














































































































































































































































































































































































Author: Петров К.М. Терехина Н.В.
Tags: география растений фитогеография флора географическое распространение растений биологические науки в целом экология россия учебник для студентов растительный покров растительные сообщества ботаническая география
ISBN: 978-5 -93808-314-1
Year: 2017




Глава 1. ИСТОРИЧЕСКИЕ УСЛОВИЯ ФОРМИРОВАНИЯ РАСТИТЕЛЬНОГО ПОКРОВА
1
К. М. Петров
Н. В. Терехина
АСТИТЕЛЬНЫЙ
ПОКРОВ
ОССИИ
Одобрено учебно-методической комиссией Института наук о Земле
Санкт-Петербургского государственного университета
в качестве учебника для студентов высших учебных заведений
в рамках подготовки специалистов естественнонаучных направлений
САНКТ-ПЕТЕРБУРГ
ХИМИЗДАТ
2017
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
2
УДК 581.9 (47+57) (075.8)
П 305
Научный редактор:
зав. кафедрой биогеографии и охраны природы Ин-та наук о Земле С.-Петер-
бургского государственного ун-та, канд. биол. наук А. А. Егоров
Рецензенты:
д.б .н., проф., зав. каф. ботаники И. И. Шамров; к.б.н., доцент каф. ботаники
И. В. Панкратова (РГПУ им. А . И. Герцена);
к.б .н., доцент каф. ботаники и дендрологии В. Ю. Нешатаев (С. -Петерб. гос.
лесотехнический ун-т им. С. М . Кирова);
д.г .н., проф. каф. страноведения и международного туризма А. А . Григорьев
(Ин-т Наук о Земле, С.-Петерб. гос. ун-т).
Петров К. М., Терехина Н. В.
П 305
Растительный покров России. Учебник. – СПб.: ХИМИЗДАТ, 2017. −
368 с.: ил.
ISBN 978-5 -93808-314-1
Излагаются базовые положения ботанической географии: исторические
условия формирования растительного покрова; учение об ареалах; учение о рас-
тительных сообществах; экологические и эколого-топологические факторы рас-
пределения растительности; географические закономерности растительного по-
крова России. Основное содержание книги посвящено описанию растительности
природных зон России: тундры, тайги, широколиственных лесов, интразональ-
ных типов растительности (суходольных лугов и болот), степей, субтропиков и
высотной поясности горных стран. Книга предназначена для обучения студентов
высших учебных заведений в рамках подготовки специалистов естественнонауч-
ных направлений, а также для практиков, занимающихся организацией особо
охраняемых природных территорий, и всех любителей природы.
Petrov K. M., Terekhina N. V.
Vegetable cover of Russia. Saint-Petersburg: Publisher...., 2017.
The basics of botanical geography are expounded: historical conditions of vege-
tation cover formation; chorology; studies of plant communities; ecological and eco-
topological factors of vegetation distribution; geographical patterns of the vegetation
cover of Russia. The main content of the book is devoted to the description of the
vegetation of the Russian natural zones: tundra, taiga, broad-leaved forests, intrazon-
al types of vegetation (meadows and bogs), steppes, subtropics, and altitudinal zones
of mountains. The book is intended for training of students with natural science spe-
cialization, as well as for practitioners involved in the organization of protected natu-
ral areas, and for all nature amateurs.
П
1805040900–034
050(01)–17
Без объявл.
Петров К. М., Терехина Н. В., 2017
ISBN 978-5 -93808-314-1
ХИМИЗДАТ, 2017
.
Глава 1. ИСТОРИЧЕСКИЕ УСЛОВИЯ ФОРМИРОВАНИЯ РАСТИТЕЛЬНОГО ПОКРОВА
3
ВВЕДЕНИЕ
5
Часть I
БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
Глава 1. ИСТОРИЧЕСКИЕ УСЛОВИЯ ФОРМИРОВАНИЯ РАСТИТЕЛЬНОГО
ПОКРОВА
8
Глава 2. УЧЕНИЕ ОБ АРЕАЛАХ
16
Глава 3. УЧЕНИЕ О РАСТИТЕЛЬНЫХ СООБЩЕСТВАХ (ФИТОЦЕНОЛОГИЯ)
24
Глава 4. ФАКТОРЫ РАСПРЕДЕЛЕНИЯ РАСТИТЕЛЬНОСТИ
34
Глава 5. ГЕОГРАФИЧЕСКИЕ ЗАКОНОМЕРНОСТИ РАСТИТЕЛЬНОГО ПОКРОВА
РОССИИ
40
Часть II
РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
Арктический и субарктический пояса
59
Глава 6. ЗОНА ТУНДР
60
6.1. Подзоны тундр
69
6.2. Ботанико-географические области тундр
81
6.2 1. Области высокоарктических тундр (полярных пустынь)
81
6.2.2. Области Арктических, Субарктических тундр и лесотундры
82
6.2.2.1. Кольская область тундр
82
6.2.2.2. Восточно-Европейская область тундр
87
6.2.2.3. Ямало-Гыданская область тундр
91
6.2.2.4. Таймырская область тундр
98
6.2.2.5. Лено-Колымская область тундр
104
6.2.2.6. Чукотская область тундр
105
6.2.2.7. Северо-Тихоокеанская островная область
113
6.3. Антропогенные нарушения ландшафтов тундр
118
Умеренный пояс
124
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
128
7.1. Подзоны тайги
135
7.2. Ботанико-географические области таежных лесов
139
7.2.1. Восточно-Скандинавская (Карельская) таежная область
139
7.2.2. Восточно-Европейская таежная область
143
7.2.3. Западно-Сибирская таежная область
154
7.2.4. Средне-Сибирская таежная область
166
7.2.5. Сахалинская таежная область
173
7.2.6. Приморская область хвойно-широколиственных лесов
178
7.3. Антропогенные нарушения ландшафтов тайги и хвойно-
широколиственных лесов
194
ОГЛАВЛЕНИЕ
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
4
Глава 8. ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
198
8.1. Материковые луга лесной зоны
198
8.2. Болота
210
8.2.1. Эколого-топологические условия образования верховых
сфагновых болот
211
8.2.2. Стадии развития верховых болот
215
8.3. Антропогенные нарушения болотных экосистем
218
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
221
9.1. Ботанико-географические области широколиственных лесов
223
9.1.1. Восточно-Европейская область широколиственных лесов
223
9.1.2. Западно-Сибирская лесостепная область
241
9.1.3. Дальневосточная область широколиственных лесов
243
9.2. Антропогенные нарушения широколиственных лесов и лесостепи
245
Глава 10. ЗОНА СТЕПЕЙ
247
10.1. Подзоны степей
252
10.2. Ботанико-географические области северных и средних степей
254
10.2.1. Восточно-Европейская (Донско-Волжская) степная область
254
10.2.2. Заволжско-Уральская степная область
258
10.2.3. Западно-Сибирская степная область
259
10.2.4. Дауро-Монгольская степная область
262
10.2.5. Прикаспийская область южных опустыненных степей
264
10.3. Антропогенные нарушения ландшафтов степей
272
Субтропический пояс
276
Глава 11. ЗОНА СУБТРОПИКОВ
276
11.1. Субтропики Южного берега Крыма
276
11.2. Субтропики Черноморского побережья Кавказа
282
Часть III
РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
290
12.1. Растительность Урала
290
12.2. Растительность Кавказа
294
12.3. Растительность Алтая
309
12.4. Растительность гор Восточного Прибайкалья
318
12.5. Растительность гор Северо-Востока Сибири
324
12.6. Растительность Камчатки
327
Литература
333
УКАЗАТЕЛЬ НАЗВАНИЙ РАСТЕНИЙ:
334
русских
334
латинских
346
Сведения об авторах
367
ОГЛАВЛЕНИЕ
.
Глава 1. ИСТОРИЧЕСКИЕ УСЛОВИЯ ФОРМИРОВАНИЯ РАСТИТЕЛЬНОГО ПОКРОВА
5
Посвящается столетию
создания кафедры ботанической географии
Санкт-Петербургского государственного университета
астительный покров – национальное достояние России, хранящее огром-
ный ресурсный и экологический потенциал, источник научного и творче-
ского вдохновения. Подготовка специалистов в области его изучения и охраны
представляется делом государственной важности. Отечественная школа бота-
нико-географов пользуется заслуженным авторитетом и мировой известно-
стью. Ее основоположник А. Н. Бекетов в 1896 г. опубликовал первый учебник
«География растений: Очерк учения о распространении и распределении рас-
тительности на земной поверхности с особым прибавлением о Европейской
России». В 1918 г. в составе географического факультета Петроградского госу-
дарственного университета была создана кафедра ботанической географии
(ныне кафедра биогеографии и охраны природы Санкт-Петербургского гос-
университета). Это первая в России кафедра, в задачи которой входило подго-
товка отечественных специалистов в области изучения растительного покрова.
На протяжении XX в. изучение тундр, лесов, болот, лугов, степей, пустынь,
растительности горных стран обогатило теорию и практику ботанической гео-
графии новыми данными. Особое место занимал опыт картографирования
растительности, создание геоботанических карт, как всей страны, так и круп-
ных регионов: Восточно-Европейской равнины, Западной Сибири, и админи-
стративных подразделений (от Ленинградской до Сахалинской областей).
Растительный покров, как единство флоры и растительности, в планетар-
ном масштабе составляет фитосферу географической оболочки Земли. Зеле-
ные растения, по словам В. И. Вернадского, – это живое вещество первого по-
рядка, исходное звено всех цепей питания в экосистеме. Растения служат не
только основанием трофической пирамиды, но и благодаря своей неподвиж-
ности они дают приют и убежище населению животного мира. Растительность
обогащает атмосферу кислородом, влияет на характер климата, формирует
ВВЕДЕНИЕ
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
6
почву с ее плодородием, выполняет водорегулирующую и противоэрозионную
функции, придает ландшафтам их неповторимый облик. Все усилия по под-
держанию на Земле условий, необходимых для жизни, не могут быть эффек-
тивными без сохранения растительного покрова и разнообразного видового
состава растений.
Ботаническая география, изучающая растительный покров как один из
компонентов современных ландшафтов и ландшафтов прошлых геологиче-
ских эпох, имеет большое практическое значение. Ее данные применяют при
сельскохозяйственном освоении земель, использовании лесных и водных ре-
сурсов, при проведении природного районирования и ландшафтного плани-
рования, при разработке программ охраны природы и сохранения биоразно-
образия.
Актуальность издания учебника «Растительный покров России» объясня-
ется тем, что последнее учебное пособие В. В. Алехина «Растительность СССР в
основных зонах» вышло в свет более полувека назад. В предлагаемом учебни-
ке излагаются базовые понятия ботанической географии и даются описания
растительности природных зон на равнинах и высотной поясности в горах
России. Зональные и региональные особенности растительного покрова ил-
люстрируются геоботаническими картами, фотографиями сообществ и видов-
доминантов. Учебник отвечает задачам подготовки специалистов, способных
решать насущные задачи изучения и охраны растительного покрова России.
Издание учебника обеспечит обучение студентов высших учебных заведений в
рамках подготовки специалистов естественнонаучных направлений и будет
стимулировать активное участие общества в природоохранной деятельности.
ВВЕДЕНИЕ
.
Глава 1. ИСТОРИЧЕСКИЕ УСЛОВИЯ ФОРМИРОВАНИЯ РАСТИТЕЛЬНОГО ПОКРОВА
7
Часть I
БАЗОВЫЕ ПОЛОЖЕНИЯ
БОТАНИЧЕСКОЙ ГЕОГРАФИИ
Описание растительности России опирается на базовые поло-
жения ботанической географии. Ботаническая география как
раздел биогеографии – наука о закономерностях географиче-
ского распределения растительности в зависимости от геологи-
ческого прошлого, современных эколого-географических усло-
вий, а также характера и интенсивности антропогенных воздей-
ствий. Объектом изучения ботанической географии является
растительный покров Земли и все его структурные подразделе-
ния, своеобразие которых тесно связано с территориальными
подразделениями географической оболочки. При изучении рас-
тительного покрова необходимо последовательно рассмотреть
следующие положения: сведения об истории формирования
растительного покрова, ареалах растений, о составе и структуре
растительных сообществ (фитоценозов), об экологических и
ландшафтных факторах и эколого-географических закономер-
ностях распределения растительного покрова.
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
8
ормирование растительного покрова России, занимающей обширные
пространства Евразии, происходило на протяжении новой геологиче-
ской эры – кайнозоя. Кайнозой (начало 60−70 млн. лет назад) подразделяется
на палеоген, неоген (раньше их объединяли в третичный период) и четвер-
тичный период – плейстоцен и голоцен.
В кайнозое наземная биота обогатилась цветковыми (покрытосеменны-
ми) растениями и коэволюционировавшими с ними членистоногими, в первую
очередь насекомыми, как необходимыми для размножения опылителями, так
и разнообразными вредителями растений. Динозавры, господствовавшие в
мезозое, уступили место млекопитающим и птицам. Большое влияние на раз-
витие органического мира в кайнозое оказали тектонические процессы и из-
менения климата.
Палеоген. Процессы горообразования затихли, уступив место действию
внешних сил – денудации. Горные сооружения были почти полностью разру-
шены, и в палеогене на их месте сформировался пенеплен – слаборасчленен-
ная равнина, где остались невысокие сопки и холмы, сложенные наиболее
устойчивыми к процессам разрушения породами. Равнинный рельеф оказал
влияние на формирование климата. Воздушные массы с океанов не встречали
препятствий, и на обширных простран-
ствах Евразии установился теплый и влаж-
ный климат. Тепло было даже за поляр-
ным кругом, так что в Гренландии и на
Шпицбергене росли субтропические леса.
На территории Евразии сформиро-
валась тропическая и субтропическая фло-
ра, получившая название полтавской. Она
характеризовалась широким распростра-
нением вечнозеленых растений: пальм,
магнолий, лавровых, миртовых, секвой, а
также каштанов, дубов и других широко-
лиственных пород (рис. 1.1).
Неоген. Это период формирования
складчатой структуры альпид и возник-
новения орографически выраженных осе-
вых частей современных горных систем
Рис. 1.1. Палеогеновый ландшафт с полтав-
ской биотой (по И. Аугуста, З. Буриану)
Глава 1
ИСТОРИЧЕСКИЕ УСЛОВИЯ ФОРМИРОВАНИЯ
РАСТИТЕЛЬНОГО ПОКРОВА
.
Глава 1. ИСТОРИЧЕСКИЕ УСЛОВИЯ ФОРМИРОВАНИЯ РАСТИТЕЛЬНОГО ПОКРОВА
9
Рис. 1.2. Ранненеогеновый ландшафт с тургайской биотой
(по И. Аугуста, З. Буриану)
Евразии, выросли Альпы, Кавказ, омолодился рельеф гор каледонской и гер-
цинской складчатости. Высокие горы стали преградой на пути западного пе-
реноса воздушных масс. К концу неогена усилились похолодание и аридиза-
ция климата.
По мере похолодания полтавская флора отступила к югу от линии: устье
Вислы в Европе – устье Янцзы в Китае. К северу от названной границы на
смену полтавской флоре пришли представители тургайской флоры, характе-
ризовавшейся развитием богатых по видовому составу широколиственных
листопадных лесов. Типичными ее представителями являются: каштан, бук,
граб, орех, ликвидамбар, береза, ольха и др.; среди хвойных – метасеквойя,
таксодиум и др. На рис. 1.2 представлен среднеевропейский ландшафт в нача-
ле неогена с лесом, образованным широколиственными и хвойными деревья-
ми. В неогене тургайская флора распространялась от высокоширотной Аркти-
ки до Кавказа и Казахстана. Из ее представителей сформировались современ-
ные неморальные широколиственные леса.
Изменения климата привели к разрыву (дизъюнкции) ареала широко-
лиственных тургайских лесов. Они полностью выпали из растительного по-
крова Сибири и заняли океанические секторы Евразии: Приатлантический и
Притихоокеанский. В это время в горах Северо-Восточной Сибири появился
пояс темнохвойной тайги. Началось иссушение внутриконтинентальных про-
странств. На севере Африки и юге Евразии впервые появились травянистые
ландшафты саванн и степей (рис. 1.3).
В истории Средиземноморья отмечен ряд катастрофических событий.
Пролив, располагавшийся в предгорном прогибе Пиренеев и соединявший
Атлантический океан со Средиземным морем, неоднократно замыкался, и то-
гда вся морская вода испарялась. В условиях сухого и жаркого климата на это
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
10
Рис. 1.3. Ландшафт саванн в Европе в конце неогена (по И. Аугуста, З. Буриану)
уходило около 5000 лет. В эти периоды впадина Средиземного моря представ-
ляла собой пустыню, покрытую толстым слоем соли. Наступала общая ариди-
зация климата побережий и формирование ландшафтов средиземноморского
типа. На территории современного Ирана и Средней Азии, в условиях жар-
кого и сухого климата на засоленных берегах и островах усыхавшего океана
Тетис возникала своеобразная флора пустынь умеренного пояса.
Четвертичный период (антропоген). Около 2,5 миллионов лет
назад в результате сильного глобального похолодания наступила ледниковая
эпоха – плейстоцен, ознаменовавшийся образованием ледовых покровов и
глубокой трансформацией растительного покрова (рис. 1.4). На протяжении
плейстоцена холодные ледниковые периоды сменялись теплыми периодами
межледниковий.
Рис. 1.4. Ледовый покров Земли в плейстоцене
.
Глава 1. ИСТОРИЧЕСКИЕ УСЛОВИЯ ФОРМИРОВАНИЯ РАСТИТЕЛЬНОГО ПОКРОВА
11
Теория оледенений в четвертичном периоде имеет как сторонников
(гляциолистов), так и противников (антигляциолистов). Последние, отрицая
покровное оледенение в Евразии, полагают, что ведущую роль в трансформа-
ции ландшафтов играли морские трансгрессии. Огромное море-озеро дей-
ствительно заливало северную низменную часть Восточно-Европейской рав-
нины, а на Западно-Сибирской низменности обширный водоем занимал ее
среднюю и южную части. Излишки вод Сибирского водоема изливались на юг
в Аральское море и далее в Каспийское, обуславливая трансгрессию, залив-
шую Прикаспийскую низменность. В результате трансгрессии воды Каспий-
ского моря через Кума-Манычскую впадину соединились с Азовским и Чер-
ным морями. На севере Восточно-Европейской равнины образовался обшир-
ный приледниковый водоем, воды которого соединялись с южными морями.
Таким образом, возникновение морей-озер объясняется не отсутствием ледни-
ков, а напротив их существованием: в Европе на Скандинавском полуострове, а
в Азии на Таймырском полуострове и севере Западно-Сибирской низменности.
Эти ледниковые щиты как огромные плотины преграждали сток рекам, теку-
щим на север, что приводило к образованию подпрудных водоемов.
Приледниковая растительность Центральной и Восточной Европы в
максимальную стадию Валдайского оледенения была представлена сочетани-
ем тундровых и степных группировок с березовым и сосновым редколесьем.
К юго-востоку они сменялись луговыми степями с сосновыми, лиственничны-
ми и березовыми лесами (перигляциальная лесостепь), которые переходили в
луговые степи с лесами из березы и сосны с участием широколиственных по-
род (рис. 1.5).
Несмотря на перестройку четвертичной биоты, вызванную похолодани-
ем, местами сохранились представители неогеновой флоры. Они находили
убежище в рефугиумах, представлявших собой благоприятные для жизни ост-
ровки среди враждебного моря холода. Главные рефугиумы Евразии были
приурочены в основном к южному поясу гор (рис. 1.6).
Послеледниковая эпоха (голоцен) началась около 10−11 тыс. лет назад.
На протяжении этого времени произошло формирование современных зо-
нальных особенностей распределения органического мира на Земле. Плане-
тарными единицами биономического районирования являются зонобиомы –
зональные подразделения растительного и животного мира, контролируемые
географической зональностью (рис. 1.7).
В течение голоцена в Евразии изменения климатических условий проис-
ходили неоднократно, но они уже не были столь резкими, как в плейстоцене.
Наиболее значительными оказались изменения, произошедшие 5−5,5 тыс. лет
назад, когда в результате длительного господства теплых и влажных климати-
ческих условий широколиственные леса продвинулись на 500−800 км север-
нее их современного распространения. В свою очередь, во время «малого лед-
никового периода» (середина XVI – середина XVII веков) на равнинах широ-
колиственные леса вновь отступили к югу, а в горах произошло повсеместное
снижение границ высотных поясов.
Важным фактором распределения растительности явились неоген-чет-
вертичные тектонические движения, обусловившие характер современного
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
12
Рис. 1.5. Растительность в максимальной стадии Валдайского оледенения
(Палеогеография Европы..., 1982, фрагмент)
.
Глава 1. ИСТОРИЧЕСКИЕ УСЛОВИЯ ФОРМИРОВАНИЯ РАСТИТЕЛЬНОГО ПОКРОВА
13
Продолжение рис. 1.5
рельефа (рис. 1.8). Соотношение равнинных пространств и гор существен-
ным образом влияет на характер климата и определяет азональные осо-
бенности подразделения арены жизни. На равнинах выражено действие
закона географической зональности. Организмы формируют свои ареалы
свободно, не встречая орографических преград. Горные сооружения усили-
вают контрастность природных условий. Распределение жизни в горах кон-
тролируется помимо широтной зональности действием закона высотной
поясности.
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
14
Рис. 1.6. Главнейшие убежища третичной биоты (Шафер, 1956):
1 – балкано-черноморское убежище; 2 – армяно-иранское убежище; 3–5
–
среднеазиат-
ские убежища; 6 – восточноазиатское убежище; 7–10 – североамериканские убежища.
Прерывистой линией обозначена область максимального оледенения в плейстоцене
Рис. 1.7. Зонобиомы Земли (по H. Walter & S.-W. Breckle):
1 – экваториальные дождевые леса; 2 – субэкваториальные сезонно-влажные леса и саванны;
3 – тропические пустыни; 4 – субтропические средиземноморского типа; 5 – влажные суб-
тропики; 6 – неморальные листопадные леса; 7 – умеренного пояса: а) пустыни, б) полупу-
стыни, в) степи; 8 – бореальные хвойные леса; 9 – арктические тундры и пустыни
.
Глава 1. ИСТОРИЧЕСКИЕ УСЛОВИЯ ФОРМИРОВАНИЯ РАСТИТЕЛЬНОГО ПОКРОВА
15
Рис. 1.8. Рельеф материков, обусловливающий подразделение арены жизни
(Мир географии..., 1984)
Вопросы к главе 1
1. Природные условия и растительность палеогена и неогена.
2. Природные условия и растительность четвертичного периода (плейстоцен, голоцен).
3. Зональные и азональные факторы подразделения арены жизни.
Рекомендуемая литература
История биогеоценозов СССР в голоцене. М.: Наука, 1976. 290 с.
Криштофович А. Н. Палеоботаника. Учебник. 4-е изд. Л.: Гостоптехиздат, 1957. 650 с.
Марков К. К., Величко А. А., Лазуков Г. И., Николаев В. А. Плейстоцен. М.: Высшая шко-
ла, 1968. 304 с.
Мейен С. В. Основы палеоботаники. М.: Недра, 1987. 403 с.
Палеогеография Европы за последние 100 000 лет (Атлас-монография). М.: Наука, 1982.
156 с.
Хотинский Н. А . Голоцен Северной Евразии. М.: Наука, 1977. 198 с.
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
16
чение об ареалах (ареалогия) является одной из основных отраслей гео-
графии растений (Толмачев, 1962, 1974). Совокупность видов растений,
встречающихся на той или иной территории, образует ее флору. В задачи бо-
танической географии входит районирование флоры, выявление неповтори-
мых флористических комплексов путем анализа особенностей географическо-
го распространения систематических таксонов (видов, родов, семейств и т. д.).
Ареал – это часть земной поверхности, в пределах которой встречается
данный таксон. Размеры, форма, внутренняя структура, границы ареала опре-
деляются историческими, географическими и экологическими факторами.
Выявление ареалов основывается на нанесении мест нахождения видов расте-
ний на карту. Чем полнее проведены наблюдения и точнее определена систе-
матическая принадлежность видов, тем точнее будет картина их географиче-
ского распространения. При выделении наиболее дробных единиц райони-
рования учитываются границы ареалов видов. Чем выше ранг таксона, тем
обширнее область его распространения. Ареалы высших таксономических
единиц охватывают территорию целых флористических царств. Новые наход-
ки, уточнение систематической принадлежности организмов подчас вносят
существенные коррективы в форму ареала, заставляют по-новому интерпре-
тировать историю его развития.
Формирование ареала связано с возникновением нового вида и завоева-
нием им пространства. Процесс формирования нового ареала монотопичен –
возникновение вида происходит в одном месте, откуда он начинает свое рас-
пространение. Новый вид происходит от одного предка или в результате ги-
бридизации. Виды, попавшие в условия географической изоляции, дают
начало ветвям филогенетического древа. Таким образом формируются непо-
вторимые черты флор, отличающихся не только по составу видов, но родов и
семейств.
В. И. Вернадский отметил огромную внутреннюю потенцию живого ве-
щества к растеканию по земной поверхности. Он назвал это явление давле-
нием жизни, энергия которого определяется быстротой размножения. Рас-
селение вида задерживается только внешними силами: неблагоприятным
режимом тепло- и влагообеспеченности, недостатком питания, конкуренци-
ей с другими видами. Если нет внешних препятствий, всякий вид в харак-
терное для него время может благодаря размножению покрыть весь Земной
шар.
Расселению организмов способствуют различные приспособления их за-
чатков (диаспóр): при половом размножении растений – спор, плодов, семян,
при вегетативном – луковиц, клубней, корневищ, ползучих побегов и т. п.
Плоды и семена водных, а также многих наземных растений легко переносят-
ся текучими водами.
Глава 2
УЧЕНИЕ ОБ АРЕАЛАХ
.
Глава 2. УЧЕНИЕ ОБ АРЕАЛАХ
17
Рис. 2.1. Плоды и семена, распространяемые ветром:
1 – одуванчик и его плоды; 2 – плоды чертополоха; 3 – семена сосны; 4 – плоды клена; 5 –
соплодие липы; 6 – плоды вяза; 7 – плод граба; 8 – плоды березы
Легкость спор и мелких семян множества видов обеспечивает возмож-
ность их переноса ветром. У растений с более тяжелыми семенами или плода-
ми возможность переноса по воздуху обеспечивается благодаря летательным
приспособлениям: волосистым придаткам, разного рода хохолкам, перепон-
чатым лопастям, «крылышкам» и т. д. (рис. 2.1).
У многих растений семена или плоды переносятся животными благодаря
разного рода прицепкам (рис. 2.2). То же может достигаться и при помощи
клейких выделений. Семена растений, плоды которых поедаются животными,
переносятся последними в кишечнике и попадают на поверхность почвы с по-
метом. Перенос семян на значительные
расстояния птицами играет особо важ-
ную роль для расселения растений.
Заметим, что вместе с экскрементами
в почву вносятся питательные веще-
ства. Отмечено, что на сфагновых бо-
лотах, почти лишенных питательных
веществ, семена голубики прорастают
только в том случае, если они попали
сюда с экскрементами птиц. Семена
многих растений обретают всхожесть
Рис. 2.2. Плоды, распространяемые жи-
вотными:
1 – соплодие лопуха; 2 – побег подмаренни-
ка цепкого с плодами; 3 – одиночный плод
череды
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
18
только после того, как пройдут через желудочно-кишечный тракт животного.
Бывает и так, что определенные животные делают для себя запасы питатель-
ных семян, но используют их не полностью или просто теряют часть своих
«складов», которые тогда и превращаются в рассадники растений соответ-
ствующего вида.
Большую роль в распространении растений играет человек. Первона-
чально растения вводились в культуру в центрах своего происхождения, а
затем начинали культивироваться широко по Земному шару. Многие вред-
ные растения и сорняки разносятся человеком по неосторожности.
На пути естественного стремления вида расширить ареал встают внешние
факторы: горные хребты, океаны, моря и т. п., а также режим тепло- и влаго-
обеспеченности. Ни один вид не образует сплошного покрова. Внутри ареала
организмы приурочены только к определенным местообитаниям. Например,
болотные мхи, травы и кустарнички (сфагнум, пушица, клюква и др.) и виды
таежного мелкотравья и кустарничков (майник, кислица, черника и др.) имеют
единый ареал, охватывающий всю зону бореальных хвойных (таежных) лесов.
Однако первые приурочены к верховым болотам, а вторые к хорошо дренируе-
мым водоразделам к травяно-кустарничковому ярусу хвойных лесов. Таким об-
разом, экологическая структура ареалов названных видов образует сложное
кружево, рисунок которого зависит от распределения верховых болот и зелено-
мошных лесов.
На пути расселения вида встают не только географические и ландшафт-
но-экологические преграды, но и сопротивление, которое встречает новосел
как со стороны уже существующих видов, так и целых сообществ. Например,
на границе леса и степи внедрению деревьев в степное сообщество препят-
ствует мощная дернина степных трав.
Первоначальную область распространения вида называют центром его
происхождения. Наряду с этим выделяют ту часть ареала, в которой наблюда-
ется максимальная изменчивость данного вида. Эту часть ареала называют
центром многообразия форм (рис. 2.3).
Границы ареала могут быть статичными, расширяющимися или сужаю-
щимися. В первом случае устойчивость границ объясняется тем, что вид до-
стиг естественных границ свое-
го ареала. Границы ареала рас-
ширяются, когда молодой вид
обладает потенцией к преодо-
лению преград. Границы ареа-
ла сужаются, когда вид теряет
завоеванную территорию.
Рис. 2.3. Количественное рас-
пределение видов коровяка
(род Verbascum) (по Ж. Лемме)
Цифры указывают количество видов,
находящихся внутри зон, ограничен-
ных линиями. Отчетливо выделяет-
ся центр многообразия форм
.
Глава 2. УЧЕНИЕ ОБ АРЕАЛАХ
19
Рис. 2.4. Основные типы ареалов на
суше (по P. Dansereau)
Объяснения в тексте
При типизации ареалов учиты-
ваются их размеры и характер рас-
пространения таксонов – непрерыв-
ный (континуальный) и разорванный
(дизъюнктивный). Выделяются ареа-
лы четырех уровней размерности:
континентальные, провинциальные,
региональные и локальные (рис. 2.4,
7В). Последние могут быть образова-
ны реликтовыми видами, то есть ви-
дами, ранее в геологической истории
широко распространенными, а те-
перь занимающими небольшие тер-
ритории, и энедемичными видами –
произрастающими только в данном
регионе и не встречающимися в других. Возможны две модели формирова-
ния ареала эндемичного вида. Первая отражает уменьшение ареала релик-
тового вида под воздействием неблагоприятных природных условий или в
результате конкуренции с другими видами (палеоэндемик); вторая связана с
формированием ареала популяции нового вида (неоэндемик).
Непрерывное распространение обозначено на рис. 2.4 нечетными но-
мерами (1, 3, 5, 7), разорванные ареалы – четными (2, 4, 6); под буквой А –
сплошные ареалы по всей площади; под буквой Б представлены ареалы, со-
стоящие из одного большого и одного или нескольких небольших участков,
связанных между собой; под буквой В – ареалы, объединяющие множество
мелких участков.
Рассмотрим примеры, иллюстрирующие разные типы ареалов. Ареал
папоротника орляк (Pteridium aquilinum) можно отнести к типу 1А (рис. 2.5).
К этому типу ареалов относятся также циркумполярные ареалы видов, при-
уроченных к арктическому поясу Северного полушария, образующие кольцо
вокруг полюса. Например, ареал крупки Draba subcapitata (рис. 2.6).
Возникновение разорванных ареалов объясняется событиями прошлых
геологических эпох: исчезновением мостов-переходов на суше, дрейфом кон-
тинентов, горообразованием, изменениями климата и т. п. Ареал вида может
быть разорванным и занимать обширные области на разных континентах –
тип 2А, например, ареал дёрена шведского (Chamaepericlymenum suecicum)
(рис. 2.7). Ареал таежного кустарничка линнеи северной (Linnaea borealis) в
основном сплошной, занимающий значительную часть Евразии. Наряду с
этим имеются его небольшие обособленные островки в различных местах
Средней и Восточной Европы, на Кавказе, на юге Западной Сибири, на
Дальнем Востоке (рис. 2.8). Следовательно, такой ареал можно отнести к ти-
пу 2Б.
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
20
Рис. 2.5. Ареал папоротника орляк (Pteridium aquilinum) (по А. И. Толмачеву)
Рис. 2.6. Циркумполярный ареал крупки (Draba subcapitata) (по А. И. Толмачеву)
.
Глава 2. УЧЕНИЕ ОБ АРЕАЛАХ
21
Рис. 2.7. Ареал дёрена шведского (Chamaepericlymenum suecicum)
(по А. И. Толмачеву)
Рис. 2.8. Ареал таежного кустарничка в пределах Евразии (Linnaea borealis)
(по А. И. Толмачеву)
Рис. 2.9. Ареал печеночницы благородной (Hepatica nobilis) (по А. И. Толмачеву)
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
22
Разрыв европейско-дальневосточного ареала (тип 4Б) спутника хвойно-
широколиственных и широколиственных лесов печеночницы благородной
(Hepatica nobilis) объясняется похолоданием климата в плейстоцене (рис. 2.9).
Особую группу образуют ареалы викарирующих видов. Викариат в пере-
воде с латинского означает замещение. В ботанической географии викаризм –
это замещение одних видов другими, образовавшимися из одного корня, т. е.
родственными, но развившимися самостоятельно в разных экологических
условиях. Викариат отражает процесс дивергентной микроэволюции, когда
обособление нового вида является следствием адаптации организмов к новым
условиям, отличным от тех, в которых обитал породивший их старый вид. Клас-
сический пример – ряд лиственниц в Азии: каяндера – гмелина (даурская) –
рупрехта – ольги – камчатская – японская, каждая из которых занимает свой
ареал (рис. 2.10).
Анализ распространения растений свидетельствует о существовании объ-
ективных флористических комплексов, формирование которых обусловлено
единством времени и места происхождения. Отображенные на карте ареалы
таксонов образуют сгущения границ в одних местах и их разрежения в других.
Рис. 2.10. Ареалы викарирующих восточносибирских и дальневосточных видов
лиственниц (по Е. Г. Боброву):
I – каяндера; II – гмелина; III – рупрехта; IV – ольги; V – камчатская; VI – японская
.
Глава 2. УЧЕНИЕ ОБ АРЕАЛАХ
23
Рис. 2.11. Фрагмент карты флористического районирования Евразии
(Тахтаджян, 1978)
Это означает, что таксоны образуют природные совокупности, каждая из
которых занимает определенную территорию, они выступают в качестве единиц
флористического районирования. Последняя работа, посвященная флористи-
ческому делению суши, принадлежит А. Л. Тахтаджяну (1978). Северную
Евразию и Северную Америку он относит к Голарктическому царству. Приве-
дем фрагмент карты флористического районирования Евразии по А. Л. Тахта-
джяну (рис. 2.11). Своеобразие флоры учитывается в дальнейшем при харак-
теристике единиц ботанико-географического районирования.
Вопросы к главе 2
1. Формирование ареалов.
2. Границы ареалов.
3. Основные типы ареалов растений.
4. Ареалы викарирующих видов.
Рекомендуемая литература
Воронов А. Г., Дроздов Н. Н., Криволуцкий Д. А, Мяло Е. Г. Биогеография с основами эколо-
гии. Учебник. 4-е изд. М.: Изд. МГУ, 2002. 392 с.
Григорьева Н. М. География растений. Учебное пособие. М.: КМК, 2014. 460 с.
Тахтаджян А. Л. Флористические области Земли. Л.: Наука, 1978. 248 с.
Толмачев А. И. Основы учения об ареалах (Введение в хорологию растений). Л.: Изд-во ЛГУ,
1962. 100 с.
Толмачев А. И. Введение в географию растений. Л.: Изд-во Ленинградского ун-та. 1974. 244 с.
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
24
астительное сообщество (фитоценоз) выступает в качестве основной ис-
ходной единицы структуры растительного покрова и представляет собой
целостную природную систему, приуроченную к определенному экотопу. В гла-
ве дается определение понятий фитоценоз, экосистема, биогеоценоз, обсужда-
ется состав сообществ, фитоценотическая роль видов, географические и гене-
тические элементы флоры, жизненные формы, экологические типы, страте-
гия видов, структура, сукцессии и границы растительных сообществ.
Фитоценозом называют сообщество растений, обитающих на относитель-
но однородном участке земной поверхности (в одном экотопе) и характеризую-
щихся определенными взаимоотношениями как между собой, так и с условиями
местообитания. Всякий естественный фитоценоз представляет собой историче-
ски сложившийся комплекс функционально связанных организмов и является
частью целостного природного образования – экосистемы (биогеоценоза).
Термин экосистема вошел в употребление в 1935 г. Он был предложен
английским ботаником А. Тенсли, который писал, что в экосистему входит не
только комплекс организмов, но и весь комплекс физических факторов, обра-
зующих условия местообитания в самом широком смысле. Биотическую и
абиотическую части экосистемы связывают непрерывные круговороты пита-
тельных веществ, энергию для которых дает солнечная радиация.
В. Н. Сукачев пришел к выводу, что растительные сообщества находят-
ся во взаимосвязи и взаимодействии с другими компонентами географиче-
ской оболочки: литосферой, атмосферой и гидросферой. Это взаимодей-
ствие на уровне фитоценозов протекает в рамках географических комплексов.
Учитывая ведущую и активную роль
живых существ в этом взаимодействии,
В. Н. Сукачев в 1945 г. предложил для
его обозначения термин «биогеоце-
ноз», близкий понятию экосистема.
Биогеоценоз – участок территории,
однородный по экологическим услови-
ям, занятый одним биоценозом. В мо-
дели биогеоценоза Сукачев выделяет
два блока: экотоп и биоценоз (рис. 3.1).
Рис. 3.1. Структура биогеоценоза и
схема взаимодействия его компонен-
тов (Сукачев, 1945)
Свойства растительного сообщества раскры-
ваются через описание его видового состава
и структуры
Глава 3
УЧЕНИЕ О РАСТИТЕЛЬНЫХ СООБЩЕСТВАХ (ФИТОЦЕНОЛОГИЯ)
.
Глава 3. УЧЕНИЕ О РАСТИТЕЛЬНЫХ СООБЩЕСТВАХ (ФИТОЦЕНОЛОГИЯ)
25
Рис. 3.2. Схема взаимодействий в биогеоценозе (по В. Н. Сукачеву)
Схема взаимодействий между организмами и средой в биогеоценозе (экоси-
стеме) показана на рис. 3.2.
Несмотря на близость понятий «экосистема» и «биогеоценоз», послед-
ний отличается тем, что имеет четкие пространственные размеры. Биогеоце-
ноз всегда приурочен к определенному участку земной поверхности, однород-
ному в экологическом отношении. Экосистема – понятие общее, в качестве
экосистемы можно рассматривать и грядку в теплице, и луг, и лес, и космиче-
ский корабль, и биосферу в целом.
Состав растительного сообщества. Основой любого описания явля-
ется список образующих данное сообщество видов. Растительные сообщества
могут быть одновидовыми, бедными или богатыми по числу видов. Степень
участия вида в сложении сообществ характеризуется его численностью (оби-
лием), биомассой, проективным покрытием, встречаемостью и т. п. Величина
названных показателей определяет фитоценотическую мощность вида.
