
дитель работы слюнных желез, дисс., 1912.
Иванов-Смоленский А. Г.. Методика исследования условных рефлексов
у человека, 1933.
Нзраильсон Ж- Г.. О тонической ДиференаировКе сочетагельяо-двигатольного
рефлекса на кожные и мышечные раздражения, дисс., Спб., 1913.
Коган Б. Л., Об иррадиации и концентрации угасателыюго торможения в коре
больших полушарий, дисс., 1914.
Коморский К). М.. иМи лл ер С. М., Условные рефлексы двигательного ана-
лизатора, Тетради физиологической лаборатории акад. И. П. Павлова, т. VI,
в. 2. 3936.
182
Э. А. АСРАТЯН
Конради Г. П., Диференцировка и взаимодействие активных условных рефлек-
сов, связанных с различными безусловными рефлексами, Труды физиологиче-
ских лабораторий акад. И. П. Павлова, т. IV, 1932.
Красногорский Н. И., ОС образовании искусственных условных рефлексов
у детей раннего возраста. Русский врач, № 36, 1907.
Крылов В. Л., О возможности образования условного рефлекса на раздражитель
через кровь, Сборник, посвященный 75-летню акад. И. П. Павлова, 1925.
Кряжев В. Я.. Характер высшей нервной деятельности гипофизэктомированных
животных, Советская неврология, т. 9—10, 1932.
Л е й б с о и Л. Г., О нервной регуляции почечной деятельности, Русский физиоло-
гический журнал, т. IX. 1926; т. X. 1927.
Майоров Ф. П., Специальный функциональный метод устранения стойкого гип-
нотического состояния у собак, Тетради физиологической лаборатории акад.
И. II. Павлова, т. V, 1933.
Павлов И. П., Двадцатилетний опыт объективного изучения высшей нервной
деятельности (поведения) животных, 1932,
Павлов И. П., Лекции о работе больших полушарий головного мозга, 2-е изд.
Медгиз, 1927.
Павлова В. И.. Образуется ли условный рефлекс при предшествовании безуслов-
ного раздражителя индиферентному? Труды физиологических лабораторий акад.
И. П. Павлова, т. V, 1933.
Петрова А. М., Об иррадиации раздражения в коре больших полушарий, Труды
Общества русских врачей, т. 80, 1913.
Подкопаев Н. А., О моменте начала иррадиации тормозного процесса. Сборник,
посвященный 75-летиго акад. И. П. Павлова, 1925.
Подкопаев Н. А., Материалы к вопросу; ЧТО Делается с клетками нндиферент-
иого и условного раздражителей во время действия безусловного раздражителя,
Труды физиологических лабораторий акад. И. П. Павлова, т. II, 1928.
Подкопаев Н. А. и С а а т ч и я и Р. Л., Условный рефлекс на иммунитет,
Журнал экспериментальной биологии и медицины, т. XI, 1929.
Протопопов В. П., О сочетательной двигательной реакции на звуковые раз-
дражения, дисс., Спб., 1909.
Райт Р. fl., Влияние безусловного рефлекса на условный рефлекс, Труды физиоло-
гических лабораторий акад. И. Г1. Павлова, т. II, 1928.
Соловейчик Д. И., Состояние возбудимости корковых клеток во время дейст-
вия безусловного раздражителя, Труды физиологических лабораторий акад.
И. П. Павлова, т. 1—11, 1928.
Ухтомский А. А., О зависимости кортикальных двигательных эффектов от по-
бочных центральных влияний, Работы физиологической лаборатории Ленин-
градского университета, т. IV—V, 1911.
Федоров Л. И.. Нарушение равновесия между процессами возбуждения и тормо-
жения у возбудимого типа собаки от повторных применений диференцировки
на частоту кожнб-механического раздражителя и восстановление равновесия
бромом. Труды физиологических лабораторий акад. И. П. Павлова, т. III, 1928.
Фурса ков Д. С., О соотношении процессов, возбуждения и торможения, Рус-
ский физиологический журнал, т. V, 1922.
Фу реи ко в Д. С., Условные рефлексы у собак без одного полушария, Архив
биологических наук, т. XXV, 1925.
Ц и т о в и ч И. С., О так называемых возомоторных психорефлексах, Русский физио-
логический журнал, т. 1, в. 3—4, 1918.
Шевалев Е. А., О процессе диференцировки сочетательных двигательных реф-
лексов на тактильное раздражение и о сегментарном типе распространенна
возбуждения, дисс., 1913.
Ш н и р м а н А. Л., К вопросу о возможности воспитания сочетательно-двигательных
рефлексов путем ретроси'гнальных сочетаний, Новое в рефлексологии и физио-
логни нервной системы, I, 1925.
Bubnoff u. Hcidcnhain, Pfltiger's Archiv, Bd. 26, 1881.
Cason, Journ. exp. psycho!.. V. 1922.
Freeman. Journ. of genet, psychol.. III.
Goltz, Pfluger's Archiv, Bd. 8, S. 460, 1874.
Rademaker, Das Stehen, 1931.
Sherrington, The integrative action of the OervoUS System, 1906.
Sherrington, At all reflex activity of the spinal cord, 1932.
НЕКОТОРЫЕ НАБЛЮДЕНИЯ НАД СОБАКАМИ, ЛИШЕННЫМИ
КОРЫ БОЛЬШОГО МОЗГА »
редставленную Гольцем (Goltz) картину жизненных отправле
ний собак, лишенных коры большого мозга, до сих пор мож-
но считать классической в физиологии поточности наблюдения,
подробности описания и по глубине анализа наблюдавшихся
явлений, хотя она пописана более 45 лет назад. Дальнейшие
исследователи этого вопроса [Ротман (Rothmann), Г. П. Зеле-
ный, Карплюс и Крейдль (Karplus и Kreidl), Тренделеибург (Trendelenburg),
Радемакер (Rademaker) и др. ] своими наблюдениями и исследованиями под-
твердили главным образом да иные Гольца. Пополнения и добавления, сде-
ланные некоторыми из них, касаются лишь деталей и больше уточняют неко-
торые стороны жизненных отправлений высших животных, лишенных коры
большого мозга. В этом плане следует особо отметить интересные работы
Г. П. Зеленого и сотрудников, посвященные изучению возможности выработ-
ки новых навыков у лишенных коры мозга собак более тонким и точным
методом, чем это сделал Гольц,—методом условных рефлексов И. П. Павлова.
Г. П. Зеленый показал, ЧТоу некоторых таких собак сохраняются половые
рефлексы и наклонность к игре. Заслуживают упоминания также работы Ра-
демакера, установившие что «магнитная реакция» (Magnetreaktion), «готов-
ность к стоянию» (Stehbercitschaft)1 2 и способность к коррекции ряда особо
ненормальных положений конечностей имеют условнорефлекторное про-
исхождение и обусловлены наличием коры большого мозга у собак.
За последние два года в процессе изучения проблемы пластичности
церв.ной системы я имел возможность сделать ряд наблюдений над доволь-
но большим числом собак с удаленной корой большого мозга (см. рису-
нок). Эти наблюдения, как мне кажется, имеют некоторую ценность и
также могут послужить в качестве дополнения и детализации к описан-
ной Гольцем картине жизненных отправлений лишенных коры собак.
Кратким описанием этих наблюдений я займусь в дальнейшем изложении.
По при меру Гольца декортикацию большого мозга я производил в два
приема: сначала удалял кору одного полушария, азатем, некоторое время
спустя после первой операции, — кору второго полушария. Благодаря
некоторым методическим нововведениям, направленным к уменьшению
кровотечения как в процессе операции, так и после нее, удалось достиг-
нуть не только большого процента выживаемости собак после удаления
коры обоих полушарий, но и сравнительно быстрой поправки собак после
1 Физиологический журнал СССР, т. XXIV, в. 1—2, 1938.
2 Под MagnctrcaKt ion Радемакер подразумевает способность животного разогнуть
согнутую конечность, когда к ее пальцам прикасаются каким-нибудь предметом, и
выпрямленной конечностью следовать за движением прикасающегося предмета;
под Siehbereitschafi — способность животного упираться в подставку конечностями
н выпрямлять нх, когда при висячем положении какая-нибудь часть его тела (хвсст
или морда) приводится в соприкосновение с этой подставкой или оно видит подставку.
184
Э А. АСРАТЯН
операции и значительного укорочения периода, названного Гольцем пе-
риодом побочных явлений (Nebencrscheinnngen). Так, почти все оперирован-
ные мной собаки на второй же день после удаления коры второго полу-
шария самостоятельно лакают молоко или воду, некоторые на второй, а
многие на третий-четвертый день стоят, делают координированные шаги.
Я проводил наблюдение над несколькими лишенными коры собаками
(см. таблицу).
С Кличка собаки Приблизи- тельный возраст в годах Промежуток между пер- вой и второй операцией в днях Продолжи-» тельность жизни после второй опе- рации п днях Примечание
1 Волчок 1J4-2 № 25 Погибла от доба- вочной операции
2 Серенада 2-3 178 107 Погибла от плохо- го ухода и исто- щен ня
3 Снежинка 1-1% 30 70 Погибла от воспа- ления легких
4 С и ипатия 1-1/2 53 250 То же
Б Пушок 1И-2 57 237 Погибла от какой- то нывыяснен- ной эпидемии
6 Красавина 4-5 77 117 То же
7 Аддис 4—5 27 37 Погибла внезапно
8 Плош 1-1 Vi 30 17 То же
9 Тузик 4-5 68 13 Погибла от наг- ноения раны
10 Шарик 2-3 38 211 Погибла от пло- хого ухода, ка- хексии
11 Усик 1 128 8 Погибла от вто* рниного крово* течения лОСлб удара головы
12 Беженка 5-6 51 Собака еще жива
13 Тахиц 7-8 36 То же
14 Белая 2—3 52 26 Погибла от про- студы
15 Радиус 2-3 37 Собака еще жива
16 Орлик 8- 9 101 — То же
Мое внимание прежде всего привлекло нарушение регуляции сердечно-
сосудистой и дыхательной систем, наблюдавшееся у собак в первые 7—13
дней после удаления коры второго полушария. В общих чертах картина
этих нарушений заключается в следующем.
Стоит этим собакам в течение 1—2 минут сделать несколько шагов по
комнате или напрячь мышцы, чтобы у них началась резко выраженная и
длительная одышка, т. е. резкое учащение и усиление деятельности сердца,
резкое учащение дыхательной ритмики, сопровождающееся высовыванием
языка, широким раскрыванием пасти и усилением саливации. При этом
собаки стоят с широко расставленными конечностями и перестают дви-
гаться. Эти сдвиги в точности совпадают со сдвигами в сердечно-сосудистой
и дыхательной системах нормальных собак после длительного бега или
после выполнения какой-нибудь другой тяжелой мышечной работы, а так-
же при перегревании.
То, что такого рода резкий сдвиг в деятельности сердечно-сосудистой
и дыхательной систем у лишенных коры собак в первые дни после операции
возникает при совершенно ничтожной мышечной работе^ а также то, что
эти сдвиги обладают большой длительностью и инерцией (иногда они длятся
20—25 минут и больше, если даже собаки уложены в мягкую люльку После
НАБЛЮДЕНИЯ НАД СОБАКАМИ, ЛИШЕННЫМИ КОРЫ БОЛЬШОГО МОЗГА
185
мышечных усилий), заставляет думать, что в регуляции деятельности
первичных центральных нервных аппаратов сердечно-сосудистой и дыха-
тельной систем кора большого мозга принимает Значительное участие,
точно так же как она принимает участие в регуляции деятельности сомати-
ческих функций организма. Говоря иначе, в результате декортикации раз-
вивается какая-то своеобразная временная астения первичных центральных
рефлекторных аппаратов как соматических, так и вегетативных систем.
Характерно, что чем короче промежуток между операциями удаления
полушарий, тем резче и ярче выступает это явление.
Препараты головного мозга собаки после удаления коры
больших полушарий
Мне кажется, что на нарушения вегетативных функции после удаления
коры большого мозга указывают также следующие наблюдения.
У большинства лишенных коры собак в течение всей их жизни наблю-
дается значительное нарушение теплорегуляции, а также некоторое из-
менение деятельности желудочно-кишечного тракта (запоры). У них от-
мечается также уменьшение силы сопротивления тканей к патогенным
факторам и ослабление силы преодоления патологических изменений (на-
ружные раны, инфекции и т. п.). Судя по всему, у этих собак изменяется
также общий обмен веществ; несмотря на усиленное кормление, все эти
собаки склонны к исхуданию. У многих из них обнаруживаются более
или менее стойкие трофические нарушения покровных тканей тела.
Осуществляется ли эта корковая регуляция функции нижележащих
отделов центральной нервной системы только путем условнорсфлекторной
деятельности хоры, что ярко выступает при образовании условных рефлек-
сов на деятельность органов с соматической и вегетативном иннервацией,
или же из специальных, точно локализованных корковых центров раздель-
но для вегетативной и соматической систем (Ландуа (Landois), В. М. Бех-
терев, Н. А. Миславский, Фултон (Fulton) и др. 1, или же она осуществ-
ляется также другими «механизмами», в частности, путем общетрофичсско-
го влияния коры на нижележащие вегетативные и соматические аппараты
(Э. Л. Асратян),— на все эти вопросы пока' не получено окончательного
ответа. Мне представляется более правдоподобным последнее предположе-
ние. Еслибы упомянутое стеническое влияние коры осуществлялось только
186
Э. А. АСРАТЯН
или преимущественно условнорефлекторным воздействием или под влия-
нием высших, специальных, точно и точечно локализованных центров, то
законно было бы ожидать таких сдвигов и после частичных экстирпаций,
чего на самом деле не бывает. Если кора осуществляет эту регуляцию не
отдельными своими составными частями, а как целостный, единый орган,
то законно предполагать, что в указанных случаях речь идет главным об-
разом об общетрофнческом, в понимании И. П. Павлова, или стеническом,
в понимании Лючиани (Luciani), влиянии коры на нижележащие соматиче-
ские и вегетативные нервные аппараты. Считаю не лишним упомянуть,
что проведенные мной несколько лет назад специальные исследования
этого вопроса позволили мне тогда еще придти к аналогичному взгляду
на общетрофическое влияние коры на центры вегетативной нервной системы.
Наблюдения над лишенными коры собаками позволили мне также дать
характеристику некоторых основных индивидуальных особенностей этих
животных до и после операции. Факты, приводимые другими исследовате-
лями, говорят о том, что основные вндреые биологические особенности
животных сохраняются и после экстирпации коры обоих полушарий: кошки
остаются кошками, собаки — собаками.
Мои наблюдения над перечисленными выше собаками с удаленной ко-
рой, проводившиеся в течение сравнительно короткого времени (что дало
возможность сделать сравнение), позволяют утверждать, что после экстир-
пации коры большого мозга у собак сохраняются не только видовые
биологические особенности, но в грубых чертах и некоторыеосновые инди-
видуальные их особенности. Мои наблюдения коснулись следующих осо-
бенностей: прожорливости собак, общей подвижности и характера общей
реакции на сторонние, преимущественно тактильные раздражения.
Прежде всего я должен сказать, что мои данные относительно самостоя-
тельного захвата, разжевывания и проглатывания пищи этими собаками,
после того как пища приводится в соприкосновение с их ртом, целиком
совпадают с данными Гольца. Я говорю об этом потому, что в более позд-
них работах по данному вопросу иногда можно встретить указания,
видимо, без достаточных оснований, будто собаки, лишенные коры, не-
способны самостоятельно брать пищу и что они проглатывают ее только
тогда, когда нм кладут пищу в рот. Однако здесь имеются индвидуальные
вариации как у нормальных, так и у оперированных собак.
Мои наблюдения позволяют сказать, что разница в общем характере
отношения собаки к пище в смысле жадности, активности захвата пищи
сохраняется и после экстирпации большого мозга. Собаки, которые не
особенно жадно ели в норме, после удаления большого мозга относились
к пище так же. Собаки же, которые в норме отличались большой жадностью
к пище, оставались жадными и после операции. Само собой разумеется,
что как в первом, так и во втором случае общая активность всех других
функций организма, и в частности реакция собак на пищу, значительно
снижается. Здесь речь идет об относительной разнице между различными
собаками, которая и сохраняется. Правда, изменения общей упитанности,
состояние здоровья и другие моменты влияют на силу реакции каждой
собаки на пищу, что может иногда запутать картину, но при приблизитель-
но равных условиях указанная индивидуальная разница сохраняет свою
силу. Из моих собак как в норме, так и после удаления большого мозга от-
личались особой жадностью Волчок, Симпатия, Аддис, Плош, Жан, Усик;
собаки Красавица, Тузик, Белая, Снежинка нс проявляли резко выражен-
ной активности к пище; остальные занимали промежуточное положение.
Представляют интерес также наблюдения над общей подвижностью
собак до и после декортикации. Я не хочу здесь заниматься выяснением
причин, почему одни собаки в норме бывают более подвижными, другие
менее ПОДвИжнымИ, Почему оДИИ Ссбаки бывают Злыми, агрессивными,
НАБЛЮДЕНИЯ НАД СОБАКАМИ, ЛИШЕННЫМИ ЦОРЫ БОЛЬШОГО МОЗГА
187
другие — сравнительно спокойными, ласковыми. Эти индивидуальные осо-
бенности, несомненно, производные очень сложного характера. Они, бес*
спорно, обусловлены как моментами врожденного характера (типовые осо-
бенности нервной системы), так и моментами приобретенного порядка
(жизненный опыт, воспитание). Несомненно, что играет роль также возраст
собаки, состояние ее здоровья, порода и ряд других моментов. Я хотел
только подчеркнуть тот факт, что существовавшая в норме разница в об-
щей подвижности собак в общих чертах сохраняется н посте декортикации,
хотя в целом степень подвижности всех собак после указанной операции
значительно снижается. Из изучавшихся мной собак до и после декорти-
кации особой подвижностью отличались Симпатия, Пушок, Уснк, Шар,
Аддис, а малой подвижностью -- Красавица, Тузик, Жан, Серенада; ос-
тальные занимали промежуточные места.
Г. П. Зеленым описал лишенную коры собаку, которая даже играла.
Он ставит эго в связь с возрастом собаки (она была лишена коры одного
полушария большого мозга, когда ей было 6 месяцев отроду, а второго
полушария, когда ей было И месяцев). Я нисколько не сомневаюсь, что
возраст, состояние здоровья, возможно, даже порода собаки имеют зна-
чение для общей Подвижности. На основании накопленного До сих пор
материала я не имею возможности входить в подробности этого вопроса,
но у меня возникают сомнения в возможности ставить общую подвиж-
ность собак в зависимость только от их возраста.
Считаю нужным сказать несколько слов о характере и силе реакции
собак на тактильные раздражения до и после удаления большого мозга.
Критерием оценки в этих случаях, правда, являлись грубые наблюдения
за общей реакцией собак, в особенности на упорные тактильные раздра-
жения; но даже при этом удалось сделать ряд точных наблюдений. Пушок,
Тузик, в значительной мере Симпатия и Красавица в норме были довольно
злыми собаками. Они не очень любили, когда их щекотали или слегка при-
щемляли какое-нибудь место кожи. Указанные черты в основном сохрани-
лись и после удаления коры обоих больших полушарий мозга. Эти собаки пос-
ле операции лаяли, ворчали, скалили зубы и даже делали попытки кусаться,
когда их брали на руки, щекотали или щипали. Остальные собаки не отли-
чались в норме особой реактивностью в этом отношении; таковыми они
остались и после операции.
На основании сделанных наблюдений я вовсе не намерен утверждать,
что указанные индивидуальные особенности целиком обусловливаются
подкорковыми нервными образованиями и что кора большого мозга не
принимает в них никакого участия или не в состоянии их переделывать.
Больше того, не исключена возможность, что в их формировании основную
роль играет именно кора, а затем только подкорковые узлы. Не исключена,
однако, также возможность, что они являются функциональными особен-
ностями этих узлов sui generis. Все это пока предположения, для уточне*
ния и конкретизации которых необходимы дальнейшие исследования.
Третье наблюдение над оперированными собаками касается характера
наркотизации. Дело в том, что в связи с моей работой по пластичности
нервной системы я некоторых собак с удаленной корой подверг одно- и
двукратной операции иного характера (Волчок, Симпатия, Пушок, Уснк).
Морфин в 1 и 2% растворе вызывал у них рвотные движения, одышку
и общее ослабление организма. При дальнейшей наркотизации эфиро-
хлороформной смесью у всех собак, хотя и в более слабо выраженном, чем
у нормальных, но в совершенно четком виде выступает фаза возбуждения,
которая дальше сменяется фазой глубокого наркоза.
Я далек от мысли на основании-сделанных наблюдений оспаривать
правильность теории Шмидеберга (Schmiedeberg) и др., объясняющей
фазу возбуждения при наркотизации как последствие ослабления задер-
188
Э А. АСРАТЯН
живающих влияний с высших корковых центров на нижележащие центры.
Я думаю, однако, что приведенные наблюдения дают основание высказать
По крайней мере два предположения даже в пределах названной теории
наркоза: 1) или фаза возбуждения нормальных животных является соче-
танным эффектом одновременного угнетения высших центров и возбужде-
ния нижележащих нервных аппаратов, 2) или же после экстирпации боль-
шого мозга происходят такие сдвиги в центральной нервной системе, что
наркоз сначала возбуждает, затем уже угнетает оставшиеся части цен-
тральной нервной системы.
В. заключение следует сказать, что лишенные коры собаки становятся
очень уязвимыми в отношении всякого рода вредоносных влияний (холод,
инфекция, недоброкачественная пища и др.) и что сопротивляемость их
организма к этим влияниям значительно понижается. Изредка на коже
у них наблюдаются дистрофические явления. Нужно еще добавить, что
как эти изъязвления, так и изъязвления механического порядка требуют
для заживления значительно больше времени, чем у нормальных животных.
Воспалительная реакция при этом бывает слабее выражена, чем в норме, а
иногда в результате ничтожных ран развивается общая инфекция организма.
Хотелось бы указать еще на одно замечательное явление, впервые на-
блюдавшееся Гольцем. Речь идет об утолщении твердой мозговой оболочки
после операции. После удаления коры полушарий я твердую мозговую
оболочку обычно сшиваю. Выясняется, что через несколько месяцев эта
тонкая оболочка сильно утолщается, достигая иногда 9—10 мм, приоб-
ретает при этом свойство хрящевой ткани и превращается в хорошую за-
щитную покрышку для нижележащих мозговых образований. Характерно,
что при удалении одного полушария часто наблюдается заметное утолще-
ние твердой мозговой оболочки и на здоровой стороне, что обнаруживается
при удалении второго полушария.
В общем характеристика изучавшихся мной собак с удаленными полу-
шариями целиком соответствует описанию, данному Гольцем около полу-
века назад. Это, в частности, относится к балансу сна и бодрствования
у этих собак. У трех собак Гольца отмечены отклонения по сравнению
с нормальными собаками, выразившиеся в том, что смена фаз бодрствования
и сна у них участилась; оперированные собак и больше нормальных бодрство-
вали, причем в состоянии бодрствования много ходили. Следует отметить,
что Гольц исхудание оперированных собак приписывает даже последнему
моменту, что едва ли можно считать правильным. Собаки же Г. П. Зеленого
ббльшую часть времени проводили во сне. Пробуждались они от голода
и для отправления своих естественных потребностей. Наблюдавшиеся
мной собаки в этом отношении походили на собак Гольца м.
ЛИТЕРАТУРА
Асратян Э. А.» К физиологии кортикальных и мозжечковых центров автономной
нервной системы, Труды V Кавказского съезда физиологов, 1935.
Зеленый Г. П., Собака без полушарий большого мозга, Труды Общества рус-
ских врачей, 1912.
Павлов И. П., Лекции о работе больших полушарий головного мозга, 1927.
Goltz F.a Pfluger’s Archiv, 51, 1892.
Karplus u. Kretdl, Zbl. f. Physiol., 25, 1912.
Ro th matin M.. Zbl. f. Physiol., 22. 1908.
Trendelenburg W., Ztschr. f. Biol., 65, 1914.
Rademaker, Das Stehen. Berlin, 1931.
1 С тех пор как данная работа была опубликована, число наших собак, лишенных
коры, резко увеличилось, достигнув 60, к тому же значительно увеличилась длитель-
ность их жизни. Описанные в данной статье наблюдения полностью подтвердились
на этом большом новом материале. —Э. А.
ПЕРЕКЛЮЧЕНИЕ В УСЛОВНОРЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ 1
дной из наиболее характерных особенностей отечественном фи-
зиологии является подлинно эволюционный подход к изучае-
мым вопросам, широкий биологический подход к объяснению
наблюдаемых явлений. Эта черта нашла наиболее яркое выра-
жение в научном творчестве нашего гениального учйтеЛЯ
И. П. Павлова.
Одним из показателей такого широкого эволюционно-биологического
подхода к изучаемым явлениям можно считать признание, наряду со спе-
цифическими, качественно особенными формами деятельности тех или
иных систем и органов, достигших высокого уровня диференциации и спе-
циализации в процессе эволюционного развития, также общности и род-
ства основных закономерностей в деятельности образований, стоящих на
различных уровнях эволюционной лестницы.
Стремление выявить «биологические корни» закономерностей деятель-
ности высших субстратов в деятельности субстратов, более низко органи-
зованных, признание специфического, высшего и поиски корней этого
высшего в низшемн общем являются, как мне кажется, одним из показа-
телей как раз эволюционного понимания сущности изучаемых явлений,
эволюционного толкования наблюдаемых фактов корифеями нашей науки—
И. М. Сеченовым, Н. Е. Введенским и особенно И. П. Павловым.
Именно И. П. Павлов, наш гениальный учитель, в течение всей своей
жизни, всем своей творческой деятельности и в особенности в период изу-
чения самых высших и сложных биологических явлений — высшей нервной
деятельности — неуклонно руководствовался этим принципом. Изучая спе-
цифические особенности, качественно новое и особенное в деятельности
коры больших полушарий, И. П. Павлов никогда не забывал, что законо-
мерности работы этого высшего органа глубоко связаны с более простыми
явлениями, с закономерностью деятельности низших этажей центральной
нервной системы и даже простых нервных образований.
С точки зрения учения И. П. Павлова, деятельность высших отделов
центральной нервной системы не только находится в неразрывной связи
с деятельностью более простых по структуре нижележащих частей цен-
тральной нервной системы и нервной системы вообще, но и имеет с ними
общие генетические корни. Многие из установленных И. П. Павловым за-
кономерностей работы больших полушарий головного мозга даже по своей
терминологической характеристике указывают на такое родство законо-
мерностей, на такую связь физиологии высших и физиологии низших от-
делов центральной нервной системы. Достаточно указать на то, как Иван
1 Обработанная стенограмма доклада на торжественном заседании Московского
общества физиологов, биохимиков и фармакологов 27. IX 3950 г», посвященном 101-й
годовщине со дня рождения И. П. Павлова, Журнал высшей нервной деятельности,
Г. III, в. I, 1951.
190
Э. А. АСРАТЯН
Петрович понимал механизм самого основного в деятельности высших
отделов, т. е. механизм образования условного рефлекса; как он понимал
возникновение, течение и многообразие форм взаимоотношения двух кар-
динальных корковых процессов — возбуждения и торможения: их ирра-
диацию, концентрацию, суммацию, взаимоиндукционные отношения и т. п.;
как он понимал и трактовал физиологическую природу гипнотических фаз,
сна и гипноза, типа и характера; как он представлял себе эволюцию охра-
нительного торможения ит. п.
Указывая постоянно на «естественную общность» закономерностей
деятельности почти всех частей нервной системы, он не забывал и другого:
что в коре больших полушарий эти закономерности достигают высшего
уровня развития, приобретают ряд специфических и качественно новых
черт. Он особенно подчеркивал это специфическое, наряду с общим, именно
в деятельности больших полушарии.
Мне кажется, что сейчас, .когда перед всеми советскими физиологами
и прежде всего перед учениками и последователями И. П. Павлова ставится
задача — многостороннее, интенсивнее, глубже и шире развивать оста-
вленное им драгоценное научное наследство, особенно учение о высшей
нервной деятельности, мы не должны забывать этот принцип, эту харак-
терную черту' его научного творчества— изучение закономерностей выс-
шей нервной деятельности в аспекте широкого эволюционно-биологиче-
ского подхода к ним. Руководствуясь этим принципом, мы должны, в част-
ности, в нашей исследовательской работе внимательно проследить историю
развития основных закономерностей высшей нервной деятельности, выявить
их «биологические корни». Более того, в целях выявления новых сторон
и специфических закономерностей деятельности коры больших полушарий
Мозга, равно как и в целях углубления наших знаний о физиологической
сущности уже известных сторон и закономерностей корковой деятельности,
мы должны смелее обращаться к общим закономерностям деятельности
нервной системы, к Истокам ЭТОЙ деятельности.
С этой точки зрения определенный интерес представляют некоторые
фактические данные, полученные мной и моими сотрудниками, правда,
давно, еще в ленинградский период нашей работы. Хотя эти данные не-
однократно нами докладывались и опубликовывались, тем не менее я осме-
ливаюсь о них доложить вновь, так как имеются все основания считать,
что они до сих пор остаются недостаточно известными не только широким
кругам отечественных физиологов, но и некоторым физиологам, которые
специально занимаются изучением высшей нервной деятельности. Иначе
трудно понять, почему они приписывают эти данные себе или же на них
должным образом не’ ссылаются (как поступали, например, некоторые
сотрудники Л. А. Орбели и П. К. Купалова, значительно позже полечив-
шие и опубликовавшие данные, аналогичные или подобные нашим).
О чем конкретно идет речь?
Давно, почти в середине прошлого столетия, гениальный основопо-
ложник отечественной физиологии И. М. Сеченов доказал на основе фактов
важное положение в физиологии, а именно: что характер рефлекторной
деятельности такого простого аппарата, как спинной мозг, вовсе не так
«машинообразен», не так статичен, не так сильно зависит от чисто анатоми-
ческих особенностей, как было принято думать в те времена. Всем хорошо
известно, что И. М. Сеченов первый показал вариабильность рефлектор-
ной деятельности под влиянием стимуляции определенных частей головно-
го мозга. Но он же впервые установил другрй факт, к сожалению, недо-
статочно известный многим физиологам, а именно, что исходное положение
конечности спинальной лягушки очень часто предопределяет и характер
рефлекторного ответа, что характер рефлекса обусловливается не только
ПЕРЕКЛЮЧЕНИЕ В УСЛОВНОРЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
191
стимулом и местом стимуляции, не только силой и характером раздра-
жителя и рецептивным полем, но и исходным положением конечности.
Например, если конечность согнута, раздражение определенного участка
кожной поверхности вызывает разгибание конечности, а ест и конечность
разогнута, то же самое раздражение того же места вызывает противо-
положный ответ—сгибание конечности. Таким образом, как показал
И. М. Сеченов, исходное положение конечности через соответствующие
чувствительные нервы отражается на функциональном состоянии спиналь-
ного рефлекторного аппарата, создает особые интрацептральные взаимоот-
ношения в спинном мозгу и тем самым до известной степени предопреде-
ляет и характер будущей реакции.
И. М. Сеченов показал, что не все зависит от характера и силы раз-
дражителя, от места стимуляции, от статических, анатомических особен-
ностей координационного аппарата, а многое в рефлекторной деятельности
зависит от состояния центральной нервной системы, от ее функциональных
особенностей, что спинальные рефлексы весьма сариабильны.
Это явление, впервые наблюдавшееся И. М. Сеченовым, было вскоре
подтверждено западноевропейскими исследователями Сандерс-Эцном, Голь-
цем — на лягушках же, Икскюлем и Иорданом — на беспозвоночных,
Шеррингтоном — на млекопитающих. Но особенно много сделал в изу-
чении этого сеченовского феномена — важного явления, открытие кото-
рого принадлежит нашей отечественной науке, — голландский физиолог
Магнус. Он экспериментально показал на млекопитающих животных, что
это явление обусловливается тоническими проприоцептивными импульсами,
исходящими из мышц конечностей, и что под влиянием потока проприо-
цептивных импульсов, исходящих из сгибателей и разгибателей при соот-
ветствующих положениях конечностей, распределяются в спинном мозгу
пути р«|>лексов (один закрываются, другие открываются), происходит
их «переключение», создается своеобразный тонический исходный функ-
циональный фон, определяется рабочая Го-товность. Если при этом проис-
ходит раздражение конечности, если аппарат будет пущен в ход, если будут
пусковые импульсы, то эти импульсы пойдут по путям, заранее предуготов-
ленным упомянутыми тоническими проприоцептивными импульсами.
Таким образом, впервые установленный И. М, Сеченовым важный
факт нашел свое дальнейшее подтверждение и развитие в работах его за-
рубежных последователей, в частности, в работах Магнуса. Но дело не
ограничилось этим. Сеченовское положение о том, что исходное состояние
органа, аппарата оказывает большое влияние на характер его ответных
реакций, развивалось, к тому же преимущественно в пашей отечественной
физиологии, также по иным линиям. Изложение этих данных отвлекло бы
меня от основной цели. Ограничусь ссылкой на классические исследования
славной ленинградской университетской физиологической школы Введен-
ского— Ухтомского, которые особенно акцентировали значение исходного
состояния субстрата, в частности, центральной нервной системы, для от-
ветной реакции, а также значение факторов, способных изменять исходное
состояние субстрата и таким путем вызывать значительные изменения
в его деятельности.
Возникает вопрос: если в низшей нервной деятельности или в деятель-
ности низших нервных образований исходное состояние играет такую важ-
ную роль, различные факторы могут изменять состояние субстрата и тем
изменять его деятельность, в частности, если проприорецепторы могут
переключать спинальные пути и предопределять характер рефлекторной
деятельности спинного мозга, создавать известную вариабильность даже
в спинальной рефлекторной деятельностй, — то не имеет ли место эта же
закономерность в высшей нервной деятельности, характерной особенностью
которой как раз и является динамичность, вариабильность, изменчивость?
192
Э. А. АСРАТЯН
Решением этого вопроса занимались я и мои сотрудники по Институту
мозга им. В. М. Бехтерева в Ленинграде в течение ряда довоенных лет.
Мои сотрудники Ф. М. Шитов, В. В. Яковлева, Я. М. Нрессмаи и В. А. За-
мятина получили ряд новых и интересных фактических данных, которые
я позволю себе привести в кратком изложении.
Но раньше несколько предварительных замечаний.
Конечно, исходя из теоретических предпосылок, можно было бы с боль-
шой долей вероятности предположить, что упомянутая выше закономер-
ность переключения путей, присущая низшим отделам центральной нервной
системы, должна быть свойственна также и высшим отделам центральной
нервной системы, в частности, коре больших полушарий.
Более того, среди богатейших фактических данных по физиологии
условных рефлексов, накопленных И. П. Павловым и его учениками на
протяжении 35 лет, имеются отдельные данные, которые косвенно указы-
вают на наличие такой закономерности в деятельности коры больших полу-
шарий. В частности, одно время в лабораториях И. II. Павлова особенно
интенсивно практиковался такой прием экспериментирования: пищевой
условный раздражитель переделывали на кислотный или наоборот, соот-
ветственно изменяя его безусловнорефлекторное подкрепление.
В этих опытах наблюдалось, что, в зависимости от некоторых обстоя-
тельств, такой условный раздражитель может стать сигналом то одного
вида деятельности, то другого, т. е. в течение определенного промежутка
времени он сохраняет оба сигнальных значения. Особенно ярко это вы-
ступало в опытах С. С. Фридемана.
Далее, каждому работающему по условным рефлексам известно, что
ситуация лаборатории, обстановка эксперимента сильно влияют на харак-
тер условнорефлекторнон деятельности животного. Есть даже такой тер-
мин: «обстановочный» рефлекс.
Итак, до наших исследований этого вопроса были известны отдельные
данные, которые косвенно указывали на то, что закономерность переклю-
чения присуща и деятельности высшнх этажей центральной нервной си-
стемы, в частности, коры больших полушарий. Но для такого заключения
недоставало еще прямых фактических данных, полученных в специальных
исследованиях. Более того, вопрос этот даже не был включен в повестку
текущей экспериментальной и теоретической работы по физиологии высшей
нервной деятельности в лабораториях И. II. Павлова и в других лабора-
ториях страны.
Вот почему необходимость специального изучения этой важной за-
кономерности нам представлялась совершенно очевидной.
Первоначальный вариант опыта сводился к следующему. Один из экспе-
риментаторов (Ф. М. Шитов) по утрам ставил опыты такого рода: метроном
120 ударов в 1 минуту’ он сделал положительным пищевым условным раз-
дражителем, а метроном60 ударов в 1 минуту—диференцировочным раздра-
жителем. После образования и закрепления этих рефлексов в той же ка-
мере, на тех же собаках, в тот же день, но позднее, ставились опыты другим
экспериментатором (В. В. Яковлевой). В последних опытах метроном
120 ударов в 1 минуту сочетался с электрическим раздражением одной из
задних лап собаки. Через несколько дней был выработан электрооборони-
тельнын условный рефлекс на этот раздражитель в вечерних опытах при
сохранении за ним значения положительного пищевого условного раздра-
жителя в утренних опытах. Интересно отметить, что после выработки
электрооборонительного условного рефлекса на M1So диференцировка
к нему, т. е. на Мвв, была налицо в первом же применении. Впоследствии
ряд других звуковых и новых раздражителей таким же путем был пре-
вращен Ф. М. Шитовым в условные пищевые раздражители, В. В. Яковле-
вой — в условные электрооборонительные раздражители.
ПЕРЕКЛЮЧЕНИЕ В УСЛОВНОРЕФЛЕКТЭРНОП ДЕЯТЕЛЬНОСТИ
193
Таким образом, эти опыты показали, что собака на один и тот же раз-
дражитель, в тон же самой камере, но у разных экспериментаторов и
в разное время дня отвечает то пищевой, то оборонительной условной
реакцией: в утреннем опыте на Mi4o она отвечала движением к кормушке,
обильным слюноотделением и всей гаммой пищевой реакции; тот же самый
М)20 У той же собаки вечером вызывал оборонительную реакцию— визг,
лай, отдергивание лапы и пр.
Итак, оказалось возможным один и тот же раздражитель, в зависи-
мости от ситуации, от обстоятельств, делать условным сигналом, вызы-
вающим то один вид деятельности организма, то другой. В этих экспери-
ментах, как показали специальные опыты, «переключателями» были экспе-
риментаторы.
Когда В. В. Яковлева, которая работала с электроюборонигельными
условными рефлексами, начала ставить опыты по утрам, а Ф, М. Шитов —
по вечерам, оказалось» что перемена времени не играет существенной роли.
Независимо от времени Дня и последовательности постановки опытов,
когда ставил опыты Ф. М. Шнтов, М121) имел пищевое значение, а когда
опыты ставила В. В. Яковлева, М,20 имел электрооборонительное значение.
Более того, если в каждом опыте необходимые для обоих опытов прибор-
чики одновременно приклеивались на различных частях тела собаки, это
также не отражалось на результатах. Но когда оба экспериментатора
одновременно присутствовали на опытах либо утром, либо вечером, это
создавало путаницу: собака реагировала наМ12а то как на пищевой сигнал,
то как на сигнал электрооборонительный.
Таким образом, при этой комбинации опытов выяснилось, что основным
переключателем было не время дня, не последовательность опытов и не те
специфические приборчики, которые в одном случае прикрепляются к ла-
пам, а в Другом Случае к щеке, а личность экспериментатора.
Но, как нами было установлено позднее, если изменять характер экспе-
риментирования, можно доказать, что таким переключателем может быть
время. Если опыты ставит один и тот же экспериментатор на одной и той же
собаке, в одной и той же камере, он может один и тот же раздражитель
утром сделать пищевым, а вечером оборонительным условным сигналом.
Роль переключателя здесь играет время дня.
Можно сделать таким переключателем к камеру. В этих целях одним
и тем же экспериментатором раздражитель в одной камере сочетается
с пищевой деятельностью, а в другой — с электрооборонительной.
Следует указать, что таким переключателем условнорефлекторной
деятельности могут быть многие факторы. Это очень важное обстоятель-
ство, и я вернусь к нему в конце своего доклада.
Возникает новый вопрос: только ли между различными по роду дея-
тельностями возможны переключения или изменения сигнального зна-
чения раздражителей или же переключение может быть в пределах, если
так можно выразиться, одной и той же деятельности?
Специально поставленные опыты показали, что возможно и это.
Я. М. Пресеман прикреплял электроды и регистрирующий движение
приборчик к левой задней лапе собаки и выработал двигательные электро-
оборонительные условные рефлексы на раздражение этой лапы. Когда
условные раздражители были выработаны, он в одном опыте прикреплял
тс же самые приборчики к противоположной правой задней лапе, т. е.
к лапе, которая никогда не раздражалась электрическим током, на кото-
рой условный рефлекс не вырабатывался. И что же оказалось? С первого
же раза, когда был дан условный сигнал, т. е. в ответ на условный сигнал,
собака дергала правую лапу, т. е. лапу, к которой были прикреплены
электроды и регистрирующие приборчики. Экстероцептивных импульсов
13. Э. А Асратян
1<М
Э. А. АСРАТЯН
с поверхности лапы, связанных с прикреплением приборчиков, было до-
статочно для того, чтсбы вызвать переключение путей, чтобы направить,
условные импульсы по иному маршруту.
В специальных опытах ситуация осложнялась дальше: к обеим задним
лапам прикреплялись электроды и регистрирующие приборчики. Здесь уже
собака запуталась. На первое применение условного раздражителя она
по очереди поднимала и опускала то одну, то другую заднюю лапу. Но
после первого раздражения одной из лап электрическим током все даль-
нейшие отсетные реакции переносились на эту лапу. Поистине эффектный
результат опыта: достаточно было однократного раздражения лапы элек-
трическим током, чтсбы переключить всю электрооборонительную двига-
тельную условнорефлекториую деятельность на эту лапу. И если в первом
варианте этих опытов переключателем служило прикрепление электродов
и приборчиков к лапе, то в этом варианте опытов эту роль выполнял пер-
гый электрический удар.
Итак, совершенно очевидно установлена возможность переключения
в пределах одного и того же рода деятельности организма, возможность
изменения сигнального значения и ответной деятельности в сфере одного
рода и знака условнорсфлекторной деятельности, имеющей важное биологи-
ческое значение.
Наконец, мы поставили перед собой еще одну задачу — попытаться
осуществить переключение или изменение физиологического знака услов-
ного раздражителя в пределах одного и того же рода деятельности орга-
низма. Говоря иначе, следовало выяснить, возможно ли переключение
одного и того же условного раздражителя с положительного на отрица-
тельный и наоборот в пределах одного рода деятельности.
Для решения этой задачи ставились опыты такого порядка (опыты
Ф. М. Шитова и В. А. Замятиной). Один и тог же экспериментатор в утрен-
них опытах в. ряду других раздражителей сочетал один раздражитель
с дачей пищи, а в вечерних опытах этот раздражитель не подкреплялся
пищей. Иначе говоря, в этой серии опытов и утром, и днем основной дея-
тельностью оставалась пищевая, все остальные условные раздражители
и утром, и вечером сохраняли свое сигнальное значение, но один раздра-
житель утром подкреплялся, а вечером не подкреплялся пищей. В резуль-
тате удалось добиться, что этот раздражитель утром действовал как поло-
жительный условный сигнал, а вечером как отрицательный.
В дальнейших опытах ситуация усложнялась. Брались два близких
раздражителя: сильный и слабый гудок. В утренних опытах сильный
гудок превратился в условный положительный раздражитель, а слабый—
в отрицательный. В дневных опытах сигнальное значение обоих раздра-
жителей менялось. Сильный гудок не подкреплялся, а слабый под-
креплялся пищей. Эта задача оказалась очень трудной, тем не менее опа
не так легко и четко, но все же некоторыми собаками решалась.
Итак, подытоживая сказанное, можно считать установленным, что
принцип переключения не только имеет место в деятельности коры боль-
ших полушарий, ио, как и следовало ожидать, здесь он достигает высшего
уровня развития и делает эту деятельность более лабильной и разнообраз-
ной, более подвижной и гибкой, делает ее средством более совершенного
приспособления организма к вечно меняющейся окружающей среде. Бла-
годаря свойству переключения, кора больших полушарий способна при-
давать разное значение одному и тому же сигналу в зависимости от ситуа-
ции, способна в широких пределах изменять свое функциональное состоя-
ние в зависимости от проприоцептивных и экстероцептпвных, возможно,
и интероцептивных импульсов, а также способна под влиянием разных
факторов создавать различные исходные функциональные фоны, разные
ПЕРЕКЛЮЧЕНИЕ В УСЛОВ1 ОРЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
195
картины распределения интрацентральных путей и тем самым готовиться
к ответу’, наилучшим образом соответствующему текущей ситуации, те-
кущим условиям существования организма. Иначе говоря, способность
переключения путей делает более совершенной, поднимает на более высо-
кий уровень деятельность коры больших полушарий н в значительной
степени обусловливает возможно точное и тонкое соответствие этой дея-
тельности конкретным условиям существования организма.
Может возникнуть вопрос относительно соотношений между законо-
мерностью переключения в высшей нервной деятельности и между неко-
торыми другими, ранее известными и близкими ему закономерностями той
же деятельности, в первую очередь образованием и протеканием разнород-
ных и комплексных условных рефлексов. Разница между переключением и
разнородными условными рефлексами совершенно очевидна; в первом слу-
чае один и тот же раздражитель сигнализирует различного рода деятель-
ности организма в зависимости ОТ ситуации, а Во втором случае разные дея-
тельности организма сигнализируются разными раздражителями. Очевидна
разница также между переключением и комплексными условными рефлек-
сами. В последнем случае все без исключения раздражители являются орга-
нической составной частью сложного условного раздражителя, пусковыми
факторами, действуют фазно, вызывают непосредственный эффект в виде
возбуждения ил» торможения той или иной деятельности организма.
В случае же переключения переключатель не является частью условного
раздражителя, не является пусковым фактором в обычном смысле, а создает
в коре больших полушарии фон определенной функциональной «окраски»,
к тому же тонического характера, предопределяет характер и род условно-
рефлекторной деятельности на определенный отрезок времени.
Из полученных нами и изложенных здесь фактических данных с пол-
ной очевидностью явствует, что принцип переключения является особой
закономерностью высшей нервной деятельности, к тому же закономер-
ностью, принадлежащей к категории универсальных для всей центральной
нервной системы.
В процессе длительного эволюционного развития простая закономер-
ность «растянутой мышцы» у беспозвоночных животных превращается
в более сложную закономерность спинального пли вообще безусловно-
рефлекторного переключения у низших и высших позвоночных животных,
а в дальнейшем достигает вершины своего развития в коре, находит свое
высшее выражение в закономерности условгорефлекторного переклю-
чения. При этом в деятельности коры больших полушарий неизмеримо
увеличивается количественный и качественный состав факторов-переклю-
чателей, а само переключение становится неизмеримо более варнабильным,
динамичным, многосторонним. Совершенно очевидно громадное биоло-
гическое значение переключения именно для деятельности коры больших
полушарии, которая как раз и характеризуется варнабилыюстыо, гиб-
костью, динамичностью, временностью.
Мне кажется, что фактические и теоретические результаты нашей иссле-
довательской работы по проблеме переключения в высшей нервной дея-
тельности, а также результаты более поздних исследований в том же на-
правлении, полученные некоторыми другими отечественными физиологами
(И. И. Лаптев, В. В. Яковлева, В. К- Федоров, Э. Г. Вацуро), не оставляют
сомнения в важности самой проблемы. Вот почему в настоящее время,
когда перед советскими физиологами ставится задача дальнейшего все-
стороннего интенсивного развития научного наследия И. П. Павлова и
в первую очередь его бессмертного материалистического учения о высшей
нервной деятельности, когда перед нами ставится важная задача более
углубленного, многостороннего изучения основных закономерностей дея-
тельности коры больших полушарии, — мы, как мне кажется, не должны
13*
]9б э А- АСРАТЯН
упускать из виду эту важную закономерность, которая впервые была
установлена для низшей нервной деятельности И. М.Сеченовым и которая,
по нашим данным, достигает наибольшего совершенства в высшей нервной
деятельности. Мы в новых, более благоприятных условиях нашей иссле-
довательской работы, наряду с другими проблемами, включаем в программу
исследований также дальнейшее всемерное изучение принципа переклю-
чения высшей нервной деятельности, считая, что Это Соответствует задачам,
поставленным партиен и правительством перед учениками и последова-
телями И. П. Павлова.
И мы с тревогой и надеждой будем ждать таких результатов нашей
дальнейшей работы по этой проблеме, которые были бы достойны дорогой
памяти любимого учителя.
-с.
К ФИЗИОЛОГИИ ВРЕМЕННОЙ СВЯЗИ 1
ессмертное творение могучего ленинского гения — «Материа-
лизм и эмпириокритицизм», в котором дано непревзойденное
по глубине и мастерству диалектико-материалистическое обоб
щен не достижений естествознания того времени, а также суро-
вая и уничтожающая критика врагов диалектического материа-
лизма, в частности, адептов «физиологического идеализма», за-
вершается знаменательными словами: «...За гносеологической схоластикой
эмпириокритицизма нельзя не видеть борьбы партий в философии, борьбы,
которая в последнем счете выражает тенденции и идеологию враждебных
классов современного общества» 2.
В переживаемое нами время в силу крайнего обострения антагонизма
между загнившим миром капитализма и цветущим миром социализма край-
него напряжения и остроты достигла также идеологическая борьба между
двумя непримиримыми лагерями современного человеческого общества —
многовековая борьба между материализмом и идеализмом. И то, что происхо-
дит в настоящее время вокруг славного имени Павлова и его гениального на-
учного творчества, в частности, вокруг его материалистического учения о
высшей нервной деятельности, является, бесспорно, частью этой борьбы,
притом частью весьма существенной, охватывающей крайне важный и
весьма значительный участок широкогофронта нашей идеологической борь-
бы. Нужно помнить, что имя Павлова является знаменем борьбы и сози-
дания не только в физиологии, по и в современной биологии и медицине.
Мы, ученики и последователи И. П. Павлова, должны отдавать себе от-
чет в том, что хотя идеализм и обречен на неминуемую гибель, но он еще
споссбен нанести значительный ущерб священному для каждого из нас делу
развития павловского научного наследия, ловко нащупать и подло исполь-
зовать отдельные слабые участки нашего единого фронта и проползти в обна-
руженные там щели, попытаться расколоть нас, задержать наше прогрессив-
ное движение, а то и вовсе приостановить его. Вот почему сейчас в большей
мере, чем когда-либо раньше, нам необходимо поднять на большую принци-
пиальную высоту нашу борьбу с зарубежными антипавловцами и с их жал-
кими эпигонами в среде советских ученых, правильно организовать и умело
вести эту борьбу, следуя во всем этом примеру великих учителей наших —
примеру Ленина и Сталина, Сеченова и Тимирязева, Павлова и Мичурина.
А между тем мы еще нс можем сказать, что в этом отношении у нас все
обстоит благополучно. Здесь нс место подробно говорить о недостатках
нашей работы в этом плане, но во имя поставленной перед нами большой,
государственной важности задачи, во имя нашей священной цели мы обя-
заны говорить об этом вообще, если только искренне хотим выявить и
* Доклад на 15-м совещании по проблемам высшей нервной деятельности, посвя-
щенном 50-летию учения акад. И. П. Павлова об условных рефлексах, 15 апреля 1952 г
Опубликован в трудах этой конференции в 1952 г.
8 В. И. Л е н и и, Соч., изд. 4-е, т. 14, стр. 343.
198
Э. А. АСРАТЯН
устранить недостатки в нашей работе по этой линии и тем самым укрепить
наши позиции, ускорить наше поступательное движение вперед.
На одном из таких недостатков ввиду его прямой связи с конкретным
научным содержанием нашего сообщения мы и остановимся. Мы имеем в виду
вербальный, декларативный и умозрительный характер многих критических
выступлений против зарубежных и отечественных антипавловцев в защиту
павловской физиологии по принципиально важным научным вопросам.
При этом мы в лучшем случае довольствуемся использованием давних фак-
тических данных павловской школы и других передовых деятелей отечест-
венной науки. А между тем известно, что научная борьба, а тем более борь-
ба в области естествознания, проводимая только или преимущественно с
помощью умозрительной критики, подкрепляемая только фактами значи-
тельной давности, не может привести к крупным и стойким успехам, как бы
остро и напряженно такая борьба ни велась.
Гениальный творец материалистического учения о высшей нервном дея-
тельности растил и развивал свое любимое детище не в оранжереях или в
безветренных долинах науки, а в суровой, непрерывной ножесточенной
борьбе со всеми черными силами реакции, со всякими лженаучными идеа-
листическими течениями и направлениями, нашедшими было приют в уют-
ном уголке «загадочных» явлений психической деятельности. Страстный и
неутомимый борец за материализм в сложнейших явлениях органической
природы, И. П. Павлов наносил разящие, сокрушительные удары по своим
идейным противникам не только л даже не столько своим острым, метким,
страстным и могучим словом, своей неотразимой логикой, своим не-
заурядном полемическим мастерством, сколько своими всегда оригина льны-
ми, вескими, капитальными и великолепными фактами, своей ьппре-
ЬЗо”<ДепИон По Совершенству и продуктивности экспериментальной работой.
В руках гениального теоретика й мыслителя эти факты имели поистине ис-
полинскую силу и неизменно обеспечивали ему полную победу в жестоких
схватках с врагами материализма. Примечательно, что сама эта борьба воин-
ствующего материалиста с его идейными противниками не только закаляла
И. П. Павлова, но «служила для него неиссякаемым источником новых
творческих замыслов, а тем самым новых экспериментов и фактов, новых и
еще более блистательных научных подвигов.
Жестокая борьба воинствующего материалиста И. П. Павлова против
старого, отживающего свои век п реакционного в пауке была борьбой не
только уничтожающей, но и созида7^ельной, двигающей науку вперед. Его
суровая и беспощадная критика антинаучных идеалистических концепций
идейных противников в кардинальных вопросах естествознания была кри-
тикой диалектической, т. е. критикой не только отрицающей, но и
творческой, конструктивной, дающей положительное решение сложнейших
и запутанных научных проблем и вопросов.
Мы — ученики и последователи И. II. Павлова — должны в порядке
самокритики сознаться, что и в этом отношении недостаточно энергично
следуем примеру своего великого учителя. Слов нет, что его бессмертное
материалистическое учение о высшей нервной деятельности, его гениаль-
ные идеи и глубокие мысли, равно как и добытый при его жизни и под его
непосредственным руководством богатейший и ценнейший фактический ма-
териал, были, есть н надолго останутся главными и решающими орудиями
нашей борьбы со всеми и всякими проявлениями идеализма в физиологии,
и не только в физиологии. Но .мы были бы плохими учениками и последова-
телями И. II. Павлова, если бы в нашей борьбе с зарубежными и отечествен-
ными антипавловцами ограничивались только использованием этих орудий,
не стремились бы своей каждодневной, систематической экспериментальной
и теоретической работой неустанно развивать дальше его великое научное
наследие и тем самым непрерывно пополнять и обогащать арсенал наших
К ФИЗИОЛОГИИ ВРЕМЕННОЙ СВЯЗИ ]9Э
•боевых средств. Необходимость творческого, а не догматического подхода
к развитию научного наследия И. П. Павлова очевидна. Эта необходимость
явствует, в частности, из того факта, что наши идейные противники посто-
янной старательно обновляют средства и формы своей борьбы с основами
материалистического учения И. П. Павлова.
Вот почему мы решили при изложении полученных нами совместно с на-
шими сотрудниками новых экспериментальных данных относительно физио-
логии временной связи и построенных на этой основе теоретических поло-
жений обратить специальное внимание па критику лженаучных концепций
зарубежных и отечественных антипавловцев по этому кардинальному воп-
росу физиологии высшей нервной деятельности.
Важнейшее значение проблемы временной связи в материалистическом
учении Н.П. Павлова о высшей нервнойдеятельностиопределяетсяцептраль-
ным положением условного рефлекса в этом учении. Вспомним, что он пи-
сал по этому поводу: «Центральное физиологическое явление в нормальной
работе больших полушарий есть то, что мы назвали условным рефлексом » *.
Пятьдесят лет назад в знаменитой мадридской речи великий ученый вы-
двинул на первым план и заложил в фундамент будущего величественного
здания этого учения только что открытое им для науки явление — факт об-
разования качественно нового рефлекса:, условного рефлекса как резуль-
тат замыкания новой временной связи между оча-
гами головного мозга, находящимися в состоя-
нии возбуждения. И. П. Павловым был установлен важный факт,
который он позже охарактеризовал словами: «нервная система представ-
ляет собой не только проводниковый, но и замыкательный прибор»2.
Спустя пять лет после мадридской речи И. П. Павлов впервые четко об-
рисовал схему образования временной связи и дал сравнительную физиоло-
гическую характеристику дуги безусловного и дуги условного рефлексов.
Он указал, что «во втором случае, следовательно, мы имели дело с вре-
мен н ы м замыканием связи как основным свой-
ством деятельности высших отделов централь-
ной нервной системы, как первым капитальным
пунктом их механики» (разрядка паша. — Э. А .)®.
А в 1927 г., характеризуя процесс образования условного рефлекса, он
писал: «Таким образом, это слияние, соединение раздражений, возникающих
в разных пунктах головного мозга, это нервное замыкание
иестьпервый нервный механизм, с к о т о р ы м м ы
встречаемся при изучении деятельности коры
больших полушарий» (разрядка наша.—Э. А.)1.
Вопрос о замыкании условной связи сохранил для И. П. Павлова свое
первенствующее значение в течение всего дальнейшего развития учения
о высшей нервной деятельности. Великин физиолог постоянно держал этот
вопрос в центре своего внимания, непрерывно изучал его Экспериментально,
неустанно продолжал разрабатывать его теоретически.
Он охарактеризовал закон замыкания временной связи как «основной
закон высшего отдел а системы», считая, что для деятельности больших полу-
шарий мозга «основное явление — образование временных связей». Ха-
рактерно, что И. П. Павлов в своих трудах иногда условные рефлексы на-
зывает «замыкательными рефлексами».
Вопросу замыкания временной нервной связи И. П. Павлов уделял
исключительное внимание не только потому, что это свойство лежит 1 2 * 4
1 И. Г1. Павлов, Полное собрание трэдов, т. III, 1949, стр. 481.
2 Т а ы же, стр. 222.
’Там же, стр. 75.
4 Т а м ж е, т. IV, 1947, стр. 46.
200
Э. А. АСРАТЯН
в основе условного реф«тскса— центрального явления во всей высшей нерв-
ной деятельности, но и потому, что в факте замыкания нервной связи как
качественно нового вида нервных явлений он видел новое, весьма важное
и веское доказательство правильности одного из развиваемых им фунда-
ментальных общих принципов рефлекторной деятельности нервной систе-
мы вообще, а именно «принципа структурности, т. е. расположения дейст-
вия силы в пространстве, приурочения динамики к структуре» \
В процессе более чем тридцатилетней напряженной экспериментальной
и теоретической разработки вопросов физиологии высшей нервной деятель-
ности, связанных с замыканием временной нервной связи, И. П. Павлов уста-
новил ряд важных закономерностей замыкания этой связи, ее упрочения
и ослабления, функционирования и задержки, выяснил роль процессов
возбуждения и торможения во всем этом и т. п. Точно установив, что «боль-
шие полушария есть орган условных рефлексов» 2, И. П. Павлов, однако,
не считал окончательно решенным вопрос,является ли замыкание временной
нервной связи исключительно функцией коры или ближайшие подкорко-
вые нервные образования также способны к замыканию подобного рода
СВЯЗИ.
Гениальному творцу учения о высшей нервной деятельности принадлежит
также ряд глубоких высказываний, уточняющих его представления отно-
сительно детален локализации и микроструктурных основ замыкания вре-
менной связи, относительно особенностей проводимости и других функцио-
нальных черт этой связи. Так, на протяжении многих лет И. П. Павлов
придерживался точки зрения, что условная связь замыкается между кор-
ковыми центрами различных анализаторов и подкорковыми центрами без-
условных рефлексов (всем известно его схематическое изображение этого
представления). В последние же годы своей творческой деятельности он
счел возможным оставить эту’ точку зрения и по существу вновь вернуться
к точке зрения, которая возникла в его лабораториях в самые первые годы
зарождения учения об условных рефлексах, а именно к допущению, что
«образование новой нервной связи, замыкательный процесс целиком
происходит в больших полушариях, т. е. в них находятся не только пункты
приложения бесчисленных индиферентных раздражений, но в них же ле-
жат и деятельные пункты, представители безусловных рефлексов, между ко-
торыми устанавливается новая связь»3. Из других высказываний И. П. Пав-
лова явствует, что он считал, что условная связь замыкается именно между
различными пунктами коры — между клетками корковых концов различ-
ных анализаторов и клетками корковых представительств безусловных
рефлексов. В приводимой ниже цитате, например, И. П. Павловне только
излагает свои соображения о «механизме» замыкания условнорефлектор-
ной связи, но уточняет и свой взгляд на затронутый выше вопрос. Он пишет:
«Основной механизм образования условного рефлекса есть встреча, совпа-
дение во времени раздражения определенного пункта коры полушарий с
более сильным раздражением другого пункта, вероятно, коры ж е, в
силу чего между этими пунктами более или менее скоро проторивается
более легкий путь, образуется соединение» 4.
Главным основанием для этих предположении И. П. Павлова служили
фактические данные его сотрудников отом, что при суммации условных реф-
лексов валовой эффект не поднимается выше определенного предела. Он счи-
тал, что это явление может быть объяснено только при допущении, чт о ус-
ловнорефлекторные связи замыкаются между корковыми клетками, харак-
теризующимися вообще низким пределом работоспособности. Если бы ус.
* И. П. П а в л о в, Полное собрание трудов, т. 111, 1949, стр. 430.
’Там же, стр. 72.
’Там же, стр. 380.
‘Там ж е, т. IV, 1947, стр. 317.
К ФИЗИОЛОГИИ ВРЕМЕННОЙ связи
201
ловиорефлекторные связи замыкались между корковыми клетками и под*
корковыми центрами, то, по И. П. Павлову, при суммировании условных
рефлексов валовой эффект был бы неизменно больше, так как предел работо-
способности подкорковых нервных центров очень высокий. Совершенно
очевидно, что' хотя аргументация эта и косвенная, но имеет большую убе-
дительную силу.
Касаясь в некоторых своих произведениях вопроса о микроструктурной
основе замыкания условнорефлекторной связи, И. П. Павлов высказал ги-
потетическое предположение, что, «вероятно, место этой замыкательной спо-
собности есть пункты сцепления нейронов (и специально коры больших полу-
шарий)» *, что «замыкание, образование новых связей мы относим за счет
функций разделительной мембраны, если она существует, или просто утон-
чающихся разветвлений между нейронами, между отдельными нервными
клетками» *.
В последние годы жизни И. П. Павлов, основываясь на некоторых факти-
ческих данных своих.лабораторий, развивал положение о том, что условно-
рефлекторная связь между двумя корковыми центрами способна проводить
нервные импульсы «туда и обратно, т. е. в противоположных направлениях»,
точнее, как бы состоит из двух связей, каждая из которых проводит нервное
возбуждение в одном направлении. Он писал по этому поводу: «Когда два
нервных пункта связаны, объединены, нервные процессы двигаются, идут
между ними в обоих направлениях. Если признать абсолютную законность
одностороннего проведения нервных процессов во всех пунктах центральной
нервной системы, то в данном случае придется принять добавочную обрат-
ного направления связь между этими пунктами, т. с. допустить существо-
вание добавочного нейрона, их связывающего» ’.
Устанавливая с помощью прямых и безупречных фактов многие законо-
мерности замыкания и функционирования временной связи, гениальный
создатель учения о высшей нервной деятельности отнюдь нс считал, что он
исчерпывающе осветил все вопросы, связанные с этой основой основ его ма-
териалистического учения. Хотя он и достиг в этой области огромных ре-
зультатов, тем не менее, благодаря присущему ему диалектическому по-
ниманию существа самого процесса исследования, ои считал, что сделанное
им является лишь этапом к достижению абсолютной истины. Его вечно
юная н беспокойная творческая натура ие могла останавливаться на достиг-
нутом. И он продолжал изучать этот вопрос до конца своих дней, считая,
что многие его стороны остаются еще недостаточно выясненными. «Услов-
ность, ассоциация по одновременности, условный рсфтекс, хотя и служит
для нас исходным фактом наших исследований, — писал он в одном из по-
следних своих произведений, — тем не менее подвергается нами дальней-
шему анализу. Перед нами важный вопрос: какие элементарные свойства
мозговой массы лежат в основании этогофакта. Этот вопрос еще не представ-
ляется нам окончательно решенным» 4. В другом месте он по тому же поводу
писал: «Сделанное до сих пор показывает, что у нас еще нет полного знания
всех тех условий, которые определяют деятельность механизма образования
условного рефлекса»5. Он сам поставил на разрешение ряд конкретных
вопросов, связанных с этой важнейшей проблемой.
Следует отметить, что многие противники материалистического учения
И. П. Павлова о высшей нервной деятельности, отдавая себе отчет в том, что
условный рефлекс является основой этогоучення, с различных позиций свое-
1 И. П. П а в л о в, Полное собрание трудов, т. 111, 1949, стр. 288.
•Там же, стр. 356.
’Там же, стр. 452.
’Там же, стр. 429.
‘Там ж е, т. IV, 1947, стр. 318.
202
Э. А. АСРАТЯН
го гнилого фронта открывали концентрированный огонь против условного
рефлекса и принципа замыкания временной связи.
Некоторые зарубежные ученые (Фултон, Лиддл) и их слепые подра-
жатели среди ученых нашей страны (Н. А. Бернштейн, В. В. Ефимов к др.),
•стремясь умалить бессмертные заслуги И. ГТ. Павлова перед наукой и зна-
чение его учения о высшей нервной деятельности, высокомерно указывали
на то, что, мол, Мейнерт, Шеррингтон, Твигмеиер или кто-то другой из ино-
странных ученых задолго до И. П. Павлова наблюдали явления типа услов-
ного рефлекса. Эти горе-критики И. П. Павлова и его учения просто
не поняли сути дела, о котором отважились высказать свои «глубоко-
мысленные» суждения. Они не поняли, что великие бессмертные заслуги
И. П. Павлова заключаются отнюдь не в том, что он установил самый факт
условного рефлекса (кстати, сам И. П. Павлов никогда не приписывал это
себе; наоборот, он неоднократно указывал, что это явление было отмечено
«уже давно и многими»), а в том, что он сумел известное с незапамятных
времен простое явление «слюнки текут», явление, привлекавшее внимание
некоторых ученых еще в XVIII столетии, но с тех пор так и непонятое и не
оцененное учеными по достоинству, сумел это простое явление наполнить
глубоким биологическим содержанием и превратить его в могущественный
ключ к раскрытию глубочайших тайн работы мозга, веками терзавших пыт-
ливый ум человека. Они не поняли того, что И. П. Павлов, впервые открыл
условный рефлекс для науки ис его помощью разрешил такие
грандиозные научные задачи, которые оказались непосильными всем его
предшественникам и современникам.
Другая группа зарубежных и наших антнпавловцев — И. С. Беритов,
П. К. Анохин, Ю. М. Коморский и др. — стала на путь дискредитации услов-
ного рефлекса, на путь низведения его до ранга примитивного, топорного,
нелабильного и негибкого вида нервной деятельности, весьма близкого по
своим свойствам к безусловным рефлексам, стали на путь принижения роли
условного рефлекса в высшей нервной деятельности. Эти ученые не видели
богатого биологического содержания условного рефлекса и важнейшей его
физиологической роли, его эволюции, многообразия, вариабильности, ла-
бильности, — не видели павловского учения об условных рефлексах во
всем его истинном величин и красе. Такое упрощенное представление об
условном рефлексе и такое принижение его значения привели одних из этих
антнпавловцев— И. С. Беритова и его последователей — в лагерь идеа-
листической зоопсихологии, к допущению существования будто бы более
высокого вида церебральной деятельности у животных, а именно так назы-
ваемой психо-нервнон Деятельности; Других ИЗ нНх — Ю. М. Конорского,
С. Миллера, Торндайка, Скиппера, Толмена и др. — к отграничению на-
выков от условных рефлексов, к ложным представлениям о том, что навыки
не могут быть сведены к условным рефлексам, а представляют ссбон белее
высокую форму поведения; наконец, третьих — П. К,- Анохина и его по-
следователей — к допущению, что «роль лобных отделов коры не можегбыть
выражена в обычных терминах, которыми мы располагаем при изучении
высшей нервной деятельности»
В довершение всего И. С. Беритов, П. К. Анохин, Ю. М. Конорский и их
единомышленники в целях маскировки этих и других своих лженаучных
и реакционных концепций, придания им видимости прогрессивного и строго
научного характера прибегают, будто по договоренности, к одному и тому
же демагогическому приему, посуществу —к клевете, а именно: к обвинению
И. II. Павлова и его бессмертного материалистического учения о высшей
нервной деятельности в отрыве от общей физиологии нервной системы, в
отсталости от современных достижений нейрофизиологии, в узости, огра
1 Сборник «Проблемы высшей нервной деятельности», 1949, стр. 9.
К ФИЗИОЛОГИИ ВРЕМЕННОЙ СВЯЗИ 203
ииченности, примитивизме и тому подобных вымышленных пороках. При
этом они, разумеется, не упускают случая выдавать себя за спасителей на-
следия великого физиолога
Наконец, выражением того же упрощенчества в понимании сущности
условного рефлекса и недооценки его важнейшей роли в высшей нервной
деятельности и еще более ярким выражением стремления низведения услов-
иогорефлехса до ранит безусловного следует считать также то, что группа
наших и зарубежных нсследов.ателей — П. К. Анохин, Н. А. Рожанский,
Шурраджер (Shurrager), Келлер (Culler) и др. — за последнее время разви-
вают концепцию о возможности образования временной связи в спинном
мозгу, образования так называемых спинномозговых условных рефлексов.
Если у наших исследователей эти предположения носят характер чисто спе-
кулятивных выдумок, лишенных какого-то ни было подобия прямой или
хотя бы косвенной фактической аргументированности, то американские
исследователи стремятся создать видимость некоего' фактического обоснова-
ния своих теоретических положении. Ниже мы увидим, насколько «солид-
ны» и «убедительны» эти их попытки.
Разумеется, что несостоятельность, порой даже абсурдность перечислен-
ных выше антипавловских концепций относительно условного рефлекса
н лежащей в его основе временной нервной связи можнз доказать гораздо
более убедительно, чем это делалось до сих пор, даже при одном только
обстоятельном критическом разборе этих концепций и лежащих в их основе
фактических данных. Мы полагаем, однако, что критика этих концепций
на основе соответствующих новых экспериментальных данных будет иметь
особенное значение. Вот почему мы считаем, что излагаемый ниже факти-
ческий материал, полученный нами и нашими сотрудниками, представляет
известный интерес и с этой точки зрения.
Наш фактический материал имеет отношение к четырем разделам пробле-
мы замыкания условнорефлекторной связи и особенностей ее функциониро-
вания, я именно: к вопросу о корковом представительстве безусловных реф-
лексов и месте замыкания временной связи, о двусторонней проводимости
условнорефлекторной связи, о функциональных состояниях временной свя-
зи при переключениях и к вопросу о возможности будто бы образования
условнорефлекторнон связи в спинном мозгу. К краткому изложению этого
материала и некоторых наших теоретических положений по соответству-
ющим вопросам мы л перейдем.
Нами уже был отмечен известный факт, что в последние годы своей твор-
ческой деятельности И. П. Павлов на основании анализа результатов сумма-
ции условных рефлексов отказался от прежних представлений о локализации
усло-внорефлекторной связи и, допуская существование корковых пред-
ставительств безусловных рефлексов, высказал предположение о замыка-
нии временной связи между корковыми пунктами. Необходимость дальней*
шей экспериментальной разработки этого важного вопроса была совершен-
но очевидна, и этим, насколько нам известно, более или менее систе\1ати-
чески занимались два ученика И. П. Павлова—П. С. Купалов с сотрудни-
ками руководимой им лаборатории и мы совместно с сотрудниками
руководимой нами лаборатории. Фактические данные и теоретические
положения П. С. Купалова по этому вопросу неоднократно излагались им
самим, и здесь нет необходимости на них останавливаться. Считаем
своим долгом лишь отметить, что из основного установленного им и его
сотрудниками факта, а именно: усиления безусловного рефлекса в
случае, когда применению безусловного раздражителя за несколько секунд
«предваряет» применение соответствующего условного раздражителя,— вы-
вод о существовании коркового представительства безусловного пищевого
рефлекса вытекает косвенно, и нельзя сказать, что он является единст-
венно возможным.
204
Э. А. АСРАТЯН
Мы подошли к изучению этого, поставленного И. П. Павловым вопроса
с иных методических позиций, а именно путем выключения коры, преимуще-
ственно хирургическим способом, стремясь таким образом получить не кос-
венное, а прямое фактическое подкрепление положения И. П. Павлова
о корковом представительстве безусловных рефлексов.
Начиная с 1934 г., мы на весьма большом числе подопытных собак наблю-
дали, что удаление коры обоих больших полушарий мозга значительно из-
меняет у них характер и течение безусловнорефлекторной деятельности.
Рис. I. Общее количество желудочного сока (в мл) у гастро-
эзофаготомнрованной собаки Тузик, выделяющегося за опыт
при мнимом кормлении м-ясом
7—непрерывная еда н течение 10 мни ; 2—прерывистая еда (по I ку-
сочку мяса через 15 сек ) в течение 10 мин (всего 40 кусков},
3—прерывистая еда в течение 5 мин (всего 20 кусков); 4—прерыви-
стая еда в течение 2‘/, мин (всего 10 кусков)
Столбики с косой штриховкой - норма, заштрихованные в клетку —
после удаления коры
Даже без специальных приемов исследования и графической регистрации,
а только при внимательном наблюдении можно легко установить, например,
что у лишенных коры собак скелетно-двигательныебезусловные рефлексы,
в особенности такие сложные, как выпрямительные и локомоторные, суще-
ственно отличаются от таковых у нормальных собак. После удаления коры
обоих полушарий большого мозга эти рефлексы делаются грубыми, негиб-
кими, инертными, несовершенными. Точно такие же изменения, но в менее
четком виде, могут быть выявлены и со стороны вегетативных безуслов-
ных рефлексов у лишенных коры собак. Все это уже давно послужило нам
основанием для внесения существенной поправки в широко распространен-
ную, почти общепризнанную формулу, согласно которой врожденная реф-
лекторная деятельность высших животных не изменяется после удаления
у них коры больших полушарий мозга. Эта формула более или менее пра-
вильна лишь в том ограниченном смысле, что у лишенных коры животных
безусловные рефлексы сохраняются; характер же этих рефлексов, их тече-
ние, степень их совершенства, как уже было отмечено, изменяются в силь-
ной степени.
Приведенные данные мы считали и считаем прямым фактическим доказа-
тельством существования коркового представительства безусловных реф-
К ФИЗИОЛОГИИ ВРЕМЕННОЙ связи
205
лекСоВ, Доказательством участия коры больших: полушарий мозга В осуще-
ствлении безусловных рефлексов, о чем мы неоднократно писали и докла-
дывали, начиная с 1934—1935 гг.
Учитывая важность этого вопроса, мы незадолго до Великой Ог-чествеи-
ной войны приступили К специальному его изучению. Эти исследования,
давшие положительные результаты, но прерванные войной, были недавно
возобновлены и позволили уже получить ценные фактические данные по ин-
тересующему нас вопросу. В весьма сжатом изложении суть этих данных
сводится к следующему.
Рис. 2. Характер желудочной секреции у гастроэзофаготомнрован-
ной собаки Тузик при мнимом кормлении мясом
Собаке закладывали в рот по одному куску каждые 15 сек,
в течение 10 мин. (всего 40 кусков)
Еще в 1938—1939 гг. наш сотрудник Я. М. Прессмаи показал, что у ли-
шенных коры собак почти в полном объеме сохраняется установленное
И. П. Павловым и сотрудниками многообразное безусловнорефлекторное
приспособление секреторной деятельности слюнных желез к роду, качеству
и количеству вводимых в рот пищевых и непищевых слюногонных веществ.
Однако эта безусловнорефлекторная регуляция функции слюнных желез
осуществляется у них менее тонко, точно и совершенно, чем у нормальных
собак.
Тоже самое было недавно установлено нашей сотрудницей Л. Л. Мар-
ковой в отношении секреторной деятельности желудочных желез. Без-
условнорефлекторная регуляция секреторной деятельности желудочных
желез после удаления коры больших полушарий значительно изменяется,
делаясь менее тонкой и совершенной. Это явствует, например, из того фак-
та, что у лишенных коры собак не так четко, как у нормальных, выступает
разница между количеством и длительностью секреции желудочного сока
при мнимых кормлениях различной длительности (рис. 1). А. Л. Маркова
установила также, что удаление коры больших полушарий влечет за собой
значительное ослабление рефлекторной секреции желудочного сока, вызван-
ной мнимым кормлением, и изменение динамики самого секреторного про-
цесса (рис. 2).
Интересный и ценный материал относительно изменения соматических
безусловных рефлексов после удаления коры больших полушарий получил
за последние годы наш сотрудник Б. Д. Стефанцов. Он не только подтвердил
давние наблюдения многих ученых, в том числе и наши, о значительном по-
вышении порога возбудимости спинальных двигательных рефлексов конеч-
ностей после удаления коры противоположного полушария большого мозга,
206
Э А. АСРАТЯН
но и установил, что рефлексы эти крайне астеничны, очень быстро утомля-
ются и долго не восстанавливаются. Характерно, что быстрая утомляемость
этих рефлексов сохраняется как явление более или менее постоянное, а из-
менения в пороге их возбудимости со временем в значительной степени вы-
равниваются. Для иллюстрации сказанного приводится кинограмма реф-
лекторного утомления сгибательного рефлекса пораженной и контрольных
задних конечностей у собаки после удаления коры полушария большого
мозга (рис. 3).
Рис. 3. Рефлекторная утомляемость задних конечностей собак на
22-й день после удаления коры левого полушария
а — кнмогрдмма собаки Альма; б-кимограмма собаки ЛеПкя Верхняя лииия-
лраа-ая конечность, средняя — левая конечность, нижняя — отметка времени в сек.
Нам кажется, что эти экспериментальные данные d сочетании с данными
наших многочисленных наблюдений, проведенных над 60 собаками, лишен-
ными коры обоих больших полушарий, представляют собой весьма веское
к тому же п р я м о е фактическое обоснование теоретического положения
II. П.” Павлова о существовании коркового представительства безусловных
рефлексов. Ия наших данных явствует также, что участие этих корковых
представительств в безусловнореф-
лекторной деятельности нижележа-
щих отделов центральной нервной
системы делает эту деятельность бо-
лее тонкой, точной, совершенной и
полноценной.
В связи с вышеизложенным мы
считаем уместным поставить сейчас
вопрос о необходимости уточнения
наших взглядов о корковых предста-
вительствах безусловных рефлексов,
а также о новой принципиальной
схеме замыкания условнорефлекторной связи между пунктами коры, т. е.
о структуре дуги условного рефлекса. Как известно, И. П. Павлов не
оставил нам подробного и четкого ответа на эти вопросы, а важность их
разрешения не подлежит сомнению.
Наиболее простым и легким решением вопроса о принципиальной схеме
замыкания условнорефлекторной связи было бы возвращение к принятой
в лабораториях И. П. Павлова самой первоначальной схеме, как предлагает
Ф. П. Майоров (рнс. 4), или же соответствующая модернизацш! общеизвест-
ной схемы И. П. Павлова (рис, 5) с тем, чтобы привести ее в механиче-
ское соответствье с последними его представлениями о замыкании времен»
нэп сеязи между различными пунктами коры (рис. 6).
К ФИЗИОЛОГИИ ВРЕМЕННОЙ СВЯЗИ
207
Нам кажется, однако, что это решение не лучшее из возможных, что эти
по существу идентичные схемы замыкания условнорефлекторной связи,
изображенные на рис. 4 и 6, нс могут нас сейчас удовлетворить даже при
больших скидках поговорках, которые обычно делаются при схематических
изображениях сложных биологических явлений. Недостатки этих двух схем,
нс в линиях, узорах и конфигурациях, а в замысле, лежащем в их основе.
Не говоря уже о чересчур
упрощенном изображении в
обеих схемах связи между ко-
рой больших полушарий и
периферическими концами
анализаторов, в них прин-
ципиально неверно указана
связь между корковым пред-
ставительством безусловного
рефлекса и между первичным
центром того же рефлекса,
а именно: она изображена как
одностронняя связь сверху
вниз.
Стремясь не догматически,
а творчески развивать идеи
И. П. Павлова о корковом
представительстве безуслов-
ного рефлекса и о замыкании
условнорефлекторной: связи
в пределах коры больших
полушарий мозга, мы еще
в 1937 г. на основании некото-
Рис. 5. Схема образования дуги условного реф-
лекса по И. II. Павлову
Временная связь замыкается между' корковыми peiten-
Тарными клвтцамм(КК) и между пищевым центром
находящимся вне коры
рых описанных в литературе
и собственных фактических
данных высказали в одной
из наших работ несколько
новых соображений по этим
вопросам и даже попытались
дать их схематическое изображение (рис. 7). Хотя мы поделились этими
мыслями и опубликовали нашу схему повторно всего несколько лет назад,
тем не менее мы считаем необходимым обратить на них внимание и сейчас.
Нам кажется прогрессивным и близким к истине предположение, соглас-
но которому корковое представительство безусловного рефлекса мыслится
в виде корковой ветви дуги этого рефлекса. Мы считаем, что уже наступила
пора расстаться с устаревшим представлением сб однолннейности дуги
безусловных рефлексов и заменить его новым, согласно которому дуга без-
условных рефлексов в своей центральной части состоит из многих ветвей,
проходящих по разным этажам центральной нервной системы. По мере подъ-
ема снизу вверх, благодаря каждой ветви такой многопучковой дуги, без-
условные рефлексы становятся все более тонким, точным и совершенным
средством нервной регуляции и объединения функций организма. На при-
веденной схеме (рис. 7) мы изобразили только первичную и корковую ветви
дуги пищевого безусловного рефлекса, пропустив, ради простоты и четкости,
все промежуточные ветви, скажем, ветви, проходящие через межуточный
мозг и ближайшие подкорковые нервные образования.
Далее, мы считаем принципиально правильным распространить это
представление также на дугу ориентировочного рефлекса, по существу
говоря, тоже безусловного рефлекса (на что указывал И. П. Павлов),
вызываемого сшдйферентньми» раздражителями. На пашей схеме по
20Я
Э. А. АСРАТЯН
Рис. 6. Модернизированная схема обра-
зования дуги условного рефлекса
К — кора; КП — корковое представительство
пищевого центра: ЗЦ — зрительный центр;
Л — пищ. си ой центр; С — слюнная железа
приведенным выше мотивам изображены также лишь первичная и корковая
ветви многоэтажной дуги ориентировочного рефлекса на звуковые раздра-
жители.
Могут спросить, а нс противоречит ли это представлсннс о корковом
представительстве безусловных рефлексов известному положению И. П.Пав-
лова о том, что «кора полу-
шарии представляет собой изо-
л ированный афферентны й отдел »
центральной нервной системы?
Конечно, нет! Во-первых, сле-
дует иметь в виду, что И. П.
Павлов относил кору боЛЬШИх
полушарий к афферентному раз-
делу центральной нервной систе-
мы при существенной оговорке,
а именно: «если ВСЮ центральную
нервную систему делить только
на две половины — афферент-
ную и эфферентную» *. При
таком подходе можно отнести
к афферентной части централь-
ной нервной системы все нерв-
ные клетки корковой ветви дуги
безусловного рефлекса, вероят-
но, даже не одной только
этой ветви. Если же допустить
существование промежуточно-
го, или вставочного, звена между
афферентными и эфферентными
отделами центральной нервной
системы (известно, что И. И. Пав-
лов не возражал и против этого
принципа классификации эле-
ментов рефлекторной дуги), то придется допустить в корковой и в других
ветвях дуги безусловного рефлекса наличие как афферентных нервных кле-
ток, так и вставочных нервных клеток, вероятно, даже многочисленных.
После этих предварительных замечаний относительно нашего понима-
ния сути коркового представительства безусловных рефлексов нетрудно
понять и нашу идею, лежащую в основе принципиальном схемы образова-
ния условнорефлекторной связи между различными пунктами коры боль-
ших полушарий мозга, рассматриваемую нами как дальнейшее развитие
тех идей И. П. Павлова по этому вопросу, которых он придерживался в по-
следние годы своей творческой деятельности. Согласно нашему представле
илю, условнорефлекторная связь замыкается между нервными клетками кор-
ковых ветвей дуги безусловного рефлекса и дуги так называемого ориенти-
ровочного рефлекса, т. е., по существу говоря, между корковыми ветвями
дуг двух безусловных рефлексов. Вот почему мы считали возможным даже
выдвинуть формулу, согласно которой у с л о в н ы й р е ф л е к с пер-
вогопорядка есть синтез двух или боль шегочис-
ла разных безусловных рефлексов, осуществлен-
ный высшим отделом центральной нервной системы, формулу, которая, как
мы понимаем, полностью гармонирует с соответствующей формулирозкой
И. П. Павлова, является дальнейшим ее уточнением и развитием. Таким
образом, с нашей точки зрения, то, что имело место в опытах М. Н Еро-
1 И. П. П а в л о в, Полное собрание трудов, т. III, 1945, стр. 38Э.
К ФИЗИОЛОГИИ ВРЕМЕННОЙ связи
209
феевой, является ле частным случаем, а основным правилом образования
условного рефлекса. На вопрос, почему же при этом обычно раздражитель
одного из рефлексов становится сигналом для другого рефлекса, можно
ответить кратко: это обусловлено в основном их силовыми соотношениями,
т. е. фактором, значение которого будет рассмотрено ниже.
Условный рефлекс высшего порядка можетбыть представлен как резуль-
тат замыкания условнорефлекторной связи между корковой ветвью без-
условного рефлекса на новый синдиферентиый» раздражитель и дугой
имеющегося условного рефлекса.
Рис. 7. Новая схема образования дуги условного
рефлекса (по Э. А. Асратяну)
УС At — дуга слухи ною подкоркопого безусловного рефлекса;
УС М — дуга слухового коркового безусловного рефлекса;
Я П‘Ж — луга слюноотделительного подкоркового безусловного
рефлекса; ЛП'Ж — дуга слюноотделительного коркового без-
условного рефлекса; П'С' — условнорефлекторная связь;
УС'Л’Я —дуга условного рефлекса
Как было сказана выше, нас интересовали также некоторые вопросы,
касающиеся функциональных особенностей условнорефлекторной связи,
в частности, поднятый И. П. Павловым вопрос о двусторонней проводимости
нервных процессов по этой условнорефлекторной связи.
Само собой разумеется, что нет ничего общего между павловским пред-
ставлением о двусторонней проводимости в корковой части условнорефлек-
торной дуги, основанным на солидных фактических данных, и спекулятив-
ными рассуждениями И. С. Беригова о так называемых «поступательной»
и «обратной» связях между корковыми центрами.
Известно, что вопросу о двусторонней проводимости временной связи
И. П. Павлов в. последний период своей творческой деятельности уделял
большое внимание в связи с назревшей необходимостью расшифровать фи-
зиологическую природу так называемых произвольных движений. Основа-
нием для допущения им существования двусторонней проводимости условно-
рефлекторной связи или, точнее, двойной условнорефлекторной связи по-
служили, с одной стороны, давно известные из практик и дрессировки живот-
ных факты выработки навыка подавать лапу, а с другой стороны, данные
специальных лабораторных экспериментов, приемы которых частично при-
ближались к приемам дрессировки (опыты М. К- Петровой, И. С.Розенталя),
а в основном были выполнены с помощью особых вариаций условнорефлек-
торной методики (опыты Ю. М. Коморского и С. Миллера, Ф. П. Майорова,
Г. В. Скипина, В. К- Федорова, И. О. Нарбутовича и Н. А. Подкопаева).
Лишь И. П. Павлов с присущей ему проницательностью смог обнаружить
в них рациональное зерно и сделать заключение о наличии двусторонней
проводимости или, точнее, о двойной временной связи, а также о том, что
14 э, А. а«г» гяи
210
Э А. АСРАГЯН
двусторонняя связь клеток Двигательного анализатора коры со всеми остал ь-
ными клетками коры и является физиологической основой гак называемых
произвольных движений. Эти фактические данные представляют весьма
пеструю карти ну, что не могло не создать противоречий и изрядной путаницы
в теоретических взглядах названных исследователей.
В нашу задачу не входит здесь обстоятельный критический обзор всех
фактов и взглядов относительно этого вопроса. Мы ограничимся лишь не-
которыми беглыми замечаниями по поводу фактических данных и теорети-
ческих положений К>. М. Конорского и С. Миллера, которым принадлежат
наиболее систематические исследования в этой области.
Работая первоначально в Варшаве, а позднее в лабораториях И. П. Пав-
лова, эти исследователи, бесспорно, получили весьма богатый, оригинальный
и ценный экспериментальный материал, что было отмечено и И. П. Павло-
вым. Однако в теоретическом освещении фактических данных они с самого
начала своей работы стояли и продолжают стоять на ошибочных позициях,
на что неоднократно указывал И. П. Павлов и что привело одного из них —
Ю. М. Конорского, — в конце концов, в лагерь ярых антипавловцев.
В условиях сильно усложненной постановки опытов по изучению услов-
норефлекторной деятельности у собак Ю. М. Конорский и С. Миллер вы-
рабатывали у подопытных животных и исследовали весьма сложные формы
временных связей путем различного рода сочетаний пассивных движений
конечностей и индиферентных раздражителей в отдельности или же в разных
комбинациях с пищевым либо оборонительными рефлексами. При этом они
не только констатировали возможность выработки положительных и отри-
цательных условных рефлексов на пассивные движения конечностей и на
инднферентные раздражители в отдельности или в разных комбинациях, т. е.
не только подтвердили в специфических условиях их опытов некоторые уста-
новленные школой И. П. Павлова закономерности условпорефлекторной
деятельности; ими были выявлены также новые интересные факты, а именно:
превращение пассивных движений, применяемых в качестве условных раз-
дражителей, в активные движения того же или противоположного рода,
вызываемые либо общей обстановкой эксперимента, либо отдельными по-
ложительными и отрицательными условными раздражителями. И вместо
того, чтобы попытаться осветить полученные ими сложные, но интересные
фактические данные с точки зрения передового материалистического учения
И. П. Павлова об условнорефлекторной деятельности, они, невзирая на
прямые указания и советы великого физиолога, стали на ошибочный путь
противопоставления этих новых фактов классическим условным рефлексам,
а в их трактовке — на скользкий путь эклектики, беспринципно заимствуя
для этой цели кое-что у субъективных психологов, кое-что у бихевиористов,
кое-что у гештальтнетов и холистов. Тем самым они загнали перспективное
дело в безвыходный тупик идеализма.
В совместных публикациях этих авторов, в особенности в последних
работах К). М. Конорского, с особым усердием и постоянством утверждается,
что полученные ими новые явления ничего общего по имеют с классическими
условными рефлексами и, П. Павлова— ни по внешним признакам, ни по
механизму, ни по особенностям и значению. IO. М. Конорский, например,
в своей последней книге написал поэтому поводу: «В течение десяти лет автор
был связан с покойным доктором С. Миллером в изучении пластических
явлений, которые не укладываются в схему павловских условных рефлек-
сов. Они представляют собой те явления, которые бихевиористская психо-
логия изучала много летпод термином «навыки» *. 11 далее он пишет, что это
такое явление, «механизм которого полностью отличен от обычного услов-
ного рефлекса» и т. п. Читая эти и подобные им высказывания, невольно
* J. Копог skI, Conditioned reflexes ar d neiron organisation, 1948, стр. 211.
К ФИЗИОЛОГИИ ВРЕМЕННОЙ СВЯЗИ
211
задаешь вопрос: почему же они называли это явление условным рефлексом,
пусть даже «второго типа»? Не является ли это выражением научного лице-
мерия?
В чем конкретно они видят эту коренную разницу между условны-
ми рефлексами, полученными И. П. Павловым, и условными рефлексами,
выработанными ими? В своих трудах они трафарстно указывают на три
таких пункта, из которых первый и самый существенный в формулировке
Ю. М. Конорского гласит так: «Условная реакция первого типа является
всегда воспроизведен нем реакции на подкрепляющий безусловный раздра-
житель, т. е. содержит только те элементы, которые присущи подкрепля-
ющему рефлексу, в то время как эффектом условного рефлекса второго
типа является любое движение животного, до сих пор пе имевшее ничего
общегос пищевой, кислотой или болевой реакцией»2. Второй пункт гово-
рит о том, что при условном рефлексе второго типа условная реакция (дви-
жение конечности) может менять свой характер в зависимости от рода
подкрепления, т. е. является детализацией первого и по существу повто-
ряет его. А третий пункт сводится к тому, что эффектом обычного услов-
ного рефлекса второго типа являются лишь соматические движения.
Оставив в стороне последний пункт как не заслуживающий внимания,
так как он по существу не говорит ни о какой разнице между двумя типами
условных рефлексов, обратимся к первым двум пунктам, толкующим о не-
сходстве условной и безусловной реакций при условном рефлексе второго
типа. Ближайшее знакомство с действительной картиной показывает, что не
существует никакой принципиальной разницы между этими двумя ти-
пами условных рефлексов, что противопоставление К). М. Конорскнм и
С. Миллером изученного ими условного рефлекса павловскому условному
рефлексу лишено всякого научного основания, что первый целиком уклады-
вается в рамки второго и является одним из его разновидностей. На это
указывал еще И. 11. Павлов; приводя соответствующие убедительные до-
воды. Мы приводим новые доводы в пользу этой точки зрения.
По поводу первого пункта можно сказать следующее. Прежде всего нуж-
но иметь в виду, что как активное движение лапы при ее раздражении элек-
трическим Током,так И пассивный ее подъем вызывают определенные пропри-
оцептивные безусловнорефлекторные реакции как деятельности организма,
на основе которых могут быть выработаны различные условные рефлексы.
Раздражителями таких условных рефлексов в числе других могут стать так-
же разные пищевые вещества. В таком случае нет ничего странного в том,
что указанные двигательные условные рефлексы могут быть вызваны и та-
кими раздражителями, которые в свою очередь являются условными для
пищевой деятельности в порядке условного рефлекса второго порядка или
цепного условного рефлекса. Правомерность этих доводов весьма четко
выступает, если исходить из общепринятой формулировки понятия услов-
ного рефлекса. Нам кажется, что она становится еще очевиднее при опреде-
лении условного рефлекса как коркового синтеза разных безусловных реф-
лексов. Для подтверждения несостоятельности второго пункта можно со-
слаться на случаи, в которых условный ответ при обычном условном реф-
лексе может менять свой характер при одном и том же подкрепляющем без-
условном рефлексе, например, при ультрапарадоксальной фазе.
Говоря об отсутствии разницы между павловскими условными рефлек-
сами и описанными Ю. М. Конорскнм и С. .Чиллером двигательными услов-
ными рефлексами, мы имеем в виду, разумеется, отсутствие принципиаль-
ной разницы. Вообще же условный рефлекс — это «центральное явление»
в деятельности больших полушарий — представлен у высших животных
1 J. Konorski, Conditioned reflexs and neuron organisation, IS48, стр. 214.
8 Успехи современной биологии, т. XV, в. 1, 1942, стр. 18.
212
Э А. АСРАТЯН
в огромном многообразии, в разных вариациях, степенях, значениях, отли-
чается рядом функциональных особенностей, С этой точки зрения, описан-
ные 10. М. Коморским и С. Миллером условные рефлексы, несомненно, ха-
рактеризуются некоторыми особенностями, точно так же как и изучаемые,
например, К. М. Быковым и сотрудниками интероцептивные условные
рефлексы или же изучаемые многими нашими учеными условные рефлексы
на так называемые автоматические раздражители. Но ни одна из этих осо-
бенностей не дает оснований рассматривать соответствующую группу
условных рефлексов как нечто принципиально отличное от классических
условных рефлексов И. П. Павлова.
Нам кажется, что некоторые функциональные особенности описанных
Ю. М. Коморским и С. Миллером рефлексов, в первую очередь двусторонняя
проводимость лежащей в их основе временной связи, обусловлены не толь-
ко тем, что в них вовлечены соматические рефлексы с их эффекторными ор-
ганами, но главным образом соотношением интенсивности возбуждения цент-
ров участвующих в них рефлексов. Более того, внимательный анализ экс-
периментального материала, полученного при изучении вопроса о двусто-
ронней проводимости условнорсфлекторной связи, привел нас к убежде-
нию, что одной из главных, если не самой главной причиной пестроты и
путаницы в относящихся к этому вопросу фактах и мнениях являются
недостаточный учет, недооценка, а то и полное игнорирование исследо-
вателями такого важного фактора, как относительная интенсивность воз-
буждения нервных центров. Известно, что И. П. Павлов всегда придавал
этому фактору исключительное значение не только в замыкании временной
связи, но и в высшей нервной деятельности вообще. Говоря, например, о
роли силовых отношений безусловнорефлекторного и индиферентиого раз-
дражителей в процессе замыкания условнорефлекторной связи, он писал;
«Силовым отношениям в деятельности коры надо приписывать, конечно,
первенствующее значение, как это уже давно дает себя если не точно
знать, то многократно замечать»1. Имея ввиду исключительную важность
вопроса, мы приведем еще одно высказывание И. П. Павлова на эту тему,
относящееся к тому периоду, когда результаты знаменитых опытов
М. П. Ерофеевой сильно занимали его ум, — высказывание, имеющее для
нас — его учеников — характер программного указания и на сегодняшний
день. «Относительная интенсивность нервного процесса,—писал он,—опре-
деляет направление нервного раздражения, определяет связь агентов с
различными деятельностями организма. Этими соотношениями интенсивно-
сти переполнена физиология условных рефлексов, и точные определения
от носитель ной интенсивности нервных процессов при действии раз-
личных раздражающих агентов составляют один из важнейших пунктов при
современном изучении нормальной деятельности больших полушарий» 2.
Одной из основных задач руководимой нами лаборатории условных реф-
лексов Института высшей нервной деятельности Академии наук СССР и яви-
лось дальнейшее систематическое многостороннее и углубленное изучение
вопроса о роли соотносительной интенсивности возбуждения нервных
центров в высшей нервной деятельности, вопрос, важное значение которо-
го И. П. Павлов, как уже было отмечено, всегда упорно подчеркивал.
Внимательно обсудив этот вопрос, мы пришли к выводу о необходимости
выделения соотносительной интенсивности возбуж-
дения нервных центров как особое правило деятельности
центральной нервной системы, в частности, коры больших полушарий мозга,
правило, в отношении которого общеизвестные принципы «очага большой
деятельности», «доминанты», «проторения путей», «переключения», «борьбы
за общий конечный путь» могли бы считаться лишь частными моментами.
1 И. П. П а в л о в, Полное собрание трудов, т. Ill, 1949, стр- 382.
’Там же, стр. 192.
К ФИЗИОЛОГИИ ВРЕМЕННОЙ СВЯЗИ
213
Основное содержание этого правила в сугубо предварительном виде мож-
но было бы изложить следующим образом: соотносительная интенсивность
возбуждения в нервных центрах определяет то или иное направление про-
ведения нервных импульсов, то или ино-е направление движения основных
нервных процессов в центральной нервной системе.
В настоящее время нами проводится ряд исследований, направленных на
дальнейшее углубление и расширение наших знаний относительно рати это-
го фактора в формировании и функционировании условнорефлекторной свя-
зи, на проверку правильности наших предположений о важной роли этого
фактора в формировании такой связи с односторонней и двусторонней про-
водимостью возбуждения и, наконец, на дальнейшее подкрепление факти-
ческими данными нашей принципиальной схемы замыкания условнорефлек-
торной связи. Некоторые новые и, как нам кажется, интересные эксперимен-
тальные данные, полученные нашей сотрудницей М. Е. Варга, будут нами
кратко изложены ниже.
Конкретная задача исследования заключается в попытке выработки и из-
учения у собак четко выраженной условнорефлекторной связи с двусторон-
ней проводимостью или двойной условнорефлекторной связи, но в таком
простом виде, чтобы полученные при этом фактические данные легко подда-
вались физиологическому анализу и объяснению. При разработке адэк-
ватной методики для разрешения этой задачи мы решили избежать принци-
пиальной методической ошибки К). М. Конорского и С. Миллера и их по-
следователей, сильно усложнивших условия и приемы эксперимента и полу-
чивших поэтому такие сложные и запутанные фактические данные, которые
на современном уровне наших знаний с большим трудом, а то и вовсе нс
поддаются удовлетворительному физиологическому анализу.
Нас не удовлетворяли также экспериментальные приемы, которыми поль-
зовались первоначально Г. П. Зеленый с сотрудниками, а в последующем
Н. А. Подкопаев и И. О. Нарбутович для решения другой, но близкой к ин-
тересующей нас, задачи. Мы Имеем в виду прием многократного сочетания
в специальных экспериментах двух так называемых индиферентных раздра-
жителей с последующей выработкой на один из них условного электрообо-
ронительного двигательного рефлекса нс испытанием после этого функцио-
нального значения второго из них. Мы считаем, что необходимость прибег-
нуть к дополнительной условнорефлекторной связи, чтобы выявить возмож-
ную временную связь между индиферентными раздражителями, значитель-
но усложняет картину и физиологический анализ полученных данных. Ска-
занное в значительной мере относится к тем приемам экспериментального
решения обсуждаемого вопроса, которыми пользовались Ф. П. Майоров
и В. К. Федоров. Не привлекали нашего внимания также дав.ние приемы
дрессировки подопытных животных на подачу лапы, которыми пользо-
вались М. К. Петрова, И.С. Розенталь идр. Хотя эти приемы и отличаются
значительной простотой, но совершенно непригодны для точного и тонкого
экспериментального анализа, для изучения динамики и особенностей ин-
тересующих явлений. Кстати, и при этом косвенно констатируется явление
двухсторонней связи.
Для дости жения нашей цели мы решили сочетать два таких биологически
значимых раздражителя, силу которых можно былобы варьировать в широ-
ких пределах и, кроме того, безусловнорефлекторные и ожидаемые условно-
рефлек торные эффекты которых могли бы быть объективно, легко- и непо
средственно учтены и точно, даже автоматически, зарегистрированы. Наш
выбор пал на приемы экспериментов, которыми в лабораториях И. П. Пав-
лова когда-то пользовались его ученцкн Н. И. Красногорский, М.Н. Ерофе-
ева и др. для решения иных важных задач, а именно: приемы сочетания пас-
сивного или вызванного электрораздражением активного движения конеч-
ности животного с пищевым (или кислотным) слюноотделительным безус
214
Э. А. АСРАТЯН
ловным рефлексом. Разработанная нами методика может считаться модифи-
кацией этой методики, измененной в соответствии со специфическими осо-
бенностями преследуемой нами цели.
В своих опытах, поставленных на свежих собаках, М. Е. Варга соче-
тала строго попеременно, т. е. то в одной, то в другой последовательности,
пищевой рефлекс сдвижением конечности, причем в одних опытах — с пас-
сивным ее поднятием, в других — с активным ее движением, вызванным
электрическим раздражением. Заметим, кстати, что это важное обстоятель-
ство, т. е. момент попеременного применения сочетаемых раздражителей,
также упускался из виду почти всеми предыдущими исследователями обсуж-
даемого вопроса, о том числе и Ю. М. Конорскнм и С. Миллером, К. С. Абу-
ладзе, Г. В. Скипиным и др., что крайне затрудняло решение поставленной
ими задачи. Мы стремились создать максимально близкие условия примене-
Рис. 8. Схема дуги [условного рефлекса 'с двойной
связью
Обозначения те же, что на рис. 7
ния двух сочетаемых раздражителен как в отношении длительности их дей-
ствия, так и по возможности в отношении их сил. Учитывая, в частности,
особую биологическую силу и главным образом инертность пищевого без-
условного рефлекса, мы для достижения указанной цели вызываем его не-
большим количеством вкусной пищи. В тех же целях конечности раздража-
ются нами индукционным током такой силы, которая немного превышает
порог возбуждения.
Не считая возможным вдаваться здесь.в детали полученных М. Е. Вар-
гой фактических результатов, остановимся на наиболее существенных из
них. М.Е.Варга установила возможность коркового синтеза этих двух без-
условных рефлексов, т. с. образования условного рефлекса с двусторонней
проводимостью его временной связи, или, точнее, двойной временной свя-
зи, к тому же в таком простом и, если можно так выразиться, чистом виде,
в каком такая связь у животных до сих пор еще никем небыла описана. Каж-
дый из сочетаемых безусловных раздражителен превратился в условный
раздражитель другого безусловного рефлекса: подача пищи вызывает л бе-
условпый пищевой рефлекс, и условнорефлекторный подъем лапы, а пассив-
ное или активное движение конечности в свою очередь вызывает и условно-
рефлекторную пищевую реакцию (рис. 8). Приведенные кинограммы до-
статочно четко иллюстрируют сказанное (рис. 9—10).
Таким образом были выработаны у сооак условные рефлексы как бы с
равноценными и равнозначными в функциональном отношении концами.
Из этого основного нашего факта, точного и проверенного, с полной очевид-
К ФИЗИОЛОГИИ ВРЕМЕННОЙ связи
215
постью вытекает несостоятельность взглядов Ю. М. Конорского и С. Мил-
лера о существовании якобы принципиальной разницы между описанными
ими условными рефлексами второго типа и классическими условными реф-
лексами И. П. Павлова. А ведь на этой мнимой, выдуманной разнице и по-
строены основные антипавловские положения Ю. М. Конорского, который
как физиолог в лагере зарубежных антипавловцев считается чуть ли не
наиболее сильным противником материалистического учения о высшей
нерв.ной деятельности.
л
б
Рис. 9. Кинограмма условных рефлексов у собаки Чернуха
а — условибр«4>лсктйрнй4 елкб>еййуд.«л«ние на паесибныП подъем лапы,-
б — услоинорефлектор ное движение лапы на подачу кормушки.
Линин сверху пни»; движения лапы, слюноотделение, отметка подачи
кормушки, отметка пассивного подъема лапы, время в секундах
Пам кажется, что этот факт является также серьезным подкреплением
предложенной нами принципиальной схемы замыкания и функционирования
условнорефлекторной связи. Из этого факта бесспорно явствует обуслов-
ленность одностороннего или двустороннего проведения процесса возбуж-
дения по условнорефлекторной дугев первую очередь соотносительной ин-
тенсивностью возбуждения сочетаемых безусловных рефлексов.
Особого внимания заслуживает динамика формирования такого услов-
ного рефлекса. Более или менее четко наблюдаются следующие четырефазы.
Во-первых, следует отметить кратковременную фазу тормозящего действия
б
'1!111Ч1П||1,||Г!Н|1 llll'IIIIMIIINIHI’l |1|1|ЛП1ГИТТТГ.*’ГТ1
<7
Рис. 10. Килограмма условных рефлексов у собаки Щ-сник
а — уелпииорефлекториев слюноотделение из электрическое раздражение
лапы; б — условнорсфлек торное движение лапы на подачу кормушки.
Линии сверху нниз=: движения лапы, слюноотделение, отметка подачи кор-
мушки, отметка электрического раздражения лапы, время в секундах
одного безусловного рефлекса на следующий за ним другой безусловный
рефлекс, очевидно, в порядке взаимной индукции. Вторая фаза характери-
зуется образованием условнорефлекторной связи с односторонним проведе-
нием возбуждения, а именно: с проведением возбуждения от менее сильного
безусловного рефлекса к более сильному. В опытах с сочетанием пассивного
движения с пищей в этой фазе подъем лапы уже начинает вызывать условно-
рефлекторное слюноотделение; а в опытах с сочетанием электрооборонитель-
ного двигательного рефлекса С пищей подача пищи в данной фазе уже начи-
216
Э. Л. АСРАТЯН
нает вызывать подъем лапы. Рефлексы противоположных направлений в этой
фазе еще отсутствуют. В третьей фазе формирования условнорефлекторной
связи наблюдается интересное явление: сильная зависимость осуществления
каждого безусловного рефлекса от следующего за ним раздражителя другого
безусловного рефлекса. В этой фазе, например, слюноотделение при подаче
пищи сильно задерживается, если вслед за подачей кормушки собака не
поднимает лапы. И, наоборот, подъем лапы в ответ на электрическое раздра-
жение задерживается, если вслед за этим раздражением не подается кормуш-
ка. Наконец, наступает завершающая фаза — фаза проведения в корковой
части условнорефлекторной дуги возбуждения и в обратном направлении
либо образования новой такой связи со способностью проводить возбужде-
ние в обратном направлении.
В заключение нужно заметить, что последовательность третьей и четвер-
той фаз неодинакова у разных подопытных собак. У иных собак обратная
проводимость условнорефлекторноп связи, или вторая связь, устанавливает-
ся очень скоро после первой, почти одновременно с ней. У этих собак фаза
резко выраженной зависимости осуществления каждого безусловного реф-
лекса от предварительного применения другого раздражителя (ставшего
для него условным) наступает после фазы двусторонней проводимости, т. е.
уже в качестве четвертой, а не третьей, как было описано выше. В этих слу-
чаях имеется как бы пятая фаза — ослабления и исчезновения этой зависи-
мости.
Нами были приведены первые точные данные, послужившие основанием
Для сделанных выше выВоДов. Но к этим выводам может быть добавлен еще
один, а именно: разработнный нами прием исследования важнейшего раз-
дела учения о высшей нервной деятельности — закономерностей замыкания
условнорсфлскторной связи и ее функционирования — является весьма
перспективным и многообещающим. И мы намерены в дальнейшем исполь-
зовать его возможно шире.
Переходим к третьему вопросу доклада.
К, особенностям функционирования временной связи имеют также близ-
кое отношение проводимые нами совместно с сотрудниками исследования
закономерностей условнорефлекторного переключения.
«Существенный признак высшей нервной деятельности, — писал
И. П. Павлов, — которой мы будем заниматься и которая в высшем живот-
ном принадлежит, вероятно, исключительно большим полушариям, состоит
не только в том, что здесь действуют бесчисленные сигнальные раздражи-
тели, ио и в том существенно, что они при опреде-
ленных условиях меняют свое физиологическое
действие» (разрядка наша.—Э. /1.) Ч Мысль о варнабильиости, измен-
чивости сигнального значения раздражителей И. П. Павлов высказывал
во многих своих произведениях, основываясь на многочисленных конкрет-
ных фактах двух основных категорий: на фактах*, свидетельствующих о пе-
ремене функционального знака сигнального раздражителя
(из положительного в отрицательный или наоборот) в пределах данного рода
условнорефлекторной деятельности, и фактах перемены рода сигналь-
ного значения раздражителя (пищевого вэлектрооборонительный или на-
оборот). Как в одном, так и в другом случае каждый раздражитель в данное
время обладает обычно только одним сигнальным значением, не считая пе-
реходных фаз в процессе переделки сигнального значения, когда раздражи-
тель в известной степени совмещает сба противоположных сигнальных зна-
чения. В случае разнородных условных рефлексов это находит свое выра-
жение в том, что при действии такого раздражителя одновременно вызы-
ваются сба рефлекса (опыты С. С. Фридемана, Г. П. Конради и др.).
1 И. П. П а в .<о в, Полное собрание трудов, т. IV, 1947, стр. 28—29.
К ФИЗИОЛОГИИ ВРЕМЕННОЙ СВЯЗИ!
217
Несомненно, к этому вопросу имеют непосредственное отношение также
отдельные наблюдения сотрудников И. П. Павлова, отметивших, что неред-
ко в силу каких-либо существенных перемен в условиях эксперимента, при-
водящих к значительному изменению обычного соотношения уровней возбу-
димости или возбуждения центров, важнейших безусловных рефлексов, ус-
ловный раздражитель быстро меняет свое сигнальное значение, вызывая
деятельность доминирующего из этих центров (опыты Г. II. Конради и др.).
Совершенно очевидно, что эта перемена рода сигнального значения раз-
дражителя по своему физиологическому механизму идентична явлению, на-
блюдавшемуся в столь яркой форме в опытах М. Н. Ерофеевой, т. е. является
переключением нервных импульсов с одних путей на другие в силу врож-
денных форм деятельности центральной нервной системы. Как раз имея
в виду это явление, И. П. Павлов писал: «Нервное возбуждение от данного
агента, ранее шедшее в один отдел нервной системы, теперь направляется в
другой. Следовательно, совершился переход нервного тока с одного путина
другой, произошло переключение нервного тока. Перед нами стоит ясный
факт, что в высшем отделе нервной системы сюда пришедшее раздражение,
смотря по условиям, проводится то в одном, то в другом направлении. Нуж-
но думать, что именно это составляет главнейшую функцию самой верхней
части нервной системы» *.
В процессе своих экспериментальных исследований Ю. М. Конорский
и С. Миллер также получили некоторый фактический материал, говорящий
о том, что при наличии у собак разнородных условных рефлексов (скажем,
пищевых и электрооборонительпых) иногда одни и те же условные раздра-
жители вызывают то рефлекс одного рода, то рефлекс другого рода, в за-
висимости о того, какой род возбудимости преобладает в больших полуша-
риях в данное время (пищевая или оборонительная). Аналогичное явление
они получали и тогда, когда в порядке экстренного изменения условии экс-
перимента повязку для регистрации движений конечностей прикрепляли
к конечностям, к которым раньше она никогда не прикреплялась: условные
раздражители часто вызывали движения этих конечностей, а не той конеч-
ности, с которой эти условные раздражители были связаны. «Из всех этих
опытов видно, — писали они, — что в. двигательном условном рефлексе
реакция не является строго приуроченной к раздражителю, на который она
была выработана, а зависит от следов предыдущих раздражителей и их ре-
акций и от всей обстановки, действующей на животное в данный «момент» *.
Эти явления были охарактеризованы авторами как «подмена», «готовность»,
«переключение» и использованы для обоснования своих антипавловских тео-
ретических положений, в частности, положения сб условных рефлексах вто-
рого типа. Этим явлениям много внимания уделяет К>- М, Конорский и в
последней своей монографии. Однако в ней, так же как и в прежних его пуб-
ликациях, они выдаются за оригинальные, новые, и ни единым словом не
упоминаются идентичные или очень близкие данные, полученные задолго
до него многими сотрудниками в лабораториях И. П. Павлова (С. С. Фри-
деман, П. Н. Васильев,М. Н. Ерофеева, А. А. Савич, Я. Е. Егоров, Г. П. Кон-
ради и др.), глубоко, всесторонне, строго научно освещенные гениальным
физиологом как раз с точки зрения соотносительной интенсивности возбуж-
дения нервных центров и охарактеризованные им как переключение нервных
импульсов с одних путей на другие.
Выявленное нами в услов-норефлекторнон деятельности переключение
существенно расширяет круг наших знаний о способах изменения сигналь-
ного значения раздражителей всоответствии с изменениями условий жизни,
в соответствии с текущими потребностям и организма. В отличие от всех пере-
1 И. П. П а в л о в, Полное собрание трудов, т. III, 1949, стр. 207.
а Труды физиологических лабораторий акад. И. П. Павлова, т. VI, в. I, 1936,
стр. 241.
218
Э. А. АСРАТЯН
численных выше способов в данном случае один и тот же раздражитель о д-
новременнои стойко сохраняет за собой два (а мо-
жет быть, и большее число) сигнальных значения, причем
выявляет их не вместе, а порознь, соответственно условиям. Кроме того,
в-данном случае переключение, как «условные рефлек-
сы, самовырабатывается. Не приходится и говорить, что эта
новая закономерность условнорефлекторной деятельности делает ее еще бо-
лее совершенной, гибкой, динамичной и вариабильной, еще более совершен-
ным средством приспособления организма к условиям существования.
Как известно, это важное явление спинальной рефлекторной деятель-
ности, установленное впервые И. М. Сеченовым, разрабатывается нами при-
менительно к условнорефлекторной деятельности коры больших Полушарий
мозга, начиная с 1936 г. В нашу задачу здесь не входит изложение факти-
ческих и теоретических результатов наших исследований в этой области.
Они, равно как и результаты исследований некоторых других наших уче-
ных (И. И. Лаптева, Э. Г. Вацуро, В, В, Яковлевой и В, К- Федорова,
И. С. Алексеевой, И. С. Беритова и др.), приступивших к изучению той
же проблемы вслед за нами, хорошо известны по соответствующим публи-
кациям. Мы сообщим лишь некоторые новые экспериментальные данные, по-
лученные нашими сотрудниками за последние годы, а также поделимся не-
которыми теоретическими соображениями поэтому вопросу, имеющими так-
же отношение к функциональным особенностям условнорефлекторной связи.
До сих пор мы совместно с сотрудниками исследовали принцип переклю-
чения в условнорефлекторной деятельности преимущественно в двух глав-
ных вариантах: либо па один и а от же раздражитель в разных условиях экс-
перимента вырабатывали два различного рода условных рефлекса (скажем,
пищевой и электрооборонительный), либо на один и тот же раздражитель
в разных условиях эксперимента вырабатывали положительный и отрица-
тельный условные рефлексы одного рода. В порядке усложнения последнего
варианта эксперимента мы иногда в одних условиях на два близких раздра-
жителя вырабатывали два условных рефлекса противоположного функцио-
нального знака, а в других условиях переделывали их функциональные
знаки в противоположные. «Переключателями» во всех этих опытах слу-
жили то замена экспериментатора, то перемена камеры, то изменение
времени или последовательности постановки экспериментов и т. д. Дру-
гие исследователи, также изучавшие эту проблему, насколько нам из-
вестно, не выходили более или менее значительно из рамок этих вариантов
э ксп ер и ментиров ания.
Возобновляя после длительного перерыва разработку этой чрезвычайно
важной и актуальной в учении о высшей нервной деятельности проблемы,
мы решили, в частности, выяснить, возможно ли выработать дв.а разнород-
ных условных рефлекса на два раздражителя (скажем, на один раздражи-
тель—пищевой, а на другой—электрооборонительный), а в других условиях
эксперимента переделать их сигнальное значение, т. е. возможно ли одно-
временное существование двух разнородных условных рефлексов с перемен-
ным сигнальным значением их раздражителей. Тщательно проведенные экс-
перименты нашего сотрудника М. И. Стручкова дали бесспорно положи-
тельный ответ на этот вопрос. В одной из камер, находящихся на третьем
этаже, М. И. Стручков вырабатывал у подопытных собак пищевые условные
рефлексы на звук зуммера и электрооборонительные — на касалку. После
выработки и значительного укрепления этих разнородных условных реф-
лексов М. И. Стручков начал ставить на тех же собаках параллельно экс-
перименты в камере, находящейся на первом этаже, сочетая теперь касалку
с пищей, а зуммер с электрическим раздражением лапы. Эти опыты не могли,
разумеется, нс оказать значительного влияния на условнорефлекторную
деятельность собак в верхней камере и вообще на их высшую нервную де-
к ФИЗИОЛОГИИ ВРЕМЕННОЙ СВЯЗИ
219
ятсльность. Болес того, решение этой трудной задачи сопровождалось раз-
витием невротических состояний у собак. Мы лишены здесь возможности
хотя бы кратко изложить поучительную историю решения этой задачи
собаками. Важно, однако, отмстить, что, проходя через фазу двойной ус-
ловнорефлекторной реакции па каждый раздражитель, через фазу некото-
рого взаимного торможения их, через фазу постепенного усиления адэкват-
ного к ослабления неадэкватного реагирования в данной камере условных
рефлексов на каждый раздражитель, собаки, в конце концов, решили задачу,
к тому же с предельной четкостью и ясностью. Приводимые ниже кимограм-
мы трех пар опытов М. И. Стручкова на одной и той же собаке достаточно
убедительно иллюстрируют сказанное (рис. 11 — 13).
пч пиinmiiiniiiнпи -------iiiiinii । in i
Касалка
_________Я ______ЛЛЛПП---------
Зуммер -------- ,--------
НИИ 1ПП П|1 II !П III 1111110 V 11НПШШ inillllHIIIII—
а 6
Рис. !1. Кимограмма условных реф-
лексов у собаки Пяткатка. с5пыт
в верхней камере
Опыт перед началом переключения. В от-
вет на раздражение зуммером выделяется
18 кашель слюны (а), в ответ на касалку
собака поднимает ногу и одновременно
с этим выделяется слюна (б).
Линии сверху вниз: а — оборонительны А
рефлекс, слюноотделение, отметка подачи
пищи, условный раздражитель, время
в секундах; б — оборонительный рефлекс,
слюноотделение, условный раздражитель,
электрический раздражитель, время в се-
кундах
пи niuian ---------
Касалка
П _____МАЮ.
Зуммер ________________
пчнянпинччииг шпини iniaiili nil luillin-
а б
Рис. 12. Кинограмма условных
рефлексов у собаки Иятнатка
12-Й опыт в верхней камере после
начала переключения (в нижней
камере переключение уже вырабо-
талось). В ответ на зуммер появ-
ляется чисто пищевая реакция (а),
в ответ на касалку — чисто обо ро-
интельная. Слюнный компонент
исчез (б).
Линии сверху вниз: а — оборони-
тельный рефлекс, слюноотделение,
отметка подачи пищи, условный
раздражитель, время в секундах;
б — оборонительный рефлекс, слю-
ноотделение, условный раздражи-
тель, электрический раздражитель,
время в секундах
Наши, совместно с сотрудниками, исследования, касающиеся переклю-
чении в условкорефлекторной деятельности/а также почти все подобные ис-
следования других ученых выполнены на традиционном подопытном живот-
ном павловских лабораторий — на собаке, если не считать некоторых на-
блюдений Э. Г. Вацуро, проведенных па обезьянах. А между тем совершен-
но очевидно, что систематическое сравнительно-физиологическое изучение
этой важнейшей закономерности высшей нервной деятельности представ-
ляло бы большой научный интерес. Мы приступили к исследованиям вэтом
направлении и уже располагаем некоторыми фактическими данными.
Наша сотрудница Г. И- Шкркова проводила на трех взрослых низших
обезьянах серию соответствующих экспериментов, по методике выработки
двигательных условных рефлексов с пищевым подкреплением. Ход опытов
регистрировался кимографическн. Опыты ставились два раза в день — ут-
ром и вечером. В утренних опытах метроном 120 ударов в 1 минуту служил
условным положительным раздражителем, метроном 60 ударов в 1 минуту—
отрицательным. После закрепления этих рефлексов в параллельных вечер-
них экспериментах Г. И. Ширкова стала изменять сигнальное значение раз-
дражителей, т. е. переключать их. Полученные ею фактические данные поз-
220
Э. А. АСРАТЯН
imunTinr.iiiMnr.il ill II. и и ||
коса/гк а
ТГ1ГПИП1И1НН11ПИ
воляют считать, что обезьяны справляются с подобной сложной задачей
значительно легче и быстрее, чем собаки, что и понятно, если иметь в виду
более высокий уровень развития и ббльшую динамичность их высшей нерв-
ной деятельности. Но даже они решают эту задачу не без известных кон-
фликтов и борьбы между старыми и новыми рефлексами. Ниже приводятся
кимограммы утреннего и вечернего опытов того же дня на одной из
подопытных обезьян после решения ею задачи адэкватного реагирования
на каждое из противоположных сигнальных значений условных раздражи-
телей соответственно условиям экс-
перимента (рис. 14).
Студенты II Московского государ-
ственного медицинского института
имени И. В. Сталина—В. Орлов,
В. Модестова и В. Тарасов—выполни-
. ли небольшую экспериментальную ра-
боту на крысах, также посвященную
этой теме. Используя модифициро-
ванную ими методику Л. И. Котля-
ревского для изучения пищевых дви-
гательных условных рефлексов у мел-
а б
Рнс. 13. Кинограмма условных рефлек-
сов у собаки Пятнатка
24-Й опыт после начала переключения r ниж-
ней камере. Опит иллюстрирует полную «м-
работку переключения. В ответ на касалку
оборонительная реакция отсутствует и раз-
вивается только пищевая (л). В ответ на
зуммер (б) пищевая реакция отсутствует,
имеется только оборонительная.
Линии сверху впил: а — оборонительный реф-
лекс, отметка подачи пищ и, слюноотделение,
условный раздражитель, время в секундах;
—оборонительный рефлекс, отметка элек-
трического раздражителя, слюноотделение,
условный раздра>китсль. время в секундах
ких грызунов, они показали на кръь
сах возможность своеобразного пе-
реключения условных двигательно-
пищевых рефлексов на один и гот
же раздражитель, а именно: образо-
вывалась то побежка к одной кор-
мушке, то побежка к противополож-
ной кормушке. Опыты показали также
что крысы способны к переключе-
нию в пределах разнородных услов-
ных рефлексов — пищевого и электро-
оборонительного, а именно: в зави-
симости от условий эксперимента,
они на один и тот же условный раздражитель давали то пищевую, тс
оборонительную реакцию. Следует, однако, заметить, что, несмотря на при-
менение в этих экспериментах ие одного, а целого комплекса переключа-
телей— экспериментатор, порядок экспериментов, специальное освеще-
ние, — крысы решали эти задачи с большим трудом и крайне медленно, во
всяком случае гораздо медленнее, чем собаки и тем более обезьяны.
Переходя к теоретическому обсуждению вопроса, мы хотели бы сначала
уточнить, идентичны лн явления, названные Ю. М. Конорским «корковым
переключением» (которые, как мы видели выше, ие представляют собой
принципиально ничего нового по ср др нению с тем, что давно было известно из
исследований лабораторий И. П. Павлова), тем явлениям, которые были по-
лучены нами и нашими сотрудниками и также названы «переключением»
в условнорефлекторной деятельности.
Нам представляется, как уже было сказано, что речь идет о совершенно
различных по своей природе и механизму явлениях и поэтому нецелесооб-
разно обозначать их одним термином. В перечисленных выше ранних на-
блюдениях сотрудников И. П. Павлова и исследованиях Ю. М. Конорского
переключение нервных импульсов с одних нервных путей па другие не яв-
ляется результатом специальной выработки, специального образования вре-
менных путей н переключателей к ним, а представляет собой безусловно-
рефлекторное изменение соотносительной возбудимости (пли интенсивности
возбуждения) разных корковых центров. Иначе говоря, в этих случаях
механизм переключения врожденный, имеет место переключение уже гото-
К ФИЗИОЛОГИИ ВРЕМЕННОЙ СВЯЗИ
221
вых условных рефлексов под воздействием различных иезусловнорефлек-
торных факторов. В нашем же случае переключение само носит истинно
условнорефлекторный характер, оно специально вырабатывается, переклю-
чатель сам является условным раздражите-
лем. Вот почему мы и думаем, что термин
«условнорефлекторное переключение» или
«переключение» в условнорефлекторной дея-
тельности следует закрепить за изучаемым
нами явлением, оставив для обозначения тех
явлении термин «подмена» или какой-либо
другой, более подходящий к их содержанию.
Интимную сторону условнорефлекторного
переключения мы представляем себе следую-
щим образом. Из факта выработки двух раз-
личного рода либо различного знака функци-
онирующих условных рефлексов на один и тот
же раздражитель можно сделать вывод, что
от коркового центра такого рефлекса к дру-
гим корковым центрам идут два УСЛОВНО
рсфлекторных пути, находящихся в фазе
четкого переключения во взаимно антагони-
стических, реципрокных отношениях. «Пере-
ключатель» как особого рода условный раз-
дражитель, а именно как тонический услов-
ный раздражитель, при своем действии соз-
дает в коре определенный функциональный
фон тонического характера, определенную
функционально-динамическую структуру от-
крытых и закрытых путей и таким образом
предопределяет характер и род условнореф-
лекторной деятельности.Этотсвоеюбразный и
специфический условный раздражитель в
одних условиях закрывает один из названных
путей от центра каждого условного раздра-
жителя и открывает другой путь, а в других
условиях изменяет картину 'в диаметрально
противоположном направлении. Действую-
щий на этом фоне обычный фазный, или пу-
сковой, условный раздражитель направляет
поток своих нервных импульсов через откры-
тый условпорсфлекторный путь и вызывает
соответствующую деятельность организма.
Нам кажется, что имеется нечто общее
между состоянием коры при действии «пере-
ключателя» и между гем ее состоянием, кото-
рое когда-то И. П. Павлов предположитель-
но считал физиологической основой сознания.
По этому повод}' он писал: «С этой точки
зрения сознание представляется мне нервной
деятельностью определенного участка боль
линия — время о секунда*
ших полушарий в данный момент, приданных условиях, обладающего из-
вестной оптимальной (вероятно, это будет средняя) возбудимостью. В этот
же момент вся остальная часть больших полушарий находится в состоя
нии более или менее пониженной возбудимости»
1 И. П. П а в л о в. Полное собрание трудов, т Ill, 1919, стр 196—197.
222
Э. А. АСРАТЯН
И. П. Павлов считал, что отдел больших полушарий с оптимальной воз-
будимостью является в данный момент «творческим» его отделом, «светлым
пятном», окруженным на всем остальном пространстве полушария более
или менее значительной тенью, что он, этот отдел, не является закреплен-
ным, а «постоя нно перемещается по всему пространству больших полушарий
в зависимости от связей, существующих между центрами, п под влиянием
внешних раздражений»1.
Нам кажется, что великий учитель с присущей ему гениальной прозор-
ливостью еще 40 лет назад необычайно четко обрисовал физиологическую
панораму коры при условнорефлекторпом переключении. Единственное
небольшое замечание, какое может быть сделано к этому описанию на ос-
новании нашего фактического материала, сводится к допущению, что опти-
мальная и пониженная возбудимость, точнее заторможенность, подвижно
охватывает не разные отделы мозга в топографическом понимании, а разные
динамические функциональные структуры коры больших полушарий.
Переходим к заключительной части доклада.
Как уже было отмечено выше, одним из проявлений антипавловского
направления в современной зарубежной и отчасти пашей физиологии
является абсурдное утверждение относительно возможности образования
условных рефлексов у высших животных даже не в ближайших или от-
даленных подкорковых нервных ганглиях, а в самом низшем отделе цен-
тральной нервной системы—в спинном мозгу.
Неглубокий смысл этих лженаучных утверждений ясен: «доказать»
ошибочность фундаментального материалистического положения И. П. Пав*
лова о том, что условный рефлекс является функцией только высшего отде-
ла центральной нервной системы и составляет сущность, содержание высшей
нервной деятельности. При этом в зарубежной периодической печати, осо-
бенно американской, этот вопрос привлекает все больше и больше внимания,
а экспериментальная его разработка и обсуждение полученных при этом
данных приняли такой характер и достигли таких пределов, что мы, ученики
и последователи И. П. Павлова, не можем пройти мимо всего этого и огра-
ничиться только выражением недоумения пли негодования. Так, например,
сели лет 8—10 назад Шуррагер (Shurragcr), Келлер (Culler) и др. на осно-
вании своих экспериментов говорили только о спинальных условных реф-
лексах у высших животных, то в последние годы Франциске (Franziskel)
выступает с утверждением, будто и спинной мозг лягушки способен к обра-
зованию условных рефлексов, а Сгонина (Sgonina) утверждает, что услов-
ный рефлекс может быть выработан даже на изолированном сердце лягушки.
Показательно и то, что американский исследователь Келлог (Kellog), ко-
торый на основании результатов своей проведенной совместно с сотрудни-
ками экспериментальной работы относится критически к фактам и взгля-
дам своих коллег по этим вопросам и придерживается более умеренной точ-
ки зрения, считает, что феномен образования специального условного реф-
лекса является самым замечательным открытием нашего времени в области
физиологии центральной нервной системы.
А между тем ближайшее ознакомление с приемами экспериментального
изучения этого вопроса американскими исследователями и с фактическими
данными убеждает в крайней несолидности, легковесности, ложности этой
основы. Мы лишены возможности разбирать здесь этот вопрос обстоятель-
но, но, чтобы дать некоторое конкретное представление о нем, укажем на
ряд характерных моментов.
Исследования Шуррагера и его сотрудников выполнены в условиях ост-
рого, вивисекционного опыта на собаках сразу после перерезки спинного
мозга, г. е. в условиях, при которых ход и характер спинальной рефлектор-
1 И. П. Павлов, Полное собрание трудов, т. III, 1949, стр. 197.
К ФИЗИОЛОГИИ ВРЕМЕННОЙ СВЯЗИ
223
ной деятельности грубо нарушены. Далее, подвергая длительному сочетан-
ному раздражению хвост и одну из задних конечностей собак электрическим
током, авторы даже не определяли порога возбудимости этих органов, что-
бы в дальнейшем исходить из него при практическом решении вопроса о
силе применяемых раздражений, как это обычно делается в физиологиче-
ских исследованиях подобного рода.
Результаты этой многолетней экспериментальной работы сводятся к
установлению факта, что после многократного (до 1000 и больше) и много-
часового сочетанного раздражения у свежеоперированных собак хвоста
слабым электрическим током и одной из задних лап — более сильным током
раздражение одного только хвоста вызывает сокращение полусухожильной
мышцы раздражавшейся ранее конечности, к тому же часто не всей мыш-
цы, а лишь отдельных ее пучков. Явление это сохраняется недолго— в пре-
делах десятка минут. Оно было названо условным рефлексом не только по-
тому, что возникает в результате сочетанного раздражения двух разных
органов животного, но и потому, что оно способно исчезнуть при дву*
троекратном раздражении одного хвоста без сочетанного раздражения
конечности. При этом на американских исследователей не подействовал
отрезвляюще и тот установленный ими факт, что в противоположность ди-
намике развития истинных условных рефлексов больших полушарий, т. е.
первоначальной генерализации и последующей концентрации, это явление
сначала возникает в виде узко локализованных сокращений отдельных
мышечных волокон, затем постепенно все больше и больше расширяет
свои границы — генерализуется. Они ограничились только констатацией
этого несоответствия.
Исследования того ж:е вопроса другими американскими учеными не вы-
ходят из описанных рамок ин в методическом, ни в каком-либо ином отно-
шении, если нс считать экспериментов Ксллога и его сотрудников, постав-
ленных в условиях хронического опыта на заблаговременно оперированных
спинальных собаках и приведших их к несколько иным результатам и более
сдержанным выводам.
Важность вопроса об изменении характера рефлекторной деятельности
спинного мозга в условиях длительных раздражений и, в частности, необхо-
димость экспериментальной критики, т. е. критики острым языком точных
фактов, изложенных вышеданных и взглядов американских исследователей
побудили нас заняться экспериментальной разработкой и этого вопроса.
11о нашей просьбе эту ответственную и нелегкую задачу взяли на себя сотруд-
ники лаборатории общей физиологии Института высшей нервной деятель-
ности Г. II. Несмеянова и Н. М. Шамарина.
В значительном количестве острых И хронических экспериментов, по-
ставленных на свежеоперированных и заблаговременно оперированных
спинальных Собаках в строгом соответствии с основными требованиями со-
временных физиологических исследований, Г, н. Несмеянова и Н. М. Ша-
марина получили весьма ценный фактический материал относительно-
изменений характера рефлекторной деятельности спинного мозга при дли-
тельных раздражениях, Этот материал, представляющий большой науч-
ный интерес, в частности, с полной убедительностью доказывает порочность
упомянутых выше фактических данных американских исследователей и
абсурдность их теоретических положений относительно возможности выра-
ботки условных рефлексов спинным мозгом.
Г. Н. Несмеяновой и Н. М. Шамариной было доказано, что в результате
многократного сочетанного раздражения хвоста и одной из задних лап элек-
трическим током возникает не однородный, неизменный и односторонний
рефлекторный ответ в виде сокращения полусухожильной мышцы при раз-
дражении хвоста, как утверждают Шуррагер и др., а целая серия разнооб-
разных и варьирующих по силе и характеру рефлекторных реакций на фоне
224
Э. А. АСРАТЯН
резкого повышения возбудимости спинного мозга, а именно: движение
хвоста при раздражении одной нз задних лап, движение лап при раздраже-
нии хвоста, явления «отдачи» при сокращении мышц хвоста и конечностей
и, что заслуживает особого внимания, рефлекторное сокращение ранее ни-
когда не раздражавшихся частей задней половины тела при раздражении
хвоста или лапы, равно как сокращение последних при раздражении пер-
вых и т. П. Еще больше подчеркивают ошибочность взглядов американских
.|ДД
\ V
исследователей следующие данные Не-
смеяновой и Шамариной. Многократное
и длительное раздражение одного только
хвоста без сочетания с раздражением
лапы или же одной только лапы без со-
четания с раздражением хвоста дает
точно такие же результаты: появление
не свойственных данным рецептивным
полям рефлекторных реакций (рис. 15),
причем эти реакции могут длиться
днями и даже неделями, если порожда-
ющие их раздражения действуют дли-
тельное время.
Из всего вышеизложенного явствует,
что при длительном изолированном
раздражении какой-нибудь части тела,
связанной со спинным мозгом ниже
места перерезки, так же как и при дли-
тельном раздражении двух различных
связанных с ним органов, в спинном
мозгу возникают и развиваются очаги
повышенной и застойной возбудимо-
сти, которые обусловливают явления
типа «проторения путей» и особенно
«доминанты», к тому же в
ванном виде, — явления,
были ошибочно оценены американски-
ми исследователями как условные реф-
лексы. Явления же типа «проторения
путей» и «доминанты», как известно, принадлежат к числу универсальных
для всей центральной нервной системы. Рызыгрываясь в коре больших полу-
шарий, они, бесспорно, играют важную роль в замыкании и функциониро-
вании условнорефлекторной св.язи; разыгрываясь же в спинном мозгу, они,
согласно взглядам И. П. Павлова о «естественной общности основных отно-
шений», могут считаться отдаленными прототипами, примитивными родо-
начальниками, как бы своеобразными биологическими корнями истинных
условных рефлексов. Метафизически мыслящие американские исследова-
Рис. 15. Рефлекторные сокраще-
ния лапы в ответ на длительное
и многократное (1250 раз) раз-
дражение хвоста
Линии сверху они»: запись движения
правой лапы, запись движения левом
лапы, запись движения хвоста, отмет-
ка раздражения хвоста, отмотка раз-
дражения лапы, время > секундах
утриро-
когорые
тели механически поставили знак равенства между этими явлениями и ус-
ловными рефлексами. В этом корень их ошибок.
Они, равно как и те из наших физиологов, которые, отдавая дань моде,
легкомысленно и голословно защищали лженаучный тезис о спинномозго-
вых «временных связях», не поняли принципиальной качественной разницы
между временностью летучей, неминуемо обреченной на исчезновение связи
при «проторении путей» или при явлении «доминанты» и временностью ус-
ловнорефлекторной связи, которая является стойкой, стабильной и может
сохраняться в течение всей жизни. Они не поняли, что условнорефлектор-
ная связь называется временной в том только смысле, что она обладает спо-
собностью при необходимости исчезать, точнее, затормаживаться. Вот что
писал И. П. Павлов по этому поводу: «В то время как суммационнын реф-
К ФИЗИОЛОГИИ ВРЕМЕННОЙ связи
225
леке есть моментальное и скоропреходящее явление, условный реф-
лекс есть постепенно укрепляющееся при вышеуказанном условии хрони-
ческое явление, представляющее характерный процесс коры» х.
Нетрудно видеть, что описанные выше состояния повышенной и застой-
ной возбудимости если не идентичны, то по меньшей мере родственны дав-
но описанным Н. Е. Введенским у лягушек явлениям «истериозиса», которое
у высших позвочных недавно было установлено Э. III. Анрапетьянцем и со-
трудниками. Важно отметить, что в тех опытахГ. Н. Несмеяновой иН.М.Ша-
мариной, в которых они длительно раздражали хвост и конечность
(изолированно или сочетанно, безразлично), это состояние повышенной
возбудимости с характерной для него рефлекторной деятельностью сохра-
нялось в течение нескольких недель, т. е. оно приобретало характер пато-
логической доминанты и в соответствии с этим придавало новым видам реф-
лекторных реакций несколько более стабильный характер. Здесь особенно
строю следует различать стабильность «временной связи» и п а т о л о г и-
ческую стабильность состояния повышенной
возбудимости (или возбуждения) спинальных центров.
Мы полагаем, что полученные нами с сотрудниками новые фактические
данные и разв.иваемые на их основе теоретические положения о физиологии
временной связи представляют собой дальнейшее развитие классических
фактических данных и глубоких идей великого учителя о замыкании вре-
менной связи между пунктами коры и ее двусторонней проводимости, об
особенностях ее функционирования в зоне «творческого отдела» коры. Мы
считаем, что приведенные факты и положения значительно укрепляют наши
позиции в борьбе с зарубежными антипавловцами них единомышленниками
в среде наших физиологов, стремящимися с помощью различных уверток
подорвать основы гениального материалистического учения И. П. Пав-
лова, повернуть вспять развитие науки о мозге, вернуть господствующее
положение идеализма в вопросах психической деятельности.
При экспериментальной и теоретической разработке вопросов физиоло-
гии высшей нервной деятельности, в частности, вопроса о временной связи,
мы еще раз убедились в глубокой истине, стань ярко сформулированной
гениальным мыслителем: «В общем же этот новый отдел физиологии поисти-
не пленителен, удовлетворяя двум, всегда рядом идущим тенденциям че-
ловеческого ума: стремлению к захватыванию всех новых и новых истин
н протесту против претензий как бы законченного где-нибудь знания»2.
1 И. П. П а в л о в, Полное собрание трудов, т. III, 1949, стр. 462.
‘Там же, т. IV, 1947, стр. 325.
15 Э. А. Асратян
О ДИАЛЕКТИКО-МАТЕРИАЛИСТИЧЕСКОМ ХАРАКТЕРЕ
УЧЕНИЯ И. П. ПАВЛОВА >
толетие со дня рождения великого русского естествоиспытате-
ля И. П. Павлова—знаменательная дата для советского на-
рода. Благодаря поддержке нашей партии, личному вниманию
В. И. Ленина и И. В. Сталина, выдающееся творчество корифея
естествознания Ивана Петровича Павлова достигло особенного
расцвета в советский период его деятельности и является гор-
достью советской науки. Учение И. П. Павлова о высшей нервной деятель-
ности должно быть по праву оценено как революционный переворот в
знании о функциях мозга.
До II. II. Павлова в процессе изучения деятельности головного мозга,
наиболее сложного и совершенного из созданий органической природы, не
было достигнуто познание того, что Ф. Энгельс и В. И. Ленин считали одной
из наиболее важных И Трудных задач естествознания: почти не были рас-
крыты протекающие в мозгу процессы; не была изучена внутренняя при-
рода и сущность работы большого мозга; не были выявлены закономер-
ности его деятельности, благодаря которой и посредством которой осу-
ществляется наиболее совершенная, тонкая и точная приспособительная
деятельность организма, т. е. высшая нервная (или психическая) деятель-
ность. Распространенные среди естествоиспытателей XIX столетня мате-
риалистические и прогрессивные взгляды и особенно воззрения И. М. Се-
ченова о рефлектор ной природе работы большого мозга оказали на
И. П. Павлова значительное влияние. Однако эта взгляды носили еще
неразвернутый характер, небыли достаточно обоснованы экспериментально
и вследствие этого были лишены необходимой действенной силы. Физиолог
Гольц, одни из крупных исследователей функций головного мозга, вы-
нужден был в конце XIX века сказать: «Каждый, кто основательно зани-
мался физиологией головного мозга, согласится со мной, что неоспоримое
знание о процессах, протекающих в этом важнейшем органе, немногим
больше наших сведений о природе планеты Марс»2. А великий И. П. Павлов,
приступая к своим классическим исследованиям в этой области на пороге
нашего столетия, с горечью констатировал, что ^физиология высшего
мозга находится сейчас в тупике», что в ней «идейно нового очень мало».
Существовавший в то время в физиологии однобокий аналитический
метод познания явлений органической природы и связанные с ним пороч-
ные приемы дробного исследования функции мозга (искусственная сти-
муляция тех или иных частей мозга в условиях грубого вивисекционного
эксперимента, повреждение или удаление отдельных его частей) оказались
непригодными для разрешения такой сложной и трудной задачи, как объ-
яснение протекающих в мозгу процессов.
1 Вопросы философии, № 3, 1949,
а Р. Гольц, Труды съезда по внутренней медицине, 1884, стр. 262 (немецкое
издание).
О ДИАЛЕКТИКО-Л1АТЕРИАЛИСТИЧЕСКОЛ! ХАРАКТЕРЕ УЧЕНИЯ И П. ПАВЛОВА 227
«Мелочная, детальная разработка, конечно, шла дальше, — говорил
И. II. Павлов, характеризуя состояние физиологии того времени, — но
основные, руководящие идеи, основные методы были исчерпаны в 70-х го-
дах. Далее идут только детальное применение и расширение их. Это уже
подражание, а не творчество; нового же за 30 лет не создано ничего, все
топчется в старых рамках» -1.
И вот на этом сером горизонте появляется сверкающий гений Павлова,
уже умудренного богатым опытом раскрытия глубоких тайн органической
природы, вооруженного совершенным синтетическим научным методом и
тонкими приемами физиологического эксперимента, с новыми, свежими,
плодотворными идеями, с новыми целевыми установками в работе.
На основе точных и достоверных естественно-научных фактов И. П. Пав-
ловым была установлена материальная природа высшей нервной деятель-
ности, была доказана детерминированность этой деятельности условиями
суш.есттов.ания организма, — Иными словами, было Доказано решающее
значение внешней среды в возникновении и формировании высшей нервной
деятельности. Путем неопровержимых фактических данных И. П. Павлов
доказал также диалектический характер процессов, протекающих в го-
ловном мозгу.
Несмотря на очевидное выдающееся значение учения И. П. Павлова
для марксистского, философского материализма, мы не можем, однако,
сказать, что научное творчество И. II. Павлова и его философские воззре-
ния в достаточной мере обобщены с точки зрения диалектического мате-
риализма. Богатейший и ценнейший экспериментальный материал и теоре-
тические концепции II. П. Павлова еще не использованы в достаточной
мере для дальнейшего развития и естественно-научного обоснования ряда
важнейших положений марксистско-ленинской философии. Это не может
считаться нормальным и терпимым, помимо всего прочего, еще и потому,
что с усилением идеологической реакции в лагере империализма весьма
активизировались враги материалистического учения И. П. Павлова,
стремящиеся отвергать учение И. П. Павлова, извращать его, противопо-
ставлять материалистическим концепциям И. П. Павлова антинаучные,
идеалистические трактовки высшей нервной деятельности. Поэтому иссле-
дование научно-теоретического содержания учения И. П. Павлова и при-
менение этого разящего материалистического оружия в борьбе с идеа-
лизмом весьма актуальны.
* ♦
Взгляды И. II. Павлова складывались и формировались под воздейст-
вием мировоззрения великих представителей русской материалистической
философии XIX века.
В годы обучения И. П. Павлова в семинарии и в университете бурно
развивалась передовая общественная мысль в России. Великие револю-
ционные демократы — Белинский, Добролюбов, Чернышевский, Герцен,
Писарев — вели напряженную, самоотверженную борьбу против реакции
в сбщестгенной жизни и в науке, за прсбуждение сознания народных
масс, за свободу, за высокие прогрессивные идеи. Много внимания уде-
ляли они горячей пропаганде идей материалистического естествознания,
в частности, биологии, и сами развивали передовые естественно-научные
воззрения. Огромно было влияние этой славной плеяды революционеров-
мыслителей на молодежь. И. П. Павлов со страстным напряжением следил
за борьбой этих властителен дум передовой общественности, с увлечением
читал их статьи в «Современнике» и в Других прогрессивных журналах,
в частности, их статьи по вопросам естествознания.
1 И. П. П а в л о в, Полное собрание трудов, т. 1, 1’940^ стр. 392.
15*
228
Э А. АСРАТЯН
В «Автобиографии» 1 И. П. Павлов писал: «Под влиянием литературы
шестидесятых годов, в особенности Писарева, наши умственные интересы
обратились в сторону естествознания, и многие из нас—в числе этих
ия — решили изучать в университете естественные науки».
Весьма сильное влияние оказало на И. П. Павлова научное творчество
отца русской физиологии И. М. Сеченова. Борясь за приоритет русской
науки, И. П. Павлов с гордостью подчеркивал роль И. М. Сеченова в раз-
витии важнейших разделов физиологии. Он писал Ленинградскому об-
ществу физиологов имени И. М. Сеченова: «Да, я рад, что вместе с Иваном
Михайловичем и полком моих дорогих сотрудников мы приобрели для
могучей власти физиологического исследования вместо половинчатого весь
нераздельно животный организм. И это —целиком наша русская неоспо-
римая заслуга в мировой науке, в обшей человеческой мысли» 2.
Особое же значение для развития воззрений И. П. Павлова имел совет-
ский период его жизни и деятельности.
И. П. Павлов оказался счастливее многих представителей славной
плеяды деятелей русской науки и культуры XIX века. Ему посчастли-
вилось не только увидеть свержение ненавистного царского режима, но
прожить после Великой Октябрьской социалистической революции почти
два десятилетия, претворить в жизнь все свои научные замыслы и стать
крупным деятелем советской науки и культуры. В условиях советского
строя, благодаря заботам нашей партии и личному вниманию В. И. Ленина
и И. В. Сталина к работам И. П. Павлова, с особой силой развернулось
творчество этого гениального ученого.
Еще в первые годы революции, когда страна переживала холод, голод
и лишения во всем и когда наш героический народ, руководимый партией
Ленина — Сталина, вел тяжелую борьбу за молодую советскую власть,
за подписью Владимира Ильича Ленина было издано специальное пра-
вительственное постановление, свидетельствующее о чрезвычайно внима-
тельном, заботливом отношении большевистской партии и советского
правительства к И. П. Павлову и его работе. В этом постановлении от-
мечались исключительные научные заслуги академика И. П. Павлова,
имеющие огромное значение для трудящихся всего мира; поручалось
комиссии во главе с Л. М. Горьким в кратчайший срок обеспечить наиболее
благоприятные условия для научной работы академика Павлова и его
сотрудников.
В дальнейшем при постоянном внимании со стороны товарища Сталина
наша партия и советское правительство неизменно содействовали созда-
нию наилучших условий для всестороннего развития научной деятельности
великого ученого.
К семидесятипятилетию И. П. Павлова был создан мощный Физио-
логический институт в системе Академии наук СССР, ныне носящий
его имя. В ознаменование восьмидесятилетия ученого в Колтушах, под
Ленинградом, был построен специальный научный институт-городок,
единственное в мире научное учреждение по изучению высшей нервной
деятельности животных. Для осуществления давнишней мечты И. II. Пав-
лова об изучении высшей нервной деятельности человека были организо-
ваны при его институтах клиники нервных и психических заболеваний.
Все руководимые им научные учреждения были оснащены новейшим обо-
рудованием.
И. П. Павлов жил и творил, окруженный всенародной любовью. Он
1 Автобиография в- сборнике «Товарищеская памятка врачей выпуска 1879 г.
бывшей Императорской медико-хирургической академии», изданная ко дню 25-летля
окончания курса, Спб., 1904.
* И. П. П а в л о в, Полное собрание трудов, т. I, 1940, стр. 27.
О ДИАЛЕК ТИКО.МАТЕРИАЛИСТИЧЕСКОМ ХАРАКТЕРЕ УЧЕНИЯ И- П. ПАВЛОВА 229
пользовался могучей моральной и материальной поддержкой советской
власти.
Важно отметить, что в советский период своей деятельности И. II. Пав-
лов испытывал благотворное влияние научного мировоззрения диалек-
тического материализма. Он читал философские произведения классиков
марксизма-ленинизма. Неоднократно, например, говорил он своим уче-
никам о внимательном чтении им книги В. И. Ленина «Материализм и
эмпириокритицизм» и заявлял о своем полном единомыслии с Владимиром
Ильичем по кардинальным вопросам философии.
