




















































































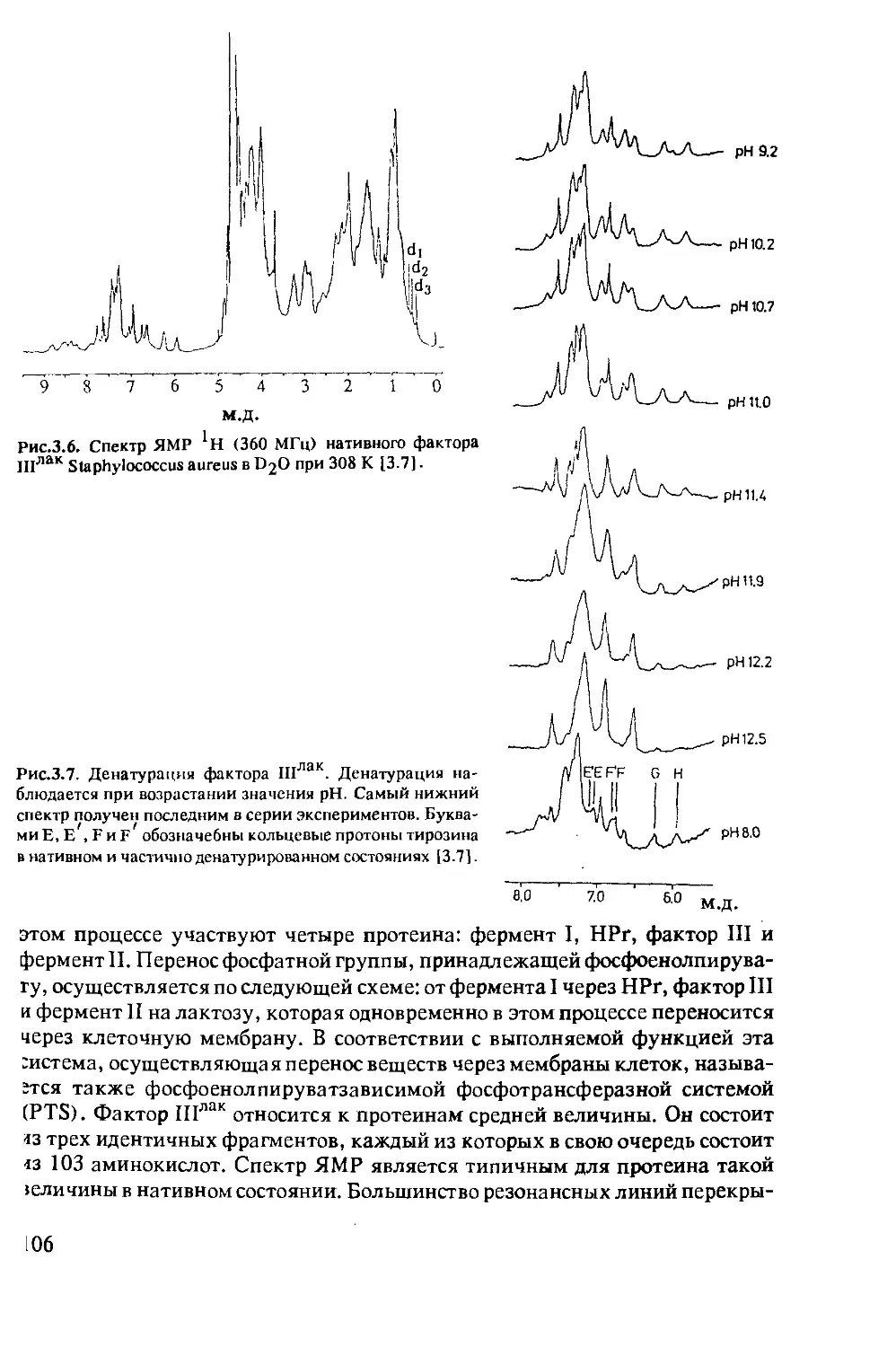



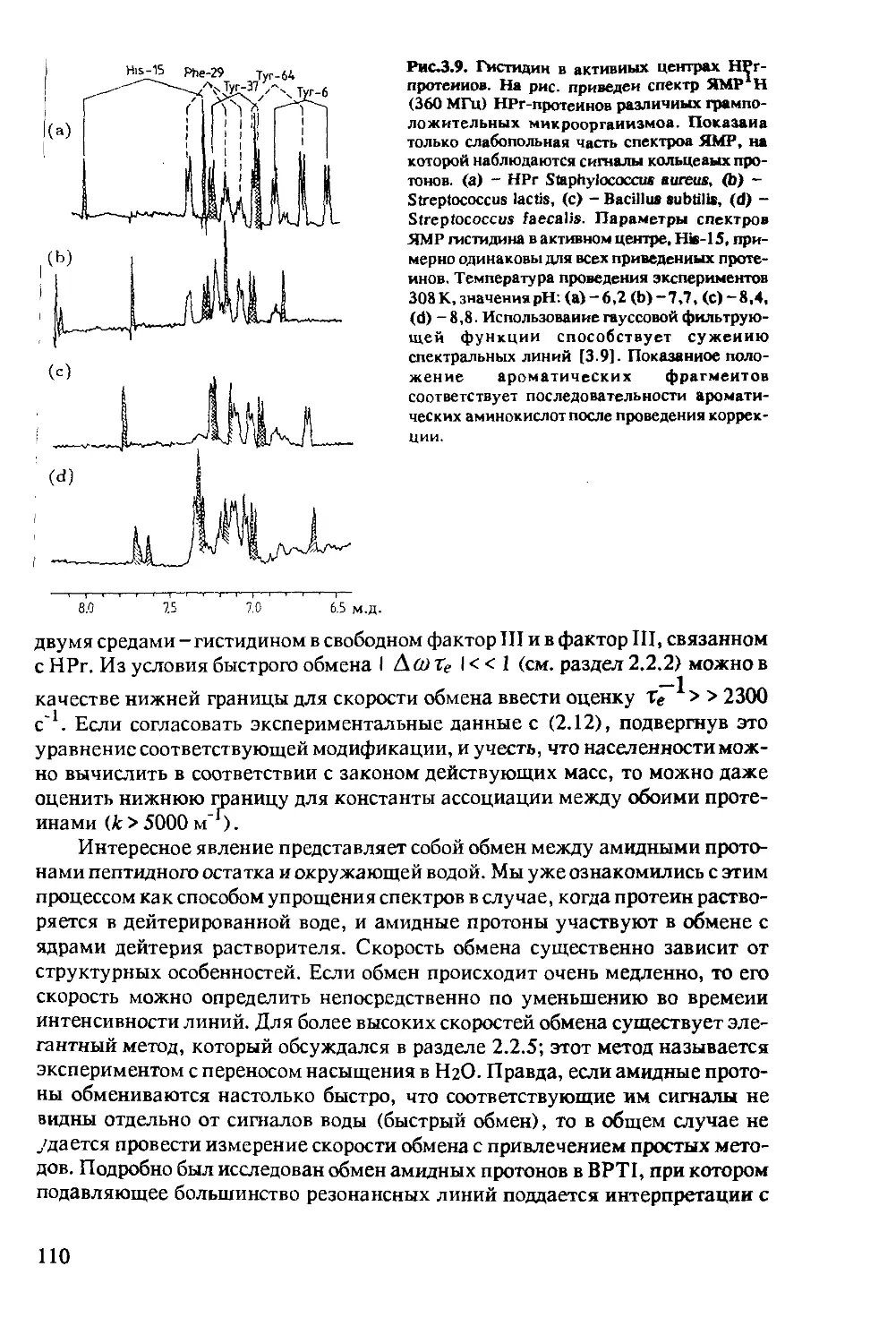
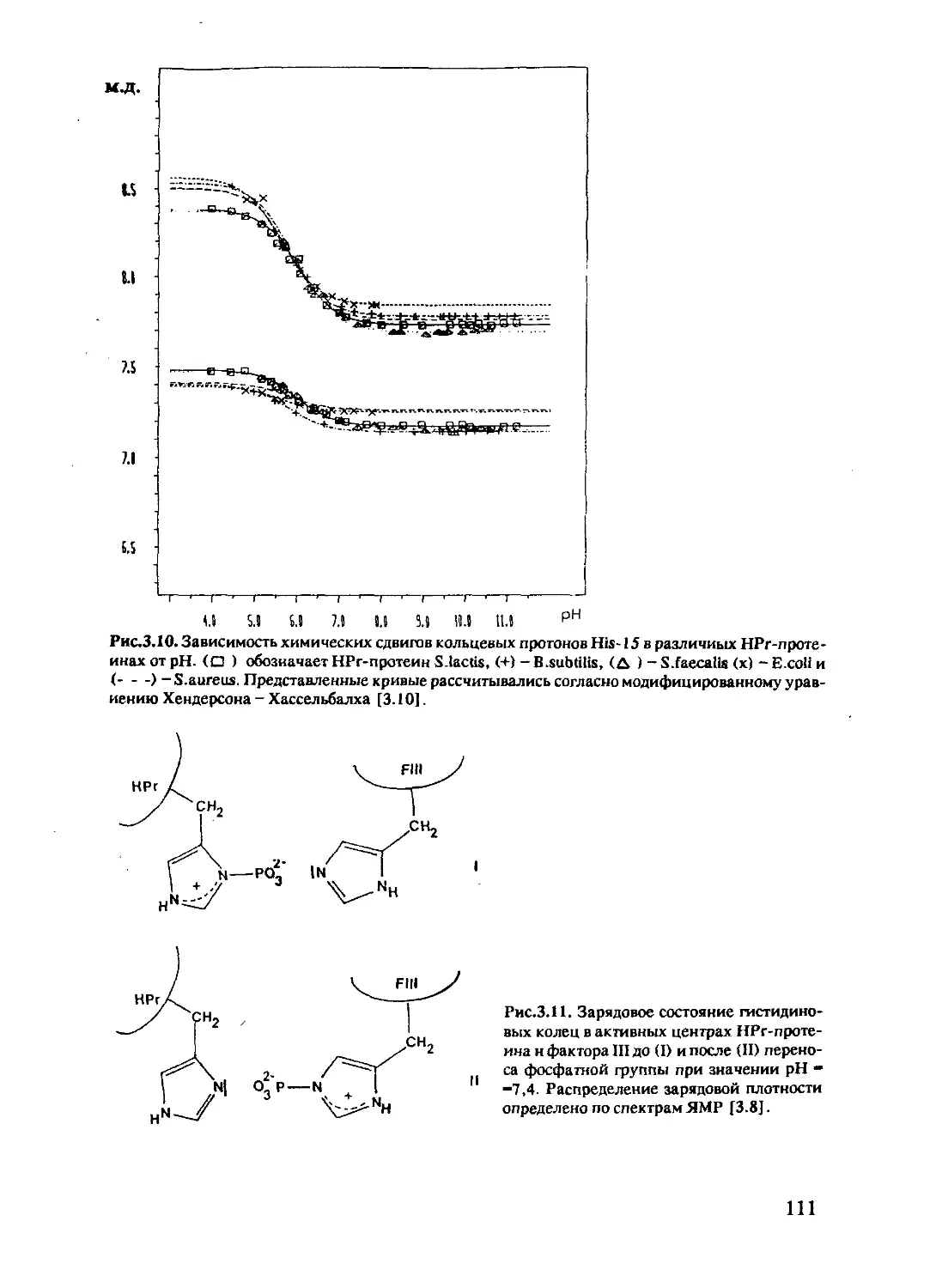

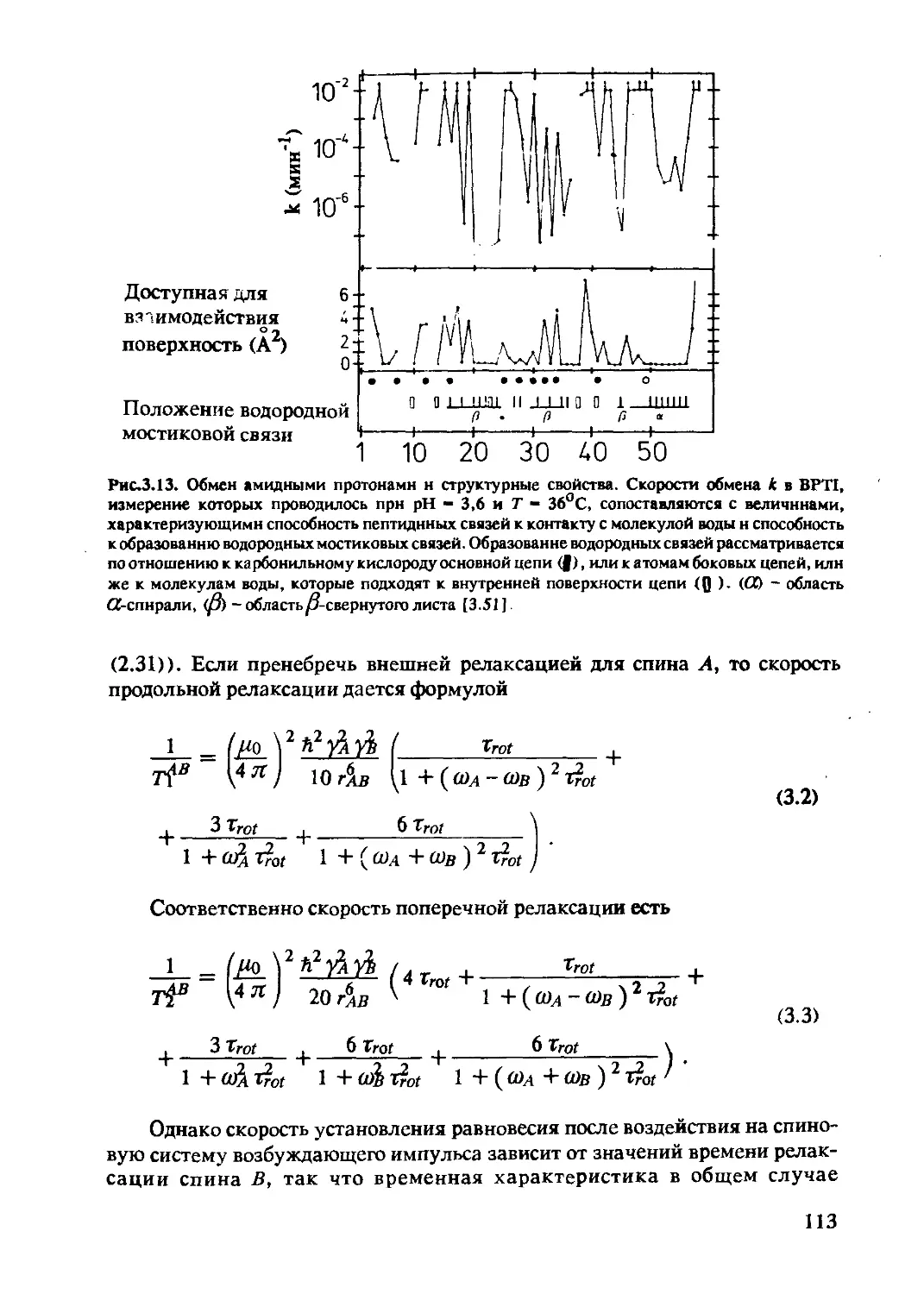



















































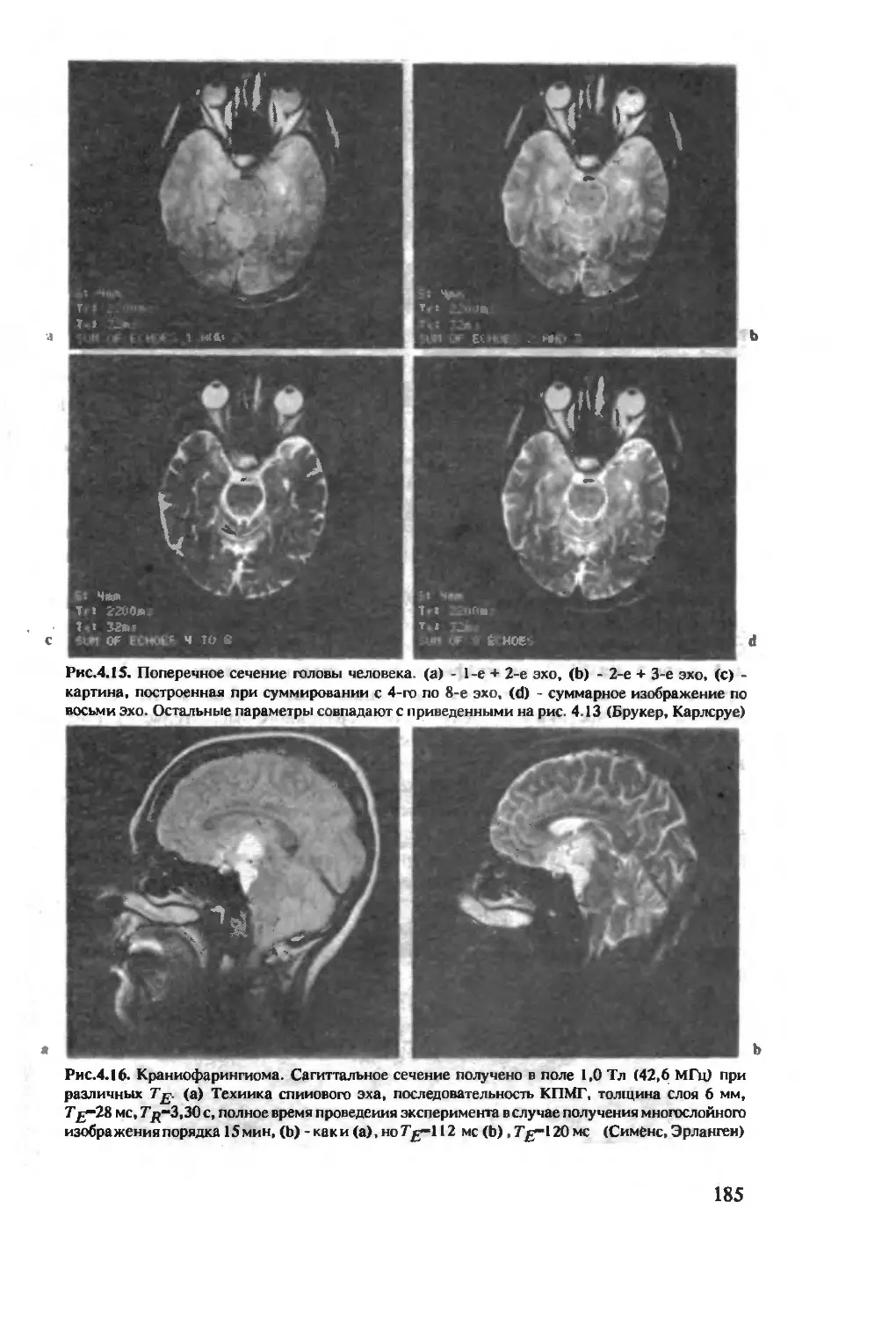
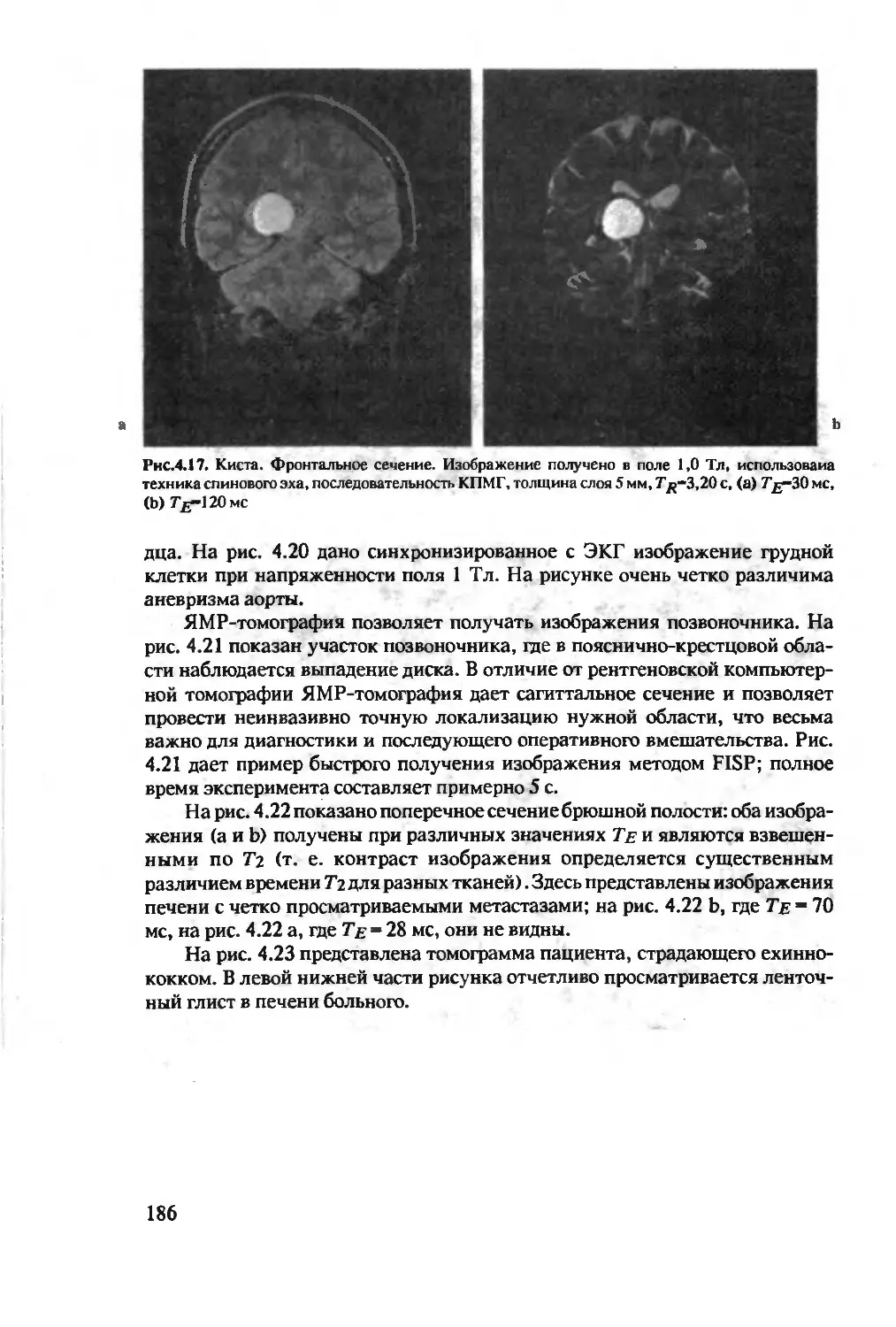
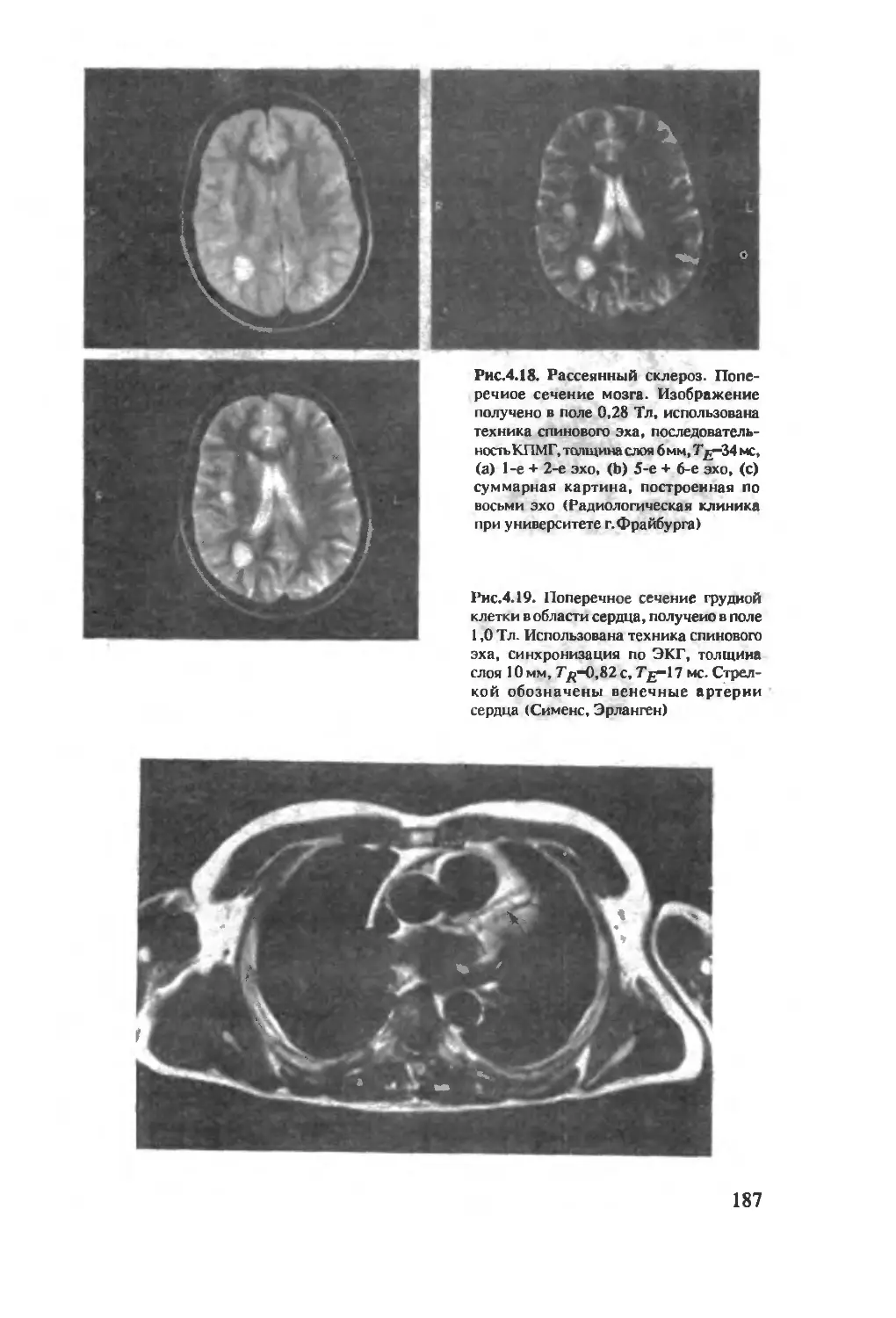
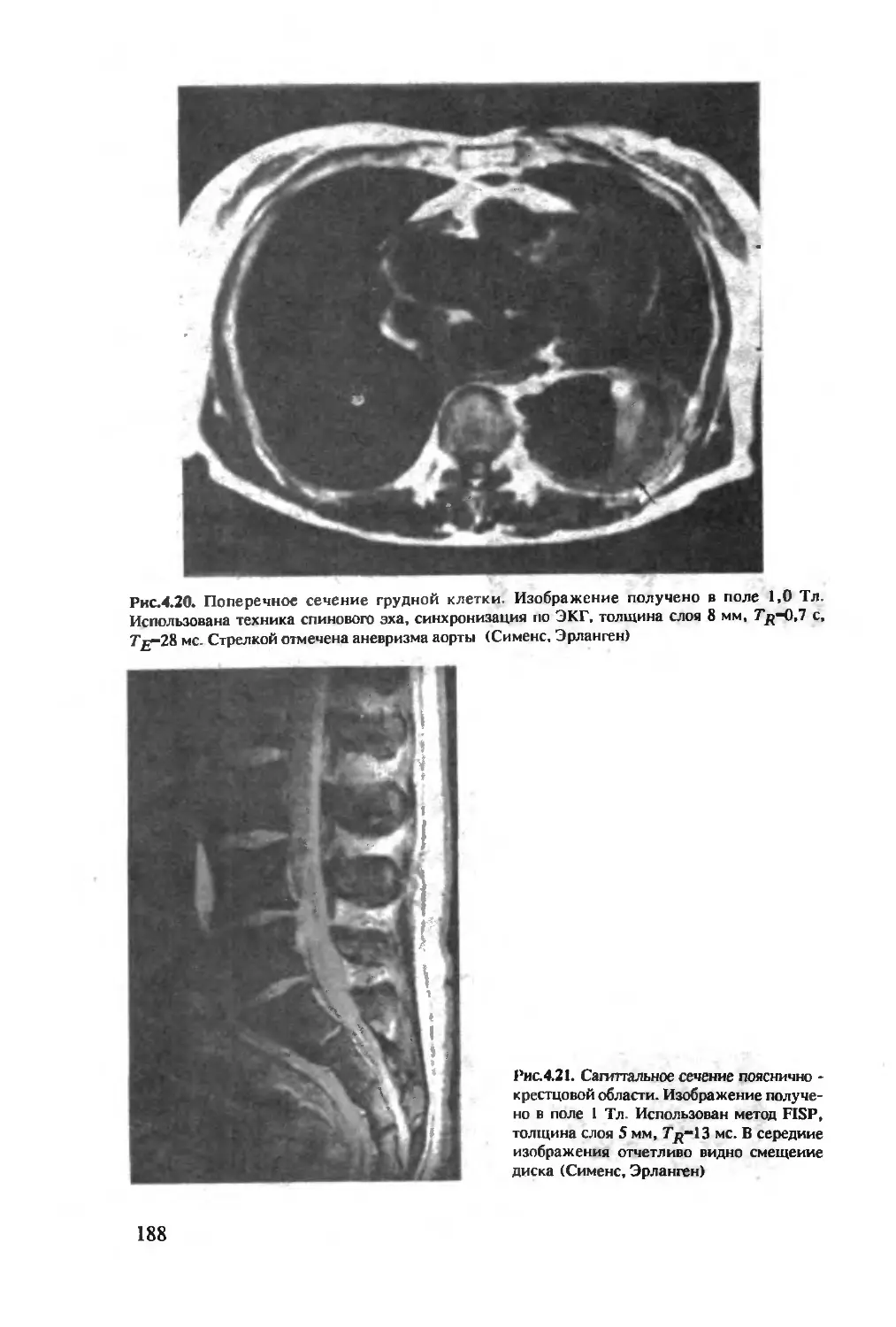
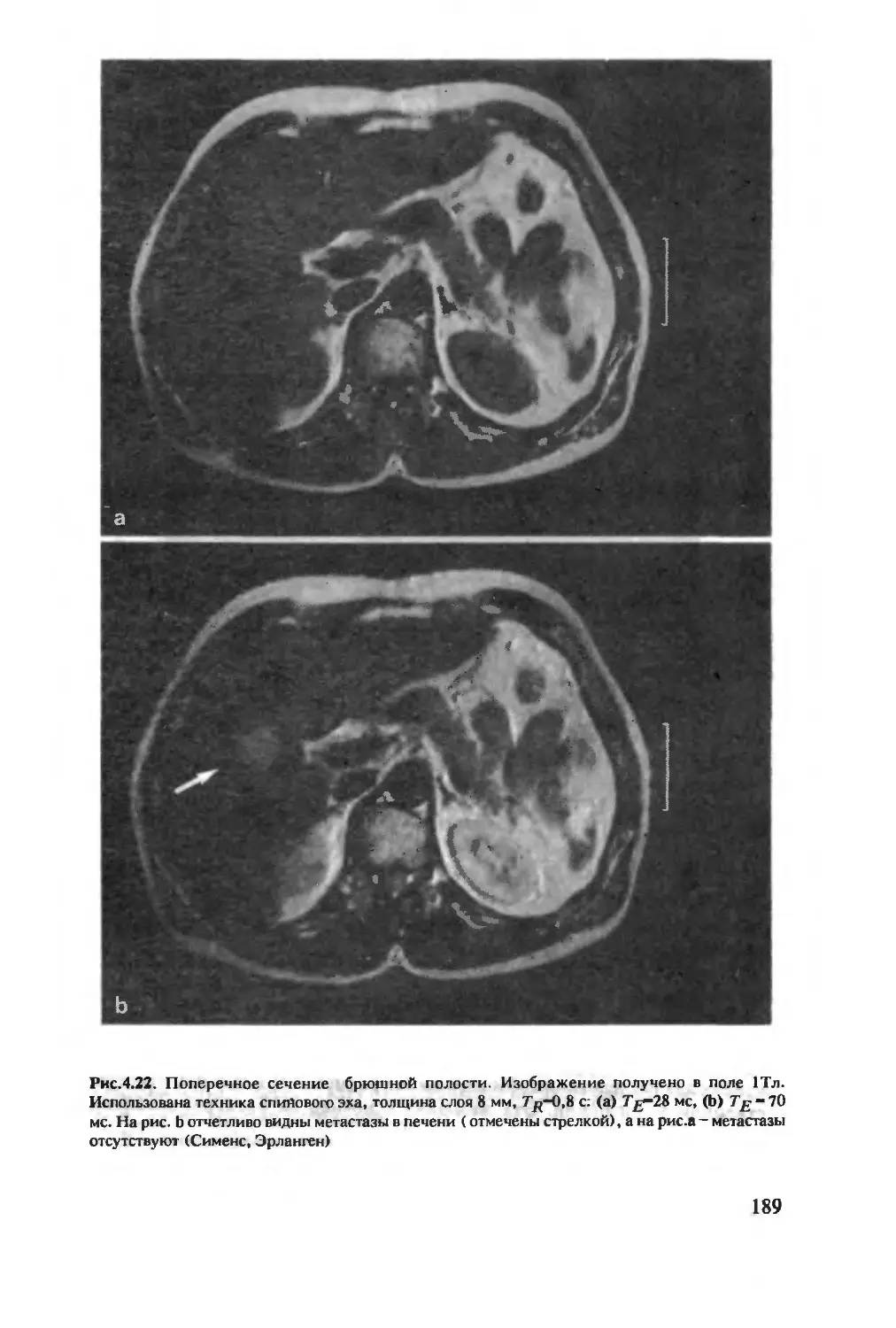
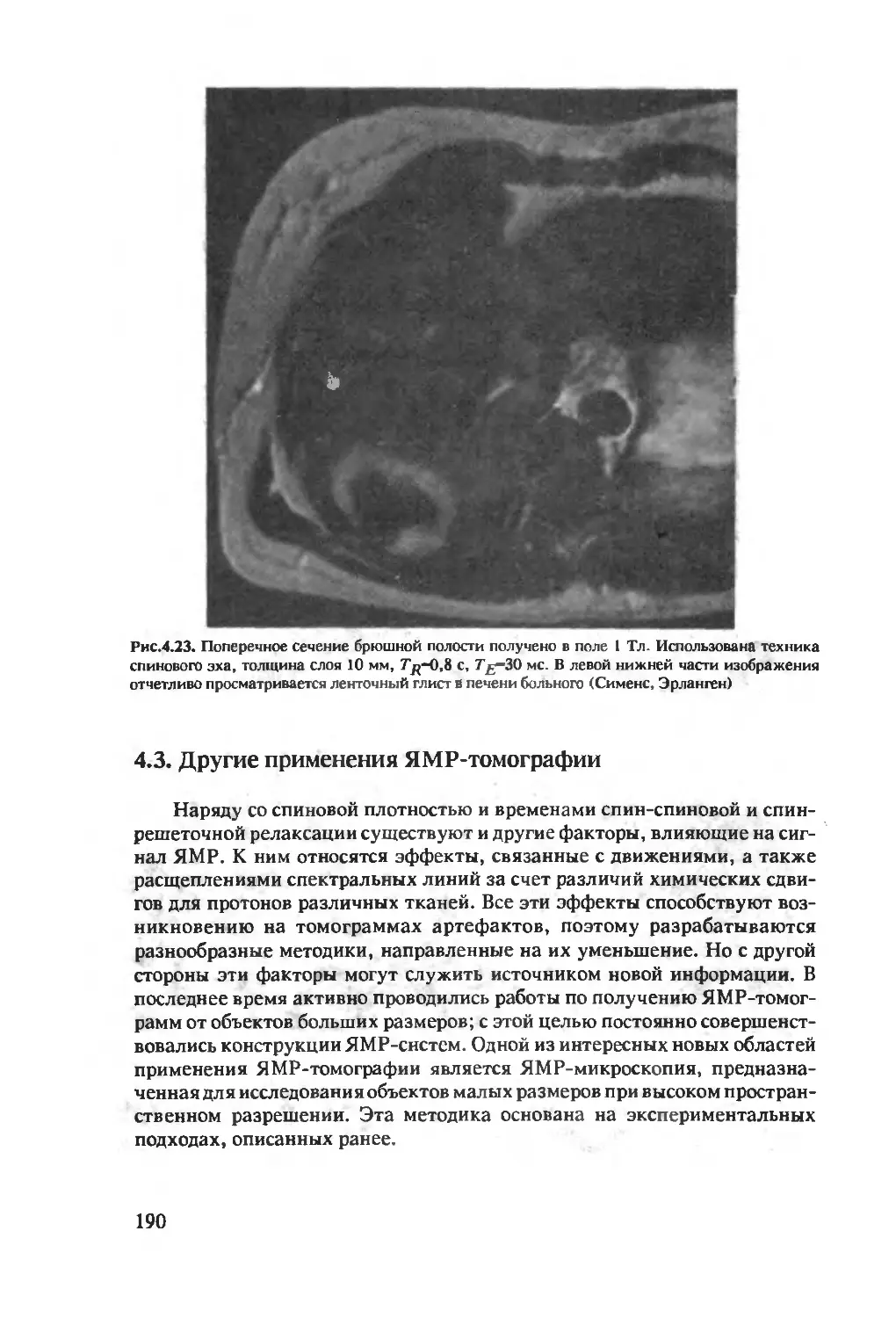






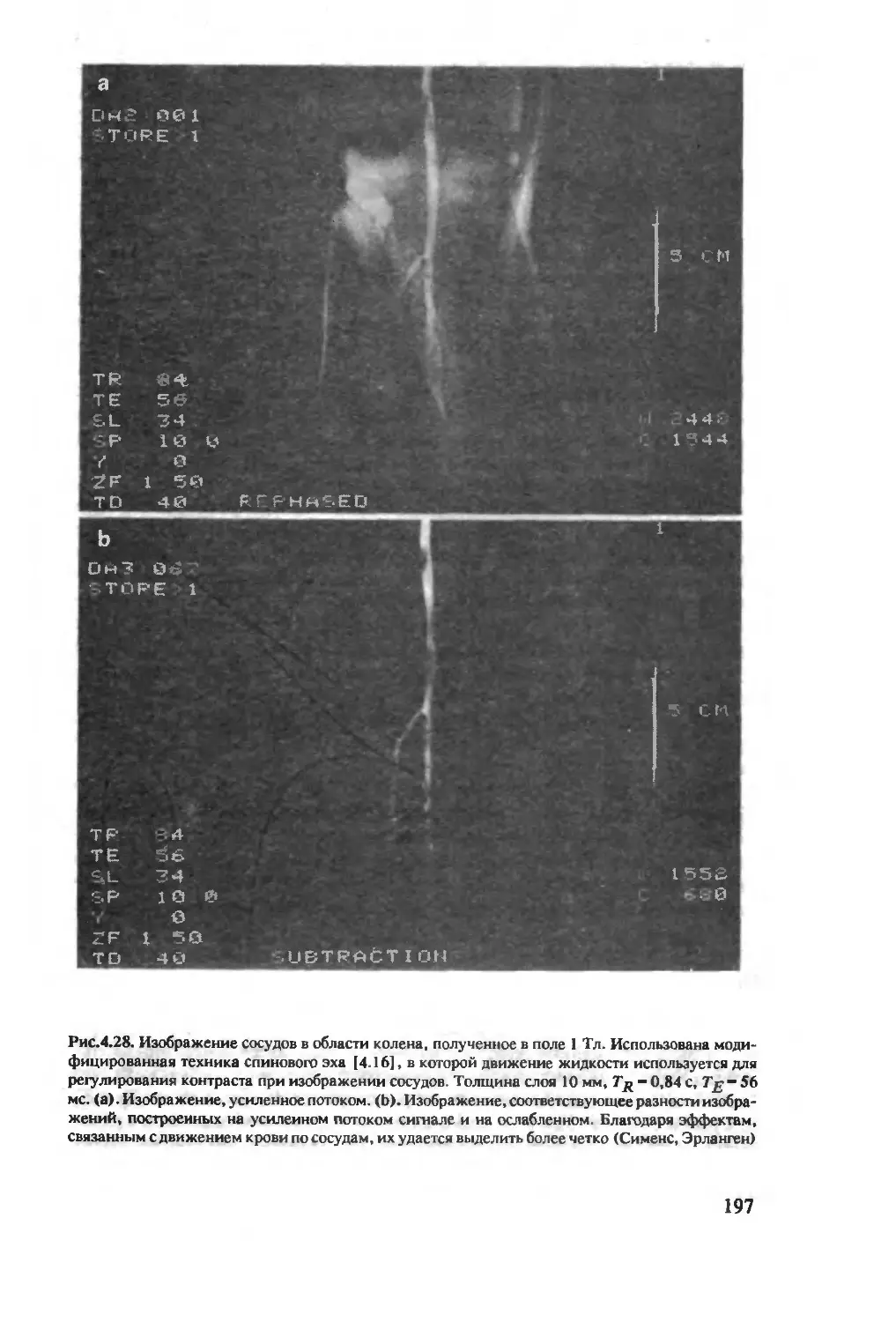




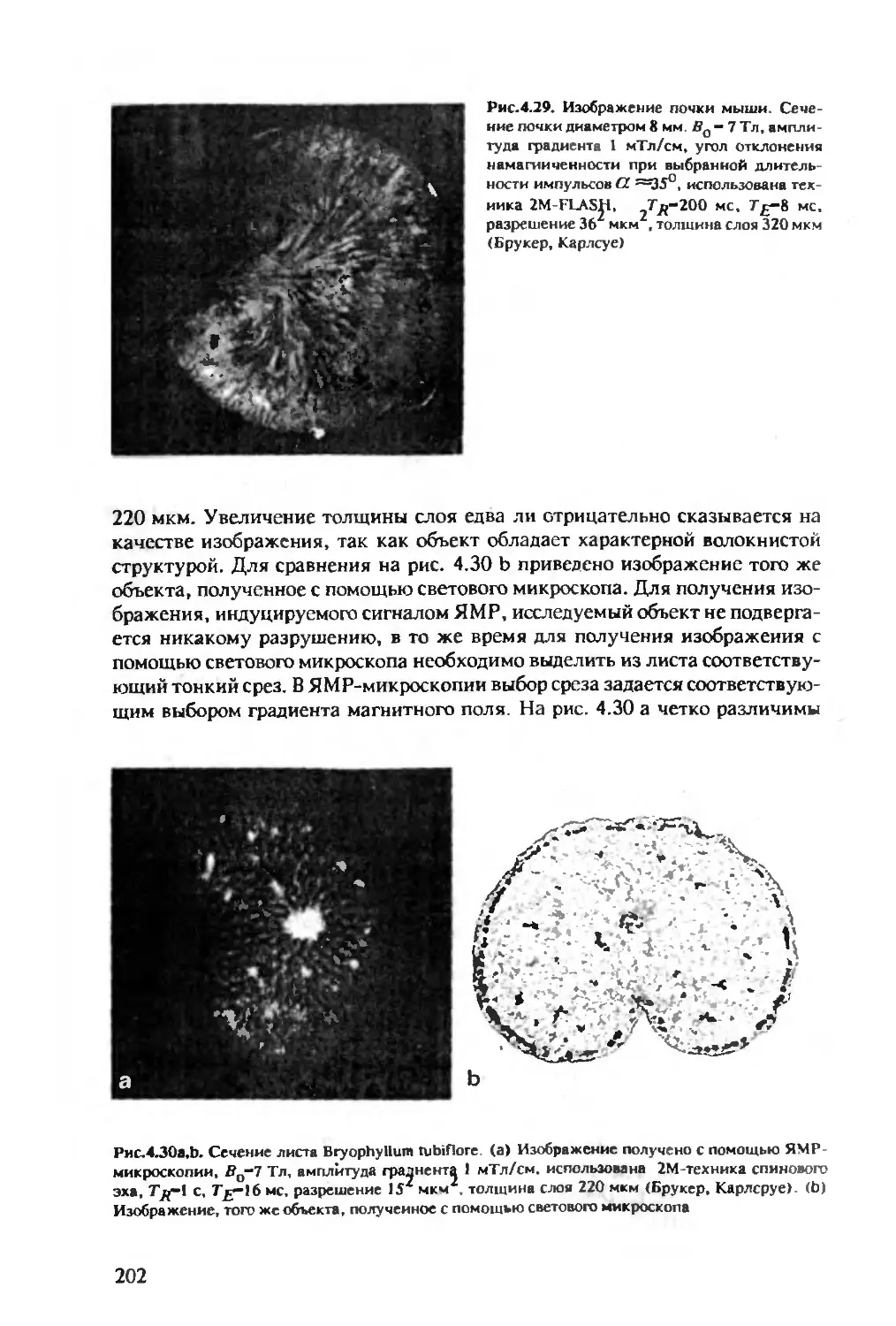
















































Author: Хуссер К.Х. Кальбитцер Х.Р.
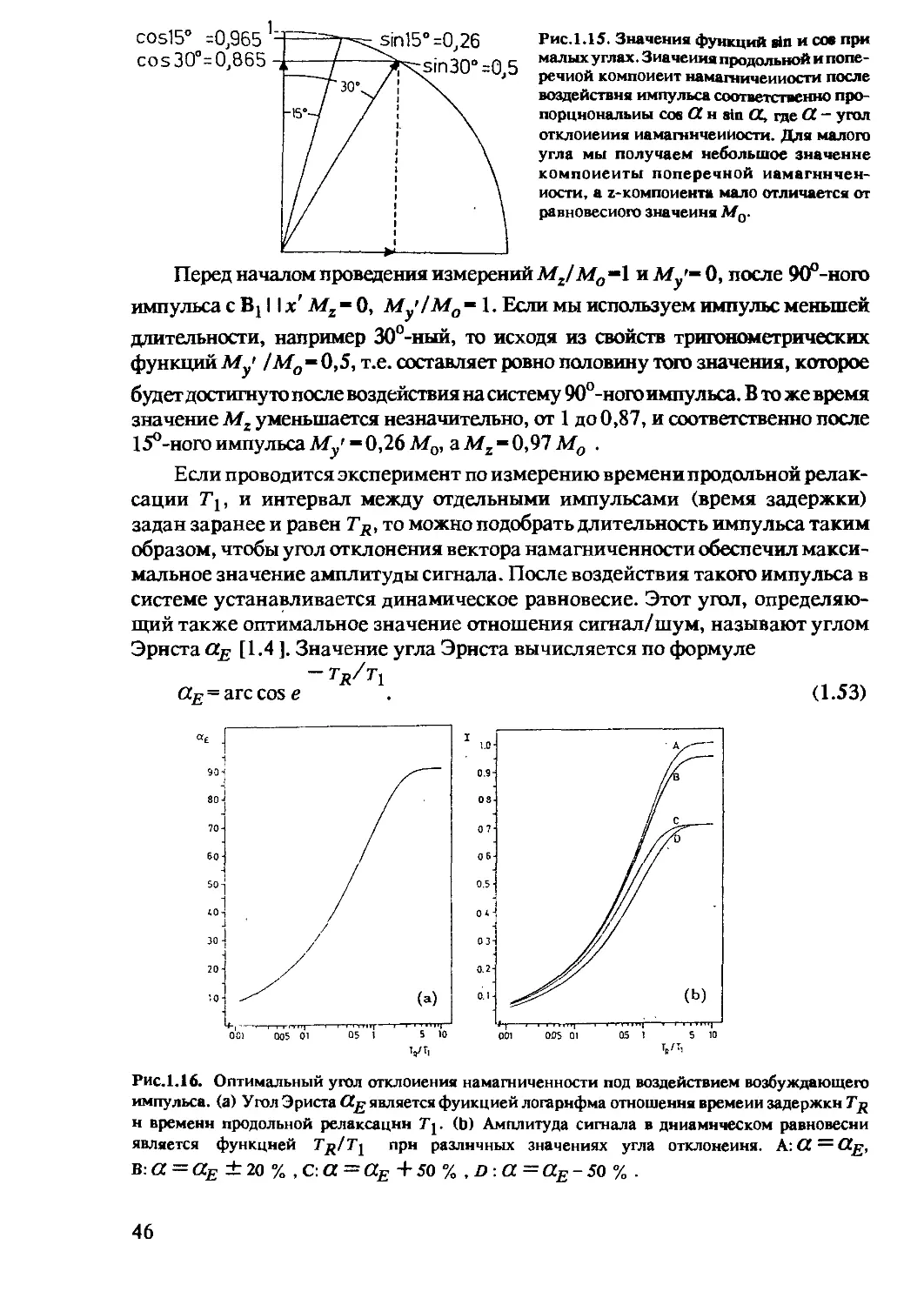
Tags: спектральные методы анализа оптические методы анализа медицина патофизиология лучевая диагностика спектроскопия томография ядерный магнитный резонанс
ISBN: 5-12-003769-0
Year: 1993




К.Х.Хауссер, Х.Р.Кальбитцер
ЯМР в медицине и биологии:
структура молекул, томография,
спектроскопия in-vivo
Перевод с немецкого
З.З.Рожковой
Под редакцией члена-корреспондента АН Украины,
доктора физ.-мат.наук С.М.Рябченко
Киев Наукова думка 1993
УДК 543.422.25
ЯМР в медицине и биологии: структура молекул, томография, спектроскопия
in-vivo/Xayccep К.Х., Кальбитцер Х.Р.; Пер. с нем. под ред. Рябченко С.М. с
предисл. Рябченко С.М., Рожковой 3.3. - Киев: Наук.думка, 1993. - 259 с.
Данная книга посвящена применению ядерного магнитного резонанса (ЯМР) к проблемам
медицины и биологии. Изложение физических основ метода ЯМР приведено в достаточно
простой форме, так что доступно для понимания широкого круга специалистов, а не только
физиков. Отличительной особенностью этой книги является то, что в ней с единой точки зрения
обсуждаются разнообразные медико-биологические приложения метода ЯМР и рассматрива-
ются связи, существующие между ЯМР высокого разрешения в жидкостях, спектроскопией
in-vivo и томографией.
Ил. 136. Табл. 13. Библиогр. в конце глав.
Редакция физики и кибернетики
Редактор А.С.Слыщенко
Издание осуществляется при финансовой поддержке фонда "Вщродження".
„ 4107000000 — 000 Х ~ А/221 ( 04 ) — 93 °°° " 93 1402010000 - 221 —94 У K.H.Hausser, H.R.Kalbitzer NMR fiir Mediziner und Biologen: Strukturbestimmung, Bildgebung, in-vivo-Spektroskopie. © Springer-Verlag Berlin, Heidelberg, 1989
ISBN 5-12-003769-0 © Перевод на русский язык, пре- дисловие к переводу, издатель- ство "Наукова думка”, 1993
Предисловие к переводу
Книга К.Х.Хауссера и Х.Р.Кальбитцера "ЯМР для медиков и биологов"
("NMR fur Medizinerund biologen") посвящена применениям метода ядерно-
го магнитного резонанса (ЯМР) к исследованиям биологических объектов:
определению структуры биомолекул, получению изображения биологиче-
ских тканей (ЯМР-томографии) и ЯМР-спектроскопии in-vivo.
Метод ЯМР, открытый и разработанный физиками, очень быстро нашел
применение в химии, и на протяжении десятилетий многие аспекты его
развития и совершенствования экспериментальной базы связывались, в ос-
новном, с проблемами исследования структуры и свойств различных хими-
ческих веществ. Разработка новых методик проведения экспериментов и
обработки данных постоянно расширяла круг решаемых с применением ЯМР
задач и позволяла исследовать все более сложные объекты. Это способство-
вало успешному применению ЯМР для исследования структуры биомолекул
и их функций в организме на уровне клеток и органов. Такая ситуация, когда
организм человека становится объектом исследования различных наук, в
настоящее время не является редкой. Это оказывается возможным благодаря
тому, что вполне сложившиеся научные дисциплины преодолевают свою
обособленность, и на их пересечении возникают новые интересные задачи.
Книга К.Х.Хауссера и Х.Р.Кальбитцера является результатом именно такого
процесса взаимодействия различных областей науки, а также примером
того, как метод, основанный на фундаментальном физическом явлении -
ЯМР, выходит за пределы чисто научных задач и выступает как метод
клинической диагностики, успешно конкурируя с рентгеновской компью-
терной томографией.
В последнее время появилось несколько прекрасных книг по современ-
ным методам ЯМР-спектроскопии, в частности, монография Р.Эрнста,
Дж.Боденхаузена и А.Вокауна "ЯМР в одном и двух измерениях", которая
была переведена на русский язык (М.: Мир, 1990). Эта книга представляет
собой энциклопедию современных методов ЯМР и исчерпывающее учебное
пособие по ЯМР-спектроскопии, хотя, возможно, несколько трудна для по-
нимания читателей, не имеющих физического образования и не владеющих
в достаточной мере современным математическим аппаратом. Браться за
перевод какой-либо монографии по ЯМР после выхода в свет такого фунда-
ментального труда, как указанная выше книга, довольно большой риск. Мы
решились на перевод и издание книги К.Х.Хауссера и Х.Р.Кальбитцера
исходя из того, что в ней, наряду с изложением основ метода ЯМР, рассчи-
танным на широкий круг читателей, рассматриваются проблемы примене-
3
ния ЯМР в медицине и биологии. Книга, несмотря на небольшой объем,
содержит обширную библиографию и знакомит читателя с самыми послед-
ними достижениями в области разнообразных приложений метода ЯМР. В
качестве дополнения к данной книге можно указать опубликованный в жур-
нале Physics Today (июнь 1992г.) обзорФ.Верлиопроблемахиперспективах
развития ЯМР-томографии.
В последнее время метод ЯМР находит все более широкое применение в
медико-биологических исследованиях, поэтому мы надеемся, что данное
издание окажется своевременным и книга найдет заинтересованного читате-
ля среди врачей, биологов, химиков, а также физиков, интересующихся
природой биологических объектов.
При переводе нами были исправлены замеченные неточности и опечатки
оригинала. Мы также решили остановиться на названии "ЯМР в медицине и
биологии", которое соответствует названию авторского перевода данной кни-
ги на английский язык: "NMR in Medicine and Biology" (Springer, 1991).
Мы благодарим доктора У.Айххоффа, который обратил наше внимание
на книгу К.Х.Хауссера и Х.Р.Кальбитцера, академиков А.А.Мойбенко,
Н.Г.Находкина и А.Г.Ситенко за поддержку идеи ее перевода, а также вы-
ражаем признательность фонду "Вщродження" за финансовую помощь.
Киев, ноябрь 1993 г. С.Рябченко,
3.Рожкова
4
Предисловие авторов
За последнее десятилетие с помощью ЯМР-спектроскопии были получе-
ны значительные результаты при исследовании макромолекул, представля-
ющих интерес для биологии, и сегодня этот метод, пожалуй, наиболее часто
используется в оригинальных биохимических работах. Возможности и успе-
хи ЯМР-томографии в настоящее время столь убедительны, что она все более
привлекает внимание со стороны медиков. Еще одна область, в которой
потенциально возможно использование пространственно селективной спек-
троскопии ЯМР, - in-vivo-спектроскопия - пока недостаточно развита и
нуждается в дальнейших технических разработках. Однако и сейчас уже
ясно, что возможности этой области ЯМР-спектроскопии чрезвычайно вели-
ки: если удастся проводить исследование биомолекул в четко локализован-
ном участке человеческого тела, т.е. осуществлять лабораторные
медицинские исследования внутренних тканей, причем неинвазивно, то это,
безусловно откроет новые перспективы в клинической диагностике.
Для того чтобы представить себе настоящие и будущие возможности
использования ЯМР в биологии и медицине, любой интересующийся этими
проблемами врач или биолог должен быть знаком хотя бы с основами явления
ядерного магнитного резонанса. Однако большинство соответствующих по-
собий написано в расчете на физиков и физико-химиков, и вследствие обилия
математических формул и специального языка малопригодно для врачей и
биологов. В настоящее время появился уже целый ряд обзорных работ по
применению ЯМР-томографии для исследования определенных участков
тела и определенных заболеваний, но они имеют, в основном, медицинскую
направленность и лишь вкратце касаются основных положений ЯМР, а воп-
росы спектроскопии в них не рассматриваются вообще. С другой стороны,
хотя для читателей с естественно-физическим образованием относительно
легко понять физические основы метода ЯМР, им часто не хватает знания
биологических и медицинских аспектов проблемы.
Мы полагаем, что основы метода, а также различные области примене-
ния ЯМР в биологии и медицине тесно связаны между собой и должны
рассматриваться в единстве. Поэтому мы старались написать доступную для
биологов и врачей книгу, в которой без громоздких математических формул
излагаются физические принципы и типичные биологические и медицин-
ские проблемы, а также пути их решения.
Книга состоит из пяти вполне самостоятельных, но глубого взаимосвя-
занных глав. Основой для понимания всей книги является глава 1. Для
читателей, интересующихся использованием ЯМР в биохимии, предназна-
5
чены главы 2 и 3. Вопросы ЯМР-томографии рассмотрены в главе 4. Те, кто
интересуется возможностями спектроскопии in-vivo, найдут соответствую-
щий материал в главе 5, убедившись при этом, насколько тесно переплета-
ются эти вопросы с материалом, приведенным в главах 2 и 4.
Мы приносим нашу искреннюю благодарность докторам М.Даймлингу,
Х.Фридбургу, профессорам У.Хеберлену и Б.Штрёбелю за просмотр рукопи-
си и сделанные устные замечания. При подготовке рукописи и рисунков
неоценимую помощь оказали нам госпожа Ц.Турм, П.Беле, Г.Ойлефельд и
Х.Кессель. Наши коллеги доктора В.Хартль, В.Кун, Е.Мюллер, П.Найдиг,
Х.Пост, Э.Р.Райнхардт и Н.Шуфф принимали участие в обсуждениях и
предоставили в наше распоряжение многие ЯМР-томограммы и другие дан-
ные. Всем им мы выражаем нашу искреннюю признательность.
Хайдельберг, март 1989 г. К.Х.Хауссер,
Х.Р.Кальбитцер
6
Содержание
Глава 1. Основы метода ядерного магнитного резонанса.....12
1.1 Основные принципы магнитного резонанса ...........13
1.1.1 Магнитный момент и ядерный спин .............13
1.1.2 Условие резонанса............................16
1.1.3 Уравнение Блоха .............................17
1.1.4 Спин-решеточна я релаксация .................22
1.1.5 Распад поперечной намагниченности и спин-спиновая
релаксация ....................................23
1.2 Спектр ЯМР .......................................27
1.2.1 Химический сдвиг ............................27
1.2.2 Диполь-дипольное взаимодействие .............28
1.2.3 Косвенное спин-спиновое взаимодействие ......29
1.2.4 Электрон-ядерное взаимодействие .............32
1.2.5 Ядерное квадрупольное взаимодействие ........33
1.3 Механизмы релаксации .............................35
1.3.1 Поперечная релаксация и ширина линий ........36
1.3.2 Временная модуляция взаимодействий ..........37
1.3.3 Дипольная релаксация ........................37
1.3.4 Релаксация за счет анизотропии
химического сдвига.................................38
1.3.5 Релаксация по механизму косвенного
спин-спинового взаимодействия .................39
1.3.6 Квадрупольная релаксация ....................40