Фитоценотипы – группы видовых популяций, характеризующиеся сход-
ной фитоценотической позицией в растительном сообществе. Доминанты –
виды, преобладающие в разных ярусах сообщества. Доминанты, слагающие
основу сообщества и играющие средообразующую роль, называются эдифика-
торами. Например, в лесу это деревья, на верховом болоте – сфагновые мхи и
т. п. Кроме того, выделяются второстепенные, редко встречающиеся, незначи-
тельные по биомассе и продуктивности виды.
Виды растений, образующие сообщество, различны по их географиче-
скому распространению и происхождению. В соответствии с особенностями
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
26
распространения видов выделяются географические элементы флоры, кото-
рые в основном формируются в пределах регионов, обладающих единством
палеогеографического развития. Любая флора состоит из многих географиче-
ских элементов. Например, А. А. Гроссгейм приводит следующее соотношение
элементов флоры Кавказа (в %): собственно кавказские – 20,0; бореальные –
38,2; средиземноморские – 13,2; колхидские – 12,7; переднеазиатские – 9,7;
понтические – 5,6; туранские – 0,1 и пришлые виды – 0,5. Данный спектр
элементов наглядно демонстрирует их роль в формировании флоры. Весьма
значительна доля собственно кавказских видов свидетельствует о том, что
Кавказ представляет собой самостоятельный центр видообразования. Широ-
кое участие бореальных элементов во флоре отражает внедрение северных
видов на Кавказ в ледниковую эпоху. Свой вклад в обогащение флоры внесли
соседние регионы.
Выяснение того, откуда происходят виды, как они попали в состав изучае-
мой растительности, предполагает выделение генетических элементов. Пре-
красным примером такого анализа является заключение известного русского
ботаника А. Н. Краснова, посетившего в конце XIX века острова Юго-Восточной
Азии, о связи тундровых кустарничков с тропической растительностью гор.
Имея списки видов, можно применить к ним методы экологического
анализа, то есть установить, какими жизненными формами и экологически-
ми типами растений образованы одни сообщества и чем они отличаются от
других.
Понятие «жизненные формы» ввел Е. Варминг. Виды, сходные по внеш-
нему облику и приспособлениям к среде, объединяют в одну жизненную фор-
му. Широко признанной классификацией жизненных форм является класси-
фикация К. Раункиера. В ее основу положена идея о том, что сходные типы
приспособлений – это сходные способы перенесения критических условий
существования. В областях с сезонной периодичностью климата таковые
наступают в зимний сезон, а в аридных областях – еще и в период летней засу-
хи. В качестве признака, выражающего приспособленность к перенесению не-
благоприятного сезона, Раункиер использовал расположение почек возобнов-
ления относительно поверхности земли. Основные жизненные формы в си-
стеме Раункиера следующие: фанерофиты – деревья и кустарники, у которых
почки возобновления расположены высоко над землей; хамефиты – невысо-
кие (20–30 см) кустарнички и полукустарнички с почками возобновления на
зимующих побегах, расположенных вблизи поверхности земли, на зиму они
укрываются слоем снега или опавшими листьями; гемикриптофиты – травя-
нистые многолетники, у которых надземные органы в конце вегетации отми-
рают, а почки возобновления находятся на уровне почвы и защищены соб-
ственными отмершими листьями и снегом; криптофиты – растения, у кото-
рых почки возобновления скрыты (у геофитов – под землей, у гидрофитов –
на дне водоема); терофиты – однолетники, переживающие неблагоприятный
период в виде семян или спор.
И. Г. Серебряковым и Т. И. Серебряковой детально разработана класси-
фикация жизненных форм, которая основывается на форме роста и длитель-
ности жизни вегетативных органов (рис. 3.3).
.
Глава 3. УЧЕНИЕ О РАСТИТЕЛЬНЫХ СООБЩЕСТВАХ (ФИТОЦЕНОЛОГИЯ)
27
Рис. 3.3. Древесные и тра-
вянистые жизненные фор-
мы, соотношение много-
летних и отмирающих ча-
стей (схема):
1 – дерево; 2 – кустарник; 3 –
кустарничек; 4 – полукустар-
ник; 5 – длиннопобеговая трава
Многолетние части показаны
черным; отмершие ранее –
двойным пунктиром; отмира-
ющие в текущем году – пунк-
тиром; цифрами с индексами
обозначен примерный возраст
отдельных скелетных осей и их
систем
Типичное дерево характеризуется тем, что у него в течение всей жизни
имеется единственный ствол, продолжительность жизни которого от несколь-
ких десятков до нескольких сотен, а изредка и тысяч лет.
Кустарники отличаются от деревьев тем, что в течение жизни особи
формируется не один единственный ствол, а несколько или много, существу-
ющих бок о бок и постепенно сменяющих друг друга.
Кустарнички – миниатюрные кустарники высотой не более 50 см, в сред-
нем 10–30 см. К типичным кустарничкам относятся господствующие растения
наших торфяных болот: багульник, мирт болотный, подбел, голубика.
Полудревесные растения характеризуются тем, что их надземные побеги
частично деревянистые, частично травянистые, т. е. верхняя, иногда значитель-
ная по размерам часть побега ежегодно отмирает, а нижняя часть с почками
возобновления, находящимися на некоторой высоте (5–15–20 см и более) над
землей, остается в качестве многолетней. К полукустарникам и полукустар-
ничкам (различаемым главным образом по величине остающейся многолет-
ней части и общим размерам) относятся, например, многочисленные виды
полыни, господствующие в растительном покрове сухих степей, многие губо-
цветные (тимьян, или богородская трава, лаванда, некоторые виды шалфея и
др.), составляющие основу растительности склонов гор в субтропиках Среди-
земноморского типа. Для полукустарников и полукустарничков в крайних усло-
виях существования характерна подушковидная форма.
Если побеги односезонные, растение называется травянистым. Среди тра-
вянистых растений выделяют однолетние и многолетние. Однолетние травы
не имеют многолетних органов и почек возобновления, а также органов веге-
тативного возобновления и отмирают после цветения и плодоношения полно-
стью, оставляя только семена. У многолетних трав надземные прямостоящие
побеги всегда однолетние, точнее односезонные, а части побегов с почками
возобновления всегда или подземные, или приземные, т. е. скрыты в подстил-
ке или плотно прижаты к почве.
Многолетние травы подразделяют на группы по характеру их многолет-
них подземных органов:
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
28
–
стержнекорневые – во взрослом состоянии имеют хорошо развитый,
часто запасающий главный корень (дудник, лопух, коровяк);
–
кистекорневые – во взрослом состоянии растения не имеют главного
корня; придаточные корни в большом количестве скучены в виде кисти на
крайне укороченной подземной стеблевой оси (подорожники, сивец луговой,
лютики едкий, купальница);
–
короткокорневищные (ирисы, гравилаты, купена, манжетки);
–
длиннокорневищные (пырей ползучий, майник);
–
дерновинные – с мощной придаточной корневой системой (мочкова-
той), к ним относятся преимущественно злаки, которые делят на рыхло-дерно-
винные (тимофеевка, овсяница луговая, мятлик дубравный) и плотно-дерно-
винные (щучка дернистая, белоус, ковыли);
–
клубнеобразующие – сборная группа, включающая растения с одним
многолетним клубнем (цикламен), со сменяющимися из года в год клубнями
(хохлатки, борцы, ятрышники), и столонно-клубневые (картофель);
–
луковичные (гусиный лук, тюльпан);
–
наземно-ползучие и наземно-столонные – травы с относительно долго
живущими (2–4 года) ползучими побегами (типа будры, лугового чая) или с
быстро отмирающими столонами-усами (земляника, костяника).
Экологические типы растений устанавливаются по отношению к отдель-
ным факторам среды. Классификация растений по приуроченности к почвам
разного уровня богатства (трофности) заключается в выделении: олиготроф-
ных видов – растения бедных почв, мезотрофных видов – растения средних по
богатству почв, эвтрофных видов – растения богатых почв. По отношению к
кислотности почвы выделяются: ацидофилы – растения кислых почв, нейтро-
филы – растения нейтральных почв, базифилы – растения щелочных почв.
Галофиты – растения, обитающие на засоленных почвах.
По отношению к увлажнению растения подразделяются на гидрофиты –
растения водоемов, гигрофиты – растения влажных местообитаний, мезофи-
ты – растения умеренно увлажненных мест, ксерофиты – растения сухих ме-
стообитаний. Ксерофиты подразделяются на гемиксерофиты – растения, не
переносящие длительного обезвоживания, и эуксерофиты – растения, перено-
сящие регулярную глубокую засуху. В качестве примера приведем экологиче-
ский ряд увлажнения и колебания обилия отдельных видов в зависимости от
условий увлажнения (рис. 3.4). Как особые типы выделяют психрофиты – рас-
тения холодных и сырых местообитаний, и криофиты – растения холодных и
сухих местообитаний.
На рисунке отчетливо выделяются зоны увлажнения, в которых опреде-
ленные виды имеют максимальное обилие. Например, к видам, характеризую-
щим лугово-степной тип увлажнения, относятся типчак, таволга обыкновенная,
подмаренник настоящий; к сухо-луговому и свеже-луговому типу увлажнения:
осока ранняя, тысячелистник обыкновенный; к влажно-луговому типу увлаж-
нения: клевер луговой, мятлик луговой, герань холмовая. Преобладание в
травостое растений определенного экологического типа позволяет судить об
условиях увлажнения данного местообитания.
Поведение вида в сообществе, выраженное в особенностях расхода жиз-
ненной энергии в целях сохранения популяции, называется жизненной страте-
.
Глава 3. УЧЕНИЕ О РАСТИТЕЛЬНЫХ СООБЩЕСТВАХ (ФИТОЦЕНОЛОГИЯ)
29
Рис. 3.4. Изменение участия в луговых травостоях отдельных видов растений
в зависимости от увлажнения (по Л. Г. Раменскому):
1 – клевер луговой; 2 – тысячелистник обыкновенный; 3 – келерия Делявина; 4 – мятлик
луговой; 5 – типчак; 6 – подмаренник настоящий; 7 – осока ранняя; 8 – таволга обыкновен-
ная; 9 – герань холмовая; 10 – короставник полевой; 11 – козлобородник коротконосиковый
гией вида. Л. Г. Раменский подразделял виды растений на виолентов (львов) –
растений, наиболее мощных по способности образовывать популяции или
стойко внедряться в сообщества; патиентов (верблюдов) – растений, побеж-
дающих в борьбе за существование, благодаря своей выносливости; и экспле-
рентов (шакалов) – растений с низкой конкурентной способностью, быстро
захватывающих свободную территорию на непродолжительное время и усту-
пающие место более конкурентно способным видам.
Американский эколог Э. Пианка выделяет два типа жизненных стратегий.
К-стратеги – это, как правило, крупные многолетние организмы, требующие
устойчивых условий существования; их жизненная энергия расходуется глав-
ным образом на прирост биомассы, а не на размножение. По классификации
Раменского это «львы». Напротив, r-стратеги, виды с непродолжительным
периодом жизни, они занимают нестабильные местообитания и характеризу-
ются высокой репродуктивной способностью. По классификации Раменского
это «шакалы».
Структура растительного сообщества характеризуется вертикаль-
ной ярусностью и горизонтальной мозаичностью. Вертикальная ярусность
свойственна всем типам наземной растительности, но особенно четко она вы-
ражена в лесных сообществах (рис. 3.5), где выделяются древесный, кустарни-
ковый, травяно-кустарничковый и мохово-лишайниковый ярусы (невысокие
деревья и кустарники иногда объединяют в подлесок).
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
30
Рис. 3.5. Вертикальная структура
растительного сообщества
Верхний ярус находится под
наибольшим воздействием внеш-
них климатических условий, под
его пологом условия существова-
ния растений значительно транс-
формируются. Ярусность может
быть не выражена в сообществах
пионеров зарастания с малой со-
мкнутостью. В моновидовых со-
обществах, особенно в полевых
агроценозах, все растения относятся к одному ярусу.
Важной количественной характеристикой хозяйственной ценности дре-
востоя является бонитет – показатель продуктивности леса. Различают 5 клас-
сов бонитета, от I – высшего до V – низшего. Определение бонитета произво-
дится по средней высоте и возрасту деревьев, с учетом происхождения древо-
стоя (семенное или порослевое) с помощью специальных таблиц. Насаждения
I–II класса бонитета называют высокобонитетными, III–IV – среднебонитет-
ными, V – низкобонитетными, Vа – непродуктивными.
Горизонтальная мозаичность, наблюдаемая в пределах почти каждого
фитоценоза, выражается в наличии сравнительно мелких групп растений,
различающихся по составу, обилию видов, их жизненности и т. п. Элементы
горизонтальной структуры фитоценозов называют микрогруппировками, или
микроценозами. Различают экотопогенную, фитогенную, зоогенную и антро-
погенную мозаичность.
Экотопогенная мозаичность, как правило, связана с неровностями мик-
рорельефа или неоднородностью почвогрунтов. Фитогенная мозаичность воз-
никает как результат взаимодействия растений в сообществе; в качестве при-
мера можно привести распределение растений под пологом леса при его не-
равномерной сомкнутости. Мозаичность может быть связана с биологией
разрастания самих видов, например, образование болотных кочек. Зоогенная
мозаичность ярко выражена в степной зоне, где вызывается деятельностью
роющих животных. Антропогенная мозаичность возникает под прямым и кос-
венным влиянием хозяйственной деятельности. Так, существенно меняется
структура растительного покрова под воздействием рубки леса, перевыпаса
скота. Благодаря мозаичности полнее используется разнообразие условий
микросреды, увеличивается устойчивость фитоценоза к неблагоприятным
факторам.
В качестве элементов пространственной структуры и сезонной ритмики
фитоценоза выделяются синузии. Они образованы определенными жизнен-
ными формами или биоморфами растений, т. е. группами видов, подчас дале-
ких в систематическом отношении, но сходных по внешнему виду, экологии и
ритму развития. Достаточно, например, назвать деревья (фанерофиты), ку-
старнички (хамефиты), травы (криптофиты), мхи и лишайники в таежном ле-
су, чтобы иметь представление об основных синузиях лесного сообщества.
.
Глава 3. УЧЕНИЕ О РАСТИТЕЛЬНЫХ СООБЩЕСТВАХ (ФИТОЦЕНОЛОГИЯ)
31
Сезонные синузии представлены разными биоморфами, развивающи-
мися на одной территории в разные сроки вегетационного периода. Свои се-
зонные аспекты характерны для травяного яруса в лесу, на лугу, в степи и т. п.
Особенно ярким примерам служат сезонные аспекты в степи. Чем больше в
сообществе синузий, тем больше его флористическое, экологическое и био-
морфное богатство.
Сукцессии растительных сообществ – это последовательная во вре-
мени смена на данной территории одних сообществ другими. Фитоценозы ме-
няются под воздействием как внутренних ценотических, так и внешних эколо-
гических факторов.
В США и в западноевропейских странах широко распространено понятие
климакса, впервые введенного Ф. Клементсом. Смена сообществ осуществля-
ются в результате того, что растения первой стадии, заселяющие обнажен-
ный субстрат, видоизменяют среду так, что она становится более благопри-
ятной для поселения растений следующей стадии. И так продолжается до тех
пор, пока растительность не достигнет климаксового состояния, стабильность
которого поддерживается равновесием между сообществом и окружающей
средой.
В. Н. Сукачев предостерегал о некритическом внедрении в отечествен-
ную науку понятия климакс. В. И. Василевич отмечал, что сохранение ста-
бильного состояния сообществ поддерживается благодаря постоянным сла-
бым нарушениям. Стабильные климаксовые сообщества в естественных ус-
ловиях на самом деле представляют собой мозаику пятен с нарушенным
растительным покровом. Например, в лесу постоянно возникают окна в со-
мкнутом древесном пологе в результате ветровалов, пожаров и т. п. Благодаря
возникающим опушкам растительный мир леса становится значительно бога-
че. Сомкнутый лес препятствует возобновлению древесных пород, поэтому он
неустойчив. В перестойных лесах наступает массовая гибель деревьев, и лес
погибает. Возобновление коренного леса начинается с опушек, поэтому слабые
нарушения сомкнутого древесного яруса – необходимое условие его устойчиво-
сти. Если деструктивная антропогенная деятельность не превышает опреде-
ленного порога, она также может способствовать устойчивому состоянию рас-
тительности. Целинная ковыльная степь сохраняет устойчивость только при
условии умеренного выпаса копытных животных, которые, скусывая и разби-
вая дернину, открывают почву для внедрения разнотравья. Субклимаксовые
сообщества, находящиеся на пути к терминальным стадиям, обладают боль-
шей способностью к восстановлению своей структуры. В настоящее время, ко-
гда влияние антропогенных факторов становится почти повсеместным, эта
категория экосистем в силу своей адаптивности является наиболее распро-
страненной.
Среди многообразных форм динамики экосистем выделяются три кате-
гории: сезонная динамика, флуктуации, сукцессии.
Под флуктуациями фитоценозов понимаются ненаправленные их изме-
нения от года к году, завершающиеся возвратом фитоценоза к исходному или
близкому к исходному состоянию. К антропогенным флуктуациям раститель-
ных сообществ можно отнести изменения, вызываемые умеренными сеноко-
шением, выпасом и т. п.
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
32
Рис. 3.6. Восстановительная сукцессия лес-
ного сообщества (по И. Н. Пономаревой)
Сукцессии, в противоположность
флуктуациям, развиваются в определен-
ном направлении, они не имеют характе-
ра колебаний около какого-то среднего
состояния. Различаются эндогенные сук-
цессии, определяемые внутриценотиче-
скими причинами, и экзогенные сукцес-
сии, вызываемые внешними факторами.
К экзогенным сукцессиям относятся как
длительные многолетние изменения рас-
тительности, связанные с глобальным по-
теплением климата, так и катастрофиче-
ские смены, являющиеся результатом
проявления природных стихий (сход ла-
вин, сели в горах, извержения вулканов,
пожары и др.). Глубокую трансформацию
растительного покрова вызывает антропо-
генная деятельность: уничтожение расти-
тельности на месте горных выработок,
при создании пахотных угодий и др.
Прекращение разрушительного воздействия внешних факторов ведет к сук-
цессиям восстановления естественной растительности (рис. 3.6).
Границы растительных сообществ. Природные системы (фитоце-
ноз, биогеоценоз) занимают определенную площадь, объем и отделены от со-
седних систем границами. Территориальные подразделения биосферы обла-
дают свойствами как прерывности (дискретности), так и непрерывности (кон-
тинуальности). Резкие границы между фитоценозами могут быть обусловлены
гранями рельефа, сменой почво-грунтов и т. п. На смену представлениям о
дискретности растительности пришла концепция о континуальной организа-
ции растительного покрова.
Рис. 3.7. Гипотетические типы рас-
пространения видов (по Р. Уитте-
керу):
A – резкие границы между сообществами
(дискретность); B – резкие границы ме-
жду сообществами обусловлены распре-
делением доминантов, однако субдоми-
нанты являются общими для соседних
сообществ; C – экотон, центр сообщества
образован специфическим набором видов,
однако на периферии сообщества виды
постепенно сменяют друг друга; D – кон-
тинуальность, резкие границы между по-
пуляциями отсутствуют, виды постепен-
но сменяют друг друга
.
Глава 3. УЧЕНИЕ О РАСТИТЕЛЬНЫХ СООБЩЕСТВАХ (ФИТОЦЕНОЛОГИЯ)
33
Растительный континуум проявляется в постепенном переходе раститель-
ных сообществ друг в друга при постепенном изменении условий внешней сре-
ды. Растительные сообщества могут быть образованы видами, экологические
амплитуды которых перекрываются в данных условиях среды. При изменении
определенных факторов этой среды постепенно уменьшают обилие и исчеза-
ют одни виды, появляются и увеличивают обилие другие, таким образом осу-
ществляется плавный переход от одного растительного сообщества к другому
(рис. 3.7). Примерами континуальных переходов могут выступать целые под-
зоны: лесотундра, хвойно-широколиственные леса, лесостепь.
Основными видами континуальности являются:
–
горизонтальный континуум – плавный переход одних сообществ в дру-
гие;
–
вертикальный континуум – постепенный переход одних ярусов фито-
ценоза в другие;
–
временной континуум – постепенное изменение состава и свойств рас-
тительного покрова во времени (при сезонных и многолетних сукцессионных
сменах).
В каждом конкретном случае границы между территориальными систе-
мами могут быть линейными или расплывчатыми, четко выраженными или
затушеванными, стабильными или подвижными, однако они объективно су-
ществуют независимо от того, обнаружены они или нет. Проведение границ
есть начало и конец каждой ботанико-географической работы.
Вопросы к главе 3
1. Понятия фитоценоз, экосистема, биогеоценоз.
2. Состав растительного сообщества (фитоценотипы, жизненные формы).
3. Структура растительного сообщества.
4. Сукцессии растительных сообществ.
5. Границы растительных сообществ.
Рекомендуемая литература
Ипатов В. С., Кирикова Л. А. Фитоценология. СПб: Изд-во С.-Петербург. ун-та, 1999.
316 с.
Корчагин А. А. Строение растительных сообществ // Полевая геоботаника. Л., 1976.
Т. 5. С. 5–320.
Онипченко В. Г. Функциональная фитоценология: Синэкология растений. М.: Красанд,
2014. 576 с.
Работнов Т. А. Фитоценология. М.: Изд-во Москов. ун-та, 1983. 296 с.
Серебряков И. Г. Жизненные формы высших растений и их изучение // Полевая гео-
ботаника / Ред. Е. М. Лавренко, А. А. Корчагин. М.–Л.: Изд-во АН СССР, 1964. Т. 3.
С. 146–205.
Сукачев В. Н. Основы лесной типологии и биогеоценологии. Избранные труды в трех
томах. / Ред. Е. М. Лавренко. Л.: Наука, 1972. Т. 1. 419 с.
Шенников А. П. Введение в геоботанику. Л.: Наука, 1964. 448 с.
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
34
аспределение растительности отражает историю формирования расти-
тельного покрова и контролируется экологическими и эколого-тополо-
гическими факторами.
Экологические факторы. Для жизни и процветания каждого орга-
низма требуется набор определенных экологических факторов: абиотических
(неживых) и биотических (живых). В число наиболее важных прямо действу-
ющих экологического факторов входят солнечная радиация, температурный
режим, количество и режим выпадения атмосферных осадков, а также физи-
ко-химические свойства почвы. Такой фактор, как рельеф, является косвенно
действующим, так как он перераспределяет потоки вещества и энергии в
ландшафте.
Соотношение тепла и влаги определяет испаряемость и в конечном счете
увлажненность местообитаний. Одним из показателей увлажненности являет-
ся гидротермический коэффициент (ГТК) Г. Т. Селянинова, вычисляемый по
формуле:
K=R⋅10/Σt
где R – сумма осадков в миллиметрах за период со среднесуточными температурами
выше 10 °С; Σt – сумма температур в градусах за то же время.
ГТК является условным выражением баланса влаги и определяет отно-
шение прихода влаги к ее расходу. Чем величина ГКТ больше 1, тем увлаж-
ненность выше; если ГКТ равно 1 – увлажнение устойчивое; ГТК ниже 1 ха-
рактеризует недостаточную увлажненность вегетационного периода; ГТК 0,5 и
ниже соответствует резкому недостатку влаги.
Для наглядной характеристики фитоклимата конкретных территорий ис-
пользуется метод построения климатограмм (рис. 4.1), предложенный Г. Валь-
тером. При построении климатограмм на график наносятся значения годово-
го хода атмосферных осадков и температур воздуха. На нижней оси абсцисс
откладываются месяцы, по левой оси ординат – температура от 0 до 50 °С с
интервалом в 10° (при отрицательных среднемесячных температурах эту ось
продолжают ниже 0 °С). По правой оси ординат откладываются осадки в мил-
лиметрах, причем шаг на левой оси 10 °С соответствует шагу 20 мм осадков на
правой, то есть соотношение должно быть 1:2 – это лучше всего передает вод-
ный и тепловой режимы растительного покрова. Если кривая осадков распо-
лагается выше термической кривой – климат влажный, гумидный, если ниже –
это засушливый период, характерный для аридного климата.
Отношение разных видов растений к условиям существования неодина-
ково: одним требуется много влаги, другие живут в сухой пустыне, одни тре-
буют много света, другие его не выносят, одни любят тепло, другие – низкие
Глава 4
ФАКТОРЫ РАСПРЕДЕЛЕНИЯ РАСТИТЕЛЬНОСТИ
.
Глава 4. ФАКТОРЫ РАСПРЕДЕЛЕНИЯ РАСТИТЕЛЬНОСТИ
35
Рис. 4.1. Климатограммы гумидного (а)
и аридного (б) районов:
----
кривая осадков; ––– температурная кри-
вая
температуры и т. п. Ход жизни орга-
низмов не нарушается, если все фак-
торы действуют в привычном для ор-
ганизма режиме. Однако картина рез-
ко меняется, если величина одного из
факторов начнет уменьшаться. Жиз-
ненность организма будет снижаться
пропорционально фактору, градиент
которого стремится к минимуму.
Идея о том, что выносливость ор-
ганизма определяется самым слабым
звеном в цепи его экологических по-
требностей, впервые была высказана
Ю. Либихом и получила название «за-
кона минимума». Действие закона ми-
нимума наглядно показано на рис. 4.2.
Высота клёпок бочки соответствует напряженности экологических факторов,
жидкость в бочке – «жизненной силе». Как видим, «жизненная сила» вытека-
ет через самую низкую клёпку, т. е. там, где напряженность экологического
фактора минимальная. Результаты опыта, иллюстрирующего закон миниму-
ма, представлены на рис. 4.3.
Ограничивающими процветание вида может оказаться не только недо-
статок (минимум), но и избыток (максимум) фактора. Существование орга-
низма ограничено областями минимума и максимума. Между ними распола-
гается наиболее благоприятная (оптимальная) экологическая область –
экологический оптимум. Представление о лимитирующем влиянии мак-
симума наравне с минимумом ввел В. Шелфорд, сформулировавший закон
толерантности (выносливости организма).
Модель толерантности, как правило, имеет
вид купола. Его центральная часть соответствует
благоприятной напряженности экологического
фактора, при котором условия существования
организма наилучшие – это область оптимума.
Края купола отвечают или слишком низкой,
или слишком высокой напряженности экологи-
ческого фактора, при которой условия суще-
ствования организмов наихудшие (рис. 4.4).
Рис. 4.2. Модель, иллюстрирующая действие за-
кона минимума
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
36
Рис. 4.3. Опыт, иллюстрирующий действие закона минимума:
вода в колбах отличается содержанием количества питательных веществ: растение, выра-
щенное в первой колбе, богатой питательными веществами, имеет максимальную продук-
тивность, в последней колбе, бедной питательными веществами, – минимальную
Существуют стенобионтные и эврибионтные виды. Первые живут в усло-
виях узкого, вторые – широкого диапазона экологического фактора (рис. 4.5).
Законы минимума и толерантности служат путеводной нитью для пони-
мания путей адаптации к экологическим факторам определенной напряжен-
ности. У организмов в процессе естественного отбора вырабатываются при-
способления, позволяющие им жить в экологических условиях, которые пона-
чалу были для них неблагоприятными.
Рис. 4.4. Купол толерантности: существование вида определяется его выносли-
востью по отношению к воздействию абиотических факторов, когда напряже-
ние фактора слишком низкое или высокое, вид гибнет
.
Глава 4. ФАКТОРЫ РАСПРЕДЕЛЕНИЯ РАСТИТЕЛЬНОСТИ
37
Рис. 4.5. Типы экологических диапазонов:
А – эврибионты; Б – стенобионты; а – эврибионты
с оптимумом в средней части градации фактора;
б, в – эврибионты с оптимумом в зонах низких и
высоких значений фактора; г, д, е – варианты сте-
нобионтов (по Ю. И. Чернову)
Биотические факторы, отражающие от-
ношения между организмами, могут иметь
разную направленность. Все многообразие
биотических отношений может быть представлено в виде матрицы
Типы
связи
+
–
0
+
–
0
++
–
+
0+
+–
–
–
0–
+0
–
0
00
У организмов, взаимно нуждающихся в совместном проживании, обе
связи положительны. Например, цветковые растения и насекомые-опылите-
ли. Связь, положительная для одного организма, отрицательна для другого –
это наиболее распространенный тип связей питания: растительноядные жи-
вотные – растения, хищник – жертва. Нейтральная для одного организма
связь может быть положительной для другого. Например, стволы и кроны де-
ревьев служат местом обитания для множества птиц, других растений, кото-
рые не наносят дереву вреда, но сами получают значительное преимущество,
находя в кроне убежище или поднимаясь ближе к свету. Обе связи отрица-
тельные у организмов, находящихся в состоянии конкуренции, как правило,
особенно сильны конкурентные отношения у особей, относящихся к одному
виду.
Эколого-топологические факторы. Внутренняя структура ланд-
шафта и растительности обусловливается разнообразием экотопов (местопо-
ложений), связанных с литологическими разностями покровных отложений, с
формами рельефа, контролирующими гидротермический режим, характер
почв, состав и структуру растительных сообществ. Л. Г. Раменский разработал
типологию местоположений, которую можно широко использовать в ботани-
ко-географических исследованиях в гумидных ландшафтах умеренного пояса
(рис. 4.6).
Важным фактором, определяющим распределение растительных сооб-
ществ по экотопам, является влагообеспеченность последних. Конкретные
условия влагообеспеченности растительных сообществ обусловлены типом
водного режима почв, который, помимо зонально-климатических факторов,
зависит от положения участков в рельефе, их дренированности, режима по-
верхностных и грунтовых вод сопредельных участков, структуры почвогрунто-
вой толщи. Приведем классификацию режимов увлажнения по В. Д. Лопати-
ну с дополнениями:
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
38
Рис. 4.6. Схема типов местоположений:
1–20 – различные типы местоположений (по Л. Г. Раменскому)
Обильный тип увлажнения, обводнение. Местообитания мокрые или сы-
рые. В растительном покрове господствуют гигрофиты. Этот тип подразделя-
ется на подтипы: обильно проточный и обильно застойный.
Устойчивый тип увлажнения. Местообитания влажные или свежие. В рас-
тительном покрове господствуют настоящие мезофиты.
Переменный тип увлажнения на плакорах характеризуется низкой вла-
гоемкостью почв. Местообитания сухие и крайне сухие. Этот тип подразде-
ляется на подтипы: умеренно переменный и резко переменный. В естествен-
ной растительности при резко переменном увлажнении господствуют ксе-
рофиты.
.
Глава 4. ФАКТОРЫ РАСПРЕДЕЛЕНИЯ РАСТИТЕЛЬНОСТИ
39
Рис. 4.7. Схема распределения
режимов увлажнения на водо-
разделе:
1 – абсолютный суходол, резко пере-
менное увлажнение; 1а – склон, пере-
менное увлажнение; 2 – нормальный
суходол, устойчивое увлажнение; 3 –
замкнутое понижение на суходоле, обильно-застойное увлажнение; 3а – ложбина, обильно-
проточное увлажнение; 4 – подножье плакора, где выклиниваются грунтовые воды (ключе-
вины), обильно-проточное увлажнение
Рис. 4.8. Схема правила предва-
рения (по В. В. Алехину):
1 – северный вид, обитающий на
плакоре, на юге переходит на се-
верные склоны и в балки; 2 – южный вид на севере встречается на южных склонах
Зависимость режима увлажнения от рельефа отражена на рис. 4.7. Ком-
плекс экологических факторов и характер растительности меняется на скло-
нах разной экспозиции. На склонах южной экспозиции приход солнечной
энергии больше, чем на плакоре, а на склонах северной экспозиции – значи-
тельно меньше. В. В. Алехин сформулировал «Правило предварения», по ко-
торому склоны северной экспозиции несут на себе растительные группиров-
ки, свойственные более северной растительной зоне, а склоны южной экспо-
зиции – сообщества, характерные для более южной зоны, что создает условия
для формирования экстразональных группировок. Например, присутствие
сообществ неморальной зоны в бореальной зоне (рис. 4.8).
Вопросы к главе 4
1. Прямо и косвенно действующие экологические факторы.
2. Закон минимума и толерантности (выносливости организма).
3. Эколого-топологические факторы.
4. Правило предварения.
Рекомендуемая литература
Березина Н. А. Экология растений (учебник). М.: Высшая школа, 2008. 546 с.
Виноградов Б. В. Основы ландшафтной экологии. М.: ГЕОС, 1998. 417 с.
Миркин Б. М., Наумова Л. Г. Основы общей экологии: Учебное пособие / Под ред.
Г. С. Розенберга. М.: Университетская книга, 2005. 200 с.
Чернова Н. М., Былова А. М. Общая экология. М.: Дрофа, 2004. 416 с.
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
40
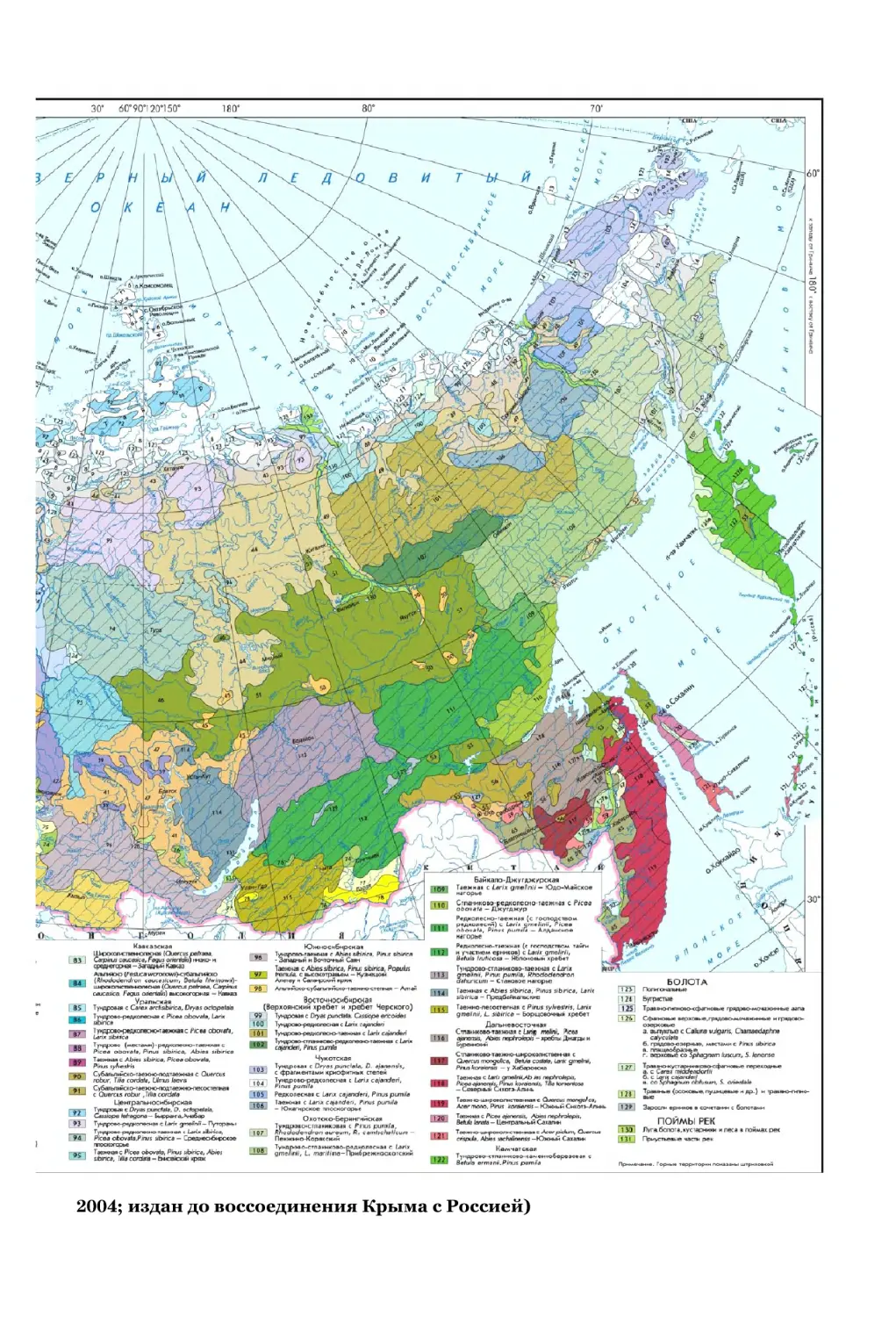
зучение географических закономерностей растительного покрова ос-
новано на двух подходах: первый – типологический, второй – рай-
онирование.
Типологический подход опирается на классификацию растительных со-
обществ и анализ их распространения путем составления геоботанических
карт. Важным событием явились «Геоботаническая карта СССР в масштабе
1:4 000 000», составленная в 1956 г. под руководством Е. М . Лавренко и
В. Б. Сочавы, и пояснительный текст к ней. В генерализованном виде эта карта
помещена в «Атласе СССР» (1985). В 1990 г. опубликована «Карта растительно-
сти СССР для ВУЗов м-ба 1:4 000 000», созданная под редакцией А. В. Белова,
С. А. Грибовой, Т. В. Котовой. В 2004 г. в «Национальном атласе России» при-
ведена карта растительности, составленная под редакцией Т. К. Юрковской,
И. С. Ильиной, И. Н. Сафроновой (рис. 5.1). В качестве единиц высшего ранга
в легенде выделяются растительность равнин и гор. Категориями следующего
ранга для равнин выступают зональные подразделения растительности: тунд-
ровая, бореальная, неморальная, степная, пустынная. Следующая ступень ле-
генды разделяет зоны по степени океаничности-континентальности. Например,
растительность бореальной зоны делится на: Приатлантическую, Восточно-
европейскую, Приуральскую (Предуральско-Зауральскую), Обь-Иртышскую,
Центральносибирскую, Восточносибирскую и Дальневосточную. Картируемой
единицей растительности гор является типичный поясной ряд, представлен-
ный чередованием высотных поясов от подножия к вершине. Особо выделяет-
ся растительность болот и речных пойм.
Второй подход к изучению географических закономерностей распреде-
ления растительности основан на ботанико-географическом районировании,
использующим геоботаническое, физико-географическое и ландшафтное рай-
онирования.
Основными критериями геоботанического районирования служат фито-
ценотические (состав и строение растительных сообществ, особенности струк-
туры растительного покрова) и флористические (видовой состав и соотноше-
ние видов, индицирующих различные экологические условия) признаки. Этот
подход был реализован в 1947 г. при составлении карты геоботанического рай-
онирования СССР и пояснительного текста к ней под редакцией Е. М. Лавренко
(рис. 5.2, табл.). Высшими единицами районирования выступают области, от-
ражающие зонально-региональные особенности растительности. Области под-
разделяются на провинции, полосы и округа, в некоторых областях выделяются
подобласти. Например, в Евроазиатской хвойнолесной области выделяются
Глава 5
ГЕОГРАФИЧЕСКИЕ ЗАКОНОМЕРНОСТИ
РАСТИТЕЛЬНОГО ПОКРОВА РОССИИ
.
Глава 5. ГЕОГРАФИЧЕСКИЕ ЗАКОНОМЕРНОСТИ РАСТИТЕЛЬНОГО ПОКРОВА РОССИИ
41
подобласти: Евросибирская темнохвойных лесов, Восточно-Сибирская светло-
хвойных лесов и Южно-Охотская темнохвойных лесов; в Евросибирской подоб-
ласти темнохвойных лесов выделяются провинции: Восточно-Европейская и
Урало-Алтайская, в последней – подпровинции: Предсибирская, Западно-Си-
бирская, Алтайско-Саянская. Провинции и подпровинции делятся на полосы,
например Восточно-Европейская провинция – на полосы осветленных темно-
хвойных лесов, зеленомошно-темнохвойных лесов и дубравно-темнохвойных
лесов. Низшей ступенью районирования служат округа.
Фитоценотические и флористические признаки использованы также в
1999 г. при составлении карты «Зоны и типы поясности растительности Рос-
сии и сопредельных территорий» под редакцией Г. Н. Огуреевой (рис. 5.3). Эта
карта выявляет зональные закономерности растительности равнинных терри-
торий и отображает типы высотной поясности в горах. Показаны единицы
разного уровня: на первом уровне выделяются зоны растительности и высот-
но-поясные системы гор, на втором − подзоны и типы поясности, на третьем
уровне – географические варианты растительности, характеризующиеся нали-
чием дифференциальных видов.
