/



















































































































































































































































































































































































































































































































































Author: Лавренко Е.М. Сочава В.Б.
Tags: ссср ан ссср растительный покров геоботаническая карта региональный вариант типологические разделы
Year: 1956




Text
ЛСЛВЕМЫЕ SCIENTMRUM URSS
INSTITUTUM BOT4NICUM ROMINE V. L.KONMROVII
DESCRIPTIO
VEGETATIONI S URSS
A D « U R S S ТА В U LA M GEOBOTANICAM » m . 1:4 000 000
ANNOTATIONS S
REMCTOHIBUS
E. M.LAVRENKO etV. B.SOCZAVA
II
£.
£
EDITIO ЛСЛПЕМ1ЛЕ SCIENTL4RUM URSS
AZ О SQ 17Л -L. E С ЛЛ />
195b
ЛКЛДЕМИЯ НЛУКСССР Ботанический институт имени БЛ.Комарова
РАСТИТЕЛЬНЫЙ ПОКРОВ СССР
ПОЯСНИТЕЛЬНЫЙ ТЕКСТ К «ГЕОБОТЛНИЧЕСКО.Й КЛРТЕ СССР»
м. 4:4 000 000
ПОД РЕДАКЦИЕЙ
Е.М. ЛАВ РЕН КО и В.Б. СОЧА ВЫ
II
к
ИЗДАТЕЛЬСТВО АКАДЕ М ИИ НАУК СССР МОСКБЛ-ЛЕНИ НГРАл]
Ответственный редактор
Е. М. ЛАВРЕНКО
Секретари редакции
В. В. ЛИПАТОВА и А. Н. ЛУКИЧЕВА
СОДЕРЖАНИЕ II ЧАСТИ
Стр.
Предисловие ............................................................ 466
Горные плодовые леса и горные кустарники. — Я. И. Рубцов ........ 467
Луга и травяные болота. — Л. А. Соколова, Е. В. Шиффере, Л. Е. Родин,
А. Н. Лукичева .................................................... 475
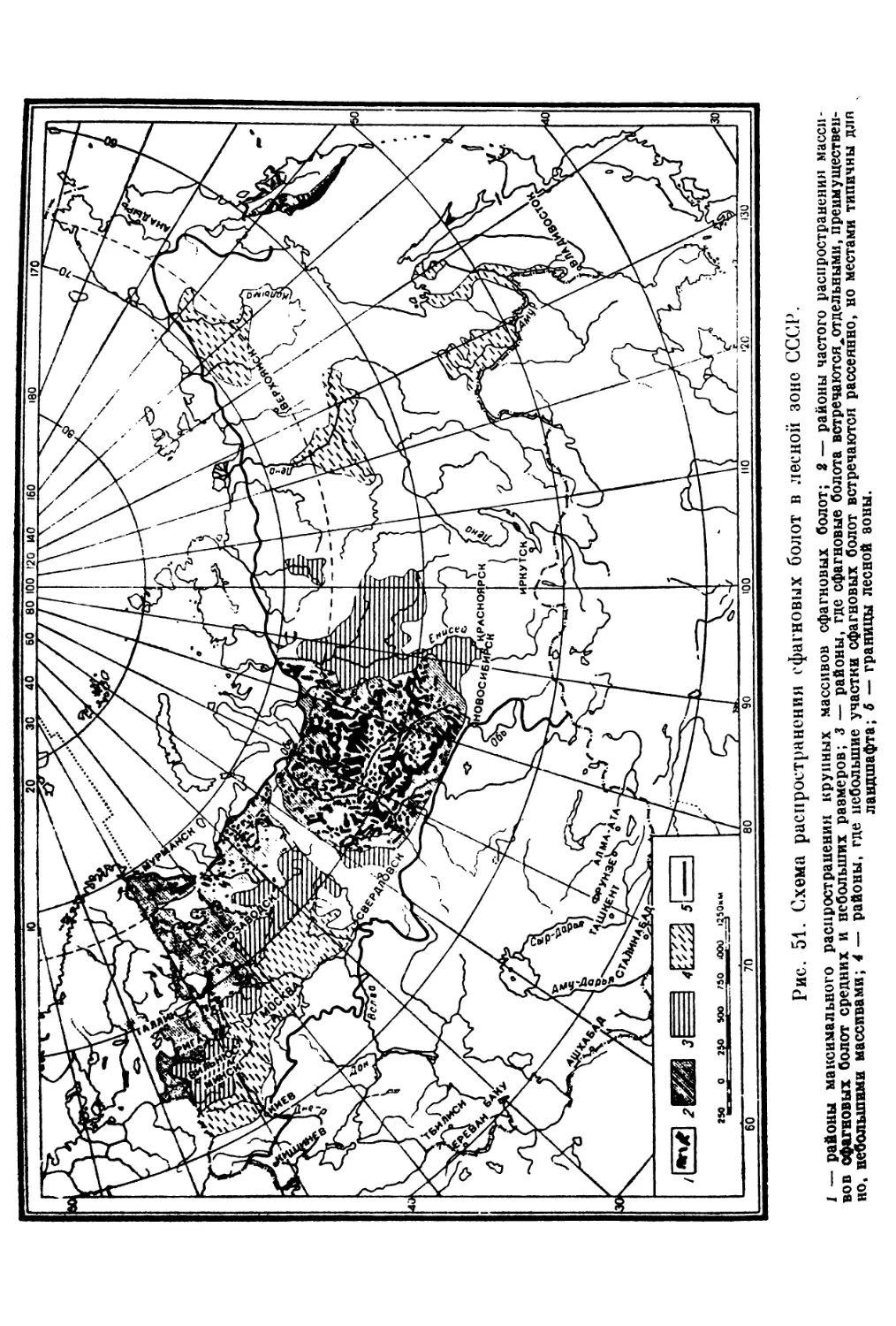
Сфагновые болота. — Е. А. Галкина ...................................... 653
Ксерофитные редколесья, нагорные ксерофиты и субтропические степи. — Н. И. Рубцов ....................................................... 573
Степи и сельскохозяйственные земли на месте степей. — Е. М. Лавренко . 595
Полукустарничковые полынные и солянковые пустыни. — Л. Е. Родин и
Н. И. Рубцов ...................................................... 731
Саксаульники и кустарниковые пустыни. — Л. Е. Родин ......... 797
Высокогорная подушковидная растительность. —Н. И. Рубцов и О. В. Заленский 825
Слоевищная растительность такыров. — Л. Е. Родин ....................... 829
Ледники и снежники. — М. М. Голлербах и В. В. Сочава ................... 835
Разреженная растительность гор (степной и пустынной зон). — Е. М. Лавренко и Л. Е. Родин....................................................... 840
Оазисы в пустынной зоне. — Е. П. Коровин ............................... 848
Литературные источники пояснительного текста к «Геоботанической карте
СССР» м. 1 : 4 000 000............................................. 859
Картографические источники «Геоботанической карты СССР» м. 1 : 4 000000 . 918
Легенда «Геоботанической карты СССР» м. 1 : 4 000 000 и указатель авторов пояснительного текста к карте ...................................... 918
Указатель латинских названий растений .................................. 927
ПРЕДИСЛОВИЕ Е
Второй частью коллективного труда «Растительный покров СССР» заканчивается публикация пояснительного текста к «Геоботанической карте СССР» м. 1 : 4 000 000, вышедшей в свет в 1955 г.
Вторая часть посвящена описанию горных плодовых лесов, лугов, травяных и сфагновых болот, ксерофитных редколесий, нагорных ксерофитов, степей, пустынь, высокогорной подушковидной растительности, растительности такыров и др.
Следует иметь в виду, что в пояснительном тексте к «Геоботанической карте» (как в первой, так и во второй части текста) характеризуются только основные обозначения, помеченные на карте цифровыми и буквенными шифрами. Дополнительные внемасштабные знаки отдельно не описываются; соответствующие сведения о дополнительных знаках нужно искать в описаниях тех основных обозначений (с цифровым и буквопным шифром), на фоне котовых показаны те или иные внемасштабные знаки.
В этом же томе публикуется список литературы к обеим частям, а также алфавитный указатель латинских названий растений, упомянутых в первой и второй 'частях «Растительного покрова СССР».
РЕДАКЦИЯ
VIII. ГОРНЫЕ ПЛОДОВЫЕ ЛЕСА И ГОРНЫЕ КУСТАРНИКИ
н. и. рубцов 1
Общий обзор
Дикорастущие горные плодовые леса и горные кустарники в виде отдельных массивов показаны на «Геоботанической карте» в южных горных системах СССР: в западных и южных предгорьях Алтая, в Тарба-гатае, в Западном Тянь-Шане, западном Памиро-Алае, Закавказье. Дополнительными внемасштабными знаками горные кустарники, а отчасти и горные плодовые леса, отмечены и в других горных системах юга СССР.
Горные плодовые леса, показанные на карте только в горах Средней Азии, представлены ореховыми лесами из грецкого ореха {Juglans regia) 1 2 и крупноплодных яблонь, главным образом киргизской {Maias kirghisorum). Грецкий орех является гималайско-переднеазиатским видом. В горах Западного Тянь-Шаня находятся, видимо, наиболее крупные массивы его естественных насаждений. Кроме последних, на «Геоботанической карте» внемасштабным знаком грецкого ореха показаны более мелкие массивы в западной части Памиро-Алая.
Киргизская яблоня, встречающаяся вместе с грецким орехом и образующая самостоятельные редкостойные насаждения, является западно-тяньшанским эндемом. Близкая к ней Сиверсова яблоня {Matus Siever-sii) образует также редкостойные насаждения в Джунгарском Алатау и Северном Тянь-Шане, что отмечено на карте внемасштабными знаками; это джунгарско-северотяныпанский вид. Наконец, в западном Памиро-Алае (к югу от Гиссарского хребта) встречается третий близкий вид — гиссарская яблоня {Malus hissarica).3
Совместно с грецким орехом и яблонями произрастает также туркестанский клен {Acer turkestanicum), являющийся среднеазиатским видом и образующий иногда самостоятельные насаждения у верхнего предела древесных пород.
Несмотря на большую удаленность от широколиственных лесов Европы, Закавказья и Малой Азии, в ореховых лесах Средней Азии встречаются некоторые типичные представители травяного покрова европейских широколиственных лесов: Brachypodium silvaticum (основное растение травяного покрова ореховых лесов) и Bromus Benekeni. Помимо весенних эфемероидов, обильны гемиэфемероиды, вегетация которых затягивается до начала лета {Allium aflatunense, Conioselinum latifolium, Ferula foliosa и др.).
1 H. И. Рубцовым составлены описания горных ореховых лесов (55) и горных кустарников|(56); общий обзор написан Е. М. Лавренко.
8 Включая Juglans fallax.
3 В настоящее время описан еще ряд видов дикой яблони из районов Северного и Западного Тянь-Шаня (Васильченко, 1951).
Наличием насаждений из грецкого ореха широколиственные леса Средней Азии резко отличаются от остальных широколиственных лесов в СССР.1
Горные листопадные кустарники [из ксеромезофильных и мезоксеро-фильных (гемиксерофильных) видов Prunus, Rosa, Spiraea, Cerasus, Cotoneaster, Crataegus, Lonicera, Abelia, Paliurus, Punica, Exochorda, Rhus, Zizyphus, Rhamnus, Berberis и др. ] показаны в виде отдельных массивов на «Геоботанической карте» в западных и южных предгорьях Алтая, в условиях низко- и среднегорного рельефа, в среднегорном поясе Тарба-гатая и гор Средней Азии и в преимущественно низкогорном поясе Закавказья. Кроме того, внемасштабным знаком шибляка эти кустарники отмечены на фоне степной растительности на Керченском и Таманском п-вах и местами в предгорьях Главного Кавказского хребта.
Вместе с зарослями степных кустарников {Spiraea hyperictfolia, S. crenata, Prunus stepposa, Amygdalus папа, Cerasus fruticosa, Caragana frutex и др.)1 2 горные кустарники, как указано далее, могут быть объединены под общим наименованием шибляка.3 От зарослей листопадных кустарников более северных лесных областей горные листопадные кустарники аридных областей (шибляк) отличаются не только большей или меньшей ксерофильностью господствующих в них видов кустарников, но и своеобразием их состава. В составе горных кустарников Кавказа и Средней Азии встречаются представители родов Paliurus, Punica, Exochorda, Rhus, Zizyphus, Abelia, отсутствующих в более северных лесных районах СССР; виды Cerasus, представленные в горах Средней Азии и Кавказа, относятся к ксерофитной секции Amygdalocerasus и т. д.
Вопрос о первичности горных кустарников (их зарослей) должен решаться дифференцированно. Несомненно, что значительная часть зарослей кустарников низкогорного и среднегорного поясов в горах западного Алтая, Средней Азии и Кавказа является вторичной, возникшей на месте уничтоженных лесов или редколесий, но часть более ксерофитных кустарников, как и заросли степных кустарников в южных частях степной зоны (области), является первичной.
Литература
М. Г. Попов (1929); Е. П. Коровин (1934); И. Т. Васильченко (1951); А. В. Калинина (1951); П. Н. Овчинников (1955); L. Adamovic (1901, 1906, 1909).
55. Горные ореховые леса, местами в сочетании с яблоневыми и кленовыми лесами
Ореховые леса из Juglans regia свойственны среднему поясу гор Средней Азии. Крупные (показанные на карте) массивы этих лесов сосредоточены на горных склонах Чаткальского и Ферганского хребтов. Более мелкими участками и фрагментами они встречаются в Пскемском и Угамском хребтах Западного Тянь-Шаня, в Гиссарском хребте и в Копет-Даге. В Северном и Центральном Тянь-Шане ореховых лесов нет.
1 На Дальнем Востоке встречается маньчжурский орех {Juglans manshurica), связанный исключительно с пойменными лесами (уремой).
2 Не следует смешивать степные кустарники, то есть их более или менее сомкнутые заросли, с кустарниковыми степями (опис. 76, 77, 78), для которых характерно' диффузное распределение тех же кустарниковых видов среди степного травяного покрова.
з Как известно, Л. Адамович (AdamoviC, 1901, 1906, 1909) к шибляку относил кустарниковые листопадные сообщества с господством Rhus coriaria, Zizyphus jujubaf Paliurus spina-Christi, Cotinus coggygria, Cerasus fruticosa, Amygdalus папа и других видов.
Ореховые леса Западного Тянь-Шаня и Гиссарского хребта лежат, примерно, в пределах высот от 1000 до 2200—2300 м над у. м., где среднегодовое количество осадков достигает иногда 1000 мм и более. Почвы под этими лесами мощные, богатые гумусом, на лёссах. Они известны под названием буроземов, причем некоторые почвоведы даже сближают эти почвы с буроземами буковых лесов Кавказа.
В последнее время эти почвы выделяются под именем черно-бурых лесных почв (Герасимов и Ливеровский, 1947).
Обычно ореховые леса представлены как чистыми, так и смешанными насаждениями; они приурочены к склонам северных экспозиций или к невысоким платообразным водоразделам, защищенным от холодных ветров горными поднятиями. Чистые насаждения грецкого ореха занимают довольно большие площади лишь в Ферганском хребте, где этот тип леса характерен для нижних, относительно пологих частей северных или западных склонов. В других горных районах чистые ореховые леса развиты в гораздо меньшей мере. Орешники указанного типа характеризуются хорошим ростом древостоя (I и II бонитеты), простотой строения и большой сомкнутостью крон (до 0.7—0.8 и более). Благодаря затенению древесным пологом, подлесок развивается довольно слабо. В подлеске изредка встречаются алыча (Prunus sogdiana), яблоня (Malus kirghisorum), жимолость (Lonicera Korolkovii, L. persica), жестер (Rhamnus cathartica). В травяном ярусе на более освещенных местах обычно доминирует лесная коротконожка (Brachypodium silvaticum), а в затененных — недотрога (Impatiens parviflora). Из других обычных компонентов орешников широко распространены некоторые зонтичные: Aegopodium tadshikorum, Archangelica decurrens, Conioselinum latifolium и др.
Смешанные кленово-ореховые и яблонево-ореховые леса приурочены к более освещенным склонам и развиваются на почвах меньшей мощности,, чем чистые орешники. В большинстве случаев это довольно изреженные, светлые насаждения, в которых вместе с грецким орехом обильно представлены клен (Acer turkestanicum) и яблоня (Malus kirghisorum), а иногда (в поймах) ясень (Fraxinus potamophila). На склонах Чаткальского хребта ореховые леса у верхнего предела своего распространения смешиваются с тяныпанской елью (Pice a Schrenkiana) и пихтой (Abies Semenovii), образуя елово-ореховые и пихтово-ореховые насаждения (19 тш).
Для всех указанных типов смешанных насаждений характерен богатый подлесок из алычи (Prunus sogdiana), жимолости (LoniceraKorolkovii, L. persica, L. Karelinii), боярышника (Crataegus turkestanica), жестера (Rhamnus cathartica), бересклета (Euonymus Semenovii), некоторых видов роз (например Rosa kokanica), кизильника (виды рода Cotoneaster). В припойменных ореховых лесах УгамаиПскема встречается виноград (Vitisvinifera).
На более сухих склонах яблоня и клен образуют небольшие участки самостоятельных яблоневых и кленовых лесков, которые сопровождаются рядом перечисленных выше кустарников. На этих же склонах в сочетании с яблонево-кленовыми лесками развиваются травяные сообщества из пушистого пырея (Agropyrum trichophorum) и луковичного ячменя (Hordeum bulbosum).
Для отмеченных на карте внемасштабным знаком яблоневых лесов Джунгарского и Заилийского Алатау, находящихся уже вне ареала грецкого ореха, характерна постоянная примесь боярышника (Crataegus altaica, С. songorica), тяныпанской осины (Populus pseudotremula), иногда урюка (Armeniaca vulgaris). Подлесок и здесь представлен кустарниками (Berberis heteropoda, Cotoneaster multiflora, C. melanocarpa, Rhamnus cathartica). В травяном покрове преобладают луговые и луговостепные виды:.
•ежа (Dactylis glomerata), крупнолистная лигулярия (Ligularia persica), душица (Origanum vulgare), володушка (Bupleurum аигеит), василистник (Thalictrum minus) и т. п. Ни пушистого пырея, ни луковичного ячменя, столь характерных спутников яблоневых лесов более южных гор Средней Азии, здесь уже совсем нет. Они замещаются типичными луговыми и степными бореальными элементами.
Плодовые леса имеют большое научное и практическое значение. Леса из грецкого ореха представляют большой научный интерес прежде всего как реликтовая формация — остаток древних (третичных) субтропических лесов Средней Азии. Принадлежность яблоневых лесов Северного Тянь-Шаня к реликтовым лесным типам тоже не вызывает сомнений. Об этом свидетельствуют явная фрагментированность их ареала, характер видового состава, географическая и флористическая связь с реликтовыми ореховыми лесами Западного Тянь-Шаня.
Народнохозяйственное значение ореховых и яблоневых лесов велико. Они играют весьма большую роль в качестве почвозащитного и водоохранного фактора, являются источником ценных пищевых продуктов (орехов, яблок) и ценной древесины. По данным С. Я. Соколова,1 один гектар орехового леса дает ежегодно урожай плодов от 120 до 350 кг. П. С. Чабан (1941) для ореховых лесов I и II бонитетов в Западном Тянь-Шане (Бостан-дыкский р-н) указывает гораздо более высокую урожайность — до 600— 800 кг с гектара. По данным этого же автора, максимум урожайности приходится на насаждение с полнотой 0.5, возраста 30—35 лет.
Грецкие орехи широко применяются в свежем (сыром) виде, в кондитерском производстве и при изготовлении высокопитательного масла. Красивая древесина орехового дерева употребляется в мебельном и фанерном производстве, а также для различных поделок. Обладающая особенно красивым рисунком древесина орехового наплыва («капа») применяется для различных архитектурно-художественных отделок. Плоды дикорастущих видов яблонь (Malus Sieversii, М. kirghisorum) используются в качестве сырья для местной пищевой промышленности.
Следует отметить, что, несмотря на крупную народнохозяйственную роль плодовых лесов Средней Азии, за ними все еще нет надлежащего ухода. Особенно это относится к яблоневым лесам Северного Тянь-Шаня, в которых до сих пор допускаются выпас скота, незаконная рубка и т. п. Организация тщательного ухода за плодовыми лесами, создание лесопосадок и специальных лесоплодовых совхозов, осуществляемые в последнее время, должны обеспечить не только сохранение ныне существующих лесных площадей из ценных дикорастущих плодовых пород, но и значительное их расширение.
Литература
С. И. Коржинский (1896); О. Э. Кнорринг и 3. А. Минквиц (1912); О. Э. Кнорринг (1915); П. А. Баранов (1924); М. Г. Попов (1929, 1934); Н. Н. Дзенс-Литовская (1930); Е. П. Коровин (1934); С. Я. Соколов (1936а); П. А. Красовский, С. Я. Соколов и Л. И. Соснин (1938); Т. Т. Трофимов (1940); П. С. Чабан (1941); И. П. Герасимов и Ю. А. Ливеровский (1947); П. П. Поляков (1948); Е. М. Лавренко и С. Я. Соколов (1949); И. Т. Васильченко (1951).
56. Горные кустарники
На «Геоботанической карте» под этим общим названием выделены контуры, объединяющие в горах растительность с преобладанием различных кустарников, образующих самостоятельные заросли или играющих крупную роль в сложении травяных сообществ (лугов и степей).
1 Соколов С. Я. 1951. Род Juglans L. В кн.: Деревья и кустарники СССР, 2, М.—Л.
Кустарниковая растительность в целом, безусловно, не представляет собою какого-либо единого ценотипа. Под этим сборным понятием объединены сообщества, эдификаторы которых относятся к кустарниковой биоморфе. Более дробное расчленение сборного кустарникового типа растительности все еще нельзя считать определившимся. Типология относящихся сюда ассоциаций представляет нередко значительные трудности вследствие слабой изученности эколого-биологических особенностей многих широко распространенных кустарников.
В ряде случаев кустарниковые сообщества являются вторичными, они возникают в результате уничтожения лесной растительности рубкой, пожарами, выпасом скота.
56 а — заросли мезофильных и полуксерофильных кустарников (шиповника, жимолости, таволги) в сочетании с луговыми степями (предгорья Алтая, горы Тарбагатая и Саура)
Под этим знаком на карте выделены заросли мезофильных и полуксерофильных кустарников, свойственные южному Алтаю, Тарбагатаю и Сауру, где кустарниковые заросли постоянно сочетаются со степными или луговыми группировками.
В юго-западном Алтае по нижнему и среднему поясу (500—1500 м над у. м.) основными эдификаторами кустарниковых сообществ являются следующие виды: шиповник (Rosa spinosissima), таволга (Spiraea hyperi-eifolia), жимолость (Lonicera tatarica), миндаль (Amygdalus папа). Постоянные спутники этих видов — луговые и степные злаки: ежа (Dactylisglomerata), коротконожка (Brachypodium pinnatum), степная тимофеевка (Phleum phleoides), типчак (Festuca sulcata), тонконог (Koeleria gracilis), вместе с которыми развивается обильное луговое и степное разнотравье. Почвы темнокаштановые или черноземы. По южным щебневатым склонам и каменистым сопкам преобладают заросли таволги, караганы (Caragana frutex) и кизильника (Cotoneaster) в сочетании с ковыльно-типчаковыми степями (Stipa capillata, S. Joannis, Festuca sulcata). По более пологим склонам северных экспозиций господствуют заросли шиповника, в сочетании с лугами из Dactylis glomerata или с луговыми степями из Phleum phleoides, Festuca sulcata, Origanum vulgare и т. д. Значительная часть степной растительности южного Алтая распахивается под зерновые и технические культуры.
Видовой состав и сложение кустарниковой растительности Саура весьма сходны с тем, что наблюдается в южном Алтае.
В Тарбагатайском хребте кустарниковые формации очень характерны для луговостепного пояса, в пределах от 1000—1200 до 1700 м над у. м. На крутых северных склонах они представлены густыми зарослями ши-повников с преобладанием Rosa spinosissima и небольшой примесью Rosa acicularis, R. Beggeriana. Среди травянистых спутников этих зарослей доминируют лесо-луговые элементы (Dactylis glomerata, Milium effusum, Brachypodium pinnatum, Vicia tenuifolia, Adenophora liliifolia, Geum ur-banum и др.). По менее крутым северным склонам в зарослях шиповников постоянно встречаются и другие мезофильные кустарники: жимолость (Lonicera tatarica), волчеягодник (Daphne altaica), смородина (Ribes Меу-eri), жестер (Rhamnus cathartica).
Предгорья южного склона Тарбагатая, в пределах от 600 до 1000— 1200 м над у. м., заняты кустарниковостепным поясом. Здесь широко распространены разнотравно-ковыльные степи с большим участием караганы
(Caragana frutex). Кустарниковые заросли данного пояса, занимающие большие площади, представлены главным образом миндальниками из Amygdalus Ledebouriana, среди которых встречаются таволга и карагана. Травяной ярус миндальников слагается луговыми и степными видами (Dactylis glomerata, Bromus inermis, Calamagrostis epigeios, Stipa capil-lata, Medicago falcata, Origanum vulgare, Ferula songorica, Verbascum son-goricum, Dipsacus azureus, Astragalus Sieversianus). Необходимо отметить, что среди миндальников постоянно встречается эндемичный для Тарбага-тая вид житняка (Agropyrum tarbagataicum).1
Районы распространения кустарниковых зарослей в Тарбагатае представляют собой пахотные и пастбищно-сенокосные угодья.
Литература
Е. П. Матвеева (1940, 1951); Н. А. Плотников (1941—1946); Е. Ф. Степанова (1950).
56 к — заросли полуксерофильныхкустарников (держидерева, таволги, гранатника, барбариса, боярышника и др.) (Кавказ)
Кустарниковые заросли Кавказа, в отличие от таковых на Алтае, Сауре и Тарбагатае, слагаются преимущественно гемиксерофильными и ксерофильными листопадными кустарниками. Подобного рода сообщества известны в литературе под именем шибляка. Понятие о шибляке впервые было установлено Л. Адамовичем (Adamovid, 1901, 1906, 1909) для Балканского п-ва. Впоследствии оно было перенесено в нашу отечественную ботанико-географическую литературу главным образом кавказскими ботаниками, широко применяющими и ныне это понятие по отношению к ксерофитным листопадным кустарниковым сообществам Кавказа.
Шибляк Кавказа приурочен к нижним и средним поясам гор, от подгорных равнин до 1500 м над у. м. Однако основными районами его развития являются главным образом предгорья в пределах (600—) 700—1000 м над у. м. На «Геоботанической карте» массивы этих кустарников показаны в Закавказье по предгорьям Карабахской системы и Щекинского нагорья, а также по предгорьям к западу от Тбилиси. Кроме того, значительные участки его выделены на юге Армении. Более мелкими участками шибляк широко распространен в целом ряде других районов Кавказа.
Основной и самой характерной формацией кавказского шибляка являются заросли чрезвычайно колючего кустарника — палиуруса или держи-дерева (Paliurus spina-Christi), развивающиеся большей частью на местах, бывших под лесом. Таким образом, как правило, палиурусо-вый шибляк — явление вторичное. Однако М. Ф. Сахокиа (1950) полагает, что значительная часть современных шибляковых сообществ представляет собою синузиальный остаток бывших лиственных редколесий с кустарниковым ярусом. Он указывает на существование и первичного
1 Этот высокий и широколистный мезофитного облика злак в качестве нового для науки вида описан недавно Н. А. Плотниковым (1941—1946). Тарбагатайский житняк весьма перспективен для культуры в качестве кормового растения. Кроме миндальников, он характерен также для разнотравно-ковыльных степей западной части предгорий южного склона Тарбагатая, слагаемых Stipa Lessingiana, S. capil-lata, Phleum phleoides и богатым луговостепным разнотравьем. В составе этих степей Amygdalut Ledebouriana также принимает значительное участие.
шибляка из Paliurus spina-Christi в безлесных районах Южного Закавказья и Дагестана.
Палиурусовый шибляк слагается целой серией ассоциаций. Наиболее распространенными являются ассоциации, в которых согосподствующим видом в травяном покрове является рыхлокустовой злак — бородач (Andropogon ischaemum).
Такой бородачево-палиурусовый шибляк особенно характерен для Восточного Закавказья. Отмеченные на карте массивы шибляка в большинстве случаев относятся именно к этому его варианту (группе ассоциаций). В составе палиурусников иногда встречаются грузинский дуб (Quercus iberica) и грабинник (Carpinus orientalis, С. caucasica), что отображено на карте соответствующим внемасштабным знаком. Необходимо отметить, что в Восточном Закавказье грабинниково-дубовые леса на нижней границе своего распространения обычно контактируют с шибляком и, в частности, с бородачевыми палиурусниками.
Из других шибляковых формаций, более или менее распространенных на Кавказе, отметим гранатники (из Punica granatum) и барбарисники (из Berberis iberica), развивающиеся по каменистым склонам сухих ущелий и по шлейфам гор, иногда в сочетании с тем же палиурусом. По мягким северным склонам предгорий и среднего пояса гор формируются тавол-жники (из Spiraea hypericifoliav. S. crenata) с участием луговостепных травяных элементов (виды Stipa, Festuca, Brachypodium). В этих же условиях встречаются заросли боярышников (Crataegus Meyeri, С. pentagyna, С. kyrtostyla) и других кустарников (Cornus mas, Cotoneaster racemiflora, Paliurus spina-Christi). Во внутреннем Дагестане и Закавказье по каменистым склонам предгорий развиты шибляковые сообщества из таких резко ксерофильных видов кустарников, как жестер Палласа (Rhamnus Pallasii), миндали (Amygdalus Fenzliana, A. zangezura, A. nairica), вишни (Cerasus incana, C. araxina,) эфедра (Ephedra procera) и т. д.
Площади, покрытые шибляком, при расчистке от кустарников могут быть использованы для плодоводства и виноградарства, а также в качестве сенокосных и пастбищных угодий. Однако при этом следует иметь в виду ,и большую почвозащитную (противоэрозионную) роль шибляковых зарослей, особенно на крутых склонах.
Литература
Н. А. Буш (1935а); А. А. Гроссгейм (1948); М. Ф. Сахокиа (1950); L. Adamovifi (1901, 1906, 1909).
56 са — заросли полуксерофильных и мезофильных кустарников (шиповника, экзохорды, боярышника и др.) в сочетании с яблоневыми редколесьями (Средняя Азия)
Кустарниковые сообщества гор Средней Азии, так же как и на Кавказе, слагаются преимущественно гемиксерофильными, а иногда и типичными ксерофильными листопадными кустарниками, и поэтому вполне могут быть причислены к категории шибляка.
Ксеромезофитным вариантом среднеазиатских кустарниковых сообществ являются розарии, то есть заросли различных ксеромезофильных шиповников (Rosa platyacantha, R. Есае, R. maracandica, R. kokanica). Они приурочены обычно к склонам северных экспозиций, но встречаются иногда и на южных склонах, если последние в достаточной степени покрыты мелкоземом.
В Северном Тянь-Шане весьма характерны розарии из желтоцветного плоскошипного шиповника {Rosa platyacantha). Южнее, в горных хребтах Западного Тянь-Шаня и Алая, розарии слагаются уже другими видами (главным образом Rosa Есае, R. maracandica). Они сопровождаются обычно травяной луговостепной растительностью {Dactylis glomerata, Origanum vulgare, О. tyttanthum, Brachypodium pinnatum, Conioselinum latifolium, Poa angustifolia, Calamagrostis epigeios, Festuca sulcata, Galium verum и т. п.).
В южном Таджикистане в средних поясах гор широко развиты розарии (из Rosa kokanica), в сочетании с которыми иногда встречаются экзо-хордники (из Exochorda Alberti). В этих зарослях постоянно присутствуют и другие кустарники {Amygdalus ulmifolia, Caragana turkestanica, виды Cotoneaster), к которым нередко присоединяются клен {Acer Regelii, A. turkestanicum) и боярышник {Crataegus pontica). Сопутствующая указанным кустарниковым сообществам травяная растительность представлена или крупным эфемероидным разнотравьем с преобладанием югана {Prangos pabularia) и комоля {Ferula Jaeschkeana), или разнотравьем луговостепного типа (из Dactylis glomerata, Vicia tenuifolia, Galium ve-rum, Glycyrrhiza glabra и др.).
Типичные, наиболее ксерофитные варианты среднеазиатского шибляка состоят из зарослей миндаля {Amygdalus spinosissima, A. bucharica). В Западном Тянь-Шане миндальники представлены Amygdalus spinosissima. Они формируются по южным щебнисто-каменистым склонам и сопровождаются травами эфемерного типа {Hordeum bulbosum, Bromus tectorum), а также полынью {Artemisia tenuisecta). Миндальники из Amygdalus bucharica свойственны главным образом южному Таджикистану, среднегорному поясу, грубым скелетным почвам. Наряду с эдификатором в них участвуют Rosa maracandica и Colutea canescens', травяной ярус слагают эфемероидные и эфемерные злаки {Hordeum bulbosum, Aegilops triuncialis, Taeniatherum crinitum, Bromus tectorum) и такое же разнотравье {Ferula Jaeschkeana, Prangos pabularia). В сочетании с этими миндальниками встречаются сумапшики (из Rhus cariaria), заросли зизифуса {Zizyphus jujuba), вишен {Cerasus sp. sp.) и других ксерофильных кустарников.
В довольно значительных контурах горных кустарников, отмененных, на «Геоботанической карте» в пределах южного Таджикистана, объединены заросли как ксеромезофильных, так и типичных ксерофильных кустарников, перечисленных выше, а также их разнообразные сочетания.
Более мелкие контуры, относящиеся к Западному Тянь-Шаню, также-представляют собою объединение ксеромезофитных и ксерофитных кустарниковых формаций, слагающихся в основном из различных видов шиповников, таволги, миндаля, иногда с примесью деревьев {Crataegus pontica, С. songorica, Acer Semenovii и др.). По склонам северных экспозиций кустарниковые формации нередко сочетаются с яблоневыми лесками из Malus kirghisorum, М. Sieversii. Такие сочетания характерны и для Джунгарского Алатау и Северного Тянь-Шаня (Заилийский Алатау).
Площади, занятые в Средней Азии под кустарниковыми зарослямиг могут быть использованы в качестве пастбищных и сенокосных угодий г а также для плодоводства и виноградарства. Очень велика почво- и водоохранная роль ЭТИХ кустарников
Л итература
С. И. Коржинскин (1896); М. Г. Попов (1929); Е. П. Коровин (1934); Н. Ф. Гон-Ж® <1936’?937)’. К- С‘ Афанасьев (1940); А. С. Королева (1940); Н. И. Рубцов Оачинииков (1948а); Е. М. Лавренко и С. Я. Соколов (1949)' А. В. Калинина (1951). '
IX. ЛУГА И ТРАВЯНЫЕ БОЛОТА
Л. А. СОКОЛОВА, Е. В. ШИФФЕРС, Л. Е. РОДИН, А. Н. ЛУКИ ЧЕВА 1
Общий обзор
В этот раздел включены те сочетания растительных сообществ, в составе которых преобладают ассоциации, принадлежащие к луговому типу растительности или к травяным болотам. К луговому типу растительности в настоящее время относят сообщества с преобладанием мезофильных многолетних травянистых растений (А. П. Щенников). Травяные болота весьма близки к лугам в таком понимании; А. П. Щенников (1938, 1941) и относит их к лугам.
В настоящую группу лугов и травяных болот включены следующие типологические единицы и их сочетания: высокогорные альпийские и субальпийские луга (57), среднегорные луга (58), травяная растительность (лугового типа) океанических островов и полуостровов побережья Тихого океана (59), приморские луга тундровой зоны (60), солончаковолуговая и лугово-болотная растительность морских побережий степной и пустынной зон (61), солончаково-луговая и лугово-болотная растительность разливов, лиманов, высыхающих плавней и побережий озер степной и пустынной зон (62), пойменные луга (63), травяные болота и луга длительно заливаемых пойм степной и пустынной зон — плавни (64),. травяные и гипново-травяные болота (65). Для части этих типологических единиц и их сочетаний с помощью буквенных обозначений ; выделены зональные варианты; это сделано для пойменных лугов (63) и травяных, и гипново-травяных болот (65). Для высокогорных (57) и среднегорных (58) лугов также с помощью буквенных обозначений отмечены их провинциальные варианты, связанные с определенными горными системами.
Растительный покров тех территорий, которые показаны на «Геоботанической карте» вышеупомянутыми обозначениями, в большинстве случаев неоднороден и не ограничивается только теми типологическими единицами, которые упоминаются в текстовой части легенды.
1 Л. А. Соколовой составлены следующие разделы: Приморские луга тундровой зоны (60), Солончаково-луговая и лугово-болотная растительность разливов, лимяпоп и т. д. (62), Пойменные луга (63, кроме 63 п), Травяные болота и луга длительно заливаемых пойм (64) и Травяные и гипново-травяные болота (65); Е. В. Шиффере составлены: Высокогорные альпийские и субальпийские луга (57, кроме 57 кп), Среднегорные луга кавказские (58 к), Солончаково-луговая и лугово-болотная растительность морских побережий (61); Л. Е. Родиным — Пойменные луга пустынной зоны (63 п) и касающиеся Средней Азии части описания Солончаково-луговой и лугово-болотной растительности разливов, лиманов и пр. (62) и Травяных болот и лугов длительно заливаемых поим (64); А. Н. Лукичевой — Травяная растительность океанических островов Тихого океана (59). Кроме того, Е. М. Лавренко составил общий обзор «Луга и травяные болота» и раздел — Среднегорные луга североказахстанские. (58 ск), Н. И. Рубцов — Среднегорные луга среднеазиатские (58 са) и В. Б. Сочава — Высокогорные альпийские и субальпийские луга карпатские (57 кп).
Так, среди высокогорных (57) и среднегорных (58) лугов значительную площадь занимают лишенные сплошного растительного покрова скалы и осыпи; кроме того, среди субальпийских лугов встречаются заросли кустарников, покрывающие часто большие площади, и участки редколесных насаждений, среди среднегорных лугов — заросли кустарников и лесные участки. Среди солончаково-луговой и лугово-болотной растительности морских побережий (61), а также разливов, лиманов, высыхающих плавней, берегов озер степной и пустынной зон (62) значительные площади заняты степной и пустынной растительностью; последняя представлена полукустарничковыми сообществами солянок и отчасти некоторых видов полыней. В поймах рек (63), помимо пойменных лугов, большие площади занимают заросли кустарников и пойменных лесов (урема), а также старицы рек с водной растительностью. В плавнях низовий рек •степной и пустынной зон (64) леса и кустарники занимают небольшую площадь, но стариц здесь также много.
Часть лугов и травяных болот, показанных на карте, является первичной, а другая часть — вторичной.
Так, к первичным лугам относятся: высокогорные альпийские луга, приморские луга тундровой зоны, галофитные (солончаковатые и солончаковые) луга и галофитные травяные болота морских побережий, а также разливов, лиманов, высыхающих плавней и берегов озер, длительно заливаемые луга и травяные болота в поймах рек, в том числе в плавнях, травяные и гипново-травяные болота тундровой зоны и частично полосы широколиственных лесов и лесостепи, а также травяная растительность островов и полуостровов Тихоокеанского побережья. При этом альпийские луга могут рассматриваться как длительно существующий коренной тип растительности в верхней части высокогорного пояса Карпат, Кавказа, гор Средней Азии и западного Алтая. Приморские луга тундровой зоны в случае даже незначительного поднятия местности переходят в зональные тундровые сообщества. Галофитные луга и травяные болота морских побережий, разливов, высыхающих плавней, побережий озер в степной и пустынной зонах в случае накопления тем или иным путем на поверхности почвы отложений (морских, аллювиальных, делювиальных, эоловых) или при поднятии местности над уровнем моря сменяются зональными степными или пустынными сообществами. Длительно заливаемые луга и травяные болота пойм рек, включая плавни степной и пустынной зон, при накоплении аллювиальных осадков или поднятии местности должны сменяться кустарниково-лесной растительностью пойм, различной в разных зонах (в лесной и степной зоне — урема, в пустынной — тугаи).1 Травяные и гипново-травяные болота тундровой зоны, приуроченные к долинам рек и к депрессиям рельефа, при накоплении торфа трансформируются в зональные тундровые сообщества. В полосе широколиственных лесов и лесостепи при накоплении торфа эти болота сменяются чер-ноолыпатниками или заболоченными березняками. Наконец, травяная растительность островов и полуостровов Тихоокеанского побережья в условиях холодного и влажного климата и сильных ветров может рассматриваться как коренная и длительно существующая. При континен-тализации климата она будет сменяться березовым редколесьем с мощным травяным покровом или зарослями кустарников (ольховника и др.).
Большинство субальпийских, среднегорных и пойменных лугов, а также часть травяных и гипново-травяных болот полосы широколи-
1 Наибольшее развитие длительно заливаемые луга получают в пойме Оби, в ее среднем и нижнем течении. Пойма этой реки могла бы быть отнесена к плавням.
ственных лесов и лесостепи являются вторичными, возникшими в резуль-тате хозяйственной деятельности человека. Субальпийские луга возникают вследствие уничтожения зарослей субальпийских кустарников (в том числе стлаников) и редколесий, среднегорные луга — на месте •сведенных лесов или кустарников, пойменные луга также возникают в результате уничтожения в поймах лесов и кустарников; последнее имеет место даже в зоне тундр, где первичным типом растительности в поймах рек являются кустарниковые заросли. Многие травяные и гипново-травяные болота лесной и степной зон возникли на месте заболоченных лесов (черноолыпатников, березняков и пр.) в условиях довольно богатого питания. В случае прекращения сенокоса и выпаса на субальпийских, среднегорных и пойменных лугах последние постепенно зарастают кустарниками, а при благоприятных климатических условиях и лесом.
Чрезвычайно велико хозяйственное значение упомянутых луговых угодий. Пойменные, а также среднегорные и солончаковые луга долин рек и котловин в степной и пустынной зонах дают бблыпую часть собираемого в стране сена. Частично выкашиваются также субальпийские луга и травяные болота. Солончаковые луга и травяные болота (заросли тростника) на юге степной зоны и в северных пустынях являются часто почти единственными здесь природными сенокосными угодьями. Альпийские, субальпийские и среднегорные луга широко используются для выпаса в летний период в условиях отгонного животноводства; многие пойменные, а также солончаковые луга в степной и пустынной зонах не только выкашиваются, но и используются как пастбища. Заросли тростника (Phragmites communis), рогоза (виды Typha) и камыша (высокорослые виды Scirpus) в поймах, а особенно в плавнях низовий рек, являются источником строительного материала.
Многие участки пойменных лугов, солончаковатых лугов в поймах, разливах, озерных котловинах, а также некоторые участки среднегорных лугов на менее крутых склонах распаханы и используются для посевов сельскохозяйственных культур, главным образом огородных, технических, отчасти и зерновых.
Литература
В. Р. Вильямс (1922); А. П. Щенников (1938, 1941); Н. Я. Кац (1941, 1948); С. Н. Тюремнов (1949); Ф. Я. Левина (1955).
57. Высокогорные альпийские и субальпийские луга, местами в сочетании с кустарниками и лесами
Высокогорные луга покрывают значительную часть вершин и склонов хребтов, пологой дугой окаймляющих с юга равнины СССР и вытянутых на громадное расстояние в широтном направлении: от 23° в. д. на западе до 90° в. д. на востоке. Сюда входят Карпаты, Кавказ, Памиро-Алай, Тянь-Шань, Джунгарский Алатау, Тарбагатай, Саур и Алтай. Формации, близкие к высокогорным лугам, встречаются небольшими участками на Урале и некоторых других горных системах. В меридирнальном направлении расстояние между крайними северными и южными точками залегания высокогорных лугов также очень велико: от 63° с. ш. на Урале до 37 ° с. ш. на Памиро-Алае.
При таком большом протяжении по широте и долготе данный растительный комплекс не может быть однообразным флористически и фитоце-
нологически; выдели, обозначенные буквами у, а, кп, к и са, имеют свои значительные отличия и свои эндемы, особенно среди альпийских лугов. И все же этот комплекс целостен на всем протяжении, будучи в основном сложен альпийскими низкотравными и субальпийскими высокотравными лугами, лишь с некоторым участием лишайниковых, мохово-лишайниковых и травяно-лишайниковых пустошей, с одной стороны, и кустарниковых и древесных сообществ, с другой.
В пределы контуров, занятых этим комплексом (57), местами входят также не выделенные по условиям масштаба карты небольшие массивы ледников и снежников. Такие массивы, так же каки более крупные (103), обычно залегают в высокогорьях выше пространства, покрытого горными лугами, и образуют так называемый нивальный (снежный) пояс. Между нивальным поясом и горными лугами часто выделяется субнивальный пояс, недавно освободившийся из-подо льда и снега, почти лишенный растительного покрова и крайне медленно заселяемый растительностью, вследствие суровых климатических условий. Здесь нередко возвышаются голые острогребенчатые хребты и скалистые вершины; большие площади покрыты глыбистыми осыпями и россыпями. Высшие растения не образуют в субнивальном поясе сколько-нибудь сомкнутых группировок. Лишь по щелям и углублениям между глыбами осыпей и по защищенным от холодных ветров ущельям подымаются сюда отдельные экземпляры растений. На Кавказе, например, отмечаются высокогорные злаки (Alopecurus glacialis идр.), камнеломки (Saxifraga flagellaris s. 1. и др.), генцианы (Gentiana Dechyana и др.), крупки (Draba hispida и др.) и некоторые другие травянистые растения. Наиболее распространены здесь лишайники, преимущественно накипные, заходящие вплоть до верхних границ субнивального пояса.
Высокогорные территории с хорошо развитым покровом из психро-мезофильных, большею частью низкорослых трав представляют собой альпийский пояс (пояс альпийских низкотравных лугов). Пояс высокотравных лугов, расположенный ниже, называется обычно субальпийским; он граничит с лесным поясом и иногда включает более или менее значительные участки заходящих сюда снизу лесных фитоценозов.
Альпийский пояс представляет собою территорию, обычно непосредственно граничащую с пространствами, покрытыми льдом и снегом или лишь недавно и неполно от них освободившимися. Здесь характерны остатки ледниковых цирков, троговые долины, моренные гряды и озера. В связи с молодостью субстрата, обнажившегося из-под ледников и часто вновь обнажающегося вследствие осыпания выветрившихся слоев, а также в связи с суровостью климатических условий высокогорий (низкие температуры, сильные сухие и холодные ветры, кратковременность вегетационного периода, резкая разница между температурами дня и ночи и т. п.), процесс завоевания субстрата растительным покровом, хотя и доходит до образования плотнодернинных травяных ассоциаций, однако все же идет медленно.
Вслед за первыми пионерами биологического выветривания горных пород (бактериями и водорослями) появляются лишайники и мхи. На моренах в верхних частях альпийского пояса они покрывают значительные площади, образуя так называемые лишайниковые и мохово-лишайниковые пустоши. С появлением среди них некоторых травянистых растений, преимущественно кобрезий (Cobresia capillifolid), а также кустарничков и полукустарничков (карликовой ивы — Salix arbuscula, водяники, брусники и др.) развиваются травяно-лишайниковые, травяно-мохово-ли-шайниковые и мохово-лишайниково-кустарничковые пустоши (с видами
Cetraria, Cladonia, Thamnolia, Polytrichum и другими), близкие по составу и строению к сообществам горных тундр (опис. 3).
Мохово-лишайниково-кустарничковые сообщества развиваются и в результате зарастания горных приледниковых озер. В их составе в таком случае обычно участвуют кобрезии (например на Кавказе СоЪ-resia schoenoides), карликовая ива, водяника, брусника, а из злаков белоус (Nardus stricta, N. glabriculmis) и луговик (Deschampsia flexuosa). Промежуточными стадиями развития растительного покрова являются при этом моховые и мохово-осоковые болотца, широко, но мелкими массивами распространенные в альпийском поясе. На первых стадиях зарастания в приледниковых бассейнах усиленно развивается Aulacomnium palustre, а затем к нему присоединяются некоторые виды сфагнов (Sphagnum Warnstorfii и др.), осоки, пушицы.
Характерными элементами растительного покрова в альпийском поясе являются открытые несомкнутые группировки скал и осыпей. Наряду с пятнами накипных лишайников и вслед за ними, здесь поселяется пестрая свита цветковых растений со своеобразным обликом, выработавшимся под влиянием природных условий высокогорных каменистых местообитаний. В большинстве это очень мелкие, низкорослые, едва возвышающиеся над почвой растения с проникающими в трещины корнями и корневищами, с крупными яркими цветами (виды Gentiana, Campanula, Poten-tilla и др.). Почти все альпийцы — многолетники; у многих травянистых растений нижняя часть стебля у основания одревесневает; много и настоящих полукустарничков; имеются в их составе также луковичные и клубневые (виды Gagea, Allium, Muscari, Crocus и др.). Часто встречаются растения-подушки с многочисленными, тесно к друг другу расположенными стеблями (Draba bryoides, Gypsophila imbricata и др.), а также суккуленты (виды Sempervivum, Sedum). У многих растений развито густое опушение всех частей или восковой налет и толстая кутикула на листьях и стеблях. Низкорослость, прижатость к земле, защищенность от воздействия низких температур, сухости, сильных ветров — все это так же характерно для растений альпийского пояса, как и наличие запасных веществ в подземных органах. Большая мобильность этих веществ позволяет альпийцам за короткий вегетационный сезон быстро достичь полного развития листьев и цветов, созревания плодов и накопления запасных веществ в соответственных органах. Значительная часть альпийских растений сохраняет зимой зеленые листья, а некоторые из них продолжают вегетацию под снегом; наблюдается также закладывание с осени зимующих почек с вполне сформировавшимися цветками.
Характерные черты альпийских растений особенно ярко выражены у обитателей скал, где наиболее резко проявляются эдафические и климатические особенности альпийских местообитаний. Менее резки эти особенности на осыпях и россыпях, где эдафические и микроклиматические условия несколько изменяются в связи с измельчанием и большей выветренностью субстрата, а также с накоплением влаги и питательных веществ между камнями и в толще осыпи. Здесь растения достигают более крупных размеров и приобретают более мезофильный облик. Зато у них имеются свои особые приспособления, связанные с подвижностью субстрата, постепенно ими закрепляемого. Типичные растения осыпей обладают длинными, крепкими, эластичными корнями и корневищами, пронизывающими толщу осыпи (виды С arum, Oxytropis, Onobrychis, Chamae-sciadium acaule и др.) или имеют стелющиеся побеги, оплетающие осыпь сверху (Arabis armena, Draba sibirica, Potentilla gelida, Sedum tenellum и др.).
Процесс почвообразования, идущий параллельно с закреплением осыпей и задернением скал, постепенно достигает такой стадии, при которой там могут поселиться некоторые мелкие рыхлокустовые или корневищнорыхлокустовые осоки и злаки (горные виды лисохвоста, кольподиума, альпийский мятлик, горолюбивая осока и др.). Пионерные открытые группировки сменяются «альпийскими коврами», вначале пестрыми от яркоцветущих растений и рыхлодернинными, а затем — по мере проникновения в них корневищных и плотнодерновинных осок и злаков (высокогорных овсяниц, грустной осоки — Carex tristis и др.) — более однотонными и плотнодернинными. Так развиваются наиболее типичные и наиболее распространенные среди альпийских низкотравных формаций альпийские плотнодернинные луга: мелкоосоково-разнотравные, разнотравно-мелкозлаковые, кобрезиево-разнотравные и другие с каменистыми, дресвяни-стыми или щебнистыми почвами, маломощными или среднемощными, большею частью торфянистыми. В травостое этих лугов подъярус с основной массой листьев обычно достигает лишь 5—8 см, а соцветия — 10 или 30 см высоты; проективное же покрытие не менее 80% и достигает 100%.
Альпийские луга по своим природным качествам являются типичными пастбищами. Их низкий плотный травостой хорошо переносит выпас и дает питательный, хорошо поедаемый «нажировочный» корм в количестве 4—8 ц/га сухой массы. Умеренный выпас способствует поддержанию хорошего состояния альпийских лугов путем разрыхления плотной дернины, естественного удобрения и предотвращения развития старики. Менее ценны в кормовом отношении пустоши. В целом, травянистые формации альпийского пояса являются высококачественными летними пастбищами и составляют основной фонд горных пастбищных угодий для многих республик и областей Кавказа, Средней Азии и Алтая.
Растительный покров субальпийского пояса характеризуется прежде всего преобладанием пышных высокотравных лугов (со средней высотой травостоя 70—90 см), но включает также группировки высокогорных кустарников и полукустарничков. Заходят сюда редколесья и криволесья (березовые, сосновые, пихтовые, буковые и др.), которые типологически должны быть отнесены к лесному поясу, но топографически настолько тесно переплетены с массивами высокотравных лугов, что не всегда могут быть выделены из пределов субальпийского пояса. Значительная часть высокотравных субальпийских лугов совершенно явственно связана с травяным ярусом того или иного типа леса, который в недавнем прошлом, очевидно, покрывал соответственные части территории субальпийского пояса. То же можно отметить в отношении травяных и кустарниковых нустошей, распространенных, как мы отмечали, по границе субальпийского и альпийского поясов.
Связь субальпийских формаций с лесными рассматривается ‘специально при описании кавказских высокогорных лугов (57 к); здесь же отметим, что значительная часть субальпийских лугов возникает в процессе зарастания осыпей и россыпей, играющих довольно заметную роль в ландшафтах субальп, хотя почвенно-растительный покров достиг здесь высокого развития и большая часть территории занята высокотравными лугами с мощными горнолуговыми почвами.
Видовой состав травостоя субальпийских лугов богат и довольно разнообразен, но основные эдификаторы и субэдификаторы преобладающих формаций в субальпах Кавказа, Алтая, гор Средней Азии и других высокогорных районов СССР немногочисленны. Некоторые из них общи для всех названных горных стран или же представлены в них близкими.
замещающими видами. Особенно велика общность видового состава в формациях, в своем генезисе явственно связанных с лесами или лесными лугами; в их составе участвуют флористические элементы с очень широкими ареалами бореального типа: евразиатского (Calamagrostis arundi-пасеа, Dactylis glomerata) и голарктического {Anthoxanthum odoratum, Deschampsia caespitosa). Немало на таких лугах крупных высоких трав, представленных в разных горных странах разными более или менее близкими видами одного и того же рода (Aconitum orientate, A. excelsum, Anthriscus nemorosa, A. silvestris и др.).
Формации, близкие к формациям альпийского пояса, более богаты эндемами, которые придают особые характерные черты субальпийским лугам отдельных, пространственно разделенных между собою горных стран.
Физиономически в травостое субальпийских лугов нередко преобладает так называемое разнотравье (виды Cephalaria, Astrantia, Beto-nica, Inula, Centaurea, Pyrethrum, Scabiosa и др.), среди которого тонут и часто остаются незамеченными листья и метелки злаков. Однако злаки так или иначе всегда присутствуют здесь. При правильном использовании луга они постепенно создают все более плотную дернину, вытесняя значительную часть видов разнотравья.
В отличие от альпийских низкотравников, пестрые злаково-разно-травные луга субальпийского пояса — типичные сенокосные угодья с высоким травостоем и значительной облиственностью той его части, которая попадает в укос; они довольно урожайны (20—30 ц/га сена), но их кормовые качества часто низки, так как в них относительно мало бобовых и злаков и много ядовитых и вредных растений (ветреницы, акониты и др.). Хозяйственные мероприятия должны быть направлены здесь на усиление злаковой и бобовой части травостоев. Количество злаков быстро возрастает при регулярном сенокосном использовании и сильно падает йод влиянием выпаса. Выпас не только вызывает выбивание части злаковой основы, но и способствует появлению и сильному разрастанию новых, сорных видов разнотравья, отсутствовавших в первоначальном составе субальпийских высокотравных лугов и быстро развивающихся на них при пастбищном использовании. В результате этого появляются вторичные разнотравные луга. Урожайность травостоя падает при этом до 8—10 ц/га, а при сильном выпасе (сбое) и до 3—4 ц/га.
Чаще всего субальпийские высокотравные луга под влиянием пастьбы превращаются в низкотравные манжетковые пастбища с очень малым участием злаков и бобовых. При прекращении пастьбы часть прежнего состава луга восстанавливается, но даже на таком «отдохнувшем» от выпаса лугу травостой значительно отличается по видовому составу и урожайности от сенокосного луга.
Сильно выбитые и истощенные луга в субальпийском поясе могут быть подвергнуты как поверхностной, так и коренной мелиорации с посевом и подсевом ценных кормовых трав. Луга могут быть заняты также под зернофуражные культуры и корнеплоды. В связи с трудностями перевозок в высокогорных условиях, желательно содержать зимою скот вблизи мест заготовок сена, а летом перегонять его на альпийские пастбища.
Как альпийские, так и субальпийские луга наиболее распространены и лучше изучены на Кавказе. Поэтому мы сравнительно детально останавливаемся на описании кавказских лугов (57 к). Субальпийские луга Карпат, принадлежащие к особому среднеевропейскому типу, также описаны несколько подробнее.
57 У — уральские
Формации, близкие к высокогорнолуговым, представлены на Урале фрагментарно.
Особенно слабо нредставлены альпийские низкотравные формации, к которым можно отнести пустошные осоково-овсяницевые луга. Основу их низкого, но довольно густого травостоя образует психрофильная горная овсяница (Festuca supina); к ней присоединяется мелкая осока (Carer hyperborea), более распространенная в горной тундре Северного Урала.
Несколько лучше развиты луга субальпийского типа, встречающиеся в сочетании с редколесьями. Горные редколесья, входящие в состав субальпийского пояса Урала, образованы преимущественно елью (Picea obovata), пихтой (Abies sibirica) и березами (Betula tortuosa, В. pubescens, реже — В. verrucosa). Для субальпийского пояса Среднего Урала особенно характерны «травянистые редколесья» со смешанным елово-пихтово-бе-резовым древесным ярусом и пышным травостоем из разных теневых трав и папоротников: Dryopteris austriaca (преимущественно на известняках), Athyrium filix-femina, Calamagrostis obtusata, C. Langsdorffii, Milium effusum, Aconitum excelsum, Oxalis acetosella, Mafanthemum bifolium, Viola biflora и др. (Игошина, 1952). На склонах, сложенных кислыми горными породами, с маломощными оподзоленными почвами часто встречаются также «луговиково-черничные редколесья»; их древесный ярус образует извилистая береза (Betula tortuosa) с примесью ели, пихты, иногда кедра; основу травяного яруса составляют луговик (Deschampsia flexu-osa) и черника (Vactinium myrtillus), иногда с примесью голубицы (V. uli-ginosum). Среди дерновин луговика кое-где пробиваются Anthoxanthum odo-ratum, Festuca supina, Anemone biarmiensis, Hieracium suberectum и др. (Игошина, 1952).
Все названные виды входят обычно в состав лугов, развивающихся на полянах между участками редколесья и относимых исследователями растительности Урала к субальпийским лугам (Игошина, 1931, 1944, 1952) или к горным пустошам (Овеснов, 1948 и др.). К. Н. Игошина (1952) различает среди субальпийских лугов Среднего Урала три группы: крупнозлаково-разнотравные, мелкозлаково-разнотравные и злаковые луговиковые.
В первой группе постоянно обильными компонентами являются Calamagrostis obtusata, С. Langsdorffii, Angelica silvestris, Cirsium heterophyl-lum, Hieracium suberectum, Polygonum bistorta, Solidago virga-aurea, Veratrum Lobelianum и некоторые другие менее высокорослые растения, как Anthoxanthum odoratum и различные виды Alchimilla. К ним присоединяются местами Digraphis arundinacea, Geranium albiflorum, Anemone biarmiensis, Aconitum excelsum и Viola biflora. В некоторых районах большие площади занимают луга с густым высоким травостоем, образованным папоротниками (Dryopteris austriaca, Athyrium filix-femina). Травостой крупнозлаково-разнотравных лугов обладает обычно густым и высоким подседом, 80—85% прективного покрытия и 50—60 см высоты; соцветия достигают 70—100 см.
Мелкозлаково-разнотравные луга образованы в разнотравной части теми же видами, но не достигающими такой большой высоты; из злаков . в них преобладают душистый колосок и волосовидная полевица (Agro-stis capillar is).
Луговиковые пустошные луга из Deschampsia flexuosa, которые Овеснов называет пустошами, имеют обычно густой и низкий травостой 10— 20 см высотой. Кроме луговика, в их травостое постоянно участвуют ду
шистый колосок, золотая розга, чемерица, ветреница пермская (Anemone biarmiensis), черника, некоторые виды осок, ожик и ситников и много видов разнотравья бореального или аркто-альппйского типа.
Литература
II. И. Крылов (1926); К. Н. Игошина (1931, 1944, 1952); А. М. Овеснов (1948, 1948а. 1950, 1951, 1952, 1952а); II. Л. Горчаковский (1955).
57 а — алтайские и североКазахстанекие
Высокогорные луга алтайского типа показаны на «Геоботанической карте» на Алтае, в Саянах и на хребтах Тарбагатай, Саур и Танну-Ола; по условиям масштаба на карте не отмечены более мелкие участки среди горных тундр (3 а) на Кузнецком Алатау, в высокогорьях Алтая и Саян.
Наибольшие массивы высокогорных лугов этого типа расположены в лучше увлажненных частях Алтая и Саян: в западных районах или на западных склонах восточных хребтов. Высотные их границы также частично связаны с различной степенью увлажнения, но преимущественно — с температурными условиями, меняющимися в направлении с севера на юг. По указаниям Б. Ф. Петрова (1952), в северных частях Кузнецкого Алатау и Восточного Саяна, на широте 54—55° субальпийские луга появляются на высоте 1200—1300 м над у. м., а на широте 52—53° они при-' урочены к высотам 1500—1600 м; в центральном же и юго-восточном Алтае, на широте 49—50°, высокогорные луга начинаются лишь с высоты 1700—1900 м над у. м. Верхние их границы доходят до 1300—1400 м на севере и до 2000—2300 м над у. м. на юге Алтайско-Саянской области. Как в западных, так и в восточных районах развиты преимущественно высокотравные субальпийские луга, низкотравные же альпийские встречаются лишь небольшими участками, редко где составляя хорошо выраженную полосу между субальпийскими лугами и тундровой растительностью.
По составу субальпийских лугов Алтая можно легко судить об их лесном происхождении. Мы видим в их высокотравных формациях такие виды, как Archangelica decurrens, Anthriscus silvestris, Aconitum excelsum, Dactylis glomerata, Calamagrostis arundinacea и другие высокие травы, свойственные алтайской «черневой тайге». Однако здесь имеются и другие формации, в которых преобладают луговые пли луговостепные растения (Dracocephalum grandijlorum, Geranium albiflorum, Ranunculus gran-difolius, Aquilegia glandule sa, Saussurea Frolowii, Rhaponticum carthamoi-des, Phlomis oreophila, Trollius altaicus). Последний вид — алтайская купальница, часто называемая за свои яркие оранжевые цветы «огоньком», распространен в более влажных районах, по границе с горной пихтовой тайгой. Фломис же, или железняк, образующий так называемые фломисо-вые луга, продвигается далеко на юг и доходит до Тянь-Шаня. В южных районах Алтая субальпийские луга остепняются; в их флористический со-став входят Helictotrichon Schellianum, Roegneria Turczaninowii, Phleum phleoides, Galium verum, Veronica spicata, Dracocephalum nutans, Gypsophila altissima и даже Artemisia sericea (Еленевский, 1938, 1940в; Келлер, 1914).
Субальпийские кустарники представлены на Алтае мощными зарослями круглолистной березы (Betula rotundifolid), образующими широкую полосу над кедрово-лиственничным редколесьем.
Собственно альпийские низкотравные луга представлены на Алтае слабее, По указанию Е. П. Матвеевой (1940), альпийские лужайкн
(с Dracocephalum altaiense,1 Aquilegia glandulosa, Trollius altaicus, Schultzia crinita и др.) вкраплены в общий фон субальпийских лугов. Местами, например на Нарымском хребте и в высокогорных котловинах юго-восточного Алтая, в альпийском поясе распространены дерновинные торфянистые осочники, образованные алтайской осокой (Carex altaica.) и кобрез-ники. Наиболее развиты здесь кобрезники из Cobresia Bellardi, С. jilt-folia, С. Smirnovii.
Своеобразным типом альпийских лугов являются алтайские разнотравные луга с Schultzia crinita', по указанию Еленевского (1940а) они занимают громадные площади верхнеальпийского пояса.
Кобрезники широко распространены и на Тарбагатае, где альпийский пояс начинается с высоты 2300 м над у. м. Е. Ф. Степанова (1950) выделяет здесь пять основных ассоциаций с преобладанием Cobresia Smirnovii* и с участием С. Bellardi («кобрезиевики» по терминологии Е. Ф. Степановой): 1) чистые, 2) моховые, 3) ивовые, 4) разнотравные и 5) типчаковые.
В широко распространенных первых двух ассоциациях среди густой щетки кобрезии ( с проективным покрытием 100% и высотой травостоя 20 см) лишь изредка встречаются единичные представители альпийского-разнотравья. Вторая ассоциация отличается бблыпим развитием мохового ковра, а третья, встречающаяся на высоте 2500 м над у. м., — участием мелкой кустарниковой ивы Salix torulosa. В четвертой ассоциации,, развивающейся по мелким понижениям рельефа в местах лучшего увлажнения, к кобрезии примешивается в значительном количестве альпийское-разнотравье: Polygonum nitens, Thalictrum alpinum, Papaver cfoceum, Rumex acetosa Swertia obtusa, Dracocephalum nutans, Alchimilla sibiricar Schultzia crinita и др. В наилучше увлажненных пониженных участках на северных склонах господство в травостое переходит полностью к разнотравью, и образуются альпийские разнотравные луга, иногда почти чисто манжетковые (с Alchimilla sibirica, A. cyrtopleura). Из злаков здесь-встречаются Роа alpina, Р. sibirica, Phleum alpinum. На южных склонах распространена типчаково-кобрезиевая ассоциация; здесь наряду с Cobresia Smirnovii большое место занимает Festuca Ganeschini, к которой примешиваются Helictotrichon asiaticus, Роа alpina, Phleum alpinum, а из разнотравья — Polygonum nitens, Minuartia biflora, Schultzia crinita, Aster alpinus, Saussurea Schanginiana, Papaver croceum и др.
Субальпийские луга занимают на Тарбагатайском хребте высоты от 1700—2000 до 2300—2400 м над у. м. на платообразных понижениях хребта и пологих склонах. В верхних частях пояса они представлены мелкотравными красочными лугами, близкими к альпийским, а в нижних — высокотравными, более типичными субальпийскими. Среди мелкотравных субальпийских лугов преобладают манжетково-разнотравные с господством Alchimilla cyrtopleura и A. rubens и со значительным участием Alchimilla sibirica, A. Krylovii, Dracocephalum altaiense s. 1., Trollius altaicus, Anemone Schrenkiana, Aquilegia glandulosa, Polygonum nitens, а из злаков — Alopecurus soongoricus, Hordeum turkestanicum, Festuca rubra, Anthoxanthum odoratum, Poa alpina и некоторых других видов.
Среди высокотравных лугов Тарбагатая преобладают злаково-разнотравные ассоциации с господством из злаков Alopecurus soongoricus: и с участием Роа pratensis, Р. relaxa, Р. dschungarica, Festuca rubra. 1 2
1 D. altaiense s. 1.
2 Прежде относилась к Cobresia schoenoides (С. A. M.) Steud.
Helictotrichon pubescens; в разнотравье обильны Alchimilla cyrtopleura, A. rubens, Hedysarum neglectum, Anemone Schrenkiana, Crepis sibirica, Aconitum soongoricum, Allium platy spathum, Dracocephalum nutans, Ligu-laria altaica, Potentilla desertorum, Serratula marginata, Saussurea Frolowii, Rhaponticum carthamoides, Linum altaicum. Как видно по этому списку, в составе тарбагатайских высокотравных лугов участвуют и низкорослые растения (виды Alchimilla, Anemone, Potentilla).
Значительное участие в растительном покрове субальпийского пояса Тарбагатая принимают степные формации (см. 83 а и 84 а), а также арчев-ники, образованные двумя видами: Juniperus sibirica и J. pseudosabina. Немалое место занимают голые скалы и осыпи с редко разбросанными экземплярами Rhodiola rosea, Luzula spicata, Roegneria Schrenkiana, R. czimganica. Попадаются небольшие болотца с пушицей — Eriophorum Scheuchzeri и с каймой из Juncus triglumis, Saxifraga hirculus, Pedicula-ris rhinanthoides.
В Западном Саяне высокогорные луга располагаются на высотах 1700—2200 м над у. м., сменяясь выше горными тундрами или каменистыми россыпями и скалами. В. В. Ревердатто (1946) различает здесь луга альпийские и субальпийские и в последних — варианты высокотравные (с травостоем до 150—180 см высоты) и «низкотравные» 1 (с травостоем, до 50—80 см).
Высокотравные субальпийские луга приурочены здесь к верхней границе темнохвойных лесов, с повышенно влажными условиями местообитания. В травостое преобладает разнотравье, среди которого в первом подъярусе обычны Rhaponticum carthamoides, Saussurea latifolia, Cirsium he-terophyllum, Aconitum excelsum, а во втором — виды Alchimilla, Cerastium dahuricum, Geranium albiflorum, Euphorbia pilosa, Lilium martagon, Polygonum bistorta, Trollius altaicus, Veratrum Lobelianum и др. Злаков и осок мало {Alopecurus brachystachyus, Роа sibirica, Carex perfusca)’, в связи с этим мало задернение. Мало также мхов, лишайники отсутствуют.
«Низкотравные» субальпийские луга, связанные с более сухими местообитаниями по границе лиственничных лесов, обычно более богаты злаками и осоками {Calamagrostis neglecta, Роа sibirica, Festuca rubra, Carex perfusca) и лучше задернены; все же и здесь явно преобладает разнотравье: Bupleurum aureum, Geranium albiflorum, Veronica longifolia, Anemone crinita, Aster flaccidus, Gentiana algida, Ptarmica Krylovii, Pedicularis compacta, Schultzia crinita, Scorzonera radiata и др.
Альпийские луга Западного Саяна характеризуются незначительным ростом растений (не выше 50 см в первом подъярусе), большим флористическим разнообразием, яркой окраской цветов, значительным покрытием почвы мхами. Яркий аспект создают здесь Aquilegia glandulosa, Dracocephalum altaiense s. 1., Trollius asiaticus, Scorzonera radiata’, кроме-них, обычны Rumex acetosa, Bupleurum aureum, Geranium albiflorum, Swer-tia obtusa, Gentiana grandiflora, Viola altaica и многие другие виды. Из злаков и осок встречаются Anthoxanthum odoratum, Festuca altaica, F. tristis, Phleum alpinum, Poa sibirica, Trisetum altaicum, Carex perfusca’, обилие злаков отмечается на «сухих альпийских лугах» по южным склонам.
В Центральном Саяне высокогорные луга располагаются на высотах 1500—1800 (2000) м над у. м. Основными компонентами субальпийских лугов в Центральном Саяне А. В. Куминова (1946а) считает высокорос-
1 Обычно низкотравными называются собственно альпийские луга, с травостоем, в 10—20 см высоты, и лишь с отдельными растениями, достигающими 40—50 см.
лые растения (90—100 см): Aconitum safanense, Rhaponlicum carthamoides и Saussurea latifolia, а также несколько менее высокое (05 см) — Veratrum Lobelianum. Средняя высота травостоя здесь равна 60 см, покрытие (проекция) 100 %, задерпованность же очень незначительная (в связи с малым участием в травостое злаков и осок). Кроме выше названных видов, здесь обычны Thalictrum minus, Cirsium heterophyllum, Bupleurum aureum, Aconitum altaicum, Geranium albijlorum, Trollius asiaticus, Viola biflora (III подъярус),Rumex acetosa, Cerastium pauciflorum и многие другие виды разнотравья. Из злаков достигает большой высоты (85 см) и часто встречается Роа sibirica f. nigra', попадаются также Triseturn altaicum (II подъярус, 25 см), Anthoxanthum odoratum (23 см), а из осок — Carex perfusca, входящая в состав I подъяруса (85 см). В самом низком III подъярусе, кроме преобладающей в нем Viola biflora (3 см), встречаются V. altiaca (10 см.), Alchimilla anisopoda (5 см), Botrychium lunaria (1 см), Veronica densiflora (7 см) и некоторые другие растения.
Почвы на этих лугах светлые горнолуговые субальпийские, небольшой мощности.
Низкотравные альпийские луга Центрального Саяна располагаются мелкими участками на стыке между субальпийскими лугами и высокогорной тундрой, часто вблизи снеговых пятен. Средняя высота травостоя с яркими крупными цветами равна здесь 10—25 см, проективное покрытие 40—50%; среди разбросанных дерновин просвечивает щебень; почвы маломощные щебнистые (светлые горнолуговые альпийские). Моховая подстилка достигает 1—2 см толщины. Наиболее распространены ассоциации с преобладанием либо Viola altaica, либо Aquilegia glandu-losa. Остальные компоненты, повторяющиеся в различных количественных соотношениях в обеих ассоциациях, следующие: Bergenia crassifolia, Callianthemum angustifolium, Campanula rotundifolia var. linifolia, Doroni-cum altaicum, Dryas oxyodontha, Gentiana grandiflora, Patrinia sibirica, Pedicularis Oederi, Polygonum bistorta, P. viviparum, Schultzia crinita, Sibbaldia macrophylla, Veronica densiflora и некоторые другие; из злаков и осок: Anthoxanthum odoratum, Festuca supina, Роа sibirica f. nigra, Phleum alpinum, Carex perfusca, C. brunnescens. Здесь же разбросаны мелкие кустарники: Rhododendron aureum, Salix reticulata, S. Turczaninowii, а местами — кусты ольховника (Alnaster fruticosus), в 2—3 м высоты.
Среди альпийских и субальпийских лугов встречаются небольшие (маломощные) болотца, основу травостоя которых образует маленький пушицевидный камыш (Trichophorum alpinum)', кроме него, характерны пушица (Eriophorum humile), осоки (Carex tripartita и С. stilbophaea *), а также соссюрея (Saussurea foliosa), сверция (Swertia obtusa) и лук (Allium schoenoprasum).
Литература
Б. А. Келлер (1914); В. И. Варанов (1926); В. А. Крюгер (1927); П. Н. Крылов (1931); Б. К. Шишкин (1937); Р. А. Еленевский (1938, 1939а, 1940а, 19406, 1940в); Е. П. Матвеева (1940); А. В. Куминова (1946а); В. В. Ревердатто (1946); Е. Ф. Степанова (1950); Ал. А. Федоров и Ан. А. Федоров (1951); Б. Ф. Петров (1952).
57 кп — карпатские
На Карпатах субальпийские луга и сочетающиеся с ними формации наиболее развиты в юго-восточной более возвышенной части хребта (Чер-ногорье, Раховский массив), где они показаны на «Геоботанической
1 В прежней литературе по Алтаю и Саянам эта осока относилась к Carex atrofusca Kuk.
карте». Нижний предел субальпийской растительности на Карпатах находится на высоте 1500—1600 м над у. м., а местами, преимущественно в результате расчистки леса, на высоте 1300—1400 м над у. м. Верхний предел леса образует чаще всего еловое редколесье (19 кп), выше которого распространены заросли кустарников, соснового стланика и луговые группировки. Местами на границе; леса развиты разреженные буковые леса; особенно обычны они там, где верхняя граница леса снижена человеком.
Отдельные деревца и кустарники местами можно встретить близ самых высоких вершин Советских Карпат; однако с высоты 1800—1900 м над у. м. обычно господствуют травяные, кустарничково-травяные и ку-старничково-лишайииково-моховые сообщества, в составе которых встречаются некоторые типичные; альпийские растении. Эта верхняя часть карпатских высокогорий, представленная на небольшой площади на самых значительных вершинах, может рассматриваться как фрагмент нижнеальпийской полосы альпийского пояса, нижняя граница которого в Татрах проходит в среднем на высоте около 1850—1950 м над у. м., а в западной части Главного Кавказского хребта на высоте около 2300 м над у. м.
Наиболее подробные сведения о растительности этой нижнеальпийской полосы сообщены М. Дейлом (Deyl, 1940), Е. М. Брадис и А. А. Запятовой (1954).
Осыпи и скалы в высокогорном поясе Карпат занимают небольшие площади и в той или иной мере характерны только для наиболее выдающихся вершин (Говерла, Пол-Иван, Петрос, Близница и др.). На них встречаются некоторые типичные скальные альпийские растения {Leontopo-dium alpinum, Antennaria carpatica, Anthyllis alpestris, Anthemis carpatica и др.); наиболее разнообразен их состав на известняках. В сходных условиях на каменистых склонах встречается изредка дриада {Dryas •octopetala) и некоторые другие альпийские растения {Bartsia alpina, Polygonum viviparum, Sesleria Bielzii и др.).
Наиболее типичны для нижнеальпийской полосы ассоциации с господством вечнозеленой осоки {Carex sempervirens), в которых, в зависимости от условий, наряду с осокой, доминируют овсяница {Festuca supina), вейник {Calamagrostis villosa), на бедных почвах черника {Vactinium туг-Hilus'), а на известняках скабиоза {Scabiosa lucida) или сон-трава {Pulsatilla alba). Широко распространены также ассоциации с господством -овсяниц {Festuca supina, или F. picta), состав флоры которых довольно разнообразен {Leontodon croceus, Carex sempervirens, Роа cenisia, Ligusti-•cum mutellina, Soldanella montana, Potentilla aurea, Sempervivum monta-num и многие другие).
Б. Павловский (Pawlowski, 1937), изучавший растительность Черно-торья, считает, что климаксом альпийских ассоциаций там является установленная по принципу характерных видов (в флористическом понимании) ass. Carex curvula—Juncus trifidus. В качестве других характерных видов упомянутой ассоциации Б. Павловский называет Festuca supina, Hieracium alpinum, Helictotrichon versicolor, Pulsatilla alba, Senecio <carpaticus, Primula minima, Sesleria Bielzii, Carex rigida {=C. Fyllae). Некоторые из них обычно не являются доминантами альпийских сообществ, в том числе и Carex curvula, в пределах Советских Карпат пока известная только в Черногорье.1
На крутых каменистых склонах (Говерла, Поп-Иван и некоторые другие вершины) развиты вейниковые {Calamagrostis villosa) луга. В них
1 Визначник рослин УРСР. Киев, 1950.
местами много овсяницы (Festuca picta) или ожики (Luzula nemorosa)-и зверобоя (Hypericum alpigenum). Во всех поименованных сообществах изредка встречаются кусты рододендрона (Rhododendron Kotschyi). На крутых каменистых склонах, обычно защищенных от ветра и зимой прикрытых снежным покровом, рододендрон Кочи на небольших участках образует ассоциации, в которых растительность нередко покрывает не более 40% поверхности субстрата. В зарослях рододендрона много черники, а местами голубицы; в напочвенном покрове распространены довольно-значительно сфагны (Sphagnum Girgensohnii, S. quinquefarium) и немногие другие мхи и лишайники (Hylocomium proliferum, Dicranum congestum, Polytrichum gracile, Lophozia alpestris, Baeomyces roseus, Cetraria islandica, Cladonia gracilis). Из трав здесь наиболее обычны ястребинка (Hieracium alpinum), вейник (Calamagrostis villosa), овсяница (Festuca-supina), ситник (Juncus trifidus), на фоне которых встречаются и другие растения (Homogyne alpina, Vaccinium vitis-idaea, Gentiana punctata, Campanula alpina и пр.). Довольно широко распространены в нижней, альпийской полосе и травяно-лишайниково-кустарничковые ассоциации, с преобладанием голубицы (Vaccinium uliginosum). В напочвенном покрове этих ассоциаций господствует исландский лишайник (Cetraria islandica), много оленьей клядонии (Cladonia rangiferina), встречаются другие лишайники и мхи (Cladonia sylvatica, С. furcata, Thamnolia vermicu-laris, Cetraria cucculata, Pleurozium Schreberi, Hylocomium proliferum, Pogonatum urnigerum и пр.). M. Дейл (Deyl, 1940) выделяет ряд ассоциаций, (социаций) этой группы: 1) Cetraria islandica—Vaccinium uliginosum— Pulsatilla alba, 2) Cetraria islandica—Vaccinium uliginosum, 3) Cetraria islandica—Juncus trifidus—Vaccinium uliginosum, 4) Cetraria islandica— Vaccinium uliginosum—Rhododendron Kotschyi, 3) Vaccinium uliginosum—Cetraria islandica—Carex sempervirens и другие. Из трав-в этих ассоциациях наиболее обычны, кроме упомянутых в названиях ассоциаций, Hieracium alpinum, Campanula alpina, Festuca supina, Heli-ctotrichon versicolor, Arnica montana и др. Кустарничковые ассоциации с рододендроном и черникой Е. М. Брадис и А. А. Запятова называют альпийскими пустошами; М. Дейл относит их к особому разряду ассоциаций альпийского пояса (Nanofruticolichenosa и Nanofruticomus-cosa).
Ниже 1800 м над у. м. на Карпатах представлено сочетание типичных субальпийских ассоциаций. Из них наиболее широко встречаются бело-усники (Nardus stricta) на бедных сухих и местами торфянистых почвах. Распространению белоуса в субальпийском поясе Карпат способствует интенсивный выпас скота. Многие белоусники развиты на месте субальпийских лугов другого состава. Выделяются собственно белоусники и разнотравные белоусники. В чистых белоусниках сравнительно немного постоянно встречающихся растений (Festuca rubra, Carex pilulifera, Luzula sudetica, L. nemorosa, Potentilla erecta, Thymus subalpestris, Vaccinium myrtillus и др.). Разнотравные белоусники развиты меньше, их сообществам не свойственно значительное задернение. Кроме белоуса, в травостое этих ассоциаций характерно участие манжеток и других трав (Ligusti-сит mutellina, Potentilla aurea, Р. erecta, Aposeris foetida, Arnica montana? Hieracium alpinum и пр.). В большей или меньшей степени белоусникам присущи мхи (Hylocomium proliferum, Pleurozium Schreberi, Polytrichum strictum) и лишайники (Cetraria islandica и пр.). Встречаются также ов-сянично-белоусовые (Festuca rubra, Festuca picta, Nardus stricta), щучково-белоусовые (Deschampsia caespitosa, Nardus stricta) и полевицево-бело-усовые (Agrostis capillaris, Nardus stricta) субальпийские луга.
Белоусовые луга на полонянах в бассейне Боржавы подробно изучены М. Малохом (Maloch, 1931); он выделяет целый ряд ассоциаций белоус ников и ассоциаций щучковых (Deschampsia caespitosa) и овсяничных (Festuca rubra) лугов, в которых белоус также широко представлен.
Из других, реже встречающихся на Карпатах ассоциаций субальпийских лугов можно назвать щучково-разнотравные, щучково овсяничные (Festuca rubra) и полевицевые (Agrostis capillaris) луга. Встречаются в субальпийском поясе и ассоциации с господством черники, возникшие на месте кустарниковых зарослей и редколесий; им обычно присущ хорошо выраженный мохово-лишайниковый ярус.
Очень типичны для субальпийского пояса Карпат асссоциации соснового стланика (Pinus mughus), в прошлом распространенные значительно шире на землях, позднее расчищенных под пастбища. В настоящее время заросли соснового стланика сохранились на крутых каменистых склонах на высоте 1500—1800 м над у. м. Местами сосновый стланик образует подлесок в ельниках на верхнем пределе лесного пояса. Заросли соснового стланика представлены несколькими ассоциациями; они бывают иногда так значительно сомкнуты, что моховой и травяно-кустарничковый покров в них не развивается. При более разреженном произрастании стланика на склонах северной экспозиции характерны сфагновые стланиковые сообщества с черникой и очень немногими травами (Homogyne alpina, Calamagrostis villosa). На менее влажных восточных и южных склонах распространены зеленомошные стланиковые заросли, в травяно-кустарничковом ярусе их также преобладает черника, но состав трав более разнообразный (Homogyne alpina, Calamagrostis villosa, Soldanella montana, Athyrium alpestre, Gentiana asclepiadea, Luzula nemorosa и др.). На глубоких и богатых почвах заросли соснового стланика почти не сохранились. В таких условиях известна ожиковая ассоциация соснового стланика (soc. Pinus mughus — Luzula silvatica', Deyl, 1940); состав трав здесь наиболее разнообразный (Luzula silvatica, Rumex arifolius, Oxalis acetosella, Calamagrostis arundinacea, Campanula abietina, Hypericum alpigenum и др.).
Еще более, чем сосновые стланики, распространены в субальпийском поясе Карпат заросли ольховника (Alnaster- viridis). Они нередко отличаются очень большой плотностью, и под их пологом произрастают немногие растения. При менее сомкнутом ярусе ольховника иногда встречается ассоциация с покровом из черники, но более обычны ассоциации с травяным покровом, развитые в местах, хорошо увлажняемых проточными водами. В зависимости от условий, в покрове этих ольховников преобладают различные виды (Rumex arifolius, Athyrium alpestre, Calamagrostis arundinacea, Luzula silvatica, Senecio nemorensis).
На склонах разной крутизны и экспозиции выше границы леса встречаются группировки можжевельника (Juniperus sibirica). В них обычно развит покров из зеленых мхов и травяно-кустарничковый ярус, общий с ельниками-черничниками. Некоторые сообщества можжевельников на высоте 1300—1400 м над у. м. возникли на месте уничтоженных ельников и елового редколесья, которому был свойствен можжевеловый подлесок.
Субальпийские пространства Карпат, а также «половины», существующие на месте бывших лесов, представляют собою основную территорию пастбищ местного скотоводства. Бессистемный выпас, имевший место в прошлом, существенно истощил эти кормовые угодья. Мероприятия по коренному и поверхностному улучшению пастбищ, а также введение загонной системы выпаса обеспечивают значительное повышение их продуктивности.
Литература
Н. Б. Вернандер (1947); II. Бугаенко и И. Безъязычный (1949); Е. М. Брадис и А. А. Запятова (1950, 1954); Е. М. Брадис (1951); А. А. Запятова (1951); Т. Sulrna (1929); М. Maloch (1931, 1932); В. Pawlowski (1937); М. Deyl (1940).
57 к — кавказские
Высокогорные альпийские и субальпийские луга в сочетании с кустарниками и лесами занимают на Кавказе весьма значительные площади (около 10 миллионов га) и образуют два явственных пояса: альпийский низкотравный (лежащий в разных районах Кавказа на высотах 2300— 3500 м над у. м.) и субальпийский высокотравный (1800—2800 м над у. м.).
В растительности высокогорных поясов Кавказа различаются: открытые, несомкнутые группировки скал, осыпей, морен (около 10% площади территории, отмеченной знаком 57 к); рыхлодернинные пестрые альпийские ковры (около 5%); плотнодернинные низкотравные луга (около 25%); приснежные луговинки (доли процента); пустоши и пустошные луга на бедных почвах (около 5—8%); болота и болотистые луга (2—3%). Около 50% занимают луга субальпийского пояса.
Открытые группировки скал, осыпей и морен являются одним из очень интересных в ботаническом отношении элементов растительного покрова. Наряду с морфологическими и биологическими особенностями, о которых говорилось выше, они интересны обилием эндемичных растений, характерных для того или иного, часто ограниченного горного района.
Среди растений, свойственных скалам кавказских высокогорий, можно, отметить: растения-подушки (Draba bryoides, D. mollissima, D. ossetica, Gypsophila imbricata, Minuartia imbricata, M. oreina, M. verna, Saxifraga j uniper if olia, S. exarata, S. moschata, Silene daghestanica и др.), суккуленты (Sedum gracile, S. tenellum, S. involucratum, Sempervivum caucasi-cum, S. pumilum и др.), луковичные и клубневые (Gagea anisanthos, G. gla-cialis, Allium oreophilum, Fritillaria lutea, Ornithogalum Schmalhausenii, Muscari coeruleum, Crocus Scharojanii и др.).
Из растений, закрепляющих осыпи и россыпи своими длинными корнями или корневищами, отметим, кроме упомянутых выше (опис. 57), Colpodium colchicum, Alopecurus Aucheri, Carex oreophila, Veronica minuta, Campanula saxifraga, Dentaria microphylla, Viola oreades, Minuartia ina-moena, Saxifraga sibirica, Pulsatilla armena, Oxyria digyna, Corydalis conorhiza, C. alpestris, Ranunculus caucasicus, Papaver caucasicum, So-bolewskya caucasica, Oxytropis cyanea, Onobrychis oxytropoides, Carum caucasicum, Symphyoloma graveolens, Nepeta supina, Scrophularia minima, Campanula Aucheri.
В видовом составе пестрых рыхлодернинных ковров Кавказа чаще всего встречаются и преобладают следующие виды: Alchimilla caucasica, Astragalus Levieri, Campanula tridens, Draba bryoides, Gypsophila tenuifo-lia, Hedysarum caucasicum, Minuartia caucasica, Oxytropis Owerini, Primula algida, Potentilla gelida, Saxifraga moschata, Taraxacum Stevenii. К ним просоединяются мелкие рыхлокустовые злаки (Alopecurus vagi-natus, Colpodium variegatum, C. versicolor, Poa alpina), а также мелкие короткокорневищные осоки (Carex rupestris, C. oreophila и др.).
Основными дернообразователями плотнодернинных низкотравных лугов являются овсяницы (Festuca supina и F. Ruprechtii) и осока (Carex tristis)', из кобрезий здесь встречаются три вида; Cobresia schoenoides, С. capillifolia (близкая к С. Bellardi) и С. persica. Овсяницы, мелкие осоки и кобрезии сочетаются с вышеперечисленными и многими другими
представителями разнотравья, образуя овсяницево-разнотравные, мелко-осоково-разнотравные и кобрезиево-разнотравные плотнодерниинные луга.
Cobresia capillifolia участвует также в составе травяно-лишайниковых пустошей, где обычно преобладают Cetraria islandica, С. nivalis, Thamno-lia vermicularis, Cladonia rangiferina, а из цветковых часто встречаются Gnaphalium supinum и Antennaria dioica. Мохово-лишайниковые пустоши образуются некоторыми видами из родов Polytrichum и Cetraria и часто включают в свой состав мелкие кустарнички и полукустарнички: Етре-trum hermaphroditum, Vactinium vitis-idaea, Salix arbuscula. Здесь часто встречается схенусовидная кобрезия (Cobresia schoenoides).
На местообитаниях с долго задерживающимся снегом развиваются приснежные луговинки с высокогорными одуванчиками (Taraxacum Stevenii, Т. porphyranthum), коллодиумом (Colpodium ponticum), тмином (Carum caucasicum). Зиббальдиевые приснежные пустоши (с Sibbaldia semiglabra и S. parviflora) выходят за пределы мелких углублений, где долго залеживается снег, и на значительных пространствах располагаются вдоль северных склонов хребтов, на западе — на высоте 2600—2800, а на востоке — на 3000—3200 м над у. м. Ниже, на высоте около 2000—2200 м, на западе широко распространены гераниевые пустошные луга с Geranium gymnocaulon и белоусники с Nardus glabriculmis.1
Болота и болотистые луга сравнительно мало распространены в высокогорьях Кавказа и встречаются изредка как в альпийском, так и в субальпийском поясах. Они развиваются преимущественно в процессе зарастания озер и реже — в порядке заболачивания лугов.
Как было указано, молодые приледниковые болота часто бывают образованы мхами (Aulacomnium palustre и Sphagnum Warnstorfii) с редким ярусом пушицы и некоторых видов осок; по окраинам зарастающего водоема развиваются влажные мелкоосоковые луга с Carex Kotschyana, С. capitellata и другими видами. Еще дальше от берега зарастающего водоема располагаются более старые осоково-гипновые и осоково-пуши-цево-сфагновые болотные комплексы с Carex dacica aEriophorum angusti-folium, co Sphagnum Warnstorfii, S. subsecundum, S. teres и др.
На выходах грунтовых вод развиваются мелкие ключевые гипново-осоковые болотца с Carex caespitosa, С. oligantha, С. dichroa, С. capillaris. Вдоль ручейков разрастаются подушки зеленых мхов (Philonotis seriata, Bryum Schleicheri, Breidleria arcuata и др.), среди которых появляются Parnassia palustris, Swertia iberica, Primula auriculata, Cardamine uligi-nosa и другие болотные растения (в том числе как редкость — Pinguicula vulgaris).
По окраинам болот развиваются болотистые и пустошные луга, преимущественно щучники с Deschampsia caespitosa, белоусники, с Nardus glabriculmis и луговиковые белоусники с Nardus и Deschampsia flexuosa', к названным эдификаторам обычно присоединяются другие злаки, свойственные бедным олиготрофным или психрофитным местообитаниям (Agrostis capillaris, Anthoxanthum adoratum, Festuca ovina),. а также некоторые виды мытников (Pedicularis condensata, P. Nordmanniana), раковые шейки (Polygonum bistorta), простой бодяк (Cirsium simplex), гравилат (Geum rivale), тмин (Carum caucasicum), валериана (Valeriana al-liariaefolia) и др.
Основными формациями субальпийских лугов на Кавказе являются: вейниково-разнотравные луга с Calamagrostis arundinacea, покрывающие-
1 Согласно А. А. Гроссгейму, Определитель растений Кавказа,. М., 1949.
значительные площади, пестрокострово-разнотравные луга с Bromus variegatus, пестроовсяничники, образованные крупной плотнодерновин-ной высокогорной овсяницей (Festuca varia) и, наконец, так называемые •субальпийские разнотравные луга и «субальпийское высокотравье», образованные целой свитой крупных, красиво цветущих трав с небольшой примесью высокорослых злаков.
Вейниково-разнотравные луга (типичные сенокосные луга с высоким, 70—90 см высоты травостоем, дающим 20—30 ц/га сена) генетически теснейшим образом связаны с травяным ярусом березовых и сосновых лесов, покрывавших значительную часть территории, занятой ныне •субальпийскими лугами, и исчезнувших частично в процессе природных •смен растительности, а частично под влиянием вырубки и выжигания их в целях расширения кормовой площади. Из современных растительных формаций, входящих в состав субальпийского пояса, вейниковоразнотравные луга связаны преимущественно с березовыми криволесьями (из Betula Litwinowii, В. Raddeana, В. Medwedewii) п с зарослями рододендрона (Rhododendron caucasicum). В видовом составе этих лугов, как и в травостое березняков и зарослей рододендрона, обычно присутствуют Anthoxanthum odoratum, Роа longifolia, Milium effusum, Aconitum nasutum, A. orientate, Anthriscus nemorosa, Betonica grandiflora, Geranium Ruprech-tii, Solidago virga-aurea, а также виды, встречающиеся в светлых высокогорных сосняках (Cephalaria gigantea, Centaurea alutacea и др.).
Некоторые ассоциации вейниково-разнотравных лугов, развивающиеся в нижних частях субальпийского пояса, в пологих горных ложбинах с хорошими условиями увлажнения, аэрации почвы и минерального питания, явно связаны с произрастающими в таких условиях кленовни-ками паркового характера, образованными высокогорным кленом Acer Trautvetteri. В их видовом составе мы встречаем, кроме вейника, бор Шмидта (Milium Schmidtianum), высокий райграс (Arrhenatherum elatius), ежу (Dactylis glomerata), длиннолистный мятлик (Роа longifolia), козлятник (Galega officinalis) и многочисленных представителей высокорослого разнотравья (Campanula latifolia, Doronicum macrophyllum, Euphorbia oblongifolia, Heracleum pubescens, Ligusticum physospermifolium, Senecio cladobotrys, Silene multifida, Telekia speciosa, Valeriana colchica и др.). Такие луга лишь особо сильным развитием вейника отличаются от так называемого «субальпийского высокотравья».
Последнее название дается горнолуговым злаково-разнотравным фитоценозам с чрезвычайно высоким травостоем, достигающим 2—3 м высоты и состоящим в основном из только что перечисленных видов злаков и разнотравья, причем обычно один-два вида (Heracleum Mante-gazzianum, Anthriscus nemorosa, Senecio platyphylloides, Telekia speciosa или др.) получают на ^некоторых участках исключительно сильное развитие. Встречаются такие фитоценозы в тех же условиях, как и парковые кленовники, являясь, возможно, как и они, реликтами третичного периода или более теплого и влажного времени послеледниковья. При регулярном сенокосном использовании как вейниково-разнотравных лугов, так и высокотравья, у них значительно усиливается злаковая основа и уменьшается разнотравность. Те и другие являются типичными сенокосными угодьями, дающими до 80—100 (высокотравье) ц/га сухой массы. Выпас действует на них губительно.1
1 Формации, обычно относимые к «субальпийскому высокотравью», по своему расположению на границе субальпийского и лесного поясов могли бы быть причислены к «среднегорным лугам». По своему строению и составу они близки, например, к высокотравным «среднегорным лугам» Средней Азии (58 са).
Пестрокостровые и пестрокострово-разнотравные луга с пестрым костром (Bromus variegatus), местами в Закавказье заменяемым ~ другим видом (Bromus adjaricus), более тесно связаны с другой группой разнотравья, широко распространенной по всему Кавказу и состоящей в основном из следующих видов: Anemone fasciculata, Campanula collina, Cepha-laria caucasica, Inula glandulosa, Linum hypericifolium, Ranunculus cauca-sicus, Scabiosa caucasica и некоторых других. В их составе обычно значительно участие бобовых (Onobrychis Biebersteinii, Anthyllis Boissieri, Trifolium canescens и др.). Из злаков обычны Koeleria caucasica, К. gracilis, Helictotrichon asiaticus.
Пестрокостровники распространены преимущественно по платообразным вершинам и пологим склонам, не имеющим следов произрастания на них леса. Их травостой несколько менее высок, чем у вейниковых лугов (50—70 см), и соответственно менее производителен (15—20 ц/га). Однако он обладает лучшими кормовыми свойствами и годится как для сенокошения, так и для умеренного выпаса.
Пестроовсяничные луга с преобладанием пестрой овсяницы (Festuca raria) — плотнокустового злака с крупными дерновинами — распространены преимущественно по южным сухим склонам и занимают на них громадные полосы до 500 м шириной (от 2300 до 2800 м над у. м.). В южных и восточных частях высокогорий Кавказа пестрая овсяница выходит на склоны других экспозиций, вплоть до северной, и поднимается высоко вверх, в альпийский и даже субнивальный пояса. В последнем она растет отдельными небольшими дерновинами; в альпийском поясе образует фитоценозы пустошного характера, а в субальпийском — близкие к горным степям или к остепненным лугам. Поселяясь в разнообразных ценозах с корневищными или рыхлокустовыми злаками, пестрая овсяница вытесняет эти злаки и образует множество разнообразных ассоциаций. Жесткая листва пестрой овсяницы охотно поедается скотом лишь весной и в начале лета, пока не загрубеет, а громадные кочкообразные дерновины затрудняют сенокошение. Вследствие этого, пестроовсяничники являются относительно плохими кормовыми угодьями, несмотря на довольно большую урожайность. В районах сильного распространения пес.троопсяния-пиков необходима борьба против дальнейшего расширения их площадей за счет лучших кормовых угодий; однако там, где пестрая овсяница за-дерняет голые скалы и осыпи, она является очень ценным растением. Ценна она также как пастбищный злак, хорошо выносящий длительный выпас с большой нагрузкой, защищающий от смыва и ос.ыпяния чрезмерно выгрызенные и вытоптанные овцами горные пастбища на крутых склонах.
Сильно распространяющимися и малополезными в кормовом отношении фитоценозами являются также белоусники, развивающиеся на Кав казе в более увлажненных районах субальпийского пояса, нижних частей альпийского пояса и верхних — лесного. На их месте желательна закладка культурных, удобряемых, высокопродуктивных лугов, вполне возможная в условиях плато и плоско-пологих склонов, с «я «ими большею частью связано распространение белоусников.
Литература
КУзаецов (1909); Я. С. Медведев (1915); Р. И. Аболин (1932); Н. А. Буш Буши Е. А. Буш (1936); Н. П. Введенский (1939); Р. А. Еленевский /<а/4 5^АсФхвдоРов (1940, 1942); В. М. Богданов (1941, 1947); А. К. Магакьян (|941, 1947, 1953); П. Д. Ярошенко (1942); А. Г. Долуханов (1946); А. Г. Долуханов г?' п‘ Сахокиа и А. Л. Харадзе (1946); А. А. Гроссгейм (1948); И.Х. Блюменталь и О. Л. Петровичева (1951); Е. В. Шиффере (1953); И. И. Абрамов (1954).
57 са — среднеазиатские в сочетании с высокогорными степями
Высокогорные среднеазиатские альпийские и субальпийские луга ( в сочетании с высокогорными степями) имеют весьма обширный ареал: от Джунгарского Алатау до горного Бадахшана. Однако пространства, показанные на «Геоботанической карте СССР» как относящиеся к этому выделу, далеко не являются сплошь луговыми и степными. В условиях Средней Азии высокогорные луга большею частью разбросаны среди каменистых склонов, осыпей, скал или узкими полосками вытянуты по вершинам горных хребтов.
Особенно фрагментарно распределение альпийских лугов, мелко-злаково-разнотравных и мелкоосоково-разнотравных, встречающихся среди каменистых обнажений, снежных полей и ледников. Основу их дернины составляют Carex stenocarpa, Festuca ovina s. 1., Роа alpina и некоторые другие осоки и злаки, а также кобрезии, но физиономически более характерны представители разнотравья с укороченными стеблями и с пестрыми яркими цветами: Primula algida, Gentiana falcata, Inula rhizocephala, Aster alpinus и др.
На моренных отложениях Алайской долины, по северным склонам на хребтах Киргизском, Заилийском и Джунгарском Алатау большие площади занимают кобрезиевые плотнодернинные пустошные луга, образованные преимущественно двумя видами — Cobresia capilliformis и С. pamiroalaica, с небольшой примесью осоки (Carex stenocarpa} и некоторых других высокогорных растений.
Значительная часть высокогорных формаций, многими ботаниками относимых к альпийским лугам, выделена на карте особым знаком (101), так как они относятся к особому типу высокогорной подушковидной растительности. Элементы этой растительности имеются и в других высокогорных областях, в частности на Кавказе, но там они не образуют крупных и обособленных массивов.
Субальпийские луга занимают большие площади, располагаясь преимущественно по северным склонам, непосредственно выше так называемых «среднегорных лугов» (58 са). Высота их очень разнообразна, нижняя граница на северных хребтах (Заилийский Алатау) расположена на высоте около 1500 м над у. м., а к югу она постепенно повышается (на Ферганском хребте — 2500—2700 м над у. м.).
Типичные субальпийские луга Средней Азии сочетаются с массивами елово-пихтовых лесов из тяныпанской ели (Picea Schrenkiana} и пихты (Abies Semenovii), с арчевниками (из Juniperus turkestanica, J. pseudosa-bina и др.), co скалистыми обнажениями и горными степями.
Злаковая основа субальпийских среднеазиатских лугов образована несколькими постоянно встречающимися видами (Trisetum sibiricum, Т. spicatum, Helictotrichon Schellianum, Brachypodium pinnatum и др.); из многочисленного пестрого разнотравья наиболее распространенными являются Aconitum rotundifolium, Doronicum altaicum, Trollius altaicus, Anemone protracta, Crepis sibirica, Anthriscus aemula, Polemonium coeru-leum s. 1., Ligularia altaica. Последняя связана с ежой (Dactylis glomerata} и более свойственна «среднегорным лугам».
Часто отмечается остепненность субальпийских лугов; к числу остепненных формаций относятся фломисовые и крестовниковые луга с эдификаторами Phlomis oreophila и Senecio soongoricus.
Остепненные луга переходят в степи с господством Stipa kirghisorum или Festuca sulcata,. с участием Helictotrichon desertorum, а также Poten-
Ulla nivea, Scabiosa songorica, Gentiana Olgae, Aster alpinus, Inula rhizo-cephala и других высокогорных растений.
Литература
П. А. Баранов (1924); Р. И. Аболин и М. М. Советкина (1930); Е. П. Коровин (1934); Р. А. Еленевский (1938, 1940а); М. М. Советкина (1938); М. Г. Попов (1941); Н. И. Рубцов (1948а); В. П. Голоскоков (1949).
58. Среднегорные луга, местами в сочетании с луговыми степями
К среднегорным лугам отнесены на «Геоботанической карте» суходольные луга лесного (пли лесо-лугового) пояса. Большая часть их лежит на высотах от 800 до 1800 м над у. м. на Кавказе и от 1000 до 2500 м над у. м. в горах Средней Азии. Лишь небольшие участки, показанные в северо-западной части Кавказа (или не попавшие на карту по масштабу), лежат значительно ниже (на высоте 200—400—600 м над у. м), но объединены со среднегорными лугами, так как типологически от них не отличаются. Верхний предел среднегорных лугов доходит на Кавказе до 2000—2400 м над у. м. (Дагестан), а в Средней Азии до 2700 м над у. м.
Большая часть среднегорных лугов, повидимому, генетически связана с лесами, а некоторые из них являются непосредственно послелесными, развивающимися на месте вырубленных и выкорчеванных лесов. Так, к послелесным лугам относятся прежде всего ассоциации вейниково-разнотравные (с Calamagrostis arundinacea.), полевицево-разнотравные (с Agrostis capillaris) и трясунково-разнотравные (с Briza media). На Кавказе они развиваются на месте буковых, дубовых и березовых лесов, преимущественно по склонам северной, северо-западной и северо-восточной экспозиций, на вышедших из-под леса бурых лесных слабооподзоленных или серых лесных оподзоленных почвах.
Не столь явно послелесное происхождение у другой группы лугов, связанных с горнолуговыми черноземовидными почвами и обычно относимых к остепненным лугам. Среди них луга, злаковую основу которых составляют перистая коротконожка {Brachypodium pinnatum), ежа (Dactylis glomerata), луговая овсяница {Festuca pratensis), луговой мятлик {Роа pratensis), белая полевица {Agrostis alba), степная тимофеевка {Phleum phleoides) и некоторые другие. Значительное место занимают на Кавказе также луга, образованные низкой осокой {Carex humilis). В составе разнотравья здесь наиболее обычны растения, часто встречающиеся на опушках лесов и в луговых степях. Характер почвы и травостоя позволяет предполагать здесь с одинаковой степенью достоверности и остепнение вышедших из-под леса лугов и олуговение степей. Возможно, что здесь перекрещивались оба эти процесса. В пределы контура входят и типично степные участки, на которых почва представляет собою чернозем, а травостой — луговые или настоящие степи. С другой стороны, часть среднегорных лугов (мелкие участки) поглощена контурами горных степей (83) или окультуренных территорий, расчищенных из-под леса (108).
Среднегорные луга большей частью являются хорошими кормовыми угодьями, преимущественно сенокосными, но частично и пастбищными? Особенно высокими качествами обладают остепненные луга, образованные прекрасными кормовыми злаками с большим количеством бобовых; собственно послелесные луга бедны в кормовом отйоше-* нии, но некоторые из них довольно высоко производительны (по грубой массе).
58 ск — североказахстанские
Североказахстанские среднегорные луга показаны на «Геоботанической карте» на северном склоне хр. Саур, в среднегорном поясе последнего. Здесь лесо-луговой пояс залегает на высоте 1650—2350 м над у. м. (Сапожников и Шишкин, 1918).
Склоны северной экспозиции заняты здесь хорошо развитыми лугами с господством злаков как луговых (Trisetum sibiricum, Deschampsia caespitosa, Роа sibirica, P. nemoralis, Alopecurus pratensis, Festuca rubra, Agropyrum repens и др.), так и луговостепных (Phleum phleoides) с большей или меньшей примесью разнотравья. У верхней границы лесо-лугового пояса появляются в составе этих лугов некоторые высокогорные виды [Роа alpina, Trisetum spicatum, Anemone narcissiflora s. 1. (видимо, A. Schrenkiana), Pachypleurum alpinum и др.]. У нижней границы лесолугового пояса в состав лугов проникают степные виды, например перистый ковыль (видимо, Stipa rubens), тырса (5. capillata) и др. По ложбинам местами развивается «алтайское большетравье», в состав которого входят высокорослые виды, главным образом из разнотравья (Aconitum excelsum, Heracleum dissectum, Bupleurum aureum, Polygonum alpinum, Sanguisorba alpina, Polemonium coeruleum s. 1., Lathyrus Gmelinii, Lilium martagon, Ligulqxia altaica, Valeriana officinalis s. 1., Allium hymenor-rhizum) с примесью злаков (Festuca pratensis и др.).
Южные склоны гор заняты степями на суглинистой почве с господством типчака (Festuca sulcata), ковылей (Stipa capillata, S. orientalis), с большей или меньшей примесью других дерновинных злаков (Koeleria gracilis, Agropyrum cristatum), степного разнотравья (Thalictrum foetidum, Potentilla acaulis, Libanotis buchtormensis, Cirsium igniarium и др.), полукустарничков, преимущественно степных (Artemisia frigida, Л. Gmelinii A. dbtusiloba, Ziziphora clinopodioides sA., Lagochilus diacanthophyllus)\ из кустарников на этих степных склонах произрастают Spiraea hypericifolia, Caragana pumila.
По крутым северным затененным склонам встречаются в большинстве случаев редкостойные насаждения сибирской лиственницы (Larix sibirica), обычно с хорошо выраженным травяным покровом с господством лугово-лесных форм. Лиственничники известны в верхних частях долин Уидене, Джеминей, Теректы, Джалпакарагай, Калмакпай, Са-рыбулак, Коржунбай, во всех истоках р. Кендерлык и по р. Чеганобо. По р. Теректы известны насаждения шренковой ели (Picea Schrenkiana).
Местные луга используются как сенокосы и пастбища.
Литература
В. В. Сапожников и Б. К. Шишкин (1918).
58 к — кавказские
Среднегорные луга развиты главным образом на Северном Кавказе: в горной части Ставропольского края, в Кабардинской АССР и в Дагестане. В Закавказье они распространены незначительно, преимущественно мелкими участками, и показаны на «Геоботанической карте» лишь в одном месте — близ г. Ахалцихе в Грузинской ССР; остальные их массивы вошли главным образом в пределы луговых горных степей (83 к), занимающих значительные пространства в Армении и Грузии.
В западной части Северного Кавказа, где обычно в связи с обилием осадков вырубаемые леса быстро восстанавливаются и вновь завоевывают территорию, сколько-нибудь значительные массивы среднегорных (вернее, здесь низкогорных) лугов развиты лишь в засушливых районах близ Геленджика и Новороссийска, да небольшими массивчиками по сухим южным склонам известняковых хребтов в других районах. Они представлены здесь преимущественно остепненными лугами, перемежающимися с горными степями.
Наиболее остепнены луговые участки на окраинных низких хребтах близ Геленджика. Мезофильные злаки (ползучий пырей, ежа) и представители луговостепного разнотравья сочетаются здесь с типчаком (Festuca sulcata), красивейшим ковылем (Stipa pulcherrima) и рядом ксерофитов и петрофитов крымско-кавказского типа (Alopecurus vaginatus, Bromus (Zerna) tomentellus, Iris laurica, Asphodeline taurica, Sideritis euxina, Trigonella cretacea и др.].
Значительно остепнены также среднегорные луга на южных склонах передовых хребтов Северного Кавказа, так называемой куэсты. Остеп-ненные луга встречаются по всему протяжению Мелового и Скалистого хребтов и особенно хорошо выражены на отрезке от р. Малой Лабы до р. Чегем, а затем в Дагестане; в лесистых районах Центрального Кавказа они занимают меньшие площади, но все же хорошо выражены. Некоторые формации остепненных лугов связаны преимущественно с южными склонами (например, сеслериево-разнотравные луга с Sesleria phleoides, осочково-костровые луга с Carex humilis и Bromus riparius, типчаково-осочковые с Carex humilis и Festuca sulcata).1 Другие (например коротконожково-разнотравные с Brachypodium pinnatum), занимая южные склоны в верхних частях лесного пояса п заходя по ним в пределы субальпийского пояса, ниже, в более теплых частях лесного пояса, переходят на склоны других экспозиций, вплоть до северной.
Коротконожковые и низкоосочковые луга — наиболее распространенные из всех остепненнолуговых формаций Северного Кавказа. Первые являются характерными представителями среднетравных лугов, используемых и под выпас и под сенокос, вторые — типичные низкотравные луга, используемые в качестве пастбищ. Те и другие — хорошие кормовые угодья с производительностью в 6—10 ц/га на низкотравных лугах и до 20—30 ц/га на среднетравных. Их кормовая ценность определяется хорошей поедаемостью и питательностью не только эдификаторов (осочки и коротконожки), но и большинства других компонентов травостоя, в состав которого часто входят такие кормовые травы, как ползучий пырей, белая полевица, береговой костер, сборная ежа, луговая овсяница, луговой овсец (Helictotrichon pratensis), луговой трищетинник (Trisetum pratense), луговая чина (Lathyrus pratensis), лядвенец (Lotus caucasicus), люцерны (Medicago glutinosa, M. falcata s. 1. и др.), эспарцеты (Onobrychis Biebersteinii, О. Cyri,O. iberica, О. inermis и др.), клевера (Trifolium ambi-guum, Т. medium, Т. montanum, Т. pratense), вики (Vicia sepium, V. va-riabilis и др.), а также многие хорошо поедаемые виды из разнотравья.
Значительно распространены остепненные луга, в которых эдифи-катором или субдоминантом является береговой костер. Они размещаются преимущественно по склонам и террасам в речных долинах; более мезо-фитные их ассоциации сближаются с коротконожковыми лугами, а более ксерофитные на каменистых склонах — с осочковыми. Некоторые их
1 Сообщества с Carex humilis, Bromus riparius и Festuca sulcata могут быть отнесены к луговым степям. (Примечание редакции).
ассоциации имеют в своем травостое большое количество типчака и ковы-лей {Stipa pulcherrima, S. capillata, S. stenophylld) и представляют собою явный переход к луговым степям, к которым они часто причисляются. Не особенно продуктивные по массе (8—12 ц/га) в природном состоянии, кострово-разнотравные луга сильно повышают свою производительность при орошении (до 20—30 ц/га); сочетание полива с удобрением может повысить их производительность до 40—50 ц/га. Такие поливные луга встречаются небольшими участками по долинам — в верховьях Кубани, Баксана, Уруха, во многих местах в Дагестане, но площади их могут быть значительно расширены.
При поливе и удобрении могли бы намного расшириться площади лугов, слабо остепненных или вполне мезофитных, в которых преобладает или играет значительную роль трищетинник {Trisetum pratense).
Собственно послелесные луга представлены на Кавказе главным образом вейниковыми (или - вейниково-разнотравными), полевицево-раз-нотравными и вейниково-полевицево-разнотравными лугами.
Вейниковые и вейниково-разнотравные луга имеют преимущественное распространение в субальпийском поясе (опис. 57); в среднегорном лесном поясе чаще встречаются вейниково-полевицево-разнотравные луга с вейниковым ярусом около 80—100 см и полевицевым — около 30—40 см высотой, с производительностью 15—20 ц/га. Кроме названных злаков-эдификаторов, здесь постоянно встречаются Anthoxanthum odoratum, Briza media, Carex pallescens, C. leporina, а из разнотравья — Achillea millefolium, Astrantia maxima, Prunella vulgaris, Campanula glomerata, Centaurea phrygia, Geranium sanguineum, Gymnadenia conopsea, Leontodon hispidus, Potentilla erecta, Ranunculus caucasicus, R. polyanthemus-, бобовых здесь мало {Trifolium hybridum, Vicia cracca и некоторые другие — единично).
На очень пологих склонах и плоских водоразделах эти луга часто заторфовываются и заболачиваются. На них поселяются Deschampsia caespitosa, Molinia coerulea, Holcus lanatus и другие растения — показатели бедных, закисленных почв. И без того невысококачественный в кормовом отношении травостой ухудшается и угодья теряют свою кормовую ценность. Этот процесс идет быстрее при выпасе и задерживается при регулярном сенокосном или комбинированном пастбищно-сенокосном использовании лугов.
Местообитания с более богатыми почвами, на которых произрастают крупнотравные луга, близкие к вышеописанному «субальпийскому вы-сокотравью» (опис. 57), в среднегорном поясе обычно распахиваются и используются под зерновые культуры, поэтому крупнотравные луга здесь имеют очень мало значения.
Литература
Т.' Ф. Пояркова (1927); Р. А. Еленевский (1939); П. Д. Ярошенко (1942а); В. М. Богданов (1941, 1947); А. А. Гроссгейм (1948); Е. В. Шиффере (1953).
58 са — среднеазиатские
В Средней Азии среднегорные луга имеют гораздо большую высотную амплитуду, чем на Кавказе. Так, в Северном Тянь-Шане они играют заметную роль в ландшафте, начиная с высоких предгорий, примерно от 1000—1200 м над у. м. Затем они весьма широко распространены в лесо-луговом поясе (до 2500—2700 м над у. м.), сменяясь выше границ этого
пояса лугами субальпийского типа. В Центральном и Западном 1 янь-Шане, а также в Памиро-Алае среднегорные луга имеют меньшее распространение. Они развиваются преимущественно по склонам северных экспозиций на почвах горнолугового и черноземного типов. В их сложении постоянное и большое участие принимают мезофильные и ксеромезофильные кустарники: шиповники {Rosa platyacantha. R. spinosissima. R. Ecae. R. Fed-tschenkoana). кизильники (Cotoneaster melanocarpa. C.multiflora). жимолости (Lonicera coerulea s. l.,L. Altmanni. L. heterophylla. L. Korolkovii). Нередко здесь встречаются небольшие участки еловых лесов (из Picea Schrenkiana) и лиственных лесов из яблони, осины, боярышника, грецкого ореха.
Невидимому, в большинстве случаев среднегорные луга являются вторичными образованиями, возникшими в результате уничтожения леса. Видовой их состав и сложение сильно меняются в зависимости от высоты над у. м., крутизны склона, экспозиции, характера почвы. Однако в них преобладает крупное мезофильное разнотравье, в котором значительную роль играют такие обычные евро-сибирские лесо-луговые виды, как Thalictrum minus. Solidago virga-aurea. Anthriscussilvestris. Galium boreale. Crepis sibirica) Geranium collinum. Aconitum excelsum. Lathyrus pratensis. L tuberosus. Lamium album. Campanulaglomerata.Polemoniumcoeruleums. 1., Nepeta pannonica. Heracleum dissectum и др. Кроме того, нередко встречаются местные горно-среднеазиатские расы, морфологически близкие к соответствующим евро-сибирским видам (например Dictamnus angu-stifolius. Dianthus Hoeltzeri. Libanotis Schrenkiana. Conioselinum latifolium). Бореальными видами особенно сильно насыщены среднегорные луга Северного Тянь-Шаня. В Центральном и Западном Тянь-Шане их значительно меньше. Совсем мало луговых бореальных элементов в системе Памиро-Алая, где горнолуговая растительность развита вообще очень слабо.
Из злаков в среднегорных лугах наиболее обычными являются Dactylis glomerata. Brachypodium pinnatum. Роа pratensis. Helictotrichon pu-bescens. Trisetum virescens. В Северном Тянь-Шане широко развиты луга с преобладанием сборной ежи (Dactylis glomerata) и коротконожки (Brachypodium pinnatum). Наблюдения показывают, что развитие этих злаков в луговом травостое в большинстве случаев связано с постоянным сенокошением определенных площадей, так как под влиянием регулярного выкашивания травостоя злаки разрастаются и получают преобладание за счет угнетаемого разнотравья. Разнотравные луга очень пестры и неустойчивы по видовому составу. Обычно в них господствует крупное разнотравье (Heracleum dissectum. Conioselinum latifolium. Lavatera thuringiaca. Ligularia persica. в Джунгарском Алатау — L. macrophylla. Dictamnus angustifolius. Inula helenium. Rumex tianschanicus. Thalictrum minus. Alcea nudiflora. Polygonum coriarium. P. alpinum.). Для Северного и Центрального Тянь-Шаня весьма характерны лигуляриевые луга с преобладанием высокого, крупнолистного сложноцветного — Ligularia persica.
Травостой среднегорных лугов густой (100% покрытия), двух-трехъ-ярусный, причем верхний ярус достигает высоты 150 см и более. При сенокошении эти луга дают от 10 до 30 ц/га хорошего сена. Значительного улучшения их можно достигнуть применением даже таких простейших мероприятий, как регулярное сенокошение и расчистка от кустарников. В условиях Северного Тянь-Шаня среднегорные луга близ нижних пределов своего распространения распахиваются под картофель и другие культуры.
В условиях сложного, сильно пересеченного, крутосклонного рельефа среднегорья, наряду с лугами, имеются и степи, которые занимают склоны южных экспозиций с почвами черноземного типа. Обычно это типчаковые или типчаково-ковыльные степи, иногда с обильным мезофильным разнотравьем, что придает им луговой облик. Так, в Джунгарском Алатау на высоте 1300—1800 м над у. м. с лугами сочетаются красноковыльноразнотравные луговые степи (на выщелоченных черноземах) с преобладанием Stipa rubens, Onobrychis ferganica, Fragaria viridis, Phleum phleoides, Polygonum alpinum, Phlomis pratensis с большим участием Bromus inermis, Poa pratensis, Galium boreale, Helictotrichon pubescens. Фрагменты подобных луговых степей встречаются также в Заилийском Алатау. На хребтах Киргизском, Кетменском, Кунгей-Алатау, Терскей-Алатау, на щебневатых черноземных почвах развиваются ковыльно-разнотравные степи с господством Stipa kirghisorum.B их видовом составе также участвует значительное число луговых, мезофильных и ксеромезофильных видов.
Горные луговые степи обладают густым травостоем и представляют собою хорошие сенокосные угодья, дающие до 10—12 ц/га сена. Они используются в качестве летних пастбищ.
Л итература
Е. П. Коровин (1934); М."М. Советкина (1938); М. Г. Попов (1941, 1941а); Н. И. Рубцов (1948).
59. Травяная растительность океанических островов (Тихий океан)
На «Геоботанической карте» травяная растительность притихоокеап-ского типа показана на Командорских о-вах, в южной оконечности Камчатки, в западной части Карагинского о-ва и на о-ве Шумшу (самый северный из Курильских о-вов). На меньшей площади в сочетании с другими формациями, преимущественно с березняками и зарослями ольховника, она встречается и в других районах Камчатки, а также на всех Курильских о-вах.
Развитию травяной растительности в названных районах очень благоприятствует морской климат с прохладным летом, мягкой зимой и большим количеством осадков в течение всего года. Зимой эти районы подвержены действию северо-восточных тихоокеанских циклонов, которые оказывают влияние также и на восточную Камчатку, принося обильные снегопады. Летом осадки приносятся юго-восточными муссонами с Тихого океана.
Сильные ветры в сочетании с очень умеренными температурами вегетационного периода и частыми туманами препятствуют развитию древесной растительности.
Из древесных пород к этим условиям наиболее приспособлена береза (Betala Ermani), а из кустарников еще в большей степени ольховник (A Inaster kamtschaticus) и некоторые ивы. Однако кустарники, по сравнению с травяными сообществами, развиваются в более защищенных местах, а рощи каменной березы там, где вегетационный период менее прохладный. В районах, где на «Геоботанической карте» показана травяная растительность притихоокеанского типа, ольховник встречается на Карагинском о-ве, о-ве Шумшу и на южной оконечности Камчатки. В последнем районе растет и каменная береза; на Карагинском о-ве известны очень небольшие рощи ее, а на о-ве Медном — отдельные деревья. На о-вах Беринга и Шумшу береза отсутствует.
Травяная растительность на тихоокеанских островах и на Камчатке нередко развивается мощно, местами образуя сплошные покровы большой высоты. Некоторые ее сообщества физиономически подобны субальпийскому высокотравыо Кавказа (Сочава, 1953). Наиболее сильно травяная растительность развивается в долинах, дренированных впадинах, на пологих склонах — там, где формируются почвы, названные на «Почвенной карте СССР» (Розов, 1954-К) дерновыми. Плодородие этих почв зависит от того, что они развиты на богатых солями вулканических породах и неоднократно обогащались вулканическим пеплом. На более бедных почвах и в местах, зимой недостаточно защищенных снежным покровом, травяные сообщества замещаются своеобразными тундроподобными верещатниками, в сочетании с которыми они обычно встречаются на островах.
На Камчатке, в результате влияния на нее азиатского антициклона в зимнее время, а также влияния холодного Охотского моря летом и, наконец, вследствие массивности самого полуострова, по сравнению с океаническими островами, создаются более континентальные условия, способствующие лучшему развитию древесной растительности. В силу этого на Камчатке шире распространены более или менее редкостойные березовые леса с мощным травяным покровом (опис. 40), нежели сообщества лугового типа. Последние на Камчатке встречаются небольшими по площади участками, в местообитаниях, где почвенно-грунтовые условия не благоприятствуют развитию древесной растительности, преимущественно в долинах рек, в подгорных ложбинах с избыточным грунтовым увлажнением. В этих условиях мощный травяной покров, в свою очередь, препятствует возобновлению деревьев. Местами в поймах Камчатки травяные сообщества развиты особенно пышно и достигают высоты до 2 м, что объясняют (Комаров, 1912, 1940) еще и обогащением почвы фосфорными и азотными соединениями в результате сгнивания большого количества рыбы, погибающей во время нереста.
На Курильских о-вах значительному развитию травяной растительности препятствует горный рельеф. Самые большие площади ее находятся на о-ве Шу мшу, который отличается от всех Курильских о-вов равнинным рельефом и выстлан толщей рыхлых пород с хорошо развитыми дерновыми почвами. На других островах Курильской гряды распространение травяных сообществ связано с речными долинами, приозерными понижениями, различными котловинами и террасами вдоль морского берега.
Развитие травяных сообществ и сходного с ними по видовому составу травяного покрова в березняках и кустарниковых зарослях явилось основанием для выделения Камчатки и прилегающих к ней островов в особую Северотихоокеанскую область (Лавренко, 1950в) или группу лесо-луговых северотихоокеанских провинций (Сочава, 1953).
О широком распространении травяных сообществ на севере Тихоокеанской области писал Пассарге (Passarge, 1921). Однако он упустил из виду распространение их на Камчатке и Курильских о-вах и ошибочно причислил к этой области слишком большие пространства на Аляске.
За пределами СССР травяные сообщества притихоокеанского типа встречаются на Алеутских о-вах и на северо-западном побережье Северной Америки. Притихоокеанская травяная растительность корреспондирует травяной растительности Приатлантической субарктики (южная Гренландия, Исландия, Фарерские и другие о-ва); последнюю Е. М. Лавренко (1950в) выделяет в особую ботанико-географическую область, «плакорным» типом растительности которой являются травяные формации
лугового типа. Сообщества высокотравья для Приатлантической Субарктики не характерны.
На о-вах Командорских, Карагинском, Шумшу и в южной части Камчатки, где на «Геоботанической карте» показана травяная растительность притихоокеанского типа, в зависимости от местных экологических условий развиваются различные группы травяных сообществ, отличающиеся по структуре травяного покрова и сочетающиеся с другими растительными группировками, которые по условиям масштаба не могли быть особо показаны на «Геоботанической карте». На богатых аллювиальных почвах речных долин с достаточным проточным увлажнением нередко развиваются сообщества высокотравья. Наиболее обычными доминантами высокотравья бывают лабазник {Filipendula kamtschatica), барапник {Senecio palmatus), вейник {Calamagrostis Langsdorfjii), дудник {Angelica refracta), кровохлебка {Sanguisorba tenuifolia). Встречаются в сообществах высокотравья и многие другие виды {Aconitum maximum, Cacalia kamtschatica, Heracleum dulce, Cirsium kamtschaticum, Chamaenerium angustifo-lium, Veratrum oxysepalum, Geum macrophyllum, G. aleppicum и др.).
В более влажных депрессиях и в поймах, где сказывается некоторое заболачивание, развиваются вейниковые сообщества {Calamagrostis Langs-dorffii), в которых разнотравье играет меньшую роль. Общее число видов в вейниковых сообществах не столь велико {Carex Gmelini, Equise-tum arvense, Equisetum Komarovii, Trollius Riederianus, Ranunculus repens, Fritillaria kamtschatcensis и др.).
Высокотравные сообщества в долинах местами сочетаются с зарослями ив и ольховника. Покров в этих кустарниковых зарослях нередко состоит из тех же представителей высокотравья, большинство которых на Камчатке растет и в березовых лесах.
Высокотравные группировки развиваются также у выходов ключей, в приозерных и других понижениях без застойного увлажнения.
На более обедненных почвах на водоразделах, часто на высоких речных террасах и морских террасах, травяной покров становится разреженнее, и высокотравье в нем не играет уже большой роли или отсутствует. Помимо разнотравья, в сложении травяных сообществ здесь принимают участие злаки, местами преобладающие на больших площадях {Calamagrostis Langsdorffii, Trisetum sibiricum, Роа pratensis, Deschampsia ber ingensis)', растут некоторые осоки {Carex subumbellata и Carex stylosa), ситник {Juncus beringensis) и ожика {Luzula parvijlora). В составе разнотравья встречаются некоторые виды, свойственные высокогорным луговым сообществам {Artemisia arctica, Arnica unalaschkensis, Anemone villa sissima), и ряд других растений {Geranium erianthum, Parageum calthifolium, Arabis Stelleri, Polygonum viviparum, Hieracium triste, Delphinium br achy centrum, Veratrum oxysepalum).
На более повышенных участках, особенно там, где ветры сдувают зимой снег, травяная растительность не образует сплошных покровов, а встречается пятнами среди сообществ горнотундрового типа, в которых нередко преобладает вороника {Empetrum nigrum) и представлены многие другие кустарнички {Diapensia lapponica, Arctous japonica, Bryanthus Gmelini, Ledum palustre, Salix arctica, Vaccinium vitis-idaea, V. uliginosum, Loiseleuria procumbens, Cassiope tetragona,C. lycopodioides) и дерновинные травы {Saxifraga bronchialiss. 1,, S. serpyllifolia), часто встречаются лишайники. Травяные сообщества среди этих тундроподобных верещатников представлены группировками, в которых господствуют горнотундровые травы {Artemisia trifurcata, Erigeron unalaschkensis, Veronica grandiflora, Coptis trifolia, Hierochloe alpina, Campanula lasiocarpa, C. Pallasiana)', растут здесь
также ожика (Luzula Kjellmanniana) и осока (Carex micropoda) и другие растения (Hierochloe odorata, Potentilla stolonifera, Primula cuneifolia, Veronica Stelleri, Pedicularis Chamissonis). Такие сочетания травяных сообществ с кустарничковыми особенно часто встречаются на о-ве Медном, в юго-восточной возвышенной части о-ва Шумшу, на столообразных возвышенностях северной половины о-ва Беринга и на сухих водоразделах западной части о-ва Карагинского.
На о-вах Карагинском и Шумшу, а также на южной Камчатке в районах повышенного рельефа, при условии снежной защиты зимой, среди травяной растительности развиваются отдельные небольшие пятна кедрового стланика и ольховника. На карте они обозначены внемасштаб-ными знаками.
Все пониженные сильно увлажненные водоразделы заняты сырыми болотного типа группировками, в которых преобладающую роль играют осоковые (Carex falcata, Carex rariflora, Eriophorum angustifolium), вейник (Calamagrostis neglecta) и ситник (Juncus Haenkei). В условиях избыточного проточного увлажнения большое участие принимают здесь представители болотного разнотравья (Iris setosa, Caltha palustris, Comarum palustre, Galium trifidum, Limnorchis convallariaefolius, Equisetum palustre и др.).
На пониженных водоразделах, где затруднен сток, развиваются участки моховых болот с типичными болотными кустарничками (Empetrum nigrum, Vaccinium uliginosum, Ledum palustre, Andromeda polifolia, Oxycoccus quadripetalus, Chamaedaphne calyculata) и травами (Rubus chamae-morus, Drosera rotundifolia и пр.). Эти группировки среди травяно-осоковых сообществ распространены повсеместно.
На о-ве Парамушире, где сырые болота обычны (Kudo, 1922), травы (Carex Middendorffii, С. crypthocarpa, Eriophorum angustifolium, Iris setosa, Cardamine pratensis, Sanguisorba tenuifolia, Juncus beringensis, Trichophorum caespitosum и многие другие, главным образом осоки и ситники) по числу видов на болотах преобладают над кустарничками (Oxycoccus quadripetalus, Andromeda polifolia). Особенностью этих болот является присутствие горнотундровых кустарничков (Salix phlebophylla, S. arctica, Sieversia pentapetala), а также остистого ятрышника (Orchis aristata).
На Командорских о-вах на болотистых замшенных местах растут пушица (Eriophorum angustifolium), морошка (Rubus chamaemorus) и другие травы (Fritillaria kamtschatcensis, Polygonum viviparum и пр.), здесь же местами встречается бузинолистная рябина (Sorbus sambucifolia).
Травяная растительность притихоокеанского типа представляет собою довольно значительный кормовой фонд, который используется в очень небольшой степени.
• Литература
Ф. Р. Челльман (1886—1887); Б. А. Федченко (1906); Е. К. Суворов(1911); В. Л. Комаров (1912, 1940); Н. В. Павлов (1936); И. К. Шишкин (1936); В. Н. Васильев (1944, 1946); А. И. Соловьев (1947); Е. М. Лавренко (1950в); В. Б. Сочава (1953); Н. Н. Розов (1954-К); Y. Yabe et К. Yendo (1904); S. Passarge (1921); V. Kudo (1922); E. Hulten (1927—1930, 1932—1933, 1937); M. Tatewaki (1928).
60. Приморские луга засоленных побережий в тундровой зоне
Приморские луга тундровой зоны формируются в особых эколого-эдафических условиях. Суровый арктический климат, засоленные почвы ' морских аллювиев, частое и систематическое орошение солеными водами
морских приливов и штормов обусловливают развитие специфической растительности.
Приморские луга расположены отдельными то крупными, то мелкими участками по всему побережью от Белого моря до Чукотского включительно. Наиболее крупные площади их отмечены на «Геоботанической карте» по побережью Янского залива (в восточной части моря Лаптевых), по побережью Омулляхской, Хромской и Гусиной губ и западнее устья Колымы — против о-вов Медвежьих (в западной части Восточно-Сиби р-ского моря). К западу и востоку от указанных массивов площади их незначительны и, по условиям масштаба, не везде могли быть показаны на карте.
Наиболее детально исследованы приморские луга тундровой зоны на западе их ареала: на Малоземельском побережье Варенцова моря (Лесков, 1936) и на побережье Мезенского залива и Чешской губы (Корчагин, 1937). А. И. Лесковым вскрыта динамика развития растительного покрова Малоземельского побережья от первоначальной стадии задер-нения до тундровой растительности. Им выделены три группы приморских лугов: луга низкого уровня, заливаемые каждым более или менее сильным приливом; луга среднего уровня, заливаемые только наиболее сильными приливами; луга высокого уровня, подвергающиеся лишь обрызгиванию при наиболее сильных приливах и штормах.
Приморские луга низкого уровня Малоземельского побережья характеризуются плотным низким ковром стелющихся и укореняющихся побегов ползучей бескильницы (Puccinellia phryganodes) с незначительной примесью Carex subspathacea, Stellaria humifusa л куртинами нитчатой водоросли. Приморские луга среднего уровня в прибрежной полосе представлены чистыми зарослями Carex subspathacea и Dupontia Fischeri, в которые местами внедряется Puccinellia phryganodes. По мере удаления от береговой линии и повышения уровня травяной покров этих зарослей обогащается внедрением Calamagrostis deschampsioides, Potentilla anseriua, Stellaria humifusa, меньше Matricaria grandiflora, Festuca rubra, Plartlago Schrenkii. В приморских лугах высокого уровня Carex subspathacea и Calamagrostis deschampsioides уже не играют роли доминантов, хотя и встречаются в значительном количестве. Для травяного покрова лугов высоких уровней характерно участие тундровых растений и мощное развитие мохового покрова. Основными ассоциациями здесь (согласно А. И. .Лескову) являются следующие: DupontiaFischeri-]-Senecio campester s. 1. — Drepanocladus uncinatus\ Salix reptans-]-Carex rariflora — Drepanocladus uncinatus\ Salix reptans -f- Carex subspathacea — Drepanocladus uncinatus. Близкие данные для приморских лугов побережья Мезенского залива и Чешской губы приводит А. А. Корчагин (1937).
По побережью Болыпеземельской тундры приморские луга более или менее значительные площади занимают в устьях рек, как, например, Пай-Яга, Хыльчу-Ю, Нерута-Вож (Андреев, 1932а), поймы которых здесь достигают 200—250 м ширины и подвержены влиянию морских приливов. На низких участках, заливаемых при каждом приливе, развивается четырехлистный хвостник (Hippuris tetraphyНа)', несколько выше расположенные, но еще часто заливаемые площади заняты зарослями дюпонции Фишера (Dupontia Fischeri) с примесью четырехлистного хвост-ника, гиперборейского лютика (Ranunculus hyperboreus), а на повышенных участках — приземистой звездчатки (Stellaria humifusa), гусиной лапчатки (Potentilla anserina),1 арктического нивяника (Leucanthemum
1 Видимо, Potentilla Egedii Wormek.
arcticum). На редко заливаемых участках с заторфовывающимися почвами развиваются заросли копьевидной ивы (Salix hastata); в травяном покрове этих зарослей господствуют редкоцветная осока (Carex rariflora), красная овсяница (Festuca rubra) 1 с примесью разнотравья (Leucanthemum arcticum, Lagotis minor, Rhodiola rosea, Potentilla Egedii, Saxifraga rivularis). Развит гипновый моховой покров (Aulacomnium palustre, Drepanocladus exannulatus, D. uncinatus).
На побережье Байдарацкой губы (Андреев, Игошина и Лесков, 1935) развиты приморские луга из галечной осоки (Carex glareosa), обертковидной осоки (С. subspathacea), щучковидного вейника (Calamagrostis deschampsioides), дюпонции Фишера (Dupontia Fischeri), темнобагряной валодеи (Vahlodea atropurpurea), песчаной овсяницы (Festuca arenaria) с примесью указанного выше разнотравья.
Для приморских лугов побережья между Леной и Колымой (Якутия) В. Б. Сочава (1933) указывает злаково-осоковые луга из Carex subspathacea и Dupontia Fischeri с участием разнотравья (Leucanthemum sibiricum, Matricaria grandiflora, Senecio congestum), сменяющиеся близ озер зарослями Carex stans и Arctophila fulva и осоково-пушицевыми болотами на равнине.
Такой же характер имеют приморские луга на побережье губы Буор-хая (Душечкин, 1937), состоящие из Carex subspathacea, С. glareosa, Stel-laria humifusa, Rhodiola borealis, Potentilla Egedii, Leucanthemum arcticum.
Для Чукотского побережья приморские луга описывает Б. Н. Городков (1939). На узкой полосе галечника поселяются отдельные экземпляры бутерлаковидной аммодении (Ammodenia peploides), приморской мертен-зии (Mertensia maritima), а на побережье Берингова моря еще ложноарникового крестовника (Senecio pseudoarnica) и пузырчатой меркни (Merckia physodes). На сильно засоленных, но уже задерненных участках господствуют Carex subspathacea и Dupontia Fischeri. На менее засоленных к ним присоединяется Calamagrostis deschampsioides, а из разнотравья — Stel-laria humifusa и темнокрасная родиола (Rhodiola atropurpurea). Повышенные участки, подвергающиеся лишь обрызгиванию солеными водами при сильных штормах, заняты бескильницево-осоковыми лугами с преобладанием тонкой бескильницы (Puccinellia tenella), морской осоки (Carex marina), обертковидной осоки (Carex subspathacea) и с участием Dupontia Fischeri, Роа arctica, Stellaria humifusa, Leucanthemum arcticum, Potentilla fragiformis, Saxifraga rivularis. В ходе дальнейшего развития приморские луга сменяются тундровой растительностью. Песчаные дюны морских побережий заняты открытыми группировками мягкого колосника (Elymus mollis), по мере задернения внедряются Calamagrostis deschampsioides, Rhodiola atropurpurea, Drepanocladus uncinatus.
Б. А. Тихомиров (1946a) связывает развитие приморских лугов Крайнего Севера Евразии со стадиями зарастания морских аллювиев. На первой стадии, в условиях активного процесса отложения и смыва морского аллювия и систематического увлажнения солеными морскими водами, развивается группа ассоциаций Carex subspathacea A-Puccinelliaphryganodes. На второй стадии при затухании аллювиального процесса и без систематического увлажнения солеными морскими водами ее сменяет группа ассоциаций Carex subspathacea-^Calamagrostis deschampsioides. На третьей стадии, в условиях прекращения аллювиального процесса и увлажнения только пресными (атмосферными) водами, развивается тундровая растительность с сохранением еще участков приморских лугов.
А. А. Корчагин (1937) в полосе побережий северных морей СССР выделяет две ботанико-географических области.
1 Видимо, Festuca arenaria Osbeck.
1. Арктическая область от Канина п-ва до Берингова пролива. Пионерами зарастания молодых аллювиев являются Puccinellia maritima, Р. phryganodesvLместами Hippuris tetraphylla. Их сменяют широко распространенные злаково-осоковые сообщества: на более засоленных участках — Carex subspathacea-\-Dupontia Fischeri, на менее засоленных участках -Carex subspathacea -|- Calamagrostis deschampsioides Dupontia Fischeri. I lo мере удаления от береговой линии и выхода из зоны затопления широко распространены осоковые сообщества — Carex discolor (в Европейской части) и С. stans (в Азиатской части). На илистых приречных и приозерных отложениях — Arctophila fulva.
2. Беломорская область (побережье Белого моря, Мезенского залива, северное и восточное побережья Кольского п-ва). Она характеризуется не только осоковыми ассоциациями с Carex subspathacea (реже С. discolor, С. stygia), но и широким развитием злаковых лугов бореального типа (Festuca rubra, Agrostis alba, Calamagrostis neglecta, Phragmites communis, Alopecurus ventricosus). Для нее свойственно также присутствие галофиль-ных растений (Plantago salsa, Triglochin maritima, Tripolium vulgare, Juncus Gerardii, из злаков — Puccinellia maritima, P. suecica, Agrostis maritima).
Приморские луга как кормовой фонд хорошего достоинства играют значительную роль на Крайнем Севере. В основном они используются для выпаса оленей. Продуктивность их различна. Наиболее широко распространенные луга из Carex subspathacea + Calamagrostis deschamp sioides-\-Dupontia Fischeri отличаются невысоким (10—15 см) травостоем и продуктивностью — 5—8 ц/га. Резко возрастает продуктивность крупноосоковых и крупнозлаковых приморских лугов в ассоциациях Carex stans, С. discolor, Arctophila fulva, Alopecurus ventricosus, Calamagrostis neglecta, развивающихся близ речек и озер с травяным покровом высотою в 60— 100 сми урожайностью от 15—20 до 25—30 ц/га. Такие луга используются под покос; они занимают сравнительно небольшие участки (полосы) близ рек и озер. Площадь их увеличивают путем искусственного спуска воды озер, днища которых зарастают Carex stans, Arctophila fulva, Calamagrostis neglecta (на востоке — Calamagrostis Langsdorffii).
Литература
В. H. Андреев (1932а); В. Б. Сочава (1933, 1934); В. Н. Андреев, К. Н. Игошина, А. И. Лесков (1935); Б. Н. Городков (19356, 1938а, 1939, 1944, 1946); А. И. Лесков (1936); В. И. Душечкин (1937); А. А. Корчагин (1937); Б. А. Тихомиров (1946а).
61. Солончаково-луговая и лугово-болотная растительность морских побережий в сочетании с солянковой, а местами с растительностью приморских песков (в степной и пустынной зонах)
Солончаково-луговая и лугово-болотная растительность морских побережий в сочетании с другими группировками на «Геоботанической карте» показана на побережьях Каспийского, Черного и Азовского морей в степной и пустынной зонах. Названное сочетание растительных сообществ развивается на морских наносах (глинистые берега и дно отступающего моря, песчаные морские косы, отмели, острова и пляжи), а также в приморской полосе речных дельт, сложенных преимущественно аллювием.
В зависимости от местообитания находится и состав растительности, всегда представленной сочетанием различных ассоциаций.
На обширной приморской полосе между Астраханью и Гурьевым, а также вдоль западного побережья Каспийского моря близ устья Кумы, Терека и Куры растительность развивается на недавно обнажившемся дне моря, покрывающемся главным образом зарослями тростника, солероса и однолетних свед Suaeda prostrata. S. confusa). Затем идут группировки татарской лебеды с солончаковой полынью, иетроси-мониями, солянками {Salsola crassa, S. dendroides). Дальше от береговой линии встречаются участки с поташником {Kalidium caspicum) и с сар-сазаном {Halocnemum strobilaceum), образующими бугристые солончаки; заросли тростника окаймлены солончаковыми лугами из прибрежии цы {Aeluropus litoralis) и бескильницы {Puccinellia dolicholepis, Р. Fominii, Р. gigantea). На песчаных дюнах, отделяющих коренной берег от бывшего морского дна, обычно растут волоснсц {Elymus giganteus) и крупная высокая полукустарниковая песчаная полынь {Artemisia Tschernieviand), а в Закавказье двулетняя A. scoparioides\ иногда здесь поселяется гре-бенчук {Tamarix ramosissima и др.) На некоторых участках берега, например близ Забурунья в Северном Прикаспии, развиты гряды бугров с кустами селитрянки {Nitraria Schoberi). Вершины более старых гряд, расположенных дальше от моря и уже менее песчанистых, покрыты группировками полукустарничковой полыни {Artemisia Lercheana. а в Закавказье — A. Meyeriana) с участием кохии, пустынного житняка и эфемеров; на еще более отдаленных от моря древних грядах развиваются полынные и ковыльно-полынные группировки с участием ковылей {Stipa capillata и S. Lessingiand).
На песчаных пляжах и островах растительность разрежена; здесь преобладают специфичные растения морских побережий. На побережье Черного и Азовского морей — обычно это разбросанные по приморскому песку экземпляры приморского синеголовника {Eryngium maritimum), морской горчицы {Cakile euxina), катрана {Crambe maritima), морковницы {Astrodaucus litoralis) и некоторых других прибрежных растений. На Каспийском побережье (в пределах Азербайджана и южного Дагестана) распространен красивый белоснежно-пушистый персидский вьюнок {Convolvulus persicus). В Азербайджане и Дагестане наблюдается, например, такая закономерность в распределении растений по песчаному пляжу, образованному прибоем морских волн. Непосредственно около воды растительность отсутствует, пляж сложен ракушняком. Дальше от воды редкими кустами растет Frankenia hirsuta, выдерживающая постоянное смачивание брызгами морской воды от прибоя. Следующая песчаная полоса густо покрыта Convolvulus persicus и Tournefortia sibirica. На месте перехода пляжа в коренной берег, на грядах песка, поселяются ситники {Juncus litoralis и J. acutus);
На песчаных побережьях Черного и Азовского морей наблюдается следующая последовательность в распределении растительности. Песчаный пляж со стороны открытого моря лишен сплошного растительного покрова. Здесь разбросаны отдельные экземпляры специфических «пляжных» растений, которые были упомянуты выше. Далее обычно следует песчаный вал, поросший волоснецом {Elymus giganteus), около которого нередко накапливаются песчаные бугры. За песчаным валом располагается также повышенная часть, сложенная песками и покрытая более или менее хорошо выраженной степной растительностью с господством Festuca Beckeri и Euphorbia Seguieriana. Кроме того, в ,составе этих степных сообществ на песках встречаются другие растения {Helichrysum arenarium, Agropyrum Lavrenkoanum, Medicago falcata ssp. erectaг
1 Medicago Kotovii Wissjul.
и др.). На сильно выбитых местах типчак исчезает. В понижениях среди такой бугристой степи на песках располагаются луга, в составе которых преобладают Agrostis alba, Calamagrostis epigeios, Holoschoenus vulgaris.
На косе Джарылгач в Каркинитском заливе наблюдается чередование ракушниковых хребтиков,покрытых тимьянниками с господством Teucrium polium- и с примесью Thymus dimorphus, Dianthus pseudarmeria и др. В понижениях между ними господствуют солончаковые луга (Agropyrum ruthenicum, Puccinellia gigantea, Aeluropus litoralis, Juncus maritimus), а также полынные (Artemisia maritima s. 1.) или солянковые группировки [Atriplex (Halimione) verrucifera, Salicornia europaea, Suaeda prostrata], с примесью Limonium Meyerii, L. caspium и других. Далее следует пониженная часть косы, примыкающая к морскому заливу (лиману). Здесь господствуют указанные выше солончаково-луговые п солончаковые группировки; нередко большие площади заняты зарослями тростника (Phragmites communis). Иногда ракушняковые валы не выражены, и тогда песчаная степь постепенно переходит в пониженную часть косы с засоленными почвами.
По низменному берегу заливов обычны валы из выброшенной во время бурь камки (Zostera minor, Z. marina), которая образует большие заросли в заливах.
В устьях высыхающих речек подъем от берега моря вглубь материка идет постепенно, но рельеф чрезвычайно расчленен многочисленными бывшими дельтовыми косами и протоками.
Последовательность смен растительного покрова здесь такова (на примере низовьев Куры): 1) непосредственно в воде — заросли тростника (Phragmites communis) с участием солероса (Salicornia europaea) и других однолетних солянок; 2) дальше от воды, но при близких грунтовых водах — солончаковатые луга из видов Puccinellia с примесью других голофитных растений; 3) еще дальше от воды на слабом повышении — заросли однолетних растений — эфемеров с преобладанием Phalaris minor и Bromus japonicus, образующих эфемеровые луга; 4) на береговых валах — пустынные сообщества с преобладанием Salsola dendroides, где участвуют полыни, эфемероид Роа bulbosa, однолетние злаки (Phleum paniculatum,Hordeum leporinum,Eremopyrum orientals и др.) и разнотравье из однолетних двудольных (Pterotheca Marschalliana и др.); 5) на коренном берегу — полынная пустыня.
Солончаково-луговая и солончаково-болотная растительность морских побережий большею частью мало производительна и представляет собой пастбища невысокой продуктивности. Местами луговые группировки иногда выкашиваются.
Литература
И. С. Косенко (1924); А. А. Гроссгейм (1926, 1932, 1948); А. Ф. Флеров (1926); А Ф Флеров и В. А. Флеров (1926); Ф. Я. Левина и М. С. Шалыт (1927); Н. А. Деся-това-Шостенко’ и Ф. Я. Левина (1928); М. И. Котов (1928а, 1938); Е. Лавренко и Н. Десятова-Шостенко (1928); Е. В. Шифферс-Рафалович (1928); Т. С. Гейдеман и И Н. Дорошко (1931); А. А. Колаковскдй (1933); А. Г. Рубцова (1940); И. Н. Бейдеман (1951,1954); Б. А. Быков (1954);. В. Г. Катышевцева (1955).
62. Солончаково-луговая и лугово-болотная растительность в сочетании с солянковыми и степными сообществами (разливы, лиманы, высыхающие плавни и озера в степной и пустынной зонах)
В степной зоне и отчасти на севере пустынной зоны, по берегам высыхающих озер, в низовьях некоторых рек, а также в плоских понижениях, куда сбрасывают свои воды бессточные реки, развивается солончаково-
луговая и лугово-болотная растительность в своеобразном сочетании с солянковыми и степными сообществами. Развитию таких сочетаний способствуют мезорельеф (чередование грив с бессточными понижениями при общей равпинности), резко выраженная переменность увлажнения (обильное длительное весеннее обводнение и сухость летнего периода), засоленность грунтовых вод и почв.
Более крупные площади солончаково-луговой и лугово-болотной растительности, сочетающейся с солянковыми и степными сообществами, показаны на карте в пределах ряда пониженных территорий степной зоны и северной части пустынной зоны: в дельте Кубани (Косенко, 1924, 1934; Богдан и Шумаков, 1925; Шифферс-Рафалович, 1928, 1928а; Флеров, 1930), по нижнему течению Куры, протекающей по Кура-Араксинской низменности Закавказья (Гроссгейм и Колаковский, 1929, 1930; Гейдеман и Дорошко, 1931; Гроссгейм, 1932, 1948; Колаковский, 1933; Бейдеман, 1951, 1953, 1954), в дельтах Сулака и Терека (Новопокровский, 1932; Шиффере, 1937; Гроссгейм, 1948). Особенно большие площади эта растительность занимает в Прикаспийской низменности — по р. Куме, в лиманных понижениях вдоль западной окраины низменности (Пачоский, 1892: Высоцкий, 1915; Захаров, 1927; Волгунов, 1928; Тереножкин, 1954), в лиманах и «разливах» междуречья Волга—Урал северной части той же низменности (Михеев, 1916; Ларин, 1926а; Лейсле, 1936; Доскач, Ларин, Левина, 1952; Иванов, 1954; Ларин, Манохина, Акимцева, 1954; Ларин, Шиффере, Левина, Замятнин, Иванина, Варивцева, Непли, Сыро-комская, 1954; Левина, 1954), в нижнем течении Урала (Пояркова и Коровник, 1931; Быков, 1954; Никитин, 1954а; Пивоваров, 1954), на востоке Прикаспийской низменности в разливах и лиманах низовий Уленты, Бул-дурты, Калдыгайты, а также в долинах Уила, Сагиза и Эмбы, протекающих по Предуральскому плато, Предустюртской равнине и Прикаспийской низменности (Никитин, 1928, 1951, 1954, 1954а; Никитин и Поярков, 1935).
Обширные площади сочетания луговой, болотной, галофитной и степной растительности занимают также в лесостепной и степной частях Западно-Сибирской низменности и в пределах наклонной равнины Павлодарского Прииртышья, главным ^образом по низменным побережьям озер Чаны, Ажбулат и Кулундинского и вдоль рек, впадающих в эти озера [Танфильев, 1902; Крылов, 1913, 1916; Кузнецов (Владимирский), 1915; Баранов и Горшенин, 1927; Куминова и Вандакурова, 1949; Куминова, 1949а; Вандакурова, 1950; Кузьмина, 1953], а также по побережьям других озер (Карасор, Улькенкарой, Теке, Селетытенгиз, Жалаулы) и низовьям рек, в них впадающих (Гордягин, 1897, 1900—1901; Городков, 1913а, 1916, Пташицкий, 1913а; Дробов, 1914; Спиридонов, 1923,1927— 1928, 1928; Баранов, 1927).
Следующий обширный район распространения этой растительности — Тургайская столовоостанцовая равнина, особенно Тургайская сквозная долина, где соответствующая растительность приурочена преимущественно к низменным побережьям озер (Короткий, 1914; Крашенинников, 1925; Павлов, 1926; Стрельников, 1938).
Значительные площади эта растительность занимает в пониженных частях Казахского мелкосопочника, в частности в Тенгизской впадине, по днищу озерных котловин, вдоль реки Нуры, а также по побережьям других озер, например Карасор (Зелинский, 1910, 1912; Капелькин, 1910; Смирнов, 1912; Рожанец-Кучеровская, 1914, 1916; Шишкин, 1914а; Косинский, 1916; Ганешип, 1917; Крашенинников, 1925; Павлов, 1949). Кроме того, массивы с соответствующей растительностью показаны на
карте в Зайсанской котловине и Алакульской впадине (Лппщиц, 1931; Борисова, 1935), в Чуйской степи в восточном Алтае (Калинина, 1948) и на Юге Борзянских степей в Забайкалье.
Для лесостепной и степной части Западно-Сибирской низменности характерна равнинность, слабое развитие речных систем, причем только наиболее крупные реки (Тобол, Ишим, Иртыш и самые крупные притоки их) не пересыхают в летний период ; большинство же степных речек пересыхает. Равнинность рельефа разнообразится невысокими увалами или грядами с крайне пологими склонами, вытянутыми с юго-запада на северо-восток и чередующимися с широкими плоскими понижениями (бывшими стоками вод), озерными котловинами и более мелкими понижениями, по которым сбрасываются весенние воды. Для понижений характерно наличие многочисленных озер, редко изолированных, чаще же в период весенних половодий соединенных в озерные системы. В летний период протоки, мелкие озера, как и степные речки, пересыхают. В связи с засоленностью почв и грунтовых вод преобладают соленые озера, но, наряду с ними, встречаются и пресные, питающиеся ключами более глубоких пресных грунтовых вод.
Наличие замкнутых понижений, озерных котловин, пересыхающих речек, засоленность грунта и грунтовых вод при резко выраженной переменности увлажнения способствуют развитию солончаково-луговой и лугово-болотной растительности в их своеобразном сочетании с солянковыми и степными сообществами. На дне котловин пресных или слабо засоленных озер и вдоль зарастающих степных речек широко развита водноболотная растительность: Phragmites communis, Scirpus Tabernaemontani, S. lacustris, Bolboschoenus maritimus, B. compactus, Typha angustifolia, T. latifolia, Scolochloa festucacea, Digraphis arundinacea, виды осок — Carex riparia, C. pseudocyperus, а в лесостепи Западной Сибири также С. 1а-siocarpa, С. diandra, с участием болотного разнотравья (Menyanthes trifoliate., Stachys palustris, Epilobium palustre, Dryopteris thelypteris и многих других). При мелководности озер и речек водно-болотная растительность часто заполняет весь водоем, занимая крупные площади («займища»). По периферии широкой полосы водно-болотной растительности развивается галофитнолуговая растительность с преобладанием злаков (Agropyrum repens, Alopecurus ventricosus, Hordeum brevisubulatum, Calamagrostis epigeios, Agrostis alba, Aneurolepidium Paboanum), а также Carex Karelinii', эти виды встречаются часто почти чистыми зарослями или более сложными смешанными сообществами. Под пологом крупных злаков и осок развит второй ярус из разнотравья с значительным участием более или менее галофильных растений (Triglochin maritima, Т. palustris, Tripo-lium vulgare, Galatella rossica, Cirsium esculentum,Limonium Gmelinii, Glaux maritima, Inula britannica и др.). Почвы под ними луговые и солончаковатолуговые. С усилением засоления почв по периферии галофитных («солончаковых») крупнозлаковых лугов развиваются галофитные бескильни-цевые луга с преобладанием Puccinellia tenuissima, часто чистыми зарослями, но обычно с примесью других галофильных растений (Aeluropus litoralis, Limonium Gmelinii, L. caspium, Juncus Gerardii. Artemisia nitrosa, Tripolium vulgare, Taraxacum bessarabicum). В более южных лесостепных и степных районах среди бескильницевых лугов пестрят пятна «злостных» солончаков с преобладанием типичных галофитов, преимущественно однолетних (Salicornia europaea, Camphorosma soongo-ricum, Petrosimonia Litwinowii, Suaeda prostrata, S. corniculata) и многолетних полукустарничковых солянок (Camphorosma monspeliacum, A triplex verrucifera), а также других галофильных полукустарничков (Frankenia
hirsuta) н галофильных видов многолетнего разнотравья {Limonium cas-pium, L. coralloides и др.).
По пологим склонам солончаковые почвы сменяются структурными столбчатыми солонцами, начиная с корковых до глубокостолбчатых с переходом на вершине склона в солонцеватые черноземные почвы. Смене почв отвечает и смена растительности: вышеописанная галофитная («солончаковая») растительность дна понижения по склонам сменяется типчаково-полынными и полынно-типчаковыми степями с преобладанием дерновин-ных злаков {Festuca sulcata, Koeleria gracilis), полукустарничковых {Artemisia nitrosa, A. rupestris) и травянистых полыней (Л. ponlica) с большим участием галофильных растений в нижних частях склона, которые выше по склону (на глубокостолбчатых солонцах и солонцеватых черноземах) уступают место степным растениям.
Такова общая схема распределения растительности в связи с мезорельефом, степенью засоления и увлажнения. Микро- и нанорельеф вносят свои коррективы: по мелким повышениям и понижениям растительность одного пояса проникает в другой пояс, что создает пестроту травяного покрова. Так, в пояс галофитных крупнозлаковых и бескильнице-вых лугов часто внедряются пятна злостных солончаков и мелкие участки па гривках типчаково-полынных сообществ, что придает комплексный характер растительному покрову.
Инос распределение растительности наблюдается у сильно соленых (горькосоленых) озер: водно-болотная растительность прибрежной полосы обычно отсутствует, прибрежная полоса покрыта выпотами солей, которые постепенно заселяются мясистыми солянками как однолетними {Sa-licornia europaea, Suaeda prostrata, S. corniculata), так и многолетними полукустарничковыми {Halocnemum strobilaceum, Atriplex verrucifera),&. также другими полукустарничковыми галофитами {Frankenia hirsuta, Limonium suffruticosum). За полосой «злостных» солончаков на менее засоленных почвах развиваются бескильницевые луга, среди которых значительную роль играют и заросли прибрежницы {Aeluropus litoralis). За бескиль-ницевыми лугами на еще менее засоленных почвах следуют крупнозлаковые галофитные луга {Agropyrum repens, Alopecurus ventricosus, Hordeum brevisubulatum), которые с переходом на склоны сменяются типчаково-полынными и полынно-типчаковыми степями на структурных солонцах. По распределению растительности по дну озерных котловин можно еще издали отличить горькосоленые от пресных и слабо соленых озер. Микро- и нанорельеф и здесь обусловливают комплексность.
В дальнейшем развитии «займищ» в лесостепной части Западно-Сибирской низменности намечается три пути: 1) смена водно-болотной растительности солончаково-луговой, а затем степной, т. е. процесс остепнения; 2) смена водно-болотной растительности сообществами дерновинных осок {Carex caespitosa) и образование кочкарников, т. е. процесс болотообразо-вания; 3) внедрение ив {Salix sibirica и др.) и позднее, по периферии, берез, т. е. путь развития березовых лесов. Последние два процесса наиболее широко наблюдаются в лесостепной полосе, а именно в северной ее части; на юге лесостепи и в степи, наоборот, господствует процесс остепнения.
Аналогичные сочетания солончаково-луговой и лугово-болотной растительности с солянковыми и степными сообществами имеют широкое распространение в пределах междуречья Волга—Урал: в Чижинских, Дюрин-ских, Балыктинских,Узеньских,Кушумских и других разливах и лиманах.
Прикаспийская низменность в междуречье Волга—Урал в северной его части, прилегающей к Общему Сырту, характеризуется обилием бес-»
сточных рек, высыхающих озер, а также крупных и мелких понижений, которые в весенний период служат местом, куда сбрасываются полые воды. Согласно А. А. Михееву (1916) и другим исследователям, весною здесь протекают реки — Дюра, Чижа, Чижа 2-я, Чижа 3-я и другие, которые теряются в Чижинских и Дюринских разливах. От оз. Балыкты к югу идет система Балыктинских разливов, южнее сливающаяся с разливами Большого Узеня и Кушума. Только Большой Узень сохраняет постоянные воды, остальные пересыхают, и на их месте остаются лишь цепи озер и высохших мелких и крупных понижений. В годы, наиболее обильные паводками, вода в лиманах застаивается до середины лета. В прибрежной полосе озер развивается прибрежноводная и водно-болотная растительность (Phragmites communis, Bolboschoenus maritimus, В. compactus, Scir-pus lacustris, S. Tabernaemontani, Butomus umbellatus, Beckmannia eruciformis, Heleocharis eupalustris, Alisma plantago-aquatica, Bhaponticuni salinum). За полосой водно-болотной растительности на луговых солонцеватых почвах идет широкая полоса пырейных лугов (Agropyrum repens). Местами развиваются почти чистые его сообщества, чаще же в состав пырейных лугов ближе к озеру внедряются водно-болотные растения, а ближе к периферии — солончаковые. Пырейные луга с усилением засоления сменяются полосой бескильницевых лугов, представленных чистыми зарослями Puccinellia dolicholepis или сообществами с примесью галофиль-ных растений (Juncus Gerar dii, Aeluropus litoralis, Atriplex verrucifera, Limo-nium caspium, L. Gmelinii). Вескильницевые луга местами прерываются зарослями Heleocharis oxylepis и пятнами пухлых солончаков с выпотами солей, зарастающих полукустарничковыми и однолетними галофитами (Halocnemum strobilaceum, Salicornia europaea, Anabasis salsa, Salsola soda, Suaeda prostrata, Atriplex litoralis, A. verrucifera,Petrosimoniaopposi-tifolia, виды Limonium и др.). Пырейные и бескильницевые луга в более глубоких понижениях ежегодно заливаются полыми весенними водами.
Выше по пологим склонам, вышедшим из ежегодного заливания, развивается растительность полупустынного или степного типа. В нижней части склонов господствуют полынные ассоциации на солонцово-солончаковых почвах с преобладанием Artemisia nitrosa и с большим участием галофильных растений. Еще выше, уже на солонцовых почвах, развиваются житняковые степи с доминированием Agropyrum pectiniforme и с примесью Роа bulbosa var. vivipara, Festuca sulcata, Koeleria gracilis и степного разнотравья. На водораздельных плакорных пространствах их сменяют полукустарничковые сообщества с господством Artemisia pauciflora и Atriplex сапа.
В более мелких понижениях, быстро освобождающихся от полых вод, или в крупных понижениях в годы слабых разливов пояс водно-болотной растительности выпадает, центр низин занимают пырейные луга, окруженные бескильницевыми лугами и солончаковой растительностью.
Поясное распределение растительности лиманов и разливов междуречья Волга—Урал’—это лишь общая схема, отражающая закономерности распределения растительности в зависимости от степени увлажнения и засоления на различных высотных уровнях мезорельефа. Необходимо отметить комплексный характер растительности в каждом поясе, что связано с условиями микро- и нанорельефа.
В древней дельте Урала заросли водно-болотной растительности (Phragmites communis и Bolboschoenus maritimus) развиты лишь в более глубоких лиманах при наличии зеркала воды. Господствуют же солончаковые бескильницевые (из Puccinellia dolicholepis, Р. gigantea) и прибрежницевые (из Aeluropus litoralis) луга с большим участием солянок. Среди бескиль-
ницевых лугов встречаются участки пырейных (из Agropyrum repens) и вострецовых (из Aneurolepidium multicaule) лугов, также с большим участием солянок.
Сочетания солоичаково-луговой и лугово-болотной растительности с солянковыми и степными сообществами широко развиты, как уже сказано выше, и по западному побережью Каспийского моря в Кура-Арак-синскоп и Терско-Сулакской низменностях. Они известны под названием «чально-луговой» растительности и описаны в работах А. А. Гроссгейма (1932, 1948), А. А. Колаковского (1933) и других. «Чалы» — бессточные понижения, в центре, в условиях постоянного переувлажнения, заняты болотной растительностью (Phragmites communis, Bolboschoenus maritimus, Scirpus lacustris, S. Taber naemontani, Cyperus longus, Typha angustifolia, T. Laxmannii, T. minima). Болотная растительность окаймлена гало-фитнолуговой (галофитночальной) растительностью с преобладанием злаков (Puccinelliagigantea, P.bulbosa, Aeluropuslitoralis, A. repens) с примесью верблюдки (Alhagi pseudalhagi), солодки (Glycyrrhiza glabra), кермека (Limonium Meyeri), полыни Шовица (Artemisia Szovitsiana,) свинороя (Cynodon dactylori), образующих и самостоятельные заросли. Здесь же встречаются заросли Imperata cylindrica и Sorghum halepense. На солончаках развивается обычная солончаковая растительность: виды Salsola, Suaeda, Petrosimonia, Atriplex, Halocnemum, Salicornia, Camphorosma. Через сообщества Artemisia Szovitsiana и Cynodon dactylon солончаковая и галофитнолуговая растительность переходит в пустынную растительность плакоров. Галофитнолуговая (чальная) растительность приурочена к темноцветным почвам, которые распахиваются под сельскохозяйственные культуры; на месте последних при прекращении распашки развиваются пырейные залежи. Пырейные луга вторичного происхождения занимают здесь большие пространства.
В восточной части Прикаспийской низменности на разливах в низовьях Булдурты, Калдыгайты и Уила распространены крупные массивы пырейных (из Agropyrum repens) лугов. В центральных, наиболее пониженных, с длительно застаивающимися водами, и заболоченных частях разливов встречаются ситняговые (из Heleocharis crassa, Н. argyrolepis, Н. euuniglumis, Н. oxylepis), бекманиевые (из Beckmannia eruciformis), камышевые (из Scirpus Tabernaemontani, S. lacustris) и тростниковые (из Phragmites communis) сообщества. На окраинных частях разливов полосами и пятнами сменяются сообщества пырейные, костровые (из Bromus inermis), бескильницевые (из Puccinellia gigantea, Р. dolicholepis), ситниковые (из Juncus Gerardii), житняковые (из Agropyrum pectiniforme). По мере нарастания засоленности почв появляются сообщества при-брежницы (Aeluropus litoralis), полыней (Artemisia nitrosa, A. mongo-lorum) и кермека (Limonium Gmelinii). В этих периферических частях разливов наблюдаются наиболее сложные комплексы и сочетания растительности, переходящие на незатопляемых территориях в полукустарничковые пустынные сообщества из черной полыни (Artemisia pauciflora), кокпека (Atriplex сапа), а также Atriplex verrucifera, Camphorosma топ-speliacum, С. Lessingii, Anabasis salsa (Никитин, 1954a).
В долине Эмбы более широко распространена галофитнолуговая и солончаковая растительность. В условиях избыточного увлажнения развиты заросли клубнекамыша (Bolboschoenus maritimus), тростника и рогозов (виды Typha). Среднеувлажненные местообитания характеризуются пырейными, ситниковыми, вейниковыми (из Calamagrostis epigeios), при-брежницевыми и бескильницевыми лугами. В обстановке недостаточного увлажнения и нарастающего засоления распространены сообщества солон-
чековой полыни, прибрежницы, скрытниц (Crypsis aculeata, С. schoenoides), кермека (Limonium Gmelinii) и верблюжьей колючки (Alhagi pseudalhagi). Значительные площади дельты Эмбы, вышедшие из сферы затопления, покрыты солончаками с сообществами однолетних солянок и полукустарничков— кзыл-кокпека (Limonium suffruticosum) и сарсазапа (llalo-спетит strobilaceum). На повышенных участках, разделяющих луга и протоки, распространены пестрые комплексы из белой полыни (Artemisia Lercheana), биюргуна (Anabasis salsa) и итсегека (Anabasis aphylla) (Никитин и Поярков, 1935; Никитин, 1954, 1954а).
В районе Тургай-Иргизских разливов также широко распространены пырейные луга, занимающие центральные, наиболее обильно увлажняемые площади.К абсолютно господствующему пырею, образующему сомкнутый травостой, в нижнем ярусе в заметном количестве прибавляется ситняг (Heleocharis palustris s. 1.). Он выступает здесь в качестве своеобразного эфемероида — быстро развиваясь с весны, заканчивает плодоношение и засыхает в июне. Характерна незначительная примесь Scorzonera parri-flora и еще реже встречающихся Juncus Gerardii, Rumex stenophyllus, Inula britannica и немногих других представителей разнотравья.
Менее увлажняемые периферические площади разливов покрыты полынно-пырейными сообществами, в которых еще сохранилась луговая основа из пырея, но травостой относительно изреживается и внедряются растения, выносящие некоторое засоление (Artemisia nitrosa, A. mongolo-rum, Limonium Gmelinii, Petrosimonia triandra).
При недостаточном увлажнении более удаленных от центра участков происходит большее переживание травостоя, исчезает луговая дернина, почвы приобретают черты глыбистых солонцов. Растительность в этих условиях слагается следующими растениями, перечисляемыми в порядке их значительности в покрове: Agropyrum pectiniforme, A. repens, Aneurole-pidium ramosum, Puccinellia gigantea, Artemisia nitrosa (или A. mongolorum), Limonium Gmelinii, Serratula cardunculus, Atriplex tatarica и некоторые другие.
Небольшие повышения, встречающиеся среди обильно орошаемых пырейных лугов, подвержены сильному засолению и на них развиваются прибрежницевые (из Aeluropus litoralis) сообщества на лугово-солончаковых почвах с примесью ряда солеустойчивых видов (Artemisia nitrosa, Puccinellia gigantea, P. dolicholepis, Psathyrostachys juncea, Petrosimonia triandra, P. brachiata, Limonium caspium).
Характерно, что в перечисленном ряду сообществ наблюдается заметное снижение урожайности сухой массы от 30 ц/га на пырейных лугах до 8—10 ц/га (и даже до 3 ц/га) на прибрежницевых с полынью сообществах. На участках пырейных лугов, которые подвержены особенно продолжительному затоплению, происходит изреживание пырея и в травостой внедряются лугово-болотные растения (Carex melanostachya, Bolboschoenus maritimus, Juncus Gerardii, Alisma plantago-aquatica, Rumex stenophyllus и др.). В общем урожае на долю пырея приходится всего 40—50% cyjfoii массы.
На топких побережьях постоянных водоемов разрастаются заросли тростника (Phragmites communis) со значительной примесью Scirpus la-custris и Typha latifolia и с меньшим участием Heleocharis palustris s. 1., Scolochloa festucacea, Alisma plantago-aquatica, Butomus umbellatus, Roripa amphibia, Scutellaria galericulata, Limosella aquatica, Carex melanostachya, C. Karelinii, Senecio paluster и др. В открытой части водоема произрастают рдесты (Potamogeton lucens, Р. pectinatus, Р. perfolialus). а также Hydrocharis morsus-ranae, Stratiotes aloides, Lemna trisulca, Ulricularia
vulgaris, Myriophyllum verticillatum и другие водные растения, которые способствуют постепенному зарастанию водоемов (Стрельников, 1938).
Значительный по площади контур рассматриваемого типа находится в Зайсанской котловине. Черный Иртыш, впадая в оз. Зайсан, образует обширную дельту, пересекаемую многочисленными рукавами, протоками, старицами. В зарастающих протоках формируются сообщества водноболотных растений (Nuphar luteum, Nymphaea Candida, Nypmphoides peltatum, Myriophyllum verticillatum, Hippuris vulgaris, Lemna trisulca). По берегам их разрастаются Heleocharis palustris s. 1., Cyperus juscus, Carex diluta, Cicuta viroa, Lythrum virgatum и другие влаголюбивые растения. Но большая часть дельты покрыта зарослями тростника (Phragmites communis), к которому примешиваются рогоз (Typha latifolia), камыш (Stirpus lacustris), сусак (Butomus umbellatus), стрелолист (Sagit-taria sagittifolia) и др.
В северо-восточной части дельты, кроме тростниковых зарослей, встречаются значительные площади злаковых лугов, слагаемых вейником (Calamagrostis ерг£его$),канареечником(£Ь‘£гaphisarundinacea),лисохвостом (Alopecurus pratensis), а также Hordeum brevisubulatum, Agropyrum repens, Agrostis alba, Bromus inermis. На более влажных участках имеется примесь Heleocharis palustris s. 1. и Bolboschoenus maritimus, а па более сухих — Oxytropis glabra, Glycyrrhiza uralensis, Sonchus arvensis и других видов разнотравья.
Незаливаемые участки, возвышающиеся над площадями тростниковых зарослей, или берега стариц заняты солончаками со специфическими для них галофитными сообществами с господством полукустарничковых и однолетних солянок (Halocnemum strobilaceum, Kalidium joliatum, Salicornia europaea, Salsola subcrassa, Suaeda proslrata, S. corniculata и др.). По краям солончаков встречаются участки чиевых сообществ (из Lasiag-rostis splendens) и группы кустов селитрянки (Nitraria sibirica)? По берегам самого русла Черного Иртыша — немногочисленные лески из белого и черного тополей (Populus alba и Р. nigra) с примесью ив (виды Salix)', участки этих поемных лесков перемежаются с луговыми сообществами.
По левому берегу Черного Иртыша наблюдаются полузаросшие, а иногда и подвижные бугристые пески. Здесь господствуют псаммофиты: Calligonum rubicundum, Elymus giganteus, Artemisia arenaria, Chondrilla pauciflora, Aristida pennata, Scorzonera ensifolia. Окраины песчаных бугров нередко бывают подвержены засолению и тогда на этих местах разрастается чий (Lasiagrostis splendens), среди которого встречаются кусты мирикарии (Myricaria alopecuroides), а в нижнем ярусе распространены Aeluropus litoralis и Limonium Gmelinii (Борисова, 1935).
В литоральной полосе оз. Зайсан, не подверженной прибою волн, наблюдаются различные сообщества прибрежноводных растений. За береговой зарослью тростника идет полоса тростника, растущего на мелководье. Далее следует узкая полоса сообщества Typha angustifolia+Nym-phoidespeltatum. За нею наблюдаются пятна сообществ с господством Nym-phoides peltatum и Ceratophyllum demersum co значительным участиемMyriophyllum verticillatum', здесь же встречаются Utricularia vulgaris и Trapa natans s. 1. В эту широкую полосу включены участки разреженной группировки Nymphoides peltatum, видов Potamogeton и Myriophyllum spicatum (Поляков, 1947).
1 К югу от оз. Зайсан, среди пустынной равнины в обширных разливах рек, стекающих с Тарбагатая, имеются обширные заросли Lasiagrostis splendens, Phragmites communis, часто с большим участием Halimodendron halodendron, не отмеченные на карте. (Примечание редакции).
Некоторым своеобразием отличается контур, показанный в депрессии между озерами Сасыкколь и Алаколь. Здесь на огромных пространствах распространены тростниковые заросли высотой 3—4 м, скрывающие от взора многочисленные протоки и старицы и образующие непроходимые «джунгли». Тростниковые заросли окаймляют берега всех озер, а также берега рек, слепо оканчивающихся на равнине Джунгарских ворот, особенно же в их устьевых окончаниях (сухих дельтах).
По берегам озер, за полосой тростниковых зарослей, па еще обильно увлажненных, но несколько засоленных участках, располагаются заросли кендыря (Trachomitum lancifolium), в примеси к которому встречаются Limonium Gmelinii, Saussurea salsa, Gypsophila trichotoma, Plantago salsa, Lactuca tatarica, Oxytropisglabra, Sphaerophysa salsula. Между сообществами с господством кендыря и тростниковыми зарослями наблюдаются переходные группировки. На более приподнятых участках с солончаковыми почвами широко распространены куртины галофитов; господствуют Atriplex verrucifera, А.сапа,а в примеси к ним встречаются Aneurolepidium angustum, Kochia prostrata, Ceratocarpus arenarius, Echinopsilon hyssopifolium, Sal-sola soda, Aeluropus litoralis, Suaeda linifolia, сочетающиеся с незадерно-ванными отакыренными участками. Заросли тростника часто выжигаются местным населением для получения более ценного пастбищного корма и для улучшения условий сенокошения. Однако наиболее ценными угодьями являются «сазоватые луга» на пониженных и слабо засоленных равнинных местах, также представленные заметными по площади участками. Травостой этих лугов слагается главным образом злаковыми сообществами из Festuca orientalis, Puccinellia gigantea, Beckmannia eruciformis, Agrostis alba, Calamagrostis pseudophragmites, C. dubia. В покрове имеется примесь осок (Carex songorica, С. melanostachya, С. diluta и др.), а также лугового разнотравья (Chartolepis intermedia, Galatella punctata s. I., Tripolium vulgare, Heteropappus altaicus, Melilotus dentatus, Mentha longifolia, Scorzonera parviflora, Vicia cracca, Euphorbia soongarica и др.). С этими лугами перемежаются тростниковые заросли.
В условиях близкого от поверхности залегания грунтовых вод развиваются заросли чия (Lasiagrostis splendens). Как было отмечено Б. А. Келлером (1911) для Кальджирской долины, а С. Ю. Липшицем (1931) для района депрессии Сасыкколь—Алаколь, имеется два варианта (группы ассоциаций) формации чиевников: 1) редкостойные зарослп, чередующиеся с участками солончаков; 2) мощные и густые заросли, сочетающиеся с луговыми сообществами.
В первом варианте между редкими дерновинами чия внедряются солянки (Atriplex verrucifera, А. сапа, Camphorosma Lessingii и др.).
Для второго варианта характерны в нижнем ярусе густых зарослей чия элементы солончаковых лугов (Sophora alopecuroides, Lepidium latifolium, Hymenophysa pubescens, Chartolepis intermedia, Galatella punctata s. 1., низкорослый Phragmites communis и др.). Небольшими пятнами попадаются группы Glycyrrhiza uralensis и Limonium Gmelinii. Нередко можно наблюдать переходы от чиевников к зарослям тростника, где сочетаются сопутствующие виды обеих формаций (Липшиц, 1931).
Солончаково-луговая и лугово-болотная растительность рассмотренных районов имеет большое значение как кормовой фонд. Пырейные, вздуто-лисохвостные, солончаковоячменные, бекманиевые, канареечниковые луга дают сено высокого кормового достоинства и в годы больших и длительных разливов отличаются высокой урожайностью: до 40—60 ц/га и выше. В годы малых разливов урожайность падает до 20—30 ц/га и ниже, т. е. продуктивность солончаковых лугов и лугово-болотной растительности
резко колеблется в зависимости от степени и продолжительности затопления. В связи с этим необходимо регулирование лиманного затопления с учетом различной реакции растений на длительность затопления. Прибреж-ницевые и переходные к плакорным полынно-типчаковые травостои используются под выпас. Солонцовые почвы пологих склонов западин и темноцветные слабозасоленные почвы под солончаково-луговой растительностью используют также под бахчи. Заросли тростника дают сырье для бумажной промышленности и строительства, заросли камышей и осок используются для выборочного сенокошения.
Литература
U.K. Пачоский (1892); А. Я. Гордягин (1897, 1900—1901); Г. И. Танфильев (1902); Ф. Ф. Зелинский (1910, 1912); В. Ф. Капелькин (1910); Б. А. Келлер (1911); В. И. Смирнов (1912); Б. Н. Городков (1913а, 1916); П. Н. Крылов (1913, 1916, 1919); М. И. Пта-шицкий (1913а); В. П. Дробов (1914); М. Ф. Короткий (1914); Н. И. Кузнецов (Владимирский) (1914, 1915); С. Е. Рожанец-Кучеровская (1914, 1916); Б. К. Шишкин (1914а); Г. Н. Высоцкий (1915); К. К. Косинский (1916); А. А. Михеев (1916); Н. В. Шип-чинский (1916); С. С. Ганешин (1917); М. Д. Спиридонов (1923, 1927—1928, 1928); И. С. Косенко (1924, 1934); В. С. Богдан и Б. А. Шумаков (1925); И. М. Крашенинников (1925); И. В. Ларин (1926а); Н. В. Павлов (1926, 1949); В. И. Баранов (1927); В. И. Баранов и К. П. Горшенин (1927); Л. 3. Захаров (1927); Д. К. Волгунов (1928); О. Э. Кнорринг (1928); С. А. Никитин (1928, 1951, 1954, 1954а); Е. В. Шифферс-Рафа-лович (1928, 1928а, 1937); А. А. Гроссгейм и А. А. Колаковский (1929. 1930); А. Ф. Флеров (1930); Т. Гейдеман и И. Дорошко (1931); С. 10. Липшиц (1931); Т. Ф. Пояркова и 3. В. Коровник (1931); А. А. Гроссгейм (1932, 1948); И. В. Новопокровский (1932); А. А. Колаковский (1933); Н. А. Иванова и И. М. Крашенинников (1934); А. Г. Борисова (1935); С. А. Никитин и В. Ф. Поярков (1935); Ф. Ф. Лейсле (1936); А. А. Генкель и П. Н. Красовский (1937); П. Г. Стрельников (1938); А. Д. Фурсаев и др. (1938); П. И. Поляков (1947); А. В. Калинина (1948); А. В. Куминова (1949а); А. В. Куминова и Е. В. Вандакурова (1949); Е. В. Вандакурова (1950); И. Н. Бейдеман (1951, 1953, 1954); А. Г. Доскач, И. В. Ларин, Ф. Я. Левина (1952); М. С. Кузьмина (1953); И. Ф. Мусаев (1953); Б. А. Быков (1954); Л. Я. Захарова (1954); В. В. Иванов (1954); И. В. Ларин, Е. В. Шиффере, Ф. Я. Левина, Б. Н. Замятнин, Л. И. Иванина, Е. А. Варивцева, Г. Н. Непли и И. В. Сырокомская (1954); И. В. Ларин, Л. А. Манохина и 3. С. Акимцева (1954); Ф. Я. Левина (1954); П. В. Пивоваров (1954); И. И. Тереножкин (1954); А. Д. Фурсаев (1954).
63. Пойменные луга, обычно в сочетании с кустарниками и лесами (урема), и сельскохозяйственные земли на их месте
Площади пойменных лугов, отвечающие масштабу карты, приурочены к поймам наиболее крупных водных артерий, принадлежащих бассейну северных морей (Северная Двина, Печора, Обь, Енисей, Лена), южных морей (Днепр, Дон, Волга, Урал, Аму-Дарья, Сыр-Дарья) и восточных морей (Амур, Анадырь) с их крупными многочисленными притоками.
Уже краткий перечень крупнейших водных артерий, пересекающих все растительные зоны СССР, от пустыни до тундры, говорит о разнообразии типов лугов равнинных областей, подразделенных на карте по зональному принципу: луга тундровой зоны (63 т), луга лесной зоны (63 л), луга степной зоны (63 ст) и луга пустынной зоны (63 п).
Зональное положение — не единственная причина разнообразия природных условий пойменных лугов. Большое значение в этом отношении имеют возраст пойм, разработанность их рельефа, высотные уровни, продолжительность поемности, характер аллювиальных и делювиальных отложений, направление почвообразовательного процесса, механический и химический состав почв, увлажнение, история развития луговой растительности, эколого-биологические и фитоценотические свойства луговых растений, влияние хозяйственной деятельности человека и т. д.
А. П. Шенников (1935, 1938, 1941) различает пять классов формаций или основных типов лугов: 1) настоящие или мезофитные луга, 2) остеп-ненвые или ксеромезофитные луга, 3) пустошные или психромезофитные луга, 4) болотные или гидромезофитные луга, 5) торфянистые или оксило-мезофитные луга. В каждом классе формаций выделяются два варианта: незасоленные (гликофитные) и засоленные (галофитные) луга.
Луга и сопутствующие им формации характеризуются ио зонам в соответствии с показом их на «Геоботанической карте». При описании зональных сочетаний растительных сообществ пойм приняты во внимание названные подразделения луговой растительности.
Литература
В. В. Алехин (1921, 1927); В. Р. Вильямс (1922); И. М. Крашенинников (1922); А. П. Шенников (1935, 1938, 1941); Р. А. Еленевский (1936, 1936а); И. В. Ларин (1936, 1950—1951); Л. Г. Раменский (1938, 1950); С. П. Смелов (1947); А. М. Дмитриев (1948).
63 т — тундровой зоны
Пойменные луга тундровой зоны показаны на «Геоботанической карте» в низовьях рр. Омы, Пеши, Печоры, Оби, Надыма, Ныды, Хадуты, Нура, Таза, Мессо-Яха, Енисея, Канчалана, Анадыря.
Суровые климатические и специфические гидрологические условия тундровой зоны, как, например, повсеместное развитие вечной мерзлоты и связанного с последней избыточного поверхностного увлажнения в вегетационный период, а также и другие физико-географические условия накладывают резкий отпечаток на растительность речных пойм тундровой зоны.
В поймах тундровой зоны на участках, вышедших из-под влияния затопления, развиваются тундровые формации. На средних уровнях пойм, в условиях краткой поемности и достаточного дренажа, широко развиты ивняковые и ерниковые формации с злаково-разнотравным или злаковоосоковым покровом лугового типа. Среди сплошных кустарниковых зарослей встречаются небольшие участки настоящих мезофильных злаковых, злаково-разнотравных и разнотравных лугов. Более характерны для пойм тундровой зоны луга низких уровней (заболоченные злаковые, злаковоосоковые и осоковые), развивающиеся в условиях более длительной поемности и избыточного увлажнения.
Большая протяженность тундровой зоны определяет и большое разнообразие растительности пойм этой зоны. Б. Н. Городков (19356,1938а) отмечает ряд основных закономерностей в распространении древесной, кустарниковой и травяной растительности речных пойм Крайнего Севера. Так, западнее Енисея лесные и кустарниковые формации, характерные для пойм южной части лесотундры и состоящие из крупнокустарниковых ив (преимущественно из Salix rossica) с примесью ольховника (Alnaster fruticosus), березы (Betula pubescens, В. tortuosa) и ели (P'icea excelsa, Р. obovata), в северной части лесотундры сменяются кустарниковыми формациями из более низкорослых ив (Salix glandulifera, S. glauca, S. hastata, S. lapponum, S. phylicifolia). Севернее, в подзоне открытых кустарниковых тундр, долинные кустарники в основном сложены ерником (Betula папа) и приземистыми ивами (Salix glauca, S. lanata, S. reptans, S. pulchra), а в подзоне лишайниково-моховых и арктических тундр сохраняются лишь Salix reptans, S. pulchra, S. lanata.
Настоящие мезофитные луга в поймах тундровой зоны занимают незначительные площади, но и здесь наблюдается определенная закономер
ность в их распространении. Если в подзоне лесотундры в составе растительного покрова лугов значительное участие принимают бореальные виды (Bromus inermis, Роа pratensis, Alopecurus pratensis и многие другие), то в подзонах открытых тундр среди кустарниковых зарослей настоящие мезофитпые луга развиты из Alopecurus alpinus, Роа alpigena, Calamagrostis groenlandica, Dupontia Fischeri и других арктических и арктоальпий-ских видов.
В приморской полосе, в сфере влияния морских приливов и засоления почв, широко развиты приморские злаково-осоковые луга из Calamagrostis deschampsioid.es, Carex subspathacea, С. discolor, С. rariflora, Dupontia Fischeri, а песчаные наносы заняты зарослями Elymus mollis.
При движении с запада на восток изменяется видовой состав растительности пойм. В лесотундре к Salix rossica на западе примешиваются Salix dasyclados, S. phylicifolia, а на востоке (восточнее Енисея) — Salix sachalinensis, S. speciosa, S. parallelinervis', Betula pubescens и В. tortuosa на востоке замещаются В. Cajanderi. На востоке тундровой зоны (восточнее Лены) местами в поймах развиты леса из тополя (Populus sua-I'eolens) и древовидной ивы — чосепии (Chosenia macrolepis). _ В подзонах открытых тундр па Дальнем Востоке господствует Salix Кгу-lovii.
Остановимся на некоторых различиях в характере луговой растительности пойм Большеземельской тундры, Обь-Енисейского водораздела и тундр, расположенных восточнее Енисея. Различия эти,наиболее ярко выражены в видовом составе настоящих мезофитных лугов и более сглажены в болотистых и торфянистых лугах. Для настоящих мезофитных лугов Большеземельской тундры (Андреев, 1932а) характерно большое участие бореальных видов (Bromus inermis, Agrostis capillaris, Trisetum sibiricum, Alopecurus pratensis, Poa pratensis, Festuca rubra, Heracleum sibiricum, Anthriscus silvestris, Achillea millefolium, Delphinium elatum, Dianthus superbus, Polygonum bistorta, Geranium silvaticum, Veratrum Lobelianum, Veronica longifolia, Equisetum arvense и др.). Арктические и аркто-альпийские виды (Hierochloe alpina, Roegneria borealis, Astragalus subpolaris, Hedysarum arcticum,Ranunculus borealis и др.) играют в составе настоящих пойменных лугов Большеземельской тундры более второстепенную роль.
По мере продвижения к востоку, с увеличением континентальности климата участие бореальных видов в луговой растительности падает, усиливается отундровение лугов и возрастает роль арктических и аркто-альпийских растений.
Б. Н. Городков (19356) отмечает, что для подзон безлесных тундр характерна кратковременная (всего несколько дней) поемность, что исключает развитие высокотравных мезофитных лугов, которые сменяются здесь пестрыми злаково-разнотравными лужайками. Он приводит описание такого луга для Обь-Енисейского водораздела (Городков, 1944). Преобладают Роа alpigena, Alopecurus borealis, Calamagrostis neglecta var. borealis, в меньшем количестве распространены Equisetum arvense, Pedicularis sudetica, Eriophorum Scheuchzeri, Deschampsia arctica, Pyrethrum bipinna-tum, Festuca cryophila. Встречается подрост Salix reptans, S. lanata. Пятнами развит тонкий моховой покров из Drepanocladus uncinatus, Pogona-tum urnigerum.
Близкие по составу вейниковые луга указаны В. Б. Сочавой (1934) для района анабарских тундр. В пойме Пенжины наиболее широко распространены вейниковые луга с господством Calamagrostis Langsdorffii и с содоминантами: Carex accrescens, Chamaenerium angustifolium, Thali-
drum sparsiflorum, Galium boreal6\ в люньшем количестве встречаются Trisetum sibiricum, Ptarmica sibirica, Geranium erianthum, Aster si-biricus, Ranunculus monophyllus, Viola repens, Trientalis europaea, Rubus ardicus. Флористический состав этих лугов говорит о бореальном характере пойменной растительности Пенжины (Тихомиров, 1935). Это сближает их с лесотундровой подзоной, что подтверждается и наличием зарослей крупнокустарниковых ив (Salix rossica, S. xerophila и др.), а также рощ тополя (Populus suaveolens), чосении (Chosenia macrolepis) и березы (Ве-tula Cafanderi). Близка к пенжинским лугам растительность поймы Анадыря (Васильев, 1936).
Настоящие (мезофитные) пойменные луга — большею частью вторичного происхождения; они развиваются под влиянием деятельности человека и животных на месте сведенных кустарниковых, преимущественно ивняковых зарослей. Меньшая часть настоящих (мезофитных) пойменных тундровых лугов — первичного происхождения. Намечается следующий путь развития первичных пойменных лугов тундровой зоны (Городков, 1944). Иловатые и песчаные речные отмели заселяются растениями-пионерами, главным образом Equisetum arvense', с закреплением субстрата хвощ сменяется зарослями Deschampsia arctica, Arctophila fulva, Eriophorum Scheuch-zeri, а последние в свою очередь сменяются злаковыми и разнотравными лугами с подростом ив. Дальнейшее развитие мелкокустарниковых ив и лишайниково-мохового покрова по мере выхода из-под влияния затопления приводит к отундровению лугов. Кустарниковые заросли с травяным покровом лугового типа являются потенциальным луговым фондом, в составе травяного покрова их постоянно присутствуют луговые растения. Так, В. Б. Сочава (1934) для анабарской тундры указывает широкое распространение на средних уровнях поймы Анабары следующих ассоциаций: Salicetum glaucae, Salicetum reptantis и Salicetum lanatae. В ассоциации Salicetum glaucae в травяном покрове доминирует Calamag-rostis neglecta, содоминантами его являются Alopecurus alpinus, Роа alpi-gena, Equisetum arvense', в меньшем количестве встречаются Saxifraga hirculus, Polygonum viviparum, Carex stans, Lagotis minor, Eriophorum Scheuchzeri, Myosotis asiatica. В ассоциации Salicetum lanatae в травяном покрове доминируют Ardagrostis latifolia и Equisetum arvense, на их фоне отмечены Alopecurus alpinus, Betula exilis, Astragalus subpolaris, A. umbel-latus, Festuca supina, F. rubra s. 1., Luzula confusa, Polygonum viviparum, Lagotis minor, Valeriana capitata, Pedicularis capitata, Saussurea Tilesii, Salix reticulata. Отмечен тонкий ковер из гипновых мхов.
По направлению к югу (к лесотундре) мелкокустарниковые формации сменяются крупнокустарниковыми с участием древесных пород.
Конкретные описания крупнокустарниковых и лесных пойменных формаций в поймах рек юга тундровой зоны имеются в работах В. Н. Андреева (1934) по Ямалу, Б. Н. Городкова (1933) по Дальневосточному краю, Ф. В. Самбука (1930) по поймам Печоры, Б. А. Тихомирова (1935) по пойме Пенжины.
Наряду с кустарниковыми зарослями в поймах тундровой зоны широко распространены болотистые луга низких уровней пойм — злаковые, злаково-осоковые и осоковые в условиях избыточного проточного увлажнения. Они представлены или чистыми зарослями (в безлесной тундре — Arctophila fulva, Carex stans, в лесотундре — Carex aquatilis, в приморской полосе — Dupo nt ia Fischeri, Carex subspathacea), или с примесью других злаков и осок и редкого болотного разнотравья. Заросли Arctophila fulva занимают все илистые отложения рек, озер, протоков, окаймляя водоемы у уреза воды. В безлесной тундре к Arctophila fulva часто примешивается
Carex stans, Dupontia Fischeri, Calamagrostis neglecta. За зарослью Arcto-phila fulva обычно идет пояс осоки — Carex stans, также с небольшой примесью Arctophila fulva, Dupontia Fischeri, Calamagrostis neglecta и редким болотным разнотравьем (Menyanthes trifoliata, Comarum palustre, Hippuris vulgaris, Polygonum viviparum и др.). В северной части лесотундры за поясом Arctophila fulva идет пояс Carex aquatilis с примесью Calamagrostis Langsdorjjii.
Для болотистых лугов, развивающихся в условиях ежегодного затопления и избыточного проточного увлажнения, характерно отсутствие накопления торфа, а также отсутствие или слабое развитие мохового покрова, состоящего из гидрофильных гипновых мхов (родов Calliergon, Drepanocladus и др.). По мере выхода из зоны затопления, но при сохранении избыточного и уже застойного увлажнения, в болотистые луга внедряются пушицы (Eriophorum Chamissonis, Е. angustifolium), мощно развивается гипновый моховой покров, в который постепенно внедряется сфагнум {Sphagnum obtusum, S. squarrosum), усиливается процесс торфо-накопления; болотистые луга переходят в травяные, гипново-травяные низинные и гипново-сфагновые переходные болота (65 т).
Арктофиловые и осоковые болотистые луга развиваются также и при зарастании или высыхании озер и при искусственном спуске последних в поймах и на водоразделах (Прянишников, 1955).
Литература
Ф. В. Самбук (1930, 1934, 1934а, 1937); В. Н. Андреев (1931, 1932а, 1934, 1935а, 1954а); А. А. Дедов (1931, 1933, 1933а); В. Б. Сочава (1932а, 1933, 19336, 1934); Б. Н. Городков (1933, 1935, 19356, 1938а, 1939, 1944, 1946, 1946а); К. Н. Игошина (1934); В. Н. Андреев, К. Н. Игошина, А. И. Лесков (1935); Б. А. Тихомиров (1935, 1946а); В. Н. Васильев (1936); В. Д. Александрова (1937); А. Н. Виноградова (1937); В. И. Душечкин (1937); И. Д. Богдановская-Гиенэф (1938); В. А. Шелудякова (1938, 1948); А. В. Прянишников (1954, 1955).
63 л — лесной зоны
Пойменные луга лесной зоны развиваются в условиях умеренного климата, который обусловливает широкое развитие мезофитной травяной, кустарниковой и древесной растительности. Естественные травостои, развивающиеся на илистых и песчаных речных отмелях, формируются в первичные пойменные луга лишь при систематическом воздействии на них человека путем сенокошения. Отсутствие сенокошения ведет к быстрому зарастанию первичных лугов кустарниками и древесными породами. Сведение кустарников и древесных насаждений обеспечивает возникновение пойменных лугов уже вторичного происхождения, но также при условии систематического сенокошения или выпаса. В зависимости от интенсивности использования под покос пойменные луга в той или иной степени покрыты зарослями кустарников, преимущественно ивняков, и островками пойменных лесов: черноольховых, осиново-березовых, еловых — на суглинистых, сосновых — на песчаных отложениях. В южной части лесной зоны и особенно в лесостепи в состав лесов пойм входят дуб, тополь, ильм и другие широколиственные породы.
Для хорошо развитых пойм характерны волнисто-гривистый рельеф, неоднородные условия увлажнения, механического и химического состава почв, режима половодий, а в связи с этим — неоднородность направления почвообразовательного процесса, что обусловливает большое разнообразие и растительного покрова пойм.
Характеристика основных типов лугов дана А. П. Щенниковым в его сводных работах (1938, 1941); ниже излагается краткая характеристика их для Европейской части в сопоставлении с типами лугов Азиатской части СССР.
В поймах рек на гривах, вышедших из зоны затопления или затопляемых, но лишенных аллювиальных наносов, с бедными почвами и недостаточным увлажнением развиваются пустошные луга и пустоши. Пустошные луга представлены мелкотравными изреженными травостоями из волосовидной полевицы (Agrostis cap Шаг is), красной овсяницы (Festuca rubra), душистого колоска (Anthoxanthum odoratum), лугового мятлика (Роа pratensis) и других мезофильных низовых злаков в угнетенном состоянии. К ним присоединяются типичные растения пустошей (психрофиты): белоус (Nardus stricta), овечья овсяница (Festuca ovina), жестковатая овсяница (Festuca duriuscula). Из разнотравья также в угнетенном состоянии принимают участие виды погремка (Rhinanthus), очанки (Euphrasia), манжетки (Alchimilla), лапчатки (Potentilla), истод (Polygala vulgaris), бед-ренец (Pimpinella saxifraga), тысячелистник (Achillea millefolium), нивяник (Leucanthemum vulgare). Характерно развитие мохового покрова (из Thuidium abietinum).
Пустошные луга крайне малопроизводительны и требуют коренных улучшений, без которых развитие психрофитов приведет к вырождению лугов в пустоши — белоусовые, овечьеовсяницевые и другие, малопригодные даже для выпаса.
Пустошные луга и пустоши значительно развиты в лесной зоне Европейской части СССР. Для Азиатской части СССР в литературе нет указаний на наличие пустошных лугов в поймах рек лесной зоны, возможно в связи со слабой освоенностью пойм, а отчасти из-за недостаточной изученности их.
В южной части лесной зоны на тех же высоких уровнях грив, что и пустошные луга, но на более теплых супесчаных и более богатых почвах развиваются остепненные луга. Для них характерно присутствие в травостое лесостепных видов: типчака (Festuca sulcata), луговой келерпи (Koeleria Delavignei), полевицы Сырейщикова (Agrostis Syreistschikovii), степной тимофеевки (Phleum phleiodes), желтой люцерны (Medicago fal-cata), горного клевера (Trifolium montanum), желтого подмаренника (Galium verum) и некоторых других. Остепненные луга особенно широко распространены в лесостепи.
В Азиатской части СССР, преимущественно на юге лесной зоны, также встречаются остепненные луга на высоких уровнях в поймах рек: в Западной Сибири — по Ишиму, Иртышу, Оби и в Средней Сибири — по Енисею и его притоку Ангаре, а также по Лене. По среднему течению Лены в районе Якутска они представлены особыми географическими вариантами; в состав их входят Festuca facutica, Linum Komarovii,1 Galium verum, Lychnis sibirica, Onobrychis sibirica, Koeleria gracilis и др. Степи и остепненные луга известны даже по Яне и Индигирке.
На средних уровнях пойм, несколько ниже пустошных и остепненных лугов, в условиях уже слабого и кратковременного затопления с незначительным наилком и с недостаточным грунтовым увлажнением развиваются мелкотравные, настоящие мезофитные луга — красноовсяничники. Невысокий разреженный травостой сложен Festuca rubra и Agrostis capil-laris с примесью Роа pratensis, Phleum pratense, Agrostis alba и обильного разнотравья (Leucanthemum vulgare, Thalictrum minus, Campanula glo-
1 Linum perenne s. 1.
merata, Galium boreale, G. rubioides, Stellaria graminea, Pimpinella saxi-fraga, Equisetum arvense, виды Rumex и др.). Из бобовых встречаются Trifolium pratense, Т. repens, Astragalus danicus, Lotus corniculatus.
Красноовсяничные луга представлены многими вариантами (ассоциациями). Располагаясь между белополевичниками и пустошными лугами, они дают или более мезофитные, или более обедненные, или остепнен-ные ассоциации. Урожайность красноовсяничников низкая — 10—12 ц/га.
В лесной зоне Азиатской части СССР в связи со слабой освоенностью пойм распространение красноовсяничников ограничено: они встречаются незначительными участками лишь в южной части лесной зоны, чаще же красная овсяница указывается только в мятликовых лугах на незаливае-мых гривах прирусловых пойм. На Дальнем Востоке А. П. Саверкин (1936) отмечает участие красной овсяницы в полевицевых (Agrostis clavata) с наземным вейником (Calamagrostis epigeios) лугах наиболее сухих местообитаний.
Из настоящих лугов, приуроченных к средним уровням пойм, широко распространены следующие луговые формации: полевичныс (Agrostis alba var. gigantea), тимофеечные(Phleum pratense), овсяничные (Festuca pratensis), лисохвостные (Alopecurus pratensis), пырейные (Agropyrum repens) и костровые (Bromus inermis). Это — урожайные, высокого кормового достоинства луга. Пырейные и костровые луга особенно характерны для более легких почв приречных пойм, остальные — для иловато-суглинистых почв центральных пойм.
Перечисленные луга редко являются монодоминантными и в этом случае занимают незначительные площади. Чаще они представлены полидо-минантными ассоциациями и с участием разнотравья. Такими смешаннозлаковыми лугами являются: луговотимофеечно-белополевицевые, луговоовсяницево-белополевицевые, луговомятликово-белополевицевыс, белополевицево-лисохвостные, болотномятликово-лисохвостные, пырейнокостровые и кострово-пырейные и т. д. Кроме злаков, большую роль часто играет мезофильное разнотравье (Achillea millefolium, Galium boreale, Geranium pratense, Ranunculus acer, Alchimilla vulgaris s. 1., Hieracium umbellatum, Rumex acetosa, R. thyrsiflorus и др.). Из бобовых характерны Trifolium pratense, Т. medium, Vicia cracca, Lathyrus pratensis. В более влажных вариантах (ассоциациях) мезофитных лугов состав разнотравья несколько иной: характерно присутствие Polygonum bistorta, Filipendula ulmaria, Sanguisorba officinalis, Veronica longifolia, Geum ri-vale, Trollius europaeus. На супесчаных почвах прирусловых пойм особенно характерны зонтичные (Heracleum sibiricum, Cenolophium Fischeri, Carum carvi, Libanotis intermedia и др.). По соотношению злаков и разнотравья различаются разнотравно-злаковые луга, в которых злаков больше 50%, и злаково-разнотравные с преобладанием разнотравья. Урожайность в нормальных условиях развития этих лугов высокая, в среднем 30—40 ц/га.
Мезофитные луга такого характера свойственны преимущественно Европейской части Союза. В таежной зоне Западной, Средней и Восточной Сибири мезофитные луга, в связи с малой освоенностью пойм, не имеют широкого распространения и приурочены преимущественно к гривам прирусловых, реже центральных пойм южной части лесной зоны. В бассейне Оби наиболее типично выражены и наиболее богаты по флористическому составу мезофитные луга лишь в Томской области (Кузнецов [Владимирский], 1915; Ильин, 1916; Куминова, 1949а; Елизарьева, 1951). Здесь они представлены преимущественно луговоовсяничными, мятликово-луговоовсяничными, тимофеевково-луговоовсяничными лугами в центральной пойме, пырейными и костровыми -г- в прирусловой, и занимают крупные
площади. Ниже по течению Оби, с увеличением поемности, площади ме-зофитных лугов резко уменьшаются, флористический состав их беднеет. Они представлены преимущественно луговомятликовыми и белополеви-цевыми лугами, приуроченными лишь к высоким уровням прирусловых пойм (Сапожников и Никитина, 1920; Бронзов, 1929; Барышников, 1933). Средние уровни центральных пойм здесь заняты вейниковыми (Calamagrostis Langsdorffii) лугами, ниже по течению сменяющимися канарееч-никовыми, а с еще большим увеличением поемности — остроосоковыми (Carex gracilis) и водноосоковыми лугами.
В бассейне Енисея настоящие (мезофитные) луга указываются по долине Ангары и ее притокам (Боровиков, 1912, 1913; Ганешин, 1912; Криштофович, 1913; Буддо, 1953), в бассейне Подкаменной Тунгуски и Нижней Тунгуски (Рубин, 1934, 1935) и по долине Енисея (Ревердатто, 1916; Зубков, 1940; Игошина, 1951; Номоконов, 1955). Долина Енисея, по сравнению с долиной Оби, менее разработана и более облесена, но и в ней отмечается определенная закономерность смен луговой растительности: по мере продвижения на север, вниз по течению, усиливается поемность, господствуют болотистые луга и одновременно внедряются арктические виды; мезофитные луга ниже Дудинки сохраняются лишь по склонам яров. В бассейне Лены настоящие (мезофитные) луга развиты преимущественно в центральной части Якутской АССР, как наиболее освоенной. Они указаны как в пойме Лены, так и в поймах крупных притоков ее — Алдана (с его притоком Амгой) и Вилюя (Дробов, 19146, 1916; Аболин, 19296; Порядин, 1930; Еленевский, Кремнев, Шахов, Работное, 1933; Работнов, 1933, 1935а; Коржевин, 1934). Мезофитные луга Лены и ее притоков представлены костровыми (Bromus sibiricus), лисо-хвостными (Alopecurus ventricosus), ячменными (Hordeum brevisubulatum), полевицевыми (Agrostis Trinii), мятликовыми (Роа pratensis) и пырейными (Agropyrum repens) лугами, чаще смешанными и с участием разнотравья и бобовых. В приленской полосе настоящие луга занимают крупные площади; в отдалении от Лены поймы ее притоков слабо освоены, сильно облесены и закустарены и здесь господствуют луга лишь первичного происхождения на долгопоемных участках — вейниковые и осокововейниковые. То же следует сказать и о пойме Лены ниже Вилюя.
Сравнивая луга Западной и Восточной Сибири с восточноевропейскими лугами, А. П. Шенников отмечает следующие различия. Разно-' травные белополевичники замещаются в Западной Сибири разнотравными луговоовсяничниками, а в Восточной Сибири (Якутия) — разнотравноячменными лугами. Лисохвостные (с Alopecurus pratensis) луга слабо ряспространрнкт в Сибири, более широко представлены солончаковатые лисохвостники (с Alopecurus ventricosus). Костровые и пырейные луга встречаются в Сибири, но не широко, а в Восточной Сибири Bromus iner-mis замещается Bromus sibiricus. To же следует сказать о Calamagrostis epigeios: в Восточной Сибири в прирусловых поймах на опесчаненных почвах его замещает Aneurolepidium dasystachys. В покрове мезофитных лугов Восточной Сибири характерно большое участие лилейных (Нете-rocallis flava, Lilium martagon, L. dahuricum).
Для Дальнего Востока, то есть для поймы Амура и его крупных притоков — Зеи (с Селемджой) и Амгуни, в литературе нет указаний не только на наличие белополевичных, пырейных, лисохвостных и костровых лугов, но даже и на участие этих злаков в составе других лугов (Доктуровский, 1909а, 1911; И. В. Кузнецов, 1911, 1913; Шишкин, 1927; Лучник, 1935, 1935а; Саверкин, 1936, 1938; Васильев, 1937; Куренцова, 1952). В работе А. П. Саверкина (1936) выделяются умеренновлажные злаково-разнотрав-
ные луга на повышенных участках пойм с господством разнотравья и особенно Sanguisorba parviflora с примесью Thalictrum simplex var. amurense, Trollius chinensis, Filipendula palmata, а из злаков — Agrostis clavata и меньше Роа pratensis, Calamagrostis epigeios. На более сухих участках преобладают злаки (Agrostis clavata, Роа pratensis, Calamagrostis epigeios v участием Koeleria gracilis, Festuca rubra). Характерно для того и другого луга наличие мезогидрофитов и гидрофитов (Digraphis arundinacea, Carex appendiculata). Близкие к ним злаково-разнотравные луга указаны для Прихапкайского района И. К. Шишкиным (1927). 3. И. Лучник (1935) для поймы Суйфуна дает характеристику разнотравно-злаковым лугам с господством разнотравья, причем злаковую основу составляют Calamagrostis Langsdorffii и С. angustifolia с примесью Festuca rubra, Роа pratensis, Р. palustris, Trisetum sibiricum, Alopecurus amurensis, Hierochloe odorata и с участием осок и тростника. Эти луга располагаются по гривкам.
К. Д. Степанова (1953) для южной части Сахалина на дренированных приречных участках также указывает вейниковые (Calamagrostis Langsdorj-fii) луга.
Низкие уровни пойм, характеризующиеся или проточным переувлажнением и аэробными почвами, или застойным переувлажнением с анаэробными почвами, заняты болотистыми и торфянистыми лугами, четкую границу между которыми трудно провести. Торфянистые и болотистые луга представлены крупнозлаковыми, крупно- и мелкоосоковыми с участием мезогидрофильного разнотравья или смешанными осоково-злаковыми сообществами. Из крупнозлаковых болотистых и торфянистых лугов наиболее широко распространены канареечниковые (Digraphis arundinacea), бекманиевые (Beckmannia eruciformis), манниковые (Glyceria aquatica, реже Glyceria fluitans), вейниковые (Calamagrostis lanceolata, C. Langsdorffii, C. neglecta), щучковые (Deschampsia caespitosa).
Наиболее близки к настоящим (мезофитным) крупнозлаковым лугам канареечники, примыкающие непосредственно к ним с понижением высотного уровня. Чаще и более крупными площадями они встречаются в прирусловой и центральной поймах, меньше в приматериковой, где приурочены лишь к берегам стариц и озерков с проточной водой. В канареечниковых лугах господствующим растением является канареечник (двукисточник) (Digraphis arundinacea), часто произрастающий чистыми зарослями с небольшой примесью разнотравья (Ranunculus repens, Caltha palustris, Galium palustre, Mentha arvensis, Agrostis stolonizans). На более сырых и заболоченных участках по границе с осочниками в канареечниковых лугах наблюдается большая примесь осок (Carex gracilis, С. caespitosa), иловатого хвоща (Equisetum heleocharis)*, формируются осоково-хвощево-канареечниковые луга. По границе с крупнозлаковыми мезофит-ными лугами наблюдается примесь лисохвоста, пырея, костра, белой полевицы, болотного мятлика, то есть развиваются переходные крупнозлаковые канареечниковые луга. Канареечники Европейской части Союза более распространены по направлению к югу лесной зоны, к лесостепи; южнее, в степной зоне, канареечник теряет роль эдификатора в связи с засоленностью почв. В Сибири канареечниковые луга особенно широко развиты в пойме Оби и Иртыша, в южной части лесной зоны и лесостепи. Ряд авторов (Сапожников и Никитина, 1920; Барышников, 1933) отмечает для пойм Западной Сибири приуроченность канареечников к различным уровням пойм. В лесостепи и южной части лесной зоны канареечники за пи-мают самые низкие уровни пойм. По мере продвижения вниз по течению Оби, с увеличением поемности, канареечники поднимаются на средние уронни, уступая низкие уровни]рстроосочникам (Carex gracilis), как, например,
5 Заказ 551
на отрезке поймы Сургут—Березово. Еще ниже по Оби, с увеличением но-емности, канареечники занимают лишь высокие уровни прирусловых пойм, Что наблюдается на отрезке Березово — Салехард. Широкого развития канареечников на Енисее иЛене нс отмечается, невидимому, в силу слабой освоенности их пойм в лесной зоне. Для Дальнего Востока есть указания лишь об участии канареечника в вейниковых лугах, хотя для южного Сахалина К. Д. Степанова (1953) выделяет и канареечниковые луга. Производительность канареечниковых лугов высокая, до 50 ц/га, но сбор сена должен быть своевременным, до огрубения стеблей.
Не менее широкое распространение имеют вейниковые луга. В южно- и среднетаежной подзонах лесной зоны Европейской части СССР они представлены вейниковыми лугами с Calamagrostis lanceolata или в форме чистых зарослей, или с примесью канареечника, манника, бекмании и незамечаемого вейника {Calamagrostos neglecta). К северо-востоку С. lanceolata сменяется С. Langsdorffii, который в среднем течении Печоры и в верхнем течении Камы уже господствует. Наиболее широко распространены лангсдорфовейниковые луга в поймах рек Западной, Средней и Восточной Сибири. На Дальнем Востоке и Сахалине вместе с осочникам и господствуют вейниковые луга с Calamagrostis Langsdorffii и узколистным вейником (С. angustifolia).
Манниковые луга с господством водного манника {Glycerin aquatica, реже G. fluitans) большого распространения не имеют и занимают незначительные площади.
Бекманиевые луга с господством обыкновенной бекмании {Вескпгап-nia eruciformis) встречаются лишь небольшими участками близ пойменных озер лесостепной зоны, заходя в южную часть лесной зоны. К бекмании примешиваются лисохвост, болотный мятлик, белая полевица. Встречаются бекманиевые луга с щучкой. Интересно отметить, что в Западной и Восточной Сибири бекмания продвигается далеко на север. М. К. Барышников (1933) отмечает участие Beckmannia eruciformis в луговой растительности поймы нижнего течения Оби (севернее с. Мужи), Б. Н. Городков (1946) указывает для пойм Индигирки (67° с. ш.) и Яны (68° с. ш.) заросли восточной бекмании {Beckmannia syzigachne), наряду с Arcto-phila fulva. О наличии бекманиевых лугов на Дальнем Востоке указаний в литературе нет.
Наиболее широкое распространение из злаковых торфянистых лугов в Европейской части СССР имеют щучковые луга с господством дернистой щучки {Deschampsia caespitosa), развивающиеся преимущественно в притеррасных поймах. Дернистая щучка обладает широкой экологической амплитудой и дает ряд ассоциаций щучковых лугов. На делювиальных дренированных наносах развиваются или чистые щучники, или влажноразнотравные щучники; на торфянистых почвах — осоковые щучники с Carex caespitosa, С.gracilis, С. acuta и ряд других.Южная граница щучни-ков проходит в лесостепной полосе. Щучковые луга еще достаточно широко развиты в Западной Сибири, но к востоку щучка постепенно теряет свое значение ценозообразователя.
Особо широким распространением из болотистых и торфянистых лугов пользуются в лесной зоне, а также в лесостепи Европейской части СССР и в Сибири остроосочники (с Carex gracilis) и дернистоосочники (с С. caespitosa). В поймах рек остроосочники приурочены к условиям проточного переувлажнения прирусловых и центральных частей пойм с богатыми минеральными почвами побережий рек, озер п стариц, дернистоосочники — к условиям застойного переувлажнения с обедненными торфянистыми почвами притеррасных и приматериковых пойм. Те и дру-
гпе представлены или чистыми осоковыми ассоциациями, или смешанными злаково- или разнотравно-осоковыми. В остроосочниках часто встречается примесь вейников (Calamagrostis lanceolata, С. neglecta, в Сибири — С. Langsdorffii), побегообразующей полевицы (Agrostis stolonizans), осок (Carex aquatilis, С. vesicaria, С. vulpina), из разнотравья распространены широколистный поручейник, калужница, иловатый хвощ и др. В дернистоосоч-никах часто наблюдается примесь дернистой щучки, собачьей полевицы, обыкновенной осоки (Carex acuta) и болотного разнотравья. В севсротаеж-пой подзоне острую осоку заменяет водная (С. aquatilis). На северо-западе Европейской части СССР в речных поймах широко развиты торфянистые луга с шершавоплодной осокой (С. lasiocarpd).
Наиболее крупные площади остроосочники занимают в пойме Оби, где они к северу также сменяются водноосочниками. Остроосочники описываются также в поймах Енисея и Лены, тогда как на Дальнем Востоке их замещают круппоосочники с Carex cryptocarpa, С. vesicata, С. Midden-dorffii, а в осоковых кочкарниках господствуют С. Schmidtii и С. Meyeriana.
Литература
А. М. Дмитриев (1904, 1905); А. Ф. Флеров (1907 —1908); Г. И. Поплавская (1908); В. С. Доктуровский (1909а, 1911); И. В. Кузнецов (1911, 1913); Г. А. Боровиков (1912, 1913); С. С. Ганешин (1912, 1925); М. Ф. Короткий (1912а); А. П. Ильинский (1913); А. Р. Какс (1913); А. Н. Криштофович (1913); А. П. Шенников (1913,1914, 1938, 1941); М. П. Григорьев (1914); Ф. Н. Дингельштедт (1914); В. П. Дробов (19146, 1916), С. М. Крылов и Н. П. Потапьевский (1914); Г. И. Ануфриев (1915, 1925, 1928); В. И. Баранов (1915); И. Г. Бейлин (1915); Н. П. Кузнецова (1915); Н. И. Кузнецов [Владимирский] (1915, 1915а); А. П. Черный и В. С. Доктуровский (1915); В. В. Алехин (1916, 1922); М. М. Ильин (1916); Г. К. Крейер (1916); В. В. Ревердатто (1916, 1924); В. С. Доктуровский и Н. Н. Жуков (1916); В. Л. Комаров (1917, 1940); В. В. Сапожников и Е. В. Никитина (1920); В. В. Зарецкий (1921); Р. А. Еленевский (1924, 1929, 19366); В. В. Алабышев (1926); В. П. Ногтев (1926); П. Н. Овчинников (1926); Е. С. Степанов (1926); И. Д. Богдановская-Гиенэф (1927); А. Я. Бронзов (1927, 1929); Р. А. Еленевский и Е. В. Еленевская (1927); К. Н. Игошина (1927, 1930, 1951); С. П. Смелов (1927, 1929, 1936); И. К. Шишкин (1927); Р. И. Аболин (19296); Н. А. Антипин и В. М. Пчелкин (1929); Ф. В. Баландин (1929, 1930); Г. Я. Бронзова (1929, 1929а); Н. А. Коновалов (1930); А. А. Корчагин (1930); А. А. Папкова (1930); В. Н. Порядин (1930); Ф. В. Самбук (1930, 19316, 1934а); Ю. Д. Цинзерлинг (1932, 19356); М. К. Барышников (1933); Р. А. Еленевский, А. И. Кремнев, А. А. Шахов и Т. А. Работ-нов (1933); Т. А. Работнов (1933, 1935а); В. С. Коржевин (1934); А. М. Рубин(1934, 1935); Я. Я. Алексеев (1935); 3. И. Лучник (1935, 1935а); О. С. Полянская (1935); И. X. Дагис (1936); В. А. Крюгер и А. А. Лютин (1936); А. П. Саверкин (1936, 1938); В. Н. Васильев (1937); Р. А. Дыдина (1940); А. И. Зубков (1940); Н. И. Темноев (1940); Д. Я. Афанасьев (1941); Б. Н. Городков (1946); М. В. Марков (1946); И. С. Буддо (1947, 1953); В. А. Крюгер (1947, 1952); Л. И. Номоконов (1947, 1951, 1955); П. Конрад (1948); Л. М. Никонов (1948); А. В. Куминова (1949а); А. М. Леонтьев (1949, 1949а); Г. С. Сабардина (1949, 1950, 1952, 1952а, 1952в, 1954); П. С. Туфанов (1949); М. Е. Баранова (1950); В. А. Михеев (1950); М. Ф. Елизарьева (1951); Г. Э. Куренцова (1952); И. С. Амелин (1953); К. Д. Степанова (1953); A. Cajander (1903, 1905).
63 ст — степной зоны
К югу от лесной зоны постепенно усиливается действие тех экологических факторов, которые обусловливают ксерофитизацию и галофити-зацию растительности пойм. Нарастают сухость климата, засоленность грунта и продолжительность паводков, увеличивается переменность увлажнения (в связи с засушливым летним периодом), в свою очередь усиливающая засоленность почв. Максимального развития эти процессы достигают в пустынной зоне, но уже в степной зоне они достаточно четко выражены. Почвы участков пойм высокого уровня развиваются по типу степных чер-5*
ноземных (долинные черноземы), а в южной части зоны — по типу каштановых почв; те и другие в той или иной мере засолены. Наряду с ними широко развиты солонцеватые и солончаковатые почвы в понижениях. Для пойменных лугов степной зоны особенно характерно совместное существование на всех высотных уровнях, но особенно на средних, растений ксеро-,гидро-игаломезофильной экологии,что обусловлено переменностью увлажнения.
Вкратце изложим характеристику основных типов лугов лесостепной и степной подзоны Европейской части СССР в сопоставлении их с лугами тех же подзон Азиатской части, в основном по А. П. Щенникову (1938 1941).
На высоких уровнях пойм лесостепи, вышедших из зоны затопления, развиваются пойменные степи и степные луга. Пойменные степи представлены типчаковыми (Festuca sulcata), реже ковыльными (Stipa Joannis) степями с небольшой примесью Роа angustifolia, Koeleria Delavignei, Medicago falcata, Artemisia austriaca и некоторых других видов. Степные луга отличаются от пойменных степей отсутствием ковыля, меньшим участием типчака, наличием Agrostis Syreistschikovii и большим участием Роа angustifolia и мезофильного разнотравья (особенно Filipendula hexapetala, Salvia stepposa, Trifolium montanum, Medicago falcata и др.). Наряду co степняками встречаются и некоторые северные мезофильные виды.
В степной зоне на тех же уровнях пойм, вышедших из зоны затопления, развиваются солонцеватые типчаковые степи и солонцеватые степные луга с участием типчака. В отличие от лесостепных они характеризуются большей ксерофитизацией и засоленностью. Для них характерно участие Limonium tomentellum, Artemisia то nogy па, A. nitrosa, Silaus Besser in других представителей галофитной растительности. Кроме указанных географически замещающих солонцеватых типчаковых степей и степных лугов, в степной зоне на более легких и незасоленных почвах широко развиты житняковые (Agropyrum pectiniforme) степи на тех же высотных не затопляемых уровнях пойм.
Несколько ниже пойменных степей и степных лугов, уже в условиях краткой поемности, в лесостепи развиваются остепненные мятликовые (Роа angustifolia) и остепненные полевицевые (Agrostis Syreistschikovii) мелкотравные луга. Краткопоемность и переменность увлажнения обусловливают сочетание в них степных и мезофильных видов. На фоне узколистного мятлика или полевицы Сырейщикова развиты ранняя осока, келерия, красная овсяница, типчак, степная тимофеевка, большое участие принимают бобовые: луговой и горный клевер, серповидная люцерна, датский астрагал, обильно представлено луговостепное разнотравье, но встречается и лугово-лесное разнотравье (нивяник, луговая герань и др.). На тех же уровнях в степной зоне развиваются солонцеватые луговомятликовые и солонцеватые полевицевые луга, причем первые известны лишь в Приднепровье и восточнее не идут. Для них также характерна примесь галомезофитов: морковника, кермека, солонцеватой полыни.
Последнее звено краткопоемных лугов на склонах высоких грив, связывающее остепненные луга с более мезофитными лугами средних уровней, в лесостепи представлено остепненными мелкозлаковыми лисо-хвостниками, замещающими здесь белополевичники северных пойм. В них наиболее ярко выражено влияние переменности увлажнения. Лисохвост дает верхний разреженный ярус, под его пологом развивается второй ярус из белой полевицы и полевицы Сырейщикова, луговой келерии, лугового и узколистного мятлика; в подседе — ранняя осока, луговой,
ползучий и шведский клевер, мышиный горошек и, наряду с ними, богатое луговостепное разнотравье.
К мелкозлаковым лисохвостникам уже на средних уровнях пойм примыкают крупноразнотравные луга с лисохвостом, развивающиеся в условиях средней поемности и увлажнения. Основу травостоя составляет крупноразнотравье сложного состава (из лугово-лесных и луговостепных видов): Sanguisorba officinalis, Filipendula ulmaria, Cenolophium Fischeri, Ly simachia vulgaris и, наряду с ними, Serratula coronata, Asparagus officinalis, Thalictrum minus, Eryngium planum, Tragopogon brevirostre s. 1., Adenophora liliifolia, Trifolium montanum, много Carex praecox. Среди крупноразнотравья теряются злаки: Alopecurus pratensis, Bromus iner-mis, Agropyrum repens, Poa palustris. В зависимости от степени увлажнения крупноразнотравные луга с лисохвостом представлены различными ассоциациями : луговомятликово-крупноразнотравными, белополевицево-крупноразнотравными, болотномятликово-крупноразнотравными, осоково-крупноразнотравными (с участием Carex vulpina, С. gracilis)', соответственно этому варьирует и состав разнотравья. Крупноразнотравные луга с лисохвостом, занимая все средние уровни центральных пойм, заходят далеко на юг в степную зону, но в южной части степной зоны они замещаются обедненными по флористическому составу крупноразнотравными лугами с пыреем (Agropyrum repens). Последние также занимают средние уровни пойм и в зависимости от степени увлажнения представлены различными ассоциациями: на более высоких и сухих участках значительную роль играет примесь Bromus inermis, на более влажных — Carex melanostachya, С. vulpina, Heleocharis eupalustris. На солонцеватых почвах развивается галофитный вариант с примесью Juncus Gerardii, Silaus Besseri, Plantago Cornutii, Trifolium fragiferum и др. Крупноразнотравные с пыреем луга, в отличие от крупноразнотравных с лисохвостом, теряют часть более мезофильного разнотравья (лабазник, кровохлебку и др.) и, наоборот, в них усиливается роль более южного лугового и луговостепного разнотравья (Ptarmica cartilaginea, Lythrum salicaria, L. virgatum, Althaea officinalis, Asparagus officinalis, Euphorbia virgata, Artemisia pon-tica и A. paniculata).
В прирусловых поймах, в условиях деятельного аллювия, в лесостепи и в степи развиваются луговые сообщества корневищных злаков: пырейные, костровые и пырейно-костровые — в лесостепи; костровые, пырейнокостровые и вейниковые (Calamagrostis epigeios) — в степной зоне. Они отличаются от пырейных, костровых и пырейно-костровых лугов лесной зоны постепенным изреживанием злакового покрова и нарастанием ксе-рофитизации в составе разнотравья. В степной зоне они развиты лишь в поймах крупных рек.
В связи с засоленностью грунтов и переменностью увлажнения, как указано выше, луговые формации лесостепи и степной зоны имеют галофитные варианты. Наиболее резко галофитизация лугов выражена в притеррасных частях пойм. В лесостепи здесь развиваются лисохвостные (Alopecurus ventricosus), бескильницевые (Puccinellia tenuissima) и осоковые (Carex disticha) луга с участием галомезофильного разнотравья (Сirsium escu-lentum, Plantago Cornutii, Taraxacum bessardbicum, Juncus Gerardii и др.).
В степной зоне бескильницевые (Puccinellia Fominii) луга распространены более широко и занимают большие площади, галофитизация их усиливается. В них присутствуют, кроме галомезофитов, и галофиты (Salicornia europaea, Atriplex verrucifera, Artemisia monogyna и др.).
Низкие уровни пойм лесостепной зоны заняты торфянистыми и болотистыми лугами. Наиболее широко из торфянистых лугов распространены
дернисто-осоковые (с Carex caespitosa), встречаются и щучпикп (с De-schampsia caespitosa) у границы своего распространения. Те п другие приурочены к приматериковой или притеррасной пойме.
Болотистые луга представлены бекманиевыми {Beckmannia erucifor-mis), манниковыми {Glyceria aquatica), канареечппковыми {Digraphis arundinacea), осоковыми {Carex gracilis, реже Carex riparia) лугами. В степной зоне осоковые луга мало распространены; сложены они преимущественно Carex riparia и С. melanostachya. Уменьшается значение и канареечника как ценозообразователя. Широкое развитие в степной зоне получают заросли водно-болотных растений; тростника {Phragmites communis), камыша {Scirpus Tabernaemontani, S. lacustris, Bolboschoenus maritimus), рогоза {Typha angustifolia, T. latifolia), ежеголовника {Sparganium ramosum s. 1.), развивающихся у уреза воды в приречной зоне, по периферии озер, стариц и протоков в центральной пойме. На юге в устьях крупных рек они известны под названием плавней. Ксерофитизация и галофитизация пойменной растительности оказывают влияние и на распределение древесной и кустарниковой растительности пойм. В лесостепи и в северной части степной зоны еще встречаются остатки лесов из Quercus robur, Populus nigra, P. alba, Ulmus scabra и U. laevis. По мере продвижения к югу состав древесных пород постепенно беднеет, и в южной части степной зоны встречаются лишь отдельные деревья Populus nigra, Salix alba, а на засоленных почвах появляются Tamarix ramosissima, Т. laxa и Elaeagnus angustifolia. В лесостепи и в северной части степной зоны в поймах местами развиты богатые по флористическому составу кустарниковые заросли: Rhamnus cathar-tica, Frangula alnus, Padus racemosa, Viburnum opulus, Crataegus mono-gyna s. 1., Prunus stepposa. Кроме них, в прирусловой пойме лесостепи развиты ивняки из Salix cinerea, S. viminalis, S. purpurea, S. triandra. G продвижением к югу кустарниковые заросли постепенно беднеют и выклиниваются. Южнее всех идет Salix triandra, которая в дельте ВолГи, например, является одним из пионеров зарастания дельтовых островов.
В Азиатской части СССР пойменные луга степной зоны развиты преимущественно в Западной Сибири.
В лесостепных отрезках пойм Иртыша и Оби на высоких уровнях развиваются типчаковый {Festuca sulcata) и ковыльно-типчаковые (со Stipa Jvannis) пойменные степи и степные луга (Спиридонов, 1927—1928; Баранов и Шелудякова, 1928; Бронзов, 1929; Бронзова, 1929а). По мере продвижения к югу, в степной полосе ксерофитизация и галофитизация усиливаются. Средние уровни пойм заняты остепненными злаково-разнотравными и разнотравно-злаковыми лугами, с богатым лугово-лесным и луговостепным разнотравьем. В северной части лесостепи они представлены остепненными овсяничниками, в которых доминирует Festuca pratensis. Наряду с луговыми компонентами {Bromus inermis, Agrostis aJba, Agropyrum repens, Poa pratensis, Filipendula ulmaria, Sanguisorba officinalis) в них принимают участие луговостепные виды {Koeleria Dela-vignei, Phleum phleoides, Agrostis Syreistschikovii, Poa angustifolia и из разнотравья: Filipendula hexapetala, Phlomis tuberosa, Galium verum, Trifolium montanum и др.).
В южной части лесостепи остепненные луга представлены мелкозлаковоразнотравными лугами, составленными Роа angustifolia, Agrostis Syreistschikovii, Phleum phleoides, Koeleria Delavignei, Festuca sulcata, богатым, «красочным» луговостепным разнотравьем и обильной Carex ргаесох. Обычна слабая засоленность.
В степной зоне в остепненных узколистномятликово-разнотравных, полевицево-разнотравных и типчаково-разнотравных лугах усиливается засоленность. Наблюдается значительная примесь Silaus Besseri, Plantago Cornutii, P. maxima, а южнее — Glycyrrhiza glarba, G. echinata, видов Artemisia.
Остепненные луга отмечены также для поймы Тобола (Иванова, 1935), Ишима (Городков, 1915), Иртыша (Спиридонов, 1927—1928; Бронзов, 1929), Оби (Баранов и Шелудякова. 1928; Бронзова,1929а; Номоконов. 1955).
Аналоги пойменных степей, стенных типчаковых лугов и остепненных мелкозлаково-разнотравных лугов развиты и в бассейне Енисея (Буддо, 1947, 1953; Номоконов, 1947, 1955). Для поймы Енисея Л. И. Номоконов описывает степные участки среди остепненных лугов: келериевые {Koele-ria gracilis), типчаковые {Festuca pseudovina), змеевковые {Cleistogenes squarrosa) и ковыльные {Stipa Joannis) ассоциации. Из остепненных мелкозлаковых лугов по Ангаре (Буддо, 1953) и по Енисею (Номоконов, 1955) отмечены житняковые {Agropyrum cristatum), типчаковые {Festuca sulcata), кистевидномятликовые {Роа botryoides), тимофеевковые {Phleum phleoides), узколистномятликовые {Роа angustifolia), полевицевые {Agrostis Trinii), волоснецовые {Aneurolepidium ramosum).1 Для них характерно присутствие среди злаково-разнотравного покрова осочки: в лесостепи — Carex ргаесох, в степи — С. duriuscula.
Восточноевропейские остепненные лисохвостники и остепненные круи-нозлаковые с лисохвостом луга, по мнению А. П. Шенникова, в Западной и отчасти в Средней (бассейн Енисея) Сибири замещаются остепненными крупноразнотравными овсяницевыми {Festuca pratensis) лугами, особенно широко развитыми в лесостепи (Кузнецов [Владимирский], 1914, 1915; Баранов и Шелудякова, 1928; Бронзова, 1929а; Куминова, 1949а; Номоконов, 1955). Восточнее (на Лене) они замещаются обедненными ячменными {Hordeum brevisubulatum) лугами. Лисохвост {Alopecurus pratensis), хотя и встречается в остепненных лугах Западной Сибири, но не играет роли эдификатора. Постепенно к югу и востоку, в силу засоленности почв, его замещает Alopecurus ventricosus. Alopecurus pratensis является эдификатором лишь в^ гидромезофитном ряду, не занимая больших площадей.
Пырейные {Agropyrum repens), костровые {Bromus inermis, а на востоке Bromus sibiricus), вейниковые {Calamagrostis epigeios) луга широко распространены в прирусловых поймах Сибири, в лесостепи и в степной зоне, где они представлены как мезофитными, так и ксеромезофитными лугами (Криштофович, 1913; Городков, 1915; Косинский, 1916; Бронзов, 1929; Стрельников, 1938: Номоконов, 1947, 1955).
Для речных пойм степной области Сибири характерно широкое развитие галофитных лугов, представленных лисохвостными (Alopecurus ventricosus), ячменными {Hordeum brevisubulatum), бескильницевыми {Puccinellia tenuissima), осоковыми {Carex disticha)\ на юге степной зоны — чиевыми {Lasiagrostis splendens) и прибрежницевыми {Aeluropus litoralis) лугами. Они развиваются в условиях засоленных почв и переменного увлажнения, в понижениях и по периферии пойменных озер и стариц центральных и особенно притеррасных пойм, где чередуются с солончаками и солонцами (опис. 62).
Более широко галофитные луга развиты в бассейне Оби (Танфильев, 1902; Городков, 1913а, 1915; Кузнецов [Владимирский], 1915; Косинский,
1 Житняковые, типчаковые и кистевидномятликовые ассоциации нужно отнести к степям. (Примечание редакции).
1916; Спиридонов, 1927—1928; Бронзова, 1929а; Куминова, 1949а; Ели-зарьева, 1951), где в речных поймах перечисленные выше галомезофиты, мезогалофиты и галофиты не только образуют самостоятельные луговые сообщества, но и внедряются в мезофитные, остепненные и болотистые луга. Хорошо выраженная в лесостепной зоне галофитизация пойменных лугов Западной Сибири возрастает в степной и резко спадает в лиственнолесной зоне.
Менее широко галофитные луга развиты в бассейне Енисея (Буддо, 1947, 1953; Номоконов, 1947, 1955). В Центральной Якутии засоленность лугов возрастает, но она ограничена долиной Лены в районе Якутска, долиной нижнего течения Амги, некоторых других рек и аласными понижениями. Галофитные луга здесь приурочены к нижним частям склонов в долины, но максимального развития достигают в районе «аласов». (Дробов, 19146; Аболин, 19296; Работнов, 1933, 1935а). Они представлены здесь также лисохвостными (Alopecurus ventricosus), ячменными (Hordeum brevisubulatum), бескильницевыми (Puccinellia tenuiflora), осоковыми (Carex disticha) лугами, чередующимися в приматериковой и притеррасной частях пойм с солончаками.
В поймах рек Дальнего Востока степные, остепненные и галофитные луга не встречаются.
Болотистые и торфянистые луга низких уровней пойм широко развиты в лесостепи Западной и Средней (бассейн Енисея) Сибири. Болотистые луга в лесостепном и степном отрезке бассейна Оби широко представлены канареечниковыми (Digraphis arundinacea), мятликовыми (Роа palustris), осоковыми (Carex gracilis, С. vesicaria, С. riparia, С. orthostachys, С. аси-tiformis, на засоленных почвах — С. disticha), вейниковыми (Calamagrostis neglecta) ассоциациями. У уреза воды рек, озер и стариц широко развиты заросли водно-болотной растительности (Phragmites communis, Scirpus lacustris, S. Tabernaemontani, Bolboschaenus maritimus, Typha latifolia, T. angustifolia и на илистых отложениях Beckmannia eruci-formis, Scolochloa festucacea). Из торфянистых лугов наиболее распространены дернистоосочники (Carex caespitosa).
К северу от лесостепи возрастает роль канареечника, вейника Лангс-дорфа и острой осоки (Carex gracilis). К югу, в степной зоне, болотистые и торфянистые луга постепенно уступают место галофитным. На южной окраине степной зоны галофитные луга развиваются в непосредственном контакте с зарослями прибрежноводной и водно-болотной растительности (тростника, камыша, рогоза).
Литература
Г. И. Танфильев (1902); А. И. Мальцев (1907); Б. Н. Городков (1913а, 1915); А. Н. Криштофович (1913); В. П. Дробов (19146); Н. И. Кузнецов [Владимирский] (1914, 1915); К. К. Косинский (1916); Д. Г. Виленский (1918, 1926); Л. Г. Раменский (1918, 1927); А. И. Бар (1919); Н. В. Властова (1919, 1919а); С. Г. Горшкова (1919); А.П.ШеииМков (1919,1930, 1938,1941); М. Д.Спиридонов (1927—1928); Н. Л. Чугунова-Сахарова (1927); В. И. Баранов и В. А. Шелудякова (1928); Р. А. Еленевский и Е. В. Еле-невская (1928); И. М. Крашенинников (1928); И. М. Крашенинников, С. Г. Горшкова и Н. А. Иванова (1928); И. М. Крашенинников и Н. А. Иванова (1928); Р. И. Або-лин (19296); А. Я. Бронзов (1929); Г. Я. Бронзова (1929а); Ф. Н. Дингельштедт (1929); Р. А. Еленевский (1929а, 19296); М. И. Котов (1930, 1931); В. Кушниренко и М. Николаев (1930, 1930а); Т. А. Работнов (1933, 1935а); М. И. Котов и В. Г. Танфильев (1934); Н. А. Иванова (1935); О. А. Елиашевич (1937, 1939); П. Г. Стрельников (1938);
Д* Шелудякова (1938, 1948); А. М. Семенова-Тян-Шанская (1940); А. Д. Алексеев (1947, 1951); Г. И. Билык (1947, 1948, 1949, 1954); И. С. Буддо (1947, 1953); Л. И. Но-
J1®47* 1955>: Д- Я- Афанасьев (1948а, 1950, 1950а, 1954); Т. Ф. Кириченко
В* Кум™°ва (1949а); М. ф. Елизарьева (1951); Л. П. Сергиевская (1951);
А. Ф. Киреев (1952); И. А. Цаценкин (1952).
63 п—пустынной зоны
Сочетания водно-болотных растительных сообществ, пойменных лугов, зарослей кустарников и лесов, называемые в пустынной зоне тугаями, показаны на карте в дельте Аму-Дарьи, а также в дельте и почти по всей долине Сыр-Дарьи. Менее крупные площади тугаев имеются в низовьях Сарысу и Чу, по долине Или и в ее низовьях, а также по долинам Лепсы, Каратала, Чарына и в самых низовьях сухих древних дельт Зеравшана, Кашка-Дарьи, Теджена и Мургаба, причем на последнем они имеются и в среднем течении, начиная от государственной границы СССР с Афганистаном.
Условия существования этого типа весьма своеобразны. Они в первую очередь определяются режимом длительного заливания тугаев паводковыми водами. При этом паводок на некоторых реках происходит дважды: весной за счет таяния снегового покрова и сброса вод поверхностного стока и летом за счет усиленного таяния ледников и снежников в горах, питающих эти реки.
Вода, поступившая в тугаи, держится в них различное время, в зависимости от положения их в общем рельефе поймы или дельты. Продолжительность периода заливания колеблется от нескольких недель до нескольких месяцев. Так, в дельте Аму-Дарьи имеются значительные площади, представляющие собою озера, которые то разливаются на десятки километров, то сокращаются до незначительных размеров, сохраняя воду только в наиболее глубоких местах. Бблыпая же часть площади таких временных водоемов бывает покрыта зарослями тростника (Phragmites communis), которые обычно с конца июня и до сентября заливаются на 0.5—1.5 м водой.