Диаметрально противоположное отношение к научной работе И. П. Пав-
лова при царизме и в социалистическом государстве не могли не
волновать великого ученого. В то время как при царском режиме он
постоянно встречал массу препятствий в своей научной деятельности,
теперь его не покидала тревога: сумеет ли он оправдать все заботы со-
ветской власти, доверие правительства и его щедрую помощь. Очень по-
казательны в этом отношении слова И. П. Павлова, сказанные им иа приеме
в Кремле советским правительством делегатов XV Международного кон-
гресса физиологов, происходившем в- 1935 г. «Мы, руководители научных
учреждений, — говорил он, — находимся прямо в тревоге и беспокой-
стве по поводу того, будем ли мы в состоянии оправдать нее те средства,
которые нам предоставляет правительство» * 4. По другому поводу И. П. Пав-
лов восторженно и взволнованно говорил: «Хочется долго жить потому,
что небывало расцветают мои лаборатории. Советская власть дала мил-
лионы на мои научные работы, на строительство лабораторий. Хочу верить,
что меры поощрения работников физиологии, а я все же остаюсь физио-
логом, достигнут цели, и моя наука особенно расцветет на родной почве» а.
Великий русский ученый И. П. Павлов, полный чувства любви к своему
народу и к своей родине, с гордостью прославлял Советскую страну и
как пламенный советский патриот жил мыслями о победоносном развитии
социалистического общества в СССР.
Незадолго до кончины он произнес волнующие слова: «Что ни делаю,
постоянно думаю, что служу этим, сколько позволяют мне мои силы,
прежде всего моему отечеству. На моей родине идет сейчас грандиозная
социальная перестройка. Уничтожена дикая пропасть между богатыми
и бедными. Я хочу жить еще до тех пор, пока не увижу окончательных
результатов этой социальной перестройки... Огромное достижение совет-
ской власти заключается в неустанном укреплении обороноспособности
страны. Хочу жить возможно дольше и потому, что за безопасность своей
родины я спокоен» 3.
И. II. Павлой ненавидел фашистских поджигателей войны. Его речь
на XV Международном конгрессе физиологов была пламенным выражением
этой ненависти, страстным призывом к непримиримой борьбе против озве-
релоГо фашизма. В последние годы своей жизни, когда все очевиднее ста-
новились агрессивные намерения фашистских вандалов и мракобесов, он
говорил о своей ненависти к ним особенно часто, говорил со священным
гневом. Он высказывал твердую уверенность в том, что наш богатырский
народ во главе с другими свободолюбивыми народами мира спасет циви-
лизацию от фашистской чумы. В войне, говорил Павлов, мы будем за-
щищать от врагов нашу настоящую родину, нашу культуру, наш}' науку.
Весь народ станет иа защиту своей страны. Выдающийся представитель
великой русской нации, И. П. Павлов глубоко воспринял идеи интерна-
ционализм;), идею дружбы народов Советского Союза и с восторгом сле-
1 И. П. II а в л о в. Полное собрание трудов, т. 1, 1940, стр. 30
4 T а м же, стр. 31.
’Там же.
230
Э А АСРАТЯН
дил за успехами в их жизни. Сожалея, что из-за своей, как говорил Павлов,
науки ему не удалось побродить по Советской стране, он постоянно рас-
спрашивал о жизни наших народов. «На-днях, — говорил И. П. Павлов,—
приезжали мои сотрудники, побывавшие в среднеазиатских республиках
и на Дальнем Востоке. Я в восторге от их рассказов; отсталые народы
теперь грамотны, просвещены, богатеют».
И. П. Павлов — великий ученый, пламенный советский патриот, строи-
тель передовой советской пауки, имя которого товарищ Сталин назвал
в числе наиболее выдающихся представителей русского народа. И. П. Пав-
лов близок и дорог всем народам Советского Союза и всему передовому
человечеству, интересам которого служила его творческая, яркая жизнь.
* *
♦
Добытый И. П. Павловым и его учениками громадный и ценнейший экс-
периментальный материал и построенное на этой основе его учение имеют
непосредственное отношение к кардинальному в философии вопросу, к воп-
росу об отношении мышления к бытию, духа — к природе. Этот вопрос
является какбы своеобразным индикатором, с помощью которого определяет-
ся философское кредо, выявляется мировоззрение того или иного ученого,
выясняется теоретический характер того или иного научного направления.
Единственно правильный, ясно и четко сформулированный ответ на этот
вопрос дает диалектический материализм: «...мышление есть продукт мате-
рии, достигшей в своем развитии высокой степени совершенства, а именно —
продукт мозга» *,
Гениальные классики марксизма-ленинизма, обобщая достижения есте-
ствознания, учат тому, что единственно правильным путем исследования
психических явлений нужно считать выведение последних из исторического
развития материи. Свойство материи — свойство простого отражения —
в процессе возникновения и развития органической природы, с усложненном
нервной системы и органов чувств, получило особое выражение; это свой-
ство в процессе развития переходит от элементарных явлений раздражимо-
сти ко все более и более совершенным видам нервной и психической деятель-
ности вплоть до сознания у человека, обусловленного общественно-исто-
рическнмч закономерностями происхождения и развития последнего.
И. П. Павлов был нс только блестящим экспериментатором, он был глу-
боким мыслителем, выдающимся теоретиком биологии и всегда очень вы-
соко ценил значение теории для естественных наук. Это он писал в свеем
мудром письме к нашей молодежи: «Изучайте, сопоставляйте, накопляйте
факты... Но, изучая, экспериментируя, наблюдая, старайтесь не оставаться
у поверхности фактов. Ие превращайтесь в архивариусов фактов. Пытай-
тесь проникнуть в тайну их возникновения. Настойчиво ищите законы, ими
управляющие» 2. Заветы, обращенные к молодежи, характеризуют путь,
пройденный в науке самим И. П. Павловым. Весьма показательна также од-
на из любимых им поговорок: «Если нет в голове идей, то не увидишь и
фактов».
На всем протяжении своего научного творчества великий мыслитель по-
стоянно прибегал к теоретическим обобщениям накопленного им и его со-
трудниками богатейшего эмпирического материала и создал ряд цельных,
монолитных и высокоидейных физиологических учений. В процессе своей
многолетней экспериментальной и теоретической работы И. П. Павлов не-
однократно высказывал глубокие мысли по многим кардинальным и по су-
ществу своему философским вопросам физиологии, биологии и естествозна-
1 История ВК.П(б). Краткий курс, стр. 107.
’ И. П. П а в л о в, Полное собрание трудов, т. 1, 1940, стр. 27.
О ДИАЛЕКТИКО-МАТЕРИАЛИСТИЧЕСКОМ ХАРАКТЕРЕ УЧЕНИЯ Н.П. ПАВЛОВА 231
ния вообще. Все это в совокупности представляет значительный материал
для суждения о его мировоззрении.
В качестве отправной точки анализа мировоззрения гениального физио-
лога необходимо дать хотя бы сжатое изложение основного содержания его
учения о высшей нервной деятельности — венца всей долголетней и исклю-
чительно плодотворной научной деятельности И. П. Павлова.
Маститый ученый, уже имевший выдающиеся достижения в развитии
физиологической науки, И. П. Павлов в начале нашего столетия приступил
к разгадке самой сложной и загадочной из тайн природы — психической
деятельности. Применяя в качестве основного орудия познания созданный
км и доведенный до высшей степени совершенства метод так называемого хро-
нического физиологического эксперимента, гениальный ученый достиг за
свою многолетнюю научную работу в этой повой области небывалых по цен-
ности результатов.
В основе величественного учения И. П. Павлова лежит простой факт,
который по-настоящему был открыт для науки лишь его проницательным
умом, оценен впервые по достоинству его гением. Этим обычным фактом яв-
ляется выделение слюны при виде или запахе пищи («слюнки текут»). Если
вспышка света, звонок или почесывание кожи несколько раз совпадают с про-
стой рефлекторной деятельностью слюнных желез во время еды, этого до-
статочно, чтобы названные посторонние для пищевой деятельности раздра-
жители приобрели для организма новое биологическое значение — стали
сигналами к деятельности слюнных желез и вообще к пищевой деятельности
организма, иначе говоря, стали раздражителями для этих органов и вызы-
вали деятельность последних. Это тоже рефлексы, но уже не врожденные,
простые рефлексы, а выработанные, более сложные рефлексы нового ка-
чества— продукт особого рода синтеза простых рефлексов, В отличие от
врожденных, такие рефлексы имеют временный характер, весьма чувстви-
тельны к изменениям в окружающей и во внутренней среде организма.
Такого рода рефлексы И. II. Павлов назвал условными, так как их образо-
вание, закрепление, осуществление и сохранение в сильной степени зависят
от условий существования организма.
Работы И. П. Павлова показали, что любое изменение во внешнем .мире
и в самом организме, воспринимаемое внешними и внутренними органами
чувств, может стать условным раздражителем или сигналом к деятельности
пищеварительных органов, равно как и к деятельности любых других органов
и систем организма и к изменению состояния организма в целом. У высших
животных и человека эти новые, высшие разнообразные и разнородные реф-
лексы обусловливаются самыми верхними отделами центральной нервной
системы. Отсюда следует, что, с точки зрения физиологической роли и био-
логического значения, условные рефлексы являются как бы средством выс-
шей регуляции со стороны коры большого мозга разнообразных функций
сложного организма в процессе его взаимодействия со средой.
И. П. Павлов оценил и осмыслил значение условнорсфлекторной дея-
тельности как истинный материалист и продолжатель материалистических
идей дарвинизма и раскрыл диалектический характер этой деятельности.
Если его прежние многолетние исследования в области физиологии крово-
обращения и пищеварения привели его к выводу о единстве организма и сре-
ды, к выводу о том, что «животный организм представляет крайне сложную
систему, состоящую из почти бесконечного ряда частей, связанных как друг
с другом, так и в виде единого комплекса с окружающей природой» ’, то его
исследования в области физиологии больших полушарий мозга привели
к расшифровке закономерностей взаимоотношений'организма с окружающей
природой. Согласно его учению, наиболее тонкое, точное и совершенное
1 И. П. Павлов, Собрание трудов, т. II, 1946, стр. 452.
2 <2
Э. А. АСРАТЯН
приспособление организма к условиям существования, самые тонкие и
совершенные взаимоотношения с окружающей средой осуществляются
пленное помощью образования (а при необходимости также и задержки)
самых разнообразных, разнородных, разностепенных и разнокачественных
углов шк рефл ексов. И. П. Павлов писал: «Первое обеспечение уравновеши-
вания, а следо вательно, и целостности отдельного организма, как и его ви-
да, составляют безусловные рефлексы... Но достигаемое этими рефлексами
уравновешивание было бы совершенно только при абсолютном постоянстве
внешней среды. Л так как внешняя среда при своем чрезвычайном разнооб-
разии вместе с тем находится в постоянном колебании, то безусловных свя-
зей как связен постоянных недостаточно и необходимо дополнение их услов-
ными рефлексами, временными связями» С этой точки зрения только и мож-
но Понять колоссальное биологическое значение физиологических особен-
ностей условных рефлексов. Их временность, изменчивость, хрупкость и обу-
словленность факторами внешней.и внутренней среды делают их гибкими,
подвижными, тонкими и совершенными средствами приспособления орга-
низма к вечно изменяющейся окружающей его среде. Сигнальный же харак-
тер условнорефлекторной деятельности позволяет организму по одним
только отдаленным сигналам (или условным раздражителям) стремиться
к благо-приятным для существования условиям и избегать неблагоприят-
ных. А в силу того, что каждая из жизненно важных деятельностей организ-
ма может быть вызвана бесчисленным количеством многообразных условных
раздражителей, сигнальная, или условнорефлекторпая, деятельность не-
измеримо расширяет диапазон восприятия предметов- и событий окружаю-
щего мира, а также диапазон деятельности самого организма.
Хотя И. П. Павлов постоянно противопоставлял условные и безусловные
рефлексы, подчеркивая разницу между ними и биологические преимуще-
ства условнорефлекторной деятельности перед безусловнорефлекторной,
но одновременно с этим он как диалектически мыслящий исследователь
всегда указывал также на относительный характер этого противопоставле-
ния, этой разницы, указывал на историческую связь между двумя назван*
ними основными видами нервной деятельности, на возможность перехода
одного вида в другой. Если условные рефлексы возн икают на основе безуслов-
ных, то, с другой стороны, «можно принимать, что некоторые из условных
вновь образованных рефлексов позднее наследственностью превращаются
в безусловные* а.
Это положение, как и постановка вопроса о единстве организма и среды,
а также ряд других положений И. П. Павлова свидетельствуют об идейно-
теоретическом родстве его учения с основными принципами мичуринской
биологии.
И. П. Павлов не ограничился безупречным экспериментальным уставов-
леннем принципиально весьма важного положения, что разнородные, много-
образные и разностепенные условные рефлексы в совокупности составляют
основной фонд психической или высшей нервной деятельности животных.
Опираясь на этот твердый фундамент и мастерски используя созданный им
метол условных рефлексов, он поднял на небывалую высоту всю физиоло-
гию большого мозга. «Для физиологии, — писал И. П. Павлов, — условный
рефлекс сделался центральным явлением, пользуясь которым можно было
все полнее и точнее изучать как нормальную, так и патологическую деятель-
ность больших полушарий» 3. Гениальный исследователь на протяжении
более 35 последних лет своей кипучей научной деятельности, и в особенности
в советский период его жизни, когда для его работы были созданы намлуч-
1 И. П. Павлов, Двадцатилетий опыт объективного изучения высшей
нервной деятельности (поведения) животных, 1938, стр. 710.
’Там же, стр. 275.
’Там же, стр. 712.
О ДИАЛЕКТИКО-МАТЕРИАЛИСТИЧЕСКОМ ХАРАКТЕРЕ УЧЕНИЯ И. П. ПАВЛОВА 233
шие условия, о которых он мог лишь мечтать при царском режиме, система-
тически и целенаправленно, всесторонне и углубленно изучал закономер-
ности формирования и осуществления высшей нервной деятельности. Он
раскрыл закономерности образования и закрепления, исчезновения и появле-
ния условных рефлексов, их взаимодействия.И. П. Павлов изучил особен-
ности протекания в коре большого мозга процессов возбуждения и тормо-
жения, закономерности целостной и локальной, синтетической и анализа-
торной деятельности коры, взаимодействия условных и безусловных реф-
лексов. Предметом исследований И. II. Павлова является также ряд таких
крайне важных вопросов, как локализация функций, физиологические
основы типа и характера, явлений сна и гипноза и т. п. Результаты много-
летней и напряженной экспериментальной работы И. П. Павлова служили
материалом для построения грандиозного и величественного здания его
материалистического учения о высшей нервной деятельности.
Изучая в течение многих лет высшую нервную деятельность в специаль-
ных опытах на собаках, И. П. Павлов в порядке «этапного» приближения
к своей исконной цели — к изучению закономерностей высшей нервной
деятельности человека — в последние годы своей жизни с увлечением зани-
мался также исследованием поведения человекообразных обезьян (шим-
панзе). И. П. Павлов придавал особенное значение изучению закономер-
ностей высшей нервной деятельности человекообразных обезьян с позиций
своего материалистического учения еще и потому, что многие зарубежные
ученые (Хобхауз, Кёлер, Перкс, Лешли и др.) довольно интенсивно изуча-
ли поведение этих животных с точки зрения идеалистической психологии,
стремясь своими лженаучными концепциями и «теориями» укрепить рас-
шатанные позиции последней. В противоположность павловскому объектив-
ному, строго научному подходу к изучению закономерностей сложного
поведения человекообразных обезьян эти ученые прибегают к субъек-
тивно-идеалистическому истолкованию наблюдаемых явлений. Они оце-
нивают внутренний мир антропоидных животных через призму субъективных
переживаний человека и приписывают им специфически человеческие фор-
мы психической деятельности: наличие идей, разума, представлений, спо-
собности к «внутреннему проникновению» в сущность предметов и явлений.
Решение этими животными сложных жизненных задач и выход их из не-
ожиданных грудных ситуаций оцениваются указанными учеными как ре-
зультат «внезапного озарения» их сознания.
Проводившаяся в течение двух-трех лет интенсивная и целеустремлен-
ная исследовательская работа И. П. Павлова и его сотрудников по изучению
Поведения человекообразных обезьян Привела к выдающимся эксперимен-
тальным и теоретическим результатам, обогащающим его материалистиче-
ское учение и в отношении частного, но весьма важного вопроса — о пове-
дений антропоидов. Верный своим давним традициям, И. П. Павлов не спе-
шил с официальной публикацией этих принципиально весьма важных
результатов, значительно дольше обычного подвергая их всестороннему
обсуждению среди своего научного коллектива — на знаменитых на-
учных конференциях по средам (павловских «средах») и в более частных
беседах в узком кругу сотрудников. Но в самые последние месяцы
своей жизни он уже счел возможным готовиться к докладу на эту тему на
предполагавшемся очередном Международном конгрессе психологов в Мад-
риде. Там же, в Мадриде, где Павлов в 1903 г. торжественно возвестил все-
му миру о рождении своего гениального учения, в 1936 г. должен был зазву-
чать могучий голос этого колосса научной мысли, страстный и беспощадный
голос боевого материалиста, направленный против приверженцев и защит-
ников тлетворного идеалистического мировоззрения. Но это его намерение
не осуществилось. Ко времени созыва конгресса И. П. Павлова уже не
было в живых.
234
Э А. АСРАТЯН
Суть добытых И. П. Павловым результатов сводится в кратких чертах
к следующему. Все поведение человекообразных обезьян, изучавшееся в ус-
ловиях своего рода естественных экспериментов, оказалось строго детер-
минированным условиями их жизни, факторами той своеобразной внешней
среды, которая создавалась для них экспериментатором. Образование у этих
обезьян сложных моторных навыков, делающих возможным добывание пи-
щи, происходит по принципу «проб и ошибок», т. е. по принципу накопления
«жизненного опыта», образования простых и сложных условных рефлексов,
а отнюдь не в силу каких-то изначально присущих им идей, представлений,
суждений, рациональных тенденций, внезапного озарения сознания или
вообще благодаря каким-то непостижимым, таинственным силам, как утвер-
ждали и продолжают утверждать сторонники реакционной, идеалистической
психологии.
Процессы возникновения и закрепления, усложнения и комплексиро-
вання, ослабления и исчезновения навыков, равно как и взаимоотношение
н взаимодействие между ними, Протекают в основном по закономерностям ус-
ловнорефлекторной деятельности. Некоторые отклонения в проявлении
этих закономерностей сравнительно с данными, наблюдаемыми у собак, не
выходят из рамок вариаций «основного мотива» и обусловливаются специфи-
ческими особенностями двигательном системы, уровнем развития и био-
логическими особенностями животного и т. п. В частности, было установ-
лено, что в образовании сложных двигательных навыков у человекообраз-
ных обезьян, как и в формировании их поведения в целом, весьма важную,
даже ведущую роль сравнительно со зрительными и другими восприятиями
играют собственные восприятия органов движения, или так называемая ки-
нестетическая рецепция. Далее, было установлено, что вырабатываемые
по принципу временных связей двигательные навыки легко и довольно ши-
роко генерализуются в коре большого мозга, что дает возможность исполь-
зовать эти навыки для решения новых задач.
Эти и ряд других достигнутых И. II. Павловым фактических и теорети-
ческих результатов ознаменовали собой очередную крупную победу мате-
риалистического понимания сложных жизненных явлений вплоть до их
высшего выражения — психических явлений. Они означали победу гениаль-
ного учения И. П. Павлова над лженаучными и беспочвенными идеали-
стическими «теориями» и «концепциями» реакционных ученых капиталисти-
ческих стран, с которыми он вел страстную научную полемику, беспощадную
идеологическую борьбу.
В последние годы жизни И. П. Павлов вплотную подошел к своей искон-
ной цели — изучению высшей нервной деятельности человека. На основа-
нии многолетней экспериментальной работы по высшей нервной деятель-
ности животных и пяти-шестилетней интенсивной работы в клинике душев-
ных и нервных заболеваний И. П. Павлов достиг выдающихся результатов
не только в выяснении сущности некоторых сложных заболеваний нервной
системы человека и их научно обоснованного лечения, но и в области по-
знания специфических особенностей высшей нервной Деятельности человека
вообще,
Великий классик ест^гтвознания обогатил свое учение об условно-
рефлекторной, или сигнальной, деятельности высших отделов центральной
нервной системы новым и исключительно ценным вкладом — развитием кон-
цепции о так называемой второй сигнальной системе. Хотя в процессе раз-
вития животного мира условнорефлекторная, или простая сигнальная,
деятельность нервной системы непрерывно развив-ается, осложняется и со-
вершенствуется, тем не менее, считал И. П. Павлов, она не претерпевает корен-
ных качественных изменений в пределах животного мира. У всех без исклю-
чения животных, на каком бы уровне эволюционного развития они ни стоя-
ли, условнорефлекторная, или сигнальная, деятельность мозга обусловли-
О ДИАЛЕКТИКО-МАТЕРИАЛИСТИЧЕСКОМ ХАРАКТЕРЕ УЧЕНИЯ И.П. ПАВЛОВА 235
вается прямым и непосредственным воздействием предметов и явлений внеш-
ней или внутренней среды организма на те или иные органы чувств. И.П. Пав-
лов считал, что «для животного действительность сигнализируется почти
исключительно только раздражениями и следами их в больших полушариях,
непосредственно приходящими в специальные клетки зрительных, слуховых
и других рецепторов организма» *. Исчерпывая почти всю высшую нервную
деятельность животных, этот вид сигнальной деятельности занимает зна-
чительное место и в психической деятельности человека. «Это то, что и мы
имеем в себе как впечатления, ощущения и представления от окружаю-
щей внешней среды как общеприродной, так и от нашей социальной,
исключая слово слышимое и видимое. Это—первая сигнальная си-
стема действительности, общая у нас с животными» 1 * * 4. Но высшая нервная
деятельность человека этим нс исчерпывается. «В развивающемся живот-
ном мире на фазе человека произошла чрезвычайная прибавка к механизмам
нервной деятельности» *. В связи с возникновением и развитием трудовой
деятельности, в условиях социальной жизни у человека «появились, раз-
вились и чрезвычайно усовершенствовались сигналы второй степени, сиг-
налы этих первичных сигналов, — в виде слов произносимых, слышимых
и видимых» *. Эта качественно новая и совершенная сигнальная система сгой-
ственна только высшей нервной деятельности человека и является «специаль-
но нашей»; она и играет исключительную роль в нашей сознательной жизни.
Эти сигналы сигналов, или слова, «представляют собой отвлечение от дей-
ствительности и допускают обобщение, что и составляет наше лишнее,
специально человеческое, высшее мышление,
создающее сперва общечеловеческий эмпиризм, а, наконец, и науку— ору-
дие высшей ориентировки человека в окружающем мире и в себе самом» 5.
Открытие И. П. Павловым речевой сигнальной системы является выдаю-
щимся вкладом в науку. Это позволило глубоко вскрыть особенности выс-
шей нервной деятельности человека. Понятно вместе с тем, что только исто-
рический материализм дает подлинную научную трактовку происхождения
и значения речи. И. П. Павлов видел уже роль труда в формировании чело-
века, но ему не пришлось завершить свои идеи о второй сигнальной системе.
Он оставался в основном в пределах непосредственно физиологического ас-
пекта этой прежде всего общественно-исторической проблемы.
♦ *
*
Все содержание учения И. П. Павлова с неоспоримостью свидетельствует,
что он был материалистом, причем материалистом не «бессознательным»,
«стихийным», а материалистом убежденным, сознательным, воинствующим.
R частности, его гениальное учение.о высшей нервной деятельности являет-
ся материалистическим не только по объективному своему содержанию, но
и по теоретическому освещению сущности и происхождения сложнейших
явлений психической деятельности. В кардинальном вопросе о соотношении
материи и психики И. П. Павлов выступает как непоколебимый в своих убеж-
дениях материалист. Для И. П. Павлова объект его исследования является
материальным, представляет собой частицу материальной природы. Все
интимные процессы, которые протекают в большом мозгу, разыгрываются на
материальной базе, «функция» и «динамика» неразрывно связаны с «види-
мым аппаратом», «приурочены к тончайшим деталям конструкции аппарата»
(И. П. Павлов). Факты, явления и процессы, которые он изучает, объективно
1 И.П. Павлов, Двадцатилетний опыт, 1938, стр. 722.
’Там же.
Л Т о »« Ж А
4 Т а м же, стр, 732.
’Там же, стр. 616.
236
Э. А. АСРАТЯН
существуют во времени и в пространстве. «Мы объясняем это чисто физиоло-
гически, чисто материально, чисто пространственно» ’» — писал он. Учение
об условных рефлексах, указывал И. П. Павлов» все время имеет дело «толь-
ко с объективными фактами, т. е. с фактами, существующими во времени
и пространстве» г. «Работа все время держится на прочном материально-
фактическом фундаменте, как во всем остальном естествознании» 3. Па кар-
динальный вопрос, что является первичным и что вторичным, производ-
ным, — материя или мышление, — И. П. Павлов всегда давал материали-
стический ответ, «...сознание, — пишет он, — представляется мне нервной
деятельностью определенного участка больших полушарий» 4.
И. П. Павлов считает, что «психическая деятельность есть результатфн-
зиологичсской деятельности определенной массы головного мозга» 5. Как
бы в ответ на снова Энгельса о том, что «материалистическое мировоззре-
ние означает просто понимание природы такой, какова она есть, без всяких
посторонних прибавлений» ®, И. П. Павлов в защиту своего материалисти-
ческого подхода к изучению мозга’еще на заре развития своего гениального
учения говорил: «Для натуралиста все — в методе, в шансах добыть непоко-
лебимую, прочную истину, и с этой только, обязательной для него, точки
зрения душа как натуралистический принцип не только не нужна ему, а
даже вредно давала бы себя знать на его работе, напрасно ограничивая сме-
лость и глубину его анализа»7. Или же: «Естествознание — это работа че-
ловеческого ума, обращенного к природе и исследующего ее без каких-
либо толкований и понятий, заимствованных из других источников, кроме
самой внешнем природы»8.
Сознательный, боевой материализм И. П. Павлова особенно ярко выяв-
ляется в его полных страсти полемических выступлениях против анимистов,
виталистов, дуалистов и зараженных другими разновидностями идеализма
психологов и физиологов, в его ожесточенной борьбе за искоренение идеа-
лизма и торжество материализма в фундаментальных вопросах биологии
и медицины, за материалистическое понимание важнейшего и сложнейшего
вопроса естествознания и философии — о соотношении материн и сознания,
Но И. П. Павлов как убежденный материалист не ограничивается при-
знанием первичности материи и вторичности психики и блестящим экспе-
риментальным доказательством правильности этого основного для материа-
листического миропонимания положения. Гениальный натуралист впер-
вые в истории естествознания экспериментально установил, что высшая
нервная, или психическая, деятельность животных детерминирована усло-
виями их жизни, факторами внешней среды, имеет «опытное происхождение»
и представляет собой совокупность многообразных, разнородных и разно-
качественных условных рефлексов, выработанных в течение их индивиду-
альной жизни. Об этом с исчерпывающей убедительностью говорят резуль-
таты многолетних его исследований высшей нервной деятельности у собак,
об этом с не меньшей убедительностью говорят результаты его исследований
высшей нервной деятельности у человекообразных обезьян, которые в крат-
ких чертах были изложены выше. Основываясь па этих результатах,
И. П. Павлов непрерывно вел ожесточенную борьбу с приверженцами и
представителями идеалистического мировоззрения в физиологии и психо-
логии. Страстность молодости, воинственность материалиста не покидали
1 И. П. Павл ов, Двадцатилетий опыт, 1938, стр. 341.
* Там же. стр. 206.
’Там же, стр. 282.
‘Там же, стр. 218.
‘Там же, стр. 706.
’ К. М ар кем Ф. Энгельс, Соч., т. XIV, стр. 651.
’И. П. Павлов, Двадцатилетии» опыт, 1938, стр. 36.
‘Там же, стр. 62.
О ДИАЛЕКТИКО-МАТЕРИАЛИСТИЧЕСКОМ ХАРАКТЕРЕ УЧЕНИЯ И. П. ПАВЛОВА 237
его до конца жизни. «Теперь от мирных дел перейдем, можно сказать, к во-
енным, к господину Кёлеру, — говорил он с присущей ему горячностью. —
С ним м.ы воюем. Это серьезная борьба... У них, повидимому, имеется жела-
ние, чтобы их предмет оставался неразъясненным; вот какая странность!
Их привлекает таинственное. От того, что можно объяснить со стороны фи-
зиологии, они отворачиваются... В этом вредном, я бы сказал, паскудном
стремлении уйти от истины психологи типа Перкса или Кёлера пользуют-
ся такими пустыми представлениями, как, например, обезьяна отошла,
«подумала на свободе» по-человечески и «решила эго дело». Конечно, это
дребедень, ребяческий выход, недостойный выход...» \ И. П. Павлов счи-
тал, что «незаконный успех этой психологии среди современных психоло-
гов можно понять только так, что среди них все еще дает себя знать дуализм
в виде анимизма, т. е. понятия о своеобразной субстанции, противопостав-
ляющейся остальной природе и обязывающей исследующую мысль держать-
ся в- отношении ее иначе, чем в отношении материальных явлений» *.
В последние годы_ своей жизни И. П. Павлов со всей силой своего могучего
гения беспощадно громил Лешли, Кёлера, Спирмана, Шеррингтона и дру-
гих зарубежных ученых за отрицание ими связи между психической деятель-
ностью и материальной структурой мозга, за отрицание ими принципа при-
чинности в высшей нервной деятельности и вообще за идеалистическое по-
нимание сущности психической деятельности, «замаскированное утвержде-
нием своеобразности психических явлений, под
которым чувствуется, несмотря на все научно приличные оговорки, все тот
же дуализм с анимизмом» а. Отрицательное отношение Шеррингтона и дру-
гих зарубежных ученых-идеалистов к материалистическому учению об ус-
ловных рефлексах И. П. Павлов совершенно справедливо объяснял тем,
что это учение своим острием направлено «против дуалистического пред-
ставления — в этом вся причина заключается; об этом говорят и прошлогод-
ние лекции Шеррингтона, где он выступил дуалистом, представляя, что
человек есть комплекс двух субстанций: высшего духа и грешного тела. Он
прямо заявляет, как это ни странно для физиолога теперешнего времени,
что, может быть, между умом и мозгом нет связи...»1 * * 4. «Кёлер заядлый ани-
мист, — с возмущением отзывался о нем Павлов, — он никак не может по-
мириться, что эту душу' можно взять в руки, взять в лабораторию, на соба-
ках разъяснить законы ее деятельности. Он этого не хочет допустить...
Кёлер — жертва анимизма. А Шеррингтон — другая жертва анимизма»- 4.
Тол ько в свете такого непримиримого отношени я материалиста И. П, Пав-
лова к идеализму, ко всем его разновидностям можно понять истинное его
отношение к психологии, вызвавшее немало- кривотолков у части наших уче-
ных. В действительности при близком знакомстве с мотивами его многолет-
него враждебного отношения к психологии нетрудно убедиться, что это —
вражда непримиримого материалиста к идеализму. Он обрушивался на субъ-
ективистскую психологию за ее антинаучную сущность, за ее индетерминизм,
за отрыв психики от материи и активно боролся за развитие научной, ма-
териал истической психологии.
Сознательный характер материализма И. П. Павлова сказывается и в дру-
гом в его признании возможности познания тайн природы. И. П. Павлов был
великим оптимистом и глубоко верил во всемогущество человеческого ума.
В произведениях И. П. Павлова на всех этапах его творчества имеется боль-
шое количество высказываний, насыщенных бодрым, материалистическим
1 «Павловские среды», т. 11, стр. 386—388.
’Там ж е, т. III, стр. 44.
•И. П. Павлов, ДвадиатилеТний опыт, 1938, стр. 548.
4 «Павловские среды», т. III, стр. 252—253.
’Там ж е, т. 11, стр. 430—431.
238
Э. А. АСРАТЯН
оптимизмом, направленных против неверия в науку, против широко рас-
пространенных среди естествоиспытателей агностицизма, релятивизма и
других разновидностей идеалистического мировоззрения. Великий мысли-
тель считал, что наука постепенно все более и более приближается к пол-
ному познанию деятельности мозга. Он писал: «Сложное берется наукой
только по частям и обрывками, но оно захватывается ею постепенно все бо-
лее н более. Следовательно, будем подеяться и терпеливо ждать, когда точ-
ное и полное знание вашего высшего органа — головного мозга — сделает-
ся нашим подлинным достоянием, а с этим и главною основою прочного че-
ловеческого счастья» х.
И. П. Павлов безгранично верил в силу научного опыта в раскрытии тайн
природы, ибо если «наблюдение собирает то, что ему предлагает
природа», то в противоположность этому «о п ы т же берет у природы то,
что он хочет».
И. П. Павлов безгранично верил в мощь естественно-научной мысли,
в то, что и физиологию «ждут такие же поражающие открытия и с ними та-
кая же чрезвычайная власть над высшей нервной системой, которые не ус-
тупят другим приобретениям естествознания» 4. Он верил в то, что его ра-
бота по высшей нервной деятельности животных окажет большую пометь
психологам в разгадке законов психической деятельности человека. Ко всему
этому следует добавить, что И. П. Павлов был строго объективным исследо-
вателем сложных явлений природы, придававшим объективным фактам
решающее значение в проверке правильности своих теоретических положе-
ний. Он считал действительность, т. е. практику, критерием истины. Среди
многочисленных упреков И. П. Павлова по адресу психологов и психологии
имеется также упрек в том, что психолог «еще не совсем отрешился от при-
страстия к философскому приему дедукции, от чисто логической работы,
не проверяющей каждый шаг мысл и со г л а с ием
с действительностью. Физиолог действует совер-
шенно обратно» (разрядка наша. — Э. А.)3. И. П. Павлов смотрел
на факты как на мощную броню против проникновения идеализма вето мате-
риалистическое учение, в его творческую атмосферу.
Некоторые наши исследователи высказывали мнение, будто материализм
великого естествоиспытателя является ограниченным, механистическим.
Подобного рода мнения являются плодом непонимания или прямого извра-
щения философских основ учения И. II. Павлова4.
Мы знаем, что отдельные высказывания И. П. Павлова обычно «крити-
ками» его воззрений трактуются как «доказательства» павловского яксбы
механицизма. Однако, исходя из отдельных, порой неудачных и нечетких
характеристик вроде того, что организм есть «физико-химическая система»,
«человек есть, конечно, система, грубо говоря,'—машина» и т. д., нельзя
характеризовать мировоззрение великого естествоиспытателя как механи-
стическое. II. П. Павлов применял понятие «механический» как синоним по-
нятия материалистический, причинно обоснованный, закономерный.
Гениальный мыслитель неизмеримо высоко стоял над уровнем механи-
стического материализма. Содержание, подлинная философская сущность
его учения носят диалектико-материалистический характер.
Великий натуралист на протяжении 35 лет объективным и точным ме-
тодом с беспримерными в нашей науке мастерством, целеустремленностью
1 И. П. П а в л о в. Лекция о работе больших полушарий головного мозга, 1927,
стр. 345.
* И. П. Павлов, Двадцатилетии^ опыт, 1938, стр. 291—292.
1 Т а м же, стр. 537.
* Эта ошибочная точка зрения высказывалась и мной в отдельпих ранних моих
работах.
О ДИАЛЕКТИКО-МАТЕРИАЛИСТИЧЕСКОМ ХАРАКТЕРЕ УЧЕНИЯ И. П. ПАВЛОВА 239
и последовательностью изучал явления и процессы, протекающие в большом
мозгу. Он выявил сложные закономерности его работы, непрерывно углуб-
лялся, говоря словами В. И. Ленина, «... от явления к сущности, от сущ-
ности первого, так сказать, порядка, к сущности второго порядка и т. д,
без кон ца»*. И неизбежным образом, повторяем, подлинная сущность,
содержание его материалистического учения являются диалектическими.
Иван Петрович Павлов является творцом мето-да так называемого хро-
нического эксперимента в физиологии. Этот правильный и плодотворный
метод физиологического эксперимента И. П. Павлов с виртуозным мастер-
ством и большим успехом применил в своих классических работах по физио-
логии пищеварительной системы; с тем же мастерством и с еще ббльшим ус-
пехом он применил его в своих исследованиях по физиологии большого моз-
га. В отличие от всех старых и подавляющего большинства новых приемов
физиологического исследования большого мозга, созданный И. П. Павло-
вым метод условных рефлексов дает возможность объективно и всесторонне
изучать функции и закономерности этого высшего интегративного органа
в его естественной, органической связи со всеми остальными органами и си-
стемами целостного организма. Он обеспечивает исследование протекающих
в мозгу процессов в их неразрывной связи и взаимодействии, в их естествен-
ной динамике возникновения и развития. Метод И. II. Павлова дал возмож-
ность изучать процессы большого мозга в их «с а м о д в и ж е и и и», в нх
спонтанейном развитии, них живой жизни»'2, осуществить многогранное
синтетическое исследование функций мозга и одновременно произвести тон-
кий анализ деятельности отдельных его органов и систем. Совершенно
бесспорно, что научно-исследовательский метод И. П. Павлова является
диалектическим по своим установкам и подходу к изучаемому объекту,
явлению.
Согласно учению И. П. Павлова, сложнейшие и многообразные явления
и процессы, протекающие в бо-льшом мозгу, как то: образованно и проте-
кание различных положительных и отрицательных условных рефлексов,
высшая анализаторная и синтетическая деятельность, явления взаимной
индукции, суммации, иррадиации и концентрации процессов возбуждения
и торможения большого мозга, находятся в постоянном взаимодействии
н взаимно обусловлены.
Неразрывно взаимосвязаны также и постоянно взаимодействуют части
большого мозга между собой, большой мозг — с другими частями нервной
системы, с органами чувств, с инкреторной системой и почти со всеми дру-
гими системами организма. Эта взаимосвязь в взаимная обусловлен несть
проявляются в усилении одних и исчезновении других групп условных реф-
лексов, в доминировании условных рефлексов одного рода над условными
рефлексами другого рода, в изменении характера течения основных нерв-
ных процессов и т. д. В конце концов, — что весьма важно, — вся деятель-
ность большого мозга тесно связана с окружающей средой и обусловлена
ею. Исследователь по своему плану может сформировать любой тип условно-
рефлекторной деятельности путем комбинации раздражителей, путем, на-
пример, создания своеобразной внешней среды в своей экспериментальной
камере. Не случайно основной вид деятельности коры большого мозга
И. П. Павловым был назван «условным рефлексом».
Приведем ряд высказываний самого И. П. Павлова по вопросу о взаимо-
связи и взаимообусловленности: «Большие полушария представляют собою в
1 В. И. Ленк и, Философские тетради, 1947, стр. 237.
я В. И. Л е и и и, Философские тетради, 1947, стр. 327.
240
Э. А. АСРАТЯН
период деятельности систему, все части которой находятся во взаимодействии
друг с другом» 1. «Кора больших полушарий представляет собой сложней-
шую функциональную мозаику из отдельных элементов, каждый из кото-
рых имеет определенное физиологическое действие — положительное или
тормозное. С другой стороны, также несомненно, что все эти элементы объ-
единены в каждый данный момент в систему, где каждый из элементов на-
ходится во взаимодействии со всеми остальными»1 2 * * * *.
В другом месте И. П. Павлов говорит о том же: «Всякое новее местное
воздействие на эту систему дает себя знать более или менее во всей, си-
стеме» *.
Характеризуя закономерности работы большого мозга, И. П. Павлов
далее пишет: «Эти явления были описаны нами отдельно, как бы вне зависи-
мости друг от друга. В действительности же. как это ясно и по смыслу дела,
они должны встречаться вместе, комбинироваться, взаимодействовать»1.
У И П. Павлова часто попадается выражение: «по общему закону взаи-
модействия нервных центров». Выбор наименования «условный» для обо-
значения нового вида рефлекса он мотивирует так: «Этим прилагательным
я желал выдвинуть характерную объективную черту' этих рефлексов, имен-
но чрезвычайную зависимость от массы условий, начиная с условности их
происхождения» ®.
Сложность условного рефлекса, говорит И. П. Павлов, «заключается не в
сложности механизма его образования, а в чрезвычайной зависимости его от
явлений как собственной внутренней среды организма, так и окружающего
внешнего мира» «.
И. В. Сталин Дал следующую яркую и глубокую характеристику пер-
вой, основной черты марксистского диалектического метода: «В противопо-
ложность метафизике, диалектика рассматривает природу не как случай-
ное скопление предметов, явлений, оторванных друг от друга, изолирован-
ных друг от друга и независимых друг от друга, — а как связное, единое
целое, где предметы, явления органически связаны друг с другом, зависят
друг от друга и обусловливают друг друга» 7
Сказанное выше с достаточной убедительностью свидетельствует о глу-
боко научном характере положений И. П. Павлова, соответствующих содер-
жанию этой черты диалектики. Диалектическим является также павловское
понимание взаимоотношений части и целого. Достаточно указать хотя бы
на его теорию о динамической локализации функций в коре большого мозга
или на его воззрения на функциональную мозаику коры большого мозга.
Теоретические положения учения И. П. Павлова полностью соответ-
ствуют и второй черте диалектики, которая рассматривает природу «не как
состояние покоя и неподвижности, застоя и неизменяемости, а как состоя-
ние непрерывного движения и изменения, непрерывного обновления и раз-
вития, где всегда что-то возникает и развивается, что-то разрушается и от-
живает свой век» 8.
Богатейший фактический материал И. П. Павлова по физиологии боль-
шого мозга является яркой иллюстрацией того, что в центральной нервной
системе вообще и в большом мозгу в особенности явления и процессы не
1 И. П. Павлов, Физиология и патология высшей нервной деятельности. 1930,
стр. 23.
’Там же, стр. 35 — 36.
* И. П. П а в л о в, Лекции о работе больших полушарий головного мозга. 1927,
стр. 202.
‘Там же, стр. 178.
* И. II. Павлов, Двадцатилетиий опыт, 1938, стр. 261.
•Там же, стр. 259.
7 История ВКП(б). Краткий курс, 1938, стр. 1QI,
•Там же.
О ДИАЛЕКТИКО-МАТЕРИАЛИСТИЧЕСКОМ ХАРАКТЕРЕ УЧЕНИЯ И.П. ПАВЛОВА 241
только протекают в тесной, неразрывной взаимосвязи и взаимодействии,
но и находятся в непрерывном движении, развитии, возникновении и исчез-
новении. Ведь одной из характерных особенностей деятельности коры боль-
шого мозга, можно сказать, даже самой характерной особенностью ее слож-
нейшей и богатейшей деятельности как раз и является исключительная под-
вижность, динамичность, изменчивость протекающих в ней процессов и яв-
лений.
Не случайно основной тип деятельности коры большого мозга
И. П. Павлов иногда называет «временным рефлексом». Часто И. П. Павлов
противопоставляет кору большого мозга другим частям центральной нерв-
ной системы именно по этому признаку ее рефлекторной деятельности. Соот-
ветственно окружающей ситуации и внутренним условиям организма в коре
большого мозга возникают, развиваются и исчезают различные положитель-
ные и отрицательные условные рефлексы, происходит сложный анализ и син-
тез, суммация, взаимная индукция, иррадиация и концентрация основных
нервных процессов и осуществляются другие виды мозговой деятельности.
Благодаря этой особенности корковой деятельности, и происходит наиболее
совершенное, тонкое, точное и быстрое приспособление организма к постоян-
но изменяющимся условиям внешнего мира, к изменениям внутри самого
организма. Этот диалектический характер явлений природы нашел свое
яркое отражение в теоретических положениях и формулировках И. П. Пав-
лова. Я приводу лишь некоторые из многочисленных его высказываний,
относящихся к этому вопросу.
Обращаясь к «внутреннему механизму работы больших полушарий»,
к основным его нервным процессам, И. П. Павлов говорит: «Первое, что
здесь прежде всего привлекает к себе внимание, — это движение этих про-
цессов...» ’.
О коре большого мозга И. П. Павлов говорит: «...деятельность ее харак-
теризуют две основные черты: чрезвычайная обусловленность и естественно
С нею связанная текучесть явлений, составляющих эту деятельность» 2,
Об особенностях условнорефлекторпой деятельноетн И. П. Павлов пи-
шет: «Изучаемые нервные явления характеризуются именно их изменяемо-
стью; каждый момент, при каждом условии они получают Новое направ-
ление» 3. Пли: «Окружающий животное внешний мир, вызывая, с одной
стороны, беспрерывно условные рефлексы, с другой стороны, также постоян-
но подавляет их» 4.
Очень характерным в этой связи является высказывание И. П. Павлова
о том, что большой мозг есть «специальный орган для беспрерывного даль-
нейшего развития животного организма» 5.
О диалектической сущности учения И. П. Павлова свидетельствует так-
же следующее: обобщив громадный материал, И. П. Павлов констатировал
«естественную общность основных отношений» в деятельности высших и низ-
ших отделов центральной нервной системы и наряду с этим неоднократно
отмечал специфический характер и новые качественные особенности основ-
ных видов деятельности коры большого мозга. Если вникнуть в сущность
процессов мозга, то нетрудно видеть, что почти все основные закономер-
ности работы большого мозга отличаются от своих аналогов в низших эта-
жах центральной нервной системы не только по степени сложности, не толь-
ко количественно, но прежде всего качественно. Это относится, например,
к различию между низшим анализом и синтезом, с одной стороны, и высшим
1 И. П. Павлов, Лекции о работе больших полушарий головного мозга,
1927, стр. 133.
’Там же, стр. 329.
’ И. П. П а в л о в. Двадцатилетний опыт, 1938, стр. 138.
‘Там же, стр. 127.
6 Т а м же, стр. 275.
16 Э, Л. Асратян
242
Э. А АСРАТЯН
анализом и синтезом — с другой; к особенностям всевозможных безуслов-
ных и условных рефлексов, а также к особенностям протекания основных
нервных процессов — возбуждения и торможения — в низших и высших
отделах центральной нервной системы и т. п.
Все это является прекрасной иллюстрацией к положению В. И. Лени-
на о поступательном развитии. На высшем витке «спирали» повторяются
примерно те же закономерности, что и на низших витках, нос новыми
качественными особенностями.
И. П. Павлов придерживался того мнения, что наука должна помочь
человеку в решении важнейших практических задач, и всегда стремился
в своей научно-творческой деятельности уделять много внимания экспери-
ментальной разработке вопросов патологии и терапии, в частности, патоло-
гии большого мозга человека, лечения его болезненных состояний. Но сме-
лый новатор науки проявлял крайнюю осторожность в вопросе о перенесе-
нии результатов лабораторного эксперимента с животных на человека как в
случае нормальной, так и в случае патологической деятельности большого
мозга. Он писал по этому поводу: «Если сведения, полученные на высших жи-
вотных относительно функций сердца, желудка и других органов, так сход-
ных с человеческими, можно применять к человеку только с осторожностью,
постоянно проверяя фактичность сходства в деятельности этих органов у че-
ловека и животных, то какую же величайшую сдержанность надо проявить
при переносе только что впервые получаемых точных естественно-научных
сведении о высшей нервной деятельности животных на высшую деятельность
человека. Ведь именно эта деятельность так поражающе резко выделяет
человека из ряда животных, так неизмеримо высоко ставит человека над
всем животным миром» *.
Наконец, следует сказать, что накопленные И. П. Павловым ценнейшее
фактические данные относительно двух основных противоречивых нервных
процессов — возбуждения и торможения, — а также его правильное теоре-
тическое освещение этих данных, в частности, его высказывания об единстве
этих активных нервных процессов, об их непрерывной борьбе и взаимном
переходе друг в друга, об их решающей роли в высшей нервной деятель-
ности, являются фундаментальными и яркими доказательствами глубокой
научности воззрений И. П. Павлова, его Диалектического подхода к оценке
физиологии мозга. Можно считать, что экспериментальные данные и теоре-
тические положения И. П. Павлова о роли и значении возбуждения и тор-
можения в деятельности мозга могут дополнить указания о специфических
противоположностях в математике, механике, физике, химии и в общест-
венных науках, какие приводятся В. И. Лениным в его гениальной рабо-
те «К вопросу о диалектике».
Можно с полным основанием словами В. И. «Ленина сказать в отношении
И. II. Павлова, что он изучил «противоречия в самой сущности предметов».
Для иллюстрации этой мысли приведу несколько высказываний Н.П. Пав-
лова: «Нервная деятельность вообщесостоит из явлений раздражения и тор-
можения. Это есть как бы две половины одной нервной деятельности» а.
«Раздражение и задерживание это—лишь разные стороны, разные прояв-
ления одного и того же процесса» «Торможение постоянно следует за воз-
буждением. ..онов некотором роде является как бы изнанкой раздражения»* 4.
«Можно было бы условно говорить о положительной и отрицательной воз-
1 И. П. П а в л о в, Лекции о работе больших полушарий головного мозга, 1927,
стр. 345.
4 И. П. П а в л о в. Двадцатилетний опыт, 1938, стр. 157.
"Там же, стр. 168
‘Там же, стр. 234.
О ДИАЛЕКТИКО-МАТЕРИАЛИСТИЧЕСКОМ ХАРАКТЕРЕ УЧЕНИЯ И. П. ПАВЛОВА 243
будимости» 1. «...синтез и анализ и исчерпывают все поведение животного...
Основные процессы, на которых основывается этот синтез и анализ, —это,
с одной стороны, раздражительный процесс, а с другой — тормозной про-
цесс, какая-то противоположность раздражительного процесса»8. Диферен-
цировка, иначе говоря, отрицательный условный рефлекс, есть, по характе-
ристике И. ГЕ Павлова, результат «борьбы между раздражением н тормо-
жением». Эта борьба имеет универсальный характер (растормаживание,
срывы, взаимная индукция, суммация, тонкий анализ и синтез, сон и бодр-
ствование и т. д.). Более того, по утверждению И. П. Павлова, «баланс меж-
ду этими процессами и колебаниями его в пределах нормы и за норму и опре-
деляют все наше поведение — здоровое и больное» ®.
Подытоживая сказанное, мы видим, что фактические данные и уче-
ние И. П. Павлова соответствуют положениям марксистского диалекти-
ческого метода о том, что предметам природы, явлениям природы свой-
ственны внутренние противоречия и что борьба между ними составляет
внутреннее содержание процесса развития.
» * *
*
И. П. Павлов — слава и гордость нашей науки, корифей естественно-
научной мысли. Он был искусным экспериментатором, виртуозным масте-
ром физиологического опыта, с необычайной силой и убедительностью рас-
крывавшим сложнейшие тайны органической природы, в изобилии добы-
вавшим достоверные, веские и красочные научные факты. На основе неоп-
ровержимых естественно-научных фактов великий ученый доказал сущест-
вование материальной основы психической деятельности и детерминирован-
ность этой деятельности условиями существования организма.
Еще на заре своей революционной и теоретической деятельности, поле-
мизируя с Михайловским, великий Ленин писал: «...Пока не умели при-
няться за изучение фактов, всегда сочиняли a priori общие теории, всегда
остававшиеся бесплодными... Нелеп тут был уже прием. Нельзя рассуждать
о душе, не объяснив- в частности психических процессов: прогресс тут дол-
жен состоять именно в том, чтобы бросить общие теории и философские по-
строения о том, что такое душа, и суметь поставить на научную почву из-
учение фактов...» 4.
Если в свете этих мыслей В. И. Ленина оценить суть изложенных выше
многолетних исследований И. П. Павлова по физиологии высшей нервной
деятельности, то можно сказать, что он блестяще претворил в жизнь то,
о чем говорил великий Ленин.
И, П. Павлов был крупнейшим теоретиком физиологии, биологии и ме-
дицины. 35-летней напряженной и целеустремленной научно-творческой
работой в области физиологии большого мозга он воздвиг величественное
здание своего бессмертного материалистического учения о высшей нервной
деятельности. И. П. Павлов был не стихийным, а сознательным материали-
стом. Он был принципиальным, непримиримым и страстным борцом за ли-
нию материализма в сложнейших и важнейших вопросах естествознания и
в сильнейшей степени способствовал его победе в этих вопросах как виртуоз-
ный мастер физиологического эксперимента и как глубокий мыслитель и тео-
ретик биологии. Трудно переоценить значение учения П. П. Павлова для
диалектического материализма. Его богатейший фактический материал по
физиологии большого мозга насыщен «объективной диалектикой», а при
1 И. П. Павлов, Двадцатилетний опыт, 1938. стр. 396.
“Там же, сгр. 410.
•Там же, стр. 11.
* В. И. Л е и и н. Собр соч., т. I, 19*47, стр. 126— 127.
16»
214
Э. Л АСРАТЯН
обобщении И осмысливании ЭТОГО материала И. П. Павлов выдвигал Диалек-
тико-материалистическую оценку сложнейшей работы большого мозга.
Учение И. П. Павлова о высшей нервной деятельности знаменует собой
крупнейшее достижение современного естествознания в области познания
глубоких тайн мозга и является могучим оружием в арсенале нашей идео-
логической борьбы со всеми черными силами реакции, со всеми и всякими
проявлениями идеализма, мракобесия в науке о жизни.
В настоящее время, когда все более и более обостряется борьба между
силами прогресса и силами реакции, наш долг — совершенствовать это ору-
жие, делать его еще более грозным и сокрушительным. Мы должны всемер-
но развивать бесценное наследство гениального ученого ио намеченным им
путям и вести наступательную борьбу со всеми врагами материалистического
учения И.П. Павлова за рубежом, а также разоблачать отдельных совет-
ских ученых, которые под влиянием враждебной буржуазной идеологии
проявляют антипатриотическое ‘ отношение в оценке воззрений И. П. Пав-
лова.
^Многочисленные ученики и последователи И. П. Павлова — ученые
Советского Союза — продолжают дело Павлова на основе единственно на-
учного мировоззрения — диалектического материализма. Они руковод-
ствуются положениями Ленина и Сталина о том, что только революционное
мировоззрение нашей партии может обеспечить победу передовой науки
и привести к разгрому ее врагов
Раздел II
КОМПЕНСАТОРНЫЕ
ПРИСПОСОБЛЕНИЯ
КОРЛ БОЛЬШОГО МОЗГА И ПЛАСТИЧНОСТЬ НЕРВНОЙ СИСТЕМЫ1
Сообщен ие I
ОПЫТЫ С АМПУТАЦИЕЙ ЛАП
риспособляемость является одной из главных основ суще-
ствования и развития живого вещества. Важнейшая роль при-
способляемости многообразно проявляется в процессе как фило-
генетического, так и онтогенетического развития организма.
Биологи до последнего времени концентрировали свое внима-
ние преимущественно на давно известных и вместе с тем наи-
более наглядных проявлениях приспособления и эволюции, а именно на-
следственных, довольно медленно протекающих изменений структуры и
функции отдельных систем организма или организма в целом. За послед-
ние годы, однако, передовые представители биологии ставят вопрос шире
и глубже; они выдвигают вопрос также о приспособительных явлениях дру-
гого порядка, имеющих важное значение для процесса эволюции в целом,
а именно о приобретенных изменениях в структуре и функции органов и
систем. Нам, нейрофизиологам, особенно радостно констатировать, что
передовая современная биология в лице академика А. Н. Северцова в
качестве важного фактора приспособления и эволюции подчеркивает зна-
чение «изменения поведения (behavior)1 животных без изменения их орга-
низации». Обозначая способность к эволюционному изменению термином
«пластичность организма», А. Н. Северцов о значении фактора ненаследст-
венного изменения поведения еще в 1922 г. писал следующее: «С биологи-
ческой точки зрения, т. е. с точки зрения приспособляемости животных, мы
имеем здесь фактор чрезвычайной важности, биологическое значение кото-
рого состоит в том, что он в весьма значительной степени повышает плас-
тичность животных по отношению к быстрым изменениям среды. При из-
менении внешних условий животное отвечает на него не изменением своей
организации, а быстрым изменением своего поведения и в очень большом
числе случаев может приспособиться к новым условиям весьма скоро» 2.
Конкретизируя свою смыль, А. II. Северцов в качестве основы этого факто-
ра указывает па «полнморфность функции органов», в особенности двига-
тельной системы, и на деятельность высших отделов центральной нерв-
ной системы, что он условно обозначает термином «деятельность разум-
ного типа».
Со своей стороны экспериментальная физиология и клиническая прак-
тика под названием викарирования, органических и функциональных
компенсаций и под другими названиями с давних пор зарегистрировали
и описали многочисленные и разнообразные проявления прижизненного
1 Успехи современной биологии, т. V, в, 5, 1936.
s А. Н. Северцов, Эволюция и психика, 1922, стр. 40,.
248
Э. А. АСРАТЯН
приспособления отдельных органов и систем, порой организма в целом,
как в условиях нормальной жизни, так и в особенности при тех или иных
грубых изменениях в структуре органов и систем, вызванных эксперимен-
тально или являющихся результатом несчастных случаев, болезней и т. д.
Общеизвестны многочисленные факты различных приспособлений дви-
гательной, пищеварительной, выделительной, дыхательной, сердечно-со-
судистой, покровной и других систем, в энзиматических процессах, гумо-
ральных средах организма и т. д. как в норме, так и при различных
болезненных состояниях организма. Их я здесь касаться не буду. Было бы
ошибкой считать, что физиология и клиника удовлетворялись лишь реги-
страцией фактов, о которых идет речь. В частности, экспериментальная
физиология в значительной степени выяснила непосредственные механизмы
отдельных проявлений приспособляемости некоторых систем. Однако мно-
гое в физиологическом механизме этих явлении до сих пор остается невы-
ясненным. В частности, до сих пор недостаточно выяснено несомненно важ-
ное значение нервной системы в целом, а также отдельных ее частей для
различных проявлений приспособления во всех названных выше органах
и системах сложного организма.
Наибольшую роль играют, однако, многообразные проявления приспо-
собляемости самой нервной системы.
Деятельность нервной системы по своему биологическому значению
является приспособительной. Еще в первоначальном гениально начер-
танном эскизе будущего величественного здания учения об условных реф-
лексах, в знаменитой мадридской речи И. П. Павлов, касаясь безуслов-
ных рефлексов, говорил: «В основании приспособления лежит простой реф-
лекторный акт», а перейдя к условным рефлексам, добавил: «Нет сомнения,
что мы имеем здесь перед собой факт дальнейшего приспособления»1. Такая
характеристика общебиологнческого значения условных рефлексов сохра-
няет свою силу в течение всего дальнейшего развития учения. Так, в
одном сообщении И. П. Павлова в 1927 г. сказано-: «Ясно, что высшее при-
способление животных, высшее уравновешивание с окружающей средой
непременно связало с этим сортом временно образующихся рефлексов» а.
А в последний период его творческой деятельности в обзорной статье
для 5СЭ, касаясь условнорефлекторном аналитической и синтетической
деятельности, И. П. Павлов писал: «Отсюда огромная широта и глубина
приспособленности, уравновешивания организма к окружающей среде»3.
Большую роль нервной системы в процессе приспособления организма
не отрицают также противники классической рефлекторной теории. Так,
Бете (Bethe) в своей совместной с Фишером обзорной статье на Эту тему
пишет: «Приспособляемость нервной системы является одним из многих
средств, которыми животный организм, если и не во всех случаях, то все
же весьма часто, может бороться с постоянной'сменой внешних и внутрен-
них условий И СО всюду его подстерегающими опасностями»4.
В этой проблеме, однако, меня особенно интересуют наиболее на-
глядные проявления приспособляемости нервной системы в виде различ-
ного рода компенсаторных приспособлении в по-вреждепном организме.
Здесь нет надобности касаться относящихся к этой категории многочислен-
ных очень интересных фактов, известных из клинической и профессио-
нально-гигиенической практики. Желающим поближе познакомиться с
этими фактами можно рекомендовать обзорные статьи Бете и Фишера
1 И. 11. Павлов, Двадпатилетний опыт объективного изучения высшей нерв-
ной деятельности (поведения) животных, 1932, стр. 25.
’Там же, стр. 395.
* И. П. Павлов, Физиологический журнал СССР, т. XIX, в. 1, 1935.
‘Bethe u. Fischer, Bethe’s Handbuch d. norm. u. pathol. Physiol., Bd. 15,
H. 2, 1931, S. 1045.
СООБЩЕНИЕ I, ОПЫТЫ С АМПУТАЦИЕЙ ЛАП
249
в «Bethe’s Handbuch», а также брошюру Н.А. Вигдорчика «Компенса-
торное профессиональное приспособление». Я хочу более подробно остано-
виться на фактах, полученных путем физиологических экспериментов.
Начиная с самого раннего периода развития экспериментальной фи-
зиологии, многие исследователи, пользовавшиеся методами разрушения
отдельных участков нервной системы, показали, что нервная система спо-
собна с различной быстротой и в разной степени компенсировать значи-
тельные отклонения от нормальной деятельности, обусловленные серьез-
ными нарушениями в се тончайшей структуре. Эго поразительное свойство
нервной системы, это замечательное проявление «своеобразного могущества
живого вещества», вызывающее истинное восхищение экспериментаторов,
И. П. Павловым образно было названо механическим иммунитетом. «В нерв-
ной системе, — писал он, — и специально в сложнейшем ее центральном
отделе, управляющем всем организмом, объединяющем все частные дея-
тельности организма, этот принцип механической самозащиты, принцип
механического иммунитета, должен был достигнуть высочайшего совер-
шенства, что действительно в массе случаев и оказывается»
Что касается фактов; проявления приспособляемости нервной системы,
то прежде всего можно указать на опыты с различного рода разрушениями
и экстирпацией частей корковой массы большого мозга у различных живот-
ных (Флуренс (Flourens), Гольц (Golz), Лучиани (Luciani), Мунк (Munk),
Гитциг (Hitzig), И. П. Павлов, В. М. Бехтерев и многие другие], а также
после удаления одного полушария собаки (Гольц). Затем можно привести
факты приспособления после частичной или полной экстирпации мозжечка
(Лучиани и др.), после перерезки задних корешков различных нервов [Би-
кель (Bickel),Магнус), после перерезки или анастомозов различных нервов
[Н. А. Миславский, Флуренс, Ленгли (Langley), Бете, Килвинпон (Kilvin-
gton) и многие другие], после перерезки ножек большого мозга и моз-
жечка [Гольц, Стар.чингср (Starl inger), Ротман (Rothman), В- М. Бехтерев
и др. ], после броун-секаровской перерезки спинного мозга и другого харак-
тера перерезок его массы [И. М. Сеченов, В. Кузьмин, Ф. Ф. Гольцингер,
Г. И Россолимо, И. В. Боровиков, В. М. Бехтерев, Вейсс (Weiss), Осава
(Osa wa), Портер (Porter) и др.]. В конце концов, сюда же можно причис-
лить факты компенсации нарушений нервной деятельности, обусловленные
повреждениями периферических эффекторных и аффекторных аппаратов,
как, например, в случаях нарушения моторики и равновесия тела в резуль-
тате ампутации конечностей (Фулд (Fold), Бете и др. ], после пересадки мышц
и сухожилий [Пертес (Perthes), Марина (Marina) и др. ], после разрушения
лабиринтов [В. М. Бехтерев,Эвальд (Ewald), Магнус и Клейн (Klein) и др. ].
Несмотря на существование огромного разнообразного и весьма цен-
ного фактического материала по затронутому вопросу, до последнего вре-
мени, однако, мы нс имели в физиологии даже попыток систематизировать
их, связать их в цельную систему, вывести общий закон. «Мне кажется, —
справедливо писал И. П. Павлов, в одной из последних своих статей, —
что до сих пор специально в физиологии нервной системы недостаточно
оценен и даже нс формулируется ясно и постоянно этот в высшей степени
важный принцип» s. .Мы не только не имели общей системы и теории, охва-
тывающей все перечисленные выше группы фактов относительно Принципа
приспособляемости нервной системы, ко до последнего времени не суще-
ствовало даже более или менее общепринятых систематизаций и теорети-
ческих концепций, которые охватывали бы явления приспособления внутри
каждой из названных групп- Многие ИЗ названных выше исследователей
в. своих теоретических обобщениях ограничивались зачастую только теми
1 И, И, Павлов, Двадцатилетии» опыт, 1932, стр. 481,
“Там же
250
Э. А. АСРАТЯН
фактами, которые ими были получены. Под названием викарирования,
замещения, органической и функциональной компенсации, способности
к механической самозащите и т. п. описывались явления приспособления
после тех или иных органических повреждений нервной системы; объяс-
нялись же они возникновением новых центров, тренировкой старых, пере-
дачей функций другим образованиям и т. д. В задачу дайной статьи но
входит систематический обзор фактов и взглядов всех хотя бы крупных
исследователей этого вопроса. Следует, однако, подчеркнуть один бесспор-
ный момент: хотя «механизмам» перечисленных разновидностей приспо-
соблений присущи специфические особенности, очевидно, что все они свя-
заны также общими закономерностями, обусловливаются тем общебио-
логическим свойством живого вещества, достигшим наивысшего развития
в нервной системе, которое в биологической и физиологической литера-
туре было названо пластичностью. Однако я считаю необходимым оста
ловиться на фактах и взглядах А. Бете, которому принадлежат не толь-
ко наиболее систематические исследования приспособительных явлений
после ампутации конечностей, по и попытка суммирования значитель-
ного литературного материала по всей проблеме и построения на этой
основе некоей общей теорий пластичности нервной системы.
Отмечая наличие способности приспособляться у всех, даже про-
стых животных и растений, Бете и Фишер пишут: «При наличии нервной
системы последняя при этом часто играет более или менее важную роль».
Авторы обзора указывают, что с классической рефлекторной теорией свя-
заны определенные достижения в изучении физиологии нервной системы,
но, по их мнению, «слишком много стало известно таких фактов, ко-
торые нельзя согласовать с этой теорией. Она была слишком механистична
и должна была быть заменена более функциональной, которую мы, однако,
еще должны ощупью отыскивать»1. По мнению Бете и Фишера, к числу
такого рода фактов относятся прежде всего факты приспособления орга-
низма к внешним и внутренним изменениям при посредстве цгнтральной
нервной системы. Исходя из этих отправных пунктов, Бете в течение ряда
лет развивал свою теорию пластичности нервной системы. Основные по-
ложения этой теории заключаются в следующем.
1. Не существует специфичности функций первичных ядерных эффе-
рентных и рецепторных нервных элементов в центральной нервной системе,
не существует также локализации центров и проводящих путей в централь-
ной нервной системе и вообще нет центров. Всю центральную нервную си-
стему можно рассматривать как «непрерывную сетчатую систему», а все
процессы в центральной нервной системе — как «переменную игру в этих
решетках». По предположению Бете и Фишера, «благодаря то более тесной,
то более дальней связи отдельных частей решетки, та или иная область
стала бы пригоднее для отдельных функций, но строго локализованных
центральных мест в этом хаосе не удалось бы обнаружить. Если при-
нять такое распределение нервной системы, то нужно было бы допу-
стить еще более широкие соотношения и во всяком случае признать воз-
можным участие в них периферических проводниковых путей и даже при-
мыкающих к ним конечных органов. Центр в периферия образовали бы
единое целее, более или менее неделимое; способности, приписываемые
обычно центральной нервной системе, распределялись бы на очень большом
протяжении, а собственно центра не было бы нигде.
Конечно, отдельные части системы участвовали бы в общей работе больше,
другие меньше, но каждая — и даже самая незначительная — часть играла
бы в целом свою роль. Каждое повреждение системы как перерыв
проводимости было бы способно влиять на все функции, на одни сильнее,
1 Beihe's Handbucb norm. u. pathol. Physiol,, Bd. 15, H. 2, 1931, S. 1050.
СООБЩЕНИЕ I. ОПЫТЫ С АМПУТАЦИЕЙ ЛАП
251
на другие слабее. Но ведь все наши операции па нервной системе ие что
иное, как в первую очередь прерывание проведения. Если мы думаем,
что при удалении, наркозе или охлаждении какого-либо центрального
участка мы сделали нечто значительно большее, чем только это, то
это является гипотезой» г. Против выводов о наличии особых центров в раз-
личных этажах центральной нервной системы, сделанных классиками
физиологии на основании многочисленных и разнообразных эксперимен-
тальных данных, авторы обзора приводят еще следующее рассуждение:
«Из всех этих целеустремленных опытов с несомненностью вытекает, что
определенные части центральной нервной системы имеют большее отноше-
ние то к одной, то к другой комбинации движений, что некоторые функции
также совершенно выпадают, когда известные области разрушены или от-
делены от остальной центральной нервной системы. Однако доказательство
того, что изучаемый процесс действительно происходит в этих частях,
почти нигде не могло быть дано. Единственно, что из этого можно вывести
определенно, это то, что та или иная область каким-то образом участвует
в процессе» г.
В более позднем произведении (1S33) Бете писал: «То, что до сих пор
представлялось нам центром, могло бы, грубо говоря, оказаться не чем
иным, как пунктами связи и прохождения между различными областями
рецепторной и эффекторной периферии»'*.
2. Хотя в совместно с Фишером написанной статье Бете допускает,
что в явлениях пластичности нервной системы анатомическим субстратом,
главным носителем этих пертурбации является центральный аппарат,
что «в основном все же придется признать, что главная роль при осуществле-
нии этих феноменов принадлежит центральным органам», — все же, по
представлению Бете, в единстве центра и периферии, в динамике нервных
процессов ведущую, определяющую роль в конечном счете играет пери-
ферия. «Периферия прямо-таки господствует над центральными явлениями:
не центральный аппарат определяет, Что должно произойти на периферии,
а периферия определяет, какую установку должен принять центральный
орган» В другом месте своей статьи Бете пишет с той же определенностью
и категоричностью: «Протекание центральных процессов, скорее, обуслов-
лено периферическими, эффекторными и рецепторными окончаниями со-
ответствующих нервов, чем анатомическими условиями в самых централь-
ных органах» 1 * * 4 5.
3. По теории Бете, в нервной деятельности, наряду с пластичностью
нервной системы и с принципом длительного регулирующего влияния
периферии на центр, важную роль играет еще третий принцип — принцип
борьбы за центральное поле, кратко названный Бете Exklusjvitat. Вот
краткая его формулировка: «Каждый процесс в нервной системе представ-
ляет собой единство, вовсе не допускающее рядом с собой никакого дру-
гого независимого от него нервного процесса или, по крайней мере, до-
пускающее его лишь в редких случаях»**.
Для обоснования своей точки зрения о принципах функционирования
нервной системы, о доминирующем регулирующем влиянии периферии на
центр и о принципе борьбы за центральное поле Бете прежде всего при-
бегает к разбору общеизвестных в физиологии старых фактов, а также
фактов из нормальной жизни животных и человека. Так, например, он
в.след за многими физиологами считает, что так называемые безусловные
1 Bethe’s Handbuch d. norm. u. pathol. Physiol., Bd. 15, II. 2, 1931, S. 1050.
a T а м же, стр. 1091.
8 Bethe A., Die Natnrwissenschaf ten, 1933.
4 Bethe’s Handbuch d. norm. u. pathol. Physiol., Bd. 15, H. 2, 1931, S. 1190.
“Таи же, стр. 1186.
•Там же, стр. 1196.
252
Э. Л. АСРАТЯН
реакции не являются абсолютно безусловными, что даже в нормальной
жизни сплошь и рядом нарушаются шаблонные координационные отно-
шения между скелетными мышцами, что одна и та же мышца, в зависи-
мости от условий, выполняет разные функции и т. д.
Хотя и заявляется авторитетно, что «перед такими фактами не устоять
классической теории, рассматривающей нервную систему как мозаичное
произведение из центров с определенными собственными функциями (Eigen-
funetjonen) \ все же Бете эти факты, видимо, не считает достаточными
для защиты позиции своей новой теории и в качестве таковой приводит
специально подобранный экспериментальный материал. Таким материа-
лом Бете считает большое количество точных и проверенных фактов,
полученных путем экспериментального нарушения целостности коорди-
национного аппарата, точнее, двигательного и сензорного аппаратов бес-
позвоночных и позвоночных животных, в виде ампутации конечностей,
пересадки мышц и сухожилий, анастомозов нервов, перерезок ннтрацент-
ральных проводящих путей, взаимного переключения моторных и сензор-
ных функций и т. д. С этой же целью приводится богатый материал хи-
рургической и неврологической клиники с аналогичными нарушениями
моторной и сензорной деятельности.
Приведенному большому экспериментальному и клиническому мате-
риалу, ярко демонстрирующему высшее проявление пластичности централь-
ной нервной системы в виде перестройки в центральном аппарате, в виде
компенсации глубоких нарушении сензорных и моторных функций
организма, Бете придает важное значение еще на том основании, что этот
материал якобы доказывает правильность некоторых частных и кон-
кретных положений, лежащих в основе изложенных выше более широ-
ких теоретических его концепций.
Указанные частные положения Бете сводятся к следующему: а) явле-
ния приспособления организма после глубоких и разнородных нарушений
в области моторной и сензорной деятельности осуществляются благодаря
быстрым перестройкам в примитивных частях центральной нервной си-
стемы; б) движущим рычагом этих перестроек в центральной нервной си-
стеме являются текущие импульсы с периферии, сигнализирующие о про-
исшедших изменениях; в) в явлениях пластичности, конкретЕЮ—в этих
перестройках примитивных частей центральной нервной системы, большой
мозг не играет роли или во всяком случае заметной роли. Перестройка
не есть процесс заучивания.
Говоря о будто менее показательных для его концепций случаях пла-
стичности (анастомозы нервов, половинная перерезка спинного мозга
и т. д.), где процесс приспособления затягивается, Бете пишет: «Таким
образом, можно сказать, что за этот период нервная система имела бы время
приспособиться к измененным условиям, и можно рассматривать все это
явление, в особенности у человека и высших животных, как процесс на-
учения. Но полученные до сих пор результаты дают нам достаточное коли-
чество примеров, которым нельзя давать подобное объяснение; это такие
случаи, когда мы вынуждены локализовать феномен пластичности в. при-
митивных частях нервной системы»2. В другом месте Бете говорит: «Пери-
ферия образует центральный орган соответственно своему составу, своему
состоянию и согласно с происходящими в ней процессами. Можно даже
сказать, что она создает для себя «центры» в центральных органах, обра-
зующиеся то сравнительно медленно и постепенно (.например, при сокра-
щении нервов и перемене мышц), то принимающие на себя новые функции
совершенно неожиданно и непосредственно (появление совершенно нз-
* Bethe's Handbuch d. norm, u. pathol, Physiol,, Bd. 15, H- 2, 1931, S. 1053.
’Там же, стр. lOiZ.
СООБЩЕНИЕ I. ОПЫТЫ С АМПУТАЦИЕЙ ЛЛП
253
мененных координаций при утрате конечностей и т. д.). Таким образом,
функции какого-либо участка центральной нервной системы определяются
не топографическим положением ил и гистологическим строением, а скорее
всего ее непосредственной связью С периферией при помощи перифери-
ческих нервов, а также непосредственными связями с ее центральными
соединениями. Но оба вида связей могут изменяться, и центральные от-
делы должны подчиняться этим изменениям»1.
Следует сказать, что теория Бете имеет как сторонников за рубежом,
так и единичных приверженцев в. Советском Союзе. Из отечественных
физиологов Следует назвать в- первую очередь П. К. АнОХИнй, работ и
воззрений которого я коснусь позже при изложении материала по анас-
томозу нервов.
Начиная с конца 1932 г., после ознакомления с обзорными статьями
Бете я стал заниматься теоретическим и экспериментальным изучением
некоторых вопросов-, имеющих отношение к пластичности нервной сис-
темы. Авторитет Бете, широко поставившего вопрос о пластичности нерв-
ной системы, не препятствовал моему критическому отношению к ряду
теоретических его установок.
Первое общее положение Бете, направленное нс только против старой
метафизической, досеченовской рефлекторной теории, но и против новой,
динамической по существу теории, — положение, отрицающее наличие
специфических центров и проводящих путей в центральной нервной си-
стеме у высших животных даже в динамическом их понимании, нельзя
считать правильным. Если старая метафизическая рефлекторная теория и
механистическое учение о специализации и локализации центров и прово-
дящих путей до сих пор крепко сидят в головах некоторых неврологов и
морфологов, то передовая физиология в лице лучших ее представителей
(И. М. Сеченов, И. П. Павлов, Гольц, Лучиани и др.), базируясь на огром-
ном количестве разнообразного и достоверного экспериментального мате-
риала, вышла из этих узких рамок и построила динамическую рефлекторную
теорию и учение о специализации и локализации центров и проводящих
путей центральной нервной системы.
Положение Бете, разделяемое некоторыми психологами и неврологами
и представляющее центральную нервную систему чем-то вроде функцио-
нально-аморфной решетки, противоречит прежде всего фактам физиологии и
клинической практики. Будучи якобы направлено против механицизма, как
утверждает Бете, это п| едставлеиие граничит с мистикой. Оно искусственно
выпячивает общее, совершенно игнорируя и растворяя в нем частное. Ме-
ханистичными следует также считать попытки Беге черпать основную
аргументацию против специфичности центров и путей у высших животных
из сравнительно-физиологического материала, причем данные о низших
беспозвоночных он механически переносит на высших животных. Бете
пишет: «Если, как было указано в предыдущей главе, уже сравнительно
физиологическое рассмотрение приводит нас к почти непреодолимым воз-
ражениям против учения о центрах (в особенности же против того воззре-
ния, что координация регулируется с определенных мест по твердо уста-
новленной схеме), то под влиянием того, чему нас учат явления пластич-
ности, начинают колебаться и последние опоры этой гипотезы» 2.
По существу концепция Бете полна противоречий. Признание нм того,
что та или иная часть центральной нервной системы принимает большее
участие в тех или иных процессах, чем другие части центральной нерв-
ной системы, уже есть отступление от явно выраженных мистических
позиций. Дальше, если согласиться с Бете, что наши опыты с экстир-
пациями говорят лишь о том, что нарушаются какие-то связи в централь-
1 Bethe’s Handbuch d. norm. u. pathol. Physiol., Bd. 15, H. 2, 1931, S. 1190.
* Там же.
254
Э А. АСРАТЯН
ном аппарате, то непонятными будут специфические отклонения от нор-
мальной деятельности, связанные с экстирпациями морфологически обо-
собленных частей центральной нервной системы. Я уже не говорю о том,
что при большей последовательности сторонники теории Бете нс должны
были допускать даже ничтожных отклонений от нормальных функций
после таких перерывов связи, ибо, согласно их взглядам, связь между
частями центральной нервной системы осуществляется почти в одина-
ковой степени многими путями.
Нельзя считать серьезным выходом из этих противоречий то, что
слово «центр» заменяется ими определениями такого порядка: та или иная
часть центральной нервной системы имеет более близкое и интимное отно-
шение к той или иной функции. Больше того, мне кажется, что если про-
являть такую категоричность, как Бете и его сотрудники, то логически
следует сделать вывод, что постановка вопроса о пластичности и пере-
стройке вообще не имеет’смысла. Раз нет центров и проводящих путей
с некоторой специфичностью функции, раз вся центральная нервная система
функционально аморфна, то о какой перестройке может идти речь?
Второе положение о важном значении периферических импульсов
для всех видов нервной деятельности является одним из бесспорных и
основных положений современной нейрофизиологии, столь общепринятым
И общеизвестным, что я на нем подробно останавливаться не буду. Огра-
ничусь напоминанием о той большой роли, которую придают перифери-
ческим импульсам со второй половины XIX века физиологи,изучающие
нервную систему, в частности, И. М. Сеченов, Магнус—в сложной к простой
координационной деятельности спинного мозга и ствола головного
мозга, И.П. Павлов—в работе большого мозга. Тем неменее нельзя согла-
ситься с Бете, когда Он периферии приписывает исключительную, всемогу-
щую роль в центральных нервных явлениях, когда он слишком упрощает
и умаляет роль самой центральной нервной системы, представляя ее в
виде как бы простого связующего звена между афферентными и эфферент-
ными нервными путями, в виде какой-то функционально аморфной пассив-
ной решетки, в которой явления формируются и разыгрываются
всемогущей периферией. Нельзя согласиться с этими воззрениями Бете,
ибо фактически все фундаментальные данные классической и современной
физиологии говорят против гиперболизирования значения пернфе-
р и и и недооценки центра. Что периферические импульсы в состоянии
изменять обычные координационные отношения во всей центральной нерв-
ной системе, нс подлежит сомнению; но как можно игнорировать массу
точных и проверенных фактов относительно изменения тех же обычных
координационных отношений в нижележащих отделах центральной нервной
системы под влиянием вышележащих ее отделов (мозжечок,большой мозг)?
В конце концов, как объяснить, если не работой большого мозга, те
факты, когда в обыденной жизни человек и животное произвольно изме-
няют деятельность координационного аппарата, когда большой мозг из
этой деятельности лепит всевозможные динамические скульптурные обра-
зы? Точка зрения Бете фактически и методологически неверна, ибо она
центральную нервную систему фактически нивеллирует, обезличивает.
Выступая внешне против механицизма в физиологии, Бете в действитель-
ности попадает в плен механистического мировоззрения.
Что же касается третьего общего принципа — борьбы за центральное
поле, то, если отбросить его метафизическую формулировку, против
существа его трудно возражать; это является одним из принципов, ко-
торый в современной нейрофизиологии не вызывает никаких существен-
ных разногласий в смысле его констатации, в особенности после класси-
ческих работ по этому вопросу И. П. Павлова, Гольца и А. А. Ухтом-
ского.
СООБЩЕНИЕ 1 ОПЫТЫ С АМПУТАЦИЕЙ ЛАП
255
Несколько замечаний по частным положениям Бете, якобы обосно-
ванным результатами его эксперментов
Никак нельзя согласиться с мнением Бете и его сторонников (П. К. Ано-
хин и др.) в Оценке роли и значения коры большого мозга в явлениях
пластичности центральной нервной системы и в характеристике возни-
кновения, развития н существа этих явлений. Бете большей частью про-
водил опыты с ампутацией конечностей у низших и высших животных
и для обоснования своих взглядов придает решающее значение резуль-
татам, полученным из этих опытов. Ввиду того что я тоже до сих пор
больше всего изучал последствия этого вида повреждений организма, я
подробнее остановлюсь на этих опытах.
Какими данными из упомянутого цикла работ аргументирует Бете
свои воззрения о значении большого мозга и о характере возникновения
компенсаторных явлений?
1. По мнению Бете, животные начинают ходить на оставшихся двух
лапах через такое короткое время после операции (в первые дни и даже
часы), что трудно допустить участие коры в этих явлениях и считать, что
они являются продуктом «научения». «Так как такие перестройки коор-
динации появляются сразу же после ампутации и таким образом не
могут быть выученными, то уже у животных с немногими конечностями
кажется невероятным, а у животных со многими конечностями является
немыслимым допущение существования заранее сформировавшегося центра
для каждой перестройки. Отпадает и та возможность, что перестройки
совершаются центром с высокой способностью приспособления [например,
путем «сознательного вчувствования (Einfiihlung)], так как удастся дока-
зать, что изменения координации не обусловлены присутствием и высших
центров» Ч
Быстроту восстановления функции состояния и ходьбы после ампута-
ции двух лап у собаки, как мне кажется, нельзя считать аргументом, иск-
лючающим предположение об участии большого мозга в этих перестройках.
Не говоря о том, что большой мозг своим трофическим влиянием может
всегда облегчать и ускорять эти перестройки, я хочу отметить возмож-
ность участия большого мозга в них также посредством условнорефлектор-
ной деятельности даже за такие короткие сроки. Ведь нельзя считать пра-
вильными взгляды, согласно которым выработка новых навыков, новых
условных рефлексов, всегда требует недель и месяцев. Не желая распро-
страняться на эту тему, укажу только, что колоссальную роль в этом
играет обстановка, сила и характер безусловной реакции, длительность
опыта и т. д. Часто бывает, что условные рефлексы, особенно оборонитель-
ные, вырабатываются через несколько сочетаний, в течение минут. В слу-
чае же ампутации лап мы имеем все благоприятствующие моменты для
быстрого возникновения условных рефлексов, которые могут привести
к новым видам передвижения животного. Однако, как показали прове-
денные мной опыты, далеко не у всех животных эта перестройка осущест-
вляется так быстро, так «молниеносно»-,как описывает Бете. Иногда это
требует десятка дней (подробное описание моих опытов см. ниже).
2. Так как экстирпация двигательных зон коры большого мозга не сни-
мает наступивших ранее перестроек (не считая длящихся несколько дней
легких нарушений, неизбежно появляющихся после экстирпации двига-
тельных зон даже у нормальных животных), Бете считает доказанным,
что кора не принимает участия в этих перестройках. «Известные наруше-
ния появлялись после каждой операции на большом мозгу (т. с. экстир-
пации двигательных зон. — Э. А.), но уже через короткий срок восста-
навливается прежний способ передвижения. Даже и после второй операции
1 Bethc’s Handbuch d. norm. u. pathol. Physiol., Bd. 15, H. 2, 1931, S. 1078.
256
Э. А. АСРАТЯН
(удаление второй зоны. — Э. Л.) животное могло прыгать со стола, не
причиняя себе вреда» к Мне думается, что этот аргумент Бете выявляет
его непоследовательность. В своих теоретических концепциях он отрицает
фиксированную локализацию и вообще локализацию функции во всей
центральной нервной системе; значит, он должен был бы отрицать и ло-
кализацию сензомоторной функции в коре большого мозга. Для доказа-
тельства же того, что кора большого мозга не участвует в процессах пере-
стройки моторных функций, он поступает так, как поступили бы ярые
сторонники метафизического представления о локализации функций, —
свои выводы базирует на результатах экстирпации только двига-
тельной зоны коры. Такая постановка вопроса, разделяемая П. К. Ано-
хиным, не выдерживает критики с точки зрения динамической теории
локализации (И. П. Павлов, Гольц, Лючиани и др.), а не только с точки
зрения теории, отрицающей всякую более или менее стойкую локализацию.
3. Ввиду того что такие компенсации моторики так же хорошо насту-
пают у «глупых животных», как и у «умных», Бете это тоже считает аргу-
ментом против участия высших центров или, говоря иначе, большого мозга
в этих явлениях. «Из параллельного опыта, который один из нас (Фишер)
многократно повторял с таким «глупым» животным, как морская свинка,
мы пришли к заключению, что большой мозг не играет при этом особой
важной роли» * *. Мне кажется, что эта аргументация против допущения
участия большого мозга в указанных компенсаторных явлениях не может
считаться научной, и потому па пей я останавливаться пе буду.
4. Для обоснования своего взгляда относительно непричастности боль-
шого мозга к описанным явлениям у высших животных Бете ссылается
на данные опытов над беспозвоночными животными, у которых экстирпа-
ция надглоточных и подглоточных нервных узлов пе оказывает никакого
влияния на появление новых видов координации движения после ампу-
тации лап. «Для теоретического объяснения всех опытов с ампутацией
имеют особенное значение опыты С DytiScuS, Так как зДеСь легко можно
доказать, что существеннейшие перестройки наступают даже в том случае,
если мозг (надглоточный ганглии) уже ранее был удален, и даже еще тогда,
когда отсутствуют и надглоточный и подглоточный ганглии. Значит, после
потери конечностей происходят изменения координации в относительно
примитивных частях нервной системы, в ганглиях брюшного мозга. Так
называемые «высшие центры» здесь участвуют мало или ВовСе не участвуют.
К аналогичным выводам, хотя и не непосредственно, пришел Будденброк
путем своих опытов над Dixippus»3.
Будучи горячим и убежденным сторонником эволюционного подхода
к разрешению любого явления в природе, в частности, в физиологии, я
не могу согласиться с Бете, когда он подобные данные физиологии нервной
системы беспозвоночных животных механически переносит в физиологию
нервной системы высших позвоночных. Хотя Бете проводит аналогию
своей теории с эволюционной теорией (в противовес рефлекторной, которую
он сравнивает с преформационной теорией), с ним никак нельзя согла-
ситься, когда он надглоточные и подглоточные узлы беспозвоночных
функционально отождествляет с корой большого мозга высших млекопи-
тающих. С точки зрения эволюционной теории мне представляется не-
правильным, механистическим также прием Бете и Фишера, заключаю-
щийся в том, что для иллюстрации явления пластичности центральной
нервной системы в опытах с ампутациями конечностей они животных
группируют не по уровню их развития, а по механическому принципу —
количеству конечностей у тех или инЬх животных. Они прямо пишут:
1 Bethels Handbuch d. norm. u. pat hoi. Physiol., Bd. 15, H. 2, 1931, S. 1072.
* T а ы же стр. 1066.
’Там же,, стр. 1077.
СООБЩЕНИЕ I. ОПЫТЫ С АМПУТАЦИЕЙ ЛАП
257
«Не считаясь с зоологической системой, мы при описании фактов перехо-
дим шаг за шагом от животных с немногими конечностями к тако-
вым с многими конечностями» *.
При критическом рассмотрении выясняется, что описанные выше част-
ные теоретические положения Бете, на которых основываются более об-
щие его теоретические концепции о нейрофизиологии, не могут считаться
обоснованными; убедительными экспериментальными данными, теоретиче-
ски строго последовательными и бесспорными, а методологически пра-
вильными.
Свое отрицательное отношение к этим данным и концепциям Бете,
возникшее после подробного ознакомления с его работами еще в 1932 г.,
я подкреплял еще следующими логическими рассуждениями. Я не мог
согласиться с его положением, что в организме высшего животного, каким
является собака, тем более в «примитивных», по выражению Бете, частях
центральной нервной системы этих животных, может происходить корен-
ная перестройка и переделка функций в результате ампутации лап, ана-
стомоза нервов, перерезки спинномозговых путей, пересадки мышц и сухо-
жилий и т. д. без деятельного участия большого мозга, без ближайшего
участия коры. Ведь современная биология и в особенности нейрофизиоло-
гия не оставляют сомнения в том, что принцип пластичности, о котором
идет речь, достигает наибольшего совершенства, наивысшего своего раз-
вития, качественно нового вида проявления в виде условнорефлекторной
деятельности именно в этом органе (Л. Н. Северцов, И. П. Павлов). Игно-
рирование значения коры большого мозга показалось мне тем более не-
правильным, что современная физиология все больше обогащается бес-
спорными данными, указывающими на явное и скрытое участие большого
мозга в регуляции работы почти всех без исключения органов и систем
высокоразвитого животного.
Большой интерес к одной из актуальнейших проблем современной
нейрофизиологии и критические отношения к теоретическим положениям
Бете побудили меня заняться исследовательской работой в этой области.
Собственные экспериментальные данные по ампутации
конечностей у собак
Я начал эксперименты В Физиологическом институте Академии наук
СССР в 1933 г., затем в 1934 г., частично в начале 1935 г. продолжал их в
ВИЭМ, а с начала 1935 г. в связи с моей работой по совместительству
в Институте мозга нм. В. М. Бехтерева я все исследования сконцентриро-
вал в этом институте, где н выполнена основная часть работ.
Я ампутировал в различных комбинациях по две лапы у 13 собак2 и
у одной кошки. Данные, полученные на кошке, неясны, так как она плохо
перенесла операцию и через несколько дней погибла. Из 13 собак со вто-
рой половины дня операции хорошо холили на двух лапах только две:
Садко и Тузик, а с 5-го дня — одна собака Золото. У первых двух собак
лапы были ампутированы крест-накрест, а у Золота ампутации подверглись
две лапы на одной стороне. Собаки были среднего и ниже среднего роста,
причем Садко была старой, Тузик— средних лет, а Золото — 6—7-месяч-
ным щенком. Две первые собаки поступили в лабораторию тренирован-
ными; они обе могли стоять и ходить на задних лапах. Точных сведений
о тренированности третьей собаки я собрать не мог. Так как из 13 со-
бак 10 научились ходить лишь через много дней и месяцев и только у
1 Bethe’s Handbuch d. norm. u. psfthol. Physiol., Bd. 15, H. 2, 1931, S. ICG®.
г В дальнейшем нами произведена ампутация лап более чем у 20 собак с теми
же результатами. Данные этих экспериментов опубликованы в «Бюллетене экспери-
ментальной биологии и медицины», т. VIII, в. 6, 1939.
17 Э. А. Асратян
258
Э. А. АСРАТЯН
3 собак (из которых 2 тренированные) компенсация наступала быстро,
именно так, как описывает Бете, у меня возникло предположение, что
большинство собак Бете было тренировано прежними хозяевами, и Бете
не обращал на этот момент должного внимания.
Нет ничего удивительного в том, что тренированные ранее собаки после
ампутации лап быстро перестраиваются и начинают пользоваться новыми
приемами передвижения. Из практики хорошо известно, что моторные
навыки способны хорошо генерализоваться. Какой-нибудь новый акроба-
тический прием не с одинаковой легкостью удается человеку, который
совершенно не занимался акробатикой, и человеку, который занимался
ею и овладевает другими акробатическими приемами.
Подчеркивая значение предварительной тренированности собаки для
быстроты наступающей перестройки, я не хочу совершенно отрицать
возможность быстрой перестройки также у предварительно нетренирован-
ных собак, в особенности при малом весе и коро1ких конечностях собаки
и при таких комбинациях ампутации лап, которые особенно тяжело от-
ражаются на функции стояния и ходьбы. Но это относится, пожалуй,
больше к исключениям, чем к правилу. Кроме того, как указывалось вы-
ше, этот момент вообще не может являться решающим для суждения о ро-
ли большого мозга в этих процессах, об их интимном характере и т. д.
Ради четкости описания я разделил собак на группы по признаку сход-
ства ампутаций лап и мозговых операций.
Собаки с ампутированными крест-накрест лапами. По утверждению
Бете, ампутация одной передней и противоположной задней лапы вызы-
вает наиболее резкие нарушения в моторике собак. Исходя из этих сооб-
ражений, я основное свое внимание сконцентрировал на этом варианте
ампутации.
У 5 преимущественно молодых здоровых собак различного роста (Хро-
мульчик, Садко, Сюита, Аллегро и Модерато) крест-накрест были ампути-
рованы одна передняя и одна задняя лапа в срединных суставах конеч-
ностей. Кроме одной из них (Садко), у остальных собак перестройка мото-
рики совершалась постепенно на протяжении длительного времени. В пер-
вые 15—20 дней все они совсем не могли вставать и при попытках передви-
гаться ползали по земле, хаотически тыкаясь в нес мордой, с помощью
здоровых и ампутированных конечностей. Когда собак ставили на две лапы,
они могли при поддержке стоять, но как только отнимали руки, собаки
сразу же падали на землю. После этой фазы как бы полной инвалидности
незаметно наступала следующая, переходная фаза, длившаяся примерно
столько же дней. Собаки начинали делать попытки вставать, прибегали
к помощи морды и культи ампутированной передней конечности. В первые
дни этой фазы попытки эти обычно кончались неудачей: собаки сплошь
и рядом снова падали на землю. Постепенно собаки совершенствовали
«технику» своего передвижения, делали два-гри прыжка, ие падая и не
прибегая к помощи культи ампутированных лап и морды. Вначале эти
успешные прыжки еще часто чередовались с падением па землю. С течением
времени, однако, у собак постепенно удлинялись периоды успешных прыж-
ков на две лапы и сокращалось количество падений на землю. Примерно
на 20—25-й день после ампутации лап собаки могли хорошо передвигаться
на расстояние нескольких метров, ни разу не падая, но только в том слу-
чае, если они прыжки совершали медленно, каждый раз как бы соразме-
ряя их. Стоило им, однако, в этом периоде участить прыжки, как они па-
дали на землю. Характерно, что в этой фазе, как правило, собаки часто
падали даже при медленных прыжках, .если их обильно кормили перед
пробами. Третья, последняя фаза, наступавшая после переходной, закан-
чивалась тем, что собаки успешно прыгали при любой частоте прыжков
даже после основательной нагрузки желудка. Следует еще отметить, что
СООБЩЕНИЕ I. ОПЫТЫ С АМПУТАЦИЕЙ ЛЛП
259
в первой фазе собаки при сталкивании их со стола калачиком падали на
землю; при той же пробе во второй фазе они хотя и не могли удержаться
на ногах, но ноги первыми достигали земли, а в третьем периоде собаки
уже прыгали и становились на здоровые лапы, как только их сталкивали
со стола.
При регулярном наблюдении за этими собаками создается впечатле-
ние, что они изо дня в день учатся, постепенно приобретают новые навыки
передвижения, непрестанно совершенствуют их. Того, что Бете называет
«молниеносной перестройкой», здесь не имеется и следа. Перестройка осу-
ществляется в процессе длительной и упорной, я бы сказал, даже мучи-
тельной для животных и экспериментатора тренировки и выучки. Живот-
ные вертятся, цепляются за что попало и делают всевозможные пробы. Из
пресмыкающихся они превращаются в двуногих животных.
Несомненно, большое значение имеют вес тела собаки, длина ее ног,
возраст и др. Эти моменты могут изменять длительность отдельных фаз
восстановления функции, соотношение между ними, общую длительность
и т. д. Но характер и существо процесса восстановления функции в основ-
ном остаются одними и теми же. Собаки делают всевозможные пробы,
учатся, тренируются.
Одного этого совершенно очевидного и бесспорного факта, мне кажется,
достаточно, чтобы оценить важную роль коры большого мозга у высших
животных в функциональных перестройках нижних отделов центральной
нервной системы, имеющих непосредственное отношение к моторике жи-
вотного. Такой вывод напрашивается на том основании, что до сих пор
кора большого мозга считается если не исключительным, то основным ор-
ганом условнорефлекторной или приобретенной деятельности.
Однако, несмотря на очевидность и бесспорность этих фактов и логич-
ность выводов, я все же не считал эти факты достаточными для оконча-
тельной оценки роли коры большого мозга в процессах перестройки низ-
ших отделов центральной нервной системы. По этой причине я не спешил
с опубликованием указанных данных, хотя они были получены еще в са-
мом начале моей работы в 1933 г., и неоднократно демонстрировались
моим товарищам по работе, а также И. П. Павлову. Я поставил перед
собой задачу экстирннровать кору одного, а затем и другого полушария
большого мозга у ампутированных собак (у которых уже завершилась
перестройка функций в центральной нервной системе), а также произ-
вести ампутацию у собак, лишенных коры одного и двух полушарий,
и проследить у них за ходом изменений моторики после этого. Экстирпа-
цию только ОДНИХ двигательных зон коры, кйк делали Бете, П. К- АНОХИН
и др., я считал неубедительной. Еще в самом начале работы я отдавал
себе полный отчет в колоссальных затруднениях, стоящих перед реализа-
цией такого дерзкого плана. Сколько нужно было иметь собак без одного
и двух полушарий, да еще подвергшихся добавочным операциям! Физио-
логам хорошо известно, с каким трудом удается получить и содержать
таких собак даже при наличии хороших условий работы. Однако, несмотря
на это, мне удалось собрать некоторый материал по затронутому вопросу.
Собаки без коры одного полушария и с крест-накрест ампутированными
лапами. Как только собаки достигали совершенной «техники» передвижения
на двух лапах, я удалял у них кору одного полушария большого мозга,
чтобы выяснить сначала влияние удаления одного полушария. Из этих
собак Хромульчмк погиб на операционном столе от наркоза. Аллегро
и Модерато погибли несколько дней спустя после операции от кровоизлия-
ния в мозг, и только Сюита жила Около 4 месяцев после удаления коры
одного полушария. Кору одного полушария я удалил также у Садко,
который, как выше было сказано, после ампутации лап быстро приспо-
собился к новому способу передвижения. Садко жил 13 дней и погиб
17»
260
Э. А. АСРАТЯН
от нагноения операционного поля. Наблюдения на Сюите и частично па
Садко показали, что после удаления коры одного полушария большого
мозга приобретенный раньше способ передвижения на двух лапах почли
целиком утрачивается. Его восстановление хотя требует в общем меньше
времени, чем приобретение (вместо! — I1/» месяцев—15—20 дней), однако
оно характеризуется почти той же Постепенностью, что и первоначальный
Рис. 1. Динамика восстановления функций стояния и ходьбы у собаки
Мальчик после ампутации передней левой и задней правой конечностей
а—через 18 пней после операции; в—iepea 22 дня: е—через 50 дней
процесс приобретения этого навыка. Характерно, что даже у Садко, перво-
начально перестроившегося «молниеносно» быстро, после удаления коры
одного полушария и после потери этой перестройки новая развивалась
уже не сразу, а постепенно. Смерть помешала собаке завершить этот про-
цесс, но развитие его протекало очень четко, в особенности в течение по-
следних 10 дней жизни.
У других собак я использовал новые варианты ампутации. У одной
молодой собаки Мальчик, среднего роста, я за один прием ампутировал
две лапы (правая передняя и левая задняя) и экстнрпировал кору левого
полушария. Мальчик жил 7 месяцев и погиб после экстирпации коры
второго полушария. Следует отметить, что у Мальчика наблюдалась та же
картина, что и у нормальных собак с ампутированными крест-накрест
лапами, с той лишь разницей, что периоды восстановления функций у
него были более затяжными. Процесс тренировки и выучки Мальчика
Заснят на кинопленке (рис. 1).
СООБЩЕНИЕ I. ОПЫТЫ С АМПУТАЦИЕЙ ЛАП
261
У двух других собак, Ту?ика и Красавицы, я удалял сначала кору
одного полушария и только спустя некоторое время (1 — Р'а месяца) ампу-
тировал лапы крест-накрест. Как уже было сказано выше, Тузик был тре-
нированной собакой и на 2-й день уже прыгал на двух лапах. Красавица же
училась ходить на двух лапах примерно в течение IVs месяцев. Разница
между этими собаками, кстати, одновременно оперированными, была
очень большой. Она опять-таки указывает на большое значение предва-
рительной тренировки даже для собак с одним полушарием, Восстапов-
ление нарушенных двигательных функций у наших собак после уда-
ления коры одного полушария не должно казаться неожиданностью, в
особенности если учесть работы Гольца, показавшие, что после удале-
ния у здоровых собак не только коры, но даже цел и ком одно го полушария,
возникшие имения выпадения со временем компенсируются.
Собаки без коры двух полушарий и с ампутированными крест-накрест
лапами. Хотя приведенный выше материал с большой отчетливостью гово-
рил о важной роли коры большого мозга в процессах функциональной
перестройки нижележащих центральных нервных аппаратов, я все же
решил получить более веские доказательства своих гипотетических поло-
жений. С этой целью я ампутировал передние И задние лапы крест-
накрест у одной молодой, среднего роста собаки Симпатия, лишенной
обоих полушарий.
5.VI 1935 г. была экстнрпнрована кора правого полушария, 28.VII —
кора левого полушария. После того как собака хорошо оправилась от
второй операции и начала хорошо ходить, 14.XI 1935 г. были ампутиро-
ваны передняя левая и задняя правая лапы. После ампутации лап Симпатия
жила примерно б1/# месяцев и за этот период не только не могла ходить
или хотя бы сделать один прыжок, но она не могла даже стоять на двух
лапах. Существенной разницы между «техникой» передвижения Симпатии
в первые дни после ампутации лап и в последние дни ее жизни не было,
кроме разве той, что безусловных попыток стоять и ходить в первые дни
она делала меньше, чем в последние. Попытки она делала непрерывно,
хаотически двигая всеми выступающими частями тела (конечности, голова,
задняя часть). Но эти попытки не привели ни к каким результатам, кроме
сильных ударов о землю, ушибов и т. д. Иногда при усердных движениях
собаке удавалось с помощью толчка встать иа две здоровые лапы, но через
миг она вновь падала на землю. Если ее поддерживали, собака опиралась
на две неповрежденные лапы. Но стоило опустить руки, и животное сразу
же падало на землю.
Кроме Симпатии, у меня была еще одна собака без двух полушарий
и без двух лап — Красавица, о которой я уже говорил. После того как
у Красавицы была удалена кора одного полушария, затем были ампу-
тированы две лапы крест-накрест, прошло 47 дней, пока она полностью
овладела «техникой» передвижения на двух лапах. Затем я экстирпировал
кору другого полушария большого мозга. После последней операции Кра-
савица жила 4 месяца без 3 дней, полностью утратя свою совершенную
«технику» передвижения на двух лапах (рис. 2). Она очень напоминала
Симпатию с той лишь разницей, что после экстирпации коры двух полу-
шарий она была спокойнее Симпатии и меньше делала безуспешных по-
пыток передвигаться.
Следует отметить, что, опираясь о стену, как Симпатия, так и в особен-
ности Красавина могли стоять в течение десятков секунд и даже минуты.
Затем они обе могли сидеть более или менее долго, когда удобно опирались
на заднюю часть, на культю амнутарованной лапы и на экстснзированную
переднюю цельную лапу. Удерживание в течение нескольких мгновений
на ногах в результате удачных прыжков вверх наблюдалось также
у Красавицы. Наблюдения за тремя другими собаками без коры двух
262
Э. А. АСРАТЯН
полушарий и без двух лап (Волчок, Мальчик и Тузик), жившими недолго
после операции (2—13 дней), хотя и не могут считатся по этой причине
полноценными, но все же целиком подтверждают данные, полученные
на Симпатии и Красавице.
Не вдаваясь пока в теоретические рассуждения, отмечу лишь, что
приведенный материал с полной очевидностью показывает, что кара
большого мозга не только имеет
Ри-е, 2- Влияние удаления коры больших
полушарий на функции стояния и ходьбы
у собаки Красавица с ампутированными
передней правой и левой задней конечностя-
ми. Через месяц после удаления коры пра-
вого полушария большого мозга были ампу-
тированы правая передняя и левая задняя
конечности
а—собака через 47 дней после второй операции:
хорошо стоит и лередпитается; б и л—через 3 ме-
сниа после удаления коры лепого полушария
большого мозга: собака на нее г да потеряла способ-
ность стоять и ходить
отношение к перестройке функ-
ции «примитивных частей» цент-
ральной нервной системы, непо-
средственно ведающих моторикой
животного, но что она играет
основную, определяющую роль
в этих перестройках.
Мой вывод подкрепляется и
другими новыми опытами с
иными вариантами ампутации
лап.
Другие варианты ампутации
лап. Прежде всего остановлюсь
на собаке Жулик, низкого рос-
та, с ампутированными у са-
мых верхних суставов передни-
ми двумя лапами. Жулик дос1иг
хорошей «техники» передвиже-
ния на двух задних лапах, на-
поминавшей движения кенгу-
ру, через месяц упорной и
мучительной тренировки. В про-
цессе этой «учебы» Жулик па-
дал на морду, на грудь, опять
поднимался, опять падал и т. д.
Так же постепенно, но в более
короткий срок, научилась хо-
дить на двух передних лапах
другая собака среднего роста
Жарик, у которой были ампу-
тированы две задние лапы в
средних суставах, К сожалению,
Жарик, как и некоторые дру-
гие собаки, погиб от случайных
причин во время моего отсут-
ствия осенью 1934 г. Не повезло
также с собакой Жулик, у ко-
торого я в один прием удалил
кору обоих полушарии, чтобы
быстро получить ответ на по-
ставленный вопрос: на второй
день после операции животное
погибло.
Интересные результаты были
получены у собаки Золото.
У этой молодой живой собаки низкого роста я ампутировал переднюю
и заднюю лапы с левой стороны. По не зависящим от меня причинам я не
мог следить за ней в первые дни после ампутации лап, а сотрудники ла-
боратории не обратили внимания на то, когда она начала прыгать на двух
сообщение г опыты с ампутацией лап
263
правых лапах, но уже на 5-й день я застал ее в совершенстве прыгающей
на этих лапах.
22 дня спустя я удалил у Золота кору левого- полушария, т. е. того
полушария, которое, благодаря превалированию перекрестных нервных
связей, имело преимущественное отношение к оставшимся целыми пра-
вым конечностям. Результаты получились, сверх ожидания, интересны-
ми. Примерно месяц собака была полностью дезориентирована и бес-
помощна. Попытки передвигаться приводили к ударам и ушибам. Месяц
спустя у Золота постепенно выработался и усовершенствовался свое-
образный прием передвижения, достойный восхищения. Собака, сильно
сгибая суставы, укорачивала здоровые лапы, выравнивала их с ампути-
рованными и на четвереньках стала ходить вначале не очень ловко, а затем
с большим мастерством. Эта (фаза у З-олота длилась примерно 2’/а—3 ме-
сяца, после чего собака стала при стоянии и ходьбе постепенно отрывать
от земли культи передней ампутированной лапы.
В последнее время Золото начала при ходьбе отрывать от земли также
культю задней ампутированной лапы.
Однако после экстирпации коры левого полушария к собаке до сих
пор так и не вернулась способность передвижения на двух лапах, кото-
рой она столь виртуозно владела до последней операции.
В настоящее время собака еще жива, и я продолжаю свои наблюдения
над ней. Это здоровая, живая, бодрая, сообразительная, игривая, полная
жизни и сил собака. Если даже не удастся сохранить Золото в живых
после удаления второго полушария (что я намерен сделать после того,
как она до-стигнет прежнего совершенства передвижения на двух целых
лапах), и полученного на ней материала достаточно для ответа на поста-
вленный вопрос и для подкрепления моих теоретических положений.
Я этому материалу придаю не менее важное значение, чем опытам на упо-
мянутых собаках без обоих полушарий.
Полученные на Золоте фактические данные больше, чем материал дру-
гих собак, дает основание предполагать, что в несомненно решающем
влиянии коры большого мозга на перестройку низших отделов централь-
ной нервной системы условнорефлекторный механизм корковой деятель-
ности преобладает над другими видами ее деятельности (трофическое
влияние и т. д.).
Выводы
Этим я заканчиваю описание опытов с ампутацией лап у собак. По-
дробно на своих взглядах я остановлюсь в другой статье после описания
результатов проведенных в том же разрезе экспериментов но анастомозу
нервов, по половинной перерезке спинного мозга и по пересадке мышц
и сухожилий, а также после критического разбора литературы о Пла-
стичности нервной системы при этих видах хирургического повреждения
организма. Здесь я бегло остановлюсь лишь на некоторых моментах.
Изучение хода перестройки у нормальных собак под влиянием удаления
коры одного полушария и в особенности двух полушарий большого мозга
с большой убедительностью говорит о решающем значении коры большого
мозга высших животных для перестройки функции в «примитивных»,
по выражению Бете, частях их центральной нервной системы. Но возни-
кает вопрос: не появились бы, в конце концов, эти перестройки, не стали бы
лишенные полушарий собаки ходить, если бы жили дольше 4 и (Р/г меся-
цев? Гольп у своей знаменитой собаки с удаленными полушариями, жившей
18 месяцев, наблюдал постоянный прогресс в некоторых отправлениях
вплоть до последних месяцев ее жизни.
264
Э. А. АСРАТЯН
Конечно, я был бы бесконечно рад и для науки было бы очень ценно,
если бы мои собаки, являющиеся воплощением мучительного труда, не
пали от неблагоприятных условии и жили дольше 4 и 6r/s месяцев
Но мне кажется, что и этого срока достаточно, чтобы можно было совер-
шенно определенно ответить на поставленный вопрос о роли коры боль-
шого мозга в явлениях пластичности нижележащих отделов центральной
нервной системы
Прежде всего у моих собак не видно было признака какого-либо поло-
жительного сдвига в восстановлении утраченных двигательных функций
в течение всего послеоперационного периода Через несколько дней, кшда
проходят ближайшие последствия наркоза, самой операции и т д , уста-
навливается какой го стандарт, который остается без существенных из-
менений в одном случае 4 месяца, в др)гом--в1/« месяцев Затем следует
напомнить, что указанный Гольцем прогресс у лишенной полушарии со-
баки не относится к моторной сфере, в особенности задних лап и вообще
задней части тела. Гольц отмечает обратное явление — через некоторое
время после удаления коры второго полушария у собак начинается по-
худание, истощение задней части тела, в частности, задних лап, вслед-
ствие чего моторные функции у них асе ухудшаются Таким образом,
едва ли можно быю бы ожидать у наших собак дальнейшего улучшения,
если бы они жили дольше, скорее нужно было ожидать дальнейшего
ухудшения моторики
Представим себе крайний, совершенно невероятный случай Если бы
мне или кому-нибудь удалось показать, что у таких собак необходимые
перестройка и форма передвижения будут, в конце концов, восстанавли-
ваться, если они после операции будут жить 1—3 года и больше, то я
в этом отнюдь не увижу отрицания того основного вывода, который
вытекает из моей работы Совершенно не исключена возможность (тому
есть даже некоторые примеры в физиологии), что за время длительного
отсутствия коры подкорковые образования в состоянии будут Хотя бы
в ничтожной степени заменить кору путем ли тренировки некоторых за-
чаточных своих свойств, как предполагал Гольц, зарождения новых функ-
циональных способностей, как думают другие, или каким-нибудь другим
способом
Полученный мной экспериментальный материал вполне позволяет
считать установки Бете и его сторонников необоснованными В То же время
он подтверждает правильность моих исходных гипотетических положений
о роли коры большого мозга высших животных в явлениях перестройки
в нижележащих отделах центральной нервной системы, обусловленной ам-
путацией двух конечностей
Какова же интимная сторона процесса восстановления нарушенных
функций и «механизм» влияния коры большого мозга на перестройку
функции центральной нервной системы’ Роль коры в этом сложном про-
цессе нельзя себе представить упрощенно, нельзя рассматривать воздей
ствие коры на эту перестройку как нечто функционально гомогенное,
однотипное Это единое и бесспорно сложное влияние коры мне представ-
ляется состоящим минимум из четырех компонентов
Во-первых, кора как высший субординационный орган оказывает на
нижележащие части центральной нервной системы трофическое влияние
(трофическое в понимании И П Павлова) С помощью этого «^механизма»
своего воздействия кора может изменять функциональное состояние ниже
лежащего нервного аппарата в широком смысле слова, в частности, лабиль-
ность его в понимании Введенского — Ухтомского И этим самым кора
может повлиять также на явления перестройки в нем
Во-вторых, кора как последний и наиболее совершенный пункт ре-
цепции может поднимать на высокий уровень, тем самым повышая их
СООБЩЕНИЕ I. ОПЫТЫ С АМПУТАЦИЕЙ ЛАП
265
функциональную ценность, все периферические импульсы, которые в ко-
нечном итоге играют большую роль как но всех видах нервной деятельности,
так и в явлениях перестройки.
Затем, мне думается, нельзя игнорировать кору как орган безусловно-
рефлекторной деятельности. В физиологии настал момент, когда мы должны
совершенно ясно поставить вопрос о том, что кора, будучи органом спе-
циального, качественно нового вида деятельности, вместе с тем не ли-
шена общих форм координационной деятельности, свойственных всей
нервной системе, что через кору, в частности, осуществляется также без-
условнсрефлекторная деятельность и, возможно, с бдльшим эффектом,
чем в нижележащем аппарате.
Наконец, кора оказывает решающее влияние на перестройку в ниже-
лежащем нервном аппарате тем видом нервной деятельности, который
является для нее основным у высших животных, т. е. условнорефлекторной
деятельностью.
Изложенное относится к взрослым высшим позвоночным. Я не вижу
противоречий между этими данными и данными, отрицающими особое
значение большого мозга в явлениях пластичности центральной нервной
системы у низших позвоночных, а также между моими данными и бесспор-
ными фактами, отрицающими значение высших отделов нервной системы
в тех же явлениях у беспозвоночных, на чем так настаивает Бете. Больше
того, не исключена возможность, что кора большого мозга не играет
решающей роли в явлениях пластичности центральной нервной системы
у эмбрионов, новорожденных, а может быть, даже у молодых высших
животных.
В заключение я хочу обратить внимание еще на один момент, чтобы
заранее предотвратить возможные недоразумения в понимании изложенных
здесь фактов и концепций. Я ие допускаю, — да было бы очень наивно
допускать, — что у взрослых высших позвоночных животных, кроме коры,
вся остальная нервная система лишена пластичности, лишена свойства
перестройки, способной привести к приспособлению, хотя бы после огра-
ниченного нарушения моторики в виде повреждения лап. Ведь лишенная
полушарий собака Гольца могла .ходить и на трех конечностях, когда
одна конечность случайно была повреждена. Радемакер специальными
опытами подтвердил это наблюдение Гольца, и я полностью присоединяюсь
к Ра дема кер у.
Однако речь идет здесь о степени совершенства пластичности, о пре-
делах ее функциональной ценности, способной обеспечивать приспособле-
ние различной степени сложности л трудности. Иначе говоря, речь идет
об историческом, эволюционном понимании явления пластичности ке
только в филогенетическом и онтогенетическом аспекте, но и в зрелом
организме. Об этом более подробно — в следующей статье.
Таким образом, оставляя более обстоятельное изложение и обосно-
вание своих взглядов до опубликования следующей статьи, в которой
будут описаны другие экспериментальные данные, здесь укажу лишь, что
в физиологическом «механизме* пластичности нервной системы, обусловли-
вающей компенсацию моторики животных с двумя ампутированными ко-
нечностями, в цепи единых по существу и многообразных по проявлению
воздействий коры основную, ведущую роль я приписываю условиорсфлек-
торной деятельности ее. С помощью этого могучего средства воздействия
кора контролирует, согласовывает, уточняет и утончает деятельность
нижележащих нервных аппаратов, создает новые виды координаций,
дающие возможность животному ходи;ь на двух лапах, более топко и
точно приспосабливаться к изменениям в окружающем мире и к измене-
ниям, произошедшим в организме
2G6
Э. А. АСРАТЯН
ЛИТЕРАТУРА
Бехтерев В. М., Основы учения о функциях мозга, т. III и IV, 1905.
Боровиков И. В., Значение задних столбов спинного мозга в проведении
мышечного чувства, дисс., 1900.
В И Г ДОрчц к Н- А., Компенсаторное профессиональное приспособление, 1934.
Ворошилов, К. В., Ludwig's Arbeiten, 1885.
Г ольцингер Ф. Ф., Чувствующие пути в спинном мозгу, дисс., 1896.
Павлов И. П., Двадцатилетний опыт объективного изучения высшей нервной
деятельности (поведения) животных, 1932.
Павлов И. П., Лекции о работе больших полушарий головного мозга, 1927.
Россолимо Г., Экспериментальные исследования по вопросу о путях, проводящих
чувствительность и движение в спинном мозгу, дисс., 1887.
Северцов А. Н., Эволюция и психика, 1922.
Сеченов И. М., Физиология нервной системы, 1866.
Bet he A. u. Fischer Е.. Handb. d. norm. u. pathol. Physiol., 15, 2, 1931.
Bickel A., Unters. fiber d Mechanismus d. nerviisen Bewegurgsregulation, 1993.
Ewald R., Physiol. Unters. fiber d. Endorgans d. Nervus Octavus, 1892.
Fl ou re ns P., Rechirches expirimentales sur les proprietfs et les functions du sys-
time nerveux dans les animaux vertebralcs, 1824.
F u 1 d, Arch. Ertwikl. Meehan., Il, 1, 1901.
Goltz F., Uber die Vcrrichtungen des Gtosshirns, 1881.
Magnus R,, Korperstallurg, 1924.
♦ •
Добавление (выдержки из статьи» опубликованной в Бюллетене
экспериментальной биологии и медицины, в. VI, т. 8, 1939). «Мои дан-
ные за последние 3 года полностью подтвердились экспериментами на
21 собаке различной породы, возраста, роста и веса, у которых дл.т
различных задач мной были ампутированы или обе передние, или обе
задние, или передняя и задняя лапы одной стороны, или ампутация
производилась крест-накрест: одна передняя и одна задняя лапа.
У нескольких собак период компенсации нарушенных функций стояния
и ходьбы длился значительно дольше, чем у собак первой серии,—21 /4—3
месяца.
У двух собак (Шарик и Хина), после того как наступила хорошая
компенсация нарушенной вследствие ампутации двух лап крест-накрест
моторики, я разрушил оба лабиринта. Разрушение лабиринтов у обоих
животных вызвало характерные резко выраженные явления со стороны
глаз, головы, туловища и конечностей. Животные совершенно утратили
способность стояния и ходьбы. С течением времени характерные явле-
ния со стороны головы и глаз постепенно исчезали (нистагм и отведе-
ние глаз, качание головы и т. д.), однако утраченная животными спо-
собность стояния и ходьбы так и не вернулась, хотя одна собака
жила 4 месяца, а вторая—около Р/2 лет после разрушения лабирин-
тов. Не касаясь теоретической стороны вопроса, можно отметить, та-
ким образом, что в компенсации моторики после ампутации лап лаби-
ринты играют исключительно важную роль.
Эксперименты над 6 другими собаками показали, что в противовес
этому зрительные рецепторы не играют почти никакой роди или иг-
рают весьма незначительную роль в этих компенсаторных явлениях.
У 3 собак (Альфа, Бета, Гамма) с ампутированными в различных ком-
бинациях лапами после наступления компенсации были крепко завяза-
ны глаза повязкой из черной толстой материи. Повязка на глазах почти
не отразилась па степени совершенства ходьбы собак на двух лапах.
На трех щенятах 4—5-месячного возраста были поставлены опыты
другого характера: у двух глаза были замазаны и затем завязаны на
операционном-столе сразу же после ампутации лап, а у одного щенка
глаза были оставлены открытыми. В дальнейшем выяснилось, что все
три щенка почти одинаково быстро и совершенно научились прыгать
на двух лапах».
КОРА БОЛЬШОГО МОЗГА И ПЛАСТИЧНОСТЬ НЕРВНОЙ СИСТЕМЫ‘
Сообщение II
ОПЫТЫ С ПОЛОВИННОЙ ПЕРЕРЕЗКОЙ СПИННОГО МОЗГА
опрос о локализации проводящих путей в спинном мозгу и
вообще в центральной нервной системе является одним из инте
реснейших и волнующих вопросов современной клиники нерв-
ных заболеваний, экспериментальной физиологии и морфоло-
гии По этому вопросу существуют разногласия не только сре-
ди представителей различных наук, но даже среди представи-
телей одной и той же дисциплины. Так, большинство клиницистов и морфо-
логов, а также значительная часть нейрофизиологов с давних пор допускает
существование множества совершенно точно локализованных, строго специ-
фических восходящих и нисходящих проводящих путей в спинном мозгу и
все больше и больше осложняет картину распределения этих путей, обна-
руживая новые и новые пучки. Ряд же представителей экспериментальной
физиологии |Шнфф (Schiff), 1896; Бете (Bethe), 1931 и др.] отрицал и отри-
цает существование специфических, тем более узко локализованных про-
водящих путей в спинном мозгу. При этом, если сторонники теории узко
локализованных, специфических проводящих спинальных путей аргу-
ментируют свои взгляды данными клиники, экспериментальной морфоло-
гий и частично данными экспериментальной физиологии, полученными
в большинстве случаев при кратковременных наблюдениях над животны-
ми с перерезанными отдельными пучками проводящей системы спинного
мозга, то сторонники теории, отрицающей такую локализацию, как пра-
вило, аргументируют своп взгляды данными более длительных наблюде-
ний над животными, оперированными указанным выше способом. Соглас-
но этим данным, всевозможные дефекты в рецепторной и моторной деяте-
льности животных, обусловленные перерезкой тех или иных частей про-
водящей системы спинного мозга, с течением времени проходят почти
бесследно. В качестве иллюстрации можно привести данные II. Вейсса
(Weiss, 1879), полученные у собак, у которых он произвел половинную пе-
ререзку спинного мозга (так называемое броун-секаровское сечение спин-
ного мозга) и спустя месяц наблюдал почти полное восстановление мо-
торных и частично сензорных функций как с пораженной, так и с проти-
воположной «^непораженной» стороны (условно назовем пораженной сто-
рону, соответствующую половинной перерезке спинного мозга, непоражен-
ной— противоположную операции сторону). Данные Вейсса были подтвер-
ждены и дополнены близкими или аналогичными опытами Г. И. Россолимо,
В. Кузьмина, Ф. Ф. Гольцингера, Я. Б. Чулковского, Осава (Osawa) на со-
баках, Г. И. Россолимо па морских свинках, Мотта (Mott), Шефера (Schafer)
* Успехи современной биологии, т. VI, в. S, 1йз7.
268
Э. А. АСРАТЯН
и др. над обезьянами и опытами Маршалла (Marschall) над кошками.
В. Кузьмин, Осава и Г. И. Россолимо осуществили более сложный экс-
перимент: они производили у собак и морских свинок поочередную дву-
кратную и даже троекратную двустороннюю половинную перерезку спин-
ного мозга на расстоянии 2—3 сегментов друг от друга Несмотря на это.
у оперированных животных через некоторое время после каждой новой по-
ловинной перерезки весьма хорошо восстанавливалась моторика и доста-
точно хорошо — чувствительность обеих задних лап. Эти данные были не-
давно подтверждены опытами Данича (Danitsch) над кроликами, а также мо-
ими опытами над собаками и кошками, о которых подробно речь будет ниже.
Если односторонние и двусторонние перерезки спинного мозга у выс-
ших позвоночных животных хотя бы временно вызывают сильные нару-
шения чувствительности и моторики и эти нарушения лишь с течением
более или менее длительного времени постепенно исчезают, то у низших
позвоночных нарушения от аналогичных перерезок очень незначительны
и проходят через очень короткое время. Об этом говорят опыты И. М. Сече-
нова с односторонней половинной перерезкой спинного мозга у ля-
гушек, а также опыты Боте с двусторонними половинными перерезками
спинного мозга у тех же животных. Аналогичные опыты, проведенные в
руководимой мной лаборатории Ф. М. Вишневецкой, показывают, что
для восстановления произвольных координированных движений лягушки
после двусторонней половинной перерезки спинного мозга требуется всего
2—3 дня. Как объяснить эти данные?
Так как современные нейрогисголот считают правильным старое
положение, что интрацентральные пути, по крайней ьсре у высокоразвитых
взрослых животных, после перерезки больше не регенерируют. то законно
допустить, что описанное выше восстановление моторики и чувствитель-
ности пораженной конечности осуществляется за счет использования ос-
тавшихся неповрежденными проводящих путей. Шифф, Бете и др. вообще
отрицают специализацию и локализацию спинальных проводящих путей.
Если одни и те же проводящие пути могут передавать различные им-
пульсы, имеют связи с различными периферическими органами и цент-
ральными аппаратами, которые по представлению узких докали зап ион истов
не должны быть связаны именно с этими путями, значит,—думают они,—
в спинном мозгу ие существует вообще специфических, тем более узко ло-
кализованных проводящих путей. Мы не говорим уже о том, что на ос-
новании опытов с повреждениями проводящих путей в других отделах
центральной нервной системы эти исследователи свой взгляд на характер
проводящих путей распространяют на всю центральную нервную систему.
Так, по их представлениям, в центральной нервной системе «каждый
пункт связан с любым другим пунктом многочисленными прямыми и не-
прямыми путями, но ни один из этих путей не существует специфически
и исключительно для какой-нибудь одной определенной связи (в крово-
обращении всякому известно такое образование коллатеральных путей)- 1
В мою задачу не входит подробное описание существующих взглядов
по затронутому вопросу. Укажу лишь, что дискуссия по этому частному
вопросу представляет собой один участок общего фронта борьбы между
различными течениями, часть общего спора о специализации и локали-
зации функций в центральной нервной системе.
ААеня 'здесь интересует частный вопрос, действительно ли в случаях
половинных одно- и двусторонних перерезок спинного мозга у высших
позвоночных животных восстановление моторики пораженных конечностей
осуществляется путем простого переключения импульсов на другие функ-
ционально готовые пути, благодаря простой, обусловлен пой внешними им-
’ Bethe’s Handbuch d. norm. u. pathol. Physiol., Bd. 15, H. 2, 1931, S. ll?0.
СООБЩЕНИЕ II. ОПЫТЫ С ПОЛОВИННОЙ ПЕРЕРЕЗКОЙ СПИННОГО МОЗГА
269
пульсами «перестройке» путей спинного мозга, без участия вышележащих
отделов центральной нервной системы, в частности, коры большого
мозга, без «научения», как это вытекает из общих концепций Бете и Фи-
шера, или же кора большого мозга принимает какое-то участие в этой
«перестройке».
Должен заранее оговориться, что прямых или косвенных эксперимен-
тальных данных, подкрепляющих их гипотетические положения по этому
вопросу, у Бете и Фишера не имеется, да и вообще, насколько мне из-
вестно, их нет в современной научной литературе. Отклонять, как это
делают Бете и Фишер, допущение, что «научение» играет роль в этих ком-
пенсаторных, восстановительных явлениях на том лишь основании, что
при односторонних повреждениях интрацентральных проводящих пу-
тей в процесс нарушения и восстановления функции вовлекается не
только оперированная сторона, но и другая половина тела, мне кажется
вовсе неубедительным. Могут указать на быстроту исчезновения нару-
шений, наступающих’ после половинной перерезки спинного мозга, у ля-
гушек для аргументации взгляда, что «научение» не играет роли в ана-
логичных процессах восстановления у высших позвоночных животных.
Но мне кажется, что такой аргумент недостаточен. Нельзя безогово-
рочно согласиться также с другим возможным допущением, направлен-
ным к отрицанию «научения» в этих восстановительных явлениях, будто
значительная задержка в ликвидации названных нарушений у высших
позвоночных животных обусловливается только или главным образом
механическими последствиями операции (шок, раневое раздражение, уще-
мление, нарушение кровоснабжения и т. д>), затушевывающими быстрое
наступление «перестройки», истинное проявление спинальной пластич-
ности. С таким допущением тоже нельзя согласиться хотя бы на том осно-
вании, что несравненно более быстрое восстановление простых спинальных
рефлексов не только после половинной, но даже после полной поперечной
перерезки спинного мозга у тех же животных уже указывает на сравни-
тельно небольшую роль этих косвенных операционных факторов в зату-
шевывании действительной картины восстановлен и я функций пораженной
и «непораженной» конечностей после половинных перерезок спинного мозга.
Таким образом, интимную физиологическую природу этих компенса-
торных процессов в центральной нервной системе, в частности, участие
большого мозга в них, можно было считать открытым вопросом в физиологии.
Исходя из некоторых теоретических установок, высказанных в моей
первой шитье, я попытался получить ответ на поставленные вопросы в спе-
циально проведенных экспериментах.
Экспериментальная часть
Работу в этом направлении я начал в 1933 г. в Академии наук СССР,
затем в 1934 г. в ВИЭМ., но в основном развернул г Институте мозга
нм. В. М. Бехтерева в течение 1935—1936 гг.
Опыты проводились главным образом над собаками. Наблюдения чад
9 собаками и одной кошкой, у которых спинной мозг был подвергнут
односторонней броун-секаровской перерезке, главным образом в облает
последних грудных и первых поясничных сегментов, а также наблюдения
над 2 собаками из этою числа, у которых спинной мозг был вторично
подвергнут перерезке с противоположной стороны на 3—4 сегмента выше
уровня первой перерезки, совершенно единодушно подтвердили данные
упоминавшихся исследователей. Правда, у большинства наших подопыт-
ных животных моторика и в особенности чувствительность пораженных
конечностей восстановились не полностью (на это указывают также многие
предыдущие исследователи), так что при внимательном наблюдении и
тщательном изучении все же можно было пораженные конечности отлгчить
270
Э. А. АСРАТЯН
от здоровых. Однако через 2 месяца после оперативных вмешательств
животные прекрасно пользовались пораженными конечностями как при
стоянии и ходьбе, так и при беге, прыжках и других сложных координи-
рованных движениях. В зависимости от возраста животных, тщательное!и
произведенной операции, ухода за животными и тому подобных моментов,
восстановление нарушенных чувствительных и моторных функций задних
конечностей требует разного времени (от 1 до I1/» месяцев для односто-
ронней и от 3 до 4 месяцев для двусторонней перерезки) и достигает раз-
личной степени совершенства.
Простое наблюдение за процессом восстановления не выявляет столь
убедительных данных, как при опытах с ампутациями лап, в пользу допу-
щения, что «научение* играет здесь столь же значительную роль, как в вос-
становлении нарушенной, вследствие ампутации лап, моторики. Однако
уже при внимательном наблюдении можно видеть, что процесс восстано-
вления нарушенной, вследствие гемнеекцни спинного мозга, моторики
имеет строго постепенный характер, что является важным моментом, уже
в какой-то мере указывающим на участие «научения» в восстановлении
этих нарушенных функций.
Спустя несколько (6—10) дней, когда исчезают ближайшие побочные
последствия самой операции — результаты нарушения кровоснабжения,
ущемления, раздавливания и т. п. — выступают непосредственные явле-
ния выпадения функций. При этом вначале животные совсем не пользуются
пораженными конечностями, хотя у них имеются как односторонние, так
и перекрестные рефлексы; непораженными же конечностями животные
в большинстве случаев пользуются свободно при всех видах движения.
В этой стадии (в разных случаях различной длительности) пораженные
конечности совсем парализованы, волочатся за телом, не принимая ника-
кою участия ни в стоянии, ни в ходьбе, ни в других видах движения.
Больше того, в этой стадии пораженные конечности обычно сильно худеют
при общей хорошей упитанности животных; на ощупь тонус мышц этих
лап заметно понижен (о том же говорят произведенные моей сотрудницей
Р. О. Барсегян специальные измерения тонуса мышц у некоторых собак).
Постепенно с течением времени упитанность конечностей и тонус их мышц
улучшаются, они все живее реагируют на рефлекторные раздражения
как их самих, так и противоположных «непораженных» конечностей, по-
являются рефлекторные реакции на раздражения передних частей тела.
Если в первой стадии парализованные конечности просто волочатся за
телом, то в этой стадии они начинают как бы пассивно принимать участие
в стоянии и даже в ходьбе. Зто пассивное участие заключается в том, что
при стоянии животное явно опирается и на пораженную лапу и более или
менее симметрично поддерживает заднюю часть тела в воздухе, а затем
при передвижении корпуса вперед оставшаяся позади пораженная экстен-
зированная конечность как бы общим сокращением мышц заднего пояса
туловища, толчком притягивается почти или совсем до уровня непора-
женной задней конечности. В этой стадии, длящейся у разных собак раз-
личное время» никогда, однако, не наблюдалось, чтобы пассивное пере-
движение экстензнрованной пораженной конечности превосходило уровень
движений непораженной задней конечности, что является обязательным
при ходьбе нормальных собак. Кроме того, в этой фазе еще ясно выражена
хромота на пораженную конечность.
Постепенно, однако, картина улучшается по всем показателям: упитан-
ность и тонус мышц повышаются, все живее и сильнее становятся рефлек-
торные движения, увеличивается размах описанных выше как бы пассив-
ных движений пораженных конечностей, появляются и постепенно уси-
ливаются уже активные разгибательные и сгибательные движения самой
пораженной конечности, движения пораженных конечностей делаются раз-
СООБЩЕНИЕ П. ОПЫТЫ С ПОЛОВИННОЙ ПЕРЕРЕЗКОЙ СПИННОГО МОЗГА
271
машистыми, но днсметричными, почему и ходьба на эту лапу отчасти на-
поминает петушиную походку. В конце концов, пораженная конечность
достигает предельное совершенства моторики, которое иногда почти не
уступает моторике здоровой конечности.
В заключение краткого описания картины компенсации этих нарушений
отмечу следующее. Наблюдения как за появлением и развитием нарушений
мотор и ю обеих задних лап собаки, обусловленных половинной перерезкой
спинного мозга, так и за наступлением и развитием компенсации этих на-
рушений убеждают в том, что причины, характер, пределы и длительность
этих нарушений для обеих лап резко различны. Об этом говорят все про-
веденные до сих пор наблюдения других исследователей, об этом говорят и
мои наблюдения.
Основной причиной сравнительно кратковременного нарушения мотор*
ных функций противоположной гемисекции конечности, безусловно, можно
считать связанные с операцией побочные факторы, а перерезке «обслуживаю-
щих» ее проводящих путей следует приписать сравнительно небольшую роль.
В то же время можно утверждать, что поражение функции односторонней
с гемисекцией конечности обусловливается в основном перерезкой «обсл у-
живаюищх»ее проводящих путей, а побочным факторам, связанным с самой
операцией, принадлежит значительно меньшая роль. В первом случае мы
имеем дело преимущественно с так называемыми побочными явлениями, ко-
торые быстро проходят, во втором случае — главным образом с действи-
тельными явлениями выпадения, которые проходят очень медленно.
Какой же вывод можно сделать на основании данных простого наблюде-
ния об интимной физиологической природе этих восстановительных про-
цессов? Мне думается, что если имеется принципиальная разница в причи-
нах нарушений функции обеих конечностей, в пределах и длительности этих
нарушений и, стало быть, в их физиологической сущности, нужно быть
последовательным и допустить существование различных физиологических
«механизмов» их компенсации. И если длительность и постепенность вос-
становления нарушенных моторных функций в какой-то мере указывают на
участие «научения» в восстановлении нарушенных функции (этого в прин-
ципе, вообще говоря, не оспаривают Беге и Фишер), то с полным правом это
допущение можно распространить и на случаи восстановления нарушенных
моторных функций конечности собаки, пораженной после гемисекции спин-
ного мозга.
На основании всего сказанного представляется совершенно неубеди-
тельной и необоснованной концепция Бете и Фишера, отрицающих значе-
ние «научения» в процессе компенсации нарушенной моторики пораженной
вследствие гемнеекции спинного мозга конечности по той только причине,
что, дескать, моторные функции противоположной лапы после этой операции
тоже временно нарушаются.
Об участии коры больших полушарий или процесса «научения» в
восстановлении функции пораженной конечности у этих собак еще ярче
свидетельствуют данные о возможности выработки двигательных услов-
ных рефлексов с этой конечности (рис. 1, стр. 273).
Однако, чтобы получить более доказательный материал в пользу до-
пущения, что «научение» участвует в названных выше компенсаторных явле-
ниях, чтобы установить пределы этого участия, а также выяснить более
общий и вместе с тем кардинальный вопрос о роли коры большого мезга
высокоразвитых взрослых позвоночных и в этих видах проявления плас-
тичнссти центральной нервной системы, мной были поставлены специаль-
ные опыты с экстирпацией коры одного и двух полушарий большого
мозга у* некоторых описанных выше собак, у которых функции пора-
женных, вследствие одно- и двусторонней броун-секаровской перерезки
272
9. А. лСРЛтЯН
спинного мозга, конечностей с течением времени восстановились и до-
стигли предельного совершенства моторики и чувствительности. Кроме
этого, у 2 других собак, ранее лишенных коры обоих полушарий боль-
шого мозга, была произведена половинная перерезка спинного мозга.
Опыты с экстирпацией коры большого мозга
У 5 из моих собак, преимущественно молодых, у которых была произ-
ведена половинная поперечная перерезка спинного мозга с левой стороны
(у одной из них на 3—4 сегмента выше первой перерезки с правой стороны
была произведена вторая половинная перерезка), через такие сроки после
спинальной операции, которые требовались для предельного восстановления
моторной функции пораженных конечностей, я сначала экстнрпировал кору
одного полушария большого мозга. Спустя некоторое время после этого
удалялась кора второго полушария большого мозга. Наблюдения над ними
проводились до конца их жизни.
Привожу сводную таблицу с отметками о времени произведенных опе-
раций и о длительности жизни собак после последней операции.
‘ U/U 1 Кличка собаки Дата спи- нальной опе- рации Дата экстир- пации коры одного полушария Дата, экстир- пации коры второго полушария Длитсл ьность жизни после последней, операции Примечания
1 Аддис 5.1 1935 г. 8.X1I 1935 г. 5.1 1936 г. 37 дней Погибла от вне- зап ного- охлажде- ния
2 Плошь 20.IX и 20.Х 1935 г. 5.1 1936 г. 4. II 1936 г. 22 дня Погибла от вос- паления легких
3 Белая 17. VII 1936 г. 28.1 1936 г. 19.XII 1936г. 26 дней То же
4 Усик 28.11! 1936 г. 20. X 1936 г. 28.Х 1 1936 г, • 8 » Погибла от вто- ричного кровоиз- лияния в резуль- тате удара голо- вой о предметы (?)
5 Чанг 28.111 1934 г. 23.V11 1934 г. 3% месяца Погибла от плохого ухода и истощения
У Аддиса, Белой, Чанга и Плоши во время первой операции была экстир-
пировала кора левого полушария, а у Усика — кора правого полушария.
Независимо от того, удалялась ли кора полушария со стороны спинальной
перерезки или кора противоположного полушария большого мозга, по-
стоянным последствием операции являлось нарушение «компенсированной»
уже моторики пораженной и частично даже «непораженной» конечности.
Хотя с первого же дня операции спинальная рефлекторная деятельность
восстанавливалась во всем своем объеме, собаки в течение 5—8 дней не
в состоянии были самостоятельно стоять или поднимать заднюю часть тела.
Передними же конечностями собаки успешно пользовались уже со второго
дня операции. Опираясь на них, они приподнимали переднюю половину
туловища и тащили за собой все тело. Когда же мы в этой фазе приподни-
мали заднюю половину тела собак и ставили их на широко расставленные
конечности, они долго не могли удерживаться в этом положении даже при
стоянии, а при попытке двигаться сразу же падали на пораженный бок.
Ближайший анализ нарушения моторики показываем, что причиной его
нельзя считать слабость задних лап, даже слабость пораженной лапы; при
рефлекторных сокращениях обе они развивали достаточную силу. Нсболь-
СООБЩЕНИЕ II. ОПЫТЫ С ПОЛОВИННОЙ ПЕРЕРЕЗКОЙ СПИННОГО МОЗГА 273
шое понижение тонуса пораженной лапы тоже нельзя было рассматривать
как причину этой дезорганизации моторики1.
Основной причиной указанного явления можно считать полный паралич
пораженной лапы, вернее, полное ее неподчинение вышележащим отделам
центральной нервной системы. Поврежденная конечность в этой фазе боль-
шей частью находится в состоянии экстензии, направляется в противопо-
ложную сторону и как бы механически мешает свободной деятельности не-
поврежденной лапы, которая уже со 2—3-го дня после удаления полуша-
рие. 1. Килограммы элеклрооборонительных двигательных услов-
ных рефлексов с пораженных лап у собаки Барс щ) и у собаки
Рыжик (б) после поперечной перерезки боковой половины спинного
мозга на уровне последних грудных сегментов
Обо з н а ч с н и Я (сверху линз): усланные и безусловные рефлексы . от-
метка ус.пятых ришражигелей, отметка безусловного раздражителя (цифры
означают расстояние между катушкалси). цремя н секундах <опы гы Л. А. Но-
виковой и Д. II. Ханутиной)
рИя Делает безуспешные попытки Совместно с передними конечностями под-
нять и двигать тело. Обычно через 5—8 дней после операции положение
постепенно улучшается. Изредка собакам удается как бы прыжком выпря-
мить тело, стать, преимущественно опираясь на непораженную лапу, и сде-
лать ею 2—3 шага; однако при дальнейших движениях они снова падают
на пораженный бок; при этом пораженная лапа обычно не принимает уча-
стия ни в стоянии, ни в ходьбе. В этой фазе, длящейся всего 2—3 дня, со-
баки в состоянии 2—3 минуты стоять, покачиваясь, когда их приподнимают
и ставят на широко расставленные задние лапы. Постепенно картина про-
должает улучшаться, и к 8—11-му дню после операции еобзки бывают
в состоянии значительное время стоять и ходить уже на четырех конеч-
1 Данные об изменении тонуса мышц, определяемого, как уже говорилось, ла
ощупь, не совсем точны. Для выяснения этого вопроса сейчас моей сотрудницей
Р. О. Барсегяи ведутся специальные исследования прибором Шпигеля.
18 э. А. Асратян
274
Э. А. АСРАТЯН
ностях, заметно прихрамывая на пораженную лапу. Вновь появляется дне-
метрия в движениях пораженной лапы. При этом из лежачего положения
собаки легко переходят в стоячее, когда лежат на «непораженном» боку.
Если же они лежат на пораженном боку, то они или совсем не могут под-
няться, или же достигают этого после длительных попыток, преимущест-
венно со стороны «непораженной» лапы. На И—15-й день моторика зад-
них конечностей достигает той степени совершенства, какая существовала
до удаления коры одного полушария. Следует сказать, что обусловленная
операцией картина нарушения моторики задних конечностей и процесс ком-
пенсации этих нарушений в общих чертах напоминают первоначальную
картину нарушений и восстановления их моторики, вызванных спинальной
операцией. В первом случае, однако, сроки более короткие.
В некоторых особенно сложных случаях нарушения и восстановления
функций этот срок резко удлиняется. В качестве примера можно при-
вести следующий случай. У собаки Чанг я сначала экстирпировал седа-
лищные нервы с обеих сторон,-что вызвало легкие проходящие нарушения
моторики задних лап (подробное описание см. в. другой статье). После
того как у Чанга восстановились нарушенные функции задних лап и он
ходил нормально, была произведена левосторонняя половинная перерезка
спинного мозга иа уровне последних грудных сегментов. Развилась обыч-
ная картина нарушения, лишь несколько более выраженная. Однако ме-
сяца через полтора у Чанга прошли и эти нарушения. Тогда я экс-
тирпировал кору левого полушария большого мозга. В результате
этого вновь возникли комбинированные нарушения, но в более резкой
форме, державшиеся больше месяца. Несмотря на постепенное улучшение,
через 31/г месяца компенсация еще не наступила. Собака погибла из-за
плохого надзора обслуживающего персонала.
Картина немного осложняется также в случаях двусторонних перерезок
спинного мозга. У собаки Плошь с двусторонними половинными перерез-
ками спинного мозга после удаления полушария были вновь поражены обе
ляпы и восстановление этих нарушений длилось 16—17 дней. Сравнение
двух задних лап показало, что на стороне последней спинальной перерезки
(справа) конечность была поражена сильнее, чем на стороне первой спи-
нальной перерезки (слева). Возможно, что эта разница обусловливалась и
тем, что у Плоти была экстирпироваиа кора левого полушария.
Како-e же влияние оказывала экстирпация коры левого или правого
полушария на тех из наших собак, у которых была произведена только
левосторонняя спинальная перерезка? Должен сказать, что путем простого
наблюдения трудно было уловить существенные различия между послед-
ствиями обоих вариантов экстирпации коры. Создалось впечатление,
что удаление коры одноименного спинальной перерезке полушария вызы-
вает будто бы более сильные и длительные нарушения, чем экстирпация
коры противоположного полушария. Мы попытались изучить этот вопрос
более точным и тонким методом — методом эдектрооборонительных услов-
ных рефлексов пока на двух собаках—Усике и Белой. У обеих собак, как
упомянуто выше, была произведена левосторонняя спинальная операция.
В процессе восстановления нарушенных моторных функций моей сотруд-
ницей Д. И. Ханутиной были успешно выработаны у обеих собак электро-
оборонительные условные рефлексы на пораженной лапе. Рефлексы выра-
батывались на зрительные, тактильные и звуковые раздражители. Когда
рефлексы хорошо закрепились, у Усика была экстирпироваиа кора правого
полушария, у Белой — кора левого полушария большого мозга. После
того как исчезли вызванные операцией нарушения функций пораженных
конечностей Д. И. Ханутина продолжала изучать условнорефлекторную
деятельность на пораженных лапах.
СООБЩЕНИЕ И. ОПЫТЫ С ПОЛОВИННОЙ ПЕРЕРЕЗКОЙ СПИННОГО МОЗГА 275
Выяснилась следующая любопытная картина. У Усика все условные
рефлексы, исчезнувшие после операции удаления полушария, быстро вос-
становились, у Белой же все условные рефлексы исчезли безвозвратно.
По зато у Белой даже после операции с большой легкостью вырабатыва-
лись условные электрооборонительпые рефлексы на остальные три конеч-
ности. В связи с необходимостью проведения второй операции на полу-
шариях мы выработку условных рефлексов на пораженную лапу у Белой
прекратили. Д. И. Ханутнна изучает этот вопрос более детально на новых
собаках. Однако уже полученные ею результаты дают основание утверж-
дать, что экстирпация коры на стороне спинального поражения оказывает
более сильное влияние на функцию пораженной лапы, чем экстирпация
коры противоположного полушария. При первом варианте операции услов-
корефлекторная деятельность пораженной конечности если не ликвиди-
руется, то во всяком случае резко ослабевает.
На основании опытов с экстирпацией коры одного полушария можно
сделать вывод, что'эта операция временно восстанавливает состояние на-
рушения функций задних лап, созданное первоначальной спинальной опе-
рацией и исчезнувшее в последующие дни.
У Плоши, Аддиса, Усика и Белой через некоторое время пссле удаления
коры одного полушария и восстановления у них моторных функций задних
конечностей я произвел экстирпацию коры второго полушария. О длитель-
ности жизни после второй операции на полушариях, а также о причине
смерти каждой собаки приведены сведения в таблице (стр. 272).
Перейду к краткому описанию последствий удаления коры обоих полу-
шарий.
У всех собак наблюдалась следующая картина, неизменно остававшаяся
в течение всей их жизни. Спинальная рефлекторная деятельность всех лап
появлялась уже с первого дня операции. Со 2-го дня она почти достигала
той же активности и силы, что и до последней операции; при этом пора-
женная лапа почти не отставала от «непораженной» задней лапы. В обеих
лапах появлялась и в дальнейшем сохранялась тоническая рефлекторная
деятельность на шейные и лабиринтные импульсы, а иногда она возникала
даже как будто «произвольно». На ощупь не очень заметно изменялся также
тонус мышц конечностей. Несмотря на все это, последняя операция на
полушариях резко нарушала моторику всей задней части тела собак, в
особенности пораженной части. В то время как передними конечностями
собаки начинали пользоваться на 2—3-й день пссле удаления второго
полушария, с помощью их поднимали переднюю половину тела и пере-
двигались, вся задняя часть их тела на пораженном боку волочилась за пе-
редней (рис. 2 в). При этом пораженная лапа обычно находилась в состоя-
нии экстензии, направлялась в противоположную сторону и вперед, почти
не производя никаких движений, «непораженная» же лапа делала ни к че-
му ие приводящие мелкие движения. Собаки утрачивали способность
самостоятельно поднимать заднюю половину тела и самостоятельно стоять
на четырех конечностях. Когда же животных ставили на широко раздвину-
тые лапы, чтобы они могли стоять, некоторые из них около полуминуты,
качаясь, могли держаться на четырех ногах, а другие сразу падали.
Картина этих глубоких нарушений, нс проявлявших никаких призна-
ков улучшения в течение всей дальнейшей жизни животных, заснята на
кинопленке. Для иллюстрации привожу несколько кадров до И после экстир-
пации коры обоих полушарий большого мозга (рис. 2 о, б, в, е).
Следует отметить, что с течением времени у лишенных коры собак со
спинальными операциями задние конечности начинают худеть, причем
особенно сильно худеет пораженная лапа. У некоторых животных заметно
уменьшается в объеме вся задняя половина тела. У них резко нарушается
по сравнению с другими собаками с удаленной корой ряд вегетативных
18*
276
3. А. ЛСРАТЯ H
функций, как то: теплорегуляция, трофика тканей, деятельность кишечного
тракта (атония кишок), иногда мочеиспускание, т. е. явления, которые у них
не наблюдались после спинальной операции, но при сохранности большого
мозга. Далее эти животные особенно чувствительны к неблагоприятным
условиям содержания. Всеми этими моментами в совокупности объясняется
сравнительно небольшая длительность их жизни по сравнению с длитель-
ностью жизни других моих собак с удаленной корой.
Для решения вопроса об участии коры большого мозга в явлениях ком-
пенсации нарушений, обусловленных половинной перерезкой спинного
мозга, я произвел операции в обратной последовательности, а именно: у
одной из собак с удаленными полушариями (Пушок), жившей около полу-
Рис 2. Влияние удаления коры больших полушарий на функции стояния н
ходьбы у собаки Аддис с поперечной перерезкой боковой половины спинного
мозга ня уровне последних грудных сегментов
4 _ через месяц после перерезки спинного мозга у собаки яосстаноюнлас* функция пора-
женной задней конечности и она пользуется ею при стоянии и. хОДЛбе; б, 9, / — та же
собака после удаления коры обоих полушарий мозга потеряла способность стоять и ходить
ня задних конечностях. Она передвигается только с помощью передних лап
года и находившейся в хорошем состоянии, я произвел половинную пере-
резку спинного мозга с левой стороны на уровне первых поясничных сег-
ментов. У собаки появилась сильная гиперестезия и гипералгезия; до нее
нельзя было дотронуться, особенно ДО пораженной конечности. Собака
жила у меня 15 дней после спинальной операции, и за этот период
признаков восстановления функций не только пораженной, но н здоровой
задней лапы не наблюдалось. Собака не могла ни стоять, ии ходить. Она
постепенно худела и пала жертвой эпидемии. Такие же результаты полу-
чены у собаки Беженки, у которой через 3 месяца после удаления коры была
произведена половинная перерезка спинного мозга. Эта собака жила 7 дней.