Ботанико-географическое районирование, основанное на характеристике
растительности природно-территориальных комплексов разной размерности,
осуществляется путем наложения геоботанических карт и схем геоботаниче-
ского районирования на карты ландшафтного районирования. Принципы
ландшафтного районирования разработаны А. Г. Исаченко. В 2001 г. им опуб-
ликована карта ландшафтно-экологического районирования России (рис. 5.4).
На высшем макрорегиональном уровне приняты система секторов, которая
отражает степень влияния океанов на ландшафты, и единицы географиче-
ской зональности – широтные ландшафтные зоны и подзоны. Сопоставление
единиц геоботанического и ландшафтного районирований показывает их
принципиальное тождество. Несмотря на различные наименования этих еди-
ниц у разных авторов, они сходны по своей сущности, поскольку обусловлены
единым естественно-историческим процессом.
Содержание настоящего учебника включает описание природных усло-
вий, растительности равнин и гор России. В качестве основных единиц описа-
ния растительного покрова предлагается рассматривать пояса, зоны, сектора и
области. Изменение гидротермического градиента в направлении север – юг
является фактором широтной поясности и зональности. На территории Рос-
сии на равнинах различаются пояса: арктический, субарктический, умерен-
ный, субтропический, которые подразделяются на зоны: тундр, бореальных
таежных лесов, неморальных широколиственных лесов, степей и субтропиков.
Изменение гидротермического градиента в направлении океан – континент
(под влиянием Западного переноса на западе Евразии, области высокого ат-
мосферного давления в Средней Сибири и муссонной циркуляции на Дальнем
Востоке) происходит разделение природных зон на сектора: Приатлантиче-
ский, Восточно-Европейский, Западно-Сибирский, Средне-Сибирский и Даль-
невосточный (Притихоокеанский).
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
42
Рис. 5.1. Карта растительности России (Национальный Атлас России,
.
Глава 5. ГЕОГРАФИЧЕСКИЕ ЗАКОНОМЕРНОСТИ РАСТИТЕЛЬНОГО ПОКРОВА РОССИИ
43
2004; издан до воссоединения Крыма с Россией)
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
44
Рис. 5.1. (продолжение). Легенда
.
Глава 5. ГЕОГРАФИЧЕСКИЕ ЗАКОНОМЕРНОСТИ РАСТИТЕЛЬНОГО ПОКРОВА РОССИИ
45
Рис. 5.1. (продолжение). Легенда
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
46
Рис. 5.1. (продолжение). Легенда
.
Глава 5. ГЕОГРАФИЧЕСКИЕ ЗАКОНОМЕРНОСТИ РАСТИТЕЛЬНОГО ПОКРОВА РОССИИ
47
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
48
Таблица
Легенда к схеме геоботанического районирования СССР
Области
Подобласти /
Группы провинций
Провинции
Подпровинции
I. Арктическая тундровая
(полоса арктических тундр;
полоса моховых и лишай-
никовых тундр)
II. Европейско-Сибирская ку-
старниковая (лесотундро-
вая) [полоса тундрово-
кустарниковая; полоса
лесокустарниковая
(лесотундра)]
а) Европейско-
Западно-
Сибирская
б) Центрально-
Сибирская
III. Берингийская кустарнико-
вая (лесотундровая)
VI. Евразиатская хвойнолесная
(таежная)
1. Европейско-
Сибирская темно-
хвойных лесов
а) Восточно-
Европейская
б) Урало-
Алтайская
1
Предсибирская
2
Западно-
Сибирская
3
Алтайско-
Саянская
2. Восточно-
Сибирская
светлохвойных
лесов
а) Средне-
Сибирская
б) Якутская
1
Вилюйско-
Хатангская
2
Витимо-
Колымская
3
Аяно-Удская
3. Южно-Охотская
темнохвойных лесов
V. Камчатская травяно-
лиственно лесная
VI. Дальневосточная хвойно-
широколиственная
VII. Европейская широколист-
венно лесная
а) Средне-
Европейская
б) Восточно-
Европейская
1
Подольско-
Бессарабская
2
Полесская
3
Средне-Русская
VIII. Средиземноморская лесная 1. Северная
а) Эвксинская
б) Дагестанская
1
Новороссийская,
2
Колхидская
3
Северо-
Кавказская
IX. Европейско-Сибирская ле-
состепная область
а) Восточно-
Европейская
б) Западно-
Сибирская
1
Средне-
Днепровская
2
Верхне-Донская
3
Закамская
.
Глава 5. ГЕОГРАФИЧЕСКИЕ ЗАКОНОМЕРНОСТИ РАСТИТЕЛЬНОГО ПОКРОВА РОССИИ
49
Продолжение
Области
Подобласти /
Группы провинций
Провинции
Подпровинции
X. Евразиатская степная
а) Восточно-
Европейская
(Понтическая)
1
Черноморско-
Азовская
2
Средне-Донская
б) Казахстан-
ская
1
Заволжско-
Уральская
2
Мугоджарско-
Тургайская
3
Восточно-
Казахстанская
в) Даурско-
Монгольская
1
Даурская
XI. Азиатская пустынная
1. Туранская группа
провинций
а) Терско-Кум-
ская
б) Северо-
Прикаспийская
в) Арало-Кас-
пийская
1
Мангышлакская
2
Усть-Урт-
Мугоджарская
3
Арало-
Балхашская
4
Кызыл-Кумская
5
Джунгаро-Тянь-
Шанская
2. Кавказско-
малоазиатская груп-
па провинций
а) Армянская
б) Атронатан-
ская
в) Кура-Арак-
синская
г) Гирканская
3. Переднеазиатская
группа провинций
а) Закаспийская
1
Восточно-
Прикаспийская
2
Кара-Кумская
б) Южно-
Туркестанская
1
Западно-Копет-
Дагская
2
Восточно-Копет-
Дагская
3
Парапамизская
4
Гиссаро-Дар-
вазская
4. Центрально-
азиатская группа
провинций
а) Центрально-
Тянь-Шанская
б) Восточно-
Памирская
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
50
Рис. 5.3. Зоны и типы поясности
.
Глава 5. ГЕОГРАФИЧЕСКИЕ ЗАКОНОМЕРНОСТИ РАСТИТЕЛЬНОГО ПОКРОВА РОССИИ
51
растительности России (1999)
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
52
Рис. 5.3. (продолжение) Легенда
.
Глава 5. ГЕОГРАФИЧЕСКИЕ ЗАКОНОМЕРНОСТИ РАСТИТЕЛЬНОГО ПОКРОВА РОССИИ
53
Рис. 5.3. (продолжение) Легенда
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
54
Рис. 5.3. (продолжение) Легенда
.
Глава 5. ГЕОГРАФИЧЕСКИЕ ЗАКОНОМЕРНОСТИ РАСТИТЕЛЬНОГО ПОКРОВА РОССИИ
55
Рис. 5.4. Ландшафтно-экологические макрорегионы России
(А. Г. Исаченко, 2001)
Условные обозначения: Восточно-Европейский сектор, зоны и подзоны: 1 – Арктическая;
2 – Субарктическая; 3 – Северотаежная; 4 – Среднетаежная; 5 – Южнотаежная; 6 –
Подтаежная; 7 – Широколиственная; 8 – Лесостепная; 9 – Северостепная; 10 – Сред-
нестепная; 11 – Южностепная; 12 – Полупустынная; 13 – Пустынная; Кавказский сектор,
зоны и подзоны: 14 – Широколиственная; 15 – Степная предсубтропическая; 16 – Сухо-
степная и полупустынная; 17 – Субсредиземноморская; 18 – Лесная предсубтропическая;
Западно-Сибирский сектор; зоны и подзоны: 19 – Тундровая; 20 – Лесотундровая; 21 –
Северотаежная; 22 – Среднетаежная; 23 – Южнотаежная; 24 – Подтаежная; 25 – Ле-
состепная; 26 – Северостепная; 27 – Среднестепная; 28 – Южностепная; Средне-
Сибирский сектор, зоны и подзоны: 29 – Арктическая; 30 – Субарктическая; 31 –
Северотаежная; 32 – Среднетаежная; 33 – Южнотаежная; 34 – Подтаежная; 35 –
Лесостепная; 36 – Степная; Восточно-Сибирский сектор, зоны и подзоны: 37 –
Субарктическая; 38 – Северотаежная; 39 – Среднетаежная; Южно-Сибирский сектор;
зона: 40 – Степная; Дальневосточный сектор, зоны и подзоны: 41 – Субарктическая;
42 – Лугово-лесная; 43 – Северотаежная; 44 – Среднетаежная; 45 – Южнотаежная; 46 –
Подтаежная; 47 – Широколиственнолесная
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
56
.
Глава 5. ГЕОГРАФИЧЕСКИЕ ЗАКОНОМЕРНОСТИ РАСТИТЕЛЬНОГО ПОКРОВА РОССИИ
57
Региональные особенности почвенно-растительного покрова, опреде-
ляемые биоклиматическими условиями, геоморфологическим строением и
характером покровных отложений, служат основанием для выделения бо-
танико-географических областей. В зоне тундр выделяются области: в подзо-
не высокоарктических тундр – две области: Баренцевоморская и Сибирская;
для подзон арктической, типичной, южной тундр и лесотундры: Кольская, Во-
сточно-Европейская, Ямало-Гыданская, Таймырская, Лено-Колымская, Чукот-
ская и Северо-Тихоокеанская островная. В пределах бореальной таежной зоны
выделяются области: Карельская, Восточно-Европейская, Западно-Сибирская,
Средне-Сибирская и Сахалинская, а также Приморская область хвойно-широ-
колиственных лесов. В зоне неморальных широколиственных лесов выделяют-
ся области: Восточно-Европейская, Западно-Сибирская (лесостепная) и Мань-
чжурская. В зоне степей в подзонах северных и средних степей выделяются
области: Восточно-Европейская (Донско-Волжская), Заволжско-Уральская, За-
падно-Сибирская и Дауро-Монгольская; в подзоне южных опустыненных сте-
пей – Прикаспийская область. В зоне субтропиков выделяются области Южного
берега Крыма и Черноморского побережья Кавказа.
Для гор характерно интегральное проявление широтных и высотно-по-
ясных особенностей распределения растительности. Здесь ботанико-географи-
ческие области соответствуют группам типов поясности. Выделяются следую-
щие основные горные страны: Уральская, Кавказская, Алтайская, Восточно-
Забайкальская, Северо-Восточно-Сибирская, Камчатская.
Дальнейшая дифференциация ботанико-географических областей равнин
связана с эколого-топологическими рядами растительных сообществ, обуслов-
ленных почвенно-геоморфологическими особенностями ландшафта. В горах
на этом уровне выделяются высотные пояса.
Ботанико-географическое районирование, отражающее основные гео-
графические закономерности растительного покрова России на макроуровне,
предлагается построить путем совмещения карт «Зоны и типы поясности рас-
тительности России и сопредельных территорий» под редакцией Г. Н. Огуре-
евой и «Ландшафтно-экологические макрорегионы России», составленной
А. Г. Исаченко. Предлагаемая схема ботанико-географических макрорегионов
представлена на рис. 5.5.
Вопросы к главе 5
1. Геоботаническое картографирование как метод познания географических законо-
мерностей растительного покрова. Анализ легенд геоботанических карт.
2. Геоботаническое районирование. Анализ карты «Зоны и типы поясности раститель-
ности России и сопредельных территорий: Карта для ВУЗов. М-б 1:8 000 000» / Под
ред. Г. Н. Огуреевой (1999).
3. Изучение закономерностей растительного покрова России на основе анализа схем
физико-географического и ландшафтного районирований.
Рекомендуемая литература
Агаханянц О. Е. Ботаническая география СССР. Учеб. пособие для пед. ин-тов по специ-
альностям 2106 «Биология» и 2107 «География». Минск: Высш. шк., 1986. 175 с.
.
Ч а с т ь I. БАЗОВЫЕ ПОЛОЖЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
58
Алехин В. В. Растительность СССР в основных зонах. Второе издание под общей редак-
цией С. С. Станкова. М.: «Советская наука», 1951. 512 с.
Заповедники СССР. Серия изданий в 10 книгах. М., 1985–2000.
Зоны и типы поясности растительности России и сопредельных территорий. М-б
1:8 000 000. Пояснительный текст и легенда к карте / Отв. ред. Г. Н. Огуреева.
М.: Изд-во центр «Интеграции», 1999. 64 с.
Исаченко А. Г. Экологическая география России. СПб.: Изд-во С.-Петерб. ун-та, 2001.
328 с.
Исаченко А. Г. Введение в экологическую географию: Учеб. пособие. СПб.: Изд-во
С.-Петерб. ун-та, 2003. 192 с.
Огуреева Г. Н. Ботанико-географическое районирование СССР. М.: Изд-во Моск. ун-та,
1991. 188 с.
Раковская Э. М., Давыдова М. И. Физическая география России. Учеб. для студ. пед.
высш. учеб. заведений: В 2-х ч. М.: Гуманит. изд. центр ВЛАДОС, 2001. Ч. 1. 288 с.
Ч.2.304с.
Растительность Европейской части СССР / Под ред. С. А. Грибовой, Т. И. Исаченко,
Е. М. Лавренко Л.: Наука, 1980. 236 с.
Растительный покров Западно-Сибирской равнины / Ильина И. С., Лапшина Е. И.,
Лавренко Н. Н. и др. Новосибирск: Наука, 1985. 251 с.
Растительный покров СССР. Пояснительный текст к геоботанической карте СССР
масштаба 1:4 000 000. Т. 1–2 / Под ред. Е. М. Лавренко и В. Б. Сочавы. М.–Л.:
Изд-во АН СССР, 1956. 972 с.
Состояние биологического разнообразия природных экосистем России / Под ред.
В. А . Орлова и А. А. Тишкова. М.: НИА-Природа, 2004. 116 с.
Сочава В. Б. Проблемы физической географии и геоботаники. Избранные труды. Но-
восибирск: Наука, Сибирское отделение, 1986. 343 с.
Шумилова Л. В. Фитогеография. Томск: Изд-во Томского ун-та, 1979. 239 с.
Юрковская Т. К. Геоботаническое картографирование и составление аналитических
карт растительности // Актуальные проблемы геоботаники. III Всероссийская
школа-конференция. Лекции, 2007. С. 43–71.
Карты
Атлас СССР. М.: Главное управление геодезии и картографии при Совете Министров
СССР, 1985. 224 с.
Геоботаническая карта СССР. М-б 1:4 000 000 / Ред. Е. М. Лавренко, В. Б. Сочава.
М.–Л.: АН СССР, 1954.
Зоны и типы поясности растительности России и сопредельных территорий: Карта для
ВУЗов. М-б 1:8 000 000 / Отв. ред. Г. Н. Огуреева. М.: «Экор», 1999. 2 л.
Карта растительности Европейской части СССР. М-б 1:2 500 000 / Отв. ред. Т. И. Иса-
ченко, Е. М. Лавренко. БИН АН СССР, 1974.
Карта растительности Западно-Сибирской равнины. М-б 1:1 500 000 / Под рук. В. Б. Со-
чавы. Иркутск: Институт географии Сибири и Дальнего востока СО АН СССР,
1976.
Национальный атлас России. Т. 1. Общая характеристика территории // Т. К. Юрков-
ская, И. С. Ильина, И. Н. Сафронова. Растительность [карта] М. 1:15 000 000 М.,
2004. С. 370–371.
.
Глава 6. ЗОНА ТУНДР
59
Часть II
РАСТИТЕЛЬНОСТЬ
ПРИРОДНЫХ ЗОН РОССИИ
От Северного полюса к югу по территории России простираются
Арктический, Субарктический, Умеренный и Субтропический
пояса, в формировании растительности которых ведущую роль
играют первичные факторы географической зональности – тем-
пературный режим и количество атмосферных осадков. По сло-
вам Александра Гумбольдта, там, где миросозерцание стремит-
ся подняться до более возвышенной точки зрения, эта картина
была бы лишена своей очаровательной прелести, если бы она
не представила нам и сферу органической жизни.
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
60
Арктический пояс охватывает Северный Ледовитый океан, характеризующий-
ся устойчивым ледовым покровом. Средняя температура самого теплого ме-
сяца здесь колеблется от 0 °C на севере до 5 °C на юге. Субарктический пояс
лежит примерно между изотермами июля 5 °С на севере и 10 °С на юге. Рас-
тительность арктического и субарктического поясов относится к зоне тундр
(при увеличении средней температуры самого теплого месяца выше 10 °C
намечается переход к зоне бореальных таежных лесов). Подразделение поя-
сов на зоны и подзоны показано на карте (рис. 6.1).
Глава 6
ЗОНА ТУНДР
риведем сведения о географическом положении, природных условиях,
флоре и способах адаптации растений к условиям обитания в зоне тундр.
Географическое положение и природные условия. Тундра (финн. tun-
turi) в переводе означает плоское безлесное пространство. Тундровая зона в
арктическом поясе занимает в основном северные острова; в субарктическом
поясе она охватывает острова Колгуев, Вайгач, южный остров Новой Земли,
о. Врангеля и материковое побережье Северного Ледовитого океана от Коль-
ского полуострова на западе до Чукотки на востоке (рис. 6.1). Бóльшая часть
зоны располагается севернее Полярного круга. На Дальнем Востоке, вслед-
ствие воздействия холодного течения, южная граница тундры смещается на юг
почти до 60° с.ш. Преобладающий тип рельефа в тундре – плоские примор-
ские низменности, которые местами осложняются возвышенностями и даже
невысокими горными хребтами.
Большая протяженность зоны тундр с запада на восток служит причиной
ее секторной неоднородности, в основе которой лежат изменения климата,
определяемые чередованием центров низкого и высокого атмосферного дав-
ления. Территория от Гренландии до п-ова Ямал находится в области низкого
давления. Циклоны, зарождающиеся над Атлантикой, несут на восток влаж-
ные и относительно теплые массы воздуха (западный перенос). В Восточной
Сибири зимой располагается центр высокого давления (Азиатский антицик-
лон). Он создает преграду на пути переноса воздушных масс с запада и опре-
деляет резко континентальный характер климата. При прогревании суши ле-
том высокое атмосферное давление сменяется низким, что вызывает начало
действия муссонных ветров с Северного Ледовитого и Тихого океанов. Поэто-
му здесь на летний период приходится 80–85 % атмосферных осадков. Центр
низкого давления между Чукоткой и Аляской порождает циклоническую дея-
тельность, смягчающую климат Дальнего Востока.
Арктический и субарктический пояса
.
Глава 6. ЗОНА ТУНДР
61
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
62
Формирование растительности тундр определяется суровыми климати-
ческими условиями. Зима продолжительная и холодная, лето короткое. Тунд-
ра оживает, как только первые лучи солнца появляются над горизонтом: веч-
нозеленые растения, доминирующие в растительном покрове, способны к
фотосинтетической деятельности даже под слоем снега. В полярный день не-
прерывный поток солнечной энергии, прогревающий землю и воздух, изоби-
лие света обеспечивают быстрое начало развития тундровой растительности,
вегетирующей около 2 месяцев. Средняя годовая температура ниже нуля, но
не зимние холода (полюс холода Северного полушария располагается, как из-
вестно, в таежной зоне, в Восточной Сибири), а холодное и короткое лето
определяет особенности условий обитания в тундре. Заморозки возможны в
любой летний день, но бывают и жаркие дни, когда температура воздуха под-
нимается до 25–30 °С. Ведущим фактором, определяющим широтную диффе-
ренциацию тундр, служат среднемесячные температуры июля (рис. 6.2).
Распределение годовых сумм атмосферных осадков меняется с запада на
восток (рис. 6.3): на побережье Кольского п-ова выпадет 600–800 мм осад-
ков, на северо-востоке Европейской части России и в Западной Сибири 400–
600 мм, в Восточной Сибири, характеризующейся резко континентальным
климатом, количество осадков минимально 200–300 мм, на Чукотском п-ове
оно увеличивается до 400–600 мм.
ГТК вегетационного периода уменьшается от 1,8–2,0 на западе до 1,0–
1,5 в Восточной Сибири, и вновь увеличивается на Чукотке до 2,0. В связи с
низкими температурами испаряемость мала, а увлажнение избыточно, что,
наряду с многолетнемерзлыми грунтами, приводит к образованию многочис-
ленных озер и болот.
Рис. 6.2. Средние месячные температура июля (Атлас Арктики, 1985)
.
Глава 6. ЗОНА ТУНДР
63
Рис. 6.3. Среднегодовое количество атмосферных осадков (Атлас Арктики,
1985)
Осадки в тундре выпадают в основном в виде снега. Продолжительность
залегания снежного покрова в Субарк-
тике до 220–280 дней. Его толщина
зависит от рельефа, а также от конти-
нентальности климата: к западу от Тай-
мыра снега больше, к востоку меньше.
Снежный покров в тундре играет роль
положительного экологического фак-
тора. В качестве хорошего теплоизоля-
тора он защищает почву, растения и
животных от морозов. Кроме того, слой
снега предохраняет растения от снеж-
ной корразии: ветви, выступающие над
снегом, срезаются или сильно повре-
ждаются зимней поземкой, поэтому
высота растений полностью зависит от
мощности снежного покрова.
Ландшафты тундры хранят па-
мять о недавнем оледенении (рис. 6.4).
Почти повсеместно распространены
Рис. 6.4. Лед под тонким корнеобитае-
мым горизонтом почвы
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
64
Рис. 6.5. Полигональная тундра
многолетнемерзлые породы – «вечная
мерзлота». Глубина сезонного оттаи-
вания колеблется от 0,3–0,4 м в под-
зоне северных тундр до 0,5–2,0 м в
южных тундрах. Постоянномерзлый
слой охлаждает почвы и замедляет
биохимические процессы, препятству-
ет фильтрации поверхностных вод и
способствует заболачиванию; он резко
ограничивает грунтовое питание рек и
их эрозионную деятельность. Морозное
растрескивание грунтов и образование
ледяных клиньев в трещинах ведут к
формированию полигонального мик-
рорельефа (рис. 6.5). Разнообразные
формы микрорельефа (рис. 6.6) обу-
словливают комплексность почвенно-
растительного покрова. При вытаива-
нии подземных льдов возникает явле-
ние термокарста – понижения округлой
формы, часто занятые озерами. Ста-
бильность многолетнемерзлых грунтов, колебания верхней границы мерзло-
ты и даже само ее существование зависят от многих обстоятельств, и в значи-
тельной степени от растительного покрова, имеющего значение важнейшего
стабилизирующего фактора. Растительная дернина затрудняет теплообмен
между мерзлым слоем и воздухом, скрепляет почву, препятствует сортировке
грунта.
В условиях сурового климата, в результате неравномерного замерзания
воды подземного источника возникает крупная ледяная линза, что приводит
к формированию положительных замкнутых форм криогенного рельефа –
бугров пучения (гидролакколитов), высота которых может достигать 80 м
(рис. 6.7.)
Недостаток тепла и обилие влаги, мерзлота, низкая емкость биологиче-
ского круговорота, небольшое количество растительного опада, его замедлен-
ная гумификация и минерализация – все это определяет главные черты гео-
химических процессов и почвообразования. Химическое выветривание проте-
кает слабо, высвобождающиеся основания вымываются из почвы, так что она
обеднена кальцием, натрием, калием, но обогащена железом и алюминием.
Широкое распространение в Субарктике, особенно в южной ее части, имеют
тундровые глеевые почвы с торфянисто-перегнойным горизонтом. В связи с
низкой продуктивностью растительности торфообразование слабое; мощность
торфа редко превышает 1 м. Почвенный покров, как и растительный, характе-
ризуется мозаичностью.
.
Глава 6. ЗОНА ТУНДР
65
Флора. Арктическая флора очень молода. Первые этапы ее развития от-
носятся к началу четвертичного периода, но полностью она сформировалась
лишь в течение послеледникового времени, т. е. за последние 10 тыс. лет. При
этом большую роль сыграла иммиграция видов, произраставших ранее вокруг
Рис. 6.6. Формы микрорельефа и почвы зоны тундр (Атлас Арктики, 1985):
Генетические горизонты: А0 – подстилка; АТ – гумусовый торфянистый; А1 – гумусо-
вый; А1С – переходный от гумусового к материнской породе; В – иллювиальный с разде-
лением на подгоризонты; G – глеевый; С – материнская порода
Комплексы почв: а – каменно-многоугольниковый (почвы: 1 – примитивно-щебнистая;
2 – перегнойно-щебнистая; 3 – почва пятна), Северная Земля – о. Большевик (высоко-
арктическая тундра); б – трещинно-нанополигональная (почвы: 1 – тундровая иллюви-
ально-гумусовая надмерзлотно- и контактно-глеевая; 2 – почва пятна; 3 – трещинно-
торфянистая), Западная Сибирь – п -ов Ямал (типичная тундра); в – комплекс почв бай-
джераховых ландшафтов (почвы: 1 – тундровая гумусная глееватая (денудированная);
2 – болотная арктическая глеевая), Новосибирские острова (высокоарктическая тундра);
г – спорадический пятнистый (почвы: 1 – тундровая глеевая; 2 – почва пятна), север
Дальнего Востока – Анадырская низменность (типичная и южная тундра)
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
66
Рис. 6 .7 . Бугор пучения, расположенный в центральной части Ямбургского
месторождения, п-ов Ямал (фото: В. И. Богоявленский)
пояса сплошного оледенения. Арктика и Субарктика входят в Арктическую
флористическую провинцию (Тахтаджян, 1978), ее флора очень бедна и
насчитывает немногим более 1500 видов. Относительно богата флора Чукот-
ки, включающая около 800 видов. Это объясняется тем, что в течение плей-
стоцена полуостров не подвергался сплошному оледенению. Эндемичных ви-
дов и подвидов в арктической флоре более 100, главным образом они произ-
растают на Чукотке. Среди них имеются лютик (Ranunculus sabinei), мак
(Papaver polare), несколько видов ивы (Salix arctica и др.), виды крестоцветно-
го рода Braya, виды мытника (Pedicularis), злаки (Puccinellia vahliana,
P. wrightii, P. angustatа) и др.
Любопытны флорогенетические связи вечнозеленых кустарничков
тундры. Их родина – горы тропических широт Юго-Восточной Азии. На это
обратил внимание русский географ А. Н. Краснов, исследовавший в конце XIX
века растительность высотных поясов островов Зондского архипелага. Приве-
дем его описание растительности субальпийского пояса одного из вулканов на
о. Ява: «... любопытно, что дошедшие до вершин формы, слагаются в ассоциа-
ции, большей частью составленные теми же родами или семействами, кото-
рые идут далеко на север и образуют зону тундры. Род Agapetes заменяет нашу
голубику. Это Ericaceae – по облику его можно назвать тропическим деревом
голубикой... Его плоды и цветы так напоминают чернику, что вы сразу при-
знаете в нем Vaccinium, а черные губы ваших спутников-малайцев напомнят
вам ваши детские прогулки по болотам и лесам родины». А. Н. Краснов делает
вывод, что «флора тундры, защищенная от мороза снежным покровом, ... есть
прямой продукт вырождения тропической растительности в полярную под
влиянием понижения температуры при сохранении прочих условий, то есть
влаги и, возможно, меньшего колебания температуры».
.
Глава 6. ЗОНА ТУНДР
67
Жизненные формы и адаптация растений. Каждая подзона тундр
характеризуется господством определенных жизненных форм растений. Для
южной тундры, особенно для многоснежных районов, характерны кустарни-
ковые сообщества. Кустарники представлены главным образом арктическими
видами ивы и березы. Очень характерны кустарнички из семейства вереско-
вых (виды Vaccinium и Arctostaphylos) и травянистые растения из семейства
осоковых – виды пушицы (Eriophorum) и осоки (Саrех). В типичной тундре
господствуют кустарничково-мохово-лишайниковые сообщества. Особенно
характерны виды куропаточьей травы (Dryas) из семейства розоцветных и не-
которые стелющиеся ивы (Salix polaris и др.), а также многолетние травяни-
стые растения из семейств мятликовых (злаковых), капустных (крестоцвет-
ных) и гвоздичных, виды камнеломки (Saxifraga) из семейства камнеломко-
вых, мытника (Pedicularis) из семейства норичниковых, лапчатки (Potentilla)
из семейства розоцветных и кисличник (Oxyria digyna) из семейства гречиш-
ных. Однолетних трав очень мало, а нередко они полностью отсутствуют. В
растительном покрове арктических тундр доминируют листостебельные, пе-
ченочные мхи и лишайники, в высокоарктических тундрах (полярных пусты-
нях) доминируют лишайники.
Для растительности тундр характерны низкорослые психрофиты и крио-
фиты, приспособленные к существованию в условиях низких температур, к ис-
пользованию тепла приземного слоя и защитных функций снежного покрова.
Наиболее распространенная жизненная форма – карликовые кустарнички и
многолетние травы (однолетники здесь – явление редкое; они не успевают за-
вершить жизненный цикл в одно лето). Стебли у них короткие, листья малых
размеров и у многих собраны в прикорневую розетку. Обычны здесь и подуш-
ковидные формы, возникающие при отмерзании верхушечных почек. В таком
случае трогаются в рост нижние пазушные почки, что приводит к обильному
ветвлению и образованию коротких стелющихся побегов. Густо расположен-
ные мелкие ветви с листьями на верхушках побегов придают кусту обтекаемую
форму, которая хорошо противостоит ураганным ветрам. Под пологом такой
подушки создается более мягкий микроклимат, а нижние отмершие листья,
постепенно разрушаясь, обогащают почву питательными веществами. Среди
полярных растений много вечнозеленых кустарничков. Таким растениям не
приходится тратить некоторую часть вегетационного периода на приобрете-
ние столь необходимых для их развития органов, как стебель и листья.
Физиологическим приспособлением растений, который защищает от гу-
бительного действия морозов, служит повышение концентрации клеточного
сока (главным образом за счет растворимых углеводов). Это предотвращает
образование внутри клетки льда, который может ее разрушить. Травянистое
растение ложечница арктическая (Cochlearia arctica) в листьях и бутонах пе-
реносит суровую зиму с температурой до –46 °С и весной продолжает свое
развитие.
Растения тундр отличаются низкой интенсивностью фотосинтеза, мед-
ленным накоплением органической массы, малой энергией роста. Годичные
кольца растений нередко состоят из одного ряда клеток. Побеги ивы полярной
удлиняются за год на 1–5 мм. Годичный прирост лишайников рода кладония
(ягель) составляет около 4 мм.
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
68
Рис. 6.8. Температура в градусах Цельсия
различных частей мака со стороны осве-
щенной солнцем и в тени (по H. Walter,
S.-W. Breckle)
Вегетационный период короткий.
Наиболее благоприятное время цветения
и плодоношения с середины июля до
конца августа, когда в основном вегета-
ция заканчивается. Такие растения, как
крупка, полярный мак, камнеломка нуж-
даются только в одном месяце, для того
чтобы начать рост, зацвести и дать семе-
на. Непродолжительность вегетационно-
го периода возмещается для растений
непрерывностью дневного освещения.
У всех, кто бы ни побывал летом в
тундре, ее аспект оставляет неизгладимое
впечатление своим светло-зеленым коло-
ритом и окраской цветков многих расте-
ний, отличающейся большей интенсив-
ностью, чем у цветков их лесных сороди-
чей. Яркое описание цветущей тундры
дал В. М. Сдобников: «Когда видишь арктическую тундру поздней осенью или
ранней весной, кажется, что здесь не может быть никаких цветов. Но вот
наступает лето. В конце июля разгар цветения растений. В это время на сол-
нечных местах, на теплых песчаных склонах по берегам озер и рек можно уви-
деть настоящие цветники. Золотистым цветом горят крупные цветы сиверсии,
полярного мака, лютиков, одуванчиков и камнеломок. Розовым цветом рас-
пускается мытник, а в теплое лето и иван-чай. Кусочками арктического неба,
упавшими в тундру, кажутся голубые цветы полярной незабудки. Цветут аст-
рагалы, паррии, куропаточья трава (дриада), калужница, звездчатка и многие
другие цветы».
Метеорологические данные о температуре воздуха летом существенно
отличаются от фактической температуры приземного слоя воздуха. Дело в
том, что земная поверхность поглощает много солнечного тепла и поэтому
температура приземного воздуха выше на несколько градусов, чем температу-
ра, измеряемая в метеорологической будке. Существенные различия темпера-
тур наблюдаются даже у разных частей одного растения (рис. 6.8). Высокая
концентрация тепла в цветке мака благоприятствует развитию генеративных
органов.
В тундре, как уже отмечалось выше, случаются дни с экстремально высо-
кими температурами порядка 30 °С. Однако обилие тепла губительно действу-
ет на растения, так как транспирация влаги ограничивается низкой темпера-
турой почвы.
.
Глава 6. ЗОНА ТУНДР
69
6.1. ПОДЗОНЫ ТУНДР
В зоне тундр, по мере продвижения на юг, тепловой режим становится
более благоприятным. В связи с этим различают подзоны: в Арктическом поя-
се – подзона высокоарктических тундр (полярных пустынь), в Субарктическом
поясе – подзона арктических (мохово-лишайниковых), типичных (кустарнич-
ково-мохово-лишайниковых), южных (кустарниковых) тундр (см. рис. 5.2.).
В пределах зоны тундр рассматривается также переходная от субарктического
к умеренному поясу подзона лесотундры.
Подзона высокоарктических тундр (полярных пустынь) имеет
циркумполярное размещение (см. рис. 6.1), она представлена прерывисто в
основном на островах. В Российском секторе Арктики занимает острова и ар-
хипелаги Северного Ледовитого океана: Земля Франца-Иосифа, Северный
остров Новой Земли, Северная Земля, Новосибирские о-ва. Бóльшая часть по-
верхности островов покрыта ледниками (рис. 6.9). Участки, свободные ото
льда, представлены каменистыми субстратами, моренными и морскими от-
ложениями, причем 75–90 % их поверхности практически лишены расти-
тельности.
Климат суровый. Зимой во время полярной ночи на западе подзоны
минимальные температуры воздуха в феврале –20–22 °С; на востоке подзоны
–25–30 °С. Лето характеризуется избытком солнечной радиации и недостат-
ком тепла. Период средних суточных положительных температур очень не-
продолжителен; средняя температура июля не поднимается выше 1–3 °С. Ат-
мосферные осадки, даже летом, выпадают в основном в виде снега. Низкие
Рис. 6.9. Ледник на о. Шпицберген
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
70
Рис. 6.10. Лишайники на каменистых скалах острова Нортбрук, Земля Франца-
Иосифа
температуры воздуха способствуют энергичному развитию морозного выветри-
вания, резко замедляют интенсивность процессов химического и биологическо-
го преобразования грунтов. В связи с этим, поверхностные отложения состоят
из довольно крупных обломков горных пород и почти лишены глинистого ма-
териала.
В растительном покрове отмечается полное доминирование лишайников
(Sphaerophorus globosus, Psoroma hypnorum, Solorina crocea, Stereocaulon
spp., Flavocetraria cucullata, Cetrariella delisei, Cetraria islandica, Thamnolia
vermicularis), мхов (Ditrichum flexicaule, Oncophorus wahlenbergii, Racomitri-
um sp., Bryum sp., Orthothecium chryseon) и водорослей (рис. 6.10). Они обра-
зуют тонкую пленку жизни с единичными экземплярами типично арктиче-
ских цветковых растений: лисохвост альпийский (Alopecurus alpinus), щучка
коротколистная (Deschampsia brevifolia), лютик серно-желтый (Ranunculus
sulphureus), камнеломка снежная (Saxifraga nivalis), мак полярный (Papaver
polare).
В связи с тем, что бóльшую площадь островов занимают ледники, расти-
тельный покров представлен фрагментарно, главным образом в береговой
зоне.
Подзона арктических тундр расположена на побережье Северного
Ледовитого океана. Под влиянием сурового климата растительный покров
здесь имеет низкое проективное покрытие. Большие пространства заняты рос-
сыпями щебня, каменными многоугольниками и т. п. (рис. 6.11). На участках,
занятых растительностью, ее сомкнутость не превышает 50–60 %. Обилие
.
Глава 6. ЗОНА ТУНДР
71
микронеровностей рельефа: бугров и впадин, пятен-медальонов, создает ха-
рактерную комплексную структуру тундровых сообществ. Растительность арк-
тической тундры представлена кустарничково-травяно-лишайниковыми, тра-
вяно-лишайниково-моховыми полигональными, пятнистыми, кочкарными
тундрами, осоково-злаково-моховыми болотами.
Из лишайников преобладают кустистые формы (Alectoria, Stereocaulon,
Flavocetraria, Cornicularia, Dactylina, Cladonia). Очень мало мхов; среди
встречающихся преобладают виды дикрановых и политриховых; также при-
сутствуют печеночные мхи. Из цветковых можно отметить: злаки (Alopecurus
alpinus, Deschampsia brevifolia, Poa arctica, Phippsia algida, Dupontia fisheri,
Arctagrostis sp. и др.), осоки (Carex bigelowii, C. rupestris), мелкие красиво цве-
тущие двудольные – полярный мак (Papaver radicatum), незабудка альпий-
ская (Myosotis alpestris), камнеломки (Saxifraga cespitosa, S. oppositifolia),
смолевка бесстебельная (Silene acaulis), незабудочник волосистый (Eritrichium
villosum); характерны пушицы (Eriophorum angustifolium, E. scheuchzeri).
На глинистых субстратах распространена пятнистая тундра (рис. 6.12).
Она состоит из слегка выпуклых округлых и овальных пятен грунта в попе-
речнике от 10–20 см до 2–3 м, лишенных растительности. Они образуются в
результате оплывания на отдельных участках верхних размокших горизонтов
почвы. В итоге появляются глинистые расплывшиеся голые пятна, окружен-
ные валиками щебня, поросшими мхами, лишайниками, травами и кустар-
ничками.
В подзоне арктических тундр начинают принимать участие прострат-
ные – стелющиеся и прижатые к почве – кустарнички, виды рр. Salix, Dryas.
В составе болотных сообществ наряду со злаками обильны осоки и пушицы.
Наличие Salix nummularia как массового фонового растения на западе подзоны
Рис. 6.11. Арктическая тундра, п-ов Таймыр
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
72
Рис. 6.12. Пятнистая тундра (по H. Walter, S.-W. Breckle):
Горизонтальная проекция и вертикальный разрез площадки 1×2 м: 1 – обнаженная почва;
2 – трещины; 3 – моховой покров в понижениях; 4 – лишайники; 5 – мятлик (Poa alpina);
6 – мак (Papaver sp.); 7 – ожика (Luzula nivalis); 8–11
–
камнеломки (Saxifraga platysepala,
S. cespitosa, S. nivalis); 12 – лисохвост (Alopecurus alpinus)
отделяет новоземельско-гыданские тундры от более восточных. В таймырско-
ленских тундрах возрастает роль Salix polaris, S. reptans, Dryas punctata,
Cassiope tetragona, Luzula confusa, Carex arctisibirica; в восточносибирских
тундрах на первое место по значению выходит Cassiope tetragona, исчезает
Dryas octopetala. Чукотские тундры выделяются распространением Salix
rotundifolia, S. phlebophylla, Dryas integrifolia, Carex lugens (последняя распро-
странена к востоку от р. Алазеи).
Подзона типичных (кустарничково-лишайниково-моховых)
тундр на северо-востоке Русской равнины занимает о. Колгуев, северную окра-
ину Малоземельской и Большеземельской тундр, на севере Западно-Сибир-
ской равнины – центральные части п-овов Ямал, Гыданский, в Средней Сиби-
ри – центральную часть п-ова Таймыр, в Восточной Сибири – узкую прибреж-
ную полосу Восточно-Сибирского моря, а на Дальнем Востоке охватывает
Чукотский п-ов и протягивается на юг по побережью Анадырской низменно-
сти. Температуры самого теплого месяца 4–6 °С. Там, где отмечается увеличе-
ние атмосферных осадков, снежный покров залегает более равномерно. По-
следний, защищая зимой растения от ветровой корразии, обеспечивает во
.
Глава 6. ЗОНА ТУНДР
73
многих случаях возможность развития довольно сомкнутого растительного
покрова, лишь местами на поверхность выступают голые пятна грунта (пятни-
стые тундры).