1.3.7 Парамагнитная релаксация ....................40
1.4 Экспериментальные методы .........................41
1.4.1 Непрерывный ЯМР .............................41
1.4.2 Импульсный ЯМР ..............................43
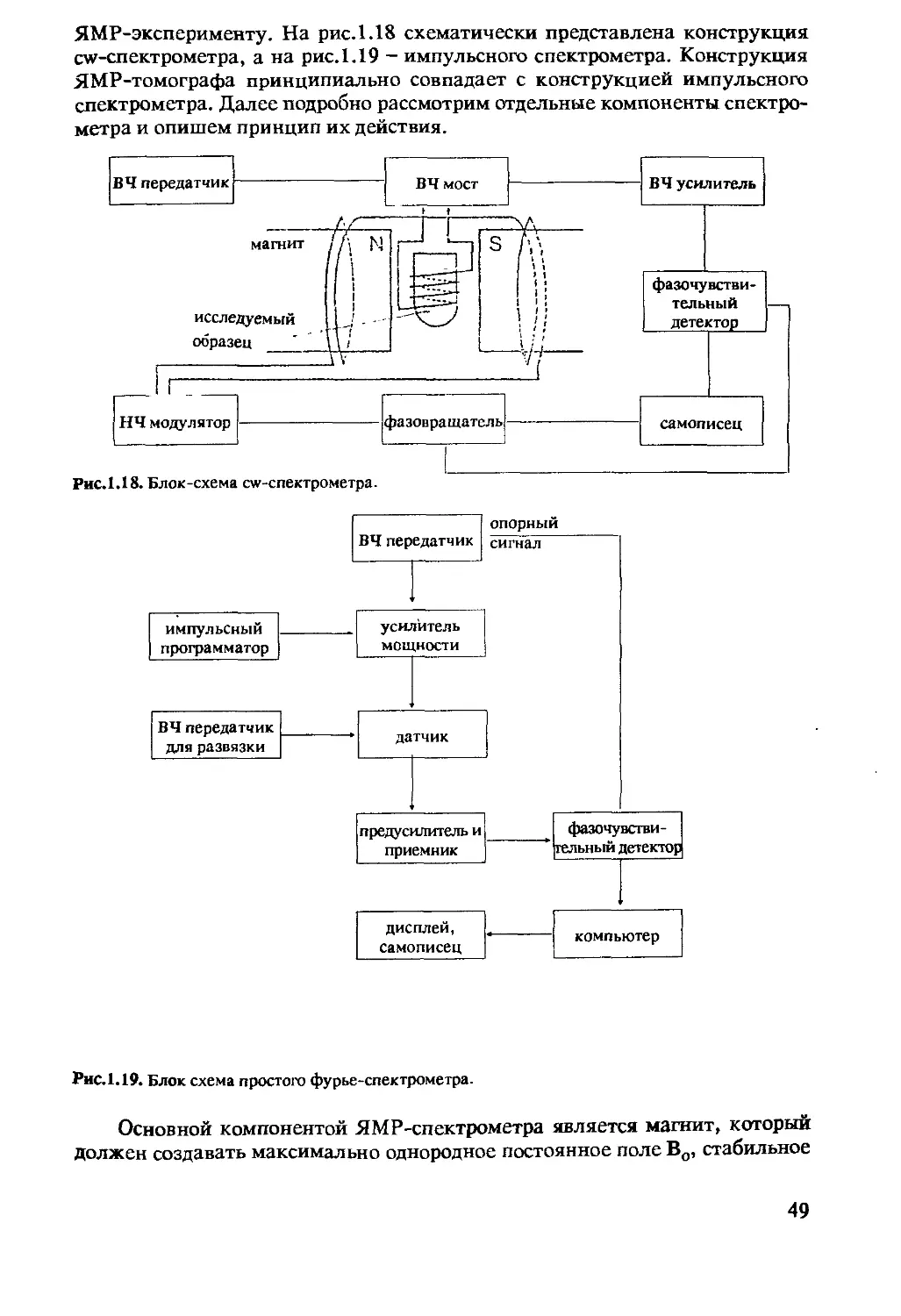
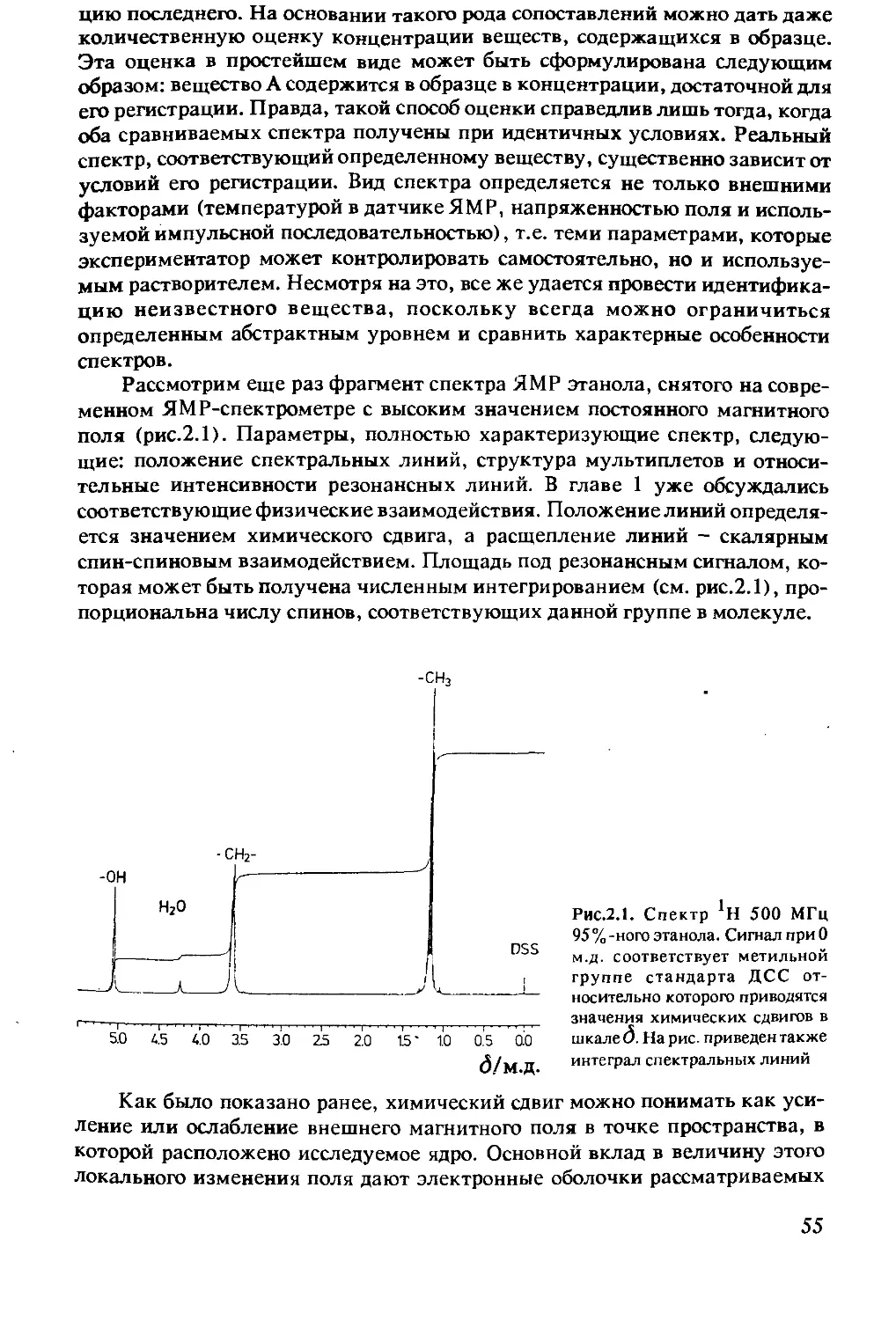
1.4.3 Спектрометр ЯМР .............................48
Литература .......................................... 52
Глава 2. ЯМР в биохимии .................................53
2.1 ЯМР как аналитический метод ......................54
2.1.1 Идентификация известных и неизвестных веществ .. 54
2.1.2 Внутренний и внешний стандарт ...............57
2.1.3 Мультиплетная структура .....................59
2.1.4 Константы скалярного взаимодействия и структура
молекул .......................................62
7
2.1.5 Определение партнера по взаимодействию .......63
2.1.6 Применение метода ЯМР для определения
концентраций ..................................65
2.1.7 Подавление интенсивного сигнала растворителя .67
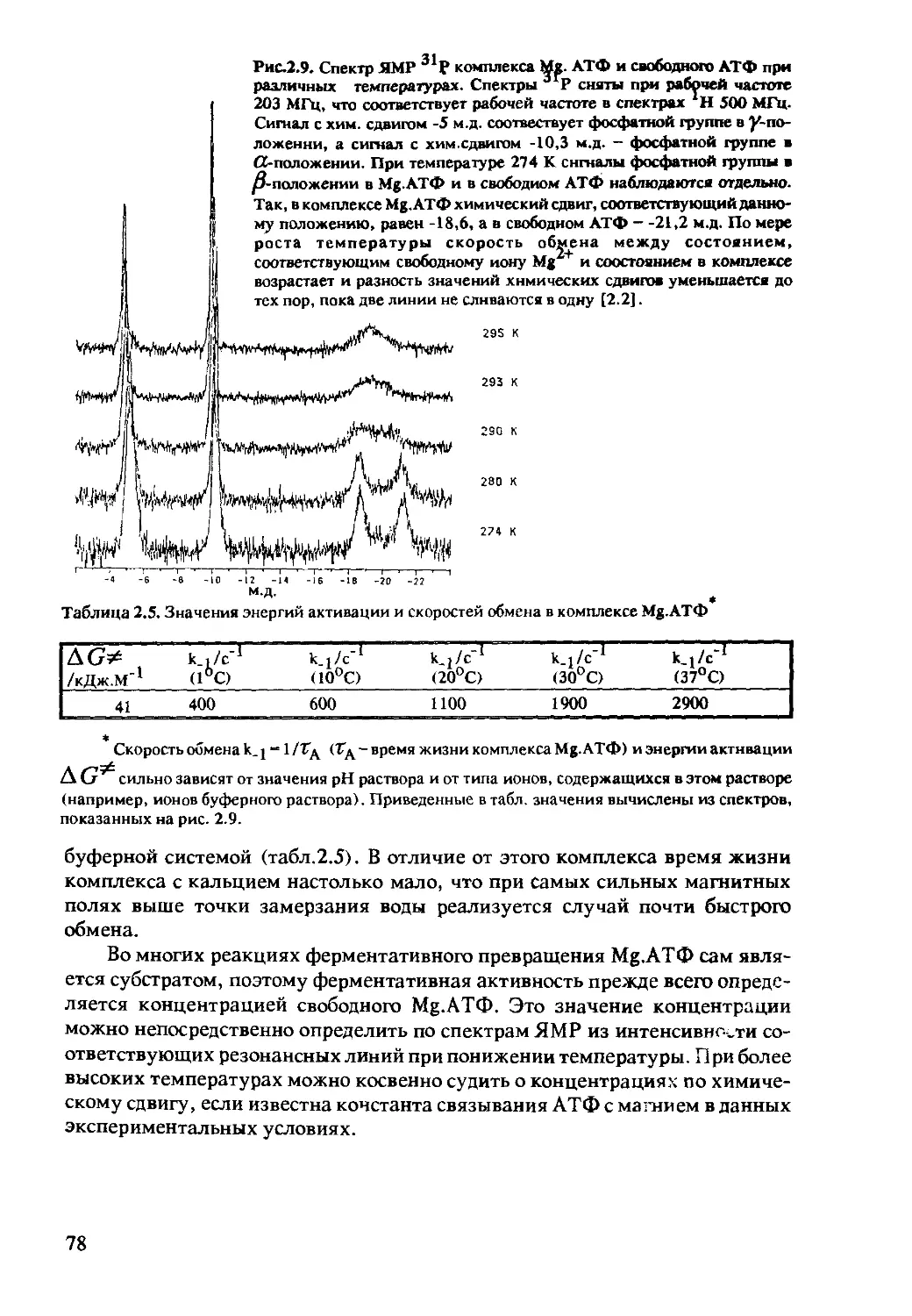
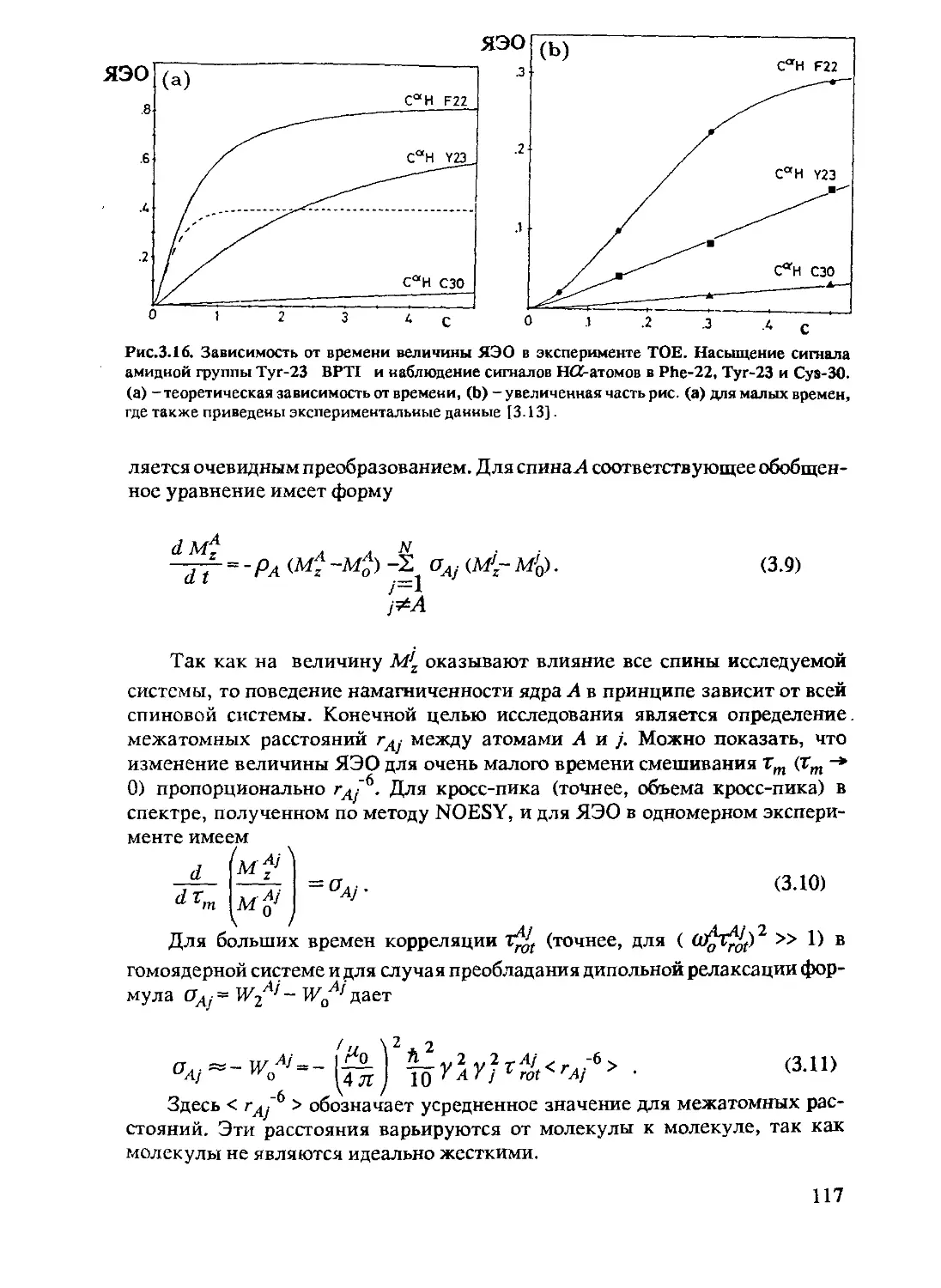
2.2 Зависящие от времени процессы в ЯМР................69
2.2.1 Временная корреляция и спектральная плотность .... 70
2.2.2 Химический обмен .............................72
2.2.3 Зависимость величины химических сдвигов от pH .... 75
2.2.4 Образование комплексов с диамагнитными
лигандами .....................................77
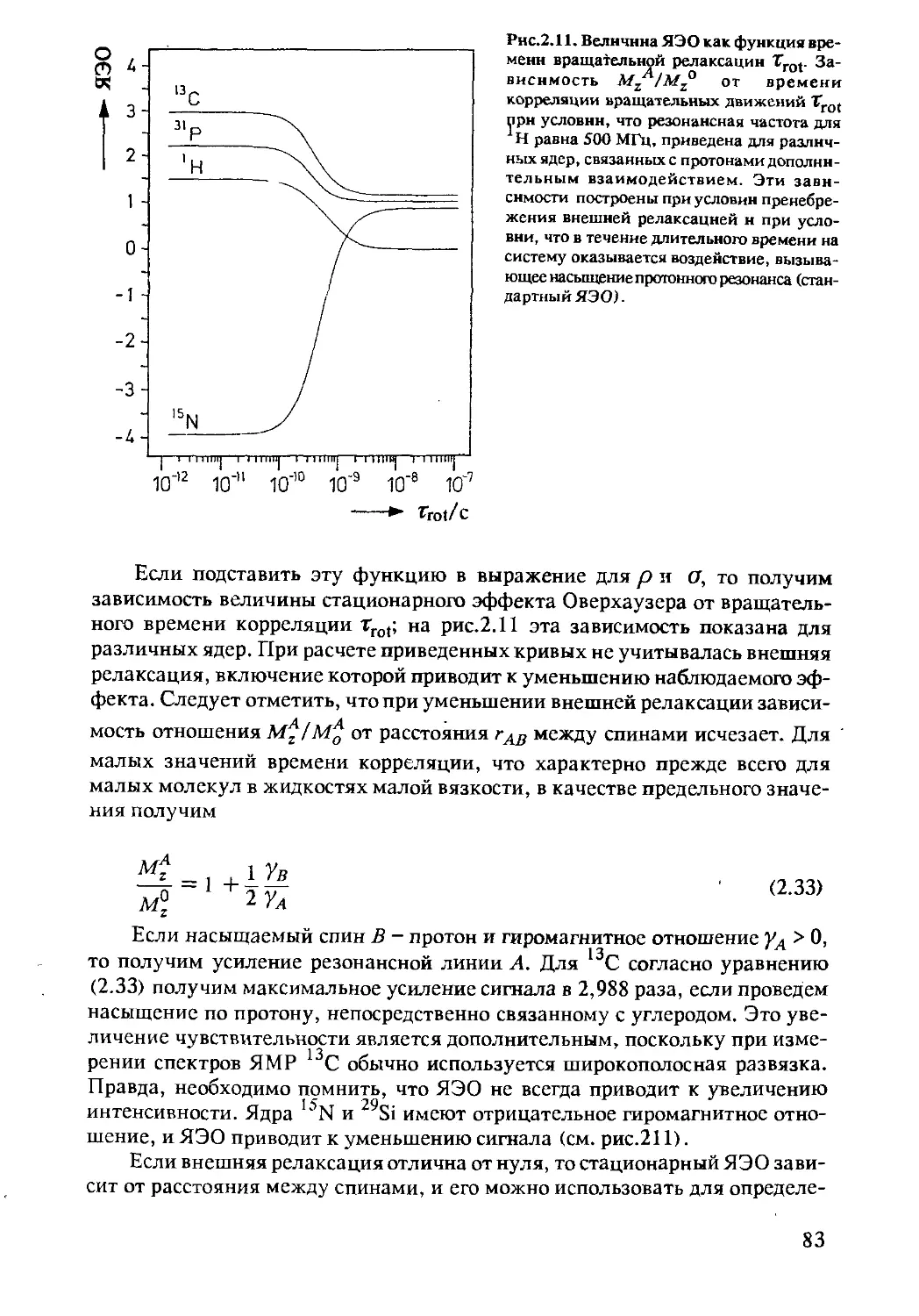
2.2.5 Перенос поляризации с насыщением .............79
2.2.6 Ядерный эффект Оверхаузера....................80
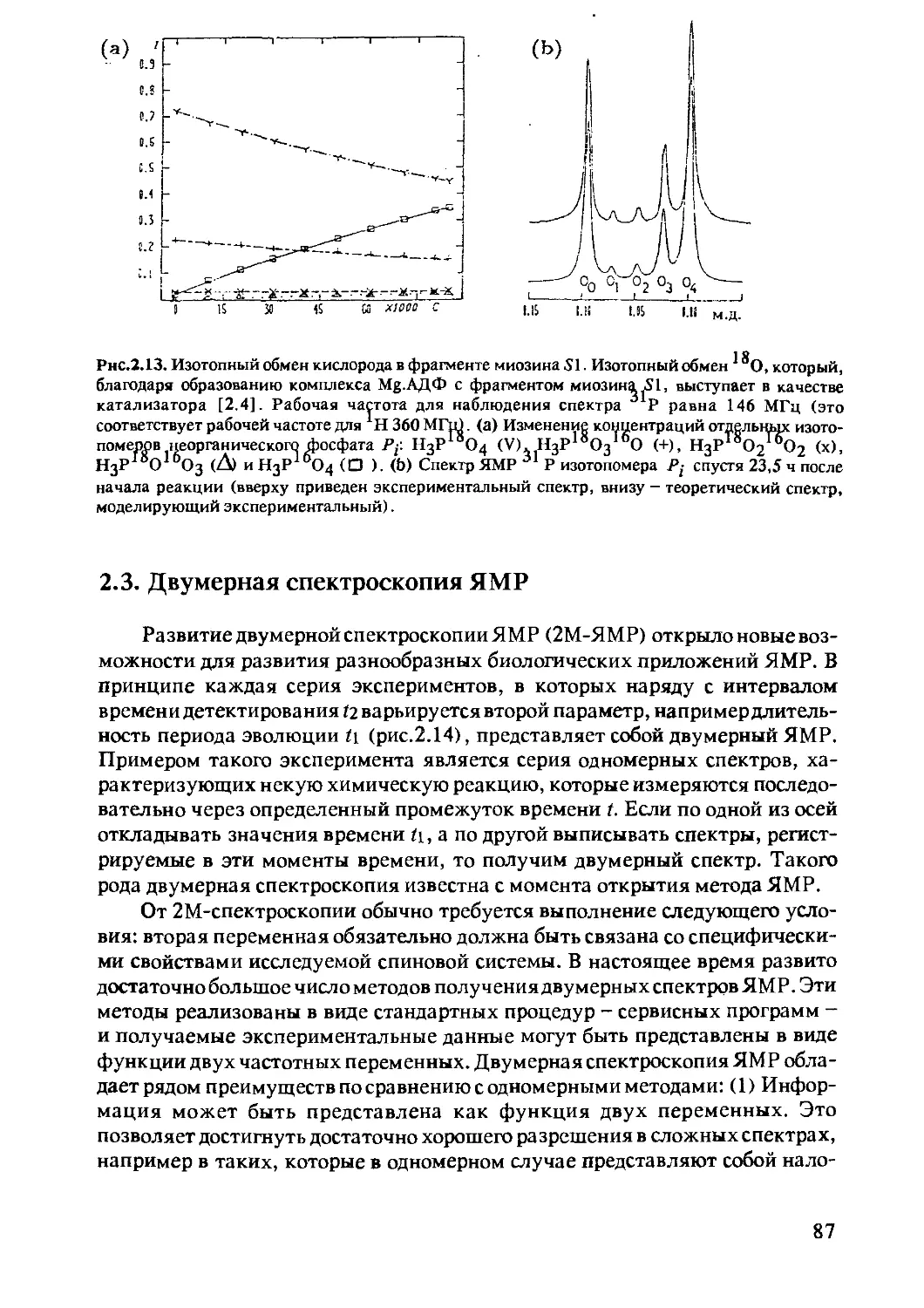
2.2.7 Использование стабильных изотопов
в качестве меток ..............................85
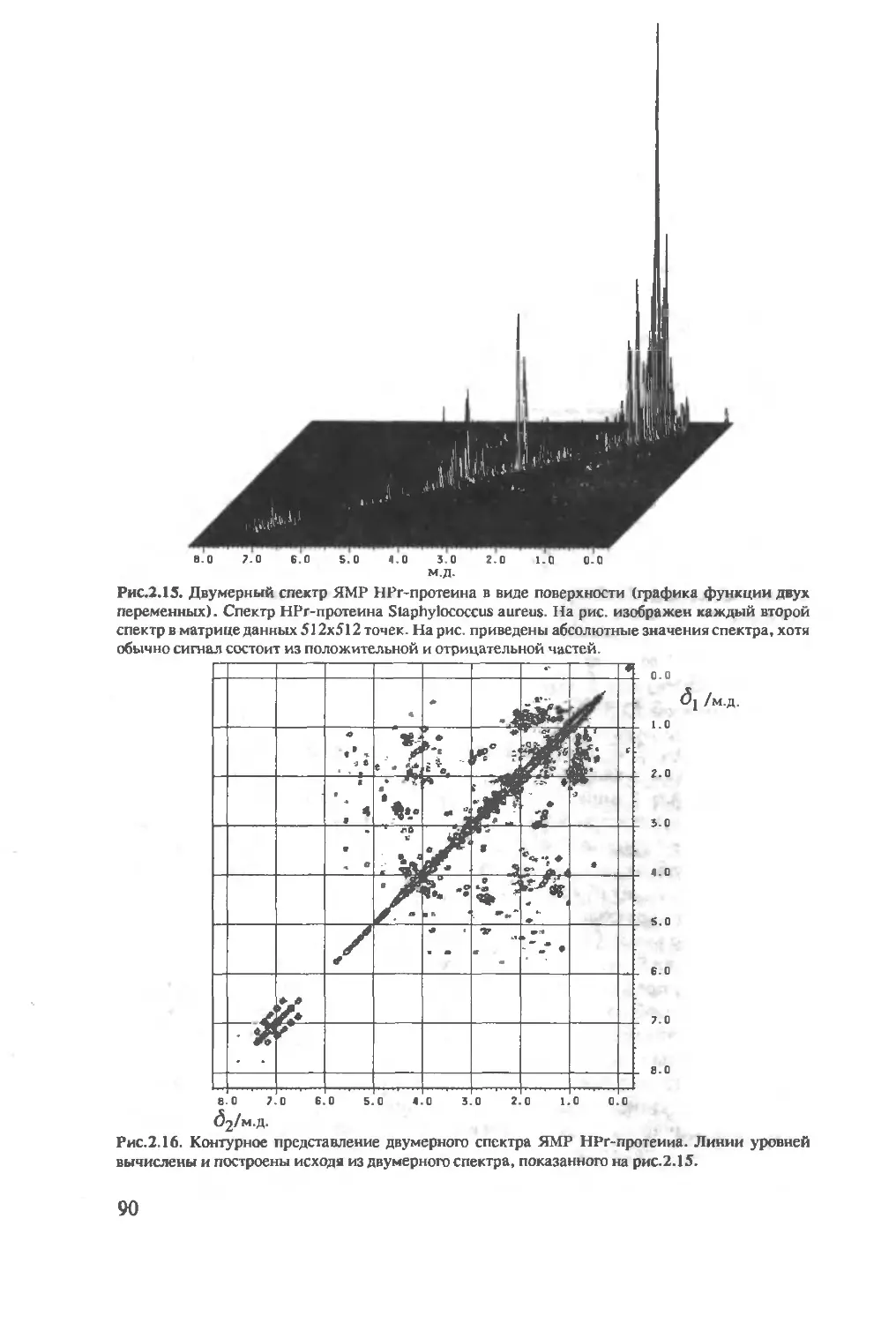
2.3 Двумерная спектроскопия ЯМР ......................87
2.3.1 Двумерный ЯМР-эксперимент ...................88
2.3.2 Обнаружение спин-спинового взаимодействия
в гомоядерном случае ..........................92
2.3.3 Измерение ЯЭО и химический обмен ............93
2.3.4 Корреляция в гетероядерном случае............94
Литература............................................95
Глава 3. ЯМР-спектроскопия биологических макромолекул .....97
3.1 ЯМР-спектроскопия протеинов ......................97
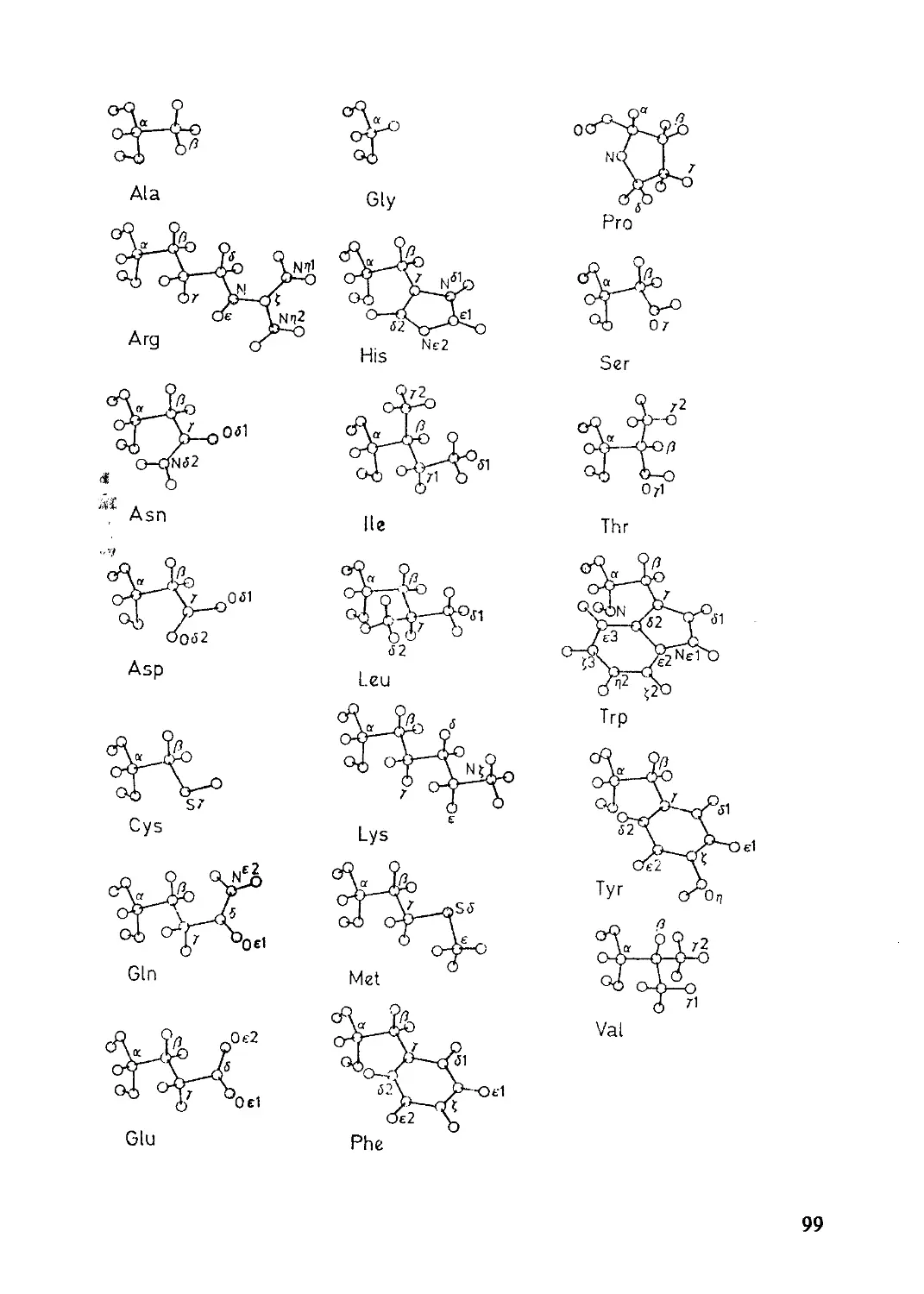
3.1.1 Строение и структура протеинов ..............97
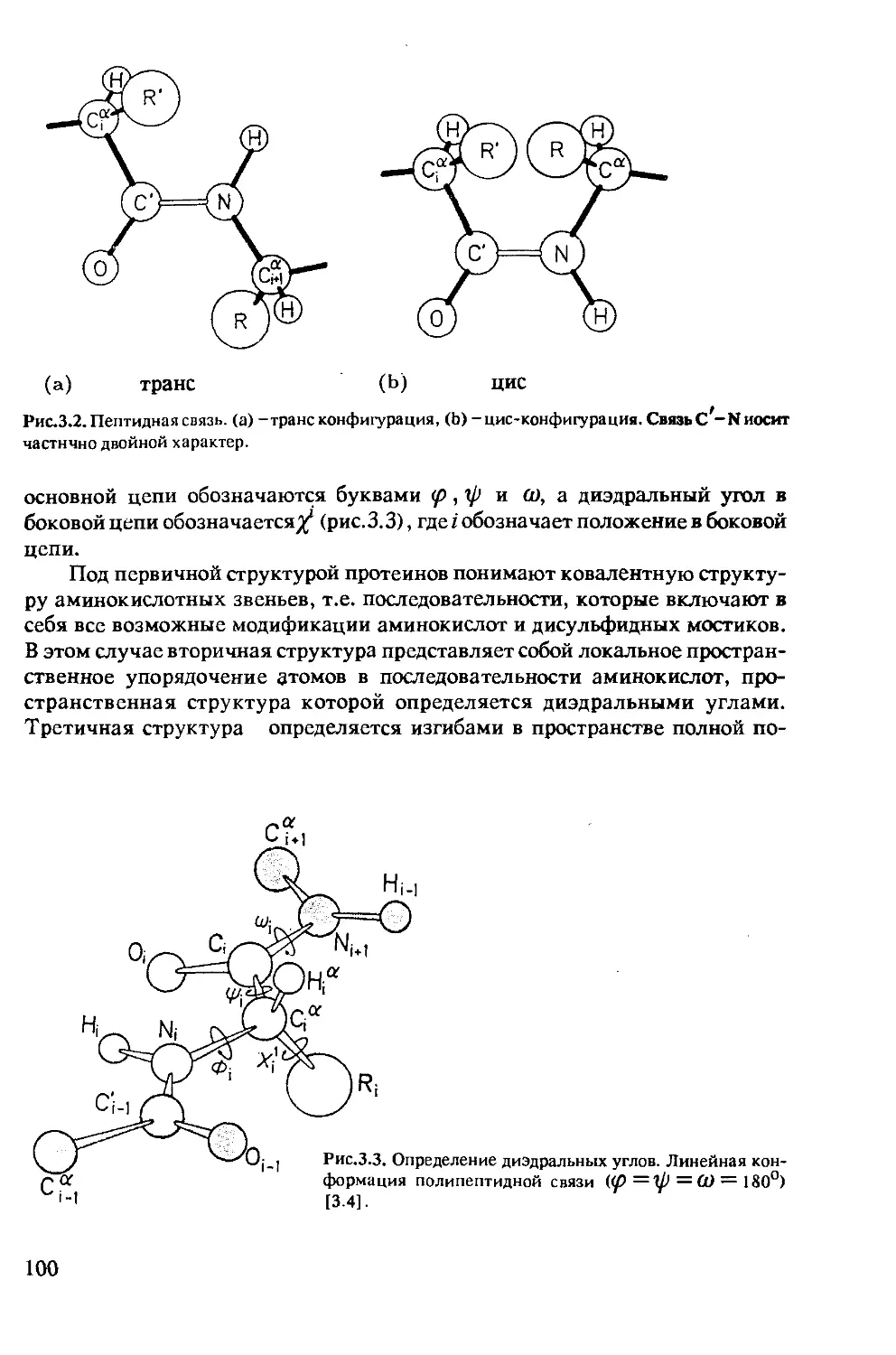
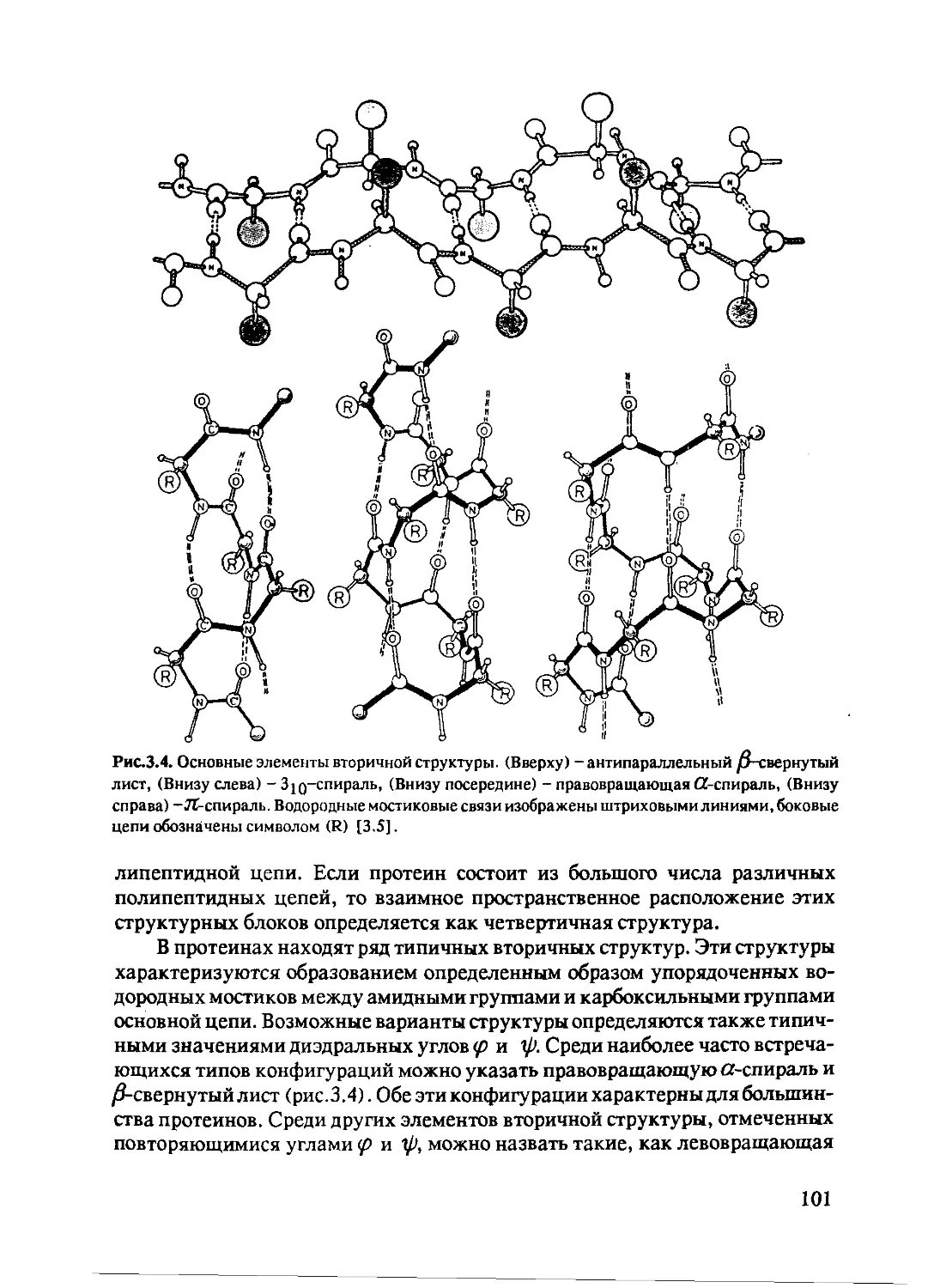
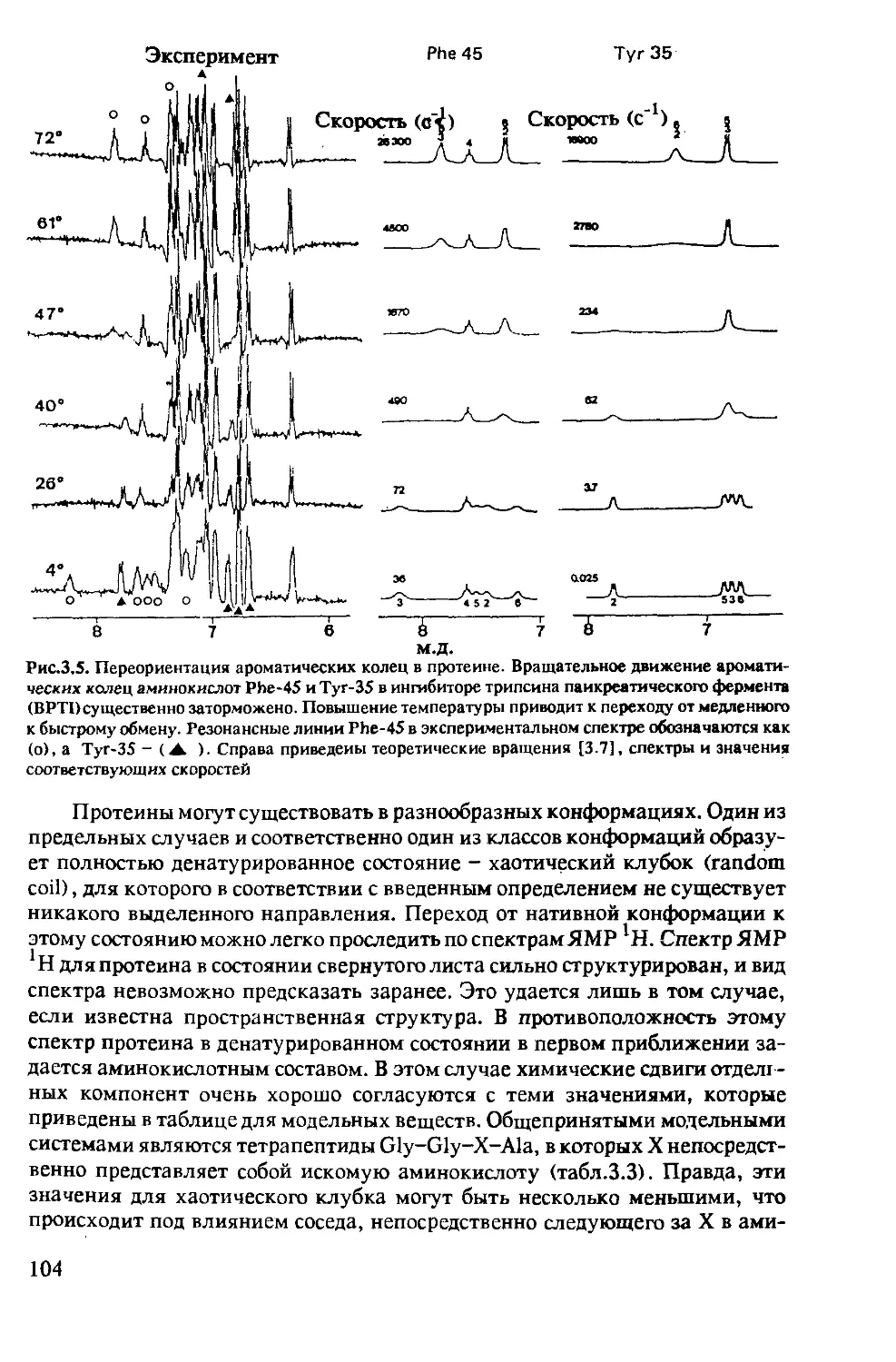
3.1.2 Динамические процессы в протеинах ...........102
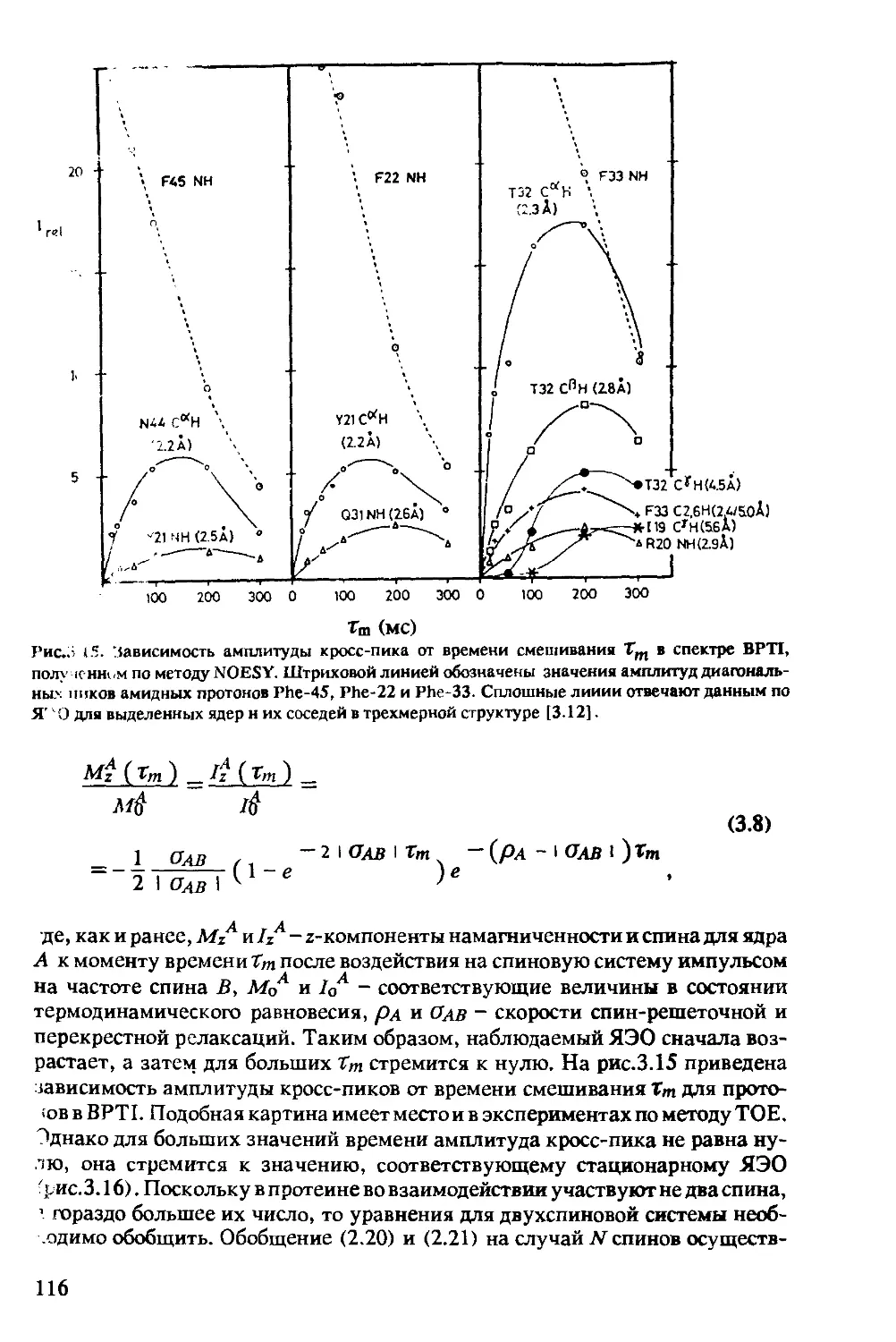
3.1.3 Определение межатомных расстояний............115
3.1.4 Отнесение резонансных линий .................128
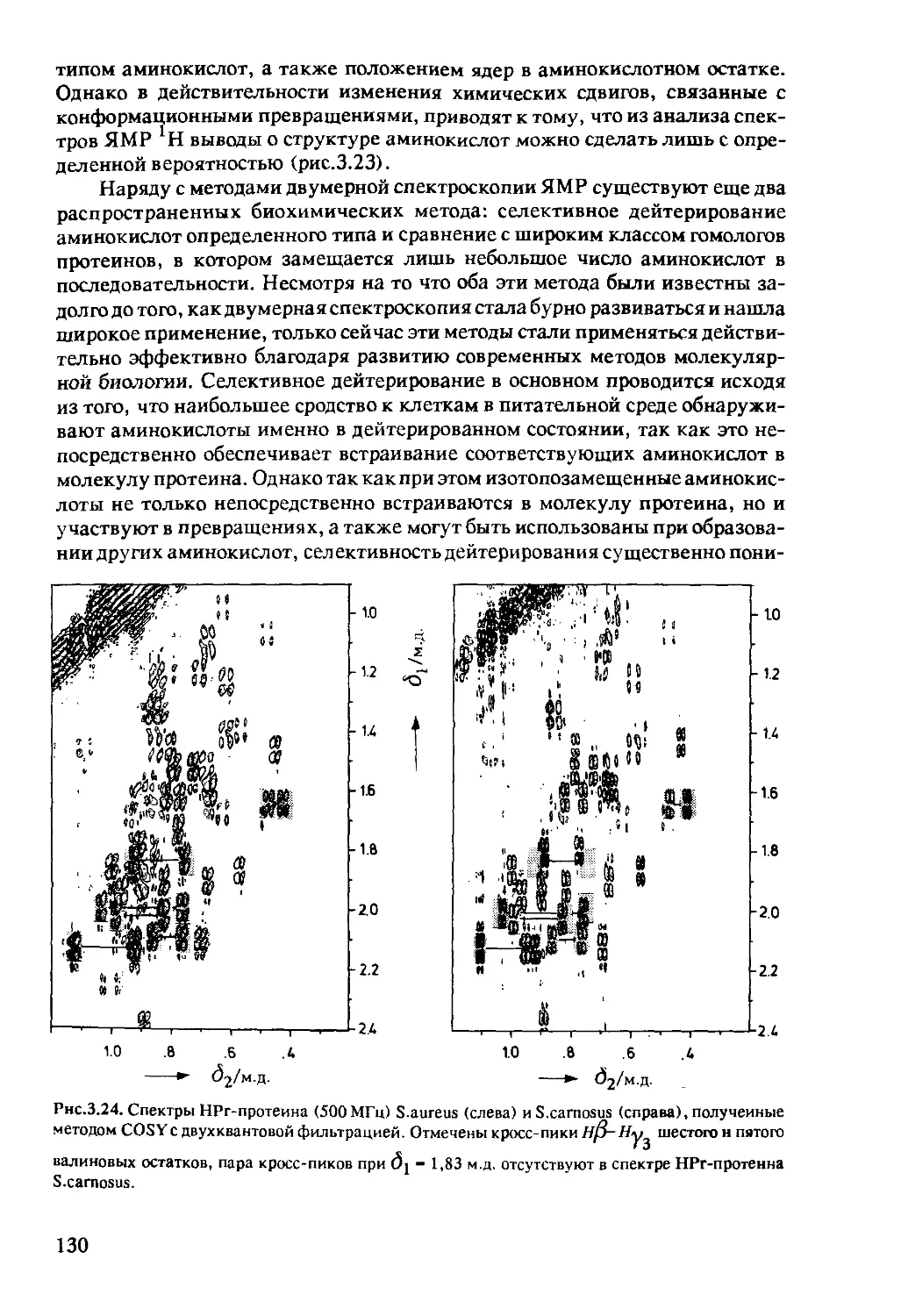
3.1.5 Интерпретация двумерных ЯМР-спектров ........135
3.1.6 Расчет структуры по данным ЯМР ..............138
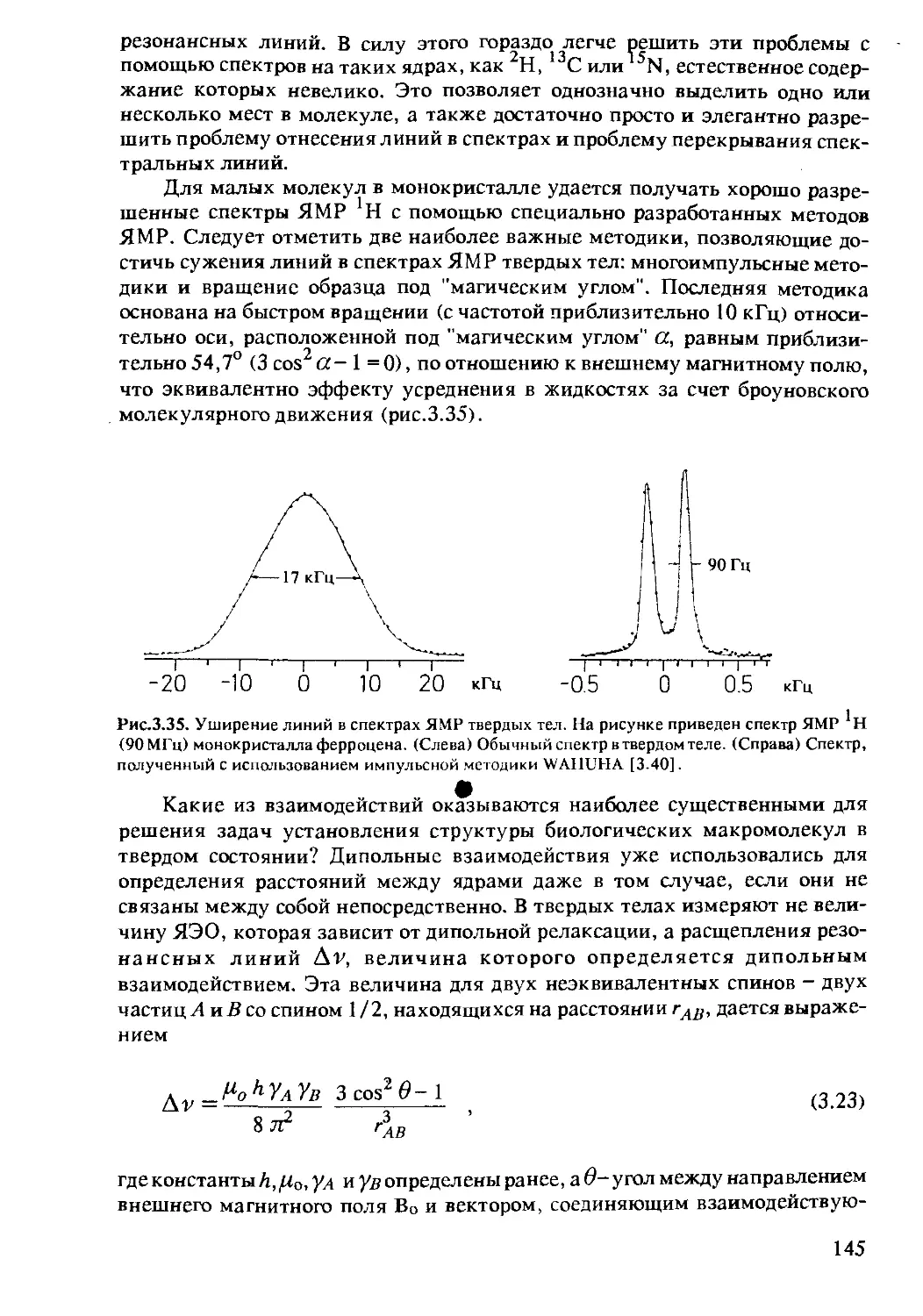
3.1.7 Твердотельный ЯМР протеинов .................144
3.2 ЯМР-спектроскопия нуклеиновых кислот,
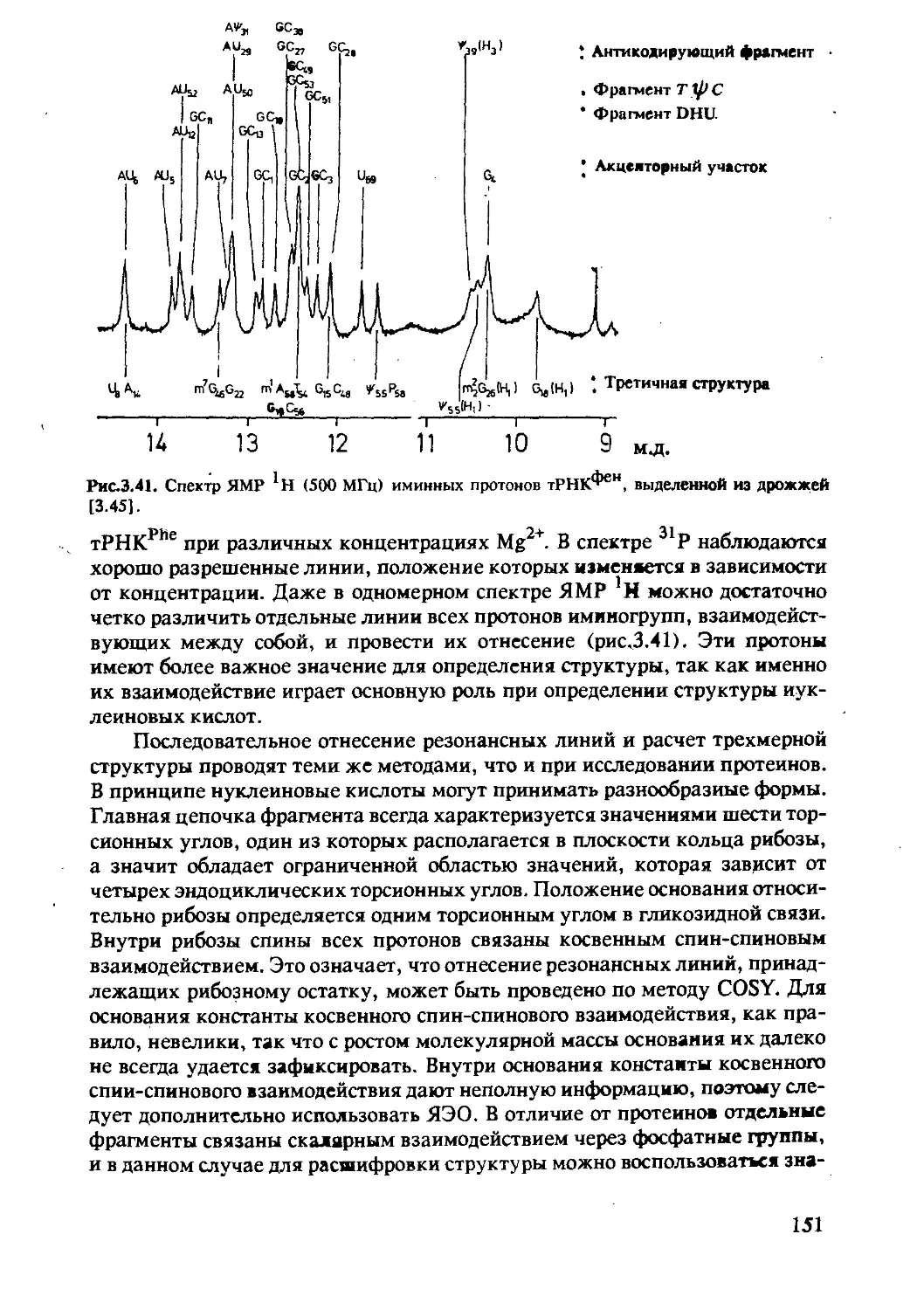
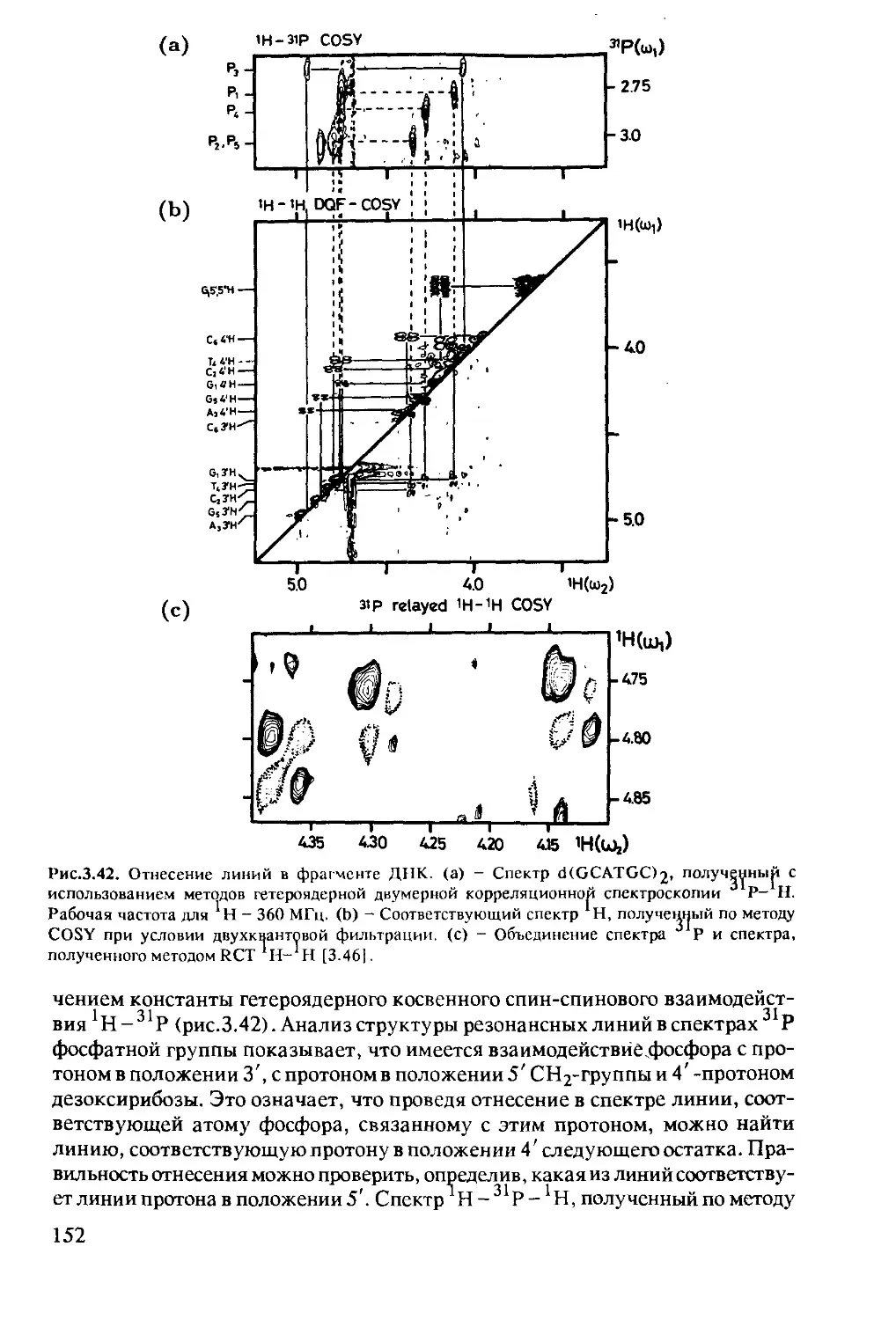
полисахаридов и липидов ..............................146
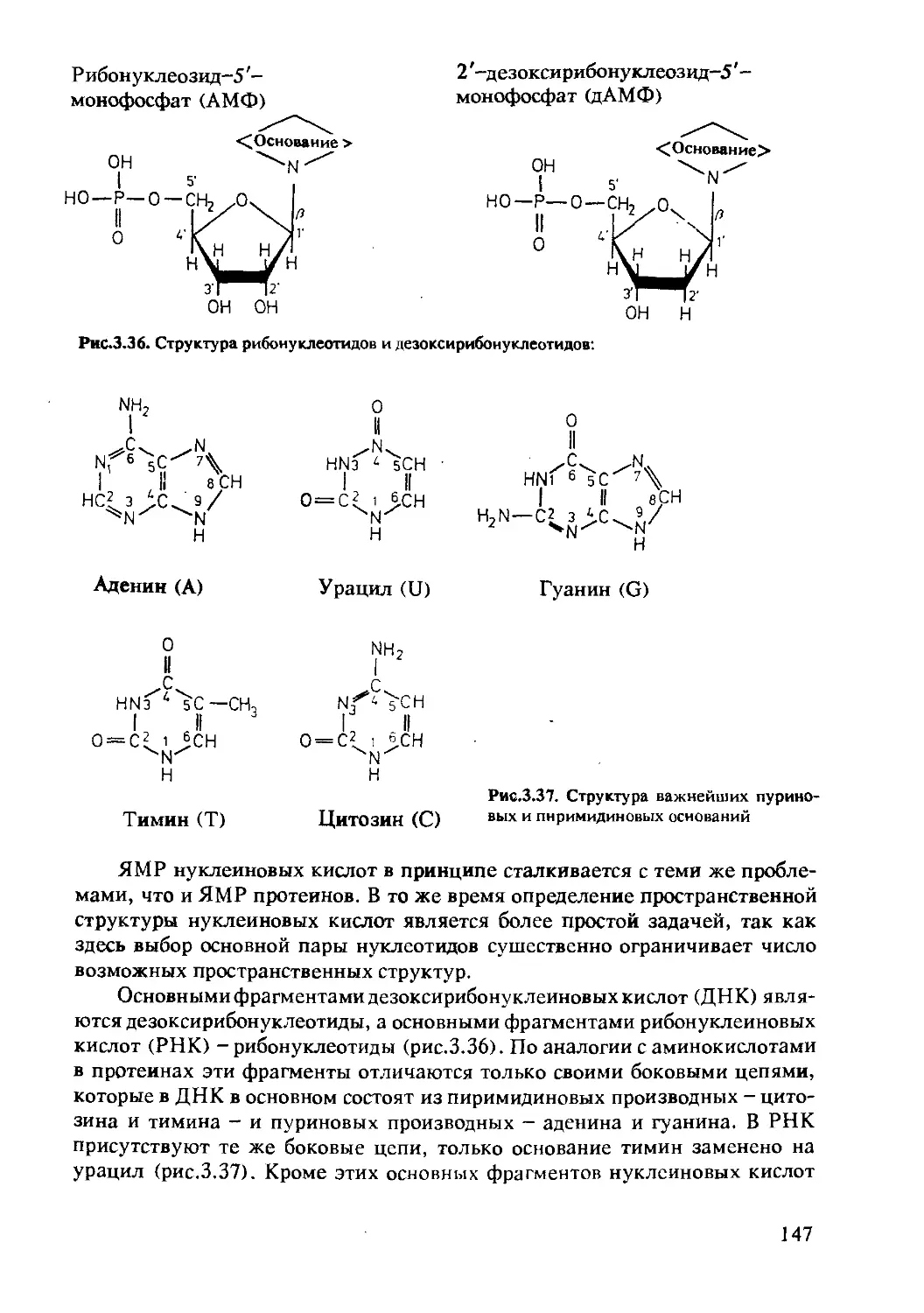
3.2.1 Состав и структура нуклеиновых кислот .......146
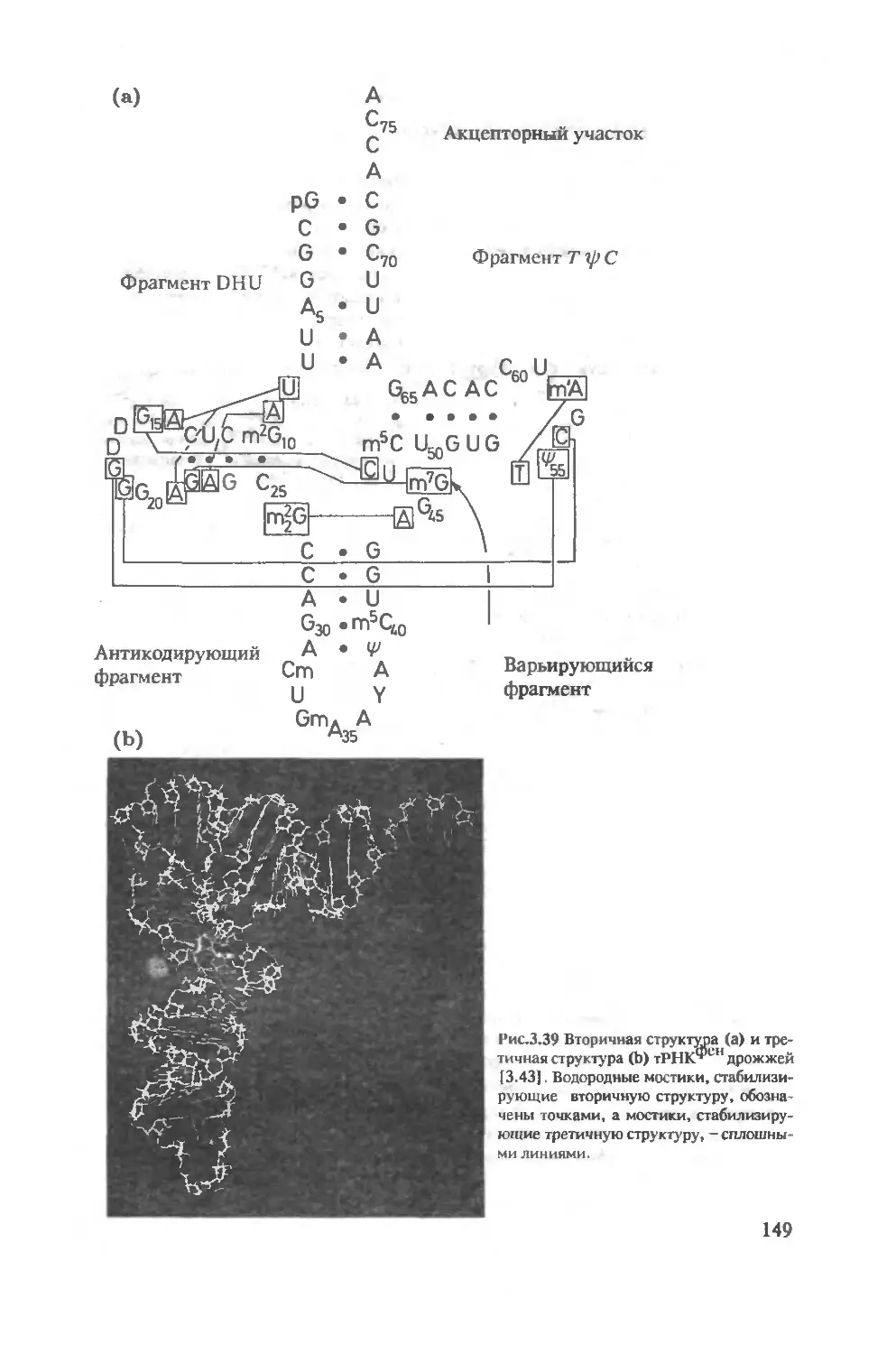
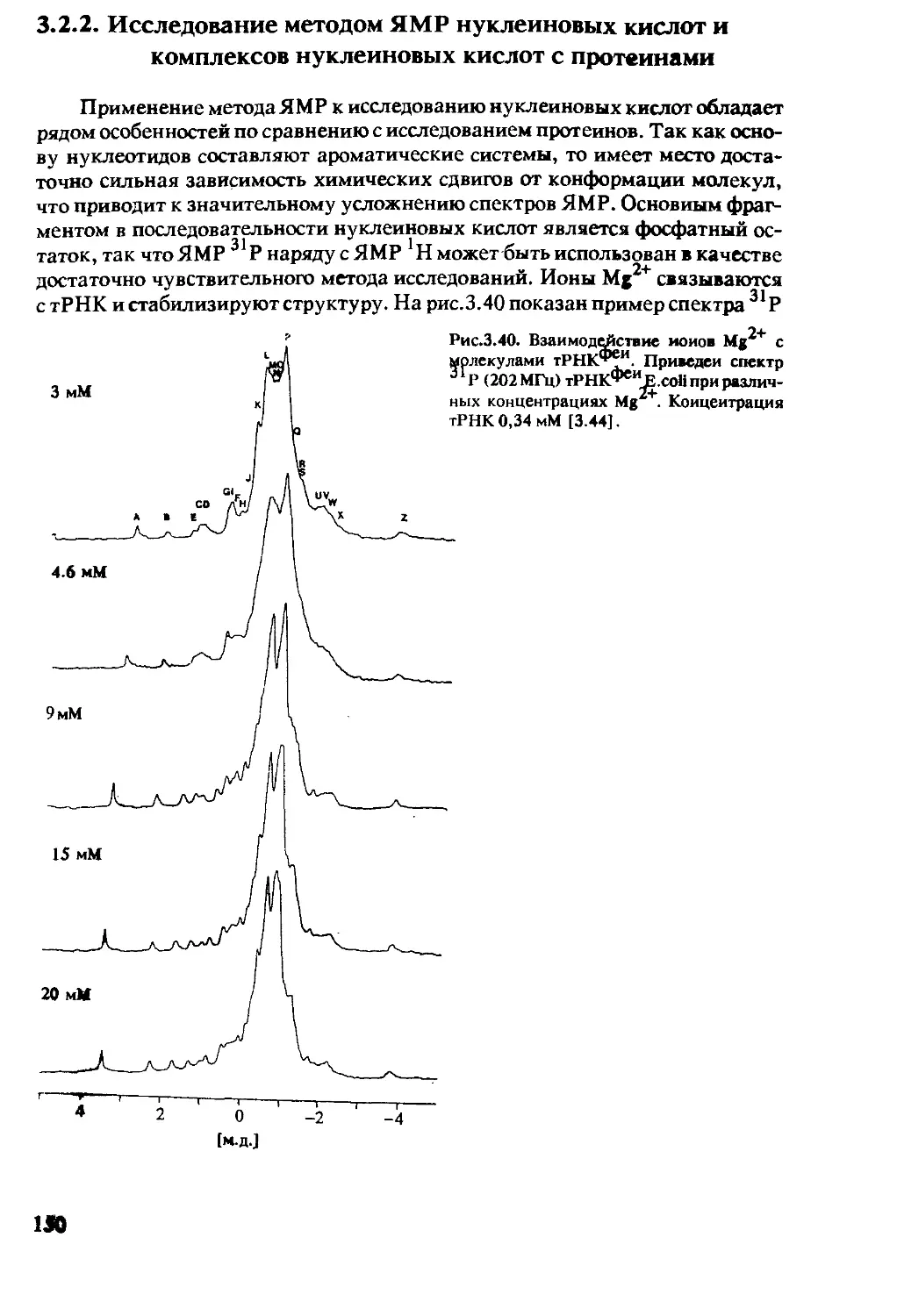
3.2.2 Исследование методом ЯМР нуклеиновых
кислот и комплексов нуклеиновых
кислот с протеинами ..........................150
3.2.3 Состав и структура полисахаридов ............153
3.2.4 Определение структуры полисахаридов..........154
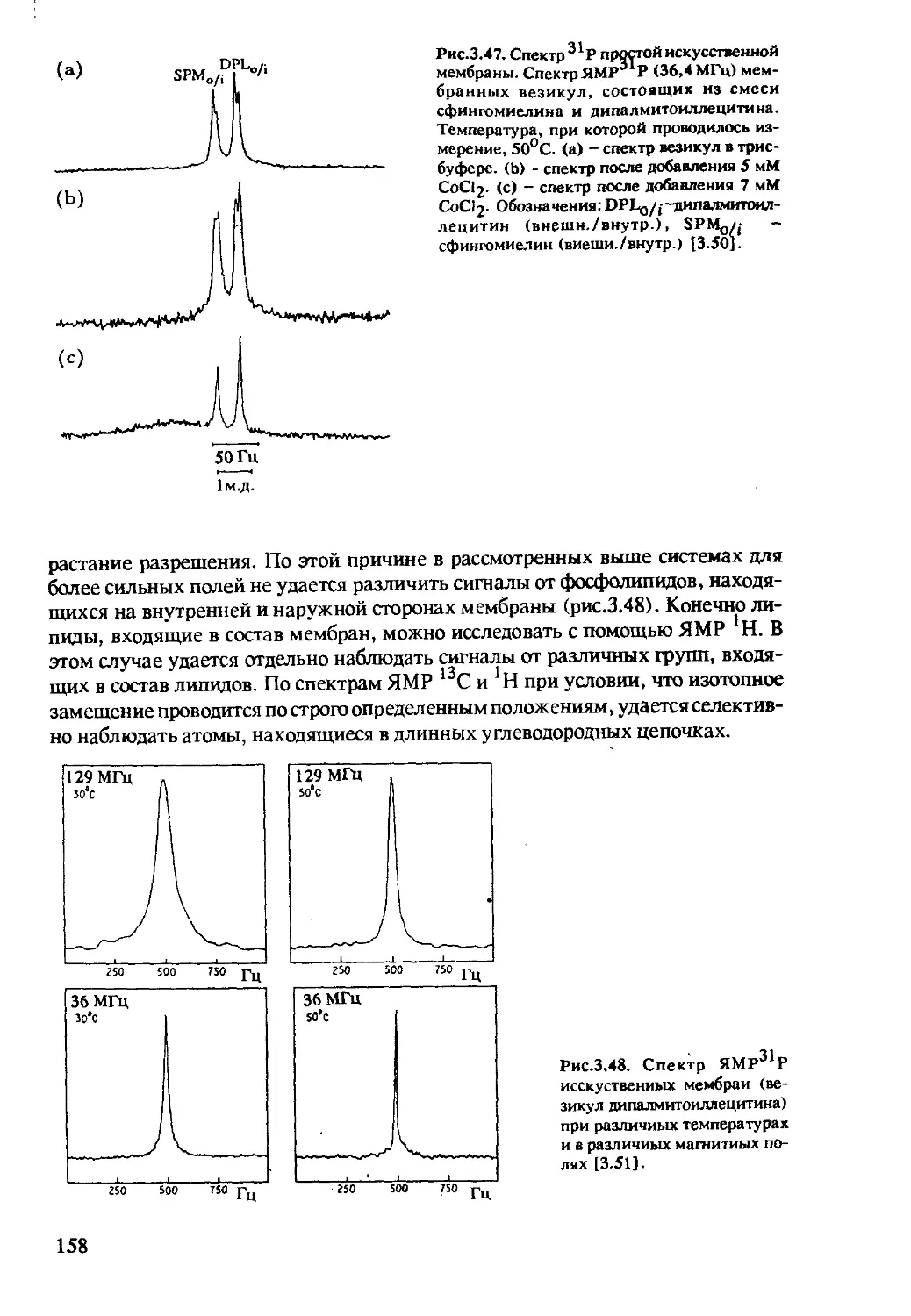
3.2.5 Исследование биологических мембран...........156
Литература............................................161
Глава 4. ЯМР-томография ...................................165
4.1 Основные принципы получения изображения ..........165
4.1.1 Двумерная проекционная реконструкция ........167
4.1.2 Методы двумерной фурье-спектроскопии ........168
4.1.3 Параметры ЯМР-томографии ....................171
8
4.1.4 Методы, позволяющие сократить время
проведения эксперимента ......................173
4.1.5 Получение трехмерных изображений ...........178
4.2 Примеры использования основных
экспериментальных методик в ЯМР-томографии ..........180
4.2.1 Изображение головы .........................180
4.2.2 Получение изображений других областей тела .183
4.3 Другие применения ЯМР-томографии ................190
4.3.1 Химический сдвиг в ЯМР-томографии...........191
4.3.2 Эффекты движения жидкостей
в ЯМР-томографии ............................193
4.3.3 Контрастные вещества в ЯМР-томографии ......198
4.3.4 ЯМР-микроскопия ............................200
4.4 Биомедицинские аспекты влияния ЯМР ..............203
4.4.1 Биомедицинские эффекты статического
магнитного поля .............................203
4.4.2 Биомедицинские эффекты
градиентных полей ............................204
4.4.3 Биомедицинские эффекты РЧполей..............204
Литература ......................................... 205
Глава 5. Пространственно селективная спектроскопия
и ЯМР-эксперименты in-vivo ...............................207
5.1 Методы повышения пространственной
селективности .......................................208
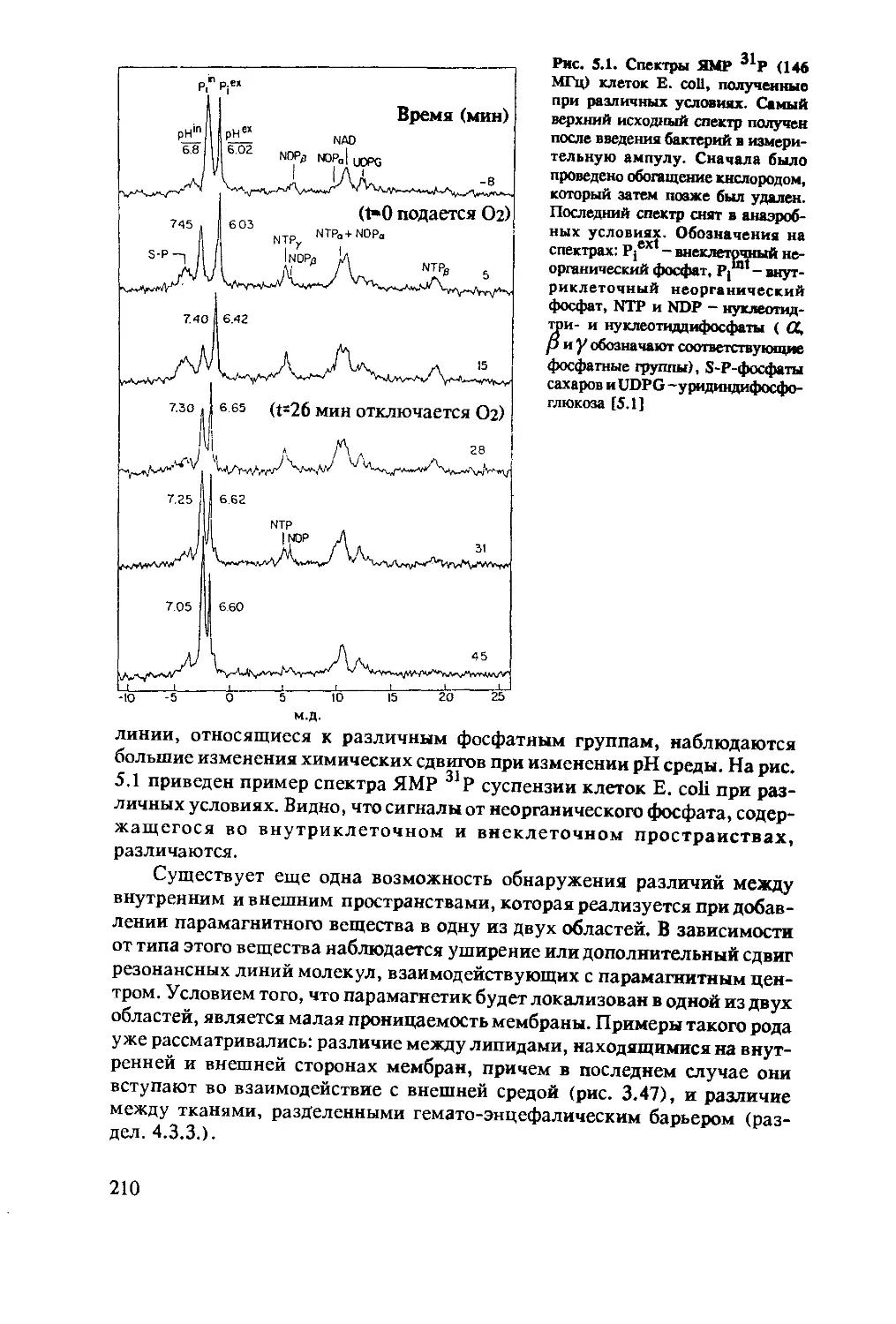
5.1.1 Пространственное разрешение для
изолированных клеток и органелл .............209
5.1.2 Пространственная селективность
при использовании РЧ полей, зависящих
от координат ................................211
5.1.3 Пространственная селекция с помощью
зависящего от координат статического
магнитного поля Во ...........................214
5.2 Типичные области применения и проблемы
ЯМР-спектроскопии живых систем ......................216
5.2.1 Сохранение естественных функций ............217
5.2.2 ЯМР-спектроскопия in-vivo на ядрах .........218
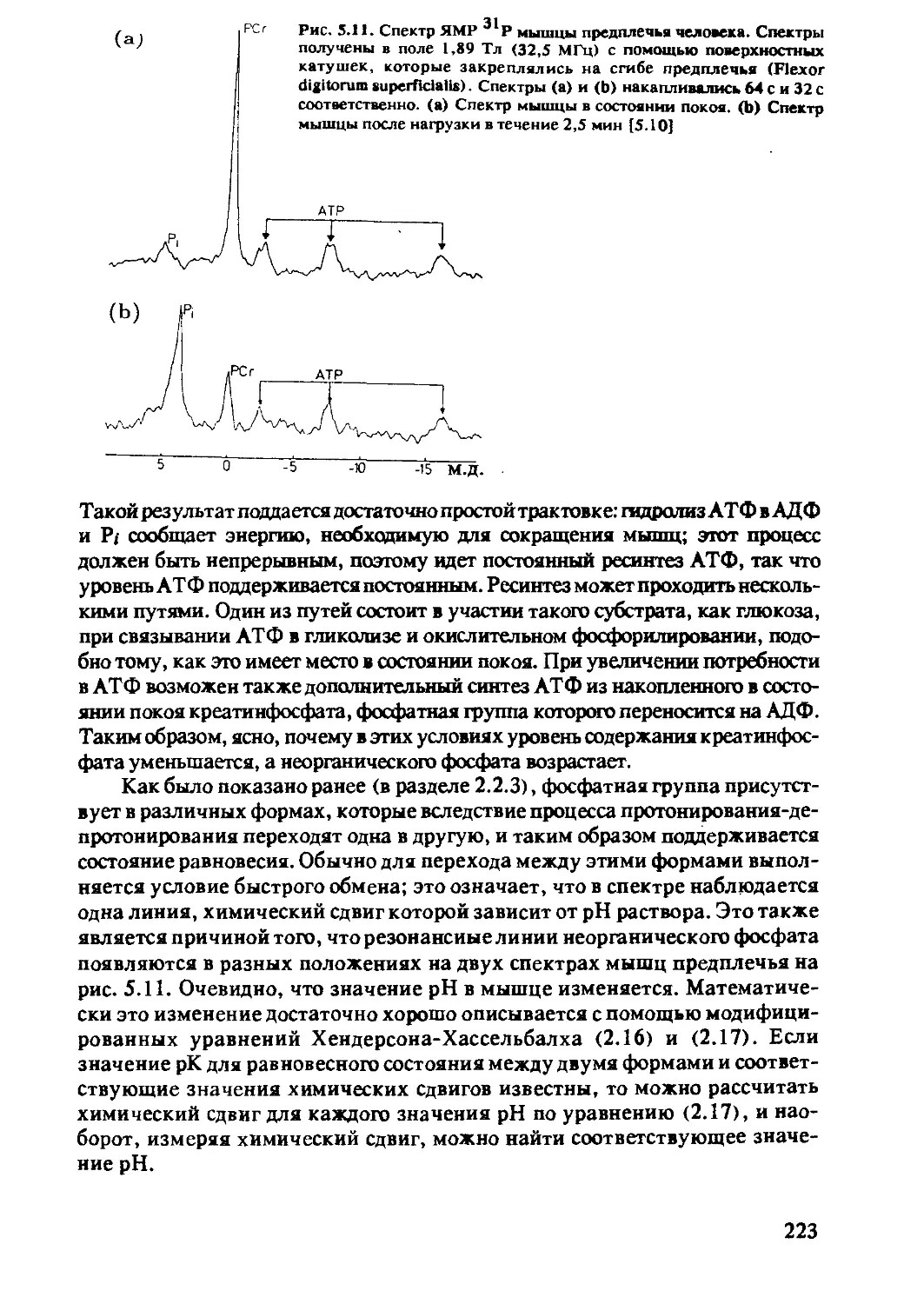
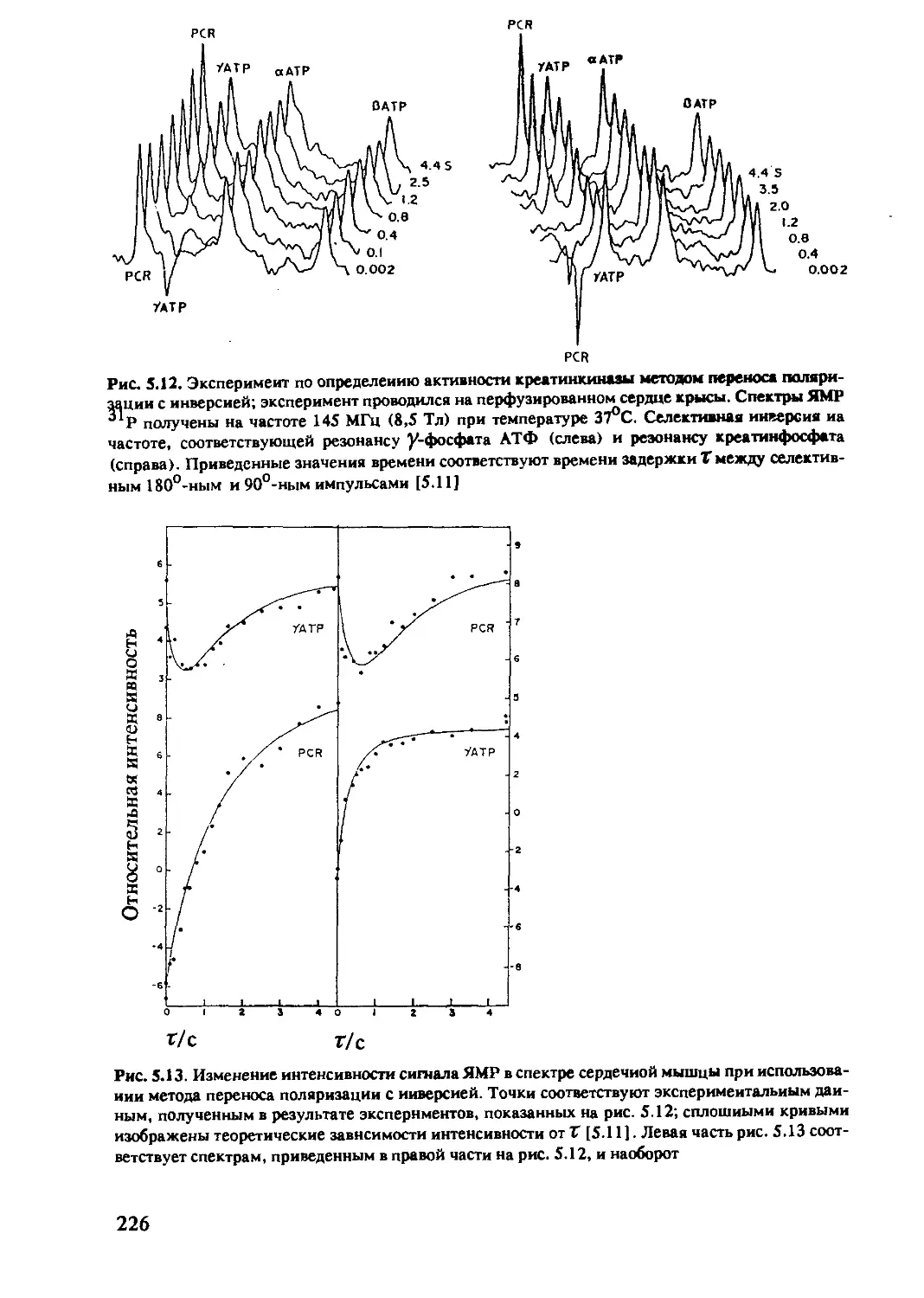
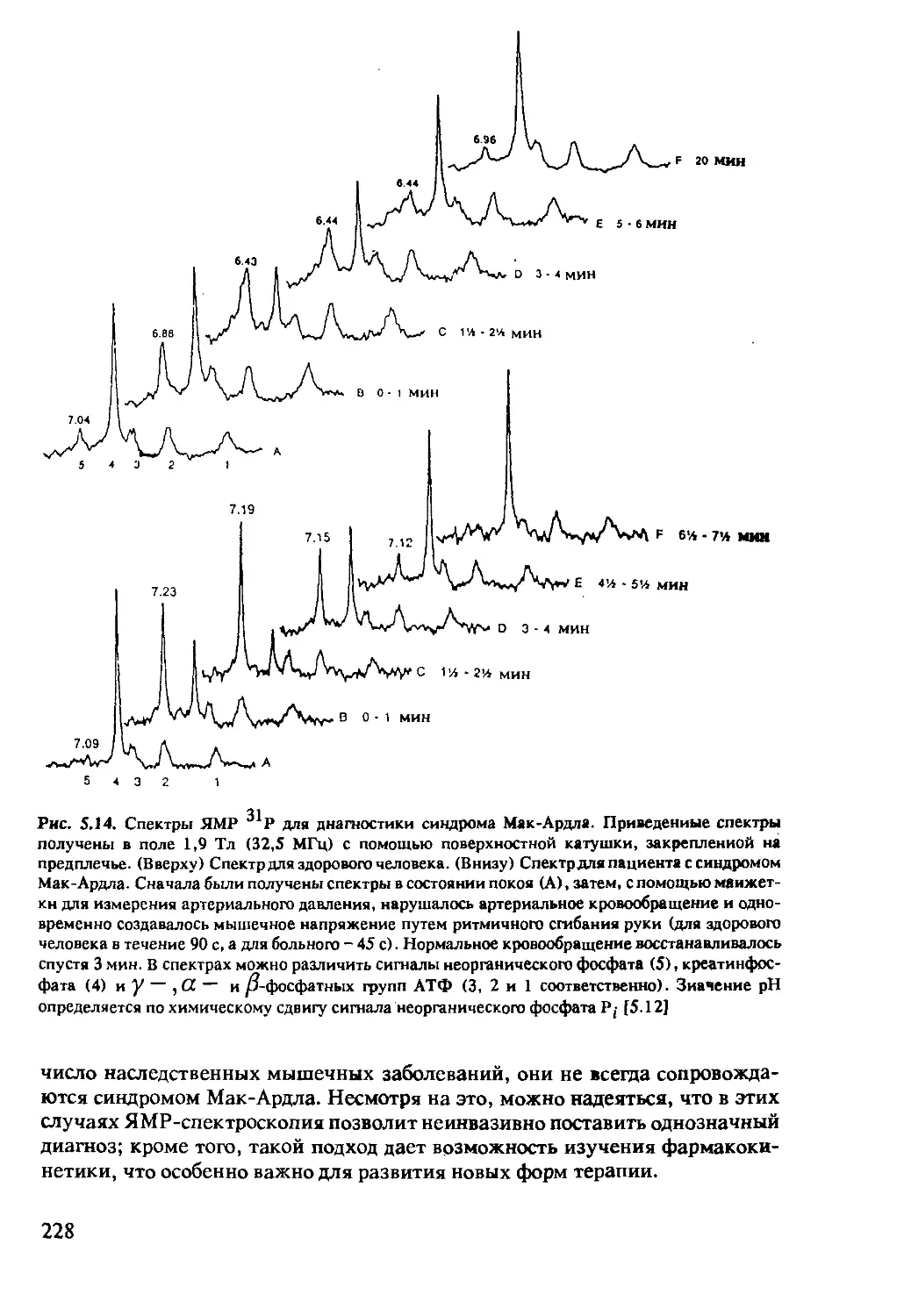
5.2.3 ЯМР-спектроскопия in-vivo на ядрах 31Р .....222
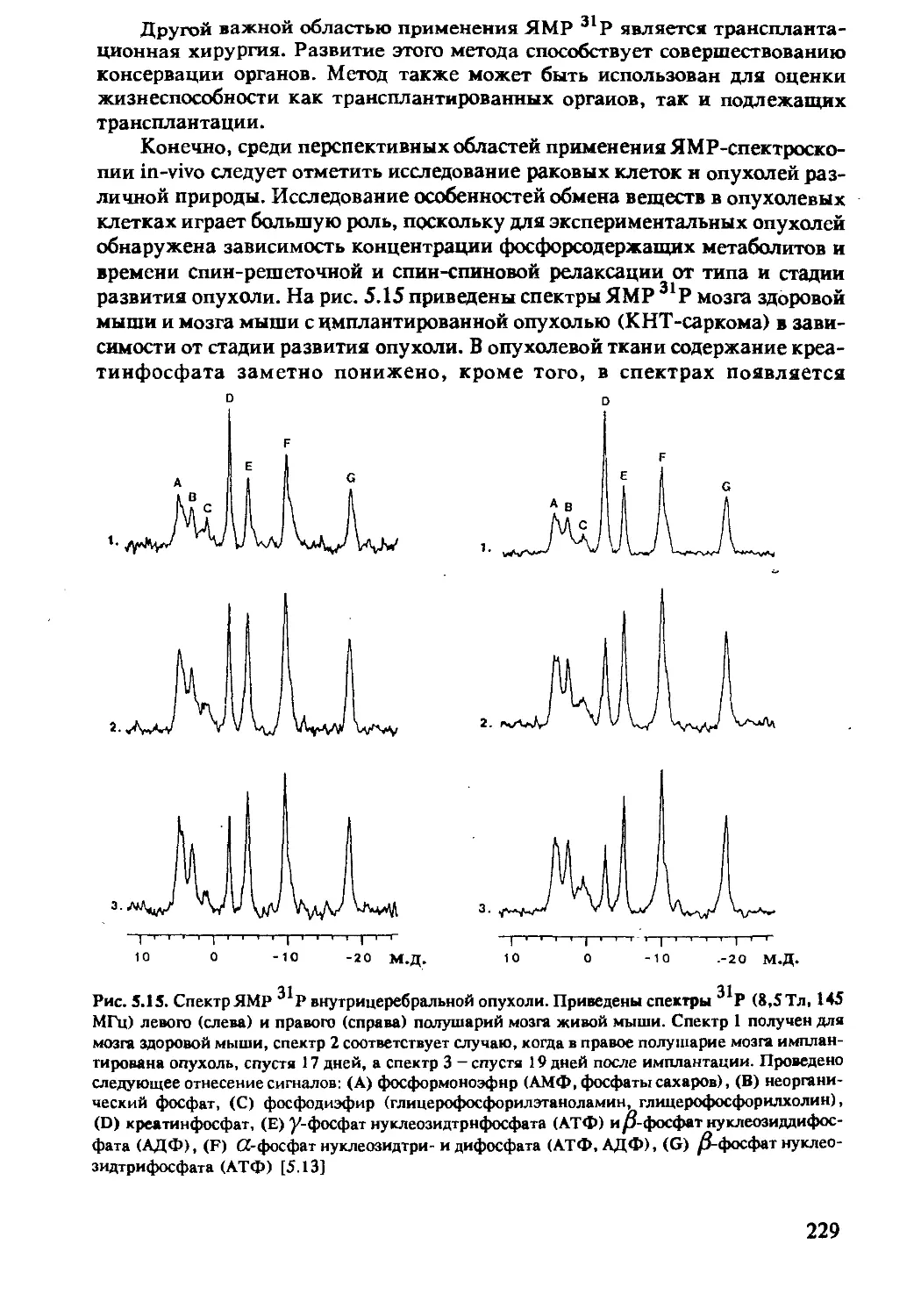
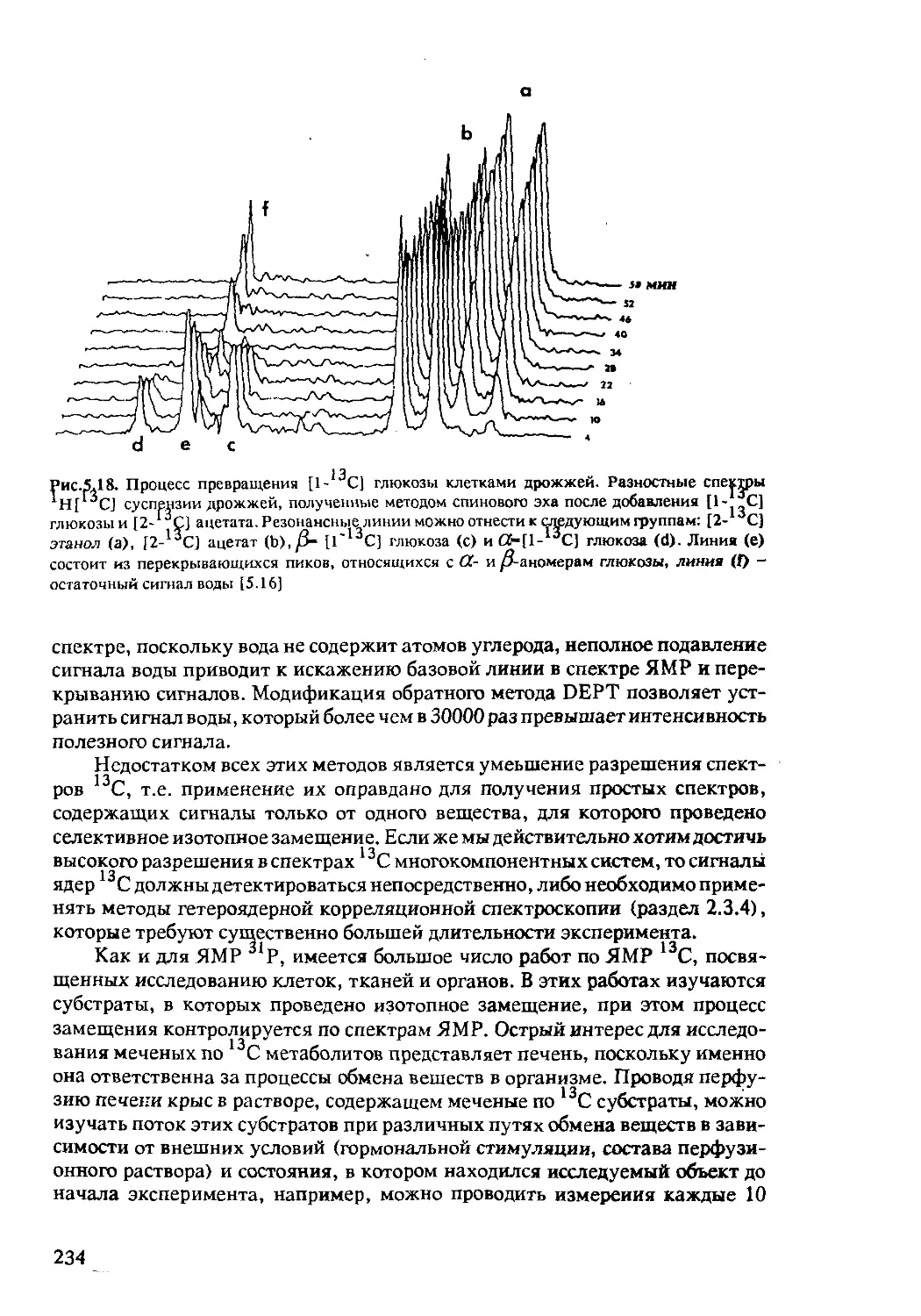
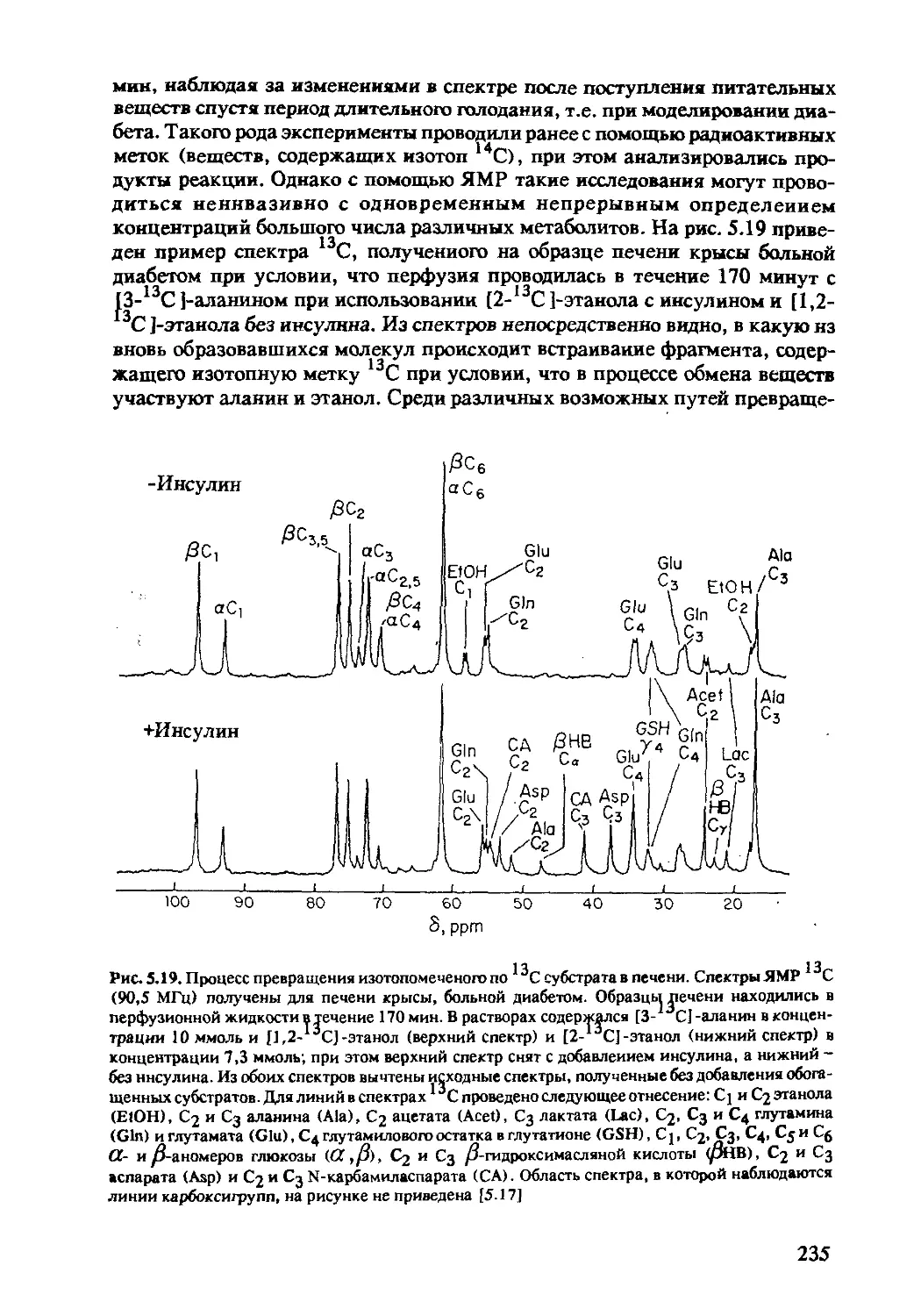
5.2.4 ЯМР-спектроскопия in-vivo на ядрах 13С .....230
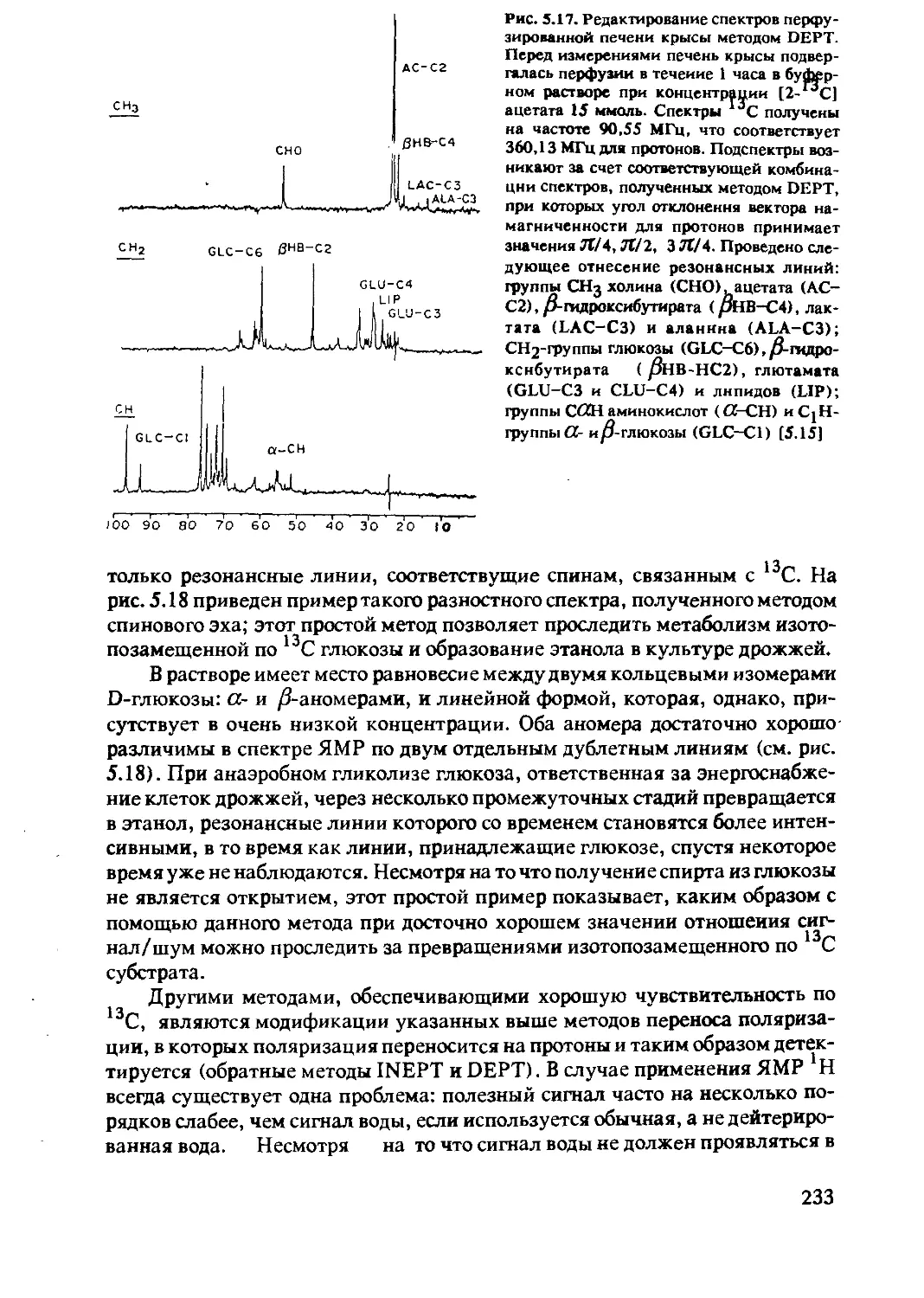
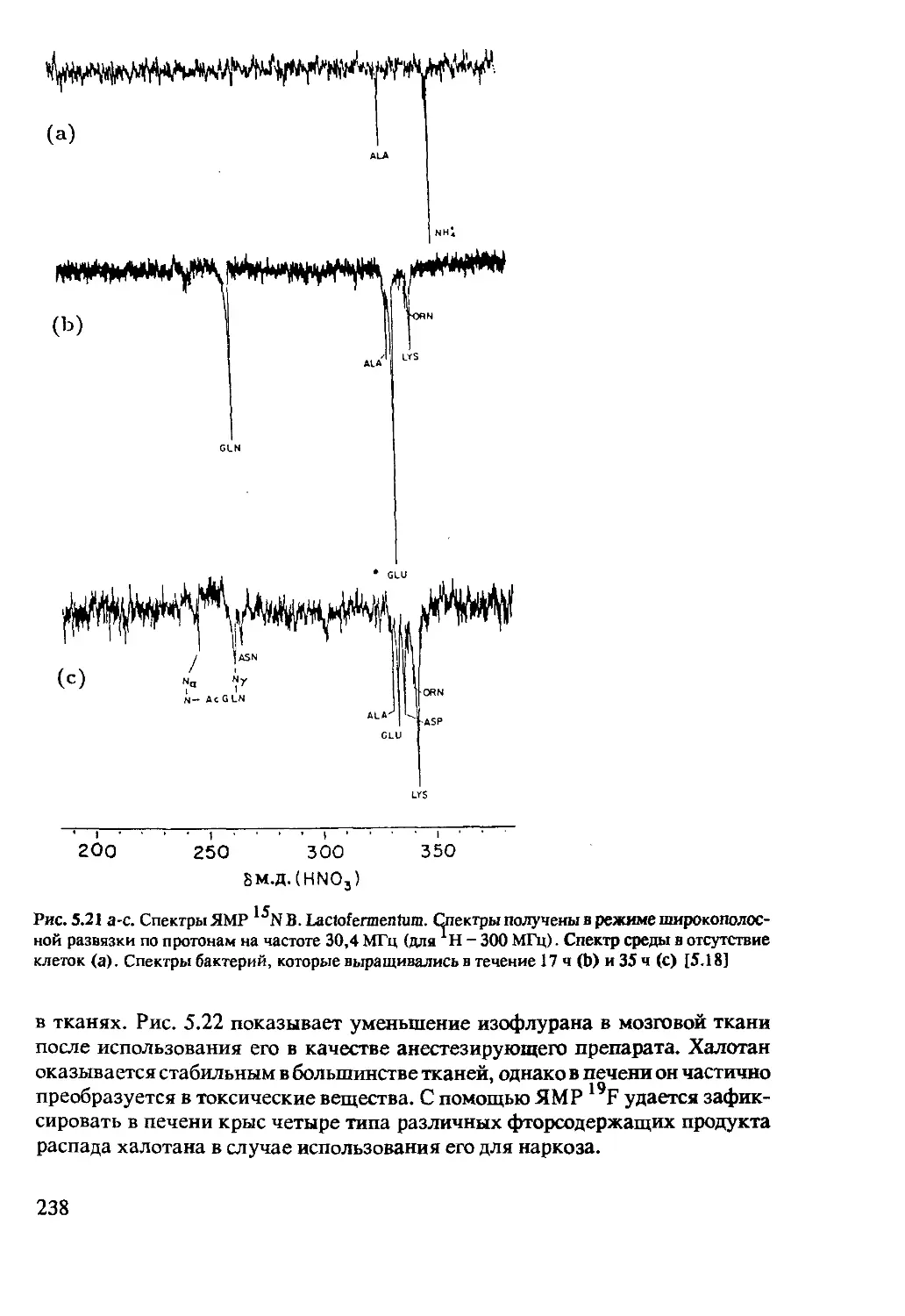
5.2.5 ЯМР-спектроскопия in-vivo на ядрах 15N 237
5.2.6 ЯМР-спектроскопия in-vivo на ядрах 19F .....237
5.2.7 Использование других ядер
в ЯМР-спектроскопии in-vivo .................239
Литература...........................................240
Предметный указатель ....................................241
9
Список сокращений и обозначений
АЦП Во, Во В1.В1 кпмг ЦП CW D е ССИ Gx, Gy, Gz аналого-цифровой преобразователь постоянное магнитное поле РЧполе, переменное магнитное поле последовательность Карра-Парселла-Мейбума-Гилла центральный процессор непрерывный ЯМР-эксперимент дейтерий, 2Н элементарный заряд (1,60219 • 10"19 А • с) спад свободной индукции градиентные магнитные поля вдоль осей декартовой системы координат
h 1i кГц РЧ Гц I постоянная Планка (6,6262 • 10'34м2 • кг • с'1) 11/271(1,0546 • 10’34 м2 • кг • с'1) 1000 Гц радиочастота (радиочастотный) герц, единица частоты ядерный спин, квантовое число, спиновое квантовое число
J к M М Мх, Му мера скалярного спин-спинового взаимодействия (Гц) постоянная Больцмана (1,3806 • 10'23 м2 • кг • с"2 • К’1) намагниченность; концентрация (моль/л) намагниченность компоненты намагниченности в плоскости, перпендикулярной внешнему магнитному полю
Mz компонента намагниченности, параллельная магнитному полю
Мо намагниченность в состоянии больцмановского равновесия; равновесная намагниченность
МГц мос ЯМР Na Pi Пиксель М.д. Q-e г 106 Гц миллион операций в секунду ядерный магнитный резонанс число Лошмидта (6,02217 • 1023моль-1) неорганический фосфат наименьший элемент плоского изображения миллионная доля рабочей частоты, 1:106 квадрупольный момент ядра радиус-вектор в сферической системе координат
10
s s S/tf T Тл Ti T2 TE спин электрона коэффициент насыщения сигнал/шум абсолютная температура; Тесла, единица напряженности магнитного поля время спин-решеточной или продольной релаксации время спин-спиновой или поперечной релаксации эффективное время поперечной релаксации время спинового эха между 90°-ным импульсом и спиновым эхо
Tr время повторения; интервал времени между последовательностями импульсов
Вексель наименьший элемент объемного изображения, соответствует пикселю при плоском изображении
a У d e Hl Hs Ho угол, угол поворота гиромагнитное отношение химический сдвиг, измеренный в м.д. азимутальный угол в сферической системе координат магнитный момент ядерного спина I магнитный момент спина электрона S магнитная проницаемость вакуума (4 • 10'7 м • кг • с’2 • А’2)
V p a T trot ftrans Tsc fs <P частота, измеряемая в Гц плотность, скорость спин-решеточной релаксации константа экранирования; скорость кросс-релаксации время корреляции время корреляции вращательных движений время корреляции трансляционных движений время корреляции при скалярных взаимодействиях время корреляции в случае парамагнитной релаксации угол в сферической системе координат; диэдральный угол в основном фрагменте пептидной цепи
X диэдральный угол в боковой пептидной цепи; магнитная восприимчивость
Ш диэдральный угол в основной пептидной цепи круговая частота (2 Л V); диэдральный угол в основной пептидной цепи
11
Глава 1. Основы метода ядерного магнитного резонанса
В первой главе настоящей книги излагаются основные положе-
ния, позволяющие понять сущность явления ядерного резонанса, в
последующих главах будет дано более глубокое и подробное его
изложение. Ядерный резонанс является одним из наиболее употре-
бительных названий метода, которому посвящена данная книга. На-
ряду с этим названием используют и такие, как ядерный спиновый
резонанс, ядерный магнитный резонанс или ЯМР (от англ, nuclear
magnetic resonance).
Первые эксперименты, в которых удалось наблюдать сигнал ядерного
резонанса в конденсированных средах, были проведены в 1945 г. независимо
Блохом и Парселлом [1.1, 1.2 ]. Следующим важным шагом было открытие
химического сдвига — величины, которая характеризует электронное окру-
жение рассматриваемого ядра. В металлах это явление (изменение резонан-
сной частоты) впервые наблюдал Найт [1.3 ], а в жидкостях — Арнольд [1.4].
Это открытие оказало колоссальное влияние на развитие не только метода
ядерного резонанса, но и других областей физики. Информация о частоте
сигнала ЯМР дает возможность получить представление об электронном
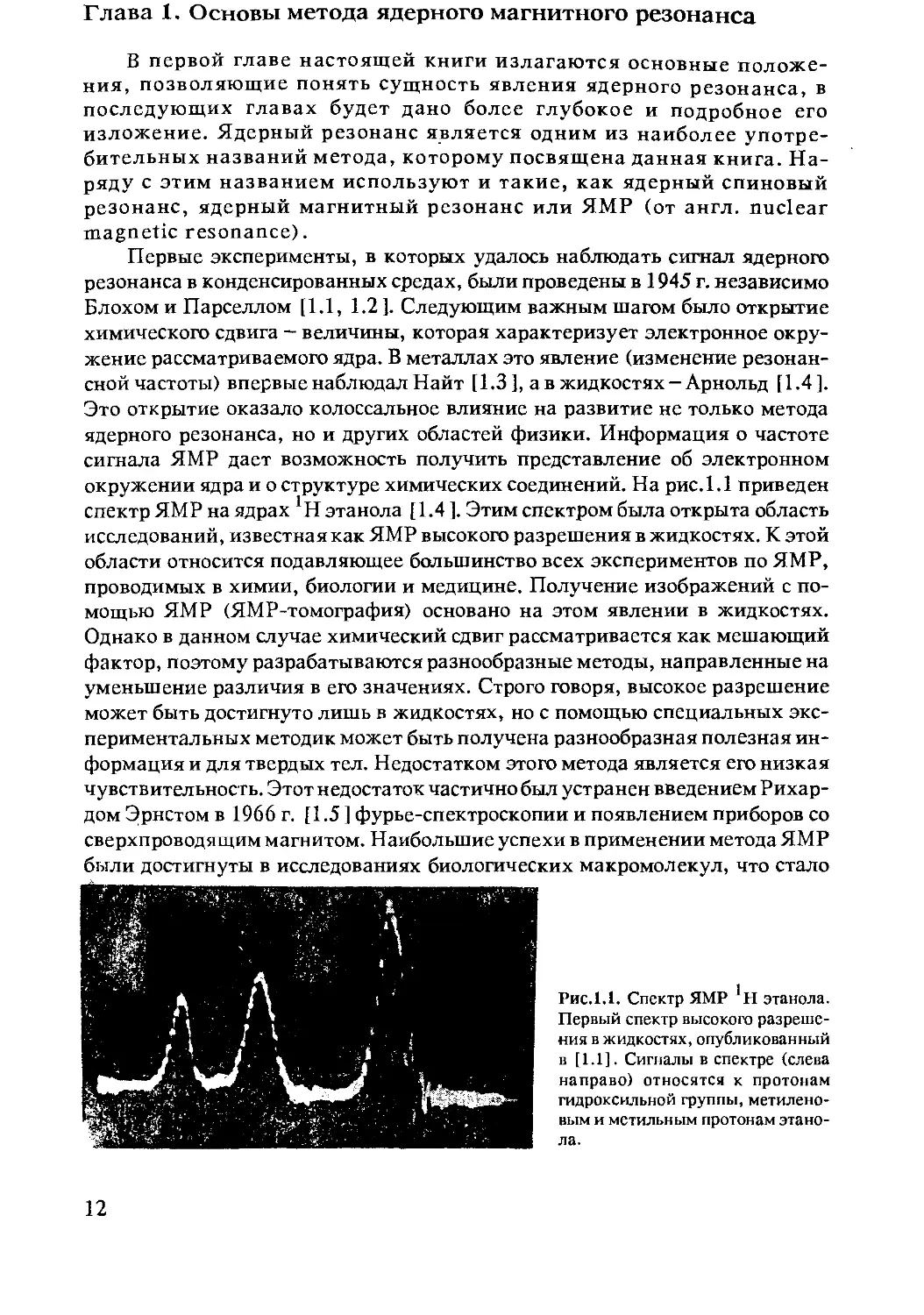
окружении ядра и о структуре химических соединений. На рис. 1.1 приведен
спектр ЯМР на ядрах 1Н этанола [1.4 ]. Этим спектром была открыта область
исследований, известная как ЯМР высокого разрешения в жидкостях. К этой
области относится подавляющее большинство всех экспериментов по ЯМР,
проводимых в химии, биологии и медицине. Получение изображений с по-
мощью ЯМР (ЯМР-томография) основано на этом явлении в жидкостях.