В условиях наиболее длительного заливания развиваются болотные почвы тяжелого (глинистого и тяжелосуглинистого) механического состава. Такие почвы характерны для чистых тростниковых зарослей.
На прирусловых валах, самых высоких элементах рельефа пойм и дельты, подверженных наиболее короткому заливанию, развиваются тугайные почвы обычно легкого (суглинистого и супесчаного) механического состава. Такие почвы типичны для тугайных лесных и древеснокустарниковых сообществ.
В связи с различиями в продолжительности заливания, значительными колебаниями уровня грунтовых вод и их минерализации, пестротой механического состава грунтов и т. п., почвенные условия и растительный покров в пределах тугайных площадей весьма разнообразны. Кроме того, на них в разной степени влияют процессы засоления, интенсивность которых может меняться под воздействием не только природных факторов среды, но и хозяйственной деятельности человека.
На значительной площади тугаи к настоящему времени сведены, и их места заняты оазисами, среди которых тугаи сохранились лишь местами в виде отдельных островков (в Хорезмском оазисе, в Ферганской котловине, в Кашка-Дарвинском и Бухарском оазисах и др.).
В узком смысле тугаями называют древесно-кустарниковую растительность. Однако здесь принято более широкое толкование. Тугаями мы именуем не только собственно древесно-кустарниковые заросли, но и заросли тростника и камышей (с кустарниками и без них) и заболоченные участки, то есть все растительные группировки низких террас, заливаемых паводковыми водами (пойменные террасы), а также и более высоких вторых террас (первых надпойменных), недавно вышедших из-под ежегод-
ного заливания и не потерявших связи с грунтовыми водами долил крупных и малых рек пустынной зоны.
В самом типичном выражении тугай представляет собою густую, трудно проходимую заросль деревьев и кустарников из видов тополей из секции Turanga {Populus pruinosa, Р. dwersifolia, Р. ariana), рослых экземпляров гребенщиков {Tamarix hispida, Т. ramosissima, Т. Karelinii, Т. laxa, Т. florida, Т. elongata и др.), видов лоха {Elaeagnus angustifolia, Е. orientalis), видов ив {Salix australior, S. songarica, S. fragilis, S. Wilhelmsiana и др.), чингиля {Halimodendron halodendron), реже облепихи {Hippophae rhamnoides). Деревья очень часто бывают оплетены лианами {Malacocar-pus crithmifolius, Clematis orientalis, Cynanchum sibiricum, Rubia dolicho-pkylla, Calystegia sepium). В сырых местообитаниях среди деревьев и под их пологом разрастается высокорослый тростник, нередко образующий участки чистых зарослей.
Деревья и кусты, оплетенные лианами, перемежаемые болотными участками густых зарослей тростника, создают действительно труднопроходимые заросли, имеющие черты сходства с джунглями (кстати, па языке туркмен тугаи именуются «джангиль»; с этим же созвучно английское «Jung]»; Кербабаев, 1954).
В менее типичных тугаях, особенно подвергшихся вырубкам деревьев и кустарников, преобладающим элементом ландшафта становятся тростниковые заросли и луговые сообщества {Calamagrostis pseudophragmites, С. epigeios, Agropyrum repens, Trachomitum lancifolium, T. scabrum, Poacy-num pictum, Cynanchum sibiricum, Typha minima, Glycyrrhiza glabra, Sophora alopecuroides, Karelinia caspica и др.), перемежаемые участками древесно-кустарниковых зарослей. В долинах Аму-Дарьи, Вахша и Пянджа, в тугаях (в широком понимании) очень типичны заросли гигантских субтропических злаков: дикого сахарного тростника — Saccharum spontaneum, Erianthus purpurascens, Imperata cylindrica, Arundo donax (Русанов, 1930).
В менее увлажненных местообитаниях, особенно на террасах, вышедших из-под ежегодного заливания (нередко на искуствепно осушенных пойменных террасах рек, воды которых разбираются выше ио течению для орошения), широко развиваются солончаковые процессы (вторичное засоление за счет подтягивания солей по капиллярам при усиленном испарении). Нередки здесь и настоящие солончаки грунтового питания. В этих условиях древесные породы выпадают и сохраняются только га-лофильные гребенщики {Tamarix hispida, Т. ramosissima, Т. laxa), чин-гиль {Halimodendron halodendron), реже лох {Elaeagnus orientalis), а к ним прибавляются поташник {Kalidium foliatum) и карабарак {Halostachys Belangeriana). Все кустарники в этих условиях угнетены; низкорослым становится и тростник, обычно сохраняющийся здесь в виде сильно разреженных зарослей с примесью типичных галофитов {Suaeda microphyНа, Limonium Gmelinii, L. otolepis, Salsola lanata, S. crassa, Salicornia europaea). В северной части пустынной области по вторым террасам речных долин нередки заросли чия {Lasiagrostis splendens), а также галофиль-ного кендыря {Poacynum pictum).
В пойме Атрека и других южных рек на луговых солончаках развиты чаирные луга (из Aeluropus litoralis и A. repens) с участием некоторых однолетних солянок {Salsola lanata, S. turcomanica) и с вкраплением сар-сазанников (из Halocnemum strobilaceum) на участках пухлых и мокрых солончаков (Нардина, 1954).
Кроме перечисленных основных районов расиространения тугаев, последние встречаются и в ряде других мест; однако они не могли быть показаны по условиям масштаба.
Отметим некоторые характерные их особенности.
И (^большие площади тугаев часто встречаются по мелким речкам, сбегающим с горных хребтов и теряющимся в пустыне у их подножья. Тугайные леса в слепых дельтах речек, оканчивающихся в депрессии Джунгарских ворот (Ргайты, Токты, Чиндалы), слагаются главным образом лавролистным тополем (Populus laurifolia), также с примесью ряда видов Salix. Выше по ущелью под полог тополевого леса проникают барбарис (Berberis heteropoda), крушина (Rhamnus cathartica), черемуха (Partus racemosa) и др. (Липшиц, 1931).
В прирусловой части ежегодно заливаемой поймы Чарына (впадающего в Или) сохранился реликтовый тугайный лес, образуемый ясенем (Fraxinus potamophila) с участием крупноплодного тополя (Populus macrocarра), лоха (Elaeagnus angustifolia) и ту райской ивы (Salix tura-nica). В подлеске произрастает ряд кустарников (Rosa laxa, Berberis ilien-sis, Lycium turcomanicum) и лиан (Cynanchum sibiricum, Clematis orientalis, Asparagus persicus), в травяном покрове господствуют злаки и некоторые виды разнотравья (Aneurolepidium multicaule, Lasiagrostis splendens. Phragmites communis, Fumaria Vaillantii, Chenopodium album, Sophora alopecuroides, Trachomitum lancifolium, hnpatiens parviflora и др.) (Быков, 1944). Наряду с ясенниками в пойме Чарына имеются еще ивняковые, лоховые и туранговыс тугаи. На регулярно заливаемых участках ясень хорошо возобновляется семенным путем, но при выходе из-под затопления наступает засоление почвы, и семенное возобновление резко сокращается (Березин, 1952).
По р. Аченахо (восточная часть хр. Кетмень) тугайные лески, по наблюдениям Л. Е. Родина, образуются ивами (Salix songarica, S. turanica, S. W ilhelmsiana и др.) с большой примесью облепихи (Hippophae rhamnoides) ; в подлеске имеется также ряд кустарников (Rosa laxa, Berberis iliensis) и особенно обильна лиана (Clematis orientalis), оплетающая кусты на опушках и забирающаяся высоко на деревья. Эти облепихово-ивняковые тугаи в пойме граничат почти непосредственно с полынными сообществами па крутых щебнистых склонах долины к руслу.
Наряду с этим в пределах Илийской долины, но сохраняющимся плесам мелких пересыхающих к лету речек, встречаются небольшие группы Tamarix laxa, Elaeagnus angustifolia, Halimodendron halodendron, а у берегов и в воде растут Scirpus lacustris, Typha angustifolia, Triglochin palustris, Haler pestes salsuginosa и водоросль—Chara уи/^ат(Липский,1906).
По Талассу, в районе выхода этой реки на равнину близ Джамбула, где она разбивается на множество протоков, пойма занята лугами (Phragmites communis, Festuca orientalis, Calamagrostis epigeios, Bolboschoenus maritimus, Trichophorum pumilum, Carex diluta, Juncus compressus, Orchis umbrosa, Iris sogdiana, Equisetum ramosissimum и др.), заболоченными зарослями тростника и рогозов, обвиваемых лианой (Calystegia sepium), и тугайными лесками из облепихи (Hippophae rhamnoides), лоха (Elaeagnus angustifolia) с примесью ив (Salix Wilhelmsiana, S. songarica) и с подлеском из Tamarix laxa, Halimodendron halodendron.
Сходная растительность имеется и на многих других речках, сбегающих с гор Средней Азии.
Не показаны на карте также туранговыс тугаи по р. Токрау (в 40 км к востоку от г. Балхаш). Древостой их сильно пострадал от вмешательства человека и в основном представлен порослью с единичными крупными деревьями туранги (Populus diversifolia) до 15 см в диаметре. Небольшие рощи этих туранговых сообществ сильно опустынены. Под их пологом, особенно по окраинам, разрастаются такие растения, как Halimodendron
halodendron, Eurotia ceratoides, Tamarix laxa, T. hispida, Salsola rigida, Ephedra lomatolepis, Camphorosma Lessingii, Kochia prostrata, Limonium suffruticosum и др. (Дмитриева, 19 м).
Площади, занятые тугаями (в широком смысле — то есть сочетаниями тростниковых зарослей, луговых сообществ и древесно-кустарниковых насаждений), используются для выпаса главным образом крупного рогатого скота и лошадей. Основным пастбищным кормом здесь является тростник, который охотно поедается только в молодом состоянии, а впоследствии быстро грубеет; однако при скусывании побегов тростник постоянно возобновляется, и выпас зачастую производится на протяжении всего вегетационного периода. Несмотря на высокую урожайность тростниковых зарослей, достигающую 115 ц/га, поедаемая масса их оказывается значительно меньше (Боровский и др., 1940).
Чаирные луга (из Aeluropus litoralis), широко распространенные на Атреке, а участками встречающиеся и в других речных долинах, представляют собою ценные пастбищные угодья для крупного рогатого скота и. особенно, для лошадей, используемые в течение года. Они дают 12 ц/га поедаемой массы, а при заготовке на сено до 16 ц га (Нечаева и др., 1943).
Литература
А. В. Каульбарс (1881); М. Н. Богданов (1882); В. И. Липский (1906); М. И. Пта-цщцкий (1910, 1913); О. Э. Кнорринг и 3. А. Минквиц (1912а, 1913); Б. К. Шишкин (1914а); И. Я. Зактрегер (1927); Ф. Н. Русанов (1930, 1934, 1947, 1949а); С. Ю. Липшиц (1931); М. И. Назаров (1932); А. В. Прозоровский (1932, 1940а); В. П. Покут-иий (1933); Е. П. Коровин (1934); И. П. Герасимов, Е. Н. Иванова и Д. И. Тарасов (1935); Н. П. Граве (1936); Г. Ф. Боровский и др. (1940); Е. А. Шингарева (1940); П. П. Поляков (1941); Н. И. Рубцов (19416, 1950); Н. Т. Нечаева и др. (1943); Б. А. Быков (1944); А. А. Дмитриева (1944); В. П. Дробов (1947, 19506); Э. Л. Березин (1952); Б. Б. Кербабаев (1954); Н. С. Нардина (1954).
64. Травяные болота и луга длительно заливаемых пойм (плавни)
На низких уровнях длительно заливаемых пойм развивается прм-брежноводная, водно-болотная и болотно-луговая растительность. Особенно широкого развития эти сочетания растительных сообществ, называемые плавнями, достигают в низовьях и дельтах наиболее крупных рек на юге СССР, в степной и пустынной зонах.
Крупные площади плавни занимают в низовьях Дуная и Днестра, главным образом в прилиманной части (Липский, 1889; Зеленецкий, 1891; Пачоский, 1914, 1927).
В низовьях Днепра, по сравнению с Дунаем и Днестром, площадь плавней резко возрастает: начинаясь от Запорожья, они доходят до устья, достигая в поперечнике 10—20 км (Пачоский, 1912а, 1927; Лавренко и Зоз, 1931; Афанасьев, 1951).
Не менее широко развиты плавни Дона и Кубани. От устья Дона, при впадении его в Таганрогский залив Азовского моря, плавни почти доходят до устья Северского Донца, достигая у Азова и в устье Маныча 30 км в поперечнике (Крашенинников и Иванова, 1928). На Кубани плавни развиты от Темрюка до Краснодара; кроме того, крупный массив плавней имеется в устье «протоки» (старого русла) Кубани, впадающей в Азовское море севернее Темрюка. Меньший массив плавней расположен в устье Ей (Косенко, 1924, 1934; Богдан и Шумаков, 1925; Шиф-ферс-Рафалович, 1928, 1928а; Флеров, 1930).
По западному побережью Каспийского моря крупные массивы плавней развиты в низовьях Терека и Сулака, где они на фоне лугово-солончаковой
и лугово-болотной растительности создают пеструю картину растительного покрова Притерской и Присулакской низменностей (Шиффере, 1937). Кроме того, по западному побережью Каспия плавни развиты в устье Куры, а также в устье Кумы, но здесь в силу большей засоленности и опу-стыненности они уступают место сочетаниям солончаковолуговой и лугово-болотной растительности (опис. 62), в связи с чем включены в контуры последних (Захаров, 1927; Волгунов, 1928; Новопокровский, 1932; Гроссгейм, 1948).
Наиболее обширны в Европейской части СССР плавни дельты Волги и ее наиболее крупного рукава Ахтубы (Коржинский, 1884; Булавкина, 1921; Доброхотова и Михайлова, 1938; Доброхотова, 1940; Фурсаев, 1940).
Слабое развитие современной дельты Урала, большая опустыненность и галофитизация растительного покрова привели к резкому уменьшению площади плавней в устье Урала (Пояркова и Коровник, 1931; Быков, 1954; Никитин, 1954а; Пивоваров, 1954).
Обширные территории плавней имеются в дельте Аму-Дарьи; меньшую площадь занимают они в низовьях Сыр-Дарьи (Назаров, 1932; Русанов, 1934; Доброхотова, 1954; Таубаев, 1954).
Прежде чем перейти к характеристике растительности плавней, следует отметить, что дельты и нижние течения большинства указанных выше рек представляют сложную систему рукавов главного русла, стариц, озер, протоков и гирл, разделенных островами, грядами, отмелями и косами, сложенными аллювиальными отложениями.
Длительная поемность, медленный спад воды, переменность увлажнения обусловливают широкое развитие, с одной стороны, водной, прибрежноводной и водно-болотной растительности, часто вызывающей зарастание водоемов, с другой — луговой и галофитной и в меньшей степени — степной и пустынной (на востоке) растительности.
Плавни низовьев Дуная, Днестра, Днепра, Дона и Кубани — рек, расположенных в степной зоне (области), по сравнению с плавнями низовьев рек, расположенных восточнее, в зоне пустынь, отличаются меньшей засоленностью. Засоление здесь усиливается по мере приближения к устью реки и наиболее резко выражено обычно в приморской части, где сказывается влияние морских вод. В связи с этим различаются плавни приречные (с пресными водами) и плавни дельтовые и приморские (с более или менее солеными водами и грунтами). В приречных плавнях водоемы (старицы, протоки, озера) отличаются богатой по флористическому составу и обильно развитой водной растительностью (Salvinia natans, Lemna minor, L. trisulca, Spirodela polyrhiza, V allisneria spiralis, Aldro-vanda vesiculosa, Trapa natans s. 1., Nymphaea alba, Nuphar luteum, Nym-phoides peltatum, Potamogetonperfoliatus, P. pectinatus, P. lucens, P. crispus, Myriophyllum verticillatum, M. spicatum, Ceratophyllum demersum, C. sub-mersum, Nafas minor, N. marina, Stratiotes aloides, Sagittaria sagittifolia, Hydrocharis morsus-ranae).
Ближе к берегу, в более мелководной полосе, водная растительность сменяется зарослями тростника {Phragmites communis), рогоза {Typha angustifolia, реже Т. latifolia), камыша {Scirpus lacustris, S. Tabernaemon-tani, S. triqueter). В пресных водах заросли тростника, рогоза и камыша достигают 4 м высоты. Под их пологом, ближе к зеркалу воды, развиваются указанные выше водные растения, а ближе к берегу — водно-болотные {Alisma plantago-aquatica, Ranunculus lingua, Butomus umbellatus, Caltha palustris, Acorus calamus). Рогоз и камыш на свободных от тростника участках с илистыми отложениями образуют самостоятельные заросли. В бо_
лее мел ко иодных участках, свободных от тростника, камыша, рогоза, развиваются заросли осок (Carer riparia, С. gracilis, ('. acutiformis, ('. melanostachya).
Заросли тростника, развивающиеся на более повышенных прибрежных участках, менее сомкнуты и в высоту не превышают 2 м; в них внедряются камыши и осоки, чистые заросли сменяются смешанными сообществами с участием болотного разнотравья (Butomus umbellatus, Sium latifolium, Oenanthe aquatica, Lythrum salicaria, L. cirgatum, Ptarniica cartilaginea, Mentha aquatica. Stachys palustris, Bidens tripartitus, Dryopteris thelyp-teris).
На береговых пологих склонах, не подвергающихся постоянному затоплению и подтоплению, среди болотных растений развиваются и луговые растения (Digraphis arundinacea, Glycerin aquatica, Hierochloe odorata, Leersia oryzoides, Calamagrostis epigeios, реже C. lanceolata, Catab-rosa aquatica).
При дальнейшем повышении рельефа подобная лугово-болотная растительность сменяется пойменными лугами низкого уровня: канарееч-никовыми, манниковыми, болотномятликовыми, осоковыми и пр.
К собственно плавням относятся лишь заросли водной, водно-болотной и лугово-болотной растительности. Однако обычно плавнями называют всю территорию низовьев и дельт рассматриваемых рек, где господствуют заросли тростника, камыша, рогоза и осок, среди которых повышенные участки заняты лугами или древесно-кустарниковыми сообществами. В связи с этим кратко остановимся на лугах низовий указанных рек.
Канареечниковые, манниковые, бекманиевые, болотномятликовые и осоковые луга низких уровней пойм и островов сменяются на средних уровнях мезофитными лугами, из которых господствуют пырейные (.4gro-pyrum repens). Меньше развиты костровые (Bromus inermis), преимущественно в прирусловой пойме, полевицевые (Agrostis alba) и мятликовые (Роа pratensis) луга. В составе этих лугов принимают участие бобовые (Vicia cracca, Lathyrus pratensis, Trifolium pratense, T. repens) и луговое разнотравье. На более дренированных участках в пырейных лугах большую роль играет Carex ргаесох, усиливается и участие лугового разнотравья. На высоких уровнях пойм, обычно с опесчаненными почвами, развиваются остепненные луга (Роа angustifolia, Agropyrum pectiniforme, Artemisia Marschalliana) с участием лугового и степного разнотравья. Однако в силу неумеренного выпаса на высоких уровнях пойм нередко господствуют сильно засоренные луга из Cynodon dactylon, Eragrostis pilosa, E. suaveolens. Лишь в притеррасных частях пойм развиты солончаковые луга и солончаки.
По мере продвижения к устью реки, особенно в приморской полосе в зоне влияния морских соленых вод, засоленность сказывается резче (Шифферс-Рафалович, 1928; Флеров, 1930). В водоемах плавней постепенно исчезают пресноводные растения и появляются наиболее солевыносливые (Ruppia spiralis, R. maritima, Zannichellia pedunculata)', в приморской полосе остаются лишь Ruppia и Chara vulgaris, а приморские заливы — лиманы совсем лишены водной растительности. Также резко беднеет прибрежноводная и водно-болотная растительность плавней. Наиболее солевыносливым является тростник; камыши и осоки постепенно отступают; рогоз не образует самостоятельных зарослей, встречаясь лишь как примесь к тростнику. В результате Phragmites communis дает почти чистые заросли с зеркалом воды под пологом или с зарослями Ruppia maritima и особенно Chara vulgaris. Заросли тростника изрежены и снижены до t.5—2 м высоты. На побережье в заросли тростника внедряются галофиль-
ные растения (Tripolium vulgare, J uncus Gerardii, Bolboschoenus maritimus, (л/nanchum acutum). На более высоких грядах заросли тростника сменяются С у perus / и sens, Acorellus pannonicus, Bolboschoenus maritimus, Puccinellia Fominii, Aeluropus litoralis, Crypsis aculeata. Вершины гряд заняты разреженными группировками полыни (Artemisia monogyna, Limonium Gmelinii, Atriplex litoralis и др.).
Ila менее засоленных участках по побережью к тростнику примешивается в большом количестве вейник (Calamagrostis epigeios). Вейниковотростниковые луга переходят выше но склону в галофнтные луга, а на высоких уровнях сменяются степными группировками.
В плавнях некоторую роль играет также древесно-кустарниковая растительность. В относительно верхних (чаще северных) частях долин рек, занятых плавнями, древесно-кустарниковая растительность представлена еще остатками лесов: осокоревых (Populus nigra), тополевых (Populus alba), ильмовых (UImus laevis), ивовых (Salix alba), реже дубовых (Quercus robur), зарослей ив (Salix triandra, S. purpurea) и других, приуроченных преимущественно к береговым валам. По мерс» продвижения к югу и понижения высоты сохраняются лишь отдельные экземпляры Salix alba и узкие разорванные полоски ивняков. На солончаковых почвах появляется Tamarix. Ближе к устью и в приморской полосе древесные и кустарниковые породы, а также луга выклиниваются, и здесь господствует исключительно растительность собственно плавней. Для Днепра, Днестра и Южного Буга, в их приустьевых частях, известны островные местонахождения лесков из черной ольхи (Alnus gluti-nosa).
Засоленность усиливается не только но направлению с севера на юг, но и с запада на восток. Если для Дуная и Днестра солончаки указываются по берегам усыхающих лиманов лишь в приустьевых частях рек, то для Днепра и Кубани они характерны и для притеррасных участков пойм.
Сопоставляя причерноморские плавни с прикаспийскими, следует отмстить, что растительные сообщества собственно плавней (водная, водноболотная и лугово-болотная растительность) сложены в основном общими ценозообразователями. Основные различия в растительности причерноморских и прикаспийских плавней объясняются более сильным засолением в Прикаспии, в связи с чем настоящие мезофитные луга здесь замещаются галофитиыми, широко развита растительность солончаков, чередующихся с участками водно-болотной и лугово-болотной растительности; при этом солончаковые растения широко внедряются и в заросли тростника. Так, в Терско-Сулакской низменности основным ценозообразователем плавней также является тростник (Phragmites communis), но роль рогоза (Typha) резко снижена, возрастает роль Bolboschoenusmaritimus. Под пологом тростника, ближе к открытой воде, среди водной растительности развиты Zannichellia pedunculata, Chara vulgaris, а по берегу — галофитная растительность (Juncus Gerardii, Tripolium vulgare, виды Atriplex). По выходе из полосы постоянного затопления, заросли тростника сменяются зарослями вейника (Calamagrostis epigeios) и осок (Carex acutiformis, С. riparia, С. melanostachya), также с участием галофитов. Выше по склону берега полоса надземного вейника сменяется полосой аржанца — Agropyrum ruthenicum с примесью A. repens и с участием Glycyrrhiza glarba, Tripolium vulgare, Galatella rossica, Gypsophila trichotoma. На внутренних склонах берегового вала распространяются солончаковатые луга с участием Puccinellia Fominii, Р. gigantea, Aeluropus litoralis, Limonium caspium, PlantagoCornutii, среди которых в западинах развиваются солончаки (Л tri-
plex verrucifera, Нaloe петит strobilaceum, Salicornia europaea). В районе приморской полосы для солончаков характерно наличие полукустарниковых солянок (Kalidium caspicum и Suaeda microphylla).
Широкое развитие солончаковолуговой и солончаковой растительности, чередующейся с растительностью собственно плавней, создает пеструю картину растительного покрова Терско-Сулакской низменности. По Куме и Куре солончаковолуговая и солончаковая растительность по площади явно преобладает над растительностью собственно плавней, и последняя не нашла отражения на карте.
В дельтовых плавнях Волги в связи с мощными потоками пресной воды засоленность плавней, по сравнению с Терско-Сулакскими, резко снижается. В плавнях Волги господствуют обширные заросли Phragmites communis с примесью Typha angustifolia, Scirpus Tabernaemontani, S. triqueter и богатая по флористическому составу и мощности развития водная растительность, среди которой еще Коржинским (1884) отмечены обширные заросли Aldrovanda vesiculosa, Trapa natans s. 1., Vallisneria spiralis, Nafas marina и лотоса—Nelumbium nuciferum, встречающегося также в низовьях Куры. Заросли тростника в плавнях Волги, кроме подводных отмелей, занимают также аллювиальные песчаные почвы островов, где тростник ассоциируется с Carex acutiformis, С. vesicaria, С. riparia, Dichostylis Micheliana, Calamagrostis epigeios, Digraphis arundinacea, Agrostis alba в разнообразных ассоциациях. На слабо солонча-коватых почвах развиты тростниково-солодковые (с Glycyrrhiza echinata), клубнекамышево-ажрековые (Aeluropus litoralis-{-Bolboschoenus maritimus) и бескильницевые луга. Наряду с тростниковыми зарослями в плавнях Волги широко представлены заросли ив, являющиеся пионерами зарастания. При этом наблюдается (Фурсаев, 1940), что ближе к морю господствуют заросли Salix triandra, а дальше от моря, с постепенным нарастанием аллювиальных отложений и поднятием высотного уровня, ее сменяет S. alba. В их травяном покрове развиваются Leersia oryzoides, Phragmites communis, Carex gracilis, Rubus caesius. Кроме того, Salix alba ассоциируется c Calamagrostis pseudophragmites, Agropyrum repens, Carex praecox. Из древесных пород, кроме Salix alba, в плавнях Волги встречаются Tamarix ramosissima, Т. laxa и Elaeagnus angustifolia, отмеченный также в плавнях у Анапы (Флеров, 1930) и в нижнем течении Терека и Су лака. Меньшую роль в плавнях Волги играют Populus nigra, Morus nigra, Ulmus laevis, встречаясь единичными экземплярами.
Восточнее Волги процесс засоления резко усиливается и уже в дельте Урала наблюдается максимальное развитие солончаковой растительности. Приморская полоса (Пояркова и Коровник, 1931) достигает ширины 10— 19 км, при этом в 4—6 км от моря проходит береговой вал. Между морем и береговым валом заросли тростника чередуются с зарослями Salicornia europaea. К северу от берегового вала растительный покров сложен кустарниковыми солянками (Halocnemum strobilaceum, виды Kalidium), к которым примешиваются Salicornia europaea, Suaeda prostrata, Limonium Gmelinii, Aeluropus litoralis, виды Salsola. На общем фоне солянковой растительности встречаются солончаковые луга: прибрежницевые (Aeluropus litoralis), бескильницевые (Puccinellia gigantea, Р. dolicholepis) и вострецовые (Aneurolepidium multicaule) с участием Salicornia europaea, Suaeda prostrata, Atriplex litoralis (Быков, 1954; Никитин, 1954a). По береговому валу Урала развиты небольшие группы ив и гребенщика (Tamarix). К югу-востоку от низовьев Урала резко возрастает опустынен-ность и засоленность. Так, в низовьях, да и по всей долине Эмбы господствует солончаковая растительность.
Рис. 48. Яблоневый лес по северному склону Заилийского Алатау близ г. Алма-Ата. На переднем плане яблоня (Malus Sieversii), (Фот. Ю. Е. Лавренко),
Рис. 49. Старица в пойме р. Днепра у г. Новогеоргиевска (Украин ская ССР). На поверхности воды листья и цветы Nymphaea alba. Над водой стебли Phragmites communis и Scirpus lacustris.
Рис. 50 Осоково-пушицевое низинное тундровое болото в окрестностях Тикси (низовье р. Лены в Якутской АССР). (Фот. Б. А. Тихомирова).
В южном Казахстане и в Средней Азии растительность плавней развита в низовьях Сыр-Дарьи и главным образом Аму-Дарьи, а также по некоторым другим рекам.
В дельте Аму-Дарьи крупный контур описываемого типа располагается в области современных разливов этой реки. Особенно значительный массив находится в левобережной части дельты. Основную площадь здесь занимают тростниковые заросли,1 котор. ч на значительном протяжении простираются от широты возвышенности Борлытау вплоть до Аральского моря (Русанов, 1934).
Однообразие тростниковых сообществ нарушается вкраплением некоторых видов, встречающихся, однако, в небольшом числе и до настоящего времени еще не полностью учтенных. В осветленных водах Аму-Дарьи на взморье Аральского моря Ф. Н. Русановым (1934) найдены такие редкие для пустынной области виды, как папоротник-щитовник {Dryopteris thelypteris) и водоплавающая сальвиния (Salvinia natans). В этих же условиях встречаются Carex pseudocyperus, Aldrovanda vesiculosa, Nymphoides peltatum.
Тростниковые заросли и сообщества прибрежноводных и погруженных растений покрывают не только область собственно дельты, но и огромные площади заливов Аральского моря. В зависимости от местных условий набор сообществ и закономерности их распределения отличаются своими особенностями.
Прибрежная полоса заливов покрыта зарослями тростника (Phragmites communis). Далее (вглубь залива) идет полоса рогоза (Typha angustifolia), которая сменяется сообществом казахстанского камыша (Scirpus kasachstanicus), образующего полосу 150—200 м. Далее следует полоса погруженной растительности с господством рдестов (главным образом Potamogeton perfoliatus, с включением участков Р. pectinatus) и с участием Scirpus kasachstanicus, вегетирующегд под водой и не достигающего поверхности. Несколько далее от берега в глубине залива наблюдаются отдельные «оазисы» водной растительности, слагаемые сообществами рдестов (Potamogeton perfoliatus и Р. pectinatus), урути (Myriophyllum spi-catum) и морской резухи (Nafas marina); нередки также участки с зарослями харовых водорослей (Доброхотова, 1954).
В приморской полосе тростник нередко образует сплавину, представляющую собой мощную толщу (Л. Е. Родин в 1953 г. наблюдал сплавину до 1 м) густо переплетенных корневищ тростника с остатками неразло-жившихся отмерших его надземных частей и примесью мелкозема. Местное население именует их «кунаками» (Доброхотова, 1954). На таких сплавинах поселяются Dryopteris thelypteris, Calystegia sepium, Lycopus euro-paeus, Stachys palustris. Молодые стебли и листья тростника идут в корм скоту, мощные взрослые стебли используются как разнообразный строительный материал для жилья, оград и т. д., а также на топливо и как подстилочный и упаковочный материал.
1 Тростник представлен в дельте Аму-Дарьи двумя видами: Phragmites communis и близким к нему Ph. Isiaca, отличающимся более крупными размерами. Тростник Изиды свойствен преимущественно приморским болотам и заболоченным берегам водоемов и распространен только в южной части СССР (основной ареал Ph. Isiaca — восточное Средиземноморье, Северная Африка, Иран и Индия), тогда как обыкновенный тростник отличается гораздо более широким экологическим и географическим ареалом. Полное отсутствие специальных исследований о взаимоотношениях обоих видов в образуемых ими сообществах лишает нас возможности говорить о них раздельно. Так как дельта Аму-Дарьи на всем протяжении изобилует протоками и замкнутыми водоемами (то есть стациями, типичными для Ph. Isiaca), можно предполагать, что тростник Изиды распространен здесь так же широко, как и тростник обыкновенный.
Среди тростниковых зарослей значительные площади приходятся на водоемы, отличающиеся большим разнообразием флоры и растительности. Т. Таубаев (1954) указывает 76 видов водно-болотных растений и 39 образуемых ими ассоциаций.
В плавнях к тростнику в незначительном количестве примешивается Lycopus europaeus, Lythrum salicaria и уже названные выше Dryopteris thelypteris, Salvinia nutans, Aldrovanda vesiculosa, Nymphoides peltatum. Carex pseudocyperus.
В пресных водоемах также господствует тростник, а с ним произрастают рогозы {Typha angustata, Т. angustifolia, Т. Laxmannii, Т. minima и др.), камыши (Scirpus Hyppolitii, S. litoralis), рдесты (Potamogeton lucens, P. crispus и др.), Ceratophyllum demersum, Salvinia natans, Nym-phaea Candida, Polygonum amphibium и др.
В солоноватых водоемах видовой состав очень обеднен. Наиболее характерны в них Ruppia maritima, Potamogeton pectinatus, Zannichellia pedunculata, а также харовые водоросли. Из последних, по личному сообщению М. М. Голлербаха, наиболее широко распространены Chara tomentosa, Nitella hyalina и Nitellopsis obtusa; в меньших количествах обнаруживаются Lamprothamnium papulosum, Chara intermedia, Ch. con-traria, Ch. delicatula, Ch. fragilis, Ch. connivens, Ch. kirghisorum и Ch. gym-nopitys.1
Все разнообразие сообществ водной растительности дельты Аму-Дарьи схематически может быть объединено в три группы формаций: 1) прибрежноводных растений (формации, слагаемые Sagittaria trifolia, Sparganium ramosum s. 1., Butomus umbellatus, Typha minima); 2) полупо-груженных растений (формации из Typha angustifolia, Scirpus lacustris. Phragmites communis, Ph. Isiaca); 3) водных или погруженных растений (формации из Nymphaea Candida, Potamogeton perfoliatus, P. lucens, P. no-dosus, P. pectinatus,Ceratophyllum demersum, Nafas marina, N. graminea, Myriophyllum spicatum, Ruppia maritima) (Таубаев, 1954).
Наряду с водоемами среди тростниковых зарослей вкраплены и островки суши, то есть участки, вышедшие из сферы заливания. Обычно они в той или иной степени засолены. На них встречаются фрагменты тугаев в виде групп гребенщиков (Tamarix laxa, Т. hispida, Т. leptostachys), луговые сообщества (из Calamagrostis pseudophragmites, Cynodon dactylon, Aeluropus litoralis), солончаки c Halostachys Belangeriana (иногда еще хранящие остатки тростника, если не на поверхности, то в почвенных разрезах), и широко распространены многочисленные переходные сообщества.
Имеется также несколько освоенных участков — оазисов (опис. 109), главным образом с культурой риса, джугары, люцерны, бахчевых.
Дельта Сыр-Дарьи также характеризуется обширными зарослями тростников, которые покрывают всю пойменную часть и подступают к самому руслу. В глубине тростников обильны озера, иловатые заболоченные низины, зарастающие старицы. Характерно и здесь отсутствие ту
1 Две последние харовые водоросли представляют особый ботанико-географи^ ческий интерес. Так, для Chara kirghisorum дельта Аму-Дарьи является самой южной частью ее эндемичного в пределах СССР ареала. Нахождение же здесь Chara gymno-pitys оказывается пока единственным для СССР, это самая северная точка тропического и субтропического ареала вида, охватывающего юго-восток Азии, Малайский архипелаг, Австралию, юг Африки и о-в Мадагаскар (М. М. Голлербах, 1951.Первая находка Chara gymnopitys А. Вг. в пределах СССР. Бот. мат. отдела споровых растений Бот. инет. АН СССР, 7).
По Аму-Дарье встречаются и другие субтропические и тропические водные и прибрежные растения, например Sagittaria trifolia и Nafas graminea.
гаев, хотя отдельные представители (Salix, Elaeagnus, Tamarix) кое-где встречаются даже вблизи Аральского моря.
М. И. Назаров (1932) указывает, что значительная часть необозримых тростниковых зарослей залита водой и бывает труднопроходимой, а в некоторых случаях и вовсе недоступной. Несмотря на исключительное однообразие этих «камышей» (наименование, часто относимое к тростниковым зарослям) и преобладание чистых, односоставных сообществ тростника, все же названный исследователь отмечает наличие более 20 видов, сопровождающих Phragmites communis. На постоянно залитых водой участках среди тростника можно изредка встретить Equisetum heleocha-ris, Typha angustifolia, T. Laxmannii, Heleocharis eupalustris, Polygonum amphtbium.
В заболоченных местах близ моря М. И. Назаров обнаружил редкую для этих южных широт бореальную орхидею (Epipactis palustris). Заросли кендыря (Trachomitum, lancifolium) весьма обычны в дельте Сыр-Дарьи в заболоченных местах. На несколько обсыхающих повышенных участках к тростнику примешиваются Calamagrostis pseudophragmites, Inula caspia. Saussurea robusta и попадаются деревца Salix cinerea, S. sibirica.
На окраинах или на возвышенных и сухих «островах» среди тростникового «моря» начинаются процессы засоления, приводящие обычно к внедрению ряда галофильных форм (Limonium otolepis, Karelinia caspica, So-phora alopecuroides, Anabasis aphylla, виды Suaeda и Petrosimonia). Здесь же иногда обнаруживаются кусты гребенщиков (виды Tamarix) и отдельные деревца Salix Wilhelmsiana и S. songarica.
Прибрежноводная флора дельты Сыр-Дарьи отличается значительно большим разнообразием. М. И. Назаров (1932) приводит список более 90 видов для нижнего течения Сыр-Дарьи, включая и дельту. В Аральском море им встречены Zostera marina,1 Nafas marina, Zannichellia palustris s. Potamogeton trichoides, В водоемах дельты обнаружено 8 видов Potamogeton (Р. crispus, Р. filiformis, Р. lucens, Р, pectinatus и др.), Sparganium minimum и S. simplex, Nymphaea Candida, Ceratophyllum demersum, Aldrovanda vesiculosa, Nymphoides peltatum и ряд других. Особенно примечательно нахождение Marsilea strigosa и бореального папоротника-щитовника — Dryopteris thelypteris (отмеченного также для амударьинской дельты).
Побережья солоноватых водоемов дают приют ряду растений: Aelu-ropus litoralis, Crypsis aculeata, Scirpus Tabernaemontani, Polygonum cor-rigioloides, Atriplex heterosperma, Salicornia europaea, Spergularia salina и др. (Назаров, 1932)
Собственно плавни в их естественном состоянии (без влияния человека) являются в ряде районов местом гнездилища саранчи, для борьбы с которой необходимо проведение системы мероприятий. Заросли тростника, камыша, рогоза и осок, хотя и отличаются высокой урожайностью, но дают сено низкого кормового достоинства. Кормовые качества сена резко повышаются при его рациональном сборе (в ранние стадии развития растений) и при силосовании. Тростник, камыш, рогоз и крупные осоки служат строительным материалом, доставляют сырье для бумажной промышленности и мелких промышленных поделок. При интенсивном ведении сельского хозяйства иловатые почвы, покрытые зарослями водно-болотной растительности, осушаются, и на их месте, с применением последующего орошения, разводят плантации риса (на юге), хлопчатника, табака, кендыря, кенафа, сеют зерновые культуры и кормовые травы. Луговые угодья в плавнях используются под покос, а растительность остепненных
1 Вероятно, Zostera minor.
грив и засоленных низин — под выпас. Следует, однако, учитывать, что урожайность лугов и пастбищ плавней резко колеблется из года в год в зависимости от степени и длительности поомности и от количества осадков. Луга и пастбища плавней в ряде случаев от поумеренного выпаса сильно изрежены и засорены. Требуется ряд мероприятий для повышения их продуктивности и кормового достоинства. Тростниковые заросли в плавнях служат мостом гнездилища для перелетной водоплавающей птицы. С целью сохранения ее необходимо уберечь крупные массивы плавней путем организации заповедников.
Литература
А. В. Каульбарс (1881); С. И. Коржинский (1884); Л. II. Краснов (1886); В. И. Липский (1889); Н. М. Золенецкий (1891); И. К. Пачоский (1912а,1914, 1927); А. И. Бар (1919); А. А. Булавкина (1921); И. С. Косенко (1924, 1934); В. С. Богдан и Б. А. Шумаков (1925); Л. 3. Захаров (1927); Д. К. Волгунов (1928); И. М. Крашенинников и Н. А. Иванова (1928); Е. В. Шифферс-Рафалович (1928,1928а, 1937); А. Ф.Фле-Ков (1930); Е. М. Лавренко и И. Г. Зоз (1931); Т. Ф. Пояркова и 3. В. Коровник (1931); [. И. Назаров (1932); И. В. Новопокровский (1932); Ф. Н. Русанов (1934, 1949а); Н. А. Буш (1935а); К. В. Доброхотова и Л. Н. Михайлова (1938); К. В. Доброхотова (1940, 19&4); А. Д. Фурсаев (1940); А. А. Гроссгейм (1948); Д. Я. Афанасьев (1951); Г. И. Билик (1954); Б. А. Быков (1954); Л. Я. Захарова (1954); И. В. Ларин, Е. В. Шиффере, Ф. Я. Левина, Б. Н. Замятнин, Л. И. Иванина, Е. А. Варивцева, Г. Н. Непли и И. В. Сырокомская (1954); С. А. Никитин (1954а); II. В. Пивоваров (1954); Т. Таубаев (1954).
65. Травяные и гипново-травяные болота
. Травяные и гипново-травяные болота развиваются в условиях сильного переувлажнения грунта и слабой проточности. Они приурочены обычно к отрицательным формам рельефа в долинах рек и на водоразделах. Общим для них является травяной покров из гидромозофильных и мезогидрофильных растений: злаков, осок, пушиц, хвощей и болотного разнотравья. Моховой покров состоит изгипновых гидрофильных мхов (виды Calliergon, Drepanocladus, Camptothecium, Meesia, Mnium и др.), но он развит не во всех ассоциациях. Травяные и гипново-травяные болота на территории СССР широко распространены от арктических тундр до степей и от западных границ страны до Дальнего Востока включительно, что обусловливает значительное разнообразие их растительного покрова. На карте выделены травяные и гипново-травяные болота тундровой зоны (65 т) и лесной зоны, включая и лесостепную (65 л).
65. т — тундровой зоны, местами в сочетании с тундровыми формациями
Более значительные площади травяных и гипиово-травяных болот тундровой зоны отмечены на карте по побережью Югорского п ва, Байда-рацкой губы, в северной трети п-ва Ямала, на побережье Гыданской и Юрацкой губы. К востоку от Енисея крупные площади гипново-травя-ных и травяных болот встречаются в долинах Агапы, Дудыпты, Хатанги, Понигая, Юели и других рек. Восточнее Лены, вплоть до Колымы, травяные и гипново-травяные болота развиты широко среди равнинной тундры. Имеется крупный болотный массив так же в устье Чауна и по побережью Чаунской губы. На Чукотском побережье травяные и гипново-травяные болота указаны восточнее м. Шмидта, включая побережье Ко-лючинской губы.
В распространении травяных и гипново-травяных болот тундровой зоны намечается ряд закономерностей, отмеченных Б. Н. Городковым (1935, 1938а, 1944, 1946). В арктической подзоне развитие болот останавливается на стадии травяно-гипновых болот, в подзоне мохово-лишайниковых и кустарниковых тундр — на стадии травя но-гиппоно-сфагновых болот. Южнее, в лесотундровой и лесной зонах, олиготрофность возрастает; гинповая стадия развития болот является кратковременной, а часто и совсем выпадает. Из осок в тундровой зоне для травяных и травяно-гипновых болот наиболее характерной является Carex stans, в лесотундре и в северной подзоне лесной зоны ее сменяет С. aquatilis. Определенная закономерность наблюдается и в распространении пушиц: для северной подзоны тундр, включая и арктические пустыни, характерны пушицевые болота с Eriophorum angustifolium, Е. Chamissonis, для южных подзон тундры и лесотундры — пушицевые кочкарники с Eriophorum vaginatum.
Наиболее обедненные травяно-гипновые болота развиты на островах Северного Ледовитого океана—Франца-Иосифа, Северной Земли, Новой Земли, Вайгач и Колгуев (Городков, 1946). На островах восточного сектора Арктики господствуют гипновые мхи (Orlhothecium chryseum, Calliergon sarmentosum, Bryum obtusifolium, Aulacomnium tur-gidum) с редкими цветковыми растениями (Caltha arctica, Ranunculus nivalis, Carex stans, Dupontia Fischeri). На островах западного сектора Арктики на фоне гипновых мхов уже развит травяной покров из Carex stans, Dupontia Fischeri, Eriophorum Chamissonis. На материковом побережье травяной покров гипново-осоковых и гипново-пушицевых болот резко обогащается. Близ м. Шмидта (Городков, 1939) травяной покров гипново-осоково-нушицевого болота представлен Eriophorum angusti-folium, Carex stans, Saxifraga cernua, Alopecurus borealis, Poa Komarovii, Arctagrostis latifolia, Caltha palustris, Polygonum viviparum, Ranunculus nivalis, Senecio frigidus, а моховой покров — Calliergon giganteum, Bryum obtusifolium, Calliergon Richardsonii, Campylium stellatum, Drepanocladus revolvens.
Для Анабарской тундры в зоне арктических тундр широко распространенными являются гипново-осоковые болота (Carex stans—Drepanocladus Sendtneri), занимающие все отрицательные формы рельефа (Сочава, 1934). При застойности и большей длительности увлажнения Carex stans сменяется С. rotundata. У основания склонов развиты пушицевые кочкарники {Eriophorum vaginatum—Dicranum elongatum). Гипново-осоковые болота с Carex stans и пушицевые кочкарники с Eriophorum- angustifolium развиты также в долине Попигая (Александрова, 1937), в долине Дудыпты (Аврам-чик, 1937; Виноградова, 1937), по побережью губы Буорхая (Душечкин, 1937). Для Обь-Енисейского водораздела Б. Н. Городков (1944) указывает ход смен растительности при заболачивании речных озер и развитии гипново-осокового болота. На первой стадии зарастания развивается луговая растительность с господством Arctophvla fulva, которая сменяется сообществом Dupontia Fischeri, Carex stans, Eriophorum angustifolium, E. Chamissonis. С нарастанием мохового покрова развивается гипново-осо-ковос болото. Доминирует Carex stans, содомин антами являются Eriophorum angustifolium, Е. Chamissonis, к ним примешиваются Carex chordor-rhiza, Comarum palustre, Pedicularis sudetica. Моховой покров развит из Drepanocladus vernicosus, D. uncinatus, D. revolvens, Calliergon Richardsonii, C. sarmentosum, Cinclidium arcticum, Camptothecium trichoides, Aulacomnium turgidum. Дальнейшее нарастание мохового покрова ведет к внедрению сфагнума (Sphagnum subsecundum) и Carex rariflorw, происходит смена гипново-осокового болота гипново-сфагновым.
Травяные и гипново-травяные болота тундровой зоны развиваются не только в долинах рек и в озерных котловинах, но широко распространены и на водоразделах, особенно в зоне полигональных тундр, а на востоке и в зоне осоково-пушицево-кочкарных тундр (опис. 7). Осоково-гипновые и гипново-осоковые болота развиты в понижениях полигонов, их широкое распространение указано в районе Янгуда—Дудыпта (Виноградова, 1937) в северной части долины Попигай (Александрова, 1937) и для арктических тундр в целом (опис. 4). Флористический состав гипново-осоковых болот полигональных тундр весьма близок к составу болот речных долин и озерных котловин. В северной части долины Попигай В. Д. Александрова для пойм и обширных надпойменных пространств в понижениях полигонов указывает широкое развитие сообщества из Carex stans, Eriophorum angustifolium, E. Chamissonis, Salix reptans, Pedicularis sudetica, Comarum palustre, Equisetum variegatum, Saxifraga hirculus с моховым покровом из Calliergon sarmentosum, Drepanocladus latifolius, Meesia triquetra. Осоково-гипновые болота подзоны арктических тундр характеризуются более мощным развитием гипновых мхов и редким обедненным травяным покровом (Carex chordorrhiza, С. rariflora, С. stans, Eriophorum angustifolium, E. Chamissonis, Dupontia Fischeri) (Городков, 1938a). Они являются последующей стадией в процессе отундровения гипново-осоковых болот и смены их моховыми тундрами.
В лесотундре, как уже указывалось выше, Carex stans сменяется С. aquatilis. Болота с господством С. aquatilis занимают большие площади как в долинах рек, так и в озерных котловинах. Та же смена осоковых болот с Carex stans водноосоковыми болотами характерна и для западного сектора тундр. На Болыпеземельских тундрах широко развиты низинные осоковые болота с Carex aquatilis близ озер или на месте бывших озер вне долин и в долинах рек (Андреев, 1932а). То же следует отметить для Ямало-Ненецкого национального округа (Самбук, 1933); лишь в северной части тундр встречаются осоковые болота с Carex stans, к югу они замещаются осоковыми болотами с С. aquatilis с примесью Caltha palustris, Comarum palustre, Polemonium acutiflorum, Cardamine dentata. Моховой покров из видов Mnium, Drepanocladus, Calliergon. Для тундровой зоны Приуралья и левобережья Обской губы также указываются водноосоковые низинные болота (Игошина, 1934; Андреев, Игошина, Лесков, 1935). Вокруг озер и протоков вслед за поясом Arctophila fulva идет пояс Carex aquatilis с примесью Calamagrostis neglecta, Eriophorum angustifolium, Comarum palustre, Saxifraga cernua, Cardamine pratensis, Caltha palustris с гипновым моховым покровом из Calliergon stramineum, Drepanocladus revolvens, D. exannulatus, Meesia triquetra. Кроме осоковых болот, на водоразделах широко развиты пушицевые болота (Eriophorum angustifolium, Е. russeolum). В южной части тундровой зоны в пределах Ямало-Ненецкого национального округа (Самбук, 1933), наряду с водноосоковыми болотами, у подножья склонов в условиях застойного увлажнения развиваются дернистоосочники с Carex caespitosa и с небольшой примесью Nardosmia frigida, Polemonium acutiflorum, Parnassia palustris.
Травяные и гипново-травяные болота широко распространены и в тундровой зоне Дальнего Востока, особенно в подзоне арктических тундр, я южнее, среди осоково-пушицево-кочкарных тундр в Анадырско-Пенжин-ской депрессии (опис. 4 и 7). По своему характеру они близки к осоково-гипновым болотам тундровой зоны Обь-Енисейского и Енисейско-Ленского водоразделов. Гипново-осоковые болота с Carex stans, Eriophorum russeolum, E. angustifolium, Carex rariflora, C. chordorrhiza, C. lugens, Hierochloe pauciflora, Betula exilis, Salix fuscescens, Pedicularis adunca,
с развитым гипновомоховым покровом указываются для бассейна Анадыря (Сочава, 1930а) и для бассейна Пенжины (Городков, 1935). В лесотундре Carex stans также сменяется С. rotundata. Наряду с гипново-осоковыми болотами развиты пушицевые кочкарники с Eriophorum angustifolium и Е. Scheuchzeri.
Травяные и гипново-травяные болота тундровой зоны частью используются как оленьи пастбища.
Литература
В. Б. Сочава (1930а, 1932а, 1934); В. Н. Андреев (1932а); Б. Н. Городков (1932, 1933, 1935, 19356, 1938а, 1939, 1944, 1946); Ф. В. Самбук (1933); К. Н. Игошина (1934); В. Н. Андреев, К. Н. Игошина, А. И. Лесков (1935); В. Н. Васильев (1936); М. Н. Ав-рамчик (1937); В. Д. Александрова (1937); А. Н. Виноградова (1937); В. И. Душеч-кин (1937); В. А. Шелудякова (1938, 1948).
65 л — лесной зоны и лесостепи
В лесной зоне травяные и гипново-травяные низинные (евтрофные) болота наиболее широко развиты в поймах рек с медленным течением. Часто евтрофные болота, не занимая больших площадей, встречаются в комплексе с верховыми и переходными болотами по периферии или в центре последних, а также небольшими массивчиками среди заболоченных лесов и лугов. В связи с этим большинство низинных болот не нашло отражения на карте, так как площади их вошли в контуры заболоченных лесов и лугов и комплексных болот. На карте отмечены лишь наиболее крупные массивы.
На крайнем севере лесной зоны, в подзоне редколесья (опис. 26 s, 26 d) травяные и гипново-травяные низинные болота показаны в нижнем течении Енисея, в бассейнах Хантайки и Дудинки и восточнее — в бассейнах Агапы, Пясины, Хеты, Попигая. На востоке они отмечены уже южнее среди горных лиственничных редколесий: в истоках Яны, Индигирки, в верховьях Колымы и, кроме того, на северо-западном побережье Охотского моря. В Европейской части Союза наиболее крупные массивы травяных и гипново-травяных болот показаны лишь в южной части лесной зоны: в Псковско-Чудской низине и в среднем течении Днепра, преимущественно в бассейне его правобережного притока Припяти (Полесье), а также по левобережным притокам, южнее Киева. В Западной Сибири на водораздельных пространствах травяные и гипново-травяные низинные болота особенно широкого развития достигают в подзоне лиственных (березовых) лесов; их крупные массивы — «займища» отмечены на водоразделах Ишим—Иртыш и Иртыш—Обь.
Наиболее северные массивы низинных болот лесной зоны, в бассейнах Хантайки, Дудинки, Хеты и других рек, развиваются в условиях высокого залегания горизонта вечной мерзлоты, что обусловливает образование многочисленных мелких водоемов — озерков и более крупных мочажин в понижениях между крупными лишайниково-моховыми буграми. В процессе зарастания водоемов развиваются осоково-гипновые и гипново-осоковые болота (Кузнецов ] Владимирский], 1932). Озерки и мочажины затягиваются сплошным гипновомоховым покровом с господством Drepanocladus exannulatus, D. fluitans, D. vernicosus, Meesia triquetra. С уплотнением гипновомохового покрова на нем развиваются Menyanthes trifo-liata, Carex rotundata, Equisetum heleocharis, Eriophorum angustifolium, Comarum palustre. Местами травяной покров обогащается Carex saxatilib, C. limosa, Eriophorum Chamissonis, Trichophorum alpinum. Осоки, пушицы, хвощ нередко дают сплошные покровы; таким образом развиваются осо-
новые, хвощевые, пушицевые, а чаще смешанные хвощево-осоковые и пушицево-осоковые гипновые болота. Площадь их обычно превышает площадь бугров. В ходе дальнейшего развития этих болот внедряются ивы (Salix hastata, S. glauca, S. arbuscula) и отдельные экземпляры березы (Betula pubescens) и лиственницы (Larix sibirica). Такого состава и близкие к ним низинные болота указываются для всей зоны лесотундры и для крайней северной полосы (подзоны редколесий) лесной зоны (Ануфриев, 1922; Городков, 1946; Кац, 1948). В пределах тех же широт, по в поймах крупных рек (нижних течений Печоры, Оби, Енисея), господствуют крупноосоковые болота с водной осокой (Carex aquatilis), которые отнесены в группу пойменных лугов (опис. 63).
Южнее, в подзонах северной, средней и южной тайги, усиливается олиготрофность. На водоразделах господствуют верховые и переходные болота.
В Европейской части СССР в зоне широколиственных лесов и лесостепи верховые болота отсутствуют или встречаются очень редко; наоборот, роль травяных и гипново-травяных болот возрастает, особенно в поймах рек. Наиболее крупные массивы низинных травяных и гипново-травяных болот имеются в среднем течении Днепра — в бассейне правобережного притока его Припяти (Танфильев, 1895; Регель, 1913; Доктуровский, Рынкевич, Копачевская, Хитрово, Деревицкий, Троицкий, 1915; Черный и Доктуровский, 1915; Доктуровский и Жуков, 1916; Тюремнов, 1931; Денисов, 1951). Широко развиты также низинные болота в поймах левобережных притоков Днепра — Сожа, Десны, Снови, Сулы и др. (Лавренко, 1928; Кушалевич, 1929; Левина, 1937; Зеров, 1938).
Низинные болота Полесья и Приднепровья представлены высокотравными, осоковыми и гипново-осоковыми болотами и черноолыпатни-ками. Очень характерными являются вьгсокотравные болота — заросли Phragmites communis, Scirpus lacustris, Typha latifolia, T. angustifolia, Equisetum heleocharis, Calamagrostis lanceolata, C. neglecta. К ним примешиваются Carex riparia, C. acutiformis, C. vesicaria, Glyceria aquatica, G. fluitans, а также водно-болотное разнотравье — Acorus calamus и др. Высокотравные болота встречаются сплошными массивами в области правобережья Припяти, захватывая и сниженные междуречья между Пиной, Припятью, Стырью, Струменом и другими южными притоками Припяти. К северу от Припяти, по ее левобережным притокам, высокотравные болота располагаются лишь узкими полосами близ уреза воды, уступая дальше от реки господство гипново-осоковым болотам (Регель, 1913).
Осоковые и гипново-осоковые болота широко представлены в северной части Полесья по низменным междуречьям рек Птич, Орессы, Тремли, на сниженном водоразделе Припять—Березина и Сож—Днепр (Тюремнов, 1931). В Приднепровье осоковые и гипново-осоковые болота наиболее широко развиты в лесостепи и на юге Полесья по левобережью Днепра. Согласно Д. К. Зерову (1938), в лесостепной полосе долины Трубежа, Недры, Супоя, Оржицы, Перевода, Ромеона, Хорола, Удоя и других рек (долины стока бывших ледниковых вод) сплошь выполнены осоковыми и гипново-осоковыми карбонатными (алкалитрофными) болотами. По фарватерной или прибрежной полосе этих долин развиты осоковые кочкарники, сложенные Carex omskiana, С. appropinquata с участием Calamagrostis neglecta, Carex diandra, C. inflata, Equisetum heleocharis, Lythrum sa-licaria, Lysimachia vulgaris, Phragmites communis и Typha latifolia', во втором ярусе — Menyanthes trifoliata, Caltha palustris, Comarum palustre, а в Воде — Lemna trisulca, Hydrocharis morsus-ranae, Nymphaea alba. Осоковые кочкарники на более обводненных участках сменяются низко
травными болотами с господством Menyanthes trifoliate., Caltha palustris, Comarum palustre, Carex limosa, C. chordorrhiza, с небольшим участием Carex inflata, C. omskiana, Calamagrostis neglecta, Equisetum heleocharis. В прибрежной полосе небольшими площадями встречаются также хвоще-вые болота с господством Equisetum heleocharis, также с участием Carex omskiana. Кроме последней, присутствуют Carex riparia, С. gracilis, С.diandra, С .vesicarian указанное выше водно-болотное разнотравье. Наиболее широко, занимая верховья долин, долинные расширения, притеррасные понижения и замкнутые низины, распространены в указанных долинах рек гипново-осоковые карбонатные болота. Моховой покров их в основном сложен Drepanocladus Sendtneri, являющимся показателем карбонатности почв. В травяном — доминируют Carex diandra, С. limosa с участием С. omskiana, Eriophorum gracile, Calamagrostis neglecta, Equisetum heleocharis, Menyanthes trifoliata, Galium palustre, Epilobium palustre, Stellaria cras-sifolia и другого болотного разнотравья. Карбонатные (алколитрофные по Д. К. Зерову, 1938) болота характерны лишь для лесостепной полосы Приднепровья, в области распространения лёссовидных суглинков.
Севернее, в Киевско-Черниговском Полесье, в области флювиогляциальных отложений, господствуют евтрофные болота, также связанные с долинами стока, со старыми руслами крупных рек и притеррасными понижениями. Из осоковых болот нечасто, по на больших площадях, развиты остроосоковые болота. Доминирует Carex gracilis, вместе с ней встречаются С. caespitosa, С. vesicarica, С. inflata, С. diandra, Роа palustris, Glyceria aquatica, Eqiusetum heleocharis, Ranunculus lingua, Acorus calamus. Из осоковых кочкарников широко развиты дернистоосоковые болота. Доминирует Carex caespitosa, к ней примешиваются Equisetum heleocharis, Scirpus silvaticus, реже — Carex appropinquata, C. inflata, C. vesicaria; второй ярус сложен болотным разнотравьем — Caltha palustris, Galium palustre и др. Встречаются гипновые мхи: Climacium den-droides, Calliergonella cuspidata, Drepanocladus aduncus, Calliergon cordi-folium. Из гипново-осоковых болот в Киевско-Черниговском Полесье наиболее широко распространены болота с травяным покровом из Carex limosa, С. diandra, С. inflata, С. caespitosa. В первом ярусе травяного покрова, наряду с доминирующими осоками, принимают участие Agrostis canina, Equisetum heleocharis, а во втором — Carex dioica, Menyanthes trifoliata. Comarum palustre, Galium uliginosum, G. palustre, Dryopteris thelypteris. Встречаются Betula humilis, Salix rosmarinifolia. Моховой покров обычно сложен Drepanocladus vernicosus, Calliergon giganteum, Calliergonella cuspidata, Camptothecium trichoides, Climacium dendroides, но отсутствуетDrepanocladus Sendtneri (показатель карбонатности болот).
В отличие от лесостепи, в Полесье, кроме гипново-осоковых болот, развиты также евтрофные сфагново-осоковые болота, приуроченные к притеррасным и приматериковым понижениям. Моховой покров их сложен гидрофильным видом сфагнума — Sphagnum obtusum. В травяном покрове этих болот также доминируют осоки — Carex limosa, С. diandra, С. inflata, к которым примешиваются Eriophorum angustifolium, Е. gracile, Calamagrostis neglecta, Equisetum heleocharis. Agrostis canina. Во втором ярусе — C ar ex chordorrhiza, C. dioica, Comarum palustre, Epilobium palustre. Scheuchzeria palustris, Galium uliginosum, G. palustre.
Под влиянием осушки осоковых и гипново-осоковых болот (Зеров, 1938) на месте их развиваются вейниковые болота с Calamagrostis neglecta, С. lanceolata, широкое развитие которых указывается и для Припятьского Полесья (Регель, 1913; Черный и Доктуровский, 1915). Значительное распространение получают также болота антропогенного (под влиянием
выпаса) происхождения — леерзиевые (Leersia oryzoides) и аирные (Acuras calamus) (Левина. 1937; Зеров, 1938).
Травяные и гипново-травяные, а из лесных черноольховые болота являются ландшафтными для долин рек лесостепи Приднепровья. Мезо-трофные осоково-сфагновые (с Carex lasiocarpa, С. limosa) болота, приуроченные к понижениям песчаных террас, ландшафтной роли не играют, их роль возрастает в лесной зоне (Лавренко, 1922).
В Азиатской части СССР крупные площади низинных болот на сниженных водораздельных пространствах указаны лишь в полосе березовых лесов и в северной лесостепи Западной Сибири (на водоразделах Обь— Иртыш и Иртыш—Ишим). Волнистый рельеф, наличие многочисленных пресных озер и понижений с богатым грунтовым водным питанием способствуют развитию высокотравных (тростниковых, камышовых, рогозовых), осоковых и гипново-осоковых болот (Кузнецов [Владимирский], 1915а; Спиридонов, 1923, 1927—1928, 1928; Барышников, 1929; Бронзов, 1936; Генкель и Красовский, 1937). Высокотравные низинные болота приурочены к периферии пресных озер или к днищам озерных котловин, а также к центральным частям крупных понижений, соединяющих озера в период весенних разливов в крупные озерные системы. Они представлены зарослями Phragmites communis, Scirpus lacustris, Typha latifolia, Scolochloa festucacea, сменяющимися по берегу крупноосоковыми болотами из Carex riparia, C. gracilis, C. vesicaria, C. orthostachys, C. melano-stachya, с участием того же водно-болотного высокотравья (тростника, камыша, рогоза) и болотного разнотравья (Polygonum amphibium, Galium palustre, Pedicularis Karoi и др.). Появляются также гипновые мхи. Такого характера высокотравные и крупноосоковые болота занимают большие площади в полосе березовых лесов и известны под названием «займищ». С выходом из зоны постоянного поверхностного увлажнения, но в условиях избыточного грунтового водного питания, высокотравные и крупноосоковые болота сменяются осоковыми кочкарниками, сложенными Carex omskiana с участием С. caespitosa и С. appropinquata. Гипновомоховой покров развит пятнами из Brachythecium rivulare, Mnium affine, M. Se-ligerii, Drepanocladus aduncus v. Kneiffii.
В южной части лесостепи развитие болот заканчивается стадией осоковых кочкарников из Carex omskiana (Генкель и Красовский, 1937). Дальнейшее развитие их идет по солончаково-луговому ряду и далее — по степному ряду (опис. 62).
В северной лесостепи и в полосе березовых лесов, кроме «займищ» и осоковых кочкарников, широко развиты гипново-осоковые болота. Для них характерно развитие сплошного гипново-мохового покрова из Drepanocladus vernicosus, D. Sendtneri, D. aduncus v. Kneiffii, Brachythecium rivulare, Calliergon trifarium, Meesia triquetra, Camptothecium tri-choides, Aulacomnium palustre. Травяной покров сложен Carex diandra, C. limosa, C. inflata, реже C. lasiocarpa. Кроме осок, в травяном покрове обычны Menyanthes trifoliata, Scheuchzeria palustris, Pedicularis Karoi, Peucedanum palustre, Equisetum heleocharis, Eriophorum angustifolium, E. gracile. Чаще гипново-осоковые болота пестрят более олиготрофными кочками или пересекаются гривками («веретьями»), занятыми осоковосфагновыми ассоциациями с Carex lasiocarpa и отдельными кустами Salix sibirica, S. pyrolifolia, S. cinerea и экземплярами Betula pubescens. Дальнейшее развитие сфагнового покрова обеспечивает превращение гипново-осокового низинного болота в осоково-сфагновое переходное.
Рассмотренные нами болота водораздельных пространств Западной Сибири широко развиты и в поймах Тобола, Ишима, Иртыша, Оби.
Севернее, в подзонах хвойных лесов Западной Сибири, низинные болота на водоразделах развиты преимущественно в комплексе с переходными и верховыми болотами (опис. 67 зс).
В Восточной Сибири в связи с орографическим строением, изрезан-ностыо рельефа и сильной облесенностью процесс болотообразования развит слабо. Более или менее крупные низинные болота имеются лишь в наиболее освоенном районе Центральной Якутии (Дробов, 19146; Або-лин, 19296; Работнов, 1939): в пойме Лены, в нижнем течении Алдана и Вилюя и в Лено-Вилюйской низине (район ал асов), где площади их вошли в контура лугов (опис. 63 л) и лиственничных среднетаежных лесов (опис. 31).
Ю. Д. Цинзерлинг (1938), рассматривая географическое распространение травяных и гипново-травяных болот, отмечает наличие географически замещающих болотных формаций. Высокотравные болота — заросли тростника, камыша, рогоза, широко распространенные в поймах и по озерным котловинам в широколиственнолесной, лесостепной и степной подзонах Европейской части Союза и в лиственнолесной и лесостепной подзонах Западной Сибири, доходят и до Дальнего Востока. Следует, однако, отметить, что на Дальнем Востоке, наряду с тростником, развиваются заросли Zizania latifolia, а из камышей — Scirpus triqueter, S. Та-bernaemontani, S. radicans, Bolboschoenus maritimus', рогоз представлен Typha latifolia (Булавкина, 1917; Комаров, 1917; Лучник, 1935; Савер-кин, 1936). Из крупноосоковых болот наибольшее распространение в поймах и озерных котловинах широколиственнолесной и лесостепной подзон Европейской части Союза имеют остроосоковые болота с Carex gracilis, широко развитые также в лиственнолесной и лесостепной подзонах Западной Сибири. Восточнее (Центральная Якутия) в аналогичных условиях местообитания развиты крупноосоковые болота из Carex orthostachys, С. aquatilis, а на Дальнем Востоке и на Камчатке остроосочники замещаются болотами из Carex cryptocarpa, С. vesicaria, С. Middendorffii (Булавкина, 1917; (Комаров, 1917, 1940; Любимова, 1940). Вейниковые болота с Calamagrostis lanceolata и С. neglecta (Европейская часть СССР) замещаются в Сибири болотами с Calamagrostis Langsdorffii, реже встречается С. neglecta. На Дальнем Востоке вместе с С. Langsdorffii широко распространен также С. angustifolia, меньше С. neglecta. Осоковые кочкарники с Carex caespitosa, наиболее распространенные в Европейской части Союза, достаточно широко распространены также в Западной и Восточной Сибири. На Дальнем Востоке из кочкообразующих осок господствуют Carex Schmidtii и С. Meyeriana. Из переходных (мезотрофных) болот наибольшим распространением пользуются в Европейской части Союза осоково-сфагновые болота с Carex lasiocarpa, С. limosa, которые доходят до Дальнего Востока включительно.
Травяные и гипново-травяные болота частично используются под покос, частично под выпас, но при интенсивном ведении хозяйства их следует расценивать как мелиоративный фонд первой очереди для организации культурных сеяных лугов и пастбищ, а также для использования их площади под зерновые, огородные и технические культуры.
Литература
Г. И. Танфильев (1895); В. Н. Сукачев (1906, 1926); В. С. Доктуровский (1911); О. И. Кузенева (1911); С. М. Филатов (1911, 1913); К. В. Регель (1913); Р. И. Аболин (1914, 1928, 19296); В. П. Дробов (19146); В. С. Доктуровский, М. И. Рынкевич, М. Н. Копачевская, В. Н. Хитрово, Н. Ф. Деревицкий, Н. А. Троицкий (1915); Н. И. Кузнецов (Владимирский] (1915а,. 1932); А. П. Черный и В. С. Доктуровский
-„ни Жуков (1916); А. А. Булавкина (1917); В. Л. Ко-(1915); В. С. Доктуровский и НН. Жуков (ig^ ig23> 1936( 1941> 1948).
маров (1917, 1928Г^1* Д Спиридонов (1923, 1927—1928, 1928); И. Д Бог-
Е. М. Лавмнко (1922 1928), М-^ пРв« 928) 1931j. м к Барышников (1929); дановская-Гаенэф (1926) С • н ^Эа, 1930, 1939); Ю.Д. Цинзсрлинг
Е. В. Кушалевич (1929), Т. А. ваоотно» v Бронзов (1936); А. П. Саверкин (1936); (1929, 1932, 1938); 3-И. Лучник (1935 , А /г ьрон^^ * л н Красов£кий С. П. Смелов (1936); Г. Н. е л Любимова (1940); Б. Н. Городков (1946);
I: S: км’• “• д“"‘“ «•«”*•А
(1951); А. Я. Вага (1953); Э. К. Вареп (1953).
X. СФАГНОВЫЕ БОЛОТА
К. А. ГАЛКИНА Общий обзор
Сфагновые болота наиболее широко распространены в лесной зоне СССР,1 особенно в тайге и в подзоне широколиственно-хвойных лесов; менее типичны они для южных районов тундровой зоны; очень не велика площадь под ними в лесостепи, а также в горных районах. Для большей части болотных массивов тундры характерно господство растительных группировок, в которых эдификаторами являются травы или гипновые мхи. В лесостепи сфагновые мхи также редко выступают в роли эдифика-торов, основная часть болотных растительных сообществ представлена травяными или лесными типами (черноольшатниками).
Размещение сфагновых растительных группировок (Sphagneta) по территории лесной зоны СССР показано в схеме на рис. 51.
Здесь прежде всего выступает зависимость между степенью обилия болотных массивов и рельефом лесной зоны. Максимальное скопление сфагновых болот наблюдается в обширной Западно-Сибирской низменности; почти полностью они отсутствуют на возвышенностях Восточной и Средней Сибири; значительное развитие сфагновых болот имеет место на Русской равнине и на крайнем востоке СССР (бассейн Амура, Колымская и Индигирская низменности).
В пределах каждой крупной ландшафтной области наблюдаются региональные закономерности распространения сфагновых болот. На относительно равнинной Европейской части СССР с севера на юг наблюдается закономерное уменьшение площадей, занятых сфагновыми болотами. Наибольшие скопления болотных массивов, в современном растительном покрове и в торфяной залежи которых господствуют сфагновые мхи, приурочены к подножьям древних террас Белого моря и к его побережье©, к крупным и средним замкнутым древним депрессиям.
В более западных депрессиях Русской равнины (Ильменской и Молого-Шекснинской) болотные массивы располагаются группами. Каждая группа состоит из ряда болотных урочищ или ландшафтов (Галкина, 1946)
1 Болотные массивы (торфяники) занимают в СССР около 71 463.75 тыс. га. Эта площадь распределяется следующим образом (в тыс. га):
РСФСР.......... 62309
Белорусская ССР . . 2900 Карело-Финская ССР 2500 Украинская ССР . . 2200
Эстонская ССР . . . 700
Латвийская ССР • . 643
Литовская ССР . . . 180
Другие республики . 31.75
(«Промышленный торфяной фонд Европейской части СССР по состоянию разведанности на 1 января 1950 г.». М., 1951).
Рис. 51. Схема распространения сфагновых болот в лесной зоне СССР.
1 — районы максимального распространении крупных массивов сфагновых болот; 2 — районы частого распространения массивов сфагновых болот средних и небольших размеров; з — районы, где сфагновые болота встречаются^отдельными, преимущественно. небольшими массивами; 4 — районы, где небольшие участки сфагновых болот встречаются рассеянно, но местами типичны для ландшафта; 5 — границы лесной зоны.
разных размеров, находящихся на различных фазах и стадиях развития. На востоке Русской равнины болотные впадины располагаются цепочкообразно по боровым террасам крупных рек (Онеги, Северной Двины, Мезени, Башки, Печоры и др.).
В Западно-Сибирской низменности самое значительное развитие сфагновых болот наблюдается в ее наиболее пониженной центральной части; площади, занятые болотными массивами, заметно сокращаются на окраинах низменности.
Болотные массивы Западной Сибири поражают своими огромными размерами и монолитностью. Местами они занимают сплошь все водораздельные пространства. На значительной части этих болотных массивов (к северу от 61° с. ш.) широко распространены мерзлые бугры.
В условиях расчлененного рельефа и резко континентального климата Восточной и Средней Сибири значение сфагновых мхов в формировании растительного покрова болот резко падает. Болотные растительные группировки, сфагновые в частности, приурочиваются к пологим, постоянно увлажняемым склонам Средне-Сибирского плоскогорья и к низменностям (Центрально-Якутской, Индигирской, Колымской и др.). На Дальнем Востоке сфагновые болотные массивы развиваются преимущественно на террасах у подножья склонов или на пологих склонах. Большей части их свойственны своеобразные гидрологический и тепловой режимы, которые способствуют процессам бугрообразования.
В зоне тундры сфагновые болотные массивы более распространены в Европейской части СССР и в Западной Сибири (опис. 66). Особенно большой процент площади занимают они в тундрах Русской равнины, где не столь суровы зимы и выпадает много осадков.
В лесостепи и в северной части степной зоны (в узком смысле) можно встретить небольшие сфагновые болотца; они приурочиваются к западинам надпойменных песчаных террас. Сфагновые мхи появились на поверхности этих массивов недавно (Лавренко, 1922, 1928; Лавренко и Левина, 1934; Зеров, 1938; Левина, 19386, 1939).
Небольшие участки сфагновых болот имеются на Кавказе, где они встречаются весьма редко и в немногих пунктах [Кобулетские и Потий-ские торфяники Рионской низменности (Терьян, 1948) и высокогорья Балкарии и Дигории (Буш, 1932) и немногие другие места]. На «Геоботанической карте» сфагновые болота Кавказа включены в контуры ольховых и грабово-ольховых лесов (53 к) и высокогорных альпийских и субальпийских лугов (57 к), среди которых они встречаются на небольшой площади. На Кавказе известно 14 видов сфагновых мхов — третья часть общего числа их в СССР (Гроссгейм, 1948).
Сфагновые мхи часто образуют сплошные покровы на лиственничных марях (опис. 34). Часть марей можно рассматривать как сфагновые болота подножий склонов, облесенные лиственницей, остальные—как заболоченные лиственничники.
Сфагновые мхи являются сильными эдификаторами болотных растительных сообществ. Они отличаются особой нетребовательностью к питательным свойствам субстрата. Чем он влажнее и беднее зольными элементами, тем больше возможности для сплошного заселения поверхности сфагновыми мхами и тем больше сказывается (до известного предела) эдификаторное значение сфагнов.
На территории нашей страны сфагновые мхи в разных районах произрастали еще, по крайней мере, в третичное время (Богдановская-Гиенэф, 1946а), но широкое распространение в растительных сообществах на болотах они получили в четвертичный период.
Всего в СССР известно 43 вида сфагновых мхов (Савич-Любицкая, 1952), значение их в сложении растительных сообществ различно.
Покровы из сфагнов секции Squarrosa {Sphagnum teres) и секции Sub-secunda (S. subsecundum и S. contortum) никогда не достигают большой плотности, а перечисленные виды сфагнов никогда не являются существенными средообразователями. В сообществах, где типичны эти сфагны, еще преобладает влияние трав или гипновых мхов.
Значительно более сильными эдификаторами являются сфагны секции Acutifolia, например Sphagnum Warnstorfii, который в ряде случаев приходит на смену сфагнов секции Subsecunda. Сфагн Барнсторфа очень широко распространен на болотах тундровой и лесной зоны. Он развивается даже на болотах в лесостепи и занимает повышения микро- и мезорельефа болотных массивов различных типов, когда эти массивы находятся в наиболее зрелых стадиях фазы богатого питания или пребывают в начальных и средних стадиях фазы смешанного питания (Галкина, 1953).
Для растительных сообществ Sphagneta Warnstorfii характерно, что их рыхлый сфагновый покров пронизан корнями осок {Carex lasiocarpa, С. appropinquata, С. dioica), вахты {Menyanth.es trijoliata), тростника {Phragmites communis) и других видов. Из деревянистых растений для этих сообществ свойственны ерник {Betula папа), болотный мирт {Chamaedaphne calyculata), а на крайнем востоке—восковница {Myrica tomentosa). Часто на покровах из Sphagnum Warnstorfii растут сосны, березы, а на востоке лиственницы. В понижениях мезо- и микрорельефа в таких массивах развиты обычно гипновые мхи и более плотные синузии трав и кустарничков.
Наиболее сильным эдификатором и мощным торфообразователем из сфагнов является Sphagnum fuscum—представитель той же секции Acutifolia. Он покрывает значительно большие площади, чем Sphagnum Warnstorfii. Этот сфагн наименее требователен к зольному питанию и занимает повышенные «сухие» элементы мезо- и микрорельефа ; как правило, он образует очень плотные ковры, особенно на выпуклых болотных массивах, переживающих зрелые стадии фазы бедного питания. В этих условиях обычно растут деревянистые микотрофные кустарнички {Andromeda polifolia, Calluna vulgaris, Empetrum nigrum, Ledum palustre, L. decumbens, Oxycoccus microcarpus, Vaccinium uliginosum) и деревья (особенно болотные сосны — Pinus silvestris ff. Litwinowii, Willkommii и pumila). Из трав здесь растут морошка {Rubus chamaemorus), пушица {Eriophorum vagina-turn) и немногие другие. Разные ассоциации группы Sphagneta fusci занимают как целые болотные массивы (или крупные участки на них), так и повышения микрорельефа (подушки, кочки, гряды, бугры), расположенные среди сфагновых болот других типов.
Sphagnum rubellum пользуется менее широким распространением, и сообщества с его господством никогда не занимают крупных пространств. Чаще всего красноватый сфагн встречается в регрессивных грядово- и озерково-мочажинных комплексах выпуклых моховиков. Здесь он приходит на смену Sphagnum fuscum или сфагнам секции Cuspidata, однако занимаемые им участки ограничены по площади, а само существование сообществ с господством S. rubellum кратковременно.
Sphagnum acutifolium, несмотря на его обширный ареал, также не преобладает в сообществах на больших пространствах. Чаще он встречается в виде примеси к Sphagnum Warnstorfii, S. fuscum, сфагнам секции Palustria (5. magellanicum, S. papillosum), а иногда и к сфагнам секции Cuspidata (например S. angustifolium).
Рис. 52. Резковыпуклый моховик (зрелая стадия развития). Аэрофотоснимок.
Рис. 53. Центральная часть полого-выпуклого моховика. Аэрофотоснимок.
Сфагны секции Palustria являются мощными эдификаторами болотных растительных группировок. Из сфагнов этой секции наиболее существенное значение имеют Sphagnum magellanicum и S. papillosum.
Sphagnum magellanicum широко распространен на болотных массивах в лесной и тундровой зонах. В чистом виде он образует небольшие куртинки на повышениях микрорельефа. Чаще же он встречается в сочетании с другими сфагнами и, в частности, со Sphagnum angustifolium. Сплошные сфагновые ковры, где сочетаются два названных сфагна, занимают целые болотные массивы, дошедшие до средних стадий фазы бедного питания, или покрывают крупные генетически однородные участки на массивах, проходящих фазу смешанного или бедного питания. Sphagnum magellanicum в чистой заросли, или в сочетании с другими сфагнами, либо с зелеными мхами (Лulacomnium palustre, Polytrichum strictum, P. commune) нередко образует повышения микрорельефа — подушки, кочки, грядки. Высота этих повышений не превышает 30—36 см.
На болотах с преобладанием Sphagnum magellanicum кустарники, кустарнички и травы, а также деревья, как правило, имеют намного лучшую жизненность, нежели в сообществах Sphagnum fuscum. Особенно это заметно на сосне, которая представлена в этих условиях формами Pinus silvestris f. uliginosa и f. Litwinowii. Влагалищная пушица образует значительно более плотные дерновины, нежели в ассоциациях Sphagnum fuscum. Там, где болотные растительные группировки лесного типа (Pi-neta, Lariceta) недавно перешли или переходят в сфагновики (Sphagneta magellanici-angustifolii), местами в большом количестве встречаются брусника, черника и осока (Carex globularis). На необлесенных или слабо облесенных болотах с преобладанием магелланского сфагна обильны четырехлепестная клюква (Охуcoccus quadripetalus), малоцветковая осока (Carex pauciflora), круглолистная росянка (Drosera rotundifolia), а на крайнем востоке СССР еще пушистая восковница (Myrica tomentosa)n осока Миддендорфа (Carex Middendorffii). Брусника, клюква, осока и росянка приурочены главным образом к болотным массивам лесной зоны.
Кроме растений, постоянно растущих в сообществах из Sphagnum magellanicum, в них можно встретить и ряд трав, оптимум развития которых находится в ассоциациях других видов сфагновых или гипновых мхов (Carex inflata, С. lasiocarpa, Phragmites communis, Menyanthes trifoliata и др.). Отмершие остатки магелланского сфагна образуют мощные слои торфа (медиум-торф).
Собщества Sphagnum papillosum пользуются значительно меньшим распространением, нежели покровы со S. magellanicum. Папиллозный сфагн приурочен главным образом к болотным массивам лесной зоны и отчасти к болотам тундровой зоны Европы и Западной Сибири. Он образует почти чистые заросли или встречается в сочетании со сфагнами секций Cuspidata (Sphagnum balticum, S. Dusenii и др.), а также co сфагнами секций Acutifolia и Palustria. Пространства, покрытые S. papillosum, характеризуются более или менее ровным микрорельефом, а чаще этот сфагн обра-зует^по душки или гряды высотой в 10—20 см. Эти повышения чередуются с мочажинами, занятыми более гидрофильными видами сфагновых мхов иных ^секций, гипновыми мхами или пленками синезеленых водорослей.
К покровам из Sphagnum papillosum приурочены главным образом травянистые растения. Из деревянистых здесь чаще всего встречаются относительно обильные кустики подбела и четырехлепестная клюква; другие кустарнички, например ерник и болотный мирт, менее обильны. В травяном ярусе ассоциаций Sphagneta papillosi преобладают осока (Carex lasiocarpa) или пухонос (Trichophorum caespitosum), либо пу
шица (Eriophorum vaginatum). Кроме названных доминантов, здесь распространены другие травянистые растения. Так, совместно с осокой (Carex lasiocarpa) растут молиния (Molinia coerulea), шейхцерия (Scheuchzeria palustris),вахта (Menyanthes trifoliata), хвощ (Equisetum heleocharis), пушица (Eriophorum angustifolium) и др. Влагалищной пушице сопутствуют, в зависимости от степени влажности, шейхцерия, малоцветковая осока (Carex pauciflord) и др.
Ассоциации Sphagneta papiilosi местами развиты по окраинам выпуклых моховиков, но чаще они занимают центральные или краевые части болотных массивов, логов и сточных котловин, находящихся в стадиях фазы смешанного питания.
Покровы из сфагнов секции Cuspidate (Sphagnum cuspidatum, S. obtu-sum, S. angustifolium, S. Dusenii и др.) пользуются широким распространением на болотных массивах тундровой, а особенно лесной зоны. В лесной зоне мхи этой секции достигают наибольшего видового разнообразия и часто покрывают обширные площади на болотах Русской равнины, Западно-Сибирской низменности и на Дальнем Востоке. В Средней и Восточной Сибири сфагны секции Cuspidata распространены меньше, так как там гидроэдафические условия более благоприятствуют развитию травяных и лесных болот. Все виды этой секции (за исключением Sphagnum lenens и S. pulchrum) гипергидрофильны.
Sphagnum obtusum имеет очень широкий ареал; он обычен в болотных сообществах тундровой и лесной зон, где совместно со Sphagnum teres и другими мхами образует то более, то менее плотные покровы. В условиях хорошей проточности и относительно богатого питания его ковры занимают обширные полосы на болотах лесной зоны. В сообществах Sphagnum obtusum или с его участием развиваются не только деревянистые, но и многие травянистые растения (Carex chordorrhiza, С. rariflora, С. stans, Eriophorum vaginatum, E. angustifolium, Comarum palustre — в тундре; Carex lasiocarpa, C. limosa, C. inflata, Menyanthes trifoliata, Equisetum heleocharis — в лесной зоне).
Sphagnum angustifolium имеет довольно широкую экологическую амплитуду по отношению к степени увлажнения, а также и зольного питания. Он образует сплошные заросли, покрывая целые болотные массивы, или крупные участки их. Ассоциации Sphagneta angustifolii местами имеют хорошо развитый ярус из сосны (Pinus silvestris f. uliginosd), березы или лиственницы. В травяно-кустарничковом ярусе чаще преобладают влагалищная пушица, морошка, болотный мирт и подбел или волосистоплодная осока (Carex lasiocarpa) с подбелом и ерником.
Не менее распространены сообщества, где эдификаторами являются более гидрофильные сфагны: Sphagnum apiculatum и 5. amblyphyllum.
Ассоциации Sphagneta apiculati, так же как и ассоциации Sphagneta amblyphylli, приурочены к топяным местообитаниям, различным по степени проточности. Среди сфагнового ковра развиваются главным образом травянистые растения (волосистоплодная, вздутая и топяная осоки, узколистная и широколистная пушицы, вахта, хвощ, шепхцерия); из деревянистых растений — клюква и подбел.
В условиях обильного застойного увлажнения водами, бедными зольными веществами,развиваются ассоциации Sphagneta ЬаШсЕОни занимают обыкновенно неглубокие, преимущественно замкнутые понижения микрорельефа. Последние чаще всего встречаются среди разнообразных Sphagneta fusci, Sphagneta magellanici или Sphagneta papiilosi. Из трав в сообществах Sphangeta baltici особенно характерна влагалищная пушица, а местами шейхцерия.
Более глубокие и влажные мочажины, увлажняемые слабо минерализованными водами, заняты ассоциациями Sphagneta Dusenii или Sphagneta cuspidati.
Sphagnum Dusenii и особенно 6’. cuspidatum достигают большого развития также в мочажинах на выпуклых моховиках северной и средней тайги Европы и Западной Сибири. В сообществе с этими сфагнами развивается ряд трав (Eriophorum I’aginatum, Scheuchzeria palustris, Carex limosa, Dro-sera anglica, в некоторых районах Rhynchospora alba и др.). Ассоциации Sphagneta Dusenii и Sphagneta cuspidati обычно встречаются в комплексе с ассоциациями Sphagneta I'usci и, несколько реже, со Sphagneta magclla-nici и Sphagneta papillosi.
В тундровой и таежной зонах, а также в притихоокеапских районах развиты ассоциации Sphagneta Lindbcrgii и Sphagneta lenensis.
Sphagnum Lindbergii, как и многие другие сфагны секции Cuspidata, гипергидрофилен. В его сообществах в тундровой зоне и на мерзлых болотах в тайге растут редкоцветная (Carex rariflora) и кругловатая (С. rotundata) осоки, пушица Шамиссо (Eriophorum Chamissonis) и другие виды.
В иных экологических условиях развивается Sphagnum lenense. На Крайнем Севере он замещает 5. juscum. Ленский сфагн — единственный из видов секции Cuspidata являющийся эдификатором «сухих» местообитаний. В лесной зоне он более вбычон на Дальнем Востоке, где в бассейне Амура и на Сахалине находится южный предел его распространения.
Развитие трав, кустарничков, кустарников и деревьев на сфагновых болотах протекает в очень специфических условиях. Корневые системы их сосредоточены в торфе, чаще всего мало- и среднсразложившемся. Корнеобитасмый слой всегда отличается недостатком кислорода и повышенной влажностью. Уровень грунтовых вод на сфагновых болотах стоит очень высоко и редко спускается ниже 30—40 см. Поверхность развивающихся болот, вследствие накопления растительных остатков, ежегодно нарастает вверх. Перечисленные особенности местообитаний сфагновых болот исключают разнообразие видового состава растений травяно-кустарничкового яруса. В этих условиях могут развиваться лишь немногие древесные породы. Травы и деревянистые растения сфагновых болот представлены древними видами с широкой областью распространения (Богдановская-Гиенэф, 1946а) и обладают рядом приспособлений. Все они многолетники; для большинства характерно преобладание вегетативного размножения над семенным. Растения сфагновых болот в той или иной мере приспособлены к нарастанию вверх торфяной массы. У сосны в этой связи сильно развит отрицательный геотропизм корней; у кустарничков, по мере обрастания стволика сфагновыми мхами, образуются обильные придаточные корни. Травы на сфагновых болотах приурочены преимущественно к более увлажненным участкам.
В листьях, стеблях, корневищах и корнях болотных трав развита система межклетников и воздушных ходов; хорошо выражен у них положительный фототропизм побегов возобновления. Многие из болотных растений, особенно деревянистых, микотрофны (Солоневич, 1954).
Сфагновые болота неразрывно связаны со всем сочетанием растительных формаций, присущим местности, где они развиваются. Главнейшие черты растительности сфагновых болот, равно как гидрологический режим и все прочие компоненты болотного биогеоценоза, обусловлены зональными особенностями природы.
На «Геоботанической карте» по совокупности признаков формации сфагновых болот объединены в два зональных типа: 1) сфагновые и дик-
раново-лишайниково-сфагновые плоско- и крупнобугристые болота тундровой зоны и полосы редколесий (опис. 66) и 2) сфагновые, местами облесенные болота лесной зоны (опис. 67). Второй тип, наиболее обширный и разнообразный, разделяется на несколько провинциальных групп формаций.
Литература
Г. И. Танфильев (1891, 1900); В. Н. Сукачев (1905, 1916, 1926); Р. И. Аболин (1914, 1915, 1928); В. Р. Вильямс (1915); Е. М. Лавренко (1922, 1928); В. С. Доктуровский (1927, 1935); О. М. Зедельмейер (1927); Д. II. Мещеряков (1927); Н. Я. Кац (1930, 1930а, 1936, 1937, 1941, 1948); С. Н. Тюремнов (1931, 1949); Н. А. Буш (1932); Ю. Д. Цинзерлинг (1932, 1938); И. Д. Богдановская-Гиенэф (1933, 1946, 1946а, 1949); Е. М Лавренко и Ф. Я. Левина (1934); Б. Н. Городков (19356); Ф. Я. Левина (1937, 19386,. 1939); Д. К. Зеров (1938); Н. И. Пьявченко (1945); Е. А. Галкина (1946, 1948, 1953, 1955); Н. В. Дылис (1946); А. А. Гроссгейм (1948); М. Н. Никонов (1948); А. Н. Терьян (1948); И. И. Тумаджанов (1948); Л. И. Савич-Любицкая (1952); Н. Г. Со-лоневич (1954); К. Metsavainio (1931).
66. Сфагновые и дикраново-лишайниково-сфагновые плоско- и крупнобугристые болота тундровой зоны и полосы редколесий
Сфагновые болотные массивы с плоско- и крупнобугристыми формами микрорельефа мерзлотного происхождения широко распространены в тундровой зоне Европейской части СССР и Западной Сибири. В восточносибирских тундрах на болотах преобладают травяные, а не сфагновые растительные группировки. Изменения в растительном покрове бугристых болот наблюдаются не только в широтном, но и в меридиональном направлениях в соответствии с климатическими особенностями, определяющими мерзлотные явления в грунте, прирост растительной массы и продолжительность вегетационного периода. На северных и восточных бугристых болотных массивах травяные и травяно-гипновые растительные группировки преобладают над сфагновыми, а на южных и западных наблюдается обратное явление. Изменяется и видовой состав сфагновых мхов (Sphagnum squarrosum, S. subsecundum, S. teres, S. Warnstorfii и S. lenense в менее суровых климатических условиях сменяются S. fuscum, S. magellanicum, S. balticum, S. Lindbergii).
Различают два основных типа бугристых болот — плоскобугристые и крупнобугристые.
На плоскобугристых болотах тундровой зоны (бугристо-полигональных) образование бугров (Пьявченко, 1949; Андреев, 1954а) связано с морозобойными трещинами, которые возникли на этих массивах в период прохождения ими травяной стадии. В процессе дальнейшего развития трещиновато-полигональных болот на них образуются плоские бугры-валики, отграниченные друг от друга мочажинами канавообразной и округлой формы. Первые возникают в морозобойных трещинах, а вторые в центральных частях полигонов. На месте этих округлых мочажин возникали и возникают озерки термокарстового происхождения. Мощность торфяной залежи колеблется в пределах 0.5—1.5 м.
Растительный покров повышений южных массивов бугристо-полигональных болот образован сфагновыми мхами (Sphagnum Warnstorfii, S. Girgensohnii, S. lenense). Местами сфагны замещаются Dicranum elon-gatum и Polytrichum strictum или лишайниками — как трубчатыми, так и кустистыми. Кустарничково-травяной ярус образован ерником, багульником, брусникой, морошкой, влагалищной пушицей и некоторыми другими. Понижения в большинстве случаев имеют покров из гипновых мхов
(виды Calliergon и Drepanocladus}. Местами среди гипновых мхов-пятнами встречаются сфагны {Sphagnum obtusum, S. subsecundum). В ./ррацяном покрове преобладают струнокорневая (Carex chordorrhizd) и редкоцветная (С. rariflora) осоки, пушица и другие травянистые растения? В еще более мягких климатических условиях, согласно Н. И. Пьявченко (1955), на буграх развиты мохово-кустарничковые группировки с лишайниками, а в напочвенном покрове этих низких бугров преобладают сфагновые мхи (Sphagnum lenense, S. balticum с примесью S. magellanicum, S. angustifolium и др.), к ним примешиваются Aulacomnium turgidum и Poly trichum strictum. На более высоких буграх сфагновые мхи замещаются Dicra-пит elongatum и лишайниками (Cladonia mitis, С. rangiferina, C<: атаи-rocrea, Cetraria cucullata,C. nivalis). Верхний ярус образован кустарцйчКами и кустарниками (Ledum palustre, Chamaedaphne calyculata,. Vqacinium uliginosum, Betula папа). Под ними чаще всего развиты Vaccinium vitis-idaea, Andromeda polifolia, Rubus chamaemorus и Eriophorum 'vaginatum. Между буграми расположены мочажины, они или широки или имеют вид мочажин — трещин; мочажины обводнены и заняты осоково-сфагновыми и осоково-гипновыми растительными группировками. Для мочажин характерны осоки (Carex chordorrhiza, С. rariflora, С. rotundata) с примесью пушицы (Eriophorum russeolum)', из сфагновых мхов там чаще всего встречаются Sphagnum balticum, S. Dusenii, S. Jensenii, S. Lindbergii, S-. obtusum, а из гипновых мхов Drepanocladus fluitans и D. vernicosus.
На одном из вариантов плоскобугристых болот в торфяной; залежи бугра с покровом из Dicranum elongatum Н. И. Пьявченко (195&) на глубине 30—40 см отметил нахождение остатков Sphagnum WarnstQrfii, составляющих основную массу (80%) торфяного слоя. Эта находка свидетельствует о возможной смене S. Warnstorfii другими, более олиготрофными видами сфагнов.
Известны и другие типы плоскобугристых болот.
Торфяная залежь плоскобугристых торфяников, как правило, мелка и редко превышает 1—2 м.
Крупнобугристые болотные массивы отличаются от бугристо-полигональных куполовидной формой бугров; склоны бугров крутые. Пространства между буграми увлажнены значительно больше аналогичных участков бугристо-полигональных болот и изобилуют озерками; кроме того, вечная мерзлота в понижениях между буграми нередко отсутствует. Среди тундровых болот крупнобугристые имеют наиболее мощную торфяную залежь (4—6 м).
Растительный покров бугров (высотой 1.5—4 м) резко отличается от растительности отделяющих их понижений. На вершинах наиболее высоких бугров, с которых сдувается снежный покров, пятнами выступает обнаженный торф, прорезанный морозобойными трещинами. Между обнажениями торфа поверхность покрыта листостебельными мхами (Dicranum congestum, D. elongatum, Polytrichum strictum и др.). Местами развиты синузии накипных лишайников (Ochrolechia tartarea, Pertusaria), бокаль-чатых лишайников, а также угнетенных кустистых (Cladonia к Cetraria) и др. Склоны бугров заняты сфагновыми мхами с куртинами кустистых лишайников (Cetraria и Cladonia). Из высших растений по склонам растут ерник, вороника, багульник, голубица, брусника, морошка, влагалищная пушица и некоторые другие. Менее высокие бугры на вершинах имеют растительный покров, сходный с растительностью склонов крупных бугров.
Понижения между буграми («ерсеи») сильно увлажнены;, их поверхность покрыта сплошь сфагнами (Sphagnum balticum, S.. Lindbergii) яля
гипновым (Calliergon cordifolium, С. stramineum, Drepanocladus ехаппи-latus) ковром. Над поверхностью мохового покрова возвышаются осоки: редкоцветная, кругловатая, струнокорневая; кроме того, узколистная пу-щица и подбел.
В Западной Сибири крупнобугристые сфагновые болота заходят дальше на юг, и их граница совпадает с южной границей подзоны северной тайги.
Литература
Р. Поле (1903); Н. И. Кузнецов [Владимирский] (1916, 1932); Г. И. Ануфриев (1922); В. Н. Андреев (1932а, 19326, 1934, 1935, 1935а, 1954а); В. Б. Сочава (1932а, 1933, 1933а, 1934); Ю. Д. Цинзерлинг (1932, 1935); Б. Н. Городков (1933, 1935, 19356, 1943, 1944, 1949); А. А. Дедов (1933); К. Н. Игошина (1934); Ф. В. Самбук (1934); В. Н. Андреев, К. Н. Игошина и А. И. Лесков (1935); П. Д. Варлыгин (1936); Н. В. Пластова (1936); А. И. Лесков (1936, 1940); Л. Н. Тюлина (1936); М. Н. Аврам-чик (1937); В. Д. Александрова (1937); А. Н. Виноградова (1937); В. И. Душечкин (1937); В. Н. Андреев и А. Л. Панфиловский (1938); И. Д. Богдановская-Гиенэф (1938); И. Я. Кац (1939, 1948); Н. Я. Кац и С. В. Кац (1948); Н. И. Пьявченко (1949, 1955); С. Н. Тюремнов (1949).
67. Сфагновые, местами облесенные болота лесной зоны
Сфагновые болотные массивы лесной зоны, как уже указывалось, различаются в зависимости от климатических и местных орографических условий. При обобщении, в соответствии с масштабом «Геоботанической карты», могут быть выделены лишь основные географические типы сфагновых болот, характеризующие обширные пространства крупных природных областей.
В Европейской части СССР характерно развитие по замкнутым котловинам выпуклых моховиков и различных типов болотных массивов подножий склонов (67 е). Большая часть этих болотных массивов уже давно вступила в фазу бедного питания. Их современный растительный покров представлен главным образом растительными группировками олиготрофно-сфагновой группы формаций, а основная часть торфяной залежи сложена растительными остатками этих же группировок. Формы микрорельефа на болотных массивах этой группы, как правило, не связаны с мерзлотными явлениями.
Скопление огромных сфагново-лишайниковых и сфагновых болотных массивов (67 бг) имеет место на севере Западно-Сибирской низменности в подзоне северной тайги. На этих болотных массивах исключительно широким развитием пользуются бугры мерзлотного происхождения и термокарстовые озерки. В средней и южной тайге Западно-Сибирской низменности широко развиты сфагновые и сфагново-гипновые болота (67 зс), для которых формы рельефа мерзлотного происхождения уже не характерны. Им присущи свои закономерности растительного покрова и свой тип сочетаний болотных урочищ. К востоку от Енисея крупные сфагновые болотные массивы встречаются спорадически. Значительные площади они занимают лишь на Камчатке и Сахалине (67 ск), где местами образуют сочетания с гипново-травяными болотами и луговыми и древесными сообществами вдоль рек. Эти болота отражают особенности сурового северотихоокеанского муссонного климата.
В соответствии со сказанным, сфагновые, местами облесенные болота лесной зоны на «Геоботанической карте» разделены на четыре основных географических типа: сфагново-лишайниковые и сфагновые, преимущественно бугристые болота (67 бг), сфагновые восточноевропейские выпуклые болота (67 е), сфагновые и сфагново-гипново-травяные западносибирские
выпуклые болота (67 зс), сфагновые и гипново-травяные сахалинско-камчатские болота (67 ск).
67 бг — сфагново-лишайниковые и сфагновые, преимущественно бугристые болота
Болотные массивы этого типа развиты на севере лесной зоны. Южный предел их в Западной Сибири определяется распространением вечной мерзлоты. На Кольском п-ве и на северо-востоке Русской равнины небольшие массивы бугристых болот распространены в условиях, где мерзлотные явления на торфяниках выражены достаточно резко. Наибольшего развития сфагново-лишайниковые и сфагновые бугристые торфяники достигают в подзоне северотаежных лесов Западно-Сибирской низменности. Здесь, в отличие от Европейской части СССР, где в лесной зоне мерзлые бугристые болота встречаются спорадически, они занимают нередко сплошь все водоразделы. Лишь вдоль западной, северной и восточной окраин низменности площади под бугристыми болотами резко сокращаются. На окраинах низменности бугристые болота встречаются только по замкнутым котловинам и очень напоминают плосковыпуклые моховики, отличаясь от последних лишь наличием мерзлых бугров с присущими им особенностями растительного покрова.
Для бугристых болот, расположенных к северу от плоского водораздела между правыми притоками Оби и реками, текущими в Обскую и Та-зовскую губы, характерно широкое развитие на буграх лишайников и листостебельных мхов (видов Dicranum). Лишайники и листостебельные мхи нередко почти нацело вытесняют прежде господствовавший Sphagnum fuscum. Из высших растений на буграх преобладают угнетенные кустарнички, свойственные для сообществ формации Sphagneta fusci. Древесная растительность отсутствует. Вершина бугров имеет уплощенную форму. Понижения между буграми заняты озерками или покровами сфагновых мхов секции Cuspidata. Здесь можно встретить сообщества типа Sphagneta Lindbergii, Sphagneta Dusenii.
К югу от упомянутого водораздела сфагновые мхи становятся эдифи-каторами как на понижениях между буграми, так и на самих буграх. Последние имеют куполовидную форму, их высота достигает 3—4 м и более. Они располагаются группами, образуя подобие колец вокруг озерков термокарстового происхождения. Пространства между буграми пересечены рядом радиально расходящихся ложбин стока. Поверхность ровных пространств и ложбин стока покрыта сплошным ковром из сфагновых мхов секции Cuspidata. На ровных пространствах господствуют более олиготрофные виды ее, а в ложбинах стока более мезотроф-ные или даже евтрофные. В зависимости от условий и вида эдификатора сфагнового ковра в травяно-кустарничковом ярусе доминируют различные растения {Eriophorum vaginatum, Scheuchzeria palustris, Carex inflata, C. lasiocarpa и др.). На крупных мерзлых буграх наблюдается закономерное расчленение поверхности на понижения и повышения. Повышения покрыты Sphagnum fuscum, а понижения более влаголюбивыми сфагнами (например S. balticum). Над сфагновым ковром возвышаются высшие растения, свойственные сообществам формации Sphagneta fusci и Sphagneta baltici. Большая часть бугров облесена сосной. На протоках между озерками и на дне только что спустившихся озерков развита травянистая растительность — главным образом осочники.
Упомянутый бугристо-озерковый комплекс занимает основную центральную часть болотного массива. Он отделяется от суходолов узкой каймой обычного грядово-мочажинного комплекса.
На повышенных водоразделах Западно-Сибирской низменности (так называемых «материках»), а также на ее западной и восточной окраинах все логовидные понижения и сточные котловины заняты сфагновыми болотами; в их торфяной толще и на поверхности мерзлота летом полностью отсутствует. По закономерностям распределения растительных группировок эти болотные массивы очень сходны с болотными массивами Карело-Финской ССР, Архангельской области и Коми АССР. Окраинные части их обычно заняты сообществами олиготрофно-сфагновой группы формаций (облесенными или безлесными).
Центр лога или сточной котловины, по сравнению с окраинами, более обводнен; он представляет собой топь. Воды, увлажняющие его, проточны, а потому и растительные сообщества, развитые здесь, более евтрофны, нежели сообщества окраин болот. В зависимости от уклонов дна торфяников, топи могут быть гладкими и грядово-мочажинными. В формировании растительного покрова топяных и притопяных местообитаний широкое участие принимают растительные группировки мезотрофно-сфагновой и евтрофно-моховой групп формаций.
В условиях Западно-Сибирской низменности болотные массивы этого типа имеют огромные размеры и местами их скопления попадают в болотный контур 67 бг.
В Средней и Восточной Сибири на пологих склонах, террасах и плато развиты сфагновые лиственничники — мари. Для их поверхности также характерен бугристый рельеф. Часть этих лиственничников, как уже указывалось, должна быть отнесена к сфагновым, облесенным лиственницей болотам, а другая — большая часть, к заболоченным лиственничникам. На «Геоботанической карте» они показаны как сфагновые лиственничные леса — мари (34)
Л итература
В. С. Доктуровский (1911); 0. И. Кузенева (1911); В. Н. Сукачев (1912а); Л. В. Шумилова (1931); Н. Я. Кац (1932, 1948); Н. И. Кузнецов (1932); Г. Н. Эйдельман (1936а); С. Л. Кушев (1939); Н. Я. Кац и С. В. Кац (1948); С. Н. Тюремнов (1949).
67 е — сфагновые восточноевропейские выпуклые болота
Восточноевропейские сфагновые болота в зоне тайги и хвойно-широколиственных лесов, объединенные в эту группу, представлены рядом типов болотных массивов, имеющих большей частью выпуклую или наклонную поверхность. Выпуклые торфяники развиваются в замкнутых котловинах, а торфяники с наклонной поверхностью — в понижениях подножий склонов или на очень пологих склонах. Все они переживают фазу бедного питания (то есть их можно причислить к группе олиготрофных верховых болот).
Современные растительные группировки и торфа различного ботанического состава распределяются по территории болотных массивов выпуклых моховиков концентрическими полосами, а у наклонных — поперечными.
Болотные массивы разных местоположений и с различной формой поверхности могут быть по особенностям растительного покрова объединены в четыре группы: 1) слабо- и сильновыпуклые облесенные моховики и облесенные моховики подножий склонов и пологих склонов; 2) резковыпуклые моховики; 3) пологовыпуклые моховики и моховики подножий склонов и пологих склонов с олиготрофными грядово-мочажинными комплек
сами; 4) плосковыпуклые моховики и моховики подножий склонов с регрессивными комплексами.
Перечисленные группы болотных массивов часто сливаются друг с другом, образуя крупные болотные системы.
1. Слабо-и сильновыпуклые моховики замкнутых котловин, облесенные сосной, а также облесенные моховики подножий склонов покрыты преимущественно сообществами формации Sphagneta magellanici, Sphagneta magellanici-angustifolii и реже Sphagneta fusci. Всегда хорошо развит древесный ярус из сосны (Pinus silvestris f. uliginosa или P. silvestris f. Litwinowii). Деревья распределяются по площади более или менее равномерно, лишь в центральных частях болотных массивов они становятся ниже и реже. По внешнему краю выпуклых моховиков почти всегда развито осоково- или пушицево-сфагновое кольцо из сообществ формации Sphagneta angustifolii или Sphagneta apiculati. На болотах с выпуклой поверхностью растительные группировки нижней части склона переходят от сфагново-моховых к лесным.
Нередко болотные массивы, мало отличающиеся по современному растительному покрову, имеют разное строение торфяной залежи. В замкнутых котловинах нижние горизонты торфяной залежи сложены преимущественно топяными (осоковыми и гипново-осоковыми) торфами, а верхние, составляющие основную часть залежи, — пушицевыми и сфагновыми (с древесными остатками) торфами. Мощность торфяной залежи — 3— 5 м, наибольшая — в центре массива.
Болота у подножий склонов имеют как топяное, так и лесное прошлое. Нижние слои торфяной залежи представлены главным образом древесными торфами. Мощность торфяной залежи меньше, а максимальные глубины ее залегают в верхней части склона болотных массивов под высоким коренным берегом.
Болотные массивы этой группы описаны в литературе под разными названиями: облесенный тип (Цинзерлинг, 1932); олиготрофно-сосновосфагновые, сосново-кустарничково-сфагновые и сосново-пушицево-кустарничково-сфагновые восточноевропейские (Кац, 1937, 1941); южнорусский облесенный тип (Богдановская-Гиенэф, 1949); притеррасные торфяные месторождения вторых надпойменных террас (Тюремнов, 1949; Тюремнов и Виноградова, 1953); болота верхового типа, переживающие стадии медиум, фускум-медиум и фускум (Галкина, 1946, 1953; Денисов, 1951).
Болотные массивы с таким растительным покровом широко распространены по всей Европейской части лесной зоны СССР, но наиболее типичны в подзоне хвойно-широколиственных лесов. Аналоги их встречаются по террасам рек, а также составляют часть водораздельных болотных систем в полосе средне- и южнотаежных лесов (так называемые рямы) в Западной Сибири.
2. Резковыпуклые моховики отличаются довольно крутыми дренированными склонами. В растительном покрове наблюдается резкое расчленение на ряд концентрических полос.
В центральной части массива сочетаются сообщества формаций Sphagneta fusci и Sphagneta baltici; древесный ярус сильно разрежен. По мере развития этой части моховика происходит смена одной экологической формы сосны другой: высокоствольная болотная сосна (f. uliginosa) уступает место более низкорослой сосне Литвинова (f. Litwinowii). Центральная часть массива окружена более или менее широкой полосой олиготрофного грядово-мочажинного комплекса, состоящего из сообществ Sphagneta fusci и Sphagneta Dusenii. Вдоль внешнего края грядово-мочажинной полосы на крутом склоне массива располагается сильно облесен
ное «кольцо» из растительных группировок Sphagneta magellanici an-gustifollii с ярусом из сосны (f. uluginosa). На особо хорошо дренированных склонах эдификаторами являются не сфагновые мхи, а древесные породы. Для мохового покрова таких участков характерно сочетание куртин сфагнов с зелеными лесными мхами. Между краем облесенного кольца и суходолом всегда имеется достаточно выраженная краевая топь (Lagg). Если вдоль нее протекает ручей или речка, то хорошо развиваются древесно-кустарниковые сообщества, в которых видное место занимают черная ольха (Alnus glutinosa) и многочисленные виды ив.
В той части краевых топей, где русло ручейка погребено в торфе или где уничтожена древесная растительность, развиты травяные или гипново-травяные (Phragmites communis, Equisetum heleocharis, Calla palustris, Comarum palustre, Carex inflata, C. vesicaria, Menyanthes trifoliata, Cicuta virosa, Peucedanum palustre, Iris pseudacoius, Naumburgia thyrsiflora, Drepanocladus exannulatus, виды Bryum), либо травяно-сфагновые (Sphagnum amblyphyllum, S. obtusum, S. riparium, S. subsecundum) сообщества.
Мощность торфяной залежи резковыпуклых моховиков достигает 5—7 м. Для нее характерно наличие прослоек сильно разложившихся пушицевых торфов, выходящих на облесенных склонах на дневную поверхность.
Томсон (Thomson, 1924) рассматривал их как «живой» пограничный горизонт, но неправильно объяснял его происхождение для центральных частей массива. В действительности этот горизонт представляет собою продукт жизнедеятельности лесной растительности, ранее покрывавшей весь массив. Развитие лесных или сильно облесенных ценозов на выпуклых моховиках средних и крупных размеров совпадало и стимулировалось климатическими и гидрологическими особенностями ксеротермического периода. Залежь на массивах этого типа стоит на грани перехода от медиум-типа к комплексному верховому.
Резковыпуклые торфяники имеют малые или средние размеры. Наиболее широко распространены они в южной тайге и в северо-западной части подзоны хвойно-широколиственных лесов. Зрелые стадии развития резковыпуклых моховиков описаны под названием болотных массивов западнорусского типа (Богдановская-Гиенэф, 1928, 1949), русского типа (Цинзерлинг, 1932), резковыпуклого типа (Галкина, 1946).
3. Пологовыпуклые моховики замкнутых котловин и моховики подножий склонов с олиготрофными грядово-мочажинными комплексами. Их центральные части в основном заняты олиготрофными грядово-мочажинными комплексами из сообществ формаций Sphagneta iusci, Sphagneta Dusenii или Sphagneta cuspidati. Группировки Sphagneta fusci занимают повышенные элементы микрорельефа — гряды, им присущ разреженный ярус из низкорослой сосны (в замкнутых котловинах — Pinus silvestris f. Willkommii, редко f. pumila; на болотах подножий склонов растут Р. silvestris f. Litwinowii, реже f. Willkommii).
Олиготрофный грядово-мочажинный центр моховиков замкнутых котловин отделяется от суходолов более или менее широкой облесенной или безлесной каймой. Сообщества, покрывающие такую кайму, относятся к формациям Sphagneta magellanici-baltici или к Sphagneta amblyphyl-lii, или Sphagneta apiculati.
Краевая топь на болотах этой группы большей частью представлена травяно-сфагновыми сообществами.
Процессы деградации сфагнового покрова на болотах с грядово-мочажинным комплексом не характерны и наблюдаются только в местах выхода
на дневную поверхность внутризалежных вод, вызывающих отмирание сфагнов.
Мощность торфяной залежи у пологовыпуклых моховиков — 7—8 м, а моховиков подножий склонов — 3—4 м. Строение залежи в замкнутых котловинах комплексно-верховое; в придонных, наиболее глубоких слоях часто залегают сапропелиты и травяно-топяные торфа. В придонных слоях торфяной залежи болотных массивов подножий склонов также обычно лежат топяные торфа.
В литературе болотные массивы этой группы именовались болотами русского типа (Цинзерлинг, 1932); болотами ильменско-западнодвинского типа (Кац, 1948); болотами среднерусского типа (Богдановская-Гиенэф, 1949); болотными массивами верхового типа олиготрофной грядово-мочажинной стадии (Денисов, 1951).
С. Н. Тюремнов и Е. А. Виноградова (1953) имеют в виду болота этой группы при описании симметричных торфяных месторождений вторых надпойменных террас размыва, торфяных месторождений зандровых равнин, месторождений склонов надпойменных террас и повышенных моренных равнин.
4. Плосковыпуклые моховики замкнутых котловин и моховики подножий склонов с регрессивными комплексами развиты главным образом вдоль побережья Белого моря и в нижнем течении крупных северных рек. В значительно меныпем количестве встречаются они в Прибалтике.
На уплощенной центральной части плосковыпуклых моховиков и на очень пологих частях поверхности болотных массивов подножий склонов широко развиты озерково-мочажинные регрессивные комплексы. В этих комплексах, кроме в различной степени гидрофитных сфагновых сообществ с господством разных видов сфагнов (Sphagnum fuscum, S. ru-bellum, S. balticum, S. cuspidatum, S. Dusenii, S. Lindbergii), развиты ассоциации с преобладанием лишайников и печеночных мхов.
На повышенных частях микрорельефа (грядах, крупных подушкообразных кочках) растут Sphagnum fuscum, кустистые и накипные лишайники. Неглубокие и обводненные понижения заняты наиболее гидрофильными сфагнами (из секции Cuspidata) и печеночниками; более глубокие мочажины залиты водой и представляют собой вторичные озерки. Развитие регрессивных комплексов на плосковыпуклых моховиках объясняется прежде всего ухудшением стока с уплощенной части (появление обильных вторичных ручьев характерно для массивов с весьма зрелыми регрессивными озерково-мочажинпыми комплексами). На болотных массивах подножий склонов развитие регрессивного комплекса обусловлено обилием вод подповерхностного стока и грунтовых, просачивающихся со склона.
Регрессивный озерково-мочажинный комплекс плосковыпуклых моховиков, находящийся в центре, окружен полосой обычного олиготрофного грядово-мочажинного комплекса. Последний занимает склоны болотного массива. На болотах подножий склонов места, занятые регрессивным комплексом, также отделяются друг от друга грядово-мочажинными участками. Однако происхождение регрессивных озерково-моча-жинных комплексов на болотах плато и подножий склонов различно. На плосковыпуклых моховиках появление в центре регрессивного комплекса всегда связано с ростом и развитием моховика и с накоплением в нем внутризалежных вод. Момент достижения этой стадии может быть ускорен или замедлен как климатическими особенностями территории, так и размерами самого массива.
Внешний край олиготрофного грядово-мочажинного комплекса на болотных массивах этого типа, как правило, окружен широкой каймой ра
стительных группировок лесных болот и заболоченных лесов. Это особенно хорошо выражено на плосковыпуклых моховиках и связано с сильным вторичным обводнением центральной части болотного массива. Сброс воды с центра торфяника осуществляется не только путем фильтрации, но и посредством весьма обильных вторичных ручьев и топей.
Эта группа болот в литературе описывалась под названиями: русско-прибалтийских и поморских болот (Цинзерлинг, 1932); советскопри-балтийских и южноприбеломорских (Кац, 1941); плосковыпуклых моховиков и моховиков подножий склонов с регрессивными комплексами (Галкина, 1946, 1955); восточноприбалтийских и южноприбалтийских болот (Богдановская-Гиенэф, 1949).
В Европейской части лесной зоны СССР, кроме названных выше групп болотных массивов, весьма широко распространены и болота, развитые в других условиях рельефа: логах, сточных котловинах, речных плесах .и пр. (Галкина, 1953). Участки таких болот, словно кружевным узором, прорезают лесные массивы главным образом северотаежной полосы. Они так же, как и мелкие выпуклые моховички подножий склонов, отмечены на «Геоботанической карте» знаком заболоченности (штриховкой) по лесным (сосняки, реже ельники) контурам.
Почти на всех болотных массивах, развитых в логах, сточных котловинах и речных плесах, сфагновые мхи лишь недавно заняли господствующее положение. Об этом свидетельствует строение торфяных залежей. Под растительными группировками евтрофно- и медотрофносфагновой групп формаций всегда залегают топяно-травяные, гипновые или реже древесные торфа. Даже под растительными группировками олиготрофносфагновой группы формаций сфагновые торфа нередко образуют лишь самые поверхностные слои залежи.
Литература
Г. И. Танфильев (1895); В. С. Доктуровский и Н. Н. Жуков (1916); Н. Я. Кац (1922, 1928, 1930, 1932а, 1937, 1941, 1948); В. В. Кудряшев (1922, 1922а); В. С. Доктуровский (1925, 1935); И. Д. Богдановская-Гиенэф (1926, 1928, 1949); Д. А. Герасимов (1926, 1929); В. В. Адамов и И. К. Ярошевич (1927); Т. А. Работнов (1929, 1929а, 1930); Н. М. Савич (1929); Ю. Д. Цинзерлинг (1929, 1929а, 1932); С. Н. Тюремнов (1931, 1949); А. А. Генкель и Е. Н. Осташева (1933); А. С. Салазкин (1934, 1936); К. И. Солоневич (1934); Н. Я. Кац, Л. В. Кудряшев, В. М. Эпштейн (1935); Г. А. Благовещенский (1936);. Е. А. Галкина (1936, 1946, 1953, 1955); Н. Я. Кац и Ц. И. Минкина (1936); П. Н. Никольский и И. И. Изотов (1936); С. П. Смелов (1936); К. И. Солоневич и Н. Г. Солоневич (1936); К. Брундза (1937); А. А. Любимова (1937); Д. К. Зеров (1938); 3. Н. Денисов (1951); Н. И. Пьявченко (1951, 1953); Н. В. Лебедева (1952); Р. П. Тихонова (1952); Л. Н. Никонов (1953); С. Н. Тюремнов и Е. А. Виноградова (1953); Г. Ф. Бачурина и Е. М. Брадис (1954); V. Auer (1922); Р. Thomson (1924, 1924а, 1929); Н Gams und Z. Ruoff (1929); I. Paasio (1939).
67 зс — сфагновые и сфагново-гипново-травяные западносибирские выпуклые болота
Сфагновые и сфагново-гипново-травяные болота развиты в южной части Западно-Сибирской низменности. Северный предел их распространения совпадает с северной границей среднетаежных лесов, а южный — с южной границей южнотаежных лесов.
В средней и южной тайге Западной Сибири болотные массивы занимают обширные водораздельные пространства. В отличие от болот северной тайги (67 бг), их края не подходят вплотную к руслам рек, а между ними и речными долинами остаются еще довольно обширные незаболоченные или слабозаболоченные лесные пространства.
Западносибирские сфагновые и сфагново-гипново-травяные болота представляют собою своеобразные болотные системы. Здесь выпуклые облесенные моховички (по местному наименованию — рямы) разбросаны среди огромных сфагновых или гипновых топяных пространств. Обширные топяные участки, характерные для этих болотных систем, пересечены рядом ложбин стока. Одни из этих ложбин непосредственно связаны с внешними водоприемниками, по другим сток вод осуществляется лишь из одной части системы в другую.
В настоящее время на поверхности западносибирских болотных систем сохранилось небольшое количество первичных озер; бблыпая часть их после понижения базиса эрозии заторфовалась и представлена сейчас рямами.
Межрямовые топяные пространства имеют местное название «галья». Современный растительный покров рямов представлен сообществами формаций Sphagneta fusci, а иногда и Sphagneta magellanici-angustifolii.
Sphagnum fuscum являлся эдификатором в болотных ценозах рямов почти с начала их возникновения. Торфяная залежь сложена слабо разложившимся фускум-торфом. Поверхность рямов сравнительно однородна (на небольших рямах) или изрезана радиально расходящимися полосами (на крупных рямах). Древостой на этих полосах очень сильно изрежен, либо отсутствует, a Sphagnum fuscum замещается более гидрофильными видами сфагновых мхов. На некоторых массивах в узлах расхождения радиальных полос имеются скопления мочажин-озерков, вероятно вторичного происхождения.
Пространства между рямами (гальи) имеют или слегка волнистую, или низкогрядовую поверхность. Повышения микрорельефа представлены сообществами формаций Sphagneta magellanici, Sphagneta papillosi или Sphagneta Warnstorfii. Состав травяно-кустарничкового яруса этих формаций различен, но весьма сходен с травяно-кустарничковой растительностью сообществ тех же формаций на болотах Русской равнины.
Понижения на гальях, в зависимости от степени проточности, заняты или сфагнами с сопутствующими им высшими растениями, или гипновыми мхами. Понижения с гипновыми мхами чаще всего примыкают к повышениям, покрытым Sphagnum Warnstorfii. Такие комплексы занимают хорошо проточные ложбины стока, сбрасывающие свои воды во внешние водоприемники.
Торфяная залежь межрямовых пространств не достигает большой мощности. Она сложена в основном не сфагновыми, а травяными торфами. Пласты травяно-топяных торфов нередко залегают на торфах древесного происхождения — ольховых или древесно-осоковых.
К группе сфагновых и сфагново-гипново-травяных западносибирских выпуклых болот (67 зс) на «Геоботанической карте» отнесены и травяно-гипновые болотные массивы подножий склонов. Их поверхность чаще всего покрыта гипновыми мхами (Drepanocladus vernicosus)', из высших растений там господствуют осоки (Carex diandra, С. limosa, С. chordorrhiza) и ряд других, преимущественно травянистых растений. Торфяная залежь сложена осоковыми и гипново-осоковыми торфами.
Литература
Н. И. Кузнецов [Владимирский] (1915а); В.И. Баранов и В.А.Шелудякова (1928); Н. В. Шипчинский (1928); М. К. Барышников (1929); А. Я. Бронзов (1930, 1936); Г. Я. Бронзова (1932); С. В. Лобачев (1934); М. И. Неиштадт(1936); Н. Я. Кац (1948); М. С. Кузьмина (1949); С. Н. Тюремнов (1949).
67 с к — сфагновые и гипново-травяные сахалинско-камчатские болота с луговыми и древесными сообществами вдоль рек
Эти болота сосредоточены главным образом по западному побережью Камчатки, встречаются они и на Сахалине. Ббльшая часть их представляет собою своеобразные болота подножий склонов, пересеченные реками, вдоль которых развиты лесные, кустарниковые (береза, ольха), а также луговые сообщества.
Поверхность болотных массивов Камчатки представлена растительными группировками травяного, евтрофномохового и мезотрофпосфаг-нового типа. Задержка многих болотных массивов в фазе богатого питания отчасти объясняется тем, что их поверхность периодически удобряется вулканическим пеплом. Для болот Камчатки характерно отсутствие древесной растительности; нет ее остатков и в торфе. Торфяные залежи сложены осоковыми, гипново-осоковыми, а иногда и сфагново-осоковыми торфами, мощность их 1.5—3.0 м, в редких случаях — 8—10 м. На ряде торфяников Камчатки наблюдаются бугры мерзлотного происхождения.
Для болотных массивов в фазе богатого питания характерны покровы гипновых мхов и хорошо развитый кустарничково-травяной ярус; в нем встречаются пушистая восковица (Myrica tomentosa), осоки (Carex ve-sicala, C. rhynchophysa), вахта (Menyanthes trifoliata) и др.
Для болотных массивов в фазе бедного питания типичны сообщества Sphagneta fusci. В травяно-кустарничковом ярусе развиты кустарнички, осока Миддендорфа (Carex Middendorffii.) и другие травы. Болес увлажненные участки сфагновых болот покрыты Sphagnum papillosum или S. Lindbergii] из трав там чаще других встречаются осоки (Carex chordorrhiza, С. li-rnosa, С. irrigua, С. pauciflora, С. rotundata.). На хорошо дренируемых участках среди покрова из Carex irrigua поселяются кустистые лишайники.
Крупный болотный массив показан на «Геоботанической карте» в По-ронайской низменности (Сахалин). Он отделяется от вод залива Терпения низким береговым уступом. Мощность торфа достигает 6 м. Поверхность большей части массива почти безлесна; на ней можно выделить три отличающиеся по растительному покрову полосы: 1) бугристо-грядовую, 2) сфагновую с угнетенной лиственницей и 3) сфагново-лиственничную.
Перечисленные участки сменяют друг друга по мере приближения к нижнему краю массива.
Бугристо-грядовый участок имеет неровную поверхность; бугры округлой или овальной формы, высотой 0.5—1.5 м, диаметром в 10—15 м. На глубине 45—65 см обнаружена мерзлота. Бугры, также как и гряды, сложены слаборазложившимся сфагновым торфом. На них растет редкая угнетенная лиственница (1—1.5 м высоты), кедровый стланик, брусника, болотный рододендрон (Rhododendron parvifolium), вороника, багульник, голубица. Сфагновый покров состоит из Sphagnum lenense, S. fuscum] на буграх сфагны местами отмирают и заменяются лишайниками. Бугры часто бывают окружены зарослями осоки Миддендорфа со сплошным сфагновым покровом (Sphagnum lenense, реже S. fuscum). Между буграми и грядами развиты понижения, покрытые S. balticum и S. Lindbergii с травяным покровом из топяной осоки. Кроме мочажин со сфагновым покровом, встречаются сильно увлажненные мочажины, заполненные вязким, почти не задернованным торфом (так называемые «римпи»). По краям и внутри таких мочажин растут отдельные экземпляры вла
галищной пущицы и некоторых других растений (llhynchospora alba, Tri-chophorum caespilosum, Carex limosa). Здесь же на небольших кочках можно встретить осоку Мнддепдорфа.
На пологих склонах болотного массива Поронайской низменности располагаются участки сфагновых сообществ с угнетенной лиственницей (высота 1 —1.5 м). Микрорельеф слегка волнистый; моховой покров сплошной (нз Sphagnum fuscum —на повышениях, и S. lenense, S. balticurn — на понижениях). В травяно-кустарничковом ярусе преобладает осока Мнддепдорфа (Carex Middendorffii), болотный мирт (Chamaedaphne caly-culata), брусника, мелкоплодная клюква и иногда багульник.
Участки сфагновых лиственничников образуют узкие полосы по краю болотного массива. Лиственница здесь достигает высоты 4—6 м. В травяно-кустарничковом ярусе господствуют кустарнички (Ledum palustre, Vactinium uliginosum, Chamaedaphne calyculata)\ среди них встречаются травы (Carex Middendorffii, liubus chamaemorus и др.). Моховой покров состоит из Sphagnum Girgensohnii и S. angustifofolium\ на кочках растут 6’. fuscum, S. magellanicum и виды Dicranum.
11о берегам многочисленных озер и вытекающих из них ручейков лиственницы достигают 15 м высоты. У края озер и вдоль протоков развита травяная растительность.
На основной части массивов верхние слои торфяной залежи до глубины 3.5—4 м сложены сфагновыми верховыми, слаборазложившимися торфами; глубже залегают низинные древесные или осоковые, довольно сильно разложившиеся торфа.
Литература
Л. И. Тихонович и II. И. Полевой (1915); Е. И. Елисеев (1932); М. И. Нойштадт (1936а); М. И. Нойштадт и М. Л. Короткииа (1936); Г. Н. Эпдольмаи (1936); Е. Л. Любимова (1940); Н. Я. Кац (1948); С. Н. Тюремнов (1949); Н. В. Пластова (1950).
Торфяная залежь сфагновых болот и их естественная растительность (в меныпей мере) используются для различных нужд народного хозяйства. Запасы торфа в СССР достигают 158 млрд, тонн, что составляет около 60% его мирового запаса. Наша торфяная промышленность располагает большим запасом торфяного сырья различного качества. Существенное значение имеют вопросы организации торфоразработок, тесно связанные с производными особенностями болотных массивов. От качества и средней мощности торфяной залежи, а также размеров болотных массивов и их конфигурации зависит возможность организации торфодобывающих предприятий того или иного назначения.
Торфяная залежь сфагновых восточноевропейских выпуклых болот (67 о) представляет собой ценный топливный фонд. Она отличается малой зольностью и пригодна для добычи топлива и получения газа. Верхние, слабо разложившиеся пласты торфа, следует использовать для изготовления изоляционных плит, заготовки стойловой подстилки, торфяной крошки для упаковки фруктов и пр.
Болотные массивы логов, сточных котловин и, отчасти, подножий склонов с низинной залежью торфа являются сырьевой базой для производства торфяных брикетов.
Сфагновые болота, переживающие евтрофную и евтрофпо-мезотрофную фазы развития, местами, где недостает сенокосных угодий, используются для сенокошения, несмотря на низкие кормовые качества присущих им трав. После мелиорации они могут быть превращены в культурные луга
или другие сельскохозяйственные угодья. Использованию болот для нужд сельского хозяйства в последнее время уделяется очень много внимания, и они в общей сложности представляют очень значительный потенциальный земельный фонд для дальнейшего развития сельского хозяйства.
Природные особенности сфагновых болотных массивов, показателем которых является их растительный покров, должны учитываться также при строительстве дорог (железных, шоссейных, проселочных и пр.), водохранилищ, плотин и прокладки газопроводов. Поверхность и недра болотных массивов всех типов и размеров в той или иной степени оказывают пагубное влияние как на ход строительства сооружений, так и на их дальнейшую эксплуатацию. Мероприятия по устранению этого влияния должны разрабатываться с учетом типа болотного массива, развития на массиве элементов водопроводящей сети и источников ее водного питания.
Важно учитывать природные особенности болотных массивов при строительстве зимних дорог, и других дорог, насыпь которых опирается на торф; На территории СССР встречаются такие пространства, на которых дорожная трасса длиною даже до десяти километров обязательно пересекает несколько болотных массивов.
При строительстве водохранилищ, как правило, затапливаются пространства, на которых сосредоточены большие скопления болотных массивов. В зависимости от строения торфяной залежи последних, через некоторое время после затопления, на поверхность водохранилищ всплывают огромные торфяные острова. Они мешают судоходству и наносят вред шлюзовому хозяйству. Чем кислее торфяные воды, тем быстрее они действуют разрушающим образом на бетонные сооружения плотин и шлюзов, а также на изолирующие -покрышки газопроводных труб. Все сказанное подтверждает необходимость комплексного исследования природы сфагновых болотных массивов и их растительного покрова, который нередко является индикатором свойств торфяной залежи и природных особенностей болота в целом. Важное значение имеет изучение закономерностей географического распространения болот и их районирование, на основе которого разрабатываются мероприятия по освоению огромной площади болот для нужд промышленности и под сельскохозяйственные угодья.
Рис. 54. Часть системы плоско-выпуклых моховиков. Аэрофотоснимок.
• <-
Рис. 55. Арчевое редколесье с травяным покровом из типчака {k\siu< <1 >иьа(а) с примесью ковылей. Темные пятна арчи (Juniperus semiglobosa nJ. turkestanica). Восточный склон ущелья по северному склону Туркестанского хребта, к югу от Шахрпстана (Таджикская ССР)- (Фот. Ю. Е. Лавренко). *
XI. КСЕРОФИТНЫЕ РЕДКОЛЕСЬЯ, НАГОРНЫЕ КСЕРОФИТЫ И СУБТРОПИЧЕСКИЕ СТЕПИ1
Н. И. РУБЦОВ
Общий обзор
Ксерофитные редколесья из древовидных можжевельников (арчи) и фисташки, нагорные ксерофиты, состоящие из ксерофильных кустарничков и полукустарничков, а также субтропические степи, слагаемые большею частью многолетними эфемероидными или близкими к последним по ритму развития травами, весьма характерны для сухих горных стран Передней и Средней Азии. В пределах СССР эти типы растительного покрова имеют широкое распространение в южных горах — в Южном и отчасти Восточном Закавказье, а также в горах Средней Азии — Копет-Даге, Памиро-Алае, Западном и отчасти Центральном Тянь-Шане. Отдельные их фрагменты проникают в более северные горные системы.
Указанные типы растительного покрова приурочены к низкогорьям, а также к средним поясам гор, поднимаясь местами до высокогорий. Связаны они с почвами сероземного типа, а выше по рельефу с коричневыми почвами; нагорные ксерофиты нередко произрастают на слабо развитых каменистых и щебнистых субстратах.
Ксерофитные редколесья, нагорные ксерофиты, колючетравники, субтропические степи — типы растительного покрова с очень различной структурой. В ксерофитных редколесьях, помимо редкостойного древесного яруса, иногда выражен кустарниковый ярус; нижний ярус в одних случаях слагается травянистыми растениями степного или эфемероид-ного типа, в других случаях полукустарничками или кустарничками. В составе нагорных ксерофитов чаще всего господствуют колючие ксеро-фильные кустарнички с большей или меньшей примесью полукустарничков и трав. Колючетравники состоят из ксерофильных колючих трав (виды Cousinia), обычно длительно вегетирующих. Субтропические степи образованы также травами эфемероидного или гемиэфемероидного типа, то есть с более или менее укороченным сроком вегетации в течение теплого сезона.
Общим для всех этих типов растительного покрова является парагене-тическое развитие, то есть сосуществование в пространстве бок-о-бок, что способствует взаимному проникновению видов из одних упомянутых выше типов растительного покрова в другие и явлениям сукцессий между этими типами растительности.
Нагорные ксерофиты, представленные группировками ксерофильных колючих кустарничков (трагакантники) или полукустарничков (тимьян-
1 Весь раздел, кроме общего обзора, написан Н.И. Рубцовым. Общий обзор составлен Е. М. Лавренко.
- нпки или томилляры), очень характерны для скелетных гор со слабо развитым почвенным покровом. По мере накопления мелкозема и вообще формирования почвы, трагакантники и томилляры сменяются травянистыми сообществами — степного или эфемероидного типа: 1) в средних и отчасти верхних поясах гор — дерновиннозлаковыми степями, преимущественно типчаково-ковыльными, сначала с большей или меньшей примесью колючих кустарничков (трагакантовые степи) и полукустарничков (тимьянниковые, а в некоторых случаях пустынные степи); 2) в нижних поясах гор — субтропическими степями.
Колючетравники, возможно, являются «вторичным» типом, возникающим в результате усиленного выпаса.
Нижние ярусы в редколесьях, как указано выше, могут быть весьма разнообразны; они состоят из кустарников, колючих кустарничков, степных дерновинных злаков (типчаки, ковыли) или эфемероидных и ге-миэфемероидных трав (Роа bulbosa, Carex pachystylis, Agropyrum tricho-phorum, виды Ferula, Inula grandis и пр.). Несмотря на в общем широкое распространение ксерофитных редколесий (арчевников, фисташников и др.) в горах Закавказья и Средней Азии, площади, занятые ими в настоящее время относительно невелики и постепенно сокращаются, почему нельзя говорить о сплошных поясах ксерофитных редколесий.
В то же время имеются основания утверждать о гораздо большем распространении в горах Средней Азии и Закавказья более или менее ксерофитных редколесий — как фисташковых, так и арчевых. В связи с ограниченным распространением настоящих лесов в этих южных горах, фисташка и арча, образующие редколесья, уже издавна истреблялись для различных целей (как топливо, строительный материал, для различных поделок и пр.). В горах Средней Азии редколесья из фисташки и арчи часто располагаются по отношению к предгорным оазисам гораздо ближе, чем участки настоящих лесов, что не могло не способствовать истреблению этих древесных пород в течение многотысячелетней культурной истории Средней Азии.
С. Н. Кудряшев (1941) высказал мысль о том, что в доагрикультурный период в низкогорном Гузаре (один из южных районов Узбекской ССР) большая часть территории, занятой в настоящее время осоково-мятликовой (Carex pachystylis, Роа bulbosa) растительностью, раньше была покрыта фисташковыми редколесьями или, как он пишет, «фисташковой саванной». Выше пояса гипотетических фисташковых редколесий в Гузаре и в настоящее время располагается пояс арчевых редколесий, где арча разбросана на фоне «крупнотравных эфемероидных андузовых лугов» (ассоциации Inula grandis).
Можно высказать весьма вероятное предположение о том, что в южной части Средней Азии (на север до Западного Тянь-Шаня) в низкогорьях на месте современных совершенно «бездревесных» осоково-мятликовых сообществ раньше существовали фисташковые редколесья с покровом из тех же видов (Роа bulbosa, Carex pachystylis), с большей или меньшей примесью полукустарничковых полыней. В современном поясе субтропических крупнотравных степей (полусаванн), занимающих высокие предгорья и частично среднегорья, располагались, вероятно, кроме фисташковых и миндальниковых (из Amygdalus bucharica), главным образом арчевые, а местами боярковые (из Crataegus pontica) редколесья с травяным покровом типа крупнотравных субтропических степей. В верхней части среднегорного пояса были широко распространены арчевые редколесья с травяным покровом степного типа (типчак и пр.) или с ярусом кустарников (типа шибляка). Редкостойность этих древесных пород обус
ловливалась, как и в настоящее время, недостатком влаги, что препят-ствует формированию сомкнутых насаждений. По северным склонам в полосе арчевых редколесий располагались сплошные заросли кустарников, арчевые, яблоневые и ореховые (Juglans regia) леса, которые в прошлом были гораздо более распространены в южной части Средней Азии, чем в настоящее время.
Литература
Е. П. Коровин (1934); А. Л. Тахтаджян (1937, 1941); С. С. Станков (1939); С. Н. Кудряшев (1941); А. А. Гроссгейм (1948); П. Н. Овчинников (1955).
68. Можжевеловые редколесья в сочетании с нагорными ксерофитами и степями
Можжевеловые редколесья (арчевники), слагаемые древовидными представителями можжевельника (род Juniperus), весьма обычны в предгорьях и горах Центрального и Западного Тянь-Шаня, Памиро-Алая и Копет-Дага. Значительное распространение они имеют в горах Кавказа, в частности в Южном Закавказье. Отдельные небольшие «рощи» арчев-ников (не показанные на карте) имеются также в горном Крыму.
По своей структуре — это настоящие редколесья, то есть редкостойные светлые насаждения паркового типа, в которых сомкнутость древесного полога обычно не превышает 0.5—0.6. Нередко она снижается до 0.1—0.2. Лишь иногда, на очень небольших площадях, среди редколесий встречаются участки настоящих густых арчевых лесов с сомкнутостью полога 0.8—0.9 и даже 1.0. Такие сохранившиеся участки густых арчевников попадаются, например, в наименее доступных частях гор Узбекистана и Киргизии. В большинстве же случаев настоящие арчевые леса уже полностью уничтожены вырубкой и последующим выпасом скота. Необходимо отметить, что и арчевые редколесья были ранее распространены гораздо шире, чем это наблюдается теперь.
68 к — кавказские
Кавказские можжевеловые (арчевые) редколесья мелкими участками представлены в Южном Закавказье (на карте не показаны). Более значительные площади арчевых редколесий (отмеченные на карте) имеются в районе Новороссийска и по среднему течению Куры.
В Закавказье арчевники слагаются из нескольких видов арчи, преимущественно Juniperus polycarpos и J. foetidissima, реже J. oxycedrus и J. excelsa.
В Южном Закавказье, также как и в Средней Азии, широко развиты арчевые редколесья в сочетании с формациями нагорных ксерофитов. Такие сочетания встречаются обычно на каменистых горных склонах, в пределах от 1000 до 2400 м над у. м. Среди нагорноксерофитных вариантов арчевников можно особо выделить трагакантовые арчевники, в нижнем ярусе которых преобладают трагакантовые (колючие) подушковидные кустарничковые и полукустарничковые астрагалы (Astragalus aureus, A. microcephalus и др.) и эспарцет (Qnobrychis cornuta). Кроме того, встречаются участки арчевых редколесий с обилием «томиллярных» элементов, т. е. различных пахучих полукустарничковых и травянистых губоцветных (например Thymus Kotschyanus, Stachys Schtschegleewii, видов Nepeta, Ziziphora, Silene).
В более высоких поясах гор, по склонам, имеющим большое количество мелкозема, на вполне сложившихся коричневых почвах, формируются степные арчевники с преобладанием степных элементов в нижнем травяном ярусе (Festuca sulcata, Stipa capillata, Koeleria gracilis).
В Южном Закавказье отмечаются также кустарниковые (шибляковые) арчевники, которые, однако, более характерны для Восточного Закавказья. В их сложении участвуют такие ксерофильные и мезоксерофиль-ные листопадные кустарники, как миндаль (Amygdalus Fenzliana, A. zan-gezura), крушина (Rhamnus Pallasii), вишня (Cerasus araxina), эфедра (Ephedra procera) и другие представители шибляка.
Арчевники предгорий Восточного Закавказья приурочены к крутым куэстообразным склонам, сложенным песчаниками, глинами и конгломератами (Сахокия, 1950у. В отличие от южнозакавказских арчевников, основу древесного яруса здесь обычно составляет острочешуйчатая арча (Juniperus foetidissima), а многоплодная арча (Juniperus polycarpos) участвует большей частью только в виде примеси. Постоянным компонентом арчевых редколесий является также кевовая фисташка (Pistacia mutica).
В районе Восточного Закавказья выделяются следующие основные типы арчевников.
1) Кустарниковые арчевники, по крутым каменистым склонам гор. Под очень светлым пологом арчи преобладают различные кустарники — держи-дерево (Paliurus spina-Christi), жимолость (Lo/iicera iberica), кизильник (Cotoneaster racemiflora), таволга (Spiraea hypericifolia), жасмин (Jas-minium fruticans), шиповники (виды Rosa).
2) Степные арчевники развиваются на более сформированных суглинистых почвах пологих склонов предгорий. Они характеризуются преобладанием в нижнем ярусе арчевников степных травянистых элементов— ковылей (Stipa Lessingiana, S. pulcherrima, 8. capillatd), типчака (Festuca sulcata), бородача (Andropogon ischaemum).
В арчевниках района Новороссийска так же, как и в арчевниках Крыма, главная роль в сложении древесного полога принадлежит уже не многоплодной арче (Juniperus polycarpos), а другим видам (Juniperus excelsa и J. foetidissima). Однако структурно они сходны с вышеуказанными арчевыми редколесьями Восточного и Южного Закавказья. Здесь можно наметить те же основные типы этих редколесий, а именно: 1) кустарниковые (шибляковые), 2) фригановые (с преобладанием в нижнем ярусе жестких, нередко колючих, подушковидных кустарничков) и 3) степные арчевники.
Литература
А. А. Гроссгейм (1948); А. В. Иванова (1949); М. Ф. Сахокиа (1950).
68 са — среднеазиатские
Среднеазиатские можжевеловые (арчевые) редколесья приурочены преимущественно к средним поясам гор (от 1200—1400 до 2000 м над у. м. и выше). Так же, как и кавказские арчевники, они встречаются преимущественно по каменистым склонам, в сочетании с нагорными ксерофитами и шибляком. По северным, более пологим склонам, с более развитыми почвами, формируются степные и даже луговые типы арчевых редколесий.
В сложении среднеазиатских арчевников участвуют иные виды арчи, чем на Кавказе (Juniperus turkestanica, J. semiglobosa, J. seravschanica.
к которым в Копет-Даге прибавляется J. turcomanica). Вообще из всех видов арчи, обитающих в Сродней Азии, имеется только один вид, свойственный также Кавказу, несмотря на многие весьма общие черты в экологической обстановке их развития.
Арчевые редколесья (реже леса) в Средней Азии приурочены преимущественно к горным системам Алая и Копет-Дага. В меньшей степени они свойственны Тянь-Шаню, главным образом Западному, где встречаются хотя и часто, но небольшими участками, которые на карте отмечены только соответствующим внемасштабным знаком. В Северном Тянь-Шане древовидные высокоствольные арчевники совсем отсутствуют, если не считать весьма небольших арчевых насаждений, встречающихся лишь в западной оконечности Киргизского хребта. Северному Тянь-Шаню свойственны только высокогорные (субальпийские) арчевые подушковидные стланики (из Juniperus sibirica и J. turkestanica). Что касается Центрального Тянь-Шаня, то и здесь высокоствольные арчевые насаждения немногочисленны и встречаются лишь в виде мелких массивов и вкраплений среди других типов растительности (нагорных ксерофитов и степей).
Среди среднеазиатских арчевых редколесий можно различить три основных структурно-экологических типа, аналогичных вышеописанным кавказским: 1) степной, 2) нагорноксерофитный и 3) луговой.
Степные арчевники (рис. 55) формируются на более или менее пологих, мягких склонах, иногда северных экспозиций. В их травяном ярусе преобладают степные злаки, преимущественно типчак (Festuca sulcata), а вместе с ним тонконог (Koeleria gracilis), степная тимофеевка (Phleum phleoides), степной мятлик (Роа relaxa) и ксероморфное разнотравье (Hypericum scabrum, Verbascum songoricum и т. п.).‘Этот вариант степного арчевника встречается главным образом в более высоких поясах гор, близ границы субальпийской растительности (1800—2000 м над у. м. и выше).
Встречаются также арчевники в сочетании с субтропическими степями из волосоносного пырея (Agropyrum trichophorum) и зонтичного — югана (Prangos pabularia). Пырейные и югановые арчевники развиваются реже типчаковых редколесий.
Нагорноксерофитные (фригановые) арчевники являются почти самыми распространенными в Средней Азии. Они широко распространены в Тянь-Шане, в системе Алая, особенно в Копет-Даге, по каменистым склонам, преимущественно южных экспозиций. Это наиболее редкостойный тип насаждений (сомкнутость крон не выше 0.2—0.3). В нижнем ярусе господствуют кустарничковые и полукустарничковые подушковидные ксерофиты (фригановая синузия). Так, для скалисто-щебнистых склонов Копет-Дага очень характерны арчевники с преобладанием жестких «подушек»/ (Acanthophyllum, Acantholimon, Onobrychis cornata, Gypsophila aretioides, Stachys turcomanica, некоторые трагакантовые астрагалы — Astragalus densissimus, A. piletocladus) (рис. 56).
Луговые арчевники распространены в гораздо меньшей степени, чем предыдущие. Они формируются в условиях повышенного увлажнения — в днищах ущелий, близ родников, у подножий северных склонов и т. п. Именно в этих условиях чаще всего встречаются настоящие густые арчевые леса. В травяном ярусе луговых арчевых редколесий довольно обычны вейник (Calamagrostis epigeios), мятлик (Роа pratensis), костер (Bromus inermis), пырей (Agropyrum repens).
Таким образом, экологический диапазон арчи в Средней Азии чрезвычайно велик — от очень сухих горноксерофитных местообитаний до мезофитных лугов; причем подобная экологическая пластичность на
блюдается нередко у одного и того же вида (например, Juniperus semigio-bosa, J. turkestanica и J. seravschanica).
Следует иметь в виду, что в контурах можжевеловых редколесий, выделенных на карте, помимо участков с преобладанием нагорных ксерофитов и степей, имеются также и участки с господством некоторых лиственных пород (например в Средней Азии Acer Semenovii, A. turcomanicum, на Кавказе Quercus pubescens), выделение которых на карте данного масштаба оказалось невозможным.
Арчевники и связанные с ними участки нагорноксерофитной или степной растительности используются преимущественно как пастбища. Зна чительная часть этих насаждений до сих пор еще служит источником для заготовок топлива, что влечет за собой обнажение горных склонов и усиление эрозии. Почвозащитная (противоэрозионная) роль арчевников совершенно несомненна, а поэтому вырубка их должна быть сведена до минимума и производиться только под строгим контролем соответствующих лесных ведомств.
Литература
О. Э. Кнорринг (1915); В. В. Огиевский (1933); Е. П. Коровин (1934); М. Г. Попов и Н. В. Андросов (1937); А. Я. Бутков (1938); М. П. Петров (1945); П. Н. Овчинников (1948а); В. П. Дробов (1950а); П. Б. Виппер (1953а).
69. Фисташковые редколесья
В пределах Советского Союза эта группа формаций слагается двумя видами — кевовой. фисташкой (Pistacia mutica) на Кавказе и в Крыму и настоящей фисташкой (Pistacia vera) в Средней Азии.
В отличие от арчевников, фисташковые редколесья занимают более низкие местоположения — предгорные равнины и сухие предгорья и никогда не создают густых сомкнутых насаждений. Это очень изреженные, светлые редколесья, в нижнем ярусе которых может развиваться самая разнообразная растительность. Таким образом, здесь, так же как и в большинстве арчевых редколесий, нет сколько-нибудь заметной связи между синузиями, входящими в состав фисташковой формации. Синузии нижних ярусов ведут себя как бы независимо от верхнего древесного полога, что, конечно, объясняется крайней изреженностью и осветленностью последнего.
Фисташники — группа формаций, весьма характерная для стран Передней и Средней Азии. Их* распространение связано главным образом с аккумулятивными формами рельефа. На твердых коренных породах они почти не встречаются. Этой ярко выраженной экологической особенностью фисташники хорошо отличаются от арчевников, которые, как было указано выше, нередко развиваются даже на скалистых местообитаниях.
69 к — кавказские из Pistacia mutica
Кавказские редколесья, слагаемые кевовой фисташкой (Pistacia mutica), наиболее типичны в Восточном Закавказье. На «Геоботанической карте» контур редколесий из кевовой фисташки показан в северо-восточных предгорьях Карабахского хребта. В других местах распространение редколесий из кевовой фисташки отмечено внемасштабным знаком. По данным А. А. Гроссгейма (1948) ксерофитные редколесья, в том числе и фисташники, характерны для Шекинского нагорья — платообразной
возвышенности, сложенной третичными отложениями. В условиях плоскохолмистого рельефа, по М. Ф.Сахокиа (1950), наблюдаются следующие основные варианты фисташников.
1) Полупустынные фисташники. Фисташка разбросана на фоне степных и пустынных покровов, образующих комплексы. В степном травостое преобладают бородач (Andropogon ischaemum), иногда ковыли (Stipa capillata, S. Lessingiana). Пустынные покровы слагаются полынью и солянками (Artemisia Meyeriana, Salsola ericoides, Kochia prostrata).
2) Бородачевые фисташники по пологим суглинисто-каменистым склонам холмов. Фисташка разбросана на фоне травостоя из бородача.
3) Ковыльные фисташники по суглинистым каштановым почвам пологих склонов развиваются на фоне степного травостоя с преобладанием ковылей (Stipa capillata, S. Lessingiana).
Кроме предгорий, фисташники на Кавказе встречаются и по речным долинам в контакте с тугайными лесами (на карте не выделены). В этих условиях формируются смешанные дубово-фисташковые насаждения с участием длинноножкового дуба (Quercus longipes) и большим или меньшим участием кустарников: держи-дерева (Paliurus spina-Christi), граната (Punica granatum),боярышника (Crataegus sp. sp.). В травяном покрове нередко преобладают типичные луговые элементы: ежа (Dactylisglomerata), пырей (Agropyrum repens), иногда галофильные растения (Puccinellia bulbosa, Limonium Meyeri).
В условиях речных долин встречаются фисташники и без кустарникового яруса; здесь фисташка разбросана на фоне галофильного злака (Aeluropus litoralis), а иногда и по фону, слагаемому сочными однолетними солянками (Petrosimonia brachiata, Salsola crassa).
Кроме горного и приречного, имеется еще третий эколого-топологический тип фисташников, встречающийся на пролювиально-делювиальных отложениях. Это пустынные редколесья из кевовой фисташки на фоне полынных, полынно-солянковых и солянковых ассоциаций (Artemisia Meyeriana, Salsola dendroides, S. ericoides).
На галечниковых террасах пересыхающих рек наблюдаются ильмовые фисташники с участием в древостое ильмов (Ulmus foliacea, U. suberosa) и с более или менее хорошо выраженным кустарниковым ярусом (Paliurus spina-Christi, Punica granatum и др.).
Литература
Н. А. Буш (1935а); А. А. Гроссгейм (1948); А. В. Иванова (1949); М. Ф. Сахокиа (1950).
69 са — среднеазиатские из Pistacia vera и Amygdalus bucbarica
Среднеазиатские редколесья из фисташки представлены на карте большим числом контуров, чем кавказские. Они встречаются в предгорьях Тянь-Шаня, Алая, Копет-Дага, на хребтах Кугитанг, Бабатаг, Актау и др., а также в южной Туркмении в районе Кушки (Бадхыз).
Настоящая фисташка (Pistacia vera), слагающая эти редколесья, представляет собою небольшое, 5—6 м высоты, обычно корявое деревцо, с шатрообразной кроной и с укороченным стволом, от которого отходят длинные ветви. Изредка встречаются экземпляры фисташки, достигающие более 10 м высоты. Обитания настоящей фисташки связаны большей частью с пустынными низкогорьями или с более высокими мягкими
предгорьями. Соответственно этому можно различать два основных эколого-топологических типа среднеазиатских фисташников — нижний и верхний.
Нижний, пустынный тип фисташников связан с пустынной полынно-эфемеровой растительностью, развивающейся на сероземах, обычно подстилаемых лёссом. Здесь фисташка растет по фону полынно-осочково-мятликовых покровов (Artemisia tenuisecta, A. scopariiformis, Carex pachystylis, Роа bulbosa), в составе которых иногда встречается более крупное эфемероидное разнотравье (Psoralea drupacea, Eremostachys labiosa, Crambe Kotschyana, Phlomis bucharica). Верхний, пустынно-степной тип фисташников, широко развитый в Таджикистане, представляет собою сочетание фисташки с полынными и фломисово-ковыльными покровами (Artemisia baldshuanica, Phlomis bucharica, Stipa capillata, S. caucasica). Подобные фисташники часто сочетаются также с миндальниками (Amygdalus bucharica) и арчевниками.
В южной Киргизии (орехосовхозы Аркит п Майли-Сай) фисташка, по данным Е. М. Лавренко и С. Я. Соколова (1949), разбросана по фону эфемерово-полынных (Artemisia tenuisecta, A. eremophila) и перовскиевых (Perovskia scrophulariifolia) покровов, а иногда по фону вишарников (Cerasus erythrocarpa, С. tianschanica).
Очень часто фисташники встречаются на обнажениях пестроцветных третичных отложений, по склонам речных долин и ущелий. В этом случае они более редкостойны. Их сопровождают виды, типичные для пестро-цветов (Zygophyllum megacarpum, Otostegia Olgae, Schraderia bucharica, Cephalorrhizum oopodum, Cirsium schakaptaricum).
По описаниям И. А. Линчевского (1935), весенний аспект кушкинских фисташников (южная Туркмения) очень красочен. Ранней весной (в конце марта) в них доминирует яркозеленый фон осочково-мятликового травостоя (Carex pachystylis и Роа bulbosa), по которому несколько позднее в большом количестве появляются яркокрасные цветы кушкинского тюльпана (Tulipa kuschkensis) и другие раннецветущие виды из лилейных, ирисовых, крестоцветных, бобовых, лютиковых. Высота травостоя (Роа bulbosa и Carex pachystylis) — 20—25 см. Задернение почвы почти полное. Уже с первой половины мая осочково-мятликовый травостой засыхает и желтеет. В это время аспект создают главным образом кусты вьюнка (Convolvulus subhirsutus), астрагала (Astragalus agameticus), иногда алтея (Alcea rhytocarpa). Летом появляются цветущие заросли псоралей (Psoralea drupacea) и гигантские (до 2 м и выше) метелки кузинии (Cousinia Raddeana), а позднее темнозеленые кусты полыни (Artemisia scopariiformis).
В фисташниках проводится сбор ценных фисташковых плодов, широко употребляемых в кондитерской промышленности. Урожайность ее плодов подвержена сильным колебаниям в зависимости от климатических условий, возраста деревьев, полноты и бонитета фисташковых редколесий. Наиболее продуктивные фисташники развиваются по глубоким сероземам на лёссе. В урожайные годы они дают в соеднем около 2 кг плодов с одного дерева. На отдельных деревьях собирается до 10—15 кг плодов (Тросько, 1947). Галлы и листья фисташки употребляются для приготовления малиновой краски, получения медицинского таннина и для дубления кож.
Все эти ценные качества фисташки вызывают необходимость организации самой строгой охраны фисташников от порубок и пожаров, рационализации сбора плодов и реконструкции этих насаждений путем создания окультуренных фисташковых лесосадов.
Литература
Е. П. Коровин (1934); И. А. Линчевский (1935); И. К. Тросько (1947); Е. М. Лавренко и С. Я. Соколов (1949); В. И. Запрягаева (1954).
70. Нагорные ксерофиты и растительность иестроцветов (фригана)
Под именем нагорных ксерофитов объединяется сильно разреженная растительность с преобладанием резко ксерофильных, большей частью подушковидных кустарничков и полукустарничков, развивающихся по каменистым склонам сухих, континентальных горных стран. По общему облику к нагорным ксерофитам очень близка изреженная растительность, обитающая по выходам пестроцветных, обычно гипсоносных, осадочных третичных пород, на которых еще нет сформированного почвенного покрова.
В нашей советской ботанико-географической литературе подобного рода растительность иногда именуется фриганой. Это народный термин, заимствованный из средиземноморских (балканских) стран, где нагорные ксерофиты также весьма широко распространены.
Кавказ и Средняя Азия принадлежат к основным очагам развития нагорноксерофитной растительности наряду с такими соседними аридными странами, как Иран, Афганистан, Турция. Нагорные ксерофиты встречаются во всех высотных поясах гор (до 3000 м над у. м. и выше), но наибольшее их развитие приурочено все же к нижним и средним поясам (от 800—1000 до 2000—2500 м), где сообщества нагорных ксерофитов нередко комбинируются с ксерофитными редколесьями.
На Кавказе главнейшими очагами формирования нагорных ксерофитов являются Армения и Дагестан. По горным склонам Большого Кавказа этот тип растительности представлен в меньшей степени (отдельные участки в верховьях Кубани, Баксана, Гильгильчая, Бугчая и др.).
В Южном Закавказье (Армения, Нахичеванская АССР) нагорные ксерофиты создают физиономию целых высотных поясов. Здесь развиваются главным образом астрагаловые трагакантники, т. е. ценозы с преобладанием колючих кустарничковых астрагалов (Astragalus microce-phalus, A. aureus, A. erinaceus, A. vedicus, A. Karjaginii, A. lagurus), вместе с которыми встречаются степные (Festuca sulcata, Koeleria gracilis) и «то-миллярные» (тимьянниковые) элементы (Thymus Kotschyanus, Scutellaria Karjaginii, Ziziphora denticulata, Z. borzhomica,1 виды рода Nepeta). В нижних поясах гор Восточного Закавказья трагакантники образованы преимущественно кавказским астрагалом (Astragalus caucasicus) в сочетании с местными видами тимьяна (Thymus tiflisiensis, Th. karamarianicus) и бородачем (Andropogon. ischaemum). В северокавказских трагакантни-ках преобладает главным образом астрагал Маршалла (Astragalus Маг-schallianus).
Кроме астрагалов трагакантники формируются также колючим эспарцетом (Onobrychis cornuta) совместно с видами акантолимона (Асап-tholimon) (рис. 59). Эти формации распространены преимущественно по известковым склонам среднего и верхнего горного пояса Восточного Кавказа (Дагестан, Азербайджан).
Следующий вариант нагорных ксерофитов, довольно широко распространенный на Кавказе, представлен тимьянниками (томиллярами), то есть ценозами с преобладанием различных полукустарничковых и тра
1 Ранее в кавказской литературе эти виды Ziziphora приводились под названием Ziziphora media.
вяных пахучих губоцветных (из родов Thymus, Nepeta, Salvia, Scutellaria, Teucrium). Тимьянники характерны преимущественно для Южного и Восточного Закавказья, а также для Дагестана. Обычно они сочетаются с трагакантниками, иногда с шибляком и ксерофитным редколесьем.
Среднеазиатская нагорноксерофитная растительность по своему общему био-экологическому складу не отличается от кавказской, но видовой состав ее здесь иной, хотя общие с Кавказом виды все же имеются (особенно в Копет-Даге, например Onobrychis cornuta).
Главными районами развития нагорных ксерофитов в Средней Азии является Копет-Даг, система Алая, Западный и отчасти Центральный Тянь-Шань. В Северном Тянь-Шане этот тип растительности почти отсутствует.
В Копет-Даге нагорные ксерофиты, преимущественно типа трага-кантников, встречаются во всех горных поясах, приурочиваясь к грубым каменисто-щебнистым субстратам, преимущественно южных склонов (рис. 57). Основными строителями относящихся сюда фитоценозов являются Gypsophilaaretioides, Onobrychis cornuta, виды родов Acantholimon, Acanthophyllum и трагакантовые астрагалы. По мере увеличения мелкозема на склонах нагорные ксерофиты постепенно сменяются степными ассоциациями. Переходным звеном к этой смене являются, например, ассоциации, образуемые типчаком и колючим эспарцетом {Festuca Вот-muelleri, Onobrychis cornuta). Ассоциации нагорных ксерофитов Копет-Дага в большей части являются вторичными, появившимися на месте вырубленных арчевников или же в результате дигрессии травяных степных ценозов под воздействием усиленного выпаса (Никитина, 1949).
В Алайской горной системе (хребты Туркестанский, Алайский, Гис-сарский и др.) нагорные ксерофиты также широко распространены. Они слагаются различными полукустарничковыми и кустарничковыми видами: Acantholimon tataricum, A. parviflorum и др., Acanthophyllum pungens, A. glandulosum, A. brevibracteatum, Onobrychis echidna, к которым примешиваются многие другие полукустарнички {Thymus serpyllum s. 1., Zi-ziphora Bungeana и др.), некоторые полукустарничковые полыни {Artemisia tenuisecta) и ксерофильные травы {Stipa caucasica, S. Hohenackeriana), а также виды родов Eremurus, Cousinia. В Туркестанском хребте и в западных отрогах Гиссарского хребта нередко встречаются участки теке-сагызников — формации с преобладанием полукустарничкового теке-сагыза {Scorzonera acanthoclada). На более мягких почвах обычным спутником текесагызников является типчак {Festuca sulcata s. 1.).
Нагорноксерофитная растительность Западного и Центрального Тянь-Шаня по своему видовому составу близка к вышеописанной алайской. Наиболее частыми ценозообразователями (кустарничками и полукустарничками) этого типа здесь являются Acanthophyllum pungens, Acantholimon alatavicum, A. Korolkovii, Onobrychis echidna, Convolvulus tragacant-hoides, Ephedra equisetina, Eurotia ceratoides' из трагакантовых астрагалов часто встречается Astragalus lasiosemius — довольно обычный компонент многих ассоциаций нагорноксерофитной растительности, иногда создающий самостоятельные ценозы. Также как и в Алае, в составе типичных нагорных ксерофитов Тянь-Шаня постоянно наблюдаются степные элементы — виды типчака, ковылей, чия {Stipa caucasica, S. Hohe-nackeriana, Lasiagrostis caragana) и более или менее развитая эфемерово-эфемероидная синузия (из Carex pachystylis, Роа bulbosa и других биологически сходных с ними видов). Столь же обычны здесь ксерофильные
или мезоксерофильные кустарники {Amygdalus spinosissima, A. Petunni-kowii, Atraphaxis pyrifolia, Cerasus tianschanica, Spiraea hypericifolia).
На рыхлых (обыкновенно гипсоносных) пестроцветных третичных толщах Средней Азии развивается сильно изреженная растительность, в которой ценотические связи крайне ослаблены или даже вовсе отсутствуют. Большей частью растения представлены здесь редкими одиночными экземплярами, рассеянными среди голых выходов горной породы. Видовой состав такого рода местообитаний в различных географических районах весьма специфичен. Он отличается обилием своеобразных эндемичных и реликтовых форм, представляющих большой интерес для выяснения истории флоры (например следующие эндемичные для этой страны виды: Otostegia Olgae, О. bucharica, Buhsea Raddeana, Cleome Gordjagini, C. Noeana, Cephalorrhizum oopodum, Chaetolimon sogdianum, Schraderia bucharica, Cousinia Sprygini, C. Badghysi, C. oopoda, Crambe Gordjagini, Ladyginia bucharica, Gaillonia bucharica и др.).
С хозяйственной точки зрения нагорные ксерофиты и растительность пестроцветов представляют собою плохие пастбища, так как большинство растений, слагающих эти ценозы, колючие и резко пахнут. Скотом они не поедаются или поедаются очень плохо. Растительность данного типа может быть использована для эксплуатации и отбора имеющихся здесь ценных эфироносных, смолоносных и камеденосных видов. Кроме того, в составе нагорных ксерофитов и на пестроцветах немало декоративных растений, которые с успехом могут быть применены в зеленом строительстве.
Литература
О. Э. Кнорринг (1915); М. Г. Попов (1923); Е. П. Коровин (1934); А. Л. Тахта-джян (1937, 1941); А. А. Гроссгейм (1948); В. Н. Никитина (1949); М. Ф. Сахокиа (1950).
71. Колючетравники (из видов Cousinia)
Эта группа формаций экологически близка к нагорноксерофитной растительности, одним из вариантов которой она в сущности и является. В своем распространении колючетравники связаны с формациями нагорных ксерофитов и нередко сочетаются вместе с последними. На карте контуры колючетравников выделены только в пределах Средней Азии. Однако мелкие фрагменты этих ценозов имеются, повидимому, также на Кавказе.
В Средней Азии ценозы с доминированием крупных колючих кузиний свойственны преимущественно верхним поясам гор Памиро-Алайскоп системы и Западному Тянь-Шаню. Довольно крупные контуры данных формаций выделены именно в пределах указанных горных систем (Туркестанский, Гиссарский и Дарвазский хребты в Алае, Чаткальский и другие хребты Западного Тянь-Шаня).
Кузинники приурочены обычно к южным щебнистым или суглинистощебнистым склонам с более или менее сформированными почвами типа коричневых луговостепных (субальпийских). Некоторые варианты этих формаций свойственны не только субальпийскому, но и альпийскому поясу.
По своему происхождению кузинники являются большей частью ценозами антропогенного (точнее зоогенного) происхождения. Их формирование в большинстве случаев обусловлено воздействием усиленного выпаса.
В горной системе Алая группа колючетравных формаций с преобладанием кузиний весьма характерна для субальпийского пояса (2500—
3000 м над. у. м.). Так, в субальпике западной оконечности Гиссарского хребта распространены кузинники с преобладанием колючей кузинии (Cousinia macilentd). В нижнем ярусе фитоценозов, образуемых этим видом, располагаются полукустарнички и ксеромезофильное мелкотравье (Arenaria Griffithii, Trigonella Popovii, Poa glabriflora, Carex duriusculi-formis). В сочетании с указанными ценозами встречаются текесагызники (Scorzonera acanthoclada) и кузинники более мезофитного типа (с преобладанием Cousinia splendida), в сложении которых участвует более крупное мезофильное разнотравье (Ligularia persica, L. Thomsonii, Nepeta podostachys, а также злаки — Dactylis glomerata, виды рода Roegneria и др.). Кроме того, в Гиссарском хребте встречаются почти чистые заросли, слагаемые кузинией Федченко (Cousinia Fedtschenkoana), с которыми по понижениям рельефа сочетаются луговидные ценозы (из Phlomis canes-cens, Ligularia persica, Poa bucharica, Roegneria ugamica, Alopecurus se-ravschanicus), а иногда ценозы с преобладанием полукустарничковой песчанки (Arenaria Griffithii).
Довольно широко представлены в горах Памиро-Алайской системы колючетравники, эдификаторами которых являются Cousinia Franche-ti, С. stephanophora, С. hissarica, С. rubiginosa, С. trachyphylla, С. pannosa и др. Среди формаций этих кузинников можно различать два типа групп ассоциаций: 1) более или менее мезофитные (луговые), с участием значительного числа луговых элементов (Geranium saxatile, Ligularia Thomsonii, L. alpigena, Poa bucharica, Nepeta podostachys и др.); 2) ксерофитные co значительным участием степных элементов (Festuca sulcata, Роа relaxa). Для этого варианта особенно характерно постоянное сочетание кузинников с группировками типичных нагорных ксерофитов из эспарцета (Ondbrychis echidna).и видов акантолимона, текесагызниками (Scorzonera acanthoclada) и тимьянниками (Thymus serpyllum s. 1., Ziziphora Bungeana, виды Scutellaria).
В субальпийском поясе Западного Тянь-Шаня (Кураминский хребет), согласно данным Г. Т. Сидоренко (1953), колючетравники представлены двумя формациями, эдификаторами которых являются Cousinia Bonva-lotii и С. egreria. Шире распространена первая из этих формаций, встречающаяся в верхней части субальпийского пояса (2700—3200 м над у. м.); она предпочитает более или менее сложившиеся почвы седловин и пологих горных склонов. На крутых склонах она развивается слабо, только в виде изреженных зарослей. В составе этой формации, выделяются две группы ассоциаций: трагакантовые и остепненные кузинники. Трагакантовые кузинники приурочены к более грубым, скелетным почвам. В их сложении значительную роль играют трагаканты (Astragalus Bornmiillerianus, Acantholimon Korolkovii) и полукустарничковые губоцветные (Ziziphora Bungeana, Lagochilus tianschanicus). Остепненные кузинники распространены шире, чем предыдущие. Они развиваются по более мягким склонам с достаточно Хорошо сложившимися почвами. В их составе имеют большой удельный вес дерновинные горностепные злаки (Festuca sulcata, Роа relaxa, Hordeum turkestanicum, Bromus turkestanicus, Piptatherum songoricum и др.).
Распространение формации с преобладанием Cousinia egreria — крупного (до 60—80 см) многолетника — приурочено к более низкому поясу гор (в пределах 1900—2600 м). В составе этой формации довольно широко распространены ассоциации с большим участием югана (Prangos pabula-ria) и ферулы (Ferula ovind). Кроме того, здесь отмечаются некоторые степ-‘ ные злаки (Роа relaxa, Festuca sulcata) и ксерофильное разнотравье (Hypericum scabrum, Allium oreophilum), а также эфемер Alyssum desertorum.
На крутых горных склонах, почти лишенных мелкозема, встречаются петрофитные варианты данной формации, характеризующиеся сильной изреженностью травостоя (20—40%) и значительной ролью типичных нагорноксерофитных элементов: трагакантовых астрагалов, колючего эспарцета, акаптолимона.
Проективное покрытие травостоя кузинииков Средней Азии колеблется в пределах от 20—30% до 60%. В большинстве случаев это плохие или посредственные пастбищные угодья с низкой кормовой производительностью из-за обилия грубых колючих непоедаемых трав. Однако некоторые варианты остепненных и луговых кузинииков представляют собою кормовые угодья более высокого качества, дающие в среднем около 4— 5 ц, а иногда до 8 ц сухой поедаемой массы с 1 га.
Литература
Е. П. Коровин (1934); А. И. Толмачев (1949); Ю. С. Григорьев (1951); Г. Т. Сидоренко (1953).
72. Субтропические низкотравные степи (полусаванны)
К субтропическим степям отнесены на карте ценозы с преобладанием в травостое эфемероидов и эфемеров, то есть растений, развитие которых заканчивается в краткий весенний период или в начале лета, хотя вегетация многих из них начинается еще с осени и фактически продолжается в течение всей сравнительно теплой зимы, характерной для Средней и Передней Азии, где эти ценозы распространены. Главной областью развития низкотравных субтропических степей в пределах Советского Союза является Средняя Азия (рис. 60). На Кавказе они выделены только в районе Апшеронского п-ва.
Согласно П. Н. Овчинникову (1940),1 указанный выше ритм развития данных фитоценозов несколько сближает их с субтропическими и тропическими саваннами. На этом основании эфемерово-эфемероидная растительность Средней Азии была выделена им в качестве особого типа, названного «полусаванной». Вообще в советской ботанико-географической литературе эфемероидная растительность описывалась под самыми различными наименованиями. Впервые ее экологическое своеобразие и особое положение среди растительного покрова Средней Азии было отмечено М. В. Культиасовым (1926—1927) и М. Г. Поповым (1927). Е. П. Коровин (1934) низкотравные эфемероидные сообщества относил к пустынному типу («эфемеровые пустыни»). Позднее (1946), в своей совместной работе с Е. Е. Коротковой, он выделил их в особый самостоятельный «тип эфемеров», а эфемероидные сообщества из пушистого пырея (Agropyrum tri-chophorum) и югана (Prangos pabularia и других видов) отнес уже к другому типу — «степей-саванн». А. В. Прозоровский (1940) низкотравные эфе-мерово-эфемероидные сообщества относил к «жаропокоящимся лугам». Н. И. Рубцов (1952) в последнее время эфемерово-эфемероидные сообщества называет «саванноидами».
Если такие термины, как «полустепь», «полусаванна», «жаропокоя-щийся луг» и «эфемеровая пустыня» нельзя считать удачными, то и название, принятое на «Геоботанической карте» — «субтропические степи»— тоже не является вполне приемлемым, так как относимые сюда растительные сообщества заметно отличаются от степного типа растительности в его типичном выражении. В них отсутствует главный характерный признак степей — преобладание в травостое многолетних травя-
нистых микротермных ксерофитов и главным образом плотподерновин-ных, узколистных злаков, свойственных умеренной зоне Евразии. Травостой эфемерово-эфемероидных сообществ слагается преимущественно из ксеромезофитов, среди которых совсем нет плотнокустовых злаков.
Низкотравные субтропические степи (полусаванны) особенно широко развиты в пределах Средней Азии, где они очень характерны для пустынных предгорных равнин и низких предгорий почти всех горных систем этой страны, за исключением Северного и Центрального Тянь-Шаня. На Кавказе относящаяся к этому, типу растительность свойственна преимущественно Апшеронскому п-ву и южной части Кура-Араксин-ской низменности (рис. 60).
72 ср —из Carex pachystylis и Роа bulbosa
Основа травостоя среднеазиатских субтропических низкотравных степей слагается двумя типичными эфемероидами — узколистной мелкой осочкой (Carex pachystylis) и луковичным мятликом (Роа bulbosa var. vivipara). Оба растения полностью заканчивают свой цикл развития к наступлению летней засухи и отмирают в надземной части уже в мае месяце. Обычными компонентами этих осочково-мятликовых сообществ являются многочисленные, большей частью мелкие травы, тоже эфемерного или эфемероидного типа. К ним относятся, например, многие однолетние астрагалы (Astragalus filicaulis, A. campylorrhynchus, A. Schmalhausenii), пажитники (Trigonella grandiflora, Т. Noeana), мелкие однолетние крестоцветные (Alyssum desertorum, A. campestre, Meniocus lintfolius, Erophila verna, Euclidium syriacum, Litvinovia tenuissima, Malcolmia turkestanica, Cryptospora falcata, Chorispora tenella), маковые (Hypecoum parviflorum, Papaver pavoninum), лютикоъые (Ceratocephalus orthoceras, Nigella integri-folia, Consolida rugulosa). Немало здесь и однолетних эфемерных злаков; особенно обычны однолетние костры (Bromus oxyodon, В. Danthoniae, В. scoparius).
Из многолетних эфемероидных видов, распространенных почти во всех районах широкого развития осочково-мятликовой растительности, укажем, например, на некоторые луковичные и клубненосные лилейные (Gagea stipitata, G. reticulata, Tulipa sp. sp., Eremurus Olgae), из зонтичных — некоторые ферулы (Ferula sp. sp.). Многолетних растений более длительной вегетации немного. Они представлены астрагалами (Astragalus тасгопух и др.), эспарцетом (Onobrychis chorassanica), некоторыми губоцветными (Eremostachys labiosa) и т. д. В общем видовой состав описываемых сообществ можно считать довольно богатым. Так, по данным Е. П. Коровина (1934), на площади 1 м2 в осочково-мятликовых сообществах района Ташкента насчитывается до 47 видов. Основной, наиболее густой подъярус, высотою 15—20 см, слагается узколистной осочкой и луковичным мятликом. Эти два доминирующих растения совместно с другими эфемерными компонентами создают полное задернение почвы. Над осочково-мятликовым подъярусом можно выделить верхний изреженный подъярус (или несколько подъярусов) более высоких трав как эфемерного типа, так и длительной вегетации.
Для осочково-мятликовых сообществ Бадхыза и Карабиля (южная Туркмения) особенно характерны гигантские, гораздо выше человеческого роста, эфемероидные зонтичные (Ferula badrakema и Dorema Aitchisonii). На предгорных равнинах и предгорьях Западного Тянь-Шаня по осочково-мятликовому фону развиваются преимущественно Pso-ralea drupacea, Phlomis salicijolia; в предгорьях Алая — Phlomis bucharica, P. thapsoides. Иногда в сложении верхнего, обычно изреженного подъяруса небольшое участие принимают некоторые полыни (Artemisia scopariiformis, A. ferganensis, A. eucina и др.).
В типичном своем выражении осочково-мятликовая растительность характерна для светлых сероземов, формирующихся на лёссах и лёссовидных суглинках. Однако существуют варианты ее, приуроченные к песчаным почвам. Таковы, например, упомянутые выше для южной Туркмении осочковые мятличники, верхний ярус которых образован гигантскими зонтичными. Вообще эфемероидное крупнотравье (виды Ferula, Prangos, Inula grandis и. др.) в порядке сочетания растительных ассоциаций нередко входит в пределы контуров, показанных на карте как низкотравные субтропические степи, что отмечено соответствующим знаком.
Среднеазиатские низкотравные осочково-мятликовые травостои используются как хорошие весенние пастбища и сенокосы. Значительная площадь, занятая ими, распахивается под поливное земледелие главным образом для культуры хлопка (например в районе Мирзачуля, Ташкента, на многих участках Ферганской котловины и т. д.). Дальнейшее развитие ирригации даст возможность освоить под земледелие большую часть плодородных земель, еще занятых в настоящее время осочково-.мят-ликовой * растительностью.
Литература
П. А. Баранов (1924); М. В'. Культиасов (1926—1927); Н. А. Базилевская (1927); М. Г. Попов (1927); Е. А. Малюгин (1931); О. Э. Кнорринг (1934); Е. П. Коровин (1934); А. П. Шенников (1938); П. Н. Овчинников (1940); А. В. Прозоровский (1940); С. Н. Кудряшев (1941); А. П. Шенников и А. Ф. Иоффе (1944); Е. П. Коровин и Е. Е. Короткова (1946); Н. И. Рубцов (1952).
72 р — из Роа bulbosa
Низкотравные сообщества с преобладанием луковичного мятлика (Роа bulbosa var. vivipara), без какого-либо участия узколистной осочки, широко развиты на Ашперонском п-ве. Встречаются они и в южной части Кура-Араксинской низменности (на карте не показаны).
На Ашперонском п-ве растительные группировки с большим обилием луковичного мятлика связаны с сухими плотными глинистыми, большею частью солонцеватыми почвами. В сложении относящихся сюда фитоценозов большую роль играют различные мелкие однолетние злаки: костры (Bromus japonicus, В. rubens, В. sterilis), ячмень (Hordeum leporinum), тимофеевка (Phleum paniculatem), лофохлоа (Lophochloa phleoides), канареечники (Phalaris minor, P. paradoxa), плевел (Lolium rigidum), эгилопсы (Aegilops squarrosa, A. cylindrica). Вместе с перечисленными злаками встречается много мелких однолетних крестоцветных (Lepidium perfoliatum, L. ruderale, Arabidopsis pumila) и бобовых (Medicago orbicularis, M. denticulate, M. arabica, Astragalus contortuplica-tus и др.).
В Кура-Араксинской низменности нередко встречается эфемеровая растительность без участия луковичного мятлика. В ней эдифика-торами являются высокие эфемерные злаки (Phalaris minor и Lolium rigidum), к которым примешивается костер (Bromus japonicus). В сочетании с этими сообществами на солончаковых луговых («чальных») почвах и по солончакам формируются сообщества с преобладанием галофильных однолетних злаков (Sphenopus divaricates, Hordeum genicula-tam, Pholiurus incurvus) и ряда других галофильных однолетних видов (Tetradiclis tenella, Frankenia pulverulenla, Psylliostachys [Sldtice] spicate).
Районы развития подобной растительности на Кавказе являются хорошими весенними и зимними пастбищами. В настоящее время они в значительной степени (особенно на Апшероне) распаханы под культуру зерновых и технических растений.,
А. А. Гроссгейм (1948).
слева) в бассейна 1СВНИКИ juniperus turcomantea) с нагорными ксерофитами (Onobrychis cornuta — внизу
Р а» на высоте 1800—1900 м над у. м. Центральный Копет-Даг (Туркменская GCP).
(Фот. Ал. А. Фсдсрова).
Рис. 57. Группировка из плотных подушек Gypsophila uretioides. 1н>пет-Да1 . (Фот. .1. К. Родина).
Рис. 58. Каменистый склон с плотными подушками Gypsophila aretioides (отмечены <g»). Горный массив Даралагез (Армянская ’ССР).
(Фот. Г. В. Аркадьева).
73. Субтропические крупнотравные степи (полусаванны) и сельскохозяйственные земли на их месте
Сюда отнесена группа формаций, эдификаторами которых являются крупные эфемероидные или гемиэфемероидные растения (Prangos pabularia, Ferula J aeschkeana, Agropyrum trichophorum,1 Hordeum bulbosum, Inula grandis). Формами эти распространены в нижнем и среднем поясах, а иногда в нижней части высокогорного пояса в Средней Азии, на черноземах и на коричневых почвах луговостепного типа (рис. 60). До сих пор в литературе подобного рода растительность известна под именем полустепи (М. Г. Попов), лугов гималайского типа и степей-саванн (Коровин, 1934; Коровин и Короткова, 1946). П. Н. Овчинников (1940) относит ее к категории крупнотравных полусаванн.1 2
73v — из Inula grandis, Prangos pabularia, видов Ferula в сочетании с ксерофильными кустарниками
Субтропические степи с преобладанием указанных крупных эфемеро-идных зонтичных: югана (Prangospabularia) и комоля (Ferula J aeschkeana, F. tenuisecta, F. ovina), а из сложноцветных крупного девясила или каран-гыза (Inula grandis) приурочены к средним, а также частично высокогорным поясам Западного Тянь-Шаня и Алая. Высотные пределы их распространения колеблются в весьма широких пределах — от 700—800 м до 3000 м над у. м., в зависимости от географического района и экспозиции горного склона. В Западном Тянь-Шане формация с преобладанием югана (Prangos pabularia) в равной степени характерна как для среднегорного, так и для субальпийского поясов (от 800 до 2800 м над у. м.). В горах Алайской системы нижняя граница распространения этой формации значительно приподнята. Она проходит примерно на высоте 1800—2000 м над у. м. По данным О. Е. Агаханянц (1953), крупнотравные субтропические степи с преобладанием югана и комоля занимают до 37%.площади субальпийского и альпийского поясов северного Дарваза.
Юганники развиваются преимущественно по южным, реже по северным горным склонам на маломощных щебнистых почвах. Средняя высота травостоя 50—60 см, проективное покрытие колеблется от 50— 60% до 100%. В составе травостоя, кроме югана, почти всегда большую, но подчиненную роль играют комоли (Ferula Jaeschkeana, F. ovina). Эти два главных растения образуют основу верхнего подъяруса. В числе второстепенных компонентов травостоя могут быть степные, нагорноксеро-фильные, луговые и даже лесо-луговые элементы, в зависимости от экспозиции горного склона, высоты над уровнем моря и характера почвы. Так, например, в составе юганников отмечены типично степные растения (Festuca sulcata, Koeleria gracilis, Nepeta pannonica), растения, характерные для фриганы (Acantholimon Korolkovii, Ziziphora Bungeana, Scorzo-nera acanthoclada, Astragalus lasiosemius, A. nigrocalyx), луговые и луговолесные виды (Dactylis glomerata, Ligularia Thomsonii, Bromus inermis, Poa nemoralis). Однако встречаются в значительном количестве (по крайней мере в низкогорных и среднегорных вариантах юганников) также эфемерово-эфемероидные (полусаванные) элементы: различные однолетние костры (Bromus macrostachys, В. oxyodon, В. tectorum, В. Dantho-niae), узколистная осочка (Carex pachystylis), луковичный ячмень (Ног-
1 Agropyrum. trichophorum не является настоящим эфемероидом. У него летом засыхает только бблыпая часть листьев; стебель же продолжает долго вегетировать.
2 Литература ко всем подразделам 73 указана в конце текста к 73 а.
9 Заказ 551
deum bulbosum), некоторые виды клубненосных зонтичных (Muretia, Scaligeria) и т. п. Вегетация юганников заканчивается при наступлении летнего сухого периода. Бблыпая часть растений уже в июле завершает свой цикл развития.
В условиях сильно защебненных (грубо скелетных) почв иногда преобладают комоли (Ferula Jaeschkeana, F. ovina). В ассоциациях с господством этих видов особенно характерны трагакантовве и вообще петрофиль-ные элементы (Scorzonera acanthoclada, виды рода Acantholimon, колючие кузинии, зизифора, некоторые колючие астрагалы — Astragalus lasiose-mius, A. nigrocalyx, виды Scutellaria и др.).
Югановые, комолево-югановые и комолевые ассоциации используются как пастбища. Иногда они скашиваются и на сено. Урожайность сухой кормовой массы достигает 8—15 ц/га. Наиболее ценными в кормовом отношении являются остепненные юганники.
К нижнегорному и среднегорному поясам (800—2000 м над у. м.) приурочены юганово-карангызовые сообщества (с преобладанием J nula grandis и Prangos pabularia). Они встречаются по склонам южных экспозиций, на темных сероземах и на коричневых почвах, формирующихся на лёссе, а иногда на элювии метаморфических сланцев. Средняя высота травостоя 50—70 см, проективное покрытие 90—100%. Первый подъярус образован карангызом и юганом. Нередко в' сложении первого подъяруса значительное участие принимает луковичный ячмень (Hordeum bulbosum). В сложении нижних подъярусов большое участие принимают мелкие эфемеры и эфемероиды (Alyssum campestre, A. desertorum, Astragalus Schmalhausenii, Роа bulbosa, Carex pachystylis, Bromus macrostachys, Are-naria serpyllifolia, Phleum paniculatam и др.).
Юганово-карангызовые сообщества используются преимущественно как весенние пастбища и частично как сенокосы. Производительность, травостоя весной (в мае) достигает 10—12 ц/га сухой кормовой массы.
В сочетании с вышеописанными крупнотравными субтропическими степями из югана, комоля и карангыза постоянно встречаются участки арчевых и кленовых (из Acer turkestanicum, A. Semenovii, A. Regelii) редколесий, а также заросли кустарников (Amygdalus spinosissima, Rosa Есае, Cerasus verrucosa, Atraphaxis pyrifolia, Berberis oblonga, виды Cotoneaster, Spiraea hypericifolia и др.). Кроме того, почти все из этих древесных и кустарниковых пород единичными экземплярами или небольшими группами рассеяны по самому фону травостоя.
73 h — из Hordeum bulbosum
Участки этих степей не велики, но встречаются они довольно часто по всем предгорьям Западного Тянь-Шаня и Алая, на более или менее пологих склонах с мелкоземистыми почвами. В большинстве случаев это сообщества вторичного происхождения, развивающиеся после распашки на старых залежах. Эдификатором их является луковичный ячмень (Hordeum bulbosum) — крупнотравный эфемероидный злак, высотою до 100—120 см, образующий густой первый подъярус. Во втором подъярусе, 20—30 см высотой, размещаются другие, преимущественно также эфемерные и эфемероидные растения: однолетние костры (Bromus tectorum, В. oxyodon), узколистная осочка (Carex pachystylis), ячменец (Taenia-therum crinitum), пырей (Agropyrum trichophorum), луковичный мятлик (Роа bulbosa), горечавка (Gentiana Olivieri), мак (Papaver pavoninum), некоторые астрагалы (Astragalus vicarius, A. filicaulis) и кузиния (Cousinia microcarpa). Наряду с эфемерами и эфемероидами в составе фитоценозов
с преобладанием луковичного ячменя встречаются также и длите льновегетирующие виды (например Glycyrrhiza glabra, Psoralea drupacea, Heteropappus canescens, Hypericum perforatum, H. scabrum, Scabiosa son-gorica, Phlomis salicifolia, Centaurea sguarrosa). Разбросаны единичные кустарники (в Западном Тянь-Шане главным образом Amygdalus communis, Cerasus verrucosa, а в горах Алайской системы —Rosa Есае, Colutea canescens и др.).
По наблюдениям В. А. Никитина, в Таджикистане имеются участки луковичных ячменников на местах, сохранившихся от распашки. Они приурочены к горным склонам со скелетными почвами типа коричневых карбонатных. В составе этих ячменников обильны душица {Origanum tyttanthum) и сборная ежа {Dactylis glomerata)', в меньшем количестве присутствуют другие виды (inula grandis, Aegilops triuncialis, Aphanop-leura capillifolia, Cousinia microcarpa, C, pulchra, Carex turkestanica, Heteropappus canescens, Hypericum scabrum, Scabiosa songorica, Phlomis fruti-cetorum). Подобные ячменники на целинных почвах встречаются и на склонах Ферганского хребта (Лавренко и Соколов, 1949; Понятовская, 1952).
Луковичный ячмень и многие другие сопровождающие его злаки, а также некоторые виды разнотравья весною (до колошения) хорошо поедаются скотом. Поэтому слагаемые им сообщества являются хорошими весенними пастбищами и сенокосами. Средняя кормовая производительность травостоя около 8 ц/га (при использовании травостоя в апреле или в начале мая). Луковичные ячменники могут использоваться также и в качестве осенне-зимних пастбищ.
73 а — из Agropyrum trichophorum и Hordeum bulbosum
Этот вариант крупнотравной эфемероидной растительности распространен в Средней Азии гораздо шире, чем сообщества с преобладанием лишь одного луковичного ячменя {Hordeum bulbosum). Следует иметь в виду, что отнесенные сюда контуры карты объединяют как смешанные ячменно-пырейные сообщества {Agropyrum trichophorum, Hordeum bulbosum), так и сочетания самостоятельных пырейных и ячменных сообществ.
Смешанные ячменно-пырейные или пырейно-ячменные травостои, так же как и чисто ячменные, формируются главным образом на старых залежах, по предгорьям всех горных систем юга Средней Азии — в Западном Тянь-Шане, в системе Алая и в Копет-Даге. Почвы под ними обычно темные сероземы или коричневые (на более высоких местоположениях). По видовому составу и сложению смешанные пырейно-ячменные сообщества почти не отличаются от вышеописанных ячменных.
Пырейники с преобладанием волосоносного пырея {Agropyrum trichophorum) развиваются и на почвах, не бывших под распашкой. Благодаря довольно большой экологической амплитуде, в этой формации можно выделить несколько групп ассоциаций. Так, на северных, более увлажненных горных склонах широко представлены луговые пырейники со значительной примесью в травостое мезофильных луговых элементов: сборной ежи {Dactylis glomerata), безостого костра {Bromus inermis), коротконожки {Brachypodium pinnatum), мятлика {Роа angustifolia), а также ряда других мезофильных или ксеромезофильных видов из группы «разнотравья».
На склонах южных экспозиций развиваются пырейники, насыщенные типичными степными элементами: типчаком {Festuca sulcata) или дерно-
винным мятликом (Роа relaxa). В типчаковых пырейниках хорошо выделяются два подъяруса: верхний — из пырея, высотою до 70—80 см, и нижний — из типчака, высотою до 30—35 см. Общее проективное покрытие травостоя — 80—90%. Из других наиболее обычных, но менее обильных компонентов типчаковых пырейников можно указать на следующие: Carex turkestanica, С. pachystylis, Роа bulbosa, Origanum tyttanthum, Anemone Tschernjaewii, Leontice Ewersmannii, Bongardia chrysogonum, Hypericum scabrum, H. perforatum, Artemisia tenuisecta, Astragalus Bornmiillerianus, Ziziphora Bungeana, Centaurea squarrosa и др. Встречаются пырейные ассоциации, в которых доминантой второго яруса является туркестанская осока (Carex turkestanica).
Пырейники со степным мятликом (Роа relaxa) свойственны более крутым южным склонам с грубыми щебнистыми почвами. Они имеют более изреженный травостой, но по видовому составу почти не отличаются от типчаково-пырейных ассоциаций.
Среди пырейников можно выделить еще третью группу ассоциаций, в которых к эдификатору — пырею — в большом обилии присоединяются главным образом эфемеры и эфемероиды. Эта группа наиболее типичных (полусаванновых) пырейников формируется в средних и нижних поясах гор, по более или менее пологим мелкоземистым склонам. В сложении ее значительное участие принимают эфемеры-однолетники (Bromus tectorum, В. oxyodon, В. macrostachys, Alyssum desertorum, Astragalus vicarius, A. Schmalhausenii), мелкие эфемероиды (Carex pachystylis и Poa bulbosa), крупнотравные эфемероиды (Inula grandis, Eremurus robustus, E. Olgae, E. Regelii, Prangos pabularia, Hordeum bulbosum, Ferula Jaesch-keana, Scaligeria hirtula и др.).
Среди пырейников всех указанных выше групп ассоциаций встречаются единичные кустарники (Amygdalus spinosissima, Cerasus verrucosa, Rosa Ecae, виды Cotoneaster, Spiraea hypericifolia), а также некоторые деревья (Crataegus turkestanica, C. pontica, Pistacia vera, Juniperus semiglobosa). Увеличение обилия кустарников на отдельных участках иногда позволяет выделять среди пырейников особые шибляковые пырейники.
Кроме Средней Азии, участки пырейников (Agropyrum trichophorum), встречаются на Кавказе, главным образом в средних поясах гор Армении. На «Геоботанической карте» они не показаны. В их сложении участвуют виды, общие с пырейниками .Средней Азии (Medicago falcata, Festuca sulcata, Hordeum bulbosum, Phleum phleoides) и специфические кавказские (Onobrychis Cyri, Scabiosa bipinnata и др.).
Растительность с преобладанием пушистого пырея используется в качестве сенокосов и весенне-осенних пастбищ. Кормовая производительность их колеблется от 6—7 до 12—14 ц/га сухой массы. Чрезмерный выпас скота приводит к изреживанию пырея, вплоть до полного выпадения этого растения из травостоя за счет разрастания других видов (Achillea micrantha, Ceratocarpus turkestanicus, некоторые кузинии, Euphorbia faxartica, Haplophyllum latifolium), в кормовом отношении менее ценных или вовсе неноедаемых.
Почвы, покрытые пырейной растительностью, во многих районах распахиваются под посевы зерновых, технических и бахчевых культур (пшеница, сахарная свекла, подсолнечник, арбузы, тыквы, дыни, иногда картофель). На залежах после распашки пырейная растительность восстанавливается примерно через 10—15, иногда 20 лет в зависимости от высоты над уровнем моря, экспозиции склона и характера почвы. Вообще в более сухих нижних поясах гор восстановление идет медленнее, чем по северным склонам средних поясов.
Литература
II. А. Баранов (1924); М. В. Кулюиасов (1926,1926- 1927); О. Э. Кнорринг (1934); Е. 11. Коровин (1934); II. Ф. Гончаров (1936, 1937); И. В. Выходцев (1937); П. Н. Овчинников (1940); М. Г. Попов (1940); Е. 11. Коровин и Е. Е. Короткова (1946); Е. М. Лавренко и С. Я. Соколов (1949); В. М. Понятовская (1949, 1952); О. Ш. Шукуров (1949); Н. И. Рубцов (1950); О. Е. Агаханянц (1953); Г. Т. Сидоренко (1953).
74. Бородачевые (из Andropogon ischaemum) субтропические степи (полусаванны) и сельскохозяйственные земли на их месте
Ценозы, слагаемые бородачом {Andropogon ischaemum), широко распространены па Кавказе и в Средней Азии, где они занимают иногда значительные площади, а поэтому выделяются на «Геоботанической карте» довольно крупными контурами (рис. 60). Однако географическое распространение этих ценозов вовсе не ограничивается указанными районами. Небольшие участки и фрагменты бородачевников (не отмеченные па карте) встречаются также и в других южных районах Советского Союза, например в Молдавии, на юге Украины, в Крыму. Вне пределов Советского Союза, судя по имеющимся литературным данным (Рубцов, 1948), бородачевые ценозы (слагаемые Andropiogon ischaemum) характерны для стран Малой и Передней Азин (Анатолия, Афганистан, Иран) и особенно для средиземноморских стран.
Бородач — многолетний рыхлокустовой злак, высотою до 80 см, обладающий исключительно большой экологической амплитудой. Он обитает в предгорьях и в средних поясах гор как на сухих, так и на влажных почвах различного механического состава — глинистых, хрящеватых, песчаных, каменистых, иногда даже на скалах. Поэтому ценозы, образуемые этим злаком, чрезвычайно разнообразны и носят то луговой, то степной облик. Интенсивно развивается бородач па вторичных (нарушенных) местообитаниях — по местам усиленного выиаса, на старых залежах, на лесных вырубках и т. д.
На Кавказе основными районами распространения бородачевников являются низкогорья по левобережью Куры, к востоку от Тбилиси (Щекинское нагорье), и северные предгорья Малого Кавказа (Армения). Кроме того, порядочные площади, занятые бородачевниками, не отмеченные на карте, имеются в Восточном Предкавказье (в пределах Терского и Сунженского хребтов), в Дагестане и ряде других мест.
В районах своего массового распространения бородачевые ценозы на Кавказе приурочены преимущественно к низкогорному холмистому, реже среднегорному рельефу, в пределах 300—800 (—900) м над у. м. Почвы обычно щебневатые, темнокаштановые, реже южные (маломощные) черноземы. Засоленных почв бородач избегает.
В наиболее сухих низкогорьях Закавказья к бородачу в качество субдоминанта нередко присоединяется полукустарничковая полынь (Artemisia Meyeriana). В таких опустыненных полынно-бородачевых сообществах встречаются и некоторые другие пустынные виды (Menio-cus linifolius, Kochia prostrata). Однако самыми распространенными и типичными вариантами описываемых степей являются почти чистые борода-чевники с безраздельным господством бородача, а также типчаково-бородачевые ценозы. Почти чистые бородачевники развиваются преимущественно на грубых, скелетных почвах. В их сложении участвует большое число видов, среди которых характерны различные однолетники (Trachynia distachya, Bromus japonicus, Sideritis montana, Minuartia Wiesneri). В типчаково-бородачевых ценозах согосподствующим видом является типчак (Festuca sulcata). Этот вариант бородачевников связан
с более мелкоземистыми почвами. Кроме главных растений — бородача и типчака, в нем участвует типичное степное ксерофильное разнотравье (Linosyris villosa, Galium verum, Achillea micrantha, Teucrium polium, Medicago falcata s. 1.).
Ковыльно-бородачевые ценозы представлены несколькими ассоциациями. В Закавказье по щебневатым почвам широко распространены ассоциации, в которых согосподствующим видом является ковыль Шовица (Stipa Szowitsiana), а в Предкавказье по темнокаштановым суглинкам — ассоциации с примесью ковыля-тырсы (Stipa capillata) и ковыля Лессинга (Stipa Lessingiana).
В Восточном Закавказье (Шнракская степь) известны сочетания бородача с солодкой (Glycyrrhiza glabra). Они формируются на плакорных участках с черноземовидными почвами. В составе солодково-борода-чевых ассоциаций отмечены как степные, так и луговые виды (Galium verum, Crucianella angustifolia, Falcaria sioides, Daucus carota, Bromus japonicus, Linum corymbulosum, Phleum paniculatum, Medicago coerulea, Polygala transcaucasica, Trifolium campestre).
По фону кавказских бородачевников часто разбросаны единичные кусты или группы держи-дерева (Paliurus spina-Christi), жестера Палласа (Rhamnus Pallasii), таволги (Spiraea hypericifolia) и других кустарников.
На Кавказе в районах распространения бородачевников встречаются небольшие (не указанные на карте) участки с преобладанием жесткого плотнодерновинного злака — золотобородника (Chrysopogon gryllus). По видовому составу и структуре этих ценозов они очень сходны с бородачев-никами.
В Средней Азии, так же как и на Кавказе, распространение бородачевых степей связано с предгорьями и низкогорьями. Основные районы их развития сосредоточены по склонам Ферганского и Чаткальского хребтов. В меньшей мере они представлены в Центральном Тянь-Шане и в горах Алая. Северной границей географического ареала бородачев-нцков (в пределах Средней Азии) следует, повидимому, считать Джунгарский Алатау (Рубцов, 1948а), где эти ценозы весьма характерны для южных щебневатых предгорий указанной горной системы.
Экологически и' структурно среднеазиатские бородачевники почти не отличаются от кавказских. В них можно различать те же основные группы ассоциаций: 1) с полным господством бородача; 2) типчаково-бородачевые; 3) ковыльно-бородачевые. Все они отличаются от соответствующих кавказских аналогов лишь флористически. В составе среднеазиатских бородачевников, наряду с общими с Кавказом видами (Festuca sulcata, Stipa capillata, S. Lessingiana, S. Szowitsiana, Kochia prostrata), имеется ряд видов, не встречающихся в кавказских бородачевниках (например Inula grandis, Psoralea drupacea, Perovskia scrophulariifolia и многие другие растения, свойственные Средней Азии).
Бородачевые степи используются главным образом для выпаса скота, реже в качестве сенокосных угодий. Бородач хорошо поедается всеми видами скота как на пастбище, так и в сене. Средняя кормовая урожайность бородачевых пастбищ весною около’ 4—5 ц/га, а осенью 6—8 ц/га сухой массы. Они могут служить и в качестве летних выпасов, так как вегетация бородача продолжается до первой половины лета.
Литература
А. А. Гроссгейм (1948); П. Н. Овчинников (1948); Н. И. Рубцов (1948, 1948а); В. В. Сохадзе и М. Ё. Сохадзе (1952).
XII. СТЕПИ И СЕЛЬСКОХОЗЯЙСТВЕННЫЕ ЗЕМЛИ НА МЕСТЕ СТЕПЕЙ
Е. М. ЛАВРЕНКО1
Общий обзор
К степям относятся растительные сообщества, состоящие преимущественно из многолетних микротермных ксерофильных (морозо- и засухоустойчивых) травяных растений, большей частью дерновинных злаков. Основные «строители», то есть господствующие растения (эдификато-ры),— дерновинные злаки степных сообществ относятся к родам: ковыль {Stipa), типчак {Festuca), житняк {Agropyrum), змеевка {Cleistoge-nes), реже мятлик {Роа), тонконог {Koeleria) и др. Значительно реже эдификаторы степей являются корневищными растениями, как, например, вострец {Aneurolepidium pseudoagropyrum) — в Забайкалье, или относятся к травяному разнотравью, как, например, сибирская пижма {Tanacetum sibiricum) — также в Забайкалье, волосистая грудница {Linosyris vil-losa), изредка господствующая в степях юга Европейской части СССР л сев'ера Казахской ССР, и некоторые другие.
Степи при определенных условиях образуют переходы к различным другим типам растительности — лугам, высокогорным пустошам, субтропическим степям (или полусаваннам), пустыням, кустарникам, трагакант-никам (или фригане), тимьянникам и пр. В этих случаях к основным степным эдификаторам в значительном количестве присоединяются в качестве согосподствующих компонентов виды, относящиеся к иным эколого-биологическим типам растений, характерным для лугов, пустошей, пустынь и пр. Во многих случаях подобные степные сообщества переходного типа занимают обширные площади.
В пределах СССР располагается наиболее обширная степная область земного шара, заходящая и в соседние с СССР страны — Румынию, Монголию и Китай. Эта область протягивается в виде более или менее широкой зоны от нижнего течения Дуная и до северо-восточного Китая. Кроме того, в горах южной части СССР, на Кавказе, в Средней Азии, в восточной части Алтая, обширные пространства заняты степями, которые в этих горных странах образуют особые степные пояса.
Степи в основном связаны, с континентальными засушливыми частями умеренного термического порса и с соответствующими по климатическим условиям поясами в горах. Для степных районов характерны холодная зима и засушливый вегетационный период (весна—лето—осень) с жарким летом. При общей засушливости вегетационного периода максимум
1 В написании этой главы принимали участие также Л. Е. Родин, составивший разделы 83 тпш, 84 цтш, 84 тшп, 84 кпд, и Н. И. Рубцов, составивпшйд>азделы 83 стш, 84 ск, 84 стш. Ф. X. Бахтеев в общем обзоре «Степи и сельскохозяйственные земли на месте степей» сделал добавления, касающиеся сельскохозяйственных культур.
осадков в степях в большинстве случаев выпадает летом — в июне— августе. Годовое количество осадков в пределах степной зоны (в широком смысле) колеблется в СССР от 600 (запад лесостепи) до 150 мм (Забайкалье).
Безлесие большинства степей является первичным явлением и связано с рядом факторов, в первую очередь с климатическими условиями — недостатком влаги для древесных пород, что усугубляется конкуренцией! последних за влагу с травяными растениями в степях. У северной границы степей хозяйственная деятельность человека содействовала их продвижению далее на север. Современная лесостепь, несомненно, была в прошлом гораздо более облесена, чем в настоящее время.
Как указано выше, в составе степей преобладают многолетние мпкро-термные ксерофилььые травы, преимущественно дерновинные злаки.1 В преобладающем большинстве случаев степные растительные сообщества являются полидоминантными с двумя или большим числом эдификато-ров. Так, особенно широко распространены на равнине и в горах типчаково-ковыльные степи с господством типчака и одного или нескольких видов ковылей. В бассейне Енисея отчасти и в Южном Забайкалье широким распространением пользуются так называемые четырехзлаковые степи с господством четырех видов дерновинных злаков: ковыля-тырсы, типчака, степного тонконога и растопыренной змеевки.
Кроме дерновинных злаков, в состав длительновегетирующих многолетних компонентов степных сообществ входят также довольно многочисленные виды так называемого разнотравья, преимущественно, из двудольных растений. Представители разнотравья большей частью относятся к стержнекорневым растениям, лишенным способности к вегетативному разрастанию (многочисленные виды Dianthus, Arenaria, Astragalus, Peucedanum, Limonium, Salvia и др.). Однако некоторые виды разнотравья в составе степей способны к вегетативному разрастанию,, так как являются или корнеотпрысковыми (например Jurinea multiflora, Euphorbia leptocaula), или корневищными (например Iris pumila, Galium ruthenicum, Achillea setacea, Artemisia glauca, Pyrethrum millefoliatum, виды Inula). Некоторое участие в типичных степях, обычно небольшое, принимают также корневищные злаки (например Роа angustifolia, Bromus' riparius, некоторые виды Aneurolepidium, виды подрода Elytrigia рода Agropyrum) и осоки (например Carex supina, С. uralensis, С. duriuscula и др.).
Выше шла речь о длительновегетирующих видах многолетних травяных растений в составе степной растительности в ее типичном выражении. Эти виды вегетируют с весны до осени. Впрочем, многие степные дерновинные злаки в степях юга Европейской части СССР и севера Казахской ССР в наиболее жаркий период лета — в июле и августе замедляют вегетацию, находятся в состоянии «полупокоят. В этот период, после плодоношения, у них засыхают листья, обычно примерно до нижней трети листа, которая остается зеленой. Осенью, поейе дождей, листья у них снова начинают отрастать.
В составе степей, особенно на юге их зонального распространения, некоторую роль играют коротковегетирующие многолетники, так называемые эфемероиды. Уже к началу лета от эфемероидов на поверхности
1 Далее дается общая характеристика главным образом типичных (настоящих)! степей.
почвы не остается никакого следа или торчат их сухие побеги с обсеменившимися плодами. Вегетация эфемероидов приурочена к весне. Большинство степных луковичных (например виды Tulipa, Hyacinthella), клубневых (например Geranium linearilobum, Valeriana tuberosa) и клубнелуковичных (например Crocus reticulatus) эфемероидов вегетируют, цветут и плодоносят только весной. Наиболее обильный эфемероид в степях — луковичный мятлик (Poa bulbosa) вегетирует, образует соцветия с луковичками, а в редких случаях цветет и плодоносит также весной; к середине мая он засыхает, но осенью, после дождей, снова отрастает. Осенью же на поверхности почвы прорастают его луковички. Наконец, весьма немногие эфемероиды (например Colchicum laetum в бассейне нижнего Дона и в Предкавказье) цветут в августе—сентябре, а плодоносят и вегетируют весной.
Некоторую роль, в общем небольшую, в составе главным образом более южных степей играют также малолетники — двулетники и однолетники. Эти малолетники иногда называются ингредиентами и противопоставляются многолетним растениям в степях, так называемым компонентам (Пачоский, 1917).
Из двулетников в степях встречаются представители родов коровяка (Verbascum), будяка (Carduus), некоторые виды полыней (Artemisia scoparia) и др. Двулетники вегетируют в течение всего вегетационного периода.
Однолетники, как и многолетники, могут быть разделены на две группы: длительновегетирующие, или летние, и коротковегетирующие, или эфемеры. К летним озимым многолетникам относятся однолетние виды качима (Gypsophila muralis, С. stepposa), виды липучки (Lappula) и немногие другие. К яровым летним (длительновегетирующим) однолетникам относится большее число видов (например однолетние виды Polygonum, Ceratocarpus arenarius, Echinopsilon sedoides). Довольно многочисленны в более южных степях так называемые эфемеры, то есть вегетирующие в основном весной, но прорастающие осенью озимые однолетники (например из ранневесенних — Eremopyrum triticeum, Erophila verna, Alyssum desertorum, Androsace elongata, A.Turczaninovii, а из поздневесенних —Bromus tectorum, B. squarrosus и некоторые другие). К яровым ранневесенним эфемерам относится Ceratocephalus orthoceras. Эфемеры и эфемероиды характерны для степей юга Европейской части СССР и Казахстана. В степях Средней и Восточной Сибири, а также в горных степях Алтая, Центрального Тянь-Шаня и Восточного Памира эти жизненные формы почти полностью отсутствуют, что связано с климатическими условиями.
В степях иногда бывает выражен и напочвенный покров из мхов, лишайников и водорослей. Так, в северных (луговых) степях нередка имеется сплошной покров из мха туидиума (Thuidium abietinum). В более-южных степях иногда в значительном количестве встречается мох тортуля (Tortula ruralis). Хорошо выражен напочвенный ярус в некоторых южных степях, представляющих переход к пустынным сообществам. В последнем случае напочвенный ярус состоит из листостебельных мхов (Tortula desertorum), кочующих кустистых и листовых лишайников (Parmelia ryssolea, Р. vagans и др.), накипных лишайников (Diploschistes scruposu? и др.), синезеленых водорослей (Stratonostoc commune и др.).
Количество однолетников значительно увеличивается на сильно сбитых скотом степных пастбищах. Малолетники (дву- и однолетники) преобладают на молодых залежах (перелогах). На более старых залежах господствуют разрастающиеся вегетативным путем степные (а отчасти
и луговые) растения. На наиболее старых залежах, как и на целинных степях, преобладают дерновинные злаки.
Растительный покров степей обычно характеризуется, помимо полидоминантности, также большой пестротой — мозаичностью, а во многих случаях и комплексностью.1
Мозаичность растительного покрова степей зависит преимущественно ют жизнедеятельности грызунов-землероев, а в некоторых случаях от влияния на условия среды отдельных более мощных видов растений. Грызуны-землерои в местах своих поселений разрыхляют почву, выносят на поверхность массу нижележащих горизонтов, удобряют почву своими экскрементами, трупами, остатками пищи. Кроме того, грызуны не только поедают надземные части растений, но вырывают и поедают подземные части вместе с почками возобновления, и тем самым нарушают нормальную структуру степных сообществ. На местах пороев грызунов обильно поселяются степные однолетники и двулетники, а также вегетативно разрастающиеся корнеотпрысковые и корневищные растения (например из последних двух групп — Artemisia austriaca, Agropyrum repens, Aneurolepidium ramosum, A. pseudoagropyrum и др.). Вообще места пороев грызунов представляют как бы миниатюрные залежи со свойственным последним процессом смен растительности.
Разбросанные по степи кусты степных кустарников или дерновины крупного злака — чия (Lasiagrostis splendens) также создают под сенью своих побегов условия, несколько отличные от условий открытых участков, и тем самым более или менее резко выраженное горизонтальное расчленение растительного покрова — мозаичность тех или иных степных ассоциаций с участием чия или кустарников.
Микрокомплексность растительного покрова также может вызываться пороями грызунов, если отдельные порой занимают более крупную площадь (в несколько кв. м или десятков кв. м), что, например, имеет место, когда по степи равномерно разбросаны холмики байбаковин или сусли-ковин. Растительный покров последних обычно представлен особыми ассоциациями, отличными от тех, которые занимают участки между холмиками грызунов. Уже в. этом случае причиной, вызывающей появление комплексности растительности, являются условия рельефа и отличия в почве, появившиеся в результате деятельности грызунов. И во многих других случаях непосредственной причиной частой смены в пространстве ассоциаций в степях бывают соответствующие смены почв и микрорельефа, обусловленные разными причинами. Далее приводятся многочисленные примеры подобной микрокомплексности растительного покрова степей.
Следует также упомянуть о том, что растительный покров степей очень изменчив в связи с изменениями метеорологических условий, главным образом в связи с чередованием более влажных и более засушливых годов или целых десятилетий, а также в связи с деятельностью грызунов-землероев. О воздействии последних на растительность степей уже сказано выше. Это воздействие бывает особенно велико и распространяется на весьма обширные площади в результате массового размножения опре- 1 * з
1 Растительный покров бывает комплексным, когда на протяжении не-
скольких метров или десятков метров одни ассоциации сменяются другими, или м о-
з а и ч н ы м, когда в составе одной и той же ассоциации наблюдается горизонтальное расчленение растительного покрова. В последнем случае отдельные мозаичные шина в ассоциации измеряются (при их пересечении) дециметрами или только немногими метрами, в связи с чем наблюдается взаимное влияние (главным образом в почве) растений, произрастающих в разных, но соседних участках мозаичной ассоциации. Между микрокомплексностью и мозаичностью растительности наблюдаются переходы.
.деленных видов грызунов, преимущественно полевок и пеструшек, что наблюдается примерно раз в 10 лет.
Как указано выше, растительный покров степей состоит в основном из многолетних растений, которые, как показали исследования последних лет, достигают в степях большого возраста — в несколько десятков лет, а иногда больше ста лет. В случае чередования сухих и влажных годов или десятилетий многолетняя часть степных сообществ обычно реагирует различным фенологическим развитием многолетних компонентов, в первую очередь дерновинных злаков. Так, например, ковыли, особенно перистые, в более или менее влажные годы могут развиваться очень пышно и обильно цвести и плодоносить; в засушливые годы ковыли развиваются гораздо слабее, плохо цветут или вовсе не развивают генеративные органы. Иначе ведут себя малолетники; количество их особей при чередовании •сухих и влажных лет резко меняется, особенно в более южных степях. Во влажные годы часто наблюдаются «волны жизни» малолетников, когда в степях в изобилии появляются двулетники (например Melilotus officinalis, Artemisia scoparia) или однолетники (например Eragrostis minor и др.).
Перейдем теперь к типологии степей. При выделении типов степей, как принято в отечественной литературе, учитываются не только господствующие, но и согосподствующие виды.
1. Типичные (настоящие) степи характеризуются господством евксерофильных и евриксерофильных степных растений; •согосподствующие виды имеют такую же экологию. Выше уже дана общая характеристика типичных (настоящих) степей. Из видов дерновинных злаков чаще всего господствуют ковыль-тырса {Stipa capillata), красный ковыль {S. rubens), украинский ковыль (S. ucrainica), Лессингов ковыль (61. Lessingiana), днепровский ковыль (S. borysthenica — осо-•бая песчаная раса Stipa Joannis s. 1.), типчак (виды из ряда Sulcatae, чаще Festuca sulcata), степной овсец {Helic to trichon deser torum), растопыренная змеевка {Cleistogenes squarrosa), гребневидный житняк {Agropyrum pectiniforme), гребенчатый житняк (Л. cristatum), а в горах Средней Азии также расползающийся мятлик {Роа relaxa), киргизский ковыль •{Stipa kirghisorum) и некоторые другие.
На «Геоботанической карте» типичные (настоящие) степи показаны дифференцировано: разнотравно-дерновиннозлаковые европейско-казахстанские степи (76), дерновиннозлаковые европейско-казахстанские степи (77), мелкодерновинные алтайско-забайкальские степи (80) и змеев-ково-тырсовые и вострецово-тырсовые енисейско-забайкальские степи (81).
Разнотравно-дерновиннозлаковые (76) и мелкодерновинные (80) степи, особенно первые, представляют собой в известно# мере переход от типичных степей к луговым степям. Следует также отметить, что европейско-казахстанские степи (76 и 77) отличаются от алтайско-забайкальских (80) и енисейско-забайкальских (81) участием в составе первых некоторого, иногда значительного количества эфемероидов и эфемеров, почти полностью отсутствующих в алтайско-забайкальских и енисейско-забайкальских степях.
2. Луговые степи менее засухоустойчивы, чем предыдущие. В составе этих степей к эдификаторам — мезоксерофильным и евриксеро-фильным злакам постоянно примешаны в значительном количестве многолетние ксеромезофильные и евмезофильные растения, главным образом из разнотравья и корневищных злаков. В составе луговых степей, которые особенно характерны для лесостепи, из дерновинных злаков и осок чаще всего преобладают ковыли {Stipa Joannis, S. stenophylla,
S. capillata), типчак (Festuca sulcata и близкие виды). В степях Забайкалья и северо-восточной Монголии в составе луговых степей часто господствует сложноцветное — пижма (Tanacetum sibiricum). В горах Средней Азии в луговых степях из дерновинных злаков преобладают, помимо Stipa Joannis, S. capillata и Festuca sulcata, также Stipa kirghisorum и Poa relaxa.
Многие согосподствующие в луговых степях виды разнотравья и корневищных злаков характерны также для остепненных лугов (например Роа pratensis, Agrostis Syreistschikovii, A. Trinii, Filipendula hexapetala, Trifolium montanum, Artemisia tanacetifolia и многие другие).
Луговые степи представляют собой переход от степей к лугам.
На «Геоботанической карте» показаны луговые степи и остепненные луга в сочетании с участками лесов (75). К луговым степям следует относить, как указано выше, также пижмовые (из Tanacetum sibiricum} забайкальские степи (79).1
3. Пустынные степи представляют собой наиболее ксеро-фитный тип степей в СССР.1 2 В составе этих степей наблюдается постоянная значительная примесь к евксерофильным и евриксерофильным дер-новинным злакам евксерофильных полукустарничков (главным образом из родов Artemisia — преимущественно подрода Seriphidium, Kochia,. Pyrethrum и др.), которые играют существенную роль в их структуре. Господствуют в травостое пустынных степей обычно следующие дерно-винные злаки: ковыли (Stipa sareptana, S. capillata, S. Lessingiana), типчак (Festuca sulcata), житняки (Agropyrum pectiniforme, A. desertorum, а на песках также A. sibiricum), а в горных пустынных степях Алтая и Средней Азии, кроме того, ряд других видов ковылей (Stipa turkestanica, S. caucasica, S. orientalis, S. glareosa).
Пустынные степи представляют собой переход от степей к пустыням.
На «Геоботанической карте» пустынные степи показаны в виде двух обозначений: полынно-дерновиннозлаковые европейско-казахстанские-степи в сочетании с пустынными сообществами (78) и ковыльковые и полу-кустарничково-ковыльковые центральноазиатские пустынные степи (82). Последние показаны в Тувинской автономной области (в Убсанурской котловине), на востоке Алтая (Чуйская степь), в Центральном Тянь-Шане и на Восточном Памире. Как указано далее, центральноазиатские пустынные степи отличаются от европейско-казахстанских не только составом господствующих и согосподствующих видов, но и почти полным отсутствием эфемероидов и эфемеров.
Пустынные степи под влиянием хозяйственной деятельности человека, а именно неумеренной пастьбы скота, несомненно расширяют свое распространение, преимущественно в северном направлении, за счет типичных дерновиннозлаковых степей.
Типичные, луговые и пустынные степи занимают преобладающие территории в пределах степной зоны (области); широко распространены они также в предгорьях и горах Кавказа (особенно Закавказья), Средней Азии, Алтая и Забайкалья.
Существует еще целый ряд типов степей; некоторые из них встречаются как в горах, так и на равнине, другие же приурочены исключительно к горным странам. На карте эти типы степей особо не выделены, так как этого не позволяет масштаб карты.
1 Ранее мы относили пижмовые степи к настоящим (типичным) степям (Лавренко, 1940а), что считаем сейчас неправильным.
2 Ранее мы называли пустынные степи «опустыненными» степями (Лавренко,
На равнине и в горах встречаются кустарниковые и тимьянниковые степи.
4. Кустарниковые степи характеризуются наличием равномерно разбросанных на фоне степного травостоя кустов степных кустарников (главным образом из родов Caragana и Spiraea). Так, на востоке Казахской ССР (в Казахском мелкосопочнике, предгорьях Тарба-гатая и южного Алтая), в Тувинской автономной области, в Бурят-Монгольской АССР, а также в Монгольской Народной Республике встречаются кустарниковые степи с Caragana pumila, С. leucophloea, С. pygmaea, С. Bungei и С. microphylla. Таким образом, структура кустарниковых степей усложнена наличием кустарниковой жизненной формы, что, кроме того, обусловливает горизонтальное расчленение растительного покрова (формирование мозаичных ассоциаций). Кустарниковые степи связаны преимущественно с легкими — щебнистыми и супесчаными почвами.
Кустарниковые степи нельзя смешивать с зарослями степных кустарников, которые представляют собою более или менее густые скопления степных кустарников из тех же родов {Spiraea, Caragana, а также Cerasus, Amygdalus, Prunus).1 Таким образом, кустарниковые степи представляют собой переход от степей к зарослям степных кустарников.
На карте кустарниковые степи и заросли степных кустарников в местах их широкого распространения показаны одним и тем же внемасштаб-ным знаком.
5. Тимьянниковые степи отличаются значительным участием в степном травостое (с преобладанием дерновинных злаков) ксерофильных полукустарничков, в большинстве случаев имеющих надземные приподымающиеся или стелющиеся по поверхности почвы побеги, образующие придаточные корни на прижатых к почве участках многолетних деревянистых побегов. Наиболее обычны полукустарнички из семейства губоцветных, особенно обилен тимьян или чебрец (весьма многочисленные виды Thymus), а также дубровник (например Teucrium ро-Нит), виды шлемника {Scutellaria, например из группы S. orientalis s. ampl.), зизифора {Ziziphora) и другие, а также и из других семейств.
Тимьянниковые степи приурочены преимущественно к более или менее каменистым склонам; иногда встречаются также на песках со слабо развитыми почвами. Эти степи довольно широко распространены в пределах степной зоны, где они приурочены к более повышенным районам, например к Средне-Русской и Приволжской возвышенностям, Донецкому кряжу, Высокому Заволжью, предгорьям Южного Урала, низко-горьям Мугоджар, Казахскому мелкосопочнику, горным степям Южного Забайкалья. Эти степи, вместе с тимьянниками, наиболее широко распространены в тех районах, где на карте показан в пределах степной зоны знак «растительности каменистых обнажений разных пород и петрофитные варианты степей и пустынь». Кроме того, тимьянниковые степи и тимьян-ники широко встречаются в горах Крыма, Кавказа и Средней Азии.
Тимьянниковые степи представляют собой переход от степей к тимьян-никам, типу растительности, свойственному лишенным почвенного покрова каменистым субстратам и отчасти пескам.1 2 В результате хозяйственной
1 Некоторые авторы, например С. И. Коржинский (1901), называют кустарниковой степью подобные заросли степных кустарников.
2Тимьянникн близки к пустыням, в составе которых также преобладают полукустарнички, но последние в большинстве случаев лишены способности укореняться своими надземными побегами на поверхности субстрата. Между тимьянниками и пустынями имеются переходы.
Ранее (Лавренко, 1940а) мы относили тимьянниковые степи к опустыненным степям.
деятельности человека, способствующей в ряде случаев развитию эрозионных процессов (смыв почвы и обнажение каменистых или песчаных субстратов), тимьянники и тимьянниковые степи несомненно сильно расширили площадь своего распространения.
Нижеследующие типы степей встречаются в СССР только в горах.
6. Полусаванновые степи отличаются значительной, примесью к степным дерновинным злакам (ковылям, типчаку и др.) видов, характерных для субтропических степей или полусаванн, в первую очередь одного из основных эдификаторов последних — волосоносного пырея {Agropyrum trichophorum), а также видов эфемероидного разнотравья (из pojsfibFerula, Eremostachys, Phlomisajip.), весьма характерных для полусаванн, широко развитых в предгорьях и низкогорьях Западного Тянь-Шаня, Памиро-Алая, Копет-Дага и частично Восточного Закавказья.
Эти степи распространены в Западном Тянь-Шане и Памиро-Алае, в полосе контакта между мезотермными субтропическими степями и микротермными дерновиннозлаковыми степями, которые в горах Средней Азии расположены по горному профилю выше первых.
7. Колючетравные степи характеризуются значительной примесью к степным дерновинным злакам (главным образом типчаку)-ксерофильных колючих многолетних трав из рода кузиния (Cousinia'). Последние образуют в горах Средней Азии, в Западном Тянь-Шане и особенно? в западной части Памиро-Алая самостоятельные группировки.
8. Трагакантовые степи отличаются значительной примесью к степным дерновинным злакам (ковылям, типчаку и др.) ксерофильных колючих кустарничков (главным образом из родов Astragalus и Acantholimon). Эти степи представляют переход от степей к трагакант-никам (фригане).
Трагакантовые степи, как и трагакантники, распространены в засушливых районах Кавказа (главным образом в Южном Закавказье — Армении, Нахичеванской АССР и др.) и в южной части Средней Азии (Памиро-Алае, Западном Тянь-Шане).
9. Пустошные (криофильные) степи характеризуются тем, что к основным степным дерновинным злакам (типчаки, мятлики, овсецы) в них примешиваются в значительном количестве компоненты высокогорных пустошей1 — дерновинные кобрезии и злаки (Cobre-sia, Ptilagrostis mongholica) и разнотравье. Таким образом, пустошные-степи являются переходом от степей к высокогорным пустошам.
Пустошные (высокогорные) степи встречаются у верхней границы, степей в восточной части Памиро-Алая и Тянь-Шаня, в Джунгарском Алатау и в восточной части Алтая. , , ' 1
Остановимся вкратце на основных географических закономерностях в распределении степной растительности в СССР.
От западных границ СССР до Алтая и Саура простирается сплошной, широкой полосой степная зона (область). На западе степная зона переходит в пределы Румынии, достигая там лесистых предгорий Карпат, а на востоке простирается далеко вглубь Монголии и Китая.
В СССР в пределах степной зоны в широком смысле можно выделить, четыре подзоны: 1) лесостепь или подзону луговых степей, 2) подзону
1 Многие авторы не отличают высокогорные пустоши от альпийских лугов..
разнотравно-дерновиннозлаковых степей, 3) подзону дерновиннозлаковых (сухих) степей и 4) подзону полынио-дерновиннозлаковых (пустынных) степей.
Рис. 61. Основные типы ареалов степных растений.
7 — паннонский вид Carex stenophylla Wahlenb.; 2—причерноморско-казахстанский вид С. uralensis С. В. Clarke; 3 — казахстанско-сибирско-монгольский вид С. dvriuscula С.А.М. (По В. И. Кречетовичу, из Д. Е. Янишевского, 1937).
Рис. 62. Основные типы ареалов степных растении: казахстанско-среднеазиатский вид Stipa kirghisorum Р. Smirn. (По Р. Ю. Рожевицу и др.).
Рис. 63. Основные типы ареалов степных растений.
1 — казахстанский (заволжско-казахстанский) вид Stipa Korshinshyi Roshev. (ориг.); 2 — даурско-монгольский олиготипный степной род Panzeria. (По Л. А. Куприяновой, Ботанические материалы Гербария Ботанического института АН СССР, т. 15, 1953).
Лесостепная подзона или подзона луговых степей географами чаще рассматривается как особая зона. В почвенном покрове этой подзоны преобладают тучные, мощные, выщелоченные и деградированные черноземы, темносерые и серые лесные
почвы, а в пониженных районах, особенно в Западной Сибири, также засоленные почвы. До распашки, в период скотоводческого (номадного) хозяйственного использования территории лесостепи, в пределах последней большая часть площади была занята луговыми степями и остепнен-ными лугами, но весьма значительную территорию занимали также лиственные леса: широколиственные — в Европейской части СССР и мелколиственные — в Западной Сибири. Местами леса даже преобладали над степями и остепненными лугами. В настоящее время леса в лесостепи сохранились на небольшой площади: территория лесостепи весьма значительно обезлесена в результате хозяйственной деятельности человека (влияния степных пожаров, пастьбы скота, рубки и корчевки лесов); особенно сильно уменьшилась лесопокрытая площадь в последний период широкого земледельческого освоения лесостепи. В настоящее время вся площадь лесостепи, кроме сохранившихся лесных массивов, часть пойменных и низинных (в озерных депрессиях) лугов распаханы и заняты культурными угодьями, главным образом зерновыми и некоторыми техническими культурами. v
Среди зерновых злаков в лесостепи преобладают озимая и яровая пшеницы, озимая рожь, яровой ячмень, овес, кукуруза; из зернобобовых наиболее распространены горох, фасоль, чечевица. В качестве крупяных широко возделываются просо, гречиха. Масличные и технические растения представлены подсолнечником, масличным льном, горчицей, озимым рапсом, рыжиком, маком, в незначительных размерах периллой. Широко возделываются картофель, сахарная свекла, кормовые корнеплоды, огородные, бахчевые, а также и плодово-ягодные. Среди последних наибольшее значение имеют яблоня, груша, вишня, слива, малина, смородина, крыжовник. Именно в этой подзоне находится Мичуринск, где И. В. Мичурин провел замечательные экспериментальные работы по созданию новых сортов яблонь, груш, вишен, винограда и других плодово-ягодных культур, которые могут выращиваться в средней полосе Европейской части Союза, в том числе в лесостепи, а также и севернее.
Подзона разнотравно-дерновиннозлаковых или, точнее, *р а з н о]т равно-типчаково-ковыльных степей так же, как и лесостепь, простирается сплошной полосой от Молдавской ССР до предгорий Алтая. В почвенном покрове этой подзоны преобладают обыкновенные и частично южные черноземы. В этой подзоне в доагрикультурный период господствовали преимущественно |разнотравно-типчаково-ковыльные степи. Леса, видимо, всегда занимали подчиненную роль и были приурочены к долинам рек и балкам. В ’настоящее время в Европейской части СССР эти степи целиком распаханы; в Азиатской^части СССР, главным образом в северной части Казахской ССР, еще местами сохранились целинные степи, преимущественно на солонцеватых черноземах; они (используются как сенокосы и пастбища.
В пределах этой подзоны расположены наиболее крупные массивы посевных площадей нашей страны, производящие ббльшую долю из общего сбора зерновых, зернобобовых и других культур. Список возделываемых сельскохозяйственных растений состоит в основном из тех же названий, которые перечислены и по отношению к предыдущей подзоне. Кроме того, в районе Северного Кавказа возделывают рис, озимый ячмень, клещевину, ляллеманцию, сорго, нут. Среди плодово-ягодных культур, кроме широкораспространенных пород, в Европейской части подзоны (к западу от Волги) имеются: абрикос, алыча, черешня, виноград и другие; растет также тутовое дерево.
Согласно постановлению ЦК КПСС и Совета Министров СССР значительные площади разнотравно-дерновиннозлаковых степей, находящиеся в Западной Сибири, северном Казахстане и на Алтае, исчисляемые миллионами гектаров, осваиваются под зерновые культуры (пшеница, кукуруза, просо, зернобобовые и др.).
Ирдзопа дерн овин и о злаковых или, точнее, г и и ч а к о в о - к о в ы л ь н ы х степе й сплошной полосой простирается от Восточного Предкавказья, нижнего Дона и Заволжья вплоть до предгорий Алтая и Тарбагатая. Кроме того, имеется большой изолированный массив этой подзоны на юге Европейской части СССР, в Причерноморье. В почвенном покрове преобладают южные черноземы и главным образом темнокаштановые и каштановые более или менее солонцеватые почвы. В Европейской части СССР эти степи почти целиком распаханы и заняты посевами озимой пшеницы, твердой и мягкой яровой пшеницы, ячменя, овса, озимой ржи, проса, кукурузы, нута, фасоли, вики, гороха, чечевицы, чины, сои, подсолнечника, масличного льна, горчицы, клещевины, суданской травы, могара, сорго, сахарной свеклы, картофеля, арбузов, дынь, огородных и плодово-ягодных культур. В Азиатской части СССР, главным образом в Казахской ССР, на значительной территории подзоны еще сохранились типчаково-ковыльные целинные степи, используемые главным образом как пастбища.
На юге подзоны местами значительно развита микрокомплексность почвенно-растительного покрова; на сильно солонцеватых каштановых почвах и на солонцах появляются пятна пустынных степей и даже пустынных растительных сообществ. Леса в этой подзоне встречаются почти исключительно в долинах рек и на песках.
Две последние подзоны можно объединить в одну — подзону типичных (настоящих) степей, как это сделано на «зональной» врезке на «Геоботанической карте».
Подзона пустынных полый но-дер и оВинно-злаковых или, точнее, полынно-типчаково-ковыльных степей тянется сплошной полосой от Восточного Предкавказья и Ергеней до предгорий Тарбагатая. На юге Украины и на севере Крыма имеется изолированная узкая полоска этих степей. В пределах подзоны на более или менее солонцеватых светлокаштановых почвах развиты пустынные, преимущественно полынно-типчаково-ковыльные и полынно-типчаковые степи. На массивах солонцов и сильно солонцеватых светлокаштановых почв преобладают пустынные растительные сообщества с отчетливо выраженными признаками остепнения. Для этой подзоны очень характерна резко выраженная микрокомплексность растительного покрова. В пределах подзоны господствуют целинные степи, используемые преимущественно как пастбища; только относительно небольшая площадь этих степей распахана.
На распаханных участках, как правило, возделываются зерновые культуры (озимая и яровая пшеницы, озимая рожь, овес, кукуруза), в качества крупяной культуры широко применяются засухоустойчивые сорта проса. Значительно меньшие площади посева занимают горох, чина, чечевица, подсолнечник, масличный лен и горчица. В качестве кормовых сеяных трав распространены суданская трава, могар, сорго, люцерна, житняк и др.
Леса в этой подзоне встречаются исключительно в долинах рек и небольшими участками в котловинах среди песков.
Вне СССР степи занимают склоны (вплоть до высокогорного пояса) и предгорные равнины Монгольского Алтая, Западную котловину боль-
10 Загса.') 551
ших озер, Гобийскую долину озер, обширное нагорье Хангая, Восточно-Монгольское плоскогорье (приподнятые равнины). Далее на восток степи, огибая Большой Хинган, уходят в северо-восточный Китай (внутреннюю Маньчжурию). Из Монголии степи заходят в пределы СССР двумя языками: 1) в Бурят-Монголию, между Хамар-Дабаном и Хэнтеем, по Селенге, и 2) к востоку от Хэнтея, в Читинскую область^по Онону и Аргуни.
На юге Сибири среди лесистых гор имеется ряд степных и лесостенных островов, приуроченных к межгорным депрессиям: Ачинско-Мину-синский лесостепной и степной, Тувинский степной, Красноярский лесостепной и степной, Канский лесостепной, Балаганско-Иркутский лесостепной, Баргузинский степной, Еравнинский лесостепной и другие, более мелкие степные и лесостепные «острова».
Система подзон в восточной части степной зоны (области) к востоку от советской части Алтая отличается от вышеописанной, характерной для европейско-западносибирско-казахстанской части степной зоны.
Облесенность Южной Сибири в значительной мере обязана горному рельефу. Вышеупомянутые лесостепные острова на юге Сибири можно рассматривать как фрагменты лесостепной подзоны. Обширное нагорье Хангая и предгорья Хэнтея заняты своеобразной горной лиственничной лесостепью с широким распространением мелкодерновинных степей (опис. 80). В пределы Селенгинского и главным образом Читинского степных районов заходят из Монголии восточные аналоги луговых степей — пижмовые степи (из Тапасе turn sibiricum) (опис. 79).
Подзона разнотравно-типчаково-ковыльных степей к востоку от Алтая отсутствует, хотя отдельные фрагменты последних имеются. К югу от Хангая и Хэнтея располагается подзона сухих дерновиннозлаковых, в основном змеевково-тырсовых степей (опис. 81), которые особенно обширные площади занимают уже вне СССР, на Восточно-Монгольском плоскогорье, откуда уходят на восток в Китай, а вдоль Селенги заходят как бы заливом в пределы Бурят-Монголии; между реками Ононом и Аргунью эти степи заходят также на юг Читинской области. Эти же стели (обычно с участием типчака — так называемые «четырехзлаковые» степи сибирских ботаников) развиты в Минусинской и Тувинской котловинах.
К югу от подзоны дерновиннозлаковых (змеевково-тырсовых) степей располагается подзона пустынных степей, которая в Монголии представлена весьма своеобразными ковыльково-луковыми и полукустарничково-ковыльковыми пустынно-степными сообществами (ковыли из ряда Ваг-batae). Подобные ковыльковые и полукустарничково-ковыльковые центральноазиатские пустынные степи (опис. 82) в пределах СССР выражены только в окраинных его частях, граничащих с Монгольской Народной Республикой и Китайской Народной Республикой: по северной окраине Убсанурской котловины, в Чуйской степи на Алтае, в Центральном Тянь-Шане ина Восточном Памире. В Монгольской Народной Республике эти степи образуют довольно широкую подзону, окаймляющую с севера пустыни Гоби. Из Монголии они заходят на восток, в пределы Китая.
Уже из вышеизложенного следует, что всю сплошную степную зону (или область) Евразии, протянувшуюся от степной части Румынии до степной части северо-восточного Китая, можно в меридиональном направлении разделить на две части (подобласти): 1) Причерноморско-Казахстанскую, охватывающую степную область до Алтая и Саура, и 2) Центрально-Азиатскую степную или Даурско-Монгольскую, включающую
восточную часть области, примерно к востоку от советской части Алтая (Лавренко, 1947, 1954).1
Как указано выше, эти подобласти отличаются набором («спектром») подзон. Кроме того, для Причерноморско-Казахстанской подобласти характерно следующее: наличие в лесостепи преимущественно лиственных лесов (сосняки только на легких почвах), преобладающая роль в травостое степей перистых ковылей (из ряда Реп-natae), значительное развитие в травостое более южных степей эфемероидов (особенно Роа bulbosa) и эфемеров. В пределах этой подобласти выделяются четыре основные провинции: в лесостепной части — Европейская лесостепная провинция с широколиственными (преимущественно дубовыми) лесами и Западно-Сибирская лесостепная провинция с мелколиственными лесами;1 2 в степной части — Причерноморская и Казахстанская степные провинции, отличающиеся различным составом эдификато-ров и других компонентов степей. Каждая из этих степных провинций состоит из трех секторов соответствующих степных подзон (см. выше).
' Для Ц.ентрально-Азиатской или Даурско-Монгольской степной подобласти характерно наличие в лесостепной части преимущественно лиственничных и сосновых лесов, преобладание в травостое степей тырсовидных (ряд Capillatae), а в подзоне пустынных степей — бородатых (ряд Barbatae) ковылей, почти полное отсутствие в степном травостое эфемероидов и эфемеров и ряд других отличий в составе и структуре степей. В пределах этой подобласти можно выделить провинции: Хангайско-Даурскую горнолесостепную, Маньчжурскую лесостепную и Монгольскую степную.3 В пределы СССР заходят только первая и частично третья провинции.
\ В настоящее время большая часть степей в пределах Европейской части СССР распахана. В Азиатской части СССР, как указано выше, также распахана значительная площадь степей, но часть последних еще сохранилась в целинном, а местами в залежном состоянии и используется как весьма ценная природная кормовая площадь.
Литература
Ф. Рупрехт (1866); В. В. Докучаев (1883, 1892); С. И. Коржинский (1888—1891, 1901); А. Н. Краснов (1891, 1894а); Г. И. Танфильев (1894а, 1896, 1898, 1902); А. Я. Гордягин (1900—1901); Г. Н. Высоцкий (1905, 1915); И. К. Пачоский (1908, 1917); В. В. Алехин (1915, 1931, 1936); П. Н. Крылов (1915, 1916); Б. А. Келлер (1916, 1923а); К. М. Залесский (1918а); И. М. Крашенинников (1923, 1925); Е. П. Коровин (1934); Г. И. Дохман (1937); И. В. Новопокровский (1937); Д. Е. Янишевский (1937); Е. М. Лавренко (1940а, 1947, 1954); Л. С. Берг (1947, 1952); П. Н. Овчинников (1947, 1955); А. А. Гроссгейм (1948); А. А. Юнатов (1950, 1954); Н. Ф. Комаров (1951).
75. Луговые степи, остепненные луга в сочетании с участками лесов (лесостепь)
Луговые степи и остепненные луга характерны для лесостепи, где они встречаются в зональных условиях, то есть на плакорах, на грунтах различного механического состава.
1 Далее при характеристике состава степей различных частей СССР указывается для части видов., преимущественно господствующих, географический тип их ареала. Это сделано для того, чтобы подчеркнуть региональные («провинциальные») различия степей в пределах степной зоны (рис. 61—63). Для горных степей это не сделано.
2 Имеются отличия также и в составе луговых степей и остепненных лугов этих двух лесостепных провинций (см. далее).
3 Провинции, выделенные А. А. Юнатовым (1950) в пределах лесостепной и степной частей Монгольской Народной Республики, следует рассматривать как подпровинции, которые выделяются и для провинций Причерноморско-Казахстанской подобласти (Лавренко, 1947).
Лесостепная зона простирается в виде то более широкой, то более узкой полосы от западных границ СССР, бассейнов Прута и Днестра, до Салаирского кряжа в Сибири. Далее к востоку, вплоть до бассейна верхнего Амура (Восточное Забайкалье), лесостепь встречается только островами. Это связано с тем, что в этих районах Южной Сибири, благодаря горному рельефу, леса далеко заходят к югу вглубь степной области. Лесостепные ландшафты здесь приурочены к внутренним горным депрессиям и долинам рек. Кроме того, отдельные небольшие участки лесостепных ландшафтов имеются и в глубине лесных континентальных областей, в Центральной Якутии и в депрессии оз. Ханка на Дальнем Востоке.
Хозяйственная деятельность человека — земледельца и особенно скотовода — в прошлом содействовала расширению территории, занятой лесостепью. Это относится даже к районам в основном реликтовой лесостепи в Центральной Якутии.