Растительность подзоны представлена кустарничково-лишайниково-мо-
ховыми, травяно-лишайниково-моховыми, бугорково-пятнистыми, пушицево-
моховыми кочкарными, низкокустарниковыми (ивовыми) тундрами, полиго-
нальными травяно-кустарничково-лишайниково-моховыми болотами. В расти-
тельном покрове доминируют зеленые мхи (рис. 6.13), лишайники (рис. 6.14),
среди кустарничков преобладают простратные формы (рис. 6.15). Важную
роль в растительности тундры играют ягодные растения (рис. 6.16).
Empetrum hermaphroditum относится к дифференцирующим видам боль-
шеземельских тундр; в ямало-гыданских тундрах его замещает E. subholarcticum;
появляется Cassiope tetragona, роль которой особенно велика в таймырских
тундрах. В этой же части подзоны произрастает Betula exilis наряду с B. nana.
Последняя исчезает в яно-колымских тундрах. В чукотских тундрах появляется
Rhododendron camtschaticum (на востоке), Salix fuscescens, Dryas integrifolia,
Carex lugens, Salix krylovii, S. richardsonii.
Подзона южной (кустарниковой) тундры характеризуется более
теплым, чем в предыдущих подзонах, летом: средняя температура воздуха в
июле составляет 9–11 °С. Среднегодовое количество атмосферных осадков на
северо-востоке Русской равнины и на Западно-Сибирской низменности со-
ставляет 400–600 мм. В Средней и Восточной Сибири, от Енисея до Колымы
количество атмосферных осадков снижается до 200–300 мм. На Дальнем Во-
стоке, в районе Анадырской низменности количество атмосферных осадков
составляет 400–600 мм.
Растительность подзоны характеризуется преобладанием ивняково-
ерниковых и кочкарных тундр, а также бугристых (в Европе) и полигональных
травяно-кустарничково-лишайниково-моховых (в Азии) болот. В раститель-
ном покрове преобладают четырехъярусные сообщества из кустарников (рис.
6.17), кустарничков, трав, мхов и лишайников. Кустарники находятся под сне-
гом, и холодные зимние ветры не уничтожают их молодые побеги и почки.
Именно вымерзание и иссушение чаще всего губят кустарники на востоке.
Для кольских тундр характерны Betula nana, Calluna vulgaris, Empetrum
hermaphroditum, Carex bigelowii. В восточноевропейско-западносибирских
тундрах фоновыми становятся Eriophorum vaginatum, Carex arctisibirica, уси-
ливается роль Betula nana; в таймырских – Betula exilis (сменяющая березу кар-
ликовую), Salix alaxensis, S. boganidensis, Duschekia fruticosa, в лено-колым-
ских – Salix richardsonii; в чукотских тундрах распространена Salix krylovii.
Лесотундра (тундролесье) является переходной и рассматривается
разными исследователями как подзона или тундр или тайги. Она ярко выра-
жена на междуречьях, где угнетенные редколесья чередуются с кустарнико-
выми или типичными тундрами. Лес и тундра, проникая друг в друга, создают
сочетания растительных сообществ, резко отличных и от тундры и от тайги.
Взаимное проникновение существенно различных типов растительности рас-
ширяется по мере нарастания континентальности климата к востоку. В преде-
лах подзоны также распространены травяно-лишайниково-моховые бугри-
стые и травяно-сфагново-гипновые аапа болота.
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
74
Aulacomnium
turgidum
Pleurozium
Schreberi
Hylocomium
splendens
Polytrichum
strictum
Рис. 6.13. Характерные виды зеленых мхов мохово-лишайниковой тундры
(www.ecosystema.ru)
.
Глава 6. ЗОНА ТУНДР
75
Cladоnia mitis
Cladonia rangiferina
Flavocetraria cucullata
Cetraria islandica
Sphaerophorns globosus
Alectoria ochroleuca
Solornia crocea
Thamnoloa vermicularis
Рис. 6.14. Лишайники типичной тундры
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
76
Дриада точечная (Dryas punctata)
Крупка волосистая (Draba pilosa)
Кассиопея четырехгранная
(Cassiope tetragona)
Диапензия лапландская
(Diapensiu lapponica)
Ива монетолистная (Salix nummularia)
Ива полярная (Salix polaris)
Фото: Б. Большакова, Л. Онищенко, Е. Глазуновой, Д. Мирина, Д. Орешкина
Рис. 6.15. Кустарнички мохово-лишайниковых (типичных) тундр
(www.plantarium.ru)
.
Глава 6. ЗОНА ТУНДР
77
Брусника (Vaccinium vitis-idaea)
Толокнянка (Arctosiaphylos uva-ursi)
Голубика (Vaccinium uliginosum)
Морошка (Rubus chamaemorus)
Водяника (Empetrum nigrum)
Клюква (Oxycoccus palustris)
Рис. 6.16. Ягодные растения типичных тундр
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
78
Заросли березы карликовой (ерник)
(Betula nana)
Ива мохнатая (Salix lanata)
Кедровый стланик (Pinus pumila)
Фото: А. Титова, Е. Берестовского
Ольховник кустарниковый
(Duschekia fruticosa)
Рис. 6.17. Кустарники южной тундры (www.plantarium.ru)
Лесотундра протягивается от Кольского полуострова до бассейна Ин-
дигирки, а восточнее – в горах Северо-Востока Сибири (см. Ч . III, гл. 13), –
занимают нижний пояс (рис. 6.18). Южная граница редколесий, разделя-
ющая субарктический и умеренный широтные пояса, является северной
границей сомкнутых лесов на водоразделах, т. е. бореальной лесной зоны
в целом.
.
Глава 6. ЗОНА ТУНДР
79
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
80
Несмотря на малое количество атмосферных осадков, для лесотундры
характерно резкое превышение увлажнения над испарением, что обусловливает
широкое распространение озер и болот, занимающих от 10 до 60 % площади.
Средние температуры воздуха в июле 10–12 °C, а в январе, в зависимости от
нарастания континентальности климата, от –10 до –40 °C. Грунты повсемест-
но многолетнемерзлые. Почвы торфянисто-глеевые, торфяно-болотные, а под
редколесьями – глеево-подзолистые.
К редколесьям (редины) принято относить древесные насаждения с со-
мкнутостью крон не более 0,3 и бонитетом не выше Vа класса. Деревья в ле-
сотундре малорослы, стволы сильно сбежисты; при высоте 4–6, иногда 10 м
диаметр ствола составляет 9–16 см (максимальный 20–22 см); их возраст мо-
жет достигать 200 лет. Крона обычно слабо развита, узкая, часто имеет флаго-
образную форму, что связано с господствующими ветрами, сухие вершины;
стволы часто изогнутые из-за солифлюкционных процессов. Иногда отмеча-
ется двухъярусное расположение ветвей на стволе (деревья в «юбках»): вни-
зу – в приземном слое и после интервала до метра – в средней и верхней ча-
стях, что объясняется действием снежной коррозии. Корни деревьев разрас-
таются в стороны от стволов до десятков метров, образуют сомкнутую систему,
сохраняющуюся вплоть до северного предела распространения деревьев.
Уменьшение корнеобитаемого сезоннотающего слоя с одновременным умень-
шением плодородия почв заставляет корни увеличивать площадь сбора пита-
тельных веществ. В южном направлении постепенно уменьшается редкостой-
ность и увеличивается сомкнутость крон. Однако вдоль долин, по бровкам
речных террас и по хорошо дренированным крутым склонам сомкнутые дре-
востои проникают далеко на север.
Разреженный древостой оказывает незначительное влияние на форми-
рование структуры и флористического состава растений в нижних ярусах.
Вместе с деревьями распространены кустарники, кустарнички и мохово-
лишайниковый покров, практически не отличающиеся от тундровых, безлес-
ных сообществ. Особенно широко распространены лишайники. Здесь их не
менее 520 видов, что связано с малоплодородными почвами. В свою очередь
лишайники вызывают обеднение флористического состава лесотундры, так
как нередко проростки высших растений погибают, не достигая корнями до
почвы, покрытой плотным слоем лишайников.
Типичным флористическим ядром лесотундры являются субарктические
элементы флоры. Некоторые бореальные виды заходят в лесотундру в силу
широты своей экологической амплитуды. Субарктические виды в лесотундре
приобретают существенную ценотическую роль благодаря своей массовости,
хотя среди них довольно разнообразны виды бореальной флоры и устойчива
небольшая примесь арктических и арктоальпийских видов растений.
В лесотундре России распространены в основном три вида кустарнико-
вых берез: береза карликовая (Betula nana), тощая (В. exilis) и растопыренная
(В. divaricata). Карликовая береза распространяется от западных границ до
бассейна Оленька. Береза тощая распространена шире – от гор Путорана до
восточных границ, простираясь от типичной тундры до Северной Монголии,
Сахалина и Камчатки. Распространение березы растопыренной близко к ареа-
лу кедрового стланика (Pinus pumila) и не выходит за пределы ареала лист-
.
Глава 6. ЗОНА ТУНДР
81
венницы даурской. Она составляет подлесок лиственничных редколесий во-
сточнее рек Лены и Алдана, замещаясь в долинах рек березой тощей. Для ле-
сотундр России также характерны кустарниковые ивы – их насчитывается не
менее 30 видов. Особенно большие площади ивняки занимают на Европей-
ском Севере и на равнинах Западной Сибири.
Непременным флороценотическим компонентом лесотундры является
род багульник (Ledum). Вид багульника болотного (L. palustre) широко рас-
пространен по верховым болотам северной окраины лесов; багульник стелю-
щийся (L. decumbens) – типичное гольцовое субарктическое растение произ-
растает восточнее Енисея. Чрезвычайно характерен для лесотундры род водя-
ники (Empetrum). По всей лесотундре и почти по всей тайге встречается
толокнянка обыкновенная (Arctostaphylos uva-ursi). На плакорах широко рас-
пространены морошка (Rubus chamaemorus), брусника (Vaccinium vitis-idaea),
голубика (Vaccinium uliginosum), пушица (Eriophorum), дриады (Dryas octo-
petala, D. integrifolia, Dryas punctata и др.), крупка (Draba nemorosa), лапчат-
ка (Potentilla), астрагал (Astragalus), смолевка (Silene acaulis) и многие другие.
Лесотундра является весьма динамичным биомом: при похолодании
климата участие древесных пород сокращается, и общий вид ландшафта ста-
новится тундровым. При повышении летних температур лесная раститель-
ность продвигается на север.
6.2. БОТАНИКО-ГЕОГРАФИЧЕСКИЕ ОБЛАСТИ ТУНДР
При характеристике тундр следует учитывать также различия, отража-
ющие их принадлежность к разным ботанико-географическим областям: для
высокоарктических тундр – к Баренцевоморской и Сибирской областям, для
арктических, типичных, южных тундр и лесотундр – к Кольской, Восточно-
Европейской, Ямало-Гыданской, Таймырской, Лено-Колымской, Чукотской
областям.
6.2.1. ОБЛАСТИ ВЫСОКОАРКТИЧЕСКИХ ТУНДР (ПОЛЯРНЫХ ПУСТЫНЬ)
Несмотря на кажущееся однообразие природных условий, в подзоне вы-
сокоарктических тундр обособляются две области: Баренцевоморская и Си-
бирская.
Баренцевоморская область, включающая архипелаг Земли Франца-
Иосифа и Северный остров Новой Земли, характеризуется повышенной цик-
лонической деятельностью и переносом сравнительно теплых атлантических
воздушных масс. Вследствие этого температура воздуха в холодный период
здесь несколько выше, чем в области, расположенной восточнее. Тем не менее,
условия для развития растительности малоблагоприятны, что объясняется
сильными ветрами и коротким вегетационным периодом.
На архипелаге Земли Франца-Иосифа насчитывается 57 видов цветко-
вых растений, 102 – мхов, 55 – печеночников, около 120 – лишайников. Рас-
тительность фрагментарно представлена травяно-лишайниково-моховыми
пятнистыми, моховыми, лишайниковыми тундрами. Характерным для данной
области является присутствие в сообществах Deschampsia alpina и Cerastium
arcticum, C. regelii ssp. caespitosa, не встречающихся восточнее, а также Papaver
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
82
polare, Saxifraga nivalis, S. cespitosa, S. cernua, S. oppositifolia, Alopecurus alpinus,
Phippsia algida.
Сибирская область, захватывающая архипелаг Северная Земля и
острова Де-Лонга, отличается устойчивой антициклональной погодой в зим-
нее время года. Летом, напротив, циклоническая деятельность несколько
усиливается. Флора Северной Земли насчитывает 61 вид цветковых растений
и около 200 видов мхов и лишайников. Из семейств наиболее широко пред-
ставлены капустные (Brassicaceae), мятликовые (Poaceae) и камнеломковые
(Saxifragaceae). Характерными видами, не встречающимися на западе, яв -
ляются: Ranunculus sabinii, Cerastium bialynickii, C. regelii, Deschampsia
brevifolia.
6.2.2. ОБЛАСТИ АРКТИЧЕСКИХ, СУБАРКТИЧЕСКИХ ТУНДР И ЛЕСОТУНДРЫ
6.2.2.1. Кольская область тундр
Кольский полуостров расположен на северо-западе России. Почти вся его
территория лежит за Северным полярным кругом. Полуостров омывается Ба-
ренцевым и Белым морями. Кольский полуостров занимает восточную часть
Балтийского кристаллического щита. Его рельеф представляет собой горы,
плато, впадины и террасы. Горные массивы полуострова возвышаются над
уровнем моря, несколько превышая 800 метров. Равнины занимают болота и
многочисленные озера. Климатические особенности Кольского п-ова опреде-
ляются его принадлежностью к Приатлантическому сектору. Климат постоян-
но влажный, океанического типа. Средние температуры января–февраля
от –8 °C на северо-западе полуострова до –14 °C в центре; июля, соответствен-
но,от8до14°C.
Ботанико-географические характеристики области представлены на
рис. 6.19. На Кольском полуострове подзоны арктических и типичных тундр
не выражены. Отмечаются южные кустарниковые тундры, занимающие рав-
нины на севере и северо-востоке, и горные тундры в центре полуострова. Они
слагаются из многих общих видов и характеризуются близкими экологиче-
скими чертами.
Кустарниковые тундры с Betula nana и Betula czerepanovii быть пред-
ставлены как зарослями из прямостоящих кустов березы до 1 м со значи-
тельной сомкнутостью, так и стелющимися формами высотой 10–15 см,
занимающими лишь депрессии микрорельефа (рис. 6.20). Ерники рас-
пространены на глинистых, песчаных и щебнистых грунтах, на глеевых,
глеево-оподзоленных и слабо подзолистых почвах, в условиях хорошего
стока. Высоту и степень развития ерника в основном определяет снеговой
режим.
При незначительной мощности снежного покрова ерники исчезают, а
при большой его мощности наблюдается значительное участие в кустарнико-
вом ярусе ив (особенно Salix glauca, S. lapponum, S. lanata, S. phylicifolia), ко-
торые, вытесняя ерник, образуют заросли тундровых ивняков. Обычное ме-
стонахождение ерниковых тундр – пологие склоны, причем высота кустов
ерника возрастает вниз по склону. Для ерниковых тундр, особенно на почвах
.
Глава 6. ЗОНА ТУНДР
83
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
84
Рис. 6.19. (продолжение) Легенда к карте растительности Кольского п-ова
.
Глава 6. ЗОНА ТУНДР
85
Рис. 6.19. (продолжение) Легенда
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
86
Рис. 6.20. Кустарниковая тундра
легкого гранулометрического состава, характерен мощный лишайниковый
покров с преобладанием Cladonia stellaris, Stereocaulon paschale, покрываю-
щий 60–80 % площади.
Кустарничковые тундры значительными массивами распространены
только в области приморского климата – на севере Кольского п-ова. Они объ-
единяют разнообразные по составу растительные группировки с господством
водяники Empetrum hermaphroditum, вереска Calluna vulgaris, дёрена Cham-
aepericlymenum suecicum, арктоуса Arctous alpina или карликовых ив, пре-
имущественно Salix herbacea. Высока степень насыщенности флоры тундр бо-
реальными видами и понижено участие арктических.
В тундровой зоне Кольского п-ова к плоходренированным понижениям
приурочены болота. Как правило, это крупнобургистые ерниковые травяно-кус-
тарничково-мохово-лишайниковые (Flavocetraria nivalis, Dicranum elongatum,
D. flexcaule, Polytrichum strictum, Ledum palustre, Vaccinium uliginosum, Vac-
cinium vitis-idaea, Rubus chamaemorus) на буграх и пушицево-осоково-гипно-
во-сфагновые (Sphagnum lindbergii, S. majus, S. riparium, Warnstorfia fluitans,
Carex rariflora, C. rotundata, C. chordorrhiza, Eriophorum russeolum) болота в
понижениях между буграми.
В приатлантической лесотундре Кольского полуострова северный предел
распространения деревьев образует береза Черепанова (Betula czerepanovii)
(рис. 6.21). Южнее, на западе полуострова доминирует сосна (Pinus sylvestris),
а восточнее нарастают площади ельников, которые состоят в основном из си-
бирской ели (Picea obovata). В изобилии встречаются различные виды кустар-
ников и мхов. В сравнительно сухих местах почву и камни толстым слоем по-
крывают лишайники рода Cladonia. Обширны ягодники (брусника, морошка,
голубика, водяника), много грибов.
.
Глава 6. ЗОНА ТУНДР
87
Рис. 6.21. Лесотундра, березовое криволесье
6.2.2.2. Восточно-Европейская область тундр
Географическое положение области показано на карте-схеме зон и под-
зон севера Восточно-Европейской равнины (рис. 6.22). Геоморфологическое
строение области определяется четвертичными ледниковыми аккумулятив-
ными формами рельефа.
Для области характерна интенсивная атмосферная циркуляция, обуслов-
ленная взаимодействием полярных атлантических воздушных масс и аркти-
ческого воздуха; с этим связана очень большая изменчивость погоды. Суще-
ственное климатообразующее значение имеет влияние относительно теплого
Баренцевого моря. В восточном и южном направлениях континентальность
климата увеличивается. Зимы здесь холодные и продолжительные, они длят-
ся с конца октября до конца апреля. Наиболее холодный месяц обычно фев-
раль – средние температуры воздуха около –20 °С. К летним месяцам относят-
ся июль и август; лето короткое, прохладное, средняя температура июля от 8
до 15 °С, в конце августа уже замечается довольно быстрый переход к осени.
Годовые осадки от 450 мм на юге до 250 мм на севере. Осадки выпадают в ос-
новном в виде снега.
В подзоне арктических тундр, которая охватывает о-в Вайгач, рас-
тительность не образует сплошного покрова и представлена простратноку-
старничково-травяно-лишайниково-моховыми, травяно-лишайниково-мохо-
выми полигональными, пятнистыми тундрами. Поверхность тундры состоит
из пятен, практически лишенных растительного покрова, и окаймляющих их
стелющихся кустарников, мхов, травянистых растений, растущих в ложбин-
ках и трещинах. Из травянистых растений характерны: смолевка бесстебельная
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
88
Рис. 6.22. Карта-схема зон и подзон севера Восточно-Европейской равнины
(http://kraeved-nao.narod.ru)
(Silene acaulis), песчанка ложнохолодная (Arenaria pseudofrigida), проломник
арктосибирский (Androsace arctisibirica), змеевик живородящий (Bistorta
vivipara), минуарция красноватая (Minuartia rubella), мак югорский (Papaver
lapponicum ssp. jugoricum), родиола розовая или золотой корень (Rhodiola
rosea).
Типичные тундры расположены на северо-востоке Малоземельской
тундры, на возвышенности Канин камень, в Большеземельской тундре при-
урочены к склонам возвышенностей Еней и Вангурей, южным склонам хребта
Пай-Хой, имеют распространение на о. Колгуев. Преобладают кустарничково-
лишайниково-моховые тундры, зачастую с болотами. В растительном покрове
доминируют зеленые мхи: Aulacomnium turgidum, A. palustre, Hylocomium
splendens, Polytrichum strictum, Rhytidium rugosum, Ptilidium ciliare,
Tomentypnum nitens, Pleurozium schreberi, Dicranum elongatum, D. flexcaule,
D. fuscescens, Warnstorfia exannulatus и др. В большей или меньшей степени
в составе напочвенного покрова присутствуют лишайники (Cladonia mitis,
С. rangiferina, Flavocetraria cucullata, Sphaerophorus globosus и др.). Травяно-
кустарничковый ярус образован кустарничками: Loiseleuria procumbens,
Phyllodoce caerulea, Arctous alpina, дриадой – куропаточьей травой Dryas
octopetala, Empetrum hermaphroditum, Vaccinium uliginosum, V. vitis-idaea,
мелкими ивами (Salix glauca, S. lanata); травами: мятликом арктическим (Poa
arctica), змеевиком живородящим (Bistorta vivpara), овсяницей приземистой
(Festuca airoides), осоки Бигелоу (Carex bigelowii), ситником трехраздельным
(Juncus trifidus). Довольно обильно представлено аркто-альпийское разно-
.
Глава 6. ЗОНА ТУНДР
89
травье: змеевик лекарственный (Bistorta officinalis), пепельник цельнолист-
ный (Tephroseris integrifolia), остролодочник грязноватый (Oxytropis sordida),
соссюрея альпийская (Saussurea alpina), копеечник альпийский (Hedysarum
alpinum), василистник альпийский (Thalictrum alpinum) и др. Во влажных ме-
стообитаниях в травяном покрове преобладают осоки (Carex bigelowii, С. con-
color). В южных районах широко развиты лишайниковые тундры с багуль-
ником (Ledum palustre). Значительные площади занимают полигональные и
бугристые травяно-кустарничково-лишайниково-моховые болота.
Южные тундры занимают бóльшую часть Восточно-Европейской об-
ласти. Полоса ерниковых тундр (Betula nana) очень широкая – она простира-
ется с севера на юг на 200–250 км. Массивы ерниковых тундр чередуются со
сфагновыми болотами и тундровыми ивняками (Salix lapponum, S. phylicifolia,
S. gmelinii, S. glauca, S. lanata). Наиболее распространены зеленомошные ер-
никовые тундры на торфяно-глеевых и торфяно-глеево-подзолистых суглини-
стых почвах. На повышениях микрорельефа встречаются некоторые лишай-
ники (Nephroma arcticum, Peltigera aphthosa, Cladonia gracilis ssp. elongata,
C. mitis и др.). В травяно-кустарничковом ярусе обычно преобладают кустар-
нички (брусника, голубика, водяника) и менее обильно представлены травы
(Carex bigelowii, С. globularis, Festuca airiodes, Petasites frigidus и др.). Зелено-
мошные ерниковые тундры встречаются обычно на склонах в условиях доста-
точного дренажа. При большей заболоченности моховой покров в ерниковых
тундрах состоит из кукушкина льна (Polytrichum commune), а при еще боль-
шем избытке застойной влаги – из сфагнов (Sphagnum russowii, S. girgen-
sohnii, S. lindbergii). При крайней степени заболачивания кустарниковый ярус
разреживается, и формируются верховые сфагновые плоские болота со сфаг-
нумом бурым (Sphagnum fuscum). В восточной части Большеземельской тунд-
ры и в Приуралье ерниковые тундры преимущественно со сплошным покро-
вом из зеленых мхов и лишайников. Лишайники покрывают до 40–50 %
площади. Широкое участие принимают Betula nаnа, Vaccinium uliginosum и
Ledum palustre. Также встречаются травяно-кустарничковые тундры; травя-
но-кустарничковый ярус образован Empetrum hermaphroditum, Vaccinium
myrtillus, V. uliginosum, V. vitis-idae, включает флористические элементы
западносибирских тундр – Avenella flexuosa, Festuca ovina, Aconitum
septentrionale, Cirsium helenioides, Trollius europaeus, T. asiaticus.
Широко распространены бугристые кустарничково-мохово-лишайнико-
вые болота. Бугры состоят преимущественно из сфагнового мха и покрыты
лишайниками, багульником, голубикой, морошкой (Rubus chamaemorus),
водяникой (Empetrum hermaphroditum), хамаедафне прицветничковой
(Chamaedaphne calyculata), березой карликовой и ивой ползучей (Salix rep-
tans). В понижениях между буграми – заросли осок (Саrех) и пушиц (Eriopho-
rum); последние в период своего плодоношения создают издали впечатление
снежного покрова.
Подзона лесотундры протягивается от горла Белого моря до подгор-
ной равнины Приполярного Урала. Ее северные пределы, представленные
изолированными островками редколесий из ели сибирской (Picea obovata)
среди описанных выше кустарниковых тундр, находятся около 67°20'–67°30'
с. ш., южные пределы расположены примерно вдоль 66 параллели с неболь-
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
90
шими отклонениями преимущественно к северу. Дальше всего на север ред-
колесья продвигаются по хорошо дренированным, всхолмленным водоразде-
лам с песчаными почвами, и особенно по склонам к долинам и террасам рек.
По заболоченным местообитаниям они проникают и в более южную подзо-
нальную полосу – в северную тайгу. Редколесья связаны с районами островно-
го распространения многолетнемерзлых пород. При этом они тяготеют к
участкам, где многолетняя мерзлота обычно отсутствует, а глубина оттаива-
ния сезонномерзлого слоя достигает нескольких метров. В лесотундре на су-
глинках формируются поверхностно-элювиально-глееватые оподзоленные
почвы, на супесчаных и песчаных субстратах – оподзоленные подбуры.
Флористическое своеобразие лесотундры заключается в согосподстве бо-
реальных и гипоарктических видов, причем это свойственно всем ярусам. Из
бореальных видов, кроме самой ели, наибольшую константность и обилие
имеет целый ряд кустарничков (Vaccinium myrtillus, V. uliginosum, V. vitis-idaea,
Ledum palustre), трав (Carex globularis, Avenella flexuosa, Equisetum sylvaticum),
мхов (Pleurozium schreberi, Polytrichum commune, Dicranum majus, D. scopari-
um). Спорадически встречаются также Rosa acicularis, Linnaea borealis, An-
dromeda polifolia, Melampyrum pratense, Solidago virgaurea, Bistorta officinalis,
Carex cinerea и др., а из мхов – Dicranum polysetum. Из гипоарктических ви-
дов постоянно, нередко в роли доминантов присутствуют Betula czerepanovii,
В. nana, Empetrum hermaphroditum, Rubus chamaemorus, менее регулярно –
Junlperus sibirica, Arctous alpina, Salix glauca и бореально-гипоарктические
Salix phylicifolia, S. lapponum.
Ели почти повсюду сопутствует береза: на севере подзоны (преимуще-
ственно к западу от Печоры) – Betula czerepanovii, на юге – В. pubescens. Бере-
за обычно сильно угнетена; ее высота 3–5 (до 7) м, диаметр ствола 4–6 (до 10)
см. Часто она представлена своеобразной формой роста, для которой харак-
терно наличие нескольких изогнутых стволиков и густой кроны в виде куста
на высоте 1,5–2 м. Кроме древесного развиты еще 3 яруса: кустарниковый
(иногда он не выражен), кустарничковый (или травяно-кустарничковый) и
напочвенный, состоящий из мхов с разной долей участия лишайников. Всем
ярусам свойственно мозаичное сложение, реже они однородны по структуре.
Еловые редколесья представлены в основном зеленомошными, лишайниково-
зеленомошными и долгомошно-сфагновыми типами. Зеленомошные редко-
лесья приурочены к средним частям пологих склонов водораздельных равнин
и речных долин, преимущественно южной экспозиции, с суглинистыми поч-
вами, местами несколько переувлажненными. По составу нижних ярусов,
структуре подлеска и древостоя хорошо различаются 2 географически заме-
щающих типа: северный (голубично-багульниковый), и южный (чернично-
вороничный). На почвах более легкого механического состава (легкосуглини-
стых и супесчаных) распространены лишайниково-зеленомошные редколесья.
На севере лесотундры встречаются также лиственнично-вороничные редколе-
сья с Larix sibirica и Empetrum hermaphroditum. В условиях избыточного за-
стойного увлажнения на болотно-подзолистых почвах распространены долго-
мошно-сфагновые еловые редколесья. Они приурочены к нижним частям по-
логих склонов, часто располагаясь по окраине депрессий, занятых болотами
или заболоченными тундрами, с которыми нередко образуют сочетания. Для
.
Глава 6. ЗОНА ТУНДР
91
травяно-кустарничкового яруса характерно повсеместное преобладание тра-
вянистых видов (Carex globularis, Rubus chamaemorus, Equisetum sylvaticum,
примесь Eriophorum vaginatum), а из кустарничков – Vaccinium myrtillus,
V. uliginosum, местами Empetrum hermaphroditum, довольно часто присут-
ствуют Oxycoccus microcarpus, Andromeda polifolia. В моховом покрове доми-
нируют сфагны (Sphagnum girgensohnii, S. angustijolium и др.) и Polytrichum
commune.
6.2.2.3. Ямало-Гыданская область тундр
Ямало-Гыданская область расположена на севере Западно-Сибирской
равнины и омывается водами Карского моря. Характер береговой линии
определяется крупными полуостровами: Ямал, Тазовский и Гыданский. С за-
пада область ограничена Уралом, с востока – р. Енисей. Суровый климат обу-
словлен неравномерным поступлением солнечной радиации в течение года,
вторжением арктических воздушных масс, близостью холодного моря. Зима
продолжительная, с длительным залеганием снежного покрова, лето холод-
ное и короткое. Средняя температура января от –24 °С на западе до –28 °С на
востоке, изотермы января вытянуты почти в меридиональном направлении.
Средняя температура июля 4 °С на севере и 8 °С на юге. Климат области уме-
ренно континентальный. Наименьшая годовая сумма атмосферных осадков
наблюдается на севере – около 300 мм, что связано с малым влагосодержани-
ем арктических воздушных масс. К югу количество осадков возрастает и со-
ставляет 350–400 мм у полярного круга. Холодный климат определяет низ-
кую испаряемость (ГТК вегетационного периода 2), что благоприятствует ин-
тенсивному заболачиванию плохо дренируемых равнин.
Почвенный покров характеризуется чрезвычайной пестротой и сложно-
стью, обусловленных особенностями биоклиматических, литолого-геоморфо-
логических и гидрологических условий. Повсеместно неглубоко залегает мно-
голетняя мерзлота, выступающая водоупором, что определяет широкое разви-
тие озер, болот, бугристого и полигонального микрорельефа. Большая часть
почвенного покрова представлена почвенными комплексами и сочетаниями
различных типов и подтипов почв, дифференцированных по микрорельефу.
Характерная черта структуры почвенного покрова – высокая заболоченность
территории (до 80 % и более). Для болотных массивов типичны полигональ-
но-валиковые комплексы болотных мерзлотных и тундрово-болотных почв.
Среди зональных почв господствуют тундровые иллювиально-гумусовые на
легких породах и тундровые глеевые – на тяжелых грунтах.
Растительность области подразделяется на подзоны арктических, типич-
ных и южных тундр, а также лесотундры (Атлас Ямало-Ненецкого..., 2004) и
представлена на фрагменте карты (рис. 6.23).
Подзона арктических тундр представлена кустарничково-моховыми
кочковатыми, осоково-пушицево-моховыми заболоченными, мохово-лишайни-
ковыми полигональными и пятнисто-полигональными тундрами. В сообще-
ствах кустарничково-моховых кочковатых тундр доминируют зеленые мхи
(Racomitrium lanuginosum, Dicranum elongatum, Aulacomnium turgidum). Ку-
старнички представлены арктоусом (Arctous alpinа), дриадой (Dryas punctata),
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
92
Рис. 6.23. Карта растительности Ямало-Ненецкого автономного округа
(Атлас Ямало-Ненецкого..., 2004)
.
Глава 6. ЗОНА ТУНДР
93
Рис. 6.23. (продолжение). Легенда к карте растительности Ямало-Ненецкого
автономного округа (Атлас Ямало-Ненецкого..., 2004)
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
94
Рис. 6.23 (продолжение). Легенда к карте
Рис. 6.23 (продолжение). Легенда к карте
.
Глава 6. ЗОНА ТУНДР
95
ивами (Salix polaris, S. nummularia), брусникой (Vaccinium vitis-idaea var. minus).
На плоских участках с полигональным микрорельефом формируются заболо-
ченные осоково-пушицево-моховые тундры. На полигонах развиты осока
(Carex arctisibirica), пушица (Eriophorum angustifolium), кустарнички (Salix
polaris, S. nummularia, Dryas punctata), мхи (Aulacomnium turgidum, Dicranum
angustum, Sphagnum fimbriatum). В трещинах встречаются осока (Carex
concolor), мхи (Scorpidium revolvens, Warnstorfia sarmentosa). На вершинах
увалов небольшими участками развиваются мохово-лишайниковые полиго-
нальные и пятнисто-полигональные тундровые сообщества, составленные ку-
старничками: арктоусом, дриадой, ивами, а также травами (Carex arctisibirica),
лишайниками (Cladoina rangiferina, C. mitis, C. macroceras, Flavocetraria
cucullata) и мхами (Racomitrium lanuginosum).
Болота в подзоне арктических тундр представлены осоково-гипновыми и
осоково-пушицевыми, которые часто сочетаются со злаково-осоково-моховыми
валиково-полигональными болотными комплексами.
Пойменная растительность образована сериями осоковых (Carex concolor)
лугов на начальной стадии, которые затем замещаются разнотравно-злаковыми
лугами, а после этого – мелкоивняковыми (Salix lanata) зарослями, сочетаю-
щимися с осоково-гипновыми болотами и травяно-моховыми тундрами.
Подзона типичных кустарничковых тундр на территории Ямало-
Гыданской области представлена на плакорах сообществами кустарничково-
травяно-моховых бугорковатых и пятнисто-бугорковатых тундр (рис. 6.24),
составленных осокой (Carex arctisibirica), разнотравьем (Luzula nivalis,
Ranunculus propinquus), кустарничками (Arctous alpina, Dryas punctata,
Vaccinium vitis-idaea var. minus), зелеными мхами (Racomitrium lanuginosum,
Aulacomnium turgidum, Dicranum angustum).
Рис. 6.24. Тундра Ямала (фото С. Тараненко)
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
96
В низинах формируются кустарничково-травяно-моховые заболоченные
тундры. В таких сообществах хорошо развиты сфагновые мхи (Sphagnum
lenense, S. lindbergii), зеленые мхи (Tomentypnum nitens, Hylocomium splendens).
Обилие трав (Luzula nivalis, Eriophorum angustifolium) и кустарничков
(Vaccinium vitis-idaea var. minus) невелико.
В южной части подзоны типичных тундр развиты травяно-кустарнич-
ково-сфагновые бугристые тундры, в растительном покрове которых обыч-
ны сфагновые мхи (Sphagnum lenense, S. lindbergii), травы (Eriophorum
angustifolium, E. vaginatum, Саrех concolor), кустарнички (Vaccinium uliginosum
ssp. microphyllum, Ledum decumbens, L. palustre, Rubus chamaemorus), низко-
рослые ива сизая (Salix glauca) и ерник (Betula nana).
Большие площади по повышенным участкам с песчаными почвами за-
няты кустарничково-зеленомошно-лишайниковыми полигональными тунд-
рами с плотным мохово-лишайниковым покровом (Cladonia rangifera, C. mitis,
C. macroceras, Flavocetraria cucullata, Polytrichum strictum).
Подзона южных кустарниковых тундр характеризуется развитием
кустарникового яруса из ерника (Betula nana), ивы мохнатой (Salix lanata),
ивы сизой (Salix glauca), ивы филиколистной (Salix phylicifolia), ольховника
(Duschekia fruticosa). Зональными типами растительных сообществ являются
ерниковые и ивняковые кустарничково-зеленомошные бугорковатые и пят-
нисто-бугорковатые тундры. На плоских слабодренируемых участках развиты
ерниковые и ивняковые, местами с ольховником травяно-кустарничково-зеле-
номошные (Sphagnum lenense, S. balticum, Dicranum angustum, Polytrichum
strictum, Vaccinium uliginosum ssp. microphyllum, Ledum palustre) заболочен-
ные тундры. На склонах увалов в южной части встречаются ивняково-ерни-
ковые с ольховником кустарничково-мохово-лишайниковые тундры, которые
часто сочетаются с лиственничными редколесьями, расположенными на скло-
нах или вершинах песчаных холмов (рис. 6.25).
Подзона лесотундры в Западной Сибири протягивается к востоку от
низовий р. Оби до Енисея. Она образована в основном редколесьем из лист-
венницы сибирской (Larix sibirica), иногда с незначительной примесью ели
сибирской (Picea obovata) и приурочена в основном к высоким террасам рек и
водоразделам, к хорошо дренируемых местам. На местности редколесья по-
стоянно чередуются с сообществами ерниковых, ивняковых и ольховниковых
тундр, а в заболоченных частях междуречий – с участками плоскобугристых и
полигональных болот.
Важнейшей особенностью горизонтальной структуры нижних ярусов
растительности редколесий является их необычайная пестрота и неоднород-
ность, выражающаяся в образовании мозаичности и комплексности. В ле-
сотундре Западной Сибири эта неоднородность проявляется повсеместно. На
дренированных участках в составе фитоценозов наблюдается постоянное чере-
дование синузий мхов, лишайников и кустарничков в зависимости от измене-
ний форм нанорельефа, режима тепла и влажности почв. В районах заболачи-
вания неоднородность структуры растительного покрова еще более выражена.
В видовом составе травяно-кустарничкового и мохово-лишайникового
ярусов преобладают бореальные и бореально-гипоарктические виды; в качестве
.
Глава 6. ЗОНА ТУНДР
97
Рис. 6.25. Лишайниковый покров редколесья
постоянной примеси присутствуют собственно гипоарктические элементы:
Arctous alpina, Ledum decumbens, Dryas punctata, Aulacomnium turgidum, Alec-
toria nigricans.
Лесотундровые лиственничные и елово-лиственничные редколесья с ер-
никово-лишайниково-зеленомошным покровом встречаются среди массивов
ерниковых тундр и приурочены к разной степени дренированным почвам
преимущественно приречных частей водоразделов. В древесном ярусе редко-
лесий этого типа абсолютно преобладает лиственница. Небольшое участие ели
обычно связано с несколько повышенной влажностью и глееватостью почв.
Примесь березы (Betula pubescens, В. tortuosa) значительна (до 3–4 единиц в
составе) и характерна как для коренных типов, так и для антропогенно-
производных редколесных березово-лиственничных древостоев.
Наиболее дренированным местообитаниям соответствует ассоциация
лиственничного лишайниково-зеленомошного редколесья, развитого на су-
глинистых среднеподзолистых глееватых почвах. Древесный ярус сомкнуто-
стью 0,1–0,2 состоит из лиственницы, имеющей высоту 6–8 м и диаметр ство-
ла 10–15 см, с единичной примесью березы (Betula tortuosa) и в некоторых
местах ели высотой до 7 м, диаметром 10–12 см. Расстояние между деревьями
10–13 м. В подлеске группы кустарников Betula nana и единичные экземпля-
ры Juniperus communis. Травяно-кустарничковый ярус представлен отдель-
ными синузиями низкорослых кустарничков, в основном: багульника, го-
лубики, водяники, среди которых часто присутствует гипоарктический вид
Ledum decumbens. Из трав обычны Carex globularis, С. bigelowii, Eriophorum
vaginatum. Хорошо развиваются группировки зеленых мхов, занимающие 50–
60 % площади фитоценоза, среди которых преобладают Pleurozium schreberi,
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
98
Polytrichum strictum, Ptilidium ciliare, и в небольшом количестве встречается
субарктический мох Aulacomnium turgidum. Синузии лишайников развиты в
более осветленных местах. Среди них обычны Cladonia stellaris, C. rangiferina,
C. uncialis, C. cornuta, а также трубчатые (Stereocaulon paschale) и листоватые
(Peltigera aphthosa, Nephroma arcticum) лишайники.
В целом четко выражена мозаичность напочвенного покрова: на повы-
шениях обычны лишайники и кусты багульника и водяники, в микропониже-
ниях – зеленые мхи и карликовая береза. Моховой покров больше развит под
низкоопущенными кронами берез и на приствольных повышениях листвен-
ниц и елей.
По окраинам болот, в местах менее дренированных, развиваются лист-
венничные зеленомошные редколесья с синузиями долгомошных и сфагно-
вых мхов и с большим участием ерника в подлеске, местами образующего от-
крытые заросли. Мозаичность и комплексность покрова в таких редколесьях
еще более характерны. Среди преобладающих по площади группировок зеле-
ных мхов (Pleurozium schreberi, Ptilidium ciliare) и кустарничков (Vaccinium
uliginosum, Empetrum nigrum, Ledum decumbens) в небольших депрессиях
концентрируются Polytrichum commune и Sphagnum girgensohnii. Древостой
становится еще более разреженным и угнетенным.