Однако в данном случае химический сдвиг рассматривается как мешающий
фактор, поэтому разрабатываются разнообразные методы, направленные на
уменьшение различия в его значениях. Строго говоря, высокое разрешение
может быть достигнуто лишь в жидкостях, но с помощью специальных экс-
периментальных методик может быть получена разнообразная полезная ин-
формация и для твердых тел. Недостатком этого метода является его низкая
чувствительность. Этот недостаток частично был устранен введением Рихар-
дом Эрнстом в 1966 г. [1.5 ] фурье-спектроскопии и появлением приборов со
сверхпроводящим магнитом. Наибольшие успехи в применении метода ЯМР
были достигнуты в исследованиях биологических макромолекул, что стало
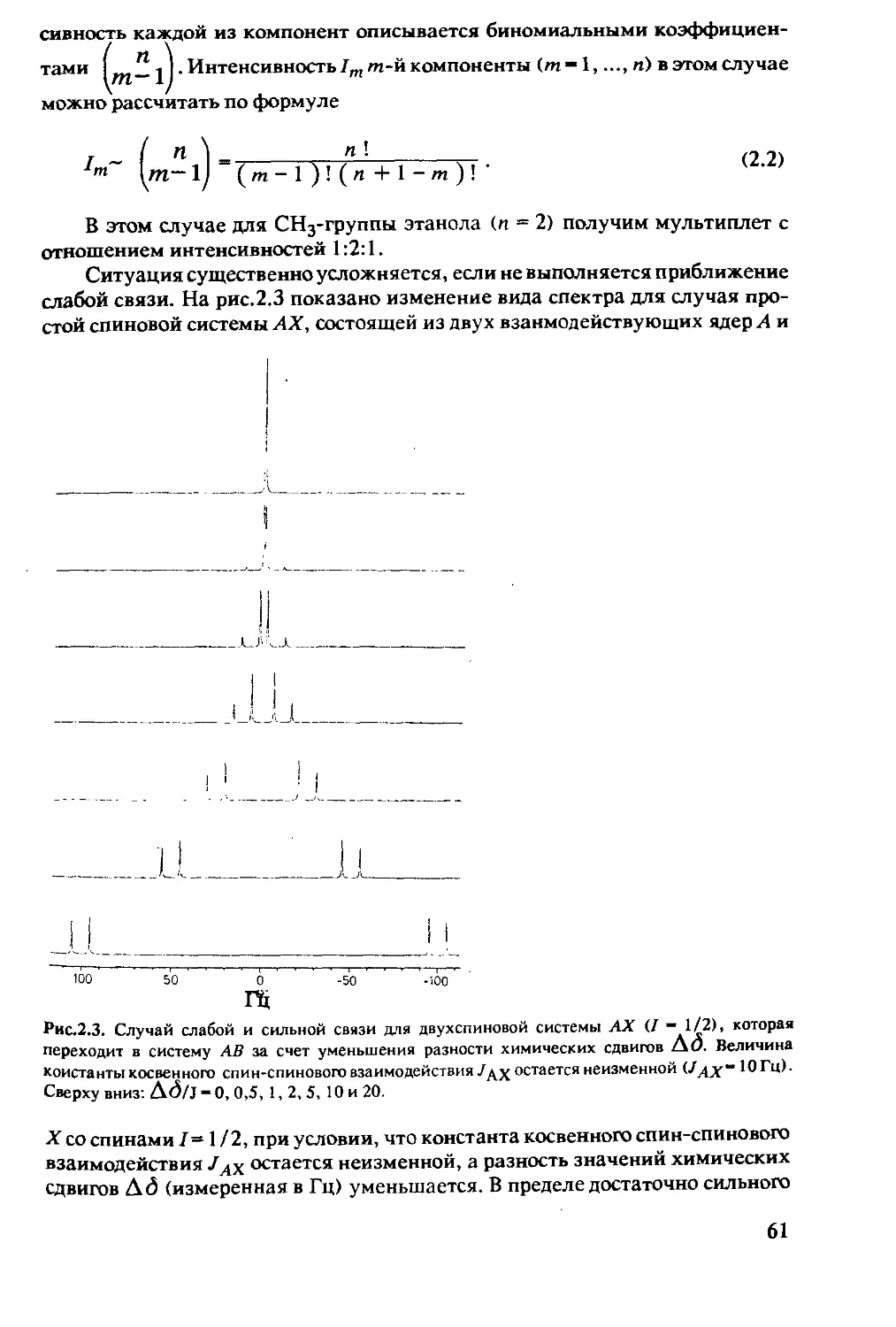
Рис. 1.1. Спектр ЯМР этанола.
Первый спектр высокого разреше-
ния в жидкостях, опубликованный
в [1.1]. Сигналы в спектре (слева
направо) относятся к протонам
гидроксильной группы, метилено-
вым и метильным протонам этано-
ла.
12
возможным в результате развития метода двумерной спектроскопии ЯМР
(2М-ЯМР), предложенного Джином Джинером [1.6]. В настоящее время
2М-ЯМР — активно развивающаяся область физики, в которой постоянно
появляются публикации о новых экспериментальных методиках. Первые
работы по применению ЯМР в биологии начались с исследования структуры
протеинов в растворе, так что для протеинов малой массы ЯМР был исполь-
зован в качестве метода, альтернативного рентгеноструктурному анализу.
Особый вклад в развитие этого направления принадлежит Курту Вютриху и
его группе.
После появления в 1971 г. работы Поля Лаутербура [1.7] и Питера
Мэнсфилда [1.8 ], посвященных изложению основ нового метода визуализа-
ции объектов (ЯМР-томографии), метод ЯМР начал использоваться в меди-
цине. Можно ожидать, что ценность спектроскопических методов для
медицинских приложений будет возрастать.
1.1. Основные принципы магнитного резонанса
Движение отдельных частиц, в частности молекул, как правило, наибо-
лее полно можно описать на языке квантовой механики - абстрактной мате-
матической теории, в которой все процессы, происходящие в природе,
выражаются с помощью операторов физических величин. При этом сами
операторы не дают наглядной физической картины, а конкретный физиче-
ский смысл приобретают только средние значения или математические ожи-
дания операторов, т.е. значения физических величин, получаемые в
результате достаточно большого числа измерений. Расчет математических
ожиданий, обычно обозначаемых парой угловых скобок, проводится соглас-
но данной теории. Например, энергия Е определяется как математическое
ожидание гамильтониана Н системы: Е = < Я >. Заметим, что во многих
случаях имеет место формальное совпадение операторных уравнений с соот-
ветствующими уравнениями для математических ожиданий, хотя их смысл,
вообще говоря, различный. Здесь, как правило, будем рассматривать мате-
матические ожидания физических величин (операторов), поэтому там, где
не возникает недоразумений, скобки, обозначающие математические ожи-
дания, для краткости будем опускать.
Основные принципы магнитного резонанса можно понять в рамках клас-
сической физики при условии, что введены дополнительные предположения,
отражающие квантовомеханические свойства системы, поэтому далее для
описания резонансных явлений часто используется квазиклассическое при-
ближение, благодаря его наглядности и простоте.
1.1.1. Магнитный момент и ядерный спин
Большинство атомных ядер обладает собственным механическим мо-
ментом вращения J, пропорциональным величине I, называемой ядерным
13
спином: J = h I, где J и I - операторы, h = hl'2.:it,h- постоянная Планка. Ядра,
обладающие собственным механическим моментом, можно рассматривать
как микроскопические гироскопы. С вращательным моментом связан ди-
польный магнитный момент ft], причем между механическим и магнитным
моментом существует простая связь
Ju7 = y7J = /i у71.
(1.1)
Коэффициент пропорциональности /у называется гиромагнитным отноше-
нием. Эта величина является константой для каждого типа ядер. Величина
спина I постоянна для каждого ядра, точнее, для каждого ядра в основном
состоянии. В физике высоких энергий наблюдаются возбужденные состояния
ядер, в которых значения вращательного момента ядер отличаются от их
значений в основном состоянии. Согласно квантовой механике, величина
ядерного спина I характеризуется максимальным собственным значением
оператора 1г - проекции оператора спина I на ось z произвольной декартовой
системы координат. Число I называют спином ядра. Собственные числа (ма-
тематические ожидания) тг оператора IL могут принимать (27+1) значений:
т7 = 7, 7-1, ...,-7,где7 > 0 может быть либо целым числом (включая нуль),
либо полуцелым (например, для ядра атома водорода Н спин 7= 1/2, а т7 =
= 1/2 и -1/2). Исходя из квантовомеханических положений можно вычис-
лить модуль момента J:
<IJ I > = Л< I I I > = й [7(7+1) ]1/2.
(1.2)
Чем больше величины у7 и 7, тем больше магнитный момент атомного
ядра, а значит, величина магнитного поля, создаваемого этим микроскопи-
чески малым магнитом.
Среди стабильных изотопов ядро атома водорода JH обладает наиболь-
шим значением гиромагнитного отношения. Так как чувствительность ме-
тода возрастает с ростом магнитного момента и большинство биологических
объектов состоит из соединений, в состав которых входят атомы водорода
(человеческое тело, например, на 60% состоит из воды), то спектры ЯМР на
ядрах *H имеют особое значение в биологии и медицине. Такие распростра-
ненные изотопы, как 1бО и 12С, обладают нулевым ядерным спином, и они
не могут быть обнаружены с помощью метода ЯМР.
Среди ядер, представляющих интерес для биологии и медицины и обла-
дающих спином I = 1/2, наряду с ХН можно назвать ядра 31Р, 13С и 15N.
Однако естественное содержание изотопа 13С составляет лишь 1 % по отно-
шению к изотопу 12С^спин которого 7= 0, а изотопа 15N - всего лишь 0,4%.
Ядра изотопов N и °Na, спин которых 7 >1, могут обладать электрическим
квадрупольным моментом, их применение в спектроскопии ЯМР весьма
ограничено. Основные свойства ядер, представляющих интерес для биологии
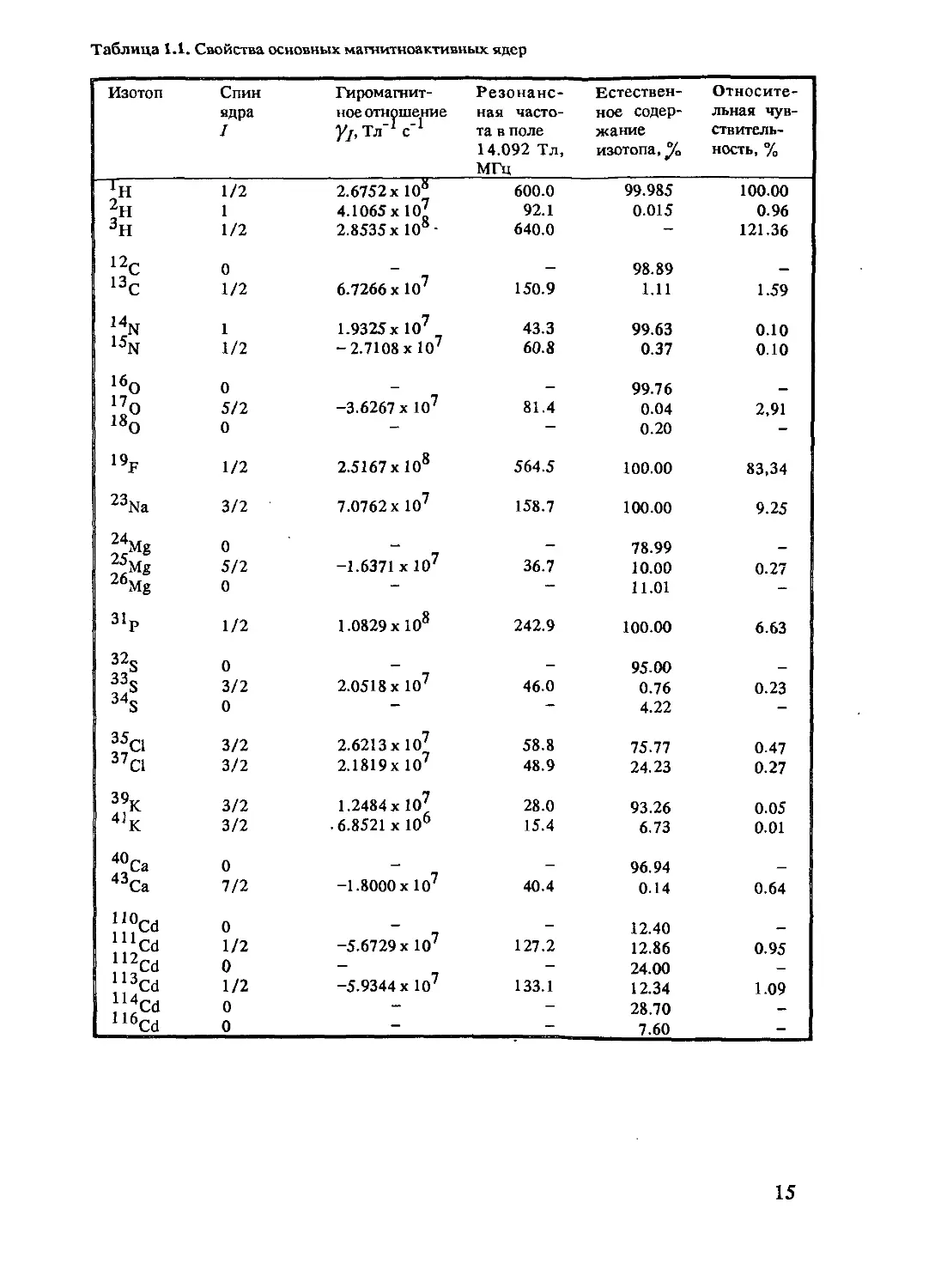
и медицины, приведены в табл. 1.1.
14
Таблица 1.1. Свойства основных магнитноактивных ядер
Изотоп Спин ядра I Гиромагнит- ное отношение Уj, Тл'1 с'1 Резонанс- ная часто- та в поле 14.092 Тл, МГц Естествен- ное содер- жание изотопа, °Ja Относите- льная чув- ствитель- ность, %
ХН 1/2 2.6752 х 108 600.0 99.985 100.00
2Н 1 4.1065 х 107 92.1 0.015 0.96
3Н 1/2 2.8535 х 108- 640.0 - 121.36
12с 0 — — 98.89 —
13С 1/2 6.7266 х 107 150.9 1.11 1.59
ltN 1 1.9325 хЮ7 43.3 99.63 0.10
15N 1/2 - 2.7108 хЮ7 60.8 0.37 0.10
16О 0 — — 99.76 —
170 5/2 -3.6267 х 107 81.4 0.04 2,91
180 0 - — 0.20 -
19f 1/2 2.5167 х 108 564.5 100.00 83,34
23Na 3/2 7.0762 х 107 158.7 100.00 9.25
24,.„ , Mg 0 - 78.99 —
5/2 -1.6371 х 10' 36.7 10.00 0.27
26Mg 0 — — 11.01 -
31p 1/2 1.0829x108 242.9 100.00 6.63
32S 0 95.00
33» 3/2 2.0518 х 107 46.0 0.76 0.23
34s 0 — — 4.22 -
35a 3/2 2.6213 х 107 58.8 75.77 0.47
37 Cl 3/2 2.1819х 107 48.9 24.23 0.27
39K 3/2 1.2484 хЮ7 28.0 93.26 0.05
4JK 3/2 • 6.8521 х 106 15.4 6.73 0.01
40Ca 0 — 96.94
43Ca 7/2 -1.8000х1О7 40.4 0.14 0.64
110Cd 0 - 12.40 —
J^Cd 1/2 -5.6729 х 10х 127.2 12.86 0.95
1 5Cd 0 — 24.00 —
Cd 1/2 -5.9344 х 10' 133.1 12.34 1.09
1 6Cd 0 — — 28.70 —
116Cd 0 — 7.60 -
15
1.1.2. Условие резонанса
В чем состоит явление ЯМР? Если наблюдать движение волчка в поле
тяжести Земли, то под влиянием этого поля ось волчка (если ее направление
отклонено от вертикали) совершает вращение вокруг направления поля тя-
жести. Такое движение называют прецессией. Частота этого вращения зави-
сит от величины поля тяжести, т.е. если представить себе, что это вращение
происходит не на Земле, а на Луне, то частота прецессии будет в шесть раз
меньше, чем на Земле. Подобное наблюдается и для магнитного диполя,
например, для атомного ядра со спином I, магнитным моментом /// и гиро-
магнитным отношением yi, помещенного в магнитное поле Во! вектор ///
совершает прецессию с угловой частотой (Di ( (1)1 = 2 Л Vf). Эта частота тем
больше, чем больше гиромагнитное отношение yi и внешнее магнитное поле
Во, а именно: (Di = yi Во.
Если же на ядерный спин I кроме статического магнитного поля Во
воздействует еще и переменное магнитное поле Bi, перпендикулярное полю
Во, то в системе может наступить резонанс. Это происходит в случае, когда
частота (О этого переменного поля равна частоте прецессии СО/ магнитных
моментов ядер. В результате наблюдаем в системе очень сильное взаимодей-
ствие, даже если переменное поле В/ мало.
Резонансный эффект - явление, широко распространенное в окружаю-
щем мире. Например, маятник можно раскачать до очень большой амплиту-
ды, если оказывать воздействие на него в определенной точке "фазы
качания".
Простая математическая формула
С0= (Di = yi Во,
(1.3)
выражающая условие резонанса, является основным уравнением ЯМР.
Рассмотрим явление ЯМР с точки зрения квантовой механики. Большое
число малых магнитных диполей/С/, связанных с ядрами атомов, в отсутствие
магнитного поля полностью разупорядочены, т.е. ориентация их статисти-
чески равновероятна. Если к этой системе ядерных спинов приложить посто-
янное магнитное поле, то эти элементарные магнитики будут
ориентироваться относительно направления постоянного магнитного поля.
Согласно фундаментальному физическому закону, открытому Штерном и
ГерЛахом в 1923 г., такие частицы со спином 7=1/2 будут располагаться
относительно магнитного поля не произвольным образом, а либо вдоль, либо
против поля, т.е. магнитное квантовое число mi, характеризующее эту ори-
ентацию, будет равным либо +1/2, либо -1/2. Этот процесс называется
квантованием.
Обычно в ЯМР положение системы координат выбирают таким образом,
что направление внешнего магнитного поля Во совпадает с направлением оси
z. Математическое ожидание z-компоненты lz оператора спина / равно
< Iz> = mi.
(1.4)
16
Энергия Ещ магнитного диполя /л во внешнем магнитном поле Во в этом
случае дается выражением
Ет = -(Ц Во) = -Я yi mi Во.
(1.5)
Рис.1.2. Расщепление уровней энергии
ядерных спинов в статическом магнитном
поле для случая 1-1/2.
Дс
---------- N" = N+e kT
hv = ДЕ =tico = "yhB0
------------------ N+
Приэтом/г выражается через I согласно уравнению (1.1). Для частиц со
спином I — 1/2, помещенных во внешнее магнитное поле, существуют два
уровня с энергиями Ej и Е2 (рис.1.2):
Ei = -Я yi • Во,
(1.6)
Е2 = Я у/ • Во.
(1.7)
Как и в любом виде спектроскопии, переходы между уровнями энергии .
могут быть индуцированы в том случае, если энергия электромагнитных
квантов Яш равна разности энергий ДЕ, и частоты радиочастотного (РЧ) поля
Bi удовлетворяют условию
Я Ш= &Е = Яу/ Во.
(1.8)
Деление на Я дает основное уравнение ЯМР (1.3), выведенное исходя из
интуитивных соображений.
1.1.3. Уравнение Блоха
Число атомных ядер в макроскопическом образце весьма велико. В ку-
бическом сантиметре воды содержится 10 ядер атомов водорода, причем
микроскопические магнитные диполи этих ядер полностью разупорядочены.
Проведем мысленно следующий эксперимент: включим на короткое время
статическое магнитное поле Во; в этом случае спины ядер Н с I - 1/2
статистически распределятся относительно направления поля Во, так что
17
одна половина их ориентируется вдоль поля, а вторая - против. Однако при
этом спиновая система не находится в состоянии теплового равновесия с
окружением. Обозначим через 7V + число спинов I, z-компонента которых Iz
ориентирована вдоль поля Во, а через N — число спинов, ориентированных
против поля, и учтем, что в состоянии теплового равновесия с окружающей
средой отношение № к 1V+ подчиняется соотношению Больцмана
м- -&Е/кТ
, (1.9)
ЛГ
где к — постоянная Больцмана, Г - температура.
В этом случае уровень с меньшей энергией населен больше и, следова-
тельно, большая часть спинов ориентирована так, что их магнитные моменты
направлены вдоль магнитного поля Во. При комнатной температуре в полях
порядка 1 Тл (10 000 Гс) относительная разность населенностей всего лишь
порядка 10'6. Однако эта небольшая величина, приводящая к тому, что
спинов, ориентированных по полю, в 1 смл вещества примерно на 1О10 боль-
ше, чем против, позволяет провести измерение макроскопической намагни-
ченности.
В состоянии термодинамического равновесия результирующая макро-
скопическая намагниченность М направлена вдоль внешнего магнитного
поля Во. Величина намагниченности Мо для комнатных температур (так
называемое высокотемпературное приближение) может быть получена из
следующего уравнения:
N h2 I (I + 1 )
мо=------(МО)
где N = + N~ - полное число ядерных спинов, находящихся в единице
объема. Из формулы видно, что макроскопическая намагниченность возра-
стает с увеличением напряженности магнитного поля Во и гиромагнитного
отношения yi и убывает с ростом температуры Т. Такое поведение намагни-
ченности М определяет большое число эффектов, наблюдаемых в ЯМР.
Время установления теплового равновесия между спиновой системой и
окружающей средой, которую даже в жидкостях принято называть решет-
кой, определяется как время спин-решеточной релаксации. Эта величина
описывает процесс установления равновесия, т.е. приближение z-компонен-
ты намагниченности М. к равновесному значению Мо, которое устанавлива-
ется в спиновой системе спустя длительный период времени. Равновесная
намагниченность устанавливается параллельно внешнему магнитному полю
Во, поэтому спин-решеточную релаксацию называют также продольной ре-
лаксацией.
Непосредственно после открытия явления ЯМР Феликс Блох на основе
классического подхода описал поведение намагниченности М, которая ха-
рактеризуется компонентами Мх, Му и Mz, с помощью системы дифферен-
циальных уравнений. Эти уравнения называются уравнениями Блоха.
18
Уравнения Блоха позволяют достаточно просто описать основные экспери-
ментальные данные: 1) Если направления намагниченности и магнитного
поля в исходный момент не совпадают, то намагниченность совершает пре-
цессию относительно направления магнитного поля. 2) Спустя достаточно
длительный промежуток времени после воздействия возбуждения в системе
устанавливается равновесная намагниченность, компонента Mz которой
вдоль направления магнитного поля равна Мо, а поперечная компонента
намагниченности, перпендикулярная направлению внешнего магнитного
поля, равна нулю. Экспоненциальное приближение Mz к равновесному зна-
чению Мо описывается уравнением
dMz Мо — Mz
(1.11)
dt I\
Постоянная Т\ называется временем продольной релаксации.
Соответственно процесс распада поперечной намагниченности описыва-
ется уравнениями для Мх и Му.
dMx Мх
~dT=~l\
dM}
dt
(1.12)
Классическое уравнение движения, описывающее прецессию намагни-
ченности в магнитном поле без учета релаксации, имеет вид
=-у7 (В X М).
(1.13)
Уравнения Блоха получаются путем феноменологического введения в
уравнение (1.13) релаксационных слагаемых в форме правых частей урав-
нений (1.11) и (1.12):
dMx
-^ = -у7(ВХМ)х
(1.14)
-^(ВХМ),-^, (1.15)
dMz
—М/(вхм)г +
m0-mz
(1.16)
В типичном ЯМР эксперименте' наряду со статическим магнитным по-
лем Во, направленным вдоль оси z, имеется еще и переменное РЧ поле с
частотой (j), магнитная составляющая которого направлена перпендикуляр-
но полю Во, например, вдоль оси х, и осциллирует с частотой V = (и/2 Л. Это
поле 2 В] cos (О t, как правило, много меньше внешнего магнитного поля Во.
Линейно поляризованное переменное магнитное поле можно представить в
19
виде разложения по двум компонентам, которые вращаются в противопо-
ложных направлениях с круговыми частотами ± (D. Взаимодействие с ядер-
ным спином той компоненты поля, которая противоположна направлению
ларморовой прецессии, пренебрежимо мало, а взаимодействие компоненты,
вращающейся в направлении, совпадающем с направлением движения ядер-
ного спина, можно рассматривать в системе координат, вращающейся вместе
с этой компонентой, как взаимодействие со статическим магнитным полем.
Таким образом, магнитное поле В, оказывающее влияние на намагничен-
ность М, имеет вид
B = Bj cos (Dti + В] sin (Dt j + B0 k, (1.17)
где i, j и k - единичные векторы в направлении осей х, у иг.
Если подставить это выражение для В в уравнения Блоха, то получим
d Мх Мх
-^ = 'YI(MyB0-MzBjS\n(Bt)--^, (1.18)
d Mv Mv
= yj(M.BJ cos (1.19)
d M. M- Mz
= y7(MySj sin (Dt— MyBj cos (Dt) + ... (1.20)
Зачастую более удобным оказывается рассмотрение поведения намагни-
ченности М во вращающейся системе координат fxz, у', z = z), частота
вращения которой относительно оси z равна Шг Соответствующим преобра-
зованием координат можно не только формально упростить уравнения, но и
преобразовать их так, что они приобретут более наглядный вид. Сложное
движение вектора намагниченности в пространстве можно разложить на два
движения: движение во вращающейся системе координат и одновременное
движение этой системы координат относительно лабораторной системы ко-
ординат, фиксированной в пространстве. Обычно частоту вращения выбира-
ют равной частоте РЧ поля, (Dr = (D, так как в этом случае поле By во
вращающейся системе координат будет неподвижным. Обозначим когерен-
тную компоненту намагниченности вдоль оси х (параллельной В у) через
М х' , а сдвинутую на 90° вдоль оси у - через М ':
М х'= Мхcos (Dt-My sin(Dt, (1.21)
М у' = +МХ sin (Dt + Му cos (D t. (1.22)
У равнения Блоха во вращающейся системе координат принимают сле-
дующий вид:
d Мх< М'
—г-----(СО7-СО) М , (1.23)
tt I j i 2
20
Рис.1.3. Магнитные поля в лабораторной и во вращающейся сис-
темах координат.
d Му' Му'
—Ty—-(a)j-a))Mx'+yJBiMz--^-, (1.24)
в
(1.25)
где 60/определена формулой (1.3).
С помощью рис. 1.3 попытаемся построить простое представление о про-
цессах, происходящих во вращающейся системе координат. Так как система
координат (х' , у', z = z) вращается с круговой частотой со, равной частоте
поля Bi, то поле Bi будет неподвижным в этой системе координат. При этом
удобно кроме суммарного поля Вг, складывающегося из полей Во и Bi,
определить еще и эффективное поле Beff, которое является векторной сум-
мой полей к (СО/ -СО) / у/и i ' Bi. Можно показать, что уравнения, описываю-
щие затухающее движение и прецессию спинов в этом эффективном
магнитном поле, имеют вид (1.14)-(1.16). Особенно простым будет вид этих
уравнений, если 60 = СО/ и эффективное поле Beff равно полю i ' В\. В этом
случае частота прецессии СО/' формально удовлетворяет условию резонанса
(1.3), в котором вместо Во используется В]:
со/'= у/В\. (1.26)
Если вначале вектор намагниченности направить вдоль оси z, и вклю-
чить кратковременно РЧ поле, например, на время 1, то вектор намагничен-
ности отклонится на некоторый угол в направлении оси у ' в плоскости у ' z,
а затем вновь возвратится к оси z. Если ВЧ поле отключается в момент
времени, когда вектор намагниченности расположится строго вдоль оси у ',
то говорят, что на систему воздействует 90°-ный или я/2-импульс. Если при
той же напряженности магнитного поля выбрать длительность РЧ импульса
такую, что вектор намагниченности отклонится от оси z в плоскости у ' z на
180°, то такой импульс называется 180°-ным или Я-импульсом (рис. 1.4). В
общем случае путем соответствующего выбора Beff и длительности импульса
можно развернуть вектор намагниченности в плоскости у' z в произвольном
направлении.
21
Рис.1.4. Прецессия вектора намагниченности, (а) Прецессия намагниченности относительно
поля В । в системе координат, вращающейся со скоростью (1) — (О[. (Ь) Прецессия намагниченно-
сти М в лабораторной системе координат после выключения радиочастотного поля В ।. Показан-
ное на рис. направление движения (прецессии) справедливо для ядерных спинов с
положительным гиромагнитным отношением.
Если под действием РЧ импульса намагниченность отклонится от оси z
(т.е. от равновесного положения), то после выключения РЧ импульса
намагниченность, в результате появления у нее поперечных компонент,
начнет прецессировать вокруг направления поля Во. Прецессия намагни-
ченности создает модуляцию во времени связанного с этой намагниченно-
стью магнитного поля. Если мы поместим образец в приемную катушку,
то изменяющееся во времени магнитное поле создаст малое индукционное
напряжение, которое может быть зарегистрировано с помощью соответст-
вующих методов. Амплитуда этого сигнала пропорциональна резонансной
частоте (O{ и намагниченности Мо; затухание сигнала во времени называ-
ют спадом свободной индукции (ССИ, free induction decay, FID).
1.1.4. Спин-решеточная релаксация
Изменение во времени намагниченности Мг может быть описано урав-
нениями Блоха [1.1 ]. Решением этих уравнений для Мг является экспонен-
циальная функция с характерным временем Tt, которое называется
временем продольной или спин-решеточной релаксации:
-t/т,
Mz(t) = М0- {Мо - Mz(o)}e . (1.27)
Если после воздействия радиочастотного импульса спиновая система
свободно эволюционирует, то она стремится к состоянию больцмановского
равновесия. В частности, после воздействия 180°-ного импульса, приводяще-
22
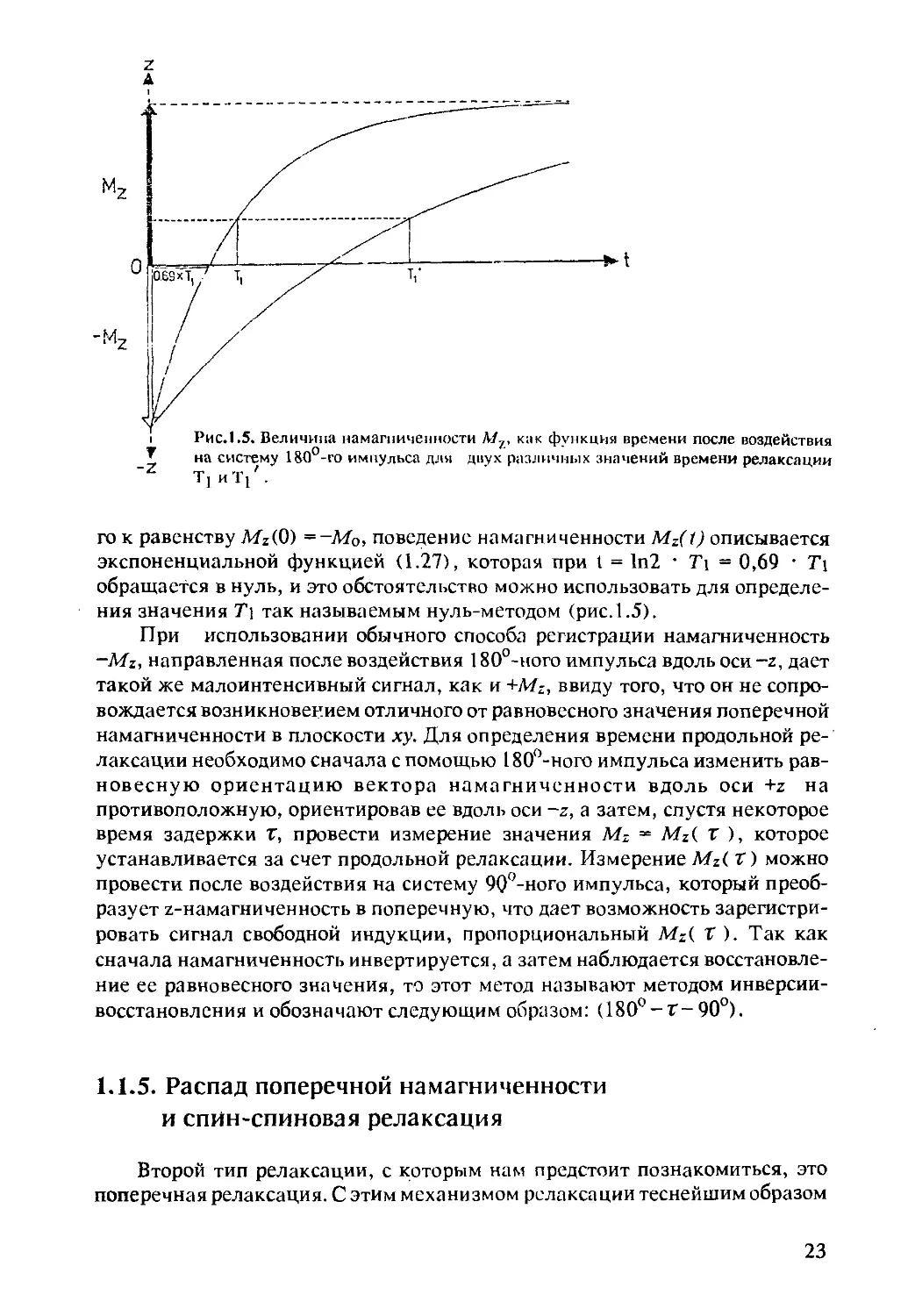
го к равенству tWz(0) = ~М0, поведение намагниченности Mz( 1) описывается
экспоненциальной функцией (1.27), которая при I = 1п2 • Т\ = 0,69 • Ti
обращается в нуль, и это обстоятельство можно использовать для определе-
ния значения Т\ так называемым нуль-методом (рис. 1.5).
При использовании обычного способа регистрации намагниченность
-Mz, направленная после воздействия 180°-ного импульса вдоль оси -z, дает
такой же малоинтенсивный сигнал, как и +MZ, ввиду того, что он не сопро-
вождается возникновением отличного от равновесного значения поперечной
намагниченности в плоскости ху. Для определения времени продольной ре-
лаксации необходимо сначала с помощью 180°-ного импульса изменить рав-
новесную ориентацию вектора намагниченности вдоль оси +z на
противоположную, ориентировав ее вдоль оси -z, а затем, спустя некоторое
время задержки Г, провести измерение значения Mz = Mz{ г ), которое
устанавливается за счет продольной релаксации. Измерение Mz( Т) можно
провести после воздействия на систему 90°-ного импульса, который преоб-
разует z-намагниченность в поперечную, что дает возможность зарегистри-
ровать сигнал свободной индукции, пропорциональный Mz( Т ). Так как
сначала намагниченность инвертируется, а затем наблюдается восстановле-
ние ее равновесного значения, то этот метод называют методом инверсии-
восстановления и обозначают следующим образом: (180°-Т-90°).
1.1.5. Распад поперечной намагниченности
и спин-спиновая релаксация
Второй тип релаксации, с которым нам предстоит познакомиться, это
поперечная релаксация. С этим механизмом релаксации теснейшим образом
23
РЧ импульс
90°
90° 180°
намагниченность М
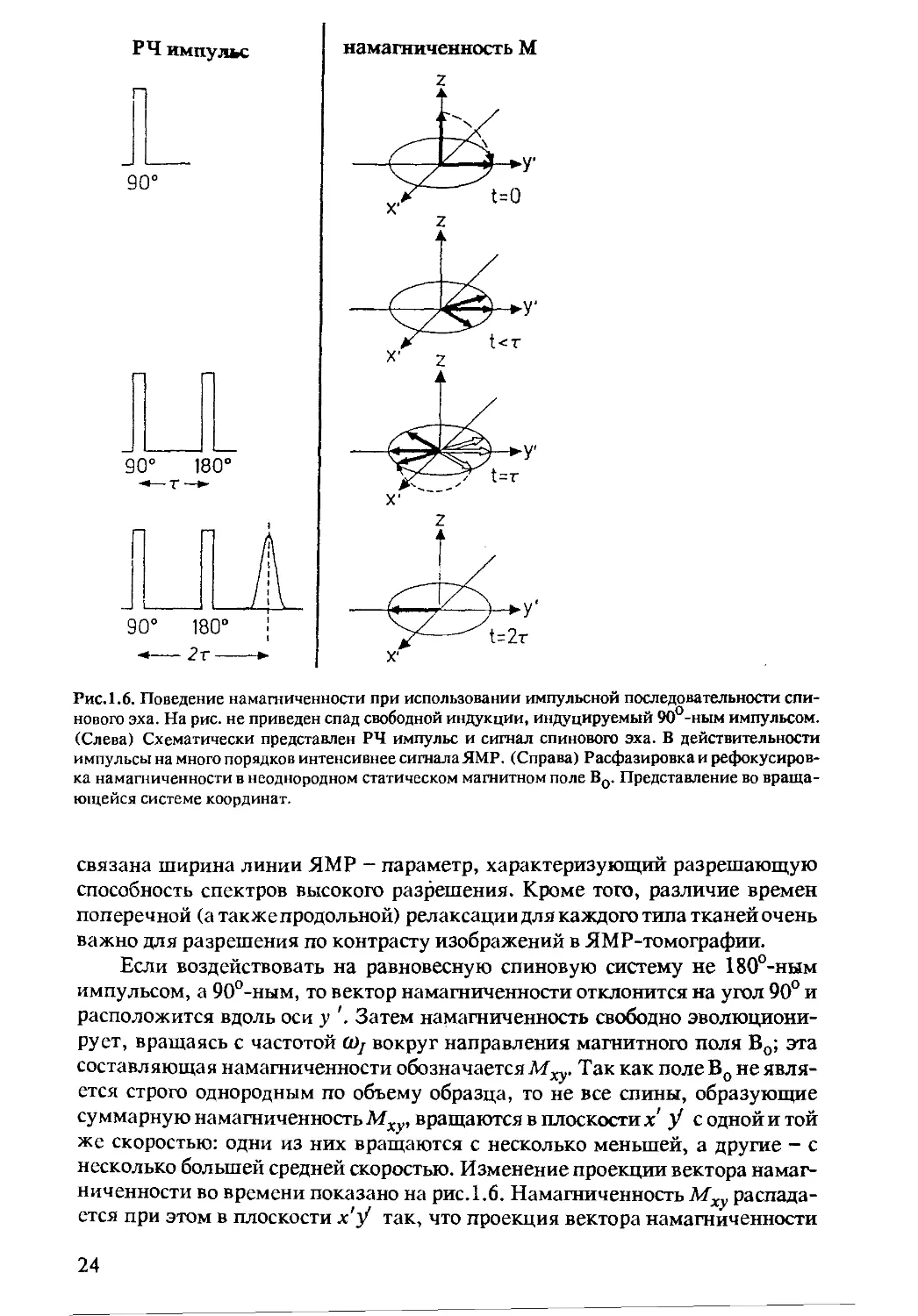
Рис.1.6. Поведение намагниченности при использовании импульсной последовательности спи-
нового эха. На рис. не приведен спад свободной индукции, индуцируемый 90°-ным импульсом.
(Слева) Схематически представлен РЧ импульс и сигнал спинового эха. В действительности
импульсы на много порядков интенсивнее сигналаЯМР. (Справа) Расфазировка и рефокусиров-
ка намагниченности в неоднородном статическом магнитном поле Во. Представление во враща-
ющейся системе координат.
связана ширина линии ЯМР - параметр, характеризующий разрешающую
способность спектров высокого разрешения. Кроме того, различие времен
поперечной (атакжепродольной) релаксации для каждого типа тканей очень
важно для разрешения по контрасту изображений в ЯМР-томографии.
Если воздействовать на равновесную спиновую систему не 180°-ным
импульсом, а 90°-ным, то вектор намагниченности отклонится на угол 90° и
расположится вдоль оси у '. Затем намагниченность свободно эволюциони-
рует, вращаясь с частотой (Dj вокруг направления магнитного поля Во; эта
составляющая намагниченности обозначается Мху. Так как поле Во не явля-
ется строго однородным по объему образца, то не все спины, образующие
суммарную намагниченность Мху, вращаются в плоскости х' У с одной и той
же скоростью: одни из них вращаются с несколько меньшей, а другие - с
несколько большей средней скоростью. Изменение проекции вектора намаг-
ниченности во времени показано на рис. 1.6. Намагниченность Мху распада-
ется при этом в плоскости х'У так, что проекция вектора намагниченности
24
в этой плоскости принимает все возможные значения, а средняя амплитуда
поперечной намагниченности и соответственно сигнал в приемной катушке
обращаются в нуль. Обычно группу сигналов, которые характеризуются
одинаковой скоростью вращения, называют спиновым пакетом. Если, спустя
время Г, на спиновую систему воздействует 180°-ный импульс, то все спино-
вые пакеты изменяют свое положение (фазу прецессии вокруг Во) таким
образом, что самые "медленно движущиеся" пакеты оказываются впереди, а
самые "быстро движущиеся" - позади. Это подобно бегу группы бегунов на
длинные дистанции, движущихся по гаревой дорожке при условии, что все они
стартуют одновременно. Спустя некоторое время бегуны распределяются вдоль
дорожки таким образом, что бегущие с большой скоростью оказываются более
удаленными от исходной точки, чем бегущие с меньшей скоростью. После
команды "кругом" бегущие с большей скоростью оказываются позади, и, если в
дальнейшем они будут двигаться с прежней скоростью, то достигнут исходной
точки одновременно с бегунами, движущимися с меньшей скоростью. Совер-
шенно аналогично вели бы себя спиновые пакеты, движущиеся с различными
скоростями, если бы направление внешнего магнитного поля мгновенно изме-
нилось. Отметим, однако, что такое мгновенное изменение направления поля
технически достаточно сложно осуществимо. Подобный эффект достигается под
действием 180°-ного импульса в момент времени X после 90°-ного импульса,
когда положение спиновых пакетов (фазы прецессии) изменится таким обра-
зом, что спустя время 27 после воздействия 90°-ного импульса или X после
180°-ного более быстрые спины догонят спины, движущиеся более медленно,
т.е. все спиновые пакеты будут иметь одинаковую фазу прецессии. Таким
образом в плоскости л/У вновь образуется результирующая намагниченность,
которая формирует сигнал, названный по имени открывшего этот эффект Эр-
вина Хана эхом Хана или спиновым эхом. Результирующее эхо имеет знак,
противоположный знаку спада свободной индукции, образовавшегося после
воздействия 90°-ного импульса, т.е. сигнал эха сдвинут на 180° относительно
сигнала, полученного после преобразования спада свободной индукции. Этого
можно избежать, сдвинув фазу 180°-ного импульса на 90°. Для этого в системе
координат, движущейся со скоростью (У/, поле В,, создающее 90°-ный импульс,
расположим вдоль оси х , а поле, создающее 180°-ный импульс - вдоль оси у'.
Тогда вращение вокруг оси у' оказывает влияние только на те спиновые пакеты,
которые совершают прецессию со скоростью большей или меньшей чем (У/, т.е.
на те пакеты, которые испытывают дефазировку под действием неоднородности
магнитного поля.
Кроме обратимого процесса дефазировки, который возникает вследст-
вие неоднородности магнитного поля Во, существует еще и необратимый
процесс, вызывающий дефазировку спинов за счет их взаимодействия между
собой. В то время, как дефазировка, возникающая за счет неоднородности
магнитного поля, является функцией неизменного во времени распределе-
ния поля, уменьшение намагниченности в плоскости х' у проходит под дей-
ствием необратимого процесса и описывается уравнением Блоха, согласно
которому убывание соответствующей проекции намагниченности описыва-
ется экспоненциальной функцией ехр(-//Т’2). Так как эта дефазировка вы-
звана спин-спиновым взаимодействием, то характерное время Т2,
25
описывающее этот процесс, называется временем спин-спиновой или попе-
речной релаксации. В эксперименте спинового эха спин-спиновое взаимо-
действие приводит к уменьшению амплитуды эха, зависящему от избранного
интервала времени Т между 90°-ным и 180°-ным импульсами, причем это
уменьшение описывается экспоненциальной функцией с характерным вре-
менем Т2- Измерение амплитуды эха как функции интервала между 90°-ным
и 180°-ным импульсами дает возможность определять Т2 в неоднородных
магнитных полях.
180°
Т"
Рис.1.7. Уменьшение амплитуды сигнала спи-
нового эха в зависимости от времени задержки
Т. Спад сигнала спинового эха определяется
временем спин-спиновой релаксации Т2- В
противоположность этому, процесс дефази-
ронки и рефокусировки отдельных спиновых
изохромат и первого Эха в основном определя-
ется неоднородностью магнитного поля.
Следует отметить, что определение значений Т2 с помощью этого метода
является достаточно трудоемкой задачей, поскольку при проведении экспе-
риментов приходится снимать множество спектров для различных значений
т (рис. 1.7). Для усовершенствования метода проведения измерений была
разработана другая импульсная последовательность - последовательность
Карра-Парселла [1.9 которая в дальнейшем была улучшена Мейбумом и
Гиллом [1.10 ]. После 90°-ного импульса на спиновую систему воздействует
серия 180°-ных импульсов, между которыми всякий раз проводится измере-
ние амплитуды спинового эха (рис. 1.8). Основным преимуществом импуль-
сной последовательности спинового эха Карра-Парселла-Мейбума-Гилла
является то, что полное измерение Т2 может быть проведено с помощью
одной импульсной последовательности. Кроме того, этот метод позволяет
проводить более точное измерение Т2, так как при использовании простой
импульсной последовательности спинового эха процессы диффузии могут
оказывать существенное влияние на точность измерений. Как будет показа-
но в главе 4, в настоящее время последовательность Карра-Парселла-Мей-
бума-Гилла приобретает особое значение для ЯМР-томографии.
26
90*х 180°у 180“у 180°у 180”у ВО”у 180"у 180”у 180“у
[t-2T +
Рис.1.8. Поведение сигнала при воздействии
импульсной последовательности Карра-
Парселла-Мейбума-Гилла (КПМГ)
1.2. Спектр ЯМР
Принцип проведения ЯМР-эксперимента можно объяснить исходя из
представлений об условии резонанса, а также о поперечной и продольной
релаксации, что в свою очередь способствует разработке специального аппа-
рата, используемого для описания эксперимента. Эти сведения достаточны
также и для того, чтобы иметь возможность описания принципов ЯМР-то-
мографии, пространственное разрешение которой определяется величиной
градиентов магнитного поля, а разрешение по контрасту - различиями в
значениях времен релаксации. ЯМР можно использовать также как анали-
тический метод, основываясь на том, что различные элементы и изотопы
обладают различными резонансными частотами. Однако для успешного при-
менения этого метода в химии и биохимии этой информации недостаточно.
Только включение дополнительных физических взаимодействий, приводя-
щих к расщеплению резонансных линий или к сдвигу соответствующих
уровней энергии ядерного спина и соответствующих частот переходов, по-
зволяет использовать ЯМР в качестве аналитического метода. В этом случае
вместо одной резонансной линии для определенного изотопа получим в спек-
тре несколько резонансных линий, положение которых в спектре связано со
свойствами молекул. В дальнейшем обсудим основные типы указанных выше
физических взаимодействий.
1.2.1. Химический сдвиг
Условие резонанса в том виде, как оно было сформулировано в уравне-
нии (1.3), строго говоря, справедливо только для ядер атомов, образующих
идеальный газ. В реальной ситуации ядра атомов находятся в веществе, как
части молекул, и окружены электронной оболочкой. Именно эта оболочка
частично экранирует атомные ядра от внешнего поля Во, так что атомные
ядра будут находиться не в поле Во, а в эффективном поле
27
В' = В0 (1 —О'),
(1.28)
где о- константа экранирования. Поэтому в основное уравнение резонанса
необходимо подставить значение эффективного поля В':
(O = 0)i = yiB' = yiB0 ( I-О'). (1.29)
Так как экранирующее действие электронной оболочки атома, а следо-
вательно, и данный сдвиг резонансной частоты зависит от химического ок-
ружения, в котором находится рассматриваемое атомное ядро, то это
изменение резонансного условия называют химическим сдвигом. Величина
химического сдвига д измеряется относительно положения линии, соответ-
ствующей стандартному веществу S, в миллионных долях (м.д.). Безразмер-
ная константа д определяется как разность констант экранирования о для
данного типа ядер в исследуемом и стандартном веществах или как относи-
тельная разность соответствующих резонансных частот (л) или V во внешнем
магнитном поле Во (с учетом того, что и<< 1):
<5 = О1 - as ~-s^~ . (1.30)
Здесь (Ji и (Js - константы экранирования, а V/ и Vs - резонансные
частоты исследуемого вещества и вещества, используемого в качестве стан-
дарта, соответственно. Положительное значение д означает, что резонанс-
ная линия ядер данного элемента в исследуемом веществе сдвинута в область
более низких частот по сравнению с резонансной частотой, на которой на-
блюдается линия стандарта.
Диапазон значений химических сдвигов для ядер 1Н, которые экрани-
рованы лишь одним электроном, достаточно мал; максимальное значение
химического сдвига равно 10 м.д. Для ядер |3С и 31Р сдвиг может составлять
сотни м.д. Несмотря на то, что для протонов эта величина мала, химический
сдвиг является важнейшим параметром ЯМР высокого разрешения: этот
метод позволяет наблюдать сигналы ЯМР ядер или группы ядер одного хи-
мического элемента, но в различном химическом окружении, так что из
величины химического сдвига можно получить информацию о химическом
окружении данного ядра. Вследствие наличия химического сдвига уже в
одном из первых спектров (см. рис. 1.1) можно было достаточно просто раз-
личить сигналы от протонов СНз-, СНг- и ОН-групп в этаноле. Однако
химический сдвиг не является единственным параметром, определяющим
вид спектра ЯМР. Спектр каждого атомного ядра также отражает взаимодей-
ствие магнитных моментов соседних ядер.
1.2.2. Диполь-дипольное взаимодействие
Взаимодействие магнитных моментов соседних атомных ядер в принци-
пе всегда приводит к расщеплению линий поглощения. Влияние магнитного
28
диполя /Л] на соседний диполь состоит в появлении дополнительного магнит-
ного поля; ядерный спин соседнего ядра ведет себя таким образом, как будто
он находится во внешнем магнитном поле Во и дополнительном локальном поле
Bloc, компонента которого, параллельная полю Во, дается выражением
Цо cos2 0- 1 )
В1ОС=4Л г3
(1.31)
Здесь г — расстояние между диполями, /zz - компонента магнитного
момента/z/, параллельнаяВ0,0-угол между магнитным полем Во и вектором
г, который соединяет оба диполя. Как видно из уравнения (1.31), диполь-ди-
польное взаимодействие убывает достаточно быстро, как третья степень
расстояния между двумя магнитными диполями. Кроме того, эта величина
анизотропна и при cos2 0= 1/3 обращается в нуль. Соответствующий угол 0,
значение которого равно примерно 55°, называют магическим углом. При
других значениях угла 0 в твердых телах, как в монокристаллах, так и в
поликристаллических материалах, наблюдается расщепление резонансных
линий. Однако поскольку кроме взаимодействия с ближайшими соседними
ядрами существует еще и более слабое взаимодействие с другими ядрами в
данной молекуле, а также взаимодействие ядер, относящихся к различным
молекулам, в общем случае в поликристаллических или аморфных телах
наблюдается не расщепление, а лишь уширение линий поглощения. Как
следует из уравнения (1.31), наличие зависимости величины диполь-ди-
польного взаимодействия от расстояния между спинами позволяет опреде-
лять расстояние между двумя ядрами, что особенно важно для ядер 1Н, так
как при установлении структуры молекул с помощью рентгеноструктурного
анализа точность определения этого параметра невелика.
В отличие от твердых тел диполь-дипольное взаимодействие в жидко-
стях не приводит к возникновению расщепления линий. Броуновское дви-
жение молекул вызывает в жидкостях быструю переориентацию молекул
относительно направления поля и усредненное по времени значение Bloc
равно нулю. Следует отметить, однако, что для молекул, представляющих
интерес с точки зрения биологии, это тепловое движение не будет доста-
точно быстрым для того, чтобы наблюдалось усреднение данного взаимо-
действия до нуля, и остаточное взаимодействие приводит к уширению
линий.
1.2.3. Косвенное спин-спиновое взаимодействие
Ранее отмечалось, что диполь-дипольное взаимодействие не вызывает
никакого расщепления резонансных линий в спектрах ЯМР жидкостей, од-
нако во многих случаях наблюдается расщепление, обусловленное непря-
мым спин-спиновым взаимодействием, которое передается через электроны
связи. Непрямое (или косвенное) спин-спиновое взаимодействие называют
еще скалярным, так как оно не зависит от ориентации спинов. Механизм
29
этого взаимодействия можно достаточно наглядно пояснить на примере пла-
виковой кислоты HF.
Если в некоторый момент времени одни из двух электронов связи «1
удерживается у протона А, то вероятность того, что спин электрона е1 ори-
ентируется антипараллельно относительно спина А, будет выше, чем веро-
ятность параллельной ориентации, поскольку электрон обладает
отрицательным магнитным моментом, а протон положительным; для ej и
ядра фтора X справедливо то же, что и для е1 я А. Однако спины обоих
электронов в соответствии с принципом Паули должны ориентироваться
антипараллельно один относительно другого. Таким образом устанавлива-
ется связь между ядрами Л и X. В такой связанной системе энергия состояния
зависит от взаимной ориентации обоих ядерных спинов; это проявляется в
расщеплении уровня энергии (рис. 1.9). Возникающее при этом различие в
значениях частот переходов приводит к наблюдению двух резонансных ли-
ний, т.е. к расщеплению линий.
В магнитном поле, В магнитном поле
В отсутствие н0 отсутствует
магнитного поля j-взаимодействие
при наличии
J-взаимодействия
•И >-А
ЕА »ЕЛ * l/4hJAX
ЕА - Ех -1/4 h JAX
— - ЕА* Ех-l/4hJAX
—-EA-EX*WhJAX
И
Рис. 1.9. Расщепление уровней энергии, возникающее под влиянием косвенного спин-спинового
взаимодействия (J-взаимодействия). Представленное на рис. дополнительное расщепление
уровней энергии за счет взаимодействия между ядрами А и X не соответствует реальному
масштабу. Обычно величина этого расщепления, намного меньше зеемановского расщепления,
возникающего под действием внешнего магнитного поля. Резонансные частоты для спинов А и X
обозначены (>-----) и ( соответственно.
Скалярное взаимодействие не зависит от величины и направления маг-
нитного поля Во, а его сила измеряется константой косвенного спин-спино-
вого взаимодействия или константой скалярного взаимодействия и
обозначается J. Эту величину, имеющую размерность энергии, как правило,
приводят в единицах частоты (Гц). Значение J убывает с ростом числа
химических связей п и при п > 3, как правило, настолько мало, что не
поддается измерению. На рис. 1.10 в качестве примера проявления косвенно-
го спин-спинового взаимодействия с константой связи J схематически при-
веден спектр несколько более сложного, чем HF, вещества PF3. В спектре
ЯМР ядра F наблюдаются две линии одинаковой интенсивности; расщеп-
ление связано с взаимодействием ядра 19F с ядром 31Р, спин которого/= 1/2.
30
Рис.1.10. СпектпЯМР для молекулы PF3. На рис. схематически представлены соответствующие
ЯМР-спектры l’F (а) и 3(Р (Ь). Значения резонансных частот для 1 г и 31Р, конечно, будут
различны, а расщепления резонансных линий совпадают друг с другом.
Квартет, наблюдаемый в спектре 31Р, возникает вследствие взаимодействия
спина ядра 31Р с суммарным спином трех эквивалентных ядер 19F, равным
3/2. Расстояние между ближайшими компонентами данного квартета будет
тем же самым, что и между компонентами дублета в спектре 1 г, и равно
константе спин-спинового взаимодействия, выражаемой в единицах часто-
ты: J = 1,44 кГц. Соотношение интенсивностей в квартете 1:3:3:1 соответст-
вует биномиальным коэффициентам. Как HF, так и PF3 являются
примерами так называемых слабосвязанных систем, для которых значение
J будет существенно меньше разности значений резонансных частот. Рас-
щепление линий и соотношение интенсивностей подчиняются довольно про-
стым правилам. Взаимодействие между ядрами одного сорта, находящихся в
химически эквивалентных положениях, подобно трем ядрам фтора в PF3
вообще не вызывает никакого расщепления резонансных линий. Если же
ядра не являются магнитно и химически эквивалентными, то мультиплет-
ность линий в спектрах легко можно предсказать, исходя из следующих
соображений: связь ядра А с ядром X со спином 1= 1 /2 приводит к появлению
в спектрах двух линий равной интенсивности, расстояние между которыми
равно JAX. Если же в спиновой системе имеется еще один спин, например М,
участвующий в спин-спиновом взаимодействии, то каждая из компонент
дублета расщепляется в дублет с константой J Особенно прост характер
расщепления в случае эквивалентных ядер, например в PF3, так как здесь
все константы равны. При взаимодействии ядра А с п эквивалентными
ядрами X в спектре системы АХп получаем (п+1) резонансную линию с
расстоянием JАХ между ближайшими линиями; распределение интенсивно-
стей внутри такого мультиплета подчиняется отношению биномиальных
31
коэффициентов. Если в спиновой системе имеются ядра со спином Z, который
больше 1/2, то вместо дублета получим мультиплет, состоящий из (2nZ+D
линий. Для дейтерия (Z = 1), например, в спектре наблюдается триплет,
причем соотношение интенсивностей в каждом конкретном случае можно
рассчитать по таблице (табл. 1.2).
Таблица!.?. Количество (2nl + 1) и интенсивности резонансных линий при взаимодейстивии п
ядер со спином I
- 1/2
13 6 7 6 3 1
1 4 10 16 19 16 10 4 1
1111
1 2 3 4 3 2 1
1 3 6 10 12 12 10 6 3 1-
1 4 10 20 31 40 44 40 31 20 10 4 1
1 - 5/2 111111
12345654321
13 6 10 21 25 27 27 25 21 15 10 6 3 1
1 4 10 20 35 56 80 104 125 140 146 140 125 104 80 56 35 20 10 4 1
Если разность значений резонансных частот ядер, связанных косвенным
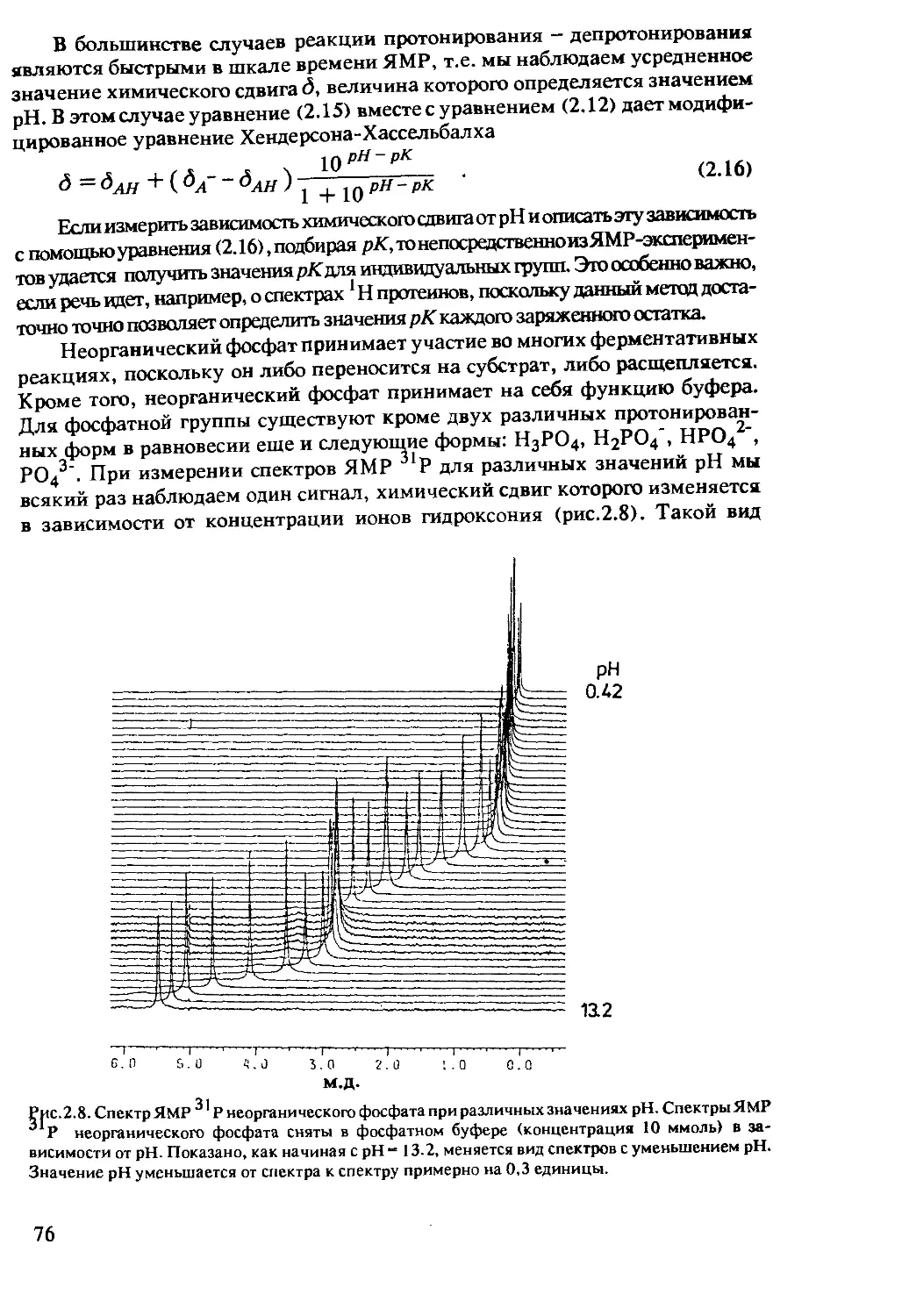
спин-спиновым взаимодействием, сравнима по величине с константой этого
взаимодействия (случай сильной связи), то мультиплетность линий в спек-
тре уже не будет подчиняться этим простым правилам. К обсуждению этого
вопроса вернемся в главе 2.
В случае PF3 взаимодействие между фосфором и фтором достаточно
велико, порядка нескольких кГц. Для ядер константа спин-спинового
взаимодействия существенно меньше. Так, в алифатическом фрагменте кон-
станта взаимодействия l3C — *Н составляет примерно 125 Гц. В этаноле, где
наблюдаются взаимодействия между протонами СН2- и СНз-групп, разде-
ленных тремя связями, константа порядка 7 Гц.
1.2.4. Электрон-ядерное взаимодействие
Магнитный резонанс атомных ядер отражает кроме взаимодействия с
соседними атомными ядрами еще и взаимодействие с магнитными момента-
ми [is неспаренных электронов, спин которых равен 1/2. В отличие от спина
ядер Iспин электронов, как правило, обозначают буквой S. Такие неспарен-
ные электроны имеются в свободных радикалах, а также в ряде ионов, кото-
рые либо входят в состав большого числа природных биологически важных
молекул, либо могут с помощью специальных химических методов вводиться
в эти молекулы (спиновые метки). В силу того, что магнитный момент
электрона Дз почти в 1000 раз больше ядерного магнитного момента Д/ (для
32
протонов/zs/pi “ 657, а для других ядер это отношение существенно больше,
поскольку их магнитные моменты jLij меньше, чем у протонов), это взаимо-
действие весьма велико. В основе этого взаимодействия лежат два механиз-
ма. Во-первых, это уже указанное выше классическое диполь-дипольное
взаимодействие, описываемое уравнением (1.31), с тем отличием, что в
числителе вместо jUi взята существенно большая величина - магнитный
момент электрона^us- Во-вторых, существует также и взаимодействие иного
типа - контактное взаимодействие Ферми. Электрон в молекуле или ионе
характеризуется определенным пространственным распределением зарядо-
вой плотности или определенным значением вероятности пребывания в каж-
дой точке пространства, характеризуемой соответствующей молекулярной
орбиталью. Если эта величина в точке, где находится ядро, имеет конечное
значение, то говорят, что между этим ядром и электроном имеет место
контактное взаимодействие. Это взаимодействие изотропно и описывается
скалярной константой а, обычно выражаемой в единицах частоты (Гц).
В случае магнитного резонанса электронного спина, электронного пара-
магнитного резонанса (ЭПР), связь спина электрона с магнитным моментом
атомного ядра приводит к весьма сложному расщеплению, которое называ-
ется сверхтонкой структурой спектра ЭПР. В ЯМР соответствующее расщеп-
ление резонансных линий, как правило, не возникает, так как вследствие
быстрой спин-решеточной релаксации электронных спинов скорость перехо-
дов между спиновыми состояниями, соответствующими ориентациям спина
по полю и против поля (т.е. между состояниями, характеризуемыми магнит-
ными квантовыми числами ms = 1/2 и -1/2), так велика, что ядерный спин
"видит" некое усредненное состояние. Однако поскольку всегда несколько
больше магнитных моментов электронов ориентировано по полю, чем против
поля, аналогично тому, как это ранее было показано для магнитных момен-
тов ядер /4/, то возникающий при этом результирующий электронный маг-
нитный момент является причиной наблюдаемых парамагнитных свойств
веществ, содержащих свободные радикалы и парамагнитные ионы: взаимо-
действие ядерного спина с электронным приводит к парамагнитному сдвигу
сигналов ЯМР, и, кроме того, включается дополнительный механизм релак-
сации, к рассмотрению которого вернемся в разделе 1.3.7.
1.2.5. Ядерное квадрупольное взаимодействие
Все ядра, спин которых J > 1, кроме магнитного момента обладают еще
и электрическим квадрупольным моментом е Q, который характеризует то,
насколько сильно распределение положительного заряда ядра отличается от
*При низких температурах, когда электронное время релаксации увеличивается, лииии
ЯМР ядер, окружающих парамагнитный центр, расщепляются. При этом зачастую а > Ябйу и
спектр усложняется. Как правило, такие спектры регистрируют методом двойного электрон-
ядерного резонанса. Другой случай проявления сверхтонкого взаимодействия в спектрах ЯМР
реализуется в магнитно-упорядоченных соединениях - ферро-, ферри- и антиферромагнетиках
(примечание редактора перевода).
33
сферического. При этом, если е — элементарный заряд, а е Q = J
г2 (3 cos2 &-1) p,roeQ>0 для продолговатых, сигарообразных ядер и е Q <
О для сплющенных, дискообразных ядер <р — зарядовая плотность, 0 - угол
между радиусом-вектором г и осью квантования спина).
В кристаллах, элементарная ячейка которых отличается от кубической,
почти во всех молекулах ядра находятся в неоднородном электрическом
поле. Соответствующий градиент поля задается второй производной потен-
циала Vпо пространственным координатам. Наибольшая компонента гради-
ента поля, которая совпадает с направлением химической связи, равна eq =
= д2 V/ д z. В таком неоднородном электрическом поле квадрупольное ядро
может принимать несколько определенных ориентаций относительно гради-
ентов поля, каждой из которых соответствует дискретное значение энергии.
Положение уровня энергии определяется как e“Qq и является произведением
квадрупольного момента eQ и градиента поля eq; это произведение непосред-
ственно измеряется в эксперименте.
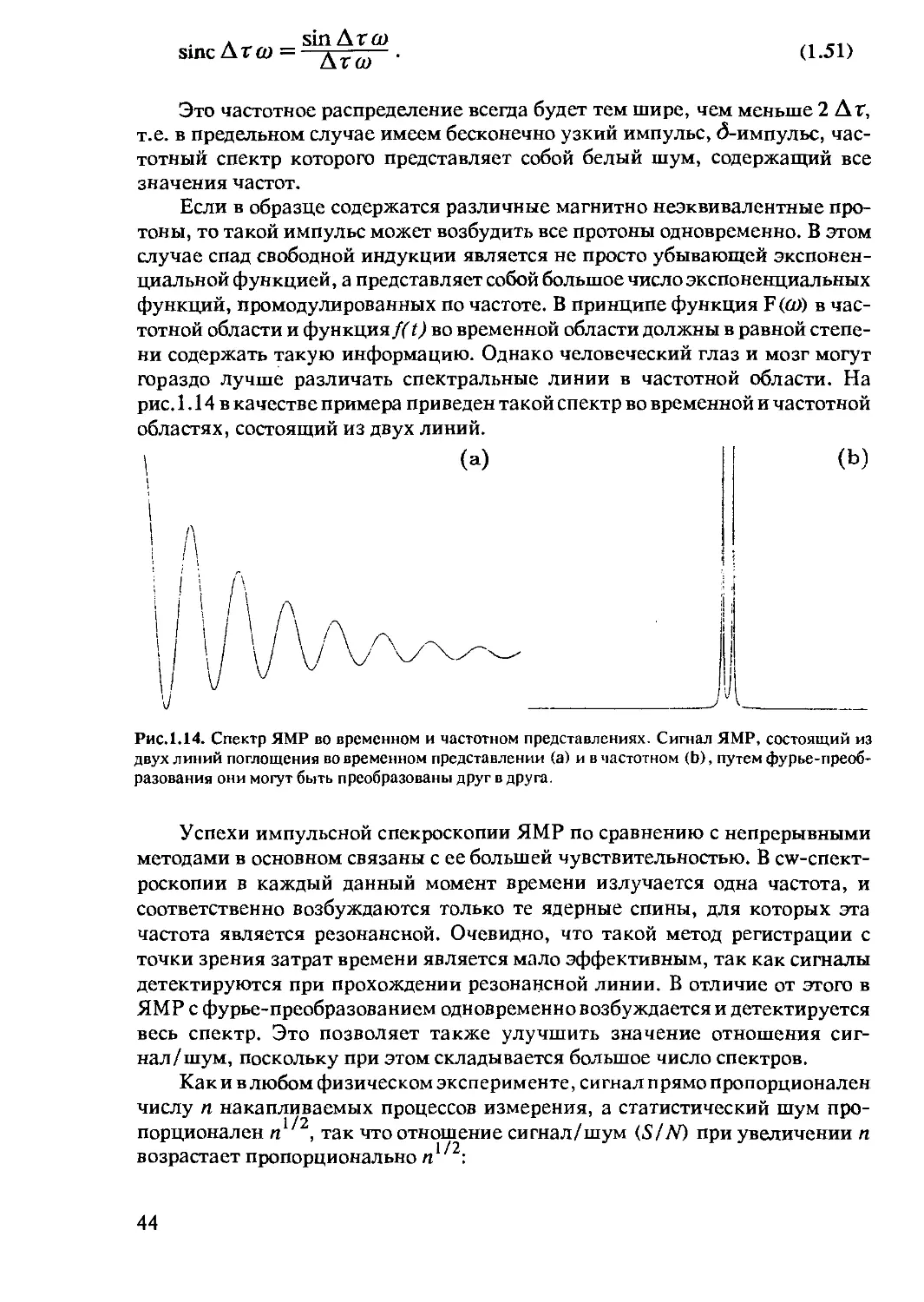
Ш (L)
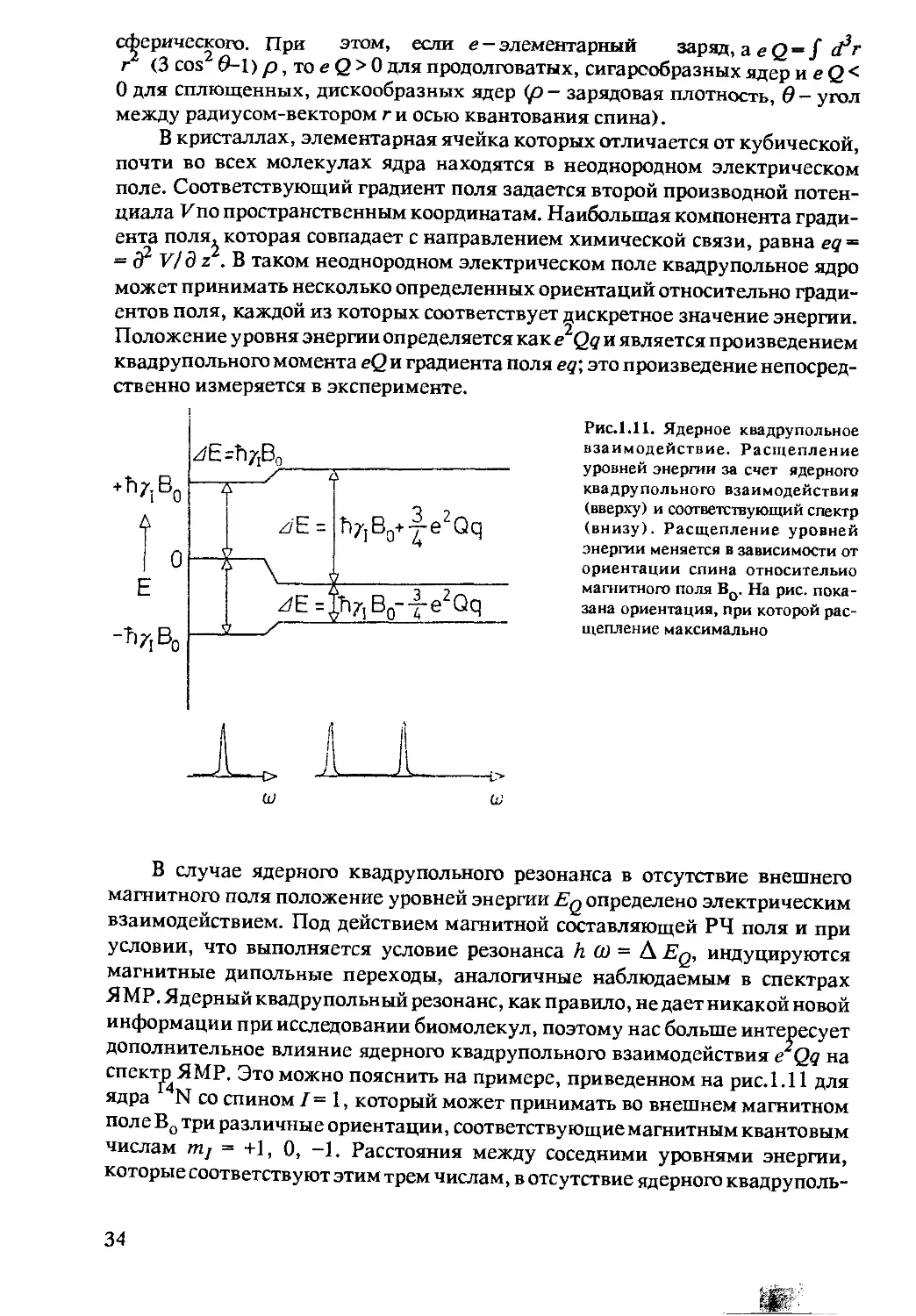
Рис.1.11. Ядерное квадрупольное
взаимодействие. Расщепление
уровней энергии за счет ядерного
квадрупольного взаимодействия
(вверху) и соответствующий спектр
(внизу). Расщепление уровней
энергии меняется в зависимости от
ориентации спина относительно
магнитного поля Во. На рис. пока-
зана ориентация, при которой рас-
щепление максимально
В случае ядерного квадрупольного резонанса в отсутствие внешнего
магнитного поля положение уровней энергии Eq определено электрическим
взаимодействием. Под действием магнитной составляющей РЧ поля и при
условии, что выполняется условие резонанса h (О = A Eq, индуцируются
магнитные дипольные переходы, аналогичные наблюдаемым в спектрах
ЯМР. Ядерный квадрупольный резонанс, как правило, не дает никакой новой
информации при исследовании биомолекул, поэтому нас больше интересует
дополнительное влияние ядерного квадрупольного взаимодействия eQq на
спектрЯМР. Это можно пояснить на примере, приведенном на рис.1.11 для
ядра N со спином I - 1, который может принимать во внешнем магнитном
пол е Во три различные ориентации, соответствующие магнитным квантовым
числам т.] = +1, 0, -1. Расстояния между соседними уровнями энергии,
которые соответствуют этим трем числам, в отсутствие ядерного квадруполь-
34
ного взаимодействия одинаковы, и в этом случае наблюдалась бы только одна
линия поглощения ЯМР. Однако поскольку имеется ядерное квадрупольное
взаимодействие, оба уровня энергии с mi = ±1 сдвигаются в одном направле-
нии (независимо от знаков !), а уровень энергии с mi = 0 сдвигается в
противоположном направлении, так что эквидистантность между уровнями
энергии нарушается, и в спектре наблюдаются две линии поглощения. В
твердых телах можно определить e2Qq из расстояния между обеими линиями
подобно тому, как это делается из наблюдения чисто ядерного квадруполь-
ного резонанса, а в жидкостях ядерное квадрупольное взаимодействие при-
водит только к нежелательному уширению линий поглощения ЯМР.
1.3. Механизмы релаксации
В предыдущем разделе мы рассмотрели взаимодействия, определяющие
положение резонансных линий в спектре ЯМР. В реальном спектре не на-
блюдаются идеально узкие линии: резонансные линии характеризуются ко-
нечной шириной. Ширина отдельных линий в отсутствие неоднородных
уширений и слабого поля/?) определяется временем поперечной релаксации
Тг. Продольная релаксация также оказывает влияние на спектр ЯМР, в
основном на интенсивность резонансных линий.
Поперечная и продольная релаксации индуцируются процессами, про-
исходящими на молекулярном уровне. Они отражают взаимодействие ядер-
ного спина с его окружением. Скорости релаксации пропорциональны
квадрату величины, характеризующей эти взаимодействия. В случае спин-
решеточной релаксации, при которой осуществляется обмен энергией с ок-
ружением, эти взаимодействия оказываются промодулированными во
времени, что происходит за счет взаимодействия спинов с флуктуирующими
магнитными полями, вызывающими переходы между стационарными состо-
яниями спиновой системы на частоте (Di. Те же процессы, которые вызывают
спин-решеточную релаксацию, ведут и к спин-спиновой релаксации, по-
скольку при спин-решеточной релаксации одновременно разрушается фазо-
вая когерентность прецессии отдельных спинов. В то же время временная
модуляция взаимодействий не является обязательным условием для разру-
шения фазовой когерентности; процессы, не модулированные во времени,
представляют собой дополнительный канал поперечной релаксации.
При исследовании спиновой релаксации в биологических системах учи-
тываются следующие пять типов взаимодействий ядерных спинов:
1) диполь-дипольное взаимодействие,
2) анизотропия химического сдвига,
3) косвенное спин-спиновое взаимодействие,
4) ядерное квадрупольное взаимодействие,
5) взаимодействие с неспаренным электроном (свободные радикалы
или парамагнитные ионы).
Еще один механизм спин-решеточной релаксации обусловлен спин-вра-
щательным взаимодействием. Этот механизм дает существенный вклад в
релаксацию для малых молекул в газах или жидкостях малой вязкости. Для
35
молекул, представляющих интерес для биохимии, этот механизм релакса-
ции, как правило, не играет никакой роли.
1.3.1. Поперечная релаксация и ширина линий
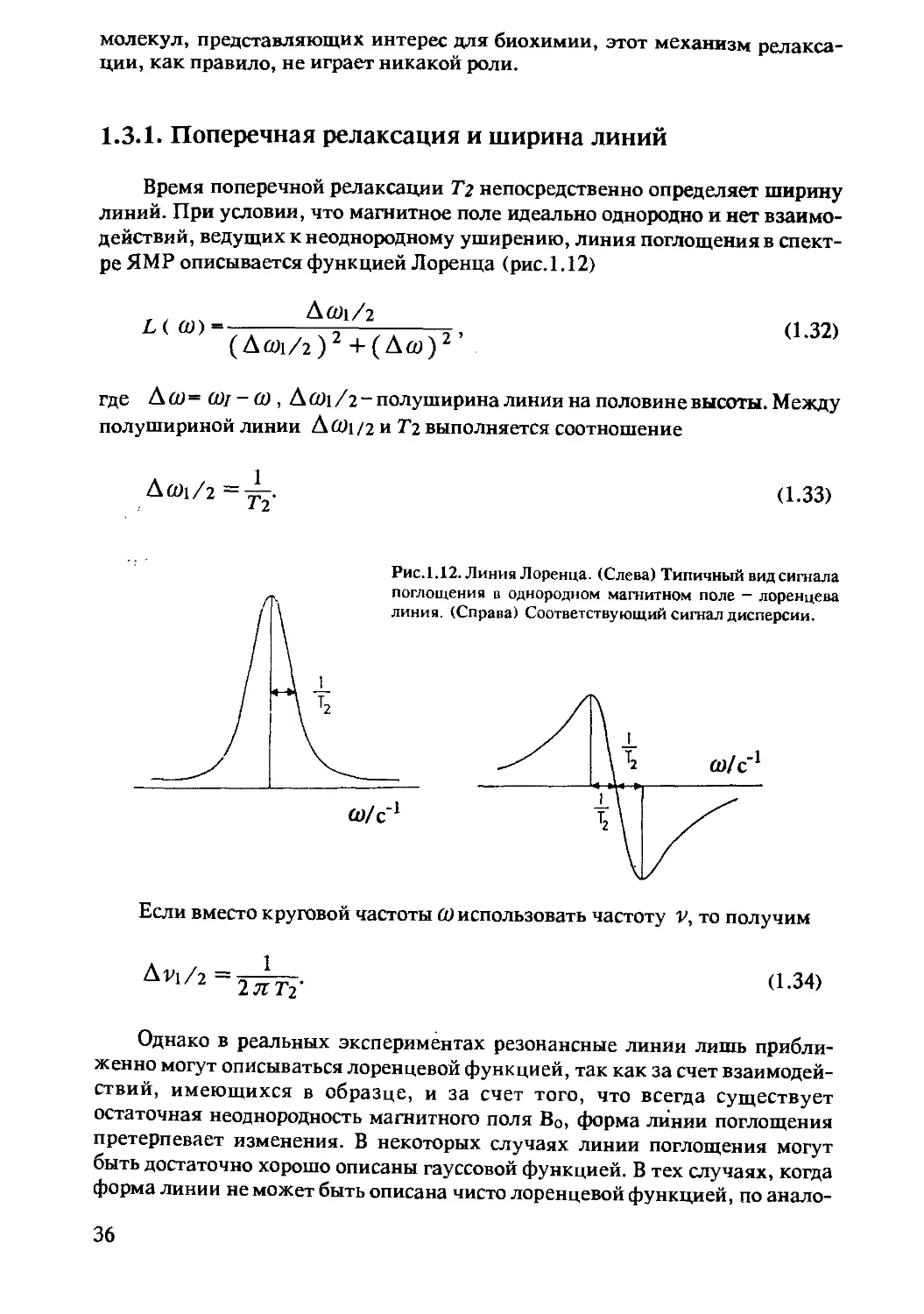
Время поперечной релаксации Т2 непосредственно определяет ширину
линий. При условии, что магнитное поле идеально однородно и нет взаимо-
действий, ведущих к неоднородному уширению, линия поглощения в спект-
ре ЯМР описывается функцией Лоренца (рис.1.12)
_______А 601/2____
L (Аал/г)2+(Д(У)2 ’
(1.32)
где А СО = 0)1 - (О , A COi /2 - полуширина линии на половине высоты. Между
полушириной линии AcOi/2 и Тт. выполняется соотношение
Аб01/2=^-. (1.33)
Если вместо круговой частоты (1) использовать частоту V, то получим
Дг?1/2 2 л: ТУ
(1.34)
Однако в реальных экспериментах резонансные линии лишь прибли-
женно могут описываться лоренцевой функцией, так как за счет взаимодей-
ствий, имеющихся в образце, и за счет того, что всегда существует
остаточная неоднородность магнитного поля Во, форма линии поглощения
претерпевает изменения. В некоторых случаях линии поглощения могут
быть достаточно хорошо описаны гауссовой функцией. В тех случаях, когда
форма линии не может быть описана чисто лоренцевой функцией, по анало-
36
гии с тем соотношением, которое описывает связь между Тз и Лй1/2, для
лоренцевой линии вводят эффективное время релаксации Тз , определяемое
соотношением
(L35>
1.3.2. Временная модуляция взаимодействий
Броуновское движение молекул в жидкостях является основной причи-
ной, определяющей зависимость от времени взаимодействий, наблюдаемых
экспериментально. Возникающие на частоте Ш/ магнитные шумы вызывают
переходы между спиновыми состояниями, обеспечивая тем самым эффек-
тивный механизм спин-решеточной релаксации. Мерой вращательной по-
движности является время корреляции вращательных движений Trot, т.е.
характерное время, за которое молекула в целом или та ее часть, которая
содержит рассматриваемый ядерный спин, повернется на угол, равный в
среднем 1 рад. В жидкостях малой вязкости для малых молекул Trot по
порядку величины равно 10’11 с, т.е. обычно выполняется следующее нера-
венство:
TrotW/<<l. (1.36)
В этом случае можно считать, что шум является белым, т.е. содержит все
частоты Ш от нуля до (точнее, частоты, для которых Ш < < Trot Ь и, кроме
того, интенсивность шумов на всех этих частотах одинакова. Однако для
биологических молекул это условие не всегда выполнимо. Значение Тг всегда
меньше Ti, за исключением нескольких специальных случаев. Это связано
с тем, что все процессы, происходящие по механизму Ti-релаксации (за счет
изменения ориентации спина при переходе из одного энергетического состо-
яния в другое), сопровождающиеся передачей или поглощением энергии в
результате взаимодействия спина с решеткой, всегда нарушают фазовую
когерентность между соседними спинами, и это приводит к возникновению
еще одного канала релаксации по механизму Т2-релаксации. При этом чем
хуже выполняется соотношение (1.36), тем больше будут различаться зна-
чения Ti и Тг, и тем лучше будет выполняться неравенство Т\ > Тг. В
последующих разделах книги мы ограничимся рассмотрением случаев, когда
справедливо неравенство (1.36) (случай максимального сужения линий и Ti
~Т2).
1.3.3. Дипольная релаксация
Для протонов основным типом взаимодействия является диполь-ди-
польное взаимодействие спинов одного сорта, поскольку эти спины обладают
37
большим магнитным моментом^ и в общем случае концентрация их велика,
кроме того, протоны всегда находятся на достаточно близких расстояниях
один от другого. Для других ядер со спином I = 1 /2 диполь-дипольное взаи-
модействие с протонами также играет определяющую роль. Количественно
скорость релаксации 1/rf^? может быть выражена через диполь-дипольное
взаимодействие между ядерным спином Лс/?1=1/2и ядерным спином X с 1Х
= 1/2 соотношением
1 _ / ^ \2
TDD / • (1^7)
Уравнение (1.37) подлежит простой интерпретации: спин А находится в
локальном магнитном поле, создаваемом спином X, и это поле согласно
(1.31) пропорционально ух/и> кроме того, зависит от угла 0между г и Во.
Зависимость от времени диполь-дипольного взаимодействия для двух ядер-
ных спинов, находящихся в данной молекуле, возникает из-за того, что в
течение длительного времени вследствие броуновского движения изменяет-
ся угол 0 относительно внешнего магнитного поля. Время корреляции вра-
щательных движений Trot является мерой скорости этого изменения. Если оба
взаимодействующих спина принадлежат различным молекулам, то под вли-
янием диффузии расстояние г также будет изменяться. Мерой этого измене-
ния является время корреляции трансляционных движений rtrans. Вклад в
скорость релаксации 1/Т^2 за счет диффузии после небольших изменений
может быть описан уравнением, подобным (1.37). В частном случае диполь-
дипольного взаимодействия двух магнитно эквивалентных ядер (ядер одного
типа, находящихся в одинаковом окружении: уА = ух, 0)А = возникает
дополнительный канал релаксации, связанный с так называемыми флип-
флоп процессами, при которых у взаимодействующих спинов происходят
взаимные изменения проекции на направление поля Во так, что суммарный
магнитный момент остается неизменным. Для учета этих процессов необхо-
димо умножить правую часть уравнения (1.37) на коэффициент 3/2.
1.3.4. Релаксация за счет анизотропии химического сдвига
Вторым типом взаимодействия, оказывающим влияние на процессы ре-
лаксации, является анизотропия химического сдвига. Электронные оболоч-
ки создают в точке расположения атомного ядра А локальное
дополнительное поле, которое почти всегда анизотропно, и в силу этого его
величина изменяется во времени под влиянием броуновского движения мо-
лекул, причем время корреляции Trot вновь является мерой этой зависимости
от времени. Так как это взаимодействие возрастает пропорционально вели-
чине поля Во, а скорость релаксации пропорциональна квадрату величины,
характеризующей силу взаимодействия, то оказывается, что вклад в скоро-
сть релаксации от временной модуляции анизотропии химического сдвига
1/7^5 для жидкостей малой вязкости в области, где выполняется уравнение
38
(1.36), в простейшем случае цилиндрической симметрии может быть вычис-
лен по формуле
(1.38)
В этом случае Д О' обозначает величину анизотропии химического сдви-
га. Для малых молекул, совершающих быстрые'вращательные движения,
Tf не равно как это имеет место для диполь-дипольной релаксации.
Здесь выполняется равенство
T2S=yTf. (1.39)
Для протонов вклад в релаксацию за счет этого механизма невелик,
поскольку ядро экранирует всего один электрон и анизотропия химического
сдвига для протона мала. Для других ядер с 1= 1/2, таких, как 13С, 15N, l9F
и 31Р, вклад в релаксацию для полей средней величины уже не является
малым. В настоящее время в спектроскопии ЯМР используются поля напря-
женностью порядка 10 Тл, и в этом случае вклад в релаксацию за счет
анизотропии химического сдвига становится определяющим.
1.3.5. Релаксация по механизму косвенного
спин-спинового взаимодействия
Рассмотренное выше косвенное спин-спиновое взаимодействие между
ядрами А и X также может давать вклад в релаксацию. Так как при враще-
нии молекулы как целого величина константы спин-спинового взаимодейст-
вия не изменяется, вращательная диффузия не является причиной
изменения релаксационных параметров во времени. Зависимость от времени
может возникать под влиянием двух различных механизмов. Во-первых,
скорость релаксации за счет косвенного спин-спинового взаимодействия мо-
жет зависеть от времени вследствие химического обмена взаимодействую-
щих ядер. Это прежде всего относится к протонам. Само явление называется
скалярной релаксацией первого рода, а соответствующее время корреляции
равно обратной скорости обмена. Во-вторых локальное поле, индуцируемое
спином X в точке, где находится спин А, модулируется релаксационными
процессами, в которых участвует спин ядра X. В этом случае говорят о
скалярной релаксации второго рода. Соответствующее время корреляции
является временем релаксации спина X.
Скалярная релаксация первого рода может давать существенный вклад
в релаксацию, в частности, для протонов и ядер дейтерия, поскольку эти
ядерные спины в большинстве случаев участвуют в быстром химическом
обмене. Скалярная релаксация второго рода особенно существенна для спина
ядра А, который связан со спином ядра X с I > 1, поскольку из-за наличия
квадрупольного взаимодействия скорость релаксации для ядра X велика.
39
Количественно скорость релаксации 1/7^с спина А за счет механизма
скалярной релаксации дается формулой
(1.40)
где 1х ~ квантовое число спина X, a TSc ~ либо время релаксации спина X,
либо обратная скорость обмена. Подобное соотношение справедливо для
скорости поперечной релаксации
^sc
---Т J АХ ^Х^Х^^ sc •
Г 3 1+(“А-“х)Чг
(1.41)
1.3.6. Квадрупольная релаксация
Для ядер со спином /2:1 квадрупольное взаимодействие вносит наибо-
лее существенный вклад в процессы релаксации. Как и в случае диполь-ди-
польного взаимодействия, возникает зависимость релаксации от времени
вследствие броуновского движения молекул. Для типичного случая, когда
симметрия градиентов поля приближается к цилиндрической, в области, где
применимо уравнение (1.36), скорости релаксации 1/Тр 2 вычисляются по
формуле
1 _ 3 2/4-3 4^.2 2 г
г?<2 Q Q rot’
(1.42)
где Trot ~ время корреляции вращательных движений молекул. В силу того,
что времена релаксации за счет квадрупольного взаимодействия малы, это
существенно уширяет резонансные линии; именно этим объясняется тот
факт, что квадрупольные ядра весьма мало используются в спектроскопии
ЯМР высокого разрешения. Исключение составляет ядро дейтерия, квадру-
польный момент которого настолько мал, что в большинстве случаев в ЯМР
спектре может быть достигнуто достаточно хорошее разрешение.
1.3.7. Парамагнитная релаксация
Ядро со спином / взаимодействует с неспаренным электроном посредст-
вом либо дипольного, либо контактного взаимодействия Ферми. В силу того,
что магнитный момент электронов p.s много больше ядерного магнитного
момента, электрон-ядерное взаимодействие является доминирующим для
ядерной спиновой релаксации. Временная зависимость релаксации в данном
случае определяется тем, что для спинов электронов время спин-решеточной
релаксации намного меньше всех других времен, т.е. соответствующее время
40
корреляции Ts равно времени релаксации электронов. Обозначим скорость
релаксации спина ядра А за счет сверхтонкого взаимодействия со спином
электрона S через l/T^J- Тогда в простейшем случае
1 7
—= a f 2 SGS+l) Ts, (1,43)
Т 1,2
причем а\ ,2 включает в себя как скалярную (фермиевскую), так и дипольную
части сверхтонкого взаимодействия. Точного количественного описания
простое выражение (1.43) не дает.