Общий вывод из этих опытов таков: удаление коры всего большого
мозга полностью и на длительное время (без малейших намеков на возвра-
щение в течение 40 дней) нарушает наступившую после половинной пере-
резки спинного мозга компенсацию опорных и локомоторных функций
задних лап.
СООБЩЕНИЕ П. ОПЫТЫ С ПОЛОВИННОЙ ПЕРЕРЕЗКОЙ СПИННОГО МОЗГА
277
Обсуждение полученных данных
Резюмируя полученные данные, следует особо подчеркнуть ряд момен-
тов, прямо или косвенно указывающих иа доминирующую, решающую
роль коры большого мозга у высокоразвитых взрослых позвоночных в ком-
пенсаторных явлениях, развивающихся после половинной поперечной пе-
ререзки спинного мозга. Такими моментами являются прежде всего мед-
ленный ход восстановления сензорных и в особенности моторных функций
пораженных конечностей, говоря иначе, медленное и постепенное исчез-
новение действительных «явлений выпадения функций». Затем следует
указать на временное возвращение этих «явлений выпадения функции»
после удаления коры одного из полушарий большого мозга, а также на
сравнительно медленное исчезновение этих функциональных нарушений.
Наконец, после удаления коры другого полушария большого мозга явления
выпадения функции возникают вновь. Момент постепенности в восстанов-
лении нарушенных функций имеет известное значение, поскольку он яв-
ляется показателем «научения», а ведь у высших животных всякое «науче-
ние» является преимущественно, если не исключительно, функцией коры
большого мозга. Следует также отметить возможность выработки дви-
гательных условных рефлексов на пораженные лапы.
Значение моментов временного и стабильного возвращения этих функ-
циональных нарушений после экстирпации коры одного или двух полу-
шарий большого мозга становится очевидным на фоне известных со времени
Гольца и многократно подтвержденных фактов, что только экстирпация
коры большого мозга у нормальных собак не нарушает ни функции стояния,
ни функции ходьбы на четырех конечностях. При этом указанные функции,
как я мог убедиться в своих опытах, в большинстве случаев появляются
уже на 2—3-й день после операции.
Еще ярче выступает значение момента постепенности на фоне следую-
щего факта; в то время как после операций на полушариях у всех описан-
ных выше собак наступала глубокая дезорганизация моторных функций
задних конечностей (в особенности поражен ной конечности), у этих живот-
ных 2—3 дня спустя после названных операций передние конечности пре-
красно выполняли свои функции опоры и локомоции. Мне думается, что
в этом отношении некоторую роль играет как кратковременное и незначи-
тельное нарушение функции «непораженной» конечности непосредственно
после удаления коры одного полушария, так и сравнительно слабое нару-
шение ее функции после второй операции на полушариях.
Как и в опытах с ампутациями, я и здесь не исключаю возможности,
что если бы собаки жили дольше, у них, в конце концов, могл и бы восста-
новиться хотя бы в слабом виде функции опоры и локомоции также
пораженных конечностей. Мне кажется, однако, это допущение мало веро-
ятным хотя бы потому, что в течение всей жизни этих ссбак я не замечал
у них какого-либо признака восстановления нарушенных функций (развив-
шаяся в.первые дни после последней операции на полушариях картина
неизменно сохранялась до момента гибели животных), а также потому,
что, как отметил еще Гольц, при длительной жизни собак с удаленкой
корой моторика задних лап не только не улучшается, но даже ухуд-
шается.
Возьмем крайний случай. Предположим даже, что это допущение в бу-
дущем получит экспериментальное подтверждение, и лишенные коры со-
баки с броун-секаровской перерезкой спинного мозга при более длитель-
ной жизни будут стоять и ходить также на задних конечностях. Я счи-
таю, что даже такой конечный исход не может явиться опровержением
тех выводов, которые сделаны мной выше о роли коры большого мозга
нормальных собак в компенсации функциональных нарушений, сбу-
278
Э. А. АСРАТЯН
словленных половинкой перерезкой спинного мозга. Такой мало вероят-
ный, но все же окончательно не исключенный исход можно было бы объяс-
нить так же, как аналогичные мало вероятные случаи в опытах с ампута-
цией: тренировкой первоначально мало выраженной и мало эффективной
пластичности и других функциональных особенностей оставшихся без по-
вреждения частей центральной нервной системы, возникновением новых
очагов, способных более или менее заменять кору в этом отношении, и тому
подобными более или менее вероятными допущениями, которые давно
имеют хождение в физиологии.
Как же представить себе интимную физиологическую природу описан-
ных выше компенсаторных явлений н участие в них коры?
Конечно, накопление нового экспериментального материала расширит
н углубит наши знания по этому вопросу, даст возможность больше вник-
нуть в сущность этих сложных и вместе с тем пленительно интересных явле-
ний пластичности нервной системы. Однако мне кажется, что уже сейчас,
па основании полученного материала, можно, не ограничиваясь конста-
тацией факта влияния коры большого мозга на явления спинальной пла-
стичности, попытаться, пока хотя бы в виде физиологической схемы, по-
строить рабочую гипотезу, способную быть, с одной стороны, ключом
К пониманию интимной природы ОПЫсЯнныХ сложных интраЦеНТралЬпыХ
взаимоотношений и взаимодействии, а с другой — быть отправным пунктом
для дальнейших исследований в данном направлении.
Само собой разумеется, что при создании физиологической рабочей
схемы я буду пользоваться не только моими данными, но, где это окажется
нужным, использую и некоторые более или менее твердо установленные
данные и положения современной нервной клиники, экспериментальной
морфологии и физиологии.
Итак, как я представляю себе на данном этапе исследований интимную
природу компенсации нарушений после половинных перерезок спинного
мозга и участие коры большого мозга в этих компенсаторных явлениях
у высших взрослых позвоночных?
Морфологические особенности центральной нервной системы вообще и
спинного мозга в частности позволяют высказать предположение, что функ-
циональная связь между какими-нибудь двумя пунктами центральной нерв-
ной системы может устанавливаться многочисленными путями: прямыми и
непрямыми, посредственными и непосредственными, малосинапсовыми и
многосинапсовымн. Однако, судя по данным клиники и экспериментальной
физиологии, в нормальных условиях, когда цельность центральной нервной
системы не нарушена, между пунктами центральной нервной системы связь
устанавливается преимущественно (если не исключительно) известными,
вполне определенными путями. Другими словами, нужно считать соответст-
вующей действительности теорию о локализации специфических проводящих
путей в центральной нервной системе вообще и в спинном мозгу в част-
ности, то, что отрицают Бете и его единомышленники и на чем упорно на-
стаивают морфологи, клиницисты и большинство физиологов.
Однако на основании экспериментов нужно допустить также, что при
нарушении целостности этих специфических л локализованных путей между
условно взятыми пунктами центральной нервной системы с течением вре-
мени, постепенно (отнюдь не сразу) устанавливается связь косвенными,
возможно, даже многосинапсовымн путями, которые до этого хотя бы в из-
вестной своей части были специфическими и служили для соединения дру-
гих областей и пунктов центральной нервной системы. Иными словами,
этим допущением я отграничиваюсь от сторонников теории узкой локали-
зации и неизменной специфичности проводящих путей в центральной нерв-
ной системе, так как допускаю относительность, вариабильность, динамич-
ность этой локализации и специфичности. Однако, как уже было сказано,
СООБЩЕНИЕ II. ОПЫТЫ С ПОЛОВИННОЙ ПЕРЕРЕЗКОЙ СПИННОГО МОЗГА
279
я не отрицаю специфичности и локализации путей, как делают Бете и его
единомышленники, и не допускаю, чтобы связь между любыми условно
взятыми пунктами центральной нервной системы в норме могла успешно
осуществляться посредством всех без исключения прямых и непрямых
морфологически проложенных путей.
«Перестройка» путей в моем представлении заключается не в простом и
быстром переключении импульсов с одних, прямых путей между центрами
на другие, окольные, существующие, по предположению Бете и его сторон-
ников, не только в виде морфологических предпосылок, но и в виде функ-
ционально полноценных, работоспособных путей. Мне кажется, что если
стать на эту точку зрения Бете и его сторонников, то не имеет смысла вообще
говорить о какой-то «перестройке» путей, ибо для того, чтобы перестраивать-
ся, нужно быть чем-то другим до этого. «Перестройку» спинальных прово-
дящих путей я рассматриваю как длительный и сложный иитрацентралъиый
процесс создания новых многосинапсовых путей, в основном из нервных
элементов, выполнявших до того иную роль, иное функциональное назначе-
ние. Повидимому, этот процесс созидания новых путей имеет многосту-
пенчатый Характер, идет от звена к звену сложной дуги нервных клеток и
их отростков, Связь- же между этими элементами, повидимому, осуществ-
ляется по принципу проторения путей, но в отличие от обычного (разного
мы здесь имеем деле со стабильным, тоническим проторением путей.
Что же приводит к ЭТОЙ «перестройке», какие факторы при этом играют
решающую роль в конкретном случае половинных перерезок спинного
мозга? На основании своих опытов я осмеливаюсь утверждать, что у высоко-
развитых взрослых животных в «перестройке» спинальных путей основную,
решающую роль играет кора большого мозга, что проявляемая в этих слу-
чаях спинным мозгом пластичность в основном представляет собой отобра-
жение пластичности коры большого мозга, наиболее пластичного по своим
функциональным особенностям и вместе с тем главного интегративного
органа центральной нервной системы высших взрослых животных. Данные
современной экспериментальной морфологии и экспериментальной физио-
логии не оставляют сомнения в том, что кора большого мозга высших жи-
вотных имеет не только морфологические предпосылки для осуществления
прямой и косвенной связи с любыми нервными элементами спинного мозга,
но что даже в нормальных условиях, при нормальных жизненных отправ-
лениях животного кора большого мозга способна настолько сильно влиять
на деятельность этих элементов, что может снять присущие им так назы-
ваемые чисто спинальные особенности работы и создать совершенно новые,
высшие, сложные и многообразные комбинации и виды их деятельности.
Таким образом, с этой точки зрения, ничего удивительного и непонятного
нет в том, что в патологических условиях, когда целостность спинного мозга
оперативно нарушается, кора большого мозга, этот господствующий в цен-
тральной нервной системе высших животных могучий орган, пользуется
всеми своими многообразными возможностями воздействия на спинальные
процессы, возможно, даже в более сильной степени, чем она это делает
в норме.
Какими же «механизмами» осуществляет кора большого мозга эту свою
сложную роль? Мне кажется, что «механизмов» этих много.
Во-первых, медленное и постепенное течение самого процесса «пере-
стройки» путей после половинных перерезок спинного мозга, в особенности
при учете тех последствий, которые вызываются операциями на полуша-
риях, уже является косвенным указанием на то, что кора участвует в этом
«механизмом» своей условнорефлекторной деятельности, что существенную
роль в этом процессе играет «научение». Как уже было указано выше,
Бете и Фишер аргументацию в пользу наличия «научения» отклоняют на
том: шатком основании, что, дескать, функции здоровой задней конечности
280
Э. Л. АСРАТЯН
тоже кратковременно нарушаются после половинной перерезки спинного
мозга. Они пишут: «У млекопитающих рядом исследователей в конечном
итоге получены одинаковые результаты (имеется в виду факт восстанов-
ления нарушенных функции, Э. Л.), но эти результаты устанавливаются
только через несколько дней и недель после операции. Эта временная за-
держка имела бы какое-то значение и могла бы быть истолкована в смысле
«научения» (Urnlernen), если бы дело касалось только оперированной сто-
роны. Но вначале, как и при полной перерезке, наступает паралич всей
задней половины тела, и только постепенно восстанавливаются и рефлексы,
и проводимость, причем большей частью на неповрежденной стороне раньше,
чем на поврежденной»1.
Некоторые критические замечания по этому поводу я уже сделал выше.
По существу говоря, я не совсем понимаю, почему кратковременное нару-
шение и быстрое восстановление функций конечности, противоположной
оперированной стороне, может служить основанием для отрицания того, что
«научение» игр-ает роль в процессе восстановления функции не только пора-
женной, но даже «непораженной» конечное^!. Если даже на минуту согла-
ситься, что «научение» никакой роли не играет в восстановлении кратко-
временно нарушенной функции «непораженной» конечности, это опять-таки
ни в коей мере не может служить основанием для отклонения такого допу-
щения в отношении восстановления функции действительно парализован-
ной конечности. В первом случае нарушение функции может оказаться по-
следствием главным образом механических воздействий операции (повреж-
дение, ущемление, раневое раздражение и т. д.), и для его восстановления
вовсе не обязательны те «механизмы» восстановления, которые необходимы
для восстановления функции парализованной конечности; в нарушении
функции пораженной конечности перечисленные механические причины
нарушения являются уже второстепенными по сравнению с перерезкой
(как таковой) основных проводящих путей.
Таким образом, имеются вес основания допускать, что «научение»,
основная функция коры большого мозга высших животных, играет важную
роль в компенсации описанных выше нарушений, в проявлении пластич-
ности центральной нервной системы при половинных перерезках спинного
мозга. Ведь, в конце концов, условнорсфлекторная и вообще приобретен-
ные виды нервной деятельности являются не чем иным, как высшим прояв-
лением пластичности, способности живого субстрата приспосабливаться
к изменениям, происшедшим внутри и вне организма. Я не считаю, однако,
что в данном конкретном случае кора большого мозга высших животных
свою задачу выполняет только через посредство могучего «механизма»
условнорефлекторной деятельности. Жизненные отправления собак, ли-
шенных коры большого мозга, в особенности неловкость и неуклюжесть
их движений, указывают на то, что кора большого мозга своеобразным,
если можно так выразиться, безусловнорефлекторным воздействием облег-
чает, делает совершеннее, тоньше и точнее деятельность нижележащих
отделов центральной нервной системы, в том числе и спинного мозга. Это
влияние коры осуществляется как тем, что через нее проходят высшие дуги
безусловных рефлексов, так и тем, что кора как могучий обобщающий
орган точно и тонко балансирует и регулирует сложные взаимоотношения
основных и вместе с тем функционально антагонистических нервных про-
цессов, формирующих всякую нервную деятельность, — взаимоотношения
процессов возбуждения и торможения.
В конце концов, можно указать на другую важную и бесспорно суще-
ственную форму воздействия коры на нижележащие нервные образования--
это трофическая функция ее. Лючиани рассматривал мозжечок как вспомога-
1 Bethe’s Handbuch d. norm. u. pathol. Physiol., Bd. 15, H. 2, 1931, S. 1127.
СООБЩЕНИЕ II. ОПЫТЫ С ПОЛОВИННОЙ ПЕРЕРЕЗКОЙ СПИННОГО МОЗГА
281
тельный орган большого мозга, а основную и единую функцию мозжечка
он видел в трофическом его воздействии на другие нервные образования,
управляющие движениями Организма. Па основании специальных ис-
следований влияния мозжечка и коры больших полушарий ла ниже-
лежащие отделы центральной нервной системы и на основании деталь-
ного анализа жизненных отправлений лишенных коры собак мы допуска-
ем что кора большого мозга как высший трофический орган своим воз-
действием изменяет функциональное состояние, функциональные свойства
(возбудимость, лабильность, общий тонус, работоспособность и т. и.)
нижележащих отделов центральной нервной системы, тем самым изменяя
и их пластичность. Отсутствие коры понижает уровень функциональной
ценности этих отделов, в том числе и спинного мозга, снижает его плас-
тические свойства, которые, судя во всему, и без того у высших взрос-
лых животных не так уже ярко представлены.
Если указанное «облегчающее», интегрирующее И Трофическое воздей-
ствие коры значительно в условиях цельности центральной нервной си-
стемы, то нет основания допускать, что значение этих разнообразных по
проявлению и, невидимому, единых по существу воздействий коры не ста-
нет более мощным фактором регуляции деятельности нижележащих отде-
лов центральной нервной системы, когда их структурная целостность на-
рушается.
Само собой разумеется, что, придавая такую решающую роль коре
большого мозга в явлениях пластичности центральной нервной системы,
я этим отнюдь не хочу умалить значение рецепторов, которые не только
постоянно сигнализируют о происшедших изменениях внутри и вне орга-
низма и тем самым помогают коре в ее сложной и ведущей анализаторной
и синтетической деятельности, но своими импульсами могут играть неко-
торую, хотя и менее важную роль без коры.
Считаю уместным здесь выдвинуть принципиально важный вопрсс
□ пластичности самого спинного мозга.
Было бы несерьезным утверждать, что спинной мозг высших взрослых
животных совершенно лишен пластических свойств, не говоря уже о спин-
ном мозге низших н высших позвоночных в первые периоды их жизни. Пла
стичность — свойство приспосабливаться к окружающей среде — обще-
биологическое свойство живого вещества; она присуща всем живым клеткам
так же, как и свойства питания, окисления, обмена веществ, возбудимости
и т. п. В процессе эволюции, одна ко, в процессе диферепциации структуры
и функций в животном организме основным и ярким носителем пластичности
становится нервная система. По ходу дальнейшего развития и диференциа-
ции самой нервной системы, когда головной мозг и в особенности высший
его отдел—кора большого мозга высших животных—приобретают домини-
рующее значение во всей центральной нервной системе, свойство пластич-
ности становится ведущим принципом деятельности, осуществляемой
в форме условнорефлекторной. В связи с этими сдвигами в центральной
нервной системе пластичность спинного мозга и других низших отделов
центральной нервной системы становится менее выраженной. Современная
физиология и клиника знают много фактов, иллюстрирующих эту' слож
ную динамику развития нервных образований и их функций. Подобно
тому как нельзя предполагать полного сходства между большим мозгом
лягушки, собаки и человека (я опускаю промежуточные звенья эволю
циониой лестницы), нельзя думать также о полной идентичности у них и
спинного мозга (то же самое можно сказать о функциональной динамике
других, морфологически более или. менее обособленных частей централь-
ной нервной системы).
Когда я на основании своих данных делаю вывод, что нарушения,
обусловленные половинной перерезкой спинного мозга у высших взрослых
2К’
Э. А. АСРАТЯН
животных, компенсируются не благодаря собственной пластичности спин
ного мозга этих животных, а главным образом благодаря пластичности
коры большого мозга, я этим хочу сказать только, что спинномозгогая
пластичность у них недостаточна для успешной компенсации таких серьез-
ных нарушении. Я не сомневаюсь в том, что при менее серьезных повреж-
дениях спинного мозга или при других видах поражений моторных
и сеизорных функций этих животных спинной мозг, в конце концов, сможет
обойтись и без значительного участия большого мозга, только ресурсами
собственной пластичности. Не исключена также возможность, что спинной
мозг у высших животных в раннем периоде их постэмбрионэльиой жизни
сможет справляться с аналогичной задачей только собственными ресурса-
ми пластичности даже при половинных его перерезках. Экспериментальная
проверка этих предположений входит в программу моей дальнейшей работы.
Сейчас, одпако, уже получено экспериментальное подтверждение другого
предположения, связанного с только что высказанными. Моя сотрудница
Ф. М. Вишневецкая показала,’ что нарушения локомоции, обусловленные
односторонней и двусторонней половинной перерезкой спинного мозга
лягушки, прекрасно и быстро компенсируются нс только при отсутствии
большого мозга у них, по даже при удалении промежуточного мозга, иначе
говоря, только собственными ресурсами пластичности их спинного мозга.
Сейчас аналогичныеолыты проводятся над птицами, и нужно ожидать, что
у них также значение спинномозговой пластичности в. компенсаторных яв-
лениях будет больше, чем у высших взрослых животных.
Во всем: этом я ничего удивительного, неожиданного или противореча-
щего моим данным, полученным на взрослых собаках, не вижу, да и никто
не увидит, если подойдет с позиций эволюционной физиологии, с позиций
диалектического материализма к пониманию развития структуры и фун кции
Зависимость восстановления . функции от эффективности, выражен-
ности пластических свойств нервного образования и от степени наносимого
повреждения заставила меня подойти к вопросу с другой стороны, — смо-
жет ли кора большого мозга высших взрослых животных компенсировать
далеко идущие нарушения, являющиеся результатом более глубоких по-
вреждений спинного мозга, чем половинная его перерезка. Для ответа на
этот вопрос я у трех собак (две взрослых, один щенок) поперечную пере-
резку спинного мозга произвел в размере больше половины. Взрослые со-
баки жили около 3 месяцев, щенок живет больше 2 месяцев, но, несмотря
на это, ни у одного животного нарушения моторики пораженных обеих
задних конечностей не компенсировались. Судя по всему, между задними и
передними половинами их тела существуют рефлекторные связи, импульсы
передаются через оставшийся «мостик» спинного мозга, однако за такой
длительный период компенсированные, интегрированные движения, ко-
торые могут обеспечить функцию стояния, не говоря уже о ходьбе, все
время отсутствуют. Что покажут дальнейшие исследования, — дело буду-
щего, но пока можно утверждать, что возможности коры большого мозга
высших животных в компенсации нарушения функций тоже не беспредельны.
На существование предела могущества коры большого мозга высших
животных указывают также наши опыты с другими вариантами операции
спинного мозга. Мы совместно с Р. О. Барсегян произвели у некоторых
собак и кошек возможно симметричное частичное продольное расщепление
спинного мозга,—то в пределах 2—3-го грудино-поясничных сегментов, то
по всей длине поясничных и сакральных сегментов в целом Эти опыты в
дальнейшем Р. О. Барсегян произвела над еще большим количеством кошек
и собак, а также над лягушками. Не дав.ая подробного описания картины
нарушений, обусловленных названными оперативными вмешательствами
(они составляют содержание двух работ Р. О- Барсегян), остановлюсь лишь
на том, что имеет непосредственное отношение к изучаемому вопросу.
СООБЩЕНИЕ II. ОПЫТЫ С ПОЛОВИННОЙ ПЕРЕРЕЗКОЙ СПИННОГС .МОЗГА 2S3
В случае частичного растепления середины спинного мозга, когда ниже
уровня расщепления сохраняются цельные спинномозговые сегменты, ин-
нервирующие задние конечности, обусловленные операцией нарушения
задних конечностей с течением времени проходят как у теплокровных,
так и у лягушек, и у животных полностью восстанавливается нарушенная
локомоция. В случае же, когда расщепление охватывает почти всю або-
ральную половину спинного мозга, когда сегментарная целостность этого
отрезка спинного мозга полностью нарушается, но сохраняются прямые
пути, связывающие верхние отделы центральной нервней системы с про-
дольно расщеп ленными половинками спинного мозга, —в этом случае глу-
бокие функциональные нарушения, обусловленные операцией, так и
не проходят ни у лягушек, ни у собак, ни у кошек, как бы долго они ни
жили (около 4 месяцев). Больше того, за все время жизни этих животных
нс наблюдается никаких признаков хотя бы смягчения этих нарушений.
Из этих опытов, а также из опытов, в которых поперечная перерезка
спинного мозга захватывает больше его половины, вытекает не только тот
вывод, что пластичность коры большого мозга и всей центральной нервной
системы имеет свои пределы; из них можно сделать другой принципиально
Важный ВыВоД, чТо нельзя центральную нервную систему рассматривать как
функционально аморфную решетку, как это делают Бете и его сторонники.
Ведь на самом деле, если бы центральная нервная система представляла
собой такую «непрерывную сетчатую систему» и вес интрацонтряльные про-
цессы—«переменную игру в этих решетках», если бы не существовало ни-
какой специализации и локализации центров и интрацентральных про-
водящих путей, если бы наши операции представляли собой в первую
очередь прерывание путей, — повторяю, если бы такой взгляд на струк-
туру и функции центральной нервной системы был верен, то мы был и бы
вправе ожидать восстановления нарушенных функций при обоих описан-
ных выше способах повреждения спинного мозга, ибо при них известные
довольно мощные столбы проводящих путей и мозговой массы структурно
не повреждаю гея и хорошо функционируют. На самом деле, как показы-
вают многочисленные эксперименты, этого не происходит.
Описанные в данной статье результаты моих экспериментов и экспе-
риментов моих сотрудников я рассматриваю как первые шаги в изучении
этой сложной и запутанной, но интереснейшей области физиологии нервной
системы. Тем не менее я осмеливаюсь сказать, что как опубликованные
уже результаты моих экспериментов по ампутации лап, так и описанные
в этой статье позволяют с чувством удовлетворения констатировать, что
высказанные мной на основании трудов корифеев отечественной науки
(И. М. Сеченов, И. П. Павлов) гипотетические положения о доминирующей
роли коры большого мозга высших животных в сложнейших проявлениях
пластичности нервной системы уже получают свое первое эксперимен-
тальное подтверждение. Об этом же говорят результаты произведенных
мной экспериментов по скрещиванию нервов и по разрушению лабиринтов,
описание которых составит содержание следующих сообщений.
ЛИТЕРАТУРА
Асратян Э. А., Кора большого, мозга и пластичность нервной системы. Вариант
с ампутацией лап, Успехи современной биологии. V, 1936.
Барсегян Р. О., К физиологии расщепленного спинного мозга. Бюллетень
экспериментальной биологии и медицины. 4, 2, 1937.
Барсегян Р. О.» Функциональные особенности продольно расщепленного спин-
ного мозга. Тезисы’диссертации,' 194Q.
Г о л ьцингер Ф. Ф., Чувствующие пути в спинном мозгу, дисс., Спб., 1896.
К узь чин В., Expert.rentelie U ntersuchungen ube.- die Leitvngsbahnen im ROken-
marks des Hundes, Medicinische Lehrbflcher, II. 2, 355, 1882.
284
Э. А. АСРАТЯН
Новикова А. А. и Ханутнна Д. И„ К вопросу об особенностях двигатель-
ных условных рефлексов у собак с половинной поперечной перерезкой спин-
ного мозга, Физиологический журнал СССР, 26, 4, 1Э39.
Россолимо Г. И., Экспериментальные исследования по вопросу о путях, прово-
дящих чувствительность и движения в спинном мозгу, дисс., М., 1887.
Сеченов И. М., Физиология нервных центров. Спб, 1891.
X а и у т и и а Д. И., Образование двигательных оборонительных условных ре-флек-
сов у собак с половинной перерезкой спинного мозга. Бюллетень эксперимен-
тальной биологии и медицины. Ill, 6, 19*37.
Чу дно вс кий Я. Б., Об изменениях чувствительности при спинной сухотке,
дисс., Спб., 1907.
B-ethe’s Handbuch d. norm. u. pat hoi. Physiologic, 15, II, 1931.
Danitsch B., Ztschr. Biol., 81, 1924.
ATar shall Ch., Proc. Roy. soc., London, 57, 1895.
Mott F., Phil, trans., 183, 1892.
Osa wa K., Untersuchungen Ober die Lcifungsbahnen im Rflckenmarks des Hundes,
Diss., 1882.
Schaffer E., Journ. physiol,, 24, 1899.
Schiff M,, Ciesammelte Beit rage zur Physiologic, 3, 1896.
Weiss N., Sitzungsber. Akad. Wiss.,’80, 1879.
♦ •
t
Добавление. Односторонняя и двусторонняя перерезка боковой по-
ловины спинного мозга на уровне последних грудных и первых пояс-
ничных сегментов была произведена нами еще у 10 собак и 1 кошки.
Полученные результаты, идентичные описанным выше, были частично
опубликованы в «Бюллетене экспериментальной биологии и медицины»,
т. VIII, в. 6, 1939. Привожу выдержку, касающуюся одной из собак,
жившей после удаления обоих полушарий более 3 лет.
«После опубликования вышеупомянутых данных я произвел одно-
стороннюю поперечную половинную перерезку спинного мозга в области
последних грудных и первых поясничных сегментов еще у ряда собак,
причем у одной из них (Бурик) после наступления компенсации я дву-
кратной операцией последовательно удалил кору одного, а затем и
другого полушария большого мозга. Удаление коры большого мозга
оказало на Бурика совершенно такое же воздействие, как и на опи-
санных мной 5собак: полностью исчезла компенсация пораженной лапы,
больше того, серьезно была задета также непораженная, т. е. противо-
положная задняя лапа. Собака утратила способность пользоваться обе
ими задними конечностями не только для ходьбы, но даже и для сто-
яния, тогда как рефлекторные и спонтанные движения обеими этими
лапами осуществлялись хорошо. Передними же лапами собака пользо-
валась нормально.
После полного удаления коры большого мозга Бурик живет уже
2 года 5 месяцев, т. е. значительно больше, чем другие собаки, под-
вергшиеся аналогичной операции, причем состояние здоровья и упи-
танность собаки хорошие. Несмотря, однако, на такое длительное время,
описанная выше картина, обусловленная удалением коры большого
мозга, остается до сих пор неизменной. Задние конечности делают хао-
тические движения, реагируют на раздражения как передней, так и зад-
ней части тела, но не в состоянии координированными движениями
хотя бы в какой-то мере способствовать поднятию задней части тела
от земли и ее передвижению.
КОРА БОЛЬШОГО МОЗГА И ПЛАСТИЧНОСТЬ НЕРВНОЙ СИСТЕМЫ* *
Сообщение 111
ВАРИАНТ ПЕРЕКРЕСТНОГО СШИВАНИЯ НЕРВОВ
.частичность нервной системы, проявляющаяся при ампута-
ции конечностей, по мнению Беге, не является очень на-
глядной, очень иллюстративной, так как она охватывает как
бы все тело,она не «локализована». В этом отношении особенно
показательны те эксперименты по проявлению пластичности
нервной системы, где искусственно изменяются исторически
сложившиеся отношения между отдельными частями тела в виде скрещи-
вания и анастомоза нервов. Недостатки этих видов нарушений (с точки зре-
ния их пригодности для изучения пластичнссти нервной системы) заключа-
ются, по мнению Бете, в том, что при них исключена возможность быстрой
перестройки, как это имеет место в опытах с ампутацией. В опытах с
локально измененными отношениями между центром и перифериен по-
средством перекрестного •сшивания нервов, которые в конечном итоге
приводят к перестройке центров, процесс регенерации, длящийся месяца-
ми, внешне значительно маскирует, по мнению Бете, характер и быстроту
наступления перестройки и может дать повод для допущения, что пере-
стройка осуществляется путем «выучивания». Бете пишет: «Таким образом,
можно сказать, что за это время нервная система имела бы возможность
приспособиться к измененным условиям и все эти явления (Vorgang), в
особенности у человека и высших животных, можно рассматривать как
процессы выучивания (Lernprozesse). Но по лученные до сих пор результаты
дают нам достаточно примеров, к которым нельзя применять подобное
объяснение. Это такие случаи, когда мы вынуждены локализовать феномен
пластичности в примитивных частях нервной системы»2.
Момент затяжки в наступлении перестройки нервных центров как при-
знак, отличающий этот процесс в случаях перекреста и анастомоза нервов
и в случае ампутации конечностей, мне кажется, теряет свое принципиаль-
ное значение после того, как мной было показано, что во многих опытах
с ампутацией перестройка тоже затягивается. Что касается характера и
существа этих перестроек, а также участия в них различных отделов
центральной нервной системы, то об этом подробно будет речь впереди.
Экспериментальные и клинические данные по скрещиванию
и анастомозу нервов
Большим теоретическим и в особенности практическим интересом, оче-
видно, следует объяснить тот факт, что перерезкой нервов, их скрещива-
нием и всевозможными анастомозами как физиологи, так и клиницисты
1 Успехи современной биологии, т. XII, в. 3, 1940.
* Bethe's Handbuch d. norm. u. pathol. Physiol., Bd. 15, H. 2. 1931, S. 1087.
286
Э. А. АСРАТЯН
занимались больше, чем какими-нибудь другими вариантами нарушения
естественной связи между периферией и центром. Первые опыты в этой
области были сделаны еще в конце XVIII века. [Фонтана (Fontana) и др.].
В дальнейшем этим вопросом занимались многие исследователи и практи-
ческие врачи. В данной статье мы лишены возможности систематически
изложить историю вопроса; желающим подробнее ознакомиться с ней можно
рекомендовать обзорные статьи Бете, Бете и Фишера, Букке (Воеке), а также
монографию Рамон-и-Кахала. Я укажу лишь на некоторые моменты.
1. С успешными результатами в смысле восстановления морфологиче-
ской структуры и функции скрещивали и анастомозировали различные
соматические нервы между собой. Наиболее интересными из этих опытов
можно считать опыты Флуранса (Flourence) с перекрещиванием главных
нервов плечевого сплетения у петухов (1828), опыты Бете с перекрещива-
нием двух седалищных нервов у собак, опыты Осборна и Килвингтона
(Osborn и Kilvington)co всевозможными вариациями новых анастомозов и
скрещиваний нервов плечевого сплетения у высших животных, опыты с за-
меной лицевого нерва акцессорным и подъязычным, а также множество
других вариантов скрещивания и замены другими черепными нервами.
2. С тем же успехом секрсщивали и анастомозировали между собой
различные вегетативные нервы — vagus sympathicus, sympathicus—
ciiorda thympani и т. д. [Н. А. Миславский, Ленгли (Langley) и др.].
3. С большим успехом в отношении восстановления морфологической
структуры и функции скрещивали и анастомозировали вегетативные нервы
с соматическими. В этом отношении представляют интерес опыты Рава
с перекрестными анастомозами пп. vagus—hypoglossus, опыты Барраго Сие-
релло (BarragoCierello), соединявшего центральный конец n. vagi с пери-
ферическим концом n. facialis, опыты Ленгли и Андерсона (Langley н Ander-
son) со сшиванием различных вегетативных нервов с соматическими нер-
вами, опыты Килвингтона с анастомозом передних корешков поясничных
сегментов с периферическими отрез ками передних корешков тазовых нервов,
опыты Эрлангера (Erlanger) с анастомозом периферического конца n. vagi
с центральным концом нервов плечевого сплетения, опыты Калюгарсану и
Анри (Calugareann и Henri) с анастомозом центрального концап. hypogios-
sis с периферическим концом n. vagi. Характерно, что как в опытах Эрлан-
гера, так и в аналогичных опытах последних двух исследователем нервы
плечевого сплетения и и. hypoglossus нс только оказывали тормозящее
влияние на работу' сердца при их раздражении (выше места анастомоза),
ко и перенимали тоническую функцию и. vagi, что Доказывается тем, что
после перерезки анастомозированного нерва учащается сердцебиение.
Наиболее интересными из этой серии опытов, повидимому, надо считать
опыты Калюгареапу и Анри с анастомозом центрального конца п. hypoglos-
sis с периферии еск им концом n. lingual is ниже места вхождения в последний
chordae thympani. В этих опытах функциональное восстановление проис-
ходило так успешно, что n. hypoglossus взял на себя регуляцию функций
желез, иннервируемых chorda thympani.
Во всех описанных случаях анастомозов и скрещиваний различных
смешанных нервов восстанавливались морфологически и функционально
как моторные, так и сензорные волокна. Многими авторами были сделаны
попытки скрестить между собой моторные и сензорные нервы (и даже ко-
решки спинного мозга), причем они получили различные результаты
[Беке (Воеке) и Др ].
Так, еще в первой половине прошлого столетия Шванн (Schwann), IIIифф
(Schiff) и др. производили такие остроумные эксперименты. Ониперсреза-
лн и сшивали различные смешанные нервы. После регенерации нервов; ис-
следователи обнажали сп инней мозг, перерезали спинальные корешки и раз-
дельно раздражали их с целью выяснить, не нарушается ли закон Бэлла-
СООБЩЕНИЕ III. ВАРИАНТ ПЕРЕКРЕСТА НЕРВОВ 287
Мажанди в результате неизбирательного врастания сензорных и моторных
волокон перерезанного нерва и возможного их смешения в процессе реге-
нерации. Все названные исследователи единодушно пришли к заключению,
что ни у холоднокровных, ни у теплокровных в описанных условиях экспе-
римента закон Бэлла-Мажанди не нарушается. В дальнейшем (1907) Бете
вновь вернулся к этому вопросу и своими экспериментами полностью под-
твердил данные и выводы указанных выше исследователей. С этими много-
кратно проверенными результатами Достоверного физиологического экспе-
римента небезынтересно сопоставить данные голландского нейрогистолога
Буккс. В серии Экспериментов Букке показал, что в результате регенера-
тивного процесса гетерогенно анастомозированных нервов центральные от-
резки чувствительных волокон не только врастают в периферические фут-
ляры моторных волокон, но, доходя до мышц, образуют в них характерные
концевые пластинки, И наоборот, центральные отрезки моторных волокон
не только врастают в периферические футляры чувствительных волокон,
но, достигая периферического органа, образуют в них характерные чув-
ствительные приборы. у1вное расхождение результатов физиологического
и гистологического экспериментов привело П. Вейсса (Р. Weiss) к новым
физиологическим экспериментам в этом направлении. На основании своих
опытов на низших позвоночных он приходит к выводу, что врастающие
в моторную периферию чувствительные волокна, в конце концов, приобре-
тают и функциональную активность: их раздражение вызывает сок ращение
мышц. Мне кажется, что опыты Вейсса еще окончательно не решают во-
проса. Ближайшее будущее покажет, насколько принципиальным и обос-
нованным является это «примирение» физиологического и гистологического
экспериментов по важнейшему и принципиальному вопросу.
Следует упомянуть о любопытных опытах Бете, в которых он сшивал
центральный конец П. maxillaris sup. С Периферическим концом И. optici
без успешного исхода не только в смысле восстановления функции, но и
морфологической структуры.
Многочисленные и разнообразные эксперименты и клинические данные
по сшиванию нервов дали возможность выявить ряд важных в теоретиче-
ском и практическом отношении моментов. Например, установлено, при
каких анастомозах получаются лучшие показатели восстановления как
морфологические, так и функциональные, какие условия способствуют и
какие препятствуют быстрому и полноценному восстановлению; выяснено
Влияние способа сшивания, возраста животных, сходства анастомозирую-
щих нервов, их длины, количества чувствительных волокон в них, наличия
так называемых сопутствующих сопряженных движений и т. д.
Совершенно точно установлено, что даже в наиболее благоприятных
случаях сшивания нервов далеко не все нервные волокна центрального
отрезка нерва врастают в футляры дегенерировавших нервных волокон
периферическо-го отрезка, если даже оба конца принадлежат одному и тому
же перерезанному нерву. Этим обстоятельством, очевидно, следует объяс-
нить также точно установленные факты, что мышцы, которые получают
новую невротизацию (в результате сшивания нервов), развивают силу,
равную примерно одной трети нормальной, и притом обычно быстро утом-
ляются. Некоторые авторы полагают, что сращение происходит немзбм-
рательно даже при перерезке и сшивании кондов одного и того же нерва.
Часто, как уже было упомянуто, врастают друг в друга афферентные и
эфферентные нервы. Далее установлено, что срастающиеся нервные во-
локна в рубце дают бифуркации и врастают в футляры различных деге-
нерировавших нервов, ведущих вачастую к совершенно разным пунктам
периферии. В литературе справедливо указывается на это как на причину
множественных аксон-рсфлексов во вновь некротизированной области.
288
Э. А. АСРАТЯН
Точный учет этих моментов дает возможность практическим врачам сде-
лать успешный выбор в случае необходимости заменить какой-нибудь
парализованный нерв Другими нервами и достигнуть значительных успе-
хов, описанием которых богата современная мировая нейрохирургическая
литература.
Следует отметить еще один момент, на который обращал внимание ряд
авторов'и который всегда надо учитывать, в особенности при физиологиче-
ских экспериментах. Дело в том, что часто при скрещиваниях и сшива-
ниях нервов, какие бы предосторожности ни принимались эксперимента-
торами, центральный конец каждого нерва, минуя всевозможные препят-
ствия, прорастает по направлению к периферическому концу и иногда
густыми пучками волокон врастает в него. В результате периферический
рабочий аппарат получает нервы как от «законного», так и от «незакон-
ного» нерва. Этот момент, часто не поддающийся точному учету в физиоло-
гических экспериментах, способен внести значительную путаницу в наши
рассуждения о перестройке центров. Во всяком случае это заставляет
избегать слишком большой категоричности даже при констатации самого
факта перестройки в центральном1 аппарате.
Меня, однако, больше интересуют неясности другого порядка, связан-
ные с восстановлением функции периферических аппаратов после сшивания
различных нервов, а именно: каков характер перестройки нервных аппа-
ратов, обусловливающей это восстановление, каково участие перифериче-
ских импульсов и коры большого мозга в этих перестройках, т. е. вопросы,
которые были подняты в моих предыдущих статьях в отношении опытов
с ампутацией конечностей и с половинной перерезкой спинного мозга. Эти
неясности, может быть, действительно усугубляются обстоятельством, на
которое указывают Бете и Фишер: затяжкой и постепенностью процесса
восстановления нарушенных функций, моментами, затрудняющими физио-
логический анализ процесса перестройки.
Как бы то ни было, после внимательного учета всех перечисленных не-
ясностей, затруднений и запутывающих моментов, связанных с изучением
проблемы пластичности нервной системы при сш ивании нервов, Бете в пони-
мании характера и существа перестройки в центральной нервной системе,
а также в оценке роли периферических импульсов и коры большого мозга
в этой перестройке придерживается техжесвоих концепций, которые выска-
зывались нм относительно опытов с ампутацией конечностей и опытов
с гемисекцией спинного мозга.
Свои выводы об интересующем меня основном вопросе — о роли коры
и о характере перестройки при скрещивании нервов Бете и Фишер аргу-
ментируют результатами некоторых опытов с раздражением двигательных
зон у собак, у которых после скрещивания нервов восстановились функ-
ции. Це говоря уже о том, что эти опыты сами по себе содержат ряд
противоречий, укажу на принципиальную сторону. В опытах, которые
ставятся под общим наркозом в условиях вивисекции, конечно, нельзя
хоть в какой-нибудь мере выявить роль коры в перестройке нижележа-
щих отделов центральной нервной системы, а тем более если такими
опытами авторы хотят опровергнуть предположение, что кора оказывает
влияние на эту перестройку механизмом «выучивания», путем условно-
рефлекторной деятельности.
Бете в своих опытах со скрещиванием седалищных нервов у Собак не
производил даже экстирпации двигательных зон (хотя он ставил вопрос
о ней еще в. 1905 г.), — эксперимента, не решающего вопроса, как указано
в моей первой статье, но более близкого к преследуемой цели, чем опыты
с раздражением. Бете придает особо важное значение опытам Осборна и
Килвингтона, которые показали, что после высокой перерезки спинного
мозга у ссбак не нарушаются вновь установленные нормальные сгиба-
СООБЩЕНИЕ III. вариант перекреста нервов
289
тельные, разгибательные и ритмические рефлексы на задние конечности,
основные нервы которых (tibialis и peroneus) заблаговременно были скре-
щены с последующим восстановлением нарушенных функций. Мне ка-
жется, что и эти опыты не являются решающими для поставленного воп-
роса. Во-первых, большая близость скрещенных нервов создает благо-
приятную почву для повой встречи обоих концов каждого нерва, а затем
нельзя отрицать, что цельные nn. fenioralis и obturatorius, а также нервы,
отходящие от ischiadicus до места его бифуркации, в состоянии в значи-
тельной мере обеспечить эти рефлексы. Наконец, нельзя пе указать ня
противоречие между этими данными и старыми данными самого Бете (1905)
о том, что при скрещивании двух седалищных нервов, когда уже проис-
ходит восстановление функции, правильная спинномозговая координация
нарушается. В этих своих экспериментах, заранее перерезая n. cruTalis,
Бете показал, что раздражение каждой лапы вызывает подъем противопо-
ложной лапы. Тогда он писал по поводу этих опытов: «Никакого привы-
кания не происходит».
Но в противовес этим фактам клиника дает огромный материал, указы-
вающий на то, что после всевозможных скрещиваний и замены одного нерва
другим, после того как появляются явные признаки восстановления мор-
фологической структуры волокон и их проводимости, перестройка надолго
задерживается, осложняясь так называемыми побочными движениями ста-
рых иннервациоипых областей, что для их преодоления необходимо боль-
шое волевое напряжение со стороны больного и что, несмотря на все это,
иногда перестройка так и не осуществляется. Некоторые клиницисты
[Шпици (Spitzy)] даже прямо указывают на связь между уровнем развития
человека и быстротой перестройки. Под явным влиянием этих фактов Бете
делает некоторое отступление от своей первоначальной категорической фор-
мулировки, в частности, для человека. По этому поводу он пишет: «Не-
смотря на то, что до настоящего времени лишь в немногих случаях при
опытах скрещивания нервов могло быть доказано, что результат сохра-
няется и после отделения высших центральных органов, все же несомненно,
что головной мозг лишь незначительно участвует в этих процессах пере-
стройки. Как вспомогательный фактор головной мозг можно принимать
во внимание только при таких перестройках, где участвуют мускулы,
«произвольно приводимые в движение»; что в таких случаях, И R особен-
ности у человека, «научение» играет большую роль, это допускают мно-
гие авторы (Шпици и др.). Мы тоже охотно соглашаемся с тем, что в
устранении сопровождающих движений участвует «произвол». Однако и
здесь остается фактом то, что иннервация через посторонний «центр» з
конечном результате осуществляется совершенно автоматически. Иначе
говоря, происходит настоящая перестройка. Однако все это не имеет зна-
чения для тех посторонних иннерваций, при которых «произвол» не иг-
рает роли, как это может быть сказано в отношении автономных областей»1.
Из приведенной цитаты видно, что механизм перестройки при сши вании
нервов Бете и Фишер представляют в таком же виде, как и при ампутации
лап. Значение большого мозга как вспомогательного фактора они непра-
вильно сводят к помощи в упразднении так называемых побочных движе-
ний, а самое главнее, т. е. перестройка центра, с их точки зрения про-
исходит «совершенно автоматически».
За последние годы П. К. Анохин и его сотрудники развернули интен-
сивную исследовательскую работу по изучению пластичности нервной
системы на объектах с анастомозами нервов. С этой целью ими были при-
менены всевозможные варианты скрещивания и анастомозов названных
выше, а также некоторых других: нервов. Так, например, П. К. Анохин и
1 Betha's ilandbach d. norm. u. pathol. Physio!., Bd. 15, H. 2, 1931, S. 1104.
19 Э. А Лсратни
290
9 А. АСРАТЯН
Д4 Иванов анастомозировали центральный конец блуждающего нерва
с периферическим концом нервов плечевого сплетения, чаще всего с n. ulna-
risun. radialis,они скрещивали n. obturatorius и n. femoralis. Если Бете
производил анастомоз п. max illaris sup. с п. opticus, то Н. И. Шумилина
анастомозировала п. vagus с п. opticus; подобно Калюгареану и Анри, осу-
ществившим анастомоз nn. hypoglossus—1 ingualis, П. К. Анохин и А. Ива-
нов анастомозировалип. vagus cn, lingualjs, а Б, Гдассон прямо повторил
опыты названных выше исследователей с темижесавершснно результатами.
В процессе многолетней систематической и целеустремленной работы
П. К. Анохина и группы сотрудников обнаружен ряд новых деталей, которые
нашли свое отражение в нашей физиологической литературе и в специаль-
ном сборнике. Из этих данных, в частности, заслуживают внимания про-
веденные П, к. Анохиным и сотрудниками наблюдения за течением функ-
ционального восстановления афферентных волокон, имеющие отношение к
проблеме специфичности возбуждения', подробно это изучено на анастомо-
зах центрального конца n. vagi с периферическими концами нервов пле-
чевого сплетения, с n. 1 ingualis, п. opticus. На этих экспериментах я здесь
подробно останавливаться небуду, так как обсуждение их не входит в за-
дачу данной статьи. Укажу только на один важный, с моей точки зрения,
момент—подробно изученный ими медленно протекающий процесс пере-
стройки функций чувствительной части ядра блуждающего нерва. Отме-
чу также сходство явлений кашля, рвоты и т. д., послуживших для них
индикаторами для суждения о перестройке центра n. v agi, с одной стороны,
и так называемых явлений сопряженных пли побочных движении, под-
робно описанных прежними исследователями, — с другой.
Большинство же фактов, полученных П. К. Анохиным и сотрудниками,
представляет известную ценность для изучения проблемы пластичное!и
нервной системы прежде всего- как повторение и подтверждение известных
уже многочисленных данных по этой проблеме, выявленных названными
выше исследователями’.
[Неоригинален П. К- Анохин также в разработке теоретических во-
просов обсуждаемой проблемы. Он находится в плену ошибочных и анти-
эволюциоиных, взглядов Бете. П. К- Анохин вслед за Бете считает, что пе-
рестройка функции происходит в низших отделах центральной нервной
системы под решающим влиянием периферических импульсов и без участия
условно рефлекторной деятельности коры больших полушарий. Идентич-
ность взглядов П. К. Анохина н Бете по всем основным вопросам проблемы
устраняет необходимость сделанные выше критические замечания в адрес
теории Бете повторять здесь внонь].
К категории опытов с нарушением естественной связи между пери-
ферическими органами и их центрами относятся и работы П. Вейсса (Weiss)
по трансплантации целых конечностей и отдельных мышц у амфибий, на-
чатые им еще в 1921 г.
Свон трансплантаты Вейсс прививал к различным пенсам тела хозяина
и обеспечивал невротизацию различными нервами этих же поясов, После
того как происходило структурное и функциональное сращение трансплан-
тата С Телом хозяина, он некоторое время еще не проявлял специфической
реакции на центральные импульсы и отвечал на всякое возбуждение цен-
тральной нервной системы. Постепенно, однако, когда по предположению
Вейсса, формируются концевые пластинки в мышцах трансплантата, эта
1 Редакция журнала «Успехи современной биологии», опубликовав статью в
1440 г. (с двухгодичным опозданием), целиком опустила весьма обстоятельную кри-
тику теоретических положений П. К. Анохина. Считая неудобным при повторном
издании полностью восстанавливать рукописный текст этой статьи, мы сочли, од-
нако, возможным в целях восполнени я явного пробела в скобках привести один аб-
зац. в СЖатон форме воспроизводя щнй основное содержание опущенной части статьи
СООБЩЕНИЕ III. ВАРИАНТ ПЕРЕКРЕСТА ПЕРВОЙ
291
неспецкфич еск ая связь заменяется специфической связью между центром и
соответствующими мышцами; наступает фаза модуляции мышц. Однако,
в зависимости от местонахождения, поведение трансплантата может быть
различным. Если конечность привита к псясу, являющемуся для нее как бы
«родным» (например, передняя конечность прививается к псясу плечевого
сплетения хозяина), то после завершения модуляции она начинает избира-
тельно реагировать на строго специфические центральные импульсы, а имен-
но: при рефлекторных и спонтанных движениях хозяина трансплантат-
конечность сокращается одновременно, однотипно и одинаково интенсивно
с одноименной конечностью хозяина. Та же картина наблюдается, если
пересаживается не цельная конечность, а отдельная мышца. Если транс-
плантат-конечнесть привита к поясу, сходному с «родным» (скажем, перед-
няя конечность привита к тазовому поясу), то не синонимные, а гомологич-
ные мышцы трансплантата-конечностн хозяина сокращаются одновременно
(например, экстензоры колена и экстензоры локтя или же флексоры тех же
суставов и т. д. сокращаются одновременно). Если же трансплантат-конеч-
ность прививается к совсем «чужому» псясу, скажем, к туловищу, то, как
показали опыты Дётуиллера, Вейсса и др., трансплантат не проявляет ни-
каких координированных рефлекторных и спонтанных движений, хотя
отдельные сокращения он осуществляет при раздражении самого туловища.
Это явление Вейсс объясняет тем, что «импульсы, специфичные для мышц
конечностей, возникают в пределах сегментов спинного мозга, соответ-
ствующих конечностям, и в- норме не выхедятза пределы этих сегментов»1.
Как мы увидим ниже, объяснение Вейсса по этому вопросу не совсем со-
ответствует основным теоретическим его концепциям. Но имеются и такие
факты, которые уже совсем не гармонируют с основными эксперименталь-
ными данными Вейсса. Несколько лет назад Николас (Nicholas) и Де-
туиллер показали, что при прививке трансплантата-ко вечности к псясу
жаберных сегментов (иннервация черепными нервами) трансплантированная
конечность осуществляет движения совместно с глоточными или жабер-
ными движениями. Этот факт настолько расходится с данными Вейсса и
так резко противоречит его основной теоретической установке (о чем речь
будет ниже), что Вейсс высказывает даже сомнение, не является ли он ар-
тефактом. Как бы там ни было, сопоставление перечисленных фактов с пол-
ной очевидностью показывает, что пояс иннервации имеет очень важное,
даже решающее значение для дальнейшей судьбы трансплантата.
С точки зрения пластичности нервной системы представляют интерес
некоторые летали экспериментов Вейсса и др. по первым двум основным
и эффективным вариантам трансплантации, именно по прививке конечно-
стей к поясам конечностей их хозяина.
Прежде всего установлено, что описанная выше картина функционирую-
щих трансплантатов неизменно сохраняется. Она не претерпевает никаких
изменений даже тогда, когда применяются самые фантастические комбина-
ции при прививке трансплантатов (когда, например, трансплантации соче-
таются с ампутациями), или даже тогда, когда, вследствие всевозможных
причудливых комбинаций этих операций, имеющиеся налицо конечности,
благодаря одновременному сокращению сипониммых и гомологичных мышц,
мешают друг другу и препятствуют нормальному выполнению других
общих моторных функций. Говоря словами Вейсса, не происходит ника-
кого «научения условной реакции, центрального приспособления». Иными
словами, никакого проявления пластичности центральной нерв ной системы
не наблюдается.
Дополн игельныеопыты Вейсса показали, что как деафферентация конеч-
ностей, так и экстирпация головного мозга не оказывают влияния на уста-
1 Р. Weiss, Biol, reviews Cambr. phil. soc., 11, № 4, 1936, p. 522.
19*
292 Э. А. АСРАТЯН
нивленьые им феномены. «Феномен гомологичного ответа, — заключает
Вейсс, — базируется на свойствах спинального механизма, и вмешатель-
ство центров, находящихся выше спинальных, пе оказывает никакого
влияния»1.
На этих данных Вейсс построил свою механистическую «резонансную
теорию», которая пользуется широкой известностью За рубежом. Согласно
этой теории, описанные выше явления объясняются наличием узко специ-
фической функциональной связи между отдельными центрами и соответ-
ствующими индивидуальными мышцами. Благодаря со настроенности
станции отправления и станции назначения, благодаря наличию «специ-
фических форм» импульсов, эти две функциональные единицы так тесно
связываются между собой, что из массы хаотических импульсов, диффузно
пробегающих по одним и тем же аналогичным путям, мышца как детектор
избирательно откликается только на те импульсы, которые адресованы ей
своим же центром. Сколько существует индивидуальных мышц и соответст-
вующих им центров, столько имеётся отдельных видов «специальных форм
импульсов».
Другими опытами Вейсс показал, что такие же узко специфические
взаимоотношения существуют между рецепторной перифериен и центром
с тон только разницей, что здесь роль станций отправления играет уже
периферия, генерирующие залпы «специальных фирм» импульсов, а детек-
торами являются соответствующие центры; «проприоцептивный импульс,
идущий до добавочного бицепса, точно узнается в центре как исходящий
от бицепса, а не от какой-либо другой мышцы». При этом сходство между
двумя парами по линии этих взаимоотношений идет так далеко, что и
здесь установившаяся раз со настроенность «неизменно сохраняется на-
всегда». «Итак,—заключает Вейсс,—мы находим между органами чувств
и центрами такие же особые взаимоотношения, какие, по нашим данным,
характеризуют отношения между центрами и мышцами, а именно: их взаи-
моотношения остаются неизменными, несмотря на произвольные изменения
в расположении периферических органов»1 2.
Здесь неместо для обстоятельной критической оценки эксперименталь-
ных данных и теоретических положений Вейсса. Нельзя не указать, однако,
на явно выраженную механистичность, схематичность, односторонность
основных его теоретических положений, в силу чего неизбежно возникает
конфликт между новыми фактами из той же области и между этими теорети-
ческими положениями. В самой теории Вейсса имеется ряд резких лротиво-
речий. С одной стороны, способность нервной системы к диффузному прове-
дению всевозможных импульсов различной «специфической формы», асдру-
гой стороны, невозможность проникновения этих импульсов через какие-то
«чужДые» им центральные районы. Концепция Вейсса о Дроблении цент-
ральной нервной системы на множество отдельных узко специализирован-
ных центров, соответственно количеству периферических индивидуальных
эффекторов и рецепторов, а также его представление о какой-то статичес-
кой, строго специфической узкой связи между соответствующими единицами
центра и периферии не только находятся в явном противоречии с основными
твердо установленными фактами и понятиями нейрофизиологии, но и носят
явную печать мистики. Далее, мне кажется противоречивым, с одной сто-
роны, резкое умаление значения структуры (диффузное проведение любых
импульсов по нервной системе), а с другой стороны, столь же резкое под-
черкивание момента крайней специализации структур — дробление нерв-
ной системы на множество узко специфических единиц. Пусть это будут
закономерности спинальной деятельности; ио ведь физиология давно из-
1 Р. Weiss, Biol. reviews Camb г. phil. soc., v. II, № 4, 1936, p. 506.
2 T а м 5K e, стр. JiOS.
СООБЩЕНИЕ 111. ВАРИАНТ ПЕРЕКРЕСТА НЕРВОВ
293
жила представления о спинном мозге как о статически функционирующем
органе. Деятельность спинного мозга очень вариабильна. Спинной мозг
эволюционирует, созревает и т. д.
Но сопсставляя данные о статическом характере установившихся
взаимоотношений центра и трансплантированной периферии у низших
животных со сходными данными, полученными у высших животных и че-
ловека, когда после изменения исторически сложившихся взаимоотношений
между сензомоторной периферией и центром, благодаря перестройке цен-
тров, мышцы меняют свои функциональные знаки в общем комплексе дру-
гих мышц и приспосабливаются к новым условиям, Вейсс в последней своей
обзорней статье высказываетпредполсжение, что это приспособление у выс-
ших животных и человека является результатом церебральной их деятель-
ности. Он пишет; «Наши эксперименты над амфибиями наводят на мысль,
что такие перестройки у высших позвоночных обусловлены деятельно-
стью головного мозга, способной замешать нарушенные спинальные шаб-
лоны; если это так, то децеребрация должна сразу же обнаружить началь-
ное нарушение. Мне неизвестно, применялся ли кем-нибудь этот способа*.
Собственные данные по анастомозу нервов
Из обзора литературы видно, что явления компенсации после всевозмож-
ных новых невротизаций, анастомозов и перекрестов нервов представляют
очень пеструю и запутанную картину, что тут гораздо больше вариаций
в нарушении функций и соответственно этому гораздо больше разнообра-
зия в «механизмах» и способах восстановления нарушенных функций, чем
в компенсации какого-либо другого вида нарушения сензорных и моторных
функций животного. Этим, очевидно, в значительной мере обусловливаются
существенные расхождения как в описаниях картин этих явлении различ-
ными исследователями, так и в теоретических положениях, высказанных
ими о факторах, путях, характере и возможностях компенсации. Мне ка-
жется совершенно бесспорным, однако, что основной причиной расхожде-
ний в констатации фактов и в теоретических концепциях является отсут-
ствие последовательного эволюционного подхода к изучаемому вопросу.
Ведь по существу для физиолога ни одна система так четко не характери-
зует генеральной линии эволюции животного мира, как центральная нерв-
ная система, в особенности в конечных звеньях развития животного мира.
А в действительности в физиологии проводилось изучение одной из карди-
нальных особенностей нейтральней нервной системы, ее функциональная
пластичность, без серьезного н последовательного учета фактора эволюции.
Отсюда одна из причин недооценки и даже игнорирования роли «научения»
и всо;бще коры большего мозга в компенсаторных явлениях <Бете и его по-
следователи), отсюда одна из главных причин указанных выше недостатков
«резонансной теории».
В согласии с исходными теоретическими положениями по пластичности
нервной системы, изложенными в предыдущих моих статьях, а также с
моими установками по экспериментальному изучению ряда вопросов этой
проблемы, в цикле опытов, поставленных мной за последние годы (1933—
1937) по перекресту нервов, я ближайшей своей целью ставил выяснение
роли коры большого мозга высокоразвитых взрослых позвоночных (собака)
в явлениях компенсации после названных оперативных вмешательств. Од-
нако прежде чем вплотную подойти к решению этой задачи, я постарался
внести некоторую ясность в один щекотливый и запутанный вопрсс.
Судя по литературным данным, последствия перерезки отдельных спин-
номозговых и черепномозговых нервов, имеющих отношение к моторике
животного, весьма различны по степейи и длительности нарушения локалъ-
1 Р. Weiss, Biol, reviewsСлтЬг. ptlil. soc., v. 11, № 4, 1936, p. 525.
294
Э. А. АСРАТЯН
ных и общих Движений. Содной стороны, бывают случаи, когдаэти наруше-
ния почти неуловимы для обычного глаза и с помощью грубых тестов, а
с другой стороны, наблюдаются и такие случаи, когда наступает длительнее
и глубокое нарушение локальных и общих моторных и опорных функций ор
ганизма. Учитывая важность всего этого для решения вопроса о роли коры
в указанных явлениях, я решил провести предварительную работу с целью
обнаружения таких нервов для перекреста, перерезка которых вызывает
серьезные и длительные последствия.
Подробная классификация и детальное изучение вопроса о последствиях
перерезок различных нервов и о путях компенсации возникших в связи
С этим нарушений — дело будущего. Однако на основании имеющегося
литературного материала, а также полученных мной предварительных дан-
ных уже можно высказать положение, что разнообразие фактов по анасто-
мозу и перекрестному сшиванию нервов можно грубо разделить минимум
на две группы. К первой группе следовало отнести те случаи, в которых
возникают ничтожные, иногда совсем незаметные нарушения, сравнительно
быстро компенсирующиеся, во. вся ком случае много раньше, чем возможно
наступление регенерации «чужого нерва и перестройки его центра. Ко
второй группе следуето гнести случаи, в которых возникают очень серьез-
ные выпадения функций и компенсация наступает с большим опозданием.
Дающим основание полагать, что ведущую роль в компенсации играет
врастание «чужого» нерва и действительная перестройка его центра. К, по-
следней группе можно отнести, например, перекрестное сшивание пп. va-
gus— modi anus и obturatorius — femoralis. Совершенно естественно, что
возможны градации как в каждой группе, так и переходные вариации между
двумя группами. Конечно, во всех случаях мы имеем дело с проявлением
пластичности центральной нервной системы, но это не мотив против по-
пытки классифицировать эти явления, ибо есть все основания допускать,
«то как по существу, так и по способам проявления и физиологической эф-
фективности пластичность пластичности рознь.
Дальнейшая экспериментальная работа, именно результаты экстирпации
коры одного или двух полушарий большого мозга, с полной очевидностью
показала принципиальную правильность такого разделения. Экстирпация
коры левого полушария у двух собак, у одной из которых справа предва-
рительно был перерезан п. radial is, а у другой n. ulnaris, не вызвала замет-
ного ухудшения в состоянии «пораженных» лап. Такой ж:е результат полу-
чен при экстирпации коры левого полушария у третьей собаки, у которой
предварительно были перерезаны сба седалищных нерва. Все три собаки
на 3-й день после операции удаления полушарий вставали, ходили и поль-
зовались «пораженными» конечностями столь же успешно, как и до опе-
рации.
Совершенно иная картина получается после декортикации большого
мозга у собак, предварительно подвергнутых операции второй группы.
Остановлюсь на этом случае подробнее.
Я решил произвести перекрестное сшивание пп. vagus — medianus. За
последние годы (1934—1937) эта операция осуществлялась у 8 собак.
У 3 из них компенсация нарушений не наступила, у остальных удовлетво-
рительная компенсация наблюдалась в среднем через 6—7 месяцев после
операции (за одним исключением). После того как компенсировались на-
рушения, я произвел декортикацию большого мозга. Две собаки жили после
названной операции недолго, к поэтому я на них останавливаться не буду.
Ценные данные получены на трех собаках—Жулике, Тахице и Радиусе
(см. таблицу).
Последствия операций на полушариях оказались неодинаковыми у всех
трех собак. Экстирпация коры левого полушария у Жулика вызвала на-
столько глубокий паралич пораженной конечности, что в первые 7—9 дней
СООБЩЕНИЕ II!. ВАРИАНТ ПЕРЕКРЕСТА НЕРВОВ
295
после операции собака при попытке опереться на эту конечность падала на
землю и не могла стоять и двигаться. Лишь на 12-й день после операции
собака могла стоять и ходить, не падая па каждом шагу. При этом она уже
опиралась на парализованную лапу, совершенно согнутую в лучезапястном
суставе и слабо согнутую в остальных суставах. Жулик прожил 16 месяцев
после удаления полушария. Небольшой сдвиг в состоянии пораженной
конечности чувствовался именно в течение 1-го месяца, в дальнейшем как бы
установился некоторый стандарт, который оставался до конца жизни со-
баки.
), Кличка 1 собаки 1 Возраст 1 Дата перекрест- ' кого сшивания п. vagus med in mis справ* Время ком- пста- 1ДЫ1 в иег. Дата удаления коры левого полушария Дат Я Уда- лен ня ко- рм право- го полу- шария Примечания
Жулик 2—3 года 25. V 1934 г. 8 19.111 1935 г. Собака по- гибла в июле 1936 г. во вре- мя эпидемии чумы
Тахиц 5—6 лет 2. VII 1936 г. 3 II. X 1936 г. 16. XI 1936 г. 1. X 1937 г. Обе собаки вне- запно погибли, по видимому, от отравления не- доброкачест- венной пищей
Радиус 2—3 года 2.VI 1936 г. 6 13.XII 1937 г. 26.1 1937 г.
Декортикация левого полушария вызвала у Тахица сильные наруше-
ния со стороны пораженной лапы; после перекрестного сшивания нервов
непривычно быстро (через 21/а— 3 месяца) наступила компенсация. Эти
явления повторной декомпенсации в общем были такого же характера, как
и у Жулика после декортикации, но в менее резкой форме. Характерно, что
у Тахица в первые дни после удаления полушария не совсем полноценно
функционировала также незатронутая передняя конечность (правая).
Правда, явлений паралича и согнутости в суставах этой конечности не
наблюдалось, но все же при попытках опереться на нее в первые дни собака
скользила и нс могла быстро подняться. Через 5—6 дней после последней
операции явления со стороны здоровой передней конечности совсем прекра-
тились, а через 14—15 дней начала несколько улучшаться и картина глубо-
ких поражений оперированной раньше передней конечности. Собака по-
степенно стала опираться на нее, не падая; при этом конечность чаше всего
сгибалась в лучезапястном суставе, так что соприкосновение с землей про-
исходило почти по всей передней поверхности ноги, начиная с лучезапя-
стного сустава. Улучшение продолжалось, и спустя месяц после операции
Тахиц часто опирался на пораженную конечность, которая уже не сгибалась.
Через 36 дней после последней операции я удалил и правое полушарие
большого мозга. Состояние пораженной конечности вновь резко ухудши-
лось. Собака опять не могла опираться на эту лапу. При стоянии она дер-
жала лапу в воздухе, когда же при ходьбе делала попытку опереться на
нее, это приводило к падению. В первые дин опять-таки была затронута
и неоперированная конечность; собака не могла делать ею четких движе-
ний, скользила. Примерно через 20 дней картина легкого нарушения функ-
ции этой конечности исчезла, а резкое нарушение функции оперированной
конечности оставалось еще в течение 2—21/» месяцев, посте чего началось
постепенное улучшение. Собака больше не держала в воздухе пораженную
296
Э. А. АСРАТЯН
конечность, лучше опиралась на нее. Па 4— 5-м месяце после последней
операции собака стала пользоваться пораженной конечностью. Тахнц жил
10 месяцев после последней операции, и все же опытный глаз легко мог об-
наружить дефект в передней правой конечности (рис. 1).
Декортикация левого полушария большого мозга у Радиуса вызвала
очень глубокое поражение функции ранее оперированной правой передней
Рис. 1. Влияние удаления коры больших полушарий на функции стояния и
ходьбы у собак Радиус и Тахиц со сшитыми перекрестно блуждающими и сре-
динными нервами с правой стороны.
Через 5—6 месяцев после перекрестного сшивания блуждающего л срединного
нервов с правой стороны функция правой передней конечности восстановилась.
После удаления коры обоих больших полушарий функция этой конечности вновь
глубоко нарушилась; конечность была сильно паретнчна и сгибалась (в особен-
ности в лучезапястном суставе) при опоре на нее
а—собака Радиус в норме,- в—с нарушенный» функциями; е—собака Тахиц п иопме; а—с на-
рушенными функциями
конечности. Во избежание повторений отмечу лишь, что по своей глубине
И стойкости это поражение занимало как бы среднее место между пораже-
ниями лап у Жулика и Тахица. К моменту' декортикации второго полуша-
рия (через 38 дней после декортикации первого полушария) лапа эта нахо-
дилась еще в Состоянии глубокого поражения. После удаления второго
полушария картина ухудшилась еще более. В отличие от Тахица, у Ра-
диуса левая передняя конечность совершенно не была затронута. Собака
прекрасно передвигалась на трех конечностях, а пользование пораженной
конечностью приводило часто к падению. Через 18—20 дней после послед-
ней операции появилось незначительное улучшение в состоянии этой лапы,
которое дли лось 30—40 дней. После чего наступил.период стандартной, почти
неизменякшейся картины весьма значительного^поражения функции. Со-
бака прожила около 9 месяцев, в очень хорошем*состоянии, прекрасно пе-
редвигалась ня трех конечностях, пользуясь п пораженной, которая хотя
СООБЩЕНИЕ III ВАРИАНТ ПЕРЕ КРЕСТА НЕРВОВ
297
и нс приводила к падению, но обусловливала весьма заметную хромоту
(рис. 1).
Несколько слов об одном факте, подробно описанном П. К. Анохиным и
сотрудниками, — о появлении у животного кашля при анастомоАг it. vap:
выше места отхождения n. rccurrentis с нервами плечевого сплетения. Как
уже сказано выше, по существу это явление принадлежит к категории явле-
ний, дав .о известных в литературе под названием сопряженных движений.
У Тах-ша и Радиуса как раз был произведен такой вариант операции.
Я не буду останавливаться иа картине появления и развития этого явления
после перекрестного сшивания нервов, ибо она изложена в работах, вы-
шедших из лаборатории П. К. Анохина. У обеих моих собак к моменту
удаления полушарий уже нельзя было вызывать рефлекторного кашля при
раздражении соответствующих конечностей. После же декортикации одного,
а затем другого полушария вновь и в очень бурной форме появился кашель.
Для его вызывания не нужно было специально раздражать ссответствующу к>
конечность; стоило собакам подвигаться или привести в движение опериро-
ванную конечность, чтобы начался приступ кашля. Это явление продол-
жалось примерно 4—5 месяцев после операции, за гем постепенно стало
ослабевать. В последний период жизни оно у обеих ссбак почти прекрати-
лось.
Экспериментальный материал, полученный па собаках Жулик, Тахип,
н Радиус, с полной очевидностью показывает исключительно важное,
я бы сказал, решающее значение большого мозга в явлениях компен-
сации у высокоразвитых взрослых позвоночных глубоких и длительных
нарушений моторных функций, вызванных перерезкой и перекрестным сши-
ванием нервов. К тому же выводу приводят данные, полученные на собаке
Чанг. То обстоятельство, что после удаления коры большого мозга у этих
животных локально поражаются именно те конечности, важные нервы ко-
торых были подвергнуты перекрестному сшиванию, что эти повторные де-
компенсации представляют собой копню первичных нарушений, на-
ступающих сразу же после перекрестного сшивания нервов, что функция
остальных конечностей у тех же собак не претерпевают заметных изменений,
достаточно убедительно говорят о том, что повторные нарушения компен-
сации нс являются результатом общих изменений в моторике животного
вследствие декортикации большого мозга.
Некоторое постепенное улучшение повторных нарушений компенсации,
наблюдающееся главным образом в первое время после удаления коры,
также не говорит против правильности заключения о важной, решающей
роли коры большого мозга в указанных компенсаторных явлениях. Прежде
всего это некоторое первоначальное улучшение можно рассматривать как
результат постепенного устранения «операционного торможения»; в 2 слу-
чаях дальнейшее улучшение было настолько незначительным, что через 6 и
7 месяцев после декортикации оно не привело к значительному ослаблению
повторных явлений декомпенсации. Но даже возможные крайние случаи
не могут быть доводом против допущения ведущей роли коры в этих компен-
саторных явлениях в норме, ибо я не исключаю возможности, что в усло-
виях отсутствия коры остальные часта центральной нервной системы, прежде
всего ближайшая подкорка, в состоянии тренировать некоторые свои функ-
циональные возможности и этим путем в той или иной мере замещать кору.
Вот основные аргументы в пользу допущения, что кора большого мозга
у высших позвоночных играет ведущую, решающую роль в компенсации
нарушений, обусловленных перекрестными анастомозами важных нервов.
При беглом ознакомлении с_ обширным и разнообразным материалом
других исследователей по анастомозам и перекрестному сшиванию раз-
личных нровов, а также с представленным в данной статье материалом по
тома же вопросу с полной очевидностью вытекает, что здесь мы не имеем
•298
Э А АСРАТЯН
дела с однородными по проявлению и сложности нарушениями функций
органов и явлениями компенсации этих нарушений. Наоборот, все говорит
о том, что здесь имеег место вереница разнообразных явлений различной
степени сложности. Точная классификация и детальная количественная и
качественная характеристика отдельных звеньев этого возрастающего по
сложности ряда явлений нарушения и компенсации функции органов —
задача дальнейших исследований, но уже сейчас небесполезно было бы
указать на существование различных механизмов компенсаторных при-
способлений соответственно степени тяжести поражения и даже рискнуть
наметить основные контуры этой классификации, указать на характерные
особенности главных звеньев этой цепи.
С другой стороны, на основании описания характера и течения насту-
пающих компенсаторных явлений, данного многими исследователями,
в частности, клиницистами, а также на основании результатов проведенных
мной экспериментов с экстирпацией коры одного и двух полушарий боль-
шого мозга у собак можно сделать заключение, что по крайней мере у выс-
ших позвоночных степень участия и роль различных отделов центральной
нервной системы в этих компенсаторных явлениях неодинаковы. Напра-
шивается предположение, что в соответствии с возрастающей сложностью
нарушения и компенсации функций органов имеется примерно восходящий
ряд доминирующих звеньев в единой цепи центральных нервных образо-
ваний, обусловливающих эти компенсаторные акты. Это допущение тоже
нуждается в детальной экспериментальной и теоретической проверке, но
мне кажется, что уже имеющийся материал позволяет не только сделать
такое допущение, но и начертать контуры будущей теории, правильно
отображающей эти сложные явления.
В самом деле, если, с одной стороны, мы имеем такие случаи, когда пе-
ререзка и перекрестное сшивание различных нервов почти никак не про-
являются или возникающие ничтожные нарушения более или менее быстро
проходят, то, с другой стороны, имеются и такие случаи, когда перерезка
и перекрестное сшивание других нервов вызывают очень глубокие и дли-
тельные нарушения. Далее, если в первой группе случаев, как показывают
предвари тельные эксперименты на высокоразвитых взрослых Позвоночных,
кора большого мозга не играет ведущей роли в компенсации, то во второй
группе 3ia ведущая роль совершенно очевидна. Можно сослаться также на
другие виды нарушения н компенсации функции моторного аппарата этих
животных, где совершенно четко выступает та же закономерность.
В этой же связи можно сослаться также на данные вышедшей из лабора-
тории Бете работы Хольста (Holst). проведенной на многоножках. Geophi-
lius, Cryptops и Lithobius.
Хольст изучал новые виды координации при ампутации различного
количества и распорядка конечностей с обеих сторон. Меня интересуют здесь
не детали этой работы, а одни замечательный, с моей точки зрения, факт.
В то время как при ампутации того или иного количества конечностей всегда
как бы мгновенно, «спонтанно» возникают новые виды координированных
движений, как бы без участия «научения», при ампутации всех конечностей
животные с помощью различных новых для них приемов действительно
«научаются» ходить и без конечностей.
С моей точки зрения, все приведенные факты, в том числе и факты
Хольста, являются иллюстрацией правильности высказанных выше сооб-
ражении. Имеется цепь нарушений функций органов различной степени
сложности; соответственно этому организм располагает массой разнообраз-
ных средств различной физиологической эффективности, иными словами, це-
лой гаммой взаимно связанных и взаимодействующих, порой различных
по характеру и по существу видов нервной деятельности, которыми он ком-
пенсирует эти нарушения.
СООБЩЕНИЕ 111. ВАРИАНТ ПЕРЕКРЕСТА НЕРВОВ
299
Если, с одной стороны, существует «механизм» дублированной иннер-
вации. простой перестройки и перекомбинацин наличных возможностей
для эффективного компенсирования этих нарушений, то, с другой стороны,
для того же «назначения» имеется возрастающая по эффективности гамма
других «средств», как то: простая тренировка функции, трофическое воз-
действие верхних этажей центральной нервной системы. Сложное безуслов-
норефлекторное интегрирование этими же частями центральном нервном
системы, условнорефлекторная деятельность. [Об этом свидетельствует уста-
Рне 2
о иичограмлл злектрооборонительных двигательных условных рефлексов
у >-обани Кочегар через 3 месяца после перекрестного сшивания бедренного
и срединного нервов, 0-килограмма электрооборонительных двигательных
условных рефлексов у собаки Радиус после перекрестного сшивания блуж-
дающего н срединного нервов (опыты В А Замятиной).
Обозначения (сверху вниз), условные и безусловные рефлексы,
отметки условного раздражителя, время в секундах, отметка безусловного
раздражителя
позленная нашей сотрудницей В. А. Замятиной Возможность выработки
условных рефлексов на деятельность конечности с анастомозированными
нервами (рис. 2)1].
Совершенно очевидно, что по крайней мере у высших позвоночных
с морфологически и функционально диференцированном, но единой нервной
системой конкретным носителем каждого из этих видов «деятельности» яв-
ляется в основном какой-нибудь отдел центральной нервной системы.
Факты показывают, что и здесь, в явлениях пластичности нервной системы,
динамика остается в связи с конструкцией и что нельзя нервную систему
1 Сведения об опытах В. А. Замятиной, опровергающих первоначальные данные
П. К. Д нохича н его сотрудников, были также опущены редакцией журнала «Ус-
пехи современной биологии» при онхбличоичнин Да1‘ЩЙ Статьи.
300
Э. А АСРАТЯН
рассматривать как функционально аморфную и равнозначную do всех
частях систему, как это пытаются делать Бете и его последователи.
В развитие высказанных в первых моих статьях предположительных
взглядов по данному вопросу я хотел сформулировать этот принцип под
названием «принцип относительной пластичности» нервной системы. Из
Приведенного мной Экспериментального материала явствует, что у высоко-
развитых взрослых позвоночных на верните этой лестницы находится кора
большого мозга с доминирующим в центральной нервной системе богатым
и могущественным арсеналом форм деятельности. Без большого риска
можно допустить, что в этой цепи другие звенья центральной нервной
системы по общей эффективности их пластических возможностей занима-
ют последовательные места, примерно соответствующие степени слож-
ности присущей нм анализаторной л синтетической деятельности.
Имеющийся в литературе экспериментальный и клинический материал,
а также собранные мной и моими сотрудниками данные по той же проблеме
позволяют предполагать, что абсолютная (возможно, даже и относитель-
ная) физиологическая эффективность каждого из этих звеньев центральной
нервной системы может изменяться как по возрастной линии, так и в эво-
люционном ряду. Не исключена также возможность индивидуальных ко-
лебаний у высших животных, в особенности у человека, поскольку речь
идет о важной роли коры большого мозга в этих явлениях, о важной роли
основного органа, обусловливающего индивидуальные различия.
С другой стороны, не подлежит сомнению, что даже в узком понима-
нии вопроса, с точки зрения свойств компенсации нарушенных функций,
общая физиологическая эффективность пластических свойств всей цен-
тральной нервной системы не представляет собой неизменную величину,
что она также развивается и увеличивается вместе с эволюцией живого
мира, возможно, даже и по возрастной линии (в некоторых пределах).
С этой точки зрения отсутствие приспособления привитых мышц и целых
конечностей к новым условиям существования у низших позвоночных,
стойкое сохранение за ними «резонансных» свойств даже в условиях цель-
ности центральной нервной системы (опыты П. Вейсса и мн. др.) в сопо-
ставлении с данными, говорящими о возможности смены функций в примерно
одинаковых случаях (пересадка мышц, природные случаи дублирования
конечностей, некоторые варианты перекрестного сшивания нервов), мне
представляются доказательством конечной низкой физиологической эффек-
тивности пластических свойств центральной нервной системы у низших
позвоночных. «Резонансное» свойство в этом понимании ие есть универ-
сальный и статический закон взаимосвязи периферии и центра, как пред’
ставляег себе автор «резонансной теории», а лишь фрагмент в общей слож-
ной и развивающейся цепи.
В пользу предположения об усилении общей физиологической эффек-
тивности пластических свойств всей центральной нервной системы по мере
подъема животного вверх по эволюционной лестнице говорят и данные
Эвальда, Бнкеля, Магнуса и его сотрудников о том, что последствия раз-
рушения лабиринтов и перерезки задних корешков у высших животных
компенсируются быстрее, чем у животных, находящихся на более низких
ступенях эволюционного ряда.
Несмотря на наличие значительного фактического материала, подтверж-
дающего основные положения развитого и сформулированного в этой ста-
тье «принципа относительной пластичности» как первого приближения
к эволюционному пониманию проблемы, я все же пока придаю ему значе-
ние лишь предварительной физиологической схемы.
сообщение hi. вариант перекреста НЕРВОВ
301
ЛИТЕРАТУРА
Анохин П. К. и др.. Проблема центра и периферии в физиологии нервной дея-
тельности, 1935.
Асратян Э. А., Кора большого мозга и пластичность нервной системы. Успехи
современной биологии, т. V, 1936, стр. 804; т. VI, 1937, стр. 45.
Корнилов Ам Ueber die Veranderungen der moforischen Functionen Beistdrungen
des Sensibilitilt, Comptes rendus d. Xll Congres international de medecine, IV,
675, 1897.
M и с л а в с к и й H. А., С. г. Soc. biol., 841, 1902.
В е t h е A., Ueber Nervenverheilung und pel arc Waclistumscrschcinungen am Ncrven,
Mtinch. med. Wschr., 25, 1967.
Bethe A., Notiz fiber die Unfahigkeit motorischer Fassern mit rezeptorischen Fas-
sern zu verhcilen, PflOg. Arch., 116, 1907.
В ethe A., Plastizitat und Zentrenlehre, Bethe's Handb. d. norm. u. path. Physiol.,
15, 2, 1931.
В ethe u. Fischer, Die Anpassuogsfdhigkoit (Plastizitat) der NcrvensyMetns,
Bethe’s Handb. d. norm. u. path. Physiol., 15, 2, 1931.
Boeke J., Nervenregeneration und verwandfe Innervationsprobleme, Ergebnisse
d. Physiol., 19. 1921.
Calugarcanu et Henri, C. r. Soc. biol., 1901, 1099.
Langley a. Anderson, The union of different kinds of fibers, J. physiol.,
31, 1904.
Langlev a. Anderson, Cn autogenetic regeneration in the nerves of the
limbs, J. physiol., 31.
Osborn u. Kilvi'ngton, Brain, 33, 1911.
R a m о n-y-Ca ja 1, The degeneration and regeneration of the nervous system, 1923.
Kava, Arch. Physiol., 1885.
W ei ss P., Selectivity controlling the central spheral relation in the nervbos system,
Biol, rev., 11, 4, 1936.
Weiss P.. Experimental innervation о-f muscles by the central ends of afferent
nerves, J. neur., 61, 1. 1936.
КОРЛ БОЛЬШОГО МОЗГА И ПЛАСТИЧНОСТЬ НЕРВНОЙ СИСТЕМЫ'
Сообщение IV
ОПЫТЫ С ДЕАФФЕРЕНТАЦИЕЙ КОНЕЧНОСТЕЙ
спинальных
пыты Бэлла и Мажанди выяснили роль передних и задник
корешков спинного мозга в сензорных и скелетно-моторных
функциях центральной нервной системы. Установленные ими
факты относительно функциональной архитектуры нервной
системы впоследствии были проверены и подтверждены многи-
ми другими экспериментаторами. Функции передних и задних
корешков изучались физиологами в условиях как острого,
так и хронического эксперимента, давшего возможность более или ме
нее продолжительного наблюдения за оперированными животными. Метод
хронического эксперимента позволил в сравнительно-физиологическом ас-
пекте исследовать картину .нарушения и динамику восстановления мо-
торных функций деафферентированных конечностей у животных, стоящих
на различных уровнях эволюционной лестницы.
Ряд авторов в многочисленных хронических опытах показал, что деаф-
ферентация конечностей у лягушек не вызывает глубоких нарушений в их
моторных функциях. При этом было установлено, что картина неглубоких
моторных нарушении деафферснтированной конечности (слабая атаксия,
вялость движении, повертывание в воде и т. д.) заметно не изменяется даже
спустя 6 месяцев после операции (Бикель (Bickel)].
Почти аналогичная картина наблюдается у птнц с деафферентирован-
ными конечностями. По данным Тренделенбурга деафферентация ног го-
лубей не вызывает существенных нарушений их моторных функций. В еще
меньшей степени нарушаются моторные функции деафферентированных
крыльев голубя. Характерно, однако, что даже эти неглубокие нарушения
компенсируются весьма незначительно. В первые 10—14 дней после опе-
рации они несколько сглаживаются, а затем устанавливается довольно
стабильная фаза, при которой дсафферентированные конечности выполняют
свои опорные, локомоторные и летательные функции почти наравне с нор-
мальными конечностями. Аналогичные данные на голубях были получены
Бальони (Baglioni).
Иную картину и динамику нарушения дает деафферентация конечно-
стей млекопитающих — коз, собак и в особенности обезьян. По данным всех
авторов, изучавших этот вопрос |Мунк (Munk), Бикель (Bickel), Мерц-
бахер (Merzbacher), Магнус (.Magnus), Орбели и Кунстман, 1924, и др. 1,
деафферентация конечностей вызывает у этих животных сильные и дли-
тельные изменения опорных и моторных.функций соответствующих конеч-
1 Физиологический ж;, риал СССР, № I, Г948. Предварительное сообщен не опуб-
ликовано в <Бюллетене; экспериментальной биологии и медицины», т. IX, в, 1, 1940.
СООБЩЕНИЕ IV. ОПЫТЫ С ДЕЛФФЕРЕНТАЦИЕЙ КОНЕЧНОСТЕЙ
303.
костей. Однако в отличие от рептилии и птиц у млекопитающих эти нару-
шения с течением времени постепенно ослабевают, и опорные и моторные
функции пораженных конечностей восстанавливаются иногда в такой сте-
пени, что неопытный глаз не сразу отличает пораженные конечности от
непораженных.
Некоторые исследователи ставили специальные эксперименты для
нения роли разных рецепторов (лабиринтов, глаз, кожных рецепторов
неповрежденных областей тела) и различных отделов головного мозга в вос-
становлении скелетно-моторных функций деафферентированных конечно-
стей низших и высших животных. Так, например, Штиллннг (Stilling}
показал, что отсутствие у лягушек серьезных моторных нарушений деаф-
ферентированных конечностей в значительной мере зависит от кожных ре-
цепторов неповрежденных конечностей. Опыты Трен дел енбурга на птицах
и опыты Балди (Baldi) на собаках выявили, что в восстановлении моторных
функций деафферентированных конечностей исключительную роль играет
афферентная иннервация симметричных конечностей. По данным Бикеля,
у лягушек, равно как и у собак, повреждение лабиринтов значительно
углубляет картину моторных нарушений деафферентированных конечно-
стей. Тренделенбург обнаружил то же самсе у голубей. Это явление Бикель
объясняет тем, что функциональные изменения, обусловленные разреше-
нием лабиринтов, суммируются с нарушениями, вызванными деафферета-
цией конечностей. Установлено, что оптические рецепторы также оказывают
значительное воздействие на картину нарушения и восстановления функ-
ций деафферентированных конечностей низших и в особенности высших
животных (Бикель, Магнус и др.).
Интересные данные были получены в экспериментах, поставленных
с целью выяснения роли верхних этажей центральной нервной системы
в этих процессах. Так, Мерцбахер показал, что удаление переднего мозга
у лягушек очень мало отражается на функциональных особенностях дегффе-
рентированных конечностей, в то время как удаление промежуточного
мозга у тех же животных значительно ухудшает состояние деафференти-
рованных конечностей. Это явление Бикель объясняет суммированием по-
следствий обоих оперативных воздействий. Он пишет: «При сочетании двух
операций, каждая из которых сама во себе способна вызвать в известной
степени кратко охарактеризованные здесь нарушения, естественно, про-
исходит суммация, простое сложение этих нарушений»1. Он экс показал,
что у лягушек перерезка спинного мозга вызывает паралич деаффтерен-
тированных конечностей.
Такне же результаты были получены на птицах. Поданным Тренделен-
бурга, удаление переднего- мозга у голубей не оказывает заметного вли-
яния на судьбу имевших место компенсаторных явлений в моторных функ-
циях деафферентированных конечностей.
Значительно бдльшую роль играют верхние этажи центральной нервной
системы в судьбе моторных нарушений деафферентированных конечностей
у млекопитающих. Согласно наблюдениям Бнкеля, у собак в стадии компен-
сации моторных нарушений деафферентированных конечностей удаление
так называемых сензомоторных зон коры головного мозга приводит к по-
явлению исчезнувших было нарушений функций пораженных конечное!ей.
С течением времени эти повторные нарушения компенсируются, но уже
с трудом и в недостаточной степени. Обратная последовательность опера-
тивных вмешательств дала по существу те же результаты. На основании
своих опытов Бикель делает вывод: «Единственно возможное объяснение
этого состоит в том, что сензомоторные зоны путем развития явлений за-
1 A. Bickel, Untersuchungen Qber den Mechanismus dor nervOsen Bewegungs-
regulation, 1903, S. 23.
304
Э. А. АСРАТЯН
мещення, путем функциональной перестройки маскируют корешковое
нарушение и приводят движения животных к норме»’.
Интересно отметить, что, несмотря на бесспорную принадлежность опи-
санных выше фактов к проблеме пластичности, Бете, автор известной теории
пластичности нервной системы, не упоминает о них ни в одной из своих
статей. А между тем эти факты, в особенности данные, указывающие на воз-
растающее в процессе филогенеза значение верхних этажей центральной
нервной системы в компенсаторных явлениях, должны были заставить
Бете и его последователей отказаться от огульного отрицания значения
верхних отделов центральной нервной системы в компенсаторных явлениях
у млекопитающих, членистоногих и лягушек.
Желая возможно полнее охватить в своей экспериментальной работе
круг фактов, так или иначе относящихся в проблеме пластичности, мы изу*
чали характер и динамику моторных нарушений деаффсрентированных
конечностей у собак. Путем более решительного оперативного воздействия
на кору головного мозга (Полной ее экстирпацией) мы пытались точнее
выяснить вопрос о значении коры больших полушарий мозга в возникаю-
щих при этом компенсаторных явлениях. Опыты нам казались тем более
необходимыми, что Бикель при решении этого вопроса ограничивался
экстирпацией ничтожно малых кусков коры головного мозга у подопытных
животных и, повидимому, по этой причине достиг лишь частичного ослаб-
ления компенсаторных явлений.
Экспериментальная часть
Для изучения поставленного вопроса нами в 1937 г. были оперированы
три собаки среднего возраста (Дунай, Рыжко и Ласточка). У собак под
эфиро-хлороформным наркозом интрадурально были перерезаны все лево-
сторонние задние корешки люмбальных и сакральных сегментов спинного
мозга (у Дуная—15.VI 1937 г., у Рыжко и Ласточки — 11.VIII 1937 г.).
Все собаки прошли описанные Бикелем стадии послеоперационного нару-
шения и восстановления функций пораженных конечностей: псевдопарали-
тическую стадию, стадию резко выраженной атаксии пораженной конеч-
ности и стадию компенсации моторных нарушений.
Псевдопаралитическая стадия у собак длилась в среднем 7—10 дней,
после чего конечности начинали четко реагиропать сначала на раздражение
симметричных непораженных задних конечностей, а затем и на раздра-
жение других частей тела. Постепенно появлялись произвольные движения
как в лежачем положении, так в особенности при стоянии и ходьбе. Эти не-
уклюжие и атактические движения постепенно усиливались, оживлялись
н совершенствовались. Процесс восстановления моторных функций пора-
женных конечностей у собак достиг более или менее стабильного уровня
соответственно через 2, 4 и 6 месяцев после операции (Бикель также отметил
большие индивидуальные колебания в сроках восстановления функции
деаффгренгированпых конечностей). Все три собаки могли опираться на
деаффгрентнрованные задние лапы, а также достаточно хорошо пользо-
ваться ими при ходьбе и беге. При внимательном наблюдении все же можно
было заметить, что движения деафферентированных конечностей несколько
размашисты и дисметричны. При стоянии собак деафферентированные ко-
нечности нередко фиксировались в позе выраженной флексии, выраженной
экстензии или в других позах.
В этой фазе восстановления моторных функций пораженных конечно-
стей у собак было произведено двухмоментное оперативное удаление всей
коры больших полушарий головного мозга.
1 A. Bickel, Untersuchungen йЬег den Mechanismus der nervSsen Bewegngs-
regulation, 1903, S. 58.
СООБЩЕНИЕ IV. ОПЫТЫ С ДЕАФФЕРЕНТАЦИЕЙ КОНЕЧНОСТЕЙ
305
9.VII 1937 г. была удалена кора правого полушария у Дуная. В пер-
вые же дни после операции наступил почти полный паралич всей задней
части тела собаки, причем это состояние по глубине превышало аналогич-
ное состояние, наступавшее после перерезки задних корешков с левой сто-
роны. Пораженная лапа совершенно потеряла способность рефлекторного
реагирования на раздражение здоровых частей тела; о спонтанных или про-
извольных движениях не могло быть и речи. Это состояние как бы полного
паралича длилось 13 дней (собака погибла вследствие инфекции). Заслу-
живает внимания то, что в первые дни после операции почти полностью без-
действовала и непораженная задняя лапа. Лишь через 4—5 дней она стала
рефлекторно реагировать на раздражения. Появились и в скором времени
достигли значительной интенсивности также «спонтанные», или произволь-
ные, движения. Однако собака до самой смерти так и не могла пользоваться
этой лапой для стояния и ходьбы, хотя спустя 10—11 дней после удаления
коры общее состояние собаки было весьма удовлетворительным и она стояла
и передвигалась на• выпрямленных передних конечностях.
У Рыжко удаление коры правого полушария головного мозга было
произведено 9.IV 1938 г., т. е. через 8 месяцев после перерезки задних
корешков. К этому времени моторные нарушения пораженной лапы собаки
хорошо компенсировались. Послеоперационная картина у Рыжко была со-
вершенно такой же, как у Дуная. В первые дни после удаления коры
одного полушария наблюдался почти полный паралич всей задней час-
ти тела собаки при хорошем функциональном состоянии передней части.
Спустя 4—5 дней после операции, началась постепенная активизация
непораженной задней конечности. На 10—11-й день собака могла уже опи-
раться на эту конечность и, благодаря этому, стоять и ходить на трех лапах.
Пораженная задняя конечность длительное время еще оставалась в состоя-
нии глубокого паралича (она неизменно находилась в состоянии контрак-
туры, в положении резкой экстензии). Лишь спустя месяц после удаления
коры, поряженная конечность проявила слабые признаки активности.
К этому времени стало постепенно ослабевать также тоническое экстен-
зорное состояние. К концу второго месяца после этой операции Рыжко
поправился. Восстановление моторных функций пораженной конечности
также достигло значительного уровня, но все же более низкого, чем до уда-
ления коры правого полушария.
7, VI 1938 г. у Рыжко была удалена кора второго, левого полушария
головного мозга. После полной декортикации задняя часть тела собаки была
совершенно парализована. Пораженная конечность почти все время нахо-
дилась в состоянии контрактуры, экстензии ц была полностью парализо-
вана. Она не проявляла никаких признаков деятельности ни спонтанно,
ин в ответ на раздражение других частей тела. Это состояние без изменения
сохранялось в течение 2 месяцев, т. е. до самой смерти животного. Не ме-
нее разительными оказались последствия удалении коры второго полуша-
рия для другой, недеаффереитированной задней конечности. Эта конечность
не потеряла рефлекторной активности, производила также довольно ожив-
ленные спонтанные движения, но уже не могла участвовать в осуществле-
нии таких сложных интегративных актов, как стояние и ходьба. Животное
могло опираться лишь на передние конечности и передвигаться только при
их помощи. Задняя же часть тела при этом пассивно волочилась за перед-
ней, а спонтанные или рефлекторные движения задней неповрежденной
конечности никакой помощи животному не оказывали. Картина функцио-
нальной активности задней неповрежденной конечности также без изме-
нения сохранялась 2 месяца (см. рисунок, стр. 306).
У третьей нашей собаки (Ласточки) кора правого полушария была уда-
лена 19.IV 1938 г., т. е. через 8 месяцев после деафферентаими задней ко-
нечности. Последствия декортикации у Ласточки были совершенно такие же.
20 э. Л. Асратян
ЗС6
Э. А. АСРАТЯН
как у первых двух собак (в особенности у Рыжко). Ласточка в послсоопс-
рационнын период поправилась настолько хорошо, что забеременела, още-
нилась и кормила своих щенят. Через 10 месяцев после удаления коры
правого полушария и к моменту удаления коры левого полушария значи-
тельно улучшилось также функциональное состояние и активность деаффе-
рентированной конечности;
Собака Рыжко. Опорные и локомоторные функции
задних конечностей, восстановленные много месяцев
спустя после перерезки задних корешков пояснич-
ных и крестцовых нервов, вновь были глубоко
нарушены после удаления коры обоих больших
полушарий мозга. Собака могла опираться только
на передние КонечноСти и передвигаться медленно
только с их помощью, волоча за собой парализо-
ван ную заднюю часть тела
она почти достигла уровня,
который существовал перед
удалением первого полуша-
рия. Последствия удаления
коры второго полушария
оказались для фуп кЦИЙ зад-
них конечностей Ласточки
также очень тяжелыми.
После этой операции Лас-
точка жила недолго — не-
многим более 2 недель. За
несколько дней ДО смерти
она была в состоянии пра-
вильно устанавливать пе-
реднюю часть тела и актив-
но пользоваться передними
конечностями для передви-
жения в полулежачем по
ложении, т. е. ползти с ме
ста на место, волоча за со-
бой заднюю часть тела,
«прикованную» к полу. При
довольно активные дви-
этом неповрежденная задняя конечность делала
жения, почти ие принимавшие, однако, участия в передвижении живот-
ного. Задняя деафферентированная конечность оставалась в тоническом
эксгензорном состоянии, не проявляя признаков какой-либо активности.
Обсуждение полученных данных
Прежде всего следует указать на одну важную особенность проведенных
опытов. Все наши собаки с деафферентироваинымн конечностями после
удаления коры головного мозга жили сравнительно недолго — от 2 недель
до 2 месяцев. Они оказались особенно чувствительными к условиям содер-
жания и погибали гораздо скорее, чем остальные собаки с удаленной корей.
При таких обстоятельствах, возможно, было бы правильнее воздержать-
ся от каких бы то ни было трактовок полученного материала, в частности,
от суждения о роли коры больших полушарий головного мозга в развитии
приспособительных явлений у собак с деафферентироваинымн конечностями,
так как трудно сказать, какова была бы финальная картина функциональ-
ного состояния и деятельности деафферентированных конечностей этих
собак, если бы они жили дольше. Однако, имея большой опыт наблюдения
над собаками, подвергавшимися другим операциям и также лишенными
коры больших полушарий, мы все же считаем возможным сделать опреде-
ленные выводы из приведенного не совсем полноценного эксперименталь-
ного материала.
Наш многолетний опыт привел нас к убеждению, что если глубокие
функциональные нарушения у лишенных коры и дополнительно опери-
рованных собак держатся упорно и без существенных изменений в течение
1—2 месяцев после удаления коры второго полушария, то, как правило,
в течение дальнейшей жизни животных такая картина претерпевает лишь
СООБЩЕНИЕ IV. ОПЫТЫ С ДЕАФФЕРЕНТАЦИЕП КОНЕЧНОСТЕЙ
307
ничтожные изменения или вовсе не изменяется. Вот почему, несмотря на
сравнительно непродолжительную жизнь наших собак, мы все же склонны
считать, что они дали весьма ценный материал для освещения сложного
физиологического вопроса, не получившего удовлетворительного решения
до последнего времени. Мы считаем, в частности, что полученные нами
данные позволяют с большей убедительностью и решительностью, чем дан-
ные Викеля, утверждать, что кора больших полушарий не только прини-
мает участие в восстановлении нарушенных моторных функций деафферен-
тированных конечностей собак, но играет при этом решающую роль.
Таким образом, устанавливается, что у собак (в отличие от лягушек и
голубей) процесс функционального переключения новых афферентных
импульсов к мотонейронам деафферентированных конечностей, процесс
восстановления моторных функций этих конечностей обусловливается
в основном деятельностью коры большого мозга. Очевидно, что важную
роль при этом играет как трофическое воздействие коры на спинной мозг,
так и в особенности её способность к сложным функциональным перестрой-
кам, к созданию новых видов интегратинной деятельности нервней си-
стемы.
Удаление коры больших полушарий отразилось в сильной степени также
на моторных функциях недеафферентированных задних конечностей собак.
Хотя при этом рефлекторная их деятельность не нарушалась, хотя они и
осуществляли разные хаотические движения значительной силы и
объема, тем не менее они не были в состоянии участвовать в таких сложных
интегративных актах, как стояние и ходьба.
Бесспорно, в данном случае дисфункция кооперированной задней конеч-
ности тесно связана с деафферентацией симметричной задней конечности.
По всей вероятности, у собак, лишенных коры больших полушарий, сим-
метричный поток афферентных импульсов является важным условием для
участия конечностей в осуществлении таких сложных безусловнорефлек-
торных интегративных актов, как стояние и ходьба. При целости коры
больших полушарий ею со временем устраняются последствия нарушений
такой симметрии, вызванных деафферентацией одной из лап.
Приведенные в этом сообщении данные полностью соответствуют другим
нашим данным о важной роли коры больших полушарии мозга в восстанов-
лении нарушенных сензорных и моторных функций организма пссле других
оперативных повреждений организма: ампутации лап, перекрестного сши-
вания нервов, половинной поперечной перерезки спинного мозга, разру-
шения лабиринтов и т. п. Тем самым еше раз подтверждаются и те выдви-
нутые нами теоретические положения о приспособительных явлениях в по-
врежденном организме, которые основаны на ряде твердо установлен-
ных общих положений нейрофизиологии (И. М. Сеченов, И. П. Павлов)
и главным образом на соответствующем собственном экспериментальном
материале.
Хотя Бете в своих обзорах и обходит молчанием данные эксперимен-
тальной физиологии относительно явлений нарушения и восстановления
функций деафферентированных конечностей, тем нс менее мы считаем себя
вправе сказать, что как те данные, так в особенности описанный в этом
сообщении новый экспериментальный материал свидетельствуют о полной
несостоятельности его основных теоретических положений о пластичности
нервной системы. Это относится, в частности, к его концепции о ничтожной,
(вернее, никакой) роли верхних этажей центральной нервной системы в про-
цессах восстановления нарушенных функций поврежденного организма
ге только у низших, но и у высших животных. Правильность наших данных
и концепций подкрепляется также данными П. К. Анохина и А. Н. Шумили-
ной относительно возможности образования «отраженных» условных реф-
лексов на деафферентированную конечность собаки. Это тем более отрадно
20*
308
Э. А. АСРАТЯН
констатировать, что, как известно, П. К,- Анохин и его сотрудники в предыду-
щих своих работах вслед за Бете отрицали возможность участия коры боль-
ших полушарий мозга в процессах восстановления нарушенных функций
поврежденного организма посредством ее специфической деятельности —
образования условных рефлексов. Тогда они (как в свое время и Бете)
признавали лишь «динамогеннос» облегчающее воздействие коры на нижеле-
жащие отделы центральной нервной системы.
Может показаться, однако, что данные Эвальда (Ewald), Бикеля, Трен-
делеибурга и др. и данные нашей лаборатории о важной роли различных
органов чувств (лабиринтов, кожных рецепторов, глаз) в этих восстанови-
тельных явлениях подтверждают правильность воззрений Бете и его после-
дователей относительно роли периферических импульсов в явлениях компен-
сации. В действительности это далеко не так. Импульсам с неповрежденных
периферических рецепторов Бете приписывает ведущую, решающую роль
в восстановлении нарушенных .функций поврежденных органов. Далее
Бете без всякого экспериментального обоснования считает, что эти импульсы
утилизируются именно примитивными частями центральной нервной си-
стемы, что под влиянием этих импульсов только они и перестраиваются
и восстанавливают нарушенные функции, что все эти процессы происходят
без всякого или в лучшем случае без значительного участия в-ысших отделов
центральной нервной Системы. По нашим же данным и воззрениям, у выс-
ших животных импульсы, направляющиеся от рецепторов, наиболее эф-
фективно утилизируются именно высшими отделами центральной нервной
системы. Эти импульсы оказывают влияние на течение восстановительных
процессов именно через посредство верхних этажей центральной нервной
системы. Мы также считаем важной роль периферических рецепторов в этих
сложных сдвигах, но она все же является подчиненной. Периферическая
сигнализация стимулирует деятельность верхних этажей центральной нерв-
ной системы, вовлекает их в сложный процесс и обеспечивает их необходи-
мым «материалом» для выполнения ими своей ведущей роли в осуществлении
наиболее трудных перестроек функции нижележащих отделов центральной
нервной системы и в выработке новых высших видов интегративной деятель-
ности.
В восстановлении моторных функций деафферентнрованных конечно-
стей птиц, а тем более рептилий, рол ь периферических импульсов, равно
как и роль нижележащих отделов центральной нервной системы, воз-
растает, а роль верхних этажей центральной нервной системы ослабевает.
Это явствует, в частности, из данных наших сотрудников А. И. Карамяна,
Б. Д. Сгефанцова, В. Д. Дмитриева (1941) и др. Эта весьма интересные и
ценны? данные экспериментальной физиологии .говорят отнюдь не в пользу
теоретических положений Бете. Наоборот, они лишний раз доказывают
йнтиэволюционную и метафизическую сущность теории, которая при оцен-
ке роли и значения тех или иных физиологических факторов в восстано-
вительных процессах поврежденного организма совершенно игнорирует
значение эволюции структуры и функции различных систем организма и
механически ставит знак равенства между значением «верхних отделов»
центральной нервной системы различных животных для этих процессов,
без учета их места в эволюционном ряду.
ЛИТЕРАТУРА
Анохин П. К. и Шумилина А. И., Новые данные к механизму условного
рефлекса. 5-е совещание по физиологическим проблемам. Тезисы докладов, 92,
1939.
Асратян Э. А., Кора большого мозга и пластичность нервной системы. Вариант
ампутации лап. Успехи современной биологии, 5, 803, 1936.
СООБЩЕНИЕ IV. ОПЫТЫ С ДЕАФФЕРЕНТАЦИЕй КОНЕЧНОСТЕЙ
309
Асратян Э. А, Кора большого мозга и пластичность нервной системы. Опыты
с половинной перерезкой спинного мозга, Успехи современной биологии, 6,
451, 1937.
Асратян Э. А., О принципе относительной пластичности нервной системы. Тетра-
ди института им. Бехтерева, 11, 172, 1939.
Асратян Э. А., Кора большого мозга и пластичность нервной системы. Вариант
с перекрестным сшиванием нервов. Успехи современной биологии, 12. 516, 1940.
Дмитриев В. Д., Новые данные по эволюции пластичности нервной системы,
Советская невропсихиатрия. б, 493. 1941.
К а рам ян А. 11., К пластичности нервной системы птиц (опыты с ампутацией
лап), Бюллетень экспериментальной биологин и медицины, 6, 1938.
Карамян А. И., К проблеме компенсаторной деятельности центральной нервной
системы, Труды Института мозга им. Бехтерева, 14, Г8, 1941.
Орбели Л. А. и Ку ист май К. И., О последствиях деафферентании задних
конечностей у собак. Известия Института им. Лесгафта, 9, 18/', 1924.
Baglioni, Arch. Anat. u. Physiol. suppL, 71. 1907.
Baldi, цит. no Luciani «Human physiology», 3, London. 1915.
Bell H., Physiologische u, pathologUche U ntersuchungcn d. Nervensystcms, Ber-
lin. 1836.
Bernard Cl., l.e;on& sur la physiologic et pathologic du systems nerveux, Paris,
1858.
Bethe's Handb. d. norm. u. pathol. Physiol., 15, 2, 1931.
Bickel A., Untersuchungen Ober d. Mechanismus d. Nervenbewegungsregulatirn,
Stuttgart, 1903.
Magendie, Vorlesungen Ober d. Nervensystem u. seine Krankheiten, Leipzig, 1841.
Magnus К., Кбгр erst el lung, Berlin, 1924.
Merzbacher, Pfltig. Arch., 88. 453. 1901. 92. 585, 1902.
Munk, SitzungsbeHchte Preus. Akad. Wissensch., 48, 1903.
Trendelenburg N., Arch. Anat. u. Physiol.. I, 1906; 499. 1907; suppl. 201
1908.
КОРА БОЛЬШОГО МОЗГА И ПЛАСТИЧНОСТЬ НЕРВНОЙ СИСТЕМЫ1
Сообщение V
ОПЫТЫ С РАЗРУШЕНИЕМ ЛАБИРИНТОВ
олее века отделяет нас от первых экспериментов и наблюде-
ний Флуранса (1830) по физиологии лабиринтов; разрушая у
голубей отдельные полукружные каналы или все вместе, он
наблюдал серьезные и длительные нарушения моторных функ-
ций, в частности, нарушение равновесия тела как в покое, так
и при движениях. С того времени разработкой физиологии
и патологии вестибулярного аппарата и полукружных каналов лабирин-
тов занималось большое число физиологов и клиницистов: И. Ф. Цион
(1873), Гольц (Goltz, 1870), Эвальд (Ewald, 1892), В. М. Бехтерев. Магнус
и Клейн (Magnus и Kleyn, 1924), Шпигель (Spigel, 1926), Лоренто де Но
(Lorento de No, 1926) и др. Благодаря этим исследованиям, наши знания
о статических и статокинетических функциях лабиринтов значительно
продвинулись вперед, накопилось много ценных данных как о лабиринтах
в целом, так и о функциях различных структурных элементов этих аппа-
ратов со сложнейшей и тончайшей архитектурой. Несмотря на все эти
фактические достижения, послужившие основанием для создания ряда
новых теорий о функциях лабиринтов, основные факты, установленные
Флурансом, и его концепции о важном значении лабиринтов для осущест-
вления и поддержания равновесия тела до сих пор остаются незыблемыми.
Из фактических данных о физиологии и патологии вестибулярного ап-
парата и полукружных каналов лабиринтов нас ближайшим образом заин-
тересовали те, которые имеют прямое или косвенное отношение к проблеме
пластичности нервной системы- К этой проблеме лабиринты имеют двоякое
отношение.
Лабиринты как рецепторы статических и статокинетических рефлек-
сов принимают, несомненно, значительное участие в компенсации нарушен-
ных моторных и сензорных функций организма, обусловленных его природ-
ными дефектами или же повреждением моторных и сензорных систем. Как
на пример укажем на важную роль лабиринтов после перерезки задних ко-
решков первых трех пар шейных нервов. (Магнус и ван Левен (Magnus
и van Leeuwen, 1924)] после деафферентацим конечностей (Бикель
(Bickel, 1903)] и их ампутации (наши данные).
Однако в данной работе нас интересовал иной род отношения лабирин-
тов к проблеме пластичности нервной системы. Исследованиями Эвальда,
Магнуса и др. было показано, что многообразные, в том числе и тяжелые
последствия разрушения одного и двух лабиринтов—отведение и нистагм
глаз, искривление шеи и туловища, нистагм и качание головы, тенден-
k Физиологический журнал СССР, т. XXX11I, №3, 1947. П редваритсльное сооб-
щение опубликовано в «Бюллетене экспериментальной биологии и медицины», т. IX,
в. 5, 1940.
СООБЩЕНИЕ V. ОПЫТЫ С РАЗРУШЕНИЕМ ЛАБИРИНТОВ
3! I
ция к круговым движениями к верчению, неправильная ориентация по от-
ношению к силе земного притяжения, нарушения функций стояния и
ходьбы и т. п. — со временем проходят, т. е. более или менее совершенно
компенсируются. В данном случае нас интересовал именно этот род отно-
шения лабиринтов к проблеме пластичности1.
Что же нам известно о природе этих компенсаторных явлений, какие
рецепторы и отделы центральной нервной системы и какое участие прини-
мают они в этих своеобразных приспособительных явлениях?
Ценные данные по этим вопросам были получены Эвальдом (1892).
Разрушая лабиринты у лягушек, голубей и собак, Эвальд наблюдал, что
хотя нарушения функций у лягушек бывают неглубокими, но они у них
почти не компенсируются; у голубей нарушения бывают глубокими, но со
временем они довольно хорошо компенсируются; у собак нарушения
бывают еще более глубокими, но зато они быстрее и компенсируются, чем
у голубей. Анализируя механизм этих «явлений замещения», Эвальд обнару-
жил, что удаление переднего мозга у таких лягушек почти не вызывает из-
менений их моторной деятельности, удаление переднего мозга у голубей
очень слабо отражается на компенсации лабиринтных нарушений. Что же
касается удаления одних только «возбудимых зон» коры больших полу-
шарий мозга, то у собак удаление этих зон вызывает почти полное исчез-
новение существовавшей компенсации лабиринтных нарушений. По данным
Эвальда, однако, эти возвратные нарушения у собак со временем вторично
компенсируются благодаря оптическим рецепторам (завязывание глаз
ослабляет эту новую компенсацию, точно так же как содержание собак
в темной комнате замедляет наступление вторичной компенсации).
Своим данным Эвальд дает следующее объяснение. Вследствие разруше-
ния лабиринтов животное лишается мышечного чувства, ослабевает сила
и точность движений и тем самым обусловливается вся известная картина
нарушений. Отсутствие мышечного чувства со временем замещается дея-
тельностью рецепторов осязания, и нарушения компенсируются. У собак
удаление «возбудимых зон» коры вызывает исчезновение компенсации, так
как эта операция выключает также и чувство осязания. Некоторое улучше-
ние состояния после этого объясняется вовлечением в процесс зри-
тельного рецептора. Ничтожные или слабые последствия удаления перед-
него мозга у лягушек и голубей Эвальд объясняет тем, что у них при этом
чувство осязания не страдает или страдает в слабой степени.
Дальнейшим детальным анализом явлений компенсации лабиринтных
нарушений занялся Магнус со своими учениками. По Магнусу, компенса-
ция лабиринтных нарушений осуществляется в основном тремя путями;
путем так называемой бехтеревской центральной компенсации, путем дуб-
лирования и «компенсаторного' усиления» шейных установочных рефлексов,
а также установочных рефлексов с тела на тело н с тела на голову и, нако-
нец, у высших млекопитающих (кошек, собак, обезьян) — путем опти-
ческих установочных рефлексов.
Бехтсревская центральная компенсация происходит очень быстро —
спустя 4 дня после операции. Когда некоторое время спустя после разру-
шения одного лабиринта разрушается второй лабиринт, у животно го прояв-
ляется ряд асимметричных функциональных нарушений, зеркально-проти-
воположных первоначальным. Иначе говоря, создается впечатление, будто
первый лабиринт вовСе не был разрушен. Первоначальные предполо-
жения, будто эту компенсацию осуществляет мозжечок, впоследствии не
нашли подтверждения. Магнус отметил, что компенсация эта происходит
1 В своих обзорных статьях о пластичности нервной системы Бете (BetJie,
1931) обходит молчанием как эту обширную область явлений, так и другие явления
нарушения и восстановления функции организма, хотя их принадлежность к про-
блеме пластичности не вызывает сомнений.
312
Э. А. АСРАТЯН
у децеребрированных животных даже после экстирпации мозжечка и дсаф-
ферентации первых трех пар шейных нервов; Шпигель и Деметриадес
(Spigel и Detnetriades, 1925) показали, что она может осуществляться даже
областью вестибулярных ядер продолговатого мозга.
Компенсация, благодаря неповрежденным шейным установочным рефлек-
сам, а также благодаря установочным рефлексам с тела на тело и с тела на
голову, имеет более медленный характер и происходит, по мнению Магнуса,
в силу того, что указанные рефлексы и лабиринтные установочные рефлек-
сы в норме частично друг друга дублируют, а также потому, что после вы-
ключения лабиринтов сохранившиеся установочные рефлексы компенсатор-
но усиливаются. Как известно, все эти установочные рефлексы осуществля-
ются средним мозгом. Происходит ли «компенсаторное усиление» ука-
занных рефлексов за счет одних только ресурсов самого среднего мозга
или же в этом принимают участие верхние этажи центральной нервной
системы, — вопрос этот не был поставлен и изучен Магнусом. У 2 кошек
с удаленными полушариями Магнус и Дюссе де Барен (Dusser de Barenne)
поочередно разрушали оба лабиринта и удовлетворились лишь констата-
цией у них установочных рефлексов с тела на тело, с тела на голову
и шейных рефлексов. Вскоре после лабиринтной операции (3 и 7 дней спустя)
животные были убиты.
Особое место занимают оптические установочные рефлексы- Благодаря
им, высшие животные компенсируют не только последствия разрушения
лабиринтов, но и дополнительные последствия перерезки задних корешков
первых трех пар шейных нервов. Эта компенсация происходит также весьма
постепенно и медленно, как отметил еще Эвальд и подтвердил Магнус.
Далее, в полном согласии с Эвальдом, Магнус наблюдал, что, чем выше стоит
животное по уровню филогенетического развития, тем быстрее и полнее
происходит компенсация лабиринтных нарушений. На основании своих
наблюдений Магнус сделал вывод, что оптические установочные рефлексы
осуществляются корой больших полушарий мозга.
Следует сказать, ЧТО и после работ Магнуса и его учеников по компенса-
ции лабиринтных нарушений многое в этой области осталось невыясненным.
МагнуС (1924) так резюмирует свои данные: «В заключение следует еще раз
подчеркнуть, что я не думаю, что этим исчерпал все компенсаторные воз-
можности; вне всякого сомнения, дело обстоит значительно сложнее, чем
до сих пор представляется»1. Представляют интерес предположения Магнуса
о том, что, помимо осуществления оптических установочных рефлексов, кора
больших полушарий мозга высших млекопитающих участвует в компенса-
торных явлениях также другими своими частями. В то же время Магнус
пишет: «Однако для этого мы до сих пор не имеем никаких доказательств».
Больше того, он вообще считает, что «анализ этих компенсаторных процес-
сов до сих пор еще не проведен до конца. До сих пор еще не выяснено,
каким образом кора большого мозга в целом и посредством каких корковых
зон действует на различные центры в стволовой части мозга, ведающие уста-
новкой тела. До сих пор такое воздействие выявилось лишь при оптиче-
ских установочных рефлексах»2.
Таким образом, вопрос о природе и факторах компенсации лабиринт-
ных нарушений, в частности, о роли коры больших полушарий мозга в этих
своеобразных приспособительных явлениях, нельзя было считать достаточно
выясненным. Фактические данные Эвальда (оставляя в стороне его неудов-
летворительные теоретические концепции), а также данные Магнуса явля-
лись только предпосылкой дин дальнейшей экспериментальной и теорети-
ческой разработки рассматриваемого вопроса. В общей связи с изучаемой
1 R. Magnus, К orperst ellung, 1J2+, S., 393.
’Там же, стр. 389.
СООБЩЕНИЕ V. ОПЫТЫ С РАЗРУШЕНИЕМ ЛАБИРИНТОВ
sis
мной и моими сотрудниками проблемой пластичности нервной системы
в условиях оперативного повреждения тех или иных систем организма мы
занялись также исследованием компенсаторных явлений, возникших после
разрушения лабиринтов, в частности, изучением роли коры больших полу-
шарий мозга в этих явлениях.
Экспериментальная часть
Эксперименты были произведены на собаках в течение 1636 — 1940 гг.
В главной серии экспериментов у собак сначала разрушали — одномо-
ментно или двухмоментно—оба лабиринта по методу Магнуса-Клей-
на, а затем, после завершения процесса компенсации лабиринтных нару-
шений, производили двухмоментную экстирпацию всей коры больших по-
лушарий мозга. В другой серии экспериментов эти разнотипные опера-
ции были произведены в обратной последовательности, т. е. сначала
декортикация большого мозга, потом—разрушение лабиринтов. После
каждой операции у животных систематически исследов.ались различные
рефлексы и функции. Характерные фазы снимались на кинопленке.
Из собак, специально оперированных для выполнения поставленной
задачи, ценный материал для наблюдений дали 5 собак, из них у 4
сначала удаляли лабиринты, а у одной — вначале производили декорти-
кацию (см. таблицу).
Кличка Сфба- КМ И При б 1И зи ТЫЛЬНЫЙ бОЭрЭСт Дата одномо- ментного раз- рушения Лабиринтов Даты дауц. ' момент кого разрушь ния ЛАбМрИМТОб Дата экстир- пации коры одного полу- шария Дата экс- тирпации коры дру- гого полу- шария Продол- житель- ность жиз- ни собак после пос- ледней о перации При мечания
Жулик 11 (молодая) Орлик (ста- рая) Лисичка (5—6 ме- сяцев) В а гр (моло- дая) Дружок (молодая) I.VI11 1936 г. IS. VII 11936г. 10. VI 3940 г. 10.III и 20.1 II 1937 г. 4.11 И 13.11 1937 г. 11. X 1936 г. 15 X 1936 г. 8. IV 1937 г. 4.111 1937 г. 19. VI 1938 г. 25.1 1937 г. 25,1 1937 г 5.V 1937 1. 5.V 1937 г. 21. V1939 г. 8 дней 10 » 25 » 580 » Около 800 дней Погибла от нагноения То же Погибла от п ролежней Погибла от нифекиии Собака жи- ла до осе- ни 1941 г.
Картина изменений после разрушения одного или двух лабиринтоя
у первой группы собак ничем особенным не отличалась ст общеизвест-
ной картины лабиринтных нарушений, описанной многочисленными ис-
следователями. Совершенно так же обстояло дело с ходом и характером
компенсации этих нарушений. Исходя из этого, считаем излишним давать
здесь описание картины лабиринтных нарушений и процесса их компен-
сации у наших подопытных собак. Хотелось только отметить ряд особен-
ностей этой картины, представляющих интерес для главной цели нашей
работы.
Компенсация лабиринтных нарушений носи г медленный и строго посте-
пенный характер- Самое поверхностное наблюдение не оставляет сомнений,
что механизм «научения» играет значительную роль в этом сложном про-
цессе. Далее, не только в различных фазах процесса компенсации, но
даже после ее завершения наблюдалось следующее явление: при быстрой
ходьбе и в особенности при беге животные постоянно смотрят то вправо
31»
Э. Л. АСРАТЯН
то влево, продвигаясь вперед по зигзагообразной траектории. Следует так-
же отметить факт дезориентации и оживления лабиринтных нарушений
у собак с законченным циклом компенсации, когда они оказываются в но-
вой, непривычной ситуации (преодоление препятствий, ходьба по ослож-
ненной дороге и т. п.). Быстрота наступления компенсации и степень ее
совершенства зависят, по нашим данным, не только от уровня развития жи-
вотного в филогенетическом ряду, как было показано Эвальдом и Магну-
сом, но и от ряда других факторов: от возраста, роста и веса животных.
Чем моложе собака (в известных пределах), чем ниже рост и меньше вес,
тем быстрее наступает компенсация я тем совершеннее бывает она. В об-
щем после разрушения одного лабиринта нарушения более или менее хо-
рошо компенсируются Спустя 4—10 Дней со времени операции, а после
двухмоментного н одномоментного разрушения обоих лабиринтов — че-
рез 8—20, а иногда и больше дней с момента операции.
Удаление коры одного полушария головного мозга у первых четырех
собак, перечисленных в таблице, было произведено после того, как у живот-
ных хорошо компенсировались нарушения. Краткая характеристика
последствий декортикации одного полушария следующая. В первые дни
после операции животные делаются очень беспокойными, у них бывают
частые приступы бурных хаотических движений всего тела, в особенности
конечностей. В слабом виде появляется нистагм глаз, нистагм и качание
головы. Животные сильнее искривляют позвоночный столб, чем нормаль-
ные животные, лишенные коры одного полушария; предпочитают лежать
на спине, иногда ложатся также на бок, противоположный той стороне,
где было оперировано полушарие. На другом боку совсем не лежат. При
укладывании на бок, соответствующий стороне операции, они сильным
рывком моментально поворачиваются на спину или на противоположный
бок. Часто во время приступа бурных движений животные совершают спи-
ралеобразные круговые движения- вокруг длинной оси тела. Долгое время
после операции они не в состоянии бывают сидеть, а тем более стоять и
ходить.
Довольно быстро затихают и исчезают нистагм головы и глаз, а также
приступы бурных движений. Животные все еще лежат на боку, противо-
положном той стороне, где было удалено полушарие, но с наклоном к
вентральной стороне. На той стороне тела, на которой была произведена
операция, они все еще не лежат. Благодаря постепенному улучшению со-
стояния, примерно через 15—20 дней, очень молодые животные (5—6 ме-
сяцев) могут сидеть и, покачиваясь, недолго стоять на широко расставлен-
ных и согнутых конечностях, сгибая туловище и шею в ту сторону, где
была произведена операция. В этой стадии они еще не могут ходить: при
попытке ходить сразу же падают на землю. У старой собаки данная стадия
наступает через 40—45 дней после операции; у собак среднего возраста —
через 20—30 дней.
Дальнейшее постепенное улучшение приводит к тому, что животные
начинают стоять все увереннее, при этом не так широко расставляют ко-
нечности и выпрямляют их, меньше качаются при стоянии; постепенно
восстанавливается симметрия тела при лежании и стоянии; животные мед-
ленно и осторожно ходят, покачиваясь из стороны в сторону. В начале этой
стадии походка у них бывает весьма шаткой; они часто падают— либо из-
за того, что конечности их цепляются одна за другую, либо из-за слабой
опорной реакции конечностей. Со временем исчезают также эти дефекты
ходьбы, но манежные движения в. сторону, где была произведена операция,
сохраняются еще долго.
Лисичка (5—6 месяцев) этот цикл завершила примерно через месяц, но
Орлик (10—12 лет) завершил его через 3 месяца, а остальные собаки мо-
лодого и среднего возраста — через Р/г—2 месяца.
СООБЩЕНИЕ V. ОПЫТЫ С РАЗРУШЕНИЕМ ЛАБИРИНТОВ
315
Таким образом, можно сказать, что экстирпация коры одного полуша-
рия головного мозга у собак с разрушенными лабиринтами и компенси-
рованными нарушениями вызывает возвращение основных лабиринтных
нарушений, которые со временем постепенно вновь компенсируются, при-
чем, как показывают наблюдения, эта вторичная компенсация происходит
также постепенно и имеет более замедленный темп, чем первичная ком-
пенсация. Судя по всему, значение оптического анализатора в этом новом
«научении» сильнее, чем в «научении» после разрушений лабиринтов. В
начальных стадиях вторичной компенсации завязывание глаз собакам
совершенно дезориентирует и дезорганизует их, а в стадии более совершен-
ной компенсации завязывание глаз хотя и не приводит собак в состояние
полной растерянности и беспомощности, но тем ие менее оказывает силь-
ное влияние на функцию стояния и в особенности ходьбы.
Экстирпация коры второго полушария головного мозга приводит к но-
вой и более резко выраженной декомпенсации лабиринтных нарушений,
чем экстирпация коры одного полушария. Животные лежат на спине, ред-
ко— на боку, противоположном той стороне тела, где в последний раз
оперировали полушарие. В лежачем положении животные изгибаются,
вернее, скручиваются в эту сторону. Вновь появляются — в слабо выра-
женном виде— нистагм глаз, нистагм и качание головы. Бурные приступы
резких динамических явлении (верчение вокруг длинной оси тела, хаоти-
ческие движения конечностей, головы, туловища) по силе и частоте зна-
чительно превосходят таковые после удаления первого полушария.
Две Собаки (Жулик и Орлик) погибли спустя 8 и 10 дней после опе-
рации от нагноения операционного поля. У третьей собаки (Лисички),
у которой приступы были особенно частыми и бурными, началось изъязвле-
ние конечностей, и она погибла примерно месяц спустя после операции
при обширных воспалительных явлениях на конечности и сильной ка-
хексии.
Четвертая собака этой группы (Вагр) жила после операции 1 год 7 меся-
цев 5 дней и погибла от случайной инфекции. Собака эта была молодой,
здоровой и живой; лабиринтные нарушения компенсировала хорошо;
удовлетворительно перенесла удаление коры одного полушария и хорошо
компенсировала также вторичные нарушения. Удаление коры второго по-
лушария вызвало все типичные нарушения, о которых шла речь выше:
бурные приступы, вращательные движения и т. д. Несмотря на царапинки,
появившиеся на конечностях в результате хаотических движений, общее
состояние этой собаки быстро улучшилось; животное стало попра-
вляться, ликвидировались побочные последствия операций. Примерно
с конца первого месяца после операции Вагр стал менее часто и бурно вер-
теться и биться, постепенно у него стало восстанавливаться нормальное по-
ложение тела при лежании; он лежал как на одном, так и на другом боку.
Примерно через два-три месяца после последней операции почти пол-
ностью исчезли нистагм глаз и головы, а также другие мелкие лабиринт-
ные нарушения. Вагр спокойно лежал в люльке, не вертясь и не совершая
хаотических движений, если не было внешнего повода. На полу, однако,
Вагр попрежнему вертелся и бился. Во время этих припадков он делал
даже попытки стоять, которые иногда ему удавались: он стоял па четырех
широко расставленных, согнутых конечностях; при этом голова собаки
была опущена вниз, позвоночник сильно сгорблен, тело покачивалось
па дрожащих конечностях. Такое состояние длилось несколько секунд
(максимум 10—15). Достаточно было собаке произвести малейшее движе-
ние конечностями, и она падала на пол, кувыркаясь через голову, вновь
начинала вертеться, биться и делать хаотические движения.
Такое состояние Багра оставалось почти неизменно до конца его жиз-
ни. Легкие и побочные явления лабиринтных нарушений миновали, но на-
316
Э А. АСРАТЯН
всегда сохранились хаотические движения, неспособность к стоянию и
ходьбе, склонность к вращательным движениям.
Как на Лисичке, так и на Вагре были сделаны специальные пробы на
наличие и живость установочных шейных рефлексов, рефлексов с тела на
тело и с тела на голову. Все эти рефлексы у данных собак были выражены
достаточно хорошо.
Пятая наша собака (Дружок) также была молодой и здоровой. Сначала
двухмоментной операцией была экстирпирована у нее вся кора больших
полушарий мозга, а год спустя был разрушен один правый лабиринт.
Последствия разрушения лабиринта у этой собаки в первые дни почти ни-
чем существенным не отличались от последствий разрушения лабиринта
у нормальной собаки. Наблюдались следующие явления: искривление
шеи и наклон темени в сторону разрушения, наклон пасти в обратную сто-
рону, понижение тонуса правых конечностей и повышение тонуса левых,
сгибание туловища в сторону разрушения, качание головы, нистагм глаз и
головы, хаотические движения; собака вертелась вокруг длинной оси тела
в направлении часовой стрелки (т. е. со стороны здорового лабиринта в сто-
рону разрушенного), неспособна была лежать на левом боку, а такж;е стоять
и ходить. Через несколько дней так называемые явления раздражения
(нистагм, бурные движения) заметно ослабели, а 10—15 дней спустя поч-
ти исчезли. Собака стала слабее и менее часто вертеться вокруг длинной
оси своего тела и меньше качать головой. Глубокие же выпадения функции
претерпевали незначительные изменения и в ярко выраженной форме
держались почти два года (собака погибла в период блокады Ленинграда).
Собака почти все время лежала. Голова была опущена книзу и находилась
в резко асимметричном положении, темя оставалось наклоненным в пра-
вую сторону, шея и туловище были согнуты, левые конечности (в особен-
ности передняя) находились большей частью в состоянии экстензии, пра-
вые— в состоянии флексии. Качание головы в слабом виде сохранилось,
изредка собака вертелась. Сна лежала либо на животе, либо на правом боку;
на левом боку совершенно не лежала; если же она случайно оказывалась
на левом боку, то резким движением моментально поворачивалась на жи-
вот или, еще чаще, минуя эту фазу, — на правый бок. Временами собака
спиралеобразно вертелась в течение нескольких минут.
Еще в конце первой недели после лабиринтной операции собака Дру-
жок делала попытки стоять; эта тенденция постепенно усилилась, в конце
месяца приняла более или менее постоянный характер и в этом виде сохра-
нилась до конца наблюдения за ссбакой. Когда Дружок вертелся, то оказы-
вался иногда на четырех неравномерно тонизированных конечностях с на-
клоном туловища вправо; при этом он несколько секунд покачивался,
кувыркался через голову и падал на левый бок, не делая пи шага либо
Делая один или два неуверенных шага только передними конечностями.
Падая иа землю, Дружок продолжал вертеться; движения собаки — до
нового перехода в позу стояния — были хаотичными.
В то время как Функции стояния и ходьбы у Дружка оставались глубоко
нарушенными, установочные рефлексы с толана голову, с тела на тело,
а также шейные и лабиринтные установочные рефлексы с левой стороны
были резко выражены. С правой же Стороны, т. е. со Стороны разрушенного
лабиринта, оставшиеся установочные рефлексы были выражены очень сла-
бо, часто их даже не удавалось обнаружить.
Некоторый материал о роли коры больших полушарий мозга в. компен-
сации лабиринтных нарушений был подучен также на 8 собаках, опери-
рованных нами для специальной работы нашего сотрудника Я. М. Прес-
смана. У этих собак была удалена кора одного полушария и на противо-
положной стороне разрушены глаз и лабиринт. Собаки жили 1—3 года,
и асимметричное положение головы (наклон в сторону поврежденных ре-
СООБЩЕНИЕ V ОПЫТЫ С РАЗРУШЕНИЕМ ЛАБИРИНТОВ
317
цепторов) сохранялось почти все время в резко выраженном виде. Конт-
рольные опыты показали, что эта асимметрия обусловливается не удале-
нием глаза, а комбинированным повреждением лабиринта и противополож-
ного полушария.
Обсуждение полученных данных
Нам кажется, что приведенным фактический материал даст достаточ
ное основание для заключения, что в компенсации глубоких лабиринтных
нарушений у собак кора больших полушарий мозга играет решающую
роль и что последняя не исчерпывается только осуществлением опти-
ческих установочных рефлексов. Догадки Магнуса о возможности более
широкого участия коры больших полушарий мозга в этих компенсатор-
ных явлениях находят экспериментальное подтверждение в наших дан-
ных. Совершенно очевидно, что возникающее после разрушения лабирин-
тов у нормальных животных явление, названное «компенсаторным усиле-
нием» неповрежденных установочных рефлексов, обусловливается в
основном также деятельностью коры больших полушарий мозга. Нашими
экспериментальными данными устанавливается Следующее: у таких жи-
вотных именно благодаря коре больших полушарий усиливается вся гамма
неповрежденных установочных рефлексов, осуществляемых средним
мозгом; благодаря коре вСе эти рефлексы в совокупности подвергаются
такой высшей и совершенной интеграции, что делается возможной почти
полная компенсация глубоких лабиринтных нарушений. Без коры сред-
ний мозг не в состоянии осуществить такой высший анализ и синтез свой-
ственных ему рефлексов- Для этого оказывается недостаточным наличие
межуточного мозга, равно как и наличие некоторых уцелевших от опе-
раций и дегенерации подкорковых нервных образовании.
Данные Эвальда относительно временного исчезновения компенсации
лабиринтных нарушений после удаления «возбудимых зон» коры большого
мозга у собак, данные Эвальда и Магнуса о роли оптических установоч-
ных рефлексов в компенсации нарушений и относительно характера их
усиления, а также предположения Магнуса о корковом происхождении
этих рефлексов явно противоречат данным и концепциям Бете о пластич-
ности нервной системы, в частности, о роли больших полушарий мозга
в. компенсации нарушений, вызванных повреждением различных систем
организма. Правда, Бете, как уже отмечалось,совершенно не упоминает
об этих фактах в своих обзорах, но это не является препятствием для
установления противоречия между его теорией и этими давно извест-
ными точными фактами.
Результаты исследований Эвальда и Магнуса не могли считаться
безупречными для решения вопроса о роли ковры больших полушарий
мозга в компенсации лабиринтных нарушений. Удаление «возбудимых
зон» коры у собак с удаленными лабиринтами вызывало лишь временное
исчезновение компенсации, а предположение Магнуса об участии коры в
компенсации лабиринтных нарушений посредством оптических установоч-
ных рефлексов и еще какими-то другими путями носили гипотетический
характер и не были строго аргументированы прямыми экспериментами.
Мы имеем, как нам кажется, основание утверждать, что наши данные
относительно полной экстирпации коры больших полушарий мозга у собак,
лишенных одного или двух лабиринтов-, позволяют сделать совершенно
определенное заключение о ведущей, определяющей роли верхних этажей
центральной нерв.ной системы в явлениях компенсации лабиринтных нару-
шений у высших взрослых животных. Эти новые данные, полученные
нами, полностью гармонируют с теми данными, которые были нами опуб-
ликованы в предыдущих сообщениях по этой проблеме. Тем самым допол
нительно подкрепляется развиваемое нами теоретическое положение о
318
Э А.АСРАТЯН
важнейшей роли верхних этажей центральной нервной системы в при-
способительных явлениях особого типа, возникающих в результате серьез-
ных повреждений различных систем организма.
Как понять наблюдавшиеся у наших собак сдвиги в сторону некоторого
смягчения картины нарушения функций после полной декортика инн го-
ловного мозга у животных с разрушенными лабиринтами или после разру-
шения лабиринтов у лишенных коры животных?
Совершенно очевидно, что значительная часть этих сдвигов является ре-
зультатом постепенного ослабления и исчезновения оперативного раздра-
жения (травма, кровотечение, дегенеративные процессы и т. д.) и вызван-
ного им торможения в центральной нервной системе. Оно обусловливает
многие из компонентов нарушения функции, наблюдаемые в первые дни
и недели после операции, и тем самым осложняет и запутывает подлинную
картину выпадения функции. Такое объяснение, вообще говоря, не являет-
ся принципиально новым, оно давно существует в физиологии. Затем, как
уже отмечалось в наших предыдущих статьях, мы допускаем, что свойство
пластичности в различной степени выраженности присуще в.сем частям
нервной системы, — в верхних этажах оно достигает лишь высшего уровня
развития. Мы допускаем также, что ничтожные нарушения функции могут
быть полностью компенсированы средними и низшими отделами цент-
ральной нервной системы. Более того, даже нарушения средней и сильной
степени в какой-то мере могут ими компенсироваться. Степень этой ком-
пенсации зависит от глубины повреждения, возраста животного, от по-
ложения животного в филогенетическом ряду и т. д. С этой именно точки
зрения мы объясняем ничтожные сдвиги в картине нарушений у наших
лишенных керы собак уже в поздний период после операции.
Эвальд удалял весь передний мозг голубей с разрушенными лабирин-
тами и не наблюдал глубоких изменений в существовавшей картине компен-
сации. Как понять расхождение между нашими описанными выше данными
и между этими данными Эвальда?
С точки зрения развиваемой нами эволюционной теории пластичности
нервной системы, мы не видим здесь никакого противоречия. Наши сотруд-
ники (Л. И. Карамяи, D. Д. Стсфанцов, В. Д. Дмитриев, Я. М. Прессман,
Г. И. Боков и др.) получили компенсацию функций после различного рода
повреждений нервной системы и других органов. По этим данным, перед-
ний мозг у голубей играет важную, но нс решающую роль в этих компен-
саторных явлениях, как, например, у собак. Это говорит об относительно
меньшем удельном весе переднего мозга и относительно большем удельном
весе нижележащих частей центральной нервной системы у голубей (по срав-
нению с удельным весом тех же частей центральной нервной системы у со-
бак) не только в нормальных приспособительных явлениях, по и в приспо-
собительных явлениях особого характера, возникающих после различного
рода повреждений тех или иных систем организма.
Во избежание повторения мы здесь не останавливаемся на нашем по-
нимании природы и механизмов этих приспособительных явлений. Мы
подробно писали об этом в уже опубликованных нами работах (Асратян,
1937—1941). Хотелось только отметить следующее. Судя по всему, основ-
ной тип деятельности верхних этажей центральной нервной системы, т. е.
условнорефлекторная деятельность, при компенсации лабиринтных наруше-
ний выявляется более отчетливо, чем при компенсации нарушений после
ампутации лап, гсмисекцнн спинного мозга, перекрестного сшивания
нервов и т. п.
Однако и в компенсации нарушенных лабиринтных функций верхние
этажи центральной нервной системы (особенно кора мозга) участвуют
также посредством трофического воздействия на нижележащие отделы и
посредством высшей безусловнорефлекторной интеграции их функций.
СООБЩЕНИЕ V. ОПЫТЫ С РАЗРУШЕНИЕМ ЛАБИРИНТОВ
319
ЛИТЕРАТУРА
Асратян Э. А., Кора большого мозга и пластичность нервной системы, Успехи
современной биологии, т. V. в. 5, 803, 193*5.
Асратян Э. А., О принципе относительной пластичности нервной снсте* и,
Груди Института мозга нм. Бехтерева, 11, 172, 1933.
Асратян Э. А., Кора большого мозга и пластичность нервной системы, Бюл-
летень экспериментальной биологии и медицины, 8, 419, 1939; 9, 27, 1940.
Асратян Э. Л., Кора большого мозга и пластичность нервной системы, Успехи
современной (пологий, 12 , 516, 1940.
Асратян Э. А., Новые данные по физиологии большого мозга, Архив биологиче-
ских наук, 61, 54, 1941.
Бехтерев В. М., Результаты перерезки слухового нерва, Меднтиекая библио-
тека. декабрь, 1882.
Бехтерев В. М., Основы учения о функциях мозга, IV, 1905.
Цион Е., Pfliig. Arch., 8, 306, 1873.
Bcthe's Handb. d. nortn. u. pathol. Physiol., 15, 11, 1931.
Bic kel A., Untersuchungen fiber den jVechanismus des nervftsen Bewegungsregula-
iion. Stuttgart, 1903.
Cam is M., 1 he physiol, of the vestibular apparattus, Oxford, 1930.
Ewald R., Physiologische Untersuchungen Ober das Endorgan des Ncrvus Octavos,
Wiesbaden. If92.
Flo lire ns M., Mem. de I’Acad. roy. des S-ci. de Paris, 9, 455, 1830.
Goltz F., Pfltig. Arch., 3, 172, 1870.
Lo rente de No R., Acta oto-lar., 9, 163, 1926.
Magnus R,, KOrperstellung, Ilerlin, 1924,
Spiegel F... Pflug. Arch., 215, 106. 1926.
Spiegel E. u. Demctriades, Pflog. Arch., 210, 215, 1925.
ОПЫТ
I Г 1
м
КОРА БОЛЬШОГО МОЗГА И ПЛАСТИЧНОСТЬ НЕРВНОЙ СИСТЕМЫ!
Сообщение VI
Ы С ПЕРЕРЕЗ.КОЙ ЗАДНИХ. КОРЕШКОВ ПЕРВЫХ ТРЕХ ПАР
ШЕЙНЫХ НЕРВОВ
ногочисленные клинические наблюдения, а также эксперимен-
тальные исследования, выполненные рядом физиологов, в част-
ности, И. М. Сеченовым (1863), Гольцем (Goltz, 1874), Шер-
рингтоном (Sherrington, 1906), И. И. Красногорским (1911),
Л. А. Орбели и К. И. Кунстман (1924), Магнусом (Magnus,
1924) и 10. М. Конорским и С. М. Миллер (1936), установили
важное значение проприоцептивных нервов не только в вызывании и под-
держании тонуса скелетных мышц, но и в осуществлении центральной
нервной системой различных видов простой и сложной координационной
деятельности. Представляют известный интерес исследования Магнуса и
его учеников относительно роли проприоцептивных, нервов в осуществ-
лении и поддержании так называемых «статических рефлексов» (рефлексы
позы или стояния и установочные рефлексы). Среди этих разнотипных
рефлексов особое место занимают выпрямительные и установочные шей-
ные рефлексы. В установлении и поддержании различных положений тела
и отдельных его частей огромную роль играют шейные мышцы. Они яв-
ляются прежде всего первичным, самостоятельным источником возник-
новения рефлексов позы и установочных рефлексов; они представляют
собой передаточное звено части лабиринтных и установочных рефлексов
на туловище и конечности, а также установочных рефлексов с туловища
на голову. Из большого материала, накопленного по этому вопросу, вы-
текает, что проприоцептивные импульсы, идущие по задним корешкам
первых трех пар шейных нервов, имеют гораздо большее значение для
упомянутых интегративных функций нервной системы, чем проприоцеп-
тивные импульсы из других частей тела.
Эту важную роль и специфические функциональные особенности зад-
них корешков первых трех пар шейных нервов Магнус и его ученики
изучали в условиях как острых, так и хронических экспериментов. Имея
в виду изучаемую нами проблему пластичности нервном системы, осо-
бый интерес представляют явления компенсаторного характера, наступаю-
щие после перерезки задних корешков первых трех пар шейных нервов
у животных.
Магнус и Ш гром ван Левен (Strom van Leeuwen. 1914) у пяти кошек про-
извели перерезку афферентных корешков первых трех пар шейных нервов.
Характерно, что такая деа(|ферентация не вызвала паралича или даже
атаксии шейных мышц. Животные способны были осуществлять разные
движения головой С первых же Дней операций. Между тем. Известно, чТо
1 Физиологический журнал СССР, т. XXXIV, № 2, 1948.
СООБЩЕНИЕ VI. ПЕРЕРЕЗКА ЗАДНИХ КОРЕШКОВ ШЕПНЫХ НнРвОВ
321
деафферептация других областей тела вызывает временный паралич соот-
ветствующих скелетных мышц, а постепенное восстановление их функций
через фазу атаксии требует длительного времени. Эта интересная осо-
бенность шейных мышц—почти нормальное функционирование с первых
же дней деафферентации, — по Магнусу, обусловливается специфиче-
ской и тесной функциональной связью лабиринтов с этими мышцами.
В нормальных условиях шейные мышцы находятся под воздействием
многих рецепторов; среди них основными являются два — собственные и
лабиринтные, которые в значительной мерс дублируют друг друга и без
длительной задержки и затруднения замещают друг Друга при выпадении
одного из них.
В то время как функциональные отправления шейных мышц заметно
не нарушаются при перерезке задних корешков I, II и III шейных нервов,
функции мышц конечностей Значительно нарушаются. Особенно ясно это
выявляется в первые дни после операции. Хотя некоторая неловкость дви-
жений передних лап кошек в опытах Магнуса проходила уже в первые дни
после операции, тем не менее у животных в специальных ситуациях на-
блюдался ряд нарушений функций передних конечностей, оставшихся
довольно долго (неделями). Так, например, при подъеме головы не вызы-
вались (или вызывались в слабой форме) разгибательные рефлексы перед-
них конечностей, а при опускании головы не вызывались (или вызывались
в слабой форме) их сгибательные рефлексы. Нарушения же функции
задних конечностей у оперированных Магнусом кошек имели более
ярко выраженный характер и длились еще дольше: конечности оставались
согнутыми как при стоянии, так и при ходьбе; задняя часть тела при ходьбе
волочилась по земле; животные шатались во время бега и прыжка, пока-
чивались и т. п. Эти дефекты, однако, со временем постепенно сглаживались
и исчезали.
Отсутствие заметных нарушений функций шейных мышц после их деаф-
ферентации Магнус объясняет наличием дублирующего сензорного меха-
низма— лабиринтов. Нарушения же функций передних и задних конеч-
ностей, обусловленные перерезкой задних корешков первых трех пар шей-
ных нервов, компенсируются, по Магнусу, усилением лабиринтных,
а также установочных рефлексов с тела на тело. На какие-нибудь другие
факторы, участвующие в компенсации этих нарушений, Магнус не указы-
вает.
Сточки зрения нашей концепции о роли коры больших полушарий мозга
в разнородных компенсаторных явлениях в организме представляло боль-
шой интерес экспериментал ыюе выяс лен ие вопроса, участвует л и кора боль-
ших полушарий мозга в явлениях компенсации нарушений, наступающих
после перерезки задних корешков первых трех пар шейных нервов, и
каков удельный вес этого участия. Некоторым основанием для допущения,
что кора больших полушарий мозга участвует в этих компенсаторных уси-
лениях, мог служить медленный и постепенный характер наступления
компенсации. Но это не могло еще считаться решением вопроса. Вероят-
ность такого предположения значительно усилилась после того, как нами
экспериментально было показано значение коры больших полушарий мозга
в компенсации нарушений, проявляющихся после других видов повреж-
дений периферических сензорных и моторных систем, а также центральной
нервной системы; ампутация лап, гемисекция спинного мозга, перекрест
нервов, разрушение лабиринтов, деафферентация конечностей. Но в дан-
ном случае установление аналогии не могло считаться решением вопроса.
Экспериментедьной разработкой этого вопроса мы занялись в 1938—
1939 гг. в Физиологическом институте им. акад. И. П. Павлова Акаде-
мии наук СССР и В Отделе физиологии центральной нервной системы
Института мозга им. В. М. Бехтерева.
21 Э. А. Асратян
322
Э. Л. АСРАТЯН
Экспериментальные данные
Вначале под нашим наблюдением было несколько кошек, о-пернрован-
ных нами большей частью без соблюдения асептики, с целью лишь практи-
ческого освоения техники операции. Перерезку задних корешков I, II и
III шейных нервов мы производили в основном по выработанному Магну-
сом способу: корешки 1 и II нервов перерезались вне спинального канала
путем экстирпации межпозвоночных узлов, а корешки III нерва— внутри
спинального канала (интрадурально). Эти кошки жили 5—8 дней. Темне
менее мы имели возможность наблюдать и познакомиться с нарушениями,
наступающими после перерезки задних корешков первых трех пар шейных
нервов, а также с наступающими за это время компенсаторными явления-
ми. На основании этих беглых наблюдений нельзя было, конечно, сделать
каких-нибудь определенных заключений. Мы могли только отметить,что
у наших кошек картина нарушений и компенсации функций в первые по-
слеоперационные дни в основном совпадает с той, которая была описана
Магнусом. Небольшое добавление, которое следует сделать к наблюдениям
Магнуса, заключается в следующем: в первый и второй день после операции
кошки ходили и стояли не только с сильно согнутыми задними конеч-
ностями, как об этом писал Магнус, но и со слабо согнутыми передними ко-
нечностями, а также с опущенной спиной. Кроме того, имело место следую-
щее интересное явление: со второго, а чаще с третьего дня кошки стали
передвигаться резко замедленными шагами, что придавало особую гра-
циозность их походке. У некоторых кошек при стоянии и ходьбе можно
было наблюдать и более широкое расставление конечностей. Уже в после-
дующие дни отмечалось исстеленное ускорение темпов ходьбы и ее
улучшение.
Основными объектами наших наблюдений были 5 беспородных ссбак,
у которых в асептических условиях перерезались по указанному выше
способу задние корешки первых трех пар шейных нервов, Так как ни Маг-
нус и его ученики, ни кто-либо другой из физиологов не наблюдали в хро-
нических условиях последствий упомянутой операции на собаках (во
всяком случае в литературе не имеется об этом никаких данных), я крат-
ко остановлюсь на описании картины функциональных нарушений и
компенсаторных явлений, наступающих ПсСЛ<& Дёйфферептации верхних
сегментов спинного мозга у собак. Следует заметить, что хотя картина нару-
шений и течение компенсаторных явлений у всех 5 собак имели в основном
один и тот же характер, однако глубина и длительность нарушений, а также
темпы компенсации у этих собак были различны. Большие вариации в этом
отношении отметил также Магнус у кошек, причем он склонен был объ-
яснить это я вление тем, что у одних кошек ярко выражены и превалируют ла-
биринтные, а у других шейные рефлексы. Отмечая тот же факт у собак,
можно к вероятным факторам, обусловливающим эти вариации, при-
бавить также общий вес тела, возраст и длину ног животных.