В долинах рек, где ближе уровень грунтовых вод и глубже или совсем от-
сутствует мерзлота, в составе лиственничных зеленомошных редколесий усили-
вается роль ели и березы (Betula pubescens), разнообразнее становятся назем-
ный покров и подлесок, в которых хорошо представлены бореальные виды.
6.2.2.4. Таймырская область тундр
Характеристика растительности области описывается на примере «Тай-
мырского» заповедника, в котором представлены сообщества от арктической
тундры до лесотундры (по: Матвеева, 1985; Исследование природы..., 2001).
Заповедник был создан 23 февраля 1979 года, а в 1995 г. ему придан статус
биосферного.
Рельеф территории заповедника – горно-равнинный. Формирование
равнин происходило под влиянием четвертичных оледенений, морских транс-
грессий и аллювиальной деятельности многочисленных рек. Вся территория
лежит в зоне сплошного распространения многолетней мерзлоты, мощность
которой достигает 500 м. Глубина сезонно-талого слоя на водоразделах ко-
леблется в зависимости от состава грунта от 15 до 100 см. Криогенные процес-
сы создают своеобразный микро- и нанорельеф и способствуют образованию
весьма неоднородного почвенного и растительного покрова. Значительная
часть территории заповедника лежит в горах Бырранга, которые представля-
ют собой самую северную в мире континентальную горную систему. Пейзаж
гор Бырранга с первого взгляда может показаться мрачным: черные однотон-
ные будто спиленные вершины, склоны ощетинились выветрелыми скальны-
ми останцами и пилообразными гребнями; до самого конца лета в распадках
белеют снежники, и лишь в долинах проглядывает зелень.
Климат резко континентальный, суровый, с долгими холодными зимами
и коротким летом. Средняя температура июля в южной части заповедника
.
Глава 6. ЗОНА ТУНДР
99
12,3 °С, в центральной части 6,5 °С, в северной, приморской 4,0 °С. Годовое
количество осадков 200–300 мм.
Основные почвообразовательные процессы – глеевый, дерновый, торфо-
образовательный. На равнинах преобладают первый и третий процессы, а в
нижнем поясе гор – второй. Зональный тип почв на Таймыре – тундровые
глеевые почвы.
На территории заповедника произрастает 429 видов сосудистых рас-
тений, 212 видов листостебельных мхов, 263 вида лишайников. Структура
растительного покрова Таймырского п-ова отображена на фрагменте карты
(рис. 6.26). Приведем краткое описание растительности.
Подзона арктических тундр. Флора цветковых насчитывает 159 ви-
дов; число видов резко возрастает по мере удаления от моря к юго-востоку.
Характер флоры – арктический, на побережье – высокоарктический; циркум-
полярных видов почти в 2 раза больше, чем азиатских, т. е. сибирская специ-
фика флоры на этих широтах теряется. Доля криофитов увеличивается до
83 %. Растительность на плакорах – ивково-моховые и лишайниково-ивково-
моховые полигонально-пятнистые тундры с ожиками, лисохвостом, мелкими
камнеломками, минуарцией крупноплодной (Minuartia macrocarpa); расти-
тельность покрывает не больше 20 % поверхности. На редких щебнистых пят-
нах распространены лишайниково-дриадовые куртинные сообщества. Со-
мкнутая растительность, разнотравно-злаковые луговины с преобладанием
мятлика альпигенного (Poa alpigena) и лисохвоста альпийского (Alopecurus
alpinus) развиваются лишь на участках южных склонов. Низкие ровные про-
странства плоских водоразделов заняты сплошными полями травяных болот.
Эти поля представляют собой заросли осоки одноцветковой (Carex concolor),
пушиц узколистной и Шейхцера (Eriophorum angustifolium, E. scheuchzeri),
дюпонции Фишера (Dupontia fisheri). Горы выше 250 м практически безжиз-
ненны, растительность представлена разреженными лишайниковыми группи-
ровками, в которых изредка попадаются некоторые крупки (Draba macrocarpa,
D. pohlei, D. pauciflora).
Подзоны типичных и южные тундр. Флора равнинных тундр
преимущественно арктическая, сибирская, в депрессиях со значительным уча-
стием субарктических видов. Всего отмечен 331 вид сосудистых растений, но
только один эндемик – тризетокелерия таймырская (Trisetokoeleria taimyrica).
Растительность плакоров представлена кустарниково-осоково-моховыми и
осоково-моховыми полидоминантными тундрами с дриадой, хамаедафне,
ивами ползучей и красивой (Salix reptans, S. pulchra), осокой (Carex arctisibiri-
ca), пушицами, брусникой, грушанкой. По долинам развиты фрагментарные
заросли кустарников, луга, разнотравно-дриадовые тундры. Территория силь-
но заболочена, поэтому обильны гигрофильно-травяные сообщества; в доли-
нах рек и спущенных озерных котловинах обширны массивы полигональных
болот (рис. 6.27).
На обнаженных щебнистых вершинах гряд, крутых склонах, озерных
и речных террасах развиты дриадово-травяные и разнотравные лугоподоб-
ные сообщества с преобладанием мятликов (Poa glauca, P. alpigena) и ов-
сяниц (Festuca brachyphylla, F. richardsonii), мытников (Pedicularis amoena,
P. verticillata) , ясколки (Cerastium maximum) , ллойдии (Lloydia serotina).
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
100
Рис. 6 .26. Карта растительности п-ова Таймыр (фрагмент карты из Атласа
Арктики, 1985)
.
Глава 6. ЗОНА ТУНДР
101
Рис. 6.27. Полигонально-валиковое болото (фото И. Поспелова)
Весьма разнообразна растительность низменных плоских озерно-аллювиаль-
ных равнин, которые сложены песками и на значительных площадях затор-
фованы. Большую часть этих равнин занимают болотные комплексы всех ти-
пов – ерниково-моховые плоскобугристые, кустарниково-осоково-моховые
плоскополигональные; полигонально-валиковые с мохово-осоковыми поли-
гонами и кустарниково-осоково-моховыми валиками, трещинно-полигональ-
ные, осоковые и злаково-осоковые.
Подзона лесотундры. На юге заповедника представлены все основ-
ные типы растительного покрова, характерные для лесотундры Таймыра –
лиственничные редколесья и редины из лиственницы даурской (Larix gmelinii),
пятнистые кустарниково-осоковые тундры, кустарниковые тундры с преобла-
данием ерника, ив красивой, сизой и ползучей (Salix pulchra, S. glauca, S. rep-
tans), в долинах – полигональные осоково-моховые болота.
В подлеске обычны ерник, багульник, голубика; в поймах – ивы, ольхов-
ник, иногда шиповник иглистый и красная смородина (Ribes rubrum); на сухих
приподнятых гривах в напочвенном покрове много кустарничков – кассиопея,
брусника, толокнянка, водяника (шикша). В напочвенном покрове более
влажных участков преобладают обычные тундровые бриевые мхи (Hylocomium
splendens var. obtusifolium, Tomentypnum nitens, Aulacomnium turgidum), пе-
ченочник Ptilidium ciliare; на сухих высоких террасах обилен ягель
(Cladonia stellaris, C . rangiferina, C. amaurocraea и др.). В пятнистых и бу-
горковых тундрах, чередующихся с редколесьями, преобладающую роль иг-
рают ивы красивая и сизая, дриада точечная, кассиопея четырехгранная (Cas-
siope tetragona), осока арктосибирская и те же виды мхов.
На речных террасах редколесья сочетаются с болотной растительно-
стью, для которой можно отметить все стадии развития полигональных болот:
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
102
Рис. 6.28. Разнотравная луговина на береговом склоне
от осоково-пушицевых и мохово-осоковых с неясно выраженной сетью рас-
трескивания до плоскобугристых. В переувлажненных понижениях моховой
покров развит слабо, здесь обильны осоки одноцветная и редкоцветковая
(Carex concolor, C. rariflora), пушицы узколистная и средняя (Eriophorum an-
gustifolium, E. medium). Разнотравные луговины речных террас и береговых
склонов в период массового цветения представляют собой сплошной цвету-
щий ковер (рис. 6.28).
Горные тундры хребта Бырранга. В состав флоры входит 340 видов
споровых и цветковых растений, из которых 38 видов произрастает только
здесь; для гор эндемичны 3 семейства и 4 рода. Эта флора – типично арктиче-
ская, континентальная, сибирская. Распределение растительности подчиняет-
ся высотной поясности, границы поясов зависят от экспозиции и крутизны
склонов. На пологих участках южного макросклона нижний пояс представлен,
в основном, ерниково-травяно-моховыми тундрами. С высоты 50–100 м они
сменяются разнотравно-мохово-дриадовыми и дриадово-моховыми тундрами.
Чем выше в горы, тем меньшие площади покрыты растениями, беднее видо-
вой состав (рис. 6.29). На менее дренированных местах дриада частично или
полностью замещается ивой полярной (Salix polaris), возрастает роль щучки
коротколистной (Deschampsia brevifolia). Верхняя граница этого пояса повы-
шается с востока на запад от 250–300 м до 400–450 м над ур.м.
Выше пояса дриадовых тундр на щебнистых грунтах обычны ивково-
мохово-сиверсиевые куртинные тундры (сиверсия ледяная или акомастилис
ледяной (Acomastylis glacialis) – таймырская роза) или эпилитно-лишайни-
ковые горные пустыни, на мелкоземистых – щучково-ивково-моховые кур-
тинные тундры. Самый верхний пояс, на плоских вершинах выше 600 м – это
разреженные фиппсиево-моховые структурные сырые тундры, сходные по ха-
.
Глава 6. ЗОНА ТУНДР
103
рактеру с арктическими пустынями. Здесь господствуют мелкие подушки мхов
и отдельные крошечные кустики фиппсии холодной (Phippsia algida), окси-
графиса ледяного (Oxygraphis glacialis), мятлика ложноукороченного (Poa
pseudoabbreviata).
Особенно богата растительность на нижней части внутригорных южных
склонов и в речных каньонах, а также на границе с ландшафтами межгорных
котловин. Здесь развиты горные луга, разнотравно-злаковые луговины, ре-
ликты более теплых эпох, на которых обильны пырейники, мятлик сизый, ов-
сяницы Ричардсона и ушковатая (Festuca richardsonii, F. auriculata), трище-
тинник колосистый (Trisetum spicatum), тонконог азиатский (Koeleria asiatica);
из разнотравья – смолевка малолистная (Silene paucifolia), копеечник аркти-
ческий (Hedysarum arcticum), арника Ильина (Arnica iljinii), пепельник тунд-
ровый (Tephroseris tundricola), лапчатка снежная (Potentilla nivea) с крупными
желтыми цветками, мак подушковидный (Papaver pulvinatum), из одной по-
душки которого иногда выходит до 50 стрелок с крупными веерообразно рас-
крытыми цветками. Местами встречается тимьян крайний (Thymus extremus).
Эти удивительные сообщества сохранились в горах Таймыра со времен терми-
ческого оптимума. Их состав свидетельствует о тесных флористических связях
Бырранга с более восточными горными системами севера Якутии и Чукотки.
На менее сухих, но хорошо прогреваемых склонах развиты пышные зла-
ково-разнотравные горные луга с преобладанием мятлика сизого, мытника
приятного (Pedicularis amoena), разнообразных бобовых – астрагалов зонтич-
ного, холодного и альпийского (Astragalus umbellatus, A. frigidus, A. alpinus),
остролодочников каргинского и Миддендорфа (Oxytropis karga, O. midden-
dorffii). Эти луга представляют собой желто-фиолетовые ковры, чередующие-
ся в небольших нишах с кустарничковыми тундрами, которые усеяны белыми
звездочками дриады, а позже обильными мелкими цветками кассиопеи.
Рис. 6.29. Горная тундра с лиственничным стлаником (фото И. Поспелова)
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
104
6.2.2.5. Лено-Колымская область тундр
Лено-Колымская область характеризуется резко выраженным континен-
тальным климатом. Средняя температура июля колеблется от 4 до 8 °С, сред-
няя температура января – от –32 до –40 °С. Дельта Лены, ограничивающая
область с запада, представляет собой веер многочисленных островов и разде-
ляющих их протоков (рис. 6.30). Преобладающая часть дельтовых островов
представляет собой участки надпойменной террасы высотой 25–30 м. Многие
острова дельты характеризуются широким развитием многолетнемерзлых
грунтов, ископаемых льдов, термокарстовых озерных котловин, а также хоро-
шо выраженных бугров пучения. Такая же высокая терраса является господ-
ствующей поверхностью в прибрежной полосе на материковом побережье
вплоть до Колымы.
Растительный покров дельты реки Лены отличается большим своеобра-
зием и включает, наряду с дельтовыми растительными группировками, при-
морские галофитные луга, болота, тундроболота, ивняки и другие тундровые
ассоциации. Преобладают широко распространенные виды равнинных тундр.
Острова дельты почти сплошь заняты полигонально-валиковыми болотами.
Полигоны заполнены водой, частично заняты зарослями осоки одноцветной
(Carex concolor), пушицы Шейхцера (Eriophorum scheuchzeri) и арктофилы
рыжеватой (Arctophila fulva); валики сформированы зелеными мхами с фраг-
ментами ивково-травяно-зеленомошной тундры.
Рис. 6.30. Дельта реки Лены (http://visibleearth.nasa.gov)
.
Глава 6. ЗОНА ТУНДР
105
По берегам многочисленных озер, а также проток на песчанистых, аллю-
виальных по преимуществу наносах, под которыми многолетнемерзлые грун-
ты залегают наиболее глубоко, появляются разнотравно-злаковые луга. Ост-
ров Тит-Ары знаменит тем, что здесь на 72° с.ш. встречаются самые северные
деревья лиственницы Каяндера (Larix cajanderi), их высота до 6 м. Среди тон-
ких стволов можно встретить огромные пни, оставшиеся от древних листвен-
ниц, что свидетельствует о прошлой более теплой климатической эпохе.
Обзорная карта растительности Лено-Колымской области представлена
на рис. 6.31. В целом за Енисеем в подзоне типичной тундры в Якутии распро-
странены алекториевые (Alectoria ochroleuca) и цетрариевые (Flavocetraria
cucullata и др.) тундры. Характерные для них лишайники переносят условия
малоснежной арктической зимы и жесткость погодных условий, губительных
для ягелевых формаций.
Восточнее дельты Лены на склонах гор распространены кустарничковые
и лишайниковые горные тундры. В кустарничковых тундрах резко преобла-
дают дриады и кассиопея. В лишайниковых тундрах господствуют на нижних
участках склонов кладонии и цетрарии, выше – алектории.
Между реками Омолой и Колыма на низменной прибрежной равнине,
сложенной рыхлыми отложениями, растительность представлена узкой при-
морской полосой арктических тундр, южнее сменяющихся широкой полосой
типичных кустарничковых тундр континентального типа. Они приурочены к
суглинистым и супесчаным, нередко щебнистым грунтам по склонам и вер-
шинам холмов, открытых действию ветров, и характеризуются пятнистым
растительным покровом. Среди разнообразных кустарничков господствуют
дриады – дриадовые тундры. Прочие цветковые растения встречаются рассе-
янно в небольшом количестве. Мхи и лишайники здесь не образуют сплошно-
го покрова и утрачивают значение эдификаторов, присущее им в мохово-
лишайниковых тундрах. На дренируемых участках в лишайниковых синузиях
преобладают Flavocetraria cucullata и Сetraria nivalis.
Особенностью растительности является отсутствие кустарниковых
тундр. Их место занимают кочкарные осоково-пушицевые тундры (рис. 6.32).
Кочкарники образованы кочками высотой до 30 см, состоящими из пуши-
цы (Eriophorum vaginatum) со мхами, лишайниками и тундровыми кустар-
ничками. Промежутки между кочками заняты мхами и лишайниками,
причем лишайники одевают также и верхушки старых, отмерших кочек
пушицы.
6.2.2.6. Чукотская область тундр
Область охватывает остров Врангеля, Чукотский полуостров и Анадыр-
скую низменность. Остров Врангеля занимает пограничное положение меж-
ду Восточно-Сибирским и Охотским морями. Рельеф острова низкогорный
сильно расчлененный. Климат формируется, главным образом, под воздей-
ствием холодного арктического воздуха. Летом с юго-востока доходит более
теплый и влажный воздух с Тихого океана, периодически поступают сухие и
нагретые воздушные массы из Сибири. Зимы продолжительные, морозные,
с сильными ветрами. Средняя температура февраля –23,5 °C. Лето холодное,
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
106
Рис. 6.31. Карта растительности Лено-Колымской области
(фрагмент карты из Атласа Арктики, 1985)
.
Глава 6. ЗОНА ТУНДР
107
Рис. 6.32. Осоково-пушицевая кочкарная тундра
случаются заморозки и снегопады, средняя температура июля +3 °C. В цен-
тре острова, отгороженном от моря горами, лето теплее. На подгорных рав-
нинах растительный покров характеризуется богатым видовым составом.
Обнаружено около 417 видов сосудистых растений, 331 – мохообразных и
310 – лишайников. Большое количество реликтов позволяет сделать вывод
о том, что исконная арктическая растительность острова не была уничтожена
ледниками. Растительный покров острова Врангеля представлен в горах по-
лярными пустынями, на подгорных равнинах – сообществами арктических и
типичных тундр (рис. 6.33). Границы зон и подзон на острове имеют субши-
ротную, а иногда и меридиональную направленность. Нижние и средние по-
яса гор заняты кустарничково-травяно-моховыми, осоково-моховыми, и тра-
вяно-лишайниковыми сообществами. Встречаются стелющиеся по земле за-
росли ивняков. Имеются болота с участием сфагновых мхов.
Климат Чукотки суровый, на побережьях – морской, во внутренних рай-
онах – резко континентальный. Годовая сумма осадков от 200 мм на севере
Чукотки до 500 мм на побережье Берингова моря. Осадки выпадают в основ-
ном летом и осенью. Зима малоснежная. Снег лежит 8–9 месяцев. Средняя
температура января от −15 °C до −39 °C. Средняя температура июля на севере
Чукотки составляет 3,8 °С, на побережье Берингова моря 7,7 °С и достигает
14 °С на Анадырской низменности.
В рельефе Чукотского п-ова преобладают горы высотой 1300–1800 м и
плоскогорья. Обширные приморские низменности покрыты мощным чехлом
рыхлых четвертичных отложений. Широко распространена многолетняя мерз-
лота и связанные с нею мерзлотные формы мезорельефа. Особо обращают на
себя внимание крупные гидролакколиты (высотой до 20–40 м) на северном
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
108
Рис. 6.33. Цветущая арктическая тундра, остров Врангеля
(http://geoman.ru/books/item/f00/s00/z0000056/st023.shtml)
побережье и солифлюкционные процессы на склонах (рис. 6.34). В условиях
развития термокарстовых процессов образуется множество озер и болот.
Большую часть территории занимают горно-тундровые почвы со значи-
тельным содержанием мелкого обломочного материала. Широко распростра-
нены полигональные грунты. На низменностях преобладают торфяно-глеевые
почвы, а в бассейне р. Анадырь встречаются торфяно-подзолистые и аллюви-
альные почвы.
Рис. 6.34. Солифлюкционные процессы на склоне, Чукотский полуостров,
окрестности поселка Провиденья (фото А. Галанина)
.
Глава 6. ЗОНА ТУНДР
109
В основу описания растительности области Чукотского полуострова по-
ложены труды по ООПТ Чукотки (по: Беликович и др., 2006). Полуостров по
богатству биоразнообразия представляет собой территорию, где на относи-
тельно небольшой площади встречаются разнообразные типы растительности
и большое количество самобытных видов. Происхождение местной флоры
связано с историей Берингийского моста суши, некогда соединявшего Азию и
Америку. Общие черты растительности Чукотской области представлены на
карте (рис. 6.35).
Подзона арктических тундр занимает северное побережье Чу-
котского полуострова. Растительность образована разреженным покровом из
арктополевицы тростниковидной (Arctagrostis arundinacea), часто с содоми-
нированием осоки траурной (Carex lugens). В целом она относительно богата
флористически; обильны кустарнички (Dryas punctata, Salix sphenophylla,
S. phlebophylla, Diapensia obovata).
Подзона типичных тундр. Кустарничково-осоково-моховые тун-
дры широко распространены на влажных склонах гор, плоских вершинах низ-
ких гор и в седловинах между острыми вершинами, а также на низких надпой-
менных террасах.
Из кустарников здесь доминирует береза тощая (Betula exilis), из кустар-
ничков – дриады (Dryas octopetala s.l., D. integrifolia (на известняках)), касси-
опея четырехгранная (Cassiope tetragona), голубика обыкновенная мелко-
листная (Vaccinium uliginosum ssp. microphyllum), брусника малая (V. vitis-
idaea var. minus), водяника черная (Empetrum nigrum), арктоус альпийский
(Arctous alpina), багульник стелющийся (Ledum decumbens); обычны рододенд-
рон мелколистный (Rhododendron parvifolium), диапенсия обратнояйцевид-
ная (Diapensia obovata); часто встречаются редкие кустики ив (Salix glauca,
S. pulchra). Среди разнотравья особенно характерны Carex lugens, С. scirpoidea,
С. vaginata, С. nesophila (в восточной Чукотке), С. podocarpa, Luzula tundricola,
L. nivalis, Hierochloe alpina, Saussurea tilesii, Acomastylis glacialis, Saxifraga
nelsoniana, Pedicularis capitata, Bistorta officinalis, Poa arctica, Hedysarum
arcticum, Anemonastrum sibiricum, Eutrema edwardsii, Endocellion glaciale,
Astragalus umbellatus. На дренированных участках шлейфов с богатым мине-
ральным питанием некоторые тундры настолько насыщены разнотравьем, что
имеют характер переходный к луговинам. В эутрофных кустарничково-осоково-
моховых тундрах на известняках с Dryas integrifolia часто содоминирует Salix
rotundifolia, а видовой состав насыщен кальцефилами: Artemisia globularia,
Carex membranacea, Claytonia tuberosa, Potentilla biflora, Silene acaulis и др.
На Чукотке имеются группировки по набору биоморф очень сходные с
«остепненными». Например, участки с доминированием Carex rupestris,
Festuca altaica или Poa glauca. Чтобы подчеркнуть их некоторое подобие степ-
ным фитоценозам, используется понятие «степоиды». Это приходится делать,
чтобы подчеркнуть, что понятие «степь» к обсуждаемой растительности не-
применимо. Среди растительности Чукотки степоиды выделяются внешним
обликом, особенно те, покрытие которых относительно велико, или те, в кото-
рых доминируют злаки и осоки. Внешний вид таких степоидов напоминает
растительность пустырей в умеренных широтах с неравномерным покрытием
и выделяющимися скоплениями полусухих листьев злаков . Флористический
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
110
Рис. 6.35. Карта растительности Чукотского п-ова (фрагмент карты из Атласа
Арктики, 1985)
.
Глава 6. ЗОНА ТУНДР
111
Рис. 6.36. Степоиды в континентальных районах Чукотки
(http://kolymintours.narod.ru/rus/ photographs.htm)
состав чукотских степоидов включает Aster alpinus, Helictotrichon krylovii,
Oxytropis campestris, O. vassilczenkoi var. substepposa, O. middendorffii, Poten-
tilla nivea, Selaginella rupestris, Thymus serpyllum, Silene repens, Eremogone
capillaris, Festuca lenensis, Eritrichium sericeum, Aconogonon ochreatum var.
laxmannii, Bupleurum triradiatum, Calamagrostis purpurascens, Elymus confu-
sus, Cnidium cnidiifolium, Pulsatilla multtfida, Lychnis samojedorum, Carex span-
iocarpa, C. obtusata, C. duriuscula, Smelowskia calycina var. porsildii и др. Сте-
поиды заходят на Чукотке севернее 68° с.ш. (рис. 6.36).
Подзона кустарниковых (южных) тундр. На плакорах Чукот-
ского полуострова растительность подзоны образована зарослями ерника
из Betula divaricata и В. exilis, ольховника из Duschekia fruticosa s. str. и
D. kamtschatica, ивняков из Salix krylovii, S. alaxensis, S. richardsonii, S. glauca,
S. pulchra s. str., S. schwerinii. Основной кустарниковой формацией являются
ольшаники на склонах гор. В состав сообщества входят Vaccinium uliginosum,
Ledum decumbens, Aconogonon tripterocarpum, Spiraea beauverdiana, Artemisia
tilesii, Calamagrostis langsdorffii, Rubus chamaemorus, Lycopodium annotinum,
L. dubium; густой напочвенный покров образуют зеленые мхи. На юге Чукот-
ки в ольшаниках встречается Rhododendron aureum.
В континентальных районах весьма обычны склоновые ивняки. Они
приурочены к различным укрытым местоположениям, или, наоборот, к вы-
ступам в нижних частях склонов, под которыми нередко лежат снежники.
В подзоне кустарниковых тундр большие площади занимают также сфагно-
вые болота. К ним тесно привязаны такие континентальные виды, как жирян-
ка лопатчатая Pinguicula spathulata, клюква мелкоплодная Oxycoccus micro-
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
112
carpus. Чукотские болота очень разнообразны, поскольку их формированию
способствует близость залегания многолетней мерзлоты и превышение коли-
чества осадков над испарением. Имеются болота олиготрофные и эутрофные,
сфагновые и гипновые, с морозообойными пятнами и без таковых, бугристые,
бугристо-мочажинные и ровные. Болота обнаруживаются на различной высо-
те, отсутствуя лишь на вершинах гор. Даже на привершинных террасах гор на
высоте 600 м они вполне обычны.
Значительная часть Чукотского побережья покрыта кочкарными пуши-
цевыми тундрами (рис. 6.37). Они поднимаются по пологим склонам гор и
плоским долинам до 200 м и простираются вдоль низменного побережья
Охотского моря далеко на юг узкой прерывистой полосой. В горах, выше
тундрового пояса располагаются гольцы, где большие пространства заняты
подвижными щебнистыми осыпями, в растительном покрове преобладают
лишайники. Особенно широко распространены горные сухие тундры со значи-
тельным развитием кустарников и кустарничков (куропаточная трава, арктоус
альпийский, на нижних частях склонов – кедровый стланик, ольховник кустар-
никовый), а также различных камнеломок. Разреженный покров образуют
дриадовые, вороничные, диапенсиевые, луазелериевые, кобрезиевые, лишай-
никово-кассиопейные, осочковые (с Carex rupestris) и т. п. сообщества. Эди-
фикатором горных тундр на Чукотке являются реликтовые виды рододендрон
камчатский Rhododendron camtschaticum и осока островная Carex nesophila.
Анадырская низменность занята преимущественно кочкарными пуши-
цевыми и кустарниковыми тундрами с большой примесью мхов и лишайни-
ков. Встречаются карликовые береза и ивы, голубика, морошка, багульник.
Роль болот здесь чрезвычайно велика, несмотря на резко континентальный
климат . Они тянутся беспрерывно на многие километры, хотя растительность
Рис. 6.37. Кочкарная тундра, Западная Чукотка (фото А. Галанина)
.
Глава 6. ЗОНА ТУНДР
113
на этом пространстве нередко существенно изменяется: участки чистых коч-
карников покрываются зарослями кустарников, которые переходят в листвен-
ничные редколесья или сменяются зарослями кедрового стланика. Состав
видов травяно-кустарничкового яруса включает: Carex concolor, С. limosa,
Eriophorum vaginatum, Betula exilis, Chamaedaphne calyculata, Ledum palustre,
Vaccinium uliginosum, Andromeda polifolia, Rubus chamaemorus, Pedicularis
lapponica.
6.2.2.7. Северо-Тихоокеанская островная область
Северо-Тихоокеанская островная область занимает Командорские о-ва,
северные и средние Курильские о-ва, а также небольшие участки Тихоокеан-
ского побережья Камчатки.
Командорские острова являются самой западной группой Алеутской
островной дуги. В состав архипелага входят два крупных острова: Беринга и
Медный, а также о. Топорков, о. Арий Камень и многочисленные мелкие ост-
ровки, небольшие камни и скалы. Они лежат на расстоянии около 175 км к
востоку от Камчатки. В 1993 г. на островах был создан Государственный при-
родный наземно-морской заповедник «Командорский».
Командорские острова – вершины крупного подводного океанического
хребта. Литогенная основа островных ландшафтов представлена мощными
вулканогенно-осадочными породами. Рельеф – низко- и среднегорный с мак-
симальными высотами до 750 м, осложненный серией морских террас разных
уровней. Берега абразионные, обрывающиеся в море крутыми уступами. В бух-
тах формируются аккумулятивные берега с песчано-галечниковыми пляжами,
серией береговых валов, иногда с дюнами (рис. 6.38).
Рис. 6.38. Береговая зона острова (вид с моря), на первом плане – каменистая
осушка, на втором – на песчаном береговом валу заросли колосняка
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
114
Климат островов – умеренный океанический: средние температуры ян-
варя –2÷0 °С, средние температуры летних месяцев не поднимаются выше
10 °С, среднегодовое количество осадков около 500 мм. Постоянный снежный
покров устанавливается в конце ноября – в декабре, а сходит в основном в се-
редине мая. Экстремальные условия для растительности создают сильные хо-
лодные ветры, дующие из Арктики. На климат островов оказывают влияние
холодные течения из Северного Ледовитого океана и ветви теплого течения
Куросио. Акватория вокруг островов не замерзает.
Гористый рельеф Командорских островов определяет формирование эле-
ментов высотной поясности. На приморских террасах и до высоты около 200 м
господствует луговая растительность, выше – комплексная растительность,
представленная кустарничково-лугово-тундровыми сообществами, выше 300 м
располагается нивально-тундровый пояс. Наиболее благоприятные условия
для формирования луговой растительности наблюдаются на низких морских
террасах и пологих склонах южной экспозиции. Под воздействием сильных
холодных арктических ветров на склонах северной экспозиции элементы
тундровых сообществ могут спускаться почти до уровня моря. Заросли из оль-
ховника кустарникового и кедрового стланика, столь обычные на Камчатке и
северных Курильских островах, на Командорских островах отсутствуют.
Флора сосудистых растений Командор включает 432 вида и подвида, от-
носящихся к 200 родам и 62 семействам. Преобладающими по числу видов
являются три семейства: Asteraceae, Poaceae и Cyperaceae. Здесь произрастают
три вида растений, занесенных в Красную книгу РСФСР: полушник азиатский
Isoetes asiatica Makino, венерин башмачок крупноцветковый Cypripedium
macranthon Sw., любка камчатская Neolindleya camtschatica (Cham. et Schlecht.)
Nevski.
В условиях влажного прохладного климата и сильных ветров у целого
ряда растений формируется комплекс морфологических адаптаций, часть из
которых свойственна растениям всех островов Северной Пацифики. К таковым
относятся: общее уменьшение высоты стеблей и укорочение междоузлий; ми-
ниатюризация растений на ветробойных склонах, особенно заметная у кустар-
ников (уменьшение размеров растения в целом, уменьшение размера листьев,
формирование плотных дерновин с более короткими стеблями); некоторое
усиление опушения стебля и листьев; более сжатый (компактный) характер
соцветия, укорочение цветоносов (но одновременно заметное увеличение числа
цветков или корзинок у сложноцветных); образование у целого ряда видов
«обвертки» соцветия из скученных верхних стеблевых листьев.
Луговая растительность Командорских островов сформирована преиму-
щественно субальпийскими и альпийскими видами бореального типа харак-
терными для Северной Пацифики. Горные тундры в наибольшей степени
сходны с сообществами высокогорий Камчатки, а также северных Курильских
островов.
Луговая растительность представлена разнотравными и крупнотравны-
ми лугами на морских террасах разного уровня и в долинах рек, а также ни-
вальными лужайками. По мере удаления от моря растительность образует от-
четливый экологический ряд. Несомкнутая галофитная растительность га-
лечных и песчано-галечных пляжей сменяется на песчаных береговых валах
.
Глава 6. ЗОНА ТУНДР
115
колосняковыми или колосняково-крестовниковыми сообществами (Leymus
mollis и Senecio pseudoarnica). Песчаные дюны и береговые валы, на которых
формируются приморские полидоминантные разнотравные луга, являются ха-
рактерными морфологическими элементами западного побережья о. Беринга и
юга о. Медного. Почти все дюны задернованы, на них развиты дерново-
песчаные почвы. На первой приморской террасе высотой 5–6 м располагают-
ся злаково-крупнотравные луга. Наиболее обычны на Командорах колосняко-
во-крупнотравные луга, в которых доминируют Leymus mollis и Heracleum
lanatum, обычны Angelica gmelinii, Arctopoa eminens, Cacalia kamtschatica,
Calamagrostis purpurea s. l., и Conioselinum chinense. Дальше от моря на лугах
уменьшается количество колосняка и увеличивается доля крупнотравья и раз-
нотравья; вместе с вышеперечисленными видами растут Aconitum maximum,
Artemisia opulenta, Carex gmelinii, Cirsium kamtschaticum, Senecio cannabifolius
и Veratrum oxysepalum. Во 2 подъярусе обычны Claytonia sibirica, Geranium
erianthum, Ligusticum scoticum, Moehringia lateriflora, Poa macrocalyx и Viola
langsdorfii. Проективное покрытие 100 %. Высота 1 подъяруса – около 1,5 м.
Крупнотравные луга встречаются по долинам рек, распадкам, ложбинам
стока на склонах, по подножьям склонов сопок и морских террас. Преоблада-
ют луга с 2–3 доминирующими видами, достигающими высоты 1,5–1,8 м.
Среди крупнотравных лугов со сложной многоярусной структурой наиболее
часто встречаются следующие типы лугов: полидоминантные крупнотравные
луга с доминированием лабазника камчатского (Filipendula camtschatica),
борщевика шерстистого (Heracleum lanatum) и крестовника коноплеволист-
ного (Senecio cannabifolius); вейниково-борщевиковые луга с доминированием
вейника пурпурного (Calamagrostis purpurea) и борщевика шерстистого; осо-
ково-дудниково-полынные луга, на которых преобладают дудник Гмелина
(Angelica gmelinii), полынь пышная (Artemisia opulenta) и осока длинноости-
стая (Carex macrochaeta).
Разнотравные и злаково-разнотравные луга существуют как в долинах
рек, так и на дренированных участках южных склонов сопок (рис. 6.39). Эти
сообщества обычно полидоминантны; в их состав входят: Anemonastrum
narcissiflorum, Carex macrochaeta, Dactylorhiza aristata, Equisetum arvense,
Geranium erianthum, Iris setosa, Avenella flexuosa, Parageum calthifolium, Poa
malacantha, Veratrum oxysepalum, разреженно встречаются представители вы-
сокотравья Cacalia kamtschatica, Conioselinum chinense, Filipendula camtschatica
и Heracleum lanatum. Во 2-м подъярусе – Coptis trifolia, Moehringia lateriflora,
Stellaria calycantha, Viola epipsiloides и Trientalis europaea. Средняя высота
травостоя в них около 0,6–0,8 м, проективное покрытие 100 %.
По корытообразным долинам рек и на равнинных участках развиты сырые
злаково-осоковые, осоково-разнотравные и осоково-вейниковые луга, где од-
ним из доминантов является Carex cryptocarpa. В их составе обычны: Angelica
genuflexa, A. gmelinii, Calamagrostis purpurea, Carex macrochaeta, Glyceria
alnasteretum, Avenella flexuosa, Comarum palustre, Polemonium acutiflorum и
Trollius riederianus. Во 2 подъярусе нередки: Bistorta vivipara, Epilobium
hornemannii, Equisetum arvense, Iris setosa, Lathyrus pilosus, Parnassia palustris,
Ranunculus repens и Rubus stellatus. Изредка встречается Salix arctica, ха-
рактерны зеленые мхи. Нередко такие луга соседствуют с монодоминантными
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
116
Рис. 6.39. Разнотравный луг на пологом приморском склоне, Командорские
острова
крупноосоковыми болотами. Торфянисто-глеевые и торфяно-болотные почвы
низинных болот формируются в условиях высокого уровня грунтовых вод.
Они наиболее широко распространены в северной низинной части о. Беринга
и на низких и средних поймах рек.
Осоково-разнотравные и кустарничково-разнотравные субальпийские
лужайки располагаются в горных цирках, на седловинах перевалов, в узких
распадках, по местам долгого лежания снега и встречаются повсюду, начиная
от уровня моря и до средней части горных склонов. На осоково-разнотравных
субальпийских лужайках доминируют: Anemonastrum narcissiflorum, Juncus
beringensis и Parageum calthifolium, обычны: Artemisia arctica, Carex riishiren-
sis, C. macrochaeta, C. micropoda, Cypripedium yatabeanum, Gentiana glauca,
Iris setosa, Lagotis glauca, Poa lanata и Polemonium acutiflorum. Проективное
покрытие 80–100 %, высота до 0,5 м.
Разреженные ивовые и ивово-рябиновые заросли распространены в рас-
падках и на склонах гор, преимущественно в защищенных от ветра местах.
Они состоят из Salix crassijulis, S. pulchra, S. lanata и Sorbus sambucifolia, под
пологом которых развит разнотравно-кустарничковый или кустарничково-
зеленомошный покров, сходный с окружающей кустарничковой тундрой. По
выположенным участкам склонов с бугристыми кустарничковыми тундрами
отмечены заросли рододендрона золотистого (Rhododendron aureum) и ряби-
ны. Ерники из Betula exilis отмечены на севере о. Беринга. Проективное по-
крытие зеленых мхов достигает 30–40 %.
Тундровые сообщества представлены различными вариантами кустар-
ничковых, разнотравно-кустарничковых и каменистых (щебнистых) тундр. На
.
Глава 6. ЗОНА ТУНДР
117
склонах северной экспозиции, испытывающих воздействие сильных холодных
арктических ветров граница тундровых сообществ спускается на более низкие
уровни. Кустарничково-рододендровые тундры распространены в нижних ча-
стях и на пологих участках склонов. Доминирует Rhododendron aureum, с ним
постоянно произрастают Chamaepericlymenum suecicum и Empetrum sibiricum.
Разнотравье содоминирует в разнотравно-кустарничковой тундре (рис. 6.40).
В напочвенном покрове преобладают зеленые мхи и лишайники.
По верхним пологим частям склонов развита шикшево-мохово-лишай-
никовая тундра, доминируют водяника сибирская, мохоцветник Гмелина
Bryanthus gmelinii и кассиопея плауновидная Cassiope lycopodioides, а из раз-
нотравья – ветреник нарциссоцветковый Anemonastrum narcissiflorum, по-
лынь арктическая Artemisia arctica, змеевик живородящий Bistorta vivipara и
лжегравилат калужницелистный Parageum calthifolium. Выше 300–350 м
располагается нивально-тундровый пояс.
Рис. 6.40. Разнотравно-кустарничковая тундра Командорских островов
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
118
На плоских и слабонаклонных приморских террасах разных уровней в
результате климатической инверсии формируются сухие приморские тундры,
представленные шикшевыми, шикшево-кустарничковыми и разнотравно-
кустарничково-шикшевыми сообществами. По составу и строению они во
многом сходны с горными кустарничковыми тундрами. В большинстве вари-
антов приморских тундр доминирует водяника, покрытие которой достигает
70–90 %. Из других видов наиболее часто отмечены дёрен шведский, щучка
берингийская Deschampsia beringensis, лжегравилат калужницелистный и мят-
лик крупночешуйный Poa macrocalyx.
6.3. АНТРОПОГЕННЫЕ НАРУШЕНИЯ ЛАНДШАФТОВ ТУНДР
Биологические природные ресурсы Крайнего Севера начали осваиваться
первобытными народами еще в доисторические времена. В XX веке важную
роль стали играть процессы, оказывающие сильное техногенное воздействие
на уязвимые тундровые экосистемы: разведка и добыча полезных ископае-
мых, строительство населенных пунктов и военных объектов.
В XIX веке тундра была мало обитаема. Местное население занималось
кочевым оленеводством (рис. 6.41) и пушным промыслом, а также рыболов-
ством и охотой на морского зверя. В начале XX века возникли немногочис-
ленные и малонаселенные поселки, расположенные в долинах рек. В послед-
ние десятилетия XX века хозяйственная деятельность в суровых условиях Се-
вера получила особенно большое развитие.