У большинства биологически важных молекул неспаренные электроны
отсутствуют. Однако при исследовании структуры молекул в определенные
положения вводят стабильные радикалы или ионы редкоземельных элемен-
тов (спиновые метки) или же проводят замещение диамагнитных ионов,
таких, например, как магний, на парамагнитные, такие, как марганец. При-
менение этих методик изложено в главе 2. Изменение значений времени
релаксации за счет введения парамагнитных веществ составляет основу ис-
пользования этих веществ в качестве контрастных средств в ЯМР-томогра-
фии (глава 4).
1.4. Экспериментальные методы
В этом разделе обсудим экспериментальные методы и принципиальную
схему ЯМР-спектрометра. Исторически более ранняя версия ЯМР - непре-
рывный ЯМР - в настоящее время в значительной степени вытеснена фурье-
спектроскопией. Следует отметить, однако, что непрерывный ЯМР
существенно проще для понимания, поскольку здесь, как в классической
спектроскопии, поглощение электромагнитных квантов является функцией
их частоты (энергии). В противоположность этому в импульсной спектроско-
пии ЯМР-сигнал зависит от времени и детектируется как спад свободной
индукции. Только математическая процедура - преобразование фурье -
превращает этот сигнал во временном представлении в сигнал в частотном
представлении, т.е. в частотный спектр, который, по крайней мере в простых
случаях, эквивалентен спектру, получаемому при непрерывной регистра-
ции. Однако фурье-спектроскопия по сравнению с методами непрерывной
регистрации значительно превосходит их по своей чувствительности и гиб-
кости. Как увидим в дальнейшем, ЯМР-спектрометры имеют много общих
свойств, несмотря на то что, например, ЯМР-томограф по своему простран-
ственному разрешению, очевидно, отличается от спектрометра высокого
разрешения.
1.4.1. Непрерывный ЯМР
В простейшем варианте ЯМР-спектроскопии, который применялся пер-
вые двадцать лет с момента открытия этого метода для измерения спектров
41
ЯМР высокого разрешения, образец помещается в однородное магнитное
поле и подвергается непрерывному воздействию РЧ поля, которое варьиру-
ется по частоте в области существования линий поглощения ЯМР для данного
образца. Этот метод называется cw-ЯМР (от англ, continuous wave).
В общем случае частота со поля Bi медленно проходит через резонанс, и
сигнал ЯМР регистрируется и вычерчивается на самописце. Обычно регист-
рации подлежит линия поглощения, т.е. компонента намагниченности Му ,
которая сдвинута по фазе относительно поля В; на 90°. Однако можно также
проводить измерение компоненты Мх' , которая совпадает по фазе с Bi и в
этом случае получаем сигнал дисперсии. Выражения для намагниченности
даются стационарными решениями уравнений Блоха (1.23)-(1.25) во враща-
ющейся системе координат (т.е., когда производные в левых частях уравне-
ний (1.23)-(1.25) равны нулю):
(1.44)
Му' — Mq
Y1 Bl Т2
1 +(со/-со)2т^ +у] в! Тх Т2
(1.45)
1 +((Dl-wyTi+yiBiTl Т2
(1.46)
Для очень слабых РЧ полей (у ;2Bi 2Ti Т2 < < 1) сигнал поглощения
описывается лоренцевой кривой (1.32). Сигналы поглощения и дисперсии
представлены на рис. 1.12.
Если скорость поглощения энергии поля Bi сравнима по величине или
превышает скорость спин-решеточной релаксации \/Т], то амплитуда сиг-
нала поглощения уменьшается, так как разность населенностей энергетиче-
ских уровней N+ - N убывает в сравнении с равновесной, определенной
законом распределения Больцмана. Одновременно возрастает ширина ли-
нии. Этот эффект называется насыщением. Конечно, степень насыщения
зависит, с одной стороны, от времен релаксации Ti и Тг, а с другой - от
величины поля Вх. Количественно степень насыщения в резонансе: (D = (В/,
может быть измерена через коэффициент насыщения s:
s= 1 +yi2Bi2 Т1Т2 = 1 + (Bt/B/opt)2.
(1.47)
Вследствие насыщения амплитуда наблюдаемого сигнала поглощения
при значении Вх выше Biopt убывает, где значение B/Opt дается выражением
Biopt-----— • (1.48)
П ^Тх Т2
Значение Biopt ограничивает область линейной зависимости амплитуды
сигнала от поля Вх (см. формулу (1.45)). С ростом Вх возникающее уширение
42
Рис.1,13. Насыщение сигнала ЯМР в cw-
спектроскопии. Полная интенсивность (а) и
амплитуда (Ь) линии поглощения ЯМР, как
функция поля В]
линий требует введения коэффициента *5 (т.е. полуширина линии А =
=s^2/T2). Полевые зависимости амплитуды и полной интенсивности (площади
под резонансной линией) сигнала поглощения представлены на рис. 1.13.
1.4.2. Импульсный ЯМР
В простейшем случае схема проведения импульсного ЯМР-эксперимен-
та выглядит следующим образом: 90°-ный импульс поворачивает вектор
намагниченности М в плоскость ху и затем проводится наблюдение спада
свободной индукции. На экране осциллографа спад свободной индукции
имеет вид функции времени f(t). Если проводится регистрация одиночного
сигнала ЯМР, например, сигнала водорода воды, и значение (D в точности
совпадает с резонансной частотой СО/, то f(t) - просто убывающая экспонен-
циальная функция. Этой экспоненциально убывающей функции можно по-
ставить в соответствие функцию F( (л)) в частотном пространстве. Форма
линии при этом является лоренцевой. Обе эти функции - во временной и
частотной областях - связаны между собой преобразованиями
оо
F(W) = J f(t) е ~ia)tdt, (1.49)
- 00
1 00
f(t) = У5- f F( Ш) eia)tda). (1.50)
00
Эти преобразования по имени французского математика Жана Батиста
де Фурье называются фурье-преобразованиями.
Типичное значение длительности 90°-ного импульса для ЯМР-спектро-
скопии высокого разрешения по порядку величины равно 10'5 с. В отличие
от непрерывного РЧ излучения, спектр такого импульса не будет монохро-
матическим, он включает в себя определенную частотную область слева и
справа от частоты заполнения импульса (О. Соответствующий частотный
спектр получают в результате фурье-преобразования этого импульса, кото-
рый для прямоугольного импульса шириной 2Ат может быть описан функ-
цией следующего вида:
43
(1.51)
A_ sin Arco
sine Атщ =
Это частотное распределение всегда будет тем шире, чем меньше 2 Аг,
т.е. в предельном случае имеем бесконечно узкий импульс, d-импульс, час-
тотный спектр которого представляет собой белый шум, содержащий все
значения частот.
Если в образце содержатся различные магнитно неэквивалентные про-
тоны, то такой импульс может возбудить все протоны одновременно. В этом
случае спад свободной индукции является не просто убывающей экспонен-
циальной функцией, а представляет собой большое число экспоненциальных
функций, промодулированных по частоте. В принципе функция F (й>) в час-
тотной области и функция f(t) во временной области должны в равной степе-
ни содержать такую информацию. Однако человеческий глаз и мозг могут
гораздо лучше различать спектральные линии в частотной области. На
рис. 1.14 в качестве примера приведен такой спектр во временной и частотной
областях, состоящий из двух линий.
Рис.1.14. Спектр ЯМР во временном и частотном представлениях. Сигнал ЯМР, состоящий из
двух линий поглощения во временном представлении (а) и в частотном (Ь), путем фурье-преоб-
разования они могут быть преобразованы друг в друга.
Успехи импульсной спекроскопии ЯМР по сравнению с непрерывными
методами в основном связаны с ее большей чувствительностью. В cw-спект-
роскопии в каждый данный момент времени излучается одна частота, и
соответственно возбуждаются только те ядерные спины, для которых эта
частота является резонансной. Очевидно, что такой метод регистрации с
точки зрения затрат времени является мало эффективным, так как сигналы
детектируются при прохождении резонансной линии. В отличие от этого в
ЯМР с фурье-преобразованием одновременно возбуждается и детектируется
весь спектр. Это позволяет также улучшить значение отношения сиг-
нал/шум, поскольку при этом складывается большое число спектров.
Как и в любом физическом эксперименте, сигнал прямо пропорционален
числу п накапливаемых процессов измерения, а статистический шум про-
порционален н1/2, так что отношение сигнал/шум (S/N) при увеличении п
возрастает пропорционально л1^2:
44
SlN~nil2. (1.52)
Так как для отдельной последовательности, состоящей из РЧ импульса
и спада свободной индукции, необходимо примерно 1 с, то за 10 000 с (2,5 ч)
можно зарегистрировать 10 000 накоплений и после фурье-преобразования
иметь 100-кратное улучшение отношения сигнал/шум по сравнению с тем,
которое достигается при одном накоплении. Правда, выигрыш в отношении
сигнал/шум, если речь идет о регистрации большого числа отдельных линий
и на регистрацию затрачивается время Та, будет не столь велик, как следо-
вало бы ожидать из приведенных выше рассуждений. При медленном накоп-
лении можно работать с передатчиком при небольшой полосе пропускания,
а в фурье-спектроскопии ширина полосы пропускания задается полной ши-
риной спектра в частотной области. Однако выигрыш в чувствительности все
еще будет значительным. Количественно он определяется отношением ши-
рины полосы пропускания в частотной области к ширине отдельной резонан-
сной линии АйД/2.
Для оценки оптимальной величины отношения сигнал/шум мы должны
еще учесть явление насыщения. Как уже отмечалось, в cw-эксперименте
амплитуда сигнала при низком уровне РЧ излучения пропорциональна ве-
личине поля Bi (т.е. корню квадратному из РЧ энергии). Однако с ростом
поля В\ вследствие уменьшения разности населенностей зеемановских уров-
ней наблюдается все большее отклонение от линейного роста, причем это
сопровождается дополнительным уширением спектральных линий, и в пре-
дельном случае, когда мощность РЧ излучения максимальна, сигнал вообще
не наблюдается.
Аналогичный эффект насыщения необходимо учитывать и в импульс-
ном ЯМР. Для того чтобы разность между населенностями двух зееманов-
ских уровней восстановилась до значения, приближенно равного исходному,
т.е. до значения, соответствующего распределению Больцмана, необходимо
выждать интервал времени, превышающий в 3-4 раза значение времени
спин-решеточной релаксации Т]. При решении задач, связанных с установ-
лением структуры биологических молекул, типичным значением Ti являет-
ся 3-5 с. Следует отметить, однако, что в фурье-спектроскопии отсутствует
эффект уширения при насыщении, который наблюдается в cw-ЯМР. Это
преимущество фурье-спектроскопии не поддается прямой оценке и потому
не столь очевидно.
Если нужно из величины относительной интенсивности резонансных
линий оценить число ядер, дающих вклад в наблюдаемую линию, то не-
обходимо поддерживать постоянным значение интервала времени между
двумя возбуждающими импульсами. Если желательно получить достаточ-
но хорошо разрешенный спектр, с хорошим отношением сигнал/шум,
причем за достаточно короткое время, то за счет сокращения длительно-
сти импульса можно существенно сократить время проведения экспери-
мента. Эта оптимизация эксперимента основана на свойствах функций sin
и cos (рис. 1.15).
45
Рис.1.15. Значения функций sin и сое при
малых углах. Значения продольной и попе-
речной компонент намагниченности после
воздействия импульса соответственно про-
порциональны сое О. и sin Ct, зде Ct - угол
отклонения намагниченности. Для малого
угла мы получаем небольшое значение
компоненты поперечной намагничен-
ности, a z- компонента мало отличается от
равновесного значения Мо.
Перед началом проведения измерений Mz/Mo -1 и Му>- 0, после 90°-ного
импульса с В] 11 х Mz - 0, Му'/Мо - 1. Если мы используем импульс меньшей
длительности, например 30°-ный, то исходя из свойств тригонометрических
функций Му> /Мо-0,5, т.е. составляет ровно половину того значения, которое
будет достигнуто после воздействия на систему 90°-ного импульса. В то же время
значение Mz уменьшается незначительно, от 1 до 0,87, и соответственно после
15°-ного импульса Му' - 0,26 Мо, a Mz - 0,97 Мо .
Если проводится эксперимент по измерению времени продольной релак-
сации Гр и интервал между отдельными импульсами (время задержки)
задан заранее и равен TR, то можно подобрать длительность импульса таким
образом, чтобы угол отклонения вектора намагниченности обеспечил макси-
мальное значение амплитуды сигнала. После воздействия такого импульса в
системе устанавливается динамическое равновесие. Этот угол, определяю-
щий также оптимальное значение отношения сигнал/шум, называют углом
Эрнста ctE [1.4 ]. Значение угла Эрнста вычисляется по формуле
— tr/t\
аЕ = arc cose . (1.53)
Рис.1.16. Оптимальный угол отклонения намагниченности под воздействием возбуждающего
импульса, (а) Угол Эриста OtE является функцией логарифма отношения времени задержки TR
и времени продольной релаксации Гр (Ь) Амплитуда сигнала в динамическом равновесии
является функцией TR/T\ прн различных значениях угла отклонения. K:Ct = CtE,
в.а = аЕ ±20 °/о ,c.ct = аЕ + so у,, d . а = аЕ -so у,.
46
На рис. 1.16 графически представлена эта зависимость оптимального
угла от времени задержки Tr и времени продольной релаксации Т\. В реаль-
ном образце можно получить несколько различных значений Т\ для каждой
из резонансных линий, а при заданном значении Tr можно оптимизировать,
отношение сигнал/шум для каждого значения Тi. На практике это оказыва-
ется не столь существенно, поскольку изменение отношения 7>/71 едва раза
вызывает лишь незначительную потерю чувствительности.
Прежде чем сигнал ЯМР, полученный при непрерывном методе регист-
рации (аналоговый сигнал), будет подвергнут обработке, он должен быть
оцифрован. Для этого в определенный момент времени измеряется значение
наведенного напряжения, и этому значению ставится в соответствие число.
Интервал A t времени между измерениями определяется по теореме Найк-
виста и зависит от значения наивысшей наблюдаемой частоты Vmax (отсчи-
тываемой от несущей частоты импульса):
А'=^Ь (154>
формулу (1.54) можно понимать следующим образом: для того чтобы
иметь возможность различать каждую частоту колебания, необходимо про-
изводить по крайней мере два измерения за период колебаний.
Перед проведением фурье-преобразования массив экспериментальных
данных подвергается фильтрации путем умножения на соответствующие
lA lA LA
75 * * 70 75 - 70 75 70
О/м.д. О/м.д. d/м.д. .
Рис.1.17. Фильтрация сигнала ЯМР. (Верхний ряд) Спектр ЯМР Н (500 МГц) НРг-протеина
в отсутствие фильтрации (слева), после экспоненциальной фильтрации путем умножения ССИ
на функцию (середина), после гауссовой фильтрации путем умножения на функцию
* (справа). ССИ состоит из 16384 (16 К) точек, которые накапливаются с разрешением
96 мкс. (Средний ряд) Спектры в частотной области Тюсле фурье-преобразования. Полная
частотная область 5208 Гц (10,4 м.д.). (Нижний ряд) Фрагмент полного спектра. Шкала в м.д.
приведена к внутреннему стандарту 2,2-диметил-2-силапентан-5-сульфоновая кислота (DSS).
47
функции. При этом преследуются две цели: с одной стороны, уменьшается
шум, а с другой - повышается разрешение за счет изменения формы резо-
нансной линии. Наиболее простой и часто используемой функцией является
•убывающая экспонента, умножение на которую приводит к небольшому
уширению линий и одновременно к уменьшению шумов (рис. 1.17). Форма
линии остается лоренцевой, поскольку умножение двух экспоненциальных
функций вновь будет экспонентой. Для сужения линий зачастую использу-
ется лоренц-гауссово преобразование. Лоренцевы линии преобразуются в
более узкие линии, а гауссовы — подобно.
Накопленный спад свободной индукции после оцифровки не будет пол-
ностью соответствовать невозмущенному спаду свободной индукции, так как
он содержит дискретный набор значений. Соответственно, фурье-преобразо-
вание спада свободной индукции, проводимое ЭВМ по алгоритму быстрого
фурье-преобразования, переводит эти данные в дискретную форму.
Несмотря на то что непрерывное фурье-преобразование может переве-
сти полный спад свободной индукции в идеальный частотный спектр, в по-
следнее время все чаще обсуждается возможность подбора наилучших
способов преобразования временного сигнала в частотное представление.
Это связано с тем, что в реальном эксперименте мы наблюдаем спад свобод-
ной индукции в течение конечного интервала времени Taq и затем повторяем
этот эксперимент, причем число повторений определяется тем значением
отношения сигнал/шум, которое нужно получить. Таким образом, в силу
конечности интервала Taq в нашем распоряжении имеется только эта диск-
ретная информация и в результате фурье-преобразования получаем частот-
ный спектр, который в точности соответствует этому усеченному спаду свободной
индукции и лишь приближенно соответствует истинному спектру.
В настоящее время развиты следующие две группы методов, позволяю-
щие более эффективно использовать имеющуюся информацию за счет того,
что спад свободной индукции продолжается за пределы интервала Taq. Один
из этих методов называется методом максимальной энтропии (ММЭ). Пер-
воначально метод ММЭ был развит для оценки данных геологоразведки,
однако в дальнейшем активно использовался при обработке изображений, и
с его помощью были достигнуты значительные результаты. Вторая группа
методов основана на линейном прогнозировании (ЛП). Методы ЛП исходят
из того, что идеальный сигнал ЯМР может быть представлен в виде произве-
дения затухающей экспоненты и косинуса. Если нам удается найти функ-
цию, которая могла бы задать наблюдаемый спад свободной индукции, то
можно было бы предсказать поведение его в любой точке на временной оси.
Оба метода обладают тем существенным недостатком, что требуют больших
затрат машинного времени, и именно этим объясняется тот факт, что в
настоящее время они используются только для решения специальных задач.
1.4.3. Спектрометр ЯМР
Конструкция всех ЯМР-спектрометров, как правило, достаточно про-
ста, и конструктивные особенности отражают требования, предъявляемые к
48
ЯМР-эксперименту. На рис.1.18 схематически представлена конструкция
cw-спектрометра, а на рис.1.19 — импульсного спектрометра. Конструкция
ЯМР-томографа принципиально совпадает с конструкцией импульсного
спектрометра. Далее подробно рассмотрим отдельные компоненты спектро-
метра и опишем принцип их действия.
Рис.1.18. Блок-схема cw-спектрометра.
Рис.1.19. Блок схема простого фурье-спектрометра.
Основной компонентой ЯМР-спектрометра является магнит, который
должен создавать максимально однородное постоянное поле Во, стабильное
49
во времени. Поле Во вызывает расщепление уровней энергии, между кото-
рыми индуцируются ЯМР-переходы. Для создания статического магнитного
поля могут использоваться три типа магнитов: электромагниты, постоянные
магниты и сверхпроводящие магниты. В настоящее время в ЯМР-спектро-
скопии высокого разрешения в основном используются сверхпроводящие
магнитные системы, поскольку лишь они могут удовлетворить высоким тре-
бованиям, предъявляемым к спектрам ЯМР. В спектроскопии ЯМР неизмен-
но сохраняется тенденция к увеличению напряженности магнитного поля,
поскольку увеличение этого параметра обеспечивает возрастание разреша-
ющей способности и выигрыш в чувствительности. В томографии наряду со
сверхпроводящими магнитами используются также и электромагниты, так
как здесь вопрос о целесообразности выбора того или иного магнитного поля
не является столь очевидным и экономические соображения не всегда явля-
ются определяющими. Постоянные магниты в настоящее время почти не
находят применения как для спектроскопии, так и для томографии.
Достижимая на практике напряженность магнитного поля зависит от
размеров исследуемого образца, которые варьируются в широких пределах:
в ЯМР высокого разрешения диаметр образца варьируется от 0,5 до 2,5 см, а
в ЯМР-томографии размеры исследуемого образца определяются размерами
человеческого тела. Для образцов малых размеров типичное значение маг-
нитного поля электромагнита - 2,5 Тл, а для больших - 0,3 Тл. Для совре-
менного уровня развития технологии напряженность поля
сверхпроводящего магнита, предназначенного для исследования образца ма-
лого объема, достигает 14 Т, а для образцов большого объема - 4 Тл.
Независимо от типа магнита при использовании ЯМР высокого разре-
шения для решения задач структурной химии к однородности поля предъяв-
ляются чрезвычайно высокие требования. Так, для ЯМР-спектрометра
высокого разрешения с рабочей частотой 600 МГц и разрешением 0,1 Гц эта
величина составляет 2 • 1О'10. Теоретически такое разрешение может быть
достигнуто при использовании сверхпроводящего магнита, однако на прак-
тике такая однородность не может быть достигнута, так как магнитная вос-
приимчивость изменяется от образца к образцу. С помощью дополнительных
резистивных катушек, которые устанавливаются между основной сверхпро-
водящей катушкой и образцом, удается провести коррекцию поля в ограни-
ченных пределах, что позволяет достичь однородности поля до 10'9 по объему
образца. Остаточную небольшую неоднородность поля в плоскости, перпен-
дикулярной Во, можно устранить путем механического вращения образца
вдоль оси.
В cw-спектроскопии величина поля Во периодически модулируется, что
позволяет наблюдать зависимость амплитуды сигнала ЯМР от времени. Эта
зависимость от времени является условием реализации метода детектирова-
ния с помощью фазочувствительного детектора (см. рис. 1.18). Однако для
фурье-спектроскопии необходимость в этом отсутствует, так как здесь спад
свободной индукции уже промодулирован во времени. Для ЯМР-томографии
необходимо создавать дополнительные градиентные поля. Они создаются с
помощью специальных градиентных катушек и воздействуют на исследуе-
мый объект одновременно с постоянным магнитным полем.
50
Для индуцирования ЯМР-переходов необходимо дополнительно пода-
вать на образец еще и РЧ поле Bi, которое поляризовано перпендикулярно
полю Во - статическому магнитному полю. РЧ поле создается передатчиком
и через катушку-резонатор подается на образец. При этом в импульсном
ЯМР передатчик создает мощные импульсы малой длительности (несколько
мкс), а в cw-спектроскопии на образец непрерывно подается сигнал малой
мощности. СигналЯМР детектируется либо той же (передающей) катушкой,
либо приемной. Этот слабый сигнал, как правило, от 10’6 до 10’1 В, перед
обработкой должен быть усилен, прежде чем будет проведена его регистра-
ция с помощью фазочувствительного детектора. В cw-спектроскопии сигнал
непосредственно подается на самописец, а в фурье-спектроскопии - на ана-
лого-цифровой преобразователь (АЦП) в ЭВМ. Этот изменяющийся во вре-
мени сигнал подвергается фурье-преобразованию и вновь подается на
устройство вывода информации - самописец или экран графического дисп-
лея.
В каждом импульсном спектрометре ЯМР ЭВМ используется для управ-
ления спектрометром, а также для накопления экспериментальных данных.
Для последующей обработки экспериментальных данных (быстрого фурье-
преобразования, реконструкции 2М- и ЗМ-изображений) необходима мощ-
ная ЭВМ. В дальнейшем мы приведем некоторые технические
характеристики, которые обеспечивают удобство эксплуатации при условии
удовлетворения современным требованиям, предъявляемым к ЯМР-экспе-
рименту.
Центральный процессор (ЦП) 32-х разрядный компьютер с быстродей-
ствием не менее 2-4 млн. операций в 1 с и памятью 8 Мбайт. Процессор
соединен с жестким диском, объем хранения информации которого состав-
ляет до 150 Мбайт. Для хранения спектральной информации может быть
использована магнитная лента или кассета емкостью до 40 Мбайт. В перспек-
тиве будут использоваться оптические устройства хранения информации -
оптические диски с гораздо более высокой емкостью. При этом существует
возможность проведения параллельной обработки данных с помощью спец-
процессора ЭВМ, который должен иметь прямой доступ к основной памяти и
к жесткому диску, что позволяет на два порядка ускорить процесс обработки
данных. Поскольку существует несколько блоков памяти, необходимо осу-
ществлять быструю связь между спектрометром и ЭВМ. Кроме того, необхо-
дим цветной графический дисплей с высоким разрешением, а также быстрая
связь с ЭВМ.
Существует альтернатива спецпроцессору. Многие фирмы предлагают
вместо него специальный быстродействующий процессор с плавающей запя-
той, поскольку для одного из основных применений, а именно, для построе-
ния двумерных спектров ЯМР мощности спецпроцессора не хватает.
Программное обеспечение, необходимое для сбора и обработки спект-
ральной информации, как правило, предоставляется фирмой-изготовите-
лем. Однако если предполагается, что одновременно к ЭВМ обращаются
несколько пользователей, например, при обработке спектральной информа-
ции одновременно несколькими операторами, то здесь необходимо подклю-
чение специального устройства - многоканальной рабочей станции. Это
51
устройство предусмотрено практически для каждой операционной системы,
если возникает необходимость трансляции на соответствующий язык про-
граммирования.
Литература
1.1. Bloch, F. (1946) Phys. Rev. 70, 460.
1.2. Purcell, E.M., Torrey, H.C. and Pound, R.V. (1946) Phys. Rev. 69, 37.
1.3. Knight, W.D. (1949) Phys. Rev. 76, 1259.
1.4. Arnold, J.T., Dharmatti, S.S. and Packard, M.E. (1951) J.Chem.
Phys. 19, 507.
1.5. Ernst, R.R. and Anderson, W.A. (1966) Rev. Sci. Instrum. 37, 93.
1.6. Jeener, J. (1971) presented at the Ampere Summer School in Basko
Polje.
1.7. Lauterbur, P.C. (1973) Nature 242, 190.
1.8. Mansfield, P., Grannell, P.K., Garroway, A.N. and Stalker, D.C.
(1973) Proc. Specialized Colloque Ampere, Krakau.
1.9. Carr, H.Y. and Purcell, E.M. (1954) Phys. Rev. 94, 630.
l.lO.Meiboom, S. and Gill, D. (1958) Rev. Sci. Instrum. 29, 688.
Дополнительная литература
Abragam, A. (1961) The Principles of Nuclear Magnetism (Oxford
University Press, Oxford)
Ernst, R.R., Bodenhausen, G. and Wokaun, A. (1990) Principles of Nuclear
Magnetic Resonance in One and Two Dimensions (Oxford University Press,
Oxford)
Poole, C.P. (1990) Theory of NMR (Wiley, London)
Shaw, D. (1976) Fourier Transform N.M.R. Spectroscopy (Elsevier,
Amsterdam)
Slichter, C.P^ (1990) Principles of Magnetic Resonance (Springer, Berlin,
Heidelberg, New York)
52
Глава 2. ЯМР в биохимии
За последнее десятилетие метод ЯМР постепенно занял ведущее поло-
жение в биохимических и биофизических исследованиях. Как в органиче-
ской химии, так и в биохимии ЯМР-спектроскопии является прежде всего
аналитическим методом, с помощью которого можно либо подтвердить, либо
опровергнуть предполагаемую структуру вновь синтезированных соедине-
ний. Кроме этой области примененияЯМР, ориентированной прежде всего
на химические приложения метода, можно получить также информацию о
пространственном расположении атомов, конфигурации биологически важ-
ных молекул и молекулярных комплексов. Такая информация позволяет
внести существенный вклад в выяснение механизмов ферментативных пре-
вращений и путей прохождения биохимических реакций (табл.2.1).
Таблица 2.1. Применение ЯМР спектроскопии в биохимии
1. ЯМР малых молекул
Отнесение всех резонансных линий и необходимое разрешение в спектрах достигается
достаточно просто
- химический анализ,
- определеиие неизвестных химических структур,
- неинвазивное определение концентраций,
- исследование путей протекания реакций и продуктов реакций,
- определение констант связи,
- конформации субстата на активном центре.
2. ЯМР макромолекул
Отнесение резонансных линий и достижение необходимого разрешения
- достаточно сложная задача
- изменение конформации молекул под влиянием внешних воздействий
(pH, температура, ионная сила, давление и т.д.),
- конформационные изменения при образовании связи с субстратом,
- структура активного центра ферментов, пространственное расположение
боковых цепей относительно субстрата, ионизация функциональных групп,
- определение временных характеристик (времени корреляции) и характера
движения при локальных и глобальных конформационных изменениях,
- взаимодействие между молекулами фермент-субстрат, протеин-протеин,
протеин-нуклеиновая кислота,
- определение вторичной структуры в растворе,
- определение третичной структуры в растворе.
Преимуществом метода ЯМР, что особенно существенно в медицинских
приложениях, является низкаяэнергия используемых РЧ квантов. Даже при
максимальных значениях магнитных полей, применяемых в настоящее вре-
мя, энергия этих квантов порядка 4 • 10 Дж, что намного меньше энергии,
необходимой для разрушения ковалентных химических связей; типичное
значение энергии химических связей по порядку равно 10 Дж. В отличие
от методов, в которых используется ионизирующее излучение, ЯМР являет-
ся неразрушаюшим методом.
53
2.1. ЯМР как аналитический метод
В этом разделе на разнообразных примерах показано, каким образом ЯМР
может быть использован в качестве аналитического метода. С этой целью более
подробно обсудим некоторые понятия, введенные в первой главе.
Применение метода ЯМР в качестве аналитического можно продемон-
стрировать на примере простейшей молекулы - молекулы этанола, посколь-
ку, как видно из рис. 1.1, протонам трех функциональных групп в этаноле:
метильной, метиленовой и гидроксильной соответствуют три различные ре-
зонансные линии, наблюдаемые в спектре. Особенность метода ЯМР прежде
всего состоит в том, что по положению резонансных линий в спектрах можно
судить о взаимном расположении отдельных атомов или групп атомов в
молекулах, причем это удается обнаружить даже для эквивалентных атомов.
ЯМР по своей информативности выгодно отличается от многих других ана-
литических методов, конкурирующих с ним.
В то же время ЯМР является довольно малочувствительным методом,
несмотря на то что в современных спектрометрах значение максимально
достижимого магнитного поля составляет 14 Тл, что соответствует рабочей
частоте для *Н 600 МГц. Минимальное количество вещества, которое может
быть зарегистрировано с помощью метода ЯМР 1Н, варьируется в широких
пределах в зависимости от условий эксперимента. Для малых молекул ти-
пичное значение, определяющее предел чувствительности, составляет
10 нмоль, т.е. то количество вещества, которое может быть проанализирова-
но простыми биохимическими методами, например, с помощью тонкослой-
ной хроматографии. При использовании других, достаточно широко
используемых методов, таких, как газовая хроматография или радиоимму-
нометрия, становятся доступными принципиально иные с точки зрения чув-
ствительности области такие, как фемто- и атоммолярная. Таким образом,
если речь идет только о регистрации спектров веществ известной структуры,
то эти методы по чувствительности превосходят ЯМР. Преимущество ЯМР
становится очевидным лишь тогда, когда возникает необходимость в получе-
нии дополнительной информации, которую может предоставить только этот
метод. Это касается тех случаев, когда структура исследуемого вещества еще
не известна. Тогда из анализа спектра ЯМР можно построить ряд возможных
структур и выбрать среди них правильную. Следует отметить также еще одну
особенность метода ЯМР: в спектрах ЯМР 1Н могут присутствовать сигналы
молекул-примесей, идентификация которых возможна только в случае при-
менения специальных методов.
2.1.1. Идентификация известных и неизвестных веществ
Использование метода ЯМР в качестве аналитического в простейшем
случае состоит в сравнении положений линий спектров образца неизвестной
структуры и стандартного образца известной структуры: совпадение спект-
ров стандартного вещества и исследуемого позволяет провести идентифика-
54
цию последнего. На основании такого рода сопоставлений можно дать даже
количественную оценку концентрации веществ, содержащихся в образце.
Эта оценка в простейшем виде может быть сформулирована следующим
образом: вещество А содержится в образце в концентрации, достаточной для
его регистрации. Правда, такой способ оценки справедлив лишь тогда, когда
оба сравниваемых спектра получены при идентичных условиях. Реальный
спектр, соответствующий определенному веществу, существенно зависит от
условий его регистрации. Вид спектра определяется не только внешними
факторами (температурой в датчике ЯМР, напряженностью поля и исполь-
зуемой импульсной последовательностью), т.е. теми параметрами, которые
экспериментатор может контролировать самостоятельно, но и используе-
мым растворителем. Несмотря на это, все же удается провести идентифика-
цию неизвестного вещества, поскольку всегда можно ограничиться
определенным абстрактным уровнем и сравнить характерные особенности
спектров.
Рассмотрим еще раз фрагмент спектра ЯМР этанола, снятого на совре-
менном ЯМР-спектрометре с высоким значением постоянного магнитного
поля (рис.2.1). Параметры, полностью характеризующие спектр, следую-
щие: положение спектральных линий, структура мультиплетов и относи-
тельные интенсивности резонансных линий. В главе 1 уже обсуждались
соответствующие физические взаимодействия. Положение линий определя-
ется значением химического сдвига, а расщепление линий - скалярным
спин-спиновым взаимодействием. Площадь под резонансным сигналом, ко-
торая может быть получена численным интегрированием (см. рис.2.1), про-
порциональна числу спинов, соответствующих данной группе в молекуле.
Рис.2.1. Спектр Н 500 МГц
95%-ного этанола. Сигнал при 0
м.д. соответствует метильной
группе стандарта ДСС от-
носительно которого приводятся
значения химических сдвигов в
шкале (5. На рис. приведен также
интеграл спектральных линий
Как было показано ранее, химический сдвиг можно понимать как уси-
ление или ослабление внешнего магнитного поля в точке пространства, в
которой расположено исследуемое ядро. Основной вклад в величину этого
локального изменения поля дают электронные оболочки рассматриваемых
55
молекул, поэтому можно утверждать, что химический сдвиг отражает
структуру молекул. Если оценивать влияние заместителей на величину хи-
мического сдвига, то с ростом числа связей между рассматриваемым атомом
и заместителем это влияние уменьшается. Однако характер электронного
распределения, а значит, и вид спектра ЯМР, зависит и от пространственной
структуры молекул, и от взаимодействия с другими молекулами в растворе,
т.е. от тех факторов, которые уже не представляются столь очевидными.
Для ЯМР *Н область значений химических сдвигов невелика (табл.2.2),
а эффекты, связанные с пространственной структурой и вызванные взаимо-
действием с другими молекулами, бывают настолько велики, что часто из
наблюдения спектров ЯМР весьма затруднительно сделать однозначный вы-
вод о химической структуре вещества.
1 13
Таблица 2.2. Химический сдвиг в спектрах Ни С
Группа Область значений химических сдвигов д в м.д. относительно ТМС
‘н-ямр 13с-ямр
-С-СН3 (обычный метил) 0,1 -1,8 5-35
-S-CH3 (тиометил) 1,5-3,8 10 20
-О-СН3 (метокси) 2,6 -4,2 45-60
-С-СН2-С (метиленовый) 0,3-3,9 15-55
-S-CHj-C (метиленовый) 1,6-4,8 25-45
-O-CHj-C (метиленовый) 3,2-4,8 40-70
-Cj-CH-C (метиновый) 0,8-2,6 30-60
-СЗ-С-С (третичный углерод) — 25 50
-СН-СН-С (алкеновый углерод) 5,0-7,0 105-145
-СН-СН-С- (ароматический) 5,0-10,0 115-145
-C-CH-N- (гетероциклический) 5,0-10,0 145-155
-С =С-С- (алкиновый) — 75-95
-С-СНО (альдегидный) 9,4-11,0 185-210
-С-СООН (кислотный) 4,6-14,0 170-185
j-C-CO-NH (пептидная связь) 6,5-11,0 170-185
Для большинства молекул в растворе конформация их при комнатной
температуре не является жесткой, онимогут существовать в виде нескольких
различных конформаций. Устанавливаемое конформационное равновесие
зависит от состава растворителя, температуры и давления. Кроме того, ис-
следуемые молекулы могут взаимодействовать между собой или с другими
молекулами и образовывать при этом более или менее стабильные комплек-
сы. Вероятность обнаружения этих взаимодействий зависит от концентра-
ции всех компонентов, присутствующих в растворе.
В отличие от спектров ЯМР !Н диапазон значений химических сдвигов
в спектрах 13С существенно больше’ (см. табл.2.2), так что положение резо-
нансных линий в спектрах ^3С определяется в основном химической струк-
турой. Значения химических сдвигов характеристичны при условии, что
влияние заместителей учитывается в виде аддитивной поправки к величине
химического сдвига.
56
2.1.2. Внутренний и внешний стандарт
Как показано в разделе 1.2.1, химический сдвиг, как правило, приводит-
ся относительно резонансной частоты (сигнала) вещества-стандарта, кото-
рое добавляется в исследуемый раствор (внутренний стандарт) или
помещается в капилляр (внешний стандарт). В этом случае, как показано
ранее, химический сдвиг зависит от резонансной частоты используемого
стандарта. Величина химического сдвига имеет смысл только в случае, если
указан стандарт, относительно которого приводится его значение. В табл .2.3
приведены основные вещества, используемые в качестве стандартов. Если же
химический сдвиг приведен относительно другого вещества, используемого
в качестве стандарта, то его значение всегда можно пересчитать относитель-
но общепринятого стандарта.
Как внешний, так и внутренний стандарты обладают рядом преиму-
ществ и недостатков. Внутренний стандарт может вступать во взаимодейст-
вие с другими молекулами, присутствующими в растворе, что приводит к
сдвигу сигнала стандарта, т.е. к сдвигу нулевой точки. Взаимодействие стан-
дарта с исследуемым веществом также может привести к изменению вида его
спектра. Поэтому основным требованием, предъявляемым к внутреннему
стандарту, является уменьшение этого эффекта. Очевидно, что внутренний
стандарт должен быть химически стабильным соединением и хорошо раство-
римым в используемом растворителе. Всем этим условиям вполне удовлет-
воряют такие вещества, как тетраметилсилан (ТМС), 2,2-диметил-2-
силапентан-5-сульфоновая кислота (ДСС) и триметилсилилпропионовая
кислота (ТСП) (см. табл.2.3), положение сигнала метильной группы для
которых используется в качестве нулевой точки в шкале химических сдвигов
и м.д. Эти вещества используются в качестве стандартов в спектрах как 1Н,
так и 13С. Для других ядер, спектры которых представляют интерес с точки
зрения биологических приложений ЯМР таких, как 15N и 31Р, пока не
установлен единый стандарт.
Внешний стандарт отделен от исследуемого вещества. Как правило, его
запаивают в капилляр и помещают в измерительную ампулу. Очевидным
преимуществом использования внешнего стандарта является то, что молеку-
лы стандартного вещества и исследуемого не взаимодействуют. Однако здесь
возникает другая проблема, связанная с тем, что магнитные восприимчиво-
сти растворителя внутри капилляра и в образе различаются. Величина маг-
нитного поля в точке расположения ядра определяется не только
экранирующим влиянием электронных оболочек, но и внешним магнитным
полем Во, величина которого зависит от магнитной восприимчивости окру-
жения. Если используется внутренний стандарт, то изменение магнитного
поля за счет восприимчивости окружения одинаково и для молекул стандар-
та, и для молекул исследуемого вещества. При использовании внешнего
стандарта это условие не выполняется. В зависимости от магнитных воснри-
имчивостей исследуемого вещества и стандарта их ядра находятся в различ-
ных магнитных полях, причем это различие в величине полей зависит не
только от магнитных восприимчивостей /РгоЬе исследуемого вещества и /Ref
вещества-стандарта, но и геометрических факторов. В ЯМР высокого разре-
57
Таблица 2.3. Стандартные вещества, используемые при исследовании биологических молекул
Спектроскопия Вещество Примечания
hi-ямр ТМС-тетраметилсилан Наиболее часто используемый в химии внутренний стандарт. Нерастворим в воде.
ДСС (2,2-диметил-2-сила- Хорошо растворим в воде, наибо-
пентан-5-сульфониевая лее часто используемый стандарт
кислота при проведении биохимических исследований. Обнаруживает ши- рокие сигналы в области 0,6 м.д., 1,73 и 2,93 м.д.
ТСП (З-(триметилсилил)- Наиболее часто используемый в
тетрадейтеропропиоиовая биохимии стандарт. Хорошо
кислота растворим в воде. Обнаружива- ет небольшую зависимость хи- мического сдвига от pH -
Тот же стандарт, что и для См. Выше.
'3С-ЯМР ЯМР *н
15М-ЯМР 15NH4+-hoh Отсутствует какой-либо "иде- альный стандарт". Часто ис-
15NOj ион пользуется в комбинации с ионом аммония как КНд nOj. Сигнал 1эМОз имеет хими- ческий сдвиг 359 м.д. относи- тельно '«Нд . +
Нитрометан 354 м.д. относительно J^NH^.
Нитробензол 349 м.д. относительно «114 .
31Р-ЯМР Фосфорная кислота (85%) Наиболее часто используемый внешний стандарт. Как прави- ло, значения химических сдвигов,определенные от- носительно других стандартов, пересчитываются на этот стандарт .
ДММФ (диметилфосфоиат) Водорастворимый стандарт, используемый в качестве внут- реннего стандарта. Его хими- ческий сдвиг составляет 39,4 м.д. относительно 85% фосфорной кислоты •
ТЭФ (триэтилфосфат) Растворим в воде, использу- ется достаточно широко в ка- честве внутреннего стандарта. Химический сдвиг составляет 0,4 м.д. относительно 85%-ной фосфорной кислоты, ХИМ. СДВИГ достаточно сильно зависит от pH.
Креатинфосфат Часто используется в качестве внутреннего стандарта в in -vivo спектроскопии ЯМР. Отличается
достаточно высоким содержани- ем в большинстве тканей. Хими-
ческий сдвиг зависит от pH (рК 4,6), в области нейтрального pH химический сдвиг составляет 2,3 м.д. относительно 85%-ной фосфорной кислоты.
58
шения образец и стандарт помещают, как правило, в цилиндрические ампу-
лы-капилляры, длинные оси которых параллельны внешнему магнитному
полю. Различие в значениях внешнего магнитного поля для образца и стан-
дарта можно рассчитать по следующей формуле:
4 тг
^B=~~T^Ref~^Probe>>B^- (2Л)
Поправка на восприимчивость оказывается особенно существенной, ес-
ли проводится сравнение спектров, полученных на спектрометре со сверх-
проводящим магнитом и с обычным электромагнитом. В последнем случае
образцы обычно располагают перпендикулярно внешнему магнитному по-
лю; согласно теоретическим расчетам коэффициент —4/3 в уравнении (2.1)
следует заменить на 2/3. Таким образом, даже для одинаковых образцов без
учета поправки на магнитную восприимчивость значения химических сдви-
гов будут отличаться.
Для образца сферической формы магнитное поле внутри и вне его оди-
наково, так что проблему, связанную с внешним стандартом, можно частич-
но обойти, если поместить стандарт в сферический капилляр.
2.1.3. Мультиплетная структура
Вторым важным свойством спектров ЯМР, позволяющим получить бо-
лее полную их характеристику, является мультиплетная структура линий.
Параметр J, количественно характеризующий расщепление резонансных
линий, называется константой связи (см. раздел 1.2.3). Величина/ называ-
ется константой гомоядерного взаимодействия, если рассматривается взаи-
модействие между ядрами одного сорта, и константой гетероядерного
взаимодействия, если взаимодействующие ядра принадлежат ядрам различ-
ных элементов. Расщепление резонансных линий, показанное на рис.2.1,
отражает гомоядерное взаимодействие между протонами, которые входят в
состав различных химических групп в этаноле. Этанол состоит не только из
атомов водорода, но и атомов углерода и кислорода, однако константа гете-
роядерного взаимодействия не может быть обнаружена по спектрам ЯМР,
поскольку изотопы 12С и 160 обладают нулевым спином. При более деталь-
ном рассмотрении спектра можно наблюдать очень слабые дополнительные
линии по обе стороны от основной - так называемые сателлиты С. Эти
линии возникают за счет взаимодействия протонов с изотопом углерода С,
естественное содержание которого составляет порядка 1 % и спин 1=1/2.
Мультиплетная структура спектров ЯМР может быть рассчитана теоре-
тически. Разработан ряд достаточно совершенных программ построения
спектров, которые позволяют по известным значениям констант спин-спи-
нового взаимодействия и химических сдвигов провести расчет теоретическо-
го спектра. Как уже обсуждалось в главе 1, в случае слабой связи характер
расщепления можно описать достаточно просто, и наоборот, исходя из харак-
тера расщепления резонансных линий, можно рассчитать, сколько спинов
59
участвует во взаимодействии и какова структура данного химического сое-
динения. При смешивании этанола с дейтерированной водой протоны гидро-
ксильной группы этанола замещаются на ядра дейтерия. В спектре исчезает
сигнал, соответствующий протонам гидроксильной группы и одновременно
исчезает взаимодействие протонов гидроксильной группы и протонов СН2-
группы (рис.2.2). Протоны, входящие в состав СН2- и СН3-групп в пределах
группы магнитно эквивалентны. Это соответствует спиновой системе А2Х3,
спектр которой приведен на рис.2.2: расщепление резонансных линий СН2-
группы - квартет, а СН3-группы - триплет.
-сн3
-сн2-
4
(Ь)
(с)
1.5 ' ' "'w
д/м.д.
Рис.2.2. Спектр этанола (концентрация 0,1 %), снятый на приборе с рабочей частотой
500 МГц. В качестве растворителя использована дейтерированная вода (99.75% обогащение по
дейтерию). Приведенный на рис. спектр получен после фурье-преобразовання ССИ для одного
прохождения. Время сбора данных 7.2 с.
(а) Фрагмент спектра, на котором представлены сигналы метиленовой и метильной групп эта-
нола; (Ь) Интеграл спектра; (с) Развязка по частоте, соответствующей резонансу метиленовой
группы при 3,65 м.д., приводит к упрощению мультиплета (триплета) при 1,18 м.д.
Интенсивность компонент мультиплета также можно рассчитать по про-
стым правилам, аналогичным правилам расчета мультиплетной структуры.
В принципе с помощью итеративной процедуры можно получить соотноше-
ние интенсивностей линий внутри мультиплета для каждой резонансной
линии. Взаимодействие данного спина с п эквивалентными ядрами со спином
1/2 вызывает расщепление резонансной линии на (п+1) компоненту, иитен-
60
сивность каждой из компонент описывается биномиальными коэффициен-
тами П . Интенсивность 1т т-тл компоненты (т — 1,..., п) в этом случае
можно рассчитать по формуле
г _ ( П \ =_______ п ___________
т (т-1) (/и- 1 )! (n + 1 -/и )! ’
(2.2)
В этом случае для СН3-группы этанола (и - 2) получим мультиплет с
отношением интенсивностей 1:2:1.
Ситуация существенно усложняется, если не выполняется приближение
слабой связи. На рис.2.3 показано изменение вида спектра для случая про-
стой спиновой системы АХ, состоящей из двух взаимодействующих ядер А и
’00 ' 50 ' ' 6 ' ' ' ' -50 ' -iOO '
Hi
₽ис.2.3. Случай слабой и сильной связи для двухспиновой системы АХ (I — 1/2), которая
переходит в систему АВ за счет уменьшения разности химических сдвигов Ад. Величина
константы косвенного спин-спинового взаимодействия /дх остается неизменной 10 Гц).
Сверху вниз: Ad/J -0, 0,5, 1,2, 5, 10 и 20.
X со спинами /= 1/2, при условии, что константа косвенного спин-спинового
взаимодействия /лх остается неизменной, а разность значений химических
сдвигов Ад (измеренная в Гц) уменьшается. В пределе достаточно сильного
61
уменьшения разности химических сдвигов реализуется приближение силь-
ной связи. В соответствии с общепринятой номенклатурой использование
букв, следующих одна за другой в алфавитном порядке, для обозначения
спиновой системы означает наличие сильной связи, например, АВ вместо
АХ, соответствующего слабой связи. Если химический сдвиг, как обычно,
измерять в м.д., то величина химического сдвига не зависит от напряженно-
сти магнитного поля. Именно на этом основано использование такой шкалы
(в м.д.). Однако если химический сдвиг измерять в Гц, то эта величина
возрастает с увеличением напряженности внешнего магнитного поля. Раз-
ность химических сдвигов, выраженная в единицах частоты, возрастает с
увеличением напряженности магнитного поля, а константа спин-спинового
взаимодействия не зависит от этой напряженности. Следовательно, при уве-
личении поля мы от приближения сильной связи переходим к приближению
слабой связи (рис.2.3 и 2.4). Следует отметить, однако, что именно зависи-
мость химического сдвига от поля осложняет сравнение спектров, снятых при
различных значениях напряженности магнитного поля.
SO 60 40 20 0 -20 -40 -60 -80
Гц
Рис.2.4. Слабая и сильная связь в системе Лз^З- Построен теоретический спектр для системы
Л2^3 Константа косвенного спин-спинового взаимодействия /дх“ 7,1 Гц, то есть соответствует
значению константы взаимодействия между метильными и метиленовыми протонами в этаноле.
Так как Д(5(измеренный в Гц) возрастает пропорционально величине внешнего магнитного
поля, то это сказывается на спектре ЯМР (за счет изменения значения рабочей частоты). Сверху
вниз: 3 МГц, 6 МГц, 15 МГц и 60 МГц.
2.1.4. Константы скалярного взаимодействия и структура
молекул
Косвенное взаимодействие двух ядерных спинов осуществляется через
электроны связи. В первом приближении можно отметить, что величина
62
этого взаимодействия убывает с ростом числа связей между ядрами. Из
анализа мультиплетности резонансных линий можно попытаться построить
альтернативную структуру для неизвестного вещества и даже выбрать наи-
более вероятную пространственную структуру.
Строго говоря, константа спин-спинового взаимодействия/ определяет-
ся не только числом связей между взаимодействующими ядрами, но также
зависит от особенностей пространственного распределения электронов. По-
скольку пространственное распределение электронов в свою очередь зависит
от диэдрального угла 0, т.е. угла на который повернуты друг относительно
друга (см. например, рис.3.3) две соседние группы ядерных спинов, связан-
ные между собой косвенным спин-спиновым взаимодействием, то и констан-
та / зависит от угла 0. Вследствие этого даже если число связей между
взаимодействующими спинами невелико, константа косвенного спии-спи-
нового взаимодействия в ряде случаев уменьшается до нуля. В дальнейшем
убедимся, что наличие зависимости константы J от угла 0дает возможность
решать достаточно сложные задачи структурной химии. Это, в частности,
позволило определить структуру такой сложной молекулы, как протеин.
2.1.5. Определение партнера по взаимодействию
Информация о химической структуре молекулы может быть получена
после того, как определено, какие из ядер данной молекулы связаны между
собой скалярным спин-спиновым взаимодействием. Величина этого взаимо-
действия позволяет сделать вывод о том, какие из ядер являются соседними
в данной химической структуре.
В рассмотренном ранее простом спектре этанола видно, что СЩ- и
СНз-группы соседствуют одна с другой. В более сложном спектре ЯМР,
состоящем из большого числа линий, достаточно сложно сделать вывод о том,
какие из взаимодействий вызывают наблюдаемое расщепление спектраль-
ных линий. В этом случае стремятся упростить спектр, применяя метод
двойного резонанса или развязку. Если в процессе детектирования на систе-
му взаимодействующих спинов подается еще одно РЧ поле, воздействующее
селективно на резонансной частоте одного из ядерных спинов, например А,
то мультиплетная структура резонансной линии, соответствующей спину
ядра X, при условии, что расщепление этой линии обусловлено спин-спино-
вой связью между спинами А и X, исчезает. Для этанола (см. рис.2.2,с)
развязка на частоте, соответствующей метиленовым протонам, приводит к
исчезновению расщепления в метильной группе. На рис.2.5 приведена схема
проведения этого эксперимента. Одновременно с возбуждающим импульсом
(поле В]) дополнительно подается импульс второго РЧ поля В2, воздейству-
ющего на частоте в течение сбора данных. Для эффективной развязки
величина поля В2 должна удовлетворять условию У > 2 ТГ/. Очевидно,
что напряженность поля развязки должна превышать напряженность поля,
создаваемого возбужденным спином. В гетероядерном случае при проведе-
нии этого эксперимента не возникает каких-либо дополнительных проблем,
поскольку разность значений частот возбуждающего поля и поля развязки
63
Передатчик
(а)
Приёмник
(Ь)
Передатчик
Приёмник
Рис.2.5. Эксперимент с развязкой от косвенного спин-спииового взаимодействия. Непрерывная
развязка (а) и развязка разделения времени (Ь). Амплитуда импульса, взаимодействующего на
частоте , используемого для регистрации, обычно много больше амплитуды импульса развяз-
ки, воздействующего на частоте ОУ^. Согласно теории, поле Шу должно включаться одновремен-
но с полем Ш2- Для того, чтобы скомпенсировать наводки, возникающие при включении поля
развязки, это поле включается уже перед включением детектирующего импульса. Приемник
включается с задержкой по сравнению с включением импульса на частоте Шу, поэтому спад
сигнала после воздействия этого импульса может быть полным.
достаточно велика. В гомоядерном случае при непрерывном облучении по-
лем В2 возникают сложности с регистрацией слабого сигнала ЯМР, посколь-
ку трудно избежать воздействия поля развязки на приемник, настроенный
на частоту детектируемого сигнала. Поэтому спиновую систему облучают
второй частотой не непрерывно, а в импульсном режиме, причем это облу-
чение синхронизировано со сбором данных (см. рис.2.5). Во время действия
импульса канал приемника остается закрытым, сбор данных осуществляется
во время интервала между импульсами (режим разделения времени), поэто-
му вовремя сбора данных значительных искажений не возникает. Несмотря
на то что облучение проводится полем В2 с напряженностью, усредненной по
всему циклу, приведенное выше условие выполняется.
В настоящее время описанные выше эксперименты с развязкой во мно-
гом утратили свой смысл. Как увидим в дальнейшем, эту же информацию
для всех партнеров по спин-спиновому взаимодействию можно получить из
одного эксперимента - двумерного ЯМР-эксперимента, требующего однако,
значительных затрат времени. Относительно больших молекул, для анализа
структуры которых необходимо определить большое число констант спин-
спинового взаимодействия, такая затрата времени вполне оправдана и, без-
условно, компенсируется получаемыми результатами. Отметим, что
существует большое число вариантов экспериментов с развязкой, которые
могут дать более полную информацию, однако используются при решении
специальных задач. Среди них можно отмстить "спин-тиклинг". Этот метод
не упрощает спектр, а наоборот, приводит к возникновению новых линий -
так называемых артефактов, которые появляются в том случае, если при
развязке мощность РЧ поля выбирается слишком малой.
64
В гетероядерном случае такие эксперименты часто направлены не на
упрощение спектров, а на получение информации о косвенном спин-спино-
вом взаимодействии, поскольку развязка приводит к полному исчезновению
мультиплетной структуры линий. С этой целью поле развязки воздействует
не селективно, на определенной частоте, а в полосе частот. Такой вариант
развязки называется широкополосной развязкой. Данный метод можно реа-
лизовать просто с помощью быстрой модуляции высокой частотой, подобно
тому, как это проводится в стандартном методе шумовой развязки. Более
эффективны современные импульсные методы (WALTZ, MLEV), которые
позволяют проводить развязку в достаточно широкой области частот и не-
посредственно учитывают свойства ядерных спинов.
Однако при использовании широкополосной развязки возникает про-
блема, состоящая в том, что образец подвергается облучению дополнитель-
ным полем в течение достаточно длительного времени. Это особенно сильно
сказывается на водных растворах и растворах электролитов, которые доста-
точно сильно поглощают РЧ излучение, что приводит к разогреву образца.
Изменение температуры сказывается на виде спектра in-vitro, а для чувст-
вительных к изменению температуры биологических молекул может приво-
дить к необратимой денатурации. При проведении исследований in-vivo это
может привести к потенциально более вредному воздействию, а именно,
разрушающему перегреву тканей.
2.1.6. Применение метода ЯМР для определения
концентраций
Задача определения концентраций с помощью метода ЯМР в идеальном
случае представляется достаточно простой. Так как площадь под резонанс-
ной линией пропорциональна числу ядерных спинов, то она пропорциональ-
на также числу соответствующих ядер. При условии, что объем образца
остается неизменным, число ядер пропорционально концентрации молекул,
содержащих эти ядра. Это позволяет по величине площади определить зна-
чение относительных концентраций, а при соответствующей калибровке —
значение абсолютных концентраций.
Рассмотрим вновь рис.2.1. Значения интегралов (площадей) под отдель-
ными резонансными линиями очевидно различаются. Соотношение площа-
дей под сигналами, соответствующими гидроксильной, метиленовой и
метильной группам, равно числу протонов в этих группах 1:2:3. Подобные
рассуждения справедливы и при определении в смеси веществ относитель-
ных концентраций присутствующих в них компонентов. При этом здесь
отсутствует необходимость в отнесении всех резонансных линий, необходи-
мо лишь установить принадлежность хотя бы одной резонансной линии к
соответствующей молекуле при условии, что в спектре отсутствует перекры-
вание спектральных линий.
Несмотря на относительную простоту такого подхода, существует, од-
нако, ряд факторов, ограничивающих точность метода. Большинство источ-
ников ошибок можно достаточно просто исключить. Поскольку методы
65
определения концентраций играют большую роль, в частности, в спектро-
скопии in-vivo, более подробно обсудим возможности устранения ошибок в
таких экспериментах.
Как было показано в разделе 1.4.2, эффекты насыщения могут сущест-
венно влиять на интенсивность резонансных линий. Так как значения вре-
мен продольной релаксации для различных групп в молекуле могут
варьироваться в широких пределах, то возможно настолько сильное насыще-
ние, что в результате будет наблюдатся полное исчезновение некоторых
резонансных линий. Это происходит в том случае, если скорость повторения
и соответственно длительность импульса, определяющая угол отклонения
вектора намагниченности, выбираются слишком большими. Если же в образ-
це присутствуют ядра с различными значениями времен релаксации и нужно
провести измерение относительной интенсивности линий с достаточно высо-
кой точностью, то необходимо построить эксперимент таким образом, чтобы
соблюдался баланс между максимальной чувствительностью и правильным
значением интегральной интенсивности резонансных линий. Максимальная
чувствительность определяется углом Эрнста, а точное определение площа-
ди под резонансным сигналом достигается тогда, когда длительность интер-
вала между импульсами выбирается из следующих соображений: спин ядра
с наибольшим значением времени релаксации должен полностью
прийти к состоянию термодинамического равновесия, прежде чем на систему
воздействует следующий импульс. Для достижения этого условия необходи-
мо, чтобы интервал времени между импульсными воздействиями на спино-
вую систему TR в несколько раз превышал максимальное из значений времен
спин-решеточной релаксации Тimax: например, TR = ЗТ[тах.
Так как возбуждающий импульс характеризуется конечной шириной,
то соответствующий спектр не для всех частот будет равноинтенсивным.
Если длительность импульса слишком велика, то не все резонансные линии
возбуждаются равномерно, что приведет, очевидно, к изменению их интен-
сивности, которая определяется разностью значений резонансной и опорной
частот.
Для улучшения отношения сигнал/шум РЧ сигнал подвергается филь-
трации с помощью аналогового фильтра (как правило, это область так назы-
ваемых промежуточных частот) .Такой фильтр позволяет пропускать только
интересующую нас область частот. Так как частотная характеристика для
аналогового фильтра не является строго прямоугольной, то сигналы по краям
спектра несколько ослаблены.
При использовании аналого-цифрового преобразователя (АЦП) сигнал,
принимающий в аналоговом представлении непрерывный ряд значений,
преобразуется в ряд целочисленных значений. В этом случае определение
площади под резонансной линией будет неточным, если на частотной оси
отсутствует достаточно число точек для хара ктеристики резонансной линии.
Точное измерение концентрации также невозможно в случае, если цифровая
фильтрация загрубляет данные о площади под резонансной линией, слишком
большой шум или перекрывание с другими сигналами затрудняет процесс
интегрирования. Если входной сигнал, поступающий на АЦП, является
очень слабым, то существляющая возможность проведения преобразования
66
позволяет получить лишь достаточно грубую информацию, так как в нашем
распоряжении имеется небольшое число значений для описания данного сигна-
ла. В предельном случае имеется только одно значение - нуль. При этом интег-
рал определяется с большой неточностью. Неточность можно устранить, если
усилить сигнал настолько, чтобы можно было располагать достаточным числом
значений. Определенные трудности возникают тогда, когда одновременно не-
обходимо проинтегрировать очень слабый и очень сильный сигналы. В этом
случае максимальное усиление входного сигнала определяется динамической
областью АЦП. В фурье-спектроскопии полный сигнал должен быть зарегист-
рирован без искажений, так как каждая точка, содержащаяся в спаде свободной
индукции, может оказывать влияние на весь спектр. Таким образом, даже
самый большой сигнал должен попадать в область значений АЦП. Для совре-
менных спектрометров с высоким значением статического магнитного поля и
соответственно высоким значением рабочей частоты, как правило, используют-
ся 12- и 16-раз^ядные АЦП, т.е. наибольший сигнал может превышать не более
чем в 4096 = 212 раз и соответственно не более чем в 65536=2Г6 раз наименьший
сигнал для того, чтобы можно было зарегистрировать эти сигналы одновремен-
но. Несмотря на то что приведенные величины весьма велики, такая ситуация
встречается на практике нередно. Если, например, нужно зарегистрировать
спектр вещества миллимолярной концентрации в воде, то сигнал воды пример-
но в 1 10000 раз интенсивнее сигнала вещества.
Если учесть все перечисленные возможности возникновения ошибок, то
при соответствующих условиях можно определять концентрацию с точно-
стью до нескольких процентов. Если используем внутренний стандарт и
добавляем строго определенное его количество, то можем достаточно точно
определить значение абсолютной концентрации. Если же условия проведе-
ния эксперимента не позволяют использовать внутренний стандарт для оп-
ределения интенсивности, то это означает, например в спектроскопии
in-vivo, что при нахождении абсолютных концентраций всегда допускается
некоторая неточность. В этом случае интенсивность резонансных линий
определяется многими внешними факторами, в частности, точным располо-
жением образца в приемной катушке и точностью настройки приемно-пере-
дающего тракта. Точность этих измерений всегда должна подвергаться
критической оценке и сопоставлениям.
2.1.7. Подавление интенсивного сигнала растворителя
Так как концентрация растворителя всегда превышает концентрацию
растворенного вещества, то проблема детектирования слабых сигналов на
фоне интенсивного сигнала растворителя возникает достаточно часто. Про-
стейший метод, позволяющий обойти эту проблему, состоит в том, что выби-
рается растворитель, не содержащий ядер, положение резонансных линий
которых совпадает с сигналами исследуемого вещества. Так, измерение
спектров на ядрах 31Р не вызывает затруднений, поскольку большинство из
используемых растворителей не содержит атомов фосфора. Однако боль-
шинство растворителей содержит протоны, так что для них наблюдаются
67
сигналы в спектрах ЯМР 1Н. Данную трудность можно обойти, используя
дейтерированные растворители, однако это также далеко не всегда примени-
мо. Наиболее часто используемый растворитель - дейтерированная вода. В
настоящее время налажен промышленный выпуск полностью дейтерирован-
ных растворителей, они нашли широкое применение. Если же по условиям
проведения эксперимента необходимо использовать протонсодержащие рас-
творители или сигнал остаточных протонов достаточно интенсивный, то этот
сигнал необходимо подавлять с использованием специальных импульсных
методов. Разнообразные методы, разработанные с этой целью, можно разде-
лить на две категории: либо в подготовительном периоде эксперимента про-
водится селективное воздействие на спиновую систему на частоте,
соответствующей сигналу растворителя, таким образом, чтобы в дальней-
шем при неселективном воздействии интенсивность сигнала от растворителя
была существенно ослаблена, либо возбуждается весь спектр, за исключени-
ем той области, в которой расположены сигналы растворителя. Наиболее
эффективен подход, в котором используется комбинация этих методов.
Рис.2.6. Методы подавления интенсивного сигнлала растворителя. На рис. схематически
представлены два типичных метода подавления интенсивного сигнала растворителя: метод
предварительного насыщения (а) ,_метод селективного возбуждения с использользованием им-
пульсной последовательности 1331 (Ь). Масштаб на рис. не соблюдается. Длительность Т
селективного импульса (воздействующего на частоте й^) , используемого для предварительного
насыщения, сравнима со значением 7"j растворителя (типичное значение Т для воды 1 с),
длительность возбуждающего импульса, воздействующего на частоте 60j, гораздо меньше
(несколько мкс). В методе (Ь) расстояние между возбуждающими импульсами определяется тем,
что в интервале Дт на расстоянии 1/227 по обе стороны от частоты опорного сигнала
располагаются максимумы возбуждения. Чтобы создать максимум возбуждения на расстоянии
1000Гц, необходимо выбрать Т-0,5 мс. (Если длительность импульса не является пренебрежимо
малой, то следует точно определить эту величину н включить ее значение в интервал Т). Дли-
тельность отдельных импульсов в последовательности соотносится, как 1:3:3:1, фаза этих им-
пульсов меняется от 0° до 180° (х и -х).
Самым распространенным является метод селективного предваритель-
ного насыщения (presaturation) сигнала растворителя, в котором использу-
ется слабый импульс большой длительности (рис.2.6). Следующий за ним
неселективный импульс возбуждает весь спектр, однако за счет предвари-
тельного насыщения интенсивность пика растворителя существенно ослаб-
ляется в 100-1000 раз. Вторую группу составляют методы, основанные на
инвертировании сигнала с помощью 180°-ного импульса и возбуждении спу-
стя время Тполного спектра, причем длительность этого интервала выбира-
68
ется такой, чтобы к моменту сбора данных намагниченность от сигнала
растворителя была равна нулю.
Недостатком описанных выше методов является то, что в процессе про-
ведения эксперимента влиянию подвергаются также и другие области спек-
тра, в частности, возбуждаются сигналы, расположенные вблизи сигналов
растворителя. В дальнейшем рассмотрим механизмы, обусловливающие, на-
пример, такие эффекты, как ядерный Оверхаузера (ЯЭО) и перенос поляри-
зации.
Методы полуселективного возбуждения обладают тем преимуществом,
что в них нежелательные эффекты отсутствуют. Полуселективные возбуж-
дающие импульсы создаются либо в виде импульсов большой длительности
и низкой интенсивности (soft pulses), либо в виде последовательности им-
пульсов большой интенсивности (hard pulses). Первая группа методов позво-
ляет получить спектр любого вещества, но реализация этих методов на
стандартном спектрометре сопряжена со значительными трудностями, так
как требует проведения специальной настройки и специального блока, кото-
рый, кстати, входит в комплект ЯМР-томографа. На практике более часто
используется простая в настройке импульсная последовательность 1331 (см.
рис.2.6). Принцип работы этой последовательности состоит в следующем:
отдельный импульс позволяет полностью "вырезать" определенную часть
спектра, а во всей остальной области сигналы компенсируются неполностью,
что позволяет провести их регистрацию. Недостатком этого метода является
сложность корректировки фаз линий в спектре.
2.2. Зависящие от времени процессы в ЯМР
Процессы спиновой релаксации за счет модуляции во времени различ-
ных взаимодействий - характерный, но далеко не единственный пример
явлений, которые могут быть изучены методом ЯМР. Эти процессы в шкале
времени ЯМР часто характеризуются либо очень малыми, либо очень боль-
шими временами. Понятие шкалы времени основано на простой концепции,
которая находит применение во многих областях. Если речь идет о медлен-
ном процессе, то предполагается, что он может быть описан последовательно,
на каждом из этапов. При рассмотрении быстрого процесса предполагается,
что наблюдаем за неким усредненным поведением системы. Здесь необходи-
мо сразу же пояснить, что понятие медленных и быстрых процессов вводится
лишь тогда, когда приводится шкала времени, в которой проводится рассмот-
рение.
Простейший путь состоит в том, чтобы задать предельные значения
времен, и в этом случае все физические явления описывать как быстрые или
медленные по сравнению с этими значениями. Очевидно, что времена, зада-
ющие масштаб, зависят также и от того, какие физические процессы описы-
ваются в данном приближении. При этом существует не одна временная
шкала ЯМР, а множество различных шкал в зависимости от типа рассматри-
ваемых взаимодействий. С теоретической точки зрения наиболее сложной
для обсуждения является промежуточная область, в которой не реализуются
69
uua предельных случая: быстрых и медленных процессов. Независимо от
используемой шкалы ЯМР физические процессы, связанные с различными
движениями, обладают своей собственной шкалой времени, которая может
быть описана временами корреляции или характеристическими частотами.
В табл.2.4 представлены типичные интервалы частот для основных молеку-
Таблица 2.4. Характерные значения частот молекулярных движений
Тип движения область значений частот (с1) |
Вибрационные и торсионные движения 101и-101л
Латеральная диффузия в мембранах 107-1010
Диффузия в растворах ю6- ю13
Вращательная диффузия в растворах 105-1012
Вращение алифатических боковых цепей в молекулах 107 - 1010
Вращение ароматических боковых цепей в молекулах 10'2- 106
Конформационные изменения протеинов 10'5- 105
Высвобождение субстратов при ферментативных реакциях 10‘3- 106
лярных движений: от очень медленных - с характеристическими временами
от нескольких минут до нескольких часов до очень быстрых - в пикосекунд-
ной области. Существуют разнообразные резонансные методы, позволяю-
щие регистрировать процессы, протекающие с характерными временами до
10 пс. Только такие сверхбыстрые процессы, как торсионные и вибрационные
движения атомов, не поддаются прямому наблюдению с помощью методов
ЯМР. Кроме того существуют макроскопические движения: потоки жидко-
стей и мышечные сокращения. Эти процессы играют второстепенную роль в
спектроскопии in-vitro, ориентированной на биохимические проблемы, од-
нако в ЯМР-томографии и в in-vivo-спектроскопии роль таких процессов
достаточно велика. Таким образом, характерные времена процессов, регист-
рируемых с помощью ЯМР, попадают в широкую область, охватывающую
несколько порядков на временной шкале, при этом каждый из ЯМР-экспе-
риментов характеризуется определенной временной областью, которая соот-
ветствует исследуемым процессам.
2.2.1. Временная корреляция и спектральная плотность
Молекулярные движения в жидкостях, например, вращения боковых
цепей, не описываются определенными частотами на больших временных
70
интервалах: мгновенная скорость вращения может варьироваться от одной
боковой цепи к другой и от молекулы к молекуле, а за счет соударений с
молекулами растворителя скорость вращения может изменяться как по ве-
личине, так и по направлению. Это означает, что молекулярные движе-
ния обладают широким спектром частот, который также присущ
флуктуирующим магнитным полям, возникающим в результате движе-
ния атомов и молекул и ответственным за релаксационные процессы в
спиновой системе.
Так как в растворе большое число одинаковых молекул находится
в состояниях, характеризуемых разнообразными движениями, то при
вычислении тех или иных величин проводят усреднение по ансамблю
молекул, а также по большому промежутку времени. Поведение во
времени какой-либо случайной величины f(t), например, флуктуиру-
ющего магнитного поля, обычно описывают с помощью корреляцион-
ной функции
G Г) >, (2.3)
где угловые скобки означают усреднение по ансамблю, а черта - усреднение
по времени. Усреднение по всем значениям времени t применимо при описа-
нии стационарного состояния, для которого с точки зрения статистики не
имеет значения, относительно какого момента времени определена корреля-
ционная функция. Для случайных процессов таких, например, как движе-
ние молекул в состоянии термодинамического равновесия, корреляционная
функция максимальна для Т = 0 и убывает во времени. Для больших Т она
должна стремиться к нулю, так как по определению для случайных процес-
сов корреляции должны уменьшаться при условии, что наблюдение за сис-
темой ведется в течение достаточно длительного времени. Ширина
корреляционной функции обычно описывается параметром, называемым
временем корреляции Тс. Эта величина характеризует определенный вид
движения и является мерой того, на каком временном интервале значения
случайной величины еще можно рассматривать как коррелированные (т.е.
G (Т) отлична от нуля).
Фурье-образ корреляционной функций G( Т ) называют спектральной
плотностью J (Ш):
00 — iWt
J(T)=f G(r) е dr. (2.4)
— 00
Точное значение времени корреляции и само определение этого парамет-
ра зависит от соответствующей корреляционной функции G<X), вид которой
определяется конкретным физическим процессом. Во многих случаях корре-
ляционную функцию можно достаточно хорошо описать экспонентой
G(r) =G(0) е
-(iri/pc)
(2.5)
71
Соответствующая спектральная плотность J (О)), которую можно по-
лучить фурье-преобразованием этой корреляционной функции, имеет ло-
ренцеву форму:
00 —(|Т|/т) -iCOT 2тг
/ (tz))= / G (0)е е dT=G(O)-----------• (2.6)
- оо 1 +аг тгс
Для случая диффузии время корреляции обычно легко поддается интерпре-
тации. Это среднее время между двумя соударениями данной молекулы с
другими молекулами. Полуширина лоренцевой функции, как видно из (2.6),
равна 1/Гс. Таким образом, уменьшение времени корреляции, очевидно,
расширяет спектральную область, на которой спектральная плотность от-
лична от нуля. Площадь под лоренцевой кривой постоянна и не зависит от
времени корреляции.
Время корреляции Гс служит характерным параметром для проведения
сравнений в шкале времени ЯМР, о которой речь шла ранее. Сама шкала
времени определяется максимальной величиной взаимодействия А О), лежа-
щего в основе наблюдаемого физического явления (амплитуда взаимодейст-
вия А (У здесь измеряется в единицах частоты). Процесс, характеризуемый
временем корреляции Гс, является медленным, если
|Ашгс | » 1, (2.7)
и соответственно быстрым, если
|Аш тс | « 1. (2.8)
2.2.2. Химический обмен
Химический обмен - один из наиболее наглядных примеров динамиче-
ских процессов. Сущность этого явления ясна из интуитивных соображений.
Под химическим обменом в общем случае понимают процессы, в которых
спин ядра в процессе релаксации может находиться в состояниях, характе-
ризуемых различным химическим окружением, что соответствует различ-
ным параметрам ЯМР. В основе изменения окружения ядерного спина может
лежать внутримолекулярный процесс, такой, например, как изменение кон-
формации, или же межмолекулярный процесс. Здесь можно рассматривать
исследуемое ядро в новой ковалентной структуре или же, наоборот, вклю-
чать в рассмотрение межмолекулярные взаимодействия тех молекул, кото-
рые содержат данное ядро, и таким образом учитывать изменение окружения
данного ядра. В простейшем случае имеется только два различных состояния,
которые отличаются по химическому сдвигу д. Время корреляции гс здесь
представлено временами жизни ТА и Гд в состояниях Ап В соотвественно, а
величина А (О непосредственно определяется разностью химических сдвигов
А <5 = дв — дА, измеренной в единицах частоты. В этом случае медленный
обмен определяется неравенством
72
|Adre|»l, (2.9)
а быстрый обмен -
|Д<5ге|«1, (2.10)
где
^1=7Л’1+ГВ'1. (2.11)
Соответствующие спектры ЯМР могут быть достаточно хорошо описаны
обобщенными уравнениями Блоха, полученными в работе Мак-Коннела
[2.1 ]. Оба предельных случая: быстрого и медленного обмена, можно объяс-
нить без привлечения серьезных расчетов, а чисто из интуитивных сообра-
жений путем предельных переходов те -*0 и соответственно те -* со. В первом
случае можно наблюдать лишь одну линию, так как ядро участвует в беско-
нечно быстром обмене между двумя состояниями с различным химическим
окружением. Во втором случае можно наблюдать две резонансные линии с
химическими сдвигами дл и <3в, интенсивности которых прямо пропорцио-
нальны относительным заселенностям (концентрациям) рл и рв, т.е. в дан-
ном случае обмен вообще отсутствует. Усреднение положений резонансных
линий при быстром обмене приводит к химическому сдвигу
д = РлдА+Рв &в- (2.12)
Для описания поведения в промежуточной области такое простое соотноше-
ние отсутствует, однако можно вычислить в явном виде положение сигнала
ЯМР как функцию времени корреляции обменного процесса Те (рис.2.7).
Условия медленного обмена приводят к уширению обеих линий, а при уско-
рении обмена - к слиянию их в одну уширенную линию при выполнении
условия! Адге I — 1. При дальнейшем возрастании скорости обмена линия
будет сужаться до тех пор, пока ее ширина не станет равной ширине линии
в отсутствие обмена.
Аналогичный процесс усреднения происходит и в случае других взаимодей-
ствий. Если окружения, соответствующие двум состояниям А и В, различаются
только константами косвенного спин-спинового взаимодействия /л и JB, то
наблюдается мультиплет с усредненной константой/' = pAJА+ рд!в до тех пор,
пока для величины А/ = JA — JB выполняется неравенство I А/ тс 1« 1
(константы J выражаются в единицах круговой частоты). Так как константы
взаимодействия/, как правило, не коррелируют с разностью химических сдви-
гов, то в ряде случаев на основе этих констант должна быть установлена шкала
времени, отличная от шкалы химических сдвигов. При наличии в спиновой
системе взаимодействия, например косвенного спин-спинового, химический
обмен не поддается описанию с помощью простых уравнений Мак-Коннела
[2.1 ]. Однако принципиально возможно описание поведения такой системы с
привлечением квантово-механических подходов.
Какую биохимическую информацию можно извлечь из изучения про-
цессов, связанных с химическим обменом? В области быстрого обмена в
73
Рис.2.7. Изменение формы линии в зависимости
от времени корреляции обменного процесса Tg.
Теоретический расчет спектра для двухпозици-
онного обмена в зависимости от I Дй)Ге1'* по
уравнению Мак-Коннела для константы равно-
весия К - рЛ/рв, равной 1 (а) и 2 (Ь).
-50 0 50 (у/с’1 Г-50 0 50 (1)!с '
принципе можно рассчитать населенности отдельных состояний при условии
наличия некоторой дополнительной информации. При этом наиболее важ-
ной является информация о числе состояний, участвующих в быстром обме-
не, так как это число присутствует в расчетной формуле в качестве
параметра. Для описания быстрого обмена между N состояниями следует
обобщить уравнение (2.12) на случай N состояний:
N
<5=2 рА
i — L
(2.13)
с учетом того, что в уравнении (2.13) необходимо экспериментально опреде-
лить значения д/. Например, в простом случае равновесной населенности
можно определить значение химического сдвига в отсутствие партнера (pi = 1,
Р2 = 0) ив случае, когда населенность второго партнера будет существенно
большей (pi = 0, р2 = 1). В общем случае в расчет следует включить еще
сведения (модельные представления) о функциональной зависимости pt.
Если найдено, что в реальном случае спектр ЯМР позволяет наблюдать
случай быстрого обмена, то отсюда непосредственно следует, что время кор-
реляции процесса обмена те меньше чем 1/1 Д д I.
74
В случае медленного обмена спектр будет более информативным. Наи-
меньшее число различных состояний можно вычислить из числа резонанс-
ных линий, а соответствующие значения населенностей можно непосредст-
венно получить из величины интеграла для каждой резонансной линии. Это
позволяет зафиксировать с помощью ЯМР различные состояния, которые
ранее удавалось зафиксировать только косвенно, например с привлечением
кинетических методов. Кроме того, здесь можно получить ограничение на
значение времени корреляции обмена это значение должно быть больше
чем 1/ I Ad I. В промежуточной области вблизи I Ад Те I — 1 обмен вызывает
существенные изменения формы линии. Это позволяет путем построения
теоретического спектра достаточно точно определять значения времени кор-
реляции обменных процессов Тх в различных состояниях X. В области уме-
ренно медленного обмена между экспериментально наблюдаемыми
значениями времени поперечной релаксации Т2Х (соответственно между
значениями ширины спектральной линии 1 / Т2Х ) > времени релаксации Г2А-
в отсутствие обмена (Тх= °°) и времени корреляции обмена Тх в состоянии х
существует следующее соотношение:
(Т2Х' У1 = (Т2Х)'1 + (ТХУ1 . (2.14)
В случае медленного обмена о числе состояний следует судить с большой
осторожностью, поскольку ЯМР дает сведения лишь о минимально возмож-
ном числе состояний системы. Можно привести многочисленные соображе-
ния, согласно которым дополнительные состояния не будут наблюдаться в
спектре. Соответствующие этим состояниям резонансные линии могут быть
сильно уширены, могут перекрываться с другими линиями или вообще не
поддаются детектированию, поскольку между некоторыми из состояний су-
ществует быстрый обмен.
2.2.3. Зависимость величины химических сдвигов от pH
Многие биологически важные молекулы содержат функциональные груп-
пы, которые в водном растворе могут находиться либо в протонированной, либо
в депротонированной форме. В общем случае в растворе имеет место равновесие
между различными формами этих молекул, что определяется концентрацией
ионов водорода. Химическое равновесие в соответствии с законом действующих
масс может быть описано константой равновесия К, которая определяет отно-
шение концентраций молекулярных форм, находящихся в равновесии. Простое
кислотно-основное равновесие для кислоты АН описывается логарифмической
функцией в соответствии с уравнением Хендерсона-Хассельбалха
pH = рК + 1g -^1 • (2.15)
Различные зарядовые состояния молекул приводят к различию в элект-
ронном распределении, что, как правило, сказывается на значениях хими-
ческих сдвигов дл_ и дАН соответствующих форм.
75
В большинстве случаев реакции протонирования — депротонирования
являются быстрыми в шкале времени ЯМР, т.е. мы наблюдаем усредненное
значение химического сдвига <5, величина которого определяется значением
pH. В этом случае уравнение (2.15) вместе с уравнением (2.12) дает модифи-
цированное уравнение Хендерсона-Хассельбалха
1 л рн ~рк
<5 = ^АН + ( дА~ ~ ^АН ) J 10 pH-рк ‘ <2' )
Если измерить зависимость химического сдвига от pH и описать эту зависимость
с помощью уравнения (2.16), подбирая рХ,тонепосредствешюизЯМР-эксперимен-
тов удается получить значения рК для индивидуальных групп. Это особенно важно,
если речь идет, например, о спектрах1Н протеинов, поскольку данный метод доста-
точно точно позволяет определить значения рК каждого заряженного остатка.
Неорганический фосфат принимает участие во многих ферментативных
реакциях, поскольку он либо переносится на субстрат, либо расщепляется.
Кроме того, неорганический фосфат принимает на себя функцию буфера.
Для фосфатной группы существуют кроме двух различных протонирован-
ных форм в равновесии еще и следующие формы: Н3РО4, Н2РО4‘, НРО4 ,
РО43'. При измерении спектров ЯМР 31Р для различных значений pH мы
всякий раз наблюдаем один сигнал, химический сдвиг которого изменяется
в зависимости от концентрации ионов гидроксония (рис.2.8). Такой вид
I - -т . . .|...7........................
с • 11 & • U 4.0 З.о 2.0 1.0 0.0
М.Д.
Ч1
^рс.2.8. Спектр ЯМР 1Р неорганического фосфата при различных значениях pH. СпектрыЯМР
Р неорганического фосфата сняты в фосфатном буфере (концентрация 10 ммоль) в за-
висимости от pH. Показано, как начиная с pH - 13.2, меняется вид спектров с уменьшением pH.
Значение pH уменьшается от спектра к спектру примерно на 0,3 единицы.
76
спектра полностью соответствует случаю быстрого обмена. Если по одной из
осей отложить значение химического сдвига, а по другой pH, то можно
построить кривую титрования, состоящую из четырех интервалов. Отсюда
следует, что вещество, участвующее в данном процессе быстрого обмена,
существует по крайней мере в четырех различных формах, которые харак-
теризуются значениями химических сдвигов <53, <32, (53 и <34, что полностью
соответствует предположениям о четырех состояниях протонирования неор-
ганического фосфата, полученным из химического подхода.
В данном случае функциональная зависимость является несколько бо-
лее сложной, чем описываемая уравнением (2.16), так как здесь для описа-
ния равновесия необходимо использовать более одного значения рК.
Используя закон действующих масс, можно расширить уравнение (2.16) на
/У значений рК{ Если кооперативные эффекты отсутствуют, то для А значе-
ний рК^ можно просто записать сумму
N pH - pKi
6 = а, + S (<Vi - а,) - ал?»
i+io’"'
Если значения р существенно отличаются друг от друга, как это имеет
место на рис.2.8, то вблизи каждого из значений р Ki можно достаточно точно
построить значения химических сдвигов, используя простое уравнение
(2.16). Этот метод применяется достаточно часто, если необходимо постро-
ить какой-либо из участков кривой титрования.
2.2.4. Образование комплексов с диамагнитными лигандами
Комплексообразование молекул с диамагнитными лигандами подчиня-
ется тем же правилам, что и процесс протонирования, зависящий от значе-
ния pH. Однако скорости обмена часто настолько малы, что условие быстрого
обмена (2.10) не выполняется. Интересными с точки зрения биологических
приложений являются комплексы с ионами металлов, которые играют роль
катализаторов в большинстве ферментативных превращений. Особенно ча-
сто встречаются комплексы двухвалентных металлов таких, как магний и
кальций. Основным веществом в биологических реакциях является адено-
зин-5-трифосфат (АТф), образующий в физиологических условиях комп-
лекс с ионом Mg .
Устойчивость Mg.ATO-KOMnneKca зависит от температуры. В больших
магнитных полях (11 Тл) переход от медленного к быстрому обмену проис-
ходит примерно при 25°С. При низких температурах можно отдельно наблю-
дать сигналы от Mg-АТФ и свободного АТФ. При повышении температуры
наблюдается усредненная резонансная линия, положение которой зависит от
концентрации реагентов (рис.2.9).
Анализ формы линии позволяет достаточно точно найти значение ско-
рости обмена и энергии активации, которые определяются используемой
77
Рис.2.9. Спектр ЯМР комплекса Mg. АТФ и свободного АТФ при
различных температурах. Спектры ЛГР сняты при рабочей частоте
203 МГц, что соответствует рабочей частоте в спектрах *Н 500 МГц.
Сигнал с хим. сдвигом -5 м.д. соотвествует фосфатной группе в у-по-
ложенни, а сигнал с хим.сдвигом -10,3 м.д. — фосфатной группе в
бХ-положеиии. При температуре 274 К сигналы фосфатной группы в
^-положении в Mg-АТФ и в свободном АТФ наблюдаются отдельно.
Так, в комплексе Mg.АТФ химический сдвиг, соответствующий данно-
му положению, равен -18,6, а в свободном АТФ - -21,2 м.д. По мере
роста температуры скорость обмена между состоянием,
соответствующим свободному иону Mg + и соостоянием в комплексе
возрастает и разность значений химических сдвигов уменьшается до
тех пор, пока две линии не сливаются в одну [2.2].
29S
293
290
280
274
К
К
К
К
Таблица 2.5. Значения энергий активации и скоростей обмена в комплексе М^.АТФ
AG* /кДж.М’1 k-i/c 4 (1бС) М/с’1 (10°С) М/с-1 (20°С) k-i/c-1 (30°С) k.j/c"1 (37°С)
41 400 600 1100 1900 2900
Скорость обмена k_ । -1 /Тд (Гд - время жизни комплекса Mg-АТФ) и энергии активации
AG* сильно зависят от значения pH раствора и от типа ионов, содержащихся в этом растворе
(например, ионов буферного раствора). Приведенные в табл, значения вычислены из спектров,
показанных на рис. 2.9.
буферной системой (табл.2.5). В отличие от этого комплекса время жизни
комплекса с кальцием настолько мало, что при самых сильных магнитных
полях выше точки замерзания воды реализуется случай почти быстрого
обмена.
Во многих реакциях ферментативного превращения Mg-АТФ сам явля-
ется субстратом, поэтому ферментативная активность прежде всего опреде-
ляется концентрацией свободного Mg-АТФ. Это значение концентрации
можно непосредственно определить по спектрам ЯМР из интенсивности со-
ответствующих резонансных линий при понижении температуры. При более
высоких температурах можно косвенно судить о концентрациях по химиче-
скому сдвигу, если известна константа связывания АТФ с магнием в данных
экспериментальных условиях.
78
2.2.5. Перенос поляризации с насыщением
В области медленного обмена используется метод, основанный на пере-
носе поляризации с насыщением. С помощью этого метода часто удается
провести более точное и однозначное определение времен корреляции об-
менных процессов по сравнению с данными, получаемыми из анализа формы
линии. Кроме того, метод переноса поляризации с насыщением позволяет
получить дополнительную информацию о том, действительно ли две наблю-
даемые в спектре резонансные линии соответствуют двум состояниям, между
которыми осуществляется обмен. Реализация этого метода достаточно про-
ста. С помощью импульса длительностью г, воздействующего селективно,
т.е. на определенной частоте, проводится насыщение линии В и измерение
обычного спектра ЯМР. Этот эксперимент является примером использова-
ния метода селективного насыщения для подавления пика растворителя.
Пусть в момент времени т положение спина, резонансная линия которо-
го подлежит насыщению, изменится и осуществится переход из положения
В в положение А. В этом случае интенсивность резонансной линии, соответ-
ствующей положению А, уменьшится. Эффективность этого метода, с одной
стороны, определяется длительностью интервала г, в течение которого осу-
ществляется обмен, и скоростью обмена в системе 1/ ТА. С другой стороны,
эффект уменьшается по мере установления равновесного распределения
Больцмана, которое характеризуется временем продольной релаксации Т1А.
Формально интенсивность 1А резонансной линии А в зависимости от времени
Т, в течение которого проводится насыщение резонансной линии В, описыва-
ется следующим выражением:
/л(г) = 1/тА + 1/г1л ехр + ] + (2Л8)
Для времени насыщения при условии, что оно существенно больше
значения Т}А, устанавливается состояние равновесия и отношение интен-
сивностей линий при наличии или отсутствии насыщения дается форму-
лой
М00)- 1
ТПоТ 1 + т1А/та
(2.19)
Существует еще одна возможность пометить спин В. Эта возможность
может быть реализована посредством инверсии намагниченности, соответст-
вующей спину В, селективным 180°-ным импульсом (inversion transfer). При
этом эффект воздействия на состояние спина ядра А может быть обнаружен
с помощью неселективного детектирующего импульса, который подается на
спиновую систему спустя время г. В этом эксперименте с ростом т прежде
всего уменьшается интенсивность сигнала от ядра А до значения, соответст-
вующего минимальному, а затем возрастает вновь до величины, соответст-
вующей значению для невозмущенной системы.
79
Источниками погрешностей, которые могут вызывать неверную интер-
претацию результатов, является ограниченная селективность инвертирую-
щих и насыщающих импульсов, которые могут искажать соседние линии, а
также вызывать ядерный эффект Оверхаузера, неявным образом влияющий
на интенсивность резонансных линий, в частности, на сигнал А.
2.2.6. Ядерный эффект Оверхаузера
Изменение интенсивности сигнала в спектрах ЯМР за счет насыщения
соответствующих резонансных состояний спина электрона впервые было
обнаружено А.В.Оверхаузером [2.2]. Впоследствии это явление было назва-
но эффектом Оверхаузера. Перенос принципов этого эксперимента на сис-
тему двух спинов, связанных между собой диполь-дипольным взаимодейст-
вием, является основой для определения пространственной структуры мето-
дом ЯМР. Эффект Оверхаузера и ядерный эффект Оверхаузера (ЯЭО) осно-
ваны на изменении поляризации, наблюдающейся в связанной системе
при условии изменения населенности одной или нескольких подсистем.
Так как диполь-дипольное взаимодействие, определяющее изменение на-
селенностей, зависит от расстояния между взаимодействующими спина-
ми, то величина ЯЭО также зависит от расстояния между ними. Эта
зависимость позволяет определить соответствующие межатомные рассто-
яния.
Экспериментальные методы, с помощью которых наблюдается ЯЭО,
подобны методам, используемым для определения скоростей обмена в слу-
чаях переноса поляризации с насыщением или инверсного переноса поляри-
зации. В данном контексте эти методы известны, как TOE (truncated driven
Overhauser effect) и переходный ЯЭО. Кроме того ЯЭО измеряется в двумер-
ных ЯМР-экспериментах, которые могут быть использованы для определе-
ния скоростей химического обмена. Обсуждению этих методов посвящен
раздел 2.3.