Для луговых степей характерно господство преимущественно мезо-ксерофильных видов с большей или меньшей примесью ксеромезофильных, евриксерофильных и евксерофильных, а для остепненных лугов — преобладание ксеромезофильных видов, отчасти с примесью овриксерофиль-ных, мезоксерофильных и евмезофильных растений. Таким образом, луговые степи и остепненные луга представляют собой переход от настоящих степей к лугам. Состав и вообще структура луговых степей и остепненных лугов сильно изменяются при движении с запада на восток, от бассейнов Прута и Днестра до верхней части бассейна Амура. В связи с этим в пределах обширной, вытянутой в широтном направлении полосы луговых степей и остепненных лугов на «Геоботанической карте» с помощью литерных обозначений выделены европейские, западносибирские, центральносибирские и забайкальские луговые степи и остепненные луга; кроме того, на карте, как указано выше, отмечены луговые степи и остепненные луга лесных областей в Центральной Якутии и в депрессии оз. Ханка.
Имеется довольно много общего в составе луговых степей и остепненных лугов Восточной Европы и Западной Сибири. Состав луговых степей и остепненных лугов юга Центральной Сибири (бассейн верхнего Енисея) и Забайкалья резко отличается от состава аналогичных сообществ более западных районов лесостепи.
75 яп — луговые степи и остепненные луга лесных областей (Центральная Якутия, Приморье)
Остепненные луга и луговые степи занимают в Центральной Якутии, в бассейне средней Лены, небольшую площадь, преимущественно по надпойменным террасам некоторых рек и по южным крутым склонам коренных берегов последних, а также по периферии приозерных (аласных) понижений. На' карте эти степи и остепненные луга показаны по левобережью Лены у Якутска и по Татте (левый приток Алдана).
С этими остепненными лугами и луговыми степями связаны черноземно-солонцеватые почвы. Часть остепненных лугов и луговых степей возникла по периферии приозерных впадин после спуска озер, а также на месте сведенных лесов.
Для центральноякутских остепненных лугов и луговых степей характерно обильное развитие степных дерновинных злаков (главным образом Festuca lenensis и Koeleria gracilis). Местами преобладают виды ко-выля-тырсы (Stipa decipiens, реже S. baicalensis1). На надпойменной тер
1 Stipa baicalensis Roshev.^5. attenuate P. Smirn.
расе Амги доминирует вострец (Aneurolepidium pseudoagropyrum). Реже по южным склонам коренных берегов рек, встречаются Helictotrichon Krylovii (эндем Центральной Якутии), Cleistogenes squarrosa, Agropyrum cristatum. Нередко обильна степная осочка (Carex duriuscula). Бобовые встречаются в небольшом количестве (более обычны Astragalus inopi-natus и Onobrychis sibirica). Богато представлено разнотравье, которое часто преобладает в составе остепненных лугов; особенно обильны полыни (Artemisia commutata — к востоку от Лены, A. pubescens — в долине Лены, A. rupestris — в привилюйских районах, а также Artemisia laciniata). Иногда на склонах встречается полукустарничковая полынь (Artemisia frigida). Часто также встречаются Pulsatilla angustifolia, Eritrichium pectinatum s. 1., Aster alpinus, Galium verum, Veronica incana, Gentiana decumbens и др. Обычно присутствуют горные растения (Cobre-sia filifolia, Papaver nudicaule и др.). Нередко развит напочвенный покров из лишайников (Parmelia vagans), синезеленых водорослей (Strato-nostoc commune) и мхов (Tortula ruralis). Эфемеры и эфемероиды отсутствуют. Часто разбросаны единичные деревья лиственницы (Larix dahuricd), березы и кустарников (главным образом Spiraea media).
Эти остепненные луга и луговые степи близки по составу и структуре к аналогичным образованиям лесостепных и степных районов Забайкалья и Монголии. Они используются как весенние и раннелетние пастбища. Значительные площади, занятые ранее остепненными лугами и луговыми степями, в настоящий момент распаханы.
Вышеописанные степные луга и луговые степи нередко сочетаются с влажными карбонатными солончаками, на которых произрастают гало-фитные луга с участием Puccinellia tenuiflora, Heleocharis euuniglumis, Alopecurus ventricosus, Beckmannia syzigachne, Carex enervis, а на сухих солончаках, частично лишенных растительности, встречаются главным образом Puccinellia tenuiflora, Agropyrum repens, Suaeda corniculata. Часто такие остепненные луга и луговые степи перемежаются с лиственничными и сосновыми лесами.
Отдельные массивы остепненных лугов и луговых степей встречаются на юге Приморского края, в пределах Ханкайско-Уссурийской равнины (впадины). На «Геоботанической карте» небольшие массивы луговых степей и остепненных лугов показаны к западу и к югу от оз. Ханка и по правобережью Суйфуна.
К верхним увалистым уровням и нижним частям делювиальных склонов возвышенностей приурочены остепненные разнотравно-злаковые луга с кустарниками, полынно-разнотравные и кустарниково-разнотравные луговые степи; из полыней очень характерна луговостепная Artemisia laciniata. Среди луговых степей в бассейне Турги, Бол. Усачи, Синтухи отмечены группировки с сибирской пижмой (Tanacetum sibiricum). К склонам и вершинам увалов и к делювиальным склонам возвышенностей приурочены также заросли разнолистной лещины (Corylus heterophylla) с небольшой примесью поросли дуба (Quercus mongolica), даурской или черной березы (Betula dahurica), осины Давида (Populus Davidiana) и др. Эти заросли возникли на месте своеобразных остепненных лещиновых черно-березово-дубовых редколесий, встречающихся в настоящее время в неизмененном виде редко. По среднему течению Синтухи (в районе с. Комиссарове) и в истоках Бол. Усачи в травяном покрове подобных дубовых редколесий господствует представитель даурско-монгольских луговых степей — сибирская пижма (Tanacetum sibiricum), здесь встречаются и другие степняки (Carex duriuscula, Cleistogenes sinensis, Scutellaria baicalensis, Saposhnikovia divaricata n др.).
По теневым склонам выше 250 м над у. м. располагается поросль монгольского дуба, даурской березы, липы (Tilia mansharica и Т. Та-queti) и мелколистного клена (Acer mono).
Южные выпуклые склоны и гребни возвышенностей заняты петрофит-ными разностями кустарниково-разнотравных и злаково-разнотравных степей, а именно змеевково-тонконогово-разнотравными (с Cleistogenes sinensis, Koeleria gracilis) и типцово-разнотравными степями с несомкнутым покровом. В подобных условиях местами встречаются группировки с тырсой (Stipa baicalensis) — долина Синтухи у с. Комиссарове. На близких местообитаниях (Ханкайский, Гродековский и Ивановский районы) растут рощи погребальной сосны (Pinus funebris).
Все эти остепненнолуговые и луговостепные группировки по составу близки к даурско-монгольским степям.
Участки с вышеописанной растительностью, приуроченной к увалам и нижним частям делювиальных склонов возвышенностей, характеризуются неустойчивым режимом увлажнения. После выпадения атмосферных осадков, особенно во влажные годы, почвы достаточно увлажнены, а местами даже временно перенасыщены влагой; в сухие сезоны (весна, осень) и в засушливые годы почти пересыхают. В последнее время почвы под этой растительностью относят к серым осолоделым и серым оподзоленным почвам; в прошлом первые, видимо, пережили луговую солонцевато-солончаковатую стадию (Ливеровский, 1946, 1947).
В большинстве случаев территории, занятые в недалеком прошлом описанными выше степями, остепненными лугами, зарослями кустарников и редколесьями, в настоящий момент распаханы и заняты пашнями, залежами, а также выпасами. Хозяйственная деятельность человека в этом районе не благоприятствует лесной растительности, но способствовала, а частью способствует и теперь остепнению местности.
Литература
Якутия: Р. И. Аболин (19296); Т. А. Работнов (1935, 1945); М. Н. Караваев (1945).
Приморье: Ю. А. Ливеровский (1946, 1947); Б. II. Колесников (1948); Г. Э. Куренцова (1950, 1951).
75 е — сельскохозяйственные земли на месте европейских луговых степей и остепненных лугов в сочетании с дубовыми лесами
Сельскохозяйственные земли на месте европейских луговых степей и остепненных лугов в сочетании с дубовыми лесами показаны на «Геоботанической карте» в виде сплошной полосы от бассейнов Прута и Днестра до Южного Урала. Это — восточноевропейская лесостепь. Кроме того, в пределах полосы, ранее занятой широколиственными (дубовыми) лесами (с остатками последних в виде отдельных массивов), сельскохозяйственные земли на месте европейских луговых степей и остепненных лугов показаны отдельными островами:1 к югу от Луцка, в районе Ровно,
1 В настоящее время многие географы и почвоведы при выделении в Европейской части СССР лесостепной зоны объединяют лесостепь в узком смысле и полосу широколиственных (дубовых) лесов; последнюю — к востоку от Полесья. Такое объединение имеет известные основания, имея в виду, во-первых, значительно бблыпую обле-сеяность лесостепи в прошлом и, во-вторых, некоторую «остепненность» опушек и полян в сохранившихся лесах и сорной растительности в полосе широколиственных лесов.
а также и на Приволжской возвышенности — к западу от Арзамаса, к югу от Сергача (так называемое «Межпьянье»), к западу от Ульяновска и т. д. (Алехин и Аверкиев, 1927, 1928; Смирнов,1927, 1940).
Луговые степи и остепненные луга, с одной стороны, и дубовые леса, с другой, в пределах европейской лесостепи (понимаемой, как и далее, в узком смысле) являются типами растительности, приуроченными к во-дораздолам (плакорам). При этом, как давно было указано исследовате-лями (Докучаев, 1891; Краснов, 1891), массивы широколиственных лесов и соответствующие им серые лесные почвы и деградированные черноземы чаще всего связаны в своем распространении с более возвышенными участками водоразделов, дренированными балками и обычно приуроченными к правым высоким берегам долин рек. Это — так называемые нагорные дубравы.
Черноземы — мощные, тучные и выщелоченные, ранее занятые луговыми степями и остепненными лугами, а теперь распаханные, приурочены большими массивами чаще всего к более пониженным и слабо дренированным участкам водоразделов, которые обычно располагаются по левобережьям рек. По занимаемой площади в настоящее время в лесостепной полосе преобладают черноземные почвы. Судя по распространению лесных почв в европейской лесостепи, последняя к настоящему времени сильно обезлесена. Больше того, весьма вероятно, что еще в доисторический период, в связи с хозяйственной деятельностью человека (рубка лесов, степные пожары, выпас домашних животных), часть лесов была уничтожена, заменена луговыми степями или остепненными лугами, а менее оподзоленные почвы при этом реградировали в черноземы (см., например, работы Авдеева, 1945, 1948).
Луговые степи и остепненные луга сохранились в настоящее время в пределах европейской лесостепи в плакорных условиях почти исключительно в заповедниках. В лесостепи Европейской части СССР имеются следующие степные заповедники: Михайловская степь в Сумской области 1 (Ширяев, 1910; Лавренкои Зоз, 1928; Зоз, 1933), Стрелецкая, Казацкая и Ямская степи в южной части Курской области2 (Алехин, 1909, 1910, 1924, 1925, 19346; Комаров и Проскоряков, 1931; Алехин и Уранов, 1933; Каден, 1940; Покровская, 1940, 1940а; Прозоровский, 1940; Зозулин, 1950). Кроме того, плакорные луговостепные и остепненнолу-говые участки, видимо, сохранились на землях Хреновского конного завода у с. Хреновое, Воронежской области (Попов, 1914; Келлер, 1931; Камышев, 1948) и на Попереченской степи в Пензенской области, принадлежащей хозяйственной организации (Спрыгин, 1923, 1925, 1930; Уранов, 1925; Лавренко, 19506).
Кроме того, небольшие участки луговых степей и остепненных лугов в лесостепной полосе сохранились местами по опушкам и на полянах в широколиственных лесах, по крутым склонам балок и пр.
Остепненные луга («разнотравно-луговые» степи Келлера, 1916, 1923а) на плакорах связаны с выщелоченными черноземами. Для остепненных лугов характерно преобладание ксеромезофильных с примесью евмезо-фильных и мезоксерофильных видов, главным образом из группы разнотравья и корневищных злаков; травостой обычно высокий и густой. В травостое особенно обильны разнотравье и отчасти бобовые (Pulsatilla patens, Filipendula hexapetala, Fragaria viridis, Trifolium montanum, Myosotis suaveolens, Salvia pratensis, Galium verum, Leucanthemum vul-
1 Степь эта подведомственна Академии наук УССР.
1 Степи эти входят в состав Центрально-Черноземного Государственного заповедника им. проф. В. В. Алехина.
gore и др.), корневищные злаки и осоки (Роа angustifolia, Calamagrostis epigeios, Agrostis Syreistschikovii, Helictotrichon pubescens, Bromus riparius, Carex praecox и др.), в меньшем количестве встречаются дерновинные луговые злаки (например Festuca pratensis). Обычна примесь более ксе-рофильных дерновинных степных злаков (типчак, тонконог, ковыли); при этом типчак и тонконог (Koeleria gracilis) располагаются в подсоде и чаще угнетены. Нередко хорошо выражен напочвенный покров из мха туидиума (Thuidium abietinum). Остепненным лугам свойственно постепенное затухание вегетации к осени, без летнего периода полупокоя, имеющего место в более южных степях.
Луговые (в узком смысле) пли «дерновинно-луговые» (Келлер, 1916, 1923а) степи характеризуются преобладанием мезоксерофильных и евриксерофильных видов с примесью ксеромезофильных и евксеро-фильных видов и связаны с мощными и тучными черноземами. Основная роль принадлежит дерновинным злакам: типчаку (Festuca sulcata), мезоксерофильным (Stipa Joannis, S. stenophylla, S. pulcherrima), и отчасти евриксерофильным ковыляй (Stipa capillata, в более южных разностях луговых степей), тонконогу (Koeleria gracilis), степному овсу {Helictotrichon (Avenastrum) desertorum], нередко значительная роль принадлежит рыхлодерновинному овсецу Шелля (Helictotrichon Schel-lianum) и корневищным злакам (особенно костру степному — Bromus riparius), а также Agrostis Syreistschikovii, реже Helictotrichon pubescens и др.). Обильно представлено луговостепное разнотравье, по составу то же, что и в остепненных лугах (см. выше), но с некоторыми более ксе-рофильными видами. Эфемероиды не обильны (Hyacinthella leucophaea, Bulbocodium versicolor, Gagea erubescens, G. pusilia). Эфемеры-однолетники встречаются только единично. Обычно бывает выражен напочвенный покров из мхов (Thuidium abietinum и отчасти Tortula ruralis)' тра-; востой густой и довольно высокий.
Как и для остепненных лугов, для луговых степей свойственно постепенное затухание вегетации к осени, без летнего периода полу покоя.
В качестве примера приведем смену аспектов для луговой Казацкой степи Курской области (Каден, 1940).
1. Предвесенний период — от момента, когда сойдет снеговой покров, до начала третьей декады апреля. Степь серовато-бурая от прошлогодней травы; к концу фазы начинается вегетация растительности; цветущих растений еще нет.
2. Конец апреля—начало мая. Период массового цветения сон-травы (Pulsatilla patens) с фиолетовыми цветами и Adonis vernalis (цветы желтые). Кроме этих основных растений, цветут еще желтоцветные виды (Gagea erubescens, Potentilla humi-fusa, Carex humilis) и фиолетово-лиловые фиалки (Viola rupestris, V. hirta).
3. Середина мая. Период массового цветения желтоцветных растений (Adonis vernalis и Primula ver is). Заканчивают цветение сон-трава (Pulsatilla patens) и фиалки. По микрозападинкам куртинами цветет золотистожелтая Draba sibirica. В конце фазы зацветают Taraxacum officinale, Lathyrus pannonicus и др.
4. Конец мая — бескрасочный период. Среди желтоватых «чашечек» первоцвета разбросаны желтые цветки Senecio integrifolius, фиолетовые Iris aphylla, белые Lathyrus pannonicus,1 Arenaria ucrainica, Anemone silvestris и др. Однако ни одно из этих растений не встречается в цветущем состоянии настолько обильно, чтобы создавать аспект.
5. Первая половина июня. Период массового цветения Salvia pratensis (цветы сине-фиолетовые), Bunias orientalis (цветки желтые), Stipa Joannis (белые «перья»). Кроме этих основных видов, значительную роль в аспекте играют белоцветные Valeriana rossica, Arenaria ucrainica hl желтоцветный Ranunculus polyanthemus. В это время цветет более 60 видов. Фаза эта соответствует максимуму развития травостоя.
1 Е. Д. Висюлина (Флора УРСР, т. 6, 1954) считает, что под названием Lathyrus pannonicus русские флористы объединили два близких вида: L. versicolor (Gmel.) Beck, (более северные степи) и L. lacteus (М. ВЛ Wissjul. (более южные степи). Таким образом, упоминаемая здесь чина должна быть отнесена к L. versicolor (6mel.) Beck.
6. Вторая половина июня. Период массового цветения сине-фиолетового шалфея (Salvia nutans) и белоцветных Leucanthemum vulgare и Filipendula hexapetala. Кроме этих основных видов, значительное участие в аспекте принимают еще виды, доминирующие в аспекте начала июня, а также Achyrophorus maculatus.
7. Конец июня—начало июля. Период массового цветения лиловоцветного Knautia arvensis и желтоцветного Galium verum. В аспекте принимают также участие Achillea setacea, отцветающий поповник и обсеменяющийся ковыль. Цветут, кроме того, рассеянные Stachys recta, Inula hirta, Phlomis tuberosa, Trifolium montanum, Onobrychis arenaria и др.
8. Середина июля. Период массового цветения Galium verum. Заметную роль в аспекте играют лиловоцветный Knautia arvensis, белоцветный Achillea setacea и желтоцветный Verbascum lychnitis.
Кроме того, цветут Campanula bononiensis, Centaurea scabiosa, Prunella grandi-flora, Veronica spuria, Helichrysum arenarium, Onobrychis arenaria и др.
9. Осенний период с конца июля до выпадения снега. Основной тон степи бурый от высохшего травостоя. Цветут отдельные разбросанные экземпляры Delphinium cuneatum, Centaurea scabiosa, Veratrum nigrum, Pieris hieracioides, Veronica incana, Helichrysum arenarium, Senecio Jacobaea и др. Под снег многие растения уходят с зелеными листьями, а иногда и с бутонами.
Состав как остепненных лугов, так и луговых степей несколько изменяется при движении с запада на восток, от западных границ СССР к Уралу.
Остановимся на распространении некоторых видов, играющих значительную роль в травостое луговых степей и остепненных лугов. Так, степной костер (Bromus riparius) встречается в значительном количестве в составе луговых степей и остепненных лугов на юге правобережной и левобережной части лесостепи в Украинской ССР, массово распространен в лесостепи более восточных районов Средне-Русской возвышенности и на Приволжской возвышенности, делается более редким в Высоком Заволжье и почти исчезает в Предуралье. Низкая осока (Carex humilis), видимо, была обычна в луговых степях правобережной части Украинской ССР, встречается в большом количестве в луговых степях и остепненных лугах Средне-Русской возвышенности, но уже отсутствует на Приволжской возвышенности, а также в Высоком Заволжье.1 Среднеевропейский вид шалфея (Salvia pratensis), обильный в составе остепненных лугов и луговых степей правобережной Украины и Средне-Русской возвышенности, доходит на восток только до Урала; уже на Средне-Русской возвышенности появляется восточноевропейско-североказахстанский вид (Salvia stepposa), который полностью замещает первый в степях Западной Сибири и северного Казахстана.
В то же время, при движении к востоку появляется ряд восточных видов. Так, степной овес (Helictotrichon desertorum) встречается только изредка на каменистых степных склонах на западе Украинской ССР, в бассейне Днестра (Покутил, Подолия), спорадически растет в составе луговых степей на меловой и реже известковой подпочве на Средне-Русской возвышенности, делается более обычным в луговых степях на пла-корах Приволжской возвышенности и, особенно, в луговых степях Высокого Заволжья Овсец Шелля (Helictotrichon Schellianum) игрьет довольно заметную роль в травостое остепненных лугов и луговых степей, начиная со Средне-Русской возвышенности (заходя только в наиболее восточную часть левобережной лесостепи Украинской ССР) и далее к востоку. На Средне-Русской возвышенности спорадически встречаются восточные виды степных корневищных полыней (Artemisia armeniaca. A. latifolia),1 2
1 Эта осока, после огромного перерыва, снова появляется в западной части Алтая.
2 Из этих видов на Украине встречается как очень редкое растение только Artemisia armeniaca (южнее лесостепи, в полосе безлесных степей).
которые делаются более обычными на Приволжской возвышенности и особенно в Высоком Заволжье. Подобные примеры можно было бы умножить.
Вообще следует указать, что состав луговых степей и остепненных лугов Приволжской возвышенности и особенно Заволжья весьма приближается к составу соответствующих травяных сообществ западносибирской лесостепи.
Для полосы распространения остепненных лугов и луговых степей (лесостепи) в Европейской части СССР, помимо часто встречающихся массивов дубовых лесов (опис. 47), характерны также заросли степных кустарников: терна (Prunus stepposa), степной вишни (Cerasus fruticosa), степного миндаля (Amygdalus папа), спиреи (Spiraea crenata), а на юге лесостепи также дерезы (Caragana frutex).1 Все эти кустарники прекрасно вегетативно разрастаются с помощью корневых отпрысков (большинство) или корневищ (дереза) и образуют более или менее густые заросли (смешанные или одновидовые), в которых обычно преобладают терн или степная вишня. В настоящее время заросли степных кустарников приурочены к опушкам лесов и к степным склонам, а в прошлом встречались, видимо, и на плакорах.
Лесостепь Европейской части СССР может быть разделена на два типа в зависимости от характера мезокомплекса почвенно-растительного покрова, в свою очередь зависящего от общих условий рельефа: возвышенную лесостепь и низменную лесостепь. Возвышенная лесостепь, преобладающая по площади в Европейской части СССР, занимает приподнятые и эродированные долинами рек и балками возвышенности — правобережную Приднепровскую, Средне-Русскую, Приволжскую и Высокое Заволжье. Низменная лесостепь, занимающая значительно меньшую площадь, приурочена к пониженным и слабо расчлененным низменностям — Приднепровской (левобережье Днепра) и Окско-Донской.
К возвышенной лесостепи приурочены крупные массивы широколиственных лесов, преимущественно дубовых или мелколиственных временников, и соответствующих лесных почв. Мелкие участки лесов показаны внемасштабными знаками соответствующих господствующих древесных пород: в пределах всей восточноевропейской лесостепи главным образом Quercus robur, а в бассейне Днестра также Q. petraea, по левобережью Прута — Q. pubescens, на севере и востоке европейской лесостепи — береза (Betula verrucosa).
Кроме того, в пределах возвышенной лесостепи по правым коренным берегам рек и по склонам балок на поверхность выходят различные горные породы, на которых поселяется растительность специфического состава. Особенно интересна растительность своеобразных луговых степей на выходах мела и известняка на Средне-Русской возвышенности.
Луговые степи на меловой и известняковой подпочве1 2 особым вне-масштабным знаком (черным кружком) показаны на карте в лесостепной
1 Д. М. Доброчаева (Флора УРСР, т. 6,1954) для лесостепи и степи Украинской ССР приводит, помимо типичной Spiraea crenata, также установленный ею близкий вид — S. Litwinowii, встречающийся на междуречье Днепр—Волга. М. И. Котов (там же) приводит Prunus spinosa только для Прикарпатья, для юго-западной части лесостепи — Р. moldavica и для большей части лесостепи и степи Восточной Европы — Р. stepposa.
2 В легенде к «Геоботанической карте» соответствующий знак охарактеризован так: «Луговые степи на меловой подпочве». Более полная формулировка должна быть иная: «Луговые степи на меловой и известняковой подпочве и растительность известняковых скал».
полосе: 1) на выходах мела — в самой верхней части бассейна Северского Донца и его левого притока Оскола, а также по Тихой Сосне и Девице и 2) на выходах девонских известняков с растительностью того же типа — в верхней части бассейна Дона — по Дону (знаменитая Галичья Гора и другие аналогичные урочища), Сосне и частично Красивой Мече (Хитрово, 1913; Козо-Полянский, 1928—1929, 1931; Виноградов и Голицын, 1948, 1954; Мешков, 1951).
На задернованных склонах, где почвообразующими породами являются мел или девонский известняк, осоково-злаковая основа травостоя этих своеобразных луговостепных сообществ состоит из рыхлодерновинной луговостепной осоки Carex humilis или из степных дерновинных злаков — Stipa Joannis, Festuca sulcata, реже Helictotrichon desertorum и др. Нередко хорошо выражен моховой покров из Thuidium abietinum. На этом фоне в большем или меньшем количестве встречаются эндемы средней части Средне-Русской возвышенности, обнаруживающие ясные родственные связи с растениями гор Монголии, юга Сибири, Южного Урала и отчасти Средней Европы. В связи с этим, указанные своеобразные луговостеп-ныо сообщества иногда называются «сниженными альпами». Эти сообщества являются реликтами так называемых «перигляциальных степей» или холодной «плейстоценовой лесостепи» (в понимании И. М. Крашенинникова).
«Сниженные альпы» разделяются на два основных варианта: северный, приуроченный к известнякам, и южный, связанный с мелами (Виноградов и Голицын, 1954). Состав горных растений в этих кальцефит-ных луговостепных группировках («сниженных альпах») неодинаков. Для северодонского известнякового варианта наиболее характерны местные эндемы: Potentilla tanaitica и Scutellaria Chitrovoi', в одном пункте обнаружен Leucanthemum sibiricum ssp. Kozo-Poljanskii. Для мелового варианта характерны: местный эндем Androsace Kozo-Poljanskii (часто встречается массово) и более широко распространенная на меловых обнажениях бассейна Дона Scutellaria creticola, а для верхнего Поосколья следующие местные эндемы: Daphne Julia, Bupleurum multinerve ssp. alaunicum, Leucanthemum sibiricum ssp. alaunicum. В обоих районах довольно обычен эндем Средне-Русской возвышенности — Schivereckia mutabilis. Последняя очень охотно поселяется на скалах известняка верхнего Дона. На скалах известняка обычен также папоротник Asplenium ruta-muraria.
Кроме того, на карте в пределах лесостепной полосы показана растительность обнажений мела по правобережью Волги, а также растительность обнажений пермских пород в Заволжье, по р. Сок. Соответствующие пояснения см. в характеристике разнотравно-дерновиннозлаковых степей (76 ч и 76 звк).
Низменная лесостепь Приднепровской низменности и Окско-Донской равнины отличается гораздо меньшей облесенностыо, чем возвышенная, а по более пониженным участкам — более или менее широким развитием комплекса засоленных почв с соответствующей растительностью. Этот комплекс особенно подробно изучен в пределах Окско-Донской равнины (Попов, 1914). На карте распространение его отмечено с помощью внемасштабных обозначений — «галофитная растительность солончаковых и солончаковатых почв» и «луговостепная, болотнолуговая и степная растительность западин в степях».
Галофитная растительность в пределах лесостепной части Приднепровской низменности приурочена к понижениям среди лёссовых террас Днепра и к притеррасовым понижениям его левых притоков (Сула).
На столбчатых солонцах и сильно солонцеватых черноземах, нередко реградированных (то есть вторично засоленных), развиты ассоциации с господством Festuca sulcata, Puccinellia convoluta s. 1. (видимо, P. Bi-lykiana), Camphorosma songoricum, иногда с примесью Plantago salsa, Tripolium vulgare. На довольно широко распространенных здесь солончаковатолуговых почвах развиты солончаковатые луга с преобладанием Carex Karelinii, Juncus Gerardii и с участием Triglochin mari-tima, Tripolium vulgare, Taraxacum bessarabicum, а по более пониженным местам — Bolboschoenus maritimus (Клеопов, 1934).
В пределах Окско-Донской равнины, между реками Воронежем и Вороной, на сильно солонцеватых черноземных почвах и глубокостолбчатых солонцах развита галофитная разность луговой степи («солонцовая степь»), в составе которой преобладают Festuca sulcata, Роа angustifolia, Limonium tomentellum, Silaus Besseri-, на столбчатых солонцах в довольно изреженном травостое — Artemisia monogyna (обычно господствует), Festuca sulcata (согосподствует), Limonium tomentellum-, на корковых солонцах доминируют Puccinllia convoluta s. I.1 и Atriplex litoralis, встречаются в значительном количестве Plantago salsa, Tripolium vulgare и др. По окраинам заболоченных котловин располагаются участки солонцоволуговых почв с господством в луговом травостое Роа angustifolia, Carex melanostachya, Polygonum pseudoare-narium, Sanguisorba officinalis, Trifolium repens, Lotus corniculatus, Silaus Besseri, Peucedanum macrophyHum,2 Leontodon autumnalis и др. («солонцовый луг»).
Вся эта растительность используется преимущественно как пастбище.
Все эти группировки в пределах Окско-Донской равнины встречаются в комплексе с лугово-болотными и луговыми западинами и осиновыми колками (Попов, 1914; Келлер, 1928).
- Луговая и лугово-болотная растительность, приуроченная к западинам, является очень характерным элементом ландшафта низинноравнинной лесостепи. Почвы в этих западинах — типа солодей. Растительность их слагается из различных луговых и лугово-болотных ассоциаций с преобладанием злаков и осок {Роа pratensis, Agropyrum repens, Agrostis stolonizans, Beckmannia eruciformis, Glyceria fluitans, G. aquatica, видов Heleocharis, Carex melanostachya, C. disticha, C. riparia, C. omskiana и др.). Часто в центре западин располагается осоковый кочкарник из Carex omskiana, а по краям — растительность лугового типа. Нередко лугово-болотные западины с кочкарником в центре окружены кольцом из кустарниковых ив, а в пределах Окско-Донской равнины — кольцом осинника. Иногда осинник заполняет всю западину (Попов, 1914); это так называемые осиновые кусты. Луговая и лугово-болотная растительность западин используется обычно как сенокосы и пастбища.
К европейской лесостепи отнесены также луговые степи и остепненные луга предгорий северного склона гор Крыма и Северного Кавказа. Лесостепные ландшафты в предгорьях Крыма и Северного Кавказа могут рассматриваться как нижний горный пояс (явление вертикальной поясностй), в большинстве случаев выше по рельефу сменяющийся поясом дубовых лесов.
Предгорная лесостепь Крыма простирается от района Севастополя почти до Феодосии, занимая южный склон третьей (внешней) гряды (куэсты) и вторую гряду (куэсту). Преобладают карбо-
1 Повидимому, ото — Р. Bilykiana Klok. из ближайшего родства Р. tenuissima Lilw. Виды рода Puccinellia в настоящем издании даны в трактовке Н. Н. Цвелева.
8 Р. macrophyllum Schischk.=P. latifolium auct.
натиые черноземы, часто щебенчатые. Коренным типом растительного покрова, видимо, являются луговые разнотравно-дерновиннозлаковые стопи с богатым разнотравьем, в состав которого входят Asphodeline taurica (крымско-кавказско-малоазиатский вид), Filipendula hexapetala, Paeonia tenuifolia, Adonis vernalis и многие другие, а из дерновиннных злаков — Stipa pulcherrima, S. pontica (крымско-закавказско-передне-азиатский вид), S. capillata и др. (Шалыт и Козлов, 1939).
Преобладающая часть территории Крымской предгорной лесостепи распахана; степные участки сохранились по каменистым склонам. Местами встречаются рощицы дуба (так называемые «дубки»), преимущественно из Quercus pubescens и Q. petraea, реже из Q. гоЬиг, в большинстве случаев очень расстроенные: разреженные, деревья низкоствольные, часто кустарниковидные. Помимо дуба, в этих «дубках» в меньшем количестве представлены Pyrus communis, Р. elaeagrifolia (изредка, крымско-малоазиатский вид), Ulmus foliacea, Crataegus monogyna s. 1., Thelycrania australis (крымско-кавказско-переднеазиатский вид), Cornus mas, Rhamnus cathartica, Corylus avellana, а по опушкам стенные кустарники (Prunus spinosa s. 1., Cotinus coggygria, Amygdalus папа), шиповники и другие. Среди этих дубовых рощ на почти ровных местах с мелкоземистыми почвами встречаются луговостепные поляны с довольно густым разнотравнозлаковым травостоем; из злаков и осок здесь больше всего растут Bromus riparius, Festuca sulcata, а также Koeleria gracilis, Stipa capillata, S. Les-singiana var. Brauneri, Carex nitida и др.; из многочисленных двудольных — Salvia nutans, Stachys recta, Paeonia tenuifolia, Plantago stepposa, Aspe-rula galioides, Polygala major, Filipendula hexapetala, Teucrium chamaedrys, Onobrychis arenaria, Convolvulus cantabrica (средиземноморский вид), Galium verum, Linum austriacum, Adonis vernalis, Helichrysum arenarium, Inula oculus-Christi и многие другие. В довольно значительном количестве представлены эфемеры (рано оканчивающие вегетацию однолетники), чем эти луговые степи отличаются от равнинных луговых степей Европейской части СССР.
По щебнистым склонам, преимущественно южной и близких экспозиций, встречаются тимьянниковые степи, с господством дерновинных злаков — Agropyrum imbricatum, A. pectiniforme, Festuca sulcata, Bromus cappadocicus (крымско-малоазиатскийвид), Andropogon ischaemum, Koeleria brevis (по более каменистым местам), в меньшем количестве— ковыли {Stipa capillata, S. Lessingiana var. Brauneri). Очень характерно большое участие полукустарничков — Thymus Callieri (крымский эндем), Convolvulus holosericeus (крымско-закавказско-переднеазиатский вид), Paronychia cephalotes, Alyssum tortuosum s. 1., Onosma tauricum (крымско-новороссийско-малоазиатский вид), Jurinea stoechadifolia, Fumana procum-bens (средиземноморский вид), Teucrium chamaedrys и ряда травянистых двудольных и однолетников, в том числе эфемеров (Дзевановский, 1925; Цырина, 1925).
В составе крымских предгорных луговых степей появляется, после перерыва, ряд видов, характерных для равнинных луговых и отчасти типичных разнотравно-дерновиннозлаковых степей юга Европейской части СССР, как, например, Stipa stenophylla, Paeonia tenuifolia, Inula ensifolia и др.1 Для крымских предгорных луговых степей, кроме того, как отчасти указано выше, характерно присутствие ряда средиземноморских, малоазиатских и переднеазиатских видов, отсутствующих в пределах равнинной лесостепи.
1 Эти виды отсутствуют на большей части территории сухих дерновиннозлаковых степей.
Предгорная лесостепьСеверного Кавказа простирается с небольшими перерывами с запада-северо-запада на восток-юго-восток, от нижнего течения Кубани до района к югу от Грозного. Кроме того, небольшие острова лесостепи европейского типа известны на наиболее возвышенных участках Ставропольской возвышенности. В основном эта предгорная лесостепь занимает высоты 500—1000 м над у. м., реже ниже. Геоморфологически это — предгорные равнины, куэстовые гряды с крутыми южными и пологими северными склонами и отчасти складчатые низкогорья. Преобладают мощные, выщелоченные и деградированные черноземы, а местами горнолесные почвы.
В предгорной лесостепи Северного Кавказа на мощных, выщелоченных или карбонатных черноземах в прошлом, видимо, были широко распространены луговые степи с узколистым ковылем и ковылем Иоанна (Stipa stenophylla, S. Joann is) (Шиффере, 1953), от которых в настоящее время остались только очень небольшие участки. Иногда в составе предкавказ-ских луговых степей из дерновинных злаков и осок преобладают Stipa pulcherrima и Carex humilis. Кроме того, более или менее обильны из злаков — Bromus riparius, Festuca sulcata, Koeleria gracilis. В составе богатого видами разнотравья обычно господствуют виды, общие с луговыми степями зональной лесостепи юга Европейской части СССР (Filipendula hexapetala. Campanula sibirica и многие другие), но встречаются также виды, отсутствующие в равнинных луговых степях и остепненных лугах и характерные для флоры Кавказа (например Psephellus leucophyllus, Veronica gentianoides и др.).
Местами на ровных и пологосклоновых участках имеются залежи; из них пырейные залежи являются хорошими сенокосами.
Если пологосклоновые степные угодья в пределах Северного Кавказа целиком распаханы, то по южным щебнистым склонам повсеместно сохранилась степная растительность, представленная чаще всего тимьяннико-выми степями. Преобладают дерновинные злаки и осоки: Andropogon ischaemum, Stipa capillata, Festuca sulcata и очень часто Carex humilis. Бородач (Andropogon ischaemum) встречается чаще как один из злаков — пионеров зарастания обнажений и на сбитых в результате выпаса местах. Из корневищных злаков в большем или меньшем количестве встречаются Bromus riparius, Agropyrum intermedium, A. trichophorum. Очень характерно обилие полукустарничков, главным образом из губоцветных — виды Thymus, Teucrium polium, Т. chamaedrys, Scutellaria polyodon (эндем предгорий Северного Кавказа). Помимо обычных широко распространенных видов степного разнотравья, здесь снова появляются некоторые луговостепные виды, отсутствующие в более засушливых степях юга Европейской части СССР (например Inula ensifolia)', встречается также ряд видов разнотравья, отсутствующих в составе равнинных степей, в том числе горные растения, как, например, Pulsatilla albana (кавказский вид), Androsace barbulata (кавказский вид) и др.
По склонам растут заросли степных кустарников: Prunus spinosa s. 1., Amygdalus папа,Rhamnus Pallasii (кавказско-переднеазиатский вид), боярышник, шиповник и др. По северным и западным склонам — остатки дубовых лесов из Quercus petraea, Q. robur, Carpinus caucasica и др. Более высокие лакколиты района Минеральных Вод также покрыты дубовыми и даже буковыми лесами, например гора Бештау (Вернандер, 1946).
Литература
Равнинная лесостепь Европейской части СССР: В. В. Докучаев (1891); А. Н. Краснов (1891); В. В. Алехин (1909, 1910, 1924, 1925, 19346); Г. И. Ширяев (1910); В. Н. Хитрово (1913); Т. И. Попов (1914); Б. А. Келлер
(1916,1923а, 1928,1931,1931г); И. И. Спрыгни (1923,1925,1930); А. А. Уранов (1925); В. В. Алехин и Д. С. Аверкиев (1927,1928); В. М. Козо-Полянский (1927, 1928—1929, 1931); П. А. Смирнов (1927, 1940); Е. М. Лавренко и И. Г. Зоз (1928); Н. Ф. Комаров и Е. И. Проскоряков (1931); В. В. Алехин и А. А. Уранов (1933); И. Г. Зоз (1933); Ю. Д. Клеонов (1933, 1934); Н. Н. Каден (1940); В. М. Покровская (1940, 1940а); Н. А. Прозоровский (1940); В. Д. Авдеев (1945, 1948); Н. П. Виноградов и С. В. Голицын (1948, 1954); Н. С. Камышев (1948, 1948а); П. П. Жудова (1950); Г. М. Зозулин (1950); Е. М. Лавренко (19506); А. Р. Мешков (1951, 1953).
Лесостепь северных предгорий Крыма: С. А. Дзевановский (1925); Т. С. Цырина (1925); М. С. Шалыт и П. К. Козлов (1939); Г. И. Поплавская (1948); Е. М. Хорват (1948).
Лесостепь северных предгорий Кавказа: И. В. Новопокровский (1927а); В. Г. Танфильев (1931); Т. Б. Вернандер (1946); Е. В. Шиффере (1953).
75 з с — сельскохозяйственные земли на .месте западносибирских луговых степей и остепненных лугов в сочетании с березовыми колками и местами
с травяными болотами
Сельскохозяйственные земли на месте западносибирских луговых степей и остепненных лугов (большая часть названных степей и лугов распахана) вытянуты в виде широкой полосы от Урала до Кузнецкого Алатау. Вплоть до Оби и Салаирского кряжа эта полоса тянется без перерыва. Леса, покрывающие Салаирский кряж, и сосняки по Оби в районе Новосибирска и далее к северу, отделяют Кузнецкую степь (лесостепь), располагающуюся между Салаирским кряжем и Кузнецким Алатау, от основной части лесостепи юга Западной Сибири. Кроме того, острова луговых степей и остепненных лугов (лесостепи) западносибирского типа имеются в северной части Казахского мелкосопочника (Казахской складчатой страны), в полосе разнотравно-дерновиннозлаковых степей и даже южнее — в полосе сухих дерновиннозлаковых (типчаковоковыльных) степей. Эти лесостепные острова на возвышенных участках среди казахстанских степей относятся к проявлениям вертикальной поясности растительного покрова.
Западносибирская лесостепь характеризуется равнинным рельефом. Западная часть западносибирской лесостепи (до долины Тобола) занимает низкохолмистый Зауральский пенеплен и главным образом равнинное Зауральское плато. Большая часть западносибирской лесостепи, на восток до левобережья Оби, то есть так называемые Ишимская и Барабин-ская степи, располагается на юге Западно-Сибирской низменности, а именно в пределах Ишимской и Барабинско-Кулундинской равнин. Лесостепная часть Кулундинской степи (левобережье Оби) занимает Приобскую повышенную равнину. Наиболее восточные части западносибирской лесостепи — Чумышская и Бийская степи — занимают увалисто-долинные предгорья Алтая и Салаирского кряжа, а Кузнецкая степь — Кузнецкую котловину с таким же рельефом.
Если сравнивать по характеру рельефа западносибирскую лесостепь’ с восточноевропейской, то бблыпую территорию западносибирской лесостепи необходимо будет отнести к низменной лесостепи. Именно таковы Ишимская и Барабинская степи (лесостепи) и лесостепь пониженноравнинного Зауральского плато. Только лесостепные районы Зауральского пенеплена, Приобской повышенной равнины (лесостепная часть Кулундинской степи) и увалистых предгорий Алтая, Салаирского кряжа и Кузнецкого Алатау, то есть Чумышскую, Бийскую и Кузнецкую степи, можно отнести к возвышенной и дренированной лесостепи.
Для низменной западносибирской лесостепи особенно характерно преобладание междуречных пространств и подчиненная роль долин, к тому же очень слабо разработанных и обычно слабо врезанных. Междуречным пространствам свойствен гривно-лощинный мезорельеф, состоящий из параллельных рядов невысоких, вытянутых в северо-восточном направлении, сравнительно узких и длинных гряд—«грив» с относительными высотами в 2—4, реже 6—10 м, и их разделяющих длинных и плоских понижений пли лощин. Поверхность грив усеяна суффозионными плоскими котловинами или блюдцами. Понижения между гривами за пяты речками, озерами, травяными болотами и более или менее засоленными лугами. Для низменной западносибирской лесостепи характерно также обилие впадин разнообразных размеров и очертаний, обычно занятых очень неглубокими озерами, солеными или пресными. Участки этой низменной лесостепи, прилегающие к долинам Иртыша и Оби, дренированы несколько лучше.
Если в восточноевропейской лесостепи господствуют выщелоченные, мощные и тучные черноземы, то в низменноравнинной западносибирской лесостепи преобладают тучные и отчасти обыкновенные черноземы, часто солонцеватые; очень большие площади занимают черноземно-луговые и засоленные почвы. Последние покрывают обширные пространства во внутренних частях недренированных междуречий, где, в зависимости от условий микро- и мезорельефа, располагаются пестрые комплексы более пли менее солонцеватых черноземов, черноземно-луговых почв, солонцов и местами солончаков с таким же комплексным растительным покровом. Под мелколиственными лесами наблюдаются деградированные черноземы, подзолистые и осолоделые почвы. В более дренированных предгорных районах «возвышенной» лесостепи Западной Сибири (см. выше) преобладают нормальные (незасоленные) черноземы, характерные для лесостепи (тучные, выщелоченные).
Флористический состав западносибирских луговых степей и остепненных лугов несколько обеднен по сравнению с их восточноевропейскими аналогами, хотя большинство видов является все же общим для тех и других. Количество эфемероидов вообще не велико в луговых степях, но и оно уменьшается в западносибирской лесостепи, по сравнению с восточноевропейской лесостепью. Так, в Западную Сибирь вовсе не заходят Hyacinthella leucophaea и Bulbocodium versicolor, являющиеся редкостью уже в Заволжье. Из эфемероидов в западносибирских луговых степях и остепненных лугах встречаются только немногочисленные виды гусиного лука. В западносибирскую лесостепь совсем не заходят следующие виды, весьма обычные в восточноевропейских луговых степях: Bromus riparius, Carex humilis? Iris aphylla, Delphinium cuneatum, Lathy-rus pallescens, Salvia pratensis, Stachys recta,Prunella grandiflora,Verbascum lychnitis и др.; из степных кустарников — Prunus stepposa. Некоторые европейские виды заходят только в западную часть западносибирской лесостепи. Так, Aster amellus s. 1. доходит лишь до Тобола, Cytisus ruthe-nicus также в основном доходит до Тобола и далее к востоку встречается только как редкое растение на водоразделе Тобол—Ишим, Inula hirta на востоке достигает Иртыша, Trifolium montanum — также (в районе Омска), но после большого перерыва снова появляется между Томском и Колыванью на Оби и т. д. Восточных видов встречается сравнительно немного. Назовем из них Geranium bifolium (западносибирский вид), Peucedanum Morisonii (западносибирско-казахстанский вид), Artemisia
1 Carex humilis снова появляется в предгорьях Алтая.
Рис. 59. Трагакантники с Onobrychis cornuta (колючие подушки). Отроги Даралагеза (Армянская ССР). (Фот. Г. В. Аркадьева).
Рис. 64. Луговая степь с березовыми колками в равнинной части Алтайского края. (Фот. Л. Е. Родина).
Рис. 65. Березовый колок в лесостепи равнинной части Алтайского края. (Фот. Л. Е. Родина).
glauca (казахстанско-южносибирский вид; на запад до Предуралья). В восточной части западносибирской лесостепи появляются некоторые восточные виды, характерные для луговых степей и остепненных лугов Средней Сибири, Забайкалья и северной Монголии, например Hemerocal-lis flava (в основном до Оби), Iris ruthenica (на запад до Иртыша), Pulsatilla Turczaninovii (на запад до долины Тобола) и др.
В составе остепненных лугов (плакорные сообщества «дернисто-луговой» подзоны лесостепной зоны в трактовке П. Н. Крылова, 1916) обычно преобладают ксеромезофильные луговые корневищные злаки—Calamagrostis epigeios, Роаpratensis, Р. angustifolia', из дерновинных злаков более обычен Phleum phleoides, а также Helictotrichon Schellianunr, остальные степные дерновинные злаки (типчак, Stipa Joannis) в небольшом количестве. Из бобовых более обильны Lathyrus pisiformis, L. pratensis, Trifolium lupinaster, Vicia craccw, из многочисленного разнотравья, которое нередко господствует над злаками, часто встречаются Achillea millefolium, Artemisia latifolia, A. pontica, A. vulgaris, Filipendula hexapelala, F. step-posa,1 Fragaria viridis, Galium boreale, G. verum, Hieracium umbellatum, Libanotis intermedia, Peucedanum Morisonii, Phlomis tuberosa, Plantago stepposa, Thalictrum simplex, Veronica longifolia, V. spicata и др.; из разнотравья особенно обильны Libanotis intermedia, Filipendula stepposa, F. hexapetala. Производительность — 18—20 ц/га (Вандакурова, 1950).
Подобные остепненные луга связаны обычно с выщелоченными или слабосолонцеватыми черноземами, а иногда и с глубокостолбчатыми солонцами, вскипающими на глубине 50—60 см. На менее глубоких («обыкновенных») солонцах, вскипающих на 30—40 см, или на сильно солонцеватых черноземно-луговых почвах встречаются галофитные разности остепненных лугов («солонцеватые» луга), в составе которых, помимо господствующих корневищных злаков (Calamagrostis epigeios, Agropyrum repens) и типчака, имеются представители галофильного лугового и луговостепного разнотравья (Plantago Cornutii, Р. maxima, Artemisia laciniata, A. rupestris, Galatella punctata s. 1. и др.) (Дохман, 1936a).
Луговые степи (плакорные группировки «разнотравно-луговой подзоны» лесостепной зоны в трактовке П.Н. Крылова, 1916)более ксерофитны. Из дерновинных злаков преобладают Festuca sulcata и Koeleria gracilis, а также Phleum phleoides', нередко доминируют также крупнодерновинные злаки-ковыли (Stipa capillata, S. Joannis, S. rubens) и степной овес (Helictotrichon desertorum); из корневищных злаков в значительном количестве встречается Роа angustifolia-, в меньшем количестве — Calamagrostis epigeios и Bromus inermis. Среди бобовых встречаются в значительном количестве Medicago falcata и Onobrychis sibirica. В составе многочисленного разнотравья более обильны Achillea millefolium, Anemone silvestris, Artemisia dracunculus, A. glauca, A. latifolia, A. Marschalliana, Campanula sibirica, Filipendula hexapetala (особенно обильна), Fragaria viridis, Galatella fastigiata, G. punctata s. L, Galium verum, Gypsophila altissima, Iris ruthenica (к востоку от Иртыша), Libanotis intermedia, Peucedanum Morisonii, Plantago stepposa, Potentilla humifusa, Pulsatilla flavescens, Salvia stepposa, Silene multiflora, Thymus Marschallianus, Veronica spicata и др. Из однолетников имеются в небольшом количестве только Androsace sep-tentrionalis, из полупаразитов — Castillefa pallida и очанка (Euphrasia).
Подобные луговые степи приурочены в более южной лесостепи к обыкновенным, иногда более или менее солонцеватым черноземам, а в северной полосе лесостепи чаще к солонцеватым черноземам. На солонцах
1 Из родства Filipendula ulmaria.
с более глубоким залеганием столбчатого горизонта (так называемые «обыкновенные» солонцы) встречаются галофитные разности луговых степей, в составе разнотравья которых в значительном количестве встречаются виды — показатели солонцеватости: Glycyrrhiza uralensis, Gala-tella punctata s. !., Artemisia pontica.
Как видно из этих списков, в составе западносибирских остепненных лугов и луговых степей преобладают виды,общие с восточноевропейскими остепненными лугами и луговыми степями. Почти все луговые степи и большая часть остепненных лугов распаханы.
Имеются все переходы между остепненными лугами, свойственными преимущественно северной части лесостепи, и луговыми степями, более характерными для южной полосы лесостепи. На юге остепненные луга встречаются по более пониженным участкам, а также по опушкам и па полянах березовых колков. На юге лесостепи часты переходы от луговых степей к типичным разнотравно-дерновпннозлаковым степям. В качестве примера таких переходных степей может быть названа «Сорочья степь», на водоразделе Тобол—Ишим, описанная еще А. Я. Гордягииым (1900—1901). Для подобных переходов от луговых к типичным степям характерно обилие Stipa rubens, Helictotrichon desertorum, обеднение видового состава разнотравья, уменьшение количества луговостепных видов и появление более южных степных видов (Seseli Ledebourii и др.).
Довольно далеко на север по южным склонам дренированных приречных участков водоразделов (например вдоль Иртыша) иропи-никают участки более ксерофитных разностей луговых степей и даже типичные степи с преобладанием ковылей, типчака, степного овса, Cleistogenes squarrosa с большим участием полукустарничковой Artemisia frigida и со сравнительно бедным разнотравьем.
На солонцах с близким к поверхности столбчатым горизонтом (не глубже 15 см от поверхности почвы) наблюдается так называемая «обедненная типчаковая степь», или «типчаковое мелкотравье» (Баранов, 1927) с господством дерновинок типчака, иногда с примесью Роа stepposa и Koeleria gracilis и с небольшим количеством довольно угнетенного разнотравья, состоящего главным образом из индикаторов резко выраженной солонцеватости почвы и близкого (к поверхности последней) залегания солей (SilausBesseri,Limonium Gmelinii,Pedicularis dasystachys, Plantago maxima, Galqtella punctata s. 1., Artemisia pontica, A. nitrosa: Peucedanum Morisonii в таких случаях дает только вегетативные побеги). Эти степи по структуре близки к типичным бедноразнотравным дерновиннозлаковым степям, что объясняется неблагоприятными почвенными условиями.
Характер сочетаний растительных сообществ в пределах западносибирской лесостепи сильно различается, с одной стороны, в пределах ее возвышенных районов, расположенных на крайнем западе полосы, в Зауралье, и на востоке, начиная с левобережья Сби, и, с другой стороны, в пределах низменных ее районов, преобладающих по площади. В пределах возвышенной лесостепи обычно отсутствуют заболоченные участки, низинные луга и болота, а также большие площади солонцов и солончаков. Солонцы встречаются небольшими пятнами. Березовые колки, характерный элемент ландшафта западносибирской лесостепи, приурочены не только к западинам, но и к вершинам холмов и их северным склонам. В Кузнецкой лесостепи, в ее южной части, по южным склонам холмов встречаются участки «каменистых» стеией на щебнистых почвах (на карте нс отмечены.)
В низменной лесостепи Западной Сибири, преобладающей по площади, растительный покров, так же как и почвенный, сильно комплексный.
Гривы, в настоящее время большей частью распаханные, в прошлом были заняты остепненными лугами п луговыми степями, а также березняками. К западинам с почвами-солодями и в настоящее время приурочены березовые леса (Betula verrucosa, В. pubescens), которые в более северных частях лесостепи иногда покрывали гривы целиком; в последнем случае под колками представлены серые лесные почвы или выщелоченные черноземы. На склонах грив и в межгрядовых понижениях, если последние узкие и неглубокие, располагаются более или менее комплексные остеп-невные луга, луговые и местами типичные дерновиннозлаковые (разно-травно-типчаково-ковыльные, типчаковые и пр.) степи на солонцеватых черноземах и солонцах; степи на солонцеватых черноземах также часто распаханы. В более широких межгривных понижениях располагаются солончаковатые луга на солончаковатолуговых почвах, нередко заболоченных, а также низинные болота, заросли кустарниковых ив, обширные заросли тростника (Phragmites communis) по берегам озер, пятна солончаков. Растительный покров подобных понижений также обычно комплексный (Баранов, 1927; Баранов и Горшенин, 1927; Иванова и Крашенинников, 1934; Дохман, 1936а).
Солончаковатые луга на солончаковатолуговых почвах преобладают также в так называемых «аллапах», обширных понижениях, в весеннее время затопляемых водой, которая в некоторых местах застаивается до середины или даже до конца лета (Баранов, 1927). В составе солончако-ватых лугов, представляющих прекрасные сенокосные угодья, господствуют в травяном покрове Hordeum brevisubulatum, Alopecurus ventricosus, Agropyrum repens, а по несколько менее влажным местам и Calamagrostis epigeios с большим или меньшим участием солончакового разнотравья и осок (Triglochin maritima, Carex Karelinii, Melilotus dentatus, Limonium Gmelinii, Plantgoa Cornutii, P. maxima, Artemisia laciniata, Cirsium escu-lentum и др.). На более засоленных солончаковых лугах преобладают виды бескильницы (Puccinellia) с участием отдельных видов солончакового разнотравья (Saussurea amara, S. salsa и др. — см. выше), полукустарничковой Artemisia nitrosa и даже сочных однолетних солянок (Suaeda corni-culata). В более южных частях низменной лесостепи (например в районе Петропавловска) появляются на мокрых солончаках по берегам озер и более южные галофиты: однолетние солянки — Salicornia еигораеа, Suaeda prostrata, Atriplex pedunculata, полукустарничковые галофиты — Atriplex verrucifera, Frankenia hirsuta и др.
В северной полосе низменной лесостепи пониженные части межречных пространств, особенно по берегам озер в Барабинской степи, заняты так называемыми «займищами». Займища представлены главным образом обширными зарослями тростника (Phragmites communis), низинными осоковыми болотами (из Carex caespitosa и др.), среди которых вкраплены гипновые и отдельные сфагновые верховые болота с низенькой сосной (так называемые «рямы»). По окраинам займищ обычно располагаются солончаковатые луга (см. выше). Часть заимищ в настоящее время осушена.
В березняках (иногда с примесью осины), столь характерных для лесостепи Западной Сибири, более или менее мощный травяной покров состоит преимущественно из лугово-лесных форм; по опушкам («прикопочное разнотравье)» к ним присоединяются некоторые луговостепные виды (опис. 42). По правым берегам крупных рек, среди довольно крупных массивов осиново-березовых лесов, встречаются, преимущественно на легких почвах, участки сосновых лесов, иногда с единичной лиственницей (Larix sibirica).
Литература
П. Н. Крылов (1898, 1915, 1916); А. Я. Гордягин (1900—1901); Г. И. Танфильев (1902); И. М. Крашенинников (1912); Б. Н. Городков (1915); Л. Ф. Ревердатто (1921)-В. И. Баранов (1923, 1927); Л. А. Петрова-Трефилова (1923, 1925); В. Ф. Семенов (1924); В. И. Баранов и К. П. Горшенин (1927); В. В. Ревердатто (1927); Н. А. Иванова и И. М. Крашенинников (1934); Г. И. Дохман (1936а); П. Н. Красовский и Г. А. Глумов (1945); Л. В. Шумилова (1947); Г. А. Глумов иП. Н. Красовский (1948); А. В. Ку-минова и Е. В. Вандакурова (1949); Е. В. Вандакурова (1950).
75цс— сельскохозяйственные землина месте центра льно сибирс к их луговых степей и остепненных лугов в сочетании сберезовыми
и с о сно в о-л и ственничны м и лесами
Центральносибирские луговые степи и остепненные луга (преимущественно сельскохозяйственные земли на их месте) показаны по окраине Минусинской котловины, в Енисейско-Чулымской впадине и в пределах лесостепных островов на юге Средней Сибири — Красноярского, Канского, Балаганско-Иркутского (по Нижней Ангаре и ее притокам), а также в виде небольших островов по другим рекам юга Средней Сибири (по Уде, Ие, Оке). Кроме того, в бассейне верхнего Чулыма отмечены луговые степи и остепненные луга, переходные между западносибирскими и центральносибирскими.
Геоморфологически все эти лесостепные острова Средней Сибири связаны или с относительно пониженными и равнинными территориями или с низкогорьями. В пределах Минусинской котловины и Енисейско-Чулымской впадины луговые степи и остепненные луга залегают по рельефу выше змеевково-тырсовых енисейско-забайкальских степей (81).
По своему составу остепненные луга и луговые степи юга Средней Сибири близки к аналогичным типам растительного покрова западносибирской лесостепи. Однако целый ряд видов, характерных для западносибирских лугов и луговых степей, отсутствует в пределах соответствующих типов растительного покрова лесостепи юга Средней Сибири или заходит только в пределы западной части последней. Так, совершенно отсутствуют здесь виды, обычно встречающиеся в составе луговых степей Западной Сибири и Европейской части СССР — Thymus Marschallianus и Salvia stepposa. Заходят как редкие растения в пределы лесостепных районов по Енисею следующие виды, общие для лесостепи Западной Сибири и Европейской части СССР: Carex ruthenica, Silene multiflora (только Енисейско-Чулымская лесостепь), Adonis vernalis, Filipendula hexapetala, Potentilla humifusa, Lavatera thuringiaca. Продвигаются несколько далее на восток, до Канской лесостепи, встречаясь здесь изредка, Stipa rubens, Plantago stepposa. Из видов, обычных в составе луговых и типичных (разнотравно-дерновиннозлаковых) степей Западной Сибири и Северного Казахстана и отсутствующих в европейских степях, проникают на восток до Енисея Peucedanum Morisonii (только Енисейско-Чулымская лесостепь) и Seseli Ledebourii (Енисейско-Чулымская и Июс-Ширинская лесостепи).
С другой стороны, в составе луговых степей и остепненных лугов лесостепных островов юга Средней Сибири имеется ряд восточных степных видов, отсутствующих в западносибирской лесостепи или заходящих только в восточную часть последней: Stipa sibirica (в основном на запад до Оби), Thalictrum petaloideum (на запад до Алтая), Bupleurum scorzonerifolium
(также), Phlofodicarpus sibiricus (на запад только до лесостепных островов вдоль Енисея)1 и др.
Остепненные луга 1 2 3 связаны обычно с выщелоченными черноземами. Чаще всего они представлены разнотравными сообществами с преобладанием ксеромезофильных и евмезофильных видов разнотравья: порезника (Libanotis intermedia), широколистной полыни (Artemisia latifolia), пижмолистной полыни (Artemisia tanacetifolia), соссюреи (Saussurea controversa), крестовника (Senecio integrifolius), зопника (Phlomis tuberosa), клубники (Fragaria viridis), кровохлебки (Sanguisorba officinalis), северного подмаренника (Galium boreale), русского касатика (Iris ruthenica) и др. Более ксерофитные ассоциации остепненных лугов характеризуются обилием желтеющего прострела (Pulsatilla flavescens). Из луговостепных дерно-винных злаков значительную роль играют Phleum phleoides, Festuca fenis-seensis (эндем степей бассейна Енисея), Роа stepposa. Эти виды в своем большинстве являются общими и для остепненных лугов и луговых степей Западной Сибири и северного Казахстана, а частью и Европейской части СССР.
Луговые степи связаны с обыкновенными, реже южными или выщелоченными черноземами. Это по преимуществу разнотравно-злаковые сообщества. Из степных (евриксерофильных и мезоксерофильных) дерновинных злаков в составе этих степей играют большую роль Helictotrichon desertorum, Festuca pseudovina? F. fenisseensis, Koeleria gracilis, Poa stepposa, реже Stipa Joannis, а из ксеромезофильных рыхлодерновинных и короткокорневищных злаков — Helictotrichon Schellianum, Phleum phleoides, Роа angustifolia. Таким образом, характерные для настоящих степей Cleisto-genes squarrosa, Stipa capillata, Poa botryoides отсутствуют в данных луговых степях или играют второстепенную роль. В подседе этих степей часто обильна осока Carex pediformis. Среди многочисленных видов ксеромезо-фильного разнотравья особенно обильны Pulsatilla flavescens и Iris ruthenica', в значительном количестве встречаются виды, упомянутые выше для остепненных лугов, а также Bupleurum multinerve, Aconitum barba-tum, Onobrychis sibirica, Trifolium lupinaster, Artemisia glauca, Galium verum, Aster alpinus и др. Указанные выше для луговых степей виды в своем большинстве также являются общими для луговых степей Западной Сибири и северного Казахстана, а частично и Европейской части СССР. Валовая урожайность луговых степей — 8.9—14.3 ц/га сухой массы.
Перейдем теперь к краткой характеристике отдельных массивов остепненных лугов и луговых степей на юге Средней Сибири. Бблыпая часть этих лугов и степей распахана и используется под посевы сельскохозяйственных культур. Следует также отметить, что хозяйственная деятельность человека, особенно скотовода, в прошлом содействовала расширению площадей, занятых остепненными лугами и луговыми степями, за счет лесов. .
В бассейне верхнего Чулыма более остепненной является Ужуро-Копь-евская лесостепь (Туркевич, 1914; Черепнин, 1953а), располагающаяся в юго-западной части бассейна этой реки. Преобладают луговостепные сообщества (с обилием желтеющего прострела и русского касатика,
1 Большинство этих видов по своему ареалу относится к сибирско-монгольской группе.
2 Далее характеристика центральносибирских остепненных лугов и луговых степей основана преимущественно на данных Л. М. Черепнина (1953а).
3 Так определяют В. В. Ревердатто, Л. М. Черепнин и др. широко распространенный в бассейне Енисея степной типчак. В широком смысле — это Festuca sulcata.
но по вторым террасам рек и южным склонам господствуют крупнополын-но-ковыльные (тырсовые) и изредка «четырехзлаковые» степи (опис. 81). По северным склонам, вершинам холмов, логам и по западинам на ровных участках встречаются березовые колки, иногда с участием сибирской лиственницы; по долинам некоторых рек — еловые насаждения. В разреженных и остепненных березовых колках развит травяной покров из Pulsatilla flavescens и Iris ruthenica ', в более густых насаждениях преобладают лугово-лесные виды (Calamagrostis arundinacea, Crepis sibirica, Heracleum dissectum, Anthriscus aemula, Trollius asiaticus, Vicia unijuga)', в этих березняках производительность травостоя — 8—12 ц/га сухой массы. На засоленных местах развиты ассоциации пикульника (Iris biglumis) ', здесь проходит северная граница распространения последнего. В пойме Чулыма — заливные луга.
Причулымская лесостепь «северного типа» (Туркевич, 1914; Черепнин, 1953а) занимает большую часть Енисейско-Чулымской котловины, которая представляет собой волнистую равнину среди отрогов гор. Господствуют остепненные луга в комплексе с березовыми колками. Засоление отсутствует. В северо-западной части лесостепи много заболоченных лугов и кустарников.
Ачинская лесостепь «северного» типа (Туркевич, 1913; Черепнин, 1953а), располагающаяся к северу от невысокого облесенного хр. Арга, отделяющего ее от Причулымской лесостепи, характеризуется волнистым рельефом, наличием березовых и осиново-березовых колков. Сохранившаяся от распашки травяная растительность представлена остекленными лугами и луговыми степями. Большая часть Ачинской лесостепи вторичного происхождения. В травостое осиново-березовых колков встречаются многочисленные виды разнотравья (Iris ruthenica, Aconitum excelsum, Thalictrum minus, Filipendula ulmaria, Bupleurum aureum, Heracleum dissectum, Geranium pseudosibiricum, Phlomis tuberosa, Galium verum, Cacalia hastata, Senecio octoglossus и др.). Таким образом, в осиново-березовых колках преобладают лугово-лесные виды при незначительном остепнении.
Степи Енисейско-Чулымской впадины и Минусинской котловины, разделенные невысоким Батеневским кряжем, окружены территориями с лесостепным ландшафтом (Ревердатто, 1925, 1954; Черепнин, 1953а). При этом при повышении местности, при переходе к горнотаежному поясу, степистость естественнр уменьшается.
Степное «ядро» Минусинской котловины, занятое типичными змеевково-тырсовыми (81) степями, окружено лесостепью «южного» типа, которая является в известной мере переходной к ландшафтам типичной степи. Л. М. Черепнин (1953а) к южной лесостепи относит следующие природные лесостепные (луговостепные) районы, окружающие с севера, востока и юга Абаканско-Минусинские степи: Боградскую степь (с запада и севера ограничена батеневским кряжем и хр. Азыр-Тала), Прибайтакскую степь (прилегает к Байтакскому хребту), Межборовую степь (к востоку от Минусинска), Иудинскую степь (вдоль Джойского хребта Западного Саяна). К южной лесостепи Л. М. Черепнин причисляет также Новосе-ловскую луговую степь, расположенную к северу от Ширинской стейи (опис. 81) и к югу от р. Чулыма. Рельеф этих в основном луговых степей довольно разнообразен: от равнинного до холмисто-увалистого низкогорного (сопочного). Почвы — обыкновенные, частью южные и выщелоченные черноземы. Большая часть луговых степей, в составе которых значительную роль играют менее ксерофильные степные дерновинные злаки (см. выше), распахана. По южным склонам довольно широко распростра-
йены крупнополыпно-ковыльные (тырсовые) и овсецово-ковыльные степи (опис. 81). По северным склонам, в логах, а иногда и в равнинных условиях встречаются березовые, лиственнично-березовые, реже сосново-лиственничные колки. И а песках (главным образом в Межборовой степи) — массивы более или менее остепненных боров. На выпасываемых первых надлуговых террасах — заросли пикульника (Iris ftigZumis), а иногда и чия (Lasiagrostis splendens). Сенокосами служат довольно частые здесь залежи и луга в поймах.
Выше (по рельефу) вышеупомянутых южнолесостепных районов, занимающих окраины Минусинской котловины, располагаются лесостепные ландшафты «северного» типа, то есть более облесенные и с луговостепной растительностью, несколько более мезофитной. Это будут следующие ле состепные районы: Беллыкская лесостепь (к югу от Беллыкского Белогорья), Курагино-Идринская лесостепь (Тубино-Сыдинское междуречье), Ермаковско-Каратузская лесостепь и Таштыпская лесостепь (предгорья Абаканского хребта). К Ширинской степи лесостепные районы, «северного» типа, по данным Л. М. Черепнина, примыкают непосредственно: Июсская лесостепь (бассейн нижнего течения Белого Июса), Батеневская лесостепь (восточная часть Батеневского кряжа) и Комская лесостепь (к северу от Беллыкского Белогорья). Рельеф — от равнинного и увалистого до низкогорного (холмистого); причем низкогорья занимают значительную площадь. Почвы — преимущественно выщелоченные и деградированные черноземы. Большинство луговых степей и остепненных лугов распахано. Крупнополынно-ковыльные (тырсовые), овсецово-ковыльные (тырсовые) и каменистые степи (опис. 81) по южным склонам занимают меньшую площадь, чем в районах «южной» лесостепи. Довольно частые залежи используются как сенокосы. Долины некоторых рек, северные склоны, а иногда и вершины холмов облесены березовыми, лиственнично-березовыми, березово-лиственничными, реже сосново-березовыми, лиственнично-сосновыми и сосновыми лесами. Ранее облесен-ность указанных районов была гораздо большей. В березовых лесах обычно преобладают Calamagrostis arundinacea и Brachypodiumpinnatum, а из разнотравья — Crepis sibirica, Pteridium aquilinum, Chamaenerium angustifolium, Thalictrum minus, Galium boreale, Rubus saxatilis, Heracleum dissectum, Sanguisorba officinalis, Trifolium lupinaster и др. Долины некоторых рек заболочены.
Красноярская островная лесостепь (И. В. Кузнецов, 1914; Черепнин, 1953а) вытянута вдоль Енисея, преимущественно влево от него, к северу от Красноярска, около которого имеется остров змеевково-тырсовых степей (81). Рельеф волнистый. Почвы—большей частью выщелоченные черноземы. В прошлом здесь преобладали луговые степи, теперь в значительной степени распаханные. В составе луговых степей доминируют Роа angustifolia. Р. stepposa, Phleum phleoides, Pulsatilla flavescens, Bupleurum multinerve и др. В составе «каменистых» вариантов этих степей встречаются Allium strictum, Veratrum nigrum, Hemerocallis flava, Lilium tenuifolium, Thlaspicochleariforme,Hedysarum neglectum, Bupleurum aureum, Gentiana decumbens, Eritrichium pectinatum, Schizonepetamultifida, Leucanthemum sibiricum, Artemisia sericea и др. На песках вдоль Енисея — заросли Thymus asiaticus, волоснецов (Elymus jenisseensis и др.), Koeleria Thoni. По северным склонам и западинам — березовые колки, количество которых увеличивается к периферии лесостепи, реже сосновые и лиственничные лески.
Канская островная лесостепь (И. В.Кузнецов, 1913а; Черепнин, 1953а) расположена в бассейне среднего течения Кана и в верховьях У солки.
Рельеф — волнистая равнина, в южной части более приподнятая и пересеченная отрогами Восточного Саяна. Преобладают выщелоченные черноземы, залегающие в комплексе со слабоподзолистыми почвами; в окрестностях Канска — обыкновенные черноземы. Ранее господствовали луговые степи с большим участием разнотравья (теперь большей частью распаханные) в сочетании с березовыми колками; последние в настоящее время окружены преимущественно пашнями и залежами. Встречаются также березово-сосновые и березово-лиственничные леса. В окрестностях Канска преобладают крупнополынно-ковыльные (тырсовые) степи (81), что на карте не отмечено. Фрагменты этих степей и участки «каменистой» степи распространены и в других частях Канской лесостепи. В центральной и особенно в северной части района распространена «бугристая степь», обязанная своим происхождением мерзлотным явлениям (Ревердатто и Буторина, 1934). «Бугристая степь» представляет собой сочетание бугров и ям между ними диаметром до 10м и больше;ямы глубиной до1—1.5 м. Бугры покрыты степной растительностью, в ряде случаев со Stipa capillata', в понижениях между ними — преимущественно более или менее остепненные луга с наличием высокогорных и аркто-альпийских растений (Allium schoenoprasum, Polygonum viviparum), а в глубоких западинах — березовые колки. Эта «бугристая степь» рассматривается как реликт ледникового периода.
В северной части Канской лесостепи на солончаковатых почвах встречаются Lasiagrostis splendens, Suaeda corniculata, Limonium Gmelinii и другие галофильные растения.
Балаганско-Иркутская островная лесостепь (Криштофович, 1910; Ревердатто, 1931; Буддо, 1953; Попов, 1953), расположенная по Ангаре и некоторым ее притокам, отличается от предыдущих участков островной лесостепи юга Средней Сибири большей ксерофитностыо степной растительности. В этом районе степная растительность приурочена к плоским надлуговым террасам рек и к южным склонам коренных берегов речных долин. Нижние («луговые») террасы рек заняты лугами и солончаками; северные склоны и прилегающие к долинам плоские возвышенности покрыты березовыми и березово-сосновыми лесами с большим или меньшим участием сибирской лиственницы.
Степи надлуговых террас носят характер типичных дерновиннозла-ковых степей, близких по составу и структуре к енисейско-забайкальским змеевково-тырсовым степям (81). Из дерновинных злаков в этих местах преобладают Festucapseudovina, Agropyrum cristatum, Stipa capillata s. 1., Poa botryoides', из разнотравья встречаются Lilium tenuifolium, Dianthus versicolor, Delphinium grandiflorum, Potentilla acaulis, P. tanacetifolia, Goniolimon speciosum, Thymus serpyllum s. 1., Veronica incana, Galium verum, Scabiosa ochroleuca, Heteropappus altaicus, Artemisia frigida и др. Степная растительность южных склонов носит более ксерофитный характер; в ее составе часто преобладают Stipa capillata s. 1. и Psathyrostachys (Elymus) juncea', встречаются Convolvulus Ammanii, Kochia prostrata и др. Под влиянием усиленного выпаса происходят следующие изменения травостоя в сторону его ксерофитизации, упрощения структуры и уменьшения растительной массы: типчаково-ковыльная (тырсовая) степь->ко-выльно (тырсово)-типчаковая степь-*змеевково-и тонконогово-типчаковая степь -*• лапчатково (Potentilla асаи/г$)-змеевковая и Tionvisao(Artemisia /п£Й/а)-тонконоговая степь-*типчаково-ланчатковая тонкойогово-полын-ная степь-> полынные (Artemisia frigida) я тимьянниковые (Thymus) группировки сбоев -*• покров из однолетних и многолетних пастбищных сорняков (Буддо, 1953).
При движении к вершинам остепненных падей, расположенных по бортам долин рек, происходит уменьшение ксерофитности (мезофитизация) степных сообществ; при этом типичные степи переходят в луговые, а последние в остепненные луга прилесных участков. Так, например, в составе луговых степей, помимо ковыля-тырсы, житняка, типчака, встречаются Phleum phleoides, Bromus inermis, Agropyrum repens, Hemerocallis flava, Aconitum barbatum, Sanguisorba officinalis, Hedysarum setigerum, Phlomis tuberosa, Galium verum, Heteropappus altaicus, Leucanthemum sibiricum, Achillea asiatica, Artemisia frigida и др. Суходольные луга среди березовых перелесков обычно только слегка остепнены; в их составе преобладает разнотравье (Geranium pratense, Anemone crinita, Sanguisorba officinalis, Vicia amoena, Aconitum barbatum, Filipendula ulmaria, Phlomis tuberosa, Hemerocallis flava, Gentiana barbata, Veratrum Lobelianum, Potentilla fragarioides и др.); иногда разбросаны также и кустарники: Cotoneaster melanocarpa и Spiraea media.
В степных сообществах в бассейне Ангары встречаются некоторые даурско-монгольские виды, не идущие далее на запад на юге Средней Сибири — Hedysarum setigerum, Tanacetum sibiricum и др.
Средняя продуктивность степных сообществ в бассейне Ангары — 5—6 ц/га, а низкотравных степных сбоев — 0.8—2.5 ц/га (Буддо, 1953).
На высоких уровнях поймы рек — остепненные луга; а в притеррасных частях поймы — солончаковые луга и солончаки с Lasiagrostis splen-dens, Suaeda corniculata, Salicornia europaea, Halerpestes ruthenica, Glaux maritima и др.
В создании открытых (безлесных) ландшафтов Балаганско-Иркутской лесостепи велика роль хозяйственной деятельности человека. И. С. Буддо (1953) считает, что луга и степи на суходолах здесь на 95% являются антропогенными и только 5% из них полностью обязаны своим происхождением природным факторам (преимущественно реликтовые степные группировки).
Литература
А. Н. Криштофович (1910); И. В. Кузнецов (1912, 1913а, 1914); С. Ю. Туркевич (1913, 1914); В. В. Ревердатто (1925, 1931, 1954); В. В. Ревердатто и Т. Н. Буторина (1934); И. С. Буддо (1953); М. Г. Попов (1953); Л. М. Черепнин (1953а).
75 зб — забайкальские луговые степи, остепненные луга и сельскохозяйственные земли на их месте в сочетании с лиственничными
и сосновыми лесами
Забайкальские луговые степи и остепненные луга показаны на карте по долинам рек (включая сюда и склоны коренных берегов) среди горных забайкальских лиственнично-сосновых и сосновых (24),- горных южносибирских лиственничных (37) и среднегорных восточносибирских среднетаежных лиственничных (36 сг) лесов. Эти степи отмечены на карте в котловине Еравнинских озер, по верхней Уде, Худану, верхнему и среднему Хилоку, верхнему Чикою, Ингоде, Онону и его левым притокам: Унде, Газимуру и др. Если забайкальские луговые степи и остепненные луга контактируют с мелкодерновинными (80) или пижмовыми (79) степями, то первые всегда располагаются по рельефу выше последних.
Состав луговых степей и остепненных лугов Забайкалья сильно отличается от аналогичных типов растительного покрова Западной Сибири; это объясняется наличием в составе луговых степей и остепненных лугов
Забайкалья большого количества даурско-монгольских, сибирско-монгольских и восточносибирско-дальневосточных видов.
Остановимся более подробно на флористическом составе этих забайкальских степей и лугов. Среди доминирующих видов представлены виды с более широким ареалом — Koeleria gracilis, Cobresia Bellardi, Aster alpinus\ казахстанско-сибирско-монгольские 1 — Artemisia laciniata', Восточносибирско-монгольские — Agrostis Trinii, Poa botryoides. Festuca lenensis, Agropyrum cristatum, Aneurolepidium pseudoagropyruni, Pulsatilla Turczaninovii, Artemisia commutata (на запад до Урала); восточносибирские (южная часть Восточной Сибири) — Phlojodicarpus sibiricus', восточносибирско-дальневосточные — Hemerocallis minor, Clematis hexapetala', даурско-монгольские — Stellera chamaejasme, Tanacetum sibiricum, Artemisia leucophylla; даурские — Oxytropis lanata (пески в низовьях Оны), Thymus eravinensis (пески в Еравнинских степях). Растения, встречающиеся менее обильно в составе забайкальских луговых степей и остепненных лугов: с широким ареалом — Helictotrichon Schellianum, Stipa capillata s. 1., Allium odorum, Veratrum nigrum, Polygonum vivipa-rum, Sanguisorba officinalis, Trifolium lupinaster, Veronica incana, Galium boreale, G. verum, Campanula glomerata, Artemisia sericea (преимущественно Сибирь), Senecio integrifolius, Serratula cardunculus, Scorzonera austrica; казахстанско-сибирско-монгольские — Potentilla acaulis, Artemisia fri-gida; сибирско-монгольские — Polygonum attenuatum, Thalictrum peta-loideum, Potentilla tanacetifolia, Astragalus adsurgens, Peucedanum baica-lense (на запад до бассейна Иртыша), Leontopodium campestre', сибирские — Bromus sibiricus, Potentilla viscosa (преимущественно Южная Сибирь), Viciamulticaulis(Южная Сибирь), Leucanthemum sibiricum', восточносибирско-дальневосточные — Lilium tenuifolium, Polygonum alopecuroides (на запад до Алтая), Bupleurum scorzonerifolium, Vicia amoena (на запад до Алтая); даурско-монгольские—Polygonum angustifolium, Delphinium gran-diflorum, Scabiosa Fischeri', даурские—Oxytropis grandif lor a, 0. myriophylla и другие виды.
Помимо состава, забайкальские луговые степи и остепненные луга отличаются от их западных аналогов менее высоким и густым травостоем и меньшей видовой насыщенностью.
Луговые степи Забайкалья изучены пока недостаточно. Далее приводится характеристика двух массивов остепненных лугов и луговых степей Забайкалья — Еравнинских степей и северной части Нерчинских степей.
Еравнинские степи приурочены к котловине Еравнинских озер, значительно приподнятой над уровнем моря и окруженной лесистыми горами. Почвы здесь более или менее засоленные (солонцеватые), суглинистые, кроме берегов озер, где развиты песчаные отложения. На приозерных песках распространены «песчаные» степи с преобладанием Pulsatilla Turczaninovii и тимьянники с господством Thymus eravinensis и с примесью Chamaerhodos grandiflora. В Еравнинских степях особенно широко распространены вострецовые степи с преобладанием корневищного вост-реца {Aneurolepidium pseudoagropyrum) и с примесью луговостепного и лугового разнотравья: Leontopodium campestre, Scabiosa Fischeri, Achillea asiatica, Artemisia laciniata, A. leucophylla, Inula britannica, Potentilla anserina, местами много Thermopsis lanceolata. По повышенным местам встречаются луговые разнотравно-дерновиннозлаковые степи.
1 Далее, поскольку речь вдет о степных растениях, эпитет «сибирский» имеет в виду преимущественно юго-восточную Сибирь (на запад обычно до Алтая); эпитет «казахстанский» подразумевает широкое распространение соответствующих видов в степной части Казахстана и в соседней части Западной Сибири.
Из дерновинных злаков преобладают Роа hotryoides и Koeleria gracilis, в меньшем количестве другие виды злаков (Agrostis Trinii, Agropyrum cristatum, Festuca pseudovina); обильно представлено разнотравье (Allium odorum. Polygonum alopecuroides, P. viviparum, Thalictrum petaloi-deum, Pulsatilla patens s. 1., Sanguisorba officinalis, Trifolium lupinaster, Vicia amoena, Astragalus adsurgens, Aster alpinus, Artemisia frigida, A. com-mutata; из осоковых встречается Cobresia Bellardi. Иногда разбросаны низкорослые кустарниковые ивы — Salix brachypoda и S. xerophila. На пятнах «влажных солонцов» (Сергиевская, 1951) господствует Puccinellia tenuiflora; встречается Artemisia anethifolia.
Северные Еравнинские луговые степи характеризуются низкорослым, сильно разреженным и бедным по видовому составу травостоем; приурочены они к ровным местам с суглинистыми трещиноватыми почвами. Господствуют из дерновинных злаков то типчак, то Agropyrum cristatum, то Роа botryoides, то Koeleria gracilis, а из разнотравья обильны полыни (Artemisia laciniata, A. commutata, A. leucophylla). Эти полыни образуют и самостоятельные ассоциации с малым участием дерновинных злаков. В качеству примеси встречаются многие из упомянутых видов луговостепного и лугового разнотравья. Интересна также кобрезиево-типчако-вая ассоциация с участием Cobresia Bellardi.
Обращает на себя внимание заметное участие в Еравнинских степях видов, характерных для высокогорий,—Cobresia Bellardi, Polygonum viviparum.
По северной окраине холмисто-равнинных Нерчинских степей (Поплавская, 1912; Сукачев, 1912) развиты разнотравные остепненные луга («разнотравные степи» в понимании В. Н. Сукачева), остепненные луга с господством Hemerocallis minor («степь лилейная» в понимании В. Н. Сукачева) и разнотравные луговые степи — ломоносовая с господством Clematis hexapetala и «зонтичная» степь с господством Phlofodicarpus sibiricus.
Разнотравные остепненные луга развиты по более возвышенным и холмистым местам на черноземной почве. Состав травостоя довольно пестрый, без значительного господства отдельных видов; в большем или меньшем количестве встречаются виды разнотравья (Bupleurum scorzonerifolium, Centaurea monanthos, Vicia multicaulis, Thalictrum peta-loideum, Tanacetum sibiricum, Oxytropis myriophylla, O. grandiflora, Ser-ratula cardunculus), злаков (Роа botryoides, Aneurolepidium pseudoagropy-rum), к ним примешиваются Veratrum nigrum, Iris ruthenica, Sanguisorba officinalis. Hemerocallis minor, Potentilla tanacetifolia, Scorzoпега austriaca и др. Имеются переходы между остепненными разнотравными лугами и вострецовой степью. Остепненные луга с господством Hemerocallis minor занимают очень плоские, широкие понижения на ровных местах, а также дно широких падей; растительный покров довольно густой. Кроме господствующего Hemerocallis minor, в меньшем количестве встречаются Vicia amoena, Artemisia laciniata, Phlomis tuberosa и др.
Ломоносовая степь с господством Clematis hexapetala имеет более ограниченное распространение, занимает более ровные места и пологие склоны к падям. В дс!вольно густом травяном покрове, помимо ломоноса, встречаются Veratrum nigrum, Hemerocallis minor, Galium verum и др.
«Зонтичная» степь с господством Phlofodicarpus sibiricus занимает участки с более развитым рельефом и более выщелоченным черноземом. Травяной покров густой; к основному растению часто примешивается в большом количестве Tanacetum sibiricum.
Вышеуказанные сообщества остепненных лугов и луговых степей местами сочетаются с растительностью столбчатых солонцов, в составе которой господствуют Aneurolepidium pseudoagropyrum и Carex duriuscula.
Северные склоны падей п вершины холмов покрыты березняками из Betula platyphylla и осины, а также кустарниками из даурско-монгольской Betula Gmelini, ив, Spiraea media, а ближе к северной границе лесостепной полосы и Rhododendron dahuricum. Местами ерники & Betula Gmelini п ивами встречаются среди ровной степи в едва заметных западинах, лишенных солонцов.
• Литература
Г. И. Поплавская (1912, 1916); В. Н. Сукаче! (1912): М. Ф. Короткий (1913); В. И. Смирнов (1914); А. В. Куминова (1938); Л. П. Сергиевская (1951).
76. Разнотравно-дерновиннозлаковые европейско-казахстанские степи
Разнотравно-дерновиннозлаковые европейско-казахстанские степи, относящиеся к типичным (настоящим степям) и связанные в почвенном отношении в основном с обыкновенными и северными разностями южных черноземов, простираются то более широкой, то более узкой полосой от р. Прут1 на западе до предгорий Алтая на востоке.
Эти степи2 характеризуются преобладанием евксерофильных и еври-ксерофильных степных растений с примесью луговостепных мезоксеро-фильных и даже луговых ксеромезофильных растений. Таким образом, более мезофитные разности (ассоциации) этих степей представляют разнообразные переходы к луговым степям. В этих степях, как и следует из их названия, господствуют плотнодерновинные злаки, преимущественно крупнодерновинные ковыли из ряда Реппаtae, и отчасти из ряда Capil-lalae (Stipa capillata), реже Pseudocapillatae (S. Korshinskyi), но при ухудшении почвенных условий (на солонцах, каменистых почвах и пр.) или при усиленном выпасе господство переходит к мелкодерновинным злакам, главным образом к типчаку (Festuca sulcata s. 1.). Более или менее обильно представлено разнотравье, в составе которого много более мезофильных видов, общих с луговыми степями (так называемое «северное степное разнотравье»), но имеется также более или менее ксерофильное разнотравье, не встречающееся в луговых степях («южное степное разнотравье»).
На каменистых почвах различной литологии развиты обычно особые «каменистые» тимьянниковые степи, в составе которых, помимо дерновин-ных степных злаков, большую роль играют полукустарнички, обычно с укореняющимися надземными деревянистыми побегами (из родов Thymus, Teucrium, Scutellaria, Artemisia и др.). На оголенном каменистом субстрате эти полукустарнички образуют и самостоятельные сообщества — тимьяниики. Тимьянниковые степи и тимьянники развиваются
1 Эти степи заходят в соседние районы Румынской Народной Республики.
4 Разнотравно-дерновиннозлаковые степи, представленные преимущественно разнотравно-типчаково-ковыльными ассоциациями, назывались также следующим образом; широколиственным ковыльником (Высоцкий, 1908—1909, 1915, 1923; частью сюда также отнесены более ксерофитные разности луговых степей); ковыльными крупнодер-новинными степями (Келлер, 1916,1923а); степями ковыльно-типцовой подзоны (Крылов, 1916); красочным типце-ковыльником (Залесский, 1918а; частью сюда такжеотне-сены луговые степи); степями зоны разнотравно-злаковой степи (Новопокровский, 1921; 1925); красочным ковыльником (Алехин, 1925, 1926, 1936); ковыльно-разнотравной степью (Крашенинников, 1932); ковыльно-луговидными степями (Клеопов и Лавренко, 1933); дерновиннозлаковыми богаторазнотравными степями (Лавренко, 1940а).
также на песках. В результате формирования почвы на каменистом субстрате тимьянники переходят в тимьянниковые степи, а последние в типичные разнотравно-дерновиннозлаковые степи.
Имеются значительные отличия в составе этих степей в бассейне Черного моря и в Заволжье, северном Казахстане и прилегающих районах Западной Сибири до предгорий Алтая. В связи с этим, разнотравно-типчаково-ковыльные степи разделены на две провинциальные разности — «причерноморские» (76 ч) и «заволжско-казахстанские» (76 звк), гра ница между которыми проходит по Волге. Конечно, эта граница не является очень резкой, так как некоторые казахстанские степные виды проникают на правобережье Волги и даже далее на запад, в бассейн Дона, а некоторые причерноморские (понтические) виды заходят в Заволжье и, реже, далее на восток.
Причерноморские разнотравно-типчаково-ковыльные степи, а также соответствующие степи в Заволжье и в Западной Сибири почти целиком распаханы. В северном Казахстане, несмотря на большую распашку в 1953—1955 гг., еще сохранилось довольно много этих степей, главным образом на солонцеватых и щебнистых почвах.
76 ч — сельскохозяйственные земли на месте причерноморских разнотравно-типчаковоковыльных степей
Сельскохозяйственные земли на месте причерноморских (понтических) разнотравно-типчаково-ковыльных степей, связанных преимущественно с обыкновенными и отчасти северными разностями южных черноземов, простираются южнее лесостепи, от долины Прута до долины Волги. Между Прутом и Днепром полоса этих степей сравнительно узкая, а восточнее Днепра вплоть до Волги она сильно расширяется; к востоку от Азовского моря эти степи обширным массивом так называемых приазовских степей спускаются к югу, к предгорьям Кавказа в бассейне Кубани. Кроме того, на предгорной равнине Крымских гор, представляющей весьма пологий северный склон внещней куэсты, имеется изолированный от основного зонального массива район разнотравно-типчаково-ковыльных степей, которые можно рассматривать как самый нижний пояс растительности Крымских гор.1
На этом обширном пространстве, ранее занятом причерноморскими разнотравно-типчаково-ковыльными степями, геоморфологические условия довольно различны. Большая часть территории, на которой расположены причерноморские разнотравно-типчаково-ковыльные степи, относится к южным частям Русской равнины и представляет собой сравнительно возвышенную и дренированную местность. Часть полосы этих степей между Прутом и Днепром является очень пологим южным склоном Приднепровской, Волыно-Подольской и Бессарабской возвышенностей, тектонически соответствующим Украинскому кристаллическому щиту. Другая часть территории этих степей, примыкающая к северному берегу Азовского моря, относится к Приазовской возвышенности, тектонически соответствующей юго-восточной части Украинского кристаллического щита. Территория к юго-западу, а отчасти и к западу от Северского Донца относится к чрезвычайно сильно пенепленизированному, но в то же время расчлененному долинами мелких рек, Донецкому кряжу (Донецкая воз
1 Другой маленький изолированный массив этих степей показан на Тарханкутском
п-ве.
вышенность). Бблыная часть бассейна среднего Дона принадлежит к южной части Средне-Русской возвышенности, сильно расчлененной балками и оврагами, а бассейн Медведицы и правый берег Волги — к южной части Приволжской возвышенности, В эту обширную, в основном возвышенную и более или менее дренированную территорию включены сравнительно узкие пониженные (низменные) участки, слабо расчлененные эрозией и связанные с древними долинами рек: Приднепровская, Донецкая, Донская и Хоперская аллювиальные равнины. К последним приурочены массивы супесчаных и песчаных почв. К югу от Дона большая часть приазовского массива разнотравно-типчаково-ковыльных степей связана с Прикубанской низменностью, представляющей восточную часть .Азово-Черноморской низины. С этой низменности разнотравно-типчаково-ковыльные степи поднимаются на наиболее возвышенную часть ('.тавропольской возвышенности; большая часть пониженных склонов этого плато занята сухими типчаково-ковыльными степями. Таким образом, на этом плато довольно отчетливо выражено явление вертикальной поясности растительного покрова.
Почти все эти степи распаханы. Более крупные их участки сохранились только в заповедниках; например, заповедными являются Стрелецкая степь Ворошиловградской области и Хомутовская степь в Сталинской области.1
В составе причерноморских разнотравно-типчаково-ковыльных степей господствуют степные дерновинные злаки, преимущественно евксерофиль-ные и евриксерофильные, частично мезоксерофильные: ковыли {Stipa rubens — только в бассейне Дона, S. ucrainica, S. stenophylla, S. capillata, реже S. dasyphylla), из мелкодерновинных злаков наиболее обилен типчак {Festuca sulcata), довольно много тонконога {Koeleria gracilis).'- Узко-листный ковыль {Stipa stenophylla) обычно приурочен к пологим северным склонам водоразделов, украинский ковыль (61. ucrainica) встречается в более ксерофитных ассоциациях, преимущественно в тех частях полосы разнотравно-типчаково-ковыльных степей, которые граничат с полосой сухих типчаково-ковыльных степей, а также в приазовских степях. Количество ковылька (б1. Lessingiana) также возрастает в составе более ксерофитных ассоциаций разнотравно-типчаково-ковыльных степей. На многих сильно выпасаемых участках среди ковылей безраздельно господствует ковыль-тырса (5. capillata), а перистые ковыли встречаются единично или вовсе отсутствуют. В более мезофитных ассоциациях этих степей, переходных к луговым, в бассейне Дона в состав домпнантов иногда входит рыхлодерновинный злак Helictotrichon Schellianum. Из корневищных злаков обилен костер {Bromus riparius), реже мятлик {Роа angustifolia)-, другие еще более мезофильные виды {Calamagrostis epigeios, Bromus inermis, Agropyrum repens) встречаются единично в составе более мезофитных ассоциаций этих степей. Обычно в небольшом количестве растет корневищная осока {Carex supina). Более или менее обильно представлено разнотравье, в составе которого наблюдается смешение «северного степного разнотравья», более мезофильного (например Filipendula hexapetala, Trifolium montanum, Vicia tenuifolia, Polygala comosa, Myosotis suaveolens и др.) и более ксерофильного (например Medicago romanica, Euphorbia stepposa, Limonium latifolium, Salvia nutans, S. austriaca, Serratula radiata и др.). Полукустарнички немногочисленны; чаще встречаются 1 2
1 Эти две заповедные степи подведомственны Академии наук Украинской ССР.
2 Из упомянутых выше дерновинных злаков к мезоксерофитной группе относятся Stipa stenophylla и S. dasyphylla.
Artemisia austriaca и Jurinea multiflora. Довольно многочисленны эфемероиды и эфемеры, что связано с относительно влажной и теплой весной причерноморских степей (по сравнению с заволжско-казахстанскими степями, где весна более сухая и холодная). Из эфемероидов упомянем о следующих, более распространенных: Роа bulbosa, виды Gagea (G. еги-bescens, G. pusilia, G. bulbifera и др.), Tulipa Schrenkii (более ксерофильные ассоциации), Т. Biebersteiniana, Ornithogalum Gussonei,Bellevaliasarmatica, llyacinthella leucophaea, Crocus reticulatus и др. Из эфемеров чаще встречаются следующие виды: Holosteum umbellatum, Arenaria serpyllifolia. Alyssum desertorum, Erophila verna, Draba nemorosa, Arabis auriculata. Viola Kitaibeliana. Androsace elongala, Veronica verna и некоторые другие. Иногда в травостое этих степей разбросаны побеги Caragana frutex, но нередко этот стенной кустарник образует более или менее компактные заросли в виде куртин, приуроченных к северным склонам водоразделов пли балок.
Причерноморские разнотравно-типчаково-ковыльные степи, вернее теперь культурные земли на их месте, занимают возвышенные и в большинстве случаев хорошо дренированные реками и балками водоразделы. В связи с этим па сохранившихся крупных участках этих степей наблюдаются характерные для балочного рельефа сочетания растительных группировок. Водоразделы заняты разнотравно-типчаково-ковыльными степями «средней» ксерофитности, по северным склонам водоразделов наблюдаются несколько более мезофитные ассоциации, а по южным склонам — более ксерофитные ассоциации этих же степей. По склонам балок северной экспозиции располагаются луговые степи, а по низу склонов — остепненные луга; те и другие наблюдаются и по дну неглубоких ложбин стока на водоразделах. На южных, обычно более крутых склонах балок, представлены различные «эдафогенные» степные ассоциации, связанные с тем, что здесь почвообразуют или даже обнажаются древние пески пли каменистые породы различной литологии. На песчаных или супесчаных почвах по склонам балок наблюдаются псаммофитные или гемппсаммофитные ассоциации разнотравно-типчаково-ковыльных или даже луговых степей со Stipa Joannis, Agrostis Syreistschikovii и псаммо-фитным разнотравьем — Potentilla arenaria, Pulsatilla nigricans 1 и др. (Келлер, 1931в; Лавренко и Дохман, 1933).
В тех случаях, когда по склонам балок, особенно в их циркообразных верховьях, к поверхности почвы подходят засоленные породы, развиваются столбчатые солонцы с растительностью типичных бедноразнотравных или даже пустынных степей. Обычно преобладает типчак, к которому в большом количестве примешивается эфемероид Роа bulbosa, а также немногочисленное степное ксерофильное разнотравье (чаще всего Linosyris villosa) или степные и пустынно-степные полукустарнички (Kochia prostrata, Artemisia austriaca, A. monogyna).
По склонам балок, преимущественно северным или западным, обычно развиваются заросли степных кустарников (Prunus stepposa, Cerasus fruticosa, Amygdalus папа, Spiraea hy pericifolia, S. crenata, Caragana frutex, Cytisus ruthenicils). Иногда в зарослях степных кустарников встречаются отдельные экземпляры древесных пород, чаще ильм (Ulmus folia-сеа), дикие груши и яблони (Pyrus communis и Malus praecox).
Остановимся вкратце на сезонном развитии разнотравно-типчаковоковыльных степей на примере целинных степей конных заводов в бассейне
1 При более узком понимании вида это — понтический вид Pulsatilla ucrainica (Ugr.) Wissjul. (Флора УРСР, т. 5, 1953).
Деркула в Ворошиловградской области (Танфильев, 1898; Лавренко ir Дохман, 1933)*
1. 2/2 III—1/2 IV. Первая (ранняя) весенняя фаза. Между побуревшими прошлогодними пучками травы местами проглядывает зелень мха Tortula ruralis и темнозеленые пластинки синезеленой водоросли Stratonostoc commune.
2. 2/2 IV. Вторая (средняя) весенняя фаза. Степь постепенно начинает зеленеть. По степи разбросаны куртины цветущего Iris pumila с желтыми, фиолетовыми и синими цветами. Цветут желтые Adonis wolgensis и гусиные луки (Gagea pusilia, G. erubescens), а несколько позже фиолетовая Viola ambigua. Из эфемеров-однолетников цветут довольно обильные Erophila verna, Holosteum umbellatum с белыми цветами, а также белоцветковая Androsace е long at а и желтоцветная Draba nemorosa. На солонцах цветут эфемероиды-тюльпаны: Tulipa Schrenkii с желтыми и красными цветами и обильный здесь желтоцветный Tulipa Biebersteiniana. По песчаным склонам цветут фиолетово-синие Pulsatilla nigricans и Р. patens.
3. 1/2 V. Третья (поздняя) весенняя фаза. На зеленом фоне подросших злаков и разнотравья цветут многочисленные двудольные: белоцветковые Lathyrus раппо-nicus, L. pallescens, Anemone silvestris, розовая Valeriana tuberosa, желтые Potentilla humifusa nP. Schurii. Цветут также степные осоки (Carex supina, С. praecox). По склонам водоразделов и балок, преимущественно северной экспозиции, цветет пунцовый Paeonia tenuifolia, встречающийся обычно большими скоплениями, а по ложбинам стока — темнорозовый Pedicularis dasystachys. Из эфемеров-однолетников цветут Alyssum deser-torum, Arabis auriculata, Viola Kitaibeliana, Veronica verna. В эту же фазу цветут степные кустарники: Prunus stepposa покрывается белыми цветами, Amygdalus папа в дерезняках — розовыми. К середине мая зацветает дереза (Caragana frutex), заросли которой становятся при этом золотисто-желтыми.
К концу фазы появляются первые «перья» (опушенные ости) у Stipa Joannis и S. rubens.
4. 2/2 V—1/2 VI. Первая (ранняя) летняя фаза, наиболее красочная. Цветут степные злаки и многочисленные двудольные. Опушенные ости цветущих перистых ко-вылей — Stipa rubens и S. Lessingiana, а по северным и песчаным склонам S. Joannis образуют волнующуюся белую пелену. Цветут также остальные злаки, менее аспек-тивные: Festuca sulcata, Koeleria gracilis, Bromus riparius. Из многочисленных цветущих двудольных выделяется обильный фиолетово-синий Salvia nutans. Из более обычных цветут также: белые Filipendula hexapetala, Trifolium montanum, Arenaria ucrai-nica, голубые Myosotis suaveolens, Campanula uralensis, красная Jurinea arachnoidea, сиренево-красная Vicia tenuifolia, темнокрасный синяк (Echium rubrum) и многие другие. Разбросаны белые шарообразные соцветия цветущего Crambe tatarica (биологический тип «перекати-поле»).
5. 2/2 VI—1/2 VII. Вторая (средняя) летняя фаза. Период полупокоя многих господствующих злаков: вышеупомянутые виды ковыля теряют свои «перья» и также, как типчак и тонконог, присыхают, в связи с чем степь принимает увядший вид. По северным склонам местами разнообразят аспект желтоватые ости цветущего в это время Stipa stenophylla.
Цветущее разнотравье менее обильно, чем в предыдущий период. Выделяются цветущие желтоцветные виды — Galium ruthenicum, Centaurea ruthenica, обильно цветет Plantago stepposa; часто раскиданы темнорозовые корзинки Serratula radiata, реже фиолетово-розовые соцветия Jurinea multiflora. В это время цветут также некоторые «перекати-поле»: выделяются одиночные розовато-сиреневые шарообразные соцветия Goniolimon tataricum, сиреневые «шары» Serratula xeranthemoides, красные Phlomis pungens, синие Limonium latifolium, а на песчаных склонах также серовато-белые «шары» Gypsophila paniculata.
6. 2/2 VII—1/2 IX (—2/2 IX). Позднелетняя и раннеосенняя фаза. Период цветения ковыля-тырсы (Stipa capillata), который зацветает то раньше, то позже, в зависимости от климатических условий. Аспект этого периода в основном определяют желто-зеленые с металлическим оттенком ости этого ковыля. Большую роль в аспекте играют также желтые и белые соцветия зонтичных: желтые*— Peucedanum ruthenicum, Р. Lubimenkoanum, Bupleurum falcatum (склоны северной экспозиции), белые — Se-seli campestre (очень часто), Libanotis intermedia, Falcaria sioides. Цветут также желтоцветные Verbascum lychnitis, V. orientale, Cephalaria uralensis, Scabiosa ochroleuca, полупаразит Orthantha lute а. В годы с влажным летом наблюдается вторичное цветение многих весенних и раннелетних растений (Anemone silvestris, Clematis integrifolia, Polygala comosa и др.).
7. 2/2 IX (1/2 X)—1/2 XI. Позднеосенняя фаза. Период предзимнего «замирания» степи. Травостой желтеет и присыхает. Доцветают только единичные экземпляры зонтичных и ворсянковых. После осенних дождей появляются всходы озимых одно-
Рис. 66. Разнотравно-типчаково-ковыльная степь с большим количеством Campanula multiflora (канделябровидное растение); ассоциация, переходная к луговой степи. Провал ьская степь на Донецком кряже Ворошиловград-ской области (Украинская ССР). (Фот. Н. А. Десятовой).
Рис. 67. Дерновиннозлаковая степь — типчаково-ковылковая (Stipa Lessingiana-г Festuca sulcata) с небольшой примесью Artemisia sublessingiana. Верхняя часть бассейна Сарысу, Казахский мелкосопочник. (Фот. В. И. Грубова).
летников (эфемеров), новые листья у Роа bulbosa и у многих дерновинных злаков (типчака, ковылей).
Таким образом, у разнотравно-типчаково-ковыльных степей уже намечается период пол упокоя части травостоя, который приурочен к наиболее жаркому времени — второй половине июня и июлю.
Урожайность травостоя этих степей в среднем около 10—12 ц/га (Куксин, 1935).
Разнотравно-типчаково-ковыльные степи на обширном пространстве между Прутом и Волгой и до предгорий Кавказа на юге, конечно, не однородны. Выделяется ряд их то менее, то более ксерофитных вариантов (описаны для Украинской ССР Ю. Д. Клеоповым и Е. М. Лавренко, 1933; для части бассейна Дона — К. М. Залесским, 1918а; см. также работы Б. А. Келлера, 1903, 1931а; И. В. Новопокровского, 1921, 1927а, 1935; Т. И. Попова, 1931; Н. Ф. Комарова, 1931; для Крыма — Н. А. Де-сятовой-Шостенко и М. С. Шалыта, 1936).
Большинство дерновинных злаков этих степей является общим для причерноморских и заволжско-казахстанских разнотравно-типчаково-ковыль-ных степей (паннонско-причерноморско-казахстанские, причерноморско-казахстанские и более широко распространенные виды): Stipa rubens (на запад до бассейна Дона включительно), S. stenophylla, S. dasyphylla, S. capillata, Festuca sulcata s. 1., Koeleria gracilis, Helictotrichon Schellianum (на запад до бассейна Дона включительно). Такое же распространение имеют многие степные корневищные злаки и осоки (Роа angustifolia, Carex supina и др.), а также многочисленные виды разнотравья: Adonis wolgensis, Anemone silvestris, Filipendula hexapetala, Potentilla humifusa, Medicago romanica, Vicia tenuifolia, Oxytropis pilosa, Libanotis intermedia, Falcaria sioides, Peucedanum Lubimenkoanum, Myosotis suaveolens, Salvia stepposa (на запад до бассейна Дона включительно), Plantago stepposa, Galium ruthenicum, Scabiosa ochroleuca, Centaurea ruthenica и др. Так же распространены полукустарнички Artemisia austriaca и Jurinea multiflora. Целый ряд видов встречается преимущественно в причерноморских степях, главным образом в разнотравно-типчаково-ковыльных, а частью также и в луговых или сухих типчаково-ковыльных степях, но заходит также в пределы заволжской части заволжско-казахстанских разнотравно-типчаково-ковыльных, а отчасти типчаково-ковыльных степей.1 К этой группе принадлежат: паннонско-причерноморские виды — Bromus riparius, Iris pumila, Paeonia tenuifolia, Crambe tatarica, Viola ambigua, Echium rubrum и др. ; кавказско-причерноморские виды — Lathyrus pallescens', причерноморские (понтические) виды — Dianthus lanceo-latus, Peucedanum ruthenicum, Limonium latifolium, Salvia nutans, Jurinea arachnoidea', причерноморско-прикаспийский вид — Serratula xeranthemoi-des 2 (несколько заходит к востоку за р. Урал) и др. Наконец, ряд степных видов вовсе не заходит к востоку за Волгу: из паннонско-причерномор-ских — Potentilla Schurii, Salvia austriaca', из причерноморских видов — Stipa ucrainica, Clematis pseudoflammula, Euphorbia stepposa и др.
Из эфемероидов общими для причерноморских и заволжско-казахстанских степей являются следующие: Роа bulbosa, Gagea pusilia, G. bulbifera, Tulipa Schrenkii, T. Biebersteiniana', заходит в Заволжье как редкое растение Hyacinthella leucophaea (причерноморский вид); Волгу не переходят Ornithogalum Gussonei (причерноморско-средиземноморский вид),
1 Эти виды заходят или только в западную часть Заволжья, или доходят до Уральских гор и р. Урала в его среднем течении.
8 Serratula xeranlhemoides и Adonis wolgensis — полуэфемероиды.
12 Заказ 551
Bellevalia sarmatica (причерноморский вид), Crocus reticulatus (панионско-причерноморский вид) и др.
Из степных кустарников общими для причерноморских и заволжско-казахстанских степей являются Amygdalus папа, Spiraea crenata, S. hy-pericifolia, Cerasus fruticosa, Caragana frutex. Причерноморский вид Prunus stepposa на восток доходит до Урала.
Очень большое количество причерноморских видов (в том числе узких эндемов) имеется в составе степных группировок на песках и каменистых склонах.
Вне заповедников целинные участки разнотравно-типчаково-ковыльных степей подвергаются усиленному выпасу, в результате чего растительный покров этих степей сильно изменяется в направлении ксерофитиза-ции, что связано с уплотнением верхних горизонтов почвы при сильном выпасе с усиленным испарением с поверхности почвы (Высоцкий, 1905; Келлер, 1916; Пачоский, 1917; Залесский, 1918, 1918а; Горшкова, 1954).
Стадии пастбищной (пасквальной) дигрессии разнотравно-типчаковоковыльных степей следующие.
1. Стадия умеренного выпаса (ковыльная), при которой сохраняется типичная структура и состав разнотравно-типчаковоковыльных степей со значительным или преобладающим участием ковылей.
2. Стадия угасания ковылей (типчаковая). При усилении выпаса ковыли начинают постепенно выпадать из травостоя. Первыми исчезают Stipa stenophylla и S. dasyphylla (в составе менее ксерофитных ассоциаций этих степей), затем S. rubens и S. Lessingiana и, наконец, S. capillata, который из всех ковылей наиболее устойчив к выпасу. Место ковылей занимается типчаком. Часто типчак в этой стадии ассоциируется с корнеотпрысковым полукустарничком {Artemisia austriaca), образуя полынково-типчаковые ассоциации, а также с молочаем и другими малосъедобными для скота видами разнотравья. Выход растительной массы сильно уменьшается по сравнению с предыдущей стадией.
3. Стадия тонконогового сбоя. При еще более усиленном выпасе типчак начинает изреживаться, и его место как преобладающего растения занимает эфемероид тонконог или живородящий мятлик {Роа bulbosa var. vivipara), игравший в предыдущих стадиях второстепенную роль. Весной развиваются также более или менее обильные эфемеры {Ceratocepkalus orthoceras, Alyssum desertorum, однолетние вероники и др.). К концу мая тонконог и эфемеры засыхают. Летом на таких Тонконоговых сбоях выделяются полукустарничковые полыни {Artemisia austriaca, а на солонцеватых черноземах — A. monogyna), нередко волосистая грудница {Linosyris villosa), ромашник {Pyrethrum millefolia-tum), а также молочаи {Euphorbia stepposa, E. Seguieriana), чабрец {Thymus Marschallianus) и др. К концу лета и осенью в годы с влажным вегетационным периодом на Тонконоговых сбоях в большом количестве развиваются некоторые летне-осенние однолетники, особенно Eragrostis minor, а на поверхности почвы — синезеленая водоросль {Stratonostoc commune) и некоторые лишайники.
4. Стадия выгона, наблюдающаяся около стойбищ скота и непосредственно у селений. Тонконог и большинство многолетников исчезают. Развиваются преимущественно летне-осенние однолетники с распростертыми стеблями — Atriplex tatarica, Ceratocarpus arenarius, Polygonum aviculare и др. После уничтожения и этих растений остается голая почва («тырло»).
Эта схема пастбищной дигрессии степного травостоя в основных чертах оказывается верной и для более южных сухих типчаково-ковыльных степей как причерноморских, так и заволжско-казахстанских.
И. К. Пачоский (1917) считает, что при недостаточном выпасе или при отсутствии последнего (при отсутствии также сенокошения) происходят изменения степного травостоя в сторону некоторой мезофитизации или «олуговения», что выражается, в основном, в угнетении степных дерновинных злаков, появлении в большом количестве более или менее мезофильных корневищных злаков (ползучий пырей, безостый костер), а также увеличении распространения некоторых видов многолетнего разнотравья, уменьшении распространения однолетников и пр. Все это, в основном, связано с накоплением на поверхности почвы относительно мощного мертвого покрова, способствующего уменьшению почвенного испарения и препятствующего семенному возобновлению большинства степных растений.
Приведем основные стадии изменения растительности залежей после прекращения распашки или, как назвал этот процесс Г. Н. Высоцкий (1915), «зацелинения» залежей (Черняев, 1865 ; Танфильев, 1898; Залесский, 1918; Высоцкий, 1923; Мальцев, 1924; Пачоский, 1927; Аврорин, 1934).
1. Стадия полевых сорняков, преимущественно одно-и двулетников (Carduus acanthoides, С. Thoermeri и др.). В тех случаях, когда поле, бросаемое под залежь, было засорено многолетними сорняками, последние могут быть обильны и в первой стадии демутации залежи. Так, на одногодичных залежах иногда преобладают корнеотпрысковые сложноцветные (Cirsium setosum, Sonchus arvensis) или корневищный злак — ползучий пырей.
2. Стадия корневищных растений, главным образом злаков, преимущественно ползучего пырея, в некоторых случаях Hiero-chloe odorata; на выпасываемых залежах в этой стадии много корнеотпрыскового полынка (Artemisia austriaca).
3. Стадия дерновинных злаков. По мере старения залежи на ней все более и более увеличивается количество дерновинных злаков, сначала типчака (Festuca sulcata) и тонконога (Koeleria gracilis), а затем и ковылей.
4. Стадия вторичной целины, когда в травостое восстанавливается типичная структура и состав разнотравно-типчаковоковыльной степи.
Подобные же стадии с некоторыми отличиями наблюдаются и при за-целинении степных залежей на нормальных (несолонцеватых) степных почвах в полосе сухих типчаково-ковыльных степей как причерноморских, так и заволжско-казахстанских.
В настоящий момент в этой полосе распахано не только большинство причерноморских целинных разнотравно-типчаково-ковыльных степей, но и залежей.
В пределах ареала причерноморских разнотравно-типчаково-ковыль-ных степей вдоль левобережий крупных рек — Днепра, Северского Донца и Дона и некоторых их притоков — находятся сравнительно широкие полосы песков и супесей с соответствующими эдафическими вариантами этих степей. Ближе к реке располагаются песчаные пространства с бугристым рельефом, а дальше от реки — супеси с более выравненной поверхностью. Вдоль левобережья Хопра, а также вдоль Чира и Ка-литвы и ее притоков располагаются главным образом супеси.1 1 2
1 На опубликованной «Карте растительности Европейской части СССР», м. 1 :
2 500 000 плкя.чяны отдельно «типчаково-ковыльные и змеевково-типчаково-ковыль-
Кроме того, в пределах более южной полосы типчаково-ковыльных (сухих) степей в низовьях Днепра, на так называемых Алешковских песках, а также на Цимлянском песчаном массиве на нижнем Дону степи на песках (но не на супесях) также относятся к разнотравно-типчаковоковыльным степям, что связано с более благоприятными условиями почвенной влажности на песках, по сравнению с соседними суглинистыми и даже супесчаными почвами.
Все эти эдафические варианты разнотравно-типчаково-ковыльпых степей на легких почвах (песках и супесях) показаны условным знаком песка.
«Супесчаные» (гемипсаммофитные) степи в большинстве случаев распаханы и используются под полевые культуры, а окультуривание песчаных пространств выражается главным образом в закреплении песков, преимущественно посадками сосны (Pinus silvestris, а на юге Украины, в полосе типчаково-ковыльных степей, местами также Р. Pallasiana). Пески используются также как пастбища. Неумеренная пастьба на песках способствует разбиванию дернины и последующей дефляции. Большие пространства песков в настоящее время находятся в стадии естественного зарастания или закрепления с помощью посадок сосны.
В составе степей на супесях (Новопокровский, 1915; Гаель, 1929,1932, 1932а; Гожев, 1929) среди дерновинных злаков преобладают, с одной стороны, Festuca sulcata, Koeleria gracilis и Stipa capillata, свойственные преимущественно степям с суглинистыми или даже глинистыми почвами, и, с другой, —Stipa borysthenica (=S. Joannis ssp. sabulosa) и Cleistogenes squarrosa (на запад до Днепра), свойственные легким почвам. Среди разнотравья встречаются как виды, приуроченные преимущественно к суглинистым почвам {Artemisia austriaca, Eryngium campestre, Taraxacum serotinum) или встречающиеся одинаково часто на легких и тяжелых почвах (например Euphorbia Seguieriana, Seseli campestre и др.), так и виды, связанные большей частью с песчаными почвами (Dianthus platyodon [ = D. polymorphus auct.], Scabiosa ucrainica, Helichrysum arenarium, Achillea Gerberi и др.).
«Песчаные» (псаммофитные) разнотравно-типчаково-ковыльные степи на почвах песчаного механического состава (песчаных черноземах или, как их часто называют, «серопесках») характеризуются весьма своеобразным составом; большинство видов, произрастающих в этих степях, свойственны только песчаному субстрату и на суглинистые почвы никогда не переходят (Новопокровский, 1915; Гаель, 1929, 1932, 1932а; Гожев, 1929; Лавренко, 1927, 1935; Лавренко и Прянишников, 1926; Лавренко и Порецкий, 1928).
Основная роль в составе этих степей принадлежит причерноморско-казахстанским (по характеру их ареала) евксерофильным и мезоксеро-фильным видам дерновинных злаков, свойственным только песчаному субстрату: Festuca Beckeri, Stipa borysthenica (=S. Joannis ssp. sabulosa), Koeleria sabuletorum,* 1 а на несколько более пониженных степных песках также Agropyrum Lavrenkoanum (причерноморский вид?); по еще более пониженным пескам к нему присоединяется плотнодерновинное осоковое — Holoschoenus vulgaris (средиземный вид). Довольно обильно
иые степи на супесях» и «разнотравно-типчаково-ковыльные степи на песках, зарастающие и голые пески».
1 М. В. Клоков («Визначник рослин УРСР», Кй1в—Харюв, 1950) отчленяет от типичной бореальной Koeleria glauca (Schkuhr) DC., которую он^рассматривает как характерную для боровых песков, две весьма близкие расы,~ свойственные степным пескам — К. sabuletorum Czern. (причерноморско-казахстанский вид) и К. borysthenica Klok. (эндем степных песков нижнего Днепра от Запорожья до гирла).
также разнотравье, из которого преобладают чаще всего стержнекорневые Euphorbia Seguieriana (причерноморско-казахстанский вид) и Artemisia Marschalliana (также), предпочитающие легкие почвы, но часто произрастающие и на суглинистых почвах. На вершинах песчаных бугров или на участках с нарушенным в результате усиленного выпаса травостоем к дсрновинпым злакам в большом количестве присоединяются полукустарнички с лежачими укореняющимися побегами — Thymus Pallasianus (причерноморский вид), Т. borysthenicus (эндем песков нижнего Днепра — от Запорожья до гирла Днепра), являющиеся облигатными псаммофитами. Подобные тимьяно-дерновиннозлаковые ассоциации песчаных степей должны быть отнесены к тимьянниковым степям. Из других видов многолетнего стержнекорневого разнотравья, приуроченных к «песчаным» степям, упомянем о следующих причерноморских псаммофитах: Dianthus platyodon, Goniolimon graminifolium (эндем долин рек южной части Украинской ССР), Asperula graveolens, а из корнсотпрыскового разнотравья о Achillea Gerberi (причерноморско-казахстанский вид), Scorzonera ensi-folia (также), Jurinea laxa (эндем песков нижнего Днепра) и др.
Растительный покров разнотравно-дерновиннозлаковых степей, а также сочетающихся с ними тимьяно-дерновиннозлаковых степей на песчаных почвах, более изрежен, чем травостой аналогичных группировок на тяжелых почвах. В связи с этим в песчаных степях в годы с более влажным вегетационным периодом встречается значительное количество малолетников, многие из которых являются облигатными псаммофитами; например среди двулетников сюда относятся Scabiosa ucrainica, Silene parviflora (паннонско-причерноморско-казахстанский вид), Syrenia ucrainica (причерноморский вид), S. dolichostylos (также), Onosma borystheni-cum (также); к двулетникам также относятся узколокальные причерноморские псаммоэндемы из рода Centaurea (например С. margaritacea и С. margarita-alba — пески по Южному Бугу у Николаева, С. konkae — пески по Днепру у Днепропетровска и Запорожья, С. breviceps— пески по нижнему Днепру и др.) и пр. Среди однолетников представлены как эфемеры, так и летне-осенние однолетники. В годы с более влажным вегетационным периодом некоторые летне-осенние однолетники — облигатные псаммофиты—развиваются в большом количестве, например Kochia lani-flora (паннонско-причерноморско-казахстанский вид), Polygonum arenarium (паннонско-причерноморский вид), Р. Bordzilovskii(причерноморский вид).
«Песчаные» разнотравно-дерновиннозлаковые степи на песчаных террасах рек в настоящее время сохранились только отдельными участками. Ранее они преобладали на большей части территории песков не только в пределах полосы (подзоны) разнотравно-типчаково-ковыльных степей, но и полосы типчаково-ковыльных (сухих) степей, сочетаясь в понижениях между песчаными буграми с древесными колками из березы,1 дуба и черной ольхи, болотами (часто кочковатыми из Carex omskiana и др.), пресными и солончаковатыми лугами и солончаками (последние в низовьях Днепра).
В настоящее время на песчаных террасах рек в результате неумеренного выпаса преобладают разбитые пески, находящиеся в различных стадиях зарастания: от голых песков до хорошо заросших, вплоть до «вторичной» песчаной степи. На песках Причерноморской степной провинции пески проходят следующие стадии зарастания (демутации).
1. Стадия голых песков, совершенно лишенных растительности или с единичными экземплярами пионеров зарастания.
1 На песках встречаются березы из группы Betula pubescens s. 1., например на песках низовьев Днепра и Южного Буга — В. borysthenica (Гринь, 19546).
2. Стадия слабозаросших песков (стадия корневищных растений). Заросли растений занимают 5—25% всей площади этих песков. Преобладают одно- или маловидовые заросли из длиннокорневищных злаков: Agropyrum dasyanthum (эндем бассейна нижнего и отчасти среднего Днепра), A. tanaiticum (эндем бассейна среднего и нижнего Дона), Calamagrostis epigeios (широко распространенный бореальный вид), Elymus sabulosus (причерноморский вид), Е. giganteus (казакстанско-центральноазиатский вид, заходящий в бассейн Дона). Местами в зарастании песков большую роль играют «вынужденнокорневищные» кустарники (Salix acutifolia, S. rosmarinifolia и причерноморско-западноказахстанский вид Cytisus borysthenicus) и полукустарнички (Artemisia arenaria s. I.).1 По более пониженным участкам зарастающих песков более или менее обильна длиннокорневищная осока Carex colchica. Однолетники обычно не играют заметной роли в зарастании перевеваемых ветром песков.
3. Стадия среднезаросших песков (стадия корневищных и стержнекорневых растений). Заросли растений занимают 25—50% от всей площади. Количество зарослей длиннокорневищных растений и «вынужденнокорневищных» кустарников увеличивается. На площадках среднего уровня (по мезорельефу) в зарослях появляются в большом количестве стержнекорневые многолетники (главным образом Euphorbia Seguieriana), двулетники (например Syrenia ucrainica, Centaurea breviceps и др.) и однолетники (например Polygonum arenarium).
4. Стадия хорошо заросших песков (стадия стержнекорневых растений). Сформированный растительный покров занимает 50—80% всей площади; остальная территория падает на еще обильные котловины выдувания. Количество зарослей длиннокорневищных растений и «вынужденнокорневищных» кустарников и полукустарничков уменьшается; преобладают стержнекорневые многолетники, главным образом Euphorbia Seguieriana и Artemisia Marschalliana, к которым примешиваются двулетники, однолетники и отдельными дерно-винами степные злаки.
5. Стадия вторичной степи (стадия дерновинных злаков). Котловины выдувания занимают не больше 5—20%. В травостое преобладают псаммофильные дерновинные злаки, а также стержнекорневые двудольные с примесью полукустарничков, корнеотпрысковых двудольных, двулетников и однолетников (Лавренко, 1927; Гаель, 1932).
Растительность каменистых обнажений разных горных пород, обычно приуроченных к правым, более высоким и крутым, берегам рек или к склонам балок, характеризуется большой пестротой: при движении по склону сверху вниз или поперек падения склона растительные ассоциации, либо чаще их фрагменты, быстро сменяют одна другую, что связано с пестротой условий существования на склонах и с их динамикой (эрозионные процессы).2
Обычно на обнажениях идет разрушение более или менее сложившихся растительных группировок, что происходит при наличии интенсивных процессов эрозии или, наоборот, формирование подобных сообществ при
1 У этих кустарников и полукустарников на засыпанных песком ветвях появляются обильные придаточные корни; эти растения, таким образом, прорастают засыпающий их песок.
* Далее растительность обнажений различной литологии описывается для причерноморских степей в целом, то есть как для полосы разнотравно-типчаково-ковыльных, так и для полосы типчаково-ковыльных степей.
зарастании смытых склонов, если процессы эрозии затухают. Развитие эрозионных процессов обусловливается как естественными факторами (большая крутизна склонов, подмыв последних рекой и пр.), так и влиянием хозяйственной деятельности человека (уничтожение лесов, усиленный выпас и т. д.).
В начальных процессах зарастания обнаженных горных пород большую роль играют ксерофильпые полукустарнички из родов Thymus, Teucrium, Hyssopus, Alyssum, Artemisia и другие, которые, обычно совместно с некоторыми злаками и видами разнотравья, образуют очень характерные для каменистых обнажений слабо сомкнутые группировки — «ти-мьянники». Эти тимьянники в дальнейшем, при некотором затухании процессов эрозии и накоплении гумуса в верхних горизонтах субстрата (начало формирования черноземной почвы), сменяются степными сообществами с господством дерновинных злаков (из родов Festuca, Koeleria, Agropyrum, Stipa), но еще с большей или меньшей примесью растений, характерных для обнажений, в том числе и упомянутых полукустарничков. Это — тимьянниковые (петрофитные) степи, которые по мере формирования почвы сменяются более устойчивыми степными сообществами — типичными или луговыми степями, в зависимости от экспозиции склона.
Выше кратко охарактеризован прогрессивный ряд растительных сообществ в процессе зарастания обнаженных горных пород в степной области. Этот ряд может с некоторыми модификациями располагаться и в обратном порядке: типичностепные ассоциациитимьянниково-степ-ные ассоциации-> тимьянниковые ассоциации. Это имеет место при интенсивно идущих процессах эрозии, например в результате усиленного выпаса, при которрм растительный покров деградирует.
Растительность обнажений используется как пастбище очень плохого качества. Ряд растений обнажений может быть использован для закрепления смытых склонов.
С обнажениями различных пород в степной Причерноморской провинции связано большое число эндемичных, часто узколокальных видов. Род Thymus представлен здесь рядом мелких, специфических для обнажений определенной литологии, видов.
На «Геоботанической карте», помимо растительности меловых обнажений, которые показаны отдельно особым внемаспггабным знаком («растительность меловых обнажений), растительность большинства обнажений, отмеченных соответствующим внемасштабным знаком («растительность каменистых обнажений разных пород...»), не дифференцирована по литологическим особенностям субстрата, что было сделано для южной части Русской равнины на «Карте растительности Европейской части СССР» м’. 1 : 2 500 000 (1948-К).
Однако существуют некоторые отличия в растительном покрове обнажений сланцев и песчаников карбона Донецкого кряжа, гранитов Украинского кристаллического массива и третичных известняков Причерноморья и Приазовья. Поэтому далее петрофитная растительность Причерноморской степной провинции характеризуется с учетом литологии обнажений.
Растительность обнажений сланцев, песчаников и реже известняков каменноугольного возраста свойственна Донецкому кряжу, где обнажения этих пород очень обычны по склонам долин рек, многочисленных балок и иногда даже на водоразделах. .
Растительность этих обнажений, как и обнажений других пород, носит очень пестрый характер, что зависит от разнообразия условий существования на склонах. Часто значительную роль играют степные дерно-винные злаки (Festuca sulcata, Agropyrum pectiniforme, Koeleria gracilis-
Stipa capillata) и другие степные растения. Обычны также растения обнажений, не привязанные к породам определенной литологии: из полукустарничков— Ephedra distachya, Silene supina, Alyssum tortuosum, Teucrium polium, Thymus dimorphus', из многолетнего травянистого разнотравья — Pimpinella titanophila, Cephalaria uralensis, Helichrysum arenarium, Pyrethrum millefoliatum, Achillea leptophylla и др.; из малолетников— Dianthus pseudarmeria, Alyssum hirsutum, Xeranthemum annuum (Клеонов, 1933a).
В составе флоры обнажений каменноугольных сланцев и песчаников мало видов, эндемичных этим обнажениям в пределах Донецкого кряжа: к последним относятся Stachys krynkensis (по р. Крынке) и Scrophularia donetzica.
Обнажения кристаллических пород — гранитов и гнейсов — с характерной растительностью встречаются в степной части Украинского кристаллического массива (щита), в двух районах: 1) к западу от Днепра, по рр. Южный Буг, Ингул, Ингулец и Бузулук; 2) в Приазовье, по рр. Молочная (верховье), Обиточная, Берда, Кальчик, Кальмиус и Груз-ский Еланчик. Таким образом, эти обнажения показаны преимущественно в полосе разнотравно-типчаково-ковыльных степей.
Растительный покров обнажений гранитов и гнейсов, как обычно, очень пестрый. На более мелкоземистых участках преобладают степные растения. На щебнистых местах, помимо растений, широко распространенных на обнажениях различных пород (из полукустарничков — Ephedra distachya, Thymus dimorphus, Teucrium polium, из дерновинных злаков — Cleistogenes maeotica, из травянистого разнотравья — Potentilla astraca-nica, Pimpinella titanophila, Cephalaria uralensis, Achillea leptophylla и др.), встречается ряд видов, свойственных только обнажениям кристаллических пород юга УССР, часто с очень ограниченными ареалами.
Особенно много таких узколокальных эндемичных видов по южной окраине приазовской части Украинского кристаллического массива: Erodium Beketovii (по Кальмпу су), Thymusgraniticus (тамже), Т. pseudogra-niticus (по Нальчику), Achillea glaberrima (гранитные сопки — Каменные Могилы в верховьях Берды), Centaurea pseudoleucolepis (там же). Из них виды рода Thymus— полукустарнички (Лавренко, 1925; Клеопов, 1933а).1
В составе флоры гранитных обнажений по южной окраине Украинского кристаллического массива, между Днестром и Днепром, такими местными эндемиками являются полукустарничковая Dianthus hypanicus-и двулетняя Silene hypanica. Здесь же имеется местный эндем — кустарничек Cytisus Skrobiczewskii, растущий не только на гранитах, но и на известняковых обнажениях, а также эфемероидное травянистое растение Leontice odessana (весьма близкое к алтайской L. altaica), которое, кроме гранитов, встречается и на других субстратах и, помимо этого района,, появляется снова в Добрудже. Кроме того, в этом районе (между Днестром и Днепром) на гранитах произрастают следующие виды с более широким ареалом, имеющие здесь островные местонахождения: Kohlrauschia рго-lifera (основной ареал в Средней Европе, Малой Азии и на Кавказе),. Alyssum saxatile (также в Приазовье; основной ареал в Средней Европе), Stachys angustifolia (Балканский п-ов, Крым) (Пачоский, 1917; Котов,. 1927а; Клеопов, 1933а).
На гранитных обнажениях встречается также ряд растений, растущих в других районах на песках (Rumex acetosella var. multifidus, Alyssum mi-
1 M. В. Клоков («Визначник рослин УРСР», Кшв—Харюв, 1950) в качестве-эндемичного вида для гранитов и гнейсов Украины приводит также Stipa graniticolcs Klok. из родства S. Joannis s. 1.
nutum, Sedum acre, Sempervivum ruthenicum, Anthemis ruthenica и др.). Кроме того, в расщелинах гранитных скал произрастает ряд более или менее широко распространенных скальных папоротников: Cystopteris-fragilis, Asplenium trichomanes, A. septentrionale, Woodsia alpina (Каменные Могилы).
Обнажения третичных известняков с характерной растительностью* встречаются в пределах полосы разнотравно-типчаково-ковыльных и полосы типчаково-ковыльных степей в Причерноморье и Приазовье г по берегам Днестра, Южного Буга, Ингула, Ингульца, Днепра, Груз-ского и Мокрого Еланчиков, Миуса и др. Кроме того, обнажения третичных известняков имеются на равнинах (Тарханкутское плато) и в предгорьях Крыма и на Ставропольской возвышенности.
На обнажениях третичных известняков обычно представлена пестрая свита растительных группировок с преобладанием степных ассоциаций и с большим участием «известколюбов». Кроме обычных степных дерновин-ных злаков, как Festuca sulcata, Stipa capillata, S. Lessingiana и другие, на этих обнажениях встречаются дерновинные злаки-известколюбы, приуроченные в степной полосе Европейской части СССР преимущественно к обнажениям известняков, — Andropogon ischaemum (переходит и на лёссовые склоны и другие обнажения), Cleistogenes bulgarica (произрастает и на гранитах). Но особенно характерен для известняковых обнажений Причерноморья и Приазовья Украины и Крымского п-ва мелкий дерновинный злак Koeleria brevis, образующий группировки по верхам («лбам») склонов. На известняковых обнажениях произрастают также многие растения, часто встречающиеся и на каменистых обнажениях других пород, преимущественно карбонатных, например мела: из полукустарничков — Ephedra distachya, Silene supina, Paronychia cephalotes. Astragalus subulatus, Onosma simplicissimum (бассейн Дона и Волги, Приазовье), Thymus dimorphus, Teucrium polium, Jurinea stoechadifolia; из многолетнего разнотравья — Potentilla astracanica, Hedysarum gran-diflorum, Pimpinella titanophila, Cephalaria uralensis, Achillea leptophylla и др.; из малолетников — Dianthus pseudarmeria и др.
Из видов, приуроченных только к обнажениям известняков юга Европейской части СССР и являющихся эндемами, укажем следующие: из кустарничков — Genista scythica (юг Украинской ССР), G. albida (Крымский п-ов); из полукустарничков — Thymus moldavicus (бассейны Днестра и Ингульца, Крымский п-ов), Pyrethrum odessanum (юг Украинской ССР, на восток до Днепра); из дерновинных злаков — Koeleria brevis (юг Украинской ССР, Крымский п-ов), К. moldavica (по р. Сухой Ягорлык в бассейне Днестра), Melica simulans (по Днестру); из разнотравья — Polygala moldavica (бассейн Днестра) (Пачоский, 1917; Котов, 1927а; Клеопов, 1933а).
Растительность меловых обнажений показана на карте в Причерноморской степной провинции, в пределах полосы разнотравно-типчаковоковыльных и отчасти полосы типчаково-ковыльных степей, в следующих районах: в южной части Средне-Русской возвышенности, в бассейне Дона и его притока Северского Донца, в средней и южной части Приволжской возвышенности (правобережье Волги)1 и к югу от Донецкого кряжа, в бассейнах Миуса и Тузлова. В этом обширном районе по правым берегам долин рек широким распространением пользуются обнажения белого пишущего мела и меловых мергелей, с которыми связана чрезвычайно своеобразная флора с большим количеством эндемов и не менее своеобразные растительные группировки — тимьянники, состоящие преимуще
1 По правобережью Волги меловые обнажения со специфической растительностью заходят довольно далеко на север, в пределы лесостепи.
ственно из ксерофильных полукустарничков, а также тимьянниковые степи. Вот неполный список этих меловых эндемов юга Европейской части СССР:1 Stipa cretacea (р. Голубая — правый приток нижнего Дона) Koeleria Talievii (бассейн средней части Северского Донца), Festuca cretacea (бассейн средней части Северского Донца и Дона), Agropyrum cretaceum (бассейн средней части Северского Донца и по р. Крынке), *Silene cretacea (широко распространенный вид, на восток до Заволжья), Diplotaxis cretacea(бассейн средней части Северского Донца), Matthiola fragrans (от бассейна Северского Донца до Предуральского плато), * Alyssum gym-nopodum (бассейн Дона), *Lepidium Meyeri (южная часть Приволжской возвышенности, бассейн среднего Дона, Заволжье, Предуральское плато), Erysimum cretaceum (бассейн Дона), Е. krynken.se (Крынка — приток Миуса), Genista tanaitica (бассейн Дона), Hedysarum cretaceum (бассейн Дона, южная часть Приволжской возвышенности и Заволжье), Н. ucrainicum (по Айдару — левому притоку Северского Донца), Poly gala cretacea (бассейн Дона), Viola cretacea (по Крынке в бассейне Миуса), *Hyssopus cretaceus (бассейн Дона), *Thymus cretaceus (бассейн Дона), *Т. Dubjanskyii (правобережье Волги, в меловых борах между Вольском и Хвалынском), Linaria cretacea (бассейны Дона, Волги и Урала), *Scrophularia cretacea (бассейн Дона и Приволжская возвышенность), *Anthemis Trotzkiana (от Медведицы до Предуральского плато), * Pyrethrum sclerophyllum (Сызранский и Хвалынский районы правобережья Волги), * Artemisia hololeuca (бассейн Дона и Миуса), Helichrysum cretaceum (бассейн Дона), Serratula tanaitica (мелы по Дону у станицы Сиротинской), Tragopogon cretaceum (правобережье Волги к югу от Вольска) и др. Многие из перечисленных двудольных относятся к биологической форме полукустарничка (эти виды отмечены звездочкой).1 2
С выходами мела связан также особый эндемичный кустарник Daphne Sophia (эндем бассейна Северского Донца, встречающийся в соседних районах Курской, Воронежской и Харьковской областей), а на мелах правого берега Дона, в излучине последнего, и по Иловле, образует заросли стланиковый казацкий можжевельник {Juniperus sabina).
На меловых обнажениях встречаются и многие другие виды, брлее или менее широко распространенные на обнажениях других пород, главным образом карбонатных (см. выше), а также многочисленные степные растения.
К чистому мелу обычно приурочена целая свита эрозиофитов, которые избегают участков обнажений с относительно сомкнутым, во всяком случае в надземной части, растительным покровом; к числу этих растений относятся эндемы меловых обнажений: Silene cretacea, Hedysarum cretaceum, Linaria cretacea, Matthiola fragrans, Scrophularia cretacea (преимущественно осыпи мела) и др. Hyssopus cretaceus на меловых обнажениях бассейна Дона особенно часто образует несомкнутые в надземной части группировки; такие же группировки образует и Thymuscretaceus, который, однако, входит и в состав сомкнутых сообществ, произрастающих на меловом субстрате с началом накопления гумуса. Характерные скопления своими подушковидными беловатыми кустиками образует на чистом мелу лбов склонов Artemisia hololeuca. Более сомкнутые сообщества формирует
1 Наиболее богата эндемами флора меловых обнажений бассейна Северского Донца и Дона. Меловые обнажения Заволжья (Общего Сырта) и Предуральского плато более бедны эндемичными видами.
2 Genista tanaitica — кустарничек; злаки относятся к группе дерновинных (Festuca cretacea имеет также корневища); Diplotaxis cretacea, Erysimum krynkense и Tragopogon cretaceum — двулетники; остальные виды из группы многолетнего травянистого разнотравья.
Artemisia salsoloides,не являющаяся собственно меловым эндемом, но все же очень характерная для молов.1 В сообщества этой полыни проникают многие степняки, в том числе типчак, житняки (Agropyrum imbricatum, Л. pectiniforme), ковыли (Stipa capillata) и др. Также меловой чебрец (Thymus cretaceus) нередко ассоциируется со степными растениями. Таким образом, при зарастании меловых обнажений полукустарничковые тимьян-пики постепенно сменяются степными сообществами — тимьянниковыми степями, в составе которых, помимо господствующих степных дерновин-ных злаков, значительную роль играют некоторые полукустарнички — Thymus cretaceus, Teucrium polium и др. (Литвинов, 1902; Дубянский, 1903, 1905; Талиев, 1904—1905; Ширяев, 1904; Кашменский, 1906; Котов, 1928, 1953; Козо-Полянский, 1931, 1931а; Семенова-Тян-Шанская, 1954).
В пределах полосы разнотравно-типчаково-ковыльных степей на низменных террасовых (лёссовых) равнинах левобережий нижнего течения левых притоков Днепра — Пела, Ворсклы, Орели и Самары, а также по левобережью излучины Северского Донца, к юго-востоку от Харькова, показана внемасштабными знаками галофитная растительность и луговоболотные фитоценозы западин. На солончаковатых солонцах здесь широко распространены ассоциации Puccinellia Bilykiana (из родства Р. tenuissima) с Camphorosmasoongoricum, а по левобережью Самары и с С. monspeliacum\ несколько выше по рельефу — ассоциации Festuca sulcata с Limonium tomen-tellum, Artemisia monogyna и Camphorosma soongoricum. Ниже по микрорельефу, на солончаковых почвах, эти ассоциации сменяются другими, в которых к господствующей бескильнице (реже Agropyrum ruthenicum) примешиваются в большом количестве галофиты — в большинстве случаев показатели хлоридо-сульфатного засоления: Salicornia europaea, Suaeda prostrata, Atriplex pedunculata, A. verrucifera (полукустарничек), Lepidium borysthenicum, Limonium caspium, Artemisia monogyna и др.1 2 Далее при снижении рельефа следуют солончаковатые луга и болота (с переменным увлажнением) — ассоциации Carex distans, Juncus Gerardii, Triglochin maritima, Bolboschoenus maritimus. Особенно богата галофитами флора солончаковых почв лёссовых террас р. Самары, где встречаются, помимо вышеупомянутых, еще Aeluropus litoralis, Frankenia hirsuta и др. (Елиашевич, 1927; Клеопов, 1934).
Гликофитная луговая и лугово-болотная растительность на лёссовых террасах рек, часто в комплексе с солонцами, встречается в западинах с осолоделыми, более или менее заболоченными почвами («степные» или «подовые подзолы»). Для подобных западин характерно резко переменное увлажнение. В наиболее глубоких частях подобных западин господствуют болотные осоки (Carex omskiana, С. vesicaria), ближе к берегам — лугово-болотные злаки (Glyceria fluitans, Beckmannia eruciformis) -, по берегу — луговая растительность. В более влажные годы, когда в этих западинах вода застаивается до осени, в них развиваются некоторые амфибийные (Elatine alsinastrum) и водяные растения (Potamogeton heterophyl-lus, Р. acutifolius, Ceratophyllum tanaiticum). Ceratophyllum tanaiticum — причерноморский эндем—распространен на запад до долины Днепра (Виленский и Лавренко, 1925; Клеопов, 1934).
В пределах полосы причерноморских (в широком смысле) разнотравпо-типчаково-ковыльных степей, особенно в северных ее частях и в возвы-
1 М. В. Клоков выделяет Artemisia salsoloides с мелов бассейна Дона в особый вид A. tanaitica Klok.
2 Atriplex verrucifera, A. pedunculata, Limonium caspium — по Днепру и его левым притокам.
шенных районах (Донецкий кряж, Калачская возвышенность, правобережья Днепра и Волги), в балках часто встречаются байрачные дубравы (из Quercus robur), а в Молдавии также лески («гырнецы») из пушистого дуба (Quercus pubescens).
Литература
Л. В. Черняев (1865); Г. И. Танфильев (1898); Б. А. Келлер (1903, 1916, 1926а. 1931а, 1931в); Г. Н. Высоцкий (1905, 1908—1909, 1915, 1923); Г. А. Боровиков (1909); И. К. Пачоский (1917, 1927); К. М. Залесский (1918, 1918а); И. В. Новопо-кровский (1921, 1927а, 1935); А. И. Мальцев (1924); Д. Г. Виленский и Е. М. Лавренко (£925); О. А. Елиашевич (1927, 1939а); ГО. Д. Клеопов (1927, 1934, 1934а); Н. Ф. Комаров (1931); Т. И. Попов (1931); А. Д. Алексеев (1933); Ю. Д. Клеонов и Е. М. Лавренко (1933); Е. М. Лавренко и Г. И. Дохман (1933); Ф. Я. Левина (1933); Н. А. Авро-Йин (1934); В. В. Алехин (19346); Н. ВЛКуксин (1935); Н. А. Десятова-Шостенко и [. С. Шалыт (1936); А. М. Левицкая (1937, 1937а); Е. И. Пестушко (1937, 1939); К. Е. Корещук (1939); А. П. Балаш (1948); Г. И. Билык (1948а); Т. С. Гейдеман (1952, 1954); М. И. Алексеенко (1954); А. А. Горшкова (1954).
Растительность песков и супесей: И. В. Новопокровский (1915); И. К. Пачоский (1922—1923); Е. М. Лавренко и А. В. Прянишников (1926); Е. М. Лавренко (1927, 1935); Е. М. Лавренко и А. С. Порецкий (1928); А. Г. Гаель (1929, 1932, 1932а, 1934); А. Д. Гожев (1929); Ф. А. Гринь (19546).
Растительность каменистых обнажений: Д. И. Литвинов (1902); В. А. Дубянский (1903, 1905); В. И. Талиев (1904—1905); Г. И. Ширяев (1904): Б. Ф. Кашменский (1906); Е. М. Лавренко (1925); М. И. Котов (1927а, 1928, 1953); Б. М. Козо-Полянский (1931, 1931а); Ю. Д. Клеопов (1933а); А. М. Семенова-Тян-Шанская (1954).
76 звк— заволжско-казахстанские разнотравно-типчаково-ковыльные степи и сельскохозяйственные земли на их месте
Заволжско-казахстанские разнотравно-типчаково-ковыльные степи на обыкновенных и главным образом на южных черноземах простираются то широкой, то более узкой полосой от левобережья Волги и до Оби. Кроме того, имеется остров этих степей в центре Кузнецкой котловины, а также ряд изолированных островов разнотравно-типчаковоковыльных степей по более высоким районам Казахского мелкосопочника в окружении более сухих типчаково-ковыльных степей. В последнем случае эти степи отражают вертикальную поясность растительного покрова в условиях низкогорий Казахского мелкосопочника. Кроме того, к разнотравно-типчаково-ковыльным степям отнесены песчаные пространства, расположенные среди боров, по правобережью Иртыша, к северо-западу от Семипалатинска.
Заволжско-казахстанские разнотравно-типчаково-ковыльные степи выступают на огромном протяжении от Волги до Оби в различных геоморфологических условиях, что определяет характер сочетаний и комплексов растительных ассоциаций в этих степях.
Бблыпая часть территории заволжско-казахстанской части полосы разнотравно-типчаково-ковыльных степей охватывает относительно возвышенные равнинные или даже низкогорные местности. Так, территория между Волгой и Уралом относится к Высокому Заволжью,, преимущественно к Заволжскому (пермскому) плато, а также к Общему Сырту (пространство между р. Самарой и р. Уралом) и к Пред-уральскому (Южно-Уральскому) плато (правобережье р. Урала к югу от Уральских гор). В Зауралье разнотравно-типчаково-ковыльные Степи занимают части Южно-Уральского и Зауральского холмистых пенепленов. Далее к востоку располагается Северо-Тургайское плато, лишенное
гидрографической сети, с большим количеством мелких соленых озер. В связи со слабым дренажем это плато по характеру растительного покрова (обилие солонцов и солонцеватых почв) приближается к «низменным» территориям. Обширный район, заключенный в огромную излучину Ишима, располагается в пределах Казахского мелкосопочника (Казахская складчатая страна), для которого характерны увалистые и волнистые возвышенные равнины, участки приречного мелкосопочника и горноостровные возвышенности. К юго-западу от Барнаула находится Приобская повышенная равнина. Остальная территория принадлежит к «низменным» степным территориям. С востока к Волге примыкает увалистая (сыртовая) равнина Низкого Заволжья (древнеаллювиальная волжская равнина, довольно сильно расчлененная в результате эрозионных процессов). К северо-востоку от Казахского мелкосопочника располагается обширная, весьма малорасчлененная Прииртышская наклонная равнина с массой соленых озер. К востоку от Иртыша простирается Барабинско-Кулундинская равнина с гривистым рельефом и также с обилием соленых озер.
В Заволжье и Приуралье подавляющая часть разнотравно-типчаковоковыльных степей распахана. В северном Казахстане до последнего времени большая часть этих степей оставалась в целинном состоянии и только в 1953—1954 гг. в связи с освоением целинных и залежных земель значительная часть этих степей была подвергнута распашке, кроме участков с большим участием солонцов или с резко выраженным сопочным рельефом. Соответствующие части Кулундинской степи в Приобье и Кузнецкой степи целиком распаханы.
Заволжско-казахстанские разнотравно-типчаково-ковыльные степи, как видно из дальнейшего, по своему составу и структуре близки к аналогичным причерноморским степям (см. для Заволжья и Приуралья работы И. И. Спрыгина, 1930; О. Э. Кнорринг, 1932; И. М. Крашенинникова, 1932; С. Е. Кучеровской, 1932; Л. Е. Родина, 1933, 19346; для юга Западной Сибири и северного Казахстана — А. Я. Гордягина, 1900— 1901; В. П. Дробова, 1914; П. Н. Крылова, 1916; М. И. Пташицкого, 1916; В. И. Баранова, 1927; А. В. Прозоровского и др., 1933-К; Е. В. Вандакуровой, 1950).1 Из дерновинных злаков доминируют такие широко распространенные виды в Причерноморско-Казахстанской степной подобласти, как Stipa capillata, S. rubens (восточнопричерноморско-казахстанский вид), Helictotrichon deser torum, F estuca sulcata s. I.,1 2 Phleum phleoides (в более мезофитных ассоциациях). В северных, менее ксеро-•фитных вариантах (ассоциациях), главным образом в Заволжье и Приуралье, в значительном количестве встречается Stipa stenophylla', наоборот, в более ксерофитных ассоциациях, особенно на юге полосы, более или менее обилен Stipa Lessingiana. На карбонатных черноземах обилен казахстанский вид Stipa Korshinskyi (рис. 63), не переходящий к западу Волги. Из корневищных злаков в более мезофитных ассоциациях встречается Роа angustifolia, реже Calamagrostis epigeios; из корневищных осок часта Carex supina. Из бобовых более обильна
1 П. Н. Крылов (1916) и другие западносибирские ботаники называют эти степи «ковыльно-кипцовыми».
2 В. И. Кречетович и Е. Г. Бобров во «Флоре СССР» (т. 2, 1934) рассматривают Festuca sulcata как паннонско-причерноморский вид, a F. pseudovina как казахстанский вид, заходящий в бассейн верхнего Енисея. П. А. Смирнов степной типчак от Украины до Казахстана относит kF. sulcata, не выделяя отдельной, pseudovina (П. А. Смирнов, К систематике и географии среднерусских представителей группы Festura ovina L. s. ampl. Бюлл. Московск. Общ. испытателей природы, отд. би-ол., т. 50, 1—2, М., 1945).
Medicago romanica; часты также Astragalus onobrychis, Onobrychis arenaria s. 1., Oxytropis pilosa. Из видов разнотравья, общих причерноморским и заволжско-казахстанским разнотравно-типчаково-ковыльным степям, более обычны: Artemisia dracunculus, A. latifolia (на запад до Средне-Русской возвышенности), И. Marschalliana, Fragaria viridis, Filipendula hexapetala,Galatella fastigiata(nu запад до Средне-Русской возвышенности), Galium verum з. 1., Hieracium echioides, Libanotis intermedia, Onosma simp-licissimum, Peucedanum Lubimenkoanum, Phlomis tuberosa, Plantago stepposa, Potentilla humifusa, Salvia stepposa (восточнопричерноморско-ка-захстанский вид), Senecio Jacobaea, Silene multiflora, Thalictrum minus, Thymus Marschallianus, Veronica incana, V. spicata s. I. Из видов разнотравья, отсутствующих в степях к западу от Волги, более обычны Aster alpinus (имеется на Жигулях), Artemisia glauca (южносибирско-казах-станский вид), Pulsatilla multifida (сибирский вид), Р. flavescens (также), Peucedanum Morisonii (казахстанский вид, на запад до р. Урала), Seseli. Ledebourii (казахстанский вид) и др. Из степных полукустарничков, широко распространенных как в причерноморских, так и в заволжско-казахстанских степях, встречаются Artemisia austriaca, Jurinea multi-flora. Эфемероиды и эфемеры немногочисленны; их несколько больше в Заволжье, и количество их уменьшается к востоку. Так, например, Роа bulbosa более или менее обилен в Заволжье в степях на южном черноземе (Родин, 19346), а в северном Казахстане луковичный мятлик встречается реже. Из луковичных эфемероидов растут Tulipa patens (волжско-казахстанский вид, на запад до Приволжской возвышенности), Т. Bie-bersteiniana (западноказахстанско-причерноморский вид), Ornithogalum Fischerianum (западноказахстанско-причерпоморский вид).
Как видно из предыдущего, имеются некоторые отличия в структуре, в первую очередь в составе, заволжско-казахстанских разнотравно-типчаково-ковыльных степей при движении с севера на юг и с запада на восток. Так, разнотравье особенно обильно в более мезофитных ассоциациях этих степей, преобладающих в северных районах соответствующей полосы; из разнотравья особенно характерны Filipendula hexapetala, Fragaria viridis, отчасти Peucedanum Morisonii, P. Lubimenkoanum и др. В Заволжье проникают некоторые причерноморские виды, не переходящие к востоку за Урал.
Общая (абсолютная) урожайность травостоя наиболее мезофитных ассоциаций в пределах Казахстана — 15—20 ц/га, при укосной массе сена около 6—7.5 ц/га; урожайность более ксерофитных ассоциаций с менее обильным разнотравьем — соответственно 15 ц/га и 5.5—7 ц/га (Прозоровский и др., см. Лавренко, 1940а). Л. Н. Соболев (1948) дает близкие показатели продуктивности для территории Казахстана; для более мезофитных ассоциаций — от 6 до 9—10 (во влажные годы) ц/га, для более ксерофитных — (4—) 5—6 (—8) ц/га.
По потяжинкам и западинам среди этих степей встречаются луговые и остепненяолуговые группировки (с преобладанием Calamagrostis epigeios, Agropyrum repens, Bromus inermis, Filipendula hexapelala, F. stepposa, Sanguisorba officinalis и др.).
Для «низменных» районов полосы разнотравно-типчаково-ковыльных степей (см. выше) с характерным для них равнинным, слабо расчлененным рельефом, а также для надлуговых террас рек с суглинистыми почвами, для бессточных котловин и вообще для участков со слабым дренажем или с близким залеганием солоносных отложений, характерно распространение почвенно-растительных миирокомплексов с большим участием солонцов и солонцеватых черноземов. На солонцах встречаются беднораз
нотравные или даже пустынно-степные группировки с господством тин-чака (Festuca sulcata), грудницы (Linosyris villosa), и некоторых полукустарничков (Artemisia austriaca, A. nitrosa, A. Lercheana, Pyrethrum achil-leifolium). Солонцовые комплексы развиты также вокруг соленых озер; ближе к берегам этих озер располагаются солончаки с галофитными сообществами из Kalidium foliatum, Atriplex verrucifera, видов Suaeda, Sali-cornia europaea и др.
Продуктивность разпотравно-типчаково-ковыльных степей на солонцеватых черноземах снижается до 5—6 ц/га, а на солонцах она гораздо ниже.
В верхней части бассейна Самары (заволжской), главным образом по правобережью Самары, располагается большой массив степей на супесях, ныне распаханных.
Стадии смен растительности на сильно выпасаемых степях и на залежах в заволжско-казахстанской части полосы разнотравпо-типчаково-ковыль-пых степей в общем те же, что и в причерноморских разнотравпо-типча-ково-ковыльных степях (76 ч).
Для большинства возвышенных районов полосы разнотравно-типча-ково-ковыльпых степей характерны обнажения коренных пород по берегам долин рек и по склонам сопок; на этих обнажениях развиты «каменистые», большей частью тимьянниковые степи, отмеченные внемас-штабным знаком („растительности каменистых обнажений. .
Растительность обнажений карбонатных пород пермской системы, главным образом пестрых мергелей татарского яруса, показана на карте в Высоком Заволжье по рр. Соку, Большой и Малой Кинели, правым притокам Самары, верховьям Демы и левым притокам (Белой.1 Обнажения этого типа имеются и по другим рекам Высокого Заволжья. Приурочены они к более крутым склонам возвышенностей и увалов, сопровождающих долины рек (Спрыгни, 1930, 1934).
В верхней части склонов и на узких плато увалов на недоразвитых карбонатных почвах, сформировавшихся на элювии указанных выше пород, развиты «каменистые» тимьянниковые степи. При этом па лбах южных склонов господствует Stipa Kor shinsky i (рис. 63), па верхних площадках водораздельных увалов преобладает S. Lessingiana, вместе с последним произрастает и «У. praecapillata (близкий вид к S. sareptana), предпочитающий, впрочем, менее сухие местообитания на этих склонах.
Для обнажений карбонатных пород татарского яруса очень характерны два эпдема Высокого Заволжья — полукустарничек Thymus baschkiri-ensis и Hedysarum liazoumovianum. Кроме того, на выходах пород пермской системы встречается и ряд других видов, свойственных обнажениям, но но приуроченных к породам определенной литологии, как, например, дериовипный злак Agropyrum desertorum и полукустарнички (Eurotia ceratoides, Alyssum tortuosum, Onosma simplicissimum), а также Pyrethrum mille/oliatum s. 1. и др.
В Заволжье, па юге Общего Сырта, имеются меловые обнажения с рядом специфических «меловых» видов (Смирнов, 1934).
В пределах Казахского мелкосопочника по склонам сопок на хрящеватых или щебнистых черноземах развиты петрофитпые варианты (ассоциации) степей с господством ковылей (Stipa rubens, S. capillata), типчака, степного овса (Helictotrichon desertorum), который часто выходит па первый план среди преобладающих видов. Ня сильно каменистых
1 Р. Сок целиком находится и приделах лесостепной полосы. Остальные местонахождения растительности обнажений пермской системы располагаются в полоса разнотраино-типчаково-ковыльных степей.
почвах крупнодерновшшые злаки выпадают и остается типчак. Очень характерно обильное развитие степного полукустарничка — холодной полыни (Artemisia frigida — казахстанско-сибирско-монгольский вид, на запад до Уральских гор включительно) и некоторых других полукустарничков: Dianthus acicularis (уральско-казахстанский вид, на запад до Волги и, кроме того, в Жигулях), Seseli eriocarpum (центральноказах-•станский вид), Thymus sp. sp., Scabiosa isetensis (волжско-казахстанский вид, на запад до правобережья Волги); встречается также листовой суккулент Orostachys spinosa (казахстанско-центральноазиатский вид) и др. Близкие по составу «каменистые» степи встречаются и в пределах Южно-Уральского и Зауральского пенепленов (на карте не отмечены).
Разнотравно-типчаково-ковыльные степи, преимущественно их петро-фитный вариант, показаны также на юго-востоке Казахского мелкосопочника, на хр. Чингизтау, а также в южных предгорьях Калбинского хребта и на северо-западных предгорьях Тарбагатая, между поселком Кызыл-кесек и станцией Жарма. Эти степи выступают здесь как высотный пояс (750—1500 м над у. м.); ниже по макрорельефу они переходят в сухие типчаково-ковыльные и даже пустынные степи (Косинский, 1916; Шип-чинский, 1916; Степанова, 1950; Быков и Степанова, 1953).
Помимо разнотравно-типчаково-ковыльных степей, в этой полосе широко распространены и кустарниковые степи, отмеченные на карте вне-масштабным знаком степных кустарников. Кустарниковые степи, приуроченные большей частью к легким почвам (щебнистым, супесчаным), характеризуются наличием кустарников, не образующих сплошных зарослей, а более или менее равномерно разбросанных среди обычного покрова из степных трав. В разнотравно-типчаково-ковыльных и кустарниковых степях здесь господствуют дерновинные злаки (Stipa capillata, S. rubens, S. kirghisorum, Festuca sulcata,1 Koeleria gracilis, реже Stipa Les-singiana, Helictotrichon desertorum). В составе многолетнего разнотравья, помимо обычных степных видов (Gypsophila paniculata, Silene wolgensis, Medicago romanica, Peucedanum Morisonii, Goniolimon speciosum, Phlomis tuber osa, Verbascum phoeniceum, Galium verum s. 1., Galatella fastigiata и др.), встречаются также виды, связывающие эти степи с Монголией и горами Средней Азии (Eremurus altaicus — восточноказахстанско-тяныпанский вид, Ferula songorica — восточноказахстанский вид, Veronica pinnata — восточноказахстанско-монгольский вид, распространенный на восток до западного Хангая). Из полукустарничков в условиях каменистых и щебнистых почв распространены петрофильные виды (Аг-iemisia frigida, в небольшом количестве A. sublessingiana — восточноказахстанский пустынно-степной вид). В составе кустарниковых степей из кустарников: чаще — Spiraea hypericifolia (причерноморско-казахстанский вид с иррадациями), Caragana frutex (причерноморско-казахстанский вид), реже — С. leucophloea (восточноказахстанско-монгольский вид), С. pumila (восточноказахстанский вид).
В горах Чингизтау и в других, в узких ущельях и местами на северных склонах, встречаются густые заросли кустарников, близкие по составу к зарослям степных кустарников западных предгорий Алтая (Rosa spi-nosissima, Lonicera tatarica, Spiraea crenata, S. hypericifolia, Caragana arbor escens).
Между Волгой и Уралом в полосе разнотравно-типчаково-ковыльных степей довольно часто встречаются дубовые лески (47 юв), приуроченные к балкам или к повышенным участкам рельефа, например к увалам Общего
1 Е. Ф. Степанова (1950) для предгорий северного склона Тарбагатая указывает Festuca Ganeschini, которую П. А. Смирнов (1945; см. стр. 649) не отличает от F. sulcata.
Сырта (Иванов, 1948). К дубу (Quercus robur) примешивается осина, реже береза. По опушкам этих дубовых рощ располагаются заросли степных кустарников (Prunus stepposa и особенно много Cerasus fruticosa, Hosa cinnamomea, Cytisus ruthenicus, Amygdalus папа, Spiraea crenata, S. hypericifolia, Caragana frutex). Стенные кустарники встречаются и независимо от дубовых рощ.
К востоку от Урала в полосе этих степей местами разбросано много березовых колков, приуроченных чаще к западинам, а в пределах Казахского мелкосопочника — также сосновых рощ на выходах кристаллических пород; по опушкам этих лесов развиты степные кустарники. Осиново-березовые лески встречаются также в ущельях среднегорных возвышенностей юго-восточной части Казахского мелкосопочника, например в горах Чингизтау и др.
Литература
А. Я. Гордягин (1900—1901); В. П. Дробов (1914); К. К. Косинский (1916); II. Н. Крылов (1916); М. И. Пташицкий (1916); Н. В. Шипчинский (1916); В. И. Баранов (1927); С. Е. Кучеровская (1927,1932); И. В. Новопокровский (1927); И. И. Спрыгни (1930, 1934); б. Э. Кнорринг (1932); И. М. Крашенинников (1932, 1939а); Е. К. Штукенберг (1932); А. В. Прозоровский, Н. И. Рубцов и А. А. Дмитриева (1933-К); JI. Е. Родин (1933, 19346); Л. А. Смирнов (1934); Е. М. Лавренко (1940ак И. М. Крашенинников и С. Е. Кучеровская-Рожанец (1941); В. В. (Иванов (1948); Л. Н. Соболев (1948); А. В. Куминова и Е. В. Вандакурова (1949); В. Е. Ванда-курова (1950); Е. Ф. Степанова (1950); Б. А. Быков и Е. Ф. Степанова (1953).
77. Дерновиннозлаковые европейско-казахстанские степи
Дерновиннозлаковые европейско-казахстанские сухие степи простираются то более широкой, то местами очень узкой полосой от Прута на западе до предгорий Алтая и Тарбагатая на востоке. Особенно суживается эта полоса в бассейне Урала, что объясняется далеким захождением здесь на север пустынных (полынно-типчаково-ковыльных и полынно-типчаковых) степей. Местами полоса типчаково-ковыльных степей прерывается, например в бассейне нижнего Дона, где разнотравно-типчаково-ковыльные степи (76) широким массивом спускаются на юг, к Кавказу, отделяя собственно причерноморский массив сухих типчаковоковыльных степей от придонско-ставропольского массива последних.
Сухие дерновиннозлаковые степи, связанные с южными черноземами (их более ксерическими разностями) и с темнокаштановыми почвами, характеризуются господством евксерофильных и евриксерофильных степных растений; мезоксерофиты, не говоря уже о евмезофитах, почти полностью отсутствуют. Господство степных дерновинных злаков еще более отчетливо, чем в разнотравно-типчаково-ковыльных степях; корневищные злаки и осоки играют незначительную роль. Разнотравье малочисленно и состоит из наиболее ксерофильных видов. Нередко заметна роль степных полукустарничков, как Artemisia austriaca, видов полыни из подрода Seriphidium, Kochia prostrata и др.; это чаще наблюдается на солонцеватых южных черноземах и на темнокаштановых почвах. Однако количество полукустарничков не достигает такого развития, как в пустынных степях. В значительном количестве встречаются эфемероиды и особенно эфемеры. В междерновинных промежутках на поверхности почвы часты лишайники.1
1 Дерновиннозлаковые сухие степи, представленные преимущественно типча-ковО-ковыльными ассоциациями, в литературе назывались также: узколистным или злаковым ковыльником (Высоцкий, 1908—1909, 1915, 1923); ковыльными и ковыльнотипчаковыми мелкодерновинными степями (Келлер, 1916,1923а); степями «безлесной лу-
13 Заказ 551
По составу и другим структурным признакам различаются причерноморские и заволжско-казахстанские типчаково-ковыльные степи.
Причерноморские и заволжские степи почти целиком распаханы. В северном Казахстане в 1954—1955 гг. в связи с освоением целинных и залежных земель под посевы пшеницы распаханы значительные площади типчаково-ковыльных степей; однако и в настоящее время здесь еще сохранились очень большие площади целинных типчаково-ковыльных степей, главным образом в районах с большим количеством солонцов или в условиях мелкосопочного рельефа.
77 ч — сельскохозяйственные земли на месте причерноморских типчаково-ковыльных степей
Причерноморские сухие степи, ныне почти полностью распаханные, а в прошлом занятые типчаково-ковыльными степными сообществами, представлены двумя большими массивами — собственно причерноморским, вытянутым в широтном направлении, и придонско-ставропольским, вытянутым в долготном направлении.
Собственно причерноморский массив типчаково-ковыльных степей занимает в основном западную часть Азово-Черноморской низменности, вовсе не расчлененной или со слабым расчленением по берегам рек (преимущественно по правым берегам), а также большую часть Тарханкут-ского плато в пределах Крымского п-ва, плато Керченского п-ва и Таманский п-ов. Придонско-ставропольский массив причерноморских сухих типчаково-ковыльных степей, как указано выше, вытянут с севера на юг, от Камышина до Моздока. На пространстве между Азовским и Каспийским морями зональность в пределах степной области имеет не широтный, а долготный характер, так как степи здесь окаймляют западную окраину пустынной области, занимающей значительную часть низменного западного Прикаспия. В границах придонско-ставропольского массива типчаково-ковыльных степей преобладают относительно возвышенные и расчлененные территории. Район между Камышином и Сталинградом представляет собою в геоморфологическом отношении южную часть Приволжской возвышенности (Приволжская расчлененная равнина). К югу от Сталинграда и до долины Маныча простирается плато Ергеней, юго-западная часть которого (среднее и отчасти нижнее течение Сала) также занята типчаково-ковыльными степями. В излучине Дона сухие степи занимают юго-восточную часть Средне-Русской возвышенности. Долина Маныча, занятая пустынными степями, прерывает долготную здесь полосу типчаково-ковыльных степей. Южная часть придонско-ставропольского массива сухих степей приурочена к пологим склонам Ставропольской возвышенности.
В этих степях господствуют евксерофильные и отчасти евриксеро-фильные дерновинные злаки, преимущественно ковыли {Stipa ucrainica, S. Lessingiana, S. capillata), а также Festuca sulcata; при этом основными («зональными») видами являются два первые. Дерновины этих злаков не сомкнуты и разделены довольно значительными промежутками. Из корневищных злаков и осок, в основном на солонцеватых почвах, встречаются Aneurolepidium ramosum и Carex uralensis. Степное разнотравье
гово-степной зоны» (Крылов, 1916); узколистным или злаковым типце-ковыльником (Залесский, 1918); степями зоны злаковой или типчаково-ковыльной степи (Ново-покровский, 1921, 1925); бескрасочным ковыльником (Алехин, 1925, 1936); ковыльными степями (Клеопов и Лавренко, 1933); дерновиннозлаковыми бедноразнотравными степями (Лавренко, 1940а).
малочисленно и в видовом отношении, и по количеству особей и представлено преимущественно наиболее ксерофильными видами: Dianthus, guttatus (на восток до Молочной), Adonis wolgensis, Medicago romanica. Astragalus Henningii, A. novoascanicus, A. dolichophyllus, Ferula orientalis, F. caspica, Cachrys odontalgica, Seseli campestre, Eryngium campestre. Goniolimon tataricum, Limonium sareptanum, L. Bungei, Achillea lepto-phylla, Pyrethrum millefoliatum (иногда обильно), Serratula xeranthemoides. Carduus uncinatus (двулетник) и др. Некоторые из этих растений имеют шарообразное соцветие, то есть относятся к биологическому типу «перекати-поле» (Seseli campestre, Eryngium campestre, Goniolimon tataricum и др.). Иногда в заметном количестве встречаются позднецветущие луковичные: Allium sphaerocephalum, A. guttatum (на восток до Павлограда и Ногайска), A. pulchellum. Нередко в заметном количестве имеются полукустарнички: Jurinea multiflora, Artemisia austriaca, реже A. taurica. В связи с относительно теплой весной и наличием значительных междер-новинных промежутков в этих степях представлено довольно много эфемероидов и эфемеров. Особенно обильны луковичный мятлик (Роа bulbosa) и тюльпаны (Tulipa Schrenkii, Т. Biebersteiniana), а также Gagea bulbifera, G. Szovitsii (только причерноморский массив этих степей) и др. Некоторые из упомянутых выше видов разнотравья относительно рано кончают вегетацию и являются переходными к эфемероидам (Adonis wolgensis, Astragalus Henningii, A. dolichophyllus, Ferula caspica, Cachrys odontalgica, Serratula xeranthemoides и др.). Из довольно многочисленных эфемеров упомянем о следующих: Erophila verna, Veronica verna, Myo-sotis micrantha, Alyssum desertorum, Androsace elongata, A. Turczaninovii, Viola Kitaibeliana, Cerastium ucrainicum (только причерноморский массив), Holosteum umbellatum и др. В теплые и относительно влажные весны озимые эфемеры развиваются обильно.
В междерновинных промежутках на поверхности почвы встречаются мхи (Tortula ruralis) и лишайники (Cladonia rangiformis, Cornicularia steppae, Parmelia ryssolea), синезеленые водоросли (Stratonostoc commune и др.).
Эти степи в пределах их собственно причерноморского и ставропольско-донского ареалов не вйолне однородны. [См. о причерноморских степях работы И. К. Пачоского (1904, 1908, 1917, 1923а); А. А. Янаты. (1913); Г. И. Поплавской (1924); М. С. Шалыта (1930, 1938); Ю. Д. Клео-пова и Е. М. Лавренко (1933); М. С. Шалыта и П. К. Козлова (1939): М. А. Альбицкой (1953); Г. И. Билыка (1953, 1954); о придонско-ставропольских типчаково-ковыльных степях—К. М. Залесского (1918,1918а); И. В. Новопокровского (1921, 1925, 1926, 1927а, 1929); Г. И. Дохмав и Т. И. Рыбаковой (1933); В. Д. Александровой, (1954)].
Для сезонного развития сухих типчаково-ковыльных степей характерно наличие отчетливого периода полупокоя, который падает на конец июня—июль. Основные фазы развития этих степей в течение вегетационного периода (Шалыт, 1938; степи Аскания-Нова) таковы.
1. III—1/2 IV. Ранневесенняя фаза. Как только оттает земля, в промежутках между желтыми полусухими дерновинами типчака и ковылей появляются зелень и первые цветущие растения. Прежде всего оживают появившиеся еще с осени листья луковичного мятлика (Роа bulbosa), обильно встречающегося на сбитых участках степи. Затем появляются отдельные золотистые звездочки гусиного лука. Чуть позже начинается массовое цветение весенней крупки (Erophila verna), а также Veronica verna, Myosotis micrantha и др.
2. 2/2 IV. Средневесенняя фаза. В дерновинах злаков между сухими и полувысох-шими листьями пробиваются свежие зеленые листочки. Очень типичен для этих степей аспект, когда на фоне уже зеленеющих дерновин степных злаков выделяются крупные
яркокрасные и желтые цветы Tulipa Schrenkii и разноцветные (желтые, фиолетовые и пр.) куртины цветущего степного ириса (Iris pumila).
3. V. Поздневесенняя — раннелетняя фаза, для которой характерен аспект цветущих и плодоносящих перистых ковылей; последние выбрасывают свои ости в первой декаде мая. В это же время цветут и другие степные злаки — типчак, тонконог. На белом фоне «перьев» ковылей выделяются яркокрасные корзинки Carduus uncinatus и фиолетовые соцветия V erbascum phoeniceum.
4. 1/2 VI. Первая летняя фаза. В первой половине июня плоды перистых ковылей и типчака осыпаются, и степь начинает присыхать. Цветут некоторые степные двудольные, как Phlomis pungens, Salvia nemorosa 1 и др.
5. 2/2 VI—1/2 VII. Вторая летняя фаза — период летнего «выгорания» степи (период полупокоя), когда степь получает соломисто-желтую окраску от высохших стеблей и листьев типчака и перистых ковылей.
6. 2/2 VII—VIII. Позднелетняя фаза. Во второй половине июля начинает выбрасывать свои соцветия и цвести Stipa capillata. В дерновинах последнего появляются новые листья. Степь снова несколько зеленеет.
7. IX—-XI. Осенняя фаза. Обсеменение тырсы происходит в конце августа, затем она высыхает, и степь снова приобретает соломисто-желтую окраску, сохраняющуюся до осенних дождей. Обычно в октябре—ноябре, после дождей, появляются проростки озимых эфемеров (Myosotis micrantha, Veronica verna и др.), розетки двулетников и многолетников (Carduus uncinatus, Verbascum phoeniceum и др.). Появляются зеленые листья у озимого эфемероида Роа bulbosa и множество молодых листочков в дерновинах ковылей и особенно типчака. Степь вступает в зиму довольно свежей — полузеленой и часто в таком виде остается до весны, покрываясь снегом только на очень короткий период.
Урожайность типчаково-ковыльных степей на юге Украины — до 10 ц/га, а степных выпасов этого типа всего 3—5—6 ц/га воздушно-сухой массы травостоя (Куксин, 1935).
Растительный покров типчаково-ковыльных степей нередко носит комплексный характер. В мелких, еле заметных западинках растительный покров более густой, увеличивается количество видов из группы разнотравья и особей последних; на пятнах солонцов развиваются ассоциации пустынных степей с господством типчака, а также полукустарничков — Kochia prostrata, Artemisia taurica (юг Украины между Скадов-ском и устьем Молочной, Ставропольская возвышенность); в бассейне Дона на солонцы заходят некоторые казахстанские полукустарнички {Pyrethrum achilleifolium, Artemisia Lercheana [=4. incana Kell.] и некоторые другие).2 По северным склонам балок, по ложбинам стока и по окраине подов встречаются менее ксерофитные ассоциации типа разно-травно-игипчаково-ковыльных степей.
По склонам Ставропольской возвышенности, между Кумой и Тереком, развиты ковыльно-андропогоновые степи, в составе которых, помимо Stipa capillata, S. Lessingiana, Koeleria gracilis, в большом количестве произрастают рыхлодерновинные злаки {Andropogon ischaemum ъ Cleistogenes serotina3) (Новопокровский, 1926).
По левобережью низовья Днепра, по Молочной, Чиру, Арче де встречаются массивы сухих степей на супесях (гемипсаммофитные ассоциации типчаково-ковыльных степей), которые, как и сухие степи на суглинистых и глинистых почвах, почти целиком распаханы. На сохранившихся целинных участках на супесях господствуют дерновинные злаки {Stipa capillata, Festuca sulcata, Koeleria gracilis), а на более легких супесях также Stipa borysthenica; характерно присутствие в значительном коли-
1 Степная раса этого вида описана теперь под названием Salvia tesquicola Klok. et Pobed. (Флора СССР, т. 21, 1954).
2 Места обилия солонцов с большим участием в их покрове полукустарничков (в бассейне Дона и др.) на карте отмечены соответствующим внемасштабным знаком.
3 Скорее это Cleistogenes maeotica или С. bulgarica.
честве змеевки (Cleistogen.es squarrosa). Разнотравье представлено в небольшом количестве факультативными псаммофитами.1
Остановимся вкратце на географическом распространении растений причерноморских типчаково-ковыльных степей.
Большинство эдификаторов — дерновинных злаков — является общим для причерноморских и заволжско-казахстанских степей (преимущественно причерноморско-казахстанские или паннонско-причерноморско-казахстанские виды): Stipa Lessingiana, S. capillata, S. borysthenica (на супесях и песках), Festuca sulcata, Koeleria gracilis, Cleistogenes squarrosa 1 2 (на супесях); к причерноморским (понтическим) видам относится Stipa uciainica.
Также общими для причерноморских и заволжско-казахстанских степей (то есть преимущественно причерноморско-казахстанскими или паннонско-причерноморско-казахстанскими видами) являются из корневищных злаков и осок: Aneurolepidium ramosum, Carex uralensis', из разнотравья: Adonis wolgensis, Medicago romanica, Limonium sareptanum, Salvia nemorosa ssp. tesquicola, Galium ruthenicum, Linosyris villosa, Pyrethrum millefoliatum', 3 из полукустарничков: Jurinea multiflora, Artemisia austriaca, из эфемероидов: Poa bulbosa, 4 Tulipa Schrenkii, T. Biebersteiniana, Gagea bulbifera и др. К восточнопричерноморско-казахстанским видам (на запад до бассейна Дона) принадлежит Dianthus leplopetalus, а к причерноморско-прикаспийским видам, редко переходящим к востоку Мугоджары, относятся из разнотравья: Astragalus doli-chophyllus, Ferula caspica, Cachrys odontalgica, Achillea leptophylla, Ser-ratula xeranthemoides. К причерноморско-средиземноморским видам, в большинстве случаев не переходящим- к востоку Волгу, относятся: Allium sphaerocephalum, Eryngium campestre, а из эфемеров: Viola Ki-taibeliana. Паннонско-причерноморским видом является Iris pumila (несколько заходит в Заволжье). К причерноморским видам, более или менее заходящим в Заволжье, относятся из разнотравья: A str agalus Неп-ningii, Seseli campestre, Goniolimon tataricum\ из полукустарничков: Artemisia taurica. К причерноморским видам, не заходящим в Заволжье — из разнотравья: Allium guttatum, A. pulchellum, Dianthus lanceolatus, D. guttatus, Astragalus novoascanicus, Ferula orientalis, Goniolimon Bes-serianum, Limonium Bungei; из эфемероидов: Cagea Szovitsii\ из эфемеров: Cerastium ucrainicum и др. Таким образом, в причерноморских типчаково-ковыльных степях много понтических видов, не заходящих или незначительно заходящих к востоку за Волгу.
Смены растительности типчаково-ковыльных степей при сильном выпасе в основных чертах проходят через те же стадии, что и в разнотравно-типчаково-ковыльных степях: ковыльную, типчаковую, мятликовую (господство Роа bulbosa) и однолетниковую (господство полусорных летне-осенних однолетников) (76 ч). Большинство сильно выписываемых целинных участков причерноморских степей и находится обычно в типчаковой или мятликовой стадии сбоя. Так, Г. И. Билык (1953, 1954) для юга Украинской ССР (полоса типчаково-ковыльных степей) указывает, что большая часть целинных степных пастбищ находится в стадии
1 Степи на песках в пределах полосы сухих типчаково-ковыльных степей отнесены к разнотравно-типчаково-ковыльным степям (76 ч).
2 Cleistogenes squarrosa — причерноморско-казахстанско-монгольский вид, a Koeleria gracilis — вид с широким голарктическим ареалом.
8 В пределах своего обширного ареала Pyrethrum millefoliatum, видимо, распадается на ряд рас.
4 Роа bulbosa обладает огромным средиземным ареалом в Евразии (в Центральной Азии отсутствует).
мятликово-полынкового или даже молочайно-полынкового сбоя 1 с урожайностью 2—7 ц/га.
Зацелиненио степных залежей в полосе типчаково-ковыльных степей также проходит через те же стадии, что и в полосе разнотравно-типчаково-ковыльных степей: 1) стадию однолетних и двулетних сорняков (с большей или меньшей примесью многолетних корнеотпрысковых и корневищных сорняков), 2) стадию корневищных (с преобладанием главным образом Agropyrum repens, а на солонцеватых почвах — Aneurolepidium ramosum) и корнеотпрысковых (Artemisia austriacd) растений, 3) стадию господства типчака, 4) стадию «вторичной» целины со строением травостоя типчаково-ковыльных степей (см. 76 ч).
В более расчлененных районах по склонам долин рек и более крупных балок встречаются каменистые обнажения со специфической растительностью (петрофитные ассоциации типчаково-ковыльных степей, тимьян-никовые степи и тимьянники). Так, в правобережной (по отношению к Днепру) более расчлененной части Азово-Черноморской низменности, на Тарханкутском плато в Крыму и местами на склонах Ставропольской возвышенности встречаются обнажения третичных известняков. По Дону, Иловле, а также по правому берегу Волги у Камышина имеются обнажения мела.2
В пределах Азово-Черноморской низменности, особенно в ее левобережной (по отношению к Днепру) наименее дренированной части, нередко встречаются обширные по занимаемой площади, но неглубокие и плоские понижения, так называемые поды. Почвы в подах — типа солодей («подовые подзолы»). Растительность подов сильно меняется в зависимости от метеорологических условий. Во влажные годы в подах развиваются группировки из луговых и болотных растений (Agropyrum pseudocaesium, Beckmannia eruciformis, Alopecurus pratensis, Heleocharis palustris s. 1., Butomus umbellatus и др.). В сухие годы поды покрываются растительностью луговостепного характера (Festuca sulcata, Phalacrachena inuloides, Inula britannica, Euphorbia virgata а др.). Урожайность травостоя до 22 ц/га воздушносухой массы (Куксин, 1935). Поды используются обычно как сенокосы (Яната, 1913; Пачоский, 1917; Шалыт, 1930, 1938).
По правобережью Дона, Волги и некоторых других рек довольно далеко на юг спускаются байрачные дубовые (из Quercus robur) леса, приуроченные обычно к балкам; на карте они показаны внемасштабным знаком. По правобережью Дона и Иловли на мелу встречаются заросли казацкого можжевельника (Juniperus sabina), также отмеченные на карте внемасштабным знаком. По балкам, склонам долин рек нередки заросли кустарников (из Prunus stepposa, Caragana frutex, видов Spiraea, Crataegus и др.).
Литература
И. К. Пачоский (1904, 1908, 1917, 1923а); А. А. Яната (1913); К. М. Залесский (1918, 1918а); И. В. Новопокровский (1921, 1925, 1926, 1927а, 1929); Г. И. Поплавская (1924); М. И. Котов (1927); М. С. Шалыт (1930, 1930а, 19306, 1938); Г. И. Дохман и Т. И. Рыбакова (1933); Ю. Д. Клеопов и Е. М. Лавренко (1933); Н. В. Куксин (1935); И. С. Шалыт и П. К. Козлов (1939); П. П. Жудова (1951); М. А. Альбицкая (1953); Г. И.^Билык (1953, 1954); В. Д. Александрова (1954); М. И. Алексеенко (1954).
1 Полынок — Artemisia austriaca, молочай—Euphorbia Seguieriana.
* Выше (76 ч) растительность этих обнажений охарактеризована как в пределах полосы разнотравно-типчаково-ковыльных, так и в полосе типчаково-ковыльных степей.
77 з в к — з а в о л ж с к о - к а з а х с т а н с к и е типчаково-ковыльные степи и сельскохозяйственные земли на их месте
Заволжско-казахстанские сухие типчаково-ковыльные степи простираются от Волги до предгорий Алтая и Тарбагатая на востоке. Они образуют то узкую, то расширяющуюся полосу, во многих районах прерываемую большими или меньшими массивами пустынных степей.
Между Волгой и Уралом сухие типчаково-ковыльные степи, здесь большей частью распаханные, занимают южную окраину увалистой (сыртовой) равнины Низкого Заволжья и частично возвышенность Общего Сырта. К югу от широтной части среднего течения Урала эти степи связаны с северной окраиной Предуральского плато. Далее к востоку полоса типчаково-ковыльных степей приурочена к южной части Зауральского пенеплена, к северной части Северо-Тургайского плато. Долиной Убагана (то есть северной частью Тургайской сквозной долины) эти степи простираются наиболее далеко на север. Степи эти занимают также северную часть Казахского мелкосопочника (Казахской складчатой страны), огибая южный выступ разнотравно-типчаково-ковыльных степей (приуроченных в основном к Кокчетавским низкогорьям и к их окраинам); далее к востоку эти степи занимают наклонную нерасчлененную равнину Павлодарского Прииртышья, где и достигают своей восточной границы по линии Кулундинское озеро—Усть-Каменогорск. Низкогорь-ями Центрального Казахстана типчаково-ковыльные степи (преимущественно их петрофитные разности), перемежаясь с массивами пустынных степей, приуроченных главным образом к более пониженным участкам территории, простираются на юго-восток вплоть до Тарбагатая. Кроме того, к повышенной части Мугоджарских гор приурочен вытянутый с севера на юг остров петрофитных разностей этих степей, сочетающихся здесь с растительностью каменистых обнажений.
В связи с земледельческим освоением целинных и залежных земель в 1954—1955 гг., значительная часть этих степей в пределах менее расчлененных территорий (Предуральское плато, Зауральский пенеплен, Северо-Тургайское плато, равнинное Павлодарское Прииртышье и др.) распахана под посевы зерновых культур, главным образом пшеницы. Типчаково-ковыльные степи низкогорий Казахского мелкосопочника, а также целинные участки в других районах этой полосы используются как весенне-летне-осенние пастбища.
В состав эдификаторов — евксерофильных и евриксерофильных дерновинных злаков — заволжско-казахстанских степей входят некоторые виды, общие с причерноморскими типчаково-ковыльными степями (пан-нонско-причерноморско-казахстанские виды): ковылок (Stipa Lessingiana), тырса (Stipa capillata), типчак (Festuca sulcata).1 Тырса часто безраздельно господствует на сильно выпасаемых участках или на легких почвах. В менее ксерофитных ассоциациях значительную роль играет восточнопричерноморско-казахстанский ковыль Stipa rubens, а в более ксерофитных ассоциациях, особенно на юге полосы типчаково-ковыльных .степей, в число эдификаторов входит иногда казахстанский вид — тырсик (Stipa sareptana).1 2 На карбонатных черноземах ;и темнокашта
1 По мнению В. И. Кречетовича и Е. Г. Боброва (Флора СССР, т. 2, 1934),гв пре-делах ареала заволжско-казахстанских типчаково-ковыльных степей степной типчак относится к виду Festuca pseudovina, который они отличают от паннонско-причерномор-ского вида F. sulcata в узком смысле.
2 Stipa sareptana на западе своего ареала заходит на Приволжскую возвышенность и в бассейне Дона, где изредка встречается на карбонатных почвах; более
новых почвах, вскипающих с поверхности, в состав эдификаторов соответствующих бедноразнотравных степных ассоциаций входит также казахстанский вид Stipa Korshinskyi (рис. 63), сочетающийся обычно со Stipa Lessingiana. Основным «зональным» видом ковыля является Stipa Lessingiana. В пределах Казахского мелкосопочника появляется горный казахстанско-среднеазиатский ковыль S. kirghisorum, особенно характерный для горных ковыльных степей Средней Азии (рис. 62). Кроме того, большую или меньшую роль в этих степях играют следующие дерновинные злаки: Koeleria gracilis (голарктический степной вид), Agropyrum desertorum (монгольско-казахстанский вид, заходящий на запад до правобережья Волги). В составе корневищных злаков и осок в небольшом количестве представлены Aneurolepidium ramosum (причерноморско-казахстанский вид), Carex supina (паннонско-причерноморско-казахстанский вид).
Виды ксерофильного разнотравья малочисленные, в большинстве случаев общие с причерноморскими степями. Упомянем, во-первых, виды с очень обширными в степной области ареалами {Medicago romanica, Galium ruthenicum, Linosyris villosa — паннонско-причерноморско-казах-станский вид — и др.), а также виды с более ограниченным распространением [Potentilla humifusa — причерноморско-казахстанский вид, Adonis wolgensis — также, Dianthus leptopetalus — восточнопричерномор-ско-казахстанский вид, заходящий на запад до бассейна Дона, Ferula caspica — причерноморско-прикаспийский вид, Serratula xeranthemoides — также, Arenaria Koriniana — казахстанский вид и др.). Из упомянутых выше Adonis wolgensis, Ferula caspica и Serratula xeranthemoides — полуэфемероиды, кончающие вегетацию в начале лета. Из полукустарничков встречаются Artemisia austriaca (паннонско-причерноморско-казахстан-ский вид), Jurinea multiflora (причерноморско-казахстанский вид), Pyrethrum achilleifolium (казахстанский вид, доходящий на запад до Дона). В составе эфемероидов: Роа bulbosa (вид с обширным средиземным ареалом), часто обильный, особенно на сбитых в результате выпаса степях, а также Tulipa Schrenkii (причерноморско-казахстанский вид), Т. Bieber-steiniana (также), Т. patens (казахстанский вид). Эфемеров меньше, чем в причерноморских типчаково-ковыльных степях.
Продуктивность казахстанских бедноразнотравных степей с тырсой и ковылком — (3—) 4 (—6) ц/га; менее ксерофитных степей со Stipa rubens, S. capillata и S. Lessingiana и бедным разнотравьем — (4.5—)5— 5.5 (—8) ц/га, а типчаково-ковыльных степей на солонцеватых почвах — 2—4 ц/га (Соболев, 1948).
По пониженным степным равнинам, в условиях слабого дренажа, а также на надлуговых террасах рек, особенно на тяжелых почвах, развиваются солонцовые комплексы с большим участием солонцов. На последних наблюдаются ассоциации пустынных степей или даже пустынные ассоциации с господством полукустарничков. В пустынно-степных ассоциациях на солонцах из дерновинных злаков господствуют типчак {Festuca sulcata) или в некоторых случаях ломкоколосник {Psathyrostachys juncea — монгольско-среднеазиатско-казахстанский вид с большими иррадиациями), из полукустарничков — Kochia prostrata и казахстанские виды — белая полынь {Artemisia Lercheana) и ромашник {Pyrethrum achilleifolium). На корковых и солончаковатых солонцах развиваются ассоциации пустынного типа с господством казахстанских видов полукустарничков — черной полыни {Artemisia pauciflora), а на юге полосы типчаково-ковыльных степей — и кокпека {Atriplex сапа).
обычен на Ергенях в составе полынно-типчаково-ковыльных степей, откуда спускается на юг до восточного Предкавказья.
В более освоенных районах полосы (подзоны) типчаково-ковыльных степей, с большим процентом распашки целины, чаще встречаются сильно сбитые в результате усиленного выпаса целинные степные участки или залежи. Так, например, в полосе типчаково-ковыльных степей на междуречье Волга—Урал на выпасываемых залежах развиты преимущественно острецово-типчаковые, типчаково-острецовые и житняково-острецовые ассоциации (острец — Aneurolepidium ramosum) с урожайностью 6—10 ц/га, а на более сильно сбитых участках — полынково-острецовые ассоциации с урожайностью 5—6 ц/га. По пониженным межсыртовым равнинам на каштановых почвах — белополынно-острецо-вые и белополынные, а на солонцах — чернополынные ассоциации (Иванов, 1954; Ларин, Шиффере, Левина и др., 1954). Следует отметить, что усиленный выпас способствует в полосе типчаково-ковыльных степей продвижению на север на солонцеватых южных черноземах или темнокаштановых почвах пустынных степей. [См. о типчаково-ковыльных степях Заволжья и Предуралья работы С. Е. Рожанец-Кучеровской (1926, 1927, 1929); И. И. Спрыгина (1930); Л. Е. Родина (1933, 19346); В. В. Иванова (1954); И. В. Ларина, Е. В. Шиффере, Ф. Я. Левиной и др. (1954); о степях юга Западной Сибири и северного Казахстана работы В. Ф. Капелькина (1910); М. Ф. Короткого (1914); С. Е. Рожанец-Кучеровской (1914, 1916); В. М. Савича (1914); К. К. Косинского (1916); П. Н. Крылова (1916); Н. В. Шипчинского (1916); В. И. Баранова (1925); И. М. Крашенинникова (1925); В. А. Крюгера (1927а, 1928); М. М. Ильина (1928); Б. А. Федченко и Н. Ф. Гончарова (1929); П. П. Полякова (1944); А. В. Куминовой и Е. В. Вандакуровой (1949); Е. В. Вандаку-ровой (1950)].
В пределах распространения заволжско-казахстанских типчаковоковыльных степей часто встречаются заросли степных кустарников, главным образом Spiraea hypericifolia (вид с очень обширным ареалом в степной области), Caragana jrutex (причерноморско-казахстанский вид) и др. Эти виды образуют компактные заросли по западинам и балкам, а также нередко произрастают в виде более или менее равномерно разбросанных экземпляров на фоне типчаково-ковыльной степи (кустарниковые типчаково-ковыльные степи); последнее наблюдается преимущественно на более легких (легкосуглинистых, супесчаных, щебнистых) почвах. В пределах Казахского мелкосопочника в составе этих кустарниковых степей на щебнистых почвах встречаются также восточные виды дерезы (Caragana pumila — восточноказахстанский вид, С. leucoph-loea — восточноказахстанско-монгольский вид) (Крюгер, 1928; Быков и Степанова, 1953).
Значительные площади типчаково-ковыльные степи, в большинстве случаев нераспаханные, занимают на легких почвах — песках и супесях, на которых развиты псаммофитные и гемипсаммофитные ассоциации этих степей. Обширные массивы типчаково-ковыльных степей на легких почвах располагаются уже в полосе пустынных степей между Прикаспийской низменностью и Мугоджарами, в пределах Предуральского плато, например по Уилу, Илеку и их притокам (Ильин, 1928; Ильин и Рожевиц, 1928; Гаель, Коликов и др., 1949; Никитин, 1954). Огромный массив типчаково-ковыльных степей на легких почвах известен на границе полосы типчаково-ковыльных степей и полосы пустынных степей в пределах Тургайского плато, между верхним течением Иргиза и оз. Са-рымоин, расположенном в Тургайской сквозной долине (Левицкий, 1938). Севернее, в полосе типчаково-ковыльных степей, располагаются, также в пределах Тургайского плато, большие массивы подобных сте
пей на легких почвах по р. Тоболу у Кустаная и южнее последнего. Имеются массивы типчаково-ковыльных степей на песках и супесях по верхнему и отчасти среднему течению Сарысу, в пределах полосы пустынных степей и даже северных пустынь (Левина, 1938). Наконец, огромный массив типчаково-ковыльных степей на легких почвах находится в пределах полосы типчаково-ковыльных степей и по границе с полосой пустынных степей вправо и влево от Иртыша, к северу и югу от Павлодара (Крюгер, 1929). В составе этих степей много общего со степями на легких почвах (супесях и песках) полосы разнотравно-типчаково-ковыльных степей (76 ч) и типчаково-ковыльных причерноморских степей (77 ч). Среди преобладающих дерновинных злаков господствуют преимущественно причерноморско-казахстанские или папнонско-причер-номорско-казахстанские виды: Stipa borysthenica( =S. Joannis ssp. sabulosa), S. capillata, Festuca Beckeri, Koeleria sabuletorum, также некоторые казахстанские виды: Stipa sareptana, Agropyrum sibiricum.1 Характерно отсутствие, помимо Stipa borysthenica, других перистых ковылей. На наиболее легких (песчаных) почвах господствуют Stipa borysthenica, Festuca Beckeri, а также Agropyrum sibiricum', обычна на песках, кроме того, Koeleria sabuletorum. На супесях преобладает Stipa capillata, а в более южных районах, преимущественно в полосе полынно-типчаково-ковыль-ных степей, Stipa sareptana, а также Agropyrum sibiricum, иногда причерноморско-казахстанско-монгольский вид Cleistogenes squarrosa. Встречаются ассоциации, в которых указанные выше эдификаторы сочетаются в различных комбинациях. На супесях, особенно тяжелых, в большом количестве представлены также злаки, характерные для тяжелых почв — Koeleria gracilis, Festuca sulcata. Из корневищных осок часто в заметном количестве встречается Carex supina (паннонско-причерноморско-казах-станский вид). Из указанных доминантов облигатными псаммофитами, никогда не переходящими на суглинистые почвы, являются Stipa borysthenica, Festuca Beckeri, Koeleria sabuletorum, Agropyrum sibiricum. Разнотравье более обильно на песчаных почвах; в этих случаях соответствующие псаммофитные ассоциации приближаются по составу к разнотравно-дерновиннозлаковым степям. На супесях гемипсаммофитные ассоциации более бедны разнотравьем. Из разнотравья и полукустарничков упомянем только более обычные в этих степях облигатные псаммофиты: причерноморско-казахстанские — Astragalus virgatus (полукустарничек), Achillea Gerberi, Scorzonera ensifoliw, причерноморско-западноказахстанские — Jurinea polyclonos, казахстанские — Chondrilla brevirostris и др. Из факультативных псаммофитов более обычны причерноморско-казахстанские виды разнотравья: Euphorbia Seguieriana и Artemisia Marschalliana.
На бугристых зарастающих песках более обильно встречаются: из кустарников, для которых характерно развитие придаточных корней на засыпаемых песком ветвях, — Salix caspica, S. acutifolia (оба эти вида ив с большим ареалом, выходящим за пределы степной области), Cytisus borysthenicus (причерноморско-западноказахстанский вид, на восток до Тургайской сквозной долины); из полукустарничков, также образующих придаточные корни на засыпаемых песком ветвях, — Artemisia arenaria s. J. (из причерноморско-казахстанско-центральноазиатской группы близких видов); из корневищных злаков — Elymus giganteus (казахстанско-центральноазиатский вид); из дерновинных злаков — Agropyrum sibiricum, из многолетнего разнотравья — Euphorbia Seguieriana.
* Agropyrum sibiricum частью заходит в бассейн Дона.
Среди песчаных массивов в понижениях с близкими грунтовыми водами встречаются березовые рощицы (местами отмечены внемасштаб-ным знаком).
Районы с широким развитием петрофитных ассоциаций типчаковоковыльных степей, сочетающихся с петрофитными пустынными и тимьян-пиковыми степями, тимьянниками и зарослями степных кустарников на выходах твердых пород, приурочены к левобережью верховьев Тобола, к наиболее высокой части (400—600 м над у. м.) Мугоджарских гор (Дохман, 1954) и главным образом к Казахскому мелкосопочнику (Косинский, 1916; Рожанец-Кучеровская, 1916; Шипчинский, 1916; Быков и Степанова, 1953).
Характер растительности на каменистых субстратах зависит от литологии горной породы, от накопления измельченных продуктов выветривания на ее поверхности и вообще от степени развития почвообразовательного процесса. Обычно растительный покров изменяется на поверхности сопок в зависимости от экспозиции склона и при движении по склону сверху вниз. Все эти явления остаются совершенно неизученными для Казахского мелкосопочника и предгорий Алтая и Тарбагатая; лучше они изучены в Мугоджарах (Дохман, 1954).
Из дерновинных злаков на щебнистом субстрате наиболее обычен типчак (Festuca sulcata), а при накоплении мелкозема также ковыли — Stipa orientalis (горный казахстанско-центральноазиатский вид, встречающийся на сильно каменистом субстрате), часто S. capillata, кроме того, S. kirghisorum, и S. Lessingiana. Очень характерно обильное развитие полукустарничковых полыней; в Мугоджарах — Artemisia Lessingiana (западноказахстанский или уральско-мугоджарский вид); в пределах Казахского мелкосопочника — A. frigida (казахстанско-сибирско-монгольский вид) и A. sublessingiana (восточноказахстанский вид); иногда встречаются также полукустарничковые виды тимьяна — Thymus mugodzharicus (западноказахстанский или уральско-мугоджарский вид) и другие виды.1 Характерно присутствие толстянкового — Orostachys spinosa (казахстанско-центральноазиатский вид). На севере полосы типчаково-ковыльных степей на каменистом субстрате встречаются виды, указанные выше для аналогичных местообитаний полосы разнотравно-типчаково-ковыльных степей (76 звк). Из кустарников в составе «каменистых» степей произрастает Spiraea hypericifolia, а в Казахском мелко-сопочнике, кроме того, Caragana pumila, С. leucophloea.
В горной области Мугоджар (Дохман, 1954) каменистые склоны заняты в основном редкостойными ассоциациями с преобладанием полукустарничковой Artemisia Lessingiana. В составе этих ассоциаций из обычных пустынно-степных полукустарничков произрастают в меньшем количестве: виды, встречающиеся на почвах разного механического состава, — Kochia prostrata, Pyrethrum achilleifolium, Artemisia Lercheana', виды, встречающиеся преимущественно на каменистых субстратах, — Ephedra distachya, Alyssum tortuosum. Из дерновинных злаков в состав Artemisieta Lessingianae, обычно в незначительном количестве, входят петрофильные виды: Stipa orientalis (казахстанско-центральноазиатский вид), встречающийся здесь нередко в заметном количестве, Koeleria sclerophylla (волжско-казахстанский вид?), Psathyrostachys lanuginosa (казахстанский вид);
1 М. В. Клоков (Флора СССР, т. 21, 1954) еще приводит для каменистых склонов и скал полосы казахстанских типчаково-ковыльных степей следующие полукустарничковые виды тимьяна: Thymus rasitatus Klok. (восточноказахстанский вид), Т. guber-linensis Iljin (Губерлинские горы), Т. roseus Schipcz. (горы Кандылатай), Т. kasak-stanicus Klok. et Shost. (западноказахстанский вид) и др.
виды, встречающиеся на почвах, как мелкоземистых, так и щебнистых, — Stipa sareptana, Festuca sulcata. Имеются петрофильные виды разнотравья, например Berteroa spathulata (казахстанский вид) и др. Разбросаны кусты Spiraea hypericifolia. По более сглаженным сопкам с начальными стадиями почвообразования распространены полынно-тырсиковые ассоциации (Stipa sareptana + Artemisia Lessingiana). Таким образом, пустынные по характеру структуры лессингиановополынные ассоциации в процессе разрушения каменистых сопок сменяются лессингиановополынно-тырсиковыми пустынными степями, а последние — ковылковыми степями. Между более высоким западным и менее высоким восточным хребтом в Мугоджарах располагается вытянутая с севера на юг центральная (водораздельная), относительно пониженная мелкосопочно-равнинная часть (высота 400—500 м над у. м.), занятая в основном в равнинных условиях степями с господством ковылка (Stipa Lessingiana), а по низким сопкам и хребтинам — полынно-тырсиковыми (Stipa sareptana + Artemisia Lessingiana) пустынными степями и редкостойными ассоциациями Artemisia Lessingiana.
На скалах, преимущественно из гранита, в Казахском мелкосопочнике встречаются петрофильные лишайники, а в расщелинах скал — кустарники (стланиковый Juniperus sabina, Lonicera microphylla, Berberis sibirica, Spiraea trilobata, Caragana pumila и др.) и многие степные петрофильные травянистые растения (см. 104 кз).
В долинах рек, по окраинам пойм и на нижних надлуговых террасах, а также по берегам соленых озер и окраинам солончаковых котловин, широко распространены группировки с преобладанием чия (Lasiagrostis splendens — казахстанско-центральноазиатский вид), с большей или меньшей примесью волоснецов (Aneurolepidium angustum — казахстанско-монгольский вид, Л. Paboanum — казахстанско-центральноазиатский вид), остреца (Aneurolepidium ramosum), а часто однолетних и полукустарничковых солянок (Бушинский, 1924; Крюгер, 1927а; Востокова, 1952). По берегам соленых озер на солончаковой почве располагаются пустынные ассоциации с господством галофильных полукустарничков (Kalidium foliatum, Halocnemum strobilaceum, Atriplex verrucifera, Limonium suf-fruticosum), а также однолетних солянок (Salicornia europaea, Suaeda prostrata, S. corniculata и др.) (Крюгер, 1913).
В условиях низкогорного рельефа в Казахском мелкосопочнике по ущельям встречаются заросли кустарников, состоящие не только из степных видов, как, например, Spiraea hypericifolia, но и более мезофильных представителей, как Lonicera tatarica, Rosa spinosissima, R. cinnamo-mea и- других, а также березовые и осиново-березовые лески с рядом бореальных растений.1 В Мугоджарах в глубоких узких долинах и лощинах также встречаются осиново-березовые лески со многими северными видами. Местами на выходах гранита в пределах Казахского мелкосопочника (Каркаралинские, Кентские и другие гранитные массивы) встречаются сильно остепненные сосняки с сосной низкого бонитета.
Литература
А. Я. Гордягин(1900—1901); Г. Н. Высоцкий (1908—1909); В. Ф. Капелькин (1910); В. А. Крюгер (1913, 1927а, 1928, 1929); М. Ф. Короткий (1914); С. Е. Рожанец-Кучеровская(1914,1916,1926, 1927,1929); В. М. Савич (1914); К. К. Косинский (1916); П. И. Крылов (1916); Н. В. Шипчинский (1916); В. П. Бушинский (1924); В. И. Баранов (1925); И. И. Крашенинников (1925); В. А. Шелудякова (1926); М. М. Ильин (1928);
1 На «Геоботанической карте» эти березовые лески показаны с помощью внемас* штабного знака березы не везде, где они имеются.
М. М. Ильин и Р. Ю. Рожевиц (1928); Б. А. Федченко и И. Ф. Гончаров (1929): И. И. Спрыгин (1930); Л. Е. Родин (1933,19346); Ф. Я. Левина (1938); С. С. Левицкий (1938); П. П. Поляков (1944); Л. Н. Соболев (1948); А. Г. Гаель, М. С. Коликов, Е. А. Малюгин и Е. С. Останин (1949);А. В. Куминова и Е. В. Вандакурова (1949); Е. В. Вандакурова (1950); Е. А. Востокова (1952); Б. А. Быков и Е. Ф. Степанова (1953); Г. И. Дохман (1954); В. В. Иванов (1954); И. В. Ларин, Е. В. Шиффере. Ф. Я. Левина, Б. Н. Замятнин, Л. И. Иванина, Е. А. Варивцева, Г. Н. Непли и И. В. Сырокомская (1954); С. А. Никитин (1954).
78. Полынно-дерновиннозлаковые европейско-казахстанские степи в сочетании с пустынными сообществами и сельскохозяйственные земли на их месте (северная полупустыня)
Полынно-дерновиннозлаковые пустынные степи распространены с некоторыми перерывами от северного побережья Черного моря на Украине и вплоть до окраин Зайсанской и Алакольской котловин в восточном Казахстане; далее на восток они уходят в пределы Китая, где приурочены к южным предгорьям Алтая. От Ергеней и Нижнего Заволжья и вплоть до Алакольской котловины эти комплексные пустынные степи образуют почти сплошную полосу, простирающуюся с запада на восток на протяжении нескольких тысяч километров. Кроме того, отдельные массивы этих степей приурочены к побережью Черного и Азовского морей, к западному Прикаспию, к северо-восточной окраине Казахского .мелкосопочника, а также к предгорьям Джунгарского Алатау, низко-горьям Чу-Илийских гор и некоторым районам предгорий Северного Тянь-Шаня.
Впервые в научной литературе растительность этих пустынных комплексных степей была подробно описана Б. А. Келлером (1907) под Красноармейском, к юго-востоку от Сталинграда. Б. А. Келлер эти степи относил к полупустыне (в более поздних работах к северной полупустыне). 1
Флористический состав этих степей в пределах их огромного ареала весьма разнообразен, но состав жизненных форм остается в основном одинаковым. Господствуют весьма засухоустойчивые евксерофильные и отчасти евриксерофильные дерновинные злаки (типчак, ковыли, житняки и др.), к которым в большом количестве примешиваются евксерофильные полукустарнички, главным образом полыни из подрода Seriphi-dium, а также ромашник (Pyrethrum achilleifolium), прутняк (Kochia prostrata) и некоторые другие. Характерно также довольно большое количество эфемероидов, из которых особенно обилен живородящий мятлик (Роа bulbosa var. vivipara), в меньшем количестве — тюльпаны, а также эфемеры (озимые однолетники). Эфемероиды и эфемеры более обильны в западных районах распространения пустынных степей. В составе пустынных степей нередко бывает выражен также напочвенный ярус из лишайников: как кустистых кочующих (Parmelia vagans, Р. rys-solea и др.), так и накипных (виды Psora и др.), синезеленых водорослей (Stratonostoc commune и др.), а иногда и мхов (Tortula).
Пустынные полынно-дерновиннозлаковые степи представляют собой переход от степей к пустыням и в своем зональном залегании образуют наиболее южную полосу (подзону) степной области (зоны).
1 Пустынные (полукустарничково-дерновиннозлаковые) степи в литературе известны под такими названиями: серый ковыльник (Stipetum cinereum) (Высоцкий, 1915), дерновиннозлаково-ромашниковые и дерновиннозлаково-полынные полупустыни (Келлер, 1923а), серый типце-ковыльник (Залесский, 1918а), полупустынная степь зоны злаково-полынной степи (Новопокровский, 1921, 1925), опустыненные (преимущественно полынно-типчаково-ковыльные) степи (Лавренко, 1940а).
В климатическом отношении пустынные степи (северная полупустыня) в их зональном залегании характеризуется очень засушливым климатом. В почвенном отношении пустынные полынно-дерновиннозлаковые степи в их зональном залегании связаны со светлокаштановыми почвами, более или менее солонцеватыми, а также с сильно солонцеватыми темнокаштановыми почвами, а в приморских районах Украинской ССР с глубокостолбчатыми солонцами. Отдельными небольшими участками, приуроченными к солонцам, пустынные степи заходят далеко на север, вплоть до южной окраины лесостепи. Пустынные степи возникают иногда и как вторичное явление. В связи с неумеренным выпасом в составе сухих типчаково-ковыльных степей может увеличиваться количество полукустарничков, а также Роа bulbosa и эфемеров.
Пустынные полынно-дерновиннозлаковые степи обычно носят комплексный характер. Характер комплексов весьма разнообразен. Как указано выше, полынно-дерновиннозлаковые степи чаще приурочены к солонцеватым светлокаштановым почвам. Западины с черноземовидными почвами заняты дерновиннозлаковыми степями с господством типчака, ковылей, житняка и с большей или меньшей примесью степного разнотравья, а иногда и степных кустарников, чаще зверобоелистной таволги (Spiraea hypericifolia). К пятнам корково- и отчасти среднестолбчатых солонцов приурочены сообщества пустынного характера с преобладанием полукустарничковых полыней,' а на солончаковатых солонцах — и солянок.
На огромном протяжении от побережья Черного моря и до Зайсан-ской и Алакольской котловин флористический состав полыней, а отчасти и степных дерновинных злаков в составе пустынных полынно-дерновин-нозлаковых степей меняется. В связи с этим на карте выделено несколько провинциальных «вариантов» пустынных степей: зч — западнопричерноморские (с Artemisia maritima s. 1.), ч — причерноморские (с A. taurica), кк — прикаспийско-казахстанские (с A. Lercheana), зк — западноказахстанские (с A. Lessingiana), вк — восточноказахстанские (с A. sublessin-giana) полынно-типчаково-ковыльные степи и кэ — прикаспийско-казахстанские полынно-эркековые степи1 (с Artemisia arenaria, A. astrachanica, Eurotia Ewersmanniana, E. ceratoides).
Учитывая такое большое количество разностей пустынных степей, остановимся вкратце на составе господствующих и согосподствующих видов. Господствуют, как указано выше, дерновинные злаки. В пределах всей полосы пустынных (полынно-типчаково-ковыльных) степей, от побережья Черного моря до Зайсанской котловины, из господствующих н пустынных степях дерновинных злаков распространены: Stipa capillata (паннонско-причерноморско-казахстанский вид с большими иррадиациями), Festuca sulcata (тот же тип ареала),2 Agropyrum pectiniforme (включая A. imbricatum; западносредиземный вид).3 Последний вид является эдификатором пустынных степей главным образом в Причерноморье. В прикаспийско-казахстанской части полосы (подзоны) полынно-типчаково-ковыльных степей (от Ергеней до Зайсанской' котловины), помимо вышеупомянутых видов, в составе дерновинных злаков-эдификаторов находим пустынно-степные казахстанские виды, только частично заходящие в бассейн Дона, — Stipa sareptana, Agropyrum sibiricum, а также монгольско-казахстанский вид Agropyrum desertorum. Часто,особенно
1 Эркек — Agropyrum sibiricum.
2 Мы в данном случае включаем в (объем Festuca sulcata Hack, и F. pseudovina Hack. В. И. Кречетович и Е. Г. Бобров (Флора СССР, т. 2, 1934) эти виды различают.
3 В странах Центральной Азии и в Восточной Сибири Agropyrum pectiniforme замещается A. cristatum з. str.
на солонцах, эдификатором является Psathyrostachys juncea, также в основном казахстанский вид (с иррадиациями).
Особенно часто встречаются пустынные степи с господством тырсика (Stipa sareptana)', этот ковыль может считаться «зональным» для полосы прикаспийско-казахстанских пустынных степей.
Согосподствующими видами в пустынных степях обычно являются полукустарничковые (пустынно-степные) полыни: в Причерноморье (к востоку от Днепра) и в Предкавказье — Artemisia taurica (восточнопричерноморский вид); в прикаспийско-казахстанских степях — A. Ler-cheana, доходящая на запад до правобережья Волги и Дона (казахстанский вид).1 В Мугоджарах и на прилегающих плато, преимущественно на каменистых и щебнистых субстратах, большую роль играет западноказахстанский пустынно-степной вид полыни A. Lessingiana, образующий и самостоятельные редкостойные группировки на каменистых склонах. На каменистых обнажениях и щебнистых светлокаштановых почвах Казахского мелкосопочника, предгорий южного Алтая, Тарбагатая и Джунгарского Алатау A. Lessingiana замещается восточноказахстанским пустынно-степным видом полыни — A. sublessingiana. Кроме того, в пределах ареала последнего вида в пустынных степях на каменистых и щебнистых склонах значительную роль часто играет более широко распространенная степная и полукустарничковая полынь A. frigida (казахстанско-сибирско-монгольский вид). О псаммофильных видах полыни сказано далее (78 кэ).
Иногда, особенно на Ергенях и в северном Прикаспии, согосподствует в пустынных степях ромашник (Pyrethrum achilleifolium — казахстанский вид). Обычно присутствуют в составе многих пустынных степей, но в меньшем количестве, полукустарнички: Kochia prostrata (средиземный вид), Camphorosma monspeliacum (средиземноморско-причерноморско-казахстанский вид).
На солонцах — корковых и солончаковатых, занимающих в полосе прикаспийско-казахстанских пустынных степей очень большие площади, господствуют полукустарнички, создавая растительные сообщества пустынного типа. По своему распространению эти полукустарнички являются в большинстве случаев казахстанскими пустынно-степными и пустынными видами (Artemisia pauciflora з. I.,1 2 Atriplex сапа, Anabasis salsa, а на щебнистых солонцах — Nanophyton erinaceum — джунгарско-казахстанский вид).
Пустынные полынно-дерновиннозлаковые степи используются преимущественно в качестве пастбищ и только в особо урожайные годы (с влажными весной и летом) — в качестве сенокосов. Полоса пустынных полынно-дерновиннозлаковых степей относится еще к зоне богарного земледелия, но крайне неустойчивого в связи с недостатком осадков. Отдельные массивы менее солонцеватых почв распаханы.
1 Artemisia Lercheana \УеЬ.=Л. i пса па Kell, et auct. Как далеко на восток идет этот вид, еще окончательно не выяснено. В полосе контакта степной и пустынной областей (зон) широко распространен в Казахстане близкий вид полыни — A. semiarida Krasch. et Lavr
2 Помимо типичной по характеру ареала пустынно-степной A. pauciflora Web., распространенной от правобережья Волги и Дона до Иртыша, отличают близкие, также казахстанские виды: A. maikara (Н. Krasch.) N. Pavl., свойственную преимущественно более южным районам, то есть северным окраинам пустынной области, и пустынностепную A. gracilescens Krasch. et Iljin, распространение которой полностью не выяснено. A. gracilescens до^сих пор достоверно известна от восточного Казахстана (Прииртышье) до Джунгарской Гоби; однако сходные с ней образцы имеются и из более западных районов, в частности из северного Прикаепия.
78 з ч — з а п а д н о п р и ч о р н о м о р с к и о п о л ы н н о-т и п ч а к о в о - к о в ы л ьп ы о и п о л ы пн о -т и п ча к о в ы е степи (с Artemisia maritima s. 1.)
Занаднопричерноморские полынно-типчаково-ковыльные и полынно-типчаковые комплексные пустынные стопи показаны па крайнем юге Украины, на низменном побережье Черного моря: на юге Измаильской и Херсонской областей. Таким образом, эти стопи приурочены к слабо-расчлененной приморской части Азово-Черноморской низменности. Лучше эти степи изучены в Херсонской области, между Ягорлыцким заливом и Скадовском.
На микроплакорах здесь преобладают полынпо-тырсово-типчаковые ассоциации. Из дерновинных злаков господствуют типчак (Festuca sulcata), тырса (Stipa capillata), житняк (Agropyrum pectiniforme), а из полукустарничков — морская полынь (Artemisia maritima s. I.),1 австрийский полынок (Artemisia austriaca)', то обильно, то в небольшом количестве встречается прутняк (Kochia prostrata) и реже камфоросма (Сат-phorosma monspeliacum). Из многолетнего разнотравья в небольшом количестве представлены Iris pumila, Dianthus guttatus, Eryngium campestre, Limonium Meyeri п др. Из эфемероидов обилен Роа bulbosa. Из летнеосенних однолетников (яровых) часто более или менее обильны Polygonum patulum, Eragrostis minor; в меньшем количестве — Polycnemum arvense, Echinopsilon sedoides, Gypsophila stepposa, Trifolium arvense. Иногда довольно хорошо выражен напочвенный покров, в составе которого преобладают то мхи (Tortula ruralis), то кочующие кустистые лишайники (Cladonia foliacea, С. rangiformis).
При усилении выпаса происходит «опустынивание» этих сообществ: уменьшается количество дерновинных злаков и увеличивается роль морской полыни (из злаков лучше противостоит выпасу житняк); увеличивается количество однолетников. На сильно сбитых участках, особенно вблизи селений, в значительном обилии встречается марь (Atriplex tata-rica)\ она же преобладает на молодых перелогах.
Подобные пустынные степи связаны с глубокостолбчатыми солонцами со столбчатым горизонтом на глубине 20—27 см. Таким образом, эти пустынные степи представляют эдафогенное явление.
Эти степи встречаются в комплексе со степными западинками и подами.
Небольшие по площади и неглубокие западинки заняты тырсово-типча-ковыми или типчаково-тырсовыми ассоциациями на обычно супесчаных или легкосуглинистых черноземовидных почвах, более или менее осолоделых, реже на осолоделых глубокостолбчатых солонцах со столбчатым горизонтом, залегающим на очень большой глубине. Полукустарничковые полыни в западинках отсутствуют или единичны. Господствуют дерно-винные злаки (типчак, тырса); более богато (но все же не обильно) представлено и степное разнотравье (Hypericum perforatum, Phlomis pungens, Galium ruthenicum и др.).
Более глубокие и несколько бблыпие по площади понижения—по-дики—заняты луговой растительностью, обычно с преобладанием темноколосовой осоки (Carex melanostachya) с примесью луговых злаков (Весктап-nia eruciformis, Agropyrum repens) и летне-осенних однолетников (Polygonum pseudoarenarium, Gypsophila stepposa, Crypsis alopecuraides. Pho-
1 Систематическое положение этой полыни остается пока не выясненным. И. В. Клоков («Визначник рослини УРСР», Ки1в— Харшв, 1950) относит ее к Artemisia Boschniakiana (Bess.) DC.
liurus pannonicus, Plantago tenuiflora, Pulicaria vulgaris и др.; первые два вида — иногда в значительном количестве)
Более обширные и глубокие поды, на дне которых выпотевает соленая верховодка, заняты солончаковой растительностью, с преобладанием аж-река (Aeluropus litoralis), солянок (Salicornia еигораеа) и др.
Вышеописанная растительность солонцового комплекса по плоскому морскому побережью переходит в растительность мокрых солончаков с преобладанием полукустарничковых солянок (Atriplex verrucifera, На-locnemum strobilaceum', оба вида ирано-туранские с иррадиациями; в степной области — причерноморско-казахстанские).
Литература
Е. Лавренко и Л. Прянишников (1926); Е. М. Лавренко и Н. А. Десятова-Шо-стенко (1928).
78 ч — причерноморские полынно-типчаково-ковыльные, полынно-типчаковые и полынно-житняковые степи
(с Artemisia taurica)
Причерноморские полынно-типчаково-ковыльные, полынно-типчаковые и полынно-житняковые комплексные пустынные степи со значительным участием таврической полыни (Artemisia taurica), по характеру распространения являющейся причерноморским видом, приурочены к северному побережью Черного моря (к северному побережью Каркинитского залива, к востоку от Скадовска), к побережью Сиваша, Керченскому п-ву (на восток от Феодосии), а также к западному Прикаспию, где массивы этих степей показаны на междуречье Кума—Терек, а также к югу от Махачкалы. Таким образом, на западе эти степи встречаются в пределах Азово-Черноморской низменности, а на востоке, в Прикаспии, — в пределах Притерек-ской наклонной равнины и в предгорьях Дагестана.
На крайнем юге Украины и в северном Крыму, по побережью Сиваша, а также Каркинитского залива и Утлюкского лимана, где эти степи лучше всего изучены, злаковую основу степи на более возвышенных равнинных участках с солонцеватыми каштановыми почвами слагают следующие дерновинные злаки: типчак (Festиса sulcata) и житняк (Agropyrum ре-ctiniforme), а также тырса (Stipa capillata), реже перистые ковыли (Stipa Lessingiana, S. ucrainica). Из полукустарничков в большом количестве встречаются таврическая полынь (Artemisia taurica), прутняк (Kochia prostrata) и полынок (Artemisia austriaca)', в небольшом количестве Jurinea multiflora. Многолетнее травянистое разнотравье — немногочисленное и необильное (Dianthus lanceolatus, Medicago romanica, Limonium czurju-kiense, Goniolimon rubellum,1 Phlomis pungens и немногие другие); иногда в значительном количестве встречается тысячелистниковый ромашник (Pyrethrum millefoliatum). Из эфемероидов обилен Роа bulbosa, довольно многочисленны тюльпаны (Tulipa Schrenkii). Много видов, а иногда и особей эфемеров (Bromus tectorum, В. squarrosus, Arenaria serpyllifolia, Cerastium ucrainicum, Erophila verna, Alyssum desertorum, Medicago minima, Trifolium arvense, T. parviflorum, Androsace Turczaninovii и др.). Из летнеосенних однолетников в составе этих степей встречаются в значительном количестве Echinopsilon sedoides и Polycnenum arvense; другие виды менее
1 М. В. Клоков отличает от волжско-западноказахстанского вида — Goniolimon rubellum (S. G. Gmel.) Klok. присивашскую pacy G. orae-syvashicae Klok. (Визнач-ник рослин УРСР, КиТв—Харюв, 1950).
обильны (Polygonum, patulum, Atriplex tatarica, Salsola ruthenica, Conso-lida paniculata и др.). Обычно выражен напочвенный ярус из лишайников, преимущественно кочующих (Parmelia ryssolea, Р. vagans, Cladonia joliacea var. convoluta, Cladonia rangiformis, C. subrangiformis, Cornicularia steppae и др.), реже мхов (Tortula ruralis). Эти степи используются как пастбища; продуктивность последних низкая (3—5 ц/га) (Куксин, 1935).
На более пониженных участках на столбчато-призматических солонцах вместе с типчаком преобладает полукустарничек Camphorosma monspeliacum.
На сильно выпасаемых полынно-типчаково-ковыльных степях ковыли исчезают, увеличивается количество Artemisia taurica и однолетников. На широко распространенных в Присивашье молодых перелогах господствуют сорные однолетники (из крестоцветных — Descurainia Sophia, Erysimum repandum, Capsella bursa-pastor is, из сложноцветных — Centaurea diffusa, из злаков — Bromus tectorum, B. squarrosus). На более старых залежах преобладает синец (Aneurolepidium ramosum), а на еще более старых — Artemisia taurica', однолетники на полынных залежах также обильны.
На фоне подобных полынно-дерновиннозлаковых степей встречаются как подчиненный элемент комплекса западинки, занятые степными дерно-виннозлаковыми ассоциациями с преобладанием типчака и ковылей, особенно тырсы, при отсутствии или при ничтожном количестве таврической полыни.
Среди этих степей разбросаны также обширные плоскодонные понижения — поды. Почвенный покров подов представлен микрокомплексами различных солонцов и солончаков. В растительном покрове преобладают в различных комбинациях следующие виды: Puccinellia Fominii (из родства Р. dolicholepis), Aeluropus litoralis, Petrosimonia oppositifolia, Atriplex verrucifera, Artemisia maritima s. 1. В выщелоченных западинках на дне подов—типчаковые или пырейные (Agropyrum pseudocaesium) ассоциации. На солончаковом днище более глубоких подов, на пухлых солончаках, господствует сарсазан (Halocnemum strobilaceum). Весной в подах развивается эфемероид Gagea Szovitsii, осенью — осенняя пролеска (Scilla autumnalis); весной также обильны эфемеры: Erophila verna, Holosteum umbellatum, Hymenolobus procumbens, Lamium Paczoskianum, Cerastium anomalum, позднее — ромашка (Matricaria chamomilla).
На Северном Кавказе массивы полынно-дерновиннозлаковых степей с участием Artemisia taurica приурочены к равнинной и предгорной части Грозненской области и Дагестанской АССР (Шиффере, 1953).
На хвалынских террасах вдоль правого берега Терека преобладают степи с господством житняка и таврической полыни; местами на повышенных участках террас — полынно-бородачевые степи (Andropogon ischaemum-}-Artemisia taurica).
Вдоль северо-восточного склона Большого Кавказа полынно-злаковые степи занимают нижние части предгорий на высотах 100—400 м над у. м. со светлокаштановыми почвами, большей частью хрящеватыми. Полыннозлаковые степи образованы преимущественно бородачем (Andropogon ischaemum) и таврической полынью (Artemisia taurica)', в значительном количестве из дерновинных злаков принимают участие Agropyrum pecti-niforme, Festuca sulcata, из эфемероидов — Роа bulbosa', кроме того, из полукустарничков — Artemisia austriaca, Kochia prostrata', из злаков — Stipa capillata, Koeleria gracilis; из разнотравья — Linosyris villosa, Achillea setacea, Peganum harmala и др.; много эфемеров — мелких зла
ков (Aegilops squarrosa, Bromus japonicus, В. squarrosus, Hordeum lepori-num), видов Alyssum, Lappula и др.
По вершинам и северо-восточным склонам мелких предгорных хребтиков здесь встречаются заросли шибляка с участием держи-дерева, таволги, боярышников, груши, а также местами остатки низкорослого дубняка из Quercus pubescens с грабинником (Carpinus orientalis).
Под влиянием усиленного скотобоя площадь распространения полынно-злаковых степей в Грозненской области и в Дагестане увеличивается за счет типчаково-ковыльных и других степей. Часть показанных на карте полынно-злаковых степей является вторичной, возникшей в результате усиленного выпаса.
В составе западнопричерноморских (78 зч) и причерноморских (78 ч) пустынных (полынно-типчаково-ковыльных и полынно-типчаковых) степей преобладают виды, общие с прикаспийско-казахстанскими пустынными степями, но имеется также много видов, отсутствующих или только незначительно заходящих в пределы ареала последних. Сюда принадлежит, во-первых, восточнопричерноморская по своему распространению таврическая полынь (Artemisia taurica). К причерноморским видам, кроме того, относятся Stipa ucrainica, Gagea Szovitsii, Iris pumila (паннонско-причерноморский вид), Cerastium ucrainicum, Dianthus guttatus, D. lanceo-latus, Limonium czurfukiense (эндем Присивашья) и многие другие. Кроме того, здесь довольно много средиземноморских видов, не переходящих или незначительно переходящих к востоку за Волгу, например Scilla autumnalis (атлантическо-средиземноморский вид), Consolida paniculata. Trifolium parviflorum, Medicago minima (средиземноморско-переднеазиатский вид) и др. В Предкавказье в пустынных степях встречаются сочетания таврической полыни с злаком Andropogon ischaemum (западносредиземный вид), который отсутствует в составе прикаспийско-казахстанских пустынных степей.
Л итература
И. К. Пачоский (1904, 1913, 1923); Ф. Я. Левина и М. С. Шалыт (1927); М. Котов и А. Прянишников (1929); Ф. Я. Левина (1929, 1938а, 1952); М. И. Котов (1935, 1949); Н. В. Куксин (1935); Н. А. Десятова-Шостенко и М. С. Шалыт (1936); Ф. Я. Попович (1938); М. С. Шалыт и П. К. Козлов (1939); М. С. Шалыт (1948); М. И. Алексеенко (1950,1950а); Н. Н. Дзенс-Литовская (1951); Н.[А. Троицкий (1951); Е. В. Шиффере (1953); М. Д. Скарлыгина (1954).
78 кк — прикаспийско-казахстанские полынно-типчаково-ков'ыльные и полынно-типчаковые степи (с Artemisia Lercheana)
Прикаспийско-казахстанские полынно-типча ково-коныл ьныр и полынно-типчаковые комплексные пустынные степи с большим участием Artemisia Lercheana занимают обширный ареал от Ергеней на западе до бассейна Тургая на востоке. К этой группе формаций условно отнесены также отдельные массивы пустынных степей в бассейне среднего Иртыша, в Павлодарской и Семипалатинской областях.1
1 Местами на «Геоботанической карте» в Казахском мелкосопочнике (Акмолинская, Карагандинская области) указан смешанный шифр — (вк-)-кк). Это означает, что для соответствующих районов характерно широкое распространение пустынных степей с участием как Artemisia sublessingiana, так и полыней родства A. Lercheana. Смешанный шифр (кк^- зк) указан и для районов к западу и востоку от Мугоджар (Актюбинская область), где, наряду с пустынными степями с участием A. Lercheana, встречаются пустынно-стенные ассоциации с А. Lessingiana (преимущественно на нистых и каменистых склонах).
Основные массивы этих степей приурочены к Ергенинскому плато (Высоцкий, 1915; Тереножкин, 1947; Жудова, 1951а; Цаценкин и др., 1952; Зворыкина, 1953; Левина, 1953; Цаценкин, 1954), к сыртовой увалистой равнине южной части Низкого Заволжья (Иванов, 1954; Ларин, Шиффере, Левина и др., 1954), северо-восточной (приуральской) части Прикаспийской низменности (Иванов, 1953; Никитин, 1954, 1954а), Предуральскому (Южноуральскому) плато и окраинным низкогорьям Мугоджар (Кучеровская-Рожанец и Рожанец, 1926; Ильин, 1928; Ильин и Рожевиц, 1928), Южно-Тургайской столово-останцевой равнине (плато) и к Тургайской сквозной равнине (Стрельников, 1938; Шахов и Земцова, 1948).
В зональном залегании на плакорах полынно-дерновиннозлаковые степи с большим участием белой полыни (Artemisia Lercheana) приурочены к светлокаштановым почвам. В пределы более северной подзоны сухих дерновиннозлаковых степей они проникают по сильносолонцеватым каштановым и темнокаштановым почвам, а иногда и по глубокостолбчатым солонцам. На юг, в область пустынь, в подзону остепненных (дерновинно-злаково-полынных) пустынь они заходят обычно по более легким почвам.
Основу травостоя этих степей образуют евксерофильные (засухоустойчивые) дерновинные злаки: типчак (Festuca sulcata), сарептский ковыль (Stipa sareptana), а также лессингов ковыль (5. Lessingiana), пустынный эркек (Agropyrum desertorum), а на более легких почвах (супесях) — тырса (Stipa capillata) и сибирский эркек (Agropyrum sibiricum). В некоторых гемипсаммофитных степных ассоциациях на супесчаных почвах Предураль-ского плато преобладает западноказахстанский ковыль Крашенинникова (Stipa Krascheninnikowii, близкий к S. rubens).
К этим господствующим дерновинным злакам в составе прикаспийско-казахстанских степей в большом количестве (в качестве субэдификаторов) присоединяются ксерофильные полукустарнички (Artemisia Lercheana, A. semiarida, Pyrethrum achilleifolium, Kochia prostrata, Camphorosma monspeliacum); обычны также степные полукустарнички (Artemisia austriaca, Jurinea multiflora). Иногда обилен корневищный злак острец или синец (Aneurolepidium ramosum — причерноморско-казахстанский вид). Ксерофильное разнотравье обычно немногочисленное, в состав его входят причерноморско-казахстанские виды (Palimbia rediviva, Limonium sarep-tanum)', восточнопричерноморско-казахстанские (Dianthus leptopetalus; Astragalus testiculatus — также горные степи Средней Азии), причерноморско-западноказахстанские (Trinia hispida), причерноморско-прикаспийские (на восток в основном до Мугоджар) (Cachrys odontalgica, Ferula caspica, Serratula xeranthemoides), казахстанские (Linosyris [Galatella] ta-tarica и др.). Как видно, большинство видов разнотравья является общим с типичными дерновиннозлаковыми (типчаково-ковыльными) степями (77 звк). Многие из видов разнотравья — гемиэфемероиды, то есть кончают вегетацию к началу засушливого лета (Astragalus testiculatus, Cachrys odontalgica, Ferula caspica, Serratula xeranthemoides и др.).
В большом количестве встречается эфемероидный живородящий мятлик (Роа bulbosa var. vivipara)', обычны тюльпаны (Tulipa Schrenkii, Т. Bieber-steiniana — причерноморско-казахстанские виды, Т. biflora — казахстанский вид). В большем или меньшем количестве представлены эфемеры, преимущественно те же, что и в типичных дерновиннозлаковых (типчаковоковыльных) степях (77). В наземном покрове нередко обильны лишайники (из родов Parmelia, Cladonia, Acarospora, Diploschistes, Psora), синезеленые водоросли (Stratonostoc), некоторые мхи.
Как видно из предыдущего, в составе прикаспийско-казахстанских пустынных степей по характеру распространен, я преобладают казахстанские и причерноморско-казахстанские виды.
Для большинства прикаспийско-казахстанских пустынных степей характерна микрокомплексность почвенного и растительного покрова, тесно связанная с комплексностью микрорельефа. Существует большое число разнообразных комплексов в полосе (подзоне) пустынных степей.
Рис. 68. Пустынно-степной микрокомплекс на главном водоразделе Ергеней.
I— ассоц. ковыльно-ромашниково-типчаковая (Festuca sulcata+Pyrethrum achilleifoli-um+Stipa capillata-]-Stipa Lessingiana) на светлокаштановой солонцеватой почве; 2 — типчаково-тырсовая (Stipa capillata-{-Festuca sulcata) на светлокаштановой несолонцеватой почве; 3—прутняково-ромашниково-типчаковая (Festuca sulcata-]-Pyrethrumach.il-leifolium+Kochia prostrata) на сильно солонцеватой светлокаштановой почве; 4 — ост-рецово-полынковая (Artemisia austriaca-]-Aneurolepidium ramosum) на светлокаштановом осолоделом солонце; 5 — полынково-острецовая (Aneurolepidium ramosum-]-Artemisia austriaca) с участием белой полыни на светлокаштановых солонцеватых почвах; б — типчаково-бел ополынна я (Artemisia Lercheana+Festuca sulcata) на глубоком и среднем солонцах; 7 — полынково-белополынная (Artemisia Lercheana-]-Artemisia austriaca) на глубоком солонце или на сильносолонцеватой светлокаштановой почве; 8— белополынно-полынковая (Artemisia austriaca-]-Artemisia Lercheana) на средних солонцах; 9— чернополынная (Artemisia pauciflora) на бугорках землероев; 10 — камфоросмовс-черкополынная (Artemisia pauciflora^-Camphorosma monspeliacum) на солончаковатом корковом солонце; 11 — камфоросмовая (Camphorosma monspeliacum) с единичной черной полынью на солончаковатом корковом солонце; 12 — камфоросмовая (Camphorosma monspeliacum) на солончаковатом корковом солонце (частые пятна голой почвы); 13 — прутняково-чернополын-ная (Artemisia pauciflora^-К ochia prostrata) на корковом солонце; 14 — молочайно-тип-чаково-тырсово-таволговая (Spiraea hypericifolia-]-Stipa capillata+Festuca sulcata-]-Euphorbia Seguieriana) на лугово-каштановой почве западинки; 15—типчаково-чабрецовая (Thymus Marschallianus+Festuca sulcata) на лугово-каштановой почве западинок; 16 — бугорки землероев (О. Я. Левина, 1953).
Лучше всего комплексность растительного покрова изучена на Ерге-нях (Димо и Келлер, 1907; Высоцкий, 1915; Левина, 1953; Зворыкина, 1953) и в северном Прикаспии (Левина, 1954).
На Ергенях, согласно исследованиям Ф. Я. Левиной (1953), на водоразделах и пологих склонах последних микроплакоры со светлокаштановыми солонцеватыми почвами заняты пустынно-степными ассоциациями с господством ковылей (Stipa Lessingiana, S. capillata), типчака и ромащника (Pyrethrum achilleifolium). Если на микроплакорах почвы относятся к глубоким солонцам, то ковыли исчезают, и с романтиком ассоциирует Agro-
pyrum desertorum. Белая полынь в этих ассоциациях встречается в небольшом количестве. К потяжинкам с более или менее выщелоченными (олуго-велыми) почвами приурочены степные ассоциации с господством ковылей и типчака и без заметного количества ромашника и полукустарничковых полыней. Сообщества пустынного типа с господством полукустарничковой белой полыни (Artemisia Lercheana), иногда с примесью типчака, приурочены к мелким понижениям диаметром в несколько метров. Эти понижения имеют обычно вид пятен различной формы (чаще округлой) с более или менее извилистыми очертаниями; почвы — глубокостолбчатые или средние солонцы. В таких же мелких понижениях, но с корковыми солонцами, развиты ассоциации пустынного типа с преобладанием полукустарничков (Artemisia pauciflora или Camphorosma monspeliacum), формирующих как чистые, так и смешанные ассоциации, совершенно лишенные степных элементов (дерновинных злаков и др.). В этих чернополынных ассоциациях, приуроченных к мелким понижениям, растительность очень изреженная, видовой состав высших растений крайне бедный, но наземный покров из лишайников и синезеленых водорослей обычно выражен очень хорошо, что объясняется не только изреженностью полыни или камфо-росмы, но и тем, что в этих западинах весной и во время дождей на короткий срок застаивается вода.
В довольно глубоких понижениях («лиманах») на Ергенях располагаются луговые сообщества, характерные для резко переменных условий увлажнения.
Близкие или несколько отличные пустынно-степные комплексы характерны для возвышенных территорий (преимущественно плато) с тяжелыми почвами полосы прикаспийско-казахстанских пустынных степей.
В настоящее время под влиянием выпаса количество полукустарничков в ассоциациях пустынных степей увеличивается. Так, по наблюдениям И. И. Тереножкина (1947), под влиянием выпаса («скотобоя») типчаковоковыльные пастбища на Ергенях превращаются в белополынно-черно-полынные, белополынно-полынково-чернополынные, то есть подвергаются опустыниванию. Больше того, в результате усиленного нерегулированного выпаса полоса пустынных степей, видимо, несколько сдвигается к северу, так как под влиянием скотобоя в состав типчаково-ковыльных степей проникают в значительном количестве полукустарничковые полыни и романтик.
В то же время по некоторым данным в полосе пустынных степей и северных (остепненных) пустынь происходит естественный процесс остепне-ния. Г. Н. Новиков (1936) считает, что в этом процессе остепнения солонцов с черной полынью и другими полукустарничками большую роль играет острец (Aneurolepidium ramosum), пронизывающий своими корневищами уплотненные столбы солонца и этим способствующий их разрушению.
Продуктивность полукустарничково-дерновиннозлаковых прикаспийско-казахстанских пустынных степей невелика. Согласно Л. Н. Соболеву (1948), общий запас растительной массы полынно-типчаково-ковыль-ных степей Казахстана весной равняется 2—3 ц/га, осенью 3—4 (5) ц/га и зимой 1—2 ц/га. Эти степи используются как выпасы, круглогодичные или осенне-зимне-весенние; выкашиваются они только в отдельные годы с большим количеством осадков, когда травостой развивается лучше.
При определенных условиях в полосе пустынных степей господствуют пустынные ассоциации с преобладанием полукустарничков. Это имеет место на выходах более или менее засоленных пород (например на Пред-уральском плато; см. Ильин и Рожевиц, 1928),1 а также на суглинистых
1 В полосе пустынных степей на каменистых или щебнистых более или менее засоленных субстратах обычно развиваются не тимьянники и тимьянниковые степи,
и глинистых верхних надлуговых террасах долин рек с преобладанием в почвенном покрове солонцов или солонцов-солончаков. В этих условиях развиваются пестрые комплексы чернополынных и солянковых ассоциаций; из солянок господствуют Atriplex сапа, Anabasis salsa, Л. aphylla, Nanophyton erinaceum, последний вид преимущественно на щебнистых субстратах.
Там, где подобные пустынные солянковые группировки особенно обильны, на карте (при шифре 78 кк) показан условный знак «пустынной растительности солонцов в степной зоне».
Участки пустынных степей в Прикаспии и в Казахстане, особенно в более северной части пустынно-степной полосы с менее солонцеватыми почвами, иногда после распашки оставляются под залежь. Изменения растительности на залежах лучше всего изучены на Ергенях (Высоцкий, 1915; Тереножкин, 1947; Левина, 1953). Здесь наблюдаются следующие стадии демутации залежей: 1) бурьяновая с преобладанием однолетников, особенно из сем. лебедовых (Salsola ruthenica, Amaranthus albus, A. ret-roflexus, Ceratocarpus arenarius, Atriplex tatarica, A. sphaeromorpha, Chenopodium album, Ch. urbicum, Echinopsilon sedoides), до 2—4 лет; 2) корне-отпрысково-корневищная с преобладанием корнеотпрысковой Artemisia austriaca, глубококорневищного Aneurolepidium ramosum, реже корневищного Agropyrum repens, с большой примесью низкорослых однолетников (Bromus, Echinopsilon и др.), до 2—6 лет; 3) стержнекорневая с господством белой полыни (Artemisia Lercheana) и с большим участием корнеотпрыскового полынка (A. austriaca), живородящего мятлика, костра (Bromus tectorum) и др., 6—10 лет, а при сильном выпасе неопределенно долгое время; 4) дерновиннозлаковая с полукустарничками стадия.
В полосе (подзоне) пустынных степей на юге Общего Сырта и в пределах Предуральского плато показана растительность меловых обнажений, приуроченная как к коренным берегам долин рек, так и к выходам мела на водоразделах 1 (Ильин, 1928; Ильин и Рожевиц, 1928; Смирнов, 1934). На этих меловых обнажениях преобладают полукустарнички; из специфических меловых эндемов представлены следующие: восточнопричерно-морско-западноказахстанские виды (Matthiola fragrans, Lepidium Meyeri, Linaria cretacea), приволжско-западноказахстанские виды (Anthemis Trolz-kiana). Кроме этих видов, на обнажениях мела Предуральского плато встречается ряд более широко распространенных, преимущественно казахстанских ( с ареалами от Заволжья или Зауралья до предгорий южного Алтая и Тарбагатая) видов, также петрофильных, но менее разборчивых в отношении литологии каменистого субстрата (Anabasis cretacea, Silene suffrutescens, Gypsophila Patrinii, Berteroa spathulata, Zygophyllum macro-pterum, Seseli glabratum, Scabiosa isetensis).
Для более низких (первых) надлуговых террас долин рек, особенно к востоку от р. Урала, характерны обширные заросли чия (Lasiagrostis splendens), сочетающегося обычно с галофитнолуговыми растениями (например видами Aneurolepidium) или даже солянками (Востокова, 1952).