Рис. 6.41. Традиционный способ ведения хозяйства – оленеводство
.
Глава 6. ЗОНА ТУНДР
119
Специальные исследования и анализ литературных источников показы-
вают, что на обширных пространствах тундр ведущую роль в формировании
их облика играло воздействие северных оленей – диких и домашних. Олене-
водство развито практически на всем протяжении тундровой зоны, лесотунд-
ры и северной тайги на площади более 3 млн. кв. км. Ягельники требуют для
перезимовки глубокого снежного покрова, почему и распространены главным
образом в западных тундрах, находящихся под влиянием влажных ветров с
Атлантического океана. Распределение лишайниковых тундр имеет большое
значение для оленеводства. Отсутствие в арктической Якутии ягельных фор-
маций не благоприятствует широкому распространению оленеводческих хо-
зяйств на побережье. Напротив, к западу от Таймыра сосредоточено наиболь-
шее поголовье оленей.
Дикие и домашние олени используют, особенно в снежный период года,
одни и те же пастбища. Они одинаково адаптированы к пастбищным услови-
ям Севера, но выпас домашних оленей имеет ряд особенностей, определяемых
системой пастбищной «стратегии» и традициями местного оленеводства.
Домашние олени круглый год содержатся в стадах по 500–2000 голов и
более. Угодья, освоенные домашним оленеводством, периодически испыты-
вают нагрузки, превышающие их биологическую емкость. В большей части
районов развития оленеводства пастбищные ресурсы используются на 80–
90 %. В структуре таких пастбищ основная роль принадлежит продукции зла-
ков, осок, разнотравья и кустарников, тогда как доля продукции мхов, лишай-
ников и кустарничков относительно невелика. Потери лишайников и зеленых
частей кустарничков могут составить в местах концентрации оленей 60–100 %
от общего запаса. В местах расположения стоянок растительность представле-
на луговинами, и ее естественный облик восстанавливается крайне медленно.
Пастбищной дигрессией в настоящее время охвачены огромные терри-
тории тундровой зоны и лесотундры. Прямое и косвенное уничтожение расти-
тельности на пастбищах, разбивание мохового и торфянистого покровов при-
водят к трансформации гидротермического режима почв и к замещению ку-
старниково- и кустарничково-моховых, лишайниковых и других тундр
злаково-моховыми, травяно-моховыми и луговинами. Лишайниковые паст-
бища на значительных территориях уже выбиты. Обращает внимание общий
процесс делихенизации – деградации лишайниковой тундры. Помимо неуме-
ренного выпаса исчезновение лишайников вызывается пожарами, а также
воздействием вездеходного транспорта.
С начала 30-х годов XX века в зоне тундр, на материке и на островах,
функционируют метеорологические станции. Для их жизнеобеспечения еже-
годно завозятся десятки бочек горюче-смазочных материалов. Пустые бочки,
десятилетиями накапливаясь вокруг жилищ людей, создают неприглядный
облик ландшафта (рис. 6.42).
Распределение промышленности, имевшее прежде очаговый характер на
Севере, в настоящее время переходит в фазу фронтального освоения террито-
рий. Существенно укрупнились контуры хозяйственной деятельности вплоть
до их слияния, включая коммуникации, промышленные объекты, населенные
пункты. Под угрозой оказались обширные районы Кольского полуострова,
низовий Печоры, Большеземельской тундры, Ямала, юга Таймыра, северной
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
120
Рис. 6.42. Антропогенное загрязнение создает неприглядный облик полярным
ландшафтам
Якутии, Чукотки и др. На этих территориях происходит интенсивное уничто-
жение, изменение, сокращение и фрагментация природных экосистем и ме-
стообитаний видов животных и растений. Большинство районов проживания
коренных народов оказалось в сфере интересов добывающих компаний, дея-
тельность которых ведет к нарушению системы традиционного природополь-
зования.
Рис. 6.43. Гусеничные вездеходы в тундре
.
Глава 6. ЗОНА ТУНДР
121
Рис. 6.44. Следы от гусениц вездехода,
превратившиеся в канавы
Глубокое, подчас невосполни-
мое разрушение ландшафтов в тунд-
рах производит техногенная деятель-
ность. Наличие в тундре многолет-
ней мерзлоты – важнейший фактор,
сдерживающий освоение территории.
Растительная дернина, в том числе
моховой покров и торфяной слой,
являются хорошим естественным
теплоизолятором. Разрушение этого
слоя способствует таянию подземных
льдов, образованию просадок и про-
валов. Поэтому необходимо исклю-
чительно бережное отношение к поч-
венно-растительному покрову. Мас-
сированное наступление на Арктику
во второй половине XX века было
связано с геологоразведкой. На дев-
ственные ландшафты тундры двину-
лись армады гусеничных вездеходов
(рис. 6.43). Достаточно вездеходу разрушить моховой покров, чтобы вскоре
следы от гусениц превратились в канавы, а затем и в глубокие промоины
(рис. 6.44). Расчищенная грунтовая площадка через несколько лет может
стать провальным термокарстовым озером.
Арктика богата запасами углеводородов. Многочисленные газоконден-
сатные месторождения (ГКМ) открыты и разрабатываются в частности на п-
ове Ямал. Строительство труб газопроводов разрушает растительный покров,
преграждает пути традиционных миграций северных оленей (рис. 6.45).
Нефтепродукты, попадающие в грунт, разлагаются очень плохо. Низкие тем-
пературы и незначительное содержание кислорода в почве способствуют дли-
тельному сохранению нефтяного загрязнения. Самоочищения в зоне много-
летней мерзлоты практически не происходит. Арктика богата также место-
рождениями различных минеральных полезных ископаемых. Их разработка
ведет к коренному разрушению тундровых экосистем (рис. 6.46).
Результатом антропогенных нарушений экосистем является разрушение
растительных сообществ. Прекращение антропогенной деятельности ведет к
восстановлению (демутации) естественного покрова. Формирование расти-
тельности на техногенных местообитаниях Крайнего Севера России детально
изучено О. И. Суминой, которая отмечает, что заселение техногенных место-
обитаний идет за счет ресурсов местной флоры. Видовой состав растительно-
сти нарушенных территорий обеднен, но при этом в каждом районе он обла-
дает значительным своеобразием. Сообщества техногенных местообитаний
очень разнообразны. От ненарушенных ценозов они отличаются не столько по
видовой насыщенности и сомкнутости покрова, сколько по составу доминантов
.
Часть II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Арктический и субарктический пояса
122
Рис. 6.45. Трубы нефтепроводов в тундре
и пространственной структуре. Следует отметить, что, несмотря на суровые
природные условия Севера, тундровая растительность обладает определенной
потенцией к демутации и восстановлению экологического потенциала ланд-
шафтов.
Рис. 6.46. Добыча полезных ископаемых
.
Глава 6. ЗОНА ТУНДР
123
Вопросы к главе 6
1. Природные условия зоны тундр; экология растений.
2. Широтные особенности растительности тундр: подзоны высокоарктических (полярных
пустынь), арктических (мохово-лишайниковых), типичных (кустарничково-мохово-
лишайниковых), южных (кустарниковых) тундр.
3. Региональные особенности растительности тундр: Кольская, Восточно-Европейская,
Ямало-Гыданская, Таймырская, Лено-Колымская, Чукотская области
Рекомендуемая литература
Александрова В. Д. Геоботаническое районирование Арктики и Антарктики. Л.: Наука,
1977. 189 с.
Атлас Арктики. М.: ГУГиК при Совете Министров СССР, 1985. 204 с.
Атлас Ямало-Ненецкого автономного округа. Омск: ФГУП «Омская картографическая фаб-
рика», 2004. 304 с.
Беликович А. В., Галанин А. В., Афонина О. М., Макарова И. И. Растительный мир особо
охраняемых территорий Чукотки. Владивосток: БСИ ДВО РАН, 2006. 260 с.
Исследование природы Таймыра. Закономерности пространственного размещения и взаи-
мосвязи климата, растительности, почв и животного мира. Ландшафты. Труды Гос.
Биосферного заповедника «Таймырский». Вып. 1. Красноярск: Восточносибирский
филиал международного ин-та леса. 2001. 274 с.
Матвеева Н. В. Зональность в растительном покрове Арктики // Труды Ботанического ин-
ститута РАН. Вып. 21. СПб., 1998. 220 с.
Мельцер Л. И. Тундровая растительность // Растительный покров Западно-Сибирской рав-
нины. Новосибирск: Наука, 1985. С. 41–54.
Мочалова О. А., Якубов В. В. Флора Командорских островов. Владивосток: БПИ ДВО РАН,
2004. 120 с.
Пармузин Ю. П. Тундролесье СССР. М.: Мысль, 1979. 295 с.
Чернов Ю. И. Жизнь тундры. М.: Мысль, 1980. 236 с.
Юрцев Б. А. Арктическая флористическая область. Л.: Изд-во: Наука, 1978. 172 с.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
124
Умеренный пояс расположен между субарктическим и субтропическим поя-
сами: от 65° до 40° с.ш. и занимает в Евразии широкую полосу от Атлантического
до Тихого океана. Для умеренного пояса характерна четкая сезонная смена тер-
мического режима от весны к лету, осени и зиме. Атмосферная циркуляция в
умеренном поясе формируется под воздействием трех центров атмосферного
давления: Атлантического (Исландского) минимума, Азиатского максимума и Ти-
хоокеанского (Алеутского) минимума. Атлантический центр определяет форми-
рование западного переноса, который, достигая Центральной Сибири, встречает
преграду в виде Азиатского антициклона. Западный перенос сопровождается
прохождением циклонов, несущих атмосферные осадки, количество которых при
продвижении на восток снижается. С удалением от Атлантического океана конти-
нентальность климата усиливается, в связи с чем в характере растительного по-
крова ярко проявляется секторность. Азиатский антициклон – холодные и плотные
массы воздуха, обусловливающие высокое атмосферное давление – формируется
в зимнее время. Действие Азиатского антициклона выражается в возникновении
зимних муссонных ветров, дующих в сторону Северного Ледовитого и Тихого оке-
анов. Режим погоды морозный, небо безоблачное. Летом, благодаря интенсив-
ной солнечной радиации, на месте Азиатского максимума подстилающая поверх-
ность нагревается, и формируются теплые и легкие воздушные массы, определя-
ющие формирование области низкого атмосферного давления – Азиатского
минимума. Направление ветров меняется, возникает летний муссон, сопровож-
дающийся вторжением влажных воздушных масс с Тихого океана и холодных
арктических – с Северного Ледовитого океана.
Как уже говорилось, при выделении природных зон первичным фактором
является температурный режим, но по мере перехода в низкие широты, где кли-
мат становится более теплым, все отчетливее проявляется роль увлажнения.
Важным экологическим фактором становится недостаток атмосферных осадков
(испарение превышает количество выпадающих осадков), ГТК вегетационного
периода становится меньше 1. В связи с этим при продвижении на юг леса сме-
няются степями, а на юго-востоке, в Казахстане и Средней Азии, степи сменяются
пустынями.
Господствующим типом растительности в умеренном поясе, где испарение
не превышает количества выпадающих осадков, являются леса (см. рис.). Для их
произрастания необходимо сочетание определенных экологических условий, в
первую очередь – достаточное количество теплоты и влаги. Испарение не должно
превышать количество выпадающих атмосферных осадков. Северная граница
распространения лесов проходит по изотерме самого теплого месяца 10 °С, а рас-
пространение лесов на юг ограничивается недостатком атмосферных осадков и
высоким испарением в летнее время – ГТК вегетационного периода несколько
больше или равно 1.
Основоположником учения о лесе является Г. Ф. Морозов. Он утверждал,
что лес есть явление географическое, существование его разнообразных типов
не может быть понято вне связи с внешней или географической средой. Настолько
Умеренный пояс
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
125
тесна и глубока эта связь, что под лесом, в сущности, мы должны разуметь не
только одну совокупность древесных растений, объединенных взаимной связью,
но и ту среду, ту арену, на которой разыгрываются те социальные процессы, кото-
рые мы все собираем, как в фокусе, в понятии лес. Лес есть стихия и, подобно
степям, пустыням, тундрам, есть часть ландшафта, часть, стало быть, земной по-
верхности, занятой в силу ее определенных биологических свойств соответствен-
ными лесными сообществами.
Леса России – ее национальное богатство, ценность которого особенно ве-
лика в связи с тем, что они относятся к возобновляемым природным ресурсам,
которые, в противоположность быстро истощаемым запасам нефти и газа, спо-
собны к самовосстановлению, если площадь и интенсивность уничтожения лес-
ной растительности вырубками и пожарами не превосходит способности лесов к
возрождению. Велико их водорегулирующее, противоэрозионное, а также рекре-
ационное значение. Дары леса – это не только древесина, но и грибы, ягоды, кед-
ровые орехи, лекарственные растения. Леса служат естественной средой обита-
ния богатой фауны – зверей и птиц, в том числе промысловых.
Леса играют огромную роль в насыщении атмосферы кислородом и погло-
щении углекислого газа. Один гектар леса в процессе фотосинтеза выделяет за год
около 1 тыс. м
3
кислорода, что удовлетворяет годовую потребность в кислороде
одного человека. Подсчитано, что кислорода, выделяемого лесами тюменского Се-
вера за год, достаточно для дыхания населения всей Тюменской области, Урала и
Казахстана. Для примера скажем, что леса США уже не могут обеспечить кислоро-
дом население этой страны. В промышленно развитые страны, где площадь лесов
сильно сокращена, кислород приносится воздушными потоками из сохранившихся
лесных ландшафтов, в том числе с территории таежной зоны России.
Площадь лесов России составляет более 800 млн. га. Лес занимает около
45 % территории нашей страны и составляет около 24 % запасов древесины всей
планеты (http://lesa-rossii.ru/). В XX–XXI вв. постепенно увеличивается тенденция
роста лесозаготовок, отчуждения таежных ландшафтов для строительства городов
и промышленных объектов. Динамика площадей основных лесообразующих по-
род России за период с 1988 по 2005 гг. отражена в таблице.
При обсуждении таблицы следует иметь в виду, что последняя карта лесов
России, основанная на данных наземного лесоустройства, была составлена в 1991 г.
Все, что создавалась позже, подготовлено на основе авиационного и космическо-
го мониторинга и имеет значительно меньшую точность. Из этого следует, что
ближе всего к истине отмечены в таблице тенденции к уменьшению площади под
елью, лиственницей, дубом высокоствольным, а также увеличение площадей под
березой, осиной, сосной обыкновенной, которые обычно представляют собой
вторичные леса. Тенденция к увеличению площадей под кедровой сосной и
буком вызывает сомнения.
Умеренный пояс подразделяется на бореальную зону хвойных лесов, не-
моральную зону широколиственных лесов, и связывающую их переходную под-
зону хвойно-широколиственных лесов (подтайги).
Умеренный пояс
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
126
Рисунок. Карта лесов России (Атлас
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
127
малонарушенных..., 2002, www.forest.ru)
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
128
Таблица
Динамика площадей основных лесообразующих пород России, тыс. га (по: priroda.ru)
Основные
лесообразующие
породы
Год учета
1988*
1993*
1998
2003
2004
2005
Хвойные
Сосна
113564,0 114326,0 116740,0 117473,0 117205,0 117295,0
Ель
78810,0 75866,3 77658,0 77198,4 76737,4 76417,7
Лиственница
277898,0 263348,0 265719,0 264287,0 263986 264269,9
Кедровая сосна
40166,0 39797,6 41033,2 40852,0 41054,6 41171,6
Твердолиственные
Дуб высокоствольный
3761,0
3808,0
3719,0
3633,7
3650,2
3611,9
Дуб низкоствольный
3198,7
2971,3
3110,3
3200,0
3169,6
3161,0
Бук
698,5
701,3
786,0
789,6
790,1
793,1
Мягколиственные
Береза
85531,0 87732,5 94170,5 97950,0 98824,8 99683,7
Осина
17711,4 18907,9 20035,0 20573,4 20682,0 20802,0
* Данные Государственного учета лесного фонда (без лесов заповедников).
Глава 7
ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
еографическое положение и природные условия. Бореальная
зона располагается на севере умеренного пояса, ее облик определяют
хвойные леса. Темнохвойная тайга – тенистый вечнозеленый лес, представ-
ляющий собой яркий пример олигодоминантной – бедной по видовому соста-
ву – растительности. Понятие «тайга», широко используемое в ботанической
и географической литературе, привнесено в науку из языков народов, прожи-
вающих в Сибири. По заключению большого знатока таежной растительности
А. И. Толмачева, во внетропических широтах северного полушария тайга яв-
ляется самым распространенным типом растительности. Ни один тип расти-
тельности на Земле не занимает столь обширного цельного пространства, как
она. Тайга является наиболее ярким выразителем зонального распределения
растительности в современную геологическую эпоху. Говоря о главнейших
чертах таежной флоры, следует обратить особое внимание на значительное
участие вечнозеленых растений в ее составе – это не только хвойные деревья,
но и вечнозеленые кустарнички и мхи. Таким образом, можно сказать, что са-
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
129
мые большие пространства суши, занятые «вечнозелеными лесами», распола-
гаются не в тропических и экваториальных широтах, а во внетропическом по-
ясе Северного полушария – это темнохвойная тайга.
Таежный тип растительности сформировался в послеледниковое время,
в голоцене, когда похолодание сменилось потеплением, и климат умеренных
широт приблизился к современному. Темнохвойная тайга умеренных широт –
молодой биом, возраст которого не превышает 10 тыс. лет. О молодости тайги
говорит и ее простая биогеоценотическая структура и бедность видового со-
става. Вместе с тем генетические корни тайги весьма древние. Наиболее веро-
ятна гипотеза происхождения тайги на равнинах Палеарктической области,
высказанная А. И. Толмачевым (1954), который полагал, что темнохвойная
тайга сложилась в горах на юге Восточной Сибири еще в неогеновый период.
Горная тайга является, таким образом, значительно более древней, чем хвой-
ные леса равнин. В свою очередь, горная тайга имеет еще более древние связи
с растительностью высотных поясов Гималаев и гор Юго-Восточной Азии,
расположенных в тропических широтах.
Темнохвойные леса широко распространены на территории России и со-
ставлены разными видами темнохвойных древесных пород, к числу которых
относятся все произрастающие в России ели, пихты и две кедровые сосны
(Pinus koraiensis, P. sibirica). Хвойные леса и сопутствующие им виды кустар-
ничков, мелкотравья, мхов и лишайников хорошо адаптированы к суровым
климатическим условиям бореальной зоны. Важным лимитирующим факто-
ром распространения древесных пород является зимняя «засуха», когда мо-
розы ограничивают доступ влаги. Хвоя – сильно редуцированная листовая
пластинка – это ксероморфное приспособление, позволяющее ограничивать
транспирацию. В Восточной Сибири, где зимы особенно суровые, даже этого
приспособления недостаточно и темнохвойные породы замещаются листвен-
ницей с опадающей на зиму хвоей. Защитой хвойных пород от мороза являет-
ся также смолистая древесина. Травяно-кустарничковый и мохово-лишайни-
ковый ярусы защищены снеговым покровом.
Типичной темнохвойной тайге свойственна не только олигодоминант-
ность древостоя, но и бедность состава таежной флоры. Локальные флоры со-
судистых растений составляют 400–700 видов. С ограниченностью видового
состава гармонирует общая скудность растительного покрова, развивающего-
ся под пологом леса. Если древостой тайги характеризуется высокой продук-
тивностью, значительностью растительной массы, сосредоточенной в верхнем
ярусе, то растения, живущие под пологом темнохвойного леса, образуют в со-
вокупности лишь ничтожную часть биомассы. Отмечается зависимость расте-
ний от тайги как среды обитания. Существование многих видов почти неотде-
лимо от темнохвойного леса – они практически не встречаются в других типах
растительности. С другой стороны, растения не таежных фитоценозов прони-
кают под полог тайги так же весьма ограничено.
Несмотря на то, что под словом тайга подразумевается темнохвойный
лес, в таежную зону входят не только леса из типичных темнохвойных пород,
но и леса из сосны обыкновенной и лиственниц, являющихся светлохвойными
породами. Кроме хвойных деревьев в тайге произрастают мелколиственные
породы: береза, осина, ольха серая, рябина, ива, которые чаще выступают как
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
130
примесь среди основных пород или формируют производные леса на месте
коренных лесов, уничтоженных вырубками или пожарами, а также возника-
ющих при зарастании заброшенных сельскохозяйственных угодий. Доля этих
лесов год от года возрастает. В целом же производными лесами занято около
30 % территории России, они сосредоточены преимущественно в центральной
части европейской России. Здесь почти не осталось первичных лесов, они есть в
основном в Якутии и на Дальнем Востоке. Большая часть коренных лесов рас-
положена в труднодоступных безлюдных местах. Около 15 % «потенциально
лесной» площади России вообще остались без леса и заняты сельхозугодьями,
поселениями, транспортной и промышленной инфраструктурой.
Климат бореальной зоны суровый, зима более морозная и более снеж-
ная, чем в зоне тундр. Средняя температура самого холодного месяца от –7 °С
(на западе) до –40 °С в Восточной Сибири, где располагается полюс холода
Северного полушария: в Верхоянске и Оймяконе зафиксирован абсолютный
минимум –68,0 °С. Лето теплое, местами жаркое. Температура самого теплого
месяца от 16 °С (на западе) до 19 °С (в Восточной Сибири). Количество атмо-
сферных осадков уменьшается с запада на восток от 600–800 мм в Восточной
Европе и Западной Сибири до 400–200 мм в Восточносибирском секторе, и
вновь увеличивается до 700 мм на Дальнем Востоке. Максимумы осадков в
теплый период года, но зимой выпадает много снега. Снежный покров играет
защитную роль. Под мощным слоем снега почва сохраняет тепло и часто не
полностью промерзает. Благодаря теплоизоляционным свойствам снега рас-
тения, а также многие животные, хорошо переносят лютые морозы. В боре-
альной зоне атмосферных осадков выпадает больше, чем испаряется. Избыток
влаги стекает в полноводные реки, пополняет грунтовые воды, способствует
образованию болот.
Для тайги зональными процессами почвообразования являются подзо-
листый и болотный. В условиях обилия влаги и кислой среды происходит глу-
бокий распад минеральной части почвы. В подзолистом горизонте сохраняет-
ся наиболее устойчивое соединение – кремнезем SiO2. Соединения алюминия,
железа, марганца и гумус выносятся в нижележащие горизонты вмывания,
гумус нередко полностью вымывается из почвенного профиля. В производных
мелколиственных лесах с развитым травяным покровом формируются дерно-
во-подзолистые почвы. Подзолистый процесс почвообразования замещается
дерновым на карбонатных материнских породах. В условиях обильного за-
стойного увлажнения идет болотный процесс почвообразования, характери-
зующийся накоплением слоя торфа.
Типы местопроизрастаний и типы еловых лесов. В бореальной зо-
не Восточно-Европейской равнины, где характерные черты темнохвойной тай-
ги, образованной в основном елью (Picea abies, P. obovata), выражены наиболее
полно, В. Н. Сукачев выделяет шесть главных типов мест произрастания:
1) водоразделы (плакоры) с суглинистыми или супесчаными средне бо-
гатыми почвами, хорошо дренированные; режим увлажнения устойчивый;
2) такие же почвы, но хуже дренированные, заболачивающиеся; режим
увлажнения переходный к обильно-застойному;
3) понижения в рельефе, почвы еще хуже дренированные, более заболо-
ченные; режим увлажнения обильно-застойный.
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
131
Эти три типа условий обитания составляют один эколого-топологичес-
кий ряд, отражающий усиление процесса заболачивания. Далее:
4) днища узких долин рек и ручьев (лога) с обильным увлажнением про-
точными водами;
5) выпуклые формы рельефа, сложенные песчаными отложениями, поч-
вы бедные, режим увлажнения резко-переменный;
6) плакоры, имеющие не свойственные бореальной зоне богатые почвы,
чаще всего связанные с близким залеганием известковых пород или с карбо-
натной мореной; режим увлажнения устойчивый.
Классическую схему эдафо-фитоценотической ординации еловых ле-
сов, связанных с названными условиями местопроизрастания, В. Н. Сукачев
(рис. 7.1) составил для зрелых лесов без учета их возрастной динамики. Схема
имеет вид креста, где центральное место представляют условия произрастания
первого типа, занятые ельниками-зеленомошниками (Piceeta hylocomiosa),
включающие сообщества ельников кисличных (рис. 7.2), черничных (рис. 7.3)
и брусничных. Направления осей указывают на последовательные смены со-
обществ, вызванные изменениями условий обитания. Вверх (ряд А) условия
приближаются к характерным для местопроизрастаний пятого типа, где ельни-
ки брусничники сменяются сосновыми борами лишайниковыми (рис. 7.4); на-
лево (ряд В) – к условиям второго и третьего типа, где ельники-долгомошники
(Piceeta polytrichosa) сменяются ельниками сфагновыми (Piceeta sphagnosa)
(рис. 7.5), а далее верховыми сфагновыми болотами; вниз (ряд D) – к услови-
ям местопроизрастаний четвертого типа, занятым приручьевыми влажно-
травяными ельниками (Piceeta uliginosa-herbosa) (рис. 7.6); вправо (ряд C) – к
условиям шестого типа, где формируются ельники сложные или неморальные
(дубравные) (Piceeta composita или P. nemorosa) – этот тип ельников характе-
ризуется участием широколиственных древесных пород, усложнением верти-
кальной структуры и заменой мохового покрова дубравным широкотравьем
(рис. 7.7).
Рис. 7.1. «Крест Сукачева» – эдафо-фитоценотическая схема еловых лесов
(по В. Н. Сукачеву)
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
132
Рис. 7.2. Ельник кисличник (фото Г. Исаченко)
Рис. 7.3. Ельник черничник (фото Д. Мирина)
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
133
Рис. 7.4. Сосняк
редкостойный
лишайниковый
(фото Г. Исаченко)
Рис. 7.5. Ельник
чернично-сфагновый
(фото Д. Мирина)
Рис. 7.6. Ельник
таволговый
приручьевой
(фото Д. Мирина)
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
134
Рис. 7.7. Ельник
травяно-дубравный
(фото М. Бекмансурова)
Схему Сукачева можно использовать для типизации как еловых, так и
сосновых лесов, экологический ареал которых во многом перекрывается с аре-
алом еловых лесов. Сосняки часто выступают в качестве производных лесов на
месте ельников, уничтоженных пожарами и вырубками (рис. 7.8).
В 1934 г. В. Н. Сукачевым была дана обобщенная схема типов леса, в ко-
торой отражена эдафическая приуроченность основных древесных пород. Она
построена в виде той же системы координат, но с нанесением на нее эдафо-
фитоценотических ареалов (рис. 7.9). Наименьшую экологическую амплитуду
имеют пихтовые леса (Abiegna), а наибольшую – сосновые (Pineta). Еловые
леса (Piceeta) идут дальше всех вниз по оси ординат, а дубравы (Querceta) за-
нимают крайние справа местообитания по горизонтальной оси.
Рис. 7.8. Основные ти-
пы условий произ-
растания еловых и
сосновых лесов и по-
ложение
сообществ
на профиле:
а – Еловые леса:
1 – ельник-брусничник;
2 – ельник-кисличник;
3 – ельник-черничник;
4 – ельник травяной;
5 – ельник-долгомошник;
6 – сфагновое болото с
низкорослой сосной;
б – Сосновые леса (боры):
1 – бор-беломошник;
2 – бор-брусничник;
3 – бор-черничник;
4 – бор-долгомошник;
5 – сфагновое болото с
низкорослой сосной
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
135
Рис. 7.9. Обобщенная си-
стема типов леса и эдафо-
фитоценотические ареалы
древесных пород
(по В.Н. Сукачеву):
Формации: Abiegna – пихтар-
ник, Cembreta – кедровники,
Lariceta dauricae – лиственнич-
ники из лиственницы даурской,
Lariceta sibiricae – лиственнич-
ники из лиственницы сибир-
ской, Piceeta – ельники, Pineta –
сосняки, Querceta – дубняк. Ти-
пы леса: Ox – кисличники (Ox-
alidosa), M – черничники (Myr-
tillosa), Pl – долгомошники (Pol-
ytrichosa), Sph – сфагновые
(Sphagnosa), Sph p – сфагновые
болота с сосной (Sphagneto-
pinosa), V – брусничные (Vaccin-
iosa), Cl – лишайниковые (Cladinosa), Ft – приручьевые (Fontinales), Til – липняковые (Til-
iosa), Q – дубравные (Quercetosa), Cr – известняковые и меловые (Calcarea et cretacea)
7.1. ПОДЗОНЫ ТАЙГИ
Зона бореальных хвойных лесов с севера на юг подразделяется на подзо-
ны: северной, средней и южной тайги, а также подтайгу. Географические ва-
рианты растительности подзон даны по: «Зоны и типы поясности...» (1999а,
1999б).
Подзона северной тайги занимает значительную часть таежной зо-
ны от лесотундры на севере до 64° с.ш. на Русской равнине, до 62° с.ш. в За-
падной Сибири и до 60° с.ш. на Среднесибирском плоскогорье. Лето холодное:
средняя температура июля от 14 до 16°, безморозный период длится около 75-
95 дней. Зональные почвы глеево-подзолистые.
Подзона образована хвойными кустарничково-лишайниково-зелено-
мошными редкостойными лесами и редколесьями. Огромные площади заняты
болотами, преимущественно сфагновыми верховыми. В Карелии раститель-
ность подзоны представлена сосновыми (Pinus sylvestris, P. friesiana), еловыми
(Picea obovata) кустарничково-лишайниково-зеленомошными (Calluna vulgaris,
Empetrum hermaphroditum, Vaccinium myrtillus, V. vitis-idaea, V. uliginosum,
Ledum palustre) лесами; грядово-мочажинными травяно-сфагново-гипновыми
аапа-болотами. В Восточной Европе – еловыми (Picea obovata, P. abies x
P. obovata), сосновыми (Pinus sylvestris) кустарничково-зеленомошными,
кустарничково-лишайниково-зеленомошными (Empetrum hermaphroditum,
Vaccinium myrtillus, V. uliginosum, Ledum palustre) лесами; грядово-мочажин-
ными сфагновыми верховыми и травяно-сфагново-гипновыми аапа болотами.
В Западной Сибири – лиственничными, лиственнично-еловыми, лиственнич-
но-сосновыми, лиственнично-елово-кедровыми (Picea obovata, Larix sibirica,
Pinus sibirica, P. sylvestris); кустарничково-лишайниково-зеленомошными
(Ledum palustre, Vaccinium vitis-idaea, V. uliginosum; V. myrtillus, Empetrum
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
136
subholarcticum, Betula nana) лесами; бугристыми кустарничково-мохово-
лишайниковыми болотами. В Восточной Сибири – лиственничными (Larix
gmelinii, L. cajanderi) зеленомошными, кустарничково-зеленомошно-лишай-
никовыми (Vaccinium uliginosum, Empetrum subholarcticum, Betula exilis) ле-
сами; моховыми марями.
Подзона средней тайги характеризуется более теплым летом, со
средней температурой июля 16–18 °С. Безморозный период увеличивается до
100–105 дней. Преобладают зеленомошные типы хвойных лесов, отличаю-
щиеся большей сомкнутостью и более высоким бонитетом по сравнению с се-
верной тайгой. Главная роль в почвенном покрове принадлежит типичным
подзолистым почвам.
Подзона образована хвойными мелкотравно-кустарничково-зеленомош-
ными (Maianthemum bifolium, Trientalis europaea, Orthilia secunda, Pyrola
spp., Luzula pilosa) лесами; болотами. В Карелии растительность представлена
сосновыми (Pinus sylvestris), еловыми (Picea abies, P. abies x P. obovata, Betula
pendula, B. pubescens) мелкотравно-кустарничково-зеленомошными (Vaccinium
myrtillus, V. vitis-idaea, Calluna vulgaris, Linnaea borealis), кустарничково-ли-
шайниковыми, кустарничково-зеленомошными лесами; грядово-мочажин-
ные сфагновые (верховые) болота. В Восточной Европе – еловыми (Picea abies,
P. abies x P. obovata, P. obovata, Betula pendula, B. pubescens), сосновыми (Pinus
sylvestris) мелкотравно-кустарничково-зеленомошными (Vaccinium myrtillus,
V. vitis-idaea, Linnaea borealis) лесами; грядово-мочажинными сфагновыми
верховыми и травяно-сфагново-гипновыми аапа-болотами. Приуральский вари-
ант – еловыми, пихтово-еловыми, сосновыми местами с кедром (Picea obovata,
Abies sibirica, Pinus sylvestris, P. sibirica, Betula pendula, B. pubescens) мелко-
травно-кустарничково-зеленомошными (Linnaea borealis, Vaccinium myrtillus,
V. vitis-idaea) лесами; грядово-мочажинными сфагновыми верховыми и тра-
вяно-сфагново-гипновыми аапа-болотами. В Западной Сибири – елово-кед-
ровыми с пихтой, лиственницей, сосновыми (Pinus sibirica, Picea obovata,
Abies sibirica, Larix sibirica, Pinus sylvestris, Betula pendula, B. pubescens) мел-
котравно-кустарничково-зеленомошными (Vaccinium vitis-idaea, V. myrtillus,
Linnaea borealis, Ledum palustre) лесами; грядово-мочажинными сфагновыми
верховыми болотами. В Восточной Сибири – лиственничными, елово-лист-
венничными, сосновыми (Larix gmelinii, Picea obovata, Pinus sylvestris, Betula
cajanderi) мелкотравно-кустарничково-зеленомошными (Vaccinium vitis-idaea,
V. uliginosum, Arctostaphylos uva-ursi, Ledum palustre), местами лимнасовыми
(Limnas stelleri) лесами с аласными лугами (Carex juncella, Calamagrostis
langsdorffii). На Дальнем Востоке – лиственничными (Larix gmelinii), пихтово-
еловыми с лиственницей (Picea ajanensis, Abies nephrolepis, Betula platyphylla)
кустарничково-моховыми (Ledum palustre, Vaccinium vitis-idaea, V. uliginosum)
лесами; лиственничными сфагновыми марями и ерниками (Betula exilis,
B. divaricata, B. fruticosa). На Сахалине – лиственничными (Larix kamtschatica)
лишайниковыми и зеленомошными лесами, часто с кедровым стлаником
(Pinus pumila), елово-лиственничными (Larix gmelinii, Picea ajanensis) с кед-
ровым стлаником багульниково-зеленомошными (Ledum palustre, Ledum
hypoleucum) лесами; темнохвойными елово-пихтовыми зеленомошными ле-
сами из ели аянской, пихты сахалинской (Abies sachalinensis), в горах – ка-
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
137
менноберезняками (Betula ermanii); мерзлыми болотами, аапа-болотами и
субокеаническими болотами (Empetrum nigrum, Myrica tomentosa).
Подзона южной тайги представлена в виде сплошной полосы толь-
ко на Русской равнине и в Западной Сибири, восточнее она занимает незначи-
тельные площади. Лето в южной тайге становится умеренно теплым, средняя
температура июля около 18–19 °С, безморозный период равен 105–120 дням.
Почвы дерново-подзолистые. Степень заболоченности ниже, а бонитет хвой-
ных лесов выше, чем в средней тайге.
Подзона образована хвойными кустарничково-травяными и травяно-
зеленомошными лесами; болотами. В кустарниковом и травяном ярусах появ-
ляются виды, свойственные широколиственному лесу, а в сосновых борах за-
метны признаки остепнения. В Восточной Европе растительность представле-
на еловыми (Picea abies, Betula pendula, B. pubescens, Populus tremula, Alnus
glutinosa), сосновыми (Pinus sylvestris) неморальнотравяными (Oxalis acetosella,
Galeobdolon luteum, Hepatica nobilis, Stellaria holostea, Pulmonaria obscura,
Convallaria majalis), кисличными лесами; грядово-мочажинныеми сфагно-
выми верховыми болотами. В Западной Сибири – кедрово-елово-пихтовыми,
елово-пихтовыми, сосновыми, с липой до р. Иртыш (Abies sibirica, Picea
obovata, Pinus sibirica, Pinus sylvestris, Betula pendula, B. pubescens, Populus
tremula, Tilia cordata) мелкотравно(Oxalis acetosella, Carex macroura, Stellaria
bungeana, Circaea alpina)-зеленомошными лесами, грядово-мочажинными
сфагновыми верховыми болотами. В Средней Сибири – лиственничными,
сосново-лиственничными, елово-пихтовыми (Larix sibirica, Pinus sylvestris,
Abies sibirica, Picea obovata, Betula pendula, B. pubescens, Populus tremula) ку-
старничково-травяно-зеленомошными (Carex macroura, Vaccinium uliginosum) с
Rhododendron dauricum лесами. На Дальнем Востоке – лиственничными (Larix
gmelinii, Betula platyphylla), сосновыми (Pinus sylvestris) кустарниковыми
(Rhododendron dauricum, Duschekia manshurica), кустарничково-травяными
(Calamagrostis langsdorfii, Vaccinium vitis-idaea) лесами; ерниками (Betula
fruticosa, B. ovalifolia), лиственничными марями и травяными болотами.
Подзона подтайги располагается между зонами тайги и широколист-
венных лесов. В Восточноевропейской области подтайга тянется хорошо вы-
раженной неширокой полосой между 53° и 60° с.ш., захватывая территории
Калининградской, Псковской, Брянской, Московской, Нижегородской обла-
стей, и далее от Казани до Перми. На юге Западно-Сибирской равнины между
56–57° и 54–55° с. ш. подтаежные леса встречаются в регионах с увлажнени-
ем от избыточного до умеренного. Климатические характеристики близки к
южнотаежным, но лето в подтайге теплее и продолжительнее, а зима мягче.
Преобладающие лесные почвы – подзолистые, дерново-подзолистые, серые
лесные.
В Европейской части России и на Дальнем Востоке подтайга состоит из
хвойно-широколиственных (смешанных) лесов, которые в Европейской части
сочетаются с черноольховыми топями, на Дальнем Востоке − с ерниково-
лиственничными марями. В условиях континентального климата Западной
Сибири хвойно-широколиственные и широколиственные леса замещаются
мелколиственными злаково-разнотравными лесами в сочетании с сосново-
кустарничково-сфагновыми верховыми и травяными низинными болотами.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
138
На Дальнем Востоке муссонные хвойно-широколиственные леса образуют
нижний пояс в горах (рис. 7.10).
В Калининградской области растительность представлена елово-широко-
лиственными, сосново-широколиственными (Quercus robur, Fraxinus excelsior,
Ulmus laevis, U. glabra, Picea abies, Pinus sylvestris; Carpinus betulus, Fagus
sylvatica) лесами; черноольшаниками (Alnus glutinosa), субатлантическими
травяными и грядово-мочажинными сфагновыми верховыми болотами с
Myrica gale. В Восточной Европе – елово-широколиственными, сосново-ши-
роколиственными, сосновыми (Quercus robur, Tilia cordata, Acer platanoides,
Ulmus laevis, U. glabra, Picea abies, Pinus sylvestris) лесами; черноольховыми
топями (Alnus glutinosa); грядово-мочажинными сфагновыми верховыми и
травяными болотами. Приуральская подтайга отличается от восточноевро-
пейской отсутствием Fraxinus excelsior и наличием Abies sibirica, а также усиле-
нием роли Tilia cordata; она представлена елово-широколиственными, липово-
Рис. 7.10. Районы распространения хвойно-широколиственных и широколист-
венных лесов (Растительный покров СССР..., 1956)
Условные обозначения: 1 – широколиственно-хвойные и хвойно-широколиственные леса;
2 – широколиственные леса; 3 – рассеянное распространение компонентов хвойно-
широколиственных формаций в таежной зоне (внемасштабный знак); I – граница распро-
странения бука в Восточной и Северной Европе; II – граница распространения граба
(Carpinus betulus); III – граница распространения дальневосточных видов граба (Carpinus
cordata и других, растущих за пределами России); IV – граница распространения сосны ко-
рейской (Pinus korainensis)
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
139
елово-пихтовыми, широколиственно-пихтово-еловыми, широколиственно-сос-
новыми (Quercus robur, Tilia cordata, Acer platanoides, Ulmus glabra, Picea
obovata, P. abies x P. obovata, Abies sibirica, Pinus sylvestris) лесами. В Западной
Сибири − березовыми, осиновыми (Betula pendula, Populus tremula) разнотрав-
но-злаковыми (Calamagrostis arundinacea, Brachypodium pinnatum, Aegopodium
podagraria), сосновыми (Pinus sylvestris) кустарничково-травяными лесами;
травяными болотами (займищами). В Приморье − хвойно-широколиствен-
ными лесами с преобладанием Pinus koraiensis, Quercus mongolica, Acer mono,
Tilia amurensis, Fraxinus rhynchophylla.