Простейшей системой, позволяющей достаточно наглядно продемонст-
рировать ЯЭО, является молекула, в которую входят два спина А и В, свя-
занные диполь-дипольным взаимодействием, причем 1А 1В - 1/2. Если
пров ести усреднение по большому числу таких молекул, то каждой молекуле
можно поставить в соответствие 4 возможных состояния, характеризуемых
определенными населенностями, т.е. установить, с какой вероятностью на-
селено каждое из состояний (рис. 2.10). В состоянии термодинамического
равновесия имеет место распределение Больцмана. Если нужно изменить
населенности, облучая спиновую систему на частоте, соответствующей ре-
зонансной частоте (1)в спина В, то вследствие диполь-дипольного взаимодей-
ствия произойдет изменение населенностей всех спиновых состояний. При
условии, что облучение проводится в течение длительного времени, в систе-
ме установится новое стационарное состояние. Возникающие при этом ре-
лаксационные процессы можно в зависимости от изменения суммарного
квантового числа т ядер А и В обозначить как нульквантовые ( Ат “ 0),
одноквантовые ( Ат = ±1) и двухквантовые ( Ат = ±2).
80
Рис.2.10. Вероятности переходов в двух-
спиновой системе. Схематическое предс-
тавление различных возможных ориен-
таций магнитного момента в двухспиновой
системе. Wy и обозначают вероят-
ности переходов между различными
состояниями. Состояния, обозначаемые
IFq обычно не обладают одинаковой энер-
гией, так что на рис. приведено условное
изображение этих уровней
Изменения населенностей приводят также к изменению z-компоненты
намагниченности спина А, величину которой к моменту времени t можно
определить с помощью 90°-ного импульса. После фурье-преобразования
найдем изменение интенсивности соответствующей резонансной линии на
частоте й>А.
Эти изменения намагниченности во времени можно описать с помощью
дифференциальных уравнений
- -рА (М* - - адв - Мв), (2.20)
б/
-jf » -рв <мв - Мв) - аВА - лф. (2.21)
В этих уравнениях рА и рв обозначают скорости спин-решеточной ре-
лаксации ядер Л и В, о^в _ скорость релаксации, описывающая обмен между
ядрами А и В, Мо - намагниченность в состоянии термодинамического рав-
новесия. Скорости релаксации определяются следующим образом:
PA=^oB+2WlA + W2B + К ext'
рв » + 2W^ + W™+ RBxt,
(TAB = (rBA = W^-W^B,
(2.22)
(2.23)
(2.24)
где Wo, Wi и Wz - вероятности нуль-, одно- и двухквантовых переходов, а
обозначает дополнительные каналы релаксации для спинов А и В.
Ядерный эффект Оверхаузера измеряется отношением интенсивности
сигнала S' после возбуждения к интенсивности сигнала So в отсутствие воз-
81
буждения или отношениемMZMO, поскольку интенсивность сигнала пропор-
циональна величине z-компоненты намагниченности. Если в течение време-
ни Т, которое существенно больше времени продольной релаксации системы,
на систему воздействует РЧ импульс на частоте й)» с мощностью, достаточ-
ной для насыщения переходов на частоте 60ц ( ув Т\Т2 >> D, то это
приведет к тому, что в стационарном состоянии MZB, dMzldt и dMB Idt в
(2.20) и (2.21) обратятся в нуль. Простым преобразованием и подстановкой
для оценки ЯЭО в стационарном состоянии получим
м* Ра м*
(2.25)
Так как равновесная намагниченность пропорциональна гиромагнитному
отношению у, то (2.25) можно переписать в виде
МА V + PA Ya
Вероятности переходов для продольной релаксации даются следующими
формулами:
(2.27)
(2.28)
И^в=|/вУ(й£), (2.29)
И^В=3/Ъ ((«£ + (/£). (2.30)
Если движение, влияющее на релаксационные процессы, изотропно, как,
например, вращательная диффузия, то коэффициент , который пропор-
ционален интенсивности взаимодействия, имеет вид
2
1
10 Utt )
(2.31)
ГАВ
где /л0 - магнитная проницаемость вакуума, гав ~ расстояние между спинами
А и В.
Соответствующая функция спектральной плотности J( ш) есть
-------- го* 7
1 )2
(2.32)
82
Рнс.2.11. Величина ЯЭО как функция вре-
мени вращательной релаксации Trot. За-
висимость Л/j от времени
корреляции вращательных движений Trot
урн условии, что резонансная частота для
Н равна 500 МГц, приведена для различ-
ных ядер, связанных с протонами дополни-
тельным взаимодействием. Эти зави-
симости построены при условии пренебре-
жения внешней релаксацией и при усло-
вии, что в течение длительного времени на
систему оказывается воздействие, вызыва-
ющее насыщение протонного резонанса (стан-
дартный ЯЭО).
Если подставить эту функцию в выражение для р и а, то получим
зависимость величины стационарного эффекта Оверхаузера от вращатель-
ного времени корреляции Trot; на рис.2.11 эта зависимость показана для
различных ядер. При расчете приведенных кривых не учитывалась внешняя
релаксация, включение которой приводит к уменьшению наблюдаемого эф-
фекта. Следует отметить, что при уменьшении внешней релаксации зависи-
мость отношения М^/М° от расстояния гАВ между спинами исчезает. Для
малых значений времени корреляции, что характерно прежде всего для
малых молекул в жидкостях малой вязкости, в качестве предельного значе-
ния получим
^£ = 1 +Ш
М* 17а
(2.33)
Если насыщаемый спин В - протон и гиромагнитное отношение уА > О,
то получим усиление резонансной линии А. Для 13С согласно уравнению
(2.33) получим максимальное усиление сигнала в 2,988 раза, если проведем
насыщение по протону, непосредственно связанному с углеродом. Это уве-
личение чувствительности является дополнительным, поскольку при изме-
рении спектров ЯМР 13С обычно используется широкополосная развязка.
Правда, необходимо помнить, что ЯЭО не всегда приводит к увеличению
интенсивности. Ядра 15N и 29Si имеют отрицательное гиромагнитное отно-
шение, и ЯЭО приводит к уменьшению сигнала (см. рис.211).
Если внешняя релаксация отлична от нуля, то стационарный ЯЭО зави-
сит от расстояния между спинами, и его можно использовать для определе-
83
3,0 7,0 6,0 м.д.
Рис.2.12. Разностный спектр HPr-протеииа, позво-
ляющий оценить величину ЯЭО. Ароматическая
область спектра *Н (360 МГц) НРг-протеина
Staphylococcus aureus. Эксперимент по методу ТОЕ
с предварительным насыщением течение 0,8 с.
(а) Внерезонансное облучение, (Ь) Насыщение на
частоте метильных протонов при-0,18 м.д.,
(с) Разность спектров (а) и (Ь) [2.3].
ния этого расстояния. Однако для количественного определения расстояний
необходимо дополнительное рассмотрение эффекта с учетом внешней релак-
сации.
Межатомные расстояния можно определить более точно, если просле-
дить за временной зависимостью ЯЭО. На рис.2.12 показано влияние пере-
носа поляризации с насыщением (ТОЕ) между метильной группой и
ароматическим кольцом в HPr-протеине, который является компонентой
фосфоенолпируватзависимой системы фосфотрансферазы. Влияние насы-
щающего импульса, который подается на частоте (Ив, можно лучше всего
оценить количественно, если кроме спектра в режиме предварительного
насыщения на частоте (Ив (режим on-resonance) снять также исходный
спектр в режиме предварительного насыщения на частоте Шв , т.е. спектр, в
котором резонансная линия на частоте (Ив отсутствует (режим off-
resonance) . Разностный спектр позволяет селективно оценить эффект Овер-
хаузера. Величина наблюдаемых эффектов часто может составлять
несколько процентов, т.е. относительно мала. Для получения хорошего зна-
чения отношения сигнал/шум необходимо провести большое число накопле-
ний. Небольшие нестабильности в работе прибора могут вносить искажения
в разностную картину, и часто именно этим объясняется необходимость
построения циклического эксперимента, причем попеременно проводится,
например, 16 экспериментов в режимах on-resonance и off-resonance, а соот-
ветствующие спады свободной индукции накапливаются отдельно. С по-
мощью этого метода можно наблюдать на современном спектрометре ЯЭО
величиной в несколько процентов. Если построить зависимость величины
84
ЯЭО от длительности насыщающего импульса (ТОЕ) или длительности интерва-
ламежду 180°-ними 90°-ным импульсами (переходныйЯЭО), то первоначальное
возрастание определится межатомным расстоянием между рассматриваемы-
ми спинами. Эта величина пропорциональна гд^6. Определение расстояний с
помощью эффекта Оверхаузера является существенным элементом алгоритма
установления структуры биологических макромолекул с помощью ЯМР. К более
детальному обсуждению этого вопроса вернемся в главе 3.
2.2.7. Использование стабильных изотопов в качестве меток
Введение радиоактивных изотопов в качестве меток - один из стандарт-
ных методов, используемых в биохимии для установления путей протекания
реакций. В ЯМР он также является основным экспериментальным методом,
однако при условии, что используются стабильные изотопы. Если ход хими-
ческих реакций известен точно, то этот метод может быть использован в
качестве вспомогательного для проведения отнесения сигналов в спектрах.
Если, например, в определенном положении протон замещается на дейте-
рий, то отсутствие соответствующей линии в спектре является точным дока-
зательством того, что именно этот протон занимает данное, определенное
положение. Для того чтобы иметь возможность проследить за ходом проте-
кания реакций, необходимо заменить изотоп, регистрируемый обычно в
спектрах ЯМР, на другой изотоп, и затем проследить за изменением интен-
сивности соответствующей линии в спектре ЯМР.
Типичным примером такого рода замещения является замена 14N на
15N, 12С на 13С и ]Н на 2Н. Это изотопное замещение наиболее интересно
при проведении исследований методом ЯМР in-vivo, что будет рассмотрено в
главе 5. Трудности возникают только для ядер 31Р, для которых отсутствуют
стабильные изотопы, при использовании которых можно было бы провести
изотопное замещение обычными методами. Здесь ситуацию можно сущест-
венно упростить, проведя маркировку по всей группе. Можно проследить за
поведением фосфатной группы в процессе реакции, если провести маркиров-
ку по изотопам кислорода 17О или 18О, естественное содержание которых
весьма невелико.
Проведение изотопного замещения по 17О, спин которого равен 5/2,
приводит к расщеплению сигнала фосфора в секстет. Наличие квадруполь-
ного момента у кислорода уменьшает время релаксации ядерного спина 17О.
Спин-спиновое взаимодействие с ядром 31Р позволяет получить представле-
ние об эффективных механизмах, дающих дополнительный вклад в релак-
сацию. Если с атомом фосфора связаны больше одного атома 17О, то линия
приобретает сложную мультиплетную структуру, так что интенсивность
сигнала уменьшается, и его уже трудно отличить от фона. Иная картина
наблюдается для изотопа 18О. Этот изотоп, как и изотоп 16О, наиболее
распространенный в природе, обладает нулевым спином и не вызывает появ-
ления дополнительной структуры в наблюдаемой спект;
Правда, замена атома 16О в фосфатной группе на атом
альной линии.
О
О приводит к
85
небольшому высокопольному сдвигу резонансной линии на 0,025 м.д., при-
чем этот сдвиг пропорционален числу связанных с атомом фосфора тяжелых
изотопов. При этом можно наблюдать сигнал ЯМР 31Р от фосфатной группы
отдельно, что позволяет установить число атомов О.
Изотопный обмен 18О может быть использован для более полного изуче-
ния реакций ферментативных превращений. При катализе большая группа
ферментов (фосфотрансфераз) расщепляет фосфаты, ответственные за энер-
гозапас, и таким образом осуществляется перенос фосфатной группы или на
другой субстрат (киназу) или на окружающую воду (гидролазу). При фер-
ментативном гидролизе нуклеозидтрифосфата (НТФ), ответственного за
энергозапас клетки, в качестве промежуточного продукта возникает такое
состояние фосфатной группы, при котором связь с ангидридом фосфорной
кислоты уже расщеплена, однако фосфатный остаток Р(- еще связан с нукле-
озиддифосфатом (НДФ) и присоединен к ферменту Е. Формально эту реак-
цию можно представить в виде следующего уравнения:
Е.НТФ +Н2О —L-.Е.НДФ - Pi _?—. Е.НДФ + Л. (2.34)
В принципе, все этапы реакции в (2.34) являются обратимыми, хотя скорость
протекания этих процессов зависит от активационной энтальпии Д G^, т.е.
от особенностей ферментативного катализа. Для многих ферментов этап (1)
повторяется многократно, прежде чем, наконец, фосфатная группа попадет
в воду. Если провести полное замещение по концевой фосфатной группе в
НТФ по изотопу 180, то при протекании реакции в обычной воде всякий раз
при прохождении первого этапа реакции атом16О встраивается в фосфатную
группу. Если же обратный процесс (1) невозможен до тех пор, пока в реак-
ционную смесь не попадет Pi, то в смеси присутствует только вещество
следующего состава: Р16О18Оз. В другом предельном случае, т.е. при усло-
вии, что первый этап реакции будет многократно повторяться, прежде чем в
реакционную смесь попадет Pt, в смеси можно обнаружить свободный Р16О4-
Использование соответствующих теоретических моделей позволяет найти
зависящее от времени распределение меток. Согласование свободных пара-
метров дает возможность предположить, что существует конечное число
обратных реакций с высвобождением Р/ и значением константы скорости к-2
для связывания Р, в комплекс Е.НДФ. На рис.2.13 приведен пример такого
моделирования для миозинового фрагмента 51. Правда, здесь проведено
рассмотрение не для меченой АТФ, а для изотопомеченого неорганического
фосфата (Р 6О4). Здесь наблюдается обратная реакция в присутствии комп-
лекса Sl.Mg.Afl®. В этой системепервый этап повторяется примерно 40 раз,
прежде чем высвобождается фосфат. Первые эксперименты по изучению
механизма ферментативного изотопного обмена в фосфатной группе прохо-
дили с привлечением масс-спектрометрии, поскольку этот метод является
весьма чувствительным для определения изотопного состава в малых моле-
кулах. Преимуществом ЯМР является то, что при определении изотопного
состава можно проводить измерения непрерывно, без дериватизации, хотя
при работе с низкими концентрациями метод масс-спектрометрии, конечно,
более эффективен.
86
18
Рнс.2.13. Изотопный обмен кислорода в фрагменте миозина 51. Изотопный обмен О, который,
благодаря образованию комплекса М£.ЛДФ с фрагментом миозина 51, выступает в качестве
катализатора [2.4]. Рабочая частота для наблюдения спектра 01Р равна 146 МГц (это
соответствует рабочей частоте для iH 360 МГг1). (а) Изменение концентраций отдельных изото-
помепов неорганического фосфата Р,: НзР18О4 (V). .НзР °Оз "О (+), НзР18О2°О2 (х),
Н3Р18О16О3 (А) и Н3Р16О4 (О ). (Ь) Спектр ЯМР 31 Р изотопомера Р; спустя 23,5 ч после
начала реакции (вверху приведен экспериментальный спектр, внизу - теоретический спектр,
моделирующий экспериментальный).
2.3. Двумерная спектроскопия ЯМР
Развитие двумерной спектроскопии ЯМР (2М-ЯМР) открыло новые воз-
можности для развития разнообразных биологических приложений ЯМР. В
принципе каждая серия экспериментов, в которых наряду с интервалом
времени детектирования ii варьируется второй параметр, например длитель-
ность периода эволюции Ц (рис.2.14), представляет собой двумерный ЯМР.
Примером такого эксперимента является серия одномерных спектров, ха-
рактеризующих некую химическую реакцию, которые измеряются последо-
вательно через определенный промежуток времени t. Если по одной из осей
откладывать значения времени Ц, а по другой выписывать спектры, регист-
рируемые в эти моменты времени, то получим двумерный спектр. Такого
рода двумерная спектроскопия известна с момента открытия метода ЯМР.
От 2М-спектроскопии обычно требуется выполнение следующего усло-
вия: вторая переменная обязательно должна быть связана со специфически-
ми свойствами исследуемой спиновой системы. В настоящее время развито
достаточно большое число методов получения двумерных спектров ЯМР. Эти
методы реализованы в виде стандартных процедур - сервисных программ -
и получаемые экспериментальные данные могут быть представлены в виде
функции двух частотных переменных. Двумерная спектроскопия ЯМР обла-
дает рядом преимуществ по сравнению с одномерными методами: (1) Инфор-
мация может быть представлена как функция двух переменных. Это
позволяет достигнуть достаточно хорошего разрешения в сложных спектрах,
например в таких, которые в одномерном случае представляют собой нало-
87
Рис.2.14. Двумерный эксперимент ЯМР. Схематическое представление 2М-ЯМР
экспериментов. Отличие представленных на рис. экспериментов наблюдается только на фазе
смешивания: в эксперименте COSY на фазе смешивания присутствует только один 90°-ный
импульс, в двухквантовом варианте COSY имеются два 90°-ных импульса, сдвинутых по фазе,
в экспериментах RCT реализована импульсная последовательность, состоящая из трех им-
пульсов (90°-180°-90°), а в эксперименте NOESY имеется два 90°-ных импульса, разделенных
достаточно большим интервалом. Здесь не приводятся сложные фазовые циклы, которые обычно
используются для того, чтобы подавлять нежелательные артефакты.
жение перекрывающихся линий. Двумерные эксперименты позволяют про-
водить надежное отнесение линий в таких спектрах. (2) Одновременно с этим
разделением по двум переменным можно провести выбор соответствующих
физических взаимодействий, что обеспечит разделение по двум измерениям.
(3) Наконец, в двумерной спектроскопии ЯМР можно достаточно просто
наблюдать многоквантовые переходы, которые в первом приближении за-
прещены правилами отбора по спиновому квантовому числу.
Путем введения других временных переменных можно осуществить пе-
реход к n-мерным спектрам ЯМР. Следует, однако, отметить, что ограниче-
ние проведения эксперимента по времени некоторым значением, обычно
реализуемым на практике, не позволяет значительно увеличить число вре-
менных переменных, так что реально достижимое число переменных — три
(ЗМ-ЯМР).
2.3.1. Двумерный ЯМР-эксперимент
В настоящее время в двумерной ЯМР-спектроскопии, как правило, ис-
пользуются методы фурье-спектроскопии. В этом случае проводится детек-
тирование сигнала S((j, 12), из которого путем фурье-преобразования
вычисляется двумерный спектр S (COi, 6O2) в частотной области. В принципе
88
двумерный ЯМР-спектр можно получить и с использованием альтернатив-
ного метода - стохастического метода, однако этот метод находится пока в
стадии разработки.
Основной 2М-ЯМР-эксперимент можно схематически представить во
временной области, разделив его на следующие 4 фазы: подготовки, эволю-
ции, смешивания и детектирования (см. рис.2.14). На фазе детектирования
сигналы, как и в одномерном случае, регистрируются через равные проме-
жутки времени Д t2, затем они подвергаются оцифровке и накапливаются.
Фаза подготовки, как правило, состоит из 90°-ного импульса, формирующе-
го поперечную намагниченность. На протяжении фазы эволюции, длитель-
ность которой равна ti, поперечная компонента намагниченности
изменяется. Затем следует период смешивания, который, вообще говоря, в
некоторых экспериментах может отсутствовать. Компоненты поперечной
намагниченности связаны между собой разнообразными взаимодействиями.
На протяжении интервала длительностью ti они подлежат детектированию
и преобразованию. Длительность периода ti постоянно возрастает от экспе-
римента к эксперименту на величину zX/'i, причем длительность интервала tj
определяется так же, как и интервала ft, теоремой Найквиста. Спектр,
соответствующий каждому значению ti, накапливается отдельно. Таким
образом строится двумерная матрица, в которой каждой паре значений (ti,
t2) соответствует сигнал амплитудой 5 (ft, ti)- Двумерное фурье-преобразо-
вание превращает сигнал во временной области S(ti, t2) в сигнал в частотной
области S((1)1,6O2) • Такое фурье-преобразование можно записать следующим
образом:
°? °? — I (Dl ft — । (1)2 Г2
S ((D1,(1)2) = J f S(ft,ft)e е dt2(lt\. (2.35)
— 00 — 00
Выражение (2.35) представляет собой следующую цепь операций: сна-
чала все сигналы свободной индукции подвергаются фурье-преобразованию
по переменной ft. Возникающая при этом новая матрица данных содержит в
строках (ось 6O2) спектры ЯМР, соответствующие значениям ft. Затем про-
водится второе фурье-преобразование по переменной ti, т.е. данные, приве-
денные в столбцах, рассматриваются как отдельные сигналы свободной
индукции, и они, как обычно, подвергаются фурье-преобразованию.
Фурье-преобразование (как и в одномерном случае) имеет действитель-
ную и мнимую части. Обычно вычисляют только действительную часть или
модуль функции 5 (COi, (1)2). Интенсивности в двумерных спектрах имеют вид
поверхностей, представленных как график функции двух переменных, т.е.
напоминают изображение земной поверхности. Графически двумерные
спектры представляют двумя способами. Первое представление - двумерная
поверхность — дает наглядную картину 2М-спектра (рис.2.15). Второе пред-
ставление выглядит как географическая карта, на которой линии уровня
соответствуют сигналам одинаковой интенсивности (рис.2.16). Такое пред-
ставление обычно используется при обработке двумерных спектров для того,
чтобы избежать перекрывания слабых сигналов сильными.
89
Рис.2.15. Двумерный спектр ЯМР HPr-протеина в виде поверхности (графика функции двух
переменных). Спектр HPr-протеина Staphylococcus aureus. На рис. изображен каждый второй
спектр в матрице данных 512x512 точек. На рис. приведены абсолютные значения спектра, хотя
обычно сигнал состоит из положительной и отрицательной частей.
Рис.2.16. Контурное представление двумерного спектра ЯМР HPr-протеииа. Линии уровней
вычислены и построены исходя из двумерного спектра, показанного на рис.2.15.
90
В табл.2.6 приведены последовательные этапы обработки данных
при построении одномерных спектров. Если попытаться перечислить
все варианты двумерной ЯМР-спектроскопии, то это была бы весьма
сложная задача, поскольку их более сотни. Однако для биологических
приложений используются лишь некоторые из них. Чтобы их упорядо-
чить, прежде всего необходимо подчеркнуть различие между гетероя-
дерной и гомоядерной 2М-спектроскопией. В гомоядерной
спектроскопии наблюдается взаимодействие ядер одного сорта, напри-
мер, протонов. В этом случае двумерная импульсная последователь-
ность состоит из импульсов, воздействующих на спиновую систему на
близких частотах. В гетероядерных экспериментах наблюдаются взаи-
модействия ядер различных типов, например, 13С и 1Н, и в импульсной
последовательности содержатся импульсы из различных частотных об-
ластей.
Таблица 2.6. Обработка данных при проведении двумерных экспериментов
1. Соответствие ССИ различным значениям инкрементов tj в матрице 5 (tj,
строк матрицы содержат отдельные сигналы свободной индукции W2 значений
в каждой строке), при этом строки упорядочены в соответствии с ростом времени /].
2. Цифровая фильтрация всех ССИ: умножение всех строк на соответствующую
фильтрующую функцию.
3. Фурье-преобразование всех ССИ, подвергнутых фильтрации; строки матрицы
содержат только спектры 5
4. Фазовая коррекция спектров 5 (Zj, <Z)p-
5. Цифровая фильтрация данных вдоль оси /|, то есть умножение столбцов на
соответствующие фильтрующие функции.
б. Дополнение матрицы нулевыми строками до тех пор, пока число
строк не станет равным числу столбцов W] -
7. Проведение фурье-преобразования данных в направлении по столбцам.
8. Коррекция фазы в направлении tp
9. Представление 2М-спектров 5 <&)| , й^).
В зависимости от типа эксперимента схема обработки данных может слегка отличаться
от представленной в таблице.
Биологические приложения 2М-спектроскопии в основном используют
эффекты, обусловленные только двумя типами взаимодействий ядериых
спинов: косвенным (скалярным) и диполь-дипольным.
91
2.3.2. Обнаружение спин-спинового взаимодействия
в гомоядерном случае
Как уже отмечалось, косвенное спин-спиновое взаимодействие, харак-
теризуемое константой взаимодействия J, тесно связано с ковалентной хи-
мической структурой. Если химическая структура известна, то можно
провести отнесение соответствующих резонансных линий. Если же структу-
ра неизвестна, то можно выбрать структуру из нескольких альтернативных.
В общем случае следует найти ответ на следующие два вопроса: (1) Какие из
ядерных спинов связаны между собой взаимодействием? (2) Насколько ве-
лико это взаимодействие? В принципе можно получить ответ на оба эти
вопроса для достаточно простых структур даже с использованием одномер-
ных методик, например, с помощью развязки или построения теоретических
спектров. В более сложном случае, когда в спектрах содержатся перекрыва-
ющиеся линии, эти методы приводят к успеху только при использовании
большого числа трудоемких и длительных экспериментов. С помощью дву-
мерных методов эту информацию можно получить из одного эксперимента.
Стандартным методом при этом является метод COSY (correlated
spectroscopy), в котором применяются два 90°-ных импульса, разделенных
временем эволюции (см. рис.2.14). Полученный спектр симметричен от-
носительно диагонали, на которой расположены так называемые диагональ-
ные пики. Эти спектры по содержащейся в них информации соответствуют
одномерному спектру. Основная информация содержится в пиках, располо-
женных вне диагонали-это так называемые кросс-пики (см. рис.2.15 и 2.16).
Именно эти пики указывают на то, между какими ядрами существует спин-
спиновое взаимодействие, т.е. они позволяют определить те константы спин-
спинового взаимодействия, которые превышают ширину линий компонент
мультиплетов. Тонкая структура кросс-пиков позволяет получить представ-
ление о величине констант спин-спинового взаимодействия.
По спектрам, получаемым по методу COSY, можно достаточно просто
установить характер связи. При этом, исходя из какого-либо кросс-пика,
находим соответствующий ему кросс-пик по горизонтали и по вертикали.
Большинство всех двумерных спектров выглядит так же, как и спектр,
полученный по методу COSY. Существует несколько вариантов эксперимен-
тов COSY. Основным является эксперимент, позволяющий существенно уп-
ростить спектры за счет «-квантовой фильтрации (п = 2,3,...). При этом
упрощение спектров может привести к потере информации. Как правило, в
основном ограничиваются получением спектров после двух- и трехкванто-
вой фильтрации, так как с ростом порядка фильтрации неизбежно понижа-
ется отношение сигнал/шум. Особую практическую ценность имеет
эксперимент, называемый эстафетным COSY-экспериментом (используется
также сокращение RELAY от relayed coherence transfer (RCT) - эстафетный
когерентный перенос). В спектре, полученном с использованием этого мето-
да, наряду с обычными COSY-пиками, наблюдаются сигналы, позволяющие
судить о наличии в спиновой системе кроме двух слабо взаимодействующих
ядерных спинов еще одного спина, с которым они оба взаимодействуют,
причем это взаимодействие достаточно велико.
92
Как правило, информация, полученная с помощью эксперимента COSY,
может быть получена и с помощью метода SECSY (spin-echo correlated
spectroscopy), в котором сбор данных задерживается по сравнению с методом
COSY на величину ti: в эксперименте используется импульсная последова-
тельность 90° - ti - 90° - ti - сбор данных. При использовании этой последо-
вательности вид спектра будет несколько иным. Одномерному спектру
соответствует спектр, расположенный на средней линии. Кросс-пики распо-
лагаются на наклонных прямых, расположенных под углом 135° по отноше-
нию к средней линии. Так как при использовании метода SECSY не удается
получить спектры с кросс-пиками, фаза которых соответствует фазе погло-
щения, то спектроскопия SECSY применяется в том случае, когда объем
памяти для накопления данных в экспериментах COSY недостаточен. Метод
супер-SECSY является более чувствительным вариантом метода SECSY.
Первым из двумерных экспериментов, нашедших практическое приме-
нение, является/-разрешенная двумерная спектроскопия, в которой исполь-
зуется следующая импульсная последовательность: 90° - ti - 180° - ti - сбор
данных, /-разрешенный спектр по информативности соответствует одномер-
ному спектру, в котором мультиплеты повернуты перпендикулярно оси (02.
Таким образом, ось (02 содержит значения химических сдвигов, соответству-
ющих положению центров мультиплетов, а по оси Wi располагаются значе-
ния констант косвенного спин-спинового взаимодействия, т.е. значения
расстояний между ближайшими линиями в мультиплете. При згом, посколь-
ку спектр не содержит информации о том, какие из ядер связаны спин-спи-
новым взаимодействием, а содержит лишь информацию о мультиплетности
спектральных линий, то в настоящее время /-разрешенная спектроскопия
имеет, пожалуй, лишь историческую ценность.
Альтернативной по отношению к спектроскопии COSY является много-
квантовая спектроскопия. Преимуществом этого метода является больший
по сравнению с COSY объем информации. Однако при этом существенно
затрудняется интерпретация спектров. Двухквантовый спектр содержит не
только информацию о том, какие из спинов связаны между собой прямым
взаимодействием, которым отвечают пики, расположенные симметрично
относительно диагонали, соответствующей двухквантовым переходам, но
также и пики, для которых симметричные относительно диагонали партнеры
отсутствуют. Спектры содержат информацию о других спинах, связанных
между собой косвенными спин-спиновыми взаимодействиями, аналогично
RCT-спектроскопии.
2.3.3. Измерения ЯЭО и химический обмен
Для определения расстояний между взаимодействующими спинами
можно использовать ЯЭО. Соответствующий 2М-эксперимент называется
NOESY (nuclear Overhauser enhancement spectroscopy). Как и в одномерном
случае, этот эксперимент можно использовать для исследования процессов
химического обмена. Соответствующая импульсная последовательность
приведена на рис.2.14. Внешний вид NOESY-спектров совпадает с видом
93
спектров, полученных по методу COSY, с той лишь разницей, что в данном
случае кросс-пики не устанавливают соответствия между взаимодействую-
щими спинами, а определяют, какие из спинов участвуют в обмене. Интен-
сивность этих кросс-пиков зависит от длительности тт выбранного периода
смешивания. С ростом Тт интенсивность кросс-пиков (объем кросс-пика)
непрерывно возрастает, достигает максимального значения, а затем снова
уменьшается до нуля. Зависимость от времени при этом соответствует на-
блюдаемой в одномерном случае при обнаружении ЯЭО. При малых значе-
ниях интервала смешивания Гт для интенсивности кросс-пика /дв между
линиями, которые соответствуют резонансу спинов А и В, справедлива фор-
мула
lAB ~ гав'6 Гт, (2.36)
где гав обозначает расстояние между ядрами, для которых наблюдаетсяЯЭО.
Более подробно зависимость величины ЯЭО от расстояния обсуждается в
главе 3.
На практике используется две модификации спектроскопии NOESY.
(1) Эксперимент NOESY с подавлением интенсивности сигналов, располо-
женных на главной диагонали. (2) Эксперимент ROESY (rotaiting frame
Overhauser enhancement spectroscopy), в котором проводится измерение ЯЭО
во вращающейся системе координат. Иногда для этого эксперимента исполь-
зуется сокращение CAMELSPIN. Эксперимент (1) полезен, если необходимо
провести качественное отнесение линий в спектре, а эксперимент (2) прово-
дят в том случае, если времена корреляции Тс таковы, что в лабораторной
системе координат ЯЭО практически не наблюдается.
2.3.4. Корреляция в гетероядерном случае
Гетероядерная корреляция, т.е. корреляция между одномерными спек-
трами ЯМР, полученными для разного сорта ядер, обладает рядом привлека-
тельных свойств. Прежде всего эти методы, как и большинство методов
двумерной спектроскопии, приводят к улучшению разрешения в спектрах,
т.е. позволяют провести отнесение линий в сложных перекрывающихся спек-
трах. Корреляция между двумя различными спектрами ЯМР, например, на
ядрах *Н (спин /) и 13С (спин S) облегчает отнесение линий в спектрах, так
как в протонных спектрах содержится дополнительная информация о спек-
трах 13С, и наоборот. Отнесение линий существенно упрощается, если про-
ведено полное отнесение в спектре хотя бы одного из ядер (JH либо 13С).
Двумерные корреляционные спектры могут быть дополнительно использо-
ваны также и для того, чтобы повысить чувствительность в спектрах ядер с
низким естественным содержанием, т.е. повысить возможности детектиро-
вания ядер с низкой чувствительностью. Простейший вариант гетероядерно-
го корреляционного эксперимента совершенно аналогичен эксперименту
COSY: используемая в этом методе импульсная последовательность также
состоит из двух 90°-ных импульсов, между которыми имеется интервал
94
длительностью Z|, причем импульсы подаются на частоте, соответствующей
резонансу ядра /, а спад свободной индукции записывается в течение времени
Различие состоит в том, что в момент, когда подается второй 90°-ный
импульс на частоте резонанса ядра I, подается дополнительный 90°-ный
импульс на частоте резонанса ядер S. Отношение сигнал/шум, достигаемое
при использовании различных методов, описывается следующим выражени-
ем:
~ Тех с /obs3/2 d-е TR/TX ) • (2.37)
Здесь уехс-гиромагнитное отношение ядерных спинов, которые возбуж-
даются в первом периоде эксперимента, до проведения наблюдения, a yobs -
гиромагнитное отношение ядер, наблюдение которых проводится в данный
момент. В экспериментах с накоплением данных необходимо учитывать и
такие параметры, как время продольной релаксакции 7\еяс возбуждаемых
спинов и интервал между импульсами TR.
Следовательно, наилучшее значение отношения сигнал/шум может
быть достигнуто в таком эксперименте, в котором сначала осуществляется
перенос поляризации от спинов с большим у к спинам с меньшим у, а затем,
на стадии детектирования, осуществляется обратный перенос поляризации.
В частности, для системы 15N-'h оценка по^ормуле (2.37) показывает, что
теоретически чувствительность в спектрах dN может быть увеличена при-
мерно в 300 раз.
Литература
2.1. McConnell, Н.М. (1958) J.Chem. Phys. 28, 430.
2.2. Sontheimer, G.M., Kuhn, W. and Kalbitzer, H.R. (1986) Biochem.
Biophys. Res. Comm. 134, 1379.
2.3. Rosch, P., Kalbitzer, H.R., Schmidt-Aderjan, U. and Hengstenberg,
W. (1981a) Biochemistry 20, 1599.
2.4. Rosch, P., Goody, R.S., Kalbitzer, H.R. and Zimmermann, H. (1981b)
Arch. Biochem. Biophys. 211, 622.
2.5. Overhauser, A.W. (1953) Phys. Rev. 92, 411.
Дополнительная литература
Brey, W.S. (ed.) (1988) Pulse Methods in ID and 2D Liquid-Phase NMR
(Academic, San Diego)
Carrington, A. and McLachlan, A.D. (1969) Introduction to Magnetic
Resonance with Applications to Chemistry and Chemical Physics (Harper and
Row, New York)
Chandrakumar, N. and Subramanian, S. (1987) Modern Techniques in
High-Resolution FT-NMR (Springer, New York, Berlin, Heidelberg)
95
Croasmun, W.R. and Carlson, R.M.K. (1987) Two-Dimensional NMR
Spectroscopy, Applications for Chemists and Biochemists (VCH, Weinheim)
Gunther, H. (1980) NMR Spectroscopy (Wiley, New York)
Harris, R.K. (1983) Nuclear Magnetic Resonance Spectroscopy (Pitman,
Marshheld)
Martin, M.L., Martin, GJ. and Delpuech, J.-J. (1980) Practical NMR
Spectroscopy (Heyden, London)
Sanders, J.K.M. and Hunter, B.K. (1987) Modern NMR Spectroscopy,
\ Guide for Chemists (Oxford University Press, Oxford)
96
Глава 3. ЯМР-спектроскопия биологических макромолекул
В этой главе обсудим проблемы ЯМР-спектроскопии биологически важ-
ных молекул. Это направление ЯМР наряду с in-vivo-спектроскопией явля-
ется одним из главных приложенийЯМР в биологии. Особенностью спектров
ЯМР таких объектов является прежде всего их сложность, возрастающая по
мере увеличения числа атомов, входящих в состав этих молекул. Кроме того,
с увеличением молекулярной массы резонансные линии все более уширяют-
ся, что, очевидно, уменьшает разрешение спектров и чувствительность ме-
тода.
Постановка задачи и пути ее решения в значительной степени опреде-
ляются особенностями исследуемого объекта. В данной главе проведено по-
следовательное изложение проблем ЯМР-спектроскопии в приложении к
каждому из классов биологических молекул: протеинов, нуклеиновых кис-
лот, полисахаридов и липидов.
3.1. ЯМР-спектроскопия протеинов
Функции полипептидов и протеинов в биологических системах весьма
разнообразны. Они играют роль структурных компонентов, активных эле-
ментов в процессах переноса, аккумуляторов метаболической энергии, био-
катализаторов и переносчиков сигналов. Несмотря на такое разнообразие
функций, строение всех протеинов в значительной степени однотипно. Как
и большинство биологических макромолекул, они состоят из отдельных
структурных фрагментов, выбор которых и связи между ними определяют
специфические свойства протеинов. Основными структурными единицами
протеинов являются аминокислоты, которые объединены в одну или не-
сколько линейных цепочек.
ЯМР протеинов относится к числу достаточно хорошо разработанных
методов и позволяет получить информацию о динамических процессах в
протеинах и о взаимодействии с субстратами и молекулярными структурами.
Эта информация едва ли может быть получена другими методами. Среди
последних достижений в этой области следует отметить определение про-
странственной структуры протеинов с помощью двумерной ЯМР-спектро-
скопии, основные принципы которой разработали Рихард Эрнст и Курт
Вютрих. Применение этих новых методов в биохимии и биофизике приобре-
тает все большую актуальность, хотя еще несколько лет назад их использо-
вание в этих областях казалось нереальным.
3.1.1. Строение и структура протеинов
Из всего многообразия аминокислот в состав протеинов входят только 20
L-аминокислот, соединение которых непосредственно программируется ге-
нетическим кодом (рис. 3.1). Название аминокислот, как правило, представ-
97
ляет собой аббревиатуру трех- или однобуквенных кодов (табл.3.1). В про-
теинах аминокислоты связаны пептидной связью (рис.3.2), которая образу-
ется из карбоксигрупп и аминогрупп двух последовательно соединенных
аминокислот.
Пептидная связь является плоской, т.е. все атомы, образующие эту
связь, расположены в одной плоскости. Пептидная связь может существовать
в двух формах: цис- и транс-форме. Подавляющее большинство всех пептид-
ных связей в протеинах находится в транс-конфигурации. Исключение здесь
составляет только пептидная связь по иминогруппе аминокислоты пролин. В
простых пептидах, содержащих пролин, обычно оба изомера присутствуют
в растворе, и константа равновесия, как правило, изменяется в пределах от
5 до 20. Если исходить из того, что длины связей и углы связей достаточно
хорошо соответствуют стандартным значениям, то следует отметить, что
пространственная структура протеина зависит только от вращения относи-
тельно одинарных связей. Соответствующие углы называются диэдральны-
ми углами и величины их зависят только от положения в аминокислотном
звене. Эти углы обозначаются греческими буквами. Три диэдральных угла
Таблица 3.1. Обозначения и свойства основных аминокислот
Наименование Трехбуквенный Однобуквен- Молекулярная массаа> рК боковой це-1 пи0'
код НЫИ КОД
Аланин Ala A 71 -
Аргинин Arg R 156 12,5
Аспаргин Asn N 114 —
Аспаргиновая кислота Asp D 115 3,9
Цистеин Cys C 103 8,3
Глутамин Gin Q 128 —
Глутаминовая кислота Glu E 129 4,3
Глицин Gly G 57 —
Гистидин His H 137 7,0 (6,2)
Изолейцин He I 113 —
Лейцин Leu L 113 —
Лизин Lys К 128 11,1
Метионин Met M 131 —
Фенилаланин Phe F 147 —
Пролин Pro P 97 —
Серин Ser S 87 —
Треонин Thr T 101 —
Триптофан Trp w 186 —
Тирозин Tyr Y 163 10,3
Валин Vai V 99 -
Молекулярная масса звена в пептидной цепи при нейтральном pH;
Значения рК цистеина и аргинина в свободных аминокислотах [3.1]. Другие значения рК
Ь)
для аминокислотного остатка X были измерены в модельном пептиде Gly-Gly-X-Ala методом
ЯМР [3.2]. Значения рК, приведенные в скобках, соответствуют наиболее вероятным значени-
ям в протеине [3.3].
Рис.3.1. Структура 20 канонических аминокислот. Представленные аминокислотные звенья
соответствуют конфигурациям, наиболее часто встречающимся в протеинах при нейтральном
pH. Атомы углерода и водорода обозначены светлыми сферами, а атомы N, О, S - темными.
98
Ala Gly
Asp
Cys
Gin
Glu
Phe
99
Hz
(а) транс
Рис.3.2. Пептидная связь. (а) -транс конфигурация, (Ь) - цис-конфигурация. Связь С*-N иосит
частично двойной характер.
основной цепи обозначаются буквами <р и О), а диэдральный угол в
боковой цепи обозначается^ (рис.3.3), где/обозначает положение в боковой
цепи.
Под первичной структурой протеинов понимают ковалентную структу-
ру аминокислотных звеньев, т.е. последовательности, которые включают в
себя все возможные модификации аминокислот и дисульфидных мостиков.
В этом случае вторичная структура представляет собой локальное простран-
ственное упорядочение атомов в последовательности аминокислот, про-
странственная структура которой определяется диэдральными углами.
Третичная структура определяется изгибами в пространстве полной по-
100
(о.
Рис.3.4. Основные элементы вторичной структуры. (Вверху) -антипараллельный /^-свернутый
лист, (Внизу слева) - Зщ-спираль, (Внизу посередине) - правовращающая «-спираль, (Внизу
справа) -7С-спираль. Водородные мостиковые связи изображены штриховыми линиями, боковые
цепи обозначены символом (R) [3.5].
липептидной цепи. Если протеин состоит из большого числа различных
полипептидных цепей, то взаимное пространственное расположение этих
структурных блоков определяется как четвертичная структура.
В протеинах находят ряд типичных вторичных структур. Эти структуры
характеризуются образованием определенным образом упорядоченных во-
дородных мостиков между амидными группами и карбоксильными группами
основной цепи. Возможные варианты структуры определяются также типич-
ными значениями диэдральных углов (р и 1р. Среди наиболее часто встреча-
ющихся типов конфигураций можно указать правовращающую «-спираль и
^3-свернутый лист (рис.3.4). Обе эти конфигурации характерны для большин-
ства протеинов. Среди других элементов вторичной структуры, отмеченных
повторяющимися углами <р и 1р, можно назвать такие, как левовращающая
101
Таблица 3.2. Основные элементы вторичной структуры в протеинах
Элемент вторичной структуры <р Водородные мостиковые связи
Довернутый лист, параллель- -119 +113 COj-.j-NHyH NH(+1-COy
ная конфигурация ИЛИ
CO,_]-NH и NHy+1-СО,
Довернутый лист, антипарал- -139 +135 COf-NH • и NHpCO .• или
лельная конфигурация иЫнД-СО.-.!
Зю-спираль -76 -5 COrNH,+3
правовращающая СХ-спираль -57 -47 CO(-NHi+4
левовращающая СХ-спираль +57 + 47 COrNH,+4
Л-спираль -57 -70 CO(—NH(+5
коллагеновая спираль -51 +153 COf-NHy.j hNHj^-CO^
-76 +127
-45 +148
Обозначения приведены согласно [3.4] и [3.6].
«-спираль, тг-спираль, Зю-спираль и коллагеновая спираль (табл.3.2).
Вследствие стерических препятствий в пептидной цепочке не все комбина-
ции диэдральных углов будут равноправны, а соответствующие энергии
могут существенно различаться. Комбинации углов <р и соответствующие
низким значениям потенциальной энергии, рассматриваются как разрешен-
ные конформации, а конформации, соответствующие высоким значениям
энергии, - как запрещенные. В простейшей модели, в которой атомы рас-
сматриваются как твердые шарики, существование конформаций с различ-
ными возможными энергиями вполне очевидно: некоторые комбинации
диэдральных углов просто невозможно реализовать по стерическим причи-
нам. Наглядно эти комбинации можно представить, воспользовавшись мо-
делью Рамачандрана, в которой каждому диэдральному углу соответствует
ось и обозначаются разрешенные и запрещенные области.
3.1.2. Динамические процессы в протеинах
Биологические полимеры в растворе не обладают жесткой структурой,
они непрерывно изменяют свою конфигурацию. Из-за большого числа еди-
ничных связей, относительно которых может совершаться свободное враще-
ние, для протеина разрешены разнообразные типы внутренних движений.
Однако большинство протеинов в том виде, в котором они встречаются в
природе, обладает вполне определенной структурой, характеризующейся
достаточно плотной упаковкой. При этом число разрешенных движений
оказывается ограниченным, поскольку многие возможные комбинации ди-
эдральных углов не могут возникнуть без нарушения естественной конфор-
мации, т.е. без денатурации. В то же время в нативном состоянии возможны
вращения боковых групп, в частности метильных, коллективные торсионные
движения основной цепи, а также переходы между различными естествен-
ными конформациями. К другим видам движений относятся процессы хими-
ческого обмена: протоны заряженных групп, амидных групп, гидроксильных
102
и сульфгидрильных остатков, а также субстраты и продукты ферментов
диссоциируют и вновь вступают в обмен. Все эти процессы также могут быть
обнаружены по спектрам ЯМР протеинов.
Плоские кольцевые системы таких аминокислот, как гистидин, тирозин,
фенилаланин и триптофан, в принципе могут совершать вращения в проте-
ине относительно осей Са-сР и - С^. Однако такие вращения часто
затруднены вследствие взаимодействия с другими атомами протеина, так что
внутри протеина эти вращения ограничены. Процесс ограничения вращений
достаточно хорошо удается наблюдать по спектрам ЯМР 1Н. Поскольку
электронные оболочки атомов соседних кольцевых систем по-разному экра-
нируют внешнее магнитное поле, то эффект экранирования можно наблю-
дать не только на данном атоме, но и на соседних атомах. Таким образом,
эффект экранировки определяется расположением протонов в пространстве
по отношению ко всей молекуле. Ароматические кольца таких аминокислот,
как тирозин и фенилаланин, симметричны относительно оси второго порядка
(см. рис.3.1), т.е. находятся в симметричном окружении, а следо-
вательно, химические сдвиги обоих (5-протонов и обоих £-протонов равны, и
в спектрах ЯМР для обеих пар протонов всегда присутствует только одна
резонансная линия. Таким образом, только нарушение симметрии окруже-
ния может приводить к различиям в значениях химических сдвигов для
симметрично расположенных протонов, а следовательно, к расщеплению
резонансных линий. Однако в случае, когда кольцо совершает быстрые вра-
щения относительно данной оси, эффекты асимметрии усредняются, и на-
блюдается "усредненная" структура, т.е. реализуется случай быстрого
движения, который характеризуется наличием в спектре одной резонансной
линии. Так как скорость вращения зависит от температуры, то иногда в
спектрах удается наблюдать переход от медленного движения к быстрому
при повышении температуры.
Прекрасной иллюстрацией изложенного может служить пример, приведен-
ный на рис.3.5, на котором показана динамика фенилаланинового и тирозинового
колец и Basic Pancreatic Trypsin Inhibitor (BPTI): при повышении температуры
(5- (2,6-) - и Е- (3,5-)-протоны всех тирозиновых и фенилаланиновых колец стано-
вятся эквивалентными. При понижении температуры спектр обоих ароматиче-
ских фрагментов изменяется (имеются ввиду Phe-45 и Туг-35), так как скорость
вращения уже не столь велика, чтобы обеспечить усреднение анизотропии. По-
строение соответствующих теоретических спектров и их сравнение с эксперимен-
тальными позволяет получить значения скоростей вращений и соответствующих
им энергий активации. Оба кольца располагаются в плотно упакованной внутрен-
ней части протеина, поэтому удивительно, что они все-таки могут достаточно
быстро вращаться. Следует отметить, однако, что в настоящее время существует
достаточно большое число доказательств того, что далеко не всегда речь идет о
равномерном вращении колец, а скорее о быстрых 180°-ных флип-движениях.
Обычные ароматические кольцевые системы в BPTI во всей области температур
не обнаруживают никаких признаков затруднения вращений; вид их спектров
остается типичным для случая быстрого движения, причем это удивительное
свойство обнаруживается для большинства протеинов.
103
М.Д.
Рис.3.5. Переориентация ароматических колец в протеине. Вращательное движение аромати-
ческих колец аминокислот Phe-45 и Туг-35 в ингибиторе трипсина панкреатического фермента
(BPTI) существенно заторможено. Повышение температуры приводит к переходу от медленного
к быстрому обмену. Резонансные линии Phe-45 в экспериментальном спектре обозначаются как
(о), а Туг-35 - (▲ ). Справа приведены теоретические вращения [3.7], спектры и значения
соответствующих скоростей
Протеины могут существовать в разнообразных конформациях. Один из
предельных случаев и соответственно один из классов конформаций образу-
ет полностью денатурированное состояние - хаотический клубок (random
coil), для которого в соответствии с введенным определением не существует
никакого выделенного направления. Переход от нативной конформации к
этому состоянию можно легко проследить по спектрамЯМР 1Н. Спектр ЯМР
Н для протеина в состоянии свернутого листа сильно структурирован, и вид
спектра невозможно предсказать заранее. Это удается лишь в том случае,
если известна пространственная структура. В противоположность этому
спектр протеина в денатурированном состоянии в первом приближении за-
дается аминокислотным составом. В этом случае химические сдвиги отдель-
ных компонент очень хорошо согласуются с теми значениями, которые
приведены в таблице для модельных веществ. Общепринятыми модельными
системами являются тетрапептиды Gly-Gly-X-Ala, в которых X непосредст-
венно представляет собой искомую аминокислоту (табл.3.3). Правда, эти
значения для хаотического клубка могут быть несколько меньшими, что
происходит под влиянием соседа, непосредственно следующего за X в ами-
104
Таблица 3.3. Химические сдвиги в спектрах 'н модельного пептида - хаотического клубка®^
Аминокис- Химические сдвиги (м.д. относительно TSP);
лотинй П на н/3 Ну другие атомы
остаток X
Ala 8,25 4,35 1,39
Arg 8,27 4,38 1,79 1,70 П8 3,32 He 7,17 fro 6,62
1,89 1,70 118 3,32
Asn 8,75 4,75 2,75 H62 6,91
2,83 H82 7,59
Asp 8,41 4,76 2,75
2,84
Cys 8,31 4,69 2,96
3,28
Gin 8,41 4,37 2,01 2,38 He2 6,87
2,13 2,38 He2 7,59
Glu 8,37 4,29 1,97 2,28
2,09 2,31
Gly 8,39 3,97
3,97
His 8,41 4,63 3,20 H82 7,14 H el 8,12
3,26
lie 8,19 4,23 1,90 1,19 Ul2 0,95 1181 0,89
1,48
Leu 8,42 4,38 1,65 1,64 116 0,90
1,65 H6 0,94
Lys 8,41 4,36 1,76 1,45 118 1,70 He 3,02 Ж 7,52
1,85 1,45 116 1,70 He 3,02
Met 8,42 4,52 2,01 2,64 He 2,13
2,15 2,64
Phe 8,32 4,66 2,99 H81 7,30 Hel 7,39 Ж 7,34
3,22 1182 7,30 He2 7,39
Prob 4,44 2,02 2,03 IIS 3,65
2,28 2,03 116 3,68
Ser 8,38 4,50 3,88
3,88
Th г 8,24 4,35 4,22 1,23
Trp 8,09 4,70 3,19 H81 7,24 Hel 10,22 11(2 7,50
3,32 Ihf2. 7,24 He3 7,65 11(3 7,17
Tyr 8,18 4,60 2,92 H81 7,15 Hel 6,86
3,13 1162 7,15 He2 6,86
Vai 8,44 4,18 2,13 0,94
0,97
а)
Ь)
Данные для тетрапептида Gly-Gly-X-Ala при 308 К в воде, pH - 7 [3.14];
Пролин в транс-конфигурации.
нокислотной цепи, а не за счет влияния вторичной или третичной структуры.
Для денатурированных протеинов ЯМР-спектр также часто отличается от
типичного спектра хаотического клубка. Это значит, что в таких состояниях
все еще сохраняется локальный порядок. Таким образом, ЯМР-спектр явля-
ется хорошим индикатором степени денатурации протеина.
На рис.3.6 приведен спектр ]Н протеина фактор Шлак, принимающего
участие в процессе фосфорилирования сахара лактозы. В этом процессе
осуществляется транспорт лактозы сквозь клеточную мембрану Bakteriums
Staphylococcus aureus, и одновременно происходит фосфорилирование. В
105
М.Д.
Рис.3.6. Спектр ЯМР ‘н (360 МГц) нативного фактора
щлак staphylococcus aureus в D2O при 308 К [3.7].
Рис.3.7. Денатурация фактора Шлак. Денатурация на-
блюдается при возрастании значения pH. Самый нижний
спектр получен последним в серии экспериментов. Буква-
ми Е, Е , F и F обозначебны кольцевые протоны тирозина
в нативном и частично денатурированном состояниях [3.7].
СЛ-А-- рН ц о
8'° ' 7'° ’ 6.0 м.д,
этом процессе участвуют четыре протеина: фермент I, НРг, фактор III и
фермент II. Перенос фосфатной группы, принадлежащей фосфоенолпирува-
гу, осуществляется по следующей схеме: от фермента I через НРг, фактор III
и фермент II на лактозу, которая одновременно в этом процессе переносится
через клеточную мембрану. В соответствии с выполняемой функцией эта
еистема, осуществляющая перенос веществ через мембраны клеток, называ-
ется также фосфоенолпируватзависимой фосфотрансферазной системой
(PTS). Фактор Шлак относится к протеинам средней величины. Он состоит
чз трех идентичных фрагментов, каждый из которых в свою очередь состоит
13 103 аминокислот. Спектр ЯМР является типичным для протеина такой
«еличины в нативном состоянии. Большинство резонансных линий перекры-
106
ваются, так что тонкая структура плохо разрешима. Только на краях спектра
(в низкопольной области, выше 6 м.д., и в высокопольной области, ниже 0,7
м.д.) можно провести отнесение сигналов от различных групп. Резонансные
линии, обозначенные di, di и d3, принадлежат к линиям различных метиль-
ных групп, причем они сдвинуты в область сильных полей. Резонансные
линии в слабопольной области спектра соответствуют протонам колец 4-ги-
стидин-, 5-фенилаланин- и 2-тирозиновых остатков. На рис.3.7 можно уви-
деть изменения в слабопольной области спектра при щелочной денатурации.
Наиболее отчетливо денатурацию удается наблюдать по двум резонансным
линиям, которые обозначены на самом нижнем спектре буквами G и Н.
Начиная со значения pH -11,4 интенсивность этих линий убывает и при
pH - 12,5 сигналы исчезают полностью. Положение резонансных линий,
соответствующих кольцевым протонам, совпадает с наблюдаемыми для кон-
формации хаотического клубка: пик при 7,7 м.д. соответствует Е1-(2)-про-
тонам гистидина, интенсивный пик при 7,3 м.д. - кольцевым протонам
фенилаланина, пик при 7,0 м.д. соответствует <52-(4)-протонам гистидина и
(5-(2,6)-протонам тирозина, а сигналы в области 6,6 м.д. -£-(3,5)-протонам
тирозина. Если же при щелочной денатурации вновь восстанавливается зна-
чение pH, близкое к физиологическому (нижний спектр на рис.3.7), то
возникает ренатурация протеина. Более тщательное сравнение спектров
показывает, что ренатурация оказывается неполной.
Конформационные изменения, возникающие при образовании связи с
субстратом, рассматриваются теориями ферментативного катализа и его
регулирования. Прекрасным примером, иллюстрирующим изложенное, яв-
ляется аденилаткиназа, которая катализирует реакцию
Mg-АДФ + АДФ - Mg-АТФ + АМФ (3.1)
и при физиологических условиях способствует тому, что при большой по-
требности в АТФ уровень содержания АДФ поддерживается низким, и одно-
временно для процесса гидролиза предоставляется дополнительное
количество АТФ. Согласно данным рентгеноструктурного анализа для аде-
нилаткиназы имеются две достаточно сильно различающиеся структуры,
которые, вероятно, соответствуют открытой форме до образования связи с
субстратом и в закрытой форме - после образования связи с субстратом.
Следует ожидать, что различным конформациям соответствуют различные
спектры ЯМР. Действительно, наибольшие изменения в спектрах *Н адени-
латкиназы человека находят в том случае, если образуется комплекс с ана-
логом бисубстрата ApsA (в Ар5А два аденозиновых остатка, которые связаны
между собой цепочкой из пяти фосфатных групп) (рис.3.8).
Так как ApsA достаточно сильно связан с ферментом, обмен оказывается
очень медленным (koff < 70 с'1), так что удается отдельно наблюдать резонан-
сные линии различных аминокислотных остатков в свободном состоянии и в
комплексе с аденилаткиназой. Например, вследствие насыщения половины
связей с активными центрами резонансная линия гистидина 36 расщепляется
на две линии равной интенсивности, которые соответствуют гистидиновому
остатку в свободном субстрате и в ферменте, связанном в комплекс. Правда,
107
Рис.3.8. Конформационные изме-
нения аденилаткииазы, индукци-
онные взаимодействия с
субстратом. Спектр ЯМР *Н (360
МГц) аденилаткииазы киназы че-
ловека в отсутствие субстрата
(вверху), после добавления Ар^А в
соотношении Г.2 (посередине),
после добавления избытка АР5А
(внизу) [3.33]. На рис отмечены
сигналы, соответствующие коль-
цевым протонам His-Зб и His-189.
и у обозначаются линии,
______________________________ соответствующие протонам Н-1,
1 1 1 1 ' 1 1 и* Н-2 и Н-8 связанной Ар гА.
9 8 7 6 5
<5/м.д.
при этом не все спектральные изменения объясняются структурной изоме-
ризацией аденилаткииазы, а сам субстрат также может отвечать за ряд этих
изменений. Оба влияния непросто разграничить, в частности, если субстрат
содержит ароматическое кольцо. Как увидим в дальнейшем, кольцевые токи
Л-электронов также могут быть причиной существенных изменений значе-
ний химических сдвигов, причем также и для ядер, находящихся достаточно
далеко от центра кольца.
В каталитических процессах зачастую решающая роль принадлежит
группам, подверженным ионизации. Основной информацией, способствую-
щей пониманию принципов, лежащих в основе каталитического процесса,
является знание величины заряда группы при физиологических условиях.
При заданных значениях pH окружающей среды заряд группы определяется
значением рК. Однако значение рК, которое характеризует равновесие меж-
ду протонированной и депротонированной формами, можно определить до-
статочно просто для каждой из групп в протеине с помощью ЯМР при
условии, что имеется возможность отнесения соответствующих линий в
спектре. Эта задача решается особенно просто в спектральной области, где
наблюдаются сигналы кольцевых протонов. В этой области резонансные
108
линии перекрываются, так как в большинстве протеинов содержание арома-
тических аминокислот невелико. Гистидин является аминокислотой, непос-
редственно участвующей в ряде каталитических процессов, сигналы которой
попадают в данную спектральную область. Сигналы гистидина достаточно
хорошо разрешимы даже для больших протеинов.
Описанные ранее компоненты протеина фосфоенолпируватзависимой
системы фосфотрансферазы в качестве интермедиата имеют фосфонатную
группу, перенос которой они осуществляют, связывая гистидиновый остаток
в активный центр. Сигналы гистидина можно идентифицировать в таких
протеинах, как НРг и фактор III. Для гистидина в активном центре гистиди-
на, входящего в состав для обоих протеинов, с помощью ЯМР было найдено
значение рК, равное примерно 6, т.е. гистидиновое кольцо при физиологи-
ческом значении pH находится в депротонированной форме. Чтобы прове-
рить, не является ли это случайным, было проведено исследование
HPr-протеинов различных микроорганизмов (рис.3.9). Несмотря на очевид-
ное различие последовательностей аминокислот, параметры ЯМР-спектров
гистидина отличаются удивительным постоянством. Однако уловить это сов-
падение, исходяиз спектров, приведенных на рис.3.9, нетакпросто, посколь-
ку эти спектры получены при различных значениях pH. Значение рК,
которое определялось титрованием pH, для всех исследованных НРг-проте-
инов было меньше, чем 6,1 (рис.3.10). Таким образом, эти эксперименты
также можно провести в фосфорилированном состоянии. Здесь для фактора
III и HPr-протеина наблюдаются необычно высокие значения рК: от 7,8 до
8,6, т.е. фосфогистидины при физиологическом значении pH протонирова-
ны. Результаты схематически представлены на рис.3.11. Следует отметить,
чтофосфонатные группы в протеине НРги факторе III связаны с различными
атомами азота в кольце. Например, в спектрах ЯМР !Н процесс фосфорили-
рования гистидина в протеине вызывает типичные изменения значений ре-
зонансных частот гистидина. Из этого можно сделать вывод не только о том,
подвергнут ли определенный гистидин процессу фосфорилирования, а также
о том, по какому из положений в кольце связана фосфонатная группа.
Взаимодействие протеинов можно также исследовать с использованием
относительно простых методов, которые позволяют обнаружить это взаимо-
действие по спектрам ЯМР !Н. Примером таких исследований является
изучение взаимодействия протеинов в фосфотрансферазной системе. При
переносе фосфонатной группы в качестве промежуточного продукта должен
образовываться комплекс из НРг и фактор III. Оба протеина независимо от
того, находятся ли они в фосфорилированном или в нефосфорилированном
состоянии, в принципе могут взаимодействовать один с другим. Следует
ожидать, что протеины могут быть обнаружены в каждой из таких комбина-
ций, однако взаимодействие будет меньшим, если обе компоненты либо
находятся в фосфорилированном состоянии, либо, напротив, в нефосфори-
лированном. Если в спектре ЯМР нефосфорилированного фактор III наблю-
дать за сдвигом резонансной линии гистидина в активном центре в
зависимости от концентрации добавляемого нефосфорилированного НРг, то
резонансная линия гистидина будет непрерывно смещаться в область силь-
ных полей. Это типичное поведение для случая быстрого обмена между
109
Рис.3.9. Гистидин в активных центрах НРг-
протеииов. На рис. приведен спектр ЯМР*Н
(360 МГц) HPr-протеинов различных грампо-
ложительных микроорганизмов. Показана
только слабопольная часть спектроа ЯМР, на
которой наблюдаются сигналы кольцевых про-
тонов. (а) - НРг Staphylococcus aureus, (b) -
Streptococcus lactis, (c) - Bacillus subtilis, (d) -
Streptococcus faecalis. Параметры спектров
ЯМР гистидина в активном центре, His-15, при-
мерно одинаковы для всех приведенных проте-
инов. Температура проведения экспериментов
308 К, значения pH: (а) -6,2 (Ь) -7,7, (с) -8,4,
(d) -8,8. Использование гауссовой фильтрую-
щей функции способствует сужению
спектральных линий [3.9]. Показанное поло-
жение ароматических фрагментов
соответствует последовательности аромати-
ческих аминокислот после проведения коррек-
ции.
8.0 7.5 7.0 6.5 м.д.
двумя средами—гистидином в свободном фактор III и в фактор III, связанном
с НРг. Из условия быстрого обмена I А О) Те I < < 1 (см. раздел 2.2.2) можно в
качестве нижней границы для скорости обмена ввести оценку Хе > > 2300
с'1. Если согласовать экспериментальные данные с (2.12), подвергнув это
уравнение соответствующей модификации, и учесть, что населенности мож-
но вычислить в соответствии с законом действующих масс, то можно даже
оценить нижнюю границу для константы ассоциации между обоими проте-
инами (к > 5000 м’ ).
Интересное явление представляет собой обмен между амидными прото-
нами пептидного остатка и окружающей водой. Мы уже ознакомились с этим
процессом как способом упрощения спектров в случае, когда протеин раство-
ряется в дейтерированной воде, и амидные протоны участвуют в обмене с
ядрами дейтерия растворителя. Скорость обмена существенно зависит от
структурных особенностей. Если обмен происходит очень медленно, то его
скорость можно определить непосредственно по уменьшению во времени
интенсивности линий. Для более высоких скоростей обмена существует эле-
гантный метод, который обсуждался в разделе 2.2.5; этот метод называется
экспериментом с переносом насыщения в НгО. Правда, если амидные прото-
ны обмениваются настолько быстро, что соответствующие им сигналы не
видны отдельно от сигналов воды (быстрый обмен), то в общем случае не
/дается провести измерение скорости обмена с привлечением простых мето-
дов. Подробно был исследован обмен амидных протонов в BPTI, при котором
подавляющее большинство резонансных линий поддается интерпретации с
ПО
М.Д.
1.5
—r~ I--1--»-1---1-]--1-1--1-1---1-1--1-J-->--]--<------
(.1 5.1 6.J 7.1 1.1 5.1 li.l U.l pH
Рис.3.10. Зависимость химических сдвигов кольцевых протонов His-15 в различных НРг-проте-
инах от pH. (□ ) обозначает HPr-протеин S.lactis, (+) - B.subtilis, (А ) - S.faecalis (х) - E.coli и
( ) - S.aureus. Представленные кривые рассчитывались согласно модифицированному урав-
нению Хендерсона - Хассельбалха [3.10].
Рис.3.11. Зарядовое состояние гистидино-
вых колец в активных центрах НРг-проте-
ина н фактора III до (I) и после (II) перено-
са фосфатной группы при значении pH -
-7,4. Распределение зарядовой плотности
определено по спектрам ЯМР [3.8].
111
помощью двумерных спектров ЯМР. Наибольшие значения скоростей, по-
рядка 2 с’1 (при pH=3,5 и Т=35°С), соответствуют значениям, наблюдаемым
для неструктурированных пептидов. Наименьшие значения - порядка 10'5
с’1, т.е. для наблюдения этого обмена необходимо ждать целый день. Рас-
смотрим подробнее, от каких факторов зависит скорость этого обмена. С
одной стороны, скорость обмена зависит от pH раствора, так как катализато-
ром обмена служат ионы НзО и ОН' (рис.3.12). С другой стороны, скорость
обмена в значительной степени определяется пространственной структурой
протеина. При этом преобладающими факторами являются доступность к
взаимодействию молекул воды и электростатическое влияние соседних
групп. Рис.3.13 иллюстрирует это на примере BPTI, для которого наряду с
данными, полученными методом ЯМР, имеются также данные рентгено-
структурного анализа. Последние можно использовать для выяснения таких
свойств, как доступность к взаимодействию молекул воды и возможность
образования водородных мостиковых связей. Следует отметить, однако, что
к этим данным следует относиться с определенной осторожностью при пере-
носе их на закономерности, наблюдаемые в растворе. Очевидно, существует
известная корреляция между доступностью к взаимодействию молекул воды
и величинами скорости обмена амидных групп, т.е. малые значения скоро-
стей обмена свидетельствуют о малой доступности пептидной связи к взаи-
модействию с протонами воды.
Строго говоря, времена релаксации протонов в протеине связаны с про-
цессами, характеризующими внутреннюю подвижность отдельных групп в
протеине, и именно поэтому они могут давать информацию принципиально-
го характера об этих процессах (см. также раздел 2.2.1). В простейшем
случае, когда речь идет о дипольной релаксации за счет вращательной диф-
фузии для двух неэквивалентных ядер А пВ со спином I = 1/2, скорость
продольной релаксации задается значением рд (уравнения (2.22), (2.27) -
нго он0
N -
Рис.3.12. Обмен амидных протонов пептидной связи, катализированный кислотой (вверху) и
щелочью (внизу) [3.11].
112
к
S
S
Доступная для
вз 1 имодействия
° 2
поверхность (А )
Положение водородной
мостиковой связи
6
2--
о:;
1СГ6
10’2
10’4
1 10 20 30 40 50
Рис.3.13. Обмен амидными протонами н структурные свойства. Скорости обмена к в BPTI,
измерение которых проводилось при pH - 3,6 и Г - 36°С, сопоставляются с величинами,
характеризующими способность пептиднных связей к контакту с молекулой воды н способность
к образованию водородных мостиковых связей. Образование водородных связей рассматривается
по отношению к карбонильному кислороду основной цепи <|>, или к атомам боковых цепей, или
же к молекулам воды, которые подходят к внутренней поверхности цепи (Q ). (CD - область
СС-спнрали, ф) - область^-свернутого листа [3.51].
(2.31)). Если пренебречь внешней релаксацией для спина А, то скорость
продольной релаксации дается формулой
1 = Лир 2 ^2^Ы (__________Trot______+
Т?в \4я/ Ю г6ав 11 +(йМ-й)в)2т?оГ
V (3.2)
। 3 Trot_।_______6 Trot_____|
1 + T?ot 1 +(a)A +O)B)2 T?ot) '
Соответственно скорость поперечной релаксации есть
1 _ Мо V 1л г .______________________Trot______
Т?3 I471/ 20г6ав ' ™ 1 +(й)л-й>в)2т?ог
। 3 Trot । 6 Trot _|________6 Trot_______\
1+йЛт?ог 1 +a&T?ot 1 +(а)А +a)B)2T^ot'
Однако скорость установления равновесия после воздействия на спино-
вую систему возбуждающего импульса зависит от значений времени релак-
сации спина В, так что временная характеристика в общем случае
113
описывается системой дифференциальных уравнений. Для двух эквивален-
тных ядер со спином 1/2 существует простая экспоненциальная зависимость,
где скорости релаксации даются следующими выражениями:
1 _ /до \2 П2/а ( 3 Trot + 12 '
\4 Лу 1Q Г^А 1 -j. 1+4 <Z)2 Tmt >
и
1 _ /до \2 й2)4 /д , 15 Trot 6TWf
|4 Л J 20 4л ' rOt 1 + ОЙ Trot 1 + 4 цД T?ot
Для эквивалентных спинов скорость продольной релаксации с ростом
О)а Trot проходит через максимум, а затем вновь убывает. Скорость попереч-
ной релаксации в отличие от скорости продольной релаксации продолжает
возрастать до тех пор, пока теория, предполагающая однородность линий,
перестает быть применимой. В случае, когда времена корреляции велики,
спектр состоит из неоднородных линий, подобных спектрам ЯМР в твердых
телах (рис.3.14).
Рнс.3.14. Зависимости продольного и по-
перечного времени релаксациии от вре-
мени корреляции. Значения Tj (А ),
Г2ЛВ(В), 1^(0 и Т-^ (D) представ-
лены как функции времени корреляции
вращательных движений Тгор Парамет-
ры, используемые для расчета: резо-
нансная частота 500 МГц, расстояние
между линиями 1 м.д., расстояние между
ядерными спинами гдд-0,175 нм
(соответствует расстоянию между двумя
10ю ю'9 юв метиленовыми протонами).
Trot/С
Времена корреляциии вращательного движения сферических молекул
можно рассчитать по уравнению Стокса-Эйнштейна
4 7Г ?? д3
Trot Зкт ,
(3.6)
где Т] — вязкость растворителя, а — эффективный радиус комплекса, к -
постоянная Больцмана, Т - абсолютная температура. Для сферической мо-
лекулы объемом V с молекулярной массой М получим
114
(37)
Для протеинов парциальный объем У*, как правило, равен приблизи-
тельно 1,2 • 10’30м3.
Уравнение (3.6) вместе с уравнением (3.7), конечно, лишь грубо позво-
ляет рассчитать значение времени вращательной релаксации, так как реаль-
ные биологические молекулы, как правило, не являются ни идеально
сферическими, ни идеально жесткими. Эти уравнения могут быть использо-
ваны для вычисления верхней границы значений времени корреляции Trot.
Первоначально предполагалось, что измерение времен релаксации мо-
жет быть испольльзовано для детального изучения процессов движения в
протеине. Тем не менее оказалось, что система обладает слишком большим
числом степеней свободы и, следовательно, слишком большим число неизве-
стных, чтобы вне зависимости от используемой модели определить характе-
ристики сложных движений в протеине. Это прежде всего касается спектров
ЯМР 1Н, где времена релаксации зависят от относительных движений боль-
шого числа протонов и от релаксационных процессов внутри молекул. В
принципе более легкой является интерпретация результатов по измерению
времени релаксации ядер13С, поскольку релаксация этих ядер, как правило,
определяется только протонами, непосредственно связанными с данным
ядерным спином.
3.1.3. Определение межатомных расстояний
В применении метода ЯМР к решению задач структурной химии важное
значение имеет определение межатомных расстояний. Основным источни-
ком получения информации о межатомных расстояниях является ядерный
эффект Оверхаузера. Дополнительная информация может быть получена из
изучения взаимодействия с парамагнитными центрами и из эффектов коль-
цевого тока. В дальнейшем подробно остановимся на рассмотрении методов,
позволяющих исходя из анализа этих взаимодействий получить информа-
цию о расстояних между атомами.
Основные сведения о ядерном эффекте Оверхаузера (ЯЭО) были изло-
жены в разделе 2.2.6. Для протеинов одномерные экспериментальные мето-
дики определения ЯЭО не имеют большого значения, так как в спектрах
ЯМР JH существует весьма ограниченная область, в которой удается достичь
разрешения, необходимого для наблюдения отдельных резонансных линий.
В общем случае необходимо провести двумерный ЯМР-эксперимент, рас-
смотренный нами ранее (NOESY), в котором перекрывание резонансных
линий не играет существенной роли. Для двухспиновой системы временную
зависимость ЯЭО можно найти путем интегрирования дифференциальных
уравнений (2.20) и (2.21). Временная зависимость амплитуды кросс-пиков в
переходномЯЭО и эксперименте NOESY описывается формулой (для случая
А АВ
медленного движения: СсЬ Trot > > 1)
115
Тщ (МС)
Рис.3 15. Зависимость амплитуды кросс-пика от времени смешивания Тт в спектре BPTI,
полученном по методу NOESY. Штриховой линией обозначены значения амплитуд диагональ-
ных пиков амидных протонов Phe-45, Phe-22 и Phe-ЗЗ. Сплошные лииии отвечают данным по
Я' О для выделенных ядер н их соседей в трехмерной структуре [3.12].
(3.8)
МА ( Тт ) ( Тщ ) _
1 ОАв / ~ 2 1 ' Тт ~ (рл ~ । Оав 1 ) Тт
=-5TS57T<1-e )'
А г А
де, как и ранее, и/г -z-компоненты намагниченности и спина для ядра
А к моменту времени Тт после воздействия на спиновую систему импульсом
на частоте спина В, М0А и 10А — соответствующие величины в состоянии
термодинамического равновесия, рл и Оав ~ скорости спин-решеточной и
перекрестной релаксаций. Таким образом, наблюдаемый ЯЭО сначала воз-
растает, а затем для больших Тт стремится к нулю. На рис.3.15 приведена
зависимость амплитуды кросс-пиков от времени смешивания Тт для прото-
ков в BPTI. Подобная картина имеет место и в экспериментах по методу ТОЕ.
Эднако для больших значений времени амплитуда кросс-пика не равна ну-
лю, она стремится к значению, соответствующему стационарному ЯЭО
'рис.3.16). Поскольку в протеине во взаимодействии участвуют не два спина,
’ гораздо большее их число, то уравнения для двухспиновой системы необ-
ходимо обобщить. Обобщение (2.20) и (2.21) на случай .V спинов осуществ-
116
Рис.3.16. Зависимость от времени величины ЯЭО в эксперименте ТОЕ. Насыщение сигнала
амидной группы Tyr-23 BPTI и наблюдение сигналов HCt-атомов в Phe-22, Tyr-23 и Cys-30.
(а) - теоретическая зависимость от времени, (Ь) - увеличенная часть рис. (а) для малых времен,
где также приведены экспериментальные данные [3.13].
лается очевидным преобразованием. Для спина Л соответствующее обобщен-
ное уравнение имеет форму
dМ, , . N . .
-тг = -Ра -м*) -2 оА} . (3.9)
“» /=1
Так как на величину MJZ оказывают влияние все спины исследуемой
системы, то поведение намагниченности ядра А в принципе зависит от всей
спиновой системы. Конечной целью исследования является определение
межатомных расстояний гА]- между атомами А и j. Можно показать, что
изменение величины ЯЭО для очень малого времени смешивания Тт (Тт -*
0) пропорционально rAj"6. Для кросс-пика (точнее, объема кросс-пика) в
спектре, полученном по методу NOESY, и для ЯЭО в одномерном экспери-
менте имеем
d
d Тт
Для больших времен корреляции (точнее, для ( (Jtifirfy)2
(3.10)
< /
гомоядерной системе и для случая преобладания дипольной релаксации фор-
мула aAj = 1У2Л/ - Дает
/„ \2*2
а . =»-jy AJ = - 1^-2- — у j у ? taL< гл~6 > . (3.11)
Д/ УГо [4Л/ IQ ' А ' J rot А!
Здесь < > обозначает усредненное значение для межатомных рас-
стояний. Эти расстояния варьируются от молекулы к молекуле, так как
молекулы не являются идеально жесткими.
117
Так как обычно неизвестны ни точные времена корреляции, ни точные
корреляционные функции (уравнения (2.27) - (2.32), строго говоря, будут
справедливы только для простой вращательной диффузии), можно попы-
таться найти их приближенно. Здесь можно реализовать следующую воз-
можность: использовать ЯЭО для двух ядерных спииов С и D в
макромолекуле, расстояние между которыми известно, для проведения со-
ответствующей калибровки. Если исходить из того, что спины Л и /описыва-
ются той же корреляционной функцией и тем же временем корреляции, что
н спины С и D, то оказывается, что для межатомных расстояний справедливо
следующее соотношение:
-6 -6 Mz]/M$
<rA/6>-rcD - z / -”д.
(3.12)
При этом определение расстояний сопряжено со следующими дву-
мя трудностями: (1) корреляционная функция является приближен-
ной, и (2) если даже данное приближение достаточно хорошее, то речь
идет только о нахождении среднего значения rAj6, а не среднего рассто-
яния rAj, которое может существенно отличаться от вычисленного из
< га/6 >.
Из изложенного следует, что определение расстояний из ЯЭО является
неточным. Однако для большинства практических приложений это не столь
существенно, как может показаться на первый взгляд. В каждом отдельном
случае ЯЭО при условии, что измерения проводятся при очень малых време-
нах смешивания, очень быстро уменьшается с ростом расстояния (пропорци-
онально шестой степени!). Следовательно, имеет смысл проводить
измерение ЯЭО для ядер, которые расположены непосредственно по сосед-
ству (это условие выполняется для большинства случаев). Это значение
достаточно хорошо согласуется как с экспериментальными данными, так и с
полученными теоретически. При измерении ЯЭО в протеинах максимальное
расстояние между протонами равно 0,5 нм.
Определение расстояния между ядрами основано на зависимости
величины диполь-дипольного взаимодействия двух ядерных спинов от
расстояния между ними. Однако диполь-дипольное взаимодействие су-
ществует не только между спинами ядер, но и между спинами ядер и
электронов. Последнее взаимодействие также зависит от расстояния
между спинами и в принципе также может быть использовано для оп-
ределения расстояний в молекулах. Это возможно вследствие того, что
гиромагнитное отношение электрона существенно больше гиромагнит-
ного отношения ядра, например протона, а значит, взаимодействие
между спинами электронов и ядер вполне поддается детектированию.
Для того, чтобы методы двумерной спектроскопии ЯМР получили ши-
рокое распространение при решении задач установления структуры моле-
кул, все надежды на определение структуры протеинов связывались с
изучением зависимости ЯЭО от расстояния.
В то же время следует отметить, что протеины являются диамагнитными
молекулами, то есть в протеине отсутствуют парамагнитные центры. А сле-
118
довательно, эти центры должны создаваться специально. Простейшим вари-
антом достижения этого является взаимодействие с другими атомами или
молекулами, обладающими неспаренными электронами, т.е. с парамагнит-
ными молекулами. Для такого рода взаимодействий подходят парамагнит-
ные ионы металлов и органические радикалы. В принципе наиболее
подходящими являются физиологические условия, при которых протеин был
бы связан с парамагнитным веществом, выполняющим роль субстрата или
дополнительного агента, так как такие вещества не вызывают каких-либо
структурных изменений. Если же такая идеальная возможность отсутствует,
то можно заменить диамагнитные лиганды на парамагнитные, которые по
своим химическим и физико-химическим свойствам наиболее хорошо соот-
ветствуют естественным условиям. Менее подходящими, нз-за трудностей в
интерпретации, являются неспецифические взаимодействия парамагнит-
ных веществ с протеином или модификация протеина органическими свобод-
ными радикалами. При неспецифическом взаимодействии возникает еще
одна проблема, связанная с тем, что отсутствует точная информация о про-
странственном расположении парамагнитной группы или групп относитель-
но протеина. Кроме того, если размеры спин-меченых молекул велики, то
нельзя быть уверенным в том, что эти молекулы не произведут такие струк-
турные изменения в протеине, что не удастся определить его структуру в
нативном состоянии.
Парамагнитные вещества могут оказывать два типа воздействий на
соседние ядерные спины. Во-первых, они могут давать вклад в значение
химического сдвига. Это так называемый парамагнитный сдвиг резонанс-
ной частоты наблюдаемого ядра. Во-вторых, наличие парамагнитных цен-
тров может вызывать уменьшение времени релаксации возбуждаемых
ядер.
Поскольку величина парамагнитного сдвига зависит от диполь-диполь-
ного взаимодействия между ядерным спином А и неспаренным спином элек-
трона S, а величина этого взаимодействия убывает как третья степень
расстояния между спинами rAS, то и величина парамагнитного сдвига про-
порциональна гА$? Так как скорость релаксации в присутствии парамагнит-
ных частиц пропорциональна квадрату взаимодействия (см. раздел 1.3), то
в первом приближении она убывает как шестая степень расстояния т.е.
пропорционально rAS6, как и в случае эффекта Оверхаузера.
Так как спины электронов, как правило, релаксируют существенно бы-
стрее, чем спины ядер, то времена корреляции, ответственные за скорость
ядерной релаксации, в основном определяются временем релаксации элект-
ронов. Скорость дипольной релаксации в области значений времен корреля-
ции тем меньше, чем меньше время корреляции. Если время релаксации
спинов электронов Т1е очень мало, то уширение линий за счет поперечной
релаксации также мало, и вид спектра в основном определяется величиной
парамагнитного сдвига (сдвиг-реагент). Противоположный эффект наблю-
дается в том случае, если значение Т1е превышает 0,1 нс, тогда уширение
линий, как правило, настолько велико, что парамагнитный сдвиг оказывает-
ся необнаруживаемым (см. табл.3.4).
119
В то время как ядерный спин можно считать жестко локализован-
ным, для спина электрона такое предположение не выполняется. В
органических свободных радикалах электронная спиновая плотность
часто распределена по большому числу ковалентных связей молекуляр-
ного остова, и даже в отсутствие ковалентных связей иона металла с
молекулой можно зафиксировать наличие отличной от нуля спиновой
плотности на соседних лигандах. Так как спиновая плотность электро-
на не убывает до нуля в точке расположения ядра, то кроме дипольного
взаимодействия существует также и контактное взаимодействие Фер-
ми. В этом случае оба взаимодействия составляют так называемое
сверхтонкое взаимодействие. Следовательно, как было отмечено в раз-
деле 1.2.4, парамагнитный сдвиг дл/, обусловленный сверхтонким вза-
имодействием, складывается из контактной дрс и чисто дипольной
частей. Дипольное взаимодействие, описываемое в рамках классиче-
ских представлений как эффект взаимодействия магнитных моментов,
создающих вокруг себя локальное магнитное поле, приводит к измене-
нию значений резонансных частот, зависящих в общем случае от угла
между радиусом-вектором, соединяющим оба ядра, и направлением
внешнего магнитного поля. Однако под действием вращательной диф-
фузии молекул в растворе этот эффект усредняется, если только маг-
нитная восприимчивость не является анизотропной, т.е. зависит от
направления. Наличие анизотропии - дополнительное требование к
веществу, которое используется в качестве сдвиг-реагента. Как прави-
ло, свободные органические радикалы не обладают достаточной анизот-
ропией, но это свойство присуще редкоземельным элементам и
некоторым переходным металлам (табл.3.4 и 3.5). Их влияние на вели-
чину химического сдвига обозначается как псевдоконтактный сдвиг дрс. На-
Таблица 3.4. Времена релаксации и ширины линий основных парамагнитных веществ
Вещество Tip/hc Ширина линий в Гц
Си 1-3 3000 - 9000
Органические свободные ради- > 10 > 20000
калы Ч: 1 -10 3000 - 20000
Mnz+ 1-10 40000 200000
Fe3+(S-|) 0,01 -0,1 400 - 5000
Gd3+ 1 -10 60000 - 400000
Nit 0,001-0,1 25-1000
Fe2+ (S-0) 0,001 70
Co2+ (5-|) 0,0001 - 0,01 50 - 200
Fe3+ (S-|) 0,0001-0,01 10-40
Dy3+ Ho“+ Yh^+ 0,001 100
0,001 100
0,001 30
Значения приведены для Во- 2,35 Тл, ^5-0,5 нм [3.4].
120
Таблица 3.5. Псевдоконтактный сдвиг, обусловленный лантаноидным сдвиг-реагентом
Ион лантаноида
Теоретическое значение для
величины псевдоконтактно-
го сдвига (м.д.)__________
6,3
100
-33
4,0
0
39
0
0
4,2
-2,0
11,0
0,7
8,6
- 53
-22
1 3+
Псевдоконтактные сдвиги, пересчитанные на эффект Dy [3.16].
блюдаемый при этом химический сдвиг д состоит из диамагнитного сдвига
ddm и сдвига, обусловленного сверхтонким взаимодействием дАу:
$ = $dm + $hf=$dm+$Fc + $pc‘ (3.13)
С точки зрения возможности получить информацию о межатом-
ных расстояниях из контактного сдвига dFc довольно сложно, однако
наличие контактного сдвига приводит к значительному увеличению
области в которой наблюдается спектр исследуемого вещества. Это
сопровождается также улучшением разрешения, и вследствие этого
часто удается проводить отнесение резонансных линий в спектрах
очень больших протеинов. Наблюдая ферми-контактный сдвиг,
можно одновременно получить дополнительную информацию о
структуре химических связей, существующих между парамагнит-
ным центром и наблюдаемым ядром.
Если для простоты предположить, что спиновая плотность сконцентри-
рована на ионе металла, то для псевдоконтактного сдвига в миллионных
долях получим выражение
dpc-F(6,<p)/r3AS. (3.14)
Здесь rAS, 6 и <р определяют положение наблюдаемого спина А относи-
тельно молекулярной системы координат, жестко связанной с парамагнит-
ным центром S (рис.3.17). Величина псевдоконтактного сдвига убывает с
расстоянием rAS (как rAS) между парамагнитным центром S и спином ядра
А, кроме того, эта величина является функцией углов 0и <р. Зависимость
F(6 ,<р) задается формулой
121
Рис.3.17. Сферическая система координат. Описание положения
точки в пространстве с помощью сферических координат г,0 и <р.
Z
F (6, <р) = - | 1 (Хх +Ху) КЗ cos2 0-1) +
+ | <%х~Ху> sin2 # cos2 2 (3-15)
где Na ~ число Авогадро. Величина химического сдвига зависит от главных
значений %х, %у и /z молярной магнитной восприимчивости %, которая явля-
ется характеристикой для соответствующего парамагнитного центра в дан-
ном комплексе. В более часто встречающемся случае аксиальной симметрии
оказывается, что/х равно Ху и зависимость от угла <р исчезает. Несмотря на
то что в зависимости от вида парамагнитного центра имеется соответствую-
щее выражение, связывающее % с наблюдаемыми величинами, основная труд-
ность состоит в точном определении компонентов каждом отдельном случае.
Однако, если проведено точное отнесение нескольких резонансных линий в
присутствии парамагнитного центра, то соответствующие химические сдви-
ги можно использовать для калибровки, а следовательно, для определения
структуры.
Основной простатической группой, входящей в состав большинства про-
теинов и обнаруживающей при этом большое число парамагнитных взаимо-
действий, является порфирин и его производные. Порфирин образует
хелатные комплексы с ионами металлов, такими, как железо, магний, цинк,
никель, кобальт, медь и редкоземельные элементы. Важнейшим среди них
является комплекс с железом - гем, который участвует не только в связыва-
нии с кислородом при образовании гемоглобина, но принимает участие также
и в других реакциях, таких, как электронный транспорт цитохрома, в ката-
литических реакциях превращения Н2О2 или в реакциях оксидирования
кислот жирного ряда в процессах, катализируемых присутствием пе-
роксидазы. В этих комплексах ион железа в зависимости от стадии окисле-
ния или типа лигандов может быть либо диамагнитным, либо парамагнитным.
Следовательно, имеются природные диамагнитные и парамагнитные комп-
лексы одной и той же молекулы, при этом параметры ЯМР-спектров этих
122
комплексов допускают достаточно простое сравнение между собой. Кроме того,
вместо иона железа в кольцевую систему можно ввести ионы других металлов.
Это может существенно упростить интерпретацию экспериментальных дан-
ных, полученных по спектрам ЯМР, например, вследствие разделения контак-
тного и псевдоконтактного сдвига. Парамагнитные свойства тема обусловлены
не только присутствием иона металла, но и наличием порфиринового радикала,
в котором неспаренный электрон распределен по всей порфириновой системе.
В этих системах результирующие химические сдвиги могут достигать значи-
тельных величин. Обычно химические сдвига в спектрах 1Н изменяются от О
мд. до 10 м.д., в данном случае резонансные линии наблюдаются в диапазоне
±100 мд. На рис.3.18 приведен спектр ’н пероксидазы хрена (Armoracia
Gaertn.), содержащей гем в качестве простетической группы.
д/м.д.
Рис.3,18. Спектр ЯМР hi (300 МГц) пероксидазы хрена, содержащей гем в качестве
простетической группы. Вверху приведем случай, когда природный гем заменяется на продукт
его разложения [3.17].
Для того чтобы выявить места связывания с ферментами таких физио-
логически важных двухвалентных ионов, как Mg и Са2 , необходимо про-
вести их замещение на парамагнитные двухвалентные ионы, наиболее
подходящими среди которых являются металлы группы лантаноидов. Обыч-
но в качестве сдвиг-реагентов используются 12 лантаноидов, за исключени-
ем Gd3+, наличие которого приводит к уширению линий, а также
диамагнитных ионов La3+ и Lu3 . Для взаимодействия ионов металлов с
молекулами растворителя часто выполняются условия быстрого обмена. На-
блюдаемый в спектрах *Н химический сдвиг усредняется по протеинам,
связанным и несвязанным с ионами металла (как было рассмотрено в разделе
2.2.2). Представителем класса Са-связанных протеинов с высоким сродством
по кальцию является парвальбумин.
123
15 10 5 0 -5 -10 (5/м.д.
Рис.3.19. Сдвиг в спектре Са-связаниого протеина, обусловленный действием лантаноидного
сдвигающего реагента. СпектрЯМР *Н (270 МГц) парвальбумина, в котором вместо имеющегося
в естественном состоянии иона Сах содержится ион Yb° . Центральная часть спектра,
содержащая сигналы протеинов и сигнал воды, представлена не полностью. ( • ) - сигналы
метильных протонов протеина, которые достаточно сильно сдвигаются под влиянием парамаг-
нитных ионов [3.18].