При сравнении данных, полученных нами на 5 собаках, с упомянутыми
выше данными Магнуса можно сделать предварительный вывод, что послед-
ствия перерезки задних корешков первых трех пар шейных нервов у ссбак
бывают значительно тяжелее и длительнее, чему кошек. Это, очевидно, го-
ворит о сравнительно большем значении шейных рефлексов для собак,
чем для кошек. Вообще картина нарушений моторных и сензорных функций
после перерезки задних корешков первых трех пар шейных нервов у со-
бак имеет очень много общего с картиной, наблюдавшейся у оперированных
аналогичным образом кошек. В первые дни после операции при хорошем
общем состоянии организма ж иво-тные, как правило, не могли стоять, они
лежали на боку, а при попытках передвигаться ползком покачивались
и падали. Голову держали симметрично; у 4 собак не выявлялось никаких
СООБЩЕНИЕ VI. ПЕРЕРЕЗКА ЗАДНИХ КОРЕШКОВ ШЕЙНЫХ НЕРВОВ
323
признаков атаксии мышц шеи, у 1 ссбаки имелись слабые проявления
атаксии в течение короткого времени. У всех без исключения собак на-
блюдались сильно выраженная атония и астения мышц конечностей, в
особенности задних. В эти дни собаки не только не могли сами стоять па
ногах, но не могли держаться на них даже тогда, когда собакам искус-
ственно придавали позу стояния. Кратковременные движения конечностей
и туловища быстро приводили к утомлению н одышке. Характерно, что
чувствительность кожи туловища и конечностей к механическим раздра-
жениям была сильно повышена.
Эта первая фаза глубоких нарушений у 2 наших подопытных собак
(Тебо и Безыменный I) длилась 3-—4 дня, у 2 собак (Шаро и Безымен-
ный П)—5 — 6 дней, а у 1 собаки (Лльмо) — 13—14 дней.
Вторая фаза менее глубоких нарушений характеризуется следующими
особенностями. Ссбакн постепенно начинают стоять, опираясь на сильно
согнутые и широко расставленные конечности, при этом задняя часть тела
опускается вниз в силу того, что задние конечности больше сгибаются и
шире расставляются. Животные при этом шатаются из стороны в сторону
и качаются взад и вперед, голову держат высоко и симметрично, движения
ее плавные, быстрые, без всяких признаков атаксии. При попытках пере-
двигаться еще белее шатается и раскачивается корпус, животные барахта-
ются, передние лапы наступают одна на другую, заплетаются; животные
падаютто наодии, то на другой бок. Особенно часто они падают при поворо-
тах или при ускорен ной ходьбе. Лапы они часто ставятнапол тыльной сто-
роной. В движениях передних конечностей наблюдаются некоторая атаксия
идисметрия, гее еще остается в ослабленной форме гиперестезия кожи ту-
ловища и конечностей, а также атония и астения мышц. Глубина моторных
и сензорных нарушений по Степени И продолжительности были различны
у каждой собаки, но у всех ссбак эти явления были четко выражены.
Постепенно, мало заметно происходит компенсация нарушений. Изо
дня в день восстанавливаются функции передних и в особенности задних
конечностей, увеличивается ИХ мышечный тонус и сила. Животные все реже
и реже падают даже при поворотах и ускоренной ходьбе, у них все больше
совершенствуется умение стоять и ходить, движения лап делаются лов-
кими, они меньше заплетаются, гиперестезия кожи постепенно исчезает.
У разных ссбак через различные промежутки времени компенсация наруше-
ний достигает такого уровня, что бывает очень трудно — без специальных
проб — уловить какие-либо следы их. Тебо и Безыменный I достигли этого
уровня на 10—12-й день операции, Шаро и Безыменный II — на 15—
20-й день, а Альмо — примерно через 1 месяц.
Медленное и постепеннее развитие компенсаторных явлений, а также
характер усовершенствования моторных функции (осторожность, заметный
оптический контроль, ухудшение движений при завязывании глаз и т. п.)
с несомненностью говорили о том, что, кроме факторов компенсации анало-
гичных нарушений у кошек, на которые указал Магнус, т. е. кроме дуб-
лирования и компенсаторного усиления неповрежденных среднемозговых
установочных рефлексов с лабиринтов н с тела, важную роль играет также
фактор «научения», т. е. корковая деятельность.
Но в целях получения более четкого и прямого ответа на вопрос о роли
коры больших полушарий мозга в описанных выше компенсаторных
явлениях мы у 3 ссбак (Тобо, Шаро и Лльмо) удалили кору одного, а затем
и другого полушария головного мозга. В приводимой ниже таблице
указываются даты произведенных операций.
Последствия удаления коры одного (правого) полушария у всех трех
ссбак оказались примерно одинаковыми, причем довольно легкими, нс
намного отличающимися от последствий удаления коры одного полушария
у нормальных ссбак. В первые дни после операции собаки лежали на левом
21*
324
Э. А. АСРАТЯН
Кличка
собак
При-
мер-
ный
воз-
раст
Дата перерез-
ки задних
Ki'peufKOB
шейных
нервов
Дата экстир-
пации коры
правого
полушария
Дата экстир-
пации коры
левого
полушария
Примечания
Тобо
Шзро
Альмо
2—3
годэ
5-6
лет
7—8
лет
6,5 29.VI 1939 г.
26. IX 1939 г.
17,0 8.1V 1910 г.
13,0
260
8,029.VI 1939 г. 26. IX 1939 г.
17,9 14.ХП939Г.
7,0 29.VI 1939 г. 26. IX 1939 г.
16,1 14.ХП939Г.
17,7
16,5
190
Погибла
notле НОВОЙ
операциипе-
рсрезки зад-
них столбов
спинного
мозга
Погибла
по неизвест-
ной причине
(отравле-
ние?)
Погибла
от истоле-
ния и ка-
хексии
5
боку (т. е. на боку, противоположном стороне, на которой была произ-
веденаоперация), выгибаясь на правую сторону; на правом боку не лежал и;
при укладывании на правый бок сразу переворачивались на левый бок через
спину; часто сами вращались вокруг длинной оси тела. Спустя 3—4 дня
после операции собаки постепенно стали лежать также на правом боку и
на животе, прекратили вращательные движения, делали попытки стоять.
Тобостоял и ходил уже на 7—8-й день после операции, Шаро —на 10—
11-й день, а Альмо — на 16—17-й день. Все три собаки вначале стояли и
ходили на широко расставленных конечностях, шатались и покачивались,
часто падали при ходьбе, вособенпости при поворотах. Вскоре у них функ-
ции стояния и ходьбы усовершенствовались, причем это наблюдалось преж-
де всего у Тобо и с опозданием на 9—10 дней у Альмо. Однако еще в про-
цессе восстановления послеоперационных нарушений у Тобо неожиданно
началось ухудшение общего состояния и в связи с этим также ухудшение
функции стояния и хоДьбы. Поправлялся Тобо постепенно, и вторая волна
послеоперационных нарушений функций стояния и ходьбы отняла у него
примерно 25—30 дней. Опередив двух других собак в восстановлении на-
рушенных функций после деафферентации нервов, а также в первый период
после удаления коры правого полушария, Тобо к полной компенсации по-
следствий операции пришел позднее, чем две другие собаки. В общем если
не принимать во внимание вмешательства Постороннего фактора, осложнив-
шего процесс восстановления функций у Тобо, то можно сказать, что
последствия удаления коры одного полушария головного мозга у собак
с перерезанными задними корешками первых трех пар шейных нервов
являются нетяжелыми и компенсируются довольно быстро.
Вскоре после удаления коры правого полушария у 2 из этих собак
(Шаро л Альмо) была удалена также кора другого (левого) полушария.
У Тобо вторая операция была произведена позднее из-за упомянутого вы-
ше ухудшения его состояния после первой операции.
Последствия удаления коры всего головного мозга у 3 собак оказались
дово-льно тяжелыми и более или менее однотипными лить в первом после-
операционном периоде. Уже в 1-й день операции, почти сразу же по
прекращении действия наркоза, собаки стали вертеться вокруг дли иной оси
СООБЩЕНИЕ VI. ПЕРЕРЕЗКА ЗАДНИХ КОРЕШКОВ ШЕЙНЫХ НЕРВОВ
325
тела. Со Следующего Дня эти движения резко участились и усилились.
Приступы такого кружения вызывали одышку и резкое учащение сердеч-
ной деятельности. После непродолжительного перерыва и отдыха начина-
лась новая волна вращательных движений и т. д. В перерывах между
приступами ссбаки лежали на спине, на левом боку, иногда в самых причуд-
ливых позах, но на правом боку не лежали. Эта фаза бурных и хаотических
движений продолжалась псслеоперации 3—4дня и привела к значитель-
ному исхуданию и ухудшению общего состояния ссбак. Креме того, из-за
резких движений и трения о стенки люльки на конечностях ссбак появились
язвы и царапины, что ещеболее ухудшило их общее сеется ним. Все это при-
вело к быстрей гибели наиболее слабую собаку Альмо. С 4—5-го дня
после операции у Шаро и Тебо приступы хаотических движений и круже-
ния стали поянляться реже, но сила их осталась прежней, а длительность
даже увеличилась (приступы дл ил ись иногда 12—15 минут). Из внешних
факторов, способствовавших появлению этих приступов, следует отметить:
наполнение мочевого пузыря и прямой кишки, кормление, ненормальное
положение тела при лежании, лежание на правом боку. Характерно, что
иногда до статен го было вертящееся животное на непродолжительное время
фиксировать руками в каком-нибудь положении, чтобы оно перестало вер-
теться. И, наоборот, стоило иногда хотя бы один раз повернуть лежащее
в спокойном ссстсянии животное или чуть-чуть поднять и опустить его го-
лову, чтобы оно начало снова бурно вертеться. Следует сказать, что как
в люльке, так и на полу с помощью этих спиралеобразных вращательных
движений ссбаки медленно передвигались вперед. Несмотря иа улучшение
общего состояния ссбак, несмотря на постепеннее восстановление сим-
метрии тела при лежании, на постепенное появление способности лежать
также на правом боку, приступы вращательных движений у ссбак сохра-
нялись еще долго, в особенности у Шаро. В этой стадии лабиринтные
установочные рефлексы, а также установочные рефлексы с тела на тело
и с тела на голову были отчетливо выражены, когда животные лежали на
правом боку. При положении на левом боку они были выражены очень
слабо, чаще их не удавалось даже обнаружить.
Еще с середины 1-го месяца после последней операции наблюдалась
небольшая разница между картиной нарушений у Шаро и Тобо. У первой
собаки приступы вращательных движений были более частыми, бурными
и длительными, чем у второй; крометого, первая менее спокойно лежала на
правом боку, чем вторая. В дальнейшем эта разница все больше и больше
увеличивалась и в итоге получились различные конечные результаты.
В конце 1-го месяца у Шаро еще продолжались бурные приступы кру-
жения. Но постепенно эти явления у него стихли ион даже пытался стоять;
чем дальше, тем чаше проявлялись у него такие попытки. Следует отме-
тить, что при попытках стоять задние конечности и вообще задняя часть
тела Шаро действовала лучше, чем передние конечности и вообще передняя
часть тела, в особенности голова. Однако эти попытки, как правило, ни
к чему не приводили: Шаро ударялся о пол, продолжал вертеться до сле-
дующей попытки встать и т. д. В конце 2-го месяца пссле операции выяс-
нилось, что Шаро на правей боку лежит почти так же, как и на левом,
и крутится как водном, так и в другом направлении. Установочные рефлек-
сы вызывались почти с одинаковой интенсивностью и живостью как при
положении на левом, так и при положении на правом боку. Примерно
с конца 2-го месяца и до ко нца жизни (6 месяцев) у Ш аро установилась бо-
лее или менее стабильная картина при хорошем ебщем ссстсянии. Он ле-
жал преимущественно на животе, придавая телу спиралеобразную форму,
лежал также на левом и правом боку. Вращательные движения стали ред-
кими и короткими. Стоять он не мог, причем не мог даже удержаться в позе
стояния, когда ему искусственно придавали эту позу.
326
Э А. АСРАТЯН
Иной была картина уТобо. Уже 20—25 дней спустя после последней
операции у Тебо исчезла асимметрия тела при лежании, он мог одинаково
хорошо лежать наобоих боках и на животе, очень редко вертелся и при этом
часто пытался стоять, При этих попытках стоять задние конечности и вооб-
ще задняя часть тела действовали хорошо: он мог поднять заднюю
половину тела, тогда как передняя часть туловища и голова оставались
как бы прикованными к полу. Если же Тобо брали на руки, то его голова
не только высоко держалась, но и свободно двигалась во все стороны.
В таком положении у собаки появлялись оживленные «спонтанные» дви-
жения также со стороны передних конечностях. Со временем Тобо не толь-
ко усовершенствовал эти своеобразные движения, но стал даже п ередвигать-
ся по полу в такой позе. Вначалеон поднимал заднюю часть тела на хорошо
выпрямленных и широко расставленных задних конечностях, прсизаоднл
«шагательные» движения вокруг «прикованной» к полу передней части тела
и головы. Далее Тебо приобрел способность передвигать тело в этой позе
по прямой линии вперед, толкая по полу переднюю часть корпуса, голову
и лежащие вытянутыми во всю длину на полу передние конечности. По-
степенно С1али активизироваться также передние конечности и шейные
мышцы; приподняв заднюю половину тела, Тобо стал постепенно отрывать
от пола и переднюю часть, а при передвижениях вначале скользил на по-
лусогнутых передних конечностях, как на санях. Постепенно все больше
и больше выпрямлялись и передние конечности, активность их движений
усилилась; постепенно выпрямилось и положение головы. В конце 2-го
месяца после операции Тобо мог стоять и ходить на четырех конечностях
с чуть опущенными головой и плечевой областью В начале этой новой
фазы он часто падал либо из-за недостаточной опоры на передние конеч-
ности, либо из-за того, что лапы цеплялись одна за другую. Позже,
однако, и этот дефект значительно уменьшился. Начиная с 3-го месяца после
операции идо конца жизни (9 месяцев) у Тобо установилась почти стабиль-
ная картина: голова и плечевая область были слегка опущены, передние
конечности при ходьбе часто цеплялись одна за другую, из-за чего собака
спотыкалась или падала; наблюдалось нарушение ритма шагания.
Обсуждение полученных данных
Если удаление коры одного полушария у всех трех собак с деафферен-
тированными шейными нервами вызывало неглубокие, но в основном одно-
типные последствия, то удаление коры второго полушария вызывало у них
более глубокие последствия, которые были однотипными лишь в первое вре-
мя после операции. У двух собак, проживших более 6 и 9 месяцев после
операции, в конечном итоге последствия полной декортикации головного
мозга по характеру оказались совершенно различными: у Шаро осуществле-
ние функции стояния и ходьбы оказалось невозможным на всю жизнь,
у Тобо после примерно 20-дневного нарушения этих функций наступила
фаза постепенного их восстановления и усовершенствования до значитель-
ных пределов.
Как понять эту' разницу в последствиях декортикации головного мозга
у Шаро и Тобо?
Прежде всего следует напомнить, что первичные нарушения, обуслов-
ленные перерезкой задних корешков первых трех пар шейных нервов, у Тобо
были менее тяжелыми и компенсировались быстрее, чем у других собак,
в том числе и у Шаро. Мы объясняем это явление у Тобо большим, чем
у Других наших собак, превалированием лабиринтных установочных реф-
лексов над шейными. Возможно, что сравнительная молодость Тобо также
играла некоторую роль. Так или иначе, уТобо первичные нарушения
были неглубокими И компенсировались значительно раньше, чем у Шарю.
СООБЩЕНИЕ VI. ПЕРЕРЕЗКА ЗАДНИХ КОРЕШКОВ ШЕЙНЫХ НЕРВОВ
327
Согласно нашим уже опубликованным данным, а также получен-
ным в последнее время данным с другими вариантами оперативного
повреждения систем организма, у взрослых собак роль и участие коры
в-компенсации различного рода нарушений Тем больше, Чем эти нарушения
тяжелее и длительнее, В случаях, когда первоначальные нарушения бывают
незаметными или неглубокими и быстро компенсируются, удаление коры од-
ного или двух полушарий Заметно не отражается на ходе компенсации.
По всем этим показателям Тобо занимает среднее место между животными с
глубокими первичными нарушениями, у которых декортикация больших по-
лушарий мозга вызывала почти поднос и безвозвратное исчезновение
имевшей место компенсации, и животными с незаметными первичными
нарушениями, у которых декортикация больших полушарий мозга не вы-
зывала заметных изменений в существующей картине. У Тобо первичные
нарушения были заметными, но не очень глубокими, и компенсировались
быстро; соответственна этому удаление коры одного, азатем и другого по-
лушария вызывало неглубокое и недлительное расстройство компенсации
этих первичных нарушении. Мы допускаем, что в компенсации описанных
нарушений относительный удельный вес неповрежденных рефлекторных
аппаратов центральной нервной системы (в частности, среднего межу-
точного мозга и недегенернрованных субкортикальных образований)
в функциональном отношении оказался у Тебо бблыним, чем у Шаро и дру-
гих собак. Таким образом, различные результаты, полученные у Шаро и
Тебо, не противоречат нашим предшествовавшим и более поздним данным,
а также нашим концепциям о решающей роли коры больших полушарий
мозга в компенсации глубоких нарушений, обусловленных серьезными по-
вреждениями тех или иных систем организма высших животных. Различны-
ми по глубине и длительности были функциональные нарушения после ОД-
НОЙ и той же операции у обеих собак; различной оказались роль и степень
участия коры больших полушарий в компенсации этих нарушений.
Это закономерное соотношение между последствиями первичных опе-
раций и удаления полушарий, как правило, отчетливо выступает уже при
удалении коры одного полушария головного мозга. Оно ясно выявилось
также у описанных выше трех собак после удаления у них коры одного
полушария. У Альмо нарушения после перерезки задних корешков
шейных нервов были самыми глубокими и длительными, у Шаро они были
менее длительными и менее глубокими, а у Тобо — еще слабее и короче.
Соответственно этому последствия удаления коры одного полушария
у Альмо были глубже и длительнее, чем у Шаро, а у Шаро — глубже и
длительнее, чем у Тобо. Однако, как было указано выше, у Тобо восстанов-
ление вторичных нарушений шло двумя волнами, в итоге затянулся ход вос-
становительного процесса и на первый взгляд создалось впечатление, что
у него не существует упомянутой выше связи между глубиной и длитель-
ностью последствий первичных и вторичных операций. Как объяснить это
явление, точнее — вторую волну ухудшения и восстановления функций
у Тобо после удаления коры одного полушария? Следует сказать, что вна-
чале для нас самих это явление было не совсем понятно, Мы знали по Преды-
дущим нашим исследованиям, что в любой стадии компенсации функций
после любых повреждений систем ухудшение состояния здоровья живот-
ных ослабляет или задерживает компенсацию. В данном случаемы склонны
были происшедшее с Тобо объяснить каким-то болезненным его состоянием,
хотя точно оценить это состоя ние не могли из-за отсутствия ясных и четких
признаков. Удаление коры второго (левого) полушария внесло ясность.
Оказалось, что у Тобо кора второго полушария претерпевала какие-то
атрсфические изменения: она была резко сморщена, васкуляризирована,
размягчена и уменьшена в объеме. Эти изменения нашли яркое выра-
жение в весе удаленного мозгового вещества (в основном вещества коры):
328
Э. А. АСРАТЯН
вес коры второго полушария был меньше веса коры первого полушария на
25% (13 г вместо 17 г). Весьма вероятно, что эта своеобразная частичная
«экстирпация» коры второго полушария была причиной второй волны ухуд-
шен ия компенсаторного процесса и ее затяжки у Тебо пссле удаления коры
первого (правого) полушария. Пользуюсь случаем заметить, что с подоб-
ным фактом атрофических изменений коры не затронутого операцией по-
лушария после удаления коры другого полушария в нашей практике мы
вообще встречались, но очень редко. Никогда, однако, таких глубоких
сдвигов, как у Тобо, мы не видели. Мы затрудняемся точно сказать, чем
обусловливается это явление; все же несомненно, что оно связано с удале-
нием коры одного полушария.
Как же понять роль верхних этажей центральной нервной системы,
в частности, коры больших полушарий мозга, в компенсаторных явлениях,
наступающих пссле перерезки задних корешков первых трех пар шейных
нервов?
Следует прежде всего отметить, что функция стояния, которая пссле
названной операции нарушается прежде всего (нарушение функций ходь-
бы является в данном случае производным), по природе является Сложным
интегративным актом и представляет собой результат комплекса разнотип-
ных установочных рефлексов, осуществляемых областью среднего мозга.
Магнус показал, что глубина и длительность нарушения этой функции пссле
выключения какого-нибудь установочного рефлекса зависят от его удель-
ного веса, процесс же компенсации этих нарушений является в основном
результатом «компенсаторного усиления» неповрежденных компонентов
указанного комплекса. В случае компенсации лабиринтных нарушений
Магнус известную роль приписывает также оптическим установочным
рефлексам коркового происхождения. Что же касается компенсации нару-
шений, обусловленных деафферецтац ней шейных нервов, то мы не находим
у Магнуса никаких указаний или даже предположении сб участии в ней
оптических установочных рефлексон и коры больших полушарий мозга
вообще. Как было уже сказано, полученные нами данные являются доста-
точным основанием для утверждения, что у ссбак при компенсации этих
нарушений, как и при компенсации лабиринтных нарушений, кора больших
полушарий мозга играет важную, иногда решающую роль.
На основании наших данных мы можем заключить: 1) что «компенсатор-
ное усиление» неповрежденных установочных рефлексов, о котором писал
Магнус, в значительной мере осуществляется благодаря коре больших по-
лушарий мозга, трофическому регуляторному влиянию коры больших по-
лушарий мозга на безусловнорефлекторный аппарат этих ре(|»лексов, а так-
же благодаря их высшей, безусловнорсфлекторной интеграции корой; оче-
видно, что в каких-то пределах такие влияния свойственны также образо-
ваниям межуточного мозга н уцелевшим подкорковым образованиям; 2) что
в компенсации этих нарушений, так же как и в компенсации лабиринт-
ных нарушений, кора больших полушарий мозга участвует путем услов-
норефлекторней интеграции неповрежденных установочных и других про-
стых спинальных, бульбарных и среднемозговых рефлексов, в частности,
путем образования оптических установочных условных рефлексов.
В тех случаях, когда шейные установочные рефлексы в сложном комп-
лексе установочных рефлексов не занимают важного .места и поэтому их
выключение не вызывает серьезного и длительного расстройства функций
стояния, эти нарушения могут быть компенсированы также d отсутствии
коры; очевидно, это про исходит благодаря более совершенной интеграции и
трофическо-регуляторному влиянию межуточного мозга И уцелевших Под-
корковых образований на рефлекторный аппарат неповрежденных устано-
вочных рефлексов, влиянию, которое обеспечивает требуемое незначитель-
ное «компенсаторное усиление» Их.
СООБЩЕНИЕ VI . ПЕРЕРЕЗКА ЗАДНИХ КОРЕШКОВ ШЕЙНЫХ НЕРВОВ
329
ЛИТЕРАТУРА
Асратян Э. А., Кора большою мозга н пластичность нервной системы. Вариант
ампутации лап, Успехи современной биологии, 5, &03, 1936.
Асратян 9. А., Кора большого мозга и пластичность нервной системы, опыты
с половинной перерезкой спинного мозга, Успехи современной биологии, 6,
451, 1937.
Асратян Э. А., Кора большого мозга и пластичность нервной системы. Вариант
перекрестного сшивания нервов, Успехи современной биологии, 12, 516, 1940.
Асратян Э. А., Но-вые данные по физиологии большого мозга, Архив биологи-
ческих наук, 61, 54, 1941.
Асратян Э. А.. Кора большого мозга и пластичность нервной системы. Вари-
ант—феномен Портера, Труды Института мозга им. Бехтерева, т. 14, 1941.
Асратян Э. А., Кора большого мозга и приспособительные явления в поврежден-
ном организме. Сообщение V. Опыты с разрушением лабиринтов, Физиологи
' ческий журнал СССР, 33, 289, 1947.
/Асратян Э. А., Кора большого мозга и приспособительные явления в поврежден-
ном организме. Сообщение IV. Опыты с деафферентацией конечностей, Физиоло-
гический журнал-СССР, 34, 1, 1948.
Конорский Ю. М. и М и л л е р С. М.. Условные рефлексы двигательного аппа-
рата, Труды физиологических лабораторий им. акад. И. П. Павлова. 6, 1. 1936.
Красногорский Н. И., О процессе задерживания и о локализации кожного
и двигательного анализатора в коре больших полушарий у собаки, дисс., Спб.,
1911.
Орбели Л. А. и Куист май К. И., О последствиях деаффереитацни задней
конечности у собаки, Труды Института иМ. Лесгафта, 9, 1924, 187.
Сеченов И. М., Собр. соч., т. 11, 1908.
Gol tz, Pflug. Arch., 8, 4450, 1874.
Magnus, Korperstellung, 1924.
Magnus u. S t г о m v a n L eeu we n, Pflflg. Arch.. 154, 157. 1914.
КОРА БОЛЬШОГО МОЗГА И ПЛАСТИЧНОСТЬ НЕРВНОЙ СИСТЕМЫ1
Сообщение VII
ОПЫТЫ С ПОВРЕЖДЕНИЕМ ДЫХАТЕЛЬНЫХ НЕРВНЫХ ПУТЕЙ
физиологии давно известно, что значительные повреждения
продолговатого мозга зачастую существенно не отражаются на
дыхательной функции. Так, например, Броун-Секар, Ланген-
дорф, Шифф и многие другие исследователи показали, что раз-
личного рода уколы, разрезы и другие умеренные хирургические
повреждения дна продолговатого мозга в области дыхательного
центра у различных млекопитающих, а также значительной силы термичес-
кие воздействия на эту область ие только не приводят к прекращению дыха-
тельных движений, но зачастую существенно пе отражаются на нормальной
их ритмике и на силе сокращения дыхательных мышц. Было установлено,
в частности, что продольно-сагиттальное сечение сегментов спинного
мозга, лежащих между продолговатым мозгом и ядрами диафрагмального
нерва, а также сегментов, дающих начало диафрагмальным нервам, не вы-
зывает сколько-нибудь серьезных и длительных расстройств дыхательной
функции. Однако из исследований упомянутых ученых явствует, что по-
перечная боковая половинная перерезка продолговатого мозга или верхних
шейных сегментов у тех же животных примерно в половине случаев вызы-
вает остановку дыхания на стороне перерезки и в Половине случаев дыха-
ние при этом сохраняется с обеих сторон. Более тщательное изучение этого
вопроса Портером на большом количестве животных показало, что при ана-
логичных операциях односторонняя остановка дыхания наступает более
чем в 90% случаев-
Лангендорф обнаружил еще одно весьма любопытное явление, которое
впоследствии стало предметом специальных исследовании Портера и ряда
других экспериментаторов. Речь идет о следующем; когда половинная пе-
ререзка шейного отдела спинного мозга на уровне первых его сегментов
вызывает остановку грудного и диафрагмального дыхания одноименной
стороны, перерезка диафрагмального нерва противоположной стороны
часто тотчас же или некоторое время спустя восстанавливает парализован-
ное диафрагмальное дыхание со стороны гемисекции спинного- мозга. Со-
гласно данным Розенблюта н Ортица, такой же результат получается и в тех
1 Первая часть статьи, посвященная перекрестному феномену диафрагмальиото
нерва (или, как мы нс совсем удачно называли его раньше, феномену Портера),
была опубликована в «Трудах Государственного института по изучению мозга
им. В. М. Бехтерева» (т. XIV, 1941). Вторая же часть.статьи, в которой излагаются
наблюдения над картиной нарушений дыхательных движений после половинной бо-
ковой поперечной перерезки спинного мозга на уровне верхних шейных еегментов,
была доложена в 1947 г. на 12-м совещании по физиологическим проблемам, пос-
вященном памяти И. П. Павлова.
СООБЩЕНИЕ VII. ПОВРЕЖДЕНИЕ ДЫХАТЕЛЬНЫХ НЕРВНЫХ ПУТЕЙ
331
случаях, когда активный диафрагмальный нерв подвергается временной и
обратимой локальной блокаде эфиром или электрическим током.
Несмотря на то, что этот интересный феномен известен в физиологии
более 60 лет и воспроизведен многими исследователями, до сих пор, однако,
отсутствует более или менее удовлетворительный физиологический анализ
его, отсутствуют точные знания о его причинах и природе. Одни исследо-
ватели причиной этого явления считали асфиксию, между тем экспери-
менты Портера показали, что асфиксия не играет существенной роли в этом
феномене. Установив, что переход дыхательных импульсов с неповреж-
денной половины шейного отдела спинного мозга на поврежденную осуще-
ствляется на уровне ядер диафрагмального нерва, Портер стремился дать
другое объяснение этому любопытному феномену. Он допускал существо-
вание перекрестной связи между мотонейронами диафрагмального нерва
двух половин шейных сегментов спинного мозга через посредство опреде-
ленной части их дендритов. По мнению Портера, в случае перерезки одного
диафрагмального нерва импульсы из мотонейронов этого нерва через на-
званные дендритные связи передаются мотонейронам противоположного
диафрагмального нерва и активируют их.
Не говоря уже о сугубо гипотетическом характере данного Портером
объяснения перекрестного феномена диафрагмального нерва, укажем, что
при таком объяснении остается совершенно открытым вопрос о подлинных
причинах перехода нервных импульсов с одних путей на другие, о факто-
рах, направляющих импульсы дыхательного центра по дендритным свя-
зям. Нельзя считать удовлетворительным сделанное Розенблютом и его
сотрудниками гипотетическое допущение, что в переходе дыхательных
импульсов с неповрежденной половины спинного мозга на поврежденную
решающую роль играет повышение чувствительности мотонейронов диаф-
рагмального нерва последней половины, обусловленное гемисекцней спин-
ного мозга, т. е. своеобразной их «денервацией».
Приходится констатировать, таким образом, что вопрос о подлинных
причинах перекрестного феномена диафрагмального нерва до сих пор
остается открытым.
Собственные экспериментальные данные
Феномен перекрестного диафрагмального нерва представляет большой
интерес со многих точек зрения. Для нас он интересен прежде всего как
яркое проявление приспособляемости нервной системы, как случай быстрого
приспособления в центральной нервной системе, возникающего после по-
вреждения определенных нервных структур.
На протяжении последних лет нами и нашими сотрудниками была
установлена решающая роль коры больших полушарий мозга лишь
в медленно протекающих и постепенно развивающихся компенсаторных
приспособительных явлениях, возникающих в организме после серьезных
повреждений структуры и функции различных частей спинного мозга,
периферических нервов, рецепторных И эффекторных органов у высоко-
развитых взрослых млекопитающих животных. В свете этих данных было
важно экспериментально выяснить, каково участие коры большого мозга
в осуществлении этого своеобразного приспособительного явления — пе-
рекрестного феномена диафрагмального нерва, при котором хотя и имеется
серьезное повреждение спинного мозга и глубокое нарушение функций
дыхательной системы, но одновременно С этим имеет место и очень быстрое
восстановление нарушенной дыхательной функции. Изучение этого во-
проса было тем более важно, что никто из предыдущих исследователей этот
феномен в указанном разрезе не исследовал.
Опыты проводились в 1940 г. на 4 собаках и И кошках в условиях
острого вивисекционного эксперимента. В ряде случаев для наркоза
332
Э. А. АСРАТЯН
мы применяли хлоролозу, а в ряде случаев легкий эфнро-хлорсформ-
ный наркоз. Во всех случаях шейный отдел спинного мозга был нами
наполовину поперечно перерезан приблизительно между 2-м и 3-м его
сегментом. Для наблюдения за диафрагмальным дыханием мы произвели
продольный разрез брюшной стенки и через это «окно» визуально конт-
ролировали движения диафрагмы.
У 5 кошек и 3 собак мы нс получили никаких данных по интересующему
нас вопросу; они частью погибли вследствие полной остановки дыхания
после гемисекции (2 кошки, 1 собака), частью вовсе не проявляли описан-
ного феномена (1 кошка и 2 собаки), а 2 кошки, у которых вначале проявился
этот феномен, погибли после деафферентации 4, 5, G и 7-го корешков с не-
перерезанной стороны спинного мозга. Только у 6 кошек и у одной собаки
с удаленными полушариями нам удалось в четком виде воспроизвести и
изучить интересующий нас феномен.
После установления у оперированных кошек своеобразного перекрестного
дыхания (реберное дыхание со стороны неповрежденной, диафрагмальное
дыхание со стороны поврежденной половины спинного мозга) мы последо-
вательно удаляли у них сначала кору больших полушарий мозга с одной,
а затем и с другой стороны. Во всех без исключения случаях удаление коры
одного и двух полушарий большого мозга заметно не отразилось на опи-
санном феномене: как реберное, так и диафрагмальное дыхание продолжа-
лось в том перекрестном виде, как и раньше. Такой же результат был полу-
чен в опыте на собаке.
У одной собаки, лишенной коры обоих полушарий за год до нашего
последнего эксперимента, мы под слабым эфиро-хлороформным наркозом
перерезали левую половину шейного отдела спинного мозга между 2-м
и 3-м сегментом. Произведенная операция вызвала точно такую же оста-
новку диафрагмального и реберного дыхания с одноименной стороны, как
у нормальной собаки. Минут через 1? после спинальной операции нами
был перерезан правый диафрагмальный нерв, что вызвало моментальное
прекращение диафрагмального дыхания и усиление реберного дыхания со-
ответствующей стороны. Через 2—3 минуты стало появляться диафраг-
мальное дыхание с левой стороны, т. е. со стороны перерезки спинного
мозга. С каждой минутой усиливаясь, диафрагмальное дыхание с этой
стороны достигло почти нормального уровня через 20—25 минут после пе-
ререзки правого диафрагмального нерва. В связи с появлением и усилением
диафрагмального дыхания с левой стороны постепенно ослабевало реберное
дыхание с правой стороны. Следует отметить, что описанное выше своеоб-
разное «перекрестное» дыхание (т. е. реберное дыхание со стороны перере-
занного диафрагмального нерва и диафрагмальное дыхание со стороны
гемисекции шейного отдела спинного мозга) очень четко выявилось и сохра-
нялось у этой лишенной полушарий собаки в течение длительного времени
(больше Р/г часов), пока не был прекращен опыт.
Таким образом, опыты на 6 кошках и одной собаке с удаленной корой
дают достаточно оснований для утверждения, что описанный выше пере-
крестный феномен диафрагмального нерва может осуществляться и без
заметного участия коры большого мозга.
В опытах на кошках нами были получены еще следующие дополни-
тельные данные, Убедившись, что удаление коры большого мозга не отра-
жается существенно на этом феномене, мы некоторое время спустя (15—
30 минут) последовательно удаляли у кошек межуточный мозг, затем сред-
ний мозг и даже мозжечок, оставив, таким образом, ТОЛЬКО продолговатый
и спинной мозг. Во всех опытах получались одни и те же результаты: по-
очередное удаление указанных частей головного мозга несколько изменяло
ритмику дыхания, его глубину и регулярность, но сам феномен сохра-
нялся во всех случаях, пока животное дышало. Характерно, что при всех
СООБЩЕНИЕ VII. ПОВРЕЖДЕНИЕ ДЫХАТЕЛЬНЫХ НЕРВНЫХ ПУТЕЙ
333
вариациях дыхания (нарушение нормального ритма, глубины) реберное
дыхание с неповрежденной стороны спинного мозга и диафрагмальное
дыхание со стороны половинной перерезки спинного мозга всегда осуще-
ствлялось синхронно.
Из приведенных данных мы делаем заключение, что описанный выше
феномен может осуществляться при наличии одного только продолговатого
мозга в тесной его связи со спинномозговыми дыхательными нервными эле-
ментами и что феномен этот не только ие обусловливается корой большого
мозга, но и всеми теми частями центральной нервной системы, которые
лежат выше продолговатого мозга.
Естественно возникает вопрос: какое место занимает этот вид приспо-
собления по своему характеру, особенностям и центральному рефлектор-
ному аппарату в комплексе других приспособительных явлений централь-
ной нервной системы, возникающих после повреждения других ее частей,
после другого рода нарушений функции нервной системы?
В опубликованных нами и нашими сотрудниками работах приведены
данные о том, что Первичные рефлекторные аппараты нервной системы мле-
копитающих даже при наличии мозжечка, среднего, межуточного мозга и
уцелевших при декортикации подкорковых нервных образований не в со-
стоянии заметно компенсировать глубокие и длительные нарушения сен-
зоркых, скелетно-моторных и вегетативных функций организма, в особен-
ности нарушений функций стояния и ходьбы, вызванных ампутацией лап,
гемисекцией спинного мозга, расщеплением спинного мозга, перекрестным
сшиванием главных нервов конечностей, разрушением лабиринтов, деаф-
фгрентацией конечностей и т. п. Для того чтобы у таких животных проис-
ходила компенсация этих функциональных нарушений, требуется наличие
коры большого мозга, этого могучего приспособительного органа с бога-
тейшими его возможностями. Однако нами было показано также, что в слу-
чаях, когда перечисленные выше органы и системы организма у тех же
животных повреждаются незначительно и возникшие в результате этого
нарушения функций бывают неглубокими и недлительными, эти нарушения
могут быть компенсированы также при отсутствии коры большого
мозга.
Совершенно очевидно, что явление приспособления в виде перекрестного
феномена диафрагмального нерва полностью не походит ни на первую из
приведенных выше групп приспособительных явлений, ни на вторую. Хотя
при этом феномене возникает тяжелое повреждение спинного мозга, в силу
чего появляются глубокие нарушения функций организма (односторонний
паралич дыхания), тем не менее восстановление функций происходит очень
быстро, значительно быстрее даже, чем при упомянутых выше легких
повреждениях органов и систем организма. Очевидно все же, что перекрест-
ный фэиомая диафрагмального нерва по характеру и по быстроте наступле-
ния компенсации ближе стоит именно к последнему* из изученных нами
типов приспособительных явлений, развивающихся при отсутствии коры
большого мозга.
Таким образом, сопоставляя полученные нами данные об отсутствии
заметной роли коры большого мозга и других высших этажей центральной
нервной системы в осуществлении приспособления типа перекрестного фено-
мена диафрагмального нерва с другими нашими данными о решающей роли
коры большого мозга в компенсации глубоких нарушений функций, воз-
никших после серьезных повреждений тех или иных частей центральной
нервной системы или же рецепторных и эффекторных органов, мы вновь
убеждаемся в правильности нашего теоретического положения о том, что
степень участия коры в компенсаторных приспособлениях обусловлена
тяжестью повреждения структур организма, глубиной нарушения компен-
сируемой функции и быстротой наступления компенсации.
334
Э А АСРАТЯН
Однако, по приведенным выше данным, перекрестный феномен диаф-
рагмального нерва осуществляется и в отсутствие мозжечка, межуточного
и даже среднего мозга, говоря иначе, является собственной функцией про-
долговатого мозга, точнее, бульбарного дыхательного центра. Как понять
это явление?
Общеизвестно, что и в нормальных условиях зависимость многих буль-
барных функций, в частности, дыхательной, от коры большого мозга и от
других высших этажей центральной нервной системы значительно слабее,
чем зависимость опорно-локомоторных функций спинального и среднемоз-
гового рефлекторного аппаратов от тех же частей центральной нервной
системы. Это, очевидно, сказывается также в случаях компенсации нару-
шений дыхательной и опорно-локомоторной функций организма, обуслов-
ленных органическим повреждением соответствующих центральных нерв-
ных образований: в первом случае роль коры большого мозга (и даже всех
других частей головного мозга) ничтожна, в другом случае она играет
огромную, решающую роль,
Длительные наблюдения за картиной нарушения функций
дыхательных мышц после гемисекции спинного мозга в области
верхних шейных сегментов
Описанные выше опыты, как и опыты всех наших предшественников
по изучению перекрестного феномена диафрагмального нерва, относятся
к числу острых. Разумеется, в условиях такого несовершенного приема
физиологического эксперимента мы могли лишь односторонне осветить
интересующий нас вопрос о роли коры больших полушарий в восстановле-
нии функций дыхательных мышц, нарушенных в результате поперечной
перерезки боковой половины спинного мозга на уровне верхних шейных
сегментов. Поэтому возникла необходимость в более обстоятельном изуче-
нии этого вопроса в условиях хронического эксперимента, т. е. наиболее
совершенного приема физиологических исследований, введенного в физио-
логию И. П. Павловым и е известным успехом использованного нами при
изучении других вопросов этой актуальной проблемы.
Исходя из полученных данных, нам представлялось необходимым прежде
всего выяснить, не удастся ли в условиях хронического эксперимента
наблюдать со временем восстановление функции дыхательных мышц,
парализованных после гемисекции спинного мозга на уровне верхних
шейных сегментов, в частности, не будет ли восстановлена функция мышц
парализованной половины диафрагмы без предварительной перерезки
диафрагмального нерва противоположной гемисекции стороны. Следует отме-
тить, что этот вопрос более или менее обстоятельно никем не исследовался.
Специальное экспериментальное изучение этого вопроса на первый
взгляд могло показаться излишним. Давно установленный факт восста-
новления опорных и локомоторных функций конечностей, пораженных
после гемисекции спинного мозга, делает весьма вероятным восстановление
также функций дыхательных мышц, пораженных в результате подобной
операции. Будучи вообще убежденными противниками решения сложных
физиологических вопросов только путем косвенных сопоставлений и меха-
нических аналогий, мы в данном случае имели еще другое основание для
прямого экспериментального изучения указанного вопроса. Мы имеем
в виду приведенные выше данные относительно перекрестного феномена
диафрагмального нерва и сделанное на их основе весьма вероятное гипоте-
тическое предположение, что из-за некоторых специфических функциональ-
ных особенностей центрального нервного аппарата дыхательном системы, в
частности, из-за относительно слабой его зависимости от коры больших
полушарий, восстановление функции пораженных после гемисекции спин-
СООБЩЕНИЕ VII. ПОВРЕЖДЕНИЕ ДЫХАТЕЛЬНЫХ НЕРВНЫХ ПУТЕЙ
335
ноп> мозга дыхательных мышц можег со временем и вовсе не произойти
или же произойдет в значительно более ограниченных пределах, чем в слу«
чае поражения локомоторных мышц после подобной же операции.
К проведению соответствующей серин экспериментов мы приступили
в 1946 г. Эксперименты были выполнены нами совместно с нашей сотруд-
ницей С. Н. Ивановом. У 10 собак при строгом соблюдении асептики и анти-
септики мы поперечно перерезали боковую половину спинного мозга на
уровне 2—3-го шейных сегментов. Большинство оперированных животных
находилось под наблюдением в течение длительного времени (значительно
больше года). Из установленных в этих экспериментах фактов особого вни-
мания заслуживают следующие.
1. Гемисекция спинного мозга на уровне верхних шейных сегментов по
сравнению с его гемисекцией на уровне грудных или поясничных сегментов
вызывает не только более обширный круг функциональных расстройств
организма, но и гораздо более глубокие нарушения двигательных, чув-
ствительных и вегетативных функций организма.
Совершенно очевидно', что эти особенности нарушения функций орга-
низма обусловливаются нс только тем, что при высокой гемисекции спин-
ного мозга гораздо большее число спинальных рефлекторных аппаратов
утрачивает естественный контакт с вышележащими отделами центральной
нервной системы, чем при низкой его гемисекции, но и тем, что зоны опе-
ративной травмы в одном и в другом случае функционально неравноценны:
как известно, шейные сегменты спинного мозга, в особенности верхние,
осуществляют гораздо более сложные рефлекторные акты, чем другие его
Сегменты (например, шейные топические рефлексы).
2, Несмотря на большом объем и глубину расстройств, большинство на-
рушенных и утраченных при высокой гемисекции спинного мозга функций
организма со временем так же медленно и постепенно восстанавливается,
как и при низких гемисекциях. Развивающиеся в поврежденном организме
разнообразные функциональные сдвиги обусловливают постепенное ис-
чезновение возникшей в результате операции асимметрии в кожной чув-
ствительности, температуре кожи, тонусе гладких мышц глаз, тонусе мышц
конечностей, туловища и шеи, а также в активности экстероцептмвных и
проприоцептивных рефлексов простого и сложного типа, что находит наи-
более яркое выражение в восстановлении нарушенных операцией функций
сидения, стояния, ходьбы и бега*.
3. Наряду с этим, возникший при высокой гемисекции спинного мозга
односторонний паралич (или очень глубокий парез) дыхательных мышц —
диафрагмы и межреберных мышц — ослабевает в ничтожных пределах или
почти неизменно сохраняется в течение длительного времени (в течение
года и больше). Это находит выражение в асимметрии дыхательных дви-
жений, которая со временем почти не претерпевает изменений или же
сглаживается в весьма ничтожных пределах в результате весьма незначи-
тельного усиления активности пораженных дыхательных мышц. Сказанное
иллюстрируется двусторонней пневмограммой: одной из оперированных
собак в различные сроки операции (см. рисунок на стр. 336).
Таким образом, нашими экспериментами установлен важный и на первый
взгляд даже несколько парадоксальный факт. Хотя после высокой геми-
секции спинного мозга наступает паралич (или очень глубокий парез) всех
1 Детальное изучение характера нарушений и динамики восстановления функ-
ций организма после односторонней и двусторонней высокой гемисекции спинного
мозга, а также выяснение роли коры больших полушарий в развитии компенсатор-
ных приспособлений нами было поручено С. Н. Ивановой, для чего было опери-
ровано еще примерно 50 собак и кошек.' С. Н. Иванова в основном эту задачу
выполнила и часть полученного материала уже опубликовала, а весь материал
в настоящее время оформляет в качестве кандидатской диссертации.
336
Э. А АСРАТЯН
соматических нервно-мышечных образований, лежащих ниже уровня пе-
ререзки, тем не менее со временем в значительных пределах восстанавли-
вается лишь функция той части этих образований, которая имеет отношение
к опорной и локомоторной функциям организма; та же часть соматических
нервно-мышечных образований, которая относится к дыхательной функции,
остается парализованной или же в лучшем случае ее активность восста-
навливается в крайне ничтожной степени.
Как объяснить это явление?
Анализ экспериментального материала показывает, что в столь резком
отставании нервно-мышечных образований дыхательной системы от нервно-
мышечных образований опорно-локомоторной системы в отношении;вос-
становления соответствующих функций организма, нарушенных в резуль-
Пневмограмма собаки Вьюн после поперечной перерезки правосторон-
ней боковой половины спинного мозга на уровне верхних шейных сег-
ментов через 5, 101 и 224 дня после операции
Верхчяя кривая—правая часть грудной клоки, нижняя—левая часть
тате высокой гемисекции спинного мозга, «неповинны» не только дыхатель-
ные мышцы, но даже иннервирующие их мотонейроны. Как явствует, на-
пример, из перекрестного феномена диафрагмального нерва, мотоней-
роны диафрагмального нерва перерезанной половины спинного мозга
способны исправно выполнять СВОЮ функцию, если для этого создаются
определенные условия путем перерезки диафрагмального нерва противо-
положной стороны. Очевидно, также обстоит делос мотонейронами межре-
берных мышц. По всей вероятности, это отставание обусловливается край-
ним затруднением в налаживании надлежащего функционального контакта
между частями центральной нервной системы, лежащими выше уровня ге-
мисекции, к между одноименными гемисекции мотонейронами диафраг-
мальных и интеркостальных нервов через «мостик» неперерезаннон поло-
вины спинного мозга.
В то же время, как следует из наших данных, такой контакт через тот
же «мостик», как правило, налаживается между теми же высшими отдела-
ми центральной нервной системы и между одноименными гемисекции
мотонейронами, иннервирующими опорно-локомоторные мышцы конеч-
ностей и туловища.
В основе такой большой разницы в восстановлении полноценного
функционального контакта между находящимися выше гемисекции цен-
СООБЩЕНИЕ VII. повреждение дыхательных нервных путей
337
тральными нервными образованиями и между находящимися ниже геми-
секции и одноименными ей мотонейронами дыхательных и опорно-локомо-
торных мышц лежит, очевидно, та же причина, что и в основе перекрестного
феномена диафрагмального нерва. Мы имеем в виду некоторые функцио-
нальные особенности дыхательного центра (точнее, констелляции дыхатель-
ных центров) и прежде всего сравнительно слабую его зависимость от выше-
лежащих отделов центральной нервной системы, в особенности от коры
больших полушарий мозга.
В то время как высокая энцефализация и, в частности, кортикализация
опорно-локомоторной функции высокоорганизованных животных обеспе-
чивает полноценное восстановление этой функции в случае повреждения
соответствующих периферических или центральных нервных структур,
благодаря главным образом условнорефлекторной перестройке функции
поврежденных центральных нервных образов-аний опор но-локомоторной
системы, относительно слабая энцефализация и, в частности, кортикалн-
зация дыхательных функций в случае повреждения в соответствующих
центральных нервных образованиях существенно ограничивает возмож-
ность эффективной условнорефлекторной перестройки функции неповреж-
денных центральных нервных структур. Возможно, что при этом извест-
ную роль играет еще одна функциональная особенность констелляции
дыхательных центров высокоразвитых животных— резкое ослабление
способности спинальных дыхательных центров (т. е. ядер диафрагмальных
и межреберцых нервов) к самостоятельному функционированию, их край-
няя зависимость от ведущего звена констелляции—бульбарного дыхатель-
ного центра. Повидимому, это обстоятельство также препятствует полно-
ценному вовлечению спинальных дыхательных центров в сферу благотвор-
ной и мощной корковой условпорефлекторной перестройки функции ниже-
лежащих центральных нервных образований, когда это диктуется необ-
ходимостью.
В заключение следует отмстить, что полученные нами данные наносят
новый сокрушительный удар по идеалистическим, антиэволюционным и
метафизическим воззрениям Бете о нервной системе как об аморфной
в функциональном отношен ин сети, лишенной всякого подобия специали-
зации и локализации функций. Если бы нервная система высокоорганизо-
ванного животного в действительности представляла собой такую сеть,
мы бы не имели столь разительного отличия в картине нарушения и восста-
новления дыхательной и опорно-локомоторной функции организма после
одного и того же повреждения определенной его части — поперечной пе-
ререзки боковой половины спинного мозга на уровне верхних шейных
сегментов.
ЛИТЕРАТУРА
Асратян Э. А., О принципе относительной пластичности нервной системы
Труды Института мозга нм. В. М, Бехтерева, 9. 1939.
Бехтерев В. М., Основы учения о функциях мозга, IV, 1905.
И нанова С. II., Восстановление функции организма после половинной перерез
кн спинного мозга на верхних уровнях, Известия АН СССР {серия биологи-
ческих наук), № 3 , 96, 195-0.
В row п • Se q u a rd, Journ. physiol., 1. 1858.
В row n>S tq u a r d, C. r. Soc. biol., 1, 1669.
I.angendorff, Arch. Physiol., S881.
Porter, Journ. physiol., 1895.
Porter, Amer, journ. physiol., IV, 1901.
Spiegel u. Demetriades, Pfliig. Archiv, 210, 1925.
Rosenbiueth A. a. Ortiz T., Amer, journ. physiol., 117, 495—513, 1936.
22 э. А. Аератян
КОРА БОЛЬШОГО МОЗГА И ПЛАСТИЧНОСТЬ НЕРВНОЙ СИСТЕМЫ
Сообщение VIII
ОПЫТЫ С ПОПЕРЕЧНОЙ ПЕРЕРЕЗКОЙ ЗАДНЕЙ ПОЛОВИНЫ
СПИННОГО МОЗГА1
нашем экспериментальной работе по систематическому изучению
проблемы компенсаторных приспособлений нервной системы мы
до сих пор использовали преимущественно^ такие виды хи-
рургического повреждения нервной системы или других систем
организма, которые до нас применялись в тех же целях другими
исследователями, а именно:ампутацию конечностей, перерезку
и перекрестное сшивание различных нервов, поперечное половинное сече-
ние боковой части спинного мозга на различных его уровнях, деафферен-
тацню конечностей или шейных мышц, разрушение лабиринтов и т. п. Но-
вое в этих исследованиях заключалось главным образом в том, что в целях
выявления роли коры больших полушарий в развивающихся в поврежден-
ном организме компенсаторных приспособлениях мы, во-первых, включили
в круг использованных нами приемов физиологического эксперимента
методику условных рефлексов и систематически применяли ее там, где
это возможно, и, во-вторых,—это мы особенно подчеркиваем — добились
успешного осуществления полного хирургического удаления коры больших
полушарий у большого числа подопытных собак с последующим сохране-
нием их жизни в течение многих месяцев, года, а иногда и больше. По-
добным же образом мы совместно с нашими сотрудниками изучали компен-
саторные приспособления у ряда позвоночных животных, стоящих на более
низких уровнях эволюционной лестницы,—у птиц, рептилий, амфибий.
В целях дальнейшего расширения наших исследований по проблеме
компенсаторных приспособлений нервной системы и дальнейшего углубле-
ния физиологического анализа этих приспособлений мы решили разрабо-
тать и новые виды хирургического или иного рода повреждения нервной
системы и других систем организма, равно как и использовать такие виды,
которые хотя и были известны в лабораторном обиходе, но не использо-
вались для изучения проблемы компенсаторных приспособлений.
Применение нами в данной работе нового вида хирургического повреж-
дения нервной системы — поперечного сечения задней половины спинного
мозга — является первой нашей попыткой в этом направлении.
Начиная с 30-х годов прошлого столетия многие ученые {ван Дин (van
Deen), Лонже (Longet), Шифф (Schiff), Ндвроцкий, К. В. Ворошилов.
1 Предварительное сообщение напечатано в сборнике «Рефераты работ учрежде-
ний Отделения биологических, наук АН СССР за 1940 г », М., 1941.
СООБЩЕНИЕ VIII ПЕРЕРЕЗКА ЗАДНЕЙ ПОЛОВИНЫ СПИННОГО МОЗГА
339
Г. И. Россолимо. Эвальд (Ewald), Бикель <Bikel) и в особенности В. М. Бех-
терев н его ученики — И. В. Боровиков, Ф. Ф. Гольцингер, Я. Б. Чуднов-
ский, а за последнее время В. П. Протопопов, Ферраро и Баррера (Ferraro а.
Barrera), М. А. Панкратов и др.] производили поперечную перерезку зад-
них столбов спинного мозга у низших позвоночных, птиц и млекопитающих
и стремились в условиях острого, а нередко и в условиях хронического
эксперимента выявить и исследовать роль этих столбов в проведении различ-
ных видов чувствительности — температурной, тактильной, проприоцеп-
тивной и болевой. При этом многие исследователи наблюдали, что у млеко-
питающих после поперечного сечения задних столбов спинного мозга,
наряду с более или менее глубоким и длительным расстройством тех или
иных видов чувствительности, во многих случаях выступают заметные на-
рушения двигательных функций пораженных конечностей, преимущест-
венно типа атаксий которые, однако, со временем постепенно и почти
полностью исчезают.
Убедившись в ряде собственных проверочных опытов на собаках, что
возникшие после поперечного сечения задних столбов спинного мозга
чувствительные и в особенности двигательные расстройства незначительны
по глубине и быстро проходят, мы решили пока воздержаться от дальней-
шего дополнительного изучения динамики нарушения и восстановления
функций организма После этой операции, а также от изучения роли коры
больших полушарий в развивающихся при этом компенсаторных приспо-
соблениях.
Как отмечалось выше, для решения этой основной задачи особен-
ный интерес представляют такие виды хирургического повреждения спин-
ного мозга или иных частей нервной системы, которые обусловливают глу-
бокие, длительные, но все же компенсируемые расстройства функций орга-
низма. С этой точки зрения наше внимание привлекли отдельные наблюде-
ния относительно глубоких последствий поперечного сечения всей задней
половины спинного мозга, попутно сделанные В. М. Бехтеревым и его уче-
никами при экспериментальном исследовании спинальных проводящих
путей, в котором они применяли, как правило, более легкие виды хирур-
гического повреждения спинного мозга — одностороннюю или двусторон-
нюю перерезку задних его столбов и т. п. В то время как у лягушек пере-
резка задней половины спинного мозга не вызывает заметных изменений
его рефлекторной деятельности, как это давно было показано II. М. Сече-
новым, Шиффом и др., у собак, как явствует из упомянутых выше единич-
ных опытов В. М. Бехтерева и его учеников, та же операция вызывает очень
глубокие и длительные нарушения чувствительных и двигательных функ-
ций организма.
С сожалением приходится отмстить не только редкое, до известной сте-
пени даже случайное применение В. М. Бехтеревым и его учениками попе-
речной перерезки всей задней половины спинного мозга у подопытных собак,
но и то обстоятельство, что последствия этой операции у своих подопыт-
ных животных они изучали, как правило, в условиях кратковременных, во
всех отношениях порочных острых—вивисекционных опытах. А это не могло
не придать полученным данным половинчатый и в известной мере даже про-
тиворечивый характер. Так, например, В. М, Бехтерев на основании постав-
ленных им еще в 1884 г. опытов пришел к выводу, что после перерезки зад-
ней половины спинного мозга в области грудных его сегментов «животное
еще вполне ясно воспринимает болевые раздражения, в чем легко убедиться
по реакции его в форме криков и явлений общего беспокойства на раздра-
жение электрическим током и уколами задней части туловища и задних
конечностей»1. Вслед за этим В. М, Бехтерев утверждает, что в подобном же
1 В. М. Бе xi грев, Основы учения о функциях мозга, т. III, 1905, стр. 832,
22*
340 9. Л. АСРАТЯН
опыте, позже поставленном его учеником Ф. Ф. Гольцингерои, «результат
t-казался тем же». Между тем из приведенного в диссертации Ф. Ф. Голь-
цннгера описания этого единственного его опыта явствует, что утверждение
В. М. Бехтерева относительно идентичности полученных им и
Ф. Ф. Гольцингером результатов лишено основания, что даже в этом
главном для их исследований пункте результаты Ф. Ф. Гольцннгера прямо
противоположны результатам подобных же опытов ею учителя. В описа-
нии этого опыта Ф. Ф. Гольцннгер указывает, что после перерезки у со-
баки задней половины спинного мозга в области грудных сегментов «при
последующем затем раздражении наведенным током с задних лап не вызы-
валось болевой реакции при надвигании катушек до 0 см»1.
Из всего вышеизложенного со всей очевидностью вытекает вывод не
только о малоцеипости этих фак t ячеек их данных для проблемы компенса-
торных приспособлений, но и о том, что на их основании трудно даже со-
ставить себе точное представление о подлинной картине выпадения функ-
ций организма после перерезки задней половины спинного мозга.
Наши критические замечания в еще большей степени относятся к резуль-
татам тех острых опытов Фултона (Fulton) и его сотрудников, в которых
они в целях изучения спинального шока у части подопытных кошек про-
извели перерезку задней половины спинного мозга.
Во всей известной нам отечественной и зарубежной литературе мы нашли
описание ЛИШЬ двух собак С поперечной перорезкой задней половины спин-
ного мозга, которые были объектами более или менее длительных наблюде-
ний и поэтому представляют значительный интерес для изучений Проблемы
компенсаторных приспособлений. Одна из них описана в диссертации Осава
(Osawa, 1882), другая — в диссертации И. В. Боровикова (1900). И та,
и другая собака были для своих хозяев как бы случайными средн много-
численных других собак, подвергавшихся иным видам хирургического
повреждения спинного мозга, стоящим в центре внимания исследователей.
Очевидно, поэтому эти две собаки не исследовались и ие описывались с
должной систематичностью и подробностью, о чем нельзя не сожалеть.
В диссертации Осава, например, вся картина нарушения и динамики
восстановления функции у собаки (опыт№ 12) исчерпывается следующими
короткими замечаниями: на второй день операции у собаки «обе задние ко-
нечности были парализованы и лишены чувствительности»; на 4-й деньона
«чуть приподнимала таз»; на 7-й день «виляла хвостом»; на 17-й день
«стояла на всех четырех конечностях и делала отдельные шатающиеся
шаги».
Через два месяца ходьба значительно улучшилась, а чувствитель-
ность полностью еще не была восстановлена. В этом состоянии собака была
оперирована вновь.
Из коротких протокольных заметок И. В Боровикова, приведенных
в его диссертации, можно узнать, что его собака (наблюдение 13-е) на второй
день после операции все время лежала с вытянутыми вперед ригидными
задними конечностями, причем чувствительность задней половины ее тела
была «слетка понижена», а сухожильные рефлексы у нее были «повсюду
сохранены».
В диссертации И. В. Боровикова отсутствуют какие бы то пи было
сведения относительно состояния собаки в последующие шесть после-
операционных дней, а в отношении дальнейших перемен отмечено следую-
щее: па 8-й день операции она пыталась ходить на передних лапах, волоча
за собой заднюю часть тела; па 18-й день стала приподнимать заднюю поло-
вину туловища; на 24-н день стала кое-как пользоваться задними конеч-
1 Ф. Ф. Гольц иигер, Чувствующие пути в спинном мозгу, дисс., Спб.,
1896, стр. 51.
СООБЩЕНИЕ VIII. ПЕРЕРЕЗКА ЗАДНЕЙ ПОЛОВИНЫ СПИННОГО МОЗГА
3’1
костями; после этого в течение месяца жизни у собаки несколько улучши-
лась двигательная активность задних лап, но все же походка оставалась
пйретическИ-атактической. К этому времени кожная чувствительность
задней половины тела собаки все еще была понижена, и, го словам
И. В. Боровикова, у нее совершенно отсутствовало «понятие о положении
задних конечностей».
Из всего этого явствует, что у описанной собаки, несмотря на тяжелое
повреждение спинного мозга, имело место постепенное и значительное вос-
становление нарушенных Функций организма, по меньшей мере двига-
тельных функций, В этом именно и заключается известное положительное
значение полученных иа этой собаке фактических данных для интересую-
щей нас проблемы.
Наряду с этим, нельзя не отметить одно досадное обстоятельство в ра-
боте И. В. Боровикова, которое для проблемы компенсаторных приспо-
соблений и вообще для науки существенно умаляет также значение факти-
ческого материала, полученного на этой собаке, единственной не только
у И. В. Боровикова» но и среди всех подопытных животных В. М. Бех-
терева и его учеников, прожившей некоторое время после порорезки задней
половины спинного мозга. Мы имеем в виду явное противоречие в харак-
теристиках, данных. И. В. Боровиковым в различных местах его диссерта-
ции последствиям поперечной перерезки задней половины спинного мозга
у этой собаки (наблюдение 13-е), что совершенно дезориентирует читателя.
Так, в протокольных записях И. В. Боровикова, кратко изложенных Ъыше,
отмечается, что на второй день операции у собаки на уколы булавкой -«чув-
ствительность задней половины слегка понижена», на 8-й день
«уколы иголкой на задн ей половине туловища с л а б е е, ч е м на пе-
редне й»1 (разрядка наша. —Э. Л.), а в дальнейшем картина тактиль-
ной и болевой чувствительности кожи как будто даже продолжает улуч-
шаться. В других же местах своей диссертации, касаясь этой темы, U.B. Бо-
ровиков дает совершенно другую характеристику состояния кожной чув-
ствительности задней половины тела этой же собаки после спинальной
операции. На стр. 142 диссертации, например, он пишет по этому поводу:
«Полная потеря кожной чувствительности тактильной и болевой
в наших наблюдениях была только в наблюдении 13-м, после перерезки
задней половины мозга» (разрядка наша. — Э. Л.). Указания на полную
потерю кожной чувствительности в задней половине туловища этой собаки
после операции встречаются также на стр. 144 и 146 диссертации.
Так и остается загадкой для читателя: какая же из двух взаимоисклю-
чающих характеристик кожной чувствительности задней половины туло-
вища оперированной собаки (наблюдение 13-е) соответствует действитель-
ности?
Если ко всему этому добавить, что самая операция поперечной перерезки
задней половины спинного мозга у этой единственной в своем роде собаки
была осуществлена несовершенно (резкая асимметрия, повреждение при-
мерно двух третей всей массы спинного мозга с задне-правебоковой сто-
роны), то станет совершенно закономерным вывод о том, что в работе
И. В. Боровикова оставалась невыясненной как истинная картина наруше-
ний двигательных, чувствительных и вегетативных функций организма
после поперечной перерезки задней половины спинного мозга, так и
динамика последующего восстановления нарушенных и утраченных
функций.
Между тем важность экспериментальной разработки этих вопросов была
очевидна.
1 И. В. Б о ровиков. Значение задних столбов спинного мазга в проведении
мышечного чувства, дисс., 1900, стр. 116.
9 42
Э А АСРАТЯН
Собственные экспериментальные данные
Мы поставили перед собой две задачи: во-первых, детально и система-
тически изучить в условиях длительного хронического эксперимента кар-
тину нарушения и динамики восстановления функций высокоразвитого
организма после перерезки всей задней половины спинного мозга, т. е.
задних столбов и задних половин боковых столбов; а во-вторых, выяснить
в. соответствии с основной целью нашей работы роль коры больших полу-
шарий мозга в ожидаемых после этой операции компенсаторных приспо-
соблениях.
Для решения этих задач нами за период 1940—1944 гг. были опериро-
ваны в общей сложности 20 взрослых собак и 3 щепка 3— 4-месячного воз-
раста. У всех 23 подопытных животных в условиях асептических и анти-
септических операций мы перерезали заднюю половину спинного мозга
либо на уровне 2—3-го шейных сегментов, либо на уровне 7—10-го грудных
сегментов.
Особенный интерес для решения первой задачи представляет материал,
полученный нами в экспериментах на 16 взрослых здоровых собаках, из
которых 6 были оперированы в 1940 г. и 10 — в 1943—1944 гг. Эксперименты
на 3 щенках интересны в несколько ином отношении и поэтому будут
описаны в отдельной статье совместно с другими данными по возраст-
ной физиологии компенсаторных приспособлений. Материал об остальных
взрослых подопытных собаках этой серии менее показателен, так как в
этих случаях спинальная операция либо комбинировалась с удалением
коры одного полушария большого мозга (у 3 собак), либо производилась
на собаке, лишенной полушарий (1 собака).
Для решения второй из интересующих нас задач мы в 1940 г. и в первой
половине 1941 г. у 10 взрослых подопытных собак и у 3 щенков спинальную
операцию в разной последовательности и комбинациях сочетали с опера-
цией удаления коры одного и двух полушарий большого мозга. Хотя, к глу-
бокому нашему огорчению» полученные в результате таких комбинирован-
ных операций данные значительно уступают результатам других вариантов
хирургического повреждения организма, тем не менее они дают достаточно
ясный ответ на поставленный вопрос.
Последствия поперечной перерезки задней половины
спинного мозга
У всех взрослых собак поперечная перерезка задней половины спинного
мозга вызывала исключительно глубокие и довольно длительные наруше-
ния чувствительных, двигательных и вегетативных функций организма,
связанных с частью спинного мозга, находящейся ниже уровня перерезки
Суть этих нарушений сводится в основном к развитию вскоре после опера-
ции глубокой депрессии соматической и вегетативной рефлекторной актив-
ности этой части спинного мозга, приближающейся к полной арефлексии,
а также к крайнему ослаблению всех видов кожной чувствительности,
приближающемуся почти к полной потере ее. Исчезали или резко ослабе-
вали также сухожильные рефлексы пораженных конечностей. При этом
нами был установлен очень интересный факт. В стадии почти полной
анестезии и арефлексии пораженных частей тела оперированные животные
довольно энергично двигали этими частями, когда им показывали мясо, по-
глаживали голову или звали их по имени, т.е. когда безусловнорефлекторно
или условнорефлекторно раздражали рецепторы головы.
Состояние глубокого спинального шока без существенных изменений
сохранялось в течение 4—6 дней, иногда и дольше, после чего начиналось
весьма медленное и постепенное восстановление угасшей рефлекторной
активности пораженного отрезка спинного мозга, а также еще более мед-
СООБЩЕНИЕ VIII. ПЕРЕРЕЗКА ЗАДНЕЙ ПОЛОВИНЫ СПИН НОГО МОЗГА
343
ленное и постепенное восстановление кожной чувствительности поражен-
ной части тела. Эти положительные сдвиги в процессе дальнейшего своего
развития приводили к восстановлению нарушенных функций стояния
(15—25-й день), ходьбы и бега (20—40-й день и позже).
У 6 собак спинальная операция была произведена на уровне 7—
10-го грудных сегментов и у 10 — на уровне 2—3-го шейных сегментов.
Последствия Поперечной перерезки задней Половины спинного мозга на
уровне верхних шейных сегментов отличаются от последствий аналогичной
операции на уровне нижних грудных сегментов не только более широким
диапазоном нарушенных функций, но и значительно большей глубиной и
длительностью поражения этих функций. Очевидно, последнее обстоятель-
ство обусловливается как топографическим положением шейных сегментов,
так и присущей им способностью к более сложной л высокой координацион-
ной деятельности по сравнению с остальными сегментами спинного мозга.
Из 16 собак у 3 поперечная перерезка задней половины спинного мозга
была сделана неудачно. У одной собаки с половинной перерезкой спинного
мозга па уровне 7—8-го грудных сегментов и у другой — на уровне
2—3-го шейных сегментов чувствительные, двигательные и вегетативные
функции были нарушены очень глубоко, процесс же восстановления
этих функций протекал у них крайне медленно, вяло и в конечном итоге
доспи невысоких уровней. Несмотря на то, что собаки жили после опера-
ции в течение многих месяцев, они так и не научились стоять и ходить.
Впоследствии было установлено, что у них было перерезано значительно
больше половины поперечника спинного мозга.
У третьей собаки последствия спинальной операции на уровне 7—8-го
грудных сегментов оказались совершенно нетипичными: она все время
кричала и визжала, возбудимость как верхней, так и нижней половины
тела была резко повышена, прикосновение к ним вызывало неистовый
визг собаки, а их рефлекторная активность не была подавлена. Спустя
несколько дней болезненность у собаки утихла, и у нес необычайно быс-
трыми темпами восстановились нарушенные функции, в том числе функции
стояния и ходьбы. Впоследствии было установлено, что у этой собаки
было перерезано значительно меньше половины поперечника спинного
мозга.
Картина нарушения функций и динамики их постановления при удач-
ной операции спинного мозга па каждом из этих двух уровней такова.
При поперечной перерезке задней половины спинного мозга на уровне
7-10-го грудных сегментов почти полностью парализуются задние конеч-
ности и хвост животных. В первые 3—4 дня операции эти части не реаги-
руют рефлекторно нн на собственное раздражение, ни на раздражение зад-
ней половины туловища, если даже в качестве раздражителя применяются
весьма сильные механические, электрические и термические воздейст-
вия—сильное сдавливание, щипок, укол, пробирки с ледяной водой
или с горячей водой 80—90°, сильный индукционный ток и т. п. При
подобного рода раздражениях нижней половины тела животных они не ре-
агируют, как правило, и движениями передних конечностей, головы, уш-
ных раковин, глаз или же какой-нибудь голосовой реакцией. Лишь в по-
рядке исключения, к тому же преимущественно у молодых собак, иногда
при сильных раздражениях хвоста, задних конечностей или задней поло-
вины туловища можно наблюдать еле заметную рефлекторную реакцию со
стороны задней или передней половины тела. В противоположность этому
даже умеренное механическое, термическое или элек трическое раздражение
передних конечностей, головы и вообще всей передней половины тела, равно
как показ мяса, свист или произнесение клички собаки вызывают не только
рефлекторные реакции со стороны органов передней части тела, но и четкие
344
Э. А. АСРАТЯН
рефлекторные сокращения нижних конечностей и в особенности хвоста.
То же наблюдается и при самостоятельных «усилиях» собак. Этот установ-
ленный нами любопытный феномен одностороннею проведения импульсов
по спинному мозгу весьма четко выявляется с первого и особенно со
второго дня после операции, т. е. еще в самом разгаре глубокого спи нал ь
ного шока.
Поперечная перерезка задней половины тела на уровне 7—10-го груд-
ных сегментов вызывает также нарушения вегетативных функций организ-
ма. Прежде всего следует отметить значительное расширение сосудов по-
раженных частей тела и в первую очередь нижних конечностей, что приво-
дит к повышению их температуры на 3—4°. Далее, у некоторых собак мы
наблюдали выраженное расслабление тонуса анальных сфинктеров, а так-
же расстройство дефекации и мочеиспускания.
Состояние такого глубокого спинального шока у собак держится
примерно 4—5 дней, после чего начинается медленное и постепенное его
ослабление и исчезновение. На- 4-й, иногда даже на 3-й день после операции
нижние конечности и хвост начинают весьма слабо реагировать на их силь-
ное механическое, термическое или электрическое раздражение, при этом
реагирует только раздражаемый орган. В следующие дни в ответ на такие
же раздражения собственная рефлекторная реакция каждой раздражаемой
части тела заметно усиливается, и, кроме того, при этом появляются слабые
рефлекторные реакции со стороны других, нераздражасмых частей задней
половины тела. Отметим, что все вызываемые в этой стадии прямые н
непрямые рефлекторные сокращения задних конечностей и хвоста в ответ
на их же раздражения имеют весьма вялый, инертный, как бы тонический
характер и быстро ослабевают, в то время как рефлекторные реакции тех же
частей тела, вызванные показом мяса, свистом, поглаживанием Головы
или произнесением клички собаки, носят характер быстрых четких сокра-
щений и ослабевают не скоро. На 6—8-й день операции восстанавливается
также проводимость спинною мозга снизу вверх. Раздражение задних ко-
нечностей и хвоста вызывает не только более четкие и несколько более быст-
рые рефлекторные их сокращения, по начинает вызывать также весьма
слабые рефлекторные реакции со стороны органов передней части тела
в виде движений передних конечностей, поднимания или поворота головы,
движений ушных раковин или глазных яблок. В этой фазе восстановления
функций собаки обычно уже могут выпрямлять переднюю часть тела и
сидеть.
В дальнейшем порог возбудимости задней половины тела все более сни-
жается, рефлекторные реакции задних конечностей и хвоста все более уси-
ливаются и убыстряются, параллельно облегчается взаимосвязь и усили-
вается взаимодействие между частями центральной нервной системы, лежа-
щими выше и ниже уровня задне-поперечной гемисекцим спинного мозга.
Все эти положительные сдвиги приводят к тому, что к 9—10-му дню после
операции собаки начинают приподниматься на широко расставленных
передних конечностях и медленно передвигаться с их помощью, волоча за
собой фактически еще бездействующую заднюю половинуг тела. Еще в этой
стадии восстановления функций собаки иногда делают попытки как бы
рывком встать на все четыре конечности, опираясь при этом на широко рас-
ставленные передние конечности и на нижнюю половину тыльной поверх-
ности сильно отогнутых назад задних конечностей. Но в этой позе они удер-
живаются не больше 10—15 секунд н затем падают. В течение ближайших
дней собаки не только чаще и успешнее делают такие попытки и не только
дольше удерживаются в такой своеобразной позе стояния, но приобретают
способность вставать на четыре конечности плавно, без рывка, а также пе-
редвигаться на четырех конечностях иа короткие дистанции. При этом зад-
ние конечности все более и более приближаются к вертикальному положе-
СООБЩЕНИЕ VIII. ПЕРЕРЕЗКА ЗАДНЕЙ ПОЛОВИНЫ СПИННОГО МОЗГА
345
нию, уменьшается площадь соприкосновения тыльной их поверхности с по-
лом и усиливается их активность.
Большинство собак через 20—25 дней после операции научается ДО-
ВОЛЬНО хорошо стоять и ходить, отличаясь от нормальных собак тем, что
у них задние конечности при этом широко расставлены, значительно- сог-
нуты во всех суставах и поэтому несколько- укорочены, при ходьбе совер-
шают атактические движения, в силу чего задняя часть тела раскачивает-
ся. Кроме того, при ходьбе собаки нередко опираются на тыльную поверх-
ность плюсны задних конечностей, временами переплетают их и падают на
землю, в особенности при быстрых передвижениях и поворотах. Через
5—10 дней сглаживаются и эти шереховатости в функциях стояния и
ходьбы оперированных животных, и становится довольно трудно отли-
чить их от нормальных собак.
Постепенно компенсируются также нарушенные вегетативные функции
организма, причем дефекация и мочеиспускание восстанавливаются в те-
чение 7—10 дней, а тонус сосудов пораженных частей тела, — насколько
об этом можно судить по уровню кожной температуры этих частей —
восстанавливается весьма медленно, в течение Р/а—2 месяцев и
дольше.
Полученный на этих собаках экспериментальный материал нельзя
считать достаточным для детальной и днференцнровочнон характеристики
динамики восстановления отдельных видов кожной н мышечной чувстви-
тельности задней половины тела, но он позволяет установить принципиаль-
но важный факт, что со временем все виды чувствительности пораженных
частей тела восстанавливаются и достигают почти прежнего уровня, при-
чем значительно позже восстанавливается болевая чувствительность.
В этой связи заслуживает внимания следующий факт: одна собака (Змейка),
спустя месяц после операции, т. е. после того как все виды чувствитель-
ности пораженных конечностей уже были восстановлены, отгрызла кон-
чики пальцев своей левой задней лапы.
При поперечной перерезке задней половины спинного мозга на уровне
2—3-го шейных сегментов, как уже было отмечено, нарушается более
обширный круг функций организма, чем при такой же перерезке на уровне
нижних грудных сегментов: парализуются все четыре конечности и хвост,
резко падает тонус наружных брюшных мышц, расширяются все поверх-
ностные сосуды (если суди ть по кожной температуре), появляется значитель-
ная брадикардия, пульс делается весьма мягким, заметно суживаются глаз-
ные щели, расслабляется третье веко, суживаются зрачки и спадают глаз-
ные яблоки, расстраиваются мочеиспускание и дефекация, животные ста-
новятся термолабильными; судя по внешним показателям, значительно
подавляется также рефлекторная активность шейных и жевательных мышц,
голосового аппарата и церебральная деятельность животных. Нами было
отмечено также, что при высокой перерезке задней половины спинного моз-
га функции организма нарушаются значительно глубже и остаются дольше,
чем при подобной же низкой его перерезке. Почти все фазы нарушения и
в особенности фазы восстановления двигательных, чувствительных и веге-
тативных функций у этих животных значительно удлинены по сравнению
с собаками с низкой перерезкой задней половины спинного мозга. При этом
если фазы соматической арефлексии, анестезии и аналгезии удлиняются
всего на несколько дней, то фазы выпрямления тела, стояния и ходьбы, а
также фазы восстановления вегетативных функции соответственно удлиня-
ются на 10—12 дней, а иногда и более.
Картина нарушений и.динамики восстановления функций у собак с вы-
сокой перерезкой задней половины спинного мозга представляет интерес
и в другом отношении. Заслуживает внимания, например, тот факт, что
функция задних конечностей нарушается глубже и восстанавливается мед-
346 э А АСРАТЯН
леннее, чем функция передних конечностей. Это находит выражение не
только в более глубокой депрессии собственной рефлекторной активности
задних конечностей по сравнению с передними, но и в том, что в стадии глу-
бокого спинального шока передние конечности легче и энергичнее задних
реагируют на раздражения рецепторов головы или на самостоятельные
«усилия» собак. Далее, особенный интерес представляют следующие факты:
в начальной стадии восстановления функции стояния собаки стоя
сильно сближают передние конечности, зачастую даже перекрещивают их,
а задние конечности, наоборот, широко расставляют. В начальной же ста-
дии восстановления функции ходьбы они наподобие кроликов передвигаются
рывками, делая при этом синхронные и однотипные прыжкообразные дви-
жения задними конечностями; только потом собаки научаются делать
раздельные ритмические шагательные движения каждой лапой. У этих со-
бак, в особенности немолодых, даже много месяцев спустя после операции,
сохраняется атактическая, шатающаяся и барахтающаяся походка, глав-
ным образом при пользовании задними конечностями, что приводит к часто-
му падению животных при ходьбе и беге Характерно также, что у собак
в течение многих месяцев так и не восстанавливается способность
фиксации кости, хлеба и других предметов передними лапами.
Наконец, заслуживает внимания то обстоятельство, что животные быст-
ро устают при ходьбе и особенно при беге, у них нередко появляется одыш-
ка, в силу чего они часто садятся отдыхать. Эта повышенная утомляемость,
обусловленная, по всей вероятности, в основном астенией спинальных цент-
ров, также сохраняется у многих собак в течение длительного времени
после спинальной операции.
В заключение краткого описания картины нарушении и динамики вос-
становления функций у собак После поперечной перерезки задней ПОЛОВИНЫ
спинного мозга на различных его уровнях приведем результаты проведен-
ных на этих собаках хронаксиметрии, тонуеометрии и исследований услов-
норефлекторной деятельности.
У 4 из этих собак до и в различные периоды после операции ме-
тодически измерялись реобаза и хроиаксия малоберцового и большебер-
цового нервов, а также реобаза и хронаксня перекрестных спинальных
рефлексов в ответ на раздражение большеберцового нерва (опыты А. И. Ка-
рамяна). В опытах было установлено, что в результате поперечной пе-
ререзки задней половины спинного мозга резко увеличивается реобаза
названных нервов (большеберцового нерва в 6—8 раз, малоберцового —
в 2—3 раза), а их хронаксня укорачивается, правда, в незначительных
пределах. Реобаза ре<|шексов при этом увеличивается в еще больших раз-
мерах, чем реобаза нервов, значительно удлиняется и хронаксня. В тече-
ние первых 15—20 дней после операции эти сдвиги в реобазе и хронаксии
двигательных нервов и особенно рефлексов пораженных конечностей умень-
шаются в незначительных пределах, а в дальнейшем, по мере восстановле-
ния нарушенных функций организма, темпы возвращения реобазы и хро-
наксии нервов и рефлексов к исходному уровню значительно ускоряются и
в конечном итоге почти достигают исходных величин.
Измерение тонуса мышц задних конечностей до и после спинальной опе-
рации показало, что тонус сгибателей после операции заметно усиливается,
а тонус разгибателей значительно ослабевает. По мере восстановления на-
рушенных функций организма сглаживаются также изменения в тонусе
мышц (опыты А. И. Карамяна).
Для нас особенный интерес представляют результаты исследования
двигательных электрооборонительных условных рефлексов у 3 из этих
собак до и после спинальной операции (опыты Г. И. Бокова и А. Л. Нови-
ковой). У 2 собак (Змейка и Бес) условные рефлексы вырабатывались
как с передней левой, так и с задней левой конечности (в последующем one-
СООБЩЕНИЕ VIII. ПЕРЕРЕЗКА ЗАДНЕЙ ПОЛОВИНЫ СПИННОГО МОЗГ А 347
рация спинного мозга у них была произведена на уровне?—8-го грудных сег-
ментов), а у одной собаки (Фокс)—с передней левой конечности (в последую-
щем операция спинного мозга у нее была произведена на уровне 2—3-го
шейных сегментов).
У 2 из этих собак (Змейка и Фокс) операция поперечной перерезки
всей задней половины спинного мозга была произведена успешно, и у них
наблюдалась описанная выше типичная картина нарушения и восстановле-
ния чувствительных, двигательных и вегетативных функций пораженных
органов и систем организма. Полученные на этих собаках данные свидетель-
ствуют о том, что поперечная перерезка задней половины спинного мозга
влечет за собой не только полное исчезновение всех двигательных электро-
обороннтел ьных условных рефлексов с пораженных конечностей на звукс-
Рнс, 1. Кинограмма электрооборонитсльных двигательных условных реф-
лексов с передней левой лавы собаки Фокс
а — в норме; <2—через 16 дней после поперечной перерезки задней половины спинного
мозга на уровне верхних шейных сегментов.
Обозначения (сверху вниз): условные и безусловные рефлексы (опыты
А. А. Нониковоя и Г. И. Бокова); отметка условных раздражителей; отметка Сезус-
.'юнного раздражителя; время в секундах
вне, световые и кожно-механические раздражители, но, как было отмечено
выше, полностью исчезают также безусловные двигательные электрообо-
ронительные рефлексы на сильнейший индукционный ток. В то же время
у собаки с половинной перерезкой спинного мозга в области грудных сег-
ментов (Змейка) из всех двигательных электрооборон и тел ьных условных
рефлексов с передней конечности исчез только условный рефлекс на кожно-
механический раздражител ь.
На рис. 1 и 2 приведены кимограммы электрооборонительных двига-
тельных условных рефлексов до и после перерезки задней половины спин-
ного мозга на высоком (рис. 1) и низком уровне (рис. 2),
Эги наши данные недавно были подтверждены Л. С. Гамбаряном.
С точки зрения основной задачи данных исследовании — выяснения
роли коры больших полушарий и особенно условнорефлекторнон ее дея-
тельности в развитии компенсаторных приспособлений — знаменателен
следующий факт. Когда спустя 4—6 дней после операции начинали вос-
станавливаться электрооборонлтельные двигательные безусловные рефлек-
сы пораженных конечностей, но их опорные и локомоторные функции
все еще оставались глубоко расстроенными, в это время восстанавливались
также и условные электрообороннтельныс двигательные рефлексы. Правда,
в течение 10—15 дней, т. е. до заметного восстановления опорных и локо-
оторных функций пораженных конечностей, эти условные рефлексы не
3»8
Э. А. АСРАТЯН
отличались особым постоянством, но самый факт их существования и осо-
бенно факт их появления раньше восстановления опорных и локомоторных
функций заслуживает внимания.
Это явление наблюдалось и у третьей нашей собаки (Бес), у которой ока-
залось перерезанным меньше половины массы спинного мозга, в силу чего
соответствующие функции организма были нарушены у нес менее глубоко,
восстанавливались более быстрыми темпами.
Последствия экстирпации коры.больших полушарий
мозга
Во второй стадии исследования мы стремились выяснить роль коры боль-
ших полушарии в развивающихся после задней поперечной гемисекцми
спинного мозга компенсаторных приспособлениях также путем удаления
коры больших полушарии у части подопытных животных. Для решения этой
задачи мы осуществили в нескольких вариантах спинальную операцию
в сочетании с операцией удаления полушарий. Во-первых, у 4 из
описанных выше собак с поперечной перерезкой задней половины спинного
мозга (Бес, Фокс, Змейка, Ру) после достижения определенного более или
менее стабильного уровня развития компенсаторных приспособлений мы
Кличка собаки и примерный возраст Уропень пере- резки задней половины спинного мозга Дата опера- ции спинного мозга Дата удале- ния коры одного г и - шария большого мозга Дата удале- ния коры второго пол упыри я большого мозга Продолжи- тельность жизни собак после послед- ней операции
Бес, 2—3 года 7—8-й ГРУДНОЙ сегмент 17. VI1940 г. 25. IX 1940 г. I7.X1I 1940г* 2 дня
Змей ха, 2—3 » То же 5.VII 13. IX 6.XII 4 мес.
Фокс, 2—3 » 2—3-й шейный сегмент 5. VII 13. IX 6.Х 11 4 дня
Р у, 8—9 мес. То же 22.Х 17. XII 6.Х I I 1941 г. —
Тобо, 3—4 года 9—10-й Груд- ной сегмент 15.1 1941 г. 26.1 X 1939 г. 18. IV1940 Г. 8 дней
Домино, 8—9 мес. 2—3-й шейный сегмент 6.II 6. II 1941 г. 19. 1V19-I1 г. Бол ьше 5 мес.
Гамбит, 3—4 года То же 6.II 6.II 23. IV 2 дня
Май. 2—3 » » » 28. IV 28. IV — Больше 5 мес.
Цыган, 3—4 » 9—10-й груд- ноЛ сегмент 13.VI 1952 г. 13.VI 1951 г. I2.X I 1951 г. 7 мес. (в на- стоящее время соба- ка жива)
удалили кору одного полушария, а через некоторое время и кору другого
полушария. Далее, у двух здоровых собак (Тобо и Цыган) первоначаль-
но удалили кору обоих полушарий, а несколько месяцев спустя перереза-
ли заднюю половину спинного мозга на уровне последних грудных сегмен-
тов. Кроме того, у 3 здоровых собак мы первоначально одномоментно
перерезали заднюю половину спинного мозга на уровне верхних шейных
сегментов и удалили кору одного полушария, с тем чтобы после развития
компенсаторных приспособлений удалить и кору второго полушария.
Как видно из приведенной выше сводной таблицы, мы достигли сравни-
тельно скромных результатов: из 9 собак с комбинированной операцией
перерезки спинного мозга и удаления коры больших полушарий более или
менее долго жили, — а значит дали удовлетворительный материал для от-
вета на поставленный вопрос,—только 3 собаки.Остальные б собак погибли
в ближайшие дин после последней операции. Животные эти погибали не
СООБЩЕНИЕ VI11 . ПЕРЕРЕЗКА ЗАДНЕЙ ПОЛОВИНЫ СПИННОГО МОЗГА
349
ИЗ-33 осложнений самой операции (большая кровопотеря во время опера-
ции, послеоперационное кровотечение, нагноение и т. п.), а в силу тяжести
сочетания указанных операций. Собаки находились в состоянии сильного
беспокойства, у многих
из них появлялись дне-
пептические явления, —
все это быстро обессили-
вало их и приводило к
гибели.
Последствия удаления
коры одного полушария
оказались по существу
одинаковыми как у собак
с поперечной перерезкой
задней половины спинного
мозга на уровне нижних
грудных сегментов (Бес и
Змейка), так и у собак
с подобной же операцией
на уровне верхних шейных
сегментов (Фокс и Ру). У
всех собак исчезали раз-
вившиеся после первой
операции компенсаторные
приспособления со стороны
сензомоторных и отчасти
вегетативных функций ор-
ганизма. После спинальной
операции рефлекторная де-
ятельность пораженных
конечностей несколько сни-
жалась, оставаясь все же в
удовлетворительном состо-
янии; но в первые дни
после операции на полу-
шариях животные не в со-
стоянии были даже сидеть,
не говоря уже о стоянии
и ходьбе. Расстраивались
также функции тазовых
органов, в ^особенности у
тех собак, у которых эти
функции раньше были глу-
Рис. 2. Килограмма электрообороннтельных дви-
гательных условных рефлексов с задней левой
лапы собаки Змейка
боко нарушены спинальной
операцией. Изменения же
со стороны функций сердеч-
но-сосудистой системы, а
также со стороны кожной н
мышечной чувствительно-
а — п норме; б — через 25 дней после поперечной пере-
резки oil дней половины спинного мозга на уровне
7—8-го грудных сегментов; о через 2 месяца после той
же операции.
Обозначения ге же. что на рис. I (опыты
А. А. Новиковой и Д. И. Ханутиной)
ста пораженных в результате спинальной операции частей тела оказались
после удаления коры одного полушария неглубокими; во всяком случае
они ненамного превышали соответствующие изменения тех же функций, воз-
никающие после удаления полушарий у нормальных собак. Помимо зна-
чительного расширения сосудов и понижения кожной и мышечной чувст-
вительности на противоположной оперированному полушарию половине
тела, что бывает и у нормальных собак после удаления коры, мы наблю-
350
Э. А. АСРАТЯН
дали подобные сдвиги также в пораженных после спинальной операции
одноименных оперированному полушарию частях тела.
Со временем у этих собак происходило повторное восстановление вто-
рично нарушенных двигательных, вегетативных и чувствительных функ
ций организма. При этом процесс повторного развития компенсаторных
приспособлений имел такой же характер, также носил явные черты «вы-
учивания», протекал постепенно, как и процесс развития компенсаторных
приспособлений после первоначальной, т. е. спинальной, операции, от-
личаясь лишь несколько более быстрым темпом. Так, например, после по-
перечной перерезки задней половины спинного мозга на уровне 7—8-го
грудных сегментов собаки Бес и Змейка могли выпрямлять тело и сидеть
к 6—7-му дню после операции, неуверенно стоять на четырех конечностях,
неуклюже передвигаться к 15—18-му дню и примерно через месяц дости-
гали почти нормального осуществления функций стояния и ходьбы. После
же удаления коры одною полушария те же собаки могли сидеть уже к 3—
4-му дню, неуверенно стоять и передвигаться на четырех конечностях к 8—
10-му дню и достигали прежнего уровня в осуществлении этих функций
к 15—20-му дню после операции. Создавалось, однако, впечатление, что
после операции на полушариях уровень компенсаторных приспособлений
оставался несколько ниже прежнего. Это явление, можетбыть, в несколько
менее выраженной форме, наблюдалось также у собак Фокс и Ру, у которых
была произведена поперечная перерезка задней половины спинного мозга
на уровне верхних шейных сегментов: соответствующие фазы развития ком-
пенсаторных приспособлений после удаления полушарии у них наступали
примерно на &—10 дней раньше, чем после спинальной операции. Не вда-
ваясь в детальный анализ этого явления, отметим лишь, что оно бесспорно
в значительной мере обусловлено наличием глубокого и длительного спи-
нального шока после спинальной операции и сравнительно неглубокого и
недлительного шока после операции на полушариях.
Кроме более быстрых темпов развития, повторные компенсаторные
приспособления отличались от первичных также некоторыми специфиче-
скими особенностями, обусловленными в основном асимметричностью
операции удаления коры полушарии. Опорные и локомоторные функции
противоположных оперированному полушарию конечностей, а также чув-
ствительность и вазомоторные функции всей этой половины тела восстанав-
ливались медленнее и хуже соответствующих функций одноименной опери-
рованному полушарию конечностей Н половины тела. Сказанное находит
отражение, в частности, в том, что у этих собак противоположные опериро-
ванному’полушарию конечности при стоянии и ходьбе отставлялись далеко
от срединной линии, были несравненно более пдретичиы, часто опускались
на пол тыльной стороной и изгибались во всех суставах (в особенности пе-
редняя лапа), при ходьбе поднимались невысоко и, скорее, скользили по
полу, чем делали нормальные движения (в особенности задняя лапа).
Удаление коры одного полушария у ссбак повлекло за собой также
исчезновение всех элоктрооборонительных двигательных рефлексов. Вновь
появлялись они в среднем через 20—25 дней после операции, т. е. на-
много позже, чем после спинальной операции (рнс. 3). Рефлексы эти не
отличались особым постоянством: они периодически то появлялись, то
вновь исчезали.
После удаления коры одного полушария у ссбак вновь произошли силь-
ные сдвиги в реобазе и хронаксии моторных нервов и в особенности рефлек-
сов, свидетельствующие о сильном повышении порога возбудимости в од-
ном и другом случае. Через 20—25 дней после операции эти изменения
сгладились.
У 3 собак мы производили одномоментно спинальную операцию и уда-
ление полушарий (Домино, Гамбит, .Май). Прежде всего отметим интерес-
СООБЩЕНИЕ VII I. ПЕРЕРЕЗКА ЗАДНЕЙ ПОЛОВИНЫ СПИННОГО МОЗГА
351
ное явление, установленное при самой операции: поперечная перерез-
ка задней половины спинного мозга на уровне верхних шейных сегментов
так резко снижала артериальное давление и церебральные функции, что уда-
ление коры одного полушария, произведенное через 15—20 минут вслед
за этой операцией, почти не требовало наркотизировання животного и про-
текало почти без кровотечения
Картина нарушения функций, а также динамика их восстановления
у этих собак, как и следовало ожидать, отражали последствия обеих опе-
раций. В первые дни после операции был налицо довольно глубокий спи-
нальный шок, арсфлексия и анестезия туловища и конечностей; в то же
время собаки могли производить движения конечностями при раздражении
головных рецепторов или при самостоятельных «усилиях»; несколько
дней спустя началось постепенное восстановление рефлекторной деятель-
ности спинного мозга, а затем кожной и мышечной чувствительности, вы-
прямительных рефлексов, функций сидения, стояния и ходьбы, как и у
Рис. 3. Кимограмма электрооборон ительных двигательных условных рефлексов
у собаки Фокс (а) и у собаки Змейка (б) через 3 месяца после удаления коры
правого полушария большого мозга
Обозначен ня (сверху вниз): условные м безусловные рефлексы (опыты Л. А. Но-
виковой и Г. Н Бокова), отметка условных раздражителей, отметка безусловного раз-
дражи геля, время в секундах
нормальных ссбак после аналогичной спинальной операции. Разница за-
ключалась лишь в том, что у собак с комбинированными операциями конеч-
ности на стороне, противоположной оперированному полушарию, заметно
отставали от симметричных конечностей в темпах и степени восстановле-
ния нарушенных функций. Кроме того, из 3 собак у 2 взрослых (Гамбит и
Май) соответствующие фазы развития компенсаторных приспособлении и
весь процесс в целом значительно затянулись—примерно в Р/а раза,[а у
очень молодой собаки (Домино) этой разницы отмечено не было. У этих
3 собак мы наблюдали некоторые любопытные явления, которые либо
отсутствовали, либо не были четко выражены у других, иначе опериро-
ванных нами собак. В стадии восстановления функции ходьбы эти собаки
не делали круговых, манежных движений, как их делают собаки, лишен-
ные коры одного полушария. Объясняется это, по всей вероятности, тем,
что поперечная перерезка задней половины спинного мозга сильно сгла-
живает асимметрию в чувствительности обеих половин тела, которая обыч-
но возникает после удаления коры одного полушария и обусловливает
манежные движения. Далее, в начальной стадии восстановления функции
стояния эти собаки (вособенности собака Май) весьма неуклюже двигались
как вперед, так и назад.
Следует отметить резкое усиление у них чесательного рефлекса, кото-
рый вызывался с обширного рецептивного поля кожной поверхности вре-
352
Э Л АСРАТЯН
менами не только со стороны одноименной задней конечности, но и со сто-
роны противоположной задней конечности.
Влияние удаления коры обоих больших полушарий мозга на картину
компенсаторых приспособлений, развивающихся после поперечной пере-
резки задней половины спинного мозга, мы могли изучать только у 2 опе-
рированных нами собак (Змейка и Домино), живших длительное время после
завершающей операции. Остальные собаки, как было отмечено, погибли
вскоре после этой операции.
Однако указанные две собаки после удаления коры обоих полушарии
дали не во всех отношениях сходные результаты.
У молодой (2—3-летней) собаки Змейка 5 июля 1940 г. была перереза-
на поперечная задняя половина спинного мозга на уровне 7—8-гО грудных
сегментов. После успешного восстановления нарушенных при этом функ-
ций 13 сентября того же года у нее была удалена кора правого полушария
большого мозга, а после вторичного восстановления возникших после этой
операции повторно нарушенных функций 6 декабря того же года у собаки
была удалена и кора левого полушария большого мозга.
У Змейки ближайший период последнем завершающей операций про-
текал гладкой рефлекторная активность всех конечностей была на сравни-
тельно высоком уровне. На 9 й день после операции она уже могла опирать-
ся на передние лапы, приподнимать переднюю часть тела и передвигаться,
волоча за собой заднюю часть тела. Задние конечности при этом делали
«холостые» и хаотические сопутствующие движения, абсолютно не содейст-
вующие передвижению животного. Собака ие могла самостоятельно стоять,
опираясь на все четыре конечности, и удерживать искусственно придан-
ную ей позу без посторонней помощи. Эта картина опорных и локомоторных
функций у Змейки сохранялась'без существенных изменений в течение
4 месяцев ее жизни за следующим исключением. В последние I1/»
месяца у Змейки несколько усовершенствовалась функция стояния и пере-
движения на передних конечностях, она приобрела способность временами
весьма недолго и неуклюже стоять на всех конечностях без посторонней
опоры, не будучи в состоянии, однако, сделать при этом ни одного шага
вперед.
У другой совсем молодой (8—9-месячной) собаки Домино 6 февраля
1941 г. была произведена поперечная перерезка задней половины спин-
ного мозга иа уровне 2—3-го шейных сегментов и сразу же удалена
кора левого полушария большого мозга. После успешного и для этой ком-
бинированной операции очень быстрого восстановления нарушенных функ-
ций 19 апреля того же года была удалена и кора правого полушария боль-
шого мозга собаки.
На 3-й день после операции собака могла выпрямлять тело и делала без-
успешные попытки стоять. Па следующий день собака уже короткое время
стояла на четырех конечностях при посторонней поддержке; без опоры она
не могла удержаться в этой позе, конечности раздвигались в сторону, сги-
бались и собака ложилась па живот. В течение следующих 6—7 дней локо-
моторные функции у собаки постепенно улучшались, и на 12-й день опе-
рации она уже могла самостоятельно стоять на широко расставленных и
полусогнутых конечностях в течение 2—3 минут, медленно покачиваясь впе-
ред и назад. При попытках ходить собака падала на живот или на бок. На
17-й день операции собака могла не только тверже и дольше стоять (к тому
же при самых причудливых положениях сильно паретичных передних ко-
нечностей: они перекрещивались, одна лапа заходила за другую, одна
или обе конечности опирались на тыльную сторону пястья), но и весьма
своеобразно передвигаться: каждая нога «ходила» сама по себе, отсутство-
вала какая-либо согласованность между движениями конечностей. При та-
кой своеобразной дробной и некоординированной походке собака почти не
СООБЩЕНИЕ VIII. ПЕРЕРЕЗКА ЗАДНЕЙ ПОЛОВИНЫ СПИННОГО МОЗГА Д5Д
двигалась с места, то и дело падая вперед, назад или на бок. Эта картина
опорных и локомоторных функций без существенных изменении сохранялась
у Домино на протяжении 5 месяцев наблюдения.
Заслуживает внимания одна особенность, появившаяся у Домино пример-
но через 1 -I’ а месяца после последней операции на полушариях: при
необходимости быстро пе-
редвигаться (он ускорял
ходьбу перед мочеиспус-
канием, дефекацией или
при длительном голодании)
Домино моментально из-
менял свою обычную ма-
неру ходить — в течение
короткого времени делал
координированные движс
ния всеми конечностями,
ставя их правильно, не
переплетая их н не падая.
После этих кратю «времен
пых«приступов» координи-
рованной и правильно"
ходьбы к собаке вновь воз-
вращались ее привычные
дробные и некоординиро-
ванные движения, причуд-
ливые комбинации скован-
ных движений паретичны-
ми конечностями, приво
дящнх к передвижению
животного большей частью
путем кувыркания.
Следует отметить повы-
шенную активность чеса-
тельного рефлекса с об-
ширной рецепторной зоной,
а также то, что у Домино,
в отличие от других наших
собак с удаленной корой,
как будто имелись некото-
рые элементы приобретен-
ной нервной деятельности
(например, он мог находить
самостоятельно пищу и
есть ее, если миска с пи-
щей была поблизости от
него).
У 2 наших взрослых
подопытных собак (Тобо и
Цыган) вначале была уда-
лена кора обоих больших
полушарий мозга и спустя
много месяцев, когда бли-
Рнс. 4. Ообакя Цыгане удаленными полушариями
после перерезки адней половины спинного мозга
(объяснения см. в тексте)
жайшие последствия операционного вмешательства прошли и когда функ-
ции стояния и ходьбы достигли $ них предельного совершенства, была
произведена поперечная перерезка задней половины спинного мозга на
уровне последних грудных сегментов.
23 Э, А. Асрагяи
354
Э. А. АСРАТЯН
Ближайшие последствия спинальной операции у этих собак оказались
в основном такими же, как у нормальных. Как у одной, так и у другой
собаки поперечная половинная перерезка спинного мозга вызвала в ниж-
нем его отрезке типичный спинальный шок, лишь незначительно уступаю-
щий по своей глубине и длительности спинальному шоку у описанных
выше собак, а также заметное угнетение рефлекторной активности верх-
него отрезка спинного мозга. Следует отметить, что и у этих собак в
стадии глубокого шока рефлекторные движения задних конечностей при
их раздражении фактически отсутствовали, ио они двигались при раз-
дражении головы или передней части туловища или осуществляли движе-
ния по неуловимым нами причинам.
Отдаленные последствия поперечной перерезки задней половины спинного
мозга МЫ могли наблюдать только У одной из этих собак (Цыгана), так
как другая собака (Тобо) погибла на 8-н день после операции.
Двигательная активность передних конечностей И ГОЛОВЫ восстанавли-
валась у Цыгана довольно быстро. На 3-й день собака могла не только под-
нимать изредка голову, но временами менять свою позу с помощью передних
лап. В последующем эти движения производились все чаще н совершен-
нее, и на 15-й день операции собака приподнималась па передние лапы
и ползала с их помощью, волоча за собой бездействующую заднюю часть
тела (рис. 4 а). В этой стадии восстановления функции собака еще не могла
держаться на четырех конечностях, когда ее приподнимали и ставили на
задние конечности. Примерно через месяц после операции Цыган уже
кратковременно удерживал искусственно приданную ему позу стояния,
раскачиваясь из стороны в сторону задней частью тела. Постепенное вос-
становление функции передних конечностей и передней части тулови-
ща привело к тому, что спустя еще 10—15 дней собака стала при быстрой
ходьбе кратковременно поднимать заднюю часть тела с помещыо мышц спи-
ны и даже опираться на сильно разогнутые, вывернутые на тыльную сторо-
ну и бездействующие при этом задние конечности, как это часто делают со-
баки с полной перерезкой спинного мозга на уровне последних грудных
сегментов и ниже (рис. 4 б и в). Способность к такого рода пассивному стоя-
нию на задних конечностях и к такого рода передвижению сохраняется
у Цыгана до сих пор (через 7 месяцев после спинальной операции) без ка-
ких-либо заметных изменений. И хотя рефлекторная активность задних
конечностей давно достигла значительного уровня, тем не менее они не при-
нимают активного участия в осуществлении функций стояния и ходьбы.
Обсуждение полученных данных
Нами экспериментально изучались два до известной степени самостоя-
тельных вопроса: последствия поперечной перерезки задней половины спин-
ного мозга у взрослых собак и последствия удаления коры одного и двух
полушарий большого мозга у этих собак после восстановления нарушенных
в результате спинальной операции функций. Установленные при этом дан-
ные в основном сводятся к следующему.
В результате поперечной перерезки задней половины спинного мозга
у собак развивается глубокий спинальный шок. Это сопровождается почти
полной потерей всех разновидностей кожной и мышечной чувствительности,
полным двигательным параличом, а также глубоким расстройством ряда
вегетативных функций организма. Однако в течение 1—2 месяцев проис-
ходит постепенное почти полное восстановление всех нарушенных чувст-
вительных, двигательных и вегетативных функций у собак. Осуществле-
ние двигательных условных рефлексов пораженными конечностями
возможно еще на ранних этапах развития компенсаторных приспособлений,
а тем более после их завершения.
СООБЩЕНИЕ VIII. ПЕРЕРЕЗКА ЗАДНЕЙ ПОЛОВИНЫ СПИННОГО МОЗГА
355
Исчезновение развившихся компенсаторных приспособлений после уда-
ления коры одного полушария большого мозга и вторичное их развитие поч-
ти до прежнего уровня осуществляются в течение 20—45 дней.
После удаления коры обоих полушарий большого мозга отмечается почти
необратимое исчезновение компенсаторных приспособлений (у двух сс-
бак) или же их исчезновение и затем весьма слабое восстановление (у одной
собаки).
Остановимся на некоторых из этих фактов. Факт развития спинального
шока у собак после поперечной перерезки задней половины спинного мозга,
а также некоторые другие описанные нами последствия этой операции с пол-
ной очевидностью показывают ошибочность господствующей в современной
нейрофизиологии теории Шеррингтона (Sherrington) о сПИНЗЛЬНОм шоке.
Согласно этой теории, спинальный шок обусловлен перерывом усиливающих
импульсов, направляющихся из церебральных центров к двигательным нерв-
ным клеткам спинального мозга, и представляет собой состояние, подобное
утомлению. Сказанное в равной мере относится и к модернизированному
варианту этой теории, разработанному Лидделом (Liddell), Фултоном
(Fulton), Мак Кучем (McCouch), Риочем (Rioch) и другими американ-
скими и английскими физиологами и допускающему, что при спинальном
шоке изменяется состояние не только мотонейронов, но и вставочных нерв-
ных клеток рефлекторных дуг.
Ошибочность теории Шеррингтона явствует из того установленного нами
факта, что в стадии глубокого спинального шока, когда никакими раздра-
жениями пораженных частей тела собак не удается вызвать какую бы то
ни было рефлекторную деятельность этих частей, последние начинают
активно и энергично двигаться при показе ссбакам пищи, поглаживании
их головы и т, п. Другие наши данные также с полной очевидностью гово-
рят о том, что спинальный шок не обусловлен изменением состояния мото-
нейронов и вставочных нейронов рефлекторных дуг. Например, когда через
несколько дней после операции спинальный шок у наших ссбак ослабевал
и в ответ на раздражение пораженных конечностей появлялись слабые реф-
лекторные их сокращения, то сокращения мышц при этом носили вялый,
растянутый, тонический характер и мышцы быстро утомлялись. В противо-
положность этому те же мышцы в те же дни в ответ на раздражения рецеп-
торов головы осуществляли быстрые и сильные сокращения и не утомлялись
быстро. Наши экспериментальные данные со всей очевидностью показывают
также ошибочность выводов Фултона, Лиддела и Риоча о том, что спи-
нальный шок якобы не развивается при перерезке задней половины и даже
задних двух третей спинного мозга. Эта допущенная ими грубая ошибка обу-
словливается, повидимому, тем, что они проводили свои исследования в ус-
ловиях острого—вивисекционного опыта, поставленного к тому же на кош-
ках, т. е. на животных, у которых, как известно, спинальный шок вообще
не отличается особой глубиной и выраженностью.
Полученный нами новый экспериментальный материал относительно
спинального шока не только позволил нам еще в 1947 г. выступить с обосно-
ванной критикой порочных данных и ошибочных взглядов упомянутых
исследователей, но и послужил основанием для развитая новых и, как нам
кажется, более передовых, связанных своими корнями с учением гениаль-
ного И. П. Павлова теоретических положений об этом в.ажном вопросе
нейрофизиологии.
Согласно нашим взглядам, спинальный шок представляет собой длитель-
ное и глубокое охранительно-целебное торможение спинного мозга, выз-
ванное грубой органической травмой его структуры, в частности, перерезкой
всей или значительной части его поперечника.
Органическая травма как сложный и многосторонне действующий болез-
нетворный фактор способна в течение короткого времени в сильной степени
23*
356 Э. А. АСРАТЯН
ослабить и истощить нервную систему, т. е. создать главные предпосылки,
при которых, согласно учению И. П. Павлова, как раз и возникает охрани-
тельно-целебное торможение. В случае полной или частичной перерезки
спинного мозга роль травмы сводится прежде всего к тому, что она преры-
вает пути положительно-трофического (в понимании 11. П. Павлова) влия-
ния церебральных нервных образований па спинальные рефлекторные
центры и тем самым значительно ослабляет их, снижает их лабильность и
работоспособность. Кроме того, перерезка как чрезмерно сильный и слож-
ный раздражитель (механическое воздействие, нарушение кровоснабже-
ния, действие сгустков крови, местный отек, воспалительные явления,
дегенеративные изменения и г. п.) вызывает сильное длительное лерераз-
дражение этих центров, в еще большей степени утомляет, ослабляет, исто-
щает их и порождает в них длительное и глубокое охранительно-целебное
торможение.
Таким образом, новое в нашем понимании происхождения и сущности
спинального шока заключается не только в том. что мы учитываем комплекс-
ное действие двух факторов —г перерыва путей и перераздражения,
раздельному действию каждого из которых Гольц (Goltz) и Шеррингтон при-
писывают возникновение спинального шока, — ио и в том, что мы сводим
сущность шока к охранительно-целебному торможению, придаем этому
состоянию определенное биологическое содержание.
С развиваемой нами точки зрения может быть удовлетворительно объ-
яснен ряд связанных с перерезкой спинного мозга явлений, которые не по-
лучил объяснения или не был «замечен» последователями существующих
односторонних теорий о спинальном шоке. Например, Шеррингтон и его
последователи отрицали и отрицают изменения в состоянии той части спин-
ного мозга, которая лежит выше уровня перерезки. Более того, они это
обстоятельство считают наиболее веским аргументом против теории Гольца
и других менее распространенных теорий о спинальном шоке. А в дейст-
вительности каждый объективный исследователь всегда может констати-
ровать наличие неглубокого шока в виде некоторой депрессии в состоянии
и деятельности верхнего отрезка спинного мозга, а в случаях высоких пе-
ререзок спинного мозга — в состоянии и деятельности церебральных нерв-
ных образований. Эта депрессия находит свое выражение в значительном
повышении порога возбудимости рефлексов., в неспособности ссбак в те-
чение некоторого времени после операций выпрямлять переднюю часть
тела и передвигаться с помощью передних конечностей (в случае полной или
частичной перерезки спинного мозга на уровне средних грудных сегментов),
во временном парезе, в параличе голосового аппарата, а также некотором
состоянии оглушения церебральных функций (в случае полной или частич-
ной перерезки спинного мозга на уровне шейных сегментов).
Мы понимаем это следующим образом: в отношении частей спинного
мезга, лежащих выше уровня перерезки, и в отношении церебральных
нервных образований спинальная травма не является фактором, прерываю-
щим идущий к ним поток благотворных ТрсфИчёСКНх импульсов, каким она
является в отношении нижнего отрезка спинного мозга, а выступает только
в роли сильного раздражителя, способного ослаблять и истощать их в сла-
бой степени и порождать в них лишь сравнительно неглубокое охрани-
тельно-целебное торможение.
Весьма вероятно, что в развитии спинального шока в вышележащем и
даже нижележащем отрезках спинного мозга, особенно при его неполных
поперечных И продольных сечениях, некоторую роль играет еще один фак-
тор или «механизм», на который в середине прошлого столетия указывал
И. М. Сеченой, при объяснении явления угнетения рефлекторной деятель-
ности спинного мозга в результате перерезки боковой его половины,
а именно: рефлекторное угнетение деятельности спинного мозга со стороны
СООБЩЕНИЕ VIII. ПЕРЕРЕЗКА ЗАДНЕЙ ПОЛОВИНЫ СПИННОГО МОЗГА
357
вышележащих отделов нейтральней нервной системы, обусловленное дли-
тельным раздражением головных концов поврежденных сп инне мозговых
восходящих проводящих путей1.
Возможность получения спинальнсто шока при так называемся «хо-
лодовой перерезке» спинного мозга [Трен дел енбург (Trendelenburg)) не
противоречит нашей точке зрения. Нам кажется, однако, что холод как
менее сильный раздражитель должен вызывать менее глубокий спинальный
шок, чем хирургическая травма. Это предположение должно быть проверено
в специальных сравнительных экспериментах.
С нашей течки зрения нетрудно объяснить также тот факт, почему при
вторичной перерезке однажды уже перерезанного и уже ссвсбсдивщегсоя
от шока спиннсго мезга не развивается шск прежней глубины. Как извест-
но, этот факт Псстсянно приводится ШеррннгТспсМ и его нс слеДс ват елям и
как аргумент в пользу их воззрений и против положений Гольца. Не го-
воря уже о том, что при повторной перерезке отсутствует фактор перерыва
потока церебральных трофических импульсов, в данном случае травма из-
за дегенерации нисходящих проводящих путей не межет уже сб у слепли-
вать такого сильного и длительного нерераздражения нижележащих
спинальных центров и вызывать в них шок такой глубины, как при
первой перерезке. А то сбстсятельство, что при этом все же развивается
шок небольшой глубины и длительности, явно противоречит установкам
Шеррингтона и подчеркивает правильность нашей течки зрения.
Паше понимание происхождения и природы спинального шока расхо-
дится со взглядами Шеррингтона него последователей еще в одном весьма
существенном пункте, о чем упоминалссь выше. На основании эксперимен-
тальных данных о возможности сокращения собаками пораженных конечнос-
тей при раздражении рецепторов головы и самостоятельных «усилиях»
животных и невозможнее!# в то же время рефлекторного сокращения этих
конечностей при их непосредственном раздражении мы считаем, что при
спинальном шоке поражаются, альтерируют нс только и даже не столько
мотонейроны И вставочные нейроны, как полагают Шеррингтон, Лндел,
Мак Куч и др,, сколько чувствительные звенья рефлекторных дуг.Этот един-
ственно возможный из описанного факта вывод подтверждается также на-
шими данными о том, что при возникновении рефлекторной лея гель нести
пораженных конечностей в ответ на их непосредственные раздражения вна-
чале рефлекторные сокращения мышц носят вялый и тонический характер
и быстро утомляются; между тем те же самые мышцы под влиянием идущих
сверху импульсов осуществляют быстрые и сильные сокращения и утомля-
ются нескоро.
Наши теоретические положения о происхождении и природе спиналь-
ного шока, в частности, положения о преимущественном поражении сен-
зорных звеньев рефлекторных дуг при этом шоке, получили подтверж-
дение в специальных исследованиях, проведенных нашей сотрудницей
В- И- Дроздовой,
1 В последнее время в частности, в статье, опубликованной в сборнике «Заболе-
вание, лечение и выздоровление» (1962), М. Г. Дурмишьян высказывает мнение,
что развитие спинального шока почти всецело обусловливается этим «механиз-
мом» — сильным рефлекторным раздражением вышележащих центральных нервных
образований. Мы лишены здесь возможности обстоятельно разбирать точку зрения
М. Г Дурмитьяна, являющуюся механическим применением указанной идеи
И. М. Сеченова (о которой, кстати говоря, он не упоминает ни единым словом
ни в рукописи своей докторской диссертации, ни в названной статье) к объясне-
нию сложнейшего явления спинального июка у высших животных без учета
достижений физиологии по этому вопросу за ’целое столетне. Эта точка зрения
М. Г. Лурмишьяна в свете современных достижений физиологии в изучении спи-
нального шока представляется крайне односторонней и примитивной.
358
Э. А. АСРАТЯН
В. Н. Дроздова установила, в частности, что в стадии максимальной для
кошек глубины спинального шока не удается вызвать рефлекторное сокра-
щенне.хвоста при его непосредственном раздражении; в то же время сокра-
щение хвоста легко наступает при раздражении боковой поверхности ниж-
ней половины туловища. При глубоком спинальном шоке у ссбак, когда не
удается вызвать рефлекторное сокращение конечностей при непосредствен-
ном их раздражении, этого легко достигнуть при раздражении области ану-
са, lie касаясь здесь причин более глубокого торможения одних рефлексов
и менее глубокого торможения других у каждого вида животных, отметим
лишь, что м3 установленных В. Н. Дроздовой фактов со всем очевидностью
вытекает, что при спинальном шоке сензорные звенья рефлекторных Дуг
страдают больше, чем вставочные и конечные звенья1.
Из всего вышеизложенного явствует, что развиваемая нами точка зре-
ния на происхождение «природу спинального шока существенно отличает-
ся также от взглядов Гольца, хотя по отдельным вопросам между ними и
имеется некоторое сходство.. В отличие от Гольца мы, во-первых, признаем
важное значение перерыва благотворного трофического влияния церебраль-
ных нервных образований на спинной мозг. Во-вторых, мы установили преи-
мущественное поражение чувствительных звеньев рефлекторных луг при
спинальном шоке. Далее, мы считаем, что торможение при спинальном шоке
не является неизбежным злом или проявлением каприза природы, а имеет
важное биологическое, охранительно-целебное значение.
Ничего общего не имеется между нашими воззрениями на спинальный
шок и между опубликованными недавно американским физиологом Хар-
ревелдом (Harreveld) спекулятивными рассуждениями о спинальном шоке
как о состоянии угнетения, обусловливаемом активностью специальных
нервных образований, или, как он говорит, «специфического тормозного
механизма» спинного мозга, освобожденного перерезкой спинного мезга от
задерживающего влияния вышележащих нервных центров. Харревелд
не утруждал себя попыткой аргументировать свои рассуждения скслько-
иибудь убедительными фактами или хотя бы доводами. Тем самым он в зна-
чительной мере освободил своих оппонентов от необходимости высказы-
ваться обстоятельно по поведу его рассуждений.
Сказанное в известной мере относится также к попытке Розенблюта
(Rosenbluth) объяснить явление спинального шока с течки зрения развивае-
мого Кеннонам (Cannon) «закона денервации». Розенблют решает эту' за-
дачу весьма своеобразно: он по существу объявляет спинальный шок не-
существующим. Он считает, что перерезка спинного мозга не изменяет воз-
будимссти нижележащих его сегментов. Слабость рефлекторной активности
этих сегменте!?, по его мнению, объясняется не их угнетением, а лишь от-
сутствием облегчающего влияния со стороны высших отделов нервной си-
стемы; последующее усиление этой активности не есть восстановление преж-
него состояния нервных клеток этих сегментов, а результат ненормально
повышенной их возбудимости вследствие их «денервации» или, точнее,
дегенерации нисходящих цереброспинальных путей. Все эти абстрактные,
лишенные всякого фактического основания рассуждения Розенблюта
представляют собой не что иное, как неудачную попытку чисто созерца-
тельного, механического перенесения сложного вопроса о спинальном шоке
в другую весьма проблематичную плоскость в физиологии.
В свете развиваемых нами на основе идей И. П. Павлова общих теоре-
тических положений о возникновении охранительно-целебного торможения
в центральной нервней системе при ееорганических повреждениях спиналь-
ный шок, равно как и многие, известные под другими названиями явления
1 Полученный В. Н. Дроздовой фактический материал о спинальном шоке
частично уже опубликован, а в целой оформляется в настоящее время в качестве
кандидатской диссертации.
СООБЩЕНИЕ VIII. ПЕРЕРЕЗКА ЗАДНЕЙ ПОЛОВИНЫ СПИННОГО МОЗГА
359
депрессии функций тех или иных поврежденных частей центральной нервной
системы (диасхизис, центральный шок и т. п.) выступают не как сбоссб-
ленные, биологически непонятные явления в поврежденной центральной
нервней системе, а как частные случаи развития охранительно-целебного
торможения с его важнейшей биологической ролью. Правильность этих
теоретических положений была доказана нами и нашими сотрудниками
в многочисленных опытах с проведением терапии снсм ближайших и
отдаленных последствий тяжелых органических повреждений тех или иных
частей нервной системы или организма в целом.
Значительный интерес представляет также установленный нами факт
постепенного и почти полного восстановления чувствительных, двигательных
и вегетативных функции организма, нарушенных вследствие поперечной
перерезки задней половины спинного мозга.
Еще в 1937 г., при трактовке собственных и литературных данных
о компенсаторных приспособлениях, развивающихся после поперечной пе-
ререзки боковой половины спинного мозга, нами была высказана мысль
о том, что специализация и локализация ннтрацентральных проводящих
путей относительна, вариабил.ьна, динамична, т. е., по существу говоря,
была высказана мысль о возможности применения к проводящим путям
спинного мозга (подразумевая под этим названием соответствующие си-
стемы нервных клеток и их отростков) теории о динамической локализации
функции, разработанной И. П. Павловым в отношении коры больших по-
лушарий. В настоящее в.ремя мы в еще большей степени убеждены в том,
что это единственно правильная точка зрения, которая спссс б на объяснить
и приведенный выше экспериментальный материал относительно восстанов-
ления нарушенных и утраченных функций организма после другого вида
массивного повреждения структуры спинного мозга.
Н а самом деле, эти явлен ня не могут быть объяснены с точки зрения тех,
которые считают, что специфичность проводящих путей спинного мозга
неизменна, а их локализация постоянна и ограничена узкими границами,
обычно изображаемыми морфологами и невропатологами на карте попереч-
ного сечения спинного мозга. Наши экспериментальные данные никак не
могут быть объяснены также с точки зрения Бете и его единомышленников,
метафизически отрицающих всякую функциональную специализацию и
локализацию в центральной нервней системе всебще и в проводящих путях
спинного мозга в частности. Против этого положения Пете и его единомыш-
леннике в говорят как специфический характер функциональных расстройств
при повреждении определенных анатомических структур спинного мозга,
так и отсутствие «молниеносного» восстановления нарушенных и утрачен-
ных при этом функций (напомним, что, согласно воззрениям Бете, быстрое
восстановление нарушенных функций свидетельствуете быстром и автома-
тическом переключении нервных импульсов от одних частей якобы
аморфной в функциональном отношении «нервней сети* к другим).
Исходя из нашего положения о существовании ядерных и рассея нных
по всему поперечнику спинного мозга запасных нервных элементов для
каждого из основных специализированных восходящих и нисходящих
проводящих систем спинного мозга, об относительности и динамичности
функциональной специализации и локализации этих путей, легко объяс-
нить не только специфичность функциональных расстройств при повреж-
дениях определенных структур спинного мозга, но и тот факт, что даже
при повреждении всей массы ядерных нервных элементов какой-нибудь
определенной системы проводящих путей спинного мозга их со временем
заменяют запасные нервные элементы, и нарушенные функции восстанавли-
ваются.
Нет необходимости подробно излагать здесь наше понимание сущности
компенсаторных приспособлений, т. е. факторов и закономерностей акти-
360
Э. А. АСРАТЯН
нации или перестройки функции неповрежденных нервных образовании
с тем, чтобы они могли более или менее полноценно замещать выведенные из
строя нервные структуры. Наш и теоретические положения по этим вопросам
достаточно подробно изложены в предыдущих наших сообщениях. Полу-
ченный в данном исследовании новый материал всецело подтверждает наши
теоретические установки по проблеме компенсаторных приспособлений и
тем самым вновь доказывает ошибочность и абсурдность антиэвелкцион
ных, идеалистических воззрений Бете и его единомышленников на эту
проблему.
Наши положения о многообразии форм физиологических процессов,
лежащих в основе компенсаторных приспособлений в повреждением орга-
низме, в частности, основное наше положение о ведущей в этом явлении
роли коры больших полушарий с ее могущественной условнорефлекторнон
деятельностью, получают также новое подтверждение. Мы имеем в виду, во-
первых, медленный н строго постепенный характер развития компенсатор-
ных приспособлений, явно носящих черты тренировки и «выучивания»;
во-вторых, возможность условнорефлекторнсго управления деятельностью
пораженных конечностей Даже в самых начальных стадиях восстановления
их функции; в-третьих, исчезновение развившихся компенсаторных приспо-
соблений после удаления коры одного полушария большого мозга и вто-
ричное их восстановление; также медленное и постепенное, с возможно-
стью осуществления условнорефлекторной регуляции функций пораженных
частей тела, как и после спинальной операции; далее, вторичное исчезнове-
ние этих компенсаторных приспособлений пссле удаления коры другого по-
лушария большого мозга, притом исчезновение необратимее (у одной ссба-
ки) либо с последующим весьма слабым восстановлением нарушенных функ-
ций (у другой ссбаки); наконец, невозмсжнссть вс установления активных
опорных и локомоторных функций задних конечностей у лишенной коры
собаки после поперечной перерезки задней половины спинного мозга.
Нам кажется, что факт ограниченного восстановления у одной из ссбак
(Домино) нарушенных спинальной операцией функций даже после удаления
коры обоих полушарий большого мозга недостаточен для того, чтобы взять
под сомнение правильность положения о ведущей роли коры в развитии
компенсаторных приспособлений у высокоразвитых взрослых животных,
г. е. правильность основного вывода всей нашей исследовательской работы
по этой проблеме,основанного на огромном экспериментальном материале.
Во-первых» нами давно было установлено, что в случаях легкого или
умеренного повреждения структур и неглубокого нарушения функций ор-
ганизма ведущая рель коры больших полушарий мозга в развитии компен-
саторных приспособлений выступает менее отчетливо. В подобных случаях
нижележащие отделы центральной нервной системы, в первую очередь
подкорковые нервные образования с их богатыми возможностями безус-
ловнорефлекторной регуляции функций организма, а значит и перестрой-
ки, способны обеспеч ить в значительных пределах восстановление нарушен-
ных и утраченных функций. Весьма вероятно также, что пссле удаления
коры сбоих полушарий большого мозга функции неповрежденных частей
головного мозга компенсаторно усиливаются, а значит увеличиваются и их
возможности в указанном направлении.
Могло случиться, что мы не полностью перерезали у Домино всюзаднюю
половину спинного мозга. Некоторым основанием для такого предположе-
ния может служить то обстоятельство, что у этой собаки быстрее, чем
у других, восстановились нарушенные пссле спинальной операции функ-
ции. Могло случиться также, что при операции на полушариях у ссбаки
уцелели более или менее значительные части коры. Это можно предпола-
гать на том основании, что собака отличалась несвойственной собакам
с удаленным полушарием высокой активностью, могла, в частности, само-
СООБЩЕНИЕ VI IJ, ПЕРЕРЕЗКА ЗАДНЕЙ ПОЛОВИНЫ СПИННОГО МОЗГА
361
стоятелык» находить и есть пищу, если пища находилась неподалеку от нее.
К сожалению, нам не пришлось путем последующей анатсмо-гистслоги-
ческон проверки уточнить полноту проделанных нами операций наспин-
ном мозгу и па больших полушариях мозга этой собаки.
Одной из причин неполного восстановления у Домино нарушенных
спинальной операцией функций после удаления коры обоих полушарий
vfor служить молодой его возраст (d момент первой операции ему было при-
мерно 8—9 месяцев). Это предположение основано не только на теорети-
ческих Параллелях и Сравнениях. В специальной серии экспериментов,
результаты которых будут изложены в следующем сообщении, нами было
установлено весьма важное значение возрастного фактора в развитии ком-
пенсаторных приспособлений, в частности, в определении участия отдель-
ных частей центральной нервней системы в этом процессе.
Не исключена, конечно, возможность, что описанная выше атипическая
картина нарушения и восстановления функций у собаки Домино обуслов-
лена своеобразной комбинацией этих трех факте рев
Как бы ни были близки к действительности наши предположения о при-
чинах известного расхождения в фактических данных, полученных на трех
лишенных коры собаках, самый факт этого расхождения, а также отсут-
ствие точных анатомических данных о размерах и локализации хирурги-
ческого повреждения спинного мезга у одной из ссбак (Цыган) не позве-
всляют считать окончательно решенным второй из поставленных в данной
работе вопрос о рели коры больших полушарий мозга в развитии компен-
саторных приспособлений взрослых ссбак пселе поперечной перерезки
у них задней половины спинного мозга. Полученный нами по этому вопросу
экспериментальный материал должен быть пспслнен дальнейшими опыта-
ми на взрослых собаках.
ЛИТЕРАТУРА
Асратян Э. А.-, Архив биологических наук, 61, 1941.
Асратян Э. А., Очерки по этиологии, функциональной патологии и терапии
травматического шока, М., 1945.
Асратян Э. А., Военно-медицинский сборник, Hi. 111, 1946.
Асратян Э. А., Тезисы и тексты докладов VII Всесоюзного съезда физиологов,
биохимиков и фармакологов, стр. 105, М., 1947 и 1943.
Бехтерев В. М., Протоколы заседаний Общества психиатров в Спб. за 1884 г.,
стр. 48, 1885.
Бехтерев В. М.. Основы учения о функциях мозга. HI, 1905.
Боков Г И. и Но ви ко на А. Л., Рефераты работ учреждений Отделения
биологических наук АН СССР за 1946 г., стр. 400.
Боровиков И. В., Значение задних столбов спинного мозга в- провсдевни
мышечного чувства, лисе., Спб., 1900.
Ворошилов К. В., Luedwig's Arbe’iten b. Jahrg. 1884, Leipzig, 1885.
Гамбарян Л. С., Условные рефлексы у собак после повреждения задних стол-
бов спинного мозга, ДАН СССР, т. 84. № 5, 1952.
Г ол ьп и н г е р Ф. Ф., Чувствующие пути в спинном мозгу, дисс., Спб. 1896.
Дроздова В Н..К механизму и природе спинального шока, Д АН СССР, т. 6, 24.
561, 1948.
Дурмишьян М. Г., О рефлекторной деятельности поврежденного спинного моз
га, Труды АМН СССР. т. 19, 1952.
Карам ян А. И., Рефераты работ учреждений Отделения биологических наук
АН СССР за 1946 г., стр. 398.
Кузьмин В., Medicinische Jahrbuch, II, 1882.
Навроцкий, Ludwig's Arbeiten b. Jatirg. 1871, Leipzig, 89. 1882.
Па икр агон M. А., Тезисы докладов Это совещания по физиологическим про-
блек.ам, 1938.
Павлов И. II.. Собрание трудов, т. Ill, IV. 1952.
Россолимо Г., Экспериментальные исследования по вопросу о путях, проводя-
щих чувствительность и движение в спинном мозгу, дисс., М., 1887.
Чудновский Я. Б., К вопросу о путях проведения волосковой чувствитель-
ности, Лисе., Спб., 1907.
362
Э. А. АСРАТЯН
Be’.hc's Handb. d. norm. u. pathol. Physiol., Bd. XV, H. 2, 1932.
Bet he u. Fischer, ibidem.
Bickel, Mtinch. med. Wschr., 37. 1898.
Cannon a. Rosenblueth, The supersensitivity of denervated structur s, 1949.
Deen van, Traite sur la physiologic de la mobile 6piniere, 1841.
Ewald, Deutsch, med.. Wschr., 30, 1898.
F er r a rc а, В e г г а гэ, Brain, 57, 1934.
Fulton, Liddell a. R ioch, Brain, 53, 1930.
F ulton. Physiology of the nervous system, 1943.
Harreveld, Amer, journ. physiol., 129, 1940.
Longet, Anatomie et physiologic du systime nervetlx de Phomme et des animanx
vertebras, Paris, 1842.
Liddell, Brain, 57, 1934.
Me Couch, Hughes a. Stewart, Journ. neurophysiol., 3, 1940 ; 4, 1941.
Ruch, Annual review of physiol., 4, 1942.
Schiff, Gesamincite Beitrage zur Physiologic, Bd. Ill, 1896.
Scherrington, Integrative action of the nervous system, 1906.
Sc herrington et a 11., Reflex activity of the spinal cord, 1932.
T rendelenburg, Pfliige-s Arch. Physiol., 133, 136, 1910.
О ПРИНЦИПЕ СООТНОСИТЕЛЬНОЙ пластичности
НЕРВНОЙ СИСТЕМЫ* 1
(Элементы эволюционной теории пластичности)
О
=<7
дним из величайших дсстижений материалистической биологии
прошлого столетия является установление того, что приспособ-
ляемость представляетссбой очень важный фактор для сущест-
вования и эволюции живого вещества. Биологи ко нстатировали,
что в процессе индивидуального приспособления и эволюции
живое вещество изменяет не только форму, но и функции, функ-
ции даже раньше формы. «Морфологические и физиологические явления,
форма и функции обусловливают взаимно друг друга»2. Это положение
Энгельса было полностью подтверждено фактами, обнаруженными био-
логами.
Биология прошлого столетня концентрировала свое внимание главным
образом на таких проявлениях пластичности, которые вырисовываются на
фоне множества поколений; она рассматривала эти явления преимуществен-
но о аспекте филогении. Передовая биология XX столетия и в первую
очередь отечественная под влиянием развития экспериментальной физио-
логии, иод растущим влиянием материалистического мировоззрения на ее
развитие эту проблему ставила шире и глубже, стала обращать внимание
на быстро развивающиеся индивидуальные приспособительные явления
и на изучение их .внутренней природы, закономерностей проявления
«самодвижения» высокоорганизованной материи по этой линии.
В частности, Крупный отечественный биолог йкаД. А. Н. Северцов в До-
советский и в особенности в советский период его плодотворной деятель-
ности обратил внимание на факт, имеющий большое значение для физио-
логов. Он указал, что в процессе эволюции животные иногда изменяют свое
поведение, не меняя своей организации, и этим путем быстро приспосаб-
ливаются к постоянно изменяющейся среде.
Называя способность к эволюционному изменению «пластичностью организма»,
А. Н. Северцов подчеркивал в своей замечательной брошюре «Эволюция и психика»,
что эта форма приспособления не была учтена биологами, что она играет большую
роль в процессе существования самого индивидуума.
А. Н. Северцов указал не только на самый факт, но и на физиологическую
природу этого вида приспособления. В качестве физиологического «механизма» он
отметил «полиморфность функции» органов, заключающуюся в том, что одни и :е же
органы, в зависимости от ситуации, выполняют различные роли,
Н. А. Северцов указал также на другой физиологический «механизм» этого
нового ьида приспособления — на высшую" нервную деятельность, на «деятельность
разумного типа».
Таким образом, в лице Л. Н. Северцова мы имеем биолога, который не
1 Доклад, прочитанный иа научной сессии Института мозга им. В- М. Бех-
терева в апреле 1938 г., Труды Государственного института по изучению мозга
им. В. J4. Бехтерева, т. IX, 1939.
1 Ф. ЭЕ1гельс, Диалектика природы, 1946, стр. 249.
364
Э. X. АСР ЛТЯН
только выдвинул на первый план проблему пластичности организма, но и
поставил вопрос о перспективном физиологическом анализе проблемы.
Общеизвестен факт, что многие системы организма приспосабливакися
к условиям и процессу труда, к климатическим условиям; ярче всего, од-
нако, эти явления приспособления человеческого организма выявляются
в случае тех или иных болезненных нарушении в организме, в частности,
в нервной системе. Практика с давних пор зарегистрировала массу фактов
приспособления организма пссле повреждения тех или иных его органов.
Практическая медицина нс только зарегистрировала такие факты, но за
последние два-три десятилетия стала вмешиваться в судьбу больного
человека, различными путями активируя приспособительные функции тех
или иных систем, чтобы помочь организму. Очень много фактов в новей*
шей медицинской литературе говорит о тем, что хирурги в случае пора
жения отдельных мышц и нервов своим вмешательством облегчают со-
стояние больного, перемещением сухожилий или перекрестом нервов
возвращают пострадавшему, человеку работоспособность.
Для физиологии проблема приспособляемости также не нога. Она воз-
никла с момента зарождения экспериментальной физиологи. Бельше чем
100 лет назад Флуране и его современники наблюдали и изучали явления
приспособляемости пссле тех или иных поражений организма. С тех пор
в физиологии накопился огромный н разнородный эмпирический материал
по этому вопросу.
Не останавливаясь на всех этих фактах, мы обратим внимание лишь
па те из них, которые относятся к области нервной системы, прежде всего
по тон причине, что замечательное свойство приспособления к механи-
ческим повреждениям, названное гениальным И. П. Павловым «мехьпи
ческим иммунитетом’, достигает наивысшего развития именно в нервной
системе. Великий физиолог считал, что функция нервной системы вообще
сводится к приспособительной деятельности, в частности, к приспособле-
нию организма к окружающей среде. Мы ограничиваемся кратким обзо-
ром фактов приспособления в области нервной системы еще и потому, "то
наш экспериментальный материал сснсван на изучении нервной системы.
Со времени Флура пса многочисленные экспериментаторы показали, что pai
личные поражения центральной нервной системы (полное и частичное удаление
коры больших полушарий, удаление хотя бы одного полушария, полная и частич-
ная экстирпация мозжечка, повреждение проводящих путей, лабиринтов, перерезка
и перекрест очень важных нервов, пересадка мышц, повреждения различных ре-
цепторных систем, ампутация конечностей и др , которые сначала вызывают очень
серьезные изменения сензомоторной деятельности организма) с течением времени
так хорошо компенсируются, что для неопытного глаза имевшиеся раньше наруше-
ния становятся неуловимыми.
Несмотря на наличие большого эмпирического материала, в физиологии
до псследиего времени отсутствовало теоретическое обоснование важной
проблемы пластичности. Многие экспериментаторы ограничивались лишь
регистрацией фактов. Те из них, которые делали попытку объяснить «меха-
низм» прнспсссбляемсстн, ограничивались лишь обсуждением полученных
ими фактов. Но общей теории, обобщающей все явления, не было создано.
И. II. Павлов, накопивший очень большой материал по этой проблеме,
характеризовал существовавшее положение следующим образом: «Мне ка-
жется, что до сих пор специально в физиологии нервной системы недостаточ-
но оценен и даже не формулируется ясно и постоя нно этот в высшей степени
важный принцип»1 (принцип прнспсссбляемсстн нервней системы.—Э. Д.).
За последние десятилетия физиологи стали больше обращать внимание
на явления пластичности. Широко был поставлен этот вопрос главным об-
разом А. Н. Северцевым и И. П. Павловым, а также Бете.
1 И. П. Павлов, Двадцатнлстний опыт объективного изучения высшей нерв-
ной деятельности (поведения) животных, 1938, стр. 560.
О ПРИНЦИПЕ СООТНОСИТЕЛЬНОЙ ПЛАСТИЧНОСТИ НЕРВНОЙ СИСТЕМЫ Зб5
Бете привлек к ней внимание экспериментаторов, собрал разнообразный
материал, сам экспериментировал в эюй области и развил общую теорию
пластичности нервной системы, в которой попытался вывести общие законы
Длй объяснения этих разнообразных явлений.
Отмечая положительные начинания Беге, нельзя не обратить внимания
и на недостатки его теории. Мы очень критически относимся к основным
положениям теории Беге о пластичности нервной системы. Неверны ос-
новные методологические установки Бете в понимании некоторых важней-
ших сторон нервной деятельности в целом и пластичности нервной системы
в частности. Отметим, что Бете рассматривает свою теорию как теорию
о нерв.ной деятельности вообще.
Основным недостатком его теории является прежде всего то, что она не
охватывает всех фактов в этой области, именно тех, которые не соответ-
ствуют представлениям автора.
Например, кто может сказать, что компенсация нарушений, которые
наступают после разрушения лабиринтов, не имеет отношения к проблеме
пластичности? Речь идет о факте, известном со времена Флуранса и под-
робно изученном многими. Однако Бете о нем не говорит ни слова.
Приспособление, которое наступает после полной или частичной экс-
тирпации мозжечка и коры больших полушарий, приспособление после
деафферентации шейных мышц и конечностей также относятся к проблеме
пластичности.
Почему эти факты были упущены Бете? Пожалуй, добрая половина их
относится к пластичности нервной системы и группируется вокруг этих
видов нарушения функции и их последующей компенсации! Из литера-
туры известно, например, что кора большого мозга в какой-то мере при-
нимает участие в компенсации нарушений сензорных и моторных функ-
ций организма. Ви всяком случае по этому вопросу имеются определенные
экспериментальные данные и довольно ясные высказывания. Бете даже
не упоминает об этом.
Недостатком теории Бете является также то, что автор отрицательно
относится к господствующей динамической рефлекторной теории нервной
деятельности. Отсюда его отрицательнее отношение даже к современной
теории специализации и локализации функций, являющейся по существу
диалектической, отсюда и его своеобразное понимание функциональ-
ной архитектуры нервной системы.
Бете слишком нмвеллирует центральный аппарат, придавая господст-
вующее значение периферическим рецепторам.
Существенным недостатком теории Бете является ее антиэво л lo-
ll и о н и ость. К эмпирическому материалу Бете относится, как истый
механист. Ему чужда идея развития, историческое понимание процессов.
Он группирует свои материал не по эволюционному принципу, а по меха-
ническим признакам, например, при классификации животных по приз-
наку количества конечностей, игнорируя уровень их развития.
Данные, полученные на беспозвоночных (результаты удаления нерв-
ных узлов у членистоногих), Бете переносит на позвоночных, ка млеко-
питающих. Он абстрактно ставит вопрос о пластичнести, опуская коли-
чественную и качественную оценку нарушений и не учитывая различных
форм проявления компенсации.
Бете и его последователи удаляют у животных только ничтожную часть
коры полушарий , а выводы делают о значении всей коры в компенсаторных
явлениях.
Абстрактность и метафизичность постановки вопроса выявляются еще
в том, что Бете ставит вопрос о значении тех или иных участков централь-
ной нервной системы в компенсаторных явлениях независимо от степени
развития животного, от того места, которое животное занимает на эвслю-
366
Э. А. АСРАТЯН
ционной лестнице. Отсюда и шаткость экспериментальной аргументации
его теоретических положений о том, что в компенсаторных процессах
у высших животных «научение» (Lernprozess) и вообще кора больших полу-
шарий не играют (или почти не играют) роли, что все процессы при этом
ограничиваются «примитивными частями» центральной нервной системы,
что ведущую, господствующую роль в этих явлениях играет периферия
(рецепторы).
За последние годы мы имели возможность не только дать теоретически
обоснованную критику этих механистических концепций Бете и его после-
дователей, но и накопить значительный экспериментальный материал, ко-
торый, соднсй стороны, подтверждает наши крнтическиезамечания, асдру-
гой — дает возможность разрабатывать новую, эволюционную теорию пла-
стичности нервней системы на основе павловского учения.
Основные аргументы и концепции Бете о том, что кора больших полу-
шарий высших животных не играет роли в явлениях компенсации, опро-
вергаются экспериментальными данными, которые были получены нами и
нашими сотрудниками.
Диапазон охвата различных видов нарушений и компенсаций функций
в наших экспериментах шире, чем у Бете и его последователей. Мы изу-
чали не только варианты ампутации конечностей, перекреста нервов, по-
ловинной поперечной перерезки спинного мозга, но и варианты разруше-
ния лабиринтов, деафферентации конечностей, продольного расщепления
спинного мозга и др.
Нами установлено, что у всех без исключения животных, подвергну-
тых указанным выше разнообразным оперативным воздействиям, пссле
наступления компенсации удаление коры даже одного
полушария снимает наступившую компенсацию
в значительной мере, а удаление коры обоих по-
лушарий приводит к полному и безвозвратному
исчезновению этой компенсации.
Нами показано также, что если животных сначала лишают коры боль-
ших полушарий, а затем подвергают указанным выше оперативным воз-
действиям, вызывающим значительные нарушения сензорной и моторной
деятельности, эти нарушения никогда не компенсируются.
Таким образом, основные положения Бете по отношению к высшим жи-
вотным (собаки) нашими Экспериментами опровергнуты.
Остановимся на некоторых наших теоретических положениях.
Мы критикуем теорию Бете за то, что он не дал классификации отдель-
ных видов пластичности нервней системы, не выяснил значения и роли от-
дельных этажей центральной нервней системы в компенсаторных явлениях,
не изучил эту проблему с точки зрения подлинной эволюции, в аспекте
фило- и онтогенетического развития. Мы сделали попытку подойти к раз-
работке поставленной проблемы с такой точки зрения, прн которой эти
недостатки были бы устранены.
Прежде всего следует не только классифицировать нарушения функции
по типу повреждения тех или иных систем, но и учесть степень сложности,
глубину нарушений при каждом типе повреждения организма и соответст-
венно этому выявить природу различных проявлений пластичности,благо-
даря которой эти нарушения компенсируются.
Приводим примеры. Тип нарушений — ампутация лап: ампутация од-
ной лапы — это нарушение известной сложности и трудности, ампута-
ция двух лап — более глубокое нарушение, ампутация трех лап — еще
более глубокое нарушение. Эксперименты показывают, что для компенса-
ции этих различных по глубине нарушений требуется пластичность раз-
личной физиологической эффективности, участие различных этажей цент-
ральной нервной системы.
О ПРИНЦИПЕ СООТНОСИТЕЛЬНОЙ ПЛАСТИЧНОСТИ НЕРВНОЙ СИСТЕМЫ
367
Удаление коры обоих полушарий не вызывает исчезновения функции
стояния и ходьбы у собак «одной ампутированной лапой (данные Гольца,
Радемакера, Э. А. Асратяна). Удаление коры обоих полушарии у ссбак,
у которых ампутированы две лапы, полностью лишает их указанных
функций и исключает возможность новой компенсации нарушенных функ-
ций. С другой стороны, ссбаки с обоими неповрежденными полушариями,
но с тремя ампутированными лапами не в состоянии стоять и передви-
гаться, хотя по физическим законам такая возможность не исключена.
Другой тип нарушения — гемисскцйЯ спинного мозга. Если попереч-
ную перерезку спинного мозга делать совсем ничтожной глубины, никаких
значительных нарушений сензомоторной функции не наблюдается, и уда-
ление коры большого мозга ничего не выявляет. Если перерезать спиннои
мозг до половины, выявляются серьезные нарушения, которые через не-
которое время компенсируются; удаление коры одного и двух полушарий
совершенно устраняет описанные компенсаторные явления. Если же пе-
ререзать больше половины спинного мозга, то даже наличие двух полу-
шарий не в состоянии компенсировать вызванные очень глубокие наруше-
ния моторных и сензорных функций.
Третий тип операции спинного мозга—продольное расщепление его.
Если произвести продольное расщепление спинного мозга в области груд-
ных сегментов, то происходят ничтожные моторные и сензорные нарушения,
которые быстро восстанавливаются. Если же расщепление производится
в области тех сегментов, которые иннервируют передние и задние конеч-
нсстн, наступает глубокое нарушение; компенсация развивается с трудом
и в ничтожных пределах.
Мы видим, таким образом, что глубина повреждения структуры и
степень тяжести возникшего при этом функционального нарушения
являются очень важными моментами, что для компенсации этих наруше-
ний требуется пластичность различной физиологической эффект ив нс ст и,
наличие совершенно различных этажей центральной нервной системы.
Единственно правильным путем для понимания интрацентральпых про-
цессов является последовательный эволюционный подход к проблеме пла-
стичности, целеустремленное экспериментальное и теоретическое изучение
приспособительных явлений на диалектико-материалистической основе.
Приспособляемость, пластичность — сбщебиологичсское свойство жи-
вого вещества, благодаря которому осуществляется постоянное приспособ-
ление животного к текущим изменениям окружающей среды. Э го свойство,
как и другие свойства живого вещества, разв-ивается, совершенствуется.
В процессе морфологической и функциональной днференциации тканей и
систем пластичность достигает высшего уровня развития и большой физио-
логической эффективности в нервной системе. Далее, в связи с эволюцией
и диференциацией самой нервной системы развивается и пластичность,
проявляющаяся в различных формах, вариантах и с различной эффектив-
ностью; при этом различаются как общие для всей нервной системы, так
и специфические для отдельных частей ее проявления пластичности. В ар-
сенале разнообразных в.идов деятельности нервней системы, благодаря ко-
торым организм в условиях нормальней жизни приспосабливается к ок-
ружающей среде, а в условиях повреждения тех иди иных систем компен-
сирует нарушенные функции, мы имеем разнообразие проявлений и ферм
пластичности. С этой точки зрения следует различать простую адаптацию
к раздражителю, торможение при очень частых или сильных раздражите-
лях (пессимум Н. Е. Введенского, запредельное торможение И. П. Павло-
ва), усвоение ритма (А. Л. Ухтомский), тренировку, викарирование, меха-
низм дублированной иннервации и запасных путей (механический имму-
нитет И. П. Павлова), трофическое воздействие, перестройку по принципу
простого переключения путей (И. М. Сеченов и Магнус), перестройку под
368
Э. А. АСРАТЯН
влиянием центров, стоящих в субординационном отношении выше,
безусловнсрефлекторное интегрирование не только нижележащими или
средними отделами центральной нервной системы, но и высшими ее от-
делами и, наконец', условнорефлекторную деятельность.
Достигая наивысшей степени развития в нервной системе, пластичнссть
не во всех частях центральной нервной системы животного определенного
вида и возраста имеет один и тот же уровень, одни и те же формы проявле-
ния, одну и ту же физиологическую эффективность. В процессе эволюции
нервной системы, морфологического и функционального ее развития
И ДВференциацин пластичность достигает различной степени совершен
ства, приобретает качественно различные формы и различную физиологи-
ческую эффективность в разных этажах периферической и в особенности
центральной нервном системы.
Имеются основания полагать, что степень совершенств.а и эффектив-
ности, а также формы проявления пластичности в каждой морфологиче-
ски и функционально обособленной части центральной нервной системы
зависят от удельного веса данной части в общей многообразной деятель-
ности нервней системы. Литературные данные, а также результаты наших
исследований позволяют высказать предположение, что степень участия
различных отделов нервной системы в компенсации упомянутых выше
нарушений неодинакова, что в соответствии с возрастанием сложности на-
рушений в процесс их компенсации вовлекаются все новые звенья единой
цепи восходящего ряда центральных нервных образований с их все воз-
растающими возможностями и силой. При этом некоторые глубокие
степени нарушений не могут быть компенсированы даже при отсутствии
повреждений всех верхних этажей центральной нервной системы. При-
веденные выше примеры можно умножить: перерезки многих второ-
степенных нервов внешне почти совершенно не изменяет функции конеч-
ностей (например, перерезка некоторых маловажных нервов плечевого
сплетения или сплетений нижних конечностей). Можно допустить, что
в этих случаях центральная нервная система очень легко и быстро пере-
страивается. Удаление коры своих полушарий почти не отражается на
картине компенсации. Но если перерезать и произвести перекрестное сши-
вание нервов, которые имеют важное значение для функций этих конеч-
ностей, происходят очень глубокие нарушения, которые компенсируются
только в том случае, если цела кора. Стоит удалить оба полушария,
и наступившая компенсация исчезает безвозвратно.