7.2. БОТАНИКО-ГЕОГРАФИЧЕСКИЕ ОБЛАСТИ ТАЕЖНЫХ ЛЕСОВ
Огромная протяженность бореальной зоны хвойных лесов с запада на
восток определяет изменение растительности. Для Восточно-Скандинавской
(Карельской) области характерны темнохвойные и светлохвойные (сосно-
вые) леса; для Восточно-Европейской области – темнохвойные таежные
леса и хвойно-широколиственные леса на юге; для Западно-Сибирской об-
ласти – темнохвойные и светлохвойные лиственничные леса; в Средне-
Сибирской области господствуют светлохвойные лиственничные леса; для
Сахалинской таежной области – темнохвойные и светлохвойные листвен-
ничные леса, а также смешанные хвойно-широколиственные леса на юге;
для Приморской области – хвойно-широколиственные леса. Приведем обоб-
щенную характеристику таежных областей с учетом их подзональных осо-
бенностей.
7.2.1. ВОСТОЧНО-СКАНДИНАВСКАЯ (КАРЕЛЬСКАЯ) ТАЕЖНАЯ ОБЛАСТЬ
Восточно-Скандинавская область таежных лесов расположена на Бал-
тийском кристаллическом щите. Занимает южную часть Кольского п-ова, се-
веро-западную часть Карелии, большую часть Карельского перешейка (Геобо-
таническое районирование..., 1989). Четвертичное оледенение здесь вырази-
лось в интенсивной деятельности ледников и сносе ледниковых отложений
флювиогляциальными потоками, следствием чего является широкое распро-
странение выхода на дневную поверхность кристаллических горных пород.
В ложбинах расположено множество озер и болот.
Климат Карелии умеренно-континентальный с чертами морского, так
как большое влияние на него оказывает положение региона на северо-западе
России, куда западный перенос с Атлантического океана несет теплые и влаж-
ные воздушные массы. Годовое количество осадков колеблется от 450–550 мм
на севере до 600–700 мм на юге. Средняя температура января –12...–13 °С
градусов на севере и –9...–10 °С градусов на юге. Средняя температура июля
в Прибеломорье 14–14,5 °С, в Северном озерном крае 14,5–15 °С, в Южном
озерном крае 15,5–16,5 °С.
Для растительности характерны сочетания сосновых, елово-сосновых и
еловых кустарничково-лишайниково-зеленомошных лесов с редкостойными
сфагновыми сосновыми лесами и с сосново-кустарничково-сфагновыми ме-
зоолиготрофными и аапа болотами. Широкое распространение имеют встре-
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
140
чающиеся только в этой области карельские кольцевые аапа болота ерниково-
молиниево-осоково-сфагновые на грядах и разнотравно-осоково-гипновые в
мочажинах с вторичными озерками.
Основные лесообразующие древесные породы: сосна обыкновенная, ели
европейская (преимущественно в подзоне средней тайги) и сибирская (в ос-
новном в северной тайге), березы пушистая и повислая (Betula pubescens,
B. pendula), осина (Populus tremula), ольха серая (Alnus incana). Ели европей-
ская и сибирская в природе легко скрещиваются и образуют переходные фор-
мы. В составе коренных лесов преобладает сосна, заметно уступает ей ель. Бе-
резняки, осинники и сероольшаники сформировались в основном под влия-
нием хозяйственной деятельности, главным образом в результате сплошных
рубок и пожаров. Преобладание сосновых лесов на севере определяется более
суровыми климатическими условиями и широким распространением здесь
бедных песчаных почв. Сосновые леса встречаются практически во всех ме-
стообитаниях – от сухих на песках и скалах до заболоченных. Наиболее рас-
пространены сосновые леса на свежих и умеренно сухих почвах – сосняки
брусничники и черничники.
В лесных сообществах отсутствуют пихта Abies sibirica и лиственница
Larix sibirica, постоянно встречающиеся далее к востоку. К видам, не распро-
страняющимся далее на восток, относятся: прострел весенний (Pulsatilla ver-
nalis), смолевка скальная (Silene rupestris), арника феноскандинавская (Arnica
fennoscandica), манжетка альпийская (Alchemilla alpina), осока Бигелоу (Carex
bigelowii), криптограмма курчавая (Cryptogramma crispa), многорядник копь-
евидный (Polystichum lonchitis) и др.
Карельская область расположена в пределах северной, средней и южной
тайги. Граница между подзонами северной и средней тайги проходит с запада
на восток несколько севернее г. Медвежьегорска. Южная тайга занимает юж-
ную окраину Балтийского щита и протягивается узкой полосой вдоль северо-
восточного побережья Финского залива.
Подзона северной тайги. На западе северная тайга занимает юг
Кольского полуострова и большую часть Карелии, в пределах Балтийского
кристаллического щита, почти доходя до Онежского озера. В северной части
подзоны господствуют сосновые редкостойные лишайниковые и зеленомош-
но-лишайниковые кустарничковые леса, чередующиеся с елово-сосновыми
редкостойными зеленомошно-лишайниковыми, сосновыми редкостойными
сфагновыми, еловыми редкостойными лишайниково-моховыми лесами, а в
понижениях рельефа – с сосново-кустарничково-сфагновыми мезооли-
готрофными и аапа болотами. В травяно-кустарничковом ярусе доминируют
Empetrum hermaphroditum, Vaccinium vitis-idaea, V. myrtillus, V. uliginosum,
Ledum palustre; в мохово-лишайниковом: Cladonia stellaris, C. rangiferina,
C. arbuscula, Cetraria islandica, Pleurozium schreberi, Hylocomium splendens.
На склонах возвышенностей и гор произрастают еловые и елово-березовые из
Picea obovata, P. abies, Betula pubescens, B. czerepanovii кустарничковые зеле-
номошные леса. Для еловых лесов даже на плакорах отмечается участие рас-
тений, свойственных болотам (водяника, багульник, голубика, морошка и др.).
На северо-западе Карелии, на уровне северного полярного круга встре-
чаются низкобонитетные еловые леса из Picea obovata, обычно представлен-
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
141
ные ассоциациями воронично-черничными лишайниково-зеленомошными.
В целом для Карелии характерно широкое распространение чернично-воро-
ничных и воронично-черничных типов ельников.
Значительную часть Карелии (к западу от Белого моря) занимает заозе-
ренное плато с небольшими абсолютными высотами 200–220 м. Здесь наибо-
лее часто встречаются сосняки лишайниковые и зеленомошно-лишайнико-
вые. Значительную долю лесов составляют сфагновые сосняки, образующие
сочетания с болотами. Преобладают аапа-болота, изредка встречаются сфаг-
новые верховые грядово-мочажинные болота.
На гранитных сельгах распространены «каменистые» типы сосняков (сос-
няки лишайниковые), в которых в условиях резко переменного режима увлаж-
нения бореальные виды соседствуют с арктоальпийскими. На выровненных
участках древних озерных террас, сложенных ленточными глинами, встречают-
ся сосновые, елово-сосновые и еловые леса, часто замещенные лугами антропо-
генного происхождения. В замкнутых бессточных котловинах формируются
сосново-кустарничково-сфагновые мезоолиготрофные болота. Широкое рас-
пространение имеют кольцевые аапа болота ерниково-молиниево-осоково-
сфагновые на грядах и разнотравно-осоково-гипновые в мочажинах с вторич-
ными озерками.
В горных районах Кольского полуострова наблюдается вертикальная по-
ясность. До высоты 300–400 метров располагается лесная растительность, вы-
соты 400–600 метров заняты березовым криволесьем и кустарниками, а выше
600–650 метров расположены горные тундры, которые занимают значи-
тельные участки на Хибинских и Ловозерских горах, в Мончетундре и неко-
торых других горных массивах, расположенных западнее. Горные тундры
характеризуются большим развитием лишайников, особенно Cladonia stellaris,
Flavocetraria nivalis, и кустарничков: дриада восьмилепестковая Dryas
octopetala, гариманелла моховидная Harrimanella hypnoides, диапенсия ла-
пландская Diapensia lapponica, ива сетчатая Salix reticulata.
Подзона средней тайги занимает центральную часть Карельского
перешейка, северо-западное Приладожье, западную часть средней Карелии,
территорию к северу от Онежского и Ладожского озер, северо-восточное побе-
режье Онежского озера. Северо-западное Приладожье, так же, как и север
Онежского озера, сильно расчленено, кристаллические коренные породы ча-
сто выходят на дневную поверхность, образуя возвышенности, именуемые
сельгами (рис. 7.11); абсолютные высоты которых равномерно повышаются с
юга на север (от 20–25 м до 150–200 м). Сосновые леса занимают большие
площади в Карелии и сильно зависят от рельефа, влажности, плодородия
почв. Самыми распространенными являются сосняки зеленомошники. Они
встречаются на хорошо дренированных участках водоразделов и в большин-
стве своем длительнопроизводные, возникшие на месте еловых лесов. Древес-
ный ярус образован сосной, иногда с примесью ели (Picea abies, P. obovata),
березы повислой, осины. Подлесок выражен слабо, хорошо развит кустарнич-
ковый ярус, мощный моховой покров.
Сосняки чернично-долгомошные сосредоточены у подножий пологих
склонов возвышенностей. В древостое обычна примесь березы пушистой и
ели . Разреженный подлесок состоит из можжевельника (Juniperus communis),
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
142
Рис. 7.11. Сельговый рельеф Карельского перешейка (фото О. Ефимовой)
рябины (Sorbus aucuparia), шиповника (Rosa acicularis), крушины (Frangula
alnus), ивы пепельной (Salix cinerea). В травяно-кустарничковом ярусе доми-
нирует черника (Vaccinium myrtillus), принимают участие брусника (Vaccinium
vitis-idaea), голубика (V. uliginosum), багульник (Ledum palustre), луговик из-
вилистый (Avenella flexuosa), вереск (Calluna vulgaris), седмичник (Trientalis
europaea) и др.
На вершинах сельг развиты сосняки лишайниковые каменистые (иногда
там растут исключительно лишайники) с луговиком извилистым, овсяницей
овечьей (Festuca ovina), тимьяном ползучим (Thymus serpyllum), купеной ду-
шистой (Polygonatum odoratum), ниже на склонах – сосняки брусничные и
бруснично-орляковые, за ними у подножий склонов – еловые черничные зе-
леномошные леса. Нередки и ельники кисличники, а местами можно встре-
тить еловые сложные леса с примесью широколиственных пород, главным
образом клена. И те, и другие развиваются на небольших неплакорных участ-
ках, наиболее обогащенных питательными веществами. В травяно-кустарнич-
ковом ярусе имеется ряд неморальных видов: сныть (Aegopodium podagraria),
печеночница благородная (Hepatica nobilis) и др. Много производных сооб-
ществ, в том числе сероольшатников. К депрессиям приурочены сосново-
кустарничково-сфагновые (Sphagnum angustifolium, S. fuscum, S. capillifolium,
Chamaedaphne calyculata, Calluna vulgaris, Vaccinium uliginosum и др.) и осоко-
во-сфагновые (Carex lasiocarpa, C. rostrata, Scheuchzeria palustris, Sphagnum
fallax, S. obtusum, S. papillosum) болота.
Среднетаежные еловые леса североевропейского типа характеризуются
простым строением: ярус кустарников отсутствует или развит слабо. Ельники
зеленомошные в Карелии больших площадей не занимают и встречаются на
почвах, близко подстилаемых коренными породами. Травяно-кустарничко-
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
143
вый ярус представлен единичными кустами черники, брусники, а также дер-
нинками луговика извилистого. Мохово-лишайниковый ярус сплошной, состо-
ит из Hylocomium splendens, Pleurozium schreberi, Rhytidiadelphus triquetrus,
Ptilium crista-castrensis, Polytrichum commune. На супесчаных и песчаных
торфяно-подзолисто-глеевых почвах заболоченные долгомошные леса пред-
ставлены бруснично-долгомошными ельниками с постоянной примесью сос-
ны. На участках с пересеченным рельефом различные типы еловых лесов и
болот образуют закономерные сочетания. Повышенные местоположения и
склоны заняты сообществами из группы зеленомошных ельников, а плоские
низины – сообществами из группы долгомошно-сфагновых и сфагновых.
Среднетаежные леса сильно видоизменены в результате хозяйственной
деятельности человека. Появление в древостое березы, осины, ольхи серой
сопровождается увеличением участия в травяно-кустарничковом ярусе марь-
янника лугового (Melampyrum pratense), костяники (Rubus saxatilis), вейника
тростникового (Calamagrostis arundinacea). Моховой покров деградирует, по-
селяются такие субнеморальные элементы, как сныть, вероника дубравная
(Veronica chamaedrys), что связано с осветлением лесов, обогащением почв за
счет поселения мелколиственных пород, а также олуговения травяно-кустар-
ничкового покрова.
7.2.2. ВОСТОЧНО-ЕВРОПЕЙСКАЯ ТАЕЖНАЯ ОБЛАСТЬ
Восточно-Европейская область расположена на Восточно-Европейской
(Русской) равнине и характеризуется относительно равнинным рельефом,
средняя высота составляет около 170 м над ур. м. Следы плейстоценовых оле-
денений выражены в виде ледниковых отложений и специфических форм ре-
льефа. Высота некоторых возвышенностей достигает 200–400 м. Климат
формируется под влиянием западного переноса, который сопровождается
прохождением циклонов, несущих атмосферные осадки. Среднее годовое ко-
личество атмосферных осадков – 600 мм. ГТК вегетационного периода от 2 на
западе, до 1,5 на востоке. Характерна умеренно холодная зима и теплое лето
со средней температурой июля от 12 °С на севере до 20 °С на юге.
Лесообразующими древесными породами коренной темнохвойной тайги
являются ель обыкновенная, ель сибирская и пихта сибирская (Abies sibirica).
На почвах легкого механического состава и на заболоченных почвах большую
роль начинают играть сосновые леса (Pinus sylvestris). Лиственница сибирская
(Larix sibirica) в качестве примеси в сосновых лесах появляется восточнее
Онеги. В результате массовых вырубок и крупных пожаров большие площади
заняты производными лесами, в которых преобладают березы (Betula pu-
bescens, B. pendula), осина, ольха серая. Производные леса также часто пред-
ставлены сосняками. Сосна, как порода с широкой экологической амплиту-
дой, выступает пионером зарастания на месте сведенных ельников. Рубки,
пожары, распашка земель с последующим восстановлением лесов – основные
факторы антропогенной динамики таежных лесов Восточной Европы.
Восточно-Европейская область характеризуется хорошо выраженным
делением на подзоны вследствие преобладания здесь плакорных местообита-
ний (хорошо дренируемых водоразделов).
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
144
Подзона северной тайги простирается на юг от лесотундры до 64°
с.ш. и представлена осветленными еловыми лесами. Характерные особенно-
сти плакорных лесов этой подзоны – ухудшение роста ели до IV–V класса бо-
нитета и разреженность древесного полога; примесь к ели светолюбивых по-
род: сосны, березы, лиственницы – как результат естественного изреживания
и осветления леса; на востоке в древесном ярусе примесь пихты и сосны кед-
ровой (Pinus sibirica). В кустарниковом ярусе встречается Betula nana. В тра-
вяно-кустарничковом распространены брусника, черника, а также Empetrum
nigrum, Vaccinium uliginosum, Ledum palustre, Carex globularis; в моховом по-
крове, кроме лесных таежных мхов, – политрихум и некоторые виды лишай-
ников. Таким образом, в северной тайге средообразующая роль темнохвойных
пород и зеленых мхов относительно низкая.
На верхних и средних частях песчаных грив, на повышенных участках
террас, на подзолистых песчаных и супесчаных хорошо дренированных почвах
встречается ельник лишайниковый. Обычно ель (Picea obovata) растет с приме-
сью березы пушистой или березы Черепанова (Betula pubescens, B. czerepanovii).
Подлесок редкий из Betula nana, Juniperus communis, Rosa acicularis, Sorbus
aucuparia. Травяно-кустарничковый ярус разрежен, чаще других встречаются
овсяница овечья (Festuca ovina), осока шаровидная (Carex globularis), кроме
них растут плаун булавовидный (Lycopodium clavatum), двурядник сплюсну-
тый (Diphasiastrum complanatum), ожика волосистая (Luzula pilosa), золо-
тарник обыкновенный (Solidago virgaurea), морошка (Rubus chamaemorus),
луговик извилистый (Avenella flexuosa). В почти сплошном лишайниковом
покрове преобладают: Cladonia arbuscula, C. mitis, C. rangiferina. По мик-
ропонижениям встречаются зеленые мхи и более влаголюбивый кукушкин
лен.
На восток от Онежского полуострова Белого моря, через верхнее течение
и дельту Северной Двины до Урала преобладает моренный плосковолнистый
равнинный рельеф. Растительный покров представлен еловыми зеленомош-
ными и сфагновыми лесами. Преобладают черничники и долгомошники.
К ели (Picea obovata, P. abies) примешиваются сосна и береза, в травяно-кус-
тарничковом ярусе – водяника, черника, брусника, голубика. Заболоченные
еловые и елово-березовые леса сочетаются со сфагновыми верховыми и пере-
ходными болотами. На междуречье Северной Двины и Мезени встречаются
массивы болотно-кустарничково-долгомошных березово-еловых лесов (голу-
бично-долгомошных, багульниково-долгомошных). Древостой разреженный.
В кустарниковом ярусе Betula nana, Salix lapponum, S. phylicifolia; в травяно-
кустарничковом ярусе, кроме упомянутых доминантов, отмечены водяника,
луговик извилистый, осока шаровидная. На песках развиты сосняки зелено-
мошные, лишайниковые, сфагновые.
К северо-востоку от Архангельска расположено Беломорско-Кулойское
плато, сложенное гипсами, известняками и доломитами верхнего палеозоя.
Наряду с характерными северотаежными сообществами здесь распространены
экстразональные среднетаежные леса – ельники черничники с кислицей, также
встречаются лиственничные леса с черникой, брусникой и с участием южнота-
ежных (вороний глаз четырехлистный Paris quadrifolia, воронец красноплод-
ный Actaea erythrocarpa и др.), неморальных (сныть Aegopodium podagraria,
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
145
медуница неясная Pulmonaria obscura и др.) и сибирских (княжик сибирский
Atragene sibirica, недоспелка копьевидная Cacalia hastata и др.) видов, что
связано с карбонатностью почв.
Уникальные природные ландшафты Среднего Пинежья охраняются на
территории Государственного природного заповедника «Пинежский». В запо-
веднике выявлено около 480 видов сосудистых растений и около 200 видов
грибов и лишайников. Во флоре заповедника много реликтовых видов: арк-
тоальпийских (дриада восьмилепестковая Dryas octopetala, бартсия альпий-
ская Bartsia alpina, арктоус альпийский Arctous alpina, ива сетчатая Salix
reticulata и др.) и южнотаежных (пион Марьин корень Paeonia anomala, един-
ственная лиана Севера княжик сибирский, латук сибирский Lactuca sibirica,
мятлик расставленный Poa remota, хохлатка дымянкообразная Corydalis
capnoides и растущая на озерах кубышка малая Nuphar pumila).
На территории заповедника встречаются эндемики Северо-Востока евро-
пейской части России: гирчовник татарский (Conioselinum tataricum), тимьян
Талиева (Thymus talijevii), который встречается еще на Урале, мятлик Танфиль-
ева (Poa tanfiljewii) и качим уральский (Gypsophila uralensis). В заповеднике
довольно обычны северные орхидеи – дремлик темно-красный (Epipactis
atrorubens), тайник сердцелистный (Listera cordata), калипсо луковичная
(Calypso bulbosa) и венерин башмачок настоящий (Cypripedium calceolus). Они
растут в местах, где близко к поверхности залегают гипсовые породы.
Основной тип растительности заповедника – леса. Преобладают ельники
из ели сибирской, меньше – сосняков, березняков и лиственничников. На
участках с песчаными грунтами преобладают брусничные сосняки и сосново-
лиственничные леса. Березняки преимущественно производные.
На Тиманском кряже господствуют еловые и лиственнично-еловые зеле-
номошные леса. Лиственницы местами образуют чистые насаждения кустар-
ничково- и травяно-зеленомошные. На выходах известняков встречаются
фрагменты кустарничково-травяных лиственничников с реликтовыми вида-
ми: спирея средняя (Spiraea media), кизильник одноцветковый (Cotoneaster
uniflorus), змееголовник Рюйша (Dracocephalum ruyschiana), ветреница лес-
ная (Anemone sylvestris). Много заболоченных еловых лесов. Ближе к Уралу
преобладают заболоченные еловые леса и еловые редколесья с участием
Betula czerepanovii. В качестве примеси присутствует сосна кедровая. Изредка
встречаются заболоченные кедрово-еловые леса. Здесь же проходит северная
граница пихты, которая растет в виде стланика.
На равнине, где воздушные массы, приносимые с запада, задерживают-
ся Уральским хребтом, осадков выпадает больше, что приводит к заболачи-
ванию. Здесь наиболее характерны в разной степени заболоченные разре-
женные леса: кедрово-еловые с березой травяно-кустарничковые долгомош-
ные и сфагновые, березово-еловые (Picea obovata, Betula pubescens) часто с
кустарниковым ярусом из Betula nana, Salix phylicifolia, S. lapponum сфагно-
вые и долгомошные. На склонах плосковершинных увалов господствуют
пихтово-еловые с примесью сосны кедровой и кедрово-еловые леса кустар-
ничково- и травяно-зеленомошные, реже кисличные и папоротниковые. Рас-
пространены также заболоченные сосновые леса. Болота – сфагновые верхо-
вые и аапа.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
146
Подзона средней тайги представлена еловыми лесами, местами в
сочетании с березово-осиновыми лесами, сосновыми борами, сфагновыми бо-
лотами и лугами. Еловые леса широко распространены на Русской равнине.
Южная граница средней тайги проходит приблизительно по 60° с.ш. На во-
стоке среднетаежные еловые леса образуют более широкую, чем на западе,
зональную полосу. Они развиваются в условиях умеренно континентального
климата с умеренно теплым и влажным летом и суровой снежной зимой.
В средней тайге, несмотря на обилие атмосферного увлажнения (когда испа-
рение меньше количества выпадаемых атмосферных осадков), уровень грун-
товых вод залегает ниже, чем на севере, а в почвах процессы оглеения выра-
жены значительно слабее.
Среднетаежные ельники особенно характерны для междуречных рав-
нинных плато, перекрытых ледниковыми отложениями, на которых форми-
руются преимущественно суглинистые подзолистые почвы, местами заболо-
ченные, с признаками оглеения. Часто их массивы разобщены сосновыми ле-
сами на флювиальных формах рельефа, сложенных песчаными отложениями.
Общими особенностями, отличающими среднетаежные еловые леса от севе-
ротаежных, являются их большая сомкнутость и постоянное участие пихты в
ассоциациях зеленомошной группы на востоке.
Общий характер еловых среднетаежных лесов на значительном протя-
жении с запада на восток не остается постоянным. Изменения касаются со-
става древостоя и нижних ярусов, но главным образом сочетаний в массивах
елового леса отдельных ассоциаций и их групп. Леса западной и восточной
части средней тайги имеют различную историю развития, и в них господ-
ствуют различные виды ели (Picea abies – на западе и Picea obovata – на
востоке).
В средней тайге наиболее полно представлены все типы ельников, выде-
ленных В. Н. Сукачевым. На дренируемых плакорах, перекрытых суглинками,
характерны леса зеленомошной группы: ельники-кисличники, ельники-чер-
ничники. На плакорах, где условия дренажа ухудшаются и повышен уровень
грунтовых вод, распространены ассоциации, переходные от зеленомошной к
сфагновой группе. В логах с проточным увлажнением и с почвами богатыми
питательными веществами формируются влажно-травяные ельники. На абсо-
лютных суходолах с резко переменным режимом увлажнения развиваются
ельники брусничники, сменяющиеся сосновыми борами лишайниковыми. На
дренируемых плакорах с богатыми почвами, особенно в южных районах сред-
ней тайги, появляются ельники субнеморальные.
Ельники черничники, кисличники, а также неморальные на всем протя-
жении среднетаежной подзоны обычно занимают наиболее плодородные
земли, и леса на них начали сводиться под пашню еще на заре развития зем-
леделия. На примере Вологодской области показан пятнистый характер ри-
сунка контуров среднетаежных ельников, постепенно сменяющихся к югу не-
моральными южнотаежными ельниками (рис. 7.12).
Подзона южной тайги на Русской равнине протягивается почти
сплошной полосой от устья Нарвы на западе до Среднего Урала на востоке. На
пространстве от Финского залива до верхней части бассейна р. Сухоны се-
верная граница южнотаежных лесов в условиях более мягкого климата захо-
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
147
дит севернее 60° с.ш. Восточнее северный предел их распространения не
идет за 59° с.ш., но южная граница подзоны проникает значительно дальше
на юг (примерно до 56,5° с.ш.) . Развитие южно-таежных ельников происходит
в достаточно влажном климате с относительно длинным вегетационным пе-
риодом. Уровень грунтовых вод вне западин, при наличии даже незначитель-
ного дренажа, находится на глубине не выше 1,5–2 м.
Южно-таежные коренные леса представлены ельниками; местами на
песчаных отложениях растет сосна. В травяно-кустарничковом ярусе многих
ассоциаций южно-таежных лесов, по сравнению с северными ельниками, за-
метно увеличивается роль трав, что способствует развитию дернового процес-
са в почве. На вырубках на смену ели обычно идет береза и часто осина. В пло-
хо дренируемых понижениях расположены сфагновые болота; поймы рек за-
няты лугами; суходольные луга сформированы и поддерживаются на месте
лесной растительности благодаря постоянному антропогенному воздейст-
вию – сенокосу и выпасу. Самые крупные массивы южнотаежных еловых ле-
сов сохранились в верховьях рек Костромы, Унжи, Ветлуги и на водоразделе
их с Северной Двиной, а также в бассейне Камы. Ряд относительно больших
участков южнотаежных ельников расположен в полосе между Вологдой и
р. Лугой (см. рис. 7.12). Основные массивы южно-таежных ельников развиты
главным образом на структурных плато, в разной степени расчлененных и по-
крытых преимущественно суглинистыми отложениями, в полосе распростра-
нения дерново-подзолистых почв, но встречаются также на подзолистых и
дерново-карбонатных почвах.
Южная тайга находится в полосе спорадического распространения ши-
роколиственных пород: липы (Tilia cordata), клена (Acer platanoides), вяза
(Ulmus sp.), дуба (Quercus robur) и др. Названные деревья, равно как и сопут-
ствующие им кустарники и травы, на северном пределе своего распростране-
ния встречаются в наиболее благоприятных условиях местообитания. Нахо-
дясь в зависимости от защиты, которую им оказывает ярус темнохвойных по-
род, широколиственные деревья в южной тайге чаще всего образуют подлесок
или произрастают в нижнем пологе. Достаточно разнообразен в южной тайге
состав неморальных трав: копытень европейский (Asarum europaeum), подма-
ренник душистый (Galium odoratum), зеленчук желтый (Galeobdolon luteum),
звездчатка ланцетовидная (Stellaria holostea), медуница неясная (Pulmonaria
obscura), папоротник щитовник мужской (Dryopteris filix-mas) и др.
В южной тайге более резко, чем в средней и северной, сказываются раз-
личия между западными и восточными темнохвойными лесами. Объясняется
это тем, что на широте южной тайги континентальность климата с запада на
восток нарастает более значительно, чем на севере. Кроме того, различна ис-
тория западных и восточных южно-таежных лесов. Первые располагаются на
территории, покрывавшейся ледником в последнюю (Валдайскую) фазу оле-
денения, вторые в плейстоцене были свободны ото льда, и темнохвойные леса
здесь поселились раньше. Западные и восточные южно-таежные леса отлича-
ются друг от друга по составу древостоя. В западных лесах господствует ель
европейская (Picea abies), в восточных – ель сибирская (Picea obovata), при
этом к востоку от р. Ветлуги большое участие в сложении древостоя принима-
ет пихта (Abies sibirica).
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
148
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
149
Рис. 7.12. (продолжение). Легенда к карте растительности Вологодской области
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
150
Государственный природный заповедник «Кологривский лес»
расположен в Костромской области. Он был образован в 2006 г. в целях со-
хранения уникальных южно-таежных коренных темнохвойных лесов Русской
равнины (http://kologrivskiy-les.ru).
На территории заповедника преобладает спокойный плосковолнистый и
плоскохолмистый рельеф; почвы дерново-подзолистые, образовавшиеся на
моренных суглинках и покровных глинах. Климат умеренно-континенталь-
ный, за год выпадает в среднем 564 мм осадков, причем большая часть – в
теплое время года. Среднемесячная температура за июль составляет 19 °С, за
январь –9 °С.
Кологривский заповедник – единственное в европейской части России
место, где сохранились небольшие «островки» коренных южно-таежных лесов
(самый большой, формирующий ядро заповедника, имеет площадь 918 га),
окруженные территорией, где такие же леса были вырублены только один раз
и сейчас идет демутация коренной растительности. Здесь представлены самые
разнообразные типы растительных сообществ: хвойные леса и хвойно-широ-
колиственные леса, заливные луга, верховые и ключевые болота с характер-
ным видовым составом флоры, вполне типичным для южно-таежных ланд-
шафтов. Ботанико-географический интерес заповедник вызывает также тем,
что на его территории смыкаются границы ареалов европейской (дуб, вяз, ме-
дуница) и сибирской (пихта, лиственница, княжик), а также бореальной и не-
моральной флор. Флора заповедника в целом типична для южной тайги, она
насчитывает 322 вида сосудистых растений, 35 видов мхов и 37 видов лишай-
ников.
Богата территория заповедника северными орхидеями. Их можно встре-
тить здесь на лесных полянах, опушках, заливных лугах, ключевых болотах.
Благодаря интенсивному и бесконтрольному освоению человеком лесов и сво-
бодных земель, осушению влажных лугов, климатическим изменениям боль-
шинство видов орхидных стали редки и сейчас находятся в Красной книге Ко-
стромской области. В настоящее время в заповеднике обнаружено и определено
13 видов орхидных: пальчатокоренники – Фукса (Dactylorhiza fuchsii), пятни-
стый (D. maculata), мясо-красный (D. incarnata), дремлик лесной (Epipactis
helleborine), гудайера ползучая (Goodyera repens), любка двулистная (Platanthera
bifolia), тайник яйцевидный (Listera ovata), гнездовка настоящая (Neottia
nidus-avis), мякотница однолистная (Malaxis monophyllos), ладьян трехнад-
резный (Corallorhiza trifida). Три вида – башмачок настоящий (Cypripedium
calceolus), надбородник безлистный (Epipogium aphyllum) и пальчатокоренник
Траунштейнера (Dactylorhiza traunsteineri) занесены в Красную Книгу РФ.
Места обитания приурочены к выходам грунтовых вод на склонах долин
малых рек и старичным понижениям, растение встречается в низинных ело-
во-сосновых и березово-сосновых травяных и осоковых болотах. Чаще можно
встретить небольшие группы по 5–10 особей или единично. В коренном ело-
вом лесу в ядре заповедника растет еще одно удивительное растение – листо-
вой лишайник лобария легочная (Lobaria pulmonaria), растение, включенное
в Красную книгу Российской Федерации. Этот лишайник крайне чувствителен
к любому типу антропогенного воздействия. В умеренном климате приурочен
к старовозрастным лесам и является признанным индикатором экологиче-
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
151
ской чистоты и ненарушенности местообитания. В коренных лесах лобарию
можно встретить на липе, рябине, клене остролистном, вязе, ели. В производ-
ных лесах – на осине и иве козьей.
Территория заповедника включает в себя 2 участка: Кологривский и
Мантуровский, сильно отличающихся друг от друга по рельефу, типам почво-
образующих пород, флоре и растительности.
Кологривский участок. Для этого участка заповедника характерно соче-
тание коренных еловых лесов и производных лесов, формирующихся на месте
естественных вывалов, вырубках различной давности и пожарищах прошлых
лет. Наибольший интерес на территории Кологривского участка представляют
субнеморальные еловые леса. Для этих лесов характерно господство в древо-
стое гибридных форм ели II класса бонитета, достигающих высоты 25–30 м
(в лучших условиях произрастания – 40–45 м высоты). В формировании пер-
вого подъяруса также принимает участие пихта и изредка, на наиболее бога-
тых почвах, липа. Во втором подъярусе присутствуют клен остролистный и вяз
гладкий (Ulmus laevis). В подлеске жимолость обыкновенная (Lonicera xylo-
steum), бересклет бородавчатый (Euonymus verrucosa), рябина (Sorbus aucu-
paria), лещина (Corylus avellana), малина (Rubus idaeus), крушина (Frangula
alnus), волчеягодник (Daphne mezereum). В травяно-кустарничковом ярусе –
бореальные и неморальные виды. В ельниках Кологривского участка растет
княжик сибирский (Atragene sibirica) – многолетняя лиана семейства лютико-
вых с нежными белыми цветками. На междуречьях, склонах и высоких поймах,
в ельниках папоротниковых весьма часто встречается еще один редкий пред-
ставитель лютиковых – воронец красноплодный (Actaea erythrocarpa). Как и
княжик, этот вид – характерный представитель таежной флоры.
Наиболее флористически богатыми являются вырубки 75–100-летней
давности. Здесь рубка носила преимущественно выборочный характер, велась
грамотно, с сохранением подроста. Эти сообщества характеризуются разнооб-
разной структурой древостоя, подлеска, подроста, травяно-кустарничкового и
мохового ярусов. Именно в таких лесах зафиксировано максимальное число
видов травянистых растений.
Наряду с субнеморальными ельниками на Кологривском участке пред-
ставлены производные сообщества на месте субнеморальных елово-сосновых
лесов: бореальные ельники, бореальные елово-сосновые леса, бореальные
сосновые леса, восстанавливающиеся гари. Небольшую площадь занимают
болота. К долинам рек приурочены фрагменты пойменных лугов.
Особое значение Мантуровского участка подчеркивается историей.
Леса здесь с XVIII века до 1917 года относились к категории казенных дач (за-
казные корабельные рощи междуречья Унжи и Ветлуги). В настоящее время
здесь господствуют сосновые леса с широким участием неморальных элемен-
тов: дуб, лещина, клен, вяз, а также почти в полном составе комплекс европей-
ского дубравного широкотравья. На участке отмечены места с единичными
взрослыми деревьями лиственницы, некогда широко распространенной на
речных и зандровых террасах и практически полностью вырубленной в ХХ
веке. Весной в сосняках Мантуровского участка появляются печеночница бла-
городная (Hepatica nobilis) и прострел или сон-трава (Pulsatilla patens). На по-
лянах сыроватых лесов встречается маленький папоротник – ужовник обык-
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
152
новенный (Ophioglossum vulgatum). На сухих песчаных травянистых склонах
и пойменных лугах растет еще один охраняемый папоротник – гроздовник
многораздельный (Botrychium multifidum).
Южно-таежные леса в той или иной степени всюду несут следы хозяй-
ственной деятельности человека. Рубки, широко практиковавшиеся в южной
тайге уже с начала XIX века, привели к значительному увеличению в южно-
таежных еловых лесах удельного веса мелколиственных пород. Как правило,
ельники в южной тайге всегда перемежаются с производными мелколиствен-
ными лесами и суходольными лугами.
Подзона подтайги (хвойно-широклиственных лесов). Ведущую
роль в формировании растительности играет положение зоны на юго-западе
лесной полосы России, в относительной близости к Атлантическому океану.
По сравнению с тайгой климат хвойно-широколиственных лесов более теп-
лый и более влажный – переходный с запада на восток от морского к умерен-
но-континентальному. На протяжении года через Калининградскую область
проходит около 50–55 циклонов. Средняя длительность безморозного перио-
да нарастает от 120 дней на северо-востоке подзоны до 165 дней на западе
Калининградской области. Годовая сумма осадков колеблется в пределах от
850 мм на западе до 600 мм на востоке. Баланс влаги положительный; ГТК
вегетационного периода около 2, на юге он приближается к 1.
Западная часть подзоны хвойно-широколиственных лесов Русской рав-
нины в четвертичный период покрывалась материковыми льдами. Следы
пребывания ледника особенно хорошо сохранились на северо-западе подзо-
ны, в полосе аккумуляции последнего (Валдайского) оледенения. Здесь широ-
ко распространен холмистый ледниковый рельеф.
С разнообразием форм и типов рельефа связана большая пестрота поч-
венного покрова хвойно-широколиственных лесов. Зональными почвами яв-
ляются дерново-подзолистые, причем наблюдается большое количество поч-
венных разностей, отличающихся одна от другой по степени оподзоленности
и гранулометрическому составу. На западе, в Калининградской области, рас-
пространены бурые лесные почвы.
На севере подзоны широколиственные виды деревьев занимают подчи-
ненное положение, а их видовой состав относительно беден. Южнее широко-
лиственные и хвойные виды деревьев занимают сходные позиции в сообще-
стве: и те, и другие достигают первого подъяруса древостоя, правда, в силу
разных причин могут преобладать то хвойные, то лиственные виды деревьев.
В травяном покрове лесов подзоны основу (по численности и биомассе) со-
ставляют неморальные и бореальные виды трав.
Близость грунтовых вод в условиях влажного климата часто вызывает
широкое развитие в подтайге процессов заболачивания. Верховые и низинные
болота покрывают большую часть низменностей и котловин, нередко они
встречаются и на высоких, но недостаточно дренированных водоразделах.
Среди болот на севере зоны преобладают верховые сфагновые торфяники,
иногда покрытые низкорослой сосной. Южнее Москвы преобладают переход-
ные и низинные болота.
Хвойно-широколиственные леса Европейской части России с запада на
восток подразделяются на следующие региональные типы:
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
153
1. Буково-грабово-еловые леса восточной Польши, Прибалтики, запада
России (Калининградская область). Древесный ярус многовидовой, в его со-
став входят Picea abies, Fagus sylvatica, Tilia cordata, Carpinus betulus, Fraxinus
excelsior, Acer platanoides, A. pseudoplatanus, Ulmus glabra и др.
2. Широколиственно-еловые леса Центра Русской равнины. Они вклю-
чают большую часть перечисленных выше видов широколиственных деревь-
ев, кроме Fagus sylvatica, Carpinus betulus и Acer pseudoplatanus. Из темно-
хвойных деревьев в лесах преобладает Picea abies.
3. Широколиственно-пихтово-еловые леса Заволжья, в которых в каче-
стве содоминантов выступают Picea obovata (включая гибридные формы),
Abies sibirica и Tilia cordata; участие дуба в древостое небольшое.
4. Липово-пихтово-еловые леса Предуралья и Урала. Здесь среди широ-
колиственных видов существенную роль играют только Tilia cordata и отчасти
Ulmus glabra, в составе сообществ постоянно присутствует Abies sibirica, хотя и
с невысоким обилием.
В качестве примера растительность подтайги приведем характеристику
растительности Национального парка Валдайский. Парк находится в
Новгородской области, в северной части Валдайской возвышенности, протя-
женность его с севера на юг – 105 км, с запада на восток – 45 км. Разнообразие
форм рельефа и почвообразующих пород предопределили чрезвычайную
пестроту и сложность почвенного покрова. Общие свойства почв – относи-
тельная обогащенность первичными минералами и наличие остаточной кар-
бонатности, слабее выраженной во флювиогляциальных и озерно-ледниковых
песках. Преобладают дерново-слабоподзолистые почвы; собственно подзолы
довольно редки. В понижениях между холмами и в лощинах распространены
дерново-подзолисто-глеевые почвы. Торфяно-болотные почвы занимают не-
большую площадь вследствие хорошей дренированности территории.