Литература
Н. А. Димо и Б. А. Келлер (1907); Г. Н. Высоцкий (1915); И. М. Крашенинников (1925); С. Е. Кучеровская-Рожанец и М. И. Рожанец (1926); М. М. Ильин (1928); М. М. Ильин и Р. Ю. Рожевиц (1928); Л. А. Смирнов (1934, 1935); Г. Н. Новиков (1936);
как в более северных подзонах степной области, а сообщества пустынные или пустынностепные.
1 На Предуральском плато выходов мела со специфической растительностью больше, чем это показано на «Геоботанической карте».
И. А. Цаценкин (1936, 1954); П. Г. Стрельников (1938); И. И. Тереножкин (1947); Л. Н. Соболев (1948); А. А. Шахов и Н. М. Земцова (1948); П. П. Жудова (1951а); Е. А. Востокова (1952); И. А. Цаценкин, В. Ф. Максимова и Т. Н. Щербиновская (1952); К. В. Зворыкина (1953); В. В. Иванов (1953, 1954); Ф. Я. Левина (1953, 1954); И. В. Ларин, Е. В. Шиффере, Ф. Я. Левина и др. (1954); С. А. Никитин (1954,1954а).
78 зк — западноказахстанские полынно-типчаково-ковыльные и полынно-типчаковые степи (с Artemisia Lessingiana)
Западноказахстанские полынно-типчаково-ковыльные степи занимают в основном предгорья Мугоджар 1 и только частью переходят на соседние участки Предуральского плато (к западу от Мугоджар) и Чаграй-ского наклонного плато (к востоку от Мугоджар). Эти степи расположены на высоте (200)250—400(500) м над у. м. (Дохман, 1954).
Для этих степей, развитых преимущественно на щебнистых почвах, характерна значительная роль в травостое, помимо более широко распространенной белой полыни (Artemisia Lercheana, казахстанский вид), также полыни Лессинга (A. Lessingiana, западноказахстанский вид).
Восточная пенепленизированная (высота ниже 400 м над у. м.) с мелкими сопками часть Мугоджар в основном занята пустынными степями; белополынно-тырсиковыми ассоциациями (Stipa sareptana-\- Artemisia Lercheana * *) на суглинистых светлокаштановых почвах; меньшую площадь занимают лессингиановополынно-тырсиковые (Stipa sareptana-\- Artemisia Lessingiana) ассоциации на почвах подобного же механического состава и маршалловополынно-тырсовые (Stipa capillata-^-Artemisia Mar-schalliana) ассоциации на дресвянистых почвах. В составе некоторых из этих ассоциаций принимает некоторое участие Spiraea hypericifolia: это — кустарниковые пустынные степи.
Значительным распространением здесь пользуются также полукустарничковые ассоциации пустынного типа. Так, на небольших сопочках нередко развиты белополынные группировки (с господством Artemisia Lercheana). Особенно большие площади пустынные ассоциации занимают в депрессиях и долинах рек, где наиболее распространены чернополын-ники (A. pauciflora), а также развиты кокпечники (Atriplex сапа) и др.
Предуральское плато, сложенное третичными и меловыми отложениями с отдельными сопочками, находится к западу от собственно Мугоджар; занято преимущественно белополынно-тырсиковыми степями (Stipa sareptana+Artemisia Lercheana)', гораздо меньшую площадь занимают здесь лессингиановополынно-тырсиковые (Stipa sareptana-}- Artemisia Lessingiana), приуроченные к сопкам, и на более легких почвах — белопо-лынно-тырсовые (Stipa capillata-^-Artemisia Lercheana) ассоциации. В долинах рек и депрессиях на солонцах и солончаках-солонцах очень большие площади занимают чернополынные ассоциации (Artemisia pauciflora), меньшую — белополынные (Artemisia Lercheana), лессингиановополын-ные (Artemisia Lessingiana), кокпековые (Atriplex сапа) и биюргуново-тас-биюргуновые (Nanophyton erinaceum-j-Anabasis salsa) группировки. Местами на песках расположены участки псаммофитных степей со Stipa borysthenica и псаммофитным разнотравьем.
1 Более высокая (400—600 м вад у. м.) собственно низкогорная часть этой невысокой горной страны на «Геоботанической карте СССР» отнесена к дерновиннозлако-вым (ткпчаково-ковыльным) степям (77 звк).
• Поводимому, помимо Artemisia Lercheana, здесь распространена также и А. зе-mlarida. В большинстве этих полынно-ковыльных ассоциаций большую роль играет также типчак (Festuca sulcata).
Валовая продукция вышеупомянутых полынно-ковыльных пустынных степей колеблется от 5.5. до 7 (9) ц/га (Дохман, 1954).
Литература
И. Г. Бортов (1865); В. А. Дубинский (1904); Г. И. Дохман (1936, 1950, 1954).
78 в к --в о с т оч н о к а з а х с т а н с ки е п о л ы н н о-типч а к о в о-
ковыльные и полынно-типчаковые степи
(с Artemisia sublessingiana)
Восточноказахстанские полынно-типчаково-ковыльные и полынно-типчаковые степи связаны в своем распространении главным образом с Казахским мелкосопочником (Казахской складчатой страной), с ее средней частью, примерно от 47 до 52° (53°) с. ш. При этом на северо-востоке мелкосопочника, от р. Селеты и до Семипалатинска, эти пустынные степи занимают относительно пониженную окраинную горно-островную равнину, с высотами обычно ниже 400 м [400—150(100 м)] над у. м. Южнее этой северной полосы пустынных степей обширные пространства занимает низко- и местами среднегорная часть мелкосопочника [(400) 500—1000(1500) м над у. м. ], по которой очень далеко на юго-восток, вплоть до Тарбагатая, спускаются сухие типчаково-ковыльные степи, что можно рассматривать как проявление вертикальной поясности растительного покрова. В пределы этой повышенной части мелкосопочника пустынные степи проникают по долинам рек и местами встречаются как пет-рогенное явление на каменистых сопках. Кроме того, пустынные степи занимают южную и восточную часть обширной Тенгизской впадины [высота (500) 400—300 м над у. м.], со всех сторон окруженной мелкосопочником. Пустынные степи Тенгизской впадины переходят в пустынные степи южной окраины средне- и низкогорной части Казахского мелкосопочника; южная полоса восточноказахстанских пустынных степей тянется с запада-северо-запада, от гор Улутау, на восток-юго-восток до южного склона Тарбагатая. На западе, до долины Сарысу, южная граница этих степей проходит примерно на высоте 500—450 м над у. м., а к востоку от Сарысу на высоте 750—600 м. Таким образом, здесь полоса пустынных степей спускается вглубь пустынь наиболее далеко на юг в связи со значительными высотами местности.
Кроме того, восточноказахстанские степи окружают почти со всех сторон пустынную Зайсанскую котловину; здесь они занимают южные предгорья Алтая и северные предгорья Тарбагатая, на высоте 750—500 м над у. м. Они имеются и в южных предгорьях Тарбагатая, на той же высоте.
Далее к югу восточноказахстанские пустынные степи выражены также в предгорьях Джунгарского Алатау и местами в предгорьях Северного Тянь-Шаня. По северным предгорьям Джунгарского Алатау, на запад до гор Конгуртау эти степи, образующие здесь хорошо выраженный нижний пояс, занимают высоты 750—600 м над у. м., а на западной и южной окраинах Джунгарского Алатау — высоты (1250)1000—750м наду. м. (Рубцов, 1948а). В Северном Тянь-Шане восточноказахстанские пустынные степи отмечены на карте в следующих местах: по восточным предгорьям Заилийского Алатау и Кунгей-Алатау; двумя маленькими массивами в предгорьях Каратау (к юго-западу от Джамбула и к юго-востоку от пос. Ачисай).
Кроме того, эти степи покрывают более повышенную часть Чу-Илий-ских гор [1250—1000 (800) м над у. м.], которые окружены с за
пада и востока пустынными территориями (Гамаюнова и Голоскоков, 1949). Для восточноказахстанских полынно-типчаково-ковыльных и полынно-типчаковых степей очень характерно более или менее обильное развитие полыней из группы Artemisia sublessingiana s. 1., ареал которой охватывает Казахский мелкосопочник, южные предгорья Алтая, предгорья Тарбагатая, Джунгарского Алатау и хребтов Северного Тянь-Шаня.1
В пределах Казахского мелкосопочника (Гордягин, 1900—1901; Зелинский, 1912; Смирнов, 1912; Семенов, 1914, 1922; Крюгер, 1927а; Павлов, 1931; Левина, 1938; Поляков, 1949), а также южных предгорий Алтая (Келлер, 1911, 1912; Борисова, 1935), северных и южных предгорий Тарбагатая (Сапожников и Шишкин, 1918; Степанова, 1950) растительный покров в полосе пустынных степей довольно сложен. В составе степей преобладают дерновинные евксерофильные злаки: ковыли {Stipa sareptana, S. Lessingiana, S. capillata), типчак {Festuca sulcata), нередко пустынный житняк {Agropyrum desertorum)-, на сильно солонцеватых почвах и солонцах нередко обилен ломкоколосник {Psathyrostachys juncea). На юге распространения этих степей из ковылей особенно обилен тырсик {Stipa sareptana). Всегда в большем или меньшем количестве встречаются полукустарничковые полыни: Artemisia semiarida (казахстанский вид), A. sublessingiana s. str. (восточноказахстанский вид, приурочен преимущественно к щебнистым и каменистым почвам), A. Lercheana (казахстанский вид).1 2 Из других полукустарничков, широко распространенных во всей полосе заволжско-казахстанских пустынных степей, встречаются Ephedra distachya, Kochia prostrata, Pyrethrum achilleifolium. Эти полукустарнички иногда представлены в небольшом количестве, и тогда подобные степи с преобладанием ковылей, связанные со слабо солонцеватыми светлокаштановыми почвами и приуроченные к межсопочным равнинам, весьма приближаются к сухим типчаково-ковыльным степям (77 звк), но обычно в этих степях полукустарничковые полыни обильны и уступают только дерновинным злакам. Разнотравье в этих степях обычно малочисленно и представлено евксерофильными и евриксерофильными видами, преимущественно общими для всей полосы заволжско-казахстанских пустынных степей, от Ергеней до Зайеанской котловины {Dianthus leptopetalus, Ferula tatarica, Phlomis tuberosa, Galium ruthenicum, Lino-syris tatarica, Artemisia dracunculus, Jurinea multiflora и некоторые другие); кроме того, встречаются некоторые восточные виды, например Allium Pallasii (восточноказахстанско-среднеазиатский вид), полупаразит Thesium multicaule (восточноказахстанский вид) и др. В предгорьях Тарбагатая имеется типичный для пустынных степей Монголии джунгарско-монгольский вид лука {Allium polyrrhizum). В заметном количестве встречаются эфемероиды; помимо видов, широко распространенных в заволжско-казахстанских пустынных степях {Роа bulbosa, Tulipa Schren-
1 Artemisia sublessingiana (Kell.) Krasch. в узком смысле широко распространена в более южных районах Казахского мелкосопочника, в предгорьях Алтая, Тарбагатая и Джунгарского Алатау. Близкие виды описаны из следующих районов: А. ка-ratavica Krasch. et Abol. (Казахстан, глинистые и щебнисто-глинистые склоны низко-горий Каратау), A. valida Krasch. (южный Казахстан, глинистые склоны и засоленные долины), A. transiliensis Р. Pol. (Казахстан, глинистые предгорья Заилийского Алатау), A. mucronulata Р. Pol. (южный Казахстан, каменистые места в верхней части гор Каратау), A. Gorjaevii Р. Pol. (Джунгарский Алатау, глинистые склоны предгорий), А. атоепа Р. Pol. (южный Алтай, гранитные низкогорья) (П. П. Поляков. Новые виды рода полыни, намеченные к описанию И. М. Крашенинниковым. Он же. Новые виды полыни из подрода Seriphidium (Bess.) Rouy. Бот. мат. герб. Бот. инет. АН СССР, т 16., 1954).
2 Типичная Artemisia Lercheana встречается, видимо, только в западных частях мелкосопочника.
kii, Т. biflora), попадается также казахстанско-среднеазиатский вид Ixiolirion tataricum. Имеются в растительном покрове этих степей также эфемеры и напочвенные лишайники.
Местами на слабо- и несолонцеватых почвах, в равнинных условиях и по шлейфам сопок, в составе пустынных степей произрастают равномерно разбросанные степные кустарники (Spiraea hypericifolia, Caragana frutex) — это кустарниковые пустынные степи (Левина, 1938; Быков и Степанова, 1953)'
Эти степи нередко являются комплексными, особенно на равнинах и шлейфах сопок и их гряд. Здесь, в связи с довольно редким травостоем пустынных степей, несмотря на незначительное количество осадков, нередко выражены эрозионные процессы — размыв поверхностных горизонтов почво-грунтов, в результате чего на поверхность выходят и захватываются процессом почвообразования засоленные породы. Возникают пестрые комплексы с большим участием солончаковатых и корковых солонцов с господством казахстанских видов солянок (Atriplex сапа, Anabasis salsa, а на щебнистых засоленных почвах также джунгарско-казахстанского вида — Nanophyton erinaceum) и черной полыни (Artemisia pauciflora s. 1.). На поверхности почвы много лишайников (Parmelia vagans, виды Aspicilia и др.). У подножья склонов останцовых гряд или горно-сопочных массивов, а также в нижней части наклонных аккумулятивных равнин (шлейфов), в местах выпотевания грунтовых вод возникают своеобразные солончаковые комплексы с фрагментами степной и сочносолянковой (Atriplex verrucifera, Camphorosma Lessingii и др.) и солончаково-луговой (Aneurolepidium angustum, Glycyrrhiza uralensis и др.) растительности.
В пределах Казахского мелкосопочника, южных предгорий Алтая и предгорий Тарбагатая широко развиты петрофитные ассоциации полынно-типчаково-ковыльных степей, а также обнажения различных пород. Растительность этих «каменистых» степей изучена пока недостаточно. В качестве примера приведем наблюдения П. П. Полякова (1949) для западной части Казахского мелкосопочника, в пределах верховьев рр. Терс-Аккан и Каракингир. Здесь на мелкощебнистых делювиальных суглинках присопочных шлейфов и наклонных равнин развиты полынноковыльные степи с господством Stipa capillata и местами S. Lessingiana и с большим участием полукустарниковых полыней (Artemisia semiarida, A. sublessingiana, а на более каменистых участках и A. frigida). При увеличении каменистости, за счет приближения к поверхности коренных пород, увеличивается количество Artemisia frigida и появляются другие индикаторы каменистости, как Ephedra distachya, Stipa orientalis (горный казахстанско-центральноазиатский вид), Berteroa spathulata(xa3ax-станский вид), Seseli glabratum (казахстанский вид, распространенный от Заволжья до китайской Джунгарии).
На вершинах сопок и по южным склонам обнажаются различные породы — граниты, песчаники, метаморфизованные сланцы, окварцован-ные мергели и др. По данным П. П. Полякова (1949), в верховьях рек Терс-Аккан и Каракингир для обнажений гранита особенно характерны и обильно встречаются Spiraea hypericifolia (причерноморско-казахстанский вид с иррадиациями) и Artemisia Marschalliana (причерноморско-казахстанский вид), характерны, но встречаются единично, Chamaerhodos erecta (двулетний казахстанско-монгольский вид), *Seseli eriocarpum (центральноказахстанский вид),1 *Onosma simplicissimum (причерноморско-
1 Далее полукустарнички отмечены звездочкой.
казахстанские вид), Thymus Marschallianus (причерноморско-казахстанский вид), Helichrysum arenarium (западнопалеарктическпй вид). На метаморфизованных сланцах довольно часто встречаются как характерные виды те же Spiraea hypericifolia и Artemisia Marschalliana, единично произрастает ♦ Artemisia semiarida, также считающаяся характерным видом для этих обнажений. На девонских песчаниках часто произрастают, по данным П. П. Полякова, характерные виды для этих пород: ♦ Artemisia semiarida и * *А. sublessingiana; характерным видом является также ★Ziziphora Bungeana. Для окварцованных мергелей являются характерными * Artemisia semiarida (встречается часто), *А. sublessingiana, Cirsium igniarium (западномонгольско-восточноказахстанскпй вид).1 ★Scabiosa isetensis (волжско-казахстанский вид),* Gypsophila Patrinii (монгольско-казахстанский вид с иррадиациями в горах Средней Азии), ★Zygophyl-lum macropterum (казахстанско-среднеазиатский, пли туранский вид), * Anabasis truncate (казахстанско-джунгарский вид), Limonium chrysocomum (восточноказахстанский вид), ★Brachanthemum. kasakhorum (восточноказахстанский вид). Одинаково часто на обнажениях разных пород отмечены Stipa orientalis, Berteroa spathulata, ♦ Artemisia frigida. На юге полосы пустынных степей на каменистых обнажениях встречается также полукустарничковая Artemisia glabella (восточноказахстанский вид). Как видно из этого, такой строгой приуроченности отдельных видов к определенным горным породам, каковая наблюдается в Причерноморской провинции степной области — к мелам, известнякам, отчасти гранитам,— здесь не имеется.
При некотором накоплении на поверхности твердых пород дресвы и мелкозема возникают редкостойные сообщества с господством полукустарничков, преимущественно полыней (Artemisia frigida, A. sublessingiana, A. semiarida, реже A. glabella). При дальнейшем формировании почвы появляются дерновинные злаки; при этом петрофитные полукустарничковые сообщества с пустынной структурой переходят в пустынные полынно-дерновиннозлаковые степи.
У подножья сопочных гряд, в долинах рек и у родников, встречаются заросли ив и рощицы березы (Betula verrucosa) с рядом бореальных видов.
С долинами рек связаны чяевники (Lasiagrostis splendens), солончако-ватые и солончаковые луга.
По низким предгорьям Джунгарского Алатау, граничащим с пустынными подгорными равнинами и характеризующимся увалистым или мелкосопочным рельефом, развиты пустынные степи 2 с преобладанием на мелкоземистых плакорах и склонах тырсика (Stipa sareptana) и. полукустарничковых полыней [восточноказахстанской (джунгарской) Artemisia sublessingiana или северотураненой A. serotina]. Господствует обычнотырсик, среди которого разбросаны пятна с преобладанием вышеуказанных видов полыни. В некоторых случаях полынь преобладает на значительном протяжении. На легких почвах, а также в слабонамеченных ложбинах доминирует тырса (Stipa capillata) с примесью тех же полыней. К этим видам примешиваются из полукустарничков Kochia prostrata, из дерновинных злаков — типчак и другие, а также некоторые эфемероиды (Роа bulbosa и др.) и эфемеры; длительновегетирующее разнотравье, как вообще в пустынных степях, представлено в незначительном количестве или почти отсутствует. На смытых склонах, вместе с теми же видами полыни, нередко преобладает Stipa Richteriana (северотуранский вид). На щебни
1 Изолированное местонахождение в Дагестане.
* Н. И. Рубцов (1948а) в своей работе о растительности Джунгарского Алатау называет пояс с этими степями «полупустынным».
стых и каменистых склонах сопочек встречаются ассоциации с господством Artemisia sublessingiana и мелких ковыльков — Stipa orientalis и +5. caucasica А По южному склону Джунгарского Алатау в этом поясе встречаются на каменистых склонах участки трагакантовых группировок (Рубцов, 1948а) с преобладанием трагакантового колючего кустарничка -\~Convolvulus tragacanthoides (центральноазиатский вид) и полукустарничка A-Helianthemum soongoricum (джунгарско-тяньшан-скнн вид). По логам — местами заросли кустов Spiraea hypericifolia. видов Atraphaxis. По пологим депрессиям в этот пустынно-степной пояс заходят пустынные серополыннпкн с господством северотуранского вида полыни (Artemisia terrae-albae). солянок (Nanophyton erinaceum и др.).
Низкогорья Чу-Илийских гор также заняты пустынными полынно-дерновпннозлаковыми, чаще полынно-ковыльными степями и растительностью щебнистых и каменистых обнажений. Чаще господствуют дерно-винные злаки с большим участием полукустарничковой северотяныпан-ской (заплийской) полыни +Artemisia transiliensis, ближайшего родства A. sublessingiana. На мелкоземистых плакорных участках и пологих склонах преобладает тырсик {Stipa sareptana). а по наиболее повышенным участкам местами киргизский ковыль (5. kirghisorum)', иногда господствует типчак {Festuca sulcata), что, возможно, объясняется влиянием выпаса. По каменистым и щебнистым склонам нередко образует очень изре-женные группировки Artemisia transiliensis с большей или меньшей примесью ковыльков — Stipa orientalis. +S. caucasica. а также Lasiagrostis caragana и типчака; иногда ковылъки господствуют над полынью. По внешним окраинам мелкосопочника на щебнисто-каменистые склоны проникает также пустынная серая полынь {Artemisia terrae-albae). Кроме того, на щебнисто-каменистых, часто оголенных склонах встречаются из низких кустарничков------}rEphedra intermedia. + Е. equisetina, -J-Cerasus
tianschanica. из полукустарничков — Artemisia juncea (восточнотуранский вид) и др. В составе этих пустынных степей много эфемероидов и эфемеров-однолетников (Гамаюнова и Голоскоков, 1949). По логам встречаются скопления кустов Spiraea hypericifolia и немногих других кустарников.
По верху шлейфов Чу-Илийских гор также распространены пустынные степи с преобладанием Stipa sareptana. S. Richteriana. Artemisia transiliensis. с большим участием эфемероидов {Poa bulbosa. луки, тюльпаны) и эфемеров-однолетников. Ниже по шлейфу преобладает Artemisia transiliensis. сменяемая на предгорных равнинах Artemisia terrae-aqbae и солянками.
В составе флоры Чу-Илийских гор имеется несколько эндемичных видов и даже эндемичный род с одним видом -\-Niedzwedzkia semiretschen-skia. произрастающим в логах.1 2
Литература
А. Я. Гордягин (1900—1901); Б. А. Келлер (1911. 1912); Ф. Ф. Зелинский (1912); В. И. Смирнов (1912); В. Ф. Семенов (191-4, 1922); В. В. Сапожников и Б. К. Шишкин (1918); В. А. Крюгер (1927а); Н. В. Павлов (1931); А. Г. Борисова (1935); Ф. Я. Левина (1938); Н. И. Рубцов (1948а); А. П. Гамаюнова и В. П. Голоскоков (1949); П. П. Поляков (1949); Е. Ф. Степанова (1950); Б. А. Быков и Е. Ф. Степанова (1953).
1 Далее крестиком (4-) обозначены виды, не встречающнеся в пределах степной зоны (области), а приуроченные в своём распространении к южным горным районам.
2 Пустынные степи Чу-Илийских гор и предгорий Джунгарского Алатау отнесены на нашей карте к восточноказахстанским пустынным степям (78 вк). Своеобразие состава и структуры (обилие эфемероидов и эфемеров) этих степей довольно сильно отличает их от более северных аналогов; их следовало бы обозначить особыми буквенными шифрами.
78 кэ — прикаспийско-казахстанские
по л ы н но-э р к е к о в ые степи (с Artemisia arenaria, A. a s tra.c h a n i с a, Eurotia Ewersmanniana, Е. ceratoides)
Прикаспийско-казахстанские полынио-эркековыс степи показаны на «Геоботанической карте» в трех местах, весьма отдаленных друг от друга: 1) в южной части массива Терских песков (Грозненская область), 2) в Рын-песках северного Прикаспия (Западно-Казахстанская область) и 3) к востоку от среднего течения Сарысу (Карагандинская область).
Все эти песчаные массивы располагаются в пределах западной или северной окраинных частей пустынной области, поэтому и растительность здесь в известной мере является переходной между растительностью песков степной области (зоны) и песков пустынной области (зоны). Характерно обилие песчаностепного казахстанского дерновинного житняка — эркека (Agropyrum sibiricum) и полукустарничковых полыней — песчаной (Artemisia arenaria) и астраханской (A. astrachanica); западноказахстанская астраханская полынь очень близка к белой полыни (A. Lerche-апа), свойственной более тяжелым почвам. Кроме того, иногда в растительном покрове значительную роль играют виды терескена (Eurotia ceratoides — средиземный вид, Е. Ewersmanniana — джунгарско-туран-ский вид).
На полузаросших бугристых или барханных песках, занимающих здесь большие площади, наблюдаются заросли однолетника — кумар-чика (Agriophyllum arenarium), корневищного кияка (Elymus giganteus) «вынужденнокорневищной» песчаной полыни (Artemisia arenaria). Здесь же растут кустарники — виды тамарикса (Tamarix), кандыма (Cdlligonum) и др. Они, так же как и полынь (Artemisia arenaria), образуют придаточные корни на засыпанных песком ветвях.
На дне котловин среди песков располагаются заросли тростника (Phragmites communis), а в Терских песках — заросли огромного субтропического злака (Erianthus purpurascens), участки лугов, иногда солончаки с солянками.
В Рын-песках имеются в котловинах рощицы тополя и осины, а в Терских песках — тополя (Populus nigra, Р. alba), лоха (Elaeagnus angustifolia), ив.
Литература
Н. В. Павлов (1931); С. Е. Рожанец-Кучеровская (1936).
79. Пижмовые (из Tanacetum sibiricum) забайкальские степи в сочетании с Вострецовыми (из Aneurolepidium pseudoagropyrum) и дерновиннозлаковыми степями и сельскохозяйственные земли
на их месте
Пижмовые степи очень характерны для горностеппого Забайкалья (для Бурят-Монгольской АССР и Читинской области), а также для северо-восточной части Монгольской Народной Республики. Особенно широко распространены пижмовые степи в Нерчинских степях и в бассейне Аргуни, Онона и Селенги, где и показаны на карте. Здесь эти степи занимают большие площади по волнистым равнинам, широким пологим увалам и склонам сопок. В бассейне Селенги пижмовые степи часто являются вторичными, развиваясь на месте уничтоженных боров. Почвы под пижмо-выми степями обычно хрящеватые, чаще всего южные, преимущественно
выщелоченные черноземы; пижмовые степи встречаются также на лесных почвах после уничтожения леса.
В пижмовых степях господствует пижма (Tanacetum sibiricum) — мезо-ксерофильное сложноцветное растение с узколинейными, иногда почти нитевидными долями сложных листьев. К ней присоединяются также другие мезоксерофильные и ксеромезофильные виды разнотравья (Stellera chamaefasme, Scutellaria baicalensis, Pulsatilla ambigua, P. Turczaninovii, Thalictrum petaloideum, Leontopodium conglobatum и др.). Указанные виды являются даурско-монгольскими или южносибирско-монгольскими видами. Благодаря малому количеству в типичных пижмовых степях дерновинных злаков, задернованность почвы незначительная — истинное покрытие всего 7—8%; проективное покрытие 60—70%. Средняя высота пижмы около 25 см; цветущие побеги до 40 см. В пижмовой степи наблюдается следующая смена аспектов (по данным А. В. Куминовой, 1938).
1) Ранней весной (вторая половина мая, до развития основной вегетативной массы травостоя) синий ковер дает ургуй (Pulsatilla Turczaninovii).
2) Для середины июня характерно цветение беловато-розовой Stellera chamaejaxme.
3) В июле аспект наиболее красочен: цветет желтоцветная Tanacetum sibiricum; к пижме в значительном количестве примешивается синецветный Scutellaria baicalensis.
4) Осенний аспект в августе образует цветущая в массе скабиоза (Scab io ха Fischeri) с яркими синими соцветиями.
5) Осенью (сентябрь) сопки, покрытые пижмовой степью, «горят заревом» отмирающей багровой листвы Tanacetum sibiricum.
Пижмовые степи по своей экологии относятся к группе луговых степей,1 которые в своем типичном выражении в Забайкалье занимают небольшие площади. Наиболее близка к обычным луговым степям группа ассоциаций пижмовых степей с обилием ксеромезофильного и евмезо-фильного лугово-степного и лугового разнотравья (главным образом Hemerocallis minor, Sanguisorba officinalis, Artemisia commutata, Potentilla tanacetifolia, а также в меньшем количестве Veratrum nigrum, Clematis hexapetala, Trifolium lupinaster и др.). В образовании растительного покрова этой разности пижмовых степей сравнительно большую роль играют дерновины мезоксерофильной стоповидной осоки Carex pediformis и типчака; в небольшом количестве встречается тырса.
Другая группа ассоциаций пижмовых степей, наоборот, тяготеет к типичным дерновиннозлаковым степям; это дерновиннозлаково-пижмо-вые степи с большой примесью дерновинных степных, преимущественно евриксерофильных злаков — ковыля-тырсы (Stipa capillata s. 1.), тонконога (Koeleria gracilis), кистевидного мятлика (Роа botryoides) и типчака. Так, по полого-увалистым степям в бассейне Онона значительное распространение имеют тырсово-пижмовые группировки. В составе этих дер-новиннозлаково-пижмовых степей многие вышеприведенные более или менее мезофильные формы разнотравья отсутствуют; разнотравье здесь (кроме пижмы) вообще менее обильно, чем в пижмовых и разнотравнопижмовых степях.
По сухим нижним частям склонов местами встречаются также востре-цово-пижмовые степи с большим количеством корневищного злака — востреца (Aneurolepidium pseudoagropyrum). Днища падей (межгорных долин) и нижние части склонов к ним заняты Вострецовыми степями с преобладанием востреца.
1 Е. М. Лавренко (1940а) ранее относил их к настоящим (типичным) степям, что неверно.
Пижмовые степи используются как пастбища; более богатые разности пижмовых степей с высоким травостоем, особенно во влажные годы, могут скашиваться на сено. Валовая урожайность пижмовых степей — около 7.6 ц/га.
Литература
В. Н. Сукачев (1912); В. И. Смирнов (1913, 1914); С. И. Данилов (1936); А. В. Ку-минова (1938); Е. М. Лавренко (1940а); А. А. Юнатов (1950, 1954); Л. П. Сергиевская (1951).
80. Мелкодерновинные (из Koeleria gracilis, Festuca sulcata s. 1., Poa botryoides, Cleistogenes squarrosa, реже ковыля — Stipa capillatas. I.) алтайско-забайкальские степи и сельскохозяйственные земли
на их месте
Мелкодерновинные алтайско-забайкальские степи распространены от Алтая до бассейна Селенги в Забайкалье. В Алтае эти степи приурочены к межгорным котловинам и долинам рек; в Тувинской автономной области — к горным склонам, спускающимся к долинам рек — притокам Кемчика и Енисея, а также к горным склонам и падям юга области по границе с Монгольской Народной Республикой; в Забайкалье, в бассейне Селенги, — к склонам и речным террасам вдоль Джиды, Уды, Нижней Селенги и Хилки. Кроме того, эти степи покрывают большую часть Баргузинской котловины, за исключением ее наиболее пониженной внутренней части, занятой более сухими змеевково-тырсовыми степями. Особенно широко эти степи распространены в Монгольской Народной Республике, в пределах Хентейско-Хангайской горной страны, а также Монгольского Алтая, для которых их описывает А. А. Юнатов (1950, 1954) под названием «горных степей» (см. также Калинина, 1954).
Эти степи по своему составу и структуре являются типичными центральноазиатскими степями. Их следует относить к настоящим степям, к мезофитным разностям последних, в известной мере переходным к луговым степям. По верхним частям горных склонов, чаще всего по опушкам лесов, мелкодерновинные степи контактируют с луговыми степями, которые, однако, в своем типичном выражении по северной окраине Центральной Азии (во внутренней части Советского Алтая, Тувинской автономной области, в Западном Забайкалье, а также в Хентейско-Хангайской горной стране) выражены только фрагментарно.
Мелкодерновинные горные степи по нижней границе своего распространения контактируют и переходят в более ксерофитные (засухоустойчивые) степи с преобладанием ковыля-тырсы, чаще всего в змеевково-тырсовые степи (81); это наблюдается, например, в Тувинской автономной области и в Западном Забайкалье — в бассейне Селенги. В восточной части Советского Алтая, на юго-востоке Тувинской автономной области и вне СССР — в Монгольском Алтае эти степи иногда непосредственно переходят в пустынные ковыльковые степи (82).
Степи эти приурочены к горночерноземным и горнокаштановым почвам, в большинстве случаев более или менее щебнистым или на террасах рек — песчаным и супесчаным.
В мелкодерновинных степях господствуют злаки с небольшими дерно-винами; типчак (Festuca sulcata s. ampl.), тонконог (Koeleria gracilis), мятлик (Poa botryoides), житняк (Agropyrum cristatum s. str.), иногда змеевка (Cleistogenes squarrosa).
Очень характерно участие в составе этих степей восточных рас степного типчака (Festuca sulcata s. ampl.), чаще всего, видимо, восточносибирско-монгольского вида — Festuca lenensis. К восточносибирско-монгольским видам относятся также Роа botryoides и Agropyrum crista-tum s. str. (последний вид с иррадиациями в горах Средней Азии). На Алтае Роа botryoides замещен западносибирско-казахстанским видом Роа stepposa; Cleistogenes squarrosa и особенно Koeleria gracilis, как известно, широко распространены в степях Евразии. Для мелкодерновинных степей характерно также отсутствие или незначительное участие крупно-дерновинных злаков — ковылей. Из последних в этих степях иногда встречается ковыль-тырса (Stipa capillata s. I.).1
К. А. Соболевская (1950) для высокогорных мелкодерновинных степей юга Тувинской автономной области приводит, кроме того, в качестве ценозообразователей тувинско-даурский (?) вид — Festuca albifolia, алтайский вид — Koeleria altaica и алтайско-даурский вид — Роа dahurica.
В составе мелкодерновинных степей обычно встречаются степные полукустарничковые полыни, чаще всего Artemisia frigida, в некоторых районах — Artemisia obtusiloba, A. subviscosa. Виды травянистого разнотравья, растущие в этих степях, в большинстве случаев относятся к сибирско-монгольским и даурско-монгольским видам.
Как и в других степях, характерных для стран северной окраины Центральной Азии, в мелкодерновинных алтайско-забайкальских степях почти полностью отсутствуют эфемероиды и эфемеры, что связано с холодной и сухой весной. Из эфемероидов встречаются только один вид тюльпана (Tulipa uniflora) и немногие гусиные луки (Gagea).
В связи с тем, что травяной покров в этих степях довольно редкий, на поверхности почвы часто довольно обильны напочвенные лишайники (Parmelia vagans и др.).
Перейдем к характеристике отдельных массивов этих степей.
В пределах внутренней части Алтая мелкодерновинные горные степи показаны на «Геоботанической карте» по Катуни и ее притокам — Урсулу, Чуе и Аргуту, то есть в тех районах, где по соседству в среднегорном поясе господствует лиственничная тайга (из Larix sibirica) или где (в верховьях Чуи) лесной пояс выпадает, и мелкодерновинные степи непосредственно контактируют с растительными группировками высокогорного пояса.
В горных степных долинах внутренней части Алтая мелкодерновинные степи приурочены в большинстве случаев к надлуговым террасам долин рек и к южным пологим склонам; к этим же позициям приурочены и переходы от горных мелкодерновинных степей к более ксерофит-ным тырсово-змеевковым или даже ковыльковым пустынным степям. По северным склонам небольших увалов в более западных и северных районах Алтая (например в долине Урсула) развиты луговые степи, а на северных склонах гор — лиственничные леса. Большую площадь занимают также обрывистые скалистые склоны.
Так, например, в Курайской степи на высоте 1500—1700 м над у. м. (по Чуе) равнинные участки с щебнистыми каштановыми почвами заняты мелкодерновинной типчаково-мятликовой степью с покрытием около 50% и преобладанием мятлика (Роа attenuata var. stepposa) и типчака.
1 Из крупнодерновинных перистых ковылей (ряд Pennatae) в районе распространения мелкодерноринных горных степей во внутреннем Алтае и в Тувинской автономной области произрастают только Stipa Joannis и kirbhisorum. Первый вид встречается в составе луговых степей.
Сизоватый фон этой степи придают кустики полукустарничковой полыни (Artemisia frigida) и дерновины Eritrichium pectinatum. Кроме того, в этой степи еще отмечены следующие виды: из дерновинных злаков — Koeleria gracilis, Stipa orientalis, из разнотравья—Allium lineare, Stellaria dichotoma, Gypsophila Patrinii (sub G. Gmelini), Dianthus versicolor (sub D. Seguieri), Stevenia cheiranthoides (sub Arabis incarnata), Medicago falcata, Astragalus 'ceratoides, A. depauperatus, A. macroceras, A. tephro-lobus, Veronica pinnata и др. Большинство видов из группы разнотравья по характеру их географического распространения относится к сибирско-монгольским или алтайским. На склонах южных и юго-восточных экспозиций развита ассоциация Stipa orientalis + Psathyr о stachys (Elymus) funcea c Caragana pygmaea и др. (Баранов и Шелудякова, 1926).
В Теньгинской степи (по Теньге — притоку Урсула), по пологим всхолмлениям равнины в долине реки (около 1000 м над у. м.), развита мелко-дерновинная типчаково-мятликовая степь (Роа attenuata var. stepposa) со значительным участием разнотравья (Thalictrum petaloideum и Thermopsis lanceolata)', из дерновинных злаков в меньшем количестве представлены Stipa capillata, Koeleria gracilis, Cleistogenes squarrosa. По соседству встречаются участки разнотравно-дерновиннозлаковой луговой степи (Келлер, 1914; Баранов и Шелудякова, 1926).
В Тувинской автономной области, согласно неопубликованным исследованиям А. В. Калининой, мелкодерновинные горные степи представлены разнотравно-мелкодерновиннозлаковыми и осочково-типчаковыми группами ассоциаций.
Разнотравно-мелкодерновиннозлаковые степи приурочены главным образом к склонам южной экспозиции. По составу преобладающих видов они имеют много общего со степями Тувинской котловины, занимающими более низкий уровень в общем рельефе (81); по данным К. А. Соболевской (1950, 1953), в них господствуют Festuca pseudovina, Koeleria gracilis, Роа botryoides, Agropyrum cristatum', роль змеевки, по сравнению со степями Тувинской котловины, значительно уменьшается, вплоть до полного ее выпадения из состава ряда ассоциации. В состав травостоя обычно в большом количестве входит стоповидная осока (Carex pediformes). К этим злакам и осокам в значительном количестве примешиваются виды из разнотравья и мелких полукустарничков. Так, К. А. Соболевская (1950) для полынно-тонконоговых степей указывает на большое развитие тех или иных видов полыни: из полукустарничковых — Artemisia frigida, A. depauperate, из травянистых — A. commutata, A. facutica. Кроме того, представлены некоторые виды травянистого разнотравья (Stellaria dichotoma, Potentilla acaulis, P. bifurca, P. sericea, Linum brevisepalum, Bupleurum pusilium, Amblynotus obovatus, Dracocephalum peregrinum, Pedicularis achilleifolia и др.). Большинство этих трав и полукустарничков по своему географическому распространению может быть отнесено к сибирско-монгольским и даурско-монгольским видам.1
Осочково-типчаковые степи покрывают верхние части холмов. Основную массу травостоя вместе с типчаком (Festuca pseudovina) составляют степные осочки (Carex pediformis и С. duriuscula). Примесь других степных растений незначительна; из более постоянных для западной части Тувинской автономной области видов разнотравья следует упомянуть ломонос Крылова (Clematis Krylovii) и для восточной части колюрию (Coluria geoides).
1 К. А. Соболевская (1950) для этих полынно-тонконоговых степей из дарпопин-ных злаков, кроме Koeleria gracilis, приводит Agropyrum cristatum, Stipa decipiens, S. orientalis, S. glareosa', последний вид сомнителен.
По опушкам лиственничных лесов, занимающих склоны северной экспозиции, эти степи контактируют с луговыми степями, характеризующимися более мощно развитым (более сомкнутым и высоким) травяным покровом. Эти луговые степи, на карте по условиям масштаба отдельно не выделенные, Л. В. Калинина считает характерными для склонов южной экспозиции средних и верхних частей лесного пояса. Основу травостоя этих степей составляют крупнодерновинные злаки (Stipa capillata, S. Joannis, Helictotrichon desertorum, Leucopoa albidw, постоянна также примесь мелкодерновинных злаков — типчака, тонконога, мятлика). В составе травостоя имеются многие виды разнотравья (Artemisia glauca, A. santolinifolia — полукустарничек, Pulsatilla Turczaninovii, Iris ruthenica и др.).
В горных степях Тувинской автономной области довольно обычны следующие кустарники: Spiraea hypericifolia, Cotoneaster melanocarpa, Grossularia acicularis, Dasiphora parvifolia. В поясе горных степей, видимо, встречаются на каменистых склонах тимьянниковые степи и тимьян-ники. К. А. Соболевская (1953) указывает для каменистых склонов и барханных песков как наиболее частый в Тувинской автономной области вид тимьяна — Thymus asiaticus.
Степи Усинской котловины (по р. Уса) представляют сочетания мелкодерновинных степей (с Роа stepposa), кустарниковых степей (с тырсой и Caragana pygmaea) и каменистых тимьянниковых степей (Куминова, 1948).
В южной части Тувинской автономной области, по южным отрогам хребтов Восточного Танну-Ола и Сангилена, а также на высоких водораздельных плато и в широких долинах юго-западной части области (Мон-гун-Тайга), на высоте 2000—2500 (3000) м над у. м., распространены высокогорные степи (Соболевская, 1950), также состоящие из мелкодерновинных злаков, но преимущественно из высокогорных видов последних (Festuca albifolia, Koeleria altaica, Poa dahurica). В составе этих степей встречаются, кроме того, злаки, характерные для горных мелкодерновинных степей среднегорного пояса (Koeleria gracilis, Agropyrum crista-tum и др.). Иногда обильны мелкие полукустарничковые полыни (Artemisia depauperata, A. frigida)', из разнотравья встречаются Potentilla sericea, Р. viscosa, Р. bifurca, Bupleurum pusilium, Allium clathratum, Leontopodium leontopodinum, Androsace Bungeana (A. Lehmanniana Sobol., 1950, non Spreng.), Arctogeron gramineus, Oxytropis oligantha, Pulsatilla Campanella, Aster alpinus, Veronica spicata, Gentiana decumbens и др.; из кустарничков — Oxytropis tragacanthoides. He входя в детали географического распространения этих видов, большинство из них по характеру ареала можно отнести к сибирско-монгольской группе. Покрытие в таких степях — около 50 %, средняя высота травостоя — около 10 см. Обращает на себя внимание обилие высокогорных форм (помимо указанных выше высокогорных видов мелкодерновинных злаков, сюда относятся Leontopodium leontopodinum, Androsace Bungeana, Pulsatilla Campanella и др.).
В юго-западной части Тувинской автономной области (Монгун-Тай-гинский природный район) вокруг озер, иногда высохших, и по надпойменным террасам рек, например Харгин-Гола (и его притоков), распространены центральноазиатские пустынные степи, на карте отдельно не выделенные. Травостой этих степей очень низкий (в среднем 5—7 см высоты); основу травостоя создают злаки (Stipa glareosa, S. orientalis, Agropyrum cristatum, Koeleria gracilis, Festuca albifolia) и мелкие полукустарничковые полыни — Artemisia frigida и A. argyrophylla. В'небольшом
количество представлены Astragalus brevifolius, Arctogeron gramineus, Bupleurum pusilium, Aster alpinus и др. По берегам озер встречается Anabasis brevifolia (Соболевская, 1950).
В Забайкалье мелкодерновинные стопи располагаются значительными массивами но окраинам стопного острова в Баргузинской депрессии и в бассейне Селенги, а именно в сродном и верхнем точении Уды, по нижнему точению Селенги и но сродному точению Джиды. Эти степи приурочены здесь к склонам гор южной и близкой экспозиции, к шлейфам склонов и к надлуговым террасам рок. Почвы — обычно легкие (щебнистые, песчаные, супесчаные, логкосуглинистые). Травостой этих стопой низкий. Преобладают чаще всего молкодерновинпыо злаки {Koeleria gracilis, Роа botryoides и Agropyrum cristatum)', иногда господствуют равномерно все эти виды («трохзлаковая степь», ио определению Л. П. Сергиевской, 1951), в других же случаях преобладает один из них или два вида в разных комбинациях. Встречаются также стопи с участием типчака {Festuca lenensis)1, однако в Баргузинских степях типчак отсутствует. К этим злакам иногда примешивается в значительном количество змеевка {Cleistogenes squarrosa)', ковыль-тырса {Stipa capillata s. 1.; чаще S. decipiens) отсутствует или играет подчиненную роль. Разнотравье обычно необильное; в более значительном количество встречаются полукустарнички и травы с более или моиоо укореняющимися надземными побегами — холодная полынь {Artemisia frigida), лапчатка {Potentilla acaulis) и тимьян {Thymus serpyllum s. 1.). Обычно co злаками комбинируется в значительном количество один или два вида из указанных полукустарничков. В меньшем количество встречаются такие виды: из мелких полукустарничков — Kochia prostrata, Arenaria capillar is, Alyssum biovulatum, A. lenense, Pti-lotrichum elongatum; из травянистого разнотравья — Allium anisopodium, A, bidentatum, Iris flavissima, Dianthus versicolor, Stellaria cherleriae, S. dichotoma, Potentilla verticillaris (южная часть Забайкалья), Oxytropis oxyphylla, Stellera chamaefasme (южная часть Забайкалья), Bupleurum scorzonerifolium, В. bicaule, Amblynotus obovatus, Schizonepeta multifida, Veronica incana, Cymbaria dahurica, Leontopodium campestre, Aster alpinus, Heteropappus altaicus, Artemisia commutata, Scorzonera austriaca и др. Иногда разбросаны кустики Caragana pygmaea. В составе напочвенного покрова имеется кочующий лишайник Parmelia vagans. Как видно из этого списка, преобладают сибирско-монгольские и даурско-монгольские виды.
В Баргузинской депрессии довольно широко распространены своеобразные и пока мало изученные группировки из полукустарничковой клейковатой полыни {Artemisia subviscosa).
В связи с усиленной пастьбой, в Забайкалье часто встречаются, особенно на песчаных и щебнистых почвах, пастбищные (дигрессионные) ассоциации с господством полукустарничков с укореняющимися надземными побегами — Artemisia frigida, Thymus serpyllum s. 1. (тимьянники)1 или некоторых видов травянистого разнотравья,' также с укореняющимися побегами — Potentilla acaulis, Veronica incana. Сюда же относятся осочковыо степи с господством молкой корневищной осоки — Carex duri-uscula. На степных «сбоях» эти виды часто встречаются в разных сочетаниях. В Баргузинских степях на песках вместе с Thymus serpyllum s. 1. иногда встречаются длиинокорневищные растения: песчаная осока {Carex sabulosa), костер Короткого {Bromus Korotkyi) и др.
На каменистых склонах в этой части Забайкалья произрастают кустарники с даурско-монгольским основным ареалом: Spiraea aquilegi-
1 Эта ассоциации ранее мы относили (Лавренко, 1940а) к опустыненным степям, что сейчас считаем неправильным.
folia, Caragana pygmaea, а у подножья каменистых склонов — группы небольших деревьев Ulmus pumila. В Баргузинских степях эти кустарники не встречаются. По северным склонам местами расположены сосновые* леса.
Мелкодерновинные степи используются под пастбища; часть их распахана. Урожайность более богатых разнотравьем мелкодерновинных злаковых степей в Забайкалье 3—8 ц/га; типчаковых и осочковых степей 2—5 ц/га, полынных 1.5—4 ц/га, и, наконец, наиболее низко-пропзводптельнымп являются тимьянниковые и лапчатковые группировки 1.5 2 ц/га (Сергиевская, 1951). Пастбища эти используются в течение круглого года.
Некоторая часть мелкодерновинных горных степей распахана; посевы в большинстве случаев орошаются. Местами возможно и богарное (неорошаемое) земледелие, но урожай без орошения не может считаться обеспеченным.
Л и т е р а т у р а
М. Ф. Короткий (1912); В. Л. Коллер (1914); Г. И. Поплавская (1914, 1916); В. И. Варанов и В. Л. Шелудякова (1926); В. К. Шишкин (1937); А. В. Куминова (1938, 1948); Е. М. Лавренко (1940а); К. А. Соболевская (195(1 1953); А. А. Юнатов (1950. 1954); Л. II. Сергиевская (1951); А. В. Калинина (1954).
81. Змеевково-тырсовые и вострецово-тырсовые енисейско-забайкальские степи и сельскохозяйственные земли
на их месте
Змеевково-тырсовые степи, относящиеся к типичным или настоящим степям, занимают значительные площади па юге Средней Сибири (бассейн Енисея) и Забайкалья, где они приурочены к межгорным депрессиям и равнинам. Эти степи занимают равнины, увалы, отлогие склоны, шлейфы гор. Вне СССР они широко распространены в Монгольской Народной Республике, главным образом в ее восточной половине (Юнатов, 1950, 1954). В Забайкалье, а также па северо-востоке Монгольской Народной Республики, помимо змоевково-тырсовых степей, довольно широко развиты также вострецово-тырсовые степи. Степи эти связаны с темнокаштановыми и каштановыми почвами, а в более северных районах — с южными черноземами, обычно более или менее легкого механического состава — легкосуглинистыми, супесчаными, хрящеватыми и щебнистыми.
Змеевково-тырсовые и вострецово-тырсовые енисейско-забайкальские стопи, среди которых имеются богаторазнотравные и бедноразнотравные группы ассоциаций, являются восточным (южносибирско-монгольским) аналогом разнотравно-дерновиннозлаковых (разнотравно-типчаково-ковыльных) и дерновинных (типчаково-ковыльных) европейско-казахстанских степей.
Отличия между ними в основном следующие: в составе европейско-казахстанских степей господствуют пористые (из ряда Pennatae), а также волосовидные или тырсовидные (из ряда Capillatae) ковыли; более или менее обильны эфемероиды и эфемеры; в составе енисейско-забайкальских степей господствуют только волосовидные ковыли; перистые ковыли или отсутствуют или, если и имеются, то играют ничтожную роль; эфемероиды и эфемеры отсутствуют или редки.
В составе упомянутых выше енисейско-забайкальских степей из дерновинных злаков преобладают: обманчивый ковыль (Stipa decipiens), воло
совидный ковыль-тырса (S. capillata s. str.),1 растопыренная змеевка (Cleistogenes squarrosa), стройный тонконог {Koeleria gracilis), типчаки {Festuca pseudovina, F. vallesiaca? F. lenensis). К этим видам в меньшем обилии примешиваются другие дерновинные злаки: гребенчатый житняк {Agropyrum cristatum) и кистевидный мятлик {Роа botryoides). В бассейне верхнего Енисея в менее ксерофитных ассоциациях этих степей местами значительную роль играет пустынный овсец {Helictotrichon desertorum). В Забайкалье нередко к тырсовидным ковылям в большом количестве примешивается корневищный злак — вострец {Aneurolepidium pseudoagropyrum), образуя таким образом вострецово-тырсовые степи. Вострец изредка встречается и в степях верхнего течения Енисея, но не играет здесь почти никакой роли в сложении травостоя степей. Из осок произрастает обычно в небольшом количестве, но значительно увеличиваясь на сильно выпасываемых участках, корневищная твердоватая осока {Carex duriuscula)', в менее ксерофитных ассоциациях иногда обильна дерновинная стоповидная осока {Carex pediformis).
Из травянистого разнотравья нередко очень обильно встречается корневищная бесстебельная лапчатка {Potentilla acaulis), а в менее ксерофитных (богаторазнотравных) степях бассейна верхнего Енисея бывает обильна корневищная сизая полынь {Artemisia glauca). Остальные виды многолетнего травянистого длительновегетирующего разнотравья обычно встречаются в небольшом обилии: из дерновинных — Allium bidentatum-, из корневищных—Fragaria viridis, Veronica incana, Galium verum; из корнеотпрысковых — Thermopsis lanceolata, Convolvulus Ammanii, Cym-baria dahurica; из по преимуществу стержнекорневых—Dianthus versicolor, Pulsatilla Turczaninovii, Delphinium grandiflorum, Erysimum altaicum, Astragalus austrosibiricus, A. inopinatus, Haplophyllum dauricum, Bupleurum bicaule,B.scorzonerifolium,Goniolimon speciosum, Artemisiacommutata, Tanacetum sibiricum, Aster alpinus, Heteropappus altaicus, Saussurea salicifolia и др. Коротковегетирующие многолетние травы (эфемероиды) почти полностью отсутствуют, только местами в небольшом количестве встречаются следующие эфемероиды из семейства лилейных: Tulipa uniflora, Gagea pauciflora, G. provisa, G. altaica. Эфемеры (коротковегетирующие однолетники) отсутствуют; из длительновегетирующих малолетников чаще других встречается Artemisia scoparia.
Из полукустарничков в этих степях произрастают Kochia prostrata, Arenaria capillaris, Alyssum lenense, Thymus sp. sp., Artemisia frigida, A. Gmelini? Из этих видов массово в енисейско-забайкальских степях встречается только холодная полынь {Artemisia frigida), а в составе тимьянников и тимьянниковых степей, на каменистых склонах и на песках, виды чабреца {Thymus). По степям местами разбросаны кусты Караган {Caragana pygmaea, С. Bungei, С. microphylla), однако никогда не образующие больших скоплений; подобные кустарниковые (карагано-вые) степи особенно характерны для бассейна Селенги (с мелколистной караганой), а также для Тувинской котловины (с караганой Бунге); имеются они и в Минусинской котловине. 1 * 3
1 Оба эти вида ковыля относятся к ряду тырсовидных или волосовидных ковы-лей, почему эти степи и названы выше «тырсовыми».
* Эти два вида типчака приводятся по В. В. Ревердатто и др. Festuca pseudovina — из группы F. sulcata s. 1.
3 Большинство этих видов — стержнекорневые; Artemisia frigida с укореняющимися надземными побегами; виды Thymus также с длинными укореняющимися надземными побегами.
На каменистых склонах и местами на песках часто встречаются тимьянниковые степи и тимьянники. Для тимьянников на каменистых склонах характерно обильное развитие видов Thymus, а также образующих подушки Chamaerhodos altaica, Androsace incana, Arctogeron gramineus и др. В составе тимьянниковых степей эти виды составляют примесь к степным дсрновинным злакам, из которых в тимьянниковых степях особенно часто преобладает типчак.
Для долин рек и окраин озерных котловин, часто встречающихся среди этих степей, очень характерны остепненные луга с господством мощного дерновинного злака, блестящего чия (Lasiagrostis splendens) и дву-чешуйного пикульника (Iris biglumis). Днища озерных котловин, а также побережья солоноватых озер заняты солончаковыми (галофитными) лугами и сообществами однолетних и реже многолетних солянок.
Виды, упомянутые выше для змеевково-тырсовых и вострецово-тыр-совых енисейско-забайкальских степей, по своему распространению являются в большинстве сибирско-монгольскими 1 или даурско-монгольскими и не заходят или только незначительно заходят в степи Западной Сибири и северного Казахстана. Так, следующие господствующие и согос-подствующие в енисейско-забайкальских степях виды могут быть отнесены к сибирско-монгольским или даурско-монгольским: Stipa decipiens, Festuca lenensis, Poa botryoides, Agropyrum cristatum, Aneurolepidium pseu-doagropyrum. Некоторые из господствующих и согосподствующих видов заходят в степи юга Западной Сибири и севера Казахстана, почему могут быть отнесены к казахстанско-сибирско-монгольским видам [Carex duri-uscula (рис. 61), Potentilla acaulis, Artemisia frigida}. Остальные из доминирующих видов более широко распространены в степной области (зоне). Из растений лугового и луговостепного типа к казахстанско-сибирско-монгольским (с иррадиациями в горах Средней Азии) видам принадлежит Lasiagrostis splendens, а к сибирско-монгольским — Iris biglumis.
Из менее обильных в этих степях растений к сибирско-монгольским и даурско-монгольским степным видам могут быть отнесены следующие: Allium bidentatum, Tulipa uniflora, Pulsatilla Turczaninovii (последний вид несколько заходит в степи Западной Сибири и северного Казахстана), Р. ambigua, Delphinium grandiflorum, Astragalus austrosibiricus, A. inopi-natus, Haplophyllum dauricum, Bupleurum bicaule, Convolvulus Ammanii, Cymbaria dahurica, Tanacetum sibiricum (эта пижма в степях бассейна Енисея отсутствует) и др.
В составе степных ассоциаций со значительным участием кустарниковых караган к даурско-монгольским и монгольским видам относятся следующие кустарники: Caragana pygmaea, С. Bungei и С. microphylla (из них С. Bungei отсутствует в Забайкалье). На каменистых обнажениях в составе тимьянников и отчасти тимьянниковых степей к числу сибирско-монгольских и даурско-монгольских видов принадлежат Chamaerhodos altaica, Androsace incana, Arctogerongramineus, некоторые виды Thymus и др.
Ряд видов, не играющих господствующей роли в травостое енисейско-забайкальских змеевково-тырсовых и вострецово-тырсовых степей, заходит далеко на запад, до Урала или даже до Волги. К таким казахстанско-сибирско-монгольским видам относятся Dianthus versicolor, Thermopsis lanceolata, Goniolimon speciosum, Saussurea salicifolia (последний вид заходит также в горы Средней Азии) и некоторые другие.
Змеевково-тырсовые и вострецово-тырсовые степи используются как пастбища; в отдельные годы, обильные осадками, они могут и выкаши
1 Как и далее, имеется в виду преимущественно Южная Сибирь.
ваться. Значительная часть этих степей в Минусинской котловине, а отчасти и в других районах их распространения, распахана.
81 ен — енисейские
(без участия Aneurolepidium pseudoagropyrum)
На юге Красноярского края, в верхней части бассейна Енисея, эти степи занимают значительные площади в Минусинской котловине (При-абаканская, Уйбатская, Сорокаозерная, Биджинская, Койбальская, Мину-синско-Тубинская, Бейская, Сабинская, Сыдинская и Саксарская степи) и в Енисейско-Чулымской впадине (Ширинская степь); небольшой участок имеется около Красноярска (Красноярская степь). Кроме того, они занимают пониженную часть Тувинской котловины. Покрывают эти степи равнины и низкогорья. Почвы — южные черноземы и темнокаштановые.
Преобладают в составе этих степей дерновинные злаки {Stipa capillata, S. decipiens, Helictotrichon desertorum, Festuca pseudovina, F. valle-siaca, Koeleria gracilis, Cleistogenes squarrosa), а часто также корневищные и рыхлодерновинные осоки (Carex duriuscula, Carex pediformis) мпълуку-старнички с укореняющимися надземными побегами (Artemisia frigida); из корневищных многолетников обычны Artemisia glauca и Potentilla acaulis.
Л. М. Черепнин (1953а)1 сообщает такие сведения о фенологии этих приенисейских степей.
В начале апреля снега в степях уже нет. Позеленение степей становится заметным в конце апреля—начале мая. Первыми в конце апреля зацветают редко разбросанные немногочисленные эфемероиды — тюльпаны (Tulipa uniflora) и слабо заметные желтоцветные гусиные луки (Gagea pauciflora, G. provisa, G. altaica и др.), а из длительновегетирующих растенией — прострел Турчанинова (Pulsatilla Turczaninovii) с яркими синими цветами.
Из преобладающих растений в степях в начале мая первыми цветут осоки — твердоватая и стоповидная, а также бесстебельная лапчатка. Все эти растения ярких аспектов не дают. Только по каменистым местам хорошо выделяются золотисто-желтые бурачки (Alyssum biovulatum, A. lenense), а несколько позже белые пятна цветущего седого проломника (Androsace incana).
Во второй половине мая степные осоки и бесстебельная лапчатка отцветают. Цветет редко разбросанный желтеющий ирис (Iris flavissima). Начинают зацветать шишковидный остролодочник (Oxytropis strobilacea), карликовая карагана (Caragana pygmaea), термопсис (Thermopsis lanceolata), альпийская астра (Aster alpinus). В местах обилия этих растений преобладают желтые и лиловые тона.
В начале июня колосятся обильные в степцх мелкодерновинные степные злаки (типчаки, тонконог). В середине июня они цветут, создавая буровато-фиолетовый аспект; по этому фону разбросаны желтые и лиловые соцветия упомянутых выше видов.
В конце июня отцветают и начинают желтеть мелкодерновинные злаки, но еще продолжается цветение разбросанных по степи термопсиса, шишковидного остролодочника, альпийской астры; к ним присоединяются другие цветущие бобовые — копеечник Гмелина (Hedysarum Gmelini), южносибирскии (Astragalus austrosibiricus) и неожиданный (A. inopinatus) астрагалы.
В июле* если лето сухое и жаркое, мелкие злаки почти совершенно высыхают, и степь принимает буроватый оттенок, но во влажные годы она сохраняет зеленые тона. Из часто встречающихся растений в это время цветут: двустебельная володушка (Bupleurum bicaule) с невзрачными желтыми зонтиками, а также лилово-синяя алтайская астра (Heteropappus altaicus) и седая вероника (Veronica incana).
Во второй половине июля обычно идут дожди, почему степь в начале августа оживает. В августе массово цветут господствующие в приенисейских степях тырсовые ковыли (Stipa capillata, S. decipiens) и полыни (Artemisia frigida, A. glauca и др.).Из ярко-Цветущих растений в это время цветут: с синими цветами — равновысокии солнечник
1 Далее использованны не только автореферат диссертации Л. М. Черепнина (1953), но и рукопись последней.
(Galatella fastigiata), крупноцветная живокость (Delphinium grandijlorum), с розовыми цветами — разноцветная гвоздика (Dianthus versicolor) и др.
В сентябре постепенно прекращается вегетация, растения подсыхают, но если заморозки начинаются поздно, то в сентябре еще сохраняется общий зеленый цвет травостоя. Отцветают разноцветная гвоздика, равновысокий солнечник, алтайская астра, крупноцветная живокость.
В октябре засыхает большинство растений. Только некотооые злаки (типчаки, тонконог), стоповидная осока, холодная полынь, астры и некоторые другие вегетируют до снега, который обычно выпадает в первой половине ноября.
Среди енисейских дерновиниозлаковых степей можно выделить как богаторазнотравные, так и бедноразнотравные ассоциации.
В составе богаторазнотравных дерновиниозлаковых енисейских степей, которые по занимаемой ими площади преобладают в степных частях Минусинской и Енисейско-Чулымской котловин, господствуют такие виды дерновинных злаков: Stipa capillata, Helictotrichon desertorum, Festuca pseudovina, Koeleria gracilis, Cleistogenes squarrosa; некоторое участие иногда принимают степные мятлики — кистевидный {Роа botryoides) и степной {Р. stepposa). Из дерновинных осок часто обильна Carex pediformis. Разнотравье также обильно. Общее проективное покрытие около 50 (60) % ; средняя высота травостоя около 20—25 см. Распространены эти степи в пределах 200—500 м высоты над у. м. Почвы под ними— южные черноземы, реже обыкновенные и темнокаштановые.
Среди богаторазнотравных дерновиниозлаковых степей бассейна верхнего Енисея наиболее распространена крупнополынно-тырсовая группа ассоциаций, довольно часто также встречается тырсово-овсецовая.1
Крупнополынно-тырсовые степи {Stipa capillata-^-Artemisia glauca) — самая распространенная группа ассоциаций типичных (настоящих) степей на юге Красноярского края. Помимо господствующегоковыля-тырсы, обильна крупная сизая полынь {Artemisia glauca), реже встречаются другие виды полыни (Л. Gmelini, A. commutata). Обильны также ложноовечий типчак, растопыренная змеевка, стройный тонконог, овсец Шелля {Helictotrichon Schellianum), стоповидная осока, а из разнотравья, кроме упомянутых выше полыней, — Potentilla acaulis, Fragaria viridis, Bupleurum scorzonerifolium, Phlomis tuberosa, Veronica incana, Galium verum, Galatella punctata s. 1., Heteropappus altaicus и др. Средняя урожайность около 7.3 ц/га сухой массы.
Тырсово-овсецовые степи {Helictotrichon desertorumA-Stipa capillata) распространены меньше, чем крупнополынно-тырсовые, и сосредоточены преимущественно в Хакассии. Помимо господствующих овсеца и ковыля-тырсы, обильны те же злаки и осоки, что и в предыдущей группе ассоциаций, а также некоторые виды разнотравья {Pulsatilla Turczaninovii, Hedysarum Gmelini, Bupleurum scorzonerifolium, Aster alpinus); встречаются также Caragana pygmaea, Cymbaria dahurica и многие другие. Средняя урожайность — около 8.8 ц/га.
Бедноразнотравные дерновиннозлаковые (сухие) енисейские степи занимают в верхней части бассейна Енисея гораздо меньшую площадь, чем вышеописанные крупнополынно-тырсовые и тырсово-овсецовые степи.1 2
1 Л. М. Черепнин (1953а) высказывает предположение, что в большинстве случаев крупнополынно-тырсовые степи являются [производными тырсово-овсецовых степей и возникают на месте последних в результате интенсивного выпаса.
В. В. Ревердатто (1954) в составе крупнополынно-тырсовой группы ассоциаций выделяет следующие «крупные ассоциации»: злаково-полынно-разнотравную, ковыльно-полынно-разнотравную и осочково-злаково-полынно-разнотравную.
2 По В. В. Ревердатто (1954), в енисейских островных степях преобладают бедноразнотравные дерновиннозлаковые степи («четырехзлаковые»). Однако, его «ковыльно-
Эти сухие степи связаны с небольшими абсолютными высотами (200— 400 м над у. м.) и с засушливым климатом, для которого характерно малое количество осадков и почти полное отсутствие снега. Почвы — каштановые, реже южные черноземы, иногда солонцеватые, часто каменистые или песчаные. Основные массивы этих степей сосредоточены в Приабаканском и Уйбатском сухостепных районах. Травостой более бедный, разреженный (общее покрытие около 35—45%), в период наибольшего развития — около 15—20 см высотой.
Наиболее типичным выражением «сухих» степей в бассейне верхнего Енисея, преимущественно на легкосуглинистых темнокаштановых почвах, является так называемая «четырехзлаковая» степь, основу которой образуют четыре дерновинных злака: обманчивый ковыль (Stipa decipiens), сизый типчак (Festuca va.llesia.ca),* 1 стройный тонконог и растопыренная змеевка. Из разнотравья постоянно встречается в большом обилии бесстебельная лапчатка (Potentilla acaulis). Остальные виды представлены в небольшом количестве (Agropyrum cristatum, Allium ani-sopodium, A. odorum, Iris flavissima, Dianthus versicolor, Potentilla bifurca Sibbaldianthe adpressa, Caragana pygmaea, Astragalus austrosibiricus, Bupleurum bicaule, Goniolimon speciosum, Convolvulus Ammanii, Veronica pin-nata, Galium verum, Heteropappus altaicus); из малолетников — Androsace septentrionalis* Artemisia scoparia и немногие другие. Средний выход сухой массы — около 4 ц/га. Иногда встречаются участки, на которых господствует один какой-нибудь вид — обманчивый ковыль, сизый типчак, стройный тонконог, твердоватая осока или холодная полынь. Господство обманчивого ковыля обычно наблюдается на старых залежах (урожайность 17—22 ц/га). Появление типчаковых, Тонконоговых, осочковых (с Carex duriuscula) и мелкополынных (с Artemisia frigida) степей связано главным образом с усиленным выпасом; при этом осочковые степи приурочены к суглинистым почвам, а мелкополынные—к супесчаным. Урожайность этих степных сбоев — 2.8—3.9 ц/га. Подобные сбои возникают и на месте вышеописанных менее ксерофитных енисейских степей.
Ассоциации «четырехзлаковых» степей, связанные с более или менее солонцеватыми темнокаштановыми почвами, характеризуются увеличением обилия таких видов, как острец (Aneurolepidium ramosum), прутняк (Kochia prostrata), алтайская астра (Heteropappus altaicus), также ситниковый волоснец (Psathyrostachys funcea), чий (Lasiagrostis splendens) и др.
На каменистых обнажениях развиты тимьянники с господством полукустарничков (Thymus minussinensis, Alyssum biovulatum, A. lenense, Gypsophila Patrinii), более или менее обильны также петрофильные травянистые растения (Eritrichium fenisseense, Potentilla acaulis, Festuca albi-folia — этот типчак только в Хакассии, Agropyrum geniculatum, а также Potentilla sericea, Dracocephalum discolor, Androsace incana, Crepis tenui-folia, Silene fenisseensis, Orostachys spinosa, Cymbaria dahurica, Bupleurum bicaule и др.).
На каменистых склонах с более или менее сформированными почвами часто встречаются также «каменистые» (тимьянниковые) степи, представляющие разнообразные переходы от «четырехзлаковых» степей к тимьян-никам. Значительный интерес представляют «каменистые» степи с большим
разнотравно-злаковые с полынью степи на четырехзлаковой основе безусловно должны быть отнесены к предыдущей (богаторазнотравной) группе степных группировок.
1 По В. В. Ревердатто (1954) —Fettuca pseudovina.
• Androsace septentrionalis является единственным эфемером в составе этих степей.
участием нитевиднолистной кобрезии (Cobresia filifolia),1 довольно часто встречающиеся на карбонатно-каменистых склонах в Ширинских степях. Эта кобрезия здесь произрастает совместно со следующими видами, представленными более или менее обильно: Festuca albifolia, Stipa decipiens, Agropyrum lolioides, Helictotrichon Schellianum, Silene fenisseensis, Potentilla acaulis, Gypsophila Patrinii, Aster alpinus, Thymus minussinensis, Crepis tenuifolia, Androsace dasyphylla. Средняя производительность каменистых степей около 3 ц/га. Каменистые степи, также как и степи на более или менее солонцеватых почвах, обычно комплексны.
В районах распространения «сухих» (бедноразнотравных) енисейских степей на хорошо заросших степной растительностью песках и легких супесях чаще встречаются ассоциации менее ксерофитных богаторазнотравных дерновиниозлаковых степей с участием таких ковылей, как Stipa Joannis, а также Calamagrostis koibalensis, Carex Korshinskyi и мелкодерновинных злаков, характерных для «четйрехзлаковой» степи. В составе обильного разнотравья — Serratula cardunculus, Potentilla acaulis и др.; из полукустарничков — Thymus minussinensis и Artemisia frigida.
На слабозакрепленных песках развиты тимьянники, главнейшие эди-фикаторы которых — полукустарнички — являются общими с тимьян-никами каменистых склонов: Thymus minussinensis, который иногда почти сплошь покрывает подобные пески, Artemisia frigida, Alyssum biovulatum, A. lenense, а из разнотравья — Potentilla acaulis и др. Имеются все переходы от подобных тимьянников к настоящим степям.
Довольно значительную площадь занимают залежи с преобладанием сорняков или корневищных злаков, или рыхлодерновинных и плотнодер-новинных злаков, в зависимости от возраста залежи.2
Бурьянистые залежи возрастом 1—2 (3) года характеризуются господством сорняков. Два основных типа этих залежей — мелко- и крупно-бурьянистые. Преобладают сорняки: Setaria viridis, Chenopodium album, Sonchus arvensis, в значительном количестве встречается также Artemisia scoparia. Средняя урожайность сухой массы — валовая 12 ц/га, кормовая 5.8 ц/га (в составе бурьянистых залежей много недоедаемых растений).
Наибольшую площадь занимают залежи, находящиеся в стадии корневищных злаков, используемые под сенокосы. Возраст этих залежей 3—5 (6) лет. Наиболее распространены залежи с господством ползучего пырея (Agropyrum r-'pens) и с примесью безостого костра (Bromus inermis) и остреца (Aneurolepidium ramosum). Менее распространены острецовые залежи (на наиболее сухих и засоленных почвах), зубровковые залежи с Hierochloe Bungeana (на легких почвах) и вейниковые залежи с Calamagrostis epigeios (в более влажных районах, на легких почвах). Средняя урожайность сухой массы пырейных залежей: валовая 19.1 ц/га, кормовая 10.9 ц/га.
Залежи в рыхлодерновинной стадии, возрастом 6—И (12) лет, представлены тимофеевко-мятликовой, мятликово-разнотравной и тимофеев-ково-дерновиннозлаковой группировками с господством Phleum phleoides, Роа angustifolia, Р. stepposa, луговостепного разнотравья, а из плотнодер-новинных злаков—Cleistogenes squarrosa, Koeleria gracilis и др. Характерно обильное развитие на некоторых залежах в этой стадии Astragalus meli-
1 По наблюдениям Л. М. Черепнина, Cobresia filifolia нередко встречается в При-абаканских и Ширинских степях, а С. Bellardi в этих же степях произрастает изредка, в составе остепненных и сырых солонцеватых лугов в котловинах. Последний вид встречается также в высокогорной мохово-лишайниковой тундре в Западном Саяне.
8 Залежи описаны по данным Л. М. Черепнина (1953).
lotoides. Урожайность сухой массы наиболее распространенных Тимофеев* ково-мятликовых залежей: валовая — 12.2 ц/га, кормовая — 9.2 ц/га.
С 9—12 лет залежь постепенно переходит к господству плотнодерновин пых злаков, и с 12—15(20) лет ее растительность мало отличается от цел и и* ной.
Упомянем еще о галофитной растительности, которая встречается в долинах рек и в озерных понижениях среди этих степей.
По окраинам озерных котловин, по надлуговым террасам рек, а иногда и по более возвышенным участкам пойм широким распространением пользуются остепненные более или менее галофитные чиевые (из Lasiagrostis splendens) и пикульниковые (из Iris biglumis) луга. По окраинам озерных котловин встречаются также галофитные луга из тонкоцветковой бискиль-ницы (Puccinellia tenuiflora). По берегам соленых озер развиты одиолет-никовосолянковые группировки (из Salicornia europaea, Suaeda cornicu-lata, Atriplex fera), а местами, например в Уйбатской степи — из полукустарничковых солянок (Kalidium foliatum), с участием ряда других галофильных растений, характерных для солончаковых почв (Saussurea papposa, Limonium Gmelinii, L. macrorrhizon, Artemisia nitrosa, Plantago salsa и др.).
Разнотравно-дерновиннозлаковые степи (крупнополынно-тырсовые и тырсово-о песцовые), по данным Л. М. Черепнина (1953а), преобладают в Енисейско-Чулымской и Минусинской котловинах. Сюда, в частности, относятся степи: Ширинская, Сыдинская, Минусииско-Тубипская, При-енисейская, Койбальская, Сорокаоэерная, Сабинская, Бейская, Биджинская и Саксарская.
В Минусинской котловине в Приабакаиской и Уйбатской степях преобладают бедноразнотравные дерновиннозлаковые степи («четырехзлаковые» или змеевково-тонконогово-типчаково-тырсовые степные группировки).
Степи Тувинской котловины, по исследованиям К. А. Соболевской (1950) и А. В. Калининой,1 также в основном могут быть отнесены к зме-евково-тырсовым степям или к переходам от последних к мелкозлаковым горным степям или к пустынным степям.
В восточной части Тувинской котловины распространены на черноземных почвах тырсовые и сизополынно-тырсовые степи. Это в основном бога-тораэнотравные степи с господством тырсы (Stipa capillata), а в некоторых случаях также и сизой полыни (Artemisia glauca); в последнем случае эти степи являются аналогом крупнополынно-тырсовых степей Енисейско-Чулымской и Минусинской котловин (см. выше). В некоторых случаях к тырсе примешиваются растения, характерные для луговых степей (Phleum phleoides, Sanguisorba officinalis, а также Medicago falcata s. 1.). Эти степи используются как сенокосы; значительная часть их распахана под богарные посевы.
К югу от хребтов Танну-Олы, к востоку от той части границы СССР с Монгольской Народной Республикой, где эта граница проходит в меридиональном направлении, имеется массив бедноразнотравных житняково-тырсовых степей (Stipa decipiens+Agropyrum < ristatum) с примесью Cleistogenes squarrosa, Stipa sibirica, Veronica pinnata, Dracocephalum discolor, Kochia prostrata и др., с разбросанными кустами караганы Бунге (Caragana Bungei) и иногда пятнами полукустарничка Nanophyton erinaceum. Эти степи используются в качестве зимних пастбищ.
1 Далее характеристика степей Тувинской автономной области в основном дана по неопубликованным данным А. В. Калининой.
Преобладают и Тувинской котлонине на каштановых почвах полынно-мелкозлаковые степи (А. В. Калинина), сложенные мелиодерновиипыми злаками: житняком (Agropyrum cristatum), тонконогом (Koeleria gracilis), растопыренной змеевкой (Cleislogenes squarrosa). Участие тырсы (Stipa capillata а. I.) в составе злаковой части травостоя не велико (поболее 5— 8% в общем покрытии травостоя). В западной части котловины в сложении этого типа степей небольшое участие принимает ковылск — Stipa orien-lalis. Кроме того, в составе этих степей в большом количестве развиваются полукустарничек — холодная полынь (Artemisia frigida) и бесстебельная лапчатка (Potentilla acaulis). Обычными, но не обильными компонентами являются корневияшая Carex duriuscula, степной суккулент Orostachys spinosa и довольно многочисленные стержнекорневые травы и полукустарнички (Arenaria capillaris, Gypsophila paniculata, Kochia prostrata, Alyssum lenense, Goniolimon speciosum и др.). Нередко примесь в подобных степях образуют кустарниковые караганы — Caragana pygmaea (15—20 см высоты) и С. Bungei (компактные кусты до 1.5—2 м высоты и примерно такого же диаметра). Кусты разбросаны на расстоянии 3—5 м и больше (карагано-полынио-мелкозлаковыо степи). На поверхности почвы довольно много лишайника Parmelia vagans. Средняя высота травостоя 15—25 см, общее проективное покрытие растительности 50—60%; урожайность сухой массы травостоя 3—5 ц/га. Используются они как пастбища для всех видов скота во все сезоны года. Эти степи в ряде случаев представляют переходы к мелкодерновинным горным степям (80).
Среди этих степей в центральной части Тувинской котловины на хрящеватых почвах, а также под влиянием усиленного выпаса на каштановых почвах встречаются сообщества пустынного полукустарничка Nano-phyton erinaceum. Пятна его зарослей порядка нескольких кв. метров обусловливают комплексность стони.
Окраины долин рек и озерных депрессий в Тувинской котловине окаймлены полосками чиевников, где вместе с чием (Lasiagrostis splendens) 9 большом количестве растут волоснецы (Aneurolepidium Paboanum и A. dasystachys) с большей или меньшей примесью галофильных и степных растений.
На берегах соленых озер в Тувинской котловине встречаются ассоциации сочных солянок: однолетних — Salicornia еигораеа и Suaeda cornicu-lata, я многолетних — Kalidium foliatum (обычно с Limonium (Statice) Gmelinii), а выше по берегам озер — ассоциации солончаковых лугов с господством Aneurolepidium dasystachys, A. Paboanum, A. angustum, Puccinellia tenuiflora, Hordeum brevisubulatum, а иногда Phragmites communis, с большим участием галофильных растений (Tripolium vulgare, Plantago salsa, Saussurea papposa я др.). Солончаково-луговые группировки используются как сенокосы и пастбища.
Литература
Б. К. Шишкин (1914); В. В. Ревердатто (1925, 1929, 1947, 1947а, 1954); В. В. Ревердатто и В. П. Голу бинцева (1930); К. А. Соболевская (1947, 1950); А. П. Самойлова (1951); Л. М. Черепнин (1953, 1953а).
81 зб — забайкальские (с участием востреца — Aneurolepidium pseudoagropyrum)
В Забайкалье змеевково-тырсовые и вострецово-тырсовые степи представлены в виде трех массивов: 1) по Селенге, на юге Бурят-Монгольской АССР; 2) по Борзе и Аргуни, на юге Читинской области; 3) небольшим изолированным массивом в центре островной Баргузинской степи
Кроме того, этими степями занята часть о-ва Ольхона на Байкале. В первых двух случаях указанные степные массивы граничат с Монгольской Народной Республикой, где эти степи занимают очень большую площадь.1 Приурочены они к плакорным позициям — невысоким увалам, отлогим, преимущественно южным и юго-западным, склонам гор и шлейфам последних, днищам сухих падей. Почвы — каштановые.
В составе степей преобладают из дерновинных злаков — тырса (Stipa decipiens) и змеевка (Cleistogenes squarrosa), из корневищных злаков — вострец (Aneurolepidium pseudoagropyrum), из разнотравья — бесстебельная лапчатка (Potentilla acaulis), из полукустарничков — холодная полынь (Artemisia frigida). Преобладают обычно один или два вида.
Чаще встречаются группы ассоциаций: змеевково-тырсовые, тырсовые, полынно-тырсовые, вострецово-тырсовые, лапчатково-змеевковые и пр. Вострецово-тырсовые степи приурочены к шлейфам гор и к днищу падей. Очень обильно вострец развивается на участках степи, потревоженных полевкой Брандта и сурками (тарбаганами). Местами, особенно на более грубых щебнистых почвах, большие площади занимают полынные группировки с господством Artemisia frigida, частью, видимо, являющиеся следствием неумеренного выпаса. В составе этих степей в значительном количестве встречаются из дерновинных злаков также тонконог (Koeleria gracilis), типчак (Festuca lenensis), кистевидный мятлик (Роа botryoides), житняк (Agropyrum cristatum). Типичные «четырехзлаковые» степи с равномерным преобладанием тырсы, змеевки, тонконога и типчака, столь характерные для сухих степей Минусинской котловины (юг Красноярской области), как выяснила Л. П. Сергиевская, для Забайкалья, видимо, не характерны.
Разнотравье — немногочисленно; встречаются Allium bidentatum, Arenaria capillaris, Erysimum altaicum, Oxytropis selengensis, Haplophyllum dauricum, Bupleurum bicaule, Goniolimon speciosum, Schizonepeta multi-fida, Veronica incana, Cymbaria dahurica, Saussurea salicifolia и др.
По более повышенным местам количество разнотравья увеличивается; в составе подобных разнотравно-дерновиннозлаковых степей (разнотравно-тырсовых, пижмово-тырсовых и пр.) появляется сибирская пижма (Tanacetum sibiricum), а также Thalictrum minus, Bupleurum scorzonerifolium, Galium verum и др.
Так как змеевково-тырсовые и близкие к ним по составу степи в Селен-гинской депрессии (в Бурят-Монголии) и на юге Читинской области занимают значительные площади, необходимо сказать несколько слов о тех типах растительного покрова, которые сочетаются с этими степями.
Среди Селенгинских степей на легких почвах (песчаных и супесчаных) местами значительные площади занимают кустарниковые степи. На фоне обычно довольно разреженного травостоя с преобладанием тех же злаков (см. выше,) а местами также Artemisia frigida, с небольшим участием разнотравья, разбросаны кусты караган (Caragana microphylla или С. pygmaea, а иногда и обоих видов вместе).
На каменистых склонах с выходами скал и нагромождениями крупного щебня и более крупных обломков камней распространены особые кустарниковые степи, где кустарники также не растут сплошными зарослями, а образуют «редкокустье» на фоне дерновиннозлаковой степи с господством
1 В Монголии эти степи (тырсовые, змеевково-тырсовые, полынно-тырсовые, вострецово-тырсовые) развиты преимущественно в восточной части страны, в пределах Восточно-Монгольского плоскогорья. Отсюда они и заходят в Читинскую область. Кроме того, степи указанного состава поднимаются, местами прерываясь, далеко на север по правобережью Орхона, а далее по Селенге.
тех же видов степных злаков, которые преобладают и в вышеописанных степях. Из полукустарничков здесь постоянно присутствуют Thymus serpyllum s. 1., Artemisia frigida, Kochia prostrata', из разнотравья обычны Allium bidentatum, A. odorum, A. anisopodium, Stellaria dichotoma, Oxyt-ropis selengensis, Schizonepeta multifida, Artemisia commutata, Crepis tenuifolia и др. На выходах скал —Orostachys spinosa, Androsace incana, Arcto-geron gramineus. На этом фоне разбросаны кустарники: Spiraea aquilegi-folia, S. media, Cotoneaster melanocarpa, Caragana pygmaea, а по низу склонов встречаются также редкостойные заросли UImus pumila.
На щебнистых склонах развиты тимьянниковые степи и тимьянники с участием полукустарничков: Thymus serpyllum s. 1., Chamaerhodos altaica, Artemisia frigida, а из травянистых форм, образующих подушки или латочки, встречаются Potentilla acaulis, Androsace incana, Arctogeron gramineus и др. Все эти полукустарнички и травы обладают способностью к вегетативному разрастанию.
В ряде случаев господство на песчано-супесчаных и щебнистых почвах тимьяна, бесстебельной лапчатки и холодной полыни является следствием усиленного выпаса.
Как в Селенгинских, так и в Борзянско-Аргуньских степях (на юге Читинской области) днища депрессий заняты солончаками и солончаковыми (галофитными) лугами. По берегам соленых озер и на гуджирных пухлых солончаках с выцветами солей растут пятнами Suaeda corniculata, Artemisia anethifolia, Limonium aureum, Puccinellia tenuiflora. На солончаковых лугах преобладают злаки: Puccinellia tenuiflora, Hordeum brevisubulatum, Alopecurus ventricosus, с примесью разнотравья — Halerpestes ruthenica, Saussurea amara и др. Окраины депрессий окружены пикульни-ково-чиевыми галофитными лугами с господством чия (Lasiagrostis splendens) и пикульника или сахилдзы (Iris biglumis).
Литература
М. Ф. Короткий (1912); В. И. Смирнов (1913); А. В. Куминова (1938); А. А. Юна-тов (1950, 1954); Л. П. Сергиевская (1951).
82. Ковыльковые и полукустарничково-ковыльковые центральноазиатские пустынные степи
Ковыльковые и полукустарничково-ковыльковые пустынные степи с господством мелких ковыл£ков из ряда Barbatae приурочены к центральноазиатским окраинам СССР — к Восточному Памиру, Центральному Тянь-Шаню, Чуйской степи на Алтае и к северным окраинам Убсунурской котловины в Тувинской автономной области, где они и показаны на карте. Таким образом, в пределах СССР они связаны в основном с горными странами (Памир, Тянь-Шань, Алтай, Танну-Ола) и в пределах последних приурочены к межгорным котловинам и широким долинам.
Эти степи сформированы двумя видами «бородатых» ковыльков: галечниковым (Stipa glareosa) и восточным (Stipa orientalis). К ним нередко примешиваются в значительном количестве некоторые полукустарнички, реже кустарнички и кустарники и в небольшом количестве некоторые злаки и разнотравье. По своей структуре эти степи характеризуются ред-котравностыо, крайне незначительным общим проективным покрытием (5—20%, реже до 40% и выше). Количество видов в ассоциациях этих степей небольшое. Эти степи должны быть отнесены в основном к пустынным.
Полукустарничково-ковыльковые и ковыльковые степи в типичном выражении весьма характерны для Центральной Азии. В плакорныхусловиях они встречаются в Монгольской Народной Республике, где образуют особую подзону пустынных степей с господством мелких «бородатых» ковыль-ков: Stipa gobica, S. glareosa, S. orientalis (последний вид только на западе Монгольской Народной Республики). Первый вид является энде-мом Монголии, второй — более широко распространенным центральноазиатским видом, заходящим в окраинную часть СССР (Памир, Центральный Тянь-Шань, Алтай и Тува), третий — казахстанско-центральноазиатским видом.1 Из других дерновинных злаков, принимающих основное участие в сложении этих степей, встречаются сибирско-центральноазитский вид Agropyrum cristatum и евразиатский степной вид Cleistogenes squarrosa.
Из полукустарничков для этих степей в СССР характерны Eurotia ceratoides — средиземный вид с обширным ареалом, Artemisia frigida — казахстанско-сибирско-монгольский вид, A. Skorniakowii — восточнопамирский вид, Anabasis brevifolia — монгольский вид, Nanophyton erina-сеит — северотуранский (джунгарско-казахстанский) вид, Keaumuria soongorica — джунгарско-монгольский. Из трагакантовидных кустарничков характерны Oxytropis aciphylla и О. tragacanthoides — монгольские виды. Из кустарников — Caragana Bungei, С. pygmaea — монгольские виды, С. spinosa — алтайско-саянско-монгольский вид.
Краткий анализ географического распространения основных господствующих и согосподствующих растений этих степей свидетельствует об их приуроченности к Центральной Азии, в основном к Монголии. Следует добавить еще, что в них вовсе отсутствуют однолетники-эфемеры и в большинстве случаев отсутствуют эфемероиды, что также характерно для центральноазиатских степей и пустынь.
Степи эти используются как круглогодовые пастбища, но преимущественно в летнее время; зимнее использование возможно в связи с отсутствием или с малой мощностью снегового покрова. Валовая урожайность травостоя этих степей очень мала: в Восточном Памире — 0.5—7 (в среднем 2—3) ц/га, в Чуйской степи — 0.7—1.2 ц/га.
Так как состав согосподствующих и второстепенных видов различен в разных районах, где представлены эти степи, то последние далее характеризуются по отдельным горным системам.
82 тв — тувинские
Пустынные полукустарничково-ковыльковые степи приурочены в Тувинской котловине к ее наиболее пониженной части, где они встречаются небольшими массивами на щебнистых смытых почвах; местами их площадь несколько увеличена под влиянием усиленного выпаса.2 В составе этих степей, поданным К. А. Соболевской (1950), преобладают из дерновинных злаков — Stipa glareosa, S. orientalis, Agropyrum cristatum, Koeleria gracilis, Cleistogenes squarrosa, а из полукустарничков —Nanophyton eri-naceum. Остальные компоненты представлены в небольшом количестве {Artemisia frigida, Oxytropis aciphylla, Convolvulus Ammanii, Caragana pygmaea, Euphorbia tshuiensis, Veronica pinnata, Potentilla acaulis, Gypsophila desertorum и др.). Высота травостоя?—15см, покрытие не болыпе40%.
1 О пустынных степях Монголии см. работы А. А. Юнатова (1950, 1954) и А. В. Калининой (1954). В составе чуйских, дентральнотяныпанских и памирских степей встречается ряд видов, отсутствующих в равнинных и низкогорных степях степной зоны (области) Евразии.
г Масштаб карты не позволил показать все мелкие массивы этих степей в Тувинской котловине, в частности, в окрестностях Кызыла.
Эти степи используются как пастбища. Полукустарничек Nano-phyton в позднеосеннее и зимнее время хорошо поедается лошадьми и верблюдами и является хорошим нажировочным кормом.
Литература
К. А. Соболевская (1950).
82 уб — убсукурские
Пустынные полукустарничково-ковыльковые степи по северной окраине Убсупурской котловины, то есть в пограничной с Монгольской Народной Республикой части Тувинской автономной области, также отличаются широким распространением подушкообразного полукустарничка Nano-phyton erinaceum, северотуранского элемента по своему ареалу, отсутствующего в соседней Чуйской степи. Редкотравные степи здесь приурочены к слабо покатым к югу, к озеру Убсу-Нур, шлейфам южных отрогов хребтов Танну-Олы. Почвы — щебнистые, переходные от каштановых к бурым. Господствуют в степи из полукустарничков — Nanophyton erinaceum, из дерновинных злаков — Stipa glareosa и Cleistogenes squarrosa; причем преобладают или указанные злаки, или нанофитон. Кроме того, в более значительном количестве представлены из полукустарничков — Artemisia frigida, Kochia prostrata ; из трав — Allium Krylovii, Scorzonera austriaca, Gypsophila desertorum. В небольшом количестве встречаются Agropyrum cristatum, Astragalus monophyllus, Stellaria dichotoma, Ptilotrichum canescens, Convolvulus Ammanii, Cymbaria dahurica, Euphorbia tshuiensis, Potentilla astragalifolia и др. Таким образом, состав этих степей близок к составу Чуйской степи; однако в пределах последней отсутствуютNa-nophyton erinaceum, Lagochilus ilicifolius и немногие другие. Высота растительного покрова — до 10 см; общее прективное покрытие до 30—40%. Местами по южным отрогам хребта Танну-Ола встречаются участки по-лынно-ковыльковых степей с частыми кустами двух видов караган (Caragana Bungei или С. pygmaea).
Литература
К. А. Соболевская (1950); А. А. Юнатов (1950, 1954); А. В. Калинина (1954).
82 а — чуйские (алтайские)
В Чуйской степи на Алтае на плоских междуречных водоразделах (на бурых галечниковых почвах) также широко развиты редкотравные ковыльковые степи. В большинстве случаев господствует галечниковый ко-вылек (Stipa glareosa); довольно значительное участие местами принимают другие виды ковыля — S. orientalis, S. decipiens (=S. Krylovii). Почти постоянно встречаются, выступая нередко в качестве согосподствующих видов, житняк (Agropyrum cristatum) и корневищная осочка (Carex durius-cula). В сложении растительного покрова этих степей принимают участие в большем или меньшем количестве кустарнички и полукустарнички (Ephedra distachya, Anabasis brevifolia, Artemisia argyrophylla, A. frigida, A. caespitosa, Reaumuria soongorica). В незначительном количестве встречаются бесстебельные астрагалы (Astragalus dilutus, A. laguroides, A. testiculatus), а также кустарничковые бобовые трагакантового типа (Oxytropis aciphylla, О. tragacanthoides). В небольшом количестве постоянно
присоединяются Dontostemon perennis, Potentilla bifurca, Convolvulus Ammanii, Goniolimon speciosum', более редко — Allium tenuissimum, Gypsophila desertorum, Ptilotrichum canescens, Euphorbia tshuiensis, Heteropappus altaicus, Potentilla astragali]olia и др. Местами разбросаны разреженные заросли караган (Caragana spinosa, С. Bungei, С. pygmaea). Высота генеративных побегов ковыльков не превышает 10—12 (—20) см, а вегетативных 3—5 см. Общее проективное покрытие в этих степях колеблется от 15—20% до 40%.
Литература
В. И. Баранов и В. А. Шелудякова (1926); А. В. Калинина (1948).
82 цтш — центра льнотяньшанские
Мелкодерновинные полынно-ковыльковые степи в Центральном Тянь-Шане более крупными массивами встречаются в бассейнах Алабуги, Сары-джаза, в верховьях Нарына, по хр. Джаманто, Байбичето и др.; здесь эти степи приурочены к горным долинам и склонам гор на высоте 2500—3000 м над у. м., то есть к контакту горных степей с «субальпийским» поясом (Выходцев, 1952). Почвы под этими степями — высокогорные светлые каштановые, обычно скелетные.
По южному борту Нарынской впадины, по данным И. В. Выходцева, в этих степях преобладают ковыльки (Stipa caucasica, S. orientalis) и полукустарничковая полынь (Artemisia tianschanica), к которым в большем или меньшем количестве примешиваются дерновинные злаки (Agropyrum cristatum, Helictotrichon desertorum, Hordeum turkestanicum), корневищные злаки (Aneurolepidium [Elymus] dasystachys), разнотравье (Leontopodium ochroleucum, Potentilla evestita, Oxy tropis globif lord). Общее покрытие около 30—40%. Из преобладающих растений достигают высоты: полынь с вегетативными побегами до 8 см высоты и генеративными до 30 см; восточный ковылек такой же высоты; кавказский ковылек — до 10 см высоты. Урожайность этих в основном весенне-летне-осенних пастбищ 3—5 ц/га.
Местами подобные степи поднимаются еще выше и встречаются в непосредственной близости к снежным и ледяным полям.
В горных долинах Центрального Тянь-Шаня по речным террасам на высоте 2000—3000 м над у. м., на каштановых, обычно скелетных почвах, встречаются полынно-типчаковые (Festuca sulcata s. 1., Artemisia tianschanica, A. serotina) степи с примесью ковыльков (Stipa caucasica, S. glareosa).
Литература
И. В. Выходцев (1952).
82 пм — памирские
В Восточном Памире редкотравные пустынные степи встречаются на высоте 3500—4500 м над у. м. Эти степи приурочены здесь к межгорным долннам, где они располагаются по днищу долин, шлейфам гор, преимущественно на легких почвах, песчаных и супесчаных. Господствуют ковыль-ки—Stipa orientalis и S. glareosa. Помимо этих дерновинных злаков и местами корневищной осочки Carex duriusculiformis, в значительном количестве встречаются ксерофильные полукустарнички (чаще Eurotia ceratoides и
Artemisia Skorniakowii, в некоторых районах также A. Kuschakewiczi и Tanacetum pamiricum), а в некоторых случаях рыхлые подушки, например Oxytropis chiliophylla.
На крайнем востоке Памира, местами на высоте 3800—4300 м. над у. м. в составе этих степей господствует дерновинный злак Ptilagrostis subses-siliflora с примесью Roegneria schugnanica, Iris tianschanica, Artemisia Skorniakowii.
В некоторых районах Восточного Памира на высоте 3900—4500 м над у. м. на песках и супесях, находящихся в начальных стадиях формирования почвенного покрова, распространены своеобразные степные редкотравные ассоциации с господством лука Allium polyphyllum и значительным участием Artemisia pamirica или Ptilagrostis subsessiliflora.
Литература
К. В. Станюкович (1949); Е. А. Варивцева (1951).
83. Горные луговые степи и остепненные луга в сочетании с зарослями кустарников и каменистыми обнажениями и сельскохозяйственные земли на их месте
Горные луговые степи и остепненные луга занимают значительные площади в большинстве горных систем южных районов СССР: на Южном Урале, на Алтае (преимущественно в предгорьях), в Тарбагатае, Джунгарском Алатау, в Северном и отчасти Западном Тянь-Шане, в северных цепях Памиро-Алая, на Кавказе и в Крыму на яйле.
По структуре и отчасти по составу горные степи и остепненные луга этих горных систем в известной мере близки к луговым степям и остепнен-ным лугам степной зоны (точнее — Причерноморско-Казахстанской подобласти Евразиатской степной области); особенно это относится к горным луговым степям Южного Урала, Алтая, Тарбагатая, Джунгарского Алатау, Северного Тянь-Шаня и Кавказа. Горные луговые степи и остепненные луга отличаются от них лишь наличием горных видов растений, отсутствующих на равнине, и широким распространением растительности каменистых обнажений (в том числе тимьянников), тимьянниковых, а местами и трагакантовых степей и т. д. С горными луговыми степями сочетаются также горные типичные (настоящие) степи, приуроченные к более сухим склонам. Позиции горных луговых степей в горах Южной Сибири и Забайкалья частично заняты мелкодерновинными степями (опис. 80).
На «Геоботанической карте» с помощью буквенных обозначений выделены: у — южноуральские, а — алтайские, ск—североказахстанские, стш — северотяныпанские, тшп — западнотяныпанские и памиро-алай-ские, к — кавказские и кр — крымские горные луговые степи и остепненные луга, краткая характеристика которых дана ниже.
83 у — южноуральские
В предгорьях Южного Урала горные луговые степи и остепненные луга щриурочены преимущественно к бассейну Сакмары и к верховьям р. Урала. В составе остепненных лугов луговые и луговостепные злаки (Calamagrostis arundinacea, С. epigeios, Helictotrichon Schellianum, Festuca sulcata) встречаются более или менее обильно, также как и некоторые осоки (Carex praecox, С. ruthenica, С. pediformis, С. montana). Но все это подчинено пестрому разнотравью из луговостепных и даже лесных форм;
из разнотравья особенно обильны Pulsatilla patens, Fragaria viridis, Trifolium lupinaster, T. montanum, Dracocephalum Ruyschiana, Galium boreale, G. verum, Inula hirta, Artemisia sericea и многие другие.
В составе луговых степей предгорий Южного Урала в значительном количестве встречаются степные злаки (Helictotrichon desertorum, Stipa Joannis, S. dasyphylla, S. stenophyIla,Festuca sulcata); из осок представлены Carex ruthenica, C. pediformis. В обильном и пышном разнотравье преобладают луговостепные виды; особенно много Pulsatilla patens, Fragaria viridis, Filipendula hexapetala, Cytisus ruthenicus (кустарник), Medicago falcata, Galium verum, Inula hirta, Artemisia armeniaca, A. sericea.
Все эти виды представлены и на равнинных степях. Только на щебйи-стых или каменистых склонах, в составе так называемых «каменистых степей», кроме растений, упомянутых выше, встречается ряд видов, не свойственных равнинным степным ассоциациям на мелкоземистых грунтах (среди этих видов имеются и горностепные): + Agropyrum reflexiaristatum (эндем Южного Урала),1 Allium rubens, Dianthus acicularis,A-Oxytropis Gmelinii (эндем Южного Урала), + О. approximata (также), Bupleurum multinerve, Scutellaria lupulina s. 1. и др.
Среди луговых степей и остепненных лугов предгорий Южного Урала встречаются заросли степных кустарников из Cerasus fruticosa, Amygda-lusnana, Spiraea crenata, Caragana frutex, Cytisus ruthenicus. По северным склонам и по днищу ложбин — березовые лески.
Литература
О Э. Кнорринг (1932); И. М. Крашенинников (1932, 1937, 1937а, 1939а); И. М. Крашенинников и С. Е. Кучеровская-Рожанец (1941); И. М. Крашенинников и Я. Я. Васильев (1949)
83 а — алтайские
Горные луговые алтайские степи показаны на «Геоботанической карте СССР» по северным предгорьям Алтая, а также в виде нескольких мелких контуров в южном и отчасти западном Алтае.
Во внутренней части западного Алтая к ним относится Абайская степь, расположенная в долине Коксу и его притока Абая, на высоте 1100— 1300 м над у. м. Невысокие горы, окаймляющие эту степь, покрыты лиственничным лесом. По окраине степи развиты заросли невысоких кустарников (-^-Sibiraea altaiensis — эндем Алтая — и Dasiphora fruticosa s. 1.). В травостое степи преобладают злаки, общие со степями равнин и низко-горий Западной Сибири и Казахстана (Роа pratensis, Koeleria gracilis, Helictotrichon Schellianum, H. desertorum, Festuca sulcata s. 1.), в небольшом количестве встречается Stipa capillata. Из разнотравья в составе этой степи представлены Gypsophila altissima, Thalictrum petaloideum, Medicago falcata, Onobrychis sibirica, Galium verum, Artemisia laciniata, Oxytropis pi-losa и др.
Горные луговые степи и остепненные луга южного Алтая остаются недостаточно изученными; ойи развиваются на различных высотах и поэтому очень разнообразны, особенно по составу.
Те массивы горных остепненных лугов, которые располагаются й низкогорной части южного Алтая (высота 400—1000 м над у. м.), занятой в основном зарослями кустарников (Rosa spinosissima, Lonicera tatarica,
1 Как к далее, крестиком (+) отмечены виды, не встречающиеся в составе равнинных в ннзкогорных степей степной зоны (области).
Spiraea hypericifolia, Amygdalus папа и др.; опис. 56а), возникли в результате уничтожения (расчистки) кустарников (Темноев, 1938,1938а). Участки остепненных лугов, используемые как сенокосы, чередуются здесь с пашнями на пологих склонах. На склонах северо-западной и западной экспозиций возникают злаково-разнотравные остепненные луга с господством разнотравья (-{-Dictamnus angustifolius, Clematis integrifolia, Lava* tera thuringiaca, Peucedanum Morisonii', в меньшем количестве-\-Bup~
leurum Krylovianum, Phlomis tuberosa, Medicago falcata, Fragaria viridis и др.); злаки (наземный вейник, ежа) встречаются как примесь; урожайность этих лугов — около 50 ц/га. По склонам северной и восточной экспозиций возникают более мезофитные разнотравно-злаковые луга с господством Brachypodium pinnatum, с примесью наземного вейника, сборной ежи, лугового мятлика, бобовых и разнотравья (Lathyrus pratensis, L. pisi-formis, L. Gmelinii, Crepis sibirica, Origanum vulgare, -{-Trollius altaicus, Iris ruthenica и др.); урожайность травостоя около 35 ц/га.
Некоторые массивы горных алтайских луговых степей граничат с высокогорными лугами, поэтому в их состав проникают высокогорные виды и формируются высокогорные криофитные (пустошные) степи.
Литература
В. В. Сапожников и Б. К. Шишкин (1918); В. В. Ревердатто (1931); Б. К. Шишкин (1937); Н. И. Темноев (1938, 1938а); Р. А. Еленевский (19406, 1940в); Е. П. Матвеева (1951).
83 ск — североказахстанские
Североказахстанские горные луговые степи показаны по западному и юго-западному склону Тарбагатая на высоте (800)1000—1300(1500) м над у. м. (Степанова, 1950).1
По пологим мелкоземистым склонам преобладают типчаковые степи из Festuca sulcata,2 к которой примешиваются дерновинные злаки (Koeleria gracilis, Phleum phleoides), разнотравье (Hypericum perforatum, Galium verum), полукустарнички (Ziziphora Bungeana) и др. Распространены также луговые степи с более разнообразным составом дерновинных злаков (Festuca sulcata, Stipa rubens, Phleum phleoides, Koeleria gracilis, Agropyrum pectiniforme,-{-A. tarbagataicum, Poa relaxa) и разнотравья (кроме упомянутых, также Thymus Marschallianus, Origanum vulgare, Veronica pinnata и др.). По степи разбросаны кустарники: Spiraea hypericifolia, а также миндаль (-{-Amygdalus Ledebouriana, близкий к А. папа). В верхней части пояса, на границе с субальпийским поясом, к типчаку в значительном количестве примешиваются овсец (-{-Helictotrichon asiaticus) или туркестанский ячмень (-{-Hordeum turkestanicum), а из разнотравья---\-Papaver
сгосеитк Myosotis silvatica s. 1., Galium verum, Veronica spicata и др.; из горностепных полукустарничков—Artemisia frigida, Ziziphora Bungeana.
Ниже по рельефу (ниже 1200—1300 м над у. м.) по пологим склонам и выравненным вершинам мелкосопочника широко распространены типчаково-ковыльные степи (типа настоящих степей) с преобладанием Festuca sulcata и ковылей (Stipa Lessingiana, S. capillata, S. kirghisorum), к которым
1 На карте контур этих степей слишком продвинут к западу за р. Каракол; здесь горные луговые степи уже сменяются более засушливыми горными степями. Горные луговые степи могли бы быть показаны узкой полосой и по северному склону Тарбагатая, ниже пояса высокогорных лугов. На карте здесь показаны североказахстанские горные степи (84 ск).
’ У Е. Ф. Степановой (1950) —Festuca Ganeschini.
примешиваются другие дерновинные злаки (Koeleria gracilis, Phleum phleoides, Psathyrostachys funcea, эфемероид Poa bulbosa) и в небольшом количестве разнотравье (Origanum vulgare, Galium verum, иногда обильная Medi-cago falcata var. tarbagataica и др.). По пологим щебнистым склонам расположены разнотравно-типчаковые кустарниковые степи (дерновинные злаки: Festuca sulcata, Роа stepposa, Stipa capillata, Agropyrum desertorum\ степное разнотравье '.Medicago falcata,Galium verum,Goniolimon specio sum., Artemisia sericea и др.; полукустарнички: Ziziphora Bungeana, Artemisia frigida-, разбросаны по степному фону кустарники: Astragalus arbuscula, Spiraea hypericifolia, Caraganafrutex, -\-CalophacaHowenii;u3 эфемеров встречаются Bromus tectoram, Alyssum campestre).
На крутых щебнистых склонах представлена редкая растительность с преобладанием ксерофильных кустарников (-\-Cerasus tianschanica, +Лг-raphaxis virgata, +Л. laetevirens, Caragana frutex) и полукустарников (Euro-tia ceratoides, Ephedra distachya, Hypericum scabrum) с примесью степного разнотравья (в том числе горностепная -\-Alcea nudiflora). По скалам разбросаны отдельные экземпляры кустарников (Lonicera microphylla, Atraphaxis virgata, Grossularia acicularis, А-Clematis songarica, A-Spiraea trilobata), полукустарников (Artemisia frigida, +Л. rutifolia) и некоторых трав; большинство этих видов встречается на скалах в пределах Казахского мелкосопочника (104 кз).
В межсопочных понижениях при благоприятных условиях увлажнения располагаются остепненные разнотравно-злаковые луга (со сборной ежой, безостым костром, ползучим пыреем). Местами по северным склонам и в ложбинах встречаются фрагменты кустарниковых зарослей (с Rosa spinosissima, Lonicera tatarica, Spiraea hypericifolia) с луговыми и степными элементами в травяной синузии. В отдельных ущельях у родников имеются заросли ивняка и березы; очень редко у осыпей встречаются небольшие группы осины (Populus tremula).
Степи западных предгорий Тарбагатая из-за недостатка водных источников используются премущественно в качестве осенне-зимних пастбищ. Возможна и заготовка зимних кормов путем сенокошения.
Литература
Е. Ф. Степанова (1950); Б. А. Быков и Е. Ф. Степанова (1953).
83 стш — северотяньшанские
Северотяныпанские луговые степи и остепненные луга свойственны Джунгарскому и Заилийскому Алатау, северным склонам Кунгей-Алатау и Киргизского хребта, где они местами и показаны на карте. Географическое распространение их определяется границами выделяемой Н. И. Рубцовым Северо-Тяньшанской провинции (Рубцов, 1950).
Луговые степи и остепненные луга в указанных хребтах местами занимают значительные площади. Основа травостоя относящихся сюда ценозов слагается как типичными ксерофильными злаками — типчаком (Festuca sulcata), ковылями (Stipa rubens, S. capillata, S. kirghisorum), так и типичными мезофильными или же ксеромезофильными видами (Dactylis glomerata, Brachypodium pinnatum, Calamagrostis epigeios, Phleum phleoides), распространенными в равнинных или мелкосопочных степях Казахстана. Столь же экологически разнохарактерна здесь и группа «разнотравья». Поэтому, в зависимости от преобладания в травостое той или иной экологической группы растений, его можно относить или к луговым степям, или к остепненным лугам.
Высотные пределы луговых степей и остепненных лугов в Северном Тянь-Шано лежат, примерно, между 1000—2000 м над у. м. Луговые степи и остепненные луга встречаются как на северных, так и на южных склонах, на черноземовидных почвах и черноземах, формирующихся на лёссовидных суглинках или на грубом каменисто-щебнистом делювии?
К категории луговых степей должны быть отнесены красноковыльно-разнотравные степи, в которых основу травостоя составляет мезофильное и ксеромозофильное разнотравье с большим участием красноватого ковыля {Stipa rubens). Такие степи встречаются, например, в высоких предгорьях Джунгарского и Заилийского Алатау, а также на хр. Кетмень. В их составе отмщены такие луговостепные виды, как Phleum. phleoides, Роа angustifolia, Fragaria viridis, -{-Onobrychis ferganica, Origanum vulgare, Vicia tenuifolia, Myosotis suaveolens, а вместе с ними и типичные луговые: Dactylis glomerata, Bromus inermis, Geranium collinum, Thalictrum simplex, Lathyrus pratensis, Ranunculus polyanthemus.
Более широко распространены типчаково-разнотравные степи. В них типчак {Festuca sulcata) полностью подчинен ксеромезофильному разнотравью, в котором нередко большая роль принадлежит типичным горносреднеазиатским элементам: -{-EremurusOlgae, -{-Ligulariapersica, -\-Eremo-stachys speciosa. Встречаются участки луговых степей со значительным участием мезофильного овсеца {-{-Helictotrichon asiaticus).
При переходе на каменистые склоны южных экспозиций травостой злаково-разнотравных луговых степей сильно изреживается. Здесь появляются литофиты вроде Sedum hybridum, 4-5. Ewersii, увеличивается обилие кустарников. Наиболее обычными кустарниками являются таволга {Spiraea hypericifolia), кизильник {Cotoneaster multiflora, С. melanocarpa), жимолость {-{-Lonicera stenantha, L. microphyIla), барбарис {-{-Berberis heteropoda), желтоцветная роза, или, по местному, «шипичка» {-{-Rosa platyacantha). Иногда среди горных луговых степей встречаются единичные деревца яблони {-{-Malus Sieversii) и урюка {-{-Armeniaca vulgaris).
Остепненные луга характеризуются явным перевесом в травостое лугового мезофильного элемента {Dactylis glomerata, Bromus inermis, Calamagrostis epigeios, Brachypodium pinnatum, -{-Helictotrichon asiaticus). Степные ксерофильные элементы здесь представлены лишь в виде незначительной примеси. Остепненные луга развиваются обычно в нижних, более увлажняемых частях склонов южных экспозиций, иногда на западных склонах. Встречаются они также и на северных склонах в сочетании с типичными (евмезофитными) лугами. Травостой их густой, высокий (до 100—120 см). В верхних подъярусах господствуют указанные выше луговые злаки или крупное луговое разнотравье {-{^Ligularia persica, -{-Rumex tianschanicus, Lavatera thuringiaca), а в нижних — степные и луговостепные виды {Festuca sulcata, Роа angustifolia, Hypericum perforatum, Origanum vulgare, Achillea millefolium, Ixiolirion tataricum). Ранней весной на этих лугах развивается довольно хорошо выраженная синузия эфемеров и эфемероидов из -{-Gagea pseudoerubescens, А-Corydalis glauces-cens, Draba nemorosa, Veronica verna.
Кустарники среди горных остепненных лугов столь же обычны, и по видовому составу они те же, что и в луговых степях. Так, например,
1 В качестве характерных признаков черноземовидных луговостепных почв Северного Тянь-Шаня М. А. Глазовская (1947) указывает большую мощность гумусового горизонта (до 100 см), высокий процент гумуса в его верхней части (17—20%), слабо выраженные морфологические признаки оподзоленности (легкая белесоватоеть структурных отдельностей в нижней части гумусового горизонта), присутствие иллювиального бурого горизонта и глубокое (130—150 см) залегание карбонатов.
здесь постоянно встречаются барбарис (A-Berberis heteropoda), кизильник (Cotoneaster) и другие перечисленные выше виды.
Густой, богатый по видовому составу травостой горных луговых степей и остепненных лугов используется как хорошее сенокосное угодье с урожайностью сена до 20 ц/га. В местах, пригодных по рельефу, черноземные почвы под этими степями интенсивно распахиваются и засеваются зерновыми или бахчевыми культурами. Местное население получает с них удовлетворительные урожаи без применения искусственного орошения (богара).
Литература
М. Г. Попов (1940); П. П. Поляков (1941а); Н. И. Рубцов (1941, 1941в, 1948а, 1950, 1954); М. А. Глазовская (1947).
83 тшп — западнотяныпанские и памиро-алайские
Горные луговые степи и остепненные луга в сочетании с зарослями кустарников и каменистыми обнажениями занимают в Западном Тянь-Шане и в горах системы Памиро-Алая особый пояс, лежащий в высотных пределах примерно от 2000 и до 3000 м над у. м. На «Геоботанической карте» показаны наиболее крупные их массивы: на южном склоне Киргизского хребта, на северном склоне Таласского Алатау, на северном и южном склонах Чаткальского хребта, в нескольких местах на Зеравшан-ском хребте и в западной части хр. Петра Первого.
На Киргизском хребте и на Таласском Алатау, по данным И. В. Выходцева (1952), наиболее широко распространены типчаковые (Festuca sulcata s. 1. и +F. Kryloviana) луговые степи. К господствующему типчаку примешиваются дерновинные злаки (Phleum phleoides, Stipa capillata, -\-Poa Litwinowiana), корневищные злаки (Роа angustifolia) и разнотравье (Hypericum perforatum, Nepeta pannonica, Galatella punctata s. 1., -\-Hedysarum songoricum, A-Gentiana tianschanica, Veronica spuria, A-Helichrysum plicatum и др.). В полусаванновых вариантах становится заметной примесь полуэфемероидных злаков и разнотравья — Agropyrum trichophorum,A-Inula grandis,-|-Prangos pabularia, A-P. uloptera, а также Andropogon ischaemum и др. В петрофитиых вариантах покров типчака изреживается, внедряются склероморфные травы и ксерофильные кустарнички. Нередки участки и с вкраплением трагакантовых кустарничков (^Onobrychis echidna, + Acantholimon alatavicum и др.). Характерно наличие в растительном покрове кустарниковых зарослей (из Spiraea hypericifolia, S. trilobata, Dasiphora fruticosa s. 1., видов Atraphaxis и др.) и разреженных арчевников (из A-Juniperus pseudosabina).
На Зеравшанском хребте и на хр. Петра Первого горные луговые степи и остепненные луга, по данным В. А. Никитина, встречаются в высотных пределах от 2000 до 2900—3200 м над у. м. Почвы — коричневые луговостепные. В сообществах этих степей преобладают степные элементы, однако присутствуют и горнолуговые, трагакантовые и тимьянниковые; они представлены различными видами разнотравья. Господствует типчак (Festuca sulcata s. 1.), к которому в заметной степени примешиваются другие дерновинные злаки (Phleum phleoides, Koeleria gracilis, -j-Poa glabriflora), представители травянистого разнотравья (Campanula glomerata, -\-Psy-chrogeton cabulicus, Achillea millefolium, Geranium collinum, -)-(?. saxatile, A-Scorzonera inconspicua, A-Inula rhizocephala), а также некоторые полукустарнички (Thymus serpyllum s. 1., -J-Arenaria Griffithii, A-Scorzonera
acanthoclada). Иногда в составе луговых степей имеется обильная примесь 4-jPoa relaxa (часто ^господствующего с типчаком) и Helictotrichon deser-torum, а также мезофильного и ксеромезофильного разнотравья (-{-Nepeta podostachys, -{-Ligularia Thomsonii, -{-Geranium saxatile -{-Erigeron alpinus и др.). Вместе с этими луговыми степями встречаются на более сухих участках разнотравные ковыльники с преобладанием киргизского ковыля или остепненные лисохвостовые (из -{-Alopecurus seravschanicus) луга с обильной примесью -\-Nepeta podostachys, а на избыточно увлажненных местах — осоковые и осоково-белополевицевые луга (из -{-Carex pseudo-foetida, -{-С. parva, Agrostis alba). Изредка встречаются небольшие заросли кустарников (-{-Ribes Meyeri и -hftosa Beggeriana).
Встречаются также трагакантовые типчаковые степи на менее мощных коричневых луговостепных и скелетных почвах (к господствующему типчаку примешиваются, а часто играют роль субэдификаторов, трагакантовые кустарнички-----\-Astragalus lasiosemius, -{-Acantholimon pamiricum,
+Я. lycopodioides, а также -{-Cousinia laetevirens и -{-C. horridula) и опустыненные типчаковые степи (с участием -{-Artemisia Lehmanniana, -{-Сои-sinia stephanophora и др.). Со всеми названными сообществами, кроме того, сочетаются обедненные типчаковые и редкостойные ковыльные степи из Stipa kirghisorum и +5. trichoides.
В ландшафте горных луговых степей характерно наличие отдельных деревьев или групп арчи (+Juniperus turkestanica и -f-J. seravschanica), изредка образующих редколесья с полнотой 0.1—0.2. Под пологом арчи появляются лугово-лесные мезофиты (-{-Ligularia Thomsonii и Роа пето-ralis).
На инсолированных склонах часты каменистые участки с разреженным и несформированным покровом, среди которого встречаются небольшие пятна кустарниковых зарослей из -{-Rosa Ecae,A~R. кокаnica,-{-Cerasus verrucosa, видов Lonicera и др.
На верхнем пределе распространения горных луговых степей покров типчака сильно изреживается, и к нему обычно примешаны высокогорные виды разнотравья: -{-Psychrogeton cabulicus, -{-Cerastium cerastoides, -{-Melandryum longicarpophorum, -{-Oxytropis Linczevskii или низенькие высокогорные полукустарнички: -{-Polygonum biaristatum, -{-Minuartia Lit-winowii, -{-Oxytropis savellanica или +<?. Michelsonii', реже встречаются следующие высокогорные травянистые виды: -{-Роа hissarica, -{-Allium monadelphum, -{-Polygonum schugnanicum, -{-Pedicular is dolichorrhiza, +/)a-paver involacratum.
С этими степями сочетаются в соответствующих местообитаниях кузинники (из -{-Cousinia Z'rancAeiu),бескильничники (из дерновинной -{-Рас-cinellia subspicata), альпийские мят лички (из -{-Роа alpina), подушечники (из -{-Oxytropis immersa или -{-Smelovskia calycina).
Литература
О. Э. Кнорринг и 3. А. Минквиц (1912); М. В. Культиасов (1926 —1927); Р. И. Або-лин и М. М. Советкина (1930); Е. П. Коровин (1934); К. С. Афанасьев (1940); А. С. Королева (1940); И. В. Выходцев (1952); К. В. Станюкович (1955).
83 к — кавказские
Горные луговые степи показаны на «Геоботанической карте» в следующих районах Кавказа: 1) на северокавказских куэстах, где они встречаются совместно со среднегорными более или менее остепненными лугами, которые в большинстве случаев преобладают; 2) во внутреннем
Дагестане; 3) в северо-западной части Армянского вулканического нагорья; 4) в юго-восточной части Армянского вулканического нагорья; 5) в Та-лышских горах.
В пределах выделенных на карте контуров горные луговые степи сочетаются с горными типичными дерновиннозлаковыми степями, занимающими более сухие позиции.
В составе горных луговых степей северокавказского куэстового района, к северу от Эльбруса, преобладают обычно Carex humilis и Stipa capillata со значительным участием Bromus riparius, Stipa pulcherrima, Festuca sulcata, Koeleria gracilis, т. e. виды, обычные и в составе равнинных степей. Обильно также разнотравье (Adonis vernalis, -\-Dianthus fragrans, Paeonia tenuifolia, A-PotentillaCrantzii, Vinca herb acea, -\-Plantago saxatilis, A-Pyrethrum parthenifolium и др.). Весной в составе этих степей обильны A-Alopecurus vaginatus, A-Pulsatilla albana и др. На щебнистых склонах преобладают Carex humilis, Andropogon ischaemum co значительным участием Bromus riparius', много также Agropyrum trichophorum, A-Muscari Szowitzianum, A-Potentilla Crantzii, часты Allium albidum, A-Astragalus Demertii, Teucrium chamaedrys, T. polium, A-Scutellaria po-lyodon, A-Artemisia caucasica. Местами разбросаны кусты A-Rhamnus Pal-lasii, Berberis vulgaris, Prunus stepposa. Ассоциации с обилием Thymus, Teucrium polium, Scutellaria polyodon должны быть отнесены к тимьян-никовым степям.
В горном Дагестане горные луговые степи занимают нижние части известняковых плато и прилегающие к ним склоны долин на высотах 1600—2000 м над у. м.; выше располагаются остепненные луга.
В составе разнотравно-типчаково-тырсовых и типчаковых степей преобладают Stipa capillata и Festuca sulcata', из других злаков — Koeleria gracilis, A-Phleum montanum, иногда встречается субальпийский костер (A-Bromus variegatus). Обильно разнотравье, в состав которого входят Thalictrum minus, Filipendula hexapetala, Salvia verticillata, Phlomis tuberosa, A-Stachys atherocalyx, Galium verum, A-Scabiosa bipinnata, А-Campanula Hohenackerii, A-Artemisia chamaemelifolia, A. Marschalliana, а из бобовых — Trifolium ambiguum, T. pratense, A-Anthyllis Boissieri. и др.
Разнотравно-перистоковыльные и кострово-осочковые степи располагаются на более или менее крутых и каменистых склонах. В их составе, наряду со Stipa pulcherrima, Bromus riparius и Carex humilis, имеется много растений, обычных для каменистых местообитаний лесного и субальпийского поясов; в более мезофитных вариантах они смыкаются с горными остепненными лугами.
В составе разнотравно-перистоковыльных степей с пестрым травостоем можно отметитьA-Bromus Biebersteinii,A~B. variegatus, А-Astragalus Biebersteinii, +4. galegiformis, A-Onobrychis Biebersteinii, A-Ferula dissecta, Dracocephalum austriacum, виды Thymus, а так же многие виды из тех, которые были выше отмечены для типчаково-тырсовых степей: Filipendula hexapetala, Phlomis tuberosa, Galium verum, -A-Scabiosa bipinnata, A-Artemisia chamaemelifolia, A. Marschalliana и др.
Кострово-осочковые луговые степи из Carex humilis и Bromus riparius обычно имеют в своем составе Phleum phleoides, Brachypodium pinnatum, Filipendulahexapetala, + Alchimillasericata, Trifolium medium, T. ambiguum, ArMedicago glutinosa, А-Lotus caucasicus, Galium verum, Achillea setacea, в меньшем количестве-----\-Pastinaca armena, Origanum vulgare, Salvia
verticillata, виды Thymus, A-Scabiosa columbaria, Leontodon hispidus и др. Костер (Bromus riparius) образует также самостоятельные группировки.
Среди горных луговых степей в Дагестане встречаются участки сухих степей, покрывающие южные склоны; для них характерны эндемичные (для Дагестана и отчасти для Северного Кавказа) эдификаторы: -{-Agropyrum gracillimum и -{-Stipa daghestanica. Кроме того, здесь обычны «нагорные ксерофиты», например -{-Bromus Biebersteinii, -{-Astragalus Вип-geanus, -{-A. daghestanicus, -{-Onobrychis radiata, -{-Nepeta cyanea, -{-Psephel-lus daghestanicus, полукустарнички: -{-Astragalus haesitabundus и -{-Medicago daghestanica, кустарничек -{-Astragalus Marschallianus и др., образующие сильно разреженные группировки, используемые под весенне-осенние и частью зимние пастбища. Эти ассоциации должны быть отнесены к трагакантовым степям.
В северо-западной части Армении и в соседней части Грузии горные луговые степи и остепненные луга, занимающие здесь большие площади, поднимаются до высоты 2300—2400 м, а в центральной и южной Армении — до высоты 2700—2800 м вад у. м. В составе злаковых луговых степей и остепненных лугов из степных злаков господствуют Festuca sulcata, Agropyrum trichophorum, -{-Роа densa, Phleum phleoides, Koeleria gracilis, из луговых — Phleum pratense, Dactylis glomerata, Festuca pratensis, из горнолуговых-----{-Bromus variegatus, -{-Koeleria caucasica. В меньшем
количестве представлены -{-Secale anatolicum, -{-Hordeum violaceum, Stipa pulcherrima, S. stenophylla и др. Обычно наличие Carex humilis. Из разнотравья встречаются -{-Lotus caucasicus, Campanula glomerata, -{-Salvia armeniaca, Myosotis alpestris, Trifolium ambiguum, T. alpestre, -{-Hypericum polygonifolium и др. Часто злаковые участки чередуются с остепнен-нолуговыми разнотравными с господством -{-Scabiosa bipinnata, Achillea setacea и -{-Prangos ferulacea или бобовых: Trifolum ambiguum, T. alpestre, -{-Vicia variabilis, -{-V. variegata. Нередко встречаются осоковые луговые степи с преобладанием приземистой осоки {Carex humilis), образующей густой, но невысокий травостой. К осоке примешиваются -{-Bromus variegatus, Koeleria gracilis, -{-К. caucasica, Phleum phleoides, -{-Poa densa, Pimpinella saxifraga, -{-Anthyllis Boissieri, -{-Medicago dzhavakhetica, Galium verum и др.
Ли тература
Н. И. Кузнецов (1910); А. К. Магакьян (1941); А. Л. Тахтаджян (1941); Е. В. Шиффере (1946, 1953); А. А. Гроссгейм (1948).
83 кр — крымские
Растительный покров Крымских яйл на высоте 1100—1540 м над у. м. носит в основном характер горных луговых степей и остепненных лугов. Такова растительность Ай-Петринской, Ялтинской, Никитской, Гурзуфской, Бабуганской (с высшей точкой Крымских гор — 1545 м), Чатыр-Дагской'яйл и Демерджи яйлы.1
Из дерновинных злаков и осок в составе горных луговых степей яйлы преобладают Carex humilis, Festuca sulcata, Bromus cappadocicus', в меньшем количестве------{-Alopecurus vaginatus, Helictotrichon Schellia-
num, Stipa pulcherrima, -{-Agropyrum strigosum и др. Из разнотравья более или менее обильны -{-Viola oreades (высокогорнолуговое растение), -{-Alchimilla taurica, +А. Stevenii, +А. jailae, +A. tytthantha, Myosotis suaveolens и др. К наиболее высоким вершинам Крымских гор приурочена
1 По условиям масштаба на «Геоботанической карте» крымские горные луговые степи показаны только на вершине Чатыр-Дага. *
характерная группировка из типчака и высокогорной -\-Draba cuspidate.. Указанные виды манжетки (большинство которых являются эндемами Крымской яйлы) образуют и самостоятельные группировки, так же как и горностепной вид + Androsace taurica (из рядг1 Villosae). К щебнистым склонам приурочены группировки из -\-Pimpinella lithophila {Р. tragium s. 1.) и полукустарничков A-Helianthemum orientate и Н. nummularium. В этих растительных группировках Крымской яйлы встречается большое количество степных растений, общих с равнинными степями:' помима типчака, низкой осоки и Helictotrichon Schellianum, также и Bromus riparius, Clematis integrifolia, Filipendula hexapetala, Betonica officinalis,. Galium verum. Inula ensifolia и др. Но здесь же много видов, отсутствующих в составе равнинных степей. Кроме ряда вышеуказанных, сюда относятся -pCerastium Biebersteinii, -^-Pulsatilla Halleri, -\-Cytisus polytrichus, -p Thymus Callieri, -\-Veronica gentianoides, -pPsephellus declinatus и др. При усиленном выпасе долго удерживаются типчак и низкая осока, затем исчезают и они, разрастаются Alchimilla jailae, Achillea millefolium, Trifolium repens. Местами на Крымской яйле выпас привел к деградации травостоя, к смыву почвы и обнажению щебнистого грунта.
Верхнюю границу леса в Крымских горах (так называемой «Первой гряды») образуют угнетенный бук {Fagus taurica) с саблевидными стволами и угнетенная сосна {Pinus hamata), формирующая на верхней границе леса редкостойные насаждения; на южном склоне верхней границы леса достигает и крымская сосна {Pinus Pallasiana). Выше границы леса местами встречаются сплошные заросли можжевелового стланца {Juniperus depressa, J. sabina). В прошлом яйлы были, несомненно, более облесены, чем в настоящее время, но вряд ли это облесение было сплошным; леса были уничтожены в результате хозяйственной деятельности человека. Вопрос о причинах безлесия яйлы породил большую литературу (Поплавская, 1948).
Литература
В. И. Талиев (1901, 1909); Г. И. Танфильев (1902а); А. Н. Криштофович (1907); А. А. Яната (1916, 1931); Е. В. Вульф (1925); Г. И. Поплавская (1930, 1933, 1948); Н. М. Чернова (1951).
84. Горные дерновиннозлаковые (типчаково-ковыльные, ковыльные, типчаковые, овсецовые и др.) степи в сочетании с каменистыми обнажениями и сельскохозяйственные земли на их месте
Горные дерновиннозлаковые степи, как и горные луговые степи и остепненные луга (83), занимают значительные площади в большинстве горных систем южных районов СССР: на Южном Урале (хр. Ирендык), в западной части горного Алтая, в северо-восточном Казахстане — на хребтах Чингиз-Tay, Тарбагатай и Саур; в Джунгарском Алатау, в Северном, Центральном и Западном Тянь-Шане, на хр. Каратау (Сыр-Дарьинском), в северных хребтах Памиро-Алая, на Западном Памире, в Копет-Даге и, наконец, на Кавказе, главным образом в Дагестане и в Южном Закавказье.
Горные дерновиннозлакбвые степи более разнообразны, чем их равнинные аналоги. Помимо типичных степей, здесь встречаются, преимущественно на щебнистых или галечниковых по механическому составу почвах, также тимьянниковые, трагакантовые, колючетравные, полу саванновые и другие степи. Полу саванновые, колючетравные и трагакантовые степи
отсутствуют в пределах равнинной и низкогорной степной зоны (области) СССР. Кроме того, в районах распространения горных степей нередки каменистые обнажения с весьма разреженным растительным покровом (опис. 104).
В составе горных степей много общих видов, в том числе и основных эдификаторов, с равнинными степями, преимущественно Причерноморско-Казахстанской подобласти; особенно это относится к горным южноуральским, алтайским, североказахстанским, северотяньшанским и кавказским степям. Но в то же время в составе горных степей много и специфических горных степных видов, отсутствующих на равнине.
На карте с помощью буквенных обозначений выделены: у — южноуральские, а — алтайские, ск — североказахстанские, стш — северо-тяныпанские, цтш — центральнотяныпанские, тпш — западнотяныпан-ские и памироалайские, кпд — копетдагские и к — кавказские горные дерновиннозлаковые степи, которые далее кратко характеризуются.
84 у — южноуральские
В Зауралье горные типчаково-ковыльные степи показаны на южной оконечности хр. Ирендык. Растительный покров здесь очень пестрый. Преобладают горные разнотравно-типчаково-ковыльные степи, и только местами, главным образом на более высоких горных ступенях, поднимающихся до 500—760 м над у. м., можно встретить островки березовых лесов, которые ниже по рельефу наблюдаются иногда в лощинах с мягкими наносами. Преобладают северные и «средние» варианты (по И. М. Крашенинникову) разнотравно-типчаково-ковыльных степей: первые — с господством Stipa stenophylla (переходящие в луговые степи), а вторые с преобладанием более южного вида ковыля (S. rubens). Довольно обильно разнотравье, частью общее с луговыми степями. В «средних» вариантах этих степей из злаков более или менее обильны Stipa capillata, Festuca sulcata, Helictotrichon, desertorum, из осок — Carex supina, из разнотравья — Trijolium montanum, Libanotis sibirica, Onosma simplicissimum, Thymus Marschallianus, Veronica incana, Plantago stepposa, Galium verum, Inula hirta, Artemisia latifolia, A. sericea и многие другие. В южной части района развит южный обедненный вариант этих степей — к Stipa rubens в большем или меньшем количестве примешаны S. capillata и S. Lessingiana. Видовой состав этих степей гораздо более ограничен, характерно наличие ряда ксерофильных видов {Dianthus leptopetalus, Linosyris [Galatella] villosa, Jurinea multiflora и др.). На склонах, где почву образуют цветные латеритные глины, обычно засоленные, развиты группировки с участием, помимо типчака и ковылей, Linosyris villosa и Artemisia maritima s. 1. на солонцеватых черноземах. На каменистых склонах появляются литофиты: -{-Agropyrum pruiniferum,1 Linaria debilis и др. Производительность (в воздушно-сухом состоянии): северного варианта 9—11 ц/га, среднего 8—10 ц/га, солонцовых полынно-типчаковых степей 6 ц/га и каменистых степей 3—4 ц/га. Эти степи используются как пастбища и сенокосы, частично они распаханы.
Литература
И. М. Крашенинников (1939а); И. М. Крашенинников и G. Е. Кучеровская-Ро-жанец (1941).
1 Здесь, как и далее, виды, помеченные крестиком (-(-), в составе ряптгиппыт и мелкосопочных степей не встречаются. Agropyrum pruiniferum, помимо Южного Урала, произрастает только в Жигулях и Мугоджарах.
84 а — алтайские
Горные дерновиннозлаковые алтайские степи показаны па «Геоботанической карте СССР» в виде двух контуров: 1) по верхнему течению Чарыша (Кырлыкская, Канская и другие степи); 2) по южным предгорьям Курчумского хребта, к западу от долины Кальджира.
Степи по верхнему течению Чарыша близки по составу и структуре к мелкодерновинным алтайско-забайкальским степям (опис. 80) и частично могли быть отнесены к последним. Так, в степи вблизи поселка Усть-Кана (высота около 1100 м над у. м.) наблюдается низкорослый травостой с преобладанием Koeleria gracilis, Festuca sulcata s. 1. В состав разнотравья входят Potentilla chrysantha, Ligularia glauca, -\-Leontopodium leontopodi-num,1 Schizonepeta multifida и др.
Степи по южным предгорьям Курчумского хребта [высота 750—1000 (1250) м над у. м.] близки по составу к казахстанским разнотравно-дер-новиннозлаковым степям, покрывающим более высокие участки Казахского мелкосопочника.
Степи эти используются как пастбища.
Литература
В. В. Сапожников и Б. К. Шишкин (1918); В. В. Ревердатто (1931); Б. К. Шишкин (1937).
84 ск — североказахстанские
Под именем североказахстанских горных дерновиннозлаковых степей на «Геоботанической карте» выделены степи Тарбагатая, а также хребтов Кан-Чингиз, Чингиз-Tay и Акшатау. По описаниям Е. Ф. Степановой (1950), в Тарбагатае они образуют пространственно хорошо выраженный степной пояс, занимающий предгорья и среднегорья от 1000 до-2000 (2200) м над у. м.
В верхних частях степного пояса, на границе с субальпийским, встречаются главным образом типчаковые степи типа луговых. К типчаку (Festuca sulcata)2 примешивается ряд других злаков (Phleum phleoides, Koeleria gracilis, -\-Roegneria czimganica). Из разнотравья наиболее обычны Galium verum, Hypericum perforatum, Thymus Marschallianus, Potentilla orientalis, Goniolimon speciosum, Dracocephalum peregrinum. У самых верхних пределов распространения типчаковых степей большое участие в сложении их травостоя принимают подушковидный кустарничек — остро-лодка иглистая (-(-Oxytropis hystrix) и копеечник (-j-Hedysarum ferganense). Е. Ф. Степанова указывает, что в период своего массового цветения копеечник нередко окрашивает склоны предгорий в красивый розовый цвет.
В типчаково-ковыльных степях преобладают типчак (Festuca sulcata), красноватый ковыль (Stipa rubens), ковылок (*S. Lessingiana), киргизский ковыль (5. kirghisorum) и тырса (Stipa capillata). Разнотравье представлено в общем теми же видами, что и в типчаковых степях. Постоянно, но в небольшом количестве, встречается полукустарничковая сублессингиановая полынь (Artemisia sublessingiana). В нижних частях степного пояса обилие этой полыни значительно увеличивается, в результате чего создаются полынно-ковыльные степи (Stipa capillata -\-Artemisia sublessingiana).
1 Возможно, это — Leontopodium campestre. !У Е. Ф. Степановой (1950) — Festuca Ganeschini.
Для степного пояса северного склона Тарбагатая весьма характерны кустарники: карагана (Caragana leucophloea), таволга (Spiraea hypericifolia'), роза (Rosa spinosissima), кизильник (Cotoneaster melanocarpa, +C. uniflora). Однако наибольшее распространение кустарники имеют не на северном, а на южном, гораздо более крутом и скалистом склоне Тарбагатайского хребта, обращенном к Балхаш-Алакольской впадине. Здесь Е. Ф. Степанова (1950) выделяет особый кустарниково-степной пояс, который слагается дерновиннозлаковыми степями с очень большим участием кустарников. Кроме того, в этом же поясе имеют место и самостоятельные кустарниковые ценозы (опис. 56а). Интересно, что, в противоположность северному склону, на южном склоне широко распространена не Caragana leucophloea, а С. frutex, весьма характерная для равнинных степей и мелкосопочника восточного Казахстана.
Кустарниковые ценозы представлены здесь главным образом миндальниками, в которых полностью господствует миндаль Ледебура (A-Amyg-dalus Ledebouriana), близко родственный алтайскому (Amygdalus папа). Из других кустарников в этих ценозах отмечены таволга (Spiraea hypericifolia) и карагана (Caragana frutex). Травяные синузии кустарниковых зарослей представлены и луговыми и степными видами: ежой (Dactylis glomerata), костром (Bromus inermis), вейником (Calamagrostis epigeios), тырсой (Stipa capillata), душицей (Origanum vulgare), ферулой (Ferula songorica) и др.
Следует отметить, что в пределах контура горных дерновиннозлаковых степей, выделенного на «Геоботанической карте» по северному склону Тарбагатая,1 имеются также и ковыльно-разнотравные степи, приближающиеся к луговым. Они приурочены к высоким межгорным долинам или равнинам, отделяющим основной хребет Тарбагатая от его предгорий. Основа травостоя луговых степей слагается красноватым ковылем (Stipa rubens), тимофеевкой (Phleum phleoides), типчаком (Festuca sulcata). Богатое по видовому составу и обилию разнотравье представлено A-Geranium albiflorum, Thalictrum minus, Veronica spicata, Phlomis tuberosa, Trifolium lupinaster, Fragaria viridis. В нижних частях склонов близ выходов родников встречаются небольшие участки злаковых лугов из лисохвоста (Alopecurus pratensis), луговой тимофеевки (Phleum pratense), щучки (Deschampsia caespitosa), иногда с небольшим участием осок.
На выходах твердых коренных пород по щебнистым склонам травостой сильно изрежен и нередко представлен лишь единичными экземплярами караганы (Caragana leucophloea), регнерии (A-Roegneria czimganica), очитка (Sedum hybridum), чабреца (Thymus Marschallianus).
Горные дерновиннозлаковые степи Тарбагатая (и территориально связанные с ними другие категории степной, кустарниковой и луговой растительности) используются в качестве сенокосных угодий и пастбищ. Луговые степи распахиваются под посевы зерновых культур.
Большой хозяйственный интерес представляет эндемичный для Тарбагатая вид житняка (Agropyrum tarbagataicum), описанный Н. А. Плотниковым. Этот житняк ютличается высоким ростом, хорошей облиствен-ностью и засухоустойчивостью (Плотников, 1941—1946).
Во флористическом отношении степи Тарбагатая являются переходными между степями Северного Тянь-Шаня и Алтая. Очень характерной особенностью этих степей является постоянное (и нередко большое) участие в их составе караганы. В этом, повидимому, сказывается уже влияние со-
1 На южном склоне этого хребта дерновиннозлаковые степи не могли быть выделены по условиям масштаба карты.
седней Центральной Азии. Не менее интересной и своеобразной чертой степей Тарбагатая является широкое развитие здесь кустарников — миндаля Ледебура {-{-Amygdalus Ledebouriana) и калофаки {-{-Calophaca Howenii). В Джунгарском Алатау эти кустарники полностью отсутствуют. Вместе с тем Тарбагатай флористически связан с Тянь-Шанем такими видами, как некоторые регнерии {-{-Roegneria Schrenkiana, 4-7?. Abolinii, 4-7?. tianschanica), и целым рядом других, свойственных тяныпанской горной системе (4- Trisetum virescens, -{-Helictotrichon tianschanicum).
Горные степи среднегорных хребтов восточной части Казахского мелкосопочника (Кан-Чингиз, Чингиз-Tay и др.) остаются до сих пор почти неизученными. Здесь, видимо, широко распространены кустарниковые степи (Косинский, 1916; Шипчинский, 1916; Быков и Степанова, 1953).
Литература
К. К. Косинский (1916); Н. В. Шипчинский (1916); В. В. Сапожников и Б. К. Шишкин (1918); Н. А. Плотников (1941—1946); Е. Ф. Степанова (1950); Б. А. Быков и Е. Ф. Степанова (1953).
84 стш — северотяньшанские
Вместе с луговыми степями дерновиннозлаковые степи в Северном Тянь-Шане (в Джунгарском Алатау, хр. Кетмень, Заилийском Алатау и Киргизском хребте) составляют особый, ландшафтно хорошо выраженный степной пояс. Они широко распространены не только в предгорьях, но и в условиях среднегорного пояса до 2500 м над у. м., а типчаковые степи достигают и высокогорий, то есть поднимаются до 3000—3500 м над у. м. Почвы под этими степями относятся к горным черноземным и каштановым.
Наиболее распространенными среди дерновиниозлаковых степей являются тырсовые степи (с преобладанием Stipa capillata). В качестве содоми-нанта в них почти постоянно выступает типчак {Festuca sulcata).1 Меньшая роль в травостое принадлежит тонконогу {Koeleria gracilis), тимофеевке {Phleum phleoides), котовнику {Nepeta pannonica), тысячелистнику {Achillea millefolium s. 1.), подмареннику {Galium verum s. 1.), чабрецу {Thymus Marschallianus) и многим другим видам, свойственным равнинным степям Евразии. Однако и горные азиатские элементы в этих степях представлены довольно значительным числом видов. Из них наиболее обычными являются -{-Carex turkestanica, -{-Rheum Wittrockii, -{-Dianthus turkestanicus, -{-D. tianschanicus, -{-Astragalus Schmalhausenii, 4~A. tibetanus, Leontopodium campestre, -{-Allium Korolkovii, 4~A. schoenoprasoides.
Средняя высота травостоя тырсовых степей равна примерно 60—70 см. Проективное покрытие колеблется от 50—60% до 80—100%. Общий флористический состав (в пределах Северного Тянь-Шаня) исчисляется 120— 130 видами (Рубцов, 1954).
Среди тырсовых степей Северного Тянь-Шаня выделяются три основные группы ассоциаций. Из них следует, прежде всего, указать на относительно высокогорную группу, формирующуюся на верхнем пределе распространения этой формации — от 1800 до 2500 м над у. м. по склонам южных экспозиций. В сложении ее травостоя (наряду с обычными степняками) большую роль играют горные и высокогорные элементы {-{-Phlomis oreophila, -{-Potentilla nervosa, -{-Carex turkestanica, -{-Ligularia Thom-
1 Некоторые авторы горный степной типчак относят к другим видам, преимущественно к Festuca Ganescnini.
sonii) и среднеазиатские виды родов Allium, Astragalus, Oxytropis. Далее выделяется среднегорная группа, представленная типчаково-тырсовыми ассоциациями, в которых роль горных азиатских видов сильно снижена. Близ нижних границ степного пояса распространена третья группа — низкогорные полынно-тырсовые ассоциации, типа пустынных степей. В их сложении в качестве субдоминанта принимает участие сублессингиа-новая полынь (Artemisia sublessingiana s. I.),1 постоянно встречается такой кссрофильный полукустарничек, как прутняк (Kochia prostrata). Травостой этих ассоциаций изреживается до. 50 % проективного покрытия.
Не менее распространенной среди дерновиннозлаковых степей является формация типчака (Festuca sulcata). Она обычна во всех высотных поясах гор, вплоть до альпийского пояса. Высокогорные ассоциации этой формации характеризуются постоянным присутствием альпийских и субальпийских растений (-\-Leontopodium leontopodinum, Aster alpinus, + Primula algida). В сложении среднегорных и низкогорных типчаковых ассоциаций обычно участвуют ковыли (Stipa capillata, S. Lessingiana, S. kirghisorum).
Типчак — превосходный пастбищный злак, особенно для лошадей и крупного рогатого скота. Горные типчаковые пастбища (а иногда и сенокосы) считаются лучшими кормовыми угодьями Тянь-Шаня. Производительность их травостоя колеблется (в зависимости от преобладания тех или иных ассоциаций) от 2—3 до 4—5 ц/га сухой массы.
Из других формаций дерновиннозлаковых степей, имеющих значительное распространение в пределах Северного Тянь-Шаня, следует указать на степи с преобладанием ковылка (Stipa Lessingiana). Они приурочены только к предгорьям и, в отличие от типчаковых и тырсовых степей, не поднимаются высоко в горы, не встречаются на песчаных почвах и избегают щебнистых почв. Типичное их обитание — темнокаштановые почвы мягких предгорий, покрытых достаточно мощным плащом лёссовидных суглинков. Ковыльковые степи флористически близки к тырсовым степям, но беднее последних, так как они развиваются в меньших высотных пределах, экологический ареал их уже. Общий видовой состав северотянь-шанских ковылковых степей определяется 70—80 видами. Средняя высота травостоя 40—45 см, проективное покрытие 70—80%, производительность от 4 до 8 ц/га сухой массы.
Среди северотяныпанских дерновиннозлаковых степей должна быть отмечена формация киргизского ковыля (Stipa kirghisorum). Она хорошо представлена только на Киргизском хребте и в западной части Кунгей-Алатау, где распространена по предгорьям и горному крутосклонному рельефу, до 2500 м над у. м. В Джунгарском Алатау, на хр. Кетмень и в Заилийском Алатау эта формация встречается сравнительно редко, небольшими вкраплениями среди степной растительности, слагаемой иными эдификаторами. Степи с преобладанием киргизского ковыля покрывают обычно щебневатые (хрящеватые) черноземы, иногда с очень близким залеганием коренных твердых пород. Грубый щебенчатый субстрат вызывает появление в видовом составе этих степей ряда петрофильных тимьянниковых элементов, например -\-Lagochilus diacanthophyllus, Pat-rinia intermedia, Ziziphora Bungeana, Thymus Marschallianus.
• Специфичными для Северного Тянь-Шаня являются красноковыльные степи (с преобладанием красноватого ковыля — Stipa rubens), которые особенно широко развиты в Джунгарском Алатау (Рубцов, 1941в,
1 В последнее время П. П. Поляков выделяет в пределах этого вида ряд более мелких — Artemisia transiliensis Р. Pol., A. Gorjaevii Р. Pol. и другие, но география их еще не вполне ясна.
1948а; Поляков, 1941а). Менее выражены они на хр. Кетмень и в Заилий-ском Алатау, где встречаются лишь в виде мелких фрагментов. В Центральном и Западном Тянь-Шане красноковыльных степей нет.
Эдификатор этих степей — Stipa rubens — является самым крупным из перистых ковылей, обитающих в Северном Тянь-Шане. Сформирован ные им степи приурочены к поясу средних и высоких предгорий (примерно до 1500—1700 м над у. м.) с достаточно мощными суглинистыми почвами черноземного типа. Вообще этот эдификатор отличается от других северо-тяныпанских ковылей-ценозообразователей повышенными требованиями не только в отношении гумусности почвы, но и к степени ее увлажнения. Можно сказать, что среди тяныпанских ковылей Stipa rubens представляет собою наиболее влаголюбивый вид. Поэтому для ценозов с преобладанием этого ковыля характерно постоянное присутствие многих растений ксеромезофильного и мезофильного типа. Красноковыльная формация в условиях Тянь-Шаня может рассматриваться как наименее ксерофитный вариант горных ковыльных степей, наиболее близкий к луговым степям. Флористически красноковыльники богаче всех других ковыльных формаций, так как вних насчитывается свыше 130 видов, из которых значительная часть представляет горноазиатский и, в частности, горносреднеазиатский элемент. Эта флористическая особенность хорошо отличает тяныпан-ские красноковыльные степи от соответствующих близлежащих равнинных степей Казахстана. Из горноазиатских флористических элементов, встречающихся в тяныпанских красноковыльных степях, можно отметить -{-Helictotrichon tianschanicum, +#. asiaticus, -{-Carex turkestanica, -{-Polygonum coriarium, -{-Ligularia persica, +L. Thomsonii, -{-Dianthus Hoelt-zeri, -{-D. turkestanicus и многие другие виды. Травостой красноковыльных степей обычно густой (нередко до 100% покрытия), до 60—70 см высотой. В его сложении можно различать 3 подъяруса. В первом (до 70 см) преобладает красноватый ковыль (Stipa rubens), вместе с которым встречаются Stipa capillata, Aneurolepidium (Elymus) angustum, а из крупного разнотравья -{-Ferula akitschkensis. Второй, наиболее густой подъярус (до 40 см) слагается Festuca sulcata, Phleum phleoides, Galium verum s. 1., G. boreale. Третий подъярус представлен более мелкими растениями (Fragaria viridis, Potentilla orientalis и др.). Иногда на фоне этих степей разбросаны -{-Rosa platyacantha («шипичка»), Cotoneaster multiflora, С. те-lanocarpa, Spiraea hypericifolia, -^-Berberis heteropoda. Красноковыльные степи используются как сенокосы, дающие не менее 10—12 ц/га сена. Почвы под этими степями интенсивно распахиваются под посевы зерновых и бахчевых культур.
В заключение характеристики северотяныпанских дерновиннозлаковых степей необходимо указать на присутствие в их составе двух формаций, образованных степным мятликом (Роа relaxa) и кавказским ковылком (-{-Stipa caucasica). Обе эти формации характерны для щебнистых почв. Первая из них небольшими участками встречается в средних и верхних поясах гор по южным склонам, вторая распространена преимущественно в низкогорьях. Хозяйственное значение этих формаций не велико. Сильно разреженный травостой всех относящихся к ним ассоциаций используется лишь в качестве летнего пастбища.
Выделение на «Геоботанической карте» северотяныпанских дерновиннозлаковых степей в качестве особой географической категории вызывается присущими им особенностями. В противоположность Центральному и Западному Тянь-Шаню, Памиро-Алаю и Копет-Дагу, в Северном Тянь-Шане полностью отсутствуют степи с доминированием или участием ковылей ряда Brevigeniculatae. Существенным отличием степной
растительности Северного Тянь-Шаня является также весьма широкое' распространение здесь тырсовых степей, которые в других частях тянь--,-шанского горного сооружения, и особенно в системе Алая, представлень/ значительно слабее, обыкновенно в виде небольших фрагментов. Красно*-ковыльные степи в пределах всей горной Средней Азии свойственны только Северному Тянь-Шаню. Все это определенно указывает на ценоти-ческое своеобразие северотяньшанские степей, что и послужило основанием особого выделения их на карте.
Литература
М. Г. Попов (1940); П. П. Поляков (1941а); Н. И. Рубцов (1941, 1941в, 1948а, 1950, 1952, 1954); К. В. Станюкович (1955).
84 цтш — центральнотяньшанские
Горные дерновиннозлаковые степи в сочетании с каменистыми обнажениями занимают обширные пространства в Центральном Тянь-Шане, и на «Геоботанической карте» показаны в следующих местах: на северном склоне хр. Терскей-Алатау (южнее оз. Иссык-Куль); этот массив на западе сливается с их крупными контурами, расположенными на хребтах-Джумголтау, Сонкёль, Байдула, Мулдатау, а на юге — с крупными массивами, занимающими северные склоны Ферганского хребта, и отходящих от него гор Джаманто, Бойбичето и хр. Атбац (окружая оз. Ча-тыркёль); три контура горных дерновиниозлаковых степей показаны на северном склоне хр. Кокшаал-Тау. Центральнотяньшанские горные дерновиннозлаковые степи представлены несколькими формациями и очень многими группами ассоциаций, которые образуют четко выраженный пояс в общем ландшафте высокогорий Тянь-Шаня. А. Н. Краснов (1888) указывал на особое богатство «сыртов» элементами степной флоры, на исключительное распространение степной растительности и на то, что степные формации Тянь-Шаня имеют тенденцию к еще более широкому распространению, в особенности на южных склонах гор, где они вытесняют сообщества других формаций.
Очень широко распространены типчаковые (из -{-Festuca Kryloviana) степи, связанные с высокогорными степными каштановидными почвами, часто скелетными и иногда щебневатыми, в высотных пределах 3000—3600 м над у. м. Эти высокогорные степи должны быть отнесены к криофитным (пустошным) степям. Мелкодерновинный типчак Крылова образует густой травостой, в составе которого присутствуют и другие мелкодерновинные злаки, достигающие также значительного обилия: Stipa orientalis, +*S. caucasica, Helictotrichon desertorum, -|-Я. tianschanicus, -{-Festuca coelestis, -\-Ptilagristismongholica, 4-P. subsessiliflora. Кроме злаков, встречаются также осоковые {-{-Carex duriusculiformis, -{-Cobresia pamiroalaica) и довольно обильные представители травянистого разнотравья {+Potentilla evestita, +Р. nervosa, -{-Leontopodium ochroleucum, +£. leontopodinum, -{-Oxytropis globiflora,{-Silenegraminifolia,-{-Astragalus nivalis,-{-Pyrethrum pyrethroides, -{-Serratula lyratifolia, -{-Bupleurum densiflorum, -{-Geranium saxatile и др.), а также некоторые специфические полукустарнички {-{-Artemisia tianschanica, -]-А. rhodantha, -{-Dracocephalum bipinnatum). Эти типчаковые степи используются как летне-осеттттие ц зимние пастбища ( давая сухой кормовой массы летом и осенью 6 ц/га, а зимой — 2 ц/га’
Более увлажненные участки характеризуются некоторым ослаблением роли типчака и усилением участия кобрезий {-{-Cobresia capilliformis
и -{-С. humilis). Для всего пояса дерновиннозлаковых степей характерно наличие стелющейся арчи {-{-Juniperus turkestanicd) (Соболев, 1952). В высокогорном поясе Центрального Тянь-Шаня распространены также криофитные таргыловые (с преобладанием A-Hordeum turkestanicum) степи, распространенные в бассейне Аксая, близ оз. Чатыркёль и других, на высотах 3600—3800 м над у. м. Степи эти занимают горные склоны, встречаясь и отдельными участками, и значительными массивами. К господствующему эдификатору {-{-Hordeum turkestanicum) примешиваются дерновинные злаки и кобрезии {-{-Festuca Kryloviana, -{-F. coelestis. -{-Pti-lagrostis mongholica, -{-P. subsessiliflora, Helictotrichon desertorum, -{-Leuco-poa Olgae, -{-Poa Albertii, -{-Cobresia pamiroalaica), полукустарничек -{-Artemisia rhodantha и др. Слепи эти также служат летними пастбищами.
Также весьма обыкновенны киргизскоковыльные (из Stipa kirghisorum) степи, распространенные во всем Центральном Тянь-Шане. Они занимают горные склоны, древние речные террасы, склоны горных долин и озерных котловин в высотных границах 2800—3500 м над у. м., т. е. в более низком поясе, чем предыдущие. Почвы — высокогорные степные каштановидные. В этих степных сообществах основным эдификатором является Stipa kirghisorum, но к нему часто в значительном количестве (иногда поднимаясь до значения субэдификаторов) примешиваются Festuca sulcata s. 1. и +F. Kryloviana, а также виды разнотравья {-{-Potentilla evestita, -{-Oxytropis globiflora, -{-Leontopodium ochroleucum, +£,. leontopodinum и др.). Киргизскоковыльные степи используются как пастбища весной и поздней осенью с запасом кормовой массы 5 ц/га.
На границе с субальпийским поясом в Центральном Тянь-Шане, в пределах 2800—3200 м над у. м. распространены также криофитные дерновиннозлаковые овсецовые степи из Helictotrichon desertorum с примесью Festuca sulcata s. 1. (или с +F. Kryloviana) и Stipa kirghisorum-, иногда встречаются участки чистых овсецовых травостоев. К названным эдификаторным и субэдификаторным видам примешиваются еще следующие растения, свойственные высокогорьям: дерновинные злаки и кобрезии {-{-Helictotrichon asiaticus, -{-Н. tianschanicus, -{-Ptilagrostis subsessiliflora, -{-Poa Albertii, -f-P. Litwinowiana, -{-Cobresia pamiroalaica), травянистое разнотравье {-{-Potentilla evestita, -{-Leontopodium ochroleucum, -{-Oxytropis globiflora, -{-Cerastium cerastoides, -{-Ligularia robusta), полукустарничек -{-Artemisia Aschurbajewii и др. Почвы под овсецовыми степями — высокогорные степные каштановидные. Овсецовые степи лучше всего выражены в котловине оз. Сонкёль, в урочище Сусамыр и в бассейне Нарыва. Используются как летние пастбища.
В пределах расположения горных дерновиннозлаковых степей широко распространены выходы скалистых пород, каменистые склоны, осыпи. На них разреженная растительность слагается различными петрофитами (104 са).
Литература
А. Н. Краснов (1888); Р. Ю. Рожевиц (1912); А. И. Михельсон (1913); О. Э. Кнор-ринг (1923); М. В. Культиасов (1926—1927); В. И. Вандышева (1948); И. В.Выходцев (1952); Л. Н. Соболев (1952, 1953).
84 тшп— западнотяныпанские и памиро-алайские
Горные дерновиннозлаковые степи g сочетании с каменистыми обнажениями чрезвычайно широко распространены в Западном Тянь-Шане и на хребтах системы Памиро-Алая, занимая здесь большие площади.
Они показаны на «Геоботанической карте» нижеперечисленными контурами: на хр. Каратау (самый крупный массив горных дерновиннозлаковых степей этого варианта), на хр. Таласском Алатау, на Чаткальском хребте (крупный массив в северной части его и два «острова» в южной), на Кураминском и Туркестанском хребтах (несколькими островными контурами на северном склоне и обширной полосой по южному склону); с последнего горные дерновиннозлаковые степи переходят на северный склон Зеравшанского хребта и на Гиссарский хребет; далее большие площади рассматриваемых степей показаны в Алайской долине, где одевают подножие южного склона Алайского и северного склона Заалайского хребтов. Южнее последнего эти степи вытянулись вдоль р. Муксу на значительном ее протяжении; еще южнее показаны небольшие «острова» на хребтах Петра Первого, Дарвазском, Шугнанском и Ваханском. Дерновиннозлаковые горные степи Западного Тянь-Шаня и Памиро-Алая весьма разнообразны: помимо типичных степей, здесь очень широким распространением пользуются криофитные, трагакантовые, колючетравные, полусаван-новые и пустынные степи.
Остановимся сначала на горных степях окраинных хребтов системы Западного Тянь-Шаня: Каратау и Кураминского хребта.
Пустынные дерновиннозлаковые степи занимают большую площадь на выдвинутой к северо-западу и внедренной в окружение пустынь оконечности гор системы Тянь-Шаня — хр. Каратау. Эти степи образуют обширный пояс в высотных пределах от 1250 до 2000 м над у. м. (Павлов, 1932; Культиасов, 1938). Для этих степей, как и для флоры всего хр. Каратау, характерно значительное количество эндемичных каратавских видов (Павлов и Липшиц, 1934), в особенности из группы ксерофитов. В частности, эндемом Каратау является субэдификатор пустынных дерновиннозлаковых (полынно-типчаковых) степей полукустарничковая полынь A-Artemisia karatavica. Эти степи распространены по плоскогорьям на основной части Каратау, а также по наиболее возвышенным местам. Основу травостоя образуют Festuca sulcata s. 1., А~ Artemisia karatavica и Stipa kirghisorum. К этим господствующим видам примешаны кустарнички (A-Astragalus neo-Lip sky anus), многолетние травы (Oxytropis pilosa, Potentilla orientalis, A-Astragalus macrotropis), эфемероиды (+A Ilium incon-spicuum, A-Tulipa turkestanica, Poa bulbosa var. vivipara) и эфемеры (Bromus Danthoniae, A-Taeniatherum crinitum, А-Nigella integrifolia, A~Lappula microcarpa). Изредка на плоскогорье встречаются небольшие группы кустарников (A-Spiraeanthus. Schrenkianus, Hulthemia berberifolia и др.). Наряду со значительной опустыненностью степей Карата^, наблюдается очень частое внедрение в степные травостои элементов субтропических степей (полусаванн) в виде Agropyrum trichophorum, A-Inula grandis, A-Prangos pabularia и многих других растений (опис. 73 а). Подобные ассоциации могут быть отнесены к полусаванновым степям.
Наконец, следует отметить еще одну характерную черту степей Каратау: в ландшафте большую роль играют кустарниковые заросли, которые не только занимают овраги и ущелья, но постоянно переходят на открытые склоны. Состав их очень разнообразен, в них входят деревца (A-Lonicera persica), кустарники (A-Rosa kokanica, Spiraea hypericifolia), кустарнички (Hulthemia berberifolia) и полукустарнички (A-Jurinea suf-fruticosa, Ziziphora Bungeana, A-Artemisia karatavica, A-Astragalus xantho-meloides и др.); характерны также весьма обильные виды разнотравья (АгAster Grimmii, А-Ferula karatavica, Hypericum elongatum, A-Astragalus Severzovii, A~Cousinia Minkwitziae, A-Leonurus turkestanicus, A-Onobrychis grandis, A-Phlomis salicifolia и др.), в большинстве своем только как ред
кость встречающиеся в травостоях дерновиннозлаковых степей, по весьма обычные в субтропических степях (полусаваннах).
В Кураминском хребте (Сидоренко, 1953) дерновиннозлаковые степи широко распространены в пределах очень большой амплитуды высот — от 1500 до 3500 м над у. м. Представлены они типчаковыми (из Festuca sulcata s. 1.) степями: как типичными, так и трагакантовыми, полусаван-новыми и опустыненными.
Типичные типчаковые степи занимают более или менее пологие склоны с развитыми суглинистыми почвами типа горностепных каштановых и черноземовидных. Эдификатором сообществ является типчак (Festuca sulcata s. 1.), обычно сильно сбитый выпасом. Наиболее часты следующие сопутствующие виды: из дерновинных злаков — Koeleria gracilis, -{-Hordeum turkestanicum (оба злака иногда становятся субэдификаторами); из травянистого разнотравья-----{-Ranunculus ajghanicus, A-Trigonella
pamirica, -{-Cousinia Bonvalotii; из эфемеров —Arenaria serpyllifolia, Veronica biloba; из полукустарничков — Ziziphora Bungeana', из трагакантовых кустарничков----{-Astragalus lasiosemius и др. Полукустарнички
и кустарнички немногочисленны.
Трагакантовые типчаковые степи Кураминского хребта приурочены к щебнисто-каменистым субстратам на крутых склонах и на более возвышенных водораздельных гребнях. Покров отличается разреженностью (проективное покрытие 60—70%). Спутниками типчака являются: из дерновинных злаков — Роа relaxa, -{-Bromus turkestanicus, -{-Leucopoa karatavica; из травянистого разнотравья--{-Cousinia Bonvalotii; из полукустарничков ----------------------------{-Arenaria Griffithii, -{-Oxytropis microsphaera, Artemisia
rutifolia',n3 трагакантовых кустарничков-{-Onobrychis echidna,
galus Bornmiillerianus, + Acantholimon Korolkovii и др.; наиболее обильны Cousinia Bonvalotii и Acontholimon Korolkovii.
Полусаванновые типчаковые степи распространены в среднегорной части Кураминского хребта. Здесь характерно сочетание двух синузий — типчака, занимающего нижний ярус, и полуэфемероидного зонтичного — югана (ArPrangos pabularia), образующего верхний ярус сообществ. Сопутствующие им виды — преимущественно выходцы из крупнотравных субтропических степей (опис. 73 v): из дерновинных злаков--{-Aneurole-
pidium alaicum, из корневищных злаков — Agropyrum trichophorum’, из длительновегетирующего разнотравья-----Н Verbascum songoricum; из эфемероидов ------------------------------{-Ferula ovina, A-Eremurus Regelii', из полукустарничков —
4--PoZygontt?n paronychioides и ряд других.
Пустынны» типчаковые степи связаны в значительной мере с деятельностью человека, способствовавшей внедрению в типчаковые сообщества полукустарничковой полыни (Artemisia tenuisecta), а также ряда других ксерофильных форм; из дерновинных злаков — Stipa capillata', из полукустарничков — A-Arenaria, Griffithii, -A-Scutellaria ramosissima, Ziziphora Bungeana', из трагакантовых кустарничков-----{-Acantholimon
Korolkovii, A~Astragalus lasiosemius, -f-Л. asaphes и др. Такие пустынные степи распространены в высоких частях хребта (до 2500 м над у. м.), главным образом на месте сведенных арчевников из A-Juniperus seravschanica.
Типчаковые степи Кураминского хребта используются под пастбища. В зависимости от состава они дают урожай сухой массы от 4 до 10 ц/га (наиболее урожайными являются полусаванновые типчаковые степи, включающие к тому же многие хорошо поедаемые травы).
- В пределах основной части системы Западного Тянь-Шаня горные дерновиннозлаковые степи занимают хорошо выраженный пояс на вы
сотах от 1800 до 3500 м над у. м.; в зависимости от местных условий высотные пределы иногда значительно колеблются. Этот пояс часто именуют поясом высокогорных степей (Кнорринг, 1915; Культиасов, 1926—1927, и др.).
Почти во всех сообществах этих степей характерна господствующая роль типчака {Festuca sulcata s. 1.), образующего плотные дерновинки, расположенные то густо (сомкнутость до 70—80%), то разреженно. Между дерновинками типчака с различными степенями обилия размещаются многие согосподствующие и сопутствующие растения. Почвы горностеппые и высокогорностепные, каштановидные, обычно суглинистые, но на склонах часто скелетные. Приведем некоторые примеры.
В горах Джебоглы-Су (Культиасов, 1926—1927) типчаковые степи представлены как трагакантовым, так и криофитным (альпийско-луговым) вариантом.
В трагакантовых типчаковых степях на фоне господствующего покрова типчака часто разбросаны подушки трагакантового (колючекустарничкового) эспарцета {А~О nobrychis echidna)', заметную роль играет колючетравная высокогорная кузиния {A-Cousinia caespitosa). Кроме того, здесь встречаются виды разнотравья {Aster alpinus, + Astragalus densiflorus, Geranium collinum, A~Oxytropis macrocarpa, Polygonum alpinum, A-Poten-tilla hololeuca), полукустарничков {A-Arenaria Griffithii), длительновеге-тирующих однолетников {A-Euphrasia Regelii) и др.
Криофитные типчаковые степи в своем составе имеют много видов, свойственных альпийским лугам. Так, в них встречаются: из дерновинных злаков-----\-Poa Litwinowiana', из разнотравья--Aconitum rotun-
difolium, A-Astragalus andaulgensis, A-Cerastium cerasioides; A~Erigeron eriocalyx, A-Gnaphalium supinum, Myosotis suaveolens, A~Oxytropis albovillo-sa, A-Pedicularis Korolkovii, A-Silene graminifolia, A-Primula algida и др.
Участки типчаковых степей перемежаются с разреженной растительностью на склонах и осыпях. В ландшафте особенно выделяется стелющаяся арча (+Juniperus turkestanica), образующая характерные круго-вины, в середине которых нередко находят приют кусты A-Lonicera Altmannii и A-Berberis heteropoda. На скалах — большой и разнообразный набор растений — хазмофитов ксерофильной и криоксерофильной экологии (опис. 104 са).
В Таласском Алатау также распространены типчаковые и ковыльнотипчаковые степи (Кнорринг и Минквиц, 1912а; Аболин и Советкина, 1930). Используются эти типчаковые степи как летние пастбища с урожайностью сухой массы 1.7—2.9 ц/га.
Помимо типичных типчаковых степей, занимающих обширные пространства плато и пологих склонов, и представляющих собою чистые травяные сообщества, в этих горах распространены еще кустарниковые типчаковые степи со значительным участием кустарниковых зарослей из A~Caragana jubata и +.С aurantiaca.
В Ташкентском Алатау, помимо широко распространенных типчаковых степей, по личному сообщению Е. П. Коровина, изредка встречаются (в частности в бассейне Ангрена) бескильницевые (из дерновинного злака А~ Puccinellia Hackeliana) степи. Бескильница образует разреженный покров, а между ее мелкими дерновинками расселяется Роа bulbosa var. vivipara, некоторые представители разнотравья {A-Ligularia altaica, Geranium collinums. 1., A-Gicer songoricum,колючелистная A-CousiniaFranchetii, мелкий полукустарничек + Polygonum rupestre) и немногие другие растения. Для бескильницевых степей характерно малое количество сопутствующих видов.
В пределах пояса дерновиннозлаковых степей широко распространена разреженная растительность скал и осыпей (опис. 104 са), занимающая в различных угодьях от 20 до 75% площади.
Дерновиннозлаковые степи хребтов системы Памиро-Алая (Туркестанский, Зеравшанский, Гиссарский и все южнее расположенные) в целом отличаются значительно большей ксерофитизованностыо, что выражается в широком распространении трагакантовых и колючетравных ассоциаций типчаковых, киргизскоковыльных, бескильницевых и других формаций.
В Гиссарском хребте в пределах 2800—3100 м над у. м. (Королева, 1940) колючетравные типчаковые степи представлены сообществами с проективным покрытием 50—70%. Господствуют Festuca sulcata s. 1. и A-Cousinia Franchetii. К этим видам-эдификаторам примешиваются: из разнотравья----\-Psychrogeton cabulicus', из мелких полукустарнич-
ков hArenaria Griffithii, А-Polygonum biaristatum и др.
Гораздо меньше участие мезофильных форм, встречающихся изредка и единично: -f-Роа bucharica, А~ Geranium saxatile.
На хр. Петра Первого типчаковые степи криофитного типа расположены на высотах более 3100 м над у. м. (Афанасьев, 1940). Сомкнутость травостоя от 40 до 70%. Господствует дерновинный алайский типчак {A-Festuca alaica), а также бухарский мятлик (А-Роа bucharica). К ним примешаны виды травянистого разнотравья {A- Arenaria turkestanica, A-Draba Olgae, А-Androsace Olgae, A-Gnaphalium supinum, A-Oxytropis immersa — образует подушки, -f-O. Michelsonii, A-Potentilla gelida, A-Psychrogeton cabulicus, A-Cerastium cerastoides, A-Melandrium longicarpophorum), некоторые полукустарнички {A-Oxytropis savellanica, + Polygonum biarista-tum, + Minuartia Litwinowii) и трагакантовые кустарнички {A~ Astragalus nigrocalyx). Как видно из этого списка, господствуют виды, свойственные высокогорным лугам и степям.
В типчаковых степях бассейна Обихингоу выше 3000 м над у. м. распространены ассоциации со значительным участием колючетравных кузиний (Гончаров, 1936). Покров их слагается типчаком {Festuca sulcata s. 1.), согосподствующим видом является -^-Cousinia stephanophora. Большое участие принимают также: из дерновинных злаков — Koeleria gracilis, Роа relaxa’, из корневищных осок — Carex duriusculiformis’, из разнотравья ---\-Geranium saxatile, A-Potentilla Lipsky ana, A-Draba Olgae’,
из трагакантовых кустарничков---Astragalus nigrocalyx’, среди менее по-
стоянных спутников ряд видов, общих с вышеперечисленными, а кроме них еще из разнотравья ^Pedicular is Olgae, A~Ligularia altaica, +-4Z-lium Winklerianum и др.
H. Ф. Гончаров (1936) указывает на последовательный ряд ассоциаций, ведущий от типчаковых степей к сообществам нагорных ксерофитов: Festucetum {Festuca sulcata s. 1.)—>Cousinieto {Cousinia stephanophora)-Festucetum-»Cousinieto-Festucetum artemisiosum (с участием A-Artemisia Lehmanniana) -» Artemisieto-Festucetum-* Festuceto-Onobrychidetum (с господством A-Onobrychis echidna).
В пределах Памиро-Алая достаточно часто встречаются также, хотя и занимают небольшие площади, ковыльные и ковыльно-типчаковые степи. Эти степи располагаются преимущественно по более сухим южным и восточным склонам, в высотных пределах 2700—3400 (3500) м над у. м. (Гончаров, 1937). Травостой густой (степень покрытия 70—80%). Эдифи-каторами являются дерновинные злаки: Festuca sulcata s. 1. и Stipa kirghisorum, а также Poa relaxa. К ним присоединяются: из разнотравья — А-Geranium saxatile, A-Nepeta podostachys, A-Ligularia altaica, A-Jurinea
hissarica, -{-Pedicularis Olgae', из трагакантовых кустарничков--HAsJra-
galus lasiosemius. Реже встречаются некоторые виды разнотравья: -{-Euphorbia serawschanica, -{-Allium Barszczewskii, -{-Pseudosedum condensatum и др.
Среди этих степей подчиненную роль играют сообщества мятликовотипчаковые (Festuca sulcata и Роа relaxa), кузиниево-мятликовые (Роа relaxa и -{-Cousinia hissarica), а на сильно затененных местах югановые (из -{-Prangos pabularia). В бассейне среднего Зеравшана, на хр. Туркестанском и Гиссарском, Ю. С. Григорьев (1941, 1944) указывает наличие типичных и опустыненных мятликово-ковыльно-типчаковых (Festuca sulcata s. 1., Stipa kirghisorum и Poa relaxa) степей. В высокогорных пустынных степях, помимо преобладающих дерновинных злаков, в качестве ^господствующего вида встречается полукустарничковая полынь -{-Artemisia Lehmanniana', в виде примеси-------{-Helictotrichon
asiaticus, -{-Scorzonera acanthoclada и др.
В Памиро-Алае (в особенности на Туркестанском хребте) встречаются сообщества трагакантовых или пустынных ковыльных степей, слагаемые в основном мелкодерновинными ковылями: Stipa orientalis, +5. caucasica, +5. trichoides. Они приурочены преимущественно к скелетным почвам, типа горных коричневых карбонатных, распространенных в нижних частях пояса дерновиннозлаковых степей или пояса остепненных арчевников. В качестве постоянных спутников отмечены следующие виды: из дерновинных злаков —Festuca sulcata s. 1. ; из полукустарничков-------\-Helian-
themum soongoricum, Acanthophyllum pungens\ из трагакантовых кустарничков \-Acantholimon compactum, +А. alatavicunr, из корневищных осок----{Carex turkestanicw, из полукустарничков — Kochia prostrata',
из эфемероидов — Роа bulbosa var. viviparw. из эфемеров-----f- Veronica
intercedens', из длительновегетирующих однолетников-------{Polygonum
polycnemoides.
Следует упомянуть еще о криофитном варианте дерновиннозлаковых степей — бескильницевых (из -{-Puccinellia subspicata) степях. Они, по личному сообщению В. А. Никитина, распространены небольшими участками на Западном Памире и Гиссаро-Дарвазской горной дуге на высотах более 3400—3500 м над у. м. Почвы — высокогорные коричневые луговостепные. В сообществах господствует мелкодерновинная бескиль-ница -{-Puccinellia subspicata, которой сопутствует низкорослое альпийское разнотравье (-{-Potentilla flabellata, -{-Cerastium cerastoides, -{-Eri-geron turkestanicus, -{-Draba Olgae, -{-Gnaphalium supinum, -{-Ranunculus turkestanicus) .Особенно характерно постоянное и обильное присутствие полукустарничка -{-Oxytropis savelianica. На более сухих участках примешиваются -{-Androsace Olgae, -{-Psychrogeton cabulicus, полукустарничек -{-Polygonum biaristatum и ряд других.
Наряду с дерновиннозлаковыми горными степями в поясе их преобладания почти повсеместно встречаются отдельные участки или хотя бы фрагменты многих других формаций. При этом особенно широко распространены весьма пестрые и разреженные сообщества скал и осыпей (опис. 104 са), которые во многих местах не могли быть показаны по условиям масштаба. Так, например, на хр. Нуратау (Кнорринг, 1934) на скалистых участках среди степной растительности встречаются отдельные подушки -{-Acantholimon alatavicum, -{-Jurinea Olgae, -{-Acanthophyllum Knorringianum. В расщелинах скал ютятся полукустарнички ^-Scutellaria orbicularis и A-Lepidolopha Komarovii. На продуктах разрушения известняков поселяются куртинками из полукустарничков — Ziziphora clinopodioides s. 1.: из разнотравья (частью эфемероидного типа)-|-Asira-
galus inaequalifolius, -{-Phlomis salicifolia, -{-Euphorbia rapulum и многие другие. Изредка встречаются одиночные кусты -{-Acer Semenovii, -{гAmygdalus spinosissima и -{-Lonicera persica.
В Кураминском хребте (Эмме-Марковская, 1940) в поясе горных дерновиниозлаковых степей встречаются кустарниковые заросли из следующих видов кустарников: +7?о$а hissarica, -[-Lonicera persica, -{-Berbe-ris oblonga, -{-Spiraea pilosa, под пологом которых развит степной травостой, близкий по составу к окружающим типчаковым степям.
В условиях более благоприятного увлажнения и богатых почв в степной ландшафт вклиниваются участки лисохвостных (из -{-Alopecurus seravschanicus) лугов, в которых эдификатор лисохвост сопровождают из злаков----f-Poa bucharica; из разнотравья----{-Nepeta podostachys,
-[•Euphorbia seravschanica, -{-Senecio soongoricus, -{-Ligularia altaica, -{-Astragalus andaulgensis, -{-Geranium saxatile и др.
Наконец, при избыточном увлажнении (берега речек и родники на склонах) развиваются болотистые осоковые луга из Carex pseudo-foetida.
Литература
G. И. Коржинский (1896); Б. А. Федченко (1903); О. Э. Кнорринг и 3. А. Минк-виц (1912, 1912а); О. Э. Кнорринг (1915, 1923, 1934); И. А. Райкова (1924); М. В. Культиасов (1926—1927, 1938); Р. И. Аболин и М. М. Советкина (1930); Н. В. Павлов (1932); Е. П. Коровин (1934); Н. В. Павлов и С. Ю. Липшиц (1934); Н. Ф. Гончаров (1936, 1937); К. С. Афанасьев (1940); А. С. Королева (1940); Л. А. Эмме-Марковская (1940); Ю. С. Григорьев (1941, 1944); Г. В. Микешин (1946); К. 3. Закиров (1947); Н. И. Рубцов (1952); Г. Т. Сидоренко (1953).
84 кпд — копетдагские
Горные дерновиннозлаковые степи в сочетании с каменистыми обнажениями имеются в наиболее высоких центральных и восточных частях Копет-Дага, а также на хр. Большой Балхан. В Копет-Даге дерновиннозлаковые степи распространены в верхнем поясе, начиная с 1600 м и до 2800 м над у. м.
Господствующей формацией являются типчаковые (из Festuca sulcata s. 1.) степи, отличающиеся в различных местоположениях набором сопутствующих видов.
Наиболее широко распространены ксерофильноразнотравные типчаковые степи, приуроченные к южным склонам и открытым выравненным участкам с щебнисто-мелкоземистыми горнокаштановыми почвами. Эти сообщества могут быть отнесены к типичным (настоящим) степям. Проективное покрытие всего 40—50%. К господствующему в покрове типчаку примешаны дерновинные ковыли {Stipa Lessingiana и +<S. caucasica}, ксерофитное разнотравье {-{-Phlomis cancellata, Centaurea squarrosa, виды Cousinia, Jurinea, Convolvulus и др.). Эти типчаковые степи характерны заметным присутствием эфемерово-эфемероидной группы растений, что является отражением зональных особенностей. Эфемероиды представлены преимущественно луковичными растениями: -{-Allium scab-rellum, +А. pseudoseravschanicum, +Л. Bodeanum, -{-Gagea dshungarica, +G. gageoides, -[-Tulipa Micheliana, -[-Hyacinthus Litwinowii, Muscari racemosum, а также Eremurus spectabilisи др. Из эфемеров обычны Alyssum campestre, -[-Bromus oxyodon, -[-Roemeria hybrida, -[-Arenaria leptoclados, Milium vernale, Eremopoa oxyglumis, -{-Minuartia Meyeri, -[-Queria hispa-nica и др.
На мелкоземистых (не скелетных) почвах заметно возрастает сомкнутость травостоя, увеличивается степень обилия типчака и падает участие
Stipa Lessingiana; нередко широко расселяется Carex pachystylis, образующая дернину; изредка встречается Роа bulbosa var. vivipara.
Мезофитные варианты всегда связаны с северными склонами и с более мощными мелкоземистыми почвами. Проективное покрытие в этих сообществах достигает 90—100%. Количество эфемеров и эфемероидов резко снижается, но внедряется ряд видов разнотравья (-[-Astragalus Jarmolen-koi, -[-Onobrychis Lipskyi, Inula oculus-Christi, Potentilla transcaspianjbp.), реже появляются примитивные полукустарнички (например -[-Astragalus brevidens).
При возрастании увлажнения (в ложбинах) дерновинные злаки резко уменьшаются в числе, сохраняясь в виде отдельных дерновинок на фоне корневищных Agropyrum repens, A. intermedium и A. trichophorum. Из других видов здесь встречаются клубнекорневые — Eremurus spectabilis и -[-Е. stenophyllus, а также стержнекорневые -[-Codonocephalum Реасо-ckianum и -^-Cousinia Smirnowii. Эти сообщества можно рассматривать как своеобразные остепненные луга.
В условиях же постоянного подпитывания талыми и дождевыми водами, на наиболее высоких местоположениях (2400—2800 м над у. м.), остепненные луга становятся еще более мезофитными. Дерновинные злаки (Stipa Lessingiana и S. capillata) отступают на задний план, и эдификаторами сообществ являются рыхлокустовые и корневищные злаки (Роа relaxa, Agropyrum repens, Dactylis glomerata, Calamagrostis epigeios) с примесью видов красочного, отчасти альпийского разнотравья, среди которых преобладают стержнекорневые (-[-Orthurus kokanicus, -[-Cephalaria subla-nata, -[-Centaurea sessilis, -[-Trachydium kopetdaghense, -[-Solenanthus circin-natus), встречаются луковичные (-[-Ixiolirion montanum, -[-Hyacinthus transcaspicus), клубнекорневые (Eremurus spectabilis, -[-Corydalis chiono-phila), а также примитивные полукустарнички (-[-Galium kopetdhagense) и однолетники (-[-Myosotis refracta и др.). Эти сообщества также могут быть отнесены к остепненным лугам.
На менее высоких вершинах, в пределах 1400—1600м над у. м., в дерновиннозлаковых степях заметную роль играют растения, являющиеся эдификаторами субтропических степей (полусаванн). Так, И. А. Линчев-ский (1935а) на вершине Сюнта отметил сообщества, слагаемые из Festuca sulcata s. 1. и Agropyrum trichophorum в равной степени обилия; на вершине же Хасардага последний даже преобладал над типчаком. Однако в обоих случаях общее участие дерновинных злаков оказывалось преобладающим, так как, помимо названного типчака, в подобных сообществах обычно присутствуют еще Stipa Lessingiana, S. pulcherrima, S. capillata, -[-S. tur-comanica, Ч-^. crassiculmis. В описываемых И. А. Линчевским типчаковых степях обильно представлены также многие другие виды: полукустарнички (-[-Stachys turcomanica), примитивные полукустарнички (-[-Onobrychis Sin-tenisii, Hypericum scabrum, -[-Crucianella Sintenisii, а также стержнекорневые виды травянистого разнотравья (-[-Convolvulus ascabadensis, Echinops ritro, -[-Salvia Linczevskii, -[-Gypsophila bicolor, -[-Phlomis kopetdagensis), клубнекорнёвые (-[-Eremostachys speciosa) и др. Эти сообщества должны быть отнесены к полусаванновым степям.
В общем ландшафте горных дерновиннозлаковых степей Копет-Дага чрезвычайно характерно наличие арчевников из -[-Juniperus turcomanica и сообществ подушкообразных нагорных ксерофитов — трагакантников (из -[-Gypsophila aretioides, -[-Onobrychis cornuta, -[-Acantholimon khoras-sanicum, +A. erinaceum, -[-Acanthophyllum glandulosum и др.). Под полог арчи внедряются и степные виды; также весьма обычны подушкообразные формы и другие виды нагорных ксерофитов. Образуемые ими
сообщества являют обширную гамму переходов между собственно арчев-никами, нагорными ксерофитами и дерновиннозлаковыми степями.
По наиболее затененным обрывистым северным склонам, под пологом высокоствольной арчи или в трещинах скал, встречаются папоротники (Cystopteris fragilis, -\-Dryopteris Komarovii, Asplenium ruta-muraria, A-Ceterach officinarum и др.) и ряд растений, совершенно отсутствующих на ровных и открытых местах (A-Dionysia tapetodes, A-Graellsia saxifragifo-lia, + Viola Sintenisii, + Valeriana ficariaefolia, + У. sisymbriifolia, + Arum Korolkowii, A-Parietaria Judaica, A-Delphinium biternatum.
Вследствие многовекового истребления арчи на топливо и для строительных целей, площади арчевников значительно сокращены, и на их месте распространяются преимущественно нагорные ксерофиты и в значительно меньшей мере — степные сообщества.
На хр. Большой Балхан горные дерновиннозлаковые степи занимают пояс в высотных пределах 1400—1800 м над у. м. Представлены они, по исследованиям Е. Г. Боброва (1931), типчаково-ковылковыми (Stipa Lessingiana и Festuca sulcata s. 1.) сообществами, весьма бедными по флористическому составу и с сомкнутостью травостоя 70—80%. По фону господствующих ковылка и типчака рассеянно и единично встречаются Galium verum, Euphorbia Seguieriana, A-Crupina vulgaris, A-Onobrychis Lyp-skyi, Centaurea squarrosa, А-Ranunculus linearilobus, + Ziziphora persica. Такого облика степи разнообразятся местами (по вершинам ущелий и склонам) пятнами кустарниковых зарослей из A~Berberis integerrima, A-Lonicera persica, и, изредка, A-Cerasus microcarpa.
Типчаково-ковылковые степи Большого Балхана приурочены к суглинистым и несколько скелетным (мелкощебнистым) горнокаштановым почвам, наиболее хорошо развитым на выровненных участках плато этого хребта. В местах же эродированных, на крутых склонах со смытым мелкоземистым плащом преобладают участки с полукустарничковыми, кустарничковыми и травянистыми представителями флоры нагорных ксерофитов (A-Stachys turcomanica, Ziziphora Bungeana, + Astragalus velatus, +Л. gaudanensis, A-Acantholimon Korovinii, A-Oxytropis Bobrovii, A-Poly-gonum paronychioides, A-Asperula balchanorum и др.).
В общем ландшафте балханских типчаково-ковыльных степей заметная роль принадлежит арчевникам (из turcomanica), кустарниковым
зарослям (A-Rhamnus Sintenisii, A-Cerasus pseudoprostrata, +C. microcarpa, A-Ephedra intermedia, А-Lonicera persica, A-Berberis turcomanica и др.), а также ^сформированным сообществам из многочисленных петрофильных (скальных) растений (A-Cousinia centauroides, +С. leptocephala, A-Zosimia absinthifolia, A-Bongardia chrysogonum, A-Allium Eugenii, A-Stachys turcomanica, A-Parietariajudaica, + Aneurolepidium kopetdaghense и др.); а в затененных местах по трещинам скал ютятся ArPeucedanum turcomanicum, A-Scrophularia turcomanica, + Valeriana sisymbriifolia, A-Orthurus heterocarpus и др. Арчевники Большого Балхана редкостойны, испорчены порубками и выпасом скота; травяной покров в них имеет преимущественно степной облик. Только в вершинах ущелий с затененными склонами, иногда увлажняемых родниковыми водами, встречаются тенелюбивые мезофиты и даже папоротники (A-Cheilanthespersica, Asplenium ruta-muraria и изредка A-Adianthum capillus-veneris).
Литература
E. Г. Бобров (1926, 1931); Е. Г. Черняковекая (1927); Е. А. Малюгин (1931);
И. А. Ливчевский (1935а); В. Н. Никитина (1954).
84 к — кавказские
На карте горные дерновиннозлаковые степи показаны по восточной окраине Большого Кавказа (в пределах Дагестанской АССР и Азербайджанской ССР) и на южнокавказском (Армянском) вулканическом нагорье (в пределах Армянской ССР и соседней южной части Грузинской ССР).
В предгорьях восточной окраины Большого Кавказа распространены сильно нарушенные выпасом степи с господством типчака {Festuca sulcata), ковыля-тырсы {Stipa capillata) и бородача {Andropogon ischaemum)', на южных склонах много полукустарничковой Artemisia taurica. По северным склонам заросли кустарников типа шибляка: неклена {Acer campestre), боярышника {Crataegus), кизила {Cornus mas), городовины {Viburnum lantana), бирючины {Ligustrum vulgare)', по южным, преимущественно каменистым склонам: держи-дерево {A-Paliurus spina-Christi), A-Rhamnus Pallasii,1 Spiraea crenata, а также сообщества нагорных ксерофитов. Урожайность 5—7 ц/га. Эти степи используются для выпаса, а частью распаханы.
Наибольшие площади горные типчаково-ковыльные и типчаковые степи занимают на вулканических нагорьях Армении, преимущественно к западу и югу от оз. Севан и на Ленинаканском плато. В Армении степи приурочены к среднегорным высотам (1000—2200 м над у. м.), где образуют широкую полосу; в южной части страны по южным склонам степи подымаются до высоты 2600—2700 м над у. м.
В составе ковыльных и типчаково-ковыльных степей Южного Закавказья по более или менее равнинным местам с мелкоземистыми почвами из ковылей чаще всего преобладает Stipa stenophylla, а местами -|-5. роп-tica и. S. Joannis. В условиях более пересеченного рельефа господствуют Stipa Lessingiana и S. pulcherrima. Из других злаков примешаны в большем или меньшем количестве Festuca sulcata, Koeleria gracilis, A-К. caucasica, Phleum phleoides, Poa bulbosa, Agropyrum imbricatum, Bromus riparius, A~B. variegatus (повыше в горах). Постоянно встречается осока {Carex humilis). Типчак часто образует самостоятельные группировки без ковылей или с небольшим их количеством, вероятно возникшие в результате усиленного выпаса. Изредка встречаются участки житняковых степей с преобладанием Agropyrum imbricatum. Из разнотравья более обычны Filipendula hexapetala, Potentilla recta, А-Onobrychis transcaucasica, Trifolium ambiguum, Euphorbia Seguieriana, Falcaria sioides, Peucedanum ruthenicum, А-Veronica orientalis, A-Linaria grandiflora, Achillea setacea л др. Как видно из этого, большинство видов закавказских типчаковоковыльных степей являются общими со степями равнин Причерноморья и даже Казахстана.
На каменистых склонах в большом количестве развиваются ковыли {Stipa capillata, +5. holosericea, S. Hohenackeriana), а из ксерофильных кустарничков типа «нагорных ксерофитов» — трагакантовые астрагалы {А-Astragalus aureus, А~А. uraniolimneus и др.). Участки с обилием последних носят название трагакантовых степей, которые часто возникают в результате усиленного выпаса. На скелетных почвах в степном поясе развиты также тимьянники (с А-Thymus Kotschyanus, +Г. eriophorus, A-Scutellaria orientalis s. 1. и др.) и трагакантники.1 2
1 Rhamnus Pallasii заходит в Ставропольскую возвышенность.
2 В Армении имеются и бородачевые степи, занимающие небольшие площади, преимущественно в Ленинаканском и Артикском районах; отдельно на карте они не показаны.
Значительная часть горных степей в Армении распахана г щие участки используются как сенокосы (на более ровных ^^ествУю выпасы. ®стах) jj ^ак
Литература
А. К. Магакьян (1941); А. Л. Тахтаджян (1941); Е В пт»жл
А. А. Гроссгейм (1943). ' h ' Шифферс (194б
’ 1®53)
XIII. ПОЛУКУСТАРНИЧКОВЫЕ ПОЛЫННЫЕ И СОЛЯНКОВЫЕ ПУСТЫНИ
Л. Н. РОДИН и Н. И. РУБЦОВ 1
Общий обзор
Полукустарничковые пустыни1 2 (полынные и солянковые) включают сообщества, состоящие из мезотермных (и микротермных в высокогорьях) и ксерофильных многолетних растений, представленных преимущественно полукустарничками.
Господствующие виды (эдификаторы, строители сообществ) полукустарничковых пустынь относятся к следующим родам: полынь (Artemisia, главным образом виды подрода Seriphidium), солянка (Salsola, исключительно многолетние виды секций Anchophyllum, Sphragidanthus, Caroxylon и Ве1ап1Ъега),3ежовник (Anabasis), саксаульник (Arthrophytum) и близкий к нему гамада (Hammada), кокпек (Atriplex), терескен (Eurotia), поташник (Kalidium), сарсазан (На1оспетит\ один вид Н. strobilaceum). Представители этих родов широко распространены в пределах пустынной области и создают сообщества, занимающие обширные пространства. Заметно меньшее значение имеют сообщества, где эдификаторами выступают тасбиюргун (Nanophyton erinaceum), карабарак (Halostachys Belangeriana), полукустарниковые шведки (Suaeda), кзылкокпек (Limonium suffruticosum), ромашник (Pyrethrum achilleifolium), прутняк (Kochia prostrata), пижма (Tanacetum xylorrhizum) и некоторые другие.
Вышеперечисленные роды распространены по всей пустынной области, но в различных районах представлены разными видами, отличающимися
1 Л. Е. Родиным составлены: Общий обзор и разделы: Злаково-полынные пустыни в сочетании с дерновиннозлаковыми степями, полынными и солянковыми пустынями (южная полупустыня) (85); Злаково-полынные пустыни на песках [из Artemisia astra-chanica или A. Massagetovii (=А. terrae-albae ssp. Massagetovii), A. arenaria, Agropyrum sibiricum и др.] в сочетании по понижениям рельефа с солянковыми и луговыми группировками и слабозаросшими бугристыми и барханными песками (86); Полынные пустыни (87); Полынные пустыни в сочетании с солянковыми пустынями (89); Эфемерово-полынные пустыни (90); Эфемерово-полынные пустыни в сочетании с солянковыми (91); Солянковые пустыни (из видов Salsola, Anabasis, Nanophyton, Reaumuria soongorica и др.) обычно в сочетании с полынными пустынями (92); Сочносолянковые пустыни [из Halocnemum strobilaceum, Halostachys caspica, (=H. Belangeriana), Kali-dium foliatum] и лишенные растительности солончаки (93); Однолетнесолянковая растительность (из однолетних видов Salsola, Suaeda, Salicornia europaea и др.) в сочетании с полукустарничковосолянковыми, галофитнолуговыми й другими сообществами (94).
Н. И. Рубцовым составлены разделы: Горные и предгорные полынные пустыни (88); Высокогорные терескеновые и полынно-терескеновые пустыни (из Eurotia сега-toides и Artemisia Skorniakowii) (95).
2 В легенде «Геоботанической карты» вместо «полукустарничковые» напечатано «полукустарниковые».
• М. М. Ильин. Ghenopodiaceae — Маревые. В кн.: Флора СССР, т. 6, 1936.
по своей характеристике. Географическое размещение их подчинено определенным закономерностям.
Так, например, эдификаторные виды полыней последовательно сменяют друг друга как в направлении с запада на восток, так и с севера на юг. Черная полынь (Artemisia pauciflora), образующая сообщества в пустынях Европейской части СССР, в Казахстане сменяется полынью майкара (Artemisia pauciflora ssp. majkara). Полыни из сборного северотуранского вида Artemisia terrae-albae s. 1. (Л. terrae-albae s. str., A. semiarida, A. ter-rae-albae ssp. Massagetovii, A. kemrudica), распространенные на севере пустынной области, при движении на юг сменяются серией видов южно-туранской полыни Artemisia herba-alba s. 1. (Л. diffusa, A. arenicola, A. bad-ghysi, A. dumosa).
Такое же явление наблюдается и среди видов боялыча (Salsola arbus-culas. 1.): в Центральном Казахстане распространен боялыч Salsola lari-cifolia, южнее, на Устюрте, он сменяется боялычем Salsola ustjurtensisA Тетыр (Salsola gemmascens), произрастающий в пределах Туранской низменности, на востоке сменяется близким видом Salsola passerina, который характерен для пустынь Монголии; в Закавказье же тетыр заменяется другим близким видом — Salsola nodulosa.
Последовательные смены видов — эдификаторов полынных пустынь наблюдаются при переходе от равнин к горам. Так, предгорья характеризуются сообществами полыней сублессинговой и туранской (Artemisia sublessingiana и Л. turanica), в среднегорных условиях они сменяются сообществами тяныпанской (Artemisia tianschanica), а в высокогорьях распространены группировки полыней Скорнякова и Лемана (Л. Scornia-kowii и Л. Lehmanniana).
Некоторые виды, широко распространенные в пределах пустынной области и даже заходящие далеко в степи, имеют очень суженный экологический ареал. Ярким примером в этом отношении служит сарсазан (Halocnemum strobilaceum), сообщества которого связаны с солончаками грунтового увлажнения, почти не выходя за пределы этих местообитаний.
На границах своего распространения, при контакте с другими типами растительности, при движении от равнин к горам и т. п. полукустарничковые пустыни образуют переходные сообщества к другим типам растительности: к лугам и тугаям (в речных долинах, прорезающих пустынную область), к степям (как в равнинных условиях по северному пределу пустынь, так и в горах на соответствующих высотных рубежах), к субтропическим степям, к высокогорной подушковидной растительности и, особенно часто, к саксаульникам и кустарниковым пустыням в пределах самой пустынной области.
Пустынная область Советского Союза занимает обширную территорию, начиная с западной части Прикаспийской низменности, где граничит со степной областью по Ергеням. Перейдя Волжско-Уральское междуречье, пустыни широким потоком вливаются в Среднюю Азию, где покрывают п-в Мангышлак и плато Устюрт, южные части Тургайской столовой страны и Казахского мелкосопочника, Бетпак-Далу, всю Туранскую низменность, на юге доходя до горных поднятий Копет-Дага, Памиро-Алая и Тянь-Шаня и на востоке Джунгарского Алатау и Тарбагатая, проникая вглубь хребтов по долинам крупных рек, и через грабены Джунгарских ворот,
1 Вид этот был намечен Е. П. Коровиным (Коровин и Гранитов, 1949), однако, В. П. Бочанцев (Chenopodiaceae — Маревые. В кн.: Флора Узбекистана, т. 2, 1953, Ташкент) не отделяет его от Salsola laricifolia и считает,что отмеченные Е. П. Коровиным отличия зависят от экологических условий.
долины Или и Кызылсу, а также через высокогорья Памира сливаясь с пустынями Центральной Азии.
Кроме того, пустыни Закавказья, где они занимают Кура-Араксин-скую низменность и долины Куры и Аракса, соединяются с пустынями Малой и Передней Азии. С последними наши пустыни смыкаются также в южной части Прикаспийской низменности и на междуречьях Теджена— Мургаба—Аму-Дарьи, а также по долине Аму-Дарьи.
Таким образом, пустыни Советского Союза составляют северную часть пустынной области Евразии и Африки.
Находясь в глубине Евразиатского материка, пустыни нашей страны отличаются резкой континентальностью климата, исключительной засушливостью вегетационного периода, определяемой ничтожным количеством осадков (в среднем 100—150 мм) и высокими температурами, достигающими в южных районах 36—40° и даже 45°. В северной части пустынной области осадки более или менее равномерно распределены по сезонам года, в то время как в южной части характерно смещение максимума осадков на период с конца осени до весны. На протяжении 5— 6 летних месяцев (V—X) дождей почти не бывает. Земледелие в пределах пустынной области может успешно развиваться только при искусственном орошении. В редкие годы с обильными осенне-зимне-весенними осадками в ограниченных условиях возможны бесполивные посевы скороспелых сортов зерновых культур (так называемая необеспеченная богара).
Почвы пустынь относятвя к типам бурых, серобурых и сероземов, в схематическом приближении сменяющихся в указанной последовательности с севера на юг. Все они отличаются низким содержанием гумуса, значительной карбонатностыо, гипсоносностыо и частым проявлением солонцеватости и солончаковатости. В депрессиях, на морских и озерных побережьях, а также в долинах и дельтах рек весьма обычны различные категории солончаков. По всей территории пустынь'Союза встречаются песчаные массивы, среди которых выделяются самые крупные песчаные пустыни Евразии — Каракумы и Кызылкумы.
Исключительная аридность условий существования растительности вызвала выработку растениями пустыни ряда морфологических и физиологических приспособлений.
Так, например, защита от испарения достигается уменьшением размеров листьев и сокращением общей листовой поверхности, что характерно не только для полукустарничков, но и для представителей других жизненных форм. В крайних случаях листовая пластинка редуцирована до ничтожной щетинки или шипика, а функция фотосинтеза перекладывается на зеленые веточки, например у саксаулов (Halozylon persicum и И. aphyl-lum), кандымов (видов Calligonum), эфедры (Ephedra strobilacea, Е. lomato-lepis) и многих других.
Широко распространены различные «анатомические» приспособления для сохранения влаги: опушенность, толстая кутикула, восковой налет, уменьшение размеров и числа устьиц, блестящая поверхность, отражающая солнечные лучи и т. п. Эта же цель достигается сбрасыванием части листьев с наступлением засушливого периода, например у полыней (видов Artemisia), у многих астрагалов (видов Astragalus), у некоторых полукустарничковых солянок (видов Salsola) и др.
У многих растений развиваются мощные корни: то уходящие вглубь на несколько метров до грунтовой воды или горизонтов с капиллярно-подвешенной влагой, то распространяющиеся неглубоко от поверхности, чтобы перехватить влагу осадков. Как правило, подземная часть пустын-
18 Заказ 551
ных полукустарничков всегда по мощности развития и массе значительно превышает их надземную часть.
Суровые условия засоленных почв привели к выработке у многих растений особых физиологических свойств, позволяющих рассматривать эти растения как особую разновидность ксерофитов, называемых галофитами. Ряд видов этой группы обладает высоким осмотическим давлением клеточного сока вследствие накапливания в организме большого количества легкорастворимых солей (например сарсазан — Halocnemum strobilaceum). Другие же галофиты, наоборот, с помощью особых железок выводят соли из организма (например кзылкокпек — Limonium suffrutico-sum и др.). Подробнее об этом говорится ниже, при описании сочносолянковых пустынь (93).
Подвижность субстрата, развевание коренных пород, перенос песка ветром вызвали также ряд специальных приспособлений: анемокарпия, свойственная большому числу видов; образование своеобразных защитных чехликов на корнях у травянистых растений и пробковых покровов — у полукустарничков (для предохранения корней от иссушения при выдувании песка).
Характерной для пустынь жизненной формой, как указано выше, являются ксерофильные полукустарнички, основные роды и виды которых уже были названы.
В жизненной форме полукустарничков наиболее ярко отражены разнообразные типы приспособления к суровым условиям существования на сухих, засоленных и бедных гумусом почвйх и на подвижных субстратах пустыни. Однако наряду с полукустарничками, выступающими в качестве эдификаторов, широко распространены и типичны для сообществ пустынь растения и других жизненных форм.
Длительновегетирующие многолетние травянистые растения представлены сравнительно ограниченным флористическим набором, причем более часты они в северной части пустынной области. Таковы виды ковылей (Stipa capillata, S. sareptana, Lasiagrostis caragana, Stipa Szowitsiana, S. Hochenackeriana),1 некоторые виды астрагалов (Astragalus), козлобородников (Tragopogon), козельцов (Scorzonera), травянистые виды колю-челистников (Лcanthophyllum), руты (Haplophyllum), кермеков (Limonium), зопников (Phlomis) и других родов. Вегетация этих растений начинается поздней весной, а плодоносят они летом и даже ранней осенью.
Почти все эти растения имеют более или менее ярко выраженную ксе-роморфную структуру — мелкие и жесткие листья, часто сведенные к колючкам, опушение и другие признаки ксерофитов.
Совсем иначе ведут себя коротковегетирующие травянистые многолетние растения — эфемероиды. Весь цикл развития они проходят в течение весны и к началу лета уже успевают принести плоды и семена. Эфемероиды используют короткий, влажный весенний период, когда еще не наступили знойные дни. Нередко можно наблюдать начало вегетации этих растений еще с осени, если дожди совпадают с теплой погодой. На зиму они замирают (но не гибнут), а весной продолжают развитие. Таким образом, у ряда эфемероидов общий период вегетации оказывается растянутым на 6—7 месяцев, так что наименование этих растений «коротковегетирующими» в значительной степени условно. Такая осенне-зимне-весенняя вегетация особенно часто проявляется в южной части пустынной области.
1 Последние два вида Stipa идут далеко на юг, особенно по щебнистым и песчаным субстратам.
В числе представителей эфемероидов выделяется значительное число луковичных растений из семейства лилейных. Это виды луков {Allium), гусиных луков {Gagea), тюльпанов {Tulipa), а также ряд видов с утолщенными корнями (стержнекорневые, клубненосные, корневищные), как, например, ревень {Rheum turkestanicum), виды ширяшей {Eremurus), касатиков {Iris), спаржи {Asparagus), несколько видов ферулы {Ferula assa-foetida, F. schair), шуманиия {Schumannia Karelinii) и др.
Эфемероиды, вегетирующие во влажное и теплое (но не жаркое) время, в подавляющем большинстве являются мезофитами, весь облик которых свидетельствует об этой их биологической природе: у них крупные листья, большая испаряющая поверхность, отсутствуют анатомические приспособления для защиты от засухи, они имеют крупные и яркие цветки, привлекающие насекомых для опыления.
К этой же биологической группе мезофитов короткого периода вегетации, но однолетних, принадлежат эфемеры. Многие из них всходят из семян весной и в течение 1.5—2 месяцев успевают завершить весь жизненный цикл. Они живут за счет влаги осадков, впитавшейся в самые поверхностные горизонты (корневые системы эфемеров развиты слабо), и едва в первые знойные дни почва просохнет, как эфемеры высыхают, ломаются ветром, крошатся и частично выносятся с мест их обитания.
Оговорка насчет условности «короткой вегетации» эфемероидов распространяется и на эфемеры: многие из них дают всходы с осени. Но есть виды, в полном объеме отвечающие понятию эфемеров, как, например, репяшок {Ceratocephalus orthoceras), действительно укладывающийся в 3—5 недель развития от всходов из семян до плодоношения.
Количество видов эфемеров очень велико; по всей пустынной области оно исчисляется многими десятками, даже сотнями. Особенно богаты эфемерами сообщества южных частей пустынной области, что в ряде случаев отмечено в самих названиях выделяемых на карте единиц растительного покрова: эфемерово-полынные, эфемерово-солянковые пустыни.1 Наиболее богато представлены семейства крестоцветных (виды Malcolmia, Lepidium, Euclidium, Tauscheria, Leptaleum, Alyssum, Meniocus, Tetracme и др.), злаков (виды Eremopyrum, Bromus, Aegilops, Schismus, Cutandia), маковых (виды Papaver, Roemeria, Glaucium, Hypecoum)', имеются также представители многих других семейств (виды Consolida, Ceratocephalus — из лютиковых, Ziziphora, Lallemantia — из губоцветных, Koelpinia, Crepis, Matricaria — из сложноцветных, Astragalus, Trigonella — из бобовых, АгпеЫа, Lappula — из бурачниковых, Plantago — из подорожниковых, Erodium — из гераниевых, Veronica — из норичниковых, Androsace — из первоцветных, Euphorbia — из молочайных и др.).
Количество видов эфемеров в одном сообществе нередко достигает 40—50, и в годы с обильными зимне-весенними осадками они образуют густой смыкающийся травостой под пологом обычно разреженных эдифи-каторных полукустарничков. В «сухие» годы эфемеры развиваются слабо и нередко погибают на ранних стадиях, не успевая принести семян.
Хорошо развившиеся эфемеры значительно повышают пастбищную ценность пустынной растительности, и в особо благоприятные годы бывает возможна даже заготовка сена.
Чрезвычайно характерна для пустынь меньшая по видовому разнообразию, но более постоянная по участию в сложении сообществ группа однолетников с длительным периодом вегетации. Это так называемые летне-осенние однолетники. Среди них больше всего представителей се-
1 В этих названиях слово «эфемерово» — трактуется широко, включая и эфемеры и эфемероиды.
мейства маревых (виды Salsola, Halimocnemis, Halotis, Gamanthus, Suaeda, Halochar is, Halogeton и др.).
Всходы этих однолетников появляются также ранней весной, и растения используют это благоприятное время для цветения (большинство маревых цветет весной). В то же время они успевают укорениться глубже, нежели это свойственно эфемерам, и, будучи ксерофитами по своей биологической природе (а часто и суккулентными ксерофитами, то есть имеющими сочные стебли и листья), они медленно развиваются в течение всего лета, а к концу его и в начале осени особенно сильно разрастаются и обильно плодоносят. Весьма вероятно, что летне-осенние однолетники используют конденсационную влагу, образование которой легче всего осуществляется к осени, когда особенно сильно сказывается разница дневных и ночных температур.
В этой группе летне-осенних однолетников наибольшим разнообразием видов отличается род Salsola, далее Halimocnemis, Halotis, Gamanthus, Halocharis и др. Виды этих родов почти все суккуленты. Однако существует значительное число видов с ксероморфной структурой. Таковы виды родов Corispermum, Ceratocarpus, Echinopsilon, Kochia, некоторые виды Salsola (S. Paulsenii, S. ruthenica).
Необходимо отметить, что в построении сообществ пустынь значительное участие принимают также споровые растения: мхи, лишайники, водоросли, грибы.
Почти все они по своей биологической природе приближаются к психрофитам, так как наиболее активное развитие их происходит в осенне-зимне-весенний период, который характеризуется прохладными температурами и значительной влажностью. Испарение влаги из поверхностных горизонтов почвы останавливает их развитие, и весь летний бездождный период они находятся в состоянии анабиоза. Однако с каждым дождем, хоть на некоторое время, их жизнедеятельность возобновляется.
Из мхов наиболее известен карахарсанг (туркм.)— Tortula desertorum, обычно встречающийся под защитой кустов в различных сообществах, но в некоторых условиях образующий сплошное покрытие поверхности почвы. В качестве реже встречающихся в пустынях могут быть названы еще виды Bryum, Grimmia, Ceratodon, Pterygoneurum, Phascum, Riccia.
Литлайники распространены гораздо более широко и представлены значительным числом видов. Их можно найти в небольших количествах на поверхности почвы в большинстве сообществ полукустарничковых пустынь. Некоторые виды поселяются на отмерших стволах и ветвях кустарников. Живущие на почве представлены двумя группами: прикрепленные к субстрату (виды Diploschistes, Acarospora, Psora, Collema, Leeidea, Caloplaca и др.) и неприкрепленные, «кочующие» виды (Parmelia, Cetraria, Aspicilia и др.).
В некоторых особых условиях лишайники столь обильно разрастаются, что сплошным и мощным слоем покрывают почву. Однако такие участки сравнительно редки, они встречаются пятнами среди различных сообществ я обычно лишены высших растений.
Водоросли, в особенности синезеленые, очень широко распространены в сообществах полукустарничковых пустынь, обитая не только на поверхности, но и проникая в почву на значительную глубину (до 1—1.5 м). В некоторых условиях они приобретают даже эдификаторное значение, например на такырах (опис. 102).
В северных районах пустынь особенно широко расселяется водоросль «кна» — Stratonostoc commune, заходящая и в степную область. В южных районах очень широко распространена синезеленая водоросль Microco-
leus vaginatus и многочисленные виды Phormidium. Реже встречаются Amorphonostoc paludosum, Л. punctijorme, Aphanothece saxicola f. nidulans, Lyngbia Margaretheana и другие виды этого рода, виды Plectonema, Апа-baena, Microcystis и очень многие другие.
Из зеленых водорослей встречаются виды Chlorella, Palmella, Protococcus, Chlorococcum, Pleurococcus и др. Отмечены также и некоторые диатомовые водоросли (Hantzschia amphioxys}.
Почвенные водоросли подчиняются характерной закономерности: их видовое разнообразие увеличивается одновременно с возрастанием обилия растительного покрова и плодородия почв (к сожалению, общая масса водорослей в различных типах почв еще до сих пор нигде количественно не определена). Но даже наиболее бедные высшими растениями солончаки содержат более 30 видов водорослей: синезеленых — 20 видов, зеленых — 8 видов, диатомовых — 2 и желтозеленых — 1 вид (Мельникова, 1953).
Несмотря на слабую изученность роли водорослей в сообществах пустынь, можно предполагать, что она весьма значительна. Водоросли во многих случаях являются первыми организмами, накапливающими гумус и, таким образом, способствуют созданию более благоприятных условий для существования высших растений.
Напочвенные грибы очень редки, по сравнению с вышерассмотренными низшими растениями. Так, шляпочные грибы попадаются редко и лишь одиночными особями, например Battarraea phalloides.
Микроскопические почвенные грибы в пустынных сообществах еще почти совершенно не изучены. Можно назвать следующие роды, представители которых были обнаружены: Stemphylium, Macrosporium, Alternaria, Aspergillus, Cladosporium, Penicillium. Как выяснено наблюдениями M. А. Литвинова (Базилевич, Голлербах и др., 1953), поселения микроскопических грибов приурочены главным образом к ризосфере цветковых растений.
Растительный покров полукустарничковых пустынь отличается значительной неоднородностью, пятнистостью, или, как это установилось называть, комплексностью.
На северной окраине пустынной области в состав комплексов оказываются включенными степные и луговые сообщества, нередко с заметным участием степных кустарников, с почвами, соответствующими этим типам растительности. Они резко отличаются от господствующих сообществ с почвами пустынного типа.
Эти комплексы, являющиеся, как бы переходными от чисто степных к настоящим пустынным типам растительного покрова, некоторые исследователи выделяли под именем полупустыни (Димо и Келлер, 1907) как особой подзоны. На нашей карте территории с господством степных сообществ в комплексе с пустынными отнесены к степям {полынно-дерновиннозлаковые степи в сочетании с пустынными сообществами (северная полупустыня) — опис. 78]. Территории с господством пустынных сообществ в комплексе со степными отйесены к пустыням [злаково-полынные пустыни в сочетании с дерновиннозлаковыми степями, полынными и солянковыми пустынями (южная полупустыня) — опис. 85].
В типичных районах пустынной области комплексный растительный покров слагается сообществами пустынного типа.
Вопросу о происхождении комплексности растительного покрова степей и пустынь посвящено значительное число работ, как геоботанических, так и почвенных. Недостатком большинства исследований было стремление объяснить происхождение комплексности одним каким-либо
фактором (рельеф и микрорельеф, влияние животных, глубина грунтовых вод, предшествующая история формирования территории, химические свойства почв или субстратов и т. п.). Теперь уже совершенно ясно, что причины, обусловливающие комплексность, многообразны, и в пределах даже ограниченной территории одни элементы комплексности произошли под влиянием иных факторов, чем другие элементы. Важно указать, что исследования последних лет, произведенные различными специалистами (ботаниками, геоморфологами и почвоведами), привели их в ряде случаев к признанию ведущей роли самой растительности в формировании многих элементов комплекса с их особой почвой и структурой сообществ (Коро* вин и Шувалов, 1948; Шувалов, 1949; Герасимов, 1951; Иванова и Левина, 1952; Момотов, 1953; Быков, 1954; Родин, 1954).
С явлением комплексности растительного покрова пустынь тесно связана и мозаичность его (см. примечание на стр. 598). причины которой также могут быть весьма различны. Так, например, к явлениям мозаичности следует отнести упомянутое выше поселение Tortula desertorum под кустами полыней или полукустарничковых солянок. Точно так же под кустами некоторых растений нередко наблюдается скопление эфемеров, отсутствующих на участках вне влияния первых. Заброшенные норы грызунов и их подземные галереи (например малого тушканчика — А1-lactaga elater) со временем оседают, и на таких местах обильно разрастаются не только эфемеры, но и многолетние формы, находящие здесь лучшие условия для своего развития. Характерные в пустынях Средней Азии колонии термитов, создающие своеобразные лишенные растительности (хотя всегда ею окружаемые) холмики—«термитники», также вызывают явления мозаичности растительного покрова, так как в результате жизнедеятельности насекомых образуются резко отличные почвы, богатые нитратами (Козлова, 1951). «Ветровая тень», создаваемая под кустами в районах развеваемых субстратов, способствует накоплению мелких частиц в виде бугорков, которые служат местами поселения растений, прежде отсутствовавших в данном сообществе. В некоторых особых условиях длительное накопление и «рост» таких бугорков приводит к образованию не только мозаичного сообщества, но даже к возникновению особых форм рельефа, отражаемых на топографических картах (чо-колаки, томмоки).
Выше уже отмечалось, что выпадение осадков в пустынной области весьма неравномерно по сезонам года, особенно в южных ее районах, что в известной мере объясняет преимущественное обилие здесь эфемеров. Однако чрезвычайно характерна не только эта сезонная, но и многолетняя периодичность (с периодом в 10—30—50 лет). В некоторые годы количество осадков и особо благоприятный температурный режим выходят далеко за пределы среднегодовых величин. Такие годы Отличаются исключительно пышным развитием эфемеров, эфемероидов, да и многолетних растений — эдификаторов пустынных сообществ (полыней, солянок и др.). Причина этого заключается не только в увеличении количества осадков и в благоприятных температурных условиях, нб и в том, что при обильном поверхностном стоке происходит выщелачивание вредных для развития растений солей, а это, в свою очередь, стимулирует особое течение микробиологических процессов в почве, изменяет ход обменных реакций, отражается на интенсивности гумификации растительных остатков и т. п. Все это в конечном итоге приводит к возникновению тех «волн жизни», к всестороннему научению которых призывал В. И. Вернадский.* 1
- - .....
1 В. И. Веряадокп. Очерка геохимии. Изд. 2-е, М.—Л., 1934.
Современный растительный покров пустынной области в целом, и рассматриваемый нами тип полукустарничковых полынных и солянковых пустынь в частности, несут на себе следы многовековой (вернее, исчисляемой тысячелетиями) человеческой культуры, хозяйственйой деятельности человека.
На самых ранних этапах своей истории человек использовал растительный покров как источник топлива, уничтожая пригодные для этой цели отлично горящие полыни и полукустарничковые солянки, что продолжается и по сей день. Прокармливаясь охотой на диких животных, человек сильно сокращал их число и тем самым нарушал естественные биоценозы, создавая в них новые формы отношений между животными и растениями. Выпас домашнего скота на следующем этапе истории внес еще более резкие нарушения в существование и развитие растительного покрова. Многие современные «эфемеровые пастбища» в прошлом несомненно представляли собою полукустарничковые эфемерово-полынные пустыни, в которых основной эдификатор (полынь) был полностью уничтожен человеком. С возникновением умения сооружать колодцы значительно возросла потребность в строительном материале, источником которого служили другие растительные сообщества (колодцы оплетаются внутри стволами песчаной акации — видов Ammodendron, не поддающейся гниению ине портящей вкусовых качеств воды). Вокруг колодцев стали возникать «тырла» и «толоки» — места с сильно или даже долностью стравленным покровом. То же самое происходило и вокруг кочевых или оседлых стоянок и поселений. На этих участках в естественном растительном покрове преимущественное развитие получили растения, не поедаемые скотом я не используемые человеком (например адраспан—Peganum harmala, каперцы — Capparis spinosa и др.), создались новые сочетания растений, новые растительные сообщества. Пасторальная дигрессия вообще вела к усилению пустынных элементов в растительном покрове (Суворов, 1949).
Мощность воздействия на весь природный комплекс, на биогеоценозы, -еще более возросла, когда человек приступил к орошаемому земледелию. На огромных площадях природная растительность была полностью уничтожена, и были созданы посевы растений, зачастую принесенных даже с других континентов. На месте многих типов растительности возникли особые новые ландшафты — оазисы. И хотя оазисы создавались в первую очередь в речных долинах и дельтах, однако искусственные каналы понесли воду и на территории, занятые полукустарничковыми пустынями.
Изменение растительного покрова в этот период происходило под воздействием вышеперечисленных факторов и под прямым и косвенным влиянием земледелия. В местах сброса оросительных вод, на участках, где грунтовые воды поднимались и т. п., возникли новые сообщества: это были и заросли гидрофитов (тростник, камыши) и солончаки с резко га-лофильными формами растений (сарсазан, карабарак и т. п.).
Упадок земледелия, происходивший порою на территориях, принадлежавших крупным государствам,1 вызывал новую перестройку растительного покрова. Огромные площади водорослевых сообществ на такырах, заросли кеурека, биюргуна, однолетних солянок и т. д., по своему строю и флористическому составу не похожие на естественные их сообщества (хотя, конечно, тоже уже изменившиеся за историческое время), покрывают территории земель древнего орошения (Древний Хорезм, Куня-Дарьинская равнина, Мешед-Мессерианская низменность и др.).
1 С. П. Толстов. Древвий Хорезм. М., 1948; По следам древне-хорезмийской цивилизации. М.—Л., 1948.
Вопрос о влиянии хозяйственной деятельности человека на растительный покров пустынной области не может быть освещен здесь достаточно подробно, и вообще он еще ждет своего исследователя. Мы считали нужным, хотя бы приблизительно, показать пути и масштабы этого влияния. Некоторые конкретные примеры будут приведены в последующем изложении.
В соответствии с видовым составом эдификаторов и согосподствующих видов, а также учитывая основные элементы, входящие в комплексы, распространенные в зональных (плакорных) условиях, полукустарничковые (полынные и солянковые) пустыни, показанные на нашей карте, разделяются на несколько типов.
1. Типичные (настоящие) полукустарничковые пустыни, в которых наиболее полно господствуют растения полукустарничковой жизненной формы, представлены у нас полынными пустынями (87), полынными пустынями в сочетании с солянковыми (89) и солянковыми пустынями (92); последние также обычно сочетаются с полынными пустынями.
В составе названных типов пустынь господствуют (в качестве эдификаторов и соэдификаторов) следующие виды полыней: белая полынь-(Artemisia Lercheana), черная полынь (Л. pauciflora и близкая к пей A. gra-cilescens), северотуранские виды полыней (Artemisia terrae-albae s. str., A. semiarida, A. terrae-albae ssp. Massagetovii, A. kemrudica, A. gurganica) и южнотуранские (Artemisia dumosa, A. badghysi, A. arenicola, A. diffusa,. A. lobulifolia), сублессинговая полынь (Л. sublessingiana), а также полукустарнички из семейства маревых: боялыч (Salsola laricifolia, S. arbus-cula), кеурек (Salsola rigida), тетыр (Salsola gemmascens), биюргуны (Anabasis salsa и Л. ramosissima), тасбиюргун (Nanophyton erinaceum), терескен (Eurotia ceratoides), прутняк (Kochia prostrata), кокпек (Atriplex сапа) и некоторые другие.
Полынные пустыни разделяются в пределах своего ареала. Так, на нашей карте выделены прикаспийско-казахстанские полынные пустыни, слагаемые белой и черной полынями (Artemisia Lercheana и Л. pauciflora), северотуранские (из форм Л. terrae-albae s. 1.) и южнотуранские (из форм Л. herba-alba s. I.).1
Полынные пустыни в сочетании с солянковыми также представлены различными географическими вариантами: прикаспийско-казахстанские (из Artemisia Lercheana, A. pauciflora, Anabasis salsa, Atriplex сапа), восточноказахстанские (из Artemisia sublessingiana, A. pauciflora ssp. maikara, Salsola laricifolia), северотуранские (из форм Artemisia terrae-albae s. 1., Salsola laricifolia, Anabasis salsa) и южнотуранские (из форм Artemisia herba-alba s. 1., Salsola arbuscula, S. gemmascens).
Солянковые пустыни в сочетании с полынными (из полукустарничковых видов Salsola, видов Anabasis, Nanophyton я др.) на карте на варианты не разделяются.
Перечисленные типы полынных и солянковых пустынь распространены преимущественно в условиях наиболее плакорных местоположений, занимая обширные пространства равнин и плато, огромной дугой опоясывающие с севера Туранскую низменность; в немногих местах они проникают и в ее пределы.
2. Остепненные пустыни характеризуются тем, что в их составе заметное участие принимают степные растения. Участие это выражается в том, что ксерофильные дерновинные злаки и степное разнотравье,
1 Последние в понимании И. М Крашенинникова.
Рис. 69. Прутняково-белополынная пустыня (Artemisia Lercheana-{-Кochia prostrata) на глубоком осолоделом солонце в полосе бурых почв северного Прикаспия (левобережье Нижней Волги). Снимок сделан 20. X. 1949, когда выпал первый снег. (Фот. Л. Е. Родина).
Рис. 70. Солянковая пустыня, преимущественно из Salsola laricifolia (на среднем и заднем плане). Юго-восточная часть пустыни Бетпак-Дала (Казахская ССР), (фот. Ю. Е. Лавренко).
Рис. 71. Полукустарничек Salsola larici folia в юго-восточной части пустыни Вотпак-Дала (Казахская ССР). Под кустом боялыча слова — полукустарничковая Artemisia turanica (Фот. Ю. Е. Лавренко).
Рис. 72. Выходы пород палеозоя, покрытые пустынным загаром, среди солянковой пустыни в юго-восточной части Бетпак-Далы (Казахская ССР). На переднем плане вправо—кусты полукустарничковой солянки Salsola laricifolia.
(Фот. Ю. Е. Лавренко).
которые здесь в виде значительной примеси присутствуют в пустынных сообществах, нередко выступают также в качестве эдификаторов, образуя самостоятельные сообщества, входящие в состав комплексного растительного покрова. Пустыни эти имеют переходный характер между типично пустынными и степными сообществами, занимая почти повсеместно выраженную полосу на границе соответствующих областей (зон). На пашей карте выделены два типа: злаково-полынные пустыни в сочетании с дер-повинпозлаковыми степями, полынными и солянковыми пустынями (85) и злаковополынпыо пустыни на песках (из Artemisia astrachanica или A. Massagetovii, A. arenaria, Agropyrum sibiricum и др.) в сочетании по понижениям рельефа с солянковыми и луговыми группировками и слабо-заросшими барханными и бугристыми песками (86).
Первый тип (85) по видовому составу эдификаторов и географическому распространению представлен несколькими вариантами: злаково-полынные пустыни предкавказские (с основным эдификатором Artemisia taurica), закавказские (из A. Meyeriana), прикаспийско-казахстанские (из Л. Lercheana и A. pauciflora), восточпоказахстапские (из A. sublessingiana и Л. gracilescens) и северотуранские (из северотуранских видов полыней — Artemisia terrae-albae s. str., A. semiarida, Л. kemrudica, A. terrae-albae ssp. Massagetovii).
Совершенно естественно, что эти разновидности отличаются и по характеру слагающих их комплексов, и по видовому составу сопутствующих растений.
3. Эфемеровые (южные) пустыни,1 распространенные в южных районах пустынной области, где в сообществах полукустарничковых пустынь значительное место принадлежит эфемерам и эфемероидам. Это следует объяснить не только климатическими особенностями (смещение осадков на осенне-зимне-весенний период), но и проникновением многих эдификаторов и сопутствующих видов из субтропических степей (полусаванн), с которыми эти пустыни во многих случаях приходят в соприкосновение. Таковы в первую очередь мятлик (Роа bulbosa var. vivipara), осока (Carex pachystylis), виды луков (Allium albanum, A. fibrosum и др.), виды мортуков (Eremopyrum orientate, E. triticeum) и многие другие виды эфемеров и эфемероидов.
Выделенные на карте эфемерово-полынные пустыни (90) и эфемеровополынные пустыни в сочетании с солянковыми (91) представлены несколькими вариантами.
Эфемерово-полынные пустыни (90) разделяются на закавказские (из Artemisia Meyeriana), среднеараксинские (из Artemisia fragrans), северотуранские (из видов Artemisia terrae-albae s. 1.)1 2 и южнотуранские (из видов Artemisia herba-alba s. I.).3
1 Сообщества эфемеровых пустынь (эфемерово-полынных и эфемерово-полынных в сочетании с солянковыми) характеризуются в принятой нами классификации эдифи-каторной ролью полукустарничков (полыней или солянок) с значительным развитием синузии эфемероидов и эфемеров. Эфемеровые пустыни, в понимании Е. П. Коровина (1934), а также глинистые пустыни с эфемерами М. Г. Попова (1925), эфемеровая злаково-осоковая формация и эфемеровая растительность М. В. Культиасова (1923,. 1927) — все это сообщества с господством Carex pachystylis и Роа bulbosa var. vivipara, сочетающихся в равных или иных соотношениях и образующих вместе с обильными, эфемерами и эфемероидами почти сплошное задернение почвы. На нашей карте эти. сообщества отнесены к субтропическим степям (полусаваннам) и частью к эфемеровополынным или эфемерово-солянковым пустыням. К этим же типам отнесены эфемеровосолянковые пустыни Е. П. Коровина (1934).
2 В группу северотуранских видов входят Artemisia semiarida, A. kemrudica,. A. terrae-albae s. str., A. terrae-albae ssp. Massagetovii, A. gurganica.
3 В группу южнотуранских видов входят Artemisia badghysi, A. dumosa, A. diffusa, A. arenicola.
Эфемерово-полынные пустыни в сочетании с солянковыми (91) разделяются на следующие варианты: закавказские (из Artemisia Meyeriana, Salsola dendroides, S. nodulosa, S. ericoides), среднеараксинские (из Artemisia fragrans, Salsola nodulosa, S. ericoides) и южнотуранские (из Artemisia herba-alba s. 1. и Salsola gemmascens).
4. Горные и предгорные полынные пустыни (88) в качестве сопутствующих видов включают элементы степей, ксерофитных кустарников (шибляка) и субтропических степей, а также сочетаются в условиях горного пересеченного рельефа с участками соответствующих сообществ. На карте различаются следующие варианты {в зависимости от видовой принадлежности эдификаторных полыней): tu — из Artemisia turanica в сочетании с ксерофитными кустарниками и t-из Artemisia tenuisecta в сочетании со степями (для обоих вариантов, приуроченных к предгорьям и частью к среднегорным условиям, характерно также развитие эфемерово-эфемероидной синузии, чем они резко отличаются от последующих); к — из A. Korshinskyi в сочетании с нагорными ксерофитами и степями и 1 — из Л. Lehmanniana (последние два варианта свойственны собственно горам и частично высокогорьям).
Горные и предгорные полынные пустыни распространены преимущественно в южных частях гор Средней Азии (в Западном Тянь-Шане и на Алае).
5. Высокогорные пустыни занимают особое место. Это терескеновые и полынно-терескеновые пустыни (из Eurotia ceratoides и Artemisia Skorniakowii) (95), распространенные главным образом на Памире. Характерно участие в их составе криофильных подушечных растений {Acantholimon diapensioides и др.). Основные эдификаторы также относятся к биологической группе криоксерофитов. Таким образом, этот тип пустынь можно рассматривать как криофилизованные пустыни.
6. Сочносолянковые пустыни (из Halocnemum strobi-laceum, Halostachys Belangeriana, Kalidium foliatum) (93) слагаются видами очень узкой экологической специализации; все они — ярко выраженные галофиты.
Помимо перечисленных и выделенных на карте типов полукустарничковых пустынь и их вариантов, есть еще некоторые типы, на карте не показанные. В речных долинах встречаются солянковые пустыни с некоторым участием галофитнолуговых растений (Aeluropus litoralis, Karelinia caspica и др.). Там же, на контакте с тугаями, в полынных и солянковых пустынях наблюдаются элементы тугайной растительности (виды Tamarix, Halimodendron halodendron, Lycium turcomanicum и др.).
В предгорьях (особенно на Копет-Даге, Большом Балхане и на хребтах системы Алая) нередко можно встретить сообщества, переходные от пу-стынь к колючетравникам и к нагорным ксерофитам. Особенно же широко распространены сообщества переходного характера между полынными и солянковыми пустынями, с одной стороны, и саксаульниками и кустарниковыми пустынями, с другой.
Все такого рода переходные сообщества, хотя они не занимают картируемых в принятом масштабе площадей, имеют большое значение для познания истории развития растительного покрова, для понимания структуры сообществ, условий формирования комплексов, изменений растительности под влиянием хозяйственной деятельности человека и т. п.
В легенде нашей карты имеется еще один тип, помещенный в общую с выше рассмотренными типами группу. Это — однолетнесолянковая растительность (из однолетних видов Salsola, Suaeda, Salicornia еигораеа и др.) в сочетании с полукустарничковосолянковыми,
галофитнолуговыми и другими сообществами (94). Сообщества этого типа на большинстве территорий — вторичные, возникшие под влиянием хозяйственной деятельности человека, и слагаются в основном однолетними растениями, так что, казалось бы, они выпадают из категории полукустарничковых пустынь. Однако условия существования однолетнесолянковой растительности чрезвычайно близки к таковым полукустарничковых пу-•стынь, п часто сообщества тех и других сочетаются вместе. Это дало основание поместить однолетнесолянковую растительность в рассматриваемую группу полукустарничковых пустынь.
Территорию пустынной области принято делить на три подзоны: остеп-ненной пустыни (полупустыни), северной пустыни и южной пустыни.
Подзона остепненной пустыни, или южной полупустыни, расположена непрерывной, сравнительно узкой полосой, пролегающей к югу от степной области (примыкая к ее подзоне пустынных степей или северной полупустыни); в эту подзону входят выделы карты 85 и 86.
Подзона северной пустыни (или подзона наиболее типичных, настоящих пустынь) занимает широкую полосу, южной границей которой является линия от северной оконечности залива Кара-Богаз-Гол через современную дельту Аму-Дарьи, несколько южнее Аральского моря, и далее на южную окраину гор Букантау (в Кызылкумах).
Подзона южных пустынь простирается на юг от этой линии до подножья гор или государственной границы.
Рассматриваемый нами тип полукустарничковых полынных и солянковых пустынь в пределах подзоны северных (настоящих) пустынь образует огромную дугу, опирающуюся на западе на северное побережье Каспийского моря, а на востоке — на предгорья Тянь-Шаня и Джунгарского Алатау. Отдельные контуры полукустарничковых пустынь встречаются средн песчаных пространств Кызылкумов и Каракумов (к подзоне се-серных пустынь относятся выделы 87 кк и 87 стр, 89 кк и 89 стр, 92 и 93 — массивы, приходящиеся на данную подзону).
Сообщества полукустарничков подзоны южных пустынь (эфемеровополынные пустыни и эфемерово-полынные в сочетании с солянковыми .пустынями) не образуют сплошной полосы, будучи приуроченными к немногим участкам местообитаний среди господствующих песчаных массивов или на контакте с ними и предгорьями, а также с оазисами (к этой 'подзоне относятся выделы 87 ютр, 89 ютр, 90, 91, 93 и 94 — массивы, приходящиеся на данную подзону).
Огромное по площади пространство пустынной области Союза ССР 'неоднородно по флористическому составу растительного покрова, что в некоторой степени было отмечено уже при кратком обзоре выделенных на карте типов. Это относится как к эдификаторам — основным видам, слагающим сообщества, так и к сопутствующим видам. Основные эди-фикаторы — полыни — закономерно сменяются при переходе от одной подзоны к другой. Точно так же ведут себя и многие сопутствующие виды. Однако и те и другие обнаруживают иные закономерности, сменяясь близкими, замещающими видами в направлении с запада на восток, причем в своем распространении они часто не ограничены пределами полосы одного типа или одной подзоны, а характерны для нескольких типов и подзон (так, например, некоторые ковыли — Stipa sareptana, S. capillata, типчак — Festuca sulcata входят в сообщества не только подзоны полупустынь и настоящих пустынь, но и степной области). С учетом такого рода закономерностей представляется возможным разделить пустынную область на ряд провинций.
Прежде всего выделяется Кавказско-Малоазиатская провинция, к которой относятся не только пустыни Закавказья в пределах Советского Союза, но и смежные с ними районы Передней и Малой Азии. Для этой провинции характерны полыни — Artemisia taurica, A. erivanica, A. fragrans, A. Meyeriana; из других полукустарничков здесь распространены Salsola dendroides, S. nodulosa, S. ericoides, Reaumu-ria hypericoides. В составе эфемеровой синузии также имеются свои характерные виды, не выходящие за пределы провинции.
Далее огромное пространство от северного Прикаспия до горных поднятий Тянь-Шаня и Джунгарского Алатау относится кТуранской провинции. На севере она граничит со степной областью, на юге граница ее почти совпадает с северной границей подзоны южных пустынь. Для этой провинции характерно исключительное господство ряда полыней, не выходящих на юг за ее пределы: Artemisia Lercheana, A. pauciflora s. 1. (заходящих на север в пределы степной области), A. semiarida, A. terrae-albae s. str., A. kemrudica, A. Lessingiana и A. sublessingiana. Большим разнообразием отличаются представители семейства маревых — виды Salsola, Anabasis, Nanophyton, Kalidium и другие, ареал многих из которых совпадает с распространением названных полыней. Точш> так же имеется много видов из других семейств с аналогичными ареалами.
Остальная, южная часть пустынной области Союза относится к П е -реднеазиатской провинции, которая простирается и за государственной границей, включая пустынные районы Передней Азии (Ирана, Афганистана и др.). В Переднеазиатской провинции характерно широкое распространение следующих видов полыни: Artemisia badghysi, A. dumosa, A. arenicola, A. lobulifolia, некоторых солянок, ареал которых не выходит за границы провинции. Чрезвычайно типично обильное участие эфемеров и эфемероидов в сложении не только пустынных, но и сообществ других типов.
Наконец, следует указать еще одну провинцию — Центральноазиатскую, в которую входят высокогорные пустыни Памира и Центрального Тянь-Шаня. Основные эдификаторы и сопутствующие виды пустынных сообществ этой провинции по своим флористическим связям едины с пустынями высокогорий Центральной Азии.
Все названные здесь провинции пустынной области СССР, которая представляет собой часть обширной пустынной области Старого Света, не являются однородными. Возможно дальнейшее деление внутри них на отдельные части, отличные от соседних (Прозоровский и Малеев, 1947).
Основное направление хозяйственного использования полукустарничковых полынных и солянковых пустынь — пастбищное хозяйство. Невысокая урожайность поедаемой кормовой массы, резко выраженная сезонность, слабая обеспеченность водопоями (редкая сеть колодцев, обусловленная бедностью близкими питьевыми грунтовыми водами), неустойчивость состояния растительного покрова в отдельные годы (связанная с резкими погодными колебаниями) и другие причины обусловили то, что на протяжении тысячелетий здесь господствовало кочевое хозяйство, полностью находившееся во власти природных стихий. В настоящее время на базе социалистической организации сельского хозяйства с применением современной техники (автомобильного и воздушного транспорта, водоподъемных механизмов, сеноуборочных машин и т. д.) использование природных кормовых ресурсов вылилось в форму отгонного животноводства. Пастбищное использование кормовых ресурсов сочетается с созданием страховых запасов кормов и с полевым кормодобыванием, устранив-
тих или ослабивших зависимость от неблагоприятных свойств природы пустынь.
Земледелие в пределах полукустарничковых пустынь возможно только на искусственном орошении, которое, как было отмечено, сосредоточено но долинам и дельтам рек. Только незначительные площади земель полукустарничковых пустынь, прилегающие к долинам, в прошлом были вовлечены в орошаемое земледелие. С применением современной техники (мощных машин для строительства каналов, электроэнергии, водоподъемных станций, дренажных устройств для борьбы с засоленными грунтовыми водами, плантажных плугов и множества сельскохозяйственных машин, химических средств борьбы с вредными свойствами грунтов, достижений агротехники колхозного производства и т. п.) открываются широкие возможности для включения новых больших площадей пустынь СССР, в том числе и земель полукустарничковых полынных и солянковых пустынь, в орошаемое высокопродуктивное земледелие.
Литература
И. Г. Борщов (1865); В. И. Липский (1891); С. И. Коржинский (1896); А. А. Еленкин (1901); Н. А. Димои Б. А. Келлер (1907); Г. Н. Высоцкий (1915); В. Н. Сукачев (1919); М. Г. Попов (1922, 1925, 1940); Б. А. Келлер (1923а); И. М. Крашенинников (1923,1925); М. В. Культиасов(1923,1926—1927); М. П. Томин(1926); Р.И. Аболин (1929, 1929а, 1929в, 1930); Е. Н. Иванова, И. П. Герасимов, С. С. Неуструев и О. Э. Кнорринг (1930); Е. П. Коровин (1934, 1935); Е. П. Коровин и Д. Н. Кашкаров (1934); Г. Н. Новиков (1936); А. В. Прозоровский (1940а); Е. П. Коровин и Е. Е. Короткова (1946); А. В. Прозоровский и В. П. Малеев (1947); Е. П. Коровин и С. А. Шувалов (1948); Е. П. Коровин и И. И. Гранитов (1949); Н. И. Суворов (1949); С. А. Шувалов (1949); Н. И. Рубцов (1950а, 1952); И. П. Герасимов (1951); А. В. Козлова (1951); Е. Н. Иванова и Ф. Я. Левина (1952); Н. И. Базилевич, М. М. Голлербах и др. (1953); В. В. Мельникова (1953); И. Ф. Момотов (1953); Б. А. Быков (1954); Л. Е. Родин (1954).
В5. Злаково-полынные пустыни в сочетании с дерновиннозлаковыми степями, полынными и солянковыми пустынями
(южная полупустыня)
Злаково-полынные пустыни сравнительно широкой полосой залегают в северной части пустынной области от подножья Ергеней на западе Прикаспийской низменности, через междуречья Волги—Урала и Урала— Эмбы, Тургайскую столово-останцовую равнину, сужаясь в центральном Казахстане под натиском степных формаций Казахского мелкосопочника, снова расширяясь на севере Бетпак-Далы, и после перерыва, образуемого степным Тарбагатайским хребтом, заканчиваются по обе стороны оз. Зайсан. Отдельные массивы злаково-полынных пустынь расположены на склонах Карабахского низкогорья в южной Армении и на Кура-Араксинской низменности.
Климатические особенности этой полосы злаково-полынных пустынь заключаются в общей недостаточности увлажнения, препятствующей развитию сомкнутого травяного (степного) покрова в плакорных условиях, в котором господствующая роль принадлежит уже пустынным полукустарничкам лишь с примесью ксерофильных степных злаков и некоторого количества видов ксерофильного разнотравья.
В связи с ярко выраженным микрорельефом происходит значите/хьное перераспределение влаги (сток поверхностных вод в микропонижения, скопление снега зимой в западинах), которое ведет к выщелачиванию солей в пониженных элементах местности и, наоборот, к подтягиванию солей к поверхности на окружающих их склонах. В понижениях (западины, падины, Лопатины) создаются условия повышенного увлажнения и осу
ществляется возможность развития порою весьма типично выраженных степных сообществ. Однако это только один из моментов сложной эволюции почвенно-растительного покрова, которую претерпевает ландшафт южной полупустыни в целом. Этому вопросу посвящена обширная литература (Димо и Келлер, 1907; Новиков, 1936; Герасимов, 1951; Левина, 1954 и др.).
Литература
Н. А. Димо и Б. А. Келлер (1907); И. В. Ларин (1926а); Е. П. Коровин (1934); Б. А. Келлер (ред.) (1936); Г. Н. Новиков (1936); А. В. Прозоровский (1940а); А. В. Прозоровский и В. П. Малеев (1947); И. П. Герасимов (1951); Е. Н. Иванова и< Ф. Я. Левина (1952); Ф. Я. Левина (1954); С. А. Никитин (1954а).