Та же закономерность выступает очень четко при разрушении лаби-
ринтов, при котором выявляется целая гамма различной сложности нару-
шений (отведение и нистагм глаз, общие нарушения тонуса мышц, глубо-
кие нарушения в статическом и динамическом равновесии тела и т. д.).
Выясняется, что различные этажи центральной нервной системы в раз-
ной степени участвуют в компенсации указанных видов нарушений. Шпи-
гель и Демитриад показали, что нарушения типа отведения глаз могут быть
компенсированы даже после удаления промежуточного мозга, благодаря
так называемой «бехтеревской компенсации». Однако глубокие нарушения,
развивающиеся после разрушения лабиринтов, могут компенсироваться
только при наличии больших полушарий (наши опыты). Аналогичные
явления наблюдаются и при нарушениях сензорной и моторной деятельно-
сти (перерезка задних корешков различных нервов).
При каждом типе повреждений мы имеем целую гамму нарушений раз-
личной градации, тяжести, сложности и величины гг соответственгго этому
целую гамму различной физиологической эффективности приспособитель-
ных процессов, пластических «свойств», присущих разным этажам цент-
ральной нервной системы, благодаря которым эти нарушения компен-
сируются. Соответственно глубине нарушений вырабатывается ряд физио-
О ПРИНЦИПЕ СООТНОСИТЕЛЬНОЙ пластичности нервной системы
3G9
логических «механизмов», с помощью которых организм компенсирует
эти нарушения. Быть может, некоторые отделы центральной нервной
системы используют с этой целью многочисленные сложные «механизмы»,
а другие — только малочисленные и элементарные. Очевидно, однако, что
в любом случае компенсации функций мы имеем дело прежде всего
с целостной деятельностью единой нервном системы, где, смотря по глу-
бине и характеру нарушения, степень участия каждого более или менее
обособленного отдела центральной нервной системы изменяется.
Можно ли считать, что пластические свойства отдельных частей
нервной системы не изменяются в филогенетическом ряду? Эксперимен-
тальный материал отвечает на этот вопрос отрицательно (А. И. Карамян,
Ф. М. Вишневецкая, Е. М. Лыхина, Мерцбахер, Бикель, Тренделен-
бург). Так, например, те дефекты, которые компенсируются у млекопита-
ющих при наличии коры больших полушарий и которые исчезают при
удалении коры, у низших животных могут быть компенсированы даже
после удаления всего переднего мозга (у черепах и лягушек довольно
быстро и совершенно, у птиц — медленнее и менее совершенно).
Значит, в эволюционном ряду степень участия и роль различных отде-
лов центральной нервной системы в компенсаторных явлениях изменяются,
и кора в компенсации сложных нарушений у высших позвоночных живот-
ных приобретает решающую роль, в то время как у других, ниже стоящих
в эволюционном ряду млекопитающих значение верхних этажей централь-
ной нервной системы меньше, у них более выражена соотносительная
физиологическая роль пластичности средних и нижележащих этажей
центральной нервной системы. Соответствующие опыты, проведенные на-
шими сотрудниками А. М. Карамяном, Е. М. Лыхиной и Ф. М. Вишневец-
кой, а также аналогичные данные в мировой физиологической литературе
свидетельствуют о том, что чем ниже стоит животное в эволюционном ряду,
гем меньше значение верхнего этажа центральной системы в приспособи-
тельных функциях организма.
Как же изменяется пластичность всей центральной нервной системы
в процессе филогенетического развития? Экспериментальные данные Пауля
Вейсса, Магнуса и Биксля, независимо от их теоретических положений,
в корне ошибочных, позволяют предполагать, что общая физиологическая
эффективность пластических свойств нервной системы повышается с ее раз-
витием, в зависимости от места, занимаемого животным в эволюционном
ряду. Экспериментальные данные, полученные Вейссом на низших позвоноч-
ных, говорят о том, что при подсадке конечностей и отдельных мышц к тем
или иным частям организма эти привитые конечности и мышцы не приспо-
сабливаются к новым условиям существования даже при целости всей
центральной нервной системы; они стойко сохраняют «резонаторные Свой-
ства» — действуют синхронно, с одинаковой силой и однотипно, с одноимен-
ными конечностями и мышцами хозяина. Если передние конечности, на-
пример, приживлять к плечевым сегментам, то эти привитые добавочные
передние конечности начинают синхронно, с одинаковой силой и однотипно,
работать с конечностями хозяина. У высших млекопитающих в аналогич-
ных случаях (пересадка мышц, перекрестное сшивание нервов, природные
случаи дублирования пальцев) возможно преодоление фазы «резонанса»,
смена функции, приспособление к новым условиям. Вейсс приводит 2 слу-
чая с дублированными пальцами на руках у людей. Эти дублированные
пальцы работали сначала, если можно так выразиться, «по теории резонан-
са»: при сгибании основного пальца сгибался и одноименный «дубликат»,
при разгибании основного разгибался его «дубликат» и т. д. Однако некото-
рое время спустя больные могли сокращать каждый из этих пальцев в от-
дельности, т. е. не по ошибочной «теории резонанса». Эти данные не только
в корне противоречат антиэволюционным, идеалистическим взглядам Вейсса
21 9. А. Аератяи
370
Э А АСРАТЯН
о якобы универсальном для всей нервной системы значении принципа
«резонанса», но вместе с тем свидетельствуют о том, что пластические
свойства центральной нервной системы у низших позвоночных выражены
слабее, чем у высших позвоночных, что физиологическая эффективность
пластических свойств усиливается в процессе эволюции.
Об этом же свидетельствуют и данные других физиологов. Так, по дан-
ным Эвальда и Магнуса, чем ниже стоит животное на эволюционной лест-
нице, тем медленнее компенсируются последствия разрушения лабиринтов.
Сравнивая Течение компенсации После разрушения лабиринтов у кроликов,
собак и обезьян, Магнус наблюдал, что у обезьян последствия разрушения
лабиринтов компенсируются быстрее, чем у собак, а у собак быстрее, чем
у кроликов. Аналогичные данные были получены Эвальдом на лягушках,
голубях и собаках.
Бикель отмечал аналогичное явление в отношении компенсации нару-
шений После Перерезки задних корешков спинного мозга у лягушек и со-
бак. У лягушек после перерезки задних корешков какой-нибудь конечно-
сти почти не наступают нарушения моторных функций, но те, которые
наступают. остаются на все время, пока лягушки живут, у собаки же пссле
такого оперативного воздействия появляются глубокие нарушения, но они
постепенно так хорошо компенсируются, что неопытный глаз не может
отличить пораженную конечность от здоровой.
Таким образом, данные других исследователей и наши данные говорят
о тем, что общая физиологическая эффективность пластичности центральной
нервной системы организма возрастаете процессе филогенетического разви-
тия, эволюции видов. А так как знаменосцем эволюции центральной нервной
системы высших животных является большой мозг или, вернее, его кора
и так как пластичность достигает наивысшего развития именно в этом органе,
можно полагать, что в центральной нервной системе человека пластичность
достигает наивысшего уровня совершенства. Многие клинические данные,
повндимому, подтверждают такое теоретическое положение. Отсюда по-
нятно, почему детальное изучение пластичности центральной нервной си-
стемы, разрешение ряда простых и сложных вопросов этой проблемы имеют
не только теоретическое, но и сугубо практическое значение.
В руководимой нами лаборатории эти явления пока изучаются только
по линии понимания внутренней природы пластичности, понимания роли
отдельных частей центральной нервной системы в компенсации в аспекте
филогенетического развития. У нас имеются отдельные данные, говорящие
о том, что участие коры большого мозга и других этажей центральной
нервной системы в компенсации различных нарушений изменяется также
в связи с возрастом.
Высказанные нами теоретические положения и наш экспериментальный
материал свидетельствуют о перспективности разработки этой важней проб*
лемы современной физиологии центральной нервной системы.
НОВЫЕ ДАННЫЕ ПО ФИЗИОЛОГИИ БОЛЬШОГО МОЗГА 1
1 протяжении более ста лет практическая медицина и экспери-
ментальная физиология накопили огромный эмпирический
материал, свидетельствующий о том, что нарушения функций,
вызванные повреждением различных рецепторных и эффекторных
органов и частей периферической и центральной нервной систе-
мы, со временем более или менее сглаживаются, компенси-
руются. В целях изучения проблемы локализации функций в коре
большого мозга И. П. Павлов и его ученики также на протяжении многих
лет производили операции частичного удаления тех или иных ее зон
у большого числа собак. Подвергая детальному наблюдению оперирован-
ных животных и объективно изучая особенности и динамику условнорсф-
лекторной деятельности у них, И. П. Павлов не только подтвердил соот-
ветствующие данные Гольца, Лючиани и др. о постепенном восстановлении
нарушенных операцией функций, но и дал детальную физиологическую
характеристику этих проявлении «своеобразного могущества живого ве-
щества» (И. П. Павлов).
По концепции I I. П. Павлова, способность к восстановлению функций,
нарушенных в результате повреждения тканей, обусловливается наличием
в организме запасных средств, его своеобразным «механическим иммуни-
тетом». При этом И. П. Павлов считал, что это общебнологическсе Сеойстео
сильно развито в нервной системе и достигает наивысшего совершенства
в больших полушариях. «Если во всем организме, — писал он, — мы по-
стоянно встречаемся с запасными средствами против частичных нарушений
его, то в нервной системе, как устанавливающей все связи и отношения ор-
ганизма, этот принцип должен обнаруживаться в высшей степени. И это
мы видим как в спинном мозгу на сложном и очень разнообразном ходе
его волокон, так и в периферической нервной системе на се широко прак-
тикуемой так называемой возвратной чувствительности, что явно значительно
нейтрализует вред механических нарушений Этих отделов. Конечно, то же
в высшей мере должно быть осуществлено в верховной инстанции аппарата,
регулирующего всю внутреннюю и внешнюю деятельность организм'’»1.
Несмотря на наличие очень богатого и разнообразного эмпирического
материала по восстановлению нарушенных функций после повреждения т?х
или иных частей организма, в частности, тех или иных частей нервной си-
стемы, несмотря иа большой теоретический интерес и практическую
важность этой обширной проблемы, ее разработкой занимались недоста-
точно систематически и целеустремленно. Отсутствовала общая теория,
очень мало было известно о природе н механизмах этих особого рода
приспособительных явлений в различных частях организма, не были
определены внешние и внутренние факторы перестроек, роль и значение
1 Архив биологических наук, т. 61, в. 3, 1941.
а И. П. Павлов, Лекции о работе больших полушарий головного мозга.
1927, стр, 281.
24
372
Э. А Ас РАТ АН
отдельных частей нервной системы в них, характер взаимодействия этих
частей и т. д. Концепции, выдвинутые рядом исследователей [Флуранс
(Flourans), Гольц (Golz), Лючиани (Luciani), Магнус (Magnus), Эвальд
(Ewald) и др.], касались узкого круга явлений и мало освещали проб-
лему в целом. Заслуга Бете (Bcthe) заключается в том, что он широко
подошел к изучению этой проблемы.
При построении своих теоретических положений Бете явно необъектив-
но использовал накопленные мировой наукой факты по изучаемой проблеме,
в частности, он совершенно обошел не совпадающий с его теорией богатый
материал, накопленный Лючиани, Гольцем, Бикелом (Bickel), И. П. Пав-
ловым и его школой. К осмыслению собственного экспериментального
материала и материала других исследователей Бете подошел не с позиций
подлинного эволюционного понимания проблемы, а как истый механист,
метафизик. Эго особенно ярко выявляется в его попытке широко обобщить
для всего животного мира данные, полученные на беспозвоночных. Ни
в коей мере нельзя считать экспериментально аргументированными, в осо-
бенности в отношении высших животных, его теоретические концепции
о решающей и императивной роли периферии или периферической рецепции
в компенсаторных явлениях; нельзя признать аргументированными и его
положения о том, будто большой мозг не играет никакой роли или играет
ничтожную роль в этих явлениях даже у высших животных. Своей метафи-
зической и неовнталистической теорией, отрывающей функцию от формы
и структуры, отрицающей функциональную специализацию и локализацию
даже в нервной системе высокоразвитых животных, недооценкой и «обез-
личкойл центральной нервной системы, в особенности высших ее этажей,
Бете и небольшая группа его единомышленников объявили войну современ-
ной динамической рефлекторной теории, категорически отрицая всякую воз-
можность объяснить описанные выше явления приспособления сточки зре-
ния последней. Нетрудно видеть, в частности, что теория Бете прямо или
косвенно направлена против учения И. П. Павлова о функциях большого
мозга — наиболее совершенного регулятора «всей внешней и внутренней
.деятельности организма» (И. П. Павлов). Пе случайно в последние годы
своей жизни И. П. Павлов с юношеской страстью в специальной статье
выступил против концепции Лешли (Lashley), единомышленника Бете, по
многим указанным выше вопросам. Ученикам И. П. Павлова хорошо из-
вестны также острые его замечания относительно упомянутых концепций
Бете.
Замечательные экспериментальные работы и глубокие теоретические
концепции И. П. Павлова по проблеме пластичности в основном охватывают
частный круг явлений — явления при частичном повреждении коры боль-
шого мозга. Что же касается его оценки состояния всей проблемы в мировой
физиологии вообще, то И, П. Павлов считал, что этой проблеме не уделяется
достаточного внимания, она целеустремленно и систематически еще не
разрабатывается, не занимает еще подобающее место в науке. В качестве
иллюстрации я приведу два высказывания его, относящиеся К начальному
и последнему периодам его работы по физиологии большого мозга.
В своей знаменитой мадридской речи (1903), касаясь только опытов
с частичной экстирпацией коры большого мозга, И. П. Павлов говорил;
«Исследования на эту тему все еще не сложились в такой специальный от-
дел, изучение которого развивалось бы неустанно и по определенному пла-
ну»1. А в сво-ей прекрасной научно-полемической статье против Лешли и
Кэтри (Cuthrie), написанной в период, когда уже было воздвигнуто величе-
ственное здание его грандиозного учения о работе большого мозга (1932),
• 1 И. П. Павлов, Двадцатилетний опыт объективного изучения высшей нерв-
ной деятельности (поведения) животных, 1938, стр. 35.
НОВЫЕ ДАННЫЕ ПО ФИЗИОЛОГИИ БОЛЬШОГО МОЗГА 373
И. П. Павлов, касаясь состояния вопроса о «механическом иммунитете»,
т. е. пластичности в центральной нервной системе вообще, писал: «Мне
кажется, что до сих пор специально в физиологии нервной системы недо-
статочно оценен и даже не формулируется ясно и постоянно этот в высшей
степени важный принцип»*.
Хотя гениальный классик естествознания и не оставил нам законченной
общей теории по затронутой проблеме в целом, но он сделал исключительно
много в этом направлении. Своим бессмертным материалистическим учением
о работе большого мозга он Создал прочный фундамент и указал правиль-
ный путь разработки проблемы пластичности.
Своевременность и важнссть систематической разработки этой актуаль-
ной проблемы были совершенно очевидны. Необходимость такой целеуст-
ремленной работы подчеркивалась еще тем, что существовавшая механи-
стическая теория Бете, как уже было сказано, не могла удовлетворить
нас ввиду се фактической веаргументирОванносТИ И Порочных Теоретиче-
ских установок, направленных против точных данных и положений со-
временной нейрофизиологии и прежде всего против учения И. П. Павлова.
Еще при жизни И. П. Павлова в Советском Союзе систематическим изу-
чением проблемы пластичности повреждений нервной системы начали за-
ниматься два его ученика — П. К. Анохин со своими сотрудниками и я со
своими сотрудниками. Я лишен возможности подробно останавливаться
здесь на работах и теоретических положениях П. К. Анохина, касающихся
главным образом нарушения и восстановления функций после анастомоза
нервов. Скажу только, что, с нашей точки зрения, теоретические установки
П. К- Анохина в этом вопросе вначале были явно ошибочными, и, несмотря
на значительный положительный сдвиг за последние годы, он до сих порете
стоит на не совсем правильных позициях. При теоретическом осмысливании
полученного им и сотрудниками фактического материала по этой проблеме,
представляющего в значительной мере повторение и подтверждение данных
других исследователей, а частью интересные новые данные, П. К. Анохин
в основном исходил из концепции Бете и его единомышленников. Доста-
точно сопоставить концепции П. К. Анохина о функциональной специализа-
ции и локализации в центральной нервной системе, о современной рефлек-
торной теории, о гиперболизированной роли периферии в интрацентраль-
ных перестройках, о недооценке роли коры большого мозга высших позво-
ночных в этих перестройках и т. д. с соответствующими концепциями Вето,
чтобы убедиться в сказанном. П. К. Анохин, так же как и Бете, стагит под
вопрос «принцип рефлекса со всеми свойственными ему чертами»; он также
утверждает, что «врядли имеет какой-либо физиологический смысл говорить
об афферентных «центрах» коры головного мозга», а двигательные центры,
по П. К. Анохину, могут быть названы «центрами только в очень условном
значении этого слова». Признавая в духе Лешли, что кора может оказать
диффузное, динамогеническое влияние на процессы перестройки в нижеле-
жащих отделах центральной нервной системы, П. К- Анохин не только отри-
цает активное влияние коры па процесс этой перестройки посредством все-
могущей условнорефлекторной деятельности, но даже отрицает запоздалое
ее «вступление» в эту перестройку, отрицает ее «условнорефлекторный конт-
роль» над деятельностью реинтегрированного участка спинного мозга,
по крайней мере в первые годы после операции. Согласно концепциям
П. К. Анохина и сотрудников, даже много времени спустя пссле заверше-
ния перестройки, «кора больших полушарий, являясь наиболее вар из би ль-
ной частью центральной нервной системы, все же не вступала в систему
спинномозговой реинтеграции»; илц же «кора — наиболее подвижный
* И. П. Павлов, Двадцатилетий опыт объективного изучения высшей
нервной деятельности (поведения) животных, I93S, стр. 5б0.
374
Э. А. АСРАТЯН
и лабильный орган — не устанавливает условной связи с реинтегриро-
ванным участком спинного мозга»1. Сходство со взглядами Бете завершается
тем. что П. К- Анохин также решающим, императивным фактором пере-
стройки считает периферию; «Именно изменения афферентных импульсов,
которые теперь бомбардируют несвойственные им центры, и приводят к
тому, что эффекторные части центральной нервной системы начинают
управлять гетерогенными органами». Или же; «Весь процесс реинтеграции
в центральной нервной системе возникает по инициативе периферичес-
ких импульсов и протекает под их постоянным контролем»2,— говорит
П. К. Анохин. Конечно, не во всех деталях концепции П. К. Ано-
хина совпадают с концепциями Бете (например, П. К. Анохин придает
очень большое значение рубцу в процессе перестройки после анастомоза
нервов), но совпадение основных теоретических положений по затронутой
проблеме является очевидным и бесспорным.
В своей экспериментальной и теоретической работе по указанной проб-
леме мы основывались на учении И. П. Павлова о работе большого мозга
и на экспериментальном материале мировой физиологии, в частности, на
данных Флуранса, Лючиаии, Гольца, Эвальда, Бикеля И Магнуса. Кроме
того, мы исходили из основных положений эволюционной теории, в част-
ности, из ряда ценных положений А. Н. Северцова об эволюционной пла-
стичности.
Полученные нами новые экспериментальные данные и развиваемые нами
теоретические положения касаются прежде всего большого мозга.
Для разностороннего и углубленного изучения проблемы пластичности
нервной системы в условиях частичного повреждения тех или иных ее частей
и других органов мы исследовали последствия не какого-нибудь определен-
ного вида оперативного повреждения отдельных частей нервной системы,
рецепторов и эффекторов, как делали почти все предыдущие исследователи,
а все основные виды повреждения наиболее- важных отделов нервной си-
стемы и рядя органов. Конкретно, в течение последних 8 лет мы изучали
и продолжаем изучать нарушение и восстановление функции после следую-
щих видов оперативных повреждений (в скобках приведены фамилии иссле-
дователей, которые раньше применяли данный вид операции): ]) ампутация
лап в различных комбинациях (Бете, Будденброк (Buddenbrock) и др.);
2) поперечная половинная перерезка спинного мозга на различных его уров-
нях (И. М. Сеченов, Вейсс (Weiss), Осава (Osawa) и др.]; 3) поперечная по-
ловинная перерезка шейного мозга почти под продолговатым мозгом (Пор-
тер (Porter)l; 4) перерезка задних столбов спинного мозга на различных его
уровнях I Фодера (Fodera), ван Дин (Van Deen), В. М. Бехтерев, М. А. Пан-
кратова и др. ]; 5) продольное расщепление спинного мозга на различных
его уровнях [Фодера, в-ан Дни, Броун-Секар (Brown-Se-quard), И. М. Се-
ченов, Тен-Катэ (Тёп-Ка(ё) и др. 1; 6) перерезка задних корешков нервов
тех или иных конечностей [Бикель, Тренделенбург (Trendelenburg) и Др.];
7) перерезка задних корешков первых трех пар шейных нервов [Магнус
и ван Левен (Van Leeuwen)]; 8) перерезка и перекрестное сшивание различ-
ных нервов [Флуранс, Осборн и Килвингтон (Osborn a. Kilwington),
Кеннеди (Kennedi), Лэнгли (Langley), Бете, П. К. Анохин и др. 1; 9) разру-
шение одного и двух лабиринтов [Флуранс, Эвальд, Магнус, Клейн (Klein)
и др. ]. Кроме того, нами был применен ряд комбинаций этих операций.
В целях изучения проблемы в эволюционном аспекте основные виды опе-
раций мы применяли на животных, стоящих на различных уровнях эволю-
ционного развития (лягушки, черепахи, птицы, млекопитающие). Собран
1 Сборник «Проблема центра и периферии в физиологии нервной деятельности»,
1935, стр. 121 и 35.
’Там же, стр. 201.
НОВЫЕ ДАННЫЕ ПО ФИЗИОЛОГИИ БОЛЬШОГО МОЗГА
375
некоторый материал также по возрастной эволюции пластичности. Каждая
серия опытов проводилась на десятках млекопитающих и на многих десят-
ках птиц и других животных. Следует сказать, что принцип эволюцион-
ного подбора подопытных животных для изучения последствий какого-
нибудь вида оперативного нарушения применялся некоторыми предыдущи-
ми исследователями (Эвальд, Бикель, Магнус, Бете и др.), однако не все
они полученный материал освещали с позиций подлинной эволюционной
теории.
В целях детального изучения картины нарушений после тех или иных
оперативных повреждений, а также в- целях изучения дальнейшей динамики
этих нарушений вплоть до их исчезновения мы пользовались не только
методом наблюдения, фотографирования и киносъемок, по и методикой ус-
ловных рефлексов и хронаксиметрии, а также тонусометрии и т. д.
Наконец, для получения точного ответа на основной и вместе с тем
спорный вопрос о роли и значении верхних этажей центральной нервной
системы в указанных выше приспособительных явлениях особого типа мы
у подопытных животных после наступления компенсации первичных нару-
шений экстирпировали не отдельные часта большого мозга, э удаляли у
млекопитающих всю кору большого мозга, у птиц и низших позвоночных—
весь передний мозг. В ряде случаев эти операции осуществлялись и в об-
ратной последовательности, т. е. у лишенных коры большого мозга собак
или у лишенных переднего мозга птиц и других животных мы производили
некоторые из перечисленных выше операций. Следует отметить, что в ука-
занных выше целях ряд исследователей также частично пли педиком уда-
лял передний мозг у низших животных (Эвальд — у голубей после разру-
шения лабиринтов; Тренделенбург—также у голубей после деафферентацни
тех или иных конечностей; Бикель и Мерцбахер — у лягушек после той
же операции; Бете — у членистоногих, удаляя надглоточные узлы после
ампутации конечностей). У млекопитающих некоторые исследователи стой
же целью удаляли лишь двигательные или сензомоторные зоны (Лючиани—
у лишенных мозжечка собак, Бикель — у собак с деафферентнрованными
конечностями, Эвальд — у собак с разрушенными лабиринтами, Бете —
у собак с ампутированными лапами, П. К. Анохин — у собак
с анастомозированными нервами). Упомянутые опыты с частичным или
полным удалением переднего мозга или верхних узлов дали много ценного
для понимания роли верхних этажей центральной нервной системы в соот-
ветствующих компенсаторных явлениях у птиц, низших позвоночных и
беспозвоночных. Следует сказать, однако, что ограниченное количество по-
допытных животных, использованных для изучения каждого варианта по-
вреждения, а также изучение лишь какого-нибудь одного из этих вариан-
тов каждым исследователем обусловливали довольно разнородные резуль-
таты и значительные неточности в выводах. Если эти разнородные данные
могли служить какой-то основой для суждения о роли переднего мозга или
надглоточных узлов в компенсаторных явлениях у беспозвоночных, низших
позвоночных н птиц, то они уже никак не могли явиться основанием для
суждения о роли коры большого мозга при тех же явлениях у млеко-
питающих.
Более ценными в этом отношении оказались данные, полученные не-
которыми исследователями, удалявшими у млекопитающих двигательную
или сепзомоторную зону. Выяснилось, что в ряде случаев после указан-
ного частичного повреждения коры большого мозга компенсаторные
явления значительно нарушаются, но со временем в слабой степени
Восстанавливаются вновь (Лючиани, Бикель, Эвальд); в других случаях
компенсаторные Процессы подвергаются ничтожным и недлительным
нарушениям (Бете, П. К. Анохин).
Совершенно очевидно, что ограниченное количество подопытных живот-
376
Э. А. АСРАТЯН
ных, исследование отдельных вариантов нарушения функций различными
исследователями, невидимому, также различные степени разрушения кор-
ковой ткани пе позволяют сделать окончательные общие выводы даже о роли
двигательных или сенэомоторных зон коры в компенсаторных явлениях,
развивающихся после различного рода повреждений нервной системы, ссн-
зорных и моторных органов. Я нс говорю уже о том, что едва ли возможно
на основании частичного удаления коры решать вопрос о ее роли в целом
в указанных выше компенсаторных явлениях. Ведь опыты Гольца, Лючиани
и II. П. Павлова ясно показали, что, благодаря эффективной пластичности
коры, последствия удаления даже очень больших ее частей компенсируются
неповрежденными ее частями.
Отсюда явствует, что для правильного решения поставленного вопроса
следовало ту или иную операцию, вызывающую первоначальные нарушения
функции, комбинировать с полной экстирпацией коры большого мозга.
А такие комбинированные операции, преследующие хотя бы иные цели, по
с более или менее удовлетворительными результатами, по данным мировой
физиологической литературы, сделаны только Радемакером (Rademaker).
Радсмакср троекратной операцией в различной последовательности уда-
лял у двух ссбак весь мозжечок и всю кору большого мозга. Одна из его
собак жила 27 дней, другая — 38 дней. Собаки Радемакера не могли ни
стоять, ни ходить; кроме того, у них была резко нарушена рефлекторная дея-
тельность неповрежденных частей центральной нервной системы. Данные
Радемакера имеют большое значение для нейрофизиологии, во всяком слу-
чае неизмеримо большее, чем наблюдения американского физиолога Фул-
тона (Fulton) и его сотрудников над несколькими недолго жившими кошками
с подобной же операцией, а также наблюдения М. А. Панкратова. Они пред-
ставляют интерес также для нас в связи с изучаемой проблемой. Однако не-
продолжительность жизни двух его собак после труднейших операций не
позволила бы сделать окончательных выводов о роли коры даже в компенса-
ции мозжечковых нарушений, если бы нс были известны данные Лючиани.
Я не говорю уже о невозможности перенесения этих данных на все другие
виды нарушений.
Для правильного решения поставленной нами перед собой трудной за-
дачи мы в последние 8 лет сочетали операцию удаления всей коры большого
мозга с какой-нибудь из перечисленных выше основных операции примерно
на трех десятках собак, которые в этом состоянии жили у нас от 2 недель до
3 лет и больше. Собак без коры одного полушария, подвергнутых одному из
указанных видов операций, было значительно больше. Собак же, у которых
производились только основные виды операций, находилось под нашим на-
блюдением значительно больше сотни. Число подопытных птиц в соответст-
вующих группах было примерно в два раза больше (голуби в опытах наших
сотрудников А. И. Карамяна, Б. О. Стефанцова, В. Д. Дмитриева,
Я. М. Прессмана). Опыты с определенными вариантами операций были про-
изведены также на большом числе лягушек и на небольшом количестве
черепах (опыты наших сотрудников Ф. М. Вишневецкой, Р. О. Барсегян,
Е. М. Лихиной и А. И. Карамяна).
Перехожу к краткому описанию полученных экспериментальных дан-
ных по отдельным вариантам повреждений нервной системы и перифериче-
ских органов.
1. Ампутации двух лап были произведены у 34 собак различной поро-
ды, возраста if роста. Детальные наблюдения за ходом компенсации, т. е.
за процессом приобретения способности стоять и передвигаться на оставших-
ся двух лапах (односторонних, обеих задних, обеих передних или крест-
накрест — передней и задней), показали, что в подавляющем большинстве
случаев компенсация наступает через 20—50, в редких случаях через 90—
100 дней после операции. Только у 2 взрослых собак и 4 щенков 4—5-ме-
НОВЫЕ ДАННЫЕ ПО ФИЗИОЛОГИИ БОЛЬШОГО МОЗГА
377
сячного возраста компенсация нарушения происходила значительно быст-
рее — через 3—5 дней. По нашим данным, быстрота наступления компенса-
ции зависит от веса, роста (длина ног), наличия предварительной трени-
ровки в ходьбе на двух лапах и от возраста. Чем меньше вес и рост и чем
моложе собаки (во всяком случае до 4-месячного возраста), тем при
прочих примерно равных условиях быстрее наступает компенсация.
Ускоряет компенсацию и предварительная тренировка (две наши взрос-
лые собаки, о которых шла речь выше, были тренированы).
Эти данные явно не согласуются с опубликованными данными Бете о
быстроте компенсации нарушений у собак с ампутированными лапами.
По данным Бете, все его собаки в первые же дни пссле операции стояли
и ходили на двух лапах. Известно, что частью на основании быстроты на-
ступления компенсации, частью па основании того, что при удалении дви-
гательных зон эта компенсация не страдает, Бете отрицает роль коры боль-
шого мозга в приспособительных явлениях у ссбак после ампутации лап
и после других видов повреждении. О том, что «научение» в описанных слу-
чаях играет весьма важную роль, говорит, по нашим данным, не только
длительность компенсации, но и строгая постепенность ее.
Удаление коры одного полушария полностью нарушало наступившую
компенсацию. Однако несколько дней спустя нарушения вновь начинали
компенсироваться и, как правило, через 15—20 Дней Достигали Прежнего
состояния. В одном случае, однако, когда у собаки были оставлены только
обе правые лапы и удалялась кора левого полушария, обратимые нару-
шения компенсировались спустя примерно 2 года. Собак с ампутирован-
ными двумя лапами и без коры одного полушария под длительным
нашим наблюдением было 7.
Удаление коры обоих полушарий у собак полностью ликвидировало
существовавшую компенсацию и лишало их способности еще раз компенси-
ровать нарушения. Собак с ампутированными лапами и без коры обоих полу-
шарий у нас было 6, из них 4 погибли в первые 2 недели после операции,
одна жила 4 месяца, одна — 6V« месяцев. У всех этих ссбак, как и следовало
ожидать, все спинальные, шейные и лабиринтные тонические рефлексы,
а также установочные рефлексы осуществлялись с достаточной ж и вестью.
Ползком передвигаясь по земле, они делали попытки стоять, иногда с по-
мощью мощного толчка они становились на ноги, но тотчас же валились на
землю. Эта картина устанавливалась через 7—10 дней после операции и со-
хранялась почти без изменения до конца жизни.
2. Половинная поперечная перерезка спинного мозга на уровне послед-
них грудных и первых поясничных сегментов была произведена у 13 со-
бак и 1 кошки (у 11 собак перерезка была односторонняя, а у 2 ссбак была
произведена также половинная перерезка с противоположной стороны сег-
мента выше уровня первой перерезки). Картина нарушений (почти полный
вялый паралич односторонней конечности, нарушение сензорной деятельно-
сти противоположной конечности ниже уровня перерезки), а также ход по-
степенной компенсации этих нарушений у животных в основном совпадали
сданными предыдущих исследователей (Вейсс, Осава, Шефер, Мотт, Дании
и др.). В общем более или менее полно эти нарушения компенсировались
через 1—1 Vr месяца после операции. Картина нарушения функции и про-
цесс компенсации у этих собак нами изучались также с помощью измерения
хронаксии моторных нервов и рефлексов и измерения тонуса сгибателей
и разгибателей (опыты Р. О. Барсегян). При этом выяснилось, что в первые
дни после операции реобаза и хронаксия моторных нервов пораженных лап
увеличиваются в небольших пределах, а реобаза и хронаксия рефлексов
с пораженной лапы — примерно в 8—10 раз. Тонус как экстензорных, так
и флексорных мышц пораженной конечности в большинстве случаев сни-
жается; в некоторых случаях, однако, наблюдается даже ничтожное усиле-
378
Э. А. АСРАТЯН
ние тонуса. Со временем, параллельно с восстановлением координационной
деятельности, восстанавливаются и эти нарушения.
У 6 оперированных собак мы исследовали также электрооборонитсльную
условнорефлекторную деятельность на пораженную лапу (данные М. И. Ха-
Путиной и А. А. Новиковой). Выяснилось, что существовавшие до операции
локальные условные рефлексы на данную конечность исчезают в первые дни
после операции, затем спустя 7—13 дней после операции они постепенно
восстанавливаются н достигают прежнего уровня.
Эти данные представляют для нас большую ценность, ибо показывают,
что корковая регуляция функции пораженной конечности является фактом,
что она осуществляется еще в процессе самой компенсации, иногда даже до
появления внешних признаков компенсации, а не только после ее заверше-
ния. У 3 из этих собак было произведено удаление коры одного полушария,
н все они находились под наблюдением длительное время. Как правило,
удаление коры одного полушария вызывало нарушение существовавшей
раньше компенсации по всем указанным выше показателям, однако со вре-
менем (примерно через 15—20 дней) вторично наступала компенсация этих
нарушений. Следует отмстить, что все эти повторные нарушения функций
пораженной конечности бывают глубже и длительнее при удалении коры
одноименного пораженной конечности полушария. Этот на первый из ляд па-
радоксальный факт в действительности легко понять, если иметь в виду, что
верхние этажи центральной нервной системы свой контроль за деятельностью
пораженной лапы могут осуществлять только через неповрежденную, т. е.
противоположную, Половину, вернее, через неповрежденный мостик спин-
ного мозга.
Удаление коры второго полушария полностью и безвозвратно ликвиди-
рует эту новую компенсацию. Под нашим наблюдением было 8 собак, ли-
шенных всей коры большого мозга и подвергнутых половинной перерезке
спинного мозга (операции у 2 собак-были произведены в обратной последо-
вательности, чем у описанных выше). Эти собаки вообще являются очень
уязвимыми и быстро погибают. Большинство из них жило после всех опера-
ций около 1 месяца или немного больше; только одна собака жила больше
3 лет. Характерно, что у этих ссбак очень глубоко нарушаются функции
не только пораженной конечности, но и противоположной задней. Хотя
как прямые, так и перекрестные рефлексы вызываются у них па обе лапы,
тем не менее собаки теряют способность активно- или даже пассивно опи-
раться на них или пользоваться ими для стояния я ходьбы. Собаки припод-
нимают переднюю часть тела, опираясь на передние конечности, и с их по-
мощью передвигаются, волоча за собой бездействующую заднюю часть тела
с окоченевшей и вытянутой пораженной лапой. Хронаксня нервов сущест-
венно не изменяется, а хрогшксия рефлексов пораженной лапы увеличивает-
ся с большим опозданием. Тонус же рефлекторных и в особенности экстеи-
зорных мышц значительно повышается.
3. Поперечная перерезка задних столбов, вернее, задней половины спин-
ного мозга па уровне последних грудных сегментов и па уровне второго
шейного сегмента нами произведена у 6 взрослых собак и у 3 щенков 3—
4-месячного возраста. Нарушения функций пораженных конечностей у этих
собак, в особенности сензорных. бывают исключительно глубокими; все
конечности парализуются, причем у взрослых собак в первые 5—10 дней;
у щенков в 1-й день никак не удается получить рефлексы как с конечности,
так и на конечности. Больше того, в это время не удается вызвать даже бо-
левую реакцию или вообще реакцию с пораженных конечностей на голову.
Характерно, однако, что в этой же фазе животные при «усилиях» или зове
их по кличке, при ласкании или показе им вкусной пиши беспорядочно дви-
гают всеми пораженными конечностями. Со временем все эти нарушения
компенсируются (у щенков через 3—6 дней, а у взрослых — через 1 —11/г ме-
НОВЫЕ ДАННЫЕ ПО ФИЗИОЛОГИИ БОЛЬШОГО МОЗГА
379
•сяца после операции). Уровень моторной деятельности достигает нормы
и почти возвращается к нормальной сензорная деятельность. Отклонения
в реобазе и хронаксии моторных нервов (резкое увеличение реобазы, ни-
чтожное — хронаксии), в реобазе и хронаксии рефлексов (резкое увеличе-
ние реобазы, значительное — хронаксии) и в тонусе мышц (увеличение
тонуса сгибателей, уменьшение тонуса разгибателей) также в это время
исчезают (опыта А. И. Карамяна).
Существенно важно то, что у собак в процессе самой компенсации, иногда
даже в ранние периоды компенсации, восстанавливаются выработанные до
операций электрооборонительные условные рефлексы на пораженные лапы.
В этом периоде удается даже вновь выработать условные рефлексы (опыты
Л. А. Новиковой и Г. И. Бокова).
Более четкие данные об участии коры большого мозга в этих компенса-
торных явлениях были получены после удаления коры одного полушария
(у 4 собак) и двух полушарий (пока у 1 собаки). Как и в предыдущих слу-
чаях, удаление коры одного полушария глубоко нарушало наступившую
компенсацию по всем перечисленным выше показателям, причем сильнее
со стороны противоположной лапы. Вторично все эти нарушения компен-
сировались спустя примерно 1 месяц, причем условпорефлекторная дея-
тельность опять восстанавливалась в процессе компенсации. Одна собака,
лишенная также коры второго полушария, живет около месяца без каких-
либо признаков компенсации утраченных функций стояния и ходьбы на
пораженных лапах.
4. Продольное расщепление спинного мозга па различных его уровнях
и с охватом различного количества его сегментов нами было произведено
у 15 собак и 5 кошек (опыты Р. О. Барсегян). Эта операция другими исследо-
вателями раньше применялась главным образом в условиях острых и полуост-
рых опытов. Как правило, ими не проводились более или менее длительные
наблюдения с целью изучения возможности компенсации возникших нару-
шений моторных И сен зорных функций.
Наши данные, полученные в условиях хронических экспериментов, по-
казали, что сензорные и моторные функции конечностей очень глубоко и
необратимо нарушаются, если продольное расщепление охватывает все
сегменты спинного мозга, иннервирующие эти конечности. Если расщеп-
ляются не все сегменты и остается хотя бы один-два из срединных или из
крайних сегментов области, то возникающие глубокие нарушения со вре-
менем (15—30 дней) компенсируются. Если же продольное расщепление
производится вне области этих сегментов, выше или ниже их, то наступают
неглубокие нарушения, которые быстро исчезают.
Нарушения эти касаются как чувствительности (от легкого понижения
До полной потери), так и моторной деятельности (от ничтожного нарушения
координации до полной арефлексии). Соответственно глубине повреждения
изменяется хронаксия моторных нервов (незначительные колебания в ту
и другую сторону) и в особенности реобаза и хронаксия рефлексов (резкое
увеличение), а также тонус мышц (ограничение диапазона колебаний) по-
раженных конечностей. В тех случаях, когда наступает компенсация, все
эти отклонения от нормы исчезают, и оказывается возможным восстановить
или выработать условные рефлексы на пораженную конечность.
У 7 из ссбак с компенсированными нарушениями была удалена кора
одного полушария. После операции описанные выше нарушения возни-
кали вновь, причем сильнее с противоположной стороны. Вторично они
компенсировались примерно с такой скоростью, как и первичные наруше-
ния (от 4—5 дней до 1 месяца и больше). Соответственным образом изменя-
лись и восстанавливались также хронаксия, тонус и условнорефлекторная
деятельность. Пссле удаления коры обоих полушарий под длительным
наблюдением находилась только одна собака, остальные по тем или иным
ЗьО
3 А. АСРАТЯН
причинам погибали в первую или вторую неделю после операции. У со-
баки, о которой идет речь, первоначально было произведено неполное
расщепление пояснично-крестцовых сегментов, и как первичные нару-
шения, так и нарушения после удаления коры одного полушария у нее
компенсировались примерно в течение 20 дней. После полней декорти-
кации большого МОЗга она целиком утрачивала Способность к компенса-
ции и около года у нее не отмечалось признаков восстановления нару-
шенных функций стояния и ходьбы, хотя как прямые, так и перекрестные
рефлексы пораженных конечностей были налицо.
5, У 3 собак нами была произведена односторонняя перерезка задних
корешков поясничных и крестцовых сегментов. Картина нарушении пара-
лизованной конечности и хода восстановления ее функций у наших собак
полностью совпадала с описанной Бикелем картиной у аналогичным об-
разом оперированных им собак. Нормальная координационная деятель-
ность пораженных лап у наших ссбак восстанавливалась через 3—6 ме-
сяцев после операции. За это время происшедшие отклонения в хронаксии
моторных нервов (небольшое увеличение) и в тонусе мышц (резкое ослаб-
ление) также постепенно выравнивались. У всех 3 собак была удалена
кора противоположного деафферентирюванной конечности полушария,
в результате чего у них не только почти бесследно исчезла наступавшая
компенсация пораженных лап, но и сильно пострадали симметричные
неповрежденные лапы. Если последние через 10—11 дней уже могли
принимать участие в стоянии и ходьбе, нарушения деафферентированных
лап вновь компенсировались только через 1 ‘/а — 2 месяца и то не в со-
вершенном виде. После удаления коры второго полушария одна ссбака
погибла в первые дни после операции, вторая — через 2 недели, третья—
через 2 месяца. Результаты наблюдения над двумя последними осбаками
говорят о том, что полная декортикация большого мозга ге только при-
водит к полней иммобилизации деаффереитиров.аннсй конечности, но и
глубоко нарушает также функции симметричной неповрежденной конеч-
ности; несмотря на наличие способности к рефлекторной деятельности,
она не принимает участия ин в стоянии, ни в передвижении тела. Живот-
ные приподнимаются на передних лапах, передвигаются с их помощью,
волоча за собой бездействующую заднюю часть тела с резко экстензл-
роканной, как бы окоченевшей пораженной лапой.
б. Перерезка и перекрестное сшивание различных нервов были произ-
ведены нами у многих ссбак, но я остановлюсь только на тех 8, у которых
эта операция касалась функционально особенно важных нервов— nn. va-
gus и modi anus. После операции сензорные и в особенности моторные
функции соответствующей передней конечности очень сильно, страдают,
животные не могут на них опираться при стоянии и ходьбе. УЗ собак
никакой компенсации не наступило, а у остальных 5 спустя б—7 месяцев
нарушения компенсировались. Оказалось, что в этой стадии можно вы-
работать электрссборонительныс условные рефлексы на пораженную лапу
(данные В. А. Замятиной). Удаление коры полушария на стороне, про-
тивоположной пораженной лапе, вызвало у 2 собак значительной глуби-
ны Декомпеисато рные явления, которые со временем, примерно через
1 месяц, вновь компенсировались. У третьей собаки после тако.'.г же
операции декомпеисаториые явления оказались очень глубокими и
в течение 16 месяцев удовлетворительного восстановления так И не про-
изошло. Удаление коры второго полушария у первых 2 ссбак опять
вызвало декомп енсаторные явления более глубокого характера, чем
в первый раз. Спустя 1—I1/» месяца после операции картина незначи-
тельно улучшилась, однако резкая функциональная недостаточность
пораженных лап сохранялась в течение всех 10 месяцев последующей
жизни собак.
НОВЫЕ ДАННЫЕ ПО ФИЗИОЛОГИИ БОЛЬШОГО МОЗГА 381
7. Из большого числа ссбак и ксшек, у которых были разрушены лаби-
ринты с обеих сторон, особенный интерес представляют 4 собаки, у которых
впоследствии была произведена декортикация большого мозга. Картина
расстройств пссле разрушения обоих лабиринтов и ход компенсации у на-
ших животных ничем существенным не отличались от картины, давно из-
вестной из данных Флуранса, Эвальда, Магнуса и др. Отмечу лишь, что, чем
моложе животное, чем меньше его вес и чем оно ниже ростом, тем быстрее
наступает компенсация нарушений.
После декортикации одного полушария лабиринтные нарушения по-
являются опять (вращательные движения, бурные приступы хаотических
движений, нистагм глаз и головы, неспособность стоять и ходить). Через
11/г — 21/а месяца эти повторные нарушения вновь исчезают. Полная Декор-
тикация большого мозга вызывает очень глубокие нарушения, которые
остаются уже навсегда. Из 4 ссбак после всех перечисленных операций
2 погибли во 2-ю неделю после операции, одна — на 4-ю, а одна жила
1 */з года. Несмотря на исключительную живость спинальных рефлексов
и на наличие ряда установочных рефлексов, собака, которая жила
iVs года, так и не могла более или менее нормально сТоЯТЬ и ходить.
Во время приступа хаотических движений она иногда оказывалась на
ногах, с искривленным корпусом делала шага два и кувырком падала
на землю. Кроме этих собак, у одной лишенной коры собаки, не подвер-
гавшейся раньше никаким другим операциям, мы разрушили лабиринты
с одной стороны. После стихания явлений раздражения (через 5—6 дней
после операции) остались только явления выпадения, и они в почти
неослабевающем виде сохранялись примерно год. Собака еще жива,
находится в хорошем состоянии, но функции стояния и ходьбы и другие
лабиринтные нарушения у нее остаются сильно выраженными.
8, Из большого числа кошек и ссбак с перерезанными задними кореш-
ками первых трех пар шейных нервов наибольший интерес представляют
3 собаки, у которых впоследствии была произведена декортикация боль-
шого мозга.
Магнус и ван Левен, первые применившие эту операцию на кошках и
сделавшие ряд интересных наблюдений в условиях хронических экспери-
ментов, показали, что после этой операции в силу исчезновения шейных
рефлексов выявляются глубокие нарушения в функциях стояния и ходьбы,
которые впоследствии довольно быстро компенсируются. Магнус полагал,
что компенсация происходит в силу компенсаторного усиления установоч-
ных лабиринтных рефлексов и установочных рефлексов с тела на тело. Зна-
чительные индивидуальные колебания в глубине нарушений и быстроте
компенсации Магнус объяснял разницей в удельном весе лабиринтных и
шейных рефлексов у различных индивидуумов. Сравнивая картину наруше-
ния и ход компенсации у наших собак (всего 5) с соответствующей картиной
у кошек, оперированных Магнусом и ван Левеном, а также нами, мы прихо-
дим к выводу, что у ссбак после перерезки задних корешков первых трех
пар шейных нервов нарушения вообще бывают глубже, чем у кошек.
Кроме того, мы у ссбак наблюдали большие индивидуальные различия в
глубине нарушений и быстроте их компенсации.
В первые дни операции собаки совершенно не в состоянии были стоять
из-за сильной атонии и астении мышц конечно стен, в особенности задних.
Передвигались они ползком, покачиваясь и падая с боку на бок. Постепен-
но они начинали стоять и ходить на широко расставленных конечностях,
покачиваясь, пошатываясь и часто падая (атактическая походка). Со вре-
менем все нарушения исчезли (у 2 собак через 10—12 дней, у 2—через 15
20 дней, у 1 — через 1 месяц пссле операции). У 3 из этих ссбак удаление
коры одного полушария вызвало лишь небольшие и кратковременные нару-
шения, немногим отличающиеся по глубине и длительности от нарушений,
382
Э. А. АСРАТЯН
появляющихся при декортикации одного полушария у нормальных ссбак.
Через 10—17 диен они могли стоять и ходить. После полного выравни-
вания картины у этих собак удалялась также кора второго полушария,
что вызывало уже глубокие изменения. Самое характерное—это по-
стоянная склонность ссбак вертеться вокруг длинном оси тела в направ-
лении оперированного в последний раз полушария. Ссбаки иногда
вертятся в течение десятков минут, пока у лих нс начинается одышка.
После некоторогоотдыхаони опять начинают вертеться и т. д. Лабиринт-
ные тонические рефлексы и установочные рефлексы хорошо выражены,
как и все виды спинальных и бульбарных рефлексов. Несмотря на это,
у ссбаки, жившей полгода, утраченные функции стояния и ходьбы так
и не восстановились. Попытки ее к стоянию или ходьбе приводили
к верчению. Передвигалась собака ползком, если так можно выразиться,
спиралеобразно, вертясь вокруг оси, что сопровождалось хаотическими
движениями всех частей тела при кратковременном нахождении в по-
ложении животом вниз,
У второй собаки, у которой первичная операция шейных нервов выз-
вала ничтожные н кратковременные нарушения, полная декортикация боль-
ших полушарий не привела к длительным и глубоким нарушениям. Резко
были выражены лишь нарушения функций шейных мышц и мышц передних
конечностей. Около 20 дней собака не могла активно менять положение
головы и спираться на передние конечности, тогда как на задние конеч-
ности она опиралась хорошо и могла даже стоять на них самостоятельно.
При этом положении она даже передвигалась, подталкивая лежащие ла
полу гслову и переднюю часть тела. Постепенно стало улучшаться со-
стояние шейных мышц и передних конечностей; собака могла приподни-
мать на небольшую высоту голову и переднюю часть тела. Координация
движений передних лап в этой фазе была нарушена. Через Р/а—2 месяца
после операции собака уже могла пользоваться передними конечностями
при ходьбе, хотя они при этом переплетались и собака часто падала.
Через 8 месяцев после операции она довольно хорошо пользуется перед-
ними ногами, хотя до сих пор при ходьбе они часто переплетаются, и жи-
вотное падает.
9. В случаях, когда первоначальные повреждения нетяжелые (ампута-
ция одной лапы, перерезка и перекрест функционально маловажных нер-
вов» поперечная перерезка и продельное расщепление спинного мозга в
ничтожных размерах и т. д.) и вызывают неглубокие и быстро проходя-
щие нарушения, декортикация не приводит к заметным декомпенсаторным
явлениям. По нашим данным, декортикация и даже децеребрация нс
оказывают заметного влияния также на быстро протекающее приспособ-
ление, известное в литературе как перекрестный феномен диафрагмаль-
ного нерва. Пссле боковой половинной перерезки спинного мозга на
уровне второго шейного сегмента прекращается одностороннее грудное
и диафрагмальное дыхание. Если же перерезается диафрагмальный нерв
противоположной стороны, то иногда сразу, иногда через некоторое
время восстанавливается диафрагмальное дыхание со стороны спинальной
перерезки. У одной из ссбак с удаленней корей этот феномен был получен
в очень ярком виде. Кроме того, у 6 кошек, у которых этот феномен по-
лучился отчетливо, пссле последовательного удаления коры, межуточ-
ного, среднего мозга и мозжечка феномен все же сохранился, хотя
ритм и глубина дыхания заметно при этом изменились.
10. При всех вариантах оперативного повреждения спинного мозга,
в особенности псясничных и крестцовых его сегментов, как правило, наряду
с соматическими расстройствами, появляются также' нарушения ряда
вегетативных функций: мочеиспускания, дефекации, температуры кожи»
отмечается образование язв и другие трофические нарушения.
НОВЫЕ ДАННЫЕ ПО ФИЗИОЛОГИИ БОЛЬШОГО МОЗГА
3.83
Эти расстройства, так же как и соматические, со временем компенси-
руются. Удаление коры одного, полушария часто вызывает ослабление
или даже временное исчезновение компенсации. После наступления вто-
ричной компенсации удаление коры второго полушария нередко спять
приводит к ее ослаблению в той или иной степени. После всего этого
иногда нарушенные вегетативные функции вновь компенсируются, ино-
гда же заметной компенсации не наблюдается. По нашим данным
(опыты Я- М. Прессмана), декортикация большого мозга значительно
отражается также на феномене появления амилазы в слюне ссбак пссле
перевода их на углеводистую диэту (В. С. Замычкина).
Резюмируя полученные нами на собаках данные, можно сказать: во
всех случаях серьезного оперативного повреждения тех или иных важных
частей нервной системы, рецепторных и эффекторных органов, вызываю’
щих глубокие и длительно компенсирующиеся нарушения соматических
н вегетативных функций организма, удаление коры одного полушария боль-
шого мозга обусловливает кратковременное, иногда длительное, а удаление
коры обоих полушарий, как правило, стойкое исчезновение компенсации
соматических функций и значительное ослабление компенсации вегетатив-
ных функций. Если же указанные серьезные оперативные повреждения
производятся на собаках, лишенных коры, то компенсации нарушений
вовсе не наступает.
Удаление коры одного и двух пслушарнй заметно не сказывается лишь
на таких компенсаторных явлениях, которые протекают быстро, возни-
кают после ничтожных оперативных повреждений и неглубоких нарушений
функций тех же систем.
Существуют, конечно, переходные формы между этими двумя крайними
положениями (как, например, оДна НЗ наших Ссбак с Деафферснтацией
шейных нервов),
Гистологические исследования мозга некоторых ссбак, произведен-
ные И. Е. Пригонниковым (в лаборатории Л. Я - Пинеса), дали следую-
щие результаты.
Привожу выписку из характеристики И. Е. Пригон ннкова: «Мик-
роскопическое изучение данного материала произведено на непрерывных
сериях препаратов, окрашенных как по методу Вейгерта (на волокна),
так и по методу Ниссли (па клетки). Па основании полученных данных
можно сказать, что цель экспериментатора — выключение коры, — не-
сомненно, достигнута. В одном случае кора полностью была удалена.
В других случаях можно было обнаружить в той или иной степени
уцелевшие пссле операции ничтожные участки коры. Чаще всего оста-
валась неудаленной кора орального отдела лобной дели и каудального
отдела затылочной доли. Однако остатки коры обнаруживали при микро-
скопическом исследовании тяжелые дегенеративные изменения. Функция
их должна была резко пострадать».
Несколько иная картина наблюдается у птиц (голубей) и тем более у че-
репах и лягушек.
По нашим данным, полученным на сотнях голубей (опыты А. И. Кара-
мяна, Б. Д. Стефанцова, В. Д. Дмитриева и Г. И. Бокова), после ампутации
одной ноги, после одно- и двусторонней поперечной половинной перерезки
спинного мозга на различных его уровнях, после перерезки задних столбов
на различных уровнях, пссле продельного расщепления спинного мезга,
охватывающего различные по числу и положению сегменты, пссле деаф-
ферентации ног, а также при применении различных комбинаций этих
операций нарушения функции бывают также достаточно глубокими, но
в большинстве случаев компенсируются значительно быстрее, чем у ссбак.
При этом у них так же, как у ссбак, в процессе компенсации вырабаты-
ваются новые и восстанавливаются старые условные рефлексы на поражен-
384
Э А АСРАТЯН
ные лапы. Удаление одного полушария переднего мозга также вызывает
декомпенсаторные явления, которые довольно быстро вновь компенсиру-
ются. И в этой фазе У голубей вырабатываются ИЛИ Восстанавливаются
условные рефлексы на пораженные лапы. Одно- или двухмоментное удале-
ние обоих полушарий переднего мозга вызывает очень глубокие декомпен-
саторные явления, которые тем не менее спустя 2—5 месяцев спять компен-
сируются, хотя и неполностью. Характерно, что когда первичные операции
производятся на голубях с удаленными полушариями, то компенсация на-
рушений принимает более затяжной характер и бывает еще менее совер-
шенной. Соответствующие данные, полученные на лягушках и черепахах
(опыты Ф. М. Вишневецкой, Е. М. Лих иной, Л. И. Карамяна, Р. О. Бар-
сегян), полностью совпадающие сданными Мерцбахера, Бикеля и др.,
говорят о том, что первоначальные нарушения у них бывают ничтожными,
иногда неуловимыми, и удаление переднего мозга существенно не отра-
жается на характере течения этих нарушений.
Исследования по возрастной физиологии пластичности нервной системы
у собак пока недостаточны дня каких-либо определенных выводов, поэтому
я на них останавливаться не буду.
Опыты с полным удалением переднего мозга у птиц и у низших
позвоночных и коры большего мозга у ссбак, бесспорно, говорят о том,
что нельзя абстрактно ставить вопрос о роли и участии верхних этажей
центральной нервной системы в компенсаторных явлениях, т. е. в; спе
циального типа приспособительных явлениях в организме, как это делает
Бете. В процессе эволюции животного мира роль и значение большого
мозга в этих особого рода приспособительных явлениях, как ив приспо-
собительных явлениях вообще, изменяются, возрастают. В то время как,
по нашим данным, у собак кора большого мозга играет решающую роль
в приспособлениях, возникающих после глубоких нарушений функций,
у птиц в аналогичных случаях передний мозг имеет менее важное значе-
ние, а у низших позвоночных — совсем ничтожное.
Таким образом, результаты наших восьмилетних исследований на боль-
шом количестве подопытных животных совершенно бесспорно иллюстри-
руют несостоятельность основного тезиса теории Бете о пластичности нерв-
ной системы, говорящего о том, будто «научение» и вообще головной мозг
не играют роли или играют ничтожную роль в приспособительных явле-
ниях, протекающих в организме после различного рода повреждений. Од-
новременно с этим наши данные полностью совпадают с классическими дан-
ными материалистической биологии и физиологии о функциях верхних
этаже)! центральной нервной системы.
Общеизвестно, что большой мозг непосредственно не связан с перифери-
ческими органами; эту связь он может осуществлять только посредством
низших отделов центральной нервной системы — через спинной, продолго-
ватый мозг, область четверохолмия л т. д.
Совершенно очевидно, что при повреждениях тех ИЛИ Иных частей низ-
ших отделов центральной нервной системы нарушенные при этом функ-
ции организма могут быть восстановлены лишь при условии, если
незатронутые части тех же отделов нервной системы в качестве пер-
вичных исполнительных аппаратов, непосредственно связанных с пери-
ферическими воспринимающими и исполнительными органами, возьмут
па себя функции поврежденных отделов.
Участие большого мозга в указанных выше приспособительных явле-
ниях в конечном итоге может заключаться только в изменении и пере-
стройке функции неповрежденных частей низших отделов центральной
нервной системы, с тем чтобы они могли замещать выпавшие из строя части
той же системы. По нашим данным и теоретическим положениям, боль-
шой мозг как самый высший и могучий орган центральной нервной си-
НОВЫЕ ДАННЫЕ ПО ФИЗИОЛОГИИ БОЛЬШОГО МОЗГА
385
стемы участвуег в перестройке функции неповрежденных частей нервной
системы различными путями, с помощью различных средств и «механиз-
мов», среди которых главными являются: 1) улучшение состояния и ра-
боте способ нести этих исполнительных частей нервной системы посред-
ством трофического» в понимании И. П. Павлова, воздействия; 2) более
совершенное восприятие и переработка сигналов периферических орга-
нов чувств о происшедших в организме изменениях и соответственно
этому более совершенная безусловнорфлекториая регуляция функции
организма; 3) выработка новых простых и сложных условных рефлек-
сов, т. с. новых высших типов церебральной деятельности, соответственно
изменившимся в организме условиям.
Судя по всему, последний механизм передачи корковой перестройки низ-
шим отделам центральной нервной системы является наиболее эффектив-
ным в решительной перестройке, изменении и приспособлении функции не-
повреждённых рабочих аппаратов нервной системы к новым условиям.
В случаях, когда компенсаторные явления приводят к успешному приспо-
соблению даже в условиях отсутствия большого мозга или коры (у птиц и
рептилий — при серьезных первоначальных нарушениях функции, у собак —
при ничтожных первоначальных нарушениях функции), перестройка нспол-
ншельных нервных приборов, очевидно, в значительной мере происходит
под влиянием неповрежденных высших интегративных аппаратов межуточ-
ного и среднего мозга, а частично в силу примитивных пластических свойств
самих исполнительных нервных центров.
Совершенно очевидно, что дальнейшее изучение интересной проблемы
пластичности нервной системы не только расширит и уточнит наши теорети-
ческие представления, но и будет иметь существенное значение для прак-
тики. Сейчас дистанция между высшим объектом наших исследований —
собакой и человеком — слишком большая, чтобы можно было без риска
впасть в механицизм на основании наших данных сделать определенные
практические выводы для человека. Мы будем стараться, однако, и в
этом вопросе идти по тому пути, по которому шел и завещал всем нам
идти наш любимый учитель Иван Петрович Павлов.
25 Э. А. Аератям
ПРЕДВАРИТЕЛЬНЫЕ РЕЗУЛЬТАТЫ И ПЕРСПЕКТИВЫ
ПРИМЕНЕНИЯ НОВОГО СПОСОБА АНЕМИЧЕСКОГО ПОРАЖЕНИЯ
ЦЕНТРАЛЬНОЙ нервной системы у высших животных1
етоднка хирургического выключения и повреждения различных
отделов мозга занимаетвидяое место средн других методик ис-
следования функций центральной нервной системы. С помощью
этой методики получено немало сведен ин относительно функций
тех или иных отделов центральной нервной системы.
Несмотря на эффективность этой методики, она обладает
рядом недостатков. Сотрясение мозга при вскрытии черепной коробки или
спинного канала; травма мозга даже при очень нежном обращении
с мозговой тканью; неизбежное нарушение циркуляции крови нс только
в тех отделах мозга, которые непосредственно подвергаются разрушению,
но и в тех его участках, которые должны были остаться нормальными,
неповрежденными; наличие сгустков крови, остающихся даже при тща-
тельной операции; рубцы, которые появляются некоторое время спустя;
отек и набухание мозговой ткани и, наконец, самое главное довольно
обширные дегенеративные изменения, которые распространяются на
большое расстояние от участка непосредственного приложения ножа
экспериментатора, — вот неполный перечень тех изменений и факторов,
которые неизбежно сопутствуют любому хирургическому вмешательству
на центральной нервной системе, искажают картину выпадения функций
и мешают экспериментатору делать более или менее определенные выводы
из своей работы.
Недостатки этого распространенного метода исследования функций цен-
тральной нервной системы побудили исследователей искать иные пути и
средства воздействия на структуру, атом самым и нафункцию Тех или иных
ее частей. И. П. Павлов всемерно поддерживал эти поиски.
Среди новых приемов воздействия на структуру и функции центральной,
нервной системы заслуживает особого внимания ос анемическое пораженно.
Многочисленные деятели практической медицины и экспериментаторы,
занимающиеся исследованием так называемой «проблемы оживления орга-
низма», давно наблюдали одно очень важное явление, а именно что наиболее
чувствительна к прекращению кровообращения в организме центральная
нервная система. Более того, эти исследователи обнаружили, что основным
фактором, препятствующим достижению их пел и—восстановлению функций
клинически умершего организма,— является как раз поражение функций
центральной нервной системы.
Многочисленные данные этих исследований, как бы собранные попутно,
и богатый материал экспериментальных работ, специально поставленных
многими нашими и зарубежными исследователями с целью изучения
___ - «
1 Доложено 28.11 1948 г. на 13-м совещании по физиологическим проблемам,
посвященном памяти И. II. Павлова. Физиологический жу риал СССР, XXXV, № 5, 1949
НОВЫЙ СПОСОБ АНЕМИЧЕСКОГО ПОРАЖЕНИЯ Ц.. Н. С.
387
последствий анемического поражения центральной нервной системы, при-
вели к другому очень важному выводу, а именно, что различные этажи
центральной нервной системы обладают неодинаковой резистентностью
к анемическому поражению, к прекращению циркуляции крови. Несколь-
ко схематизируя, можно сказать, что, по данным этих исследователей,
наиболее чувствительными к анемизации являются филогенетически наи-
более молодые образования центральной нервной системы — мозжечок,
кора головного мозга, а последовательно все более и более резистент-
ными к прекращению циркуляции крови — менее молодые части; наибо-
лее же резистентными являются бульбарные центры и спинной мозг
[И. Р. Петров, Л. Л. Андреев, С. С. Брюхоненко, В. Л. Неговский,
Купер (Cooper), Гейманс (Heymans), Хилл (Hill) и др.].
Результаты экспериментов, полученные различными исследователями,
неоднородны; быть может, это объясняется тем, что разные исследователи
применяли различные способы выключения или прекращения кровоснабже-
ния. Каждый же из этих способов, обладая определенными преимущест-
вами в одних отношениях, страдал недостатками в других, и в итоге созда-
валась довольно пестрая картина последствий анемического поражения.
Так, например, очень распространенным способом анемизации мозга
является перевязка питающих его сосудов: либо четырех крупных сосудов,
питающих головной мозг, либо брюшной аорты — в целях апемизирования
спинного мозга [Купер, Броун-Секар (Brown-Sequard)]. Выяснено, что эти
приемы не приводят к цели. Очень часто различными окольными путями
и головной мозг, и спинной все же снабжаются кровью, хотя и не в доста-
точном количестве. В целях устранения или хотя бы ослабления путей
окольного кровоснабжения мозга возникла масса вариантов этого способа
анемизации (П. К- Анохин и др.).
Другой прием, также довольно распространенный, заключается в сле-
дующем: либо выпускают полностью кровь из организма, вызывая как бы
клиническую смерть вплоть до прекращения работы сердца с тем, чтобы
последующим вливанием крови восстановить работу сердца и функции
организма; либо прекращают циркуляцию крови путем остановки сердца
(электрическим ударом, механическим путем, наркозом или каким-нибудь
другим способом) или же путем зажатия каких-нибудь крупных сосудов,
например, аорты или легочных артерий. Этот способ тоже страдает суще-
ственным недостатками. Лишается крови не только центральная нервная
система, но и весь организм, а если учесть, что капилляры сосудов
по своей чувствительности к аноксии не уступают многим нервным клет-
кам, то легко понять, что при выпускании" крови или полном прекра-
щении ее циркуляции вызывается не только анемизации центральной
нервной системы, но и сильное поражение всей капиллярной сети, в том
числе и мозговых капилляров. Л это не может, конечно, не отразиться
на всей последующей картине нарушения и восстановления функций
организма, в том числе и функции центральной нервной системы.
Существующие способы анемизации центральной нервной системы пе
дают возможности точно дозировать наносимое поражение. Креме того,
эти способы не позволяют в момент анемического поражения централь-
ной нервной системы точно, графически регистрировать динамику изме-
нений основных функций организма.
Вот почему, несмотря на то, что с помощью указанных способов было
накоплено много новых и важных сведений относительно близких и
отдаленных последствий анемического поражения центральной нервной
системы, необходимость дальнейшего усовершенствования этого приема
не отпала. Представлялось важным усовершенствовать этот своеобразный
«анемический нож» и сделать ею пригодным к более тонким вмешательствам
в структуру центральной нервной системы, к более тонким поражениям
25*
ЗВ8
Э А АСРАТЯН
различных ее отделов, которые могут сравниваться между собой и по-
служить основанием для точных научных выводов.
Убедившись на основании мноюлстнего применения в недостатках
хирургического способа поражения центральной нервной системы, я за-
нялся изысканием способа ее анемизацни; эта работа проводилась мной
совместно с Г Т. Сахиулиной
Мы разработали способ анемизацни центральной нервной системы собак,
который, как мнекажстся. выгодно отличается от всех перечисленных выше.
В основу его положен прием искусственного повышения ликворного
давления, предложенный Лейденом (Leiden) и в последующем в раз-
личных целях использованный клиницистами. Суть его заключается
Кнмограыма опыта с анемизапией центральной нервной системы у собак
Обозначснкм (смсрху вниз) дыхание, артериальное д.-шление нулевая линия давле-
ния, прямоугольник — период повышенного давления (ЗОО мм) м сублуральноэ» пространстве
в следующем: производят субдуральную пункцию, иглу соединяют с ре-
зервуаром, содержащим рингеровскии раствор. Резервуар ставят на такой
Высоте, чтобы развиваемое жидкостью Давление равнялось 300—380 мм
ртутного столба, Т. е. было почти в 2 раза выше нормального арте-
риального давления. Организм в целом, если можно так выразиться,
живет, так как сердце работает, а путем искусственного дыхания
он снабжается кислородом; лишается крови только центральная нервная
система (в стадии анемизацни введенные в ток крови краски не окраши-
вают мозга). При этом, как показали наши контрольные опыты, вся
центральная нервная система может быть полностью анемизирована
сразу же. После прекращения этого давления немедленно возобновляется
циркуляция крови в центральной нервной системе. Это делает возможным
точную дозировку времени анемизацни или степени травмы.
В случае необходимости можно анемизировать всю центральную нерв-
ную систему или отдельно спинной и отдельно головной мозг. Техника
анемизацни очень проста и дает возможность как до, так и в момент
анемизацни и после ее прекращения производить графическую регистра-
цию артериального давления и дыхания, а также вести другие наблюде-
ния. На рисунке приведена кнмографическая запись опытов в момент
анемизацни.
Вскоре после начала анемизацни прекращается дыхание, а вслед за тем
НОВЫЙ СПОСОБ АНЕМИЧЕСКОГО ПОРАЖЕНИЯ Ц. Н. С
3S9
в известной последовательности ослабевают и исчезают спинальные и буль-
барные рефлексы. Отчетливо изменяется артериальное давление.
После прекращения анемизации постепенно, через определенные интер-
валы и в известной последовательности, восстанавливаются исчезнувшие
рефлексы — бульбарные, спинальные, среднемозговые, межуточно-мозго-
вые, а некоторое время спустя и функции коры больших полушарий, если
анемизации была не так длительна, чтсбы вызвать сильное повреждение
нервных клеток или смерть.
Нам удалось наблюдать в течение длительного времени большое число
животных, центральная нервная система которых подвергалась анемизации
в течение 6—20 минут. У нас имеются две собаки, одна из которых живет
уже 2 месяца после 20-мннутной анемизации всей центральной нервной си-
стемы, а вторая живет больше месяца.
Известно, что другим исследователям, пользовавшимся другими прие-
мами анемизации центральной нервной системы, удавалось сохранить жизнь
животных и вести наблюдения за ними в течение длительного времени лишь
после?—10-минутной анемизации. Мы с Г. Т. Сахиуловон почти удвоили
время анемизации этой системы с последующим длительным сохране-
нием Жизни животных.
Каковы отдаленные, последствия такой анемизации? Ограничусь лишь
некоторыми краткими сообщениями предварительного характера. Следует
сказать, что основное положение, которое подчеркивалось многими авто-
рами, а именно что филогенетически более молодые образования централь-
ной нервной системы (в частности, кора больших полушарий) страдают от
анемизации раньше и глубже филогенетически более древних (спинальные,
бульбарные, среднемозговые центры), в условиях наших более точных экс-
периментов в основном подтвердилось. Сохранение жизни собак после
более длительной анемизации позволило нам выяснить, что при углубле-
нии анемического поражения можно постепенно вывести из строя
вслед за корой и менее молодые образования: базальные, меж у точно-мозго-
вые и среднемозговые. Собака, которая была подвергнута 20-мннутной ане-
мизации, 2 месяца лежит на боку, простые рефлексы и основные жизненные
функции протекают у нее более или менее нормально, но сложные рефлек-
торные акты, как стояние и ходьба, остаются для нее невозможными. У нее
отсутствуют даже такие среднемозговые рефлексы, как выпрямительные.
При 16-минутной анемизации эти рефлексы сохраняются, и такие животные
очень напоминают собак, лишенных коры хирургическим способом. У собак,
у которых анемизации производится в течение 12минут и меньше, рефлексы
восстанавливаются более полно, а после 6—7-минутной анемизации — на-
столько полно, что при обычном наблюдении трудно выявить какую-ни-
будь разницу между ними и нормальными собаками.
Нас особенно интересовали многие проявления специфических рефлек-
торных актов, свойственных тем или иным уровням центральной нервной
системы, и главным образом условнорефлекторная деятельность. Г. Т.Са-
хиулина занимается специальным изучением этого вопроса. Она
выяснила, например, что у собак, подвергнутых анемизации в течение
10—12 минут, вырабатываются условные рефлексы, хотя, судя по литера-
турным данным, корковые элементы у таких собак должны быть
сильно повреждены. Более того, нечто очень напоминающее условные реф-
лексы имеется и у собаки, подвергнутой 16-минутной анемизации.
Нужно все же подчеркнуть, что эти рефлексы занимают какое-то погра-
ничное положение между условными рефлексами и явлениями типа доми-
нанты или проторения путей.
Имеются некоторые основания предполагать, что примитивные реакции
временного характера, иначе говоря, примитивные условные рефлексы,
39»)
Э. А. АСРАТЯН
по крайней мере до уровня развития собаки, могут осуществляться также
базальными ганглиями.
Что касается динамики восстановления нарушенных анемизацией функ-
ции, то хотя она н варьирует в зависимости от длительности анемизации,
от возраста, состояния здоровья и некоторых других особенностей подопыт-
ных собак, тем не менее можно четко выявить н исследовать ряд более или
менее определенных фаз пли стадий этой динамики. Вслед за первоначальной
фазой восстановления жизненных функций организма наступает фаза воз-
буждения, затем фаза более или менее глубокого угнетения функций выс-
ших отделов центральной нервной системы, которая иногда нарушается
вспышками возбуждения.
Таким образом, некоторое время фаза угнетения (явно охранительной
природы) и фаза возбуждения довольно часто сменяют друг друга. Нако-
нец, наступает фаза медленного восстановления моторных и сензорныхфунк-
ций организма в тех или иных пределах. Сензорные функции как будто
страдают глубже и восстанавливаются позже моторных. Следует заметить,
что снотворные средства ускоряют темпы восстановления нарушенных функ-
ций организма.
Предложенный нами способ, несомненно, является наиболее совершен-
ным из существующих способов анемизации. Но результаты его применения
тогда только приобретут большую убедительность, когда препараты мозга
животных будут подвергаться гистологическому исследованию. Лишь при
этом условии можно надеяться, что многие вопросы физиологии центральной
нервной системы, в Особенности коры больших Полушарий мбзгй, смогут
быть успешно разрешены.
ЛИТЕРАТУРА
Андреев Л. А., Деятельность головного моога собаки и церебральное кроно-обра’
теине, Архив биологических наук, т. 45, в. 2, 1937; Arch, neuro), a. psych., 34,
699, 1935.
Анохин II. К., Способ получения хронической анемии головного мозга, Русский
физиологический журнал, 13, 3, 395, 1930.
Брюхоненко С. С., Применение метода искусственного кровообращения для
оживления организма. Сб. трудов Научно-исследовательскогоинститута экспери-
ментальной физиологии и терапии, 1937.
Неговский В. А., Восстановление жизненных функций организма, находящегося
в состоянии агонии или в периоде клинической смерти, М,, 1943.
Петров И. Р., Кислородное голодание мозга. М., 1948.
Павлов И. П., Двадцатилетии it опыт объективного изучения высшей нервной
деятельности (поведения) животных, 222, 1938.
С ах и ул и на Г. Т., Влияние снотворных веществ на восстановление функций
анемизированного спинного мозга собаки. Рефераты работ учреждений биологи-
ческого отделения ЛИ СССР за 1945 г.
В rown-Sequard, J. de la physiologic de I'homine et des animaux, 1,95,729, 1858.
Cooper, Guy’s hosp. rep., 1, 457. 1836.
Cushing 11., Brit. i. surg., 5. 558, 1917—1918.
Leyden, Virch. Archiv. 37, 519, 1866.
Hi I!, The cerebral circulation, 1896.
Heymans u Bouckaeri, Ergebnisse der Pnysiologie, Bd. 41, 28, 1939.
НЕКОТОРЫЕ ОБЩИЕ ЧЕРТЫ НАРУШЕНИЯ И ВОССТАНОВЛЕНИЯ
НЕРВНОЙ ДЕЯТЕЛЬНОСТИ>
В
К_Л
ся творческая деятельность И. П. Павлова была проникнута
принципом единства теории и практики, стремлением познать
явления природы, чтобы управлять ими, поставить науку на
службу человеку.
Считая правомерной и полезной связь физиологии с практи-
кой жизни и со многими научными дисциплинами — психоло-
гией, педагогикой и т. д., — великий Павлов был особенно горячим сторон-
ником союза между физиологией и медициной. Он считал даже, что «пони-
маемые в глубоком смысле физиология и медицина неотделимы». Этой теме,
т. е. союзу физиологии и медицины, И. П. Павлов посвятил много блестя-
щих выступлений и докладов, а также много ярких страниц своего
гениального научного творчества. Он считал, что этот союз полезен как для
медицины, так н для физиологии. Для физиологии потому, что все болез-
ненные состояния организма, которые изучаются врачами, представляют
как бы своего рода эксперименты, поставленные «невидимой рукой» при-
роды, эксперименты, которые зачастую выявляют такие стороны течения
жизненных процессов в организме, которые в обычном эксперименте на
животных так ярко не выявляются. Но особенно выгоден этот союз, по
мнению И. П. Павлова, для медицины, ибо способ, которым пользуется
медицина для познания тайн природы, а именно наблюдение,
является пассивным и недостаточно эффективным для глубокого и многосто-
роннего познания происхождения и течения жизненных процессов. Физио-
логия же в качестве основного способа познания этих закономерностей
пользуется более эффективным средством — опыта м физиологическим
экспериментом, искусным мастером которого был сам Павлов. Он говорил:
«Наблюдениесобираетто, чтоему предлагает природа опыт же берет у при-
роды то, что он хочет. И сила биологического опыта поистине колоссальна»2.
И. П. Павлов считал, что вся медицина должна пройти через «огонь
эксперимента», что эксперимент сделает медицину сознательной; он считал,
что «чем полнее будет проделан опыт на животных, тем менее часто придется
больным быть в положении опытных объектов со всеми печальными послед-
ствиями этого». Исходя из всего этого, он считай целесообразным учредить
в медицинских высших учебных заведениях три специальные эксперимен-
тальные кафедры физиологии: нормальной, патологической и терапевтиче-
ской физиологии.
Выражением такого правильного понимания принципа единства теории
и практики, а также успешного претворения этого принципа в жизнь было
1 Доклад, прочитанный на сессии, посвященной 100-летню со дня рождения
И. П. Павлова, в 1949 г. Известия Академии наук СССР, серия биологическая,
№ 6, 1949.
8 И. П. Павлой. Полное собрание трудов, т. II, 1946, стр. 357.
392
Э. Л АСРАТЯН
то, что И. П. Павлов, кроме специальных физиологических исследований
в различных важнейших областях физиологии, имеющих сами по себе гро-
маднее значение для различных отделов медицины, занимался на протяже-
нии многих лет также систематическим изучением этиологии, функциональ-
ной патологии и терапии болезненных состояний некоторых важнейших
систем организма, а именно: сердечно-сосудистой, пищеварительной системы
и главным образом больших полушарий мозга.
Особенно следует отметить работы гениального физиолога го
экспериментальной патологии и терапии высшей нервной деятельности —
исследования, которые являются поистине классическими, создавшими
новую эпоху в ряде разделов медицины, в частности, в психиатрии и невро-
патологии.
Чтобы оценить все громадное значение этих исследований И. П. Павлова
для медицины, следует учесть, что в такой важнейшей медицинской дисцип-
лине, как общая патология, или, как теперь более принято называть,
патологическая физиология, до указанных исследований фактически
отсутствовала глава о функциональной патологии нервной системы. Не-
богатый материал, попутно накопленный в физиологических исследова-
ниях, не был обобщен с этой точки зрения, а специальных систематиче
ских и методических исследований в этой области, как известно, не про-
водилось. Достаточно сказать, что ие только во всех учебных, но и в спе-
циальных капитальных руководствах по патологической физиологии от-
сутствовал (и до сих пор, можно сказать, еще отсутствует) этот важней-
ший раздел патологии, именно патологии нервной системы.
Таким образом, исследования И. П. Павлова по патологической деятель-
ности больших полушарий мозга вообще являются первым, к тому же бес-
ценным вкладом в эту важнейшую область общей патологии. Классические
исследования гениального мыслителя заложили гранитный фундамент но-
вой главы общей патологии, открыли перед ней новые широкие горизонты
и предначертали пути дальнейших исследований в этой важнейшей области
экспериментальной медицины. По этим путям шли многие ученики
И. П. Павлова, в их числе и мы.
На протяжении 15 лет в руководимой нами лаборатории проводится
экспериментальное изучение проблемы нарушения и восстановления функ-
ций нервной системы. Изложению полученных нами данных и посвящено
настоящее сообщение.
Постоянно подчеркивая не только высокий уровень, но и качественно
специфический характер закономерностей работы коры больших полушарий
мозга по сравнению с закономерностями низших отделов центральной нерв-
ной системы, И. П. Павлов как диалектически мыслящий исследователь од-
новременно отмечал общность основных отношений, родство основных
закономерностей работы больших полушарий и низших отделов централь-
ной нервной системы.
И. П. Павлов неоднократно указывал, что изучение закономерностей
условнорефлекторной деятельности может оказать большую помощь
в познании закономерностей безусловнорефлекторной деятель нести, так
как в первом случае мы имеем дело с процессом возникновения и становле-
ния нервной деятельности, а во втором — с готовыми соотношениями,
установленными в отдаленном прошлом, на раннем этапе развития централь-
ной нервной системы. Как известно, с этим связаны и идеи И. П. Павлова
о возможности в определенных условиях наследственного закрепления
условных рефлексов, т. е. об их превращении в безусловные.
Логично было думать, что такая же общность существует между зако
номерноегями патологической деятельности коры больших полушарий,
обнаруженными и изученными И. П. Павловым, и между закономерностями
патологической деятельности низших отделов центральной нервной системы,
ОБЩИЕ ЧЕРТЫ НАРУШЕНИЯ И ВОССТАНОВЛЕНИЯ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ 393
к исследованию которых научно-исследовательская мысль только присту-
пает.
Иначе говоря, логично было думать, что подобно тому, как установлен-
ные И. П. Павловым закономерности нормальной условнорефлекторной
деятельности в настоящее время по праву считаются закономерностями ста-
новления и протекания рефлекторной деятельности вообще (со специфи-
ческими особенностями этой деятельности для каждой морфологически
и фунцкионально более или менее обособленной части центральной
нервной системы), подобно этому определенные И. П. Павловым зако-
номерности патологической деятельности и закономерности восстановления
нарушенной деятельности высших отделов, центральной нервной системы
также имеют универсальный характер для всей центральной нервной сис-
темы, иначе говоря, имеют значение и для патологической деятельности
низших отделов центральной нервной системы.
Результаты экспериментальной работы нашей лаборатории по вопросу
о нарушении и восстановлении нервной деятельности с отчетливостью
показывают, что это действительно так.
Вызывая патологические состояния различных отделов центральной
нервной системы—Головного мозга, периферических нервных образований—
различными патогенными воздействиями (хирургическим, термическим,
анемическим, грубомеханическим повреждением, сильной электрической
травмой, повреждением симпатической нервной системы и тому' подобными
патогенными воздействиями) и изучая возникновение и протекание патоло-
гических процессов в этих образованиях, мы убедились, что закономерности
патологической деятельности низших отделов центральной нервной системы
в своей основе родственны закономерностям патологической деятельности
больших полушарий головного мозга, выявленным и изученными. П. Пав-
ловым и его сотрудниками.
В дальнейшем я на конкретных примерах проиллюстрирую правиль-
ность этого положения.
Прежде всего о закономерностях возникновения патологических со-
стояний. И. П. Павлов неоднократно отмечал, что при так называемых
функциональных патогенных воздействиях на организм патологическая
деятельность коры больших полушарий возникает очень часто не тотчас,
а некоторое время спустя, как бы через некоторый длительный «латентный
период». Речь идет только о функциональных патогенных воздействиях,
а не об органических, при которых явно нарушается структурная целост-
ность коры больших полушарий мозга.
Наши эксперименты показали, что такую же картину мы имеем в воз-
никновении патологического состояния низших отделов центральной нерв-
ной системы.
Опыты с анемизацией изолированного спинного мозга и всей централь-
ной нервной системы у собак, а также опыты с термическим воздействием
на спинной мозг собак и кошек выявили, что нарушения деятельности
высших и низших отделов центральной нервной системы ярко выступают
не сразу после этих своеобразных (правда, не совсем функциональных,
но в известной мере к ним близко стоящих) патогенных воздействий, а спу-
стя много часов.
Далее, одной из характернейших особенностей патологической деятель-
ности коры больших полушарий, вызванных не только функциональными,
но и органическими патогенными воздействиями, И. П. Павлов считал асте-
нию, ослабление этой деятельности, Это само собой понятно. Во всяком слу-
чае клиницистам это явление известно очень хорошо. Наши эксперименты
как с анемизацией и термическим поражением низших отделов центральной
нервной системы, так и с органическими повреждениями их показали, что
394
Э. А. АСРАТЯН
та же самая черта характеризует патологическую деятельность и этих
отделов центральной нервной системы.
Если перерезать спинной мозг у собаки и произвести десимпатизацию
такого изолированного спинного мозга, то его рефлекторная активность
сильно понижается — рефлексы делаются слабыми, неустойчивыми, быстро
исчезают и, что особенно характерно, исчезнувшие рефлексы восстанавли-
ваются крайне медленно. То же явление наблюдалось и у холоднокровных
животных. Эта закономерность очень четко выявляется и при патологиче-
ских состояниях, вызванных анемизацией, термическим поражением и дру-
гими воздействиями наспинной мозг и на другие частя центральной нерв-
ной системы. Конечно, тот факт, что одной из характернейших черт болез-
ненных состояний высших и низших отделов центральной нервной системы
является астения, не может считаться новым. Мы упоминаем о нем лишь
для того, чтобы облегчить понимание сути следующей особенности пато-
логической деятельности центральной нервной системы.
По И. П. Павлову, очень важной особенностью патологической деятель-
ности больших полушарий является развитие в истощенных послабленных
болезнетворным воздействием нервных клетках коры полушарий охранитель-
но-целебного торможения, этого своеобразного природного средства само-
защиты и самолечения нервной системы, развившегося в процессе много-
вековой эволюции.
И. II. Павлов изучал закономерности развития охранительно-целебного
торможения в коре большого мозга при так называемых функциональных
патогенных воздействиях на организм; при действии чрезмерно сильных раз-
дражителей, при столкновении процессов возбуждения и торможения и при
всякого рода грудных и конфликтных ситуациях для организма. В тот пе-
риод, когда Он Систематически изучал последствия органических поврежде-
ний коры больших полушарий — последствия удаления тех или иных ее
частей, концепция об охранительно-целебной роли торможения не была еще
им развита. Возвращаясь позднее к этим исследованиям, И. П. Павлов
отмечал охранительный характер торможения, быстро развивающегося
в поврежденной коре под воздействием адэкватных в поврежденной зоне
условных раздражителей. Но - нам представляется, что можно с точки
зрения этой концепции И. П. Павлова осветить и осмыслить полученный
им же экспериментальный материал, относящийся к прямым послед-
ствиям органических повреждений коры больших полушарий. Даже при
беглом обзоре этого богатого и разнообразного экспериментального
материала И. П. Павлова и его сотрудников мы убеждаемся, что тормо-
жение, которое развивается в коре больших полушарий в результате
органических ее повреждений как таковых, представляет собой охра-
нительно-целебное торможение. Все эти данные говорят о том, что почти
все условные рефлексы после более или менее значительных поврежде-
ний любых частей коры большого мозга сильно ослабевают и даже исче-
зают на более или менее длительный срок. А по сути дела это есть не что
иное, как универсальное охранительное торможение ослабленной травмой
коры больших полушарий. Если такая кора и способна к условнорефлек-
торной деятельности, то только в ответ на слабые раздражители. Но эти
условные рефлексы отличаются крайней хрупкостью, быстро исчезают
(«раздражительная слабость» по И. П. Павлову).
В настоящее время имеются все основ.ання считать, что как эксперимен-
тальный материал И. П. Павлова и других исследователей относительно
последствий органических повреждений коры большого мозга, так и данные
нейрохирургических клиник относительно гак называемого центрального
шока, или диасхизиса, при повреждениях коры мозга свидетельствуют
о том, что охранительное торможение возникает в коре больших полу-
шарий не только при функциональных, но и при органических патоген-
•ОБЩИЕ ЧЕРТЫ НАРУШЕНИЯ И ВОССТАНОВЛЕНИЯ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ 395
ных воздействиях па кору, что явления центрального шока, или
диасхизнса, по своей природе суть не что иное, как павловское охрани-
тельно-целебное торможение. На это мы указывали еще 1941 г., и, судя по
всему, эта точка зрения получила признание. Кроме Того, наша эксперимен-
тальная работа позволила установить, что охранительно-целебное торможе-
ние представляет собой универсальное явление для всей нервной системы,
т. е. присуще не только коре больших полушарий, но и всем нижележа-
щим частям центральной нервной системы и даже периферическим нерв-
ным образованиям. В этих частях нервной системы, так же как и в коре боль-
ших полушарии, при различного рода болезненных состояниях, вызванных
органическими или функциональными патогенными воздействиями, рефлек-
торная деятельность, как правило, ослабевает, а часто временно исчезает
совсем. Было установлено, что развившееся в данном случае диффузное
торможение по природе является охранительным, т. е. играет такую же
важную биологическую роль, как и при патологических состояниях коры
больших полушарий. Об этом свидетельствует, в частности, то обстоятель-
ство, что если в таких случаях и возникает какая-нибудь рефлектор-
ная деятельность, то в основном на слабые раздражители. Положительное
влияние снотворных на темпы восстановления нарушенных при этом функ-
ций организма является другим убедительным доказательством правиль-
ности высказанного положения.
По данным 14. П. Павлова, одной из характернейших особенностей пато-
логического состояния коры больших полушарии является нарушение тон-
кой и совершенной координационной ее деятельности. Это выступает доста-
точно четко при болезненных состояниях, вызванных функциональными
патогенными воздействиями, но особенно ярко при органических патоген-
ных воздействиях на кору больших полушарий. Известно, что после того,
как проходит фаза универсального торможения уцелевших отповреждения
частей коры (мы можем сказать: охранительно-целебного торможения),
постепенно восстанавливаются положительные условные рефлексы, но тор-
мозные условные рефлексы длительное время остаются расторможенными,
нарушенными. Бесспорно, это является следствием ослабления условного
торможения (я предлагаю назвать его торможением координационным в от-
личие от охранительно-целебного торможения). Ослабление координацион-
ного торможения может считаться одной их характерных черт патологиче-
ской деятельности коры больших полушарий. Очевидно, этим обусловливает-
ся и некоторое усиление положительных условных рефлексов, и инертный
характер их протекания в начальной фазе восстановления нарушенной
условнорефлекторной деятельности поврежденной коры больших полуша-
рий мозга.
Эти черты — ослабление координационного торможения, инертность,
зачастую некоторая экзальтированность процесса возбуждения — харак-
терны также для низших отделов центральной нервной системы в периоды
патологической их деятельности, в особенности в начальные фазы восста-
новления утраченных или нарушенных функций. Тонический характер со-
кращения скелетных мышц и нарушение координации движений являются
типичной чертой патологической деятельности низших отделов центральной
нервной системы.
Согласно воззрениям И, П. Павлова, волнообразное течение условнореф-
лекторной деятельности является одной из типичных черт патологического
состояния и патологической Деятельности коры больших полушарий. Он
объяснял это явление слабостью коры и борьбой между процессом торможе-
ния (охранительным по природе) и возбуждением. Слабые корковые нервные
клетки быстро истощаются от более или менее значительной работы и охва-
тываются охранительным торможением. После некоторого укрепления в ре-
зультате торможения они вновь развивают временную деятельность и т. д.
396
Э. А. АСРАТЯН
Та же к артина наблюдается и при болезненных состояниях низших отде-
лов центральной нервной системы и, очевидно, имеет такой же механизм.
Я не буду ссылаться на наш специальный экспериментальный материал.
Можно указать на ряд явлений подобной волнообразности в деятельности
нервной системы, давно известных в физиологии и медицине, до сих пор
по-настоящему не расшифрованных, но могущих быть удовлетворительно
объясненными с этой точки зрения. К ним относятся дыхание Чейн-Стскса,
волны Траубе-Герингса в артериальном давлении. Здесь, как и в случаях
волнообразности в деятельности коры больших полушарий, речь идет,
очевидно, о борьбе между торможением охранительного порядка и воз-
буждением. Имеются все основания считать, что эта борьба и обуслов-
ливает волнообразность в деятельности ослабленных патологическим со-
стоянием клеток спинного, продолговатого мозга и т. п.
И. П. Павлов указывал, что в картине болезненного состояния коры
больших полушарий очень существенную роль играют типологические
особенности больного животного. Мы получили экспериментальный ма-
териал, говорящий о том, что, во-первых, эти типологические особенности
свойственны не только коре больших полушарий, как склонен был
думать И, П, Павлов, а что они являются, по всей вероятности, универ-
сальными, т. е. присущи всем отделам центральной нервной системы;
во-вторых, они сказываются на характере и течении болезненного состоя-
ния всех отделов, а не только коры.
В моей работе, опубликованной еще в 1938 г.1, был приведен достаточно
убедительный материал о том, что типологические особенности в какой-то
степени присущи всем отделам центральной нервной системы. У подопыт-
ных ссбак, оперативным путем лишенных коры больших полушарии го-
ловного мозга (их было около 50), многие индивидуальные особенности
нервной деятельности оставались в примитивной форме и после удаления
коры. Из этих ссбак пссле операции игривые оставались в общем игри-
выми, обжоры — обжорами, смирные—смирными, злые*—злыми и т. д.
Вызывая анемизацию спинного мозга или оказывая на него хирургическое
или иного рода патогенное воздействие, мы убедились, что очень часто
как глубина нарушения функций, так и характер и течение их обуслов-
ливаются далеко не только характером и тяжестью патогенного воздейст-
вия, возрастом, состоянием здоровья и упитанностью подопытного
животного, но очень часто определяются, говоря осторожно, инди-
видуальными особенностями нервной системы животных. Имеется доста-
точно оснований считать, что здесь речь идет главным образом о типо-
логических сссбенностях нервной системы.
Наиболее важным в нашей работе я считаю вывод о большей уязви-
мости чувствительных элементов рефлекторных дуг к болезнетворным
факторам. И. П. Павлов считал, что кора больших полушарий сплошь
состоит из чувствительных анализаторных, афферентных нервных элемен-
тов, осуществляющих наиболее совершенный анализ и синтез, создающих
«главную сложность и даже главную славу» деятельности коры больших
полушарий. Далее, И. П. Павлов неоднократно указывал, что эти клетки
являются наиболее уязвимыми к патогенным воздействиям, обладают наи-
более низким пределом, или «потолком», работоспособности. Хотя он и
не указывал на какую бы то ни было связь между этими двумя обстоя-
тельствами, тем не менее их сопоставление не лишено было бы определен-
ного значения.
Мне неизвестно, чтобы клиницисты отмечали большую уязвимость чув-
ствительных элементов рефлекторных Д\Т В различных чЗСгях централь-
ной нервной системы. Но наш экспериментальный материал говорит, что
Физиологический журнал, т. XXIV, в. I. 1938.
ОБЩИЕ ЧЕРТЫ НАРУШЕНИЯ] И ВОССТАНОВЛЕНИЯ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ 397
эго именно так, что при патогенных воздействиях общего характера пер-
выми нарушаются сензорные функции, первыми выводятся из строя
сензорные элементы рефлекторных дуг. Это относится ко всем частям
центральной нервной системы, начиная от коры больших полушарий
и кончая спинным мозгом. Первые доказательства правильности этого
положения были нами получены еще перед Великой Отечественной
войной. При резком ухудшении состояния спинного мозга (его перерезка,
анемизацни, десимпатизация и т. п.) он после кратковременной активной
работы прекращает рефлекторную Деятельность конечностей при их
раздражении, которые, одиако, долго сохраняют способность сокра-
щаться под влиянием импульсов из высших отделов центральной нервной
системы или же при раздражении других рефлексогенных зон. Та же
картина наблюдается при перерезке у собак на высоком уровне только
задней половины спинного мозга. Спинальная рефлекторная деятельность
исчезает, но под влиянием импульсов, исходящих из вышележащих
отделов центральной нервной системы, конечности сокращаются»
Полученный нами и нашими сотрудниками разнообразный фактический
материал, относящийся к различным отделам центральной нервной сис-
темы, показывает, что спинальный шок, центральный шок и многие
явления центрального паралича и пареза обусловливаются главным
образом нарушением чувствительных функций, выведением из строя сен-
зорных элементов рефлекторных дуг на соответствующих этажах цент-
ральной нервной системы.
Нам кажется, что и это положение, если не прямо, то косвенно гармони-
рует с концепцией И. П. Павлова о патологической деятельности коры
больших полушарий головного мозга.
Быть может, такая высокая чувствительное™, уязвимость чувствитель-
ных звеньев рефлекторных дуг обусловливаете! тем, что они, согласно
существующим воззрениям об эволюции нервной деятельности, являются
наиболее молодыми образованиями. Правильно ли это предположение или
нет, покажет будущее. Наш экспериментальный материал, относящийся
к этому вопросу, говорит о том, что не только экспериментаторам, но и
клиницистам надлежит пересмотреть под развиваемым нами новым углом
зрения как материал о разнообразных центральных параличах и парезах,
так и взгляды на их происхождение и природу. Центральные параличи
и парезы часто без достаточного основания приписывают нарушению
двигательных элементов центральной нервной системы. Они же большей
частью обусловлены именно нарушением функции сензориых элементов.
Мы видим, таким образом, что первые более или менее достоверные све-
дения о закономерностях возникновения и протекания патологической дея-
тельности низших отделов центральной нервной системы и о физиологиче-
ской природе этой деятельности свидетельствуют о значительной близости,
о родстве этих закономерностей с теми, которые для патологической
деятельности коры больших полушарии были выявлены и изучены
гениальным И. П. Павловым. Это в значительной мере относится и к за-
кономерностям восстановления нарушенной деятельности высших и низ-
ших отделов центральной нервной системы. В качестве иллюстрации
сказанного приведем несколько примеров.
Прежде всего следует отметить, что близкие и отдаленные последствия
различного по глубине оперативного повреждения проводящих путей и
серого вещества в различных отделах спинного и головного мозга приводят
к заключению, что основные принципы павловской теории динамической
локализации функций в коре больших полушарий могут быть отнесены так-
же ко всем нижележащим центральным’ нервным образованиям и массив-
ным проводящим путям, иначе говоря, носят универсальный характер.
В массивных центральных нервных образованиях и проводящих путях,
398
Э Л. АСРАТЯН
так же как и в коре, отдельные специализированные зоны не отделены друг
от друга четкими границами, а состоят из сконцентрированных ядерных
и рассеянных периферических элементов и своими краями перекрывают
друг друга. Далее, эти данные свидетельствуют о том, что и в этих
низших отделах центральной нервной системы процесс восстановления
нарушенной или утраченной функции после их хирургического повреж-
дения в значительной мере может рассматриваться как постепенная
активизация уцелевших от разрушения элементов, что степень и размеры
замещения утраченной функции зависят от объема уцелевших запасных
или рассеянных элементов, что для темпов и качества этой активизации
(иначе говоря, восстановления функции) большое значение имеют частота
осуществления функции, отдых, усиление охранительного торможения
фармакологическими и иными средствами и т. п.
Наши экспериментальные данные, полученные в опытах на живот-
ных, а также значительный клинический материал довольно подробно
освещены нами в ряде статей. В последние годы нами и нашими
сотрудниками получены убедительные экспериментальные данные отно-
сительно того, что внутривенное введение значительных доз препаратов
кальция является мощным терапевтическим средством при тяжелейших
повреждениях низших и высших отделов центральной нервной системы,
что при некоторых видах нарушения деятельности последней весьма
положительный терапевтический эффект дает также аденозннтрифосфор-
ная кислота. Имеются известные основания считать, что положитель-
ное терапевтическое действие препаратов кальция обусловливается их
свойством регулировать проницаемость клеточных мембран, ослаблять
отек и набухание патологически измененных нервных образований и
способствовать развитию в них охранительно-целебного торможения.
Положительное целебное действие аденаз интрнфосфор ной кислоты
может быть предположительно объяснено в свете современных данных
и взглядов В. А. Энгельгардта и других об ее громадной роли в биоэ-
нергетике клеток.
Резюмируя изложенные в этой статье данные, мы считаем необходи-
мым еще раз подчеркнуть, что исследования И. П. Павлова по эксперимен-
тальной патологии и терапии больших полушарий головного мозга явля-
ются органической и весьма важной составной частью его научного творче-
ства, величественным памятником новаторской мысли и титанического
труда.
Эти исследования, как яркий маяк, освещают и пути изучения патологии
и терапии всей нервной деятельности.
Они ярко отражают характерную черту’ творчества гениального фи-
зиолога, а именно утверждение принципа единства теории и практики,
необходимости союза физиологии и медицины.
И сейчас, когда партия, правительство, советский народ требуют от
нас — учеников и последователей И. П. Павлов-а — всемерно развивать его
бесценное и многогранное научное наследие, создав для этого все необхо-
димые условия,— мы должны уделять больше внимания развитию работ
и идей И. П. Павлова по экспериментальной патологии и терапии, укреп-
лению этого мостика между физиологией и медициной и тем самым обес-
печить еще более действенное участие науки в строительстве величест-
венного здания коммунизма на нашей священной советской земле.
£г^—-Xi
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В СВЕТЕ УЧЕНИЯ
И. Н. ПАВЛОВА1
Эти черты
ногогранное научное творчество И. П. Павлова, который нахо-
дился под влиянием передовых идеи славной плеяды русских ре-
вол юц ио иных демо к р -атов -п ро светител ей ( Ге рце н а, Пел и н с ког о,
Чернышевского, Добролюбова, Писарева), характеризует его
как убежденного, боевого и непоколебимого материалиста, как
гениального физиолога с широким биологическим кругозором,
научного творчества великого ученого нашли свое яркое вы-
ражение, в частности, в его воззрениях на приспособляемость как на
фундаментальное свойство живого вещества, в его воззрениях на детер-
минированность функции организма условиями существования, факторами
внешней среды, в его воззрениях относительно целостности организма
и его единства со средой, в его подлинно эволюционном подходе к изу-
чаемым сложным физиологическим явлениям.
Для И. П. Павлова как биолога-материалиста приспособляемость ж иных
существ! является важной основой биологических явлений. При этом
И. П. Павлову как биологу-физиологу в высокой степени присущи тезаме-
чательные черты передовых деятелей отечественной биологии И. М. Сече-
нова, И. И. Мечникова, К. А. Тимирязева, А. Н. Северцева и особенно
В. И. Мичурина и Т. Д. Лысенко, которые ставят их на голову выше
зарубежных ученых, именующих себя последователями Дарвина. Эти уче-
ные подняли проблему приспособляемости на новый, более высокий уро-
вень и обеспечили нашей биологической науке авангардную роль в ми-
ровой биологии. Речь идет в первую очередь о новаторском, творческом
подходе славной плеяды наших биологов и физиологов к изучению много-
образных явлений приспособления в органической природе в целях глубо-
кого познания их и активного управления ими, а нс созерцательной,
пассивной регистрации их, столь характерной для подавляющего боль-
шинства зарубежных биологов. Речь идет об утверждении нашими
биологами и физиологами важнейшего для эволюционного процесса
значения быстро развивающихся индивидуальных приспособительных
явлений В функциях н в строении организма в ответ на быстрые же из-
менения в окружающей среде, а также о многостороннем и углубленном
изучении их в противовес значению, придаваемому зарубежными буржуаз-
ными учеными крайне медленным, тысячелетиями протекающим видо-
вым приспособительным явлениям морфологического и функционального
характера в ответ на медленные изменения среды.
И. П. Павлов, искусный мастер физиологического эксперимента, преи-
мущество опыта как средства познания и овладения биологическими
явлениями над наблюдением видел в том, что «наблюдение ссби-
1 «Павловское чтение» на торжеств-енном собрании Академии наук УССР и
Общества физиологов, биохимиков и формакологов УССР 30 ноября 1951 г , прове-
денное согласно постановлению Научною совета го проблемам физиологического
учения акад. И. II. Павлова при Президиуме Академии наук СССР.
+00
Э. А. АСРАТЯН
рает то, что ему предлагает природа, опыт же берет у природы то, что он
хочет. И сила биологического опыта поистине колоссальна*1.
Как это созвучно указанию К. А. Тимирязева, что биологические
явления нужно исследовать «не в страдательной роли наблюдателя,
а в деятельной роли испытателя», а также девизу И. В. Мичурина:«Мы не
можем ждать милос!ей от природы; взять их у нее — наша задача».
Эта яркая черта творчества выдающихся деятелей передовой отечествен-
ной биологии характеризует их не только как убежденных, боевых материа-
листов и великих оптимистов, но и диалектически мыслящих деятелей
передовой науки.
Вся многолетняя кипучая и исключительно плодотворная научно-
творческая деятельность великого натуралиста И. П. Павлова может
быть охарактеризована как непрерывное и целенаправленное исследование
различного рода приспособительных явлений в высокоразвитом животном
организме. Доминирующий принцип многогранного научного творчества
И. П. Павлова— нервизм является не только выражением его идей о веду-
щей роли нервной системы в регуляции многообразных функции организма.
Одновременно этот принцип подчеркивает и то положение, что все твор-
чество великого ученого насквозь пронизано идеей приспособлен ня, что
приспособляемость является своеобразным биологическим стержнем, вокруг
которого вращаются все творения его могучего ума—все эксперимен-
тальные и теоретические его исследования по физиологии кровообраще-
ния, по физиологии пищеварения и особенно обессмертившие его имя
исследования по фиЗиолоГНН И патологии Высшей нервной Деятельности.
Гениальный мыслитель все многообразие функций нервной системы
снизу доверху сводил по существу к приспособительной деятельности. Еще
в. знаменитой мадридской речи —в этом первоначальном гениальном
эскизе будущего величественного здания его материалистического учения
о высшей нервной деятельности—И. П. Павлов, касаясь врожденных при-
способительных явлений в деятельности органов и систем высокоразвитого
организма, говорил, что в этих случаях «в основании приспособления лежит
простой рефлекторный акт»; перейдя к характеристике биологической
роли условных рефлексов, он добавил: «пет сомнения, что мы имеем здесь
перед собой факт дальнейшего приспособления»2. Такая характеристика
ебщебиологического значения безуслов-норефлекторной и условпорефлек-
торной деятельности, т. е. всей нервней деятельности в целом, сохраняет
свою силу во всех последующих произведениях И. П. Павлова по физио-
логии высшей нервной деятельности, в которых он так или иначе касается
этой темы. Утверждая, что «высшее приспособление животных, высшее
уравновешивание с окружающей средой непременно связано с этим сортом
временно образующихся рефлексов»3, и характеризуя высшую нервную дея-
тельность как деятельность, которая обеспечивает сложнейшие взаимо-
отношения высокоразвитого животного организма с окружающей средой,
осуществляет его наиболее тонкое, точное и совершенное приспособление
к условиям существования, И. П. Павлов постоянно подчеркивал крайнюю
динамичность, вариабильность, пластичность этой деятельности. «Главней-
шее, сильнейшее и постоянно остающееся впечатление от изучения высшей
нервной деятельности нашим методом, — писал он, — это чрезвычайная
пластичность этой деятельности, ее огромные возможности: ничего несстает-
ся неподвижным, неподатливым, а все всегда может быть достигнуто, из-
меняться к лучшему, лишь бы были осуществлены соответственные усло-
вия»*.
1 И. П. Павлов, Полное собрание трудов, т. 11, 1946, стр. 357.
! Там же. т. Ill, 1949, стр. 30.
’Там же, стр. 371.
'Тан ж е, сгр 454.
компенсаторные приспособления в свете УЧЕНИЯ И. П. ПАВЛОВА
401
Выражением такого подлинно эволюционного понимания И. П. Павло-
вым важнейшей биологической роли высших отделов центральной нервной
системы в постоянно развивающейся и совершенствующейся приспособи-
тельной деятельности организма являются также его положения об отно-
сительном характере противопоставления двух основных видов приспосо-
бительной деятельности нервной системы — условных и безусловных реф-
лексов, о возможности превращения условных рефлексов в безусловные,
если это диктуется большой биологической необходимостью, и о том, что
большие полушария мозга с этой точки зрения представляются в роли спе-
циального органа дальнейшей непрерывной эволюции организма. Великий
мыслитель по этому поводу писал: «Таким образом, большие полушария
являются органом анализа раздражений и органом образования новых
рефлексов, новых связей. Они — орган животного организма, который
специализирован на то, чтсбы постоянно осуществлять все более и более
совершенное уравновешивание (И. П. Павлов этот термин употребляет
как синоним термина,«приспособление». — Э. Л.) организма с внешней
средой,—орган для соответственного и непосредственного реагирования
на различнейшие комбинации и колебания явлений внешнего мира, в из-
вестной степени специальный орган для беспрерывного дальнейшего раз-
вития животного организма.
Можно принимать, что некоторые из условных вновь образованных реф-
лексов позднее наследственностью превращаются в безусловные»1.
В этой связи мы хотели бы отметить также полное созвучие принципов
И. П. Павлова о детерминированности приспособительной деятельности
нервной системы условиями существования организма и о возможности на-
следственного закрепления приобретенных в индивидуальной жизни рефлек-
сов и передачи их потомству в качестве выработанного многочисленными
предками ценнейшего фонда приспособительной деятельности основным
принципам мичуринской биологии, в частности, принципу наследственного
закрепления индивидуально приобретенных свойств, ярко и ляконичею
сформулированному Т. Д. Лысенко: «Наследственность есть эффект кон-
центрирования воздействий условий внешней среды, ассимилированных
организмами в ряде предыдущих поколений»2.
* *
♦
Среди развивающихся в животном организме многообразных приспо-
собительных явлений совершенно особое место занимают те, которые воз-
никают после его повреждения, после значительного нарушения структуры
и функций тех или иных его органов и систем под воздействием механиче-
ских, термических, химических или иных болезнетворных воздействий и
в конечном итоге приводят к частичному или полному восстановлению на-
рушенных или утраченных функций поврежденного организма.
В окружающей животный организм внешней среде имеется немало вредо-
носных факторов, в частности, факторов, способных вызвать механическое
повреждение, нарушить анатомическую целостность тех или иных отделов
организма, тем самым нарушить соответствующие его функции и ослабить
его жизнеспособность в тех или иных пределах. Разумеется, последствия
такой механической травмы организма в сильной степени зависят от объема,
глубины и локализации нанесенного повреждения. По степени тяжести они
могутбыть гибельными дчя организма или могутбыть такими ничтожными,
что не вызовут сколько-нибудь значительных изменений в функциях орга-
низма, не скажутся на уровне его жизнеспособности. Механические по-
1 И. П. Павлов, Полное собрание трудов, т. Ill, 1949-, стр. 216—217.
’ О положении в биологической науке. Стенографический отчет сессии Все-
союзной академии сельскохозяйственных наук им. В. И. Ленина, 1948, стр. 33.
26 Э. А. Асратян
402
Э. А. АСРАТЯН
вреждения организма, занимающие по степени тяжести промежуточное
место между этими крайними степенями, способны вызывать многообразные
по характеру, объему и глубине нарушения структуры и функций организ-
ма. При этом функциональные нарушения в отличие от структурных имеют
не только региональный характер, но и в. той или иней степени отражаются
на организме в целом. Нарушение же нормального течения важнейших
жизненных процессов целостного организма не может не ухудшать пер-
спективы дальнейшего его существования. В таких именно драматиче-
ских для организма случаях с особой яркостью вырисовывается важней-
шее биологическое значение приспособляемости, с особой силон выяв-
ляется это «своеобразное могущество живого вещества» (И. П. Павлов).
Врачам, физиологам и биологам хорошо известно, что в подобных слу-
чаях организм различными путями исредствами неустанно борется спослед-
ствиями повреждения целостности его структуры, стремится разнообраз-
ными способам и восстановить свои нарушенные или утраченные функ-
ции, приспособиться к новым -условиям’ жизни, улучшить Предпосылки
к дальнейшему благополучному существованию.
Общеизвестно, что организм многих беспозвоночных и низших позвоноч-
ных животных справляется с этой задачей сравнительно престо и легко:
он довольно быстро восстанавливает не только поврежденные, но даже
утраченные части тела, тем самым восстанавливая и нарушенные или утра-
ченные функции. Известно, например, что у кишечнополостных, червей,
морских звезд восстанавливается целость организма из ничтожных
его остатков, а из низших позвоночных аксолотли, тритоны, саламандры
и др. способны восстанавливать отрезанные пальцы и даже конечности,
хвост, челюсти и даже глаза. В отношении высокоразвитых позвоночных
животных до недавнего времени принято было думать, что у них лишь
некоторые примитивные ткани — эпителиальные и соединительные —
способны к размножению, росту и регенерации, в силу чего-у высших жи-
вотных и человека последствия механического повреждения организма
ликвидируются лишь в ограниченных пределах, главным образом в виде
гнстогепетической регенерации. В качестве примера можно сослаться на
общеизвестные явления заживления ран кожного покрова, мышц, костей и
многих внутренних органов и на давно установленный знаменитым нашим
патологом В. В. Исдвыссцким факт регенерации печени и других желез
у млеко питающих.
Благодаря новейшим достижениям советских биолотов и гистологов
О. Б. Лепешинской, Л. Н. Студитского, И. И. Рампан и др., стало известно,
что псе ткани высокоразвитых органов, в том числе мышечная и нервная,
способны восстанавливать нарушенные структуры как путем размножения
соответствующих клеточных образований, так и путем образования клеток
из неклеточного живого вещества. К сожалению, в действительности
эта чудесная способность тканей реализуется у высокоразвитых организ-
мов опять-таки в крайне скромных размерах, преимущественно в пределах
гистогенеткческоЙ регенерации, в виде заживления ран. Объем же наших
знаний не позволяет еще активно- управлять этим процессом и перевести
его на путь органссбразовапия.
В силу ли непрерывно возрастающей в процессе эволюции специализа-
ции и днференцнации структуры и функций тканей, непрерывного услож-
нения структуры и функций органов и систем высокоразвитых животных
и человека, как считали раньше; в силу ли редкой утраты органов на про-
тяжении бесчисленных поколений или возникшей в процессе эволюции
необходимости быстрого преграждения путей проникновению микробе в
в организм посредством быстрого гистогснетйческого процесса — заживле-
ния ран, как считает А. Н. Студитский; в силу ли каких-либо других
обстоятельств, — это в настоящее время сказать трудно, ь:0 потеря ТыССЮ-
КОМПЕНСАТОРНЫЕ. ПРИСПОСОБЛЕНИЯ В СВЕТЕ УЧЕНИЯ И. П. ПАВЛОВА
4U3
развитым организмом способности к полней или хотя бы частичной реге-
нерации поврежденных и утраченных органов является непреложным
фактом. Затруднительность и даже невозможность органогеиетической
регенерации как коренная перемена в условиях жизни организмов на
определенном этапе их эволюционного развития, очевидно, и направили
их на другой путь борьбы с тяжелыми последствиями грубого повреждения
структуры тех или иных органов и систем—на путь развития компен-
саторных приспособлений.
В подобных, па первый взгляд как будто совершенно безнадежных и
безвыходных ситуациях.поврежденный высокоразви1ый организм добивает-
ся значительного, а нередко даже полного восстановления нарушенных или
утраченных функции либо путем мобилизации запасных возможностей и
усиленней тренировки уцелевших остатков поврежденных органов и си-
стем, либо путем сложной перестройки нервных центров и путей, т. е. лом-
ки старых и создания новых форм координационной деятельности не-
поврежденных органов и систем, либо путем каких-нибудь других функ-
циональных изменений’ в нервней системе и в других системах организма.
Явления подобного рода функционального приспособления у травми-
рованных, раненых или оперированных животных и человека привлекают
внимание клиницистов и экспериментаторов, начиная с XVII1 столетня.
Особенно энергично ими стали интересоваться некоторые выдающиеся дея-
тели медицины и физиологии прошлого и текущего столетий — И. М. Се-
ченов, И. П. Павлов, В. М. Бехтерев, Г. II. Россолимо, В. Кузьмин, А. Кор-
нилов, Флуранс, Броун-Секар, Гольц, Лючиани, Эвальд, Бикель, Магнус
и др. Под названием компенсации, механического иммунитета, тренировки,
замещения, привыкания, внкарирования, перестройки к под другими наз-
ваниями они описали много разнообразных и ярких случаев восстановления
нарушенных и утраченных функций у высокоразвитых животных и у чело-
века пссле хирургического повреждения органов движения и других испол-
нительных органов, после повреждения периферических рецептаров
или же нервной системы в различных ее частях. Конкретнее, ими изуча-
лись явления нарушения и восстановления функций после следующих
повреждении организма; ампутации конечностей, пересадки сухожилий
мышц конечностей; после повреждения различных внутренних органов,
в частности, органов пищеварительной системы, сердца, легких, печек,
эндокринных желез; после повреждения лабиринтов, глаз или других
органов чувств или чувствительных нервных корешков различных нервов,
иннервирующих конечности или шейные мышцы; после перерезки и пере-
крестного сшивания различных нервов, иннервирующих наружные или
внутренние органы; пссле повреждения различных отделов центральной
нервной системы, в частности, спинного мозга, мозжечка и коры боль-
ших полушарий, и, наконец, после повреждения мощных путей внутри
центральной нервной системы и на различных ее уровнях.
Следует отметить, что этисссбого рода приспособительные явления или,
как мы предпочитаем теперь их называть, компенсаторные приспсссбления,
достигают особой яркости и выраженности именно при повреждении нерв-
ной системы, т. е. той системы, роль которой, по И. П. Павлову, и сводится
к приспособительной деятельности, к осуществлению согласованной ра-
боты различных органов и систем организма, к созданию целостной его
деятель нести, к обеспечению сложнейших взаимоотношений организма и
внешней среды. Сбразно называя свойства приспсссбления организма
к механическим повреждениям «механическим иммунитетом», И. П. Пав-
лов по этому поводу писал: «Ведь система организма слагалась среди всех
окружающих ее услов-нй: термических, электрических, бактерийных и
других и между ними также механических условий и должна была их всех
уравновесить, к ним приспособиться, возможно предупредить илиограни-
26*
404
Э Л АСРАТЯН
чить разрушительное их на себе действие. В нервной системе и специально
в сложнейшем ее центральном отделе, управляющем всем организмом, объ-
единяющим все частные деятель нести организма, этот принцип механиче-
ской самозащиты, этот принцип механического иммунитета должен был до-
стигнуть Высочайшего Совершенства, что Действительно в массе случаев и
оказывается»1. Имея в виду именно удивительные по своей выразительности
компенсаторные приспособления после грубого повреждения высших от-
делов центральной нервной системы, он, искусный мастер физиологической
хирургии, говорил; «Такова специальная особенность, таково своеобраз-
ное могущество живого вещества»2.
В завершение чисто внешней характеристики разнообразных компенса-
торных приспособлений в поврежденном организме можно сказать еще сле-
дующее. Если хирургия внутренних органов давно оставила роль пассив-
ного регистратора этих компенсаторных приспособлений и активно и це-
леустремленно способствует развитию таковых путем создания новых,
искусственных связей и соотношений между органами и даже путем созда-
ния новых «органов», то за последнее время на тот же путь все смелее ста-
новятся также нейрохирургия и ортопедическая хирургия. Так, например,
пользуясь чудесным свойством нервных центров перестраивать свои функ-
ции, нейрохирурги нередко со значительным успехом активируют функцию
парализованных скелетных мышц человека и возвращают их встрой путем
сшивания периферических концов атрофированных нервов этих мышце
центральными концами здоровых нервов, расположенных недалеко от
них. Используя то же свойство перестройки нервных центров в случаях
переделки функции скелетных мышц с пересаженными сухожилиями, совре-
менная ортопедическая хирургия добивается немалых успехов в создании
различного рода искусственных двигательных «органов» из уцелевших от
повреждения костей и мышц конечностей человека (пальцы Крукенберга,
мышечные каналы и т. п.). Эти «органы» дают ему возможность искусно
использовать различные механические устройства для выполнения много-
образных сложных функций, осуществлять весьма сложные бытовые и
трудовые манипуляции — ходить, писать, бриться, пользоваться вилкой,
ножом, пилой, молотком и другими предметами.
В этой связи нельзя не отметить также определенные успехи современ-
ной сурдопедагогики и невропатологии. Путем ли методической и упорной
тренировки, путем ли мобилизации резервных возможностей организма,
путем ли «заимствования» чувствительных импульсов из неповрежденных
рецепторов и выработки новых навыков нередко удается восстанавливать
в значительном объеме различные простые и сложные функции двигатель-
ной системы и устную речь, нарушенные либо в результате разных заболе-
ваний, либо в результате механического повреждения тех пли иных частей
нервной системы и рецепторов. Можно сослаться на пример талантливой
советской девушки Ольги Скороходовой, воспитанницы проф. И. А. Соко-
лянского, которая, будучи слепоглухонемой, обучилась устной и письмен-
ной речи и достигла весьма высокого уровня умственного развития.
Как же обстоит вопрос с пониманием сущности этих замечательных
явлений, с теоретическим освещением обсуждаемой проблемы?
Необходимо с сожалением отметить, что, несмотря на совершенно оче-
видную теоретическую и практическую важность проблемы, несмотря на
давний интерес к пен со стороны значительного числа биологов, физиоло-
гов и клиницистов, тем не менее в течение продолжительного времени явле-
ния компенсаторных приспособлений не привлекали к себе должного вни-
мания, не стали в центре внимания хотя бы ограниченной группы исследо-
1 И. П. Павлов, Полное собрание трудов, т III, 1949, стр 445—446.
‘Тач же. 1 IV, 1947, стр 266.
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В СВЕТЕ УМЕНИЯ И. П ПАВЛОВА
405
вателей, не подвергались более иди менее систематическому эксперимен-
тальному, клиническому или теоретическому изучению. Имеющий к этой
проблеме отношение богатый И разнообразный экспериментальный И кли-
нический материал, о котором упоминалось выше, был накоплен не в спе-
циально проведенных систематических и целенаправленных лабораторных
или клинических исследованиях, а в основном собран попутно при изу-
чении других проблем и задач, что не могло не придать этому материалу
отрывистый, разношерстный и в известной мере даже разноречивый ха-
рактер. Не делались даже попытки, если можно так выразиться, чисто
механического суммирования и систематизации относящегося к этой акту-
альной проблеме разнородного, разбросанного по страницам различных
журналов эмпирического фактического материала. Указанные данные ио
нашли отражения не только на страницах учебников по физиологии, пато-
физиологии или специальных медицинских дисциплин, но и в соответству-
ющих многотомных капитальных изданиях типа руководств пли обозрений.
Можно ли в таком случае удивляться отсутствию общей научно обосно-
ванной теоретической базы, отсутствию теории этой весьма важной и ак-
туальной биологической и медицинской проблемы! Полная неясность и
разнобой, отсутствие более или менее общепринятой научной точки зрения
но вопросу о приспособляемости были характерны не только для пред-
ставителей различных теоретических и клинических дисциплин, по и для
специалистов одной и той же дисциплины. Известное распространение сре-
дн значительной группы физиологов и клиницистов получило лишь мнение
о важной роли нервной системы во многих компенсаторных приспособле-
ниях. Но и это представлялось в. весьма общей и туманной форме; ничего
конкретного, ясного, а тем более фактически обоснованного не было ив этом
вопросе. В литер.атуре высказывались различные, подчас противоречивые
предположения, соображения и догадки относительно факторов и механиз-
мов частных случаев или узкого круга явлений из области компенсаторных
приспособлений. Суть этих предположений л догадок, сделанных к тому же
в виде мимолетных замечаний, нашла свое выражение в упомянутых выше
названиях «замещение», «тренировка», «привыкание», «викарирование»,
«перестройка» и др., предложенных разными физиологами и клиницистами
для обозначения наблюдавшихся ими частных случаев компенсаторных
приспособлений.
Таким образом, до недавнего времени проблема компенсаторных при-
способлений не изучалась целенаправленно, систематически ни в. аспекте
экспериментальном и клиническом, ни в аспекте теоретическом. Говоря
иначе, эта важная во всех отношениях проблема не была по-настоящему
включена в число актуальных проблем физиологии и медицины, не стала
предметом специального исследования хотя бы со стороны ограниченного
числа ученых. Вот почему И. II. Павлов, касаясь этой темы незадолго до
своей кончины, в 1932 г., писал: «Мне кажется, что до сих пор специально
в физиологии нервной системы недостаточно оценен и даже не форму-
лируется ЯСНО и постоянно этот в высшей степени важный принцип»1.
На фоне такой неутешительной картины с экспериментальной, клини-
ческой И особенно теоретической разработкой проблемы нельзя неотметить
попытку немецкого физиолога Альбрехта Бете собрать воедино значитель-
ный экспериментальный и клинический материал по проблеме компенса-
торных приспособлений, дополнить его результатами собственных лабора-
торных исследований некоторых частных вопросов и развить некую общую
теорию пластичности нервной системы, нашедшей приверженцев не только
среди зарубежных ученых (Фишер, Гольдштейн, Лешли, Хольст и др.),
но и среди некоторых ученых нашей страны (П. К- Анохин и группа его
1 И. П. Павлов, П-олное собрание трудов, т. III, 1949, Стр. 445.
4П6
Э А АСРАТЯН
сотрудников). Бете тем самым привлек внимание исследователей к пробле-
ме, стимулировал исследовательскую работу в этой области.
Однако значение заслуг Бете в сильной степени умаляется ошибочно-
стью идейных основ развиваемой им теории и другими коренными недостат-
ками проведенной им литературной и экспериментальной работы. Мы ог-
раничимся лишь отдельными замечаниями, отсылая интересующихся этим
вопросом к другим нашим работам по рассматриваемой проблеме, в кото-
рых дана обстоятельная критика теории и работ Бете.
Взгляды Бете иа физиологию компенсаторных приспособлений выте-
кают из его ошибочных, антиэволюционных и по существу своему виталисти-
ческих общетеоретических положений относительно принципов строения и
деятельности нервной системы, находящихся в полном противоречии с со-
временной материалистической рефлекторной теорией нервной деятель-
ности, в частности, с вершиной этой теории — сучением И. П. Павлова сб
условнорсфлскториом деятельности больших полушарии. Согласно мета-
физической теории Бете, не существует принципиальной разницы между
структурными и функциональными особенностями нервной системы у жи-
вотных, стоящих на различных ступенях эволюционной лестницы. Как
у высокоразвитых животных, так и у примитивных многоклеточных живот-
ных нервная система представляет собой в функциональном отношении
аморфную однородную сеть нервных клеток и проводников. Как у одних,
так и у других животных части этой «непрерывной сетчатой системы» мо-
гут отличаться друг от друга лишь в морфологическом отношении; в функ-
циональном же отношении они принципиально равноценны, эквипотен-
циальны; в ней не существует в сколько-нибудь заметной степени специа-
лизации и локализации центров и проводящих путей. Бете и Фишер писа-
ли; -«То, что до сих пор представлялось нам центром, могло бы, грубо
говоря, оказаться не чем иным, как пунктом связи и проведения между
различными областями рецепторной и эффекторной периферии»1.
Категорически отрицая возможность объяснить явления компенсатор-
ных приспособлений на основе рефлекторной теории, поднятой И. М. Се-
ченовым И И. II. Павловым на небывалую высоту'— на уровень глубоко
эволюционного и строго материалистического учения о деятельности нерв-
ной системы в целом, Бете и его единомышленники утверждали, что эти
явления могут быть удовлетворительно освещены лишь с позиций раз-
виваемых ими антинаучных, механистических и виталистических концеп-
ций, фактически отрицающих эволюцию нервной системы, отрывающих
функцию от структуры, обезличивающих центральную нервную систему и
принижающих ее значение.
С точки зрения Бете и его единомышленников, при механическом по-
вреждении какой-нибудь части центральной нервной системы, анатоми-
чески тесно связанной с определенными органами, другие части этой эк-
випотенциальной «сети» весьма быстро и без особого труда перестраиваются
и берут на себя функции поврежденной части. Точно такая же быстрая и
легкая перемена функций происходит в отдельных центральных узлах этой
«сети» и в тех случаях, когда повреждаются иннервируемые другими узла-
ми периферические рабочие органы, когда нарушается естественная нерв-
ная связь между этими узлами и органами и т. п. Толчком и решающим фак-
тором подобных функциональных перемен в центральной нервной системе
является периферия, импульсы с периферических органов чувств. По сло-
вам Бете, «периферия образует себе центральный орган... создает себе
«центры» в центральной нервной системе», «периферия прямо-таки господ-
ствует над центральными явлениями: ие центральный аппарат определяет,
что должно произойти на периферии, а периферия определяет, какую уста-
1 Bethe’s Handbuch d. norm. u. pathol. Physiol,, Bd. XV, H. 2, 1931, S. 1190.
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В СВЕТЕ УЧЕНИЯ И. П. ПАВЛОВА
407
новку Должен принять центральный орган»1. Достаточна информация пе-
риферия о происшедших в организме повреждениях, чтобы произошла мол-
ниеносная и как бы автоматическая перестройка соответствующих цент-
ральных звеньев «нервной сети», т. е. возникли соответствующие компен-
саторные приспособления.
Важею отметить еще один момент. Беге считает, что процесс перестройки
функций происходит в примитивных частях, т. е. низших отделах централь-
ной нервной системы, и но своей природе не есть «выучивание» (Unilernen).
Он считает, далее,— на это следует обратить особенное внимание, — что
кора больших полушарий в этих особого рода приспособительных явле-
ниях не играет сколько-нибудь существенной роли или вовсе не играет
роли.
Нетрудно видеть, что теория Бете находится в явном противоречии с
наиболее достоверными экспериментальными и теоретическими данными
современной биологии, физиологии и медицины относительно функций нерв-
ной системы, в особенности с материалистическим учением И. П. Павлова
о деятельности высших отделов центральной нервной системы, являющим-
ся вершиной этих достижений. В то время как И. II. Павлов наосновании
громадного, точного, многократно проверенного и исключительно ценного
фактического материала установил, что кора больших полушарий осуще-
ствляет наиболее тонкую, точную и совершенную приспособительную дея-
тельность высокоразвитого организма, является наиболее мощным и со-
вершенным регулятором «всей внешней и внутренней деятельности организ-
ма», верховным «распределителем и распорядителем всей деятельности ор-
ганизма..., держит в своем ведении все явления, происходящие в теле»,—
Бете и его единомышленники отрицали важную роль коры в развитии
компенсаторных приспособлений, причем отрицали голословно, на осно-
вании крайне малоценного, неубедительного фактического материала.
Это отчетливо видно даже на примере наиболее обстоятельно разработан-
ного им вопроса —о развитии компенсаторных приспособлений после
ампутации конечностей у низших и высших животных,
Свой ошибочный взгляд на роль коры больших полушарий в компен-
саторных приспособлениях Бете, как это ни странно, аргументирует прежде
всего тем, что эти приспособления не исчезают после удаления’ окологло-
точных нервных узлов у жука Dytiscus, а также после удаления двигатель-
ных зон коры больших полушарий у ссбак. Не говоря уже о недопустимости
на современном уровне развития биологических наук вульгарного отож-
дествления роли окологлоточных узлов у жуков и коры больших полуша-
рий у высших млекопитающих, укажем на неубедительность фактических
данных Бете относительно удаления двигательных зон у собак, равно
как и на его непоследовательность в этом вопросе. После того, что стало
известно из опытов Гольца, Лючиани и особенно И. П. Павлова с частич-
ной экстирпацией коры больших полушарий мозга, крайне наивно со
стороны Бете на основании результатов подобных опытов судить о роли
всей коры. Нельзя не заметить также, что самым фактом постановки таких
опытов с удалением двигательных зон коры он явно вступает в неприми-
римое противоречие со своими же ошибочными теоретическими установ-
ками. Ведь он фактически отрицает специализацию и локализацию функ
ций в центральной нервной системе!
Бете свои ошибочные выводы о роли процесса «выучивания» вообще и
роли коры больших полушарий мозга в компенсаторных приспособлениях
аргументирует еще тем, что, согласно его опытам (заметим — весьма ма-
лочисленным), не только у беспозвоночных, но якобы и у млекопитающих
1 Bethe's Handbuch <1. norm. u. pathol. Physiol., Bd. XV, H. 2, 1931, S. 118-6.
408
Э. А. АСРАТЯН
эти приспособления наступают очень быстро пссле нанесения поврежде-
ния, в частности, после ампутации конечностей. При этом он обращает осо-
бенное внимание на то крайне «важное» с его точки зрения обстоятель-
ство, что в этом отношении «умные» млекопитающие (таковыми он считает
собак) ничем не отличаются от «глупых» (таковыми он считает морских
свинок). Как мы увидим ниже, при изложении нашего экспериментального
материала, эти фактические данные Бете в отношении собак являются
плодом недоразумения. На самом деле, у заранее нетренированных собак
компенсаторные приспособления развиваются весьма постепенно и мед-
ленно— неделями, а в некоторых случаях даже месяцами. Рассужде-
ния же Бете об «умных» и «глупых» животных в этом аспекте слишком
несолидны, чтобы их стоило сбсуждать.
В заключение критической оценки работы Бете по пластичности нервной
системы хотелось обратить внимание еще на одно любопытное обстоя-
тельство.
Когда мы приступили к йзученню обсуждаемой проблемы и в связи
с этим стали тщательно собирать литературные данные, мы выявили весь-
м а богатый и ценный экспериментальный материал о компенсаторных при-
способлениях, который странным сбразом ускользнул от внимания Бете.
Мы считаем этот факт странным потому, что этот материал представлен в
основном в немецкой научной литературе и в значительной части в про-
дукции той лаборатории, Воспитанником которой является сам Бете,—
лаборатории известного физиолога Гольца. В частности, ни водной из ста-
тей и обзоров Бете и его единомышленников не получили отражения инте-
ресные данные Эвальда о последствиях разрушения лабиринтов у птиц и
собак, о характере и темпе восстановления нарушенных при этом функций
и о последствиях удаления двигательных зон коры больших полушарий
у Ссбак и всего переднего мозга у птиц. Бете и его единомышленники обхо-
дят полным молчанием также ценный фактический материал Бикел я отно-
сительно последствий деафферентация конечностей у ссбак, динамики вос-
становления нарушенных при этом функций и относительно последствий
удаления двигательных зон у них. Это же относится к богатейшему и цен-
ному экспериментальному материалу по физиологии компенсаторных при-
способлений, полученному в лабораториях И. П. Павлова, а также Лючиа-
ни, Магнусом и др. Не потому ли Бете «не замечал» этот материал, что
он находился в явном и непримиримом противоречии с его антинаучны-
ми виталистическими взглядами? Впрочем, подобные случаи — пе такое уж
редкое явление среди ученых буржуазных стран, в особенности в США.
Недостаточность одной только словесной полемики с Бете и его сторон-
никами, равно как и своевременность в важность систематической экспери-
ментальной разработки обсуждаемой актуальной в теоретическом и прак-
тическом отношении проблемы с позиций материалистического учения
гениального И. П. Павлова были совершенно очевидны еще примерно два
десятка лет назад, когда мы приступили к целенаправленной исследова-
тельской работе в этой области. Как марксисты-ленинцы и ученики
И. П. Павлова мн не могли безразлично отнестись к тому обстоятельству,
что в одной из важнейших областей физиологии и медицины в те годы
почти безраздельно господствовала реакционная, неовиталистическая и
антиэволюционная теория Бете. Переход от словесной критики теории Бете
к более вескому в области естествознания языку достоверных и точных
экспериментальных данных, к изучению проблемы по-павловски стал на-
сущной необходимостью и потому, что эта энтипавловская теория Беге
в силу ли его авторитета, в силу ли заманчивости новизны, по идеоло-
гическим ли мотивам или по другим причинам за сравнительно короткое
время приобрела, как уже было отмечено, значительное число сторонни-
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В СВЕТЕ УЧЕНИЯ И. П. ПАВЛОВА 409
ков не только среди ученых буржуазных стран, но, как эго ни странно,
и среди наших ученых (П. К- Анохин).
Каково же было отношение самого И. П. Павлова к этой проблеме? Сле-
дует отметить, что И. П. Павлов в первый период своей исследовательской
работы по физиологии условных рефлексов, занимаясь систематическим изу-
чением проблемы специализации и локализации функций в коре больших
полушарий и произведя в этих целях многочисленные операции частичной
экстирпации различных ее зон, попутно накопил весьма ценный материал
также относительно компенсаторных приспособлений при повреждении
больших полушарий. Хотя ои специально не изучал этой проблемы и поэ-
тому не создал соответствующей теории, тем не менее на основании анализа
попутно накопленного специфического материала гениальный физиолог
высказал весьма глубокую идею относительно активирования запасных
нервных образований и замещаемости как ближайшей физиологической!
основы компенсаторных приспособлений в нервной системе вообще. Он
писал: «Если во всем организме мы постоянно встречаемся с запасными
средствами против частичных нарушений его, то в нервной системе, как
устанавливающей все связи и отношения организма, этот принцип должен
обнаруживаться в высшей степени. И это мы видим как в спинном мозгу на
сложном и очень разнообразном ходе его волокон, так и в периферической
нервной системе на ее широко практикуемой, так называемой возвратной
чувствительности, что явно значительно нейтрализует вред механических
нарушений этих отделов. Конечно, то же самое в высшей мере должно быть
осуществлено в верховной инстанции аппарата, регулирующего всю внут-
реннюю и внешнюю деятельность организма»1.
Великий физиолог до конца своей жизни живо интересовался пробле-
мой компенсаторных приспособлений, но, будучи поглощен изучением дру-
гих акту'альных проблем физиологии и патологии высшей нервной деятель-
ности, он, к сожалению, так и не приступил к ее специальному, углублен-
ному лабораторному исследованию.
Значение сделанного И. П. Павловым для проблемы компенсаторных
приспоссбленнй далеко не исчерпывается накопленным им эксперимен-
тальным материалом относительно последствий хирургического повреж-
дения коры больших полушарий и высказанными нм глубокими идеями,
имеющими отношение к этой проблеме. Его гениальное материалистиче-
ское учение о высшей нервной деятельности, послужившее твердой
научной основой для решения многих актуальных и сложных проблем
современной биологии и медицины, пролило яркий свет на проблему
компенсаторных приспоссбленнй, указало единственно правильный путь
ее дальнейшей успешной экспериментальной и теоретической разработки.
В нашей стране, кроме нас, систематическим изучением проблемы
компенсаторных приспособлений занимался также другой ученик
И. П. Павлова — П. К- Анохин. Случилось так, что мы и fl. К- Анохин
пошли разными путями. Он следовал по пути неовиталиста и гештальтиста
Бете, а мы без колебаний встали на павловский путь в физиологии.
И неизбежно мы и П. К. Анохин пришли к разным итогам.
П. К. Анохин и его сотрудники более или менее обстоятельно исследова-
ли лишь компенсаторные приспособления после перерезки, перекрестного
сшивания одного из блуждающих нервов с каким-нибудь нервом плечевого
сплетения. Они выполнили отдельные исследования, посвященные компен-
саторным приспособлениям, развивающимся после деафферентации конеч-
ностей или же после пересадки сухожилий их мышц. Для выявления физио-
логической природы компенсаторных приспособлений у подопытных
животных (почти исключительно ссбак) П. К. Анохин и его сотрудники вре-
1 И. П. Па вл ов, Полное собрание трудов, т. JV, 19-47. стр. 268-.
•110
Э А. АСРАТЯН
менами использовали методику двигательных электросборонительных ус-
ловных рефлексов, а в единичных случаях прибегали также к экстирпации
двигательных зок коры больших полушарий у них.
Переходя к оценке результатов этой работы, следует отметить, что по-
лученный П. К. Анохиным и его сотрудниками Л. Ивановым, В. Матве-
евым, И. Лаптевым, Б. Глассоном, С. Балакиным, Н. Шумилиной и др.
фактический материал по обсуждаемой проблеме не отличается ориги-
нальностью и в основном представляет собой повторение и подтверждение
ранее известных данных с небольшим добавлением новых фактов.
Особенно ДсСаДно То, что при теоретическом освещении фактических
данных и всей проблемы в целом П. К,- Анохин полностью примкнул к оши-
бочным, идеалистическим, антинаучным воззрениям Бете, Лешли (Lashley),
II. Вейсса (Р. Weiss), Когхилла (Koghill), Вейцзекера (Weizsacker), Гольд-
штейна (Goldstejn) и др. на коренные вопросы нейрофизиологии, перешел
в. лагерь этих прожженных антипавловцев, стал их убежденным единомыш-
ленником л подобострастным проповедником их идей, теоретических ус-
тановок и положений в отечественной физиологии, правда, опять-таки часто
выдавая эти идеи, положения и установки за собственные и оригинальные.
Это относится, в частности, к ошибочным н антинаучным взглядам Вете и
его сторонников на материалистическую рефлекторную теорию, на спе-
циализацию и локализацию функций в центральной нервной системе, на
локализацию и сущность перестройки в центральных нервных образова-
ниях, на роль периферических импульсов и на роль коры больших полуша-
рий в этих интрацентральных перестройках.
П. К. Анохин вслед за Бете и его единомышленниками и в духе его
взглядов брал под удар «принцип рефлекса со всеми свойственными ему
чертами»; он считал, что«вряд ли имеет какой-либо физиологический смысл
говорить об афферентных «центрах» коры головного мозга», что Двигатель-
ные центры могут быть названы «центрами только в очень условном зна-
чении этого слова» и что «когда обычно говоря т: «центр управляет», «центр
посылает импульсы» и т. д., то в этих выражениях по сути дела нет ни-
какого физиологического смысла»1. Он отрицал также специфичность внут-
рицентральных проводящих путей.
Имея в виду материалистическую рефлекторную теорию нервной дея-
тельности, П. К. Анохин в полном согласии со взглядами названных выше
ученых-идеалистов считал, что «установки старой нейрофизиологии долж-
ны быть коренным образом изменены». Нельзя в этой связи не указать на
дерзкий и возмутительный упрек П. К. Анохина, явно направленный по
адресу И. П. Павлова и его учеников и надменно облеченный в слова:
«Вместо того, чтобы немедленно взяТЬСЯ за. исправление фундамента, дав-
шего катастрофическую трещину, нейрофизиологи продолжают беспечно
украшать верхние этажи этого несовершенного здания»2. П. К. Анохин
вслед за Бете считал, что перестройка (или, по его терминологии, реин-
теграция) функций происходит в низших отделах центральной нервной
системы, точнее, в пределах спинного мозга. Новое, добавленное им, сво-
дится к явному антипавловскому допущению, что при этом вырабатываются
спинномозговые связи типа условнорефлекторных. «Из всего сказанного
вытекает один важнейший вывод, — писал П. К- Анохин, — что установка
новых связей в области спинномозговых отношений происходит по прин-
ципу временных связей, т. е. по принципу условного рефлекса. Вог почему
мы и высказали предположение, что нервный анастомоз может быть мето-
1 Сборник «Проблема центра и периферии в физиологии нервной деятельно-
сти», 1935, стр. 203.
“Там же. стр. 70.
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В СВЕТЕ УЧЕНИЯ И. П. ПАВЛОВА |
дом изучения образования временных Связей на уровне спинного мозга»1.
Далее П. К. Анохин в духе Лешли— известного американского единомыш-
ленника Бете, подвергшегося острой критике И. П. Павлова, — считал,
что кора больших полушарий способна лишь оказывать диффузное не-
спец ифическое, «эквипотенциальное»облегчающее влияние ца процесс пе-
рестройки в низших отделах центральной нервной системы. Это допущение
он аргументировал тем, что в его опытах удаление двигательной зоны
коры больших полушарий соответственно стороне перекреста нервов ко-
нечности вызывало нарушение функций этой конечности лишь в первые
дни после операции; спустя несколько дней, нарушенные функции вновь
восстанавливались.
Этим, с точки зрения П. К. Анохина, и исчерпывается участие коры
больших полушарий в развитии компенсаторных приспособлений.
П. К. Анохин отрицал не только активное участие коры с ее всемогущей
условнорсфлекторной деятельностью в процессе этой перестройки, он от-
рицал даже исследующее ее «вступление» в эту перестройку спинальных
центров, отрицал ее «условнорефлекторный контроль» над деятельностью
уже «реинтегрированных участков спинного мозга» по крайней мере в те-
чение длительного времени — Г/г года спустя пссле произведенной опе-
рации и пссле нескольких сот сочетаний инднферентных раздражителей
с безусловным рефлексом. По этому поводу П. К.Анохин и А.Иванов писали:
«Из всего вышесказанного следует, что переучивание центров на уровне
спинномозговых сегментов происходит в системе целостного нервного ком-
плекса, и только благодаря ему. Отсюда понятным становится тот на первый
взгляд парадоксальный факт, что кора больших полушарий, являясь наи-
более вариабельной частью центральной нервной системы, все же не всту-
пила в систему спинномозговой реинтеграции»4. В подобного рода утвержде-
ниях в работах П. К. Анохина нет недостатка. В другом месте, например,
он писал: «Кора — наиболее подвижный и лабильный орган — не устанав-
ливает условной связи с реинтегрированным участком спинного мозга»3.
Невозможность выработки двигательных и электросборонительных
условных рефлексов на конечность с перекрестно сшитыми нервами и при-
мерно год спустя пссле операции, когда двигательные функции конечности
хорошо восстановлены, привела П. К. Анохина и А. Иванова к выводу;
«Все факты говорят за то, что у животного исчезла способность управлять
с ко’ры локализованными двигательными актами в области реинтегриро-
ванных спинных центров»1. А когда у одной из собак с анастомозом цент-
рального конца блуждающего нерва с периферическими отрезками трех
корешков плечевого сплетения П. К. Анохин и А. Иванов получили,
вопреки основным своим экспериментальным данным, электрооборони-
тельный двигательный условный рефлекс, они поспешили объяснить это
«сохранившейся пирамидной иннервацией» конечности и «большей дифг-
ренцированноетъю» ее иннервации, т. е. поспешили свести эти данные
к случайным и подчеркнуть правильность основного положения о том,
что кора больших полушарий не участвует в Восстановлении функции
конечностей с анастомозированными нервами и теряет способность к их
управлению.
Наконец, П. К. Анохин так же, как и Бете, основным, решающим фак-
тором перестройки считает периферические импульсы, периферию. Qh вмес-
те с А. Ивановым писал: «Так как во всех случаях окончательным результа-
том такого постепенного реинтегративного процесса является функция,
1 Сборник «Проблема центра и периферии в физиологии нервной деятельно-
сти», 1935, стр. 66.
* Т а м же, стр. 121.
’Тан же, стр. 35.
‘Там же, стр. 118.
412
Э. А. АСРАТЯН
адэкватная именно периферическим образованиям, а не нервным центрам,
связанным с ними, то мы имеем полное основание говорить о том, что весь
процесс реинтеграции в центральной нервной системе возникает по ини-
циативе периферических импульсов и протекает под их постоянным конт-
ролем»1.
В другой статье П. К. Анохина и его сотрудников говорится: «Нам
кажется, что наши результаты дают возможность высказать предположе-
ние о механизме «переучивания» и поставить прогноз его при каждом
отдельном анастомозе гетерогенных нервов. Несомненно одно, что пере-
становка в нервных центрах при гетерогенном анастомозе происходит под
влиянием афферентной сигнализации. Именно изменения афферентных
импульсов, которые теперь бомбардируют не свойственные им центры,
и приводят к тому, что эффекторные части центральной нервной системы
начинают управлять гетеро!енными органами»2.
Указывая на полное совпадение взглядов Бете и П. К. Анохина по ко-
ренным вопросам физиологий нервной системы и, в частности, по вопросу
о компенсаторных приспособлениях, мы не отрицаем наличия определен-
ных расхождении между ними по второстепенным вопросам, по деталям.
Например, II. К. Анохин в отличие от Бете придавал нервному рубцу ис-
ключительно большое значение в процессах перестройки, приписывая ему
какую-то мифическую функцию активного трансформатора и перераспре-
делителя нервных импульсов.
Небезынтересен и тот факт, что П. К. Анохин, как и Бете, в своих про-
изведениях обошел полным молчанием те из упомянутых выше ценных фак-
тических данных Бикеля, Лючиани, Эвальда, Магнуса и др. по пробле-
ме компенсаторных приспособлений, которые противоречат его установкам.
В заключение следует отметить, что хотя П. К. Анохин в дальнейшем,
в особенности за последние годы, несколько отошел от многих своих
ошибочных теоретических установок, как мы полагаем, не без известного
влияния нашей многолетней неустанной и суровой критики его ошибок
И Р основном ПОД влиянием критики его ошибок на' исторической Объ-
единенной сессии Академии наук СССР и Академии медицинских наук
СССР в 1950 г., тем не менее До Сих пор отсутствуют еще явные и бес-
спорные доказательства его искреннего и действительного отказа от своих
ошибочных антипаьловских взглядов в целом
Мы в своей работе не только стремились твердо и непоколебимо стоять
на позициях материалистической рефлекторной теории Сеченова — Пав-
лова, целиком исходить из учения нашего гениального учителя о высшей
нервной деятельности, ио и руководствоваться павловскими принципами
организации н ведения научно-исследовательской работы, т. е. стремились
разработать пленительно интересную проблему целенаправленно, последо-
вательно, многосторонне, многократно проверяя полученные данные.
Разработкой проблемы компенсаторных приспособлений мы со своими
сотрудниками занимаемся уже на протяжении 20 лет, не считая небольшого
перерыва в период Отечественной войны. Работа в этом направлении
была начата нами в 1932—1933 гг. с одобрения И. П. Павлова и в руко-
водимом им Институте физиологии Академии наук СССР, затем в силу
ряда обстоятельств в 1934 г. была перенесена в руководимый И. П. Пав-
ловым же Отдел физиологии Института экспериментальной медицины,
а, начиная с 1935 г., после назначения меня на должность заведующего
Отделом физиологии центральной нервной системы в Институте мозга
им. Бехтерева, вся эта работа была сконцентрирована в этом отделе
и проводилась там до войны.
’ Сборник «Проблема центра и периферии в физиологии нервной деятельности»,
1935, стр. 101.
‘Там же, стр. 201.
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В СВЕТЕ УЧЕНИЯ И. П. ПАВЛОВА .ЦЗ
После некоторого перерыва, обусловленного переключением на разра-
ботку вопросов терапии сном последствий органических повреждений,
в 1944 г. работа в этом направлении была возобновлена в лаборатории
восстановления функций Академии наук СССР.
В работе принимало и принимает участие значительное число на-
ших сотрудников — Р. О. Барсегян, Г. И. Боков, Ф. М. Вишневецкая,
В. Д. Дмитриев, В. II. Дроздова, В. А. Замятина,С. Н. Иванова, Е.М. Лы-
хина, Л. И. Карамян, Л. Л. Новикова, Е. А. Романовская, Н. И. Санкова,
Г. Т. Сахиулина, Б. Д. Стефанцов, Я- М. Пресеман, К. М. Ротарь, Т. Урган-
джян, Д. И. Хаиутнна, которым, пользуясь случаем, выражаю свою сер-
дечную благодарность.
Несколько слов о принципах проведения нашей исследовательской ра-
боты.