На рис.3.19 представлен спектр, который демонстрирует эффект заме-
щения диамагнитных ионов кальция парамагнитными ионами Yb . Наблю-
даемые сдвиги, возникающие за счет сверхтонкого взаимодействия, не столь
велики, как в рассмотренном ранее примере (см. рис.3.18), но тем не менее
значительны. С помощью рентгеноструктурного анализа определена струк-
тура парвальбумина карпа (Cyprinus carpio L.). Этими данными о структуре
можно воспользоваться для определения в первом приближении положения
иона металла относительно протеина и для оценки компонент восприимчи-
3+
Рнс.3.20. Положение Yb в парвальбумине.
Изображенная поверхность определяет поло-
жение ядерных спинов вблизи иона иттербия;
эти спины испытывают парамагнитный сдвиг
27 м.д. [3.18]. Ион металла расположен в цен-
тре симметрии поверхностной структуры; на
рис. приведено объемное изображение.
124
вости в (3.15). На рис.3.20 показано, как на основании этих данных можно
построить поверхность, на которой расположен протон, так, чтобы получить
величину псевдоконтактного сдвига, равную 27 м.д. Форма этой поверхности
задается выражениями (3.14) и (3.15).
Влияние парамагнитных ионов на время релаксации, как и на величину
химического сдвига, обусловлено контактным и дипольным взаимодействи-
ями. Поскольку скорость релаксации квадратично зависит от энергии взаи-
модействия, то дипольный вклад пропорционален гд/. Чтобы оценить, как
связаны между собой скорости релаксации и значения расстояний до пара-
магнитного центра, то в общем случае следует исходить из того, что вклад
контактного слагаемого пренебрежимо мал, что, как правило, достаточно
хорошо выполнимо, если между парамагнитным центром и наблюдаемым
ядром отсутствует ковалентная связь, и лиганд не находится в непосредст-
венной близости от парамагнитного центра. Скорости продольной l/T1Af и
поперечной 1/Т^ релаксации в парамагнитном комплексе можно предста-
вить таким образом, чтобы в них выделить парамагнитный вклад 1/Т
связанный с неспаренным электроном парамагнитного иона, и диамагнитный
вклад 1/Tj 2д, наблюдаемый в отсутствие парамагнитного центра. Это можно
осуществить приближенно, если рассмотреть парамагнитные лиганды, близкие
по своей структуре к диамагнитным. Парамагнитные ионы с достаточно боль-
шими временами электронной спиновой релаксации такие, как Мп2+ и Gd3*
(табл.3.4), оказывают настолько большое влияние на время поперечной релак-
сации, что резонансные линии, соответствующие ядрам, удаленным от пара-
магнитного центра на сравнительно большие расстояния (порядка 1,5 нм),
уширяются настолько сильно, что их вообще не удается наблюдать.
Для определения меньших расстояний рассматривается усреднение, со-
ответствующее быстрому обмену, характерному для нековалентно связан-
ных лигандов. Наблюдаемые при этом скорости релаксации 1/Tj 2obs
являются статистическими средними скоростей релаксации в свободном
1 /Ту if и связанном 1 /Ту 2М состояниях лиганда:
_J_=_±£_+_PL_.
Tl,2t>Ar Г1,2/ Т1,2М
(3.16)
Усреднение здесь проводится с вероятностями /у и характеризующи-
ми соответственно свободное и связанное состояния лиганда (pf+Рь ж D • Эти
простые соотношения выполняются в том случае, если среднее время жизни
парамагнитного комплекса Тм существенно меньше времен релаксации
Тj 2м и обратной разности химических сдвигов для свободного и связанного
состояний 1/1 Adi. Варьируя условия проведения эксперимента, необходи-
мо подобрать такие времена релаксации, которые позволят наблюдать хоро-
шее разрешение спектральных линий.
Наряду с парамагнитным влиянием существует и другой эффект - диа-
магнитная анизотропия. Этот эффект зависит от расстояния, поэтому суще-
ствует принципиальная возможность использования его для определения
125
структуры. Кроме того, существует еще одни достаточно хороший с точки
зрения качественного понимания эффект, который наблюдается в аромати-
ческих системах, - эффект кольцевого тока (ring current shift). В зависимости
от относительного расположения рассматриваемого ядра и ароматического
кольца резонансная частота наблюдаемого спина может либо возрастать,
либо уменьшаться. Данное явление допускает следующее простое и нагляд-
ное объяснение: подобно тому, как это происходит в электрической цепи, по
системе 7Г-орбиталей кольца происходит перенос электронов, т.е. течет
электрический ток, индуцируемый внешним магнитным полем, при этом
вторичное поле, создаваемое этим током, ослабляет внешнее магнитное поле
[3.22 ]. Для описания диамагнитной анизотропии ароматических кольцевых
систем имеются следующие четыре основные модели: классическое диполь-
ное уравнение, которое приближенно описывает магнитное поле, индуциро-
ванное магнитным диполем в центре кольцевой системы [3.23], двойное
дипольное уравнение, описывающее магнитное поле, создаваемое двумя
диполями, один из которых расположен над кольцом, а другой - под кольцом
[3.24], квазиклассическое уравнение Джонсона-Бови [3.25 ] для магнитного
поля двух замкнутых проводников с током, один из которых находится над
кольцом, а другой - под кольцом, и квантово-механическое уравнение Хая-
Маллиона [3.26 ]. Сравнительные исследования, проведенные для протеи-
нов, структура которых определена по данным рентгеноструктурного
анализа, показали, что для ядер, не принадлежащих ароматической спино-
вой системе, подходят после соответствующей калибровки все модели, кото-
рые с достаточной степенью точности описывают "кольцевой сдвиг", т.е.
химический сдвиг, обусловленный кольцевыми токами. В частности, это
справедливо для простых кольцевых систем, таких, как тирозин, фенилала-
нин и гистидин. Для более сложных кольцевых систем, которые обнаружены
для триптофана и гема, наиболее подходящим является уравнение Джонсо-
на-Бови. Простейшим уравнением, описывающим магнитное поле кольце-
вого тока, является дипольное уравнение, которое определяет кольцевой
сдвиг дя, зависящий от характерной для данной кольцевой системы констан-
ты С, а также от расстояния г от центра кольцевой системы и угла 9 (г и О
определены на рис.3.17, где ось z перпендикулярна плоскости кольца):
dR—C —/"Л (3.17)
Г
Константа С определяется следующим выражением:
3Na ’ (ЗЛ8)
где г-коэффициент, определяющий интенсивность кольцевого тока 0 = 1 для
фенилаланина), Na ~ число Авогадро, %l = 49,5 • 10'12 м^ -моль"1.
На рис.3.21 показан сдвиг за счет кольцевого тока, который можно было
бы ожидать согласно уравнениям Джонсона-Бови и Хая-Миллиона для гекса-
126
Рис.3.21. Диамагнитная анизотропия в ароматических системах. Изменение химического сдвига
3g (в м.д.) вблизи ароматического кольца (в плоскости кольца), (а) и (с) рассчитаны по
уравнению Джонсона-Бови, (Ь) и (d) - по уравнению Хая-Маллиона [3.19]. В формуле (3.17)
Og имеет противоположный знак.
1-.__._,_____
0.0 0.5
НМ
Рис.3.22. Сдвиг, наблюдаемый под влиянием кольцевых токов в плоскости, перпендикулярной
кольцевой системе и проходящей через ее центр. Сдвиг (5^ (в м.д.) рассчитан согласно уравне-
нию Джонсона-Бови. Жирная линия обозначает размер ароматического кольца; обозначенные
на рис. точки соответствуют экспериментальным данным для метильных групп лизоцима [3.19].
(Знак соответствует определению, введенному на рис. 3.21.).
127
и пентагональных колец. Оказывается, что для больших расстояний г оба
расчета дают очень похожие картины. И, наконец, на рис.3.22 показан коль-
цевой сдвиг в плоскости, перпендикулярной плоскости кольца при условии,
что эта плоскость проходит через его центр. Трудность, возникающая при
использовании кольцевого сдвига для структурных расчетов, связана с суще-
ствованием других механизмов изменения химического сдвига, зависящих
от конформаций. Отметим, что значение дл определяется с большой погреш-
ностью, поскольку, как правило, разделить все эти влияния достаточно
сложно.
3.1.4. Отнесение резонансных линий
Отнесение резонансных линий в спектрах ЯМР к определенным атомам
в макромолекулах является одним из наиболее важных условий надежного
определения молекулярной структуры. Хотя в принципе для получения оп-
ределенной информации о структуре достаточно провести отнесение неболь-
шого числа резонансных линий, точность структурной информации
возрастает с увеличением числа расшифрованных линий, так как в этом
случае можно провести дополнительное сравнение результатов. С ростом
надежности расшифровки спектров существенно возрастает также и надеж-
ность следующих из этого выводов, которые часто весьма чувствительны к
ошибкам, допущенным при отнесении резонансных линий. Расшифровка
спектров ЯМР протеинов, как правило, проводится в две стадии: сначала
пытаются выяснить, какие резонансные линии относятся к аминокислотно-
му остатку, и при этом не заботятся о том, какое положение занимает данная
аминокислота в протеиновой последовательности. Таким образом во многих
случаях выясняют, о какой из аминокислот идет речь. Второй шаг состоит в
том, чтобы определить место этой аминокислоты в протеиновой последова-
тельности. Как правило, эта проблема решается с помощью независимых,
биохимических методов: либо с использованием биохимического аминокис-
лотного анализа, либо косвенным путем - по восстановлению соответствую-
щей последовательности в ДНК. Поскольку такая расшифровка спектров
предполагает определение параметров, характерных для каждой последова-
тельности аминокислот, то очевидно, что эффективность расшифровки спек-
тров зависит от того, насколько правильно определена эта первичная
структура. Неточности, допущенные в определении первичной структуры,
могут быть обнаружены в результате сравнения с данными, полученными
методом ЯМР. Эта расшифровка основана на сравнении расстояний, которые
являются типичными для аминокислот, следующих одна за другой в после-
довательности. Одновременно соотношения между расстояниями использу-
ются для упорядочения элементов вторичной структуры, так что при таком
последовательном отнесении резонансных линий можно получить информа-
цию о вторичной структуре.
Как указывалось ранее, отнесение резонансных линий к определенному
типу аминокислот можно было бы считать простой процедурой, если бы
существовала простая зависимость между значениями резонансных частот и
128
6 5 4 3 2 1 д/м.д.
1 1 " Ч 1 ' 1 1 Ala 1 1 Arg Дсп! "41 ! vpr-rii-г a I 1 1 I 1 l РЧ V f >r I 1 1 1 1 1 1 1 I 1 1 1 1 I 1 1 1 1 I 1 1 1 1 I 1 U ! ^=3, t , 1
a
рфаг-} ,—।— i ।
= o.
»r—
— - p 1
।
ASPi «4™==, P'^-Нь -J i I
— Cys oti i —г । । । —, i i i, p I f
Gin ai— , I 1 1 , 1 1 Jb3k=i 1
Glu Gly , 1* ” 1 1 vl 1 JW=-4—k— i ! !
His 1 1 1 1 1 । , 1 1
lie Leu a Г ижш. >=, 1— р^йКЙт^—i
Lys Met 81 |V 1 jt , j » c 1 1 1 1
Phe :»! 1 | I
ai •дммг^т
Pro | 55 >1 r , 4 ,1
Ser ar= L 1 ! p 1 ।
Thr | at= p— 1 1 1 ’ 1
Trp Туг 1 «£== Hl й====™ пр: д 1 ! 1 i 1 1 I SSFL-, | j |
Vai p 1 j 1 n Im, , !
MlHlI.I p' ‘T ' r '“ 'i i t 1 i 1 i , , 4 l i i i 1 i i > x. 1 , 1 I , 1 1
Рис.3.23. Химические сдвиги (5 в спектрах ЯМР протеинов (относительно внутреннего
стандарта DCC). Приведены медианы, квартили, 5-е и 95-е доли процентов значений хими- .
ческих сдвигов [3.3].
129
типом аминокислот, а также положением ядер в аминокислотном остатке.
Однако в действительности изменения химических сдвигов, связанные с
конформационными превращениями, приводят к тому, что из анализа спек-
тров ЯМР выводы о структуре аминокислот можно сделать лишь с опре-
деленной вероятностью (рис.3.23).
Наряду с методами двумерной спектроскопии ЯМР существуют еще два
распространенных биохимических метода: селективное дейтерирование
аминокислот определенного типа и сравнение с широким классом гомологов
протеинов, в котором замещается лишь небольшое число аминокислот в
последовательности. Несмотря на то что оба эти метода были известны за-
долго до того, как двумерная спектроскопия стала бурно развиваться и нашла
широкое применение, только сейчас эти методы стали применяться действи-
тельно эффективно благодаря развитию современных методов молекуляр-
ной биологии. Селективное дейтерирование в основном проводится исходя
из того, что наибольшее сродство к клеткам в питательной среде обнаружи-
вают аминокислоты именно в дейтерированном состоянии, так как это не-
посредственно обеспечивает встраивание соответствующих аминокислот в
молекулу протеина. Однако так как при этом изотопозамещенные аминокис-
лоты не только непосредственно встраиваются в молекулу протеина, но и
участвуют в превращениях, а также могут быть использованы при образова-
нии других аминокислот, селективность дейтерирования существенно пони-
1.0 .8 .6 .4
----д2/м.д.
1.0 .8 .6 .4
----► д2/м.д.
Рнс.3.24. Спектры НРг-протеина (500 МГц) S.aureus (слева) HS.camosus (справа), полученные
методом COSY с двухквантовой фильтрацией. Отмечены кросс-пики Н/3-Ну$ шестого и пятого
валиновых остатков, пара кросс-пиков при dj - 1,83 м.д. отсутствуют в спектре НРг-протенна
S.camosus.
130
Рис.3.25. Использование методов COSY и RCT для установления спин-спинового взаимо-
действия, Спектр валинового остатка, полученный по методу COSYh RCT, содержит диагональ-
ные пики ( ) и кросс-пики («, о ); ( • )-кросс-пик, полученный методом COSY, а (о )-
методом RCT. Обычно кросс-пики, которые наблюдаются методом COSY, наблюдаются и мето-
дом RCT. Мультиплетная структура кросс-пиков не показана.
жается. Для повышения селективности можно выбрать ауксотрофные му-
танты, которые сами не приспособлены для синтеза соответствующей ами-
нокислоты и используют аминокислоту, получаемую в побочном синтезе
прежде всего при синтезе пептида. Сначала этот метод ограничивался в
основном только ферментами бактерий, так как только бактерии могут вы-
ращиваться в больших объемах. Для того чтобы распространить этот метод
на высокоорганизованные системы, можно было бы в принципе отклониться
от метода, использующего культуры соответствующих клеток, поскольку
этот метод сопряжен с большими затратами, и соответственно практическая
ценность его невелика.
Эта проблема может быть решена также при выделении исследуемых
протеинов на культуре ткани и на клонах бактерий. К этому следует также
добавить, что можно было бы подобрать условия, при которых бактерии
будут продуцировать именно тот протеин, который необходим. Это, конечно,
существенно повышает эффективность использования дорогостоящей дейте-
рированной аминокислоты.
Интерпретация полученных спектров проста: если, например, ввести
дейтерированный лейцин, то ядра дейтерия в лейцине вообще не дадут сиг-
нала в спектрах ЯМР 1Н. Следовательно, все отсутствующие в спектрах
резонансные линии (по сравнению с обычным спектром, полученным для
недейтерированного соединения) соответствуют линиям лейцина. Подобная
информация может быть получена из сравнения спектров протеинов с близ-
131
кими последовательностями аминокислот. В этом случае отсутствующие в спек-
трах линии соответствуют аминокислотам, участвующим в дейтерообмене. Та-
ким образом можно установить либо природные мутанты, либо
туденаправленно создавать новые мутанты. На рис.3.24 показан пример, кото-
рый может быть использован для сравнения природных гомологов: НРг-проте-
ина Staphylococcus aureus и HPr-протеина Staphylococcus camosus. Обе эти
структуры отличаются только по двум положениям в последовательности: НРг-
протеин Staphylococcus aureus содержит в положении 7 валин и в положении 71
серин, Staphylococcus camosus содержит в этих положениях треонин и аланин.
Соответствующие кросс-пики в двумерном спектре НРг-протеина
Staphylococcus camosus исчезают. Если же в этих спектрах отсутствуют сигна-
лы только одной из аминокислот, то одновременно можно получить дополни-
тельную информацию о положении аминокислоты в последовательности. Это
условие выполняется всегда при проведении целенаправленного мутагенеза.
Иная картина наблюдается при дейтерировании, так как в этом случае отсутст-
вуют сигналы, соответствующие всем аминокислотам одного типа.
В принципе для отнесения всех резонансных линий не существует более
элегантного метода, чем методы двумерного ЯМР. Однако эффективность
этого метода существенно понижается с ростом молекулярной массы.
Отнесение резонансных линий к определенному типу аминокислот ос-
новывается на том, что в аминокислотных остатках большинство протонов
связаны между собой косвенным спин-спиновым взаимодействием. В то же
время спин-спиновое взаимодействие между протонами двух соседних ами-
нокислот очень слабое, поскольку между ближайшими парами протонов:
На-протоном и амидным, имеются четыре связи (см. рис.3.3), т.е. каждый
аминокислотый остаток протеина можно рассматривать как изолированную
спиновую систему. Так что для каждой аминокислоты имеет место типичная
картина спин-спинового взаимодействия, наблюдаемая в двумерных спект-
рах ЯМР. Рис.3.25 дает схематическое представление о косвенном спин-спи-
новом взаимодействии для валинового остатка, соответствующее методам
COSY и RCT. Такая же картина должна наблюдаться и в реальном экспери-
ментальном спектре (рис.3.26). Интерпретация спектра осложняется не
только тем, что неизвестны точные значения химических сдвигов для иско-
мых резонансных линий, но и тем, что не может быть проведено надежное
отнесение отдельных кросс-пиков в спектре. Это может быть также связано
и со слишком большой шириной резонансных линий кросс-пиков, так как
уширение линий сопровождается также уменьшением их амплитуды, и часто
рассматриваемые линии сливаются с фоном. Поскольку ширины линий, ко-
торые в основном определяются временем поперечной релаксации и скоро-
стью химического обмена, заранее неизвестны, то отсутствует уверенность
в том, что проведено правильное отнесение линий. Особенно существенно на
отнесении линий сказывается ширина линий в спектрах, полученных по
методу COSY, в которых пики в подспектре расщепляются на пики с отрица-
тельными и положительными знаками, так что полный интеграл пиков
кросс-мультиплета равен нулю. Чем больше ширины линий, тем менее за-
метны эти линии в спектре. Это проявляется тем нагляднее, чем ближе
распол ож ены одна к другой линии различных знаков, что происходит в том сл у ча е,
132
.5
dj/ м.д.
Рис.3.26. Картина связей валинового фрагмента HPr-протеина. Спектр (500 МГц) НРг-протеи-
на S.aureus в D2O, полученный методом COSY после двухквантовой фильтрации.
Концентрация протеина 4 ммоль, эксперимент проводился в течение двух дней.
если величины констант спин-спинового взаимодействия ядер, которым со-
ответствует кросс-пик, уменьшаются. При J = 0 кросс-пики исчезают. Так
как величины констант косвенного спин-спинового взаимодействия зависят
от диэдральных углов, то они зависят также от конформации, а значит,
возможны случаи, когда отдельные кросс-пики исчезают вследствие конфор-
мационных изменений.
Вторая проблема связана с перекрыванием искомого кросс-пика с дру-
гими сигналами, что не позволяет провести однозначное отнесение соответ-
ствующих сигналов в спектре. Решение этой проблемы может быть найдено
при использовании других методов двумерной спектроскопии. Если же, на-
пример, вследствие перекрывания сигналов невозможно провести отнесение
кросс-пиков в спектре, полученном с использованием метода COSY, по их
принадлежности к соответствующей спиновой системе, то часто удается
провести отнесение с использованием метода RCT, так соответствующие
картины кросс-пиков, полученные в этих двух экспериментах, различаются:
кросс-пики оказываются в разных местах спектра, и перекрывание отсутст-
вует.
133
Наряду с уже известными биохимическими методами имеются другие
методы, предназначенные для модификации боковых цепей молекул, что
позволяет обнаружить корреляцию наблюдаемых изменений в спектрах
аминокислот с изменениями, наблюдаемыми в последовательности. Эти ме-
тоды в основном используют тот факт что, в нативном состоянии протеина
реакционная способность аминокислот в зависимости от их положения в
молекуле может различаться достаточно сильно. Это становится понятным,
если представить, что аминокислота находится во внутренней части протеи-
на, которая труднодоступна для взаимодействия с химическими веществами.
В этом случае месторасположение модификации в последовательности мо-
жет быть установлено с помощью химических методов. Распространены в
основном методы H-D-обмена Е1 -протонов гистидина или нитрование тиро-
зина. Правда, все эти методы обладают тем недостатком, что здесь имеется
опасность неверной интерпретации изменений, наблюдаемых в спектрах, в
случае, если эти изменения возникают не в результате введения специфиче-
ских меток, а, напротив, связаны с неспецифическими процессами денату-
рации.
Двумерная спектроскопия в данном случае является самым элегантным
методом исследования. Если известно, какая из аминокислот анализируется,
и где она расположена в последовательности, то можно охарактеризовать тип
спиновой системы, соответствующей данному фрагменту, адля каждого типа
подобрать наиболее подходящий метод. Если удается однозначно устано-
вить, какие из аминокислот (одна или несколько) расположены по соседству
с данной аминокислотой и таким образом определить фрагмент последова-
тельности, то можно провести согласование данного фрагмента с фрагментом
в известной последовательности. Простейший вариант реализуется тогда,
когда аминокислота встречается в протеине только один раз. В этом случае
для определения последовательности отсутствует необходимость в получе-
нии данных о соседних аминокислотах.
Каким образом можно получить информацию о соседних аминокисло-
тах? Так как величины констант косвенного спин-спинового взаимодействия
не дают сведений о пептидных связях, то их значения не могут рассматри-
ваться как источник информации. Остается еще ядерный эффект Оверхау-
зера, который можно наблюдать для протонов соседних аминокислот при
условии, что они расположены достаточно близко в последовательности.
Правильность этих рассуждений следует из рис.3.3 и рис.3.4. Следует отме-
тить, однако, что расстояния между взаимодействующими протонами зави-
сят от диэдральных углов. На рис.3.27 представлены основные обозначения
для расстояний в соответствии с общепринятой номенклатурой. Несмотря на
то что пролиновый остаток не содержит амидных протонов, модельные тео-
ретические расчеты показывают, что при произвольных поворотах устанав-
ливаются такие углы <р и 1р, которые обеспечивают расстояния, не
превышающие 0,3 нм, что позволяет наблюдать ЯЭО. В принципе в трехмер-
ной структуре протоны могут быть расположены на таких расстояних, даже
если они не принадлежат аминокислотам, непосредственно следующим одна
за другой. Из статистического анализа кристаллической структуры протеи-
нов следует, что это менее вероятно: 88 % всех расстояний daN и <1nn, а также
134
Vai 65
Gly 67
Рис.3.27.Расстояния в последовательности, получаемые по данным ЯЭО. На рис. указаны наи-
более важные межъядерные расстояния между протонами для наиболее важных соседних ами-
нокислот в последовательности
определены из спектров ЯМР
первичной структуры HPr S.faecalis.
и приведены значения ЯЭО, которые в принципе могут быть
Н. Показанные аминокислоты представляют собой фрагмент
76 % всех расстояний d^N, которые в этих структурах протеинов меньше или
равны 0,3 нм, принадлежат к аминокислотам, следующим одна за другой,
так что, согласно [3.5], данные, полученные при измерении ЯЭО, могут
вполне достоверно отражать структуру аминокислотной последовательно-
сти. Возможную неоднозначность исключают сравнением с реально сущест-
вующей аминокислотной последовательностью.
Расстояния dxN и dOCN в последовательности зависят от диэдральных
углов <р и основной цепи, причем по определению величины этих углов
определяют вторичную структуру. Следовательно, величины, измеряемые
при наблюдении ЯЭО, характеризуют вторичные структуры.
3.1.5. Интерпретация двумерных ЯМР-спектров
Расшифровка двумерных спектров биологических макромолекул
"вручную" - деятельность, требующая достаточно больших затрат времени
и далеко не всегда интересная. Например, для небольшой молекулы, такой,
как HPr-протеин Staphylococcus aureus, в состав которой входят 87 амино-
кислот, в спектре, полученном по методу COSY, наблюдается примерно 600
кросс-пиков, а по методу RCT- порядка 1000. В этих спектрах прежде всего
должны быть измерены координаты кросс-пиков, а затем с учетом всех
кросс-пиков следует провести идентификацию аминокислот в последова-
тельности. После этого необходимо провести расшифровку спектров, по-
лученных по методу NOESY, чтобы полностью расшифровать всю последо-
вательность. Обычно при этом проводят расшифровку массива спектров,
снятых в НгО, а затем повторяют то же самое для спектров, снятых в D2O. В
последнем случае спектры отличаются лучшим качеством вследствие малой
135
интенсивности сигнала воды. Правда, эти спектры не содержат информации
о протонах, обменивающихся с ядрами дейтерия растворителя. Очевидно,
что расшифровка спектров — самый трудоемкий процесс при использовании
двумерной спектроскопии ЯМР для установления структуры протеинов.
Именно этим объясняются попытки по возможности автоматизировать об-
суждаемый процесс.
Существуют два принципиально различных подхода, позволяющих
провести расшифровку спектров. Сюда следует отнести возможность исполь-
зования всей имеющейся информации для расшифровки эксперименталь-
ных спектров. Если известны физические взаимодействия, то существует
возможность проведения моделирования любого двумерного эксперимента в
случае, если достаточно хорошо проведено отнесение спиновой системы к
определенному типу, правда, при этом релаксационные процессы описыва-
ются лишь весьма приближенно. Спиновая система определяется исходя из
принципиальной структуры аминокислот. Соответствующие константы
спин-спинового взаимодействия и химические сдвиги являются при это сво-
бодно варьируемыми параметрами, которые согласовываются со значения-
ми, определяемыми экспериментально, и затем могут быть использованы
для полного описания спектров. Однако на практике этот метод требует
слишком много времени. Можно несколько улучшить этот подход, если
провести серию специальных дополнительных экспериментов. На рис. 3.28
показано, что таким методом, по крайней мере для небольших молекул,
Рис.3.28. Наглядное представление двумерного спектра 2,3-ди-бромпропионовой кислоты, пол-
ученного по методу COSY-45 при двухквантовой фильтрации, (а) рассчитанный спектр, (Ь) -
фрагмент, (с) - соответствующий экспериментальный спектр [3.20].
можно получить даже из очень слабых сигналов в спектре достаточно полную
информацию.
В качестве альтернативного можно использовать подход, аналогичный
поэтапной оценке, проводимой без применения ЭВМ. Используя этот метод,
можно извлечь из двумерного спектра все основные признаки структуры, а
затем перейти к более полному уровню описания (рис.3.29). При таком
способе анализа структуры используется не вся информация, но расчеты при
этом менее сложны и применимы к спектрам больших молекул.
Проблема, возникающая при автоматической обработке спектров, со-
стоит в том, что в спектрах возникают артефакты, которые зачастую можно
136
Рис.3.29. Упрощенная схема расшифровки
двумерных спектров ЯМР.
Рис.З.ЗОа.Ь. Способы уменьшения артефактов в двумерных спектрах. Спектр ЯМР Н <360
МГц), полученный по методу NOES Yдля НРг-протеинаЗ.ГаесаПв (время смешивания Тт-100 мс).
(а) - экспериментальный спектр, демонстрирующий типичные артефакты: (j-шумы (верти-
кальные полосы в спектре) и антидиагональные полосы (полосы, расположенные слева-направо
вниз). (Ь) — спектр после соответствующей обработки, в результате которой артефакты
существенно ослаблены и уменьшена интенсивность диагональных пиков [3.21].
137
достаточно просто увидеть в спектрах.без привлечения специальных мето-
дик, т.е. "на глаз". Упростить возникающие в связи с артефактами проблемы
можно только специальным образом построив эксперимент.
В качестве следующего этапа в каждом случае можно попытаться путем
соответствующего преобразования матрицы данных, полученных в резуль-
тате двумерного эксперимента, по возможности уменьшить артефакты,
прежде чем перейти к расшифровке спектров (рис.3.30). В приведенных
спектрах, полученных по методу NOESY, присутствуют два типичных экс-
периментальных артефакта: сигналы, расположенные слева направо вниз
(зеркальное отражение нормального спектра) и/1-шумы (интенсивные вер-
тикальные сигналы). Оба артефакта можно уменьшить путем соответствую-
щей обработки. Одновременно уменьшится и интенсивность диагонального
пика, который не содержит какой-либо полезной для проведения оценки
информации, но при этсм кросс-пики, находящиеся вблизи диагонали, могут
перекрываться.
На следующем уровне отнесения пиков и контурных диаграмм, как
правило, удается распознать и по возможности уменьшить большую часть
оставшихся артефактов. При этом для надежности все-таки следует сравни-
вать между собой результаты, полученные на различных этапах расшифров-
ки, и подвергать их контролю.
3.1.6. Расчет структуры по данным ЯМР
После полного отнесения резонансных линий, что не имеет большого
значения с биологической точки зрения, следует определить структуру про-
<^ак<|.н-4) СХ-спираль Зц^-спираль Вращение! Вращение!! Вращение l' Врапдавек' Полуоборот
^«.Й-З) Й.Н-3) dNN^>^2) <«,Н-2) <1^(/,Н-1) dCXN 3jON <Hz) 1 1 > яшЛЯЛ 11 1
4444444 444444 4 9 4 5 7 5 7 9 4 9
1234567 123456 12 3 4 12 3 4 12 3 4 12 3 4 12 3 4
Рис.3.31. Расстояния, полученные по данным ЯЭО, и константы косвенного спин-спинового взаи-
модействия которые следует ожидать для вторичной структуры в стандартной геометрии.
Самая нижняя строка в таблице обозначает относительное положение аминокислоты в полипеп-
тидной цепочке. Толщины линий пропорциональны предполагаемой величине ЯЭО [3.27].
138
Рис.3.32. Зависимость вицинальных коиствнт
косвенного спин-спинового взаимодействия
в пептидах от величины диэдрального угла
Кривые приведены для случая нвилучшего
соответствие экспериментальных данных (с ука-
занием допустимого интервала погрешностей)
уравнению Карплуса (3.19) [3.28]. Угол допре-
делен как 0-6О°-(р.
теина исходя из межатомных расстояний, которые вычисляются согласно
данным по ЯЭО. Не применяя специальных расчетов, можно получить све-
дения о вторичной структуре протеина из данных по ЯЭО. Типичным при-
мером этого являются данные поЯЭО для протонов протеинового фрагмента,
о которых речь шла ранее. Для каждого элемента вторичной структуры
исходя из известных стандартных значений координат можно рассчитать
относительные значения величин, описывающих ЯЭО, при условии, что для
них имеет место простая зависимость типа 1/А В этом случае для каждого
типа вторичной структуры получают типичные комбинации данных по ЯЭО,
что позволяет сформулировать гипотезы о вторичной структуре исходя из
экспериментальных данных по ЯЭО (рис.3.31). Предположения о структуре
можно также подтвердить дополнительно путем измерения значений кон-
стант косвенного спин-спинового взаимодействия J между амидным прото-
ном и Н Ct-протонами в зависимости от диэдрального угла <р. Количественно
зависимость константы спин-спинового взаимодействия J от диэдрального
угла 0 (0 60° - 0) дается уравнением Карплуса
J-Л cos2 д-7? cos d+С, (3.19)
где А, В и С - эмпирические константы, величина которых зависит от типа
исследуемой молекулы. Для диэдрального угла <р в пептидах эта зависи-
мость, согласно [3.13 ], может быть достаточно хорошо описана для значений
А, В и С, равных соответственно 5,41, 1,27 и 2,17 (рис.3.32), Как видно из
рис.3.32, уравнение Карплуса может иметь более одного решения. Другая
трудность состоит в том, что при быстром вращении относительно одинарной
связи наблюдается усреднение величин констант косвенного спин-спинового
взаимодействия (случай быстрого движения). Однако для глобулярных про-
теинов следует ожидать, что значения констант косвенного спин-спинового
взаимодействия достаточно хорошо передают значения диэдральных углов в
основной цепи и во внутренней части протеина, так как здесь, вероятно,
отсутствует значительное усреднение по углам. С помощью этого усредне-
ния можно надежно идентифицировать ct-спираль и ^-свернутый лист. На
рис.3.33 приведен такой пример - большой антипараллельный ^-свернутый
139
Рис.3.33. Структура^-свернутого листа для HPr E.coli [3.29] HS.faecalis [3.21], полученная по
спектрам ЯМР. Сплошной линией показаны связи, для которых удалось наблюдать ЯЭО для
обоих протеинов. Штриховой линией показаны связи, для которых удалось наблюдать ЯЭО для
одиого из протеинов.
лист, идентификация которого была проведена с помощью метода ЯМР в
HPr-протеине E.coli и S.faecalis. Хотя в этой области значений диэдральных
углов только 9 из 25 аминокислот идентичны, в обоих протеинах данные по
ЯЭО характерны для структуры ^-свернутого листа. Кроме того, эта струк-
тура представляет также интерес как пример того, когда структура молекул,
находящихся в кристаллическом состоянии, принципиально отличается от
структуры в растворе, определяемой по спектрам ЯМР. Структура в кристал-
лическом состоянии содержит два ^-свернутых листа, которые отделены
один от другого лежащей между ними СС-спиралью [3.30 ].
Другим примером, представляющим собой иллюстрацию подобного раз-
личия структуры протеина в кристаллическом состоянии и в растворе, явля-
ется металлотионин-2 - протеин, связанный с ионом металла. Конфигурация
связей для ионов Cd, полученная поданным рентгеноструктурного анализа,
существенно отличается от полученной из анализа спектров ЯМР [3.31-
3.32 ]. Это не означает, что одна из двух структур определена неверно, а лишь
подтверждает, что найденная кристаллическая структура отличается от
структуры, преобладающей в растворе. Можно предположить, что и для
других протеинов будет найдено подобное свойство. В частности, это пред-
положение может быть справедливым для протеинов с малой массой и по-
липептидов, для которых внутримолекулярные взаимодействия в кристал-
лическом состоянии являются достаточно сильными. Следовательно, для
определения структуры малых протеинов в растворе может быть использован
метод ЯМР.
Для того чтобы построить поданным ЯМР полную трехмерную структу-
ру протеинов, необходимо скомбинировать межатомные расстояния, опреде-
ленные экспериментально, с возможными конформациями, которые может
140
принимать полипептидная цепочка в пространстве, и с учетом возникающих
при этом ограничений провести расчет структуры, основываясь на макси-
мально возможной совместимости данных. Методы, которые, как правило,
используются в настоящее время и хорошо себя зарекомендовали, реализо-
ваны в так называемом алгоритме расстояний [3.33 ], [3.34 ] и методе огра-
ниченной молекулярной динамики [3.35 ].
Алгоритм расстояний является одним из наиболее эффективных. Он
основан на том, что полная структура молекул описывается согласно данным
о межатомных расстояниях. Возможная степень неоднозначности, связанная
с вращениями и, как следствие, с изменениями диэдральных углов, задается
в таких системах указанием максимальных и минимальных расстояний. Так
как ЯЭО позволяет определять максимальные межатомные расстояния, то в
этом методе можно использовать данные, полученные с помощью ЯМР. В
ряде случаев метод расстояний оказывается достаточно эффективным. Воз-
можность проверки правильности выбранной структуры состоит в том, чтобы
исходя из данных рентгеноструктурного анализа рассчитать данные по ЯЭО.
С этой целью прежде всего необходимо определить расстояния, которые соот-
ветствуют ожидаемым из данных поЯЭО. Из результатов рентгеноструктурно-
го анализа можно выбрать все пары протонов, которые находятся на более
близких расстояниях, чем те максимальные расстояния, которые можно в прин-
ципе определить из данных по ЯЭО. Для того чтобы максимально приблизить
оценкик реальным условиям, необходимо провести дополнительное разделение
ожидаемых значений интенсивности по классам и определить, например, силь-
ный, средний и слабый ЯЭО для расстояний менее 0,25 нм, от 0,25 до 0,3 нм и
от 0,3 до 0,4 нм соответственно. Для того чтобы надежно смоделировать струк-
туру, необходимо располагать достаточным числом экспериментальных дан-
ных. В противном случае, если из эксперимента получены не все данные по
ЯЭО, то провести полный расчет не удается.
Кристаллическая структура BPTI принадлежит к наиболее хорошо под-
дающимся расшифровке и поэтому была выбрана для проверки в [3.36].
Результат приведен на рис.3.34. Жирная линия обозначает положение
С (Z-атомов в кристаллической структуре, тонкие линии обозначают различ-
ные рассчитанные структуры, которые согласуются с данными по ЯЭО. Итак,
для BPTI по данным ЯЭО были получены 508 расстояний, которые разделены
затем на несколько классов, согласно введенным ранее определениям, затем
из них выбраны 356 значений, используемых в дальнейшем для расчета, и
получено отличное согласие между данной структурой и расчетной
(рис.3.34,а). Даже если ограничиться 170 наибольшими значениямиЯЭО и
использовать их для расчета, то это позволяет определить основную плоско-
сть сгиба для молекулы протеина (рис.3.34). Доказательством надежности
метода ЯМР служит также и то, что получено достаточно хорошее совпаде-
ние для ранее неизвестной структуры ингибитора CZ-амилазы тендамистата
при проведении исследований одновременно и независимо двумя группами
исследователей, одна из которых проводила рентгеноструктурный анализ
[3.37 ], а другая использовала метод ЯМР [3.38 ]. Им удалось впервые пока-
зать, что ЯМР и рентгенострукурный анализ могут независимо друг от друга
приводить к близким результатам.
141
Рис.3.34а-Ь. Расчет структуры по данным ЯЭО. Приведенные структуры рассчитаны с помощью
алгоритма расстояний с учетом данных по ЯЭО (представлена только основная цепочка). Жир-
ной линией обозначена структура, построенная по данным рентгеноструктурного анализа, и эта
структура сравнивается с альтернативными, построеннымы на основании 356 (а) и 170 (Ь)
межатомных расстояний [3.34].
Число варьируемых переменных в принципе можно сократить, если
исходить из стандартной геометрии и рассматривать только те диэдральные
углы, которые совместимы с простыми модельными построениями. Учет
меньшего числа степеней свободы позволяет применить метод для протеинов
большего размера. Эффективность этого метода была показана на примере
BPTI [3.39].
Методом, который в принципе лучше использует информацию и не
зависит от экспериментальных данных, является метод ограниченной моле-
кулярной динамики. Этот метод основан на использовании по возможности
наиболее хорошей исходной структуры, параметры которой вводят в качест-
ве коэффициентов в N классических уравнений движения Ньютона для
отдельных атомов, входящих в макромолекулу. Решения уравнений движе-
ния находят численным интегрированием. Ускорение zZ2rz/z/Z2 z-ro атома
массой mi определяется производной по пространственным координатам по-
тенциала взаимодействия между атомами V:
d2rt
= -grad V (rz,..., rN).
(3.20)
Если известны значения пространственных координат rz всех атомов i и
значения их скоростей vz к моменту времени t0, то можно рассчитать поведе-
ние всех атомов в молекуле в некоторый момент времени t путем решения
уравнения (3.20) при условии, что известна потенциальная энергия V. Урав-
нение (3.20) может быть решено численно, при этом dt в выражении для
производной заменяется конечным интервалом Д/, а все величины в момент
142
времени t + At вычисляются в линейном приближении. Например, прибли-
жение для скорости к моменту времени to + Az имеет вид
л
vz (("*0 + ^ = Аг + у((г=г0) •
(3.21)
Эмпирический потенциал V состоит из энергетических вкладов за счет
ковалентных связей и из вкладов за счет электростатического взаимодейст-
вия, в принципе в расчете должны быть учтены все пары атомов, входящих
в молекулу. Оказывается, что расчеты, проводимые по методу молекулярной
динамики для макромолекул, кристаллическая структура которых известна,
моделируют структуры, которые выглядят вполне правдоподобными, но при
этом, как правило, не соответствуют нативным структурам. Этого и следует
ожидать, так как в расчеты введены существенныеприближения. Два из этих
приближений совершенно очевидны. Во-первых, в большинстве расчетов
пренебрегают взаимодействием с естественным окружением, например, с
растворителем, и проводят расчет в вакууме, хотя из биохимии достаточно
хорошо известно, что конформация протеина сильно зависит от растворите-
ля. Во-вторых, в расчетах принимается, что продолжительность некоторых
событий, например сворачивание протеина, составляет несколько пикосекунд,
в то время как в действительности это событие длится несколько секунд. Даже
при проведении расчетов с помощью больших компьютеров невозможно учесть
большие временные интервалы, поскольку длительность отдельных шагов Аг,
которые выбираются для интегрирования уравнений движения, должна быть
исключительно мала нравна, какправило, нескольким фемтосекундам. Однако
при проведении расчетов структур по данным ЯМР выбор этого параметра
играет второстепенную роль, так как в данном случае в систему вводится допол-
нительная информация. Межатомные расстояния, которые получают из дан-
ных по ЯЭО, вводят в уравнения (3.20) с помощью псевдопотенциалов и таким
образом определяют изгиб в определенном направлении. Типичное выражение
для такого псевдопотенциала имеет вид
Кяэо —
для гу > гу ,
для Гу < Гу .
(3.22)
Здесь Ci и С2 - соответствующим образом выбранные константы, гу° —
расстояния между атомами i и у, определенные согласно данным по ЯЭО, г у -
фактическое значение расстояния между двумя атомами. Конечно, подо-
бные псевдопотенциалы можно построить и для других величин, например,
таких, как диэдральные углы, и затем ввести их в расчет.
Проблема развития методов расчета трехмерных структур протеинов в
настоящее время еще не исчерпана. Оптимальный метод определения струк-
туры использует наряду с данными по ЯЭО все многообразие дополнитель-
ной информации. Подбор этой разнообразной информации, по-видимому,
143
наиболее целесообразно проводить с использованием методов, развитых в
моделях искусственного интеллекта. Однако эти методы только тогда пре-
восходят обычные методы, если они объединяют подходы, позволяющие
получить надежные физические данные, содержащиеся в расчетах по методу
молекулярной динамики.
3.1.7. Твердотельный ЯМР протеинов
Большинство ЯМР-экспериментов в биологических системах являются
типичными примерами приложения ЯМР к исследованию жидкостей, где
большая часть всех взаимодействий, величина которых зависит от ориента-
ции соответствующих связей относительно магнитного поля, усредняется до
нуля за счет быстрой вращательной диффузии молекул в растворе, и эти
взаимодействия могут рассматриваться лишь в качестве источника релакса-
ции. Преимущество метода ЯМР в жидкостях очевидно: в ряде случаев
водный раствор воспроизводит естественное физиологическое окружение
исследуемой молекулы. Ширина линий растворителя достаточно мала и
поэтому часто спектры растворенных веществ поддаются интерпретации на
атомарном уровне.
В то же время существуют неоспоримые аргументы в пользу применения
твердотельного ЯМР в биологии: величины, отражающие зависимость от
направления, которые усредняются за счет быстрых движений в растворите-
ле, содержат весьма важную и часто однозначно интерпретируемую допол-
нительную информацию о структуре исследуемых веществ. Кроме того, в
биологических системах содержатся компоненты, нерастворимые в воде. В
основном они образуют надмолекулярные структуры. К ним относятся мем-
браны, рассмотрение которых будет проведено нами в дальнейшем, волок-
нистые протеины, строение которых напоминает структуру коллагена.
Коллаген является компонентой клеточного остова. К ним относятся также
большие системы, состоящие из большого числа отдельных компонент, каж-
дая из которых является водорастворимой, таких, как актомиозиновая сис-
тема мышечных клеток или фрагментов, входящих в состав сложных
вирусов. Эти системы иногда могут кристаллизоваться, и в этом случае,
конечно, они могут достаточно эффективно анализироваться с использова-
нием методов рентгеноструктурного анализа. В ряде случаев эти системы
можно ориентировать в сильных постоянных магнитных полях за счет нали-
чия у них магнитных дипольных моментов, что существенно упрощает про-
блемы, возникающие в ЯМР-спектроскопии.
В то же время ЯМР в жидкостях - преобладающий метод, который
находит свое применение при анализе биологически важных молекул имен-
но потому, что ширины резонансных линий оказываются порядка несколь-
ких герц, что является несомненным преимуществом по сравнению со
значениями, наблюдаемыми в спектрах ЯМР твердых тел. Поскольку в мак-
ромолекулах содержится большое число протонов, и ширина соответствую-
щих им резонансных линий в спектрах ЯМР в твердой фазе достаточно
велика, это в высшей степени затрудняет разделение и отнесение отдельных
144
резонансных линий. В силу этого гораздо легче решить эти проблемы с
помощью спектров на таких ядрах, как 2Н, 12С или ^N, естественное содер-
жание которых невелико. Это позволяет однозначно выделить одно или
несколько мест в молекуле, а также достаточно просто и элегантно разре-
шить проблему отнесения линий в спектрах и проблему перекрывания спек-
тральных линий.
Для малых молекул в монокристалле удается получать хорошо разре-
шенные спектры ЯМР 'Н с помощью специально разработанных методов
ЯМР. Следует отметить две наиболее важные методики, позволяющие до-
стичь сужения линий в спектрах ЯМР твердых тел: многоимпульсные мето-
дики и вращение образца под "магическим углом". Последняя методика
основана на быстром вращении (с частотой приблизительно 10 кГц) относи-
тельно оси, расположенной под "магическим углом" а, равным приблизи-
тельно 54,7° (3 cos2 CL - 1 = 0), по отношению к внешнему магнитному полю,
что эквивалентно эффекту усреднения в жидкостях за счет броуновского
молекулярного движения (рис.3.35).
Рис.3.35. Уширение линий в спектрах ЯМР твердых тел. На рисунке приведен спектр ЯМР Н
(90 МГц) монокристалла ферроцена. (Слева) Обычный спектр в твердом теле. (Справа) Спектр,
полученный с использованием импульсной методики WAI1UHA [3.40].
Какие из взаимодействий оказываются наиболее существенными для
решения задач установления структуры биологических макромолекул в
твердом состоянии? Дипольные взаимодействия уже использовались для
определения расстояний между ядрами даже в том случае, если они не
связаны между собой непосредственно. В твердых телах измеряют не вели-
чину ЯЭО, которая зависит от дипольной релаксации, а расщепления резо-
нансных линий Av, величина которого определяется дипольным
взаимодействием. Эта величина для двух неэквивалентных спинов - двух
частиц А и В со спином 1/2, находящихся на расстоянии гАВ, дается выраже-
нием
3 cos2 0-1 > (3.23)
8 л2 г3АВ
где константы Л, До, Уа и ув определены ранее, а 0- угол между направлением
внешнего магнитного поля Во и вектором, соединяющим взаимодействую-
145
щие ядра. В неупорядоченном образце угол 0 принимает все возможные
значения. Таким образом получаем спектр, который представляет собой
суперпозицию спектров, соответствующих всем возможным ориентациям,
т.е. спектр порошкообразного вещества. В ориентированном образце 0 при-
нимает вполне определенное значение, и в этом случае в спектре наблюда-
ется дублет. Так как в общем случае расстояние между атомами известно при
условии, что ядра связаны между собой ковалентной связью, то получаем
простое соотношение между величиной расщепления резонансных линий и
ориентацией ковалентной связи в пространстве. Для структуры протеина
особое значение приобретает ориентация основной цепи. В этом случае мож-
но определить константы диполь-дипольного взаимодействия для пептидных
связей *5N - 'Ни 15N- Си для С «-группы.
Другим типом ориентационно зависящего взаимодействия является
анизотропия химического сдвига, которая играет существенную роль в спек-
трах i5N и 13С. Величины экспериментально определяемых химических
сдвигов зависят от ориентации главных осей соответствующего тензора от-
носительно магнитного поля. Здесь полное число параметров должно быть из-
вестно (по крайней мере в первом приближении для модельных структур).
Аналогии ная ориентационная зависимость имеет место для квадруполь-
ного взаимодействия. В этом случае также существует возможность опреде-
ления ориентации связей.
С помощью этих методов можно успешно определять трехмерную структуру
протеинов, подобно тому, как это было сделано для плазматического протеина
/tZ-вируса [3.41 J. Правда, в настоящее время еще окончательно неясно, какое
практическое значение имеет твердотельный ЯМР в данной области, поскольку
этот метод сопряжен со значительными экспериментальными трудностями, кото-
рые связаны, например, с необходимостью введения изотопных меток.
3.2. ЯМР-спектроскопия нуклеиновых кислот,
полисахаридов и липидов
Наряду с протеинами существуют и другие макромолекулы, которые
выполняют важные биологические функции. Большинство методов ЯМР,
используемых для исследования протеинов, могут быть при этом непосред-
ственно перенесены на другие макромолекулы. В этом разделе рассмотрим
характерные особенности применения метода ЯМР для исследования нукле-
иновых кислот, полисахаридов и липидов.
3.2.1. Состав и структура нуклеиновых кислот
Нуклеиновые кислоты занимают ключевую позицию в процессах хра-
нения и передачи информации о наследственных свойствах организма. Как
и протеины, они являются линейными макромолекулами, состоящими из
небольшого числа различных фрагментов, нуклеотидов.
146
2 '-дезоксирибонуклеозид-5'-
монофосфат (дАМФ)
Рибонуклеозид-5 '-
монофосфат (АМФ)
ОН ОН
Рис.3.36. Структура рибонуклеотидов и дезоксирибонуклеотидов:
О
II
HN3 4 5СН
I II
0=С2 1 6.СН
Н
Аденин (А) Урацил (U) Гуанин (G)
НИз^'бС— СН,
I II 3
0 = 0? 1 6СН
Н
NH2
N^4>CH
I, »
0 = С2 1 6 CH
H
О
Тимин (Т) Цитозин (С)
Рис,3.37. Структура важнейших пурино-
вых и пиримидиновых оснований
ЯМР нуклеиновых кислот в принципе сталкивается с теми же пробле-
мами, что и ЯМР протеинов. В то же время определение пространственной
структуры нуклеиновых кислот является более простой задачей, так как
здесь выбор основной пары нуклеотидов сушественно ограничивает число
возможных пространственных структур.
Основными фрагментами дезоксирибонуклеиновых кислот (ДНК) явля-
ются дезоксирибонуклеотиды, а основными фрагментами рибонуклеиновых
кислот (РНК) - рибонуклеотиды (рис.3.36). По аналогии с аминокислотами
в протеинах эти фрагменты отличаются только своими боковыми цепями,
которые в ДНК в основном состоят из пиримидиновых производных - цито-
зина и тимина - и пуриновых производных - аденина и гуанина. В РНК
присутствуют те же боковые цепи, только основание тимин заменено на
урацил (рис.3.37). Кроме этих основных фрагментов нуклеиновых кислот
147
А-ДНК В-ДНК Z-ДНК
Рис.3.38. Трехмерная структура дезоксирибонуклеиновой кислоты. Модели основных форм
двойной спирали ДНК [3.42].
имеется еще небольшое количество производных этих оснований-так назы-
ваемые редкие основания.
ДНК представляет собой двойную спираль, образованную двумя пере-
плетающимися (комплементарными) нитями. Она может существовать в
нескольких различных формах, значение которых для процессов регуляции
и контроля передачи наследственной информации в дальнейшем будет под-
робно обсуждаться (рис.3.38). РНК, как правило, существует в виде мономе-
ра. Основными формами РНК являются матричная РНК (мРНК),
транспортная РНК (тРНК) (рис.3.39) и рибосомная РНК (рРНК). Как ДНК,
таки РНК вступаете разнообразные взаимодействия с протеинами, в резуль-
тате чего происходит либо считывание информации, либо ее регуляция.
Дополнительно ДНК в эукариотических клетках образуют ассоциаты с про-
теинами: гистонами, которые участвуют в строительстве хромосом. Подо-
бные взаимодействия демонстрируют РНК, например рРНК связывается с
протеинами, из которых состоят рибосомы. В процессах расшифровки и
передачи информации мРНК и тРНК вступают в разнообразные взаимодей-
ствия с ферментами, участвующими в этих процессах. Поэтому ясно, что
исследование взаимодействий между протеинами и нуклеиновыми кислота-
ми является одной из самых интересных областей применения метода ЯМР
к биологическим задачам.
148
A
C75
c
Акцепторный участок
pG • C
C • G
G • C70 Фрагмент Tl])C
Фрагмент DHU G U
A • U j
G30 *m5Q0
Антикодирующий A • W
фрагмент Cm А Варьирующимся
U у фрагмент
(b)
GmA A
A35
Рис.3.39 Вторичная структура (а) и тре-
тичная структура (b) тРНК*’>ен дрожжей
[3.43]. Водородные мостики, стабилизи-
рующие вторичную структуру, обозна-
чены точками, а мостики, стабилизиру-
ющие третичную структуру, — сплошны-
ми линиями.
149
3.2.2. Исследование методом ЯМР нуклеиновых кислот и
комплексов нуклеиновых кислот с протеинами
Применение метода ЯМР к исследованию нуклеиновых кислот обладает
рядом особенностей по сравнению с исследованием протеинов. Так как осно-
ву нуклеотидов составляют ароматические системы, то имеет место доста-
точно сильная зависимость химических сдвигов от конформации молекул,
что приводит к значительному усложнению спектров ЯМР. Основным фраг-
ментом в последовательности нуклеиновых кислот является фосфатный ос-
таток, так что ЯМР 31Р наряду с ЯМР 1Н может быть использован в качестве
достаточно чувствительного метода исследований. Ионы Mj2+ связываются
с тРНК и стабилизируют структуру. На рис.3.40 показан пример спектра 31Р
Рис.3.40. Взаимодействие иоиов Mg* с
молекулами тРНК*®". Приведен спектр
d 1Р (202 МГц) тРНкФенЕ.соИ при различ-
ных концентрациях Mg . Концентрация
тРНК0,34 мМ [3.44].
3 мМ
4.6 мМ
9мМ
15 мМ
[М.Д.]
ISO
*’,39IH3I ; Антикодирующий фрагмент •
Фрагмент Tty С
' Фрагмент DHU
AV3, ССЯ
GC„ GCj,
| «с»
АОщ AUs)
I GC, GC»
AOe GCu
Рис.3.41. Спектр ЯМР 'н (500 МГц) иминных протонов тРНК*^6”, выделенной из дрожжей
[3.45].
тРНК₽Ке при различных концентрациях Mg2+. В спектре 31Р наблюдаются
хорошо разрешенные линии, положение которых изменяется в зависимости
от концентрации. Даже в одномерном спектре ЯМР !Н можно достаточно
четко различить отдельные линии всех протонов имииогрупп, взаимодейст-
вующих между собой, и провести их отнесение (рис.3.41). Эти протоны
имеют более важное значение для определения структуры, так как именно
их взаимодействие играет основную роль при определении структуры нук-
леиновых кислот.
Последовательное отнесение резонансных линий и расчет трехмерной
структуры проводят теми же методами, что и при исследовании протеинов.
В принципе нуклеиновые кислоты могут принимать разнообразные формы.
Главная цепочка фрагмента всегда характеризуется значениями шести тор-
сионных углов, один из которых располагается в плоскости кольца рибозы,
а значит обладает ограниченной областью значений, которая зависит от
четырех эндоциклических торсионных углов. Положение основания относи-
тельно рибозы определяется одним торсионным углом в гликозидной связи.
Внутри рибозы спины всех протонов связаны косвенным спин-спиновым
взаимодействием. Это означает, что отнесение резонансных линий, принад-
лежащих рибозному остатку, может быть проведено по методу COSY. Для
основания константы косвенного спин-спинового взаимодействия, как пра-
вило, невелики, так что с ростом молекулярной массы основания их далеко
не всегда удается зафиксировать. Внутри основания константы косвенного
спии-спинового взаимодействия дают неполную информацию, поэтому сле-
дует дополнительно использовать ЯЭО. В отличие от протеинов отдельные
фрагменты связаны скалярным взаимодействием через фосфатные группы,
и в данном случае для расшифровки структуры можно воспользоваться зна-
151
5.0 4.0 ’Н(ш2)
3i р relayed ’Н-’Н COSY
435 430 425 420 415 ’HCWj)
Рис.3.42. Отнесение линий в фрагменте ДНК. (а) — Спектр dlGCATGC^, получ^ныр с
использованием методов гетероядерной двумерной корреляционной спектроскопии Р— Н.
Рабочая частота для - 360 МГц. (Ь) - Соответствующий спектр Н, получегшый по методу
COSY при условии двухквантовой фильтрации, (с) - Объединение спектра Р и спектра,
полученного методом RCT 1Н-1Н [3.46].
чением константы гетероядерного косвенного спин-спинового взаимодейст-
вия 1Н - 31Р (рис.3.42). Анализ структуры резонансных линий в спектрах 31Р
фосфатной группы показывает, что имеется взаимодействие фосфора с про-
тоном в положении 3', с протоном в положении 5' СН2-группы и 4' -протоном
дезоксирибозы. Это означает, что проведя отнесение в спектре линии, соот-
ветствующей атому фосфора, связанному с этим протоном, можно найти
линию, соответствующую протону в положении 4' следующего остатка. Пра-
вильность отнесения можно проверить, определив, какая из линий соответству-
ет линии протона в положении 5'. Спектр ' Н - 31Р - 1Н, полученный по методу
152
RCT, позволяет отнести линии, соответствующие протонам в положениях 3'
и 4', которые принадлежат двум соседним кольцам дезоксирибозы.
Информация, получаемая из значений констант спин-спинового взаи-
модействия и ЯЭО, отчасти является избыточной, так что спиновая система
нуклеозида может быть определена достаточно надежно. Наряду с этим
имеются зависящие от пространственной структуры данные по ЯЭО для
последовательности, исходя из которых можно идентифицировать соседние
нуклеотиды, а также данные по сильному ЯЭО для спаренных оснований
комплементарных нитей.
Расчет структуры для двойной спирали ДНК относительно прост, так
как вследствие спаривания оснований и регулярности структур (А-, В- и
Z-ДНК) уже имеются достаточно надежно подтверждаемые структурные
гипотезы, среди которых необходимо провести выбор наиболее вероятной и
затем уточнить структуру. Подобное справедливо также и для ДНК с одной
спиралью или для РНК, еслиони обладают такой структурой, как, например,
тРНК.
Если структура фрагмента ДНК известна и проведено отнесение резо-
нансных линий, то можно изучить взаимодействие ДНК со специфическими
протеинами. Примером этого может служить взаимодействие С протеином,
который является лак-репрессором и подавляет транскрипцию лактозоспе-
цифических ферментов, так что лактоза практически отсутствует в клетках.
Структура головной части этого протеина, которая ответственна за связыва-
ние с ДНК, недавно была установлена методом ЯМР и опубликована в [3.35 ].
По спектрам комплекса, полученным по методу NOESY, можно определить
положение протеина относительно ДНК в том случае, если известно лишь
несколько расстояний из измерений ЯЭО. Экспериментально по данным
ЯЭО для !Н для трех различных ароматических колец лак-репрессора (Туг-
7, Туг-17 и His-29) найдены структуры, соответствующие определенным
нуклеотидам лак-оператора. По этим оценочным значениям расстояний
получена структура комплекса, в котором лак-репрессор развернут на 180°
в сравнении со структурой, полученнной из модельных представлений
(рис.3.43).
3.2.3. Состав и структура полисахаридов
Третью группу биологических макромолекул, состоящих из простых
фрагментов, образуют олиго- и полисахариды. Они состоят из простых мо-
носахаридных фрагментов, связанных между собой. Функции полисахари-
дов весьма разнообразны. Они играют роль резервных веществ, например
крахмал, и структурных элементов, например целлюлоза. Важную функ-
цию распознавания клеток, а также роль рецепторов выполняют поверхно-
стные элементы: олигосахариды и малые полисахариды, связанные с
липидами, свойства которых определяет их первичная структура.
Основные фрагменты полисахаридов - моносахариды - обнаруживают
большее разнообразие, чем аминокислоты. Правда, моносахариды с пятью
атомами углерода (пентоза) и с шестью атомами углерода (гексоза) встреча-
153
G®C 12
G-C II
OG 10
G°C 9
ДоТ 8
G°C 7
To A 6
G°C 5
ToA 4
To A 3
A°T 2
A° T I
G°C -I
G°C -2
Рис.3.43. Модель, построенная по
данным ЯМР для лак-репрессора в
комплексе с лак-оператором. Схема-
тически представлено положение го-
ловной части лак-репрессора
относительно двойной спирали ДНК
лак-оператора [3.47].
Рис.3.44. Строение полисахаридов. Структура дисахарид мальтозы (а), которая состоит из двух
молекул глюкозы и сахарозы, и сахарозы (Ь), состоящей из одной молекулы глюкозы и одной
молекулы фруктозы.
ются чаще, чем другие углеводы. Моносахаридные фрагменты обычно связа-
ны между собой гликозидными связями (рис.3.44). В отличие от протеинов
для них в основном характерно наличие разветвленных цепей, так как для
сахаридов каждая из гидроксигрупп может образовывать точку ветвления.
3.2.4. Определение структуры полисахаридов
Химический анализ полисахаридов достаточно сложен, поэтому многие
исследования сахаридов направлены на определение их химической струк-
туры. Кольцо молекулы сахаридов может принимать различные конформа-
ции: наиболее часто встречающиеся среди них - кресло и ванна. Обе
154
конформации характеризуются типичными значениями констант косвенно-
го спин-спинового взаимодействия и могут достаточно просто определяться
с помощью ЯМР Н. В обеих конформациях расстояния между отдельными
кольцевыми протонами также будут различать совершенно определенным
образом. Следовательно, и данные по ЯЭО также должны существенно отли-
чаться. В противоположность тому, что было получено при рутинном поста-
дийном определении структуры протеинов, в химии сахаридов отсутствуют
простые методы последовательного определения структуры.
Для полного описания структуры необходимо знать не только картину
связей моносахаридов, но и характер их связывания, т.е. необходимо допол-
нительно определить то, какой является связь: CL- или /3-гликозидной (см.
рис.3.44). Здесь самым надежным является метод двумерного ЯМР. Если же
нас интересует только картина связей моносахаридов, то отнесение сигналов
в спектрах может быть существенно упрощено в случае проведения проце-
дуры дериватизации (рис.3.45).
R
V IV III II I 1,1* 2 3
Galal —4GloHAoB1 —ЗО«1В1 — 4GloB1 — СН2-СН-СН( OAc)-CH=CH-Alk
HHCOAlk
м.д.
Рнс.3.45. Спектр ЯМР (500 МГц), полученный по методу COSY для перацетилированного
гликосфинголипида, выделенного из эритроцитарной мембраны [3.48].
155
С точки зрения биологии значительный интерес представляет трехмер-
ная структура полисахаридов, которая определяется конформацией кольце-
вых систем и вращением относительно простых связей в нециклических
частях молекул. Принципиально структуру полисахаридов можно опреде-
лить путем измерения констант связи и ЯЭО, хотя здесь могут возникнуть
существенные трудности при оценке биологического соответствия определя-
емых структур. Связывание полисахаридов с липидами и протеинами, а
также характер упаковки их в клеточные мембраны, по-видимому, оказыва-
ет сильное влияние на их пространственную структуру, так что измерения,
проводимые в изолированных полисахаридах, даже при условии, что они
проводятся в водной среде, необязательно должны давать реальное представ-
ление об интактной структуре. Но с другой стороны, такие измерения, по-
видимому, дают наилучшее представление о структуре полисахаридов.
3.2.5. Исследование биологических мембран
Липиды образуют большой гетерогенный класс органических молекул,
которые могут быть экстрагированы неполярными растворителями. Наряду
с изолированными молекулами, такими, как жирорастворимые витамины и
стероидные гормоны, для которых ЯМР успешно применяется для установ-
ления химической структуры, важны также фосфолипиды, которые образу-
ют плоские, надмолекулярные структуры - двойные липидные мембраны. В
соответствии с той ролью, которую играют мембраны, они были исчерпыва-
юще исследованы с использованием всех имеющихся в распоряжении био-
физических методов, включая ЯМР.
При исследовании мембран всегда имеют дело с большим числом моле-
кул. Мембраны, встречающиеся в природе, представляют собой очень слож-
ные системы, состоящие из большого числа различных липидов и протеинов,
встроенных в липидную мембрану (рис.3.46). В силу этого исключительно
сложно получить детальное представление об индивидуальных молекулах
подобно тому, как это было сделано для протеинов в растворах. Только для
некоторых модельных систем, таких, как мицеллы и липосомы, которые
состоят из вполне определенных компонентов, можно надеяться на то, что
будут получены надежные ответы на принципиальные вопросы, касающиеся
их организации и движения. В дальнейшем результаты, полученные для
модельных мембран, могут быть перенесены на мембраны, встречающиеся в
природе.
В зависимости от температуры и состава мембраны могут существовать
в различных физических фазах. При понижении температуры мембраны
обнаруживают свойства твердых тел, при повышении температуры они пе-
реходят в жидкокристаллическое состояние, которое характеризуется боль-
шей подвижностью молекул в плоскости мембраны. В жидкокристаллическом
состоянии найдено, что коэффициенты латеральной диффузии почти так же
высоки, как и в воде. Как правило, в таком состоянии находятся биологиче-
ски активные мембраны при физиологических условиях. Ограничение дви-
жения в одной плоскости приводит к тому, что в спектрах ЯМР наблюдаются
156
Рис.3.46. Модель плазматической мембраны. В биологической мембране, которая в основном
построена из фосфолипидов и холестерола, протеины и гликолипиды встраиваются в мембрану
и могут свободно диффундировать в плоскости. Остатки олигосахаридов в основном
располагаются на внешней стороне мембраны [3.49].
достаточно широкие линии, так как в данном случае изотропное движение
невозможно. Ниже температуры перехода ширина спектральных линий
очень велика и составляет несколько миллионных долей, что соответствует
значениям, наблюдаемым для спектров ЯМР в твердых телах.
Так как фосфолипиды содержат фосфатные группы, с помощью ЯМР 31Р
можно наблюдать фосфорсодержащие липосомы. Выше температуры фазо-
вого перехода при благоприятных условиях в искусственных мембранных
везикулах можно наблюдать сигналы от различных фосфолипидов
(рис.3.47). В малых везикулах удается различить линии, соответствующие
фосфолипидам, находящимся на внутренней и внешней сторонах мембраны
(химические сдвиги отличаются на несколько Гц). Для более надежного
отнесения соответствующих резонансных линий фосфолипидов на внутрен-
нюю или внешнюю поверхность мембраны, необходимо добавить парамаг-
нитное вещество, для которого проницаемость мембраны невелика, и в
основном будет наблюдаться связывание этого вещества с фосфолипидом,
находящимся на одной из сторон поверхности. Резонансные линии липидов,
связанных с парамагнитным веществом, в этом случае сильно уширяются и
практически не наблюдаются в спектре. Спектры ЯМР 31Р липосом также
являются подтверждением сделанного ранее вывода о том, что увеличение
напряженности магнитного поля далеко не всегда обеспечивает более высо-
кое разрешение, так как для ядер фосфора вклад в релаксацию за счет
анизотропии химического сдвига будет значительным. В этом случае скорость
релаксации возрастает как квадрат напряженности магнитного поля (см.
формулу (1.38)),а разность значений химических сдвигов увеличивается с
ростом поля линейно, поэтому уширение линий может компенсировать воз-
157
Рис.3.47. Спектр З^Р простой искусственной
мембраны. Спектр ЯМР3 (36,4 МГц) мем-
бранных везикул, состоящих из смеси
сфингомиелина и дипалмитоиллецитина.
50 Гц
1м.д.
Температура, при которой проводилось из-
мерение, 50°С. (а) - спектр везикул в трис-
буфере. (Ь) - спектр после добавления 5 мМ
СоС<2- (с) - спектр после добавления 7 мМ
СоС12- Обозначения: DPL0/(--дипалмитоил-
лецитин (внешн./внутр.), ЗРМдД- -
сфингомиелин (виеши./внутр.) [3.50].
растание разрешения. По этой причине в рассмотренных выше системах для
более сильных полей не удается различить сигналы от фосфолипидов, находя-
щихся на внутренней и наружной сторонах мембраны (рис.3.48). Конечно ли-
пиды, входящие в состав мембран, можно исследовать с помощью ЯМР 1Н. В
этом случае удается отдельно наблюдать сигналы от различных групп, входя-
щих в состав липидов. По спектрам ЯМР 13С и при условии, что изотопное
замещение проводится по строго определенным положениям, удается селектив-
но наблюдать атомы, находящиеся в длинных углеводородных цепочках.
Рис.3,48. Спектр ЯМР31Р
исскуствениых мембран (ве-
зикул дипалмитоиллецитина)
при различных температурах
и в различных магнитных по-
лях [3.51].
158
Липидные мембраны обладают тем характерным свойством, что моле-
кулы липидов определенным образом упорядочены. В одном из предельных
случаев все молекулы полностью упорядочены, как в кристаллическом со-
стоянии. Другим предельным случаем является статистический порядок
(беспорядок), который в большей или меньшей степени наблюдается для
жидкостей и порошкообразных сред. Для количественного описания степени
упорядочения, называемого жидкокристаллическим, используется понятие
параметра порядка. Эта величина является мерой макроскопического упоря-
дочения некоторой величины в образце: например, упорядочения направле-
ния вектора, соединяющего два выделенных атома в молекуле, относительно
некоторой оси. Параметр порядка может быть определен как величина, ко-
торая принимает значение, равное единице, если соответствующие величи-
ны, измеряемые в молекулах (например, направление выделенной связи),
являются одинаковыми для всех молекул (что отвечает полному упорядоче-
нию данной величины), и значение, равное нулю, если эти величины прини-
мают все возможные значения (что соответствует отсутствию какого-либо
упорядочения в пространстве выделенных связей в молекулах). На практике
параметр порядка не всегда допускает такую простую и наглядну.ю интер-
претацию, а также не всегда удается установить непосредственную связь с
величинами, которые могут быть измерены по спектрам (см. ниже).
Важные результаты, необходимые для установления структуры мемб-
ран, могут быть получены для ориентированных мембран с помощью резо-
нанса на ядрах дейтерия. Ориентация создается следующим образом.
Мембрану помещают между двумя тонкими стеклянными пластинами, каж-
дая из которых задает ориентацию прилежащего слоя, т.е. получается двой-
ной слой. При проведении экспериментов необходимо прежде всего решить
проблему улучшения отношения сигнал/шум. Здесь существует достаточно
простой вариант решения: для этого необходимо сложить несколько стеклян-
ных пластинок с мембранами стопкой и получить спектр в таком образце.
При наличии ориентации спектры мембран переходят из области резонанса,
характерного для жидкости, когда вследствие изотропного движения боль-
шая часть взаимодействий полностью усредняется, в область резонанса, ха-
рактерного для твердых тел. В ориентированных системах наблюдается
расщепление резонансных линий Av, которое зависит от ориентации систе-
мы относительно внешнего магнитного поля и характеризуется константой
квадрупольного взаимодействия, отличной от нуля для дейтерия, спин ядра
которого равен единице. В жидкокристаллическом состоянии вращательная
диффузия относительно оси, перпендикулярной поверхности мембраны, как
правило, происходит с достаточно высокими скоростями, поэтому различия
в спектрах, наблюдаемых для молекул, находящихся в этих плоскостях,
усредняются до нуля. Величина квадрупольного расщепления А V зависит от
угля 0 между направлением связи 13С - 2Н и нормалью к плоскости мембра-
ны, а также от угла, который составляет эта нормаль по отношению к внеш-
нему магнитному полю. Соответствующий параметр порядка SqD
определяется по следующей формуле:
SCD = ^< 3 cos2 6- 1 >. (3.24)
159
I___I I----------1
20 кГц 50 кГц
7
Рис.3.49. Спектр ЯМР на ядрах Н (46,1 МГц) реконструированных саркоплазматических
везикул, в которых естественные липиды заменены на 1,2-диелайдоил-глицерол-З-фосфохо-
лин, в котором в положении 9 н Юпроведено замещение на дейтерий, (Вверху) Спехтр мембраны
без протеина. (Внизу) Спектр мембраны, в которой содержатся естественные мембраны проте-
ина. Измерения при 25°С (а) и при 4°С (Ь) [3.52].
Если рассмотреть не ориентированную мембрану, а липосому и провести
усреднение по всем возможным направлениям нормали к поверхности мем-
браны, то найдем максимум, соответствующий перпендикулярной ориента-
ции. Следовательно, в полном спектре будут наблюдаться в основном
дублетные сигналы от этих молекул, и получим два максимума. Наблюдае-
мое расщепление дается формулой
Av = SCD,
4 h CJJ
(3.25)
где h - постоянная Планка, eQ - квадрупольный момент дейтерия и eq -
градиент электрического поля в направлении связи 13С — 2Н. На рис.3.49
приведен спектр ЯМР 2Н реконструированных саркоплазматических вези-
кул ниже и выше температуры фазового перехода. Жирные кислоты мемб-
ранных липидов селективно помечены дейтерием в положениях 9 и 10. При
встраивании саркоплазматической АТФазы в мембрану наблюдаемая кон-
станта квадрупольного взаимодействия убывает, т.е. параметр порядка
уменьшается. Ниже температуры перехода наблюдается уширенный спектр,
типичная ширина линий которого характерна для состояния геля. Встраива-
ние протеина в мембрану еще более сильно сказывается на виде спектра
ЯМР.
160
Литература
3.1. Lehninger, A.L. (1983) Biochemie (VCH, Weinheim).
3.2. Bundi A. and Wuthrich, K. (1979) Biopolymers 18, 285.
3.3. Gross, K.-H. and Kalbitzer, H.R. (1988) J.Magn. Res. 76, 87.
3.4. ШРАС-ШВ (1970) J.Biol. Chem. 245, 6489.
3.5. Schulz, G.E. and Schirmer, R.H. (1979) Principles of Protein
Structure (Springer, Berlin, Heidelberg, New York).
3.6. Ghelis, C. and Yon, J. (1982) Protein Folding (Academic, New York).
3.7. Wuthrich, K. and Wagner, G. (1978) TIBS 3, 227.
3.8. Kalbitzer, H.R., Deutscher, J., Hengstenberg, W. and Rosch, P.
(1981) Biochemistry 21, 6178.
3.9. Kalbitzer, H.R., Marquetant, R., Rosch, P. and Schirmer, R.H.
(1982a) Eur. J.Biochem, 126, 531.
3.10. Kalbitzer, H.R., Hengstenberg, W., Rosch, P., Muss, P., Bemsmann, P.,
Engelmann, R., Dorschug, M. and Deutscher, J. (1982b) Biochemistry
21,2879.
3.11. Tuchsen, E. and Woodward, C. (1985) J. Mol. Biol. 185, 421.
3,12. Wagner, G. (1983) Quart. Rev. Biophys. 16, 1 .
3.12. Kumar, A., Wagner, G., Ernst, R.R. and Wuthrich, K. (1981) J. Am.
Chem. Soc. 103, 3654.
3.13. Dubs, A., Wagner, G. and Wuthrich, K. (1979) Biochem. Biophys.
Acta 577, 177.
3.14. Wuthrich, K. (1986) NMR of Proteins and Nucleic Acids (Wiley,
New York).
3.15. Bertini, I., Luchinat, C. and Messori. L. (1987) in Metal Ions in
Biological Systems, Vol. 21, edited by H. Siegel (Marcel Dekker, Basel).
3.16. Bleaney, B., Dobson, C.M., Levine, B.A., Martin, R.B., Williams,
R.J.P. and Xavier, A.V. (1972) J.C.S. Chem. Comm., 791.
3.17. Morishima, I., Shiro, Y. and Nakajiama, K. (1986) Biochemistry 25, 3576.
3.18. Lee, L. and Sykes, B.D. (1983) Biochemistry 22, 4366.
3.19. Perkins, S.J. and Dwek, R.A. (1980) Biochemistry 19, 245.
3.20. Meier, B.U., Bodenhausen, G. and Ernst, R.R. (1984) J. Magn. Res.
60,161.
3.21. Glaser, S. and Kalbitzer, H.R. (1986) J.Mag. Res, 68, 350,
Glaser, S. (1987) Doctoral Thesis, University of Heidelberg
3.22. Pauling, L. (1936) J.Chem. Phys. 4, 673.
3.23. Pople, J.A. (1956) J. Chem. Phys. 24, 1111.
Robbins, A.H., McRoe, D.E., Williamson, M., Collet, S.A., Xuong,
N.H., Furey. W.F., Wang, B.C. and Stout, C.D. (1990) J. Mol. Biol,
submitted
3.24. Abraham, R.J., Fell, S.C.M. and Smith, K.M. (1977) Org. Magn.
Res. 9, 367.
3.25. Johnson, C.EJr. and Bovey, F.A. (1958) J.Chem. Phys. 29, 1012.
3.26. Haigh, C.W. and Mallion, R.B. (1972) Org. Magn. Res. 4, 203.
3.27. Wagner, G., Neuhaus, D., Worgotter, E., Vasak, M., Kagi, J.H.R.
and Wuthrich, K. (1986) J. Mol. Biol. 187, 131.
161
3.28. DeMarco, A., Minas, M. and Wuthrich, K. (1978) Biopolymers 17, 637.
3.29. Klevit, R.E. and Waygood, E.B. (1986) Biochemistry 25, 7774.
3.3O. E1-Kabbani, O.A.L., Waygood, E.B. and Delbaere, L.T.J. (1987) J.
Biol. Chem. 262, 12926.
3.31. Frey, M.H., Wagner, G., Vasak, M., Sorensen, O.W., Neuhaus, D.,
Worgotter, E., Kagi, J.H.R., Ernst, R.R. and Wuthrich, K. (1985) J.
Am. Chem. Soc. 107, 6847.
3.32. Furey, W.F., Robbins, A.H., Clancy, L.L., Winge, D.R., Wang, B.C.
and Stout, C.D. (1986) Science 231, 704.
3.33. Braun, W., Bosch, C., Brown, L.R., Go, N. and Wuthrich, K. (1981)
Biochem. Biophys. Acta 667, 377
3.34. Havel T.F., Kuntz, I.D. and Crippen\ G.M. (1983) J.Theor. Biol. 104,359.
3.35. Kaptein, R., Zuiderweg, E.R.P., Scheek, R.M., Boelens, R. and van
Gunsteren, W.F. (1985) J.Mol. Biol. 182, 179.
3.36. Havel, T.F. and Wuthrich, K. (1985), J.Mol. Biol, 182, 281.
3.37. Pflugrath, J.W., Wiegand, G., Huber, R. and Vertesy, L. (1986) J.
Mol. Biol. 189, 383.
3.38. Kline, A.D., Braun, W. and Wuthrich, K. (1986) J.Mol. Biol. 189, 377.
3.39. Braun, W. and Go, N. (1985) Mol. Biol. 186, 611.
3.40. Waugh, J.S,. Huber, L.M. and Haberlen, U. (1968) Phys. Rev. Lett.
20,180.
3.41. Opella, S.J., Stewart, P.L. and Valentine, K.G. (1987) Quart. Rev.
Biophys. 19, 7.
3.42. Seelig, J. and Seelig, A. (1980) Quart. Rev. Biophys. 13, 19.
3.42. Dickerson, R.E. (1983) Sci. Am. 249 (6) 100.
3.43. Kim, S.H., Suddath, F.L., Quigley, G.J., McPherson, A., Sussman,
J.L., Wang, A.H.J., Seeman, N.C. and Rich, A. (1974) Science 185, 435.
3.44. Hyde, E.I. and Reid, B.R. (1985) Biochemistry 24, 4315.
3.45. Heerschnap, A., Haasnot, C.A.G. and Hilbers, C.W. (1983) Nucl.
Acid Res. 11, 4501.
3.46. Frey, M.H., Leupin, W., Sorensen, O.W., Denny, W.A., Ernst, R.R.
and Wuthrich, K. (1985) Biopolymers 24, 2371.
3.47. Boelens, R., Scheek, R.M., van Boom, J.H. and Kaptein, R. (1987)
J.Mol. Biol. 193, 213.
3.48. Dabrowxki, J., Dabrowski, U, Hanfland, P., Kordowicz, M. and
Hull, W.E. (1986) Maqn. Res. Chem. 24, 729.
3.49. Bretscher, M.S. (1985) Sci Am. 253 (4) 86.
3.50. Berden, J.A., Barker, R.W. and Radda, G.K. (1975) Biochem.
Biophys. Acta 375, 186.
3.51. Berden, J.A., Cullis, P.R., Hoult, D.I., McLaughlin, A.C., Radda,
G.K. and Richards, R.E. (1974) FEBS Lett, 46, 55.
3.52. Billeter, M., Braun, W. and Wuthrich, K. (1982) J.Mol. Biol. /55,321.
3.53. Wuthrich, K. (1976) NMR in Biological Research: Peptides and
Proteins (North-Holland, Amsterdam).
162
Дополнительная литература
Jarde tzky, О. and Roberts, G.C.K. (1981) NMR in. Molecular Biology
(Academic, New York)
Neuha us, D. and Williamson, M.P. (1989) The Nuclear Overhauser Effect in
Structural and Conformational Analysis (VCH, Weinheim)
Wuhrich, K. (1976) NMR in Biologial Research: Peptides and Proteins
(North-Holland, Amsterdam)
Wuhrich, K. (1986) NMR of Proteins and Nucleic Acids (Wiley, New York).
163
Глава 4. ЯМР-томография
В этой главе речь пойдет еще об одном приложении ЯМР в биомедицине -
о получении изображений, индуцированных сигналом ЯМР. Этот метод на-
зывают ЯМР-томографией или магнитно-резонансной томографией. Суще-
ствует также еще одно, менее часто употребляемое название -
ЯМР-цойгматография. В данном случае измеряемой величиной является
намагниченность ядер определенного типа в выделенном элементе объема
(точнее, спиновый отклик на переменное магнитное поле). В этом состоит
принципиальное отличие данного метода от рентгеновской компьютерной
томографии, где измеряемой величиной является коэффициент поглощения
рентгеновского излучения различными тканями.
Как известно, спиновый отклик, регистрируемый в ЯМР-эксперименте,
зависит не только от спиновой плотности, но и от других параметров. Важ-
нейшими среди них являются времена продольной и поперечной релаксации,
которые так же, как и спиновая плотность, содержат информацию о струк-
туре и составе исследуемой ткани. Таким образом, в отличие от рентгено-
вской компьютерной томографии можно выделить по крайней мере три
параметра, которые могут быть использованы для характеристики исследу-
емых тканей. Вследствие этого в тканях, однородных по одному из парамет-
ров, можно, путем вариации параметров соответствующей импульсной
последовательности усилить контраст по другим параметрам. В дополнение
к тому, что в ЯМР-томографии не используется ионизирующее излучение,
возможности разрешения по контрасту делают ЯМР-томографию более
предпочтительной по сравнению с рентгеновской компьютерной томогра-
фией, хотя отношение сигнал/шум в рентгеновской томографии может быть
значительно выше, чем в магнитно-резонансной.
4.1. Основные принципы получения изображения
При использовании метода ЯМР высокого разрешения для установления
структуры биомрлекул стремятся построить эксперимент таким образом, чтобы
сигналы, регистрируемые от всех элементов объема исследуемого образца, по
возможности были бы одинаковыми, и этим объясняются высокие требования,
предъявляемые к однородности постоянного магнитного поля Во. В противопо-
ложность этому при исследовании гетерогенных образцов, например, тела че-
ловека, эксперимент стремятся построить так, чтобы сигналы, регистрируемые
от различных элементов объема образца, были бы максимально различимыми.
С этой целью в объеме исследуемого образца создается градиент магнитного
поля Во, и протоны, выбор которых определяется их высоким естественным
содержанием и большим магнитным моментом, в разных участках образца
вступают в резонанс на различных частотах.
В основу метода пространственно-селективного ЯМР положен тот факт,
что за счет приложения градиента магнитного поля осуществляется выбор
тонкого плоского слоя в исследуемом образце. Принцип селективного воз-
165
Рис.4.1. Принцип селективного возбуждения.
Поскольку на образец одновременно воздействует
статическое магнитное поле Во и постоянный гра-
диент ноля Gz, то селективное по частоте
высокочастотное поле возбуждает только ядерные
спины, находящиеся в тонком слое
буждения наглядно иллюстрирует рис. 4.1. В методе селективного возбуждения
создается градиент постоянного поля вдоль определенного направления, так что
величина постоянного поля вдоль этого направления* линейно изменяется с
расстоянием, а его направление остается неизменным . На рис. 4.1 в качестве
направления градиентов выбрана ось z, однако в принципе можно выбрать
произвольное направление в пространстве. Если образец облучается РЧ полем
в узкой полосе частот, то условие резонанса (1.3) выполняется только для
спинов ядер, находящихся в некотором тонком слое (в связи с наличием гради-
ента поля), и поэтому только ядра этого слоя дают вклад в измеряемый сигнал.
Для того чтобы получить двумерное изображение данного слоя исследу-
емого объекта, необходимо разбить слой на (п х л) частей, т. е. построить
матрицу Ap,q, состоящую из (и х и) элементов изображения или пикселей
(picture elements или pixels), при этом п выбирается исходя из желаемого
значения отношения сигнал/шум или из каких-либо соображений, связан-
ных с техникой обработки сигналов, например п = 256. В эксперименте
определяются значения амплитуд сигналов от этих пикселей Арл и затем на
основании этих данных строится изображение, например, на экране. Для
построения изображений можно реализовать следующие варианты: последо-
вательно выделить л2 элементов и провести измерение от каждого из них.
Этот метод называется методом чувствительной точки. Визуализация изо-
бражений с помощью этого метода требует значительных затрат времени.
Очевидно, что время проведения эксперимента будет существенно меньшим,
если получать изображение по линиям (одновременно от и элементов в ряду
или в столбике) или, что еще лучше, если одновременно получать сигналы
ЯМР от всех л2 элементов, расположенных в слое. Последний метод называ-
ется методом чувствительной плоскости. Часто на практике достаточно рас-
смотреть одну или несколько таких плоскостей, но можно также получить
изображение от всего трехмерного образца, рассматривая слой за слоем.
В литературе встречается не очень точный термин: "градиенты линейного поля”, который
подразумевает, что поле линейно зависит от пространственных координат, а градиенты являются
константами/
166
И, наконец, вместо того, чтобы ограничиться некоторым слоем, можно
получить одновременно изображение от всего трехмерного объема. Следует
отметить, однако, что для большинства способов получения ЯМР-изображе-
ний объектов требуются значительные затраты времени. Именно поэтому
методы, направленные на сокращение времени проведения экспериментов,
приобретают особое практическое значение.
4.1.1. Двумерная проекционная реконструкция
Среди всего многообразия методов получения плоских изображений
практическое значение имеют два: метод двумерной проекционной реконст-
рукции и двумерной интроскопии. Впервые изображение, индуцированное
сигналом ЯМР, получил Поль Лаутербур [4.1 ] именно проекционно-рекон-
структивным методом, который в настоящее время представляет скорее ис-
торический интерес. Принцип этого метода состоит в следующем: сначала в
исследуемом образце выделяется слой, затем в плоскости этого слоя создает-
ся градиент поля и проводится измерение амплитуды сигнала ЯМР. Таким
образом получаем одномерную проекцию протонной плотности вдоль на-
правления этого градиента. Направление, вдоль которого формируется гра-
диент, поворачивается в плоскости п раз на одинаковый угол, равный 180°/и,
так что получаем серию п одномерных проекций протонной плотности в этой
плоскости. На следующем этапе подобно тому, как это проводится в рентге-
новской компьютерной томографии, восстанавливается картина протонной
плотности в выделенной плоскости (рис. 4.2).
Рис.4.2. Методы проекционно-реконструктивной
томографии. Схема реконструкции изображения
объекта по данным ЯМР; в качестве объекта
использовались две пробирки, наполненные водой
[4.1]
167
У
90°
Рис.4.3. Двумерная фурье-интроскопия. После выбора слоя (см. рис.4.1.) в течение интервалов
tx и ty, в направлениях хи у, соответственно формируются градиенты поля Gx и Gy
4.1.2. Методы двумерной фурье-интроскопии
Используемые в настоящее время методы получения плоских изображе-
ний являются разнообразными вариантами двумерной фурье-интроскопии.
Эти методы тесно связаны с методами двумерной ЯМР-спектроскопии, раз-
работанными Рихардом Эрнстом с сотрудниками [4.2 ]. Остановимся на ос-
новных принципах фурье-интроскопии. Сначала в образце создается
градиент магнитного поля, что позволяет выделить определенный слой. За-
тем образец подвергаетссявоздействию РЧ импульса, например 90°-ного или
180°-ного, так что возбуждаются или инвертируются спины, лежащие в
выделенном тонком слое. На рис. 4.3 показана схема эксперимента. Для
выделения определенного слоя в плоскости (ху) прикладывается градиент
поля вдоль оси z. После воздействия первого возбуждающего импульса вклю-
чается градиент поля Gx вдоль оси х на время tx, а затем градиент поля Gy —
на время ty вдоль оси у. Затем сигнал ЯМР подвергается оцифровке и запи-
сывается для п эквидистантных значений /у, и таким образом получают п
экспериментальных точек. Затем эксперимент повторяется для п эквиди-
стантных значений tx. Таким образом формируется матрица п эксперимен-
тальных точек. Двумерная картина получается после двумерного
фурье-преобразования.
Остановимся на этом вопросе несколько подробнее: обозначим протон-
ную плотность в некоторой точке (х, у) в слое черезр (х, у). Если пренебречь
релаксационными эффектами, то сигнал ЯМР ds, регистрируемый от эле-
мента dxdy, в точке (х, у) может быть выражен следующим образом:
' Y1 ( tx + В2 ty )
ds-С pdxdy е , (4.1)
где С — коэффициент пропорциональности. Магнитные поля В[ и В? имеют вид
Bi~B0 + xGx, (4.2)
B2 = Bo+yGy, (4.3)
где Во - значение напряженности магнитного поля в точке (0,0).
168
После подстановки (4.2) и (4.3) в (4.1) сигнал ЯМР от элемента dxdy на
частоте резонанса (о = у, Во и при условии, что используется режим фазо-
чувствительного детектирования с частотой со7, равен
, п „ I I i’Yl{xGxtx +yGyty)
ds-С р dxdy е , (4.4)
а для всего слоя
. . ‘Vi (d)xtx + a>vtv)
s(tx> ty) =C f f p (x, y) e dxdy**
г r ‘ ( Шх *x + fy )
= CA Jjp(a)x,(oy)e 7 d(iixd(Dy, (4.5)
W
<DX = Yl*Gx’ O)y = yjyGy,A=( y2GxGy)-1 . (4.6)
Протонная спиновая плотность p(x, у), распределение которой нужно
реконструировать, вследствие свойств фурье-преобразования является
фурье-образом функции s(tx, ty), которая измеряется экспериментально.
Следует отметить, что наличие в формулах произведения Gxtx позволя-
ет модифицировать эксперимент. Вместо того, чтобы варьировать tx, как
было указано выше, можно при постоянном tx задавать последовательность
п значений Gx. Этот подход обладает рядом достоинств и часто используется
на практике [4.3 ].
В настоящее время развито столь большое число методов, являющихся
комбинацией градиентных и импульсных методов, что в рамках данного изло-
жения мы вынуждены ограничиться лишь основными принципами и рядом
простых примеров. Простейшая импульсная последовательность состоит из
одного 90°-ного импульса и периода наблюдения сигнала в сочетании с подхо-
дящим выбором градиентов магнитного поля. Импульсную последовательность
можно дополнить, вводя сначала еще один 90°-ный насыщенный импульс.