85 пк — предкавказские (из Artemisia taurica)
Предкавказские злаково-полынные остепненные пустыни отличаются от их более северного варианта (85 кк) не только преобладающим видом полыни (таврическая полынь — Artemisia taurica), но и некоторыми другими компонентами. В составе их редко играют сколько-нибудь существенную роль ковылек (Stipa Lessingiana) и типчак (Festuca sulcata) и гораздо большее значение имеют гребневидный житняк (пырей) (Agropyrum pecti-ntforme) и пустынный житняк (A. desertorum)', заметное участие в травостое принимают также свинорой (Cynodon dactylon), верблюжья колючка (Alhagi pseudalhagi) и солодка (Glycyrrhiza glabra). Отличается этот вариант и строением растительного покрова. Комплексность имеет характер мезокомплекса, связанного не с мелкими неровностями поверхности почвы, а с более крупными элементами рельефа — террасами, ложбинами, западинами. Особенно сказывается это отличие в дагестанской приморской полосе.
Злаково-полынные остепненные пустыни из Artemisia taurica (85 пк)* показаны на карте в следующих местах: на Терско-Кумской равнине, сложенной лёссовидными суглинками, залегающими поверх флювиогляциальных песков и галечников; в дельте Терека (в северной ее части и по южной окраине); на узкой полосе Прикаспийской низменности к югу от Махачкалы до р. Самур.1 В пределах последней полосы выделяется, несколько морских террас, сложенных верхнетретичными известняками, конгломератами, глинами и песками и подымающихся в предгорья Дагестана до высоты около 200 м над у. м. Вдоль берега моря отлагаются* современные осадки Каспия в виде ракушечных песков.
Массив Терско-Кумской равнины с каштановыми и светлокаштановыми почвами почти полностью распахан, и остепненные пустынные формации представляют собой толоки (иногда, правда, обширные) среди полей хлопчатника и пшеницы. На толоках преобладает таврическая полынь (Artemisia taurica), к которой присоединяются житняки (Agropyrum desertorum-и A. pectiniforme), кровельный костер (Bromus tectorum), луковичный мятлик (Роа bulbosa var. vivipara), свинорой (Cynodon dactylon), полынок (Artemisia austriaca) и веничная полынь (A. scoparia), молочай (Euphorbia Seguieriana), тысячелистники (Achillea nobilisst A. setacea), верблюжья колючка (Alhagi pseudalhagi), гармала (Peganum harmala) и другие пустынно-степные ксерофиты.
На приморских террасах Дагестана собственно злаково-полынная* пустынная растительность чередуется с поливными полями и сенокосными*
1 Как показали исследования Е. М. Лавренко, Ф. Я. Левиной и Е. Ф. Шиффере В 1949 г., Artemieia taurica в значительном количестве встречается гораздо севернее-долины Кумы и Маныча, где она произрастает совместно с А. Lercheana и А. paucif lora..
поливными залежами; последние представляют собою преимущественно пырейно-солодковые луговые ценозы из пырея и солодки (Agropyrum repens и Glycyrrhiza glabra) с участием морской полыни (Artemisia maritima s. 1.), мелких солянок (Echinopsilon sedoides и др.), а также свинороя (Су-nodon dactylon), голубой люцерны (Medicago coerulea), обыкновенного мятлика (Роа trivialis), -ползучей лапчатки (Potentilla reptans) и других представителей луговой и лугово-солончаковой флоры. В более пониженных частях террас имеются нераспахиваемые болотистые солончаковатые луга, образованные восточной овсяницей (Festuca orientalis и русским, пыреем (Agropyrum ruthenicum), с участием осок, ситников, тростника и др.
Старые залежи, редко поливаемые и используемые под зимний выпас, теряют луговые элементы и приближаются к злаково-полынным пустыням; на них большое развитие получает Camphorosma Lessingii, к которой присоединяются Artemisia taurica, Agropyrum pectinijorme, Poa bulbosa', на почве много лишайников и мелких пустынно-степных мхов (Tortula ruralis), а также синезеленой водоросли (Stratonostoc commune).
Наиболее высокие участки террас, не поливаемые вообще и используемые под зимний выпас, покрыты злаково-полынной пустыней, образованной таврической полынью с участием морской полыни, полынка, гребневидного житняка (пустынный житняк здесь уже не имеет ценотического значения), типчака, верблюжьей колючки, прутняка (Kochia prostrata), луковичного мятлика, колосовидного подорожникоцветника (Psylliosta-chys spicata), однолетней тимофеевки (Phleum paniculatum) и других однолетников — эфемеров.
Литература
И. В. Новопокровский (1925, 1927а); Е. В. Шиффере (1953).
85 к — закавказские (из Artemisia Meyeriana)
Нанесенные на карте контуры закавказских злаково-полынных остепненных пустынь до некоторой степени условны. В связи с быстрой сменой экологических условий на крутых склонах, с одной стороны, а также-с сильной распаханностью пологих склонов, с другой, контуры эти включают как собственно злаково-полынные пустынные сообщества, так и по--лынно-злаковые опустыненные степи (в верхних частях склонов), а также окультуренные земли — поля, сады, залежи.
К полыни (Artemisia Meyeriana)1 здесь присоединяются типчак (Festuca sulcata), ковыль Щовица (Stipa Szowitsiana) и гребневидный житняк (Agropyrum pectiniforme). По крутым каменистым склонам с неразвитыми почвами распространены ксерофитные фриганоидные группировки с Astragalus calycinus, Stocky s fruticulosa, Ephedra procera и др.; в нижней части.. . юго-восточного массива, на переходе к эфемерово-полынным пустыням. и эфемерово-полынным в сочетании с солянковыми (опис. 90 к и 91 к), на плотных глинистых склонах разбросаны редкие кустики Reaumuria hype-ricoides (=R. cistoides) и Kochia prostrata.
Литература
А. А. Гроссгейм (1932, 1948); А. К. Магакьян (1941); А. Л. Тахтаджян (1941);. M. П. Богданов (1954).
1 В понимании А. А. Гроссгейма. Определитель растений Кавказа. М., 1949..
85 кк — прикаспийско-казахстанские (из Artemisia Lercheana и A. pauciflora)
Прикаспийско-казахстанские злаково-полынные пустыни из белой и черной полыней (Artemisia Lercheana и A. pauciflora) распространены в пределах Северного Прикаспия, в бассейне Эмбы и на равнинах Тургай-ской столово-останцовой страны, в восточной части которой они смыкаются с восточно-казахстанскими злаково-полынными пустынями.
Злаково-полынные пустыни занимают плоские или слабо волнистые равнины с резко выраженным микрорельефом, обусловливающим’ характерную для них пестроту (комплексность) почвенного и растительного покрова. Преобладающими являются почвы тяжелого механического состава — глинистые и суглинистые светлокаштановые в комплексе с лугово-каштановыми западинными почвами и различными разновидностями солонцов.
Наиболее типичной чертой растительного покрова является сочетание сообществ степного типа с сообществами пустынными, что и создает характерную картину пятнистости (комплексности).
В западной части ареала этих злаково-полынных пустынь (Сарпин-ская низменность и низменность между Волгой и Уралом) наиболее распространены следующие основные члены комплекса.
1) Типчаково-белополынные (из Artemisia Lercheana и Festuca sulcata), житняково-белополынные (из Artemisia Lercheana и Agropyrum desertorum) и прутняково-белополынные (из Artemisia Lercheana и Kochia prostrata) пустыни на светлокаштановых солонцеватых почвах, занимающих микроплакорные, ровные участки. Во всех этих сообществах главное значение имеют пустынные полукустарнички: белая полынь (Artemisia Lercheana) и другие (Artemisia pauciflora, Pyrethrum achilleifolium), с заметным, а иногда большим участием степных трав (Stipa Lessingiana, S. sareptana, Koeleria gracilis). Очень характерно наличие некоторого числа эфемероидов и эфемеров (Роа bulbosa var. vivipara, Carex uralensis, Tulipa biflora, Alyssum desertorum, Ceratocephalus orthoceras, Astragalus testicula-tus и др.), ранней весной покрывающих открытые плешинки, свободные от полукустарничков и многолетних степных трав. Характерно также значительное количество на самой поверхности почвы водорослей (Stratonostoc commune, Microcoleus vaginatus), лишайников (Parmelia ryssolea, P. vagans), мхов (Bryum caespitosum) и некоторых других видов споровых растений.
Сообщества подобного остепненно-пустынного облика в составе комплексов занимают от 10 до 40 (60)% площади.
2) Чернополынные (из Artemisia pauciflora), рояашниково-чернополын-ные (с участием Pyrethrum achilleifolium), камфоросмово-чернополынные {с участием Camphorosma monspeliacum), острецово-чернополынные (с участием Aneurolepidium ramosum) пустыни на столбчатых, корково-столбчатых, солончаковатых и других разновидностях солонцов на едва повышенных элементах рельефа. К перечисленным господствующим растениям иногда примешиваются биюргун (Anabasis salsa) и прутняк (Kochia prostrata). Число видов цветковых растений в отдельных сообществах около 8— 10, редко достигает 12. В небольшом количестве здесь иногда встречаются живородящий мятлик (Роа bulbosa var. vivipara) и немного эфемеров (Alyssum desertorum, Lepidium perfoliatum); более заметны летне-осенние однолетники (Ceratocarpus arenarius и Echinopsilon sedoides). Очень значительную роль играют споровые растения, особенно водоросли (Stratonostoc commune. Microcoleus vaginatus) и лишайники (Parmelia vagans, P. ryssolea,
Collema crispum, Psora decipiens, Acarospora Schleicheri, виды Placodium), а также мхи {Bryum caespitosum, Tortula ruralis и др.). Видовой состав споровых растений здесь наиболее богат.
Эти типично пустынные сообщества занимают в комплексах от 25 до 50—60%.
3) Типчаковые (из Festuca sulcata), тырсово-типчаковые (из типчака и Stipa capillata), ковылково-типчаковые (из типчака и Stipa Lessingiana), житняково-типчаковые (из типчака и Agropyrum pectini-forme) степи па лугово-каштановых почвах, занимающих наиболее пониженные места в микрорельефе (блюдца, западины, лопатины). Обилие степных дерновинных злаков в этих сообществах придает последним типичный степной облик, резко выделяя их на фоне остальной растительности в ландшафте.
В качестве примеси к основным названным растениям встречаются тонконог {Koeleria gracilis), красный ковыль {Stipa rubens), острец {Aneurolepidium ramosum) и значительное количество видов степного —главным образом «сухого» — разнотравья: Linosyris villosa, Jurinea multiflora, Potentilla bijurca, Achillea nobilis, Galium ruthenicum, Phlomis tuberosa, Medicago falcata, Melandrium viscosum, Artemisia austriaca и многих других. Травяной покров достигает иногда почти полной сомкнутости, и, хотя здесь имеются и эфемеры и некоторые споровые растения, роль их крайне ничтожна. Во многих западинах, особенно в северной части полосы злаково-полынных пустынь, встречаются степные кустарники: таволожка {Spiraea hypericifolia), бобовник {Amygdalus папа) и астрагал {Astragalus brachylobus), которые, в местах, слабее затронутых влиянием выпаса, образуют иногда густые заросли.
Степные западины в среднем занимают от 10 до 40% площади в комплексах. По мере движения с севера (где процент западин наиболее высок) на юг происходит постепенное ослабление их роли в комплексах, наряду с обеднением видового состава, уменьшением густоты покрова и степени участия кустарников.
Соотношение площадей этих основных членов комплексов значительно колеблется: появляются достаточно хорошо очерченные переходные между ними сообщества (особенно на различных разновидностях солонцов); комплекс часто может усложняться до четырех-пяти членов (лишь в наиболее простом случае он состоит всего из двух).
Двучленные комплексы наиболее типичны для северных районов описываемых пустынь, где сочетаются тырсово-типчаковые степи в западинах с чернополынными пустынями на корково-столбчатых солонцах. Их площади примерно равны, но на самом севере степные западины иногда занимают 60—70%.
Четырехчленные комплексы состоят из следующих сообществ: типчаково-белополынные (из Artemisia Lercheana и Festuca sulcata) на солонцеватых светлокаштановых почвах; прутняково-белополынные (из Artemisia Lercheana и Kochia prostrata) на глубокостолбчатых солонцах; чернополынные (из Artemisia pauciflora) и камфоросмово-чернополынные (из Artemisia pauciflora и Camphorosma monspeliacum) на корковых и корковостолбчатых солонцах; тырсово-типчаковые (из Festuca sulcata и Stipa capillata) или тырсиково-типчаковые (из Festuca sulcata и Stipa sareptana) на лугово-каштановых почвах западин. Соотношения между названными отдельными членами комплекса сильно меняются, но, во всяком случае, на долю степных, сообществ приходится всего 10—30%, остальная же площадь занята пустынными сообществами.
Исследования взаимоотношений различных сообществ в комплексах северной части Волжско-Уральского междуречья показали, что в настоящее время здесь происходит процесс активного разрушения солонцов под воздействием наступающей на них степной растительности. Чернополынные, белополынные, прутняково-белополынные и другие сообщества пустынного типа приобретают характер остаточных, и они сменяются степными сообществами различного состава (в зависимости от данных конкретных условий). В целом, эволюция почвенно-растительного покрова ведет к ослаблению комплексности и направлена к формированию степных и остепненных сообществ более однородного состава (Левина, 1954).
В Прикаспийской низменности очень характерным элементом ландшафта являются частые сусликовые холмики. Молодые холмики покрыты разреженной растительностью, в которую входят черная полынь (Artemisia pauciflora), белая полынь (Artemisia Lerheana), камфоросма (Campho-rosma monspeliacum), прутняк (Kochia prostrala), итсегек (Anabasis aphyIla), однолетние солянки, немногие виды эфемеров (Lepidium perfoliatum, Alyssum desertorum и др.) и редкие комочки кны (Stratonostoc commune). Многие холмики, вследствие естественной мелиорации, осуществляемой под влиянием почвенного гипса, выброшенного землероями на поверхность (Формозов и Воронов, 1935), активно зарастают степной растительностью: ковылком (Stipa Lessingiana), тырсиком (Stipa sareptana), типчаком (Festuca sulcata), грудницей (Linosyris villosa) и другими видами.
Сусликовые холмики занимают от 5 до 15% площади, еще более усложняя и без того пестрый комплексный покров.
Точно так же усложняют общую картину часто встречающиеся «лиманы» и «разливы», вызывающие появление ряда сообществ переходного порядка от га л офитно луговой (а иногда еще и лугово-болотной) растительности к пустынным сообществам.
Необходимо отметить, что злаково-полынные пустыни северного При-каспия, особенно на Волжско-Уральском междуречье, в силу некоторых местных условий (сравнительно недавнее освобождение от морской трансгрессии, остаточный рельеф морского дна, значительная эрозионная моделировка водами временных весенних рек и поверхностного стока, развитие процессов суффозии и др.) отличаются от других районов этих пустынь сугубой комплексностью, выраженной в очень яркой форме. Эта комплексность растительного покрова, процессы формирования которой не закончились и в настоящее время, неоднократно привлекала внимание ряда исследователей (Б. А. Келлера, В. А. Ковды, И. П. Герасимова), работы которых позволили дать более подробную характеристику остепненных пустынь в этом районе.
Пустыни, лежащие далее на восток, изучены значительно менее детально, однако даже те недостаточные данные, которые опубликованы в литературе, позволяют наметить некоторые их отличия.
Прежде всего следует указать на значительно менее яршГ проявленную комплексность.
Так, на междуречье Урала и Эмбы, по данным С. А. Никитина (1954а), характерны главным образом двучленные комплексы с преобладанием белополынных сообществ (из Artemisia Lercheana) и с участием чернополынных (из Artemisia pauciflora). Для белополынников очень типично участие злаков: пустынного житняка (Agropyrum desertorum), сибирского житняка (Agropyrum sibiricum), живородящего мятлика (Роа bulbosa var. vivipara) и тырсика (Stipa sareptana).
Представлены злаково-белополынные сообщества (житпяково-бело-полынные (из Artemisia Lercheana и Agropyrum desertorum), тырсиково-житняково-бслополыппые (из Artemisia Lercheana, Agropyrum desertorum и Stipa sareptana)], особенно характерные для этих пустынь к востоку от р. Урала на светлокаштановых почвах. В них довольно богатое «сухое» разнотравье (Serratula xeranthemoides, Linosyris tatarica, Jurinea multiflora, Dianlhus lep topetalus), эфемероиды и эфемеры (Allium paniculatum, A. in-deriense, Tulipa biflora, Poa bulbosa var. vivipara, Tauscheria lasiocarpa, Veronica biloba, Descurainia Sophia, Ceralocephalus orlhoceras), много летне-осенних однолетников (Salsola tamariscina, Ceratocarpus arenarius), а также довольно обильны лишайники. Для этих районов характерна также частая встречаемость татарского ревеня (Rheum tataricum).
Черпополынвые пустыни (белополынно-чернополынные (из Artemisia pauciflora и A. Lercheana), итсегеково-чернополынлыс (с участием Anabasis aphylla), камфоросмово-чернополынные (с участием Camphorosma топ-speliacum), коклеково-чернополынные (с участием Atriplex сапа)] распространены не только в виде пятен среди господствующих злаково-белополынных сообществ, по встречаются на больших площадях на солонцовых почвах низовьев бессточных рек (Сагиза, Кушума, Джаксыбая и др.).
На обширных пространствах Подуральского плато господствуют своеобразные итсегеково-бслополынные сообщества (из Artemisia Lercheana и Anabasis aphylla), на однородном сером фоно которых выделяются темнозеленые крупные кусты итсегека. Флористический состав этих комплексов сходен с ранее описанными (здесь встречаются Kochia prostrata, Agropyrum desertorum, Stipa sareptana, Serratula xeranthemoides, Cachrys odontalgica, Ferula tatarica, Poa bulbosa и тс же эфемеры и летне-осенние однолетники). Однако видовая насыщенность отдельных участков меньше, также как и высота растений и степень покрытия. На участках с разреженным травостоем особенно обильны лишайники.
В Мугоджарах на щебнистых почвах характерны сообщества лессин-гианополыпников (из Artemisia Lessingiana). Помимо формирования самостоятельных сооб1цеств в общем комплексном покрове, Artemisia Lessingiana в качестве субэдификатора входит здесь в злаковые сообщества.
Для комплексов зауральской части (к востоку от р. Урала) характерно выпадение кустарниковых сообществ в западинах и исчезновение типчака в злаково-белополынных группировках.
Злаково-полынные пустыни западной части Тургайской столово-останцовой равнины представлены комплексами типчаково-полынных (Artemisia terrae-albae в. str. и Festuca sulcata), белополынных (Artemisia Lercheana, A. astrachanica), чернополынных (Artemisia pauciflora s. I.),1 биюргунных (Anabasis salsa) и изредка тасбиюргунных (Nanophyton eri-naceum) сообществ (Крашенинников, 1925). *
Обширные пространства южных полупустынь используются главным образом под пастбищное животноводство. Выпас скота на пастбищах производится с середины апреля до конца октября—середины ноября. Средняя урожайность пастбищ 4—5 ц/га поедаемой сухой массы. В отдельных районах и в благоприятные годы возможен частичный выпас и в зимний период.
В стойловый период животноводство может быть полностью обеспечено сеном, заготовляемым на лиманах и разливах. Однако здесь необходимо перераспределение сена из районов избытка его в районы с недостатком
1 Включая Artemisia pauciflora ssp. maikara.
сенокосных угодий (например в Волжско-Уральском междуречье необходима переброска сена из района Чижинскнх разливов в западную часть).
При комплексном сложении почвенно-растительного покрова земледелие в южной полупустыне имеет выборочный характер. Распахиваются в первую очередь степные падины с темноцветными почвами, а также участки территории с преобладанием в комплексе злаковых сообществ и малой долей участия чернополынных на солонцах.
В наибольшей мере освоена Волжско-Уральская часть южной полупустыни. В северной полосе, где много падин, ее освоение под земледелие достигает 50%, в средней же и южной — процент распашки сокращается, хотя там имеются резервы пахотнопригодных падинных со степными злаками почв. Рост технического оснащения сельского хозяйства, особенно в связи со строительством Сталинградской гидроэлектростанции, открывает широкие перспективы для дальнейшего освоения под земледелие значительных территорий целинных земель южной полупустыни, пока что используемых в основном как пастбища.
Литература
Г. Н. Высоцкий (1915); И. М. Крашенинников (1925); II. В. Ларин (1926а); Н. В. Павлов (1926); М. М. Ильин и Р. Ю. Рожевиц (1928); О. Э. Кнорринг (1928); А. Н. Формозов и А. Г. Воронов (1935); Г. Н. Новиков (1936); Е. М. Лавренко (1940а); А. В. Прозоровский (1940а); И. И. Тереножкин (1947); А. В. Прозоровский и В. П. Малеев (1947); И. П. Герасимов (1951); Ф. Я. Левина (1954); С. А. Никитин (1954а); Б. А. Быков (1955).
85 вк — восточноказахстанские (из Artemisia sublessingiana и A. pauciflora)
Восточноказахстанские злаково-полынные пустыни распространены к востоку от прикаспийско-казахстанских, начиная от восточной части Тургайской столовой страны, далее встречаются в южной части Казахского мелкосопочника, на северной окраине Бетпак-Далы и в северном Прибалхашье. В западной части ареала они сочетаются с прикаспийско-казахстанскими, а в восточной — с северотуранскими (на некоторых контурах это показано индексами кк -|- вк и вк -f- стр).
Для этого варианта злаково-полынных пустынь характерно некоторое участие представителей собственно туранской флоры. В особенности существенно появление здесь северотурайской полыни (Artemisia semiarida), .сменяющей произрастающую на западе белую полынь (A. Lerche-апа). На щебнистых почвах (Казахский мелкосопочник и другие районы) распространены сублессингианополынники (из A. sublessingiana).
Наряду с этим, восточноказахстанский вариант отличается несколько меньшей комплексностью, а также значительно меньшим участием в сложении растительного покрова степных сообществ на лугово-каштановых почвах западин. В то же время здесь особенно характерно присутствие представителей солянковых пустынь: кокпека (Atriplex сапа), биюргуна (Anabasis salsa), тасбиюргуна (Nanophyton erinaceum) как в виде примеси в полынных сообществах, так и в виде самостоятельных группировок, включенных в злаково-полынные пустыни небольшими площадями.
Литература
Н.В.Павлов(1931); Ф. Я. Левина (1938); И. В. Ларин (1941); Н. И. Рубцов (1950а).
85 стр—северотуранские (из северотуранских видов полыни)
Северотуранские злаково-полынные пустыни показаны небольшими контурами на Мангышлаке (горы Каратау), а основной их массив лежит к востоку от долины Сарысу в северной части Бетпак-Далы и в северном Прибалхашье, где, как отмечено выше, они обозначены общим индексом вк + стр.
Этот вариант злаково-полынных пустынь характеризуется большим участием элементов туранской флоры. Среди типчаково-полынных (полынь -{-Festuca sulcata), тырсиково-тинчаково-нолынных (полынь-|-Fe-stuca sulcata-)-Stipa sareptana), ковылково-типчаково-полынных (полынь -{-Festuca sulcata-{-Stipa Lessingiana) ассоциаций эдификаторную роль играет Artemisia semiarida, A. t rrae-albae s. str., а также A. sublessingiana.
Комплексность растительного покрова снова становится более сложной, особенно в районах мелкосопочника, где на равнинах и в депрессиях рельефа заметное значение приобретают степные кустарники: таволожка {Spiraea hypericifolia) и карагана {Caragana balchaschensis), а на южных щебнистых склонах мелкосопочника — сублессингианополынники (из Artemisia sublessingiana) и боялычники (из Salsola laricifolia). Кроме того, небольшими участками встречаются группировки из Nanophyton erinaceum и Anabasis truncata. В долинах характерны сообщества кокпека {Atriplex сапа), иногда занимающие большие площади, и биюргуна {Anabasis salsa). Вдоль речных русел господствуют заросли различных галофитов и широко распространены чиевники (из Lasiagrostis splendens).
Литература
С. С. Неуструев и И. М. Крашенинников (ред.) (1930); Ф. Я. Левина (1938); А. В. Прозоровский (1940а); А. В. Прозоровский и В. П. Малеев (1947); Н. И. Рубцов (1950а).
86. Злаково-полы иные пустыни на песках (из Artemisia astrachanica или A. Massagetovii, 1 A. arenaria, Agropyrum sibiricum и др.), в сочетании по понижениям рельефа с солянковыми и луговыми группировками и слабозаросшими барханными и бугристыми
песками
Злаково-полынные пустыни на песках распространены на равнинах северного Прикаспия и Приаралья. Наиболее крупные массивы лежат в западной части Прикаспия (между Ергенями, Волгой и Каспийским морем, а также на междуречье Волги и Урала — крупный песчаный массив Рын-пески); к востоку от р. Урала эти пустыни занимают отдельные песчаные массивы: в низовьях сухих дельт Уила и Сагиза, Булдурты, Калды-гайты, Джаксыбая и др.; в северном Приаралье встречаются небольшими пятнами на северной оконечности Мангышлака (п-ов Бузачи), на песках Большие Барсуки и Приаральские Каракумы, а кроме того, изолированно от названных районов — в южном Прибалхашье (в южной части песков Сары-Ишикотрау и на песках Таукум).
Господствующий элемент ландшафта — плоскохолмистые или весьма пологоувалистые, иногда мелкобугристые и даже совсем плоские равнины. Среди них почти всегда имеются мелкие, а иногда и довольно крупные
1 В монографии подрода Seriphidium П. П. Поляков этот вид рассматривает как Artemisia terrae-albae ssp. Massagetovii.
массивы подвижных голых или зарастающих песков, возникших в результате хищнической эксплуатации «песчаных пастбищ» в середине прошлого столетия. В районе песков, примыкающих к дельте Волги, характерны хорошо выраженные, вытянутые в широтном направлении бэровские бугры. В некоторых зауральских песчаных массивах, а также в песках Приаралья, встречаются участки с развитым грядовым рельефом.
Почвы здесь песчаные, супесчаные, реже глинисто-песчаные, слабо развитые (примитивные) типа бурых. Между бэровскими буграми, а также между грядами и в котловинах (при бугристом рельефе) часто встречаются сильно солонцеватые и солончаковые почвы; плоские солончаковые понижения обычны и на равнинах.
Будучи распространенными в пределах полосы остепненных пустынь (южных полупустынь), рассматриваемые злаково-полынные пустыни на песках находятся в тех же климатических условиях, что и «зональный тип» ранее описанных комплексных злаково-полынных пустынь (85). Однако условия их существования значительно отличаются в силу особенностей песчаного субстрата, обеспечивающего гораздо более благоприятный водный режим. Последний обусловлен наличием подповерхностного горизонта влажности, сохраняющегося в течение почти всего вегетационного периода. Этим в основных чертах объясняется более далекое продвижение по пескам на юг ряда форм, являющихся типичными для плакорных условий на территориях, лежащих на значительном расстоянии к северу (в частности это относится к злакам, например к Agropyrum sibiricum, Stipa Joannis и др.). В то же время существование на нередко подвижных субстратах привело к выработке своих «псаммофильных» форм, наиболее яркими представителями которых являются виды Calligonum, а также такой злак, как Aristida pennata или песчаная осока — ранг Carex physo-des, широко распространенные в пустынях, расположенных значительно южнее.
Флористический состав сообществ злаково-полынных пустынь на песках в пределах обширного ареала несколько изменяется, так как долины Волги и Урала являются рубежом для некоторых восточных элементов. Так, например, ранг (или илак, Carex physodes), будучи обычным растением к востоку от Урала, лишь как исключительная редкость встречается в песках Волжско-Уральского междуречья и совершенно отсутствует к западу от Волги. Точно так же не переходит через Волгу джузгун {Calligonum aphyllum), который не часто находится в Рын-песках и весьма обычен восточнее Урала, дополняемый здесь несколькими другими видами этого рода. Не идут далее Волги на запад и астраханская полынь {Artemisia astrachanica), селин {Aristida pennata), кулан-гуйрюк {Eremo-sparton aphyllum) и др.
На задерненных равнинных, увалистых и мелкобугристых уплотненных песках растительность представлена мятликово-белополынными {Artemisia Lercheana или Artemisia astrachanica и Роа bulbosa), еркеково-белополынными (те же полыни и Agropyrum sibiricum) и осочково-белопо-лынными {Artemisia astrachanica и Carex uralensis) пустынями в сочетании с белополынными {Artemisia Lercheana или A. astrachanica) и песчанополынными {Artemisia arenaria) пустынями на рыхлых песках.
В составе названных сообществ встречаются следующие виды: прутняк {Kochia prostrata), молочай {Euphorbia Seguieriana), хвойник {Ephedra distachya), кровельный костер {Bromus tectorum) и ряд других эфемеров, как, например, Eremopyrum triticeum, Alyssum desertorum, Lepidium perfo-liatum, Descurainia Sophia. Также весьма характерен и часто очень обилен эбелек {Ceratocarpus arenarius) — обычный показатель влияния
выпаса скота. Урожайность этих весьма ценных пастбищ равна 5—7 ц/га сухой массы.
На бугристых и грядово-бугристых песках восточнее Урала распространены еркековые (из Agropyrum sibiricum), еркеково-белополынные (Artemisia astrachanica и Agropyrum sibiricum), илаково-белополынные (Artemisia astrachanica и Carex physodes) и янтаковые (из Alhagi pseudalhagi) сообщества с примесью некоторых псаммофильных полукустарников — видов Calligonum, Eurotia ceratoides, видов Astragalus из секции Ammo-dendron. Для названных сообществ характерны следующие виды: Achillea Gerberi, Syrenia siliculosa, Salsola pellucida, Kochia laniflora, Helichrysum arenarium, Allium sabulosum, Lappula semiglabra, большинство которых типично для степей на песчаных почвах (Никитин, 1954, 1954а). Урожай сухой массы 6—10 ц/га.
Равнинные и волнисто-холмистые пески в южной части Прибалхашья покрыты еркеково-прутняково-полынными (из Artemisia terrae-albae ssp. Massagetovii, Kochia prostrata и Agropyrum sibiricum) и полынными (из Artemisia terrae-albae s. 1.) сообществами на песчаных уплотненных серо-бурых почвах. По более крутым склонам распространены заросли терескена (Eurotia ceratoides), а на подвижных песках (на вершинах гряд и бугров и на участках, разбитых выпасом) разреженная растительность из видов джузгунов (Calligonum aphyllum, С. leucocladum и др.) с селином (Ari-stida pennata), гелиотропом (Heliotropium dasycarpum), хондриллой (Chond-rilla ambigua) и др.
Песчаный массив Таукум несколько отличается от других. По низинам между гряд для него характерны полынные (из Artemisia terrae-albae ssp. Massagetovii) и прутняково-полынные (тоже с Kochia prostrata) сообщества, а на склонах гряд — сообщества из джунгарской полыни (Artemisia songorica) и борджока (Ephedra lomatolepis) с еркеком (Agropyrum sibiricum), терескеном (Eurotia ceratoides), Salsola pellucida, Eremurus inderiensis и др. На разбитых и подвижных бугристых и грядово-бугристых песках широко распространены сообщества песчаной полыни (Artemisia arenaria), отличающиеся довольно пестрым видовым составом сопровождающих ее растений: Agropyrum sibiricum, Bromus tectorum, Lappula semiglabra, Artemisia scoparia, Ceratocarpus arenarius, Salsola ruthenica, S. pellucida, Chondrilla brevirostris, C. ambigua, Agriophyllum arenarium, Eremosparton aphyllum и др.
На барханных песках встречаются крайне изреженные сообщества, вернее — фрагменты сообществ, представленные групповыми зарослями песчаного овса (Elymus giganteus), песчаной полыни (Artemisia arenaria), селина (Aristida pennata), кумарчика (Agriophyllum arenarium), Heliotropium arguzioides, Chondrilla ambigua, Salsola pellucida и некоторых других. Из этих растений особую кормовую ценность имеет песчаный овес.
В песках Таукум на солончаковатых почвах, связанных с различными понижениями в рельефе, господствуют солянково-полынные и солянковые пустынные сообщества (Artemisia Schrenkiana, Atriplex сапа, A. verruci-fera, Anabasis aphylla, A. salsa, однолетние солянки и шведки — виды Salsola и Suaeda), а на солончаках, подпитываемых близкими засоленными грунтовыми водами, характерны сообщества сарсазана (Halocnemum strobilaceum), поташника (Kalidium foliatum), солероса (Salicorniaeuropaea) и однолетних солянок (Salsola nitraria, S. crassa, Suaeda confasa и др.), в которых также встречаются Camphorosma Lessingii, Plantago salsa, Limonium suffruticosum, Frankenia hirsuta, Tournefortia sibirica и другие галофильные растения.
В приморских участках песков в северном Прикаспии, а иногда и в пониженных котловинах среди песчаных массивов обычны группы гребенщиков {Tamarix ramosissima, Т. laxa, Т. gracilis, Т. Ewersmannii) и селитрянки {Nitraria Schoberi).
Среди песчаных массивов Зауралья с действующими протоками и озерами по депрессиям рельефа встречаются участки лугов из вейника (Calamagrostis epigeios), солодки {Glycyrrhiza glabra), пырея {Agropyrum repens), а на засоленных местах луга из чаира {Aeluropus litoralis) и полыни {Artemisia nitrosa и A. mongolorum); кроме этих сообществ, распространены также заросли тростника {Phragmites communis) и клубнекамыша {Bolboschoenus maritimus).
В целом, злаково-полынные пустыни на песках представляют собой ценные пастбищные угодья, частью используемые, помимо вегетационного периода, также и зимой, с урожайностью кормовой массы от 4 до 12 ц/га.
Литература
А. Н. Краснов (1886а); В. М. Савич (1910); В. А. Дубинский (1911); А. И. Михельсон (1913); С. А. Никитин (1928, 1935, 1954, 1954а); Б. А. Борнеман и М. Д. Спиридонов (1929); И. И. Томашевский (1931); М. А. Орлов (1940); Н. И. Рубцов (1941а); И. И. Тереножкин (1947); А. Г. Гаель и др. (1949, 1950); А. А. Трушковский (1952); Б. А. Быков (1955).
87. Полынные пустыни
Полынные пустыни распространены относительно небольшими площадями в северной половине пустынной области от Ергеней на западе до предгорий Тянь-Шаня на востоке, будучи обычно расположенными по соседству (или включенными в них) со злаково-полынными пустынями, сочетающимися с дерновиннозлаковыми степями и полынными, а также солянковыми пустынями (85) и злаково-полынными пустынями на песках (86).
Экологические условия существования данного типа пустынь характеризуются летним дефицитом влаги, что вызывает частичное ослабление вегетации основных компонентов этих пустынь (полыни и представители маревых), маломощным снеговым покровом, глубоким залеганием уровня грунтовых вод и широким распространением солонцеватости почв, то есть явлений, типичных для северной полосы пустынной области. Почвы (зональные, плакорные) относятся к типу бурых и частью серо-бурых почв.
Расчлененность микрорельефа выражена значительно слабее, чем в злаково-полынных пустынях (85). В частности, здесь западины меньше по размерам и менее глубоко врезаны; соответственно ослабляется степень промачивания их. Почвы западин часто несут признаки солонцеватости, а в летнее время испытывают глубокое иссушение. Для полынных пустынь характерны следующие особенности: менее четко выраженная комплексность, по сравнению со злаково-полынными пустынями; более или менее безраздельное господство полыней {Artemisia Lercheana, A. ter-rae-albae s. sir., A. turanica, A. kemrudica, A. gurganica, A. diffusa и др.); появление на более засоленных почвах кокпека {Atriplex сапа) или биюр-гуна {Anabasis salsa)', резкое сокращение количества и размеров западин со степными сообществами. В более северных районах к полыням примешивается небольшое количество злаков.
87 кк — прикаспийско-казахстанские (из Artemisia Lercheana)
Прикаспийско-казахстанские полынные пустыни, иногда с примесыо степных дерновинных злаков, довольно широко распространены в север
ном Прикаспии, а также к востоку от Урала, в бассейне Уила и Сагиза, занимая сниженные равнины, обычно слабо расчлененные, а также нижние пологие части шлейфов незначительных возвышенностей и увалов, причем и для тех и других характерно слабое развитие микрорельефа. Преобладающие почвы — бурые пустынно-степные, легкосуглинистые и супесчаные.
Для характеристики этих пустынь можно привести данные по одному из типичных и хорошо изученных районов к югу от оз. Баскунчак. Здесь-растительный покров представлен следующими комплексами.
1) Житняково-белополынные (из Artemisia Lercheana и Agropyrum desertorum) и, реже, типчаково-белополынные (из Artemisia Lercheana и Festuca sulcata) пустыни на бурых слабосолонцеватых почвах. К этим основным растениям примешиваются ромашник {Pyrethrum achilleifolium), прутняк {Kochia prostrata), мятлик {Роа bulbosa var. vivipara), острец. {Aneurolepidium ramosum), изредка ковылок {Stipa Lessingiana), грудница {Linosyris villosa) и некоторые другие виды ксерофильного разнотравья. Имеются здесь и эфемеры {Alyssum desertorum, Veronica biloba, Descurainia Sophia и др.), однако количество их значительно меньше, чем в прикаспийско-казахстанских злаково-полынных пустынях (85 кк). Встречаются также и напочвенные лишайники.
2) Белополынные (из Artemisia Lercheana) и прутняковые (из Kochia prostrata) пустыни на столбчатых и корковых солонцах (рис. 69). В качестве сопутствующих растений здесь встречаются кокпек {Atriplex сапа), камфоросма {Camphorosma monspeliacum), кзылкокпек {Limonium suffru-ticosum), биюргун {Anabasis salsa) и др. Довольно обычными являются мятлик {Роа bulbosa var. vivipara), немногие эфемеры и летне-осенние однолетники, а также напочвенные водоросли (особенно кна — Stratonostoc commune) и лишайники.
3) Типчаковые (из Festuca sulcata), тырсиково-типчаковые (типчак и Stipa sareptana) степи на слабо выраженных лугово-каштановых почвах мелких плоских западин. В составе этих степных сообществ чаще примешаны элементы пустынные, нежели степные. Так, здесь встречаются прутняк {Kochia prostrata), ромашник {Pyrethrum achilleifolium), грудница {Linosyris tatarica), мятлик {Poa bulbosa var. vivipara), изредка ковылок {Stipa Lessingiana) и некоторые другие. Степные кустарники {Spiraea hypericifolia, Amygdalus папа) отсутствуют.
4) Неопределенные сочетания белой полыни {Artemisia Lercheana, местами A. astrachanica), прутняка {Kochia prostrata), камфоросмы {Camphorosma monspeliacum), изредка черной полыни {Artemisia pauciflora) на молодых (свежих) сусликовых холмиках.
Пустыни Урало-Кушумского водораздела на почвах более тяжелых, чем предшествующие, слагаются комплексами следующего состава.
1) Типчаково-белополынные (из Artemisia Lercheana и Festuca sulcata) и типчаково-ромашниково-белополынные (из Artemisia Lercheana, Pyrethrum achilleifolium и Festuca sulcata) пустыни на бурых почвах.
2) Белополынные (из Artemisia Lercheana), чернополынно-белополын-ные (из Artemisia Lercheana и A. pauciflora) и белополынно-кокпековые (из Atriplex сапа и Artemisia Lercheana) пустыня на различных разновидностях солонцов.
3) Тырсиково-типчаковые (из Festuca sulcata и Stipa sareptana) и ост-рецово-типчаковые (из Festuca sulcata и Aneurolepidium ramosum) сообщества в едва приметных западинках с лугово-каштановыми слабосолонцеватыми почвами.
4) Пестрые сочетания кокпека (Atriplex сапа), камфоросмы (Campho-rosma monspeliacum), белой и черной полыни (Artemisia Lercheana is. А. pauciflora) и биюргуна (Anabasis salsa) на сусликовых холмиках.
На междуречье Джаксыбая и Уила и в других местах Зауралья житняко-во-белополынные (из Artemisia Lercheana и Agropyrum desertorum) и тырси-ково-житняково-белополынные (тоже и Stipa sareptana) пустыни с участием ксерофильного разнотравья (Serratula xeranthemoides, Ferula tatarica, Jurinea multiflora) и небольшого числа эфемеров (Alyssum desertorum, Tauscheria lasiocarpa, Leptaleum filifolium и др.) сочетаются с белополынными (из Artemisia Lercheana), чернополынно-болополынпыми (из Arte-misih Lercheana и Л. pauciflora), итсегеково-белополынпыми (из Artemisia Lercheana и Anabasis aphylla), биюргуново-белополыпными (из Artemisia Lercheana и Anabasis salsa) и кокпековыми (из Atriplex сапа) сообществами (Никитин, 1954а).
Литература
И. М. Крашенинников (1925); О. Э. Кнорринг (1928); С. С. Неуструев и И. М. Крашенинников (ред.) (1930); Е. П. Коровин (1934); Б. А. Келлер (ред.) (1936); А. В. Прозоровский (1940а); С. А. Никитин (1954а); Б. А. Быков (1955).
87 стр — северотуранские (из северотуранеких видов полыни)
Этот вариант полынных пустынь встречается маленькими контурами на Мангышлаке, в Центральном Казахстане, в бассейне Сарысу, к северу от оз. Алаколя, в нескольких местах у северных подножий Каратау (Сыр-дарьинского), Чу-Илийских гор и Тянь-Шаня, а также по северной окраине останцовых возвышенностей в Центральных Кызылкумах.
Характерным для них является приуроченность к шлейфам небольших поднятий, останцовых плато и предгорий. Почвы серо-бурые, глинистые, реже суглинистые и супесчаные.
Повсюду на своем ареале полынные пустыни контактируют с контурами полынных пустынь, сочетающихся с солянковыми пустынями (89).
Северотуранские полынные пустыни в наиболее яркой степени несут •черты «настоящих пустынь» (в понимании А. В. Прозоровского, 1940а). Наименьшее во всей пустынной области количество осадков (всего 150— 100 мм и менее) распределяется более или менее равномерно в течение года с небольшим максимумом весной, когда растительный покров разнообразится эфемерами и эфемероидами. Летний же период, с мая по октябрь, • отличается малым количеством осадков, а в силу этого заметным дефицитом влаги. В этом сезоне даже полыни сбрасывают значительную часть листьев, вступая в фазу крайне ослабленной жизнедеятельности.
На протяжении почти всего года, за исключением короткого весеннего •периода, преобладает однообразный серый аспект, обязанный господствующему фону полукустарничков, главным образом полыни. Развивающиеся весной эфемеры и эфемероиды не образуют сплошного покрытия поверхности почвы; повсюду видны плешинки субстрата, на которых характерно наличие заметного количества лишайников, мхов и водорослей.
Основу растительного покрова слагают северотуранские виды полыни (Artemisia terrae-albae s. str., A. semiarida, A. kemrudica, A. turanica и др.), к которым примешиваются в незначительном обилии некоторые другие пустынные полукустарнички: прутняк (Kochia prostrata), кеурек (Salsola rigida), терескен (Eurotia ceratoides) и еще реже боялыч (Salsola laricifolia, S. arbuscula).
Изредка встречаются и злаки; иногда (при более легком механическом составе почв) образуются даже довольно сомкнутые куртинки; однако
это уже не степные злаки, примесь которых характерна для злаково-полынных пустынь, а пустынные. Это пустынные виды ковылей (Stipa Szowitsiana, S. Richteriana), обычно встречающиеся отдельными дернинами и только изредка, как отмечено, более или менее сомкнутыми куртинками. Более обычны эфемероиды: мятлик (Роа bulbosa var. vivipara) и осочка (Carex stenophylloid.es), а также в незначительном обилии виды Allium, Tulipa, Gagea. Эфемеры хотя и не образуют сомкнутого покрова, однако представлены большим числом видов: Alyssum desertorum, Bromus teclorum, Tauscheria lasiocarpa, Roemeria hybrida, Microlonchus minimus, Astragalus tribuloides, Lepidium perfoliatum, Tetracme quadricornis, Trigo-nel/a arcuata, Veronica biloba и др.
Характерна также примесь ксерофитных летне-осенних однолетников — видов из семейства маревых (Chenopodiaceae): Ceratocarpus arenarius, Gir-gensohnia oppositiflora, Salsola brachiata, Halimocnemis sclerosperma, Kochia Schrenkiana и др. Эти растения также встречаются одиночно или рассеянно, однако количество их возрастает на солонцеватых и солонча-коватых разновидностях почв. Лишайники представлены следующими видами: Diploschistes scruposus, Psora decipiens, P. Saviczii, Aspicilia desertorum, Lecanora aspera, виды Collema, Squamaria и др.
Степень покрытия растениями в сообществах северотуранских полынных пустынь колеблется в пределах 30—40%, а нередко и ниже.
Будучи распространенными в пределах северной части пустынной области, полынные пустыни характеризуются некоторыми отличиями по флористическому составу.
Так, например, на Мангышлаке, где полынные пустыни представлены небольшими контурами, характерно преобладание в качестве эдификатора (или соэдификатора, наряду с Artemisia terrae-albae s. str.) гурганской полыни (Artemisia gurganica), близкой к крымской полыни (Artemisia taurica). Кроме того, здесь характерно наличие, наряду с пустынными ковы-лями (Stipa Szowitsiana, Lasiagrostris caragana) —степных ковылей (Stipa Lessingiana, S. sareptana), далее на восток уже исчезающих. В этом же географическом районе типичны следующие коротковегетирующие растения-эфемероиды: Роа bulbosa var. vivipara, Colpodium humile, Rhinopeta-lum Karelinii и др.
В Центральном Казахстане и в северной части Бетпак-Далы полынные пустыни (из Artemisia terrae-albae s. str., A. turanica) в своем составе имеют пустынные полукустарнички, как, например, терескен (Eurotia ceratoides), прутняк (Kochia prostrata), кеурек (Salsola rigida)', иногда встречается ковыль (Stipa Hohenackeriana). Эфемероиды и эфемеры представлены ограниченным числом видов: Colpodium humile, Tulipa Behmiana, Rheum tataricum, Gagea reticulata, Tauscheria lasiocarpa, Leptaleum fili~ folium, Alyssum desertorum и некоторые другие (Павлов, 1931; Коровин и Миронов, 1935).
Полынные пустыни Чу-Илийских гор сменяются различными сообществами в зависимости от экспозиции. На северных склонах господствует Artemisia transiliensis (из ближайшего родства A. sublessingiana) со значительной примесью тырсика (Stipa sareptana), с участием эфемероидов (Роа bulbosa, var. vivipara, Ixiolirion tataricum, AlliumBarszczewskii, иногда Eremurus altaicus) и эфемеров (Eremopyrum orientate, Rochelia Bungei, Consolida rugulosa, Trigonella orthoceras и др.).
На южных склонах преобладают сообщества прутняка (Kochia prostrata) и той же полыни (Artemisia transiliensis). Здесь покров заметно изрежен (проективное покрытие 60%), и участие эфемеров и эфемероидов резко ослаблено. На засоленных почвах под пологом Artemisia transili-
ensis основной фон составляют эфемер Ceratocephalus orthoceras и летнеосенний однолетник Ceratocarpus turkestanicus, а другие эфемеры и эфемероиды редки и угнетены (Гамаюнова и Голоскоков, 1949).
На предгорных равнинах к северу от Каратау полынные пустыни слагаются сообществами с значительным покрытием (до 80%) Artemisia kara-tavica с примесью Astragalus mucidus и Hulthemia {Rosa} persica. Между кустами этих растений встречаются как многолетние травы, так и некоторые эфемероиды и эфемеры. Травы представлены следующими видами: Stipa Hochenackeriana, Lasiagrostis caragana, Agropyrum pectinijorme, Korovinia tenuisecta, Potentilla soongorica, Bungea vesiculif ra, Centaurea squarrosa и др. Из эфемероидов отмечены Ixiolirion tataricum, Tulipa Greigii, Eranthis longistipitata, Poa bulbosa. Эфемеры немногочисленны: Bromus tectorum, B. Danthoniae, Hordeum leporinum, Roemeria rejracta, Ziziphora tenuior, Euphorbia inderiensis, Scabiosa Olivieri, Alyssum desertorum, Ceratocephalus orthoceras, Nigella integrifolia (Культиасов, 1938).
Полынные пустыни из тех же видов полыней (Artemisia terrae-albae s. str., A. turanica) на шлейфах Букантау (в Кызылкумах) имеют примесь кеурека (Salsola rigida), тасбиюргуна (Nanophyton erinaceum). Здесь отмечено небольшое количество эфемеров (Schismus arabicus, Eremopyrum orientale, Veronica biloba, Thuspeinantha persica) и однолетних видов маревых (Girgensohnia oppositiflora, Halimocnemis mollissima, H. villosa, Salsola foliosa, S. hispidula, Halotis pilosa, Gamanthus gamocarpus). Характерно, что довольно обычным растением является гигантское зонтичное — чомуч (Ferula assa-foetida), особенно при наличии на поверхности песчаного плаща (Русанов, 1936).
Литература
И. Г. Борщов (1865); Б. А. Федченко и И. М. Крашенинников (1910); А. И. Михельсон (1913); И. Д. Спиридонов (1918,1930); И. М. Крашенинников (1925); Б. А. Борнеман и М. Д. Спиридонов (1929); С. С. Неуструев и И. М. Крашенинников (род.) (1930); Н. В. Павлов (1930, 1931, 1935); Е. П. Коровин (1934); Е. П. Коровин и Б. А. Миронов (1935); Ф. Н. Русанов (1936); М. В. Культиасов (1938); Ф. Я. Левина (1938); А. В. Прозоровский (1940а); А. П. Гамаюнова и В. П. Голоскоков (1949); Н. И. Рубцов (1950а, 1952); С. А. Никитин (1954, 1954а).
87 ютр — южнотуранские (из Artemisia herba-alba s. 1.)
Полынные пустыни южнотуранского варианта в виде мелких контуров на карте показаны преимущественно в Западном Копет-Даге, где они включены в господствующие сообщества эфемерово-полынных пустынь. Однако полынные пустыни этого типа (87 ютр) широко распространены по всему югу пустынной области в виде внемасштабных включений в солянковые пустыни (92) и полынные пустыни в сочетании с солянковыми (89).
Полынные пустыни этого типа в Копет-Даге представлены сообществами полыней: бадхызской (Artemisia badghysi) и тернистой (A. dumosa} с примесью Salsola Aucheri и S. laricifoliw, пустыни эти приурочены к засоленным и гипсоносным примитивным почвам на бедлендах. Очевидно' неблагоприятные свойства субстрата (подвижность, засоленность, гипсо-носность) ограничивают возможность развития здесь эфемерового покрова, и травянистые растения представлены преимущественно летне-осенними однолетними маревыми (Salsola turcomanica, S. lanata, S. stellulata, Halimocnemis Karelinii, Halocharis hispida, Halotis pilosa), Herniaria hir~ suta и немногими другими.
Характерными для этих условий являются некоторые многолетники, которые обладают мощной корневой системой, способствующей к удерживанию их в оползающем субстрате бедленда, как, например, Asparagus Irichophyllus, Zygophyllum macropterum, Noaea mucronata, Zosimia absinthi-folia, Crambe edentula, Eremostachys Regeliana, Scutellaria Litwinowii, La-gochilus cabulicus, Haplophyllum acutifolium, Rindera turkestanica и др. Однако эти растения встречаются единичными' экземплярами и успешно развиваются лишь в годы с обильными осадками.
Литература
С. И. Коржинекий (1896); Е. Г. Черняковская (1927); Е. П. Коровин (1934); 11. А. Линчевский (1935а); А. В. Прозоровский (1940а); Н. Т. Нечаева и др. (1943); А. В. Прозоровский и В. 11. Малеев (1947).
88. Горные и предгорные полынные пустыни
Основания горных систем Средней Азии всюду окружены пустынной растительностью. Для наклонных подгорных равнин, шлейфов гор и предгорий особенно характерны полынные пустыни, которые по своему видовому составу и структуре заметно отличаются от прилегающих к ним полынных пустынь па плакорах. Здесь эдификаторами выступают особые виды полукустарничковых полыней подрода Seriphidium, географическое распространение которых связано с горами, вне которых они не встречаются или же встречаются редко. Сюда относятся, например, такие виды полукустарничковых полыней, как Artemisia tenuisecta nA. baldshua-nica. Для предгорий Северного Тянь-Шаня весьма характерной является Artemisia sublessingiana s. I.1 (Рубцов, 1950a), хотя она встречается и в пла-корных пустынях (Бетпак-Дала и др.). Типично горными видами являются Artemisia Korshinskyi и A. Lehmanniana, распространение которых связано главным образом с высокими холодными нагорьями Центрального Тянь-Шаня и Памира.
Для большей части предгорных пустынь Средней Азии весьма характерна выраженная синузия из эфемеров и эфемероидов, пышно развивающихся в весеннее время. В высокогорных полынных сообществах Центрального Тянь-Шаня и Памира эфемерово-эфемероидная синузия отсутствует. Таким образом, горные полынные пустыни грубо можно разделить на две экологические и структурно резко отличные группы формаций: 1) группу низкогорную эфемерово-полынную и 2) группу высокогорную криофильно-полынную, совершенно лишенную синузии эфемеров.
Литература
Е. П. Коровин (1934); М. Г. Попов (1940); А. В. Прозоровский (1940а); Н. И. Рубцов (1950а, 1952).
88 tu — из Artemisia turanica в сочетании с ксерофиль-ными кустарниками
Полынники из туранской полыни {Artemisia turanica) сколько-нибудь значительных площадей не занимают. Они встречаются обычно в сочетании с различными солянковыми группировками, а иногда [по склонам Каратау (Сырдарьинского) ] в сочетании с ксерофильными кустарниками.
1 В последнее время П. П. Поляков в пределах Artemisia sublessingiana s. 1. выделил ряд мелких видов, в частности для предгорий Заилийского Алатау A. transilensis, а для Джунгарского Алатау Л. Gorjaevii (см. Вотан, матер. Гербария Вотан, инет. АН СССР, т. 16, 1954).
Основным районом распространения туранополынников является юго-восточная Туркмения, где они приурочены к предгорным равнинам и предгорьям Паропамиза. Здесь они развиваются на песчаных и супесчаных, иногда засоленных сероземах в комплексе с солянковой растительностью солончаковых понижений. В составе туркменских туранополынников солянки также играют существенную роль; наиболее обычны здесь полукустарничковые (Aellenia subaphylla и Salsola arbuscula) и однолетники (Halocharis hispida и Salsola carinata). Весною в этих полынниках развивается эфемеровая синузия, главным образом из различных крестоцветных (Malcolmia, Leptaleum), маковых (Papaver pavoninum, Нуресоит parviflorum) и других весенних однолетников, вместе с которыми встречаются и такие эфемероиды, как Carex physodes, С. pachystylis, Роа bulbosa var. vivipara.
В юго-восточной Туркмении (Бадхыз) в сложении туранополынников принимает значительное участие бадхызская полынь (Artemisia badghysi). Такого рода смешанные полынные сообщества постоянно встречаются по подножьям каменистых сопок в районе Ер-Ойланской впадины (Линчев-ский, 1935). Для них характерно присутствие крупного кустарникового парнолистника (Zygophyllum atriplicoides) и некоторых специфических петрофильных видов, как, например, Plantago lachnantha, Acantholepis orientalis, Dipterocome pusilia. Из крупных полукустарниковых солянок здесь встречаются Salsola arbuscula и Aellenia subaphylla.
Туранополынники, отмеченные на карте в районе северо-восточных склонов Каратау (Сырдарьинского) имеют несколько иной характер. Они развиваются на опустыненных щебнистых предгорьях, где нет никаких признаков засоления. Поэтому солянки, столь обычные для туркменских туранополынников, здесь отсутствуют. В составе каратавских сообществ из Artemisia turanica видную роль играют ксерофильные и петро-фильные кустарники: Atraphaxis spinosa, A. frutescens, Ephedra dista-chya, Acantholimon alatavicum, Acanthophyllum pungens. Особо следует отметить, что в пределах описываемого контура встречается замечательный, реликтовый кустарник — Spireanthus Schrenkianus. Из травянистых компонентов каратавских туранополынников наиболее обычными являются Stipa caucasica, Lasiagrostis caragana, Festuca sulcata, а также эфемеры и эфемероиды. Выше по склонам Каратау туранополынники сменяются зарослями ксерофильных (на южных экспозициях) и мезофильных (на северных экспозициях) кустарников.
Растительность с преобладанием туранской полыни используется в качестве весенних и осенне-зимних пастбищ. Кормовая производительность-их незначительная — не более 2—2.5 ц/га.
Литература
.Н. А. Базилевская (1929); И. А. Лиичевский (1935).
88 t — из Artemisia tenuis'-eta в сочетании со степями
Формация с преобладанием тонкорассеченной полыни (Artemisia tenui-secta) очень широко распространена по предгорьям Западного Тянь-Шаня и Алая. Особенно характерна она для предгорий, спускающихся в Ферганскую котловину, где относящиеся сюда сообщества образуют хорошо выраженный пояс предгорных полынных пустынь.
Благодаря своему широкому географическому распространению и большому экологическому диапазону, формация тонкорассеченной полыни представлена очень многими ассоциациями. Наиболее типичной ассоциа
цией являются эфемеровые полынники, формирующиеся на щебнистых сероземах лёссовых предгорий в пределах от 600—700 до 1000 м над у. м. Для них очень характерна хорошо выраженная синузия из весенних эфемеров и эфемероидов, состоящая главным образом из осочки (Carex pachystylis) и луковичного мятлика (Роа bulbosa var. vivipara). К этим двум главным компонентам весенней синузии присоединяются многочисленные эфемеровые злаки (Phleum paniculatum, Bromus tectorum, В. macrosta-chys, В. Danthoniae, Eremopyrum orientale) и столь же многочисленное эфемеровое мелкое разнотравье (Alyssum desertorum, Meniocus linifolius, виды Trigonella, Hypecoum, Astragalus, Papaver и т. д.). По зеленому фону весенних растений разбросаны кусты полыни, слагая довольно изреженный ярус. С наступлением летних засух (к концу мая) эфемеры выгорают, стравливаются скотом, и от них иногда не остается даже и следов. В это время в растительном покрове резко преобладает полынь, уже выбрасывающая свои цветоносы. Вместе с ней развиваются другие компоненты летнего травостоя: дерновинные злаки (Stipa Hohenackeriana, Lasiagrostis caragana), зопники (Phlomis salicifolia, P. bucharica), псоралея (Psoralea drupacea) и др.
Нередко в сложении эфемеровых полынников участвуют некоторые солянки: Aellenia subaphylla, Salsola rigida, S- arbuscula, Girgensohnia oppositiflora. При некотором засолении почвы обилие их увеличивается, а участие эфемеров при этом снижается. Таким образом, эфемеровые полынники замещаются галофитными вариантами. Галофитные полынники также довольно широко распространены. Они нередко сочетаются как с эфемеровыми полынниками, так и с настоящими солянковыми пустынями с господством боялыча (Salsola arbuscula, S. laricifolia), биюргунов (Anabasis salsa, A. eriopoda, A. ramosissima).
Вместе с тем, в районах распространения эфемеровых полынников постоянно встречаются участки субтропических степей из крупных эфемероидных злаков: волосоносного пырея (Agropyrum trichophorum), луковичного ячменя (Hordeum bulbosum) и эфемера — тонкоостника (Taenia-therum crinitum) (Лавренко и Соколов, 1949). Таким образом, контуры, обозначенные на карте шифром 88 t, объединяют, в сущности, не только сочетания сообществ Artemisia tenuisecta со степями, но и сочетания их с солянковыми пустынями. Это замечание в особенности относится к контурам, выделенным в пределах Таджикистана.
Следует отметить, что контуры полынников, обозначенные на карте-шифром 88 t, в пределах Таджикистана объединяют собой сообщества не только с преобладанием Artemisia tenuisecta, но и других близких к ней видоц полыней, а именно A. baldshuanica, A. glandulifera, A. Korshinskyi. Полынники, слагаемые этими видами, фитоценотически мало чем отличаются от сообществ, слагаемых A. tenuisecta. Они формируются на более грубых почвах. В их сложении иногда значительное участие принимают юган (Prangos pabularia), Rheum Maximowiczii, Ferula gigantea, Echinops maracandicus, а также растения, распространяющиеся при усиленном выпасе скота, — Peganum harmala и Ceratocarpus turkestanicus.
В составе формации Artemisia tenuisecta, кроме указанных выше эфемеровых и галофитных ассоциаций, может быть выделена еще петрофитпая группа ассоциаций, связанная в своем развитии с грубыми скелетными почвами смытых склонов предгорий и среднегорного пояса (до 1500 — 2000 м над у. м.). Для нее характерно обилие ксерофильных полукустарничков и кустарников, как, например, Eurotia ceratoides, Convolvulus-subhirsutus, Cerasus erythrocarpa, C. verrucosa, Amygdalus spinosissima, A. bucharica. Главным образом к этому варианту полынников, слагаемых.
Artemisia tenuisecta, приурочено распространение фисташки (Pistacia veraj, что отображено на карте соответствующим внемасштабным знаком.
Полынники из Artemisia tenuisecta, и экологически тесно связанная с ними группа горных полынных формаций с эдификаторами из того же подрода Seriphidium, используются в качестве весенних и осенне-зимних пастбищ. Наилучшими кормовыми качествами обладают эфемеровые полынники. Весною их кормовая продуктивность достигает до 5 ц/га сухой массы, но осенью, после исчезновения эфемеров, она снижается до 1.5— 2 ц. Галофитные и петрофитные варианты полынников представляют собой преимущественно осенне-зимние пастбища; их кормовая продуктивность обычно не превышает 2—3 ц/га. Летом Artemisia tenuisecta и близкие к ней виды почти не поедаются скотом, повидимому, из-за наличия в них большого количества эфирных масел. Осенью, после заморозков, полыни становятся вполне поедаемыми. Часть площади, занимаемая по-лынниками, в настоящее время распахивается под поливные зерновые и технические культуры, особенно в Ферганской котловине.
Литература
О. Э. Кнорринг (1915); Е. П. Коровин (1934); Н. Ф. Гончаров (1937); Е. М. Лавренко и С. Я. Соколов (1949); Г. Т. Сидоренко (1953).
88 к — из Artemisia Korshinskyi в сочетании с нагорными ксерофитами и степями
Формация с преобладанием полыни Коржинского не занимает сколько-нибудь значительных площадей. Относящиеся сюда на карте очень небольшие контуры имеются лишь в пределах Таджикистана: один контур относится к высокой долине Зеравшана, другой лежит вдоль западных склонов хребтов Бадахшана 1 (Язгулемского, Рушанского, Шугнанского, Вахан-ского).
Artemisia Korshinskyi — вид, близкий к A. tenuisecta, которую он сменяет на более высоких местоположениях, а именно средних и высоких поясах гор. Экологический ареал A. Korshinskyi в верхней своей границе смыкается с ареалом типичной высокогорной полыни — A. Lehmanni-апа, а в нижней границе — с A. tenuisecta. Таким образом, полынь Коржинского в экологическом отношении занимает как бы промежуточное место -между этими двумя видами.
Сообщества, образуемые полынью Коржинского, встречаются в условиях сильно пересеченного рельефа, на грубоскелетных почвах по крутым каменистым склонам, на высотах от 2000 до 3000 м над у. м. В их сложении большое участие принимают типичные пустынные полукустарнички и нагорные подушковидные ксерофиты (Kochia prostrata, Eurotia ceratoi-des, Onobrychis echidna, виды Acantholimon). Этот вариант полынников, слагаемых Artemisia Korshinskyi, является наиболее характерным и распространенным. Кроме указанных полукустарничков, в нем встречаются некоторые злаки: чаще всего кавказский ковыль (Stipa caucasica) и типчак {Festuca sulcatas. 1.), реже Stipa turkestanica и S. kirghisorum. В условиях более мягких склонов гор указанные злаки приобретают господствующее значение, формируя степные ассоциации. Наоборот, увеличение скелет-ности почвы ведет к преобладанию нагорных ксерофитов. Таким образом, контуры карты, обозначающие полынники из Artemisia Korschinskyi, включают не только эти сообщества, но и сочетания их со степями, а также с нагорными ксерофитами.
1 В трактовке К. В. Станюковича (1952).
В Бадахшане, в пределах контура, показанного на карте, встречаются колючетравники с господством видов Cousinia.
Благодаря сильно пересеченному рельефу, значительная часть площади, занятая полынниками из Artemisia Korshinskyi, представлена каменистыми склонами, осыпями и скалами, почти совсем лишенными растительности. В хозяйственном отношении вся относящаяся сюда территория являет собою скудное, с низкой производительностью пастбище, преимущественно для мелкого рогатого скота.
Литература
Н. Ф. Гончаров (1937).
88 1 — из Artemisia Lehmanniana
Полынь Лемана (Artemisia Lehmanniana) — ксерофитный полукустарничек 25—35 см высоты — является типичным высокогорным видом. Нижняя граница распространения ее редко спускается ниже 2000 м над у. м. Сообщества, образованные этим видом, встречаются в высоких нагорьях Центрального Тянь-Шаня, в высокогорных поясах Алая, но в наиболее развитом виде они представлены на Памире.
Согласно данным К. В. Станюковича (1949), памирские полынники из Artemisia Lehmanniana располагаются в субальпийском и в нижней части альпийского поясов, на высоте от 3800 до 4300 м над у. м. Их развитие связано главным образом с западными районами Памира, лежащими на границе с Бадахшаном, а также с южными частями Восточного Памира. Эти районы лучше обеспечены атмосферными осадками и, в частности, зимним снеговым покровом. Широко распространены указанные полынники и в субальпийском поясе горного Бадахшана. Почвы под ними — хрящевато-суглинистые или супесчано-хрящеватые высокогорные сероземы, переходные к типу бурых. Проективное покрытие растительностью в среднем около 30—40%, высота покрова 25—30 см. В покрове господствуют кустики Artemisia Lehmanniana, к которым, в зависимости от различных условий среды (главным образом от степени увлажнения), примешиваются различные содоминанты. Для наиболее сухих местообитаний субальпийского пояса Памира характерна терескеново-полынная ассоциация (из Artemisia LehmannianaA-Eurotia ceratoides). В менее сухих условиях развивается полынная ассоциация с Tanacetum pamiricum. По северным склонам, лучше обеспеченным почвенной влагой, формируются остепненные злаково-полынные сообщества, в которых к Artemisia Lehmanniana примешиваются некоторые степные дерновинные злаки (Stipa orientalis, S. turkestanica, Poa relaxa), степная осочка (Carex duriusculiformis), различное ксерофильное разнотравье (Cousinia rubiginosa, Lindelofia stylosa и др.). На склонах гор, у верхних границ субальпийского пояса, встречаются полынные ассоциации со значительным участием подушковидных кустарничков и полукустарничков: куртковника (Acantholimon diapen-sioides), остролодки (Oxytropis Poncinsii, О. melanotricha), лапчатки (Potentilla pamiroalaica).
В горном Бадахшане, наряду с широко распространенными остепнен-ными полынниками с участием Festuca sulcata s. 1., Stipa orientalis, S. turkestanica, Poa relaxa, широко развиты колючеподушечные их варианты с Acanthol imon pamiricym, в составе которых постоянными компонентами являются Arenaria Griffithii, Cousinia rubiginosa, Carex duriusculiformis и другие ксерофильные виды.
В общем формация, слагаемая полынью Лемана, как и многие другие пустынные формации, развитые на высоких нагорьях Азии, флористически
очень бедна. Судя по имеющимся данным, в ней насчитывается всего липп» Ю—15 видов. При использовании ее в качестве пастбища для мелкого рогатого скота, она дает незначительную кормовую массу. Бблыпая часть площади в районах распространения этой формации приходится на крутые, каменистые трудно доступные склоны.
Литература
И. А. Райкова (1930); Е. П. Коровин (1934); Н. Ф. Гончаров (1937); К. В. Станюкович (1949, 1952, 1955); Г. Т. Сидоренко (1953).
89. Полынные пустыни в сочетании с солянковыми
Пустыни этого типа наиболее широко распространены в пределах пустынной области. На карте они показаны самостоятельными крупными контурами на обширных пространствах равнин от низовьев Эмбы до северного Устюрта и Мангышлака, в южной части Тургайской столовой страны, в Центральном Казахстане, занимают почти всю Бетпак-Далу, северное побережье Балхаша, покрывают подгорные равнины Каратау и северных цепей Тянь-Шаня, а также останцовых плато центральных Кызылкумов. Мелкие массивы полынных пустынь в сочетании с солянковыми пустынями встречаются на огромном протяжении от побережья оз. Челкар на северо-западе пустынной области до Джунгарских ворот на востоке, достигая на юге Красноводского плато и депрессий Унгу за. В этих пределах они контактируют с различными типами пустынь (злаково-полынными, полынными, эфемерово-полынными, солянковыми и др.), и, кроме того, очень часто встречаются в пределах последних, не будучи показаны на карте по условиям масштаба карты.1
В своем распространении полынные пустыни в сочетании с солянковыми связаны с областями третичных отложений и, реже, с отложениями четвертичного периода.
Господствующий равнинный рельеф, сопровождаемый к тому же в большинстве случаев горизонтально залегающими коренными породами, обусловливает бессточность занятых сообществами этого типа территорий. Как правило, почвообразующие коренные породы богаты гипсом и другими солями. Все это, влияя на почвенные процессы, в значительной мере определяет и основные черты растительного покрова. Почвы этого типа пустынь серо-бурые, часто солонцеватые и с выраженным гипсовым горизонтом.
Грунтовая вода повсеместно залегает на большой глубине, за исключением впадин и других глубоких депрессий. Незначительные колебания в строении поверхности (наличие слабо выраженного микрорельефа), либо в механическом составе почв и почвообразующих пород, глубина залегания скоплений гипса и т. п. отражаются на характере растительного покрова, который отличается достаточно четкой комплексностью, одна ко менее ярко выраженной, чем это наблюдается в пределах злаковополынных пустынь.
Количество осадков в пределах ареала рассматриваемых пустынь является наименьшим для пустынной области СССР, что и накладывает отпечаток на растительный покров, характеризующийся ярким проявлением черт ксерофитности и угнетенности, а также исключительным преобладанием в сложении его представителей полукустарничковой жизненной
1 Ареал полынных пустынь в сочетании с солянковыми лежит в пределах Туран-ской группы провинций и заходит в северную часть Переднеазиатской группы провинций (Прозоровский и Малеев, 1947).
формы, а именно видов полыни из подрода Seriphidium (Artemisia terrae-albae s. str., A. turanica, A. gurganica, A. kemrudica) и ряда видов Salsola (Salsola arbuscula, S. laricifolia, S. rigida) и других видов семейства маревых (Anabasis salsa, Nanophyton erinaceum и др.).
Полынные сообщества связаны с наиболее «пресными», а потому и наиболее благоприятными экологическими условиями. В них растительный покров достигает наибольшей сомкнутости (до 40—50% покрытия) и наибольшего видового разнообразия за счет сопутствующих видов, в частности эфемеров (вообще бедно представленных в этом типе пустынь). Сообщества же кеурека (Salsola rigida), боялыча (Salsola laricifolia, S. arbuscula), биюргуна (Anabasis salsa) и других полукустарничков из семейства маревых всегда связаны с сильно солонцеватыми или солончако-ватыми вариантами серо-бурых почв и отличаются разреженностью растительного покрова; между отдельными кустами остаются большие открытые проплешины, на которых нередко развиваются лишайники, мхи и водоросли.
Наиболее типичный (часто встречающийся) комплекс представлен тремя сообществами: полынным, боялычно-полынным и биюргуно-вым.
Сообщества полыни (Artemisia terrae-albae s. str., A. turanica, A. kemrudica и др.) всегда приурочены к слегка повышенным местам и к почвам более легкого механического состава. Нередко на поверхности почвы и в верхних горизонтах ее присутствует небольшое количество мелкой щебенки, грубого хряща и гравия. Основной фон образует полынь, кусты которой дают до 40—50% покрытия. В качестве незначительной примеси встречается кеурек (Salsola rigida), очень редки курчавка (Atra-phaxis spinosa), прутняк (Kochia prostrata) и некоторые другие. Из эфемероидов наиболее обычны тюльпаны (Tulipa Greigii, Т. Behmiana, Т. Albertii, Т. turkestanica) и луки (Allium Schubertii, A. Borszczowii), гусиные луки (Gagea reticulata, G. stipitata), Rhinopetalum Karelinii, Colpodium humile-, из эфемеров — Lepidium perfoliatum, Koelpinia linearis, Litvino-via tenuissima, Tauscheria lasiocarpa, Ziziphora tenuior, Heteracia Szowitsii, Trigonella orthoceras, Lappula spinocarpos, Leptaleum filifolium, Alyssum desertorum, Pachypterygium multicaule и др. Иногда встречаются участки полынников со значительным участием шайра (Ferula ferulaeoides).
Биюргунники (сообщества Anabasis salsa) занимают незначительно пониженные, с глинистыми почвами, участки. Растительный покров отличается своей изреженностью, степень покрытия 15—20%; нередко с биюр-гуном растет тасбиюргун (Nanophyton erinaceum)-, другие полукустарнички встречаются крайне редко: боялыч (Salsola laricifolia), кеурек (Salsola rigida), но чаще других виды Anabasis (A. eriopoda, A. truncata). Эфемероиды в биюргунниках редки и бедны по видовому составу, что объясняется, повидимому, такыровидностью почвенной корки и засоленностью почвы; здесь изредка можно встретить эфемероиды (Tulipa Albertii, Carex pachystylis, Colpodium humile, Eremurus inderiensis, Megacarpaea megalocarpa, Rheum tataricum, Eremostachys affinis) и эфемеры (Lepidium perfoliatum, Veronica biloba и др.); кроме того, обычны единичные однолетние маревые (виды Halimocnemis, Salsola, Gamanthus).
Полынно-боялычные (а часто и чисто боялычные) сообщества занимают промежуточные между двумя названными позиции, встречаясь также на пологих склонах сопок и бессточных впадин. Кусты боялыча, даже с примесью полыни, обычно покрывают менее 50% поверхности. Здесь также встречаются единичные экземпляры терескена (Eurotia ceratoides), курчавки (Atraphaxis spinosa) и единичные или рассеянные, но более раз-20*
нообразные, чем в биюргунниках, эфемероиды (Rheum turkestanicum, Eremurus inderiensis, Ferula Syreitschikovii, виды Allium a Gagea) и эфемеры ‘(Alyssum desertorum, Lepidium perfoliatum и многие другие виды, уже ранее перечисленные). В центральной части Бетпак-Далы в различных сообществах отмечено около 20—25 видов эфемероидов и 50 видов эфемеров. Однако они нигде не образуют сомкнутого травостоя, а встречаются отдельными экземплярами или группками (Рубцов, 1943; Рубцов и Кубанская, 1949). Таков облик сообществ, составляющих комплексы в Бетпак-Дале, которая может служить образцом наиболее типичного проявления этого типа пустынь (Коровин и Миронов, 1935).
Однако, на столь огромной площади распространения, как было отмечено в начале этого раздела, полынные пустыни в сочетании с солянковыми имеют значительные отличия по флористическому составу и набору входящих в комплексы сообществ, определяемые местными природными особенностями, и представлены поэтому различными вариантами.
Литература
И. М. Крашенинников (1923, 1925); Е. П. Коровин (1934, 1935); Е. П. Коровин и Б. А. Миронов (1935); А. В. Прозоровский (1940а); Н. И. Рубцов (1943, 1952); А. В. Прозоровский и В. П. Малеев (1947); Н. И. Рубцов и 3. В. Кубанская (1949); Н. Т. Агеева и А. 3. Соломченко (1951).
89 кк — прикаспийско-казахстанские (из Artemisia pauciflora s. 1., A. Lercheana, Anabasis salsa, Atriplex сапа)
Прикаспийско-казахстанские полынные пустыни в сочетании с солянковыми простираются от р. Урала до Тургайской столовой страны, доходя на юг не далее сухой дельты Эмбы.
Для широкой полосы Прикаспийской низменности в низовьях Урала— Эмбы характерен (Никитин, 1954а) сильно расчлененный мезорельеф: увалы (гривы 7—15 м высотой) чередуются с понижениями, обычно занятыми солончаками (сорами); количество последних возрастает по мере приближения к Каспийскому морю. Для этого обширного района характерен следующий комплекс. На увалах и гривах распространены белополынные (из Artemisia Lercheana) и биюргуново-полынные (из Artemisia Lercheana и Anabasis salsa) сообщества на солонцеватых бурых и серо-бурых почвах. В годы с благоприятным тепловым режимом и обильными ' осадками весной развиваются луковичный мятлик (Роа bulbosa var. vivipara) и довольно обильные эфемеры: Leptaleum filifolium, Rochelia disperma, Goldbachia laevigata, Sterigmostemum tomentosum, Arnebia decumbens, Lappula patula, L. spinocarpos, Alyssum desertorum, Trigonella arcuata, MalcoUnia taraxacifolia, Plantago minuta, Lepidium perfoliatum, Chorispora tenellavijvp. Одиночными экземплярами встречаются эфемероиды: Tulipa biflora, Ferula nuda, F. canescens, Cachrys odontalgica, Scorzonera pusilia. Характерны здесь и летне-осенние однолетники: Halimocnemis sclerosperma, Salsola brachiata, S. aralensis, S. lanata, S. tamariscina, а также весьма обычны лишайники. В годы, бедные осадками, эфемеры развиваются очень слабо.
В нижних частях склонов преобладают биюргунники (из Anabasis salsa), отличающиеся крайне изреженным покровом биюргуна с ничтожным участием немногих наиболее галофильных эфемеров с однолетними маревыми (виды Halimocnemis, Salsola).
По краям солончаковых депрессий господствуют сообщества сарса-зана (Halocnemum strobilaceum) с примесью полукустарничковых галофитов: поташника (Kalidium foliatum), кзылкокпека (Limonium suffru-ticosum), кокпека (Atriplex сапа), солончаковой полыни (Artemisia топо-gyna), кустарничковой шведки (Suaeda dendroides) и некоторых однолетних маревых (Salicornia europaea, Salsola brachiata, Petrosimonia triandra, P. brachyphylla).
Характерным элементом ландшафта в предустюртской части Прикаспийской низменности являются останцовые возвышенности; плоская поверхность их покрыта биюргуново-серополынными (из Artemisia terrae-albae s. 1. и Anabasis salsa) сообществами с участием боялыча (Salsola ar-buscula) и кеурека (Salsola rigida), сходными с пустынями северотуран-ского варианта, с которыми этот район приходит в соприкосновение.
Литература
И. М. Крашенинников (1925); Н. В. Павлов (1926); О. Э. Кнорринг (1928); Б..А. Борнеман и М. Д. Спиридонов (1929); М. Д. Спиридонов (1930а); Е. П. Коровин (1934); А. В. Прозоровский (1940а); И. В. Ларин, Е. В. Шиффере и др. (1954); С. А. Никитин (1954, 1954а).
89 стр — северотуранские (из северотуранских видов полыни, Salsola laricifolia, Anabasis salsa и др.)
Северотуранский вариант полынных пустынь в сочетании с солянковыми распространен наиболее широко — от Каспийского моря до Джунгарских ворот. В общей характеристике уже были очерчены основные, слагающие его комплексы на примере растительности Бетпак-Далы. Ознакомимся еще с некоторыми географическими районами для более подробного освещения пустынь этого типа.
В этом отношении интересен район северо-западного Устюрта, Мангышлака и п-ва Бузачи.
Широко распространенные здесь сообщества северотуранских видов полыни (Artemisia terrae-albae s. str., A. terrae-albae ssp. Massagetovii, A. diffusa, A. turanica), зачастую связанные переходными формами между собой и с Artemisia Lercheana (особенно по северной границе распространения этого варианта), отличаются значительным флористическим разнообразием сопутствующих видов и входят в различные комплексы.
Обычны здесь двучленные комплексы, которые (например на п-ве Бузачи) состоят из полынников и боялычных полынников на серобурых «пресных» (нормальных) или солонцеватых почвах и биюр-гунников на такыровидных солонцах; эти последние сообщества занимают 30—40% площади (Борнеман и Спиридонов, 1929; Агеева и Соломченко, 1951; Момотов, 1953).
Биюргунники, как обычно, бедны сопутствующими растениями; кроме биюргуна (Anabasis salsa), здесь единично встречаются полукустарнички: итсегек (Anabasis aphylla) и кеурек (Salsola rigida), летне-осенний однолетник Ceratocarpus arenarius и эфемеры: Eremopyrum orientale, Meniocus linifolius, Leptaleum filifolium, Lepidium perfoliatum, Tetracme quadricornis, Veronica amoena, Lappula spinocarpos. Обычны здесь лишайники (Parmelia vagans, P. ryssolea, виды Collema, Aspicilia desertorum и др.) и водоросли.
Полынники (в широком понимании, то есть чистые полынники, боялыч-ные полынники. биюпгуновые полынники, кеурековые полынники и др.)
отличаются довольно богатым составом входящих в них растений, причем количество их всегда возрастает при опесчанивании субстрата и расчленении рельефа. В полынниках этого района встречаются следующие виды полукустарничков: Artemisia gurganica, A. terrae-albae s. str., A. kemrudica (и большое число связующих их переходных форм), итсегек (Anabasis aphylla), прутняк (Kochia prostrata), боялыч (Salsola laricifolia), тасбиюргун (Nanophyton erinaceum). Изредка попадаются кустарнички: карагана (Caragana grandiflora) и вьюнок (Convolvulus fruti-cosus). На песчаных и щебнистых участках встречаются единично и небольшими группами пустынные ковыли (Stipa sareptana ч&г. kasakorum, S. Szowitsiana, Lasiagrostis caragana) и еркек (Agropyrum sibiricum). Эфемероиды и эфемеры в «средние» по осадкам годы немногочисленны, но во влажные образуют иногда даже почти сплошное покрытие почвы. Здесь обычны следующие виды: из эфемероидов — Gagea reticulata, Роа bulbosa var. vivipara, Colpodium humile, Astragalus erioceras; из эфемеров — Eremopyrum orientate, Alyssum desertorum, Meniocus linifolius, Leptaleum fili-folium, Veronica amoena, Androsace Turczaninovii, Astragalus tribuloides, A. oxyglottis, A. commixtus, Lappula spinocarpos, Ceratocephalus ortho-ceras, Tetracme quadricornis, Roemeria hybrida, Diptichocarpus strictus, Eragrostis minor, Trigonella arcuata и некоторые другие. Из летне-осенних однолетников наиболее обычен Ceratocarpus arenarius, на сбитых выпасом местах образующий часто сплошной покров; кроме того, встречаются Halogeton glomeratus, Salsola crassa, S. foliosa S. nitraria, Halimocnemis scterosperma, H. Karelinii, Horaninowia ulicina я. некоторые другие.
Этот обширный список является сводным по многим описаниям для различных сообществ с господствующей ролью полыни (Борнеман и Спиридонов, 1929; Русанов, 1930а); обычно же в конкретных сообществах количество входящих в них видов колеблется в пределах 10—15, редко достигая 20.
Помимо перечисленных высших растений, широко распространены лишайники, которые присутствуют почти во всех сообществах. Количество и видовое разнообразие их возрастает в менее сомкнутых группировках. Характерны следующие виды: Diploschistes scruposus, Squamaria peltata, S. muralis, Lecanora aspera, L. frustulosa, Aspicilia desertorum, Parmelia ryssotea, P. prolixa, Placodium elegans и др.
Необходимо отметить, что двучленные комплексы только в схеме характеризуют растительный покров наиболее однородных плакорных условий; на этом основном фоне всегда отмечается присутствие еще нескольких сообществ, вносящих значительное разнообразие в общую картину. Так, например, на п-ве Тюб-Карагане среди господствующего фона полынников и биюргунников встречаются злаковые полынники (на склонах с песчаным или дресвяно-песчаным плащом), сообщества еркека (на песках), тасбиюргунники (на каменистых и щебнистых склонах), сарсазанники (по солончаковым депрессиям) и луговые сообщества из Aelu-ropus litoralis (по менее засоленным понижениям), а также многие переходные между перечисленными сообщества.
На юго-восточной окраине Тургайской столовой страны, по данным М. Д. Спиридонова (1918), широко распространены боялычево-полынные (Artemisia terrae-albae s. str. -|- Artemisia turanicaA-Salsola laricifolia) и полынные (Artemisia terrae-albae s. str. и A. turanica) комплексные сообщества на серо-бурых почвах. Боялычно-полынники занимают бблыпую часть территории. Видовой состав этого сообщества (в целом) характеризуется большим разнообразием (свыше 100 видов учтено М. Д. Спиридоновым на основе 32 описаний), включая полукустарнички (Salsola rigida.
Kochia prostrata, Ephedra di st achy а), длительновегетирующие многолетники (Stipa Szowitsiana, S. capillata, S. caucasica, Lasiagrostis caragana, Eremostachys tuberosa), эфемероиды (Poa bulbosa, var. vivipara, Allium caesium, Colpodium humile, Asparagus Breslerianus, Tulipa Borszczowii, Ferula ferulaeoides, Oedibasis apiculata), длительновегетирующие однолетники (Ceratocarpus arenarius, Girgensohnia oppositiflora, Salsola tamariscina, Echinopsilon sedoides, Kirilowia eriantha) и эфемеры (Eremopyrum orientate,
Рис. 73. Проекция комплекса на площади 1 га северной части полынно-солянковой пустыни
Устюрт.
1 — мелкий биюргун (Anabasis salsa) на солончаковатосолонцеватой серо-бурой почве; 2 — крупный биюргун (Anabasis salsa) на слабосолонцеватой серо-бурой почве;
3— полынь (Artemisia terrae-albae и A. maikara) на слабосолонцеватой серо-бурой почве; 4 — боялыч (Salsola laricifo-На), полынь (Artemisia terrae-albae и A. maikara) на слабосолонцеватой серо-бурой почве; 6 — полынь (Artemisia terrae-albae) с ковылем (Stipa Korshinskyi) и кустарниками (Atraphaxis spinosa) на промытой почве западин; 6—-биюргун с полынью на слабосолонцеватой серо-бурой почве; 7— боялыч без Польши; 8 — холмики землероев; 9 — кусты курчавки (Atraphaxis spinosa) (И. Ф. Мрмотов, 1953).
С er otocephalus orthoceras, Schismus arabicus. Alyssum desertorum, Chorispora tenella, Veronica biloba, Leptaleum filifolium, Tauscheria lasiocarpa, Senecio coronopifolius я др.). Характерно постоянное и почти всегда обильное присутствие лишайников: Aspicilia alpino-desertorum и, реже. Psora decipiens.
Среди господствующих боялычно-полынников в виде островных включений встречаются еще сообщества биюргуново-полынные (Artemisia terrae-albae s. str. + Artemisia turanica-}-Anabasis salsa), биюргуново-тас-биюргуновые (Anabasis salsa-}-Nanophyton erinaceum) и карагановые (Caragana balchaschensis)', последние приурочены к небольшим замкнутым понижениям. Флористический состав зарослей караганы весьма богат; наряду с некоторыми типичными пустынными видами в них входит большое ко* '
личество степных растений. Биюргуново-полынники и биюргуново-тас-биюргунники слагаются незначительным числом видов.
Общий запас растительной массы боялычно-полынников М. Д. Спиридонов оценивает в 1.5—2.3 ц/га (в воздушно-сухом состоянии).1
Весьма сходную картину дает и И. В. Ларин (1941) для Центрального Казахстана (район между Карсакпаем и Джезказганом). Здесь господствуют: 1) полынники (из Artemisia semiarida) на суглинистых сероземах (светлобурые почвы) в комплексе с 2) боялычниками (из Salsola laricifolia с примесью полыней, чаще всего Artemisia turanica) на сероземах, нередко щебневатых (рис. 71). Кроме того, в комплексе часто присутствуют еще следующие сообщества: 3) биюргунники (из Anabasis salsa} на такыровидных почвах и корковых солончаках; 4) чернополынники (из Artemisia pauciflora) на солонцах; 5) тасбиюргунники (из Nanophyton erinaceum) на сильнощебневатых, с близким гипсовым горизонтом, почвах.
В равнинных условиях в северной части плато Бетпак-Далы распространены комплексы почвенно-растительного покрова, также слагаемые 4—5 членами: полынные сообщества (из Artemisia terrae-albae s. str., иногда с примесью A. turanica); биюргуново-полынные (из Artemisia terrae-albae s. str. и Anabasis salsa); биюргуновые или биюргуново-тасбиюр-гуновые (из Anabasis salsa или из Nanophyton erinaceum и Anabasis salsa); полынно-боялычевые (из Salsola laricifolia и Artemisia terrae-albae s. str.); карагановые сообщества (из Caragana balchaschensis с участием степных злаков). Господствующими среди них являются полынные сообщества (из Artemisia terrae-albae s. str.) на серо-бурых почвах, которые рядом исследователей рассматриваются как выражение зонального типа растительности (Спиридонов, 1918; Павлов, 1931 и др.).
Литература
Г. С. Карелин (1883); Б. А. Федченко и И. М. Крашенинников (1910); А. И. Михельсон (1913); М. Д. Спиридонов (1918, 1930а); И. Ми Крашенинников (1925); Н. В. Павлов (1926, 1930, 1931, 1935); М. М. Ильин и Р. Ю. Рожевиц (1928); Б. А. Борнеман и М. Д. Спиридонов (1929); Г. И. Доленко (1930); С. С. Неуструев и И. М. Крашенинников (ред.) (1930); Р. Ю. Рожевиц (1930); Ф. Н. Русанов (1930а); Е. П. Коровин (1934, 1935); Б. С. Закржевский и Е. П. Коровин (1935); Е. П. Коровин и Б. А. Миронов (1935); Е. П. Коровин и В. А. Селевин (1935); П. Г. Стрельников (1938); И. В. Ларин (1941); М. Г. Попов (1941); Н. И. Рубцов (1941а, 1944, 1950а); Е. П. Коровин и С. А. Шувалов (1948); 3. В. Кубанская (1949); Н. И. Рубцов и 3. В. Кубанская (1949); Н. Т. Агеева и А. 3. Соломченко (1951); И. Ф. Момотов (1953); Ш. И. Коган (1954); Н. Н. Пельт (1955); Н. Н. Пельт и Л. Е. Родин (1955).
89 ютр— южнотуранские (из Artemisia herba-alba s. I., Salsola gemmascens, S. arbuscula)
Южнотуранский вариант полынных пустынь в сочетании с солянковыми представлен на карте немногими контурами. Наиболее крупные из них лежат на Красноводском плато, на западных склонах южного Устюрта (обращенных к заливу Кара-Богаз-Гол), в центральной и южной части Кызылкумов, а также вдоль Унгуза. Мелкие контуры пустынь этого типа приурочены к останцам в районе Сарыкамышской дельты Аму-Дарьи
1 Н. Т. Агеева и А. 3. Соломченко (1951) оценивают валовую производительность комплексных биюргуновых и полынных пастбищ в 7.9 ц/га, в то время как поедаемая масса равна в среднем всего 2.9 ц/га (последняя цифра сильно колеблется в зависимости от вида скота, выпасаемого на пастбище: для крупного рогатого скота — 1.4 ц/га, для овец и коз — 1.6 ц/га, для верблюдов — 5.6 п/га).
(из них по условиям масштаба показаны только некоторые), где они лежат среди солянковых пустынь и черносаксаульников.
Это — южный вариант пустынь данного типа. Основные строители сообществ — виды южнотуранских полыней: кемрудской и бадхызской (Artemisia kemrudica, A. badghysi и формы, переходное между A. kemrudica и A. gurganica),1 образующие то почти чистые сообщества (с незначительной примесью других полукустарничков), то тетырово-полынные (полынь и Salsola gemmascens), то боялычно-полынные (полынь и Salsola arbuscula) на серо-бурых почвах, отличающихся солончаковатостью (в случаях с тетыром) или некоторой песчанистостью (в случаях с боялы-чом).
На Устюрте наиболее типичны трехчленные комплексы: полынники на повышенных элементах рельефа и верхних частях склонов увалов с серо-бурыми почвами; тетырники на солончаковатых почвах нижних частей склонов2 или плоских и сточных депрессий; биюргунники в замкнутых понижениях с застоем вод поверхностного стока на такыровидных солонцеватых почвах. Полынники занимают более значительные площади и заметно разнообразнее по флористическому составу. Помимо основного строителя сообществ — полыни (представленной зачастую несколькими формами), обычно присутствуют и другие полукустарнички и кустарнички: боялыч (Salsola arbuscula), кеурек (Salsola rigida), вьюнок (Convolvulus fruticosus), курчавка (Atraphaxis spinosa). Изредка встречаются многолетние травы — пустынные ковыли (Stipa Szowitsiana, S. Richteriana). Эфемероиды и эфемеры здесь также обычно не образуют сомкнутого покрова (за исключением весен с особо благоприятными условиями по осадкам и тепловому режиму, когда они достигают значительного развития). Первые представлены следующими видами: Allium caspium, А. Вог-szczowii, Tulipa Borszczowii, Т. Buhseana, Т. Biebersteiniana, Carex physodes (отнюдь не на песчаных почвах, а на глинистых; единично, рассеянно). Из эфемеров характерны: Eremopyrum orientals, Matricaria lamellata, Koelpinia linearis, Matthiola chenopodiifolia, Amberboa turanica, Zollicoferia (Launea) Korovinii, Heteroderis pusilia, Acantholepis orientalis, Tetracme quadricornis, Senecio coronopifolius, Lappula spinocarpos, Leptaleum filifolium, Scorzonera pusilia, Hypecoum pendulum, Meniocus-linifolius, Lallemanthia Royleana, Alyssum marginatum, A. desertorum, I satis-trachycarpa, Nonea turcomanica, Heterocaryum Szovitsianum, Londesia eriantha и др. Летне-осенние однолетники немногочисленны: Halimocnemis-Karelinii, Н. sclerosperma, Salsola lanata, Halocharis hispida, Petrosimonia glauca, Gamanthus gamocarpus.
Видовой состав биюргунников самый бедный, как и в других, уже описанных вариантах. К господствующему биюргуну чаще других полукустарничков примешивается кеурек (Salsola rigida), в меньшей степени тасбиюр-гун (Nanophyton erinaceum) и совсем редко тетыр (Salsola gemmascens} и боялыч (Salsola arbuscula). Эфемероиды отсутствуют, эфемеры крайне-редки, летне-осенние однолетники встречаются чаще.
Сообщества тетыра (Salsola gemmascens) хотя и несколько богаче, но' все же очень ограничены по видовому составу. Зато флористическое разнообразие их резко возрастает в местах выхода на поверхность известняков, песчаников и мергелей и, особенно, на обрывах и в оврагах, а также на песчаных плащах, где сразу появляются многие скальные формы и виды,
1 Эта виды ранее включались в Artemisia herba-alba s. 1.
9 По наблюдениям Ф. Н. Русанова (1930а), тетыр (Salsola gemmascens) занимает* по преимуществу склоны с более или менее размытой поверхностью.
связанные с дополнительным увлажнением. На всех этих местообитаниях встречаются следующие виды: Caragana grandtflora, Rhamnus Sintenisii, Cousinia onopordioides, Jurinea persimilis, Crambe edentula, Schumannia Karelinii, Stipa Szowitsiana, S. Hohenackeriana, Seseli eriocephalum, Zygo-phyllum Eichwaldii, Z. turcomanicum, Allium sabulosum, A. caspium, Hap-lophyllum Bungei, H. obtusifolium, Calligonum junceum, Capparis spinosa и еще ряд других, а также очень многие эфемеры и летне-осенние виды маревых (Русанов, 1930а).
Следует отметить, что в нагорных местах среди полынников и боялыч-ников встречается черный саксаул (Haloxylon aphyllum) в виде низкорослых и крайне редко разбросанных экземпляров.1
Полынные пустыни в сочетании с солянковыми на Унгузе развиваются в основном под влиянием сильной солончаковатости почв этого своеобразного района. Сообщества комплексов Унгуза разнообразятся заметным распространением там саксаульчика (Arthrophytum Lehmannianum), а также участием боялыча (Salsola arbuscula), при наличии небольшого количества галофильных эфемеров (Matricaria lamellata, Streptoloma desertorum, Astragalus tribuloides, Schismus arabicus, Amberboa turanica) и летне-осенних однолетников (Salsola carinata, S. sclerantha, Halotis pilosa, Halimocnemis sclerosperma). Солянковые пустыни представлены сообществами те-тыра (Salsola gemmascens), боялыча (Salsola arbuscula) is. весьма разреженных сообществ из видов Anabasis (Л. eriopoda, A. brachiata, A. truncate) (Шингарева, 1940).
Чрезвычайно пеструю картину образуют полынные пустыни в комплексе с солянковыми на останцовых возвышенностях в центральных Кызылкумах. Широкое распространение щебнистых, дресвяных и песчаных -субстратов обусловливает внедрение в состав полынных и солянковых -сообществ многих видов, в особенности скальных и псаммофильных: Aellenia subaphylla, Astragalus squarrosus, Artemisia santolina, Acantho-phyllum pungens, Convolvulus hamadae, Cousinia affinis, Ferula assa-foe-tida, Iris songarica, Aristida pennata, Stipa Hohenackeriana, Carex physo-des и другие, присутствие которых маскирует границы между отдельными сообществами в комплексах.
Для названного района характерно взаимное наложение ареалов северотуранских и южнотуранских видов полыни (Artemisia terrae-albae •8. str., A. terrae-albae ssp. Massagetovii, A dijjusa, A. arenicola и др.) и почти постоянное участие, наряду с названными, еще и A. turanica. Характерно также совместное произрастание Роа bulbosa var. vivipara и Carex physodes. Точно так же весьма обычным является присутствие единичных и очень сильно рассеянных экземпляров белого саксаула (Haloxylon persicum) и некоторых видов джузгуна (Calligonum microcarpum, С. alatiforme и др.) (Порецкий и др. 1936; Гранитов, 1950).
Обширные площади полынных пустынь в сочетании с солянковыми используются как пастбища, преимущественно осенне-зимние, когда становятся поедаемыми полукустарничковые солянки и главным образом ' полынь, и лишь в годы с обильным развитием эфемеров — как весенние.
Урожайность поедаемой массы на лучших полынных пастбищах достигает 4.3 ц/га; она падает с ослаблением участия полыни (полынно-боялычные, полынно-тетыровые и другие дают всего 1.5—2 ц/га поедаемой массы), в биюргунииках снижаясь до 1—1.6 ц/га.
Полынники представляют весьма ценные (нажировочные для овец) пастбища, однако степень использования их очень незначительна, так
1 Некоторые исследователи считают, что здесь распространен зайсанский саксаул (Haloxylon ammodendron) (Коровин и Гранитов, 1949; Нечаева, 1951).
как в районах их распространения очень редкая сеть колодцев и, как правило, грунтовые воды залегают глубоко и труднодоступны.
Литература
Е. П. Коровин (1927, 1927а, 1934); Ф. Н. Русанов (1930а); В. Н. Минервин (1935); А. С. Порецкий и др. (1936); Е. А.. Шингарева (1940); Н. Т. Нечаева и др. (1943); Ш. И. Коган (1949); Е. П. Коровин и И. И. Гранитов (1949); И. И. Гранитов (1950); Н. Т. Нечаева (1951); Н. Н. Пельт (1955); Н Н. Пельт и Л. Е. Родин (1955).
90. Эфемерово- полынные пустыни
Эфемерово-полынные пустыни показаны на карте несколькими группами контуров: на крайнем западе в Закавказье в долинах Аракса и Куры с прилегающими низкогорьями, на подгорных равнинах и в низкогорьях Копет-Дага и на подгорных равнинах останцового плато в центральных Кызылкумах. Кроме того, отдельные небольшие массивы этих пустынь показаны на подгорной равнине Каратау (Сырдарьинского), а также к северу от Таласского и Заилийского Алатау. Самый восточный массив показан на подгорной равнине Чу-Илийских гор.
Таким образом, эфемерово-полынные пустыни лежат почти исключительно в пределах Переднеазиатской группы провинций (в понимании А. В. Прозоровского; Прозоровский и Малеев, 1947).
Как показывает перечень районов распространения этих пустынь, они всегда связаны с подгорными равнинами и низкогорьями, в пределах -абсолютных высот от 150 до 400—500 м над у. м. (в редких случаях, как, например, в Копет-Даге, поднимаясь и выше).
В этих высотных* пределах распространены светлые и типичные сероземы, отличающиеся значительной карбонатностью и часто солончакова-тостью, залегающие на лёссах и лёссовидных породах. Однако обычное положение их на склонах и шлейфах предгорий сообщает им во многих случаях заметную скелетность, а иногда даже защебненность.
Экологические условия существования этого типа пустыни определяются в основном режимом осадков, наибольшее количество которых приходится на осень—зиму—весну (с X по IV—V). Такой режим благоприятствует тому, что многие растения начинают вегетировать (или прорастать из семян) с осени. Это в особенно яркой форме проявляется в наиболее южных районах распространения эфемерово-полынных пустынь.
Господствующими в растительном покрове являются полыни из подрода Seriphidium — как эндемичные для пустынь Средней Азии, так и связанные с другими флористическими центрами. К среднеазиатской группе относятся Artemisia terrae-albae s. str., A. kemrudica, A. arenicola, A. diffusa, A. badghysi, A. turanica, A. gurganica; к переднесреднеазиатской — A. dumosa, A. lobulifolia, A. santolina и, наконец, в причерноморско-закавказскую группу входят A. taurica, A. erivanica, A. fragrans, A. Meyeriana.
В летнее время эти пустыни представляют собою однообразные, серые по окраске заросли полыни, кустики которых покрывают 20—30% площади, оставляя между кустов открытую или покрытую побуревшими остатками эфемероидов почву. Но уже с осени намечается появление молодых побегов мятлика (Роа bulbosa var. vivipara) или осочки (Carex pachystylis), а также всходов эфемеров. Ранней весной развивается богатый покров эфемероидов и эфемеров, который зачастую, особенно в годы с благоприятным водно-тепловым режимом, достигает высоты 20—30 см и в зна
чительной степени маскирует голубовато-серые кусты полыни, энергичное развитие побегов которой несколько запаздывает.
Зеленый ковер мятлика и осочки расцвечен яркими красками многочисленных цветущих эфемероидов и эфемеров, отдельные виды которых, развиваясь в разные сроки, образуют красочные, сменяющие один другой аспекты.
В эту пору эфемерово-полынные пустыни создают иллюзию луговых травостоев, одевающих склоны и предгорья. В весенний период — это-ценнейшие пастбища, в некоторые годы используемые даже как сенокосы.
Литература
Е. П. Коровин (1934); А. С. Порецкий и др. (1936); А. В. Прозоровский (1940а);. А. В. Прозоровский и В. П. Малеев (1947).
90 к — закавказские (из Artemisia Meyeriana)
На равнинах в предгорьях Восточного Закавказья, в Кура-Араксин-ской низменности и прилегающих низкогорьях, на сероземах, бурых пустынно-степных и отчасти светлокаштановых почвах широко распространены эфемерово-полынные пустыни с господством Artemisia Meyeriana (в понимании А. А. Гроссгейма, 1948).
Эти пустыни представлены почти чистыми полынными сообществами; при сильном преобладании полыни к ней примешиваются следующие многолетники: Agropyrum pectintforme, Kochia prostrata, Euphorbia Marschal-liana, Allium pseudoflavum, A. rubellum, виды Cousinia и др.; из длительновегетирующих однолетников часто встречается Ceratocarpus arenarius. Характерна сезонность развития растительного покрова. Осенью и ранней весной, а частью и зимой, в эфемерово-полынной пустыне Закавказья цветет и вегетирует большое количество эфемероидов (Роа bulbosa, Colpodium humile, виды Gagea, Muscari, Bellevalia, Tulipa и др.) и эфемеров (Androsace Turczaninovii, Koelpinia linearis, Ziziphora tenuior, Erophila verna, Papaver Belangeri, виды Valerianella и Roemer ia). Эфемеры и эфемероиды размещаются на открытых пятнах почвы между кустами полыни, которая также усиленно вегетирует ранней весной, а летом замедляет свое развитие; вторая вспышка развития приурочивается к периоду осенних дождей (в сентябре—октябре), когда она цветет и плодоносит; семена созревают в октябре—ноябре.
Эфемерово-полынная пустыня в Восточном Закавказье дает переходы в сторону различных формаций солянковых пустынь и вместе с ними служит в качестве зимних пастбищ для обширных стад, главным образом овечьих, со средней продуктивностью от 2 до 5 ц/га сухой массы. В западной части Мильской степи на сильно солонцеватых почвах в комплексе с полынными сообществами распространены группировки Camphorosma Lessingii. Значительная часть территории закавказских полынных пустынь распахивается под поливные культуры, главным образом под хлопчатник. В Кобыстане (плоскогорье и небольшие возвышенности юго-восточной оконечности Главного Кавказского хребта), по данным М. П. Богданова (1954), распространены эфемерово-полынные, эфемерово-солянково-полынные и эфемерово-солянковые пустыни. Эфемерово-полынные пустыни на серо-бурых слабо солонцеватых почвах слагаются из господствующей здесь Artemisia Meyeriana с обильным покровом Роа bulbosa и со значительной примесью Colpodium humile, образующих задернение-почвы до 85% в период массового весеннего развития. Кроме этих основных растений травяного покрова, из эфемероидов встречаются Allium rubellum
и Tragopogon tuberosus. Но гораздо более заметную роль играют многочисленные эфемеры, из них главнейшие: Eremopyrum orientate, Е. triti-ееит, Erodium cicutarium, Aegilops cylindrica, Medicago minima, Bromus japonicus, Filago spathulata, Alyssum desertorum, Helianthemum salicifo-lium, Torularia contortuplicata и многие другие.
На солонцеватых и солончаковатых разновидностях серо-бурых почв распространены солянково-полынные (Artemisia Meyeriana, Salsola den-roides, S. nodulosa, S. ericoides, Suaeda dendroides) и полынносолянковые (Salsola nodulosa, S. dendroides, S. ericoides с примесью Artemisia Meyeriana) пустыни. Эфемерово-эфемероидовая синузия в этих пустынных сообществах заметно ослаблена. Задернение, образуемое Роа bulbosa и Colpodium humile, обычно около 40—50%. Количество видов эфемеров значительно меньше, чем в полынных пустынях, и здесь характерно появление таких солеустойчивых видов, как Hordeum leporinum, Lepidium perfoliatum, а также заметное внедрение летнеосенних однолетников (Salsola crassa, Petrosimonia brachiata, Filago spathulata, Gamanthus pilosus).
В условиях сильно расчлененного рельефа Кобыстана, изобилующего глубокими оврагами, балками, крутыми склонами и т. п., перечисленные сообщества, хотя и являются наиболее распространенными, однако далеко не исчерпывают встречающихся здесь формаций. К ним еще добавляются формации сочносолянковых пустынь (сарсазановые из Halocne-тит strobilaceum, поташниковые из Kalidium foliatum, сведовые из Suaeda dendroides и др.) на солончаках, образующие пестрые сочетания в соответствующих условиях. В масштабе нашей карты они не могли быть показаны.
Литература
А. А. Гроссгейм (1930, 1948); М. П. Богданов (1954).
90 ар — среднеараксинские (из Artemisia fragrans)
Распространены на Араксинской межгорной равнине, в среднем течении Аракса и прилегающих предгорьях.
Сообщества эфемерово-полынных пустынь с преобладанием Artemisia fragrans занимают как равнинные участки, так и нижние склоны холмов и предгорий на суглинистых и глинистых сероземных, слабо засоленных почвах; довольно часты и каменистые участки. Эдификаторами сообществ здесь является полукустарничковая Artemisia fragrans, которую сопровождают травянистые и полукустарничковые многолетники: Euphorbia Seguieriana, Acantholimon Karelinii, Kochia prostrata, Capparis spinosa, Helichrysum rubicundum, Verbascum saccatum, Centaurea squarrosa, Noaea mucronata и другие, сохраняющие свою роль в аспекте весь вегетационный период. Весной же пустыни покрываются сплошным зеленым ковром эфемероидов, что особенно красочно в годы, обильные зимне-весенними осадками. Здесь характерны следующие эфемероиды: луковичный мятлик (Роа bulbosa var. vivipara), кольподиум (Colpodium humile), узколистная осочка (Carex stenophylloides), птицемлечник (Ornithogalum Gussonei), мерендера (Merendera trigyna), герань (Geranium tuberosum). Эфемеры — злаки весьма разнообразны по видовому составу: Bromus tectorum, В. Danthoniae, В. squarrosus, В. japonicus, Eremopoa persica, Eremopyrum orientate, Aegilops triunciaiis, Taeniatherum, crinitum. Эфемеровое разнотравье представлено следующими видами: Helianthemum salicifolium,
Erodium oxyrrhynchum, Koelpinia linearis, Filago arvensis, Pterotheca Marschalliana, Androsace Turczaninovii, Alyssum dasycarpum, A. strictum, Ceratocephalus falcatus, Erysimum persicum, Trigonella radiata, Medicago minima, Rochelia disperma, Meniocus linifolius, Lepidium vesicarium, Astragalus commixtus, A. oxyglottis, Roemeria hybrida, Minuartia sclerantha и многие другие. В общем списке растений, входящих в эту полынную формацию, насчитывается более 110 видов (Прилипко, 1939).
Эфемерово-полынные пустыни Армении — ценные пастбища, используемые весной всеми видами животных (в некоторые годы возможно сенокошение), а зимой — овцами и верблюдами.
Литература
А. А. Гроссгейм и Д. И. Сосновский (1927); А. А. Гроссгейм (1928, 1932); Л. И. Прилипко (1939); А. К. Магакьян (1941); А. Л. Тахтаджян (1941).
90 стр — северотуранские (из северотуранских видов полыни)
Северотуранский вариант эфемерово-полынных пустынь показан на карте в виде нескольких контуров: на подгорной равнине Каратау (Сырдарьинского) на стыке с долиной Сыр-Дарьи, по левобережью Сыр-Дарьи (в ее так называемом Чардаринском отрезке); ряд мелких контуров расположен к северу от Таласского и Заилийского Алатау и, как уже указывалось, самый восточный массив приурочен к предгорной равнине Чу-Илийских гор.
В пределах так называемого Чардаринского участка местами характерно сочетание двух видов полыни: Artemisia terrae-albae s. str. и A. eu-cina. На большей же части территории господствует только первая; в редких местах встречается вместе с серой полынью еще и туранская полынь (Artemisia turanica). Повсеместно под пологом кустов полыни и между ними развивается заметный покров эфемероидов и эфемеров, представленный следующими видами: из эфемероидов — Carex pachystylis, Роа bulbosa var. vivipara (эти эфемероиды образуют основу травостоя), Allium Schubertii', из эфемеров — Eremopyrum prostratum, Bromus tectorum, В. Danthoniae, Veronica biloba, Ceratocephalus falcatus, Meniocus linifolius, Hoiosteum umbellatum, Nigella integrifolia. В летнее время, после засыхания эфемеров, становятся заметными развивающиеся в эту пору однолетние маревые: Halocharis hispida, Gamanthus gamocarpus, Ceratocarpus arenarius, Salsola crassa. Количество их увеличивается на засоленных почвах; в этих условиях полынь имеет несколько угнетенный облик, и среди ее кустов внедряется в большем или меньшем количестве биюргун (Anabasis salsa) (Культиасов, 1926).
На подгорной равнине к югу от западной части Каратау эфемеровополынные пустыни отличаются своеобразным флористическим составом. Среди кустиков полыни (Artemisia terrae-albae s. str.), Astragalus mucidus и Hulthemia (Rosa) berberifolia довольно обильны склероморфные многолетние травы: Cousinia mollis, С. syrdariensis, Astragalus turkestanus, A. Severzovii, A. macrotropis, которые сохраняют ландшафтное значение в течение почти всего вегетационного периода. Однако гораздо более обильны эфемероиды и эфемеры, создающие весной богатый и часто почти сомкнутый покров. Здесь наиболее характерны из эфемероидов: Allium caesium, Tulipa Greigii, T. turkestanica, Ixiolirion tataricum, Poa bulbosa -var. vivipara, Carexpachystylis', из эфемеров: Alyssum desertorum, Astragalus filicaulis, A. commixtus, Trigonella Noeana, Bromus Danthoniae, Papaver
pavoninum, Scabiosa Olivieri, Ziziphora tenuior, Malcolmia africana, Koel-pinia linearis, Veronica campylopoda, Heteracia Szowitsii, Eremopyrum orientale, Heteroderis pusilia, Nigella integrifolia и др. (Культиасов, 1938).
Самый восточный массив эфемерово-полынных пустынь приурочен к Чу-Илийским горам. Здесь, в высотных пределах 200—250 м над у. м., на склонах гор господствуют эфемерово-полынные пустыни, представленные двумя формациями. Эфемерово-прутняково-полынные (Artemisia terrae-albae s. str., Kochia prostrata и эфемеры) сообщества отличаются хорошо развитым ярусом полукустарничков, образующих 25—55% покрытия и дающих 60—90% всей надземной массы. Из эфемероидов и. эфемеров встречаются следующие виды: Carex pachystylis, Роа bulbosa var. vivipara, Seseli sessiliflorum, Alyssum desertorum, Ziziphora tenuior, Koelpinia linearis, Garhadiolus papposus, Bromus faponicus и др.
Эфемерово-ковыльно-полынные (Artemisia transiliensis из родства A. sublessingiana, Stipa capillata и эфемероиды) сообщества отличаются заметным участием Stipa capillata или S. caucasica. Эти сообщества приурочены к северным склонам и лежат на большой абсолютной высоте. Участки со Stipa caucasica связаны всегда с несколько щебнистыми почвами; растительный покров в этих местах обычно сильно изрежен, всего 20— 25% проективного покрытия. В более же распространенных сообществах с участием ковыля-тырсы (Stipa capillata) степень покрытия в среднем 35—40%, а запас кормов 3—4 ц/га, не считая весенних эфемеров, масса которых сильно колеблется по годам. Видовой состав эфемероидов и эфемеров следующий: Роа bulbosa var. vivipara, Carex pachystylis, Ixiolirion tataricum, Trigonella arcuata, Nonea caspica, Aphanopleura capilltfolia, а также виды, перечисленные выше для первой формации.
Как общую характерную черту Чу-Илийских эфемерово-полынных пустынь, следует отметить наличие в составе сообществ ряда степных элементов. Помимо уже названных, типичны еще и такие степные, злаки: Festuca sulcata, Stipa Lessingiana и Koeleria gracilis', они встречаются далеко не на всех участках, обычно рассеянно или единично. Характерно-также единичное участие летне-осенних однолетников, представленных только эбелеком (Ceratocarpus arenarius) (Блюменталь, 1937).
Литература
М. В. Культиасов (1926, 1938); И. X. Блюменталь (1937).
90 ютр — южнотуранские (из Artemisia herb а-alba s. 1.)
Южнотуранский вариант эфемерово-полынных пустынь распространен в южной части пустынной области, на подгорной равнине гор Большого Балхана и Копет-Дага (занимая особенно значительные площади в Западном Копет-Даге), а также на подгорных равнинах и шлейфах останцового плато в центральных Кызылкумах. Эфемерово-полынные пустыни контактируют здесь с полынными пустынями в сочетании с солянковыми (89) и эфемерово-полынными пустынями в сочетании с солянковыми (91), будучи зачастую вкрапленными среди них мелкими площадями, не укладывающимися в масштаб карты.
В Копет-Даге строителем сообществ является бадхызская полынь . (Artemisia badghysi), которая сопровождается еще другими видами полы-
ней (Л. kopetdagensis, A. dumosa). Обильные эфемероиды представлены следующими видами: Allium scabriscapum, A. xiphopetalum, Eremurus Olgae, Codonocephalum Peacockianum, Gagea stipitata. Все это крупные растения, заметно возвышающиеся не только над основным осочково-мятликовым (Роа bulbosa var. vivipara и Carex pachystylis) с эфемерами покровом, но и над кустами полыни. Из эфемеров особенно обильны здесь: Leptaleum filifolium, Ziziphora tenuior, Hypecoum pendulum, Goldbachia laevigata, Holosteum glutinosum, Fumaria Vaillantii, Eruca sativa, Veronica campy-lopoda, V. didyma, Alyssum desertorum, Adonis aestivalis, Papaver pavo-ninum, Roemeria hybrida, Spinacia tetrandra, Androsace Turczaninovii и др. Среди многолетних трав встречаются Cousinia eryngioides, С. bipinnata, С. Komarovii, Jurinea derderioides, Eremostachys labiosa, Teucrium polium и другие, вегетация и цветение которых продолжаются еще в начале сухого периода, после отмирания эфемеров. Показательно, что обильная весенняя влага позволяет развиваться некоторым видам мхов (Pterygoneu-rum ovatum, Tortula ruralis, Phascum cuspidatum). В районе Кара-Калы, по И. А. Линчевскому (1935а), эфемерово-полынные пустыни распространены по низким предгорьям, слагаемым соленосными палеогеновыми глинами, на светлых сероземах. Почвы здесь своеобразны, они «сплываются» после дождей, и на них образуется поверхностная корка. Этому району свойственен характерный ландшафт бедленда.
На северных склонах здесь господствует бадхызская полынь (Artemisia badzhysi),1 под пологом которой развит покров Carex pachystylis и Роа bulbosa var. vivipara с примесью эфемероидов (Gagea reticulata, Allium albanum и др.) и эфемеров (Epilasia hemilasia, Diarthron vesiculosum, Koelpinia linearis, Medicago rigidula, Onobrychis micrantha, Leptaleum filifolium, Ziziphora tenuior, Helianthemum salicifolium, Meniocus linifolius, Bromus Danthoniae, Eremopyrum Buonapartis, Aegilops triuncialis и др.). Изредка встречаются здесь кустики Zygophyllum atriplicoides и Noaea mucronata, а также многолетние травы — Stipa Hohenackeriana и др.
На южных крутых склонах поверхность почвы обнажена; здесь встречаются разреженные группки немногих эфемеров (Arnebia decumbens, Eremopyrum Buonapartis, Koelpinia linearis, Bromus tectorum, Malcolmia turkestanica) и летне-осенних однолетников (Salsola crassa, S. sclerantha и др.).
В пределах этого пустынного ландшафта характерны заросли держидерева (Paliurus spina-Christi) с примесью многих более или менее ксеро-фильных кустарников (Ephedra equisetina, Colutea Buhsei, Celtis cauca-sica, Cerasus microcarpa, Punica granatum)', однако площадь их ничтожна, и в масштабе нашей карты они не могут быть показаны.
Пестрота условий существования в пределах сильно расчлененного ландшафта бедленда определяет значительную неоднородность покрова эфемерово-полынных пустынь и его продуктивности. Общий запас сухой растительной массы, по определениям И. А. Линчевского (1935а), колеблется в пределах от 11 до 26 ц/га; поедаемая кормовая масса значительно меньше, всего 3—5 ц/га, а на изреженных участках снижается до 0.5— 1.5 ц/га (Нечаева и др., 1943).
В пределах распространения эфемерово-полынных пустынь на Копет-Даге очень часто выходят на поверхность мергели, известняки и рухляки нижнемелового возраста. В подобных местах растительность резко отличается преобладанием скальных форм, так называемых «нагорных ксеро
1 И. А. Линчевский указывает Artemisia herba-alba в.?1. Можно, однако, полагать, что здесь встречаются три вида полыни: A. badghysi, A. dumosa и A. kopetdagensis.
фитов»: Scutellaria Litwinowii, Stachys turcomanica, Dendrostellera turcmeno-rum, Convolvulus fruticosus, C. subhirsutus, Zygophyllum eurypterum, Acantholimon pterostegium, Jurinea Antonowii. Buhsea coluteoides, Amygdalus spinosissima, Ephedra intermedia, виды Astragalus и многие другие растения, относящиеся к этой экологической группе. Здесь эфемеры, хотя и встречаются, но преимущественно под пологом других растений, редко выходя на открытые участки.
Эфемерово-полынные пустыни останцовых Кызылкумов приурочены к хплейфам и полого-волнистым равнинам с суглинисто-супесчаными сероземами. Среди кустов песчаной полыни (Artemisia arenicola) 1 встречается небольшая примесь туранской полыни (A. turanica), а также единичные кусты кеурека (Salsola rigida) п боялыча (S. arbuscula); еще реже встречаются чогон (Aellenia subaphylla). Эфемероиды и количественно и по видовому составу немногочисленны: Carex, physodes, Роа bulbosa var. vivipara, Allium filidens, Rhinopetalum Karelinii, Tulipa lehmanniana. Зато эфемеры представлены очень большим количеством видов. Наиболее часто здесь встречаются Eremopyrum orientale, Bromus tectorum, Consolida soongorica, C. rugulosa, Malcolmia Karelinii, Koelpinia linearis, Streptoloma desertorum, Arnebia decumbens, Psammogeton setifolium, Euphorbia Turczaninowii, Ziziphora tenuior, Roemeria hybrida, Ceratocephalus orthoceras, Silene папа, Heteroderis pusilia, Hemilasia acrolasia, Astragalus corrugatus, Plantago lagocephala и многие другие, всего более 55 видов (Порецкий и др., 1936). Обычно присутствует и небольшое количество летне-осенних однолетников: Ceratocarpus turkestanicus, Salsola iliensis, Girgensohnia oppositiflora, Kirilowia eriantha, Halotis pilosa.
Эфемерово-полынные пустыни рассматриваемого района дают около 6 ц/га сухой массы, причем в наиболее пышно развитых зарослях это количество повышается до 10 ц/га. Большую часть этой продукции составляет полынь, а на долю эфемеров приходится в среднем около 1 ц/га сухой массы. Разреженные и обедненные по составу сообщества (с ослабленной ролью полыни и возросшей — полукустарничковых маревых) дают 3.5 ц/га, из которых на эфемеры приходится 0.5 ц/га.
При опесчанивании поверхности происходит заметная перестройка сообществ: изреживается полынь и за ее счет внедряются боялыч (Salsola arbuscula) или терескен (Eurotia ceratoides) и другие полукустарнички; изредка попадается низкорослый черный саксаул (Haloxylon aphyllum); появляются Stipa Szowitsiana, Iris tenuifolia, Acanthophyllum elatius, Allium sabulosum, более пышно развивается Carex physodes, а эфемеры убывают в количестве и также значительно падает их видовое разнообразие.
Литература
С. И. Коржинский (1896); Е. Г. Черняковская (1924, 1925, 1927); Е. П. Коровин (1927а); И. А. Линчевский (1935а); А. С. Порецкий и др. (1936); Н. Т. Нечаева и др. (1943); А. П. Шенников и А. Ф. Иоффе (1944); Г. В. Микешин (1946а); И. И. Гранитов (1950); В. Н. Никитина (1954); Н. Н. Пельт (1955); Н. Н. Пельт н Л. Е. Родин (1955).
91. Эфемерово-полынные пустыни в сочетании с солянковыми
На нашей карте эфемерово-полынные пустыни в сочетании с солянковыми самостоятельными контурами показаны в Закавказье: по Араксу на границе с Ираном, по долине Куры в ее среднем течении, а особенно
1 Ранее этот вид включался в Artemisia herba-alba s. I. (Порецкий и др., 1936).
в дельте Куры и Аракса, в нескольких местах достигая побережья Каспийского моря; в Средней Азии: обширный контур на южном Устюрте, ряд мелких контуров в пределах юго-западной Туркмении, затем в западном Копет-Даге, протягиваясь узкой и почти непрерывной полосой вдоль Копет-Дага на восток до 59° в. д. и, наконец, по Унгузу, в восточной его части, нигде не выходя за пределы Переднеазиатской группы провинций в смысле А. В. Прозоровского (Прозоровский и Малеев, 1947).
Пустыни этого типа контактируют по большей части с эфемерово-полынными (90) и солянковыми пустынями (92), однако в ряде случаев они приходят в соприкосновение и с другими типами пустынь (89, 99, 100, 102 и др.). Кроме того, в пределах перечисленных типов встречаются участки эфемерово-полынных пустынь в сочетании с солянковыми, которые не могли быть показаны на карте по условиям масштаба.
Таким образом, описываемые пустыни распределены в южной части пустынной области, переходя незначительно к северу от 40° с. ш.
Эфемерово-полынные пустыни в сочетании с солянковыми в своем распространении связаны с глинистыми долинными и дельтовыми речными наносами, наиболее пониженными частями подгорных шлейфов, а также с выходами более древних засоленных и гипсоносных пород.
Климатические условия существования идентичны с эфемерово-полынными пустынями, основное же отличие лежит в почвенных условиях. Почвы характеризуются широким распространением солонцеватых разновидностей светлых сероземов и, местами, серо-бурых почв.
Растительный покров слагается из сочетания преобладающих (а иногда участвующих в равной степени) эфемерово-полынных пустынных сообществ и сообществ солянковых пустынь.
Различается несколько географических вариантов.
4
Литература
Е. П. Коровин (1927а); А. А. Гроссгейм (1930, 1948); А. В. Прозоровский (1940а); Н. Т. Нечаева и др. (1943); А. В. Прозоровский и В. П. Малеев (1947).
91 к — закавказские (из Artemisia Meyeriana, Salsola dendroides, S. nodulosa, S. ericoides)
Эфемерово-полынные пустыни в сочетании с солянковыми распространены в Кура-Араксинской низменности в Закавказье, на засоленных разновидностях сероземов и светлокаштановых почв.
Наиболее распространены карганные пустыни из Salsola dendroides (=S. verrucosa). Они покрывают большие пространства в низовьях Куры и характерны для засоленных почв аллювиального происхождения, сравнительно недавно вышедших из стадии поймы. Salsola dendroides встречается как в виде густых и высоких (80—90 см) чистых зарослей, так и в сочетании с полынью {Artemisia Meyeriana),1 эфемероидными злаками (Роа bulbosa var. vivipara, Colpodium humile), однолетними солянками (Salsola crassa, Petrosimonia brachiata, Suaeda altissima), верблюжьей колючкой (Alhagi pseudalhagi) и другими видами галофильного разнотравья (Glycyrrhiza glabra, Limonium Meyeri и др.). Многочисленные эфемеры {Eremopyrum orientate, Hordeum leporinum, H. geniculatum, Bromus rubens, B, japonicus, Hemiaraia hirsuta, Medicago minima, Filago spathulata и др.) образуют рано развивающиеся и отцветающие весенние синузии.
* В ш—м—« А. А. Гроесгейма. Определитель растений Кавказа. М., 1949.
Эфемерово-полынные пустыни (из Artemisia Meyeriana) занимают второе место по их значению в Закавказье. Они свойственны светлокаштановым и каштановым почвам, преимущественно незасоленным или слабозасоленным. Под покровом полыни, к которой примешиваются иногда полукустарничковые солянки Salsola dendroides и S. nodulosa (что обычно свидетельствует о некотором засолении почв), всегда развита синузия эфемероидов и эфемеров. Главная роль здесь принадлежит мятлику (Роа bulbosa var. vivipara), образующему постоянное и заметное задернение поверхности почвы, в большей степени, чем в эфемерово-солянковой (кар-ганной) пустыне. Среди эфемеров более широко распространены представители разнотравья, нежели злаков. Особенно характерны такие растения, как Helianthemum salicifolium, Medicago minima, Torularia contortupli-cata и другие, встречающиеся и в сообществах солянковых пустынь Закавказья (Гроссгейм, 1932).
В случае вторичного засоления почвы или на почвах, первично засоленных (но сильнее, чем под Salsola dendroides), развивается формация полукустарничковой мелколистной шведки (Suaeda microphylla), очень обычная и широко распространенная на низменности Восточного Закавказья. Эта формация часто замещает сообщества каргана при отмирании последнего в случае прогрессирующего засоления почвы.
Вересковидная солянка (Salsola ericoides) — низкорослый полукустарничек, обычно произрастает в смеси с полынью Мейера (Artemisia Меуе-riana) на сухих склонах предгорий Большого Кавказа и возвышенностей среди Кура-Араксинской низменности (Баба-Занан, Кюровдаг, Боздаг). В этом сообществе часто доминирует полынь Мейера; ей сопутствует эфе-мероидный луковичный мятлик, и имеется примесь эфемеров: Bromus Danthoniae, Koelpinia linearis, Medicago minima и др.
Пустынные сообщества горной солянки (Salsola nodulosa) развиваются на шлейфах гор и на низких возвышенностях, на выходах третичных гипсоносных пород. Особенно большие площади они занимают в Восточном Закавказье к западу от Баку — в Кобристане. В их составе-принимают значительное участие также другие полукустарнички: Artemisia Meyeriana, Salsola ericoides, Reaumuria hypericoides. Весною же здесь развивается обильный покров эфемероидов («эфемеретум» кавказских ботаников): Роа bulbosa var. vivipara, Medicago minima, Aegilops squarrosa, Trachynia distachya п многие другие. Встречаются также ксерофильные кустарнички — Astragalus, aureus и др. В эфемерово-полынных сообществах, по мере поднятия на предгорья, появляются дерновинные злаки (виды Stipa, Koeleria и др.), а значение эфемеровой синузии ослабляется.
Литература
А. А. Гроссгейм (1930, 1932, 1948); Л. И. Прилипко (1939).
91 ар — среднеараксинские (из Artemisia fragrans, Salsola nodulosa, S. ericoides)
Этот вариант эфемерово-полынных пустынь в сочетании с солянковыми распространен только на Араксинской равнине к северо-западу от Нахичевани. Здесь характерно сочетание участков полынных, эфемеровополынных и солянковых сообществ. В первых двух сообществах господствует Artemisia fragrans-,1 причем на участках с щебнистыми, сильно уплотненными почвами она занимает особо доминирующее положение,
1 В понимании А. А. Гроссгейма. Определитель растений Кавказа. М., 1949.
а на более лугких суглинистых почвах значительно развиваются эфемеры.
На участках полынной пустыни наиболее характерны следующие сопутствующие виды: а) полукустарнички — Acantholimon Karelinii, Kochia prostrata, Ephedra distachya, Teucrium polium', б) длительновеге-тирующие многолетние травы — Chondrilla juncea, Centaurea squarrosa, Euphorbia Mar chailiana', в) эфемероиды — Poa bulbosa var. vivipara, Allium rubellum, Iris caucasica', г) эфемеры — Taeniatherum crinitum, Koelpinia linearis, Meniocus linifolius, Eremopyrum orientals, Erodium Hoejjtianum. Длительновегетирующих однолетников почти нет, кроме отчасти рудерального Ceratocarpus arenarius (Прилипко, 1939).
Эфемерово-полынные (из той же Artemisia jragrans) сообщества выделяются высокой степенью обилия и видового разнообразия эфемероидов (Роа bulbosa var. vivipara, Colpodium humile, Carex stenophylla s. I.,1 Merendera trigyna, Ornithogalum Gussonei) и эфемеров (Bromus tectorum, B. Danthoniae, B. japonicus, Eremopoa persica, Eremopyrum orientale, Fi-lago arvensis, Androsace Turczaninovii, Alyssum hirsutum, Taeniatherum crinitum, Erysimum persicum, Medicago rigidula и др.).
Солянковые сообщества слагаются главным образом вересковидной солянкой (Salsola ericoides) с участием других полукустарничковых маревых: Salsola glauca и Kochia prostrata. В сообществе этих растений принимают некоторое участие галофильные эфемеры и длительновегетирующие однолетники: Sphenopus divaricatus, Lepidium versicarium, Psylliosta-chys spicata, Nepeta micrantha, Seidlitzia florida и другие однолетние маревые.
В предгорной полосе на многих склонах изредка встречаются эфеме-рово-полынно-солянковые сообщества, в Которых Artemisia fragrans, Salsola nodulosa и S. glauca равномерно смешаны. Здесь также очень богатый состав эфемероидов и эфемеров, однако характерно несколько большее количество галофильных осенних однолетников (Магакьян, 1941).
Литература
А. А. Гроссгейм (1932); Л. И. Прилипко (1939); А. К. Магакьян (1941).
91 ютр — южнотуранение (из Artemisia herba-alb a s. 1., Salsola gemmascens, S. arbuscula)
Этот вариант эфемеропо-полтлтных пустынь в сочетании с солянковыми на южном Устюрте характеризуется растительным покровом, слагаемым из двух формаций: формации полыни (чаще всего Artemisia dijfusa, A. kem-rudica и A. badghysi', в самой северной части могут встречаться формы, близкие к A. terrae-albae s. str.) и формации тетыра (Salsola gemmasens), (или же формации других полукустарничковых солянок — S. arbuscula, S. rigida). Они занимают самостоятельные участки в сочетании, но иногда встречаются смешанные сообщества из полыни и тетыра.
Приурочены эти пустыни к светлым сероземам, обычно солонцеватым, суглинистым, реже супесчаным; полынные сообщества обычно приурочены к более выщелоченным разновидностям почв, нередко связанным с едва приметными понижениями.
Основную массу растительности в этих сочетаниях составляет полынь.
В полынных сообществах обычна незначительная примесь кеурека {Salsola rigida) и боялыча (Salsola arbuscula), растущих группами; кроме того, единичными экземплярами встречается низкорослый черный саксаул
1 Возможны Carex pachystylis и С. stenophylloides.
(Haloxylon aphyllum). Сомкнутость полукустарничкового яруса незначительная, не более 30—40%. Между кустами поселяются эфемероидная осочка (Carex pachystylis), некоторые луковичные эфемероиды (Tulipa sogdiana, Allium sabulosum, A. fibrosum, Gagea reticulata, G. divaricata) и эфемеры (Eremopyrum orientale, Leptaleum filifolium, Astragalus arpilo-bus, A. oxyglottis, Amberboa turanica и др.). Характерно также присутствие летне-осенних однолетних маревых, например Girgensohnia minima.
Эфемерово-полынные пустыни в сочетании с солянковыми в основном представляют собою пастбища для верблюдов, но весной они могут быть использованы и овцами.
Во влажные годы эфемеры и эфемероиды хорошо развиваются, образуя густой травостой. В такие годы урожайность поедаемой массы достигает 4—4.3 ц/га, в засушливые — снижается до 0.8—1 ц/га.
Основное затруднение при использовании этих пастбищ заключается в плохом обеспечении водопоями, в силу чего в осенний и зимний периоды, когда корма становятся сухими, выпас овец не производится. При более густой сети колодцев было бы возможно производить круглогодичный выпас (Нечаева и др., 1943).
В пределах Копет-Дага эфемерово-полынные пустыни в сочетании с солянковыми отличаются некоторыми особенностями. Они связаны со светлыми, часто солончаковатыми сероземами подгорной равнины и низких предгорий. Растительный покров отличается неоднородностью, пятнистостью: наряду с участками эфемерово-полынных сообществ встречаются и эфемерово-солянковые, или же полукустарничковые солянки внедряются среди кустов полыни. И в тех и других сообществах характерно наличие синузии эфемероидов и эфемеров. На подгорной равнине восточной части Копет-Дага описываемые эфемерово-полынные пустыни слагаются следующими видами: а) полукустарнички — Artemisia badghysi, Salsola gemmascens, S. rigida, S. dendroides; б) эфемероиды — Poa bulbosa var. vivipara, Carex pachystylis (образует дерновину только латками, покрывая около 50% почвы); в) эфемеры — Papaver pavoninum, Koelpinia linearis, Arnebia transcaspica, Astragalus filicaulis, Lallemantia Royleana, Trigonella Noeana, Bromus Danthoniae (на солончаковатых почвах перечисленные виды эфемеров изреживаются или. даже выпадают, и преимущественное значение приобретают солеустойчивые виды — Matricaria lamellata, Lepidium perfoliatum, Consolida rugulosa, Amberboa odorata, Psylli-ostachys spicata, Spergularia salina, Plantago coronopus и др.); г) летнеосенние однолетники, приуроченные преимущественно к сообществам с господствующей • ролью полукустарничковых солянок — Halocharis hispida, Halimocnemis mollissima, Girgensohnia oppositiflora, Salsola lanata (в полынниках тоже встречаются, но редко и единичными особями) (Коровин, 1927а).
В западном Копет-Даге эфемерово-полынные пустыни распространены в весьма своеобразных условиях. Они приурочены к пролювиям, отлагающимся в нижних частях чрезвычайно эродированных соленосных глинистых отложений, преимущественно палеогеновых и плиоценовых, образующих характерный ландшафт бедленда. Наряду с господствующими здесь в сообществах полынями (Artemisia dumosa, A. badghysi, A. kop et daghensis), распространены полукустарничковые солянки (Noaea mucronata, Salsola Aucheri, Aellenia subaphylla), некоторые полукустарнички (Convolvulus subhirsutus, Scutellaria Litwinowii, Acantholimon pterostegium), некоторые многолетние длительновегетирующие травы (Stipa Hohenackeriana, Euphorbia sogdiana) и весьма обильные эфемеры (Diarthron vesiculosum, Koelpinia linearis, Leptaleum filifolium, Onobrychis micrantha, Hypecoum
trilobum, Erodium cicutarium, Scandix stellata, Roemeria refracta, Epilasia hemilasia, Ziziphoratenuior, Senecio vernalis, LallemantiaRoyleana и многие другие виды) и некоторые эфемероиды {Carex pachystylis, Роа bulbosa var. vivipara, Tulipa Hoogiana, Allium fibrosum, A. Borszczowii, Iris kopetda-gensis, Geranium Kotschyi, Ixiolirion tataricum, Dorema Aitchisonii)', число летне-осенних однолетников незначительно {Salsola sclerantha, Girgen-sohnia oppositiflora, Ceratocarpus turkestanicus).
Особые черты имеет растительный покров гор Малого Ба л хана, на нашей карте также отнесенный к этому типу. По недавно вышедшей работе Р. П. Тарасова (1954) две трети площади Малого Балхана представляют собою обнажения горных пород, различных по возрасту и литологии. На скалах, на щебнистой коре выветривания, на известняках и мергелях, на глинах и песчаниках, чрезвычайно характерны разнообразные сообщества нагорных ксерофитов и скальных растений, представленных более чем 50 видами {Rhamnus Sintenisii, Astragalus Marschallianus, A. pileto-cladus, A. deserti, Ephedra intermedia, Pennisetum orientale, Parietaria judaica, Antitoxicum pumilum, Zygophyllum atriplicoides, Z. portulacoides и др.). Многие сообщества характеризуются приуроченностью к определенным по литологии и химизму отложениям.
Только в предгорьях, на более обогащенных мелкоземом субстратах с почвами, близкими к сероземам, встречаются эфемерово-полынные пустыни в сочетании с полынно-солянковыми и солянковыми. Однако и здесь наблюдается примесь нагорных ксерофитов, а эфемероидно-эфемеровая синузия {Роа bulbosa var. vivipara, Carex pachystylis, Allium rubellum, Iris longiscapa, Matricaria lamellata, Roemeria hybrida, Erodium oxyrrhynchum, Amberboa suaveolens, Eremopyrum orientale, Lachnoloma Lehmannii и др.) несколько ослаблена.
Контуры эфемерово-полынных пустынь в сочетании с солянковыми, расположенные по Унгузу, чередуются с белосаксаульниками на песчаных грядах. В сообществах описываемого варианта господствуют полыни: Artemisia arenicola, A. kemrudica, A. terrae-albae ssp. Massagetovii. Значительно участие полукустарничков: боялыча {Salsola laricifolia) и саксаульника {Arthrophytum Lehmannianum) и, реже, Reaumuria oxiana, Salsola rigida, Anabasis ramosissima, A. truncata, Haplophyllum obtusifolium. Характерно заметное количество эфемероидов {Asparagus turkestanicus, Epilasia hemi-lasia), эфемеров {Malcolmia Karelinii, Isatis emarginata, Arnebia transcaspica, Streptoloma desertorum, Astragalus tribuloides, Garhadiolus papposus, Matricaria lamellata) и длите льновегетирующих однолетников {Ceratocarpus turkestanicus, Horaninowia anomala, Halimocnemis Karelinii, Salsola sclerantha). Запас поедаемой растительной массы в пределах этого контура очень низкий — в среднем около 0.9 ц/га (Шингарева, 1940).
Литература
Е. П. Коровин (1927а); И. А. Линчевский (1935а); А. В. Прозоровский (1940а); Е. А. Шингарева (1940); Н. Т. Нечаева и др. (1943); Г. В. Микешин (1946а); В. Н. Никитина (1954); Р. П. Тарасов (1954); Н. Н. Пельт (1955); Н. Н. Пельт и Л. Е. Родин (1955).
92. Солянковые пустыни (из Reaumuria soongorica, видов Salsola, Anabasis, Nanophyton и др.) обычно в сочетании с полынными пустынями
Пустыни этого типа широко распространены, занимая по площади следующее место после полынных пустынь.
Наиболее крупные площади их сосредоточены на Устюрте, в южной части Мангышлака, в южной части Бетпак-Далы, в Прибалхашье, вдоль
староречий Жана-Дарьи, в низовьях Сыр-Дарьи и северном Приаралье, в Сарыкамышской котловине.
Таким образом, солянковые пустыни распространены, с одной стороны, на древних платообразных поверхностях, а с другой, на древних аллювиальных равнинах и участках речных долин, давно вышедших из сферы влияния грунтовых вод. Полукустарничковосолянковые пустыни в основном лежат в подзоне северных пустынь и связаны преимущественно с серо-бурыми пустынными почвами или с почвами близкого к ним типа, встречающимися в комплексе с солонцами, солончаками и такырами (или в той или иной степени отакыренными почвами).
Почти на всем пространстве полукустарничковосолянковые пустыни встречаются в комплексе с полынными пустынями; отдельные, иногда довольно значительные площади последних обычны среди этих комплексных солянковых пустынь, и контуры тех и других чередуются между собой.
На столь обширном пространстве своего ареала солянковые пустыни достаточно разнообразны, а в еще большей мере разнообразны их комплексы, в зависимости от видового состава основных эдификаторов (рис. 73).
Так, в общих чертах можно отметить, что в северных частях ареала преобладают боялычники (из Salsola laricifolia), которые комплексируются с биюргунниками (из Anabasis salsa) и полынниками (из Artemisia semiarida, A. terrae-albae s. str., A. kemrudica и др.). Южнее распространены боялычники (из Salsola arbuscula) в комплексе с кеуречниками (из Salsola rigida) и полынниками (из Artemisia kemrudica, A. diffusa).
Наконец, в южной (особенно юго-восточной) части Устюрта комплексы слагаются из преобладающих сообществ биюргунников (из Anabasis salsa и A. ramosissima) с боялычниками (из Salsola laricifolia), полынниками (из Artemisia kemrudica, заменяемой A. kemrudica var. karabugasica в юго-западной, прикарабогазской части).
В отдельных частях ареала солянковых пустынь комплексы еще более усложняются, так как, помимо названных сообществ, к ним прибавляются фрагменты черносаксауловых ассоциаций, сообщества водорослей и лишайников на такырах, а также значительно возрастает роль эфемеровой синузии. Кроме того, на гипсовых и солончаковых почвах и солончаках, которые часты среди обширных пространств полукустарничковосолянковых пустынь, встречаются фрагменты сообществ различных видов Anabasis {A. eriopoda, A. brachiata, A. truncata), Nanophyton erinaceum, а в отдельных случаях и типичных представителей растительности солончаков — Halocnemum strobilaceum, Kalidium caspicum и др.
Нередко, особено на Устюрте, встречаются участки со щебнистыми, а иногда только лишь с хрящеватыми почвами, на которых появляется ряд довольно типичных петрофитов, как тасбиюргун {Nanophyton erinaceum), Acanthophyllum pungens, Haplophyllum obtusifolium, Atraphaxis spinosa, Anabasis brachiata, Convolvulus fruticosus и др. Часто в этих местах особенно заметно возрастает количество боялыча Salsola laricifolia. На щебне и на камнях в таких случаях обычно довольно значительно разрастаются накипные лишайники: Diploschistes scruposus, Teloschistes brevior, Lecanora aspera, Aspicilia desertorum, и др., а в некоторых случаях, особенно на так называемых «бозынгенах» (сухие гипсовые солончаки), колонии лишайников иногда образуют сплошной покров на почве.
Полукустарничково-солянковые пустыни — это малопродуктивные осенне-зимние пастбища, используемые главным образом верблюдами,
с урожайностью 1.5—2.5 ц/га поедаемой кормовой массы. При участии в сложении сообществ полыни, а также в сочетании с полынными сообществами, запас поедаемой массы возрастает до 3.5—4.5 и в хорошие годы даже до 5 ц/га.
Ввиду крайне слабой обводненности территорий солянковых пустынь (очень редкие колодцы, слабые возможности использования вод поверхностного стока) пастбищное использование их очень не велико.
Литература
Г. С. Карелин (1883); Б. А. Федченко и И. М. Крашенинников (1910); О. Э. Кнор-ринг и 3. А. Минквиц (1913); М. Г. Попов (1915, 1922, 1940); И. И. Спрыгин и М. Г. Попов (1915); М. Д. Спиридонов (1918, 1921, 1930); И. М. Крашенинников (1923, 1925); Г. И Доленко (4930); С. С. Неуструев и И. М. Крашенинников (ред.) (1930); Н. В. Павлов (1930, 1935); Р. Ю. Рожевиц (1930); Ф. Н. Русанов (1930); Е. П. Коровин (1934); Б. С. Закржевский и Е. П. Коровин (1935); Е. П. Коровин и В. А. Селевин (1935); В. Н. Минервин (1935); Е. Г. Победимова (1935); А. С. Порецкий и др. (1936); П. Г. Стрельников (1938); А. В. Прозоровский (1940а); Л. Е. Родин (1940, 1948); Н. И. Рубцов (1941а, 1943, 1944, 1950а, 1952); Ш. И. Коган (1949, 1954); 3. В. Кубанская (1949); Н. И. Рубцов и 3. В. Кубанская (1949); С. А. Никитин (1954, 1954а); Н. Н. Пельт (1955); Н. Н. Пельт и Л. Е. Родин (1955).
93. Сочносолянковые пустыни (из Halocnemum strobilaceum, Halostachys caspica,1 Kalidium foliatum) и лишенные растительности солончаки
Сочносолянковые пустыни в своем распространении связаны с четырьмя группами местообитаний: 1) солончаками морских и озерных побережий; 2) дельтовыми окончаниями рек, как впадающих в моря и озера, так и не достигающих их; 3) речными долинами; 4) внутриматериковыми солончаками (рис. 74).
Наиболее крупные площади сочносолянковых пустынь сосредоточены по восточному побережью северной части Каспийского моря. Это — соры Мертвый Култук и Кайдак, представляющие собой недавно осушенное дно мелководных заливов Каспия, и примыкающая к ним береговая часть п-ва Бузачи.
В южной части Каспийского моря сочносолянковые пустыни занимают значительно меньшие по площади пространства, будучи вытянуты относительно узкой полосой как вдоль побережья самого Каспия, так и его залива Кара-Богаз-Гола. Меньшие площади сочносолянковых пустынь расположены на южном побережье оз. Балхаш и в озерной депрессии между последним и оз. Алаколь и по его берегам, а также в западной части оз. Зайсан.
Вторая группа местообитаний сочносолянковых пустынь — солончаки дельтовых окончаний современных рек: Куры, Урала, Эмбы, Тургая, Сыр-Дарьи, Аму-Дарьи, Или, Зеравшана и Атрека, обязанные эрозионноаккумулятивной деятельности этих рек.
К этой же группе местообитаний надо отнести обширный солончак Хаки на Прикаспийской низменности, который, очевидно, связан с деятельностью рек Большого и Малого Узеней, имевших в этих местах свою дельту (Ковда, 1950), а также солончаки вдоль Сарпинской дельты на западной части Прикаспийской низменности. Сюда же следует отнести и сор Балханский, представляющий собою одновременно высохший залив Каспийского моря и дельту Узбоя.
1 В настоящее время по номенклатурным соображениям этот вид именуется Halostachys Belangeriana.
неясная по происхождению депрессия которого граничит с Заунгузскими и Низменными Каракумами.
Рис. 74. Схема распространения сочносолянковых пустынь и лишенных растительности солончаков в Средней Азии и на павказе. основные массивы сочносолянковых пустынь и лишенных растительности солончаков (93).
УЗ. Сочносолянковые пустыни и лишенные растительности солончаки 789
Третья группа местообитаний сочпосолянковых пустынь связана с речными долинами крупных рок, обычно в их среднем течении. 'Гаковы массивы сочпосолянковых пустынь в среднем точении Или, Сыр-Дарьи (в пределах Ферганской котловины) и Аму-Дарьи (вблизи Керков и выше). Эти местообитания по существу очень близки но генезису ко второй группе, так как они почти повсеместно приурочены к низовьям мелких притоков названных крупных рек, сбегающих с соседних хребтов и часто (хотя бы часть года) по доносящих своих вод до главной реки.
Четвертая группа местообитаний сочпосолянковых пустынь — впутри-материковые солончаки, обычно залегающие в глубоких депрессиях рельефа. В наибольшей мере они распространены на Устюрте и на Мангышлаке, где занимают нередко большие площади. Таковы, например, соры Барса-Кольмес, Каратюлой, Чумыштыколь, Алтай, Сам, Карын-Ярык и многие другие, разбросанные в депрессии, примыкающей к северному чинку Устюрта. Некоторые из солончаков лежат во впадинах, имеющих дно ниже уровня океана: таковы, например, впадины Карагие (Батыр) и Каунды на Мангышлаке, Карашор на южной окраине Устюрта (под чинком Капланкыр), впадина Акчакая под обрывом останца Эшок-Апкрон-кыр, впадина Ер-Ойлан-Туз между горами Дузенкыр и Бадхызом.
Несколько особое место занимает Сарыкамышская котловина, которая в недавнее геологическое время была заполнена водами и представляла собой озеро. Сейчас на дне этой впадипы имеются значительные площади солончаков с пустынной сочносолянковой растительностью.
Среди впутриматериковых местообитаний следует особо выделить сочносолянковые пустыни, разбросанные мелкими пятнами среди песчаных массивов Низменных Каракумов в их западной части, где они, вероятно, связаны с древней аллювиальной равниной Пра-Аму-Дарьи.
Нужно отметить, что по типу засоления почв в пределах СССР выделяются четыре провинции (Ковда, 1946).
Провинция хлоридного соленакопления охватывает низменности, окружающие Каспийское море (Прикаспийскую низменность, Кура-Арак-синскую и Терско-Сулакскую низменность, юго-западную Туркмению и низовья Атрека). Для нее характерно сложное сочетание дельт крупных рек и приморских равнин, образованных трансгрессией Каспия. Наиболее крупные массивы солончаковых пустынь попадают в эту провинцию.
Провинция сульфатно-хлоридного соленакопления охватывает обширные пространства Туранской низменности (дельта Сыр-Дарьи, долины Сыр-Дарьи и Аму-Дарьи, долина Вахша, дельты и низовья Мургаба и Тед-жена), связанные как с процессами континентального соленакопления, так и с многовековым воздействием на процессы засоления орошаемого земледелия. К ней относятся и низовья Кубани и Дона, побережье залива Сиваша и причерноморские лиманы. В этой провинции сочносолянковые пустыни занимают значительные площади, а также встречаются многочисленными мелкими контурами, не показанными на карте.
Провинция хлоридно-сульфатного засоления охватывает дельту Аму-Дарьи, низовья Зеравшана и Ферганскую котловину, а в основном к ней относится обширная территория Центрального Казахстана. Засоленные почвы этой провинции приурочены к речным и приречным террасам и к дельтам рек. Кроме перечисленных районов, сочносолянковые пустыни встречаются здесь мелкими вкраплениями, не показанными на карте по условиям масштаба.1
1 Провинция сульфатио-содового засоления, по данным В. А. Ковды (1946), охватывает равнины лесостепной и степной зоны, где засоленные почвы приурочены
Помимо сочное,оливковых пустынь, выделенных особыми контурами на нашей карте, они вкраплены среди других типов пустынь в виде мелких пятен, которые но могли быть изображены в масштабе карты.
Распространенно их также подчиняется определенной закономерности. 'Гак, они отмечены на контурах полынных пустынь (87) в низовьях Большого и Малого У зоной; среди злаково-полынных пустынь (86) в южной части песчаного массива между Волгой и Уралом (1’ын-нески); среди полынных пустынь в сочетании с солянковыми (89) в низовьях Сагиза; па контурах эфемерово-полынных пустынь (90) и эфемерово-полынных пустынь в сочетании с, солянковыми (91) по долине Куры и Лракса в Закавказье; среди черкезпиков (99) на песках в пониженной части древней дельты Лтрека, па контакте ее с приморскими песками, в юго-западной Туркмении; в котловинах барханных песков (100) на п-ве Челекене; на обширной площади Низменных Каракумов в котловинах среди грядовых песков, покрытых белосаксаулышками (98); и, наконец, среди смешанносаксаульников (97) в юго-восточных Каракумах, в зоне Келифского Узбоя.
Как видно из этого перечисления, мелкие участки сочносолянковых пустынь, не показанные в масштабе карты, повсюду связаны в своем распространении с сухими дельтами современных рек, слепо оканчивающихся в пустыне, либо же с древними (реже четвертичными) дельтами, перекрытыми позднейшими аллювиально-пролювиальными (и эоловыми — Низменные и юго-восточные Каракумы) наносами. Совершенно очевидно, что приуроченность сочносоляпковых пустынь к этим районам объясняется теми активными процессами соленакоплепия, которые в широких масштабах свойственны сухим дельтам.
Во всех отмеченных группах местообитаний основным фактором, определяющим условия жизни растений, является близкое залегание к поверхности засоленных грунтовых вод.
Во многих случаях, когда солончаки лежат в глубоких депрессиях, последние являются водосбором вод поверхностного стока. Нередко на дне таких депрессий в периоды после выпадения осадков образуются озера, которые высыхают в последующее жаркое и сухое время года. Обычно центральные части таких депрессий покрыты лишенными высших растений мокрыми и пухлыми солончаками, а растительность развивается преимущественно в краевой полосе депрессий.
Солянковые пустыни слагаются немногими растениями, приспособленными к условиям крайне высокого засоления почв и грунтовых вод.
Таковы представители типичных галофитов: сарсазан (Halocnemum strobilaceum), поташники (Kalidium foliatum и К. caspicum), карабарак {Halostachys Belangeriana), кзылкокпек (Limonium suffruticosum), мелколистная шведка (Suaeda microphyНа), карган (Salsola dendroides), а также травянистые однолетние виды семейства маревых: Salsola crassa, S. soda, S. lanata, S. turcomanica, Salicornia europaea.
Следует отметить, что среди галофитов выделяются две группы растений. Одни из них накапливают в своих ассимилирующих органах большое количество легкорастворимых хлористых и сернокислых солей натрия и калия. Вследствие этого, их клеточный сок обладает очень высоким осмотическим давлением; это приспособление не только позволяет данным растениям существовать на крайне засоленном субстрате, но и придает им большую устойчивость против высоких температур и сухости воздуха. Показательно, что обычные анатомические приспособления для защиты листьев от испарения отсутствуют у них: кутикула тонкая, устьица открытые и не
к речным и приозерным террасам. В пределах этой провинции сообщества сочносолянковых пустынь встречаются незначительными участками.
погруженные, нет защитных волосков. Таковы, например, солерос (Salicornia europaea), сарсазан (Halocnemum strobilaceum), нетросимония (Pet-rosimonia oppositifolia), Ofaiston monandrum, Suaeda prostrata, Salsola brachiata, S. mutica, Atriplex сапа, Л. verrueijern.
Вторая группа — растения, которые содержат значительно меньшее количество легкорастворимых солей и обладают особыми железками, с помощью которых соли выводятся наружу, отлагаясь на поверхности листьев, а впоследствии попадают в почву (непосредственно с растений, а также вместе с опадом). При этом осуществляется как бы перекачка солей из глубоких слоев почвы на ее поверхность. Сюда относятся некоторые виды гребенщика (Tamarix ramosissima, Т. gracilis, Т. laxa, Т. leptosta-chys и др.) и кермека (Limonium Gmelinii, L. caspium, L. coralloides, L. suffruticosum), виды чаира (Aeluropus litoralis, Л. repens), однолетняя франкения (Frankenia pulverulenta) и др.
Сообщества сочносолянковых пустынь отличаются крайней пестротой и, как правило, сильной *изреженностыо растительного покрова. На особенно «злостных» солончаках преобладают обнаженные от растительности пространства, порою занимающие огромные площади (например солончак Соленые Грязи или Хаки в Заволжье, сор Барса-Кельмес на Устюрте, солончак Карашор в западной Туркмении и др.).
Рассмотрим некоторые наиболее широко распространенные формации сочносолянковых пустынь.
Сарсазанники (из Halocnemum strobilaceum) связаны с так называемыми мокрыми солончаками или солончаками грунтового питания. Условия существования растительности здесь наиболее тяжелые в силу крайне высокой степени концентрации почвенных растворов. Сарсазан нередко образует чистые, одновидовые сообщества. Растения разбросаны на 2—3 м одно,от другого. Куст сарсазана имеет характерную лепешковидную форму, веткй достигают всего 20—30 см высоты. При удалении грунтовых вод от поверхности и при наличии хотя бы временного нисходящего тока, к сарсазаиу примешиваются некоторые другие растения: Limonium suffruticosum, Halostachys Belangeriana, Frankenia hirsuta и немногие однолетние солянки (Salicornia europaea, Salsola incanescens, S. turcomanica, S. lanata, Petrosimonia glauca, Halimocnemis sclerosperma). Изредка в виде мелких угнетенных экземпляров встречается Tamarix Karelinii.
Поташники (из Kalidium caspicum) распространены на солончаках с грунтовыми водами на глубине 3—4 м. Среди резко разбросанных кустов поташника встречаются единичные растения каргана (Salsola dendroides), чогона (Aellenia subaphylla), тетыра (Salsola gemmascens) и также редкие однолетние солянки (Salsola turcomanica, S. lanata, Halochar is hispida, Petrosimonia glauca и др.).
Карабарачники (из Halostachys Belangeriana) очень часто бывают приурочены к вторичным солончакам, возникшим в орошаемых районах, в том числе и на территориях земель древнего орошения. Наряду с крупными кустами карабарака, здесь встречаются (одиночно или рассеянно) угнетенные кустики гребенщиков (Tamarix ramosissima, Т. laxa), однолетние солянки (Salsola turcomanica, Halogeton glomeratus й изредка чахлые дернинки чаира (Aeluropus litoralis).
Сообщества солероса (Salicornia europaea) расположены в виде узких полосок по мокрым солончакам с грунтовыми водами у самой поверхности, а также по прибрежной части соленых озер. Они же часто образуют полосу вдоль низменных побережий Каспийского моря.
Рассмотренные формации — только наиболее широко распространенные; обрисованы они лишь в своих основных чертах. Обычно же сочно-
солянковые пустыни представлены пестрыми сочетаниями фрагментов этих сообществ с многими переходами между ними. Пестрота видового состава усиливается всякий раз, когда происходит какое-либо, даже небольшое, изменение эдафических условий. Так, например, при наносе на поверхность солончака песка, последний накопляется в виде небольших бугорков под кустами сарсазана, поташника и других полукустарничков и в этих случаях здесь поселяются некоторые солеустойчивые эфемеры (Matricaria lamellata, Lepidium, perfoliatum, Alyssum desertorum, Koelpinia linearis, Veronica campylopoda), наряду с более пышно разрастающимися солянками.
При наличии проточных грунтовых вод внедряются луговые галофиты, вроде чаира (Aeluropus litoralis), свинороя (Cynodon dactylon), бескиль-пицы (виды Puccinellia), солодки (Glycyrrhiza glabra), акбаша (Kareli-nia caspica) и др.
Сочносолянковые пустыни не имеют пастбищной ценности и обычно значатся в категории неудобей и бросовых земель. Однако некоторые растения используются населением для кустарного получения соды и поташа (Halocnemum strobilaceum, Kalidium caspicum, Halostachys Belanger iana и др.). Кроме того, они содержат инсектисиды и являются сырьем для промышленного получения этих продуктов (например итсегек — Anabasis aphylla).
Литература
Г. С. Карелин (1883); В. С. Арцымович (1911); М. Д. Спиридонов (1918, 1930); М. Г. Попов (1922); Б. А. Келлер (1923а, ред. 1936, 1940); М. М. Ильин (1927); С. С. Неуструев и И. М. Крашенинников (ред.) (1930); Ю. А. Скворцов (1930); Е. П. Коровин (1934); В. Н. Минервин (1935); Е. Г. Победимова (1935); А. В. Прозоровский (1940а); Н. Т. Нечаева и др. (1943); В. А. Ковда (1946, 1950); М. П. Богданов (1954); Н. С. Нардина (1954); С. А. Никитин (1954, 1954а); Н. Н. Пельт (1955); Н. Н. Пельт и Л. Е. Родин (1955); Л. Е. Родин и Н. И. Базилевич (1955).
94. Однолетнесолянковая растительность (из однолетних видов Salsola, Suaeda, Salicornia herbacea 1 и др.) в сочетании
с полукустарничково-солянковыми, галофитно-луговыми и другими сообществами
Однолетнесолянковая растительность встречается небольшими пятнами по окраинам орошаемых оазисов в нижней части долины Куры, в дельте Аму-Дарьи, а также небольшими участками в ее среднем течении, в нижних частях сухих дельт Теджена и Мургаба, Кашка-Дарьи, Зеравшана, в районе ст. Урсатьевской (левобережье Сыр-Дарьи).
Однолетнесолянковая растительность повсюду очень четко приурочена к окраинам крупных оазисов, где она встречается на землях, недавно вышедших из-под орошения и заброшенных по причине вторичного засоления.
Растительный покров обычно разреженный; повсюду выступает обнаженный субстрат, часто изобилующий выцветами солей на поверхности.
Наиболее распространены здесь однолетние виды Salsola (Salsola turcomanica, S. crassa s. 1.), Halimocnemis К arelinii, H. sclerosperma, H. mol-lissima и др.), виды Suaeda, Petrosimonia, Gamanthus (G. gamocarpus), Salicornia europaea и некоторые другие.
1 В настоящее время согласно номенклатурным правилам этот вид именуется Salicornia europaea.
Наряду с господствующими здесь однолетними формами, обычно на более давно заброшенных землях, встречаются и многолетние полукустарнички (Halocnemum strobilaceum, Halostachys Belangeriana, Anabasis salsa), а на землях, недавно вышедших из-под орошаемого земледелия, сорные виды (Alhagi persarum, A. kirghisorum, Karelinia caspica и др.). Реже, на участках с близкими грунтовыми водами, среди однолетнесолянковой растительности попадаются небольшие пятна галофитных лужков, слагаемых чаиром (Aeluropus litoralis и A. repens).
Пастбищное значение этих сообществ ничтожно; скот неохотно поедает солянки лишь в осенне-зимний сезон.
Л и тература
М. Д. Спиридонов (1921); А. А. Гроссгейм (1930, 1932); Е. П. Коровин (1934); Ф. Н. Русанов (1934); Б. А. Келлер (1940); А. В. Прозоровский (1940а); Н. Т. Нечаева и др. (1943); И. И. Гранитов (1950); Н. И. Рубцов (1952).
95. Высокогорные терескеновые и полынно-терескеновые пустыни (из Eurotia ceratoides, Artemisia Skorniakowii)
В пределах СССР эти пустыни свойственны главным образом Памиру, в меньшей степени высокогорным сыртам Центрального Тянь-Шаня.
По данным К. В. Станюковича (1949), терескенники являются на Памире господствующей формацией. Они встречаются в пределах от 3500 до 4300 м над у. м., на склонах всех экспозиций и на почвах самого различного механического состава — от песков и грубощебенчатых разновидностей до суглинков.
Характернейшей особенностью высокогорных терескенников является их крайняя разреженность и исключительная бедность видами. Проективное покрытие в среднем не превышает 10%, а число видов, участвующих в данной формации, обычно не достигает даже и десятка. Обычно относящиеся сюда ассоциации слагаются всего лишь тремя-пятью видами. Сам терескен представлен угнетенной низкорослой формой, в виде кустиков 15—25 см высоты.
На песках и песчаных почвах, в условиях пониженного минерального питания, формируются сильно разреженные терескенники с участием Christolea pamirica и Stipa glareosa, Zygophyllium Rosovii.
По дну долин, по песчаным террасам и древним конусам выноса обычны разреженные ассоциации Eurotia ceratoides с участием Artemisia Skorniakowii, Stipa glareosa, S. orientalis, Macrotomia euchroma.
На мелкоземистых, но в различной степени захцебненных почвах конусов выноса, по шлейфам и склонам гор, обычным компонентом является полынь Скорнякова (Artemisia Scorniakowii), которая часто создает и самостоятельные сообщества, без участия терескёна. Проективное покрытие полынно-терескеновой ассоциации достигает 25%. В составе ее обычны следующие виды: из злаков — Stipa glareosa, S. orientalis, Poa Litwino-wiana, из криофильных подушечных форм — Acantholimon diapensioides. При лучшем увлажнении в этой ассоциации появляются Hordeum turkesta-nicum, Aneurolepidium dasystachys, Oxytropis Poncinsii, Artemisia pamirica.
В наиболее благоприятных условиях увлажнения — у подножий северных склонов и по сухим руслам весенних водотоков — развивается ячменно-терескеновая ассоциация, где содоминантом является Hordeum turkestanicum. Проективное покрытие травостоя здесь достигает максимальной величины — 30—40%. В сложении этой ассоциации, кроме
указанных двух главных видов, участвуют криофильные растения-подушки— Acantholimon diapensioides, A. Hedinii, злаки — Роа Litwino-wiana и Aneurolepidium dasystachys, рыхлые подушки Smelovskia calycina.
На засоленных почвах по днищам долин и по речным террасам весьма широкое распространение имеют терескенники с большим участием парнолистника (Zygophyllum Rosovii), а по пологим склонам гор с участием куртковника (Acantholimon diapensioides). Эти ассоциации обыкновенно связаны с некоторым засолением субстрата, но по своему видовому составу они все же мало отличаются от терескенников, описанных выше. Из растений, не отмеченных ранее, здесь встречаются такие травянистые ксерофиты, как Dracocephalum heterophyllum и Christolea pamirica, а из ксерофильных полукустарничков—Tanacetum xylorrhizum и Ephedra Fedtschenkoi.
В наиболее высоких местоположениях, уже в пределах «альпийского» пояса Памира (выше 4000 м над у. м.), терескенники слагаются особенно сильно угнетенной формой Eurotia ceratoides в виде приземистых кустиков не более 15—20 см высоты. Редкостойность относящихся сюда сообществ достигает максимальной величины, так что корневые системы растений перестают быть связанными друг с другом. В этих условиях к терескену в качестве согосподствующего вида присоединяется Tanacetum xylorrhizum, в результате чего создаются пижмовые терескенники — ассоциация, чрезвычайно широко распространенная в «альпийском» поясе Памира. В сложении «альпийских» терескенников большую роль играют также криофильные высокогорные подушки — Acantholimon Hedinii и Oxytropis immersa.
Как было указано выше, терескеновые пустыни свойственны и высокогорьям Центрального Тянь-Шаня. На карте они не показаны в качестве самостоятельных контуров. Терескенники Центрального Тянь-Шаня вошли частью в контур 101 тш и частью в контур 104 са. По характеру своего сложения они сходны с памирскими терескенниками, несколько отличаясь от последних главным образом лишь видовым составом (Кашка-ров, Жуков, Станюкович, 1937).
Полынные пустыни из Artemisia Scorniakowii, как сообщает К. В. Станюкович (1949), распространены менее широко, чем терескеновые и полынно-терескеновые. Полынь Скорнякова избирает легкие, хорошо аэрированные почвы на песчаных или щебнистых склонах и шлейфах, преимущественно в верхней части субальпийского пояса, на высотах от 3900 до 4300 м над у. м. Характерно частое наличие на небольшой глубине вечной мерзлоты.
Видовой состав этих полынников богаче, в одной ассоциации встречается около 7 видов. Значительно выше проективное покрытие — в среднем 20—25%.
В большинстве долин типичны полынники из Artemisia Skorniakowii с доминантом Ptilagrostis subsessiliflora, а также с участием Роа Lit-winowiana, Braya pamirica, Oxytropis Poncinsii и единичными Tanacetum xylorrhizum, Roegneria czimganica и Eurotia ceratoides.
В центральных районах Памира характерна ассоциация полыни Скорнякова со Stipa orientalis при участии Acantholimon diapensioides, Artemisia pamirica, Hedysarum cephalotes, Eurotia ceratoides и единичными Oxytropis sp. и Potentilla Moorcroftii.
Кроме того, встречаются сообщества формаций Artemisia rutifolia и A. Kuschakewiczii, имеющие ограниченное распространение.
В пределах альпийского пояса, на высотах от 4300 до 4700 м лад у. м. и даже выше, наиболее типичными становятся сообщества фор
мации Tanacetum xylorrhizum. Они занимают самые сухие и холодные местообитания, — куда уже не заходит терескен,—на песках, галечниках, скоплениях щебня, такыровидных и засоленных почвах. Сообщества пижмы настолько изрежены, что К. В. Станюкович (1949) не устанавливает сложившихся ассоциаций; при раскопке корневых систем часто обнаруживалось отсутствие смыкания корневых систем. Спутниками Tanacetum xylorrhizum являются Parrya ехсара (наиболее постоянный вид в сообществах), Oxytropis Poncinsii, Braya pamirica, Pyrethrum pyrethraides’, реже других отмечены .Hordeum turkestanicum и Aneurolepidium dasystachys\ в верхней части альпийского пояса появляются Melandrium apetalum, Rhodiola pamiroalaica, R. heterodonta. Сообщества пижмы в хозяйственных целях не используются.
Высокогорные терескеповые и полынно-терескеновые пустыни используются в качестве пастбищного угодья и в качестве источника топлива. Несмотря на низкую кормовую продуктивность, значение этих угодий (на Памире) все же достаточно велико, так как они занимают большие площади и могут эксплуатироваться в течение круглого года. Заготовка терескена на топливо производится в больших количествах, без учета его крайне замедленного роста и плохого возобновления, что может привести к полному оголению горных склонов в районах распространения этого растения (Райкова, 1930; Станюкович, 1949).
Литература
О. Э. Кнорринг (1923); И. А. Райкова (1930); П. А. Баранов (1936); Д. Кашкаров, А. Жуков, К. Станюкович (1937); К. В. Станюкович (1949, 1955); Е. А. Варивцева (1951); Н. И. Рубцов (1952).
XIV. САКСАУЛЬНИКИ И КУСТАРНИКОВЫЕ ПУСТЫНИ
л. JJ. годин
Общий обзор
Саксаульники и кустарниковые пустыни включают сообщества, слагаемые крупными ксерофильными полукустарниками и кустарниками (часто древовидными), по биологической природе относящимися к мезотерм-ным растениям. •
Господствующие виды — эдификаторы (строители сообществ) относятся к следующим родам: саксаул (Haloxylon), основная площадь ареала которого приходится на Советскую Среднюю Азию, но один вид уходит на восток в пределы пустынь Центральной Азии, а другие близкие виды встречаются в Африке; солянки (Salsola) — древовидные полукустарники черкезы и чогон, в основном эндемичные для Средней Азии и только несколько заходящие в Иран; кандым, или джузгун (Calligonum), насчитывающий в СССР более 70 видов, большинство которых (57 видов) являются эндемами пустынной области нашей страны и только два уходят в Иран и пять в Центральную Азию; хвойник, или борджок (Ephedra), представленный двумя вида'ми, эндемичными для Средней Азии (один — Ephedra strobilacea — заходит и в Иран);песчаная акация, или сюзен(Атто-dendron), виды которой формировались в Средней Азии, являются ее эндемами (только один вид уходит несколько на восток за ее пределы в Джунгарию и Кашгарию); астрагал (Astragalus), некоторые из видов которого типичны для описываемых кустарниковых пустынь (они относятся к особой секции рода — секции Ammodendron Bge.); ряд монотипных родов (Smirnovia, Eremosparton) и другие роды и виды, также эндемичные для наших пустынь.
Представители перечисленных родов, являясь эдификаторами либо сопутствующими видами саксаульников и кустарниковых пустынь, распространены не только в их пределах, но проникают и в другие типы растительности в качестве сопутствующих или редких растений. Так, черный саксаул (Haloxylon aphyllum) нередко встречается в виде незначительной примеси в некоторых сообществах полынных и солянковых пустынь (например на Устюрте); иногда он поднимается довольно высоко в горы до границы с арчевниками (например в Большом Балхане). Некоторые виды кандымов (Calligonum) внедряются в полынные пустыни. Зайсанский саксаул (Haloxylon ammodendron) на северной границе своего ареала иногда может быть найден среди степных сообществ и т. п. В саксаульники, в свою очередь, нередко (особенно в речных долинах) проникают элементы тугайной растительности. В сообществах кустарниковых пустынь встречается примесь некоторых видов, характерных для колючетравников и нагорных ксерофитов.
Поэтому нередко наблюдаются сообщества переходного характера между названными типами.
Саксаульники и кустарниковые пустыни в основном свойственны песчаным массивам Средней Азии и в меньшей степени древним дельтам (некоторые саксаульники и черкезники), где они приурочены как к песчаным, так и к глинистым почвам.
Отмеченная преимущественная связь описываемых сообществ с песками явилась основанием к выделению саксаульников и кустарниковых пустынь в качестве особого типа — «песчаных пустынь», принятого некоторыми авторами, наряду с выделявшимися ими «глинистыми», «щебнистыми», «гипсовыми» и «солончаковыми» пустынями.
Наиболее крупная площадь саксаульников и кустарниковых пустынь — это массивы Каракумы и Кызылкумы, занимающие значительную часть Туранской низменности, далее следуют значительно меньшие по площади Прикаспийские пески, Приаральские Каракумы, Муюнкумы и пески Сары-Ишикотрау (пески южного Прибалхашья). Кроме того, имеется значительное количество мелких песчаных массивов, разбросанных по всей территории пустынной области Советского Союза и обычно приуроченных к древним долинам и дельтам рек, часто уже угасших к настоящему времени. Эти небольшие площади кустарниковых пустынь и саксаульников расположены среди полынных и солянковых пустынь и субтропических степей (полусаванн).
Климатические условия саксаульников и кустарниковых пустынь в основных чертах повторяют общие особенности климата пустыни: засушливость, континентальность, высокие летние температуры при отсутствии осадков и т. п. Однако в песках, особенно в таких гигантских песчаных массивах, как Каракумы или Кызылкумы, возникают некоторые своеобразные и очень важные для жизни растений отличия.
Так, в пределах Каракумов количество осадков снижается до 100— 120 мм в год, а в некоторых местах даже до 75 мм. Слабо одетая растительностью песчаная поверхность, во-первых, очень сильно нагревается солнцем (до 60°), в связи с чем чрезвычайно быстро испаряется влага в поверхностных горизонтах. Во-вторых, пески обладают высокой отражательной способностью. Это приводит к тому, что растения испытывают как бы двойной нагрев: прямых солнечных лучей и отраженной лучистой энергии. Средняя суточная температура воздуха доходит до 30—35°, а в дневные часы в некоторых районах (например в юго-восточных Каракумах) достигает 49—50° (правда, дни с такими температурами бывают )редко). В связи с этим относительная влажность воздуха, обычно составляющая в Каракумах менее 30%, падает в дневные часы до 2— 3%.
Высокий йагрев почвы создает сильные восходящие токи горячего воздуха. Этим объясняется возникновение в «песчаных пустынях» уже вскоре после восхода солнца ветра, достигающего в последующие часы наибольшей силы и постепенно затихающего к вечеру. Характерно также полное отсутствие облачности на протяжении нескольких месяцев. Снеговой покров, как правило, отсутствует. Снег держится только несколько дней, а часто только и несколько часов, стаивая под влиянием прямого солнечного нагрева даже при отрицательных температурах воздуха. Только в очень редкие суровые и многоснежные зимы снеговой покров удерживается в течение двух-трех недель.
Таким образом, в саксаульниках и кустарниковых пустынях климатическая обстановка и, в особенности, климат приземного слоя воздуха отличаются еще большей аридностью, еще большей резкостью проявления отдельных экологических факторов, имеющих жизненно важное значение для растений. 4
Под саксаульниками и кустарниковыми пустынями формируются примитивные, слабо развитые песчаные почвы, относящиеся к типам серобурых и сероземов (в южной части). Под сообществами черного саксаула, приуроченными к древним дельтам и к понижениям среди песков, обычно образуются солончаковые или солопчаковатые почвы, в некоторых случаях отакырепныс.
Резко выраженная аридность (пустынность) климата определила целый ряд морфологических и экологических особенностей основных строителей — эдификаторов сообществ. Все это — ксерофиты и галоксе-рофиты, то есть растения крайне засушливых условий существования.
Для многих растений характерна слабая облиственность, часто сочетающаяся с уменьшением листовой пластинки до размеров, сходных с хвоей (например у черкеза — Salsola Richteri). У ряда растений выработалась большая или меньшая афиллия (безлистность) с передачей функции фотосинтеза зеленым веточкам (например у видов саксаула — Haloxylon, у кандымов — Calligonum и др.). В результате крайнего дефицита влаги в летний период у некоторых распространенных здесь растений наблюдается частичный летний листопад (и веткопад — у афилльных форм). Растения сбрасывают некоторую часть листьев или зеленых веточек, что сокращает транспирацию в наиболее жаркое время (например виды Calligonum, Ammodendron, Astragalus и др.). У многих растений существуют различные анатомические приспособления для сокращения испарения (защитные волоски, погруженные устьица, утолщенная кутикула, блестящая поверхность листьев и т. п.). В «поисках» влаги все эдификаторы сообществ развивают мощную корневую систему, углубляющуюся до горизонта капиллярно подвешенной влаги, а иногда и непосредственно до грунтовых вод.1 Многие растения образуют корни, распространяющиеся горизонтально, придерживаясь подповерхностного влажного горизонта. Таковы многие виды Calligonum, длина горизонтальных корней которых достигает 30 м. У некоторых растений, существующих в условиях подвижных песков, протянутые далеко в стороны корни, кроме того, выполняют функцию своего рода «якорей», удерживающих растение на месте при развевании песков. Это особенно хорошо выражено у селинов (Aristida), а также у кандымов (Calligonum), причем корни их одеты пробковидной корой, предохраняющей от высыхания. Виды Calligonum в то же время обладают способностью образовывать придаточные корни на засыпанных песком частях стволов, что значительно содействует улучшению водоснабжения растения (Дубянский, 1928). Подобные же придаточные корни образуются у эремоспартона (Eremosparton flaccidum), черкеза (Salsola Richteri), белого саксаула (Haloxylon persicum) и др.
Плоды и семена многих растений имеют приспособления к переносу их ветром (анемокарпия). Имеются и другие приспособления, обеспечивающие растениям успешное существование в своеобразных и трудных условиях песчаных, нередко подвижных субстратов и в экстра аридной климятп-ческой обстановке. По ряду таких специфических признаков большинство эдификаторов и очень многие сопутствующие виды, входящие в сообщества саксаульников и кустарниковых пустынь, относят к особой категории — псаммофитам, то есть к растениям, приспособленным к жизни в песках.
1 Распространение саксаульников и большинства сообществ кустярникляит пустынь, по э. Н. Благовещенскому (1940), связано с образованием в песках влажного конденсационного горизонта на глубине 2—5 м, возникающего за счет внутри-почвенного испарения от грунтовых вод, при условии залегания их не глубже 28 и; при большей их глубине конденсационный горизонт не образуется, и в таких условиях саксаульники и другие сообщества отсутствуют.
Наиболее характерной жизненной формой описываемого типа растительности являются непрофильные полукустарники и кустарники.
К жизненной форме полукустарников относятся растения, генеративные побеги которых ежегодно отмирают. Это виды саксаула (черный саксаул — Haloxylon aphyllum, белый саксаул — И. persicum, зайсапский саксаул — Н. ammodendron), черкеза (Salsola Richteri и 5. Paletzkiana). чогоп (Aelle-nia subaphylla). Очевидно к этой же группе следует относить и некоторые астрагалы (виды Astragalus из секции Ammodendron). Возможно, что к полукустарникам могут быть причислены и виды кандымов (Calligonum), однако биологические свойства этих растений изучены еще недостаточно.1
Кустарники представлены видами песчаной акации (Ammodendron Conollyi, A. Lehmannii, A. Eichwaldii, A. Karelinii). Взрослые экземпляры песчаной акации, не подвергавшиеся порубкам, имеют древовидную форму. К группе кустарников следует отнести и единственные вечнозеленые растения наших пустынь: борджок (Ephedra strobilacea) и близкий к нему Е. lomatolepis (распространенный в песках Прибалхашья и Муюнкумах). Названным выше эдификаторам сопутствует некоторое число полукустарничков. Это виды полыней (Artemisia terrae-albae ssp. Massagetovi, A. are-nicola, A. santolina, A. eriocarpa, A. Dimoana, A. Kelleri} и вьюнков (Convolvulus Korolkovii, C. erinaceus, C. fruticosus). Сравнительно редко, но все же встречаются полукустарничковые солянки: боялыч (Salsola arbuscula), кеурек (5. rigida).
Травянистые многолетние растения, сопутствующие вышеперечисленным в образуемых ими сообществах, представлены здесь двумя группами: длительновегетирующими и коротковегетирующими (эфемероидами). Длительновегетирующие немногочисленны: селин (Aristida pennata), еркек-селин (Aristida Karelinii), гелиотроп (Heliotropium arguzioides), хрозофора (Chrozophora gracilis), турнефортия (Tournefortia sogdiana), руты (Haplophyllum Bungei и H. pedicellatum), Astragalus chivensis, Cousinia bipinnata и некоторые другие. Большинство их является ксерофитами; у них имеются соответствующие анатомические приспособления, они слабо облиствены, склероморфны и т. п. У некоторых из них корешки одеваются как бы чехликом из склеенных корневыми выделениями песчинок, который предохраняет корни от высыхания при обнажении их ветром.
Очень богато представлены эфемероиды. Таковы, например, виды Ferula (F. assa-foetida, F. oopoda, F. badrakema и др.), часто играющие заметную роль в облике некоторых саксаульников и кустарниковыхх пустынь. Довольно обильно участие луков (Allium caspium, A. sabulosum). Почти всюду встречаются ширяш (Eremurus inderiensis) или другие виды этого рода, украшающие весной пустыню высокими красочными стрелками соцветий. Весьма обычен выделяющийся огромными листьями, до 70—80 см длиной при почти такой же ширине, ревень (Rheum turkestanicum). Менее часто встречаются Eremostachys Regeliana, Leontice Ewersmannii, Eminium Lehmannii, Iris songarica, Gagea stipitata, Tulipa sogdiana, Bongardia chrysogonum и др. К группе эфемероидов относится илак, или песчаная осока (Carex physodes). Она своими сильно ветвящимися корневищами скрепляет песчаную поверхность, предохраняя ее от развевания. Илак — лучшее кормовое растение в пустыне, хорошо поедаемое скотом даже в сухом состоянии (своеобразное «сено на корню»). Все эфемероиды по своему облику (строению листьев, их размерам и т. п.)
1 Возможно, что Знды Calligonum образуют особую жизненную форму, по некоторым признакам сближающуюся с полукустарниками, а по другим — с кустарниками.
являются настоящими мезофитами. Они начинают развиваться с ранней весны (а иногда и с осени) и заканчивают весь цикл развития, включая плодоношение, к моменту наступления засушливого периода.
Такой же быстрый цикл развития проходят и эфемеры — однолетние травянистые растения, большое количество видов которых весной одевает склоны песчаных гряд иногда сплошным зеленым ковром, украшенным множеством цветков (особенно в годы с особо обильными осадками). Все они также по биологической природе являются мезофитами. Они представляют собою своеобразный «ярус во времени», образуя эфемеровую синузию в сообществах растений весной. К ним относятся Bromus tectorum, В. Danthoniae, Eremopyrum orientals, Papaver pavoninum, Malcolmia gran-difloia, Cutandia memphitica, Schismus arabicus, Streptoloma desertorum, Spirorhynchus sabulosus, Koelpinia macrantha, Consolida camptocarpa, Silene папа, Lappula semiglabra, Erodium oxyrrhynchum, Cousinia annua, Cithareloma vernum и множество других.
Довольно разнообразны и однолетники летне-осеннего цикла развития, однако количественное их участие в построении сообществ не столь велико. К ним относятся следующие растения: Agriophyllum latijolium, Corispermum aralo-caspicum, C. Lehmannianum, Salsola aperta, S. cari-nata, S. praecox, Horaninowia ulicina, Euphorbia Turczaninowii, Kochia iranica, Atriplex dimorphostegia и некоторые другие.
Из споровых растений в сообществах саксаульников и кустарниковых пустынь заметное место принадлежит пустынному мху — Tortula desertorum. Лишайники, если и встречаются, то в малом количестве и только на выходах щебня и скалах. Водоросли имеются и здесь, однако они до настоящего времени еще не изучены.
Растительный покров описываемых пустынь не однороден. Для него характерна комплексность, однако она значительно отличается от обрисованной в предшествующем разделе (XIII). Комплексность саксаульников и кустарниковых пустынь обусловлена в основном мезорельефом песчаных массивов; последние почти на всей территории характеризуются гря-довым или бугристым сложением. Растительность склонов гряд, межгрядовых понижений (или котловин в бугристых песках) и вершин гряд (или бугров) всегда представлена сообществами различных ассоциаций, нередко весьма отличных не только по составу сопутствующих видов, но и по эдификаторам. В качестве примера укажем, что в Кызылкумах очень характерны следующие элементы комплекса: 1) илаковый белосаксауль-ник (сообщество из Haloxylon persicum с покровом из Carex physodes)1 на вершинах гряд с хорошо закрепленной поверхностью песка; 2) илаковый джузгунник (виды Calligonum—Carex physodes) на склонах гряд с закрепленной поверхностью песка; 3) моховой черносаксаульник (Haloxylon aphyllum—Tortula desertorum) на дне межгрядовых понижений с сильно уплотненным (оглиненным) песком (Порецкий и др., 1936).
В Каракумах очень часто встречаются комплексы из следующих ассоциаций: 1) селиновый сюзенник (Ammodendron Conollyi—Aristida реп-nata) на вершине гряды с подвижной песчаной поверхностью; 2) илаковый белосаксаульник (Haloxylon persicum—Carex physodes) на склоне гряды с закрепленной поверхностью песка; 3) эфемеровый черносаксаульник (Haloxylon aphyllum—эфемеровое разнотравье) на глинисто-песчаных почвах межгрядовых понижений.
1 Флористический состав сообществ гораздо богаче, но здесь указываются только основные эдификаторы и основные сопутствующие виды.
Для многих районов Низменных Каракумов (например для района древних дельт Теджена и Мургаба) характерно комплексирование различных сообществ саксаульников и кустарниковых пустынь на песках с водорослевыми сообществами на такырах. В некоторых местах (например в песках между Узбоем и Копет-Дагом) часто наблюдается комплекс различных сообществ саксаульников на песках с сообществами сочносолянковых пустынь на сильно засоленных почвах и солончаках. Разнообразие подобных комплексов очень велико.
Было отмечено выше, что комплексы растительного покрова в «песчаной пустыне» обусловлены строением мезорельефа. Это верно только в первом приближении. Истинные причины оказываются значительно глубже и сложнее, и в большинстве случаев ведущая роль принадлежит самому растительному покрову. Так, например, сообщества, занимающие склон гряды (или какое-либо иное положение), отличаются особыми условиями просачивания метеорных вод, степенью засемененности почв, а следовательно, и возможностями возобновления растений. Более густой покров эфемеров и эфемероидов определяет количество ежегодно поступающих в почву зольных элементов и степень обогащения ее органическими веществами,' что не только обусловливает направление почвообразующих процессов, но и существенным образом меняет и многие другие факторы среды и сказывается на последующем развитии сообщества и т. д. В условиях подвижного песчаного субстрата постоянно наблюдается деградация одних сообществ и становление других,1 и роль растительности в этих процессах выступает с наибольшей яркостью.
В ряде случаев наблюдаемые в настоящее время комплексы имеют сложную историю. Так, в речных долинах, на плато и других местах саксаульники или сообщества каких-либо кустарниковых пустынь внедрились значительно позже, когда на указанных территориях уже существовал растительный покров, прошедший длительную эволюцию. В этих сообществах уже были сформированы особые, им присущие почвы, свой ритм развития, своя сезонность и т. п. В результате наложения новых сообществ (возникших вследствие эоловых или аллювиальных процессов в ходе общего развития ландшафта) формировались новые комплексы, в которых не механически сочетались старые и юные формы, а созидались новые сообщества, постоянно развивающиеся и взаимообусловленные. Этим объясняется значительная неустойчивость, подвижность комплексов.
Так, например, в древней дельте Аму-Дарьи можно наблюдать наступание элементов саксаульников и кустарниковых пустынь на водорослевые сообщества такыров. Простой комплекс — черносаксаульник на песчаных грядах и водорослевый такыр — становится более сложным. Появляются переходные сообщества между названными. При заселении такыра растения накапливают песок, существенно меняя гидрологический режим такыра. Водоросли отмирают, и на недавнем такыре возникает новое сообщество, которого не было вначале в указанном простом комплексе, из сочетания двух сообществ: это может быть серополынник (сообщество из Artemisia terrae-albae) или кеуречник (из Salsola rigida) и др. Под полынником и кеуречником развиваются уже иные почвы (в данном случае близкие к серо-бурым), в профиле которых, однако, вскрываются следы еще недавнего такырного прошлого.
В некоторых местах в Сарыкамышской котловине под влиянием растительности произошло столь резкое изменение условий существования,
1 Кстати, именно эти последние явления привлекали наибольшее внимание исследователей, и стадии смен растительности в процессе разбивания и зарастания песков освещены во многих работах (В. А. Дубянскнй, М. П. Петров и др.).
что это привело к полному отмиранию эдификаторов на больших территориях, к полной замене одних сообществ другими (например на месте отмерших гребенщиковых тугаев возникли черносаксаульники), и к возникновению новых отношений между имевшимися там элементами комплексов.
Наряду с комплексностью растительного покрова существует еще и мозаичность (см. примечание на стр. 598). В описываемых здесь пустынях это явление изучено слабо. Известно, что мозаичность вызывается различными причинами. Так, например, под кустами черного саксаула (в сообществах его с покровом однолетних солянок) наблюдается развитие синузии эфемеров, представленной рядом видов {Matricaria lamellata, Veronica campylopoda, Lepidium perfoliatum, Eremopyrum orientate, Bromus tectorum, Malcolmia grandiflora, Consolida camptocarpa, Alyssum desertorum, Amberboa turanica и др.). Это явление некоторыми авторами отмечалось под наименованием «бугра вегетации»1 (Шингарева, 1940а; Зонн и Леонтьев, 1942 и др.).
Установлено, что почвы таких бугров отличаются большим содержанием гумуса (за счет разложения стволов и корней саксаула), и условия для возобновления саксаула здесь более благоприятны вследствие повышенного содержания питательных веществ — азота и фосфора (Зонн и Леонтьев, 1942).
Очень часто причиной, вызывающей мозаичность, является ветер. Несомые ветром песчинки задерживаются под кустами (в «ветровой тени») и, накапливаясь, образуют бугор. Бугор как бы цементируется органическими веществами, поступающими с опадом растений. Постоянно приходится встречать такие бугры, достигающие иногда больших размеров (до 1—1.5 м высоты), особенно под кустами борджока {Ephedra strobilacea). Подобные же бугры образуются и под кустами саксаула и других растений. Бугры эти оказываются настолько прочными, что и после отмирания куста они долго сохраняются даже среди сильно развеваемых песков, предоставляя приют многим растениям.
Нередки случаи мозаичности в плановых белосаксаульниках, среди которых на склонах гряд часто наблюдаются колонии большой песчанки (Rhombomys opimus). Устраивая многочисленные норы, соединенные под землей сложной сетью ходов, песчанки служат причиной образования сообщества, отличного от исходного. Влияние песчанок связано не только с их роющей деятельностью, но и с тем, что они, поедая одни растения, устраняют конкурентов у других, развивающихся более обильно. Песчанки вносят в почву большое количество органических веществ в виде несъедобных остатков растений, а также своих экскрементов. Объедая молодые ветки саксаула, они придают кусту такую своебразную форму, по которой безошибочно определяется наличие по соседству колоний песчанок.
Мозаичные сообщества возникают и под влиянием чрезмерного выпаса скота, приводящего к разбиванию скрепленной илаком поверхности и к последующему развеванию участка («язвы дефляции»), на котором поселяются «пионерные» виды, отсутствовавшие в первоначальном покрове.
Необходимо все же указать здесь, что слабая дефляция, в виде отдельных очагов на фоне илакового белосаксаульника (или какого-либо иного сообщества), создает мозаичное сообщество, в котором сохранен ярус
1 Очевидно, в связи с тем, что под кустами саксаула за счет скопления частиц мелкозема, приносимых ветром, и обильного опада однолетних ассимиляционных побегов образуется плоский бугор, на котором развивается эта синузия эфемеров.
эдификаторов п меняется лишь состав сопутствующих видов. Но дефляция на большой площади и заходящая более далеко приводит к смене растительности., возникновению нового сообщества с ипымл эдифпкаторными и сопутствующими видами. В этом случае уже приходится говорить о комплексе сообществ плановых белосаксаульников {Haloxylon persicum— Carex physodes) на закрепленных песках и, например, гелиотроповых сю-зенников {Ammodendron Conollyi—Heliotropium arguzioides) на подвижных песках.
Точно так же, как и в полукустарничковых (полынных и солянковых) пустынях, в саксаульниках и кустарниковых пустынях проходят «волны жизни»: эфемеры и все остальные растения более пышно развиваются в годы с особо обильными осадками. Однако следует отметить, что здесь это явление не столь контрастно. Дело в том, что легкая водопроницаемость песков обусловливает образование на некоторой глубине подвешенного горизонта увлажнения, обладающего значительным постоянством (хотя он и сокращается в «сухие» годы и становится мощнее в годы «влажные»). Наличие этого постоянного горизонта обеспечивает и в «сухой» год развитие саксаула и других кустарников, хотя и ослабленное в сравнении с благоприятным годом, однако не в такой резкой форме, как в такой же год в полукустарничковых пустынях с их главным образом глинистыми почвами. Не следует упускать из виду и другое важное обстоятельство — накопление в песках конденсационной влаги. Последняя несомненно играет существенную роль в жизни растений на песках (Благовещенский, 1954). На развитии саксаула, кандымов и остальных эдификаторов это, конечно, менее заметно, чем на эфемерах и эфемероидах. «Пески» весной зеленеют почти всегда (правда, период вегетации может быть очень коротким), в то время как полынные и солянковые пустыни часто бывают лишены весенней эфемеровой синузии.
Очевидно, с этой же периодичностью благоприятного увлажнения связаны «семенные годы» у саксаулов, черкеза, кандымов и других растений, когда плодоношение их значительно выше, чем в средние годы. За годами с обильным плодоношением следуют годы и более обильного семенного возобновления (если не наступит катастрофически сухой год, когда происходит массовая гибель всходов). Это легко обнаруживается при внимательном изучении растений в сообществах, в частности, их возраста. Всегда четко вычленяются разновозрастные группы подроста (однолетние, четырехлетние, восьмилетние) и гораздо труднее обнаружить особи, которые по возрасту распределились бы в последовательный ряд (1, 2, 3, 4, 5 и т. д. лет).
Хозяйственная деятельность человека на протяжении предшествующих тысячелетий в сильнейшей степени отразилась на состоянии растительности, на формировании определенных сообществ, на пространственном размещении отдельных типов саксаульников и кустарниковых пустынь.
Подвижность песчаных субстратов, при уничтожении связывающего их растительного покрова, обусловила быструю и почти всегда необратимую смену сообществ, часто приводящую к возникновению новых ландшафтов. Таковыми, например, являются барханные пески и заросли псаммофиль-ных кустарников (опис. 100). Распределение их на территории Средней Азии совершенно четко обнаруживает приуроченность этих образований к современным оазисам или к землям древнего орошения (то есть к бывшим оазисам, заброшенным на том или ином этапе истории). Чрезмерный выпас, вырубки на топливо, движение караванов и стад — вот причины, породившие этот ландшафт. Фрагменты типа разреженной псаммофитной расти
тельности имеются и на песчаных массивах, нетронутых или сравнительно слабо затронутых воздействием человека, но огромные барханные поля в физико-географических условиях Средней Азии могли возникнуть лишь в результате стихийной хозяйственной деятельности человека.
Уничтожение растительности (прямое и косвенное) стихийно совершалось вплоть до самого последнего времени. С 90-х годов прошлого столетия начались пескоукрепительные работы в Средней Азии в связи с заносами подвижными песками Закаспийской (ныне Ашхабадской) железной дороги. Но лишь в советское время вопросам борьбы с подвижными песками был придан государственный плановый характер, и в этом деле имеются серьезные достижения.
На ранних этапах своей истории человек очень охотно поселялся в песках, среди разнообразной растительности, обеспечивавшей охотника топливом, а впоследствии скотовода круглогодичными пастбищами и также топливом.
Изобретение гончарного круга было прогрессивным шагом в истории материальной культуры человека, но последовавшее за этим массовое сооружение керамических печей, поглощавших большое количество топлива., явилось губительным для окружающих саксауловых зарослей (или других сообществ), служивших источником топливной древесины.
Умение строить глубокие колодцы в песках имело большое значение для развития кочевого скотоводства. Но для укрепления колодезного ствола требовалась высококачественная древесина: прочная, стойкая к сырости, не портящая вкусовых достоинств воды и т. п. Таким явился сюзен (Ammodendron Conollyi и другие виды этого же рода).
Для сооружения колодца глубиной 20 м (это более или менее средняя глубина колодцев Каракумов) требуется более 5000 отрубков сюзена. Из одного экземпляра этого деревца получается не более двух отрубков. На один колодец надо вырубить около 2500 особей сюзена. Для боле( глубокого колодца требуется гораздо большее количество материала: в колодец глубиной в 60 метров надо уложить около 30000 отрубков, то есть уничтожить сюзен на площади 600 га. Легко представить, какие количества этого (часто встречающегося, но разреженно растущего) отличного пескоукрепляющего растения потребовалось для тысяч колодцев Каракумов, которые выкапывались, забрасывались и вновь сооружались на протяжении многих веков или даже тысячелетий.
Ближайшие окрестности колодца вскоре бывают окружены оголенными от растительности и вздыбленными выше окружающих грядовых песков барханами. «Акчайга» — белые пески (то есть лишенные растительности) — называет их население) и эти пески в караванных переходах стали надежным ориентиром колодца в «песчаном море». Когда колодец забрасывается, «тырло» зарастает растительностью, но это уже будет новое сообщество, резко отличное от того, которое было ранее; для него будет характерным внедрение ряда сорняков, которые пришли со стадами и человеком.
Выше уже немного говорилось о влиянии выпаса доматных животных. Выпас диких животных также сказывается на состоянии растительности: между растениями и животными существовали тесные биоценотические отношения. Однако эти отношения только в редких случаях приводили к полному уничтожению растительного покрова. Имеются основания полагать, что выпас диких животных имел серьезное значение в разбивании песков (Станюкович, 1950). И сейчас, в отдаленных от населенных пунктов и колодцев районах, где нет выпаса домашних животных, можно наблюдать сложную сеть троп, выбитых джейранами (Antilope subgutturosa), участ-
ков, стравленных дикими кабанами (Sus scrofa), и т. п. Но лишь выпас домашних животных, производившийся хищнически, обусловил резкие смены сообществ саксауловой и кустарниковой пустыни на больших площадях. Так совершалось веками.
Только с установлением новой системы планового отгонного животноводства представилась возможность рационального использования богатейших кормовых ресурсов «песчаных пустынь» (загонная система пастьбы, регулирование выпаса по нагрузке скота на единицу площади, правильное использование по сезонам, более густая сеть колодцев и иных источников водопоя и т. п.). Однако в этом деле много недоработанного, нередки нарушения рациональной системы и т. д., что также не бесследно проходит для современного растительного покрова.
Если в деле регулирования выпаса скота, установления рационального использования пастбищ и повышения их продуктивности имеются большие достижения, то в использовании топливных ресурсов еще нет необходимого порядка. Лет 15—20 тому назад основным транспортным средством был верблюд. На верблюдах вывозили заготовленный на топливо саксаул к железнодорожным станциям. «Радиус действия» равнялся суточному переходу верблюда, т. е. 30—35 км. Но и при этом заготовки саксаула осуществлялись в огромных размерах. Так, только из бывшего Перовского у. (ныне Кзыл-Ординская область Казахской ССР) за три года (1905—1907) было вывезено с семи железнодорожных станций в пределах этого уезда 161.7 тысяч тонн саксаула, а всего было выбрано из зарослей 196.6 тысяч тонн (Верховский, 1911). Если считать, что на 1 га в среднем приходится 5 тонн топливной древесины саксаула, то окажется, что за эти три года был выбран весь ценный саксаул на площади 40 тысяч га. Учитывая, что возобновление древостоя в саксаульниках требует 15—20 лет, можно ясно представить, какие огромные площади саксауловых зарослей были вовлечены в хозяйственный оборот. При этом следует учесть, что (по данным того же автора) со времени окончания постройки железной дороги Оренбург—Ташкент происходило ежегодное значительное увеличение вывоза саксаула (за 1905—1906 гг. было вывезено по железной дороге 65.6 тысяч тонн, а за один 1907 год было вывезено уже 96.1 тысяч тонн).
В настоящее время с развитием автотранспорта «радиус действия» многократно увеличился — до 120—150 км. Лишь на ббльшем расстоянии от крупных населенных пунктов можно теперь встретить добротные полнодревесные заросли саксаула. Только при наличии труднопроходимых препятствий, в виде барханных цепей или топких солончаков, сохранившиеся заросли встречаются более близко.
Типичными («настоящими», в наиболее яркой степени несущими черты этих ксерофитных сообществ) среди описываемых саксаульников и кустарниковых пустынь являются сообщества, в которых заметное участие принимают в качестве сопутствующих растений виды полыней {Artemisia semiarida, A. terrae-albae s. str., A. diffusa, A. arenicola и др.): полынные саксаульники, полынные черкезники, полынные кандымники и т. п. Они связаны с уплотненными и оглиненными песками или с глинистым почвами, одетыми песчаным плащом. В этих случаях проявляются признаки зональных черт почвообразования (серо-бурого и сероземного). Однако такие «настоящие» пустынные типы встречаются на сравнительно небольших площадях (в северных и центральных частях Кызылкумов, в Заунгузских Каракумах и в некоторых других местах).
Гораздо более широко развиты различные варианты, отклоняющиеся от этих «зональных» типов (в той мере, насколько можно сближать их с подлинными зональными, занимающими плакорные местообитания).
Наиболее широко распространены полусаванновые (то есть со значительным участием элементов субтропических степей или полусаванн) саксаульники и кустарниковые пустыни. Для них характерна богатая эфе-мерово-эфемероидная синузия, представленная большим количеством видов. Это в первую очередь Carex physodes, Rheum, turkestanicum, виды Ferula, Allium, Iris и т. п. многочисленное эфемероидное разнотравье.
Остепненные саксаульники и кустарниковые пустыни характеризуются участием в их составе степных растений в качестве сопутствующих видов в сообществах. Таковы еркек {Agropyrum sibiricum), ковыли {Stipa Lessingiana, S. capillata, S. Szowitsiana, Lasiagrostis caragana), широко распространенные в степях полыни {Artemisia Lercheana, A. astrachanica, A. scoparia, A. arenaria), молочай {Euphorbia Seguieriana) и другие виды «сухого» (ксерофильного) разнотравья, обычные в пределах южных частей степной области. Такие остепненные сообщества встречаются только на наиболее северных песчаных массивах (пески Сам, Приаральские Каракумы, пески в северной Бетпак-Дале, мелкие массивы в бассейне Эмбы и др.).
Известны и луговые, и тугайные варианты саксаульников, в сложении которых участвуют галофитнолуговые злаки, как чаир {Aeluropus litoralis), урук {Calamagrostis dubia), тростник {Phragmites communis), а также такие травы, как, например, Karelinia caspica, Goniolimon speciosum и др. Наиболее типично наличие в этих саксаульниках видов гребенщика {Tamarix sp. sp.), дерезы {Lycium turcomanicum), чингиля {Halimodendron halodendron). Подобные саксаульники встречаются в древних дельтах и на окраинах речных долин. Площадь, ими занимаемая, не велика.
Крайне редки варианты кустарниковых пустынь с элементами растительности нагорных ксерофитов. Они встречаются только в виде недостаточно четких фрагментов на фоне господствующих типичных или полуса-ванновых вариантов кустарниковых пустынь.
Все названные нами варианты не могут быть показаны в масштабе нашей карты.
В предшествующем разделе говорилось о том, что в размещении полукустарничковых пустынь имеется определенная закономерность, и могут быть выделены полосы остепненных пустынь (или полупустыни), настоящих и эфемеровых пустынь. Примерно так же могут быть разделены саксаульники и кустарниковые пустыни, с той лишь разницей, что территориально они размещены отдельными участками и поэтому нельзя выделить более или менее определенных полос. Кроме того, особые свойства песчаного субстрата облегчают продвижение на юг северных растений и, наоборот, проникновение к северу южных. При этом «зональные» границы как бы смазываются, становятся менее четкими и определенными, чем это наблюдается на растительности в плакорных условиях.
Однако можно отметить, что наиболее северные песчаные массивы покрыты растительностью со значительным участием степных флористических элементов, что позволяет и здесь говорить о подзоне остепненных саксаульников и кустарниковых пустынь (выше были отмечены их отличительные особенности). Часть песчаных массивов несет растительность с преобладанием типичных («настоящих») вариантов этих групп пустынь. Для них характерно преимущественное развитие синузии полукустарничков (главным образом видов Artemisia) с очень незначительным участием эфемеров. В эту подзону попадают песчаные массивы или их часть (например северная часть Кызылкумов), лежащие в пределах той климатической подзоны, для которой характерно более или менее равномерное распределение осадков по сезонам.
Подзона южных саксаульников и кустарниковых пустынь выражена более определенно. К ней относятся Каракумы, большая часть Кызылкумов, Муюнкумы, часть песков Прибалхашья, приморские пески восточного побережья Каспийского моря и некоторые другие. Эта подзона очень обширна, она занимает большую часть всего ареала саксаульников и кустарниковых пустынь. Основная характерная черта южной подзоны — исключительное обилие эфемеров и эфемероидов во всех формациях.
Помимо такого подразделения ареала саксаульников и кустарниковых пустынь на подзоны, обусловленного главным образом климатическими особенностями, возможно разделение на провинции с учетом флористических связей, отражающих некоторые общие моменты генезиса растительного покрова.
Значительная часть ареала описываемых пустынь относится к Туран-ской провинции. Для нее характерно большое разнообразие кандыма (Calligonum), представленного здесь более чем 40 видами (преимущественно из секции Pterococcus) из 65, встречающихся в кустарниковых пустынях Средней Азии. Точно так же в этой провинции весьма многочисленны виды полыней, в качестве сопутствующих видов входящие в эти сообщества (Artemisia arenaria, A. Marschalliana, A. Lercheana, A. astrachanica, A. terrae-albae s. str., A. turanica п др.). Отличительной чертой является наличие многих видов степного происхождения, которые, будучи обычными в северных районах провинции, проникают в центральные и даже южные ее части (виды Stipa, Agropyrum sibiricum, A. desertorum). Кроме того, очень показательно большое разнообразие видов Chondrilla и Tragopogon и наличие ряда эндемичных для этой провинции видов (Eremosparton aphyllum, Ephedra lomatolepis, Haloxylon ammodendron и др.).
Переднеазиатская провинция выделяется по исключительному обилию видов эфемеров и эфемероидов. Разнообразие полыней (Artemisia) здесь не столь велико, причем имеются виды, эндемичные для этой провинции (A. badghysi, A. Dimoana, A. Kelleri, A. persica). Виды Calligonum также обильны, однако количество их почти вдвое меньше, чем в пределах Туранской провинции (около 20 видов), и они относятся к секциям Pte-rigobasis и Eucalligonum этого рода. В большом числе представлены виды Ferula, порою имеющие ландшафтное значение в растительном покрове (F. oopoda, F. badrakema, F. assa-foetida и др.). Характерно также большее, чем в Туранской провинции, видовое разнообразие сингренов (видов Astragalus из секции Ammodendron).
Здесь отмечены только наиболее существенные отличия провинций между собой. Каждая из провинций может быть в свою очередь разделена на подпровинции (Прозоровский и Малеев, 1947).
Саксаульники и кустарниковые пустыни представляют огромную народнохозяйственную ценность. Это лучшие пастбища для каракульских овец, составляющих существенную долю народного богатства республик Средней Азии. Большое разнообразие видового состава, включающего виды различной фенологии, позволяет почти всюду использовать эти пастбища на протяжении всего года. Средняя продуктивность поедаемой массы от 0.8 до 1.9 ц/га. Кроме того, заросли саксаула и псаммофиль-ных кустарников служат источником топливной древесины. Особенно ценными являются черносаксаульники, дающие наибольший выход древесины с более высокой калорийностью, приближающейся к бурому углю (в среднем от 5 до 12 т/га). Белосаксаульники менее продуктивны [от 0.5 до 1.5 (2) т/га]. Многие города республик Средней Азии снаб
жаются саксауловыми дровами, заготовка которых производится в значительном от них отдалении. Заросли псаммофильных кустарников имеют чисто потребительское значение на месте и лишь там, где уже уничтожены саксаульники. Во флоре саксаульников и кустарниковых пустынь имеется много растений, используемых для получения поташа, дубильных веществ, красок, эфирных масел, лекарственных средств и т. п.
Литература
И. Г. Борщов (1865); С. И. Коржинский (1896); А. А. Еленкин (1901);Н. П. Верховский (1911); В. А. Дубинский (1911, 1928, 1929); В. П. Дробов (1922, 1929); М. II. Томин (1926); В. М. Савич (1927—1928); В. И. Аболин (1930); М. II. Петров (1933, 1946, 1950); Е. И. Коровин (1934); Е. II. Коровин и Д. Н. Кашкаров (1934); А. С. Порецкий и др. (1936); А. В. Прозоровский (1936, 1940а); Э. Н. Благовещенский (1940, 1954); Г, Ф. Боровский и др. (1940); М. Г. Попов (1940); Л. Е. Родин (1940, 1948); Е. А. Шингарева (1940, 1940а); С. В. Зоны и В. Л. Леонтьев (1942); Н. Т. Нечаева и др. (1943); Е. П. Коровин и Е. Е. Короткова (1946); О. И. Морозова (1946); А. В. Прозоровский и В. П. Малеев (1947); Б. С. Виноградов (1948); В. Н. Кунин (1950); К. В. Станюкович (1950); М. С. Шалыт (1951); Н. И. Рубцов (1952); Б. И. Тарабаева (1953, 1953а); В. Л. Леонтьев (1954); Н. Т. Нечаева (1954, 1954а); О. Paulsen (1911).
96. Черносаксаульники из Haloxylon aphyllum и саксаульники из Н. ammodendron
Черносаксаульники, в наибольшей степени отвечающие представлению о «лесах пустыни», имеются только в очень немногих местах. Таковы черносаксаульники Кос-Кудукской дачи (левобережье Чу), Баканасской дачи (правобережье Или), Нидербслентской и Яраджинской дачи (в юго-восточных Каракумах), между котловиной Чакпакты и колодцем Узун-Кую на Устюрте, в некоторых местах по долине Сыр-Дарьи.
В этих и очень немногих других местах черносаксаульники представляют собою густые, иногда даже смыкающиеся насаждения, в которых кусты и деревца черного саксаула достигают высоты 4—5 м. Площади перечисленных насаждений сравнительно не велики; не все массивы даже могут быть отображены в масштабе нашей карты и, к сожалению, до сих пор отсутствуют достаточно полные описания этих редких по своему мощному развитию насаждений.
Вообще же сообщества черносаксаульников или комплексов, в которых им принадлежит господствующее значение, достаточно велики и широко распространены в пределах Средней Азии (рис. 75). Наиболее северные массивы черносаксаульников расположены в западной части Бет-пак-Далы и в среднем течении Сарысу. Далее черносаксаульники имеются в правобережной части нижнего течения Или (Баканасская дача); небольшие пятна есть по обоим берегам в долине среднего течения Или; крупные массивы черносаксаульников лежат между р. Чу и Муюнкумами, а также в нижнем течении Чу, теряющейся на равнине. Далее к югу черносаксауловые сообщества протянулись узкой полосой по нижнему течению Сыр-Дарьи и по древнему ее руслу Жана-Дарья, почти смыкаясь с крупными площадями их у подножья возвышенности Бельтау на северо-западной окраине Кызылкумов; на той же широте небольшие пятна черносаксаульников встречаются на Устюрте и по нему идут с перерывами на юг до чинка.
На Устюрте черносаксаульники покрывают впадину Ассаке-Аудан и разбросаны небольшими островками к северу, а также к западу от этой впадины; в нескольких местах примыкают к солончаку Барса-Кель-мес и, наконец, самый северный участок черносаксаульников примыкает с юга к пескам Сам.
Менее крупные по площади контуры черносаксаульников показаны на карте в древней (Куня-Дарьинской) дельте Аму-Дарьи, а также в Са-рыкамышской котловине; узкая полоса черносаксаульников окаймляет западную окраину песков Кумсебшен. В этой западной части Каракумов черносаксаульники, хотя и встречаются, однако лишь в комплексах, и не могут быть показаны в масштабе карты. В юго-восточных частях Кызылкумов и Каракумов черносаксаульники представлены только мелкими пятнами.
Общий обзор ареала черносаксаульников показывает преимущественную приуроченность этих сообществ к современным и древним долинам и дельтам рек Средней Азии, а также к депрессиям среди песчаных мас-
Рис. 75. Схема распространения черносаксаульников в Средней Азии. Основные массивы сообществ Haloxylon aphyllum и Н. ammodendron.
сивов. Исключение составляют черносаксаульники Бетпак-Далы и Устюрта. Возможно, что черный саксаул здесь представлен особой экологической формой, а, может быть, даже более правильно относить его к виду Haloxylon ammodendron (Нечаева, 1951).
В общей форме можно считать, что местообитания черного саксаула связаны с относительно близкими грунтовыми водами или с некоторым дополнительным увлажнением, хотя бы водами поверхностного стока.
Почвы черносаксаульников — примитивные, часто отакыренные, сероземы и серо-бурые, обычно солончаковые и солончаковатые, иногда гипсоносные, большей частью глинистые, сравнительно редко супесчаные или песчаные.
Общей характерной чертой сообществ черного саксаула является значительно большая простота строения их, чем, например, белосаксаульников.
Примесь полукустарников очень ограничена и по видовому составу и количественно. Значительнее роль низких полукустарничков; боялыча {Salsola arbuscula), кеурека (5. rigida), биюргуна {Anabasis salsa) и полыней {Artemisia terrae-albae s. str., A. kemrudica и других — на севере или A. arenicola и A. diffusa — на юге). Названные виды обычно входят в разные сообщества черного саксаула.
Травянистых растений в черносаксаульниках всегда мало. Больше других заметно участие группы однолетних солянок (Halimocnemis longi-jolia, Н. villosa, Ceratocarpus turkestanicus, Salsola lanata, S. turcomanica, S. brachiala, Horaninowia anomala, H. ulicina, Suaeda Lipskyi, Kochia Schrenkiana, Londesia eriantha, Kirilowia eriantha), и в более редких случаях к ним примешиваются эфемеры (виды Eremopyrum, Matricaria lamellata, Euphorbia densa, Tauscheria lasiocarpa, Leptaleum filifolium, Acantho-lepis orientalis, Schismus arabicus, Lallemantia Royleana, Senecio subdenta-tus, Chrozophora gracilis) и еще реже эфемероиды (Rhinopetalum Karelinii).
Довольно часто встречаются сообщества моховых черносаксаульников, которые отличаются крайней изреженностью, плохим ростом черного саксаула (всего 1—1.5 м высоты) и значительным количеством отмерших кустов, которые четко выделяются на почти сплошном покрове пустынного мха — Tortula desertorum.
Как большая редкость встречаются лишайниковые (главным образом с Psora decipiens) черносаксаульники на такырных почвах. Здесь саксаул также изрежен и имеет высоту кустов 1—1.5 м. Такие черносаксаульники отмечены в Баканасской даче (Тарабаева, 1953а).
Сравнительно редки черносаксаульники, в которых развит богатый покров эфемеров и эфемероидов, главным образом илака (Carex physodes). Такие илаковые черносаксаульники врсгда бывают приурочены к нижним частям склонов пологих гряд или к котловинам и межгрядовым понижениям в грядовых, бугристых и котловинных песках. Илаковые черносаксаульники обычно встречаются в комплексе с другими сообществами (белосаксаульники, смешанносаксаульники).
В сильно изреженных черносаксаульниках Устюрта и его останцов в северных частях Каракумов с редкостойным покровом биюргуна (Anabasis salsa) флористический состав поражает исключительной бедностью и ничтожным количеством особей. Черный саксаул отличается низкорослостью и обилием сухостоя. Почвы в этих местообитаниях приближаются к такырам, на их поверхности наблюдается развитие сообществ водорослей и лишайников.
Черносаксаульники используются как пастбища для верблюдов; урожайность кормовой массы (поедаются главным образом зеленые веточки саксаула; участие остальных растений в общем кормовом запасе крайне ничтожно) исчисляется в 1.2—1.4 ц/га. Сезонные колебания сравнительно не велики.
В современных речных долинах, а также по временным руслам черносаксаульники обычно примыкают к тугаям, часто образуя переходные к ним сообщества. Генетическая преемственность черносаксаульников от тугаев в настоящее время не вызывает сомнений. Наиболее часто встречаются сообщества черного саксаула с ббльшим или меньптим участием видов джингила (гребенщика). На нашей карте они выделены внемас-штабным знаком последнего.
Необходимо все же отметить, что названные переходные от тугаев к черносаксаульникам сообщества распространены гораздо шире, чем это позволяет отобразить масштаб карты. Гребенщиковые черносаксаульники известны в нижних частях Куня-Дарьинской дельты, по Узбою, в нижней части дельты Мургаба и Теджена, в Сарыкамышской котловине, по староречьям Куня-Дарьи и Жана-Дарьи.
Литература
Б. К. Шишкин (1914а); И. И. Спрыгни и М. Г. Попов (1915); В. Н. Шнитников (1925); Н. А. Базилевская (1927); Е. П. Коровин (1927); В. М. Савич (1927—1928);
В. А. Дубинский (1928); М. П. Петров (1934); В. Н. Минервин (1935); С. А. Никитин (1935); А. С. Пороцкий и др. (1936); А. Г. Гаель и Е. С. Останин (1939); Л. Е. Родин (1940, 1948); Э. Н. Благовещенский (1941); Ю. С. Новиков и В. JI. Леонтьев (1942); Е. П. Коровин и И. И. Гранитов (1949); И. И. Гранитов (1950); II. И. Рубцов (1950а, 1952); В-Л. Леонтьев (1951, 1952, 1954); Н. Т. Нечаева (1951); II. Н. Пельт (1951, 1955); Б. И. Тарабаева (1953а); Н. Н. Пельт и Л. Е. Родин (1955).
97. Смешанные саксаульники (из Haloxylon persicum и Н. aphyllum)
Смешанные саксаульники в виде небольших участков постоянно встречаются среди обширных площадей белосаксаульников, нередко входя постоянным членом в состав комплексов растительного покрова песчаных массивов. В этих случаях они занимают промежуточные позиции между типичными обитаниями белого саксаула на склонах бугров и гряд и обитаниями черного саксаула на дне котловин и межгрядовых понижений.
Однако смешанные саксаульники встречаются не только мелкими участками в комплексе с другими сообществами, но и довольно значительными площадями, господствуя в растительном покрове.
Так, в пределах Средней Азии известны большие контуры смешанных саксаульников в Муюнкумах, где они соприкасаются с массивами черносаксаульников, мелкие пятна в среднем течении Сыр-Дарьи и более значительные площади в ее нижнем течении. Особенно велики площади смешанных саксаульников по северо-западной окраине Кызылкумов; небольшие контуры их показаны в центральной части этой пустыни, где они примыкают к полынникам кызылкумских останцовых возвышенностей. Крупные массивы смешанных саксаульников находятся между староречьем Жана-Дарьи и Аральским морем, а также в самом нижнем течении Сыр-Дарьи; узкая полоса показана по долине Узбоя. Далее эта формация представлена на значительной площади в восточной части Заунгузья, и, наконец, в крайней юго-восточной части Каракумов отмечен обширный массив между Аму-Дарьей и Мургабом.
Таким образом, довольно четко обрисовывается приуроченность смешанных саксаульников к внешним краевым частям древних аллювиальных равнин и сухих дельт и к аллювиальным отложениям древней долины Аму-Дарьи (в среднем ее течении — массив к югу от Дарганаты и между Аму-Дарьей и Мургабом).
В целом, для смешанных саксаульников характерен несколько обедненный видовой состав сравнительно с формацией белосаксаульников, отличающейся среди саксаульников наибольшим флористическим богатством. Видовой состав полукустарников небогат, так же как невелико и количественное их участие в построении фитоценозов (черкез, чогон, джузгуны — Calligonum eriopodum, С. undulatum, С. microcarpum и немногие другие — встречаются единично). Незначительна и роль полукустарничков, если не считать полыней (Artemisia terrae-albae ssp. Massagetovii, A. arenicola, A. diffusa), при близком подстилании коренных пород (особенно песчаников), принимающих большое участие в сложении сме-шанносаксаульников. Селин встречается сравнительно редко. Участие эфемероидов и эфемеров значительно, хотя непостоянно (из них назовем Carex physodes, Allium caspium, Ferula assa-foetida, Dorema sabulosum, Haplophyllum Bungei, Amberboa turanica, Eremopyrum orientals, Cryptodiscus ammophilus, Matricaria lamellata, Schismus arabicus, Erodium Litvino-vii, Euphorbia cheirolepis, Koelpinia linearis и др.) так же, как и некоторых однолетних маревых (Salsola carinata, S. sclerantha, S. turcomanica, Hora-
ninowia ulicina, Н. anomala, Halimocnemis longifolia и др.). Как и в бело-саксаульниках, при уплотнении субстрата развивается напочвенный покров из Tortula desertorum.
Сообщества смешанных саксаульников относятся ко многим ассоциациям, образующим корреспондирующие ряды с ассоциациями белосак-саульников (полынные, плановые, селиново-илаковые, моховые).
Смешанные саксаульники встречаются в различных комплексах и сочетаниях с сообществами других формаций. На нашей карте показаны следующие.^
1) Смешанные саксаульники в сочетании с массивами слабозаросших и подвижных песков. Массив их находится на левобережье Аму-Дарьи между Керками и Чарджоу; он расположен за полосой барханных песков, прилегающей к оазису. Находящиеся под воздействием хозяйственной деятельности человека эти сообщества во многих местах подвергались сильному выпасу. Пески пришли в подвижное состояние, что и обусловило возникновение данного сочетания сообществ илаковых смешанных саксаульников на грядовых песках с пятнами селиновых сюзенников на разбитых песках. К черному и белому саксаулу (Haloxylon aphyllum и Н. persicum) примешаны черкез (Salsola Richteri), борджок (Ephedra strobilacea), виды кандымов (Calligonum), изредка чогон (Aellenia subaphylla). Травяной ярус состоит из негустого покрова илака (Carex physodes) со значительным участием эфемеров (Bromus tectorum, Papaver pavoninum, Malcolmia africana, M. turkestanica, Arnebia decumbens, Nonea caspica и др.) и летнеосенних однолетников (Salsola carinata, S. lanata, Gamanthus gamocarpus, Horaninowia ulicina и др.). На разбитых песках — разреженные сообщества сюзена (Ammodendron Conollyi) с селином (Aristida pennata) и еркек-селином (Aristida Karelinii).
2) Смешанные саксаульники в сочетании с сообществами джингильников (гребенщиков). Они окаймляют с северо-западной стороны песчаный массив Кызылкумов, с другой стороны примыкая к древнеаллювиальной равнине староречья Жана-Дарьи с ее солянковыми пустынями с примесью черного саксаула. Здесь характерно сочетание илаковых и полынных смешанных саксаульников; и те, и другие занимают 30—40% площади на невысоких песчаных грядах. Разреженные сообщества гребенщика (Tamarix hispida) приурочены к понижениям или котловинам. В пределах этого контура имеются остатки древней оросительной сети и полей. Во многих местах встречаются разреженные заросли биюргунников (из Anabasis salsa) на такыровидных почвах.
3) Смешанные саксаульники с преобладанием сообществ с господством эфемерово-эфемеро-идной синузии, выраженной в развитии главным образом илакового покрова. Небольшие пятна этих сообществ имеются в западной и центральной частях Кызылкумов, примыкая к останцовым возвышенностям. Бугристые и пологоволнистые пески занимают здесь равнинные пространства на контакте останцовых возвышенностей и песчаных массивов собственно Кызылкумов. К повышенным элементам песчаного рельефа приурочены белосаксаульники (из На1оху1оц persicum), к пониженным элементам (котловинам и межбутровым понижениям) — черносаксаульники (из Haloxylon aphyllum); однако характерны участки, где совместно произрастают оба вида. Песчаная поверхность хорошо закреплена густым покровом илака (Carex physodes) со значительным количеством эфемеров. Здесь обычны Eremopyrum orientals.
Schismus arabicus, Erodium Litvinovii, Leptaleum jilifolium, Streptoloma desertorum, Euphorbia cheirolepis, E. Turczaninowii, Matricaria lamellata, Chrozophora gracilis, Nonea caspica, Tetracme quadricorn is, Lappula semiglabra. Встречаются также и летне-осенние однолетники: Ceratocarpus arenarius, Salsola pellucida, S. praecox, Horan inowia anomala и др.
4) Смешанные саксаульники с с о л я н к о в о й растительностью на солончаках. Они показаны на. нашей карте только по долине Узбоя в его широтной части (начиная от Ак-Яйлинской петли и заканчиваясь несколько ниже оз. Ясхан). Смешанные саксаульники здесь покрывают небольшие участки светлых (серых) песков, расположенных на второй и третьей террасах Узбоя, перемежаемые участками солончаков и засоленных такыровидных почв с фрагментами сообществ сарсазана (Halocnemum strobilaceum), карабарака (Halostachys Belangeriana), поташника (Kalidium foliatum и К. caspicum) и однолетних маревых (Salsola lanata, S. turgaica, S. turcomanica, Halimocnemis longifolia, Gamanthus gamocarpus, Horaninowia anomala, Anabasis micradena и др.).
Растительность долины Узбоя, конечно, гораздо богаче и разнообразнее, чем это допускает показать масштаб карты. Наличие во многих местах долины серии террас, каждая из которых отличается своим почвенно-растительным покровом, выходы то соленых, то пресных грунтовых вод, озера, старицы, заполненные ключевыми водами или водами поверхностного стока, песчаные скопления на разных террасах и т. п. создают чрезвычайную пестроту растительного покрова по Узбою.
Так, в районе колодца Джингильды отмечена следующая картина по поперечному профилю от дна русла: 1) лишенный растительности сор (на дне древнего русла); 2) сообщество сарсазана (Halocnemum strobilaceum) на низком террасовидном уступе; 3) сообщество лугово-гребенщикового тугая (заросли Tamarix с покровом из Aeluropus litoralis) на второй террасе с солончаковыми остаточно-тугайными почвами; 4) однолетнесолянковый черносаксаульник (Haloxylon aphyllum с покровом однолетних маревых) на третьей террасе на солончаковых почвах с погребенным гумусовым горизонтом (древние тугайные почвы).
В верхнем отрезке долины Узбоя в районе колодца Бала-Ишем схема несколько иная: 1) солончак, лишенный растительности на дне древнего русла; 2) пестрое сочетание нескольких сообществ (боялычник из Salsola arbuscula, кеуречник из S. rigida, кзылкокпечник из Limonium suffruti-cosum и др.) на солончаковых почвах второй террасы; 3) серополынный черносаксаульник на серо-бурых солончаковых почвах третьей террасы; 4) илаковый белосаксаульник на волнистых песках, лежащих на четвертой террасе Узбоя.
Пастбищное значение территории Узбоя с его пестрым растительным покровом не велико и регулярного выпаса здесь нет. Но так как вдоль Узбоя расположено значительное количество колодцев, то при прогоне на водопой скот всех видов непродолжительное время выпасается и в самой долине. В целом же смешанные саксаульники представляют собой ценные пастбища круглогодового использования. Урожайность поедаемой массы близка к тому, что дают белосаксаульники, и колеблется в пределах от 1.5 до 3 ц/га в зависимости от степени развития полыни, илака и эфемеров.
Литература
О. Э. Кнорринг и 3. А. Минквиц (1913); Н. А. Базилевская (1927); В. П.Дробов 1928); А. С. Порецкий и др. (1936); Л. Е. Родин (1940, 1948); Е. А. Шингарева (1940,
1940а); Э. Н. Благовещенский (1941, 1949а); 10. С. Новиков и В. Л. Леонтьев (1942); И. Т. Нечаева и др. (1943); Н. Н. Пельт (1951, 1955); В. Л. Леонтьев (1952, 1954); Н. II. Пельт и Л. Е. Родин (1955).
98. Белосаксаульники (из Haloxylon persicum)
Формация болосаксаульников образована белым саксаулом {Haloxylon persicum'), образующим наиболее типичные для песчаных массивов Средней Азии редкостойные заросли, своеобразные «леса пустыни». Белый саксаул — крупный полукустарник, иногда приобретающий форму и размеры деревца высотой 2—3 м (изредка до 3.5 м).
Белосаксаульники занимают большую площадь в песках Каракумов и Кызылкумов, а также менее значительные площади в других песчаных массивах: Чильмамедкум, Кумсебшен, Учтагап, Муюнкумы, Таукум, Сары-l 11пикотрау и др.
На огромном пространстве своего ареала — от Каспийского моря до Джунгарии — белосаксаульники представлены многими ассоциациями, отличающимися как по структуре сообществ, так и по флористическому составу, сохраняя, однако, многие общие черты. В наиболее полном развитии белосаксаульники — сложные сообщества, слагаемые из нескольких ярусов.
1) Ярус белого саксаула, к которому примешиваются крупные полукустарники: черкез {Salsola Richteri), чогон {Aellenia subaphylla), борджок {Ephedra strobilacea, в северных районах заменяемая Е. lomatolepis) и некоторые рослые джузгуны {Calligonum arborescens, С. eriopodum). Число кустов белого саксаула колеблется в пределах около 120—180 на 1 га, а всех кустов около 200—300 на 1 га.
2) Ярус низкорослых полукустарников и полукустарничков, представленный видами джузгунов {Calligonum turkestanicum, С. microcarpum, С. rotula, С. setosum и др.), вьюнков {Convolvulus erinaceus, С. divaricatus, С. Korolkovii), полыней {Artemisia santolina, A. arenaria, A. terrae-albae ssp. Massagetovii и др.), терескена {Eurotia Ewersmanniana, E. ceratoides) и др.
3) Ярус травянистых эфемероидов и длительновегетирующих многолетников. Среди них наиболее часты Aristida pennata, Rheum turkestanicum, Eremurus inderiensis, Allium caspium и A. sabulosum, Cryptodiscus ammophilus, Dorema sabulosum, Schumannia Karelinii и многие другие.
4) Ярус эфемеров, представленный большим видовым разнообразием: Matricaria lamellata, Bromus tectorum, Eremopyrum orientale, Euphorbia densa, Centaurea pulchella, Corispermum papillosum, Amberboa turanica, Cutandia memphitica, Crepis tectorum, Koelpinia linearis, Isatis emarginata, Ceratocephalus orthoceras, Descurainia Sophia и многие другие. К этому же ярусу следует отнести и илак {Carex physodes), который является наиболее постоянным спутником белого саксаула. Илаковые белосаксаульники принадлежат к наиболее распространенным сообществам в этой формации. Флористический состав илаковых белосаксаульников отличается большим видовым разнообразием. Точно так же и структура этой ассоциации, широко распространенной в Кызылкумах, Каракумах и других песчаных массивах, оказывается наиболее сложной, так как в ней обычно представлены растения различных ярусов и разных биологических групп.
Однако сообщества белого саксаула, в которых были бы все перечисленные ярусы, относительно редки, и большей частью наблюдаются такие, где под пологом эдификатора развиты один—два яруса или же сочетания растений из нескольких вышеназванных ярусов. В особых условиях, по днищам котловин в бугристых и грядовых песках, исчезают почти
полностью цветковые растения, и господствующее значение приобретает пустынный мох (Tortula desertorum), сплошным покровом одевающий поверхность почвы.
Белосаксаульники распространены по грядовым, бугристым, ячеистым и другим пескам названных ранее песчаных массивов Азии. Размеры кустов саксаула, состав и количество видов, слагающих сообщества белосаксаульников, сильно изменчивы в зависимости от различных условий: мощности песчаного наноса, нахождения на склоне или в котловине, ме~ ханического состава песка, состояния песчаной поверхности под влиянием ветров, степени воздействия хозяйственной деятельности человека (выпас скота, вырубка на топливо саксаула и других полукустарников) и т. п.
Так, например, в Кызылкумах А. С. Порецкпй, Ф. Н. Русанов и К. С. Афанасьев (1936) установили следующие ассоциации белосаксаульников.
Белосаксаульники боялычные (Haloxylon persicum—Salsola arbuscula), приуроченные к волнистым пескам на близколежащих щебнистых третично-меловых породах. Характерно развитие яруса боялыча под пологом саксаула; последний несколько угнетен и низкорослый. К обильному боялычу примешиваются еще некоторые полукустарнички : Astragalus squarrosus, Ammothamnus Lehmannii, Artemisia terrae-albae s. str., A. terrae-albae ssp. Massagetovii. В нижнем ярусе покров и лака неравномерный и разреженный. Из однолетников более заметны длительновегетирующие растения (Salsola praecox, Ceratocarpus arenarius, Horaninowia ulicina и др.), нежели эфемеры.
Белосаксаульники полынные (Haloxylon persicum—Artemisia terrae-albae s. str. и A. terrae-albae ssp. Massagetovii) характерны для маломощных песчаных субстратов с близким подстиланием коренных пород. Белый саксаул здесь также разрежен и угнетен. Полынь преобладает в ярусе полукустарничков; кроме нее, здесь единично встречаются Astragalus squarrosus, Salsola rigida, Ammothamnus Lehmannii. Из крупных трав обычны (также единичные) Ferula assa-foetida и Stipa Szowitsiana. Характерен покров илака с немногими длительновегетирующими однолетниками и эфемерами.
Белосаксаульники джузгуновые (Haloxylon persicum—Calligonum sp. sp.) распространены в бугристых и бугристо-грядовых песках на склонах бугров, преимущественно в верхней их части. Саксаул здесь хорошего роста, однако немногочислен. Большая роль в сложении сообщества принадлежит джузгунам (Calligonum membranaceum, С. microcarpum, С. alatum и др.). Полукустарнички почти отсутствуют. Довольно хорошо развит покров илака с большим количеством видов длительновегетирующих однолетников и эфемеров (Salsola praecox, S. pellucida, Horaninowia ulicina, Corispermum Lehmannianum, Kochia Schrenkiana, Centaurea pulchella, Erodium Litvinovii, Arnebia decumbens, Scabiosa Olivieri, Veronica biloba и др.).
Белосаксаульники илаковые (Haloxylonpersicum—Carexphysodes) наиболее широко распространены в грядовых и бугристо-грядовых песках, занимая их склоны (преимущественно средние части и пологие верхние). Белый саксаул отличается хорошим ростом и значительным обилием. В одном ярусе с саксаулом встречается черкез (Salsola Richteri) и, реже, борджок (Ephedra strobilacea), а под их пологом — небольшое количество кустов джузгунов (Calligonum leucocladum, С. membranaceum, С. aralense и др.). Из полукустарничков, немногочисленных по обилию, отмечены Astragalus unifoliolatus, Artemisia eriocarpa. Обычно присутствуют здесь единичные Dorema sabulosum и Ferula assa-foetida, а также
небольшое количество дерновинок Aristida pennata. Единично встречаются длительновегетпрующие травы и мелкие полукустарнички: Helio-tropium arguzioides, Convolvulus divaricatus, Acanthophyllum elatius и др. Илак (Carex physodes) образует сплошной покров, однако на его фоне, хотя и в незначительном обилии, наблюдаются многие длительновегетирующие однолетники (Corispermum Lehmannianum, Horaninowia ulicina, Salsola praecox) п особенно эфемеры (Centaurea pulchella, Spirorhynchus sabulosus, Erodium Litvinovii, Leptaleum filifolium, Arnebia decumbens, Koelpinia linearis, Streptoloma desertorum, Schismus arabicus и др.). Таким образом, из приведенного неполного перечня видов, характерных для илаковых белосаксаульников, можно заметить, насколько разнообразны их флористический состав и набор представителей различных жизненных форм и биологических групп.
Бело саксаульник и мятликово-илаковые (Haloxylon рег-sicum — Carex physodes-}-Роа bulbosa var. vavipara) распространены по пологим северным склонам широтно ориентированных гряд, на уплотненных песках, обогащенных пылеватыми частицами и с признаками почвообразования. Вобщем облике сообществфизиономически выделяются полыни (Artemisia terrae-albae s. str. и Л. terrae-albae ssp. Massagetovii), тогда как белый саксаул очень угнетен и изрежен.1 Примесь полукустарников ничтожна, чаще всего встречаются угнетенные экземпляры Calligonum leu-cocladum. Характерен почти сплошной покров илака и мятлика, по которому разбросаны в небольшом обилии, но довольно разнообразные по видовому составу, длительновегетирующие однолетники и эфемеры (Ceratocarpus arenarius, Salsola praecox, Scabiosa Olivieri, Alyssum desertorum, Veronica biloba, Consolida songorica и др.).
В Каракумах Л. Е. Родин (1948, а также по исследованиям 1951— 1953 гг.) выделяет следующие, наиболее часто встречающиеся ассоциации белосаксаульников.
Белосаксаульники реомюриевые (Haloxylon persicum — Reaumuria oxiana), распространенные только по маломощным песчаным плащам на вершинах заунгузских останцов, сложенных песчаниками (песок крупнозернистый, и в нем всегда имеется примесь песчаниковой щебенки). Белый саксаул хорошего роста. Большую роль в сложении сообщества играет реомюрия (Reaumuria oxiana), образующая хорошо выраженный ярус. Полукустарники единичны (Astragalus paucijugus, Salsola arbuscula, Calligonum gracile) так же, как и полукустарнички (Artemisia are-nicola, A. eriocarpa). Характерно присутствие здесь Aristida pennata, менее характерна Ferula assa-foetida. Илак также присутствует, но не образует заметного покрова. Однолетников (длительновегетирующих и эфемеров) незначительное количество видов, и обилие их ничтожно.
Белосаксаульники черкезовые (Haloxylon persicum-}-Salsola Richteri) приурочены к грядовым пескам в древних дельтах Аму-Дарьи, Мургаба и Теджена; пески характеризуются некоторой засоленностью. В одном ярусе с хорошо развитым саксаулом представлен черкез (Salsola Richteri), играющий большую роль в сообществе; иногда к нему присоединяются Tamarix leptostachys и Aellenia subaphylla. Полукустарничков почти нет. Травянистых растений мало, это преимущественно эфемероиды (Allium sabulosum, Dorema sabulosum, Rheum turkestanicum, HaplophyUum Bungei). Однолетники главным образом длительновегетирующие (Salsola carinata, S. lanata, Atriplex dimorphostegia, A. moneta, Gamanthus gamocarpus.
1 А. С. Порецкий и др. (1936) данное сообщество относят к формации полынни-ков; однако, иак нам представляется, правильнее отнести его к оелосаксаульникам.
Halotis pilosa), а эфемеров мало (Eremopyrum orientale, Amberboa turanica и др.).
Белосаксаульники полынные (Haloxylonpersicum — Artemisia arenicola) приурочены к уплотненным пескам, покрывающим тонким плащом останцы Устюрта, лежащие восточнее Сарыкамышской котловины. Сак’ саул здесь угнетен и редкостойный. Полынь образует хорошо выраженный ярус; среди кустов полыни встречаются редкие экземпляры Salsola arbus^ cula, Calligonum setosum, Convolvulus subhirsutus. Очень характерны здесь эфемероиды Allium caspium и Ferula assa-foetida; илак (Carex physodes) всегда имеется, но связного покрова не образует; вместе с ним встречаются в незначительном количестве немногие длительновегстирующие однолетники (Salsola carinata, Horaninowia anomala) и эфемеры (Eremopyrum orientate, Senecio subdentatus и др.).
Белосаксаульники ковыльные (Haloxylon persicum — Stipa Szowitsiana) распространены преимущественно в Заунгузье, где они приурочены к равнинным, иногда слабо волнистым пескам, подстилаемым песчаниковыми породами, на плоских вершинах останцов (кыров), на пологих их склонах, а также в межкыровых понижениях. Саксаул здесь изрежен, преимущественно низкорослый. В одном ярусе с ним встречаются некоторые виды Calligonum (единичные особи). Из полукустарничков характерен Convolvulus Korolkovii. Наиболее же типично большое участие ковыля Шовица (Stipa Szowitsiana), дерновинки которого образуют проективное покрытие до 30%. Свободные участки покрыты илаком; оба эти растения хорошо скрепляют поверхность песка. Примесь однолетников (длительновегетирующих и эфемеров) очень незначительна (Horaninowia anomala, Се-ratocarpus turkestanicus, Eremopyrum orientale, Centaurea pulchella и др.).
Белосаксаульники селиново-илаковые (Haloxylon persicum—Carex physodes A-Aristida pennata) отмечены на пологих склонах грядовых песков в верхнем отрезке Узбоя. В господствующем ярусе хорошо развитого белого саксаула почти отсутствует примесь других крупных кустарников (единичные AmmodendronLehmannii, Ephedra strobilacea). Немногочисленны виды полукустарничков, встречающихся единично (Artemisia eriocarpa, Salsola arbuscula, Acanthophyllum elatius), но зато чрезвычайно ярко выражен травяной покров, слагаемый илаком и селином, которые почти равны по обилию (илака несколько больше). Остальные травы (многолетние и однолетние) играют малозаметную роль, будучи представлены немногими видами при ничтожной степени обилия (Heliotropium arguzioides, Cousinia bipinnata, Dorema sabulosum, Haplophyllum Bungei, Eremopyrum orientale и еще несколько видов эфемеров).
Белосаксаульники илаковые (Haloxylon persicum — Carex physodes) широко распространены в Каракумах по пологим склонам гряд. Белый саксаул здесь обычно хорошо развит, достигая крупных размеров и большого обилия. Заметная доля в этом ярусе принадлежит черкезу (Salsola Richteri). Единично встречаются еще многие полукустарники (Calligonum turkestanicum, С. microcarpum, С. gracile, С. undulatumu др., Ephedra strobilacea, Aellenia subaphylla) и некоторые полукустарнички (Artemisia eriocarpa, Astragalus unifoliolatus, Convolvulus Korolkovii, C. erinaceus, Haplophyllum obtusifolium). Многолетних длительновегетирующих трав мало (Aristida pennata, Jurinea derderioides), эфемероидов больше (Rheum turkestanicum, Dorema sabulosum, Ferula assa-foetida, Allium sabulosum, Eremurus inderiensis и др.); особенно выражено участие в сложении сообществ илака (Carex physodes), прочно скрепляющего почву от развевания. По илаковому ковру весной разбросано большое количество видов эфемеров (Papaver psooninum, Arnebia decumbens, Koelpinia linearis, Streptoloma desertorum,
Ziziphora tenuior, Valerianella dufresnia, I satis emarginata, Chartoloma pla-lycarpum, Cithareloma vernum, Senecio subdentatus, Consolida camptocarpa, Euphorbia Turczaninowii и др.). Участие длительновегетирующих однолетников здесь незначительно (Horaninowia ulicina, Salsola carinata, S. sclerantha и др.).
Белосаксаульники моховые (Haloxylon persicum—Tortula desertorum) в Каракумах встречаются редко; они обычны лишь в Заунгузье, где занимают днища межгрядовых котловин, весьма редко заходя на нижние части склонов гряд. Саксаул здесь крайне угнетен, изрежен, много мертвого валежника. Полукустарники чрезвычайно редки и тоже угнетены. Многолетние травы почти не встречаются. ПоЪерхность почвы покрыта подушками мха Tortula desertorum. На его фоне разбросаны однолетники (Matricaria lamellata, Eremopyrum orientale, Bromus tectorum, Senecio subdentatus, Amberboa turanica, Horaninowia ulicina, H. anomala, Euphorbia densa, Salsola carinata, Halimocnemis longifolia). Характерно наличие ряда видов лишайников: Placodium Tominii, Psora decipiens, виды Collema и др.