Желая возможно многостороннее и обстоятельнее изучить основные,
наиболее общие закономерности возникновения и развития компенсатор-
ных приспособлений, мы изучали их не при каком-нибудь одном частном
виде хирургического повреждения двигательных и чувствительных органов
или различных отделов нервной системы, а при целой серии разновидностей
подобных повреждений, при многих формах хирургического повреждения
различных систем организма и в первую очередь нервной системы. Говоря
конкретно, компенсаторные приспособления мы изучали при ампутации
конечностей, при перерезке и перекрестном сшивании нервов, перерезке
задних корешков нервов шейных мыши, разрушении лабиринтов, при бро-
ун-секаровской перерезкеСПИнноГомозга, ПроДоЛЬноМ расщепленииспИнно-
го мозга на различных участках, при перерезке задней и передней половицы
спинного мозга, экстирпации мозжечка, при половинной поперечной пере-
резке ствола мозга. Временами мы комбинировали эти виды повреждения.
Таким образом, наносимые нами повреждения включали почти все ва-
рианты хирургических операций, применяющихся теми или иными ис-
следователями этой проблемы, а также некоторые заново разработанные
нами варианты.
Далее, предполагая, что возрастные, типологические и иные, не всегда
поддающиеся точному учету' особенности организма животных могут иг-
рать известную роль в развитии компенсаторных приспособлений, и желая
точнее выявить общие закономерности их возникновения и течения даже на
фоне этих осложняющих обстоятельств, мы значительно увеличили число
подопытных животных в каждой группе по сравнению с тем, которое при-
меняется обычно. По каждому из указанных вариантов операций число
подопытных животных иногда в десятки раз превышало таковое у наших
предшественнико-в и современников по изучению этой проблемы. В общей
сложности под нашим наблюдением находилось около 300 оперированных
ссбак и небольшое число кошек.
Особенность нашей работы заключается также в том, что мы поставили
перед собой задачу изучать проблему приспособления в аспекте эволюцион-
ного, прежде всего филогенетического развития. В число наших подопыт-
ных животных входятлягушки и черепахи, бараны, разные птицы и млеко-
питающие, главным образом собаки.
Мы изучаем динамику нарушения и восстановления функций у подо-
пытных животных не только путем простого наблюдения, но и с помощью
большого числа современных точных физиологических методик исследова-
ния — измерения тонуса мышц конечностей, специального изучения осо-
бенностей разного рода безуслов-ных рефлексов, изучения хронаксии нер-
вов, мышц и рефлексов конечностей, двигательных электросборонительных
условных рефлексов. Все это дает нам возможность полнее охарактеризовать
особенности возникновения и течения компенсаторных приспособлении и,
в частности, выяснить, в какой степени кора больших полушарий участвует
414
Э Д ЛСРАГЯЯ
в весстановлении нарушенных функций поврежденного организма. Мы до-
вольно широко используем также (фотографию и кинематографию.
Для выяснения роли верхних этажей центральной нервной системы
в развитии компенсаторных приспособлений мы у низших позвоночных
животных — амфибий, рептилий и птиц—удаляем весь передний мозг,
как это в тех же целях и на тех же животных делали многие наши пред-
шественники (Эвальд, Бикель, Мерцбахер, Тренделенбург и др.).
. Наконец, наиболее характерной чертой нашей работы является то, что
в целях выяснения роли и участия высших отделов центральной нервной
системы в компенсаторных приспособлениях мы удаляли и удаляем у собак
нс какую-нибудь изолированную зону или часть коры больших полушарий,
как это делали все наши предшественники и современники (в том числе Бете
и 11. К. Анохин), которые вообще считали возможным иногда прибегать
к этой операции, а экстирпируем всю кору больших полушарий у жи-
вотных. При этом в большинстве случаев кору больших полушарий мы
удаляем после максимального развития компенсаторных приспособлений.
Значительно реже мы практикуем обратный порядок операции — сначала
удаляем кору больших полушарий, а затем производим другие пере-
численные выше операции.
Важность такой радикальной операции для решения центрального
вопроса проблемы совершенно очевидна и не нуждается в особых разъяс-
нениях. Выше мы уже указывали, что в старой физиологической литерату-
ре имеется некоторый фактический материал, свидетельствующий о том.
что удаление двигательных зон коры больших полушарий у собак устраняет
в известном мере компенсацию нарушений, обусловленных первоначаль-
ным разрушением лабиринтов (Эвальд), деафферентацией конечностей
(Бикель) и удалением мозжечка (Лючианн). Тогда же мы указали, что эти
ценные данные странным образом выпали из поля зрения Бете и П. К. Ано-
хина; точно так же выпали из поля их зрения и более новые данные голланд-
ского физиолога Магнуса относительно того, что у кошек и собак нарушен-
ные после повреждения лабиринтов и после деафферентация шейных нер-
вов двигательные функции восстанавливаются благодаря оптическим уста-
новочным рефлексам, которые, по его гипотетическим предположениям,
осуществляются корой больших полушарии.
Таким образом, еще до наших исследовании имелись кое-какие данные,
говорящие об участии коры больших полушарий в развитии компенса-
торных приспособлений. Однако они были лишены необходимой доказатель-
ной силы, и вот почему. Хотя,согласно этим данным, удаление двигатель-
ных зон у собак и расстраивает компенсаторные приспособления, возник-
шие у них пссле удаления мозжечка, после разрушения лабиринтов и деаф-
ферентации лап, но со временем эти приспособления в большей или меньшей
степени появляются вновь, т. с. вторично. Следует добавить, что эти данные
были получены в весьма ограниченном числе опытов и характеризовались
значительной пестротой, если не противоречивостью. Например, в то время
как, по данным Лючиани, Бикел я и Эвальда, удаление двигательных зон у
собак вызывает глубокое и длительное расстройство компенсаторных при-
способлений,— по данным Бете и II. К. Анохина, последствия этой опе-
рации весьма кратковременны и неглубоки. Чем это обусловлено — осо-
бенностями ли первоначальной операции, величиной удаленной массы двига-
тельных зон, промежутками между операциями,—было неясно.
Решение этой задачи и было главной целью наших исследований.
Чтобы составить представление об исключительной трудности задачи,
которую мы дерзнули поставить перед собой, — задачи комбинирования
разнородных сложных хирургических операций, вызывающих первона-
чальные нарушения функций организма, со сложнейшей операцией полного
хирургического удаления коры больших полушарий у значительного
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В СВЕТЕ УЧЕНИЯ И. И. ПАВЛОВА -||5
числа собак, — следует вспомнить, что до нашей работы в этой области
никому из отечественных или зарубежных ученых еще не посчастливилось
в целях изучения компенсаторных приспособлений с успехом осуществить
хотя бы одну такую операцию у высших млекопитающих. А в иных целях,
насколько нам известно, только голландскому физиологу Радсмакеру
(Radetnaker) удалось более или менее успешно комбинировать полную экс-
тирпацию мозжечка с полным удалением коры больших полушарий у двух
собак, одна из которых прожила после этого 38 дней, а другая и того мень-
ше — 27 диен. Высоко оценивая полученные Радемакером на этих собаках
данные относительно характера спинальной, бульбарной, среднемозго-
вой и межуточномозговой рефлекторной деятельности, мы вынуждены вмес-
те с тем отметить, что непродолжительность существования этих собак,
осложненного к тому же кйкОЙ-ТО тяжелой Инфекцией, не позволяет быть
категоричным даже и выводах относительно истинных последствий упомя-
нутой выше комбинированной операции, не говоря уже о картине возмож-
ных при этом компенсаторных приспособлений.Сказанное еще в большей
степени относится к научной ценности наблюдений Л. А. Орбели и
М. А. Панкратова над несколькими одномоментно оперированными кошками
с удаленным мозжечком и корон, равно как и к наблюдениям, проведенным
американскими физиологами Фултоном, Лндделом и Риочем (Fulton, Lid-
dell (Rioch) на некоторых трехмоментно оперированных кошках: как
у тех, так и других исследователей большинство кошек прожило всего
несколько дней после операции, и только единичные из них жили нем-
ногим более 10 дней.
В этой связи следует отметить, что научный мир уже 20 лет тщетно ждет
выполнения данного Фултоно-м и его сотрудниками твердого обещания
успешно осуществить эту комбинированную операцию у кошек, собак и
даже обезьян, сохраняя их жизнь на неопределенно долгое время. Это обе-
щание было дано в докладе о результатах наблюдений на упомянутых вы-
ше кошках, сделанном на заседании Нью-Йоркской медицинской академии в
ноябре 1931 г. и опубликованном в 1932 г.‘.Авторы поместили эту статью как
«сообщение первое». Остальные сообщения так и не последовали до сих пор.
Будучи лишены возможности подробно изложить экспериментальные и
теоретические результаты выполненной нами работы по проблеме компен-
саторных приспособлений, мы остановимся лишь на тех из них, которые
имеют непосредственное отношение к основной цели этих исследований,
а именно: к выяснению роли высших отделов центральной нервной системы,
роли «выучивания», т. е. условнорефлекторной деятельности, в развитии
компенсаторных приспособлений у низших и высших позвоночных живот-
ных.
Наши данные говорят прежде всего о том, что процесс восстановления
нарушенных и утраченных функций, в особенности после значительной
тяжести хирургических повреждений, не иосит автоматического характера
и не осуществляется стремительно быстро, как утверждал Бете, а проте-
кает весьма медленно и постепенно и имеет характер упорной тренировки
и «выучивания». Это особенно наглядно выступает копытах с ампутацией
конечностей, т. е. при основном варианте эксперимента Бете на собаках. Сле-
дует отметить, что у него было всего несколько собак с двумя ампутирован-
ными конечностями, у нас же около 40 собак разных пород и возрастов
с ампутациями двух лап в различных комбинациях. Из этого числа толь-
ко у 3 взрослых собак функции стояния и ходьбы па двух лапах восстано-
вились быстро, в течение 2—3 дней, т. е. как у собак Бете. Болес же чем
у 90% подопытных животных этой группы восстановление функции стоя-
ния и ходьбы происходило путем методической, упорной и очень длитель-
1 Arch, neural, a. psichiatr., 28, № 3, 1932, р. 542—570.
416
3. А. АСРАТЯН
ной тренировки (см. рис. 1, стр. 260). Большинство собак научилось стоять
и ходить только через 1 — 1 */> месяца, а
Рис. 1. Динамика восстановления функций
стояния и ходьбы у собаки Асланка после
поперечной перерезки передней половины
спинного моз!а на уровне последних груд-
ных сегментов
а через 10 дней, 6 —через 13 пней и в—через
30 дчей ппсле one-рации
некоторые даже через 3 месяца
и позже.
Следует отметить, что ею
быстроту наступления компен-
сации в сильной степени влияют
возраст, вес и длина конечно-
стей собаки.
В чем же, однако, заклю-
чается секрет того, что только
у 3 наших собак и у всех 4 со-
бак Бете с ампутированными
конечностями так быстро вос-
становилась утраченная способ-
ность стоять и ходить? Этот
секрет раскрыла одна из на-
ших подопытных собак, котс-
рая, по нашим наблюдениям,
до ампутации лап умела стоять
и ходить на задних лапах. Как
эта собака, так и две другие
специально подобранные нами
собаки были предварительно
тренированы своими прежними
владельцами и могли «служить»,
т. е. стоять и ходить на задних
лапах. После ампутации лап эти
3 собаки, как уже отмечалось,
быстро начали стоять и ходить,
по всей вероятности, в силу
генерализации имевшихся ра-
нее сложных двигательных на-
выков. На основании этих дан-
ных мы даже предполагали, что
упомянутые выше подопытные
собаки Бете случайно оказались
тренированными в этом смысле
прежними их владельцами. Это
предположение подтверждается
тем,что, судя по фотографиям,
приведенным в работе Бете,
все они — домашние собаки
(шпицы).
По нашим данным, молние-
носной перестройки функции
центров и восстановления на
рушенных функций у собак нс
наблюдается не только п ри ампу-
тации лап, но и при всех других
вариантах повреждения орга-
низма: при различного рода повреждениях спинного мозга, нервов и орга-
нов чувств (рис. 1 и 2). Наши более достоверные факты позволяют сделать
вывод, что у собак именно «научение», т. е. выработка условных рефлек-
сов, а следовательно, кора больших полушарии играет решающую роль
в восстановлении нарушенных функций. Мы имеем и другие косвенные на-
блюдения, говорящие в пользу того же вывода. Если мы создавали опери-
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В СВЕТЕ УЧЕНИЯ И П ПАВЛОВА
417
Рис. 2. Динамика восстановления функций стояния
и ходьбы у собаки Цербер после перерезки боковой
половины спинного мозга на уровне 2—3-го шейных
сегментов
рованным животным такую обстановку, которая лишала их возможно-
сти тренироваться (например, держали их в люльке), восстановление
нарушенных и утраченных функций задерживалось надолго. Если же, нао-
борот, создавалась об-
становка для усиленной
тренировки животных
(скажем, давали им воз-
можность много дви-
гаться), это резко уско-
ряло темпы восстанов-
ления функций. Следует
отметить, что,это отно-
сится как к животным
с ампутированными ла-
пами, так и к животным
с другими оперативны-
ми повреждениями.
Далее, вопреки оши-
бочным утверждениям
П. К. Анохина, нами
было показано, что у
собак в случае наруше-
ния функций какой-
нибудь конечности в
результате оперативного
повреждения спинного
мозга или соответству-
ющих нервов на пора-
женной конечности мо-
гут быть выработаны
двигательные электро-
оборонительные услов-
ные рефлексы со всех
анализаторов, причем
образование и упроче-
ние условных рефлексов
происходят, как прави-
ло, параллельно вос-
становлению функций
этих конечностей, а
иногда даже раньше,
чем ПОЯВЛЯЮТСЯ другие
признаки восстановле-
ния функций. Это явст-
вует из кимограммы ус
ловных рефлексов конеч-
ности, пораженной в ре-
зультате боковой,задней
или передней половин-
ных поперечных перерезок спинного мозга (рис. 3; см. также рисунки
на стр. 273, 299, 347, 349, 35) и 424),
Если же электрооборонительные условные рефлексы с задних лап вы-
рабатываются у собак до спинальной операции, то после операции они ис-
чезают, но вскоре появляются вновь, во всяком случае, как правило, задол-
го до возникновения и развития опорных и локомоторных функций пора-
женных конечностей.
27 3. А. Асратян
418
Э. А. АСРАТЯН
___[ Нас (______________, 39 (________—, СО ,-------------
I । । । 1 । । । 111 । । । । । । 11 м । । । । 1 | 11 11 । | 111 111 1-1 1 11 11 111 । I 11 1111 I >
Рис. 3. Кинограмма электрооборонительных двигательных условных
рефлексов
а—у собаки Фикс (поперечная перерезка задней половины спинного мозга в обла-
сти 2—3-го шейных сегментов); б-у собаки Валет (поперечная перерезка боковой
половины спинндго моэга на том же уровне); а—у собаки Шоколадка (поперечная
перерезка передней половины спииного мо-зга на уровне последних грудных сег-
ментов).О бозначення (сверху вниз): условные и безусловные рефлексы.отметка
условных раздражителей, отметка безусловных раздражителей, время в секундах
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В СВЕТЕ УЧЕНИЯ И. П ПАВЛОВА 419
Эти данные являются новым веским доказательством правильности на-
шего положения о роли условнорефлекторной деятельности коры в восста-
новлении нарушенных функций.
Еще более показательны наши данные относительно последствий уда-
ления коры одного из больших полушарий мозга. Во всех без исключения
случаях более или менее значительного хирургического повреждения тех
или иных рецепторных и эффекторных органов, нервов или важных в функ-
циональном отношении отделов центральной нервной системы у собак,
при котором закономерно возникают глубокие и долго компенсирующиеся
нарушения соответствующих функций организма, последующее удаление
коры одного большого полушария мозга, как правило, влечет за собой рас-
стройство уже развившихся компенсаторных приспособлений. Со временем,
однако, эти компенсаторные приспособления возникают вновь, т. е. вто-
рично. При этом наблюдается одно интересное и важное явление.
Если после восстановления нарушенных и утраченных функций у ссбак
мы удаляем кору одного полушария, то получаем различные последствия,
в зависимости от того, каким образом мы сочетаем эту' операцию с первона-
чальным операционным повреждением конечностей или их нервов, спин-
ного мозга или органов чувств. Допустим, например, что мы ампутиро-
вали у собак передние и задние конечности с левой стороны, а после вос-
становления у них нарушенной функции стояния и ходьбы у одних под-
опытных животных удаляли кору одноименного, т. е. левого, полушария,
а у других — кору противоположного, т. е. правого, полушария. Если
бы операция, являющаяся тяжелой травмой, сама по себе имела решающее
значение и обусловливала все или основную часть возникающих при
этом последствий, то результаты в обоих случаях должны быть одни и
те же. В действительности же получаются разные результаты. В первом
случае, когда операция производилась на одной стороне, удаления коры од-
ного полушария было достаточно для того, чтобы функции стояния и ходьбы
нарушились весьма длительно и глубоко и вторично полностью не вос-
станавливались в течение длительного времени, хотя одно полушарие оста-
валось нетронутым, (У одной нашей собаки с ампутированными двумя левы-
ми лапами и с экстирпированной корой левого же полушария повторное
восстановление функций длилось более двух лет.) В другом случае, когда
лапы ампутировались на одной стороне, а кора одного полушария уда-
лялась на противоположной стороне функции стояния и ходьбы не нару-
шались сколько-нибудь глубоко и длительно, восстанавливаясь примерно
на 20—25-й день. Следует отметить, что повторное восстановление нару-
шенных после удаления полушарий функций стояния и ходьбы опять-таки
происходило постепенно и в результате методической тренировки.
Из сказанного явствует, что удаления коры полушария, одноименного
стороне повреждения, т. е. функционально больше соответствующего здо-
ровым конечностям, благодаря преобладанию перекрестных нервных свя-
зей, достаточно для того, чтобы животное почти полностью и надолго потеря-
ло способность стоять и ходить, хотя практически оно здорово, питается
нормально и внешне не отличается от здоровых собак. Удаление коры полу-
шария, которое функционально соответствует поврежденным конечностям,
также не остается без последствий, очевидно, благодаря наличию прямых
нервных связей; однако последствия эти неглубокие и быстро компенсиру-
ются.
Приведенный выше материал касается собак с ампутированными ла-
пами. То же происходит при одностороннем повреждении центральной или
Периферической иннервации конечностей, например, при половинном боко
вой перерезке спинного мозга на тех Или иных его уровнях. Допустим, что
произведена поперечная перерезка боковой половины спинного мозга с левой
стороны и на уровне последних грудных сегментов, а возникшие вследствие
27*
420
Э. А. АСРАТЯН
этого нарушения двигательных и чувствительных функций нижних конечно-
стей со временем хорошо восстанавливаются. Для выяснения роли коры боль-
ших полушарий в развитии компенсаторных приспособлений и в данном
случае небезразлично, кору какого полушария мы удалим в дальнейшем.
Как показали наши опыты, при удалении коры одноименного пораженной
конечности полушария развиваются более тяжелые и длительные послед-
ствия, чем при удалении коры противоположного пораженной конечности
полушария.
Еще более убедительные данные мы получили при осуществлении глав-
ной и наиболее трудной части наших исследований — при экстирпации коры
обоих полушарий у большого числа подопытных собак с предварительными
оперативными повреждениями конечностей, спинного мозга, органов чувств,
нервов, лабиринтов, мозжечка, нервных корешков и т. л., у которых в пос-
ледующем наступала максимальная компенсация нарушений, вызванных
этими повреждениями.
Экстирпацию коры больших полушарий мозга мы осуществляем двух-
моментно, т. е. первоначально удаляем у собак кору одного полушария и,
спустя некоторое время, кору другого полушария. В такой последователь-
ности мы успешно оперировали ОКОЛО 40 собак. Исследования, проведенные
на этих собаках, с особой ясностью показали правильность нашего теорети-
ческого положения о важнейшей, ведущей роли коры больших полушарий
в компенсаторных приспособлениях поврежденного организма, полностью
гармонирующего с основными принципами учения И. П. Павлова о функ-
циях коры больших полушарий, о высшей нервной деятельности. Нами
было установлено, что у всех без исключения животных навсегда исчезают
ранее восстановленные функции, как бы долго собаки ни жили (2—3 года)
(рис. 4; см. также рисунки на стр. 262, 276, 296, 306 и 353).
Нами были поставлены также опыты с операциями, производившимися
в обратной последовательности, а именно: у ряда других собак сначала
удалялась кора больших полушарий, а затем после исчезновения ближай-
ших последствий операции, т. е. после того, как собаки значительно по-
правлялись и начинали хорошо стоять и ходить, осуществлялась ампутация
лап, половинное боковое сечение спинного мозга и т. д. Нами было выяв-
лено, что у этих собак функции, нарушенные в результате последних опе-
раций, больше не восстанавливаются.
Таким образом, опыты с хирургическим повреждением организма, про-
изводившиеся в различной последовательности, привели к одним и тем же
результатам: к исчезновению компенсаторных приспособлений или к исклю-
чению возможности их развития после удаления коры обоих полушарий.
Все сказанное до сих пор относилось к так называемым соматическим
функциям организма, нарушение и восстановление которых вследствие
повреждения соответствующих структур выявляется особенно рельефно.
К сожалению, мы не смогли до последнего времени заниматься системати-
ческим изучением динамики нарушения и восстановления так называемых
вегетативных функций в поврежденном организме, в частности, изучением
роли коры больших полушарий при этом. По этому вопросу мы пока рас-
полагаем небольшим экспериментальным материалом, накопленным попут-
но в процессе исследования компенсаторных приспособлений в сфере сома-
тических функций. Разумеется, единство соматических и вегетативных функ-
ций целостного организма и их подчиненность коре больших полушарий иск-
лючают возможность каких-либо существенных расхождений в закономер-
ностях возникновения и развития компенсаторных приспособлении в одном
и другом случае. Но вместе с тем нельзя отрицать и известной специфично-
сти протекания этих процессов в поврежденном организме. Об этом свиде-
тельствует, в частности, имеющийся в нашем распоряжении предваритель-
ный материал о развитии компенсаторных приспособлений в сфере вегета-
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В СВЕТЕ УЧЕНИЯ И. II. ПАВЛОВА 42!
тивных функций поврежденного организма. В дальнейшем мы намерены
подвергнуть этот важный вопрос специальному изучению.
Приведем некоторые дополнительные данные, подкрепляющие основное
наше теоретическое положение о роли коры в развитии компенсаторных
приспособлений.
Следует отметить, что в тех случаях, когда повреждение спинного мозга
или нервов в силу своего характера и тяжести вызывает преимущественно
локальные нарушения функций организма, потом компенсирующиеся,
Рис. 4. Последствия удаления коры обоих больших полушарий у собак Шарик
и Эльма после перерезки боковой половины спинного мозга на уровне 2—3-гошейных
сегментов
а-собаиа Эльма через 229 дней после половинной перерезки спинного мозга и через 179 дней
после удалении коры левого полушария; в—через 56 пней поСле Удаления коры правого полу-
шария; л -собака Шарик через 265 дней после поперечной перерезки боковой половины спинного
мэзга на уровне 2-3-го шейных сегментов и 175 лней после удаления коры правого полу-
шария; г— через 202 дня после удаления коры левого полушария
удаление коры обоих полушарий устраняет локальные компенсаторные
приспособления, не отражаясь заметно на общем состоянии организма.
Так, например, удаление коры обоих полушарий у собак с односторонней
гемисекцией спинного мозга на уровне последних грудных сегментов сильно
нарушает опорные и локомоторные функции одноименной нижней и соответст-
вующие функции задней конечности, но заметно не изменяет функции перед-
них конечностей. Такиесобаки способны приподниматься на передних конеч-
ностях и активно передвигаться на них, волоча за собой бездействующую
422
Э. А. АСРАТЯН
заднюю часть тела с окоченелой и вытянутой пораженной лапой. То же
наблюдается при удалении коры обоих полушарий у собак с деафферента-
йней какой-нибудь конечности, перерезкой и перекрестным сшиванием
нервов конечностей и т. и. Пораженные конечности при этом страдают очень
глубоко, а остальные—весьма слабо (см. рисунки на стр. 276, 296,306 и 353).
Одно маленькое, но существенное замечание. Мы удаляем у собак кору
больших полушарий головного мозга, следовательно, ОНИ становятся
«бескорковыми». Между тем в литературе имеется некоторая неясность по
поводу обозначения таких собак, которых чаше всего называют «бесполушар-
нымн». Это название неправильно потому, что, как правило, удаляются не
полушария целиком, а только кора. В то же время, как показывают после-
дующие гистологические исследования препаратов мозга оперированных со-
бак, оставшаяся часть больших полушарий головного мозга и прежде всего
ближайшие подкорковые узлы вслед за хирургическим вмешательством под-
вергаются значительным дегенеративным изменениям. В этой связи мож-
но указать, что гистологические исследования препаратов мозга 12опериро-
ванных нами собак, произведенные в лабораториях проф. Л. Я. Пинеса,
проф. В. С. Дойникова иМ. М. Александровской, подтвердили полноту про-
изведенной экстирпации у большинства из них и констатировали фактдеге-
неративных изменений в подкорковых образованиях. Таким образом, наши
собаки по роду операций являются «бескорковыми», а по отдаленным послед-
ствиям этой операции в значительной степени «бесполушарными».
Следует отметить, что последствия не всякого повреждения организма
могут быть удовлетворительно компенсированы, если даже кора больших
полушарий совершенно не повреждается. С другой стороны, при некоторых
очень легких повреждениях последствия их у высших животных могут быть
компенсированы и без решающего участия коры больших полушарий. Чтобы
это стало яснее, приведем ряд примеров. Если перерезать немного больше
боковой или дорзальной половины спинного мозга у собак, нарушенные
при этом функции, как правило, не восстанавливаются в более или менее
значительной степени, хотя при этой операции остается, казалось бы, мате-
риальная основа — «мостик» для того, чтобы верхние отделы центральной
нервной системы могли сообщаться с нижним отрезком спинного мозга и со-
действовать восстановлению его функций. Или если у собак продольно и
сагиттально расщепить 2—3 сегмента спинного мозга в тех его частях,
которые непосредственно не иннервируют конечности (грудные или копчи-
ковые сегменты), то хотя это весьма сильно отражается на функциях конеч-
ностей, в особенности лежащих ниже уровня расщепления, но нарушенные
функции со временем восстанавливаются. При подобном же расщеплении
в области люмбальных или нижних шейных сегментов, т. о. сегментов, кото-
рые непосредственно иннервируют конечности, функции их нарушаются
весьма глубоко и, как правило, больше нс восстанавливаются, хотя голов-
ной мозг, в том числе и кора больших полушарий, при этом сообщается
с каждой половиной расщепленного спинного мозга.
Доказательством правильности обратного положения является то, что
некоторые легкие нарушения функции организма, вызванные неглубокими
повреждениями тех или иных его структур, могут быть компенсированы и
без решающего участия коры больших полушарий, т. е. в основном ресур-
сами подкорковых узлов и низших отделов центральной нервной системы.
Например, если ампутировать у собак только одну конечность, перерезать
нс половику спинного мозга, а одну треть, или же перерезать какие-ни-
будь маловажные нервы конечности и наложить анастомоз, то нарушения
функции организма при Этом будут неглубокими и быстро восстановятся.
Последующее удаление коры больших полушарий также существенно не
отразится на процессе восстановления функций.
Следует указать, что исход зависит не только от степени тяжести нале-
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ 3 СВЕТЕ УЧЕНИЯ И. П. ПАВЛОВА
423
сенной организму травмы, но и от глубины нарушений его функций. Мы
располагаем фактическими данными, свидетельствующими о том, что значи-
тельную роль играют и такие моменты, как возраст животных, их вес,
длина конечностей, их индивидуальные особенности (мы употребляем этот
собирательный термин, так как у нас нет еще точных данных о значении
типологических особенностей животных, их породы и т. п.), а также степень
зависимости поврежденных нервных структур и нарушенных при этом функ-
ций организма от коры больших полушарий мозга. Вобщем из наших дан-
ных явствует, что, чем моложе животное, чем меньше его вес и чем короче
его конечности, тем быстрее и полнее компенсируются нарушенные функции
организма и гем значительнее участие низших отделов центральной нерв-
ной системы в развитии компенсаторных приспособлений.
Значение степени зависимости нарушенных функций от корковой регу-
ляции для развития компенсаторных приспособлений иллюстрируется сле-
дующими данными. Нами совместно с С. Н. Ивановой было установлено,
что поперечная перерезка боковой половины спинного мозга па уровне
2—3-го шейных сегментов влечет за собой глубокое нарушение опорных
и локомоторных функций одноименных конечностей и почти полный пара-
лич одноименных дыхательных мышц. Но опорные и локомоторные функции
пораженных конечностей через 30—40 дней почти полностью восстанав-
ливаются, нарушенные же функции дыхательных мышц остаются без за-
метных изменений в течение многих месяцев, года и больше (см. рисунок
на стр. 336).
С этой точки зрения весьма показательны наши данные относительно
роли коры больших полушарий мозга в осуществлении так называемого
перекрестного феномена диафрагмального нерва. При поперечной пере-
резке боковой половины спинного мозга» в которой берут начало диафраг-
мальные нервы (на уровне 1—2-ю и даже 3-го шейного сегмента), соот-
ветствующая половина диафрагмы, как и следовало ожидать, парали-
зуется. Но если перерезать диафрагмальный нерв с противоположной
СТОрОНЫ, парализованная (вследствие гемиселекЦИИ спинного мозга) поло-
вина диафрагмы начинает функционировать. Значит, происходит весь-
ма быстрая, как бы молниеносная, перестройка функций спинальных цент-
ров. В специально проведенных нами опытах было установлено, что в. этом
весьма своеобразном приспособительном феномене кора больших полу-
шарий не принимает значительного участия: этот феномен наблюдается у со-
бак с удаленными полушариями.
Разумеется, эти данные ни в коей мере не противоречат данным павлов-
ской школы, в особенности классическим исследованиям К. М. Быкова,
доказавшим безграничность корковой регуляции функции организма, но
отнюдь не отрицающим наличия различных градаций в силе этой регуля-
ции.
Значение индивидуальных особенностей животных для участия коры
в развитии компенсаторных приспособлений достаточно четко выступает
из следующих данных. Нами было установлено, что у собак последствия
перерезки задних корешков первых трех пар шейных нервов сильно варь-
ируют по степени тяжести и длительности. У одних собак при этом на-
ступали очень глубокие нарушения функций стояния и ходьбы, причем
нарушенные функции восстанавливались через месяц и позже, у других на-
рушения эти были неглубокими и исчезали через 10—12 дней. После-
дующая экстирпация коры обоих полушарий и в этом случае приводила
к разным последствиям: у первых собак компенсаторные приспособления
исчезали безвозвратно, а у вторых собак эти приспособления хотя и исчезали,
но со временем в слабой степени вновь’восстанавливались.
Несколько слов о собаках, лишенных целиком и мозжечка, и коры боль-
ших полушарии. У 4 из собак с удаленным мозжечком, у которых макси-
424
Э. А. АСРАТЯН
мально компенсировались мозжечковые расстройства, мы удаляли кору
одного полушария. Последствия удаления коры даже одного полушария
оказались для всех них крайне тяжелыми, во всяком случае гораздо тяжелее,
чем у собак с другими первоначальными хирургическими повреждениями
периферических органов и центральной нервной системы. Но и здесь ВЫ-
ЯВИЛИСЬ индивидуальные различия. У двух собак (Злюки и Лебедя) после
удаления коры одного полушария способность стоять и неуклюже перед-
вигаться в значительной степени восстановилась, хотя на это и потребо-
валось 4—6 месяцев; другие же две собаки (Шарик и Веселый) вот уже
Рис. 5. Последствия удаления коры
больших полушарий у лишенных моз-
жечка собак
а—собака Лебедь после удаления мозжечка
и коры одного полушария; (5—та нее собака
после удалении коры второго полушария;
•—собака Злюка после удаления коры обоих
полушарий
7—10 месяцев после удаления коры одного полушария находятся в удов-
летворительном состоянии, но у них не проявляется никаких признаков
восстановления функции стояния и ходьбы. Нечего и говорить уже о том.
что удаление коры второго полушария у собак Злюка и Лебедь вызвало пол-
ное и безвозвратное исчезновение всех компенсаторных приспособлений
(рис. 5), причем Лебедь прожил после этого Немногим больше месяца, Злюка
же жила 1 >/а года, что во много раз превышает продолжительность
жизни 2собакРадемаксра, лишенных коры обоих полушарий н мозжечка,
не говоря уже об оперированных таким же образом кошках Фултона.
Разумеется, научная ценность нашей уникальной подопытной собаки
далеко не исчерпывается полученными у нее данными о компенсаторных
приспособлениях, но мы лишены здесь возможности останавливаться
на этом вопросе.
Таким образом, можно утверждать, что не каждое нарушение функций
организма может быть компенсировано удовлетворительно при наличии
всей коры или даже коры одного полушария; 'с другой стороны, возможны
случаи восстановления функции и без существенного участия коры. Однако
приведенные для иллюстрации этого положения фактические данные пред-
ставляют все же исключения, которые ни в коей мерс не опровергают уста-
новленного нами основного положения о ведущей роли коры больших по-
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В- СВЕТЕ УЧЕНИЯ И П. ПАВЛОВА
425
лушарнй мозга о развитии компенсаторных приспособлений в высокораз-
витом организме после значительных механических повреждений струк-
туры двигательных и воспринимающих органов, периферической нервной
системы или различных отделов центральной нервной системы.
Следовательно, и в отношении изученного нами нового класса физио-
логических явлений подтверждается положение И. П. Павлова, что кора
больших полушарий, этот верховный орган высокоразвитого организма,
держит в своем ведении, является верховным распределителем и распоря-
дителем всех происходящих в организме процессов.
Несколько слов о наших экспериментальных данных относительно ком-
пенсаторных приспособлений у низших позвоночных животных.
У низших позвоночных нами получены несколько иные результаты в от-
ношении темпа и характера компенсаторных приспособлений, а также роли
переднего мозга при этом, чем у собак. У птиц, например, удаление перед-
него мозга почти полностью устраняет компенсаторные приспособления,
развившиеся после повреждения нервов, задних корешков спинного мозга,
конечностей или после удаления мозжечка. Восстановленные функции вновь
надолго нарушаются. Но у птиц, в отличие от собак, эти функции со вре-
менем в слабой степени вновь восстанавливаются.
У лягушек и черепах нарушение и восстановление двигательных функ-
ций после повреждения нервов или спинного мозга выступают недостаточ-
но рельефно. Более или менее убедительным можно считать то, что соот-
ветствующее хирургическое повреждение периферической или центральной
нервной системы у них не вызывает такого глубокого нарушения функций,
как у высших животных. Да это и понятно, потому что диференциация,
специализация и локализация функций центральной нервной системы у них
стоят на весьма низком уровне. Интересно, однако, что те ничтожные
нарушения функций, которые наступают у этих животных в результате
повреждения спинного мозга или деафферентации конечностей, компен-
сируются в ничтожных пределах и крайне медленно, а удаление переднего
мозга вовсе не оказывает влияния.
В настоящее время мы приступаем к проверке этих, пока предвари-
тельных данных и выводов на более подходящих подопытных животных
из класса рептилий, а именно на крупных пустынных ящерицах-варанах.
* *
*
Переходим к заключительной части нашего доклада — к общим вы-
водам и краткому теоретическому обобщению полученного нами факти-
ческого материала.
Прежде всего отметим, что не только анализ собственных эксперимен-
тальных данных, но и критический анализ достоверного фактического ма-
териала наших предшественников по разработке обсуждаемой проблемы
приводят пас к твердому убеждению, что сложные и многообразные ком-
пенсаторные приспособления в поврежденном организме не являются, как
это утверждал Бете, выражением какой-то переменной игры перифериче-
ских нервных импульсов в якобы пассивной и аморфной центральной Чести
«нервной сети», результатом каких-то однородных и монотонных сдвигов
в «примитивных» частях центральной нервной системы, будто бы совер-
шенно идентичных по принципу у всех животных, независимо от их место-
положения в эволюционном ряду.
Наш обширный материал позволяет утверждать, что компенсаторные
приспособления в поврежденном организме, осуществляемые благодаря
комплексу тесно связанных между собой факторов, представляют собой слож-
ный синтез многообразных, взаимодействующих в организме и в особен-
ности в нервной системе процессов. К этим процессам относятся, в частно-
сти, регенерация поврежденных структур, активирование дублирующих
426
Э. А. АСРАТЯН
механизмов, мобилизация и активирование запасных возможностей, трени-
ровка неповрежденных частей и их функциональное и структурное вика-
рированне. улучшение функционального состояния не разрушенных трав?
мой, но сильно альтерированных ею нервных образований путем развития
в них запредельного (охранительно-целебного) торможения, простая или
безусловнорефлекторная перестройка функций нервных центров и путей
и, наконец, условнорефлекторная перестройка функций нервных центров
и путей, являющаяся вершиной этой вереницы своеобразных защитных мер
и средств организма против Последствий ёГО механического повреждения.
Факты говорят о том, что и здесь живая природа проявила срою нелюбовь
к стандарту, шаблону. Против различных по характеру, тяжести и локали-
зации механических повреждений организма, неизбежно порождающих
различного рода, объема и глубины расстройства его функций, организм
в процессе своей эволюции развивал массу многообразных защитных мер
и средств различной физиологической эффективности, в совокупности или
в отдельных комбинациях обеспечивающих развитие спасительных ком-
пенсаторных приспособлений.
Многими отечественными и зарубежными исследователями еще до нас
в общих чертах было установлено важное значение нервной системы,
а также значение отдельных примитивных форм ее деятельности в развитии
компенсаторных приспособлений. На основании нашего эксперименталь-
ного материала не только вы явлено значение некоторых новых примитивных
форм нервной деятельности для подобных приспособлений, по и твердо
установлено важнейшее для всей проблемы положение, полностью про-
тиворечащее метафизической теории Бете, а именно что в процессе
филогенетического развития животного мира
роль высших отделов центральной нервной
системы в компенсаторных приспособлениях
непрерывно возрастает, достигая у высших жи-
вотных значения доминирующего, решающего
фактора.
Из нашего экспериментального материала относительно разных послед-
ствий удаления высших отделов центральной нервной системы у ссбак,
птиц, рептилий и амфибий с одинаковыми предварительными повреждения-
ми, относительно разных последствии удаления коры больших полушарий
у собак с разными степенями предварительных повреждении организма,
а также относительно слабого участия коры в восстановлении нарушенной
травмой дыхательной функции явствует следующее: хотя при развитии ком-
пенсаторных приспособлений нервная система действует как единое целое,
тем не менее степень и характер участия различных ее отделов изменяются
в зависимости как от уровня развития организма, от места, занимаемого им
на эволюционной лестнице, так и от характера, тяжести и локализации на-
несенного повреждения. При этом было также установлено, что в большин-
стве случаев имеет силу одно правило, а именно: чем тяжелее
повреждение организма, глубже поражение его
функций и выше занимаемое им место в эволю-
ционном ряду, тем важнее роль и активнее учас-
тие высших отделов центральной нервной
системы в развивающихся при этом компенса-
торных приспособлениях.
С этим тесно связана еще одна крайне интересная и важная закономер-
ность. На обширном сравнительно-физиологическом материале мы убеди-
лись в правильности факта, давно установленного многими учеными,
а именно: примерно одинаковые по характеру, тяжести и локализации
повреждения центральной нервной системы (например, гемисекция боковой
половины спинного мозга на определенном его уровне) вызывают неоди-
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В СВЕТЕ УЧЕНИЯ И. П. ПАВЛОВА
427
наковые по глубине, объему и длительности нарушения функций у живот-
ных, стоящих на разных уровнях эволюционной лестницы; чем выше стоит
животное, тем эти нарушения глубже и обширнее. Каждому непредубеж-
денному врачу и биологу ясно, что этот факт выявляет полную несостоя-
тельность антиэволюционных взглядов Бете и его единомышленников
относительно функциональной аморфности и эквипотенциальности, пред-
ставляющих якобы общий принцип функциональной архитектуры цент-
ральной нервной системы у всех животных, обладающих нервной системой.
В противоположность взглядам Бете этот факт доказывает правильность
положения о возрастающей в процессе эволюции дифференциации, специа-
лизации и локализации структуры и функций в центральной нервной
системе.
Но нас сейчас интересует не этот факт как таковой, а сделанное нами
существенное добавление к нему, у низших позвоночных незначительные по
своему объему и глубине нарушения функций организма, возникшие в ре-
зультате половинной перерезки спинного мозга или в результате других
подобных повреждений спинного мозга, рецепторных органов и нервов,
почти не компенсируются, а у высших позвоночных глубокие нарушения,
возникшие от аналогичных же операций, со временем почти полностью ком-
пенсируются. Эго относится не только к последствиям повреждения спин-
ного мозга, но в значительной мере распространяется м на последствия мно-
гих видов повреждений центральной нервной системы у высших и низших
позвоночных. Из этих фактов логично вытекает следующий вывод: более
глубокое нарушение функций организма при многих повреждениях цент-
ральной нервной системы как своего рода биологический «минус», обу-
словленный прогрессирующей специализацией и локализацией в централь-
ной нервной системе в процессе эволюции, нейтрализуется другим, одновре-
менно протекающим (быть может, даже взаимосвязанным) изменением в
центральной нервной системе, а именно возрастающей в процессе эволю-
ции ролью ее высшего отдела в компенсаторных приспособлениях, в осо-
бенности бурно растущей мощью этого отдела на верхних ступенях эволю-
ционной лестницы.
На основе этих взглядов получают удовлетворительное объяснение мно-
гие непонятные явления в физиологии и медицине. В порядке иллюстрации
сошлемся на установленный П. Вейссом и другими зарубежными исследо-
вателями, но ложно истолкованный ими факт отсутствия приспособления
пересаженных мышц и целых конечностей у низших позвоночных к новым
местам приживления, к новым условиям существования, стойкое сохране-
ние этими прижившими частями так называемых «резонансных» свойств
в условиях полной невредимости центральной нервной системы. А ведь
известно, что у высших животных и человека в подобных примерно случаях,
скажем, при пересадке сухожилий мышц, при перекрестном сшивании нер-
вов и даже в случаях врожденного дублирования пальцев руки, происхо-
дит полная перестройка функций, т. е. приспособление к новым условиям
существования; функционирование же этих частей по принципу «резонанса»
представляет лишь кратковременную начальную фазу этого приспособи-
тельного процесса. Таким образом, в свете наших данных и теоретических
установок «резонансное» свойство выступает не как универсальный, аб-
солютный и статический закон взаимосвязи периферии и центра, как оши-
бочно представляет Пауль Вейсс, автор широкоизвестной лженаучной «ре-
зонансной» теории, а лишь как фаза, как звено в развивающейся цепи при-
способительных явлений.
Считая излишним детальное изложение нашего понимания физиологи-
ческой сущности всех перечисленных выше разнородных сдвигов и процес-
сов в организме, участвующих в развитии компенсаторных приспособле-
ний, коротко осветим лишь взгляд на внутреннее содержание основного
428
Э. Л. АСРАТЯН
установленного нами факта—ведущей роли коры больших полушарий в ком»
пенсаторных приспособлениях у высокоразвитых животных.
Общеизвестно, что у высших животных кора больших полушарий ана-
томически непосредствен но не связана с периферическими исполнительными
и воспринимающими органами; эту связь она осуществляет только посред-
ством нижележащих отделов центральной нервной системы — базальных
ганглиев больших полушарий, промежуточного и среднего мозга и в ко-
нечном итоге посредством продолговатого и спинного мозга и исходя-
щих из них нервов. Совершенно очевидно, что в случае повреждения тех
или иных органов или систем, нервов или тех или иных отделов централь-
ной нервной системы нарушенные при этом функции организма могутбыть
восстановлены в конечном итоге лишь при обязательной перестройке функ-
ций неразрушенных частей нижележащих отделов центральной нервной
системы, имеющих непосредственную связь с периферией. При таком по-
ложении участие коры больших полушарий как верховного «распределителя
и распорядителя» в организме, как решающего фактора в формировании и
осуществлении компенсаторных приспособлений в конечном итоге сводится
к изменению, перестройке функций этих нижележащих и подчиненных ей
исполнительных нервных образований, уцелевших от разрушения частей
низших отделов центральной нервной системы с тем, чтобы они оказались
в состоянии замещать вышедшие из строя части того же самого исполнитель-
ного аппарата.
На основании полученных нами экспериментальных данных мы при-
ходим к заключению, чТо эту важную Преобразующую, созидательную
роль в отношении функций нижележащих неразрушенных нервных обра-
зований кора больших полушарий осуществляет с помощью комплекса
взаимосвязанных воздействий на эти образования. Наиболее важное зна-
чение имеют следующие три рода влияния коры на нижележащие отделы
центральной нервной системы или следующие три стороны единой дея тель-
ности коры, которые различными путями с помощью разных механиз-
мов властно изменяют деятельность всех остальных нервных образова-
ний.
Мы имеем в виду прежде всего трофическое (в павловском понимании)
влияние коры больших полушарий на нижележащие части центральной
нервной системы и на связанные с ними периферические нервные образо-
вания и органы, установленное нами в специально поставленной серии ост-
рых опытов еще в 1932—1933 гг. и выявленное в процессе многолетних на-
блюдений над большим числом лишенных коры полушарий собак (в общей
сложности под нашим наблюдением находилось более 60 таких собак).
О трофическом влиянии коры свидетельствуют весьма интересные и ценные
исследования Б. Баяндурова и недавно полученные в нашей лаборатории
новые данные Б. Д. Стефаицова.
Кора больших полушарий как высший трофический орган способна
сильно изменять функциональное состояние — возбудимость, лабильность,
тонус, работоспособность — нижележащих отделов центральной нервной
системы и тем самым создавать благоприятные предпосылки к их функци-
ональной перестройке. Отсутствие же коры резко ухудшает функциональное
состояние этих образований со всеми вытекающими отсюда последствиями
и, в частности, для перестройки их функций.
Далее, мы имеем в виду участие коры в безусловнорефлекторной: дея-
тельности нервной системы. Сказанное многим может показаться непонят-
ным, так как распространено мнение, что кора больших полушарий являет-
ся лишь органом условнорсфлекторной деятельности и не имеет отношения
к безусловнорефлёкторной деятельности. А между тем это мнение по су-
ществу ошибочно. Ведь высказывания И. П. Павловао корковом предста-
вительстве безусловных рефлексов нельзя рассматривать иначе как допу-
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В СВЕТЕ УЧЕНИЯ И. П. ПАВЛОВА 429
щение участия коры больших полушарий в безусловнорефлекторной дея-
тельности. Наблюдения за собаками, лишенными коры больших полушарий,
приводят к твердому убеждению, что хотя у них и сохраняются простые и
сложные безусловные рефлексы, в том числе и такие сложные рефлектор-
ные функции, как стояние и ходьба, тем нс менее эти рефлексы, в особен-
ности сложные из них, не обладают тем совершенством, тонкостью, точ-
ностью и быстротой, как у нормальных собак.
В настоящее время этот вопрос является темой наших специальных ис-
следований, и мы надеемся, что в ближайшее время сможем представить
более Точную качественную и количественную характеристику этих изме?
нений в безусловнорефлекторной деятельности лишенных коры собак.
Но самый факт упрощения и обеднения безусловиорефлекторной деятель-
ности у собак с удаленной корой мы считаем установленным.
Этот факт, рассматриваемый нами как прямое подтверждение идеи
И. П. Павлова о корковом представительстве безусловных рефлексов, по-
служил основанием для развития положения о том. что через кору больших
полушарий проходит высшая ветвь рефлекторной дуги любого безуслов-
ного рефлекса, которая, благодаря более совершенной рецепции и более
совершенному объединению (интеграции) протекающих в центральной
нервной системе процессов., и придает безусловным рефлексам предельное
для них совершенство, тонкость, точность и лабильность.
На основании всего изложенного считаем, что одним из мощных прие-
мов воздействия коры больших полушарий на перестройку функций нераз-
рушенных нервных образований нижележащих отделов центральной нерв-
ной системы является участие коры в осуществлении безусловнорефлектор-
ной деятельности. Удаление коры делает их деятельность примитивной,
малоценной со всеми вытекающими для перестройки их функций послед-
ствиями.
Третий и главный вид деятельности коры больших полушарий, играю-
щий ведущую роль в перестройке функций неразрушенных исполнитель-
ных нервных образований нижележащих отделов центральной нервной
системы, а тем самым и в создании и осуществлении компенсаторных при-
способлений в поврежденном организме, — это ее условнорефлекторная
деятельность, ее способность к образованию в соответствии с произо-
шедшими в организме коренными сдвигами новых простых и сложных услов-
ных рефлексов, к созданию качественно новой и более высокой интегратив-
ной деятельности, к осуществлению более тонкого и совершенного анализа
и синтеза в тяжелые для организма периоды жизни. Об этом свидетельствует
медленный темп и постепенное развитие компенсаторных приспособлений
при значительных по своей тяжести повреждениях организма. Об этом же
говорит характер развития этих компенсаторных приспособлений, характер
выработки новых сложных двигательных условных рефлексов типа разно-
образных навыков с прогрессивным их усовершенствованием путем упор-
ной и методической тренировки.
Все эго было нами доказано, вопреки необоснованным утверждениям
Бете, на большом разнообразном и абсолютно достоверном эксперименталь-
ном материале.
Наконец, об этом свидетельствуют наши да иные о способности животных
к образованию двигательных электрооборонительных условных рефлексов
на деятельность пораженных — в результате нарушений иннервации —
конечностей не только после восстановления их функции (что первона-
чально отрицал П. К. Анохин), но даже в процессе развития самых пер-
вых признаков компенсаторных приспособлений.
Да и могло ли быть иначе, если условнорефлекторная деятельность по
своей сущности является высшим выражением приспособительной дея-
тельности живой материи вообще!
4 ЗА
Э Л. АСРАТЯН
Только закоренелые идеалисты и реакционеры в науке могли отрицать
столь очевидные факты и то, конечно, голословно.
Несколько слов о нашем понимании роли периферических импульсов в
в развитии компенсаторных приспособлений.
Как уже было отмечено выше, Бете и его последователи приписывают
периферическим импульсам значение решающего фактора в развитии ком-
пенсаторных приспособлений в поврежденном организме. При этом он умо-
зрительно делает заключение, что эти импульсы «утилизируются» именно
примитивными частями центральной нервном системы, что перестройка
функций в последних осуществляется под прямым и непосредственным воз-
действием периферических импульсов, без всякого или значительного уча-
стия высших отделов центральной нервной системы. По нашим данным,
периферические рецепторы, периферические импульсы действительно
играют важную роль в развитии компенсаторных приспособлений. В от-
личие от Бете мы экспериментально показали задерживающее влияние вы-
ключения зрения и разрушения лабиринтов на динамику компенсаторных
приспособлений при ампутации лап и другого рода повреждениях ор-
ганизма.
Однако в полном соответствии с материалистическим учением И. М. Се-
ченова и II. П. Павлова о важнейшей роли периферических чувствительных
импульсов и чувствительности вообще в нормальной и патологической дея-
тельности нервной системы мы иначе, чем Бете, понимаем значение этих
импульсов в развитии компенсаторных приспособлений. В нашем понима-
нии Эти чувствительные импульсы участвуют в перестройке функций нераз-
рушенных исполнительных нервных образований в основном посредственно,
через высшие отделы центральной нервной системы, где они достигают наи-
большей своей выраженности и «утилизируются» с максимальным для этой
цели эффектом.
Подытоживая все сказанное, мы должны отметить, что не только наш
фактический материал, но и наши выводы и теоретические положения о
компенсаторных приспособлениях коренным образом расходятся сданными,
выводами и теоретическими положениями Бете и его последователен.
Одновременно с этим мы с особым удовлетворением констатируем, что
наши данные и теоретические положения находятся в полном соответствии
с основными принципами материалистического учения великого И. П. Пав-
лова о высшей нервной деятельности, в частности, с фундаментальным поло-
жением этого учения о том, что кора больших полушарий мозга является
наиболее могучим органом в осуществлении сложнейших взаимоотношений
высокоразвитого здорового организма с окружающей средой, органом наи-
более тонкого, точного и совершенного' его приспособления к условиям
существования. Наши исследования с полной убедительностью доказывают,
что такую же крайне важную роль кора больших полушарий мозга играет
и в тех специфических приспособительных явлениях, которые возникают
в высокоразвитом поврежденном организме, т. е. в компенсаторных приспо-
соблениях. Таким образом, полученные нами точные экспериментальные
данные позволили нам применительно к специальной области биологичес-
ких явлений в известной мере расширить границы использования этого уче-
ния и распространить его на новый весьма важный в биологии класс приспо-
собительных явлений, который привлекает все большее внимание физиоло-
гов и клиницистов.
Развиваемая нами на основе учения И. П. Павлова эволюционная тео-
рия компенсаторных приспособлений нервной системы рассматривает роль
высших отделов центральной нервной системы, различных внутренних и
наружных рецепторов и других факторов в развитии этих приспособлений
не метафизически и абстрактно, а диалектически, в зависимости от уровня
филогенетического развития организма, его возраста, характера, глубины
КОМПЕНСАТОРНЫЕ ПРИСПОСОБЛЕНИЯ В СВЕТЕ УЧЕНИЯ И. П. ПАВЛОВА
431
и локализации повреждения, условий существования организма и т. п.
Мы рады констатировать, что полученные нами вместе с группой со-
трудников экспериментальные данные и построенные на нх основе теорети-
ческие положения не только удовлетворительно разрешают некоторые слож-
ные вопросы важнейшей биологической и медицинской проблемы в свете
материалистического учения гениального И. П. Павлова, но и расширяют
перспективы дальнейшей экспериментальной, клинической и теоретиче-
ской ее разработки в отечественной науке.
♦ •
♦
Мне была оказана высокая честь — поручено возобновить прерванный
Великой Отечественной войной цикл «Павловских чтений», к тому же в
прекрасной столице цветущей Советской Украины.
В этот радостный для меня день я с глубоким волнением вспоминаю
своего любимого учителя — великого И. И. Павлова, который горячо
поддерживал мои начинания по разработке пленительно интересной проб-
лемы компенсаторных приспособлений в свете его гениального учения и не-
задолго до своей кончины на одной из знаменитых «сред» (6 ноября 1935 г.)
уделил значительное внимание тогда еще первым ощутимым итогам моих
исследований в этой области. С сожалением констатируя факт, «в какой сте-
пени прочно сидит дуализм в ученых головах», и причисляя к ним Бете,
великий физиолог говорил: «Я думаю, воюя с условными рефлексами,
он сделал довольно грубую ошибку в своей работе, хотя он, надо сказать,
малый очень сообразительный,—теперь Эзрас Асратович его поправляет.
Это ему, конечно, послужит, к порядочному стыду, напоминанием, чтсбы
не очень впутывать в научное мышление свое общее миросозерцание».
Подвергая, далее, уничтожающей критике фактическое содержание экс-
периментальной работы Бете по ампутации конечностей у ссбак, И. П. Пав-
лов продолжает: «Был известнейший всем факт, что искалечение с те-
чением времени выравнивается, а он прибавил к этому слова: это надо от-
нести на «таинственное» (потому что без дальнейшего анализа) свойство спин-
ного мозга. Вот вам все его работы и весь разговор о пластичности. Я опять
готов понимать это как действие того же дуализма. Но что же он такое сде-
лал своими совершенно ненужными опытами? И тем не менее он нашел себе
последователей. Тут никакого приобретения нет. Тут одни пустые слова
брошены. Дуалистический пыл против монизма, который выступает в наших
условных рефлексах, до такой степени затемнил господина Бете, что ему
и в голову не пришло обратить внимание на условные рефлексы. Но, поз-
вольте, все, что вы говорите о пластичности спинного мозга, — все это при-
ложимо и к нашим условным рефлексам в коре. Следовательно, первое, что
он должен был бы сделать, если бы он не был во власти дуалистического
миросозерцания, а немножко обратил бы внимание на наши условные реф-
лексы, когда эти собаки обучались, наконец^,двигаться, возвратили себе
локомоцию, он должен был бы спросить себя, не потеряют ли они эту спо-
собность, если у них отнять большие полушария. Тогда все дело свелось
бы на условные рефлексы коры. Это он не сделал, а сделал это Эзрас Асра-
тович и оказался совершенно прав. Все эти собаки восстанавливают ло-
комоцию при помощи больших полушарий, T. е. При НОМощи условных
рефлексов. Если собаку, которая после искалечения обучалась двигаться,
лишить больших полушарий, то она делается неисправимой калекой.
Вот вам, что значит игра слов, довольствование словами, что значит
бросить слово «пластичность» и на этом закончить и успокоиться.
Это очень поучительная история»1.
Если гениальное материалистическое учение о высшей нервной деятель-
’ Павловские среды, т. 3, стр. 19*19, 253—254.
432 Э. А. АСРАТЯН
ности было идейным истоком моих исследований по физиологии компенса-
торных приспособлений, то положительное отношение учителя к этим иссле-
дованиям и к первым их результатам послужило мощной моральной под-
держкой и источником НОВЫХ сил В последующей моей 16-летней исследова-
тельской работе в том же направлении, протекавшей, как известно, в жесто-
кой борьбе с антипавловцами и далеко не всегда ГЛМко, но все же Давшей
достаточный фактический материал для провозглашения победы павловских
идей еще в одной из запутанных областей биологии и медицины.
Вот почему я сегодня не только преисполнен чувства искренней приз-
нательности и глубокой благодарности к Павловскому научному совету’
при Президиуме Академии наук СССР, руководству Академии наук
Украинской ССР и руководству Общества физиологов, биохимиков и
фармакологов Украинской ССР за оказанную мне высокую честь, но
н полон неудержимого желания продолжать дальнейшую разработку
важной проблемы современной биологии и медицины по столбовой до-
роге, указанной нашим великим .учителем—И. П. Павловым.
J3 а з 8 e л III
ОХРАНИТЕЛЬНО-ЦЕЛЕБНОЕ
ТОРМОЖЕН И E
ОХРАН И ТЕЛЬНАЯ И ЦЕЛЕБНАЯ РОЛЬ ПРОЦЕССА ТОРМОЖЕНИЯ1
ради многочисленных и многосторонних достижений отечест-
венной физиологии достижения в области изучения рели и при-
роды процесса торможения занимаютоссбоемссто. Общеизвест-
но, что в этом вопросе наши физиологи играли ведущую рсЛЬ
в мировой физиологической науке.
Открытие интрацентрального торможения И. М. Сеченовым, его
мысли и высказывания о роли процесса торможения при разнообразных ви-
дах координационном деятельности нервной системы явились отправной точ-
кой для дальнейших исследований простых и сложных форм интегративной
деятельности различных отделов центральной нервной системы, исследо-
ваний, которые легли в основу современной физиологии нервней системы
[И. П. Павлов, Н. Е. Введенский, Гольц (Goltz), Магнус (Magnus) и мн.
др.]. Оригинальные, глубокие, с мастерством художника выполненные ис-
следования Н. Е. Введенского, направленные на разгадку тайн внутренней
природы процесса торможения, на раскрытие его сущности, уже много де-
сятков лет стоят в центре внимания ведущих физиологов мира, работавших
и работающих в этом направлении. А в работах И. П. Павлова по физиоло-
гии и патологии большого мозга, в которых он достиг вершины физиологи-
ческой мысли, были не только глубоко изучены указанные стороны про-
цесса торможения при самой высшей, совершенной и сложной анализа-
торной и синтетической деятельности нервной системы, но и была рас-
крыта новая сторона, новая роль процесса торможения в деятельности боль-
шого мозга, а именно охранительно-целебная роль торможения.
Многолетние исследования И. П. Павлова и его сотрудников показали,
что, благодаря своевременному развитию торможения, устраняется снгснссть
истощения наиболее реактивных клеток, клеток коры больших полушарий,
угрожающая им при действии на организм сильных раздражителей или при
длительном действии раздражителен умеренной силы. В двинем случае
роль возникшего торможения заключается не в формировании координа-
ционной деятельности корковых нервных клеток, а в их охране, в их зашите
от вредоносного действии раздражителя. Известно, что не только так назы-
ваемое запредельное торможение, но и естественный периодический сон
с его переходными фазами И. П. П авлов рассматривал как проявление охра-
нительного торможения. Сб охранительной роли торможения при нормаль-
ной деятельности большого мозга он писал: «С одной стороны, постоянно,
наравне с процессом раздражения, участвуя в разнообразной деятельности
животного во время бодрого состояния, торможение, сдругой стороны, так-
же постоянно является в роли охранителя реактивнейших клеток организма,
клеток коры больших полушарий, защищая как специально против чрез-
вычайного напряжения их деятельности при встрече с очень сильными
1 Вводная глава монографии «Очерки по этиологии, патологии и терапии трав
маткческого шока»', 1945.
28*
436
Э. А. АСРАТЯН
раздражениями или при длительном повторении хота бы и несильных раз-
дражений, так и обеспечивая им необходимый покой после ежедневной
нормальной работы в виде сна»1.
Классические исследования И. П. Павлова и его учеников показали,
что эта специфическая и с биологической точки зрения очень важная роль
торможения еще ярче выступает при разнообразных патологических состоя-
ниях большого мозга. Одной из важнейших предпосылок для развития пато-
логических состояний большого мозга И. П. Павлов считал слабость его
нервных клеток, обусловленную различными наследственными или инди-
видуальными факторами. «Такая нервная система при встрече с трудностя-
ми,—писал он,—или пссле непосильного возбуждения неизбежно перехо-
дит вссстояние истощения. А истощение есть один из главнейших физиоло-
гических импульсов к возникновению тормозного процесса как охранитель-
ного процесса»2. Это охранительное торможение И. П. Павлов рассматривал
как «нормальный прием физиологической борьбы против болезнетворного
агента», как «физиологическую меру», предохраняющую слабые нервные
клетки от «угрожающего разрушения вследствие непосильной работы».
При этом охранительное торможение имеет более стойкий характер, чем
в условиях нормальной деятельности большого мозга. В зависимости от
преимущественной локализации торможения в массе большого мозга, от
его интенсивности, от вызванных им явлений растормаживания, индукции,
иррадиации и других функциональных сдвигов в больших полушариях,
а также в зависимости от типологических особенностей животного или че-
ловека, от степени слабости их нервной системы, от характера, силы и дли-
тельности действия предрасполагающих и этиологических факторов и т. д.
создается тот или другой тип нарушения деятельности большого мозга.
Однако, по концепции И. П. Павлова, прнболезненных состояниях боль-
шого мозга роль торможения нс ограничивается только охраной или заши-
той ослабленных и истощенных нервных клеток от дальнейшего истощения.
В отношении этих клеток торможение играет Другую, исключительно важ-
ную роль—целебную или, как говорил И. П. Павлов, целительную
роль, роль своеобразного природного лекарства. Следует сказать, что об
этом свойстве торможения многие из наших ученых обычно забывают,
излагая взгляды И- П. Павлова на указанный вопрос, и сбычно упоминают
только сб охранительной роли торможения, или об охранительном
торможении.
Данные эксперимента и клиники о положительных терапевтических
результатах естественного торможения, усиленного или не усиленного ле-
карственными средствами, нельзя удовлетворительно объяснить только ох-
ранительной ролью торможения. А. Г. Иванов-Смоленский и некоторые дру-
гие наши ученые положительный эффект сонной терапии при патологиче-
ских состоя ниях большого мозга трактуют как результат действия двух фак-
торов: фактора охранительного торможения, которое пассивно защищает
нерв-ные центры от дальнейшего вредоносного действия патогенного начала,
и активно-оборонительно действующего фактора в виде своеобразной «ве-
гетативной мобилизации» (сдвиги в деятельности вегетативной нервной си-
стемы, в составе крови, в теплорегуляции и т. д.). А. Г. Иванов-Смоленский
пишет по этому поводу: «Терапевтическое действие длительного наркоза
представляет собой единство двух тесно сплетенных и неразрывно связан-
ных между собой компонентов: пассивно-защитного (охранительное тормо-
жение) и активно-оборонительного (вегетативная самозащита)»3.
Совершенно очевидно, что при таком понимании торможению отводится
* И. П, Павлов, Двадцатилетний опыт объективного изучения высшей
нервной деятельности (поведения) жннотных, 1938, стр. 510.
“Там же, стр. 514.
’ Труды психиатрической клиники акад. И. П. Павлова, сб. 11, 1940, стр. 32.
ОХРАНИТЕЛЬНАЯ И ЦЕЛЕБНАЯ РОЛЬ ТОРМОЖЕНИЯ
437
роль пассивного, вспомогательного, второстепенного фактора, роль какого-
то придатка к более эффективному и активному фактору, играющему веду-
щую рель в восстановлении функций истощенных н ослабленных нервных
образований. Мне кажется, что такое понимание роли торможения является
односторонним и половинчатым и едва ли полностью гармонирует со взгля-
дами И. II. Павлова. В описанных выше восстановительных явлениях
И.П. Павлов придавал торможению не только роль «пассивно-защитного»
фактора, но и роль активно действующего целебного фактора, роль веду-
щего, а не придаточного фактора в системе Других возможных факторов. В
пользу такого понимания взглядов И. П. Павлова говорит ето трактовка ле-
чебного действия брома как фактора, углубляющего и усиливающего тор-
можение. Говоря о действии брома при патологических состоя пнях боль-
шого мозга, вызванных перенапряжением тормозного процесса и действием
сильного раздражителя, он писал: «В качестве целительного мо-
мента в обоих Случаях резко подчеркивается сущест-
веннейшее значение восстановления и усиления
тормозного процесса, так как на основании многих других
наших опытов, помимо только что описанных, брому должно быть припи-
сано непосредственное отношение к тормозному процессу, именно как вос-
станавливающему и усиливающему его агенту»1 (подчеркнуто нами.— Э. Д).
В целях внесения большей четкости в терминологию и более точной
передачи сущности павлогского понимания охранительной и целебной
роли торможения в нормальной и патологической деятельности большого
мозга я предложил говорить сб охранительной роли торможения, или об
охранительном торможении, когда оно возникает в усло-виях нормальной
деятельности большого мозга, и сб охранительно-целебней роли торможе-
ния, или сб охранительно-целебном торможении, когда оно появляется при
патологических состояниях большого мозга.
Павловская концепция сб охранительной и охранительно-целебной роли
процесса торможения является новым словом в биологии и ценнейшим вкла-
дом в физиологию и в медицину. В частности, эта концепция лучше других
ранее существовавших и ныне существующих освещает механизмы и
природу действия так называемой сонно-наркозной терапии психических
и нервных заболеваний, является лучшей теоретической основой этого ста-
ринного эмпирического терапевтического направления иоткрываетширокие
перспективы для его дальнейшего развития. Эта глубокая и оригинальная
концепция гениального мыслителя представляет ценнейшую часть его мно-
гостороннего научного творчества, а значит, и оставленного нам, его учени-
кам и последователям, драгоценного научного наследия. И наш долг перед
страной, перед народной наукой, перед бессмертным учителем — не только
беречь это наследство, но и творчески развивать его дальше, уделяя при
этщя должное внимание важной органическом его части — идее сб охрани-
тельной и охранительно-целебной роли процесса торможения.
* *
*
Существует еще много вопросов, связанных сфмзиологией и патологией
охранительной и целебной роли торможения, которые нуждаются в даль-
нейшей теоретической и экспериментальной разработке. К числу их относит-
ся, например, вопрос сб интимной природе этого торможения; вопрос об
идентичности или неидентичности его координационному торможению;
вопрос об особенностях обмена веществ и энергии в клетках, охваченных
этим торможением; вопрос о месте возникновения этого торможения в мак-
ро- и микроструктурных образованиях нервной системы, а быть может,
и в пределах всех возбудимых тканей организма и г. д.
1 И. П. Павлов, Двадцати летний опыт, 1938, стр. 622.
438
Э. А. АСРАТЯН
Имеется немало неразрешенных вопросов и в практическом применении
этой концепции И. П. Павлова, т, е. в практике терапии сном патологи-
ческих состояний нервной системы К ним относится, например, вопрос об
уточнении круга патологических состоянии большого мозга, при которых
допустимо и даже показано применение терапии сном; кардинальный воп-
рос о возможности применения этой терапии при патологических состояниях
других отделов центральной нервной системы; вопрос об оптимальной до-
зировке, глубине и длительности лечебного сна в случаях, где применение
его показано; вопросы выбора наиболее действенных снотворных веществ,
их комбинаций с другими терапевтическими факторами и т. д. Совершенно
бесспорно, что всесторонняя разработка этого комплекса взаимно связан-
ных актуальных вопросов дала бы многое как для теоретического освещения
проблемы, так и для практического применения терапии сном. Это не
означает, однако, что углубленное и систематическое изучение отдельных
частных вопросов указанного комплекса лишено смысла и не в состоянии
дать ощутимые результаты для дальнейшего развития теории и практики
терапин сном. Я за последние Годы занимался теоретической и экспери-
ментальной разработкой только одного из поставленных мной вопросов.,
и полученные результаты в основном оправдали мои надежды.
Известно, что экспериментальные данные павловских лабораторий об
охранительной и целебной роли торможения, а также соответствующие тео-
ретические концепции И. П. Павлова относятся лишь к деятельности боль-
шого мозга: нормальной или патологической, обусловленной преимущест-
венно функциональными поражениями. Далее известно, что до сих пор так
называемая сонно-наркозная терапия в клинике применялась также в основ-
ном для лечения патологических состояний большого мозга, обусловленных
опять-таки преимущественно его функциональными поражениями.
Возник, однако, вопрос; ограничивается ди охранительно-целебная роль
торможения только пределами большого мозга при нормальной его дея-
тельности и при его функциональных поражениях?
В процессе систематической теоретической и экспериментальной работы
в области физиологии и патологии нервной системы у меня постепенно сло-
жилось впечатление, что охранительное н целебное значение процесса тор-
можения выявляется: 1) не только в деятельности большого мозга, но и
в деятельности всей нервной системы (возможно, даже в деятельности всех
возбудимых тканей организма) и 2) не только при функциональных пора-
жениях большого мозга и различных других отделов нервной системы (или
возбудимых тк-аней вообще), но и при органических их поражениях. Соот-
ветственно этому у меня созревала идея о возможности дальнейшего раз-
вития концепции И. П. Павлова об охранительной и целебной роли процесса
торможения: во-первых, полиции значительного расширения границ при-
ложения этой концепции, хотя бы в пределах нервной системы, и, во-вто-
рых, по линии ее приложения не только к функциональным, но и к органи-
ческим— травматическим поражениям различных отделов нервной системы
и организма вообще. Мысли в этом направлении в осторожной форме я вы-
сказывал в опубликованных мной работах, начиная с 1933 г., но более кон-
кретное выражение они приобрели лишь в предвоенные годы.
Что послужило основанием для этих гипотетических положений? Глав-
ным основанием было общепризнанное теоретическое положение о близости,
сродстве или тождестве основных закономерностей деятельности всех
разделов нервной системы. И. П. Павлов это положение охрактеризовал
как «естественную общность основных отношений» нервных процессов
и явлений. В качестве же иллюстрации этой «естественной общности» от-
дельных закономерностей деятельности и функциональных особенностей
коры большого мозга, межуточного, среднего, продолговатого и спинного
мозга, периферических нервных образований и даже слюнных желез идру-
ОХРАНИТЕЛЬНАЯ И ЦЕЛЕБНАЯ РОЛЬ ТОРМОЖЕНИЯ
439
г их рабочих органов (иррадиация, суммация, проторение путей; индукция,
фазы и градации возбуждения и торможения, истощения и т. д.) И. П. Пав-
лов и его ученики, а также многие ученые других школ и направлений
представили многочисленныефактичсскиедаиные[И. М. Сеченов, Н. Е. Вве-
денский, К. М. Быков, А. А. Ухтомский, А. Д. Сперанский, Д. С. Фурсн-
ков, И. П. Разенков, Ю. В. Фольбюрт, Н. А. Подкопаев, Л. А. Орбели, Лу-
чиани (Luciani), Гольц (Goltz), Магнус (Magnus) и др.]. У И. П. Павлова
встречаются отдельные высказывания о том, что и запредельное торможе-
ние имеет свой аналог в низших отделах центральной нервной системы. Он
писал: «Надо принимать, что этому торможению находится аналоги в
низших отделах центральной нервной системы»1. В развитие этой его
мысли я еще лет 10 назад указал на сходство и даже тождество «явления
извращения» или торможения спинальных рефлексов при сильных раз-
дражениях чувствительных нервов,«пессимума»Н. Е. Введенского и«запрс-
дельного (т. е. охранительного) торможения» И. ГЕ Павлова.
Основанием же для предположения, что охранительная и целебная
роль торможения выявляется не только при функциональных поражениях
различных частей нервной системы, но и при органических—травматических
их поражениях, послужил громадный лабораторно-экспериментальный и
клинический материал. При оперативном или иного рода травматическом
воздействии на центральную нервную систему выпадают или нарушаются
не только функции непосредственно поврежденных участков, по и функции
таких ее участков, которые находятся топографически близко или даж:е зна-
чительно удалены от очага прямого органического повреждения субстрата.
На протяжении более 100 лет это явление наблюдалось н изучалось
большим числом экспериментаторов и клиницистов. Именовали это явление
центральным шоком, спинальным шоком, торможением, диасхизисом и.т. п.
п трактовали его как результат сотрясения мозгового субстрата, циркуля-
торных нарушений, локального или общего отека мозговой массы, как ре-
зультат рубцеваний, нарушений субординационных отношений, косвенной
травмы, дегенерационного раздражения со стороны травмированных участ-
ков и т. д.
Работая в течение ряда лет над проблемой пластичности нервной системы,
я имел возможность совместно со своими сотрудниками (А. И. Кар амин,
А. А. Фадеева, Я. М. Пресеман, И. В. Введенская, Р. О. Барсегян,
Д. И. Ханутина, Б. Д. Стсфанцов, Ф. М. Шитов, В. Д. Дмитриев, Г. И. Бо-
ков и Др.) наблюдать и исследовать эти явления в. самых различных со-
стояниях, вызванных различного рода оперативными воздействиями на
большой мозг, мозжечок, продолговатый и спинной мозг, на органы чувств
и периферические нервы рептилий, птиц и млекопитающих. Анализ обшир-
ного литературного материала и собственные наблюдения привели меня
к заключению, что среди комплекса взаимно связанных факторов, обуслов-
ливающих нарушение функций непосредственно травмой не задетых частей
нервной системы, важную роль играет процесс торможения и что в отноше-
нии этих нервных образований он играет роль охранительного и целебного
фактора. Если учесть, что действие всякого органического повреждения
нервной системы прямо или косвенно включает две основные, в понимании
И. П.Павлова, предпосылки для возникновения охранительно-целебного
торможения — ослабление нервной системы и ее перераздражение, —то
нельзя отрицать правдоподобность сделанного мной выше предположения.
Следует отметить, что в трудах И. П. Павлова встречаются близкие к
этому предположению высказывания. Он считал, что «после повреждения
определенных мест коры больших полушарий» раздражение соответствую-
1 И. П. Павлов, Двадцатилетпий опыт, 1938, стр. 608.
440
Э. А. АСРАТЯН
щих поврежденным зонам рецепторов вызывает охранительное тормо-
жение.
Вот в кратком изложении те фактические данные и теоретические пред-
посылки, которые породили мысль о возможности и необходимости дальней-
шего развития концепции И. П. Павлова. Они легли также в основу выска-
занных мной гипотетических положений об универсальности охранитель-
ной и целебной роли торможения для всех частей нервной системы и не
только в условиях нормальной их деятельности, не только при патологиче-
ской их деятельности, вызванной функциональными воздействиями на
них, НО и при патологической деятельности, обусловленной их непосредст-
венной органической травмой.
Я глубоко сожалею, что слишком поздно — только к началу Великой
Отечественной войны — приступи.’! к специальной экспериментальной про-
верке этих гипотетических положений и к выяснению возможности исполь-
зования их в практике здравоохранения нашей страны.
На первых этапах исследовательской работы по этой проблеме мне
удалась более или менее обстоятельно изучить лишь некоторые весьма ак-
туальные вопросы, связанные с этиологией, патологией и терапией травмати-
ческого иожогового шока, развивающегося при таких травматических воз-
действиях на организм, при которых поражения различных отделов цент-
ральной нервной системы, за редким исключением, носят в основном функ-
циональный характер.
Широкий план теоретической, экспериментальной и клинической работы,
намеченный мной в указанном выше направлении, далеко не полностью ине
реализован. Но те скромные результаты, которые мной были получены по
травматическому пожоговому шоку и будут кратко изложены в этой работе,
а также данные, полученные мной и моими сотрудниками — Г. Т. Сахиули-
ной, Е. В. Гуровой, Е. А. Романовской и Л. В. Симуковой об органических
поражениях различных отделов нервной системы, все больше убеждают
в правильности высказанных мной гипотетических положений и в возмож-
ности успешного-приложения мыслей и концепций великого Павлова к част-
ным областям или даже отдельным вопросам физиологии и медицины, ис-
следованием которых он ие занимался.
ПРОФИЛАКТИКА И ТЕРАПИЯ ШОКА»
чхроблема шока, еще до войны занимавшая внимание многих
Я советских ученых, в настоящее время стала одной из централь-
1 пых тем наших научных исследований Я попытался разрабо-
тать новый способ борьбы с шоком, в особенности с наиболее
/2) тяжелыми и грозными его формами. Речь идет прежде всего
о глубоких стадиях вторичного травматического шока.
Поздние стадии его представляют собой многостороннее и сложное па-
тологическое состояние, в структуре которого ярче всего вырисовываются
три главные взаимодействующие черты. Это, во-первых, истощение и ослаб-
ление центральной нервной системы, затем глубокие изменения в составе
крови, в эффективном ее объеме, в функциях сосудистой системы, в особен-
ности капилляров, и, наконец., наводнение организма токсическими продук-
тами гнстолигнчсского происхождения и продуктами нормального н нару-
шенного обмена веществ. Соответственно с этим я считал, что для успешной
борьбы с шоком следует пользоваться таким комплексным средством, ко-
торое включало бы адэкватные терапевтические факторы для каждого из
этих основных элементов функциональной патологии позднею шока. Нельзя
согласиться с исследователями, которые обращают внимание па какую-
нибудь одну сторону патогенеза шока (скажем, на изменения в нервной си-
стеме, в гемодинамике или в составе крови и тканевой среды и т. п.) и соот-
ветственно с этим для его лечения применяют какой-нибудь один терапевти-
ческий фактор, например, соду (Кэннон, Кунз и др.), хлористый каль-
ций (Лапонте), фосфорнокислый калий (Г. Я. Хголес и др.), симпатомиметин
(И. П. Чукичев), отдельные снотворные или наркотические вещества (Са-
Еори, М. А. Топчнбашев, И. Р. Петров и др.), различные гормоны, витамины,
к ровоза мешающие растворы и т. д. Эта экспериментальные и клинические
исследования оставляли в стороне данные многолетней практики борьбы с
шоком, гоЕорящие именно о преимуществе комбинированной терапии шока.
Те же экспериментаторы, которые рекомендуют врачам в борьбе с шоком
комбинировать те или иные известные противошоковые средства, сами в ус-
ловиях лабораторного эксперимента на животных предварительно не дока-
зали действительного преимущества или хотя бы фактическую возможность
сочетания этих средств (И. Р. Петров и др.), а исходили из опытов, постав-
ленных с каждым ингредиентом в отдел ьности. Вот почему такие предложе-
ния также нельзя считать удовлетворительными.
Для определения подлинной ценности любого нового противошокового
средства необходимым условием является предварительная строгая и адэк-
ватная проверка его эффективности в условиях лабораторного эксперимен-
та на животных.
К этому вопросу не все наши экспериментаторы подходят с требуемой
строгостью. Некоторые из предложенных нашими исследователями
(Г. Я. Хвюлес, И. П. Чукичев) противошоковых средств (фосфорнокислый
1 «Медицинский работник», № 24 (502), 17.VI 1943 г.
442
Э А. АСРАТЯН
калий, симпатомиметин) в условиях лабораторного эксперимента испыты-
вались лишь при раннем, или так называемом первичном, шоке. Между тем
эту стадию шока по патогенезу, практической значимости и по трудности
лечения никак нельзя сравнить с поздней стадией, или так называемым вто-
ричным, шоком, в. состоянии которого больные, как правило, попадают в
лечебные учреждения как в условиях войны, так и в условиях мирного вре-
мени, Мне кажется, что именно это является главной причиной значитель-
ного расхождения между данными экспериментальной терапии животных,
полученными названными авторами, и данными врачей, применявших эти
средства у шоковых больных. Нельзя также считать строгими испытания,
проведенные М. А. Топчибашевым и И. Р. Петровым с предложенными ими
противошоковыми средствами, так как М. А. Топчнбашев и его сотрудники
эфиро-масляную смесь назначали больным, находящимся в состоянии сред-
него и легкого шока (примерно с максимальным кровяным давлением 90 мм),
а И. Р. Петров и его сотрудники гексенал, амитал-натрий с паральдегидом,
хлоролозу, 10% спирт и 10% глюкозу, 10 % раствор бромистого натрия раз-
дельно вводили кроликам также при легких передних степенях эксперимен-
тального шока (вещества эти испытывались при введении per os, рог rectum
и внутримышечно; 54% контрольных животных выживало без всякой
помощи; выживаемость же леченных животных была всего на 24% выше).
Наконец, нельзя считать полноценным испытание новых противошоковых
средств при экспериментальном шоке, вызванном различными химическими
средствами, манипуляциями на кишечнике и т. д. (Г. Я. Хволес, А. Ф. Лепу-
калн и др.), так как эти виды шока не могут считаться адэкватными общеиз-
вестному и распространенному виду травматического шока у человека.
Исходя из своих теоретических установок о борьбе с шоком путем комби-
нации некоторых взаимодействующих терапевтических факторов соответ-
ственно основным патогенетическим особенностям шока, я попытался при-
менить в качестве фактора для восстановления истощенных и астеничных
центров снотворные вещества и бромиды, вопреки господствующей в настоя*
шее время тенденции пользоваться возбуждающими средствами. Это решение
исходило в основном из концепции И. П. Павлова об охранительной и целеб-
ной роли торможения — сна для истощающихся и истощенных нервных цент-
ров.
Совершенно естественно было ожидать, что такие средства, как хлоро-
форм, эфир, гексенал, амитал-натрий, введенные в организм тяжелого шо-
кового больного в виде паров или концентрированных растворов, резко
ослабляют и без того ослабленные до критического уровня жизненно важные
функции организма (кровяное давление, дыхание, окислительные процес-
сы, теплорегуляция и др.) и приведут, скорее, к ухудшению общего состоя-
ния организма, чем к восстановлению истощенных нервных центров. Наши
опыты показали, что даже наиболее подходящие для этой цели снотворные—
гедонал и уретан — в подобных случаях шока и при том же способе приме-
нения дают примерно такие же неутешительные конечные результаты, прав-
да, в несколько затяжной и мягкой форме. Выход из положения был найден
в полном отказе от неподходящих наркотиков и сильных снотворных и в ис-
пользовании наиболее пригодных для нашей цели препаратов (гедонал, уре-
тан, в меньшей мерс паральдегид, веронал и некоторые другие), а главное —
в сочетании этих средств с такими терапевтическими факторами, которые
были бы способны не только препятствовать развитию угнетающего дейст-
вия снотворных на жизненно важные функции организма, но даже активизи-
ровать их.
В результате экспериментальной работы, проведенной со строгим учё-
том особенностей изменений в сосудистой системе, в составе крови и тканей
при позднем шоке, а также фармакодинамических свойств ряда химиче-
ских препаратов, была Составлена жидкость для внутривенного вливания.
ПРОФИЛАКТИКА И ТЕРАПИЯ ШОКА
443
Грубая характеристика ее такова. По сравнению с рингер-локковским раст-
вором жидкость примерно в три раза гипертоничнее (главным образом за
счет двукратного увеличения дозы поваренной соли и двадцатнкрагного
увеличения глюкозы), это помогает возможно дольше удерживать жидкость
в крови, ослабить отек тканей и набухание эритроцитов, усилить жажду
и водно-солевой обмен. Доза двууглекислого натрия увеличена в 10 раз для
нейтрализации ацидоза. Более чем в 20 раз увеличена н доза хлористого
кальция,чтобы тонизировать и уплотнить капилляры и сбалансировать вред-
ную гиперкалиемию, возникающую в поздних стадиях шока. По тем же
соображениям в настоящее время совсем изъят из состава жидкости
хлористый калий. Для углубления торможения в нервной системе и раз-
вития сна в качестве главных лечебных средств в противошоковую жид-
кость введены бромиды и снотворные вещества. В тех же целях, а главное
для консервации органических веществ в жидкость введен алкоголь. Мы
добиваемся также увеличения вязкости жидкости путем прибавления к ее
составу безвредных коллоидных веществ.
Противошоковые терапевтические качества нашей жидкости были под-
вергнуты строгому испытанию при участим Л. В. Симуковой более чем
в 200 опытах, поставленных частично на кроликах, а главным образом на
кошках и собаках. Шок вызывался у ненаркотизированных животных силь-
ной травматизацией лап, сочетаемой в ряде опытов с кровопусканием. Па
фоне глубоких степеней вторичного шока, когда кровяное давление падало
в три-четыре раза по сравнению с исходным уровнем, внутривенно медленно
вливалась жидкость. В то время как все бел исключения контрольные жи-
вотные погибали вскоре после наступления глубокого необратимого вторич-
ного травматического шока, более 80 животных, получивших жидкость
внутривенно, были выведены из шока и спасены.
Экспериментальная часть работы была начата в сентябре 1941 г., закон-
чена и доложена в марте 1942 г. После этого на пунктах скорой помощи., а в
основном B.Q фронтовых санитарных учреждениях мной при участии воен-
врача И. В. Каменской жидкость была введена более чем 100 больным, на-
ходившимся в состоянии тяжелого вторичного травматического шока, ВЫЗ-
ВАННОГО городской или военной травмой (апрель—октябрь 1942 г.). В слу-
чаях неосложненного травматического шока даже очень большой глубины
(при максимальном кровяном давлении 50—60 мм и ниже) жидкость давала
даже более эффективные результаты, чем в опытах на животных.
Вот общая картина действия жидкости. Немедленно после вве-
дения начинается повышение общего и пульсового кровяного давления,
которое через 15—20 минут достигает двух третей нормального уровня
у животных и почти нормального у человека. Быстро наступает улучшение
дыхания по глубине, ритму и частоте, улучшается цвет кожи, а также цвет
артериальной крови и ее свертываемость; прекращается потоотделение, сужи-
ваются зрачки. Вскоре после начала вливания происходит оживление дви-
гательной и чувствительной а|>еры: люди начинают жаловаться на боли
в пораженных частях тела, оживленно двигаются, охотно разговаривают,
констатируют улучшение своего состояния, становятся бодрее. Через 15—
20 минут после начала введения на фоне дальнейшего развития этих сдви-
гов незаметно появляется и усиливается дремотное состояние, переходящее
потом в сон, который очень напоминает естественный, может быть легко прер-
ван и восстановлен. Длительность его в среднем равна 3—4 пасам. В стадии
глубокого сна описанные выше положительные сдвиги бывают несколько
ослаблены, но вскоре после пробуждения вновь выравниваются и даже
усиливаются. Резко возрастают жажда и усвояемость воды. Люди
многократно и обильно пьют воду еще.до наступления сна. Заснув они
часто просыпаются, пьют воду и опять засыпают.
Травматический шок по своей функциональной патологии очень близок
4)4
Э. А. АСРАТЯН
к ожоговому. Поэтому я при участии лаборантов Л. В. Снмуковой и
П. В. Станкевича в условиях лабораторного эксперимента на животных
провел испытание жидкости для лечения ожогового шока. Результаты
оказались такими же, как и при травматическом шоке.
Физиологический анализ действия жидкости и конечных результатов
ее применения позволяет считать, что под влиянием гласным образом сна
происходит постепенное функциональное восстановление истощенных и
астеничных жизненно важных нервных центров, в результате чего они
в свою очередь, несомненно, усиливают нервную регуляцию трофики и функ-
ций пострадавших от шока органов и тканей, как бы завершая этим самым
круг взаимодействующих восстановительных процессов.
Руководящие теоретические установки данной работы, основанные на
глубоких павловских концепциях об охранительной и целебной роли тор-
можения, поднимают на более высокую ступень старую забытую практику
терапии шока наркотическими веществами.
В свете этих концепций становится понятным не только профилактиче-
ское, но и терапевтическое действие седативных, снотворных и наркотиче-
ских веществ при травматическом шоке. Описанные же выше экспери-
ментальные И клинические данные получены в результате попытки внести
новый момент в практику терапии шока сном путем сочетания более
удачно подобранных снотворных веществ с факторами, направленными
на восстановление нарушенной при шоке гемодинамики, а такж:е химизма
и состава крови и тканей.
УЧЕНИЕ И. П. ПАВЛОВА ОБ ОХРАНИТЕЛЬНОЙ’И ЦЕЛЕБНОЙ
РОЛИ ТОРМОЖЕНИЯ И ТРАВМЫ НЕРВНОЙ СИСТЕМЫ1
нашей физиологической и клинической литературе обычно
упоминается только концепция И. П. Павлова об охранитель-
ном значении торможения,а при трактовке результатов терапии
сном психических и нервных заболевании обычно учитывается
только охранительная сторона торможения (см., например,
работы проф. А. Г. Иванова-Смоленского и сотрудников). Та-
кое понимание взглядов И. П. Павлова является односторонним и не-
полным. При таком понимании роль торможения в восстановлении функции
пораженных нервных центров ограничивается только охраной, блокирова-
нием или ограждением истощающихся и истощенных нервных центров от
дальнейшего вредного воздействия патогенных стимуляций, т. е. тормо-
жению отводится значение вспомогательного «пассивно-защитного», косвен-
ного и второстепенного фактора ла фоне каких-то других более эффектив-
ных и активных факторов, играющих ведущую роль в восстановлении
функций истощенных и ослабленных нервных центров (сдвиги в вегетатив-
ной нервной системе, в составе крови и т. д.). Если же учесть не только
охранительную, но и особую целебную роль торможения, т. е. приписать
ему значение активного, эффективного, самостоятельного терапевтического
фактора, играющего при восстановлении функции пораженных нервных
центров в комплексе других факторов не второстепенную, а ведущую
роль, то этр, с моей точки зрения, полнее и вернее отразит взгляды
И. П. Павлова по обсуждаемому вопросу.
В соответствии с таким пониманием взглядов И. II. Павлова на Охрани-
тельную и целебную роль торможения я позволяю себе сделать одно пред-
положение. Мне кажется, что в целях установления терминологической
четкости и более точной передачи сущности концепции И. П. Павлова было
бы целесообразнее говорить об охранительно и роли торможения,
или об охранительном торможении, когда речь идет о его возникновении
в условиях нормальной деятельности большого мозга, и сб о х р а н и -
тел ь но-цсл еб ной роли торможения, или об охранительно-целеб-
ном торможении, когда речь идет об его возникновении при патологиче-
ских состояниях большого мозга.
Было бы легкомысленно утверждать, что все вопросы, связанные с ох-
ранительной и в особенности с целебной ролью торможения, уже исчерпы-
вающе ясны даже в ч исто теоретич еском аспекте. 11 асборот, многие вопросы,
в том числе и вопрос об интимной природе охранительного и целебного дей-
ствия торможения, вопрос об особенностях обмена веществ и энергии в
клетках, охваченных торможением, вопрос о месте возникновения и локали-
зации этого торможеиия в м икро и макроструктурах нервной системы,вопрос
1 Доклад, прочитанный 3,Х 1Q4? г, на конференции, посвященной проблемам
высшей нервной деятельности, созванной в Москве в связи с 94-й годовщиной
рождения акад. И. П. Павлова. «Невропатология и психиатрия», № 4. 1944.
446
Э. А. А6РАТЯН
о рамках применения этой концепции и многие другие до сих пор еще не вы-
яснены. Четких и ясных ответов на эти и подобные вопросы, относящиеся
к интимной природе основных нервных процессов, к сожалению, не имеется
не только в трудах И, П, Павлова (который, кстати говоря, не ставил себе
задачей исследование нервной деятельности в этой плоскости), но и в трудах
тех исследователей, которые проводили и проводят Специальные исследова-
ния в этом направлении. Однако едва ли кто станет отрицать возможность
оперирования бесспорными фактами, явлениями и понятиями, относящимися
к охранительной и целебной роли торможения (как и вообще к торможению,
возбуждению и другим элементарным свойствам нервной системы), лишь
на том основании, что интимная природа этих процессов наукой еще не рас-
шифрована. Отрицание такой возможности и необходимости противоречило
бы всему ходу развития современной физиологии и науки вообще, противо-
речило бы диалектическому пониманию процесса познания. Разбирая воп-
рос о возможных плоскостях изучения жизненных явлений, И. П. Павлов,
например, одну из них характеризовал так: «Охватывая деятельность слож-
ных конструкций во всем их действительном объеме, можно отыскивать стро-
гие правила этой деятельности, пли, что то же, констатировать все те усло-
вия, которые точно определяют течение деятельности во всех ее моментах
и вариациях»1. Указывая далее, что в своих исследованиях он сам стоял eia
такой именно плоскости, И. П. Павлов продолжал: «Наэтой плоскости наше
внимание не занимал вопрос о том, чтб такое раздражение и торможение в их
последнем глубоком основания. Мы брали их как два фактических данных...
Мы даже не останавливались подробно на вопросе о возможном размещении
двух основных явлений этой деятельности—раздражения и торможения—по
двум элементам нашей конструкции: нервным клеткам и соединительным
пунктам или путям между нервными клетками, удовлетворяясь временным
предположением, что то и другое суть функции нервных клеток»2.
Имеется очень много неразрешенных вопросов и в практическом приме-
нении концепции И. П. Павлова об охранительно-целебной роли торможе-
ния, т. е. в практике терапии сном патологических состояний нервной си-
стемы. К ним относится, в частности, вопрос о границах применения
терапии сном при всевозможных патологических состояниях большого моз-
га, вопрос о возможности проведения этой терапии при патологических со-
стояниях различных других частей центральной нервной системы, вопросы
дозировки лечебного сна по глубине и длительности в тех или иных пока
занпых случаях, вопрос о выборе снотворных веществ, их комбинации
с другими лекарственными средствами и т. д.
Совершенно очевидно, что разрешение перечисленных выше и других
теоретических и практических вопросов, составляющих важную часть гро-
мадного и многогранного научного наследия И. П. Павлова, потребует
интенсивного научно-творческого труда его учеников и последователей.
Считая своим долгом ученика перед памятью незабвенного учителя Ивана
Петровича принять возможно активное участие в разработке и дальнейшем
развитии его научного наследия, я в последние годы занимался теоретиче-
ским и экспериментальным изучением ряда вопросов, которые органически
Связаны с изложенными выше концепциями И. П. Павловаоб Охранитель-
ной и вособенности об охранительно-целебной роли торможения. В весьма
сжатом изложении результаты этой работы будут приведены ниже.
• •*
* •
Известно, что свои концепции сб охранительной и охраннтельно-целеб-
ной роли торможения И. П. Павлов развил применительно к нормальным
1 И. П. Павлов, Лекции о работе больших полушарий головного мозга,
1927. стр. 328.
® Та м же.
УЧЕНИЕ И. П, ПАВЛОВА ОБ ОХРАНИТЕЛЬНОЙ И ЦЕЛЕБНОЙ РОЛИ ТОРМОЖЕНИЯ 447
и патологическим состояниям большого мозга. Известно также, что к боль-
шому мозгу относится и терапия сном психических и нервных заболева-
ний.
Еще задолго до начала Великой Отечественной войны, в особенности в
тотпериод, когда я совместно со своими сотрудниками занимался исследова-
нием некоторых сторон проблемы пластичности нервной системы на основе
учения И. П. Павлова о функциях большого мозга, у меня возникла идея
о возможности расширения рамок теоретического и практического приме-
нения концепции И. П. Павлова сб охранительной и охранительно-целеб-
ной роли торможения, о возможности распространения этих концепций
на всю центральную нервную систему.
Эта идея своими корнями тесно связана с теоретическим положением о
генетическом сродстве, о внутренней связи и сходстве основных закономер-
ностей длительности всех важненш их отделов центральной нервной системы.
Давая общую характеристику своей многолетней работы по физиологии
большого мозга, И. II. Павлов писал: «Нельзя не видеть очень много пунк-
тов, где наши факты, полученные при исследовании больших полушарий,
совпадают с фактами физиологии спинного мозга, что свидетельствует сб
естественной общности основных отношений и там и здесь»1. Известно,
что эта «естественная общность» подчеркивалась И. П. Павловым
и в отношении гипнотических фаз коры большого мозга и изученных
Н. Е. Введенским парабиотических стадий простых нервных образований.
В своих исследованиях по физиологии большого мозга и центральной
нервной системы вообще я постоянно руководствовался этим основным
принципом даже при изучении специфических особенностей той или иной,
структурно и функционально обособленной части центральной нервной си-
стемы. В частности, руководствуясь положением И. П. Павлова об «есте-
ственной общности основных отношений» и следуя примеру некоторых стар-
ших учеников И. П. Павлова, каждый из которых в своей области проводил
и проводит эту установку учителя, я еще лет 10 назад указал на сродство
«запредельного торможения» И. П. Павлова, пессимума Н. Е. Введенского
и явления торможения спинальных рефлексов при сильных раздражениях
чувствительных нервов, а несколько позже применил принцип «относитель-
ной функциональной подвижности (лабильности)» Н. Е. Введенского к уче-
нию И. П. Павлова о работе большого мозга. Если я раиыие указывал на
пессимум и торможение рефлексов как на проявления охранительного тор«
можения в пределах нормальной деятельности низших нервных образований,
то по ход}' дальнейших исследовании проблемы пластичности нервной си-
стемы, как уже было сказано, у меня появилась идея о возможности приме-
нить павловскую концепцию об охранительно-целебной роли торможения к
патологическим состояниям низших отделов центральной нервной системы.
Непосредственным толчком к мысли о возможности более широкого
распространения этих павловских концепций послужило следующее. В це-
лях изучения явлений пластичности нервной системы я и мои сотрудники
на протяжении ряда лет производили различные операции на нескольких
сотнях животных (броун-секаровское сечение спинного мозга, перерезка
задней его половины, частичное его расщепление, деаффереитация конеч-
ностей, перекрестное сшивание различных нервов, разрушение лабирин-
тов, ампутация отдельных конечностей, частичная или полная экстирпация
коры большого мозга или мозжечка и т. д.). Наблюдения за этими животны-
ми привели меня к заключению, что торможение, которое развивается
в близких и отдаленных от очага поражения частях центральной нервной
системы и которое в системе других факторов травмы участвует в создании
* И. П. Павлов, Лекции о работе больших полушарий головного мозга,
1927, стр. 329.
448
Э. А. АСРАТЯН
сложной картины функциональных нарушений центральной нервной систе-
мы, имеет в значительной мере ту же физиологическую природу и значение,
что и при различных хронических гипнотических состояниях большого
мозга, обусловленных его функционал ьными поражениями,т. е. имеетхарак-
тср охранительно-целебного торможения. Такое предположение было тем
вероятнее, что по своему прямому и косвенному действию травма включает
две основные предпосылки для возникновения и развития охранительно-
целебного торможения: ослабление нервной системы и ее перераздражение.
Для экспериментальной проверки этих предположении я решил испы-
тать влияние снотворных веществ на течение компенсаторных процессов
и возникновение новых форм координационной деятельности у оперирован-
ных упомянутыми выше способами животных. Я рассчитывал на то, что
при правильности исходных теоретических предпосылок эти средства
путем углубления охранительно-целебного торможения (а в иных случаях
путем его создания) ускорят восстановление функции истощенных клеток,
темпы компенсации и образования новых форм координационной деятель-
ности и, возможно, поднимут их на более высокий уровень.
При проведении терапии сном травматических поражений различ-
ных отделов нервной системы я считал необходимым строго Соблюдать
следующие правила: 1) избегать шаблона в применении этой терапии
и Дозировать лечебный сон по его глубине и длительности соответственно
локализации, давности и характеру поражения нервной системы; 2) по-
добрать такие снотворные вещества, которые вызывают сон, возможно
близкий к естественному, и которые максимально свободны от побочных
вредных воздействий на жизненно важные функции организма, в част-
ности, на нервную систему; 3) кроме истощения нервной системы, выз-
нанного травмой, точно учесть другие основные патогенетические черты
каждого вида болезненного состояния и соответственно этому сочетать
снотворные вещества как фактор лечения истощенных центров с другими
лечебными факторами, направленными против других сторон патогенеза
заболевания.
Травматический шок. Результаты моей теоретической, экс-
периментальной и клинической работы по лечению травматического шока
более или менее подробно освещены в ряде моих других докладов и статей,
поэтому здесь я привожу лишь основные данные.
В соответствии с изложенными выше принципами в качестве основного
лечебного фактора для истощенных и ослабленных центров я использовал
снотворные вещества, тогда как многие ученые предлагают для этой цели
возбуждающие средства. Вообще говоря, применение снотворных и нарко-
тических средств в борьбе с шоком не ново. Не говоря уже о двухвековой
давности применения опия и его препаратов-для обезболивания больных,
находящихся в состоянии шока, укажу на то, что, начиная со второй поло-
вины прошлого столетия, многие ученые с той же целью эпизодически при-
меняли наркотические или сильные снотворные средства (хлороформ, эфир,
ими тал-иа трий, гексенал и др.).
Вводя в ослабленный при шоке организм больного вещества, облада-
ющие выраженным угнетающим действием на жизненные функции орга-
низма, к тому же применяя их в чистом или концентрированном виде,
эти ученые, естественно, не могли добиться благоприятных результатов
при тяжелом шоке, а могли получать положительные результаты лишь
при предупреждении шока или при лечении легких и в лучшем случае
средних его степеней.
В своей работе я исходил из иных теоретических предпосылок и со-
ответственно этому преследовал иную цель, а именно: добиться углубления
охранительно-целебного торможения в центральной нервной системе шоко-
вых больных, при этом не подавляя, а повышая уровень жизненных фунК-
УЧЕНИЕ Н.П. ПАВЛОВА ОБ ОХРАНИТЕЛЬНОЙ И ЦЕЛЕБНОЙ РОЛИ ТОРМОЖЕНИЯ 449
ций организма. Исходя из сказанного, я, во-первых, отказался от пере-
численных Еыше средств, а использовал снотворные вещества из группы
карбаминовой кислоты (резонал и уретан), которые, поданным фармаколо-
гии и клиники, почти лишены вредных побочных свойств. Далее, я вводил
их в организм не в чистом или концентрированном виде, а в составе комп-
лексной жидкости, обладающей и другими лечебными свойствами.
В результате мне удалось достигнуть более эффективных результатов
не только при профилактике шока или при лечении легких и средних его
степеней, но, что особен но важно, и при лечении глубоких степеней позд-
него шока.
Кроме снотворного, как указано, предложенная мной жидкость содержит
ряд других лечебных факторов, которые представлены резкой ее гипертони-
чностью (для ослабления отека тканей, набухания эритроцитов, усиления
жажды и усвояемости воды, увеличения объема и тока крови и т. д.), большой
дозой хлористого кальция (Для Тонизирования И уплотнения капилляров,
для нейтрализации вредной гиперкалиемии л стимуляции сердечно-сосуди-
стой системы вообще) и'бикарбоната натрия в ее составе (для нейтрализации
ацидоза и увеличения щелочного резерва). По замыслу перечисленные
сдвиги на фоне наступающего сна должны были обеспечить резкое и до-
статочно стойкое повышение кровяного давления, значительное усиление
органического, водно-.сслевого и газового обмена и в результате этого
косвенно ослабить аноксию, азотемию, токсемию и другие изменения,
произошедшие в тканях и крови.
Ожоговый и тепловой шок. Исходя из общепризнанного
положения, что ожоговый шок по патогенезу и симптоматологии очень
близок к травматическому (некоторые ученые говорят даже об их идентич-
ности), я и мои лаборанты Л. В. Самукова и II. В. Станкевич испытали те-
рапевтическое действие жидкости также в отношении ожогового шока на
двух десятках ссбак. Выяснилось, что жидкость не выводит животных из
теплового шока, и они, как и контрольные, все без исключения вскоре
погибают. Тогда я изменил состав жидкости, учтя одну важную особенность
патогенеза теплового шока, упущенную мной вначале, — резкое обезвожи-
вание организма. Новый состав жидкости по компонентам не отличался от
основного состава; в нее, как и в основную жидкость, входили снотворные
вещества и бром, а также хлористый кальций и сода в резко увеличенных
дозах по сравнению с раствором Рингера. От основной жидкости опа, од-
нако, отличалась тем, что не была гипертоничной, что позволило нам
вводить ее в организм животного в дозах, в три-четыре раза больших, чем
основную жидкость. Новая серия опытов с измененной жидкостью по-
казала ее высокую эффективность и при лечении теплового шока.
Органические поражения центральной нерв-
ной системы. Травматический шок и некоторые виды шока, близко сто-
ящие к нему по своему патогенезу, симптоматологии, а также по практиче-
скому значению, были до сих пор главной темой моей исследовательской ра-
боты^ проводимой с целью практической проверки теоретического положения
о возможности приложения концепции И. П. Павлова об охарннтельно це-
лебной роли торможения к травматическим поражениям нервной системы.
Те же виды органического, операционно-травматического поражения нерв-
ной системы, которые по перво начально намеченному плану должны были
быть основными объектами моей исследовательской работы с упомянутой
выше точки зрения, в силу изменившихся условий изучались мной до сих
пор скорее ориентировочно, чем в плане систематической работы. И если,
несмотря на это, я кратко излагаю здесь результаты, то только потому,
что полученные предварительные данные являются очень ободряющими.
Примерно у десятка животных я вызывал различного рода операцион-
ные травмы СпинноГо мозга (броун-секаровскоё сечение, перерезка задней
29 Э. А. Асратяи
450
Э. А. АСРАТЯН
половины и полная перерезка спинного мозга). Для однотипной операции
животных (собаки и кошки) подбирали попарно с возможно близкими пока-
зателями веса, возраста, упитанности и роста. Послеоперационный уход за
животными был по возможности одинаков, за исключением того, что конт-
рольные не получали снотворных, а подопытные три раза в день получали
дозу снотворных и бромидов, достаточную для развития и поддержания
длительного сна оптимальной глубины. В даче снотворных через каж
дые 2—3 дня делался перерыв для проведения сравнительных функцио-
нальных проб и наблюдений. В общей сложности снотворные давались в те-
чение 6—10 дней. Во всех случаях у подопытных животных рефлексы
появлялись раньше и восстановление функции пораженного спинного
мозга происходило быстрее, чем у контрольных животных.
Аналогичные результаты были получены и при применении снотворных
в комбинации с хлористым кальцием в случаях экспериментально вызван-
ного отека — набухания головного мозга у ссбак.
Сопоставляя симптоматологию отека— набухания мозга у человека
с последствиями экстирпации верхних шейных симпатических узлов у жи-
вотных, я высказал предположение, что в генезе отека — набухания мозга
важную рель играет ослабление тонуса симпатической нервной системы,
которое могло возникнуть как результат травматического поражения
высших отделов центральной нервной системы. Это допущение показа-
лось мне заманчивым не только потому, что в свое время мне удалось по-
казать известное трофическое влияние симпатической нервной системы
на состояние большого мозга, но в основном и потому, что снятие или ос-
лабление регуляции то нуса симпатической нервной системы и, как я пола-
гал, трсфики эндотелия капилляров создает Ситуацию, близкую к той, ко-
торая в свете современных данных сб этиологии и патогенезе отека— на-
бухания мозга является важной предпосылкой для развития этого болез-
ненного Состояния. С целью проверки.указанных предположений я еще в
Ленинграде, азатем в Ташкенте удалил у 14 ссбак верхние шейные симпа-
тические узлы для специального наблюдения в сравнения картины функци-
ональных расстройств большого мозга этих животных с картиной функци-
ональных расстройств того же органа у двух других ссбак, у которых
отек— набухание мозга был вызван хроническом супердуральной комп-
рессией мозга стерильными зернами фасоли. Как данные этих наблюдений,
так и анагомо-гистслогические исследования мозга четырех из симпатэкто-
мированных мной ссбак, произведенные сотрудниками Г. Н. Терехова, пока
залп, что мои предположения о важной роли снижения тонуса симпатичес-
кой нервной системы в генезеотека—набухания мозга в основном оправда-
лись.
Кроме этих предварительных экспериментальных данных на животных,
я располагаю весьма ограниченным числом предварительных наблюдений
о благоприятном действии снотворных и бромидов на острые и хронические
последствия спинальной и церебральной травмы у людей (коммоция —
контузия, ранение спинного и головного мозга, каузалгня к т. п.). Часть
наблюдений проводилась на фронте н часть в клиниках Л. Г. Шаргород-
ского и Г. М. Фрейдовича (Ташкент). За исключением одного больного
с каузалгией, у всех больных лишь удлинение и углубление ночного сна
в течение ряда дней путем систематической дачи снотворных и бромидов
вызывали заметнее улучшение их состояния и самочувствия.
* *
♦
Резюмируя свое сообщение о проведенной работе по терапии сном
шока ио предварительных наблюдениях над результатами терапии снем
некоторых видов травматических поражений центральной нервной системы,
я должен сказать следующее. Высказанное мной несколько лет назад
УЧЕНИЕ И.П. ПАВЛОВА ОБ ОХРАНИТЕЛЬНОЙ И ЦЕЛЕБНОЙ РОЛИ ТОРМОЖЕНИЯ 451
теоретическое положение о возможности расширения области теоретичес-
кого и практического применения концепции И. II. Павлова сб охрани-
тельно-целебной роли торможения, о возможности распространения этой
концепции на всю центральную нервную систему, в частности, на ее пато-
логические состояния, вызванные травматическими воздействиями, в
настоящее время уже имеет известное экспериментальное и даже клиничес-
кое обоснование. Совершенно очевидно, что проведенная мной с этой точки
зрения работа по шоку поднимает на более высокий уровень теорию и прак-
тику старого эмпирического направления в области применения седатив-
ных, снотворных и наркотических препаратов при различных видах
шока. Что же касается результатов работы по органическим травмам
центральной нервной системы, то хотя они носят предварительный
харак тер и потому лишены необходимой доказательности и убедительности,
тем не менее имеют большое значение для правильной ориентации
в будущей работе в этом направлении. Я счастлив буду сознавать, что
в какой-то мере сумел использовать идеи гениального учителя при раз-
решении теоретических вопросов и практических задач, столь актуаль-
ных в переживаемые нашей родиной дни.
29*
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКОГО ШОКА'
современной медицине существует много направлений в изу-
чении шока, а еще больше различных мер и средств, рекомен-
дованных для профилактики и терапии различных его видов, в
ч астности, травматического шока. Размеры статьи не позволяют
дать хотя бы сжатый критический анализ и оценку всех этих
направлений и. средств. Можно сказать только, что, несмотря
на большие достижения медицинской науки в распознавании патогенеза
шока и в области борьбы с ним, несмотря на наличие значительного коли-
чества старых и новых средств, способных более или менее успешно пре-
дупреждать и лечить те или иные разновидности шока, борьба с ним до сего
времени все езде остается в основном в пределах симптоматической терапии.
Особого внимания заслуживает одно направление в профилактике
и терапии шока, которое, несмотря на свою большую давность, привлекало
к Себе Сравнительно меньше внимания теоретических и практических
работников и поэтому развивалось менее интенсивно, чем другие. Мы имеем
в виду борьбу против шока, в частности, против травматического, путем
применения различных седативных, снотворных и наркотических средств.
Сравнительно слабое внимание, уделяемое этому направлению профилак-
тики и в особенности терапии шока, тем бол ее досадно, что с точки зрения
неврогенной теории патогенеза травматического шока именно это направ-
ление имеет больше научных оснований считаться приближением к кау-
зальной терапии и профилактике данного страдания, чем иные из сущест-
вующих и более распространенных видов борьбы с шоком, базирующихся
на других патогенетических теориях.
Включаясь в разработку весьма актуальной и вместе с тем сложной
проблемы шока, мы прежде всего преследовали сугубо практическую цель:
найти новый способ профилактики и терапии травматического шока, в осо-
бенности его тяжелых степеней. Отправной точкой для наших исканий,
а также основной научной базой для нашей теоретической и эксперимен-
тальной работы в указанном направлении являлось учение великого
И. П. Навловаоб охранительной и целебной роли процесса торможения для
истощенных высших нейтральных нервных образований. В процессе работы
наше руководящее направление слилось со старым направлением борьбы
с шоком, о которой мы говорили выше, и, как нам кажется, значительно
усилило последнюю. В результате проведенных нами за последние два
года (1941—1942) теоретических, экспериментальных и клинических ис-
следований с настоящее время можно предложить еще один новым способ
профилактики и терапии травматического шока, который в основной своей
части может рассматриваться как дальнейшее развитие упомянутого выше
направления борьбы с шоком, освещенного с позиций учения II. П. Пав-
лова.
1 Военно-медицинский сборник, № 2, 1945. Печатается со значительными сок-
ращениями.
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКОГО ШОК К
•153
Прежде чем приступить к краткому описанию разработанного нами
споссба и к изложению полученных экспериментальных и клинических
данных по борьбе с травматическим шоком, мы дадим сжатый критический
обзор известных нам литературных данных о результатах применения
различных седативных, снотворных и наркотических средств в целях про-
филактики и терапии этого страдания. Необходимость хотя бы неполного
обзора истории и современного ссстоянвя упомянутого направления борь-
бы с шоком диктуется тем, что в современной монографической и периоди-
ческой литературе по шоку они совсем нс освещены; имеются лишь упомина-
ния о применении морфина и подобных препаратов. Яркой иллюстрацией
сказанного являются монографии Кэннона (Cannon), Блэлока (Blelcck),
Скуддера (Scudder) и др., посвященных проблеме шока. Болес того, с со-
жалением приходится констатировать, чго известные нам современные
авторы частных работ, посвященных применению тех или иных обезболи-
вающих наркотических или снотворных средств для профилактики или
терапии шока, почти совершенно умалчивают сб истории вопроса и не
ссылаются даже на аналогичные новые работы других исследователей.
* *
♦
На заре возникновения учения о шоке, когда врачи имели лишь смут-
ное представление о его природе и очень мало знали Даже его Внешнюю
картину, возникли два диаметрально противоположных направления
лекарственного лечения шока. Для «ремонта» изношенной при шоке «жиз-
ненней машины» [ле Драп (le Dran)], для ессстансвления истощенных при
этом сил организма одни рекомендовали применять стимулирующие в воз-
буждающие лекарственные средства, другие—успокаивающие и боле-
утоляющие. Со временем параллельно с углублением и расширением зна-
ний о природе шока не только выдвигались новые принципы и меры борьбы
с различными его разновидностями (трансфузия цельной крови и ее отдель-
ных компонентов, введение различного рода кровозамеияюших жидкостей,
применение кислорода, углекислоты, гормонов, витаминов и т. д.), но и
продолжалось развитие двух первоначальных направлений, и особенно
интенсивно первого. Бурное развитие этого направления привело к резкому
увеличению числа противошоковых возбуждающих и стимулирующих
средств., появились даже предложения ввести некоторые из этих веществ
непосредственно в жидкую среду центральном нервной системы (ле Гран
и др.). А вялое и зигзагообразное развитие второго направления, осново-
положником которого следует считать ле Драна, привело от применения
опия и препаратов его группы к использованию бромидов и различного
рода снотворных и наркотических средств большим числом клиницистов
и экспериментаторов XIX и XX столетий [Н. И. Пирогов, Иордан (Iordan),
Савори (Savory), Гренинген (Groeningen), Нуссбаум (Nussbaum), Н. Н. Бур-
денко, А. В. Вишневский, At А. Топчибашев, В. И Попов, И. Р. Петров,
Б. П. Абрамсон, Г. Образцов, М. Н. Ахутин, П. Л. Сельповским, П.П. Ве-
селкин, Г. Н. Ханукой, И. А. Труханов, А. М. Трубенко, И. М. Григоров-
ский и др.]. Следует указать, что почти все экспериментаторы и клини-
цисты, применявшие седативные, наркотические и снотворные средствадля
профилактики или терапии травматического шока, исходили нз тех теоре-
тических предпосылок, что в генезе этого страдания решающую или важную
роль играютболевые и иного рода импульсы, поступающие из травмирован-
ных областей тела и приводящие к нерераздражению и истощению централь-
ной нервной системы или жизненно важных центров. Применением на-
званных веществ преследовалась главным образом одна цель—путем вы-
зывания аналгезии, сна или общей, спиральной и региональной анестезии
или же блокады нервов добиться ослабления или прекращения потока
этих нервных импульсов в центральную нервную систему.
454
Э. А. АСРАТЯН
Для профилактики и терапии тр авматического шока применял ись седатив-
ные, снотворные и наркотические вещества в том виде, в каком их обычно
применяют в медицинской Практике, Т. е. в виде чистых препаратов или
концентрированных растворов. При этом способы введения указанных
веществ в организм в подавляющем большинстве случаев также оставались
трафаретными, т. с. такими, какими обычно пользуются в медицинской
практике для введения больным без явлений шока. В виде исключения мо-
жно назвать практикуемое за последнее время внутривенное введение кон-
центрированных растворов морфина, алкоголя и бромидов [Функ-Брентано
(Funck-Brentano), Фразир (Frazier), П. Н. Веселкин]. Далее, в своей прак-
тике борьбы с травматическим шоком врачи применяли эти вещества или
самостоятельно, или дополнительно к другим противошоковым средствам
(горячие и спиртные напитки и стимулирующие средства, внутривенное
введение различных жидкостей, цельной крови и ее компонентов, кислород,
углекислота, гормоны, витамины и т. д.). Такого рода последовательное
или одновременное применение разнородных, иной раз даже противопо-
ложно действующих факторов, направленных против тех или иных симпто-
мов травматического шока, напоминает, скорее, арифметическую суммацию,
чем сочетание органически связанных факторов, одновременно действу-
ющих на основные патогенетические черты травматического шока. Экспе-
риментаторы же, как правило, влияние седативных, наркотических и сно-
творных средств на течение травматического шока изучали изолированно
от других противошоковых факторов. И если некоторые из них тем не
менее высказываются в пользу желательности комбинирования этих ве-
ществ с другими общеизвестными противошоковыми средствами при ле-
чении травматического шока у человека, то это носит в основном умозри-
тельный и декларативный характер, так как нс подкреплено эксперимен-
тально, что явствует из опубликованных до сего времени данных. Так,
например, И. Р. Петров совсем недавно рекомендовал при терапии травмати-
ческого шока комбинировать наркотические и снотворные вещества с не-
которыми общеизвестными противошоковыми средствами (с цельной и
разбавленной кровью, аскорбиновой кислотой, кислородом и углекисло-
той, алкоголем и т. дЛ, не приводя, однако, каких-либо эксперименталь-
ных данных о возможной и действительной пользе хотя бы простого сумми-
рования этих факторов при лечении даже легких и средних степеней трав-
матического шока. Б. II. Абрамсон рассматривает магнезиальную кровь
как сочетание двух противошоковых терапевтических факторов— наркоти-
ческого вещества (сернокислая магнезия) и крови. Он не приводит однако,
никаких экспериментальных данных о самостоятельной терапевтической
ценности сернокислой магнезии при травматическом шоке.
Подавляющим большинством этих авторов Получены хорошие резуль-
таты от применения седативных, снотворных и некоторых наркотических
средств, когда они использовали их в целях предупреждения травматичес-
кого шока или в целях прекращения Дальнейшего развития и углубления
его начальных стадий. Более или менее удовлетворительные результаты
отмечены при терапии эректнлыюй фазы, а также легких и средних степе-
ней торпидной фазы травматического шока. К этому выводу привели дан-
ные ученых XIX века.Об этом же говорят новые данные. Как пример можно
привести клинические наблюдения М. А. Топчибашева с сотрудниками и
экспериментальные данные И. Р. Петрова с сотрудниками. Нужно отметить,
что названные авторы не проводят в своих работах этой диференцнации
в отношении эффективности примененных ими средств при тех или иных
степенях травматического шока. Мы даем эту диференциальную опенку на
основании представленного ими фактического материала. М. А. Топчиба-
шев, например, пишет, что аналгезии «у травматиков резко ослабляет уже
развивающиеся явленна шока», а из приведенных М. Л. Абиевым выдержек
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКОГО ШОКА
455
историй болезни видно, что у всех больных, выведенных из состояния трав-
матического шока до инъекции аналгезина, максимальное кровяное давле-
ние составляло 80—95 мм, т. е. было характерно для легких и средних
степеней травматического шока. Авторы не приводят данных относительно
эффективности применения аналгезина при тяжелом травматическом шоке,
когда максимальное кровяное давление падает до 60 мм и ниже. Мы апри-
ори допускаем, однако, что в этих случаях трудно ожидать положитель-
ного терапевтического эффекта от анелгезина хотя бы на том основании,
что при этих степенях травматического шока всасывание веществ не только
из подкожных слоев (куда вводится аналгезии), нои изжелудочпо кишеч-
ного тракта сильно затрудняется. Возможно, что частично этим объяс-
няется резкое противоречие между результатами применения аналгезина
М. А. Тончибашевым с сотрудниками, с одной стороны, нВ. А. Парамоно-
вым — с другой.
И. Р- Петров с сотрудниками, как и многие авторы до них, изучали влия-
ние гексенала, хлоралозы, амнтал-иатрия в комбинации с паральдегидом,
мединалом и бромидами на течение и исход травматического шока у кро-
ликов. Эти вещества они использовали или в виде чистых препаратов, или
в виде концентрированных растворов и вводили их в организм подопытных
животных принятыми в медицинской практике способами. Новым, пожалуй,
является только то, что П. Н. Веселкин вводил 10% раствор бромистого нат-
рия внутривенно. Благодаря применению этих веществ, выживаемость трав-
мированных животных увеличилась на 12—25% по сравнению с контроль-
ными животными. Отсутствие кИмоГрафИчеСкой записи кровяного давления
или каких-либо других объективных показателей степени глубины шока не
позволяет нам делать заключение о фазе и степени тяжести травмати-
ческого шока у подопытных животных И. Р. Петрова и его сотрудников.
Однако при анализе имеющихся косвенных данных можно придти к вы-
воду, что авторы вызывали у животных в основном травматический шок
легкой п средней степени. Об этом говорят, во-первых, данные о том, что
значительное большинство контрольных животных жило больше одних
суток, тогда как общеизвестно, что животные, в частности, кролики, с тя-
желым шоком живут всего несколько часов, во всяком случае в пределах
одних суток. Об этом говорят и другие данные: введенные per os, per rectum
и внутримышечно снотворные и наркотические вещества всасывались и
оказывали заметное терапевтическое действие. Касаясь клинических дан-
ных М. А. Топчибашева с сотрудниками, мы уже указывали, что эффектив-
ное всасывание веществ при ректальном, оральном, внутримышечном н под-
кожном их введении в организм само по себе уже является показателем
отсутствия глубокой степени шока. В основе этого положения лежат м ново-
численные клинические и экспериментальные данные [Мэммори (Мишптогу),
Сайме (Symes), Витинг (Wieeting), Мун (Moon) и др.]. Достаточно вспом-
нить, что при тяжелых стадиях шока подкожная инъекция даже заведомо
летальных доз стрихнина не оказывает никакого действия на организм
[Андерхилл (Underhill), Блэлок (Blalock)]. Для определения степени
тяжести травматического шока у подопытных животных, а также для оцен-
ки эффективности избранного И. Р. Петровым и сотрудниками способа
применения наркотических и снотворных средств весьма показателен еще
один факт. Фразг.р (1935) и другие внутривенным введением 10% раствора
глюкозы и 10 % алкоголя достигли удовлетворительных результатов при
лечении травматического шока. Точно пользуясь рекомендованным Фрази-
ром составом, а такжеспоссбом его применения, без указания фамилии это-
го автора, И. Р. Петров в серии своих экспериментов добился несравненно
лучших результатов, чем в аналогичных же опытах, проводившихся нм
с наркотическими и снотворными средствами, вводимыми в организм per
os, per rectum или внутримышечно в виде чистых препаратов или концен-
45$
Э, А. АСРАТЯН
трированных растворов (в первые сутки 100% выживаемость при введении
алкоголя и глюкозы вместо 54% выживаемости контрольных и 70—78%
выживаемости при наркотических и снотворных средствах).
Обзор старых и новых клинических и экспериментальных данных при-
водит к заключению, что при тяжелых стадиях травматического шока при-
нятые шаблонные способы введения в организм седативных, снотворных
и в особенности наркотических веществ, к тому же в виде чистых препара-
тов НлИ концентрированных растворов не только не дают положительного
эффекта, поскорее вызывают отрицательный. Одержанное и даже отрица-
тельное отношение старых и современных авторов к применению этих
веществ, в особенности наркотических, в глубоких стадиях травматиче-
ского шока [ле Гросс Кларк (Le Gross Clark), Н. И. Пирогов, Рихтер (Kich-
tcr), Фишер (Fischer), Кэннон и др.] и при травматическом шоке вообще
[Гренинген (Groeningen), Гендерсон (Henderson), Мун и др.], в этом от-
ношении так же показательн2, как и тут факт, что, будучи одно время
предметом увлечения, это направление профилактики и терапии так в не
было внедрено в практику; насборот, постепенно и на продолжительное
время оно было предано забвению как практиками, так и теоретиками.
Исключение было сделано для препаратов группы опия, и то преиму-
щественно в целях профилактики или терапии легких и средних стадий
травматического шока.
Профилактическое и терапевтическое действие седативных, снотворных
и наркотических веществ при травматическом шоке почти все перечислен-
ные выше авторы приписывают главным образом тому, что эти вещества,
снижая чувствительность, создавая локальную или общую аналгезию или
анестезию, прекращают или ослабляют доступ интенсивного потока нерв-
ных импульсов, в особенности болевых, из области травмы в истощенные
или истощающиеся нервные центры или центральную нервную систему
вообще. Некоторые авторы полагают, что под влиянием названных веществ
усиливается стойкость нервных центров к недостаточному снабжению кисло-
родом [Крайл (Crile), Кэннон и др.]. Совершенно очевидно, что оба эти
объяснения, достаточные Для понимания механизма предохранения нерв-
ных центров от истощения и приостановки начавшегося процесса истоще-
ния, совершенно недостаточны для понимания механизма восстановления
функции уже истощенных нервных центров.
Некоторые авторы тем или иным из этих веществ приписывают допол-
нительное, как бы производное терапевтическое влияние на те или иные
органы и системы организма. Считают, например, что препараты группы
опия при травматическом шоке снимают депрессию, уравновешивают и ре-
гулируют кровообращение (Моррис и др.), увеличивают резервную щелоч-
ность крови [Кэттел (Cattell), Кэннон и др.]. Даже в отношении хлоро-
форма было сделано Допущение, что он при травматическом шоке усили-
вает работу сердца и регулирует его ритм [Листер (Lister), Скрайв (Strive)
и др.]. Небез интерес но отметить, что Моррис, Дюбуа (Du Bois) и многие
американские клиницисты положительное профилактическое и терапевти-
ческое влияние препаратов группы опия и алкоголя при травматическом
шоке необоснованно объясняют стимулирующим Их влиянием на цент-
ральную нервную систему.
В заключение следует сказать, что сторонники ограниченного приме-
нения седативных, снотворных и наркотических веществ при глубоких
степенях травматического шока, а также противники их применения при
любых его степенях свои позиции аргументируют тем, что указанные
средства вызывают депрессию медуллярных нервных центров, уси-
ливают аноксемию, ослабляют метаболические процессы в организме,
снижают температуру и т. д. [Гренинген, Кендрик (Kendrick) и др.].
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКОГО ШОКА
457
Прежде чем перейти к описанию полученных нами экспериментальных
и клинических данных по те-p ап ии травматического шока, мы изложим наши
руководящие 'теоретические представлений о функциональной патологии
тяжелых степеней вторичного травматического шока, основанных на ли-
тературных данных по этому вопросу, некоторых данных современной
физиологии нервной системы и частично на наших собственных исследо-
ваниях как по физиологии нервной системы, так и специально по шоку.
Мы исходим из старых представлений о том, что ведущим моментом
патологии вторичного травматического шока является нарушение функций
нервной системы, обусловленное в основном мощным и длительным пото-
ком разнородных нервных импульсов из травмированных частей тела
[Траверс (Travers), II. И. Пирогов, Фишер, Митчелл (Mitchell), Гренинген,
Крайл, Г. В. Алипов, 11. Н. Бурденко и др.] и другими этиологическими
факторами.
Известно, что неврогенная теория патогенеза травматического июня
имеет несколько вариантов. Из них вариант Гренингена—Крайла нам пред-
ставляется наиболее близким к истине. Мы принимаем его с некоторыми
поправками и дополнениями, наиболее существенными из которых яв-
ляются следующие. Еще несколько лет назад мы высказали мысль, что
так называемые «пусковые» и «трофические» функции нервных центров
суть две стороны единой их функции, что они переходят одна в другую,
взаимно связаны, взаимодействуют и т. д. Исходя из этих концепций при-
менительно к патогенезу травматического шока, мы считаем возможным
предполагать, что при истощении и астении комплекса соматических и ве-
гетативных центральных образований страдают как «пусковые», так и
«трсфические» их регуляторные функции. Если, например, обратиться
к сосудодвигательному центру, функциональное нарушение которого при
травматическом шоке играет основную роль среди поражений других
центров, то, согласно нашим представлениям, в результате его истощения
и астении нарушается нс только нервная регуляция моторной деятельности
сосудов, но и нервная регуляция трофики и функционального состояния
тканевых элементов сосудистой системы, в частности, эндотелия капилля-
ров. И такой важный момент функциональной архитектуры травматиче-
ского шока, как увеличение проницаемости капилляров, сбусловливается,
с нашей точки зрения, не только стазом крови в них и изменением их хи-
мической среды, но н в значительной степени ослаблением регуляции их
трофики и функционального состояния со стороны истощенного сосудо-
двигательного центра.
Мы считаем, что сложная картина патологии вторичного травмати-
ческого шока создается не одним фактором, каким бы он ни был важным,
а комплексом взаимодействующих факторов, составляющих основные
звенья в так называемых «порочных кругах». Новые литературиыеданные,
а также результаты специальных наших экспериментов на животных
позволяют считать, что, наряду с нарушением функций нервных центров
как основной причины, в патогенезе глубоких стадий вторичного травмати-
ческого шока, как правило (а не как исключение, как склонны думать упомя-
нутые выше авторы), существенное место занимают также токсемия (про-
дукты аутолиза тканей, нарушенного и нормального метаболизма «бакте-
риальные токсины, плохо выводимые из организма из-за олигурии при
шоке) и местная и общая плазмо-кровопотеря. К этим трем факторам могут
присоединяться также нарушения функций эндокринной системы, жировая
эмболия и др. ’.
1 В первом издании статьи мы писали о практической равноценности этих трех
факторов, чю по существу неправильно.
458
Э Л АСРАТЯН
На рис. 1 схематически изображена архитектура патологических про-
цессов при позднем травматическом шоке, сгруппированных нами не по
принципу последовательности их возникновения, а по функциональному
признаку.
В соответствии с теоретическими представлениями и целью наших
исканий — разработкой нового СпсСсба терапии Тяжелых степеней вто-
ричного травматического шока — мы решили сочетать минимум три дей-
ственных и взаимно друг друга дополняющих и усиливающих терапевти-
ческих фактора: 1) фактор для восстановления функции истощенных и ас-
Рис. I. Схематическое изображение функциональной патологии
при позднем травматическом шоке
гемических нервных центров, 2) фактор для ослабления или ликви-
дации нарушений в циркуляторной системе (атония мелких сосудов, ато-
ния и порознсстъ капилляров, уменьшение количества крови и т. д.),
3) фактор для усиления газового, органического и водно-солевого обмена
с целью освобождения организма от токсических продуктов, а также ослаб-
ления или ликвидации нарушений в составе и химизме циркулирующей
крови (нарушение ионного равновесия, аноксемия, азотемия, ацидоз, гемо-
концентрация, отек эритроцитов и тканей и т, д.).
Нам кажется, что такой подход к терапии шока находится в соответст-
вии с лучшими традициями векового опыта практической медицины, а
с научной точки зрения выгодно отличается от одностороннего подхода
тех исследователем, которые при разработке новых спсссбов лечения шока
обращают внимание на какую-либо одну сторону патогенеза шока и соот-
ветственно с этим применяют какой-нибудь один адэкватный терапевтиче-
ский фактор простого или сложного состава.
Не излагая исходных теоретических концепций и принципов терапии
шока многих отечественных и зарубежных исследователей, укажем, на-
пример, на предложения применять раствор соды [Маршалл, Кэннон,
Кунсе (Coonse)], хлористого кальция [Лапента (Lapenta), Калъцолари
(CaUolari), Эпингер, В. Бабук и др.], фосфорнокислого калия (Е. М. Бер-
кович, Г. Я. Хволес и др.), отдельные снотворные и наркотические ве-
щества [Неи дор ([юр (Neudcrfer), Савори, Эрихзен (Erichsen), Иордан,
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКОГО ШОКА
459
М. Л. Топчибашев и др.], раствор эфедрина [Чен (Chen), Джонсон (Johnson),
А. Ф. Лепукалн и др.], симпатсмиметнн (И. П. Чукичев и Иванов), стрих-
нин {Лейден (Leyden), Шнейдер (Schneider), Гренинген и др.], раствор
алкоголя и глюкозы [Фразир (Frasier), Сбразцов и др.], большое число так
называемых кровозаменяющих жидкостей (нермазол, укрмнфузнн, тутсфу-
зин, жидкости В. И. Попова, ЛИПК, ЦИПК, П. Л. Сельповского и др.),
различные возбуждающие и стимулирующие вещества, гормоны, витамины,
газы и т. п. (кофеин, камфора, бензедрин, питуитрин, инсулгн, кортин, ас-
корбиновая кислота, витамин В, кислород, углекислый газ и др.). Следует
сказать, что практика вносит существенные коррективы в подобные пред-
ложения. Перечисленные вещества в большинстве применяются не в каче-
стве самостоятельных противошоковых средств, а в качестве дополнитель-
ных к таким испытанным противошоковым средствам, как цельная или
разбавленная кровь, нормальные плазма и сыворотка или растворы сухой
плазмы и сыворотки.
Мы не ограничились теоретическими соображениями о преимуществе,
желательности и необходимости применения комплексного противошоко-
вого средства, представляющего комбинацию трех терапевтических фак-
торов, адэкватно направленных против трех основных сторон патологии
позднего шока, а попытались разрешить эту трудную задачу эксперимен-
тально. Ключом к разрешению первой задачи явилось учение И. П. Пав-
-лова об охранительной и, что еще важнее, целебной роли процесса торможе-
ния для истощенных или истощающихся высших нервных центров, учение,
которое было обосновано богатейшим экспериментальным материалом и
подкреплено многолетним опытом терапии сном психических и нервных
заболеваний. В соответствии с поставленной целью мы считали возможным
несколько расширить область применения этого учения и распространить
его также на низшие центры, преимущественно истощающиеся при травма-
тическом шоке. Исходя из этого, в качестве первого терапевтического
фактора мы решили применить бромиды и снотворные.
В пользу такого решения говорили также литературные данные о сдвиге
реакции крови в сторону щелочности во время сна {Динст (Dienst) 1, старые
данные о положительном влиянии покоя и естественного сна па состояние
больного при шоке (ле Дран, Н. И. Пирогов, Крайл и др.), данные о ре-
зультатах применения старых противошоковых средств — алкоголя и
препаратов группы опия и, наконец, материалы других исследователей
по терапии легких и средних степеней травматического шока с помощью
наркотических и снотворных средств.
Однако, в отличие от предыдущих исследователей, мы еще в самом
начале своей работы считали недостаточным и даже опасным применять
для лечения тяжелых степеней вторичного травматического шока одни
только снотворные и бромиды, а тем более наркотические средства в чистом
виде или в виде концентрированных растворов. Наши опасения, основан-
ные на данных физиологии медикаментозного сна, в общем совпадали
с доводами против применения этих веществ при травматическом шоке,
выдвинутыми Гренингеном и др. Мы считали, что снижение кровяного
давления, аноксемия, ослабление метаболизма, падение температуры и дру-
гие сдвиги, обусловленные действием этих веществ, могли быть еще тер-
пимы при начальных и средних степенях травматического шока, но они
могли оказаться грозными и даже фатальными при глубоких его степенях,
к значение положительного их действия на истощенные центры свелось бы
к нулю, если таковое в подобных условиях вообще имело бы место.
На основании этих соображений мы решили, во-первых, пользоваться
такими снотворными веществами, которые нс оказывают сильного депрес-
сорного действия иа жизненно важные функции организма, прежде всего
на функции бульбарных центров, а во-вторых, сочетать снотворный фактор
4 60
Э А. /ЧСРАТЯН
с двумя другими терапевтическими факторами в надежде на то, что они
смогут компенсировать отрицательные стороны их действия на организм
и даже создать противоположные им сдвиги в организме больного, нахо-
дящегося в состоянии шока. Учитывая необходимость оказания быстрой
помощи находящемуся в тяжелом шоке больному, мы решили соче гать эти
лечебные факторы в комплексной жидкости и рекомендовать ее вводить
нс per os, per rectum или подкожно, а внутривенно.
Проведенная нами теоретическая и экспериментальная работа привела
к удовлетворительному решению этих задач. Была составлена жидкость
следующего состава: NaCl 16 г, СаС12 3 г, NaBr 1,5 г, NaHCO3 1,2 г, глю-
козы 30 г, снотворное (доза в зависимости от его вида), дважды дестилли-
роваиной воды 1 л. По нашим данным, дня указанной цели лучшим сно-
творным является гедонал (1,2—1,5 г на 1 л) и уретан (2,5—3 г на 1 л);
значительно менее эффективны паральдегид (4—5 мг на 1 л) и в особен-
ности веронал (0,7—0,8 т на 1 л). Можно комбинировать первые с послед-
ними. В противоположность данным предыдущих исследователей (А. И. Ме-
лентьева, И. Р. Петров и Др.), мы не могли получить положительных ре-
зультатов от применения амитал-натрия и гексенала даже при введении
их в организм в составе нашей жидкости. Причиной расхождения является,
очевидно, разница между исходной глубиной купированного шока у нас
и у них. В небольшом количестве опытов мы не получили положительных
результатов также от применения эфира и мединала.
В начале нашей экспериментальной работы к этому составу жидкости
мы добавляли еще небольшую дозу спирта. Желая, однако, выявить по-
длинную терапевтическую ценность нашей жидкости в «чистом виде»,
без участия этого давнишнего противошокового средства, в дальнейшей
экспериментальной работе и в клинике мы исключили из состава жидкости
спирт. В настоящее время, когда эту задачу можно считать в основном
решенной в положительном смысле, по техническим соображениям (кон-
сервирование снотворных и глюкозы) мы вновь прибавляем к состав;у
пашей противошоковой жидкости чистый спирт в объеме 25—35 мл на
I л жидкости. Следует также указать, что наши представления о меха-
низме лечебного действия спирта при шоке этому не препятствовали.
Дозируется жидкость соответственно весу больного (примерно 10 мл
на 1 кг веса). Краткая характеристика ее следующая
Жидкость примерно в три раза гипертоиичнее рннгер-локковского
раствора. В ней приблизительно в 7—10 раз увеличена доза бикарбоната,
в 15—20 раз доза хлористого кальция, в 11/3—2 раза доза хлористого натрия,
в 20—30 раз доза глюкозы; из состава жидкости исключен хлористый
калий. Прибавлены бромиды, снотворное вещество и спирт. Жидкость
прозрачная, бесцветная, имеет слабощелочную реакцию (в целях улучше-
ния осмотических свойств жидкости в настоящее время проводится работа
по изысканию какого-нибудь доступного и безвредного коллоидного
вещества для прибавления к ее составу).
Проведенная нами экспериментальная работа (более 200 опытов на
кроликах, в основном на кошках и собаках) и клинические наблюдения
(110 шоковых больных) свидетельствуют об эффективном терапевтическом
действии нашей жидкости и тем самым о правильности исходных поло-
жений и оправдывает в основном наши расчеты, ожидания и надежды.
Прежде чем перейти к краткой характеристике клинических и экспери-
1 Жидкость изготовляется в виде двух отдельных растворов. Детали изготов-
ления, дозировки и применения жидкости описаны в специально составленной
инструкции. Более подробное обоснование терапевтических свойств жидкости н от-
дельных ее химических компонентов дано в напей монографии «Очерки по этиоло-
гии, функциональной патологии и терапии травматического шока» (1945).
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКОГО ШОКА
461
ментальных данных, скажем несколько слов о нашем понимании принци-
лов испытания нового противошокового средства.
Исключительно важным и серьезным является вопрос о предваритель-
ной тщательной, многосторонней и адэкватной экспериментальной про-
верке всякого нового лечебного средства, в частности, противошокового
средства, предназначенного для внедрения в практику здравоохранения.
К. этому вопросу, однако, не все ваши исследователи проблемы шока под-
ходят с требуемой осторожностью. В качестве иллюстрации можно при-
вести несколько примеров.
Предложенные нашими исследователями несколько лет назад новые
противошоковые средства — раствор фосфорнокислого калия и симпато-
миметин — в стадии экспериментального изучения на животных были
испытаны авторами лишь при раннем, или так называемом первичном,
травматическом шоке. Нам кажется, что в свете современных данных о па-
тологии и терапии шока такая экспериментальная проверка новых про-
тивошоковых средств не может считаться полноценной и адэкватной. Во-
первых, как в гражданской жизни, так и в условиях войны исключительно
редко приходится встречаться С первичным, ИЛи ранним, шоком из-за
его кратковременности; как правило, врачи застают пострадавших в
стадии позднего, или вторичного, шока. Далее, совершенно бесспорно, что
лечение первичного шока является несравненно более легкой задачей,
чем лечение вторичного шока. Достаточно напомнить, что один из широко
известных лозунгов борьбы с шоком гласит: «Начни лечить шок воз-
можно раньше».
По данным большинства современных исследователей, эксперименталь-
ный первичный шок, например, без большого труда может быть устранен
вливанием изотонических растворов и даже простым питьем воды; в то же
время общеизвестно, что для лечения вторичного шока эти средства со-
вершенно недостаточны. И .в этом нет ничего удивительного, так как вто-
ричный шок со своей сложной и запутанной патологией, со своими «по-
рочными кругами» разнообразных патологических процессов коренным
образом отличается от первичного шока. С практической точки зрения,
проблема шока почти целиком исчерпывается проблемой вторичного шока,
и борьба с шоком — это прежде всего борьба со вторичным шоком. Это
обстоятельство и упускается из виду многими нашими экспериментаторами.
Нам кажется, в частности, что в основе расхождений между результатами
терапии экспериментального шока у животных авторами этих средств и
шокового состояния человека при применении этих же средств лечащими
врачами лежит именно этот момент, т. е. то, что в первом случае эти
средства были испытаны при раннем, или первичном, шоке, а во втором
случае они, как правило, применялись при позднем, или вторичном, шоке.
Нельзя считать также достаточноадэкватным испытание новых противо-
шоковых средств многими нашими исследователями, если оно произво-
дится при легких и средних степенях позднего шока или при экспери-
ментальном шоке, вызванном различными химическими средствами и ма-
нипуляциями над кишечником и висцеральными нервами, т. е. при таких
видах экспериментального шока, которые по этиологии и патогенезу далеки
от типичного травматического шока у человека.
Экспериментальная проверка нашей противошоковой жидкости была
проведена со строгим учетом всех перечисленных выше моментов. Опыты
ставились главным образом на ненаркотизированных собаках и кошках,
а частью на кроликах при ближайшем участии лаборанта Л. В. Симу-
ковой. Как правило, мы вызывали у животных шок очень глубокой сте-
пени путем сильной травматизации задних конечностей (100—1000 ударов
молотком или металлической палкой), а иногда травматизацией в сочетании
с кровопусканием (10—30% крови). В стадии вторичного шока через
462
Э А АСРАТЯН
1—7 часов после травматизации кровяное давление у животных снижалось
в три-четыре раза и больше по сравнению с исходным уровнем, резко падала
температура тела, и вообще четко выступали все характерные признаки
глубокого вторичного шока со стороны дыхательной, двигательной и чув-
ствительной систем, со стороны зрачков и т д В этом состоянии глубокого
Л
р.
Рас 2 Лечение жидкостью позднего травматического шока у животных
Обозначения (сверху вниз> ди ха ине (стрелка показывает начало и конец вЛияа-
ння) артериальное давление путевая линия время п секундах
А Опыт > 142 Кошка / — через 3 часа после травматнзшии плипанис жидкости
1J через 4 часа после окончания вливания Б Опыт № 149 Кошка Мере:» 2 часа 20 мин
после траемятизации вливание жидкости при наличии симптомов паралича дыхательного
центра (/> й Олы г .S'- 135 Собака / — через 2 часа После трЛбМ1Тизации ьлинаиие Жид-
кости U — через 23 часа после вливания жидкости
шока все животные без исключения погибали, если им не оказывалось
никакой помощи, они погибали также в том случае, если им производи-
лось внутривенное вливание физиологического иЛИ рннГёр-ЛокКОВСКОГО
раствора', составленной нами жидкости, но без снотворных, снотворных
без нашей жидкости или снотворных в составе рингер-локковской жидкости
Не Помогли этим животным согревание и питье Смерть, как правило,
наступала через 1—5 часос. редко через ’10—15 часов
Внутривенное введение нашей противошоковой жидкости в обычном
составе давало совершенно другие результаты Более 80% животных уда-
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКОГО ШОКА
463.
лось вывести из состояния глубочайшего, фактически необратимого шока
путем внутривенного введе.ния жидкости в полном составе и организации
соответствующего ухода (согревание и питье) (рис. 2—3).
По данным точной графической регистрации и объективных наблюде-
ний, картина действия нашей жидкости такова: немедленно после начала
Рнс. 3. Лечение жидкостью позднего травматического шока у животных
A. Onur NV 123. Собака, / — через в часов после трввматиз&ции вливание жидкости;
/У—через сутки после влиаания. Б — опыт № til. Кошка. У — через 2 часа пссле трав-
натизации влнваные жидкости; //—через сутки после вливания. В — опыт № 42/1 11.
Кошка. I — через 1 час 45 мин. послетравматизаиии вливание жидкости; //-через 4 часа
после вливания. Обозначения те же. что и на рис. 2
ее вливания начинается подъем кровяного давления и в дальнейшем оно
продолжает повышаться; улучшается работа сердца и дыхательной системы,
снижается гемокОНцентрация, кровь приобретает алый цвет, увеличи-
вается ее свертываемость; повышается чувствительность и двигательная
активность животных; через 10—15 минут после начала вливания жи-
вотные постепенно погружаются в дремотное Состояние и наступает Сои
иа фоне удовлетворительной деятельности сердечно-сосудистой, дыхатель-
ной системы и обменных процессов в организме. Просыпаясь через 1—2 часа
после введения жидкости, животные с большой жадностью пьют, стоят,
ходят, бегают и вообще мало чем отличаются от нормальных животных.
В этой стадии кровяное давление и температура тола достигают у них
нормального уровня. В первые два дня после опыта температура у живот-
464
Э. А. АСРА1ЯН
пых повышается на 1—2° по сравнению с нормой» затем, спустя 2—3 дня,
выравнивается. После выведения из шока животные находились под нашим
наблюдением в течение 3—7 месяцев. На некоторых из них по два-три раза
ставились опыты с шоком и они с одинаковым успехом выводились из шока
нашей жидкостью. Длительное наблюдение не выявляло заметных изме-
нений в состоянии здоровья животных даже после неоднократного введения
им жидкости (рис. 4).
• ♦
*
В настоящее время мы располагаем данными применения жидкости
более чем у НО раненых, в подавляющем большинстве находившихся
в состоянии тяжелейшего торпидного шока, с максимальным кровяным
Рис. 4. Излеченные от смертельного шока животные
давлением 60 мм и ниже (эти данные получены нами на фронтах Оте-
чественной войны при участии врача И. В. Каменской).
Краткая характеристика клинических результатов такова.
Прежде всего следует сказать, что имеется поразительное сходство
между данными, полученными нами в экспериментах на животных, и дан-
ными клиники. Больше того, мы должны с удовлетворением констатиро-
вать, что терапевтический эффект нашей противошоковой жидкости у че-
ловека оказался значительно выше, чем полученный в экспериментах на
животных. Это обстоятельство мы склонны объяснять в значительной
мерс тем, что при выведении из шока люди находятся в несравненно более
благоприятных условиях, чем подопытные животные; во время введения
жидкости животные остаются привязанными к станку, лишены возмож-
ности пить и т. д. За исключением двух случаев, наша антишоковая жид-
кость весьма быстро и эффективно выводила больных из шока. Одиннад-
цать больных с тяжелым шоком до нашего вмешательства не могли быть
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКОГО ШОКА
465
выведены из него ни переливанием крови, ни инфузией различных других
противошоковых жидкостей, ни применением обычных противошоковых
мер; инфузия же нашей жидкости быстро выводила больных из шока.
В тех сравнительно редких случаях, когда у больных кровопотеря была
очень велика, мы производили им дополнительно переливание крови спустя
4—10 часов после того, как они были выведены из шока. В данном случае
трансфузией крови мы преследовали цель ослабить анемию у больных,
находящихся в основном уже вне шока.
Конечный результат применения нашей жидкости (мы имеем в виду
длительное выздоровление больных) зависел от характера ранения и на-
личия тяжелых осложняющих моментов в виде перитонита, газовой флег-
моны, повреждения важных органов, пневмо-гемоторакса, внутричерепных
кровотечений и т. д. Если шок не давал осложнений или если осложнение
заключалось только в умеренной кровопотере, то процент выздоровления
доходил до 100. Несколько показательных случаев приведено на рис. 5
и в таблице.
Рис. 5 Уровень максимального кровяного давления до (пёрВый
стозбик) и после (следующие столбики) введения жидкости.
Близко расположенные столбики показывают давление в течение
одних суток, остальные столбики — в следующие дни. Пунктир-
ный столбик—отсутствие пульса на лучевой артерии
I - осколочное ранение голени, повреждение сосудов, кровопотеря, шок.
выздоровление. Н — осколочное ранение коленного сустава, кровопотеря,
шок, иыздоривленне. ИГ - травматическая ампутация стопы, ушиб
голени, шок. выздоровление. IV — проникающее ранение живота, оско-
лочное ранение бедра, множественные осколочные ранения стопы и го-
лени. кровотечение, шок. выздоровление (до введения жидкости транс*
фузия крови не помогала). V — осколочное ранение голени, раздробление
костей, ушиб брюшной стенки и кровотечение, шок, выздоровление що
введения жидкости трансфузия кроем не помогала)
Остановимся кратко на картине изменений в состоянии больного, на-
ходящегося в шоке, при введения жидкости. Через 3—5 минут после начала
введения у больных появляется пульс (если его раньше не было) и в даль-
нейшем он все время улучшается как по наполнению, так и по напряжению
и частоте. Очень быстро повышается артериальное давление, достигая в кон
це введения почти нормального уровня (повышение в- пределах 50—60 мм и
больше). Постепенно улучшается цвет лица и слизистых покровов, прекра-
щается потоотделение, суживаются зрачки. Очень быстро наступает также
улучшение дыхания по глубине, ритму и частоте. Постепенно восстанавли-
вается чувствительная и двигательная сфера больных; они начинают жало-
ваться на боли в раненых частях тела, становятся активнее движения, по-
является интерес к окружающему. До наступления дремотного состояния
больные охотно вступают в разговор, сообщают об улучшении самочувствия,
у них улучшается настроение и т. д. Через 15—20 минут больные постепенно
погружаются в дремотное состояние, а некоторое время спустя и
в сон. Глубина и длительность сна обусловливаются не только дозой сно-
творного и типологическими особенностями больных, но к условиями,
30 Э А. Асратян
46G
Э. А. АСРАТЯН
Дата Инициалы и возраст больного Диагноз Характер и нремя опера- тивного вме- шательства Время введения ЖИДКОСТИ
до операции после операции
VIII 3. и., 28 лет Сквозное осколочное проникающее ранение живота. Повреждение пенен и. Внутреннее кро- вотечение. Перитонит. Шок Лапарото- мия, Эфир- ный наркоз 19 ч. 25 м.