Варьируя интервал времени t между этими импульсами, получаем изображе-
ние, которое называют взвешенным по Т\. Этот метод известен как метод
насыщения-восстановления (saturationrecovery). Еслисначалавместо90°-ного
импульса ввести 180°-ный, то получим импульсную последовательность (180°-
(-90°), описанную в разделе 1.1.4 и предназначенную для измерения времени
спин-решеточной релаксации Тj; время I здесь также используется в качестве
варьируемого параметра. Поскольку под действием 180°-ного импульса спино-
вые изохроматы располагаются вдоль оси - z, а затем происходит их восстанов-
ление к равновесному положению - вдоль оси z, то эта последовательность
называется последовательностью инверсии-восстановления (inversion
recovery). Изображение, получаемое с помощью этой последовательности, за-
висит от t и является сильно взвешенным по Ту.
Отметим, однако, что в большинстве экспериментов используется метод
спинового эха. Сигнал эха формируется либо с помощью соответствующей
169
последовательности РЧ импульсов, либо посредством градиентных импуль-
сов с варьирующимися знаками. Наиболее распространенной последователь-
ностью РЧ импульсов является последовательность Карра-Парселла-Мей-
бума-Гилла (рис. 1,8), описанная в разделе 1.1.5. Для получения достаточно
хорошего значения отношения сигнал/шум необходимо провести усредне-
ние, например, по восьми сигналам эха после восьми 180°-ных импульсов
(рис. 4.4). Для получения изображения, называемого взвешенным по Тг,
можно использовать только одни определенный сигнал эха, например, пер-
вый или восьмой.
Импульсная последовательность Карра-Парселла-Мейбума-Гилла
(КПМГ) в зависимости от числа сигналов эха длится от 100 до 300 мс.
Поскольку время продольной релаксации обычно порядка 2 с, интервал
времени перед включением следующей импульсной последовательности
должен быть не менее 2 с (рис. 4.4), так что если для получения изображения
необходимо, например, сформировать 256 градиентов поля Gx, то продолжи-
тельность такого эксперимента составляет порядка 8,5 мин. Гораздо более
эффективным является эксперимент, в котором во время ожидания изменя-
ется частота передатчика, и таким образом выделяются дополнительные,
следующие один за другим слои, для которых проводятся измерения (см. рис.
4.4). Эта методика называется техникой мульти-эхо/мульти-слой и наибо-
лее часто используете:. в настоящее время. Экономия времени при,использо-
вании этой методики достигается за счет того, что во время накопления
информации от одного слоя происходит запись информации сразу от не-
скольких слоев. При этом время, необходимое для накопления информации
от одного слоя, остается неизменным.
90° 150° 100° 180° 180°
О ‘ 50 100 150 200
2000 Время t/мс
О
I* 50 100 150 200 ’ 250 300 350 .',00 050
1 -ый слой
2000 Время t/мс
следующий
градиент
2-ой слой
3-ий слой
Рис.4.4. Схематическое представление метода Карра-Парселла-Мейбума-Гилла - модифициро-
ванного метода спинового эха. (Вверху) Последовательность мульти-эхо. (Внизу) Последова-
тельность мульти-эхо / мульти-слой
170
4.1.3. Параметры ЯМР-томограмм
Качество Я MP-томограмм и возможность использования их в диагности-
ке определяются следующими характеристиками:
1. Разрешение по контрасту.
2. Пространственное разрешение.
3. Качество изображения, зависящее от отношения сигнал/шум.
4. Отсутствие артефактов.
5. Время, необходимое для проведения экспериментов.
Разрешение по контрасту - отличительная особенность ЯМР-томогра-
фии.
Несмотря на то что все ткани человеческого организма состоят из разно-
образных молекул, содержащих атомы водорода, наблюдаемый сигнал ЯМР
в основном определяется протонами воды - внутри- и внеклеточной. Это
связано с тем, что вода присутствует в организме в очень высокой концент-
рации. Кроме того, для очень больших молекул или молекул, входящих в
состав макроструктур, соответствующие резонансные линии оказываются
сильно уширенными, что существенно сокращает возможности их детекти-
рования. Поэтому наряду с водой вклад в сигнал ЯМР дают малые водород-
содержащие молекулы, а также молекулы, обладающие большой
подвижностью, такие, как липиды, входящие в состав жировых тканей, при
этом вклад в суммарный сигнал пропорционален их концентрации. Очевид-
но, что содержание воды в различных тканях будет сказываться на интенсив-
ности сигнала ЯМР этих тканей. Патологические изменения в тканях также
сказываются на содержании воды. Еще более существенным параметром
является время спиновой релаксации для протонов воды в различных тканях,
поскольку эта величина отражает взаимодействие молекул воды с молеку-
лярными компонентами клетки. Как правило, амплитуда сигнала зависит от
всех трех параметров: спиновой плотности р, времени спин-решеточной
релаксации Т\ и времени спин-спиновой релаксации Ti. Используя описан-
ные ранее методы, можно учесть вклад этих факторов в величины, опреде-
ляющие качество изображения, что позволяет варьировать контраст
изображения в широких пределах.
Остальные параметры, определяющие качество изображения, как пра-
вило, связаны между собой. Если задать определенное желаемое значение
отношения сигнал/шум, то величина пространственного разрешения, как и
во всяком физическом эксперименте, будет зависеть от соотношения между
точностью, которая может быть достигнута в данном эксперименте, и време-
нем, необходимым для проведения данного эксперимента. Например, про-
странственное разрешение по одной из координат - линейное разрешение -
можно увеличить в два раза, удвоив амплитуду градиента. При этом интен-
сивность сигнала на пиксель, т. е. отношение сигнал/шум, уменьшится в
четыре раза. Если же нужно провести измерение с постоянным значением
отношения сигнал/шум, то в данном случае необходимо провести измерение
16 раз и усреднить результат по всем измерениям, так что время проведения
измерения при увеличении линейного разрешения в два раза при постоянной
толщине слоя увеличивается в 16 раз. Если требуется увеличить в два раза
171
разрешение в третьем измерении, например при получении трехмерного или
двумерного изображения, сократив в два раза толщину выделенного слоя при
сохранении отношения сигнал/шум, то время регистрации возрастаете 4 раза,
т. е. при удвоении линейного разрешения в каждом из трех пространственных
измерений полное время проведения эксперимента возрастает в 64 раза.
На практике при матрице изображения 256 х 256 пикселей в зависимо-
сти от величины градиента достигается пространственное разрешение поряд-
ка 1 мм; если одновременно накапливать информацию от восьми слоев, то в
этом случае эксперимент продлится 5-10 мин. Поскольку такая продолжи-
тельность эксперимента слишком велика для практических целей, большин-
ство разрабатываемых методик направлено на сокращение времени
проведения эксперимента. Несмотря на то что метод ЯМР-томографии ус-
пешно применяется в дифференциальной диагностике, следует отдавать себе
отчете том, что сокращение времени проведения эксперимента всегда влечет
за собой либо уменьшение пространственного разрешения, либо ухудшение
отношения сигнал/шум, либо появление артефактов, либо комбинацию всех
этих негативных моментов.
Принципиальная возможность улучшения отношения сигнал/шум со-
стоит в повышении напряженности поля Во, поскольку интенсивность сигна-
ла пропорциональна В^ (1 < Л < 2). Однако здесь следует отметить ряд
ограничений принципиального характера. Эти ограничения не только тех-
ническиелибофинансовые, нои чисто физические. Последние связаны с тем,
что с ростом напряженности магнитного поля увеличивается значение рабо-
чей частоты, а следовательно, растет и обычное (неядерное) поглощение
микроволнового излучения тканями тела человека. Правда, на практике
оказывается, что данный эффект возрастания нерезонансного поглощения
оказывается несколько меньшим, чем можно было бы ожидать исходя из
теоретических оценок. В общем случае можно сказать, что для in-vivo-спек-
троскопии, о которой речь пойдет в главе 5, где разрешение зависит от
величины химического сдвига, пропорционального Во, напряженность маг-
нитного поля должна быть максимально возможной. Для визуализации изо-
бражения критерий выбора наиболее эффективного значения поля Во еще не
сформулирован окончательно, значение поля, как правило, попадает в ин-
тервал от 0,02 до 2 Тл. В области полей порядка 0,3 Т разработаны томогра-
фы, предназначенные для исследования всего тела, в которых магнитное
поле создается резистивными катушками; для более высоких полей необхо-
димо использовать магниты со сверхпроводящими катушками. Приобрете-
ние таких приборов сопряжено со значительными затратами, а их
эксплуатация также требует больших средств, поскольку необходим жидкий
гелий. Однакоэти приборы обладают и рядом преимуществ, в частности, они
практически не потребляют электроэнергии. Наряду с меньшей стоимостью
основное преимущество приборов, снабженных слабопольными магнитами,
состоит в том, что для них невелико поле рассеяния, кроме того, в экспери-
ментах используются РЧ импульсы с относительно малой мощностью, иа
томограммах отсутствуют артефакты за счет химических сдвигов, а также неве-
лико время спин-решеточной релаксации Т\, поскольку, как известно, эта вели-
172
чина для воды, содержащейся в тканях, существенно возрастает с увеличе-
нием напряженности магнитного поля. Последнее позволяет увеличивать
скорость повторения импульсов, что частично компенсирует возрастание
необходимого для проведения эксперимента времени при уменьшении на-
пряженности магнитного поля. Основное преимущество приборов с более
высоким значением напряженности магнитного поля - при заданном разре-
шении для проведения эксперимента необходимо меньшее время вследствие
лучшего отношения сигнал/шум.
4.1.4. Методы, позволяющие сократить время проведения
экспериментов
Как уже отмечалось, основным недостатком ЯМР-томографии является
длительность каждого отдельного измерения. Это обусловлено в основном
тем, что между двумя импульсными последовательностями необходимо вво-
дить время задержки, равное нескольким значениям времени спии-решеточ-
иой релаксации, для восстановления равновесного значения
намагниченности. Для линейного разрешения 256 пикселей при условии, что
время задержки между импульсами выбрано равным примерно 1 с, для
одноимпульсной последовательности необходимо 256 с, а при усреднении по
двум наборам последовательностей необходимо уже 512 с, т. е. примерно 8,5
мин для получения изображения от одного слоя. Следовательно, изображе-
ния грудной клетки или брюшной полости искажаются артефактами, связан-
ными с движениями (движениями сердца и легких, перистальтикой
кишечника). В случае периодических движений этого можно избежать с
помощью стробоскопических методов. Недавно были разработаны специаль-
ные градиентные импульсные последовательности (MAST и EXORCIST),
которые компенсируют нежелательные эффекты расфазировки, связанные
с движениями. В каждом конкретном случае при сокращении времени про-
ведения экспериментов руководствуются следующими соображениями:
уменьшением нагрузки на организм больного, что особенно важно прн об-
следовании тяжело больных, а также более эффективным использованием
аппаратуры. Кроме того, сокращение времени проведения эксперимента
позволяет наблюдать изменение некоторых процессов во времени.
Методы, направленные на сокращение времени измерений, которые
рассматривались различными коллективами авторов, в принципе можно
разделить на следующие две группы: методы быстрого получения изображе-
ний с использованием техники градиентного эха и методы, основанные на
различном фазовом кодировании сигналов (эха), регистрируемых в преде-
лах одной импульсной последовательности (обычно КПМГ) после однократ-
ного возбуждения спиновой системы. К первой группе методов,
позволяющих существенно уменьшить время проведения эксперимента, от-
носится метод, разработанный Питером Мэнсфилдом [4.4 ], называемый ме-
тодом планарного эха (echo planar imaging). Отметим, что этот метод в
известном смысле содержит элементы обеих групп методов, указанных вы-
173
ше. Данный метод не нашел достаточно широкого применения, так как в нем
используются мощные градиенты, которые включаются на очень короткое время.
Собственно говоря, техника градиентного эха применяется достаточно
давно. Эта техника, подобно 180°-ному импульсу, позволяет получать по-
следовательность сигналов спинового эха, варьируя градиенты в соответст-
вующих временных интервалах. Здесь следует отметить существенное
отличие последовательности КПМГ от градиентного эха. Изменение в после-
довательности КПМГ фазы РЧ импульса позволяет ликвидировать возник-
шую за счет неоднородности поля /?0 (см. раздел 1.1.5) расфазировку
спиновых изохромат, что дает возможность провести регистрацию сигналов
спинового эха. При использовании инверсии градиентов фокусировка спино-
вых изохромат может быть достигнута только в том случае, если их расфа-
зировка была вызвана воздействием соответствующего градиента. Именно
этим объясняются столь высокие требования к однородности магнитного
поля До- Существенным ограничением в данном случае является неоднород-
ность магнитной восприимчивости в объеме исследуемого образца.
Начиная с 1985 г. группа исследователей в Гёттингене занималась раз-
работкой методов быстрого получения изображений на основе метода гради-
ентногоэха [4.7 ]. Основной идеей этого метода, названного авторами FLASH
(Fast Law Angle Shot - быстрый малоугловой снимок), является то, что
спиновая система возбуждается импульсом, длительность которого соответ-
ствует углу отклонения спиновых изохромат порядка 15-30°, вместо исполь-
зуемого обычно 90°-ного импульса, вследствие чего скорость повторения
импульсов в последовательности может быть существенно увеличена, так
как время задержки между импульсами можно выбрать меньшим времени
спин-решеточной релаксации Т\. Обоснование применимости этого метода
приведено в разделе 1.4.2.
Техника градиентного эха используется для считывания информации об
изображении. На рис. 4.5 представлены детали импульсной последователь-
ности FLASH. Одновременное градиентом, с помощью которого осуществля-
ется выбор слоя, подается РЧ импульс, длительность которого соответствует
углу отклонения спиновых изохромат от 15 до 30°; на втором этапе реализа-
ции последовательности достигается фокусировка за счет градиента, осуще-
ствляющего выбор слоя, и предварительная расфазировка, возникающая под
действием считывающих градиентов. Одновременно на образец воздейству-
ют п различных фазокодирующих градиентов, амплитуда которых возраста-
ет линейно. После изменения знака считывающих градиентов сигнал
наблюдается в виде сигнала градиентного эха. Остаточная поперечная на-
магниченность разрушается затем с помощью градиентного импульса
(spoiler gradient). Затем этот цикл вновь повторяется без введения дополни-
тельной задержки. Выбор наиболее предпочтительных углов отклонения
спинов (или длительностей импульсов) и времени задержки зависит от вре-
мени Т\. Подбирая значения длительности импульсов в последовательности,
а также варьируя времена задержки, можно получить различные изображе-
ния, взвешенные по 74. Амплитуда сигнала в методе FLASH пропорциональ-
на значению равновесной намагниченности Мо, а также следующей
величине:
174
Р Ч-импульс
Сигнал
Градиент выбора
слоя
Фазокодирующий
градиент
Считывающий
градиент
Разрушающий
градиент
Рис.4.5. Импульсная и гра-
диентная последовательнос-
ти в методе FLASH
(1-е 7^/7'1 ) sin а -Те/т*
1- е~ TR^Ti cos а
(4.7)
где Tr - период повторения последовательности, Те - время наблюдения эха,
Т2 - эффективное время поперечной релаксации, а- угол поворота намаг-
ниченности. Оптимальным углом поворота вектора намагниченности явля-
ется угол Эрнста данный формулой (1.53). При этом значении угла в
методе FLASH удается получить максимальное значение амплитуды сигна-
ла. Время повторения в данном случае порядка 10-20 мс; это соответствует
для 2562 пикселей полному времени проведения эксперимента порядка не-
скольких секунд. Такое сокращение времени проведения эксперимента по
крайней мере частично компенсирует ухудшение отношения сигнал/шум.
Метод, также основанный на последовательности градиентного эха, был
разработан А. Оппелем с сотрудниками [4.5 ] и назван ими FISP (Fast Imaging
with Steady Precession - быстрое изображение при стационарной прецессии).
Схема проведения эксперимента с использованием этого метода приведена
на рис. 4.6. Особенность этого метода состоит в том, что градиент, осуществ-
ляющий выбор слоя, фазокодирующий градиент и считывающий градиент
включаются вовремя проведения эксперимента дважды, продолжительность
их воздействия одинакова, но при этом при повторном включении градиента
его знак меняется на противоположный, так что к моменту окончания после-
довательности все спины находятся в исходном положении, т. е. в фазе.
Вследствие этого для продольной и поперечной намагниченностей достига-
ются значения, соответствующие стационарному состоянию. Если интенсив-
ность сигнала высока, то интервал между импульсами должен быть меньше
времен релаксации Т\ и Т2 . Амплитуда сигнала, кроме того, что она пропор-
циональна равновесной намагниченности Мо, пропорциональна также
-----------s5---------------------, (4.8)
( 1 + Г1/Т* ) + ( 1 - 7’1/7’* ) cos а
что для 90°-ного импульса и при условии, что в жидкостях Т\ = Т2, соответ-
ствует установившемуся значению намагниченности, равному половине Mz.
175
Рис.4.6. Импульсная и градиентная последовательности в методе FISP
В общем случае оптимальное значение угла отклонения, зависящее от отно-
шения Т\/Тг , определяется соотношением
ctopt = arc cos ——. (4.9)
Ti + Г*
Как следует из рис. 4.5 и 4.6, существенное отличие между методами FLASH
и FISP состоит во временной зависимости градиентов. Несимметричная за-
висимость градиентов от времени в методе FLASH разрушает фазовую коге-
рентность на каждом этапе выполнения последовательности; однако
остаточная поперечная намагниченность в плоскости (ху) разрушается соот-
ветствующим импульсным градиентом (spoiler gradient). После прохожде-
ния нескольких циклов последовательности устанавливается стационарная
намагниченность Mz. С помощью последовательности FLASH получаются
изображения, которые в основном передают распределение спиновой плот-
ности. Соответствующим выбором параметров: угла отклонения вектора на-
магниченности а и периода повторения Tr, можно получить изображение,
взвешенное по Ту. Поскольку фазовая когерентность так или иначе наруша-
ется, последовательность FLASH не слишком чувствительна к движениям,
которых не удается избежать при исследовании, например, в абдоминальной
области. В противоположность этому последовательности в методе FISP сим-
метричны относительно центра эха. При этом поперечная намагниченность
в плоскости (ху) полностью восстанавливается до значения, начиная с кото-
рого она убывает под влиянием спин-спиновой релаксации с постоянной
времени Т2 После прохождения нескольких последовательностей, как и для
поперечной намагниченности в плоскости (ху), стационарное состояние ус-
танавливается для продольной намагниченности. Это и определяет тот факт,
что с помощью последовательности FISP получается изображение, взвешен-
176
Рис.4.7. Последовательности эхо: а) - последовательность, используемая в обычной технике
"мульти-эхо" и (Ь) последовательность, используемая в методе RARE. Величина столбика на
рис. характеризует степень расфазировки спиновых изохромат, возникающей под действием
фазокодирующих градиентов
иое как по Ti, так и по Тг. Несмотря на то что эта последовательность
позволяет существенно уменьшить время проведения эксперимента, ЯМР-
томограммы, полученные с помощью этой последовательности, обладают
достаточно высокой контрастностью. Следует отметить, что в методе FISP
предъявляются очень высокие требования к однородности магнитного поля.
На практике оба метода в известном смысле дополняют одни другого. При-
менения этих методов изложены в следующем разделе.
Иной подход, также преследующий своей целью сокращение времени
проведения эксперимента, предложен в методе RARE (Rapid Acquisition with
Relaxation Enchancement - быстрый сбор с релаксационным усилением) [4.9 ].
Основная идея этого метода состоит не в том, чтобы сократить время прове-
дения эксперимента за счет уменьшения длительности периода повторения,
а в том, чтобы зарегистрировать ряд значений фазокодирующих градиентов
при одном возбуждении спиновой системы за счет различного фазового ко-
дирования различных сигналов эха. Рис. 4.7 поясняет различие между обыч-
ным методом мульти-эхо и методом RARE. В обоих методах используется
последовательность КПМГ, и после каждого возбуждения спиновой системы
одновременно регистрируется, например, восемь сигналов эха. В технике
мульти-эхо фазовое кодирование для всех восьми эхо одинаково, так что в
зависимости от соотношения между длительностью интервала между им-
пульсами и величиной Тг получаем восемь различных эхо-изображений,
взвешенных по Тг (рис. 4.7а). В методе RARE все восемь эхо содержат
различную информацию о фазе, так что для получения полной картины,
содержащей фазовую информаци о 256 точках, необходимо подать лишь 32
возбуждающих импульса вместо 256, т. е. коэффициент экономии времени
равен 8 (см. рис. 4.7b). Полученные таким образом изображения обнаружи-
вают средний контраст, наблюдаемый для эхо-изображений 4 и 5 в обычной
серии мульти-эхо-изображений. Метод RARE позволяет получить изображе-
177
ние, контраст которого является взвешенным по Тт. и может изменяться
определенным образом в зависимости от числа сигналов эха. Этот метод
подходит для многослойной техники, например, для исследования мозга.
Однако он совершенно не годится для исследования подвижных частей тела,
например, грудной клетки и брюшной полости, поскольку движения разру-
шают фазовую когерентность в последовательности КПМГ. Кроме того, по-
скольку для высокополевых приборов необходимо вводить в
последовательность достаточно большое число 180°-ных импульсов, в тече-
ние короткого времени будет достигнуто максимально допустимое значение
РЧ нагрузки.
Как отмечалось выше, эхо-изображение, полученное данным методом,
сильно зависит от Тт. Это влечет за собой интересное приложение модифи-
цированного метода RARE. Так, последовательность эхо можно построить
таким образом, что все сигналы из областей с Тт < 500 мс исчезают, т. е.
наблюдаются только области, содержащие воду в высокой концентрации,
например, ликворную жидкость в спинно-мозговом канале. Так как в этом
случае отсутствует опасность перекрывания различных областей исследуе-
мого объекта (области с большим содержанием воды встречаются достаточно
редко), можно исключить градиент, осуществляющий выбор слоя, и соответ-
ственно построить проекцию исследуемой области тела на проходящую через
эту область плоскость [4.6 ]. Такая методика эффективно используется при
исследовании шейных участков позвоночника при получении миелограммы
без применения контрастных средств. В этом случае фиксируется не только
тонкий слой, а получается проекция на плоскость, что позволяет наглядно
изобразить анатомические пропорции подобно тому, как это наблюдается на
рентгеновском снимке, при этом спинно-мозговой канал всегда остается в пло-
скости изображения. Такое изображение может быть получено за несколько
секунд, например, с помощью 128 сигналов эха на одно возбуждение при усло-
вии, что временной интервал между сигналами эха составляет 20-30 мс.
4.1.5. Получение трехмерных изображений
Впервые идея получения трехмерного фурье-изображения, индуциро-
ванного сигналом ЯМР, была предложена Эрнстом с сотрудниками [4.2].
Преимущества этого метода состоят в следующем.
1. Максимально возможное значение отношения сигнал/шум, посколь-
ку при каждом воздействии импульса возбуждается и соответственно дает
вклад в сигнал весь объем образца. В то же время при построении двумерного
изображения вклад в сигнал дают только спины, находящиеся в выделенной
плоскости, а спины всего объема дают вклад в шум.
2. Оптимальное разрешение по всем трем направлениям в пространстве.
3. Возможность исследования любой поверхности (даже не плоской) в
объеме образца.
Несмотря на эти преимущества методы получения трехмерных изобра-
жений до последнего времени не находили широкого применения. Прежде
всего это связано с тем, что проведение таких экспериментов требует значи-
178
тельных по сравнению с методами получения двумерных изображений за-
трат времени. Если, например, для последовательности КПМГ выбраны сле-
дующие параметры сбора данных: Tr = 1 с и число накоплений А - 2, для
которых строится матрица 1283 векселей (voxel - термин, обозначающий
элемент объема), то в этом случае для проведения эксперимента потребуется
9 часов! Существует, кроме того, еще одна техническая проблема. Это объем
памяти ЭВМ, который необходим для обработки матрицы экспериментальных
данных и последующего построения изображения. Для приведенного выше при-
мера - 1283 векселей - требуется объем памяти, равный 16 Мбайт. С развитием
вычислительной техники эта проблема становится скорее финансовой, чем тех-
нической. Методы, рассмотренные в предыдущем разделе, направленные на со-
кращение времени проведения экспериментов, позволяют построить трехмерное
изображение за время, вполне удовлетворяющее практическим требованиям.
Пример, приведенный выше, т. е. изображение 1283 векселей с использованием
импульсной последовательности 3M-FLASH, требует примерно 5-10 минут.
Методы быстрого получения трехмерных изображений являются естест-
венным развитием методов, описанных в предыдущем разделе: FLASH и
FISP, в основу этих методов положена техника градиентного эха. Для пол-
учения трехмерного изображения в последовательность вместо градиента,
осуществляющего выбор слоя, вводят второй фазокодирующий градиент,
который осуществляет независимо от первого градиента кодирование в п
стадий [4.7 ]. Если второй фазокодирующий градиент так же, как и первый,
осуществляет кодирование в 128 стадий, то получаем однородный массив
данных с одинаковым пространственным разрешением по всем трем направ-
лениям. На основе этих данных с помощью ЭВМ для диагностических целей
Селективный РЧ-импульс
и градиентное эхо
Первый фазокодирующий
градиент
Разрушающий
градиент
Второй фазокодирующий
градиент
Считывающий
градиент
Рис.4.8. Импульсная и градиентная последовательности в методе 3 М -1 ; ASH.
179
Селективный РЧ-импульс
и градиентное эхо
Первый фазокодирующий
градиент
Второй фазокодирующий
градиент
Считывающий
градиент
Рис.4.9. Импульсная и градиентная последовательности в методе 3M-FISP.
можно построить изображение любой поверхности, проходящей через иссле-
дуемую область. На рис. 4.8 и 4.9 представлены схемы методов 3M-FLASH и
3M-FISP. Различие между этими двумя методами такое же, как и в двумер-
ном их варианте.
4.2. Примеры использования основных экспериментальных
методик в ЯМР-томографии
В данном разделе основные особенности различных методов визуализа-
ции объектов иллюстрируются рядом конкретных примеров . Основное вни-
мание уделяется изложению методик, а не деталям диагностики, которые,
конечно, имеют значение для клинической практики.
4.2.1. Изображение головы
Среди наиболее важных областей примененияЯМР-томографии прежде
всего следует указать получение изображения головы. Это связано с тем, что
кости черепа не являются препятствием для ЯМР-томографии, в отличие от
рентгеновской компьютерной томографии, так как кости практически про-
зрачны для РЧ излучения. Кроме того, изображения, взвешенные по Т\ нТ2,
обеспечивают оптимальное разрешение по контрасту мозга и мягких тканей.
Приведенные томограммы получены на приборах фирм Брукер (Карлсруе) и Сименс
(Эрланген) н в радиологической клинике при университете г.Фрайбурга. Марки приборов:
BrukerBNT 1100 (рис.4.10.-4.13,4.18),BrukerMedspec20/100 (рис. 4.13-4.15), Siemens Tomikon
(рис. 4.16, 4.17, 4.19-4.23, 4.28).
180
Исследованиям головного мозга не мешают внутренние движения, которые
затрудняют получение ЯМР-томограмм грудной клетки (движение сердца и
легких) и брюшной полости (перистальтика кишечника). Для иллюстрации
на рис. 4.10-4.15 приведены изображения головы одного из авторов
(К.Х. Хауссера) в слабом (0,28 Тл) и сильном (2,0 Тл) полях с использова-
нием разнообразных методик.
На рис. 4.10 показана томограмма сагиттального сечения головы при
напряженности поля 0,28 Тл (12 МГц). Следует отметить, что такое изобра-
жение не может быть получено с помощью рентгеновской компьютерной
томографии. Эксперимент проводился с использованием техники мульти-
эхо/мультислой для последовательности КПМГ. На рис. 4.10 a-d показаны
изображения, взвешенные по Тг (в зависимости от выбора отдельного эха).
На рис. 4.11 дана суммарная картина по всем восьми эхо-изображениям,
полученным в этом эксперименте. На рис. 4.12 (а) и 4.12 (Ь) представлены
два различных изображения, взвешенных по Тг, на рис. 4.12 (с) - суммарное
(по восьми эхо) изображение, а на рис. 4.12 (d) - изображение, полученное
методом RARE. Как видно, метод RARE, несмотря на то что эксперимент
требует существенно меньшего времени, позволяет получить достаточно
хорошее изображение.
Для сравнения следующие изображения получены в поле 2 Тл (85 МГц).
На рис. 4.13 приведены четыре различных, взвешенных по Тг, изображения
сагиттального сечения головы, а на рис. 4.14 - суммарное изображение. На
рис. 4.15 показано поперечное сечение головы на уровне глаз для различных
значений времени наблюдения эха.
На следующих рисунках показаны различные примеры патологий голо-
вного мозга. Во всех примерах матрица данных от 2562 пикселей записыва-
ется для слоя толщиной 5-10 мм (на каждом рисунке приводится точное
значение этого параметра). Это соответствует линейному разрешению по-
рядка 1 мм.
На рис. 4.16 изображена краниофарингиома, сагиттальное сечение по-
лучено методом спинового эха на приборе с напряженностью поля 1 Тл.
Сопоставление эхо-изображений для двух различных значений времени на-
блюдения эха Те наглядно демонстрирует возможности разрешения по
контрасту, т. е. возможности получения изображений, взвешенных по Тг.
Сравнение этих изображений позволяет увидеть различие между опухолью
и отеком тканей перифокальной области.
На рис. 4.17 показано изображение кисты мозга, полученное методом
спинового эха при напряженности поля 1 Тл. Как и в предыдущем случае,
имеем изображение, взвешенное по Тг. Изменение разрешения по контрасту
достигается варьированием значения Те. Так как киста содержит жидкость,
для которой значение времени релаксации Гг велико, то при увеличении
времени Те контраст между опухолью и окружением усиливается.
На рис. 4.18 представлено поперечное сечение головы пациента, страда-
ющего рассеянным склерозом. Более светлые области, соответствующие бо-
лее интенсивным сигналам, - это области очаговых процессов; наиболее
четко они видны на втором изображении. Изображения, взвешенные по Т2,
получены методом КПМГ.
181
Рис.4.10. Сагиттальное сечение головы че-
ловека. Изображение получено в поле на-
пряженностью 0,28 Тл (12 МГц) с
использованием техники мульти-эхо /
мульти-слой по восьми сигналам эха в
последовательности Карра-Парселла-
Мейбума-Гилла. Толщина слоя 6 мм, время
повторения 7р-2с, изображение построено
по 25(> пикселям, время проведения
эксперимента 8,5 мин, 7£-34 мс. В за-
висимости от номера эхо построены изо-
бражения, взвешенные по Т^: (а) 1-е эхо:
Т£]“34 мс, (Ь) 3-е эхо.’Т£3-102 мс, (с) 5 е
3Xo:Tgg-l70 мс, (d) 8-е 3xo.‘7’£g_272 мс.
(Брукер, Карлсруе)
Рис.4.11. Сагиттальное сечение головы че-
ловека. Суммарная картииа построена по
восьми сигналам эха (см. рис.4.10)
182
•,/Н & 8 ECHOES
• 3 I. -ЗЧ«и-..2’1 .Л.Ч1СНГ 4 л>»
я
Чг Т. - ЗЧв» SCHIOH й>
Ь
L НС .
ECHO fc
Т« » 294»» SCHICHT tea
Рис.4.12. Поперечное сечение головы человека. Тд-3 с, полное время эксперимента порядка 13
мин. (а) - второе эхо Т£2“68 мс, (Ь) - шестое эхо: Тg^-204 мс. (с) - изображение построено по
восьми эхо, (d) - изображ ние получено методом RARE, Tg-Ь с, сигналы эха соответствуют
различным кодированиям фазы, полное время проведения эксперимента порядка 100 с (Брукер,
Карлсруе)
d
4.2.2. Получение изображений других областей тела
ЯМР-томография находит разнообразные применения для получения
изображений не только головы, но и других частей тела, однако при возра-
стании диаметра исследуемой области и неизменной матрице данных 25б2
пикселей достигается несколько меньшее пространственное разрешение -
от 1 до 2 мм. При проведении экспериментов возникают проблемы, связан-
ные с движениями сердца при исследовании области грудной клетки и с
перистальтикой кишечника при исследовании брюшной полости. Если дви-
жения периодические, то удается получить изображение с достаточно хоро-
шим разрешением, синхронизируя сбор данных с частотой этих движений.
На рис. 4.19 приведено изображение сердца при напряженности поля 1 Тл и
синхронизации сбора данных с ЭКГ. На этом рисунке достаточно четко
видны все анатомические детали; стрелкой отмечены венечные артерии сер-
183
Рис.4.13. Сагиттальное сечение головы че-
ловека. Изображение получено в магнитном
поле напряженностью 2,0 Тл (85 МГц). Пара-
метры: техника мульти-эхо / мульти-слой,
последовательность КПМГ, толщина слоя 4
мм, время повторения Т^—2,2 с, 256г пиксе-
лей, время проведения эксперимента поряд-
ка 9 мин, Тут—32 мс, при использовании раз-
личных эхо получены изображения,
взвешенные по Т^. (а) - суммарное изобра-
жение, построенное по 1-му + 2-му эхо, (Ь) -
по 3-му + 4-му эхо, (с) - по 5-му + 6 му эхо
(d) - по 7-му + 8-му эхо. Изображения, при-
веденные на рис.4.13 - 4.15, получены на
томографе со сверхпроводящим магнитом
(Брукер, Карлсруе)
Рис.4.14.»Сагиттальноое сечеиие головы че-
ловека. Изображение построено по восьми
эхо-экспериментам, приведенным иа
рис.4.13 (Брукер, Карлсруе)
184
Рис.4.15. Поперечное сечение головы человека, (а) - 1-е + 2-е эхо, (Ь) - 2-е + 3-е эхо, (с) -
картина, построенная при суммировании с 4-го по 8-е эхо, (d) - суммарное изображение по
восьми эхо. Остальные параметры совпадают с приведенными на рис. 4.13 (Брукер, Карлсруе)
Рис.4.16. Краниофарингиома. Сагиттальное сечение получено в поле 1,0 Тл (42,6 МГц) при
различных Tg. (а) Техника спинового эха, последовательность КПМГ, толщина слоя 6 мм,
Т£”28 мс, 7р“3,30с, полное время проведения эксперимента в случае получения многослойного
изображения порядка 15мин, (Ь) - каки (а), но Tf-l12 мс (Ь), Т£-120 мс (Сименс, Эрланген)
185
РиС.4.17. Киста. Фронтальное сечение. Изображение получено в поле 1,0 Тл, использована
техника спинового эха, последовательность КПМГ, толщина слоя 5 мм, 7^-3,20 с, (а) Т^т-ЗО мс,
(Ь) Tj^ 120 мс
дца. На рис. 4.20 дано синхронизированное с ЭКГ изображение грудной
клетки при напряженности поля 1 Тл. На рисунке очень четко различима
аневризма аорты.
ЯМР-томография позволяет получать изображения позвоночника. На
рис. 4.21 показан участок позвоночника, где в пояснично-крестцовой обла-
сти наблюдается выпадение диска. В отличие от рентгеновской компьютер-
ной томографии ЯМР-томография дает сагиттальное сечение и позволяет
провести неинвазивно точную локализацию нужной области, что весьма
важно для диагностики и последующего оперативного вмешательства. Рис.
4.21 дает пример быстрого получения изображения методом FISP; полное
время эксперимента составляет примерно 5 с.
На рис. 4.22 показано поперечное сечение брюшной полости: оба изобра-
жения (а и Ь) получены при различных значениях Те и являются взвешен-
ными по Тг (т. е. контраст изображения определяется существенным
различием времени Тг для разных тканей). Здесь представлены изображения
печени с четко просматриваемыми метастазами; на рис. 4.22 Ь, где Те “ 70
мс, на рис. 4.22 а, где Те “ 28 мс, они не видны.
На рис. 4.23 представлена томограмма пациента, страдающего ехинно-
кокком. В левой нижней части рисунка отчетливо просматривается ленточ-
ный глист в печени больного.
186
Рис.4.18. Рассеянный склероз. Попе-
речное сечение мозга. Изображение
получено в поле 0,28 Тл. использована
техника спинового эха, последователь-
ность КПМГ, толщина слоя 6мм,Т£-34мс,
(а) 1-е + 2-е эхо, (Ь) 5-е + 6-е эхо, (с)
суммарная картина, построенная по
восьми эхо (Радиологическая клиника
при университете г.Фрайбурга)
Рис.4.19. Поперечное сечение грудной
клетки в области сердца, получено в поле
1,0 Тл. Использована техника спинового
эха, синхронизация по ЭКГ, толщина
слоя 10 мм, 7yj-O,82c, Tf—17 мс. Стрел-
кой обозначены венечные артерии
сердца (Сименс, Эрланген)
187
Рис.4.20. Поперечное сечение грудной клетки. Изображение получено в поле 1,0 Тл.
Использована техника спинового эха, синхронизация по ЭКГ, толщина слоя 8 мм, Т^-0,7 с,
Tg-2% мс. Стрелкой отмечена аневризма аорты (Сименс, Эрланген)
Рис.4.21. Сагиттальное сечение пояснично -
крестцовой области. Изображение получе-
но в поле 1 Тл Использован метод FISP,
толщина слоя 5 мм, 7^-13 мс. В середиие
изображения отчетливо видно смещение
диска (Сименс, Эрланген)
188
Рис.4.22. Поперечное сечение брюшной полости. Изображение получено в поле 1Тл.
Использована техника спинового эха, толщина слоя 8 мм, 7р-0,8 с: (а) 28 мс, (Ь) Тр - 70
мс. На рис. Ь отчетливо видны метастазы в печени (отмечены стрелкой), а на рис.а - метастазы
отсутствуют (Сименс, Эрланген)
189
Рис.4.23. Поперечное сечение брюшной полости получено в поле 1 Тл. Использована техника
спинового эха, толщина слоя 10 мм, 7^-0,8 с. Тут—30 мс. В левой нижней части изображения
отчетливо просматривается ленточный глист в печени больного (Сименс, Эрланген)
4.3. Другие применения ЯМР-томографии
Наряду со спиновой плотностью и временами спин-спиновой и спин-
решеточной релаксации существуют и другие факторы, влияющие на сиг-
нал ЯМР. К ним относятся эффекты, связанные с движениями, а также
расщеплениями спектральных линий за счет различий химических сдви-
гов для протонов различных тканей. Все эти эффекты способствуют воз-
никновению на томограммах артефактов, поэтому разрабатываются
разнообразные методики, направленные на их уменьшение. Но с другой
стороны эти факторы могут служить источником новой информации. В
последнее время активно проводились работы по получению ЯМР-томог-
рамм от объектов больших размеров; с этой целью постоянно совершенст-
вовались конструкции ЯМР-снстем. Одной из интересных новых областей
применения ЯМР-томографии является ЯМР-микроскопия, предназна-
ченная для исследования объектов малых размеров при высоком простран-
ственном разрешении. Эта методика основана на экспериментальных
подходах, описанных ранее.
190
4.3.1. Химический сдвиг в ЯМР-томографии
Как уже обсуждалось, сигнал ЯМР, регистрируемый от тканей челове-
ческого организма, в основном определяется концентрацией содержащейся
в них воды. Вообще говоря, все протоны, входящие в состав подвижных
молекул, также вносят свой вклад в сигнал ЯМР, однако концентрации этих
протонов, а значит, и интенсивности сигналов от них намного порядков ниже
концентрации и интенсивности сигналов воды. Исключение составляют ли-
пиды, входящие в состав клеток жировой ткани, так как концентрация
липидов может быть очень высокой.
Резонансные линии, соответствующие метиленовым протонам липи-
дов, сдвинуты примерно на 3,5 м. д. в сторону сильного поля относительно
линии воды. Абсолютный химический сдвиг, измеряемый в единицах час-
тоты, пропорционален напряженности внешнего магнитного поля Во (см.
(1.3)). В слабых полях, напряженность которых порядка 0,2 Тл, сигналы воды и
жиров сдвинуты один относительно другого на 30 Гц, поэтому их достаточно
трудно различить. Совершенно иная картина наблюдается для поля порядка 2 Тл.
Здесь резонансные линии отстоят одна от другой примерно на 300 Гц, что
может существенно сказаться на качестве получаемого изображения. Как
правило, это проявляется в том случае, когда при проведении эксперимента
необходимо с помощью селективного по частоте импульса выделить опреде-
ленный слой или же четко отделить один слой от другого. В настоящее время
развит ряд методов, позволяющих сгладить влияние этих различий в значе-
ниях химических сдвигов. Но в то же время возможности разделения сигна-
лов от жиров и воды могут быть использованы для построения изображений
жировой ткани и воды, например, для получения отчетливых изображений
лимфатических узлов и прилежащей к ним жировой ткани при диагностике
новообразований молочной железы.
В принципе существуют два совершенно различных пути разделения
протонов жиров и воды на основе различия их химических сдвигов. Один из
методов, названный его авторами методом CHESS (chemical shift selective -
селективный по химическому сдвигу), основан на том, что импульсная по-
следовательность, используемая для получения изображения, возбуждает
только один из двух типов протонов. Принципы этого метода изложены в
работе [4.6 ]. Один из способов достижения необходимого результата состоит
в насыщении спинов одного из двух типов протонов с помощью селективного
РЧ импульса без приложения градиента поля, а затем, как обычно, подается
импульсная последовательность. При этом получаем изображение, постро-
енное по сигналам протонов только одного типа, поскольку намагниченность
протонов второго типа (находящихся в другом химическом окружении), для
которых выполняется условие насыщения, равна нулю [4.8 ]. В этом методе
однородность магнитного поля Во во всем исследуемом объеме должна быть
значительно лучше чем 3,5 м. д. Так как используемый обычно в ЯМР-то-
мографии магнит обеспечивает однородность в исследуемом объеме не хуже
1 м. д., то это условие выполняется.
В другом варианте этого метода протоны одного сорта не исключаются
за счет насыщения, а нужные резонансные линии выбираются селективным
191
90’,CHESS 90’ 90’
РЧ-импульс —ty---------ty---------------------
Сигнал эха Ф ф
Градиент выбора слоя ---------------L---1-----—— -П------ТР----
Фазокодирующий ____Ц_ ____________________________
градиент S___ - _______
Считывающий ____I___L___1_______|____I______I__
градиент
Рис.4.24. Импульсная и градиентная последовательности в методе CHESS
возбуждением, которое действует вместе с РЧ импульсом для выбора плоско-
сти, изображение которой следует получить. Этот вариант CHESS-томогра-
фии можно реализовать с помощью трехимпульсной последовательности. В
частности, это вариант стимулированного эха (4.9), описанного Ханом еще
в 1950 г. [4.10 ]; в дополнение к обычным сигналам эха в результате воздей-
ствия импульсной последовательности, состоящей из трех РЧ импульсов:
90° - И - 90° - Ц - 90° - /з, к моменту времени /з = d возникает сигнал
дополнительного эха, называемого стимулированным.
На рис. 4.24 представлена последовательность РЧ импульсов и градиен-
тная последовательность. Для получения двух первых эхо-сигналов исполь-
зуются селективные по частоте импульсы, которые могут быть заданы
разными способами: например в виде РЧ импульса гауссовой формы длитель-
ностью порядка 10 мс, что соответствует гауссовому распределению по час-
тоте с полушириной порядка 100 Гц. Импульсы в пределах
последовательности можно достаточно произвольно менять местами. Напри-
мер, для выбора слоя можно использовать третий импульс вместо второго,
селективного по частоте. Преимущество этого метода состоит в следующем.
1. Отсутствуют проблемы, связанные с вихревыми токами, поскольку
перед первым импульсом не выключаются никакие градиенты.
2. Качество изображения не зависит от точности выбора длительности
возбуждающего импульса и от однородности поля В\, так что подобный
эксперимент можно проводить даже с поверхностными катушками.
3. Эта техника позволяет получать изображения одновременно
по большому числу резонансных линий, причем независимо от
принадлежности этих линий одному или нескольким различным
веществам.
Один из вариантов метода CHESS представлен в работе [4.11 ]; в этом
варианте одна из двух спиновых изохромат инвертируется под действием
180°-ного импульса, используемого в последовательности CHESS, и затем
проводится обычный двумерный ЯМР-эксперимент, например, с использо-
ванием импульсной последовательности КПМГ. При этом целесообразно
построить эксперимент таким образом, чтобы инвертировалась намагничен-
192
ность с большим значением времени 7\; это позволит уменьшить ошибку при
определении амплитуды в процессе релаксации. И, наконец, эксперимент
повторяется без инвертирующего импульса. Сумма результатов обоих экс-
периментов позволяет построить изображение только по сигналам протонов,
входящих в состав жировой ткани, а разность — только по протонам воды.
Совершенно иной метод, предложенный Диксоном [4.12] и названный
методом CSI (chemical shift imaging - изображение по химическому сдвигу),
основан на том, что спиновые изохроматы, соответствующие протонам воды
и протонам жиров, после воздействия на спиновую систему 90°-ного импуль-
са вращаются в плоскости (ху) с различными круговыми частотами (if и (if.
При этом разность фаз линейно возрастает со временем. Однако при исполь-
зовании обычной импульсной последовательности спинового эха 90° - Т-180°
после 180°-ного импульса и к моменту регистрации сигнала эха эта разность
фаз вновь будет равна нулю, так что намагниченности, соответствующие
протонам воды ЛГ и протонам жировой ткани MF опять находятся в фазе, и
сумма их модулей: М™+ MFдает вклад в спиновое эхо. Однако если в период
рефокусировки на спиновую систему дополнительно воздействовать гради-
ентом поля, то обе компоненты намагниченности не будут фокусироваться к
одному и тому же моменту времени. При этом, если градиентное и спиновое
эха совпадают, то получается такая же суммарная картина, как и в отсутст-
вие биполярного градиента: Mw + MF, а если наблюдается разность фаз,
равная половине периода Л/ ((if - (if), то получится разностная картина.' MF -
Mw. Сложение этих двух изображений дает томограмму, построенную толь-
ко по сигналам протонов жировой ткани, а вычитание - томограмму, постро-
енную по сигналам протонов воды.
4.3.2. Эффекты движения жидкостей в ЯМР-томографии
Человеческий организм не является неподвижной жесткой системой.
Даже в состоянии покоя в организме происходит ряд процессов, связанных с
внутренними макроскопическими движениями. Особое значение имеют не-
прерывные процессы движения жидкостей, без которых невозможно нор-
мальное функционирование организма. Важнейшим среди этих процессов
является движение крови по кровеносной системе. К ним относится движе-
I 41
(а)
(Ь)
V г
Рис.4.25, (а) Однородный поток. Элемент объема дли-
ной /, двигаясь со скоростью v, за время t проходит путь
Л/ (t>) Ламинарный поток. Профиль скоростей пара-
болический
193
ние лимфы по лимфатической системе, перенос мочи по мочеточникам и
поток ликворной жидкости в спинно-мозговом канале.
Для направленного движения жидкости в системах имеющих форму
трубок, рассматривают три типа потоков: однородный (plug flow-поршневой
поток), ламинарный и турбулентный. В случае однородного потока скорость
течения по всему сечению сосуда одинакова (рис. 4.25). Однородное движе-
ние жидкости является наиболее простым для описания, поэтому часто ис-
пользуется для моделирования потока, хотя далеко не в полной мере
отвечает процессам, протекающим в человеческом организме. Ламинарное
течение часто имеет параболический профиль скоростей (рис. 4.25 Ь) и
является типичным для потока, движущегося с низкой скоростью в сосуде
большого диаметра. Эти условия выполняются для движения венозной кро-
ви. Турбулентное движение является типичным для движения с достаточно
высокой скоростью, что может иметь место для движения крови по артериям.
Если сравнить интенсивности сигналов от движущейся и покоящейся жидко-
сти, то эффекты, обусловленные течением жидкости, могут сказываться на
ЯМР-томограммах таким образом, что это может приводить как к уменьше-
нию, так и увеличению интенсивности сигнала.
В эксперименте с накоплением для того, чтобы оптимизировать отноше-
ние сигнал/шум, период повторения последовательности обычно выбирается не
меньше трехкратного значения времени спин-решеточной релаксации. Ис-
пользование более коротких 90°-ных импульсов для выбора слоя приводит к
частичному насыщению сигнала в жидкости. При проникновении в выбранный
слой новых потоков жидкости насыщение частично снимается и в результате
интенсивность сигнала будет несколько возрастать. Ситуация оказывается бо-
лее сложной, если последовательность состоит из нескольких импульсов,
Рис.4.26. Влияние движения потока
жидкости при использовании техники
спинового эха. (а) Используемая им-
пульсная последовательность. (Ь)
Эксперимент при одиовремеииом по-
токе жидкости по трубке, (с) График
зависимости интенсивности сигнала от
vt при однородном потоке
194
а слой выбирается после первого возбуждающего импульса. Предположим,
что после первого селективного по частоте 90°-ного импульса в последова-
тельности 90°-180° - эхо возбуждается цилиндр длиной I (рис. 4.26). При
этом возбуждается объем жидкости V-Лг2/. Если за время Т- ТЕЦ., где Т-
интервал между 90° и 180°-ным импульсом, жидкость, движущаяся со ско-
ростью v, пройдет путь Д/ - vTg/2, то объем жидкости, находящейся под
действием обоих импульсов, уменьшится на величину Ду - л^2 А/ и станет
равным У* =°17Г r\l — ДГ), а сигнал эха соответственно уменьшится в отноше-
нии, равном У* / Упо сравнению с сигналом в неподвижной жидкости (см. рис.
4.25). Отсюда следует, что сигнал в таких условиях при достаточно высокой
скорости потока v может уменьшиться до нуля, если &1—nTe/'2. > L
Прямая пропорциональная зависимость амплитуды сигнала от скорости
справедлива только тогда, когда отсутствует насыщение, т. е. в том случае,
если период повторения TR много больше времени релаксации. Для меньших
значений TR следует дополнительно учесть описанный выше эффект увели-
чения сигнала за счет дополнительного включения в возбуждаемый объем
спинов, для которых не выполняется условие насыщения. Например, если
вторая импульсная последовательность подается через время ТЕЦ. после
формирования сигнала эха {TR - ЗТ£/2), то объем У" - г2 Л ЗД/ будет
включать новые спины, для которых не выполняется условие насыщения.
Следовательно, интенсивность сигнала от объема V" равна интенсивности
сигнала после первой последовательности, уменьшенной в V"/V1 раз, при
этом оставшийся объем
У- У" = г2ТГ(/-3 Д/)
вследствие частичного насыщения даст существенно меньший вклад в сум-
марный сигнал. Очевидно, что при определенной скорости потока v в зави-
симости от степени насыщения, т. е. от отношения Ti/Tr, оба эти эффекта
могут компенсировать друг друга, и можно получить такой же по интенсив-
ности сигнал, как и для неподвижной жидкости.
Если же необходимо независимо от скорости потока жидкости v пол-
учить максимальную интенсивность сигнала, то объем Уцо, который воз-
буждается под действием 180°-ного импульса, смещается за счет сдвига
частоты в присутствии градиентов в направлении потока на расстояние а по
отношению к объему У90, возбуждаемому 90°-ным импульсом (см. рис.
4.26). В этом случае для а > I в неподвижной жидкости не наблюдается
никакого сигнала. Для движущейся.жидкости интенсивность сигнала явля-
ется функцией v/; до vt = а -1 интенсивность сигнала равна нулю, а затем она
линейно возрастает до vt=а и при vt- а+l вновь уменьшается до нуля подобно
тому, как это представлено на рис. 4.26.
Ранее отмечалось, что предположение о равенстве скоростей всех спинов
в потоке нереалистично. В действительности поток крови в кровеносной
системе является либо турбулентным, либо ламинарным. По сравнению со
случаем, приведенным на рис. 4.26, более реальным является случай лами-
нарного течения (рис. 4.27). Для ламинарного потока скорость непрерывно
возрастает от стенок к центру, и профиль скоростей близок к параболическо-
195
му. В примере, показанном на рис. 4.27, после воздействия как 180°-ного, так
и 90°-ного импульса возбуждаются только те спины, которые расположены
внутри объема, обозначенного двойной штриховкой, и именно они дают
вклад в сигнал.
Рис.4.27. Эффекты, возникающие при лами-
нарном потоке
Как было видно, сигнал отсутствует, если в импульсной последователь-
ности 90° - Т - 180° либо 90°-ный, либо 180°-ный импульс воздействует на
разные объемы V и V'. Однако, если часть спинов, находящихся в объеме V,
в промежуток времени ТЕ/2 между 90°-ным и 180°-ным импульсом перено-
сится из объема V в объем У', например, движущейся кровью, то получим
сигнал ЯМР. Этот подход можно использовать в различных вариантах, на-
пример, для селективного изображения на ЯМР-томограммах кровеносных
сосудов (рис. 4.28).
Скорость v, с которой однородная жидкость протекает по жесткой труб-
ке, можно измерять методом ЯМР по наблюдаемому сокращению времени
спин-спиновой релаксации. Если жидкость движется по трубке по скоростью
v, то измерение времени спин-спиновой релаксации Т2 в слое, т. е. в неболь-
шой области цилиндра, с помощью последовательности КПМГ дает меньшее
значение ^, , чем в случае покоящейся жидкости. Между Т^иТ^ существует
количественная связь:
Л=у-+7 , <4.Ю)
тг *2 I
где I— длина цилиндрического слоя. Так как Тг,Тг тл1 можно измерить независи-
мо, то скорость v можно определитьиз соотношения (4.10). Можно также провести
измерение Тг для различных Z, и в этом случае определить скорость v из формулы
(4.9), не зная при этом значения Тг. Если известны скорость потока v и площадь
поперечного сечения трубки Q, то можно рассчитать количество Q v жидкости,
которое протекает через трубку в единицу времени.
До сих пор предполагалось, что жидкость движется с постоянной скоро-
стью v. Это предположение плохо выполняется для движения потока крови
в венах и еще менее реалистично для артерий, поскольку потоки крови в этих
сосудах пульсируют синхронно с сокращениями сердца. Применение им-
пульсной последовательности КПМГ, синхронизированной с ЭКГ, напри-
мер, на аорте, позволяет определить время Т2 в различных фазах работы
сердца, что в свою очередь дает возможность по значениям Т2 определить
скорость потока v в соответствующих фазах сердечного цикла.
196
Рис.4.28. Изображение сосудов в области колена, полученное в поле 1 Тл. Использована моди-
фицированная техника спинового эха [4.16], в которой движение жидкости используется для
регулирования контраста при изображении сосудов. Толщина слоя 10 мм, Tr - 0,84 с, Tg - 56
мс. (а). Изображение, усиленное потоком. (Ь). Изображение, соответствующее разности изобра-
жений, построенных на усиленном потоком сигнале и на ослабленном. Благодаря эффектам,
связанным с движением крови по сосудам, их удается выделить более четко (Сименс, Эрланген)
197
Чтобы в этом случае определить количество крови, которое протекает
через кровеносный сосуд в единицу времени, то вместо произведения Qy
необходимо вычислить интеграл
п t2
S ^(t)dt, (4.11)
‘2 Ч ti
где длительность импульсного цикла. Однако количественная оценка
таких измерений весьма проблематична, поскольку она основана на несколь-
ких упрощениях, которые, как правило, не выполняются для циркуляции
крови в организме. Кровь - неоднородная жидкость, она представляет собой
двухфазную систему, состоящую из эритроцитарной массы и плазмы. Зна-
чения Т\ и Т2 зависят от ряда параметров, таких, как вязкость, температура,
pH и содержание кислорода. Кроме того, разность давлений в кровеносных
сосудах, стенки которых эластичны, распространяется волнообразно. Эти
причины определяют значительные погрешности измерений количества и
скорости движения крови в сосудах.
До сих пор мы ограничивались рассмотрением потока в направлении,
перпендикулярном плоскости изображения, так как в этом случае течение
однородной жидкости в жесткой трубке сравнительно легко описать. Более
сложными оказываются случаи, когда поток распространяется в плоскости
изображения, и особенно, когда направление распространения потока явля-
ется произвольным. Этот случай также был подробно рассмотрен как теоре-
тически, так и экспериментально, однако его изложение выходит за рамки
настоящей книги. Поэтому сошлемся на литературу; достаточно подробный
обзор, содержащий свыше 100 ссылок, дан в работе [4.13 ]. Проблема, возни-
кающая при изучении движения крови в артериях и венах, состоит в появ-
лении интенсивного фонового сигнала от окружающих неподвижных тканей.
Однако этот сигнал можно эффективно подавить с помощью методов, опи-
санных в [4.14].
Несмотря на довольно сложную теорию, исследования по движению
крови в настоящее время имеют важное диагностическое значение. С по-
мощью ЯМР можно исследовать те области, которые принципиально недо-
ступны ультразвуковым исследованиям, а также проводить неинвазивную
диагностику нарушений циркуляции крови, вызванных частичным или пол-
ным перекрыванием крупных сосудов, например, при образовании тромбов.
4.3.3. Контрастные вещества в ЯМР-томографии
Успех ЯМР-томографии во многом связан с тем, что различные ткани
могут быть эффективно исследованы этим методом без применения контр-
астных веществ и неинвазивно с высоким разрешением по контрасту за счет
вариации параметров, зависящих от различия времен релаксации Т] и Tj
для разных тканей. Тем не менее возникает вопрос о необходимости приме-
нения контрастных веществ. Оказывается, что их применение полезно в тех
198
случаях, когда указанные различия между Т\ и Т2 для соседних тканей так
малы, что за счет вариации параметров импульсной последовательности не
удается достаточно четко выявить эти различия. Использование контраст-
ных веществ позволяет, в частности, увидеть различие между опухолевой
тканью и отеком перифокальной области.
Общим свойством всех контрастных веществ, применяемых в ЯМР-то-
мографии, является то, что они парамагнитны, и следовательно, приводят к
уменьшению значений Tj и Т2 (см. раздел 1.3.7). В подавляющем большин-
стве случаев контрастные вещества являются хелатами или относятся к
соединениям, включающим в себя элементы редкоземельной группы. Среди
этих веществ наиболее часто используемым является комплекс Gd-DTPA
(гадолинийдиэтилентриаминпентауксусная кислота). Правда, в настоящее
время уже имеются данные по применению и других контрастных веществ,
таких, в частности, как микроскопические ферриты и стабильные свободные
нитроксидные радикалы.
Очевидное ограничение, существующее для применения Gd-DTPA, со-
стоит в том, что это вещество не может проникать сквозь клеточную мемб-
рану, например, через гемато-энцефалический барьер. Однако это свойство
находит важное применение в диагностике: поскольку, если вследствие трав-
мы, инфаркта, развития опухоли или в результате применения некоторых
лекарственных препаратов гемато-энцефалический барьер становится про-
ницаемым для контрастного вещества, то оно может, например, накапли-
ваться в опухоли. Повреждения гемато-энцефалического барьера часто
сопровождаются отеками, которые затрудняют выявление небольших мета-
стаз на обычных ЯМР-томограммах. Здесь необходимый контраст может
обеспечить Gd-DTPA. Особенно эффективно применение Gd-DTPA в слу-
чае, если необходимо точно определить размеры опухоли и ее локализацию.
Контрастное вещество накапливается именно в опухоли, а не в прилежащих
тканях, что усиливает контраст между опухолью и ее окружением.
С помощью Gd-DTPA могут исследоваться и другие органы такие, как
почки, печень и селезенка. В сочетании с методами быстрого получения
изображений (такими, как FLASH) можно изучать зависимость от времени
процесса накопления Gd-DTPA в опухоли.
Стабильные нитроксидные радикалы достаточно давно нашли примене-
ние в биологии, однако в ЯМР-томографии возможности их использования
изучены в меньшей степени. Тем не менее они представляют потенциальный
интерес для ЯМР-томографии по двум причинам: во-первых, эти радикалы
легко связываются с молекулами-носителями, которые могут избирательно
накапливаться в определенных тканях; во-вторых, эти вещества могут про-
никать сквозь наружные мембраны клеток, что позволяет надеяться на по-
лучение более подробной информации о метаболических процессах,
протекающих внутри клеток. Следует отметить, однако, что применение
нитроксидных радикалов в ЯМР-томографии находится на начальном этапе.
Решающим моментом при выборе контрастного вещества является воп-
рос его токсичности. Несмотря на то что сам гадолиний как элемент токси-
чен, в настоящее время отсутствуют данные о каких-либо отрицательных
побочных явлениях, связанных с применением Gd-DTPA; сообщалось лишь
199
о небольших обратимых изменениях концентрации сывороточного железа, а
также об уменьшении степени связывания железа. Тот факт, что длительность
полупериода выведения Gd-DTPA через почки составляет 20 мин, свидетель-
ствует о том, что этот препарат практически безвреден для организма.
4.3.4. ЯМР-микроскопия
В этом разделе мы обсудим проблемы максимально достижимого про-
странственного разрешения при получении дву- и трехмерных изображений
объектов малых размеров. Термин "ЯМР-микроскопия" или "визуализация
микрообъектов" используется в том случае, если достигаемое в эксперименте
разрешение превышает разрешение человеческого глаза, которое составляет
порядка 50 мкм. При таком разрешении выбору 1282 пикселей на плоскость
или 1283 векселей в трехмерном объеме соответствуют объекты, линейные
размеры которых меньше 5 мм. Для исследования объектов больших разме-
ров с меньшим пространственным разрешением говорят о визуализации
миниобъектов (mini-imaging) в отличие от визуализации макрообъектов
(macro-imaging). Не будем останавливаться на этом более подробно, по-
скольку нет принципиального различия между макро- и микровизуализа-
цией. Достижимое значение разрешения в ЯМР-микроскопии ограничено
максимально допустимым значением градиента поля и прежде всего отноше-
нием сигнал/шум [4.15, 4.16].
Необходимая для достижения заданного разрешения величина градиен-
та зависит от ширины линии и соответственно от области частот, в пределах
которой с учетом разброса значений химических сдвигов лежат соответству-
ющие резонансные линии исследуемых ядер. Ограничимся рассмотрением
ядер 1Н, поскольку они имеют наибольшую чувствительность. Ширина ли-
нии без учета факторов, зависящих от формы линии, по порядку величины
равна I/T2. В жидкостях значения Т2 изменяются в пределах от 10’2 до 1 с,
так что для достижения разрешения 10 мкм величина градиента должна быть
порядка нескольких мТл/см. При современном уровне развития техники
достижение таких значений вполне реально. В твердых телах типичные
значения порядка 10'5 с, и реализация условий ЯМР-микроскопии в этом
случае является весьма серьезной проблемой (вращение под магическим
углом и вращающиеся градиентные поля).
Вследствие разности значений химических сдвигов, а следовательно,
разности резонансных частот, вообще говоря, нецелесообразно получать
изображения по всем ядрам 1Н. Поэтому, как правило, изображение строит-
ся по сигналам ядерных спинов определенного сорта, например, протонам
воды, находящимся в одинаковом окружении, для которых область значений
химических сдвигов не превышает 0,5 м. д. Изменение поля за счет градиента
в области одного пикселя должно быть больше поля, соответствующего ука-
занному значению химического сдвига. Например, для поля 11,75 Тл (рабо-
чая частота для протонов 500 МГц) и разрешения 10 мкм это соответствует
величине градиента порядка 6 мТл/см, что в настоящее время вполне дости-
жимо для ЯМР-микроскопии.
200
Иначе обстоит дело со значением отношения сигнал/шум, так как вслед-
ствие малой разности заселенностей различных спиновых уровней при ком-
натной температуре и малого магнитного момента ядер метод ЯМР обладает
сравнительно малой чувствительностью. При измерениях в магнитном поле
11,7 Тл (500 МГц) удается достичь отношения сигнал/шум 10:1 при комнат-
ной температуре при условии, что в образце содержится 1015 ядер водорода.
Для большинства биологических тканей это соответствует кубику с длиной
ребра 20 мкм, и такая величина относится к достижимому разрешению. Если
по третьей переменной ограничиться несколько меньшим разрешением, на-
пример, в два раза меньшим, то это позволит выделить вексель размером 20х
х 20 х 40 мкм . Размер образца по одной из пространственных переменных
часто делают существенно большим, например, от нескольких сотен мкм до
1 мм.
Рассуждения, приведенные в разделе 4.1 для выбора параметров опти-
мизации отношения сигнал/шум при проведении трехмерных эксперимен-
тов, справедливы также и для ЯМР-микроскопии. Если же мы хотим достичь
более высокого разрешения чем 20 мкм, то любое улучшение разрешения
требует значительных усилий. Так, для увеличения линейного разрешения
в два раза в трехмерном пространстве необходимо увеличить отношение
сигнал/шум в 23 = 8 раз, т. е. почти на порядок. Поскольку образец мал, то и
шум также мал, поэтому можно попытаться уменьшить шум на входе уси-
лителя за счет охлаждения приемной катушки до гелиевой температуры.
Однако это очень сложно сделать при исследовании биологических объектов
из-за большого различия в температурах приемного контура и образца.
Даже в том случае, если удается дополнительно повысить отношение
сигнал/шум, то необходимо учесть еще один фактор, влияющий на макси-
мально достижимое разрешение, а именно молекулярную диффузию, кото-
рая ограничивает пространственное разрешение диффузионным смещением
для рассматриваемой молекулы. Эта величина для чистой воды при исполь-
зовании последовательности спинового эха за время ТЕ = 10-20 мс достигает
максимум 2-3 мкм; в биологических условиях эта величина еще меньше. В
простейшем варианте использования последовательности спинового эха
диффузия не создает каких-либо серьезных ограничений для разрешения.
Для более сложной импульсной последовательности часть пространственной
информации может теряться.
Ниже приведены два примера микроскопических ЯМР-исследований.
Изображения микрообъектов были получены при 300 МГц (7 Тл) при значе-
нии градиентов 1 мТл/см, матрица изображений состоит из 256 пикселей.
На рис. 4.29 представлено сечение почки мыши диаметром около 8 мм.
Изображение получено с помощью импульсной последовательности FLASH.
Разрешение в плоскости сечения составляет 362 мкм2, толщина слоя 320 мкм,
т. е. разрешение по пространственной переменной в направлении, перпенди-
кулярном плоскости слоя, почти на порядок меньше. Внутренние структуры
ясно видны даже при таких малых размерах объекта.
На рис. 4.30 а представлено сечение листа Bryophyllum tubiflore; диаметр
объекта около 3 мм. Изображение получено с использованием техники спи-
нового эха, разрешение 152 мкм2 в плоскости изображения, толщина слоя
201
Рис.4.29. Изображение почки мыши. Сече-
ние почки диаметром 8 мм. Во - 7 Тл, ампли-
туда градиента I мТл/см, угол отклонения
намагниченности при выбранной длитель-
ности импульсов CL —35°, использована тех-
ника 2M-FLASH, 7jj-200 мс, 7^—8 мс,
разрешение 36х мкм2, толщина слоя 320 мкм
(Брукер, Карлсуе)
220 мкм. Увеличение толщины слоя едва ли отрицательно сказывается на
качестве изображения, так как объект обладает характерной волокнистой
структурой. Для сравнения на рис. 4.30 b приведено изображение того же
объекта, полученное с помощью светового микроскопа. Для получения изо-
бражения, индуцируемого сигналом ЯМР, исследуемый объект не подверга-
ется никакому разрушению, в то же время для получения изображения с
помощью светового микроскопа необходимо выделить из листа соответству-
ющий тонкий срез. В ЯМР-микроскопии выбор среза задается соответствую-
щим выбором градиента магнитного поля. На рис. 4.30 а четко различимы
Рис.4.30а,Ь. Сечение листа Bryophyllum tubiflore (а) Изображение получено с помощью ЯМР-
микроскопии, Во-7 Тл, амплитуда градиента 1 мТл/см. использована 2М-техника спинового
эха, Tjj-! с, 7^—16 мс, разрешение 152 мкм2. толщина слоя 220 мкм (Брукер, Карлсруе). (Ь)
Изображение, того же объекта, полученное с помощью светового микроскопа
202
светлые главные направления волокон, на рис. 4.30 b они выглядят темными.
Именно по этим каналам в листе осуществляется транспорт воды.
4.4. Биомедицинские аспекты влияния ЯМР
Применение ЯМР-томографии в медицине имеет то очевидное преиму-
щество по сравнению с другими диагностическими методами, что здесь пол-
ностью исключается влияние ионизирующего излучения. Однако, как и для
любого физического или фармацевтического воздействия на организм чело-
века, в этом случае также необходимо учитывать разнообразные побочные
воздействия. Особое внимание при этом следует уделить вопросу о том,
насколько серьезны эти воздействия по сравнению с той полезной информа-
цией, которая может быть получена при использовании данного метода.
Необходимо также оценивать, какова вероятность возникновения неблагоп-
риятных последствий в результате оказываемых воздействий. Для того, что-
бы оценивать степень риска, необходимо исследовать вопрос о том, какие из
физических взаимодействий наиболее существенны для человеческого орга-
низма при проведении ЯМР-томографического обследования, и какие био-
логические эффекты могут при этом возникать. С физической точки зрения
имеет смысл обсудить влияние статического магнитного, градиентного и РЧ
полей.
4.4.1. Биомедицинские эффекты статического магнитного поля
Статическое магнитное поле Во оказывает существенное влияние на
парамагнитные и ферромагнитные вещества. В однородном магнитном поле
магнитные диполи упорядочиваются, а при наличии градиентов магнитного
поля упорядочение диполей оказывается неоднородным в направлениях,
вдоль которых создается градиент магнитного поля. Эти магнитные поля
оказывают влияние на макроскопические объекты, которые могут представ-
лять собой серьзную опасность для здоровья. Подвижные ферромагнитные
предметы, такие, как ножницы или скальпели, могут достаточно сильно
притягиваться к магниту, и это может нанести серьезные травмы пациентам.
Кроме того, если у пациента имеются ферромагнитные имплантанты (регу-
лятор сердечного ритма, гемостатические зажимы), то под действием маг-
нитного поля может изменяться их положение, что также представляет
определенную опасность.
Можно оценить неблагоприятное воздействие статического магнитного
поля и на молекулярном уровне, ито в основном сказывается на молекулах,
обладающих дипольным магнитным моментом. Известно, что в сильных
магнитных полях липиды мембран ориентируются (см. раздел 3.2.5), что
используется в биофизике как один из наиболее известных способов получе-
ния ориентированных структур. Очевидно, что такая ориентация молекул
может приводить к нарушениям их естественных функций. Следует отме-
203
тить, однако, что ориентирующее воздействие, как правило, компенсируется
влиянием тепловой диффузии, причем оба эти взаимодействия по порядку
величины примерно одинаковы.
Так как в настоящее время отсутствует серьезный опыт в оценке опас-
ного воздействия побочных факторов, то рекомендуемые предельные значе-
ния напряженности магнитного поля лежат в диапазоне от 2,0 до 2,5 Тл.
Однако эти предельные значения могут быть несколько сдвинуты в сторону
увеличения значения напряженности магнитного поля, если это диктуется
медицинскими показаниями.
Среди возможных вредных факторов можно также указать на искаже-
ния показаний механических и электронных приборов, находящихся побли-
зости ЯМР-магнитов.
4.4.2. Биомедицинские эффекты градиентных полей
Изменяющиеся во времени градиентные магнитные поля играют суще-
ственную роль в ЯМР-томографии. Эти поля гораздо мейьше поля Во, однако
сильно неоднородны. Статическое влияние этих полей такое же, как и поля
Во. Следует отметить особое влияние переменной во времени составляющей
градиентных полей: переменные магнитные поля создают в проводниках, а
также в человеческом теле индукционные токи, которые могут приводить к
деполяризации мембран. Наряду с электрическими эффектами могут возни-
кать также зависящие от времени механические деформации. По-видимому,
они являются причиной возникновения известных достаточно давно фосфе-
нов, которые возникают при переключении градиентных полей и восприни-
маются пациентами как световые вспышки.
Величина индукционного тока пропорциональна скорости изменения
магнитного поля dB/dt, поэтому рекомендуется ограничивать максимальное
изменение магнитного поля в единицу времени интервалом от 3 до 20 Тл/с.
Эти предельные значения существенно зависят от интервала времени, в
течение которого изменяется напряженность магнитного поля (обычно этот
интервал порядка мс).
4.4.3. Биомедицинские эффекты РЧ полей
Как обсуждалось ранее, энергия РЧ квантов на много порядков ниже
энергии, необходимой для разрушения ковалентных химических связей.
Основной эффект воздействия РЧ излучения - разогрев тканей, который
проявляется как диатермия. Поглощение РЧ-излучения зависит от частоты
и увеличивается с ее ростом. Кроме того, поглощение зависит от состава
ткани и возрастает при увеличении ионной силы среды.
В организме человека даже в состоянии покоя постоянно выделяется
определенное количество теплоты (примерно 1 Вт/кг), так что поглощение
энергии, которая значительно меньше этой величины, может рассматривать-
204
ся как безвредное. Как правило, система терморегуляции человека может
справиться с существенно большими значениями тепловой нагрузки, напри-
мер, возникающей в результате повышенной мышечной активности или
подводящейся извне (под воздействием солнечного излучения, температуры
воздуха). Поэтому предельно допустимые значения тепловой нагрузки оп-
ределяются, с одной стороны, значением выделяемого тепла, а с другой стороны -
максимальным его количеством, с которым может справиться система теп-
ловой регуляции организма без существенного повышения температуры те-
ла. Типичные значения допустимого тепловыделения для тканей составляют
несколько Вт/кг или такие, которые повышают температуру тела не более
чем на 1°С.
Литература
4.1. Lauterbur, Р. (1973) Nature 242,190.
4.2. Kumar, A., Welti, D. and Ernst, R.R. (1975) J.Magn.Reson. 18,69.
4.3. Edelstein, W.A., Hutchison, J.M.S., Johnson, G. and Redpath, T.
(1980) Phys.Med.Biol. 25, 751.
4.4. Mansfield, P. (1977) J.Phys.C 10, L55.
4.5. Oppelt, A., Graumann, R., Barfuss, H., Fischer, H., Hartl, W. and
Schajor, W. (1986) Electromedica 54,15.
4.6. Hennig, J., Friedburg, H. and Strobel, B. (1986) J.Comp.
Ass.Tomography 10, 375.
4.7. Johnson, G., Hutchison, J.M.S., Redpath, T.W. and Eastwood, L.M.
(1983) J.Magn.Reson. 54, 374.
4.8. Haase, A., Frahm, J., Hanicke, W. and Mathaei, D. (1985) Phys.
Med.Biol. 30, 341.
4.9. Haase, A. and Frahm, J. (1985) J.Magn.Reson. 64,94.
Haase, A., Frahm, J., Mathaei, D., Hanicke, W. and Merboidt, K.-D.
(1986) J.Mag.Reson. 67, 258
4.10. Hahn, E.L. (1950) Phys.Rev. 80, 580.
4.11. Hennig, J. and Friedburg, H. (1986) Magn.Reson.Med. 3. 844.
4.12. Dixon, W.T. (1984) Radiology 153,189.
4.13. Hennig, J., Nauerth, A., Friedburg, H. and Ratzel, D. (1984)
Radiologe 24, 579.
4.14. Deimling, M., Muller, E., Lenz, G., Barth, K., Fritschy, P., Seiderer,
M. and Reinhardt, E.R. (1986) Diagn.Imag.Clinic.Med. 55,37.
4.15. Laukien, F.H. (1984) B.Sc.Thesis, Massachusetts Institute of Tech-
nology, Cambridge MA.
4.16. Kuhn, W. (1990) Ang.Chem. 102, 1.
4.17. Hennig, J., Mueri, M., Brunner, P. and Friedburg, H. (1988) Radio-
logy 166,237
4.18. Laub, G. and Kaiser, W. (1988) J.Comp.Ass.Tomography 12, 377
4.19. Moran, P.R. (1982) Magn.Reson.Imag. 1, 197
4.20. Muller, E., Deimling, M. and Reinhardt, E.R. (1986) Magn.Reson.Med.
3,331
205
4.21. Petersen, S.B., Muller, R.N. and Rinck, P.A. (1985) An Introduction
to Biomedical Nuclear Magnetic Resonance (Thieme, New York).
Дополнительная литература
Berquist, Т.Н., Ehman, R.L. and Richardson, M.L. (1987) Magnetic
Resonance of the Musculosketal System (Raven, New York)
Bradley, W.G. and Bydder (1990) MRI Atlas of the Brain (Deutscher
Arzteverlag, Koln)
Brant-Zawadzky, M. and Norman, D. (1986) Magnetic Resonance Imaging
of the Central Nervous System (Raven, New York)
Gerhardt, P. and Frommhold, W. (1988) Atlas of Anatomic Correlations in
CT and MRI (Thieme, Stuttgart)
Higgins, C.B. and Hricak, H. (eds.) (1987) Magnetic Resonance Imaging of
the Body (Raven, New York)
Huk, W.J., Gademann, G.F.and Friedmann, G. (1989) Magnetic Resonance
Imaging of the Central Nervous System Diseases (Springer, Berlin, Heidelberg,
New York)
Kazner, E., Wende, S., Grumme, T., Stochdorph, O., Felix, R. and
Claussen, C. (1989) Computed Tomography and Magnetic Tomography of
Intacranial Tumors. A Clinical Perspective (Springer, Berlin, Heidelberg, New
York)
Mansfield, P. and Morris, P.G. (1982) NMR Imaging in Biomedicine
(Academic, New York)
Matwiyoff, N.A. (1989) MagneticResonanceWorkbook (Raven, New York)
Morris, P.G. (1986) Nuclear Magnetic Resonance Imaging in Medicine and
Biology (Oxford University Press, Oxford)
Partain, C.L., James, A.E., Rollo, F.D. and Price, R.R. (1983) Nuclear
Magnetic Resonance Imaging (Saunders, Philadelphia)
Partain, C.L., Price, R.R., Patton, J.A., Kuikami, M.V. and James, A.E.Jr.
(1988) Magnetic Resonance Imaging (Saunders, Philadelphia)
Petersen, S.B., Muller, R.N. and Rinck, P.A. (1985) An Introduction to
Biomedical Nuclear Magnetic Resonance (Thieme, New York)
Schulthess, G.K. von (1989) Morphology and Function in MRI,
Cardiovascular and Renal Systems (Springer, Berlin, Heidelberg, New York)
Stark, D.D. and Bradley, W.G. (1988) Magnetic Resonance Imaging
(Mosby, St.Louis)
Wehrli, F.W., Shaw, D. and Kneeland, J.B. (1988) Biomedical Magnetic
Resonance Imaging: Principles, Methodology, and Applications (VCH,
Weinheim)
Young, S.W. (1988) Magnetic Resonance Imaging. Basic Principles (Raven,
New York)
206
Глава 5. Пространственно селективная спектроскопия
и ЯМР-эксперименты in-vivo
ЯМР-спектроскопия в живых системах принципиально отличается от
спектроскопии ЯМР высокого разрешения в однородных средах, изложению
принципов которой посвящены главы 2 и 3. Характерной особенностью жи-
вых систем является чрезвычайно сложная пространственная структура, и
поэтому большинство методов ЯМР, используемых для исследования столь
сложных объектов, с необходимостью требует учета зависимости спектраль-
ной информации от пространственных координат. В качестве простейших
моделей таких систем можно рассматривать фрагменты клеток и клеточные
органеллы, хотя, строго говоря, только с переходом к интактным живым
клеткам мы переходим в область собственно спектроскопии in-vivo. Живые
клетки могут рассматриваться как фрагменты тканей, и затем на следующей
степени организации они объединяются в органы. В медицине ЯМР-спектро-
скопию in-vivo понимают как исследование живых высокоорганизованных
систем. В этом случае реализация пространственной селективности оказы-
вается технически более сложной, чем в методе ЯМР, предназначенном для
исследования изолированных клеток. Однако проблемы, которые могут быть
решены с помощью ЯМР-спектроскопии in-vivo и методы, используемые для
решения такого рода задач, не представляют собой ничего принципиально
нового по сравнению с проблемами и методами исследований живых клеток
или их фрагментов.
Живые системы являются многокомпонентными, поэтому можно ду-
мать, что спектры ЯМР состоят из перекрывающихся линий от большого
числа различных молекул и едва ли поддаются расшифровке. Однако реаль-
ная ситуация не столь безнадежна, поскольку концентрация лишь неболь-
шого числа компонент, присутствующих в этих системах, достаточно велика
для того, чтобы соответствующие им сигналы ЯМР существенно отличались
от фона. Этим объясняется тот факт, что эти спектры выглядят обычно
достаточно просто и вполне поддаются расшифровке.
Если же наряду с пространственной информацией требуется получить
дополнительную спектральную информацию, то необходимо ввести еще од-
ну переменную и изучить распределение интенсивности сигнала по этой
величине. Это сопряжено зачастую с возникновением проблемы увеличения
чувствительности при детектировании. Если уменьшить пространственное
разрешение за счет усреднения по большему объему, то можно обнаружить
даже малые концентрации метаболитов. Таким образом, любые эксперимен-
ты, за исключением тех, которые основаны на использовании внутренних
свойств биологических систем для получения грубого пространственного
разрешения, всегда представляют собой компромисс между пространствен-
ным разрешением и пределом чувствительности. В то время как дискуссия
по вопросу об оптимальной величине напряженности магнитного поля в
ЯМР-томографии продолжается, для спектроскопических целей необходима
максимально возможная величина магнитного поля, поскольку именно этим
определяется максимальное разрешение в спектрах и максимальная чувст-
207
вительность. Правда, при таком подходе возникает чисто техническая про-
блема - создание магнитов с очень высоким значением напряженности маг-
нитного поля и большим внутренним диаметром. В настоящее время для
исследования методом ЯМР высокого разрешения образцов диаметром 10 мм
имеются промышленно изготавливаемые магниты с напряженностью поля
14 Тл (рабочая частота для наблюдения резонанса иа ядрах *Н равна
600 МГц), в то же время максимальное значение напряженности магнитного
поля при диаметре образца 1,2 м практически ограничено значением 4 Тл
(170 МГц для 1Н).
5.1. Методы повышения пространственной селективности
В ЯМР-спектроскопии in-vivo имеются два вида качественно различной
пространственной информации. С одной стороны, это информация об абсо-
лютном положении в лабораторной системе координат, которую мы получа-
ем в ЯМР-томографии, например, как информацию о распределении
спиновой плотности протонов молекул воды. С другой стороны, это инфор-
мация об относительном положении соответствующих спинов в определен-
ной биологической структуре, т.е. о том, находится ли рассматриваемая
молекула внутри или вне определенной замкнутой структуры, такой, как
клетка или ее фрагменты.
Все методы пространственно селективной ЯМР-спектроскопии для до-
стижения необходимого значения отношения сигнал/шум требуют про-
странственного усреднения. Как правило, оно проводится по большому числу
клеток или их фрагментов. Поэтому необходимо с большой осторожностью
относится к интерпретации спектроскопических данных, если они рассмат-
риваются на морфологическом уровне. Это особенно важно, если на основа-
нии измерения концентрации субстрата в гетерогенных тканях мы делаем
выводы о функциональном состоянии отдельных клеток. Даже для суспензий
одинаковых клеток следует учитывать, что усреднение проводится по боль-
шому числу различных областей, для которых условия резонанса могут быть
совершенно различными.
Основной проблемой пространственно селективной спектроскопии in-
vivo является улучшение отношения сигнал/шум. Наряду с указанным выше
ограничением на практически достижимые значения напряженности маг-
нитного поля Во эта проблема связана в основном с коэффициентом запол-
нения, поскольку речь идет о магните с большим зазором, предназначенным
для исследования всего тела. Под коэффициентом заполнения понимают
отношение объемов исследуемого образца и катушки. Этот коэффициент
должен быть максимально близким к единице. Если, например, измеряется
образец объемом 1 см3 в катушке, предназначенной для исследования всего
тела диаметром примерно 60 см и длиной порядка 60 см, то коэффициент
заполнения по порядку величины равен 10’5.
Существуют две принципиальные возможности достижения основных
целей пространственно селективной спектроскопии: либо на образец по-
даются РЧ поля, зависящие от координат, которые, как правило, создаются
208
поверхностными катушками, либо выделяется определенный объем, как и в
случае ЯМР-томографии, где наряду с постоянным магнитным полем Во в
образце формируются три градиента магнитного поля вдоль трех взаимно
перпендикулярных направлений. Оба метода можно также скомбинировать,
приложив, например, вдоль оси поверхностной катушки еще один градиент.
Другая возможность состоит в том, что возбуждение производится с по-
мощью катушки, предназначенной для исследования всего тела при наличии
соответствующих градиентов поля, а сигнал детектируется с помощью по-
верхностной катушки. Преимущество поверхностной катушки состоит в том,
что коэффициент заполнения в данном случае будет на много порядков
превышать коэффициент заполнения для катушки, предназначенной для
исследования всего тела. Однако следует отметить также существенный не-
достаток поверхностных катушек, состоящий в том, что они предназначены
только для исследования органов, расположенных вблизи поверхности тела,
и совершенно не пригодны для исследования внутренних органов. Можно
надеяться, что в дальнейшем будут разработаны катушки для эндоскопи-
ческих исследований.
5.1.1. Пространственное разрешение для изолированных
клеток и органелл
Сигналы от молекул, относящихся к различным областям, удается наи-
более просто различить, если соответствующий спектр ЯМР, относящийся
хотя бы к одной из этих областей, можно измерить отдельно. Например, в
случае суспензии клеток образец центрифугируется, а затем отдельно сни-
мают спектры центрифугата (внутриклеточной массы) и внеклеточной мас-
сы. При проведении экспериментов in-vivo для поддержания
жизнедеятельности клеток среда должна постоянно обновляться с помощью
специальной аппаратуры. Это дает возможность отдельно определять кон-
центрацию субстрата в растворе.
Конечно, эта проблема может быть решена более корректно, если в
одном эксперименте будут отдельно наблюдаться сигналы внеклеточной и
внутриклеточной сред. Это возможно в том случае, если обе среды оказывают
различное влияние на наблюдаемые спектральные свойства, например, раз-
личие магнитных восприимчивосгей внутриклеточной и внеклеточной сред
обусловливает для этих сред различие химических сдвигов, а также может
создавать в переходной области сильную локальную неоднородность, что
приводит к различию в значениях скоростей релаксации.
Довольно часто внутриклеточные и внеклеточные компоненты среды
различаются по значению pH и ионному составу, и оба эти параметра могут
влиять на значения химических сдвигов для содержащихся в среде веществ.
Особенно велика разность значений химических сдвигов для веществ, для
которых значения рК группы, по которой проводится титрование, находится
вблизи значения pH. Как было показано для неорганического фосфата в
разделе 2.2.3 (см. рис. 2.8), в спектрах ЯМР 31Р, содержащих резонансные
209
Рис. 5.1. Спектры ЯМР 31Р (146
МГц) клеток Е. coll, полученные
при различных условиях. Самый
верхний исходный спектр получен
после введения бактерий в измери-
тельную ампулу. Сначала было
проведено обогащение кислородом,
который затем позже был удален.
Последний спектр снят в анаэроб-
ных условиях. Обозначения на
спектрах: Р,ех* - внеклетр<шый не-
органический фосфат, Р| - внут-
риклеточный неорганический
фосфат, NTP и NDP - нуклеотид-
три- и нуклеотиддифосфаты ( (X,
р и у обозначают соответствующие
фосфатные группы), S-Р-фосфаты
сахаров hUDPG -уридиндифосфо-
глюкоза [5.1]
линии, относящиеся к различным фосфатным группам, наблюдаются
большие изменения химических сдвигов при изменении pH среды. На рис.
5.1 приведен пример спектра ЯМР 31Р суспензии клеток Е. coli при раз-
личных условиях. Видно, что сигналы от неорганического фосфата, содер-
жащегося во внутриклеточном и внеклеточном пространствах,
различаются.
Существует еще одна возможность обнаружения различий между
внутренним и внешним пространствами, которая реализуется при добав-
лении парамагнитного вещества в одну из двух областей. В зависимости
от типа этого вещества наблюдается уширение или дополнительный сдвиг
резонансных линий молекул, взаимодействующих с парамагнитным цен-
тром. Условием того, что парамагнетик будет локализован в одной из двух
областей, является малая проницаемость мембраны. Примеры такого рода
уже рассматривались: различие между липидами, находящимися на внут-
ренней и внешней сторонах мембран, причем в последнем случае они
вступают во взаимодействие с внешней средой (рис. 3.47), и различие
между тканями, разделенными гемато-энцефалическим барьером (раз-
дел. 4.3.3.).
210
5.1.2. Пространственная селективность при использовании
РЧ полей, зависящих от координат
В каждом традиционном ЯМР-эксперименте, в котором длина образца
превышает длину РЧ катушки, в известном смысле проводится пространст-
венная селекция, так как детектируются только сигналы, поступающие от
внутренней области катушкн. Это происходит по двум причинам: (1) РЧ
возбуждение во внешней области незначительно, и (2) возможность детек-
тирования сигналов из внешней области также очень мала.
РЧ поле Bi однородно только внутри катушки, а вне ее оно очень быстро
убывает. Поэтому спины, находящиеся за пределами передающей катушки, под
действием РЧ импульса возбуждаются не полностью, и их вклад в суммарный
сигнал невелик. Сигнал ЯМР регистрируется путем измерения напряжения,
индуцируемого в катушке магнитным полем прецессирующих ядерных спинов.
Чем больше величина этого наведенного напряжения, тем выше чувствитель-
ность данного эксперимента. Пространственную зависимость чувствительности
можно легко оценить по принципу взаимности: наведенное напряжение, кото-
рое создается в точке Р внутри катушки намагниченностью, изменяющейся во
времени, пропорционально полю Bi, которое создается в этой точке при усло-
вии, что через катушку течет ток соответствующей частоты. Так как поле Bi
внутри катушки, как правило, достигает максимального значения, а вне катуш-
ки быстро убываете расстоянием, то чувствительность для спинов, находящихся
внутри катушки, максимальна и в соответствии с принципом взаимности быс-
тро убывает вне катушки.
Первые эксперименты по пространственной селекции с животным in-
vivo действительно проводились с помощью обычной приемной катушки
спектрометра высокого разрешения, причем эту часть тела животного, кото-
рую требовалось исследовать, помещали в приемную катушку и получали
спектр ЯМР. Важным этапом в развитии этого метода была разработка такой
конфигурации катушки, для которой характерно сильно неоднородное рас-
пределение магнитного поля; такая катушка называется поверхностной
(рис. 5.2). Большинство всех экспериментов in-vivo в настоящее время про-
водится с помощью таких простых устройств.