Белосаксаульники вообще, а в особенности илаковые, являются наиболее ценными пастбищами для каракульских овец, причем во многих районах они используются круглогодично. Илак поедается овцами не только в свежем зеленом виде весной, но и в высохшем состоянии летом и осенью, так как сохраняет свои кормовые качества. Осенью и зимой, и частично и летом, овцы поедают и саксаул. Однако урожайность кормовой массы в белосаксаульниках не велика, всего 1.5—2 ц/га, причем сильно колеблется в зависимости от погодных условий, прежде всего от количества и времени выпадения осадков.
В настоящее время все белосаксаульники несут на себе следы хозяйственной деятельности человека, и только в районах, удаленных от населенных пунктов и колодцев и в стороне от дорог и караванных троп, сохранились сообщества белого саксаула с ненарушенным растительным покровом. Однако, как показали наблюдения многих исследователей, при полном отсутствии выпаса, в первую очередь в котловинах, а впоследствии и на склонах, а также на равнинных участках песков, происходит уплотнение поверхностных горизонтов песка, в результате чего изре-живается покров илака, уменьшается количество эфемеров и многолетних травянистых растений, и происходит отмирание полукустарников, в том числе и самого саксаула. Очевидно, в этих условиях (при уплотнении субстрата) затрудняется естественное возобновление, полукустарники же отмирают в силу естественного старения. Внешне эти участки производят удручающее впечатление кладбищ с массой валяющихся мертвых •стволов на почве, поверхность которой занята подушками пустынного мха (Tortula desertorum). Отмечено, что при нарушении мохового покрова здесь разрастаются илак и другие травянистые растения, и при последующем умеренном выпасе создаются вновь более благоприятные условия для развития сообщества белого саксаула в его нормальном облике. Очевидно, умеренный выпас, помимо разрыхления плотных подушек Tortula desertorum, благоприятствует заделке семян, устраняет накопление в дернинах селина мертвых остатков и т. п., что и создает более разнообразные условия для поселения растений.
Необходимо все же отметить, что распространение моховых белосак-саульников связано не только с районами слабого выпаса (пески Учта-ган), но и с теми районами, где пески залегают маломощным слоем на 'близких плотных коренных породах (западные Кызылкумы, пески Учта-ган).
На нашей карте особым значком показаны белосаксаульники, в построении сообществ которых особенно велико участие эфемеров и эфемероидов, прежде всего илака (Carex physodes). Если субтропические степи опоясывают предгорья Средней Азии, то белосаксаульники с ила-ковым покровом примыкают к ним на песчаных равнинах, разливаясь широким потоком в южной части пустынной области и постепенно суживая свой ареал при движении к северу, где место эфемерово-эфемероидной синузии занимает синузия полукустарничковая (полыни).
Белосаксаульники, как большинство других формаций пустыни, встречаются в комплексах и сочетаниях с сообществами других формаций. Однако масштаб карты не позволяет показать эти детали строения растительного покрова. Исключение могло быть сделано только для обширных районов южной части Низменных Каракумов, где небольшие песчаные массивы с господством белосаксаульников сочетаются с мелкими участками водорослевых сообществ на такырах. Для выделения их на карте принят немасштабный условный знак такыров.
Литература
О. Э. Кнорринг и 3. Л. Минквиц (1913); А. И. Михельсон (1913); Б. К. Шишкин (1914а); М. Г. Попов (1915); И. И. Спрыгни и М. Г. Попов (1915); М. Д. Спиридонов (1924, 1930); В. Н. Шнитников (1925); Н. А. Базилевская (1927, 1929); Е. П. Коровин (1927); В. А. Дубянский (1928); С. С. Неуструев и И. М. Крашенинников (ред.) (1930); М. П. Петров (1933, 1935); В. Н. Минервин (1935); С. А. Никитин (1935); А. С. Порецкий и др. (1936); А. Г. Гаель и Е. С. Останин (1939); Л. Е. Родин (1940, 1948); Е. А. Шингарева (1940, 1940а); Э. Н. Благовещенский (1941); Ю. С. Новиков и В. Л. Леонтьев (1942); Н. Т. Нечаева и др. (1943); И. И. Гранитов (1950); В. Л. Леонтьев (1954); Н. Н. Пельт (1955); Н. Н. Пельт и Л. Е. Родин (1955).
99. Черкезники (из Salsola Richteri), джузгунники (из видов Calligonum), кзылчевники (из Ephedra strobilacea, Е. lomatolepis)
Сообщества этой группы формаций значительными площадями расположены в юго-западной Туркмении (пески Хан-Баагыкум, пески п-ва Дарджа, пески Гейрджаны и др.), на Красноводском п-ве, во впадине Карынжарык (западная окраина южного Устюрта), в центральных Кызылкумах, в юго-восточных Каракумах (между Аму-Дарьей и Тедженом), по южной окраине Низменных Каракумов, в северной и южной частях Муюнкумов, в песках Таукум и небольшими контурами на песках верхних отрезков долины Аму-Дарьи и по Сыр-Дарье. Кроме того, эти сообщества мелкими участками довольно часто встречаются среди всех песчаных массивов Средней Азии.
Из названных формаций наиболее широко распространены сообщества джузгунников, слагаемые многочисленными видами Calligonum. Эти сообщества характерны для песчаных массивов с относительно мало развитым, разреженным травяным покровом, в силу чего поверхность песчаных гряд и бугров слабо закреплена и отличается некоторой подвижностью песка.
Кустарниковый ярус джузгуновых сообществ обычно состоит не только из немногих видов Calligonum, но к ним часто примешаны Salsola Richteri, реже S. arbuscula, виды Astragalus из секции Ammodendron, а изредка и белый саксаул — Haloxylon persicum.
На маломощных песках с близким подстиланием глинистых или каменистых пород и на уплотненных песках у подножья гряд в джузгун-никах бывает развит ярус полукустарничковых полыней (Artemisia ter-
Рис. 76. На переднем плане такыр с редкостойной ассоциацией Anabasis salsa* на заднем плане — пылевато-супесчаная гряда с черносаксаульником (Haloxylon aphyllum). Юго-восточная часть пустыни Бетпак-Дала (урочище Саксаул-Дала). (Фот. Ю. Е. Лавренко).
Рис. 77. Черный саксаул (Haloxylon aphyllum) в пустыне Саксаул-Д ал а (в юго-восточной части пустыни Бетпак-Дала). (Фот. Л. Е. Родина).
ic. 78. Разбитые пески с кустами Ephedra strobilacea вблизи колодца. На заднем гане слева — юрты туркмен у колодца. Южная часть пустыни Каракумы (Туркменская ССР). (Фот. А. В. Прозоровского).
Рис. 79. Барханные сыпучие пески по окраине долины Аму-Дарьи, к югу от Хорезмского оазиса (Туркменская ССР). (Фот. Е. А. Шингаревой).
rae-albae s. str., A. terrae-albae ssp. Massagetovii, A. santolina, A. eriocarpa,. A. arenicola}.
Травяной покров, как сказано, развит слабо: разреженный илак (Carex physodes}, редкие дернинки селина (Aristida pennata} и немногочисленные, хотя и разнообразные по видовому составу, эфемеры и эфемероиды (Allium sabulosum, A. caspium, Senecio subdentatus, Tetracme quadricornis, Spirorhynhus sabulosus, Scabiosa Olivieri, Londesia eriantha, Erodium oxyr-rhynchum, Acantholepis orientalis, Matthiola chenopodiifolia, Cithareloma vernum и др.). Как исключение, можно встретить джузгунники с густым покровом и лака, хорошо скрепляющим склоны гряд. Весьма вероятно, что такие джузгунники являются производными на месте белосаксаульников, где белый саксаул выбран на топливо.
Джузгунники очень часто наблюдаются в сочетании с сообществами белого саксаула, занимая всегда различные позиции по признаку закрепленности песчаной поверхности (джузгунники всегда на слабо закрепленных и иногда даже на подвижных песках, а белосаксаульники — на хорошо закрепленных песках), что и дало повод некоторым исследователям считать во многих случаях, что джузгунники — сообщества вторичные, производные от уничтоженных человеком белосаксаульников. Очевидно, в определенных случаях такое предположение имеет под собою реальную-почву, но в то же время во многих районах (Кызылкумы, пески юго-западной Туркмении, юго-восточные Каракумы) джузгунники, видимо, являются естественными образованиями.
Фрагменты сообществ джузгунников весьма часто сопровождают вершины песчаных гряд, почти всегда отличающиеся некоторым пере-веванием песка.
Черкезники, сообщества из крупных древовидных полукустарников (Salsola Richteri и, реже, S. Paletzkiana}, имеют более ограниченное распространение. Значительные массивы имеются лишь в нескольких местах (пески Хан-Баагыкум в юго-западной Туркмении, в юго-восточных Каракумах по границе с Мургабским оазисом), где они представляют естественные сообщества.
Сомкнутость кустарникового яруса в них незначительная (0.2—0.4); помимо господствующего черкеза, имеется обычно примесь двух-трех видов джузгунов (Calligonum}, реже сюзена (Ammodendron Karelinii или A. Lehmannii); травяной ярус представлен иногда густым покровом илака с эфемеровым разнотравьем (например в юго-восточных Каракумах), а чаще негустым покровом илака и селина с небольшой примесью эфемеров (Bromus tectorum, В. macrostachys, Eremopyrum orientale, E. Buo-napartis, Papaver pavoninum, Malcolmia africana, Leptaleum filifolium, Senecio subdentatus}. Встречаются также сообщества черкеза с значительным участием полыни (Artemisia diffusa, A. arenicola} и эфемеров.
Черкезники могут быть не только первичными, но и вторичными, антропогенными образованиями, представляя собою дериват белосаксаульников и смешанных саксаульников, в которых саксаул был уничтожен на топливо. Такие черкезники обычно встречаются вблизи колодцев, являвшихся местом зимовки значительного скотоводческого населения-(например у колодца Бала-Ишем на Узбое, в некоторых местах на равнинах Куня-Дарьинской дельты и др.), однако площади их не велики и в масштабе нашей карты не могут быть показаны. Обычно в этих вторичных черкезниках характерно более заметное участие видов Calligonum и, вследствие некоторой разбитости песка, внедряются виды, характерные для подвижных песков, как ffeliotropium arguzioides, Chrozophora graci-
Us, Aristida pennata, Corispermum aralo-caspicum, Agriophyllum latifolium и некоторые другие.
Кзылчевники (из видов хвойника — Ephedra strobilacea и Е. lomato-lepis) встречаются лишь небольшими участками, будучи вкрапленными в основной фон джузгунников или черкезников. Они характеризуются очень близким к этим сообществам видовым составом с господством в кустарниковом ярусе одного из видов Ephedra [в южных районах Средней Азии встречается Е. strobilacea (рис. 78), а в северных — Е. lomatolepis]. В качестве небольшой примеси эти виды Ephedra встречаются также там, где отсутствует хорошее задернение песчаной поверхности.
Пастбищная ценность этой группы формаций значительно ниже, чем, например, белосаксаульников. Они служат для выпаса в тех местах, где хорошо развит покров илака и эфемеров; в этих условиях максимальный поедаемый запас приходится на весенний сезон и колеблется в пределах 1—2.3 ц/га.
Литература
Г. С. Карелин (1883); В. П. Дробов (1922); М. Д. Спиридонов (1924); Н. А. Базилевская (1927, 1929); В. А. Дубянский (1928); С. С. Неуструев и И. .М. Крашенинников (ред.) (1930); С. А. Никитин (1935); Е. Г. Победимова (1935); А. С. Порецкий и др. (1936); А. Г. Гаель и Е. С. Останин (1939); Л. Е. Родин (1940, 1948); Н. Т. Нечаева и др. (1943); Н. И. Рубцов (1952); Н. Н. Пельт (1955); Н. Н. Пельт и Л. Е. Родин (1955).
100. Разреженные псаммофильные кустарники (из видов Ammodendron, Eremosparton, Astragalus sect. Ammodendron, Calligonum и Convolvulus fruticosus) в сочетании с барханными песками
Разреженные псаммофильные кустарники всегда сочетаются с барханными песками. Во многих случаях следует говорить о массивах подвижных и барханных песков с участием разреженной растительности.
Этого рода сочетания крупными массивами расположены в восточной части Приаральских Каракумов, в юго-западной Туркмении, по северной окраине Каракумов, в юго-восточных Каракумах и в южных Кызылкумах, а также небольшими пятнами в среднем течении Аму-Дарьи (в районе Термеза).
Все эти массивы почти везде примыкают к современным оазисам или к землям древнего орошения. Такое расположение названных сообществ и барханных песков указывает на их вторичное происхождение, связанное с длительным (на протяжении сотен лет и даже тысячелетий) воздействием хозяйственной деятельности человека, приведшей к полному уничтожению былой растительности, вследствие чего пески пришли в подвижное состояние, образовав барханы иногда на весьма крупных площадях.
Неустойчивость и сыпучесть субстрата ограничивает возможность поселения растений; на такого рода массивах существуют только специфические псаммофильные растения, обладающие рядом приспособлений к жизни в этих условиях. Одни виды имеют громадную по охватываемой площади поверхностную корневую систему для улавливания влаги осадков (Aristida Karelinii), другие обладают активной способностью образовывать придаточные корни при засыпании растений песком (виды Ammodendron, некоторые виды Tamarix), третьи снабжены мощными •и длинными корнями, удерживающими растения при выдувании песка (виды Calligonum), многие имеют семена и плоды, легко разносимые Петром, и другие приспособления к жизни в условиях подвижных песков.
Наиболее обычны и широко распространены сюзенники — сообщества из видов Ammodendron (Л. Conollyi, A. Karelinii, A. Lehmannii)-, кусты этих пород разбросаны на значительных расстояниях; иногда примешиваются виды Calligonum (чаще всего С. caput-Medusae} и Eremosparton flaccidum, еще реже Salsola Richteri, с единичными травянистыми растениями, также характерными для подвижных песков, как, например, Ari-stida Karelinii, Tournefortia sogdiana, Horaninowia ulicina, Heliotropium arguzioides и некоторые другие.
Фрагменты формации сюзенников нередко встречаются среди белосаксаульников, на участках с разбитыми песками, и обычно сопровождают барханные пески вблизи колодцев (возникшие от неумеренного выпаса скота); однако эти участки в масштабе карты не могут быть показаны.
Спнгренники (сообщества из видов Astragalus sect. Ammodendron) занимают ограниченные площади слабо закрепленных и подвижных песков, отличающихся примесью грубозернистых (гравийных) фракций и часто залегающих на коренных породах плато (или кыров), сильно подвергнутых дефляции. В Кызылкумах сингренники состоят из Astragalus squarrosus с примесью Zygophyllum Eichwaldii и Convolvulus hamadae с травяным покровом редких эфемеров и солянок. Более редки сингренники с развитым илаковым покровом, либо с полынью, или с боялычом.
Такие небольшие площади сингренников имеются на песках юго-западной Туркмении и в юго-западных Каракумах, где состоят из Astragalus unifoliolatus и других видов из секции Ammodendron, с небольшой примесью видов Calligonum и редким покровом эфемеров {Malcolmia, Senecio, Leptaleum, Tetracme и др.). Покров илака встречается редко.
Вьюнковники (сообщества из Convolvulus hamadae} известны только в юго-западных Кызылкумах, где обитают вперемежку с сингренниками, па равнинах с маломощным плащом супесей с примесью гравия и хорошо выраженным гипсовым горизонтом. Наряду с эфемерами во вьюнковниках появляются многолетние растения (виды Stipa, Iris, Cleome и др.).
В непосредственной близости к орошаемым землям на разбитых песках, чаще в их котловинах, наряду с сюзеном, наблюдается примесь янтака {Alhagi persarum и A. kirghisorum). Иногда встречаются и чистые заросли янтака.
Подвижные барханные пески приносят большой вред народному хозяйству заносами железных дорог, промышленных и жилых строений, полей и садов, соляных промыслов, колодцев, запылением воздуха, порчей машин и механизмов и т. п. В России борьба с подвижными песками начата с конца XVIII столетия. История пескоукрепительных работ изложена у М. П. Петрова (1950). В пустынях Средней Азии пескоукрепительные работы стали осуществляться с 80-х годов прошлого столетия. Наиболее эффективные результаты приносит фитомелиорация, то есть посев и насаждение на подвижных песках растений. В качестве пескоукрепителей используется большой ассортимент дикорастущих пустынных растений, главным образом псаммофитов: многие виды джузгу-нов {Calligonum}, белый и черный саксаул {Haloxylon persicum и Н. aphyllum}, виды сюзена {Ammodendron}, черкезов {Salsola Richteri и S. Paletz-kiana), гребенщиков {Tamarix), сингренов {Astragalus paucijugus, A. unifoliolatus и др.), а также многие травянистые растения {Elymus giganteus, Aristida Karelinii, A. pennata, Agropyrum sibiricum, Agriophyllum arena-rium, A. latifolium, Horaninowia ulicina и др.).
Борьба с подвижными песками ведется также с помощью механических защит и нанесением на поверхность песка битумной эмульсии.
_ пяйонах распространения подвижных песков достигнуть,
Во многих Раи°н pap £остановлены и поросли насаждениями успешные результат . других пескоукрепляющих пород. Та«
саксаула, “дытоТко в Узбечкой ССР и в Туркменский
ССРРИзакреплено св^ше 66 000 га песков, покрытых сейчас зарослями, дающими* топливо населению и подножный корм для скота (Петров, 195Важнейшее значение для предотвращения возникновения новых площадей подвижных песков имеют меры по сохранению существующих на песках насаждений путем рациональной системы использования их под выпас, заготовку топлива и т. п.
Литература
М. Г. Попов (1915); В. А. Дубинский (1928); Н. А. Базилевская (1929); С.А Ни китин (1935); М. П. Петров (1935, 1950); Е. Г. Победимова (1935); Т. Ф. Якубов (19Чч\? А. С. Порецкий и др. (1936); А. Г. Гаель (1938); Л. Е. Родин (1940, 1948); Е. А. Шип’ гарева (1940, 1940а); Н. Т. Нечаева и др. (1943); А. Г. Гаель и др. (1950); В. Н Rv" нин (1950); Н. Г. Захаров и др. (1954); Н. Н. Пельт и Л. Е. Родин (1955).
XV. ВЫСОКОГОРНАЯ ПОДУШКОВИДНАЯ РАСТИТЕЛЬНОСТЬ
Н. И. РУБЦОВ и О. В. ЗАЛЕНСКИЙ1
Различные виды растений-подушек, распространенные на Восточном Памире и в Тянь-Шане, образуют в этих районах своеобразный тип растительности, названный К. В. Станюковичем (1948) «высокогорные (криофильные) подушечники». Характерной особенностью этого типа, хорошо отличающей его от других высокогорных формаций (криофитных субальпийских и альпийских полукустарничковых пустынь, степей, лугов, пустошей и т. п.), является доминирование растений-подушек. Последние представляют собой особую жизненную форму растений, возникшую в результате редукции процессов роста надземных органов, вызванной неблагоприятными условиями. Редукция роста надземных органов достигает у растений-подушек крайней степени своей выраженности и обусловливает у них появление ряда морфологических признаков (укороченный рост побегов; радиальный тип ветвления; шаровидная или плоская форма растения; сомкнутость ветвей, между которыми накапливаются отмершие листья, а также почва, образующие материал, наполняющий подушку, и т. д.). Перечисленные признаки характерны только для растений-подушек и могут быть использованы для их более детальной классификации (см. Hauri, 1912; Rauch, 1940 и др.).
Всего на земном шаре зарегистрировано более 200 видов цветковых растений-подушек (Schroter und Hauri, 1914). Из них более 50% встречается в Южной Америке особенно в Андах, в Патагонии и на Фалькленд-ских о-вах), в Новой Зеландии и на о. Кергуелен. Следующее место (около 25%) занимают Гималаи, нагорья Центральной Азии, Средняя Азия, Малая Азия и Кавказ. Именно в континентальных районах Евразии растения-подушки имеют наиболее широкое распространение.
Комплексное эколого-физиологическое изучение растений-подушек в последние годы осуществляется коллективом сотрудников Памирской биологической станции АН Таджикской СССР (Заленский, 1948; Станюкович, 1948; Зайцева, 1949, 1949а; Свешникова, 1952 и др.). В результате стационарных исследований морфологии различных видов рода Acantholimon и других растений было показано, что характерная для но-душек компактность возникает вследствие крайнего торможения роста побегов в длину при незначительном темпе их ветвления. Развитие определенной формы (от вогнутой до полусферической) определяется соотношением скоростей роста побегов периферии и центра подушки, а также особенностями их отмирания и возобновления. Были разработаны методы определения возраста растений-подушек и показано, что для них характерна исключительно высокая продолжительность жизни. Так, возраст крупных экземпляров Acantholimon diapensioides достигает нескольких
1 Общий очерк написан О. В. Заленским; остальное — Н. И. Рубцовым.
сотен лет. Высокая продолжительность жизни сочетается у растений-подушек с рядом своеобразных физиологических особенностей, а именно, с низкой интенсивностью и большой равномерностью изменений различных процессов обмена веществ (фотосинтез, транспирация, осмотическое давление, углеводный и азотный обмен). О. В. Заленский и К. В. Станюкович предполагают, что растения-подушки следует рассматривать как жизненную форму, возникшую в условиях высокогорий под влиянием специфического для них режима факторов внешней среды, изменяющего многие важные для формообразования особенности биосинтеза.
Литература
О. В. Заленский (1948); К. В. Станюкович (1948); М. Г. Зайцева (1949, 1949а); В. М. Свешникова (1952); Н. Hauri (1912); С. Scbroter tind Н. llauri (1914)-О. Paulsen (1920); W. Rauch (1940).
101 им — памирская (из Acantholimon diape nsioides, Tanacetum xylorrhizum, Oxytropis Poncinsii, 0. tianschanica, Potentilla p a m i г о a 1 a i c a)
На высокогорьях Памира весьма характерной формацией подушковидной растительности являются куртковники (с господством куртков-ника — Acantholimon diapensioides). Эта формация встречается преимущественно по северным склонам па мелкоземисто-щебневатых почвах, на высоте от 3800 до 4500 м над у. м. Основу ее составляют плотные подушки куртковника, диаметром от 20—40 до 100 см. В качестве содо-минанта обычно выступает терескен (Eurotia ceratoides). К этим главенствующим растениям иногда примешиваются Dracocephalum Paulsenii, Роа relaxa, Potentilla pamiroalaica, Potentilla Moorcroftii, Oxytropis immersa и некоторые другие. Общее покрытие в сообществах, относящихся к данной формации, обыкновенно не превышает 15—20%.
Из других формаций подушковидной растительности, характерных для высокогорий Памира, отметим остролодочники, слагаемые подушковидными формами рода Oxytropis (О. Poncinsii, О. tianschanica). Они формируются на песчаных и щебнистых почвах на высоте свыше 4000 м над у. м. Кроме указанных господствующих видов, в составе остролодочников участвуют Braya pamirica, Oxytropis immersa, Tanacetum xylorrhizum, Macrotomia euchroma, Aneurolepidium dasystachys, Allium poly-phyllum, Roegneria schugnanica, Lindelofia stylosa, Astragalus heterodontus. Общее проективное покрытие в остролодочниках не более 20—25%.
Довольно распространенными на Памире являются формации из подушковидных лапчаток (Potentilla pamiroalaica, Р. pamirica).
Формация Potentilla pamiroalaica встречается во всех районах альпийского пояса Памира на песчано-каменистых почвах. В ее сложении принимают участие Tanacetum xylorrhizum, Oxytropis immersa, О. Poncinsii, Dracocephalum heterophyllum, Saussurea sorocephala, Smelovskia calycina, Allium polyphyllum, Thalictrum alpinum, Roegneria schugnanica, Androsace akbaitalensis. Проективное покрытие достигает 50%.
Формация Potentilla pamirica приурочена преимущественно к самой верхней части альпийского пояса, выше 4500 м над у. м., где встречается по щебнистым склонам гор. Общее проективное покрытие около 30%. В ее состав входят некоторые из видов, указанных для предыдущей формации. Кроме того, в ней отмечены Роа Litwinowiana, Astragalus char-guschanus.
В числе формаций подушковидной растительности следует отметить также формацию с преобладанием Oxytropis immersa, фрагментарно пред-
ставленную в альпийском поясе Памира (4300—5000 м над у. м.) и формацию с господством Dryadanthe tetrandra, характерную лишь для наиболее увлажняемых местообитаний альпийского пояса (подножья северных склонов, днища долин возле снеговых пятен).
Литература
О. Э. Кнорринг (1923); И. А. Райкова (1930); П. А. Варанов (1936); П.Н.Овчинников (1948); К. 13. Станюкович (1948, 1949, 1955).
101 т ш — т я п ь ш а н с к а я (из Sibbaldia let г and га, Thylacospermum rupifragum1 и др.)
Подушковидная растительность, развивающаяся на высоких, холодных и сухих сыртах Центрального Тянь-Шаня, в противоположность Памиру, представлена главным образом дриадантниками (из Dryadanthe tetrandra). Значительно распространены здесь также ассоциации мохо-видки (Thylacospermum caespitosum).
Подушки Dryadanthe ietrandra имеют обычно округлую, плоско-выпуклую форму. Высота их над поверхностью почвы не превышает 6—7 см, а в диаметре они достигают 60—70 см. Подушки эти настолько плотные, что свободно выдерживают тяжесть человека, почти не деформируясь. Плотность их создается за счет весьма многочисленных укороченных побегов, неоднократно переплетающихся между собою, причем рост побегов в горизонтальном направлении идет быстрее, чем в вертикальном, что и создает плотную подушку.
Проективное покрытие дриадантников Центрального Тянь-Шаня колеблется от 50 до 60%, причем на долю самой Dryadanthe приходится иногда не менее 30—35%. Можно различить два основных варианта этой формации: пустошный и луговой. Первый из них связан с более сухими местообитаниями и характеризуется значительным участием кобрезии, а вместе с ней таких ксероморфных видов, как Leontopodium ochroleucum, Potentilla hololeuca, Oxytropis chionobia. Второй вариант связан с более влажными местообитаниями и отличается большой примесью к эдифика-тору преимущественно мезоморфного альпийского разнотравья лугового типа: Papaver croceum, Ranunculus gelidus, Taraxacum pseudoalpinum, Viola tianschanica, Potentilla gelida, Trisetum altaicum.
Ассоциации, образуемые моховидкой (Thylacospermum caespitosum), в Центральном Тянь-Шане распространены в гораздо меньшей степени, чем дриадантники. Небольшими фрагментами они встречаются в верхней части альпийского пояса (выше 3000 м над у. м.) по щебнистым почвам горных склонов и на днищах речных долин.
Подушки моховидки иногда достигают 150 см в диаметре и 15—20 см высоты. На участках, ценотически наиболее выравненных, они покрывают до 40% поверхности почвы. Между подушками остаются промежутки голой земли, которые иногда заселяются отдельными экземплярами высокогорных растений. Интересно, что и на самих подушках поселяются различные виды цветковых растений, например Trisetum spi-catum, Crepis Karelinii, Poa alpina, P. relaxa.
1 Согласно номенклатуре, принятой во «Флоре СССР», Sibbaldia tetrandra Bge. должна называться Dryadanthe tetrandra (Bge.) Juz. (Флора СССР, т. 10,1941, стр. 229), а приоритетным названием Thylacospermum rupifragum Schrenk является бином Thylacospermum caespitosum (Camb.) Schischk. (Флора СССР, т. б, 1936, стр. 546), как эти растения в тексте и именуются.
В связи с этим, на них формируются своеобразные микробиоценозы, которые Гейнис (Heinis, 1925, 1937) описал для Альп. На Памире к аналогичным микробиоценозам должны быть отнесены «агрегаты», описанные И. А. Райковой (1930).
Thylacospermum caespitosum представляет собою монотипный род, ареал которого заходит из высокогорий Центральной Азии и Гималаев в горы Средней Азии только своим северо-западным краем. На Памире этот род отсутствует.
Следует иметь в виду, что ассоциации подушковидной растительности, формирующиеся на высокогорьях Памира и Центрального Тянь-Шаня, являются весьма изреженными. Кроме того, они перемежаются большими участками голых каменистых россыпей и скал. Поэтому пастбищное значение подобного рода растительности, в сущности, ничтожно.
Литература
М. Г. Попов (1928); И. А. Райкова (1930); Д. Кашкаров, А. Жуков и К.]|Ста-шокович (1937); Г. G. Сабардина (1951); F. Heinis (1925, 1937).
Рис. 81. Водорослевый такыр на подгорной равнине Копет-Дага (Туркменская ССР). (Фо!1. Л. Е. Родина).
Рис. 82. Лишайниковый такыр на Мешед-Мессерианской равнине (Турк менская ССР). (Фот. Л. Е. Родина).
XVI. СЛОЕВИЩНАЯ РАСТИТЕЛЬНОСТЬ ТАКЫРОВ
Л. Е. РОДИН
102. Водорослевые и лишайниковые сообщества такыров
Такыры — характерный элемент ландшафта южных пустынь. Они широкой полосой окаймляют Коиет-Даг, занимая подгорную равнину его в западной и восточной частях хребта. Эти такыры расположены на пролювиальных отложениях, слагающих подгорную равнину в области развития силовых потоков и активного поверхностного стока. Кроме того, такыры широко распространены в Низменных Каракумах и в крайней западной части Заунгузья. Они вкраплены небольшими площадями среди белосаксаульников, занимая котловины в песках с грядовым или котловинногрядовым рельефом (рис. 80).
Такыры Низменных Каракумов залегают на окончании силевых потоков, стекающих с Копет-Дага, а также в зонах растекания паводковых вод, связанных с деятельностью Мургаба и Теджена, сухие дельты которых в значительной мере перекрыты песчаными наносами. Очевидно, все эти такыры приурочены к поверхности древней аллювиальной равнины Пра-Мургаба и Пра-Теджена, которая впоследствии была перекрыта песками также аллювиального происхождения. Такыры распространены здесь в широкой полосе; для нее характерны не только показанные па карте более крупные по площади такыры, но и огромное количество мелких, некартируемых при масштабе нашей карты.
Крупные массивы такыров имеются в пределах Куня-Дарьинской древнеаллювиальной равнины. Они расположены здесь преимущественно на землях древнего орошения, начиная от сухого русла Даудана на севере до развалин крепости Гяур-Кала на юге.1
Небольшие площади такыров имеются также в низовьях Кашка-Дарьи.
Кроме того, такыры небольшими участками, не показуемыми в масштабе нашей карты, широко распространены в различных типах пустынь; для ‘этих случаев предусмотрен особый немасштабный знак. Такие мелкие такыры особенно часто встречаются на Устюрте, будучи приуроченными там к мелким местным, большей частью бессточным понижениям рельефа. Они имеются в Низменных Каракумах, в песках Учтаган и Чильмамедкум
1 На нашей карте данный такырцый массив не показан, так как в момент ее составления по этой территории отсутствовали достоверные материалы; по имевшимся в то время материалам экспедиции Агролеспроекта на нашей карте показаны черносаксаульники с участием такыров. В результате работ Аму-Дарьинской экспедиции Ботанического института им. В. Л. Комарова и Совета по изучению производительных сил АН СССР в 1953 г. удалось получить более точное представление о площадях, занимаемых такырами в пределах Куня-Дарьинской равнины, однако уже не было возможности внести необходимые исправления на карте, в то время находившейся в печати.
и в западной части Заунгузья; точно также они распространены в широкой полосе, примыкающей к среднему и нижнему отрезкам Дарьялыка, а также в восточной и южной частях Сарыкамышской котловины. Кроме того, такие мелкие такыры встречаются в низовьях Сарысу, в южной
части Бетпак-Далы, в нижней части древнего протока Жана-Дарьи (рис. 80).
До недавнего времени считалось общепринятым, что такыры — это территории, совершенно лишенные растительного покрова (Герасимов, 1931,1933; Коровин, 1934; Успанов, 1940). Однако после работ Н. Н. Болышева (Болышев и Евдокимова, 1944; Болышев и Манучарова, 1946, 1947) и углубленных исследований такыров Ботаническим институтом
им. В. Л. Комарова и Институтом почвоведения им. В. В. Докучаева АН СССР в 1930—1953 гг. (Родин, 1952, 1954; Базилевич, Голлербах п др., 1953; Базилевич, Родии и др., 1953; Родин и Голлербах, 1954 и др.) составилось ясное представление о растительном покрове этих почвенных образований.
Такыры — это области развития растительно!'© покрова, слагаемого сообществами си незеленых водорослей и лишайников при ничтожном участии эфемеров и однолетних солянок, часто слаборазвитых и в неблагоприятные годы не выходящих из стадии всходов.
Сообщества водорослей (водорослевые такыры) характерны для геологически молодых областей современной аккумуляции континентальных осадков и хорошо выраженного поверхностного стока атмосферных вод, обусловливающего периодическое затопление территории. В результате этого па равнинных пространствах создается совершенно своеобразный режим кратковременных мелких водоемов (глубиной всего в несколько сантиметров), на опресненном дне которых развиваются сообщества синезеленых водорослей, главным образом из семейства Oscillato-riaceae (виды Phormidiumn Microcoleus). Общий видовой состав водорослей различных разновидностей такыров оказывается весьма богатым. Так, в северной части Туранской низменности (древняя дельта Аму-Дарьи, северный отрезок Каракумов и прилегающие отроги южного Устюрта) на водорослевых такырах Н. В. Сдобниковой (1956) выявлено 92 вида водорослей: 43 вида синезеленых (Cyanophyta), 35 — зеленых (СЫого-phyta), 5 — желтозеленых (Xanthophyta), 1— эвгленовых (Euglenophyta) и 8 видов диатомовых водорослей (Bacillariophyta). По данным же Л. И. Новичковой (1955), на подгорной равнине Копет-Дага на такырах и связанных с ними сообществах, входящих в общий с ними ландшафт равнины, обнаружено 186 видов, разновидностей и форм водорослей 95 видов синезеленых, 54 — зеленых, 2 — желтозеленых, 2 — эвгленовых и 33 вида диатомовых. Водоросли образуют на поверхности почвь мощные пленки (достигающие нескольких миллиметров толщины) что и составляет наиболее характерную черту этих водорослевых такыров. Количество сухой органической массы водорослей составляем 0.5—1.0 т/га, а по определениям Л. Н. Новичковой (1955) иногда 1.4 т/га. Вегетация водорослей происходит не только в период затопления поверхности и образования временных мелководных «озер», но в при увлажнении атмосферными осадками.
Лишайниковые такыры распространены на территориях, лишенных современной аккумуляции, с крайне слабым поверхностным стоком, и обычно увлажняемых только влагой атмосферных осадков, без затопления (Базилевич, Голлербах и др., 1953). Лишайниковые сообщества представлены колониями главным образом из видов Diploschistes albissimus, D. scruposus и Squamaria lentigera, а также видов Collema, Aspicilia, Psora и др. Сухой органической массы, накапливаемой лишайниками, также около 0.5—1.0 т/га.
На такырах широко распространены почвенные микроскопические грибы. Как показали специальные исследования М. А. Литвинова (1954), во флоре почвенных микромицетов Преобладающими являются грибы семейства Dematiaceae (из родов Stemphylium, Macrosporium, Alternaria, Cladosporium и др.). Степень распространения и флористический состав почвенных микромицетов обнаруживают специфические особенности на различных категориях такыров и определенную взаимосвязь с сообществами водорослей, расселением высших растений, физико-химическими свойствами почв и стадией развития ландшафта в целом.
Флористический состав сообществ различных разновидностей такыров отличается заметным разнообразием, причем выделяются группы видов, играющие роль эдификаторов.
Так, например, Л. Н. Новичковой (1955) установлены различные группы эдификаторов в разных частях исследованной ею подгорной равнины Копет-Дага. В нижней части равнины, характеризующейся такырами начальных стадий развития, сообщества водорослей слагаются Phormidium autumnale с прпмесью Microcoleus vaginatus. В средней части, с наиболее типичными такырами, в сообществах доминирует Microcoleus vaginatus в сочетании с рядом видов рода Phormidium (Ph. autumnale, Ph. corium, Ph. foleolarum, Ph. inundatum, Ph. molle и др.). Кроме водорослей, в сообществах на этих такырах принимают участие лишайники, главным образом Collema minor, С. granulatum, Psora lurida, Diploschistes albis-simus, Aspicilia desertorum и др. Иногда они достигают столь заметного обилия, что такие такыры именуют лишайниково-водорослевыми. В верхней части равнины характерны так называемые зарастающие такыры, на которых намечается поселение некоторых цветковых растений (эфемеров и эфемероидов), а в покрове водорослей хотя и господствуют синезеленые (преимущественно представители рода Phormidium), но видовой состав их заметно сокращается, и характерно внедрение зеленых (виды Chlorococcum, Macrochloris, Chlorella) и некоторых диатомовых водорослей.
Водорослевые, лишайниково-водорослевые и лишайниковые такыры в пределах показанных на «Геоботанической карте» контуров, не занимают всей их площади. Территории эти характеризуются широко развитой комплексностью почвенно-растительного покрова.
Так, на подгорных равнинах, особенно в их средней части, очень характерны падины («ойтаки», турки.) — понижения округлой формы, глубиной в 30—60 см и более, размером 20—50 м в поперечнике. Осенью и весной они заполняются водами поверхностного стока, которые промачивают почву на глубину 3—4 м и более и создают в ней большие запасы влаги, что обеспечивает развитие растительности лугового типа. Здесь образуются сообщества с господством чаира (Aeluropus litoralis), лисохвоста (Alopecurus ventricosus), вики (Vicia peregrina) и ряда эфемеровых злаков (Bromus Danthoniae, В. squarrosus, В. macrostachys, Eremopyrum orientale, Aegilops triuncialis, Pholiurus pannonicus, Sclerochloa dura) и многих видов эфемерового разнотравья. По внешней окраине падин очень характерна полоса из луковичного мятлика (Роа bulbosa var. vivipara) с небольшой примесью полыни (Artemisia badghysi).
Вследствие усиленного испарения воды, поступающей по капиллярным ходам, по периферии падин образуется солончаковое кольцо, на котором наблюдаются разреженные заросли каргана (Salsola dendroides), мелколистной шведки (Suaeda microphylld) с однолетними маревыми (виды Salsola, Halimocnemis, Halocharis и др.).
Помимо таких хорошо развитых падин с луговым покровом, характерно наличие мелких западин, имеющих небольшую глубину и незначительные размеры, с еще несформировавшимся луговым травостоем. В них можно встретить (в зависимости от размеров и глубины западин) сообщества эфемеровые, мятликово-эфемеровые, мятликовые, полынномятликовые и другие, знаменующие различные стадии развития западин и, соответственно, их растительного покрова (Родин, 1954).
Среди такыров обычны разливы временных водотоков, в которых растительность представлена луговидными сообществами почти чистых травостоев арпагана (Eremopyrum orientale) с участием заячьего ячменя (Hordeum leporinum). Эти эфемерово-злаковые сообщества развиваются
только в годы со значительным количеством осадков или с обильным поверхностным стоком. Тогда они достигают высоты 35—50 см и дают высокую урожайность. В засушливые же годы наблюдаются лишь бедные разреженные заросли однолетних солянок с ничтожным участием эфемеровых злаков. Под влиянием обильного увлажнения и богатого луго-видного травостоя почвы разливов приобретают характер луговых, на которые, однако, наложена значительная солончаковатость (Родин, 1954).
Характерным элементом комплекса является растительность вдоль русел временных потоков, которые в изобилии пересекают подгорную равнину. В основных чертах растительный покров здесь представлен пестрыми сочетаниями фрагментов луговидно-эфемеровых (арпагановых, ячменных), эфемеровых (эфемеровое разнотравье), полынных (из Artemisia badghysi), мятликовых (из Роа bulbosa) сообществ с редкими группами каргана (Salsola dendroides) и частыми пятнами однолетних солянок (Salsola lanata, S. turcomanica, Halimocnemis Karelinii, H. mollissima, Halocharis hispida и др.).
Наиболее выраженные и наименее комплексные такыры расположены далее всего от предгорий. Именно здесь на больших площадях распространены водорослевые такыры, незначительно испещренные участками сообществ высших растений.
Чем ближе к горам, тем более усложняется общая картина растительности подгорной равнины: наряду с участками водорослевых сообществ на такырах появляются лишайниковые такыры и переходные между первыми и вторыми. Все большую площадь занимают сообщества высших растений и, наконец, появляются зарастающие такыры со значительным покровом из эфемеров и эфемероидов, среди которых вкраплены пятна эфемерово-мятликовых полынников на почвах, близких к светлым сероземам. Эта верхняя полоса предгорной равнины является переходной к зональным почвенно-растительным сообществам предгорий с мятликовыми полынниками на светлых сероземах.
На небольших такырах среди Каракумов растительность не имеет такой ярко выраженной комплексности. Замкнутые котловины каракумских такыров более равномерно заливаются водой и таким образом достигается большее однообразие условий местообитания. В Каракумах отмечены только сообщества синезеленых водорослей (Microcoleus vagi-natus, Phormidium ambiguum, P. corium, P. dimorphum, P. lividum)', лишайниковые такыры не встречены. По краям такыров наблюдается поселение некоторых высших растений (Salsola arbuscula, S. rigida, S. gemmascens, Haloxylon aphyllum, Artemisia arenicola).
При наличии на такырах песчаных наносов на последних развивается растительный покров, сходный с растительностью ближайших песков, однако несколько обедненного флористического состава (всегда выпадают наиболее типичные псаммофиты).
Такыры и такыровидные образования (имея в виду переходные сообщества между типичными водорослевыми и лишайниковыми такырами и более примитивными сообществами, с одной стороны, и более сложными — с участием цветковых растений — с другой стороны) широко распространены не только в пределах Туранской низменности, но также в Афганском Туркестане и на пустынных равнинах Ирана и Джунгарии. Однако соответствующие исследования их в этих странах еще не осуществлены. Повидимому, к этой же категории следует отнести «тойримы» Гоби в Центральной Азии, а также «playas» пустынь Большого Бассейна в Северной Америке и, возможно, «dry lakes» западной Австралии.
Литература
М. Г. Попов (1922); И. П. Герасимов (1931, 1933); И. И. Гранитов (1934)- Е П и* Йовин (1934); У. У. Успанов (1940); Н. Н. Болышев и Т. И. Евдокимова Но/у?” L Н. Болышев и Е. А. Манучарова (1946, 1947); М. Л. Пархоменко (1949)- И Ь к А лышев (1952, 1955); Л. Е. Родин (1952, 1954); Н. И. Базилевич, М. М. Голлообау i, (1953); Н. И. Базилевич, Л. Е. Родин и др. (1953); М. А. Литвинов (1954)- Л р рР-дин и М. М. Голлербах (1954); Н. И. Базилевич и Л. Е. Родии (1955) Л н и'1 °* нова (1955); Н. В. Сдобникова (1956). «ович-
103. ЛЕДНИКИ И СНЕЖНИКИ
ИГ. М. ГОЛЛЕРБАХ и В. Б. СОЧАВА
Ледники и снежники на территории СССР наибольшую площадь покрывают в Арктике (54 000 км1 2 — 78% всей площади оледенения СССР).1 Значительные скопления льда имеют место на Новой Земле, Северной Земле и Земле Франца-Иосифа, где ледниковые языки местами спускаются к берегу мори. Эти крупнейшие ледниковые районы находятся в при-атлаптической провинции зоны полярных пустынь (опис. 2) с крайне бедным и разреженным растительным покровом. По краю ледников, например на базальтовых скалах и осыпях Земли Франца-Иоснфа, увлажняемых водами тающих ледников, растительность бывает несколько богаче, чем на сухих скалах, за счет разрастающихся моховых подушек и некоторых цветковых растений {Raniulculus nivalis, Cochlearia arctica и др.). Сходное явление наблюдается в местах, орошаемых ледниковыми водами, на северном острове Новой Земли. Выступающие среди ледников обнаженные каменистые поверхности (нунатаки) характеризуются очень немногими цветковыми растениями и небольшим числом видов мхов и лишайников.
В бореальном поясе значительную площадь ледники занимают в Верхоянско-Колымской горной стране (около 600 км2), главным образом в бассейне Индигирки.2
Они показаны на «Геоботанической карте» в окружении осыпей и скал высокогорий умеренного пояса с очень разреженной растительностью. Самые крупные долинные ледники здесь спускаются до высоты около 1800 м над у. м. Приледниковая растительность этого района пока не изучена.
Вторым крупным центром оледенения в бореальном поясе является Алтай, где ледники по последним данным3 покрывают площадь около 590 км2. Значительные скопления ледников на Алтае наблюдаются на хр. Катуньские и Чуйские белки, много ледников в северной части Шапшальского хребта. Остальные ледники Алтая рассеяны по высокогорьям (снеговая линия на западе на высоте 2400—2600 м, в центральном Алтае — 2400—3100 м и в южном Алтае — 3100—3500 м над у. м.) и включены в контуры разреженной растительности высокогорий умеренного пояса (опис. 1), а частью альпийских лугов (57 а) и горных тундр (3 а).
1 П. А. Шумский. Современное оледенение Советской Арктики. Вопросы географии, 4, 1947.
* Л. Л. Берман. Современное оледенение верховьев р. Индигирки. Вопросы гео» графин; 4, 1947. По другим данным площадь оледенения Верхоянско-Колымской горной страны значительно меньшая (около 250 км1).
з ft. В. Тронов. Современное оледенение Алтая. Томск, 1949.
О немногих высших растениях, встречающихся на Алтае у края ледников и фирновых полей, упоминалось при описании разреженной растительности высокогорий (опис. 1).
На Кавказе под ледниками занято около 2000 км2 и наибольшая площадь оледенения находится в центральной части Главного Кавказского хребта, где ледники только и показаны на «Геоботанической карте» среди контуров альпийских лугов. Снеговая линия на Главном Кавказском хребте между верховьями Кубани и истоками Андийского Койсу проходит на высоте от 3000 до 3500 м над у. м. Вокруг снежных пятен и у края ледников нередко выделяются по распространению лютики (Ranunculus trisectilis, R. oreophilus) и некоторые первоцветы (Primula algida). Непосредственно к ледникам и снежникам часто подходит особый тип альпийских ковров; в их составе растут колокольчик (Campanula tridentata), горечавка (Gentiana djimilensis), манжетка (Alchimilla sericea), часто зибальдии (Sibbaldia parviflora, S. semiglabra) и некоторые другие растения. По краю угасающего карового ледника нередко развивается полоса несомкнутой пригляциальной растительности, которая ниже переходит в сообщества альпийских ковров. Растительность альпийских ковров на Кавказе, как утверждает Ан. А. Федоров (1942), имеет много общего с околоснежной растительностью и в своем происхождении связана с ней. В Верхней Сванетии на высоте 4000 м над у. м. А. Л. Харадзе (1944) наблюдала только два вида камнеломок (Saxifraga junipertfolia, S. то-schatd).
На Малом Кавказе фирновые снега на небольшой площади развиты в области вершины Арагаца. У самой кромки снегов здесь местами обильно разрастается своеобразный ромашник (Chamaemelum melano-lepis), по экологии типичный пригляциальный криофит (Федоров, 1945); кроме того, растут немногие другие виды (Oxyria digyna, Alopecurus glacialis, Colpodium fibrosum).
Наибольшая площадь высокогорного оледенения в СССР сосредоточена на границе умеренного и субтропического поясов, в горах Средней Азии (около 11 000 км2).1 Снеговая граница там проходит на высоте около 3500 м на западе и достигает 5500 м над у. м. на Памире. Значительные скопления ледников и фирновых полей в горах Средней Азии совпадают с основными орографическими центрами и горными узлами. Ледники окружены огромными, преимущественно щебнистыми пространствами, которым присуща разреженная растительность среднеазиатских высокогорий, ниже по склону с небольшими участками степных, пустынных и нагорноксерофитных формаций (опис. 104). Огромные высоты, значительная расчлененность местности и большие углы падения способствуют обнаженности склонов. Воды, стекающие с ледников и снежников, смывают мелкозем и вместе с другими факторами физического выветривания способствуют сползанию каменных россыпей. При таких условиях в полосе вечных снегов и ледников встречаются очень немногие высшие растения, растущие преимущественно между камнями в защищенных местах. Более разнообразен видовой состав растений, встречающихся по моренам по краю тающего ледника (Parryа excapa, Didymophysa Fed-
1 С. В. Калесник. Горные ледниковые районы СССР. Л., 1937. По подсчетам Р. Д.Забирова(ОледенениеПамира.М.,1955) площадь современного оледенения в горах Ж- дей Азии больше. Только на Памире она, поданным названного автора, превышает км*. Судя по картосхеме современного оледенения Памира, составленной Р. Д. Заборовым, пространства под ледниками Памира на «Геоботанической карте» преуменьшены за счет преувеличения территории под разреженной растительностью высокогорий.
czenkoana, Chorispora macropoda, Astragalus Beketovii, Oxytropis pagobia, Waldheimia Korolkovii, Erigeron Olgae и др.), среди них некоторые относятся к числу типичных пригляциальных растений. На нунатаке ледника Федченко (4200 м над у. м.) на небольших участках, орошаемых водами тающих снегов, развито сообщество дриаданты (Dryadanthe [Sib-baldia] tetrandra), в состав которого входят немногие виды (Saxifraga hirculus, Leontopodium ochroleucum, Potentilla pamiroalaica, Cobresia stenocarpa и др.). В Восточном Памире в нивальном поясе выше других растений (5250 м над у. м.) подымаются Melandrium apetalum и Pachypleurum alpinum (Станюкович, 1949).
В арктической и бореальной зонах для нриледниковой полосы наиболее характерны виды горнотундрового флористического комплекса, а на Кавказе и в Средней Азии — растения, генетически связанные с флорой пустынных высокогорий. Мезофильный элемент флоры типичных альпийских лугов у края ледников и снежников представлен более скудно.
В Арктике и в высокогорных областях, преимущественно по склонам гор и по краям ледников, на огромных пространствах, казалось бы, безжизненного снега, могут в большом количестве развиваться низшие растительные организмы, окрашивающие его в свойственный им цвет. Наибольшую известность получил «красный снег».
В пределах СССР «красный снег» представляет довольно обычное явление в Арктике, у подошвы снеговых гор в юго-западной части Новой Земли, во многих местах на Земле Франца-Иосифа и в других местах. На Кавказе он наблюдался в горах Главного Кавказского хребта (в истоках Теберды, в Балкарии, Сванетии и Дигории на высоте от 2050 до 3400 м над у. м.); широко распространен он в зоне вечных снегов Камчатки; находили его также на восточном склоне Северного и Полярного Урала, в Сибири (Алтай, Становой хребет), в Джунгарском Алатау и в других местах.
В зависимости от степени развития организмов, вызывающих эту окраску, красный цвет снега может меняться в интенсивности от бледно-розового до кровянокрасного и темномалинового и нередко распространяется на огромные пространства. Так, например, в горах Карачая на перевале Басса, на высоте около 3000 м над у. м., в 1929 г. было найдено на снегу красное поле площадью в несколько квадратных километров. Толщина окрашенного снега измерялась сантиметрами. Однако более обычными являются сравнительно небольшие окрашенные пятна снега; площадь их составляет десятки квадратных метров и толщина измеряется миллиметрами. Известен случай розовой окраски снега на глубине 10—20 см при неокрашенной поверхности снега.
Организмом, вызывающим «красный снег», является прежде всего один из видов одноклеточных подвижных водорослей хламидомонад (Chlamy-domonas nivalis—Sphaerella nivalis), относящийся к типичным зеленым водорослям (Chlorophyta), но вырабатывающий в своих клетках еще и красный пигмент — гематохром. Большую часть времени эта хламидомонада находится в состоянии покоящихся неподвижных шаровидных клеточек, густо заполненных гематохромом, но при оттаивании верхних слоев снега она начинает очень быстро размножаться, образуя неподвижные мелкие клетки и типичные подвижные хламидомонады. Розовую и красную окраску снега могут вызывать также несколько видов дрожжевых грибков (из родов Torula и Torulopsis) и некоторые другие организмы, сопровождающие водоросли.
Кроме «красного снега», известно еще много других случаев, когда снег становится окрашенным от развития в нем водорослей и, вместе с ними, грибов и бактерий. Окраска снега может быть еще зеленой, желтой, коричневой, бурой и даже черной, в зависимости от преобладания в нем тех или иных видов. Все же чаще других встречается зеленое «цветение» снега, вызываемое различными видами зеленых водорослей, из которых весьма обычной является нитчатая Rhaphidonema nivale. Коричневую окраску снега вызывают чаще всего диатомовые водоросли (Bacillario-phyta) и некоторые дрожжи. Совершенно черным цветом отличается дрожжевидный грибок (TVadsoniella nigra), выделенный из окрашенного снега, наблюдавшегося на Кавказе (Аделсу, Буго, Баш-Кара).
В итоге, в пределах СССР в окрашенном снеге было найдено следующее количество видов низших растений.
Водоросли
Тип Chlorophyta (Зеленые водоросли) — 19 видов Тип Bacillariophyta (Диатомовые водоросли) — 29 »
Тип Cyanophyta (Синезеленые водоросли) — И » Тип Rhodophyta (Красные водоросли) — 1 вид
Всего 60 видов
Грибы (в том числе дрожжи и дрожжевые грибки)
Класс Phycomycetes (Грибы-водоросли) — 7 видов
Класс Ascomycetes (Сумчатые грибы) — 23 вида
Класс Fungi imperfect! (Несовершенные грибы) — 35 видов
Всего 65 видов
Класс Actinomycetales (Актиномицеты) — 2 вида Бактерии — 12 видов
Таким образом, общее количество низших растений, найденных в окрашенном снеге в пределах СССР, составляет значительную цифру — 139 видов. Однако следует думать, что не все они являются специфически приспособленными к обитанию в верхних слоях снега, так как многие из них были найдены и в других местообитаниях. Наличие в установленном списке некоторого процента занесенных форм вряд ли подлежит сомнению. Тем не менее, существование типично «снежных» низших растений (Chlamydomonas nivalis, Rhaphidonema nivale, Nadsoniella nigra и др.) столь достоверно, а распространенность их массового развития на больших пространствах снежных полей в горах столь велика, что вполне уместно выделять особо зону низших растений окрашенного снега в высокогорных областях СССР.
Жизненные условия этой растительной группировки весьма своеобразны. Большую часть времени, вследствие преобладания отрицательных температур, все снежные организмы проводят в покоящемся состоянии, проявляя исключительно высокую жизненную стойкость при промерзании. Но как только под влиянием сильной солнечной радиации поверхность снега начинает хотя бы слегка подтаивать, снежные организмы размножаются с исключительно высокой интенсивностью.
Как и во всех других биотопах, первичным звеном здесь являются Хлорофиллоиосные автотрофные организмы — водоросли, способные фотосинтезировать в этих условиях, довольствуясь минимальным количеством минеральных веществ, содержащихся в снеге. Органические вещества, прижизненно выделяемые водорослями или поступающие в окружающую
среду после их отмирания, в свою очередь представляют тот питательный субстрат, который создает возможность поселения гетеротрофных растений — грибов, актиномицетов и бактерий.
Ледники и снежники, имеющие большое значение в питании рек в Арктике и в особенности в высокогорьях, являются также существенным экологическим фактором. Влияя на климат прилегающих мест, а также определяя режим увлажнения грунтов высокогорных районов, ледники и снежники тем самым воздействуют на состав и строение высокогорных растительных группировок. С деятельностью ледников связано развитие форм высокогорного рельефа, в зависимости от которых находится и распределение растительности.
Литература
В. В. Сапожников (1906, 1916); А. А. Еленкин (1914); Н. Н. Воронихин (1930); А. Ф. Казанский (1932); Е. П. Коровин (1934); Г. С. Филиппов (1934); А. Г. Салимов-ская-Родина (1936); Ан. А. Федоров (1942, 1945); А. Л. Харадзе (1944); К. В. Станюкович (1949); Л. И. Леонов (1953).
104. РАЗРЕЖЕННАЯ РАСТИТЕЛЬНОСТЬ ГОР СТЕПНОЙ И ПУСТЫННОЙ ЗОН
Е. М. ЛАВРЕНКО и Л. Е. РОДИН'
Разреженная растительность гор показана на «Геоботанической карте» в аридных областях — в Казахском мелкосопочнике (Казахской складчатой стране) и в горах Средней Азии, где она занимает большие площади в средне- и высокогорном поясах. Имеется она и в Восточном Закавказье, но занимает здесь меньшие площади, почему на карте не отмечена.
Несмотря на суровость условий существования, каменистые склоны и осыпи создают большое разнообразие экологических ниш, в которых находят приют многие виды. Здесь встречаются не только собственно литофиты (петрофиты, хазмофиты), но и растения, широко распространенные в луговых, степных и других сообществах высокогорий, отличающихся значительно меньшей флористической пестротой. Естественно, что исследователи высокогорий всегда уделяли особое внимание флоре скал, каменистых россыпей и т. п. местообитаний, крайне разреженная растительность которых выделялась флористическим богатством.
104 кз—Казахской складчатой страны с фрагментами степной, луговой и лесной растительности
Разреженная растительность гор, преимущественно на выходах гранита, в пределах Казахской складчатой страны показана на карте в горном массиве Улутау (Зелинский, 1912; Павлов, 1931), к северу от Караганды и, главным образом, к югу от линии Караганда—Каркаралинск. В последнем районе разреженная растительность занимает выходы кристаллических пород, преимущественно гранитов, в горах Кос-агач, Конур-Кызылтау, Манатау, Бес-Чеку, Кызыл-Караган, Конур-Кульджа, Бер-кара и др. (Рожанец-Кучеровская, 1916; Крюгер, 1927а). Высота этих горных массивов — 750—1250 (1500) м над у. м.
Растительность высоких каменистых сопок очень разреженная. Поверхность скал обычно покрыта лишайниками. Очень характерно присутствие довольно многочисленных видов, свойственных каменистому субстрату (из полукустарничков: Ephedra distachya, Gypsophila Patrinii, Ziziphora Bungeana, виды Thymus, Artemisia frigida, A. glabella, A. sublessingiana', из петрофильного разнотравья: Thalictrum foetidum, Libanotis buchtormensis, Patrinia intermedia', суккулент Orostachys spinosa и др.). Встречаются участки с более или менее сформированными степными ассоциациями с преобладанием дерновинных злаков (Festuca sulcata, Koeleria gracilis, Stipa capillata) с большей или меньшей примесью вышеупомянутых видов полукустарничковых полыней и степных видов разнотравья. В затененных расщелинах скал произрастают папоротники
1 Е. М. Лавренко составлен общин очерк и текст к 104 кз, Л. Е. Родиным — текст к 103 са.
(Cystopteris fragilis, Woodsia ilvensis, Polypodium vulgare, Asplenium septentr ionale). В каменистых расщелинах скал и на осыпях встречаются кустарники, то единичными экземплярами, то зарослями: Juniperus sabina (стланик), Berberis sibirica, Spiraea hypericifolia, S. trilobata (восточная часть мелкосопочника), Dasiphora parvifolia, Caragana pumila, Lonicera microphylla и др. По каменистым ущельям образуют заросли Rosa spinosissima, R. cinnamomea, Caragana frutex и др.
В ущельях обычны березовые и осиново-березовые рощи (из Betula verrucosa и Populus tremula) с рядом бореальных форм, заросли ив, фрагменты лугов. В районе Каркаралинска на скалах местами встречаются единичные сосны и редкостойные насаждения их.
Литература
Ф. Ф. Зелинский (1912); С. Е. Рожанец-Кучеровская (1916); В. А. Крюгер (1927а); Н. В. Павлов (1931).
104 са — Средней Азии с фрагментами степной, пустынной и нагориоксерофитной растительности
Разреженная растительность гор пустынной зоны распространена чрезвычайно широко в Средней Азии, занимая наиболее высокие хребты и на верхнем своем пределе соприкасаясь с ледниками и снежниками (опис. 103).
На «Геоботанической карте» она представлена то крупными контурами, то мелкими островками.
Соответствующие контуры показаны на Заилийском Алатау, далее они имеются на Киргизском хребте, в Таласском Алатау, Терскей-Алатау, Кокшаалтау, в Туркестанском и Зеравшанском хребтах, на хр. Алайском и Заалайском и на всех горных хребтах Памира. На последнем площади, занятые этой растительностью, особенно велики.
Условия местообитания растений в высокогорьях весьма разнообразны. Здесь характерны не только собственно скалы, но и гребни вершин, нагромождения выветривающихся пород и продуктов их разрушения, осыпи, каменистые россыпи, моренные галечники и валунники и т. п. Наряду с инсолированными участками, сухими и нагреваемыми солнцем, столь же часты и затененные, увлажняемые сочащейся по трещинам грунтовой влагой, а также дождями и росами, и почти не попадающие на прямое солнечное облучение. •
Особенно же важна для жизни растений резкая смена температур воздуха и почвы, в больших пределах изменяющаяся здесь как в годовом, так и суточном ходе. В целом температурный режим суров, вегетационный период краток и нередко прерывист, почвы примитивны и бедны органическим веществом.
В силу этих условий большинство растений мелки и низкорослы, часто прижимаются к субстрату, или же погребаются им и от стелющихся под осыпью стеблей выставляют на поверхность лишь концы побегов. Однако их корневые системы, как правило, мощны, энергично проникают в трещины и грубый субстрат, часто удерживая растение даже на ползущей вниз осыпи.
Общими чертами всех стаций являются разомкнутость растительного покрова, невыработанность сообществ, агрегатность группировок, пестрота видового состава и .преобладание среди растений биологических типов криофитов, криоксерофитов, часто образующих формы плотных дерновин и разнообразных подушек.
Характерно также, что большинство растений является многолетниками, и близкие виды, представленные на равнине однолетними формами, в высокогорьях на скалах имеют многолетних родичей (Краснов, 1888).
Наряду с этим на скалах и осыпях находят приют и типичные влаголюбивые растения, которые, как пишет П. А. Баранов (1925), в тени окружающих скал, чувствуют себя нисколько не хуже, чем на берегу горного ручейка. Такова, например, Impatiens parviflora, обычнейший, спутник как каменистых осыпей Чимгана, так и берегов горных ручьев
и скал около водопадов.
Таким образом, скалы и осыпи как субстрат для развития растительного покрова при преобладании условий, благоприятных ксерофитам, представляют возможности для поселения и других растений весьма различной экологии. Среди растений скал, осыпей и т. п. М. Г. Попов (1940) разделяет хазмофиты на облигатные и факультативные. Кроме того, им обращено внимание на определенную связь многих растений с химизмом субстрата и физическими особенностями выветривания различных горных пород (гранитов, известняков, сланцев и т. д.), что накладывает заметный отпечаток на флористический состав растительности. М. Г. Попов приводит для Тянь-Шаня сравнительные списки видов одноименных родов из многих семейств, из которых одни связаны в своем распространении с гранитами, другие — с известняками. К сожалению, подобных сведений нет по остальным горным системам Средней Азии, в силу чего нет возможности дать сводные данные.
В процессах первичного почвообразования на скалах особо важная роль принадлежит мхам и лишайникам (см., например, работы М. А. Гла-совской, 1950 и И. Н. Антипова-Каратаева, 1951), однако недостаточная
флористическая изученность этих важных для становления арены жизни в высокогорьях групп растений не позволяет изложить сведения о них сколько-нибудь достоверно.
В ряде случаев по условиям масштаба соответствующие контуры на «Геоботанической карте» не могли быть показаны. Так, например, в Джунгарском Алатау скалы и каменистые россыпи занимают в общем не менее половины площади в альпийском поясе. Для этих местообитаний Н. И. Рубцов (1948а) указывает многие растения, ютящиеся в трещинах, на карнизах, каменистых склонах и т. п., часто едва приметные среди нагромождений породы, чуть затронутой процессами почвообразования. На скалах характерны Paraquilegia grandiflora, Р. dnemonoides, Potentilla orientalis, Saxifraga oppositifolia, Dracocephalum imberbe, Parrya pinnatifida, P. ste-nocarpa, Chorispora Bungeana, Allium caricoides, Pachypleurum mucrona-tum, Valeriana caespitosa, Rhodiola coccinea.
На скоплениях мелкозема по карнизам скал встречаются обычные представители альпийских лугов -.Polygonum nitens, Р .viviparum, Dryadanthe tetrandra, Cerastium tianschanicum, Trisetum altaicum, Melandrium ape-talum. На теневых и влажных склонах скал характерны Saxifraga hircu-lus, S. Komarovii, S. sibirica, а на теплых южных склонах встречаются кустарники: Lonicera humilis и Dasiphora fruticosa. На каменистых россыпях Джунгарского Алатау флора значительно беднее (а многие россыпи совершенно лишены растений). Здесь отмечены Cerastium lithospermifo-lium, Veronica macrostemon, Dracocephalum stamineum, Rheum rhizostachyum, Delphinium dasyanthum, Erysimum altaicum, Polygonum rupestre, Allium platyspathum, A. oreophilum, Androsace dasyphylla, Valeriana caespitosa.
В Заилийском Алатау разреженная растительность скал и осыпей, по В. П. Голоскокову (1949), занимает около половины площади ялытий-
ского и нивального поясов. Названными автор различает здесь несколько стаций.
На скалах нивального и альпийского поясов, лишенных мелкозема, обильны лишайники (виды Lecidea, Rhizocarpon, Gyrophora) и встречаются в небольшом количестве мхи (из семейств Grimmiaceae, Orthotrichaceae и др.), подготовляющие субстрат для поселения цветковых, единичные особи которых появляются лишь изредка в трещинах с накопившимся мелкоземом (Potentilla biflora, Rhodiola coccinea, R. gelida, Saxifraga op-positifolia, Thylacospermum caespitosum, Dryadanthe tetrandra, Paraquile-gia grandiflora, Rosularia alpestris, Oxytropis atbaschi; в нивальном поясе отмечены лишь первые шесть видов). В субальпийском поясе наряду со многими из вышеперечисленных растений встречаются еще Valeriana caespitosa, Viola biflora, Allium schoenoprasoides, A. caricoides, A. filifo-lium, Silene lithophila, Minuartia Kryloviana, Dracocephalum nodulosum, Aster limonifolius, Pyrethrum pyrethroides, Poa Albertii, P. relaxa.
На осыпях нивального пояса встречаются очень немногие растения: Cerastium litospermifolium, Thylacospermum caespitosum, Pastinacopsis glacialis. В альпийском поясе видовое разнообразие возрастает. Здесь для осыпей характерны некоторые низкорослые распластанные кустарнички: Dasiphora phyllocalyx, Spiraea tianschanica, Lonicera Olgae, L. humi-lis, Caragana fubata. Из травянистых растений обычны следующие: Роа Lipskyi, Leucopoa Olgae, Dracocephalum imberbe, Allium platyspathum, Corydalis Fedtschenkoana, Cerastium lithospermifolium, Chorispora songorica, Ch. Bungeana, Pastinacopsis glacialis и др. Осыпи субальпийского пояса отличаются особым своеобразием: по периферии они окаймлены полосой кустарников. На северных склонах кайму образуют Salix alatavica и S. hastata, а на южных — Juniperus turkestanica, Spiraea ferganensis, Ribes Meyeri, Lonicera Karelinii, L. hispida, Dasiphora phyllocalyx, Spiraea tianschanica, Caragana fubata.
На самих же осыпях встречаются немногие травянистые формы: Allium polyphyllum, A. platyspathum, Роа relaxa, Р. Albertii, Dracocephalum imberbe и др.
Растительность каменистых склонов, уже заметно обогащенных мелкоземом, значительно богаче во всех поясах, включая большинство растений скал и осыпей. Для экономии места упомянем лишь растения, ранее не названные. Так, в нивальном поясе прибавляются Astragalus densiflorus, A. Beketovii, Leonthopodium ochroleucum. В альпийском — число таких видов больше: Astragalus nivalis, A. lithophilus, Oxytropis Severzovii, Dracocephalum Paulsenii, Veronica macrostemon, Pa-paver tianschanicum, Ranunculus gelidus, Callianthemum alatavicum, Angelica brevicaulis, Draba oreades и ряд других. В субальпийском поясе на каменистых склонах видовое разнообразие особенно велико, так как конкуренция между разреженными растениями еще не возникает, а общие условия существования значительно благоприятнее. Помимо большинства растений скал и осыпей, здесь появляются следующие виды: Polygonum rupestre, Ligularia robusta, Serratula lyratifolia, Sedum Ewersii, Saxifraga setigera, Adenophora hymalayna, Plantago arachnoidea, Gypsophila microphylla, Oxytropis assiensis, Taphrospermum altaicum и многие другие.
На современных моренах уже наблюдаются куртинки с преобладанием одного-двух видов растений. Так, на ровных мелкоземистых участках куртинки образуют Saussurea sorocephala, Pyrethrum leontopodium, Waldhei-mia tridactylites, Erigeron Schmalhausenii. На склонах северной экспозиции группировки слагают Carex Griffithii, Сhamaenerium latifolium, Cerastium
lithospermifolium, Colpodium leucolepis, Papaver tianschanicum', на южных склонах — Trisetum spicatum и подушечные Thylacospermum caespitosum, Dryadanthe tetrandra.
Кроме того, на моренах единичными особями встречаются Erigeron Olgae, Crepis Karelinii, Melandrium apetalum, Pastinacopsis glacialis, Chorispora songorica, Ch. Bungeana, Luzula spicata, Leontopodium ochroleu-cum, Saxifraga setigera, S. cernua, Draba oreades, Cerastium cerastoides, Aster flaccidus.
Галечники и каменисто-песчаные водно-ледниковые наносы речек, рождающихся из ледников, не имеют такого специфического набора видов, свойственного некоторым из ранее рассмотренных стаций. В. П. Голоскоков подчеркивает, что здесь встречается «конгломерат» видов, занесенных водными потоками с окружающих ближайших склонов со скалами и осыпями, а нередко принесенных ими издалека из других поясов. Так, на галечниках альпийского пояса встречаются следующие типичные «альпийцы»: Thylacospermum caespitosum, Oxytropis chionobia, О. Severtzovii, Astragalus densiflorus, A. lithophilus, Chamaenerium latifolium, Smelowskia calycina, Draba oreades, Papaver tianschanicum, Erigeron turkestanicus-, совместно с ними произрастают такие растения морен, как Saussurea sorocephala, Pyrethrum leontopodium, Waldheimia tridactylites, Carex Griffithii, Crepis Karelinii и многие другие.
В субальпийском поясе галечники обогащаются новыми видами: кроме всех названных выше, здесь встречаются Sedurn Ewersii, Pachypleu-rum mucronatum, Festuca rubra s. 1., Draba lanceolata, Leontopodium campestre, Polygonum rupestre и др. Характерно здесь еще и наличие ряда кустарничков: Dasiphora phyllocalyx, Salix alatavica, Juniperus turkestanica, Ephedra Fedtschenkdi, Comarum Salesovianum, Caragana fubata.1
На Киргизском хребте и Таласском Алатау О. Э. Кнорринг и 3. А. Минквиц (1912а) указывают только 24 вида на скалах и осыпях в высокогорьях. Здесь встречены ими следующие кустарнички и полукустарнички: Oxytropis Poncinsii, Lonicera humilis, Dasiphora fruticosa, Artemisia rutifolia. Из травянистых растений найдены Allium oreophilum, Melandrium apetalum, Paraquilegia grandiflora, Chorispora Bungeana, Parrya excapa, Rosularia alpestris, Potentilla hololeuca, P. nivea s. 1., Euphorbia humilis, Androsace septentrionalis, A. dasyphylla, Dracocephalum Paulsenii, D. integrifolium, D. nutans, D. oblongifolium, Plantago ara-chnoidea, Tanacetum megalocephala, T. Herderi, Pyrethrum pyrethroides.
Несомненно, список этот не полон, как это следует из материалов Р. И. Аболина и М. М. Советкиной (1930) по альпийскому поясу Таласского Алатау и Сусамырского хребта. Для скал, каменистых склонов, каменистых осыпей и т. п. этого пояса, занимающих в его пределах более 70% площади, они приводят список из 92 видов растений; к сожалению, нет указаний о приуроченности растений к определенным стациям. Укажем, что ими встречены следующие кустарнички, полукустарнички и подушковидные формы: Sibbaldia macrophylla, Dryadanthe tetrandra,
1 Мы привели данные В. П. Голоскокова с сокращениями, имея целью показать значительное разнообразие видового состава рассматриваемой растительности. К сожалению, по другим горным хребтам нет подобных тщательных исследований. В то же время можно предполагать, что лучше увлажняемый Северный Тянь-Шань выделяется ббльшим богатством флоры высокогорий, нежели Центральный и хребты си* стемы Памиро-Алая. Еще А. Н. Краснов (1888) подчеркивал особое богатство на Тянь-Шане флоры скал, осыпей и т. п. Однако это можно оудет установить лишь при налички более подробных данных, которые будут получены новыми исследователями. Поэтому в последующем изложении неизбежна значительная схематичность и краткость.
Рис. 83. Бухарский оазис. Г. Бухара. В центре — средневековый мавзолей; влево на переднем плане — остатки глинобитной городской стены с отдельными кустами Alhagi spar si folia] вправо — заброшенный участок с зарослями Alhagi sparsifolia, (Фот. Ю. Е. Лавренко).
Astragalus lithophilus, Oxytropis humifusa, О. chionobia, О. microsphaera, 0.Lehmannii, Lonicera glauca, Pyrethrum pulchrum, Tanacetum megalocephala, Pyrethrum pyrethroides, Artemisia Aschurbajewii. Из обширного списка травяных форм, встреченных названными авторами, укажем только чаще всего наблюдавшиеся в разных точках изученной ими территории: Dracocephalum oblongifolium, Aster alpinus, Pyrethrum pulchrum, Waldheimia Korolkovii, Crepis multicaulis, Trisetum spicatum, Papaver nudicaule, Draba lanceolata, Saxifraga sibirica, Potentilla hololeucaviP. gelida, Aulacospermum simplex, Carum atrosanguineum.
В субальпийском поясе скалы и сходные местообитания распространены преимущественно на южных склонах и занимают сравнительно небольшую площадь по отношению ко всему поясу. Р. И. Аболин и М. М. Советкина (1930) указывают 18 видов кустарников и полукустарничков {Juniperus sabina, J. turkestanica, J. semiglobosa, Ribes Meyeri, Rosa platyacantha, Cotoneaster melanocarpa, Lonicera humilis, L. Altmannii, L. heterophylla, Artemisia rutifolia, Thymus serpyllum s. 1. и др.). Травянистые растения чаще всего встречались следующие: Phleum phleoides, Роа Litwinowiana, Festuca sulcata s. 1., Carex turkestanica, Eremurus altaicus, Phlomis oreophila, Pedicularis pycnantha, Rheum Wittrockii, Eremostachys speciosa, Galatella punctata s. 1. Другие растения встречались реже. Всего названные авторы приводят список из 62 видов.
На вершине Большой Чимган (в горах Чимган) П. А. Баранов (1924) на скалах и осыпях обнаружил низкорослую арчу, подушки Onobrychis echidna, несколько кустарничковых видов Oxytropis, Astragalus Olgae, а также следующие травянистые и полукустарничковые растения: Erigeron andryaloides, Pyrethrum pyrethroides, Crepis multicaulis, Allium oreophylum, Minuartia lineata, Arenaria Griffithii, Macrotomia ugamen-sis, Parry a albida, P. asperrima, Aulacospermum turkestanicum, Valeriana caespitosa, Echinops tschimganicus, Rubia tibetica, Astragalus Baranovii, Oxyria digyna, Rhodiola Semenovii.
В Центральной части Гиссарского хребта (высота 3200 м, а особенно выше 3400—3500 м над у. м.) ландшафт скал и щебнисто-каменистых склонов имеет преобладающее распространение. А. С. Королева (1940) указывает, что здесь очень типичны Polygonum hissaricum, Cousinia Fran-chetii Dracocephalum oblongifolium, Pyrethrum pyrethroides, встречаются немногие злаки {Calamagrostis laguroides, Poa bucharica), а также следующие растения: Arenaria Griffithii, Psychrogeton cabulicus, Parrya steno-carpa, Draba Olgae, Geranium saxatile, Aulacospermum simplex.
В западной части Гиссарского хребта Ю. С. Григорьев и др. (Григорьев, Королева и Никитин, 1936) в пределах альпийского пояса (выше 3000 м над у. м.) указывают на широкое распространение по скалистым и щебнистым склонам южных экспозиций сообществ Astragalus lejosemius, Polygonum hissaricum и Cousinia acicularis. Сообщества эти сильно изрежены, однако на участках с большим количеством мелкозема достигают 50—70% проективного покрытия, обычно же гораздо меньше. В первом сообществе в качестве примеси встречаются Oryzopsis molinioides, Euphorbia seraw-schanica, Alopecurus himalaicus и др. В сообществе с господством Cousinia acicularis примешиваются Adonis turkestanicus, Astragalus lejosemius, Oxytropis immersa, Scorzonera acanthoclada, Arenaria Griffithii, Psychrogeton cabulicus, Lagotis Korolkowii, Trigonella Popovii и др.
В субальпийском поясе названные авторы отмечают преобладающее распространение разнотравно-камолевых {Ferula Jaeschkeana -f-разнотравье) сообществ, имеющих ничтожное покрытие (часто лишь 20%, иногда больше) и приуроченных к сухим щебнистым и каменистым склонам южной и воо
точной экспозиций. В составе этих сообществ обычно встречаются Cousinia commix ta, Euphorbia serawschanica, Oryzopsismolinioid.es, A donis lurkestanicus, Hypericum scabrum, Senecio soongoricus. Реже могут быть найдены Роа bulbosa var. vivipara, Polygonum bucharicum, Astragalus lejosemius, Artemisia persica, Cicer songoricum, Ziziphora Bungeana, а на особо каменистых субстратах—RheumFedtschenkoi, Hyssopus seravschanicus, Angelica ternata. В нижней части субальпийского пояса встречаются единичные особи арчи и жимолости (Lonicera serawschanica), шиповник (Rosa kokanica) и некоторые другие кустарники.
Растительность скал, осыпей и т. п. в Алае была впервые описана С. И. Коржинским (1896). В нижней части альпийского пояса он указывает наличие на скалистых склонах ущелий стелющейся арчи (Juniperus pseudosabina), ряда низкорослых кустарников: Cerasus pseudopro-strata, Ephedra equisetina, Cotoneaster racemiflora, Spiraea trilobata, Rosa spinosissima, Lonicera Olgae, Caragana jubata, а также подушечных растений — Dryadanthe tetrandra и Oxytropis humifusa.
На высотах более 3500 м над у. м. С. И. Коржинским были собраны Saxifraga hirculus, Calamagrostis anthoxanthoides, Potentilla sericea, Eutrema Edwardsii, Smelowskia calycina, Chorispora macropoda, Lloydia serotina, Leontopodium leontopodinum, Aneurolepidium alaicum и Hedysarum Semeno-vii. Два последних растения характерны для осыпей, играя важную роль в задержании осыпающегося щебня; особенно выделяется в этом отношении Aneurolepidium alaicum.
На меньших высотах, на скалах и тому подобных местах встречаются Dracocephalum Paulsenii, D. heterophyUum, Allium oreophilum, Rheum Fedtschenkoi, Ixiolirion tataricum, Parrya stenocarpa. Встречались здесь и некоторые другие из видов, упомянутых ранее.
На Памире, по опубликованным данным О. Э. Кнорринг (1923) и К. В. Станюковича (1949), по гребням хребтов и их склонам близ снеговой линии на скалах и осыпях встречаются разрозненные пятна — фрагменты лугов (из Carex melanantha, С. oxyleuca, Cobresia capilliformis) и отдельные подушки Androsace akbaitalensis, Oxytropis melanotricha, Dryadanthe tetrandra, Potentilla pamirica. Нередко подушки этих растений служат приютом для других видов, обитающих также и непосредственно на щебнисто-скальных субстратах: Aster alpinus, Melandrium apetalum, Leontopodium ochro-leucum, Poa Litwinowiana, Rhodiola pamiroalaica, Lloydia serotina и др. Наряду с ними очень широко распространены криоксерофильные полукустарнички и травы: Tanacetum xylorrhizum, Dracocephalum heterophyllum, Potentilla Moorcroftii, Erigeron andryaloides, Pyrethrum pyrethroides, Ephedra Fedtschenkoi.
На моренных галечниках, а также на песках единичными особями разбросаны Christolea pamirica, Braya pamirica, Parrya excapa, Poa Litwinowiana.
На осыпях альпийского пояса очень типичны специально приспособившиеся к подвижному субстрату ползучие и засыпаемые щебнем растения: Waldheimia tridactylites, Christolea flabellata, Didymophysa Fedczenkoana, Saussurea sorocephala.
На моренах О. Э. Кнорринг (1923) отмечала присутствие Artemisia rupestris, A. Skorniakowii, Acantholimon diapensioides, Potentilla Moorcroftii.
Характерна приуроченность к гипсовым солончакам (например на перевале Акбайтал) следующих, единично встречающихся растений: Calamagrostis anthoxanthoides, Pyrethrum pyrethroides, Lagotis decumbens, Braya rosea var. albiflora, Hymenolobus procumbens, Acantholimon diapensioides.
В субальпийском поясе на скалах и осыпях возрастает общее количество растений и намечается тенденция к формированию более или менее выраженных сочетаний. Здесь уже .встречаются фрагменты терескенников (из Eurotia ceratoides) и полынников (из Artemisia rutifolia), наряду с которыми растут одиночные особи или небольшие группы дерновинных злаков (Stipa glareosa, S. orientalis), а также криоксерофильные кустарнички (Comarum Salesovianum, Dasiphora fruticosa s. 1. и др.) и травянистые многолетники (Dracocephalum heterophyllum, Arnebia guttata и др.).
В Шугнане, в отличие от Восточного Памира, растительность скал, каменистых склонов и т. п. отличается особенно богатым флористическим составом. Неудивительно, что Б. А. Федченко (1909) в своих исследованиях уделил особенное внимание именно этим местообитаниям. На перевалах Койтезек, Вранг, Ямг, Кара-горум и др. Б. А. Федченко чаще всего обнаруживал следующие виды (перечисляемые в алфавитном порядке): Arenaria Griff ithii, Calamagrostis anthoxanthoides, Cerastium cerastoides, Corydalis Gortschakovii, Erigeron Poncinsii, Draba alpina, Hedysarum cephalotes, Primula nivalis, Pyrethrum pyrethroides, Ranunculus alpigenus, Rosularia alpestris, Saxifraga hirsulis, Smelowskia calycina, Linde-lofia stylosa, а также подушечные растения (Acantholimon diapensioides, A. alatavicum, A. parviflorum, Astragalus lasiosemius, Oxytropis immersa) я низкорослые полукустарнички и кустарнички (Artemisia rupestris, А. persica, Dasiphora fruticosa s. 1., Lonicera Semenovii).
Реже, лишь в некоторых из помещенных Б. А. Федченко мест, были встречены Allium monadelphum, A. polyphyllum, Aster flaccidus, Astragalus alatavicus, A. schugnanicus, A. Scheremetevianus, Cicer songoricum, Delphinium Brunonianum, Dracocephalum Paulsenii, Erithrichium pami-ricum, Braya oxycarpa, Geranium collinum, Heracleum Olgae, Paraquilegia grandiflora, P. anemonoides, Lloydia serotina, Macrotomia euchroma, Mala-curus lanatus, Nepeta podostachys, Oxytropis platonychia, Papaver croceum, Poa Litwinowiana, Polygonum molliiforme, Pyrethrum Stoliczkai, Ranunculus rufocepalus, Rhodiola gelida, Senecio bucharicus, Swertia marginata, Winklera silaifolia, Ziziphora Bungeana.
На осыпях обычно встречаются Waldheimia tridactylites и Saxifraga Komarovii; на моренах — Saussurea pamirica я на цесках, отложенных ледниковыми речками, — Paracaryum hymalayense.
По ледниковым ручьям и речкам изредка встречаются группы Myri-caria alopecuroides, Salix oxycarpa (я другие виды этого рода), Hippo-phae rhamnoides, Betula schugnanica, Rosa nanothamnus.
В заключение необходимо отметить, что в пределах контуров 104 са распространены также отдельными участками или фрагментами высокогорные альпийские и субальпийские луга (57), высокогорная подушковидная растительность (101), горные пустыни (88) и горные степи (84).
Литература
А. Н. Краснов (1888); С. И. Коржинский (1896); Б. А. Федченко (1903, 1909, 1912а); О. Э. Кнорринг и 3. А. Минквиц (1912а); О. Э. Кнорринг (1923); П. А. Баранов (1924, 1925); И. А. Райкова (1924, 1930); Р. И. Аболин и М. М. Советкина (1930); Ю. С. Григорьев, А. С. Королева и В. А. Никитин (1936); А. С. Королева (1940); М. Г. Попов (1940); Н. И. Рубцов (1948а); В. П. Голоскоков (1949); К. В. Станюкович (1949); М. А. Глазовская (1950); И. Н. Антипов-Каратаев (1951).
109. ОАЗИСЫ В ПУСТЫННОЙ ЗОНЕ (ЕСТЕСТВЕННЫЙ РАСТИТЕЛЬНЫЙ ПОКРОВ БОЛЕЕ ИЛИ МЕНЕЕ ЦЕЛИКОМ ЗАМЕНЕН КУЛЬТУРНЫМ)1
Е. П. КОРОВИН2
Растительность оазисов, то есть районов современного орошаемого земледелия, отличается большим богатством, и внешний облик их резко контрастирует с окружающими пустынными ландшафтами. В оазисах орошаемые поля с множеством различных сельскохозяйственных культур (хлопчатник, пшеница, сорго, рис, сахарная свекла, кукуруза, ячмень, подсолнечник, клещевина, зерновые бобовые, обширный ассортимент бахчевых и овощных культур и др.) сочетаются с плодовыми садами и виноградниками, насчитывающими тоже огромное количество видов и особенно сортов, а также с парками и древесными посадками вдоль каналов й арыков, дорог и на приусадебных участках.
Существование и территориальное распределение в Средней Азии оазисов сопряжено в первую очередь с наиболее крупными реками: Аму-Дарьей, Сыр-Дарьей, Зеравшаном, Тедженом, Мургабом, Чирчиком, Сурхан-Дарьей, Кашка-Дарьей, Вахшем, Арысыо и другими, а также многими мелкими реками, сбегающими с горных хребтов Средней Азии от Джунгарского Алатау на северо-востоке и до Копет-Дага на юго-западе. Исключением являются небольшие оазисы в подгорных районах, бедных поверхностным стоком, — существование их поддерживается грунтовыми водами, выводимыми наружу с помощью так называемых кяризов. Подобного рода оазисы находятся кое-где вдоль Копет-Дага и некоторых других горных хребтов. Лишь отдельные малые оазисы питаются водами напорными и ключевыми, как, например, Тамдынский оазис в центральных Кызылкумах (по малым своим размерам на карте не показан).
Зависимость существования оазисов от водоносных рек определяется тремя главным факторами: водным балансом реки, условиями стока и ирригацией. С развитием гидротехники роль ирригации в жизни оазисов приобретает первостепенное значение. Доказательством этого является громадный прирост поливных земель в Средней Азии, происшедший в советское время благодаря плановому развитию ирригации.
Задачей ирригации является выведение воды из речного русла каналами и распределение ее по оросителям на орошаемой территории. Достигается это самотеком или, в случаях низкого уровня речного потока, с помощью машинного подъема воды. Устройство водоемов, плотин и дру
1 На нашей карте оазисы показаны только в пределах среднеазиатских республик, так как в момент составления карты отсутствовали необходимые материалы по другим районам.
* Очерк, написанный Е. П. Коровиным, с согласия автора несколько сокращен применительно к данному изданию Л. Е. Родиным.
гих гидротехнических сооружений имеет целью управление режимом речного стока и регулирование распределения поливной воды на территории оазиса. Ирригационная сеть каналов забирает из русла реки нередко значительную часть воды, направляя ее на земли, вышедшие из сферы влияния вод речной долины.
Растекание воды по каналам и оросителям, особенно в больших оазисах, вносит весьма существенные изменения в природные условия охваченной ими местности. Действующую в оазисах ирригационную сеть следует рассматривать в качестве своеобразного составного элемента бассейна данной реки (Шульц, 1949).1 Значение ее в режиме таких многоводных рек, как Аму-Дарья и Сыр-Дарья, может быть, мало ощутимо, но в гидрологии малых рек оно сказывается в сильной степени, например в гидрологии рек Ферганской котловины. Ирригационный расход воды бывает настолько велик, что многие реки, питающие оазисы, уже не доходят до естественного места впадения.
Однако на орошаемых территориях изменяется весь комплекс природных условий.
Ирригационная сеть во много раз увеличивает поверхность испарения, а в связи с этим оказывает глубокое влияние на естественный водно-тепловой баланс атмосферы, особенно в ее приземном слое. Изменяются в оазисах все элементы климата. Орошение, особенно на больших площадях, действует умеряющим образом на климат: становятся ниже температуры, увеличивается влажность воздуха. Сравнительное изучение теплового обмена в оазисах и пустынях показывает, что наиболее заметные различия наступают в летние месяцы, достигая по средней месячной температуре 3.1°, а по среднему минимуму — 1.4°. Весной и осенью тепловые различия сглаживаются, и разница выражается в десятых долях градуса. В связи с этим в оазисах сокращаются периоды с температурой, например выше 10 и 15°, на шесть-семь дней, период же с температурой выше 25° — на 26 дней. Вместе с тем в оазисах удлиняется на несколько дней безморозный период. При всем этом сумма эффективных температур, например вычисленная по хлопчатнику, в оазисах меньше, чем в окружающей пустынной местности. Для оазиса в низовьях Аму-Дарьи она на 500° меньше, чем рядом в пустыне.
Влажность воздуха в крупных оазисах превышает, по крайней мере в полтора раза, влажность в пустыне. Однако эта разница возникает за счет главным образом летнего времени, когда относительная влажность увеличивается на 20% по сравнению с пустыней. Весной и осенью относительная влажность воздуха в пустыне и оазисах значительно выравнивается. Следует также отметить, что влажность воздуха заметно возрастает в оазисах лишь по утрам и вечерам, в середине же дня она остается довольно низкой. Все это сказывается на продолжительности периода воздушной засухи, который с 200 дней в пустыне уменьшается в оазисах до 150 (Погосян, 1952).
Орошение, как видно, смягчает сезонные контрасты климата, столь характерные для пустынных районов Средней Азии, подчеркивая тем самым их своеобразие. Еще резче проявляется действие этого фактора в приземном слое воздуха и в почве. К физическому обмену влажности и теплоты здесь прибавляется влияние транспирации мощного покрова хлопчатника и люцерны, занимающих в оазисах Средней Азии массивы в сотни тысяч гектаров. Установлено еще ранее, что среди растений на поле в дневные часы температура на 8—12° ниже, а влажность на 40—60%
1 В. Л. Шульц. Реки Средней Азии. М., 1949.
выше, чем над обнаженной почвой (А. А. Скворцов и Ю. А. Скворцов, 1927). Большие различия зарегистрированы в нагревании поверхности почвы под хлопчатником и без него. В середине июля они достигают почти 100%; так, например, при 64° на свободной поверхности почвы, температура в поле не повышается более 34е. По-разному прогревается почва. В указанном случае температура в 34' была прослежена на обнаженной площадке до глубины 20 см. Наибольшие температурные контрасты бывают, естественно, во время максимального развития хлопчатника, когда растения полнее затеняют поверхность почвы. Сильное влияние на температурные различия на поле, занятом посевом или свободном от него, оказывает испарение растений (Бабушкин, 1953).
Большое значение для характеристики микроклимата хлопкового поля имеют более точные данные о тепловом балансе. В наиболее жарком месяце (июле) тепловой баланс на хлопковом поле на 0.2 кал. см8/мпн. превышает баланс прилежащей пустыни. Объясняется эта разница различной степенью отдачи тепла, излученном поверхностью хлопкового поля и пустыни. Вследствие пониженной температуры поверхности хлопкового поля излучение тепла снижается, а поэтому возрастает и радиационный баланс. Таким образом, хлопковое поле в июльские дни имеет запас тепла значительно больший, чем в пустыне.1
Все изложенное свидетельствует о том, что в оазисах под влиянием полива и агрикультуры формируется свой климат, заслуживающий особого названия — агроклнмата.
Вместе с тем в оазисах остаются сезонные контрасты климата, характерные для окружающей пустыни, правда в несколько смягченном виде. Однако, благодаря искусственному обводнению, они, естественно, обусловливаются только тепловым режимом. В этом отношении в структуре климата оазисов Средней Азии выделяются три хорошо выраженных сезона: микро-, мезо- и макротермические сезоны. В пустыне высокая солнечная радиация летом, сочетаясь с наступающим дефицитом влаги, ограничивает вегетацию растений, а то н исключает ее возможность; в оазисах же она вызывает лишь воздушную засуху в дневные часы. Последняя не влияет на продолжительность вегетации большинства растений, но временно, в часы дневного максимума температуры, вызывает нарушения в водном хозяйстве растений и некоторые отрицательные явления (например опадение бутонов и завязей у хлопчатника, торможение роста побегов у деревьев я др.).
Ирригация вносит глубокие изменения в режим верхних толщ почвогрунта. Если разветвленная сеть каналов является своеобразным элементом речной сети, то она вызывает в почвогрунтах орошаемой местности процессы, характерные для природного режима речной долины.
Многие оазисы и орошаемые земельные массивы расположены в Средней Азии на первых и вторых террасах современной речной системы. Эти террасы в процессе развития речных бассейнов вышли за пределы влияния текучих речных вод и до орошения находились в большей своей части в субаридных условиях. Лишь отдельные освоенные ирригацией территории еще не оторвались в полной мере от непосредственного влияния. речной долины. Это почти целиком Нижнеамударьинский оазис — периодические разливы Аму-Дарьи в высокие паводки покрывают обширные пространства, вода заполняет старые озерные понижения, вследствие
1 В. А. Айзенштат Т. В. Кириллова. Сравнительная характеристика составляющих радиационного баланса полупустыни и хлопкового поля. ДАН УзССР, 1954, 2.
чего восстанавливается характерный для поймы гидроморфный режим. То же самое происходит п в приамударьпнской культурной полосе.
Иные явления наблюдаются в оазисах более высокого по отношению к современным речным уровням положения. Так, Голодностепскнй оазис формируется на землях межгорной долины, потерявшей связь с сырдарьнн-скнм стоком к концу четвертичного периода. В естественных почвогрунтах неосвоенных частей Голодной степи не улавливаются черты ее гидроморфного прошлого, они подвергались позднее процессу пустынного почвообразования. Действительно, почвы на этой равнине представлены пустынными, хорошо промытыми и свободными от солей сероземами, а растительность — покровом из пустынной осоки (Carex pachystylis) и мятлика (Роа bulbosa var. vivipara). Таково же природное состояние земель Ташкентского оазиса, развернувшегося на следующей по уровню террасе, которая в геологическом прошлом вступила ранее Голодной степи в фазу континентального режима. Погребенные в толщах лёсса, образующего эту террасу, прослойки перегнойных почв свидетельствуют об ее далеком лугово-аллювиальном прошлом. На уцелевших целинных участках Ташкентского оазиса можно встретить растительность и почвы, соответствующие эфемерондной формации с типичными сероземами.
Изменения в почвогрунтах под влиянием ирригации имеют один характер, одно направление — в сторону восстановления гидроморфных процессов, столь характерных в развитии речной поймы.
Орошение повышает зеркало грунтовых вод. Прп избыточном поливе последние выходят наружу, восстанавливая тем самым в почвогрунте типичный режим поймы; почвы заболачиваются и требуют особой мелиорации. Важно отметить,что с подъемом уровня грунтовых вод устанавливается физическая связь их с поверхностью почвы и с приземным слоем воздуха. Благодаря испарению и капиллярности в почвогрунте возникают восходящие токи воды, которые транспортируют к поверхности растворенные в ней вещества. Таким путем происходит миграция солей снизу вверх и их накопление в поверхностных горизонтах почвы. Засоление почв — это вторая тенденция в изменениях почвогрунта под влиянием ирригации. То и другое направление — заболачивание и засоление — свойственно не всем оазисам, о чем будет сказано дальше.
В оазисах Средней Азии наблюдается также луговой процесс почвообразования как вторичное явление, связанное с орошением земель.
Указанные изменения в характерном для них виде проявляются лишь в условиях неорганизованного водного хозяйства. Хозяйство, поставленное научно, предусматривает установление определенных поливных норм, предупреждающих заболачивание и засоление почв. Оно направлено к поддержанию почвы на уровне, удовлетворяющем требования культурных растений.
В оазисах Средней Азии в течение многих веков интенсивной агрикультуры сформировались особые почвы. Они не находят места в классификации естественных почвенных типов и выделяются в особый тип оазисно-культурных или культурно-поливных почв, которые образованы мощным слоем агроирригационных наносов. В древних оазисах, например в Бухарском, мощность культурного слоя достигает трех метров, а в бывшем Мешед-Мессерианском оазисе (юго-западная Туркмения) обнаружены культурно-поливные почвы мощностью до четырех метров.
Для оазисных почв характерны следующие особенности: более равномерное распределение по горизонтам гумуса и карбонатов, а также и азота, бблыпее количество валового фосфора. Плодородие оазисно-культурных почв заметно убывает с глубиной, но не столь резко, как в целинных
почвах: даже на глубине двух метров оно выше, чем в подпахотном горизонте целины. Можно говорить об остаточном плодородии оазисных почв, отмечая этим прогрессивное возрастание элементов пищи растений по мере образования агроирригационных наносов.
Общий облик оазисов определяется культурной растительностью, то есть сельскохозяйственными культурами и лесопарковыми насаждениями. Первые имеют доминирующее значение в экономике оазисов, а вторые придают оазисам характерный и своеобразный внешний вид, резко контрастирующий с окружающими и почти весь год сухими равнинами и холмами.
Древесно-кустарниковая растительность, представленная в оазисах рощами и полезащитными полосами, а в городах и селениях — парками и садами, вносит в природу оазиса лесной элемент.
Избыток воды, обилие тепла, длинный вегетационный период, относительно теплые зимы — все это благоприятствует росту и развитию древесных и кустарниковых пород и акклиматизации разнообразной дендрофлоры. Собственных, автохтонных древесных пород в оазисах Средней Азии насчитываются единицы, и они являются выходцами из тугаев речных долин. Наиболее характерными представителями местной оазисной дендрофлоры являются карагачи, встречающиеся в нескольких архитектурно выраженных формах (Ulmus Androssowii, U. densa). Карагачи не так давно были обычными деревьями в любом селении, но в последние годы их стало меньше, в ряде же мест они совершенно исчезли. Широко распространены в оазисах ива (Salix Daviesii, S. acmophylla), шелковица (Morus alba, M. nigra), пирамидальный тополь (Populus pyramidalis), туранга (Populus pruinosa, P. diversifolia), джида (Elaeagnus angustifolia), ясень (Fraxinus excelsior), облепиха (Hippophae rhamnoides). Реже в городах и селениях встречаются местные же породы, но вышедшие из гор Средней Азии: грецкий орех (Juglans regia s. 1.), чинар (Platanus orientalis), арча (виды Juniperus), боярка (виды Crataegus) и некоторые другие.
Вообще же лесопарковые насаждения характеризуются значительным разнообразием, и этим они обязаны иноземной, интродуцированной в разное время флоре. Последняя сосредоточена главным образом в городах, меньше видов встречается в селениях. Интродукция древесно-кустарниковой флоры в Средней Азии имеет давнюю историю, равную, повидимому, истории культуры ее народов. Время появления в оазисах ряда иноземных видов деревьев установить не удается. Основная же масса растущих в республиках Средней Азии иноземных деревьев и кустарников введена в советское время и, отчасти, в конце прошлого столетия. Однако к настоящему времени еще только немногие представители иноземной флоры успели широко распространиться по городам и селам Средней Азии. В быт населения республики вошли белая акация (Robinia pseudoacacia), некоторые тополи (Populus BoUeana, Р. nigra, Р. deltoides, Р. balsamifera и др.), гледичия (Gleditschia triacanthos), айлант (Ailanthus altissima), ильмы (Ulmus scabra, U. pumila), некоторые клены (Acer campestre, A. velutinum, A. pseudoplatanus) и др.
Основной же ассортимент экзотов сосредоточен в парках и садах городов среднеазиатских республик, особенно в Узбекистане. Здесь растут вполне акклиматизировавшиеся выходцы из влажных субтропиков: болотный кипарис (Taxodium distichum), магнолия (Magnolia obovata и др.), тюльпанное дерево (Liriodendrontulipiferum), мыльное дерево (Koel-reuteria paniculate,), японская софора (Sophora japonica), пекан (С ary а pecan), эвкоммия (Eucommia ulmoides), альбицция (Albizzia Julibrissin), катальпа (Catalpa Bungei), керрия (Kerria japonica), парротия (Parrotia
persica), дзельква (Zelcova carpinifolia) и многие другие. В садах и парках оазисов Средней Азии зарегистрировано около 900 видов древесных и кустарниковых пород, включая представителей отечественной флоры (Шипчинский, 1953). Среди них преобладают вполне прижившиеся к оазисам растения. Разнообразие дендрофлоры оазисов свидетельствует о больших возможностях обогащения растительных ресурсов в Средней Азии.
Создаваемые поливной культурой условия способны удовлетворить требования самой разнообразной интродуцируемой флоры. Это подтверждается многолетним опытом ботанического сада в Ташкенте, в котором собрано около 1700 видов деревьев и кустарников (Русанов, 1953). Следует обратить внимание на то, что Ташкентский оазис расположен в теплой, но не жаркой зоне Средней Азии.
На долю хвойных в дендрофлоре оазисов Средней Азии приходится 49 видов: мамонтово дерево, болотный кипарис, ель (7 видов), сосна (13 видов), кедр (3 вида), туя (2 вида), можжевельники и др. Среди лиственных наиболее разнообразно представлены тополь, ива, береза, ильм, клен (22 вида), ясень, жасмин, барбарис, боярышник, виноград. Опыты по культуре пальмы в южных широтах Средней Азии пока не дали положительных результатов; испытанные пальмы из родов хамеропс, вашингтония, трахикарпус, а также финиковая пальма, не выдерживают низких температур в особо суровые зимы и в такие годы обмерзают, а некоторые погибают совершенно. Только на крайнем юге, в Каракале и Кизыл-Атреке, названные пальмы более или менее прижились.
Изучение интродуцированной флоры Средней Азии с точки зрения ее происхождения показывает, что наиболее успешно поддаются акклиматизации деревья и кустарники восточной и юго-восточной Азии и Северной Америки. Представители тропической флоры приживаются в виде исключения, например некоторые виды стеркулии, мелии, кассии, цезальпинии. Вечнозеленые формы, как правило, вымерзают.
Перспективы интродукции вообще и дендрофлоры, в частности, в оазисы Средней Азии чрезвычайно велики, и к ней предъявляются высокие требования со стороны народного хозяйства. Необходимо иметь в виду, что облесение оазисов, окруженных знойными пустынями, имеет громадное, еще не вполне понятое и оцененное значение. Помимо ландшафтноархитектурной роли, древесные насаждения в оазисах несколько умеряют температурные контрасты воздуха. Кроме того, они влияют на влажность воздуха и вообще играют большую роль в мелиорации земель оазисов. Нельзя представить себе оазис в Средней Азии, лишенный древесных насаждений, они необходимы для него.
Лесопарковая растительность в оазисах перемежается с полевыми и садовыми культурами. Состав тех и других в Средней Азии исключительно разнообразен. Первое место среди сельскохозяйственных растений по экономическому значению и размерам занимаемой площади принадлежит хлопчатнику. Оазисы Средней Азии, в частности расположенные на территории Узбекской ССР, являются основной базой производства хлопкового сырца в СССР и одной из важнейших баз мирового хлопководства. Хлопчатник представлен сортами советской селекции, принадлежащими к типам упланда («hirsutum») и египтянина («barbadense»).
Кроме хлопчатника, в оазисах Средней Азии возделываются в поливной культуре на волокно кенаф (Hibiscus cannabinus) и джут (Corchorus capsularis). Обе культуры представлены высокоурожайными и скороспелыми сортами. На стадии выхода из испытания находится юкка (У исса), прекрасно выживающая во всех оазисах и дающая большой урожай листьев и семян. Второстепенное значение имеет лен и еще меньшее —
конопля. Хорошо растет в поливных условиях китайское волокнистое растение — рами (Boehmeria nivea), не вошедшее еще в производство.
Прочие возделываемые в оазисах сельскохозяйственные растения распределяются в следующие группы: хлебные, зерновые бобовые, огородные, бахчевые, кормовые, плодовые и ягодные культуры.
Наибольшее значение из хлебных культур имеют кукуруза и рис, дающие громадные урожаи во всех оазисах Средней Азии. В Кара-Калпакии, кроме того, издавна разводится хлебное сорго, или джугара (Sorghum сегпиит). Помимо перечисленных культур, следует назвать пшеницу, ячмень, просо (все они имеют в поливном хозяйстве республики второстепенное значение) и гречиху (последняя еще не вошла в хозяйство, хотя имеет шансы занять в нем определенное место).
Широко культивируются в оазисах представители различных бобовых: нут (Cicer arietinum), маш (Phaseolus aureus) и вигна (Vigna sinensis). Менее характерны обыкновенная фасоль, чечевица, горохи, которые встречаются лишь на приусадебных участках.
Масличные представлены древней культурой кунжута (Sesamum indicum) и относительно новой культурой клещевины (Bicinus communis). Кунжут распространен преимущественно в южных районах Средней Азии. Хорошие урожаи дает в поливных условиях подсолнух.
Огородные и бахчевые культуры обладают исключительным разнообразием видов и сортов. Из огородных культур на первом месте стоит картофель. Эта культура введена в Средней Азии сравнительно недавно. К относительно молодым огородным культурам также относятся помидоры, овощной перец, баклажаны, кабачки, сахарная свекла; последняя разводится преимущественно в оазисах Киргизской ССР. На пороге к внедрению находятся культуры крахмалистых корнеплодов: ямса (Dioscorea batatas) и батата (Ipomaea batatas). Более широким признанием пользуется топинамбур (Helianthus tuberosus), дающий высокие урожаи. Выше не были упомянуты обычные овощные, возделываемые в Средней Азии: огурцы, морковь, редис, редька, капуста, петрушка, лук, укроп и другие — все они в поливных условиях высоко продуктивны.
Бахчевые культуры заслуживают особого внимания, тем более, что оазисы Средней Азии являются одним из важнейших центров происхождения бахчевых. Большое экономическое значение имеют дыни, арбузы, тыквы. По данным К. И. Пангало (1928),1 в Средней Авии находится родина нескольких видов дыни; сортовой состав их исключительно разнообразен. Богата по составу и тыква. Население Средней Азии с давних пор занимается разведением этой культуры, употребляя плоды в пищу и на корм скоту. Менее разнообразен состав арбузов, насчитывающих немало эндемичных форм. Дыни, тыквы, арбузы — это излюбленные культуры у народов Средней Азии, поэтому они распространены широко и повсеместно во всех малых и больших оазисах.
Кормовые растения имеют также большое значение. Первое место занимает люцерна, разводимая в оазисах в качестве сменной культуры в хлопковом севообороте; местами ее заменяет клевер, рекомендуемый для каир-ных (долинных) земель, а в люцерновой травосмеси — райграс, сборная ежа, красная овсяница и некоторые другие злаки..
Плодовым культурам принадлежит большой удельный вес в растениеводческих ресурсах оазисов Средней Азии. Разводимые в садах промышленного значения плодовые деревья участвуют вместе с тем в составе зеленых насаждений в парках городов и селений. В общем аспекте оази
1 К. И. Пангало. Дывн. Л., 1928.
сов роль плодовых нс меньше, нежели лесных и декоративных пород. Среди них имеются все основные группы: косточковые/ орехоплодные п ягодные. Имеющие промышленное значение культуры представлены в большинстве формами, вышедшими из народной селекции далекого прошлого Средней Азии (Попов, 1929). Мало того, такие культуры, как абрикос, миндаль, грецкий орех, фисташка, гранат, инжир, некоторые яблони и груши, имеют центр своего происхождения в Средней Азии, где они растут в диком состоянии. Ряд сортов привлечен из иных очагов древнего земледелия: Китая, стран Средиземноморья и др. В последнее время успешно интродуцируются сорта яблони, груши, вишни и сливы европейской селекции.
Наибольшее разнообразие плодовых сосредоточено в южных и средних широтах Средней Азии. Для представления о богатстве плодовых в оазисах Средней Азии достаточно указать, что только в Ферганской котловине зарегистрировано около 685 сортов, из них абрикоса — 320, миндаля —185. Второй пример: в Кара-Калпакском и Хорезмском оазисах описано 97 сортов абрикосов, более 20 сортов груш и 20 сортов яблонь. Сортовые фонды плодовых в одном только Узбекистане превышают 3300 сортов.
Семечковые представлены яблоней, грушей, айвой, гранатом, инжиром; косточковые — абрикосом, персиком, миндалем, сливой, алычой, черешней, вишней, кизилом; ягодные — малиной, смородиной, крыжовником и виноградом. Среднеазиатские абрикосы по своей сахаристости, а миндали по содержанию жира — превосходят все известные сорта.
Виноградарство принадлежит к числу древних автохтонных отраслей сельского хозяйства в Средней Азии. Сортовые фонды винограда чрезвычайно своеобразны и связаны с древнейшей культурой страны.
Из орехоплодных в оазисах, кроме грецкого ореха, обладающего множеством различных форм, прекрасно растут лещина, фисташка и пекан. Промышленное значение имеет только грецкий орех.
Подводя итоги растениеводческим ресурсам оазисов Средней Азии, мы еще раз убеждаемся в громадном, далеко еще не исчерпанном потенциале возможностей, скрытых в самой природе оазисов — в климате и почвах. Глубокая самобытность состава сельскохозяйственных культур, их разнообразие и непревзойденные во многих отношениях хозяйственные качества свидетельствуют о древности земледельческой культуры в среднеазиатских оазисах. Однако нельзя не видеть в этом и отражения специфических условий оазисов, создаваемых природой и трудом человека.
В культурной флоре оазисов Средней Азии за время многовековой культуры собраны представители различных флор мира, связанные своим происхождением со многими древними центрами земледелия. В составе перечисленных выше растений хорошо различаются выходцы стран Средиземноморья, Индии, Китая, Центральной и Южной Америкии, отчасти Абиссинии. Важно не только разнообразие культурной флоры Средней Азии, но и большая биологическая неоднородность самих переселенцев.
С некоторым приближением к действительности можно представить это богатство числом около 7—7.5 тысяч различных форм. В распределении культурной флоры в пределах Средней Азии по оазисам проявляется определенный порядок, объясняемый отчасти природными особенностями отдельных оазисов.
При всей глубине влияния культуры на естественно-исторические условия, последние, однако, продолжают сказываться на оазисах, обусловливая те или иные различия в их режимах. В этих различиях проглядывают региональные особенности климата, геологии и почвообразо
нательных процессов в соответствии с географическим положением оазиса. Очень хорошо различаются следующие группы равнинных оазисов: южная (Атрекский, Тедженский, Мургабский, Вахшский, Ширабадский, Кзылаякский), центральная (Бухарский, Голодностепский, Чарджуй-ский) и северная (Хорезмский, Каракалпакский).
Вместе с тем оазисы каждой из названных групп имеют свои характерные геолого-почвенные особенности, сильно влияющие на всю организацию земледелия: на подбор сортов сельскохозяйственных культур, на мелиорацию, на агротехнику и т. д.
Развитие земледелия, в частности хлопководства, во многом зависит от зональных особенностей климата и, прежде всего, от теплового режима. В земледелии южных оазисов доминирующую роль играет культура тонковолокнистого хлопчатника типа «barbadense», в центральных оазисах рентабельна культура среднеспелых сортов типа «hirsutum», в северных оазисах успешно разводятся скороспелые сорта того же типа. В связи с изменением теплового режима находится состав других сельскохозяйственных культур и приемы их возделывания. Понятно, что климатические различия еще далеко не исчерпывают индивидуальную характеристику оазисов выделенных групп.
Особое место занимают подгорные оазисы (Ташкентский, Самаркандский, Ферганский, Сурхандарьинский, Джамбульский, Алмаатинский и др.). Повышенное положение относительно аллювиальных равнин объясняет многое в режиме этих оазисов: по-иному складываются условия стока, как поверхностного, так и подземного, исключающие застой грунтовых вод; по-иному протекает водно-солевой обмен в почвогрунтах; иначе формируется агроклимат оазиса.
Не менее важно для естественно-исторической характеристики подгорных оазисов наличие толщ лёсса, на которых они расположены. Лёсс обладает, как известно, хорошими фильтрационными свойствами; кроме того, он не содержит легкорастворимых солей. В силу указанных причин — благоприятных условий стока и опресненного грунта—почвы подгорных оазисов не испытывают, как правило, ни вторичного заболачивания, ни вторичного засоления, столь характерных для равнинных оазисов.
В подгорных оазисах, помимо того, относительно более умеренные климатические условия. Например, в Ташкентском и Самаркандском оазисах выпадает осадков в 1%—2 раза больше, чем в Бухарском или Голодностепском оазисах; зато они уступают последним по сумме эффективных температур.
Все это значительно расширяет возможности подбора, акклиматизации и культуры растений в оазисах описываемого типа.
Большинство оазисов (исключая Самаркандский, Ташкентский и ряд менее крупных подгорных оазисов) расположено на древних аллювиальных равнинах. Последние немногим превышают уровень моря, а в отдельных случаях, например в низовьях Аму-Дарьи, находятся почти на уровне Аральского моря. История формирования аллювиальных равнин в Средней Азии характеризуется двумя процессами: прогрессивным соленакоп-лением, по мере отложения речного аллювия, и последующей аккумуляцией солей в нижних горизонтах аллювия под влиянием их вымывания. Неумеренные поливы в оазисах этого равнинного типа вызывают вторичную подвижность солей — к естественному горизонтальному перемещению их в грунте прибавляется вертикальная миграция, обусловливающая засоление почв. Земледелию в равнинных оазисах приходится постоянно сталкиваться с засолением на участках полей, где до .сего времени не было его признаков. Указанное обстоятельство накладывает глу<-
бокий отпечаток навесь природный режим оазиса. Естественное почвообразование в таких условиях находится все время на рубеже, за которым начинается солончаковый ряд развития (Ковда, Егоров и др., 1954).
Почвы равнинной дельты характерны тем, что в своем развитии они проходят обязательно луговую стадию почвообразования. Однако эта стадия чрезвычайно кратковременна и не оставляет заметного следа. Такой вывод, относящийся к низовьям Аму-Дарьи, может быть на основании геоботанических данных распространен вообще на развитие почв речных долин Средней Азии. Систематически увлажняемые почвы оазисов как будто имеют все условия для проявления лугового почвообразования. В действительности здесь, как и в естественных почвах речной долины, луговая фаза нигде не проявляется в ее типичном виде. Поэтому в равнинных оазисах, как правило, отсутствуют луговые почвы.
Высказанными выше соображениями объясняется характер спонтанной растительности, возникающей в оазисах на залежах, по обочинам каналов, дорог, и т. д. Встречающиеся в этих условиях группировки растений обладают большим разнообразием, маскирующим естественные отношения между ними. Состоят они, в основном, из элементов тугайной местной и адвентивной флоры. Рассматривая эти группировки по составу и во взаимодействии с почвами, можно их объединить в типы: болотный, солончаковый и тугайный.
Болотная растительность в оазисах сосредоточена в местах с близкими или поверхностными грунтовыми водами. Это бывает в понижениях, заполняемых сбросными водами, вдоль каналов и коллекторов. Растительность в этих случаях образована густыми зарослями тростника {Phragmites communis). В виде примеси наблюдаются рогозы {Typha Laxmannii, Т. minima, Т. elephantina),cin:BJiKa{Juncus lampocarpus, J. Ge-rardii), камыши {Scirpus Tabernaemontani, S. litoralis и др.) и некоторые другие; в южных оазисах можно встретить гигантский злак арундо {Arundo donax).
На засоленных болотах удерживается дольше всех растений тростник, обладающий значительной солеустойчивостью. По мере концентрации солей заросли его становятся разреженными, растения перестают цвести, подавляется их рост и возобновление, и болото превращается в мокрый солончак. Заросли тростника на мокрых сильно засоленных почвах заменяются группировками солероса {Salicornia europaea). Солерос отмечает одну из фаз развития солончаковой растительности в оазисах, представленной на перелогах и залежах весьма разнообразно, в соответствии с характером и степенью засоления почв. Наиболее обычны на молодых перелогах группировки однолетних солянок (из цикла Salsola crassa), шведок {Suaeda arcuata, S. paradoxa), кохии {Kochia scoparia), летроси-монии {Petrosimonia sibirica), лебеды {Atriplex heterosperma, A. tatarica и др.), кермека {Limonium otolepis) и др.
На старых перелогах, подверженных вторичному засолению, образуется густой солянковый покров, поддерживающий и даже усиливающий поверхностное засоление почв. Одной из фаз изменения солончаковой растительности в этих условиях является образование группировок из ажрека {Aeluropus litoralis). Этот корневищный злак плотно задер-новывает почву, он развивает мощную систему корневищ и корней и формирует густой травостой. Ажрек выделяет на поверхности листьев поваренную соль, вследствие чего задернение им почв не способствует рассолению, хотя и улучшает их структуру.
Весьма характерны для залежей и перелогов в равнинных оазисах заросли янтака (виды Alhagi; рис. 83), карелинии {Karelinia caspica)
и парнолистника {Zygophyllum fabago, Z. oxianum). Первые два растения, благодаря корнеотпрысковому размножению, быстро покрывают взрыхленную после обработки почву. Каждое из jinx доминирует в покрове в зависимости от степени засоленности почвы и положения грунтовых вод. Между ними могут быть пятна пальчатника {Cynodon dactylon), солодки {Glycyrrhiza glabra), лебеды (виды Atriplex), донника {Melilotus officinalis), астры {Tripolium vulgare), однолетних злаков и других трав, свидетельствующих о поверхностном засолении почвы. В сходных условиях на залежах появляются крупный дерновинный злак эриантус {Erianthus ригри-rascens), кустики гребенщика (виды Tamarix), чингиля {Halimodendron halodendron) и даже деревца туранги {Populus diversifolia, Р. pruinosa). Это уже показатели изменений почвогрунтового режима, имеющих явно тугайное направление.
В растительности оазисов играет заметную роль адвентивная флора. Элементы ее дольше сохраняются на перелогах, по обочинам дорог, вдоль каналов, откуда они переходят на поля. На хлопковых полях находят благоприятные условия для размножения многие сорняки. К ним относятся сыть {Cyperus rotundas), гумай {Sorghum halepense), вьюнок {Convolvulus arvensis), додартия {Dodartia orientalis), пальчатник {Cynodon dactylon), а из однолетников: паслен {Solanum nigrum и близкие виды), лебеда (виды Atriplex), гибискус {Hibiscus trionum), абутилон {Abutilon Theophrasti), портулак {Portulaca oleracea), щирица {Amaranthus retrofle-xus), щетинник {Setaria viridis, S. glauca) и др. На поля люцерны, джута, кенафа переходят виды повилик {Cuscuta).
На рисовые поля попадают через оросительную сеть водолюбивые сорняки: крупноплодное и куриное просо {Echinochloa macrocarpa, Е. crus-galli), морской клубнекамыш {Bolboschoenus maritimus), рогоз {Typha Laxmannii) и ряд других болотных растений.
Состав сорной растительности изменяется в зависимости от состояния земледелия в оазисе и от его природных условий.
В пределах контуров оазисов, показанных на карте, имеются залежи, а также земли, временно вышедшие из-под орошения по причине засоления или несовершенства оросительной сети. Точно так же в пределах отдельных оазисов весьма различен коэффициент земельного пользования, имеющий свои физико-географические и агротехнические причины. В силу этого в пределах оазисов встречаются участки тугайных лесов, тростниковых зарослей, а также мелкие массивы песков или участков с пустынной растительностью. Эти сообщества претерпели значительное изменение не только в результате непосредственной хозяйственной деятельности человека, но и под влиянием преобразованной им среды. Последняя настолько видоизменена, что в пределах оазисов существует свой температурный режим, особый режим влажности, определенный ритм многих растений и т. п., что позволяет говорить об оазисах как о совершенно особом ландшафте.
Литература
А. Ф. Миддендорф (1882); О. Э. Кнорринг и 3. А. Минквиц (1913); О. Э. Кнорринг (1915); А. А. Скворцов и Ю. А. Скворцов (1927); Н. И. Вавилов (1929); М. Г. Попов (1929, 1940); Ю. А. Скворцов (1930); Е. А. Малюгин (1931); Е. П. Коровин (1934, 1934а); Ф. Н. Ру-санов (1934, 1947, 1949, 1953); В. П. Дробов (1935); А. В. Прозоровский (1940а); Л. Н. Бабушкин и Е. П. Коровин (1947); Н. В. Шипчинский (1950, 1953); Л. Н. Бабушкин (1952, 1953); X. П. Погосян (ред.) (1952); В. А. Ковда, В. В. Егоров и др. (1954).
ЛИТЕРАТУРНЫЕ ИСТОЧНИКИ ПОЯСНИТЕЛЬНОГО ТЕКСТА К «ГЕОБОТАНИЧЕСКОЙ КАРТЕ СССР» м. 1 : 4 000 000 1
Аболин Р. И. 1910. Некоторые данные о лесных и других растительных формациях Жигулевских гор Симбирской губ. Лесн. ж., 40, 3. — 1912. Тайга между рр. Нерчей и Куэнгой в Забайкальской обл. В кн.: Предварит, отчет об организ. и исполн. работ по иссл. почв Аз. России в 1911 г., СПб. — 1913. В тайге Лено-Вилюйской равнины. В кн.: Предварит, отчет об организ. и исполн. j)a6oT по иссл. почв Аз. России в 1912 г., СПб. — 1914. Опыт эпигенологическои классификации болот. Болотоведение, 3—4.— 1915. Болотные формы Pinus silvestris L. Тр. Бот. муз. АН, 15. — 1917. Лено-Вилюйская равнина Якутской обл. (Опыт эпигенологической монографии). Тр. Почв.-бот. экспед. по иссл. колониз. районов Аз. России в 1912 г., ч. 1, 2. [Тираж целиком утрачен. Текст этой работы в несколько измененной редакции см.: Аболин, 19296]. — 1927. Вертикальные почвенные пояса в Центральном Тянь-Шане. Бюлл. почвовед., 1—2. — 1928. К вопросу о классификации болот Северо-Западной обл. Мат. по опытн. мелио-рат. делу, 2. — 1929. Южная часть Алма-Атинского окр. Казанской АССР в естественно-историческом отношении. Тр. Инет, почвовед, и геобот. САГУ, Казахстанская сер., 1. — 1929а. Восточная часть Сыр-Дарьинского окр. Казанской АССР в естественно-историческом отношении. Тр. Инет, почвовед, и геобот. САГУ, Казахстанская сер., 2. — 19296. Геоботаническое и почвенное описание Лено-Вилюйской равнины. Тр. Комиссии по изуч. Якутск. АССР, 10. — 1929в. Основы естественно-исторического районирования Советской Средней Азии. Тр. САГУ, сер. ХП-а, геогр., 2. — 1930. От пустынных степей Прибалхашья до снежных вершин Хантенгри. Тр. Инет, почвовед, и геобот. САГУ, Казахстанская сер., 5. — 1932. Краткая характеристика основных типов естественных угодий горного Дагестана. Махачкала. — 1933. Естественные кормовые угодья Киргизии и очередные задачи их исследования. В кн.: Проблемы растениеводческого освоения пустынь, 7, Л.
Аболин Р. И. и М. М. Советкина. 1930. Горные пастбища Та лас-Сус амырского района в Киргизской АССР. Мат. Комиссии экспед. иссл. АН СССР, 27.
Абрамов И. И. 1954. Рододендроновые верещатники высокогорий Юго-Осетии. Уч. зап. Ленингр. унив., сер. геогр. наук, 9.
Абрамова А. И. 1951. Природные ресурсы Томской обл. и пути их освоения. Тр. Томск, унив., 114.
Авдеев В. Д. 1945. История леса и степи в западном Закамье. Изв. ВГО, 67,1—2. — 1948. Возникновение степей в Закамье. Казань.
Аверкиев Д. С. 1929. Растительность Заветлужья Красно-Ваковского у. Производительные силы Нижегородск. губ., 13.
Аврамчик М. Н. 1937. Геоботаническая и пастбищная характеристика района р. Ду-дыпты. Тр. Арктич. инет., 63.
Аврорин Н. А. 1933. Геоботаническая карта Хибинского района. В кн.: Хибинские апатиты, 6, Л. — 1933а. Материалы о типах лесов горного Алтая. Тр. Инет, по изуч. леса АН СССР, 7. — 1934. Растительность разновозрастных залежей Каменной степи. (К вопросу о восстановлении степной растительности из-под пашни). Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 7.
1 Список составлен В. В. Липатовой при участии авторов текста. Включены все работы ботанико-географического содержания, упомянутые в тексте. Таким образом, библиография заканчивается первой половиной 1954 г. включительно и является наиболее полной из всех имеющихся, хотя и далеко не исчерпывает литературу вопроса, достигающую, вероятно, не менее 10 000 названий. Литература, опубликованная в 1954—1955 гг., учтена только отчасти.
Библиографические ссылки проверены по книгам (de visu) в библиотеке Ботанического института им. В. Л. Комарова АН СССР и частично в других книгохранилищах Ленинграда.
Аврорин Н. А., М. X. Качурин и А. А. Коровкин. 1936. Материалы по растительности Хибинских гор. Тр. СОПС АН СССР, Кольская сер., 11.
Агаханянц О. Е. 1953. Полусаванновая растительность Северного Дарваза. Изв. АН ТаджССР, отд. естеств. наук, 5.
Аггеенко В. Н. 1890. Флора Крыма, 1. Ботанико-географический очерк Таврического полуострова. Тр. СПб. оощ. естествоиспыт., 21, отд. бот. — 1897. Обзор растительности Крыма с топографической и флористической точки зрения. СПб. Агеева Н. Т. и А. 3. Соломченко. 1951. Растительность пастбищ урочища Сам. Уч. зап. Казахск. унив., 13, 1.
Адамов В. В., И. К. Ярошевич. 1927. Обзор растительности Белорусского Полесья. Мат. по изуч. растит. Белоруссии, 2.
Акинфпев И. Я. 189ч. О растительных и преимущественно лесных зонах в Центральном Кавказе. Русск. лесн. дело, 14, 15. — 1894а. Северный Кавказ. 1. Ботаническое исследование Ставропольской губ. 1889 г.; 2. Верховья Калауса и Ставропольское поднятие. Зап. Кавказ, отд. РГО, 16. — 1897. Альпийские растения Центрального Кавказа. Зап. Кавказ, отд. РГО, 19. — 1899. Северный Кавказ. 2. Ботаническое исследование Кубанско-Терского водораздела и Эльбруса. Тр. Тифл. бот. сада, 3.
Алабышев В. В. 1926. Очерк растительности поймы правого берега р. Волхова от д. Слутки до р. Пчевжи. Мат. по иссл. р. Волхова и его бассейна, 9.
Алгазин В. С. 1939. Растительность Приобья. Новосибирск.
Александрова В. Д. 1937. Тундры правобережья р. Попигай. Тр. Арктич. инст.,0<3.— 1945. Краткий очерк растительности Новой Земли по линии пересечения залив Рогачев—бухта Савина. Проблемы Арктики, 5—6. — 1946. Растительный покров острова Междушарского. Проблемы Арктики, 5. — 1950. О содержании понятия «полярная пустыня» как тип растительности. (Природа, 9. — 1953. О границах растительных подзон в низовьях р. Яны. Изв. ВГО, 85, 1. — 1954. Краткий очерк растительности вдоль Дона от Воронежа до Ростова-на-Дону. Тр.’ Бот. инет. АН СССР, сер. 3, геоботаника, 9.
Александрова В. Д. и А. И. Зубков. 1937. Физико-географический очерк Новой Земли. В кн.: Новоземельская экскурсия, 1, Л.
Алексеев А. Д. (О л е к с i 1 в). 19оо. Степов! щлини Грипшпського району на Артем!в-щиш. Ж. Б!о-бот. циклу ВУАН, 5—6. — 1947. Пойменные луга Харьковской обл. Зап. Харьковск. с.-х. пнет., 6. — 1951. Достижения в изучении пойменных лугов УССР. Зап. Харьковск. с.-х. инет., 7.
Алексеев Е. В. (ОлексНв). 1926. Типи nicy по мокрих грунтах. Зап. Кшвськ. с.-г. !нст., 1. — 1928. Типы украинского леса. Правобережье. Киев.
Алексеев Я. Я. 1935. Луга Западной обл. В кн.: Я. Я. Алексеев, А. В. Жуковский, Г. Л. Граве, Растительность Западной обл., Смоленск.
Алексеенко М. И. 1950. О растительности засоленных почв Приазовья. Тр. Н.-иссл. инет. биол. Харьковск. унив., 13. — 1950а. Пастбища и сенокоЗы овцеводческих совхозов югаУсСР. Тр. Н.-иссл. инет. биол. Харьковск. унив., 13.—1954. К во-просу о динамике растительности песков Нижнего Днепра в связи с проблемой их закрепления и освоения. В кн.: Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР, М.—Л.
Алехин В. В. 1909. Очерк растительности и ее последовательной смены на участке «Стрелецкая степь» под Курском. Тр. СПб. общ. естествоиспыт., 40, 1. — 1910. Казацкая степь Курского у. в связи с окружающей растительностью. Тр. СПб. общ. естествоиспыт., 41, 3. — 1915. Типы русских степей. Изв. Бот. сада Петра Великого, 15, 3—4. — 1916. Растительность лугов р. Цны и нижнего течения р.Мокши. Тамбов.—-1921. Луга как зональное явление. В кн.: Дневн. Первого Все-росс, съезда русских ботаников в Петрограде в 1921 г., 3, Пг. — 1921а. Основные черты в распределении растительности Европейской России. М. — 1922. Растительность лугов р. Вороны. Ж. Моск. отд. Русск. бот. общ., 1. — 1924. Зональная и экстрозональная растительность Курской губ. в связи с разделением губернии на естественные районы. Почвоведение, 1—2. — 1925. Растительный покров степей Центрально-Черноземной обл. Воронеж. — 1925а. Флора и растительный покров Московского края. В кн.: Московский край, М. — 1926. Современное состояние вопроса о классификации русских степей. В кн.: Дневн. Всесоюзн. съезда ботаников в Москве в январе 1926 г., М. — 1927. Луга и методы их исследования. Бюлл. МОИП, отд. биол., 36, 1—2. — 1929. По лесам от верхнего Приветлужья до р. Волги. Производительные силы Нижегородск. губ., 13. — 1929а [редактор]. Предварительный отчет о работах Нижегородской геоботанической экспедиции в 1928 г. Производительные силы Нижегородск. губ., 13. — 1930. Карта растительности Азиатской части СССР. (Краткий объяснительный текст). В кн.: Атлас промышленности СССР, 5, Л. — 1930а. Карта растительности Европейской части СССР. (Краткий объяснительный текст). В кн.: Атлас промышленности СССР, 5, Л. — 1931. Русские степи и методы
исследования их растительного покрова. Бюлл. МОИП, отд. биол., 40, 3—4. — 1934. Геоботанические карты Московской обл. (Краткий объяснительный текст к картам). В кн.: Доклады сов. делегации на Междунар. Геогр. конгр. в Варшаве, М. — 1934а. Геоботанические карты б. Нижегородской губ. Вкн.: Доклады сов. делегации на Междунар. Геогр. конгр. в Варшаве, М. — 19346. Центральночерноземные степи. Воронеж. — 1935. Объяснительная записка к геоботаническим картам (современной и восстановленной) б. Нижегородской губ (в м. 1 : 500 000). Л.— 1936. Растительность СССР в ее основных зонах. В кн.: Г. Вальтер — В. Алехин, Основы ботанической географии, М.—Л. — 1946. История русской фитоценологии и ее особенности. Уч. зап. Моск, унив., 103, 2, кн. 1. — 1947. Растительность и геоботанические районы Московской и сопредельных областей. М. — 1951. Растительность СССР в основных зонах. Изд. 2-е. М.
Алехин В. В. и Д. С. Аверкиев. 1927. Растительность степного Заалатырья. Производительные силы Нижегородск. губ., 6. — 1928. Смены растительности от Запо-чинья до р. Волги. Производительные силы Нижегородск. губ., 9.
Алехин В. В. и А. А. Ура нов. 1933. Методика исследовании степей. Сов. бот., 2.
Альбицкая М. А. 1953. Очерк растительности Мелитопольского и Акимовского районов Запорожской обл. Материалы исследования искусственных лесов в районе р. Молочной и Молочного лимана. Научн. зап. Днепропетр. унив., 38.
Альбов Н. М. 1892. Леса Абхазии. Зап. Общ. сельск. хоз. Южн. России, 4. — 1894. Ботанико-географические исследования в западном Закавказье в 1893 г. (Наблюдения над флорой юрских известняков). Зап. Кавказ, отд. РГО, 16. — 1896.Ботанико-географические исследования в западном Закавказье в 1894 г. Зап. Кавказ. отд. РГО, 18.— 1896а. Очерк растительности Колхиды. Землеведение, 3, 1. Амелин И. С. 1953. К характеристику лугов Львовской обл. Об. научн. тр. Львовск. вет.-зоот. инет., 6.
Андреев В. Н. 1931. Растительность тундры северного Канина. (Геоботаническое исследование оленьих пастбищ). В кн.: В. Н. Андреев, А. А. Дедов и Ф. В. Самбук, Оленьи пастбища Северного края, Архангельск. — 1932. Подзоны тундры Северного края. Природа, 10. 1932а. Типы тундр запада Большой Земли. Тр.
Бот. муз. АН СССР, 25. — 1933. Кормовые ресурсы оленеводства в западной части Болыпеземельской тундры. В кн.: Оленьи пастбища Северного края, 2, Л. — 1934. . Кормовая база Ямальского оленеводства. Сов. оленеводство, 1. — 1935. О растительном покрове юго-восточного Канина. Тр. Полярн. комиссии АН СССР, 20. — 1935а. Растительность и природные районы восточной части Болыпеземельской тундры. Тр. Полярн. комиссии АН СССР, 22. — 19356. Лесная растительность Южного Тимана. (Геоботаническая характеристика района рр. Ухты и Ижмы в Печорском крае). Тр. Полярн. комиссии АН СССР, 24. —1952. Применение аэрометодов для геоботанического картирования и инвентаризации кормовых площадей. Бот. ж., 37, 6. — 1954. Продвижение древесной растительности в тундру в связи с защитными свойствами лесопосадок на севере. Бот. ж., 39, 1. — 1954а. Растительный покров восточноевропейской тундры и мероприятия по его использованию и преобразованию. Л.
Андреев В. Н., К. Н. Игошина и А. И. Лесков. 1935. Оленьи пастбища и растительный покров Полярного Приуралья. Сов. оленеводство, 5.
Андреев В. Н. и А. Л. Панфиловский. 1938. Обследование тундровых пастбищ с помощью самолета. Тр. Инет, полярн. землед., животн. и промысл, хоз., сер. оленеводство, 1.
Антипин Н. А. и В. М. Пчелкин. 1929. Заливные луга по р. Нерли. Тр. Иваново-Возн. научн. общ. краевед., 6.
Антипов-Каратаев И. И. 1951. Выветривание и почвообразование на Восточном Памире. Тр. АН ТаджССР, 1, (Инет, почвовед, мелиор. и ирриг.^.
Антонов А. А. 1890—1892. О растительных формациях Закаспиискои обл. Бот. зап. Бот. сада СПб. унив., 3, прилож.
Ануфриев Г. И. 1915. Сенокосные угодья юго-восточной части Новоржевского у. Мат. по организ. и культуре корм, площади, 11. — 1922. О болотах Кольского п-ова. Работы организованного Геогр. инст-ом в 1920 г. Кольск. почв.-бот. отр. Сев. н.-промысл, экспед., 3. — 1925. Краткий очерк растительности поймы оз. Ильменя и нижнего течения рек Ильменского бассейна. Мат. по иссл. р. Волхова и его бассейна, 4. — 1928. Стационарные геоботанические наблюдения в пойме р. Волхова. Мат. по иссл. р. Волхова и его бассейна, 22.
Арманд Д. Л. 1950. Опыт математического анализа связи между типами растительности и климатом. Изв. ВГО, 82, 1.
Архипов С. С. 1932. Заболачивание и типы лесов Котласского леспромхоза Северного края. М.
Арцымович в. С. 1911. Мокрые солонцы окрестностей Баскунчакского озера. Опыт ойкологического исследования растительности мокрых солонцов. Тр. Общ. испыт. прир. при Харьковск. унив., 44.
Афанасьев Д. Я. (Афанасьев) 1941. Заплавн! луки р. Десни. КиГв. — 1948 (1947). Наслщю i перспективи геоботашчних досл!джень на У крапп. Бот. ж. УРСР, 3—4. — 1948а (1947). Короткий нарис грунтчв та рослинност! заплавини р. Дьоми. Бот. ж. УРСР, 4, 3—4. — 1950. Заплавш луки середнього Дшпра та заходи lx полшшення. КиГв. — 1950а. Основы типи лук заплави середнього лкостепового Дншра. Бот. ж. АН УРСР, 7, 3. — 1951. Геоботашчний нарис Нижньодншровських плавшв. Бот. ж. АН УРСР, 8, 2. — 1954. Природы кормов! упддя Украшсько! РСР, ix полшшення та рашональне використання. Бот. ж. АН УРСР, Р, 1. F
Афанасьев К. С. 1940. Очерк растительности Гармского и смежных частей Ромит-ского, Комсомолабадского и Тавильдаринского районов Таджикистана. Тр. Тадж. базы АН СССР, 8.
Бабушкин Л. Н. 1952. О температуре и влажности воздуха среди хлопчатника и люцерны в условиях орошаемого хозяйства Узбекистана. Тр. Ташкент, геофиз. обсерв., 7 (8). — 1953. Метеорологические факторы и растение. Тр. САГУ, нов. сер., 35 (биол. науки, 13).
Бабушкин Л. Н. и Е. П. Коровин. 1947. К вопросу районирования хлопковой зоны в Средней Азии. Изв. АН УзССР, 1.
Бажанов С. С. 1926. Поемные луга в Самарской губ. Пзв. Самарск. с.-х. инет., 3. Базилевич Н. И., М. М. Голлербах, М. А. Литвинов, Л. Е. Родин и Д. М. Штейнберг.
1953. О роли биологических факторов в образовании такыров на трассе Главного Туркменского канала. Бот. ж., 38, 1.
Базилевич Н. И. и Л. Е. Родин. 1955. К вопросу о генезисе и эволюции такыров и принципах их мелиорации. Тр. Арало-Каспийск. комплексн. экспедиции АН СССР, 4.
Базилевич Н. И., Л. Е. Родин, Е. И. Рачковская, Р. А. Калашникова и Н. Е. Бека-ревич. 1953. Изменение почв такыров под влиянием растений. Почвоведение, И.
Базилевская Н. А. 1927. Очерки растительности юго-восточных Каракумов. Изв. Главн. бот. сада, 26, 2. — 1929. Растительность юго-восточных Каракумов: грядовые пески и холмистые предгорья Бадхыза. Изв. Главн. бот. сада, 28, 5-6.
Баландин В. Н. 1936. Кормовые угодья Азово-Черноморья и их использование. Ростов-на-Дону.
Баландин Ф. В. 1929. Сенокосы Ленинградской обл. и Карелии. Изв. по с.-х. опытн. делу Ленингр. обл., 10. — 1930. Кормовой вопрос в молочных районах Ленинградской обл. и пути его разрешения. М.—Л.
Балаш А. П. 1948. Смена аспектов и фенологический спектр Аксайской степи (под г. Ростовом-на-Дону). Уч. зап. фак. естествозн. Ростовск.-на-Дону пед. и учит, инет., 1.
Бар А. И. 1919. Растительность дельты р. Дона. Мат. по изуч. растит, и лугов долины р. Дона в пределах Донского края, 1.
Баранов В. И. 1915. Нижнее течение р. Свияги и р. Волга близ г. Свияжска. Мат. по исслед. лугов Казанской губ., 4. — 1923. Очерк растительности Калачинского у. Омской губ. Тр. Сиб. с.-х. акад., 2. — 1925. Южная граница черноземных степей в Кустанайской губ. (по данным исследования 1923 г.). Тр. Общ. изуч. Казахстана, 6. — 1926. Вертикальная зональность растительности в горах Юго-Восточного Алтая. В кн.: Дневн. Всесоюзн. Съезда ботаников в Москве в январе 1926 г., М. — 1927. Растительность черноземной полосы Западной Сибири. Зап. Зап.-Сиб. отд. ГГО, 39, 3. — 1928. Почвы и растительность Демьян-Иртышского водораздела. Иссл. колониз. фондов, 1, сер. Б, Новосибирск. — 1931. Высокогорная тундра в юго-восточном Алтае. В кн.: 25 лет научно-пед. и общ. деятельности Б. А. Келлера, Воронеж. — 1948. Кулигаш. Тр. Казан, фил. АН СССР, сер. биол. и с.-х. наук, 1.
Баранов В. И. и К. П. Горшенин. 1927. К познанию солонцовых комплексов черноземной полосы Западной Сибири. Тр. Сиб. инет, сельск. хоз. и лесовод., 7, 1.
Баранов В. И. и П. П. Поляков. 1936. Геоботанические исследования в Восточном Алтае. Уч. зап. Казанск. унив., 96, 6.
Баранов В. И. и М. Н. Смирнов. 1931. Пихтовая тайга на предгорьях Алтая. Уч. зап. Пермск. унив., 1.
Баранов В. И. и В. А. Шелудякова. 1926. К изучению степей юго-восточного Алтая. Тр. Сиб. с.-х. акад., 5. — 1928. Материалы к познанию лугов и болот долины р. Оби. Тр. Сиб. инет, сельск. хоз. и лесовод., 10, 5.
Баранов П. А. 1924. Очерк растительности Чимгана. Изв. Туркестан, отд. РГО, 17. — 1925. К познанию растительности горных каменистых осыпей. Бюлл. САГУ, 9. — 1936. Биологическая наука на службе освоения высокогорного Памира. Природа, 2.
Баранов П. А. и И. А. Райкова. 1928. Дарваз и его культурная растительность. Изв.
Общ. для изуч. Таджикистана и иранских народностей за его пределами, 1.
Баранова М. Е. (Баранава) 1950. Натуральный сенакосы i пасб!шчы БССР, ix выкарыстанне i палепшанне. Мшск.
Барышников М. К. 1929. Осоково-гипновые болота Западного Васюганья. Бюлл. Инет. луг. и болотн. культуры, 2. — 1933. Луга Оби и Иртыша Тобольского Севера. М.
Бачурина Г. Ф. и Е. М. Брадис (Бачур1на та Брад!с). 1954. Торфов! болота Украшенного Полшся та шляхи ix використання в сельскому господарств!. Ки1в.
Бейдеман И. Н. 1951. Смены растительного покрова по берегам и дну залива им. Кирова в связи с отступанием Каспийского моря. Бот. ж., 36, 1. — 1953. Эколого-биологические основы смен растительного покрова (на примере низменности восточного Закавказья). Бот. ж., 38, 4. — 1954. Развитие растительности и почв в низменности восточного Закавказья. В кн.: Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР, М.—Л.
Бейлин И. Г. 1915. Пойма р. Свияги в среднем ее течении. Мат. по обслед. лугов Казанской губ., 5.
Бекетов А. Н. 1874—1877. Примечания в кн.: А. Гризебах, Растительность Земнога шара согласно климатическому ее распределению, 1 (1874), 2 (1877), СПб. — 1884» Фитогеографический очерк Европейской России. В кн.: Э. Реклю. Земля и люди. Всеобщая география. 5, 2 (дополнения), СПб. — 1896. География растений. СПб.
Бельгард А. Л. 1947. Основные закономерности растительного покрова юго-востока УССР. В кн.: Растительный и животный мир юго-востока УССР, 1, 1, Днепропетровск. — 1950. Лесная растительность юго-востока УССР. Киев.
Бельский П. С. 1950. Природа и типы леса горной тайги Саяно-Сибирской естественно-исторической провинции. Охрана природы, 11.
Берг Л. С. 1913. Опыт разделения Сибири и Туркестана на ландшафтные и морфологические области. В кн.: Сб. в честь 70-летия Д. Н. Анучина, М. — 1928. Зона тундр. Опыт ландшафтной характеристики. Изв. Ленингр. унив., 1. — 1938. Природа СССР. Изд. 2-е. М. — 1947. Географические зоны Советского Союза. Изд. 3-е. М. — 1952. Географические зоны Советского Союза, 2. М.
Березин Э. Л. 1952. Естественное возобновление ясеня влаголюбивого в Чарьшской лесной даче. Алма-Ата.
Билык Г. И. (Biлик). 1938. Рослиншсть засолених грунт!в л!вобережжя середнього Дшця. Геобот. зб. АН УРСР, 2. — 1940. Еколопчш ряди галофшьних фгго-ценозхв приморсько! смуги УРСР. Бот. ж. АН УРСР, 1, 3—4. — 1947 (1946). Геоботан!чний опис басейну р!ки Молочно! i Молочного лиману. Бот. ж. АН УРСР, 3, 1—2. — 1948 (1947). Заплавш луки р. Вовчо! (л!ва притока р. П!в-нхчного Дшця). Бот. ж. АН УРСР, 4, 3—4. — 1948а. Рослиншсть Дшстро-Бузького байрачного степу. Бот. ж. АН УРСР, 5, 2. — 1949. Заплавш луки р. Швшчного Дшця. Бот. ж. АН УРСР, 6, 4. — 1953. Природш кормов! уг!ддя та заходи по ix полшшенню. В кн.: Природа i господарство швденних райошв УРСР, Кшв. — 1954. Естественная кормовая база юга Украинской ССР и пути ее реконструкции. В кн.: Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР, М.—Л. — 1954а. Лучна рослиншсть При-тисенсько! низовини та г!рсько-л!сового пояса. В кн.: Рослиншсть Закарпатсь-ко1 обл., Кихв. — 19546. До класифшацп галоф!льно! рослинност! Украшьско! РСР. Бот. ж. АН УРСР, 11, 4.
Биркенгоф А. Л. 1932. Лесной покров и лесные ресурсы северо-восточного края Якутской АССР. В кн.: Якутская АССР, 3, Л. — 1932а. Краткий предварительный отчет о лесоисследовательских работах, произведенных Индигирским гидрологическим отрядом Якутской экспедиции АН СССР, 1929—1930 гг. Тр. СОПС АН СССР, Якутская сер., 6. — 1935. Заметка о распространении некоторых деревьев и кустарников в бассейне р. Бурей (ДВК). Сов. бот., 2. — 1936. Сведения о границах ареалов некоторых хвойных пород на Дальнем Востоке. Бюлл. МОИН, отд. биол., 45, 2. — 1938. Леса центральной части п-ва Камчатки. Тр. СОПС АН СССР, Камчатская сер., 6.
Битрих А. А. 1908. Очерки лесов Усть-Сысольского у. Леей, ж., 38, 4—5.
Благовещенский В. В. 1951. (напер. 1950). Лесная растительность Южноульяновского водораздела в связи с ее водоохранной ролью. Уч. зап. Ульяновск, пед. штст,к 3. — 1951а (на пер. 1950). О лесных ассоциациях с осокой волосистой в Ульяновском Заволжье. Уч. зап. Ульяновск, пед. инет., 3. — 1954. Ясень обыкновенный на восточной границе своего распространения. Бюлл. МОИП, отд. биол.^ 4. — 1955. Ассоциации сосновых лесов Ульяновского правобережья Волги-1 Сосновые леса зеленомошники. Уч. зап. Ульяновск, пед. тает-, 6.
Благовещенский Г. А. 1936. Эволюция растительного покрова болотного массива «1007 км» у ст. Лоухи (Карелия). Тр. Бот. инет. АН СССР, сер. 3, геоботаника. 3.
Благовещенский Э. Н.—-1940. Новые данные о внутрипочвенной конденсации. Метеорология и гидрология, 3.-—1941. Саксаульники низменных Кара-Кумов. Иав. ВГО, 73, 2. — 1949. Кустарниковые пустыни Азии. В кн.: Тр. Второго Всесоюзн. геогр. съезда, 3, М. — 1949а. Некоторые данные по экологии корневых систем деревьев и кустарников в песчаной пустыне. Изв. Туркм. фил. АН СССР, 2. — 1954. О задачах Каракумской станции в связи с лесомелиоративными мероприятиями в пустынях. В кн.: Пустыни СССР и их освоение, 2, М.—Л.
Блюменталь И. X. 1937. Геоботанический очерк западной оконечности Заилийского Ала-Тау и Чу-Илийских гор. Л.
Блюменталь И. X. и О. Л. Петровичева. 1951. Геоботанический очерк высокогорных мелкоосоковых пастбищ северо-западного Кавказа. Уч. зап. Ленингр. унив., сер. биол., 30.
Бобров Е. Г. 1926. К вопросу о поясах растительности Среднего Копет-Дага. Вкн.: Сб. Геогр.-эконом, иссл. инет., Л. — 1929. Очерк растительности юго-западного Приуралья. Изв. Главн. бот. сада, 28. — 1931. Растительность гор Большие Балханы. Тр. Бот. сада АН СССР, 44.
Боброва Л. И. и М. X. Качурнн. 1936. Очерк растительности Монче-тундры. Тр. СОПС АН СССР, Кольская сер., 11.
Богдан А. В. 1936. Кормовая площадь горной части Азово-Черноморского края. Тр. Краснодарск. с.-х. инет., 4.
Богдан В. С. и Б. А. Шумаков. 1925. Закубанские плавни в почвенно-ботаническом и мелиоративном отношениях. Тр. Кубанск. с.-х. инет., 3.
Богданов В. М. 1941. О растительном покрове горной области Северо-Осетинской республики. Тр. Горек, с.-х. инет., 4 (12). — 1947. Сенокосы и пастбища бассейна р. Ардона (в его среднем течении). Тр. Сев.-Осет. с.-х. инет., 7 (14).
Богданов М. Н. 1871. Птицы и звери черноземной полосы Поволжья и долины средней и нижней Волги. Тр. Общ. естествоисп. при Казанск. унив., 1. — 1875. Обзор экспедиций и естественно-исторических исследований в Арало-Каспийской обл. с 1720 по 1874 г. Тр. Арало-Касп. экспед., 1. — 1882. Очерки природы Хивинского оазиса и пустыни Кызыл-Кум. В кн.: Описания Хивинского похода 1873 г., 12, Ташкент.
Богданов М. П. 1954. Зимние пастбища Кобыстана и основные пути их рационального использования и улучшения. Тр. Инет. бот. АН АзССР, 18.
Богдановская-Гиенэф И. Д. 1926. Ключевые болота Кингисеппского у. Ленинградской губ. Ж. Русск. бот. общ., 11, 3—4.—1927. Луга долины р. Луги и ее притоков в Кингисеппском у. Ленинградской губ. Зап. Ленингр. с.-х. инет., 4. — 1928. Растительный покров верховых болот русской Прибалтики. Тр. Петер-гофск. естеств.-научн. инет., 5. — 1933. О распространении некоторых видов сфагнов на Полистовских болотах. Бот. ж. СССР, 18, 1—2. — 1936. Образование и развитие гряд и мочажин на болотах. Сов. бот., 6. — 1938. Природные условия и оленьи пастбища о. Колгуева. Тр. Н.-иссл. инет, полярн. землед. животн. и промысл, хоз., сер. оленеводство, 2. — 1946. О некоторых основных вопросах болотоведения. Бот. ж. СССР, 31, 2. — 1946а. О происхождении флоры бореальных болот Евразии. Мат. по ист. флоры и растит. СССР, 2. М.—Л.— 1949. Типы верховых болот СССР. В кн.: Тр. Второго Всесоюзн. геогр. съезда, 3, М.
Болотова В. М. 1953. Распространение и условия произрастания липы сердцевидной в Коми АССР. Тр. Коми фил. АН СССР, 1.
Болышев Н. Н. 1952. Происхождение и эволюция почв такыров. Почвоведение, 5. — 1955. Происхождение и эволюция почв такыров. М.
Болышев Н. Н. и Т. И. Евдокимова. 1944. О природе корочек такыров. Почвоведение, 7—8.
Болышев Н. Н. и Е. А. Манучарова. 1946. О растительности такыров. Вести. Моск.
Ь увив., 3—4. — 1947. Распределение водорослей в профиле некоторых почв пустынной зоны. Вести. Моск, унив., 8.
Борисова А. Г. 1935. От озера Зайсан к Нарымскому хребту. Изв. ВГО, 67, 4.
Борнеман Б. А. и М. Д. Спиридонов. 1929. Очерк почв и растительности полуостровов Мангышлак и Бузачи. Мат. Комиссии экспед. иссл. АН СССР, 12.
Боровиков Г. А. 1909. Очерк растительности юго-западной части Области Войску Донского. Зап. Новоросс. общ. естествоиспыт., 33. — 1912. Очерк растительности Восточного Заангарья. Тр. Почв.-бот. экспёд. Переселенч. упр. по иссл. колониз. районов Аз. России в 1909 г., ч. 2, 7. — 1913. Очерк растительности Западного Заангарья. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1910 г., ч. 2, б.
Боровский Г. Ф., Г. А. Зиновьев, В. Н. Минервин, Н. А. Мордвинов, И. А. Мосолов, Н. Т. Нечаева и Н. Н. Пельт. 1940. Кормовые растения равнинной Туркмении. Тр. Туркменок, респ. опытн. станции по животн., 1.
Борщов И. Г. 1865. Материалы для ботанической географии Арало-Каспийского края. Зап. Ими. Акад, наук, 7 (прилож. 1).
Брадис Е. М. (Брад! с). 1951. Пол^нини Закарпатсько! обл., !х використання та шляхи полшшення. Ки!в.
Брадис Е. М. и А. А. Запятова. (С. М. Брадш та О. О. Зап’ятова). 1950. Высокопрна рослишпсть Закарпатсько! обл. Бот. ж. АН УРСР, 7, 1. — 1954. Загальний характер високопрно! рослинност’1 i вивчення !! кормово! цшностГ В кн.: Рос-линнхсть Закарпатсько! обл. УРСР, Ки!в.
Бронзов А. Я. 1927. Типы лугов по р. Мологе. Тр. Луг. инет., 1. — 1929. Зональные явления в пойме р. Иртыша. Изв. Луг. инет., 4—6. —1930. Верховые болота. Нарым-ского края (бассейн р. Васюгана). Тр. Н.-иссл. торф, инет., <3. — 1936 Гипновые болота на южной окраине западносибирской равнинной тайги. Почвоведение, 2.
Бронзова Г. Я. 1926. Работа экспедиции по изучению лугов и болот Сибири. Изв. Луг. инет., 4. — 1929. Естественные луга в ландшафте таежной поймы р. Васю-гапа. Бюлл. Инет. луг. и болотн. культуры, 3. — 1929а. По Обским лугам. (Предварит, сообщ.). Изв. Луг. инет., 4—6. — 1932. Торфяные болота Сибирской части Урало-Кузнецкого комбината. Торф, дело, 5.
Брундза К. И. (К. Brundza). 1937(1936) Kamanos. Aufbau und Pflanzendecke. Jahrb. d. Landwirtsch. Akad. in Dotnuva, 3—4.
Бугаенко П. и И. Безъязычный. 1949. Полонины Закарпатья. Ужгород.
Буддо И. С. 1947. Луга Иркутской обл. и пути их улучшения. Иркутск. — 1953. Луга Приангарья. Л.
Будищев А. Ф. 1867. Ботаническое описание древесных ^ кустарных пород, произрастающих в Амурском, Уссурийском и Зауссурийском краях. Зап. Сиб. отд. РГО, 9, 10.
Булавкина А. А. 1917. Растительность Сучана и острова Путятин в Южно-Уссурийском крае. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1913 г., ч. 2, 2 (прилож. 1). — 1921. Астраханский край. Растительность. В кн.: Россия, 12, Пг.
Бутков А. Я. 1938. Растительность гор Ходжа-Гургур-Ата. Ташкент.
Буш Н. А. 1904. По горам и ущельям Хевсурии и Тушетии. Тр. СПб. бот. сада, 23, 3. — 1905. Ботаническое путешествие по западному Дагестану. Тр. СПб. бот. сада, 24, 3. — 1909. О ботанико-географических исследованиях Кубанской обл. в 1908 г. Изв. РГО, 46, 4—6. — 1913. О делении Сибири на ботанико-географические области. Изв. Акад. Наук, сер. 6, 7, 1. — 1915. К ботанической карте западной половины северного склона Кавказа. Изв. РГО, 51. — 1923. Ботанико-географический очерк России. Европейская Россия. Естеств. произвол. силы России, 5, Растительный мир, отд. 1, Пг. — 1923а. Ботанико-географический очерк России. Кавказ. Естеств. произвол, силы России, 5, Растительный мир, отд. 1-а, Пг. — 1930. Обзор работ по флоре и растительности Крыма, Кавказа и Закавказья с 1920 по 1929 г. Ж. Русск. бот. общ., 14, 3. — 1932. О болотах озерного происхождения в Балкарии и Дигории (Центральный Кавказ) Тр. Бот. муз. АН СССР, 25. — 1935. Ботанико-географический очерк Европейской части СССР. Изд. 4-е. М.—Л. — 1935а. Ботанико-географический очерк Кавказа. М.—Л. — 1937. Ботанические исследования Кавказа за 20 лет советской власти. Природа, 11. ,
Буш Н. А. и Е. А. Буш. 1936. Высокогорные луга Юго-Осетии. Изв. Юго-осет. н.-иссл. инет., 3. — 1936а. — Растительный покров восточной Юго-Осетии и его динамика. Тр. СОПС АН СССР, Закавказская сер., 18.
Бушинский В. П. 1924 (1922—1923). Природные условия роста чия (Lasiagrostis splendens Kuntb.) и возможность введения его в культуру. Тр. по прикл. бот. и селекц., 13, 2.
Быков Б. А. 1944. Реликтовый ясеневый лес поймы р. Чарын. Изв. Казахет. фил. АН СССР, сер. бот., 1. — 1950. Еловые леса Тянь-Шаня, их история, особенности и типология. Алма-Ата. — 1954. К вопросу о происхождении комплексности растительности в Прикаспии. 6 кн.: Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР, М.—Л. — 1955. Растительность и кормовые ресурсы Западного Казахстана. Алма-Ата.
Быков Б. А. и Е. Ф. Степанова. 1953. Кустарниковые степи как тип растительности. Изв. ВГО, 85, 1.
Бычкова Е. X. 1950. Флора и растительность юго-востока Европейской части СССР. Библиография Саратовской обл., 2. Саратов.
Бэр К. М. (С. Б. Baer) 1838. Expedition a Novai’a Zemlia et en Laponie. 3. V6g6-tation et climat de Novaia Zemlia. Bull. Sc. de Г Acad. des sc. ae St. P6tersb.. 3, 11—12.
Вавилов H. И. 1929. Возделываемые растения Хивинского оазиса. Тр. по прикл. бот., генет. и селекц., 20.
Вага А. Я. 1953. Фитоценозы болот Эстонской ССР. В кн.: Научная сессия по вопросам биологии и сельского хозяйства. Рига, 22—26 окт. 1951 г., М.
Вандакурова Е. В. 1950. Растительность Ку^ундинской степи. Новосибирск.
Вандышева В. И. 1948. Сырты Иссык-Кульской области. Тр. Киргизск. н.-иссл. инет, животн., 8.
Варварин Б. Г. 1950. Пастбища и сенокосы Тувинской авт. обл. Тр. Тувинск. с.-х. опытн. станц., 2.
Варен Э. К. (Е. V а г е р). 1953. Eesti USV laaneosa soode geobotaaniline iseloomust.us (Геоботаническое исследование болот западной части Эстонской ССР). В кн.: Eesti HSV fauna ja floora uurimise kiisimusi, Tallinn.
Варивцева E. A. 1951. Краткая характеристика пастбищ Восточного Памира. Тр. Тадж. фил. АН СССР, 28.
Варлыгин П. Д. 1936. Торфяные болота Кольского п-ова. Тр. Центр, торф, опытн. станции, 1.
Василевич П. П. 1954. Пойменные луга окрестностей г. Кирова и пути их улучшения. Киров.
Васильев В. Н. 1936. Оленьи пастбища Анадырского края. Тр. Арктич. инет., 62. — 1937. Растительный покров Малого Хингана. Тр. Дальневост. фил. АН СССР, сер. бот., 2. — 1941. Каменная береза (Betula Ermani Cham. s. 1.). (Экология и ценология). Бот. ж. СССР, 26, 2—3. — 1944. Растительность северной части вулканического кольца Тихого океана. Изв. ВГО, 76, 5. — 1946. Краткий очерк растительности Курильских островов. Природа, 6.
Васильев В. Ф. 1931. К характеристике можжевеловых лесов Крыма. Ж. Русск. бот. общ., 16, 4.
Васильев Я. Я. 1929. Естественно-исторический очерк лесов северной части Залаир-ского кантона Башкирской республики. Лесовед. и лесовод., 7. — 1933. Леса и лесовозобновление в районах Братска, Илимска и Усть-Кута. Тр. СОПС АН СССР, Сибирская сер., 2. — 1935. Объем понятия типа леса и схема классификации типов леса. Сов. бот., 1. — 1938. Леса из цельнолистной пихты. Природа, 9. — 1938а. Лесные ассоциации Супутинского заповедника. Тр. Горнотаежной станции Дальневост. фил. АН СССР, 2.
Васильченко И. Т. 1951. Дикорастущие виды яблони в Средней Азии. В кн.: Мат. Первого Всесоюзн. совещ. ботаников и селекционеров 24—27 марта 1950 г., 2, М.-Л.
Введенский Н. П. 1939. Растительность пастбищного массива горы Большой Бамбак и ее кормовое значение. Тр. Кавказ, заповеди., 2.
Вернандер Н. Б. 1947. Почвы закарпатской обл. УССР. Почвоведение, 6.
Вернандер Т. Б. 1929. Анализ растительности на степях б. Орловской губ. Изв. Сев.-Черноземн. (б. Шатиловской) обл. с.-х. опытн. станции, 3, 2. — 1946. Растительным покров Бештаугорского лесопарка. Уч. зап. Моск, унив., 97.
Верховский,Н. П. 1911. Саксаул — дерево-кустарник песчаных степей Средней Азии. Сельское хоз. и лесовод., 237, 10.
Веселовский В. П. и Н. С. Плотников. 1930. Материалы по изучению типов леса Майкопского окр. Тр. Сев.-Кавказ, ассоц. н.-иссл. инет., 64.
Викторов С. В. 1951. Растительность как индикатор литологических условий на Северном Усть-Урте и в западноказахстанских степях. Бюлл. МОЙП, отд. биол., 66, 1.
Виленский Д. Г. 1918. Из наблюдений над растительностью естественных кормовых угодий Новоузенского у. Самарской губ. Бюлл. Отд. прикл. бот. Саратовск. обл. опытн. станции, 1. —- 1926* Растительность заливных лугов Республики немцев Поволжья. В кн.: Сб. статей и мат. Госплана АССР немцев Поволжья, 1, Покрове к.
Виленский Д. Г. и Е. М. Лавренко (В i ленський i Лавренко). 1925. Про умовв, в яких расте на Харвдвщин], в околицях с. Змпвського Лиману. Ceratophyllum tanaiticum Sapjegin. BicTi Харювськ. с.-г. шст., 2—3.
Вильямс В. Р. 1915. Типы болот с точки зрения почвообразовательного процесса. Мат. по организ. и культуре корм, площади, 13. — 1922. Общее земледелие. II. Естественно-исторические основы луговодства или луговедение. М.
Виноградов Б. С. 1948. Влияние животных на почвы и растительность пустынь. В кн.: Животный мир СССР, 2, М.—Л.
Виноградов И. С. 1940. Система экологических рядов ассоциаций заливных лугов.
Виноградов Н. П. и С. В. Голицын. 1948. Послевоенное состояние наиболее интересных местонахождений реликтовых растений Верхнего Поосколья и Северо-Донского реликтового района. Тр. Воронежск. унив., 16. — 1954. Сниженные альпы и тимьянники Средне-Русской возвышенности. Бот. ж., 39, 3.
Вивогоадова А. Н. 1937. Геоботанический очерк оленьих пастбищ района р. II ясины. Тр. Арктич. инет., 63.
Виппер П. Б. 1953. Леса Юго-Западного Хентея. Тр. Монг, комиссии АН СССР, 54. — 1953а. Арчевники Средней Азии как лесной тип растительности. Бот. ж., 38, 3. — 1954. Горные леса Средней Азии. В кн.: Вопросы лесоведения и лесоводства. М.
Вл а сто на 11. В. 1919. Растительность долины р. Дона между станицами Мариинской и Аксайской. Мат. по изуч. растит, и лугов долины р. Дона в пределах Донского края, 1. — 1919а. Растительность нижней части долины р. Донца. Мат. по изуч. растит, и лугов долины р. Дона в пределах Донского края, 1.—1933. Растительный покров переходного болота Орехово-Зуевского района и некоторые технические свойства залежи. Тр. Всесоюзн. Инет, торфа, 3. — 1936. Торфяные болота нижнего течения р. Оби. Тр. Центр, торф, опытн. станции, 1. — 1956. Материалы к познанию растительности торфяников Южного Сахалина. Геогр. сб. ВГО, 8. М.— Л.
Воейков А. И. (W о о i к о w). 1871. Beitrage zur Kenntnis der Wald- und Regenzonen des Kaukasus. Zeitschr. d. osterreich. Ges. f. Meteorol., d, 14.
Волгунов Д. К. 1928. К познанию растительности плавней р. Кумы» В кн.: Мат. по флористич. и фаупистич. обслед. Терского окр., Пятигорск.
Воробьев Д. В. 1953. Типы лесов Европейской части СССР. Киев.
Воробьев Д. В. и П. С. Погребняк. (Воробйов i По гр е б н я к). 1929. Л1со-вий типолопчний визначник Украшського Пол!сся. Тр. з Л1сово! досв^дноТ справи па Укр., 11.
Воробьев Д. П. 1935. Растительный покров южного Сихотэ-Алиня и дикорастущие плодово-ягодные растения в нем. Тр. Дальневост. фил. АН СССР, сер. бот., 1. — 1937. Растительность южной части побережья Охотского моря. Тр. Дальневост. фил. АН СССР, сер. бот., 2. — 1947. Растительность Курильских островов. Изв. ВГО, 79. 4.
Воронихин Н. Н. 1930. Водоросли Полярного и Северного Урала. Тр. Ленингр. общ. естествоиспыт., 60. 3.
Воронов Ю. Н. 1905. Краткий отчет о ботанических экскурсиях в Абхазии летом 1902 г. Тр. СПб. общ. естествоиспыт., 34. отд. бот. — 1908. Очерк растительности Абхазии. I. Растительность морского побережья от Сухума до устья р. Кодора. Вести. Тифл. бот. сада, — 1910. Очерк лесных областей и распространение главнейших древесных пород на Кавказе. Кавказ, хоз., 8, 10. — 1911—1912. Батумская область. Изв. Кавказ, отд. РГО, 21. 1.
Воропанов П. В. 1950. Ельники Севера. М.—Л.
Востокова Е. А. 1952. Чиевники Западного Казахстана. Бюлл. МОИП, отд. биол., 57. 1.
Вульф Е. В. 1925. Растительность восточных яйл Крыма, их мелиорация и хозяйственное использование. М. — 1929. Керченский полуостров и его растительность в связи с вопросом о происхождении флоры Крыма. Зап. Крым. общ. естествоиспыт., 11.
Высоцкий Г. Н. 1899. Почвенные зоны Европейской России в связи с соленосностью грунтов и характером лесной растительности. Почвоведение, 1. 1. — 1904. О карте типов местопроизрастании. В кн.: Современные вопросы русского сельского хозяйства, СПб. — 1905. Степи Европейской России. В кн.: Полная энциклопедия русского сельского хозяйства, 9. СПб. — 1906. Почвенно-ботанические исследования в южных Тульских засеках. Тр. опытн. лесничеств, 4. -г 1908—1909. О лесорастительных условиях Самарского удельного окр. Почвенно-ботанический лесоводственный очерк, 1 (1908), 2 (1909), СПб. — 1909. О фито-топологических картах, способах их составления и их практическом значении. Почвоведение, 11. 2. — 1912. Лесные культуры степных опытных лесничеств с 1893 по 1907 г. Тр. по лесн. опытн. делу в России, 41. — 1913. О дубравах в Европейской России и их областях. Леей, ж., 43. 1—2. — 1915. Ергеня. К ул ьтурно-фито логический очерк. Тр. Бюро по прикл. бот., 8. 10—И. — 1915а. Природные растительные условия и результаты лесоразведения на Ергенях Астраханской губ. Ежегодн. лесн. департамента, 1. — 1923 (1922—1923). О перспективах нашего степного полеводства и скотоводства. Тр. по прикл. бот. и селекц., 13. 3.
Выходцев И. В. 1937. Переднеазиатские пырейные степи с преобладанием Agropyrum trichophorum в Тянь-Шане и Памире-Алае. Тр. Киргизск. н.-иссл. инет, животн., 3. — 1945. Геоботанические ландшафты Киргизии. Изв. Киргизск. фил. АН СССР, 2—3. — 1946. Ботаническая наука в Киргизии. В кн.: Наука в Киргизии за 20 лет, 1926—-1946 гг., Фрунзе. — 1952. Растительность пастбищ и сенокосов Киргизской ССР и ее кормовое значение. Л.
Гаель А. Г. 1929. Пески нижнего Дона. Цымлянско-Донской и Романовский песчаные массивы, их естественно-исторические особенности и пути хозяйственного использования песков. Тр. ио лесн. опытн. делу, 4. — 1932. Пески верхнего Дона.
Изв. ГГО, 64, 1—3. — 1932а. Донецко-Кундрючский песчаный массив. Изв. ГГО. 64, 4—5. — 1934. К вопросу о генезисе и эволюции песков СССР. В кн.: Тр. Первого Всесоюзн. геогр. съезда, 3, Л. — 1938. Некоторые особенности водного режима почвогрунтов в песчаных пустынях. Изв. ГГО, 70, 4—5.
Гаель А. Г., М. С. Коликов, Е. А. Малюгин и Е. С. Останин. 1949. Пески Урало-Эмбенского района и пути их освоения. Тр. Пнет, пустынь АН КазССР, 1. — 1950. Песчаные пустыни северного Приаралья и пути их освоения. Тр. Инет, пустынь АН КазССР, 2.
Гаель А. Г. и Е. С. Останин. 1939. Южно-Казахстанский песчаный массив Муюнкум. В кн.: Освоение пустынь, полупустынь и высокогорий, М.
Гайдук А. и А. Борисов. 1916. Леса Витимского края Иркутской губ. Пг.
Галиновский В. И. 1938. Березовые колки Западно-Сибирской низменности. Леей, индустрия, 2.
Галкина Е. А. 1936. Типы болот Тунгудского района Автономной Карельской ССР. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 3. — 1946. Болотные ландшафты и принципы их классификации. В кн.: Сборник научи, работ [Бот. инет. АН СССР], выполненных в Ленинграде за три года Великой Отечественной войны (1941—1943), Л. — 1948. Применение аэросъемки при изучении болотных массивов. В кн.: Тр. Второго Всесоюзн. геогр. съезда, 2, М. — 1953. Пути использования аэрофотосъемки в болотоведении. Бот. ж., 38, 6. — 1955. Болотные ландшафты лесной зоны. Геогр. со. В ГС), 7.
Гамаюнова А. П. и В. П. Голоскоков. 1949. Материалы к флоре и растительности Чу-Илийских гор. Изв. АН КазССР, 74, сер. бот., 4.
Ганешин С. С. 1912. Растительность Ангаро-Илимского края Иркутской губ. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1909 г., ч. 2, 5. — 1917. Ботанико-географический очерк средней части Акмолинской обл. Тр. Почв.-бот. экспед. Переселенч. Унр. по иссл. колониз. районов Аз. России в 1914 г., ч. 2, 1. — 1925. Растительность поймы р. Волхова. Мат. по иссл. р. Волхова и его бассейна, 4. — 1927. Краткая история и результаты исследования флоры и растительности .Чужского у. Ленинградской губ. Тр. Ленингр. общ. изуч. местного края, 1. — 1927а. О реликтовом характере лесов некоторых естественно-исторических районов Лужского у. Ленинградской губ. В кн.: Юбилейный сборник, посвящ. И. П. Бородину, Л.
Гейдеман Т. С. 1940. Краткий очерк растительности Закатало-Белоканской низменности. Тр. Бот. инет. Азерб. фил. АН СССР, 9. — 1950. Ксероморфные дубравы «гырницы» южной части Молдавской ССР. Научн. зап. Молд. н.-иссл. базы АН СССР, 2. — 1952. Краткий очерк растительного покрова Молдавской ССР. Изв. Молд. фил. АН СССР, 4—5 (7—8). — 1954. Растительность оползней, оврагов и склонов, подвергшихся смыву на территории Молдавской ССР. Изв. Молд. фил. АН СССР, 2 (16).
Гейдеман Т. С. и И. Н. Дорошко. 1931. Очерк растительности Сальянской степи. Тр. по геобот. обслед. пастбищ АзССР, сер. А, зимние пастбища, 8.
Генкель А. А. и П. Н. Красовский. 1937. Материалы по изучению озер, займищ, болот и торфяников западно-сибирской лесостепи. Уч. зап. Пермск. унив., 3, 1.
Генкель А. А. и Е. И. Осташева. 1933. Висячие болота окрестностей горы Яман-Тау на Южном Урале. Изв. Пермск. биол. н.-иссл. инет., 8, 6—8.
Геоботаническое районирование СССР. 1947. Тр. Комиссии по естеств.-ист. районированию СССР, 2, 2.
Герасимов Д. А. 1926. Геоботаническое исследование торфяных болот Урала. Торф, дело, 3. — 1929. Полистовский болотный район. Торф, дело, 1.
Герасимов И. П. 1931. О такырах и процессах такырообразования. Почвоведение, 4. — 1933. О «такырах», их генетической сущности и о процессе такырообразования. Почвоведение, 5. — 1945. Мировая почвенная карта и общие законы географии почв. Почвоведение, 3—4. — 1951. Географические наблюдения в Прикаспии. (К проблеме обводнения и орошения Прикаспийской низменности). Изв. АН СССР, сер. геогр., 4.
Герасимов И. П., Е. Н. Иванова и Д. И. Тарасов. 1935. Почвенно-мелиоративный очерк дельты и долины р. Аму-Дарьи. Тр. СОПС АН СССР, Каракалпакская сер., 6.
Герасимов И. П. и Ю. А. Ливеровскии. 1947. Черно-бурые почвы ореховых лесов Средней Азии и их палеогеографическое значение. Почвоведение, 9.
Герасимов И. П. и К. К. Марков. 1939. Ледниковый период на территории СССР. Тр. Инет, геогр. АН СССР, 33.
Глваовская М. А. 1947 (на пер. 1949). Почвы горных областей Казахстана. Изв. АН КазССР, 52, сер. почв., 4. — 1950. Выветривание горных пород в нивальном поясе центрального Тянь-Шаня. Тр. Почв. инет. АН СССР, 34.
Глуедавов С. И. 1953. Луга Западной Сибири. Новосибирск.
Глумов Г. А. 1948. К вопросу о морфологической структуре почвенно-растительных комплексов. Тр. Молотовск. с.-х. инет., 12.
Глумов Г. А. и Красовский П. Н. 1948. Основные черты зацелинения залежей Троицкого лесостепного заповедника. Изв. Естеств.-научн. инет, при Молотовск. унив., 12, 8.
Глушков Н. Н., И. II. Долбилин, В. И. Венгров и Ф. С. Тимашев. 1948. Леса Урала. Свердловск.
Гмелин И. Г. (G m е 1 i п). 1747. Flora sibirica sive historia plantarum Sibiriae, 1. СПб. — 1751—1752. Reise (lurch Sibirien von den Jahr 1733 bis 1743. 1 (1751), 2 (1752), 3 (1752), 4 (1752). Gottingen.
Гоби X. Я. 1876. О влиянии Валдайской возвышенности на географическое распространение растений в связи с очерком флоры западной части Новгородской губ. Тр. СПб. общ. естествоиспыт., 7.
Говорухин В. С. 1929. Растительность бассейна р. Ылыча (Сев. Урал). Тр. Общ. изуч. Урала, Сибири и Дальнего Востока, I, 1. — 1933. Очерк растительности летних пастбищ северного оленя в тундрах Обско-Тазовского п-ова. Землеведение, 35, 1. — 1936. Пятнистая тундра в горах Северного Урала. Землеведение, 38, 2. — 1940. Лесотундра Старого Света. Уч. зап. геогр. фак. Моск. обл. пед. инет., 3, 1.
Говорухин В. С. и Ф. Н. Мильков. 1951. Леса восточной части Великолукской обл. Уч. зап. Моск. обл. пед. инет., 17.
Годнев Е. Д. 1949. Ергени и опыт их облесения. М. — Л.
Гожев А. Д. 1929. Типы песков области Среднего Дона и их хозяйственное использование. Тр. по лесн. опытн. делу, Центр, лесн. опытн. станции, 3. — 1934. Болота Удского района. Тр. СОПСАН СССР, Дальневосточная сер., 3. — 1934а. Леса Удского района. Тр. СОПС АН СССР, Дальневосточная сер., 3.
Голгофская К. Ю. 1953. Типы байрачных лесов севера Ворошилошрадской обл. в связи с мероприятиями по рационализации в них лесного хозяйства. М.
Голицин С. В. 1935. Опыт ботанико-географического картирования юго-западного Закавказья с дендрологической точки зрения. Тр. Воронежск. унив., 7.
Голоскоков В. П. 1949. Флора и растительность высокогорных поясов Заилийского Алатау. Алма-Ата.
Голубинский С. С. 1934. Ленточные боры Западно-Сибирского края. Тр. Лебяжинск. зон. лесн. опытн. станции, 1.
Гончаров Н. Ф. 1936. Очерк растительности центрального Таджикистана. Тр. Таджикско-Памирской экспед. 1932 г., 24. — 1937. Районы флоры Таджикистана и их растительность. В кн.: Флора Таджикистана, 5, М.—Л.
Гордягин А. Я. 1897. Геоботанические исследования в южной полосе Тобольской губ., произведенные в 1896 году. Ежегодн. Тобольск, губ. муз., 7. — 1897а. О кокчетавских лесах. Зап. Зап.-Сиб. отд. РГО, 22. — 1900—1901. хМатериалы для познания почв и растительности Западной Сибири. Тр. Общ. естествоиспыт. при Казанск. унив., 34, 3 (1900); 35, 2 (1901). — 1921 (на пер. 1922). Растительность Татарской республики. В кн.: Географическое описание Татарской республики, 1, Казань.
Городков Б. Н. 1912. Очерк растительности низовьев р. Конды. Ежегодн. Тобольск, губ. муз., 20. — 1913. Поездка в Салымский край. Ежегодн. Тобольск, губ. муз., 21. — 1913а. Растительность Ишимского у. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1912 г., СПб. — 1915. Подзона лиственных лесов в пределах Ишимского у. Тобольской губ. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1912 г., ч. 2, 3. — 1916. Опыт деления Западно-Сибирской низменности на ботанико-географические области. Ежегодн. Тобольск, губ. муз., 27. — 1924. Западносибирская экспедиция Российской Академии Наук и Русского географического общества. Природа, 7—12. — 1924а (1922). Тезисы к докладам об основаниях ботанико-географического районирования и картографирования России. Ж. Русск. бот. общ., 7. — 192о. Полярный Урал в верхнем течении р. Соби. Тр. Бот. муз. АН СССР, 19. — 1928. Крупнобугристые торфяники и их географическое распространение. Природа, 6. — 1929. Безлесье тундры. Природа, 3. — 1929а. Полярный Урал в верховьях рр. ВоДкара, Сыни и Ляпина. Мат. Комиссии экспед. иссл. АН СССР, Уральская сер., 7. — 19296. Тундра и альпийский пояс гор. Природа, 11. — 19о2. Почвы Гыданской тундры. Тр. Полярн. комиссии АН СССР, 7. — 1933. Естественные пастбищные угодья тундровой зоны ДВК. Сов. оленеводство, 2. — 1935. Геоботанический и почвенный очерк Пенжинского района Дальневосточного края. Тр. Дальневост. фил. АН СССР, сер. бот., 1. — 1935а. Материалы для познания горных тундр Полярного Урала. Тр. ледниковых экспед., 4. — 19356. Растительность тундровой эоны СССР. М.—Л. — 1936. Растительность Полярного и Северного Урала. В кн.: Природа Урала, Свердловск. — / 1938. Изучение растительности Советской Арктики за последние 20 лет. Сов.
бот., 1. — 1938а. Растительность Арктики и горных тундр СССР. В кн.: Растительность СССР, 1, М.—Л. — 1939. Ботанико-географический очерк Чукотского
побережья. Уч. зап. Ленингр. пед. инет. им. А. И. Герцена, 21. — 1943. Полярные пустыни о. Врангеля. Бот. ж. СССР, 28, 4. — 1944. Тундры Обь-Енисейского водораздела. Сов. бот., 3—5. — 1946. Ботанико-географический очерк Крайнего Севера и Арктики СССР. Уч. зап. Ленингр. пед. инет, им. А. И. Герцена, — 1946а. Движение растительности на севере лесной зоны Западно-Сибирской низменности. Проблемы физ. геогр., 12. — 19466. Опыт классификации растительности Арктики. Сов. бот., 14, 1, 2. —1947. Полярные пустыни и арктические тундры. (Сравнительное почвенно-ботаническое исследование). Тезисы доклада. Бот. ж. СССР, 32, 1. — 1949. О почвенно-растительных комплексах тундровых и полярно-пустынных ландшафтов. В кн.: Тр. Второго Всесоюзн. геогр. съезда, 3, М. — 1952. Происхождение арктических пустынь и тундр. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 8. — 1954. Материалы для познания четвертичных ландшафтов Арктики. Бот. ж., 39, 1. — 1956. Растительность и почвы о. Котельного (Новосибирские о-ва). В кн.: Растительность Крайнего Севера и ее освоение, 2, М.—Л.
Горчаковский П. Л. 1949. Сосновые леса Приобья как зональное ботанико-географическое явление. Бот. ж., 34, 5. — 1949а. Таежные и лесостепные березняки Приобья. (Ботанико-географический очерк). Сб. тр. по лесн. хоз., 1, Свердловск. — 1950. Высокогорная растительность заповедника «Денежкин камень». Свердловск. — 1954. Высокогорная растительность Яман-Тау — крупнейшей вершины Южного Урала. Бот. ж., 39, 6. — 1954а. Лесная растительность подгольцового пояса Урала. Сб. тр. по лесн. хоз., 2, Свердловск. — 19546. Пихтовая тайга Среднего Урала. Зап. Уральск, отдела Геогр. общ. СССР, 1. — 1955. Луга высокогорных районов Урала. Свердловск.
Горшкова А. А. 1954. Материалы к изучению степных пастбищ Ворошиловградской обл. в связи с их улучшением. Тр. Бот. инет. АН СССР, сер. 3,^ геоботаника, 9. — 1954а. Растительность необлесенной части Еравинского аймака Бурят-Монгольской АССР. Мат. по изуч. производит, сил. Бурят-Монг. АССР, 1.
Горшкова С. Г. 1919. Растительность долины р. Дона от станицы Казанской до станицы Семикаракорской. Мат. по изуч. растит, и лугов долины р. Дона в пределах Донского края, 1.
Грабарь В. А. 1951. Очерк растительности бассейна р. Тересвы. Наук. зап. Ужго-родск. ушв., сер. б!ол., 4.
Граве Н. П. 1936. Тугайные джунгли низовьев Аму-Дарьи. М.
Гранитов И. И. 1934. К вопросу о взаимоотношениях в эфемерном покрове пустыни. В кн.: Хозяйственное освоение пустынь Средней Азии и Казахстана, М.— Ташкент. — 1950. Карта растительности юго-западных Кызыл-Кумов. Тр. САГУ, нов. сер., 19 (биол. науки, 8).
Грибанов Л. Н. 1954. Ленточные боры Алтайского края и Казахстана. М.
Григорьев А. А. 1924. Полярная граница древесной растительности в Большеземельской и некоторых других тундрах; факторы ее обусловливающие и колебания ее в ближайшую к нам эпоху. Землеведение, 26, 1—2. — 1946. Некоторые итоги разработки новых идей в физической географии. Изв. АН СССР, сер. геогр. и геофиз. 10, 2. — 1946а. О географических радиационных рубежах и характеристиках радиационных условий горизонтальных физико-географических зон. Проблемы физ. геогр., 12. — 1947. Субарктика. М.—Л.
Григорьев М. П. 1914. Второй краткий предварительный отчет о работах 1913 г. по исследованию лугов Владимирской губ. Владимир.
Григорьев Ю. С. 1941. Основные черты растительного покрова бассейна Верхнего Ягноба. Изв. Тадж. фил. АН СССР, I. — 1944. Очерк растительности бассейна Среднего Зеравшана. Изв. Тадж. фил. АН СССР, 7. — 1948. О растительности окрестностей Сталинабада в связи с вопросом о вертикальных поясах южного Таджикистана. Сообщ. Тадж. фил. АН СССР, 3. — 1951. Материалы к истории трагакантовой флоры Памиро-Алая. Тр. Тадж. фил. АН СССР, 18.
Григорьев Ю. С., А. С. Королева и В. А. Никитин. 1936. Очерк растительности западной части южных склонов Гиссарского хребта. Тр. Тадж. базы АН СССР, 2.
Гриневецкий Б. Б. 1913. Восточная граница бука в Европе. Тр. Тифл. бот. сада, 12, 2. Гринь Ф. А. (Ф. О. Гринь). 1940. Рослиншсть Хенсько! лкотундри. Бот. ж. АН
УРСР, 1, 1. — 1950. Нарис лково! рослинност! Терношльсько! обл. Бот. ж. АН УРСР, 7, 1. — 1954. Дубов! л!си. В кн.: Рослиншсть Закарпатсько! обл. УРСР, Ки!в. — 1954а. Хвойш л!си. В кн.: Рослиншсть Закарпатсько! обл. УРСР, Ки!в. — 19546. Про минуле i сучасне лкових гайк!в на Нижньодш-провських шсках. Бот. ж. АН УРСР, 11, 1.
Гроздов Б. В. 1950. Типы леса Брянской, Смоленской и Калужской обл. Брянск. Гроссгейм А. А. 1915. Очерк растительности Араздаянского имения. Тифлис. — 1926. Краткий очерк растительного покрова Азербайджана. Мат. по районированию АзССР, 1, 2. — 1926а. Флора Талыша. Тифлис. — 1928. Краткий очерк растительного покрова ССР Армении. Мат. по районированию АрмССР, 2. —'
1928а. Опыт классификации растительности на засоленных местообитаниях Закавказья. Изв. Азерб. унив., отд. естествозн. и медиц., 7. — 1929. Введение в геоботаническое обследование зимних пастбищ ССР Азербайджана. Тр. по гео-бот. обслед. пастбищ ССР Азербайджана, сер. А, зимние пастбища, 1. — 1930. Очерк растительного покрова Закавказья. Тифлис. — 1932. Очерк растительности Кура-Араксинской низменности. В кн.: Мат. к общей схеме использования водных ресурсов Кура-Араксинского басе., 4, Тифлис — 1939. К географии хвойных Кавказа. Тр. Азерб. унив., 1. — 1940. Развитие ботанической науки в Азербайджане за 20 лет. Изв. Азерб. фил. АН СССР, 2. — 1948. Растительный покров Кавказа. Мат. к познанию фауны и флоры СССР, отд. бот., нов. сер., 4 (12). Гроссгейм А. А. и А. А. Колаковский. 1929. Геоботанический очерк зимних пастбищ левобережной части Казахского у. Тр. по геобот. обслед. пастбищ ССР Азербайджана, сер. А, зимние пастбища, 3. — 1930. Очерк растительности зимних пастбищ правобережной части Казахского у. Тр. по геобот. обслед. пастбищ ССР Азербайджана. Сер. А, зимние пастбища, 5.
Гроссгейм А. А. и Л. И. Прилипко. 1931. Очерк растительности территории Ленкоранской лесной опытной станции. Баку.
Гроссгейм А. А. и Д. И. Сосновский. 1927. Опыт ботанико-географического районирования Кавказского края. Изв. Тифл. политехи, инет., 3.
Гроссгейм А. А., М. Ф. Сахокиа, Д. И. Сосновский и А. Л. Тахтаджян. 1945. Опыт построения классификационной схемы растительного покрова Кавказа. ДАН АрмССР, 2, 3.
Гроссет Г. Э. 1928. Материалы к изучению лесов Воронежской губ. I. Леса Валуй-ского у. Тр. Н.-иссл. инет, при Воронежск. унив., 2. — 1929. Материалы по изучению лесов Центрально-Черноземной обл. II. Некоторые данные о дубравах юго-западной части б. Воронежской и юго-восточной части б. Курской губ. Тр. Н.-иссл. инет, при Воронежск. унив., 3. — 1930. Лес и степь в их взаимоотношениях в пределах лесостепной полосы Восточной Европы. Воронеж.
Грубов В. И. 1940. О современном состоянии липового острова в предгорьях Кузнецкого Алатау. Сов. бот., 1.
Грудзинская И. А. 1953. Широколиственные леса предгорий северо-западного Кавказа. В кн.: Широколиственные леса северо-западного Кавказа, М.
Гудошников С. В. 1953. Растительность бассейна р. Джиды в пределах Бурят-Монгольской АССР. Томск.
Гулисашвили В. 3. 1940. Некоторые экологические особенности ели восточной (Picea orientalis) и пихты кавказской (Abies Nordmanniana) и климатические границы их распространения. Сообщ. Груз. фил. АН СССР, 1, 1. — 1955. Закономерности в распространении лесной растительности и главных древесных пород в Закавказье. Бот. ж., 40, 1.
Гуман В. В. 1911. Почвы и типы насаждений Заволжской дачи Казанской губ. Тр. по лесн. опытн. делу в России, 31.
Гурский А. В. 4945. Несколько слов о степях, полупустынях и пустынях. Бот. ж. СССР, 30, 6.
Гусельников Е. П. 1936. К вопросу об изучении растительности утугов. Тр. Вост.-Сиб. унив., 3.
Гуторович И. И. 1912. Краткое описание типов насаждений, встречаемых в Вятской и Пермской губерниях, в северных их частях. Лесн. ж., 42, 4—5.
Дагис И. X. (J. D a g у s). 1936. Apascios up6s pievij mazieji viksvyuai. Vytauto Did-ziojo Univ. Mat.-Gamt. Facult. Darbai, 11.
Дадыкин В. П. 1952. Особенности поведения растений на холодных почвах. М. Данилов С. И. 1936. Пал в Забайкальских степях и его влияние на растительность.
Вести. Дальневост. фил. АН СССР, 21.
Данилова М. М. 1948. Болота долины реки Камы. Изв. Ест.-научн. инет, при Моло-товск. унив., 12, 6.
Двораковский М. С. 1948. Грабовые леса вблизи восточной границы распространения граба (Carpinus betulus L.). Вестн. Моск, унив., 5.
Дедов А. А. 1931. Оленьи пастбища Тиманской тундры. В кн.: В. А. Андреев, А. А. Дедов, Ф. В. Самбук, Оленьи пастбища Северного края, Архангельск.— 1933. Летние оленьи пастбища восточной части Малоземельской тундры. В кн.: Оленьи пастбища Северного края, 2, Л. — 1933а. Материалы к характеристике кормовой площади Таймырского окр. Сов. оленеводство, 2.
Денисов А. К. 1954. Пойменные дубравы лесной зоны. М.—Л.
Денисов 3. Н. 1951. Естественно-историческая классификация болот БССР. Сб. научн.
трудов Инет, мелиор., водного и болотного хоз. АН БССР, 1.
Десятова-Шостенко Н. А. и Ф. Я. Левина (Н. О. Десятова-Шостенко та Ф. Я. Л е в i н а). 1928. Ботатчне дослщження Чорноморських Kic та остров!в: Тендера, Джарилгача, Орлова та Довгого. Мат. охорони природи на УкраТш, 1.
Десятова-Шостеико Н. А. и М. С. Шалыт (Н. О. Д есятова-Шостенко та М. G. Шалит) 1936. Матер1али до вивчення цытинних стетв швшчного Криму. Тр.Н.-дослщч. шст. бот. Харювськ. унив., 1.
Дзевановский С. А. 1925. Ботаническая экскурсия в «Осминские дубки». Зап. Крым, общ. естествоиспыт. и любит, природы, 8.
Дзенс-Литовская Н. Н. 1930. Типы горных лесов Джаляль-Абадского кантона Киргизской АССР. В кн.: Сб. Геогр.-эконом, иссл. инет, при геогр. фак. Ленингр. университета за 1928 г., Л. — 1933. Материалы для изучения еловых лесов Киргизской АССР. Тр. Инет, по изуч. леса АН СССР, 1. — 1951. Почвы и растительность Присивашья. Изв. ВГО, 83, 5.
Димо Н. А. и Б. А. Келлер. 1907. В области полупустыни. Саратов.
Дингельштедт Ф. Н. 1914. Некоторые черты растительности долины р. Свири. Изв. Общ. изуч. Олонецкой губ., 3. — 1929. Очерк пойменных лесов долины Дона. В кн.: Очерки по фитосоциологии и фитогеографии, М.
Дмитриев А. М. 1904. Луга Холмогорского района. СПб. — 1905. Луга Поволжья и низовьев Шексны и Мологи в Ярославской губ. Сельское хоз. и лесовод., 217, 5, 6. — 1948. Луговодство с основами луговедения. Изд. 2-е. М.
Дмитриева А. А. 1944. Материалы по флоре и растительности северного Прибалхашья. Изв. Казахст. фил. АН СССР, сер. бот., 1.
Доброхотова К. В. 1940. Ассоциации высших водных растений как фактор роста дельты Волги. Тр. Астрах, заповеди., 3. — 1954. Значение водной растительности в эволюции нерестовых водоемов дельты Аму-Дарьи. Тр. Лаборат. озеровед. АН СССР, 3.
Доброхотова К. В. и Л. Н. Михайлова. 1938. Материалы к изучению фитоценозов приморской части дельты Волги в пределах Астраханского заповедника. Тр. Астрах, заповеди., 2.
Добрынин Б. Ф. 1925. Ландшафтные (естественные) районы и растительность Дагестана. Мем. геогр. отд. Общ. любит, естествозн. антропол. и этногр., 1.
Добрынин Г. М. 1938. Влияние сроков сенокошения на величину и качество урожая лугов «Присухонской низины» Вологодской обл. в 1935 г. Зап. Пушкинск. с.-х. инет., 8.
Доктуровский В. С. 1907. Очерк растительности хМинского Полесья. Тр. студ. кружка для иссл. русской природы при Моск, унив., 3. — 1909. Амурская обл. Район рр. Норы и Мамына. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1908 г., СПб. — 1909а. Растительность бассейна рр. Норы и Мамына в Амурской обл. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1908 г., ч. 2, 1. — 1911. Растительность Тырминско-Б у рейнского района и Амурской обл. вообще. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1909 г., ч. 2, 3. — 1925. О торфяниках Пензенской губ. Тр. по изуч. заповеди. Отд. охраны природы Главнауки НКП РСФСР, 3. — 1927. О границах сфагновых болот и о болотных районах. Торф, дело, 8. — 1935. Торфяные болота. Изд. 2-е. М.—Л. — 1936. Материалы по изучению торфяников Закавказья. Почвоведение, 2.
Доктуровский В. С. и Н. Н. Жуков. 1916. Ботанические исследования по среднему течению р. Птичь. Мат. по иссл. рек и речи, долин Полесья, I, Киев.— 1916а. Ботанические исследования в пойме р. Брагинки. Мат. по иссл. рек и речи, долин Полесья, 1. Киев.
Доктуровский В. С., М. И. Рынкевич, М. Н. Копачевская, В. Н. Хитрово, Н. Ф. Дере-вицкий, Н. А. Троицкий. 1915. Исследования болот Волынской губ. Пг.
Докучаев В. В. 1883. Русский чернозем. СПб. — 1891. К вопросу о соотношениях между возрастом и высотой местности, с одной стороны, характером и распределением черноземов, лесных земель и солонцов — с другой. Вести, естествозн.г 1—3. — 1892. Наши степи прежде и теперь. СПб. — 1899. К учению о зонах природы. СПб.
Доленко Г. И. 1916. Части Лено-Вилюйского водораздела Якутской обл. В кн.: Предварит. отчет об организ. и исполн. работ по иссл. почв Аз. России в 1914 г., СПб. — 1930. Краткое описание ландшафтных районов западного Усть-Урта и равнинного Мангышлака. Мат. Комиссии экспед. иссл. АН СССР, 26, Казахстанская сер.
Долуханов А. Г. 1932. Верхние пределы леса в горах восточной части Малого Кавказа. Тр. геобот. обслед. пастбищ ССР Азербайджана, сер. Д, 3. — 1938. Геоботанический очерк лесов ущелья р. Чхалты. Тр. Тбил. бот. инет. Груз. фил. АН СССР, 5. — 1938а. Очерк лесной растительности бассейна р. Тала-чай Зака-тальского района. Тр. Бот. инет. Азерб. фил. АН СССР, 4. — 1940. К вопросу о взаимоотношении темнохвойных и сосновых лесов Кавказа. Бот. ж. СССР. 25, 4—5. — 1941. Краткий очерк растительности Лагодехского заповедника, В кн.: Лагодехский заповедник, 1, Тбилиси. — 1946. Верхние пределы альпийской растительности Аварского Койсу (Дагестан). Тр. Тбил. бот. инет. АН
ГрузССР, 9. — 1949. Леса Зангезура. (Типологический очерк). Тр. Бот. инет. АН АрмССР, 6. — 1952. О природе реликтов субальпийской мезофильной дендрофлоры Колхиды. Тр. Тбил. бот. инет. АН ГрузССР, 14. — 1953. Каштановые леса Грузии. Тр. Тбил. бот. инет. АН ГрузССР, 15.
Долуханов А. Г. и М. Ф. Сахокиа. 1941. Опыт геоботанического районирования Закавказья. Сообщ. АН ГрузССР, 2, 4.
Долуханов А. Г., М. Ф. Сахокиа и А. Л. Харадзе. 1942. К вопросу о высокогорных растительных поясах Кавказа. Тр. Тбил. бот. инет. АН ГрузССР, 8. — 1946. Основные черты растительного покрова Верхней Сванетии. Тр. Тбил. бот. инет. ЛИ ГрузССР, 9.
Доскач Л. Г. 1937. Физико-географический очерк Зейско-Буреинской равнины. Тр. 11 нет. геогр. АН СССР, 27.
Доскач А. Г., И. В. Ларин и Ф. Я. Левина. 1952. Пути улучшения хозяйственного освоения области бессточных рек Прикаспийской низменности. Сообщ. Комплекс, научн. экспед. по вопросам полезащ. лесоразвед. АН СССР, 1.
Достойнова Е. Я. 1925. Влияние сенокошения на состав и строение луговых сообществ. Мат. Вологодск. обл. с.-х. опытн. станции, 2.
Дохман Г. И. 1930. Фитосоциологический анализ растительного покрова Старобель-ских целинных степей. Изв. Ассоц. н.-иссл. инет, при физ.-мат. фак. 1-го Моск, унив., 3, 2-А. — 1935. Подзона лиственных лесов или северная лесостепь в пределах б. Ишимского окр. Землеведение, 37, 2. — 1936. О некоторых классификационных единицах комплексов. Землеведение, 38, 3. — 1936а. Опыт эколого-генетической классификации растительности Ишимской лесостепи. Бюлл. МОИП, отд. биол., 45, 3. — 1937. Вопросы классификации степей. Землеведение, 39, 4—5. — 1940. О картировании комплексов. Уч. зап. Моск, унив., 36. — 1950. Растительные формации Мугоджар, их взаимоотношение и динамика. Землеведение, нов. сер., 3 (43). — 1954. Растительность Мугоджар. М.
Дохман Г. И. и Т. И. Рыбакова. 1933. Очерк растительности степей Сало-Манычского водораздела (б. Сальский окр. Северокавказского края). Бюлл. МОИП, отд. биол., 42, 2.
Дробов В. П. 1909. Растительные формации Мариинско-Чулымской тайги. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1908 г., ч. 2, 2. — 1910. Растительные формации Лено-Киренгского края Верхоленского у. Иркутской губ. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1909 г., ч. 2, 1. — 1914. Из степей Акмолинской обл. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1913 г., СПб. — 1914а. Материалы к изучению типов лесных насаждений Вытегорского у. Олонецкой губ. Изв. Общ. изуч. Олонецкой губ., 3, 1—2. — 191чб. — Растительность в районе Якутско-Усть-Майского тракта Якутской обл. и окр. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1912 г., ч. 2, 1. — 1916. Общий очерк растительности в бассейне рр. Нижней Тунгуски и Вилюя. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1914 г. СПб. — 1922. Генезис и эволюция песков западной Ферганы. Изв. Туркест. отд. РГО, 15. — 1923 (1921). Загадки саксаула. Ж. Русск. бот. общ., 6. — 1927. Краткий очерк растительности Ленско-Алданского плато. Мат. Комиссии по изуч. Якут. АССР, 8. — 1928. Пески у ст. Репетек и их растительность. Тр. САГУ, сер. VIII-в, бот., 1. — 1929. Материалы к познанию растительности песчаных пустынь Средней Азии. В кн.: Очерки по фитосоциологии и фитогеографии, М. — 1935. Древесная и кустарниковая растительность Ташкентского оазиса. Бюлл. САГУ, 21. — 1947. Тугайная растительность Кара-Калпакской АССР. Изв. АН УзССР, 5. — 1950. Экология некоторых древесных и кустарниковых форм растений Узбекистана. Изв. АН УзССР, 2. — 1950а. Леса Узбекистана. Ташкент. — 19506. Тугайная древесная и кустарниковая растительность Кара-Калпакской АССР. Мат. по произвол, силам Узбекистана, 1, Ташкент. — 1952. Растительность песчаных пустынь Узбекистана. Ташкент.
Дроздов И. П. 1938. Кормовые растения естественных сенокосов и пастбищ Бурят-Монголии. Улан-Удэ.
Дубинский В. A. J903. О характере растительности меловых обнажении (по исследованиям в Воронежской губ.). Изв. СПб. бот. сада, 3, 7. — 1904. Краткий очерк поездки в Тургайскую и Уральскую области. Изв. СПб. бот. сада, 4, 7. — 1905. Характер растительности меловых обнажений в бассейне р. Хопра. Изв. СПб. бот. сада, 5, 3. — 1911. Растительность русских песчаных пустынь. В кн.: И. Вальтер, Законы образования пустынь в настоящее и прошлое время, СПб. — 1928. Песчаная пустыня юго-восточные Кара-Кумы, ее естественные районы, возможности их сельскохозяйственного использования и значение для ирригации. Тр. по прикл. бот., генет. и селекц., 19, — 1929. Песчаные пустыни
Туркмении. В кн.: Туркмения, 3, Л. — 1932. Песчано-пустынные пространства
Центрального Казакстана. В кн.: Казакстан, М. Л. — 1947. Работа Русского географического оощества по изучению сыпучих песков. Вопросы геогр., 3. Дунин-Горкавнч А. А. 1904—1910. Тобольский Север. Общий обзор страны, ее естественных богатств и промышленной деятельности населении. 1 (1904), 2 (1910). СПб.
Душечкнн В. И. 1937. Оленьи пастбища в Хараулахскнх горах (Якутия). Тр. Арктич. мнет., W.
Дыдмна Р. А. 1940. Урожай и фактический сеиосбор па лугах Молого-Шекспинского междуречья. Тр. Бот. инет. АП СССР, сер. 3, геоботаника, •/.
Дылнс 11. И. 1939. Редколесные березняки и безлесные ерпиковые заросли Печорской тайги как отголоски ландшафта ледникового времени. Бот. ж. СССР, 24, 4. — 1940. Тиша лиственничных лесов Южного Тимана. Тр. Бот. пнет. АП СССР, сер. 3, геоботаника, 4. — 1946. Материалы к географии сфагновых мхов Коми АССР. Бот. ж. СССР, <П, 1. — 1947. Сибирская лиственница. Мат. к познанию фауны и флоры СССР, отд. бот., нов. сер., 2 (Ю). 1947а. Реликты ледниковой
флоры на р. Верхней Лоне. Бюлл. МОПП, отд. биол., 52, 4. — I960. К характеристике лиственничных лесов Алданского нагорья. Тп. Пнет. леса АП СССР, 3. — 1953. Еловые леса Сродного Сихотэ-Алиня. В кн.: II. В. Дылиси П. Б. Виппер, Леса западного склона среднего Сихотэ-Алиня, М.
Дылио Н. В. и 11. Б. Виппер. 1953. Леса западного склона сродного Сихотэ-Алиня.М.
Елагин И. II. 1951. Грушевые леса северо-западного Кавказа. М.
Еленевскнй Р. А. 1924. Ънекие луга. Перспективы мелиорации основных типов кормовой площади на фоно генезиса долины р. Оки. М. — 1929. Пермский луговой массив. Пэв. Луг. инет., 4-6. — 1929а. Пойма р. Сулы от г. Л убей до с. Жов-нин (Лувенского и Кременчугского око.). В кн.; Тп. совощ. по*вопросам луговедения и опытного луговодства, 2, М. —* 19296. Пойма р. Суры от истоков до Пензы. Изв. Луг. инет., 1—3. - 1936. Вопросы изучения и освоения пойм. М. — 1936а. К вопросу о происхождении лугов. Учен. зап. Горьк. унив., 5. — 19366. Пойма р. Оки в пределах Московской обл. Уч. зап. Горьк. унив., 5. — 1938. Динамика ландшафтных смен от Алтая к Тянь-Шаню. Бюлл. МОИП, отд. биол., 47, 3. — 1939. Горнолуговыо этюды Кавказского заповедника. Тр. Кавказск. заповеди., 2. — 1939а. Прошлое лугов Южного Алтая. Природа, 9к _ 1940. Высокогорный продольный ландшафтный профиль северного склона Западного Кавказа. Бюлл. МОИП, отд. биол., 49, I. — 1940а. Горные луга Евразии как ландшафтно-географическое явление. Землеведение, нов. сер., 1 (41). — 19406. Кормовые ресурсы Рудного Алтая. В кн.: Сельское хозяйство Рудного Алтая, М.—Л. — 1940в. Основные орографические и ландшафтные черты Рудного Алтая. В кн.: Сельское хозяйство Рудного Алтая, М.—Л.
Еленевский Р. А. и Е. В. Еленевская. 1927. Тетеревская пойма на фоне Днепровско-Припятского пойменного ландшафта. Изд. Полесск. с.-х. опытн. станции, отд. полевод., 14/28, — 1928. Геоботанический очерк заливных лугов Козаровичской поймы р. Днепра. Козаровичская опытно-мелиор. луг. станция, 1, Киев.
Еленевский Р. А., А. И. Кремнев, А. А. Шахов и Т. А. Работноп. 1933. Природные ресурсы южной Якутии в связи с социалистической реконструкцией сельского хозяйства. М—Л.
Еленкин А. А. 1901. Кочующие лишайники пустынь и степей. Изв. СПб. бот. сада, 2, 1. — 1914. Пресноводные водоросли Камчатки. В кн.: Тр. Камчатской экспедиции Ф. 11. Рябушинского, бот. отд., 2, М.
Елиашевич О. А. (Ел1ашович). 1927. Матер1али до флори долини р. Самари. Тр. с.-г. боташки, 1, 3. — 1937. (1936). Луки середнього Дншра (Курил1вськ1 плавш). 36. 5об1т Дншропетр. бот. саду, 1 — 1939. Растительность речных долин Приазовья.
б. праць Дншропетр. бот. саду, 4.—1939а (1938). Степова рослинтсть Л1вобе-8ежжя середнього Дншра (р1знотравно-типчинно-ковилов! степи). 36. праць [шпропетр. бот. саду,
Елмзарьева М. Ф. 1951. Луговая растительность Томской обл. как кормовая база животноводства. Тр. Томск, унив., 114.
Елисеев Б. Н. 1932. Геологическое описание пересечения о. Сахалина по линии рр. Вал и Тенги. Отчет по исследованиям 1930 г. Тр. Нефтяного геол.-развед. инет., сер. А, 14.
Еремнн Г. Г. 1947. Географический очерк Костромской обл. Кострома.
Еремин Г. П., Н. С. Конюшков, С. П. Смелов. 1950. Улучшение и использование сенокосов и пастбищ нечерноземной полосы. М.
Жадовспй А. Е. 1929. Растительность Заветлужья Ветлужского у. Производительные силы Нижегородок. губ., 13.
Жудова П. П. 1950. О поли Stipa pulcberrima С. Koch на степях Башкирского Пред-уралъя. Бюлл. МОИП, отд. оиол., 55, 2. — 1951. Геоботанический очерк север-
ноги отрезка государственной лесной полезащитной трассы Камышин -Сталинград. Бюлл. МОП 11, отд.биол., 56, 2. — 1951а. Растительность Волжско-Кумско-Герского междуречья и прилегающих склонов Ергенинскои и Ставропольской возвышенностей. Вести. Моск, уннв., 11.
Жуков А. Б. 1969. Дубравы УССР и способы их восстановления. В кн.: Дубравы СССР, 1. (Всесоюзн. 11.-иссл. инет. лесн. хоз., 28), М.
Зайцев Б. Д. 1933. О распределении сосновых и еловых насаждении в Северном крае в зависимости от почвенных условий. В кн.: Мат. Второй конференции по изуч. производит, сил Северного края, 2, Архангельск.
Зайцева М. Г. 1949. Об определении возраста растений-подушек. Сообщ. Тадж. фил. АН СССР, 17. 1949а. О развитии формы у растений-подушек. Сообщ.
Тадж. фил. АН СССР, 17.
Закиров К. 3. 1939. Полынная и вьюнковая растительность 11 шпанской равнины («стони») и их экологические особенности. Тр. Узбекист. унив., 22, 8. —1939а. Растительность предгорий (адыров) южного Узбекистана в пределах Каршин-ского района. Тр. Узбекист. унив., 12, 8. — 1947. К проблеме зональности и терминологии ботанической географии в Средней Азии. Высотные пояса долины Зеравшана. Вюлл. САГУ, 25.
Закржевскнй Б. С. и Е. 11. Коровин. 1935. Экологические особенности главнейших растении Ветнак-дала. Тр. САГУ, сор. Vlll-в, бот., 23.
Зактрегер И. Я. 1927. Тугайные леса нижнего течения р. Аму-Дарьи. Мат. Особого комитета АН СССР по иссл. союзы, п авт. республик, 10, Казахстанская сер.
Залензкпй О. В. 1948. О температурном п водном режиме растений-подушек. Вот. ж., 33, 6. - 1949. Краткие итоги ботанических исследований Памира. Бот. ж., 34, 4. — 1950. Пути растениеводческого освоения высокогорий Памира. В кн.: Пустыни СССР и их освоение, I, М.—Л.
Залесский К. М. 1918. Залежная и пастбищная растительность Донской обл. Ростов-на-Дону — 1918а. Материалы к познанию растительности Донских степей. Ростов-на-Дону.
Запри гнева В. И. 1948. Березняки Горного Зеравшана. Сообщ. Тадж. фил. АН СССР, 10. — 1954. Биологические особенности фисташки в связи с ее культурой в Таджикистане. Бот. ж., 39, 3.
Запятова А. А. (О. О. 3 а п’я т о в а) 1951. Полонини Перечинсьского окр. Закарпат-сько1 обл. Бот. ж. АН УРСР, 7, 4.
Зарецкий В. В. 1921. Пойма р. Десны в окрестностях г. Чернигова, ее флора и хозяйственное значение. Чернигов.
Захаров Л. 3. 1927. Плавни низовьев р. Кумы, как гнездилища перелетной саранчи. (Естеств.-ист. очерк). Изв. Сев.-Кавказ, краев, станции защ. раст., 3.
Захаров Н. Г., И. Б. Ревут, В. Л. Леонтьев, В. П. Дубровский и Л. С. Доценко. 1954. Новый способ закрепления подвижных песков. М.— Л.
Захарова Л. Я. 1954. Ландшафты Волго-Ахтубинской поймы и пустынной степи западного Прикаспия в пределах землепользования колхоза им. Сталина. Уч. зап. Моск, унив., 170.
Зворыкина К. В. 1953. Фенология и подземные части основных растительных ассоциаций Ергеней. Тр. Комплекс, научн. экспед. по вопросам полезащ. лесораз-вед. АП СССР, 2, 5.
Зедельмейер О. М. 1927. Распространение торфяных болот и сфагновых мхов на Кавказе. Торф, дело, 7.
Зеленецкпй 11. М. 1891. Отчет о ботанических исследованиях Бессарабской губ. 1. Одесса.
Зелинский Ф. Ф. 1910. Растительность Кушмурунской вол. Петропавловского у. Акмолинской обл. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1909 г., ч. 2, 2. — 1912. Очерк растительности средней части Атбасарского у. Акмолинской обл. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1910 г., ч. 2, 1.
Зеров Д. К. 1930. Ол1готро^н1 сфагнов! болота швшчно-захщньо! частили Коростен-ськоТ окр BicTi Укршсторфу, 1. — 1938. Болота УРСР. Рослитпсть 1 стра-тиграф!я. Кшв.
Зов И. (1. Зоэ). 1933. Рослинтсть Михайл1всько! шлини на Сумщиш. Ж. Bio-бот. циклу ВУАН, 5-6.
Зозулин Г. М. 1950. «Стрелецкая степы. Центрально-черноземный заповедник им. проф. В. В. Алехина. Курск.
Золотарев С. А. 1950. Взаимодействие между почвой и аянским хвойным лесом. Хабаровск. — 1953. О влиянии главнейших хвойных пород Дальнего Востока на почву. Почвоведение, 2.
Золотовскпй М. В. 1938. Очерк растительности Алтайского Государственного заповедника. Тр. Алтайск. заповеди., 2.
Зонн С. В. 1950. Горно-лесные почвы северо-западного Кавказа. М.—Л. — 1954. Влияние леса на почвы. М.
Зонн С. В. и В. Л. Леонтьев. 1942. О почвообразовательном значении саксаула на песках пустыни Кара-Кумы. Почвоведение, 8.
Зонов Б. В. 1931. Описания бассейна р. Омолона. Мат. партии по иссл. р. Коломы, 4, Иркутск.
Зубков А. И. 1932. Тундры Гусиной Земли. Тр. Бот. муз. АН СССР, 25. — 1935. Продолжительность вегетационного периода на северном острове Новой Земли. Арктика, 3. — 1940. Геоботаническая характеристика растительности окрестностей Игарки в связи с с.-х. освоением. Тр. Н.-иссл. инет, полярн. землед., животн. и промысл, хоз., сер. растениеводства, 1.
Иваненко Б. И. 1925. Типы насаждений Государственного Крымского заповедника. Тр. по изуч. заповеди. Отд. охр. природы Главнауки НКП РСФСР, 1. — 1952. Дубравы Крыма. В кн.: Дубравы СССР, 4. (Всесоюзн. Н.-иссл. инет. лесн. хоз., 31), М.
Иванов В. В. 1948. Колки и дубравы южных отрогов Общего Сырта. Изв. ВГО, 80, 6. — 1953. К изучению лесорастительных условий долины р. Урала и прилежащих территорий. Геогр. сб. ВГО, 2. — 1953а. Черная ольха в долине р. Урала. Природа, 2. — 1954. Кормовая база и пути ее реконструкции в северной части междуречья Волга—Урал. В кн.: Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР, М.—Л.
Иванов Д. А. 1953. Луга и пастбища. М.—Л.
Иванов Н. Н. 1948. Л андшафтно-климатические зоны земного шара. Зап. ВГО, нов. сер., 1.
Иванова А. В. 1946. Можжевеловые редколесья Южной Армении. Тр. Бот. инет. АН АрмССР, 4. — 1949. К вопросу возобновления фисташника в Армении. ДАН АрмССР, 10, 3. — 1949а. Перспективы использования можжевеловых редколесий в Армении. Изв. АН АрмССР, биол. и с.-х. науки, 2, 2.
Иванова Е. Н. и Ф. Я. Левина. 1952. Солонцовые комплексы Прикаспия. Почвоведение, 10.
Иванова Е. Н., И. П. Герасимов, С. С. Неуструев и О. Э. Кнорринг-Неуструева. 1930. Почвенные и ботанико-географические исследования в Кара-Калпакской автономной области. Тр. Почв. инет. АН СССР, 3—4.
Иванова Н. А. 1935. Растительность поймы Тобола в б. Курганском окр. (юго-запад-
. ная Сибирь). Бот. ж. СССР, 20, 5.
Иванова Н. А. и И. М. Крашенинников. 1934. К истории развития растительных ландшафтов Западной Сибири. Землеведение, 36, 1.
Ивашкевич Б. А. 1926. Леса Советского Сахалина. Экономическая жизнь Дальнего Востока, 4. — 1927. Леса восточного побережья Советского Сахалина. Экономическая жизнь Дальнего Востока, 4. — 19z7a. Леса северного Сахалина. В кн.: Сахалин. Горно-геол, экспед. 1925 г. (Мат. по общей и прикл. геологии, 112), Л. _ 19276. Типы лесов Приморья и их экономическое значение. В кн.: Производительные силы Дальнего Востока, 3, Растительный мир, Хабаровск—Владивосток. — 1929. Девственный лес, особенности его строения и развития. Лесн. хоз. и лесн. пром., 10 (73). — 1933. Дальневосточные леса и их промышленная будущность. М.—Хабаровск.
Игошина К. Н. 1927. Растительные сообщества на аллювиях Камы и Чусовой. Тр. Биол. н.-иссл. инет, при Пермск. унив., 1, 1. — 1930. Растительность северной части Верхнекамского окр. Уральской обл. Тр. Биол. н.-иссл. инет, при Пермск. унив., 3, 2. — 1931. Высокогорная растительность Среднего Урала. Ж. Русск. бот. общ., 16, 1. — 1934. Ботаническая и хозяйственная характеристика оленьих пастбищ в районе Обдорской зональной станции. Сов. оленеводство, 1. —1935. Оленьи пастбища Полярного Урала в верховьях рр. Лонгот-Юган и Щучьей. Сов. оленеводство, 5, (прилож. 1). — 1944. Растительность Среднего Урала. Сов. бот., 6. — 1951. К изучению растительности Енисейского кряжа. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 7. — 1952. Растительность субальп Среднего Урала. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 8.
Ильин М. М. 1914. Абаканская степь. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1913 г, Пг. — 1916. Растительность водораздельной полосы Чулым Чичкаюл. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1914 г., Пг. — 1927. Растительность Эльтонской котловины. Изв. Главн. бот. сада, 26, к. — 1928. Очерк растительности Хобдинского района. Мат. Комиссии экспед. иссл. АН СССР, 5, Казахстанская сер. — 1936. Растительность Среднего Урала. В кн.: Природа Урала, Свердловск.
Ильин М. М. и Р. Ю. Рожевиц. 1928. Очерк растительности районов рр. верхней Эмбы, Темира и Чегана. Мат. Комиссии экспед. иссл. АН СССР, 5, Казахстанская сер.
Ильинский А. П. 1913. Геоботаническое исследование лугов Тверской губ. Тверь. — 1929. Методы и задачи изучения географического распространения древесных пород в СССР. Тр. по прикл. бот., генетике и селекц., 21, 3. — 1936. (А. Р. II-j i n s k i). Zur Phytocoenologie der Larchenwaldcr der Ilmengcbirges. Ber. Schweiz. Bot. Ges., 46. — 1937. 20 лет советского геоботанического картирования. Сов. бот., 5. — 1939. К вопросу о северной границе хвойно-широколиственных лесов в Европейской части РСФСР. В кн.: Президенту АН СССР акад. В. Л. Комарову к 70-летию со дня рождения и 45-летию научн. деятельности, Л. — 1945. Растительность советских Карпат. Вюлл. МОИН, отд. биол., 50, 3—4.
Ильинский А. П. и Е. В. Шиффере. 1936. О фитогеографическом картировании в СССР. Сов. бот., 5.
Ильинский Н. В. 1915. Луга юго-западной части Вологодской губ. в с.-х. отношении. 1. Долинные луга. Вологда.
Иорданская И. Н. и И. Г. Серебряков. 1954. О морфогенезе жизненной формы кустарника на примере бересклета бородавчатого—Evonymus verrucosa Scop. Бот. ж., 39, 5.
Кабанов Н. Е. 1935. Библиографическая сводка материалов по растительному и почвенному покрову Дальневосточного края за последнее десятилетие. Тр. Дальневост. фил. АН СССР, сер. бот., 1. — 1937. Типы растительности южной оконечности Сихотэ-Алиня. Тр. Дальневост. фил. АН СССР, сер. бот., 2. — 1940. Лесная растительность Северного Сахалина. Владивосток. — 1946. Растительность о-ва Петрова на Дальнем Востоке. Бюлл. МОИП, отд. биол., 51, 4—5. — 1950. Ботанико-географические районы Сахалина. Сообщ. Дальневост. фил. АЙ СССР, 1.
Каден Н. Н. 1940. Очерк растительности Казацкой степи под Курском. Тр. Центр.-черноземн. заповеди., 1.
Казанский А. В. 1952. Возрастные изменения почв и луговых угодий озерно-речной поймы в пустынных эндореических водосборах Казахстана. М.
Казанский А. Ф. 1932. К микрофлоре Новой Земли. Тр. Полярн. комиссии АН СССР, 7.
Какс А. Р. 1913. Луга по нижнему течению р. Меши. Тр. по обслед. лугов Казанской губ., 1.
Калашников Л. Н. 1927. К характеристике боровой растительности Кададинской лесной дачи Кузнецкого у. Саратовской губ. Изв. Саратовск. инет, сельск. хоз. и мелиор., 3. — 1929. К характеристике сосновых ассоциации Кузнецкого окр. Средне-Волжской обл. Изв. Саратовск. инет, сельск. хоз. и мелиор., 5.
Калинина А. В. 1948. Растительность Чуйской степи на Алтае. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 5. — 1951. Кустарниковые заросли в районе распространения ореховых лесов на склоне Ферганского и Чоткальского хребтов Западного Тянь-Шаня. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 7. — 1954. Стационарные исследования пастбищ Монгольской Народной Республики. Тр. Монг, комиссии АН СССР, 60.
Камышев Н. С. 1948. (на пер. 1949) Аспекты Хреновской степи и их происхождение. Тр. Воронежск. унив., 15. — 1948а. Растительность Воронежской обл. Воронеж.
Капелысин В. Ф. 1910. Очерк растительности окрестностей оз. Денгиз и р. Терс-Аккан, Атбасарского у., Акмолинской обл. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1908 г., ч. 2, 5.
Караваев М. Н. 1945. Краткий анализ флоры степей Центральной Якутии. Бот. ж. СССР, 30, 2.
Карандина С. Н. 1949. Растительность стационара в липово-дубовом лесу. Уч. зап. Ленингр. унив., сер. биол. наук, 17. — 1950. Корневые системы древесных пород широколиственных лесов лесостепи. Уч. зап. Ленингр. унив., 134, сер. биол. наук, 25. — 1954. Распределение подземных частей растений в некоторых ассоциациях Хибинской тундры. Бот. ж., 39, 6.
Карасев И. И., И. П. Минина, Л. Г. Раменский, С. П. Смелов и И. А. Цаценкин. 1941. Сенокосы и пастбища. М.
Карелин Г. С. 1883. Экспедиция для осмотра восточных берегов Каспийского моря в 1832 г. Зап. РГО; 10.
Карпов В. Г. 1952. О лесной растительности предгорий Лабинско-Тебердинского окр. северо-западного Кавказа. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 8. — 1953. Современное состояние естественных и искусственных лесных насаждений в районе трассы государственной лесозащитной полосы. [Сталинград—Степной— Черкесск]. Тр. Комплекс, научн. экспед. по вопросам полезащ. лесоразвед., 2, 7.
Карягин И. И. 1930. Очерк древесной растительности Кубинского у. Баку.
Катышевцева В. Г. 1955. Растительность побережья Каспийского моря в междуречье Волга—Урал. Алма-Ата.
878
Каульбарс А. В. 1881. Низовья Аму-Дарьи, описанные по собственным исследов^ в а7310Г»23ОэтетГоб исследовании болот Богородского у. Московской губ. Ма,
Кац Н. Я. 1922. Отчет оо ис д _ Материалы к геоботаническим исслп ‘ по опытн. Делу Моск. iyo.,^- в ^19 и 1920 гг Изв н "ед0.
ваниям болот Ива™»^ 0 типах олиготрофных сфагновых болот ЕврОпеа^ торф. Инуи-3-4„тной имеридиональной^зональности. Тр. Бот. н.-Лл ' Д’* России и их широт рд в _ 1929 Растительность правобепп^' при физ.-мат. фак. 1 НижегопОдокой губ. Производительные силы НюиТЬя Красно-Баковского уч Нижегородско ^Р^ N^^H^ero-
P°^K- ^Rot Centralbl бб, 2. — 1930а. Die grundlegenden pesetzmassigkeE' л v r^Hon iind der Begriff der Assoziation. Beitr. zur Biologie der PfIanZen ЙГ2 - 1932 Болота Биробиджанского района Дальневосточного края. Мел^ 50’ 2‘ 1 1932а К вопросу о районировании болот Средней ПечоЛ;
^птИЯж ЖФР 17 5—6 - 19326? Я JК а Л) Zur Kenntnis der MoorX Fernen bstens Вег’. d. Deutsch. Bot. Ges., 50. - 1936. Болота Европейской части Союза ССР 2 Водные и болотные растительные ценозы и закономерности «5 Структуры Бот. ж СССР, 21, 4. - 1937. Типы болот и их размещение на терри* тппии Евпопейской части СССР. Землеведение, 39 4-5. — 1939. Болота ни-аовьев р Оби В кн.: Президенту АН СССР акад. В. Л. Комарову к 70-летию со дня рождения и 45-летию научи, деятельности, Л. — 1941. Болота и торфя. ники. М.__ 1948. Типы болот СССР и Западной Европы и их географическое
Кац ^^яГн^В. Кац. 1948. Стратиграфия торфяников Приобского Севера. Тр. Ко-миссии по изуч. четвертичн. пер. АН СССР, 7, 1.
Кац Н. Я., Л. В. Кудряшев и В. М. Эпштейн. 1935. О торфяниках дельты Северной Двины. Землеведение, 37, 4. „
Кац Я. Я. и Ц. И. Минкина. 1936. Торфяные болота северной части бассейна р. Печоры. Тр. Центр, торф, опытн. станции, 1,
Кашкаров Д. Н., А. Жуков и К. В. Станюкович. 1937. Холодная пустыня Центрального Тянь-Шаня. Л. о
Кашменский Б. Ф. 1906. Растительность меловых обнажении Старобельского у. Харьковской губ. (бассейн р. Айдара). Тр. СПб. бот. сада, 26, 1.
Квецинский В. 1917. О типах лесонасаждений в Изосимовской, Нагорной и Заречной дачах Костромского у. Костромской губ. Тр. Костромск. научн. общ. по изуч. местного края, 6.
Келлер Б. А. 1903. Из области черноземно-ковыльных степей. Ботанико-географические исследования в Сердобском у. Саратовской губ. Тр. Общ. естествоиспыт. при Казанск. унив., 37, 1. — 1907. Очерки и заметки по флоре юга Царицынского у. В кн.: Н. А. Димо и Б. А. Келлер, В области полупустыни, Саратов. — 1911. Ботанико-географические исследования в Зайсанском у. Семипалатинской обл. Очерк растительности «Кальджирской долины». Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1908 г., ч. 2, 10,— 1912. Ботанико-географические исследования в Зайсанском у. Семипалатинской обл., ч. 2. Тр. Общ. естествоиспыт. при Казанск. унив., 44, 5. — 1914. По долинам^ и горам Алтая. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1910 г., ч. 2, 6.—1916. К вопросу о классификации русских степей. Русский почвовед, 16/18. — 1921. Опыты и некоторые общие выводы по экологии солончакового растения Salicornia herbacea L. Вести, опытн. дела, 1—-2, Воронеж. 1923. Значение и направления геоботанических работ. Ьюлл. Всеросс. конф, по изуч. естеств. производит, сил страны, 6. — 1923а. астительныи мир русских степей, полупустынь и пустынь. Очерки экологи-
И Фитосо^иол®1ические. Вып. 1. Введение. Общая часть. Тр Гос. солон-Лневн ЛвХпюаСТ’’ ~ 1^6‘ Геоботаническая карта Воронежской губ. В кн.: т™ ЮЗН- сЭезда Ваников в Москве в январе 1926 г., М. - 1926а. нии Р8Боа stOTonhvlln0raHH4eCKH« в экологические заметки. I. О распростране-Avena“ б'гСаРатовск°й губ. II. Степь с преобладаем 5. — 19266. Экологичргки^п^ У' Саратовской губ. Зап. Воронежск. с.-х. инет., почвах полупустынь по т!ияыЫ Раститольных ассоциаций на резко засоленных 5-7. - ЖУ(В“кпочвенных анализов. Бюлл. почвоведа, pean Russia, 'journ. of Ecology 'fetation of the plains of Ewj
почв полупустыни. В кн • ЮбпттО.“.'т/k J927a- Из ЖИЗНИ растении засоленных Л. - 1928. К ха^вдет^ике сборник’ посвяЩенный И. П. Бородину, ного чернозема. Зап Одесск ₽?^втвльиости и почв солонцов в подзоне мош степи. Типчаково- и ковшьип-л^ Твствоиспыт-’ 44 ~ 1931. в Хреновскои земной обл., М.—Л. — 1931а УкпЫв степи- в кн.: Степи Центра л ьно-Черя®" в балке Таловой около ДокучяАи,Лп'-1ЬНЫв (лоссингианново-тырсовые) степ® «по Докучаевой опытной станции и в балке Гия»*
в районе Вейделевки. В кн.: Степи Центрально-Черноземной обл., М.—Л. — 19о1б. Степь как производительная сила природы и значение степных заповедников. В кн.: Степи Центрально-Черноземнойобл., М.—Л.— 1931в. Степи на рендзинном черноземе и черноземовидной супеси. В кн.: Степи Центральночерноземной обл., М.—Л. — 1931г. [Редактор.) Стени Центрально-Черноземной обл. (Степные сенокосы и пастбища). М.—JI. — 1936. [Редактор]. Растительность Каспийской низменности между рр. Волгой и Уралом. Тр. СОПС АН СССР, Волжско-Каспийская сер., 11. — 1938. Главные типы и основные закономерности в растительности СССР. В кн.: Растительность СССР, I, М.—Л. — 1940. Растительность засоленных почв СССР. В кн.: Растительность СССР, 2, М,—Л.
Кеппен Ф. П. 1885. Географическое распространение хвойных дерев в Европейской России и на Кавказе. Зап. Акад. Наук, 50, (прилож. 4.).
Кербабаев Б. Б. 1954. Джангильная растительность поймы Аму-Дарьи. Тр. Инет, биол. АН ТуркмССР, сер. бот., 1.
Кепдовели Н. II. 1935. [Основные типы растительного покрова Грузии]. Тбилиси. (На груз. яз.).
Киреев А. Ф. 1952. Ботанический очерк лугов в среднем течении р. Иловли. Тр. Ста-лингр. с.-х. инет., 2.
Кириченко Т. Ф. 1948. Луга среднего течения р. Самары. Научн. зап. Днепропетр. унив., 32.
Клеопов Ю. Д. 1927. Хомутовський степ (Мар1юп!льська окр.). Охорона пам’яток природи на УкраТш, 1. — 1933. Залишки степово! рослинностх на Ки1всько! височиш. Журн. Bio-бот. циклу ВУАН, 5—6. — 1933а. Рослиние вкриття швденно-захщно! частини Донецького кряжа (кол. Сталшсько! окр.). Bicfl. Ки!в. бот. саду, 15. — 1934. Геоботашчний есюз л!вобережжя середньо! Над-дшпрянщини. Ж. 1нст. бот. ВУАН, 2 (10). — 1934а. Рослишпеть Йарл1вського стенового заповщника ВУАН. Bien. Ки!’в. бот. саду, 17. — 1935. Про геомор-фогенетичш мотив! розвитку рослинного вкриття УССР. Ж. 1нст. оот. УАН, 5 (13). — 1941. Основные черты развития флоры широколиственных лесов Европейской части СССР. Мат. по ист. флоры и растит. СССР, 1.
Клеопов Ю. Д. и Е. М. Лавренко. 1933. Сучасний стан класифшацп украхнських стешв. Ж. Bio-бот. циклу ВУАН, 5—6. — 1938. Легенда до карти рослинносп УРСР. Геобот. зб. 1нст. бот. АН УРСР, 2.
Клопотов Б. Н. 1910. Кузнецкий у. Томской губ. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1909 г., СПб.
Кнорринг О. Э. 1915. Ботанико-географический очерк Намаганского у*. Тр. Почв.-бот. эксщд. Переселенч. упр. по иссл. колониз. районов Аз. России в 1912 г., ч. 2, 5. — 1923. Ботанико-географические наблюдения на Памире в 1913 г. Изв. Главн. бот. сада, 22, 1. — 19*8. Очерк растительности районов рр. Сагида и нижней Эмбы в пределах Адаевского и Темирского уездов Актюбинской губ. Мат. Комиссии экспед. иссл. АН СССР, Казахстанская сер., 5. — 1932. Растительность западного склона Южного Урала в бассейне р. Большого Ика. Тр. СОПС АН СССР, Башкирская сер., 2. — 1934. Растительность Нуратаусских гор. Изв. ГГО, 66, 5.
Кнорринг О. Э. и 3. А. Минквиц. 1912. Растительность Андижанского у. В кн.: Предварит. отчет о бот. иссл. в Сибири и Туркестане в 1911 г. СПб. — 1912а. Растительность Аулие-Атинского у. Сырдарьинской обл. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1909 г., ч. 2, 6. — 1913. Растительность Перовского у. Сыр-Дарьинской обл. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1910 г., ч. 2, 2.
Ковда В. А. 1946. Происхождение и режим засоленных почв, 1. М.— Л. — 1950. Почвы Прикаспийской низменности (северо-западной частиц. М.—Л.
Ковда В. А., В. В. Егоров, А. Т. Морозов и Ю. П. Лебедев. 1954. Закономерности процессов соленакопления в пустынях Арало-Каспийской низменности. Тр. Почв. инет. АН СССР, 44.
Коган Ш. И. 1949. Эфемеры Южного Усть-Урта. Изв. Туркм. фил. АН СССР, 2. —1954. Растительность Южного Усть-Урта. Тр. Инет. биол. АН ТуркмССР, бот. сер., 2.
Кожевников А. В. 1935. Материалы по экологии буковых лесов Западного Закавказья. Сов. бот., 5. — 1936. Материалы по динамике ярусности луговых травостоев. Бюлл. МОИП, отд. биол., 45, 3. — 1937. Наблюдения 8а сезонной и годовой изменчивостью растительного покрова колхидского дубового леса. Бюлл. МОИП, отд. биол., 46, 5.
Кожевников П. П. (К о ж е в н i к о в). 1931. Типи nicy та nicoBi асоц1яцн Под1лля. Всес. н.-досл. 1нст. л1сового госп. та агролшомелюр., сер. наук, видань, 10. —1936. Естественно-исторические районы и типы.лесов Украины. В кн.: Лесные культуры (Укр. н.-иссл. инет. лесн. хоз., 18), Киев. •— 1939. Дубовые леса лесостепи Европейской части СССР. Тр. Всесоюзн. Н.-иссл. инет. лесн. хоз., 1.
Козлова А. В. 1951. О накоплении нитратов в термитниках Туркмении. Почвоведение, 10.
Козо-Полянский Б. М. 1925. Сводная геоботаническая карта Центрально-Черноземной обл. Мат. по районированию Центр.-Черноземн. обл., 2, Воронеж. — 1927. Черноземный рододендрон, след древней растительности на Тимской гряде. Воронеж, краевед, сб., 4. — 1928—1929. (В. М. К о s о - Р о 1 j a n s k i) Gla-ziale Pflanzenrelikte auf dem Orel-Kurskiscben Plateau in Siiden der Mittelriissi-schen Hochebene, I—II. Vegetationsbilder, 19, 1—2, 7—8. — 1931. В стране живых ископаемых. Очерк из истории горных боров на степной равнине Центрально-Черноземной обл. М. — 1931а (В. М. Koso-Poljanski). Хего-therme Relikten am Flusse Tichaja Ssosna (Siidrussland). Vegetationsbilder, 22, 2. Колаковский A. A. 1933. Очерк растительности Ширванской степи. Тр. по геобот. обслед. пастбищ ССР Азербайджана, сер. А, зимние пастбища, 10. — 1937. Растительность Бзыбского известнякового хребта как кормовая база для животноводства. Тр. Инет, абхазской культуры, 9. — 1947. Фито ландшафты Абхазии и история их развития. Тбилиси. — 1948. Основные фитоландшафты приморской низменности Южной Абхазии. Тр. Тбил. бот. инет. АН ГрузССР, 12.
Колесников Б. П. 1936. Заповедник охотской флоры и фауны. Зап. Приморск, фил. ГГО, 6 (23). — 1937. Растительность хребта Тачинчжан. Вестн. Дальневост, фил. АН СССР, 24. — 1937а. Чозения [Ghosenia macrolepis (Turcz.) Кот.] и ее ценозы на Дальнем Востоке. Тр. Дальневост. фил. АН СССР, сер. бот., 2. — 1938. Растительность восточных склонов среднего Си1&тэ-Алиня. Тр. Сихотэ-Алинского заповеди., 1. — 1945. Обыкновенная сосна (Pinus silvestris L. s. 1.) на юго-восточной границе своего ареала. Бюлл. МОИП, отд. биол., 50, 5—6. — 1946. К систематике и истории развития лиственниц секции Pauciseriales Patschke. Мат. по ист. флоры и растит. СССР, 2. — 1947. Лиственничные леса Средне-Амурской равнины. Тр. Дальневост, базы им. акад. В. Л. Комарова АН СССР, сер. бот., 1. — 1948. О характере ландшафта Суйфуно-Ханкайского геоботанического округа. Мат. к изуч. природных ресурсов Дальнего Востока, 1. — 1951. Кедровые леса Приморского края. Владивосток. — 1954. Корейский кедр на советском Дальнем Востоке. Комаровские чтения Дальневост. фил. АН СССР, 4.
Комаров В. Л. 1897. Ботанико-географические области бассейна Амура. Тр. СПб. общ. естествоиспыт., 28. 1. — 1912. Путешествие по Камчатке в 1908—1909 гг. В кн.: Камчатская экспедиция Ф. П. Рябушинского, снаряженная при содействии РГО, бот. отд., 1, М. — 1917. Типы растительности Южно-Уссурийского края. Тр. Почв.-бот. экспед. Переселенч. ^гпр. по иссл. колониз. районов Аз. России в 1913 г., ч. 2, 2. — 1922. Краткий очерк растительности Сибири. Мат. для изуч. естеств.-производ. сил России, 46. — 1926. Введение в изучение растительности Якутии. Тр. комиссии АН СССР по изуч. ЯкАССР, 1. — 1928. Библиография к флоре и описанию растительности Дальнего Востока. Зап. Южн.-Уссур. отд. РГО, 2. — 1934. Ботанико-географический очерк хвойных деревьев (Gymnospermae) СССР. Бюлл. Тихоок. комитета АН СССР, 3. — 1940. Ботанический очерк Камчатки. В кн.: Камчатский сборник, 1, М.—Л.
Комаров Н. Ф. 1931. Степи юго-востока Россошанского окр. В кн.: Степи Центральночерноземной обл., М.—Л. — 1951. Этапы и факторы эволюции растительного покрова черноземных степей. Зап. ВГО, нов. сер., 13.
Комаров Н. Ф. и Е. И. Проскоряков. 1931. Западные степи Центрально-Черноземной обл. В кн.: Степи Центрально-Черноземной обл., М.—Л.
Коновалов Н. А. 1928. Лесные ассоциации северо-западной части Вартемягской дачи Парголовского учебно-опытного лесничества. В кн.: Природа и хозяйство учебно-опытн. лесничества Ленингр. лесн. института, М. — 1929. Типы леса подмосковных опытных лесничеств. Тр. по лесн. опытн. делу. Центр. Лесн. опытн. станции, 5. — 1930. Очерк растительности дельты р. Селенги. Тр. Комиссии АН СССР по изуч. оз. Байкал, 3. —1936. Очерк типов хвойно-широколиственных лесов в Брянском лесном массиве. Тр. Ленингр. общ. естествоиспыт., 65, 3. — 1949. Очерк широколиственных лесов центральной лесостепи. Уч. зап. Уральского унив., 10, биол.
Конрад П. 1948. Пойменные заболоченные луга Земгалии и повышение их урожайности. Изв. АН ЛатССР, 11.
Корещук К. Е. (К. 6. К о р е щ у к). 1939 (1938). Нарнс рослинност! долини р. Вов-чо1 (в межах: гнрло р. Вовчо!—с. Василыавка). 36. праць Дншропетр. бот. саду, 3.
Коравевин В. С. 1934. Растительность долины р. Алдана. Тр. СОПС АН СССР, Якутская сер., 16.
Коржинекий С. И. 1884. Предварительный отчет о ботанической экскурсии в дельту Ь Волги. Тр. Общ. естествоиспыт. прн Казанск. унив., 13, i. — 1888—1891. верная граница черноземно-степной области восточной полосы Европейской'
России в ботанико-географическом и почвенном отношении. 1—2. Тр. Общ. естествоиспыт. при Казанск. унив., 1 (1888) —18, 5; 2 (1891) — 22, 6. — 1892. Отчет об исследованиях Амурской обл. как земледельческой колонии. Изв. Вост.-Сиб. отд. РГО, 23, 4—5. — 1896. Очерки растительности Туркестана. Зап. Акад. Наук, сер. 8, 4. — 1898. (К о г s h i n s k y). Tentamen florae Rossiae orientalis. Зап. Акад. Наук, сер. 8, 7, 1. — 1899. Россия. Растительность. Вкн.: Энциклопедический словарь Брокгауза и Ефрона, 27 А, СПб. — 1901. Степи. В кн.: Энциклопедический словарь Брокгауза и Ефрона, 31А, СПб.
Кормовые ресурсы тундровой зоны СССР. 193о—19о4. Ч. 1, Сов. оленеводство, 1 (1934); ч. 2, Сов. оленеводство, 2 (1933).
Коровин Е. П. 1927. Заметка о растительности центральных Кара-Кум. Изв. Инет, почвовед, и геобот. САГУ, 3. — 1927а. Основные черты в строении растительного покрова горной и подгорной части Копет-Дага, преимущественно заключенной между станциями Гяуре—Келята. Изв. Инет, почвовед, и геобот. САГУ, 3. — 1934. Растительность Средней Азии и Южного Казахстана. М.—Ташкент. — 1934а. Сорная растительность хлопковых полей Средней Азии и меры борьбы с ней. Ташкент, инет, удобр. и агропочвовед., сер. научн., 1.—19346. Экологические типы пустынь Средней Азии и Казахстана в перспективах их хозяйственного освоения. В кн.: Хозяйственное освоение пустынь Средней Азии и Казахстана, М.—Ташкент. — 1935. Бетпак-дала как особый тип пустыни. Тр. САГУ, сер. VIII-в, бот., 27. — 1935а. Динамика растительности Бетпак-дала. Тр. САГУ, сер. VIII-в, бот., 26. — 1945. Итоги изучения ботанической географии Узбекистана и ее очередные задачи. Бюлл. АН УзССР, И—12. — 1950. Геоботаническая карта Узбекистана. В кн.: Юбилейный сборник, посвящ. 25-летию УзССР, Ташкент. [На титульном листе год издания не указан, на выпускных знаках помечено «подписано к печати 12-ХП 1949 г.»].
Коровин Е. П. и И. И. Гранитов. 1949. Растительный покров [Кара-калпакского Усть-урта]. В кн.: Усть-урт Кара-калпакский, его природа и хозяйство, Ташкент.
Коровин Е. П. и Д. Н. Каппсаров. 1934. Типы пустынь Туркестана. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 1.