11.V1II О и Слепое осколочное ранение мягких тканей левой голени. Повреж- дение суставов. Крово- потеря. Шок Обработка раны. Мест- ная анесте- зия — 15 > 20 »
I7.VIII Е. В., 29 лет Сквозное осколочное ранение левого колен- ного сустава с раздроб- лением костей. Крово- потеря. Шок Резекция сустава. Местная анестезия 22 ч. 20 м. —
23.VII1 В. И., 36 лет Травматическая ам- путация левой стопы. Ушиб голени. Шок Ампутация нижней тре- ти голени — 6 ч. 25 м, 23.VIII
24. VIII Б. Л., 18 лет Слепое осколочное проникающее ранение живота. Сквозное оско- лочное ранение нижней трети бедра с повреж- дением кости. Множеств венное осколочное ра- нение правой и левой стопы и голени. Крово- потеря. Шок. Лапарото- мия. Обра- ботка ран. Эфирный наркоз —я 8 ч СО м.
27.VIII G. А., 22 лет Осколочное ранение левой голени с разд- роблением обеих костей и большим дефектом мягких тканей. Ушиб брюшной стенки. Кро- вопотеря. Шок. Обработка раны голе- ни. Местная анестезия 23 » 00 >
1 .IX С. В.. 6 лет Множественное оско- лочное ранение головы (повреждены правый глаз, кости носа, раз- рез верхней губы), ту- ловища и конечностей. Шок Перевязка без. анесте- зии, 23 ч. ОО м. 14 ч. 30 м.
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКОГО ШОКА
467
Клинические показатели до п не дени Я жидкости Клинические показатели после введения жидкости
лул1 с дыхание артери- альное давление В JAM рТ. СТ. темпе- ратура время и дата исследования 2 * В дыхание артери- альное давление в мм рт. ст. темпе- ратура Примечания
126 34 38/62 — 20 ч. 00 м. 11.VIII 102 1'4 20 24 64/118 74 — Смерть от пери тонига L1.VIH
134 34 43/64 35,5 16 ч. 00 м. 21 » 00 > 12.VIII 96 96 124 18 20 28 80/110 80/И2 72/108 36,1 37,2 39,0 Выздоровление
132 38 40 макс. — 23 ч. СО м. 18.VIII 3 ч. 20 м. 12 > 00 » 105 108 IOO 22 18 20 67/118 68/114 70/110 38,0 То же
120 Не про- щупы- вается 27 36 42/62 Не Опре- делялось — 7 > 00 > 11 » 15 » 24. VI11 J2 ч. 30 м. 25. VI11 J0 ч. 40 м, 8 » 30 » 14 » 30 » 22 > 00 » 25.VII1 11 ч. 00 м. 90 108 108 105 108 96 106 109 120 2G 18 19 21 21 24 21 24 64/110 60/102 64/108 60'102 58/106 58/102 62/108 68/110 ОЭ СР & ® 1 1 1 > г* II о о о » > Выздоровление. До введения жид- кости переливание крон и без эффекта
120 14 37/35 — 23 ч. 30 м. 28. VIII 16 ч. 00 м. 90 102 18 18 65/112 70/114 — Г ыздоровление. До введения жид- кости переливание крови и ЖИДКОСТИ Сельповского без эффекта
147 27 43 макс. —— 15 ч. 30 м. 22 » 10 » 2. IX 12 ч. 00 м. 120 120 120 16 20 28 — 37.6 Выздоровление
30*
468
Э. А. АСРАТЯН
1 Характер и время опера- тивного вме- шательства Время введения ЖИДКОСТИ
Пата Инициалы и возраст Сального Диагноз до операции после операции
<к IX Л. Ф-. 21 года Травматическая ам- путация верхней трети правого предплечья. Множественное мелко- осколочное ранение грудной кдеткн, при- чем одно проникаю- щее (закрытый пневмо- торакс). Множествен- ное ранение лица, че- репа, левой кисти, сто- пы, легкая контузи-я Ампутация. Обработка ран. 16 ч. 30 м. 15 ч. 15 м. —
16 IX С. И., 38 лет Множественное оско- лочное ранение верх- ней трети левой голени и сустава. Осколочное ранение левого бедра с переломом кисти; большая кровопотеря. Шок Обработка ран. Мест- ная анесте- зия 19 ч 15 м. 20 » 10 »
17.IX Б. Н., 23 лет Сквозное пулевое ра- нение грудной клетки Открытый пневмогемо- торакс. Шок Закрытие пневмото- ракса, Мест пая анесте- зия 15 ч. 30 м. 14 ч. 00 м.
21.IX К.Н.. 34 лет 1 Осколочное р?некие бедра с переломом ко- сти. Кровопотеря. Шок Ампутация. Эфирный наркоз. 16 я. 50 м. 15 » 00 —
19. IX с н., 19 лет Травматическая ам- путация средней трети левого бедра. Большая кровопотеря. Шок Ампутаци я Хлорэтил. Эфирный наркоз 14 30 » —
23. V И. Ш., 20 лет Семь ножевых ране- ний на различных участках тела. Одно — проникающее в груд- ную клетку. Глубокий травматический шок Обработка ран. Мест- ная анесте- зия 2 ч. 20 и. 1 » 25 » —
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКОГО ШОКА
469
П родолжсние
Клинические показатели до введения жидкости Клинические показатели после введения жидкости Примечании
л улье дыхание артери- альное давление н мм рт. СТ. ГСМ гм- мт ура время и дата нсследона имя пульс дыхание артери- альное давление в мм рт. ст. темпе- ратура
96 1*0 Не про- щу- пы- вает- ся 136 145 116 26 31 39 42 21 27 Чейн- Сток- са 56 макс. 50/72 Не опре делял ось 56 макс 36/53 36/55 34,0 35.4 36,2 15 ч. 50 м. 17 » 51 » 17.1.Х 12 ч. 50 ы. 21 » 00 » 24 » 00 » 17.1Х 13 ч. 00 м. 18.1 X 14 ч, 00 м. 14 » 55 » 18 э СО » 22 » 40 » 18.1Х 14 ч. 00 м. 1 9.1 X 12 ч. 10 м. 15 » 55 * 16 » 40 » 15 ► 50 » 16 » 50 » 22 » 10 » 20. IX 9 ч. 20 м. 14 » 00 » 2 » 00 » 2 » 40 » 8 » 30 > 2.1Х 58 6G 75 100 105 99 93 10О 105 99 87 99 |0‘2 105 120 136 120 140 120 100 100 93 20 22 24 27 24 21 21 21 24 24 21 33 16 18 18 18 18 21 21 18 112 102 64/118 65/110 63/106 67/112 75/124 73/113 67/102 70/114 70/110 68/112 66 115 68/118 R5/I32 60/104 64/112 65/114 64/112 68/115 105 75/110 35,0 37,0 36,4 36,7 38,5 Выздоровление Выздоровление. До введения жид- кости переливание Крови без эффекта Выздоровление То же Выздоровление. Дополнительное переливание 450 мл крови из-за анемии Выздоровление
470
Э. А. АСРАТЯН
в- которых они находятся, дальнейшими лечебными манипуляциями, ухо-
дом и т. д. В общем сон очень напоминает естественный к длится 2—4 часа.
В стадии глубокого сна кровяное давление иногда падает па 10—15 мм
и несколько ухудшается пульс. После того, однако, как больные просы-
паются, эти сдвиги вновь выравниваются. Характерным и важным является
одно обстоятельство: как правило, через 10—15 минут после введения
жидкости у больных резко усиливается жажда; они неоднократно пьют
воду еще до наступления сна, затем часто просыпаются, пьют воду и опять
засыпают. У многих жажда остается в течение нескольких часов и после
прекращения сна. Иные в течение 4—5 часов, выпивают по 2—3 л жид-
кости и больше. Увеличивается диурез. С прекращением сна состояние
больных продолжает улучшаться по всем перечисленным показателям.
Характерно, что многие больные молодого возраста, проснувшись, про-
сят есть.
Очень важным показателем усиления окислительных процессов и об-
мена веществ у больных является tq, что в случаях выраженной гипотер-
мии (34—35°) температура тела повышается на 1—2° еще во время пере-
ливания жидкости. Вообще, как правило, температура больных в тече-
ние первых двух дней после выведения из шока повышается на 1—2° по
сравнению с нормальной, а затем постепенно выравнивается.
В условиях фронта мы могли следить за состоянием здоровья вы-
веденных из шока больных лишь на протяжении 5—7 дней. На пунк-
тах же скорой помощи Ташкента мы имели возможность наблюдать за состоя-
нием здоровья выведенных нами из шока больных в течение 1—3 месяцев.
На основании этих наблюдений можно сказать, что наша жидкость не вызы-
вает каких-либо уловимых нарушений в состоянии здоровья больных как
в первые дни после выведения из шока, так и в последующие дни и недели.
Имеющиеся у нас немногочисленные экспериментальные и клиниче-
ские данные свидетельствуют о еще большей эффективности противошоко-
вой жидкости при применении се в более легких стадиях травматического
шока и с целью профилактики, т. е. при таких шоковых и предшоковых
состояниях, борьба с которыми представляет значительно меньшие труд-
ности, чем борьба с тяжелыми состояниями шока. Далее, наши новые
эксперименты выявили весьма эффективнее терапевтическое действие
предложенной жидкости при ожоговом шоке. Эти данные будут опубли-
кованы в отдельной статье.
Физиологический и клинический анализ наших экспериментальных и
клинических данных позволяет сделать следующие аргументированные,
частью гипотетические предположения о природе действия разработанной
нами противошоковой жидкости на организм больного. Благодаря своим фи-
зико-химическим свойствам И фармакодинамическим особенностям входящих
в ее состав компонентов, она дольше удерживается в кровяном русле, чем
изотонические растворы и простые гипертонические растворы кристаллои-
дов, а также способствует возвращению в кровяное русло интерцеллю-
лярной и интрацеллюлярной отечной жидкости из тканей и эритроцитов,
ослабляет гемоконцентрацию, усиливает жажду и активирует водно-соле-
вой обмен; жидкость тонизирует стенки капилляров и уменьшает их по-
розность (СаСЬ), нейтрализует ацидоз (NaIlCO3) и ослабляет гипогликемию
(глюкоза). Далее жидкость увеличивает количество циркулирующей крови,
повышает кровяное давление путем значительного улучшения кровообра-
щения в организме, резко ослабляет аноцссмню и аноксию, активирует
все основные метаболические процессы в организме, повышает темпера-
туру тела, увеличивает диурез и т. д. Наконец, на этом фоне она вызывает
ослабление'болев.ых ощущений, дремотное состояние и сон. Имеется веское
основание для гипотетического предположения, что под совокупным воз-
действием перечисленных сдвигов в организме происходит постепенное
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКОГО ШОКА
471
функциональное восстановление истощенных низших и в особенности выс-
ших нервных центров, а они, постепенно усиливая свой контроль над про-
текающими в организме процессами, в свою очередь оказывают на них бла-
гоприятное влияние. Таким образом, вместо разорванного «порочного кру-
га» патологических процессов при травматическом шоке, как бы создается
круг взаимно усиливающих положительных сдвигов в организме, свое-
образный благотворный круг процессов восстановления.
Выводы
1. Учение И. П. Павлова об охранительной и целебной роли процесса
торможения относится не только к нормальным и патологическим состоя-
ниям большого мозга, оно имеет более широкую область применения.
Можно полагать, что это учение имеет универсальное значение для всей
центральной нервной системы и может быть применено ко всем тем забо-
леваниям организма, при которых истощение и астения нервной системы
играют в патогенезе заболевания более или менее существенную роль.
Пам кажется, в частности, что сделанная нами попытка применить учение
И. П. Павлова к патогенезу и терапии травматического шока поднимает
на более высокий уровень теорию терапии сном этого заболевания, так
как она объясняет и охранительное, и целебное действие седативных и сно-
творных веществ, тогда как существующие до сих пор представления
способны были объяснить лишь охранительное действие этих веществ.
2. Терапия сном нс должна быть однотипной, шаблонной при всех
показанных для ее применения видах заболеваний. Необходимо в каж-
дом отдельном случае строго учитывать ие только нарушения в деятель-
ности нервной системы, но и другие важные черты патогенетической архи-
тектуры данного заболевания и соответственно с этим комбинировать
терапию сном с другими адэкватиыми терапевтическими факторами. Далее
необходимо учитывать функциональные особенности центральных нервных
образований, которые нарушаются при каждом отдельном виде заболева-
ния, и соответственно с этим устанавливать оптимальную дозу назначае-
мого сна как по длительности, так и глубине, подбирать для этой цели
наиболее подходящие по своим фармакодинамическим свойствам медика-
менты, определять лучший способ их введения в организм и т. д.
Руководствуясь этими правилами при разработке нашего способа тера-
пии травматического шока сном, мы сочетали снотворный фактор с факто-
рами, регулирующими изменения в крови, в сосудистой системе, в мета-
болктических процессах и т. п., определили вид лучших снотворных, их
дозировку, способ введения в организм и т. д.
Нам кажется, что достигнутые нами в эксперименте и клинике резуль-
таты лечения таких тяжелых степеней вторичного травматического шока,
в отношении которых предложенные до сих пор способы седативной, сон-
ной и наркозной терапии оказывались неэффективными, обусловли-
ваются именно этими новыми моментами, внесенными нами в теорию и
практику терапии травматического шока.
ЛИТЕРАТУРА
Абрамсон Б. П.. О длительности периода «готовности к шоку» после
кровопотери, Архив биологических наук, 1, 4, 51—57, 58, 1940.
Абрамсом Б. П., Переливание магнезиальной крови при экспериментальном
шоке, Вестник хирургии, 56, кн. 5, 659, 1938.
Бурденко Н. Н., Шок, Центральный медицинский журнал, 12, 4, 393, 1933.
Веселкин П. Н., Экспериментальные данные по вопросу о патогенезе
травматического Шока, Вестник хирургии, 51, 211, 1937.
В и шве некий А., Шок как нервно-дистрофический комплекс, Новый хирур-
гический архив, 49, 26, 1941.
Г р и г о р о в с к и й И. М., Послеоперационный шок, Вестник хирургии, 55,
472
Э. А. АСРАТЯН
Мелентьева А. И., Опыт применения гексенала в экспериментальной хирур-
гии, Вестник хирургии, 59, № 1—3, 35, 1940.
Образцов Г. Д., Экспериментальные и клинические материалы к обоснова-
нию алкогольной терапии травматического шока. Вестник хирургии, 53, 123, 1937.
Парамонов В. А., Новый хирургический архив, 47, кн. 3, 1940.
Петров И. Р., О патогенезе и терапии травматического шока. Военно-сани-
тарное дело. 9, 3, 1942.
Петров И. Р., О терапии травматического шока, Клиническая медицина, 21),
10, 3-9, 1942.
Попов В. А., сб. «Шок», 1938.
Пирогов Н. И., Основы военно-полевой хирургии, 1866.
Топчибашев М. А., Травматический шок и меры борьбы спим, Сборник
по эфирно-масляиому инъекционному наркозу, 1939.
Троянов А., О влиянии обширных ожогов кожи на животный организм,
дисс., 1882.
Хамуков Г. Н., О травматическом шоке, сб. «Материалы военно-полевой
хирургии», 94, 1949.
Agnew D., Principl pract. of surgery, 1881.
Blalock A., Principl of surgical care, 1940.
Bergmann G., Ztsenr. f. Aertzl. Forbildung, 35, 192, 1938,
Cannon \V., Traumatic shock.
Cowel I E., J. Amer. med. assoc., 70 607, 1918.
Grille G., Keen’s surgery, 1, 922, 1914; 6, 145, 1915.
Dienst C„ Klin. Wschr., 11, 3=0. 1938.
Erich sen J., Practischcs Handl), f. Chirurgie, I, 1864.
Fischer H., Ueber d. Schock, 1870.
Fischer H. Handb. d. Kriegschr., 1, 1882.
Frazier Ch., J. Amer. med. assoc., 105, 1731, 1935.
FunckBrentano, Mem. Acad, chin, 66, 615, 1940.
Groeningen G., Ueber d. Shock, 1885.
Krisc liner M., Die Chirurgie, 10,314, 1938.
I. eveuf J., De chirurgie, 55, 403, 1949.
Moon V., Shock 8. related capillary phenomena, 1938.
Mummery a. S у m e s, Brit, med. j., 2, 786, 1908.
Neudorfer E., Handb. d. Kriegschir., 1, 1864.
Sender J. S„ Shock, 1940.
Seeley Essax a. Matin, Ann. surg., 104, 332, 1936.
S 1 о m e D. a. O’S haugnessy Z., Brit. j. surg., 22 , 589, 1935.
Wi eting J., L'rgebnisse d. Chir. u. Orihop., 14, 617, 1921.
Wiggers C. J., Amer. med. assoc., 117, 1143, 1941.
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКИХ ПОРАЖЕНИЙ
НЕРВНОЙ СИСТЕМЫ1
бщеизвестно, что крупные достижения нашей отечественной 4п-
знологип в области изучения роли и природы процесса тормо-
жения в нервной системе неразрывно связаны с Петербургским
государственным университетом, с именами двух его профес-
соров—И. М. Сеченова и Н. Е. Введенского—и с именем егс
воспитанника—И.П. Павлова. Работы этих гениальных корифеев
нашей пауки явились идейным истоком для дальнейшего многостороннего
и углубленного изучения роли и природы процесса торможения многочис-
ленными их учениками и последователями как у нас, так и за рубежом.
Так как тема моего краткого сообщения также связана с изучением
роли процесса торможения в функциях организма и так как в процессе
своей исследовательской работы по этой теме я постоянно ощущал благо-
творное влияние университетской физиологической школы на образ и ход
моих мыслей, я счастлив был любезно предоставленной мне возможностью
поделиться на этой сессии результатами проведенной мной и моими со-
трудниками работы и этим самым принести дань глубокого уважения слав-
ному юбиляру — Ленинградскому государственному университету — мо-
гучему очагу развития нашей отечественной науки и культуры.
Основой и руководящим началом моей работы были данные и концеп-
ции моего учителя И. П. Павлова сб охранительно-целебной роли про-
цесса торможения в нормальной и патологической деятельности большого
мозга. Известно, что эти данные и концепции гениального мыслителя от-
крыли новую и с биологической точки зрения весьма важную роль про-
цесса торможения в деятельности большого мозга, а именно’: рель охрани-
теля его нервных клеток от угрожающей им опасности истощения при их
перевозбуждении и роль своеобразного естественного целительного на-
чала, способствующего восстановлению функции истощенных но той или
иной причине нервных клеток коры большого мезга.
Эти идеи И. П. Павлова как ценнейшая часть его богатейшего науч
него наследия положены в основу исследовательской работы многих его
учеников и последователей (Ю. В. Фольборт, Н. А. Подкопаев, А, Г. Ива-
нов-Смоленский, А. О. Долин и др.). В своем кратком сообщении я воздер-
жусь от обзора результатов этих исследований и ограничусь в основном
эскизным изложением результатов проведенной мной и моими сотрудни-
ками теоретической, экспериментальной и клинической работы в этой
области.
На основании проведенной мной литературно-теоретической и экспери-
ментальной работы по физиологии и экспериментальной патологии нервной
системы у меня возникла мысль о возможности и необходимости значитель-
1 Доклад на юбилейной сессии Ленинградского государственного университета
в 1945 1. Труды юбилейной научной сессии ЛГУ, 1946.
474
Э. Л. АСРАТЯН
ного расширения рамок теоретического и практического приложения кон-
цепций И. П. Павлова об охранительно-целебной роли торможения, мысль
о возможности развития этих концепций до степени общебиологического
принципа. Несколько лет назад мной были сформулированы два органи-
чески связанных между собой положения: 1) процесс торможения играет
роль охранительно-целебного фактора не только в деятельности большого
мозга, но и в деятельности всей нервной системы (быть может, в деятель-
ности всех возбудимых тканей вообще); 2) охранительно-целебная роль
процесса торможения выявляется не только при функциональных пораже-
ниях большого мозга (и нервной системы вообще), но н при травматических,
органических повреждениях всех главных отделов нервной системы. Осно-
ванием для первого положения я считал наличие большого числа общепри-
знан пых и точных фактических данных, свидетельствующих о близости,
сродстве или тождестве основных нервных процессов и об «естественной
общности» основных закономерностей деятельности всех отделов нервной
системы (фазы и градации возбуждения и торможения, суммация, прото-
рение путей, переключение путей, иррадиация, доминанта, индукция —
электротонические явления, псссимум— извращение — запредельное тор-
можение и т, д,). Одним из оснований для второго положения я считал
то, что всякая органическая травма нервной системы представляет собой
сочетание двух существенных факторов, являющихся, по воззрениям
И. П. Павлова, главными предпосылками для возникновения и развития
охранительно-целебного торможения, а именно: свойства перераздражать
нервную Систему И вызывать ее Ослабление и истощение.
Исходя из приведенных положений, можно было преследовать двоя-
кую цель: во-первых, цель теоретическую — попытаться с новой точки зре-
ния осветить и осмыслить ряд важных явлений нормальной и патологиче-
ской деятельности нервной системы; во-вторых, цель практическую — пу-
тем искусственного усиления охранительно-целебного торможения в по-
раженной нервной системе попытаться улучшить ее функциональное со-
стояние и тем самым ускорить и усовершенствовать процессы восстановле-
ния ее нарушенных функций.
Совершенно естественно, что меня как физиолога первая цель привле-
кала больше, чем вторая. Руководствуясь, однако, запросами военного
времени и желая внести свою лепту в дело обеспечения здоровья солдат
и офицеров героической Красной Армии, я основное внимание сконцент-
рировал на второй цели — на разработке интересовавшей меня проблемы
с точки зрения практических интересов здравоохранения страны.
Соответственно проведенной работе теоретических вопросов в своем
кратком сообщении я коснусь лишь мимоходом и, скорее, в порядке их по-
становки, чем разрешения.
Одним нз таких вопросов является много лет назад поднятый мной во-
прос о сродстве и даже тождестве феноменов «пессимума» Н. Е. Введен-
ского, установленного И. М. Сеченовым явления торможения спинальных
рефлексов при сильных раздражениях чувствительных нервов (позже обоз-
наченного Шеррингтоном как «извращение») и «запредельного торможения»
И. П. Павлова. Известно, что классическим объектом изучения каждого
из этих феноменов соответственно были: нерв с его периферическим рабо«
чим прибором, спинной и головной мозг. До сих пор эти феномены иссле-
довались главным образом с точки зрения уточнения условий их возник-
новения, развития и исчезновения, с точки зрения их многосторонней ха-
рактеристики, определения их места и значения в координационной дея-
тельности тех или иных отделов нервной системы и т. д. В добавление
к этому И. П. Павлов обнаружил и изучил новую и биологически весьма
важную сторону «запредельного торможения» в деятельности большого
мозга—охранительную, а в известных случаях и целительную его
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКИХ ПОРАЖЕНИЙ НЕРВНОЙ СИСТЕМЫ
475
роль. Естественно возникает вопрос: не заключается ли биологическое
значение «пессимума* и «извращения» хотя бы в известных случаях в ох-
ране соответствующих субстратов от «порчи», повреждения, которое могло
возникнуть в результате сильных, частых или длительных стимуляций?
Приведу лишь некоторые фактические данные в пользу такой поста-
новки вопроса.
Много лет назад некоторые физиологи высказали мысль, что развитие
торможения играет значительную роль в так называемом феномене утом-
ления рефлексов. Ряд данных, полученных моими сотрудниками до войны
в Институте мозга им. Бехтерева (А. А. Новикова, Д. И. Ханутнна,
Н. Л. Соловьева и др.), позволил не только убедиться в правильности этой
мысли, но также констатировать парабиотическую природу этого торможе-
ния и охранительное его значение. Особенно показательны в этом отноше-
нии данные, полученные Д. И. Ханутиной. Сильные ритмические раздраже-
ния кожной поверхности предплюсны одной из задних лап довольно быстро
приводят к утомлению сгибательного его рефлекса у животных с перерезан-
ным спинным мозгом. В то время как рефлексы на сильные раздражения
•совершенно исчезают, переход к слабым раздражениям восстанавливает
рефлекторную деятельность изолированного спинного мозга. Эти данные
с несомненностью говорят как о развитии торможения в спинальных цент-
рах при утомлении рефлексов, так и о парабнотическом характере этого
торможения. Охранительное же значение торможения ярко выступает
в других опытах. Экстирпация люмбальных и сакральных симпатических
узлов значительно изменяет функциональное состояние изолированной
части спинного мозга. Одним из выражений этого является резкое увели-
чение его работоспособности; при ритмических раздражениях лапы даже
относительное рефлекторное ее утомление наступает с большим опозда-
нием и трудом. Этот «неутомимый» аппарат, однако, после такой более или
менее значительной рефлекторной деятельности впадает в состояние край-
него истощения и в течение длительного времени (иногда днями) теряет спо-
собность к полноценной рефлекторной деятельности умеренной длитель-
ности и интенсивности. Вероятное объяснение механизма описанного яв-
ления заключается в том, что после упомянутых выше оперативных вме-
шательств ослабевает способность центрального рефлекторного аппарата
к своевременному развитию охранительного торможения достаточной ин-
тенсивности, могущего предотвратить истощение и ослабление нервной
системы.
Мне кажется, что некоторые данные II. Е. Введенского о положитель-
ном действии пессимума на возбудимость и работоспособность мышцы,
а также недавно опубликованные данные А. Н. Магницкого и сотруд-
ников о влиянии пессимума на метаболизм мышц служат лишним
основанием для сделанного нами допущения, что «извращение» и «несси-
мум», хотя бы в известных случаях,’ являются выражением охранитель-
ного торможения и представляют собой защитно-приспособительные про-
цессы важного биологического значения.
Еще несколько слов о группе явлений из области патологии нервной
деятельности, имеющих отношение к обсуждаемому вопросу.
Известно, что многие загадочные патологические состояния большого
мозга И, П, Павловым были блестяще освещены на основе развиваемых мм
концепций об охраннтельно-целебной роли торможения. В этом свете стал
понятен также механизм благоприятного Действия Так называемой СОННОЙ
терапии при некоторых видах психических и нервных заболеваний. Воз-
никает вопрос: нельзя ли в свете концепции об охранительно-целебной
роли торможения понять также природу ряда патологических состоянии
других частей неравной системы?
Приведу несколько примеров, иллюстрирующих такую возможность.
476
Э А. АСРАТЯН
Многие авторы считали и считают, что в этиологии и патогенезе трав-
матического шока торможение сосудодвигательного центра играет важную
и даже ведущую роль (Мельтцер, Рише, А. В. Леонтович, И. П. Чукичев,
И. А. Аршавский, Н. В. Голиков и др.). Будучи несогласен с тем, что тор-
можением сосудодвигательного и других центров при травматическом шоке
исчерпывается вся сложность его этиологии и функциональной патологии,
я тем не менее не только признаю его наличие и приписываю ему важную
роль, но считаю, что оно играет роль охранительно-целебного фактора
для этих истощенных центров.
Такую же охранительно-целебную роль я приписываю торможению,
которое развивается при так называемых локальных шоках или рефлек-
торных параличах.
Наконец, в свете концепции об охранительно-целебной роли торможе-
ния я склонен рассматривать также ряд явлений из сложного и запутан-
ного комплекса патологических Сдвигов в дятельности центральной нерв-
ной системы при ее непосредственных органических — травматических по-
вреждениях. Причинно-следственные отношения при этих нарушениях
очень сложны и запутаны. Грубо схематически можно выделить следующие
моменты в функциональной патологии поврежденной нервной системы:
1) выпадение функций разрушенных или же резко альтерированных
травмой клеточных образований и проводящих путей (разрушение, раз-
давливание, кровоизлияние, ирритация, воспаление и т. д.);
2) выпадение или нарушение функции из-за изменения функциональ-
ного состояния и деятельности близких к очагу повреждения и отдален-
ных от него отделов центральной нервной системы, непосредственно не за-
детых травмирующим фактором, но функционально тесно связанных с по-
врежденным очагом (межцентральпая функциональная связь в виде субор-
динации, трофического действия, индукции и г. д.).
Как известно, при органических травмах тех или иных отделов цент-
ральной нервной системы сензорные и моторные нарушения производного
характера занимают значительное место в общей картине нарушений ее
состояния и деятельности и часто даже преобладают над очаговыми наруше-
ниями. Более ста лет эта явления знакомы клиницистам и экспериментато-
рам. Они были описаны и изучены под различными названиями (шок,
диасхизис, сотрясение и т. дЛ. Я также имел возможность в течение мно-
гих лет наблюдать и изучать эти загадочные явления в самых различных
вариациях, градациях и фазах их возникновения, развития и исчезновения
при различного рода хирургических повреждениях большого мозга,
мозжечка, продолговатого или спинного мозга различных высших и низ-
ших позвоночных. Критический анализ обширных литературных данных
и собственные исследования привели меня к заключению, что в числе
факторов, обусловливающих перечисленные выше функциональные нару-
шения, а также исчезновение или ослабление этих нарушений, весьма
важную роль играет охранительно-целебное торможение.
Я глубоко убежден, что многие из не поддающихся удовлетворительному
объяснению явлений нормальной и патологической деятельности нервной
системы со временем смогут быть более удовлетворительно объяснены и ос-
мыслены на основе развиваемых здесь положений сб универсально-биоло-
гическом характере охранительно-целебной роли процесса торможения.
Некоторым подтверждением правильности этих положении могут считаться,
в частности, результаты, которые за последние 3—4 года получены мной
и моими сотрудниками в процессе изучения терапии сном травматиче-
ских поражений организма вообще и нервной системы в частности.
К краткому изложению этих данных я и перехожу.
Прежде всего я позволю себе сделать несколько предварительных за-
мечаний об общих установках, которыми мы руководствовались в свсей
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКИХ ПОРАЖЕНИЙ НЕРВНОЙ СИСТЕМЫ
477
работе м которые во многих отношениях отличаются от установок других
исследователей, также занимающихся разработкой вопросов терапии
сном.
1. Основной нашей задачей мы считали возможно детальную разработку
поставленных вопросов в экспериментах на животных. При этом мы стре-
мились к тому, чтобы вызванные нами экспериментальные патологиче-
ские состояния были возможно близки к тем, которые наблюдаются при
травме человека. Как правило, к работе в клинике мы переходили лишь
после предварительной детальной разработки вопросов в лабораторных
экспериментах на животных.
2. Учитывая то обстоятельство, что при многих патологических состоя-
ниях травмированного организма, кроме функциональной или органиче-
ской травмы нервной системы, происходят также нарушения функций
многих других систем, мы стремились сочетать фактор терапии сном (пред-
назначенной для вызывания или усиления охранительно-целсбного1 тор-
можения в нервной системе) с другими терапевтическими факторами,
адэкватно направленными против других черт архитектуры функциональ-
ной патологии заболеваний.
3. Для вызывания или усиления охранительно-целебного торможения
в. пораженной нервной системе мы стремились применять такие снотворные
средства (гедонал, уретан, бромиды, паральдегид и др.), которые вызывают
сон, возможно близкий к естественному, не действуют вредно на жизненно
важные функции организма даже при значительных дозах и не засоряют
организм вредными продуктами своего распада. Следует напомнить, что
большинство других исследователей, применяющих терапию сном для
лечения различных психических и нервных заболеваний, не только не
избегает такого засорения организма, но приписывает этому даже важное
терапевтическое значение.
4. Мы стремились значительно углубить и удлинить естественный сон
пораженного организма, а не ввергать его в состояние глубокого и дли-
тельного наркоза, как делает большинство исследователей, применяющих
терапию сном.
5. Наконец, мы стремились к значительному расширению области при-
менения терапии сном. Мы охватили ие только такие виды травмати-
ческого поражения организма, при которых с давних пор эмпирически
применяют те или иные наркотические или снотворные препараты (шок,
каузалгня и т. д.), но и такие, при которых терапия сном до последнего
времени обычно не применялась (рефлекторные параличи, парезы и конт-
рактуры, органическая травма спинного и головного мозга, их анемизация,
термическая травма и т. д.).
Наиболее обстоятельно мы изучали терапию сном травматического
шока и близких к нему Состояний. Нами была разработана противо-
шоковая жидкость, которая представляет собой сочетание трех терапевти-
ческих факторов, адэкватно направленных на углубление охранительло-це-
лебного торможения, па улучшение состояния гемодинамики и на регуля-
цию нарушенного при шоке химизма тканей и крови. Испытание терапев-
тической ценности этой жидкости в большом числе экспериментов на со-
баках и кошках (ввергнутых в смертельный поздний шок грубой механи-
ческой травматизацией одной или двух задних лап) показало, что она обла-
дает высокими целебными свойствами.
Л. В. Симукова, П. В. Станкевич и я в эксперименте на животных до-
бились удовлетворительных результатов также при терапии сном ожо-
гового шока и солнечно-теплового удара.
Весьма утешительные результаты были нами получены также при тера-
пии сном экспериментально вызванного отека—набухания мозга у со-
бак (экстирпация верхних шейных симпатических узлов, локальная ме-
478
Э А АСРАТЯН
химическая травма большого мозга). Особенно положительными оказа-
лись эти результаты в тех случаях, когда снотворные вещества в течение
ряда послеоперационных дней сочетались с хлористым кальцием, предназ-
наченным для дегидратации мозга и для ослабления воспалительных про-
цессов в нем.
В значительном числе экспериментов на собаках нами производилась
оперативная травма спинного мозга (броун-секаровское сечение, перерезка
задних столбов и т. д.Е Как известно, в таких случаях вслед за фазой вы-
падения функции наступает фаза медленного их восстановления до значи-
тельных уровней. Мы рассчитывали, что путем искусственного усиления
охранительно-целебного торможения нам удастся ускорить процессы нор-
мализации функций пораженных нервных элементов спинного мозга и
тем самым создать благоприятные предпосылки для быстрого возникнове-
ния и развития всей гаммы процессов восстановления нарушенных функ-
ций (компенсация в силу тренировки, перестройка функции под влиянием
периферических импульсов и под влиянием коры большого мозга и т. д.).
Наш расчет полностью оправдался. Применение снотворных веществ в те-
чение ряда послеоперационных дней значительно ускоряло темпы восста-
новления нарушенных функций, сделало это восстановление более совер-
шенным и стойким, чем у контрольных животных.
Большую и интересную работу' с терапией сном анемнзированного
спинного мозга па лягушках и собаках проводила моя сотрудница
Г. Т. Сахиулина. В опытах на лягушках Г. Т. Сахиулина перевязывала
аорту’ на длительное время (от нескольких десятков минут до нескольких
часов). После восстановления циркуляции лягушкам подопытной группы
она вводила раствор снотворных средств и получила поразительные резуль-
таты: введение снотворных веществ в два-три раза ускоряло восстановле-
ние утраченной рефлекторном деятельности спинного мозга, придавало
восстановленным рефлексам более яркий характер и делало их значительно
более стойкими, чем у контрольных лягушек. Хронические же наблюде-
ния показали, что при систематическом введении снотворных лягушки
с описанными травмами жили значительно дольше контрольных.
Подобное же положительное действие снотворных средств на течение
восстановительных процессов Г. Т. Сахиулина получила после локальной
анемизации люмбальных и сакральных сегментов спинного мозга у ссбак.
Моя сотрудница Е. В. Гурова изучала влияние снотворных веществ
при остром рефлекторном истощении изолированного путем перерезки
спинного мозга лягушки. Выяснилось, что и при этом виде острой рефлек-
торной недостаточности спинного мозга снотворные оказывают благотвор-
нее влияние.
В другой серии интересных экспериментов, поставленных на лягушках
и кошках, Е. В. Гурова вызывала длительные рефлекторные параличи и
парезы одной из задних лап животных посредством длительного их раздра-
жения сильным электрическим током (несколько ослабленный ток город-
ской сети). Электрическая травма, помимо локальных изменений в рефлек-
торной деятельности и трофике пораженной лапы, вызывала ухудшение
также общего состояния животных. Полученные Е. В. Гуровой данные
свидетельствуют о том, что прием снотворных веществ и течение ряда
дней значительно ускоряет процесс восстановления функций пораженной
лапы и вообще оказывает благотворное влияние на общее состояние и дли-
тельность жизни животных.
Моя сотрудница Е. А. Романовская изучает характер и течение восста-
новления рефлекторной деятельности у лягушек после локальной термиче-
ской травмы спинного мозга. По предварительным данным Е. А. Романов-
ской, снотворные средства оказывают положительное влияние на течение
восстановительных процессов при этом виде дисфункции спинного мозга.
ТЕРАПИЯ СНОМ ТРАВМАТИЧЕСКИХ ПОРАЖЕНИЙ НЕРВНОЙ СИСТЕМЫ
479
Совершенно естественно, что по мере накопления положительных экспе-
риментальных лабораторных данных мы должны были делать осторожные
попытки применить разработанные нами методы терапии сном при соот-
ветствующих болезненных состояниях организма человека. Физиологам
и биологам должно быть понятно, что это решение было принято нами не
без тревоги, сомнений и волнения. Я рал сообщить, что результаты при-
менения разработанных нами способов терапии сном при ряде патологи-
ческих состояний человеческого организма не только успокоили нас, но
даже превзошли самые лучшие наши ожидания.
Еще в начале 1942 г. в ряде клиник Ташкента, а затем на одном из
фронтов Великой Отечественной войны мной совместно с Л. В.Симуковой,
И. В. Каменской и большим числом военных врачей противошоковая -
жидкость была применена при тяжелых формах травматического шока
у раненых. Полученные нами результаты показали высокую терапевтиче-
скую эффективность этой жидкости. Позже, когда было организовано
фабричное производство жидкости для нужд фронта, такие же данные в
большом объеме были получены в ряде клиник Москвы и в особенности
на многих фронтах Отечественной воины.
Весьма положительные результаты от применения снотворных веществ
были получены мной в ряде случаев коммоцнонно-контузионного синдрома.
Аналогичные данные были получены А. О. Долиным и А. Г. Ивановым-
Смоленским.
В конце 1942 г., находясь в Ташкенте, я приступил к проведению те-
рапии спсм больных с огнестрельными ранениями слиннсго и головного мез-
га, каузалгней, рефлекторными параличами, контрактурами нт.д. Несмотря
на весьма благоприятные предварительные результаты, я вынужден был
прервать эту работу по техническим причинам. За последнее время я и
мои сотрудники (Е. В. Гурова, Г. Т. Сахиулина) возобновили применение
терапии сном при указанных поражениях в двух клиниках Москвы.
Наши клинические наблюдения в общей сложности охватывают несколько
десятков больных с каузалгней, ранениями головного и спинного мозга
и с рефлекторными контрактурами. Наиболее четкие и стойкие положи-
тельные результаты от применения терапии сном мы получили пока при ка-
узалгии и ири ранениях головного мозга. Должен сказать, однако, что
как бы хороши ни были эти клинические результаты, мы их рассматриваем
как предварительные, ибо количество наблюдений у нас еще невелико
В настоящее время нами приняты меры к расширению клинической работы
в этом направлении. С тревогой я жду, когда закон больших цифр скажет
свое веское слово по поводу этого наиболее интересного и значительного
этапа нашей работы, наших исканий и наших надежд.
В заключение я хочу затронуть еще один вопрос: как понять природу
охраиительно-целебного торможения.
Для ответа на этот вопрос исключительной важности я не располагаю
никакими прямыми фактическими данными. В поисках временной рабочей
гипотезы я вынужден был дать свободу полету своей фантазии.
Было бы анахронизмом при современном уровне развития физиологии
прибегать к концепциям Геринга, Ферворна и Гаскелла о природе тормо-
жения как сб ассим иля торном или нутритивном процессе и попытаться
с этой точки зрения понять охранительно-целебное действие торможения.
Имеются, однако, некоторые достоверные данные, которые позволяют
предполагать, что в нервных центрах, охваченных торможением, проис-
ходят какие-то интенсивные процессы, но иного характера, чем те, которые
происходят в них при возбуждении. Более того, на основании этих данных
можно полагать, что активная деятельность нервных центров при тормо-
жении ограничивается только пределами самих центров, обращена «внутрь»
центров. Я имею в виду данные В интерштейна, Усуи (из лаборатории
480
3. А. АСРАТЯН
Варбурга), Мориса, Леба и др. о том, что применение значительных доз
наркотических и в особенности снотворных веществ почти не отражается
на количестве поглощаемого нервными центрами кислорода. Из этих дан-
ных можно как будто заключить, что, ввергая нервные центры (клетки)
в глубокое торможение, наркотические и снотворные вещества в умерен-
ных дозах прекращают лишь внешнюю их деятельность и как бы Переводят
активность нервных клеток на другие рельсы, направляют эту* активность
внутрь центров (клеток). На основании всего этого я и предполагаю, что
охранительно-целебное торможение — эго особое деятельное состояние,
которое оказывает благоприятное целебное действие на истощенную и
ослабленную по различным причинам нервную клетку.
Я осмеливаюсь искать подтверждение этих предположений в могучем
арсенале славной физиологической школы Ленинградского университета,
где рождалось и созревало классическое учение о парабиотическом тор-
можении.
Гениальный Н. Е. Введенский впервые охарактеризовал торможение
как «неколеблющееся состояние своеобразного возбуждения», как деятель-
ное, активное состояние нервных элементов.
Не являются ли приведенные мной выше фактические данные дополни-
тельным подкреплением фактов и учения Н. Е. Введенского? Далее,
в своих предположениях о природе охранительно-целебного действия тор-
можения не могу ли я основываться также на богатейшем фактическом
материале школы Введенского—Ухтомского и научении о парабиозе?
Я склонен дать положительный ответ на тот и другой вопрос.
^Физиологический синтез совпадает,
отождествляется с медициной; с этой
тучки зрения он может быть по npapj
назван экспериментальной терапией».
И. П. Павлов
К ЭТИОЛОГИИ, ФУНКЦИОНАЛЬНОЙ ПАТОЛОГИИ И ТЕРАПИИ
ПОСЛЕДСТВИЙ ОРГАНИЧЕСКИХ ПОРАЖЕНИЙ
НЕРВНОЙ СИСТЕМЫ »
Введение
течение ряда довоенных лет я совместно со своими сотрудни-
ками (А. И. Карамян, Я. М. Прсссман, Б. Д. Стефанцов,
1 Р. О. Барсегян, А. А. Фадеева, И. В. Введенская, Д. И. Хану-
| тина, В. Д. Дмитриев, К. М. Ротарь, В. А. Замятина и др.)
вел теоретическую и экспериментальную работу в области
одной из актуальных проблем современной биологии и меди-
цины— проблемы приспособляемости или пластичности нервной системы.
В этих исследованиях мы концентрировали свое внимание главным
образом на выявлении и изучении физиологических факторов, обусловли-
вающих восстановление различных моторных и сензорных функции
организма, нарушенных оперативной травмой различных этажей цен-
тральной нервной системы, нервов, а также повреждением рецепторных
и эффекторных органов. Нами было установлено, в частности, что среди
множества факторов различного характера и эффективности (адаптация,
тренировка, викарирование, запредельное торможение, дублированная
иннервация, простая перестройка нервных центров под влиянием пери-
ферических импульсов и т. п.) главную роль играет высшая интегратив-
ная деятельность большого мозга (у высших животных кора большого
мозга). Далее, по вашим данным, эта важная роль верхних отделов
центральной нервной системы в процессе восстановления утраченных или
нарушенных функций организма неуклонно возрастает по мерс восхожде-
ния по эволюционной лестнице, приобретая у высших животных значение
решающего фактора.
Наши данные позволили также сделать вывод, что эту важную биоло-
гическую роль большой мозг осуществляет благодаря своей трофической,
безусловнорефлекторной и высшей анализаторной и синтетической дея-
тельности, т. е. условнорефлекторной деятельности, способности образо-
вывать новые навыки, осуществлять новые виды координационной деятель-
ности, сложные и трудные перестройки функций низших отделов централь-
ной нервной системы.
Следует указать, что наши экспериментальные данные и воззрения
о решающей роли коры большого мозга в этих особого рода приспособи-
тельных явлениях организма, в противовес господствовавшим тогда в фи-
зиологии данным и воззрениям, находятся в полном соответствии с факти-
ческими данными и учением И. П. Павлова-о работе большого мозга.
1 Военно-медицинский сборник, № 3, 1946.
31 Э. А. Асра1ян
482
Э А. АСРАТЯН
Направление наших довоенных исследований этой проблемы все больше
побуждало нас перейти к разработке актуальных вопросов практической
медицины. Задачи военного времени, несомненно, побудили ускорить
этот переход. Я и мои сотрудники продолжали изучение проблемы приспо-
собляемости поврежденного организма, но уже под новым углом зрения,
в свете актуальных практических запросов переживаемого момента. Мы
ставили перед собой задачу искать новые пути, средства и возможности
для ускорения и усовершенствования приспособительных процессов, для
активного воздействия на течение и исход напряженной борьбы организма
за восстановление нарушенного здоровья. Хотя ив новой фазе своей работы
мы широко использовали достижения мировой медицины и биологии по
Интересующим нас вопросам, но исходной точкой и путеводной звездой для
наших новых изысканий попрежнему являлись работы и идеи гениального
И. П. Павлова. При этом особое внимание нами было уделено трудам и
концепции учителя об охранительной и целебной роли процесса торможе-
ния — сна.
И. П. Павлов показал, что в деятельности большого мозга роль про-
цесса торможения не ограничивается его участием в создании высших
форм координационной деятельности нервной системы, или высшей нервной
деятельности. Многочисленными работами И. П. Павлова и его сотрудни-
ков было установлено, что в нормальной и патологической деятельности
большого мозга торможение играет и другую, в биологическом отношении
весьма важную роль — роль природного охранительного и целебного
фактора, т. е. фактора, способного при необходимости охранять нежнейшие
нервные клетки большого мозга от болезнетворного действия чрезмерно
сильных или длительных раздражении, а также оказывающего благо-
творное целебное влияние на эти клетки, если они под воздействием не-
благоприятных обстоятельств в какой-то мере истощаются, ослабевают
и ввергаются в болезненное состояние.
Руководствуясь этой концепцией, И. П. Павлов не только по-новому
осветил природу ряда психических и нервных заболеваний, но подвел
также твердую научно-теоретическую базу под старое эмпирическое тера-
певтическое направление — лечение психических и нервных заболеваний
лекарственным сном или наркозом, поднял это направление на более вы-
сокий уровень и открыл блестящие перспективы для дальнейшего его
развития.
Обширные экспериментальные и клинические данные школы И. П. Пав-
лова и ряда других исследователей о некоторых функциональных осо-
бенностях травмированной нервной системы, а также наши прежние дан-
ные, полученные в этом направлении, позволили выдвинуть вопрос о воз-
можности и необходимости значительного расширения рамок теоретиче-
ского и практического применения упомянутой концепции И.П. Павлова.
В качестве руководящей рабочей гипотезы мной были развиты следую-
щие теоретические положения: 1) процесс торможения играет роль охра-
нительно-целебного фактора не только в нормальной и патологической
деятельности большого мозга, но и в нормальной и патологической дея-
тельности всей нервной системы; 2) процесс торможения выполняет эту
важную роль не только при функциональных поражениях всех отделов
нервной системы, но и при органических ее повреждениях. Таким образом,
в порядке дальнейшего развития идей И. П. Павлова и в. соответствии
с его высказываниями о существовании «аналога» запредельного тормо-
жения в низших отделах центральной нервной системы мной было сделано
допущение, что способность к развитию охранительно-целебного тормо-
жения принадлежит к категории универсальных и кардинальных свойств
всех нервных элементов (быть может, к категории общебиологических
свойств всех возбудимых тканей), зарождается в них в определенных уело-
ПОСЛЕДСТВИЯ ОРГАНИЧЕСКИХ ПОРАЖЕНИЙ НЕРВНОЙ СИСТЕМЫ
483
виях функциональных и органических их поражений и играет важную
роль в динамике возникших при этом патологических процессов.
Я не буду в данной статье разбирать литературный материал, равно
как и наши прежние экспериментальные данные, лежащие в основе этих
теоретических положений. Я не буду также затрагивать те вопросы физио-
логии и патологии нервной системы, которые на основе этих положений
получают, как мне кажется, новое освещение. Ограничусь лишь кратким
изложением своего взгляда на отдельные стороны функциональной пато-
логии близких и отдаленных последствий некоторых видов органических
поражении нервной системы, послужившей отправной точкой для поис-
ков новых путей их лечения1.
К функциональной патологии и терапии последствий некоторых форм
органических поражений нервной системы
1. Стойкое угнетение центрального происхож-
деи и я. Давно известно, что различные острые и хронические послед-
ствия непосредственной механической травмы тех или иных частей цен-
тральной нервной системы (параличи, парезы, сензорные и трофические
нарушения, вызванные размозжением, ушибом, компрессией и другими
травмами мозговой ткани) обусловливаются не только разрушением или
грубой и необратимой травматической альтерацией определенных ее очагов,
но и глубоким угнетением состояния и функции клеточных масс обширных
ее областей, находящихся поблизости и даже вдали от мест повреждения.
При этом степень участия диффузного угнетения не разрушенных травмой
нервных элементов в создании пестрой картины расстройств тех или иных
функций организма зависит как от тяжести, характера и локализации
травмы, так и от возрастных, типовых и индивидуальных особенностей
пострадавшего, от его состояния в момент поражения и т. д. И если при
наличии гнездных повреждений мозговой ткани диффузное угнетение
нервных образований играет роль важной, но все же дополнительной
причины расстройств функций организма, то бывают та кис формы травмы
организма, при которых оно играет роль единственной или во всяком
случае основной причины этих расстройств (различные виды сотрясения
головного и спинного мозга).
Таким образом, угнетение функций уцелевших от травматического
разрушения обширных масс нервных образовании занимает важное место
в архитектуре функциональной патологии близких и отдаленных послед-
ствий многих видов органического поражения любых отделов централь-
ной нервной системы. И если учесть, что патологическая альтерация цент-
ральных нервных образований при этом в большинстве случаев имеет
в той или иной степени обратимый характер, то станут попятными как
исключительная важность многостороннего исследования ее генеза и при-
роды, так и тот неослабевающий интерес, который проявляют к этому воп-
росу многие исследователи и клиницисты на протяжении трех столетий L.'C’
ти (Petit), Холл (Hall), Н. И. Пирогов, Гольц (Goltz), Лючиани (Luciani),
И. П. Павлов и многие его ученики, А. Л. Поленов, Н. Н. Бурденко, Леваи-
довский (Lewandowski), Кушинг (Gushing), Монаков (Monakow) и др, 1.
Обзор истории и современного состояния обширных экспериментальных
1 Моя и моих сотрудников теоретическая, экспериментальная и клиническая
работа по этиологии, функциональной патологии и терапии травматического,
ожогового и теплового шоков является органической частью всего комплекса
наших исследований по проблеме травматических повреждений организма, прово-
димых нами на основе упомянутых выше теоретических положений, начиная с 1941 г.
Эта часть нашей работы достаточно подробно освещена в ряде статей и поэтому
здесь не излагается.
31*
484
Э. А. АСРАТЯН
и клинических данных об этом явлении, а также многочисленных теорети-
ческих воззрений на его происхождение и сущность отвлек бы меня от
основной задачи. Ограничусь лишь некоторыми общими замечаниями.
Явление травматического угнетения состояния и функций центральных
нервных образований разными авторами обозначалось различными термина-
ми: церебральный шок, спинальный шок, угнетение, сотрясение, диасхизнс,
торможение и т. и. Относительно его генеза и природы были высказаны
многочисленные противоречивые и в большинстве своем односторонние гипо-
тезы. Главными или единственными причинами этого явления разные авторы
считали и считают молекулярные или микроструктурные изменения в нерв-
ных клетках, какие-то невыясненной природы функциональные изменения
в клетках и синапсах, нарушение кровоснабжения мозга, нарушение субор-
динационных отношений или системных связей между отдельными очагами
центральной нервной системы, составляющими определенные функцио-
нальные констелляции, и т. п.
Совершенно очевидно, что трудно действием одного только фактора
удовлетворительно объяснить всю сложность картины патологических
изменений, возникающих при органической травме тех или иных отделов
центральной нервной системы. Существовавшие по этому вопросу одно-
сторонние аналитические концепции за последнее время все смелее сме-
няются более или менее стройными синтетическими концепциями.
Высокая чувствительность нервных клеток к недостаточности снабже-
ния кислородом и питательными продуктами, их крайняя бедность соб-
ственными запасами питательных средств и буферных систем [Крайль
(Crile)], исключительная их чувствительность к механическим, химиче-
ским, температурным и иного рода факторам, а также ряд других струк-
турных и функциональных особенностей этих нежнейших образований
обусловливают их чрезмерную уязвимость и ранимость как при самых
разнотипных сильных воздействиях извне, так и многообразных резких
изменениях внутри организма. А ведь органическая травма и представляет
собой многосторонне действующий патогенный фактор, вернее, комплекс
таких факторов. Даже чисто механический компонент этого комплекса
(удар, контрудар, компрессия, сильное движение ликвора и т. п.) один
способен вызвать глубокие и стойкие изменения функционального состоя-
ния обширных участков мозговой ткани, независимо от того, вызывает он
гнездные разрушения или нет. Зона и глубина поражения, вызванного
действием одного этого фактора, вскоре могут быть расширены и углуб-
лены вовлечением в действие целой гаммы других разнообразных факторов
первичного и производного характера; перерыв межсистемных связей,
кровоизлияние, анемия механического и рефлекторного происхождения,
действие гистолитических, а часто и бактериальных токсинов, изменения
химизма жидких сред, нарушение ликворообращен и я и т. л. Совокупного
действия только этих факторов уже достаточно для замыкания «порочного
круга» патологических процессов, который в дальнейшем может еще больше
осложниться вовлечением в цепь новых патогенных факторов. К числу
последних можно отнести образование рубцов, развитие отека — набуха-
ния, нарушение состояния и деятельности вегетативной нервной системы,
з частности, симпатического ее компонента. Будет ли нарушение функции
вегетативной нервной системы вызвано вторично или, что еще хуже, будет
обусловлено первичным травматическим поражением центральных обра-
зований этой системы, расположенных на различных уровнях центральной
нервной системы, — последствия включения этого звена в замкнутую
цепь «порочного круга» могут быть очень тяжелыми. Прежде всего это
может вызвать дисфункцию многих жизненно важных систем организма
и косвенным образом углубить патологическую альтерацию пораженных
образований центральной нервной системы.
ПОСЛЕДСТВИЯ ОРГАНИЧЕСКИХ ПОРАЖЕНИЙ НЕРВНОЙ СИСТЕМЫ
485
Вовлечение симпатической нервной системы в «порочный круг» может
вызвать также далеко идущие последствия иного характера. Совершенно
бесспорно, что нарушение функции симпатической нервной системы спо-
собно углубить степень патологической альтерации центральных нерв-
ных образований. Существенную роль при этом могут играть также измене-
ния сосудистого русла мозга и проницаемости капиллярного эндотелия,
который, во моему предположению, контролируется симпатической нерв-
ной системой, так же как и моторные элементы сосудов и капилляров.
Я считаю принципиально важным, что все эти первичные и производ-
ные патогенные факторы, как бы они ни отличались друг от друга по своему
характеру и природе, как бы они разнотипно ни действовали на централь-
ные нервные образования, — все они в конечном итоге способны вызвать
резкое ослабление и истощение этих образований, вызвать в них близкие
или даже идентичные состояния патологической альтерации той или иной
глубины.
Нет необходимости, чтобы во всех случаях органической травмы цен-
тральной нервной системы были вызваны на арену действия все перечис-
ленные выше внешние факторы и внутренние сдвиги, все названные звенья
замкнутой цепи «порочного круга». В зависимости от тяжести, характера
и локализации трав.мы, от возраста и состояния пострадавшего, а также
от ряда других моментов, из комплекса патогенных факторов могут от-
сутствовать или, наоборот, доминировать в нем то одни, то другие, архи-
тектура функциональной патологии болезненного состояния организма
может иметь различный характер, динамика патологического процесса
может иметь неодинаковое течение. Однако при всех этих условиях почти
всегда быв.аст достаточно предпосылок для такой степени ослабления, исто-
щения и перераздраженни центральных нервных образований, которые
находятся обычно за пределами нормы.
Если интимная природа патогенного действия перечисленных выше
внешних и внутренних факторов на состояние центральных нервных обра-
зований все еще остается неизвестной, то внешняя сторона или грубый
механизм этого действия в свете современных данных о физиологии и пато-
логии нервной системы представляется более или менее понятным и не
нуждается в особых разъяснениях.
Несколько труднее понять хотя бы грубый механизм участия альтери-
рованной симпатической нервной системы в возникновении патологического
состояния или в углублении патологической альтерации центральных нерв-
ных образований при органической травме последних. Этот частный,
но важный вопрос привлекает внимание многих клннпцистов н являет-
ся предметом острых дискуссий. Он ждет еще своей детальной экспери-
ментальной и теоретической разработки. В настоящее время можно выска-
зать лишь более или менее вероятное предположение.
Можно допустить, что возникшая вследствие травмы центральной
нервной системы патологическая альтерация подчиненной ей симпатиче-
ской нервной системы обусловливает либорезкее повышение, либо рез-
кое снижение нормального уровня ее активности, т. е. вызывает сдвиги,
которые сами по себе прямым и окольным путем могут оказать сущест-
венное влияние на динамику патологического состояния в центральных
нервных образованиях, изменять его течение в том или другом направле-
нии. В настоящее время среди экспериментаторов и клиницистов довольно
распространено мнение, что именно повышение или понижение активности
симпатической нервной системы является характерной чертой многих
патологических состояний организма.
Возможны также другие предположения о характере действия симпа-
тического нерва на патологический процесс. Можно представить, напри-
мер, такую возможность: уровень активности симпатического нерва су-
486
Э. Л. ЛСРАТЯН
щественно не отклоняется от нормы, но в условиях патологической аль-
терации иннервируемого им субстрата его импульсы вызывают не обычные
положительные трофические сдвиги, а сдвиги иного характера, способные
углубить имеющуюся патологическую альтерацию. Во всяком случае
некоторые намеки на такую возможность имеются. Наконец, не лишено
основания предположение, что значительная патологическая альтерация
самого симпатического нерва извращает природу его импульсов и вызы-
вает в иннервируемых им субстратах изменения, в корне отличные от
таковых в норме, вызывает изменения дистрофического характера. Из-
вестным основанием для такого предположения могут служить те на-
блюдения И. П. Павлова, из которых он сделал вывод о существовании
двух противоположно действующих родов трофических нервов: подни-
мающих и ослабляющих «жизненность ткани». И. II. Павлов говорил о двух
видах трофических нервов, но мыслимо, что и один и тот же нерв, в зави-
симости от его состояния, сможет оказывать противоположные трофи-
ческие действия на иннервируемый субстрат. О возможности изменения
характера трофического действия нервов, говорят также весьма интересные
экспериментальные данные А. Д. Сперанского и его сотрудников.
Чтобы не быть неправильно понятым, в заключение скажу, что пере-
численные выше предположения о характере влияния симпатического
нерва на динамику патологического состояния в травмированной централь-
ной нервной системе не рассматриваются мной как взаимоисключающие.
Наоборот, весьма вероятно, что в одних случаях травмы имеет место один
род влияния симпатическою нерва на патологические явления, в других
случаях — другой. Возможны также случаи, когда в процессе развития
патологического состояния вовлеченный в «порочный круг* симпатический
нерв меняет характер своего участия, переходя от одного типа действия
на патологическое состояние к другому.
Мы видели, таким образом, что механическая травма центральной нерв-
ной системы представляет собой комплекс разнородных первичных и вто-
ричных болезнетворных факторов, которые, действуя в полном или даже
неполном составе, способны не только быстро разрушать или медленно
некротизировать определенные очаги центральной нервной системы, но
в той или иной степени угнетать и функцию обширных ее участков, про-
изводить своеобразную функциональную экстирпацию участков со всеми
вытекающими отсюда последствиями для организма в целом.
В чем сущность этого явления угнетения? В чем заключается биологи-
ческий смысл развития этого состояния5
Современная теоретическая и практическая медицина пе дает более
или менее удовлетворительного ответа на первый вопрос, а второй вопрос
даже и не поставлен. Называя явление угнетения шоком, диасхизисом,
сотрясением, асинапсией и т п., физиологи и клиницисты стремились вы-
яснить условия его возникновения, уточнить круг обусловливающих его
причин, проследить динамику его развития и т. д. Но сущность явления
стаегся неразгаданной. А между тем это вэпрос не только большого
теоретического значения, но и огромной практической важности, ибо от
степени познания сущности этого весьма распространенного при травмах
центральной нервной системы явления зависит правильный выбор и при-
менение соответствующих терапевтических средств и мероприятий.
Мне казалось, что для того, чтобы сделать хотя бы первые шаги в этом
направлении и подойти к раскрытию сущности этого сложного явления,
необходимо применить концепцию И. П. Павлова об охранительно-целеб-
ной роли торможения и отчасти родственную ей теорию Н. Е. Введенского
о парабиозе.
В свете теоретических установок двух классиков нашей науки развитие
и сущность обсуждаемого явления мио представляются в следующем виде.
ПОСЛЕДСТВИЯ ОРГАНИЧЕСКИХ ПОРАЖЕНИЯ НЕРВНОЙ СИСТЕМЫ
•187
Едва ли может вызвать у кого-нибудь сомнение, что перечисленные
выше патогенные факторы в комплексе или раздельно способны вызвать
ослабление и истощение центральных нервных образований путем их стой-
кого и длительного раздражения и перераздражения, ослабления их кро-
воснабжения, изменения состава и реакции их жидких сред н т. д. Было $5ы
неправильно, однако, считать, что сущность функционального угнетения
центральных нервных образований сводится целиком к их ослаблению
и истощению. Если вспомним, что И. ГТ. Павлов считал слабость и исто-
щенность нервных клеток ближайшими предпосылками для развития
в них охранительно-целебного торможения, и одновременно учтем, что
в комплексе факторов органического поражения нет недостатка в таких,
которые способны обеспечить постоянное присутствие «генератора» охра-
нительно-целебного торможения, т. е. перераздражения, то логично будет
предположить, что в ослабленных и истощенных органической травмой
центральных нервных образованиях развивается охранительно-целебное
(или парабиотическое) торможение. Мне кажется, в этом и заключается
суть травматического угнетения функции указанных образований. Я далек
от мысли считать, что этим полностью раскрывается природа обсуждаемого
явления — стойкого центрального угнетения. Однако было бы неправильно
думать, что сведение названного угнетения в основном к охранительно-
целебному торможению не является шагом вперед, что это лишь иное сло-
весное выражение одного и того же понятия. Хотя интимная природа
охранительно-целебного торможения (как торможения и возбуждения
вообще) все еще остается невыясненной, ио оно, несомненно, относится
к более конкретным, изученным и биологически более понятным явлениям,
чем угнетение, центральный шок, диасхизис и т. п. Это станет очевидным,
если обратимся ко второму из поставленных выше вопросов.
Если не стать на развиваемую мной точку зрения, то одна только по-
становка вопроса о биологическом значении явления травматического
угнетения центральных нервных образований может показаться абсурдной.
Физиологический процесс, лежащий в основе такого угнетения, большин-
ством исследователей оценен как явление отрицательного биологиче-
ского значения. Более того, даже с точки зрения Гольца, трактовавшего
центральный шок как явление торможения, а также с точки зрения
Н. Е. Введенского, рассматривавшего торможение как особое «неколеблю-
щееся состояние своеобразного возбуждения», или как перевозбуждение,
нельзя правильно понять биологическое значение торможения, развиваю-
щегося при травматических, в частности, при органических поражениях
нервной системы. Лишь с точки зрения концепции И. П. Павлова стано-
вится понятным огромное биологическое значение торможения при этих
поражениях. Исходя из учения И. П. Павлова, я расцениваю возникно-
вение охранительно-целебного торможения в травмированных централь-
ных нервных образованиях как явление положительного биологиче-
ского значения, явление приспособления, как меру предупреждения даль-
нейшего углубления патологической альтерации, меру естественной само-
защиты и самолечения.
Совершенно естественно, что нервные образования, в той или иной
степени охваченные стойким торможением, тем самым выводятся из строя
со всеми вытекающими отсюда последствиями для внутрицентральной и
в нецентральной деятельности нервной системы в целом. Но ведь такое
своевременное функциональное выведение пораженных нервных образо-
ваний из строя является лучшей физиологической гарантией их возможно
быстрого возвращения в строй в более иди менее здоровом состоянии;
в противном случае им угрожало бы несколько позднее, но уже необрати-
мое выведение из строя, а быть может, и окончательная гибель.
Против такого понимания существа явления не может считаться веским
48»
Э. А. АСРАТЯН
также возражение, что охранительно-целебное торможение истощенных
и ослабленных нервных образований как процесс естественной самозащиты
и самолечения не всегда приводит к удовлетворительному восстановлению
их функций. Известно ведь, что многие другие защитные реакции орга-
низма также часто не приводят к желательным результатам, в особенности
если существует большая разница между степенью поражения организма
и потенциальными возможностями его защитных сил. В таких именно слу-
чаях особенно ценна умелая помощь врача.
2. Стойкое угнетение рефлекторного происхо-
ждения . Клинике и экспериментальной патологии хорошо знакомы
также состояния стойкого очагового или более или менее генерализованного
угнетения функций центральной нервной системы, обусловленного травма-
тическим повреждением не мозгового вещества, а периферических нервов
и рецепторов.
К таким состояниям относится, в частности, так называемый локальный
шок или ступор, по терминологии старых клиницистов III. II. Пирогов,
Фишер (Fischer)I, и сходное явление, привлекавшее особеннее внимание
врачей со времен первой мировой войны и известнее в современной меди-
цинской литературе под названием рефлекторных параличей [Бабинский
и Фроман (Babinski et Fromen), М. II. Асгвацатуров, С. Н. Давнденков].
Существуют разные взгляды на механизм возникновения рефлекторных
локальных параличей, парезов, сензорных, сосудодвигательных и трофи-
ческих нарушений. Их считают результатом сотрясения центрального
рефлекторного прибора или же рефлекторного нарушения кровоснабжения
пораженной области, указывают на их физиопатическую природу и на их
отличие от истерических и истеротравматических параличей и т. п. Неко-
торые клиницисты (М. II. Аствацатуров и др.) подчеркивают близость
отдельных видов рефлекторных и истерических параличей.
Литературные данные, а также собственные наблюдения над неболь-
шим числом больных с рефлекторными параличами привели меня к заклю-
чению, что, как бы истерические и рефлекторные параличи, парезы и сен-
зорныс нарушения пи различались по своей этиологии, динамике развития
и симптоматологии, все же они весьма близки друг к другу по природе
функциональной патологии пораженных центральных образований. В свое
время И. П. Павлов дал блестящую физиологическую трактовку генеза
и природы истерических параличей, парезов и сензорных расстройств.
11. П. Павлов считал, что эти нарушения обусловливаются развитием
охранительного торможения в определенных очагах слабой и истощенной
центральной нервной системы.
Мне казалось не лишенным основания предположение, что и при трав-
матических рефлекторных параличах основная сущность функциональной
патологии пораженных центральных нервных образований заключается
в их ослаблении, истощении и охранительно-целебном торможении.
Весьма вероятно, что травматические рефлекторные параличи развиваются
именно у лнц, которые отличаются природной слабостью нервной системы,
пли у которых нервная система ослаблена под влиянием неблагоприят-
ных Внешних условий. Быть может, этим и объясняется то не понятое до
сих лор обстоятельство, что одинаковое примерно повреждение рецепторов
и нервов вызывает локальные рефлекторные расстройства функций только
у небольшого числа лиц. Но и без этой важной предпосылки сильнее трав-
матическое рефлекторное возбуждение центральных нервных образований
само по себе способно вызвать острое их ослабление и истощение, а вслед
за этим и более или менее стойкое охранительно-целебное торможение
в них, т. е. процесс естественной самоохраны и самолечения.
3. Стойкое повышение возбудимости централь-
ного и периферического происхождения. Следствием
ПОСЛЕДСТВИЯ ОРГАНИЧЕСКИХ ПОРАЖЕНИЯ НЕРВНОЙ СИСТЕМЫ 48g
органического поражения центральных и периферических нервных обра-
зований может быть не только стойкое угнетение нервной деятельности,
ио и состояние диаметрально противоположного характера — состояние
стойкой повышенной активности, развитие и доминирование повышенной
возбудимости или возбуждения разной экстенсивности и интенсивности.
К последствиям второго типа органических поражении центральной и
периферической нервной системы относятся каузалгия, фантомные ощу-
щения и боли, проводниковые и корешковые боли, рефлекторные кон-
трактуры, различного рода гиперкинезы и т. п.
Большинство современных клиницистов совершенно правильно указывает
на то, что в генезе каузалгических, фантомных, корешковых и проводни-
ковых болей играет роль целая гамма факторов, связанных с механической
травмой и возникающих вторично. К числу таких факторов относятся:
перерыв, размозжение, сотрясение и компрессия проводников; механическо-е,
рефлекторное л, возможно, аксон-рефлекторное нарушение кровоснабжения
периферических и центральных частей пораженной области, инфекция,
нарушение химизма жидких сред пораженных тканей, гормональные сдвиги,
местные воспалительные явления, отек — набухание, рубец и т. п.
Но современным данным, динамика развития патологического про-
цесса рисуется в следующем виде. Часть первично действующих факторов
вызывает сильную и длительную механическую и химическую ирритацию
пораженного субстрата; постепенно во-влекаются в действие остальные
фзкторы производного характера, которые в комплексе с первыми со
временем создают целую систему тесно связанных патологически альтери-
рованных очагов в периферической и центральной нервной системе: в районе
локализации травмы, в районе периферической проекции травмированны*
проводников, в районах центральной их проекции на спинальном, среднс-
и межуточномозговых и кортикальном уровнях. При этом, как правильно
считают некоторые авторы (М. И. Аствацагуров, II. И. Руссцкий и др.),
между этими альтерированными центральными и периферическими оча-
гами устанавливается порочное взаимодействие, приводящее к взаимному
углублению патологической альтерации отдельных звеньев и системы
в целом. Таким образом, создается «порочный круг» патологически альте-
рированных нервных образований, в который на каком-то еще не выяс-
ненном этапе развития вовлекается симпатическая нервная система.
Несколько слов об одном важном теоретически и практически вопросе.
Обширные литературные данные наших и зарубежных клиницистов позво-
ляют считать, что если роль измененной периферии в возникновении пато-
логического состояния придает периферическим звеньям «порочного круга»
некоторое доминирующее значение в начальных стадиях заболевания,
то со временем значение доминирующего фактора приобретают централь-
ные звенья «порочного круга». В большинстве случаев каузалгнй и
фантомных болей происходит именно такое своеобразное «перекочевы-
вание» патологической доминанты с периферии в центры. Об этом го-
ворит не только развитие реперкуссии или сннсстезалгии. Это явствует
также из данных многих клиницистов о том, что лечебные мероприятия
физиотерапевтического, медикаментозного и хирургического характера,
направленные на периферические звенья «порочного круга», дают более
или менее удовлетворительные результаты лишь в начальных стадиях
этих болезненных состояний; в поздних же стадиях они в большинстве
случаев оказываются неэффективными, в особенности если страдание
имеет выраженный характер. Я имею в виду рентгенотерапию, УВЧ-тера-
пию, грязелечение, ионофорез с различными средствами, инфильтрацион-
ную и проводниковую блокаду аналгезйрующими средствами, невролиз,
невроэктомию, невротомию, периартериальную невротомию, хордотомию,
перерезку задних корешков и т. п.
490
Э. А. АСРАТЯН
Как бы в противоречии с этим положением стоят результаты одного
оперативного воздействия. В целях избавления больных от каузалгических
болен хирурги прибегают также к довольно смелой операции — к удале-
нию симпатических узлов и рамисекции н нередко получают хорошие
результаты, если даже каузалгня имеет большую давность [Лериш (I.e-
riche), Тиноль (Tinel), А. Л. Поленов, В. А. Бондарчук, В. Н. Шаме в,
И. И. Орлов, Б. Г. Егоров, Ф. М. Лямперт, С. Ю. Минкин, А. Н. Баку-
лев]. Целебный эффект объясняют различного рода сосудодвигательными
изменениями в области поражения, перерывом путей болевых ощущений,
ослаблением повышенной чувствительности рецепторов и т. п. Все эти
разнородные объяснения говорят о том, что как будто бы и на поздних
этапах развития каузалгического синдрома доминируют изменения в
периферических звеньях «порочного круга*.
Мне кажется, что положительный лечебный эффект от упомянутых
операции обусловливается не только и не столько перечисленными выше
сдвигами в сосудах и рецепторах, а в основном тем, что соответствующие
альтерированные спинальные сегменты избавляются от патогенного воз-
действия альтерированных симпатических узлов *. С этой точки зрения
исключительный интерес представляют следующие факты. Описаны случаи,
когда односторонняя ганглиоэктомия снимает одновременно каузалгические
или иного происхождения боли как с одноименной, так и с противополож-
ной конечности (П. Е. Бабицкий и др.).
Сказанное о доминировании центральных звеньев «порочного круга»
в функциональной патологии поздних стадий каузалгии, фантомных болей
и т. п. не является, конечно, универсальным правилом. Бывают случаи,
когда периферические звенья «порочного круга» стойко сохраняют зна-
чительную роль на всех этапах болезненного процесса (обширные рубцы
или стойкие воспалительные явления в районе поражения нервов, грубое
нарушение кровоснабжения из-за повреждения крупных сосудов и т. д.).
Бесспорно, однако, что и в теории, и в особенности в лечебной практике
до сих пор недооценивается важное значение центральных звеньев «пороч-
ного круга» в функциональной патологии каузалгии и подобных ей бо-
лезненных состояний.
В своем кратком обзоре я попытался с некоторой новой точки зрения
осветить природу наиболее типичных последствий отдельных видов орга-
нической травмы центральной и периферической нервной системы: стойкое
угнетение и стойкую повышенную возбудимость центральных нервных
образований. Я не коснулся всего многообразия патологических процессов,
которые возникают в нервной системе и в организме вообще как При опи-
санных выше, так и при других видах органической травмы нервной си-
стемы и которые далеко не укладываются в рамки очерченной мной простой
схемы. Это и не входило в мою задачу. Имеется, однако, один вопрос,
о котором я хотел сказать несколько слов.
Чем объяснить то явление, что при некоторых формах органической
травмы нервной системы патологический процесс характеризуется доми-
нированием либо стойкого угнетения, либо стойкой повышенной возбуди-
мости? На современном уровне наших знаний трудно дать исчерпывающий
ответ на этот вопрос. Совершенно бесспорно, что важную роль играют
характер, тяжесть и локализация травмы, возраст пострадавшего, состоя-
ние его организма в момент травмирования и т. д. Мне кажется, что, как
и при функциональных заболеваниях нервной системы, здесь важную роль
играет тип нервной системы пострадавшего. Возможно, этим хотя бы ча-
1 Весьма показательны в этом отношении данные Е. Г. Егорова, Л. Я. Пннеса
и др. относительно гистопатологических изменений в нервных элементах симпати-
ческих узлов, удаленных но поводу каузалгии.
ПОСЛЕДСТВИЯ ОРГАНИЧЕСКИХ ПОРАЖЕНИЙ НЕРВНОЙ СИСТЕМЫ
491
стично Объясняется не разгаданное до сих пор явление, почему при оди-
наковых примерно поражениях одних и тех же нервов и при более или
менее одинаковом течении местных процессов в них у ОДНИХ раненых раз-
виваются явления вялого рефлекторного паралича, у других — контрак-
туры, у третьих — каузалгня и т. п. Известное основание для такого пред-
положения дают материалы лаборатории И. П. Павлова и его учение о
значении типа нервной системы для характера невроза, вызванного функ-
циональными воздействиями на организм.
4. Вопросы терапии. В основной части работы я и мои со-
трудники поставили перед собой скромную задачу: попытаться экспери-
ментально разработать новый способ терапевтического воздействия на
пораженную органической травмой центральную нервную систему с
целью ослабить или снять патологическую альтерацию (в виде ли стой-
кого угнетения или стойкого повышенного возбуждения) неразрушенных
ее элементов.
Если мое понимание физиологической природы угнетения функции
центральных нервных образований при определенных видах органиче-
ской травмы нервной системы было близко к истине, то следовало ожи-
дать положительного терапевтического эффекта от применения средств,
способных углубить охранительно-целебное торможение в этих образова-
ниях, создать для них еще более эффективный физиологический покой.
Такими средствами могли оказаться удачно подобранные, рационально
дозированные и умело примененные снотворные препараты. Применение
терапии сном в указанных выше случаях логически вытекало из всего
изложенного о роли естественным путем возникшего охранительно-целеб-
ного торможения в явлениях центрального и рефлекторного угнетения
травмированной нервной системы.
На первый взгляд может показаться нелогичным другое наше решение:
применить терапию сном также в случаях, когда посттравматическая
патологическая альтерация в центральных нервных образованиях характе-
ризуется преобладанием стойкого повышения возбудимости или возбужде-
ния. Однако, если вспомнить, что, по данным лаборатории И. П. Павлова,
средства, вызывающие охранительно-целебное торможение, оказывают бла-
гоприятное терапевтическое действие и при тех формах нарушения высшей
нервной деятельности, которые вызываются функциональными воздействия-
ми на собак возбудимого типа, т. е. при нарушениях, которые также харак-
теризуются доминированием стойкой повышенной возбудимости в боль-
шом мозге, то трудно будет в нашем решении видеть что-либо нелогичное.
По данным лаборатории И. П. Павлова, охранительно-целебное тормо-
жение является как бы своеобразным универсальным нормализатором,
способным оказывать целебное действие на деятельность большого мозга,
нарушенную под влиянием патогенных функциональных воздействий.
Мы рассчитывали, что при применении терапии сном нам удастся в одних
случаях углублением имеющегося в нервных образованиях охранительно-
целебного торможения, а в других случаях — созданием в них такового
восстановить в тон или иной мере их нормальное состояние и функцию.
Исходя из наших теоретических предположений, что развитие охрани-
тельно-целебного торможения свойственно всем нервным элементам, мы
надеялись также, что терапия сном окажет благотворное влияние одно-
временно если нс на все, то на большинство звеньев «порочных кругов»,
т. е. окажет своеобразное комплексное терапевтическое действие- Более
того, мы рассчитывали, что нормализация состояния этих нервных обра-
зований даст им возможность не только вернуться в строп и вновь при-
нимать прежнее участие во внутрицентральной и внецентралыюй дея-
тельности нервной системы в целом, но и, в порядке функциональной пере-
стройки и организации новых видов координации, взять на себя в какой-то
•192
Э Л АСРАТЯН
мере и функцию тех нервных образований, которые были разрушены или
необратимо альтерированы травмой. Поэтому мы считали, что терапия
сном создаст лучшие предпосылки также для действия стимулирующих
лечебных мер и средств (лечебная физкультура, трудовая терапия, физи-
ческие и лекарственные средства), к которым так часто прибегают лечащие
врачи при травматических нарушениях функций нервной системы и кото-
рые, будучи применены'преждевременно, т. е. в стадии, когда нервнье
образования еще очень слабы и истощены, способны лишь «муштровать»
их, оказать лишь кратковременный и преходящий терапевтический эффект,
а порой вызвать и вредные последствия.
К мысли о разработке нового способа терапии сном последствий орга-
нических поражений нервной системы я пришел на основании изложен-
ных выше теоретических соображений.
Однако в моем решении некоторую роль сыграли и следующие обстоя-
тельства.
Общеизвестна многолетняя практика эпизодического применения раз-
личных обезболивающих, снотворных и наркотических препаратов в борьбе
с отдельными симптомами травмы — с бессонницей, возбужденным со-
стоянием, болями и т. п. В этих целях пользовались и пользуются бро-
мидами, опием и препаратами его группы, пирамидоном, новокаином,
вероналом, люминалом, хлоралгидратом, сернокислой магнезией и другими
препаратами, применяя их либо отдельно, либо в сочетании с другими
лечебными средствами. Ограниченная цель применения этих препаратов
и отсутствие более или менее точного представления о природе их действия
не могли, конечно, способствовать правильному выбору лечащими вра-
чами препаратов, их дозировки, способа и длительности их применения,
а также установлению показаний и противопоказаний к их назначению.
Естественно, что при таких условиях эмпирическая медицина не могла
извлечь всю возможную пользу от применения этих препаратов для борьбы
с тяжелыми последствиями органических поражений нервной системы.
II хотя в литературе больше проявлялось отрицательное, чем положитель-
ное отношение к использованию снотворных и седативных препаратов при
указанных болезненных состояниях, тем не менее при анализе и крити-
ческой оценке литературных данных я находил определенное обоснование
для своего решения о методическом проведении терапии сном при орга-
нических поражениях нервной системы
Для реализации подобного решения можно было пользоваться двумя
путями: либо механически перенести в клинику органических поражений
нервной системы один из существующих способов сонно-наркозной тера-
пии, применяемых в клинике психических и нервных заболеваний, либо
разработать принципиально новый способ терапии сном с учетом не-
которых особенностей последствии органических поражений нервной систе-
мы. В первом случае имелись бы даже известные исторические и научные
основания Известно, что именно давнишняя практика эпизодического при-
менения снотворных препаратов для успокоения буйных психически
больных со временем привела к появлению и развитию методической
сонно-наркозной терапии лиц с функциональными заболеваниями боль-
шого мозга. Более 20 лет наши и зарубежные клиницисты с опре-
деленным успехом пользуются смесью Клоэтта и другими составами сно-
творных и наркотических препаратов для лечения некоторых видов пси-
хических и нервных заболеваний, а за годы этой войны — и постгравма-
тнческих психоневрозов.
Если при этом учесть то обстоятельство, что учение И. П. Павлова об
охранительно-целебной роли торможения—сна является бесспорно лучшей
теоретической основой для сонно-наркозной терапии психических и нерв-
ных заболеваний, то мысль о перенесении этого способа терапии в клинику
ПОСЛЕДСТВИЯ ОРГАНИЧЕСКИХ ПОРАЖЕНИЙ НЕРВНОЙ СИСТЕМЫ
493
органических поражений нервной системы покажется не только логичной,
но и заманчивой.
Однако по некоторым теоретическим и практическим соображениям
я решил отказаться от этого пути и избрал путь разработки принципиально
нового способа терапии сном. Дело в том, что существующий способ сонно-
няркозной терапии предполагает применение таких препаратов (хлорал-
гидрат, амитал-натрнн, люминал, сернокислая магнезия и др.) и в таких
дозах, что больные ввергаются почти в круглосуточный наркоз. Более
того, посте нескольких суток сонно-наркозной терапии в организме боль-
ных происходит ряд таких серьезных изменений (ацидоз, азотемия, ток-
семия, гипертермия и т. п.), которые приводят к значительному ослаб-
лению витальных функций организма, а нередко и к коллапсу. В отлично от
многих психиатров я не считал, что эти сдвиги могут иметь положитель-
ное целебное значение. Наоборот, по моему мнению, они способны только
ослабить целебное действие собственно сна и поэтому должны быть по
возможности устранены. Я считал также, что наркотическая, или песси-
мальная, глубина торможения, которая достигается в этих случаях,
едва ли является наилучшей для достижения искомой цели. Меня не удов-
летворяли также некоторые сложности техники применения сонно-нар-
козной терапии, грубое нарушение режима больных при лечении и т. д.
Следует сказать, что ряд советских ученых за последние годы
стал на путь более систематического применения седативных снотвор-
ных и наркотических препаратов при органических поражениях нервной
системы. Так как некоторые из э-.их ученых свою работу в этом направле-
нии начинали одновременно и независимо от нас, а большинство из них —
после нас, то я их коснусь в- конце статьи, чтобы иметь возможность прове-
сти параллель и дать сравнительную оценку.
Экспериментальная терапия сном последствий органических
поражений нервной системы животных
Исследования Клода Бернара, И. П. Павлова и других классиков
физиологии являются яркой иллюстрацией того, что экспериментальная
разработка вопросов терапии различных заболеваний относится к одной
из существенных задач физиологических исследований.
Приступая к серии опытов по экспериментальной терапии животных,
я преследовал двойную цель: во-первых, всесторонне проверить правиль-
ность своих теоретических положении о возможности расширения диапа-
зона теоретического и практического применения концепций И. II. Павлова
об охранительно-целебной роли торможения—сна; во-вторых, в случае
подтверждения правильности этих положении попытаться использовать их
для нужд практической медицины — разработать новый способ терапии
последствии органических поражений нервной системы
Работа была начата мной в 1941 г. Со временем в нее были вовлечены
мои сотрудники Л. В. Сммукова, Г. Т. Сахиулина. Е. В, Гурова, Е- А, Ро-
мановская, С. Н. Иванова и П. В. Станкевич. Опыты ставились на собаках,
кошках, крысах и лягушках. Оперативной, механической, электричь?кгй
и термической травмой, а также длительной анемизацией тех или иных
частей нервной системы мы вызывали у подопытных животных различного
рода глубокие нарушения двигательных, чувствительных, сссудодвпга-
тельных и трофических функций. Вслед за этим травмированных живот-
ных делили на две группы; животные одной (подопытной) группы в течение
определенного времени систематически получали снотворные вещества,
а животные Другой (контрольной) группы снотворных не получали. Ре-
жим животных той и другой групп в остальном был одинаков. Мы внима-
тельно следили, чтобы для параллельных наблюдений были взяты животные
494
Э. А. АСРАТЯН
с возможно одинаковыми показателями веса, возраста, упитанности и
роста, а также с близкими показателями характера, объема и глубины
поражения функций. Если же из попарно взятых животных одно более
или менее заметно отличалось от другого по этим показателям, то мы, как
правило, животного с худшими показателями включали в подопытную
группу. Динамика восстановления функций у животных нами контролиро-
валась разными способами; тщательное наблюдение за общим состоянием
и картиной нарушенных функций; определение уровня возбудимости
в ответ на механическое, химическое и электрическое раздражение;
тесты на способность к стоянию, прыжкам, к выравниванию нарушенного
равновесия и т. д.; тесты на наличие рефлексов и на их активность; тесты
на утомляемость; иимографическая регистрация двигательных рефлексов.
Кроме того, в случае глубоких нарушений жизненноважных функций,
приводящих, в конце концов, к гибели животных, учитывалась длитель-
ность жизни животных после травмы. К сожалению, мы были лишены
возможности пользоваться такими ценными приемами физиологических
исследований, как хронаксиметрия, тонусометрия и изучение биоэлектри-
ческих потенциалов. Однако в процессе работы хны убедились, что для ре-
шения поставленных практических задач можно временно обойтись без
этих исследований и ограничиться грубыми, но достаточно показательными
тестами. В целях же выбора наиболее эффективных снотворных и седатив-
ных препаратов, определения их оптимальной терапевтической дозы,
уточнения вопросов о наиболее рациональных способах и сроках их при-
менения, о возможносш их комбинации между собой н с другими медика-
ментозными средствами мы в процессе работы пользовались различными
препаратами в разных дозах, давали их разными способами, в различных
комбинациях и в различные сроки.
В одной статье невозможно подробно описать результаты проведенной
мной и моими сотрудниками обширной и многосторонней эксперименталь-
ной работы. Ограничусь кратким изложением наиболее важных и характер-
ных данных.
Для нарушения двигательных, чувствительных, сосудодвигательных
и ряда других функций мы подвергали подопытных животных следующим
патогенным воздействиям: производили экстирпацию верхних шейнь'х
симпатических узлов и компрессию коры большого мозга, полную пере-
резку и броун-секаровское сечение спинного мозга, перерезку задних
столбов спинного мозга, вызывали длительную анемизацию центральной
нервной системы, сильную электротравму отдельных конечностей, острее
истощение перерезанного спинного мозга, коммоцию—контузию спинного
мозга, органическую травму продолговатого мозга и т. п.
Вообще мы применяли травмы значительной интенсивности, чтобы
вызвать глубокие функциональные расстройства подопытных животных,
не отличающихся, как известно, особенно высокими показателями струк-
туры и функции центральной нервной системы. В общей сложности под
нашим наблюдением было около 150 млекопитающих и много сотен холод-
нокровных животных.
Большинство указанных видов травматических воздействий приме-
нялось и к теплокровным, и к холоднокровным животным.
Общий вывод из нашей работы по экспериментальной терапии сном
последствий описанных выше видов травматических поражений нервной
системы животных таков: снотворные препараты, как правило, оказывают
благотворное влияние на процесс восстановления нарушенных при этом
функций животного организма.
Приведу весьма краткое описание полученных нами эксперименталь-
ных данных.
ПОСЛЕДСТВИЯ ОРГАНИЧЕСКИХ ПОРАЖЕНИЙ НЕРВНОЙ СИСТЕМЫ 495.
У 6 собак и 2 кошек был перерезан спинной мозг на уровне послед-
них грудных сегментов. Все явления спинального шока (подавление
рефлекторной деятельности задних конечностей и хвоста, расстройсша
функции тазовых органов и т. п.) у животных подопытной группы проте-
кали значительно менее интенсивно и проходили на 3—4 дня раньше, чем
у животных контрольной группы.
Благоприятное влияние снотворных на течение восстановительных про-
цессов очень ярко выявилось также в других моих опытах (8 собак), в ко-
торых я производил броун-секаровское сечение спинного мозга также
на уровне последних грудных сегментов. У собак подопытной группы пара-
литические и паретические явления со стороны пораженной конечности
исчезали значительно быстрее, и собаки на 5—6 дней раньше животных
контрольной группы стали пользоваться своими конечностями для осу-
ществления функций стояния и ходьбы. Следует напомнить, что эти явле-
ния у собак и кошек вообще проходят довольно быстро, примерно через
12—15 дней.
Несколько менее четко было влияние снотворных на состояние живот-
ных, у которых путем экстирпации верхних шейных симпатических узлов
и компрессией коры большого мозга был вызван отек—набухание мозга.
Комбинация снотворных с хлористым кальцием давала более эффективные
результаты (ослабление депрессивного состояния, вегетативных рас-
стройств, улучшение аппетита и общею состояния, большая выжнвас
мость). Всего под наблюдением было 14 ссбак.
Очень четкие результаты были получены у 10 других собак, у которых
я перерезал заднюю половину спинного мозга на уровне 11 шейного сег-
мента. Нами давно было установлено, что эта операция вызывает очень
глубокие и длительные (примерно 30 — 40-дневные) расстройства двига-
тельных и чувствительных функций всех конечностей и хвоста (полный
паралич и анестезия, вслед за тем усиленная болевая чувствительность
и т. п.), а также глубокие нарушения функций вегетативной нервной
системы. Снотворные вещества в среднем почти наполовину сокращали
срок восстановления утраченных или нарушенных при этом функций, в
частности, функции стояния, ходьбы и бега.
Пользуясь методикой повышения гидростатического давления в суб-
дуральном пространстве, Г. Т. Сахи улина у 26 собак и кошек вызывала 20—
40-минутную анемию спинного мозга ниже уровня последних грудных
сегментов. Закономерным следствием такого воздействия были очень глу-
бокие нарушения моторных и сензорных функций задних конечностей,
функций тазовых органов, а также сосудодвигательные и трофические
расстройства, которые в большинстве случаев со временем постепенно
ослабевали и даже полностью исчезали, если поражение спинальных нерв-
ных элементов не имело необратимого характера. После некоторого коли-
чества ориентировочных и менее показательных опытов Г. Т. Сахиулина
в ряде последующих опытов обнаружила, что снотворные препараты резко
стимулируют процесс восстановления нарушенных функций по всем по-
казателям. Они иногда почти в 2 — 2*/8 раза сокращают сроки восстанов-
ления функций стояния, ходьбы и бега у собак.
В серии других опытов Г. Т. Сахиулина в течение ll/t—5 часов анс-
мизировала лягушек путем перевязывания аорты у выхода ее из сердца.
Динамика исчезновения и появления рефлекторной деятельности у части
животных регистрировалась кимографически, у другой части — простыми
тестами. На обширном экспериментальном материале Г. Т. Сахиулина
весьма четко выявила положительное действие снотворных и в этих опытах:
они примерно в 2 раза ускоряют восстановление утраченных рефлексов.
Исключительный интерес представляет одно явление, регулярно наблю-
даемое у лягушек, подвергшихся 4—5-часовой анемизацни. Когда день-два
493
Э. А. АСРАТЯН
спустя после анемизации рефлекторная деятельность у лягушек контроль-
ной группы восстанавливается в такой степени, что они в состоянии плавать
в аквариуме, то после более или менее энергичных движений лягушки очень
часто впадают в состояние кататонии, длящейся несколько минут. Даже
сильными раздражениями не удается вывести их из этого состояния. У ля-
гушек подопытной группы никогда не наблюдалось развития кататони-
ческого состояния, как бы долго и энергично они ни плавали в аквариуме.
Е. В. Гурова в серии хронических опытов на кошках и лягушках вы-
зывала рефлекторный паралич одной из задних лап путем длительного ее
раздражения сильным электрическим током (ток от городской сети, ослаб-
ленный до 80—100 V). Парализованные лапы обычно отекали; часто наблю-
дались н трофические расстройства (изъязвления, выпадение шерсти ит. д.).
Систематическое применение снотворных значительно ускоряло исчезно-
вение как рефлекторного паралича, так и трофических расстройств иотека.
В то время как полное восстановление функций пораженной лапы имело
место лишь у 33% животных контрольной группы, у подопытной группы
этот процент достигал 76. В серии других опытов Е. В. Гурова перерезала
спинной мозг лягушек на уровне 111—IV сегментов и вызывала острое
его утомление и истощение путем длительного раздражения задних лап
или нервов сильным фарадическим током. И в этих опытах снотворные
ускоряли восстановление функций истощенного спинного мозга, к тому же
изолированного от верхних отделов центральной нервной системы,
Е. А. Романовская достигала глубокого нарушения рефлекторной
деятельности спинного мозга у крыс и лягушек путем термической его
травмы (орошение того и иного участка мозга физиологическим раствором,
подогретым до ,0э и выше). И в этих опытах снотворные препараты оказы-
вали благоприг иное влияние на течение и исход восстановительных процес-
сов. Так, например. глубокие нарушения моторной функции (параличи
и парезьй проходили у 45,4% животных подопытной группы и лишь у 17,6%
животных контрольной группы, а глубокие нарушения кожной чувстви-
тельности (анестезия, гнпестезия)—у 40% животных подопытной группы
и лишь у 17,3% контрольной группы.
С. Н. Иванова уколами в продолговатый мозг у крыс и лягушек вы-
зывала значительные нарушения вестибулярных рефлексов и локомоции,
а также ухудшение общего их состояния. Применение снотворных и в этих
опытах ускоряло воссганоиленпе нарушенной рефлекторной деятельности,
но особенно ярко сказывалось на общем состоянии и выживаемости подопыт-
ных животных. Достаточно сказать, что продолжительность жизни жи-
вотных подопытной группы в среднем более чем в 2 раза превышала тако-
вую у животных контрольной группы.
В опытах Л. В. Симуковон синдром коммоции—контузии головного или
спинного мозга у крыс и лягушек вызывался ударом металлического шара,
брошенного с определенной высоты. Вскрытие показало, что в травми-
рованных частях центральной нервной системы при этом происходит более
или менее значительное кровоизлияние. Вообще травма вызывает глубокое
нарушение двигательных и чувствительных функций организма, а также
функций витальных систем. Хотя эти опыты являются ориентировочными,
но полученные данные создают впечатление, что снотворные препараты
< в этих случаях значительно ускоряют темпы восстановления этих функ-
ций и увеличивают выживаемость животных.
Таким образом, наши данные в. совокупности показывают, что при всех
примененных нами вариантах экспериментального нарушения функций
нервной системы снотворные препараты значительно сокращают период
выпадения или нарушения функций, ускоряют'появление и темпы развития
процессов восстановления функций, улучшают общее состояние, ускоряют
процессы заживления ран и увеличивают выживаемость животных. Мы неод-
ПОСЛЕДСТВИЯ ОРГАНИЧЕСКИХ ПОРАЖЕНИЯ НЕРВНОЙ СИСТЕМЫ 497
некратно имели возможность убедиться также в том, что снотворные уве-
личивают прочность восстановленных чувствительных и двигательных
функций. Мной и моими сотрудниками еще раньше было сделано наблюде-
ние, что ухудшение общего состояния животных (из-за случайных инфек-
ций, плохого ухода и г. п.) не только отрицательно влияет на темпы вос-
становления нарушенных функций, но может в той или иной степени спо-
собствовать возвращению этих нарушений, если они успели уже исчезнуть.
Это наблюдение было подтверждено в описанных экспериментах. При
этом выяснилось, что подопытные животные обладают значительно боль-
шей резистентностью к отрицательному действию упомянутого выше фак-
тора, чем контрольные.
Существуют, однако, значительные вариации благоприятного терапев-
тического действия снотворных препаратов на течение и исход процессов
восстановления нарушенных травмой функции животного организма. Об-
щий обзор наших данных показывает, что степень этого благоприятного
действия зависит от многих факторов. Очень важную роль играет уровень
филогенетического и онтогенетического развития нервной системы жи-
вотных. При однотипных травмах нервной системы применение снотвор-
ных дает более быстрые и эффективные результаты у собак и кошек, чем
у крыс и в особенности у лягушек. Точно так же этот эффект на молодых
животных сказывается ярче, чем на старых. Как и следовало ожидать,
важное значение имеют характер, экстенсивность, интенсивность и локали-
зация травмы. Снотворные препараты оказывают более эффективное дей-
ствие при травмах головного, чем при травмах спинного мозга. Далее,
положительное действие снотворных ярче проявляется при травмах сред-
ней, чем при травмах большой интенсивности и экстенсивности; при лег-
ких же травмах восстановление нарушенных функций и без снотворных
происходит быстро, поэтому положительное действие последних в этих
случаях не всегда четко выступает.
Исключительно важное значение имеет фармакодинамическое свойство
снотворных препаратов, их дозировка и сроки применения. В начальных
стадиях нашем работы хотя и были получены положительные результаты,
но они отличались пестротой и меньшей выраженностью. По мере накопле-
ния опыта мы начали получать все более однородные и лучшие результаты.
Ввиду того что названные вопросы длительное время стояли в центре
наших исканий, я кратко изложу результаты работы в этом направлении.
Это тем более необходимо, что на них мы строили основные принципы
нового способа терапии сном последствий органических поражении нерв-
ной системы.
Лучшие терапевтические результаты нами получены:
а) при применении снотворных препаратов карбаминовой группы —
гедонала и уретана (они и продукты их расщепления нетоксичны даже
в значительных концентрациях). За ними по эффективности следуют бро-
миды, паральдегид и веронал. Менее эффективны люминал, хлоралгид-
рат и вообще сильнодействующие снотворные и наркотические препараты;
б) при применении таких доз снотворных, которые вызывают сон дли-
тельностью 10—15 часов в сутки, по характеру близкий к естественному,
но глубже его. Дозы, обусловливающие менее глубокий и менее длительный
сон или же сон, близкий к наркозу, действуют менее эффективно;
в) при систематическом применении снотворных препаратов от несколь-
ких дней до 1—P/s месяцев, в. зависимости от характера поражения
и его глубины;
г) при комбинации снотворных препаратов с другими терапевтическими
факторами, если в функциональной патологии болезненного состояния
четко выступают нарушения функций других систем организма (в част-
ности, с хлористым кальцием — в случаях воспалительных явлений
32 Э. А. Асратян
498
Э А АСРАТЯН
или увеличения проницаемости капилляров, с витаминами Вх и С—при
соответствующих гиповитаминозах, с биологическими и химическими
антисептиками — при наличии инфекции и т. д).
Было- установлено также, что при перечисленных выше болезненных
состояниях организма энтеральный способ введения снотворных веществ
имеет ряд преимуществ (простота техники введения в организм, медленный
темп поступления в кровь и т. д.).
Терапия сном последствий органических поражений
нервной системы человека
Положительные результаты лабораторно-экспериментальной разработки
и применения терапии сном близких и отдаленных последствий органи-
ческих поражений нервной системы животных создали естественно-научные
предпосылки для перенесения нашей работы в клинику соответственных
заболеваний человеческого организма. Не без известного волнениям трево-
ги мы приступили к проведению терапии сном органических пораже-
ний нервной системы человека. Нами строго учитывался более высокий
уровень развития и организации нервной системы человека, главным обра-
зом верхних ее этажей, специфические условия и особенности генеза и те-
чения патологических состояний, а также исключительное многообразие
форм поражения нервной системы у человека и ряд других моментов. Мы
и не думали разработанный нами способ терапии сном травмированных
животных механически переносить в клинику человека. На работу в кли-
нике мы смотрели до известной степени как на продолжение нашей экспери-
ментальной работы в новых условиях и на новом объекте. Хотя мы, физио-
логи, тревожились в начале нашей работы в клинике, мы не расценивали
наш поступок как необоснованный риск. Вопреки распространенному
средн многих ученых стилю работы — начинать проверку своих гипо-
тетических положений о лечении того или иного болезненного состояния
прямо с клинического исследования, минуя фазу предварительных экспе-
риментов на животных, мы приступили к клинической работе после того,
как все основные вопросы были решены в экспериментах на животных.
Возникшие в клинике новые вопросы мы пытались опять-таки предвари-
тельно решать в параллельно проводимом лабораторном эксперименте на
животных.
Ориентировочная наша клиническая работа была начата еще в 1942 г.
на одном из фронтов Великой Отечественной войны, а затем в двух клини-
ках Ташкента. В основном же эта работа была выполнена мной и мои-
ми сотрудниками Е. В. Гуровой, Г. Т. Сахиулнной, А. М. Джавадян,
Р. С. Орловой и С. II. Ивановой за последние два года в ряде эвакогоспита-
лей и клиник Москвы при ближайшем участии лечащих врачей этих учреж-
дений (Институт неврологии ВИЭМ, клиники Института протезирования).
В порядке общей оценки результатов нашей клинической работы можно
сказать, что они полностью подтвердили нс только правильность исходных
наших теоретических положений, но также и правильность основных прин-
ципов экспериментально разработанного нами способа терапии сном.
Однако в процессе работы в клинике, в особенности в начальном периоде,
мы много внимания уделяли поискам лучших путей для приспособления
и применения этого способа к лечению последствии тех или иных частных
и специфических форм органических поражений нервной системы чело-
века, зачастую не воспроизводимых в условиях эксперимента на живот-
ных, Выяснению вопроса О Показаниях и противопоказаниях к его при-
менению И т. п.
Я не стану в данной статье описывать весь ход нашей работы в клинике,
все наши «зигзаги», удачи и неудачи в процессе этих поисков. Могу лишь
ПОСЛЕДСТВИЯ ОРГАНИЧЕСКИХ ПОРАЖЕНИЙ НЕРВНОЙ СИСТЕМЫ
499
сказать, что, к счастью, все возникшие затруднения были в основном пре-
одолены в первые же месяцы работы. Фактически в клинике мы внесли
только одно существенное изменение в разработанный нами способ терапии
сном, заключающееся в добавлении анталгических средств к снотворным
препаратам при лечении больных с выраженным болевым синдромом. Мы
прибегли к этому потому, что установленные нами оптимальные терапев-
тические дозы снотворных не вызывали у указанных больных сна доста-
точной глубины и длительности, а от применения слишком больших доз
снотворных мы воздерживались по теоретическим и практическим соображе-
ниям. Включение обезболивающих средств в состав предложенной нами
снотворной смеси в данном случае удовлетворяло требованиям комплексной
терапии об одновременной борьбе против всех или основных звеньев
«порочного круга» заболевания. При лечении каузалгни, например, этим
принципом руководствуются многие наши клиницисты (И. И. Русецкий
и др.). В качестве наиболее подходящего обезболивающего препарата
мы избрали эйкодал (текодин), который во многих отношениях выгодно
отличается от морфина, пантопона и других подобных препаратов (меньшая
токсичность, меньшая опасность привыкания к нему больных и т. гг.).
Накопленный в первом периоде клинических исследований опыт дал
нам возможность выработать несколько основных правил применения тера-
пии сном последствий органических травм нервной системы человека
и в последующем руководствоваться ими. Прежде всего мы стремились
возможно упростить технику применения нашего способа терапии. В этих
целях был разработан стандартный рецепт снотворных препаратов. За не-
имением в своем распоряжении достаточного количества гедоилла (наибо-
лее эффективного в данном случае препарата) мы включили в этот рецепт
уретан. А ввиду того, что уретан является слабым снотворным, мы вынуж-
дены были присоединить к нему также веронал. Учтя специфическое от-
ношение препаратов брома к нервной системе, мы включили в состав ре-
цепта также бромиды. -Этот рецепт нами предложен в двух вариантах.
Кроме указанных трех препаратов, входящих в состав обоих вариантов
рецепта в одинаковых дозах, в состав одного рецепта включен аналге-
зирующий препарат эйкодал, а в состав другого—пирамидон. Первый ре-
цепт предназначен для терапии сном посттравматических нарушений
нервной деятельности с выраженной гнпералгезией (каузалгия, фантом-
ные боли, корешковые боли и т. п.), а второй рецепт — для других форм
посттравматических нарушений.
1. Urethani ......... Г ,0
Verona И.............0,25
Natrii brotnati.......0,2
Eucodali.............0,01
2. Urethani..................1,0
V eronali.................0,25
Katrii bromati............0,2
Pyramidoni ...............0,05
Считаю нужным вновь подчеркнуть, что уретан и веронал с большей
пользой могут быть заменены гедоналом (0,7), а также бромуралом или
люминалом (менее эффективный).
Условно- мы называем приведенные составы «стандартными единицами»
терапии сном. В вопросе о назначении суточной дозы «единиц» мы руко-
водствовались следующим принципом: дать ту минимальную общую дозу
снотворных, которая обеспечивает в среднем 12—13-часовой крепкий ноч-
ной сон. Клинические наблюдения подтвердили правильность ранее сде-
ланного нами вывода о том, что менее длительный и глубокий сон, как
и очень длительный сон, приближающейся к наркозу, не дает необходи-
мых терапевтических результатов. В последнем случае у больных очень
скоро развивается общая слабость, появляются признаки грозного ток-
сикоза и т. п. Рабочая формулировка нашего принципа дозировки сно-
32*
500
Э. А. АСРАТЯН
творных такова: следует лечить не дозой снотворных, а дозой хорошего,
крепкого сна. Исходя из этого принципа, а также учитывая возраст, вес
и общее состояние больных, как и длительность естественного сна у них,
мы давали одну, две или три «единицы» {при необходимости приме-
няли и дробные ее части) перед ужином и после него с перерывами при-
мерно в 20—30 минут между приемом каждой единицы. Кроме того, боль-
ным с каузалгней (как правило) и больным с фантомными н корешковыми
болями (как исключение) мы давали по одной единице смеси 1 также днем.
Чтобы избежать опасности развития значительного токсикоза и сла-
бости больного при систематическом применении даже умеренных доз
снотворных и чтобы дать организму возможность освободиться от нако-
пившихся продуктов распада снотворных, мы в течение курса лечения
эпизодически устраивали перерывы («отдых»). На основании нашего опыта
в клинике мы пришли к заключению, что самым целесообразным и удоб-
ным вариантом «отдыха» является однодневный перерыв после шестиднев-
ного приема смеси. Мы пришли также к выводу, что полезно в период всего
курса лечения, в особенности при назначении состава 1, улучшать пита-
ние больных и давать им витамины Bi и С.
Нам долго не удавалось установить длительность курса лечения при
тех или иных формах нарушения нервной деятельности. Особенно сильно нас
дезориентировали те больные, у которых в начале лечения нашим способом
имели место неплохие сдвиги, но затем, по неизвестным причинам, темпы
этих сдвигов постепенно снижались. Стремясь добиться удовлетворитель-
ных конечных результатов, мы в первый период нашей работы лечили та-
ких больных 2—2’7* месяца и больше и, к нашему огорчению, без оссбого
у’спеха. Часто, несмотря на перерывы, больные ослабевали от затянувше-
гося курса лечения. В нескольких случаях мы, наоборот, очень быстро
достигали хороших результатов и сразу же прекращали лечение. У части
этих больных через короткое время появлялись рецидивы, которые не во
всех случаях поддавались быстрой ликвидации, в особенности если мы
опаздывали с возобновлением лечения. На основании дальнейшего опыта
мы пришли к определенным выводам, правильность которых была полно-
стью подтверждена последующей нашей работой, а именно: курс лечения
должен проводиться не меньше 30—35 дней, какие бы ни были достигнуты
хорошие результаты, и не дольше 40—50 дней независимо от достигнутых
результатов. Кроме того, мы пришли к выводу о целесообразности посте-
пенного уменьшения дозы снотворных в течение последней недели лече-
ния (на половину, на треть и па четверть обычной дозы).
Противопоказанием к применению нашего способа терапии мы счи-
таем болезни печени, почек и выраженные нарушения сердечно-сосуди-
стой системы. Во всяком случаемы избегали лечить больных с подобными
заболеваниями.
Разработанный нами способ терапии сном был применен всего у 143
больных с общими и местными последствиями: коммоцией—контузией го-
ловного мозга (14 больных); проникающими ранениями головного мозга
(14 больных); коммоцией—контузией, компрессией и проникающими ране-
ниями спинного мозга (30 больных); рефлекторными параличами и паре-
зами (3 больных); каузалгней (48 больных); фантомными болями (31 боль-
ной); гиперкинезами различной этиологии (3 больных). У 22 больных ле-
чение Сном ПО Той ИЛИ иной причине было Прекращено Через 3—10 дней
после его начала- Хотя эти больные не могут быть приняты do внимание
при общей оценке эффективности нашего способа терапии сном, тем не
менее следует отметить, что у многих больных, которые лечились не меньше
6—7 дней, намечались явные положительные сдвиги. Из 121 больного
прошедшего более или менее полный курс лечения сном (в среднем 30—
45 дней), хорошие результаты были получены у 60 (50%), удовл створи тел ь
ПОСЛЕДСТВИЯ ОРГАНИЧЕСКИХ ПОРАЖЕНИЙ НЕРВНОЙ СИСТЕМЫ 50!
ные — у 34 (28%), незначительное улучшение состояния отмечено у 16
(13%) и безрезультатным оказалось лечение у 11 больных (9%).
Приводимая ниже таблица дает представление о результатах лечения
последствий отдельных видов травматических поражений нервной системы.
. Вид поражения Общее число боль- ных Результаты лечения Преждевре- менное пре- кращение ле- чения
хоро- шие удов - лство- ри- тсль- ныс не-зна- читесь ныс от- сутст- вуют
Каузалгия Фантомные боли Специальные травмы Коммоция—контузия головного мо-зга . . Проникающие ранения черепа ...... Рефлекторные параличи—парезы - - . Гиперкинезы 43 31 30 14 14 3 3 23 13 6 10 6 2 13 4 II 3 2 1 3 6 2 1 3 1 4 5 2 9 4 6 3
Всего , 143 60 34 16 11 22
Многие больные после продолжения курса лечения находились под
наблюдением в течение 1—3 месяцев, а значительная часть — в течение
5—7 месяцев. Судя по данным этих наблюдений, достигнутые результаты
имеют стойкий характер. Более того, нередки случаи, когда после прекра-
щения курса лечения восстановленные функции продолжают прогрессивно
улучшаться.
Привожу несколько выдержек из историй болезни некоторых леченных
по предложенному нами способу больных.
Больные, леченные в госпиталях Г. Т. Сахиулиной при участии вра-
чей Артемьевой, Мирзоевой, Михайловой, А. М. Джавадян и Р. С. Орловой.
1. Больной Г-в. Проникающее ранение височной области; осколок находится
на глубине 6 см. Жалобы на упорные головные боли, особенно в области ранения.
Сильное нарушение речи и слуха; резкая гипотония и астения мышц. Ходит очень
плохо, находится в состоянии депрессии; пассивен, слаб; плохой аппетит. Лечение
продолжалось 45 дней. Больной выписан со следующим заключением: «В резуль-
тате лечения сном по методу проф. Асратяна наблюдалось значительное улучшение
в состоянии больного: больной стал активен, хорошо слышит, говорит, значительно
лучше ходит. Головной бо.чи нет; жалуется на боль только в области раны».
2. Больной Д-х. Пулевое ранение поясничной области < повреждением кореш-
ков конского хвоста. Жалуется на сильные постоянные боли в ногах. Резкое
ограничение движений во всех суставах ног. Не стоит, не ходит. После 40-днев-
ного курса лечения наши?! способом боли полностью исчезли; больной ходит па
костылях.
3. Больной Х-С. Пулевое ранение нижней трети правого плеча с частичным
повреждением срединного нерва. Резко выраженные каузалгические боли в руке.
Синестезалгня, алгическая контрактура, слабость, бледность, бессонница. После
44-дневиого лечения сном боли полностью исчезли, в результате чего резко увели-
чился и объем движений во всех суставах, Значительное улучшение общего
состояния.
4. Больной Г-й. Ранение разрывной пулей в левое плечо с повреждением кости
и плечевого сплетения. Резкие каузалгические боли. После 26-дневного лечения
боли полностью прекратились.
5. Больной P-в. Пулевое ранение в левое предплечье, правое бедро и правую
ягодицу. Рефлекторный спастический парез •-паралич правой ноги: отсутствие
движений в пальцах и голеностопном суставе, резкое ограничение движений
в коленном суставе, заметное —в тазобедренном. Зоны анестезии и гипестезии.
Стоять и ходить не может. В конце курса -лечения сном (40 дней) полностью вос-
станавливают!: я движения й пальцах и В голеностопном суставе, резко увеличи-
вается объем движений в коленном и тазобедренном суставах. Восстанавливается
Чувствительность. Больной стоит н ходит, хромая на правую н-огу.
502
Э. Л. АСРАТЯН
G. Больной К-н. Сквозное осколочное ранение в шейную область без поврежде-
ния мозга. Сильная контузия головного мозга. Полный паралич и анестезия правой
руки; сильный парез и гнпестезия правой ноги; головные боли. Стоит и ходит
с большим трудом. После курса лечения сном почти совсем исчез парез ноги,
больной стал хорошо стоять' и ходить; исчезли головные боли, восстановилась
чувствительность в. правой руке, а также умеренное движение в плечевом и лок-
тевом суставах и в меньшей степени — в пальцах,
7. Больной П-к. Осколочное ранение правого плеча. Сильно выраженная
каузалгня. Слнестезалгня, алгнческая контрактура, отек кисти. После 40-дневного
курса лечения сном исчезновение болей и алгичесхой контрактуры, спадение отека.
Больные из клиник Института протезирования. Лечащий врач Иванова
при участии врачей Орлов-ой и Иваненковой.
1. Больной Т-в. Осколочное ранение левой стопы и правой голени. Ампутация
и реампутация левой голени и правого- бедра. Резкие и постоянные фантомные
боли с обеих сторон. Лечение физиотерапевтическими процедурами не давало ощу-
тимых результатов. Лечение сном (39 дней) полностью сняло фантомные боли.
В дождливую погоду иногда ощущает тупые, слабые, скоро- проходящие боли.
2. Б-ольная П-ва. Травматическая ампутация левой голени и правого бедра.
Реампутация. Резко выраженные стойкие фантомные боли с обеих сторон. Бессон-
ница. Лечение сном (41 день) снимает фантомные боли н ощущения. Лишь време-
нами в дождливую погоду ощущает пальцы и легкую боль.
а. Больной Т-х. Пулевое ранение правой стопы и левого бедра. Реампутация.
Сильные и постоянные фантомные боли. Физиотерапевтические методы лечения пе
дают ощутимых результатов. Лечение сном (35 дней) почти полностью снимает
фантомные боли и ощущения.
4. Больной Ш-й. Осколочное ранение обеих голеней. Реампутация. Резко вы-
раженные и стойкие фантомные боли. Из многочисленных физиотерапевтических
процедур только УВЧ терапия оказала некоторое кратковременное положительное
действие. Лечение сном (38 дней) привело к ликвидации фантомных болей и ощу-
щений. Лишь в дождливую погоду появляются кратковременные слабые боли.
Больной из клиники Института неврологии АМН СССР. Лечащие вра-
чи Е, В, Гурова и Д, И. Лунев,
Больной К-н. Ушиб головы с кровоизлиянием в области основания черепа.
Перелом основания черепа (?). Ушиб левого голеностопного сустава. Нарушение
функций ряда черепномозговых нервов, головные боли, головокружение, тошнота,
бессонница, приступы резчайших болей в зоне иннервации левого тройничного
нерва. Положение в постели вынужденное. Не может стоять и ходить. Больной по-
лучил очень короткий курс лечения сном—всего 10 дней. Тем не менее состояние
больного значительно улучшилось: приступы стали весьма редкими и слабыми, резко
уменьшилась гиперпатия в области выхода тройничного нерва, исчезло вынуж-
денное положение; больной поправился, стал стоять и ходить.
Выше я уже указывал, что у определенной группы больных мы не могли
добиться каких-либо положительных терапевтических результатов или же
достигали весьма незначительного улучшения. У нас имеются основания
считать, что у некоторого числа этих больных мы не получили ожидаемых
результатов, потому что в первый период нашей работы в клинике мы не
знали еще многих важных деталей применения нового способа терапии.
Однако неудачи бывали и в последний период нашей работы. Чем их объ-
яснить? Одну из причин мы знаем довольно точно: это — хронический
алкоголизм больных. Нам не удавалось вызвать у них глубокий и длитель-
ный сон даже при назначении значительных доз снотворных. Это делает
понятным отсутствие у них желательных терапевтических результатов.
Однако нс все наши неудачи могут быть объяснены алкоголизмом боль-
ных. Нс исключена возможность, что в некоторых случаях наших неудач
в лечении каузалгии и фантомных болей причинами являлись грубые изме-
нения в периферических звеньях «порочного круга» (обширные невромы
и спайки нервов, их компрессия, воспалительные явления в них, грубое
нарушение кровообращения и т. д.). Очевидно,’ в таких случаях хорошие
результаты могут быть получены лишь при сочетании терапии сном с хи-
рургическими и иного рода воздействиями на эти «звенья». Возможно
ПОСЛЕДСТВИЯ ОРГАНИЧЕСКИХ ПОРАЖЕНИЙ НЕРВНОЙ СИСТЕМЫ
503
также, что часть наших неудач обусловливалась глубокими, необратимыми
изменениями в травмированных центральных нервных образованиях.
Могли быть, конечно, и другие причины.
Я закончу краткий обзор нашего клинического материала несколькими
дополнительными замечаниями общего характера.
В подавляющем большинстве положительных случаев через б—7 дней
после начала лечения появляются первые ощутимые признаки облегчения
основного страдания больного. В дальнейшем темпы восстановления на-
рушенных функций все ускоряются и через 36—45 дней приводят к ста-
бильным хорошим или удовлетворительным результатам.
Еще во время курса лечения у больных, принимавших состав 2, улуч-
шались, как правило, общее состояние, самочувствие и аппетит. В ряде
случаев отмечалось ускорение процесса заживления ран, а у некоторых
больных гипертонией—значительное снижение артериального давления.
Те же сдвиги наблюдались у определенной части больных, принимавших
состав 1. Однако у другой части больных последней группы в процессе ле-
чения появлялась некоторая общая слабость и ухудшение аппетита.
После окончания курса лечения эти явления быстро’ проходили.
За все время работы в клинике не было ни одного несчастного случая
или серьезного осложнения у леченных нами больных. Следует, однако,
отметить, что в начальном периоде нашей работы в клинике, т. е. в период
поисков оптимальных доз снотворных и рациональной смены дней лече-
ния и отдыха, у отдельных больных с каузалгнен и с фантомными болями
мы наблюдали проходящие явления тошноты и рвоты (состав 1).
Мы не наблюдали также более или менее заметного привыкания к сно-
творным смесям. Тем не менее должен заметить, что у части лечащих
врачей возникало подозрение относительно привыкания некоторых боль-
ных к снотворным. В этой связи следует указать, что после прекращения
дачи снотворных у небольшого числа больных в течение первых 2—3 дней
нарушался сон. Вскоре Эго, однако, проходило, и нормальный сон восста-
навливался даже у тех, кто раньше страдал выраженной бессонницей.
* «
Я»
В теории и практике охранительно-целебного торможения имеется еще
очень много невыясненных вопросов. К числу их относится важный вопрос
о механизме и природе целебного действия этого торможения. Современ-
ная физиология и клиника почти не располагают прямыми фактическими
данными по этому вопросу. Поэтому до поры до времени приходится удо-
влетворяться лишь догадками. При современном уровне развития нейро-
физиологии концепции Геринга (Hering), Гаскелла (Gaskell) и Ферворна
(Verworn) о торможении как о нутритивном или ассимилятор ном процессе
не могут считаться правильными. Классические исследования Н. Е. Вве-
денского и И.П. Павлова дают достаточно оснований считать процесс тор-
можения таким же активным процессом, как и процесс возбуждения. Воп-
рос заключается, очевидно, в природе этой активности и точке ее приложе-
ния. Мне кажется, что в связи со сказанным представляют значительный
интерес некоторые экспериментальные данные Вннтерштейна (Winterstein),
Усуи (Usui), Леба (Loeb), Жовстта (Jowctt) и других о том, что действие
на центральные нервные образования значительных доз некоторых сно-
творных средств, достаточных для прекращения внешней деятельности этих
образований, почти не отражается на количестве поглощаемого ими кисло-
рода. Создается впечатление, что активность нервных образований при
этом не прекращается и даже не ослабевает существенно, а переключается
на новые рельсы, приобретая новый характер и направление. Не исклю-
чена возможность, что подобная своеобразная активность охранительно-
504 Э. Л. АСРЛТЯН
целебного торможения и оказывает благотворное целебное действие на
истощенные и ослабленные нервные образования.
Я уже упоминал о том,что за последние годы в практической медицине
значительно усиливается тенденция к более систематическому применению
седативных, снотворных и наркотических препаратов в борьбе, с отдель-
ными последствиями травм нервной системы, а именно с последствиями
коммоции — контузии головного мозга (J1. Ю. Шпнрт и А. А. Кислицин,
А. О. Долин, А. Г. Иванов-Смоленский) и с каузалгней (Б. И. Серафимов,
В. В. Зикеев, Е. М. Стеблов, В. Л. Грингот, И. М. Сливко, И. И. Русецкий,
С. Д. Каминский, Ф. А. Андреев) Снотворные применялись также
для стимуляции регенеративных процессов в пораженной коже человека
(К. Л. Поляков).
Чем отличаются наши исследования и предложенный нами способ терапии
сном: от исследований и предложений перечисленных выше авторов, зани-
мавшихся изучением указанной проблемы частью одновременно и независимо
от вас (А. О. Долин, Л. Ю. Шпирт, К. Л. Поляков), а большей частью
позже нас?
Не касаясь существенных расхождений в теории изучаемой проблемы,
укажу вкратце на некоторые практически важные моменты, отличающие
нашу работу от работ перечисленных выше авторов. Эти моменты следу-
ющие:
а) обширные п возможно адэкватпые экспериментальные исследования
животных как обязательный предварительный этап к переходу в клинику;
б) выбор, проверка и включение в практику терапии сном более эф-
фективных и безвредных препаратов (гедонал, уретан, бромурал, эйкодал),
ранее не применявшихся для этой цели;
в) лечение сном, близким к естественному по характеру и отчасти по
глубине и длительности, а не почти круглосуточным наркозом или же сном,
близким к наркозу (А. О. Долин, А. Г. Иванов-Смоленский,Б. И.Сера-
фимов, В. В. Зикеев, Е. М. Стеблов, Ф. А. Андреев, С. Д. Каминский);
г) наш способ терапии сном является в основном самостоятельным
(при комбинации с другими лечебными мерами—основным), а не дополне-
нием к другим, более радикальным лечебным воздействиям (В. В. Зикеев,
Е. М. Стеблов и др.);
д) мы лечили и предлагаем лечить нашим способом не один или два
частных вида последствий травматических поражений нервной системы
(коммоция—контузия или каузалгия), а обширную группу близких и от-
даленных последствий различного рода травм нервной системы;
е) наш способ терапии сном выгодно отличается от существовавших
и вновь предлагаемых способов сонно-наркозной терапии физмологич-
ностыо, простотой и доступностью техники применения, безвредностью
и, судя по всему, большей эффективностью.
Бесспорно, предложенный нами способ терапии может быть в дальней-
шем усовершенствован. Но я уверен, что и в настоящем виде он в состоя-
нии принести немалую пользу в борьбе с последствиями боевых травм,
т. е. в разрешении одной из основных послевоенных задач теории и прак-
тики нашей медицинской науки. Я допускаю также, что новый способ
терапии сном может быть применен для лечения последствий функциональ-
ных поражений нервной системы.
Подытоживая результаты пятилетних экспериментальных и клиниче-
ских исследований нашего маленького коллектива по разработке нового
способа терапии сном последствий органических травм нервной системы,
я не могу не констатировать с удовлетворением, что эти результаты в об-
щем подтверждают исходные наши теоретические положения, в основу ко-
торых положены гениальные идеи бессмертного учителя И. П. Павлова.
Этим самым одна из преследуемых нами целей в тон или иной мере уже
ПОСЛЕДСТВИЯ ОРГАНИЧЕСКИХ ПОРАЖЕНИИ НЕРВНОЙ СИСТЕМЫ
505
достигнута. В отношении второй цели мы еще переживаем период тревож-
ных надежд. Я уже знаком с горечью переживаний авторов-теоретиков
по поводу печальной судьбы нх практических предложений, любовно раз-
работанных ими иной раз в трудных условиях. Моей радости не было бы
границ, если бы разработанному нами новому способу терапин посчаст-
ливилось возможно безболезненно и быстро войти в арсенал средств, с по-
мощью которых наши врачи ведут неустанную борьбу за восстановление
здоровья и трудоспособности лучших сынов нашей великой Родины.
Для общей же характеристики планов нашей дальнейшей исследова-
тельской работы я не нахожу лучших слов, чем слова, сказанные
И. II. Павловым об экспериментальной терапии почти полвека назад:
«Таким образом, эта только что народившаяся экспериментальная терапия
является лишь грандиозным результатом предшествующей физиологиче-
ской работы, который дает современному физиологу полную уверенность
в надежности его пути и необыкновенно возбуждает его энергию для бу-
дущих работ»1.
Приношу сердечную благодарность проф. II. И. Гращенкову и Е. К. Мо-
лодой, а также докторам Шейхаметову, Голованову, Мелкумовой и др.
за помощь и поддержку в нашей работе в клиниках и госпиталях.
литература
Асратян Э. Л., Новые данные ПО физиологии большого мозга, Архив био-
логических наук, 3, 54, 1941.
Асратян Э. Л., Учение Павлова об охранительной и целебной роли тормо-
жения и травмы нервной системы, Невропатология н психиатрия, 4, 34, 1944.
Ас то а цату ров М., Сборник избранных трудов, 1939.
Бурденко Н. Н., Ампутация как нейрохирургическая операция, 1942.
Да виден ков С., Травма периферических нервов, Вопросы нейрохирургии,
2, 56, 1944.
Долин А. О., Здродопе ка я И. И., Резников С. И., Сперан-
ская М. Н., Экспериментально-лабораторное и клиническое изучение сотрясений
н контузий головного мозга и их лечение на физиологической основе, Тезисы до-
кладов научной сессии по Павлову, 1942.
Иванов-Смоленский А. Г., О нарушениях нервной деятельности копту-
зионно-комыоиионного происхождения, Военно-медицинский сборник АН СССР, 2,
193 » 205, 1945.
Каминский С. Д. и Резников Л. Б., К патогенезу и терапии каузалгии,
Невропатология и психиатрия, 6, 30, 1944.
Орбели Л. А», Лекции по физиологии нервной системы» 1935.
Павлов И. П., Полное собрание трудов, 1, 1940.
Поленов А. Л., Повреждение костей черепа, мозга, оболочек и черепных
нервов, Основы практической нейрохирургии, 1943.
Русецкий И. И., Боль, ее формы и патогенез, 1944.
Серафимов Б, И,, Опыт лечения каузалгии длительным сном. Невропато-
логия и психиатрия, 1, 15, 1944.
Сперанский А. Д., Нервная система в патологии, 1930.
Членов Л. Г., О патогенезе каузалгии, Вопросы нейрохирургии, 3, 7, 1914.
Bost г oern A., Mohr's Handbuch ftir innere Medizin, 15, 631, 1943.
Cushi ng H., Brit, journ. surg., 5, 558, 1917—1918.
Denny-Brown, New England, journ. med., 227, 771, 1941.
Denny-Brown, Brain, 64, 223, 1941.
Doupe-Cullen, Journ. neurolog., neurosurg. a. psychiatr.. 7, 33, 1944.
F. ri ch sen I.. On concussion of the spine, nervous shock etc., 1882.
Goltz Е.» Pfliiger's Arch. ges. Physiol., 51, 570, 1892; 76, 411, 1899.
Groat, Magoun, Amer, journ. physiol., 141, 117, 1944.
Grant ahi, Ueber die Erkennung der traurnatischen Hirnverletzung, 1936.
G u r d j i a n a. Webster, J. surg., gynecol. a. obstetr., 76, 623, 1943.
Gurdjiana. Webster, Amer, journ. surg., 63, 236, 1944.
Head a. Holmes, Brain, 34, 102, 1911.
И. П. Па в лов, Полное собрание трудов т. I, 1940, стр. 364.
506
Э. А. АСРАТЯН
J ас к son J., Brit. med. journ., 1, 591, 188-4.
Ler iche R., La chirurgie de la douleur, 1937.
Lewandowski M., Die Funktionen des Zentralnervensystems, 1907.
Lewis T., Pain, 1942.
Lucian i L., Human physiology, HI, 1915.
Marburg a. Helf a nd, Injuries of the nervous system, 1939.
Mecluria a. Crawford, Arch. surg.. 16, 451, 1528.
Minkowsky M., PflUger’s Arch., 141, 171, 1911.
Mitchell \V., Amer, journ. med. sci., 73, 305, 1877.
Moore Ch., Journ. Amer. med. assoc., 108, 859, 1937.
Peab a. Raffler. Bull, war med., 5, 98, 1944.
Reichardt M., Uethe's llandb, d. norm. u. path. Physiol., 10, 193, 1927.
Sherrington Ch.. The integration, action of the nervous system, 1906.
Sherrington a. Brawn, G r a h a tn, Journ. physiol., 13, 209, I9ll.
Takats, Arch, neurol. a. psychiatr., 50, 318, I943.
Tine I, Le systtme nerveux vegetatif, 1967.
Trendelenburg W., Zschr. f. Biol., 65, 103, 1915.
К ТЕОРИИ И ПРАКТИКЕ ПАВЛОВСКОГО
ОХРАНИТЕЛЬНО-ЦЕЛЕБНОГО ТОРМОЖЕНИЯ*
течение 8 довоенных лет я и мои сотрудники проводили
экспериментальные и теоретические исследования в одной из
актуальных областей современной биологии и медицины —
по проблеме приспособляемости, или пластичности, нервной
системы. Мы изучали не обычные приспособительные явления
здорового организма, а приспособительные явления особого
типа, возникающие в организме после органического поражения различ-
ных частей центральной или периферической нервной системы, рецепторов
или эффекторных органов.
Экспериментальные и клинические исследования в этой области велись
давно, но вяло и несистематично. Начиная с 30-х годов нашего столетня
внимание экспериментаторов к этой проблеме значительно усилилось, и
ее исследованием занялся ряд советских и зарубежных ученых. Наши
исследования существенно отличались от исследовании П. К. Анохина,
Бете, Фишера и др. Наиболее характерные особенности нашей работы
заключались в следующем.
1. В своей работе мы руководствовались основными положениями уче-
ния И, П. Павлова о роли и о рефлекторной деятельности коры головного
мозга.
2. Проведенная нами работа по объему значительно превышала работу
других ученых. Наши исследования осуществлялись не на десятках, а
на сотнях животных, притом стоящих на различных ступенях эволюцион-
ной лестницы.
3, Для изучения приспособительных явлений в организме мы подвер-
гали подопытных животных не одному-двум видам оперативного воздей-
ствия (в целях нарушения тех или иных функций), а десяткам различных
зидов оперативных воздействий.
4. Для изучения характера последствий травм нервной системы и дру-
гих органов, а также динамики приспособительных явлений мы пользо-
вались возможно большим числом современных методик физиологического
исследования; мы не ограничивались только наблюдениями и фото-кинэ-
сьемкой подопытных животных, но применяли также методики Для Точ-
ного измерения тонуса скелетных мышц, для оценки состояния рефлек-
торной деятельности, для измерения хронаксии нервов и рефлекторных
дуг, для ныработки оборонительных условных рефлексов на пораженные
конечности и т. д.
5. И, самое главное, для ответа на основной и спорный вопрос о роли
верхних отделов центральной нервной системы в этих приспособительных
явлениях мы нс ограничивались только локальными поражениями отдель-
1 Труды Объединенной сессии, посвященной 10-летию со дня смерти И. П. Пав-
лова, 1946.
508
Э. Л. АСРАТЯН
них частей высших отделов центральной нервной системы, как делали дру-
гие ученые, а практиковали полную экстирпацию этого органа, т. е. пол-
ное удаление переднего мозга у низших позвоночных и коры большого
мозга у высших позвоночных (главным сбразом у ссбак).
Наш систематический, разнссбразный и, как нам кажется, более полно-
ценный экспериментальный материал позволил придти к выводам и теоре-
тическим положениям, которые шли вразрез с выводами и теоретиче-
скими положениями других исследователей; эти выводы, однако, полностью
гармонировали с основными установками И. П. Павлова о функциях коры
большого мозга и о закономерностях ее деятельности. Паши эксперимен-
тальные исследования нового к.часса приспособительных явлений, кото-
рые возникают после органических повреждений нервной системы и дру-
гих частей организма, позволили значительно расширить круг биологи-
ческих явлении, охватываемых учением И. II. Павлова.
Основные выводы сводятся к следующему.
1. По нашим данным, основная роль среди многочисленных факторов,
участвующих в создании приспособительных явлений в поврежденном ор-
ганизме (тренировка, мобилизация запасных путей и центров, функцио-
нальное внкарирование, адаптация, запредельное торможение, простая пе-
рестройка функций под воздействием периферических импульсов и т. п.),
принадлежит деятельности высших отделов центральной нервной системы,
а у высших млекопитающих — деятельности коры большого мозга,
2. Роль большого мозга в явлениях приспособления тем больше, чем
Выше стоит животное на эволюционной лестнице. У высших животных Кора
большого мозга приобретает значение решающего фактора,
3. Роль большого мозга в явлениях приспособления осуществляется
не только с помощью трофического воздействия на низшие отделы цен-
тральной нервной системы; на это были сделаны указания л до нас
(динамогенные или сблегчающие воздействия, по данным других авторов).
Влияние большого мозга осуществляется благодаря безусловнорефлек-
торной и, самое главное, условнорефлекторной деятельности — благода-
ря неограниченным возможностям перестраивать деятельность низших
отделов центральной нервной системы, организовывать новые виды инте-
гративной деятельности и т. д.
Работы многих клиницистов о лечении пострадавших в Отечественной
войне воинов показали, что наши выводы и положения были в основном
правильны.
Полученные нами фактические данные и высказанные нами теорети-
ческие положения расширяли диапазон наших исследований в этой сб*
ласти и одновременно с этим они все больше приближали нас к задачам
практической медицины.
В дни Великой Отечественной войны мы начали изучать органические
поражения нервной системы человека. По существу мы продолжали ра-
ботать над той же проблемой, но с совершенно иной точки зрения. Мы
искали новых путей и споссбов ускорить и усовершенствовать эти приспо-
собительные явления, оказать воздействие на течение и исход напряжен-
ной борьбы больного организма за восстановление нарушенного здоровья.
И на этот раз путеводной звездой для нашего исследования явились кон-
цепции нашего гениального учителя И. II. Павлова.
По какому пути шли наши поиски? Уже давно было известно, что по-
следствия органических поражений центральной и периферической нерв-
ной системы обусловливаются пе только тем, что травма разрушает или
необратимо изменяет какие-то нервные очаги и пути, но также и тем, что
она косвенно изменяет и функциональное состояние других нервных эле-
ментов на обширной территории, ввергает их в своеобразное патологиче-
ское состояние. При этом указанная косвенная, или отраженная, функцио-
О ПАВЛОВСКОМ ОХРАНИТЕЛЬНО-ЦЕЛЕБНОМ ТОРМОЖЕНИИ 509
нальная патологическая альтерация охватывает преимущественно области,
лежащие как поблизости от очага прямого и первичного поражения, так
и вдали от него, но находящиеся в тесной функциональной связи с послед-
ним. Плохо нс только то, что вовлечение обширных неразрушенных ча-
стей мозгового вещества в. патологический процесс расширяет объем на-
рушения функций организма, но и то, что такой патологической функцио-
нальной альтерацией из строя прежде всего выводятся уцелевшие от раз-
рушения нервные клетки и пути; последние в дальнейшем должны взять
на себя функции нарушенных очагов и заполнить брешь в функциональ-
ном ансамбле, образовавшуюся вследствие уничтожения или необрати-
мой альтерации определенной группы нервных образований.
Наше внимание было привлечено именно к этим нервным образованиям.
Следовало ожидать, что если бы удалось возможно скорее вернуть в строй
эти нервные клетки я их пути, то тем самым удалось бы не только быстро
восстановить значительную часть нарушенных функции организма, обу-
словленных патологическом альтерацией, но и одновременно создать не-
обходимые предпосылки для стимулирования приспособительных процес-
сов (тренировки, мобилизация запасных возможностей, простой и сложной
перестройки функций и т. д.), направленных на восстановление нарушен-
ных функций и в той части организма, поражение которой обусловлено раз-
рушением или необратимой альтерацией известной группы клеток и путей.
Для достижения этой цели существует много путей, и практическая ме-
дицина эмпирически пользуется некоторыми из них. Нам казалось, что
на один из таких новых путей ускорения и усовершенствования процесса
нормализации функционального состояния этих патологически альтери-
рованных клеток и центров указывает учение И. П. Павлова сб охрани-
тельной н целебной роли торможения в патологической деятельности боль-
шого мозга, вызванной функциональными патогенными воздействиями.
Запредельное торможение и раньше причислялось нами к таким физиоло-
гическим факторам, которые при органических поражениях нервной си-
стемы играют существенную роль в восстановлении нарушенных функций.
Но теперь в наших новых исследованиях охранительно-целебная роль
торможения стала в центре нашего внимания. Мы сочли возможным рас-
пространить концепции И. П. Павлова об охранительно-целебной роли
торможения и на обширную область органических поражений нервной си-
стемы. Кроме того, мы распространили ее на всю нервную систему — снизу
доверху, условно даже иа все возбудимые ткани организма. Мы были убеж-
дены, что концепция И. П. Павлова в таком расширительном понимании, а
также его учение о типах нервной системы помогут нам понять многие явле-
ния, давно изученные физиологами, но недостаточно глубоко ими оценен-
ные с точки зрения биологической их роли. Мы надеялись,что они помогут
нам понять многие неясные стороны природы разных болезненных состоя-
ний организма, вызванных органическими травмами нервной системы и
других систем организма; мы рассчитывали, что они помогут найти пути
рациональной терапии этих болезненных состояний.
Органические травмы являются не простым патогенным фактором, а
представляют собой комплекс патогенных факторов, как бы гамму сопут-
ствующих и вторично возникающих факторов (инфекция, интоксикация,
спайки, рубцы и т. п.); эти факторы, действуя совместно или отдельными
группами, способны резко истощать и ослаблять нервную систему, дли-
тельно и сильно псрераздражать сс; таким образом, они способны созда-
вать все предпосылки для возникновения и развития в ней охранительно-
целебного торможения. Поэтому не может быть сомнения в том, что уче-
ние И. П. Павлова об охранительной и целебной роли торможения может
быть распространено и на последствия органической травмы нервной си-
стемы.
510
Э А. АСРАТЯН
Есть ли основание распространять эту концепцию на всю нервную си-
стему — на головной, спинной мозг, на периферические ганглии и г. д.?
Прежде всего мы имеем в виду общее положение современной физиологии,
которое с особой силой было подчеркнуто И. П. Павловым. Согласно этому
положению, все явления и процессы, протекающие в коре большого мозга,
своими корнями генетически связаны с явлениями и процессами в низших
отделах центральной нервной системы. Кроме того, у И. П. Павлова
имеются указания, что запредельное торможение, т. е. одна из разновид-
ностей охранительного торможения, имеет свою аналогию в процессах,
протекающих в нижележащих частях нервной системы. В порядке даль-
нейшего развития и конкретизации этих мыслей И. П. Павлова мы более
10 лег назад указывали на то, что явления пессимума Н. Е. Введенского
и явление угнетения спинальных рефлексов при сильном раздражении
чувствительных нервов, установленное впервые И. М. Сеченовым, могут
рассматриваться как выражение охранительного торможения. Следует
отметить, что ни у И. Е. Введенского, ни у И М. Сеченова не имеется ука-
Рис. 1. Рефлекторное сокращение конечности в ответ на электрическое раз-
дражение ее (объяснение в тексте)
заний на эту биологически важную сторону изученных ими явлений. Л с
точки зрения концепции И. П. Павлова эти явления приобретают другое
значение, биологически осмысливаются глубже и полнее.
Доказательством того, что охранительное торможение является уни-
версальным для всех частей нервной системы, может служить также сле-
дующее. Шеррингтон давно отметил, что в феномене утомления спиналь-
ных рефлексов существенную роль играет развитие торможения. Однако
он не оценил этого явления в достаточной степени, На основе учения
И. II. Павлова оно приобретает большое биологическое содержание. Еще
ЗаДолГо До Войны мы уделяли значительное внимание изучению этого явле-
ния, но, к сожалению, работа была прервана войной. Особенно ценные
данные, ярко иллюстрирующие охранительную роль торможения в дея-
тельности спинного мозга, были получены нашей сотрудницей Д. И. Хану-
тиной.
У подопытных ссбак оперативным путем травмировался спинной мозг.
Сильное ритмическое электрическое раздражение одной из задних лап
очень быстро вызывало «утомление» ее рефлексов (на кимограмме видно,
что они почти исчезают). Если пссле этого сразу же переходили к более
слабым ритмическим раздражениям, рефлекторная деятельность вновь во-
зобновлялась почти в полном объеме (рис. 1). Совершенно бесспорно, что
сильное раздражение вызывало пар абиотическое или, лучше сказать, охра-
нительное торможение в ослабленных и истощенных спинальных нервных
образованиях. Имеется еще много данных (о них ниже), которые также
позволяют считать, что охранительно-целебное торможение не является
специфической особенностью нервных образований коры большого мозга,
а универсальным свойством всех нервных элементов.
Можно ли на основе учения И. П. Павлова по-новому осветить некото-
рые неясные явления в болезненных состояниях организма, возникших
в. результате органической травмы центральной и периферической нервной
системы, рецепторов и т. п.? Если подойти к этим явлениям не только.
О ПАВЛОВСКОМ ОХРАНИТЕЛЬНО-ЦЕЛЕБНОМ ТОРМОЖЕНИИ
511
с точки зрения концепции И. П. Павлова об охранительно-целебной роли
торможения, ио и с точки зрения его учения о типах нервной системы, то
многие из этих явлений могут быть освещены и объяснены более удовлет-
ворительно, чем это имело место до сих пор в экспериментальной невро-
логии животных и в клинике органических поражений нервной системы
человека. Этот вопрос детальнее нами ссвешен в ряде работ. Здесь мы
ограничимся лишь обсуждением природы патологических состояний цен-
тральных звеньев «порочных кругов» при тех болезненных состоя ниях, ко-
торые возникают в результате органической травмы нервной системы.
Мы присоединяемся к мнению авторов, считающих, что по крайней
мере в поздних стадиях болезненных состоянии именно эти звенья являются
ведущими; поэтому на них мы и концентрировали внимание. Нам кажется,
что физиологическая сущность явления центрального угнетения, которое
развивается при органических поражениях тех или иных частей централь-
ной нервной системы или периферических нервов и в той или ином степени
лежит в основе параличей- и парезов центрального и рефлекторного про-
исхождения, сводится в основном к развитию и доминированию стойкого
охранительно-целебного торможения в ослабленных и истощенных цент раль-
ных нервных образованиях. Мы имеем в виду физиологическую природу
явления, которое давно известно в литературе под названием центрального
шока, диасхизиса и т. п. То, что до сих пор считалось неизбежным и не-
желательным отрицательным следствием органическом травмы, в новом по-
нимании представляется как проявление целесообразной биологической ре-
акции природной самозащиты: развитие охранительно-целебного тормо-
жения предохраняет клетки от дальнейшего истощения и ослабления,
устраняет опасность их гибели и оказывает на них целебное действие,
С этой точки зрения физиологическую сущность интрацентральных про-
цессов, возникающих при фантомных болях, каузалгии, спастических ре-
флекторных параличах и т. д.,-можно видеть в возникновении и доминиро-
вании стойкой повышенной возбудимости (или стойкого процесса возбуж-
дения) в соответственных альтерированных нервных образованиях.
Почему органическая травма нервной системы дает в одних случаях одни
результаты, в других — другие, в одних случаях — развитие и доминиро-
вание стойкого охра китель но-иелебного торможения, в других — развитие
стойкой возбудимости или возбуждения? Известно, что это зависит от мно-
гих факторов: от характера, локализации и тяжести травмы, от сопутст-
вующих осложняющих факторов и т. п. Мы полагаем, что в этом вопросе
существенную роль играет также тип нервной системы пострадавшего.
Этот вопрос — один из сложных в клинике органических поражений, и
в его разрешении существенную помощь может оказать учение И. П. Пав-
лова о типах нервной системы.
И. П. Павлов показал, что принадлежность животных к тому или иному
типу может явиться фактором, определяющим характер последствий па-
тогенных функциональных воздействий на организм. Пам кажется, что
типовая принадлежность нервной системы может играть большую роль
также в характере последствий органической травмы организма, в част-
ности, органической травмы нервной системы. Мы допускаем, например,
что природная или приобретенная слабость нервной системы с нарушенным
балансом между процессом возбуждения и торможения в пользу послед-
него является благоприятной почвой для развития и доминирования стой-
кого охранительно-целебного торможения. Если же при наличии сильной
или слабой нервной системы баланс процесса возбуждения и торможения
нарушен в пользу первого (природно или под воздействием предраспола-
гающих факторов), это создает благоприятную почву для развития и до-
минирования стойкого процесса возбуждения или повышенной возбуди-
мости.
512
Э. А. АСРАТЯН
Исходя из представлений о природе этих болезненных состоянии и об
универсальном значении охранительно-целебного торможения, мы сде-
лали следующее предположение. Если придти на помощь природе в тех
случаях, когда охранительное торможение развивается, но недостаточно
интенсивно, и в известных пределах усилить это торможение медикамен-
тозными снотворными средствами, то, быть может, нам удастся ускорить
процесс нормализации этих поврежденных клеток и тем самым создать
благоприятные предпосылки для развития приспособительных явлений
в организме. Мы ожидали такой же целебный результат от применения
снотворных средств и в тех случаях, когда охранительно-целебное тормо-
жение не развивается вовсе или развивается в очень слабой степени, т. е.
в тех случаях, когда в результате органической травмы в центральной нерв-
ной системе превалирует стойкая повышенная возбудимость или возбуж-
дение. Ведь в лабораториях И. П. Павлова подобные целебные результаты
от применения снотворных средств были получены как в случаях, когда
под воздействием патогенной функциональной травмы организма в коре
большого мозга возникает и превалирует охранительное целебное тормо-
жение, так и в тех случаях, когда под таким же воздействием в коре боль-
шого мозга возникает и превалирует процесс возбуждения. По аналогии
мы ожидали, что и здесь искусственно вызванное охранительно-целебное
торможение окажет положительное действие как в одних, так и в других
случаях последствий органической травмы нервной системы.
Для проверки правильности наших предположений нами совместно с
сотрудниками были поставлены специальные эксперименты.
При проведении терапии сном для борьбы с разнообразными послед-
ствиями органической травмы центральной и периферической нервной си-
стемы мы отказались от трафаретного пути переноса в эту новую область
тех способов наркозной терапиц, которыми пользуются клиницисты для ле-
чения некоторых психических и нервных заболеваний. Мы учитывали ток-
сичность применяемых при этом снотворных и наркотических препаратов,
употребление их в чрезмерно больших дозах, парабиотическую или нарко-
тическую глубину сна — торможения, а также такие побочные и, с нашей
точки зрения, вредные сдвиги в организме больного, как азотемия, аци-
доз, токсемия, гипертермия. Поэтому мы считали целесообразным огра-
ничиться глубиной сна, приближающегося к естественному; такой сон вы-
зывался небольшими дозами нетоксических снотворных средств.
От экспериментальных исследований на животных мы ожидали не
только подтверждения наших положений о полезности терапии сном
при органических травмах нервной системы, по и подтверждения или от-
рицания наших соображений о характере и особенностях такой терапии.
Иначе говоря, мы должны были экспериментально разработать основы
нового способа терапии сном.
Мы далеки от мысли, что в условиях эксперимента на животных мы
сможем воспроизвести во всем объеме и деталях те последствия органиче-
ских травм, которые наблюдаются у человека при травматизации его на
войне или в быту. Тем не менее эксперименты на животных оказывают
большую помощь в разрешении вопросов, подобных тем, которые мы поста-
вили перед собой; они ориентируют нас, они, пусть даже только в основ-
ных и грубых чертах, дают правильный ответ на интересующие физиолога
и клинициста вопросы.
Мы опускаем изложение результатов своей и своих сотрудников экспе-
риментальной и клинической работы по этиологии, функциональной па-
тологии н терапии сном травматического шока — они достаточно хо-
рошо известны по нашим докладам и публикациям. Мы кратко приведем
только те экспериментальные и клинические данные, которые относятся
к последствиям органической травмы нервной системы и организма вообще.
О ПАВЛОВСКОМ ОХРАНИТЕЛЬНО-ЦЕЛЕБНОМ ТОРМОЖЕНИИ
513
Мы вызывали у наших животных: 1) отек и набухание головного мозга
путем непосредсгвенной его травмы или путем удаления шейных симпа-
тических узлов; 2) травму спинного мозга путем перерезки задних его стол-
бов на уровне шейных сегментов или путем броун-секаровского его се-
чения на разных уровнях (Э. А. Асратян); 3) анемизацию центральной
нервной системы или отдельных ее частей (Г. Т. Сахиулина): 4) термиче-
скую травму спинного мозга (Е. А. Романовская); 5) органические пораже-
ния продолговатого мозга путем уколов или перерезки (С. Н. Иванова);
6) рефлекторные параличи (Ё. В. Гурова); 7) коммоцию—контузию голов-
ного или спинного мозга (Л. В. Симукова) и т. п. При этом наши экспери-
Рис. 3. Восстанув-
ление рефлектор-
ной деятельности
у контрол ьны х
(столбики с двой-
ной штриховкой) и
леченных сном
(столбики с орди-
нарной штрихов-
кой) лягушек пос-
ле длительной ане-
мнзации их цен-
тральной нервной
системы:
Рис. 2. Восстановление дви-
гательных функций у со-
баки после анемизацни
спинною мозга
Столбики с двойной штрихов-
кой — вртлая <в днях), через-
которое контрольные животные
стоят холят, бегают. Столбики
с ординарной штриховкой—вре-
мя, через которое те же функ-
ции осуществляются животны-
ми, леченными сном I, //„
III — отдельные животные
менты охватывали представителей как высших, так и низших позвоноч-
ных. Последствия этих и некоторых других разновидностей органиче-
ской травмы различных отделов нервной системы в виде различного рода
моторных, сензорных, сосудистых и трофических расстройств нам хорошо
были известны как ПО литературным Данным, так и по прежним нашим
исследованиям. Тем це менее мы изучали их вновь. Животных разделили
на две группы: контрольных и подопытных; часть животных (контрольные)
с меньшими поражениями не подвергалась лечению, а другая часть (под-
опытные) с более тяжелыми поражениями получала снотворные вещества
в течение 10—20 дней и больше.
Во всех случаях, когда терапия сном подопытных животных прово-
дилась рационально, период выпадения функций был более кратковремен-
ным, быстрее возникали приспособительные явления, уровень их был бо-
лее высоким и, что особенно важно, достигнутые результаты отличались
большей стойкостью, чем в случаях, когда снотворные вещества не приме-
нялись. Некоторые примеры приведены на рис. 2, 3, 4 и 5.