(а) (Ь) (с)
Рис. 5.2. Простые приемные катушки, (а) Седловая катушка (модифицированная катушка
Гельмгольца), (Ь) Соленоид, (с) Поверхностная катушка
211
Рнс.5.3. Распределение значений угла отклонения намагниченности для поверхностных кату-
шек. Приведенные на рисунке линии соответствуют равным значениям угла отклонения вектора
намагниченности для плоскостей, проходящих через ось поверхностной катушки. Поверхност-
ная катушка радиусом R - 10 мм расположена в плоскости (x,z). Длительность импульса
выбирается такой, чтобы при (х,у) - (0, R/2) угол отклонения вектора намагниченности ОС был
равен (а) 90° и (Ь) 180° [5.3]
Располагая на поверхности тела плоскую, как правило, круглую катуш-
ку, рассчитанную на соответствующую мощность и настроенную на резонан-
сную частоту исследуемого ядра в магнитном поле Во, получаем, без
соблюдения каких-либо специальных условий, только сигналы от элементов
объема, находящихся вблизи катушки, при этом сигнал максимален в центре
катушки и непрерывно убывает вдоль оси катушки перпендикулярно повер-
хности (рис. 5.3). Отметим, что существуют различные приемы, позволяю-
щие еще более ограничить исследуемый объем. Это можно сделать
вследствие того, что РЧ поле В\ сильно неоднородно по объему образца. При
этом ограничение по объему легко достигается для приповерхностных уча-
стков, поскольку, как следует из геометрических соображений, именно эти
участки дают наибольший вклад в сигнал. Предположим, что мы выбрали
импульсную последовательность, которая начинается с 90°-ного импульса;
если поле В\ таково, что в центре катушки угол отклонения вектора намаг-
ниченности равен 135°, то на расстоянии от поверхности вдоль оси катушки,
равном примерно одному радиусу катушки, величина поля соответствует
углу отклонения 45°, при этом вклад от более удаленных объемов пренебре-
ги
жимо мал. Область, обеспечивающая достаточную чувствительность детек-
тирования сигналов в этом простом случае соответствует примерно полусфе-
ре с радиусом поверхностной катушки 7? и центром, совпадающим с центром
катушки. Для более глубоко расположенной области величина поля В1 дол-
жна достигать такого значения, чтобы импульс на глубине 0,7 - 1,5 7? соот-
ветствовал отклонению намагниченности на 90° ± 45°, что, безусловно,
сопряжено с существенными трудностями. Действительно, поле 7?j такой
напряженности в областях, Примыкающих к поверхности, соответствует
значению угла отклонения намагниченности 270 ± 45°, что вызывает эф-
фект, аналогичный воздействию на спиновую систему 90°-ного импульса,
т.е. намагниченность отклоняется в плоскость (х,у). Так как в приповерхно-
стной области определенной величины РЧ ток создает в три раза большее
поле Bj, чем в глубине образца, то согласно принципу взаимности сигнал от
приповерхностной области соответственно индуцирует в РЧ катушке в три
раза большее напряжение, и его вклад в суммарный сигнал оказывается
большим, чем от нужного объема.
Таким образом, с помощью поверхностной катушки проводится двойная
пространственная селекция: при возбуждении и при детектирования. Обе эти
функции можно разделить. В ЯМР-экспериментах определенного типа для
возбуждения необходимо использовать однородное поле В]. Это поле созда-
ется катушкой, предназначенной для исследования всего тела, а сигнал де-
тектируется с помощью дополнительной поверхностной катушки.
Пространственную селективность возбуждающего импульса можно по-
высить с помощью соответствующей импульсной последовательности, осо-
бенностью которой является зависимость угла поворота вектора
намагниченности от пространственных координат. Лишние сигналы, т.е. сигна-
лы, детектируемые от всех других областей, кроме исследуемой, можно устра-
нить организовав определенным образом фазовые циклы; это приводит к тому,
что нежелательные сигналы уничтожаются, а сигналы, подлежащие регистра-
ции, сохраняются. В настоящее время известны разнообразные импульсные
последовательности, способствующие достижению этой цели; к наиболее рас-
пространенным принадлежат, пожалуй, последовательности "глубоких" им-
пульсов (depth pulses). Простейшая из импульсных последовательностей
состоит из импульса длительности ta и непосредственно следующего за ним
импульса длительностью 2 ta, фаза которого циклически изменяется на 90°. В
областях, в которых длительность импульса соответствует 90°-ному импуль-
су, импульсе удвоенной длительностью приводит к инверенн намагниченности,
в то время как в других областях сигнал ослабляется [5.2 ].
Несовершенство всех этих методов состоит в том, что исследуемый объем
не имеет определенной формы, поэтому весьма затруднен точный выбор
точек в пространстве. Однако для структур, лежащих вблизи поверхности,
детектирование с помощью поверхностных катушек, обладает тем сущест-
венным преимуществом, что в данном случае высокая чувствительность
обеспечивается большим коэффициентом заполнения. Поверхностные ка-
тушки иногда используются в томографии для получения изображений
структур, лежащих вблизи поверхности.
213
5.1.3. Пространственная селекция с помощью зависящего
от координат статического магнитного поля Во
Метод пространственной селекции путем формирования в образце поля
Во, изменяющегося в зависимости от координат за счет приложенных гради-
ентов поля, является технически более сложным по сравнению с описанным
выше методом, но имеет более широкую сферу применения. Основной про-
блемой остается увеличение отношения сигнал/шум. Можно выделить сле-
дующие две группы методов: методы, позволяющие проводить локализацию,
в которых, как и в обычной спектроскопии, спектры ЯМР высокого разреше-
ния получаются от отдельных, строго определенных элементов объема, и
томографические методы, в которых спектроскопические данные одновре-
менно накапливаются от большого числа элементов объема. Методы локали-
зации идейно соответствуют методам чувствительной точки, которые
раньше использовались в ЯМР-томографии и в настоящее время для нее
представляют лишь исторический интерес, так как требуют больших затрат
времени для получения полного изображения. Для спектроскопии данное
обстоятельство не оказывается столь важным, поскольку в большинстве слу-
чаев речь идет только о спектрах в отдельной области, в которой, например,
расположена опухоль, а информация об остальной области ие представляет
интереса.
Большинство методов, используемых в томографии, достаточно просто
модифицировать для получения дополнительной информации о химическом
сдвиге. Для того чтобы уширение линий было небольшим, т.е. для обеспече-
ния высокого разрешения, необходимо, чтобы во время сбора спектральной
информации магнитное поле было по возможности максимально отнород-
ным. Поэтому при сборе данных все градиенты отключаются. Выключение
градиентов непосредственно перед началом сбора данных существенно нару-
шает однородность магнитное поля за счет возникновения вихревых токов в
металлических деталях датчика и в материале магнита, поэтому необходимо
предусмотреть небольшой интервал задержки (длительностью порядка 10
мс) между выключением градиентов и началом сбора данных.
К идеальной технике должны предъявляться дополнительные требова-
ния, которым до настоящего времени не удовлетворяет ни один из методов:
кодирование пространственной информации не должно никоим образом ска-
зываться на спектральной информации, и кроме того, любая схема проведе-
ния эксперимента, известная из спекроскопии высокого разрешения, должна
применяться без каких-либо изменений.
Среди всего многообразия экспериментальных методик, применяемых в
данной области, остановимся на одной, называемой LOCUS (localization of
unaffected spins - локализация невозбужденных спинов) (рис. 5.4). При
использовании этой методики процедура селекции оставляет спины, находя-
щиеся в определенной области, невозбужденными, так что затем в отсутст-
вие градиентов в этой области может быть проведен любой ЯМР-эксперимент.
С этой целью включают градиенты, а намагниченность окружения под дейст-
вием селективных по частоте 90°-ных импульсов, спектр которых не содержит
214
90° 90° 90° 90° 90° 90° 90° 90° 90° 90° 90° 90° 90°
РЧ-нмпульс —{}-(} {) {} {}-(} {} {) {}-------------------j-----------
Сигнал
Gx
Выбор объема
Детектирование
Рис.5.4. Схематическое представление эксперимента LOCUS. Fj ,...,Fj 2 изображают селектив-
ные по частоте 90°-е импульсы с различными значениями несущей частоты; эти импульсы
воздействуют на образец в присутствии градиентов Gx, Gy и Gz [5.4]
резонансных частот для выбранной области, поворачивается и располагается
в плоскости (х,у), в которой происходит быстрая расфазировка спинов в
градиентном поле. Эффективность этой процедуры легко проверить при
записи обычной томограммы после процедуры селекции; в идеальном случае
на томограмме должна быть изображена только выделенная область. Селек-
тивность этого метода ограничивается продольной релаксацией спинов ок-
ружения, которая снимает насыщение.
Преимущество спектроскопических методов визуализации по сравне-
нию с методом локализации состоит в том, что необходимую область
можно выбрать после проведения эксперимента и по крайней мере в прин-
ципе можно получить спектр любой желаемой области. На рис. 5.5 приве-
дена модификация эксперимента "спиновое эхо", известного из
томографии (см. рис. 4.4), который позволяет получать спектральную
информацию из предварительно выделенной плоскости. Единственное
различие по сравнению с обычным получением изображения состоит в
том, что постоянный считывающий градиент в направлении х, который
обычно приложен во время сбора данных, теперь включается в течение
периода эволюции. Для пространственного кодирования градиенты вдоль
направлений х и у должны изменяться дискретно от эксперимента к экс-
перименту. Соответствующий трехмерный эксперимент можно получить
заменой селективного 90°-ного импульса неселективным, а также диск-
ретным изменением z-градиента одновременно с другими градиентами.
При этом, однако, значительно увеличивается минимальное число от-
дельных экспериментов, и следовательно, возрастает время получения
трехмерного изображения.
215
180"
^Спиновое эхо
V
Рис. 5.5. Схема эксперимента для получения спектроскопического изображения с помощью
спинового эх*. Двумерная версия спинового эха; градиент Gz, воздействующий одновременно с
селективным 90°-ным импульсом, используется для выбора плоскости. Величины градиентов Gx
и Gy варьируются независимо друг от друга и от эксперимента к эксперименту (5.5)
5.2. Типичные области применения и проблемы
ЯМР-спектроскопии живых систем
В этом разделе показано, какая биохимическая информация может быть
получена с помощью ЯМР-спектроскопии in-vivo. При выборе примеров не
отдавалось предпочтения тому или иному методу пространственной селек-
ции, так как для большинства примеров отсутствует необходимость в про-
странственной селекции по условиям проведения эксперимента, либо в
простых случаях пространственная селекция проводится с помощью повер-
хностных катушек. Это отражает сегодняшний уровень развития исследова-
ний, которые сопряжены с рядом спектроскопических или технических
проблем, поэтому всегда, если это возможно, стараются максимально упрощать
методы пространственной селекции при применении их к таким сложным сис-
темам, как человек, хотя существует принципиальная возможность более пол-
ного применения этих методов к исследованию человеческого организма.
Критика, часто высказываемая в адрес спектроскопии in-vivo, состоит в
том, что этот метод лишь подтверждает уже известные результаты, найден-
ные ранее с помощью других методов. Это действительно во многом верно и
свидетельствует не против in-vivo-спектроскопии как таковой, а скорее под-
тверждает богатый арсенал возможностей биохимиков, которые, используя
классические биохимические методы, смогли получить правильные резуль-
таты. Несмотря на это, in-vivo-спектроскопия позволяет получить и нечто
новое по сравнению с тем, что может быть получено другими методами:
неинвазивное и непрерывное наблюдение метаболических процессов в клет-
ках на молекулярном уровне. Дополнительно во многих случаях этот метод
позволяет проводить непосредственно измерение величин, которые очень
сложно получить другими методами.
216
5.2.1. Сохранение естественных функции
Важным условием получения данных с помощью спектроскопии in-vivo
является поддержание исследуемого объекта в определенном и легко воспро-
изводимом состоянии. В большинстве случае это состояние должно быть
близким к интактному физиологическому состоянию. Поскольку биологиче-
ские системы, как правило, не являются равновесными, а гомеостаз устанав-
ливается только за счет изменений во всей системе, состоящей из
биологической системы и ее окружения, то эти изменения прежде всего
должны приниматься в расчет при достаточно длительных экспериментах.
Здесь имеются различные решения, адаптированные к специфическим
проблемам. Если речь идет о простых системах, в которых наблюдаются
только субстраты н продукты в открытой среде, то достаточно предотвратить
разделение частиц и использовать субстраты в большом избытке. Часто вра-
щением образца, а возможно, в комбинации с перемешиванием удается
предотвратить разделение частиц. Еще проще это сделать при изучении
обмена веществ в образцах, представляющих собой суспензию клеток и
тканей с низкой метаболической активностью. Для этого можно плотно
упаковать клетки и соответственно надлежащим образом зафиксировать
исследуемую ткань в измерительной ампуле. Однако в общем случае суще-
ствует необходимость следить за концентрациями субстратов и метаболитов
в открытой среде посредством ее обновления. На рис. 5.6 приведена схема
типичной экспериментальной установки. Исключительно эффективный метод,
значения pH
Кислородные
электроды
Рис. 5.6. Установка для перфузии при
проведении эксперимента in-vivo на
изолированных клетках и тканях. На
схеме показано, что парциальное дав-
ление кислорода и значение pH в среде
регулируется, а сама среда в процессе
проведения эксперимента обновляет-
ся. Температура регулируется с по-
мощью специального температурного
блока, входящего в комплект каждого
спектрометра ЯМР высокого разреше-
ния; температура среды может также
дополнительно стабилизироваться с
помощью внешнего устройства [5.6].
217
позволяющий предотвратить разделение клеток и обеспечивающий обмен с
окружающим раствором так, чтобы клетки не вымывались из объема образ-
ца, состоит в том, что клетки упаковываются в тонкие нити агара. В зависи-
мости от требований, предъявляемых к эксперименту, могут быть
использованы многочисленные другие экспериментальные подходы, напри-
мер, такие, при которых обмен веществ осуществляется через полупроница-
емые мембраны для диализа, или такие, при которых клетки закрепляются
на жестких носителях с большой площадью поверхности. В случае более
крупных клеточных образований опасность их вымывания перфузионным
раствором уменьшается. Правда, при этом возникает проблема, состоящая в
том, что для слишком крупных частиц тканей большие диффузионные длины
препятствуют эффективному обмену веществ. При благоприятных условиях
можно из отдельных клеток вырастить так называемые сфероиды, т.е. груп-
пы клеток, размеры которых достаточно малы (порядка 100 мкм) для того,
чтобы существенно не препятствовать днффузни, и в то же время достаточно
велики для того, чтобы предотвратить их вымывание. При исследовании
органов целиком процесс их перфузии может прводиться с помощью подхо-
дящего обогащенного кислородом питательного раствора. Следует отметить,
однако, что эта проблема в биохимии и физиологии не нова. Имеющиеся
методики должны быть адаптированы к конструктивным особенностям
ЯМР-спектрометров.
Несмотря на то что при проведении экспериментов с животными физи-
ологические функции в значительной степени являются саморегулирующи-
мися, здесь может возникнуть проблема, связанная с тем, что во время
проведения эксперимента животные находятся под наркозом. Для того, что-
бы получить достоверные результаты, необходимо проводить тщательный
контроль всех жизненных функций. Несмотря на то что ЯМР-спектроско-
пию можно рассматривать как относительно безвредный метод в применении
к человеку (см. раздел 4.4), тем не менее здесь необходимо соблюдать суще-
ственно более жесткие условия при проведении исследований.
5.2.2. ЯМР-спектроскопия in-vivo на ядрах *Н
На первый взгляд ЯМР-спектроскопия на ядрах 1Н - наиболее пригод-
ный метод для целей исследования in-vivo, так как, во-первых, ЯМР *Н
обладает исключительно высокой чувствительностью, а во-вторых, высокое
естественное содержание ядер JH не требует изотопного замещения. С дру-
гой стороны, поскольку область значений химических сдвигов, в которой
наблюдаются резонансные линии биологически важных молекул, достаточ-
но мала (см. разделы 1.2.1 и 2.1.1), это существенно затрудняет интерпрета-
цию спектров, а наличие интенсивного пика воды затрудняет наблюдение
сигналов, представляющих интерес метаболитов. Селективное возбуждение
или селективное насыщение, как правило, позволяет существенно ослабить
сигнал воды (см. раздел 2.1.7).
Часто в спектрах ЯМР lH in-vivo наблюдается существенное перекрыва-
ние спектральных линий. Поэтому можно попытаться извлечь необходимую
218
Рис. 5.7. Спиновое эхо в системе с косвенной спин-спиновой связью. Схематическое представ-
ление спиновых изохромат во вращающейся системе координат после воздействия на спиновую
систему импульсной последовательности вида 90х° - Т- 1&Оу° - Т. Намагниченность после
воздействия импульса 90х° (а), спустя времяТ/2 (Ь), к моменту времениТ- 1/2J (с); намагни-
ченность после воздействия импульса 180у° (d), спустя время 3/2 Г (е) и к моменту времени
2Т - 1 /7(f). В случае (а) обе компоненты дублета находятся в фазе, в случае (с) и (d) - в
противофазе, а в случае (f) - вновь в фазе, однако по сравнению со случаем (а) знак меняется
на противоположный.
информацию за счет целенаправленного упрощения спектров. Эта процеду-
ра обычно называется редактированием спектров. Ранее уже отмечалось, что
для этого может быть эффективно использована простая импульсная после-
довательность (90х° - Т - 180у° - т), которая приводит к ^-взвешиванию, а
также изменяет фазы сигналов, зависящие от мультиплетной структуры.
Поэтому спектры, полученные по методу спинового эха, содержат сигналы
10 8 6 4 2 0 (5/м.д.
Рис. 5.8. Спектр ЯМР Н (270 МГц), получен-
ный методом спинового эха для эритроцитов,
(а) Обычный спектр ЯМР. Спектр, получен-
ный методом спинового эха при Т мс (Ь) и
при Т мс (с). В области слабых полей
наблюдаются сигналы ароматических колец
нуклеотидов и гистидиновых остатков гемогло-
бина, в сильном поле - сигналы иона глутатио-
на, пирувата и лактата [5.7]
219
как с положительными, так и с отрицательными фазами. Это явление может
быть описано с помощью классической векторной модели (рис. 5.7), 180°-ный
импульс приводит к рефокусировке вследствие неоднородности поля или
различия в значениях химических сдвигов, но расфазнрование индивиду-
альных компонентов мультиплета, обусловленное косвенным спин-спино-
вым взаимодействием, сохраняется. Спустя время 2г, равное 1/7 (где J
измеряется в Гц), спиновые изохроматы, соответствующие компонентам
дублета, располагаются в направлении -х и, таким образом получается
сигнал с инвертированной фазой (отрицательный сигнал), в то время как
спиновые изохроматы, соответствующие синглетам или центральной компо-
ненте триплета, располагаются вдоль положительного направления оси х и
получается сигнал с нормальной фазой (положительный сигнал).
На рис. 5.8 приведены спектры эритроцитов, полученные по методу
спинового эха; эритроциты являются наиболее удачным объектом исследо-
вания, поскольку они обнаруживают незначительную анаэробную метабо-
лическую активность. В силу этого эритроциты можно без особых
предосторожностей поместить максимально плотно в измерительную ампу-
лу, подобрав соответствующий буфер, и снять спектр. В спектре, снятом по
методу спинового эха, широкие сигналы макромолекулярных компонент в
основном подавляются. Вследствие коротких времен спин-спиновой релак-
сации для данных спинов их намагниченность к моменту наблюдения сигна-
ла эха приближается к нулю, однако сигналы от целого ряда молекул можно
наблюдать. Несмотря на то что сигналы от малых молекул, полученные по
этой методике, значительно усиливаются, обычно в спектре присутствуют
также остаточные сигналы гемоглобина, концентрация которого в эритроци-
тах очень велика. Если подойти к интерпретации спектров со всей тщатель-
ностью и учесть взвешенность по Tj, то с помощью этого метода можно
определить внутриклеточную концентрацию метаболитов и проследить за
их изменениями в зависимости от условий эксперимента.
Рис. 5.9. Спектр ЯМР Н (360 МГц) мозга мыши в
состоянии гистидинемии, полученный методом
спинового эха. Спектр получен с помощью поверх-
ностных катушек для времени задержки 20 мс (а) и
60 мс (Ь). В спектре четко наблюдаются сигналы
кольцевых протонов гистидина (7,83 м.д. и 7,08
м.д.), метильных протонов холина (3,20 м.д.), кре-
атина и креатинфосфата (3,03 м.д.), атакжеМ-аце-
тиласпартата (2,01 м.д.) [5.8]
220
Гистидинемия - это врожденное нарушение обмена веществ, связан-
ное с отсутствием гистидазы — фермента, участвующего в расщеплении
такой аминокислоты, как гистидин. Это сопровождается тем, что в крови
и тканях наблюдается повышенная концентрация гистидина. У людей
этот дефект проявляется в нарушении нормального умственного разви-
тия. В спектрах, полученных для лабораторных животных (гистидинемия
мыши) по методу спинового эха, можно наблюдать сигналы от гистидина
в тканях мозга. На рис. 5.9 приведены спектры, полученные этим методом
на живой мыши с помощью поверхностных катушек. Кроме резонансных
линий, относящихся к протонам гистидинового кольца с химическим
сдвигом 7,83 и 7,08 м.д., наблюдаются также интенсивные сигналы от
метильных протонов холина, креатина, креатинфосфата и N-ацетилас-
паргиновой кислоты.
Были развиты разнообразные методы, позволяющие эффективно
уменьшить интенсивность тех мультиплетов в спектре, которые не пред-
ставляют интереса. Если, например, спектр, полученный методом спино-
вого эха, в котором компоненты дублета находятся в противофазе,
сложить с нормальным спектром, то получим спектр, в котором дублетные
линии с константой косвенного спин-спинового взаимодействия J подав-
лены.
Используя различные варианты этого разностного метода, можно
провести селекцию резонансных линий, т.е. выяснить, какие из спинов
связаны между собой взаимодействием. В этом случае два различных
образом полученных спектра вычитаются один из другого: нормальный
спектр, полученный методом спинового эха, в котором Г выбирают так,
что интересующий сигнал является отрицательным, и спектр, также по-
лученный методом спинового эха, но при включенном в течение времени
Г мощном поле развязки на резонансной частоте партнера по косвенному
взаимодействию. Развязка приводит к тому, что резонансные линии соот-
ветствующих спинов, связанных косвенным спин-спиновым взаимодейст-
вием, в отличие от всех остальных линий, наблюдаемых в спектре, не
изменяют фазы, следовательно, при использовании разностного метода
остается только этот сигнал. На рис. 5.10 показан пример действенности
этого метода или исследования функционирования биологических тка-
ней; в мышце, находящейся в рабочем состоянии с высокой нагрузкой,
когда кислородный обмен недостаточен, накапливается значительное ко-
личество молочной кислоты (СН3-СНОН-СООН). С помощью описанно-
го метода можно селективно наблюдать сигнал метильной группы
молочной кислоты с химическим сдвигом 1,33 м.д. при условии, что про-
ведена развязка на частоте, соответствующей резонансу протонов СН-
группы - 4,1 м.д. (см. рис. 5.10). Примечательно, что такое
редактирование сигналов в спектре удается провести даже в том случае,
когда вследствие неоднородности поля мультиплеты в спектрах недоста-
точно хорошо разрешены.
221
Рис. 5.10. Спектр ЯМР 'н (360МГц) мышцы
крысы, полученный методом спинового эха с
использованием техники двойного резонан-
са (360 МГц). (а) Фрагмент обычного спект-
ра, полученного методом спинового эха при
времени задержки Т - 60 мс, (Ь) Спектр,
полученный методом спинового эха с ис-
пользованием техники двойного резонанса;
этотспектр представляет собой разность двух
спектров: спектра, полученного при регист-
рации каждого второго спада свободной ин-
дукции с развязкой иа частоте,
соответствующей резонансу при 4,1 м.д., и
спектра без развязки [5.9]
• • 31
5.2.3. ЯМР-спектроскопия in-vivo на ядрах Р
31
Характерными свойствами ЯМР-спектроскопии на ядрах Р являются
большой диапазон значений химических сдвигов, составляющий для биоло-
гически важных молекул порядка 30 м.д., достаточно высокая чувствитель-
ность (порядка 7% по отношению к протонам) и 100%-ное естественное
содержание данного изотопа. Поскольку в большинстве исследований одно-
временно присутствует ограниченное число фосфорсодержащих молекул,
проблемы, связанные с перекрыванием резонансных линий, возникают не
так часто, как в спектрах ЯМР на ядрах 'Н. Все эти свойства определяют то,
что большинство всех исследований in-vivo проводятся с использованием
ЯМР-спектроскопии на ядрах 31Р.
В биологически активных системах в Зависимости от функционального
состояния с помощью спектров ЯМР 31Р могут быть зафиксированы разно-
образные фосфорсодержащие метаболиты. Правда, в том случае, когда их
концентрация ниже 0,1 ммоль они, как правило, не поддаются детектирова-
нию. Вследствие физиологического значения уровень внутриклеточного со-
держания АТФ достаточно высок, поэтому в спектрах живых клеток почти
всегда удается наблюдать три сигнала от фосфатных групп АТФ. Кроме того,
в большинстве случаев наблюдаются резонансные линии, соответствующие
неорганическому фосфату Р/ и креатинфосфату.
Все эти сигналы, как правило, наблюдаются в спектре мышц. На рис.
5.11 приведен пример спектра 31Р, полученного с помощью поверхностных
катушек для огибающей мышцы предплечья человека. Сильное напряжение
приводит к уменьшению сигнала креатинфосфата и к возрастанию интенсив-
ности сигнала неорганического фосфата. Это означает, что при таких усло-
виях концентрация креатинфосфата уменьшается, концентрация
неорганического фосфата растет, а концентрация АТФ остается неизменной.
222
(а)
31
Рис. 5.11. Спектр ЯМР Р мышцы предплечья человека. Спектры
получены в поле 1,89 Тл (32,5 МГц) с помощью поверхностных
катушек, которые закреплялись на сгибе предплечья (Flexor
digitorum superficialis). Спектры (а) и (Ь) накапливались 64 с и 32 с
соответственно, (а) Спектр мышцы в состоянии покоя. (Ь) Спектр
мышцы после нагрузки в течение 2,5 мин [5.10]
5 0 -5 -Ю -15 М.Д.
Такой результат поддается достаточно простой трактовке: гидролиз АТФ в АДФ
и Р/ сообщает энергию, необходимую для сокращения мышц; этот процесс
должен быть непрерывным, поэтому идет постоянный ресинтез АТФ, так что
уровень АТФ поддерживается постоянным. Ресинтез может проходить несколь-
кими путями. Один из путей состоит в участии такого субстрата, как глюкоза,
при связывании АТФ в гликолизе и окислительном фосфорилировании, подо-
бно тому, как это имеет место в состоянии покоя. При увеличении потребности
в АТФ возможен также дополнительный синтез АТФ из накопленного в состо-
янии покоя креатинфосфата, фосфатная труппа которого переносится на АДФ.
Таким образом, ясно, почему в этих условиях уровень содержания креатинфос-
фата уменьшается, а неорганического фосфата возрастает.
Как было показано ранее (в разделе 2.2.3), фосфатная группа присутст-
вует в различных формах, которые вследствие процесса протонирования-де-
протонирования переходят одна в другую, и таким образом поддерживается
состояние равновесия. Обычно для перехода между этими формами выпол-
няется условие быстрого обмена; это означает, что в спектре наблюдается
одна линия, химический сдвиг которой зависит от pH раствора. Это также
является причиной того, что резонансные линии неорганического фосфата
появляются в разных положениях на двух спектрах мышц предплечья на
рис. 5.11. Очевидно, что значение pH в мышце изменяется. Математиче-
ски это изменение достаточно хорошо описывается с помощью модифици-
рованных уравнений Хендерсона-Хассельбалха (2.16) и (2.17). Если
значение рК для равновесного состояния между двумя формами и соответ-
ствующие значения химических сдвигов известны, то можно рассчитать
химический сдвиг для каждого значения pH по уравнению (2.17), и нао-
борот, измеряя химический сдвиг, можно найти соответствующее значе-
ние pH.
223
Таким образом, неорганический фосфат можно использовать в качест-
вае естественного индикатора pH. Для значений pH, лежащих вблизи эффек-
тивного значения рК (рК' порядка 6,9), малые изменения значения pH
приводят к очень большим изменениям химического сдвига. В силу этого
изменения значений pH в этой области очень точно могут быть определены
по наблюдаемым изменениям химических сдвигов в спектрах 31Р; наблюда-
емые изменения значений pH составляют около 0,02 единиц pH. Для исполь-
зования зависимости pH от величины химического сдвига для точного
определения pH необходимо учитывать, что в принципе все константы рав-
новесия, а также все значения рК фосфатных групп зависят от температуры.
Кроме того, ионная сила оказывает влияние на установление ионного равно-
весия, а специфические взаимодействия такие, как связывание ионов Mg и
Са, могут приводить к изменению в процессе титрования.
На практике для соблюдения высокой абсолютной точности прежде все-
го рекомедуется построить градуировочную кривую с учетом внешних усло-
вий. Существуют дополнительные возможности для проверки, если
одновременно проводятся наблюдения за изменением химического сдвига
другого ионизируемого фосфатного остатка. Воспроизводимость измерений
может быть существенно повышена при использовании внутреннего стан-
дарта. Естественным внутренним стандартом является креатинфосфат, со-
держащийся во многих клетках. Если для простоты предположить, что в
области значений рК для описания функциональной зависимости достаточно
только одного значения рК, то после преобразования (2.16) можно получить
значение pH как функцию химического сдвига d сигнала неорганического
фосфата:
, d-dj
рН = рК +log5^3 • (5.1)
Величины рК', di и d2 варьируются от ткани к ткани, при этом численные
значения параметров, полученных разными авторами для одной и той же
ткани, могут различаться. Вообще говоря, с точки зрения оценки этих дан-
ных для практических приложений это не имеет столь большого значения,
как может показаться на первый взгляд, поскольку изменения значений pH,
представляющие интерес, как правило, достаточно хорошо воспроизводимы.
Типичными значениями для скелетных мышц теплокровных животных яв-
ляются рК' = 6,75, di = 3,27 м.д. и d? = 5,69 м.д. [5.18 ].
Использование зависимости химических сдвигов от pH также обладает
рядом недостатков: если анализируется смесь неизвестных веществ, то не-
возможно однозначно провести отнесение резонансных линий к веществам
определенного типа, не выяснив при этом точно условий проведения экспе-
римента. Если же воспользоваться литературными данными и на основании
этого попытаться провести анализ спектров, то очевидно, что такой метод
неизбежно повлечет за собой ошибки. Кроме того, при получении спектров
Р, как правило, используется внешний стандарт, что в зависимости от
геометрии и магнитной восприимчивости образца может вызвать существен-
ные различия в значениях измеряемых величин (см. раздел 2.1.2), поэтому
здесь также по возможности следует использовать внутренний стандарт.
224
Процессы обмена веществ в живых клетках регулируются с помощью
ферментов. Ферментативная активность может регулироваться разнообраз-
ными способами: концентрация ферментов может изменяться как на стадии
синтеза, так и на стадии расщепления; ковалентная модификация и взаимо-
действие с другими компонентами системы, со специфическими регулятора-
ми, с субстратами или продуктами определяют реальную активность
каждого из ферментов в системе регулирования, внутри которой существуют
разнообразнейшие связи. В силу этого невозможно ожидать, что фермента-
тивные характеристики, определенные из экспериментов in-vitro, дадут ис-
черпывающую информацию© процессах, протекающих в клетках. Несмотря
на то, что в клетках постоянно осуществляется синтез и одновременно распад
различных веществ - в организме человека, например, ежедневно подверже-
но превращению примерно 70 кг АТФ - в целом в организме поддерживается
достаточно стабильное равновесие. Следовательно, важно подобрать метод,
который позволит определить при этих условиях ферментативную актив-
ность в интактных клетках. Такую возможность дает перенос спиновой по-
ляризации (раздел 2.2.5), который можно количественно оценить с помощью
методов селективного переноса с насыщением, селективного инверсного пе-
реноса или с использованием методов обменной двумерной спектроскопии.
Среди реакций, исследуемых в различных тканях с помощью данного
метода, следует указать реакцию креатинкиназы, которая способствует бы-
строй регенерации АТФ при большом потреблении АТФ из креатинфосфата
и катализует ресинтез креатинфосфата (РКг) из креатина (Кг) в случае
низкого уровня потребления АТФ. Обе реакции можно представить следую-
щим образом:
*1
РКг2’ + Mg2+ . ADP3' + Н+ XКг + Mg2+ . ATP4’. (5.2)
fc-1
Таким образом фосфатная группа участвует в быстром обмене между двумя
состояниями с разным химическим окружением - креатинфосфатом и АТФ.
На рис. 5.12 показаны спектры перфузированного сердца крысы, полученные
методом переноса инверсии для различных значений времени задержки Г.
Зависимость интенсивности резонансных линий креатинфосфата и /-фосфа-
та АТФ как функция Т (рис. 5.13) наглядно демонстрирует эффект переноса
поляризации. Экспериментальные зависимости, данные точками, можно
описать уравнениями, приведенными в разделе 2.2.5. Из параметров, входя-
щих в эти уравнения, можно рассчитать значения скоростей для реакций,
протекающих в сердечной мышце. Основной проблемой в такого рода экспе-
риментах является зависимость полученных результатов от выбранной мо-
дели. Клетки - неоднородные системы, разделенные на компартменты.
Внутри каждого из компартментов, которые в той или иной степени связаны
между собой, протекают различные метаболические процессы, однако в
ЯМР-эксперименте проходит усреднение по всем компартментам. Большая
часть АТФ образуется в митохондриях, в то время как потребляется АТФ в
225
PCR
Рис. 5.12. Эксперимент по определению активности креатинкиназы методом переноса поляри-
зации с инверсией; эксперимент проводился на перфузированном сердце крысы. Спектры ЯМР
ЭТР получены на частоте 145 МГц (8,5 Тл) при температуре 37°С. Селективная инверсия на
частоте, соответствующей резонансу ^-фосфата АТФ (слева) и резонансу креатинфосфата
(справа). Приведенные значения времени соответствуют времени задержки Тмежду селектив-
ным 180°-ным и 90°-ным импульсами [5.11]
Рис. 5.13. Изменение интенсивности сигнала ЯМР в спектре сердечной мышцы при использова-
нии метода переноса поляризации с инверсией. Точки соответствуют экспериментальным дан-
ным, полученным в результате экспериментов, показанных на рис. 5.12; сплошными кривыми
изображены теоретические зависимости интенсивности от Г [5.11]. Левая часть рис. 5.13 соот-
ветствует спектрам, приведенным в правой части на рис. 5.12, и наоборот
226
основном в цитозоле для таких специфических функций, как сокращение
мышц. Креатинкиназа и креатинфосфат также находятся в цитозоле и ми-
тохондриях; креатинфосфат может проникать сквозь мембраны митохонд-
рий по специальной транспортной системе - креатинфосфатному депо.
Константы скоростей в формуле (2.18) правильно описывают индивидуаль-
ные процессы только в случае быстрого установления равновесия между
цитозолем и компартментами.
До настоящего времени почти все органы исследовались с помощью
метода ЯМР на ядрах 31Р. Эксперимент по получению спектров 31Р доста-
точно прост, н также достаточно просто может быть проведена расшифровка
спектров 31Р. Ценность результатов, полученных из спектров 31Р in-vivo,
зависит от правильности постановки задачи с биохимической и медицинской
точек зрения.
Исследование физиологии и патологии мышц в настоящее время явля-
ется одной из главных областей применения спектроскопии ЯМР 31Р in-vivo;
в основном это определяется тем, что поверхностные мышцы могут быть
достаточно эффективно исследованы с помощью поверхностных катушек. В
качестве типичного примера можно было бы привести исследование пациен-
та с синдромом Мак-Ардла [5.13 ]; этот пример из-за его наглядности цити-
руется достаточно часто и приведен здесь на рис. 5.14. Спектры мышц
пациента в состоянии покоя не отличаются от соответствующих спектров
здорового человека и содержат сигналы от неорганического фосфата, креа-
тинфосфата и АТФ. Значения pH, определенные по величинам химических
сдвигов, в пределах установленной погрешности совпадают в обоих случаях.
Как показано ранее, в здоровых тканях при нагрузке уровень креатинфос-
фата убывает, а уровень неорганического фосфата возрастает. Дополнитель-
но из гликогена, содержащегося в мышцах, высвобождается глюкоза,
которая участвует в регенерации АТФ. Если подача кислорода нарушена, то
имеет место процесс анаэробного гликолиза, конечным продуктом которого
является молочная кислота, накапливаемая в мышцах и приводящая к рез-
кому понижению pH в мышцах. Тогда на фазе регенерации вновь устанав-
ливается прежнее состояние равновесия. У пациентов с синдромом
Мак-Ардла наблюдается отсутствие фосфорилазы в мышцах, что приводит к
нарушению процесса высвобождения глюкозы из гликогена. Следовательно,
нет возможности для регенерации АТФ путем анаэробного гликолиза, и
молочная кислота не образуется. В результате уровень креатинфосфата
уменьшается знаачительно быстрее, а уменьшения внутриклеточного pH не
происходит (нет образования молочной кислоты). Наоборот, наблюдается
небольшое увеличение pH, что можно было бы ожидать для реакции креа-
тинкиназы (5.2), при которой происходит связывание протонов при регене-
рации АТФ. Фаза восстановления опять не наступает. Наблюдаемые
спектральные изменения можно хорошо объяснить известным проявлением
такой патологии, как синдром Мак-Ардла; наблюдение такого рода проявле-
ний может быть использовано в качестве неинвазивной диагностики синдро-
ма Мак-Ардла.
Следует отметить, однако, что практическая ценность такого рода ис-
следований уменьшается в связи с тем, что хотя существует значительное
227
qi
Рис. 5.14. Спектры ЯМР Р для диагностики синдрома Мак-Ардла. Приведенные спектры
получены в поле 1,9 Тл (32,5 МГц) с помощью поверхностной катушки, закрепленной на
предплечье. (Вверху) Спектр для здорового человека. (Внизу) Спектр для пациента с синдромом
Мак-Ардла. Сначала были получены спектры в состоянии покоя (А), затем, с помощью манжет-
ки для измерения артериального давления, нарушалось артериальное кровообращение и одно-
временно создавалось мышечное напряжение путем ритмичного сгибания руки (для здорового
человека в течение 90 с, а для больного - 45 с). Нормальное кровообращение восстанавливалось
спустя 3 мин. В спектрах можно различить сигналы неорганического фосфата (5), креатинфос-
фата (4) и у — , Ct — и /^-фосфатных групп АТФ (3, 2 и 1 соответственно). Значение pH
определяется по химическому сдвигу сигнала неорганического фосфата Р, [5.12]
число наследственных мышечных заболеваний, они не всегда сопровожда-
ются синдромом Мак-Ардла. Несмотря на это, можно надеяться, что в этих
случаях ЯМР-спектроскопия позволит неинвазивно поставить однозначный
диагноз; кроме того, такой подход дает возможность изучения фармакоки-
нетики, что особенно важно для развития новых форм терапии.
228
Другой важной областью применения ЯМР 31Р является транспланта-
ционная хирургия. Развитие этого метода способствует совершествованию
консервации органов. Метод также может быть использован для оценки
жизнеспособности как трансплантированных органов, так и подлежащих
трансплантации.
Конечно, среди перспективных областей применения ЯМР-спектроско-
пии in-vivo следует отметить исследование раковых клеток н опухолей раз-
личной природы. Исследование особенностей обмена веществ в опухолевых
клетках играет большую роль, поскольку для экспериментальных опухолей
обнаружена зависимость концентрации фосфорсодержащих метаболитов и
времени спин-решеточной и спин-спиновой релаксации от типа и стадии
развития опухоли. На рис. 5.15 приведены спектры ЯМР 31Р мозга здоровой
мыши и мозга мыши с имплантированной опухолью (КНТ-саркома) в зави-
симости от стадии развития опухоли. В опухолевой ткани содержание креа-
тинфосфата заметно понижено, кроме того, в спектрах появляется
о 1
Рис. 5.15. СпектрЯМР 31Р внутрицеребральной опухоли. Приведены спектры Р (8,5Тл, 145
МГц) левого (слева) и правого (справа) полушарий мозга живой мыши. Спектр 1 получен для
мозга здоровой мыши, спектр 2 соответствует случаю, когда в правое полушарие мозга имплан-
тирована опухоль, спустя 17 дней, а спектр 3 - спустя 19 дней после имплантации. Проведено
следующее отнесение сигналов: (А) фосформоноэфнр (АМФ, фосфаты сахаров), (В) неоргани-
ческий фосфат, (С) фосфодиэфир (глицерофосфорилэтаноламин, глицерофосфорилхолин),
(D) креатинфосфат, (Е) у-фосфат нуклеозидтрнфосфата (АТФ) и р-фосфат нуклеозиддифос-
фата (АДФ), (F) (X-фосфат нуклеозидтри- и дифосфата (АТФ, АДФ), (G) уЗ-фосфат нуклео-
зидтрифосфата (АТФ) [5.13]
229
малоинтенсивный пик с химическим сдвигом -12,25 м.д., который можно
отнести к уридиндифосфоглюкозе - промежуточному продукту биосинтеза
гликогенов, который не может быть обнаружен в нормальной ткани мозга. В
зависимости от стадии развития опухоли спектр ЯМР 31Р будет изменяться,
поскольку на поздних стадиях развития опухоли нарушается питание сосу-
дов. При этом значение pH уменьшается за счет возрастания анаэробного
гликолиза, что отражается на величине химического сдвига линии, соответ-
ствующей неорганическому фосфату. На поздней, некротической стадии
развития в спектре не наблюдаются сигналы, соответствующие фосфатам,
ответственным за энергозапас клетки.
Несомненный интерес представляет также и такое приложение ЯМР 31Р
в онкологии, как исследование обмена веществ по концентрации фосфорсо-
держащих метаболитов после применения различных терапевтических
средств. Это один из возможных путей получения новой информации о
механизмах реакций. Однако еще более значительные результаты могут
быть получены из спектров ЯМР 31Р в будущем, если появится возможность
контролировать эффективность терапии в онкологии. В настоящее время уже
получены первые результаты в этом направлении.
13
5.2.4. ЯМР-спектроскопия in-vivo на ядрах С
Углерод является одним из элементов, входящих в состав почти всех
биологически важных молекул. Наиболее распространен в природе изотоп
12С — ядро с четным числом протонов и четным числом нейтронов. В основном
состоянии такие ядра обладают нулевым спином. Явление ЯМР наблюдается
для изотопа 13С, который обладает спином 1/2; естественное содержание
этого изотопа очень мало, приблизительно 1,1%. Так как гиромагнитное
отношение также невелико, то удается получать спектры 13С за разумное
время только для веществ, концентрация которых достаточно велика (при
напряженности поля 11 Тл и объеме образца 2 мл концентрация должна быть
порядка 0,1 М). Правда, при использовании изотопного замещения и увели-
чении объема образца это предельное значение по концентрации можно
существенно увеличить. При этом следует отметить различие между селек-
тивным и неселективным изотопным замещением: при неселективном заме-
щении за счет косвенного спин-спинового взаимодействия с соседними
ядрами возникает расщепление резонансных линий, что приводит к умень-
шению интенсивности отдельных линий. Следовательно, если изотопное
замещение проводится только для общего увеличения чувствительности,
например, при росте бактерий в питательной среде, обогащенной 13С, то
сначала по мере обогащения, чувствительность повышается, а затем при
более высоких концентрациях опять падает, так как увеличивается вероят-
ность того, что два атома 13С окажутся в одной молекуле. В соответствии с
биномиальным распределением можно оценить, что в данном случае наибо-
лее благоприятным будет обогащение от 10 до 20%. Это ограничение, конеч-
но, не относится к случаю селективного обогащения, когда проводится
изотопное замещение по определенному положению в молекуле.
230
Как правило, большинство атомов углерода, входящих в биомолекулы,
связзно с одним или несколькими атомами водорода. Наблюдаемое при этом
расщепление линий в спектрах 13С в основном дает информацию о числе
атомов водорода, непосредственно связанных с атомами углерода. Посколь-
ку расщепление линий одновременно приводит к уменьшению их интенсив-
ности, то часто этой информацией пренебрегают и во время сбора данных
включают поле развязки от протонов, подавляя тем самым расщепление
линий. Если же при этом в течение достаточно длительного периода времени
проводить облучение спиновой системы на частоте протонного резонанса, то
можно осуществить перенос спиновой поляризации от протонов как от более
чувствительных ядер к менее чувствительным - ядрам 13С. Для малых мо-
лекул усиление сигнала за счет ядерного эффекта Оверхаузера может дости-
гать значения, равного 2,98, что сопровождается значительным повышением
чувствительности при детектировании. Правда, реально достижимое увели-
чение интенсивности зависит от многих факторов, которые определяют ве-
личину ЯЭО. Отметим также, что в таких условиях невозможно достаточно
надежно определить относительную концентрацию исходя из площадей под
резонансными линиями.
В качестве достоинства метода ЯМР на ядрах 13С можно указать боль-
шую область значений химических сдвигов (более чем 200 м.д.). Правда, для
сложных систем это не исключает иногда зна чительного перекрывания спек-
тральных линий, несмотря на то что столь большая область значений хими-
ческих сдвигов должна была бы гарантировать достаточно хорошее
разрешение. Поэтому с целью упрощения спектров часто приходится ис-
пользовать методы, разработанные в спектроскопии 1Н. Вследствие этого
большинство импульсных последовательностей, описанных ранее для ЯМР
1Н, может быть непосредственно перенесена на ЯМР 13С. С целью повыше-
ния чувствительности ЯМР 13С разработаны специальные методы: DEPT
(distortionless enhancement by polarization transfer - усиление без искажений
за счет переноса поляризации) и INEPT (insensitive nuclei enhanced by
polarization transfer - усиление сигналов малочувствительных ядер за счет
переноса поляризации). Методы DEPT и INEPT, а также разнообразные их
модификации предназначены для увеличения чувствительности в ЯМР 1JC
за счет переноса поляризации от протонов, непосредственно связанных с
соответствующими атомами углерода (рис. 5.16и5.17).В реальных условиях
часто не удается полностью реализовать теоретические возможности увели-
чения чувствительности, так как в такого рода экспериментах по исследова-
нию биологических тканей из-за высокой концентрации электролита не
обеспечивается необходимая однородность поля Вр
Константы косвенного спин-спинового взаимодействия углерода с не-
посредственно связанными с ним протонами могут достигать достаточно
больших значений, так что соответствующие линии хорошо разрешимы в
спектре. Взаимодействие спина протона со спином ядра С приводит к
дублетному расщеплению резонансной линии *H, величина которого более
чем 100 Гц, т.е. присутствие в молекуле ядра 13С может быть обнаружено
неявным образом по соответствующему протонному сигналу. Существует
очень простой метод, который является модификацией разностного метода
231
Рис. 5.16. Методы переноса поляризации ^Н-^С. Поляризация спинов/ (^Н) на спины S (^3С)
переносится посредством изображенных импульсных последовательностей [5.14]
спинового эха. С помощью неселективных импульсов последовательности
(90х° - Т - 180у° - Й создается сигнал протонного спинового эха, при этом
время 1 выбирается таким образом, что мультиплеты, возникающие за счет
косвенного спин-спинового взаимодействия с 13С, наблюдаются в противо-
фазе (Т = 1/2 J). 180°-ный импульс, подаваемый по каналу 1Н, приводит к
рефокусировке намагниченности вдоль оси у к моменту времени 2 Т. Если же
спиновая система одновременно облучается 180°-ным импульсом как по
каналу 1Н, так и по каналу 13С, то, как и в гомоядерном случае, возникает
рефокусировка вдоль оси -у тех компонент, которые связаны с °C, и наблю-
дается сигнал поглощения с отрицательной фазой. Если найти разность спек-
тров, полученных с помощью последовательности, содержащей и не
содержащей импульс, возбуждающий 13С, то в разностном спектре остаются
232
CH2 GLC-C6 бнв-с2
Рис. 5.17. Редактирование спектров перфу-
зированной печени крысы методом DEPT.
Перед измерениями печень крысы подвер-
галась перфузии в течение 1 часа в буфер-
ном растворе при концентрации [2-гзС]
ацетата 15 ммоль. Спектры 13С получены
на частоте 90,55 МГц, что соответствует
360,13 МГц для протонов. Подспектры воз-
никают за счет соответствующей комбина-
ции спектров, полученных методом DEPT,
при которых угол отклонения вектора на-
магниченности для протонов принимает
значения 7Г/4, Ttlt, 3 7174. Проведено сле-
дующее отнесение резонансных линий:
группы CHj холина (СНО), ацетата (АС-
С2), Дгидроксибутирата <рНВ-€4), лак-
тата (LAC-C3) и аланина (ALA-C3);
СН2-группы глюкозы (GLC-C6), уЗ-гидро-
кснбутирата ()ЗНВ-НС2), глютамата
(GLU-C3 и CLU-C4) и липидов (LIP);
группы ССМ аминокислот ((Х-СН) и CjH-
группыО!- и Д-глюкозы (GLC-C1) [5.15]
IЬо' 90 ' а'о 70 60 50 40 30 2 0 10
только резонансные линии, соответствущие спинам, связанным с 13С. На
рис. 5.18 приведен пример такого разностного спектра, полученного методом
спинового эха; этот простой метод позволяет проследить метаболизм изото-
позамещенной по 13С глюкозы и образование этанола в культуре дрожжей.
В растворе имеет место равновесие между двумя кольцевыми изомерами
D-глюкозы: а- и Д-аномерами, и линейной формой, которая, однако, при-
сутствует в очень низкой концентрации. Оба аномера достаточно хорошо
различимы в спектре ЯМР по двум отдельным дублетным линиям (см. рис.
5.18). При анаэробном гликолизе глюкоза, ответственная за энергоснабже-
ние клеток дрожжей, через несколько промежуточных стадий превращается
в этанол, резонансные линии которого со временем становятся более интен-
сивными, в то время как линии, принадлежащие глюкозе, спустя некоторое
время уже не наблюдаются. Несмотря на то что получение спирта из глюкозы
не является открытием, этот простой пример показывает, каким образом с
помощью данного метода при досточно хорошем значении отношения сиг-
нал/шум можно проследить за превращениями изотопозамещенного по С
субстрата.
Другими методами, обеспечивающими хорошую чувствительность по
13С, являются модификации указанных выше методов переноса поляриза-
ции, в которых поляризация переносится на протоны и таким образом детек-
тируется (обратные методы INEPT и DEPT). В случае применения ЯМР Н
всегда существует одна проблема: полезный сигнал часто на несколько по-
рядков слабее, чем сигнал воды, если используется обычная, а не дейтериро-
ванная вода. Несмотря на то что сигнал воды не должен проявляться в
233
a
13
Рис.5.18. Процесс превращения [1- С] глюкозы клетками дрожжей. Разностные спектры
^Нг3С) суспензии дрожжей, полученные методом спинового эха после добавления [1-JzC]
глюкозыи [2- С] ацетата. Резонансные линии можно отнести к следующим группам: [2- С]
этанол (a), [2- С) ацетат (Ь),/3- [Г13С] глюкоза (с) и(2-[1- С] глюкоза (d). Линия (е)
состоит из перекрывающихся пиков, относящихся с СС- и /3-аномерам глюкозы, линия (О “
остаточный сигнал воды [5.16]
спектре, поскольку вода не содержит атомов углерода, неполное подавление
сигнала воды приводит к искажению базовой линии в спектре ЯМР и пере-
крыванию сигналов. Модификация обратного метода DEPT позволяет уст-
ранить сигнал воды, который более чем в 30000раз превышает интенсивность
полезного сигнала.
Недостатком всех этих методов является умеьшение разрешения спект-
ров 13С, т.е. применение их оправдано для получения простых спектров,
содержащих сигналы только от одного вещества, для которого проведено
селективное изотопное замещение. Если же мы действительно хотим достичь
высокого разрешения в спектрах13С многокомпонентных систем, то сигналы
ядер 13С должны детектироваться непосредственно, либо необходимо приме-
нять методы гетероядерной корреляционной спектроскопии (раздел 2.3.4),
которые требуют существенно большей длительности эксперимента.
Как и для ЯМР 31Р, имеется большое число работ по ЯМР 13С, посвя-
щенных исследованию клеток, тканей и органов. В этих работах изучаются
субстраты, в которых проведено изотопное замещение, при этом процесс
замещения контролируется по спектрам ЯМР. Острый интерес для исследо-
вания меченых по 13С метаболитов представляет печень, поскольку именно
она ответственна за процессы обмена веществ в организме. Проводя перфу-
зию печени крыс в растворе, содержащем меченые по 13С субстраты, можно
изучать поток этих субстратов при различных путях обмена веществ в зави-
симости от внешних условий (гормональной стимуляции, состава перфузи-
онного раствора) и состояния, в котором находился исследуемый объект до
начала эксперимента, например, можно проводить измерения каждые 10
234
мии, наблюдая за изменениями в спектре после поступления питательных
веществ спустя период длительного голодания, т.е. при моделировании диа-
бета. Такого рода эксперименты проводили ранее с помощью радиоактивных
меток (веществ, содержащих изотоп 14С), при этом анализировались про-
дукты реакции. Однако с помощью ЯМР такие исследования могут прово-
диться неннвазивно с одновременным непрерывным определением
концентраций большого числа различных метаболитов. На рис. 5.19 приве-
ден пример спектра 13С, полученного на образце печени крысы больной
диабетом при условии, что перфузия проводилась в течение 170 минут с
[3-13С [-аланином при использовании [2-13С [-этанола с инсулином и [1,2-
3С [-этанола без инсулина. Из спектров непосредственно видно, в какую нз
вновь образовавшихся молекул происходит встраивание фрагмента, содер-
жащего изотопную метку 13С при условии, что в процессе обмена веществ
участвуют аланин и этанол. Среди различных возможных путей превраще-
14 13
Рис. 5.19. Процесс превращения изотопомеченого по °C субстрата в печени. Спектры ЯМР С
(90,5 МГц) получены для печени крысы, больной диабетом. Образцу ^1ечени находились в
перфузионной жидкости в течение 170 мин. В растворах содержался [3- JC] -аланин в концен-
трации 10 ммоль и [1,2- ^С] -этанол (верхний спектр) и [2- с]-этанол (нижний спектр) в
концентрации 7,3 ммоль; при этом верхний спектр снят с добавлением инсулина, а нижний -
без инсулина. Из обоих спектров вычтены исходные спектры, полученные без добавления обога-
щенных субстратов. Для линий в спектрах1JC проведено следующее отнесение: Ci и С2 этанола
(EtOH), С2 и С3 аланина (Ala), С2 ацетата (Acet), С3 лактата (Lac), С2, С3 и С4 глутамина
(Gin) и глутамата (Glu), С4 глутамилового остатка в глутатионе (GSH), Cj, С2, С3, С4, С5И Cg
CL- и ^3-аномеров глюкозы (CJ ,/3), С2 и С3 /3-гидроксимасляной кислоты (рНВ), С2 и С3
аспарата (Asp) и С2 и С3 N-карбамиласпарата (СА). Область спектра, в которой наблюдаются
линии карбоксигрупп, на рисунке не приведена [5.17]
235
ния [3-13С]-аланина с образованием [3-13С ]-пирувата один путь приво-
дит к глюконеогенезу, т.е. к новому синтезу глюкозы, а другой — к ресин-
тезу исходных продуктов (рис. 5.20). В процессе трикарбонилкислотного
цикла атом 13С из [3-13С ]-аланина встраивается в положение 2 фосфое-
нолпирувата и, пройдя через несколько стадий циклических превраще-
ний, встраивается также в положение 1 фосфоенолпирувата. В
соответствии с этим изменяется также распределение атомов 1ЛС в про-
дуктах реакции: глюкозе и аланине. Анализ этого распределения позво-
ляет исследовать процесс замещения по отдельным путям обмена веществ.
В этих экспериментах было обнаружено, что в печени крыс при условии
голодания их в течение 24 часов содержание аланина совпадает с имею-
щимся в исходном состоянии, т.е. весь образовавшийся аланин расходует-
ся на восстановление исходного продукта (futile cycle), а значит, вновь
используется для синтеза глюкозы.
ЦИТОЗОЛЬ МИТОХОНДРИЯ|
Рис. 5.20. Упрощенная модель глюконеогенеза. Звездочками и жирными буквами выделены
места первоначального замещения по 1 °C в аланине (Ala), а черные точки обозначают положе-
ния. которые после многократного прохождения трикарбоксилкислотиого цикла занимают ато-
мы 1,5С. Используемые сокращения: трикарбоксилкислотный цикл (ТСА), оксалацетат (ОАА),
пируват (PYR), малат (MAL), фумарат (F(JM), фосфоенолпируват (РЕР) [5.17]:
1 - цитозоль, 2 - митохондрии
Получение информации о том, проведено ли в данной молекуле одно-
кратное или многократное изотопное замещение, сопряжено со значитель-
ными трудностями при использовании классических методов. Метод ЯМР
позволяет получить эту информацию достаточно просто, так как в спектре
удается наблюдать возникающее в результате косвенного спин-спинового
взаимодействия расщепление резонансных линий. Этот метод уже приме-
нялся в описанном выше эксперименте (рис. 5.19) по наблюдению разложе-
ния этанола. Этанол при изотопном замещении по двум положениям,
[ 1,2-13С ]-этанол, можно наблюдать по дублетной линии на верхнем спектре
на рис. 5.19. Оба атома встраиваются в ацетат и глютамат, что сопровожда-
ется появлением в спектрах соответствующих дублетных сигналов. Таким
236
же образом можно детально проследить за многими превращениями, кото-
рыми сопровождаются процессы обмена веществ.
5.2.5. ЯМР-спектроскопия in-vivo на ядрах 15N
Наиболее распространенным в природе изотопом азота является 14N,
для которого спин ядра равен 1. Наличие у ядер14N квадрупольного момента
обусловливает настолько сильное уширение спектральных линий, что дан-
ный изотоп практически не используется для исследования биологически
важных молекул. Совершенно иная ситуация имеет место для изотопа 15N,
спин которого 1 = 1/2. Хотя естественное содержание этого изотопа всего
0,37 %, ядра 15N в качестве изотопных меток являются даже более подходя-
щими, чем ядра13С, естественное содержание которого составляет 1,1%, так
как возмущающий сигнал фона для ЯМР 15N соответственно слабее.
Вследствие того, что гиромагнитное отношение для изотопа 15N отрица-
тельно, применение широкополосной развязки по протонам не приводит к
достижению желаемого результата - увеличению чувствительности, по-
скольку ядерный эффект Оверхаузера вызывает полное исчезновение сигна-
ла (см. раздел 2.2.6). Для повышения чувствительности метода ЯМР на ядрах
15N подходят все методы, которые были рассмотрены нами при обсуждении
ЯМР 13С. Так как область значений химических сдвигов очень велика, то
здесь можно ожидать хорошей разрешимости спектров.
На рис. 5.21 приведен пример спектра 15N Brevibacterium
lactofermentum, выращенного в среде, содержащей 15NH4C1 в качестве ис-
точника азота. С помощью этого приема можно достаточно хорошо просле-
дить внедрение в клетку изотопной метки. Спустя некоторое время меченая
изотопом15N аминокислота аланин (Ala) может быть обнаружена за преде-
лами клетки, т.е. происходит ее передача от бактерии к среде, в то время как
другие аминокислоты остаются внутри клеток.
19
5.2.6. ЯМР-спектроскопия ш-vivo на ядрах F
Несмотря на то, что подходящий с точки зрения ЯМР изотоп фтора 19F
характеризуется 100%-ным естественным содержанием, в этом случае так-
же необходимо проводить изотопное замещение веществ, поскольку в боль-
шинстве систем, встречающихся в природе, в растворимых фазах фтор
практически отсутствует. Большим преимуществом этого ядра со спином 1 =
1/2 является, наряду с большой областью значений химических сдвигов,
высокая чувствительность, близкая к чувствительности протонов. Вещества,
содержащие фтор, часто являются лекарственными препаратами, и одной из
областей применения ЯМР 19F является исследование обмена веществ в
присутствии этих препаратов.
Анестезирующие средства такие, как халотан и изофлуран, содержат
фтор; это позволяет проследить за процессами их накопления и превращения
237
200 250 300 350
8М.Д.(НЬЮ3)
Рис. 5.21 a-c. Спектры ЯМР 15N В. Lactofermentum. Спектры получены в режиме широкополос-
ной развязки по протонам на частоте 30,4 МГц (для ХН - 300 МГц). Спектр среды в отсутствие
клеток (а). Спектры бактерий, которые выращивались в течение 17 ч (Ь) и 35 ч (с) [5.18]
в тканях. Рис. 5.22 показывает уменьшение изофлурана в мозговой ткани
после использования его в качестве анестезирующего препарата. Халотан
оказывается стабильным в большинстве тканей, однако в печени он частично
преобразуется в токсические вещества. С помощью ЯМР 19F удается зафик-
сировать в печени крыс четыре типа различных фторсодержащих продукта
распада халотана в случае использования его для наркоза.
238
19
Рис. 5.22. Спектр ЯМР F анестезирующего
средства, введенного в мозговую ткань.
Спектры ’г (75,5 МГц) нзофлурана
(CFjCHCIOCFjH) в мозгу живого кролика,
которому вводился нзофлуран перед проведе-
нием измерения как анестезирующее средство.
Спектры показывают уменьшение содержания
анестетика из ткани мозга. Сигнал прн 0 м.д.
принадлежит внешнему стандарту, в качестве
которого взят дибромтетрафторэтан. Измере-
ние проводилось с помощью поверхностной ка-
тушки [5.19]
Подобные эксперименты можно провести также с антиметаболитами,
используемыми в качестве противоопухолевых препаратов, такими, как
дифторметилонитин, который является ингибитором орнитиндекарбоксила-
зы; его избирательно поглощают некоторые виды опухолей. Такие исследо-
вания имеют как теоретическое, так и чисто практическое значение,
поскольку результаты, полученные методом ЯМР, могут быть использованы
для клинических исследований пациентов. Так, по спектрам ЯМР 19F соот-
ветствующих лекарственных препаратов можно определять их концентра-
цию в очаговой области, например, в опухоли.
5.2.7. Использование других ядер в ЯМР-спектроскопии
in-vivo
В спектроскопии ЯМР in-vivo, конечно, могут быть использованы все
ядра, спин которых отличен от нуля. Среди них прежде всего следует указать
на такие квадрупольные ядра, как 23Na и 3^К, содержание которых в биоло-
гических системах достаточно велико. Дейтерий, естественное содержание
которого невелико, в принципе можно использовать в качестве метки в
биологических молекулах. Однако более предпочтительным является пря-
мое наблюдение сигналов *H, в этом случае отсутствие соответствующего
сигнала в спектре 1Н свидетельствует о том, что именно в этом положении
проведено изотопное замещение.
Ширины резонансных линий, наблюдаемых в спектре квадрупольных
ядер, часто столь велики, что при проведении исследований in-vivo разность
значений химических сдвигов оказывается недостаточной для разделения
резонансных линий, принадлежащих к различным молекулам. Ионы натрия
239
и калия не образуют ковалентных связей в биологических молекулах, и
следовательно, невозможно указать характерные значения химических
сдвигов их резонансных линий. Взаимодействие ионов натрия и калия с
макромолекулами, входящими в состав клеток, в основном приводит к
уменьшению времени релаксации. В случае, если ионы находятся в различ-
ном химическом окружении, иногда удается наблюдать многоэкспоненци-
альный спад свбодной индукции.
Литература
5.1. Ugurbil, К., Rottenberg, Н., Glynn, Р. and Shulman, R.G. (1982)
Biochemistry 21,1068.
5.2. Bendall, M.R. and Pegg, D.T. (1985) Megn. Reson. Med. 2, 91
5.3. Haase, A., Hanicke, W. and Frahm, J. (1984) J. Magn. Reson. 56, 401.
5.4. Haase, A. (1986) Magn. Reson. Med. 3. 963.
5.5. Maudsley, A.A. and Hilal, S.K. (1985) Magn. Reson. Med. 2. 218.
5.6. Lyon. R.C., Faustino. P J. and Cohen. J.S. (1986) Magn. Reson. Med. 3.663.
5.7. Brown, F.F., Campbell, I.D., Kuchel, P.W. and Rabenstein, D.C.
(1977) FEBS Leu. 82, 12.
5.8. Gadian, D.G., Proctor, E., Williams, S.R., Caby, E.B. and Gardiner,
R.M. (1986) Magn. Reson. Med. 3, 150.
5.9. Routhman, D.L., Arias-Mendoza, F., Shulman, G.J. and Shulman.
R.G. (1984) J. Magn. Reson. 60, 430.
5.10. Arnold, D.L., Matthews, P.M. and Radda, G.K. (1984) Magn. Reson.
Med. 1, 307.
5.11. Degani, H., Laughlin, M., Campbell, S. and Shulman, R.G. (1985)
Biochemistry 24, 5510.
5.12. Ross. B.D., Radda, G.K., Gadian, D.G., Rocker, G., Esiri, M. and
Falconer-Smith, JU. (1981) New Engl. J. Med. 304, 1338.
5.13. Ross, B.D., Higgins, R.J., Conley, F.K. and True, N.S. (1987) Magn.
Reson. Med. 4, 323.
5.14. Sorensen, O.W. and Ernst, R.R. (1983) J. Magn. Reson. 51, 477.
Taylor, D J., Styles, P., Matthews, P.M., Annond, D.A., Gadian, D.G.,
Bore, P. and Radda, G.K. (1986) Magn. Reson. Med. 3, 44
5.16. Foxall, D.L., Cohen, J.S. and Tschudin, R.G. (1983) J. Magn. Reson.
51, 330.
5.17. Cohen, S. (1987) Biochemistry 26, 573.
5.18. Haran, N., Kahana, X.E. and Lapidot, A. (1983) J. Biol. Chem. 258,
12929.
5.19. Bendall, M.R., den Hollander, J.A., Arias-Mendoza, F., Rothman,
D.L., Behar, K.L., and Shulman, R.G. (1985) Magn. Reson. Med. 2, 56.
5.20. Wyrwicz, A.M., Conboy, C.B., Ryback, K.R., Nichols, B.G. and
Eisile, P. (1987) Biochem. Biophys. Acta 927, 86.
240
Предметный указатель
А
АДФ, см. аденозиндифосфат
АМФ, см. аденозинмонофосфат
АТФ, см. аденозинтрифосфат
АЦП, см. аналого-цифровой преобразователь
АЦ-преобразователь, см. аналого-цифровой преобразователь
Абдоминальная область 148, 154, 159
Аденилаткиназа 89
Аденин 125
Аденозиндифосфат 69, 89, 193
Аденозинмонофосфат 89, 125, 199
Аденозинтрифосфат 61, 69, 89, 193, 205
Азот 15
Аланин 80, 87, 109, 202, 207
Алгоритм расстояний 119
Алкены 42
Алкины 42
Амидные протоны 84, 93, 111, 113, 117
Аминокислоты, см. также отдельные аминокислоты 80
Аналого-цифровой преобразователь 35, 51
Аневризма 159
-аорты 159
Анизотропия 16, 123
- химического сдвига 22, 24, 134
- магнитной восприимчивости 100
Аорта 159, 170
Аргинин 80_87, 109
Ароматические углеводороды 96
Артефакты 115, 146, 163
- связанные с движениями 196
Аспаргиновая кислота 80, 87, 109, 207
Аспаргин 80,87,109,114, 204, 207
Атом водорода, см. 'н
Ацетат 202
Ацетиласпаргиновая кислота 191
Б
БФП (быстрое Фурье-преобразование) 33
Биномиальные коэффициенты 17, 46
241
Больцмановское равновесие 16, 27, 30, 51
Броуновское движение молекул 16, 23, 54, 123
В
Валин 98, 102, 129
Везикулы, саркоплазматические 159
Венечные артерии сердца 189
Вероятность перехода 81
Взаимодействие
— дипольное 28,33, 38, 40, 118, 145
- сверхтонкое 33, 41, 118
- электрон-ядерное 32, 33
— ядерное квадрупольное 33
-скалярное, см. также J-взаимодействие 30, 39, 59, 73, 93, 133,139,152
Взаимодействие, см. связь
Внеклеточное
-пространство 146, 181
- масса 146, 181
Возбуждаемый объем 142, 167, 182
Возбуждающий импульс 28, 51, 53, 142, 167, 182
Воксель, см. элемент объема
Восприимчивость магнитная 56, 209, 225
Вращательная диффузия 36, 83, 113, 144
Вращение образца 50
Вращение под магическим углом 29, 145, 200
Временная область, временной интервал 43, 89
Временная зависимость 34,37, 50, 53,68,79,86,103, 142, 143, 222, 238
Временная корреляция 70, 144
-обмена 72, НО, 144
- вращательных движений 40, 80, 114, 144
- трансляционных движений 39
Временная шкала, см. шкала ЯМР
Время измерений, см. измерения время 88, 171, 179
Время повторения 46, 66, 175, 179, 194
Время релаксации
-эффективное 37, 175, 196
-поперечной (спин-спиновой) 17,23,37,42,46,66,79,94,113,170,199,215
- электронной 42, 120
-спин-решеточной (продольной) 17, 23, 37, 42, 66, 79, 94,113,170,199, 215
Время смешивания 89, 92
Время спин-решеточной релаксации, см. время релаксации
Время формирования эха 12, 145, 150, 167, 186, 189, 200
Вторичная структура 101, 114,138
Выбор слоя 166, 174, 191
242
Высокое разрешение 12, 50
Вязкость 144, 196
Г
Гадолиний DTPA 171
Гамильтониан 12
Гауссова форма импульса 165
Гауссов фильтр 32
Гелике 83,94,117, 126
Гемато-энцефалический барьер 159
Гем 102
Гетероцикл 56
Гиромагнитное отношение 14, 21, 38, 42, 63, 82, 83, 95, 113, 114, 117
Гистидинемия 221
Гистидин 80, 84, 87, 90, 126, 129, 219
Гликолиз 193,197, 203
Глицин 80, 87, 109, 114
Глюкоза 132, 182, 197, 199, 202
Глюконеогенез 197
Глютаминовая кислота 80, 87, 97, 109, 202, 204, 207
Глютамин 80, 109, 204, 207
Глютат-ион 190, 204
Градиента амплитуда 148, 172
Градиентное эхо 148, 165
Градиент
- электрического поля 20, 25
- магнитного поля 141, 166, 174, 191
Градиент выбора слоя 35, 166, 174, 191
Грудная клетка 174, 183
Гуанин 125
д
ДНК 124
ДСС 32, 41, 43, 109
ДТПА, см. Gd-DTPA
Двумерная спектроскопия ЯМР, см. 2В-ЯМР
Двумерное изображение 36, 142, 172
Двумерное преобразование Фурье 51, 88, 168
Дезоксирибоза 125, 129
Дейтерий,см. 2Н
Деполяризация мембран 159
Детектирование фазочувствительное 34, 169
243
Дефазировка 11,23, 148
Дефекты изображения
— за счет движений 148, 154
- за счет разности химических сдвигов 164
— за счет течения жидкостей 166
Диагностика 205
Диатермия 176
Дисперсии сигнал 22, 27
Диффузия 14, 23, 54, 94, 134, 173, 189
Длительность импульса 19, 28, 53
Е
Естественное содержание 13
Ехиннокок163
Ж
Железо, см. ион железа 100, 103
Жировая ткань 146, 164
3
Зеемановский уровень 29, 46
Значение pH 53, 58, 75, ПО, 224, 227
И
Изображение
-трехмерное 36, 142, 147,153, 172
- головы 154
- спектральное 185
- двумерное 36, 142, 172
Изображение микрообъектов 165, 172
Изображение, восстановленное по химическому сдвигу (CSI) 164
Изображения макрообъектов 165, 172
Изолейцин 80, 87, 97, 109
Изотопная метка 68, 108, 135, 202
Изотопное обогащение 135, 200, 206
Изотопный эффект 18О 85
Изотоп 13
Изофлуран 206
Имплантант 175
Импульсная последовательность Карра-Парселла-Мейбума-Гилла 27, 171,
173,177,181,192,196
Импульсная последовательность спиновое эхо 26, 27,171,173,177,181,192,196
244
Импульс (РЧ-импульс)
-широкополосный 50, 66, 200, 206
-селективный по частоте 142, 148, 165
Импульс, осуществляющий выбор слоя 142, 148, 165
Инверсия градиента 148
Индукционный ток 176
Инсулин 204
Инфаркт 171
Ионная сила 39, 176, 194
Ионы Cd2+118
Ион Са2+ 100
Ион Се3+ 101
Ион Со2+100, 102, 135
Hoh15NH4+61
Ион 15NO3' 58
Ион Еи3+ 101
Ион Fe2+ 100, 103
Ион Fe3+ 100, 103
Ион La3+ 101
Ион Lu3+ 101
Ион Mg2+ 100
Ион Мп2+120
Ион Nd3+120
Ион Ni2+120
Ион Рш3+ 121
Ион Рг3+ 121
Ион Sm3+ 121
Ион ТЬ3+ 121
ИонТш3+ 121
ИонУО2+121
ИонУЬ3+ 121
HoHGd3+100
К
КПМГ - см. Карра-Парселла-Мейбума-Гилла последовательность
Карбамиласпарат 202
Катушка
-резистивная 49, 173
-сверхпроводящая 49, 173
Катушка Гельмгольца 211
Катушка градиентная 49, 173
Качество изображения 146
Квадрупольный момент 14, 34, 148, 160, 237, 239
245
Квадрупольный момент ядра 14, 34, 148, 160, 237, 239
Квантовая механика 13, 16, 72
Квантовое число, магнитное 14,34,37,40,82,113, 145, 159
Квант взаимодействия Планка 14, 34, 37, 40, 82, 113, 145, 159
Кислород 14, 86, 177, 221, 227
Клетка 201, 217, 233
Клетки крови, красные, см. Эритроциты
Количество крови 170
Комплекс нуклеиновая кислота-протеин 150
Константа взаимодействия 30, 32
Константа спин-спинового взаимодействия 30, 39
Константа экранирования 16
Контрастные вещества 41, 199
Контраст 165, 171, 176, 199
Контраст изображения 11, 141, 146, 150, 171
Конформационные изменения 53, 56, 72, 108
Конформация 53, 56, 104
Корреляционная функция 71
Коэффициент заполнения 180, 185
Коэффициент насыщения 42, 46, 66, 175, 194
Краниофарингиома 154, 158
Креатинкиназа 221, 225
Креатинфосфат 61, 69, 89, 193, 205, 220, 223
Креатин 221
Кристалл 144
Кровеносные сосуды 159, 166, 171
Кросс-пик 74, 97, 110, 116, 128, 130, 133
Кросс-релаксация 63, 96
Круговая частота 16
Л
Лактат 233, 236
Лак-репрессор 120, 153, 199
Лантаниды 121, 123, 199
Ларморова прецессия 16
Лейцин 98, 105, 129
Ленточный глист 163
Лизин 98, 105, 129
Ликвор 178
Линия
- поглощения 36, 42
- дисперсии 42
- гауссова 36
246
— однородная 42
- неоднородная 36
- лоренцева 36
Линия гауссовой формы 36
Липиды 146, 155, 203
Липосомы 155
Локальное поле 29
Лоренцева функция 36
М
Магический угол 29, 145
Магнитное поле
- эффективное 21, 28
-однородное 49, 174, 214
- неоднородное 28, 49, 174
- в лабораторной системе координат 18
- локальное 29
- зависящее от координат 214
- в вращающейся системе координат 18
- статическое 49
- переменное поле 17, 42, 63, 64, 166, 204, 208
Магнит 172
электро- 49, 50,172
постоянный- 49, 50, 172
сверхпроводящий 49, 50, 172
Максимальное сужение 23
Мак-Ардла синдром 227
Мак-Коннела уравнение 73
Малтоза 154
Математическое ожидание 12, 55, 136
Межклеточное пространство 146, 181
Мембрана 156
Метаболизм 207, 221,227
Метастазы
- в печени 189
Метионин 98, 105, 129
Методы получения двумерных изображений 143, 167
Методы получения изображений
- с помощью градиентного эха 174
- спинового эха 175
Метод инверсии-восстановления 53, 145, 165
Метод максимальной энтропии 48
Метод переноса поляризации 79
Метод последовательной точки 166
247
Механизмы релаксации 36
- за счет анизотропии химического сдвига 36, 39, 157
- за счет диполь-дипольного взаимодействия 36, 82, 113
- за счет сверхтонкого взаимодействия 36, 41, 113
- за счет квадрупольного взаимодействия 36, 40
— за счет скалярного взаимодействия 36, 39
— за счет спин-вращательного взаимодействия 36
Миелограмма 178
Микроскопия 200
Миозин 98
Многослойная техника 170, 177
Мозг 151, 154, 177, 199, 226
Молекулярная диффузия 37, 70, 112, 155
-динамика 141
Молочная кислота (см. лактат)
Момент, магнитный 13, 16, 20
Моча 157, 166
Мультиплет, см. J-взаимодействие
Мышца 222
Н
Наведенное напряжение 40, 45
Намагниченность
-продольная 19, 42, 46, 116, 175
- макроскопическая 18
-поперечная 19, 42, 46, 175
Направления волокон 203
Напряженность поля 13, 40, 45, 147, 173
Насыщение 42, 46, 66, 175, 194
Нитробензол 58
Нитроксидный радикал 120
Нитрометан 58
Нуклеиновая кислота 146
О
Обмен
-амидных протонов 84, 93, 111, 113, 117
- химических 25, 56, 75, 84, 89, 105, 194
Обмен веществ, см. метаболизм
Обработка изображений 33
Оператор 13
Определение концентраций 54, 207, 209, 220, 230
Определение расстояний 16, 63, 75, 96, 123
248
Определение структуры 16, 63, 75, 96, 126
-протеинов 13, 115, 138, 144
Опухоль 180,181,183,189, 198, 229, 238
- в печени 189
Органелла 207, 209, 217, 235
Орнитин 237
Отек 181,186,199
Отношение сигнал/шум 27, 30, 70
Отношение, гиромагнитное 13
Оцифровывание 27, 30
Очаговые процессы 156, 160
П
Парамагнетизм 33, 40, 118, 156, 199
Параметр порядка 160
Парвальбумин 126
Пептидная связь 97
Передатчик 22, 55, 200, 211
Переносимость контрастных веществ 198
Перенос поляризации 63, 196
Перенос поляризации, см. также ЯЭО 115, 138
Переход 17, 34, 81
Перистальтика 174, 183
Перистальтика кишечника 148, 154
Пероксидаза хрена 123
Перфузия 217, 226, 233, 234
Печень 189,199,233
Пиксель, см. элемент изображения
Пируват 219, 236
Плавиковая кислота 16
Плазма крови 170
Плотность, спектральная 27, 195
Побочные явления 199
Поверхностная катушка 192, 209, 211, 216, 220, 223, 227, 239
Поглощение
- энергии 199
- высокочастотное 199
Подавление сигнала растворителя 67, 218
Полисахариды 153
Полуширина линии 22
Поперечное сечение 180
Порфирин 121
Последовательность эхо 12, 145, 150, 167, 186, 189, 200
249
Постоянная Больцмана 16, 95
Потенциал, электрический 34
Потоковый эффект 166
Поток
-поршневой 166
— ламинарный 166
— турбулентный 166
Поток крови 166
Почка 202
Пояснично-крестцовая область 186
Предварительное насыщение 68
Прецессия 16, 19, 21, 34
Приемная катушка 34, 173, 182
Принцип Паули 30
Приращение времени 47, 88
Программное (математическое) обеспечение 51
Проекционная реконструкция 167
Проекция 167
Пролин 20, 105, 129
Пространственная селекция
- с помощью РЧ-поля 210
— с помощью статических магнитных полей 165,200, 207
- с помощью градиентов 210
Пространственное разрешение 165, 200, 207
Протеин 97, 150
Протонная плотность 167
Протоны
- метиленовые 12, 28, 55, 60, 233
-метильные 12, 28, 55, 60, 233
Протоны жировой ткани 146, 164
Протон, см.’Н 12, 54, 55, 56, 58, 60, 68, 105, 123, 145, 151, 155, 218
Профиль скоростей, параболический 166
Прохождение медленное 27, 30
Процессор быстродействующий с плавающей запятой 30
Прямоугольный импульс 43
Псевдоконтактный сдвиг 121
Р
РНК 147
Равновесие
- больцмановское 16, 27, 30, 51
-динамическое 31, 149
-температурное 16, 27, 30, 51, 63, 96, 149
Равновесная намагниченность 17, 27, 30, 51, 63, 96, 149
250
Радикал 32, 41, 118, 198
Радиочастотный (РЧ)
— импульс, см. импульс
- полоса пропускания РЧ поля 30
- передатчик 34
Развязка 49,192
- широкополосная 55, 66, 200, 206
- многоимпульсная 50
- шумовая 50
Разностное изображение 166, 169
Разностные методы 192, 220, 223
Разность заселенностей 16, 27, 30, 63
Разность фаз 192
Разогрев
-тканей 50, 176
Разрешение
-пространственное 142, 146, 159, 172
- спектральное 32, 34, 39, 70, 79, 123, 134, 203
Разрушающий градиент 175
Рамачандрана модель 102
Рассеянный склероз 227
Расфазировка 175
Расщепление 14, 16, 41, 45, 200
Расщепление энергетического уровня 15, 33
- за счет дипольного взаимодействия 16, 19
- за счет J-взаимодействия 16
- за счет ядерного квадрупольного взаимодействия 20
Регулятор сердечного ритма 203
Редактирование спектров 218, 230
Резонансная частота 15, 21, 27, 43
Резонатор 50
Рентгеновская томография 165, 180, 183
Рентгеноструктурный анализ 13, 121, 126, 141
Рефокусировка 24, 192, 219, 231
Рибоза 147
С
Сагиттальное сечение 181
Сахарид 153
Сахароза 154
Свернутый лист 83, 117
Сверхтонкая структура 19
Сверхтонкое взаимодействие 19, 26, 99
Сдвиг
— химический 12, 28, 36, 55, 75, 105, 109, 129, 190, 209, 214, 219, 222,
232,237,239
— контактный 32, 120
-парамагнитный 120
— псевдоконтактный 121
- за счет кольцевого тока 109, 125
Сдвиг-реагент 120
Седловая катушка 211
Сердечный ритм 189
Серин 98, 105, 123
Сигнал воды 55, 68, 192, 218
Сигнал поглощения 22, 27
Сигнал протонов жировой ткани 146, 164
Сигнал, см. сигнал ЯМР 22, 43, 47
Система координат 14
-сферическая 122
-вращающаяся 14, 24, 42
Склероз рассеянный 227
Скорость потока 166
Скорость релаксации 36
Сложение спектров 189, 201
Смещение диска 159, 161
Собственный возбуждающий импульс 13
Соленоид 211
Спад свободной индукции (ССИ) 12, 28, 32, 51, 70, 145
Спектрометр
- cw- 27, 35
- FT- 48
Спектрометр ядерного резонанса, см. ЯМР-спектрометр
Спектр шумов 38
Спецпроцессор 36
Спинномозговой канал 151, 166
Спиновая метка 33, 41, 118
Спиновая плотность 165, 171
Спиновое квантовое число 14
Спиновое эхо
-КПМГ (Карра-Парселла-Мейбума-Гилла) 27,171, 173, 177,181,192,196
-Хана 26
Спиновый пакет 24
Спин-вращательное взаимодействие 36
Спин-решеточная релаксация 18, 22, 37, 42, 46, 65, 78, 94, 113, 170,
196,215
Спин-спиновое взаимодействие, см. J-взаимодействие 30, 39
Спин-тиклинг 65
Стандарт 28, 57
252
— внешний 57, 225
- внутренний 28, 57
Суммарное изображение 180
Сферическая система координат 122
Считывающий градиент 148
Т
ТМС (тетраметилсилан) 57
Температурная реакция тканей 176
Температурная регуляция 204, 217
Температурное равновесие 17
Теорема Найквиста 47
Тепло
- количество, 204
-регуляция 204, 217
Техника мульти-эхо/мульти-слой 170, 177
Тимин 147
Тирозин 98, 102, 105, 116, 126, 129
Токсичность 200
Томография 165
-сосудов 183, 193
- химический сдвиг в томографии 191
-сердца 174, 183
- контрастные вещества в томографии 41, 178, 198
-головы 180
-печени 189, 200, 233
- микрообьектов 200
- почек 189, 200
-опухолей 181, 183, 186, 189, 200
Томограф для всего тела 26, 35, 141
Травма 200
Треонин 98, 105, 116, 129
Третичная структура 53, 100, 139
Триптофан 98, 102, 105, 116, 126, 129
Трифторид фосфора (PF3) 30
т-РНК 147
Ti-взвешенное изображение 170, 175
Тг-взвешенное изображение 170, 175, 217
У
13
Углерод,см. С
Угол
-диэдральный 63, 100, 139
— магнитный 29, 145, 200
Угол Эрнста 31, 51, 149
Угол поворота 30, 51, 148, 165, 183
- в методе FISP 150
- в методе FLASH 149
-Эрнста 31, 51, 149
Ультразвуковое исследование 198
Упорядочение 126
Уравнение Джонсона-Бови 106
Уравнение Карпл уса 117
Уравнение Стокса-Эйнштейна 114
Уравнение Хая-Маллиона 126, 127
Уравнение Хендерсона-Хассельбалха 76, 111, 223
Уравнения Блоха 16,27
Урацил 147
Уретра 193
Условие резонанса 16,21,27
Уширение линии 36, 74
Уширение сигналов 36, 42, 74
Ф
Фазовая когерентность 34, 176
Фазовое кодирование 172, 215
Фенилаланин 98, 102. 116, 126, 129
Ферми-контактное взаимодействие 19, 26, 100
Ферриты 171
Ферромагнитные вещества 175
Ферроцен 123
Фильтрующая функция 32, 73
Фосфат, неорганический 76, 85, 212, 223, 227
Фосфены 204
Фосфоенолпируват 105, 236
Фосфорилаза 227
Фосфорная кислота, см. фосфат неорганический
Фосфор, см. 31Р
Фосфотрансферазная система 104, 109
Фронтальное сечение 159
Фруктоза 132
Функция sine 44
Фурье
- интроскопия 143
- спектроскопия 12, 26, 28, 70
— спектрометр 34
254
Фурье " преобразование
- одномерное 26, 28
- двумерное 36, 70
- трехмерное 70, 152
X
Халотан 237
Хаотический клубок 104
Хелаты редкоземельных элементов 171
Холин 191
ц
Цветной графический дисплей 36
Циркуляция, см. кровообращение
Цистеин 98, 102, 105, 116, 129
Цитозин 98, 102, 105, 116,129
Цифровая фильтрация 32, 73
Ч
Частота Лармора, см. ларморова частота 16, 35
Частотная область 28
-спектр 26,28,70,144, 152
Чувствительность 13, 29, 34, 40, 76, 141, 179, 185, 192, 200, 206
Чувствительность детектирования, см. чувствительность
Ш
Шиммирующая катушка 50
Ширина линии
- однородной 36
- неоднородной 36
Широкополосная развязка 50, 66, 200, 206
Шумы 44, 47, 170, 200, 208, 214, 234
Э
ЭКГ-синхронизация 148, 159
ЭПР 19
Эквивалентные ядра 18, 24, 45
Электронная оболочка 15, 24
Электронное распределение 12
255
Электронный
— спин 19, 26, 99
— спиновый резонанс 19
Электрон, неспаренный 19, 26, 99
Элементарный анализ 14
Элементарный магнит 15
Элемент изображения 142, 172
Элемент объема 142, 172
Энергия
- поглощения 27, 48
— уровень 15, 17, 64
- активации 62, 86
Энзим 39, 69, 79, 195, 203
Эритроцит 170,190
Этанол 13, 15, 18, 40, 50, 202
Эффект Оверхаузера 80, 93, 115, 134, 141, 151, 231
Эффект кольцевого тока 109, 125
Эхо
- Хана 12
— градиентное 148, 165
— стимулированное 164
Эхо-планарное изображение 148
Я
ЯМР
- cw27, 35, 42, 49
-трехмерный 70, 87, 88
-твердотельный 12, 16, 29, 34, 144
- in-vivo 179
- импульсный 43, 49
- высокого разрешения 18, 53, 97
-многоимпульсный 145
-пространственноселективный 165, 207
-двумерный 87, 88
ЯМР 1Н, 2Н 12, 54, 55, 56, 58, 60, 68, 105, 123, 145, 151, 155, 208, 218, 239
ЯМР inCd 12
ЯМР 113Cd 12
ЯМР 13С 12, 18, 24, 42, 66, 122, 136, 200
ЯМР 15N 15, 85, 145, 237
ЯМР 17О 12, 18, 85
ЯМР 19F 14, 17, 24, 206
ЯМР 23Na 208, 239
ЯМР 31Р 14, 30, 39, 57, 76, 82, 85, 150, 158, 209, 222
ЯМР 33S 12, 18
256
ЯМР35С1 12, 18
ЯМР 37С112, 18
ЯМР39К, 41К208, 239
ЯМР 43Са 12
ЯМР-импульс, см. импульс
ЯМР-микроскопия 200
ЯМР-спектрометр 48
-томография 12, 24, 50, 165, 214
ЯМР-спектроскопия в твердых телах 12, 16, 29, 34, 144
ЯМР-спектроскопия in-vivo 207 - 239
- 13С 230
-19F237
-’Н218
- 2Н 239
-39К239
- 15N 237
- 23Na 239
-31Р222
ЯЭО (ядерный эффект Оверхаузера) 80, 93, 115, 134, 141, 151, 231
- стационарный 116,237
-средний 141
-слабый 141
- нестационарный 116, 237
— с насыщением, при переносе поляризации 84
Явление ядерного резонанса 14
Ядерная спиновая томография, см. ЯМР-томография
Ядерное квадрупольное взаимодействие 33, 40, 160, 237
Ядерно-электронное взаимодействие, см. сверхтонкое взаимодействие
Ядерный квадрупольный резонанс 33
Ядерный резонанс, см. ЯМР
Ядерный спин 14
Ядерный эффект Оверхаузера, см. ЯЭО
3M-FISP 152
3M-FLASH 152
BPTI 86, 93, 97, 120
Bryophyllum tubiflore 174
CAMELSPIN 94, 76
CHESS-томография 164
COSY 87, 130, 150, 155
CSI - см. изображение, восстановленное по химическому сдвигу
DEPT 207
DFMO 207
EXORSIST 148
FISP 150-152
257
FLASH 150-152
Faktor III 86, 90
Gd-DTPA 100, 171
НРг-протеин 32, 67, 72, 86, 90, 110, 116, 119
INEPT 200, 203
J-взаимодействие 16, 24, 41, 45, 57, 74, 111, 117, 128, 132, 189, 200, 205
LOCUS 214
MAS, см. вращение под магическим углом
MAST 173
MLEV 65
N-ацетиласпаргиновая кислота 221
NOESY89, 93, 115, 135, 152
PF3, см. трифторид фосфора 30
Pi, см. фосфат неорганический
RARE 177
RCT89, 92, 131, 152
ROESY 94
SECSY 92
TOE 81, 84
WAHUHA 129
WALTZ 65
cw-ЯМР 27, 35, 42, 49
- спектрометр 33
pH-зависимость 53, 58, 75, ПО, 224, 227
258
Наукове видання
Хауссер Карл X.,
Кальбкцер Ханс Р.
ЯМР в медицин! та бюлогн
(росшською мовою)
Оформления художника Сергеева Г.О.
Здано до наборуЮ.11.93. Пщп. до друку 29.12.93. Формат 60x84/16. Обл.-вид. арк. 14,5.
Офс. друк. Ум.-друк. арк.16,1. Ум. фарбо-вщб. 15,8. Зам. 3076. Тираж 1000
Видавництво "Паукова думка",
252601, КиТв-4, вул. Терещенювська, 3.
Виробничо-видавничий комбшат "Патент", 294013 м.Ужгород, вул. Гагар1на, 101
Отсканировал Семенкяенко Владимир
chem_vova@mail.univ.kiev.ua; vova2002@mail.ru
К.Х.Хауссер, Х.Р.Кальбитцер
ЯМР
в медицине
и биологии:
структура молекул,
томография,
спектроскопия in-vivo
НАУКОВА
ДУМКА
Карл X. Хауссер является профессором
Хайдельбергского университета, членом
Общества Макса Планка и рада международ-
ных научных организаций. Степень доктора
философии получил в Тюбингенском
университете в 1950 году. Имеет многочислен-
ные работы в области ЯМР, ЭПР и динамичес-
кой поляризации адер.
Ханс Р. Кальбитцер изучал медицину и
физику в университетах Геттингена и Хайдель-
берга. Имеет докторские степени по медицине
(1976 г.) и физике (1981 г.). Является профессо-
ром Хайдельбергского университета. Область
научных интересов — ЯМР- и ЭПР-исследования
структуры и динамики биомолекул.