Коровин Е. П. и Е. Е. Короткова. 1946. Типы растительности Средней Азии. Тр. САГУ, нов. сер., 8 (биол. науки, 2).
Коровин Е. П. и Б. А. Миронов. 1935. Главнейшие растительные ассоциации восточной Бетпак-дала и их распределение в зависимости от рельефа и почв. Тр. САГУ, сер. VIII-в, бот., 21.
Коровин Е. П. и В. А. Сел евин. 1935. Физико-географический очерк Бетпак-дала. Тр. САГУ, сер. ХП-а, геогр., 13.
Коровин Е. П. и С. А. Шувалов. 1948. О биогенной комплексности почвенно-растительного покрова в аридной зоне. Бюлл. МОИП, отд. биол., 53, 1.
Коровкин А. А. 1934. Геоботанический очерк Хибинского массива. Изв. ВГО, 66, 6. Королева А. С. 1940. Очерк растительности центральной части южного склона Гис-сарского хребта и ее естественно-кормовые ресурсы. Тр. Тадж. базы АН СССР, 8.
Короткий М. Ф. 1912. Баргузинские степи. В кн.: Предварит, отчет об организ. иисполн. работ по иссл. почв Аз. России в 1911 г., СПб. — 1912а. К вопросу о распределении лугов и лесов в зависимости от почв. Мат. по изуч. растит. Псковской губ., I. — 19126. Очерк растительности Зейско-Буреинского района Амурской обл. Тр. Амурской экспед., 16, бот. иссл. 1910 г., 3. — 1913. Еравинские степи. В кн.: Предварит, отчет об организ. и исполн. работ по иссл. почв Аз. России в 1912 г., СПб. — 1914. Кустанайские степи. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1913 г., СПб. — 1916. Экспедиция на р. Мую. В кн.: Предварит, отчет об организ. и исполн. работ по иссл. почв Аз. России в 1914 г., СПб. — 1916а. Степные явления в Баргузинской тайге. (Экспедиция на р. Мую). В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1914 г., СПб.
Корчагин А. А. 1926. Взаимоотношения травянистых и лесных ассоциаций в районе учебно-опытного Парголовского лесничества. Ж. Русск. Бот. общ., 11, 1—2.— 1929. К вопросу о типах леса по исследованиям в Тотемском у. Вологодской губ. В кн.: Очерки по фитосоциологии и фитогеографии, М. — 1932. Поемные луга р. Северной Двины в Черевковском районе и их хозяйственная оценка, тр. Бот. муз. АН СССР, 25. — 1933. Мезенско-Тиманский отряд. В кн.: Экспедиции Всесоюзн. Акад. Наук 1932 г., Л. — 1933а. Об основных понятиях тун-дроведения. Сов. бот., 2. — 1937. Растительность морских аллювиев Мезенского залива и Чешской губы. (Луга и луговые болота), тр. Бот. инет. АН СССР, сер. 3, геоботаника, 2. — 1940. Растительность северной половины Печорско-Ылычского заповедника. Тр. Печорско-Ылычск. заповеди., 2.— 1945. Происхождение темнохвойных лесов Евразии В кн.: Научная сессия Ленингр. университета 16—30 ноября 1945 г. Тезисы докладов по секции географ, наук, Л. — 1946»
К вопросу о принципах классификации лесных группировок. В кн.: Сборник научн. работ [Бот. инет. АН СССР], выполненных в Ленинграде за три года Великой Отечественной войны (1941 —1943), М.—Л. — 1954. Влияние пожаров на лесную растительность и восстановление ее после пожара на европейском Севере. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 9. — 1956. Еловые леса западного Притиманья в бассейне р. Мезенской Пижмы. Уч. зап. Ленингр. унив., сер. геогр. наук, 11.
Корчагина М. В. и А. А. Корчагин. 1931. Растительность Хибинских гор. В кн.: Путеводитель по Хибинским тундрам, Л.
Корякина В. Ф. 1951. Динамика роста, накопления массы и химического состава травостоя северодвинских пойменных лугов. Тр. Бот. инет. АН СССР, сер. 4, эксперим. бот., 8.
Косенко И. С. 1924. К познанию растительности лиманов и плавней Приазовского побережья Кубанского края. Тр. Кубанск. с.-х. инет., 1, 2. — 1928. Природные кормовые угодья бассейна р. Лабы. Тр. Северо-Кавказ. ассоц. н.-иссл. инет., 46. — 1930. Ботанико-географическая характеристика районов табаководства Кубанского и Майкопского округов. Гос. Пнет, табаковед., 75. — 1934. Материалы к познанию эколого-географической природы приазовских плавень. Тр. Всесоюзн. Центр, станции рисового хоз., 5.
Косец Н. И. (М. I. Косець). 1948 (1947). Буков! л!си Зах!дного Под!лля. Бот. ж. АН УРСР, 4, 3—4. — 1949. Нарис рослинност! НрськоТ частини Закарпатсько! обл. УРСР. Бот. ж. АН УРСР, 6, 1. — 1953. Нарис л!сово! рослинност! Льв!всько! обл. УкраТнсько! РСР. Бот. ж. АН УРСР, 10, 4. — 1954. Буков! л!си. В кн.: Рослиншсть Закарпатсько! обл. УРСР, Кшв.
Косинский К. К. 1916. Растительность юго-западной части Семипалатинского у. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1914 г., Пг.
Котов М. И. 1927. Ботанико-географические исследования в Причерноморских степях. В кн.: Наук. зап. по б’юл., Харюв. — 1927а. Ботан!чно-географ!чний нарис долины р. 1нгульця. Тр. с.-г. бот., 1, 3. — 1928 (1927). Ботанико-географический очерк растительности меловых обнажений по р. Осколу и его притокам. Ж. Русск. бот. общ., 12, 3. — 1928а. Новые материалы к растительности севера Арабатской стрелки. Ж. Русск. бот. общ., 13, 3—4. — 1930. Ботанико-географический очерк долины низовьев р. Самары. Тр. Гос. Ихтиол, опытн. станции, 6, 1, Херсон. — 1930а. Геоботанический очерк буковых лесов по р. Збруч. Ж. Русск. бот. общ., 15, 1—2. — 1931. Материалы к растительности долины р.Оскола между городами Купянском и Валуйками. Ж. Русск. бот. общ., 16, 5—6. — 1931а. Растительность расположенного близ Сиваша соленого оз. Оверьянов-ского и его берегов. В кн.: 25 лет научн.-пед. и общ. деятельности Б. А. Келлера, Воронеж. — 1933. Засоленные почвы и их растительность в Донецком бассейне. Природа, 8—9. — 1935. Рослиншсть щлини на п-ов! Чонгар. (Над-сивашшя). Ж. 1нст. бот. ВУАН, 3 (11). — 1938. Генезис солонцово-солончакового комплекса в Закавказье. Сов. бот., 3. — 1940. Геоботашчний нарис шв-денно! частини Вшницько! обл. Бот. ж. АН УРСР, 1, 2. — 1947. Высокогорная флора растительности Южного Урала. Сов. бот., 15, 3. — 1947а. Прсыи соснов! бори як проблема вторично! ооташчно! географ!!. Бот. ж. АН УРСР, 4, 1—2. — 1949. Рец. на ст. М. С. Шалыт «О растительности Присивашья. (К вопросу о существовании полынных степей на юге Украины и в Северном Крыму)», Бот. ж. УРСР, 6, 3. — 1953. Флора крейдяних вщелонень в басейн! С!верського Д!нця в межах УРСР у звязку з геолончним в!ком та л!толог!чним складом пор!д. Бот. ж. АН УРСР, 10, 2.
Котов М. и А. Прянишников (М. Котов та О. Прян!шн!ков). 1929. Мате-р!али до вивчення рослинности Понадсивашсько! смуги Украгни. Мат. досл!дж. земель мел!офонду на Мелиюшлыциш, 1. Харк!в.
Котов М. И. и В. Г. Танфильев. (М. I. К о т о в та В. Г. Т а н ф i л ь е в). 1934. Боташчно-географ!чний нарис долили р. 1нгула. Ж. Iнет. бот. ВУАН, 2 (10). Кочкин М. А. 1952. Леса Крыма. Симферополь.
Крайнев В. П. 1951. Дубравы Заволжья. В кн.: Дубравы СССР, 3, (Всесоюзн. Н.-иссл. инет. лесн. хоз., 30), М.—Л.
Красильников П. К. 1937. Почвы и растительность Горемыкского сельского совета Северо-Байкальского района. Тр. СОПС АН СССР, Восточно-Сибирская сер., 4.
Краснов А. Н. 1886. Геоботанические исследования в Калмыцких степях. Изв. РГО., 22,1. — 1886а. Заметки о растительности Калмыцких степей вообще и растительности песков в особенности. Изв. Геол, комитета, 5, 5. — 1886—1887. О зависимости между почвой и растительностью в черноземной полосе Европейской России. Мат. по изуч. русских почв, 2 (1886), 3 (1887). — 1888. Опыт истории развития флоры южной части восточного Тянь-Шаня. Зап. РГО, 19. — 1891. Материалы для флоры Полтавской губ. Тр. Общ. испытат. природы при Харьковск. унив., 24. — 1893. Рельеф, растительность и почвы Харьковской губ.
Харьков. — 1894. Ботанико-географический очерк Полтавской губ. В кн.: Мат. к оценке земель Полтавск. губ., СПб. — 1894а. Травяные степи северного полушария. Изв. Общ. любит, естествозн., антропол. и этногр., 83 (ошибочно вместо 81), (Тр. Геогр. отд., 1). — 1895. К флоре бассейна р. Чаквы. Тр. общ. испит, природы при Харьковск. унив., 28. — 1899. Основы землеведения, 4, 1, География растений, Харьков. — 1906. Очерки растительности и животного мира ближайших окрестностей г. Батума. В кн.: Батум и его окрестности, Батум.
Красовский П. А., С. Я. Соколов и Л. И. Соснин. 1938. Ореховые леса Южной Киргизии. Тр. Бот. инет. АН СССР, сер. 5, растит, сырье, 1.
Красовский П. Н. и Г. А. Глумов. 1945. Основные черты зацелинения залежей Троицкого лесостепного заповедника. Уч. зап. Молотовского унив.. 4. 2.
Красюк А. А. 1927. Почвы Ленско-Амгинского водораздела (Якутекий окр.). Мат. Комиссии по изуч. Якутск. АССР АН СССР, 6.
Крашенинников И. М. 1909. Тургайский у. Вторая Наурзумская вол. В кн.: Предварит. отчет о бот. иссл. в Сибири и Туркестане в 1908 г., СПб. — 1910. Бассейн р. Аргуни. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1909 г., СПб. — 1912. Пз наблюдений над,формациями леса и степи в Челябинском у. в 1910 г. Пзв. СПб. бот. сада, 12, 1. — 1913. К характеристике ландшафтов восточного Забайкалья. Землеведение, 20, 1—2. — 1919. Ботанико-географические группировки и геоморфология Южного Урала в их взаимной связи. Ж. Новочеркасск. отд. Русск. бот. общ., 1, 1. — 1922. Цикл развития растительности долин степных зон Евразии. Изв. Геогр. инет., 3. — 1923. Киргизские степи как объект ботанико-географического анализа и синтеза. Изв. Главн. бот. сада, 22, 1. — 1925. Растительный покров Киргизской республики. Оренбург. — 1928. Ботанико-географический очерк долины Дона от устья Донца до станицы Мелеховской. Тр. Главн. бот. сада, *41, 2. — 1932. Организация работ Южного геоботанического отряда Башкирской экспедиции АН СССР. Тр. СОПС АН СССР, Башкирская сер., 2. — 1936. Растительность Южного Урала. В кн.: Природа Урала, Свердловск. — 1937. Анализ реликтовой флоры Южного Урала в связи е историей растительности и палеогеографией плейстоцена Сов. бот., 4. — 1937а. Взаимоотношения леса и степи на южной окраине Уральской возвышенности. Землеведение, 39, 6. — 1939. Основные пути развития растительности Южного Урала в связи с палеогеографией северной Евразии в плейстоцене и голоцене. Сов. бот., 6—7. — 1939а. Физико-географические районы Южного Урала. I. Предгорья восточного склона и прилегающие части пенепленов. Тр. СОПС и Пнет, геогр АН СССР, Уральская сер., 7. — 1946. Опыт филогенетического анализа некоторых евразиатских групп рода Artemisia L. в связи с особенностями палеогеографии Евразии. Мат. по ист. флоры и растительн. СССР, 2.
Крашенинников И. М. и Васильев Я. Я. 1949. О лесостепи западного склона Южного Урала. Тр. Почв. инет. АН СССР, 30.
Крашенинников И. М., С. Г. Горшкова и Н. А. Иванова. 1928. Ботанико-географическая характеристика районов элементарных ландшафтов долины Нижнего Дона от устья до станицы Мелеховской. Тр. Главн. бот. сада, 41, 2.
Крашенинников И. М. и Н. А. Иванова. 1928. Краткий анализ фитосоциальных отношений среди ассоциаций долины Нижнего Дона от устья до станицы Мелеховской. Тр. Главн. бот. сада, 41, 2.
Крашенинников И. М. и В. М. Крашенинникова. 1905. Сосновые боры Челябинского у. Изв. СПб. бот. сада, 5, 4.
Крашенинников И. М. и С. Е. Кучеровская-Рожанец. 1941. Растительность Башкирской АССР. В кн.: Природные ресурсы Башкирской АССР, 1, М.—Л.
Крашенинников С. П. 1755. Описание Земли Камчатки, 1, 2. СПб.
Крекер Г. К. 1916. Луга и болота бассейна р. Лахвы Могилевской губ. но иссл. 1912— 1914 гг. 1. Луга. Юрьев.
Криштофовнч А. Н. 1907. К вопросу о растительности Крымской Яйлы. Изв. СПб. бот. сада, 7, 5—6. — 1910. Ботанико-географические исследования в области Березового хребта и Балаганской степи в Иркутской губ. Тр. Почв.-бот. экспед. Переселенч. упр. по’иссл. колониз. районов Аз. России в 1908 г., ч. 2, 3, — 1913. Очерк растительности Око-Ангарского края (Иркутской губ.). Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1910 г., ч. 2, 3. — 1941. Каталог растений ископаемой флоры СССР. В кн.: Палеонтология СССР, 12 (приложение), М.—Л.
Круберг Ю. К. 1937. О географическом распространении сосны Pinus silvestris в Азиатской части Союза ССР. Уч. зап. Ленингр. пед. инет. им. А. И. Герцена, 4, 2.
Крылов Г. В. 1953. Березовые леса Томской обл. и их типы. Новосибирск.
Крылов Г. В. и Н. Г. Салатова. 1950. Леса Западной Сибири. Новосибирск.
Крылов П. Н. 1878—1885. Материал к флоре Пермской губ. I—IV. Тр. Общ. естество-
испыт. при Казанск. унив., 6, 6 (1878); 9, 6 (1881); 11, 5 (1882); 14, 2 (1885). — 1891. Липа в предгорьях Кузнецкого Ала-Тау. Изв. Томск, унив., 3, отд. 2. — 1898. Очерк растительности Томской губ. В кн.: Научные очерки Томского края, Томск. — 1898а. Тайга с естественно-исторической точки зрения. В кн.: Научные очерки Томского края, Томск. — 1902. Краткий очерк флоры Томской губ. и Алтая. Изв. СПб. бот. сада, 2, 3. — 1913. Растительность в Барабинской степи и смежных с ней местах. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1912 г., СПб. — 1915. К вопросу о колебании границы между лесной и степной областями. Тр. Бот. муз. АН, 14. — 1916. Степи западной части Томской губ. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1913 г., ч. 2, 1 — 1919. Очерк растительности Сибири. Стат.-эконом, бюлл., 17, Томск. — 1926. Вишерскии край. Исторический и бытовой очерк Северного Приуралья. Урал, 8, Свердловск. — 1931. Фито-статистический очерк альпийской области Алтая. Изв. Томск, отд. Русск. бот. общ., 3, 1—2.
Крылов С. М. и Н. П. Потапьевский. 1914. Притоки р. Меши: р. Нурма, Иинка, Упшя. Среднее и верхнее течение р. Меши. Камская пойма. Мат. по иссл. лугов Казанск. губ., 2.
Крылова И. Л. 1953. Очерк растительности верхней границы леса в горах Крыма. Тр. Крымск, фил. АН СССР, 8,
Крюгер В. А. 1913. Ботанические и географические наблюдения в Семипалатинской обл. Тр. Общ. естествоиспыт. при Казанск. унив., 45, 3. — 1927. Главнейшие* черты растительности и почв южной части Чингистайской вол. Бухтарминского у. Семипалатинской губ. Тр. Общ. изуч. Казахстана, 8, 1. — 1927а. Фито-геогра-фические этюды (Каркаралинский у. Семипалатинской губ.). Зап. Семипалатинск. отд. ГРГО, 16, 2. — 1928. Растительность Семипалатинского опытного* поля. Зап. Семипалатинск, отд. ГРГО, 17, 2. — 1929. Геоботанические исследования в южной части Павлодарского у. (Семипалатинской губ.). Тр. Семипалатинск. окр. муз., 2. — 1935. Изменение растительности засоленных почв в некоторых районах различных фитогеографических зон. (Западная Сибирь и Северный Казахстан). Уч. зап. Пермск. унив., 1, 4. — 1947. К изучению флоры поймы Камы и ее формирования. Уч. зап. Молотовск. унив., 6, 1. —1952. О зональном распределении растительности поймы р. Камы. Бот. ж.г 37, 3.
Крюгер В. А. и А. А. Лютин. 1936. Почвенно-ботанические исследования в долине р. Камы и ее притоков в связи с построением Левшинской плотины. Изв. Пермск. биол. н.-иссл. инет, при Пермск. унив., 10, 9—10.
Кубанская 3. В. 1949. Растительность западной части пустыни Бетпак-Дала. Изв. АН КазССР, 74, сер. бот., 4.
Кудряшев В. В. 1922. Оршинский мох Тверской губ. Вестн. торф, дела, 1—2Г (прилож.). — 1922а. Отчет по исследованию болот по линии Московско-Вин-давской ж. д. Вестн. торф, дела, 1—2.
'Кудряшев С. Н. 1941. Растительность Гузара. (Опыт ботанико-растениеводческой региональной монографии). Ташкент.
Куэенева О. И. 1911. Эволюция болотных формаций Амурской обл. и опыт их класси-tHKaqHH. Почвоведение, 13, 2. — 1914. Палы таиги Приамурья. Лесн. ж., 44г -10.
Кузнецов И. В. 1911. Растительность Болонь-Оджальского района. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1910 г., СПб. — 1912. Растительность Канского у. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1911 г., СПб. — 1913. Растительность Амгунского района. Мат. к иссл. колониз. районов Аз. России, 2. — 1913а. Растительность Красноярского у. В кн.: Предварит. отчет о бот. иссл. в Сибири и Туркестане в 1912 г., СПб. — 1914. Минусинский у. Енисейской губ. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1913 г., Пг. — 1916. Растительность Обь-Енисейского водораздела в южной части Енисейской губ. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1914 г., Пг.
Кузнецов Н. И. 1891. Элементы Средиземноморской области в Западном Закавказье. Зап. РГО, 23, 3. — 1909. Принципы деления Кавказа на ботанико-географические провинции. Зап. Акад. Наук, сер. 8, 24, 1. — 1910. Нагорный Дагестан и значение его в развитии флоры Кавказа. Изв. РГО, 46, 6—7. — 1912. Опыт деления Сибири на ботанико-географические провинции. Изв. Акад. Наук,, сер. 6, 6,14. — 1913. В дебрях Дагестана. Изв. РГО, 49, 1—3. — 1923. Ботанико-географический очерк Рионской низменности. Изв. Н.-мелиор. инет., 3—4. — 1924. Ботанико-географическая карта Европейской части СССР. Почвоведение г 3—4. — 1927. Юго-западная, южная и юго-восточная границы распространения лиственницы (Larix sibirica Led.) в пределах Европейской России. В кн.: Юбилейный^ сборник, посвящ. И. П. Бородину, Л. — 1928. Геоботаническая карта Европейской части СССР, I, л. 14-и. Краткая пояснительная записка (Казан-
ский край), Л. — 1928а. Ботанико-географическое картирование Европейской России. Зап. Одесск. общ. естествоиспыт., 44.
Кузнецов Н. И. [Владимирский]. 1910. Бассейн левых притоков р. Шилки — Черной, Горбичанки и Желтуги. В кн.: Предварит, отчет о оот. иссл. в Сибири и Туркестане в 1909 г., СПб. — 1911. Растительность Лено-Киренского края. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1910 г., СПб. — 1912. Растительность бассейна левых притоков р. Шилки (Забайкальской обл.). Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России* в 1909 г., ч. 2, 4. — 1914. Очерк растительности Барнаульского у. В кн.: Предварит. отчет о бот. иссл. в Сибири и Туркестане в 1913 г., СПб. — 1915. Материалы по исследованию почв и растительности в средней части Томской губ. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России^ в 1912 г., ч. 2, 2. — 1915а. Очерк растительности Нарымского края Томской губ. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1911 г., ч. 2, 1. — 1916. Растительность Енисейской лесотундры. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1914 г., Пг. — 1932-Лайды в низовьях р. Енисея, их строение, образование и место в классификационной системе болотно-лесных образований. Тр. Полярн. комиссии АН СССР, 12.
Кузнецова Н. П. 1915. Сенокосные угодья средней части Опочецкого у. Мат. по организ. и культуре корм, площади, 11.
Кузьмина М. С. 1949. Эволюция растительности болот Барабинской низменности в связи с их осушением и естественным усыханием. В кн.: Тр. Юбилейной сессии, посвящ. столетию со дня рождения В. В. Докучаева, М.—Л. — 1953. Растительность Барабы. Тр. Почв. инет. АН СССР, 36.
Кузякин А. П., А. С. Корина, В. А. Николаев, М. Н. Першина, Г. И. Дохман, А. Н. Желоховцев, А. П. Шапошников, Ф. И. Левин и П. П. Жудова. 1951-Государственная защитная лесная полоса Камышин—Сталинград. Тр. Комплекс. научн. экспед. по вопросам полезащ. лесоразвед., 1, 1.
Куксин Н. В. (М. В. Кукс in). 1935. Природш кормов! вг!ддя УСРР. Харюв. Культиасов М. В. 1923. Очерк растительности гор Пистали-Тау. Тр. Туркестан, научн. общ., 1. — 1926. Очерк растительности Чардаринского района. Изв. Инет, почвовед, и геобот. САГУ, 2. — 1926—1927. Вертикальные растительные зоны в Западном Тянь-Шане. Бюлл. САГУ 14 (1926), 15 (1927). — 1938. Тау-сагыз-и экологические основы введения его в культуру. М.—Л.
Куминова А. В. 1936. Очерк растительности Алданского района Якутской АССР. Тр. Томск, унив., 90. — 1938. Степи Забайкалья и их место в ботанико-географическом районировании Даурии. Тр. Биол. инет, при Томск, унив., 5. — 1946. Растительность Кизир-Казырского междуречья. Изв. Зап.-Сиб. фил. АН СССР, сер. биол., 1, 1. — 1946а. Альпийская область Центрального Саяна. Изв. Зап.-Сиб. фил. АН СССР, сер. биол., I, 2. — 1948. Степи бассейна р. Уса в Центральных Саянах. Изв. Зап.-Сиб. фил. АН СССР, сер. биол., 2, 1 (бот.). — 1949. Весенняя фаза развития липового леса в Кузнецком Ала-Тау. Изв. Зап.-Сиб. фил. АН СССР, сер. биол., 3, 1 (бот.). — 1949а (на пер. 1950). Растительность Кемеровской обл. Новосибирск.
Куминова А. В. и Е. В. Вандакурова. 1949. Степи Сибири. Новосибирск.
Кунин В. Н. 1950. О причинах исчезновения кустарников в Южных Каракумах. Бот. ж., 35, 3.
Куренцова Г. Э. 1939. Монгольский дуб и его участие в фитоценозах бассейна р. Супу-тинки. Тр. Горно-таежн. станции Дальневост. фил. АН СССР, 3. — 1950. Остепненные дубовые и сосново-дубовые леса бассейна р. Сиитухи. Сообщ. Дальневост. фил. АН СССР, 1. — 1951. Остепненные ассоциации Суйфунской равнины. Сообщ. Дальневост. фил. АН СССР, 2. — 1951а. Типчаковые кедрово-дубовые леса и их производные в восточных приханкайских районах Приморского края. Сообщ. Дальневост, фил. АН СССР, 2. — 1952. Луга восточной части Прихан-кайской равнины и их хозяйственное значение. Тр. Дальневост. фил. АН СССР, сер. растениевод., I. — 1952а. Порослевые древесно-кустарниковые заросли юго-западной части Приморского края. Тр. Дальневост. фил. АН СССР, сер. растениевод., 1. — 1955. К происхождению растительности Приханкайской равнины Приморского края. Бот. ж., 40, 2.
Кучеровская С. Е. — см. Кучеровская-Рожанец С. Е.
Кучеровская-Рожанец С. Е. 1912. Растительность Ангаро-Ленского края близ Илимского тракта. (Б. Мамырь—Усть-Кут). В кн.: Предварит, отчет о бот. иссл. в Сибири и в Туркестане в 1911 г., СПб. — 1913. Растительность Мариинского у. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1912 г., СПб. — 1914. Растительность Павлодарского у. В кн.: Предварит, отчет о бот. иссл. в Сибири и в Туркестане в 1913 г.» СПб. — 1916. Очерк растительности района* Баян-аул—Каркаралы. В кн.: Предварит, отчет о бот. иссл. в Сибири и в Туркестане в 1914 г., Пг. — 1926. Очерк растительности Оренбургской губ. Вести»
просвещенца, 10, Оренбург. — 1927. Естественные и культурные растительные ландшафты Предуралья в пределах Спасско-Петровского района Оренбургской губ. Тр. Оренбургск. почв.-бот. бюро, 3. — 1929. Растительность района Оренбургской сельскохозяйственной станции. Тр. Оренбургск. почв.-бот. бюро, 4. — 1932. Растительность башкирской части Общего Сырта. Тр. СОНС АН СССР, Башкирская сер., 2. — 1936. Геоботанические ландшафты южной части Терско-Кумского песчаного массива. Изв. ГГО, 68, 4. —
Кучеровская-Рожанец С. Е. и М. И. Рожанец. 1926. Почвенно-ботанические условия участка Кумсай и прилегающих местностей. Тр. Общ. изуч. Казахстана, 7, 1. Кушалевнч Е. В. 1929. (Кушалевгч). Кабылянская балота. Наш край, 6—7 (45—46), Менск.
Кушев С. Л. 1939. Морфология и генезис бугристых марей и их географическое распространение. Тр. Комитета по вечной мерзлоте АН СССР, 8.
Кушниренко В. и М. Николаев (В. Кушн1ренко та М. Hi ко лае в). 1930. Кузьминський лучний масив Грунського району. Зап. Полтавськ. с.-г. шст., 4, 1. — 1930а. Попередне повщомлення про nepe6ir робк експедицп по дослщ-женню луюв Полтавсько! округи в 1930 р. Зап. Полтавськ. с.-г. шст., 4, 1.
-Лаасимер Л. Р. 1950. Экология альварного леса. Тарту.
Лавренко Е. М. 1922. Болота Харьковской губ. С.-х. жизнь, 4—8, Купянск. — 1925. Ботанико-географические исследования между рр. Миусом и Кальмиусом. Тр. с.-х. опытн. учреждений Дона и Сев. Кавказа, Ростов-на-Дону. — 1926. Леса Донецкого кряжа. Почвоведение, 3—4. — 1927. Пастбищная дигрессия на Нижнеднепровских песках в связи с задачами работ Алешковской опытной мелиоративной песчаной станции. С.-х. опытн. дело, 3 (И), Харьков. — 1927а (Е. L a v-г е n к о) The flora of Ucraina. В кн.: Contributions to the study of the soils of Ucraina (Мат. дослщж. груипв Украши, 6), Харьков. — 1928. Болота Украши. Bien, природознавства, 3—4. — 1930. Лесные реликтовые (третичные) центры между Карпатами и Алтаем. Ж. Русск. бот. общ., 15, 4. — 1931. Некоторые данные о степной растительности северных черноземов левобережной Украины. В кн.: 25 лет научн.-пед.и общ. деятельности Б. А. Келледа, Воронеж. — 1935. Некоторые наблюдения над корневой системой, экологией и хозяйственным значением псаммофитов нижнеднепровских песков. В кн.: Проблемы растениеводческого освоения пустынь, 3, Л. — 1936. О генезисе сфагновых болот в пределах степной зоны в бассейнах рр. Буга, Днепра и Дона. Сов. бот., 3.— 1938. История флоры и растительности СССР по данным современного распространения растений. В кн.: Растительность СССР, 1, М.—Л. — 1940. Обзорные карты растительности СССР. Изв. ВГО, 72, 3. — 1940а. Степи СССР. В кн.: Растительность СССР, 2, М.—Л. — 1942. О провинциальном расчленении Евр-азиатской степной области. Бот. ж. СССР, 27, 6. — 1943. Региональные исследования и картография растительности за 25 лет (1917—1942). Изв. ВГО, 75, 5. — 1945. Советская геоботаника (фитоценология). В кн.: Успехи биол. наук в СССР за 25 лет (1917—1942), М.—Л. — 1947. Евразиатская степная область. Тр. Комиссии по естеств.-истор. районированию СССР, 2, 2 (Геобот. районирование СССР).— 1950. Вопрос о взаимоотношениях степи и леса на новом этапе. Проблемы ботаники, 1. — 1950а. Изучение растительности степей и пустынь. (Советская геоботаника на службе социалистического народного хозяйства СССР в Сталинскую эпоху). Бюлл. МОИП, отд. биол., 55, 1. — 19506. Некоторые наблюдения над влиянием пожара на растительность северной степи. (Попереченская степь Пензенской обл.), Бот. ж., 35, 1. — 1950в. Основные черты ботанико-географического разделения СССР и сопредельных стран. Проблемы ботаники. 1. — 1954. Степи Евразиатской степной области, их география, динамика и история. В кн.: Вопросы ботаники, 1, М.—Л.
-Лавренко Е. М. и Н. А. Десятова-Шостенко. (С- Л а в р е н к о та Н. Д е с я-това-Шостенко). 1928. Рослиншсть засолених грунт!в Ягорлицького п-ва (Херсоне ько! окр.). Мат. дослщж. груннв Украши, 2.
Ляврспко Е. М. и Г. И. Дохман. 1933. Рослиншсть Староошьских стешв. Ж. Bio-бот. циклу ВУАН, 5—6.
Лавренко Е. М. и И. Г. Зоз. (С. Лавренко та I. 3 о з). 1928. Рослиншсть цглнни Михайл1вського юнного заводу (кол. К ап Hi ста), Сумсько! окр. Охорона пам’яток природи на Укра1ш, 2, Харю в. — 1931. Растительность конкских плавень р. Днепра («Великого луга») Запорожского окр. Мат. по пробл. нижи. Днепра, 2.
Лавренао Е. М. и Ф. Я. Левжна (6. Лавренко та Ф. Л е.в i н а). 1934. Остр1вне находище бореально! рослинност! в околицях ВшницЕ Bien. Кшв. бот. саду ,17.
Лаврами» Е. и А. Порецкий (6. Лавренко та А. II о р е ц ь к и й). 1928. Рослиншсть Челбаського 1 1ван1вського масив!в та Кшбурнсько? коси Нижнв-Дшщювських nicKie. Мат. охоронн природи на УкраТш, 1, Харюв.
Лавренко Е. М. и А. В. Прозоровский. 1939. Растительность Европейской части СССР. В кн.: Почвы СССР, 1, М.-Л.
Лавренко Е. М. и А. В. Прянишников (С. Л а в р е н к о та О. П р я н i ш-н i к о в). 1926. Рослишпсть Нижнедшпровських (Олешювських) шск!в та швденного району, що з ним мсжуе (по дослщженню 1925 року). Мат. по дослщж. грунт1в Украши, 1, 3.
Лавренко Е. М. и С. Я. Соколов. 1949. Растительность плодовых лесов и прилегающих районов Южной Киргизии. В кн.: Плодовые леса Южной Киргизии и их использование (Тр. Южно-Киргиз. экспед., 1), М.—Л.
Лавренко Е. М. и В. Б. Сочава [редакторы]. 1950. Карта растительности Европейской части СССР. М. 1 : 2 500 000. Пояснительный текст. М.—Л.
Лазаренко А. С. 1936. Очерк бриофлоры заповедника Горнотаежной станции. Тр. Горнотаежной станции Дальневост. фил. АН СССР, 1.
Лапшина Е. И. 1952. Карта растительности Сибири (южной части) в м. 1 : 2 500 000. Новосибирск.
Ларин И. В. 1926. Опыт определения по растительному покрову почв, материнских пород, рельефа, сельскохозяйственных угодий и других элементов ландшафта средней части Уральской губ. Тр. Общ. изуч. Казахстана, 7, 1. — 1926а. Растительность, почвы и сельскохозяйственная оценка Чижинских разливов. Мат. Особого комитета АН СССР по иссл. союзн. и авт. республик, Казахстанская сер., 2.— 1927. Растительные зоны средней части Уральской губ. Ж. Русск. бот. общ., 12, 1—2. — 1930. Почвы и растительность. Естественные районы [Уральского окр.]. Вкн.: Уральский округ него районы, 3, 2, Уральск.— 1931. Естественные кормовые ресурсы Западной Сибири. Новосибирск. — 1936. Материалы по динамике растительной массы и химических веществ травостоев в течение вегетационного периода в различных зонах СССР. Тр. Инет. физ. геогр. АН СССР, 21. — 1941. Предварительные итоги стационарных исследований Центрально-Казахстанского пустынного отряда Бот. инет. им. В. Л. Комарова АН СССР. Сов. бот., 1—2. — 1950—1951 [редактор]. Кормовые растения сенокосов и пастбищ СССР. 1 (1950), 2 (1951). М.—Л.
Ларин И. В., Л. А. Манохина и 3. С. Акимцева. 1954. Влияние лиманного орошения на естественную растительность. В кн.: Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР, М.—Л.
Ларин И. В., Е. В. Шиффере, Ф. Я. Левина, Б. Н. Замятнин, Л. И. Иванина, Е. А. Варивцева, Г. Н. Неплии И. В. Сырокомекая. 1954. Основные закономерности распределения растительности и геоботаническое районирование Северного Прикаспия в пределах междуречья Волга—Урал. В кн.: Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР, М.—Л.
Лебедев Д. В. 1956. Введение в ботаническую литературу СССР. Пособие для геоботаников. М.—Л.
Лебедев Н. Н. 1941. Географический очерк Талыша. Тр. Почв. инет. АН СССР, 26, 1. Лебедева Н. В. 1952. Связь растительного покрова с внутренней водопроводящей сетью болотных массивов. Л.
Левина Ф. Я. (Л е в i н а). 1929. Матер1али до вивчення рослинност! Понадсиваш-сько! смуги Украши. Мат. дослщж. земель мелюфонду на Мел1тополыциш, 1, Харюв. — 1933. Залишки ц!линно1 степово! рослинности на Куп'янщин! у Велико-Вурлуцькому райош. Ж. Bio-бот. циклу ВУАН, 6—6. — 1937. Болота Черниговского Полесья. Бот. ж. СССР, 22, 1. — 1938. Растительность долины р. Сары-Су в зоне полупустыни. Бюлл. МОИП, отд. биол, 47, — 1938а. Весняна рослиншсть Надсивашшя. Геобот. зб. 1нст. бот. АН УРСР, 2. — 19386. Болота Черниговского полесья. Уч. зап. Ленингр. пед. инет. им. А. И. Герцена, 9 (фак. естеств. наук, 4). — 1939. Материалы к стратиграфии торфяников Черниговского Полесья. Изв. Крым. пед. инет., 8. — 1952. Полынь таврическая (Artemisia taurica Willdenow) в пределах пустынно-степного юга и юго-востока Европейской части СССР. Тр. Бот. инет. АН СССР, сер. 3, геоботаника 8. — 1953. Растительный покров [района лесной полосы Сталинград—Степной—Черкесск]. Тр. Комплекс, научн. экспед. по вопросам полезащ. лесораэвед., 2, 7,— 1954. Микрокомплексы области бессточных рек междуречья Волга—Урал, основные черты их структуры и развития. В кн.: Вопросы улучшения кормовой базы в степной, полупустынной и пустынной эонах СССР, М,—Л. — 1955. К вопросу о генезисе луговой флоры и растительности юго-востока Европейской z равнины. Бот. ж., 40, 4.
Левина Ф. Я. и М. С. Шалыт (Л е в i н а та Шалит). 1927. Про рослиншсть ocTpoBiB Чурюка та Чурюк-Тюба на Сиваш! Мел1топольсько1 окр. Охорона пам'яток природи на УкраТш, 1, Харюв.
Левицкая А. М. (Л е в и ц ь к а). 1937 (1936). Геоботатчний нарис друго! тераси Дншра. Зб. робгг Д Hi пропето, бот. саду, 1. — 1937а. Матергали до монографИ типчатниюв (Festuca sulcata). Зб. праць Дншропетр. бот. саду, 2.
Левицкий А. П. 1910. Верхнезейская экспедиция. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1909 г., СПб. — 1910а. К вопросу об эволюции болот Амурской обл. Почвоведение, 12. 1.
Левицкий С. С. 1938 (1937). Материалы к изучению флоры и растительности Наур-зумского заповедника. Тр. Наурзумск. заповеди., 7.
Лейсле Ф. Ф. 1936. Лиманы. В кн.: Растительность Каспийской низменности между рр. Волгой и Уралом, 1 (Тр. СОПС АН СССР, Волжско-Каспийская сер., 2).
Леонов Л. И. 1953. В высоких широтах. (Записки натуралиста). М. [Глава «Красный снег»].
Леонтьев А. М. 1937. Геоботанические районы Беломорско-Кулойской части Северного края. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 2. — 1949. Основные закономерности распространения растительности Молого-Шекснинского междуречья до образования Рыбинского водохранилища. Тр. Дарвинск. заповеди, на Рыбинском водохр., 7. — 1949а. Пустошные, мелкозлаковые и осоковые луга Молого-Шекснинского междуречья до образования Рыбинского водохранилища. Тр. Дарвинск. заповеди, на Рыбинском водохр., 1.
Леонтьев В. Л. 1951. О создании саксауловых насаждений в связи со строительством Главного Туркменского канала. Изв. ВГО, 83. 3. — 1952. Развитие подроста и формирование древостоя черного саксаула [Haloxylon aphyllum (Minkw.) Iljin] в пустыне Кара-Кум. Тр. Бот. инет. АН СССР, сер. 3. геоботаника, 8.— 1954. Саксауловые леса пустыни Кара-Кум. хМ.—Л.
Леонтьев Ф. С. 1948. Северный предел распространения лиственных древесных пород на Омолоно-Чукотском Дальнем Востоке. Бюлл. МОИП, отд. биол., 53. 4.
Лепехин И. И. 1771—1805. Дневные записки путешествия по разным провинциям Российского государства. 1 (1771), 2 (1772), 3 (1780), 4 (1805). СПб.
Лесков А. И. 1932. Верхний предел лесов в горах Западного Кавказа. Бот. ж. СССР, 77, 2. — 1936. Геоботанический очерк приморских лугов Малоземельского побережья Варенцова моря. Бот. ж. СССР, 27, 1. — 1940. Очерк растительности долины р. Полуя. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 4. — 1940а. Фито-ценологический очерк редколесий бассейна р. Полуя. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 4. — 1943. Принципы естественной системы растительных ассоциаций. Бот. ж. СССР, 28. 2.
Ливеровский Ю. А. 1946. О ландшафте равнин южного Приморья и Приамурья и его генезисе. Проблемы физ. геогр., 72. — 1947. Почвы Амурской лесостепи. Почвоведение, 7.
Ливеровский Ю. А. и Б. П. Колесников. 1949. Природа южной половины Советского Дальнего Востока. М.
Линчевский И. А. 1935. Растительность Бадхыза. В кн.: Растительные ресурсы Туркменской ССР, 7, Л. — 1935а. Растительность западного Копет-Дага. В кн.: Растительные ресурсы Туркменской ССР, 7, Л.
Липпмаа Ф. (Т. L i р р m а а). 1935. Eesti geobotaanika pohijooni. Act. Inst, et Horti Bot. Univ. Tartuensis, 4. 3—4. — 1938 (на nep. 1939). Areal und Altersbestimmung einer Union (Galeobdolon — Asperula — Asarum — U.) sowie das Problem der Charac-terarten und der Konstanten. Act. Inst, et Horti Bot. Univ. Tartuensis, 6. 2—3.
Липский В. И. 1889. Исследования о флоре Бессарабии. Зап. Киевск. общ. естествоиспыт., 10. 2. — 1891. Ботаническая экскурсия за Каспий. Зап. Киевск. Общ. естествоиспыт., 77, 2. — 1899. Флора Кавказа. Свод сведений о флоре Кавказа за двухсотлетний период ее исследования, начиная от Турнефора и кончая XIX в. Тр. Тифл. бот. сада, 4. — 1902—1903. Флора Средней Азии. 1. Литература по флоре Средней Азии. 2. История ботанического исследования Средней Азии. Тр. Тифл. бот. сада, 7, 1 (1902), 2 (1903). — 1906. По горным областям Русского Туркестана (Тянь-Шаня). Изв. РГО, 42. 1. — 1911. Лесная растительность в Туркестане. Тр. по опытн. лесн. делу в России, 30. — 1915. Ботанические исследования в Закаспийской обл. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1912 г., ч. 2, 4.
Липшиц С. Ю. 1929. Предварительный отчет о ботанико-географических работах в Южном Урале в 1927 г. Бюлл. МОИП, отд. биол., 38. 3—4. — 1931. Растительность Чулакского плато и Джунгарских ворот. Бюлл. МОИП, отд. биол., 40. 1—2. — 1936. К познанию флоры и растительности горячих источников Камчатки. Бюлл. МОИП, отд. биол., 45. 2. — 1947. Систематика, флористика и география растений. В кн.: Л. П. Бреславец, Б. Л. Исаченко и др., Очерки по истории русской ботаники, М. — 1947—1952. Русские ботаники. Биографо-библиографический словарь. 7 (1947), 2 (1947), 3 (1950), 4 (1952). М.
Липшиц С. Ю. и Ю. А. Ливеровский. 1937. Почвенно-ботанические исследования и проблема сельского хозяйства в центральной части долины р. Камчатки. Тр. СОПС АН СССР, Камчатская сер., 4.
Литвинов Д. И. 1891 (1890). Геоботанические заметки о флоре Европейской России.
Бюлл. МОИП, нов. сер., 4. 3. — 1902. О реликтовом характере флоры камени-
стых склонов в Европейской России. Тр. Бот. муз. АН, 1. — 1909. Библиография флоры Сибири. Тр. Бот. муз. АН, 5. — 1927. О некоторых ботанико-географических соотношениях в нашей флоре. Л.
Литвинов М. А. 1954. О путях формирования и расселения почвенной микроскопической флоры на такырах. Тр. Бот. инет. АН СССР, сер. 2, споровые растения, 9.
Лобачев С. В. 1934. Экспедиция на р. Вах. (Предварит, краткий отчет Нарымской экспед.). Изв. ГГО, 66, 1.
Лоспцкий К. Б. 1949. Дубравы северной лесостепи и зоны смешанных лесов. В кн.: Дубравы СССР, 2 (Всесоюзн. Н.-иссл. инет. лесн. хоз., 29), М.—Л.
Лупинович И. С. 1947. Описание зон и провинций по естественно-историческим стра-намДТр. Комиссии по естеств.-ист. районированию СССР, 1, Естеств.-истор. районирование СССР.
Лучник 3. И. 1935. Поемные луга и болота Нижнего Суйфуна. Тр. Дальневост. фил. АН СССР, сер. бот., 1. — 1935а. Фенологические наблюдения над растительностью лугов р. Супутинки. Тр. Дальневост. фил. АН СССР, сер. бот., I.
Любимова А. А. 1937. Растительность и почвы побережья оз. Ловозера (Коль-i * ский п-ов). Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 2.
Любимова Е. Л. 1940. Некоторые данные о болотах западного побережья Камчатки. Камчатск. сб., 1, М.—Л.
Магакьян А. К. 1941. Растительность Армянской ССР. М.—Л. — 1947. Этапы развития высокогорных лугов Закавказья. Ереван. — 1953. Рододендроновые заросли в ’Армянской ССР. Тр. Тбил. бот. инет. АН ГрузССР, 15.
Маидель Г. А. 1894. Путешествие по северо-восточной части Якутской обл. в 1868— 1870 гг., I. Зап. Акад. Наук, 74 (прилож. 3).
Максимович К. И. (М а х i m о w i с z). 1859. Primitiae florae amurensis. Versuch einer Flora des Amurlandes. Mem. a 1’Acad. d. Sc. de St. Petersb., 9. — 1883. Очерк растительности восточной Азии, преимущественно Маньчжурии и Японии. Вестн. садовод., плодовод, и огороди., 1—7.
Малеев В. П. 1927. Пицундская сосновая роща. Тр. Абхаз, научн. общ., 1, 2. — 1931. Растительность района Новороссийск—Михайловский перевал и ее отношение к Крыму. Зап. Никитск. бот. сада, 13, 2. — 1933. Можжевеловый лес на мысе Мартьян в Крыму. Бот. ж. СССР, 18, 6. — 1936. Дубняки Абхазии. Тр. СОПС АН СССР, Закавказская сер., 19. — 1936а. Флора и растительность Абхазии. Тр. СОПС АН СССР, Закавказская сер., 19. — 1937. Исследования флоры и растительности Кавказа и Крыма за 20 лет после Великой Октябрьской Социалистической революции. Сов. бот., 5. — 1940. Растительность причерноморских стран (Эвксинской провинции Средиземноморья), ее происхождение и связи. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 4. — 1948. Растительность Южного Крыма. Тр. Никитск. бот. сада, 25, 1—2.
Мальцев А. И. 1907. Растительность лугов (по наблюдениям в Корочанском у. Курской губ.). Тр. Бот. сада Юрьевск. унив., 8, 1. — 1924. Фитосоциологические исследования в Каменной Степи. Тр. по прикл. бот. и селекц., 13, 2.
Малюгин Е. А. 1931. Кара-Калинский район. Естественно-исторические условия и опыты культуры древесных пород. Тр. Туркм. лесокульт. опытн. станции, 1. Ашхабад.
Марголина Д. Л. 1941. Флора и растительность Таджикистана. Библиография. Тр. Тадж. фил. АН СССР, 15.
Марков М. В. 1935—1939. Лес и степь в условиях Закамья. 1, 2. Уч. зап. Казанск. унив., 95, 7 (1935); 99, 1 (1939). — 1940. Природные условия развития растительности в пойме. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 4. — 1946. Луга Татарской АССР (сенокосы и пастбища). Казань. — 1948. Растительность Татарии. Казань.
Мартыненко М. А. 1938. О новом для Алтая папоротнике Dryopteris oreopteris (Ehrh.) Maxon. Тр. Ленингр. общ. естествоиспыт., 67, 3. — 1941. Пихтовая тайга Алтая. Л.
Матвеева Е. П. 1940. Основные растительные богатства Алтая и Саян. Природа, 9. — 1950. Геоботанические районы Восточно-Казахстанской обл. Изв. АН КазССР, сер. бот., 5. — 1951. Геоботаническое районирование Восточно-Казахстанской обл. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 7. — 1953. Сенокосы и пастбища и меры их улучшения. М.—Л.
Матренинский В. В. 1917. Леса Кологривского у. в естественно-историческом отношении. Тр. Костромск. научн. общ. по изуч. местн. края, 6.
Махатадзе Л. Б. 1941. Леса Северной Армении. (Типы лесов, лесовозобновление, хозяйственные мероприятия). Тр. Кировоканск. опытн. станции, 1.
Медведев Я. С. 1882. Очерки закавказских лесов. Лесн. ж., 4, 7—8. — 1915. Растительность Кавказа. 1. Тр. Тифл. бот. сада, 18, 1. — 1919. Деревья и кустарники Кавказа Изд. 3-е. Тифлис.
Мелехов И. С. 1944. О взаимоотношениях между сосной и елью в связи с пожарами в лесах Европейского Севера СССР. Бот. ж. СССР, 29, 4.
Мельникова В. В. 1953. О составе и распространении водорослей в некоторых почвах Вахшской и Гиссарской долин южного Таджикистана. Л.
Мешков А. Р. 1951. Районы флоры меловых и известняковых обнажений Средне-Русской возвышенности. Бот. ж., 36, 3. — 1953. Схема геоботанических районов черноземного центра. Вопросы геогр., 32.
Мещеряков Д. П. 1927. Типы болот, как отражение особенностей геоморфологических районов. В кн.: Тр. совещ. по вопросам луговедения и опытного луговодства,!, М.
Миддендорф А. Ф. 1867. Путешествие на север и восток Сибири, ч. 1, отд. 4. Растительность Сибири. СПб. — 1882. Очерки Ферганской долины. СПб.
Микешин Г. В. 1946. Ковыльно-типчаковые степи Копет-дага. Бюлл. МОИП, отд. биол., 51, 3. — 1946а. Полынные пустыни Копет-дага. Бюлл. МОИП, отд. биол.. 51, 6.
Милованович Д. А. 1928. Типы лесов Среднего Урала. Пермь.
Мильков Ф. Н. 1950. Лесостепь Русской равнины. Опыт ландшафтной характеристики. М. — 1950а. О черноольшатниках Среднего И лека. Землеведение, нов. сер., 3 (43). — 1953. Воздействие рельефа на растительность и животный мир. М. — 1953а. Среднее Поволжье. М.
Минервин В. Н. 1935. Объяснительная записка к карте типов пастбищ равнинной части Туркменской ССР. (В ст.: Пути развития кормовой базы Кара-Кумов). В кн.: Проблемы Туркмении, 2, М.—Л.
Минквиц 3. А. и О. Э. Кнорринг. 1910. Растительность Чимкентского у. Сыр-Дарьин-ской обл. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1908 г., ч. 2, 4.
Минкина Ц. И. и Р. В. Федорова. 1936. Смена лесов и история развития торфяников Дальневосточного края по данным ископаемой древесной пыльцы. Сов. бот., 4. Михайличенко В. С. 1936. Есюз рослинносп швшчносхщно! окра!ни п-ва Ямалу. Ж. 1нст. бот. УАН, 7 (15).
Михайлова М. Г. 1938. Флора та рослиншсть УРСР. Б1блюграфия. Ки!в. — 1941. Доповнення. Ки!в.
Михеев А. А. 1916. В полынных равнинах и разливах Приуралья (Уральское казачье войско). Изв. Бот. сада Петра Великого 16 (прилож. 1).
Михеев В. А. 1950. Естественные сенокосы и пастбища Башкирии. Уфа.
Михельсон А. И. 1913. Очерк растительности части Пржевальского и Джаркент-ского у. (Семиреченской обл.). Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1910 г., ч. 2, 4.
Мишкин Б. А. 1953. Флора Хибинских гор, ее анализ и история. М.—Л.
Молчанов А. А. 1953. Сосновый лес и влага. М.
Момотов И. Ф. 1953. Растительные комплексы Усть-Урта. Ташкент.
Морозов Г. Ф. 1926. Учение о лесе. Изд. 3-е. Л. — 1931. Учение о типах насаждений. М.—Л.
Морозова О. И. 1938. Влияние выпаса на пастбища пустынной зоны. В кн.: Сельскохозяйственное освоение пустынь Средней Азии, Ташкент. — 1946. Пастбищное хозяйство в каракулеводстве Средней Азии. М.
Мусаев И. Ф. 1953. Растительность кормовых угодий Присулакской низменности (Дагестанская АССР). Л.
Мусатова К. М. 1939. Динамика массы, питательность веществ и отавность растительности основных типов пастбищ и сенокосов лесостепной зоны Западно-Сибирского края. Бот. ж. СССР, 24, 2.
Мушкетов И. В. 1886. Туркестан. (Геологическое и орографическое описание), 1. СПб.
Назаров М. И. 1932. Кендырь среди растительности долины р. Сыр-Дарьи. Тр. Инет, нов. лубяного сырья, 3. — 1934. Основные типы растительности Бурят-Монгольской АССР и их кормовое значение. Бот. ж. СССР, 19, 1. — 1935. Очерк растительности Окинского края в Восточном Саяне. Изв. ГГО, 67, 1.
Напалков Н. В. 1951. Дубравы Центральной лесостепи и Нижнего Поволжья. В кн.: Дубравы СССР, 3, (Всесоюзн. Н.-иссл. инет. лесн. хоз., 30), М.—Л.
Нардина Н. С. 1954. Очерк растительности нижнего течения Атрека. Тр. Инет. биол. АН ТуркмССР, сер. бот., 2.
Нат С. Г. 1915. Леса и воды Печорского края. Лесн. ж., 45, 4.
Неврли И. 1912. Леса севера Европейской России. Изв. [СПб.] Лесн. инет., 22.
Недригайлов С. Н. 1928. Лесные ресурсы Ленско-Алданского плато и Заалданско-Верхоянского горного района. Мат. Комиссии АН СССР по изуч. Якут. АССР, 12. — 1928а. Лесные ресурсы центральной части Южного Урала. Тр. Геогр. отд. КЕПС АН СССР, 1. — 1932. Лесной покров и лесные ресурсы комплекса лесных районов лесного края Якутской АССР. В кн.: Якутская АССР, 3, Л.— 1936. О росте леса в Анадырском крае. Тр. Арктич. инет., 40.
Нейштадт М. И. 1936. Торфяные болота Барабинской лесостепи. Тр. Центр, торф, опытн. станции, 1. — 1936а. Торфяные болота западной Камчатки. Тр. Центр, торф, опытн. станции, 1.
Нейштадт М. И. и М. Я. Короткина. 1936. Торфяные болота юго-восточной Камчатки. Тр. Центр, торф, опытн. станции, 1.
Некрасова Т. П. 1938. Растительность альпийского и субальпийского поясов Чуна-тундры. Тр. Лапландск. заповеди., 1.
Неуструев С. С. и И. М. Крашенинников [редакторы]. 1930. Картапочвенно-ботани-ческих районов Усть-урта и п-ва Мангышлак. Мат. Комиссии экспед. иссл. АН СССР, 26, Казахстанская сер.
Нечаева Н. Т. 1951. Материалы к биологии Haloxylon ammodendron Bge. Бот. ж., 36, 1. — 1954. Влияние выпаса на пастбища Каракумов как основа пастбшце-оборота. В кн.: Пустыни СССР и их освоение, 2, М.—Л. — 1954а. Засемененность пастбищ юго-восточных Каракумов и влияние выпаса на заделку семян. В кн.: Пустыни СССР и их освоение, 2, М.—Л.
Нечаева Н. Т., Н. А. Мордвинов и И. А. Мосолов. 1943. Пастбища Кара-Кумов и их использование. Ашхабад.
Никитин С. А. 1928. Очерк растительности песков Кокджида, Джага-булак и Кара-тугай по рр. Темиру и Эмбе. Мат. Комиссии экспед. иссл. АН СССР, 5, Казахстанская сер. — 1934. Типы песков Каракалпакских Кизил-кумов и пути их хозяйственного использования. В кн.: Кара-Калпакия, 2, Л. — 1935. Пески западного Прибалхашья. Тр. Почв. инет. АН СССР, 11. — 1951. Государственная защитная лесная полоса гора Вишневая—Каспийское море. Тр. Комплекс, научн. экспед. АН СССР по вопросам полезащ. лесоразвед., 7, 1. — 1954. Растительность восточной части Прикаспийской низменности. В кн.: Пустыни СССР и их освоение, 2, М.—Л. — 1954а. Растительность междуречий Ку-шума, Урала и Эмбы и ее кормовые ресурсы. В кн.: Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР, М.—Л.
Никитин С. А. и В. Ф. Поярков. 1935. Луговые районы пустынного Зауралья. Тр. Почв. инет. АН СССР, 11.
Никитина В. Н. 1949. Флора и растительность восточного Копет-Дага. Л. — 1954. Растительность восточного Копет-Дага в связи с ее поясностью. Тр. Инет. биол. АН ТуркмССР, сер. бот., 1.
Никитина Е. В. 1927. Материалы к изучению торфяников Западной Сибири. «Рямы» Барабинского окр. по исследованиям 1926 г. Изв. Томск, отд. Русск. бот. общ., 2, 1—2.
Николаева М. Г. 1941. Кустарниковый тип растительности южной части Большого и Малого Ямала. Бот. ж. СССР, 26, 1. |
Никольский П. Н. и И. И. Изотов. 1936. Очерк растительности полосы вдоль Паран-дово-Ругозерского тракта (Карелия). Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 3.
Никонов Л. Н. 1948. Геоботаническое описание лугов с определением их хозяйственного значения в окрестностях Витебска. Уч. зап. Белорусск. унив., 7.
Никонов М. Н. 1948. О роли познания современных торфяных отложений для выяснения генезиса ископаемых углей. Бюлл. МОИП, отд. геол., 28, 6. — 1953. Торфяники Средней Печоры. Тр. Инет, леса АН СССР, 13.
Новиков Г. Н. 1936. Растительно-почвенные комплексы северной части Каспийской равнины, их типы и происхождение. Тр. СОПС АН СССР, Волжско-Каспийская сер., 2.
Новиков Ю. С. и В. Л. Леонтьев. 1942. Пути восстановления древесно-кустарниковой растительности Кара-Кумов. Тр. Туркм. фил. АН СССР, 2.
Новичкова Л. Н. 1955. Сообщества низших растений такыров подгорной равнины Копет-Дага. Л.
Новопокровский И. В. 1909. Забайкальская обл. Долина рр. Нерчи, Куэнги, Белого Урюма и Унгура. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1908 г., СПб. — 1910. Растительность района Амурской ж. д. от Нерчинска и ст. Куэнги до низовьев Белого Урюма. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1908 г., ч. 2, 9. — 1912. Ботаникогеографические исследования в Нерчинском и Читинском округах Забайкальской обл. летом 1908 г. Землеведение, 19, 1—2. — 1915. Растительность войсковых песчаных лесничеств Донской обл. Изв. Бот. сада Петра Великого, 16, (прилож. 1). —1921. Растительность Донского края. (Ботаникохгеографический очерк). Новочеркасск. — 1925. Растительность Северо-Кавказского края. Изв.-Донск. инет, сельск. хоз. имелиор., 5 (прилож.). —1926. Растительность Моздокской степи. Тр. Северо-Кавказ. ассоц. н.-иссл. инет., 18.—1927. Некоторые данные о растительности степей и речных долин Южного Предуралья. В кн.: Тр. со-вещ. по вопросам луговедения и опытного луговодства, 1, М. — 1927а. Растительность Ставрополья. Тр. Северо-Кавказ. ассоц. н.-иссл. инст.г22.—1929.. Матери. .
алы к познанию растительности Приманычской степи. Тр. Северо-Кавказ. ассоц. н.-иссл. инет., 70. — 1931. Материалы для познания растительности Южного Предуралья. Стерлитамакский и Уфимский кантоны. М.—Л. — 1932. Результаты геоботанического обследования района канала Октябрьской революции и Присулакской низменности. Изв. Бот. сада АН СССР, 30, 1—2. — 1935. Растительность учебно-опытного совхоза № 2 Зернотреста (Азовско-Черноморского края). Тр. Краснодарск. с.-х. инет., 1. — 1937. Зональные типы степей Европейской части СССР. (Опыт классификации русских степей). Землеведение, 39, 3. Ногтев В. П. 1926. Поемные луга Нижегородской губ., предварительные итоги и перспективы их геоботанического изучения. Производительные силы Нижегородск. губ., 3.
Номоконов Л. И. 1947. Пойменные луга р. Енисея в пределах Минусинской котловины. Бюлл. Сиб. бот. сада, 1. — 1951. Луговая растительность Валдайского района. Уч. зап. Ленингр. унив., 143 (сер. биол. наук, 30). — 1955. Пойменные луга р. Енисея. Л.
О климатических различиях России, в связи с местными обстоятельствами по видам сельского хозяйства. 1834. Земледелья, газ., 1, прибавл. (Статья без подписи; Траутфеттер (1880) приписывает ее Е. Ф. Канкрину).
Обручев С. В. 1931. Колымско-Индигирский край. Географический и геологический очерк. Тр. СОПС АН СССР, Якутская сер., 1.
Овеснов А. М. 1948. Горные луга Вишерского Урала. Тр. Естеств.-научн. инет, при Молотовск. унив., 10, 1. — 1948а. Заметки об олуговении горных тундр на Северном Урале. Изв. Естеств.-научн. инет, при Молотовск. унив., 12, 8. — 1950. Луга долины р. Сосьвы (Средний Урал). Изв. Естеств.-научн. инет, при Молотовск. унив., 12, 10. — 19Й. Естественные луга в верховьях р. Вишеры (Северный Урал). Изв. Естеств.-научн. инет, при Молотовск. унив., 13, 2—3. — 1952. Горные луга Западного Урала. Молотов. — 1952а. Горные луга Лопьинского камня и хребта Оше-ньер (Северный Урал). Изв. Естеств.-научн. инет, при Молотовск. унив., 13, 4—5.
Овсянников В. Ф. 1930. Лиственные породы. Изд. 2-е Хабаровск. — 1930а. Поездка в долину р. Анадырь летом 1929 г. Зап. Владивост. отд. РГО, о (22).
Овчинников П. Н. 1926. Очерк растительности поймы р. Волхова от д. Завижье до с. Пчевы. Мат. по иссл. р. Волхова и его бассейна, 9.—1940. К истории растительности Средней Азии. Сов. бот., 3. — 1941. Sibbaldia tetrandra Bge. и вопрос о происхождении криофильной растительности Средней Азии. Сов. бот., 1—2. — 1947. О принципах классификации растительности. Сообщ. Тадж. фил. АН СССР, 2. — 1948. Основные моменты происхождения горных степей. Сообщ. Тадж. фил. АН СССР, 3. — 1948а. О главнейших типах древесной растительности Таджикистана. Сообщ. Тадж. фил. АН СССР, 6. — 19486. О типологическом расчленении травянистой растительности Таджикистана. Сообщ. Тадж. фил. АН СССР, 10. — 1951. Ботанические исследования Таджикистана. Тр. Тадж. фил. АН СССР, 27. — 1955. Основные направления видообразования в связи с происхождением типов растительности Средней Азии. Тр. АН Тадж. ССР, 31.
ЮгиевскиЗ В. В. 1933. Туркменский можжевельник (арча) и его возобновление. В кн.: В. П. Покутний и В. В. Огиевский, Леса Туркмении, Ашхабад.
Окиншевич Н. А. 1908. Леса Бессарабии и их отношение к рельефу местности и почвам. Зап. Новоросс. общ. естествоиспыт., 32.
Орлов А. Я. 1951. Темнохвойные леса Северного Кавказа. М. — 1953. Буковые леса северо-западного Кавказа. В кн.: ПГироколиств. леса северо-западного Кавказа, М.
Орлов М. А. 1940. Пески Астраханской полупустыни, методы их укрепления и хозяйственного использования. М.
Орлов Ф. Б. 1947. Об изменении живого напочвенного покрова на гарях. Сб. н.-иссл. работ Архангельск, лесотехнич. инет., 9.
Ососков П. А. 1909—1912. Зависимость лесной растительности от геологического состава коренных пород. Лесн. ж., 39, 2—5; 8—9 (1909); 41, 3—4 (1911); 42, 4-5 (1912).
Павлов Н. В. 1925. Островные леса Тургайской обл. и некоторые вопросы их биологии. Изв. Главн. оот. сада, 24. — 1926. Ботанико-географическии очерк северо-восточной части Тургайской обл. Ж. Русск. бот. общ., 11, 1—2. — 1930. Введение в растительность Карсакпая. Изв. Ассоц. н.-иссл. инет, при 1 МГУ, 3, 1-А. — 1931. Комплексные степи и пустыни Карсакпая. (Опыт био-географической монографии^. М. — 1932. К истории исследования и географии сыр-дарьинского Кара-тау. Сов. каучук, 10. — 1935. Растительность западной Бетпак-дала и Карсакпайского плато. Тр. САГУ, сер. VIII-в, бот., 22. — 1936. Березовые леса западного побережья Камчатки. Бюлл. МОИП, отд. биол., 45 , 2. — 1940.
Литературные источники по флоре и растительности Казахстана. Тр. Казахст. фил. АН СССР, 19. — 1945. Идеи и достижения советской ботаники в Казахстане за 25 лет. Изв. АН КазССР, сер. бот., 2. — 1948. Ботаническая география СССР. Алма-Ата. — 1949. Основные закономерности в растительном покрове Казахстана. В кн.: Тр. Третьей сессии АН КазССР, Алма-Ата. «
Павлов Н. В. и С. Ю. Липшиц. 1934. Эскиз флористических элементов Сырдарышского Каратау. Сов. бот., 1.
Павлов II. В. и П. Н. Чижиков. 1937. Природные условия и проблема земледелия на юге Большерецкого района Камчатки. Тр. СОПС АН СССР, Камчатская сер., 3.
Палибип И. В. 1903. Ботанические результаты плавания ледокола «Ермак» в Северном Ледовитом океане летом 1901 г. Изв. СПб. бот. сада, 3, 2, 3, 5.
Паллас П. С. 1773—1786. Путешествие по разным провинциям Российской империи. 1 2 (1786), 3 (1786), СПб. — 1796. (Р. S. Pallas) Tableu physique
et topographique de la Tauride tire du journal d’un vojage fait en 1794. SPb.
Панютин П. C. 1942. Болота Колхиды. Высокогорные болота. Бот. ж. СССР, 27,5. Папкова А. А. 1930. Растительность лугов поймы р. Луги в пределах Лужского района Ленинградского окр. Тр. Бот. муз. АН СССР, 22.
Пархоменко М. Л. 1949. К вопросу об эволюции некоторых такыров. Почвоведение, 9. Пачоский И. К. 1892. Флорографические и фитогеографические исследования Калмыцких степей. Киев. — 1904. Очерк растительности Днепровского у. Таврической губ. Зап. Новоросс. общ. естествоиспыт., 26. — 1908. Причерноморские степи. Ботанико-географический очерк. Зап. Общ. сельск. хоз. Южн. России, 78, 7—9. — 1910. Основные черты развития флоры юго-западной России. Зап. Новоросс. общ. естествоиспыт., 34, (прилож.). — 1912(1911—1912). Материалы для флоры Бессарабии. Тр. Бессарабск. общ. естествоиспыт., 3. — 1912а. Плавневые солонцы в низовьях Днепра. Тр. Бот. сада Юрьевск. унив., 13, 1. — 1913. Ботаническая экскурсия в Аскания-Нова и на Сиваш. Зап. Крым. общ. естествоиспыт. и любит, природы, 2. — 1914. Очерк растительности Бессарабии. Кишинев.— 1915. Описание растительности Херсонской губ. I. Леса. Мат. по иссл. почв и грунтов Херсонской губ., 8. — 1917. Описание растительности Херсонской губ. II. Степи. Херсон. — 1922—1923. По пескам Днепровского у., 1, 2. Изв. Гос. Степного заповеди. Аскания-Нова, 1, (1922), 2 (1923). — 1923. Ботаническая экскурсия в Крым. Изв. Гос. Степного заповеди. Аскания-Нова, 2. — 1923а. Наблюдения над растительным покровом степей Аскания-Нова в 1922 г. Изв. Гос Степного заповеди. Аскания-Нова, 2. — 1926. Наблюдения над целинным покровом в Аскании-Новой в 1923 г. BicTi Держ. стенового заповщн. «ЧаплЬ», 3. — 1927. Описание растительности Херсонской губ. III. Плавни, пески, солончаки, сорные растения. (Мат. по иссл. почв и грунтов Херсонской губ.). Херсон.— 1930. (I. Paczoski). Lasy Bialowiezy. (Die Waldtypen von Bialo-
wieza). Pahstw. Rada ochrony przyrody. Monogr. naukowe, 1, Poznaii.
Пашковский К. A. 1951. Возобновление сосны в ленточных борах Прииртышья. Алма-Ата. — 1952. Саксаульники Коскудукского лесного массива. Алма-Ата.
Пельт И. Н. 1951. Земли древнего орошения Джанадарьинской древне-аллювиальной равнины. Изв. ВГО, 83, 3. — 1955. Краткая характеристика пастбищ Западной и Северной Туркмении. Тр. Арало-Каспийск. комплёксн. экспедиции АН СССР, 4.
Пельт Н. Н. и Л. Е. Родин. 1955. Карта пастбищ Западной Туркмении и прилегающих частей Узбекистана и Казахстана. Тр. Арало-Каспииск. комплексн. экспедиции АН СССР, 4.
Перфильев И. А. 1928. Материалы к флоре островов Новой Земли и Колгуева. Архангельск.
Пестушко Е. И. (6.1. П е с т у ш к о). 1937. Рослиншсть засолених грунпв долили середнього Дшпра (елисавет1всько-Курил!вський масив). Зб. праць Дшпропетр. бот. саду, 2. — 1939 (1938). Геоботашчний нарис долини р. Самары-Дншрянсь-ко! (вщ гирла р. Вовчо! до гирла р. Самари). Зб. праць Дшпропетр. бот. саду, 3.
Петров Б. Ф. 1934. Наносы и почвы бассейна р. Конды (приток Иртыша). Изв. ВГО, 66, 5. — 1948. Ландшафты и почвы центральной части Восточных Саян. Землеведение, нов. сер., 2 (42).— 1952. Почвы Алтайско-Саянской обл. Тр. Почв, инет. АН СССР, 35.
Петров М. П. 1933. Корневые системы растений песчаной пустыни Кара-Кумы, их распределение и взаимоотношения в связи с экологическими условиями. Тр. по прикл. бот., генет. и селекц., сер. 1,7. — 1934. К вопросу о происхождении растительности пустыни Каракумы. В кн.: Хозяйственное освоение пустынь Средней Азии и Казакстана, М.—Ташкент. — 1935. Экологический очерк растительности Репетекскогб песчано-пустынного заповедника в юго-восточных Каракумах. В кн.: Проблемы растениевод, освоения пустынь, 4, Л. — 1944. Исследование аэросъемочного материала при геоботаническом обследовании пустынь
Средней Азии. Тр. Туркм. фил. АН СССР, 5. —• 1945. Древесно-кустарниковая растительность юго-западного Копет-Дага и ее связь с лесной растительностью Северного Ирана. Изв. Туркм. фил. АН СССР, 1. — 1946. Роль растительности в эволюции рельефа песчаных пустынь. Изв. Туркм. фил. АН СССР, 3—4. — 1950.»Подвижные пески пустынь Союза ССР и борьба с ними. М.
Петрова-Трефилова Л. А. 1923. Почвы и растительность сродней части Барабинской степи. Изв. Биол. н.-иссл. инет, при Пермск. унив., 2, 2. — 1925. Растительность солонцов и солончаков Барабинской степи. Пзв. Биол. н.-иссл. инет, при Пермск унив., 3, 8.
Пивоваров П. В. 1954. Пастбища колхозов Гурьевского междуречья Волга—Урал. В кн.: Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР, М.-Л. J
Плеске Ф. Д. 1887. Критический обзор млекопитающих и птиц Кольского п-ова. СПб. Плотников Н. А. 1941—1946. Житняки Балхашско-Алакульской котловины и их внутривидовое разнообразие в условиях обитания. Тр. Омск. с.-х. инет., 20.
Победимова Е. Г. 1935. Растительность прибрежных песков и пустынь залива Кара-бугаза. Тр. Почв. инет. АН СССР, 11.
Поварницын В. А. 1929. Произрастание лиственницы и ее возобновление в Ваковском лесничестве Нижегородской губ. В кн.: Очерки по фитосоциологии и фитогеографии, М. — 1931. Типы буковых лесов Джалабетского лесного массива Юго-Осе-тии. Тр. СОПС АН СССР, Закавказская сер., 2. — 1932. Леса лено-алданского водораздела по Саныяхтахской тропе. В кн.: Якутская АССР, 3, Л. — 1932а. Леса долины р. Алдана от г. Томмота до устья р. Ноторы. В кн.: Якутская АССР, 3, Л. — 1933. Леса долины р. Алдана от г. Томмота до устья р. Учура. Тр. Инет, по изуч. леса АН СССР, 1. — 1934. Леса и лесовозобновление в бассейне р. Б. Белой в Восточных Саянах. Тр. СОНС АН СССР, Сибирская сер., 7. — 1936. Типы лесов Абхазии. Тр. СОПС АН СССР, Закавказская сер., 19. — 1937. Почвы и растительность бассейна р. Верхней Ангары. Тр. СОПС АН СССР, Восточно-Сибирская сер., 4. — 1940. Типы лесов Черноморского побережья между рр. Сукко и Пшадой. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 4. — 1941. Типы лесов сибирской лиственницы СССР. В кн.: Со. тр. Сибирского лесотехн, института, Л. — 1944. Кедровые леса СССР. Красноярск. — 1949. Леса даурской лиственницы СССР. Бюлл. МОИП, отд. биол., 54, (3). — 1950. (В. О. П о в а р н i ц и н). Л1си Закарпаття. Бот. ж. АН УРСР, 7, 3.___
1954. Вклад росшських та украТнських вчених у вивченпя рослиннрсп УРСР. Бот. ж. АН УРСР, П, 2.
Погосян X. П. [редактор]. 1952. Изменение климата в связи с планом преобразования I» природы засушливых районов СССР. Л.
Погребняк П. С. 1927. JIico-рослинш умови Правобережного Пол1сся УкраТни. Тр. з лк. доев, справи на Украшь 7. — f929. (Р. S. Pogrebnjak) Uber die Methodik der Standortsuntersuchungen in Verbindung mit Waldtypen. В кн.: Verb. II Int. Kongr. forstl. Versuchsanstalten. — 1931. ЛИсорослинщ умови Под1лля. Н.-досл. 1нст. лков. госп. та arponicoMeniop., сер. наук, вид., 10. Харюв. — 1951. Исторический анализ развития лесоводственной типологии. В кн.: Тр. совещания по лесной типологии, М.
Поздняков Л. К. 1955. Леса бассейна р. Олёкмы. Природа, 1.
Покровская В. М. 1940. Описание растительности Ямской степи. Тр. Центр.-черноземн. заповеди, им. пр оф. В. В. Алехина, 1. — 1940а. Стрелецкая степь в аспективных картинах. Тр. Центр.-черноземн. заповеди, им. проф. В. В. Алехина, 1.
Покутний В. П. 1933. тугайные леса (верхней и средний части бассейна реки Мур-габа). В кн.: В. П. Покутний и В. В. Огиевский. Леса Туркмении. Ашхабад.
Поле Р. 1903. (R. Pohl е). Pflanzengeographische Studien fiber die Halbinsel Kanin und das angrenzende Waldgebiet. Тр. СПб. бот. сада, 21. 1. — 1906. О лесах северной России. СПб. — 1910. Программа для ботанико-географического исследования тундры. В кн.: Программы для бот.-геогр. исследований, изданные бот.-геогр. подкомиссией при почв, комиссии Вольного экономии, общ., 2, СПб. — 1919. (R.) Р h о I е. Beitrage zur Kenntnis der westsibirischen Tiefebene. Zeitschr. d. Ges. fur. Erdkunde zu Berlin, 9/10.
Пешков П. П. 1929. Типы лиственничных лесов Хакассии. Тр. Сиб. инет. с.-х. лесоводства, 12, 1—3. — 1934. Ботанико-географические очерки Кузнецкой котловины, Салаира и западной Предсалаирской полосы. Тр. СОПС АН СССР, Сибирская сер., 13. — 1934а. Краткий очерк растительности северо-восточных отрогов Холзунского хребта на Алтае. Тр Бот. инет. АН. СССР, сер. 3, геоботаника, 1. — 1941. Растительность дельты р. Или. Тр. Казахст. фил. АН СССР, 20. — 1941а. Растительность Джунгарского Алатау в оайоне рр. Саркана и Баскана. Тр. Казахст. фил. АН СССР, 20. — 1944. Растительность бассейна средней части р. Нуры в Северном Казахстане. Изв. Казахст. фил. АН СССР, сер. бот., 1. — 1947. Прибрежно-водная расти-
тельность оз. Зайсан. Вестн. АН КазССР, 1—2 (22—23). — 1948. Лиственные леса Заилийского Алатау. Тр. Алмаатинск. заповеди., 7. — 1949. Растительность водораздела рок Терс-Аккан—Кенгир. Изв. АН КазССР, 74 (сер. бот., 4). Полянская О. С. 1929. Растительность Белоруссии. Природа, И. — 1935. Луга и пастбища Ленинградской обл. Л. — 1936. Об инверсии поясов растительности в Хибинских горах. Сов. бот., 4.
Полянская О. С., М. Ф. Корнилов и А. Г. Трутнев. 1937. Почвенно-геоботанические исследования на территории колхозов Крестецкого района Ленинградской обл. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 2.
Пономарев А. II. 1948. Лесостепной комплекс северной окраины Кунгурской лесостепи. Изв. Естеств.-научн. инет, при Молотовск. унив., 12, 6.
Пономарев 11^ А. 1933 (на пер. 1932). Березы СССР. М.—Л. — 1934. Лиственницы Понятовская В. М. 1949. Сезонный ритм жизни травяных фитоценозов лесного пояса Южной Киргизии. В кн.: Плодовые леса Южной Киргизии и их использование, М.—Л. — 1952. Стационарное изучение травянистых сообществ лесного пояса Южной Киргизии. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 8.
Поплавская Г. И. 1908. Материалы к изучению луговой растительности Псковской губ. Тр. СПб. общ. естествоиспыт., о7, 4. — 1912. Лесная полоса в Нерчинских степях. В кн.: Предварит, отчет об организ. и исполн. работ по иссл. почв Азиат. России в 1911 г., СПб. — 1912а. Материалы по изучению растительности северо-восточной части Забайкальской обл. Тр. Амурской экспед., 16 (бот. иссл. 1910 г., 2). — 1913. Бассейн р. Верхней Ангары. Южные открытые-склоны. В кн.: Предварит. отчет об организ. и исполн. работ по иссл. почв. Азиат. России в 1912 г., СПб. — 1914. Исследования в Верхнеудинском у. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1913 г., Пг. — 1916. На северной окраине Селенгинской Даурии. (Ботанико-географический очерк). Тр. Бот. муз. АН, 15. — 1924. Опыт фитосоциологического анализа растительности целинной заповедной степи Аскания-Нова. Ж. Русск. бот. общ., 9. — 1925. К вопросу о характере верхней границы бука в Крыму. Ж. Русск. бот. общ., 10, 1—2. — 1925а. Материалы по изучению растительности Крымского Государственного заповедника. Тр. по изуч. заповеди. Отд. охр. природы Главнауки НКП РСФСР, 2. — 1928. О березе в Крыму. Ж. Русск. бот. общ., 13, 1—2. — 1929. О некоторых взаимо-замещающих буковых ассоциациях в Крыму. В кн.: Очерки по фитосоциологии и фитогеографии, М. — 1930. К растительности Чатырдага. Ж. Русск. бот. общ., 15, 1—2. — 1933. Некоторые данные по изучению горных лугов в геоботаническом и кормовом отношениях. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 1. — 1948. Растительность горного Крыма. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 5.
Попов В. В. и Б. П. Тихомиров. 1940. Лиственничные леса бассейна рр. Маны и Кана в Восточных Саянах. В кн.: Лиственница сибирская, Красноярск.
Попов М. Г. 1915. Ботанико-географический очерк гор Султан-Уиз-Даг. В кн.: Почвенные исследования в бассейне рр. Сыр-Дарьи и Aky-Дарьи, 1, М. — 1922. Ботанико-географический очерк северной Хивы. Изв. Туркестан, отд. РГО, 15. — 1923. Флора пестроцветных толщ (краснопесчаниковых низкогории) Бухары. (Фрагмент к истории флоры Туркестана). Тр. Туркестан, научн. общ., 1. — 1925. Экологические типы растительности пустынь южного Туркестана. Изв. Главн. бот. сада, 24. — 1927. Основные черты истории развития флоры Средней Азии. Бюлл. САГУ, 15. — 1928. Растительно-высотные поясы в горах Средней Азии. В кн.: Дневн. Всесоюзн. съезда ботаников в Ленинграде в январе 1928 г., Л. — 1929. Дикие плодовые Средней Азии. Тр. по прикл. бот., генет. и селекц., 22, 3. — 1934. Дикие плодовые леса Узбекистана. В кн.: Узбекистан, Тр. и мат. Первой конф, по изуч. производительных сил Узбекистана 19—28 де-/ кабря 1932 г., 3, Л. — 1940. Растительный покров Казахстана. Тр. Казахст. фил.
АН СССР, 18. — 1941. Ботаническая характеристика верховьев Чилика. Тр, Казахст. фил. АН СССР, 20. — 1941а. Высотные пояса Заилийского Алатау. Тр. Казахст. фил. АН СССР, 20. — 1949. Очерк растительности и флоры Карпат. Мат. к познанию фауны и флоры СССР, отд. оот., нов. сер., 5 (13). —1950. Растительность о-ва Монерон (Японское море). Бот. ж., 35, — 1953. О взаимо
отношении леса (тайги) и степи в Средней Сибири. Бюлл. МОИП, отд. биол., 58, 6. Попов М. Г. и Н. В. Андросов. 1937. Растительность заповедника Гуралаш и Заамин-ской лесной дачи. Ташкент.
Попов Т. И. 1914. Происхождение и развитие осиновых кустов в пределах Воронежской губ. (Геоботанический очерк). Тр. Докучаевск. почв, комитета, 2. — 1931. Краснянские степи Борисоглебского окр. В кн.: Степи Центрально-Черноземной обл., М.—Л.
Попович Ф. Я. 1938. Новые данные к флоре и растительности района Присивашья. Сов. бот., 1.
Порецкий А. С. 1936. Геоботанические районы юго-западной части Кара-Калпакских Кызыл-Кумов. Тр. СОПС АН СССР, Кара-Калпакская сор., 4.
Порецкий А. С., Ф. Н. Русанов и К. С. Афанасьев. 1936. Растительные группировки Кара-Калпакских Кызыл-Кумов. Тр. СОПС АН СССР, Кара-Калпакская сер., 4.
Порфирьев В. С. 1947. К познанию пихтовых ценозов Татарской АССР. Уч. зап. Ка-занск. пед. инет., фак., естествозн., 5. — 1950. Темнохвойпо-широколиственные леса северо-востока Татарии. Уч. зап. Казанск. пед. инет., фак. естествозн., Р.
Порядин В. Н. 1930. Луговодство в Якутском окр. В кн.: Мат. комиссии АН СССР по изучению Якутской АССР, 29, Л.
Пояркова Т. Ф. 1927. Очерк растительности хребта Маркотх на Северном Кавказе. Изв. Главн. бот. сада, 26, 3.
Пояркова Т. Ф. и 3. В. Коровник. 1931. Почвы и растительность дельты р. Урала. В кн.: Баксайский район, 7, Уральск.
Пояснительный текст к карте растительности СССР в м. 1 : 5 000 000. 1941. М.—Л. Прасолов Л. И. 1927. Южное Забайкалье. Почвенно-географический очерк. Мат. Особ, комитета АН СССР по иссл. союзн. и авт. респуб., 12, Бурят-Монгольская сер.
Прилипко Л. И. 1939. Растительные отношения в Нахичеванской АССР. Тр. Бот. инет. Азерб. Лил. АН СССР, 7. — 1944. Заповедник гирканского низового леса. Изв. Аэерб. фил. АН СССР, 7. — 1954. Лесная растительность Азербайджана. Баку.
Прозоровский А. В. 1932. Илийский кендырь. Материалы по систематике и экологии кендыря в долине Или. Казахст. инет, удобр. и агропочвовед., 1, Алма-Ата. — 1935. О зональных типах пустынь Советской Средней Азии. Изв. ГГО, 67, 3. — 1936. О биологических типах растений пустыни. Бот. ж. СССР, 21, 5. — 1937. Геоботаническое изучение полупустынь, пустынь и гор Сродней Азии за 20 лет. Сов. бот., 5. — 1940. Изучение биоценотических взаимоотношений между травянистыми растениями мезофильного и ксерофильного типов. Сов. бот., 5—6. — 1940а. Полупустыни и пустыни СССР. В кн.: Растительность СССР, 2, М.—Л.
Прозоровский А. В. и В. П. Малеев. 1947. Азиатская пустынная область. Тр. Комиссии АН СССР по естеств.-ист. районированию СССР, 2, 2. Геобот. районирование СССР.
Прозоровский Н. А. 1929. К изучению растительности Ямской степи. Курск. — 1930. На северном пределе степей. I. Алгасовские степи. Исслод. и мат. Рязанск. обл. Средне-Окского музея, 6. — 1940. Изменение растительности Стерлецкой степи при отсутствии пастьбы скота и сенокошения. Тр. Центр.-Черноземн. заповеди, им. проф. В. В. Алехина, 1. — 1949. Очерк растительного покрова центрально-черноземных областей. Вопросы геогр., 13.
Протопопов Г. Ф. 1954. Естественное семенное возобновление ели Шренка. Тр. Инет, бот. и растениевод. Киргиз, фил. АН СССР, 1 (5).
Прохоров Н. И. 1905. Ботанико-географические письма из русской лесостепи. Почвоведение, 4.
Прянишников А. В* 1954. Залужение тундры. Бот. ж., 39, 1. — 1955. Образование лугов на дне осушенных озер в тундре. Бот. ж., 40, 3.
Пряхин М. И. 1940. О вертикальных растительных зонах южного Таджикистана. Изв. ВГО, 72, 6.
Пташицкий М. И. 1910. Растительность бассейна среднего течения р. Или. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1908 г., ч, 2, 8. — 1913. Растительность бассейна нижнего течения р. Или в пределах Верненского у. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1909 г., ч. 2, 8. — 1913а. Растительность Ишим-Иртышского водораздела в пределах Тюкалинского и Тарского уездов. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1912 г., СПб. — 1916. Опыт методологического изучения растительности Акмолинских степей. В кн.: Предварит, отчет о бог. иссл. в Сибири и Туркестане в 1914 г., Пг.
Пьявченко Н. И. 1945. Болото и торфяник. Сов. бот., 1. — 1949. О генезисе бугристого рельефа торфяников в северо-восточной части Европейской России. Почвоведение, 5. — 1951. Некоторые данные о торфяниках долины Волги. ДАН СССР, 78, 1. — 1953. Зоринские болота Курской обл. Тр. Инет, леса АН СССР, 13. — 1955. Бугристые торфяники. М.
Работнов Т. А. 1929. Болота б. Никольского у. Северо-Двинской губ. Изв. Луг. инет., 4—-6. — 1929а. О болотах Калужской губ. Торф, дело, 1. — 1930. О болотах Архангельского окр. Торф, дело, 5. — 1933. Природные условия сельского хозяйства долины р. Амги. В кн.: Р. А. Еленевский, А. И. Кремнев, А. А. Шахов и Т. А. Работнов, Природные ресурсы южной Якутии, М. — 1935. Ландшафты песчаных образовании в низовьях Вилюя. Землеведение, 37, 4. — 1935а. Природные районы южной части Якутской АССР. Бот. ж. СССР,
20, 2. — 1936. Леса из Betula Ermani Cham. в южной Якутии, Природа, 4. — 1936а. Обзор работ по изучению растительности Якутской АССР. Бот. ж. ,СССР, 21, 6. — 1937. Растительность высокогорного пояса бассейна рр. Алдана иТимп-топа. Изв. ГГО, 6’9, 4. — 1939. О болотах центральной Якутии. Изв. ГГО, 71, 4. — 1945. О степях центральной Якутии. Природа, 2. — 1950. Жизненный цикл многолетников травянистых растений в луговых ценозах. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 6.
Радде Г. И. 1901. Основные черты растительного мира на Кавказе. Зан. Кавказ, отд. РГО, 22, 3.
Райкова И. А. 1924. Материалы к ботанико-географической характеристике Памира. Изв. Турксст. отд. РГО, 17. — 1930. Растительные ландшафты Памира. Тр. САГУ, сер. VIП-в, бот., 12. — 1936. Климат и растительность Памира. В кн.: Конференция по сельскохозяйственному освоению Памира, М.—Л.
Раменская М. Л. 1954. Улучшение сенокосов и пастбищ Карело-Финской ССР. Петрозаводск.
Раменский Л. Г. 1918. Исследование лугов Воронежской губ. Мат. по естеств.-ист/ иссл. Воронежск. губ., отд. 1, 1. — 1924. Основные закономерности растительного покрова. Вестн. опытн. дела. Воронеж. — 1927. Сравнительный географический очерк лугов Среднечерноземной обл. и Второго Донского окр. Сталинградской губ. В кн.: Тр. Совещ. по вопросам луговедения и опытного луговодства, /, М. — 1938. Введение в комплексное почвенно-геоботаническое исследование земель. М. — 1941. Типы материковых лугов и болот лесной зоны. В кн.: И. И. Карасев, И. И. Минина, Л. Г. Раменский, С. II. Смелов, И. А. Цаценкин, Сенокосы и пастбища, М. — 1950. Вопросы классификации, типировки и характеристики пастбищ и сенокосов при их паспортизации. Бот. ж., 35, 3.
Растительность СССР. 1938—1940. 1 (1938), 2 (1940). М.—Л.
Ревердатто В. В. 1916. Растительность прибрежной зоны р. Енисея в Туруханском крае. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1914 г., Пг. — 1921. Основные формации высокогорно-тундровой зоны северо-восточного Алтая. Изв. Томск, отд. Русск. бот. общ., 1, 1—2. — 1924. Луговая растительность долины р. Томи в пределах Кузнецкого и Щегловского у. Изв. Томск, унив., 74. — 1925. Растительные зоны Абаканской степи. Изв. Томск, унив., 75. — 1927. О происхождении растительности Бийской степи. (Опыт фитогеографического анализа). Томск. — 1929 (1928). Приабаканские степи и орошаемые земли в системе р. Абакана (в пределах Минусинского и Хакасского окр. Сибирского края). Изв. Томск, унив., 81. — 1931. Растительность Сибири. В кн.: Естественно-исторические условия сельскохозяйственного производства Сибири, 3, Новосибирск. — 1931а. Растительность Сибирского края. Изв. ГГО, 63,1. — 1946. Очерк Гастительности Западного Саяна. Изв. Зап.-Сиб. фил. АН СССР, сер. биол., I. — 947. Некоторые замечания об «островных» степях Сибири. Сов. бот., 15, 6. — 1947а. О флористическом составе некоторых ассоциаций Хакасских степей. Ивв, Зап.-Сиб. фил. АН СССР, сер. биол., 2, 1.—1954. Степи Хакасии. Ивв. ВГО,£б, 3.
Ревердатто В. В. и Т. Н. Буторина. 1934. Бугристые степные ассоциации в среднесибирских степях. Бот. ж. СССР, 19, 3.
Ревердатто В. В. и В. П. Голубинцева. 1930. Сорная растительность орошаемых и неорошаемых полей и залежей южносибирских степей. М.—Л.
Ревердатто Л. Ф. 1921. Очерк растительности Кузнецкой степи. Изв. Томск, отд. Русск. бот. общ., 1, 1—2. — 1924. Очерк растительности юго-восточной части Томской губ. Тр. Томск, унив., 74.
Регель К. В. 1913. Растительность болот северного Полесья и влияние на нее осушки и орошения. Тр. Бюро по прикл. бот., 6, 9.
Роговской П. А. 1950 (1949). К процессу демутации в Прикаспийской степи. Уч. зап. Кабардинск. пед. и учит, инет., 2.
Родин Л. Е. 1933. Типы степей Нижнего Заволжья. Бот. ж. СССР, 18, 4. — 1934. Материалы к познанию лесов Тянь-Шаня. Ельники Кетмень-Тау. Изв. ГГО, 66, 1. — 1934а. Материалы к познанию лесов Тянь-Шаня. Ельники северного склона Джунгарского Алатау. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 1. — 19346. Типы зональной растительности [сыртовой части Нижне-Волжского края в области проектированного орошения]. Тр. Комиссии по ирриг. АН СССР, 2. — 1940. Основные черты растительного покрова северных (Заунгузских) Кара-кумов. В кн.: Природные ресурсы Кара-кумов, 1. М.—Л. —1948. Материалы по изучению растительности Северных и Заунгузских Кара-кумов. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 5. — 1952. Работы Ботанического института им. В. Л. Комарова АН СССР на трассе Главного Туркменского канала. Бот. ж., 37, 6. — 1954. Роль растительности в образовании такыров и их комплексов. В кн.: Пустыни СССР и их освоение, 2, М.— Л.
Родин Л. Е. и Н. И. Базилевич. 1955. О круговороте зольных элементов и азота в некоторых пустынных биогеоценозах. Бот. ж., 40, I.
Родин Л. Е. ж М. М. Голлербах. 1954. Биогеоценозы такыров и их генезис. В кн.: Вопросы ботаники 2, М,— Л.
Рожанец-Кучеровская С. Е. см. Кучеровская-Рожанец С. Е.
Рожевиц Р.^Ю. 1912* Растительность западной части Пржевальского и южной части Пишпекского уездов Семиреченской обл. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1908 г., ч. 2, J2. — 1930. Очерк растительности восточного Усть-Урта. Мат. Комиссии экспед. иссл. АН СССР, 26, Казахстанская сер.
Рослиншсть ЗакарпатськоТ облает! УРСР. 1954. Кихв.
Рослякова А. Ф. 1952. Алданское нагорье в пределах Алдано-Тимптонского междуречья. (Опыт физико-географической характеристики). М.
Рубин А. М. 1934. Краткий очерк растительности нижнего течения р. Нижней Тунгуски. Тр. Полярн. комиссии АН СССР, 15. — 1935. Растительность долины g. Чуни, притока Подкамеиной Тунгуски. Тр. Полярн. комиссии АН СССР, Рубцов Н. И. 1941. Геоботанические исследования в бассейне р. Малой Алмаатинки.
Тр. Казахст. фил. АН СССР, 20. — 1941а. Геоботанический очерк окрестностей г. Балхаша. Тр. Казахст. фил. АН СССР, 20. — 19416. Заметка о растительности дельты р. Пли. Тр. Казахст. фил. АН СССР, 20. — 1941в. Растительность Джунгарского Алатау в районе рр. Аксу и Биень. Тр. Казахст. фил. АН СССР, 20. — 1943. Весенние ботанические наблюдения в пустыне Бетпак-дала. Сов. бот., 1. — 1944. Растительность урочища Кокашик в пустыне Бетпак-Дала. Изв. Казахст. фил. АН СССР, сер. бот., 1. — 1945. Обзор геоботанических исследований в Казахстане за 25 лет. Изв. АН КазССР, сер. бот., 2. — 1946. Тяныпанская осина. Сов. бот., 14. 5. — 1948. К познанию бородачевых ценозов СССР. Бюлл. МОИП, отд. биол., 53, 4. — 1948а. Растительный покров Джунгарского Алатау. Алма-Ата. — 1950. О геоботаническом районировании Тянь-Шаня. Бюлл. МОИП, отд. бнол., 55. 4. — 1950а. Пустыни Северного Тянь-Шаня. Изв. АН КазССР, сер. бот., 5. — 1952. Растительный покров Казахстана. В кн.: Очерки по физической географии Казахстана, Алма-Ата. — 1954. Степи Северного Тянь-Шаня. Изв. АН КазССР, сер. биол., 7.—1955. Геоботаническое районирование Северного Тянь-Шаня. Изв. АН КазССР, сер. биол., 10. — 1955а. Луга Северного Тянь-Шаня. Тр. Инет, ботаники АН КазССР, 1. — 19556. О типах растительности Тянь-Шаня. Бюлл. МОИП, отд. биол., 60. 5.
Рубцов Н. И. и 3. В. Кубанская. 1949. Флора пустыни Бетпак-дала. Изв. АН КазССР, 52 (сер. бот., 3).
Рубцова А. Г. 1940. Растительный покров Кызыл-Агачского заповедника им. С. М. Кирова. Тр. Бот. инет. Азерб. фил. АН СССР, 11.
Рупрехт Ф. 1866. Геоботанические исследования о черноземе. Зап. Акад. Наук, 10 (при лож. 6).
Русанов Ф. Н. 1930. Место кендыря в зарослях гигантских злаков—тугаях долины р. Пянджа и его притоков. Кендырь—Рами, 6. — 1930а. Очерк растительности западного Усть-Урта и равнинного Мангышлака. Мат. Комиссии экспед. иссл. АН СССР, 26. Казахстанская сер. — 1934. Растительность дельты Аму-Дарьи и ее хозяйственное значение. В кн.: Каракалпакия, 2. Л. — 1936. Геоботанические районы юго-восточной части Кара-Калпакских Кызыл-Кумов. Тр. СОПС АН СССР, Кара-Калпакская сер., 4. — 1947. Некоторые особенности среднеазиатских речных долин и их растительности. Бюлл. АН УзССР, 7. — 1949. Какие деревья выращивать в лесозащитных и полезащитных полосах Узбекистана. Ташкент. — 1949а. Среднеазиатские тамариксы. Ташкент. — 1953. Амурская дендрофлора и поведение ее видов в условиях Ташкентского оазиса. Тр. Бот. сада АН УзССР, 3.
Сабардина Г. С. 1949. Естественные луга Рижско-Елгавской низменности. Изв. АН ЛатССР, 3 (20). — 1950. Геоботаническое обследование естественных лугов Латвийской ССР. Бот. ж., 35. 5. — 1951. Высокогорные пустыни южной части Центрального Тянь-Шаня. Уч. зап. Ленингр. унив., 143 (сер. биол. наук, 30). — 1952. Естественные луга северо-западного берега Лубанского озера. Сб. тр. Инет, зоотехн. и зоогигиены АН ЛатССР, 1. — 1952а. Луга бассейна р. Абуле в среднем ее течении. Сб. тр. Инет, зоотехн. и зоогигиены АН ЛатССР, 1. — 19526. Дикорастущие кормовые растения Латвийской ССР. Сб. тр. Инет, зоотехн. и зоогигиены. АН ЛатССР, 2. — 1952в. Естественные луга в районе нижнего течения р. Венты. Сб. тр. Инет, зоотехн. и зоогигиены АН ЛатССР, 2. — 1954. Луговая растительность Латвийской ССР. Л.
Саверкин А. П. 1936. Естественная кормовая база Приморья ДВК. Вестн. Дальневост. фил. АН СССР, 17. — 1938. Итоги изучения естественной кормовой базы — лугов и пастбищ ДВК. Вестн. Дальневосточ. фил. АН СССР, 30 (3).
Савич В. М. 1910. Очерк флоры западной части заволжских песков Астраханского края с хозяйственной точки зрения. СПб. — 1910а. Тургайская экспедиция. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1909 г.» СПб. — 1914. «Борбас». Наурзумские ковыльные степи Арало-Ишимского водораздела. Изв. Бот. сада, 14, 1—2. — 1927—1928. Саксауловые рощи Туркмении. Лесовед. и лесовод., 4—о. — 1928. Типы растительного покрова севера Приморья. Мат. по изуч. колониз. районов ДВК, 1.
Савич Н. М. 1926. Результаты геоботанических исследований в б. Рогачевском уезде летом 1923 г. Минск. — 1929. Вынш! геабаташчных дасьледваньняу у Mari-леускай акрузе клетку 1925 г. Матер* ялы да вывучэньня флёры i фауны Беларусь 3, Менск. — i930. Результаты геоботанических исследований в Бельском у. Смоленской губ. летом 1926 г. Тр. Общ. изуч. природы Смоленск, края, 5.
Савнч-Любпцкая Л. И. 1952. Сфагновые (торфяные) мхи. Флора споровых растений СССР, 7, Листостебельные мхи (1). Л.
Савкина 3. П. 1953. Кормовая база животноводства Крайнего Севера и основные пути ее укрепления. В кн.: Доклады VI расширенной сессии Уч. совета Пнет, полярн. земледелия, житотновод. и промышл. хоз., 2, Л.
Сажинов Г. И. 1949. Улучшение и использование сенокосов я пастбищ в условиях Вологодской обл. Вологда.
Салазкин А. С. 1934. Естественные кормовые угодья Мурманского окр. Сов. оленеводство, 1. — 1936. Очерк растительности бассейна р. Умбы. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 3.
Салазкин А. С., Ф. В. Самбук, О. С. Полянская и М. И. Пряхин. 1936. Оленьи пастбища и растительный покров Мурманского окр. Тр. Арктич. инет., 72.
Салимовская-Родина А. Г. 1936. К микрофлоре цветного снега. Архив биол. наук, 43, 2/3.
Самбук Ф. В. 1927. Растительные ассоциации на желтоподзолистых почвах Коношской дачи, Вологодской губ. Ж. Русск. бот. общ., 12, 1—2. — 1930. Ботаникогеографический очерк долины р. Печоры. Тр. Бот. муз. АН СССР, 22. — 1931. Геоботаническая характеристика зимних пастбищ у устья р. Печоры. В кн.: В. Н. Андреев, А. А. Дедов, Ф. В. Самбук, Оленьи пастбища Северного Края, Архангельск. — 1931а. Методика маршрутного исследования тундровых пастбищ. Тр. Полярн. комиссии АН СССР, 6. — 19316. Основные типы лугов в пойме средней Печоры. Тр. Бот. муз. АН СССР, 23. — 1932. Печорские леса. Тр. Бот. муз. АН СССР, 24. — 1933. Пастбищные угодья первого ненецкого оленеводческого колхоза. В кн.: Оленьи пастбища Северного края, 2, Л. — 1934. Кормовые угодья тундр Ненецкого окр. Северного Края. Сов. оленеводство, 1. — 1934а. Поемные луга бассейна р. Печоры. Сыктывкар. — 1937. Краткий очерк растительности Таймыра. Проблемы Арктики, 1. — 1937а. О классификации растительности тундровой зоны. Сов. бот., 2.
Самбук Ф. В. и А. А. Дедов. 1934. Подзоны Припечорских тундр. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 1.
Самойлова А. П. 1951. Растительность берегов некоторых соленых озер Тувинской автономной обл. Тр. Томск, унив., 118.
Самойлович Г. Г. 1948. Новые методы (авиаметоды) для изучения географического распространения лесов. В кн.: Тр. Второго Всесоюзн. геогр. съезда, 2, М.
Сапожников В. В. 1906. Очерки Семиречья. Джунгарский Алатау и одна экскурсия в Заилийском Алатау. Изв. Томск, унив., 28. — 1911. Монгольский Алтай в истоках Иртыша и Кобдо. Томск. — 1913. Экспедиция в Джаркентский и Пржевальский уезды. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1912 г., СПб. — 1916. У верхней черты растительности. В кн.: Сборник статей, посвящ. К. А. Тимирязеву его учениками, М.
Сапожников В. В. и Е. В. Никитина. 1920. Поездка в низовье Оби и Обскую губу в 1919 г., Изв. Инет, исслед. Сибири, 2.
Сапожников В. В. и Б. К. Шишкин. 1918. Растительность Зайсанского у. Томск.
Саркисова-Федорова О. В. 1929. К биологии травяного покрова еловых лесов. В кн.: Очерки по фитосоциологии и фитогеографии, М.
Сахаров М. И. 1939. Типы леса Кавказского Государственного заповедника в верховьях р. Мзымты. Тр. Кавказ, заповеди., 2.
Сахокиа М. Ф. 1950. Ксерофитные редколесья. В кн.: Е. М. Лавренко и В. Б. Сочава [ред.], Карта растительности Европейской части СССР. М. 1 : 2 500 000. (Пояснительный текст), М.—Л. — 1950а. Смешанные широколиственные леса колхидские. В кн.: Е. М. Лавренко и В. Б. Сочава (ред.], Карта растительности Европейской части СССР. М. 1:2 500 000. (Пояснительный текст), М.—Л.
Свешникова В. М. 1952. Корневые системы растений Памира. Тр. Инет. бот. АН ТаджССР, 4.
Сдобникова Н. В. 1956. Почвенные водоросли такыров северной части Ту раненой низменности. Л.
Северцев Н. А. 1873. Вертикальное и горизонтальное распределение туркестанских животных. Изв. Общ. любит, естествозн., антропол. и этногр., 8, 2.
Семенов В. Ф. 1914. Ботанические работы в Акмолинской обл. в 1912 и 1913 годах (предварительный отчет). К материалам по флоре киргизских степей. Изв. Томск, технологич. инет., 32, 4. — 1922. От Омска до Перовска через Акмолинскую обл. К материалам по фитогеографии Киргизского края. Тр. Сиб. с.-х. акад., 7.— 1924. Краткий очерк растительности Омской губ. Изв. Зап.-Сиб. отд. РГО, 4, 1. — 1926. О растительности Бухтарминского края и хребта Холзун. Тр. Сиб. с.-х. акад., 6‘, 9.
Семенов П. П. 1867. Поездка из укрепления Верного через горный перевал у Суок-Тюбе 'и ущелье Буам к западной оконечности оз. Иссык-Куль в 1856 г. Зап. РГО, 1.
Семенов-Тян-Шанский М. Д. 1948. К вопросу о ландшафтном районировании севера Восточно-Европейской равнины. Уч. зап. Ленингр. пед. инет. им. М. Н. Покровского, 6 (геогр., 2).
Семенова-Тян-Шанская А. М. 1940. Краткий очерк лугов и пастбищ долины р. Свияги в районе г. Ульяновска. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 4. — 1954. Биология растений и динамика растительности меловых обнажений по р. Деркул. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 9.
Сергиевская Л. П. 1951. Степи Бурят-Монголии. В кн.: Памяти П. Н. Крылова в связи со столетием со дня рождения. (Тр. Томск, унив., 116).
Серебряков И. Г. 1945. Анализ высотного распределения растений горных ельников Заилийского Алатау. Бюлл. МОИП, отд. биол., 50, 5—6.
Серебряков И. Г. и В. Б. Куваев. 1951. Материалы о высотном распространении растений в условиях Хибинских гор. Уч. зап. Моск. гор. пед. инет. им. В. П. Потемкина, 19.
Серебрянников П. П. 1913. О типах насаждений и их значении в северном лесном хозяйстве. Лесн. ж., 43, 1—2.
Сидоренко Г. Т. 1953. Растительность и кормовые ресурсы Кураминского хребта. Тр. Инет. бот. АН ТаджССР, 9.
Синская Е. Н. 1933. Основные черты эволюции лесной растительности Кавказа в связи с историей видов. Бот. ж. СССР, 18, 5—6.
Скарлыгина М. Д. 1954. Основные черты растительного покрова Крымского При-сивашья. Л.
Скворцов А. А. и Ю. А. Скворцов. 1927. Искусственное орошение, климат и почва оазисов. Л.
Скворцов Ю. А. 1930. Почвенно-географический очерк нижнего течения р. Атрек. Тр. САГУ, сер. VII-а, почвовед., 1.
Скворцова А. В. 1952. Орошаемые луга южных районов Хакасской автономной обл. Новосибирск.
Скородумов А. С. 1954. Почвы Черного леса. Киев.
Смелов С. П. 1927. Луга Заволжья Ярославской губ. Тр. Луг. инет., 2.—1929. Луга и болота в области Волго-Северо-Двинского водораздела. В кн.: Тр. Совещ. по вопросам луговедения и опытного луговодства, 2, М. — 1936. Лугово-болотный комплекс водораздела Волги, Днепра и оз. Ильмень. Бот. ж. СССР, 21* 2. — 1947. Биологические основы луговодства. М.
Смелов С. П. и Т. А. Работнов. 1929. Материалы к изучению реакции луговых почв и распределения в связи с ней луговой растительности. Изв. Луг. инет., 1—3.
Смиренский А. А. 1951. Болота Северного Казахстана. Вопросы геогр., 26.
Смирнов В. И. 1903. Ботанико-географические исследования в северо-восточной части Саратовской губ. Тр. Общ. естествоиспыт. при Казанск. унив., 37, 4. — 1910-Долина р. Абакана в Енисейской губ. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1909 г., СПб. — 1912. Растительность в области рр. Сары-су и Кон (Акмолинская обл.). Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1908 г., ч. 2, 11. — 1913. Растительность Троицко-савского и Селенгинского уездов. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1912 г., иПб. — 1914. Растительность западной части Акшин-ского у. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1913 г.г СПб.
Смирнов Л. А. 1934. О флоре меловых выходов в Заволжье. Бюлл. МОИП, отд. биол., 43, 1. — 1935. Комплексы растительного покрова Заволжья и пути их эволюции. Сов. бот., 5.
Смирнов П. А. 1927. Растительность Межпьянья Сергачского и Арзамасского уездов. Производительные силы Нижегород. iy6., б. — 1940. Флора и растительность Центрально-промышленного района. Мат. к познанию фауны и флоры СССР, нов. сер., отд. бот., 1, (9).
Смирнова А. Д. 1936. К вопросу о структуре еловых лесов Приветлужья. Уч. зап. Горьк. унив., 5. — 1943. Типы еловых лесов крайнего севера Кировской обл. Бот. ж. СССР, 28, 5. — 1951. Типы еловых лесов крайнего севера Кировской обл., ч. 1. Уч. зап. Горьк. унив., 19. — 1954. Типы еловых лесов крайнего севера Кировской обл., ч. 2. Уч. зап. Горьк. унив., 25.
Смирнова 3. И. 1928. Лесные ассоциации северо-западной части Ленинградской обл. Тр. Петергофск. естеств.-научн. инет., 5. — 1928а. Материалы к флоре сфагновых мхов Урала. Изв. Биол. н.-иссл. инет, при Пермск. унив., 6, 2. — 1938. Растительные ассоциации о-ва Колгуев. Бот. ж. СССР, 23, 5—6.
Соболев Л. Н. 1948. Естественные кормовые угодья Казахстана. (Ботанико-географическое и кормовое описание). Тр. экспед. по изуч. земельных фондов Казахской ССР, 9, Алма-Ата. — 1950. Растительность и почвы как элемент ее место— обитания. Физико-географическое районирование. В кн.: Казахстан. (Общая физ.-геогр. характеристика), М.—Л. — 1952. Краткий очерк растительности района работ Тяныпанской физико-географической станции. Тр. Инет, геогр. АН СССР, 49. — 1953. Степи средней части подгорной равнины и северных предгорий Терскей-Алатау. Тр. Инет, геогр. АН СССР, 56.
Соболевская К. А. 1941. Реликтовые ассоциации ледниковой эпохи в Хакассии. Изв. ВГО, 78, 3. — 1946. К вопросу о реликтовой флоре восточных склонов Кузнецкого Алатау и Хакасских степей. Изв. Зап.-Сиб. фил. АН СССР, сер. биол., I, 2. — 1947. О флористическом соотношении степей Тувинской обл. Изв. Зап.-Сиб. фил. АН СССР, сер. биол., 2, 1. — 1949. Растительность горных пастбищ Уленьской котловины (восточный склон Кузнецкого Алатау). Изв. Зап.-Сиб. tnn. АН СССР, сер. биол., 3, 1. — 1950. Растительность Тувы. Новосибирск. — 953. Конспект флоры Тувы. Новосибирск.
Советкина М. М. 1930. Растительность юго-западной части Центрального Тянь-Шаня в пределах Нарынского кантона Киргизской АССР и ее кормовые запасы. Ташкент. — 1938. Пастбища и сенокосы Средней Азии. Ташкент.
Соколов С. Я. 1926. Рекогносцировочное исследование типов леса Лисинского лесничества Ленинградской губ. Лесовед. и лесовод., 3. — 1928. Лесные растительные-ассоциации и торфяники Осинорощинской дачи Парголовского учебно-опытного-лесничества Ленинградского лесного института. В кн.: Природа и хозяйство учебно-опытных лесничеств Ленингр. лесн. института, М. — 1929. К вопросу о классификации типов еловых лесов. В кн.: Очерки по фитосоциологии и фитогеографии, М. — 1931. Общий естественно-исторический и лесоводственный очерк Сочинского района. Тр. и иссл. по лесн. хоз. и лесн. пром. 14. — 1931а. Типы леса восточной части Баково-Варнавинского учебно-опытного леспромхоза. В кн.: Природа и хозяйство учебных леспромхозов Лесотехнической академии, 2, М.—Л. — 1936. Классификация типов леса Абхазии. Тр. СОПС АН СССР и Бот. инет., Закавказская сер., 19. — 1936а. Плодовые леса Киргизии и перспективы их использования. В кн.: Проблемы Киргизской АССР, 2, Л. — 1937—1938. Успехи советской лесной геоботаники. Сов. бот., 6 (1937), 1 (1938). 1951. Лесохозяйственное значение типов леса таежной полосы. В кн.: Тр. совещ. по лесной типологии. 3—5 февраля 1950 г., М. — 1952. Некоторые ценные древесные и кустарниковые породы северной части Черноморского побережья Кавказа. Тр. Бот. инет. АН СССР, сер. 5, растит, сырье, 3.
Соколова Л. А. 1937. Материалы к геоботаническому районированию Онего-Северо-двинского водораздела и Онежского п-ва. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 2. — 1951. Основные черты растительности западного склона (северной части) Южного Урала. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 7.
Соловьев А. И. 1947. Курильские острова. Изд. 2-е. М.—Л.
Соловьев К. П. 1935. Материалы по изучению растительного покрова п-ва Муравьева-Амурского. Тр. Дальневост. фил. АН СССР, сер. бот., 1. — 1951. Еловопихтовые леса Южного Сахалина и их естественное возобновление. Сб. работ Дальневост, н.-иссл. инет. лесн. хоз., 3.
Солоневич К. И. 1934. Геоботанический очерк района западной части Кемь-Ухтин-ского тракта (Карелия). Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 1. — 1936. К растительности Ловоэерских гор. (Кольский п-ов). Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 3. — 1940. О регрессии ареала сосны на Кольском п-ве. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 4.
Солоневич К. И. и Н. Г. Солоневич. 1936. Геоботанический очерк района между станциями Кивач и Кяппесельга Кировской ж. д. (Карелия). Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 3.
Солоневич Н. Г. 1954. Эколого-биологические особенности основных компонентов травяно-кустарничкового яруса болотных растительных сообществ. Л.
Сортовое районирование сельскохозяйственных культур на 1954 г. 1954. М.
Соснин Л. И. 1939. Типы леса Кавказского Государственного заповедника. Тр. Кавказ, заповеди.,2.—1949. Восточная ель в Кавказском заповеднике. Охрана природы, 7.
-ОмшавсшЬ Д. И. 1915. Очерк растительности Верхней Сванетии. Вести, русск. флоры, 2, 3.
Сохадзе Е. В. и М. Е. Сохадзе. 1952. О биоэкологических особенностях бородача и структуре бородачевых сообществ Кавказа. Бюлл. хМОИП, отд. биол., 57, 3.
♦Сочава В. Б. 1927. Ботанический очерк лесов Полярного Урала от р. Нельки до р. Хулги. Тр. Бот. муз. АН СССР, 21. — 1927а. Северная граница кедра (Pinus sibirica Маут.) на Урале. Изв. АН СССР, сер. 6, 21. — 19^9. О пределе лесов на крайнем северо-востоке Азии. Природа, 12. — 1930. К фитосоциологии темнохвойного леса. Ж. Русск. бот. общ., 15, 1—2. — 1930а. О пятнистых тундрах Анадырского края. Тр. Полярн. комиссии АН СССР, 2. — 19306. Пределы лесов в горах Ляпинского Урала. Тр. Бот. муз. АН СССР, 22. — 1930в. (S о-с z a w а V.) Das Anadyrgebiet. Botanisch-geographische Beobachtungen in ausser-sten Nordosten Asiens. Zeitschr. d. Ges. f. Erdkunde zu Berlin, 7/8. — 1931. Некоторые основные понятия и термины тундроведения. Ж. Русск. бот. общ., 16, 1. — 1932. Высокогорная флора Дуссэ-Алиня. Бот. ж. СССР, 17, 2. — 1932а. По тундрам бассейна Пенжинской губы. Изв. РГО, 64, 4—5. — 1933. Естественные кормовые угодья тундровой зоны Якутии. Сов. оленеводство, 2. — 1933а. На истоках рр. Шугора и Северной Сосвы. Изв. ГГО, 65, 6. — 19336. Тундры бассейна р. Анабары. Изв. ГГО, 65, 4. — 1934. Растительные ассоциации Анабарской тундры. Бот. ж. СССР, 19, 3. — 1934а. Растительный покров Буреин-ского хребта к северу от Дульниканского перевала. Тр. СОПС АН СССР, Дальневосточная сер., 2. — 1940. О безлесья тундр. Тр. Ленингр. общ. естествоиспыт., 68. 3. — 1944. О генезисе и фитоценологии аянского темнохвойного леса. Бот. ж. СССР, 29, 5. — 1944а. Причины безлесья гольцов Восточной Сибири и в Приамурье. Природа, 2. — 19446. О происхождении флоры северных полярных стран. Природа, 4. — 1945. Фрагменты горной степи на Среднем Урале. Сов. бот., 3. — 1945а. Фратрии растительных формаций СССР и их филоценогения. ДАН СССР, 45, 1. — 19456. Элементы растительного покрова Северного Сихотэ-Алиня и их взаимоотношения. Сов. бот. 1. — 1946. Вопросы флорогенеза и фило-ценогенеза маньчжурского смешанного леса. Мат. по ист. флоры и растит. СССР, 2. — 1946а. Некоторые ботанико-географические результаты экспедиции Академии Наук СССР на Северо-Западный Кавказ в 1945 г. Сов. бот., 14, 3. — 19466. Тайга и гольцы Северного Сихотэ-Алиня. Уч. зап. Ленингр. пед. инет, им. А. И. Герцена, 49. — 1947. Геоботанические наблюдения в горах и ущельях Мало-Аджарского хребта. Сов. бот., 15, 5. — 1948. Географические связи растительного покрова на территории СССР. Уч. зап. Ленингр. пед. инет, им. А. И. Герцена, 73. — 1949. О происхождении буковых лесов Кавказа. Изв. АН СССР, сер. биол., 2. — 1950. Новейшие вертикальные движения земной коры и растительный покров. Землеведение, нов. сер., 3 (43). — 1953. Растительность лесной зоны. В кн.: Животный мир СССР, 4, М.—Л.
Сочава В. Б., Т. И. Исаченко и А. Н. Лукичева. 1953. Общие черты географического распространения лесной растительности Западно-Сибирской низменности. Изв. ВГО, 85, 2.
Сочава В. Б. и В. В. Липатова. 1952. Распространение бука в лесах Молдавии. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 8.
Сочава В. Б. и А. Н. Лукичева. 1953. К географии кедрового стланика. ДАН СССР, 90. 6.
Спиридонов М. Д. 1918. Очерки растительности Киргизских пустынных степей. Изв. Главн. бот. сада, 18. 2. — 1921. Голодная степь. Тр. Главн. бот. сада, 35. — 1923. Схема естественно-исторических образований западной половины Омской губ. Омск. — 1924. Характеристика естественно-исторических образований юго-восточной части песков Кызыл-Кумы. Изв. РГО, 55, 2. — 1927—1928. Материалы к изучению растительных ландшафтов в Западной Сибири. Изв. Главн. бот. сада, 26, 5 (1927); 27, 1 (1928). — 1928. Материалы к изучению озер и болот в южной части Западной Сиоири. Изв. Зап.-Сиб. муз., 1. — 1930. Очерк растительности западной части песков Сам. Мат. Комисе, экспед. иссл. АН СССР, 26, Казахстанская сер. — 1930а. Очерк растительности района северо-западного и западного Усть-Урта. Мат. Комиссии экспед. иссл. АН СССР, 26. Казахстанская сер.
Спрыгни И. И. 1900. Почвенные и ботанические исследования в Пензенском и Горо-дищенском уездах в 1896—1899 гг. Тр. Общ. естествоиспыт. при Казанск. унив., 33, 5. — 1915. Новая работа из области северных степей. Тр. Пензенск. общ. лк)бит. естествозн., 2. — 1922. Борьба леса со степью в Пензенской губ. Пенза. — 1923. Материалы к описанию степи около д. Поперечной Пензенского у. и заповедного участка на ней. В кн.: Работы по изуч. пензенских заповедников, 1. Пенза. — 1925. Из области Пензенской лесостепи. 1. Травяные степи Пензен-<ской губ. Тр. по изуч. заповедников Отд. охраны природы Главнауки, 4. — 1930. Растительный покров Средне-Волжского края. Самара. — 1934. Выходы пород
татарского яруса пермской системы в Заволжье как один из центров видообразования в группе калькофильных растений. Сов. бот., 4. — 1936. О некоторых лесных реликтах Приволжской возвышенности. Уч. зап. Казанск. унив., 9о, 6.
Спрыгни И. И. и М. Г. Попов. 1915. Ботанико-географические исследования в Туркестане. В кн.: Почвенные исследования в бассейне рр. Сыр-Дарьи и Аму-Дарьи, I, М.
Срединский Н. К. 1874. Очерк растительности Рионского бассейна. Зап. Новоросс. общ. естествоиспыт., 2, 3.
Ссорин В. А. 1951. Пихтовые леса Верхней Сванетии (по среднему течению р. Ингур). Бот. ж., 36, 5.
Станков С. С. 1926. Южный берег Крыма. Н. Новгород. — 1928. Растительность Заволжья Нижегородского и Городецкого уу. и суглинисто-супесчаного района Заочья. Производительные силы Нижегородск. губ., 9. — 1929. Растительность юго-западного правобережья Ветлужского у. Производительные силы Нижегородск. губ., 13. — 1933. Основные черты и распределение растительности Южного Крыма. (Севастополь—Феодосия). Бот. ж. СССР, 18, 1—2. — 1939. О нагорных и степных ксерофитах Южного Крыма в связи с географической изменчивостью можжевеловых лесов между Ласпи и К ар ад атом. Бот. ж. СССР, 24, 5—6.
Станюкович К. В. 1948. Альпийские подушечники как своеобразный тип высокогорной растительности. Природа, 12. — 1949. Растительный покров Восточного Памира. Зап. ВГО, нов. сер., 10. — 1950. О роли диких копытных в образовании подвижных песков. Сообщ. Тадж. фил. АН СССР, 24. — 1952. Еще раз о том, что называть Памиром. Изв. ВГО, 84, 4. — 1955. Основные типы поясности в горах СССР. Изв. ВГО, 87, 3.
Стариков Г. Ф. и П. Н. Дьяконов. 1952. Леса полуострова Камчатка. М.—Л. Старк С. К. 1933. Леса Верхнеколымского района. М.
Стеблин-Каменский Н. 1917. Типы насаждений Погонно-Лосиноостровской лесной дачи Московской губ. и у. Изв. Пгр. лесн. инет., 31.
Степанов Е. С. 1926. Очерк растительности поймы правого берега р. Волхова от г. Новгорода до д. Слугки и левого берега от г. Новгорода до р. Водосьи. Мат. по иссл. р. Волхова и его бассейна, 9.
Степанов Н. А. 1952. Дубравы Северного Кавказа. В кн.: Дубравы СССР, 4 (Всесоюзн. Н.-иссл. инет. лесн. хоз., 31), М.—Л.
Степанова Е. Ф. 1950. Растительность западного Тарбагатая. Изв. АН КазССР, 98 (сер. бот., 5).
Степанова К. Д. 1953. Луга южной части Сахалина. Л.
Степунин Г. 1914. Леса Ставропольской возвышенности. Лесн. ж., 44, 8. Стрельников П. Г. 1938. Тургай-Иргизская полупустыня. Л.
Строгий А. А. 1934. Деревья и кустарники Дальнего Востока. Их лесоводственные свойства, использование и техническое применение. М.—Хабаровск.
Струмилин С. Г. 1947. Итоги естественно-исторического районирования СССР. Тр. Комиссии по естеств.-ист. районированию СССР, 1. Естеств.-ист. районирование СССР, М.—Л.
Стуков Г. А. 1907. Очерк флоры восточного Забайкалья. Изв. Читинск. отдел. При-амурск. отд. РГО, 8.
Суворов Е. К. 1911. Из поездки на Командорские о-ва. Изв. РГО, 47, 6.
Суворов Н. И. 1949. О пасторальной дигрессии растительности в пустынях южного Прибалхашья. Изв. АН КазССР, 52 (сер. бот., 3).
Сукачев В. Н. 1903. Ботанико-географические исследования в Грайворонском и Обоян-ском уездах Курской губ. Тр. Общ. испыт. природы при Харьковск. унив., 37. — 1903а. О болотной и меловой растительности юго-восточной части Курской губ. Тр. Общ. испыт. природы при Харьковск. унив., 37. — 19036. Очерк растительности юго-восточной части Курской губ. Изв. СПб. лесн. инет., 9. — 1904. О ботанико-географических исследованиях в Бузулукском бору Самарской губ. Тр. опытн. леснич., 2. — 1905. О болотной сосне. Лесн. ж., 35, 3. — 1906. Материалы к изучению болот и торфяников озерной области. Тр. преснов, биол. станции СПб. общ. естествоиспыт., 2. — 1908. Лесные формации и их взаимоотношения в Брянских лесах. Тр. по лесн. опытн. делу в России, 9. — 1911. К вопросу о влиянии мерзлоты на почву. Изв. Акад. Наук, сер. 6, 5, 1. — 1912. Нерчинские степи. В кн.: Предварит, отчет об организ. и исполн. работ по иссл. почв Аз. России в 1911 г., СПб. — 1912а. Растительность верхней части бассейна р. Тунгира Олекминского окр., Якутской обл. (Фитосоциологический очерк). Тр. Амурской экспед., 16 (бот. иссл. 1910 г., 1). — 1913. Бассейн р. Верхней Ангары. (Общий очерк). В кн.: Предварит, отчет об организ. и исполн. работ по иссл. почв Аз. России в 1912 г., СПб. — 1914. О Betula pubescens Ehrh. и близких к ней видах в Сибири. Изв. Акад. Наук, сер. 6, 8, 3. — 1916. О «теории дернового процесса» проф. В. Р. Вильямса. Почвоведение, 2. — 1919. Общие
задачи, программа и организация изучения растительных сообществ в долине-p. Чу, Пишпекского у. Сомиреченской обл. В кн.: Растительность долипы р. Чу, 7, Пг. — 1921. К вопросу о ближайших задачах изучения растительности Кольского п-ва. В кн.: Работы организованного Геогр. инст-ом в 1920 г. Кольского’ почв.-бот. отряда Сев. н.-промысл, экспед., 7, Пг. — 1923 (1921). Фитосоциоло-гические очерки. I. Ж. Русск. бот. общ., 6. — 1924. К истории развития лиственниц. В кн.: Лесное дело, Л. — 1926. Болота, их образование, развитие и. свойства. Изд. 3-е. Л. — 1927. Краткое руководство к исследованию типов леса. М. (3-е изд. в 1931 г.). — 1928. (Sukachev). Principles of classification of the spruce communities of European Russia. Journ. of Ecology, 76, 1. — 1931.. Типы леса Бузулукского бора. Тр. и иссл. по лесн. хоз. и лесн. пром., 13. — 1934. Дендрология с основами лесной геоботаники. Л. (2-е изд. 1938). — 1938. Главнейшие понятия из учения о растительном покрове. В кн.: Растительность СССР, 7, М.—Л. — 1938а. История растительности СССР во время плейстоцена. В кн.: Растительность СССР, 7, М.—Л. — 1948. Кокчетавские горные леса. Землеведение, нов. сер., 2(42). — 1949. Задачи стационарного фитоценотического изучения дубового леса и некоторые общие результаты его. Уч. зап. Ленингр. унив., сер.оиол. наук, 17.
Сукачев В. Н. и Г. И. Поплавская. 1914. Ботанические исследования северного побережья Байкала в 1914 г. Изв. Акад. Наук, сер. 6, 8, 17. — 1927. Растительность Крымского Государственного заповедника. В кн.: Крымский Гос. заповедник, его природа, история и значение, М.
Сукачев В. И., А. И. Савенкова и Е. В. Наливкина. 1916. Княжедворский луговой стационарный пункт в 1914—1915 гг. Мат. по организ. и культуре корм, площади, 14.
Сумгин М. и Б. Демчинский. 1940. Область вечной мерзлоты. М.—Л.
Талиев В. И. 1901. Флора Крыма и роль человека в ее развитии. Тр. Общ. испыт. природы при Харьковск. унив., 35. — 1904—1905. Растительность меловых обнажений южной России. Тр. Общ. испыт. природы при Харьковск. унив., 39, 1 (1904); 40 (1905). — 1909. О растительности Крымской Яйлы. Тр. Общ. испыт. природы при Харьковск. унив., 42. — 1913. Введение в ботаническое исследование Харьковской губ. Харьков.
Танфильев В. Г. 1931. Описание естественных кормовых угодий УЗОС. Изв. Урупск. зоотехн. опытн.-племен, станции, 4, Пятигорск.
Танфильев Г. И. 1891. Болота. В кн.: Энциклопедический словарь Брокгауза и Ефрона, 4, СПб. — 1892. О связи между растительностью и почвой, по наблюдениям в Воронежской губ. Тр. СПб. общ. естествоиспыт., 22. — 1894. По тундрам тиманских самоедов летом 1892 г. Изв. РГО, 30, 1. — 1894а. Пределы лесов на юге России. СПб. — 1895. Болота и торфяники Полесья. СПб. — 1896. Доисторические степи Европейской России. Землеведение, 3, 2. — 1897. Физико-географические области Европейской России. Тр. Вольн. экон, общ., 1. — 1898. Ботанико-географические исследования в степной полосе. Тр. экспед. снаряж. Лесным департаментом под рук. проф. Докучаева, Научн. отдел, 2, 2. — 1900. Болота и торфяники. В кн.: Полная энциклопедия русского сельского хозяйства, 7, СПб. — 1900а. География растений. (С ботанико-географической картой Российской империи). В кн.: Полная энциклопедия русского сельского хозяйства, 2, СПб. — 1902. Бараба и Кулундинская степь в пределах Алтайского окр. Тр. Геол, части кабинета Е. И. В., 5, 1, СПб. — 1902а. К вопросу о причинах безлесия Крымской яйлы. Изв. СПб. бот. сада, 2, 1. — 1903. Главнейшие черты растительности России. (С ботанико-географической картой Российской империи). В кн.: Е. Варминг, Распределение растений, 3, СПб. — 1904. Очерк главнейших районов Черноморского побережья Кавказа. В кн.: Современные вопросы русского сельского хозяйства, СПб. — 1911. Пределы лесов в полярной России по исследованиям в тундре тиманских самоедов. Одесса. — 1915. Галиция и Буковина. Одесса. — 1923. Очерк географии и истории главнейших культурных растений. Одесса.
Тарабаева Б. И. 1953. К редукции листовой поверхности у саксаула. Уч. зап. Казахск. SiHB., 14, 4. — 1953а. Растительность приилийских саксаульников. Уч. зап. азахск. унив., 14, 4.
Тарасов Р. П. 1954. Растительность Малых Балхан. Тр. Инет. биол. АН ТуркмССР, Сер. бот., 2.
Таскаева Н. Я. 1943. К познанию лесов средней части Молотовской обл. Бот. ж. СССР, 28, 6.
Таубаев Т. 1954. Водная растительность низовьев Аму-Дарьи. Ташкент.
Тахтаджян А. Л. 1937. Ксерофильная растительность скелетных гор Армении. Тр. Армянск. фил. АН СССР, сер. биол., 2. — 1941. Ботанико-географический очерк Армении. Тр. Бот. инет. Армянск. фил. АН СССР, 2.
Темноев Н. И. 1938. Луга низкогорной части Казахстанского Алтая. Сов. бот. 4—5. — 1938а. Некоторые флористические особенности лугов горностепной части юго-западного Алтая. Бот. ж. СССР, 23, 2. — 1940. Очерк растительного покрова верхнего отрезка долины Волги от д. Иваньково, Кимрского района, до д. Н. Каменец, Мышкинского района. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 4.
Тереножкин И. И. 1934. Скотобой, его роль и значение для пастбищного хозяйства в сухо-степных и полупустынных районах Нижнего Поволжья. Сов. бот., 4. — 1947. Пастбища Сталинградской обл. (Характеристика основных типов пастбищ и перспективы их улучшения). Сталинград. — 1954. Растительность и кормовая база Волго-Донского междуречья и пути их улучшения. В кн.: Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР,
Терьян А. II. 1948. Торфяные месторождения Грузии. Торф, промышленность, 1. Тимофеев В. П. 1947. Лиственница в культуре. М.—Л.
Тиханович Н. и II. Полевой. 1915. Геоморфологический очерк русского Сахалина. Тр. Геол, комитета, нов. сер., 120.
Тихомиров Б. А. 1935. Краткий очерк долинной растительности Пенжинского района. Тр. Дальневост. фил. АН СССР, сер. бот., 1. — 1941. К вопросу о динамике полярного и вертикального пределов лесов в Евразии. Сов. бот., 5—6. — 1946. К происхождению ассоциаций кедрового стланика (Pinus pumila Rgl.). Мат. по ист. флоры и растительн. СССР, 2. — 1946а. К происхождению лугового типа растительности в арктической Евразии. В кн.: Сборник научн. работ, выполненных [Бот. Инет. им. В. Л. Комарова АН СССР] в Ленинграде за три года Великой Отечественной войны (1941—1943), Л. — 19466. Пути формирования растительного покрова арктической Евразии в четвертичном периоде. Сов. бот., 14, 5. — 1948. К характеристике флоры западного побережья Таймыра. Тр. Карело-Финск. унив., 2. — 1949. Кедровый стланик, его биология и использование. Мат. к познанию фауны и флоры СССР, нов. сер., отд. бот., 6 (14). — 1953. Безлесье тундры него преодоление. Бот. ж., 38, 4.
Тихомиров Н. К. 1927. Очерк растительности о-ва Ольхона на оз. Байкал. Тр. Комиссии АН СССР по изуч. оз. Байкал, 2.
Тихонова Р. П. 1952. К вопросу использования индикаторных свойств растительного покрова для сельскохозяйственного освоения болотных массивов Карело-Фин-скои ССР. Л.
Ткаченко М. Е. 1911. Леса севера. Тр. по лесн. опытн. делу в России, 25. — 1939. [редактор] Общее лесоводство. Изд. 1-е, Л. (Изд. 2-е, 1952).
Толмачев А. И. 1923. Ботанико-географические работы в районе Югорского Шара в 1921 и 1922 гг. Изв. Главн. бот. сада, 22, 2. — 1929. (Т о J m а б е у) Beitrage zur Kenntnis des Gebietes von Matotschkin Schar und der Ostkiiste Nowaja Semlia’s. Изв. АН СССР, сер. VII, отд. физ.-мат. наук. — 1930. Предварительный отчет о работе Таймырской экспедиции Академии Наук СССР в 1928 г. Тр. Полярн. комиссии АН СССР, 1. — 1931. О распространении древесных пород и о северной границе лесов в области между Енисеем и Хатангой. Тр. Полярн. комиссии АН СССР, 5. — 1939. О некоторых закономерностях распределения растительного покрова в Арктике. Бот. ж. СССР, 24, 5—6. — 1948. Основные пути формирования растительности высокогорных ландшафтов северного полушария. Бот. ж., 33, 2. — 1949. Наблюдения над Cousinia Fedtschenkoana С. Winkl. и некоторые вопросы генезиса нагорно-ксерофитной растительности Средней Азии. Бот. ж., 34, 1. — 1950. О высокогорной флоре горы Лопатина (о. Сахалин). Бот. ж., 35, 4. — 1954. К истории возникновения и развития темнохвойной тайги. М.—Л. — 1956. Вертикальное распределение растительности на Сахалине. Геогр. сб. ВГО, 8.
Томашевский И. И. 1931. Пески Астраханской степи. Тр. по лесн. опытн. делу Центр, лесн. опытн. станции, 10.
Томин М. П. 1910. Очерк растительности Манзурской возвышенности и отрогов Березового хребта в Верхоленском у. Иркутской губ. Тр. Почв.-бот. экспед. Пепе-селенч. упр. по иссл. колониз. районов Аз. России в 1908 г., ч. 2, 6. — 1926. Лишайники, встречающиеся на солонцеватых почвах в пустынной области юго-востока. В кн.: Б. А. Келлер, Растительный мир русских степей, полупустынь и пустынь, 2, Воронеж.
Траутфеттер Р. Э. 1849—1851. (Е. R. Trautvetter). Die pflanzengeographischen Verhaltnisse des Europaischen Russlands. 1 (1849), 2 (1850), 3 (1851). Riga. — 1851. О растительно-географических округах Европейской России. В кн.: Тр. Комиссии при Унив. св. Владимира для описания губерний Киевского учебного округа, Киев. —1880. Florae rossicae fontes. Тр. СПб. бот. сада, 7, 1.
Троицкий В. Д. 1941. Роща камчатской пихты и физико-географические условия района, ее окружающего. Изв. ВГО, 73, 3.
Троицкий Н. А. 1939. Остатки лесов в верхнем течении р. Дзорагет. Научи, тр» Ереванск. унив., 9. — 1951. Ботаническая и производственная характеристика пастбищ каракулеводческих совхозов Крыма. Изв. Крым. отд. Геогр. общ. СССР, 1.
Троицкий Н. Д. 1929. Дубовые леса Крымского Государственного заповедника (Quer-cetum sessiliflorae). Тр. по изуч. заповеди., 10. — 1936. О пицундской сосне-в Южном Крыму. Природа, 9.
Тросько И. К. 1947. Реконструкция фисташковых рощ и культура фисташки в Средней Азии. Ташкент.
Трофимов Т. Т. 1940. Растительность ореховых лесов южной Киргизии. Бюлл. МОИП„ отд. биол., 49, 5—6.
Трутнев А. Г. 1953. Целинные земли Европейского севера СССР и их освоение. М—Л. Трушковский А. А. 1952. Пески пустынного Заволжья как пастбищный фонд и оценка естественного зарастания песков. Бот. ж., 37, 6.
Тумаджанов И. И. 1938. Геоботанический очерк сосновых лесов Атенского ущелья. Тр. Тбил. бот. инет. Груз. фил. АН СССР, 2. — 1938а. Леса горной Тушетии. Тр. Тбил. бот. инет. Груз. фил. АН СССР, о. — 19386. Типы лесов бассейна р. Белокан-Чай. Тр. Бот. инет. Азерб. фил. АН СССР, 4. — 1938в. Типы лесов, бассейна р. Киш-Чай. Тр. Бот. инет. Азерб. фил. АН СССР, 4. — 1938г. Краткий очерк лесной растительности бассейна р. Курмух-Чай. Тр. Бот. инет. Азерб. фил. АН СССР, 4. — 1947. Лесная растительность долины Теберды в свете послеледниковой истории развития фитоландшафтов. Тр. Тбил. бот. инет. АН ГрузССР, 11. — 1948. Очерк болотной растительности долины Теберды. Тр. Тбил. бот. инет. АН ГрузССР, 12.
Туркевич С. Ю. 1913. Растительность Ачинского у. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1912 г., СПб. — 1914. Очерк растительности южной части Ачинского и северной части Минусинского у. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1913 г., Пг.
Туфанов П. С. 1949. Луга и пастбища Ивановской обл. В кн.: Растительные ресурсы Ивановской обл., 1, Иваново.
Тюлина Л. Н. 1924 (1922). К фитосоциологии елового леса. Ж. Русск. бот. общ., 7. — 1929. Из высокогорной области Южного Урала. В кн.: Очерки по фитосоциологии и фитогеографии, М. — 1929а. К эволюции растительного покрова предгорий Южного Урала. В кн.: Гос. Ильменский минералогический заповедник, 1, Златоуст. — 1930. Материалы по изучению перелогов Госзаповедника «Чапли» (б. Аскания-Нова). BicTi Держ. Степ, заповщн. «ЧаплЬ>, 7. — 1931. Материалы по высокогорной растительности южного Урала. Изв. ВГО, 63, 5—6. — 1936. О лесной растительности Анадырского края и ее взаимоотношениях с тундрой. Тр. Арктич. инет., 40. — 1937. Лесная растительность Хатангского района у ее северного предела. Тр. Арктич. инет., 63. — 1948. Материалы по высокогорной растительности Баргузинского хребта. Землеведение, нов. сер., 2 (42). — 1949. Очерк растительности Баргузинского заповедника. Научн.-методич. зап. Главн. упр. по заповеди., 12. — 1950. Из истории растительного покрова северо-восточного побережья Байкала. Проблемы физ. географ., 15. — 1954. Лиственничные леса северо-восточного побережья Байкала и западного склона Баргузинского хребта. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 9.
Тюремнов С. Н. 1928. Геоботаническое исследование болот восточной части Центрально-Черноземной обл. Торф, дело, 7. — 1931. Болота Белорусской республики. Торф, дело, 1. — 1949. Торфяные месторождения и их разведка. Изд. 2-ег М.—Л.
Тюремнов С. Н. и Е. А. Виноградова. 1953. Геоморфологическая классификация торфяных месторождений. Тр. Моск. торф, инет., 2.
Тюрин А. В. 1916. Еловые насаждения в северной и северо-восточной России. Тр. по лесн. опытн. делу в России, 58.
Уранов А. А. 1925. Материалы к фитосоциологическому описанию заповедной степи Пензенской губ. в связи с законом константности. Тр. по изуч. заповеди. Отд-охр. природы Главнауки НКП РСФСР, 7.
Усков С. П. 1930. Типы лесов Карелии. Петрозаводск.
Уснанов У. У. 1940. Генезис и мелиорация такыров. Тр. Почв. инет. АН СССР, 19, 1-Уткин Л. А. 1935. Ботанико-географический очерк Причулымского края. Бот. ж. СССР, 20, 6.
Федоров Ал. А. 1949. Каштанолистный дуб (Quercus castaneifolia С. А. М.) в Талыше! и его использование. Тр. Бот. инет. АН СССР, сер. 5, растит, ресурсы, 2.
Федоров Ал. А. и Ан. А. Федоров. 1951. Два года в Саянах. М.
Федоров Ан. А. 1940. Капуджих. Изв. Армянск. фил. АН СССР, 4—5. — 1942. Альпийские ковры Кавказа и их происхождение. Изв. Армянск. фил. АН СССР*
.9—10. — 1945. Околоснежная растительность горы Арагац (Алагез) в Армении. Сов. бот., 13, 4.
Федченко Б. А. 1902. Очерк растительности Памира, Шугнана и Алая. Тр. СПб. Общ. естествоиспыт., 33, 1. — 1903. В Западном Тянь-Шане летом 1902 г. Изв. РГО, 39, 5. — 1906. (В. F е d t s с h е n k о). Flore des lies du Commandeur. Cra-covie. — 1909. Шугнан. Тр. СПб. бот. сада, 30, 1. — 1912. К схематической ботанико-географической карте Азиатской России. В кн.: К. Д. Глинка и Б. А. Федченко, Краткая характеристика почвенных и растительных зон Азиатской России, СПб. — 1912а. Растительность Алая и Памира. В кн.: Предварит. отчет о бот. иссл. в Сибири и Туркестане в 1911 г., СПб. — 1925. Изучение растительности Сибири за последние годы (1918—1925). Сев. Азия, 5—6. — 1925а. Очерки растительности Туркестана. Л. — 1937. Успехи советской флористики (1917—1937). Сов. бот., 5.
Федченко Б. А. и Н. Ф. Гончаров. 1929. Очерк растительности восточной части Оренбургской губ. Тр. Главн. бот. сада, 41, 1.
Федченко Б. А. и И. М. Крашенинников. 1910. Растения Тургайской обл. Тр. Почв.-бот. экспед. Переселенч. упр. по иссл. колониз. районов Аз. России в 1908 г.,
Федченко Б. А. и А. Ф. Флеров. 1907—1911. Растительность России — Russlands Vegetationsbilder (на русск. и нем. яз.). I, 2 (1907), 3 (1908), 4 (1911). СПб.
Фигуровский И. В. 1916. Деление Кавказа на физико-географические области и районы. Изв. Кавказ, отд. РГО, 24, 2.
Филатов С. М. 1911. Цевельское болото. Мат. по изуч. вост, болотного района Псковской губ., 1. — 1913. Болото между озерами Полисто и Цевло. Мат. по изуч. вост, болотного района Псковской губ., 2.
Филиппов Г. С. 1934. Микрофлора красного снега некоторых районов Кавказа. Изв. АН СССР, сер. 7, отд. матем. и естеств. наук, 7.
Флеров А. Ф. 1907—1908. Окская флора. 1 (1907), 2 (1908). СПб. — 1926. Растительность северо-черноморского побережья Кавказа. II. Анапский район. Тр. Сев.-Кавказ. ассоц. н.-иссл. инет., 12. — 1930. Типы растительности дельты и низовьев р. Кубани и р. Анапки. Тр. Сев.-Кавказ, ассоц. н.-иссл. инет., 83. — 1936. Растительность Кобулетских болот. Почвоведение, 2. — 1951. Растительность Колхидской низменности. Бюлл. Всесоюзн. н.-иссл. инет, чая и субтропич. культур, 1.
Флеров А. Ф. и В. А. Флеров. 1926. Растительность северо-черноморского побережья Кавказа. I. Растительность п-ва Абрау и побережья Анапа—Новороссийск. Тр. Сев.-Кавказ, ассоц. н.-иссл. инет., 8.
Фокин А. Д. 1929. Краткий очерк растительности Вятского края. В кн.: Вятский край за 1929 г. (В помощь учителю), Вятка. — 1930. Три года работы геоботанического отряда Вятской почвенной экспедиции. Вятское хоз., 2, 3.
Формозов А. Н. и А. Г. Воронов. 1935. Основные черты деятельности грызунов на пастбищах и сенокосных угодьях. ДАН СССР, о, 8.
Форет А. 1917. Типы насаждении Екатерининской дачи Ветлужского у. Тр. Костромск. научн. общ. по изуч. мести, края, 6.
Фурсаев А. Д. 1934. О географической зональности в распределении флоры и растительности нижней Волги. Уч. зап. Саратовск. унив., 11, 2. — 1940. Материалы к вопросу о сукцессиях лесных ассоциаций в дельте Волги. Тр. Астрах, заповеди., 3. — 1952. Естественные леса в пределах трассы Государственной лесной полосы. Саратов—Камышин. Уч. вал. Саратовск. унив., 29. — 1954. Растительность Волго-Ахтубинской поймы и перспективы ее улучшения. В кн.: Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР, М.-Л.
Фурсаев А. Д., Н. Г. Басов, В. В. Гришанин, М. П. Кирсанов и В. И. Князевская.. 1938. Сукцессии приморской полосы дельты Волги. Уч. зап. Саратовск. унив., 1 (14), (биол. сер., 2).
Харадзе А. Л. 1944. Очерк флоры субнивального пояса Верхней Сванетии. Заметки по систематике и геогр. растений Тбил. бот. инет., 12. — 1946. К изучению пе-ригляциальной растительности Центрального Кавказа. Сообщ. АН ГрузССР,. 7, 9—10.
Хитрово А. А. 1907. Казанские нагорные дубравы. Лесн. ж., 37, 5.
Хитрово В. Н. 1913. Путеводитель по Галичьеи горе. Иэв. Общ. для иссл. природы! Орловск. губ., 3. — 1925. Растительность [Орловского* края]. В кн.: Природа» Орловского края, Орел.
Хорват Е. М. 1948. Растительность крымских «дубков». Тр. Никитск. бот. сада,.. 25, 1—2.
Цаценкин И. А. 1936. Естественные кормовые угодья Калмыцкой АССР. Изв. ГГО, 68, Ь. — 1940. Типологический состав естественных сенокосов и пастбищ по*
природным зонам СССР. Вестн. с.-х. науки. Кормодобывание, 4. — 1940а. Угодья кормового пользования в СССР. Вести, с.-х. пауки. Кормодобывание, 5. — 1947. Природные кормовые угодья СССР. В кн.: Вопросы кормодобывания, М. — 1952. Геоботаническая классификация пастбищ и сенокосов правобережной части Прикаспийской низменности и Ергоной. Уч. зап. Моск, унив., 160 (геогр., о). — 1954. Классификация пастбищ и сенокосов западного Прикаспия и вопросы их комплексного изучения. В кн.: Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР, М.—Л.
Цаценкин И. А. и II. А. Антипин. 1951. Типы природных кормовых угодий. Вкн.: Многолетние травы в лугопастбищных севооборотах, М.
Цаценкин И. А., В. Ф. Максимова и Т. II. Щсрбиновская. 1952. Геоботаническое районирование пастбищ и сенокосов Западного Прикаспия — Ергоной, Сарпинской низменности и Черных земель. Вюлл. МОИП, отд. биол. 57, 1.
Ципзерлинг Ю. Д. 1929. Очерк растительности по среднему точонию р. Печоры. Изв. Главн. бот. сада, 28, 1—2. — 1929а. Результаты исследования болот и некоторых других геоботанических наблюдений в районе оз. Имандра. В кн.: Очерки по фитосоциологии и фитогеографии, М. — 1932. География растительного покрова северо-запада Европейской части СССР. Тр. Геоморф, инет., сер. физ.-геогр., 4. (Часть тиража выпущена в 1934]. — 1935. Материалы по растительности северо-востока Кольского п-ва. Тр. СОПС АН СССР, Кольская сер., 10. — 1935а. Очерк растительности массива Сабля. Тр. Ледниковых экспед., 1. Урал. — 19356. Растительность поймы среднего течения р. Печоры от с. Усть-1Цугор до с. Усть-Уса. Тр. Полярн. комиссии АН СССР, 24, — 1938. Растительность болот. В кн.: Растительность СССР, 1, М.—Л.
Цымек А. А. 1950. Главнейшие лиственные породы Дальнего Востока. Хабаровск. — 1951. Роль лиственных пород в сложении лесного покрова на Дальнем Востоке. Сб. работ Дальневост, н.-иссл. инет. лесн. хоз., 3.
Цырина Т. С. 1925. Ливенские дубки. Зап. Крым. общ. естествоиспыт. и любит, природы, 8.
Чабан П. С. 1941. Казахстанские субтропики — Бостандыкский район Казахской ССР. Тр. Казахст. фил. АН СССР, 20.
Чекановский А. Л. 1896. Дневник экспедиции по рр. Нижней Тунгуске, Оленеку и Лене в 1874—1875 гг. Зап. РГО, 20, 1.
Челльман Ф. Р. 1886—1887. О явнобрачной флоре Командорских островов. Бот. зап. при бот. саде СПб. унив., 1.
Черепнин Л. М. 1953. Растительность залежей южной части Красноярского края. Уч. зап. Красноярск, пед. инет., 2. — 1953а. Флора и растительность южной части Красноярского края. Л.
Чернов В. Н. 1940. Геоботанический очерк Окского Государственного заповедника. Тр. Окского заповеди., 1.
Чернова Н. М. 1951. Растительный покров западных яйл Крыма и их хозяйственное использование. Тр. Никитск. бот. сада, 25, 3.
Черный А. П. и В. С. Доктуровский. 1915. В области Полесья. Исследование болот и лугов в долине р. Лани. Мат. по организ. и культуре корм, площади, 10.
Черняев В. М. 1858. О лесах Украины. М.
Черняев Л. В. 1865. Очерки стопной растительности. Сельск. хоз. и лесовод., 88, 2.
Черняховская Е. Г. 1924. Весенняя растительность Каракалинского района Закаспийской обл. I. Изв. Главн. бот. сада, 23, 2. — 1925. Весенняя растительность Ка-?акалинского района Закаспийской обл., II. Изв. Главн. бот. сада, 24, 1. — 927. Очерк растительности Копет-дага. Изв. Главн. бот. сада, 26, 3.
Чижиков П. Н. 1951. О березовых лесах юга Камчатки. Бюлл. МОИП, отд. биол., 56,
Чугунов Л. А. 1951. Луговодство. Изд. 2-е. М.—Л.
Чугунова-Сахарова Н. Л. 1927. Материалы по изучению дельты Волги и прилегающей предустьевой части Каспийского моря. Наш край, 6, Астрахань.
Шалыт М. С. (Шали т). 1930. Великий Чанельсышй шд в Асканп-Нова та його Еослиншсть року 1927—1928. BicTi Держ.Степ. заповщн. «ЧаплЬ (кол. Аскашя-1ова), 7. — 1930а. Геоботанический очерк Государственного степного заповедника «Чапли» (б. Аскания-Нова). Бюлл. Фитотехн. станции Гос. степн. заповеди. «Чапли» (о. Аскания-Нова), 1. — 19306. Деяк! вщомост! запроцес вщновлювання степово! рослинност! на перелогах Державного Стенового заповщника «ЧаплЬ (кол. Аскашя-Нова). BicTi Держ. Степ, заповщн. «ЧаплЬ (кол. Аскан1я-Нова), 7. — 1938. Растительность степей Аскания-Нова. Изв. Крым. пед. инет., 7. — 1948. О растительности Присивашья. (К вопросу о существовании «полынных степей» на юге Украины и в Северном Крыму). Бюлл. МОИП, отд. биол., 53, 6. — 1951. Дикорастущие полезные растения Туркменской ССР. М.
Шалыт М. С. и II. К. Козлов. 1939. Геоботаническое районирование Крыма. Изв. ГГО, 71, 3.
Шаханип 11. И. 1945. Ботанико-географическая характеристика Ярославской обл. Уч. зап. Ярославок, пед. инет., 6 (16).
Шахов Л. Л. 19ч5. Экологические и фитоценотические области солончакового фитоценоза. Бюлл. МОП 11, отд. биол., 50, 3—4. — 1947. Борьба за существование в ценозе галофитов. Сов. бот., 15, 4.
Шахов Л. Л. и II. М. Земцова. 1948. Динамика кормового запаса и химизма растений полупустынных пастбищ. Бюлл. МОИП, отд. биол., 53, в.
Шелковинкой Л. Б. 1906. Экскурсия в Талыш. Вести. Тифл. бот. сада, 3.
Шелудякова В. А. 1926. Атбассарские степи. (Растительность северо-западной части Атбассарского у.). Изв. Зан.-Сиб. отд. РГО, 5. — 1938. Растительность бассейна р. Индигирки. Сов. бот., 4—5. — 1948. Растительность северо-востока Якутии. В кн.: Доклады на первой научной сессии Якутской базы АН СССР, Якутск.
Шенников А. II. 1913. Аллювиальные луга в долинах рр. Сев. Двины и Сухоны в пределах Вологодской губ. Мат. по организ. и культуре корм, площади, 6. — 1914. Материковые и озерные луга Олонецкой губернии. Мат. по организ. и культуре корм, площади, 9. — 1919. Луга Симбирскои губернии, 1. Симбирск. — 1930. Волжские луга Средневолжской обл. Л. — 1935. Принципы ботанической классификации лугов. Сов. бот., 5. — 1937. Теоретическая геоботаника за последние 20 лет. Сов. бот., 5. — 1938. Луговая растительность СССР. В кн.: Растительность СССР, 1, М.—Л. — 1940. К ботанической географии лесного северо-востока Европейской части СССР. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 4. — 1941. Луговедение. Л. —1948. Успехи советской геоботаники. Вести. Ленингр. унив., 9.
Шенников А. П. и А. Ф. Иоффе. 1944. К биологии пустынных злаков-эфемеров. Бот. ж. СССР, 29, 1.
Шимашок А. П. 1931. Опыт изучения северных лесов. (Типы лесов Чарондского края). М.—Л.
Шингарева Е. А. 1940. Кормовые ресурсы Северных Кара-кумов. В кн.: Природные ресурсы Кара-кумов, 1, М.—Л. — 1940а. Растительность и кормовые ресурсы Низменных Кара-кумов. В кн.: Природные ресурсы Кара-кумов, 2, М.—Л.
Шипчинский И. В. 1916. Растительность юго-восточной части Семипалатинского у. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1914 г., Пг. — 1928. Краткий очерк растительности восточной части Нарымского края. Изв. Главн. бот. сада, 27, 4. — 1950. К озеленению города Балхаша. Тр. Бот. инет. АН СССР, сер. 6, интродукц. и зел. строит., 1. — 1953. Материалы по интродукции деревьев и кустарников в равнинах Средней Азии (для озеленения). Тр. Бот. инет. АН СССР, сер. 6, интродукц. и зел. строит., 3.
Ширяев Г. И. 1904. Материалы для флоры южной части Старобельского и восточной части Купянского уу. Харьковской губ. Тр. Общ. испыт. природы при Харьковск. унив., 38, 1. — 1910. Флора долины р. Пела в Лебединском у. Харьковской губ. Тр. Общ. испыт. природы при Харьковск. унив., 43.
Шиффере Е. В. 1928. Таманский п-ов и сев.-вост, часть Керченского. Изв. Главн. бот. сада, 27, 2. — 1928а. Приазовские лиманы и плавни нижнего течения р. Кубани. Изв. Главн. бот. сада, 27, 5—6. — 1937. Сорная растительность дельты р. Терека и других плоскостных районов Дагестана в ее связи с местными естественными фитоценозами. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 2. — 1941. Геоботаническое районирование и исследование природных кормовых угодий северных склонов Б. Кавказа. Природа, 3. — 1946. Природная кормовая растительность горного Дагестана. В кн.: Сельское хозяйство Дагестана, М.—Л. — 1951. К характеристике растительных природных кормовых угодии северо-западной части Кавказа. Тр. Бот. инет. АН СССР, сер. 5, геоботаника, 7. — 1953. Растительность Северного Кавказа и его природные кормовые угодья. М.—Л.
Шифферс-Рафалович Е. В. см. Шиффере Е. В.
Шишкин Б. К. 1914. Очерки Урянхайского края. Изв. Томск, унив., 60. — 1914а. Растительность западной части Копальского у. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1913 г., Пг. — 1937. Растительность Алтая. В кн.: Ойротия, Тр. сессии СОПС АН СССР по изуч. производит, сил Ойротской авт. обл., М.—Л.
Шишкин И. К. 1927. Типы лугов Приханкайского района и их хозяйственная ценность. В кн.: Производительные силы Дальнего Востока, 3, растительный мир, Хабаровск—Владивосток. — 1933. К познанию ольгинской лиственницы. (Larix olgensis A. Henry). Бот. ж. СССР, 18, 3. — 1933а. Сосна на юге Уссурийского края. Вести. Дальневост. фил. АН СССР, 1—3. — 1934. Лиственница Любарского (Larix Lubarskii Suk.) в Уссурийском крае. Вести. Дальневост. фил. АН СССР, 9. — 1935. Microbiota decussate Кош. как элемент растительного покрова Уссурийского края. Тр. Дальневост. фил. АН СССР, сер. бот., 1. —> 1936.
Материалы к флоре Карагинского о-ва. Вестн. Дальневост. фил. АН СССР, 20.
Шмидт Ф. Б. 1874. Труды Сибирской экспедиции РГО, физ. отд., 2, бот. часть. СПб. Шнитаиков В. Н. 1925. Растительность Джетысу. В кн.: Джетысу (Семиречье), Ташкент.
Штукенберг Е. К. 1932. Растительность Мурапталовского зерносовхоза Мелеузовского района Башкирской АССР. Тр. СОПС АН СССР, Башкирская сер., 2.
Шувалов С. А. 1949. К вопросу о комплексности почвенно-растительного покрова Усть-Урта. В кн.: Тр. Юбилейной сессии, посвящ. столетию со дня рождения В. В. Докучаева, М.—Л.
Шукуров О. Ш. 1949. Эгилопсовая формация в Таджикистане и ее происхождение. Сообщ. Тадж. фил. АН СССР, 20.
Шумилова Л. В. 1931. О бугристых торфяниках южной части Туруханского края. Изв. Томск, отд. Русск. бот. общ., 3, 1—2. — 1933. Материалы по изучению оленьих пастбищ в районе оз. Пясино и Норильских гор в Туруханском крае. Мат. по изуч. Сибири, 4, Томск. — 1947. Схема ботанико-географических районов Ба-рабы. Уч. зап. Томск, унив., 7. — 1949. О расчленении Сибири на ботанико-географические провинции. В кн.: Вопросы географии Сибири, I, Томск. — 1949а. Растительность Центрально-Сибирского плоскогорья. В кн.: Тр. Второго Всесоюзн. геогр. съезда, 3, М.
Щербаков И. П. 1951. Естественно-исторические условия районов дубрав промышленного значения. Тр. Комплекс, научн. экспед. по вопросам полезащ. лесоразвед., 7, 2.
Эмме-Марковская Л. А. 1940. Материалы по растительности южных склонов Кураминского хребта. Тр. Тадж. базы АН СССР, 8.
Эйдельман Г. Н. 1936. Торфяные болота Дальне-Восточного края. Тр. Центр, торф, опытн. станции, 1. — 1936а. Торфяные болота Енисейского Севера. Тр. Центр, торф, опытн. станции, 1.
Эттинген Г. Г. 1910. Нижне-Амурская экспедиция. В кн.: Предварит, отчет о бот. иссл. в Сибири и Туркестане в 1909 г., СПб.
Юдин Ю. П. 1937. Северная граница пихты и кедра в пределах Ижмо-Печорского и Ижмо-Вымского междуречий. Изв. ГГО, 69, 1. — 1940. Распространение и характер произрастания липы в бассейне р. Конды. Тр. Бот. инет. АН СССР, сер. 3, геоботаника, 4. — 1950. Очерк растительности бассейнов рек Шугора и Подче-рема. Бот. ж., 35, 5. — 1951. Вертикальная зональность и верхняя граница леса в горах Шугорского Урала. Изв. Коми фил. ВГО, 1, 1. — 1953. Основные группы типов леса Коми АССР. Тр. Коми фил. АН СССР, 1.
Юнатов А. А. 1950. Основные черты растительного покрова Монгольской Народной Республики. Тр. Монг, комиссии АН СССР, 39. — 1954. Кормовые растения пастбищ и сенокосов Монгольской Народной Республики. Тр. Монг, комиссии АН СССР, 56.
Юрьев М. М. 1912. Болота и леса северной части Великолуцкого у. Мат. по изуч. растит. Псковской губ., 2.
Яковлев Ф. С. 1946. Классификация и основные закономерности распространения ольшаников и дубняков в поймах рр. Дона и Нижней Волги. Научн. зап. Воронежск. лесохоз. инет., 9.
Якубов Т. Ф. 1935. Пески Урдинского района Западного Казахстана. Тр. Почв. инет. АН СССР, 9.
Яната А. А. 1913. Флора степи Мелитопольского и юго-западной части Днепровского уездов, Таврической губ. Тр. Естеств.-ист. муз. Таврическ. губ. земства, 2. — 1916. О природе и хозяйстве Крымской яйлы в связи с влиянием ее на водный режим горного Крыма. В кн.: По Крыму, 3, Симферополь. — 1931. Яйла. Экономика и культура Крыма, 3.
Янишевский Д. Е. 1937. К характеристике осок засушливых областей Союза ССР. Сов. бот., 4.
Яровой М. И. 1939. Растительность бассейна р. Яны и Верхоянского хребта. Сов. бот., 1.
Ярошенко Г. Д. 1929. Сосна и дуб в Армении. Эривань.
Ярошенко П. Д. 1942. Опыт классификации травянистой высокогорной растительности Кавказа. Изв. Армянск. фил. АН СССР, 1—2 (15—16). — 1942а. О сменах растительности в лесной области Закавказья. Изв. Армянск. фил. АН СССР, 7 (21). — 1946. О взаимоотношениях лугов и некоторых других фитоценозов в высокогорьях Кавказа. Изв. АН АрмССР, естеств. науки, 1. — 1947. Нариси рос-линносп Закарпатсько! обл. Наук. зап. Ужгородськ. ушв., 1.
Adamovic L. 1901. Die Sibljak-Fonnation, ein wenig bekanntes Buschwerk der Bal-kanlander. Engler’s Bot. Jabrb., 31. — 1906. Ober eine bisher nicht unterschie-dene Vegetationsformation der Balkanbalbinsel, die Pseudomacchie. Verh. der Zool.-Bot. Ges. in Wien, 56, 4. — 1909. Die Vegetationsverhaltnisse der Balkan-lander (Mosische Lander), umfassend Serbian, Altserbien, Bulgarian, Ostrumelien, Nordturakien und Nordmazedonien. Die Vegetation der Erde, II, Leipzig.
Auer V. 1922. (1923) Suotutkimuksia Kuusamon ja Kuolajarven vaara-alueilta. Common. ex Inst. Quaest. Forest. Finlandiae, 6.
Borza Al. 1937. Cercetari fitosociologice asupra pSdurilor basarabene. Phytosociologi-cal studies on the forests of Basarabia. Bui. Grad. Bot. si al Muz. Bot. dela Univer. din Cluj, 17, 1—2.
Braun-Blanquct J., G. Sissingh und J. Vlieger. 1939. Klasse der Vaccinio-Picee-tea. Prodromus der Pflanzengesellscbaften, 6.
Cajander A. 1903. Beitriige zur Kenntnis der Vegetation der Alluvionen des nordlichen Eurasiens. I. Die Alluvionen des unteren Lena-Thales. Act. Soc. Sc. Fenn., 32, 1. — 1904. Studien uber die Vegetation des Urwaldes am Lena-Fluss. Act. Soc. Sc. Fenn., 32, 3. — 1905. Beitrage zur Kenntnis der Vegetation der Alluvionen des nordli-cben Eurasiens. II. Die Alluvionen des Onega-Thales. Act. Soc. Sc. Fenn., 33, 6.
Deyl M. 1940. Plants, soil and climate of Pop Ivan. Opera Bot. Cechica, 2.
Dornin K. 1930. Budiny Podkarpatske Rusi s hlediska sociologickeho. Sp. Pfirodov. Fac. Karlovy Univ., 107.
Du Rietz G. E. 1925. Die regionale Gliederung der skandinawischen Vegetation. Sven. Vaxtsociol. Sallskap. Handl., 8.
Gams H. und S. Ruoff. 1929. Geschichte, Aufbau und Pflanzendecke des Zehlaubruches. Schriften der Phys.-Okon. Ges. zu Konigsberg, 66, H. 1, T. 1.
Hauri H. 1912. Anabasis aretioides Moq. et Coss.—eine Polsterpflanze der algerischen Sahara. Beih. Bot. Centralbl., 38, 1.
Heinis F. 1925. Uber die Mikrofauna alpiner Polster und Rosettenpflanzen. Festschr. zur Feier des 60 Geburtstages von Friedrich Zschokke. Basel. —1937. Beitrfige zur Microbiocoenose in alpinen Pflanzenpolstern. Ber. Geobot. Inst. Rubel in ; urich f. 1936.
Hulten E. 1925. (Г у л ь т е н). Некоторые географические заметки о карте Южной Камчатки. Изв. РГО, 57, 1. — 1927—1930. Flora of Kamtchatka and the adjacent islands. I (1927), 2 (1928), 3 (1929), 4 (1930). Stockholm. — 1932—1933. Siid-Kamtschatka. 1—12. Vegetationsbilder, 23, 1—2. — 1937. Flora of the Aleutian islands. Stockholm.
Kihlman A. 1890. Pflanzenbiologische Studien aus Russisch Lappland. Act. Soc. pro Fauna et Flora Fenn., 6, 3.
Kirstein K. 1929. Lettlands Waldtypen. Act. For. Fenn., 34.
Kostyniuk M. i R. Wieczorek. 1937. Zespoly lesne okolicy Morszyna. Kosmos, 62, 3, Lwow.
Kudo Y. 1922. Flora of the Island of Paramushir. Joum. of the Coll, of Agricult.» Hokkaido Univ., 9, 2.
Kujala V. 1926. Untersuchungen fiber Waldvegetation in Slid- und Mittelfinnland. Gommun. ex Inst. Quaest. Forest. Finlandiae, 10. — 1929. Untersuchungen fiber Waldtypen in Petsamo und in angrenzenden Teilen von Inari-Lappland. Commun. ex Inst. Quaest. Forest. Finlandiae, 13.
Kupffer K. R. 1925. Grundzuge der Pflanzengeographie des Ostbaltischen Gebietes. Abhandl. des Herder—Instituts zu Riga, I, 6.
Linkola K. 1929. Zur Kenntnis der Waldtypen Eestis. Act. Forest. Fenn., 34.
Maloch M. 1931. Borzavske poloniny v Podkarpatsk£ Rusi. Sb. vyzk. ustavfi zemedel. RCS., 67. — 1932. Agrobotanicka studie о Nardetech boriavskych polonin na Podkarpatske Rusi. Sb. vjfzk. ustavfi zemedel RCS., 83.
Metsavainio K. 1931. Untersuchungen uber das Wurzelsystem der Moorpflanzen. Ann, Bot. Soc. Zool-.Bot. Fenn. Vanamo, I, 1.
Miklaszewski J. 1928. Lasy i leSnictwo w Polsce. I. Warszawa.
Ostenfeld С. H. and C. Syrach Larsen. 1930. The species of the genus Larix and their |eo|raphical distribution. Kgl. Danske Videnskab. Selskab. Biol. Meddelelser, Paasio I. 1939. Zur Vegetation der eigentlichen Hochmoore Estlands. Ann. Bot. Soc. Zool.-Bot. Fenn. Vanamo, II, 2.
Passarge S. 1921. Vergleichende Landschaftskunde. 2. Kaltewusten und Kaltesteppen. Berlin.
Paulsen O. 1911. Track af Vegetationen i Transkaspiens Lavland Kebenhavn. — 1920. Studies in the vegetation of Pamir. (The second Danish Pamir expedition). Copenhagen.
Pawlowski В. 1937. Einfiihrung in die Pflanzenwelt der Czamohora in den Ostkarpathen. Krakdw. — 1947 (1946). Caract4ristique geobotanique generale des Monts de Gzywc-zyn. Bull, de ГАс. Polonaise des Sc. et aes Lettres, ser B, Sc. Natur., (1).
Pawlowski B. et J. Walas. 1949. (1948). Les associations des plantes vasculaires des Monts de Czywczyn. Bull, de ГАс. Polonaise des Sc. et des Lettres, s6r. B, Sc. Natur., (1).
Podpera J. 1925. Versuch einer epiontologischen Gliederung des europaischen Wal-des. Verb, des Geobot. Inst. Rubel in Zurich., 3.
Rauch W. 1940. Die Wuchsformen der Polsterpflanzen. T. 1, 2. Botanisches Archiv, 40, 2, 3.
Rubel E. (Red.). 1932. Die Buchenwalder Europas. Verb, des Geobot. Inst. Riibel in Zurich., 8.
Rubner K. 1934. Das natiirliche Waldbild Europas. Zeitschr. fur Weltforstwirtschaft, 2, 1/3. — 1953. Die pflanzengeographischen Grundlagen des Waldbaues. Berlin.
Scbroter C. und H. Hauri. 1914. Versuch einer Uebersicht der siphonogamen Polsterpflanzen. Engl. Bot. Jahrb. 50, (Suppl. Fest. Band fiir Engler).
Srodoh A. 1947 (1946). The upper limit of forest in the Gzarnohora Mountains and the Czywcryn Mountains. Bull, de ГАс. Polonaise des Sc. et des Lettres, ser. B, Sc. Natur., (1).
Steffen H. 1936. OstpreuBens Eichenwalder. Beih. Bot. Centralbl., 55, B, 1/2.
Sulma T. 1929. Kosodrzewina i jej zespoly w Gorganach. Act. Soc. Bot. Poloniae, 6,2. Svoboda P. 1952. 2ivot lesa. Praha. — 1953. Lesni dreviny a jejich porosty. 1. Praha. Szafer W. 1935. Las i step na zachodniem Podolu. Polsk. Ak. Um. Roz. wydz matemat.-przyrod., 71, B. 2. — 1936. Rezerwaty lefine w Szutromincach na Podolu. Ochrony przyrody, 16.
Tatewaki M. 1928. On the plant-communities in the middle part of the island of Urup in the Kuriles. Bot. Mag., 42, 501, Tokyo.
Tengwall T. A. 1920. Die Vegetation des Sarekgebietes. Naturwissenschaft. Untersuch. des Sarekgebirges in Swedisch—Lappland, 3, Bot. Lif. 4.
Thomson P. 1924. 1st der Grenzhorizont im Sphagnumtorflager eine synchrone Bildung? Bot. Labor, der Estland. Moorversuchsst. Thoma bei Wagewa. — 1924a. Vor-laufige Mitteilung uber neue Fundorte und Verhreitungsgehiete einiger Moorpflan-zen in Estland. Sitzungsber. der Naturforsch. Ges. Univ. Dorpat., 31, 3—4. — 1929. Die regionale Entwicklungsgeschichte der Walder Estlands. Act. et Comment. Univ. Dorpat., 17, 2.
Vineent J. 1936. Rachovsko. В кн.: Topografie lestt v Ceskoslovensk£ republice Sb. v^zk. ustavd zemedel. CSR, 150, 2.
Yabe Y. et K. Yendo. 1904. Plants of Shimushu Island. Bot. Mag., 18, 212, Tokyo.
Zlatnlk A. 1935. V^voia slozeni prirozenych lesd na Podkarpatske Rusi a jejich vztah ke stanovisti. Studie о statnich lesfch na Podkarpatske Rusi, 3. Sb. vyzk. listavfi zemidel. CSR, 127. — 1938. Vegetace a stanoviSte reservace Stuiica, Javornik a Pop Ivan. В кн.: Prozkum prirozenych lesflna Podkarpatske Rusi, 1.Sb. vVzk.iistavd zemedel. CRS, 152.
КАРТОГРАФИЧЕСКИЕ ИСТОЧНИКИ «ГЕОБОТАНИЧЕСКОЙ КАРТЫ СССР М. 1:4 000 000, УПОМЯНУТЫЕ В ПОЯСНИТЕЛЬНОМ ТЕКСТЕ1
Аболин Р. И. 1929-К. Почвенно-ботаническая карта южной части Алма-Атинского округа Казакской АССР, м. 1 : 600 000. См. Аболин Р. И. 1929. *
Аврамчик М. Н., Е. Г. Бобров, А. В. Вазингер, Е. А. Кондратьева, Д. В. Лебедев, Г. А. Мельвиль, Л. Е. Родин, Л. А. Смирнов. 1933-К. Геоботаническая карта области проектируемого орошения северной части Нижневолжского края, м. 1 : 840 000. В работе: Аврамчик М. Н. Геоботаническая карта. Тр. Комиссии по ирриг. АН СССР, 2, 1934, Л.
Аггеенко В. Н. 1890-К. Геоботаническая карта Крыма, м. 1:840 000. См. АггеенкоВ. Н. 1890.
Алехин В. В. 1921-К. Растительные зоны Европейской России, м. 1 : 32 000 000. См. Алехин В. В. 1921а. — 1930-К. Карта растительности Азиатской части СССР, м. 1 : 16 000 000. См. Алехин В. В. 1930. — 1930-Ка. Карта растительности Европейской части СССР, м. 1 : 7 000 000. См. Алехин В. В. 1930а. — 1934-К. Карта современного растительного покрова Московской области, м. 1 : 1 500 000. См. Алехин В. В. 1934. — 1936-К. Карта растительности СССР, м. 1 : 25 000 000. В кн.: Вальтер Г. — Алехин В. Основы ботанической географии. 1936. М.—Л.
Андреев В. Н. 1934-К. Карта природных типологических районов Ямальского округа, м. 1 : 4 000 000. См. Андреев В. Н. 1934.
Атлас Мира. 1954-К. Изд. Главн. упр. геодезии и картографии. М.
Берг Л. С. 1913-К. Схематическая карта ландшафтных зон России, м. 1 : 30 000 000. См. Берг Л. С. 1913.
Богдановская-Гиенэф И. Д. 1938-К. Схематическая карта растительности о-ва Колгуева, м. 1 : 300 000. См. Богдановская-Гиенэф И. Д. 1938.
Большой Советский атлас мира. 1937-К. 1. М.
Борнеман Б. А. и М. Д. Спиридонов. 1929-К. Карта почвенно-ботанических районов Усть-Урта и п-ва Мангышлак, м. 1 : 2 000 000. См. Борнеман Б. А. и М. Д. Спиридонов. 1929.
Боровиков Г. А. 1912-К. Схематическая карта растительности Заангарья, м. 1 :820000. См. Боровиков Г. А. 1912.
Борщов И. Г. 1865-К. [Ботанико-географические карты Арало-Каспийского края], 2 карты, м. 1 : 4 200 000 им. 1:8 400 000. См. Борщов И. Г. 1865.
Буш Н. А. 1918-К. Карта ботанико-географических провинций Европейской России и Сибири, м. 1 : 16 500 000. См. Буш Н. А. 1923.
Вульф Е. В., П. К. Козлов и П. П. Кожевников. 1939-К. Геоботанические районы Крыма, м. 1 : 2 200 000. См. Шалыт М. С. и П. К. Козлов. 1939.
Геоботаническая карта Крыма. 1947-К. м. 1 : 1 100 000. В приложении к кн.: Добрынин Б. Ф. Физическая география СССР, Европейская часть и Кавказ. М. 1948.
Геоботанические районы Карельской АССР и Мурманского округа Ленинградской области. 1934-К. м. 1 : 2 000 000. Растительность, л. III. Атлас Ленинградской обл. и Карельской АССР. Изд. Геогр.-экон. инет, при ЛГУ.
Геоботанические районы Ленинградской обл. 1934-К. м. 1 : 1 000 000. Растительность, л. I—II. Атлас Ленинградской обл. и Карельской АССР, Изд. Геогр.-экон. инет, при ЛГУ.
Гипсометрическая карта Европейской части СССР. 1941-К. м. 1 : 1 500 000. Под ред. Т. Н. Гунбинои. Изд. Главн. упр. геодезии и картографии. М.
Гипсометрическая карта СССР. 1949-К. м. 1 : 2 500 000. Под общей ред. И. П. Заруц-кой. Изд. Главн. упр. геодезии и картографии. М. 1 2
1 Описание карт составлено А. Н. Лукичевой.
2 Данная и аналогичные ссылки в дальнейшем означают, что карта опубликована при работе, которая указана в списке литературных источников (стр. 859—912).
Гоби X. Я. 1876-К. (Карты распределения некоторых растений на русской равнине], 3 карты, м. 1 : 20 000 000. См. Гоби X. Я. 1876.
Гордягин А. Я. 1900—1901-К. Карта западной части бассейна р. Иртыша, м. 1 : 2 100 000. См. Гордягин А. Я. 1900—1901.
Городков Б. Н. 1915-К. Схематическая карта распределения растительности Ишимского у., м. 1 : 820 000. См. Городков Б. Н. 1915. — 1916-К. Карта ботанико-географических областей Западно-Сибирской низменности, м. 1:9 000 000 См. Городков Б. Н. 1916.
Гроссгейм А. А. 1930-К. Карта растительного покрова Закфедерации: Азербайджана, Армении и Грузии, м. 1 : 420 000. Изд. Закавказ. опытн.-иссл. инет, водного хоз. Тифлис.
Добрынин Б. Ф. 1924-К. Схематическая геоботаническая карта Дагестана, м. 1 : 1 600 000. См. Добрынин Б. Ф. 1925.
Доктуровский В. С. 1909-К. Маршрутная ботаническая карта Норско-Мамынского водораздела, м. 1 : 420 000. См. Доктуровский В. С. 1909. — 1911-К. Ботаническая карта (схематическая) Амурской обл., м. 1 : 3 486 000. См. Доктуровский В. С. 1911. — 1911-Ка. Маршрутная карта района Тырминско-Буреинской экспедиции 1909 г., м. 1 : 420 000. См. Доктуровский В. С. 1911.
Дохман Г. И. 1954-К. Карта растительности средней и северной частей Мугоджарских гор. См. Дохман Г. И. 1954.
Дробов В. П. 1909-К. Карта части Мариинско-Чулымской тайги Томской губ. Мариинского у., м. 1 : 420 000. См. Дробов В. П. 1909.
Золотовский М. В. и М. С. Хомутова. 1937-К. Схематическая карта растительности Алтайского заповедника, м. 1 : 420 000. В кн. Дмитриев В. Вч М. В. Золотовский, С. С. Фолитарек, М. С. Хомутова и П. В. Юргенсон, Алтайский Гос. заповедник. 1937. М.
Ивашкевич Б. А. 1927-К. Карта лесов юго-западной части Советского Сахалина, м. 1 : 400 000. См. Ивашкевич Б. А. 1927а.
Ивашкевич Б. А., А. Н. Иванов и Л. В. Канунников. 1923-К. Карта лесов Приморской губ., м. 1 : 840 000. Изд. Приморского губ. лесного отдела. Владивосток.
Игошина К. Н. 1951-К. Карта растительности Енисейского кряжа, м. 1 : 5 000 000. См. Игошина К. Н. 1951.
Карта геоботанического районирования СССР. 1947-К. м. 1 : 10 000 000. См. Геоботаническое районирование СССР. 1947.
Карта геоморфологического районирования СССР. 1947-К. м. 1 : 10 000 000. В кн.: Геоморфологическое районирование СССР. Тр. комиссии по естеств.-истор. районированию СССР, 2, 1, М.—Л. 1947.
Карта лесов Азиатской России. 1914-К. м. 1 : 12 600 000. Атлас Азиатской России. Изд. Переселенч. упр. Главн. упр. землеустройства и земледелия. СПб.
Карта лесов Европейской России, находящихся в ведении казенного лесного управления. 1909-К. м. 1 : 1 680 000. Изд. Отдела статистики и картографии Мин. путей сообщ. СПб.
Карта лесов Европейской части СССР с распределением по породам. 1931-К. м. 1 : 1 800 000. Изд. Отчетно-эконом. сектора ПЗУ Наркомлеса. М.
Карта лесов Западносибирского края, Бурят-Монгольской АССР, Тувинской народной республики и Восточносибирского края. 1932-К. м. 1 : 2 000 000. Изд. упр. промышл. освоения лесов «Промлеса» Запсиблестреста, Томск.
Карта лесов Иркутской обл. и Бурят-Монгольской АССР. 1948-К. м. 1 : 500 000. Мин. лесной и бумажной пром. СССР.
Карта лесов Красноярского края. 1946-К. м. 1 : 1 000 000. Трест лесной авиации. Сиб. авиалесоустр. контора.
Карта растительности Азиатской России. 1914-К. м. 1 : 12 600 000. Атлас Азиатской России. Изд. Переселенч. упр. Главн. упр. землеустройства и земледелия. СПб.
Карта растительности Европейской части Союза ССР. 1937-К. м. 1 : 7 500 000. Большой Советский атлас мира, 1. М.
Карта растительности Европейской части СССР. 1948-К. Под ред. Е. М. Лавренко и В. Б. Сочавы. м. 1 : 2 500 000. Изд. Бот. инет. АН СССР. Л.
Карта растительности Союза Советских Социалистических республик. 1939-К. Отв. ред. Е. М. Лавренко, м. 1 : 5 000 000. Изд. Бот. инет. АН СССР. Л.
Карта растительности Союза ССР (растительные зоны, подзоны и округа). 1937-К. м. 1 : 15 000 000. Большой Советский атлас мира, I. М.
Кеппен Ф. П. 1885-К. Карта Европейской России [границы лиственницы и кедра], м. 1:16 800 000. См. Кеппен Ф. П. 1885. — 1885-Ка. Карта Европейской России, распространение ели, м. 1 : 16 800 000. См. Кеппен Ф. П. 1885. — 1885-Кб. Карта Европейской России, распространение сосны, м. 1 : 16 800 000. См. Кеппен Ф. П. 1885.
Кворрянг О. Э. 1915-К. Ка рта распределения растительности Наманганского у. Ферганской обл., м. 1 : 820 000. См. Кнорринг О. Э. 1915.
Кнорринг О. Э. и 3. А. Минквиц. 1910-К. Схематическая карта распределения растительных областей Чимкентского у., м. 1:1680 000. См. Минквиц 3. А. и О. Э. Кнорринг. 1910. — 1912-К. Карта растительности Андижанского у. Ферганской оол., м. 1 : 840 000. См. Кнорринг О. Э. и 3. А. Минквиц. 1912. — 1912-Ка. Схематическая карта растительных областей Аулие-Атинского у., м. 1 : 820 000. См. Кнорринг О. Э. и 3. А. Минквиц. 1912а. — 1913-К. Карта растительности районов Перовского у. (Сыр-Дарвинской обл.), м. 1 : 820 000. См. Кнорринг О. Э. и 3. А. Минквиц. 1913.
Колесников Б. П. 1938-К. Карта растительности южной части Тернейского района Приморской обл. Приморского края, м. 1 : 500 000. См. Колесников Б. П. 1938.
Коликов М. С. и Е. А. Малюгин. 1950-К. Схематическая карта типов песков северо-западной части Приаральских Кара-Кумов. См. Гаель А. Г., М. С. Коликов, Е. А. Малюгин и Е. С. Останин. 1950.
Коржевин. В. С. 1934-К. Карта растительности долины р. Алдана между устьями рр. Учура и Ноторы, м. 1 : 2 500 000. См. Коржевин В. С. 1934.
Коржинский С. И. 1898-К. Rossiае Europaeae orientalis tabula Geograpica in regiones pbytogeograpbicas divisa. См. Коржинский С. И. (Korshincky). 1898. — 1899-K. Карта ботанических областей Российской империи, м. 1 : 23 100 000. См. Коржинский С. И. 1899. — 1899-Ка. Карта распространения некоторых древесных пород по Европейской России, м. 1 : 10 500 000. См. Коржинский С. И. 1899. — 1899-Кб. Карта Российской империи с обозначением реликтов третичных лесов, центров распространения и путей переселения деревьев, м. 1 : 23 100 000. См. Коржинский С. И. 1899.
Коровин Е. П. 1932-К. Карта растительности Среднеазиатских республик, м. 1 : 3 000 000. См. Коровин Е. II. 1934.
Краснов А. Н. 1888-К. [Карта части Центрального Тянь-Шаня и прилегающих районов], м. 1 : 2 800 000. См. Краснов А. Н. 1888.
Крашенинников И. М., С. Ё. Кучеровская-Рожанец и И. В. Новопокровский. 1941-К. Карта ботанико-географических районов Башкирской АССР, м. 1:3 000 000. См. Крашенинников И. М. и С. Е. Кучеровская-Рожанец. 1941.
Криштофович А. Н. 1910-К. Ботаническая карта Балаганского и Верхоленского у. Иркутской губ., м. 1 : 630 000. См. Криштофович А. Н. 1910. — 1913-К. Карта растительности части Нижнеудинского и Балаганского у. (Иркутской губ.), м. 1 : 420 000. См. Криштофович А. Н. 1913.
Крылов П. Н. 1913-К. Карта Барабы и смежных мест, м. 1 : 2 100 000. См. Крылов П. Н. 1913. — 1916-К. Карта флористических областей и их зон и подзон в западной части Томской губ., м. 1 : 2 100 000. См. Крылов П. Н. 1916.
Кузнецов Н. И. 1891-К. Карта Черноморского окр. Границы распространения древесных пород, м. 1 : 1 260 000. См. Кузнецов Н. И. 1891. — 1909-К. Карта ботанико-географических провинций Кавказского края, м. 1 : 5 000 000. См. Кузнецов Н. И. 1909. — 1910-К. Ботанико-географическая карта нагорного Дагестана, м. 1 : 1 080 000. См. Кузнецов Н. И. 1910. (редактор). — 1927— 1932-К. Геоботаническая карта Европейской части СССР, м. 1:1 050 000. л. 5 (1930); л. 6 (1930); л. 7 (1932); л. 9 (1932); л. 10 (1929)+плюр (1930); л. 14 (1927)4-плюр (1927); л. 15 (1928); л. 16 (1928). Изд. Главн. бот. сада. Л. — 1928-К. Карта растительности Европейской части СССР, м. 1:4 000 000. Изд. Всесоюзн. Инет, прикл. бот. и Гос. Инет, опытн. агрон. НКЗ. РСФСР. Л. — 1930-К. Map of vegetation of the European part of the USSR, m. 1 : 10 000 000. В кн.: Kusnetzow N. I. 1930. Vegetation in the European part of the USSR. Guidebook for the excursion of the second international congress of soil science, 1, Moskow.
Кузнецов [Владимирский] И. И. 1912-К. Схематическая карта распределения главнейших растительных формаций в районе работ Н. И. Кузнецова в Забайкальской обл., м. 1 : 370 000. См. Кузнецов [Владимирский] Н. И. 1912. —- 1915-К. Нарымский край в области течения левобережных притоков Оби — рр. Чаи, Парабели и нижней части Васюгана, м. 1 : 1 680 000. См. Кузнецов [Владимирский] Н. И. 1915а.
Куминова А. В. 1934-К. Карта растительности Даурской провинции, м. 1 : 5 500 000. См. Куминова А. В. 1938. — 1949-К. Карта растительности Кемеровской обл., м. 1 : 900 000. См. Куминова А. В. 1949а.
Кучеровская-Рожанец С. Е. 1914-К. Схематическая карта растительных районов (зон) Павлодарского у. Семипалатинской обл. и смежной части Тюкалинского у. Тобольской губ., м. 1 : 1 680 000. См. Кучеровская-Рожанец С. Е. 1914.
Леонтьев А. М. 1937-К. Карта геоботанических районов Беломорско-Кулойского края, м. 1 : 1 000 000. См. Леонтьев А. М. 1937.
Леса и лесное хозяйство Карельской АССР и Мурманского окр. Ленинградской обл., 1934-К. М. 1 : 3 000 000 и 1 : 10 000 000» Леса, л. IV. Атлас Ленинградской обл. и Карельской АССР. Изд. Геогр.-экон. инет, при ЛГУ. Л.
Леса и лесное хозяйство Ленинградской обл. 1934-К. м. 1 : 1 000 000 и 1 : 5 000 000. Леса, л. I—II. Атлас Ленинградской обл. и Карельской АССР. Изд. Геогр.-экон. инет, при ЛГУ. Л.
Липпмаа Ф. (L i р р m а а Т.). 1935-К. Eesti geobotaaniline kaart, m. 1 : 1 000 000. См. Липпмаа Ф. 1935.
Лобаев И. И. 1950-К. Схематическая карта геоботанических комплексов песчаного массива Малые Барсуки, м. 1 : 800 000. См. Гаель А. Г., М. С. Коликов, Е. А. Малюгин и Е. С. Останин. 1950.
Лобова Е. В. 1946-К. Почвенная карта Казахской СССР, м. 1 : 2 500 000. Изд. Почв инет. АН СССР. Л.
Любимова А. А. 1937-К. Геоботаническая карта побережья оз. Ловозера, м. 1 : 400 000. См. Любимова, А. А. 1937.
Медведев Я. С. 1907-К. Карта фитогеографических областей Кавказа, м. 1 : 1 680 000. В кн.: Медведев Я. С. Об областях растительности на Кавказе. Вести. Тифл. бот. сада, 8, 1907.
Цедригайлов С. Н. 1925-К. Схематическая карта лесных районов по маршрутам Якутской экспедиции Академии Наук СССР, м. 1 : 600 000. См. Недригайлов С. Н. 1928.
Неуструев С. С. и И. М. Крашенинников. 1930-К. Карта почвенно-ботанических районов Усть-Урта и п-ва Мангышлак, м. 1:2 000 000. См. Неуструев С. С. и И. М. Крашенинников (род.). 1930.
Новопокровский И. В. 1925-К. Геоботаническая карта района капала Октябрьской революции и прилегающей части Прикаспийской низменности, м. 1 : 210 000. См. Новопокровский И. В. 1932. — 1925-Ка. Схематическая карта ботаникогеографических районов Северокавказского края, м. 1:3 000 000. См. Новопокровский И. В. 1925.
Новопокровский И. В. и С. Ю. Туркевич. 1927-К. Схематическая карта ботаникогеографических районов Ставрополья, м. 1 : 1 050 000. См. Новопокровский И. В. 1927а.
Останин Е. С. 1950-К. Схематическая карта геоботанических комплексов песчаного массива Большие Барсуки, м. 1 .*800 000. См. Гаель А. Г., М. С. Коликов, Е. А. Малюгин и Е. С. Останин. 1950.
Поварницын В. А. 1934-К. Карта растительности бассейна р. Б. Белой, м. 1 : 400 000. См. Поварницын В. А. 1934.
Поварницын В. А. и П. К. Красильников. 1937-К. Карта растительности бассейна р. Верхней Ангары и северо-западного побережья оз. Байкал, м. 1 : 500 000. См. Поварницын В. А. 1937.
Почвенная карта Европейской части Союза ССР. 1947-К. Под ред. Л. И. Прасолова и И. П. Герасимова, м. 1 : 2 500 000. Изд. почв. инет. АН СССР. М.
Прозоровский А. В., Н. И. Рубцов и А. А. Дмитриева при участии М. Н. Аврамчика. 1933-К. Геоботаническая карта Казахстана, м. 1 : 2 000 000. Изд. Казахст. н.-иссл. инет, удобрений и агропочвоведения. Алма-Ата.
Прозоровский Н. А. 1949-К; Карта современной и восстановленной растительности Курской, Орловской, Тамбовской и Воронежской областей, м. 1:2 500 000. См. Прозоровский Н. А. 1949.
Работнов Т. А. 1935-К. Схематическая карта природных районов южной части Якутской АССР, м. 1 : 10 000 000. См. Работнов Т. А. 1935.
Рожевиц Р. Ю. 1912-К. Схематическая карта распределения растительности части Пржевальского и Пишпекского уездов по иссл. 1908 г., м. 1:1 680 000. См. Рожевиц Р. Ю. 1912.
Розов Н. Н. (при участии Е. В. Лобовой). 1954-К. Почвенная карта СССР для высших учебных заведений, м. 1 : 4 000 000. Под общей ред. И. П. Герасимова. Изд. Главн. упр. геодезии и картографии. М.
Самойлович Г. Г. 1948-К. Схематическая карта кедровых лесов в бассейнах нижних притоков р. Оби Ямальского лесхоза. См. Самойлович Г. Г. 1948. — 1948-Ка. Схематическая карта сосновых лесов в бассейнах нижних притоков р. Оби Ямальского лесхоза. См. Самойлович Г. Г. 1948.
Сибирцев Н., А. Ферхмин и Г. Танфильев. 1901-К. Почвенная карта Европейской России, составленная по почину и плану проф. В. В. Докучаева, м. 1 : 2 520 000. СПб.
Спиртов В. И. 1912-К. Схематическая карта растительности района между рр. Сары-Су и Коном в Акмолинском у., м. 1 : 420 000. См. Смирнов В. И. 1912.
Соболев Л. Н. 1950-К. Карта физико-географических районов Казахстана, м. 1 : 10 000 000. См. Соболев JI. Н. 1950.
Соболевская К. А. 1950-К. Схематическая карта поясно-зонального расчленения растительности Тувы. См. Соболевская К. А. 1950.
Соколова Л. А. 1937-К. Карта геоботанических районов Онего-Северодвинского водораздела и Онежского п-ва, м. 1 : 1 000 000. См. Соколова Л. А. 1937.
Сочава В. Б. 1953-К. Схема геоботанического районирования лесной зоны. См. Сочава В. Б. 1953.
Сочава В. Б., Т. И. Исаченко и А. Н. Лукичева. 1952-К. Геоботаническая карта лесной зоны Западно-Сибирской низменности, м. 1 : 9 000 000. См. Сочава В. Б., Т. И. Исаченко и А. Н. Лукичева. 1953.
Спрыгин И. И. 1930-К. Схематическая карта растительного покрова Средневолжского края, м. 1 : 2 000 000. См. Спрыгин И. И. 1930.
Старк С. К. 1933-К. Карта приречной части бассейна верхней Колымы с указанием лесонасаждений, обследованных лесоэкономической экспедицией Якутской АССР, 1930—1931 гг., м. 1 : 1 000 000. См. Старк С. К. 1933.
Сукачев В. II. 1938-К. Схематическая геоботаническая карта СССР, м. 1 : 15 000 000. В кн.: Сукачев В. Н. Дендрология с основами лесной геоботаники. Изд. 2-е. 1938. Л.
Схематическая карта лесов Главзапсиблеса. 1943-К., м. 1 : 1 000 000. Трест лесной авиации. Сиб. авиалесоустроительная контора.
Схематическая карта лесов Молотовской обл. 1946-К., м. 1 : 750 000. Изд. Модотов-ского Облнитолеса.
Танфильев Г. И. 1894-К. Карта распределения лесов на юге Европейской России, м. 1:6 000 000. См. Танфильев Г. И. 1894а. — 1897-К. Карта физико-географических областей Европейской России, м. 1 : 11 550 000. См. Танфильев Г. И. 1897. — 1900-К. Ботанико-географическая карта Российской империи, м. 1 : 25 200 000. См. Танфильев Г. И. 1900а. [Эта карта без всяких изменений была опубликована в приложении к работе: Танфильев Г. И. 1903].
Томин М. П. 1910-К. Схематическая карта растительных сообществ Верхоленского и Балаганского уездов Иркутской губ., м. 1 : 420 000. См. Томин М. П. 1910.
Туркевич С. Ю. 1914-К. Схематическая карта растительности южной части Ачинского и северной части Минусинского уездов Енисейской губ., м. 1 : 1 008 000. См. Туркевич С. Ю. 1914.
Федченко Б. А. и О. Э. Кнорринг. 1926-К. Карта растительности Туркестана, м. 1 : 4 200 000. См. Федченко Б. А. 1925а.
Фокин А. Д. 1929-К. Подзоны лесной полосы в пределах Вятского края, м. 1 : 4 500 000. См. Фокин А. Д. 1929.
Цинзерлинг Ю. Д. 1931-К. Карта геоботанических районов северо-запада Европейской части СССР, м. 1 : 2 500 000. См. Цинзерлинг Ю. Д. 1932.
Шелудякова В. А. 1938-К. Геоботаническая схематическая карта бассейна р. Индигирки, м. 1 : 2 000 000. См. Шелудякова В. А. 1938.
Шенников А. П. 1940-К. Схематическая карточка подзон и провинций северо-востока Европейской части СССР, м. 1 : 6 000 000. См. Шенников А. П. 1940.
Шипчинский Н. В. 1916-К. Карта восточной части Семипалатинского у., м. 1 :1 680 000. См. Шипчинский Н. В. 1916.
Шиффере Е. В. 1937-К. Геоботаническая карта Кавказа, м. 1 : 4 000 000. В кн.: Большая Советская энциклопедия, 30. 1937. — 1953-К. Карта растительности и природных кормовых угодий Северного Кавказа, м. 1 : 2 500 000. См. Шиффере Е. В. 1953.
Шумилова Л. В. 1949-К. Ботанико-географические страны Сибири, м. 1 : 25 000 000. См. Шумилова Л. В. 1949.
ЛЕГЕНДА «ГЕОБОТАНИЧЕСКОЙ КАРТЫ СССР» М. 1 : 4 000 000 и УКАЗАТЕЛЬ АВТОРОВ ПОЯСНИТЕЛЬНОГО ТЕКСТА К КАРТЕ
Стр. Введение в обзор растительного покрова СССР на основе «Геоботанической
карты» м. 1 : 4 000 000. Составил В. Б. Сочава.......................I, 9
I. Арктические пустыни и тундры
Общий обзор. Составил В. В. Сочава ...................................: 61
1. Разреженная растительность осыпей и скал гор Арктики и высокогорий
умеренного пояса. Составил Б. Н. Городков ........................... 76
2. Арктические пустыни. Составил Б. Н. Городков 1...................... 79
2 тм — травяно-моховые. 2 окм — обедненные кустарничково-моховые 2 км — кустарничково-моховые
3. Горные тундры, местами в сочетании с кустарниками и лугами. Составил
В. Б. Сочава......................................................... 82
Общий очерк......................................................... »
3 ко — Кольские.................................................... 92
3 у — уральские.................................................... 95
3 с — среднесибирские ............................................. 98
За — алтайско-саянские ........................................... 101
Зв — восточносибирские ........................................... 105
3 тх — притихоокеанские........................................... 110
4. Арктические тундры в сочетании с гипново-травяными болотами. Составил
Б, Н. Городков ..................................................... 113
5. Моховые и лишайниковые тундры в сочетании с гипново-травяными и ди-краново-лишайниково-сфагновыми болотами. Составил В. Б. Сочава . . . 115
6. Кустарничковые тундры в сочетании с моховыми и лишайниковыми, а также гипново-травяными и дикраново-лишайниково-сфагновыми болотами. Составил В. В. Сочава .......................................... 124
7. Осоково-пушицевые кочкарные тундры в сочетании с гипново-травяными
болотами. Составил Б. Н. Городков................................... 129
8. Тундровые ивняки в сочетании со сфагновыми и дикраново-лишайни-ково-сфагновыми болотами. Составил В, Б. Сочава....................... 130
9. Ер пиковые тундры, в сочетании со сфагновыми и дикраново-лишайни-ково-сфагновыми болотами и местами тундровыми ивняками. Составил В. Б. Сочава.......................................................... 132
II. Темнохвойные леса
Общий обзор. Составил В. В. Сочава...................................... 139
10. Предтундровые редколесья из сибирской ели в сочетании с тундрами и болотами (лесотундра). Составил В. Б. Сочава .......................... 152
11. Притихоокеанские и восточносибирские заросли кедрового стланика (Pinus pumila) и ольхово-кедровые заросли. Составил В. Б. Сочава ......... 155
12. Европейские северотаежные еловые леса, местами в сочетании с березня-
ками и сфагновыми болотами. Составил В. Б. Сочава................... 159
Общий очерк...................................................... »
12 е — из Picea excelsa........................................... 160
12 о — из Picea obovata........................................... 162
1 В тех случаях, когда пояснительный текст к выделам на карте, имеющим добавочное буквенное обозначение, написан не одним автором, указывается авторство отдельно по каждому выделу.
Стр-
13. Европейские среднетаежные еловые леса, местами в сочетании с березово-осиновыми лесами, сфагновыми болотами и лугами. Составил В. Б. Сочава...........................................................If 164
Общий очерк....................................................... »
13 е — из Picea excelsa........................................... 166
13 о - из Picea obovata .......................................... 168
14. Европейские южнотаежные еловые леса, местами в сочетании с березово-
осиновыми лесами, сфагновыми болотами и лугами. Составил В. Б. Сочава 172 Общий очерк............................................................ >
14 е — из Picea excelsa........................................... 174
14 о — из Picea obovata........................................... 176
15. Сибирские северотаежные лиственнично-елово-кедровые леса, местами в сочетании со сфагновыми болотами. Составил В. Б. Сочава ............. 179
16. Сибирские среднетаежные елово-пихтово-кедровые леса, местами в соче-
тании со сфагновыми болотами и березняками. Составил В, В. Сочава . . . 182
17. Сибирские южнотаежные елово-кедрово-пихтовые леса, местами в сочетании со сфагновыми болотами, оерезняками и осинниками. Составил В. Б. Сочава .......................................................... 185
18. Сибирские горнотаежные темнохвойные леса. Составил В. Б. Сочава . . . 188
Общий очерк....................................................... >
18 см — преимущественно смешанные елово-кедрово-пихтовые, елово-
пихтовые и елово-кедровые................................. 190
18 пх — с преобладанием сибирской пихты (черневая тайга) .... 194
18 кд — с преобладанием сибирского кедра........................... 197
19. Горные темнохвойные леса, местами в сочетании с хвойно-широколиственными и широколиственными лесами. Составил В. Б. Сочава 201 Общий очерк............................................................... »
19 кп — карпатские из Picea excelsa, местами с участием Abies alba. 203 19 к — кавказские из Abies Nordmanniana и Picea orientalis........ 206
19 тш — тяныпанские из Picea Schrenkiana, местами с участием Abies
Semenovii ................................................. 210
19 дв — дальневосточные из Picea jezoensis, местами с участием Abies nephrolepis и A. sachalinensis .................................... 212
III. Сосновые леса
Общий обзор. Составила А. М. Семенова-Тян-Шанская........................ 217
20. Европейско-западносибирские северотаежные сосновые леса, местами
в сочетании со сфагновыми болотами. Составила А. М. С еменова-Тян-Шанская 224
21. Европейско-западносибирские средне- и южнотаежные сосновые леса, местами в сочетании со сфагновыми болотами. Составила А. М. Семенова-Тян-Шанская............................................................ 226
22. Казахстанско-западносибирские остепненные травяно-кустарничковые и
травяные сосновые леса. Составила А. М. Семенова-Тян-Шанская . . . 231
23. Сибирские травяно-кустарничковые и кустарничковые лиственнично-сосновые и сосновые леса в сочетании с лишайниково-кустарничковыми сосняками. Составила А. М. Семенова-Тян-Шанская........................... 234
Общий очерк.......................................................... »
23 s — с Larix sibirica............................................. 235
23 d — с Larix dahurica, местами в сочетании с багульниково-моховыми лиственничниками.................................................... 239
24. Забайкальско-амурские травяно-кустарничковые лиственнично-сосновые и сосновые леса, на юге остепненные и в сочетании со степями. Составила А. М. Семенова-Тян-Шанская............................................. 240
25. Горные крымско-кавказские сосновые леса. Составила А. М. Семенова-Тян-Шанская............................................................ 244
IV. Лиственничные леса
Общий обзор. Составил В. Б. Сочава ....................................... 249
26. Предтундровые лиственничные редколесья, в сочетании с тундрами и болотами (лесотундра). Составил В. Б. Сочава............................. 265
Общий очерк.......................................................... »
26 s — иэ Larix sibirica ......................................... 266
26 d — из Larix dahurica....................................• . . 267
27. Западносибирские северотаежные мохово-кустарничковые и лишайниково-кустарничковые лиственничные, елово-лиственничные и сосноволиственничные леса. Составил В. Б, Сочава ;............................ 269
Стр.
28. Восточносибирские северотаежные кустарничково-моховые и лишайниковые лиственничные леса, местами в сочетании с болотами. Составил В. Б. Сочава .......................................................I, 271
29. Восточносибирские северотаежпые лиственничные леса в сочетании с частыми гинново-травяными болотами и приозерными травяными сообществами (Колымская и Индигирская низменности). Составил В. Б. Сочава 274
30. Центральносибирскио сроднетаежные кустарничковые и травяно-кустарничковые лиственничные леса борового типа. Составил В. Б, Сочава . . 277
Общий очерк ...................................................... »
30 а — из Larix sibirica, часто с участием сосны................ 278
30 d — из Larix dahurica, часто в сочетании с багульниково-моховыми лиственничными лесами , . ...................................... 281
31. Центральносибирские среднетаежные леса из даурской лиственницы в сочетании с приозерными злаково-осоковыми (аласными) группировками. Составил В. Б. Сочава .............................................. 286
32. Центральносибирские среднетаежные леса из даурской лиственницы в сочетании с гинново-травяными болотами, орниками и лугами. Составил В. Б. Сочава ....................................................... 287
33. Дальневосточные багульниковые лиственничные леса, местами в сочетании с травяно-болотными и сфагновыми лиственничными лесами (марями). Составил В. В. Сочава .............................................. 290
Общий очерк ......................................................... »
33 ш — то же, в сочетании с травяными лиственничниками, в полосе рассеянного распространения широколиственных пород. Составил В. В. Сочава................................................ 294
34. Дальневосточные травяно-болотные и сфагновые лиственничные леса (мари), местами в сочетании с багульниковыми лиственничными лесами и болотами. Составил В. Б. Сочава .................................... 296
35. Горные северотаежные редкостойные лиственничные леса. Составил В. Б. Сочава . .................................................... 298
Общий очерк.................................................... »
35 s — из Larix sibirica и Larix Sukaczowii..................... 299
35 d — из Larix dahurica.............................•.......... 801
36. Горные восточносибирские среднетаежные лиственничные леса. Составил В. Б. Сочава...................................•.................... 804
Общий очерк....................................................... »
36 сг — среднегорные ............................................ 806
36 р — подгольцовые редкостойные................................. 808
37. Горные южносибирские лиственничные леса. Составил В. Б. Сочава ... 311
38. Горные южносибирские кедрово-лиственничные леса, в сочетании с кедровыми лесами. Составил В. Б. Сочава ............................... 815
V. Березовые и осиновые леса
Общий обзор. Составила Г. И. Исаченко ............................... 819
39. Предтундровые березовые редколесья, в сочетании с тундрами и болотами (лесотундра). Составила А. Н. Лукичева ........................ 827
40. Редкостойные березовые леса из березы Эрмана, местами в сочетании с травяными и кустарниковыми группировками. Составила А. Н. Лукичева 881
41. Березовые в осиновые леса (в хвойнолесной зоне, местами в сочетании с хвойными лесами, лугами и болотами). Составила Т. И. Исаченко . . . 885
42. Березовые и осиновые леса лесостепного типа в сочетании с лугами и местами с гипново-травяными болотами. Составила Г. И. Исаченко .... 842
VI. Хвойно-широколиственные леса
Общий обзор. Составил В. В. Сочава .................................. 846
43. Европейские широколиственно-еловые леса. Составила А. М. Семенова-Тян-Шанская .............. .......................................... 851
Общий очерк...................................................... »
43 е — с Picea excelsa.......................................... 852
43 о — с Picea obovata и Abies sibirica ....................... 855
44. Европейские широколиственно-сосновые леса. Составила А. М. Семенова-Тян-Шанская '. .......................... 856
45. Дальневосточные хвойно-широколиственные леса. Составил В. Б. Сочава 859
Стр.
Общий обзор. Составил В. Б. Сочава ................... I, 865
46. Прикарцатско-предкавказские дубовые и дубово-грабовые леса с участием ряда других широколиственных пород. Составила А. М. Семенова-Тян-Шанская ....................................................... 880
Общий очерк..................................................... »
46 кн — прикарпатские.......................................... 881
46 к — предкавказские.......................................... 888
47. Восточноевропейские дубовые леса. Составила А. М. Семеноеа-
Тян-Шанская.................................................... 886
Общий очерк..................................................... »
47 дн — приднепровские........................................ 889
47 ср — среднерусские......................................... 891
47 зв — среднезаволжские...................................... 894
47 юв — юговосточные.......................................... 895
48. Предуральскио и приволжские дубово-липовые и липовые леса с ильмом и кленом. Составила А. М. Семенова-Тян-Шанская .................. 896
49. Присредиземыоморские дубовые леса из пушистого дуба. Составил В. Б, Сочава..................................................... 898
50. Горные дубовые леса. Составил В. В. Сочава ............. 401
Общий очерк................................................... »
50 к — северокавкаэско-крымские (из Quercus potraea) и закавказские
(из Quercus iberica).................................. 408
50 г — гирканские (из Quercus castaneifolia).................. 409
51. Буковые леса. Составил В. Б. Сочава ................. 411
Общий очерк................................................... »
51 кп — карпатские (из Fagua silvatica)....................... 412
51 крп — крымско-подольские (иэ Fagua taurica)................ 415
51 к — кавказские (из Fagua orientalis)....................... 417
52. Закавказские смешанные широколиственные леса. Составил В. Б. Сочава 421 Общий очерк......................................................... »
52 кл — колхидские (иэ Fagus orientalis, Castanea sativa, Quercus Hart-wissiana, Carpinus caucasica)............................ 422
52 г — гирканские (иэ Quercus castaneifolla, Parrotia persica, Carpinus caucasica)............................................. 425
53. Низинные лиственные леса. Составил В. В. Сочава................ 427
Общий очерк..................................................... »
53 е — европейские ольховые (из Alnus glutinosa) леса......... 428
53 к — кавказские ольховые (иэ Alnus barbate), грабово-ольховые и дубово-ольховые леса с лианами...................../. . 480
53 кд — кавказские ольховые (из Alnus barbata) и грабово-ольховые леса, в сочетании с низинными лесами из Quercus longlpes и других лиственных пород............................................. 483
54. Дальневосточные широколиственные леса из монгольского дуба, амурской липы и других лиственных пород. Составил В. В. Сочава ........... 484
. . VIII. Горные плодовые леса и горные кустарники
55. Горные ореховые леса, местами в сочетании с яблоневыми и кленовыми лёсами. Составил Н. И. Рубцов ................... 468
56. Горные кустарники. Составил Н. И. Рубцов .............. 470
Общий очерк......................................... »
56 а — заросли мезофильных и полуксерофильных кустарников (шиповника, жимолости, таволги!, в сочетании с луговыми степями (предгорья Алтая, горы Тарбагатая и Саура)........ 471
56 к — заросли полуксерофильных кустарников (держи-дерева, таволги, гранатника, барбариса, боярышника и др.) (Кавказ). 472 56 са — заросли полуксерофильных и мезофильных кустарников (шиповника, экзохорды, боярышника и др.), в сочетании с яблоневыми редколесьями (Средняя Азия).................. 478
JX. Луга и травяные болота
Общий обзор. Составил Е. М. Лавренко .................. 475
57. Высокогорные альпийские и субальпийские луга, местами в сочетании с -кустарниками’и лйвса^и’ ‘.......................... 477
Стр;
Общий очерк. Составила Е. В. Шиффере ...........................11,477
57 у —уральские. Составила Е. В. Шиффере .........................*482
57 а — алтайские и североказахстанские. Составила Е. В. Шиффере 483
57 кп — карпатские. Составил В, Б. Сочава ........................ 486
57 к — кавказские; Составила Е. В. Шиффере ....................... 490
57 са — среднеазиатские в сочетании с высокогорными степями. Составила Е. В. Шиффере ..................................... 494
58. Среднегорные луга, местами в сочетании с луговыми степями ........ 495
Общий очерк. Составила Е. В. Шиффере ............................. »
58 ск — североказахстанские. Составил Е. М. Лавренко ............. 496
58 к — кавказские. Составила Е. В. Шиффере ....................... >
58 са — среднеазиатские. Составил Н. И. Рубцов ................... 498
59. Травяная растительность океанических островов (Тихий океан). Составила А. Н. Лукичева .................................................. 500
60. Приморские луга засоленных побережий в тундровой зоне. Составила Л, А. Соколова ....................................................... 503
61. Солончаково-луговая и лугово-болотная растительность морских побережий в сочетании с солянковой, а местами с растительностью приморских песков (в степной и пустынной зонах). Составила Е. В. Шиффере .... 506
62. Солончаково-луговая и лугово-болотная растительность в сочетании с солянковыми и степными сообществами (разливы, лиманы, высыхающие плавни и озера в степной и пустынной зонах). Составили Л. А. Соколова и Л. Е. Родин (последний в части Средней Азии и Казахстана) .......... 508
63. Пойменные луга, обычно в сочетании с кустарниками и лесами (урема) и сельскохозяйственные земли на их месте ............................... 517
Общий очерк. Составила Л. А. Соколова ............................ »
63 т — тундровой зоны. Составила Л. А. Соколова .................. 518
63 л — лесной зоны. Составила Л, А. Соколова ..................... 521
63 ст — степной зоны. Составила Л, А. Соколова ................... 527
63 п — пустынной зоны. Составил Л. Е. Родин^ . . . . ........ 533
64. Травяные болота и луга длительно заливаемых пойм (плавни). Составили
Л. А. Соколова и Л. Е. Родин (последний в части Средней Азии и южного Казахстана)......................................................... 536
65. Травяные и гипново-травяные болота. Составила Л. А. Соколова .... 544
Общий очерк...................................................... »
65 т — тундровой зоны, местами в сочетании с тундровыми формациями. » 65 л — лесной зоны и лесостепи................................ 547
X. Сфагновые болота
Общий обзор. Составила Е. А. Галкина .................................. 553
66. Сфагновые и дикраново-лишайниково-сфагновые плоско- и крупнобугрис-
тые болота тундровой зоны и полосы редколесий. Составила Е. А. Галкина 560
67. Сфагновые, местами облесенные болота лесной зоны. Составила
Е. А. Галкина....................................................... 562
Общий очерк ........................................................ >
67 бг — сфагново-лишайниковые и сфагновые, преимущественно бугристые болота................................................ 563
67 е — сфагновые восточноевропейские выпуклые болота............... 564
67 зс — сфагновые и сфагново-гипново-травяные западносибирские выпуклые болота.............................................. 568
67 ск — сфагновые и гипново-травяные сахалинско-камчатские болота с луговыми и древесными сообществами вдоль рек............. 570
XI. Ксерофитные редколесья, нагорные ксерофиты и субтропические степи
Общий обзор. Составил Е. М. Лавренко................................... 573
68. Можжевеловые редколесья в сочетании с нагорными ксерофитами и степями Составил Я. И. Рубцов ....................... . . 575
Общий очерк......................................................... »
68 к — кавказские.................................................. >
68 са — среднеазиатские.......................................... 576
69. Фисташковые редколесья. Составил Н. И Рубцов...................... 578
Общий очерк......................................................... >
69 к — кавказские из Pistacia mutica............................... »
69 са — среднеазиатские из Pistacia vera и Amygdalus bucharica .... 579
70. Нагорные ксерофиты и растительность пестроцветов (фригана). Составил Н. И. Рубцов......................................................... 581
Стр.
71. Колючетравники (из видов Cousinia). Составил Н. И Рубцов.....II, 683
72. Субтропические низкотравные степи (полусаванны). Составил Н. И. Рубцов ......................................................... 585
Общий очерк ....................................................... »
72 ср — из Carex pachystylis и Роа bulbosa...................... 587
72 р — из Роа bulbosa............................................. 588
73. Субтропические крупнотравные степи (полусаванны) и сельскохозяйствен-
ные земли на их месте. Составил Н. И. Рубцов...................... 589
Общий очерк.................................................... >
73 v — из Inula grandis, Prangos pabularia, видов Ferula в сочетании с ксерофитными кустарниками.............................. >
73 h — из Hordeum bulbosum...................................... 590
73 а — из Agropyrum trichophorum и Hordeum bulbosum............. 591
74. Бородачевые (из Andropogon ischaemum) субтропические степи (полуса-
ванны) и сельскохозяйственные земли на их месте. Составил Н. И, Рубцов 593
XII. Степи и сельскохозяйственные земли на месте степей
Общий обзор. Составил Е. М. Лавренко при участии Ф. X. Бахтеева . . . 595
75. Луговые степи, остепненные луга, в сочетании с участками лесов (лесостепь). Составил Е. М. Лавренко .................................... 607
Общий очерк ....................................................... »
75 яп — луговые степи и остепненные луга лесных областей (Центральная Якутия, Приморье)............................................ 608
75 е — сельскохозяйственные земли на месте европейских луговых
степей и остепненных лугов в сочетании с дубовыми лесами . . 610
75 зс — сельскохозяйственные земли на месте западносибирских луговых степей и остепненных лугов в сочетании с березовыми колками и местами с травяными болотами................................. 619
75 цс — сельскохозяйственные земли на месте центральносибирских луговых степей и остепненных лугов в сочетании с березовыми и сосново-лиственничными лесами.................................. 624
75 зб — забайкальские луговые степи, остепненные луга и сельскохозяйственные земли на их месте в сочетании с лиственничными и «сосновыми лесами.................................................. 629
76. Разнотравно-дерновиннозлаковые европейско-казахстанские степи. Соста-
вил Е. М. Лавренко................................................ 632
Общий очерк........................................................ >
76 ч — сельскохозяйственные земли на месте причерноморских разно-
травно-типчаково-ковыльных степей......................... 633
76 звк — заволжско-казахстанские разнотравно-типчаково-ковыльные" степи и сельскохозяйственные земли на их месте........... 648
77. Дерновиннозлаковые европейско-казахстанские степи. Составил
Е. М. Лавренко ............................. 653
Общий очерк ..................................................... »
77ч — сельскохозяйственные земли на месте причерноморских типчаково-ковыльных степей.......................................... 654
77 звк — заволжско-казахстанские типчаково-ковыльные степи и сельскохозяйственные земли на их месте............................. 659
78. Полынно-дерновиннозлаковые европейско-казахстанские степи в сочетании с пустынными сообществами и сельскохозяйственные земли на их месте (северная полупустыня). Составил Е. М. Лавренко........................ 665
Общий очерк ...................................................... »
78 зч — западнопричерноморские полынно-типчаково-ковыльные и
полынно-типчаковые степи (с Artemisia maritima s. 1.)....668
78 ч — причерноморские полынно-типчаково-ковыльные, полыннотипчаковые и полынно-житняковые степи (с Artemisia taurica). 669 78 кк — прикаспийско-казахстанские полынно-типчаково-ковыльные и
полынно-типчаковые €тепи (с Artemisia Lercheana)........ 671
78 зк — западноказахстанские полынно-типчаково-ковыльные и полынно-типчаковые степи (с Artemisia Lessingiana)............... 676
78 вк — восточноказахстанские полынно-типчакрво-ковыльные и по-
лынно-типчаковые степи (с Artemisia sublessingiana)..... 677
78 кз — прикаспийско-казахстанские полынно-эркековые степи (с Arte-
misia arenaria, A. astrachanica, Eurotia Ewersmanniana, E. ceratoides) ................................................ 682
Стр.
79. Пижмовые (из Tanacetum sibiricum) забайкальские степи, в сочетании с Вострецовыми (Aneurolepidium pseudoagropyrum) и дерновиннозлаковыми степями и сельскохозяйственные земли на их месте. Составил Е. М. Лавренко............................................................Ц, 682
80. Мелкодерновинные (из Koeleria gracilis, Festuca sulcata s. 1., Poa botryoides, Cleistogenes squarrosa, реже ковыля — Stipa capillata s. 1.) алтайско-забайкальские степи и сельскохозяйственные земли на их месте. Составил
Е. М. Лавренко.................................................... 684
81. Змеевково-тырсовые и вострецово-тырсовые енисейско-забайкальские
степи и сельскохозяйственные земли на их месте. Составил Е. М. Лавренко 689 Общий очерк ......................................................... »
81 ен — енисейские (без участия Aneurolepidium pseudoagropyrum). 692
81 зб — забайкальские (с участием востреца — Aneurolepidium pseud )-agropyrum)....................................................... 697
82. Ковыльковые и полукустарничково-ковыльковые центральноазиатские пустынные степи. Составил Е. М, Лавренко ......................... 699
Общий очерк......................................................... »
82 тв — тувинские................................................ 700
82 у б — убсанурские............................................. 701
82 а — чуйские (алтайские)........................................ *
82 цтш — центральнотяныпанские . •........................... ... 702
82 пм — памирские.................................................. »
83. Горные луговые степи и остепненные луга в сочетании с зарослями кустарников и каменистыми обнажениями и сельскохозяйственные земли на их месте............................................................. 703
Общий очерк. Составил Е. М. Лавренко ............................ »
83 у — южноуральские. Составил Е. М, Лавренко.................... »
83 а — алтайские. Составил Е. М. Лавренко........................ 704
83 ск — североказахстанские. Составил Е. М. Лавренко ............ 705
83 стш — северотяныпанские. Составил Н. И. Рубцов ............... 706
83 тшп — западнотяныпанские и памироалайские. Составил Л, Е. Родин 708
83 к — кавказские. Составил Е. М. Лавренко при участии Е. В. Шиффере.................................................... 709
83 кр — крымские. Составил Е. М. Лавренко........................ 711
84. Горные дерновиннозлаковые (типчаково-ковыльные, ковыльные, типчаковые, овсецовые и др.) степи, в сочетании с каменистыми обнажениями и сельскохозяйственные земли на их месте.......................... . . 712
Общий очерк. Составил Е. М. Лавренко............................... »
84 у — южноуральские. Составил Е. М. Лавренко ................... 713
84 а — алтайские. Составил Е. М. Лавренко ....................... 714
84 ск —североказахстанские. Составил Н. И. Рубцов ............. »
84 стш — северотяныпанские. Составил Н. И. Рубцов ............... 716
84 цтш — центра л ьнотяньшанские. Составил Л. Е. Родин .......... 719
84 тшп — западнотяныпанские и памиро-алайские. Составил Л. Е. Родин 720
84 кпд — копетдагские. Составил Л. Е. Родин...................... 726
84 к — кавказские. Составил Е. М. Лавренко при участии
Е. В. Шиффере ....................... 729
XIII. Полукустарничковые полынные и солянковые пустыни
Общий обзор. Составил Л. Е. Родин .......................................... 731
85. Злаково-полынные пустыни в сочетании с дерновинно-злаковыми степями, полынными и солянковыми пустынями (южная полупустыня)...................... 745
Общий очерк. Составил Л. Е. Родин ...................................... »
85 пк — предкавказские (из Artemisia taurica). Составил Л. Е. Родин при участии Е. В. Шиффере............................................. 746
85 к — закавказские (из Artemisia Meyeriana). Составил Л. Е. Родин 747 85 кк — прикаспийско-казахстанские (из Artemisia Lercheana и A. pau-
ciflora). Составил Л. Е. Родин................................ 748
85 вк — восточноказахстанские (из Artemisia sublessingiana и A. pauciflora). Составил Л. Е. Родин.......................................... 752
85 стр — северотуранские (из северотуранских видов полыни). Составил Л. Ё. Родин ...................................................... 753
86. Злаково-полынные пустыни на песках (из Artemisia astrachanica или
A. Massagetovii, A. arenaria, Agropyrum sibiricum и др.) в сочетании по понижениям рельефа с солянковыми и луговыми группировками и слабо заросшими барханными и бугристыми песками. Составил Л. Е. Родин ... »
Стр.
87. Полынные пустыни. Составил Л. Е. Родин.........................II, 750
Общий очерк..................................................... »
87 кк — прикаспийско-казахстанские (из Artemisia Lercheana) ... »
87 стр — северотуранские (из северотуранских видов полыни) .... 768
87 ютр — южнотуранские (из Artemisia-herba alba s. 1.).......... 760
88. Горные и предгорные полынные пустыни. Составил Я. И. Рубцов .... 761
Общий очерк .................................................... »
88 tu — из Artemisia turanica в сочетании с ксерофильными кустарниками .................................................... >
88 t — из Artemisia tenuisecta в сочетании со степями........... 762
88 к — из Artemisia Korshinskyi в сочетании с нагорными ксерофитами и степями......................................................... 764
88 1 — из Artemisia Lehmauniana....................... 766
89. Полынные пустыни в сочетании с солянковыми. Составил Л. Е. Родин 766 Общий очерк................................................ »
89 кк — прикаспийско-казахстанские (из Artemisia pauciflora s. 1.,
A. Lercheana, Anabasis salsa, Atriplex сапа).............. 768
89 стр — северотуранские (из северотуранских видов полыни, Salsola laricifolia, Anabasis salsa и др.)............................. 769
89 ютр — южнотуранские (из Artemisia herba-alba s. 1., Salsola gem-mascens, S. arbuscula)............................................ 772
90. Эфемерово-полынные пустыни......................................... 775
Общий очерк. Составил Л. Е. Родин .............................. »
90к — закавказские (из Artemisia Meyeriana). Составил Л. Е. Родин
при участии Е. В. Шиффере............................... 776
90 ар — среднеараксинские (из Artemisia fragrans). Составил
Л, Е. Родин при участии Е. В. Шиффере .................. 777
90 стр — северотуранские (из северотуранских видов полыни). Составил Л. Е. Родин ......................................... 778
90 ютр — южнотуранские (из Artemisia herba-alba s. 1.). Составил Л. Е. Родин.............................................. 779
91. Эфемерово-полынные пустыни в сочетании с солянковыми............. 781
Общий очерк. Составил Л. Е. Родин .............................. »
91 к — закавказские (из Artemisia Meyeriana, Salsola dendroides, Salsola nodulosa, Salsola ericoides). Составил Л. E. Родин при участии E. В. Шиффере.................................... 782
91 ар — среднеараксинские (из Artemisia fragrans, Salsola nodulosa, Salsola ericoides). Составил Л, E. Родин при участии E. В. Шиффере ........................ 783
91 ютр — южнотуранские (из Artemisia hcrba-alna s. 1., Salsola gem-mascens, S. arbuscula). Составил Л. E. Родин............. 784
92. Солянковые пустыни (из Reaumuria soongorica, видов Salsola, Anabasis, Nanophyton и др.) обычно в сочетании с полынными пустынями. Составил Л. Е. Родин................................................. 786
93. Сочносолянковые пустыни (из Halocnemum strobilaceum, Halostachys caspica, Kalidium foliatum) и лишенные растительности солончаки. Составил Л. Е. Родин................................................... 788
94. Однолетнесолянковая растительность (из однолетних видов Salsola, Suaeda, Salicornia herbacea и др.) в сочетании с полукустарничково-солянковыми, галофитнолуговыми и другими сообществами. Составил Л. Е. Родин . . 793
95. Высокогорные терескеновые и полынно-терескеновые пустыни (из Euro-tia ceratoides, Artemisia Skorniakowii). Составили H. И. Рубцов и Л. Е. Родин.......................................................... 794
XIV. Саксаульники и кустарниковые пустыни
Общий обзор. Составил Л. Е. Родин.................................... 797
96. Черносаксаульники из Haloxylon aphyllum и саксаульники из Н. ammodendron. Составил Л. Е. Родин...................................... 809
97. Смешанные саксаульники (из Haloxylon persicum и Н. aphyllum). Соста-' вил Л. Е. Родин .............................................., . 812
98; Белосаксаульники (из Haloxylon persicum). Составил Л. Е. Родин .... 815
99. Черкезники (из Salsola Richteri), джузгунники (из видов Calligonum), кзыл-
чевники (из Ephedra strobilacea, Е. lomatolepis). Составил Л. Е. Родин 820
100. Разреженные псаммофитные кустарники (из видов Ammodendron, Eremo-sparton, Astragalus sect. Ammodendron, Calligonum и Convolvulus fruti-cosus) в сочетании с барханными песками. Составил Л. Е. Родин .... 822
30 Заказ 551
Стр.
XV. Высокогорная подушковидная растительность
101. Высокогорная подушковидная растительность ..........................11,825
Общий очерк. Составил О, В. Заленский............................... *
101 пм — памирская (из Acantholimon diaponsioides, Tanncetum ху-lorrhizum, Oxytropis Poncinsii, О. tianschanica, Potentilla pa-miroalaica). Составил //. И. Рубцов......................... 82ft
101 тш — тяныпанская (из Sibbaldia tetrandra, Thylacospermum rupifra-gum и др.). Составил H. И. Рубцов............................ 827
XVI. Слоевищная растительность такыров
102. Водорослевые и лишайниковые сообщества такыров. ('оставил Л. Е. Родин 829
103. Ледники и снежники. Составили М. М. Голлербах и В. Б. Сачава . . . 835
104. Разреженная растительность гор стенной и пустынной зон» .... 840»
Общий очерк. Составил Е. М. Лавренко................................... »
104 кз — Казахской складчатой страны с фрагментами стопной, луго-
вой и лесной растительности. Составил Е, М. Лавренко ... »•
104 са — Средней Азии с фрагментами степной, пустынной и нагорно-ксерофитной растительности. Составил Л. Е. Родин..................... 841
105. Сельскохозяйственные земли на месте таежных лесов, ('.оставили
Т. И. Исаченко и Ф. X, Бахтеев ...................................... 1, 442
106. Сельскохозяйственные земли на месте хвойно-широколиственных лесов. Составили Т. И. Исаченко и Ф. X. Бахтеев................................... 449
107. Сельскохозяйственные земли на месте лиственных лесов, ('оставили Т. И. Исаченко и Ф. X. Бахтеев............................................. 461
108. Сельскохозяйственные земли на месте широколиственных и хвойных ле-
сов горных областей. Составили Г. И. Исаченко и Ф. X. Бахтеев; .... 458
109. Оазисы в пустынной зоне (естественный растительный покров более или
менее целиком заменен культурным). Составил Е. П. Коровин .... II, 848
Литературные источники пояснительного текста к «Геоботанической карте СССР» м. 1 : 4 000 000. Составила В. В. Липатова при участии авто-
ров текста............................................................... 859
Картографические источники «Геоботанической карты СССР» м. 1 : 4 000 000, упомянутые в пояснительном тексте. Составила А. И. Лукичева . , „ 918
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ РАСТЕНИЙ1
Abella В. Вг. 468.
-- corymbosa Hgl. et Schmalh. 212.
Abies alba Mil). 144, 201- 203, 415, 910. gracilis Korn. 145, 215.
- - holophylla Maxim. 145, 348, 360, 363. Mayriana Miyabe et Kudo 145. nephrolepis Maxim. 144, 145, 202, 212, 214, 360, 919.
N ordman nian a (Stev.) Spacli 143, 144, 202, 206, 871, 919.
sachalinensis Mast. 144, 145, 212, 214, 293, 919.
Semenovii B. Fedtach. 144, 202, 210, 212, 469, 494, 919.
- sibirica Ldb. 96, 141, 143, 174, 188, 210, 310,- 348, 355, 482, 920.
Abutilon Theophrasti Mod. 858.
Acantholepis orientalis l/OSfl. 762, 773, 811, 821.
Acantholimon Boiaa. 577, 581, 582, 590, 602, 764, 825.
qlatavicum Bge. 582, 708, 725, 762, 847.
- compactum E. Kor. 725. diapensioides Boiaa. 742, 765, 794, 795, 825, 826, 846, 847, 926.
erinaceum (Jaub. et Spach) Lincz. 727. Hedinii Oaten f. 795.
- Karelinii (Stachegi.) Bgc. 777, 784.
- khorassanicum Czerniak. 727.
- Korolkovii (Rgl.) E. Kor. 582, 584, 589, 722.
Korovinii Czerniak. 728.
- lycopodioides (Girard) Boise. 709.
- pamiricum Czerniak. 709, 765.
- parvijlorum Rgl. 582, 847.
- pterostegium Bge. 781, 785.
lataricum Boiaa. 582.
Acanthophyllum С. A. M. 577, 582, 734.
— brevibractealum Lipaky 582.
elatius Bgo. 781, 817, 818.
glanduloBum Bge. 582, 727.
- Knorringianum Schischk. 725.
- pungens (Bgo.) Boiaa. 582, 725, 702, 774, 787.
Acarospora Mass. 77, 672, 736.
— Schleicher! (Ach.) Maae. 749.
Acelidanthus anticleoides Trautv. et Mey. 158 215.
Acer oampestre L. 371, 390, 391, 393, 400, 403, 409, 431, 729, 852.
— ginnala Maxim. 365, 371.
1 Составила В. В. Липатова.
Acer laetum С. A. M. 384, 40®, 421, 425.
— manshuricum Maxim. 213, 360.
— mono Maxim. 360, 363, 371, 435, 436, 610.
pic turn Tliunb. 438.
platanoidcs L. 165, 178, 192, 207, 245, 336, 354, 371, 382, 389, 390, 394, 396, 397, 423.
pseudo platan us L. 371, 382, 389, 413, 414, 852.
pseudosieboldianum (Рлх) Koni. 213, 360, 362, 436.
Hegelii Pax 474, 590.
Semenov i i Hgl. et Hord. 2L2, 474, 578, 590, 726.
Stevenii Pojark. 405.
lataricum L. 365, 371, 383, 392 , 393, 395, 399.
tegmentoeum Maxim. 213, 360, 371. Traulvetteri Medw. 207, 245, 371, 419, 492.
turcomanicum Pojark. 578.
turlteetanicum Pax 212, 467,469,474,590. ukurunduense Trautv. et Mey. 88, 360, 363, 371.
- velutinum Boles. 408, 409, 421, 427, 432, 433, 852.
Achillea aeiallca Sorg. 629, 630.
— Gerberi M. B. 640, 641, 662, 755.
— glaberrima Klok. 644.
- leptophylla M. B. 644, 645, 655, 357.
- • micranlha M. B. 592, 594.
millefolium L. 395, 445-447, 498, 519, 522, 523, 621, 707, 708, 712. - millefolium s. 1. 716.
- nobllie L. 746, 749.
- cetacea W. et K. 596, 613, 671, 710, 711, 729, 746.
Achyrophorus maculatue (L.) Scop. 613, Aconitum altaicum Steinb. 486.
— barbatum Pers. 448, 625, 629.
- Ciekanovtkyi Steinb. 316.
— delphlnlfoltum DC. 112.
— excehum Rchb. 164, 167, 168, 170, 185, 187. 191, 192, 194, 198, 197, 236, 238, 260, 280, 301, 312, 313, 316, 355, 356, 397, 481—483, 485, 496, 499, 626.
Pitcher I Rchb. 111.
— maximum Pall. 333, 502.
— Napellur s. ]. 211, 316. .
— naeutum Fisch. 325, 492.
Aconitum orientate Mill. 209, 325, 481,492. — Raddeanum Rgl. 361.
— rotundifolium Kar. et Kir. 494, 723. — sajanense Kumin. 486.
- soongoricum Stapf 485.
- umbrosum (Korch.) Korn. 307.
— volubile Pall. 306, 311.
Acorellus pannonicus (Jacq.) Palla 539.
Acorus calamus L. 537, 548—550.
Actaea erythrocarpa Fisch. 174.
— spicata L. 170, 208.
Actinidia arguta (Sieb. et Zucc.) Planch. 363.
— kolomikta Maxim.214, 215, 360,374,436. — polysama (Sieb. et Zucc.) Maxim. 363. Adenophora hymalayana Feer. 843.
— latifolia Fisch. 305, 341.
— liliifolia Ldb. 192, 471, 529.
— sublata Kom. 435.
Adianthum capillus-veneris L. 728.
— pedatum L. 361.
Adonis aestivalis L. 780.
— turkestanicus (Korsh.) Adolf 845, 846.
— vernalis L. 338, 343, 390, 612, 617, 624, 710.
— wolgensis Stev. 636, 637, 655, 657, 660.
Adoxa moschalellina L. 179, 211.
Aegilops L. 735.
— cylindrica Host 588, 777.
— squarrosa L. 588, 671, 783.
— triuncinalit L. 474, 591, 777, 780, 832.
Aegopodium alpestre Ldb. 200, 211, 212. — podagraria L. 167, 168, 175, 178, 187, 191, 192, 227, 232, 336-340, 344, 350, 354, 355, 357, 376, 381, 382, 384, 388, 390—395, 397, 415, 416, 446.
— tadshikorum Schischk. 469.
Aellenia (Salsola) subaphylla (G. A. M.) Aell. 762, 763, 774, 781, 785, 792, 800, 813, 815, 817, 818.
Aeluropus litoralis (Gouan) Pari. 507, 508, 510—516, 531, 534, 536, 539, 540, 542, 543, 579, 647, 669, 670, ‘ 742, 756, 770,' 792—794, 807, 814, 832 857
- repens (Desf.) Pari. 513, 534, 792, 794.
Agrimonia eupatoria L. 381, 390, 395, 454.
— pilosa Ldb. 448.
Agriophyllum arenarium M. B. 682, 755, 823
— latifolium F. et M. 801, 822, 823.
Agropyrum Gaertn. 595, 596, 643.
— cretaceum Klok. et Prokud. 646.
— cristatum (L.) Gaertn. 496, 531, 599, 609, 628, 630, 631, 666, 684—688, 690, 691, 694, 696—698, 700—702.
— dasynathum Ldb. 642.
desertorum (Fisch.) Schult. 600, 651, 660, 666, 672, 674, 678, 706, 746, 748, 750, 751, 757, 758, 808.
— geniculatum (Trin.) Korsh. 694.
— gracillimum Nevski 711.
—* imbricatum (M. B.) Roem. et Schult.
F 617, 647, 666, 729.
—* intermedium (Host) P. B. 400, 618, \ 727.
AgropyrumLavrenkoanum Prokud. 507, 640. — lolioides (Kar. et Kir.) Roshev. 695. — pectiniforme Room, et Schult. 512— 514, 528, 538, 599, 600, 617, 643, 647, 666, 668—670, 705, 746, 747, 749, 760, 776.
• - pruiniferum Nevski 713.
- - pseudocaesium Prokud. 658, 670.
— reflexiaristatum Nevski 704.
- repens (L.) P. B. 390, 396, 399, 446, 447, 496, 510—515, 523, 524, 529,
530, 531, 534, 538—540, 577, 579, 598, 609, 616, 621, 623, 629, 634, 650,
658, 668, 675, 695, 727, 747, 756.
- ruthenicum (Grisb.) Prokud. 508, 539, 647, 747.
— sibiricum (Willd.) P. B. 600, 662, 666, 672, 682, 731, 741, 750, 753—755, 770, 807, 808, 823, 924.
— strigosum (M. B.) Boiss. 711. tanaiticum Nevski 642.
- tarbagataicum Plotn. 472, 705, 715. — tianschanicum 211.
— trichophorum (Link) Richt. 469, 574, 577, 585, 589—592, 602, 618, 708,
710, 711, 721, 722, 727, 763, 923.
Agrostis alba L. 340, 343, 446, 453, 454, 460, 495, 506, 508, 510, 515, 516,
522, 530, 538, 540, 709.
— alba L. var. gigantea (Roth) Mey. 523. — canina L. 95, 460, 549.
— capillaris L. 408, 445, 454, 460, 482, 488, 489, 491, 495, 519, 522.
clavata Trin. 523, 525.
- maritima Lam. 506.
— stolonizans Bess. 460, 525, 527, 616.
— Syreistschikovii P. Smirn. 452, 522, 528, 530, 600, 612, 635.
— Trinii Turcz. 286, 288, 524, 531, 600, 630, 631.
Ailanthus altissima (Mill.) Swingl. 852. Albizzia Julibrissin Durazz. 425—427, 852. Albovia tripartita (Kalenicz.) Schischk. 403. Alcea nudiflora (Lindl.) Boiss. 499, 706. — rhyticarpa (Trautv.) Iljin 580.
Alchimilla L. 482, 485, 522.
— anisopoda Juz. 486.
— caucasica Bus. 490.
— cyrtopleura Juz. 484, 485.
— jailae Juz. 711, 712.
— Krylovii Juz. 484.
— rubens Juz. 484, 485.
— sericata Rchb. 710.
— sericea Willd. 836.
— sibirica Zamelis 484.
— Stevenii Bus. 711.
— taurica Juz. 711.
— tytthantha Juz. 711.
— vulgaris s. 1. 445, 450, 523.
Aldrovanda vesiculosa L. 537, 540—543.
Alectoria nigricans Nyl. 80, 94, 119. — ochroleuca (Hoffm.) Mass. 69, 70, 78, 94, 97, 100, 103, 104, 107, 108, 111, 119, 121, 122, 126, 158.
Alfredia acantholepis Kar. et Kir. 211.
Alhagi Adans. 857.
— kirghisorum Schrenk 794, 823.
— persarum Boiss. et Bushe 794, 823.
Alhagi pseudalhagi (М. В.) Desv. 513. 514, 746, 755, 782.
Alisma plantago-aquatica L. 432, 512. 514, 537.
Alliaria officinalis Andrz. 390.
Allium L. 479, 717, 735, 759, 768, 807.
- aflatunense B. Fed lech. 467.
- - albanum Grossh. 741, 780.
— albidum Fisch. 710.
- - anisopodium Ldb. 688, 694, 699. Barszczewskii Lipsky 725, 759.
- bidentatum Fisch. 688, 690, 691, 698, 699.
- - Bodeanum Rgl. 726.
— Borszczowii Rgl. 767, 773, 786.
- - caesium Schrenk 771, 778.
— caricoides Rgl. 842, 843.
— caspium (РаП.) M. B. 773, 774, 800, 812, 815, 818, 821.
— clathratum Ldb. 687.
— Eugenii Vved. 728.
— fibrosum Rgl. 741, 785, 786.
— filidens Rgl. 781.
— filifolium Rgl. 843..
— gutattum Stev. 655, 657.
— hymenorrhizum Ldb. 496.
— inconspicuum Vved. 721.
— inderiense Fisch. 751.
— Korolkovii Rgl. 716.
— Krylovii Sobolevsk. 701.
— lineare L. 686.
— monadelphum Less. 709, 847.
— odorum L. 630, 631, 694, 699.
— oreophilum С. A. M. 490, 584, 842, 844—846.
— Pallasii Murr. 678.
— paniculatum L. 751.
- platyspathum Schrenk 485, 842, 843.
— polyphyllum Kar. et Kir. 703, 826, 843, 847.
— polyrrhizum Turcz. 678.
— pseudoflavum Vved. 776.
— pseudoseravschanicum M. Pop. et Vved. 726.
- pulchellum Don. 655, 657.
— rubellum M. B. 776, 784, 786.
— rubens Schrad. 704.
— sabulosum Stev. 755, 774, 781, 785, 800, 815, 817, 818, 821.
— scabrellum Boiss. et Bubse 726.
— scabriscapum Boiss. et Ky. 780.
— schoenoprasoides Rgl. 716, 843.
— schoenoprasum L. 486, 628.
— Schubertii Zucc. 767, 778.
— sphaerocephalum L. 655, 657.
— strictum Schrad. 627.
— tenuissimum L. 702.
— ursinum L. 375, 417.
— victorialis L. 195, 196, 236.
— W inkier ianum Rgl. 724.
— xiphopetalum Aitch. et Baker 780. Alnaster (Alnus) fruticosus Ldb. 90, 97, 99, 100, 130, 138, 153, 158, 182, 215, 221, 235, 236, 239, 240—242, 258, 267, 268, 273, 274, 279, 282, 284, 289, 290, 301—303, 305, 310, 447, 486, 518.
— (Alnus) kamtschaticus (Kom.) J arm.
ex Czerep. 91, 111, 112, 155, 158, 159, 334, 500.
— (Alnus) Maximowiczii (Call.) J arm. 332, 334.