Положительное действие терапии сном было отмечено нами у жи-
вотных после броун-секаровского сечения, спинного мозга, после пере-
резки задних столбов спинного мозга, после органической травмы про-
долговатого мозга, после тепловой или «анемической» травмы спинного
мозга, при экспериментальном рефлекторном параличе и парезе задних ко-
33 Э. А. .Асратян
514
Э А АСРАТЯН
вечностей, а также при лечении последствий ряда других видов органи-
ческих поражений нервной системы у низших и высших позвоночных.
Полученные данные привели нас к заключению, что снотворные веще-
ства при всех экспериментально вызванных поражениях функций нервной
системы укорачивают период их выпадения и ускоряют восстановление на-
рушенных или утраченных функций. Более того, при грубых поражениях,
представляющих смертельную опасность для животных, снотворные ве-
щества увеличивают выживаемость животных или удлиняют их жизнь.
Рис. 4. Кинограмма тех же опытов, что на рис. 3
а — контрольная лягушка (опыт 11 III 1944 г), начало
перфузии жидкости Рингер-Локка 13 ч 05 м., перевязка
аорты 13 ч 35 м . стрелка указывает момент снятия с аорты
лигатуры, в-лягушка леченная сном (опыт 10 IV 1Й44 г )
начало перфузии жидкости Рингер-Локка I I ч 15 м , перевяз-
ка аорты 11ч 45 м . стрелка указывает момент снятия с
аорты лигатуры и введения уретана
Наилучшие результаты мы получали в тех случаях, когда применяли
не такие токсические снотворные вещества, как хлоралгидрат, амитал-нат-
рий, люминал и т. п., а такие, как гедоиал и уретан отдельно или в сочета-
нии с вероналом и бромидами. Далее, эти результаты были особенно яр-
кими, когда снотворные давались в дозах, вызывавших не наркоз или
очень глубокий сон, а сон, близкий к естественному по глубине, харак-
теру и длительности (т. е. 10—12 часов в сутки). В тех случаях, когда
мы применяли эти снотворные вещества в дозах, которые вызывают
глубокий сон, близкий к наркозу, иначе говоря углубляли торможе-
ние до глубины парабиоза, мы имели, как правило, значительно более
слабый, нередко даже отрицательный эффект, т. е. ухудшение состояния
подопытных животных по сравнению с контрольными.
Благоприятное действие оптимальных доз известной группы снотвор-
ных веществ на течение восстановительных процессов при органических
поражениях центральной и периферической нервной системы у эксперм-
О ПАВЛОВСКОМ ОХРАНИТЕЛЬНО-ЦЕЛЕБНОМ ТОРМОЖЕНИИ
515
ментальных животных дало нам основание осторожно применить эти веще-
ства в клинике органических поражений нервной системы человека. Пер-
вые шаги были сделаны еще в 1942 г. на фронте, а также в Ташкенте; более
систематически мы занимались этим начиная с 1943 г. Наши клинические
наблюдения охватывают около 150 больных с каузалгией,фантомными бо-
лями, со спинальными церебральными трав-
мами, контузиями головного мозга и т. д. Эти
данные уже опубликованы, поэтому останавли-
ваться на них подробно мы не будем. Укажем
лишь, что примерно у 75% больных были до-
стигнуты удовлетворительные или хорошие
терапевтические результаты стойкого харак-
тера.
Как представить себе «механизм» целебного
действия торможения? Мы не будем сейчас об-
суждать вопрос, локализуется ли это торможе-
ние в клетках или в синапсах, заключается ЛИ
его сущность в химических, физиологических
или каких-либо других изменениях. Мы выс-
кажем только соображения, возникшие у нас
при знакомстве с некоторыми данными других
исследователей. Винтерштейн, Леб, У суя, Жовет
и некоторые другие зарубежные исследователи
в процессе своих экспериментальных исследова-
/ П Ш
Рис. 5. Восстановление дви-
гательной функции у собак
с перерезанными на уровне
П шейного сегмента задни-
ми столбами спинного мозга
Обозначения те же, что на
рис. 2
иий, преследующих разные цели, отмстили один
важный факт, который.
с нашей точки зрения, может быть очень полезен для понимания Природы
целебного действия охранительного торможения на альтерированные цен-
тральные нервные образования. Они подвергали воздействию наркотиче-
ских веществ изолированные части центральной нервной системы в до-
бивались прекращения внешнего выявления их деятельности; при этом
количество поглощаемого нервной тканью кислорода существенно не умень-
шалось.
Нам кажется, эти данные позволяют высказать следующее суждение:
при охранительно-целебном торможении происходит не ослабление жиз-
ненных процессов в нервной ткани, а какая-то перестройка, изменение на-
правленности и характера этой активности. Клетки как бы замыкаются
в себе, всю свою жизненную активность направляют на структурное
и функциональное восстановление тех нарушений, которые в них про-
изошли под влиянием тех или иных факторов. Прямых доказательств этого
еще не получено, но такое предположение весьма вероятно. В настоящее
время мы вернулись к детальному систематическому изучению этого и
ряда других актуальных вопросов обсуждаемой проблемы, чего мы не
имели возможности делать в годы войны, когда были заняты в основном
практическим решением этой проблемы.
ИДЕИ И. П. ПАВЛОВА И КЛИНИКА ОРГАНИЧЕСКИХ
ПОРАЖЕНИЙ НЕРВНОЙ СИСТЕМЫ 1
дной из характерных. чертИ. П. Павлова как ученого был по-
стоянный интерес его к вопросам функциональной патологии
и терапии изучаемых им органов и систем. Эта черта великого
экспериментатора и теоретика нашла особенно яркое отраже-
ние в его классических исследованиях, большого мозга.
Разработке вопросов функциональных поражений большого
мозга И. П. Павлов посвятил почти 25 лет своей творческой деятельности.
Гениальный мыслитель искусно синтезировал богатейшие результаты своих
исследований в учении о патологических состояниях больших полушарий
и отчасти в учении о природе сна, в учении о типах нервной системы.
Результаты «физиологических экскурсов» И. П. Павлова в важную об-
ласть функциональной патологии мозга человека—в область нервных и
психических болезней—всем известны.
Одновременно следует отметить, что вопросы органических поражений
большого мозга занимали И. П. Павлова преимущественно в начальном пе-
риоде создания им учения об условных рефлексах в связи с исследованием
проблемы локализации функций в коре большого мозга. В поставленных им
опытах было получено много ценных фактических данных об общих законо-
мерностях деятельности большого мозга при органических его поражениях.
Однако на той стадии развития учения об условных рефлексах И. П. Пав-
лов не мог так же полно развить и углубить данные об органической па-
тологии. как это было им впоследствии сделано в отношении функциональ-
ных поражений большого мозга. В то время многие закономерности нор-
мальной деятельности большого мозга не были еще известны, учение
И. П. Павлова о сне только зарождалось и еще не было речи об охранитель-
ной и целебной роли торможения, о физиологических основах типов нерв-
ной системы, о последствиях функциональных воздействий на большой
мозг и т. и. Возвращаясь к этой теме много лет спустя, И. П. Павлов с до-
садой говорил, что, вероятно, при этих «сведениях утилизация материала
с экстирпациями была бы шире и глубже».
Совершенно естественно, что ученики И. П. Павлова, работающие над
дальнейшим развитием его богатейшего научного наследия, не могли пре-
дать забвению этот важный раздел научного творчества учителя. Во время
Великой Отечественной войны была начата разработка на основе учения
И. П. Павлова вопросов, связанных с органическими поражениями нерв-
ной системы. Результаты проведенных в этом направлении работ Л. А. Орбе-
ли, А. Г. Иванова-Смоленского, А. О. Долина и др. известны.
1 Доклад, прочитанный на 11-м совещании по физиологическим проблемам, по-
священном 10-летию со дня смерти И, П. Павлова, 23 HI 1946 г. Сборник «Клини-
ческая физиология органов чувства, изд. АМН СССР, 1949.
ИДЕИ ПАВЛОВА И КЛИНИКА ОРГАНИЧЕСКИХ ПОРАЖЕНИИ НЕРВНОЙ СИСТЕМЫ 517
Цель данного сообщения — показать возможность извлечения из со-
кровищницы фактических данных и всего учения И. П, Павлова значи-
тельной пользы для клиники органических поражений нервной системы
человека.
Анализ богатейших фактических данных лабораторий И. П. Павлова,
а также клинических наблюдений, собранных нами и нашими сотрудни-
ками, позволяет осветить ряд неясных вопросов этиологии, функциональной
патологии и терапии некоторых болезненных состояний, вызванных ор-
ганическими поражениями центральной и периферической нервной си-
стемы. В качестве иллюстрации коснемся некоторых из них.
Данные экспериментальной и клинической неврологии свидетельствуют,
что характер, тяжесть, локализация и длительность последствий травма-
тических поражений нервной системы зависят не только от характера,
локализации и тяжести самой травмы, от сопутствующих факторов (крово-
течение, инфекция, воспалительные явления, рубцевание, спайки и т. п.),
от возраста и общего состояния пострадавшего, от характера и эффектив-
ности лечебных мероприятий и т. и., но также от каких-то еще недостаточно
выясненных факторов. В клинике органических поражений нервной си-
стемы довольно обычными являются случаи самых разнообразных функцио-
нальных расстройств при примерно одинаковом течении ранения и уходе.
В качестве примера можно привести последствия ранения или контузии
тех или иных периферических нервов. В значительном большинстве слу-
чаев эти последствия сводятся к чувствительным, двигательным, сосуди-
стым и трофическим нарушениям, адекватным травме по топике, тяжести
и т. п. Отраженных явлений при этом либо не бывает, либо они выражены
слабо. Нередки, однако, и другие случаи, когда при таких же типах травмы
на первое место выступают обширные отраженные яв.ления в виде каузал-
гических и фантомных болей, контрактур, спастических и вялых парали-
чей, сосудистых и трофических расстройств, не адэкватных травме. Нам
кажется, что к разрешению этого важного вопроса необходимо применить
учение И. П. Павлова о типах нервной системы и о значении типологи-
ческих особенностей нервной системы для характера реакций большого
мозга при патогенных воздействиях на него.
Какие имеются основания для использования концепций И. П. Пав-
лова при анализе последствий органических повреждений большого мозга,
а тем более последствий повреждений других частей нервной системы?
Всякое, даже ограниченное органическое поражение центральной или
периферической нервной системы более или менее сказывается на всей
системе, во всяком случае на тех областях ее, которые топографически
близки или функционально тесно связаны С очагом поражения. Следователь-
но, органическая травма определенных участков нервной системы (независи-
мо от того, обратимы ли возникшие при этом очаговые расстройства или нет)
является одновременно причиной функционального поражения других зон
этой же системы. В отношении коры большого мозга правильность этого
положения не вызывает сомнения. Сам И. П. Павлов неоднократно ука-
зывал на производные или косвенные изменения функции одних частей
коры большого мозга при ограниченных органических поражениях дру-
гих его частей. По нашему мнению, нс может вызвать особых возражений
и допущение, что органическая травма периферических нервов, проводя-
щих путей или клеточной массы низших отделов центральной нервной
системы также способна обусловить закономерные изменения функцио-
нального состояния клеток коры большого мозга.
Если кора большого мозга вовлекается в патологический процесс при
органической травме любой части центральной или периферической нерв-
ной системы, то имеются все основания полагать, что при патогенных воз-
действиях на нервную систему, вызывающих в последней органические на-
518
Э. А. АСРАТЯН
рушения, типологические особенности нервной системы должны играть
существенную роль в определении характера последствий поражения. При
этом следует отметить, что наши давние экспериментальные наблюдения
на собаках, лишенных коры больших полушарии, позволяют сделать
вывод, что типологические особенности свойственны не только нервным
элементам коры большого мозга, но и более низко расположенным обра-
зованиям мозга, в первую очередь базальным ганглиям. Опыты показали,
что после декортикации мозга собаки в примитивной форме сохраняют
основные черты темперамента.
Мы полагаем, что типологические особенности нервной системы играют
важную роль в формировании характера последствий органических
трамв нервной системы даже тогда, когда поражение локализуется в пери-
ферических нервах и в низших отделах центральной нервной системы,
не затрагивая непосредственно коры большого мозга.
Возможно, что в развитии патологического синдрома известную роль
играет функциональное состояние нервной системы в момент травмы.
Клиницисты и экспериментаторы давно оценили значение таких пред-
располагающих ко многим болезненным состояниям факторов, как утомле-
ние, истощение, голод, холод, интоксикации, бессонница, психическое
напряжение и т. п. Пам кажется вероятным предположение, что эти пред-
располагающие факторы, изменяя основные типологические функциональ-
ные свойства нервной системы, оказывают влияние на патогенез болезнен-
ных состояний, вызванных органической се травмой.
Значение типовых особенностей нервной системы при этих состояниях
сводится к изменению ее силы, равновесия и подвижности основных нерв-
ных процессов — возбуждения и торможения.
На основе учения И. П. Павлова об охранительно-целебной роли тор-
можения можно понять также физиологическую природу некоторых пато-
логических состояний, развивающихся в центральной нервной системе
после органической травмы. С нашей точки зрения, физиологическую сущ-
ность функционального угнетения, вызванного органической травмой раз-
личных отделов центральной нервной системы, можно свести в основном
к развитию стойкого охранительно-целебного торможения в соответствую-
щих альтерированных нервных образованиях.
Положение И. П. Павлова о сходстве основных процессов во всей нерв-
ной системе позволяет допустить, что не только нервные элементы коры
большого мозга, но и все вообще нервные элементы способны при соответ-
ствующих ситуациях развивать охранительно-целебное торможение.
Физиологическую сущность ннтраиентральных явлений при каузалгии,
фантомных болях, .рефлекторных параличах и других подобных состоя-
ниях можно свести в основном к развитию и доминированию стойкого цент-
рального возбуждения в соответствующих нервных образованиях. Боль-
шую роль играет при этом наличие значительного нарушения баланса
между процессами центрального торможения и возбуждения в пользу
последнего (при этом нервная система может быть как сильной, так и сла-
бой) либо аналогичное функциональное состояние, созданное внешними
факторами.
Сформулировав основные положения, мы должны ответить на вопрос,
соответствуют ли высказанные нами теоретические соображения точно
установленным данным клиники органических поражений нервной си-
стемы человека.
Мы думаем, что наши представления о важной роли типовых особен-
ностей и функционального состояния нервной системы в момент ранения
не противоречат представлениям клиницистов о существенном значении
других факторов в генезе и развитии этих болезненных состояний.
Все эти ранее выявленные и перечисленные нами выше факторы создают
ИДЕИ ПАВЛОВА И КЛИНИКА ОРГАНИЧЕСКИХ ПОРАЖЕНИЙ НЕРВНОЙ СИСТЕМЫ 519
в совокупности «порочные круги» патологических состояний, взаимодей-
ствующие звенья которых располагаются частью на периферии, частью на
различных уровнях центральной нервной системы. При этом наше внима-
ние было сосредоточено преимущественно на центральных звеньях этих
«порочных кругов» как ведущих звеньях в цепи патологически альтери-
рованных нервных образований.
Может возникнуть вопрос, ие переоцениваем ли мы роль центральных
звеньев «порочных кругов» в патологии подобных болезненных состояний.
Нам кажется, что нет. Многие передовые невропатологи давно указывали,
что периферические звенья «порочных кругов» играют ведущую роль лишь
в начальных стадиях развития каузалгий, фантомных болей и подобных
состояний: со временем, однако, центральные звенья как факторы, опреде-
ляющие динамику процесса, берут верх. В последнее время это положение
было подкреплено многими убедительными данными (И. И. Русецкий и др.).
Необходимо учесть, правда, что за последние десятилетня нейрохирур-
гия обогатилась весьма ценными данными о положительном целебном
эффекте оперативного воздействия на симпатическую нервную систему
при ряде болезненных состояний, которые клиницисты оценивают как до-
казательство постоянного превалирования периферических звеньев «по-
рочных кругов» над центральными. Обнаружение изменений в структуре
удаленных симпатических узлов рассматривается как веское подкрепле-
ние этой точки зрения. Не противоречат ли эти клинические данные нашим
представлениям? Нам кажется, что нет. Ближайшее ознакомление с этими
данными современной нейрохирургии показывает, что роль симпатиче-
ской нервной системы в этнологии и функциональной патологии каузалгии,
фантомных болей, рефлекторных параличей и подобных состояний боль-
шинство клиницистов понимает односторонне. Односторонним является
не только представление о том, что положительный эффект десимпатиза-
ции артерий, рамисекции или ганглноэктомии обусловливается якобы
только сосудодвигательными изменениями на периферии или же перерез-
кой болепроводящих путей, идущих по симпатической цепочке, но и те
сравнительно новые представления, на основании которых этот эффект
связывают со снижением уровня чувствительности периферических рецеп-
торов.
Если ограничиться приведенными выше трактовками результатов ган-
глиоэктомии и рамисекции, можно сказать, что при этом учитывается отно-
шение симпатической нервной системы только к периферическим образо-
ваниям — к сосудам, нервам, рецепторам и т. п. А между тем многочислен-
ные данные свидетельствуют о том, что симпатическая нервная система,
находясь в подчинении высших отделов головного мозга, в свою очередь
оказывает влияние на функциональное состояние самой центральной
нервной системы снизу доверху.
Патологическая альтерация тех или иных частей симпатической нервной
системы не может быть безразличной для центральной нервной системы.
И в таких случаях положительный терапевтический эффект ганглиоэкто-
мии и рамисекции можно трактовать в значительной мере как результат
прекращения поступления патогенных симпатических импульсов в цент-
ральную нервную систему.
Таким образом, мы видим, что данные нейрохирургии о положитель-
ном действии ганглноэктомии и рамисекции при последствиях органиче-
ской травмы периферических нервов ле только не противоречат допущению
о превалировании центральных звеньев «порочных кругов» в функци-
ональной патологии этих болезненных состояний, но даже подкрепляют
это допущение. Более того, становится весьма вероятным также пред-
положение, что изменение состояния симпатической нервной системы
может играть Известную роль в развитии патологических состояний
520
Э А. АСРАТЯН
при органических травмах различных отделов центральной нервной си-
стемы. Это предположение в настоящее время получило эксперименталь-
ное подтверждение. В частности, нами и нашими сотрудниками —
Д. И. Ханутиной, II. А. Соловьевой, Р. О. Барсегян и Б Д. Стефанцо-
вым — за последние 10 лет были получены экспериментальные данные,
показывающие, что после экстирпации брюшной симпатической цепочки
резко ухудшается функциональное состояние перерезанного спинного
мозга у лягушек. У собак после симпатэктомии также резко ухудша-
ется состояние травмированного спинного мозга, в частности, ослабевает
его способность к своевременному развитию охранительного торможения
при интенсивной рефлекторной деятельности.
Кроме этих данных, лег 5 назад нами были получены эксперименталь-
ные данные, показывающие, что повреждение симпатической нервной
системы играет определенную роль и в развитии отека и набухания
головного мозга.
В заключение нужно отметить следующее. За годы Великой Отечест-
венной войны нами и нашими сотрудниками — Л. В. Симуковой,
Г. Т. Сахиулиной, П. В. Станкевичем, Е. В. Гуровой, Е. А. Романовской
и С. II. Ивановой — была выполнена экспериментальная работа на хо-
лоднокровных и теплокровных животных, результаты которой нами рас-
сматриваются как существенное подкрепление формулированных выше по-
ложений о том, что охранительно-целебное торможение может развиваться
не только в пределах большого мозга при функциональных его поражениях,
но и во всей нервной системе и при любом типе поражений, г. е. при страда-
ниях как функциональных, так и органических. Исходя из этих положений,
мы предложили противошоковую жидкость, которая оказалась достаточно
эффективной как в экспериментах на животных, так и при широком приме-
нении ее в клинике и на фронтах Отечественной войны. Кроме того, на-
ми был разработай новый способ терапии сном применительно к послед-
ствиям органических поражений нервной системы, который выгодно от-
личается от существующих способов наркозной терапии своей доступ-
ностью, физиолотичностью, безвредностью и эффективностью. Он ПОЛНО-
СТЬЮ оправдал наши надежды как при экспериментальной терапии жи-
вотных с органическими травмами нервной системы, так и при предва-
рительной клинической проверке на людях (под нашим наблюдением на-
ходилось около 150 больных с каузалгней, фантомными болями, последст-
виями ранения или контузии спинного и головного мозга и др.).
ОБ ОХРАНИТЕЛЬНОМ ТОРМОЖЕНИИ И ЕГО ЗНАЧЕНИИ
В МЕДИЦИНЕ1
орошо известно, что нашей отечественной физиологии принад-
лежит громадная заслуга в разработке проблемы торможения.
Классическое открытие интрацентрального торможения
И. М. Сеченовым, а также его важное теоретическое положение
о роли торможения в деятельности нервной системы, по сущест-
ву говоря, открыли новую страницу в истории физиологии вооб-
ще и в истории физиологии центральной нервной системы в частности.
Вся последующая научно-исследовательская деятельность наших и за-
рубежных физиологов, занимающихся изучением центральной нервной си-
стемы. явилась дальнейшим развитием фактических данных и идей слав-
ного отца нашей отечественной физиологии. По существу все работы этих
ученых о координационной деятельности спинного мозга и других этажей
центральной нервной системы, о роли процессов возбуждения и торможе-
ния в координационной деятельности центральной нервной системы могут
рассматриваться как последующее развитие классических исследований
И. М. Сеченова,
Исследования ученика И. М. Сеченова Н. Е. Введенского, посвященные
выяснению природы торможения, несомненно, являются непревзойденными
ПО своей глубине. Никто из мировых Физиологов не сделал так много для
познания сущности процесса торможения, как Н. Е. Введенский.
Наконец, И. П. Павлов своими выдающимися исследованиями по физио-
логии больших полушарий мозга не только установил важную роль процес-
сов возбуждения и торможения в создании самой совершенной и высшей
координационной деятельности нервной системы, самого высшего прояв-
ления мозговой деятельности, но и сказал совершенно новое слово в науке,
открыл совершенно новую роль процесса торможения в деятельности боль-
ших полушарий мозга — охранительную и целительную его роль.
Однако, если нам, физиологам и патофизиологам, хорошо известны ве-
ликие заслуги И. П. Павлова в разработке и выяснении роли торможения
в координационной деятельности коры больших полушарий, то необходимо
признать, что мы не так хорошо знаем и недостаточно высоко оцениваем
грандиозное по своему значению открытие И. П. Павловым этой принци-
пиально новой роли торможения.
В чем же заключается сущность открытия И. П. Павлова?
Акад. И. П. Павлов впервые в истории науки (не в истории физиологии,
но вообще .в истории науки) показал, что в деятельности коры больших по-
лушарий мозга при определенных трудных ситуациях, при определенных
условиях перевозбуждения возникает торможение, значение которого со-
стоит не в том, чтобы, наряду с процессом возбуждения, создавать тот или
1 Стенограмма лекинн, (прочитанной 15. I 1951 г. на декаднике ЦПУ.
’ Сборник «Учение И. П. Павлова в теоретической и практической медицине»,
1951.
522
Э. А. АСРАТЯН
иной вид координационной деятельности, тот или иной вид высшей нервной
деятельности,—торможение возникает в целях охраны или защиты реактив-
нейших и нежнейших нервных образований.
Речь идет о торможении, которое защищает клетки коры больших по-
лушарий от опасности повреждения, а то и гибели, предотвращает серьез-
ную угрозу, возникающую при перевозбуждении клеток, в случаях, когда
они вынуждаются к выполнению непосильных задач, при катастрофиче-
ских ситуациях, при истощении и ослаблении их под влиянием различных
факторов. В этих случаях торможение возникает не для того, чтобы ко-
ординировать деятельность клеток этого высшего отдела нервной системы,
а для того, чтобы охранять, защищать их.
Торможение здесь имеет физиологическое значение естественного само-
охранения, самозащиты. И. П. Павлов говорил, что в этих случаях тормо-
жение выступает как «.нормальный прием физиологической борьбы» про-
тив болезнетворных факторов, как «физиологическая мера», предотвращаю’
щая угрозу разрушения нервных клеток коры вследствие непосильной
работы.
Какие же предпосылки, условия и факторы, по экспериментальным дан-
ным И. П. Павлова, способствуют развитию этого охранительного торможе-
ния? Чтобы понять это, необходимо остановиться на некоторых структур-
ных и функциональных особенностях нервных клеток и главным образом
клеток коры больший полушарий.
Нервные клетки, в особенности корковые нервные клетки, отличаются
значительной реактивностью, уязвимостью, чувствительностью ко всевоз-
можным изменениям во внешней и внутренней среде организма. Хорошо
известно, что многие клетки других тканей организма не только зависят от
централизованного снабжения необходимыми питательными продуктами
в широком смысле и так называемыми буферными веществами, но и сами
содержат некоторое количество тех и других. В этом отношении нервные
клетки, особенно нервные клетки коры больших полушарий, крайне бедны.
Другая особенность нервных клеток, которую следует учесть, чтобы
понять всю важность роли охранительного торможения, заключается в их
исключительно интенсивном функционированни-
Многочисленными биохимическими исследованиями установлено, что
окислительная активность многих нервных клеток гораздо выше, чем ряда
других клеток. При этом потребление кислорода нервными клетками в де-
сятки раз превышает потребление его другими клетками и тканями орга-
низма.
Эта крайняя чувствительность, уязвимость, эта непомерная «жадность»
к питательным продуктам, в особенности к кислородному и углеводному
питанию, и, наряду с этим, крайняя бедность собственными питательными
к буферными ресурсами делают нервные клетки очень хрупкими, подат-
ливыми ко всевозможным колебаниям как во внутренней, так и во внешней
среде организма.
Ближайшими предпосылками и стимуляторами, порождающими охра-
нительное торможение в клетках коры больших полушарий, согласно ис-
следованиям И. П. Павлова, являются их ослабление и истощение.
Если какие-нибудь сильные внешние агенты, действуя на организм,
ставят перед нервными клетками коры непосильную, как говорил
И. П. Павлов, задачу, вызывают их перевозбуждение и тем более длитель-
ное, ослабляют и истощают их, то в этих случаях и возникает «физиологи-
ческая мера» защиты в виде охранительного торможения.
При этом слабая по своей природе цервная система является особенно
благоприятной почвой для возникновения охранительного торможения.
«Такая нервная система,— писал И. П. Павлов,— при встрече с трудно-
стями ... или после непосильного возбуждения неизбежно переходит в со-
ОБ ОХРАНИТЕЛЬНОМ ТОРМОЖЕНИИ И ЕГО ЗНАЧЕНИИ В МЕДИЦИНЕ
523
стояние истощения. А истощение есть один из главнейших физиологиче-
ских импульсов к возникновению тормозного процесса как охранительного
процесса»1.
Охранительное торможение, естественно, может возникать и в сильной
нервной системе, если находятся такие раздражители, которые визы
ваюг длительное ее перевозбуждение. Это важно подчеркнуть, так как
при изложении этих концепций И. II. Павлова иногда представляют
дело таким образом, что возникновение охранительного торможения при-
суще только слабой нервной системе. На самом же деле удается вызвать
(хотя иногда и с трудом) истощение, ослабление и охранительное торможе-
ние в любой нервной системе. Разница лишь в том, что в одних случаях
это торможение возникает при одной степени и длительности перевозбуж-
дения, а в других случаях —при другой интенсивности и длительности
перевозбуждения. Предел, после которого возникает торможение, может
быть разный, но такой предел существует для нервной системы любой
силы.
В эксперименте на животных И. П. Павловым и его сотрудниками было
установлено, что в клетках коры больших полушарий мозга охранитель-
ное торможение закономерно возникает при применении сверхсилыюго
условного раздражителя. Известная закономерность силовых соотношений,
можно сказать, универсальная закономерность в деятельности нервной
системы вообще (т. е. закономерность, которая проявляется и в деятель-
ности примитивных нервных образований, и в деятельности коры больших
полушарий и согласно которой величина ответа субстрата в каких-то пре-
делах соответствует силе его стимуляции) извращается, когда усиление
условного раздражителя переходит известные границы.
Если постепенно усиливать условный раздражитель, например, звонок,
то при известных пределах усиления этот звонок как положительный услов-
ный раздражитель вызывает все более и более сильный условный эффект.
Однако наступает момент, когда дальнейшее усиление этого положитель-
ного условного раздражителя не только не обусловливает дальнейшего
нарастания условного рефлекса, но, наоборот, вызывает более слабый ус-
ловный рефлекс, который, в конце концов, может совершенно исчезнуть.
Таким образом, усиление условного или сигнального раздражителя выше
определенного предела может вызвать торможение, названное И. П. Пав-
ловым «запредельным торможеннемд
Это одно из проявлении новой, открытой И. П. Павловым физиологиче-
ской роли или функции охранительного торможения в нормальной высшей
нервной деятельности организма.
Другим проявлением охранительной роли торможения в нормальной
деятельности больших полушарий является периодический сон. Если поды-
тожить все фактические данные и все высказывания И. П. Павлова о физио-
логической природе нормального сна, Т, е, периодического естественного
сна, то мы можем сказать, что, по И. П. Павлову, сон как универсальное
торможение больших полушарий является выражением именно охрани-
тельного торможения. Ослабление и истощение нервных клеток, т. е. бли-
жайшие физиологические предпосылки и стимуляторы к возникновению
охранительного торможения, согласно учению Й. П. Павлова, возникают
во время сна не из-за кратковременного действия сверхсилыюго раздражи-
теля, а благодаря тому, что в течение длительного периода днев-
ного бодрствования различные раздражители, действуя непрерывно на
центральную нервную систему и, в частности, на корковые клетки,
вызывают длительную работу их, а. в результате этого ослабление
л истощение.
1 И. П. Павлов, Полное собрание трудов, т. III. 1949, стр. 410.
524
Э. А. АСРАТЯН
Возникшее при этом в каком-нибудь очаге охранительное торможение
быстро распространяется на всю кору, ослабленную и истощенную в основ-
ной своей массе длительным бодрствованием, а тем более интенсивной ра-
ботой. Затем это торможение распространяется вниз на подкорковые нерв-
ные образования, а в дальнейшем в какой-то степени и на нижние отделы
центральной нервной системы, обусловливая тем самым состояние естест-
венного сна, «обеспечивая им необходимый покой после ежедневной нор-
мальной работы», прекращая «дальнейшее функциональное разрушение
клетки» и способствуя восстановлению их нормальной работоспособности.
Я хочу обратить внимание на одну существенную принципиальную
черту учения И. П. Павлова о сне, которая, как правило, упускается из
виду при изложении современного состояния проблемы сна и при сопостав-
лении этого передового учения с другими современными теориями о сне,
принадлежащими в основном зарубежным ученым, а именно с токсеми-
ческой и сосудодвигательной теориями сна, с теориями, допускающими
наличие специальных центров сна в различных частях промежуточного
мозга, с теорией, объясняющей возникновение сна прекращением проприо-
цептивных импульсов, и т. п.
Все эти теории не могут быть поставлены в один ряд с учением
И. П. Павлова о сне не только потому, что они менее аргументированы до-
стоверными фактами и наблюдениями, что они менее убедительны по своей
идейной основе, что они односторонни и т. д., но и по той причине, что ни
одна из этих теорий, ни один из авторов или сторонников этих теорий не
ставит перед собой задачи освещения физиологической природы и сущ-
ности явления сна. Все без исключения теории сна толкуют лишь о том,
какие причины предрасполагают или порождают сон. какие факторы спо-
собствуют и какие противодействуют наступлению сна (и к тому же делает-
ся это односторонне, упрощенно), какова внешняя картина и динамика
сна ит. и., но ни одна из них нс может ответить на вопрос, в чем же
заключается сущность сна. Только наиболее передовая, наиболее аргу-
ментированная теория И. П. Павлова, точнее, учение его о сне, ставит
перед собой эту задачу и блестяще ее разрешает.
Исходя из учения И. П. Павлова, можно не только шире, глубже и
истинно научно осветить все вопросы, связанные с возник новен нем, раз-
витием и природой сна, но и оценить и осмыслить истинное значение от-
дельных достоверных научных данных относительно сна, полученных дру-
гими исследователями этой проблемы и неправильно истолкованных ими
в свете односторонних и ошибочных теоретических концепций. Таким обра-
зом, в настоящее время единственно научной теорией сна является уче
пне И. П. Павлова, согласно которому сущность сна сводится к торможе-
нию. мы можем сказать больше—к охранительному торможению.
Необходимо отметить далее, ЧТО И. П. Павлов выявил и исследовал
важнейшую роль охранительного торможения при патологической деятель-
ности коры больших полушарий. При этом он не ограничился накопле-
нием огромного в крайне ценного фактического материала относительно
экспериментальной патологии высшей нервной деятельности животных, ил-
люстрирующего роль и значение охранительного торможения в функцио-
нальной патологии мозга. Умудренный многолетним и богатейшим опытом,
маститый физиолог в последний период своей научной деятельности весьма
активно включился в разработку ряда весьма актуальных и сложных во-
просов патологии высшей нервной деятельности человека преимуществен-
но под этим углом зрения.
В своих блестящих экскурсах в область психиатрии и невропатологии,
проведенных на основании непосредственного изучения многих больных,
страдающих различными формами психических и нервных болезней,
И. П. Павлов дал блестящую физиологическую трактовку многих не-
ОБ ОХРАНИТЕЛЬНОМ ТОРМОЖЕНИИ И ЕГО ЗНАЧЕНИИ В МЕДИЦИНЕ
525
разгаданных, сложных болезненных состояний больших полушарий мозга
животных и человека. Он объяснял эти болезненные состояния главным
образом возникновением охранительного торможения в коре больших полу-
шарий и ближайшей подкорке, охранительного торможения хронического
характера, различной интенсивности, различной экстенсивности и различной
локализации. Если учесть, что при этом нарушается нормальный баланс
между двумя основными нервными процессами —возбуждения и торможе-
ния, нарушается нормальная картина их взаимоотношений и взаимодействии,
к тому же в разных случаях по-разному, то нетрудно понять самый факт
многообразия форм болезненных состояний, обусловленных в основном
наличием охранительного торможения в тех или иных частях больших
полушарий.
Все, что сделано И. П. Павловым в этой области, не только весьма ори-
гинально, весомо, но и крайне ценно для клиники, в особенности для пси-
хиатрической и невропатологической. И. П. Павлов считал, что при раз-
личных болезненных состояниях больших полушарий головного мозга,
в частности, при шизофрении, истерии и др., мы имеем дело со своеобраз-
ным, можно сказать, двойственным состоянием. С одной стороны, здесь
налицо физиологическое состояние, потому что болезнетворный фактор,
ослабляя и истощая нервную систему, прежде всего кору больших полу-
шарий, порождает в ней охранительное торможение как меру физиологи-
ческой самозащиты с ее благотворной функцией. А с другой стороны,
торможение как бы частично экстирпнрует кору больших полушарий пли
подкорку и в той или иной степени выводит из строя крупные или мел-
кие участки мозговой коры или ближайшей подкорки, тем самым нару-
шая нормальную целостную деятельность этого верховного органа, этого
верховного «распределителя и распорядителя* функций во всем организ-
ме. Своеобразное сочетание патологии и физиологии!
И. П. Павлов нс только дал блестящую физиологическую трактовку
многих болезненных состояний, не только раскрыл природу, физиологи-
ческую основу этих патологических состояний и тем наметил пути для
дальнейшей работы в этом направлении, но, верный своим давним и слав-
ным традициям, он стремился извлечь из всего этого практическую пользу,
стремился разработать меры борьбы с ними, научно обоснованные приемы
профилактики и лечения этих болезненных состояний.
Гениальный ученый проводил интенсивную работу' не только по экспе-
риментальной патологии высшей нервной деятельности, но эксперимен-
тально разрабатывал и ряд вопросов терапии, установив, в частности, что
средства, способные усиливать охранительное торможение, значительно
ускоряют восстановление нарушенной деятельности больших полушарий
мозга. Здесь выступает вторая сторона физиологической функции охра-
нительного торможения, а именно целебная, или, как говорил И. П. Пав-
лов, целительная функция торможения, функция восстановления нарушен-
ной деятельности коры больших полушарий и ближайших подкорковых
нервных образований.
Таким образом, еще в лабораторных исследованиях И. П. Павлова, на-
ряду с охранительной ролью торможения, была Показана и целебная его
роль. Это открытие имеет величайшее теоретическое и практическое зна-
чение. Оно лежит в основе того направления современной медицины, ко-
торое известно под названием лечения сном или сонной и наркозной те-
рапии различных болезненных состояний организма и прежде всего пси-
хических и нервных заболеваний.
К этому великому открытию И. П. Павлов пришел через лабораторию,
через лабораторный эксперимент. Свои теоретические положения он по-
строил на основании достоверных экспериментальных данных по патоло-
гии и терапии высшей нервной деятельности животных.
526
Э. А. АСРАТЯН
Известно, что эмпирическая медицина пользовалась, хотя и в весьма
ограниченных пределах, снотворными средствами для лечения некоторых
психических и нервных заболеваний человека. Однако эта практика, как
всякая практика, лишенная передовой теории, была слепа, и поэтому при
применении снотворных веществ для лечения психических и нервных за-
болеваний было очень много неудач. В силу этого эмпирическое направ-
ление лечения снотворными и наркотическими препаратами переживало
кризис, зашло в безвыходный тупик и неизбежно прекратило бы свое су-
ществование, если бы не слилось с мощным научным потоком, берущим
начало в лабораториях И. II. Павлова, с передовым и оригинальным
учением гениального физиолога об охранительной и целебной роли про-
цесса торможения.
Можно смело сказать, что научной основой терапии сном в целом,
этого интересного и перспективного направления современной медицины,
которое сейчас расширяется И распространяется на все новые виды болез-
ненных состояний организма, несомненно, является учение И. 11. Павлова,
об охранительной и целебной роли торможения, независимо от тсго, что
делалось до пего или одновременно с ним отдельными представителями
эмпирической медицины.
Л\ожно ли сказать, что И. П. Павлов успел исчерпывающе разрабо-
тать все вопросы, связанные с открытой им новой важной ролью торможе-
ния— охранительной и целебной его ролью? Бесспорно, нет. Как многие
другие разделы его величайшего материалистического учения о высшей
нервной деятельности, так и его учение об охранительно-целебном тормо-
жении должны стать предметом дальнейшей экспериментальной, теорети-
ческой и клинической разработки. Многие вопросы этой столь важной для
теории и практики проблемы являются нерешенными. Разрешить их, бес-
спорно, возможно, лишь следуя по пути, намеченному И. П. Павловым.
Неясно, в частности, что представляет собой охранительное торможение.
Идет ли речь о каком-то новом виде процесса торможения или это особая
сторона единого процесса торможения? Идет ли речь сб особой роли тор-
можения, наряду с координационной его ролью (мы предлагаем этоттер-
мин для внесения большей ясности), или же, наряду с координационным
торможением, существует также торможение охранительное? Если это два.
разных торможения, то какова связь между ними? Если это две функции
единого процесса торможения, то каковы взаимоотношения между ними?
У И. П. Павлова мы не находим четкого ответа на эти вопросы, да на ос-
новании того фактического материала, которым он располагал, он и не
мог его дать.
Неясно также, в каких микроструктурных образованиях нервной си-
стемы генерируется охранительное торможение,— в самих ли клетках,
в синапсах ли или в клетках и синапсах одновременно.
Мы не знаем также, что происходит с обменом веществ при охранитель-
ном торможении нервных клеток. А ведь этот вопрос имеет не только теоре-
тическое, но и сугубо практическое значение. Пока мы экспериментально
не выясним характера обмена веществ в нервных центрах, охваченных
охранительным торможением, мы не поймем по-настоящему природу охра-
нительного, целебного действия торможения,
Далее, при жизни И. П. Павлова не был достаточно ясен (да и сейчас
нуждается в дальнейшем уточнении) вопрос о Том, только ли клеткам боль-
ших полушарий присуще генерирование охранительного торможения или
же эта функция свойственна и нервным клеткам других образований,
Других отделов центральной нервной системы и нервной системы вообще.
Наконец, при жизни И. П, Павлова неясно было, возникает ли охрани-
тельно-целебное торможение в клетках больших полушарий тольков при
так называемых функциональных патогенных воздействиях млн же оно-
ОБ ОХРАНИТЕЛЬНОМ ТОРМОЖЕНИИ И ЕГО ЗНАЧЕНИИ В МЕДИЦИНЕ
527
возникает в этих клетках (и клетках других отделов нервной системы),
также при токсических и иного рода патогенных воздействиях, ведущих
к органическим изменениям в нервных клетках.
Вот далеко не полный перечень вопросов, которые были н в значитель-
ной степени остаются неясными, нуждались и нуждаются в дальнейшей
экспериментальной и теоретической разработке.
Несомненно, что если на эти вопросы будут даны четкие и убедительные
ответы, этим мы не только углубим и расширим круг наших знаний о при-
роде охранительного торможения, о закономерностях его возникновения
и течения, но еще больше вооружим нашу медицину, еще больше укрепим
то перспектив.ное направление в современной медицине, которое связана
с именем великого И. П. Павлова, — направление терапии сном различ-
ных болезненных состояний организма.
Что же сделано после смерти И. П. Павлова его учениками и исследова-
телями в развитие этого учения?
Можно сказать определенно, что кое-что нами сделано в этом направле-
нии, но пока крайне мало н в значительной мере не так, как следовало бы,
как было указано И. П. Павловым.
Говоря о более или менее значительных результатах работы наших
физиологов и клиницистов по линии дальнейшего развития учения
И. П. Павлова об охранительно-целебной роли торможения, следует прежде
всего указать на интересные и ценные лабораторные и клинические иссле-
дования М. К. Петровой, А. Г. Иванова-Смоленского, А. С Долина,
Ф. А. Андреева и других исследователей, в том числе и наши совместно
с сотрудниками, а также на исследования значительной группы наших
клиницистов по сонно-наркозной терапии психических заболеваний с по-
мощью ранее известных и новых методических приемов.
Разумеется, мы не можем дать здесь полный обзор результатов много-
летней работы большой группы наших ученых. Поэтому мы коснемся
результатов тех работ, которые представляют принципиальный интерес.
Остановимся прежде всего на достижениях в области теории обсужда-
емого вопроса. Известно, что экспериментальные данные лабораторий
И. II. Павлова и его теоретические положения относительно охранительно-
целебной роли торможения относились лишь к деятельности больших по-
лушарии головного мозга, а из болезненных состояний последних охваты-
вали лишь те, которые возникают при функциональных, патогенных воз-
действиях на организм.
13 плане дальнейшего творческого развития этих положений учителя
нами выдвинуты (и па протяжении 10 лет защищаются) теоретические по-
ложения о том, что, во-первых, охранительно-пслебноеторможениеприсуще
не только большим Полушариям мозга, но является универсальным свой-
ством всей нервной системы, и, во-вторых, о том, ч то это торможение воз-
никает в нервной системе не только при функциональных, но и при орга-
нических изменениях в ней.
Основанием для первого положения явились высказывания И, П, Пав-
лова относительно «естесгвенной общности» основных отношений и законо-
мерностей деятельности различных отделов нервной системы, в частности,
его указания относительно существования аналога запредельного тормо-
жения в низших отделах центральной нервном системы, некоторые косвен-
ные литературные данные относительно отдельных форм периферического
и спинального торможения и в основном значительный фактический ма-
териал, полученный в специальных экспериментах нами и нашими сотруд-
никами в предвоенные годы.
Из литературных данных, говорящих в пользу этого положения, можно
указать на феномен пессимума знаменитого нашего физиолога II. Е. Вве-
денского и на феномен торможения спинальной рефлекторной деятельности
528
Э. А. АСРАТЯН
при сильных раздражениях, описанный отцом нашей физиологии И. М. Се-
ченовым, а вслед за ним и другими отечественными и зарубежными физио-
логами (А- С. Дмитриев, Шеррингтон).
Мы глубоко убеждены в том, что истинное биологическое значение этих
явлений можно понять в полном объеме лишь на основе учения И. П. Пав-
лова об охранительной роли торможения. Необходимо отметить, что
II. Е. Введенский, открывший феномен пессимума, в полном объеме не
осветил, нс раскрыл биологического значения этого явления. Он лишь де-
тально охарактеризовал его природу, условия возникновения, течения
и т. п. И только в свете учения И. П. Павлова нам стало ясно биологиче-
ское значение этого явления, и мы даже можем использовать самый факт
пессимума для обоснования положения о том, что ох ранителыгое торможе-
ние является универсальным во всей нервной системе.
То же самое можно сказать относительно торможения спинальных реф-
лексов при сильных раздражениях. Эго было подтверждено в специальных
экспериментах наших сотрудников,-в частности, в экспериментахД. И. Ха-
пугиной, поставленных на собаках до и после удаления у них брюшных
симпатических цепочек.
Можно утверждать, что открытое И. П. Павловым в больших полуша-
риях охранительное торможение имеет характер универсального биоло-
гического принципа для всей нервной системы, а возможно, и для всех
возбудимых тканей. Конечно, способность к возникновению охранитель-
ного торможения, степень его выраженности, его динамика и некоторые
особенности его возникновения и течения могут быть разными для раз-
личных этажей центральной нервной системы, для нервных систем, стоя-
щих на различных уровнях эволюционного развития. Однако ясно, что
это явление присуще всем отделам нервной системы. Это положение может
считаться в значительной степени доказанным как нашими данными,
так и данными проф. А. Н. Магницкого и проф. А. С. Дмитриева.
Основанием для второго нашего положения относительно того, что охра-
нительно-целебное торможение возникает не только при воздействиях на
кору больших полушарий мозга, вызывающих функциональные изменения
в ней,—что изучалось экспериментально и клинически И. II. Павловым,—
но и при болезненных состояниях, сопровождающихся органическими из-
менениями в нервной системе, послужили прежде всего проведенные много
лет назад экспериментальные и клинические данные других исследовате-
лей, не оцененные ими с этой точки зрения и не понятые ими по-настоя-
щему. В качестве примера можно сослаться на Давно известные явления,
возникающие при механическом повреждении различных отделов централь-
ной нервной системы, в особенности высоко развитой, и вошедшие в лите-
ратуру под названием центрального шока, диасхизиса и под другими
названиями. Специальные исследования, поставленные в нашей лаборато-
рии, дают все основания утверждать, что длительное торможение, возни-
кающее при хирургических повреждениях тех или иных частей центральной
нервной системы, является охранительным торможением.
Аналогичные данные получены нами также при анемизацни и терми-
ческих повреждениях центральной нервной системы. С этой точки зрения
весьма интересны данные А. Г. Иванова-Смоленского и сотрудников отно-
сительно того, что охранительное торможение возникает и при различного
рода нейротоксикозах.
Нашими учеными достигнуты также некоторые успехи в области прак-
тического использования охранительно-целебного торможения. Прежде
всего следует отметить определенные достижения в применении экспери-
ментальной терапии сном и наркозом ряда болезненных состояний ор-
ганизма, преимущественно нервной системы. Напомню знаменитые опыты
М. К. Петровой, в которых она показала благотворное терапевтическое
ОБ ОХРАНИТЕЛЬНОМ ТОРМОЖЕНИИ И ЕГО ЗНАЧЕНИИ В МЕДИЦИНЕ
529
влияние физиологического сна и медикаментозного наркотического сна не
только ла течение экспериментальных неврозов у собак, но также на тро-
фические расстройства кожи и других органов (язва, экзема, папиллома,
облысение и т. п.), вызванных, как она считала, болезненным состоянием
больших полушарий мозга.
Весьма интересные и ценные данные были получены А. Г. Ивановым-
СмоленСКИМ И его сотрудниками относительно благотворного Действия дли-
тельного наркотического сна на течение нейротоксикозов у морских свинок
и у белых крыс, вызванных мескалином, тетраэтилсвинцом и другими хи-
мическими веществами. Определенных положительных результатов до-
бился А. О. Долин при терапии сном (наркозом) экспериментальной
коммоции и контузии у собак.
Обширные экспериментальные исследования, проведенные в руково-
димой нами лаборатории, с большой убедительностью показали благотвор-
ное влияние терапии сном при травматическом, ожоговом и тепловом
шоках, а также при поражениях, возникающих в результате анемизации
центральной нервной системы или в результате хирургических, механи-
ческих, тепловых и электрических воздействий на различные отделы цен-
тральной нервной системы ссбак, кошек, крыс и лягушек.
Клинически терапия сном и наркозом получила особенно широкое
применение в годы Великой Отечественной войны и послевоенные годы.
Если до войны сон но-наркозную терапию проводили лишь при ограничен-
ном числе психических и нервных заболеваний, то в настоящее время об-
ласть ее применения значительно расширилась. Если раньше этим зани-
мались единичные врачи, к тому же почти исключительно психиатры
(В. П. Протопопов, А. Г. Иванов-Смоле некий, Е. К. Краснушкин и др.),
то сейчас терапия сном и наркозом различных заболеваний применяется
значительным количеством хирургов, терапевтов, невропатологов, психиа-
тров, дерматологов, педиатров и др.
Несколько слов о недостатках нашей работы в этой области. Мы обра-
щаемся ко всем советским физиологам и патофизиологам, возглавляющим
эти дисциплины, потому что именно от них требуется вскрыть корни,
причины недостатков, принять все зависящие меры, чтобы устранить их
и тем самым способствовать дальнейшему успешному развитию важнейшей
части научного наследия И. П. Павлова — учения об охранительно-це-
лебной роли торможения.
В чем же заключаются основные недостатки нашей работы в данной
области?
Прежде всего — это недооценка учения И. П. Павлова об охранитель-
ной и целебной роли торможения нашими физиологами и патофизиологами.
Ведь не секрет, что это учение должным образом не освещается в курсах
наших лекций и что соответствующие гемы почти не фигурируют в наших
научных планах. Если бы мы отнеслись к этому серьезно и занялись ин-
тенсивной экспериментальной разработкой учения обохраниТелЬнО-Целеб-
ной роли торможения, то не было бы так много нерешенных вопросов, о чем
мы говорили выше (характер связи между координационным и охранитель-
ным торможением, сущность обмена веществ в нервных центрах при охра-
нительном торможении, установление структурных единиц, в которых оно
возникает, природа его и т. д.). А между тем удовлетворительное решение
этих вопросов представляет большой теоретический интерес и имеет ис-
ключительное значение для практической медицины.
Особенное внимание следует обратить на следующий недостаток. Как
было указано выше, в период Великой Отечественной войны у многих на-
ших исследователей и клиницистов резко возрос интерес к учению
И. П. Павлова сб охранительном торможении в чисто практическом ас-
пекте, т. е. многие стремились применять терапию сном или наркозом
34 Э А. Асратян
530
Э. А. АСРАТЯН
ДЛЯ лечения последствий боевых травм или ИНЫХ болезненных состояний.
Однако все это делалось и делается далеко не так, как следовало бы. Не
только клиницисты, но Даже физиологи и ПаТсфнЗиолоГИ, работающие
в указанном направлении, начинают применять терапию сном непосред-
ственно на больном человеке, без предварительной разработки соответст-
вующих вопросов в лабораторных экспериментах на животных.
Это является крупнейшим недостатком в работе некоторых наших уче-
ных, стремящихся внедрить в те или иные разделы медицины учение
И. П. Павлова об охранительно-целебной роли торможения. Подобные
приемы работы не имеют ничего общего со стилем и принципами работы
самого И. П. Павлова. К сожалению, мы часто забываем завет И. П. Пав-
лова, что, «чем полнее будет проделан опыт на животных, тем менее часто
придется больным быть в положении опытных объектов со всеми печаль-
ными последствиями этого»1.
Мы забываем завет И. П. Павлова, что все такие важные вопросы
должны быть предварительно' изучены, разрешены в экспериментах на
животных, что окончательная победа медицины, а значит и окончательная
победа нового терапевтического направления — терапии сном должна
придти через лабораторный эксперимент. Поэтому, если мы решили за-
няться дальнейшей разработкой учения И. П. Павлова об охранительно
целебной роли торможения и приложить его к тем или иным разделам прак-
тической медицины, мы должны прежде всего позаботиться об устранении
этого недостатка, сб исправлении допущенной нами крупной ошибки.
Многие вопросы терапии сном и наркозом остаются нерешенными,
ide решен, в частности, вопрос о том, какие снотворные наиболее эффек-
тивны и, значит, должны преимущественно применяться в этих целях.
Среди наших клиницистов по указанному поводу существуют большие
разногласия. Далее, нерешенными остаются и вопросы о дозировке сно-
творных, а также о наиболее эффективной глубине сна. До войны и в тече-
ние войны подавляющее большинство наших клиницистов лечило больных
длительным, 23 —24-часовым, глубочайшим сном, весьма близким
к наркозу. А между тем мы убеждены, что торможение такой глуби-
ны и длительности едва ли особенно эффективно в терапевтическом отно-
шении. Мы с сотрудниками, а вслед за нами и некоторые другие исследо-
ватели стали пропагандировать для этой цели торможение умеренной
глубины и длительности, максимально приближающееся ко сну. Хотя
понемногу эта идея и распространяется среди клиницистов, однако мож-
но утверждать, что вопросы выбора наиболее эффективных снотворных,
дозировки их, а также глубины сна остаются до сих пор нерешенными
удовлетворительно ни экспериментально, ни теоретически.
Удивительно лИ поСлё ЭТоГо, что До исторической Павловской сессии
среди наших клиницистов, преимущественно психиатров-невропатологов,
появилось отрицательное отношение к терапии сном? Известно, что мно-
гие из них считали, что на практике она себя не оправдала, и поэтому об-
ратились к другим способам борьбы с болезненными состояниями боль-
ших полушарий, в том числе к лоботомии.
Мы, физиологи и патсфизИологИ, как Представители Эксперименталь-
ной медицины должны в порядке самокритики признать себя виновными
атом, что наша медицина не извлекает всей возможной пользы из учения
И. П. Павлова об охранительно-пелебной роли торможения. Пора покон-
чить с ошибочным мнением и связанной с ним порочной практикой, будто
физиологам надлежит разрабатывать лишь вопросы, представляющие
чисто теоретический интерес, дающие «пишу для души*. Пора нам спус-
титься на землю, как это делал великий pf П. Павлов, и занято ся экспс-
1 И. П. Павлов, Полное собрание трудов, т. II, 1946, стр. 363.
ОБ ОХРАНИТЕЛЬНОМ ТОРМОЖЕНИИ И ЕГО ЗНАЧЕНИИ В МЕДИЦИНЕ аЗ|
риментальной и теоретической разработкой важнейших вопросов, непосред-
ственно связанных с учением И. П. Павлова об охранительно-целебной
роли торможения и имеющих важное значение для лечебной практики.
Очень важным является еще один вопрос, решение которого может
значительно усилить эффективность терапии сном. Речь идет о показа-
ниях и противопоказаниях к этому виду лечения.
Недавно нам прислали Из оДнон весьма ответственной организации
проект инструкции по терапии снсм. Там бь'ло перечислено около 60
нозологических единиц, рекомендованных для терапии сном, начиная
от травматического шока и кончая гипертонией. И для лечения всех этих
болезненных состояний рекомендовался единственный трафаретный прием
лечения сном или, точнее, применения снотворных средств.
Это является грубой сшибкой, И мЫ ДОЛЖНЫ Внести яСнсСТЬ В этот во-
прос, Необходимо точно установить, при каких болезненных состояниях
допустимо и полезно применение терапии сном, при каких оно недопу-
стимо. Необходимо разработать отдельные приемы применения снотвор-
ных веществ соответственно сссбеннсстям болезненных ссстся! й и т. в.
Это особенно важно сейчас, когда имеются признаки легксмысль кого от-
ношения к данной терапии, когда иные врачи готовы без особого размы-
шления, без предварительной лабораторной проверки применять терапию
снсм при самых различных заболеваниях, выдавая это своеобразное лиха-
чество в науке за внедрение идей И. П. Павлова в медицину. От такси
профанации ничего хорошего ждать нельзя.
Очень вджен также вопрос о комбинации терапии снсм с другими
лечебными средствами. Ведь ясно, что при воздействии многих болезне-
творных факторов на организм поражаются многие его системы, нару-
шаются многие его функции. Даже при как бы изолированном поражении
нервной системы, при различных как бы сбсссбленных локальных пато-
логических ее состояниях неизбежно, прямо или косвенно, нарушаются
функции многих других систем. Можно ли при таких условиях пользо-
ваться одними и теми же снотворными средствами и дозировками их,
одними и теми же трафаретными приемами терапии снсм при разнород-
ных болезненных состояниях? Конечно, нельзя!
Таким образом, мы считаем, что необходима тщательная и всесторонняя
разработка и экспериментальное решение физиологами и патофизиологами
вопросов о показаниях и противопоказаниях к терапии сном, наиболее
подходящих для этого снотворных средствах, их дозировках, сочетании
их с другими медикаментозными средствами и т. д. Эго решение должно
быть достигнуто путем лабораторного эксперимента на животных, как
это завещал нам великий И. П. Павлов, а не путем клинического экспери-
мента на больных.
Заканчивая наше сообщение, мы вновь подчеркиваем необходимость
усиления внимания к учению И. П. Павлова сб охранительней и целебней
роли торможения. Мы должны впредь в своей преподавательской работе
уделять Должное внимание этому учению, полнее ссвсщать его значение.
В своей исследовательской работе мы также должны больше заниматься
вопросами охранительно-целебной роли торможения, в том числе теми,
на которых мы остановились выше и которые имеют особое значение для
здравоохранения нашей страны,
Этим самым мы не только увеличим долю нашего участия в дальнейшем
развитии ценнейшего научного наследия Великого И. П. Павлова, важ-
ной частью которого является его учение об охранительной и целебней
роли торможения, но и примем деятельное участие во внедрении его ге-
ниальных идей в различные области теоретической и практической меди-
цины. _______
34»
йр--
ЭКСПЕРИМЕНТАЛЬНАЯ ТЕРАПИЯ СНОМ ПОСЛЕДСТВИЙ
ОРГАНИЧЕСКИХ ПОВРЕЖДЕНИЙ
ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ1
роблема торможения1 является одной из центральных проб-
лем современной физиологии нервной системы и физиологии
вообще. А нашей отечественной физиологии принадлежит не
только честь открытия внутрицентрального торможения, ио
и громадные заслуги в экспериментальной и теоретической
разработке наиболее сложных и важных вопросов этой акту-
альной проблемы. Выдающиеся достижения корифеев нашей отечественной
науки И,М,Сеченова, Н. Е. Введенского и особенно И. П. Павлова в экс-
периментальной и теоретической разработке проблемы торможения были
и по сей день остаются основным идейным истоком новых исследований
отечественных и зарубежных физиологов, направленных па расширение и
углубление наших знаний о природе процесса торможения и его роли в
деятельности различных отделов центральной нервной системы.
Вслед за открытием И. М. Сеченовым внутрицентрального торможения
и определения им важнейшей роли этого процесса в деятельности нервной
системы Н. Е. Введенский достиг выдающихся результатов в познании
внутренней природы этого процесса, установил его единство с процессом воз-
буждения, а также его важную роль в координационной деятельности спин-
ного мозга. Это направление творческой работы отечественных физиологов
было поднято на небывалую высоту гением И. П. Павлова. Он не только
выявил и детально изучил роль торможения в координационной деятель-
ности самых высших отделов центральной нервной системы, но и открыл
совершенно новую, весьма важную роль торможения в деятельности
нервных клеток коры больших полушарий мозга — охранительную и це-
лительную. Громадный фактический материал по этому вопросу, получен-
ный И. ГТ. Павловым и его сотрудниками в процессе многолетней экспери-
ментальной работы, послужил для него основанием для создания физио-
логического учения об охранительной и целебной роли торможения, являю-
щегося одним из самых замечательных достижений современного естество-
знания и имеющего огромное значение для теоретической и практической
медицины.
Согласно воззрениям И. П. Павлова, в условиях нормальной деятель-
ности корковых клеток, отличающихся от других нервных клеток гораздо
большей реактивностью, низкой работоспособностью и быстрой утомляе-
мостью, охранительная роль торможения выявляется в виде нестойкого
запредельного торможения при сверхснльных раздражениях или же в виде
каждодневного сна после длительного бодрствования и деятельности.
В том и другом случае возникшее торможение прекращает деятельность
истощенных нервных клеток, ограждает их от вредного влияния внешних
* Журнал высшей нервной деятельности, т. I, в 6, 1951
ТЕРАПИЯ СНОМ ПОСЛЕДСТВИЙ ОРГАНИЧЕСКИХ ПОВРЕЖДЕНИИ Ц. Н. С.
533
факторов, предотвращает опасность дальнейшего их истощения, создает
для них наиболее полноценный физиологический покой и способствует вос-
становлению их состояния и работоспособности.
Охранительная, защитная роль торможения в отношении клеток коры
больших полушарий мозга еще ярче выступает при их разнообразных бо-
лезненных состояниях. Одной из важных предпосылок для развития по-
добных состояний является природная или приобретенная слабость самих
клеток. Такие клетки под воздействием сверхсильных раздражении или
иного рода трудностей утомляются, истощаются и впадают в состояние
более или менее длительного торможения. Хотя в этих случаях торможе-
ние и выводит из строя значительные массы корковых клеток и тем самым
нарушает целостную деятельность мозга, тем не менее в отношении ослаб-
ленных и истощенных нервных клеток оно н здесь выступает в качестве
охранительного процесса, в качестве «физиологической меры*, способной
предохранять слабые нервные клетки от дальнейшего истощения, от угро-
жающего разрушения-вследствие непосильной работы, выступает «как нор-
мальный прием физиологической борьбы против болезнетворного агента»
(И. П. Павлов.).
Однако в отношении ослабленных и истощенных корковых нервных
клеток торможение выступает не только в роли природного- фактора само-
защиты, но и в качестве природного фактора само-лечення. Речь идет о це-
лебной или, как говорил И, П. Павлов, целительной роли торможения.
Торможение способствует нормализации состояния и восстановлению
функций ослабленных и истощенных нервных клеток, а тем самым и вос-
становлению нормальной, целостной деятельности больших полушарий
мозга.
Трудно переоценить значение фактических данных и теоретических по-
ложений И. П. Павлова об охранительной и целебной роли торможения не
только для физиологии, Но и для медицины, по крайней мере для основных
ее разделов. В научном творчестве великого физиолога эти труды прина-
длежат к тем звеньям, которые особенно тесно связывают физиологию с ме-
дициной. Достаточно сказать, что это учение об охранительно-целебной
роли торможения является научной основой одного из самых актуальных
и перспективных терапевтических направлений современной медицины —
терапии сном или наркозом болезненных состояний организма, прежде
всего психических и нервных заболеваний.
Гениальный ученый не успел исчерпывающе разработать все вопросы,
касающиеся теоретического и практического значения открытой им новой
роли торможения.
К ним относятся, в частности, вопросы об идентичности или неидентич-
ности охранительного торможения «координационному торможению»1, об
особенностях обмена веществ и энергии в нервных клетках при охрани-
тельном торможении, о границах распространения способности (хотя бы
в пределах нервной системы) к развитию охранительного торможения,
о составе факторов, способных порождать охранительно-целебное торможе-
ние в корковых и других нервных клетках, и т. п. Можно утверждать, что
эти и некоторые другие актуальные в теоретическом отношен ин вопросы
о природе и закономерностях возникновения и течения охранительного
торможения остаются нерешенными до сих пор, хотя проблема охрани-
тельного торможения и привлекала внимание ряда учеников и последова-
телей И.П. Павлова, в том числе и наше. А это в сильной степени обязы-
вает нас всех.
1 Термин «координационное торможёнис» был нами введен в «физиологический
обиход» в 1944 г. для удобства различения координационной роли торможения и
охранительно-целебной Позже наше предложение было поддержано А. Н. Магницким
и А. С. Дмитриевым.
534 Э А АСРАТЯН
Наше отставание в разработке этого важнейшего раздела научного на-
следия И. П. Павлова особенно значительно по линии экспериментальной
и теоретической. Мы можем указать лишь на несколько интересных ис-
следований .
Это, во-первых, работы М. К. Петровой по физиологической и медика-
ментозной терапии сном различных трофических расстройств кожи, под-
кожных тканей и внутренних органов у собак (язвенные и экзематозные
процессы, доброкачественные и злокачественные новообразования, облы-
сение, фурункулез и т. п.), обусловленных, как опа считала, нарушением
нормального состояния и деятельности коры больших полушарий мозга.
К этого рода работам относятся также проводимые с 1917 г. исследования
А. Г. Иванова-Смоленского и сотрудников по лечению длительным нарко-
тическим сном тяжелых нарушений высшей нервной деятельности у крыс
и морских свинок, вызванных различными токсическими веществами и от-
дельными инфекциями. Начиная с 1941 г. нами и нашими сотрудниками
проводятся исследования по терапия сном различного рода болезненных
состояний у высших и низших животных, вызванных органическим, ане-
мическим и тепловым повреждениями различных частей центральной и пе-
риферической нервных систем. Отдельные наблюдения о наркозной тера-
пии коммоции — контузии у собак имеются у А. О. Долина (1942). Этим
исчерпывается список экспериментальных лабораторных исследования про-
блемы охранительной и целебной роли торможения. Нельзя, конечно, не
радоваться тому, что клиническая разработка этой проблемы в нашей
стране проводится более широко и интенсивно. Однако это не может пол-
ностью компенсировать наше отставание по линии лабораторно-экспери-
ментальной. разработки проблемы, т. е. по той линии, которую
И. П. Павлов считал решающей в. подобных случаях. Не может быть двух
мнений о том, что после исторического перелома на физиологическом
фронте —Объединенной сессии Академии наук СССР и Академии медицин-
ских наук СССР — должна быть резко улучшена и наша эксперименталь-
ная работа по проблеме охранительно-целебного торможения,
Это обстоятельство, а также наблюдаемый за последнее время усиленный
интерес широких кругов физиологов и врачей нашей страны к вопросам те-
ории и практики терапии сном делают, как нам кажется, своевременным
и необходимым обмен опытом работы в этом области каждым из нас. Ру-
ководствуясь этими соображениями, мы и решили воспользоваться пред-
ставившейся возможностью не только для изложения полученного нами све-
жего, еще не опубликованного материала по этой проблеме, но и для сжа-
того обзора прежнего нашего материала и некоторых принципиальных на-
ших установок по многим важным теоретическим И практическим вопро-
сам терапии сном.
К исследованию охранительной и целебной роли торможения мы пришли
в ходе нашей многолетней работы по изучению проблемы компенсаторных
приспособлений, закономерностей нарушения и восстановления функций
поврежденного организма. Мы решили использовать охранительно-целеб-
ное торможение в качестве фактора, способного, по нашим предположе-
ниям, содействовать процессу нормализации альтерированных нервных кле-
ток поврежденной коры больших полушарий или других частей централь-
ной нервной системы и тем самым создавать благоприятные предпосылки
для ускорения темпов и улучшения развивающихся в поврежденном ор-
ганизме компенсаторных приспособлений.
Как известно, четкие и предельно ясные формулировки концепций
И. П. Павлова об охранительной и целебной роли торможения относи-
лись только к клеткам больших полушарий мозга, к тому же лишь в усло-
виях их нормальной деятельности и в условиях тех видов патологической
деятельности, которые обусловлены функциональным поражением. В со-
ТЕРАПИЯ СНОМ ПОСЛЕДСТВИЙ ОРГАНИЧЕСКИХ ПОВРЕЖДЕНИЙ Ц. И. С. 535
ответствии с этим" терапия сном или наркозом применялась в его лабо-
раториях и клинике лишь для лечения функциональных болезненных со-
стояний больших полушарий. Вопрос о применимости этих концепции
к клеткам других отделов центральной нервной системы в условиях нор-
мальной и патологической их деятельности, равно как и к патологической
деятельности нервных клеток коры больших полушарий, обусловленной
их органическими поражениями, был оставлен открытым, если не счи-
тать отдельных высказываний и догадок И. П. Павлова по этому
п золу.
На основании тщательного анализа громадного экспериментального ма-
териала лабораторий И. П. Павлова и других отечественных и зарубежных
исследователей относительно глубокой и длительной центральной депрес-
сии (центральный шок, диасхизис и т. п.), вызванной органической трав-
мой коры и подкорковых образований больших полушарий и других ча-
стей центральной нервной системы, анализа открытых И. М. Сеченовым
и Н. Е. Введенским феноменов торможения в примитивных центральных
и периферических нервных образованиях при сильных или частых
раздражениях, а также на основании значительного и разнообразного но-
вого материала нашей лаборатории относительно центрального торможе-
ния при различного рода травмирующих воздействиях на центральную
и периферическую нервную систему мы считали возможным расширить
рамки теоретического применения концепций учителя. Нами были раз-
виты положения, что: а) открытая И. П. Павловым охранительно-целеб-
ная роль торможения принадлежит к категории универсальных явлений
и присуща всей нервной системе (и даже всем возбудимым тканям); б) тор-
можение как процесс естественной самозащиты и самолечения возникает
в нервной системе не только при функциональных, но и при ее органи-
ческих повреждениях, Мы ограничиваемся здесь только перечнем основа-
ний для этих теоретических положений, отсылая интересующихся указан-
ным вопросом, а также нашим пониманием значения этих положений для
теории физиологии нервной системы к ранее опубликованным нашим рабо-
там. Укажем лишь, что эти теоретические положения являются дальнейшим
развитием идей гениального учителя, в частности, его идеи об «естествен-
ной общности основных отношений», о родстве основных закономерностей
нервной деятельности, а также его положения о том, что истощение и сверх-
сильное раздражение нервных клеток являются основными предпосылками
для развития охранительного торможения.
В соответствии со значительным расширением границ теоретического
применения гениальных концепций учителя об охранительной и целебной
роли торможения мы стремились расширить также рамки практического их
использования, а именно: попытались проводить терапию сном при бо-
лезненных состояниях организма, обусловленных органическим повреж-
дением как больших полушарий, так и нижележащих частей центральной
и периферической нервной системы. Проведенная нами совместно с сотруд-
никами Л. В. Симуковой, ГТ. В. Станкевичем, Г. Т. Сахиулнной, С. Н. Ива-
новой, Б. Д. Стефанцовым, П. Ф. Минаевым, Е. А. Романовской, Е. В. Гу-
ровой, В. Н. Дроздовой большая экспериментальная работа не только пол-
ностью подтвердила правильность наших предположений и оправдала наши
надежды, по и позволила установить ряд теоретически и практически важ-
ных положений относительно выбора и дозировки снотворных средств для
достижения оптимальных эффектов терапии сном, относительно комби-
нирования их между собой и с другими терапевтическими факторами, спо-
собов их применения, длительности терапии сном, показаний и про-
тивопоказаний к ее применению и т.п.
Подопытными животными служили собаки (более 500) и кошки (около
200); в известном числе опытов нами использовались также кролики,
530
Э. Л. АСРАТЯН
крысы и лягушки. Как правило, мы ставим хронические эксперименты;
острые опыты ставятся весьма редко.
Л^еханической, операционной, термической и электрической траьмой,
а также длительной анемнзацией тех или иных частей центральной и пе-
риферической нервной системы или организма у подопытных животных вы-
зываются различного рода глубокие нарушения двигательных, чувстви-
тельных и вегетативных функций. Вслед за этим травмированных живот-
ных делят на две группы: животных одной группы (контрольной) обеспе-
чивают обычным лабораторным уходом за больными животными; живот-
ные другой (подопытной) группы вдобавок получают снотворные (одно-
кратно или систематически в течение определенного времени, в зависимости
от характера, степени и длительности нарушения функций). При разделе-
нии животных на две группы обращают строгое внимание на то, чтсбы для
параллельных наблюдений были взяты животные с возможно одинаковыми
показателями веса, возраста, упитанности, роста, а также характера, глу-
бины и объема поражения функций. Если же одно из парных подопытных
животных более или менее значительно отличается от другого по перечи-
сленным выше показателям, то животное с худшими показателями вклю-
чается в подопытную группу. В целях возможно полной и точной характе-
ристики динамики нарушения и восстановления функций у подопытных
животных мы используем различные современные приемы физиологического
эксперимента для выявления качественной и количественной характери-
стики, а при возможности и объективной графической регистрации дея-
тельности интересующего нас органа, системы органов или той или иной
деятельности организма в целом.
В случае глубоких нарушений жизненно важных функций организма,
приводящих, в конце концов, к гибели подопытных животных, учитывается
длительность их жизни после травмы. В опытах, как правило, приме-
няются травматические повреждения значительной интенсивности, чтсбы
вызвать у подопытных животных глубокие и длительные расстройства
функций.
Эффективность терапии сном нами исследовалась и исследуется при
следующих специфических патогенных воздействиях: тяжелой механиче-
ской травме нижних конечностей (для вызывания травматического шока);
жогс значительной части кожной поверхности (для вызывания ожого-
вого шока); сильном перегревании всего организма (для вызывания тепло-
вого шока); экстирпации верхних шейных симпатических узлов и ком-
прессии больших полушарий мозга (в том и другом случае для вызывания
отека — набухания мозга); полном поперечном сечении спинного мозга
(для вызывания спинального шока и нарушения соответствующих двига-
тельных, чувствительных и вегета тивных функций); раздельном поперечном
сечении боковой, передней или задней половины спинного мозга; ло-
кальной термической травме спинного мозга, коммоции —контузии спин-
ного и головного мозга; локальной органической травме продолговатого
мозга и больших полушарии; остром истощении перерезанного спинного
мозга, вызванном длительным раздражением нервов; длительной анеми-
зациеи всем центральной нервной системы; близких и отдаленных тяжелых
последствиях органической травмы спинного мозга (приступы бурных дви-
жений, эпилептические припадки, глубокие дистрофические измене-
ния и т. п.).
Весьма сжатая характеристика полученного нами фактического мате-
риала по экспериментальной терапии сном вызванных нами болезнен-
ных Состояний у животных такова.
1. В течение 1941—1942 гг. нами при участии лаборантов Л. В. С.'иму-
ковой и П. В. Станкевича было выполнено весьма обширное эксперименталь-
ное исследование по разработке на основе учения И. П. Павлова ряда слож-
ТЕРАПИЯ СНОМ ПОСЛЕДСТВИИ ОРГАНИЧЕСКИХ ПОВРЕЖДЕНИЙ Ц. Н. С. 53“
нейших вопросов этиологии, функциональной патологии и терапии травма-
тического шока (иод опытом было более 250 собак и кошек). В этом иссле-
довании, в частности, был впервые разработан способ терапии сном травма-
тического шока, который сводился к внутреннему введению довольно
сложной по составу противошоковой жидкости, включающей три взаимо-
связанных терапевтических фактора, направленных против трех взаимодей-
ствующих сторон функциональной патологии позднего травматического шо-
ка: а) прежде всего снотворные и седативные вещества для восстановления
состояния и функции истощенных и ослабленных нервных центров путем
усиления в них охранительно-целебного торможения; б) средства для вос-
становления функций капилляров и нарушенной гемодинамики вообще;
в.) средства для нормализации нарушенного химизма крови и организма
вообще. При стопроцентной гибели контрольных (нелеченных) животных
от вызываемого экспериментального травматического шока разработан-
ный нами способ терапии сном спасал от шока такой же тяжести около 85%
подопытных животных, которые после опыта жили под нашим наблюде-
нием месяцами.
За последние годы наш сотрудник П. Ф. Минаев добился дальнейшего
повышения эффективности разработанного способа терапии сном травма-
тического шока, прибавляя к противошоковой жидкости также небольшие
дозы аденозпнтрифоСфорной кислоты.
2. В 1942—1943 гг. нами совместно с лаборантами Л. В. Симуковой и
П. В. Станкевичем в экспериментах на 50собаках были разработаны способы
терапии сном ожогового шока или, как принято называть, солнечнотепло-
вого удара. Суть этих весьма эффективных способов сводится к сочетанию
снотворных и седативных средств с другими терапевтическими средствами,
подобранными соответственно специфическим особенностям функциональ-
ной патологии ожогового и теплового шока.
3. Еще в 1942 г. у 6 взрослых собак и 2 взрослых кошек мы перерезали
спинной мозг на уровне последних грудных сегментов и вызывали у них
спинальный шок значительной глубины (подавление рефлекторной актив-
ности задних конечностей и хвоста, расстройства функции тазовых орга-
нов и т. п.). Затем с помощью введения .медикаментов достигали у подопыт-
ных животных углубления и удлинения сна в течение 4—5 дней. Оказа-
лось, что спинальный шок у них исчезал на 3—4 дня раньше, чем у жи-
вотных контрольной группы.
За последние годы наша сотрудница В. Н. Дроздова получила новые и
более веские факты относительно положительного влияния снотворных
на течение спинального шока у 8 собак, у которых она перерезала спинной
мозг на уровне последних шейных сегментов. Для оценки глубины спи-
нального шока В. Н. Дроздова пользовалась также другими показате-
лями— состоянием рефлекторной активности сосудодвигательных центров
перерезанного спинного мозга, быстротой наступления утомления сгиба-
тельного рефлекса задних конечностей. По данным В. Н. Дроздовой, у со-
бак подопытной группы соматические регексы и рефлекторная регуляция
мочеиспускания восстанавливаются почти в два раза быстрее, чем у Смбак
контрольной группы. Рефлекторная активность спинальных сосудодвига-
тельных центров у собак подопытной группы восстанавливалась на 25—
30-й день, а у собак контрольной группы — на 50—70-й день. У собак под-
опытной группы утомление сгибательного рефлекса наступало почти в два
раза медленнее, чем у собак контрольной группы.
4. В 1942—1944 гг. у 8 взрослых собак мы перерезали боковую поло-
вину спинного мозга на уровне X—XI сегментов спинного мозга (так на-
зываемое броун-секаровское сечение). У собак подопытной группы, полу-
чавших в течение 7—8 дней снотворные, паралитические и паретические
явления со стороны пораженной задней конечности исчезали значительно
538 э А- АСРАТЯН
быстрее, чем у контрольных. Собаки подопытной группы в среднем на 5—
6 дней раньше собак контрольной группы начали пользоваться поражен-
ными конечностями для стояния и ходьбы.
За последние годы наша сотрудница С. Н. Иванова отметила благотвор-
ное влияние снотворных препаратов на динамику восстановления двига-
тельных, чувствительных, вегетативных функций у 10 собак после боковой
половинной перерезки спинного мозга на уровне верхних шейных сегментов
спинного мозга, при которой функциональные расстройства отличаются
значительно большим объемом, глубиной и длительностью, чем при по-
добной перерезке на уровне грудных сегментов спинного мозга. По данным
С. Н. Ивановой, собаки подопытной группы ползали на 9—10-й день, на-
чинали ходить на 13—18-й, хорошо ходили на 20—31-fi день; собаки же
контрольной группы ползали на 14—21-й день, начинали стоять на 19—
25-й, хорошо ходили на 38—44-й день.
5. В 1943—1944 гг. у 10 взрослых собак мы перерезали заднюю попе-
речную половину спинного мозга' на уровне верхних шейных сегментов.
Как было показано нами раньше, эта операция вызывает длительное (30—
40-дневное) и глубокое расстройство двигательных, чувствительных и ве-
гетативных функций организма, а именно: вначале очень глубокую деп-
рессию рефлекторной активности спинного мозга1 (почти полный паралич
и анестезия конечностей, туловища и хвоста), длящуюся несколько дней,
и длительную утрат}' способности стоять и ходить после того, как глу-
бокий спинальный шок проходил. У собак подопытной группы, получав-
ших снотворные в течение 12—15 дней, почти наполовину сокращался срок
восстановления утраченных и нарушенных двигательных, чувствительных
и вегетативных функций, в частности, функции стояния, ходьбы и бега.
Собаки подопытной группы в среднем стояли уже на 12-й день, ходили на
14-й, бегали на 17-й день; собаки >ке контрольной группы в среднем стояли
на 23-й день, ходили на 27-й, бегали на 41-й день.
6. В 1945—1946 гг. С. И. Иванова уколами в продолговатый мозг у зна-
чительного числа крыс и лягушек вызывала серьезные нарушения вести-
булярных рефлексов локомоции, а также существенное ухудшение общего
их состояния, приводившее многих к гибели. Применение снотворных и
в этих опытах оказывало благоприятное влияние на темпы восстановле-
ния нарушенной ре4>лекторной деятельности и особенно на темпы улуч-
шения общего состояния и па выживаемость подопытных животных.
Животные подопытной группы в среднем живут в два раза дольше живот-
ных контрольной группы.
7. В 1944—1945 гг. В. Л. Симукова вызывала коммоцию— контузию
спинного или головного мозга у крыс и лягушек при помощи локального
удара дозируемой силы. Эта травма вызывает глубокое нарушение двига-
тельных, чувствительных и вегетативных функций организма и частую
гибель животных (по данным вскрытия, в травмированных частях нервной
системы происходит значительное кровоизлияние). Снотворные вещества
и в этих опытах значительно ускоряли темпы восстановления нарушенных
и утраченных функций и заметно увеличивали выживаемость подопытных
животных.
8. В 1943—1944 гг. Е. В. Гурова вызывала у кошек и лягушек рефлек-
торный паралич одной из задних лап путем длительного раздражения силь-
ным и прерывистым электрическим током. Травмированные конечности
обычно отекали; часто возникали также дистрофические явления типа изъ-
язвления и выпадения волос. Применение снотворных препаратов не только
1 Мы с недоумением узнали, что М. Г, Дуомишьян в своей докторской диссер-
тации чуть ли не себе приписывает приоритет открытия этого явления. Па делеон
лишь подтвердил, и то с опозданием примерно на 10 лет, полученный нами и не-
однократно описывавшийся факт.
ТЕРАПИЯ СНОМ ПОСЛЕДСТВИЙ ОРГАНИЧЕСКИХ ПОВРЕЖДЕНИЙ Ц Н С 53.9
резко ускоряло и улучшало восстановление рефлекторной активности по-
раженных конечностей (у животных подопытной группы полное восстанов-
ление парализованной конечности имело место в 76% случаев, а у живот-
ных контрольной группы лишь в 33% случаев), но весьма благоприят-
но влияло также на темпы исчезновения дистрофических явлений.
В другой серии опытов Е. В. Гурова вызывала острое утомление и исто-
щение перерезанного спинного мозга у лягушек путем длительного раздра-
жения задних конечностей или нервных стволов сильным фарадическим
током. И в этих опытах снотворные значительно ускоряли восстановление
рефлекторной деятельности истощенного спинного мозга.
9. В течение 1944—1948 гг. Е. А. Романовская выполнила обширную
экспериментальную работу но локализованной и дозированной тепловой
травме спинного мозга у лягушек, крыс, кошек и собак. Обстоятельные
исследования Е. А. Романовской показали, что снотворные препараты
весьма благоприятно влияют на процесс восстановления нарушенных при
этой травме двигательных,- чувствительных и вегетативных функций под-
опытных животных, а в случае очень глубоких травм и на выживаемость
животных. По данным Е. А. Романовской, это благотворное влияние осо-
бенно заметно при травмах умеренной интенсивности. Так, например,
глубокие нарушения двигательной функции (параличи, парезы) проходили
у лягушек подопытной группы в 45,4%, а контрольной группы—лишь
в 17.6%; глубокие нарушения чувствительности (анестезия, гипестезия) —
соответственно у 40% и 17,3% животных. Собаки подопытной группы сто-
яли на 5—6-й день, ходили на 11-й, а собаки контрольной группы стояли
на 10—13-й день, ходили на 13— 16-й день.
10. В течение 1943—1946 гг. Г. Т. Сахиулина весьма обстоятельно ис-
следовала влияние снотворных препаратов на динамику восстановления
рефлекторной активности спинного мозга или всей центральной нервной
системы у лягушек и собак, нарушенной дозированной анемнзацией. Много-
численные тщательно поставленные острые и хронические опыты Г. Т. Са-
хиулинон с большой убедительностью показали, что снотворные препа-
раты существенно ускоряют (примерно в два раза) и улучшают процесс
восстановления нарушенных и утраченных двигательных, чувствительных
и вегетативных функций как у лягушек, так и у собак. Например, если
в условиях острого опыта у лягушек контрольной группы рефлекторная
активность спинного мозга после 3-часовой анемизации организма восста-
навливается примерно через 2 часа после возобновления циркуляции, то
у лягушек подопытной группы восстановление происходит через 20 минут.
У собак подопытной группы после 20-минутной анемизации спинного мозга
функции тазовых органов восстанавливались через 3—4 дня, у собак кон-
трольной группы — через 7—10 дней. Первые стояли через 8—10 дней,
а вторые — через 20—25; первые ходили через 18—20 дней, вторые — че-
рез 35—40 диен. Из полученных Г. Т. Сахиулиной данных заслуживают
особого внимания два факта. Если анемизации перерезанного спинного
мозга лягушки в условиях острого опыта приводит к исчезновению его
рефлекторной активности, то это имеет необратимый характер; рефлек-
торная активность никогда больше не восстанавливается после возобнов-
ления перфузии препарата питательной жидкостью; препарат гибнет. При-
бавление же снотворных веществ к жидкости, как правило, приводит
к восстановлению угашенной рефлекторной активности перерезанного
спинного мозга. У тотально анемизированных в течение 4—5 часов лягушек
контрольной группы в условиях хронического опыта наблюдается любо-
пытное явление: когда у них рефлекторная активность восстанавливается
в такой степени, что они в состоянии плавать в аквариуме, то после более
или менее энергичных движений они очень часто мгновенно впадают в со-
стояние своеобразного оцепенения, длящегося до 1 минуты. Из этого со-
510
Э А АСРАТЯН
стояния их не удается вывести даже сильными раздражениями. У лягушек
же подопытной группы никогда не наблюдалось это кататоническое состоя-
ние, как бы долго и энергично они ни плавали в аквариуме.
11. В 1942—1944 гг. нами у 14 собак был вызван отек — набухание
мозга: у части животных путем одновременной экстирпации обоих верхних
симпатических узлов, у части — путем компрессии коры больших полу-
шарий. Применение снотворных в сочетании с хлористым кальцием в боль-
шей степени улучшало состояние животных и повышало их выживаемость,
чем применение одних снотворных.
12. За последние годы в- процессе работы, связанной с осуществлением
весьма сложных мозговых операций, к тому же зачастую в различных ком-
бинациях (экстирпация коры больших полушарий и мозжечка, половин-
ная перерезка шейных сегментов мозга и удаление коры больших полуша-
рий мозга и т. п.), нами и нашими сотрудниками накоплен значительный
экспериментальный материал, свидетельствующий об исключительно благо-
творном терапевтическом влиянии снотворных препаратов при близких
и отдаленных, в большинстве крайне тяжелых последствиях этих серьез-
ных органических повреждении высших отделов центральной нервной си-
стемы у высших животных (преимущественно у собак). Имеется в виду
в первую очередь усиленное двигательное беспокойство в первые дни пссле
перечисленных операции, глубокие и обширные дистрофические явления
в позднем послеоперационном периоде и эпилептические припадки в близ-
ких и отдаленных послеоперационных периодах.
Не желая отклоняться от основной темы, мы не будет здесь вдаваться
в анализ причин этих явлений. Отметим лишь, что они большей частью
возникают при комбинации тяжелых операций и имеют сложное проис-
хождение, говоря конкретнее, обусловливаются комплексом многообраз-
ных сдвигов не только в поврежденных частях центральной нервной си-
стемы, но и во всей нервной системе, сдвигов порядка выпадения функции,
механических изменении, нарушения кровоснабжения и ликворообраще-
ния, дегенеративных изменений, нарушения компенсаторных приспособле-
ний и т. п. В соответствии с основным содержанием данной статьи мы
ограничимся лишь кратким изложением результатов, достигнутых нами
по терапин сном этих тяжелейших и опаснейших последствий органи-
ческих поражений мозга.
Л. У многих подопытных животных в первые дни после тяжелых и осо-
бенно комбинированных мозговых операций возникают приступы двига-
тельных реакций в виде сложных двигательных автоматизмов, чаще всего
своеобразного «бега на месте», вращательных движений вокруг длинной
осн тела, жевательных движений и различного рода движений головой.
Изменяясь в своей интенсивности, частоте и продолжительности, в зави-
симости от различных обстоятельств, эти приступы зачастую принимают
характер «двигательной бури», сопровождаются стоном, визгом и лаем под-
опытных животных и, как правило, приводят их к гибели, если не пред-
принимаются энергичные и эффективные терапевтические меры. Мы
должны признать, что на начальном этале наших исследований даже при
значительном опыте ухода за животными с мозговыми операциями нам
редко удавалось спасти жизнь собак при таких катастрофах, А в настоя-
щее время мы можем с удовлетворением констатировать, что своевременным
применением Снотворных в комбинации с некоторыми простыми терапев-
тическими мероприятиями, направленными на поддержание питания со-
бак, мы, как правило, спасаем животных от смерти. В этих случаях сно-
творные мы применяем в течение 4—10 дней, в зависимости от обстоя-
тельств.
Б. У некоторых собак с тяжелыми и особенно комбинированными моз-
говыми операциями со временем возникают разнообразные дистрофические
ТЕРАПИЯ СНОМ ПОСЛЕДСТВИЙ ОРГАНИЧЕСКИХ ПОВРЕЖДЕНИЙ Ц. Н. С
541
явления типа отеков отдельных частей тела, подкожных циститов, изъяз-
влений, выпадения волос, экземы и т. д. Нами накоплен небольшой еще
экспериментальный материал относительно результатов терапии сном
этих дистрофических изменений тканей, что пока лишает нас возможности
быть категоричными в. своих выводах. Этот предварительный материал
оставляет, однако, весьма благоприятное впечатление: 10—15-дневное при-
менение снотворных оказывает благотворное терапевтическое влияние при
упомянутых тяжелых последствиях органических повреждений мозга.
В. Одним из частых, крайне тяжелых и в большинстве своем отдален-
ных последствий осуществленных нами сложных мозговых операций, в осо-
бенности комбинированных, являются эпилептические припадки у под-
опытных ЖИВОТНЫХ, представляющие Собой сложное сочетание тониче-
ских и клонических судорог, а также различных двигательных автома-
тизмов. Эти эпилептические припадки в большинстве случаев протекают
весьма бурно, повторяются часто и, как правило, приводят к быстрому
ослаблению, истощений» И гибели животных. Нами накоплен большой фак-
тический материал, ясно свидетельствующий о том, что систематическим
применением снотворных препаратов в течение 10—20 дней МОЖНО совер-
шенно освободить собак от этих припадков и спасти их жизнь. В качестве
примера исключительно благоприятного действия терапии сном при
этих припадках можно привести следующий случай. У собаки Шарик
24.Х 1950 г. удален мозжечок, a 25.VI 1951 г. — кора правого полушария
большого мозга. Ближайший послеоперационный период протекал у
нее гладко. 1. VII у собаки внезапно начались бурные эпилептические при-
падки, которые после 2—3-кратного повторения привели к острому ослаб-
лению и истощению организма животного и остановке дыхания. Некото-
рое время жизнь животного поддерживалась искусственным дыханием.
С трудом удалось спасти собаку от гибели. Состояние было очень тяжелым.
На следующий день эти припадки повторились. Не дожидаясь дальнейших
осложнений, мы применили снотворные. Терапия сном продолжалась
в течение 2 недель и привела к исчезновению припадков. С тех пор прошло
5 месяцев, и собака находится в удовлетворительном состоянии.
Таким образом, полученные нами и нашими сотрудниками богатые и
разнообразные фактические данные показывают, что при всех осуществлен-
ных нами вариантах экспериментального нарушения функций нервной си-
стемы у животных применение снотворных веществ ускоряет появление про-
цессов восстановления нарушенных и утраченных функций пов.режденного
организма, ускоряет темпы и повышает стойкость этих процессов, улуч-
шает общее состояние организма, а в случаях опасных для жизни травм
увеличивает выживаемость животных. Мы имели неоднократную возмож-
ность убедиться также в том, что снотворные вещества ускоряют процесс
заживления ран.
Накопленный нами фактический материал показывает, что эффектив-
ность применения снотворных препаратов при органических поражениях
нервной системы зависит от многих факторов. Важное значение имеет, на-
пример, уровень филогенетического н, повидимому, онтогенетического раз-
вития животных. При близких по характеру и интенсивности травмах нерв-
ной системы животных применение снотворных дает более быстрые и яр-
кие результаты у собак и кошек, чем у крыс л особенно лягушек. Точно
так же эффект терапии сном у молодых животных более выражен, чем
у старых. Весьма важное значение имеют характер, интенсивность
и локализация самой травмы. Снотворные вещества оказывают более эф-
фективное терапевтическое действие, при повреждениях высших отделов
центральной нервной системы, чем при повреждениях низших ее отделов.
Положительное терапевтическое действие терапии сном ярче прояв-
ляется при травмах средней интенсивности, чем при травмах большой глу-
542
Э А АСРАТЯН
бины и размера; при легких же травмах это действие не всегда четко высту-
пает, так как восстановление нарушенных функций происходит быстро
и без снотворных.
Исключительно важное значение имеют фармакодинамические свойства
снотворных препаратов, дозировка и длительность их применения, а
также способ введения их в организм. В начальных стадиях нашей экспе-
риментальной работы по этой проблеме, когда мы еще недостаточно хо-
рошо ориентировались в указанных вопросах, результаты проведенной
нами терапии сном хотя н были положительными, но характеризовались
пестротой и в общем были Mtliee показательными:. По мерс обогащения и
уточнения наших знаний в этой области мы стали получать все более одно-
родные и лучшие результаты. Мы считаем необходимым поделиться опытом
нашей экспериментальной работы в этой области не только потому, что
рассматриваемые вопросы имеют первостепенное теоретическое и практиче-
ское значение, но и потому, что они почему-то не привлекают должного
внимания наших экспериментаторов и врачей, занимающихся изучением
проблемы терапии сном.
Наша многолетняя и обширная экспериментальная работа позволила
сделать несколько выводов по этим вопросам.
1. Наилучшими из снотворных веществ для проведения терапии снем
являются препараты карбаминовой группы — годовал и отчасти уретан.
В отличие от других снотворных, в частности, снотворных из группы бар-
битуратов, которыми, как правило, пользуются наши исследователи и кли-
ницисты для терапии сном и наркозом, гедонал и уретан, а также продукты
их расщепления не токсичны и в больших дозах, не нарушают деятельности
жизненно важных систем и характера метаболизма в организме даже при
длительном применении.
2. При лечении поледствнй органических повреждений нервной системы
и организма вообще лучшие результаты получаются от таких доз снотвор-
ных, которые вызывают сон длительностью 10—15 часов в сутки, т. е. близ-
кий по характеру к естественному, физиологическому сну, но несколько
глубже него. Гораздо менее эффективны дозы, вызывающие менее глубо-
кий и менее длительный сон, а также более глубокий и более длителънын
сон, и особенно дозы, вызывающие сон, близкий к наркозу. В этом отноше-
нии также наши выводы не совсем совпадают с практикой других наших
экспериментаторов и клиницистов (Ф. А. Андреев и др.), отошедших от
наркозной терапии, но все же предпочитающих дозы снотворных, вызываю-
щие 18—20-часовой сон, т. е. «длительный наркотический сон», по харак-
теристике этих исследователей, который лишь с большой натяжкой может
считаться близким к естественному, физиологическому сну.
Указывая на некоторые расхождения между нами и другими исследо-
вателями в оценке эффективности отдельных снотворных и степени глу-
бины и длительности сна для оптимального терапевтического эффекта,
мы хотим подчеркнуть необходимость дальнейших специальных экспери-
ментальных исследований этих важнейших вопросов терапин сном.
3. Длительность применения снотворных препаратов, споссб их вве-
дения в организм, комбинация их с другими терапевтическими средствами
н мерами должны определяться характером, интенсивностью и особенно-
стями болезненного состояния. Мы пришли к выводу, что в этом вопросе
не должно быть шаблона, как. впрочем, и во всех серьезных вопросах те-
рапии. Если при травматическом шоке потребуется однократный соч
длительностью в несколько часов, внутривенное введение снотворных ве-
ществ в комбинации с одними терапевтическими средствами, то при орга-
нических повреждениях центральной нервной системы необходима много-
дневная терапия сном с введением снотворных веществ энтерально (и.'и
в особых случаях подкожно) и р комбинации с другими терапевтическим»
ТЕРАПИЯ СНОМ ПОС.Г ЕДСТВИЙ ОРГАНИЧЕСКИХ ПОВРЕЖДЕНИЙ Ц Н С 543
средствами. На необходимость избегать шаблона при терапии сном мы
указываем потому, что, судя по имеющимся данным, среди некоторых кру-
гов наших клиницистов имеется ошибочная и вредная тенденция к трафа-
ретному применению одного более или менее удачного приема терапии сном
при самых разнообразных, порой весьма отдаленных друг от друга бо-
лезненных состояниях организма.
4. Мы позволим себе привести еще один вывод из опыта многолетнего
нашего изучения актуальных вопросов теории и практики терапии сном.
Этот вывод касается не существа дела, а относится к характеру организа-
ции самой исследовательской работы, к сложному вопросу взаимоотно-
шений лабораторных и клинических исследований по функциональной па-
тологии и терапии. Как явствует из всего изложенного, во всей своей ра-
боте по теоретическому и практическому изучению обсуждаемой проб-
лемы мы постоянно и неуклонно придерживались важного павловского
принципа о необходимости решения сложных вопро-
сов патофизиологии и особенно терапии прежде
всего при помощи лабораторных экспериментов
п а животных и только вслед за тем критического использования по-
лученных результатов в медицинской практике.
Нет надобности излагать здесь результаты применения разработанных
нами способов терапии сном травматического шока и ряда близких и от-
даленных последствий тяжелых органических повреждений нервной си-
стемы у раненых на фронтах Великой Отечественной войны и в эвакогоспи-
талях/ Интересующихся отсылаю к нашим прежним публикациям. Следует
лишь указать, что каждый раз, когда по ходу нашей исследовательски й
работы возникала необходимость во внедрении разработанных нами спо-
собов лечения в практику здравоохранения, мы использовали предоставлен-
ную нам возможность не без понятной тревоги, но зато со спокойной
совестью, так как, следуя завету великого И.П. Павлова, мы предвари-
тельно пропускали эта способы «через огонь эксперимента»—эксперимента
лабораторного.
А между тем, многие врачи и физиологи, в том числе даже ученики
И. П. Павлова, разрабатывающие вопросы терапии сном применительно
к различным болезненным состояниям организма, отходя от этого важного
принципа И. П. Павлова, свою работу, свои поиски начинают пря^о
с исследования на человеке, без соответствующих предварительных экспе-
риментов на животных. Эти врачи и физиологи забывают указание
И. II. Павлова, что окончательная лебеда медицины при разрешении слож-
нейших вопросов патофизиологии и терапии может придти только через
лабораторный опыт, через эксперимент на животных и что, «чем полнее
будет проделан опыт на животных, тем менее часто придется больным быть
в. положении опытных объектов со всеми печальными послед-
ствиями этого»1.
Чтсбы не быть голословным, сошлюсь на известную работу Ф. А. Ан-
дреева по терапии сном различного рода внутренних и иных заболева-
ний человека, в том числе гипертонии, язвы желудка, язвы двенадцати-
перстной кишки. Хотя, судя по некоторым данным, эта работа проводится
не без определенного успеха, хотя в настоящее время Ф. А. Андреев имеет
значительное число последователей среди врачей, тем не менее мы счи-
таем совершенно ненормальным и непростительным, что вся эта работа
проводилась и до сих пор проводится исключительно на людях без соот-
ветствующего предварительного или хотя бы параллельного эксперимента
на животных. Можно ли сомневаться в. том, что полноценный лаборатор-
ный эксперимент на животных, эксперимент в высоком павловском понн-
1 И. П. Павлов, Полное собрание трудов., т. 1, 1946, стр. 363.
544
Э. Л. АСРАТЯН
мании, может оказать неоценимую пользу в разрешении таких практически
важных вопросов при терапии сном этих болезненных состояний, как вы-
бор снотворных, определение их оптимальной дозировки, комбинирова-
ние с другими терапевтическими средствами, уточнение показаний и про-
тивопоказаний к их применению и т. п.
В годы Великой Отечественной войны, кроме нас, многие другие наши
ученые также применяли различные способы терапии сном и наркозом
разнообразных последствий боевых травм (А. О. Долин, Л. Г. Иванов-
Смоленский, Г. Е. Стеблов, С. Д. Каминский, Б. А. Серафимов,
Ф. А. Андреев и др.). А между тем никто из них не использовал богатых
возможностей лабораторного эксперимента на животных для предваритель-
ного решения многих важных вопросов терапии сном упомянутых болез-
ненных состояний. Единственным исключением является А. О. Долин,
который, судя по одной его заметке, провел несколько ориентировочных
наблюдений подобного рода. Разумеется, на современном уровне наших
знаний и методических возможностей весьма трудно, а пожалуй, и не-
возможно добиться удовлетворительного решения многих вопросов пато-
физиологии и терапии, в том числе и терапии сном, исключительно в усло-
виях лабораторного эксперимента на животных, и осторожное клиниче-
ское применение в этих случаях неизбежно. Но весь вопрос в том, чтобы
такая тактика, такой стиль являлся исключением, а не правилом.
Мы и раньше неоднократно указывали на ошибочность такого подхода
к разработке важных вопросов теории и практики терапии сном, на опас-
ность отклонения от намеченных И. П. Павловым путей. Пользуясь, слу-
чаем, мы еще раз обращаем внимание научной и медицинской обществен-
ности на указанный существенный недостаток в проводимой нашими физио-
логами и врачами работе по терапии сном различных болезненных со-
стояний организма. Крайне необходимо изжить этот недостаток в нашей
общей работе именно сейчас, когда в связи с резким расширением исследо-
вательской и клинической работы в пашен стране по вопросам теории и
практики терапии сном значительно возросла опасность профанации и
вульгаризации, которые, как известно, всегда являются нежелательными
спутниками в любом серьезном и важном деле.
литература
Андреев Ф. А., Функциональная церебропатология внутренних заболеваний
и терапия их длительным сном, Труды Объединенной сессии, посвященной 10-летню
со дня смерти И. П. Павлова, 1948.
Асратян Э. А., Новый способ терапии травматического шока, Военно-сани-
тарное дело, № 8, 1943.
Асратян Э А., Новое в профилактике и терапии шока. Достижения совет-
ской медицины, № 2, 1944.
Асратян Э. А., Сонная терапия травматического шока, Военно-медицинский
сборник, АН СССР, № 2, 1945.
Асратян Э. А., Очерки по этиологии, патологии и терапии травматического
шока, 1945.
Асратян Э. А., Учение И. П. Павлова об охранительной и целебной роли
торможения и травмы нервной системы. Невропатология и психиатрия, № 4, 1944.
Асратян Э. Л., К этиологии, функциональной патологии и терапии послед-
ствий органических поражений нервной системы, Военно-медицинский сборник
АН СССР, № 3, 1946.
Асратян Э. А., К теории и практике охранительной к целебной роли тор-
можения, Физиологический журнал СССР, 32, № I, 1946.
Асратян Э. А., Сонная терапия травматического поражения нервной системы,
Труды объединенной научной сессии Ленинградского университета. 1946.
Асратян Э. А., К теории и практике павловского охранительно-целебного
торможения. Труды Объединенной сессии, поен я щен ной 10-летию со дня ечерти
И. П. Павлова, 1948.
Асратян Э. А., К профилактике и терапии теплового шока. Рефераты работ
Биологического отделения АН СССР, 1943—3944
Введенский В. Е., Собрание сочинений, т. 4, 19Ут
ТЕРАПИЯ СНОМ ПОСЛЕДСТВИЙ ОРГАНИЧЕСКИХ ПОВРЕЖДЕНИЙ Ц. Н. С. 545
Гурова Е, В,, Влияние снотворных на восстановление рефлекторной дея-
тельности истощенного изолированного спинного мозга, ДАН СССР, -51, № 9, 1916.
Гурова Е. В., Влияние снотворных на восстановление функций спинного
мозга при рефлекторных параличах, ДАН СССР, 51, № 7, 1946.
Долин А. О. и др., Сотрясение и контузия головного мозга и их лечение
сонным торможением, Невропатология и психиатрия, № 3, 19-16.
Иванов-Смоленский А. Г.3 О нарушениях нервной деятельности кончу
.«ионно-ко.ммоционного происхождения, Военно-медицинский сборник АН СССР,
№ 2, 1945.
Иванов-Смоленский Л. Г., Экспериментально-клинические исследования
в области охранительного торможения и длительного сна, Журнал высШён нервной
деятельности, 1, в. 3, 1951.
Иванова С. Н., Сонная терапия фантомных болей, Рефераты работ Биологи-
ческого отделения АН СССР, 1945.
Иванова С. Н., Восстановление функций организма после половинной по-
перечной перерезки спинного мозга на верхних уровнях, АН СССР, серия биоло-
гических наук, № 3, 1950.
Иванова С, Н,, Сонная терапия последствий оперативного повреждения
продолг овагого мозга животных, Рефераты работ Биологического отделения
АН СССР, 1945.
Минаев П. Ф., Лечение травмгатического шока у животных аденозинтрифос-
формой кислотой в сочетании с различными веществами, ДАН СССР, 74, № 2 1950.
Павлов И. Г!., Полное собрание трудов, т. Ill, 1949.
Павлов И. П.. Полное собрание трудов, т. IV, 1947.
Петрова М. К., Сонное наркотическое, сонное гипнотическое, а также
запредельное торможение и их терапевтическое значение, Труды Объединенной
сессии, посвященной 10-летию со дня смерти И. П. Павлова, 1948.
Романовская Е. А., Тепловая травма спинного мозга лягушек и поиски
способов стимуляции процессов восстановления нарушенных функций, Рефераты
работ Биологического отделения АН СССР, 1945.
Романовская Е. А., Сонная терапия последствий тепловой травмы спин-
ного мозга, автореферат дисс., 19.я2.
Сахиулина Г. Т., Влияние снотворных веществ на восстановление функций
анечизнрованной центральной нервной системы лягушки, ДАН СССР, 52, № I, 1916.
Сахиулина Г. Т., Влияние снотворных веществ на восстановление функций
анемнзированной центральной нервйой системы лягушки, ДАН СССР, 52, № 9, 19-46.
Сах нули на Г. Т., Влияние снотворных и стимулирующих нервную систс"”
веществ на восстановление функций анечизнрованной центральной нервной системы
лягушки, ДАН СССР, 52, № 3, Р946.
Сахиулина Г. Т., Сонная терапия каузалгий. Рефераты работ Биологиче-
ского отделения АН СССР, 1945.
Сахиулина Г. Т., Ближайшие и отдаленные последствия анемизации цент-
ральной нервной системы, дисс., 1919.
Сахиулина Г. Т., Анемическая травма спинного мозга еобак и физиологи-
ческая стимуляция процессов восстановления функций, Известия АН СССР, серия
биологических наук, № 3. 1951.
Сеченов И, АС, Избранные труды, 1935
Э5 9. А. Асратян
У ЧЕНИЕ И. П. ПАВЛОВА —ОСНОВА МЕДИЦИНЫ1
^объединенная сессия Академии наук СССР и Академии медицин-
ских наук СССР, посвященная проблемам физиологического
учения акад. И..П. Павлова, была выдающимся событием не
только в физиологии, но и во всей советской биологии и медм-
/jp цине. Сессия, как известно, была созвана вскоре после опубли-
кования гениального научного произведения товарища Сталина
о языкознании и протекала в атмосфере суровой и деловой критики и
самокритики, была как бы первым отзвуком наших биологов и врачей на
руководящее указание вождя, что «никакая наука нс может развиваться
и преуспевать без борьбы мнения, без свободы критики».
В единодушно принятом постановлении сессии подчеркивается, что
«созданный великим русским физиологом И. П. Павловым естественно-
научный объективный метод изучения жизненных функций организма, а
также его оригинальные исследования По физиологии кровообращения,уче-
ние о пищеварении и трофической иннервации, в особенности же его
гениальное учение о высшей нервной деятельности знаменуют новую эпоху
в развитии творческого естествознания».
Далее в резолюции подчеркивается особенное значение научного ме-
тода и научного творчества И. П. Павлова для медицины как области на-
учной и практической деятельности человека, органически связанной
с естествознанием, в известной мере являющейся частью естествознания.
Отметив, определенные достижения советских ученых в дальнейшем раз-
витии учения II. П. Павлова и во внедрении его идей в медицину, сессия
констатировала, что «в целом фактические н теоретические результаты
работы по развитию научного наследия И. П. Павлова, в особенности ис-
следования по высшей нервной деятельности, далеко не соответствуют за-
дачам, поставленным перед учениками и последователями великого уче-
ного, и условиям, созданным для этой цели советским правительством,
партией» и что по вине некоторых наших ученых: академиков Л. А. Ор-
белн, И. С. Бсриташвили, А. Д. Сперанского, преф. П. К. Ано-хина и дру-
гих, длительное время занимавших руководящее положение в нашей физио-
логии и патофизиологии, «разработка’ научного наследия И. П. Павлова во
многих отношениях не шла по столбовой дороге развития его идей». Сес-
сия отметила также слабое внедрение идей и направлений II. П. Павлова
в медицину.
В своем постановлении сессия со всей силой подчеркнула необходимость
коренного улучшения в области дальнейшей разработки научного насле-
дия И. П. Павлова, всемерного развития его идей и направления, равно
как и необходимость смелого внедрения его идей и научных принципов в ме-
1 Доклад, прочитанный 26 октября 1951 г.'на объединенной сессии Отделения
биологических наук Академии наук СССР и Академии наук Латвийской, Эстонской
и Литовской ССР в Риге. Советская книга, ,V° 12, 1951.
УЧЕНИЕ И. ГТ. ПАВЛОВА - ОСНОВА МЕДИЦИНЫ
547
дицину, необходимость перестройки медицины на созданных им новых на-
учных основах.
Как явствует из названия моего доклада, он имеет отношение как раз
к последнему вопросу, к вопросу о перестройке медицины на твердом есте-
ственно-научном фундаменте многогранного творчества гениального уче-
ного.
Прежде всего остановимся на одном вопросе — на павловском понима-
нии задач физиологии как естественно-научной дисциплины и на его пони-
мании взаимоотношений физиологии и медицины. Этот вопрос на первый
взгляд может показаться простым и малозначительным; на деле же он
принадлежит к разряду принципиальных и исключительно важных во-
просов.
В традиционном понимании не представляет особой трудности опреде-
ление задач физиологии и ее взаимоотношении с медициной. -Физиология
экспериментально и теоретически изучает закономерности нормальной дея-
тельности отдельных органов, отдельных систем органов и организма в це-
лом, изучает природу и динамику протекающих в них процессов в различ-
ных условиях существования. Нетрудно понять важное для медицины зна-
чение точных фактических данных и правильных теоретических положений
передовой материалистической физиологии. Эту важную задачу физиоло-
гии и этуг сторону взаимоотношений физиологии и медицины И. П. Павлов
охарактеризовал весьма красочно. Он писал: «Понимаемые в глубоком
смысле физиология и медицина неотделимы. Если врач в действительности
и тем более в идеале есть механик человеческого организма, то всякое но-
вое физиологическое приобретение рано или поздно непременным образом
увеличивает власть врача над его чрезвычайным механизмом, власть —
сохранять и чинить этот механизм»1.
Однако, как бы это все не было важно, этим, по И, П, Павлову,
далеко не исчерпывается задача физиологии и ее взаимоотношения с
медициной. Ярый противник тезиса, согласно которому паука яв-
ляется самоцелью для ученого, убежденный и горячий сторонник
принципа, согласно которому наука служит могучим средством реше-
ния важнейших практических Задач, воЗНикакщих Перед человеком,
И. П. Павлов признавал за физиологией еще более активную, еще более
действенную роль. Согласно воззрениям гениального мыслителя, физио-
логия должна стремиться познать явления, процессы, закономерности дея-
тельности организма с тем, чтобы можно было управлять, командовать
ими, поставить науку на службу жизни, активно содействовать разреше-
нию актуальных ее задач и в первую очередь задач медицины. Он считал,
в частности, что одной из главнейших задач физиологических исследований
является экспериментальная разработка и теоретическое освещение воп-
росов возникновения и течения, а также природы и лечения различных
болезненных состояний организма, и придерживался мнения, что со време-
нем такого рода исследования должны быть положены в основу всех важ-
нейших разделов медицины. По этому поводу он писал: «И только какой-
нибудь неисправимый схоласт мог бы сказать, что это не наше дело. На-
против, именно физиолог с его компетенцией — в методических и логи-
ческих приемах исследования жизни — является здесь самым законным
работником»* 2.
Более чем шестидесятилетия я кипучая и исключительно плодотворная
научная деятельность великого ученого, выдвинувшая нашу отечествен-
ную физиологию на первое место в мире, насквозь проникнута этим стрем-
лением. Наряду со своими классическими исследованиями по физисло-
* И П. Па влов, Полное собрание трудов, т. III, 1949, стр. 70.
2 Там же, т. II. 1946, стр. 345.
35*
548
Э. А. АСРАТЯН
гии сердечно-сосудистой системы, пищеварительного тракта и больших по-
лушарий головного мозга, создавшими новую эпоху в физиологии и имею-
щими огромное значение для медицины, И. П. Павлов специально занимался
экспериментальной и теоретической разработкой ряда актуальных во-
просов функциональной патологии и те pan и ив сфере деятель-
ности тех же систем организма.
Эту новую черту физиологии и ее взаимоотношений с медициной
И. П. Павлов весьма ярко характеризует следующими словами: «Механик
кончает свое изучение той или иной машины тем, что подвергается экза-
мену, состоящему в сборе разобранной и спутанной машины. То же должно
быть и с физиологом. Только тот может сказать, что он
изучил жизнь, кто сумел вернуть нарушенный
ход сек норме» (разрядка наша.—3. А.)1. В течение своей многолет-
ней научно-творческой деятельности И. П. Павлов высказывает эту мысль
неоднократно и по разным поводам. Так, например, за два года досмерги,
касаясь своих исследований по экспериментальной патологии и терапии
высшей нервной деятельности у животных, гениальный физиолог с закон-
ным удовлетворением писал: «Конечно, наша власть знания над нервной
системой должна выявиться еще в большей степени, если мы будем уметь
не только портить нервную систему, но потом и поправлять по желанию.
Тогда уже доподлинно будет доказано, что мы овладели процессами и ими
командуем. Это так и есть: во многих случаях мы не только производим
заболевание, но и устраняем его, так сказать, по заказу совершенно точно»2.
Таким образом, мы видим, что II. П. Павлов резко расширил традици-
онный диапазон применения физиологических исследований, охватил ими
также вопросы экспериментальной патологии и экспериментальной тера-
пии. Совершенно очевидно, что в этом новом, павловском понимании за-
дача физиологии, значение ее для медицины неизмеримо возрастают. Фи-
зиологический эксперимент в его высоком,павловском понимании приоб-
ретает значение могущественного фактора также в разрешении сложней-
ших вопросов патофизиологии и терапии, т. е. сугубо медицинских во-
просов. Великий физиолог считал, что «только пройдя сквозь огонь экспе-
римента, вся медицина станет тем, чем должна быть, т. е. сознательной
а следовательно, всегда и вполне целесообразно действующей»3. Это поло-
жение И. П. Павлов аргументировал тем, что медицина в познании слож-
ных процессов в человеческом организме почти исключительно пользуется
одним приемом — наблюдением, т. с. пассивным и сравнительно примитив-
ным приемом, достаточным лишь для изучения простых явлений, но абсо-
лютно непригодным для выявления, изучения и познания сущности возни-
кающих и протекающих в сложном организме процессов. Медицина по
вполне понятным причинам не в состоянии использовать для этой цели
активное, действенное, могучее орудие современного естествознания —
опыт, эксперимент, который в области физиологии был поднят И. П. Павло-
вым на небывалую высоту. Поэтому для своего прогресса медицина должна
возможно шире и Смелее опираться па физиологию. Должна всемерно ис-
пользовать беспредельную возможность лабораторного физиологического
эксперимента. «Наблюдение, — писал II. П. Павлов. — собирает то.
что ему предлагает природа, опыт же берет у природы
то, что он хрчет, И сида биологического опыта поистине
колоссальна» (разрядка наша.—3. Л.)4.
Чтобы полностью понять важное Значение, которое придавал И. П. Пав-
лов данным лабораторного эксперимента на животных для медицины,
1 И. П. Павлов, Полное собрание трудов, т. II, 1946, стр. 354.
4 Там же, т. III, 1949, стр. 545.
'Там же, т. П, 1945, стр. 360.
‘Там же, стр. 357.
УЧЕНИЕ И П. ПАВЛОВА — ОСНОВА МЕДИЦИНЫ 549
необходимо отметить еще следующее. Великий физиолог был противником
механического перенесения на человека экспериментальных данных и све-
дений, полученных на животных относительно деятельности различных
органов и систем, особенно относительно деятельности больших полушарий,
которая «так поражающе резко выделяет человека из ряда животных». На-
ряду с этим, он считал не только возможным, но и необходимым критическое
использование данных лабораторного эксперимента для понимания слож-
нейших закономерностей деятельности человеческого организма в условиях
нормы и патологии, и притом в отношении всех систем организма,
не исключая и высшие отделы нервной системы, ибо, говорил он: «Самые
общие основы высшей нервной деятельности, приуроченной к большим
полушариям, одни и те же как у высших животных, так и у людей,
а потому и элементарные явления этой деятельности должны быть одина-
ковы у тех к у других как в норме, так и в патологических случаях»1.
Отношения между физиологией и медициной И. П. Павлов рассматри-
вал нс в виде односторонней связи, а как союзнические отношения, по-
лезные не только для медицины, но и для физиологии. Считая полезной и
закономерной связь физиологии со многими отраслями знания и практи-
ческой деятельности человека, И. П. Павлов был особенно горячим сторон-
ником тесного, неразрывного союза между физиологией и медициной. Ге-
ниальный физиолог посвятил этой теме много ярких страниц своих науч-
ных трудов по физиологии и патофизиологии кровообращения, пищеваре-
ния, трофики тканей и высшей нервной деятельности, а также ряд специ-
альных докладов, блестящих по форме и содержанию, в которых с такой
страстью говорится о союзе между физиологией и медициной.
Мы уже говорили о павловском понимании значения физиологии для
медицины. В чем же видел великий ученый пользу от этого союза для фи-
зиологии? «Мир патологических явлений,—писал он,—представляет собой
бесконечный ряд всевозможных особенных, т. е. не имеющих места
в нормальном течении жизни, комбинаций физиологических явлений. Это,
бесспорно, как бы ряд физиологических опытов, делаемых природой и
жизнью, это часто такие сочетания явлений, которые бы долго не пришли
в голову физиологам и которые иногда даже не могли бы быть нарочно вос-
произведены техническими средствами современной физиологии. Отсюда
клиническая казуистика остается навсегда богатым источником новых фи-
зиологических мыслей и неожиданных физиологических фактов. Поэтому-то
физиологу естественно желать более тесного союза физиолог ин с ме-
дициной»*.
Нет необходимости что-либо добавлять к сказанному гениальным мы-
слителем, чтобы понять всю великую правду его положения о том, что «по-
нимаемые в глубоком смысле физиология и медицина неотделимы».
Попутно обратим внимание на следующие важные обстоятельства.
Из приведенной цитаты И. П. Павлова явствует, что он в отличие от не-
которых наших физиологов и патофизиологов (А. Д. Сперанский и его по-
следователи) патологические явления рассматривает как особенные
комбинации физиологических явлений, а не как особенные яв-
ления или процессы, отличные от физиологических.
Далее, И. П. Павлов проводит сравнение между механиком и врачом,
а также употребляет слово «механизм» для обозначения организма и т. п.
Это не должно служить поводом к ошибочным заключениям, как бывало,
будто И. П. Павлов iro своим воззрениям на кардинальные вопросы естество-
знания и философии был механистическим или вульгарным материали-
стом. В действительности он пользовался этими и подобными им сравне-
1 И, П. Павлов, Полное собрание трудов, т. IV, 1947, стр. 326.
! Та м ж е, т. 11, 19491, стр. 57.
550 Э. А. АСРАТЯН
ниями и словами лишь в целях, если можно так выразиться, риторических,
лишь для яркого, образного выргокения своих глубоких мыслей.
Чтобы не отклоняться сильно в сторону от основной нашей темы, мы не
можем обстоятельно осветить здесь важный вопрос о мировоззрении
И. П. Павлова. Интересующихся этим вопросом отсылаем к соответствую-
щим литературным источникам, в частности, к нашей специальной статье
на эту тему*. Здесь же отметим, что великий мыслитель был материалистом-
диалектиком, а не механистом. Об этом свидетельствует, в частности, его
понимание вопроса о задачах физиологии и об ее взаимоотношениях с ме-
дициной, т. е. по существу говоря, правильное диалектико-материали-
стическое понимание единства теории и практики, а также его понимание
сущности другого обсуждаемого здесь вопроса—относительно возможности
и характера применения к человеку фактов и сведении, полученных в
экспериментах на животных.
Итак, нам кажется, что павловское понимание задач физиологии и ее
взаимоотношений с медициной является существенной частью того креп-
кого научного физиологического фундамента, на который уже опирается,
а в будущем должна в еще большей степени опираться советская медицина.
Но наиболее важной частью этого фундамента, его краеугольным камнем
являются конкретные методические, фактические и теоретические резуль-
таты, достигнутые гением Павлова в процессе его многолетней кипучей и
напряженной научно-творческой работы, придавшие всей физиологической
науке современный, павловский облик, создавшие новую, павловскую эру
в физиологии, вошедшие в золотой фонд нашей отечественной науки.
Несколько слов о созданном И. П. Павловым физиологическом методе,
отражающем его основные научные принципы. Речь идет не о каких-то
частных методиках, приемах л способах физиологического эксперимента,
а о ведущих принципах, о широком методологическом подходе к экспери-
ментальной разработке важнейших вопросов физиологии и медицины.
И. П. Павлов придавал научному методу первостепенное значение. «Для
натуралиста — все в методе», говорил он.
В допавловской физиологии почти безраздельно господствовал крайне
односторонний: и упрощенный подход к изучению сложнейших функций
организма, получивший отражение в методе острого—вивисекционного
эксперимента. Экспериментаторы, пользовавшиеся вивисекцией, произ-
водили всевозможные разрезы на теле наркотизированного или ненарко-
тмзированного животного, грубо нарушали целостность организма, обна-
жали нужные органы, а то и вырывали их из тела, разрушали или нарушали
естественную связь и взаимодействие между ее частями, искажали нормаль-
ный характер и течение жизненных процессов в организме и стремились
при столь запутанной картине выявить и изучить закономерности работы
тех или иных органов, систем или организма в целом путем их стимуляции
или угнетения различного рода электрическими, механическими, химиче-
скими, термическими и другими воздействиями.
Виртуозно владея техникой вивисекционного эксперимента, И. П. Пав-
лов тем не менее первым обрушился на этот порочный, топорный и мало-
продуктивный метод исследования сложнейших функций организма и пер-
вым ввел ю систему хронические физиологические эксперименты, т. е.
эксперименты на неповрежденных животных или на животных, заранее
оперированных по всем правилам хирургической антисептики и асеп-
тики и доведенных до нормального состояния тщательным послеопераци-
онным уходом. Эти эксперименты дают возможность детально, тонко и
многосторонне исследовать функции тех или и.ных органов и систем в це-
лостном организме, в живой связи этих органов и систем со всеми осталь-
1 Э. А. Асрат ян. Вопросы философии, Ni 3, 1949.
УЧЕНИКИ П ПАВЛОВА - ОСНОВА МЕДИЦИНЫ 551
ними, в естественных условиях кх работы, исследовать физиологические
процессы в их нормальном виде, в динамике, «изучать деятельность орга-
низма в целом и его частей в строго нормальных условиях и в связи с этими
условиями» (И. П. Павлов).
Искусный мастер филигранного анализа сложнейших функций орга-
низма, И. П. Павлов первым в мировой физиологии стал на путь синтети-
ческого изучения этих функций, точнее, сочетал аналитический и синтети-
ческий подход к их изучению и создал единый и объективный научный метод
познания глубоких тайн органической природы,— метод Павлова, по суще-
ству своему диалектический, самый совершенный и плодотворный метод в
мировой физиологии. Зародившись в самый ранний период научной дея-
тельности И. П. Павлова, этот метод в Процессе дальнейших исследований
великого ученого непрерывно совершенствовался, обеспечивая богатство
достоверных и ценнейших фактических данных при экспериментальной раз-
работке всех тех вопросов физиологии, патофизиологии и терапии, к кото-
рым он приложил свой могучий ум и чудодейственные руки.
Из всего сказанного явствует, что не каждое резание животного есть
совершенный физиологический эксперимент. В павловском понимании фи-
зиологический эксперимент имеет весьма высокое содержание, является
действенным и плодотворным орудием раскрытия закономерностей функ-
ционирования организма в различных его состояниях. Говоря о перво-
степенной важности эксперимента для медицины, великий физиолог имел
в виду именно такой эксперимент.
R научном методе И. П. Павлова не в меньшей стелена, чем в остальном
его научном творчестве, сказывается его мировоззрение, его принципиаль-
ные установки на кардинальные вопросы биологии и естествознания во-
обще, установки, которые имеют фундаментальное значение также для всей
медицинской науки. Речь идет прежде всего о воззрениях великого физио-
лога относительно неразрывной целостности организма и единства орга-
низма с окружающей средой, — воззрениях, которые родият его с выдаю-
щимися представителями передового советского дарвинизма — К. А. Ти-
мирязевым, И. В. Мичуриным и Т. Д. Лысенко. «Как ясно каждому, —
писал И. П. Павлов, — животный организм представляет крайне слож-
ную систему, состоящую из почти бесЕСонечного ряда частей, связанных
как друг с другом, так и в виде единого комплекса с окружающей средой
и находящихся с ней в равновесии»1.
Разработанный И. П. Павловым метод хронического эксперимента не
только обеспечивает объективное изучение функций организма в условиях
его ненарушенной целостности, но и сохраняет единство организма со сре-
дой, пусть порой искусственной, лабораторной. Именно это обстоятельство
позволило гениальному натуралисту' установить детерминированность про-
стых и сложных функций организма условиями их существования, выя-
вить во всем величии и красе роль нервной системы в высокоразвитом слож-
ном организме, всесторонне и глубоко изучить эту' роль и развить на этой
основе ведущий принцип его многогранного научного творчества — прин-
цип нервизма. Этот плодотворный и передовой научный принцип, прохо-
дящий красной нитью через всю многолетнюю научно-творческую работу
И. П. Павлова по различным проблемам физиологии и утверждающий ве-
дущую роль нервной системы в регулировании функций высокоразвитого
животного организма, в обеспечении его целостности и его единства со
средой, в приспособительной деятельности органов, систем и организма
в целом,— этот принцип привел великого натуралиста, в конце концов,
к изучению физиологии больших полушарии мозга, где достиг своего апо-
гея. И. П. Павлов установил фундаментальное для всей медицины и био-
1 И. П. Павлов, Полное собрание трудов, т. II, 1946, стр. 452.
552
Э А АСРАТЯН
логин положение, что большие полушария мозга являются верховным рас-
порядителем и распределителем всего, что происходит в организме, и одно-
временно обеспечивают самое совершенное, точнее и топкое приспособле-
ние организма к среде, регулируют его сложнейшие и тончайшие взаимо-
отношения с окружающим миром.
Перейдем к краткому обзору тех конкретных фактических и теорети-
ческих достижений И. П. Павлова, которые имеют особенное значение для
меди цины.
В начальном периоде своей научной деятельности И. П. Павлов в тече-
ние примерно 15 лет интенсивно изучал ряд актуальных вопросов физио-
логии кровообращения и обогатил эту важнейшую область физиологии
ценнейшими вкладами экспериментального и теоретического характера,
которые в то же время имеют первостепенное значение для многих раз-
делов медицины, в первую очередь для терапии и хирургии. Им было уста-
новлено, в частности, что собственные чувствительные «приборы» в
сердечно-сосудистой системе имеют несравненно более широкое распростра-
нение, чем принято было думать в то время, и что они способны улавли-
вать изменения не только в уровне артериального давления, но и в хими-
ческом составе крови. Им было установлено также, что в совокупности эти
воспринимающие приборы через соответствующие рефлексы осуществляют
саморегуляцию сердечно-сосудистой деятельности, приспосабливают эту
деятельность к текущим потребностям организма, поддерживают артериаль-
ное давление в известных средних пределах, наиболее благоприятных для
кровоснабжения основных жизненно важных органов и систем организма.
Впоследствии И. П. Павлов на основании новых фактических данных раз-
вил более широкое теоретическое положение, согласно которому во всех
органах и тканях организма имеются разнообразные и специфические чув-
ствительные нервные образования, «приспособленные каждое к своему
своеобразному раздражителю механического, физического или химиче-
ского характера» и играющее очень важную роль в рефлектор ной регуля-
ции разнообразных функций организма.
Как известно, эти данные, лежащие в их основе идеи и все это направле-
ние исследовательской работы И. П. Павлова, представляющие исключи-
тельный интерес для всех основных разделов медицины, в наше время
с успехом развиваются выдающимся его учеником акад. К. М. Быковым и
сотрудниками, а также другими отечественными и зарубежными учеными.
К раннему периоду исследовательской работы II. П. Павлова относится
также установление нм факта, что среди центробежных нервов сердца
имеются такие, которые способны усиливать сердечные сокращения без изме-
нения их частоты, и такие, которые способны ослаблять сердечные сокра-
щения без изменения их частоты. Детальный экспериментальный ана-
лиз этих фактов привел проникновенного теоретика к мысли о существо-
вании особого, в те времена неизвестного, рода нервного влияния на
ткани организма, а именно нервной регуляции интимных химических
процессов или «питания» (трофики) тканей и органов.
Позднее при изучении И. П. Павловым физиологии пищеварения эти
первоначальные факты были существенно подкреплены им новыми факта-
ми того же рода и положены в основу оригинального и глубокого учения
о трофической иннервации тканей, являющегося одним из выдающихся
достижений отечественной физиологии и имеющего первостепенно важное
значение для всей медицины.
Исследования И. П. Павлова, посвященные физиологии пищеварения,
по новизне и ценности теоретических и экспериментальных результатов,
а также по оригинальности и мастерству выполнения являются поистине
классическими. Они и выдвинули И. II. Павлова в первые ряды славной
плеяды отечественных физиологов и снискали ему мировую известность.
УЧЕНИЕ И П ПАВЛОВА - ОСНОВА МЕДИЦИНЫ
553
Не приходится говорить о колоссальном значении результатов этих иссле-
дований для медицины, особенно для терапии и хирургии. Передовые дея-
тели советской медицины по достоинству оценили важное значение экспе-
риментальных и теоретических достижений И. П. Павлова в физиологии
пищеварения и немало способствовали их проникновению в клинику.
ДопавловСкая физиология пищеварения была одним ИЗ самых отсталых
разделов физиологии. Старания большого числа выдающихся отечественных
и зарубежных физиологов ликвидировать это отставание или хотя бы зна-
чительно ослабить его оказались тщетными. Лить гению И. П. Павлова
суждено было вывести физиологию пищеварения из застоя и поднять ее на
небывалую высоту. Началось с разработки и блестящего осуществления
целон серии остроумных и утонченных хирургических операций над упря-
танными вглубь тела органами пищеварительной системы, операций, де-
лающих доступной для наблюдения и исследования деятельность этих ор-
ганов без нарушения их кровоснабжения, иннервации и вообще условии
их нормального существования и работы.
Многолетние напряженные и целеустремленные исследования И. П. Пав-
лова и его учеников по физиологии пищеварительной системы увенчались
блестящими результатами. Вопреки распространенным в то время оши-
бочным взглядам, И. П. Павловым было установлено, что основные пище-
варительные железы (желудочные и поджелудочная) имеют секреторную
иннервацию, т. е. нервы, способные вызывать и тонко регулировать сек-
реторную деятельность желез. Было показано, что, благодаря нервной
регуляции состояния н функции главных пищеварительных желез, ди-
намика их секреторной деятельности, равно как и ферментативные, хими-
ческие и физические свойства выделенных ими пищеварительных соков
поразительно топко и точно приспосабливаются к качеству, количеству
и роду принятых пищевых продуктов, а также введенных в пищеваритель-
ный тракт несъедобных веществ. Было обнаружено и изучено явление
«психического возбуждения» пищеварительных желез, установлено суще-
ствование весьма тонкой специфической возбудимости слизистой оболочки
в разных участках пищеварительного тракта, выявлены и изучены другие
стороны и закономерности деятельности пищеварительного тракта в уело
виях нормы и патологии, в условиях голода и насыщения и т. п.
Полученный И. П. Павловым и его учениками богатейший и ценней-
ший фактический материал по физиологии пищеварения был обобщен и
освещен им с позиций материалистического миропонимания и положен им
в сснову его монументального учения о физиологии пищеварения. Без
преувеличения можно сказать, что фактически И. П. Павлов заново соз-
дал этот важный раздел физиологии и тем самым создал твердую естест-
венно-научную основу для важнейшего раздела медицины, изучающего
этиологию, функциональную патологию и терапию заболеваний желудочно-
кишечного. тракта. Его классические исследования создали твердый фун-
дамент для дальнейших экспериментальных и клинических исследовании
в этом направлении как у нас, так и за рубежом (И. П. Разенков, К. М. Бы-
ков, Ю. В. Фольборт и др.).
На пороге XX столетия маститый физиолог переключился на разра-
ботку физиологии больших полушарий мозга — самого высшего и слож-
ного создания земной природы. В течение 35 лет напряженной, целенаправ-
ленной и исключительно продуктивной научно-исследовательской работы
И. П. Павлов создал свое гениальное материалистическое учение о выс-
шей нервной деятельности, являющееся вершиной его научного творчества,
одной из наиболее крупных побед человеческой мысли над природой и
подтверждающее правильность разрешения марксистско-ленинской теорией
кардинального философского вопроса — отношения материи и психики.
Если учесть при этом, что в течение многих лет веЛИКИЙ ученый проводил
554 э- А. АСРАТЯН
весьма интенсивное и целеустремленное экспериментальное и клиническое
изучение важнейших вопросов патологии и терапии высшей нервной дея-
тельности, то станет совершенно очевидно колоссальное значение достиг-
нутых им теоретических и экспериментальных результатов, его напряжен-
ного творческого труда по физиологии и патологии больших полушарий
для всей медицины, в первую очередь для психиатрии и невропатологии.
Прежде всего следует отметить, что при изучении сложнейших законо-
мерностей деятельности больших полушарий мозга И. П. Павлов реши-
тельно и бесповоротно стал на испытанный путь натуралиста, на путь точ-
ного и объективного физиологического эксперимента и строго научного
мышления, ибо, как он считал, «при этом работа все время держится на
прочном, материально-фактическом фундаменте, как во всем остальном
естествознании, благодаря чему поистине неудержимым образом накоп-
ляется точный материал и чрезвычайно ширится горизонт исследования»1.
Далее, И. П. Павлов установил, что выработка условных рефлексов
является основным и наиболее характерным видом деятельности больших
полушарий, что разнородные и разностепенные условные рефлексы в со-
вокупности и составляют основной фонд высшей нервной или, говоря иначе,
психической деятельности, что большие полушария посредством образова-
ния (а при необходимости и подавления) условных рефлексов обеспечи-
вают наиболее совершенное, точное и тонкое приспособление организма
к условиям существования, осуществляют наиболее тонкие взаимоотно-
шения организма со средой, наиболее совершенную анализаторную и син-
тетическую деятельность, а также наиболее тонкую и совершенную регу-
ляцию функций наружных и внутренних органов.
И. П. Павлов доказал строгую детерминированность высшей нервной
деятельности животных условиями их существования. Он установил наличие
у человека еще особого, более высокого и совершенного вида высшей нерв-
ной деятельности — второй сигнальной системы, или речи, обусловленной
трудовой деятельностью человека и социальной жизнью.
Следует указать еще на важное значение для медицины теоретических
и экспериментальных исследований И. П. Павлова относительно динамиче-
ской локализации функций в коре больших полушарий, физиологических
основ типа и характера, физиологической природы сна и гипноза.
Не приходится и говорить о прямом отношении к медицине, и специально
к психиатрии и невропатологии, исследований И. П. 11авлова по экспери-
ментальной патологии и терапии высшей нервной деятельности животных
и в особенности по патологии и терапии высшей нервной деятельности че-
ловека.
Выдающиеся исследования И. П. Павлова по этим вопросам по существу
открыли новую яркую страницу в истории невропатологии и психиатрии,
а их значение выходит далеко за пределы указанных двух важнейших раз-
делов медицины, охватывая все основные ее разделы.
В процессе своих многолетних исследований по физиологии и патофи-
зиологии больших полушарий мозга И. П. Павлов нс только выявил и де-
тально изучил роль процесса торможения в координационной их деятель-
ности, но и открыл совершенно новую, в биологическом отношении крайне
важную роль этого процесса в деятельности нервных клеток коры больших
полушарий мозга, а именно: охранительную и целебную роль про-
цесса торможения. Об охранительной роли торможения в условиях норма-
льной деятельности больших полушарий он писал: «С одной стороны, посто-
янно, наравне с процессом раздражения участвуя в разнообразной дея-
тельности животного во время бодрого состояния, торможение, с другой
стороны, также постоянно является в роли охранителя реактивнейших кле-
1 И. П. Павлов, Полное собрание трудов, т. III, 1949, стр. 221.
УЧЕНИЕ И. П. ПАВЛОВА - ОСНОВА МЕДИЦИНЫ
555
ток организма, клеток коры больших полушарий, защищая как специально
против чрезвычайного напряжения их деятельности при встрече с очень
сильными раздражениями или при длительном повторен ии хотя бы и не-
сильных раздражений, так и обеспечивая им необходимый покой после
ежедневной нормальной работы в виде сна» \
Таким образом, согласно воззрениям Н.П. Павлова, в условиях нор-
мальной деятельности корковых клеток, отличающихся от других нервных
клеток гораздо более высокой реактивностью, низкой работоспособностью
и быстрой утомляемостью, охранительная роль торможения выявляется
либо ft виде нестойкого запредельного торможения при сверхсильных
раздражениях, либо в виде каждодневного сна после длительного бодр-
ствования и деятельности. В том и другом случае возникшее торможение
прекращает деятельность истощенных нервных клеток, ограждает их от
вредного влияния внешних факторов, предотвращает опасность дальней-
шего их истощения, создает для них наиболее полноценный физиологи-
ческий покой и способствует восстановлению нх состояния и работо-
способности.
Охранительная, защитная роль торможения в отношении клеток коры
больших полушарий мозга еще ярче выступает при разнообразных болез-
ненных состояниях последних. Одной из важных предпосылок для разви-
тия подобных состояний является природная или приобретенная слабость
самых клеток. Такие клетки под воздействием сверхсильиых раздражений
или иного рода трудностей утомляются и истощаются и впадают в состоя-
ние более или менее длительного торможения. Хотя в этих случаях тормо-
жение выводит из строя значительные массы корковых клеток и тем самым
нарушает нормальную целостную деятельность мозга, тем не менее в от-
ношении ослабленных и истощенных нервных клеток оно и здесь выступает
в качестве охранительного процесса, в качестве «физиологической меры»,
способной предохранить слабые нервные клетки от дальнейшего истощения,
от угрожающего разрушения вследствие непосильной работы, выступает
«как нормальный прием физиологической борьбы против болезнетворного
агента» (И. II. Павлов).
Однако в отношении ослабленных и истощенных корковых нервных кле-
ток торможение выступает не только в роли природного фактора самоза-
щиты, но и в качестве природного фактора самолечения. Речь
идет о целебной или, как говорил И. П. Павлов, целительной роли
торможения. Торможение способствует нормализации состояния и восста-
новлению функций ослабленных и истощенных нервных клеток, а тем са-
мым и восстановлению нормальной, целостной деятельности больших полу-
шарий мозга.
Трудно переоценить значение фактических данных и теоретических
положении И. II. Павлова об охранительной и целебной роли торможения
не только для физиологии, но и для медицины в целом, в особенности для
психиатрии и невропатологии. В научном творчестве великого мыслителя
они принадлежат к тем звеньям, которые особенно крепко связывают фи-
зиологию с медициной.
Напоминаем, что в своих блестящих «физиологических экскурсах» в об-
ласть психиатрии и невропатологии, сделанных на основании непосредст-
венного наблюдения психически и нервных больных, И. П. Павлов объяс-
нил многие из этих болезненных состояний (шизофрения, истерия и т. и.)
главным образом как результат возникновения стойкого охранительного
торможения в больших полушариях мозга. Легко понять, что многообра-
зие форм болезненных состояний можёт быть обусловлено различной экс-
тенсивностью, интенсивностью и локализацией возникшего охраннтель-
* И. П Павлов, Полное собрание трудов» т. Ill, 1949, стр. 407.
556
Э. А. АСРАТЯН
кого торможения. Следует указать, что это учение является научной осно-
вой одного из самых актуальных и перспективных терапевтических
направлений современной медицины — терапии сном пли наркозом бо-
лезненных состояний организма и прежде всего психических и нервных за-
болеваний.
Накопленные И. П. Павловым и его сотрудниками экспериментальные
и клинические данные, а также его учение об охранительной и целебной
роли процесса торможения послужили основанием для дальнейших лабо-
раторных и клинических исследований в этом направлении со стороны зна-
чительной группы его учеников и последователей: М. К. Петровой,
Л. Г. Иванова-Смоленского, Ф. Л. Андреева, А. О. Долина, Э. А. Асра-
тяна и др.
В конце доклада мы коснемся некоторых трудностей и шероховатостей
в нашей работе по этой линии.
Большевистская партия и советское правительство поставили перед
нашими учеными трудную и ответственную задачу: всемерно развивать на-
учное наследие гениального И. П. Павлова и внедрять его идеи в различ-
ные области нашей социалистической практики, в первую очередь в меди-
цину. Не касаясь всего комплекса вопросов, связанных с реализацией
этой задачи, мы соответственно содержанию нашего доклада коснемся лишь
некоторых вопросов, имеющих отношение к внедрению идей и направления
И. П. Павлова в медицину, к перестройке советской медицины на началах
павловской физиологии.
На пути успешной реализации этой задачи стояло и продолжает стоять
несколько серьезных опасностей, о которых мы никогда не должны за-
бывать. Это, во-первых, опасность недооценки громадного значения на-
учного творчества И. П. Павлова для медицины со стороны убежденных
противников материалистического учения Й. II. Павлова, со стороны мало-
численной группы клиницистов —-М. О. Гуревича, А. С. Шмарьява,
А. О. Эдельштейна и их единомышленников, зачастую примыкающих к тем
или иным модным или устарелым течениям реакционной буржуазной
медицины.
Существует и другая опасность на пути успешного внедрения идей
И. П. Павлова в медицину — опасность профанации, вульгаризации, упро-
щенчества, опошления крайне ответственного и серьезного дела формаль-
ным подходом, склонностью к дешевым сенсациям и рекламе.
Далеко не так уж редки уродливые факты декларативной, искусствен-
ной, а то и насильственной «увязки» учения ил и не которы х идей И. П. Пав-
лова с теми или иными специфическими, зачастую мелкими вопросами ме-
дицины. Нужно ли доказывать, что подобного рода поступки и побужде-
ния, чем бы они ни обусловливались, всегда свидетельствуют о незнании
учения И. П. Павлова или в лучшем случае о крайне поверхностном его
знании, не имеют ничего общего ни с подлинной наукой, ни с ее правиль-
ным внедрением в практику.
Наконец, еще об одной серьезной опасности. Некоторые ученые, раз-
рабатывающие те или иные важные вопросы патофизиологии и терапии
в. свете учения И. П. Павлова, уклоняются от указанного И. П. Пав-
ловым пути, от его принципа предварительной разработки этих вопросов
в лабораторных экспериментах на животных и только затем критического
использования полученных результатов в клинике. Разумеется, на совре-
менном уровне ваших знании и при теперешнем арсенале наших методи-
ческих возможностей крайне затруднительно или даже невозможно до-
биться удовлетворительной разработки многих важных вопросов пато-
физиологии и терапии только в- условиях 'лабораторного эксперимента
на животных. В подобных случаях осторожное клиническое исследование
неизбежно.
УЧЕНИЕ И. П. ПАВЛОВА — ОСНОВА МЕДИЦИНЫ
5.57
Но ученые, о которых мы говорим, такой прием разработки вопроса
патофизиологии и терапии превратили в основное правило, забывая ука-
зание И. П. Павлова о важной роли эксперимента для медицины, а также
его слова о том, что «чем полнее будет проделан опыт на животных, тем ме-
нее часто придется больным быть в положении опытных объектов, со всеми
печальными последствиями этого» ’.
Было бы непростительно, конечно, игнорировать или замазывать эти
опасности и не предпринимать энергичных мер борьбы с ними в соответ-
ствии с источниками, характером и степенью опасности. Эти и другие труд-
ности на путях перестройки советской медицины на началах павловской
физиологии могут и должны быть преодолены эффективными научными и
организационными мероприятиями, среди которых, с нашей точки зрения,
одно из первых мест должна занимать организация систематического, ме-
тодического и углубленного изучения гениального научного творчества
великого Павлова всеми нашими физиологами, патофизиологами, клинн-
цистами и врачами-различных специальностей. Можно не сомневаться, что
каждый передовой ученый, каждый мыслящий врач, который будет добро-
совестно осваивать его гениальное учение, идеи, направление, будет оча-
рован и пленен величием ученого и его научным творчеством. Каждый из
них, несомненно, будет стремиться связать учение И. П. Павлова или те
или иные его идеи .'со спецификой своей научной или практической деятель-
ности по внутреннему убеждению, с любовью, со знанием дела и, конечно,
с большой пользой.
Многолетняя научно-творческая деятельность И. П. Павлова может
быть охарактеризована как триумфальное шествие его сверкающего ге-
ния на поприще науки. Бесспорно, в его выдающихся научных дости-
жениях, помимо его огромных природных дарований, весьма важную роль
сыграли высокая культура, совершенный стиль его научных исследований и,
что особенно следует подчеркнуть, его безграничная и беззаветная любовь
к народу, его пламенный патриотизм в самом высоком и благородном
смысле этого слова.
Можно не сомневаться, что если мы не только в научном, но и в этих
отношениях будем достойными его учениками и последователями, то успех
будет нашим непременным спутником на путях разрешения трудной и от-
ветственной задачи, поставленной перед нами большевистской партией, со-
ветским правительством и мудрым вождем нашим великим Сталиным.
J И. П. Павлов. Полное еоГраиие трудов, т. II, 1946, стр. 363.
СОДЕРЖАНИЕ
С гр
От автора............................................................. 5
Раздел I. Условные рефлексы
Новая методика изучений условных рефлексов у черепах (совместно с Алек-
саняном и Барсегян)............................................. И
Матерна.-ы по условным рефлексам у черепах (совместно с Алексаняном) . . 14
Условнореф-екториая деятельность у черепах без больших полушарий и без
промежуточного мозга (совместно с Алексаняном)................. 19
Системность работп» больших полушарий головного мозга.................24
Влияние пищевого безусловного рефлекса на соответствующне условные реф-
лексы .................................. ...... ......... ...... 38
Влияние посторонних и условных раздражителей на величину безусловного
рефлекса................................................... 46
Влияние экстирпации верхних шейных симпатических узлов ка пищевые
условные рефлексы у собаки..................................... 58
Влияние одновременной перерезки обоих шейных симпатических нервов на
пищевые условные реф|ексы у собаки ................. 77
О влиянии симпатической нервной системы на условнорсфлектооную деятель-
ность собаки .............................................. ........ 95
Связь между дтительностью действия условного раздражителя и величиной
условного рефлекса..........................’. .................96
К физиологии иррадиации и концентрации процессов в коре больших полу-
шарий , , 106
К учению о физиологической лабильности высших центральных этажей . . . ПО
Влияние условного оборонительно-двигательного ]>еф.чекса на безусловную
болевую реакцию собаки ......................... 128
К вопросу о локализации центральной части рефлекторной дуги двигательного
оборонительного условного рефлекса........................... 13-1
Влияние питуитрина на условнее пищевые слюноотделительные рефлексы
(предварительное сообщение)................................... 139
О прочности условных рефлексов .................................. 145
К вопросу об анатомо-гистологической основе условнорефлекторнон деятель-
ности высших животных............................................ . 147
Принцип «переключения» в условнорефтекторной деятельности............162
К физиологии двигательных оборонительных условных рефлексов ...... 168
Некоторые наблюдения над собаками, лишенными коры большого мозга . . . 183
Перектючение в усювнорефтскторнай деятельности.......................189
К физиологии временной связи....................................... 197
О диалекгико-материзлистическом характере учения И. П. Павлова .... 226
СОДЕРЖАНИЕ
559
Стр.
Раздел II. Компенсаторные приспособления
Кора большого мозга и пластичность нервной системы. Сообщение I. Опыты
с ампутацией лап ....................................................... 247
Кора большого мозга и пластичность нервной системы. Сообщение II. Опыты
с половинной перерезкой спинного мозга..............................267
Кора большого мозга и пластичность нервной системы. Сообщение III. Вари-
ант перекрестного сшивания нервов...................................285
Кора большого мозга и пластичность нервной системы. Сообщение IV. Опыты
с деафферентацией конечностей ......................................302
Кора большого мозга и пластичность нервной системы. Сообщение V. Опыты
е разрушением лабиринтов ......................................... 310
Кора большого мозга и пластичность нервной системы. Сообщение VI. Опыты
с перерезкой задних корешков первых трех пар шейных нервов..........320
Кора большого мозга л пластичность нервной системы. Сообщение VII. Опыты
с повреждением дыхательных нервных путей ......................... 330
Кора большого мозга' и пластичность нервной системы, ('.сообщение VIII.
Опыты с поперечной перерезкой задней половины спинного мозга .... 338
О принципе соотносительной пластичности нервной системы................363
Новые данные по физиологии большого мозга ..............................371
Предварительные результаты и перспективы применения нового способа ане-
мического поражения центральной нервной системы у высших животных . 385
Некоторые общие черты нарушения и восстановления нервной деятельности 391
Компенсаторные приспособления в свете учения И. П. Павлова ....... 399
Раздел III. Охранительно-целебное торможение
Охранительная и целебная роль процесса торможения ................... 435
Профилактика и терапия шока............................................441
Учение И. П. Павлова об охранительной и целебной роли торможения м
травмы нервной системы ................. ........................... 445
Терапия сном травматического шока .....................................452
Терапия сном травматических поражений нервной системы .................473
К этиологии Функциональное патологии и терапии последствий органических
поражений нервной системы......................................... 481
К теории и практике павловского охранительно-целебноп> торможения . . . 507
Идеи И. П. Павлова и клиника органических поражений нервной системы . 516
Об охранительном торможении к его значении в медицине ............... . 521
Экспериментальная терапия сном -последствий органических повреждений
центральной нервной системы .......... ............................. 532
Учение И. П. Павлова—основа медицины ..................................546
Печатается по поС1ан0И.ч£Н1т10
Редакционно-издательского совета
Академии медицинских наук СССР
Редактор Г. В. Архшылыкий
Технический редактор Н. Л. Кирсанова
Г03415 Подп. к печати 15.IV 19л4 г
Изд. № 68 Зак. 622
Форм, бумаги 70Х1081/и Бум. л. 17,5
Печ. .1 47,95. Уч.-изд. л. 49,33
Цена 34 о. 55 к. по прейскуранту 1952 г
Переплет I р.Тираж 10000
1-я тип Трансжелдорнздата МПС
ЗАМЕЧЕННЫЕ ОПЕЧАТКИ
Стра- ница Строка Напечатано Следует читать
6 21 сверху ошибочность экспери- ошибочность части
ментальных экспериментальных
234 4—3 снизу коренных качественных коренных, качественных
237 10—9 снизу в доугом в его прмзна- в другом—в его
НИИ признании
243 10 сверху котсбания ми колеба ния
247 11 снизу смыль мысль
374 ] » стр. 201 стр. 201 и 101
387 21 > существенным существенными
389 17 сверху Сахнулевой Сахнулиной
411 20 снизу спинных спинальных
413 11 снизу бараны вараны
449 27 сверху Самукова Симукова
4рб 1 сверху историй из историй
Э. А. Асратян Зак. 622-
35 р. 55 к.