(Alnus) viridis (Chaix) Spach. 205, 489.
Alnus barbala С. A. M. 207, 208, 378, 405, 423, 425, 428, 430, 432, 433, 921. glutinosa (L.) Gartn. 168, 348, 357, 378, 383, 388, 405, 428, 430, 539, 566, 921. hirsuta Turcz. 289, 297, 332.
incana (L.) Moen ch 161, 167, 172,
383, 444.
japonica Sieb. et Zucc. 438, 458.
— kolaensis Orlova [A, incana (L.) Mo-ench var. virescens Wahlb.] 328, 330.
- subcordata С. A. M. 409, 427, 432. Alopecurus L. 112.
- alpinus Sm. 77—79, 81, 82, 114, 123, 519, 520.
- - amurensis Kom. 525.
Aucheri Boiss. 490.
- borealis Trin. 519, 545.
- brachyslachyus M. B. 485.
- glacialis C. Koch 478, 836.
- glaucus Less. 238.
— himalaicus Hook. f. 845.
— pratensis L. 123, 300, 336, 451, 496» 515, 519, 523, 524, 529, 531, 658, 715.
— seravschanicus Ovcz. 584, 709, 726.
— soongoricus (Roshev.) V. Petr. 484. — vaginatus Pall. 490, 497, 710, 711.
— ventricosus Pers. 506, 510, 511, 524, 529, 531, 532, 609, 623, 699, 832.
Alternaria Nees ex Wallr. 737, 831.
Althaea officinalis L. 529.
Alyssum L. 643, 671, 735.
— biovulatum N. Busch 97, 688, 692, 694, 695.
— campestre L. 587, 590, 706, 726.
— dasycarpum Steph. 778.
— desertorum Stapf 584, 587, 590, 592, 597, 635, 636, 638, 655, 669, 748, 750, 754, 757—760, 763, 767, 768, 770, 771, 773, 777—780, 793, 803, 817.
— gymnopodum P. Smim. 646.
— hirsutum M. B. 644, 784.
— lenense Adams 688, 690, 692, 694, 695, 697.
— marginatum Steud. 773.
— minutum Schlecht. 644.
— saxatile L. 644.
— strictum Willd. 778.
— tortuosum W. et K. 644, 651, 663. . — tortuosum s. 1. 617.
А тагаnthus a lb us L. 675.
— retroflexus L. 675, 858.
Amberboa odorata II jin 785.
— suaveolens (Willd.) Iljin 786.
— turanica Iljin 773, 774, 785, 803, 812, 815, 818, 819.
Amblynotus obovatus (Ldb.) Johnst. 686, 688.
A mblystedium serpens (Hedw.) Br. et Ammodendron Fisch. 739, 797, 799, 822, 823, 925.
Ammodendron Conollyi Biro. 800. 801. 804, 805, 813, 823.
— Eichwaldii Ldb. 800.
— Karelinii F. et M. 800, 821, 823.
— Lehmannii Bge. 800, 818, 821, 823.
Ammodenia peploides (L.) Rupr. 505.
Ammothamnus Lehmannii Bge. 816.
Amorphonostoc paludosum (Kutz.) Elenk. 737 •
— punctiforme (Kutz.) Elenk. 737.
Amygdalus L. 601.
— bucharica Korsh. 474, 574, 579, 580, 763> 922
— communis L. 591.
— Fenzliana (Fritsch) Lipskv. 473, 576.
— Ledebouriana Schlecht. 472, 705, 715, 716.
— nairica Fed. et Takht. 473.
- папа L. 358, 390, 393, 395, 399, 468, 471, 614, 617, 618, 635, 636, 638, 653, 704, 705, 715, 749, 757.
— Petunnikowii Litw. 583.
— spinosissima Bge. 474, 583, 590, 592, 726, 763, 781.
— ulmifolia (Franch.) M. Pop. 212, 474.
— zangezura Fed. et Takht. 473, 576.
Anabaena Bory 737.
Anabasis L. 731, 740, 744,’774, 786, 787, 925.
— aphylla L. 514, 543, 675, 750, 751, 755, 758, 769, 770, 793.
— aretioides Moq. et Coss. 911.
— brachiata F. et M. 774, 787.
— brevifolia С. A. M. 688, 700, 701.
— cretacea Pall. 675.
— - eriopoda (Schrenk) Ben th. 673, 767, 774, 787.
— micradena Iljin 814.
— ramo sis sima Minkw. 740, 763, 786, 787.
— salsa (С. A. M.) Benth. 512—514, 667, 675, 676, 679, 740, 748, 751—753, 755—758, 763, 767—769, 771, 772, 778, 787, 794, 810, 811, 813, 925.
— truncate (Schrenk) Bge. 680, 753, 767, 774, 786, 787.
Andromeda polifolia L. 125, 219, 266, 296, 330, 503, 556, 561.
Andropogon ischaemum L. 339, 473, 576, 579, 581, 593, 617, 618, 645, 656, 670, 671, 708, 710, 729, 923.
Androsace L. 735.
— akbaitalensis Derg. 826, 846.
— barbulata Ovcz. 618.
— Bmgeana Schischk. et Bobr. 687.
— dasyphylla Bge. 695, 842, 844.
— elongate L. 597, 635, 636, 655.
- incana Lam. 691, 692, 694, 699, 712.
— Koso-Polfanskii Ovcz. 615.
— Lehmanniana Sobol, non Spreng. 687.
— Olgae Ovcz. 725.
— eeptentrionalis L. 621, 694, 844.
— taurica Ovcz. 712.
— triflora Adams 126.
— Turczaninovii Freyn 597, 655, 669, 770, 776, 778, 780, 784.
Anemone L. 384, 485.
- altaica Fisch. 174, 191, 195.
— biarmiensis Juz. 98, 482, 483.
Anemone crinita Juz. 108, 236, 485, 629.
— dichotoma L. 279, 288, 289, 305.
— fasciculata L. 493.
- narcissiflora s. 1. 496.
— nemorosa L. 175, 414.
— protracta (Ulbr.) Juz. 494.
— ranunculoides L. 375, 382, 411, 430.
— reflexa Steph. 279, 280.
— Schrenkiana Juz. 484, 485, 496.
— sibirica L, 108, 111, 112.
— silvestris L. 448, 612, 621, 636, 637.
— T scher njaewii Rgl. 592.
— udensis Trautv. et Mey. 438.
— villosissima (1) C.) Juz. 502.
Aneurolepidium Nevski 596, 675.
— alaicum (Korsh.) Nevski 722, 846.
— angustum (Ttin.) Nevski 516, 664 679, 697, 718.
— dasystachys (Trin.) Nevski 103, 524, 697, 702, 794-796, 826.
— kopetdaghense (Roshev.) Nevski 728.
— multicaule (Kar. et Kir.) Nevski 513 535, 540.
— Paboanum (Claus) Nevski 510, 664 697.
— pseudoagropyrum (Trin.) Nevski 595 598, 608, 630—632, 682, 683, 690— 692, 697, 698, 924.
— ramosum (Trin.) Nevski 514, 531, 598 654, 657, 658, 660, 661, 664, 67o' 672—675, 694, 695, 748, 749, 757. ’
Angelica brevicaulis (Rupr.) B. Fedtsch 843.
— dahurica (Fisch.) Benth. et Hook 342.
— Maximowiczii (Fr. Schmidt) Benth 294, 297.
— refracta Fr. Schmidt 502.
— silvestris L. 167, 194, 429, 482.
— ternata Rgl. et Schnialh. 846.
— ursina (Rupr.) Rgl. et Schmalh. 332.
Anomodon Hook, et Tayl. 411.
— attenuatus (Schreb.) Huben. 377, 411.
Antennaria carpatica (Wahlb.) R. Br 487.
— dioica (L.) Gaerth. 201, 231, 491.
Anthelia Ji^ratzkana (Limp.J Spruce 82.
Anthemis carpatica Willd. 487.
— dumetorum D. Sosn. 408.
— ruthenica M. B. 645.
— Trotzkiana Claus 646, 675.
A nthox anthum odoratum L. 94. 108, 191 195, 245, 300, 316, 324, 325, 445* 481, 482, 484—486, 491, 492, 498, 522.'
Anthriscus aemula (Woron.) Schischk 494, 626.
— nemorosa (M. B.) Spreng. 481, 492.
— silvestris (L.) Hoffm. 313, 390, 447 481, 483, 499, 519.
Anthyllis affinis Britt. 460.
— alpestris Rchb. 487.
— Boissieri Sag. 493, 710, 711.
A ntitoxicum ( Vincetoxicum) officinale
(Moench) Pobed. 228.
— (Vincetoxicum) pumilum (Dcnl.) Pobed. 786.
— (Vincetoxicum) scandens (Somm. et-Lev.) Pobed. 404, 406.
Aphanopleura capillifolia (Rgl. et Schmalh.) Lipsky 591, 779.
Aphanothece saxicola Nag. f. nidulans (P. Richt.) Elenk. 737.
Aposeris foelida (L.) Cass. 488.
Aquilegia glandulosa Fisch. 108, 483— 486.
— Karelinii (Baker) O. et B. Fedtsch. 211.
— parviflora Ldb. 436.
— sibirica Lam. 236, 313.
Arabidopsis pumila (Steph.) N. Busch 588.
Arabis armena N. Busch 479.
— auriculata Lam. 635, 636.
— incarnata Pall. 686.
— septentrionalis N. Busch 78.
- Stelleri DC. 502.
Aralia rnanxkurica Rupr. et Maxim. 360, 436.
Arbutus andrachne L. 400.
.Archangelica decurrens Ldb. 469, 483.
Arctagrostis latifolia (R. Br.) Griseb. 70, 97, 112, 117, 118, 121, 126, 137, 230, 267, 520, 545.
Arctogeron gramineus (L.) DC. 687, 688, 691, 699.
Arctophila fulva (Trin.) Anders. 68, 114, 276, 505, 506, 520,521, 526, 545, 546.
Arctostaphylos uva-ursi (L.) Spreng. 94, 181, 220, 221, 224, 230, 236, 239, 241, 259, 270, 282, 301, 329.
Arctous alpina (L.) Niedenzu 69, 94, 100, 107, 108, 112, 121, 125, 136, 153,
158, 181, 191, 259, 266, 269, 270,
272, 280, 282, 300, 301, 306, 307,
309, 329.
— japonica Nakai 502.
Arenaria L. 596.
— capillaris Poir. 688, 690, 697, 698.
— formosa Fisch. 92.
— Griffithii Boiss. 584, 708, 722-—724, 765, 845, 847.
— funcea M. B. 439.
— Koriniana Fisch. 660.
— leptoclados Guss. 726.
— serpyllifolia L. 590, 635, 669, 722.
— turkestanica Schischk. 724.
— ucrainica Spreng. 612, 636.
Aristella bromoides Bert. 400, 410.
Aristida L. 799.
— Karelinii (Trin. et Rupr.) Roshev. 800, 813, 822, 823.
— pennata Trin. 515, 754, 755, 774, 800, 801, 813, 815, 817, 818, 821—823.
Aristolochia clematitis L. 395.
Armeniaca manshurica (Koehne) Skvortz. 439.
— vulgaris Lam. 469, 707.
Arnebia Forsk. 735.
— decumbens (Vent.) Coss, et Kral. 768, 780, 781, 813, 816—818.
— guttata Bge. 847.
— transcaspica M. Pop. 785, 786.
Arnica frigida С. A. M. 111.
— Lessingii Gr. 111.
— montana L. 204, 488.
— unalaschkensis Less. 502.
Arrhenatherum elatius (L.) M. et K. 492.
Artemisia L. 531, 600, 632, 643, 731, 733, 807, 808, 883.
— amoena P. Pol. 678.
- anethifolia Web. 631, 699.
— annua L. 433.
— arctica Loss. 111, 502.
- arenaria DC. 515, 666, 682, 731, 741, 753—755, 807, 808, 815, 923, 924. arenaria s. 1. 642, 662.
- arenicola Krasch. 732, 740, 741, 744, 774, 775, 781, 786, 800, 806, 810, 812, 817, 818, 821, 833.
- argyrophylla Ldb. 687, 701. armeniaca Lam. 229, 613, 704.
— Aschurbajewii C. Winkl. 720, 845.
- astrachanica P. Pol. 666, 682, 731, 741, 751, 753—755, 757, 807,808,923, 924.
— austriaca Jacq. 376, 396, 528, 598, 635, 637—640, 650, 651, 653, 655. 657, 658, 660,668—670, 672, 673, 675, 746, 749.
— badghysi Krasch. et Lincz. 732, 740, 741, 744, 760, 762, 773, 775, 779, 780, 784, 785, 808, 832, 833.
— baldshuanica Krasch. et Lavr. 580, 761, 763.
— Boschniakiana (Bess.) DC. 668.
— caespitosq Ldb. 701.
— caucasica Willd. 710.
— chamaemelifolia Vill. 710.
— commutata Bess. 286, 609, 630, 631, 683, 686, 688, 690, 693, 699.
— depauperata Krasch. 686, 687.
— diffusa Krasch. 732, 740,741,756, 769, 774, 775, 784, 787, 806, 810, 812, 821.
— Dimoan a M. Pop. 800, 808.
— dracunculus L. 286, 621, 650, 678.
— dumosa P. Pol. 732, 740, 741, 744, 760, 775, 780, 785.
— eremophila Krasch. et Butk. 580.
— eriocarpa Bge. 800, 816—818, 821.
— erivanica (Bess.) P. Pol. 744, 775.
— eucina P. Pol. 587, 778.
— ferganensis Krasch. 587.
— fragrans Willd. 741, 742, 744, 775, 777, 783, 784, 925. •
— frigida Willd. 103, 496, 609,
622, 628—631, 652, 663, 667, 679, 680, 685—688, 690—692, 694, 695, 697—701, 705, 706, 840.
— glabella Kar. et Kir. 680, 840.
— glandulifera Krasch. 763.
— glauca Pall. 596, 620, 621, 625, 650, 687, 690, 692, 693, 696.
— glomerata Ldb. 111.
— Gmelinii Web. 436, 439, 496, 690, 693.
— Gorfaevii P. Pol. 678, 717, 761.
— gracilescens Krasch. et Iljin 667, 740, 741.
— gurganica Krasch. 740, 741, 756, 759, 767, 770, 773, 775.
— herba-alba s. I. 732, 740—742, 760, 772, 773, 779—781, 784, 925.
— hololeuca M. B. 646.
— incana Kell, et auct. 656, 667.
— facutica Drob. 686.
Artemisia juncea Kar. et Kir. 681.
— karatavica Krasch. et Abol. 678, 721, 760.
— Keiskeana Miq. 437.
— Kelleri Krasch. 800, 808.
— kemrudica Krasch. 732, 740, 741, 744, 756, 758, 767, 770, 773, 775, 784, 786, 787, 810.
— kemrudica Krasch. var. karabugasica Krasch. 787.
— kopetdagensis Krasch. 780, 785.
— Korshinskyi Krasch. 742, 761, 763— 765, 925.
— Kuschakewiczii C. Winkl. 703, 795.
— laciniata Willd. 242, 259, 281, 305, 315, 436, 456, 457, 609, 621, 623, 630, 631, 704.
— latifolia Ldb. 259, 261, 613, 621, 625, 650, 713.
— Lehmanniana Bge. 709, 724, 725, 732 742, 761, 764, 765, 925.
— Lercheana Weh. 396, 507, 514, 651, 656, 660, 663, 666, 667, 671—676, 678, 682, 740, 741, 744, 746, 748— 752, 754, 756—758, 768, 769, 807, 808, 923—925.
— Lessingiana Bess. 663, 664, 666, 667, 671, 676, 744, 751, 923.
— leucophylla Turcz. 630, 631.
— lobulifolia Boiss. 740, 744, 775.
— maikara (Krasch.) N. Pavl. 667, 771.
— maritima s. 1. 508, 666, 668, 670, 713, 747, 923.
— Marschalliana Spreng. 538, 621, 641, 642, 650, 662, 676, 679, 680, 710, 808.
— Massagetovii Krasch. 731, 741, 753, 924.
— Maximovicziana Krasch. 435.
— Meyeriana Bess. 507, 579, 593, 741, 742, 744, 747, 775—777, 782, 783, 924, 925.
— mongolorum Krasch. 513, 514, 756.
— monogyna W. et K. 528, 529, 539, 616, 635, 638, 647, 769.
— mucronulata P. Pol. 678.
— nitrosa Web. 510—514, 528, 622, 623, 651, 696, 755.
— norvegica Fries. 126.
— obtusiloba Ldb. 496, 685.
— pamirica C. Winkl. 703, 794, 795.
— paniculate Lam. 529.
— pauciflora Web. 512, 513, 660, 667, 673, 674, 676, 732, 740, 741, 746, 748—752, 757, 758, 772, 924.
— pauciflora s. 1. 667, 679, 744, 751, 768, 925.
— pauciflora Web. ssp. maikara Krasch. 732, 740, 751.
— persica Boiss. 808, 846, 847.
— pontica L. 511, 529, 621, 622.
— pubescens Ldb. 221, 240, 609.
— rhodantha Rupr. 719, 720.
— rupestris L. 511, 609, 621, 846, 847.
— rutifolia Steph. 706, 722, 795, 844, 845, 847.
— salsoloides Willd. 647.
— santolina Schrenk 774, 775, 800, 815, 821.
Artemisia santolinifolia Turcz. 687.
— Schrenkiana Ldb. 755.
— scoparia W. et K. 597, 599, 690, 694 695, 746, 755, 807.
— scopariiformis M. Pop. 580, 587.
— scoparioides Grossh. 507.
— semiarida Krasch. et Lavr. 667, 672 676, 678—680, 732, 740, 741, 744, 752, 753, 758, 772, 787, 806.
— ser ice a Web. 232, 242, 312, 315, 483. 627, 630, 704, 706, 713.
— serotina Bge. 680, 702.
— Skorniakowii C. Winkl. 700, 703,
731, 732, 742, 794, 795, 846, 925.
— songorica Schrenk 755.
— stolonifera Maxim. 435—437.
— sublessingiana (Kell.) Krasch. 652, 663, 666, 667, 671, 677—681, 714, 732, 740, 741, 744, 752, 753, 759, 779, 840, 923, 924.
— sublessingiana s. 1. 678, 717, 761.
— subviscosa Bess. 685, 688.
— Szovitsiana Bess. 513.
— tanacetifolia L. 221, 240, 286, 600r 625.
— tanaitica Klok. 647.
— taurica Willd. 655—657, 666, 667, 669—671, 729, 741, 744, 746, 747, 759, 775, 887, 923, 924.
— tenuisecta Nevski 474, 580, 582, 592, 722, 742, 761—764, 925.
terrae-albae Krasch. 681, 732, 740, 741, 744, 751, 753, 756, 758—
760, 767, 769—772, 774, 775, 778, 779, 784, 787, 802, 806, 808, 810, 816, 817, 820.
— terrae-albae s. 1. 732, 740, 741, 755, 769.
— terrae-albae Krasch. ssp. Massagetovii Krasch. 731, 732, 740, 741, 753, 755, 769, 774, 786, 800, 812, 815— 817, 821.
— tianschanica Krasch. 702, 719, 732.
— Tilesii Ldh. 111.
— transiliensis P. Pol. 678, 681, 717, 759, 761, 779.
— trifurcate Steph. 79, 100, 107, 108, 502.
— Tschernieviana Bess. 507.
— turanica Krasch. 732, 742, 756, 758— 762, 767, 769—772, 774, 775, 778, 781, 808, 925.
— valida Krasch. 678.
— vulgaris L. 621.
Arthrophytum Schrenk 731.
— Lehmannianum Bge. 774, 786.
Arum Korolkowii Rgl. 728.
Aruncus asiaticus Poiark. 215, 342, 361-— kamtschaticus Ryab. Ill, 112. — vulgaris Raf. 205.
Arundinella anomala Steud. 439, 448, 457.
Arundo donax L. 534, 857.
Asarum europaeum L. 170, 173, 175, 179, 191, 192, 195, 205, 208, 340, 354, 355, 357, 375, 382, 388, 390, 392, 397, 413.
— Sieboldii Miq. 375.
Asparagus L. 735, 808.
— Breslerianus Schult. 771.
— officinalis L. 396, 529.
— persicus Baker 535.
— trichophyllus Bee. 761.
— turkestanicus M. Pop. 786.
Aspergillus Fr. 737.
Asperula balchanorum Bobr. 728.
— galioides M. B. 617.
— graveolens M. B. 641.
— Maximoviczii Kom. 435.
— odorata L. 174, 177, 178, 191, 192,
194, 195, 197, 204, 206—208, 337,
340, 365, 375, 382, 390, 393, 395,
397, 404, 405, 408, 410, 413—416, 418, 419.
Asphodeline taurica (Pall.) Kunth 497, 617.
Aspicilia (Mass.) Koerb. 679, 736, 831.
— alpino-desertorum (Kremplhb.) Elenk. 771.
— desertorum (Kremplhb.) Mer. 759, 769, 770, 787, 832.
Asplenium ruta-muraria L. 615, 728.
— septentrionale (L.) Hoffm. 234, 645, 841.
— trichomanes L. 645.
Aster alpinus L. 245, 484, 494, 495, 609, 625, 630, 631, 650, 687, 688, 690, 692, 693, 695, 717, 723, 845, 846.
— Amellus s. 1. 620.
— consanguineus Ldb. 111.
— flaccidus Bge. 102, 485, 844, 847
— Grimmii (Rgl. et Schmalh.) B. Fedtsch. 721.
— limonifolius (Less.) B. Fedtsch. 843.
— scaber Thunb. 436, 457.
— sibiricus L. 520.
Astragalus L. 596, 602, 717, 733—735, 755, 763, 781, 797, 799, 800, 808, 820, 822, 823, 925.
— adsurgens Pall. 630, 631.
— agameticus Lipsky 580.
— alatavicus Kar. et Kir. 847.
— andaulgensis B. Fedtsch. 723, 726.
— arbuscula Pall. 706.
— arpilobus Kar. et Kir. 785.
— asaphes Bge. 722.
— aureus Willd. 575, 581, 729, 783.
— austrosibiricus Schischk. 690—692,
694.
— Baranovii M. Pop. 845.
— Beketovii (Krassn.) B. Fedtsch. 837, 843.
— Biebersteinii Bge. 710.
— BornmUllerianus B. Fedtsch. 584, 592, 722.
— brachylobus DC. 749.
— brevidens Freyn et Sint. 727.
— brevifolius Ldb. 688.
— Bungeanus Boiss. 711.
— calycinus M. B. 747.
— campylorrhynchus F. et M. 587.
— caucasicus Pall. 581.
— ceratoides M. B. 686.
— charguschanus Freyn 826.
— chivensis Bge. 800.
— commixtus Bge. 770, 778.
Astragalus contortuplicatus L. 588-
— corrugatus Bert. 781.
— daghestanicus Grossh. 711.
— danicus Retz. 343, 523.
— Demertii Charadze 710.
— densiflorus Kar. et Kir. 723, 843,. 844.
- - densissimus Boriss. 577.
— depauperatus Ldb. 686.
— deserti M. Pop. 786.
— dilutus Bge. 701.
— dolichophyllus Pall. 655, 657.
— erinaceus F. et M. 581.
— erioceras F. et M. 770.
— filicaulis F. et M. 587, 590, 778, 785
- frigidus (L.) Bge. 112.
— galegiformis L. 710.
— gaudanensis B. Fedtsch. 728.
— glycyphylloides DC. 408.
— glycyphyllus L. 381.
— haesitabundus Lipsky 711.
— Henningii (Stev.) Boriss. 655, 657-
— heterodontus Boriss. 826.
— inaequalifolius N. Basil. 726.
— inopinatus Boriss. 609, 690—692.
— Jarmolenkoi Gontsch. 727.
— Karfaginii Boriss. 581.
— laguroides Pall. 701.
— lagurus Willd. 581.
— lasiosemius Boiss. 582, 589, 590, 709». 722, 725, 847.
— lefosemius (Lipsky) M. Pop. 845, 846..
— Levieri Freyn 490.
— lithophilus Kar. et Kir. 843—845.
— macroceras С. A. M. 686.
— macronyx Bge. 587.
— macrotropis Bge. 721, 778, 780.
— Marschallianus Fisch. 581, 711, 786..
— melilotoides Pall. 695.
— microcephalus Willd. 246, 575, 581.
— monophyllus Bge. 701.
— mucidus Bge. 760, 778.
— neo-Lipskyanus M. Pop. 721.
— nigrocalyx Slob. 589 590, 724.
— nivalis Kar. et Kir. 719, 843.
— novoascanicus Klok. 655, 657.
— Olgae Bge. 845.
— onobrychis L. 399, 650.
— oxyglottis Stev. 770, 778, 785.
— paucifugus С. A. M. 817, 823.
— piletocladus Freyn et Sint 577, 786.
— Scheremetevianus B. Fedtsch. 847.
— Schmalhausenii Bge. 587, 590, 592, 716..
— schugnanicus B. Fedtsch. 847.
— Severzovii Bge. 721, 778.
— Sieversianus Pall. 472.
— squarrosus Bge. 774, 816, 823.
— subpolaris Boriss. et Schischk. 118».
123, 519, 520.
— subulatus M. B. 645.
— tephrolobus Bge. 686.
— testiculatus Pall. 672, 701, 748.
— tibetanus Benth. 716.
— tribuloides Delil 759, 770, 774, 786..
— turcomanicus Bge. 780.
— turkestanus Bge. 778, 786.
— umbellatus Bge. 118, 520.
Astragalus unifoliolatus Bge. 816, 818, 823.
__ uraniolimneus Boiss. 729.
— vedicus Takht. 581.
— velatus Trautv. 728.
— vicarius Lipsky 590, 592.
— virgatus Pall. 662.
— xanthomeloides E. Kor. et M. Pop. 721.
Astrantia L. 209, 481.
— maxima Pall. 498.
Astrodaucus litoralis (M. B.) Drude 507.
.Athyrium Roth. 414.
— acrostichoides (Sw.) Diels 361.
— alpestre (Hoppe) Rylands 159, 196, — 325, 489.
— crenatum (Sommerf.) Rupr. 170, 186, 194, 356.
— -filix-femina (L.) Roth 168, 175, 186, 191, 192, 194, 196, 197, 204, 206,
207, 215, 356, 376, 382, 388, 414,
418, 424, 432, 482.
Atractylodes ovata Thunh. 436.
A tr a gene ochotensis Pall. 214
— sibirica L. 211.
Atraphaxis L. 681, 708.
— frutescens (L.) Ewersm. 762.
— laetevirens (Ldb.) Jaub. et Spach 706.
— pyrifolia Bge. ‘583, 590.
— spinosa L. 762, 767, 771, 773, 787.
— virgata (Rgl.) Krassn. 706.
Atrichium undulatum (Hedw.) P. B. 377.
Atriplex L. 513, 539, 731, 858.
— сапа C. A. M. 512, 513, 516, 660, 667, 675, 676, 679, 740, 751—753, 755—758, 769, 792, 925.
— dimer pho stegia Kar. et Kir. 801, 817.
— (Obione) fera (L.) Bge. 696.
— *heterosperma Bge. 543, 857.
— litoralis L. 512, 5394 540, 616.
— moneta Bge. 817.
— (Obione) pedunculate L. 623, 647.
— sphaeromorpha II jin 675.
— tatarica L. 514, 638, 668, 670, 675, 857.
— (Obione) verrucifera M. B. 508, 510— 513, 516, 529, 539, 623, 647, 651, 664, 669, 670, 679, 755, 792.
Aulacomnium acuminatum (Lind, et Am.) Par. 260, 282, 303.
— palustre (Hedw.) Schwaegr. 102, 108, 109, 116—118, 169, 260, 267, 280, 284, 288, 294, 296, 300, 303, 310, 330, 479, 491, 505, 550, 557.
— turgidum (Wahlb.) Schwgr. 69, 77, 80, 97, 102, 107, 113, 116—119, 121, 128, 129, 137, 138, 260, 268, 274, 282, 284, 300, 302, 303, 545, 561.
Aulacospermum simplex Rupr. 845.
— turkestanicum (French.) Schischk. 845.
Avena desertorum Less. 878.
Bacidia*(De-Notar.) Th. Fr. 78.
Baeomyces rose us Pers. 488. /
Bartsia alpina L. 95, 487.
Battarraea phalloides Pers. 737.
JBeckmannia eruciformis (L.) Host. 512, 513, 516, 525, 526. 530. 532, 616, 647, 658, 668.
Beckmannia syzigahne (Steud.) Fernaid 286, 526, 609.
Bellevalia Lapeyr. 776.
— sarmatica (Pall.) Woron. 635, 638. Berberis L. 468.
— amurensis Rupr. 360.
— heteropoda Schrenk 469, 535, 707, 708 718, 723.
— iberica Stev. et Fisch. 473.
— iliensis M. Pop. 535.
— integerrima Bge. 728.
— oblonga (RgL) G. K. Schn. 590, 726.
— sibirica Pall. 664, 841.
— turcomanica Karel. 728.
— vulgaris L. 710.
Bergenia crassifolia (L.) Fritsch 196, 198 201, 241, 318, 486.
— pacifica Kom. 334.
Berteroa spathulata (Steph.) G. A. M. 664 675, 679, 680.
Betonica L. 481.
— grandiflora Willd. 245, 404, 492.
— officinalis L. 381, 395, 399, 403, 454, 712.
Betula ajanensis Kom. 320.
— alajica Litw. 319.
— baicalensts Sukacz. 320.
— borysthenica Klok. 320, 641.
— Cajanderi Sukacz. 273, 274, 320, 519 520.
— costata Trautv. 213, 322, 360, 362 435, 437.
— dahurica Pall. 295, 322, 341, 435, 436 439, 448, 457, 609. ’
— Ermanii Cham. 106, 108, 157, 214 215, 310, 322—324, 332, 438, 500, 897’.
— Ermanii s. 1. 331, 866.
— exilis Sukacz. 70, 71, 90, 99, 101 106 107, 128, 129, 133, 268, 271, 272* 274’ 276, 287, 288, 293, 296, 303, ’ ЗОб’ 520, 546.
— fruticosa Pall. 134, 240, 272, 273 280 288, 289, 296, 303, 307.
— Gmelinii Bge. 632.
— grandiflora Litw. 320.
— humilis Schrank 316, 549.
— ircutensis Sukacz. 320.
— japonica Sieb. 322, 342.
— Kelleriana Sukacz. 322.
— Korshinskyi Litw. 322.
— Krylovii G. Kryl. 320, 338.
— lanata (Rgl.) Vassil. 332.
— Litwinowii A. DoL 245, 322, 323. 325 419, 492. r
— manshurica (Rgl.) Nakai 322, 342 436 437, 460.
— Medwedewii Rgl. 325, 492.
— megrelica D. Sosn. 322.
— microphyIla Bge. 322.
— Middendorffii Trautv. et Mey. 70 91 106, 108, 130, 134, 138, 158, 241, 261’
274, 290, 292, 293, 296, 302, 307, 310’
— папа L. 70, 71, 90, 93, 94, 96, 97* 115
117, 118, 120, 121, 131, 133, 153, 161’
162, 181, 190, 220, 224, 225, 227, 266
267, 270, 300, 301, 329, 330, 518, 556
561.
— ovalifolia Rupr. 294, 296, 297, 456.
— pamirica Litw. 322.
Betula paraermani Vassil. 332.
- platyphylla Sukacz. 280, 290, 303, 313, 320, 341, 448, 632.
— Prochorowii Kuzen. et Litw. 332.
— procurva Litw. 322.
— pubescens Ehrh. 161, 167, 195, 224, 230, 269, 320, 324, 329, 336, 338—340, 344, 345, 354, 394, 482, 518, 519, 548, 550, 623, 903.
- pubescens s. 1. 641.
— Raddeana Trautv. 245, 322, 492.
— Rezniczenkoana (Litw.) Schischk. 322. — rotundifolia Spach. 90, 103—105, 312, 314, 316, 318, 483.
— Saposhnikovii Sukacz. 322.
— Schmidtii Rgl. 322, 360, 363.
— schugnanica (B. Fedtsch.) Litw. 322, 847.
— Sukaczewii Soczava 134.
— tianschanica Rupr. 322.
— tortuosa Ldb. 90, 93, 95—97, 153, 154, 224, 267, 300, 319, 322—324, 327, 482, 518, 519.
— turkestanica Litw. 322.
— ulmifolia Sieb. et Zucc. 332.
— verrucosa Ehrh. 224, 230, 245, 311, 320, 325, 336, 338, 344, 354, 394, 414, 419, 482, 614, 623, 680, 841.
Bidens tripartitus L. 431, 538.
Blechnum spicant With. 424.
Boehmeria nivea (L.) Gaud. 854.
Bolboschoenus compactus (Hoffm.) Drob. 510, 512.
— maritimus (L.) Palla 510, 512—515, 530, 532, 535, 539, 540, 551, 616, 647, 756, 858.
Bongardia chrysogonum (L.) Boiss. 592, 728, 800.
Boschniakia rossica (Cham, et Schlecht.) Hult. 158.
Botrychium lunaria (L.) Sw. 486.
Brachanthemum kasakhorum Krasch. 680.
Brachypodium P. B. 473.
— pinnatum (L.) P. B. 178, 195, 211, 229,
232, 233, 238, 246, 337, 338, 340, 344,
358, 376, 384, 385, 392, 393, 395, 403,
404, 406, 454, 471, 474, 494, 495, 497,
499, 591, 627, 705—707, 710.
— rupestre (Host) Roem. et Schult. 246, 400, 406, 407.
— silvaticum (Huds.) P. B. 192, 212, 246, 247, 381, 384, 385, 397, 400, 403, 406, 408—410. 413, 415, 424, 427, 447, 467, 469.
Brachythecium Br. eur. 108, 411, 429. — curtum (Lindb.) Lindb. 354.
- - Mildeanum (Schimp.) Milde 430.
— populeum (Hedw.) Br. et Sch. 206.
— rivulare Br. et Sch. 550.
— rotabulum (Hedw.) Br. et Sch. 377.
— salebro sum (Web. et Mohr.) Br. et Sch. 377.
— velutinum (Hedw.) Br. et Sch. 206, 377. Braya oxycarpa Hook. f. et Thoms. 847. — pamirica (Korsh.) O. Fedtsch. 795, 796, 826, 846.
— rosea (Turcz.) Bge. var. albiflora Maxim. 846.
Breidleria arcuata (Lindb.) Loeske 491.
Briza media L. 445, 460, 495, 498.
Bromus L. 675, 735.
— adjaricus Somm. et Lev. 493.
— Benekenii (Lge.) Trimen 376, 467.
— Biebersteinii Roem. et Schult. 710, 711.
— cappadocicus Boiss. et Bal. 617, 711.
— Danthoniae Trin. 587, 589, 721, 760, 763, 777, 778, 780, 783—785, 801, 832.
— inermis Leyss. 343, 396, 447, 472, 500, 513, 515, 519, 523, 524, 529—531, 538, 577, 589, 591, 621, 629, 634, 650, 695, 707, 715.
— japonicus Thunb. 508, 588, 593, 594, 671 777, 779, 782, 784.
— Korotkyi Drob. 688.
— macrostachys Desf. 589, 590, 592, 763, 821, 832.
— oxyodon Schrenk 587, 589, 590, 592, 726.
— riparius Rehm. 245, 246, 497, 596, 612, 613, 617, 618, 620, 634, 636, 637, 710, 712, 729.
— rubens L. 588, 782.
— scoparius L. 587.
— sibiricus Drob. 260, 281, 286, 313, 524, 531, 630.
— squarrosus L. 597, 669—671, 777, 832. — sterilis L. 588.
— tectorum L. 474, 589, 590, 592, 597, 669, 670, 675, 706, 746, 754, 755, 759, 760, 763, 777, 778, 780, 781, 784, 801, 803, 813, 815, 819, 821.
— tomentellus Boiss. 497.
— turkestanicus Drob. 584, 722.
— variegatus M. B. 492, 493, 710, 711, 729.
Brunnera macrophylla (M. B.) Johnst. 420. Bryanthus Gmelinii D. Don. 110, 502.
Bryopogon (Link.) Koerb. 163.
— calybreiforme (L.) Elenk. 169.
Bryum Hedw. 81, 566, 736.
— caespitosum Schimp. 748, 749.
— obtusifolium Lindb. 545.
— Schleicheri Schwagr. 491.
Buhsea coluteoides (Boiss.) Bge. 781.
— Raddeana (Trautv.) Boriss. 583.
Bulbocodium versicolor (Кег.-Gawl.) Spreng. 612, 620.
Bungea vesicultfera (Herd.) Schischk. 760.
Bunias orientalis L. 612.
Bupleurum aureum Fisch. 193, 194, 316, 344, 356, 470, 485, 486, 496, 626, 627.
— bicaule Helm. 688, 690—692, 694, 698.
— densiflorum Rupr. 719.
— falcatum L. 636.
— Krylovianum Schischk. 705.
— multinerve DC. 232, 238, 625, 627, 704.
— multinerve DC. ssp. alaunicum K.-Pol. 615.
— pusilium Kryl. 686—688.
— scorzonerifolium Willd. 242, 439, 448, 457, 624, 630, 631, 688, 690, 693, 698.
Butomus umbellatus L. 512, 514, 515, 537, 538, 542, 658.
Buxus colchica Pojark. 423, 431.
— sempervirens L. 420.
Cacolia hastata L. 164, 187, 192, 260, 307.
312, 332, 342, 356, 397, 436, 457, 626.
Cocalia kamtschatica (Maxim.) Kudo 502. Cachrys odontalgica Pall. 65a, 657, 672, 751, 768.
Cakile euxina Pobed. 507.
Calamagrostis angustifolia Kom. 525, 526, 551.
— anthoxanthoides (Munro) Rgl. 846, 847. — arundinacea (L.) Roth 192, 221, 229, 232, 233, 235, 238, 245, 246, 261, 279, 311, 313, 325, 338, 339, 343, 344, 376, 384, 385, 394, 395, 403, 404, 408, 414, 446, 447, 481, 483, 489, 491, 495, 626, 627, 703.
— deschampsioides Trin. 504—506, 519.
— dubia Bge. 516, 807.
— elata Blytt 187.
— epigeios (L.) Roth 195, 225, 233, 238, 270, 339, 343, 344, 385, 407, 446, 447, 454,457, 472, 474,508,510,513, 515, 523—525, 529, 531, 534, 535, 538— 540, 577, 612, 621, 623, 634, 642, 649, 650, 695, 703, 706, 707, 715, 727, 756.
— groenlandica (Sohr ad.) Kunth 118, 120, 137, 519.
— Holmii Lang] 132.
— koibalensis Reverd. 695.
— laguroides Rgl. 845.
— lanceolata Roth 168, 345, 525—527, 538, 548, 551.
— Langsdorffii (Link) Trin. 68, 111, 131, 159, 168, 185, 191, 193, 195, 196, 214, 215, 225, 241, 260, 261, 274, 282, 286, 288, 289, 291, 292, 294, 296—298, 301, 305, 307, 330, 332, 333, 336, 339, 341, 344, 436—438, 457 , 482 , 502 , 506, 519, 521, 524—527 551.
— lapponica (Wahlbg.) Hartm. 96, 97, 112, 159.
— neglecta (Ehrh.) P. B. 131, 153, 162, 296, 330, 485, 503, 506, 520, 521, 525—527, 532, 546, 548, 549, 551.
— neglecta (Ehrh.) P. B. var. borealis LdesL 519
— obtusata Trin. 187, 191, 194, 196, 197, 241, 311, 312, 325, 338, 340, 341, 343, 344, 482.
— pseudophragmites (Hall, f.) Koeler 516, 534, 540, 542, 543.
— purpurascens R. Br. 129.
— Turczaninowii Litw. 236.
— villosa Mutel. 487—489.
Calamintha grandiflora (L.) Moench 208. — officinalis Moench 407, 426.
Calla palustris L. 176, 429, 430, 566. Callianthemum alatavicum Freyn. 843. — angustifolium Witasek 104, 486.
Calliergon Kindb. 132, 521, 544, 546, 561.
— cordifolium (Hedw.) Kindb. 168, 330, 429, 549, 562.
— cuspidatum (Hedw.) Kindb. 430.
— giganteum (Schimp.) Kindb. 78, 114, 118, 545, 549.
— Richardsonii (Mitt.) Kindb. 545.
— same n to sum (Wahlenb.) Kindb. 113, 114, 118, 131, 545, 546.
— str ament urn (Brid.) Kindb. 546, 562.
— trifarium (Web. et Mohr.) Kindb. 550.
Calliergon turgescens (Th. Jens.) Kindb. 78, 102.
Calliergonella cusp i data (Hedw.) Loeske 549.
Calligonum L. 682, 733, 754, 755, 797, 799, 800, 801, 808, 813, 818, 820, 821—823, 925.
— alatiforme N. Pavl. 774.
— alatum Litw. 816.
— aphyllum (Pall.) Giirke 754, 755.
— aralense Borszcz. 816.
— arborescens Litw. 815.
— caput-Medusae Schrenk 823.
— eriopodum Bge. 812, 815.
— gracile Litw. 817, 818.
— junceum (F. et M.) Litw. 774.
— leucocladum (Schrenk) Bge.755, 816, 81’7.
— membranaceum (Borszcz.) Litw. 816.
— microcarpum Borszcz. 774, 812, 815, 816, 818.
— rotula Borszcz. 815.
— rubicundum Bge. 515.
— setosum Litw. 815, 818.
— sp. sp. 816.
— turkestanicum (E. Kor.) N. Pavl. 815, 818.
— undulatujn Litw. 812, 818.
Calluna vulgaris (L.) Hill 95, 125, 219, 224, 227, 228, 329, 357, 556.
Calophaca Th. Fr. 78, 736.
— Howenii Schrenk 706, 716.
Caltha arctica R. Br. 545.
— palustris L. 184, 330, 503, 525, 537, 545, 546, 548, 549.
Calystegia sepium (L.) R. Br. 374, x408, 429, 534, 535, 541.
Campanula L. 479.
— abietina Grisb. 204, 414, 489.
— alliariaefolia Willd. 246, 384.
— alpina Jacq. 488.
— altaica Ldb. 201.
— Aucheri DC. 490.
— bononiensis L. 343, 613.
— collina M. B. 493.
— glomerata L. 286, 454, 498, 499, 522F 630, 708, 710.
— Hohenackeri F. et M. 710.
— lactiflora M. B. 209.
— lasiocarpa Cham. Ill, 502.
— latifolia L. 208, 492.
— Pallasiana Vest. 103, 108, 502.
— rapunculoides L. 384, 407.
— rapunculus L. 406.
— rotundifolia чат. linifolia Wahlb. 486-
— saxifraga M. B. 490.
— sibirica L. 343, 618, 621.
— tridens Rupr. 490.
— tridentata Schreb. 836.
— uralensis Nevski 636.
Camphorosma L. 513.
— Lessingii Litw. 513, 516, 536, 679». 747, 755, 776.
— monspeliacum L. 510, 513, 647, 667r 668, 670, 672—674, 748—751, 757.
— soongoricum Bge. 510, 616, 647.
Camptothecium Br. et Sch. 544.
— trichoides Broth. {Tomenthypnum" ni-tens (Schreb.) Loeske] 77, 80, 81, 97,102r
107, ИЗ, 116, 118, 128, 129, 268, 274, 282, 300, 303, 545, 549, 550.
Campylium stellatum (Hedw.) J. Lange et C. Jens. 114, 545.
— Zemliae G. Jens. 77.
Capparis spinosa L. 739, 774, 777.
Capsella bursa-pastor is (L.) Med. 670.
Caragana Lam. 601.
— arborescens Lam. 186, 194, 231, 313, 652.
— aurantiaca Koehne 723.
— balchaschensis (Kom.) Pojark. 753, 771,
Bungei Ldb. 601, 690, 691, 696, 697, 700—702.
— frutex (L.) G. Koch 231, 232, 358, 393, 395, 468, 471, 472, 614, 635, 636, 638, 652, 653, 658, 661, 679, 704, 706, 715, 841.
- grandiflora (M. B.) DC. 770, 774.
— fubata (Pall.) Poir. 212, 314, 723, 843, 844, 846.
— leucophloea Pojark. 601, 652, 661, 663, 715.
- microphyIla (Pall.) Lam. 601, 690, 691, 698.
- pumila Pojark. 496, 601, 652, 661, 663, 664, 841.
— pygmaea (L.) DC. 601, 686—694, 697— 702.
— spinosa (L.) DC. 700, 702.
— turkestanica Kom. 474.
Cardamine bellidifolia L. 78, 82.
— dentata Schultes 546.
— impatiens L. 424.
— macrophyIla Willd. 96.
— pratensis L. 503, 546.
— uliginosa M. B. 491.
Carduus L. 597.
— acanthoides L. 639.
— Thoermeri Weinm. 639.
— uncinatus M. B. 655, 656.
Carex L. 288.
— accrescens Ohwi 198, 294, 519.
— acuta L. 445, 451, 526, 527. acutiformis Ehrh. 532, 538—540, 548.
— algida Turcz. 98, 125, 168.
— altaica Gorodk. 102, 484.
— alticola Popl. 108.
— appendiculata (Trautv.) Kilk. 293, 525.
— appropinquata Sebum. 548—550, 556.
— aquatilis Wahlb. 131, 276, 330, 520, 521, 527, 545, 546, 548, 551.
— atrofusca Kiik. 486.
— atrofusca Schkuhr 98.
— brevicollis DC. 376, 389.
— brizoides Juslen. 383.
— brunnescens (Pers.) Poir. 162, 329, 486.
— Buschiorum V. Krecz. 245.
— caespitosa L. 131, 168, 286, 344, 445, 491, 511, 525, 526, 530, 532, 546, 549, 550, 551, 623.
— campylorhina V. Krecz. 214, 362, 376.
• — Canescens L. 330.
— capillaris L. 97, 98, 491.
— capitata L. 288.
— capitellata Boiss. et Bal. 491.
— chordorrhiza Ehrh. 267, 344, 545, 546, 549, 558, 561, 569, 570.
Carex colchica Gay 642.
— contiqua Hoppe 426.
— cryptocarpa С. A. M. 503, 527, 551.
— curvula All. 487.
— dacica Heuff. 491.
— diandra Schrank 344, 510, 548—550, 569.
- dichroa Freyn 491.
- digitata L. 204, 246, 357, 390, 413, 415.
- dduta M. B. 515, 516, 535.
dioica L. 549, 556.
discolor Nyl. 506, 519.
— disperma Dew. 168, 176.
- distans L. 647.
— disticha Huds. 345, 529, 531, 532, 616.
— divulsa Stokes 410, 427.
- duriuscula С. A. M. 273, 286, 531, 596, 603, 609, 631, 686, 688, 690—692, 694, 697, 701.
— duriusculiformis N. Krecz. 584, 702, 719, 724, 765.
— elata Bell. 344.
— elongata L. 394, 430.
— enervis С. A. M. 609.
— ensifolia Turcz. 112.
— ericetorum Poll. 270.
— falcata Turcz. 282, 503.
— flava L. 451.
— Fyllae Holm. 487.
— glacialis Mack. 111.
glareosa Wahlb. 505.
— globularis L. 96, 136—158, 153, 159, 162, 164, 166, 167, 169, 170, 180, 181, 198, 227, 269, 284, 288, 289, 291—294, 301, 338, 339, 557.
— Gmelinii Hook, et Arn. 502.
— gracilis Curt. 394, 524—526, 529, 530, 532, 538, 540, 549, 550, 551.
— Griffithii Boott 843, 844.
— Grioletii Roem. 406.
— humilis Leyss. 408, 495, 497, 612, 613, 615, 618, 620, 710, 711, 720, 729.
— humilis s. 1. 246.
— hyperborea Drej. 69, 78, 94, 97—100, 115, 117—119, 121, 125, 126, 136— 138, 268, 300, .303, 329, 482.
— Ilfinii V. Krecz. 92, 108, 198.
— inf lata Huds. 287, 344, 429, $48—550, 557, 558, 563, 566.
— irrigua Wahlb. 95, 570.
— Karelinii Meinsh. 510, 514, 616, ^23.
— koraginensis Meinsh. 111.
— koreana Kom. 438.
— Кor shinsky i Kom. 695.
— Kotschyana Boiss. et Hohen. 491.
— lanceolata Boott 439.
— lasiocarpa Ehrh. 184, 339, 344, 457, 510, 527, 550, 551, 556—558, 563.
— lenaeensis Kiik. 138.
— leporina L. 498.
— limosa L. 184, 267, 344, 547, 549—551, 558, 559, 569—571.
— lithophila Turcz. 288, 289.
— lugens Holm. 69, 107, 112, 129, 138, 159, 546.
— macrogyna Turcz. 99, 108.
— macroura Meinsh. 196, 238, 281, 286, 313, 339.
Carex manshurica Meinsh. 214.
— marina Dew. 505.
— melanantha С. A. M. 846.
— melanocarpa Cham. 67, 112, 126.
— melanostachya M. B. 392, 396, 514, 516, 529, 530, 538, 539, 550, 616,
668.
— melanostoma Fisch. 111.
— Meyeriana Kunth 296, 297, 307, 527, 551.
— Michelii Host 381, 382, 384.
— micropoda С. A. M. 113, 503.
— Middendorffii F. Schmidt. 293, 503, 527, 551, 557, 570, 571.
— minuta French. 260, 276, 292, 294, 297.
— misandra R. Br. 70, 107, 112, 126.
— montana L. 381, 703.
— nanella Ohwi 363.
— nitida Host 617.
— obtusata Liljebl. 102, 286.
— oligantha Stend. 491.
— omskiana Meinsh. 548—550, 616, 641, 647.
— oreophila С. A. M. 490.
— orthostachys С. A. M. 345, 532, 550, 551.
— oxyleuca N. Krecz. 846.
— pachystylis Gay 574, 580, 582, 587, 589, 590, 592, 727, 741, 762, 763, 767, 775, 778—780, 784, 785, 786, 851, 923.
— pallescens L. 498.
— panicea L. 450, 451.
— parva Nees 709.
— pauciflora Lightf. 176, 557, 558, 570.
— pauxilla V. Krecz. 108.
— pediformis С. A. M. 242, 259, 272, 280—282, 312, 313, 340, 625, 683, 686, 690, 692, 693, 703, 704.
— perfusca V. Krecz. 485, 486.
— phyllostachys С. A. M. 427.
— physodes M. B. 754, 755, 762, 773, 774, 781,800, 801, 804, 807, 811—813, SISSIS, 820, 821.
— pilosa Scop. 206, 337, 350, 354, 355, 357, 376, 382, 386, 390—392, 394, 395, 398, 413, 414, 416.
— pilulifera L. 488.
— praecox Schreb. 39*0, 529—531, 538, 540, 612, 636, 703.
— pseudo-cyperus L. 431, 510, 541, 542.
— pseudo-foetida Kuk. 709, 726.
— quadriflora (Kuk.) Ohwi 362.
— rariflora Wahlb. 503—505, 519, 545, 546, 558, 559. 561.
— Redowskiana С. A. M. 268, 280.
- remota L. 432.
— rhynchophysa С. A. M. 570.
— rigida Good. 487.
— rigidioides Gorodk. 107—109, 158, 309.
— riparia Curt. 345, 429, 430, 510, 530, 532, 538—540, 548—550, 616.
— rotundata Wahlb. 130, 545, 547, 550, 561, 570.
— rupestris Bell. 69, 97, 98, 104, ill, 126, 300, 490.
— ruthenica V. Krecz. 624, 703, 704.
— sabulosa Turcz. 688.
— sabynensis Less. 104.
— saxatilis L. 547.
Carex Schmidtii Meinsh. 260, 286, 288r 289, 296, 297, 307, 527, 551.
— scirpoidea Mchx. 112.
— sempervirens Vill. 487, 488.
— setina (Christ) V. Krecz. 126.
— siderosticta Hance 362, 376, 436.
— silvatica Huds. 204, 376, 384, 426, 427.
— songorica Kar. et Kir. 516.
— spaniocarpa Steud. 273.
— stans Drej. 99, 114, 117, 119, 125, 130r 132, 137, 505, 506, 520, 521, 545— 547, 558.
— stenocarpa Turcz. 102, 104, 314, 494.
— stcnolepis Less. 330.
— stenophylla Wahlb. 603.
— stenophylla s. 1. 784.
— stenophyllaides V. Krecz. 759, 777, 784. stilbophaea V. Krecz. 102, 486.
— strigosa Huds. 432.
stygia Fries 506.
- stylosa С. A. M. 502.
-- subspathacea Wormskj. 504—506, 519r 520.
-- subumbellata Meinsh. 502.
- supina Willd. 233, 596, 634, 636, 637r 649, 660, 662, 713.
- tripartita All. 96, 98, 111, 123, 486.
— tristis M. B. 480, 490.
— turkestanica Rgl. 591, 592, 716, 718r 725, 845.
— uralensis С. B. Clarke 596, 603, 654r 657, 748, 754.
— ussuriensis Kom. 214, 362, 376, 436.
— Vanheurckii Muell. Agr. 261, 282.
— vesicaria L. 330, 429, 430,. 527, 532r 540, 548—551, 566, 647.
— vesicata Meinsh. 527, 570.
- vulpina L. 394, 527, 529.
— wiluica Meinsh. 131, 240,. 260, 286r 288, 430.
— xyphium Kom. 214.
Carpinus betulus L. 352, 357, 367, 370r 381, 389, 390, 392, 413, 871.
— caucasica Grossh. 370, 378, 381, 384r 403, 407, 408, 417, 419, 422, 423, 425„ 426, 473, 618, 921.
— cordata Blume 213, 351, 360, 362, 367r 370, 436, 438.
— orientalis Mill. 247, 370, 399, 400r 405, 407, 408, 424, 473, 671.
— schuschaensis H. Winkl. 370, 408, 410.
Carum L. 479.
— atrosanguineum Kar. et Kir. 845.
— carvi L. 460, 523.
— caucasicum (M. B.) Boiss. 490, 491.
Carya pecan (Marsh.) Engl, et Graebn. 852..
Cassiope ericoides (Pall.) D.. Don 67, 107,. 108, 111, 158, 303, 309.
— lycopodioides (Pall.) D. Don 91, 110r 502.
— Redowskii (Cham.. et Schlecht. )i
G. Don 108.
— tetragona (L.) D. Don 69, 70, 100, 108r 118, 119, 121, 126, 128, 309, 502.
Castanea sativa Mill. 209, 366, 370, 378r 417, 420, 422, 423, 921.
Castillefa pallida (L.) Kunth 621.
Catdbrosa aquatica (L4 P. B. 538.
Catalpa Bungei С. А. М. 852.
Calocarpon (Koerb. et Th. Fr.) Stein. 78.
Caulophyllum robuslum Maxim. 375.
Celaslrus jlagellaris Rupr. 360.
Celtis caucasica Willd. 780.
Cenolophium Fischeri (Spreng.) Koch 523, 529.
Centaurea L. 481, 641.
— alutacea Dobrocz. 492.
— breviceps Iljin 641, 642.
— diffusa Lam. 670.
— hyrcana J. Bornm. 410.
— konkae Klok. 641.
— margarita-alba Klok. 641.
— margaritacea Tenore. 641.
— M arschalliana Spreng. 357.
— monanthos Georgi. 631.
— phrygia L. 498.
— pseudoleucolepis Kip. 644.
— pulchella Ldb. 815—818.
— ruthenica Lam. 636, 637.
— scabiosa L. 446, 613.
— sessilis Willd. 727.
— sibirica L. 229.
— squarrosa Willd. 591, 592, 726, 728, 760, 777, 784.
Centrostis abortiva (L.) Sw. 421.
Cephalanthera caucasica Kranzlin 421.
— longifolia (Huds.) Fritsch. 407.
— rubra (L.) L. C. Rich. 421.
Cephalaria Schrad. 481.
— caucasica Litw. 493.
— gigantea (Ldb.) E. Bobr. 492.
— sublanata (Bornm.) Szabo 727.
— uralensis Schrad. 636, 644, 645.
Cephalorrhizum oopodum M. Pop. et Kor. 580, 583.
Cephaloziella divaricata (Franc.) Schiffn. 82 Cerastium alpinum s. 1. 80.
— anomalum W. et K. 670.
— arvense L. 233.
— Beeringianun Cham, et Schlecht. 78, 112, 126.
— Bialynizckii A. Tolm. 11, 81, 82.
— Biebersteinii DC. 712.
— cerastoides Britt. 709, 720, 723—725, 844, 847,
— dahuricum Fisch. 485.
— Fischerianum Ser. 91.
— lithospermifoHum Fisch. 842—844.
— maximum L. 111.
— pauciflorum Stev. 312, 486.
— pusilium Ser. 108.
— Regelii Ostenf. 77, 81.
— tianschanicum Schischk. 842.
— ucrainicum Paez. 655, 657, 669, 671.
Cerasus Juss. 468, 601.
— araxina Pojark. 473, 576.
— avium (L.) Moench 381, 384, 389, 414.
— erythrocarpa Nevski 580, 763.
— fruticosa (Pall.) Woron. 343, 344, 358, 390, 393—395, 399, 468, 614, 635, 638, 653, 704.
— incana (Pall.) Spach 473.
— mdhaleb (L.) Mill. 212.
— Maximowiczii (Rupr) Kom. 360.
— microcarpa (С. A. M.) Boiss. 728, 780.
— pseudoprostrata Pojark. 728, 846.
Cerasus sachalinensis (Fr. Scbm.) Kom. et Klob.-Alis. 363.
— sp. sp. 474.
— tianschanica Pojark. 580, 583, 681, 706.
— verrucosa (Franch.) Nevski 590, 591, 592, 709, 763.
Ceratocarpus L/ 736.
— arenarius L. 516, 597, 638, 675, 748, 751, 754, 755, 759, 769, 770, 771,’ 776, 778, 779, 784, 814, 816, 817.
— turkestanicus Sav.-Rycz. 592, 760, 763, 781, 786, 811, 818.
Ceratocephalus Moench 735.
— falcatus Pers. 778.
— orthoceras DC. 587, 597, 638, 735, 748, 751, 760, 770, 771, 781, 815.
Ceratodon Brid. 736.
Ceratophyllum demersum L. 515, 537. 542, 543.
— submersum L. 537.
— tanaiticum Sapjeg. 647, 866.
Cesia corallioides (Nees) Carruth. 82.
Ceterach officinarum Willd. 728.
Cetraria (Ach.) Th. Fr. 296, 479, 491, 561, 736.
— chrysantha Turk. 78, 97, 100, 111.
— crispa Nyl. [C. tenuifolia (Retz.) Vain.] 94, 100, 102, 103, 267, 328.
— cucullata (Bell.) Ach. 69, 94, 97, 100,
102—104, 116, 118, 121, 122, 126, 128,
130, 138, 153, 158, 181, 195, 266, 267,
270, 287, 300, 302, 303, 328, 488, 561.
— hiascens (Fr.) Th. Fr. 77, 80, 98, 123, 128
— islandica (L.) Ach. 77, 80, 81, 94, 102, 114, 116, 121, 136, 138, 158, 180, 198, 219, 224, 229, 239, 309, 310, 330, 488, 491.
— nivalis (L.) Ach. 93, 94, 97, 100, 102, 103, 107, 120, 125, 126, 136, <38, 158, 267, 270, 300, 328, 330, 491, 561.
— Tilesii Ach. 78, 97, 102, 111. Chaerophyllum maculatum Willd. 407.
Chaetolimon sogdianum Lincz. 583.
Chamaedaphne calyculata (L.) Moench 180, 184, 219, 230, 231, 284, 292, 294, 296, 297, 503, 556, 561, 571.
Chamaemelum melanolepsis Boiss. et Buhse 836.
Chamaenerium angustifolium (L.) Scop. 195, 241, 261, 293, 325. 340, 502, 519, 672.
— latifolium (L.) Th. et Lange 843, 844. Chamaepericlymenum (Cornus) canadensis (L.) Graebn. 214, 293.
— (Cornus) suecicum (L.) Graebn. 125, 161, 329.
Chamaerhodos altaica (Laxm.) Bge. 691,
— erecta (L.) Bge. 679.
— grandiflora (Pall.) Ldb. 630.
Chamaesciadiuni acaule (M. B.) Boiss. 479. Chandonanthus setiformis (Ehrh.) Lindb. 77. Chara connivens Salzm. 542.
— contraria A. Br. 542.
— delicatula Ag. 542.
— fragilis Desv. 542.
— gymnopitys A. Br. 542.
Chara intermedia А. Вг. 542.
— kirghisorum Lessing Л emend Нollerh. 542. — tomentosa L. (Ch. ceratophylla Wallr.) 542.
— vulgaris L. 535 , 538, 539.
Chartolepis intermedia Boiss. 516.
Chartoloma platycarpum Bge. 819. Chsilanthes persica (Bory) Mett. 728. Chenopodium album L. 535, 675, 695. — urbicum L. 675.
Chimaphila (Pyrola) umbellata (L.) Nutt. 214, 233, 357.
Chlamydomonas nivalis Wille 837, 838. Chlorella Beyer. 737, 832.
Chlorococcum Fr. 737, 832.
Chondrilla L. 808.
— ambigua Fisch. 755.
— brevirostris F. et M. 662, 755.
- funcea L. 784.
— pauciflora Ldh. 515.
Chorispora Bungeana F. et M. 78, 842, 843, 844.
— macropoda Trautv. 837, 846.
— songarica Schrenk 843, 844.
- tenella (Pall.) DC. 587, 768, 771.
Chosenia macrolepis (Tures.) Kom. 272, 273, 332, 437, 519, 520, 880.
Christolea flabellata (Rgl.) N. Busch 846. — pamirica Korsh. 794, 795, 846.
Chrotophora gracilis F. et M. 800, 811, 814, 821.
Chrysopogon gryllus (L.) Trin. 594.
Chrysosplenium tetrandrum (Lund) Th. Fries 132.
Cicer arietinum L. 854.
— songoricum Steph. 723. 846, 847.
Cicerbita all9ina (L.) Wallr. 205.
— asurea (Ldh.) Beauv. 211.
— Gmelinii Beauv. 356, 397.
Ciouta virosa L. 286, 429, 515, 566, Cimicifuga foetida L. 236.
Cinclidium arcticum (Br. eur.) Schim. 545. Circaea alpina L. 195, 214, 358.
— lutetiana L. 207, 413, 432.
Cirsium esculentum С. A. M. 510, 529, 623.
- heterophyllum (L.) Hill 93, 94, 192— 194, 196, 197, 233, 301, 329, 339, 482, 485, 486.
— igniarium Spreng. 496, 680.
— kamischaticum Ldb. Ill, 332, 502.
- - pendulum Fisch. 307.
— schakaptaricum B. Fed tach. 580. setosum M. B. 639.
- simplex С. A. M. 491.
Cistus tauricus Presl 246, 247.
Cithareloma vernum Bge. 801, 819, 821.
Cladonia (Hill.) Hoffm. 102, 161, 181, 287, 296, 479, 561, 672.
- alpestris (L.) Rabh. 69. 78, 93, 94, 100, 103, 104, 107, 108, 112, 120,
122, 123, 125, 136, 137, 153, 158,
163, 181, 195, 198, 219, 224, 225 , 233, 239, 266, 289, 270, 303, 309 , 316, 328, 330.
— amaurocrea (Floerk) Scbaer. 103, 107, 130, 138, 181, 195, 561.
— bellidlflora (Ach.) Schaer. 94.
Cladonia chordalis Flk. 94.
— elongate Jacq. 94, 118, 136.
— foliacea (Huas.) Schaer. 668.
— foliacea (Huds.) Schaer. var. convolute (Lam.) Wain. 670.
— jurcata (Huds.) Schrad. 488.
— gracilis (L.) Willd. 97, 121, 130, 138, 163, 180, 270, 330, 448.
— mitis Sandst. 94, 116, 118, 120, 121, 136, 137, 153, 266, 303, 316, 328, 330, 561.
- pyxidata (L.) Fr. 77, 80, 94, 267.
— rangiferina (L.) Web. 69, 78, 94, 100,
116, 120, 121 -123, 125, 130, 137,
138, 158, 160, 163, 180, 198, 219,
224, 229 , 233, 266, 267, 267 , 270, 300, 303, 309 , 328, 330, 488, 491, 561.
— rangiformis Hoffm. 655, 668, 670.
- subrangiformis Sandst. 670.
— sylvatica (L.) Hoffm. 69 , 97, 100, 104, 112, 122, 123, 125, 128, 130, 138, 158, 160, 163, 180, 219, 225, 229, 232, 233 , 239 , 267 . 269 , 270, 300 , 309 , 328, 488.
— uncialis (L.) Web. 94, 97, 121, 128, 138 328.
Cladosporium Link et Fr. 737, 831.
Claytonia acutifolia Pall. 107.
— arctica Adams 100, 107, 112, 118, — Esehscholtsii Cham. 91.
— Joanneana Roem. et Schult. 102.
Cleistogenes Kong 595.
— bulgarica (Bornin.) Kong 645, 656.
— maeotica Klok. et. Zoz 644, 656.
— serotina (Link) Keng 656.
— sinensis (Hance) Keng 609, 610.
— squarrosa (Trin.) Keng 531, 599, 609, 622, 625, 640, 657, 662, 684-686, 688, 690, 692, 693, 695—698, 700, 701, 924.
Clematis hexapetala Pall. 630, 631, 683.
— integrifolia L. 636, 705, 712.
— Krylovii (Kom.) Reverd. 686.
— orientalis L. 534, 535.
— pseudoflammula Schmalh. 637.
— songarica Bge. 706.
— vitalba L. 374, 408, 432, 433.
— viticella L. 432.
Cleome L. 823.
— Gordfaginii M. Pop. 583.
— Noeana Boiss. 583.
Climacium dendroides (Hedw.) Web. et Mohr. 186, 192, 549.
Clinelymus (Blymus) dahuricus (Tures.) Nevski 313.
— (Elymus) sibiricus (L.) Nevski 286. CHxo^odt'um umbrosum (M. В.) C. Koch — vulgare L. 382, 399, 406.
Clintonia udensis Trautv. et Mey. 214.
Cnldium afanense (Rgl. et Til.) Drude 158, 215, 306, 309.
Cobresia Willd. 602.
— Bellardi (All.) Degl. 102—104, 108, 112, 201, 484, 490, 630, 631. 695.
— c&pillljolia (Decne.) С. B. Clarke 478, — oapilliformis Iv. 494, 719, 846.
Coptig trifolia (L.) Sallsb. 214, 293, 502. Corchorus capgularig L. 853. Coriepermum L. 736. — aralo-caspicum Iljin 801, 822. — Lehmannianum Bge. 801, 816, 817. — papillosum (Ktzej Iljin 815. Cornicularia divergene Adi. 78. 97. 100. 112, 119, 121, 126. — sieppae Sav. 655, 670.
Cornus mag L. 248, 373, 381, 382, 384, 400, 403, 405, 406, 473, 817, 729.
Coronilla varia L. 403.
Corydalis Med. 375, 384, 411. — alpestris С. A. M. 490. — bracteata (Steph.) Pera. 195. — cava (Mill.) Scfrweigg. et KOrte 382. — chionophile E. Czerniak. 727, — oonorhisa Ldb. 490, — Fedtschenkoana Rgl. 843. — glaucescens Rgl. 707. — Gortschakovii Schrenk 847. — Halleri Willd. 390, 430. — Marschalliana (Pall.) Pers. 427. Corylus avellana L. 205, 245, 338, 355, 373, 381, 383—385, 389, 394, 397, 403, 404, 423, 434, — heterophylla Fisch. 295, 341, 435, 437, 448, 458—458, 809.
— iberica Wittm. 208. — manehurioa Maxim. 213, 295, 435, 437.
Cotinus coggygria Scop. 247, 373, 400, 404, 488, 617.
Cotoneaster Med. 488, 469, 471, 474, 590, 592, 708.
— integerrima Med. 313.
— melanocarpa Lodd. 231, 233, 318, 340, 469, 499, 629, 687, 699, 707, 715, 718, 845.
— multiflora Bge. 343, 469, 499, 707, 718.
— raoemiflora (Desf.) C. Koch 473, 578, 846.
— uniflora Bge. 715.
Cousinia Cass. 573, 582, 583, 802, 765, 776, 922.
— acicularis French. 845. — affinis Schrenk 774. - annua C. Winkl. 801.
> — badghysi Kult. 583. — bipinnata Boiss. 780, 800, 818. — Bonvalotii French. 584, 722. — caespitosa C. Winkl. 723. — eentauroidee F. et M. 728. — oommiata Kult. 846. — egreria Jut. 584. — eryngioides Boiss. 780. — Fedtschenkoana C. Winkl. 584, - Franohetii C. Winkl. 584, 709, 724, 845.
- hissarica C. Winkl. 584, 725.
— horridula Jus. 709. — Komarovil C. Winkl. 780. — laetevirens C. Winkl. 709. — leptooephala F. et M. 728 - maoilenta C. Winkl. 584. — miorocarpa Boiss. 590, 591. — MinMtgiao Bomm. 721. — mollis Schrenk 778.
et
831
491.
Cobresia filijolia (Turcz.) С. B. Clarke 102, 307, 484, 609, 695.
— humilis (С. A. M.) Serg. 720.
— pamiroalaica Iv. 494, 719, 720.
— persica Ok. et Bomm. 490.
~ *choenoides (С. A. M.) Steud. 479, 4УО, 491.
— sibirica Turcz. 97, 98, 107, 112.
— Smirnovii Iv. 102, 484.
— stenocarpa (Kar. et Kir.) Steud. 837. Gochlearia arctica Schlecht. 78, 81, 835. — groenlandica L. 82.
Codonocephalum Peacookianum Aitch Hemsl. 727, 780.
Colchioum laetum Stev. 597.
Collema Wigg. 736, 759, 769, 819, — crispum Wifi». 749.
— granulatum (L.) R6hl. 832.
— minor (Pach.) Tomin 832.
Colpodium altaicum Trin. 104
— colchicum (Albov) Woron. 490, — fibrosum Trautv. 836.
~ B.) Griseb. 759, 767, 770,
771,776, 777, 782, 784.
leucolepis Nevski 844.
— ponticum (Bal.) Woron. 491.
—• variegatum Boiss. 490.
— versicolor (Stev.) Schmalh. 490.
Colwia geo ides (Pall.) Ldb. 688.
Colutea Buhsei (Boiss.) Shap. 780.
— canescens Shap. 474, 591.
c°marum palustre L. 99, 162, 330, 339, 503, 521, 545-549, 558, 566.
““««omenum (Steph.) Aschers. et Gr. 844, 847.
Conioselinum latifolium Rupr. 467, 469, 474, 499. F
Conocephalum conieum (L.) Dum. 204.
Consolida (Delphinium) s. F. Gray 735. 801^*803 elo camPtw>^rPa Nevski (^Ip^inium) panioulata Schur 670,671. (fielphinium) rugulosa (Boiss.) SchrOd. 587, 759, 781, 785.
Nevskin,Wm) (Kar. et Kir.)
Convallarle miialie ’ L. 167, 175, 206, 305, 357, 376, 382, 384, 395, 396, 436, 448, 457.
Convolvulus L. 726»
— Ammanii Desr. 628, 690, 691, 694, 700—702.
— arvensis L. 858.
— asoabadensis Bornm. et Sint. 727.
— cantabrioa L. 617.
— divaricatus Rgl. et Schmalh. 815, 817. — erinaeeus Ldb. 800, 815, 818.
— fruticosug Pall. 770, 773, 781, 800, 822, 925.
— hamadas (Vved.) V. Petr. 774, — holoeerioeus M. B. 617.
— Rgl. et Schmalh. 800,
354, 390, 617.
373,
363,
399,
728,
905. 723,
787, 823.
815,
— hamadas (Vved.) — holoeerioeus M. B.
818.
— perstoug L. 507.
— subhirsutue Rgl. et Schmalh. 763, 785, 818.
— tragacanthoides Turcz. 582, 681.
580,
Cousinia onopordioides Ldb. 774.
— oopoda Jus. 583.
— pannosa C. Winkl, 584.
— pulchra C. Winkl. 591.
— Raddeana C. Winkl. 580.
— rubiginosa Kult. 584, 765.
— Smirnowii Trautv. 727.
— splendida C. Winkl. 584.
— о prygin ii Kult. 583.
— ste^hanophora C. Winkl. 584, 709, — syrdarfensis Kult. 778.
— trachyphylla Juz. 584.
Crambe edentula F. et M. 761, 774.
— Gordfaginii Spryg. et Pop. 583.
— К о techy ana Boiss. 580.
— marifima L. 507.
— tatarica Sebeok 636, 637.
Crataegus L. 373, 383, 400, 401, 407, 425, 468, 658, 729, 852.
— altaica Lge. 469.
— kyrtostyla Fingorh. 389, 390, 392, 403, 409, 410, 423, 426, 433, 434, 473.
— Maximowicsii G. K. Selin. 364, 437.
— Meyeri A. Pojark. 473.
— microphy Ila C. Koch 426.
— monogyna Jacq. 205, 373, 381, 382, 389, 390, 392, 399, 413, 452.
— monogyna s. 1. 530, 617.
— oxyacantha L. 382.
— pentagyna W. et K. 433, 434, 473.
— pontica C. Koch 474, 574, 592.
— . sanguinea Pall. 394.
— songorica C. Koch 469, 474.
— sp. sp. 579.
— turkestanica A. Poiark. 469, 592.
Cratoneurum curvicaule Broth. 77.
Crepis L. 735.
— chrysantha (Ldb.) Turcz. 103, 118.
— Karelinii M. Pop. et Schischk. 827, 844.
— lyrata (L.) Froel. 193.
— multicaulis Ldb. 845.
— paludosa (L.) Moench 167, 168.
— rumicifolia Boiss. et Bal. 325.
- sibirica L. 164, 193. 194, 211, 236, 260, 312, 485, 494, 499, 626, 627, 705.
— tectorum L. 286, 815.
— tenuifolia Willd. 242, 694, 695, 699.
Crocus L. 479.
— Heuffelianus Herb. 417.
— reticulatus Stev. 597, 635, 638.
— Scharofanii Rupr. 490.
Crucianella angustifolia L. 594.
— Sintenisii bornm. 727.
Crupina vulgaris Cass. 728.
Crypsis aculeata (L.) Ait. 514, 539, 543.
— atogecuroides (Pill, et Mitt.) Schrad. — schoenoides (L.) Lam. 514.
Cryptodiscus ammophilis Bge. 812, 815.
Cryptospora falcata Kar. et Kir. 587.
Ctenidium molluscum (Hedw.) Mitt. 204. Cuscuta L. 858.
Cutandia Willi. 735.
— memphitica (Spreng.) Richt. 801, 815. Cyclamen abchasicum (Medw.).Kolak. 412. — elegant Boiss. et Buhse 427.
Cymbaria d ah uric a L. 688, 690, 691. 693, 694, 698, 701.
Cynanchum acutum L. 539.
— sibiricum Willd. 534, 535.
Cynodon dactylon (L.) Pers. 513, 538 542, 746, 747, 793, 858.
Cynosurus cri status L. 460.
— echinatus L. 410.
Cyperus fuse us L. 515, 539.
— longus L. 513.
— rotundas L. 858. —
Cysto^teris^ragilis (L.) Bornh. 211, 645, — mon tana (Lam.) Bornh. 318.
— sudetica A. Br. et Milde 186.
Cytisus borysthenicus Grun. 642, 662.
— caucasicus Grossh. 247.
— polytrichus M. B. 712.
— ruthenicus Fisch. 357, 358, 388, 394, 395, 620, 635, 653, 704.
— Skrobiszewskii Paez. 644.
— Zingeri (Nenuk.) V. Krocz. 228, 229.
238, 388.
Dactylis glomerata L. 195, 211, 238, 246, 260, 311, 355, 376, 381, 382, 392, 395, 396, 399, 400, 406—408, 410, 427, 446, 451, 470—472, 474, 481, 483, 492, 494, 495, 499, 579, 584, 589,
591, 706, 707, 711, 715, 727.
Danae racemosa (L.) Moench 421, 426. Daphne altaica Pall. 471.
— Julia K.-Pol. 615.
— mezereum L. 177, 178, 192, 195, 205 373, 413.
— pontica L. 419, 423.
— Sophia Kalen. 646.
Dasiphora (Potentilla) fruticosa (L.) Rydb. 280, 289, 312, 316, 842, 844.
— fruticosa s. 1. 704, 708, 847.
— parvifolia (Fisch.) Juz. 687, 841.
— phyllocalyx Juz. 843, 844.
Daucus carota L. 594.
Delphinium bitematum Huth 728.
— brachycentrum Ldb. 502.
— Brunonianum Royle 847.
— cheilanthum Fisch. 70, 112.
— crassifolium Schrad. 314.
— cuneatum Stev. 613, 620.
— dasyanthum Kar. et Kir. 842.
— elatum L. 131, 164, 192, 194, 196 519.
— ^n^^orum k- 628, 630, 690, 691, Dendrostellera turkmenorum Pobed. 781. Dentaria bulbifera L. 204, 207, 384, 417, — glandulosa W. et K. 411, 417.
— microphylla Willd. 490.
— quinquefolia M. B. 411, 416.
— tenuifolia Lbd. 118.
Deschampsia arctica (Spreng.) Schischk. 79, 81, 82, 97, 98, 117, 123, 519, 520.
— beringensis Hult. 502.
— borealis (Trautv.) Roshev. 126.
— caespitosa (L.) P. B. 330, 445, 481, 488, 489, 491, 496, 498, 525, 526, 530, 715.
— flexuosa (L.) Trin. 94, 95,120, 125, 136,
153, 164, НИ), 219, 245, 301, 306, 329, 419, 479, 482, 491.
Descurainia Sophia (L.) Schur. 670, 751, 754, 757, 815.
Deutz in amurensis (Hgl.) Airy-Show 436.
Dianthus L. 596.
— acicularis Fisch. 652, 704.
— arenarius L. 228, 357.
— fragrans Adams 710.
— guttatus M. B. 655, 657, 668, 671.
- - Hoeltzeri Winkl. 499, 718.
— h у panic, us Aiidrz. 644.
— lanccolatus Stov. 637, 657, 669, 671.
— leptopetalus Willd. 657, 660, 672, 678, 713, 751.
— platyodon Klok. 640, 641.
— polymorph us auct. 640.
— pseudarmeria M. B. 507, 644, 645.
— Seguieri Ldb. 686.
— superbus L. 201, 316, 519.
— tianschanicus Schischk. 716.
— turkestanicus Preobr. 716, 718.
— versicolor Fisch. 104, 628, 686, 688, 690, 691, 693, 694.
Diapensia lapponica L. 69, 70, 126, 502.
Diarthron vesiculosum (F. et M.) С. A. M. 780, 785.
Dicentra peregrina (Rudolphi) Fedde 91, 109.
Diehostylis M icheliana (L.) Nees 540.
Dicranoweisia cirrata (Hedw.) Lindb. 126. — crispula (Hedw.) Lindb. 80.
Dicranum Hedw. 197, 296, 300, 563, 571.
— acutifolium (Lindb. et Am.) C. Jens. 102.
— angustum Lindb. 118.
— Bergeri Bland. 300, 305.
— Bonjeanii De Not 330.
— congestum В rid. 116, 120, 305, 307, 309, 488, 561.
— elongatum Schleich. 69, 97, 102, 109, 114, 116, 118, 119, 120, 129, 136, 138, 300, 303, 545, 560, 561.
— fuscescens Turn. 70, 116, 137, 300.
— mafus Turn. 153.
— scoparium Hedw. 125, 211, 329, 414.
— spadiceum Zett. 97, 128.
— undulatum Br. eur. 233, 236, 310, 357, 429.
Dictamnus albus L. 381.
— angustifolius G. Don. 499, 705.
— caucasicus Fisch. 384.
Didymophysa Fedczenkoana Rgl. 836, 846.
Diervilla Middendorffiana Garr. 334. — suavis Kom. 215.
Digitalis grandiflora Mill. 395.
Digraphis arundinacea (L.) Trin. 345, 482, 510, 515, 525, 530, 532, 538, 540.
Dionysia tapetodes Bge. 728.
Dioscorea batatas Decne. 854.
— caucasica Lipsky 374, 407, 423.
Diospyros lotus L. 421, 423, 425, 427, 431.
Diploschistes Norm. 672, 736.
— albissimus (Ach.) Dalia Torre et Parnth. 831.
— scruposus (Schreb.) Norm. 597, 759, 770, 787, 831.
Diplotaxis cretacea Kotov 646.
Dipsacus azure us Schrenk 472 — laciniatus L. 433.
Diptcrocome pusilia F. et M. 762.
Diptychocarpus str ictus (Fisch.) Trautv. 770.
Distichium capillaceum (Hedw.) Br. eur. 77, 118, 211.
— Hagenii Ryan 81.
Ditrichum flexicaule (Scbleiph.) Hampe 77, 80, 81, 119.
Dodartia orientalis L. 858.
Dontostemon perennis С. A. M. 702.
Dorema Aitchisonii E. Kor. • 587, 786. — sabulosum Litw. 812, 815—818.
Doronicum altaicum Pall. 92, 108, 211, 486, 494.
— macrophyllum Fisch. 492.
Dorycnium graecum (L.) Ser. 247, 403, 405.
- - intermedium Ldb. 246.
Draba L. 80, 82.
— alpina L. 81, 847.
- bryoides DC. 479, 490.
- cuspidata M. B. 712.
— hispida Willd. 478.
— lactea Adams 126.
— lanceolata Hoyle 844, 845.
— macrocar pa Adams 80.
— micropetala Hook. 126.
— niollissima Stev. 490.
— nemorosa L. 635, 636, 707.
— Olgae Rgl. et Schmalh. 724, 725, 845.
— oreades Schrenk 843, 844.
— ossetica (Rupr.) Somm. et Lev. 490.
— pilosa DC. 126.
— sibirica (Pall.) Thell. 479, 612.
Dracocephalum altaiense s. 1. 484, 485.
— austriacum L. 710.
— bipinnatum Rupr. 719.
— discolor Bge. 694, 696.
— grandiflorum L. 104, 316, 483. .
— heterophyllum Ben th. 795, 826, 846, 847.
— imberbe Bge. 842, 843.
— integrifolium Bge. 844.
— nodulosum Rupr. 843.
— nutans L. 483—485, 844.
— oblongifolium Rgl. 844, 845.
— Paulsenii Briq. 826, 843, 844, 846, 847.
— peregrin urn L. 686, 714.
— nuyschiana L. 228, 233, 376, 704. — stamineum Kar. et Kir. 842.
Drepanocladus (C. Mull.) Roth 80, 132, 197, 287, 288, 429, 521, 544, 546, 561.
— aduncus (Hedw.) Moenkem. 549.
— aduncus var. Kneiffii (Schimp.) Moenkem. 550.
— badius (Hartm.) Roth 303.
— exannulatus (Br. eur.) Warnst. 102, 116, 330, 505, 546, 547, 562, 566.
— jluitans (Hedw.) Warnst. 384, 547, 561.
— intermedius (Lindb.) Warnst. 102, 267.
— latifolius (Lindb. et Am.) Warnst. 546.
— revolvens (Turn.) Warnst. 78, 113, 114, 118, 545, 546.
— Sendtneri (Schimp.) Warnst. 102, 344, 545, 549, 550.
— uncinatus (Hedw.) Warnst. 80, 81, 102,
108, 113, 114, 118, 123, 131, 211, 303, 330, 504, 505, 519 , 545.
Drepanocladus vernicoaus (Lindb.) Wamst. 287,344,545, 547, 549, 550, 501, 569.
Droaera anglica Hude. 559.
— rotundifolia L. 503, 557.
Dryadanthe (Sibbaldia) tetrandra (Bge.) Jus. 827, 837, 842—844, 846.
Dryaa ajananaia Juz. 108.
— grandia Juz. 105.
— kamtachatioa Juz. 79, 110.
— octopetala L. 94, 97, 98, 487.
— oxyodonta Jus. 102—105, 486.
— punctata Jus. 69, 78, 80, 81, 97, 98, 100, 103, 105, 107, 111, 115, 118, 119, 125—127, 138, 287, 268, 303.
— viaoosa Jus. 308.
Dryopteria Adans. 432.
— amurenaia Christ 214, 381.
— austriaca (Jacq.) Woynar 159, 170, 178, 191, 192, 204, 214, 482.
— Borreri (Rouy) V. Krecz. 424.
— Buaohiana Fom. 214, 361, 362.
— orlatata (L.) A. Gray 170.
— filix-maa (L.) Schott 174, 175, 177, 178, 192, 194, 195, 204, 207 , 208 , 211, 341, 354, 365, 378, 382, 388, 397, 413, 414, 418, 418, 421.
— Komarovii Koss. 728.
— Linnaeana Christens. 161, 166, 167, 170, 178, 180, 184, 191, 204, 214, 307, 318, 329, 333, 338, 355, 356.
— oreopteria (Ehrh.) Maxon 196, 889.
— phegopteria (L.) Christens. 178, 191, 204, 214, 215, 355.
— Robertiana (Hoffm.) Christens. 240.
— apinulosa (Mull.) Ktze. 186, 170, 178, 186, 194, 196, 198,
— thelypterie (L.j A. Gray 429 , 430, 510, 538, 541-543 , 549.
Dufurea arctica Hook. 78, 81, 97.
Duponlla Fischeri R. Br. 88, 70, 81, 114, 504-506 , 519, 521, 545 , 546. t
Bchtnochloa crus-galli (L.) Roem. et Schult. 858.
— macrocarpa Vaslng. 858.
Echinopanax datum Nakai 215.
Bohtnopa maraoandicua Bge. 763.
— ritro L. 727.
— taohimganicua B. Fedtsch. 845. BoMnopaUon Moq. 675, 738.
— hyaaopifolium (Pall.) Moq. 516.
- eadoides (Pall.) Moq. 59?, 668, 669, 675, 747, 748, 771.
Bchium rubrum Jacq. 638, 637.
Blaeagnua L. 543.
— angustifolia L. 433, 530, 534, 535, 540, 682, 852.
— orientalis L. 534.
Blatlne alelnastrum L. 647.
Bleutherococoua eentiooaue (Rupr. et Ma* xim.) Maxim. 213, 341, 3ft0.
Blymus giganteua Vahl 507, 515, 642, 662, 682. 755, 823.
— fenleaeenait Tures. 627.
— mollis Trin. 505, 519.
— eabulosus M. B. 642.
Elymue villosisaimus Seri bn. 106.
Eminium Lahmannii (Bge.) Ktze. 800.
Empetrum hermaphroditum (Lange) Hager. 491.
— nigrum L. 69, 93, 94, 96—98, 100, 107, 108, 110, 117, 119 — 121, 125,
138, 153, 160, 161, 181, 190, 195, 219
220, 224, 225, 230, 236, 241, 245, 266,
267, 269, 270, 272, 296, 300, 302, 309,
314, 316, 329, 330, 502, 503, 556.
Encalypta oiliata Hedw. 211.
Entodon orthocarpus (La Pyl.) Lindb. 211. Ephedra L. 797, 822.
— distachya L. 644, 645, 663, 678, 679, 701, 708, 754, 762, 771, 784, 840.
— equisetina Bge. 582, 881, 780, 848. — Fedtschenkoi PauJa. 795, 844, 846. — intermedia Schr. et С. A. M. 681, 728, 781, 786.
lomatolepix Schrenk 536, 733, 755, 800, 80S, 815, 820, 822, 925.
- procera F. et M. 473, 576, 747.
— strobilacea Bge. 733, 797, 800, 803, 813, 815, 818, 818, 820, 822, 925.
Epilaaia hemilaaia (Bge.) С. B. Clarke 780, 786.
Epilobium alpinum L. 111.
— Hornemannii Rchb. 111.
— montanum L. 204, 245.
— palustre L. 510, 549.
— aertulatum Hausskn. 111.
Bpimedium colchicum (Bolaa.) Trautv. 406.
Epipactia palustris (L.) Crantz. 543.
Equisetum arvense L. 448, 502, 519, 520. 523.
— heleocharis Ehrh. 162, 280, 296, 330, 344, 525, 543, 547—550, 558, 586.
— Komarovii Iljin 333, 502.
— majua Gars 432.
— palustre L. 96, 99, 184, 503.
— pratense Ehrh. 125, 168, 180, 236, 274, 280, 282, 283, 338, 339.
— ramoeiasimue Desf. 535.
— aoirpoides Michx. 180, 282, 305.
— silvaticum L. 162, 164, 168, 167, 169— 171, 176, 180, 181, 184, 214, 267, 282, 291, 297, 446.
— variegatum Schlelch. 118, 546. Eragrostis minor Host 599, 838, 688, 770. — plloea (L.) P. B. 538.
— auaveolena Beck. 538.
Branthia longiatipitata Rgl. 780.
Bremopoa oxyglumis (Boiss.) Roshev. 728. — persica (Trin.) Roshev. 777, 784.
Eremopyrum Jaub. et Spach. 735, 811, — Buonapartis (Spreng.) Nevski 780, 821. — orientale (L.) Jaub. ot Spach 508, 741, 759, 760, 783 , 789 , 770, 771, 773 , 777, 779, 781, 782, 784—786, 801, 803, 812 813, 815, 818, 819, 821, 832.
— prostratum Podpira 778.
— tritloeum (Gaertn.) Nevski 597, 741, 754, 777.
Eremosparton F. et M. 797, 822, 925. — aphyllum (Pall.) F. et M. 754, 755, 808. — flaccidum Litw. 799, 823.
Bremostachye Bge. 802.
— affinle Schrenk 787.
Ere most achy 8 labiosa Bge. 580, 587, 780.
— Regeliana Aitsh. ot Hemsl. 761, 800.
- • speciosa Rupr. 707, 727, 845.
— tubeгоза (Pall.) Bge. 771.
Eremurus M. B. 582, 735.
— altaicus (Pall.) Stev. 652, 750, 845.
— inderiensis (M. B.) Rgl. 755, 767, 768, 800, 815, 818.
— Olgae Rgl. 587, 592, 707, 780.
- - Regelii Vved. 592, 722.
robuBtus Rgl. 592.
spedabilis M. B. 726, 727.
— stenophyllus (Boise, et Buhse) Baker 727.
Erianthus purpuraacens Anderss. 534, 682, 858.
Erigeron a Ip in us L. 709.
— andryaloidea (DC.) Booth. 845, 846.
- eriocalyx (Ldb.) Vierh. 102, 723.
— Olgae Rgl. et Schmalh. 837, 844.
- Poncinsii (French.) Botsch. 847. Schmalhausenii M. Pop. 843.
— turkestanicua Vierhap. 725, 844.
— unalaschkensie (DC.) VierhaD. 502.
Eriophorum angustifolium Roth ol, 114, 118, 130, 132, 268, 276, 330, 491, 503, 521, 545—547, 549, 550, 558.
— brachyantherum Trautv. 118.
Cha mis son is С. A. M. 521, 545—547, 559.
— gracile Koch 95, 549, 550. humile Turcz. 102, 486.
- - japonicum Maxim. 109. latifoliuni Hoppo 102.
• rusaeolum Fr. 132, 546, 561. Scheuchseri Hoppe 114, 130, 485, 519, 520 547
— vaginatum L. 69, 106, 118, 121, 122, 126, 129, 132, 159, 162, 260, 268, — 276, 288, 291, 292, 296, 307, 545, 556, 558, 559, 561, 563.
Eritrichium fenissecnse Turcz. 694.
— pamiricum B. Fedtsch. 847.
— peotinatum (Pall.) DC. 627, 686.
— pedinatum s. 1. 609.
- villosum (Ldb.) Bge. 69, 102, 118.
Erodium L’Il6r. 735.
— Beketov it Schmalh. 644.
— cicutarium (L.) L’H4r. 777, 786.
— Hoefftianum С. A. M. 784.
— Litvinovii Woron. 812, 814, 816, 817.
— oxyrrhynchum M. B. 778, 786, 801, 821
Erophila verna (L.) Besser 587, 597, 635, 636, 655, 669, 670, 776.
Eruca sativa Lam. 780.
Eryngium campestre L. 640, 655, 657, — maritimum L. 507.
— planum L. 529.
Erysimum altaicum С. A. M. 690, 698, 842.
— cretaceum (Rupr.) Schmalh. 646.
— krynkense Lavr. 646.
— Pallasii (Pursch) Femald 107.
— persicum Boiss. 778, 784.
— repandum L. 670.
Erythronium sibiricum (F. et M.) Kryl. 195.
Euclidium R. Br. 735.
Euolidium ayriacum (L.) R. Br. 587.
Eucommia и Imo idea Oliv. 852.
Euonymus europaea L. 354, 374, 382, 384, 388-393.
— папа M. B. 373.
— pauciflora Maxim. 341, 436.
— Semenovii Rgl. et Herd. 212, 469.
— verrucosa Scop. 178, 247, 337, 354, 355, 374, 376, 382, 388-390, 392, 394, 395, 397, 877.
Eupatorium cannabinum L. 429.
Euphorbia L. 735.
- a™^loide8 L. 246, 404, 406, 410,
— angulata Jacq, 381.
cheirolejpis F. et M. 812, 814, — densa Schrenk 811, 815, 819.
- - humilis С. A. M. 844.
— inderiensis Less. 760.
— jaxartica Prokh. 592.
— leptocaula Boiss. 596.
— macroceras F. ot M. 420.
- - Marschalliana Boiss. 776, 784.
— oblongifolia C. Koch 492,
— pilosa L. 485.
- - rapulum Kar. et Kir. 726. semivillosa Prokh. 394.
— serawschanica Rgl. 725, 726, 845, 846. - SeguierianaNeck.396.507,638,640—642, 658, 662, 673, 728, 729, 746, 754, 777, 807.
sogdiana M. Pop. 786. soongarica Boiss. 516. stepposa Zoz 634, 637, 638.
- tshuiensis (Prokh.) Serg. 700—702.
— Turczaninowii Kar. et Kir. 781, 801, 814, 819.
— virgata W. ot K. 529, 658.
Euphrasia L. 522, 621.
— Regelii Wettst. 723.
Eurhynchium striatum (Hedw.) Schlmp. 204, 206.
— Swartsii (Turn.) Curn. 377.
Eurotia Adans. 731.
— ceratoides (L.) С. A. M. 536, 582, 651, 666, 682, 700, 702, 706, 731, 740, 742, 754. 755, 758, 759, 763—765, 787, 781, 794, 795, 815, 828, 847, 923, 925.
— Ewersmanniana Stschegl. 666, 682, 815, 923.
Eutrema Edwardeii R. Br. 102, 112, 846. Bxochorda Lindl. 468.
— Albertii Rgl. 474.
Fagus crenata Blume 370.
— orientalis Lipsky 378, 380, 384, 385, 412, 417, 418, 422, 423. 921.
— silvatica L. 352, 378, 380, 412, 414, 416, 460, 921.
— silvatica L. ssp. moesiaoa (Maly) Czecz. 415.
- taurica Popl. 378, 380, 412, 415, 416, 418, 712, 921.
Falcaria sioides (Wib.) Aschers. 594, 636, 637, 729.
Ferula L. 574, 587, 589, 602, 800, 808, 923.
_ 4„лЛ™78К8-Р..1. 587 , 800, 808.
Z SS“nUB. 6S5, 057, 560, 672.
~ jStoA Е. К»,. 757, 77«.
—. foliosa Lipsky 467.
__sisantea В. Fedtsch. 763.
- ShLna Vatke 474, 589, 590, 592,
— karatavica Rgl. et Schmalh. 721.
— nuda Spreng. 768.
— oopoda (Boiss. et Buhse) Boiss. 800, 808.
— orientalis L. 655, 657.
— ovina Boiss. 584, 589, 590, 722.
— schair Borszcz. 735.
— songorica Pall. 472, 652, 715.
— sp. sp. 587.
— Syreitschikovii K.-Pol. 768.
— tatarica FiSch. 678, 751, 758.
— tenuisecta E. Kor. 589.
Festuca L. 473, 595, 643.
— alaica Drob. 724.
— albifolia Reverd. 685, 687, 694, 695.
— altaica Trin. 102, 104, 112, 485.
— arenaria Osheck 505.
— Beckeri Hack. 507, 640, 662.
— Bornmuelleri (Hack.) V. Krecz. et Bobr. 582.
— brevifolia R. Br. 69, 100, 118,126,267.
— coelestis (St.-Yves) V. Krecz. et Bobr. 719, 720.
— cretacea Czern. 646.
— cryophila V. Krecz. et Bobr. 519.
— duriuscula L. 522.
— Ganeschinii Drob. 212, 484, 652, 705, 714, 716.
— gigantea (L.) Vill. 195, 197, 365, 376, 407.
— jacutica Drob. 259, 281, 282, 284, 286, 288, 522.
— jenisseensis Reverd. 625.
— Kryloviana Reverd. 98, 102, 708, 719, 720.
— lenensis Drob. 221, 240, 259, 273, 608, 630, 685, 688, 690, 691, 698.
— montana M. B. 204, 208, 384
408, 418.
— orientalis Kerner 516, 535, 747.
— ovina L. 445, 491, 522, 649.
— ovina s. 1. 494.
— picta Kit. 487, 488.
— pratensis Huds. 406, 446, 451
460, 495, 496, 523, 530, 531, 612,
— pseudo-sulcata Drob. 364, 439,
— pseudovina Hack. 531, 625, 628
649, 659, 666, 686, 690, 692—694’
~ 484- 485- 488, 489*
496, 504—506, 519, 522, 525.
— rubra s. 1. Hl, 520, 844.
- -Buj>recAt«(Boise.)V. Krecz.etBobr.490 Ж'»7,(4Й! ’41У‘П' *92’ l95’
— eulcata Hack’ 221, 228, 231, 245,
385,
454, 711. 448,
631,
246, 338, 357, 393, 396, 399, 400 4чч 471, 474, 496, 497, 511, 512 ’ 529’
528, 530, -531, 576, 577, 581’ кя?
589, 591—594, 599, 600, 612*
615—618, 621, 625, 634, 636, 63?
640, 643, 645, 647, 649, 651, 652 654 656—660, 662—664, 666, 668—670
672, 673, 676, 678, 681, 684, 685
703—707, 710, 711, 713—718, 725
729, 743, 746—751, 753, 757, 762*
779, 840, 887.
Festuca sulcata s. 1. 582, 632, 637 649 684, 690, 702, 704, 708, 714, 720-1
728, 764, 765, 845, 924.
— supina Schur 72, 94—97, 104, 105 117, 120, 136, 153, 181, 270, 300, 301’ 306, 482, 486—488, 490, 520.
— tristis Kryl. et Ivanitzk. 485.
— vallesiaca Schleich. 454, 690, 692 694.
— varia Haenke 492, 493.
Ficaria verna Huds. 384.
Ficus colchica Grossh. 431.
— hyrcana Grossh. 425, 427.
Filago arvensis L. 778, 784.
— spathulata Presl 777, 782.
Filipendula denudata (Presl) Fritsch 383. — hexapetala Gilib. 246, 337, 338, 394
452, 454, 528, 530, 600, 611, 613, 617’ 618, 621, 624, 634, 636, 637, 650, 704,’ 710, 712, 729.
— kamtschatica (Pall.) Maxim. 332, 333 502.
— palmata (Pall.) Maxim. 307, 525.
— stepposa Juz. 621, 650.
— ulmaria (L.) Maxim. 131, 168, 170 233, 330, 339, 343, 376, 388, 398, 429, 436,454,523,529,530, 621, 626,629.
Fissidens taxtfolius Hedw. 377.
Fragaria vesca L. 233, 404.
— viridis Duch. 399, 500, 611, 621, 625, 650, 690, 693, 704, 705, 707, 715, 718.
Frangula alnus Mill. 168, 177, 178, 192, 357, 373, 390, 423, 429, 432, 530.
Frankenia hirsuta L. 507, 510, 511, 623, 647, 755, 792.
— pulverulenta L. 588, 792.
Fraxinus coriaritfolia Scheele 371.
— excelsior L. 348, 354, 365, 371, 382, 384, 389, 391, 392, 396, 409, 414, 432, 433, 852.
— manshurica Rupr. 360, 364, 365, 371, 437, 460.
— ornus L. 371.
— oxycarpa Willd. 371, 400.
— potamophila Herd. 212, 469, 535.
— rhynchophylla Hance 222, 360, 371, 438.
Fritillaria kamtschatcensis (L.) Fisch. 502, 503.
— lutea Mill. 490.
Fu/nanaprocu/ndens (Dunal)Gr. et Godr.617. Fumaria Vaillantii Loisl. 535, 780.
Gagea Salisb. 375, 382, 411, 479, 635, 685, 735, 759, 768, 776.
— altaica Schischk. et Sumn. 690, 692. — anisanthos C. Koch 490.
Gagea bulbifera (Pall.) Roem. et Schult. 635, 637, 655, 657.
— divaricata Rgl. 785.
— dshungarica Rgl. 726.
— erubescens Bess. 612, 635, 636.
— gageoides (Zucc.) Vved. 726.
— glacialis C. Koch 490.
— pauciflora Turcz. 690, 692.
— provisa Pascher 690, 692.
— pseudoerubescens Pascher 707.
— pusilia (Schmidt) Roem. et Schult. 612, 635—637.
— reticulata (Pall.) Roem. et Schult. 587, 759, 767, 770, 780, 785.
— stipitata Merklin 587, 767, 780, 800.
— Szovitsii (Lang) Bess. 655, 657, 670, 671.
Gaillonia bucharica B. Fedtsch. et N. De-sjat. 583.
Galanthus nivalis L. 389.
Galatella fastigiata (Ldb.) Nees 621, 650, 652, 693.
— punctata s. 1. 343, 516, 621, 622, 693, 708, 845.
— rossica Novopokr. 510, 539.
Galega officinalis L. 492.
— orientalis Lam. 408.
Galeobdolon luteum Huds. 174, 175, 204, 337, 354, 357, 375, 391, 413, 430.
Galium L. 232.
— boreale L. 289, 339, 340, 457, 499, 500, 520, 523, 621, 625, 627, 630, 704, 718.
kopetdaghnse Pobed. 727.
— Krylovii Iljin 196, 312.
— mollugo L. 247.
— palustre L. 525, 549, 550.
— rubioides L. 390, 523.
— ruthenicum Willd. 596, 636, 637, 657, 660, 668, 678, 749.
— Schultesii Vest 204, 205.
— trifidum L. 503.
— uliginosum L. 289, 549.
— vernum Scop. 381.
— verum L. 286, 311, 312, 394, 395, 452, 454, 474, 483, 522, 530, 594, 609, 611, 613, 617, 621, 625, 626, 628—631, 690, 693, 694, 698, 704—706, 710—714, 728.
— verum s. 1. 650, 652, 716, 718.
Gamanthus Bge. 736, 767, 793.
— gamocarpus (Moq.) Bge. 760, 773, 778, 793,813, 814, 817.
— pilosus (Pall.) Bge. 777.
Garhadiolus papposus Boiss. et Buhse 779, 786.
Genista albida Willd. 645.
— angustifolia Schischk. 403.
— scythica Paez. 645.
— tanaitica P. Smirn. 646.
— tinctoria L. 228, 357, 388.
Gentiana L. 479.
— algida Pall. 102—105, 108, 112, 485.
— asclepiadea L. 489.
— barbata Froel. 238, 629.
— Dechyana Somm. et Lev. 478.
— deoumbens L. 609, 687.
— djimilensis G. Koch 836.
Gentiana falcata Turcz. 494.
— glauca Pall. Ill, 113.
— grandiflora Laxm. 102, 104, 485, 486.
— nipponica Maxim. 113.
— Olgae Rgl. et Schmalh. 495.
— Olivieri Griseb. 590.
— punctata L. 488.
— tenella Rottb. 111.
— tianschanica Rupr. 708.
— turkestanorum Gand. 211.
Geranium albiflorum Ldb. 96, 108, 191,
194, 196, 300, 312, 325, 482, 483,
485, 486, 715.
— bifolium Patrin 620.
— collinum Steph. 211, 499, 707, 708,
723, 847.
— collinum. s. 1. 723.
— erianthum DC. Ill, 502, 520.
— eriostemon Fisch. 438.
— gymnocaulon DC. 491.
— jacutense Drob. 281.
— Kotschyi Boiss. 786.
— linearilobum DC. 597.
— Maximowiczii Rgl. et Ma ack 435.
— pratense L. 523, o29.
— pseudosibiricum J. Меуёг 188, 238, 279, 312, 395, 626.
— Robertianum L. 195, 204, 207, 245.
— Ruprechtii Woron. 325, 492.
— sanguineum L. 404, 498.
— saxatile Kar. et Kir. 584, 708, 709, 719, 724, 726, 845.
— silvaticum L. 93,167,168, 329, 330, 519.
— tuberosum L. 777.
— Vlassovianum Fisch. 294, 436.
Geum aleppicum Jacq. 502.
— macrophyllum Willd. 502.
— rivale L. 491, 523.
— urbanum L. 471.
Girgensohnia minima E. Koi*. 785.
— oppositiflora (Pall.) Fenzl 759, 760, 763, 771, 781, 785, 786.
Glaucium Adans. 735.
Glaux maritima L. 510, 629.
Glechoma heder acea L. 354, 390, 393.
— hirsuta W. et K. 413, 414.
Gleditschia caspia Desf. 427.
— triacanthos L. 852.
Glyceria alnasteretum Kom. 159.
— aquatica (L.) Wahlb. 287, 430, 525, 526, 530, 538, 548, 549, 616.
— fluitans (L.) R. Br. 525, 526, 548, 616, 647.
Glycyrrhiza echinata L. 531, 540.
— glabra L. 474, 513, 531, 534, 539, 591, 594, 746, 747, 756, 782, 793, 858.
— uralensis Fisch. 515, 516, 622,
679.
Gnaphalium norvegicum Gunn. 95.
— silvaticum L. 197.
— sup i num L. 123, 491, 723—725.
Goidbachia laevigata (M. B.) DC. 768, 780.
Goniolimon Besserianum (Schult.) Kusn. 657.
— graminifolium (Ait.} Boiss. 641.
— rubellum (S. G. Gmel.) Klok. 669.
— orae-syvashicae Klok. 669.
— speciosum (LJ Boiss. 628, 652, 690,
691, 694, 697, 698, 702, 706, 714, 807. Goniolimon tataricum (L.) Boiss. 638, 655, 657.
Goodycra repens (L.) R. Br. 147, 208, 245. Graellsia sax ifragifolia (DC.) Boiss. 728. Grimm ia Hedw. 736.
— ovalis (Hedw.) Lindb. 102.
Grossularia acicular is (Smith) Spach 687, 706.
Gymnadenia conopsea (L.) R. Br. 498. Gypsophila altissima L. 232, 483, 621, 704. — aretioides Boiss. 577, 582, 727.
— bicolor Freyn et Sint. 727.
— desertorum (Bge.) Fenzl 700—702.
— Gmelinii Bge. 686.
— imbricata Rupr. 479, 490.
— microphylla (Schrenk) Fensl 843.
— muralis L. 597.
— paniculata L. 232, 636, 652, 697. — PatriniiSer. 675, 680, 686,694, 695,840. — stepposa Klok. 597, 668.
— tenuifolia M. B. 490.
— trichotoma Wend. 516, 539.
Gyrophora (Ach.) Koerh. 78, 843.
Halerpestes ruthenica (Jacq.) Ovcz. 103, 629, 699.
— salsuginosa (Pall.) Green 103, 535. Halimocnemis С. A. M. 736, 767, 768, 832.
— Karelinii Moq. 760, 770, 773, 786, 793 833.
— longifolia Bee. 811, 813, 814, 819. — mollissima Bge. 760, 785, 793, 833. — sclerosperma (Pall.) С. A. M. 759, 768,-770, 773, 774, 792, 793.
— villosa Kar. et Kir. 760, 811.
Halimodendron halodendron (Pall.) Voes. 515, 534, 535, 742, 807, 858.
Halocharis Moq. 736, 832.
— hispida (С. A. M.) Bge. 760, 762, 773, 778, 785, 792, 833.
Halocnemum M. B. 513, 731.
— strobilaceum (Pall.) M. B. 507, 511, 512, 514, 515, 534 , 539, 540, 664 , 669, 670, 731, 732, 734, 742, 755, 769, 777, 787, 788, 791—794, 814, 925.
Haloeeton С. A. M. 736.
— elomeratus (M. В.) С. A. M. 770, 792. Halostachys Belangeriana (Moq.) Botsch.
534, 542, 731, 742, 788, 791—
794, 814. ’
- caspica (Pall.) С. A. M. 731, 788, 925.
Halotis Bge. 736.
- pilosa (Moq.) Iljin 760, 774, 781, 818.
Haloxylon Bge. 797, 799.
— ammodendron (С. A. M.) Bge. 774, 797, 800, 808, 809, 810, 892, 925.
— aphyllum (Minkw.) Iljin 733, 774, Tel, 785 , 797, 800, 801, 809, 810, 812-814, 823, 833, 889, 925.
— persicum Bge. 733, 774, 799, 800, 801, 804, 812, 813, 815—820, 823, 925.
Hammada Iljin (A rthrophytum Schrenk) 731.
Hantsschia amphioxys (Ehr.) Grun. 737.
Haplophyllum A. Juss. 734.
— acutifollum (DC.) G. Don 761.
— Bungei Trautv. 774,800,812,817, 818.
Haplophyllum dauricum (L.) G. Don 69f) 691, 698.
— latifolium Kar. et Kir. 592.
— obtusifoilum Ldb. 774, 786, 787, 818
— pedicellatum Bge. 800.
Harimanella (Cassiope) hypno ides (L.) Cov. 78, 94, 95, 98.
— (Cassiope) Stelleriana (Pall.) Cov. Ho. Hedera caucasigena Pojark. 375.
— colchica C. Koch 374, 375, 407, 420 423, 432.
— helix L. 247, 353, 374, 382, 411, 417 433.
— Pastuchovii Woron. 375, 408, 421, 426 433.
— taurica Carr. 375.
Hedysarum alpinum L. 117.
— arcticum B. Fedtsch. 519.
— caucasicum M. B. 490.
— cephalotes French. 795, 847.
— cretaceum Fisch. 646.
— ferganense Korsh. 714.
— Gmelinii Ldb. 233, 692, 693.
— grandiflorum Pall. 645.
— neglectum Ldb. 485, 627.
— Rasoumovianum Fisch, et Helm 651.
— Semenovii Rgl. et Herd. 846.
— setigerum Turcz. 629.
— songoricum Bone. 708.
— ucrainicum B. Kasehm. 646.
Heleocharis R. Br. 616.
— argyrolepis Kjerulff 513.
— crassa F. et M. 513.
— eupalustris Lindb. f. (H. palustris var. eupalustris Syr.) 512, 529, 543.
— euuniglumis Zinserl. (H. uniglumis auct.) 286, 513, 609.
— oxylepis (Meinsh.) B. Fedtsch. 512, 513. — palustris s. 1. 514, 515, 658.
Helianthemum nummularium (L.) Mill. 712.
— orientale (Grosser) Juz. et Pozd. 712. — salicifolium (L.) Mill. 777, 780, 783. — soongoricum Schrenk 681, 725.
Heliantnus tuberosus L. 854.
Helichrysum arenarium (L.) Moench 221, 239, 357, 358, 507, 613, 617, 640, 644, 680, 755.
— cretaceum P. Smirn, 646.
— plica turn DC. 708.
— rubicundum (C. Koch) D. Sosn. 777.
Helictotrichon asiaticus (Roshev.) Grossh. 105, 484, 493, 705, 707, 718, 720, 725.
— desertorum (Less.) Pilger 103, 494, 599, 612, 613, 615, 621, 622, 625, 649, 651, 652, 687, 690, 692, 693, 702, 704, 709, 713, 719, 720.
— Krylovii N. Pavl. 609.
— mongolicum (Roshev.) Henrard 108.
— pratensis (L.) Bess. 497.
— pubescent (Huds.) Pilger 211, 358, 485, 499, 500, 612.
— Schellianum (Hack.) Kitagava 338, 358, 447, 483, 494, 612. 613, 621, 625, 630, 634, 637, 693, 695, 703, 704, 711, 712.
tianschanicum (Roshev.) Henrard 716, 718—720.
Helictotrichon versicolor (Vill.) Klok. 487» 488.
Heliotropium arguzioides Kar. et Kir. 755, 800, 804, 817, 818, 821, 823.
— dasycarpum Ldb. 755.
Hemerocallis flava L. 446, 524, 621, 627, 629.
— minor Mill. 630, 631, 683.
Hemilasia acrolasia (Bge). C. B. Clarke 781. Hepatica nob i I is Garsault 174, 354, 365, 414. Heracleum dissectum Ldb. 194, 447, 496, 499, 626, 627.
— dulce Fisch. Ill, 332, 502.
— Mantegazzianum Somm. et Lev. 492.
— Olgae Rgl. et Schmalh. 847.
— pubescens M. B. 492.
— sibiricum L. 233, 519, 523.
Herniaria hirsute L. 760, 782.
Heteracia Szowitsii F. et M. 767, 779.
Heterocaryum Szovitsianum (F. et M.) DC. 773.
Heteroderis pusilia Boiss. 773, 779, 781.
Heteropappus altaicus (Willd.) Novopokr. 516, 628, 629, 688, 690, 692—694, 702.
— canescens (N. ah E.) Novopokr. 591. Hibiscus cannabinus L. 853.
— trionum L. 858.
Hieracium alpinum L. 487, 488.
— echioides Lumn. 650.
— Krylovii Nevski 192.
— pocuticum Wol. 205.
— rotundum Kit. 203, 414.
— suberectum Schischk. et Steinb. 325, 482.
— transsilvanicum Heuff. 203.
— triste Willd. 502.
— umbellatum L. 404, 523, 621.
Hierochloe alpina (Lilj.) Roem. et Schult. 97, 100, 103, 105, 107, 108, 121, 158, 215, 300, 309, 502 519.
— Bungeana Trin. 695.
— odorata (L.) Wahlb. 503, 525, 538, 639.
— pauciflora R. Br. 68, 99, 114, 546.
Hippophae rhamnoides L. 247, 534, 535, 847, 852.
Hippuris tetraphylla L. f. 504, 506.
— vulgaris L. 515, 521.
Holcus lanatus L. 454, 498.
Holoschoenus vulgaris Link 508, 640.
Holosteum glutinosum (M. B.) F. et M. 780.
— umbellatum L. 635, 636, 655, 670, 778.
Homalia (Brid.) Br. et Sch. 411.
— trichomanoides (Hedw.) Br. et Sch. 377.
Homalothecium Philippeanum (Spruce) Br. eur 211.
Homogyne8д^Па Cass. 2^4, 414,
Horaninowia anomala (С. A. M.) Moq. 786, 811, 813, 814, 818, 819.
— ulicina F. et M. 770, 801, 811—813, 816, 817, 819, 823.
Hordeum brevisubulatum (Trin.) Link 286, 510, 511, 515, 524, 531, 532, 623, 697, 699.
— bulbosum L. 469, 474, 589, 590, 591, 592, 763, 923.
Hordeum geniculatum All. 588, 782.
— leporinum Link 508, 588, 671, 760, 777, 782, 832.
— turkestanicum Nevski 484, 584, 702, 705, 720, 722, 794, 796.
— violaceum Boiss. et Huet. 711.
Hulthemia berberifolia (Pall.) Dumort. 721, 778.
— persica (Michx.) Bornm. 760.
Humulus lupulus L. 374, 423, 429, 432.
Hyacinthella Schur 597.
— leucophaea (C. Koch) Schur 612, 620, 635, 637.
Hyacinthus Litwinowii E. Czerniak. 726.
— transcaspicus Litw. 727.
Hydrangea petiolaris Sieb. et Zucc. 214. Hydrochar is morsus-ranae L. 514, 537, 548. Hylocomium alascanum (Lesq. et James) Kindb. 80, 81.
— proliferum (L.) Lindb. (Я. splendens (Hedw). Br. et Sch.] 69, 93, 96, 97, 103, 116—120, 125, 128, 131, 136, 137, 150, 153, 161, 162, 166, 168, 180, 181, 184, 186, 191, 192, 195, 200, 205, 208, 214, 219, 225, 227, 229, 233, 236, 241, 266, 268—270, 274, 280, 290, 296, 300, 302, 303, 305, 307, 310, 312, 329, 330, 354, 357, 429, 488.
— pyrenaicum (Spruce) Lindb. 310.
Hylomecon vernalis Maxim. 362.
Hymenolobus procumbens (L.) Nutt. 670, 846.
Hymenophysa pubescens С. A. M. 516. Hypecoum L. 735, 763.
— parviflorum Kar. et Kir. 587, 762.
— pendulum L. 773, 780.
— trilobum Trautv. 786.
Hypericum alpigenum Kit. 488, 489.
— androsaemum L. 426.
— bupleuroides Griseb. 420.
— elongatum Ldb. 721.
— inodorum Willd. 406.
— perforatum L. 192, 246, 403, 591» 592 668, 705, 707, 708, 714.
— polygonifolium Rupr. 711.
— scabrum L. 577, 584, 591, 592,1706, 727, 846.
Hypnum arcuatum Lindb. 310.
— В amber ger i Schimp. 126.
— cupressiforme Hedw. 206, 384.
Hyssopus L. 643.
— cretaceus Dub. 646.
— seravschanicus (Dub.) Pazij 846.
Ilex colchica Pojark. 208, 209, 420, 423, 431, 432.
— hyrcana Pojark. 409, 410, 421, 426.
— rugosa Fr. Schmidt 214.
— stenocarpa Pojark. 208.
Impatiens noli-tangere L. 168, 382, 429.
— parviflora DC. 469, 535, 842.
Imperata cylindrica (L.) P. B. 513, 534.
Inula L. 481, 596.
— britannica L. 510, 514, 630, 658.
— caspia Blume 543.
— ensifolia L. 247, 617, 618, 712.
— glandulcsa Willd. 493.
Inula grandis Schrenk 574, 587, 589,
590—592, 594, 708, 721, 923.
— helenium L. 499.
— hirta L. 343, 613, 620, 704, 713.
— oculus-Christi L. 617, 727.
— rhizocephala Schrenk 494, 495, 708.
Ipomoea batatas (L.) Lam. 854.
Iris L. 735, 807, 823.
— aphylla L. 612, 620.
— biglumis Vahl. 626, 627, 691, 696, 699.
— caucasica Hoffm. 784.
— flavissima Pall. 688, 692, 694.
— kopetdagensis Vved. 786.
— longiscapa Ldb. 786.
— pseudacorus L. 429, 430, 432, 566.
— pumila L. 596, 636, 637, 656, 657, 668, 671.
— ruthenica Ker.-Gawl. 221, 232, 236, 279, 280, 313, 340, 446, 621, 625, 626, 631, 687, 705.
— setosa Pall. Ill, 503.
— sibirica L. 289, 296.
— sogdiana Bge. 535.
— songarica Schrenk 774, 800.
— taurica Loddiges 497.
— tenuifolia Pali. 781.
— tianschanica (Maxim.) Vved. 703.
— uniflora Pall. 315, 438, 439.
I satis emarginata Kar. etKir. 786, 815, 819.
— trachycarpa Trautv. 773.
Isopyrum thaliqtroides L. 382, 417.
Isottyecium myurum (Brid.) В rid. 414.
Ixiolirion montanum (La Bill.) Herb. 727.
— tataricum (Pall.) Roem. et Schult. 679, 707, 759, 760, 778, 779, 786, 846.
Jasminium fruticans L. 404, 576.
Jeffersonia dubia (Maxim.) Benth. et Hook 362, 375.
Juglans L. 470.
— fallax Dode 467.
— manshurica Maxim. 360, 364, 437, 460, 468.
— regia L. 212, 467, 468, 575.
— regia s. 1. 852.
Juncus acutus L. 507.
— beringensis Buch. Ill, 502, 503,
— compressus Jacq. 535.
— effusus L. 430, 431.
— Gerardii Lois. 506, 510, 512—
514, 529, 539, 616, 647, 857.
— Haenkei E. May. 503.
— lampocarpus Ehrh. 857.
— litoralis С. A. M. 507.
— maritimus Lam. 508.
— trifidus L. 78, 98, 487, 488.
— triglumis L. 485.
Juniperus L. 575, 852.
— communis L. 190, 224, 227, 233, 235, 245, 300, 301, 316, 339.
— depressa Stev. 245, 712.
— excetsa M. B. 247, 399, 400, 460, 575, 576.
— foetidissima Willd. 405, 575, 576.
— oblonga M. B. 245, 246.
— oxycedrus L. 246, 247, 400, 575.
— polycarpos C. Koch 575, 576.
Juniperus pseudosabina F. et M. 198, 316, 485, 494, 708, 846.
— sabina L. 646, 658, 664, 712, 841, 845.
— semiglobosa Rgl. 576, 578, 592, 845.
— seravschanica Kom. 576, 578, 709, 722.
— sibirica Burgsd. 94, 163, 205, 239, 272, 279, 310, 329, 333, 485, 489, 577.
— turcomanica B. Fedtsch. 577, 727, 728.
— turkestanica Kom. 212, 494, 576—
578, 709, 720, 723, 843—845.
Jurinea Gass. 726.
— Antonowii G. Winkl. 781.
— arachnoidea Bge. 636, 637.
— cyanoides (L.) Rchb. 357.
— derderioides C. Winkl. 780, 818.
— hissarica Iljin 724.
— laxa (Fisch.) Korsh. 641.
— multiflora (L.) B. Fedtsch. 596, 635— —637, 650, 655, 657, 660, 669, 672, 678, 713, 749, 751, 758.
— Olgae Rgl. et Schmalh. 725.
— persimilis Iljin 774.
— polyclones DC. 662.
— stoechadifolia (M. B.) DG. 617, 645.
— suffruticosa Rgl. 721.
Kalidium Moq. 540, 731, 744.
— caspicum (L.) Ung.-Sternb. 507, 540, 787, 791—793, 814.
— foliatum (Pall.) Moq. 515, 534, 651, 664, 696, 697, 731, 742, 755, 769. 777, 788, 791, 814, 925.
Kalonymus (Euonymus) latifolia (Mill.) Prokh. 384, 409.
— (Euonymus) leiophloea (Stev.) Prokh. 373.
— (Euonymus) macroptera (Rupr.) Prokh. 213, 374.
— (Euonymus) sachalinensis (F. Schmidt) Prokh. 214, 334.
Kalopanax septemlobum (Thunb.) Koidz. 214, 360, 362, 438.
Karelinia caspica (Pall.) Less. 534, 543, 742, 793, 794, 807, 857.
Kerria japonica (L.) DC. 852.
Kirilowia eriantha Bge. 771, 781, 811.
Knautia arvensis (L.) Coult. 613.
— tatarica (L.) Litw. 356, 397.
Kochia Roth. 600, 736.
— iranica (Hausskn. et Bomm.) Litw. 801.
— laniflora (S. G. Gmel.) Borb. 641, 755.
— prostrata (L.) Schrad. 103, 516, 536, 579, 593, 594, 628, 635, 653, 656,
660, 663, 665, 667—670, 672, 673,
678, 680, 688, 690, 694, 696, 697,
699, 701, 717, 725, 731, 740, 747— 751, 754, 755, 757—759, 764, 767,
770, 771, 776, 777, 779, 784.
— Schrenkiana (Moq.) Iljin 759, 811, 816.
— scoparia (L.) Schrad. 857.
Koeleria Pere. 595, 643, 783.
— altaica (Dornin) Kryl. 104, 685, 687.
Koeleria borysthenica Klok. 640.
— brevis Stev. 617, 645.
— caucasica Dom. 493, 711, 729.
— Delavignei Czern. 452, 454, 522, 528, 530.
— glauca (Schruhr) DC. 228, 357, 640.
— gracilis Pers. 221, 232, 286, 338, 358, 393, 439, 457,471, 493, 496, 511, 512, 522, 525, 531, 576, 577, 581,
589, 608, 610, 612, 617, 618, 621,
622, 625, 630, 631, 634, 636, 637,
639, 640, 643, 652, 656, 657, 660,
662, 670, 683—688, 690, 692, 693, 695, 697, 698, 700, 704—706, 708,
7Ю, 711, 714, 716, 722, 724, 729, 748, 749, 779, 840, 924.
— Luerssenii Dom. 245.
— moldavica M. Alex. 645.
— sabuletorum Czern. 640, 622.
— sclerophylla P. Smirn. 663.
— seminuda (Trautv.) Dom. 67, 221, 240. '
— Talievii Lavr. 646.
— Thonii Dom. 627.
Koelpinia Pall. 735.
— linearis Pall. 767, 773, 776, 778—781, 783—786, 793, 812, 815, 817, 881. macrantha C. Winkl. 801.
Kohlrauschia prolifera (L.) Kunth. 410, 644.
Koelreuteria paniculata Laxm. 852.
Korovinia tenuisecta (Rgl. et Schmalh.) Nevski et Vved. 760.
Kummer о via striata (Thunb.) Schindl. 457.
Lachnoloma Lehmannii Bge. 786.
Lactuca muralis (L.) Fresen. 204, 406, 413.
— sibirica (L.) Benth. 188, 288, 293.
—- tatarica (L.) С. A. M. 516.
Ladyginia bucharica Lipsky 583.
Lagochilus cabulicus Benth. 761.
— diacanthophyllus (Pall.) Benth. 496, 717.
— ilicifolius Bge. 701.
— tianschanicus Pavl. 584.
Lagotis altaica (Willd.) P. Smirn. [Z. integ-rifolia (Willd.) Shischk.] 102.
— decumbens Rupr. 846.
— Gmelinii (Cham, et Schletd.) Rupr [L. glauca Gaertn.] 111.
— Korolkowii (Rgl. et Schmalh.) Maxim. 845.
— minor (Willd.) Standi. 97, 118, 123, 300, 505, 520.
Lallemantia F. et M. 735.
— Royleana Benth. 773, 785, 786, 811.
Lamium album L. 499.
— barbatum Sib. et Zucc. 375.
— Paczoskianum Vorosch. 670.
Lamprothamnium papulosum Groves (Z. alopecuroides A. Br.) 542.
Lappula Gilib. 597, 671, 735.
— microcarpa (Ldb.) Giirke 721.
— patula (Lehm.) Aschers. 768.
— semiglabra (Lab.) Giircke 755, 801, 814.
Lappula spinocarpos (Forsk.) Aschers. 767—770, 773.
Lapsana communis L. 382, 407.
Larix Mill. 912.
— Cajanderi Mayr 302.
— Czekanowskii Szafer 250.
— dahurica Turcz. 70, 90, 106, 130, 201, 221, 234, 239, 241, 242, 250, 265, 267, 271, 276, 281, 290, 299, 301,437, 609, 919, 920.
— decidua Mill. ssp. carpatica Dom. 251.
— Komarovii B. Kolesn. 251.
— kurilensis Mayr 251.
— laricina C. Koch 250.
— Lubarskii Sukacz. 251, 437, 909.
— maritima Sukacz. 251.
— Middendorffii B. Kolesn. 251.
— ochotensis B. Kolesn. 251.
— olgensis A. Henry 250, 437, 909.
— polonica Racib. 251.
— sibirica Ldb. 90, 96, 106, 154, 193, 220, 221, 234, 235, 242» 250, 265, 266, 269, 278, 299, 348, 496, 548, 623, 685, 884, 919, 920.
— Sukaczewii Djil. 154, 164, 221, 225, 229, 238, 251, 265, 299, 920.
— trilobum (L.) Borkh. 393, 408.
Laser trilobum (L) Borkh. 393, 408.
Lasiagrostis caragana Trin. et Rtipr. 582, 681,734,759,760,762, 763, 770,771,807.
— splendes (Trin.) Kunth 515, 516, 531, 534, 535, 598, 627—629, 664, 675, 680, 691, 694, 696, 697, 699, 753, 865.
Lathyrus alatus (Maxim.) Kom. 435.
— Gmelinii (Fisch.) Fritsch 196, 238, 312, 344, 446, 496, 705.
— humilis Fisch. 229, 236, 239, 242, 280, 281, 284, 313.
— lacteus (M. B.) Wissjul. 612.
— laxiflorus (Desf.) Ktze. 247,406,407,410.
— niger (L.) Bernh. 382.
— pallescens (M. В.) C. Koch. 620, 636, 637.
— pannonicus (Kramer) Garcke 612, 636.
— pisiformis L. 229, 232, 621, 705.
— pratensis L. 232, 450, 497, 499, 523, 538, 621, 705, 707.
— quinquinervis (Miq.) Litw. 297.
— roseus Stev. 407, 408.
— tuberosus L. 499.
— vernus (L.) Bernh. 170, 229, 232, 344, 354, 395, 397, 446.
— versicolor (Gmel.) Beck. 612.
Laurocerasus officinalis Roem. 208, 419, 420, 423, 432.
Laurus nobilis L. 424.
Lavatera thuringiaca L. 396, 499, 624, 705, 707.
Lecanora Ach. 78.
— aspera (Mer.) Tom. 759, 770, 787.
— epibryon Ach. 81.
— frustulosa (Dicks.) Ach. 770.
Lecidea (Ach.) Mass, et Th. Fr. 78, 736, 843.
Ledum decumbens (Ait.) Small 69, 97, 107, 108, 115, 125, 129, 138, 158, 309, 310, 556.
— dilatatum Wahlb. 292—294.
Ledum hypoleucum Kom. 292, 294.
— palustre L. 134, 153, 159—162, 184, 198, 219—221, 224, 225, 227, 230,
231, 236, 241, 266, 267, 269, 284,
289, 294, 300, 302, 305, 314, 330.
343, 502, 503, 556, 561, 571.
Leersia oryzoides (L.) Swartz. 431, 538,
450, 550.
Lemna minor L. 537.
— trisulca L. 514, 515, 537, 548.
Leontice altaica Pall. 644.
— Ewersmannii Bge. 592, 800.
— odessana Fisch. 644.
Leontodon autumn a Us L. 616.
— croceus Haenke 487.
— hispidus L. 445, 498, 710.
Leontopodium alpinum Cass. 487.
— campestre (Lab.) Hand.-Mazz. 630, 688, 714, 716, 844.
— conglobatum Turcz. 683.
— leontopodinum (DC.) Hand-Mazz. 687, 714, 717, 719, 720, 846.
— ochroleucum Beauv. 702, 719, 720, 827, 837, 843, 844, 846.
— sibiricum Cass. 286.
Leonurus turkestanicus V. Krecz. et Kupr. 721.
Lepidium L. 735.
— borysthenicum Kip. 647.
— latifolium L. 516.
• л- Meyeri Claus 646, 675.
— perfoliatum L. 588, 748, 750, 754, 759, 767—769, 777, 785, 793, 803.
— ruderale L. 588.
— vesicarium L. 778, 784.
Lepidolopha Komarovii C. Winkl. 725. Leptaleum DC. 735, 762 , 823.
- filifolium (Willd.) DC. 758, 759, 767— 771, 773, 780, 785, 786, 811, 814, 817, 821.
Leptobryum pyriforme (Hedw.) Wils. 211.
Lespedeza bicolor Tuzcz. 341, 363, 437, 456—458.
Leucanthemum arcticum (L.) DC. 504, 505. — sibiricum DC. 315, 505, 627, 629, 630. — sibiricum DC. ssp. alaunicum K.-Pol.
615.
— sibiricum DC. ssp. Kozo-Poljanskii Golitc. 615.
— vulgare Lam. 445, 446, 460, 522, 611, 613.
Leucopoa albida (Turcz.) V. Krecz. 687. — karatavica (Bge.) V. Krecz. et Bobr. 722. — Olgae (Rgl.) V. Krecz. et Bobr. 720, 843.
Libanotis buchtormensis (Fisch.) DC. 496, 840.
— intermedia Rupr. 523, 612, 625, 636, 637, 650.
— monstrosa (Willd.) DC. 102.
— Schrenkiana С. A. M. 499.
- sibirica (L.) С. A. M. 233, 395, 454, 713.
Ligularia alpigena Pojark. 584.
— altaica DC. 485, 494, 496, 723, 724, 726.
— glauca (L.) 0. Hoffm. 714.
— macrophylla (Ldb.) DC. 499.
Ligularia persica Boiss. 470, 499, 584, 707, 718.
— robusta (Ldb.) DC. 720, 843.
— sibirica (L.) Cass. 215, 310.
— Thomsonii (Clarke.) Pojark. 584, 589, 709, 716, 718.
Ligusticum mute Ilina (L.) Crantz 487, 488.
— physospermifolium Alb. 492.
Ligustrina amurensis Rupr. 360.
Ligustrum vulgare L. 247, 384, 392, 433, 729.
Lilium avenaceum Fisch. 111.
— dahuricum Ker.-Gawl. 524.
— martagon L. 193, 316, 485, 496, 524.
— tenuifolium Fisch. 627, 628, 630.
Limnas Stelleri Trin. 259, 261, 281, 282, 305, 307.
Limnorchis convallariaefolius (Fisch, ex Lindb.) Rydb. 503.
Limonium Mill. (Statice L.) 512, 596, 734.
— aureum (L.) Hill 699.
— Bungei (Claus) Gamajun. 655, 657.
— caspium (Willd.) Gams 508, 510—512, 514, 539, 647, 792.
— chrysocomum (Kar. et. Kir.) Ktze. 680.
— congestum (Lab.) Ktze. 103.
— cor allo ides (Tausch) Lincz. 511, 792. - czurjukiense (Klok.) Lavr. 669, 671.
— Gmelinii (Willd.) Ktze. 510, 512— 516, 534, 539, 540, 622, 623, 628, 696, 697, 792.
— latifolium (Sm.) Ktze. 634, 636, 637.
— macrorrhizon (Ldb.) Ktze. 696.
— Meyeri (Boiss.) Ktze. 508, 513, 579, 668, 782.
— otolepis (Schrenk) Ktze. 534, 543, 857.
— sareptanum (Becker) Gams 655, 657, 672.
— suffruticosum (L.) Ktze. 511, 514, 536, 664, 731, 734, 755, 757, 769, 791, 792, 814.
— tomentellum (Boiss.) Ktze. 528, 616, 647.
Limosella aquatica L. 514.
Linaria Bungei Kupr. 759.
— cretacea Fisch. 646, 675.
— debilis Kupr. 713.
— grandiflora Desf. 729.
Lindelofia stylosa (Kar. et Kir.) Brand 765, 826, 847.
Linnaea borealis L. 158, 166, 167, 169, 178, 180, 182, 186, 190, 191, 196,
198, 200, 208, 214, 230, 236, 245,
260, 269, 279, 282, 305, 310, 316,338, 354, 358, 429.
Linosyris tatarica (Less.) С. A. M. 672, 678, 751, 757.
— villosa (L.) DC. 594, 595, 635, 638, 651, 657, 660, 670, 713, 749, 750, 757.
Linum altaicum Ldb. 485.
— austriacum L. 617.
— brevisepalum Juz. 686.
— corymbulosum Rchb. 594.
— hypericifolium Salish. 493.
— Komarovii Juz. 522.
— perenne s. 1. 522.
Liriodendron tulip if erum L. 852.
Listera cordata (L.) R. Br. 147, 208.
— major Nakai 215.
Lithospermum purpureo-coeruleum L.
381, 382, 399, 408.
Litvinovia tenuissima (Pall.) N. Busch 587, 767.
Lloydia serotina (L.) Rchb. 82, 97, 98, 103, 111, 112, 846, 847.
Lobaria Schreb. 163.
— pulmonaria Hoffm. 169.
Loiseleuria procumbens (L.) Desv. 67, 69, 94, 110, 125, 136, 160, 309, 502.
Lolium rigidum Gaud. 588.
Londesia eriantha F. et M. 773, 811,821. Lonicera L. 468, 709.
— altaica Pall. 196—198, 235, 301, 311— 313, 316, 338, 343, 344, 447.
— A Itmann ii Rgl. et Schmalh. 211, 212, 499, 723, 845.
— caucasica Pall. 245.
— Chamissoi Bge. 333, 334.
— chrysantha Turcz. 341, 360.
— coerulea L. 245, 499.
— edulis Turcz. 241, 307, 333.
— glauca Hook, et Thoms. 845.
— heterophylla Decaisne 499, 845.
— hispida Pall. 211, 212, 843.
— humilis Kar. et Kir. 842—844, 845.
— iberica M. B. 576.
— Karelinii Bge. 211, 212, 469, 843.
— Korolkovii Sapf. 469, 499.
— Maximoviczii Rupr. 215, 360, 437.
— micro phy Ila WilldL 664, 706, 707,841.
— nigra L. 204, 413.
— Olgae Rgl. et Schmalh. 843. 846.
— persica Jaub. et Spach 469, 721,726, 728.
— sachalinensis (Fr. Schm.) E. Wolf. 334.
— Semenovii Rgl. 847.
— serawschanica Pojark. 846.
— stenantha Pojark. 707.
— tatarica L. 336, 343, 394, 471, 652, 664, 704, 706.
— xylosteum L. 167, 168, 177, 178, 184, 186, 192, 206, 245, 354, 355, 373, 391, 394, 397.
Lophochloa phleoides (Vill.) Rchb. 588, 780.
Lophozia alpestris (Schleich.) Evans 488 — Binsteadii (Kaalaas) Evans 126.
— lycopodioides (Wallr.) Gogniaux. 131, 310.
Lotus caucasicus Kupr. 497, 710, 711.
— corniculatus L. 450, 451, 523, 616.
Luzula arcuata Wahlb. 78.
— confusa Lindb. 77, 81, 82, 97,
118, 121, 126, 520.
— Forsteri (Sm.) DC. 426, 427.
— kamtschadalorum (Sam.) Gorodk. 111.
— Kfellmanniana Miyabe et Kudo 503.
— nemorosa E. Mey. 414, 488, 489.
—'nivalis Laest. 70, 81, 82, 114, 115.
— parviflora (Ehrh.) Desv. 201, 316, 502.
— pilosa (L.) Willd. 167, 184, 219, 236, 280, 354, 384.
— silvatica (Huds.)Gaud.205,208,414,489.
— spicata (L.) DC. 485, 844.
Luzula sudetica (Willd.) DC. 488.
Lychnis fulgens Fisch. 457.
— sibirica L. 522.
Lycium turcomanicum Turcz. 535, 742, 807.
Lycopodium anceps Wallr. 184, 241, 259, 269 293
— annotinum L. 147, 153, 158, 167, 180, 184, 190, 195, 200, 208, 214, 338, 339, 415.
— appressum (Desv.) V. Petr. 78.
— chinense H. Christ 215.
— c lava turn L. 184, 195.
— pungens La Pylaie 329.
Lycopus europaeus L. 431, 541, 542.
— exaltatus L. f. 376.
Lyngbia Margaretheana G. Schmid 737.
Lysimachia d ah uric a Ldb. 436. — vulgaris L. 376, 529, 548. Lythrum salicaria L. 431, 529, 538, 542, 548.
— virgatum L. 515, 529, 538.
MaacKia amurensis Rupr. et Maxim. 295, 360, 436.
Macrochloris Korschik. 832. Macrosporium Fr. 737, 831. Macrotomia euchroma (Royle) Pauls. 794, 826, 847.
— ugamensis M. Pop. 845
Magnolia obovata Thunb. 852.
Majanthemum bi folium (L.) F. Schmidt 147, 161, 166, 167, 176, 177, 184,
186, 187, 191, 195, 196, 198, 214,
219 , 225 , 236 , 239 , 241, 260, 269,
279, 280, 293, 305, 335, 338, 339,
354, 482.
— d Hat a turn (Wood.) Nels, et Mach. 159, 293.
Malacocarpus crithmifolius (Retz.) G. A. M. 534.
Malocurus lanatus (Korsh.) Nevski 847.
Malcolmia R. Br. 735, 762, 823.
— africana (L.) R. Br. 778, 813, 821.
— grandiflora (Bge.) Ktze. 801, 803.
— Karelinii Lipsky 781, 786.
— taraxacifolia Balb. 768.
— turkestanica Litw. 587, 780, 813.
Malus hissarica S. Kudr. 467.
— kirghisorum Al. et An. Theod. 467, 469, 470, 474.
— manshurica (Maxim.) Kom. 295, 364.
— orientalis Uglitzk. 384, 431.
— praecox (Pall.) Borkh. 635.
— Sieversii (Ldb.) M. Roem. 212, 467, 470, 474, 707.
Marsilea strigosa Willd. 543.
Matricaria L. 735.
— chamomilla L. 670.
— grandiflora (Hook.) A. Tolm. 504,
— lamellata Bge. 773, 774, 785, 786, 793, 803, 811, 812, 814, 815, 819.
Matthiola chenopodiifolia F. et. M. 773, fragrans Bge. 646, 675.
Medicago arabica All. 588.
— coerulea Less. 594, 747.
Medicago daghestanica Rupr. 711.
— denticulate Willd. 088.
— dzhavakhetica E. Bordz. 711.
— falcata L. 343, 394, 447, 454, 472, 522 528, 592, 621, 686, 704—706, 749.
— falcata s. 1. 497, 594, 696.
— falcata L. ssp. erecta Kotov 507.
— falcata L. var. tarbagataica Sumn. 706.
— glutinosa M. B. 497, 710.
— Kotovii Wissjul. 507.
— minima Grufberg 669, 671, 777, 778, 782, 783.
— orbicularis All. 588.
— rigidula Desr. 780, 784.
— ro manic a Prod. 634, 637, 650, 652, 655, 657, 660, 669.
Meesia Hedw. 544. e
— triquetra (Hook, et Tayl.) Angstr. 344, 546, 547, 550.
Megacarpaea megalocarpa (Fisch.) Schischk. 767.
Melampyrum arvense L. 403.
— pratense L. 225, 269.
— setaceum (Maxim.) Nakai 437.
Melandrium apetalum (L.) Fenzl 795,837, 842, 844, 846.
— longicarpophorum Kom. 709, 724.
— visco sum (L.) Cel. 749.
Melica nutans L. 167, 184, 187, 245, 279, 329,339, 341, 354, 355, 376, 392.
— picta C. Koch 376.
— simulans Klok. 645.
— uniflora Retz. 382.
Melilotus dentatus (W. et K.) Pers. 516, 623.
— officinalis (L.) Desr. 599, 858.
Meniocus Desv. 735.
— linifolius (Steph.) DC. 587, 593, 763, 769, 770, 773, 778, 780, 784.
Mentha aquatica L. 538.
— arvensis L. 525.
— longifolia (L.) Huds. 516.
Menyanthes trifoliata L. 162, 170, 339, 344, 429, 430, 510, 521, 547—549, 550, 556—558, 566, 570.
Merckia physodes (DC.) Fisch. 505.
Mercurialis perennis L. 175, 178, 204— 206, 337, 350, 354, 355, 375, 382, 390, 391, 393, 398, 406, 413—415, 416.
Merendera trigyna (Adams) Woron. 777, 784.
Mertensia kamczatica (Turcz.) DC. 112.
— maritima (L.) S. F. Gray 505.
Mespilus germanica L. 247, 409, 410, 426, 433, 434.
Microbiota decussata Kom. 159, 334, 909.
Microcoleus Desmaz. 831.
— vaginatus (Vauch.) Gom. 736, 748, 832, 833.
Microcystis (Kutz.) Elenk. 737.
Microlonchus minimus Boiss. 759.
Micromeles anifolia (Sieb. et Zucc.) Koehne 360, 362, 438.
Microetylis monophyllos (L.) Lindl. 234.
Milium effusum L. 178, 187, 196, 204, 211, 236, 313, 325, 329, 330, 339, 355, 376, 397, 471, 482, 492.
Milium Schmidtianum C. Koch 492. — vernale M. B. 726.
Minuartia arctica (Stev.) Asch, et Gr. 78, 97, 103, 111, 118, 300.
— biflora (L.) Schinz et Thell. 103, 484.
— caucasica (Ad.) Mattf. 490.
— imbricata (M. B.) Woron. 490.
— inamoena (С. A. M.) Woron. 490.
— Kryloviana Schischk. 843.
— lineata (С. A. M. ) Bornm. 845.
— Litwinowii Schischk. 709, 724.
— macrocarpa (Pursh) Ostenf. 118, 126.
— Meyeri (Boiss.) Bornm, 726.
— oreina (Mattf.) Schischk. 490.
— rubella (Wahlb.) Graebn. 78, 111.
— sclerantha (F. et M.) Thell. 778.
— verna (L.) Hiern. 490.
— Wiesneri (Stapf) Schischk. 593.
Miscanthus purpurascens Anderss. 439 r 457, 458.
Mitella nuda L. 200, 214, 279, 282, 307.
Mnium Hedw. 132, 192, 411, 544, 546.
— affine Bland. 330, 377, 384, 550.
— cinclidioides (Blytt.) Hiib. 186, 330, 429.
— cuspidatum Hedw. 377.
— longirostre Brid. 377.
— pseudopunctatum Br. et Schimp. 96.
— punctatum Hedw. 204.
— Seligerii (Jur.) Limpr. 550.
— spinosum (Voit) Schwaegr. 211.
— stellare Hedw. 377.
Molinia coerulea (L.) Moench 95, 384» 498, 558.
— litoralis Host. 246, 403, 431.
Moneses (Pyrola) uniflora (L.) A. Gray 192, 245.
Morus alba L. 852.
— nigra L. 540, 852.
Muretia Boiss. 590.
Muscari Mill. 479, 776.
— coeruleum A. Los. 490.
— racemosum (L.) Mill. 726.
— Szowitzianum Baker 710.
Myosotis alpestris Schmidt 711.
— asiatica Schischk. et Serg. 78, 98,. 118, 520.
— Krylovii Serg. 196.
— micrantha Pall. 655, 656.
— refracta Boiss. 727.
— silvatica s. 1. 705.
— suaveolens W. et K. 611, 634, 636» 637, 707, 711, 723.
Myrica tomentosa (DC.) Asch, et Graebn. 296, 297, 556, 557, 570.
Myricaria alopecuroides Schrenk 515, 847.
Myriophyllum spica turn L. 515, 537, 541» 542.
— verticillatum L. 515, 537.
Nadsoniella nigra Issatsch. 838.
Najas graminea Del. 542.
— marina L. 537, 540, 541—543.
— minor All. 537.
Nanophyton Less. 701,731,740,744,786,925.
— erinaceum (Pall.) Bge. 667, 675, 676, 679, 681, 696, 697, 700, 701, 731»
740, 751—753, 760, 767, 770—773, 787.
Nardosmia frigida (L.) Hook. 99, 118, 136, 137, 288, 546.
— glacialis Ldb. 107.
Nardus L. 491.
— glabriculmis Sakalo 479, 491.
— stricta L. 95, 445, 479, 488, 522.
Naumburgia thrysiflora (L.) Rchb. 566.
Nelumbium nuciferum Gartn. 540.
Neottia nidus-avis (L.) L. C. Rich. 415.
Nepeta L. 575, 581, 582.
— cyanea Stev. 711.
— micrantha Bge. 784.
— pannonica L. 454, 499, 589, 708, 716.
— podostachys Benth. 584, 709, 724, 726, 847.
— supina Stev. 490.
Nephroma arcticum (L.) Torss. 125, 136, 160, 161, 181, 225, 269.
Niedzwedzkia semiretschenskia B. Fedtsch. 681.
Nigella integrifolia Rgl. 587, 721, 760, 778, 779.
Nitella hyalina (DC) Ag. 542.
Nitellopsis obtusa (Desv.) Groves [Tolypel-lopsis stelligera (Bauer) Mig.] 542.
Nitraria Schoeberi L. 507, 756.
— sibirica Pall. 515.
Noaea mucronata (Forsk.) Asch, et Schweinf. 761, 777, 780, 785.
Nonea caspica (Willd.) G. Don 779, 813, 814.
— turcomanica M. Pop. 773.
Novosieversia (Sieversia) glacialis (Adams)
F. Bolle 97, 100, 107, 126, 127, 303.
Nuphar luteum (L.) Sm. 515, 537.
Nymphaea alba L. 537, 548. — Candida Presl 515, 542, 543. Nymphoides peltatum (S. G. Gmel.) Ktze.
515, 537, 541—543.
Ochrolechia androgyna Am. 82.
— frigida (Sw.) Lynge 77, 81.
— inaequatula A. Zahlh. 77, 80, 81.
— tartarea Mass. 125, 561.
Oedibasis apiculata (Kar. et Kir.) K.-Pol. 771.
Oenanthe aquatica (L.) Poir. 538.
Ofaiston monantdrum (Pall.) Moq. 792.
Oncophorus Wahlenbergii Brid. 77, 80.
Onobrychis Adans. 479.
— arenaria (Kit.) DC. 338, 452, 613, 617. — arenaria s. 1. 650.
— Biebersteinii Sirjaev 493, 497, 710.
— chorassanica Bge. 587.
— cornuta (L.) Desv. 575, 577, 581, 582, 727.
— Cyri Grossh. 497, 592.
— echidna Lipsky 582, 584, 708, 722— 724, 764, 845.
— ferganica (Sirjaev) Grossh. 500, 707.
— grandis Lipsky 721.
— iberica Grossh. 497.
— inermis Stev. 497.
— Lipskyi E. Kor. 727, 728.
— micrantha Schrenk 780, 786.
— oxytropoides Bge. 490.
Onobrychis radiata M. B. 711.
— sibirica Turcz. 522, 609, 621, 625, 704.
— Sintenisii Bomm. 727.
— tanaitica Spreng. 228.
— transcaucasica Grossh. 729.
Onosma borysthenicum Klok. 641.
— simplicissimum L. 645, 650, 651, 679, 713.
— tauricum Pall. 617.
Oplismenus undulatifolius (Ard.) P. B. 424, 426, 431, 432.
Orchis aristata Fisch. 503.
— triphylla C. Koch 421.
— umbrosa Kar. et Kir. 535.
Origanum tyttanthum Gontsch. 474,591,592.
— vulgare L. 246, 344, 376, 381, 390, 394, 403, 470—472, 474, 705—707, 710, 715.
Ornithogalum Fischerianum Krasch. 650. — Gussonei Ten. 635, 637, 777, 784. — Schmalhausenii N. Alb. 490.
Orostachys spinosa (L.) С. A. M. 652, 663r 694, 697, 699, 840.
Orthantha lutea (L.) Kern. 636.
Orthothecium chryseum (Schwaegr.) Br. et Sch. 77, 545.
Orthurus heterocarpus (Boiss.) Juz. 728. — kokanicus (Rgl. et Schmalh.) Juz. 727. Oryzopsis molinioides (Boiss.) Hack. 845, 846.
Osmorhiza aristata (Thunb.) Mak. et Yabe 194, 365.
Osmunda cinnamomea 214, 215, 294, 295, 436, 438.
— regalis L. 431.
Otostegia bucharica B. Fedtsch. 583.
— Olgae (Rgl.) Korsh. 580, 583.
Oxalis acetosella L. 147, 150, 161, 167, 170, 175—178, 184, 186, 187, 191, 192, 195—197, 204, 205, 207, 208, 214, 230, 245, 260, 307, 318, 335, 337, 338, 354, 358, 362, 414—416, 419, 429, 482, 489.
— corniculata L. 424, 427.
Oxycoccus microcarpus Turcz. 159, 180, 266, 296, 556.
— quadripetalus Gilib. (O. palustris Pers.) 170, 219, 227, 292, 296, 297, 503, 557.
Oxyria digyna (L.) Hill. 77, 78, 81, 82, 123, 490, 836, 845.
Oxytropis DC. 479, 717, 826, 845.
— aciphylla Ldb. 700, 701.
— alb о-villosa B. FedtSch. 723.
— alpina Bge. 102.
— approximata Less. 704.
— assiensis Vass. 843.
— atbaschi Saposhn. 843.
— Bobrovii B. Fedtsch. 728.
♦ — chiliophylla Royle 703.
— chinobia Bge. 827, 844, 845.
— cyanea M. B. 490.
— glabra (Lam.) DC. 515, 516.
— globiflora Bge. 702, 719, 720.
— Gmelinii Fisch. 704.
— grandiflora (Pall.) DC. 630, 631.
— humifusa Kar. et Kir. 845, 846.
— hystrix Schrenk 714.
112.
617,
422,
701.
846.
128.
Oxytropis immersa (Baker) Bge. 709j 795, 826, 84$847.
- lanata (Pall.) DC 630.
— Lehmannii Bge. 845.
__ Linczevsku Gontsch. 709.
_ longines Fisch, ill.
— macrocarpa Kar. et Kir.. 72d.
— melanotricha Bge. 765, 846.
__ Michelsonii B. Fedtsch. 709, 724.
—- microsphaera Bge. 722, 845.
— Middendorffii Trautv. 99.
__ mvriophylla (Pall.) DC. 630, 631.
- nig^cens (Pall.) Fisch. 100, 107, 111, 128.
— oligantha Bge. 687.
— Owerinii Bge. 490.
— oxyphylla (Pall.) DC. •688.
— pagobia Bge. 837.
- pilosa (L.) DC. 228, 637, 650, 704, 721.
— platonychia Bge. 847.
- Poncinsii French. 765, 794—796, 826, 844, 926.
— revoluta Ldb. 111.
— savellanica Bge. 709, 724, 725.
— selengensis Bge. 698, 699.
— Severzovii Bge. 843, 844.
— sordida (WiDd.) Pers. 117, 300.
— sp. 795.
— strobilacea Bge. 692.
— tianschanica Bge. 826, 926.
— tragacanthoides Fisch. 687, 700,
Pachyphragma macrophyllum (Hott.) N. Busch 208, 418.
Pachypleurum alpinum Ldb. 70, 97, 102, 123, 300, 496, 837.
— mucronatum (Schrenk) Schischk. 842, 844.
Pachypterigium multicaule (Kar. et Kir.) Bge. 767.
Padus Maackii (Rupr.) Kom. 213, 360.
— racemosa (Lam.) Gilib. 168, 186, 194 195, 197, 236, 338, 339, 354, 390, 397* 530, 535.
Paeonia anomala L. 340.
— obovata Maxim. 215.
— tenuifolia L. 617, 636, 637, 710.
— triternata Pall. 246.
Palimbia rediviva (Pall.) Thell. 672.
Paliurus Mill. 468.
— spina-Christi Mill. 400, 401 404 407 468’ 472’ 473> 576’ 579’ 594, /Zu, ZoU.
Palmella Lyndb. 737.
Pal^lla^quarrosa (Hedw.) Brid.
Panzeria Moench 603 Papaver L. 735, 763.
— Belangeri Boiss. 776.
— caucasicum M. B. 490
- croceum Ldb. 484, 705, 827 847
— involucratum M. Pon. 709 ’
— nudicaule L. 609 845
" % % ™2'
— polare A. Tolm. 79 81 so
- ndicatum R3ttb. 77,112^ is - iiansehanicum M. Pop 843 844.
96,
846
Paracaryum himalayense (Klotzschi г г» Clarke 847. ' В.
Parageum calthifolium. (Menzies) Nakai
Paraquilegia anemonoides (Kar. et Ifi. i N. Schipcz. 842, 847. Л1г<)
— grandiflora (Fisch.) Drum, et Нпы.
78, 842—844, 847. tch-
Parietaria judaica Strand. 728, 786.
Paris incompleta M. B. 404.
— quadrifolia L. 184, 187, 195, 197 onz 245, 260, 280, 339, 344, 354. ’ ’
Parmelia (Ach.) De Not. 78, 102 Чях 672, 736. ’ °*’
— conspersa (Ehrh.) Ach. 103.
— molliuscula Ach. 102.
— prolixa (Ach.) Nyl. 770.
— ryssolea (Ach.) Nyl. 597, 655, 665 670 748, 769, 770. ’ ’
— vagans Nyl. 597, 609, 665, 670 679 685, 688, 697, 748, 769. ’ ’
Parnassia Kotzebuei Cham, et Schlecht. — Laxmannii Pall. 314.
— palustris L. Ill, 131, 491, 546.
Paronychia cephalotes (M. B.) Bess. 645.
Parrotia persica (DC) С. A. M. 378 425, 426, 852, 921.
Parrya albida M. Pop. 845.
— asperrima M. Pop. 845.
— exscapa С. A. M. 796, 836, 844,
— nudicaulis (L.) Rgl. 121, 126, — pinnatifida Kar. et Kir. 842.
— stenocarpa Kar. et Kir. 842, 845, Pastinaca armena F. et M. 710.
Pastinacopsis glacialis Golosk. 843, Patrinia intermedia Roem. et Schult. 840.
— scdbiosaefolia Link. 448, 457.
— sibirica (L.) Juss. 102, 104, 108, Pedicularis achilleifolia Steph. 686. — Adamsii Hulten 107, 126, 128. — adunca M. B. 546.
— amoena Adams 99, 313.
— capitata Adams 111, 520.
— Chamissonis Stev. 503.
— compacta Steph. 485.
— condensata M. B. 491.
— dasystachys Schrenk 622, 636.
— dolichorrhiza Schrenk 709.
— elata Willd. 316.
— grandiflora Fisch. 296.
— hirsuta L. 121.
— Karoi Freyn 550.
— Korolkovii Rgl. 723.
— Kusnetzovii Kom. 215.
— labradoricd Wirsing. 241, 280, 291.
— lapponica L. 118, 137, 159, 296.
— Nordmanniana Bge. 491.
* — Oederi Vahl 97, 98, 111, 486.
— Olgae Rgl. 724, 725.
— pycnantha Boiss. 845.
— resupinata L. 280, 448, 457.
— rhinanthoides Schrenk 485.
— septrum-carolinum L. 131, 288.
— sudetica Willd. 99, 114, 117, 519» 545, 546.
— tristie L. 314.
844.
717,
486.
Pedicularis verticillata L, 104, 1Ц.
Peganum harmala L. 671, 739, 74'5 733
Peltigera aphthosa (L.) Willd.’ 102* 11 я’ 136, 180, 181, 225, 269. ’
— canina (L.) Willd. 114, 384.
— malacea (Ach.) Funck. 181.
— scabrosa Th. Fr. 180.
Penicillium Fr. 737.
Pennisetum orientale Rich. 786.
Periploca graeca L. 374, 423, 432, 434
Perovskia scrophulariifolia Bge. 580’ 594
Perlusaria DC. 561. ’
Petasites albus (L.) Gaertn. 205, 206 414
Petrosimonia Bge. 513, 543, 793. ’
— brachiata (Pall.) Bge. 514, 579, 777, 782. ’
— brachyphylla (Bge.) Iljin 769.
— glauca (Pall.) Bge. 773, 792.
— Litwinowii Korsh. 510, 543.
— oppositifolia (Pall.) Litw. 512, 670,
— sibirica (Pall.) Bge. 857.
— triandra (Pall.) Simonkai 514, 769.
Peucedanum L. 596.
— baicalense (Redow.) C. Koch 630.
— latifolium auct. 616.
— Lubimenkoanum Kotov 636, 637, 650.
— macrophyllum Schischk. 616.
— Morisonii Bess. 620—622, 624, 650, 652, 705.
— palustre (L.) Moench 429, 550, 566.
— ruthenicum M. B. 636, 637, 729.
— terebinthaceum Fisch. 448, 457.
— turcomanicum Schischk. 728.
Phalacrachena inuloides (Fisch.) Iljin 658.
Phalaris minor Retz. 508, 588.
— paradoxa L. 588.
Phascum Hedw. 736.
— cuspidatum Hedw. 780.
Phaseolus aureus Roxb. 854.
Phellodendron amurense Rupr. 360, 364. — sachalinense (Fr. Schm.) Sarg. 214, 438, Philadelphus caucasicus Koehne 423.
— tenuifolius Rupr. et Maxim. 213, 360, 374, 436.
Philonotis seriata (Mitt.) Lindb. 491.
Phippsia R. Br. 68.
— algida (Soland.) R. Br. 70, 77, 79, 80, 82 123. *
Phleum alpinum L. 70, 484—486.
— montanum G. Koch 710.
— paniculatum Huds. 508, 588, 590, 594, 747, 763.
— phleoides (L.) Simk. 408, 447, 448, 454, 471, 472, 483, 495, 496, 500, 522; 530, 531, 577, 592, 621, 625, 627, 629, 649, 695, 696, 705—708, 710, 711, 714— 716, 718, 729, 845.
— pratense L. 446, 451, 454, 522, 523, 711, 715.
Phlofodicarpus sibiricus (Steph.) K.-Pol. 625, 630, 631.
Phlomis L. 602, 734.
— bucharica Rgl. 580, 587, 763.
— cancellata Bge. 726.
— canescens Rgl. 584.
— • fruticetorum Gontsch. 591.
— kopetdagensis Knorr. 727.
K„. „ Klt 483. 4M
— pratensis Kar. et Kir. 500
~ pungens Willd. 636, 656 rrr aaq salicifolia Rgl. 587, 591 721 726 763*
— thapsoides Bge. 587 ’ /6“’
— tuberosa L. 260, 312, 343, 344 392 454, 530, 613, 621, 625, 526 629 Ml’ 650, 852, 678, 693’, 705’, 715’, 715’, 749*
Pholuirus incurvus (L.) Schinz et Thell’ □00.
— pannonicus (Host) Trin. 668, 832 Phormidium Kutz. 737, 831, 832.
— ambiguum Gom. 833.
— autumnale (Ag.) Gom. 832
— corium (Ag.) Gom. 832, 833.
— dimorphum Lemm. 833.
— foleolarum (Mont.) Gom. 832.
— inundatum Kutz. 832.
— lividum Nag. 833.
— molle (Kiitz.) Gom. 832.
Phragmites communis Trin. 233, 286. 296 345, 429—431, 477, 506, 508, 510* 512—516, 530, 532, 535, 537—543* 548, 550, 556, 557, 566, 623, 682. 697* 756, 807, 857.
— Isiaca (Del.) Kunth 541, 542.
Phyllitis scolopendrium (L.) Newm. 421. 424, 426, 427, 432.
Phyllodoce aleutica (Spreng.) A. Heller 91,110.
— coerulea (L.) Bab. 78, 94, 110, 160, 191, 215.
Phyteuma argutum Rgl. 212.
Picea ajanensis Fisch. 212.
— excelsa Link 141, 143, 160, 166, 174, 201—203, 328, 329, 348, 352, 354, 398, 415, 460, 518, 918—920.
— fennica Rgl. 141, 160.
— Glehnii Mast. 145.
— fezoensis (Sieb. et Zucc.) Carr. 106, 108, 143, 144, 202, 212, 293, 307, 310, 341, 360, 363, 919.
— koraiensis Nakai 141, 215, 360, 364.
— obovata Ldb. 70, 96, 141, 143, 152, 160, 166, 168, 174, 176, 188, 201, 215, 266, 289, 307, 328, 348, 352, 355, 482, 518, 918—920.
— orientalis (L.) Link 144, 202, 871 919.
— Roberta P. Vipp. 212.
— Schrenkiana F. et M. 143, 144, 469, 494, 496, 499, 919.
— Wilsonii Mast. 202.
Pieris-hieracioides L. 613.
Pimpinella lithophila Schischk. 712. — saxifraga L. 445, 446, 522, 523, — titanophila Woron. 644, 645. — tragium s. 1. 712.
Pineuicula vulgaris L. 491.
Pinus cembra L. 139, 144, 203. ___ funebris Kom. 217, 222, 439, 610.
Z tt D. Sosn. 217, 222, 244, 246,
- toraienws Slab, et Zucc. 143, 144, 213, 348, 367.
— lapponica Mayr 217, 328.
___ mughus Scop. 205, 217, 489.
206,
210,
711.
Указатель
«“‘иг"' 222' Ш'
~ ^3^ 141—143, 155, 158, 215, 239, 241, 242 * 244 , 272, 303, 308, 310, 334, 905,
__ sibirica (Rupr.) Мауг 90, 96, 106, 139, 141, 143, 1Й8, 197, 220, 225, 242, 310, 348* 902.
__sibirica Мауг f. turjosa Gorodk. 180.
— sibirica Мауг var. humistrata Litw. 142.
- silvestris L. 154, 217, 348, 640, 883.
— silvestris s. 1. 880.
— silvestris L. f. Litwinowii Sukacz. 227, 556, 557, 565, 566.
— silvestris L. f. pumila Abolin 227, 556.
— silvestris L. f. uliginosa Abolin 227, 557, 558, 565, 566.
— silvestris L. f. Willkommii Sukacz. 227, 556, 566.
— Stankeviczii (Sukacz.) Fom. 247, 400.
Piptatherum songoricum (Trin. et Rupr.) Roshev. 584.
Pistacia mutica F. et M. 400, 460, 576, 578, 922.
— vera L. 578, 579, 592, 764, 922.
Placodium (DC.) Vain. 479.
— elegans (Link.) Ach. 770.
— Tominii Sav. 819.
Plagiochilla asplenioides (L.) Dum.
Plantago L. 735.
— arachnoidea Schrenk 843, 844.
— canescens Willd. 286.
— Cornutii Gouan 529, 531, 539, 621, 623.
— coronopus L. 785.
— lachnantha Bge. 762.
— lagocephala Bge. 781.
— major L. 447.
— maxima Juss. 531, 621—623.
— media L. 445.
— minuta Pall. 768.
— salsa Pall. 506, 516, 616, 696
— saxatilis M. B. 710. ’
— Schrenkii C. Koch 504.
~ 65(^713 KUpr‘ 617, 621 ’ 624’ 636> 637>
— tenuiflora W. et K. 669.
Platantherabifolia (L.) L. C. Rich.
— chlorantha Cust. 415, 421.
Platanus orientalis L. 852
Plectonema Thur. 737.
Pleurococcus Wild. 737
V-,™. »<•
STS, fit и' j£‘u?3>
32»: 3$ m?: «3 «»’
РоЛлл 1111 112> 595
— Albertii Rg], 720, 843
~ alptgena (Fr.i .
217, 222,
433, 434,
204.
697, 755.
184.
93,
150,
mW, «UU, 61*1,
229, 233, 236, 241, 266, 269, 280, 290, 296, 300, 305, 310, 312, 329, 339, 354, 357, 488.
Ill, 112, 595.
iioertii Rgl. 720, 843.
- „Ipigena (Fr.) Lindm. 122, 519, 520. — alpina L. 102, 122, 484, 490, 494, 496, 709, 827.
595.
Poa altaica Trin. 104.
__ angustifolia L. 338, 396, 399, 448. 452 474, 528, 530, 531, 538, 591, 596, 612 616, 621, 625, 627, 634, 636, 649, 695' 707, 708.
_ arctica R. Br. 107, 115, 118, 120, 126 132, 158, 505.
— attenuate Trin. var. stepposa Kryl. 685 686,
__ botryoides Trin. 221, 240, 286, 531 625, 628, 630, 631, 683—686, 688“ 690, 691, 693, 698, 924.
__ bucharica Roshev. 584, 724, 726, 845.
__ bulbosa L. 508, 574, 580, 582, 587, 588, 590, 592, 597, 607, 635, 637, 650 655—657, 660, 666, 668—670, 679-681, 706, 729, 747, 751, 754, 760, 776, 777, 833, 923.
__ bulbosa L. var. vivipara Koeler 512, 587, 588, 638, 665, 672, 721, 723, 725, 727, 741 746, 748, 750, 751, 757, 759, 762, 763’, 768, 770, 771, 774, 775, 777-786, 817, 832, 846, 851.
__ cenisia All. 487.
__ Chaixii Vill. 205.
__ dahurica Trin. 108, 273, 685, 687.
— dense Troitzky 711.
— dschungarica Roshev. 484.
— glabriflora Roshev. 584, 708.
__ hissarica Roshev. 709.
__ Komarovii Roshev. 79, 545.
__ Lipskyi Roshev. 843.
__Litwinowiana Ovcz. 708, 720, 723, 794, 795, 826, 845—847.
__ longifolia Trin. 325, 492.
__ masenderana Freyn et Sint. 426.
__ nemoralis L. 211, 212, 245, 376, 381,382, 406—408, 410 , 416, 432, 496 , 589 , 709.
__ palustris L. 286, 390, 525, 529, 532, 549.
__ naucispicula Scribn. et Merr. 82.
__ pratensis L. 246 , 281, 289 , 312 , 340, 343, 376, 448, 451, 454, 484, 495, 499, 500, 502, 519 , 522, 524 , 525, 530, 538, 577, 600, 616, 621, 704.
__ relaxa Ovcz. 484, 577, 584, 592, 599, 600, 705, 709, 718, 722, 724, 725, 727, 765, 826, 827, 843.
— sibirica Roshev. 201, 236, 288, 311, 312, 316, 339, 484, 485, 496.
— sibirica Roshev. f. nigra Roshev. 486.
— stepposa (Kryl.) Roshev. 622, 625, 627, 685, 687, 693, 695, 706.
— sterilis M. B. 406.
— trivialis L. 747. ,
Poacynum (Apocynum) pictum (Schrenk)
BaiH. 534.
Pogonetum urnigerum (Hedw.) P. B. 4oo, 519.
Polemonium acutiflorum Willd. 131, 310, 546.
— boreale Adams 123. ...
— coeruleum L. 188, 211, 297, 312, 316.
— coeruleum s. I. 494, 496, 499. Polyblastia Lonnr. 78.
Polycnemum arvense L. 668, 669. —
Polygala anatolica Boiss. et Heldr.
— comosa Schkuhr 634, 636.
— cretacea Kotov 646.
Polygala major Jacq. 617.
— moldavica Kotov 645.
— sibirica L. 457.
— transcaucasica Tamamsch. 594.
— vulgaris L. 522.
Polyganatum acuminatifolium Kom. 457
— latifolium Desf. 399.
— multiflorum (L.) All. 384, 396.
— officinale All. 357, 376, .395, 396.
Polygonum L. 597.
— alopecuroides Turcz. 630, 631.
— alpinum All. 496, 499, 500, 723.
— amphibium L. 542, 543, 550.
— angustifolium Pall. 630.
— arenarium W. et K. 641, 642.
— attenuatum V. Petr. 630.
— aviculare L. 638.
— biaristatum Ait ch. et Hemslev 709 724,- 725. J
— bistorta L. 96—98, 102, 105, 117, 131 267, 300, 312, 316, 482, 485, 486 491 519, 523.
— Bordzilovskii Klok. 641.
— bucharicum Grig. 846.
— coriarium Grig. 499, 718.
— corrigioloides Jaub. et Spach 543.
— ellipticum Willd. 118.
— hissaricum M. Pop. 845.
— hydropiper L. 432, 433.
— molliiforme Boiss. 847.
— nitens (F. et M.) V. Petr. 484, 842.
— nodosum Pers. 432.
— paronychioides С. A. M. 722, 728.
— patulum M. B. 668, 670.
— polycnemoides Jaub. et Spach 725.
— pseudo arenarium Klok. 616, 668.
— rupestre Kar. et Kir. 723,
— schugnanicum Kom. 709.
— songoricum Schrenk 211.
— tripterocarpum A. Gray 129, — viviparum L. 102, 104, 111, 132, 486, 487, 502, 503, 520, 521', 545i 628, 630, 631, 842.
Polypodium lineare Thunb. 360.
— serratum (Willd.) Futo 424.
— vulgare L. 234, 432, 841.
Polystichum Braunii F6e 195, 197, 214, 414.
— lobatum (Sw.) Presl 204, 414, 421, 426, 427.
— Woronowii Fom. 424, 426.
Polytrichum Hedw. 197, 219, 266, 479, 491. — alpinum Hedw. 77, 80, 81, 98, 118, 123. — commune Hedw. 70, 98, 104, 116, 118, 120, 136, 138, 153, 160, 163, 166, 169, 180, 181, 184, 190, 191, 192, 195, 200, 220, 225, 227, 236, 261, 267, 284, 291, 301, 329, 338, 339, 557.
— formosum Hedw. 414.
— gracile Sm.^88.
— hyperboreum R. Br. 97, 120, 121,
— Jensenii Hag. 129.
— juniperinum Willd. 239, 2-70.
— norvegicum Hedw. 98.
— piliferum Hedw. 107, 120, 239, 270, 309. n
— strictum Sm. 70, 102, 103, 113, 118, 121, 125, 138, 160, 162,169 181, 296, 300, 330, 488, 557, 560, 561.
842—844.
274, 310.
118, 123,
123.
261,
116,
P°P682* aU>a L' 382> 390’ 515’ 53°- 539,
— ariana Dode 534.
— balsamifera L. 852.
— Bolleana Lauche 852.
— Oavidiana Dode 609.
— deltoides Marschall 852.
— ^^sifolia Schrenk 534, 535, 852, 858
— hybnda M. B. 433.
— hyrcana Grossh. 432.
— koreana Rehder 364.
— laurifolia Ldb. 535.
macrocarpa (Schrenk) N. Pavl. et Lipsch. 535.
— Maximowiczii A. Henry 364, 437
— nigra L. 515, 530, 539, 540, 682* 852
— pseudotreniula N. Rubtz. 323, 469.
— pruinosa Schrenk 534, 852, 858.
— pyramidalis Rozier 852.
— Sieboldii Miquel 323.
— suaveolens Fisch. 272, 273, 437. 519 520.
— tremula L. 240, 320, 322, 325, 336.
338, 354, 435, 460, 706, 841.
Portulaca oleracea L. 858.
Potamogeton L. 515, 543.
— acutifolius Link 647.
— crispus L. 537, 542, 543.
— filiformis Pers. 543.
— heterophyllus Schreb. 647.
— lucens L. 514, 537, 542, 543.
— nodosus Poir. 542.
— pectinatus L. 514, 537, 540—543.
— perfoliatus L. 514, 537, 541, 542.
— trichoides Cham, et Schlecht. 543.
Potentilla L. 479, 485, 522.
— acaulis L. 496, 628, 630, 686, 688, 690—695, 697—700.
— anserina L. 447, 504, 630.
— arenaria Borkh. 635.
— argentea L. 376.
— astracanica Jacq. 644, 645.
— astragalifolia Bge. 701, 702.
— aurea Tom. 487, 488.
— biflora Willd. 843.
— bifurca L. 103, 686, 687, 694, 702,J49.
— chrysantha Trev. 714.
— Crantzii (Grantz) Beck 710.
— desertorum Bge. 485.
— Egedii Wormsk. 504, 505.
— elegans Cham, et Schlecht. 107, 108.
— emarginata Pursh 82, 99, 126.
erecta (L.) Hampe 488, 498.
—/evestita Th. Wolf 702, 719, 720.
— flabellata Rgl. et Schmalh. 725.
— fragarioidesL, 629.
— fragiformis Willd. 505.
— gelida С. A. M. 479, 490, 724, 827, 845,
— hololeuca Boiss. 723, 827, 844, 845.
— humifusa Willd. 612, 621, 624, 636, 637, 650, 660.
— Lipsky ana Th. Wolf 724.
— micrantha Ram. 412.
— Moorcroftii Wall 795, 826, 846.
— multifida L. 103.
___ nervosa Juz. 716, 719.
- nivea L. 100» 103» 112, 495.
— nivea s. 1. 844.
Potentilla orientalis Juz. 714, 718, 721, 842.
— pamirica Th. Wolf 826, 846.
— pamiroalaica Juz. 765, 826, 837, 926.
— recta L. 729.
— reptans L. 747.
— Schurii Fuss 636, 637.
— sericea L. 686, 687, 694, 846.
— soongorica Bge. 760.
— stolonijera Lehm. 503.
— tanacetifolia Willd. 628, 630, 631, 683.
— tanaitica Zing. 615.
— transcaspia Th. Wolf 727.
— verticillaris Steph. 688.
— viscosa J. Don 446, 630, 687.
Prangos Lindl. 587.
— ferulacea (L.) Lindl. 711.
— pabularia Lindl. 474, 577, 584f 585, 589, 590, 592, 708, 721, 722, 725, 763, 923.
— uloptera DC. 708.
Primula acaulis (L.) Hill. (P. vulgaris Huds.) 246, 247, 406.
— algida Ad. 102, 490, 494, 717, 723, 836.
— auriculata Lam. 491.
— cuneifolia Ldb. Ill, 112, 503.
— heterochroma Stapf 410, 426.
— macrocalyx Bge. 247, 407.
— maegaseifolia Boiss. et Bal. 420.
— minima L. 487.
— nivalis Pall. 78, 847.
— veris L. 612.
Protococcus Agardh. 737.
Prunella grandiflora (L.) Jacq. 613, 620.
— vulgaris L. 446, 498.
Prunus Mill. 468, 601.
— divaricata Ldb. 212.
— moldavica Kotov 399, 614.
— sogdiana Vass. 469.
— spinosa L. 205, 381, 614.
— spinosa s. 1. 617, 618.
— stepposa Kotov 389, 390, 392, 393, 395, 452, 468, 530, 614, 620, 635, 636, 638, 653, 658, 710.
Psammogeton setifolium Boiss. 781.
Psathyrostachys juncea (Fisch.) Nevski 514, 628, 660, 667, 678, 686, 694, 706.
— lanuginosa (Trin.) Nevski 663.
Psephellus daghestanicus D. Sosn. 711.
— de alb at us (Willd.) Boiss. 408.
— declinatus (M. В.) C. Koch 712.
— > leucophyllus (M. В.) G. A. M. 618.
Pseudosedum condensatum Boriss. 725.
Psora (Hall.) Koerb. 665, 672, 736, 831.
— decipiens (Ehrh.) Hoffm. 749, 759, 771, 811, 819, 831.
— lurida (Sw.) DC. 832.
— Saviczii Tom. 759.
Psoralea bituminosa L. 247, 403.
— drupacea Bge. 580, 587, 591, 594, 763.
Psoroma hypnorum S. Gray 77.
Psychrogeton cdbulicus Boiss. 708, 709, 724, 725, 845.
Psylliostachys (Statice) spicata (Willd.) Nevski 588, 747, 784, 785.
P tar mica bisserata DC. 384,* 403r 404, 407.
— cartilaginea Ldb. 529, 538.
— Krylovii Serg. 485.
— sibirica Ldb. 306, 520.
Pteridium aquilinum (L.) Kuhn 175, 194, 227, 228, 232, 237, 238, 247, 293—295, 333, 337, 338, 340, 341, 344, 357, 363, 384, 388, 395, 436, 627.
— tauricum (Presl) V. Krocz. 246, 403, 404, 406, 424, 425.
Pteris cretica L. 420, 424.
Pterocarya pterocarpa (Michx) Kunth 424, 427, 432, 433.
Pterotheca Marschalliana (Rchb.) Grossh. 508, 778.
Pterygoneurum Juz. 736.
— ovatum (Hedw.) Dix. 780.
Ptilagrostis mongholica (Turcz.) Griseb. 101, 103—105, 602, 719, 720.
— subsessiliflora (Rupr.) Roshev. 703, 719 720 795.
Ptilidium ciliare (L.) Hanipe 69, 112, 113, 116, 118, 121, 128, 129, 158, 180, 260, 268, 281, 300, 302, 303.
Ptilium crista-castrensis (Hedw.) De Not 150, 162, 180, 181, 184, 195, 200, 233, 236, 269, 305, 310, 312.
Ptilotrichum canescens С. A. M. 701, 702.
— elongatum С. A. M. 688.
Puccinellia Pari. (Atropis Rupr.) 508, 616, 623, 793.
— angustata (R. Br.) Band et Redf. 77.
— Bilykiana Klok. 616, 647.
— bulbosa Grossh. 513, 579.
— convoluta s. 1. 616.
— dolicholepis V. Krecz. 507, 512—514, 540, 670.
— Fominii Bilyk 507, 529, 539, 670.
— gigantea Grossh. 507, 508, 512—514, 516, 539, 540.
— Hackeliana V. Krecz. 723.
— maritima (Huds.) Pari. 506.
— phryganodes (Trin.) Scribn. et Merr. 81, 504—506.
— subspicata V. Krecz. 709, 725.
— suecica Holmb. 506.
— tenella V. Krecz. 505.
— tenuiflora (Turcz.) Scribn. et Merr. 103, 286, 532, 609, 631, 696, 697, 699.
— tenuissima Litw. 510, 529, 531, 616, 647.
— vulgaris Gaertn. 669.
Pulmonaria Filarszkyana Javorka 204.
— mollissima Kern. 187, 238, 344, 382.
— obscura Dumort. 170, 178, 179, 337, 386, 388, 392, 397.
Pulsatilla alba Rchb. 487, 488.
— albana (Stev.) Bercht. 618, 710.
— ambigua (Turcz.) Juz. 313, 683, 691.
— angustifolia Turcz. 609.
— armena (Boiss.) Rupr. 490.
— Bungeana G. A. M. 103.
— campanella Fisch. 687?
— flavescens (Zuccar.) Juz. 221, 621. 625—627, 650.
— Halleri (All.) Willd. 712.
— multifida (Pritzel) Juz. 281, 284, 65Q.
— nigricans Storck 635, 636.
— patens (L.) Mill. 611, 612, 636, 704.
— patens s. 1. 221, 231—233, 236, 259. 305, 307, 315, 340, 446, 448, 631.
— pratensis (L.) Mill. 228.
Pulsatilla Turczaninovii Kryl. et Serg. 242, 621, 630, 683, 687, 690—693.
— ucrainica (Ugr.) Wissjul. 635.
Punica L. 46o.
— granatum L. 401, 473, 579, 780.
Pyrethrum Gaertn. 481, 600.
— achilleifolium M. B. 651, 656, 660, 663, 665, 667, 672, 673, 678, 731, 748, 757.
— bipinnatum (L.) Willd. 519.
— corymb о sum (L.) Willd. 384, 390, 392, 394.
— leonlopodium (C. Winkl.) Krasch. 843, 844.
— millejoliatum (L.) Willd. 596, 638, 644, 655, 669.
— millejoliatum s. 1. 651, 657.
— odessanum Klok. 645.
— parthenijolium Willd. 710.
— pulchrum Ldb. 845.
— pyrethroides (Kar. et Kir.) C. Winkl. 719, 796, 843—847.
— sclerophyllum Krasch. 646.
— Stoliczkai Clarke 847.
Pyrola incarnata Fisch. 147, 241, 281, 293, 305, 307.
— japonica Sieb. 362.
— media Sw. 233, 245, 316.
— minor L. 245, 316.
— rotundifolia L. 166, 200, 234, 236, . 239, 245, 279, 280, 316, 337, 357.
— inrescercsSchweigg.233,236,246,247,279.
Pyrus Balansae Decaisne 431.
— Boissieriana Buhse 409.
— caucasica An. Fed. 384, 403.
— communis L. 399, 617, 635.
— elaeagrijolia Pall. 617.
— Grossheimii A. Fed. 409.
— Korshinskyi Litw. 212.
— salicifolia Pall. 401.
— ussuriensis Maxim. 364.
Quercus araxina (Trautv.) Grossh. 369, 408.
— castaneifolia С. A. M. 369, 378, 402, 404, 408, 409, 425, 426, 459, 906, 921.
— cerris L. 369.
— dentata Thunb. 369, 438, 439, 458.
— dschorochensis C. Koch 369.
— erucifolia Stev. 369.
— Hartwissiana Stev. 369, 378, 403, 422—425, 458, 921.
— hypochrysa Stev. 369.
— iberica Stev. 246, 369, 378, 402—404, 406, 408—410, 423, 424, 459, 473, 921.
— imeretina Stev. 369, 423, 425, 431, 458.
— longipes Stev. 369, 378, 428, 432—434, 579, 921.
— macranthera F. et M. 245, 325, 369, 408—410.
— Medwedewii D. Sosn. 369.
— mongolica Fisch. 222, 341, 360, 363,
— 366, 369, 435, 438, 448, 456, 458, 460, 609.
— petraea Liebl. 246, 352, 369, 378, 381—385, 399, 402—404, 413, 415, 459, 614, 617, 618, 921.
Quercus pontica G. Koch 369.
— pubescens Willd. 246, 247, 369, 398; 404, 459, 460, 578, 614, 617, 648. 671.
— robur L. 173, 348. 354, 357, 368, 381, 384, 386, 389, 392, 396, 397, 399, 403, 414, 530, 539, 614, 617, 618, 648, 653, 658.
Queria hispanica L. 726.
Ramischia (Pyrola) secunda (L.) Garcke 147, 153, 233, 245, 246, 280, 293, 357, 388.
Ranunculus acer L. 445, 450, 523.
— ajghanicus Aitch. et Hemsley 722.
— alpigenus Kom. 847.
— altaicus Laxm. 111.
— auricomus L. 445, 450.
— borealis Trautv. 325, 519.
— caucasicus M. B. 490, 493, 498.
— gelidus Kar. et Kir. 827, 843.
— glabriusculus Rupr. 98.
— grandifolius С. A. M. 196, 483.
— hyperboreus Rottb. 504.
— lanuginosus L. 205.
— linearilobus Bge. 728.
— lingua L. 537, 549.
— monophyllus Ovcz. 520.
— nivalis L. 70, 114, 123, 545, 835.
— oreophilus M. B. 836.
— platanijolius L. 205.
— polyanthemus L. 498, 612, 707.
— propinquus С. A. M. 96, 108, 300, 312.
— pygmaeus Wahlb. 82.
— repens L. 502, 525.
— rufocepalus Francb. 847.
— Sabinii R. Br. 82.
— sulphureus Soland. 77, 78, 80, 82.
— trisectilis Ovcz. <836.
— turkestanicus Franch. 725.
Reaumuria cistoides Adam 747.
— hypericoides Willd. 744, 747, 783.
— oxiana (Ldb.) Boiss. 786, 817.
— soongorica (Pall.) Maxim. 700, 701, 731, 786, 925.
Rhacomitrium hypnoides Lindb. 78, 80, 97, 107, 113, 120, 121.
Rhamnus L. 468.
— cathartica L. 390, 395, 399, 452, 469, 471, 530, 535, 617,
— imeretina Booth 420.
— Pallasii F. et M. 401, 473, 576, 594, 618, 710, 729.
— Sintenisii Rech. f. 728, 774, 786.
Rhaphidonema nivale Lagerh. 838.
Rhaponticum carthamoides (Willd.) Iljin 483, 485, 486.
— salinum (Spreng.) Less. 512.
Rheum Fedtschenkoi Maxim. 846. *
— Maximowiczii A. Los. 763.
— rhizostachyum Schrenk 842.
— tataricum L. f. 751, 759, 767.
— turkestanicum Janisch. 735, 768,
800, 804, 815, 817, 818.
— Wittrockii Lund str. 716, 845.
Rhinanthbs L. 522.
Rhinopetalum Karelinii Fisch. 759, 767, 781, 811.
Rhizocarpon (Ram.) Stein. 78, 843.
Rhodiola atropurpurea (Turcz.) Trautv. et May. 505.
— borealis Boriss. 505.
— coccinea (Royle) Boriss. 842, 843.
— gelida Schrenk 843. 847.
— neterodonta (Hook, et Thoms.) Boriss. 795.
— pamiroalaica Boriss. 795, 846.
— quadrtf ida [Pall.) F. et M. 97, 108.
— rosea L. 485, 505.
— Semenovii (Rgl. et Herd.) Boriss. 845.
Rhodobryum roseum (Hedw.) Limpr. 196.
Rhododendron Adamsii Rehd. 101, 105, 107, 108, 303.
— aureum Georgi 90, 104, 105, 108— 110, 158, 198, 215, 306, 310, 333, 334, 486.
— caucasicum Pall. 209, 245, 325, 419, 492.
— dahuricum L. 221, 236, 241, 244, 259, 282, 290, 295, 305, 318, 447, 632.
— hypopitys Pojark. 215.
— kamtschaticum Pall. 91, 112, 333.
— Kotschyi Simk. 488.
— Ledebourii Pojark. 313.
— luteum Sweet 246, 325, 373, 385, 403, 419, 423—425.
— mucronulatum Turcz. 435.
— parvtfolium Adams 101, 105, 108, 258, 259, 296, 303, 307, 314, 570.
— ponticum L. 208, 406, 419, 420, 423, 424, 432.
— Redowskianum Maxim. 108.
Rhus L. 468.
— coriaria L. 247, 400, 468, 474.
Rhynchospora alba (L.) Vahl 559, 571.
Rhytidiadelphus squarrosus (Hedw.) Warnst. 196.
— triquetrus (Hedw.) Warnst. 161, 168, 178, 184, 186, 191, 192, 195, 196» 208, 211, 214, 307, 354, 429.
Rhytidium rugosum (HedwJ Lindb. 69, 97, 102, 107, 109, 112, 116—119, 260, 267, 281, 300, 302, 305.
Ribes alpinum L. 413.
— atropurpureyfn С. A. M. 280.
— Biebersteinii Berl. 208, 245, 325.
— carpaticum Kit. 206.
— hispidulum Pojark. 195—197.
— » Meyeri Maxim. 471, 709, 843, 845.
— » nigrum L. 187, 339, 343, 429.
— pallidiflorum Pojark. 307.
— rubrum L. 168, 236, 268, 289, 301, .. 330, 343.
Ricca Mich. 736.
Ricinus communis L. 854.,
Rindera ^turkestanica Kuan. 761.
Robinia pseudoacacia L. 852.
Rochelia Bungei Trautv. 759.
— • disperma (L. f.) G. Koch 768, 778.
Roegneria C. Koch 584.
— Abo I in ii (Drob.) Nevski 716.
— angustiglumis Nevski 313.
— borealis (Turcz.) Nevski 286, 519.
— caucasica C. Koch 407.
— czimganica (Drob.) Nevski 485, 714, 715, 795.
Roegneria macroura (Turcz.) Nevski 286.
— Schrenkiana (F. et M.) Nevski 485, 716.
— schugnanica Nevski 703, 826.
— tianschanica (Drob.) Nevski 716.
— Turczaninovii (Drob.) Nevski 286, 483.
— ugamica (Drob.) Nevski 584.
Roemeria Med. 735, 776.
— hybrida (L.) DC. 726, 759, 770, 778, 780, 781, 786.
— refracta (Stev.) DC. 760, 786.
Roripa amphibia (L.) Bess. 514.
Rosa L. 400, 452, 468, 576.
— acicularis Lindl. 158, 161—163, 182, 184, 190, 224, 230, 232, 235, 239, 241, 281, 301, 305, 307, 311, 313, 338— 341, 343, 344, 447, 471.
— Alberti Rgl. 211.
— amblyotis G. A. M. 333, 334.
— Beggeriana Schrenk 211, 471, 709.
— canina L. 205.
— cinnamomea L. 232, 233, 330, 340, 344, 374, 394, 653, 664, 841.
— dahurica Pall. 341.
— Ecae Aitch. 473, 474, 499, 590—592, 709.
— Fedtschenkoana Rgl. 499. .
— hissarica Slobodov 726.
— kokanica Rgl. 469, 473, 474, 709, 721, 846.
— laxa Retz. 535.
— maracandica Bge. 473, 474.
— nanothamnus Bouleng. 847.
— pendulina L. 204, 206.
— platyacantha Schrenk 473, 474, 499, 707, 718, 845.
— rugosa Thunb. 438, 458.
— spino sis sima L. 233, 471, 499, 652, 664, 704, 706, 715, 841, 846.
Rosularia alpestris (Kar. et Kir.) Boriss. 843, 844, 847.
Rubia dolychophylla Schrenk 534.
— tibetica Hook 845.
Rubus arcticus L. 131, 288, 289, 293, 338, 339, 520.
— caesius L. 540.
— caucasicus Focke 425.
— chamaemorus L. 114, 136, 153, 158— 160, 162, 180, 184, 291, 296, 330, 339, 503, 556, 561, 571.
— hirtus W. et K. 413, 425, 432.
— humiltfolius G. A. M. 167.
— idaeus L. 184, 192, 195, 198, 211, 245, 325, 394, 397, 447.
— Raddeanus Focke 426.
— sanguineus Friv. 247, 432.
— saxatilis L. 167, 184, 186, 192, 227, 339, 344, 354, 627.
Rumex L. 523.
— acetosa L. 484—486, 523.
— acetosella L. var. multtfidus (L.) DC. 644.
— arcticus Trautv. 70.
— artfolius All. 131, 489.
— gramintfolius Lamb. 120.
— stenophyllus Ldb. 514.
— thyrstflorus Fingerh. 108, 523.
— tianschanicus A. Los. 499, 707.
I
Ruppta L. 538.
— maritima L. 538, 542.
— spiralis L. 538.
Ruscus hypophyllum L. 209, 420, 423, 424, 432.
— hyrcanus Woron. 426, 432.
— ponticus Woron. 247, 420, 423.
Saccharum spontaneum L. 534.
Sagittaria sagittifolia L. 515, 537.
— trifolia L. 542.
Salicornia L. 513, 793.
— europaea L. 508, 510—512, 515, 529,
534, 539, 540, 543, 623, 629, 647,
651, 664, 669, 696, 697, 731, 742,
755, 769, 791—793, 857.
— herbacea L. 793, 878, 925.
Salix L. 515, 535, 543.
— acmophylla Boiss. 852.
— acutifolia Willd. 642, 662.
— alatavica Kar. et Kir. 211, 843, 844.
— alba L. 390, 530, 539, 540.
— arbuscula L. 316, 478, 491, 548.
— arctica Pall. 79, 80, 97, 98, 100, 110, 128, 133, 502, 503.
— aurita L. 227.
— australior Anderss. 433, 534.
— berberifolia Pall. 104, 105.
— brachypoda (Trautv. et Mey.) Kom. 281, 289, 292, 456, 631.
— caprea L. 178, 229, 230, 325, 339, 343, 397, 454.
— caspica Pall. 662.
— Chamissonis Anderss. 110, 114.
— cinerea L. 168, 236, 343, 345, 430, 444, 454, 530, 543, 550.
— cuneata Turcz. 108, 110.
— dasyclados Wimm. 519.
— Daviesii Boiss. 852.
— fragilis L. 390, 534.
— fumosa Turcz. 105, 138.
— fuscescens Anderss. 546.
— glandulifera Floder. 518.
— glauca L. 70, 81, 90, 93, 96, 99, 101, 103, 117, 118, 121, 125, 131, 133,
134, 136, 268, 301, 303, 328, 518,
548.
— hastata L. 96, 99, 117, 211, 268, 274, 301, 505, 518, 548, 843.
— herbacea L. 94, 120, 121, 125.
— Integra Thunb. 437.
— Krylovii E. Wolf 92, 103, 106, 107, 519.
— lanata L. 70, 90, 93, 96, 99, 101, 117, 131, 132, 134, 137, 301, 303, 307, 310, 328, 518, 519.
— lapponum L. 125, 134, 136, 137, 162, 227, 328, 518.
— livida Wahlb. 444.
— myrsinites L. 93.
— myrtilloides L. 162, 227, 296, 307.
— nigricans (Sm.) Enand. 301, 444.
— nummularia Anderss. 102, 104.
— orbicularis Anderss. 110.
— oxycarpa Anderss. 847.
— Pallasii Anderss. 110.
— parallelinervis Floder. 274, 332, 519.
— pentandra L. 168, 339, 345, 444.
Salix phlebophylla Anderss. 91, 503.
— phylicifolia L. 90, 93, 117, 131, 132, 134, 136, 137, 162, 190, 268, 300, 301, 328, 444, 518, 519.
— polaris Wahlb. 69, 78—80, 81, 97, 100, 114, 115, 118, 120.
— pulchra Cham. 70, 71, 96, 97, 99, 101, 114, 117, 118, 121, 128, 518.
— purpurea L. 530, 539.
— pyrolifolia Ldb. 279, 307, 550.
— reptans Rupr. 81, 98, 114, 118, 128, 133, 504, 518, 519, 546.
— reticulata L. 94, 98—100, 119, 120, 486, 520.
— rhamnifolia Pall. 105.
— rosmarinifolia L. 549, 642.
— rossica Nas. 518—520.
— rotundifolia Trautv.69, 99, 115, 120,126»
— sachalinensis F. Schmidt 332, 519.
— saxalilis Turcz. 105.
— sibirica Pall. 343, 345, 511, 543, 550.
— songarica Anderss. 534, 535, 543.
— speciosa Hook, et Am. 519.
— sp. sp. 268, 429.
— Starkeana Willd. 437.
— stipulifera Floder. 137.
— torulosa Trautv. 108, 484.
— triandra L. 530, 539, 540.
— turanica Nas. 535.
— Turczaninowii Laksch. 486.
— viminalis L. 530.
— Wilhelmsiana M. B. 534, 535, 543.
— xerophila Floder. 240, 241, 273, 281, 290, 341, 343, 454, 456, 520, 631.
Salsola L. 513, 540, 731, 733, 736, 740, 742, 744, 755, 767—768, 786, 793,
797, 832, 925.
— aperta Iljin 801.
— aralensis Iljin 768
— arbuscula Pall. 740, 758, 762, 763, 767, 769, 772—774, 781, 784, 785, 787, 800, 810, 814, 816—818, 820, 833, 925.
— arbuscula s. 1. 732.
— Aucheri Bge. 760, 785.
— brachiata Pall. 759, 768, 769, 792, 811.
— carinata С. A. M. 762, 774, 801, 812, 813, 817—819.
— crassa M. B. 507, 534, 579, 755, 770, 777, 778, 780, 782, 791, 857.
— crassa s. 1. 793.
— dendroides Pall. 507, 508, 579, 742, 744, 777, 782, 783, '785, 791, 792, 832, 833, 925.
— ericoides M. B. 579, 742, 744, 777, 782, 783, 784, 925.
— foliosa (L.) Schrad. 760, 770.
— gemmascens Pall. 732, 740, 742, 772— 774, 784, 785, 792, 833, 925.
— glauca M. B. 784.
— hispidula Bge. 760.
— iliensis Lipsky 781.
— incanescens С. A. M. 792.
— lanata Pall. 534, 760, 768, 773, 785, 791, 792, 811, 813, 814, 817, 833.
— laricifolia (Turcz.) Litw. 732, 740, 753, 758, 760, 763, 767, 769—772, 786, 787, 925.
Salsola mutica С. А. М. 792.
— nitraria Pall. 755, 770.
— nodulosa (Moq.) Iljin 732, 742, 744, 777, 782—784, 925.
— Paletzkiana Litw. 800, 821, 823.
— passerina Bge. 732.
— Paulsenii Litw. 736.
— pellucida Litw. 755, 814, 816.
— praecox Litw. 801, 814, 816, 817.
— Richteri Karel. 799, 800, 813, SISSIS, 820, 821, 823, 925.
— rigida Pall. 536, 740, 758—760, 763, 767, 769, 770, 773, 781, 784—787, 800, 802, 810, 814, 816, 833.
— ruthenica Iljin 670, 675, 736, 755.
— sclerantha G. A. M. 774, 780, 786, 812, 819.
— soda L. 512, 516, 791.
— stellulata E. Kor. 760.
— subcrassa M. Pop. 515.
— tamariscina Pall. 751, 768, 771.
— turcomanica Litw. 534, 760, 791—793, 811, 812, 814, 833.
— turgaica Iljin 814.
— ustfurtensis E. Kor. 732.
— verrucosa M. B. 782.
Salvia L. 582, 596.
— armeniaca (Bordz.) Grossh. 711.
— austriaca Jacq. 634, 637.
— glutinosa L. 204, 208, 382, 407, 413.
— bine zev ski i Kudr. 727.
— nemorosa L. 656.
— nemorosa L. ssp. tesquicola (Klok. et Pobed.) Lavr. 657.
— nutans L. 613, 617, 634, 636, 637.
— pratensis L. 452, 611, 613, 620.
— stepposa Schost. 452, 528, 613, 621, 624, 637, 650.
— tesquicola Klok. et Pobed. 656.
— verticillata L. 710.
Salvinia natans (L.) All. 537, 541, 542.
Sambucus ebulus L. 433.
— nigra L. 208, 384, 390.
— sibirica Nakai 187, 230, 339.
Sanguisorba alpina Bge. 496
— officinalis L. Ill, 236, 289, 294, 297, 300, 313, 340, 341, 454, 523, 529, 530, 616, 625, 627, 629—631, 650, 683, 696.
— parviflora (Maxim.) Takeda 525.
— sitchensis С. A. M. 113.
— tenuifolia Fisch. 296, 341, 502, 503.
Sanicula europaea L. 194, 207, 208, 382, 408, 410, 416, 418.
Saposhnikovia divaricata (Turcz.) Schischk. 609.
Sasa kurilensis (Rupr.) Makino et Shi-bata 215, 334.
Saussurea alpina (L.) DC. 97, 98, 117, 120, 201, 267.
— amara (L.) DC. 623, 699.
— amurensis Turcz. 297.
— controversa DC. 229, 238, 625.
— foliosa Ldb. 486.
— Frolowii Ldb. 483, 485.
— Kitamuraria Miyabe et Tatew. 109. — latifolia Ldb. 196, 197, 312, 485, 486. — nuda Ldb. 79.
Saussurea pamirica C. Winkl. 847.
— papposa Turcz. 696, 697.
— robusta Ldb. 543.
— salicifolia (L.) DC. 690, 691, 698.
— salsa (Pall.) Spreng. 516, 623.
— Schanginiana (Wydl.) Fisch. 112, 484..
— sorocephala Schrenk 826, 843, 844,. 846.
— Tilesii Ldb. 126, 520.
Saxifraga L. 69, 82.
— bronchialis L. 67, 126.
— bronchialis s. 1. 502.
— caespitosa L. 81, 82, 107.
— cernua L. 77, 80, 114, 118, 126, 545r 546, 844.
— exarata Vill. 490.
— flagellaris Willd. 107, 126.
— flagellaris s. 1. 478.
— foliolosa R. Br. 70.
— hirculus L. 78, 102, 118, 314, 485, 520, 546, 837, 842, 846, 847.
— juniperifolia Adams 490, 836.
— Komarovii A. Los. 842, 847.
— melaleuca Fisch. 104.
— moschata Wulf. 490, 836.
— nivalis L. 77, 78, 80, 126.
— oppositifolia L. 77, 78, 80, 81, 107,. 126, 842, 843.
— punctata L. 118, 325.
— rivularis L. 505.
— serpyllifolia Pursh 82, 502.
— setigera Pursh 843, 844.
— sibirica L. 201, 316, 490, 842, 845.
Scabiosa L. 481.
— bipinnata C. Koch 592, 710, 711..
— caucasica Willd. 493.
— columbaria L. 710.
— Fischeri DC. 630, 683.
— isetensis L. 652, 675, 680.
— lucida Vill. 487.
— ochroleuca L. 232, 628, 636, 637.
— Olivieri Coult. 760, 779, 816, 817r 821.
— songorica Schrenk 495, 591.
— ucrainica L. 640, 641.
Scaligeria DC. 590.
— hirtula (Rgl. et Schmalh.) Lipsky 592.
Scandix stellata Soland. 786.
Scheuchzeria palustris L. 549, 550, 558.
559, 563.
Schismus P. B. 735.
— arabicus Ness 760, 771, 774, 801, 811 r 812, 814, 817.
Schistidium gracile (Schleich.) Limpr. 77r 80.
Schivereckia mutabilis Alex. 615.
Schizandra chinensis (Turcz.) Baill. 360..
Schizonepeta multifida (L.) Briq. 627. 688, 698, 699, 714.
Schraderia bucharica (M. Pop.) Nevski 580, 583.
Schultzia crinita (Pall.) Spreng. 102—105. 484-486.
Schumann ia Karelinii (Bge.) E. Kor. 735, 774, 815.
Scilia L. 375.
— autumnalis L. 670.
— bifolia L. 382, 389, 390.
Scilla Hohenackeri F. et M. 427.
— sibirica Anderws 384.
Scirpus L. 477.
— (Schoenoplectus) Hyppolitii V. Krecz. 542.
— kasachstanicus Dobroch. 541.
— (Schoenoplectus) lacustris L. 510, 512— 515, 530, 532, 535, 537, 542, 548, 550.
— (Schoenoplectus) litoralis Schrad. 542, 857.
— radicans Schkuhr 551.
— silvaticus L. 429, 549.
— (Schoenoplectus) Tabernaemontani Gmel. 286, 510, 512, 513, 530, 532, 537, 540, 543, 551, 857.
— (Schoenoplectus) triqueter L. 537, 540, 551.
Sclerochloa dura (L.) P. B. 832.
Scolochloa festucacea Link. 286, 510, 514, 532, 550.
Scorzonera L. 734.
— acanthoclada French. 582, 584, 589, 590, 708, 725, 845.
— austriaca Willd. 630, 631, 688, 701.
— ensifolia M. B. 515, 641, 662.
— inconspicua Lipsch. 708.
— parviflora Jacq. 514, 516.
— pusilia Pall. 768, 773.
— radiata Fisch. 305, 313, 485.
Scrophularia cretacea Fisch. 646.
— donetzica Kotov 644.
— minima M. B. 490.
— Scopolii Hoppe 205.
— turcomanica Bornm. et Sint. 728.
Scutellaria L. 582, 584, 590, 601, 632.*
— baicalensis Georgi 609, 683.
— Chitrovoi Juz. 615.
— creticola Juz. 615.
— galericulata L. 431, 514.
— Karjaginii Grossh. 581.
— Litwinowii Bornm. et Sint. 761, 781 785.
— lupulina s. 1. 704.
— orbicularis Bge. 725.
— orientalis L. 601.
— orientalis s. 1. 729.
— polyodon Juz. 618, 710.
— ramosissima M. Pop. 722.
— Tournefortii Benth. 410.
— ussuriensis (Rgl.) Kudo 375.
Secale anatolicum Boiss. 711.
Sedum L. 479.
— acre L. 645.
— cyaneum Rud. 107.
— Ewersii Ldb. 707, 843, 844.
— gracile С. A. M. 490.
— hybridum L. 233, 707, 715.
— involucratum M. B. 490.
— tenellum M. B. 479, 490.
Seidlitzia florida (M. B.) Boiss. 784.
Selaginella sibirica (Milde) Hieron. 111.
Sempervwum L. 479.
— caucasicum Rupr. 490.
— montanum L. 487.
— pumilum M. B. 490.
— ruthenicum (Koch) Schnittsp. et Lehm. 357, 645.
Senecio L. 823.
— bucharicus C. Winkl. 847.
— campester s. 1. 504.
— carpaticus Herb. 487.
— caucasicus (M. B.) DC. 209.
— cladobotrys Ldb. 492.
— congestum (R. Br.) A. Tolm. 505.
— coronopifolius Desi. 771, 773.
— frigidus Less. 545.
— Fuchsii Gmel. 205.
— integrijolius (L.) Clairv. 117, 120 r. 286, 612, 625, 630.
— Jacobaea L. 613, 650.
— Jacquinianus Rchb. 208, 325.
— kamtschaticus Kom. Ill, 332.
— nemorensis L. 489.
— octoglossus DC. 164, 192, 215, 312,-626.
— palmatus Pall. 293, 332, 502.
— paluster (L.) Hook. 514.
— platyphylloides S. et L. 492.
— pseudoarnica Less. 505.
— resedifolius Less. 98, 112, 118.
— soongoricus Fisch. 211, 494, 726, 846.-
— subdentatus Ldb. 811, 818, 819, 821.
— vernalis W. et K. 786.
Serratula cardunculus (Pall.) Schischk. 514, 630, 631, 695.
— toronata L. 529.
— lyratifolia Schrenk 719, 843.
— marginata Tausch. 485.
— quinquejolia M. B. 384, 407, 408.-
— radiata M. B. 634, 636.
— tanaitica Smirn. 646.
— xeranthemoides M. B. 636, 637, 655, 657, 660, 672, 751, 758.
Sesamum indicum L. 854.
Seseli campestre Bess. 636, 640, 655, 657.
— eriocarpum (Schrenk) Вк Fedtsch. 652t. 679.
— eriocephalum (Pall.) Schischk. 774.
— glabratum Willd. 675, 679.
— Ledebourii G. Don 622, 624, 650-
— sessiliflorum Schrenk 779.
Sesleria anatolica Deyl. 247.
— Bielzii Schur 487.
— coerulea (L.) Ard. 450, 451.
— phleoides Stev. 497.
Setaria glauca (L.) P. B. 858.
— viridis (L.) P. B. 695, 858.
Sibbaldia macrophyIla Turcz. 486, 844-
— parvijlora Willd. 491, 836.
— procumbens L. 98, 123.
— semiglabra С. A. M. 491, 836.
— tetrandra Bge. 827, 892, 926.
Sibbaldianthe adpressa (Bge.) Juz. 694.
Sibiraea altaiensis (Laxm.) С. K. Schn. 704-
Sideritis euxina Juz. 497.
— montanb L. 593.
Sieversia pentapetala (L.) Greene 503.
— pusilia (Gaertn.) Hulten 158.
Silaus Besseri DC. 528, 529, 531, 616, 622-
Silene L. 575.
— acaulis L. 69, 78, .97, 125.
- r bupletirbides L. 410.
— chamarensis Turci. 104.
— chlorantha (Willd.) Ehrh. 228.
— compacta Fisch. 246.
SUene cretacea Fisch. 646.
— daghestanica Rupr. 490.
— graminifolia Otth. 719, 723.
— hypanica Klok. 644.
— jenisseensis Willd. 242, 456, 694, 695.
— lithophilla Kar. et Kir. 843.
— multi] ida (Ad.) Rohrb. 209, 492.
— multiflora (Ehrh.) Pers. 621, 624, 650.
— папа Kar. et Kir. 781, 801.
— nutans L. 233.
— parviflora (Ehrh.) Pers. 641.
— paucifolia Ldb. 98.
— Ruprechtii Schischk. 245.
— stenophylla Ldb. 67, 107.
— suffrutescens M. B. 675.
— supina M. B. 644, 645.
— wolgensis. (Willd.) Bess. 652.
Sium latifolium L. 538.
-Skimmia repens Nakai 214.
. Smelov ski a alba (Pall.) Rgl. 108.
— calycina (Steph.) С. A. M. 103, 709, 795, 826, 844, 846, 847.
Smilacina dahurica Turcz. 294, 297.
— trifoliata Dest. 292, 297.
.S mil ax excelsa L. 374, 407, 408, 420, 423, 426, 432—434.
.Smirnovia Bge. 797.
.Smyrnium perfoliatum L. 403, 406.
.Sobolewskya caucasica (Rupr.) N. Busch 490.
Solanum dulcamara L. 374, 429.
— nigrum L. 858.
.Soldanella montana Willd. 204, 487, 489.
.Solenanthus circinnatus Ldb. 727.
.Solidago virga-aurea L. 68, 94, 131, 184, 187, 197, 225, 233, 236, 288, 293, 300, 306, 310, 329, 330, 482, 492, 499.
Solorina crocea Ach. 94.
Sonchus arvensi# L. 515, 639, 695.
Sophora alopecuroides L. 516, 534, 535, 543.
— faponica L. 852.
.Sorbaria sorbifolia (L.) A. Br. 187, 298, 307, 437.
.Sorbus amurensis Koehne 215.
— aucuparia L. 161, 177, 178, 182, 190, 227, 325, 336, 354, 357, 397.
— caucasigena Kom. 245.
— glabrata Hedl. 329.
— sambucifolia Roem. 110, 158, 292, 333, 334, 503.
— Schneideriana Koehne 215, 334.
— sibirica Hedl. 186, 187, 194, 195, 197, 198, 200, 236, 269, 279, 289, 338, 339.
— tianschanica Rupr. 212.
— torminalis (L.) Grantz 247, 409.
. Sorehum cernuum Host. 854.
— halepense (L.) Pers. 513, 858.
.Sparganium minimum Hill 543.
— neglectum Beeby 431.
— polyedrum Asch, et Gr. 432.
— ramosum s. 1. 530, 542.
— simplex Huds. 543.
Spergularia salina J. et C. Presl 543, 785.
-Sphaerella nivalis (Bauer.) Sommerf 837.
Sphaerophorus globosus Vain. 78, 94, 97, 111, 116, 120, 121.
^Sphaerophysa salsula (Pall.) DC. 516.
Sphagnum L. 266.
— acutifolium Ehrh. 163, 179, 180, 181, 290, 556.
— amblyphyllum Russ. 184, 558, 566.
— Angstroemii Hartm. 306.
— angustifolium G. Jens. 96, 153, 158, 169, 180, 291, 296, 306, 310, 330, 556—558, 561, 571.
— apiculatum H. Lindb. 178, 179, 296, 558.
— balticum Russ. 129, 138, 303, 557, 560, 561, 563, 567, 570, 571.
— centrale G. Jens. 169, 296, 330, 429, 431.
— compactum DG. 107, 125, 138, 296, 330.
— contortum Schultz. 556.
— cuspidatum Ehrh. 431, 558, 559, 567.
— Dusenii G. Jens. 557—559, 561, 567.
— fimbriatum Wils. 114.
— fuscum (Schimp.) Klinggr. 136, 180, 287, 296, 297, 330, 339, 556, 557,
559, 560, 563, 567, 569, 570, 571.
— Girgensohnii Russ. 125, 129, 136—138, 158, 163, 164, 166, 168, 169, 178, 179, 180, 205, 214, 290, 303, 310, 339, 488, 560, 571,
— Gravetii Russ. 431.
— Jensenii H. Lindb. 561.
— lenense H. Lindb. 138, 260, 558—561, 570, 571.
— Lindbergii Schimp. 136, 306, 559— — 561, 567, 570.
— magellanicum Brid. 109, 137, 180, 287, 291, 296, 297, 556, 557, 560, 561, 571.
— obtusum Warnst. 521, 549, 558, 561, 566.
— papillosum Lindb. 431, 556, 557, 570.
— platyphyllum (Sull, ex Lindb) Warnst. 268.
— pulchrum (Lindb.) Warnst. 558.
— quinquefarium (Lindb.) Warnst. 488.
— riparium Angstr. 184, 330. 429, 566.
— rubellum Wills. 330, 556, 567.
— Russowii Warnst. 136, 137, 162,169, 330.
— squarrosum Crome 114, 169, 303, 339, 429, 521, 560.
— subsecundum Nees 114, 297, 491, 545, 556, 560, 561, „566.
— teres (Schimp.) Angstr. 268, 303, 310, 491, 556, 558, 560.
— Warnstorfii Russ. 93, 107, 129, 137, 158, 169, 179, 268, 303, 306, 330, 479, 491, 556, 560, 561, 569.
— Wulfianum Girg. 339.
Sphenopus divaricatus (Gouan) Rchb. 588, 784.
Spinacia tetrandra Stev. 780.
Spiraea L. 468, 601, 658.
— alpina Pall. 104, 280.
— aquilegifolia Pall. 688, 699.
— betulifolia Pall. 292, 334.
— chamaedryfolia L. 311, 313,
— crenata L. 233, 343, 394, 468, 473,
614, 635, 638, 652, 653, 704, 729.
— ferganensis Pojark. 843.
— hypericifolia L. 407, 468, 471, 473,
496, 576, 583, 590, 592, 594, 635, 638,
652, 653, 661, 663, 664, 666, 673,
676, 679—681, 687, 705—708, 715,
718, 721, 749, 753, 757, 841.
Spiraea Litwinowii Dobrocz. 614.
— media Schmidt 194, 197, 231, 232,
235, 236, 281, 307, 313, 316, 338, 340, 341, 344, 447, 456, 609, 629, 632, 699.
— pilosa Franch. 726.
— salicifolia L. 288, 289, 298, 307, 316, 437.
— tianschanica Pojark. 843.
— trilobala L. 664, 706, 708, 841, 846.
— ulmifolia Scop. 204.
— ussuriensis Pojark. 435.
Spiraeanthus Schrenkianus (F. et M.) Maxim. 721, 762.
Spirodela polyrhiza (L.) Schleid. 537.
Spirorhynchus sabulosus Kar. et Kir. 801, 817, 821.
.Spodiopogon sibiricus Trin. 439.
Squamaria DC. 759.
— lentigera Nyl. 831.
— muralis (Schreb.) Elenk. 770.
— peltata DC. 770.
-Stachys angustifolia M. B. 644.
— atherocalyx C. Koch 710.
— fruticulosa M. B. 747.
— krynkensis Kotov 644.
— palustris L. 510, 538, 541.
— recta L. 613, 617, 620.
— Stchtschegleewii Sosn. 575.
— sylvatica L. 194, 195, 197, 397.
— turcomanica Trautv. 577, 727, 728, 761.
Staphylea colchica Stev. 423.
— pinnata L. 373.
Stellaria chamaejasme L. 630, 683, 688.
— cherleriae (Fisch.) Williams 688.
— crassifolia Ehrh. 549.
— dichotoma L. 686, 688, 699, 701.
— Edwardsii R. Br. 77, 78, 80, 82, 118.
— Eschscholtziana Fenzl 112.
— graminea L. 446, 523.
— holostea L. 174, 175, 178, 191, 227, 337, 354, 357, 382, 386, 388, 390— 393, 395, 397.
— humifusa Roth 81, 504, 505.
— nemorum L. 413.
— petraea Bge. 103.
Stemphylium Wallr. 737, 831.
Stereocaulon alpinum Laur. 77, 266.
— paschale (L.) Fr. 94, 103, 120, 121, 123, 125, 136, 153, 181, 195, 266, 270, 300, 301, 309, 328.
— rivulorum Magn. 80.
— tomentosum Fr. 120.
S ter igmo sternum tomentosum M. B. 768. Stevenia cheiranthoides DC. 686.
Stipa L. 473, 595, 643, 734, 783, 808, 823.
— attenuata P. Smirn. 608.
— baicalensis Roshev. 608, 610.
— borysthenica Klok. 599, 640, 656, 657, 662, 676.
— capillata L. 376, 396, 399, 456, 471, 472, 496, 498, 507, 576, 579, 580, 594,
599, 600, 612, 617, 618, 621, 625, 628,
632, 634, 636—638, 640, 643, 645, 647,
649, 651, 652, 654, 656, 657, 659, 660,
662, 663, 666, 667, 668—670, 672, 673,
676, 678—680, 686, 687, 690, 692, 693, 696, 704—706, 708, 710, 713— 718, 722, 727, 729, 734, 743, 749, 771, 779, 807, 840.
Stipa capillata s. 1. 628, 630, 683— 685, 688, 697, 924.
— caucasica Schmalh. 580, 582, 600, 681, 702, 718, 719, 725, 726, 762, 764, 771, 779.
— crassiculmis P. Smirn. 727.
— cretacea P. Smirn. 646.
— daghestanica (Westb.) Grossh. 711.
— dasyphylla Czem. 634, 637, 638, 704.
— decipiens P. Smirn. 608, 686, 688, 689, 691, 692, 694—696, 698, 701.
— glareosa P. Smirn. 600, 686, 687, 699, 700—702, 794, 847.
— gobica Roshev. 700.
— graniticola Klok. 644.
— Hohenackeriana Trin. et Rupr. 582, 729, 734, 759, 760, 763, 774, 780, 785.
— holosericea Trin. 729.
— Joannis Cel. 221, 231, 312, 357, 358, 393, 471, 528, 530, 531, 599, 600, 612, 615, 618, 621, 625, 635, 636, 685, 687, 695, 704, 729, 754.
— Joannis s 1. 599, 644.
— Joannis Cel.ssp. sabulosa Paez. 640, 662.
— kirghisorum P. Smirn. 494, 500, 599, 600, 603, 652, 660, 663, 681, 685, 705, 706, 709, 714, 717, 720, 721, 724, 725, 764.
— Korshinskyi Roshev. 603, 631, 649, 651, 660, 771.
— Krascheninnikowii Roshev. 672.
— Krylovii Roshev. 701.
— Lessingiana Trin. et Rupr. 393, 472, 507, 576, 579, 594, 599, 600, 634, 636, 638, 645, 649, 651, 652, 654, 656, 657, 659, 660, 663, 664, 669, 672, 673, 678, 679, 705, 713, 714, 717, 726—729, 746,748—750,753,757,759, 779, 807.
— Lessingiana Trin. et Rupr. var. Brau-neri Paez. 617.
— orientalis Trin. 496. 600, 663, 679— 681, 686, 687, 697, 699—702, 719, 725, 765, 794, 795, 847.
— pontica P. Smim. 617, 729.
— praecapillata Alech. 651.
— pulcherrima C. Koch, 400, 497, 498, 576, 612, 617, 618, 710, 711, 727, 729, 874.
— Richteriana Kar. et Kir. 680, 681, 759 773
— rubens P. Smim. 496, 500, 599, 621, 622, 624, 634, 636—638, 649, 651, 652, 659, 660, 672, 705—707, 713— 715, 717, 718, 749.
— sareptana Becker 600, 651, 659, 662, 664, 666, 667, 672, 676, 678, 680,
681, 734, 743, 748—751, 753, 757,
758, 759.
— sareptana Becker var. kasakorum Ro-shev. 770.
— sibirica (L.) Lam. 624, 696.
— stenophylla Gzern, 498, 599, 612, 617, 618, 634, 636—638, 649, 704, 711, 713, 729, 878.
Stipa Szowitsiana Trin. 594, 734, 747, 759, 770, 771, 773, 774, 781, 807, 816, 818.
— trichoides P. Smirn. 709, 725.
— turcomanica P. Smirn. 727.
— turkestanica Hack. 600, 764, 765.
— ucrainica P. Smirn. 599, 634, 637, 654, 657, 669, 671.
Stratiotes aloides L. 514, 537.
Stratonostoc Elenk. 672.
— (Nostoc) commune (Vauch.) Elenk. 78, 113, 114, 597, 609, 636, 638, 655, 665, 736, 747, 748, 750, 757.
Streptoloma desertorum Bge. 774, 781, 786, 801, 814, 817, 818.
Streptopus amplexifolius (L.) DC. 159. Struthiopteris filicastrum All. 187, 194, 208, 341, 355, 388, 392, 395, 418, 427, 432, 437.
Suaeda Forsk. 513, 543, 651, 731, 736, 742, 755, 793, 925.
— altissima (L.) Pall. 782.
— arcuata Bge. 857.
— confusa Iljin 507, 755.
— corniculata (С. A. M.) Bge. 510, 511, 515, 609, 623, 628, 629, 664, 696, 697, 699.
— dendroides (С. A. M.) Moq. 769, 777.
— linifolia Pall. 516.
— Lipskyi Litw. 811.
— microphylla Pall. 534, 540, 783, 791, 832.
— paradoxa Bge. 857.
— prostrata Pall. 507,508, 510, 511, 512, 515, 540, 623, 647, 664, 792.
Swertia iberica F. et M. 491.
— marginata Schrenk 847.
— obtusa Ldb. 102, 484—486.
Symphyoloma graveolens С. A. M. 490.
Symphytum asperum Lepech. 209.
— cordatum W. et K. 204, 414.
— grandiflorum DC. 207, 208, 384.
— tauricum Willd. 400.
Syrenia dolichostylos Klok. 641.
— siliculosa (M. B.) Andrz. 755. — ucrainica Klok. 641, 642.
Syringa robusta Nakai 215.
Taeniatherum crinitum (Schreb.) Nevski 474, 590, 721, 763, 777, 784.
Tamarix L. 539, 540, 543, 682, 742, 814, 822, 823, 858.
— elongata Ldb. 534.
— Ewersmannii Presl 756.
— florida Bge. 534.
— gracilis Willd. 756, 792.
— hispida Willd. 534, 536, 542, 813.
— Karelinii Bge. 534, 792.
— laxa Willd. 433, 530, 534—536, 540, 542, 756, 792.
— leptostachys Bge. 542, 792, 817.
— ramosissima Ldb. 507, 530, 534, 540, 756, 792.
Tamus communis L. 374, 407, 423, 432.
Tanacetum Herderi Rgl. et Schmalh. 844. — megalocephaia (Rupr.) P. Pol. 844, 845. — pamiricum* O. Hoffm. 703, 765.
— sibiricum L. 242, 439, 456, 595, 600,
609, 629—631, 682, 683, 690, 691, 826, 698, 924.
Tanacetum xylorrhizum Krasch. 731, 795r 796, 826, 846, 926.
Taphrospermum altaicum G. A. M. 843.
Taraxacum bessarabicum (Hora.) Hand.-Maz. 510, 529, 616.
— officinale s. 1. 447.
— officinale Web. 612.
— porphyranthum Boiss. 491.
— pseudoalpinum Schischk. 827.
— serotinum (W. et K.) Poir. 640'.
— Stevenii (Spreng) DC 490, 491.
Tauscheria Fisch. 735.
— lasiocarpa Fisch. 751, 758 , 759, 76T, 771, 811.
Taxodium distichum (L.) Rich. 852.
Tax us baccata L. 203, 348, 353, 423.
Teleckia speciosa (Schreb.) Baumg. 209r 492.
Teloschistes brevior Nillm. 787.
Tetr acme Bge. 735, 823.
— quadricornis (Steph.) Bge. 759, 769, 770, 773, 814, 821.
Tetradiclis tenella (Ehrenb). Litw. 588.
Teucrium L. 582, 632, 643.
— chamaedrys L. 246, 617, 618, 710.
— polium L. 508, 594, 601, 618, 644, 645, 647, 710, 780, 784.
Thalictrum alpinum L. 98, 102, 103, 111, 112, 117, 300, 484, 826.
— contortum L. 341, 436.
— filamentosum Maxim. 362.
— flavum L. 436.
— foetidum L. 279, 496, 840.
— minus L. 187, 188, 279, 280, 293, 311, 312,313, 333, 344, 470,486,499, 522, 529, 626, 627, 650, 698, 710, 715.
— petaloideum L. 624, 630, 631, 683, 686, 704.
— simplex L. 288, 289, 621, 707.
— simplex L. var. amurense Maxim. 525.
— sparsiflorum Turcz. 519.
Thamnolia Schaer. 479.
— vermicularis Ach. 77, 80, 81, 97, 111, 114, 128, 158, 488, 491.
Thelycrania (Cornus) alba (L.) Pojark. 187, 339, 343, 394.
— (Cornus) australis (G. A. M.) Sanadze* 381, 384, 400, 403, 425, 433, 617.
— (Cornus) sanguinea (L.) Fourr. 381, 383, 390—392.
Thermopsis alpina (Pall.) Ldb. 103.
— lanceolata R. Br. 630. 686, 690—692-
Thesium chinense Turcz. 439.
— multicaule Ldb. 678.
Thlaspi cochleariforme DC. 627.
Thuidium Br. et Sch. 192, 377.
— abietinum Br. et Sch. 107, 211, 522, 597, 612, 615.
Thuspeinantha persica (Boiss.) Briq. 760-
Thylacospermum caespitosum (CamJb.) Schischk. 827, 828, 843, 844.
— rupifragum Schrenk 827, 926.
Thymus L. 221, 358, 582, 601, 618, 628, 632, 643, 644, 690, 691, 710, 840.
— asiaticus Serg. 627, 687.
— baschkiriensis Klok. et Shost. 651.
Thymus borysthenicus Klok. et Shost. 641.
— Callieri Bobr. 617, 712.
— cretaceus Klok. et Shost. 646, 647.
— dimorphus Klok. et Shost. 508, 644, 645.
— Dubjanskii Klok et Shost. 646.
— eravinensis Serg. 630.
— eriophorus Ronn. 729.
— graniticus Klok. et Shost. 644.
— guberlinensis Iljin 663.
— karamarianicus Klok. et Schost. 581.
— kasakstanicus Klok. et Shost. 663.
— Kotschyanus Boiss. et Hohen. 575, 581, 729.
— Marschallianus Willd. 228, 621, 624, 638, 650, 673, 680, 705, 713—717.
— minussinensis Serg. 694, 695.
— moldavicus Klok. et Shost. 645.
— mugodzharicus Klok. et Shost. 663.
— Pallasianus H. Braun 641.
— pseudograniticus Klok. et Shost. 644.
— pulegioides L. 460.
— rasitatus Klok. 663.
— roseus Schipcz. 663.
— serpyllum s. 1. 233, 306, 582, 584, 628, 688, 699, 708, 845.
— sp. sp. 652, 690.
— subalpestris Klok. 488.
— tijlisiensis Klok. et Shost. 581.
Tilia amurensis Rupr. 213, 365, 370, 435—438.
— begoniijolia Stev. 371.
— caucasica Rupr. 371, 384, 417, 419, 423, 424, 433.
— cor data Mill. 165, 173, 177, 186, 192, 336, 339, 340, 348, 354, 365, 366,
370, 382, 389, 390, 392, 394, 396,
397, 413, 414, 417.
— cordijolia Bess. 371.
— dasystyla Stev. 371.
— manshurica Rupr. 364, 371, 610.
— multiflora Ldb. 371, 423.
— platyphyllos Scop. 371.
— rubra DC. 371.
— sibirica Fisch. 195, 370.
— Taqueti С. K. Schn. 213, 360, 362, 364, 370, 610,
— tomentosa Moench 371, 382.
Timmia bavarica Hessl. 211.
Tofieldia cernua Smith 306.
— palustris Huds. 125.
Tortella fragilis (Drum.) Limpr. 126.
'Tortula Hedw. 665.
— desertorum Broth. 597, 736, 738, 801, 811, 813, 816, 819.
— ruralis (Hedw.) Schwaegr. 211, 377, 597, 609, 612, 636, 655, 668, 670, 747, 749, 780,
T^orula Pers, ex Tr. 837.
Torularia contortuplicata (Steph.) O. Schulz 777, 783.
Torulopsis Berl. 837.
Tournefortia sibirica L. 507, 755.
— sogdiana (Bge.) M. Pop. 800, 823.
Trachomitum (Apocynum) lancifolium (Russan.) Pobed. 516, 534, 535, 543.
— 5^£осУпит) scabrum (Russan.) Pobed.
Trachydium kopetdaghense E. Kor. 727. Trachynia distachya (L.) Link. 593, 783.
Trachystemon orientale (L.) D. Don 208, 406, 418, 424.
Tragopogon L. 734, 808.
— brevirostre s. 1. 529.
— cretaceum Nik. 646.
— major Jacq. 454.
— tuberosus C. Koch 777.
Trapa natans s. 1. 515, 537, 540.
Trichophorum alpinum (L.) Pers. 486, 547.
— caespitosum (L.) Hartm. 95, 503, 557, 571.
— pumilum (Vahl) Schinz et Thell. 535. Trientalis europaea L. 158, 161, 166, 167, 177, 178, 180, 184, 187, 191, 219, 225, 241, 269, 280, 288, 318, 329, 338, 339, 354, 520.
Trijolium alpestre L. 246, 408, 711.
— ambiguum M. B. 497, 710, 711, 729.
— arvense L. 668, 669.
— campestre Schreb. 594.
— canescens Willd. 493.
— fragiferum L. 529.
— hybridwh L. 451, 498.
— lupinaster L. 229, 231, 233, 242, 260, 305, 313, 316, 339, 446, 621, 625, 627, 630, 631, 683, 704, 715.
— medium L. 497, 523, 710.
— montanum L. 394, 452, 497, 522, 528— 530, 600, 611, 613, 620, 634, 636, 704, 713.
— parviflorum Ehrh. 669, 671.
— pratense L. 446, 450, 451, 497, 523, 538 710
— repens L. 450, 451, 523, 538, 616, 712.
— spadiceum L. 445.
— trichocephalum M. B. 245.
Triglochin maritima L. 506, 510, 616, 623, 647.
— palustris L. 510, 535.
Trigonella L. 735, 763.
•— arcuata С. A. M. 759, 768, 770, 779.
— cretacea (M. B.) Grossh. 497.
— grandiflora Bge. 587.
— Noeana Boiss. 587, 778, 785.
— orthoceras Kar. et Kir. 759, 767.
— pamirica Boriss. 722.
— Popovii E. Kor. 584, 845.
— radiata (L.) Boiss. 778.
Trinia hispida Hoffm. 672.
Tripolium {Aster) vulgare Nees. 506, 510, 516, 539, 616, 697, 858.
Trisetum altaicum (Steph.) Roshev. 104, 485,' 486, 827, 842.
— pratense Pers. 454, 497, 498.
— sibiricum Rupr. Ill, 201, 316, 494, 496, 502, 519, 520, 525, 844.
— spicatum (L.) Richt. 70, 79, 100, 102, 111, 112, 126, 494, 496, 827, 845.
— virescens (Rgl.) B. Fedtsch. 499, 716.
Trollius altaicus С. A. M. 301, 483—485, 494, 705.
— asiaticus L. 108, 236, 306, 313, 485, 486, 626.
— chinensis Bge. 525.
— europaeus L. 93, 94, 167, 330, 523.
— Ledebourii Rchh. 341.
Trollius Riederianus F. et M. Ill, 112, 502.
Tulipa L. 597, 735, 758, 776.
— Albertii Rgl. 767.
—4 Behmiana Rgl. 759, 767.
— Biebersteiniana Schult. 635—637, 650, 655, 657, 660, 672, 773.
— biflora Pall. 672, 679, 748, 751, 768.
— Borszczowii Rgl. 771, 773.
— Buhseana Boiss. 773.
— Greigii Rgl. 760, 767, 778.
— Hoogiana B. Fedsch. 786.
— kuschkensis B. Fedtsch. 580.
— Lehmanniana Merckl. 781.
— Micheliana Hoog 726.
— patens Agardh. 650, 660.
Schrenkii Rgl. 635—637, 655—657, 660, 669, 672, 679.
— sogdiana Bge. 785, 800.
— sp. sp. 587.
— turkestanica Rgl. 721, 767, 778.
— uniflora (L.) Bess. 685, 690—692.
Typha L. 477, 513, 539.
— ahgustata Bory et Ghaub. 542.
— angustifolia L. 510, 513, 515, 530, 532, 535, 537, 540—543, 548.
— elephantina Roxb. 857.
— latifolia L. 431, 510, 514, 515, 530, 532, 537, 548, 550, 551,
— Laxmannii Lepech. 513, 542, 543, 857 858
— minima Funk 513, 534, 542, 857.
Ulmus Androssowii Litw. 852.
— densa Litw. 852.
— el lip tic a C. Koch 207, 423, 424.
— foliacea Gilib. 348, 382, 389, 390, 392, 393, 396, 399, 400, 423, 425, 433, 579, 617, 635.
— laciniata (Trautv.) Mayr 213, 364.
— laevis Pall. 173, 178, 348, 354, 384, 388, 390, 392, 394, 396, 397, 530, 539 540.
— propinqua Kidz. 360, 364, 437.
— pumila L. 689, 699, 852.
— scabra Mill. 165, 178, 192, 207, 348, 354, 385, 392, 396,397,419,530,852.
— suberosa Moench 348, 400, 433, 434, 579.
Urtica dioica L. 388, 392, 397, 398, 429, 433.
Usnea (Dill.) Ach. 163.
— barbata (L.)Hoffm. 169.
Utricularia vulgaris L. 514, 515.
Vaccinium arctostaphylos L. 406, 419, 423, 424.
— myrtillus L. 93, 94, 96, 103, 125, 153, 161, 162, 167, 169, 178, 184, 195, 198, 205, 215, 219, 224, 225, 227, 230, 231, 235, 236, 245, 269, 279, 300, 318, 329, 338, 354, 357, 358, 415, 419, 482, 487, 488.
— ovalifolium Smith 214, 215, 292, 297. — uliginosum L. 93, 94, 97, 98, 117, 121, 125, 126, 129, 138, 153, 158, 160—162, 219, 220, 224, 225, 227, 230, 235,
236, 241, 266, 269, 272, 279, 282,
284, 289, 291—293, 300, 301, 307,
316, 318, 329, 330, 482, 488, 502v 503, 556, 561, 571.
Vaccinium vitis-idaea L. 69, 94, 96, 103,. 108, 111, 115, 117, 118, 120, 121,. 125, 129, 137, 138, 153, 158, 169, 180,.
190, 198, 200, 219, 221, 224, 225, 227,
230, 231, 233, 235, 236, 241, 245, 266,.
267, 269, 279, 282, 284, 293, 294, 300,
302, 307, 309, 310, 316, 329, 343, 354,. 357, 358, 415, 488, 491, 502, 561, — vulcanorum Kom. 111.
Vahlodea atropurpurea (Wahlbg.) Fries 505.. Valeriana alliariaefolia Vahl 208, 49K — caespitosa Rupr. 842, 843, 845.
— capitata Pall. 111,117,118,267,300,520. — colchica Utk. 492.
— ficariaefolia Boiss. 728.
— officinalis s. 1. 496.
— rossica Smirn. 612.
— sisymbriifolia Desf. 728.
— tuberosa L. 597, 636.
Valerianella Mill. 776.
— dufresnia Bge. 819.
Vallisneria spiralis L. 537, 540.
Veratrum Lobelianum Bernh. 159, 187r 193, 196, 301, 482, 485, 486, 519, 629.
— Maackii Rgl. 448, 457.
— nigrum L. 613, 627, 630, 631, 638. — oxysepalum Turcz. Ill, 297, 332, 502. Verbascum L. 597.
— lychnitis L. 613, 620, 636.
— orientale M. B. 636.
— phoeniceum L. 652, 656.
— saccatum G. Koch 777.
— songoricum Schrenk 472, 577, 722. Veronica L. 735.
— amoena Stev. 769, 770.
— biloba L. 722, 751, 757, 759, 760„ 767, 771, 778, 816, 817.
— campylopoda Boiss. 779, 780, 793, 803.
— chamaedrys L. 382.
— densiflora Ldb. 486.
— didyma Ten. 780.
— gentianoides Vahl 618, 712.
— grandiflora Gaerth. Ill, 502.
— incana L. 233, 260, 286, 312, 609, 613. 628, 630, 650, 688, 690, 692, 693, 698, 713.
— intercedens Bornm. 725.
— longifolia L. 454, 485, 519, 523,621-— macrostemon Bge. 842, 843.
- maxima Mill. 204, 205, — minuta G. A. M. 490. — olgensis Kom. 435. — orientalis Mill. 729.
— pinnata L. 652, 686, 694, 696, 700, 705.
— prostrata L. 396.
— sibirica L. 436, 457.
— spicata L. 228, 233, 483, 621, 687, 705, 715.
— spicata s. 1. 650.
— spuria L. 613, 708.
— Stelleri Pall. 503.
— verna L. 635, 636, 655, 656, 707. Verrucaria Th. Fr. 78.
Viburnum lantana L. 245,373,381, 390,729-— opulus L. 167, 192, 195, 383, 390.
429,-432, 530.
Viburnum orientale Pall. 423.
— pubinerve Blume. 437.
— Sargentii Koehne 360, 436.
Vicia amoena Fisch. 281, 448, 629—631
— amurensis Oett. 435.
— baicalensis (Turcz.) B. Fedtsh. 2R0
— cracca L. 232, 281, 446, 448 451 доя’ 516, 523, 538, 621. ’ ’
— crocea (Desf.) B. Fedtsch. 208 407
— multicaulis Ldb. 630, 631.
— peregrina L. 832.
— sepium L. 232, 311, 497.
— silvatica L. 184.
— tenuifolia Roth 471, 474, 634. 636
637, 707. ' ’
— unijuga A. Br. 232, 446, 448, 626.
— variabilis Freyn et Sint. 497, 711.
— variegata Willd. 711.
Vigna sinensis Endl. 854.
Vinca herbacea W. et K. 390, 400, 710.
Viola alba Besser 406.
— altaica Ker-Gawl. 92, 104, 108, 485 486.
— ambigua W. et K. 636, 637.
— bijlora L. 113, 195, 312, 482, 486, 843.
— cretacea Klok. 646.
— hirta L. 612.
— Kitaibeliana Roem. et Schult. 635, 636, 655, 657.
— mirabilis L. 178, 381, 392, 395,397.
— oreades M. B. 490, 711.
— repens Turz. 520.
— rupestris F. W. Schmidt 612.
— sacchalinensis De Boiss. 293.
— Selkirkii Pursh 191.
— Sieheana W. Bckr. 410, 426, 427.
— Sintenisii W. Bckr. 728.
— sp. sp. 384.
— suavis M. B. 396.
— sylvestris Lam. 415.
— tianschanica Maxim. 827.
— tricolor L. 395.
— uniflora L. 195, 200, 236, 312, 339.
Vitis amurensis Rupr. 214. 360, 374, 436.
— silvestris Gmel. 432, 433.
— v in if er a L. 408, 469.
Waldheimia Korolkovii Rgl. er Schmalh. 837, 845.
Waldheimia tridactylites Kar. et Kir. 78, 843, 844, 846, 847.
Waldsteinia ternata (Steph.) Fritsch 295.-Winklera «ilaifolia Koren. 847.
Woodsia alpina (Bolton) Gray 645.
— ilvensis R. Br. 841.
Yucca L. 853.
Xeranthemum annuum L. 644.
Zannichellia palustris s. 1. 543
— pedunculala Rchb. 538, 539, 542.
Ze z°O7a carPinif°lia (Pall.) Dipp. 425,. 427, 853.
— hyrcana Grossh. et A. Jarm. 425 427 Zizania latifolia Turcz. 551. ’
Ziziphora L. 575, 581, 601, 735. — oorzhomica Juz. 581.
— Bungeana Juz. 582, 584, 589, 592. 680, 705, 706, 717, 721, 722,728.840. 846, 847.
— clinopodioides 8. 1. 496, 725.
— denticulata Juz. 581.
— media Link. 581.
— persica Bge. 728.
— tenuior L. 760, 767, 776, 779—781,. 786, 819.
Zizyphus Mill. 468.
— jufuba Mill. 468, 474.
Zollicoferia (Launea) Korovinii M. Pop.-773.
Zosimia absinthifolia (Vent.) Lunk. 728v 761.
Zostera marina L. 508, 543.
— minor (Cavol.) Nolte 508, 543.
Zygadenus sibiricus (L.) A. Grey 238..
Zygophyllum atriplicoides Fisch. 762, 780F 786.
— Eichwaldii С. A. M. 774, 823.
— eurypterum Boiss. et Buhse 781.
— fabago L. 858.
— macropterum С. A. M. 675, 680, 761-
— megacarpum Boriss. 580.
— oxianum Boriss. 858.
— portulacoides Cham. 786.
— Rosovii Bge. 794, 795.
— turcomanicum F. et M. 774.
Утверждено к печати Ботаническим институтом Академии Наук СССР
♦
Редактор Издательства Д. В. Лебедев Технический редактор Р. С. Певзнер Корректоры Т. А. Пострелова и К. Н. Феноменов
♦
РИСО АН СССР № 10®—38В. Подписано к печати 30 мая 1956 г. М-09347. Бумага 70ХЮ81/1в. Бум. л. 16. Печ. л. 43,84. Уч.-изд. л. 47,43 + +10 вклеек + карта на 8 листах (15,09-уч.-изд. л.) Тираж 5000 экз. Заказ № 551. Цена книги с приложением геоботанической карты 48 руб.
1-я тип. Издательства АН СССР, Ленинград, В. О., 9 линия, д. 12.
ИСПРАВЛЕНИЯ И ОПЕЧАТКИ В 1 И П ЧАСТЯХ
Страница Строка I |__ Напечатано | _ 1 __ Должно быть
88 77 104 126 15S 21 сверху । 12 снизу 2 сверху 1 27 снизу ' 21 сверху 1 данных) Kununcu/us Ptilagrosti, ospicatuin, Roschniacia ! данных. Ranunculus Ptilagrostis, spica^um, Roschniakia
184 6 снизу повышених повышениях
210 24 сверху 58 с a 57 са
210 25 сверху 57 са 58 са
32В 4 сверху Populus. Populus
В ВО 9 сверху Preurozium Pleurozium
ВВ4 16 сверху Scheideriana Schneideriana
874 10 снизу vitabla vitalba
384 24 снизу ranunculoides rapunculoides
407 2В снизу ranunculoides rapunculoides
408 11 сверху vitabla vitalba
419 20 снизу защищенные ущелья защищенных ущельях
4В2 18 сверху vitabla vitalba
4ВВ 20 снизу vitabla vitalba
489 7 сверху Spodiopogon Gmelini S podiopogon sibiricus
507 колон-титул лугово-болотистая j лугово-болотная
507 1 сверху Гурьевым | Гурьевом
508 И сверху Meyerii I Meyeri
525 19 снизу примесью разнотравья (Яапил-i примесью Agrostis stolnnizans и
cuius repens, Caltha palustris. разнотравья (Ranunculus re-
Galium palustre, Mentha ar- pens, Caltha palustris, Galium
vensis, Agrostis stolonizans). palustre, Mentha arvensis).
520 18 сверху осочникам и ос очниками
540 3 снизу югу-востоку юго-востоку
546 16 сверху Франца-Иосифа Земли Франца-Иосифа
558 21 сверху lenens lenense
562 9 сверху В. Н. Андреев (1932а, 19326, В. Н. Андреев (1932, 1932а,
580 18 снизу rhytocarpa rhyticarpa
600 22 снизу Убсанурской Убсунурской
606 11 снизу Убсанурской Убсуну рекой
610 13 снизу сплошной почти сплошной
688 14 сверху выпасе с выпасе и с
642 649 649 658 666 6 снизу 2 снизу 8 снизу 22 сверху 2 сверху полукустарников отд. би-ол. Festura В. Е. Вандакурова характеризуется полукустарничков отд. биол. Festuca Е. В. Вандакурова характеризуются основе»
694 4 снизу основе мятличники
709 719 786 800 11 снизу 20 сверху 17 сверху 8 снизу мятлички Бойбичето ксероморфной Tulipa sogdiana, Bongardia Байбичето склероморфной Tulipa sogdiana и др.
chrysogonum и др. 14,
859 10 сверху 15, usv NSV
866 8 сверху