Флора национального парка Валдайский довольно богата: список сосу-
дистых растений насчитывает не менее 750, а список мхов (включая печеноч-
ники) – не менее 126 видов. Здесь произрастают 17 видов растений, занесен-
ных в Красную книгу Российской Федерации: полушник щетинистый (Isoetes
setacea), меч-трава обыкновенная (Cladium mariscus), венерин башмачок насто-
ящий (Cypripedium calceolus), пальчатокоренник длиннолистный (Dactylorhiza
baltica), глянцелистник Лёзеля (Liparis loeselii), ятрышник шлемоносный
(Orchis militaris) и др. В парке обнаружено немало видов, которые довольно
редки на всем Северо-Западе европейской части России и/или в Новгородской
области: пухонос альпийский (Trichophorum alpinum), лилия саранка (Lilium
martagon), тайник сердцевидный (Listera cordata), гнездовка настоящая
(Neottia nidus-avis), ладьян трехнадрезанный (Corallorrhiza trifida), мякотни-
ца однолистная (Malaxis monophyllos), хаммарбия болотная (Hammarbya
paludosa), бородник шароносный (Jovibarba globifera), зимолюбка зонтичная
(Chimaphila umbellata), шалфей мутовчатый (Salvia verticillata), мытник
царский скипетр (Pedicularis sceptrum-carolinum), Петров крест чешуйчатый
(Lathraea squamaria), лобелия Дортманна (Lobelia dortmanna), бузульник си-
бирский (Ligularia sibirica) и др. Наибольшее число редких видов обнаружено
в неморальных лесах, особенно дубравах, на безлесных мезотрофных болотах,
окруженных сфагновыми сосняками, и в богатых заболоченных лесах.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
154
Располагается парк на границе двух растительных подзон южной тайги и
хвойно-широколиственных лесов. Здесь преобладают подтаежные леса, глав-
ным образом вторичные осиновые и березовые с участием широколиственных
пород – липы, клена, дуба, и сельскохозяйственных земель на их месте (всего
более 60 %). Однако довольно часто (более 10 %) встречаются участки с расти-
тельностью южнотаежного типа: еловые леса с кислицей (Oxalis acetosella),
майником (Maianthemum bifolium) и др. и с участием неморальных трав, не-
редки также вторичные южнотаежные березовые, осиновые, сероольховые
леса. На повышениях, сложенных песками, встречаются сосняки брусничные,
вересковые, иногда лишайниковые. Выровненные впадины на песках и супе-
сях заняты в основном ельниками с участием сосны, а на тяжелосуглинистых
дерново-слабоподзолистых почвах, прежде покрытыми широколиственными
лесами, сохранились местами рощицы из угнетенного дуба с примесью бере-
зы, рябины, ясеня, лещины, черемухи (Padus avium), волчеягодника в подлес-
ке, в травяном покрове преобладают неморальные виды. В долинах рек часто
встречаются таволговые ельники с Filipendula ulmaria и черноольшатники. На
обширных плоских низинах иногда наблюдаются долгомошные и сфагновые
березово-еловые леса.
Благоприятные климатические условия и относительно плодородные
почвы обусловили сведение коренных лесов подтайги и формирование на их
месте сельскохозяйственных угодий.
7.2.3. ЗАПАДНО-СИБИРСКАЯ ТАЕЖНАЯ ОБЛАСТЬ
Западно-Сибирская область простирается от Урала до Енисея, 85 % ее
площади занимает Западно-Сибирская низменность. Она представляет собой
обширную, сильно заболоченную низину, с абсолютными высотами 80–120 м,
слабо наклоненную к северу. В четвертичный период ледник покрывал север-
ную часть равнины. На фоне низменности резко выделяются образованные
ледником возвышенности: Сибирские Увалы (до 200 м), Аганский увал (до
150 м), Белогорский материк (до 200 м) и др. Похолодание в плейстоцене
привело к глубокому промерзанию грунта. В наши дни многолетнемерзлые
породы захватывают территорию подзоны северной тайги.
Положение Западной Сибири в умеренных широтах, удаленность от оке-
анов, наличие Уральских гор, стоящих преградой на пути воздушных масс,
открытый доступ холодных воздушных масс с севера обусловили формирова-
ние здесь умеренно-континентального и довольно сурового климата. Лето
умеренно теплое, средняя температура июля на севере около 14 °С, на юге
19 °С. Зима снежная, холоднее, чем в тундровой зоне, где смягчающее воздей-
ствие оказывает океан. Средняя температура января –30 °С на северо-востоке
и –15 °С на юго-западе. Осадков выпадает около 400–500 мм в год. Вслед-
ствие малого испарения создаются условия высокого увлажнения (ГТК веге-
тационного периода 2), что способствует высокой заболоченности и заозерен-
ности Западно-Сибирской низменности. На дренированных участках форми-
руются подзолистые и дерново-подзолистые почвы, в то время как на
недренированных междуречьях – глеево-подзолистые, болотно-подзолистые,
лугово-болотные и болотные.
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
155
Область характеризуют темнохвойные леса из ели, сосны кедровой и
пихты сибирской на увалах и террасах рек. Значительную роль в растительном
покрове играют лиственница сибирская и сосна обыкновенная. Из широко-
лиственных пород в южной подзоне встречается липа, причем редко и в не-
большом количестве, и вяз гладкий (Ulmus laevis), произрастающий лишь на
юго-западе. Почти всем лесным сообществам западно-сибирской тайги свой-
ственно участие мелколиственных пород.
Слабая дренированность Западно-Сибирской низменности сильно огра-
ничивает распространение темнохвойных лесов, огромные площади занима-
ют верховые сфагновые болота, поэтому данную территорию называют лесо-
болотной зоной. Историю формирования болот рассмотрим на примере Сур-
гутского Полесья, которое расположено в центральной части Западно-
Сибирской равнины к северу от долины р. Аган. В начале атлантического пе-
риода (7860±70 лет) первичными очагами заболачивания являлись наиболее
глубокие депрессии рельефа, в которых в условиях более богатого водно-
минерального режима образовались эвтрофные травяные и травяно-моховые
болота, сложенные низинным осоковым, гипновым или осоково-гипновым
торфом мощностью не более 0,5 м. Эвтрофная стадия в развитии болот была
очаговой и сравнительно непродолжительной. Завершающим этапом было
формирование верховых болот с грядово-мочажинными и озерково-грядово-
мочажинными комплексами.
Ландшафт Сургутского Полесья представляет собой низменную равнину,
сформированную на зандровых песках. Равнина пересекается реками, теку-
щими почти в меридиональном направлении. К приречным песчаным гривам
приурочены редкостойные сосновые лишайниковые леса, площадь которых
занимает не более 2–3 % от площади Полесья. В их напочвенном покрове пре-
обладают лишайники (Stereocaulon paschale, виды Cladonia, Nephroma arcticum,
Peltigera malacea, P. aphthosa, Flavocetraria cucullata и др.). Травяно-кустар-
ничковый ярус образован брусникой (Vaccinium vitis-idaea), толокнянкой
(Arctostaphylos uva-ursi).
Общий облик территории определяют заболоченные междуречные про-
странства с многочисленными озерами. В сплошных болотных массивах до-
минируют грядово-озерковые, грядово-мочажинно-озерковые и грядово-озер-
ковые болотные комплексы с сосново-кустарничково-сфагновыми (рямовы-
ми) сообществами по грядам, осоково-пушицево-сфагновыми мочажинами и
многочисленными озерами (рис. 7.13). Значительное распространение имеют
грядово-мочажинные комплексы, представляющие собой чередование гряд
разнообразной формы и топких мочажин (рис. 7.14). Характерными элемен-
тами озерно-болотных комплексов Сургутского Полесья являются хасыреи –
плоские днища бывших озер округлой формы, находящиеся на разных стади-
ях развития болотообразовательного процесса от остатков акваторий мелко-
водных водоемов и мелкозалежных низинных осоково-гипновых топей до ме-
зотрофных осоково-сфагновых сплавин и верховых сфагновых и сосново-
кустарничково-сфагновых участков.
Особенно высокой заболоченностью отличается пространство к северу от
Широтного Приобья (включающего территорию, где течение реки Оби при-
обретает широтное направление до слияния ее с рекой Иртыш) . На отдельных
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
156
Рис. 7.13. Космическое изображение ландшафта Сургутского Полесья
(по Google Earth)
Рис. 7.14. Грядово-мочажинный комплекс
(по С. М. Горожанкиной, В. Д. Константинову)
Растения: 1 – Rubus chamaemorus, 2 – Eriophorum vaginatum, 3 – Ledum palustre, 4 –
Drosera rotundifolia, 5 – Betula nana, 6 – Chamaedaphne calyculata, 7 – Andromeda polifolia,
8 – Sphagnum fuscum, 9 – Cladonia stellaris, 10 – Cladonia arbuscula, 11 – Pleurozium
schreberi, 12 – Dicranum polysetum, 13 – Sphagnum balticum
Виды торфа: 14 – фускум; 15 – мочажинный
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
157
участках, например, в Васюганье, заболоченность достигает 70 %. Характер-
ными элементами грядово-озерковых комплексов являются термокарстовые
озера округлой формы. Они возникают в результате протаивания и проседа-
ния мерзлых грунтов.
По окраинам болот встречаются рямы – сосново-сфагновые сообщества с
угнетенными тонкоствольными деревьями. Кустарниковый и кустарничковый
ярусы здесь обычно сомкнуты, они состоят из багульника, болотного мирта
(Chamaedaphne calyculata) и ерника (Betula nana). В травяно-кустарничковом
ярусе встречаются Rubus chamaemorus, Vaccinium vitis-idaea, Carex globularis.
Подзона северной тайги граничит с лесотундрой, а ее южная грани-
ца проходит по широте Сибирских Увалов. Болота этой подзоны по площади
преобладают над лесами. Среди болот наиболее обычны сфагновые плоско- и
крупнобугристые торфяные. Первые развиты преимущественно на севере,
вторые господствуют на юге.
Крупнобугристые комплексные болота характеризуются чередованием
крупных куполообразных торфяных бугров с мерзлым минеральным ядром и
обводненных вытянутых мочажин, в которых обычны термокарстовые озера.
Растительность бугров образована лишайниками, бриевыми мхами (Dicranum
flexcaule, D. elongatum), кустарничками (багульник, брусника, голубика, морош-
ка, ерник). Встречаются отдельные деревья сосны (Pinus sylvestris), листвен-
ницы (Larix sibirica), березы (Betula pubescens). В мочажинах обычны осоки и
пушицы. Моховой покров составлен в основном сфагновыми и гипновыми
мхами (Drepanocladus, Calliergon).
Леса подзоны представлены редкостойными лиственничными и лист-
веннично-еловыми зеленомошно-лишайниковыми, кустарничково-сфагновы-
ми сообществами. Основой этих сообществ являются лиственница сибирская
(Larix sibirica) и ель сибирская (Picea obovata) Широко распространены гипо-
арктические кустарнички: водяника, багульник, голубика, карликовая береза,
брусника. Кроме того, характерны осока шаровидная (Carex globularis), пла-
ун годичный (Lycopodium annotinum), линнея (Linnaea borealis), седмичник
(Trientalis europaea), голокучник обыкновенный (Gymnocarpium dryopteris),
хвощи (Equisetum pratense, E. sylvaticum, E. scirpoides). В напочвенном покрове
значительную роль играют лишайники: Cladonia stellaris, С. rangiferina. Зеле-
ные мхи (Pleurozium schreberi, Hylocomium splendens, Ptilium crista castensis,
Polylrichum commune), характерные и для других подзон тайги, занимают
здесь обычно второстепенное место. При заболачивании увеличивается роль
сфагновых мхов. Здесь, особенно в южной части подзоны, более или менее
значительны вкрапления кедровых и сосновых лесов. На междуречьях в ред-
костойных лесах кедр и сосна встречаются преимущественно в виде примеси,
причем обычно доля кедра меньше, чем сосны. Сомкнутые лесные насажде-
ния развиты только по долинам рек, придолинным склонам междуречий и на
небольших участках на более дренированных увалах.
С севера на юг роль лиственницы ослабевает, местами попадаются чистые
ельники лишайниково-зеленомошные, ельники со значительной примесью
сосны и березы, лиственнично-елово-кедровые лишайниково-зеленомошно-
кустарничковые, приуроченные к глеево-подзолистым и подзолисто-болотным
почвам. Послепожарные сообщества на месте коренных лесов представлены
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
158
березовыми (Betula pubescens) с сосной сибирской (Pinus sibirica), елью (Picea
obovata), лиственницей (Larix sibirica) зеленомошно-кустарничковыми лесами,
которые являются продолжительной стадией в восстановлении коренных лесов.
Леса Верхнетазовской возвышенности носят экстразональный характер.
Они имеют черты среднетаежных лесов и представлены: елово-кедровыми с
лиственницей кустарничково-зеленомошными, сосновыми кустарничково-
зеленомошными и сосновыми с елью и сосной сибирской кустарничково-
сфагновыми лесами. На этой территории основан Верхне-Тазовский государ-
ственный природный заповедник. Заповедник расположен в восточной части
Северных Увалов, почти полностью в пределах Верхне-Тазовской возвышен-
ности. Это довольно крупная равнина, приподнятая на высоту до 200 м и бо-
лее, с характерным моренно-холмистым рельефом, формы которого сложены
главным образом песчанистыми валунными суглинками, перекрытыми лес-
совидными суглинками, и впоследствии сильно расчлененными эрозией. Бо-
лот на высоких междуречьях мало, почти отсутствуют и торфяники.
В плакорных условиях развиваются подзолистые и таежно-поверхност-
но-глеевые типы почв. Подзолистые почвы представлены песчаными подзо-
лами, развивающимися под светлохвойной тайгой – сосновыми и листвен-
нично-сосновыми лесами. Таежно-поверхностно-глеевые почвы формируются
на глинистом и суглинистом субстратах и развиваются под елово-кедровыми
темнохвойными лесами.
По данным 1997 г. в Верхне-Тазовском заповеднике произрастает 310 ви-
дов сосудистых растений, относящихся к 166 родам и 60 семействам; 111 ви-
дов листостебельных мохообразных, принадлежащих к 26 семействам; 91 вид
лишайников 36 родов и 20 семейств.
Растительность представлена почти исключительно таежными лесами.
Леса с преобладанием сосны составляют 59,4 % лесопокрытой площади запо-
ведника; леса с преобладанием сосны сибирской – 16,6 %, лиственницы –
11,0 %, ели – 2,8 %, березы (повислой и пушистой) – 7,1 %, осины – 0,4 %. Из
сосняков наиболее распространены сухие бруснично-лишайниковые боры –
самые светлые и разреженные, с древостоями паркового типа; они распро-
странены на самых бедных песчаных почвах высоких надпойменных террас и
на песчаных холмах увалов. Единично в таких лесах встречается кедр. Подле-
сок отсутствует. Среди кустарничков доминирует брусника. Хорошо развит ли-
шайниковый покров из кладина звездчатая, оленья, лесная (Cladonia stellaris,
C. rangiferina, C. arbuscula).
Темнохвойные леса занимают, по сравнению с сосняками, небольшие
площади, но они весьма разнообразны. Эти леса образованы преимущественно
кедровой сосной и елью, небольшое участие в них принимает пихта. Темно-
хвойные леса различны по составу и возрасту. Это связано с тем, что после ча-
стых здесь пожаров коренной лес отмирает не полностью. В результате образу-
ется смешанная темнохвойная тайга с большим участием березы и осины. Чи-
стых кедровников (из сосны сибирской) мало, они распространены отдельными
небольшими массивами, в основном по хорошо дренированным берегам рек.
Моховые ельники, напротив, распространены по более сырым местам, часто в
высокой пойме. Основной зональный тип темнохвойных лесов в заповеднике –
елово-кедровая (часто смешанная) кустарничково-зеленомошная тайга. Со-
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
159
мкнутость древостоя – 0,6–0,7. Подлесок развит слабо; обычна рябина. Рассе-
янно растут кусты шиповника, можжевельника. В наземном покрове домини-
руют бореальные кустарнички и мхи. Среди кустарничков преобладают брус-
ника и черника, среди мхов – гилокомиум блестящий и плевроциум Шребера.
Моховой покров часто сплошной или почти сплошной. Постоянно встречают-
ся «пятна» листоватых лишайников – пельтигеры пупырчатой и собачьей,
нефромы арктической.
Южная граница подзоны совпадает с южной границей широкого рас-
пространения лиственничных редкостойных лесов и крупнобугристых болот,
а на междуречье Оби и Енисея – близка к северной границе распространения
пихты.
Подзона средней тайги представлена елово-пихтово-кедровыми ле-
сами в сочетании со сфагновыми болотами и березняками. К хорошо дрениру-
емым моренным грядам и возвышенностям приурочены темнохвойные леса.
В этих лесах преобладает сосна сибирская, которая наряду с елью играет суще-
ственную роль. Пихта становится почти постоянным компонентом древостоя
сначала в лесах речных долин, а затем и междуречий. В Западной Сибири
темнохвойные леса из ели, сосны сибирской и пихты называются урманами
(рис. 7.15). В напочвенном покрове здесь часто господствуют зеленые боре-
альные мхи, а в травяно-кустарничковом ярусе обильны брусника, багульник,
местами черника.
Рассмотрим особенности темнохвойных лесов на примере Аганского ува-
ла, расположенного в междуречье р. Аган и среднего течения р. Обь (рис. 7.16).
Для Аганского увала характерны елово-кедровые с пихтой, местами кедрово-
еловые, мелкотравно-бруснично-зеленомошные леса. Почвы под ними подзо-
листые элювиально-глееватые на средних и легких суглинках. Для тяжелосу-
глинистых почв более характерны елово-кедровые мелкотравно-чернично-зе-
леномошные леса . Подлесок развит слабо . Единично, реже группами (по 2–3
Рис. 7.15. Темнохвойная тайга (урман) Западной Сибири
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
160
Рис. 7.16. Темнохвойный лес (урман) на Аганском увале
Урманы выделяются темно-зеленым цветом по сравнению с окружающими их болотно-
озерными комплексами, получившими светлое изображение (по Google Earth)
экземпляра), рассеянно встречаются рябина сибирская (Sorbus sibirica), ши-
повник иглистый (Rosa acicularis), ива козья (Salix caprea), малина, реже жи-
молость (Lonicera altaica).
В травяно-кустарничковом покрове преобладают бореальные кустарнич-
ки – брусника, черника, линнея северная. Характерной особенностью являет-
ся присутствие группы видов таежного мелкотравья – кислицы, майника дву-
листного, седмичника европейского, гудайеры ползучей (Goodyera repens),
ортилии однобокой (Orthilia secunda). Из других травянистых видов следует
отметить большое участие плаунов (Diphasiastrum complanatum, Lycopodium
annotinum), папоротников (Gymnocarpium dryopteris), хвощей (Equisetum syl-
vaticum). Единичными экземплярами встречаются лугово-лесные виды: вей-
ник притупленный и Лангсдорфа (Calamagrostis obtusata, C. langsdorffii), ма-
рьянник луговой (Melampyrum pratense), золотарник обыкновенный (Solida-
go virgaurea). Общее покрытие травяно-кустарничкового яруса 60–85 %.
Моховой покров сплошной из Hylocomium splendens и Pleurozium schreberi
с постоянным включением небольших латок Ptilium crista-castrensis, Dicranum
polysetum, Polytrichum commune. Все микропонижения заняты сфагновыми
мхами, преимущественно Sphagnum russowii, S. fallax.
Большие площади занимают вторичные леса. Восстановительные смены
елово-кедровых мелкотравно-бруснично-зеленомошных лесов представляют
темнохвойно-сосновые и темнохвойно-мелколиственные зеленомошные со-
общества. Наиболее распространены елово-березовые и березово-осиновые с
пихтой и сосной сибирской леса.
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
161
Подзона южной тайги отличается широким распространением вто-
ричных темнохвойно-мелколиственных травяных и, несколько реже, мелко-
травно-зеленомошных лесов. Характерным признаком коренных ассоциаций
темнохвойных лесов является широкое участие в елово-кедровых насаждени-
ях пихты. В травяном покрове северной части подзоны преобладает мелкотра-
вье, в южной – большую роль играют виды широкотравья, свойственные ши-
роколиственным лесам европейской части России. В подлеске или во втором
пологе, реже в первом, встречается липа (Tilia sibirica); в подлеске снытьевых
березово-осиновых лесов ее участие более значительно. Местами липа пред-
ставляет чистые сообщества. Песчаные гривы заняты сосновыми борами (Pinus
sylvestris). Южная граница этой подзоны более или менее совпадает с южной
границей распространения еловых лесов.
Фрагменты темнохвойных лесов подзоны сохранились на сравнительно
небольшой площади, главным образом в восточной части южной тайги За-
падно-Сибирской низменности, например, в Кемеровской области. Неболь-
шие массивы этих лесов встречаются среди производных березовых и березо-
во-осиновых насаждений на Обь-Иртышском междуречье, а также на мень-
шей площади в приуральской части южной тайги Западной Сибири.
Одной из причин сокращения площади южно-таежных сибирских тем-
нохвойных лесов является быстрое олуговение вырубок и пожарищ. Чем даль-
ше на юг, тем сильнее становится конкурентная способность травянистых со-
обществ. Мощный травяной покров во вторичных березняках и осинниках
задерживает возобновление темнохвойных пород, но полностью не исключает
его. Поэтому, наряду с производными мелколиственными лесами, в южной
тайге нередко встречаются темнохвойно-березово-осиновые леса как стадия
восстановления темнохвойной южной тайги.
В южнотаежных темнохвойных лесах подлесок обычно состоит из не-
многих видов: черемуха (Padus avium), рябина (Sorbus sibirica), жимолость
(Lonicera xylosteum), карагана (Caragana arborescens). Моховой покров пред-
ставлен разобщенными куртинами зеленых мхов (Rhytidiadelphus triquetrus,
Climacium dendroides, Hylocomium splendens, Pseudobryum cinclidioides и др.).
В травяном покрове обычно таежное мелкотравье: кислица (Охаlis acetosella),
майник (Maianthemum bifolium), костяника (Rubus saxatilis) и линнея (Linnaea
borealis). Рассеянно встречаются папоротники (Athyrium filix-femina, Diplazium
sibiricum, Rhizomatopteris sudetica, Dryopteris carthusiana), злаки (Calamagrostis
obtusata, Melica nutans, Milium effusum) и представители разнотравья – сныть
(Aegopodium podagraria), медуница (Pulmonaria mollis).
Для южнотаежных темнохвойных лесов Западной Сибири характерны
три группы ассоциаций. Ассоциации темнохвойно-зеленомошной группы
приурочены к повышенным и дренированным местоположениям. На сырых
участках пихта утрачивает господствующее положение в древостое, в моховом
покрове среди зеленых мхов появляются сфагны. Видовой состав травяно-
кустарничкового яруса более однообразный, в нем выделяются черника, брус-
ника и лесной хвощ. На плохо дренируемых водоразделах по окраинам болот
намечается переход к темнохвойно-сфагновой группе ассоциаций. Сообщества
елово-пихтовой группы с участием неморальных видов в травяном ярусе на
легких дерново-подзолистых почвах по склонам речных долин характеризу-
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
162
ются разнообразным травяным покровом (рис. 7.17). В нем преобладает вей-
ник тростниковидный (Calamagrostis phragmitoides), а на прогалинах – высо-
котравье (Aegopodium podagraria, Aconitum septentrionale, Pleurospermum
uralense, Thalictrum minus, Solidago virgaurea и др.). Наряду с разнотравьем в
южно-таежных темнохвойных лесах присутствуют типичные представители
таежного мелкотравья (Trientalis europaea, Maianthemum bifolium, Oxalis ace-
tosella, Paris quadrifolia).
В подзоне южной тайги, наряду с возрастанием щелочности грунтов, па-
дает общая заболоченность территории, уменьшаются размеры болотных мас-
сивов, преобладание переходит к гипново-осоковым болотам, а еще южнее
господствующим типом становятся болота травяные, приуроченные к меж-
гривным лощинам и периферии зарастающих озер. Среди травяных болот
наиболее обычны кочкарные осоковые и кустарниково-осоковые, и лишь в
неширокой полосе вдоль южной окраины подзоны получают распростране-
ние тростниково-осоковые и тростниковые «займища».
Эвтрофные болота, особенно кочкарные осоковые и кустарниково-
осоковые, повсеместно встречаются в долинах Оби, Иртыша и их крупных
притоков в виде мелких массивов, рассеянных среди лугов и пойменных ку-
старников. В группу эвтрофных торфяных болот входят также долинные со-
гры, т. е кочкарные осоково-древесные (осоково-березовые, осоково-еловые и
т. п.) болота, нередко сочетающиеся с небольшими по площади облесенными
мезотрофными осоково-сфагновыми болотами, которые занимают узкие по-
лосы по линии контактов согр, заболоченных лесов и олиготрофных болот.
Подзона подтайги характеризуется наиболее оптимальным сочетани-
ем увлажнения и радиации по сравнению с тайгой и степями; высокой степе-
нью гидроморфности и широким развитием процессов – оглеения, заболачи-
вания, осолодения и засоления почв. В почвенном покрове под лесами развиты
Рис. 7.17. Темнохвойный лес с участием неморальных видов
(фото Геблеровского экологического общества)
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
163
зональные почвы – дерново-подзолистые и выщелоченные черноземы, серые
и темно-серые лесные почвы. В западной, приуральской, части большие пло-
щади занимают почвы легкого механического состава.
Основная лесообразующая порода в подтайге Западной Сибири – береза
повислая (Betula pendula). На засоленных в разной степени почвах, вплоть до
структурных солонцов, из всех древесных пород способна развиваться только
эта. Чем глубже находится засоленный горизонт, тем лучше развивается ее
поверхностная корневая система. В условиях постоянного переувлажнения и
заболачивания на окраинах болот и в понижениях береза повислая уступает
доминирующее положение березе пушистой (Betula pubescens). Эта порода
более требовательна к влаге и плохо переносит засоление почвогрунтов. Оси-
на на юге равнины встречается в березовых лесах в качестве примеси, ее уча-
стие в древостоях увеличивается на богатых и хорошо увлажненных почвах,
чистые насаждения осина образует редко.
Своеобразие березовых лесов Западно-Сибирской равнины проявляется в
экологическом составе доминантных видов травяного яруса. На границе с юж-
ной тайгой в лесах из Betula pendula наряду с основными синузиями подтаеж-
ных лесных злаков (Calamagrostis arundinacea) и разнотравья (Aegopodium
podagraria, Pulmonaria mollis) представлены: южно-таежное мезофильное раз-
нотравье (Delphinium elatum, Aconitum septentrionale) и таежное мезофильное
мелкотравье (Maianthemum bifoliит, Trientalis europaea). На юге подтайги и в
лесостепи в сложении фитоценозов большую роль играют ксеромезофильные
растения, травостой составляют лесные мезофильные злаки (Brachypodium
pinnatum), лугово-лесные ксеромезофильные злаки (Calamagrostis epigeios,
Phleum phleoides), луговолесное ксеромезофильное разнотравье (Filipendula
vulgaris, Seseli libanotis).
В северной части подзоны на дренируемых плакорах распространены
березовые и осиново-березовые (из Betula pendula), местами с липой, злаково-
разнотравные леса. Почвы под северными березняками дерново-подзолистые.
Кустарниковый ярус слагают шиповник майский (Rosa majalis), малина (Rubus
idaeus), боярышник (Crataegus sanguinea), виды ив (Salix caprea, S. cinerea),
рябина (Sorbus sibirica), калина (Viburnum opulus), черная смородина (Ribes
nigrum), спиреи средняя и иволистная (Spiraea media, S. salicifolia), жимолость
(Lonicera xylosteum). Травяной покров высокий и густой. Доминируют вейники
Calamagrostis arundinacea, С. epigeios, С. langsdorffii, С. obtusata, С. canescens.
Большую роль в травостое сообществ играют лесные виды Rubus saxailis, Carex
macroura, Pulmonaria mollis, Aegopodium podagraria, Pleurospermum uralense,
Kadenia dubia, Angelica sylvestris, Crepis sibirica, Delphinium elatum, Aconitum
septentrionale, Rubus arcticus, Paris quadrifolia и многие другие, а также луго-
во-лесное разнотравье – Geranium pseudosibiricum, Galium boreale, Hieracium
umbellatum, Vicia cracca, V. sepium, Lathyrus pratensis, Gentiana pneumonanthe,
Achillea millefolium, Stellaria graminea. Характерно присутствие в составе сооб-
ществ видов таежного мелкотравья (Trientalis europaea, Maianthemum bifolium,
Pyrola rotundifolia, P. minor, Ortilia secunda) и папоротников (Gymnocarpium
dryopteris, Dryopteris carthusiana, Athyrium filix-femina, Phegopteris connectilis,
Matteuccia struthiopteris, Pteridium aquilinum). Примером состава подобных
сообществ могут служить разнотравно-вейниковая, разнотравно-злаковая,
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
164
разнотравно-коротконожковая, разнотравно-костяничная ассоциации Приир-
тышских березовых лесов. В нарушенных сообществах увеличивается видовое
разнообразие злаков, в значительном количестве появляются: Brachypodium
pinnatum, Elytrigia repens, Phleum phleoides, Poa pratensis, P. trivialis, Festuca
pratensis, Agrostis gigantean.
В понижениях рельефа, где высокое стояние уровня грунтовых вод, вызы-
вающее заболоченность почвы, широко развиты березовые (из Betula pubescens)
и осиново-березовые осоковые, осоково-вейниковые леса в сочетании с осо-
ковыми и тростниково-осоковыми болотами. Преобладающие виды осок:
двурядная, дернистая и Буксбаума (Carex disticha, C. cespitosa, C. buxbaumii).
В травостое разнотравно-осокововых сообществ доминируют гигрофильный
Calamagrostis canescens и ксеромезофильный С. epigeios. Подтаежное лесное
разнотравье состоит из мезофилов (Rubus saxatilis, Aegopodium podagraria,
Lathyrus pisiformis, Serratula coronata, Chamaenerion angustifolium, Crepis
sibirica) и гигромезофилов (Filipendula ulmaria, Kadenia dubia, Aconitum
volubile, Angelica silvestris, Heracleum sibiricum).
В результате неумеренного выпаса в березовых лесах развиваются разно-
травно осоковые сообщества. Они представляют собой кочковатые гигрофиль-
ные сообщества, состоящие из Carex brunnescens, C. cespitosa, С. vaginata,
С. rostrata, Calamagrostis neglecta, Agrostis gigantea, Comarum palustre, Kadenia
dubia, Vicia cracca.
Для южной части подтайги, на дренируемых плакорах, характерны березо-
вые и осиновые вейниковые и вейниково-высокотравные леса. Почвы под вей-
никовыми и вейниково-высокотравными березняками серые лесные. В кустар-
никовом ярусе встречаются Ribes nigrum, Padus avium, Rosa majalis, Cotoneaster
melanocarpus, Sorbus sibirica, Viburnum opulus. Травостой высокий (до 1 м) и
густой. В травяном покрове доминируют злаки: вейники – Calamagrostis
epigeios, С. arundinacea, коротконожка – Brachypodium pinnatum, ежа –
Dactylis glomerata; менее обилен перловник – Melica nutans. Отличительной
особенностью фитоценозов является ярко выраженный и богато представлен-
ный ярус высокотравья. Его слагают зонтичные (Angelica silvestris, Heracleum
dissectum, H. sibiricum, Kadenia dubia, Pleurospermum uralense, Aegopodium
podagraria, Сarum carvi), сложноцветные (Crepis sibirica, Centaurea scabiosa,
Serratula coronata, Cirsium heterophyllum, Cacalia hastata, Tanacetum vulgare),
розоцветные (Agrimonia pilosa, Filipendula ulmaria) и представители других
семейств: Rumex crispus, Lilium martagon, Polygonatum odoratum, Thalictrum
simplex. Сообщества богаты мезофильными видами лесного (Pulmonaria mollis,
Viola mirabilis, Equisetum sylvaticum, Origanum vulgare, Rubus saxatilis, Vicia
megalotropis, V. sylvatica, Polemonium caeruleum) и лугово-лесного (Hieracium
umbellatum, Vicia sepium, Achillea millefolium, Sanguisorba officinalis) разно-
травья.
Примером подобных березовых лесов может служить березняк вейнико-
во-коротконожково-разнотравный; характерны также ассоциации осинника
костянично-разнотравно-перловникового. Ксеромезофильно-разнотравно-зла-
ковые остепненные березовые леса представлены ассоциациями с доминиро-
ванием вейника наземного или пырея ползучего. Древостой преимущественно
из Betula pendula (рис. 7.18).
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
165
Рис. 7.18. Березовые леса на юге Западной Сибири
(http://21region.org/2010/07/19/yekskursiya-v-omsk.html)
В восточной приобской части области на суглинках структурно-денуда-
ционных плато в составе березовых лесов обычна примесь темнохвойных по-
род – кедра, ели, пихты.
В полосе южной подтайги березовые леса на сухих открытых участках
чередуются с массивами лесных суходольных лугов. На хорошо дренируемых
территориях луга занимают 35–50 % площади. Они богаты по видовому со-
ставу. Доминантами и субдоминантами луговых сообществ являются Dactylis
glomerata, Роа pratensis, Festuca pratensis, F. rubra, Phleum pratense, Ph. phleoides,
Brachypodium pinnatum, Calamagrostis epigeios, Vicia sepium, Trifolium pratense,
T. repens, Lathyrus pisiformis, Galium boreale, Centaurea scabiosa, Thalictrum
simplex, Th. minus, Geranium sylvaticum, Filipendula ulmaria, Hylotelephium
triphyllum, Aegopodium podagraria, Pulmonaria mollis, Kadenia dubia, Seseli
libanotis, Phlomoides tuberosa, Solidago virgaurea, Tanacetum vulgare, Achillea
millefolium, Hieracium umbellatum и многие другие. В нарушенных местооби-
таниях травостой низкий 15–20 см, степень проективного покрытия 40–50 %,
на выбитых участках поселяется Festuca valesiaca.
В понижениях рельефа с неустойчивым водно-солевым режимом на
оглеенных и оторфованных солодях развиваются гигрогенные сообщества бе-
резняков. Они составлены березой пушистой с ивами, представлены раз-
нотравно-вейниковыми (с Calamagrostis canescens) и разнотравно-осоковыми
(с Carex disticha) сообществами.
На боровых террасах Пышмы, Туры, Тобола, Иртыша сосредоточены ос-
новные массивы подтаежных сосновых лесов из Pinus sylvestris. Для этих тер-
риторий характерен волнисто-холмистый или равнинный рельеф с дюнны-
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
166
ми всхолмлениями, сложенными мелкозернистыми песками и супесями.
В древесном ярусе подтаежных лесов присутствуют темнохвойные породы:
ель, пихта, кедровая сосна, что несвойственно остепненным сосновым лесам.
На западе подзоны в составе сосновых лесов широко распространена липа
сердцевидная, которая произрастает преимущественно в подлеске. В группе
подтаежных лесов преобладают различные варианты травяных и травяно-
кустарничковых лесов. Подлесок в этих лесах редкий из единичных экзем-
пляров шиповника иглистого и собачьего, рябины, жимолости, крушины
ломкой, можжевельника, черемухи. Переходный характер подтаежных со-
обществ сосновых лесов особенно ярко проявляется в составе травяно-
кустарничкового яруса: наряду с таежными растениями здесь отмечено ши-
рокое участие лугово-лесных, а на юге подзоны появление лугово-степных
растений. Доминируют Vaccinium vitis-idaea и V. myrtillus. Наиболее харак-
терные лугово-лесные и боровые виды: Chimaphila umbellata, Maianthemum
bifolium, Rubus saxatilis, Galium boreale, Pulmonaria obscura, Geranium sylvati-
cum, Calamagrostis arundinaceae, Pyrola minor, P. rotundifolia, Stellaria ho-
lostea. Зеленые мхи встречаются фрагментарно (Pleurozium schreberi, Ptilium
crista-castrensis, Dicranum polysetum, Rytidiadelphus triquetrus). Нижние ча-
сти склонов и выположенные, хорошо дренированные местоположения по-
крыты травяными, преимущественно вейниковыми лесами. На площади под-
таежных боров, ранее подвергашихся выборочным рубками или низовым
пожарам, широко распространены производные березовые и сосново-березо-
вые леса.
Сосновые лишайниковые леса встречаются небольшими участками среди
других типов подтаежных лесов. Древесный ярус распределен неравномерно;
подлесок редкий из можжевельника (Juniperus communis) и рактиничка рус-
ского (Chamaecytisus ruthenicus) в лесах Притоболья и Caragana arborescens,
Rosa acicularis, Spiraea media в лесах восточных районов. Травяной покров
разрежен, беден по видовому составу. Наиболее обычны: Antennaria dioica,
Chimaphyla umbellate, Melampyrum pratense, Hieracium umbellatum,
Trommsdorffia maculata, Orthilia secunda. Лишайники пятнами покрывают
40–50 % почвы. Преобладает более ксерофильный вид Cladonia arbuscula с
участием Cl. stellaris, Cl. rangiferina. Из мхов – единичные куртины Dicranum
polysetum, Pleurozium schreberi.
7.2.4. СРЕДНЕ-СИБИРСКАЯ ТАЕЖНАЯ ОБЛАСТЬ
Западная граница Средне-Сибирской области проходит по склону Сред-
несибирского плоскогорья, с востока она ограничена Верхоянским хребтом.
Основные черты рельефа определяются положением области на Среднесибир-
ском плоскогорье и на Центральноякутской и Ленской низменностях.
Нахождение в глубинной части огромного континента почти исключает
влияние влажных и теплых морских воздушных масс. Западный перенос не
достигает Средней Сибири, а влияние Тихого океана ослаблено горными барь-
ерами. Зимой здесь господствует холодный континентальный воздух (Азиат-
ский антициклон). Резкая континентальность климата проявляется в больших
амплитудах температур (годовые до 100 °С, а в теплый период амплитуды су-
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
167
точных температур нередко достигают 20–30 °С) и в малом количестве атмо-
сферных осадков (200–400 мм). В результате здесь средние месячные зимние
температуры воздуха намного ниже средних температур соответствующих
широт Восточно-Европейской и Западно-Сибирской областей, а летние, наобо-
рот, выше. ГТК вегетационного периода от 1,3 до 1, а местами 0,8. Тем не ме-
нее, несмотря на засушливость, для Средне-Сибирской области характерны
лесные, озерные и болотные ландшафты. Причиной этому служит повсемест-
но распространенная (кроме прирусловых полос крупнейших рек) многолет-
няя мерзлота грунтов, мощность которой достигает 200–350 м. На ее холод-
ном водоупорном слое конденсируется влага, питающая нижние горизонты
почвы вместе с деятельным слоем, мощность которого достигает 0,6–3,5 м.
Мерзлые грунты содержат большое количество жил, линз и клиньев ископае-
мого льда. Процессы образования и таяния ископаемого льда создают запа-
динно-озерный рельеф, а интенсивные мерзлотные процессы в рыхлых грун-
тах – кочковато-мелкобугристый микрорельеф поверхности.
Подзолистые почвы в Средней Сибири теряют свое доминирующее зна-
чение, их место занимают совершенно особые почвы: буротаежные и таежно-
мерзлотные ожелезненные под лиственничными лесами. В связи с изменени-
ем растительного покрова при вырубках, пожарах, а иногда и естественным
путем, нарушается термический режим грунтов и усиливаются термокарсто-
вые процессы. В результате чего все низменные поверхности испещрены озе-
рами и аласами – плоскими неглубокими понижениями с лугово-степной рас-
тительностью, нередко на месте высохших озер.
Представление о растительном покрове Средне-Сибирской области дает
карта растительности Якутии (рис. 7.19). Центральноякутская низменность
и Лено-Алданское плато заняты лиственничной тайгой с включением сос-
новых боров, остепненно-луговых березняков (чаранов), а на карбонатных
субстратах южной части области наблюдаются примесь темнохвойных по-
род и небольшие участки еловых лесов. Повсюду среди тайги, особенно в
бассейне р. Вилюя и в низовьях р. Алдана, распространены безлесные луго-
во-степные участки, низинные болота, заросли кустарников, а в северо-вос-
точной части тайги – участки с развеваемыми песками. Лесная раститель-
ность сочетается с аласами (рис. 7.20 и 7.21) и со степными ковыльно-по-
лынными ассоциациями. Под чаранами и лугово-степными полянами
обычны луговые черноземы, в той или иной степени засоленные, солоди –
под лугами.
Подзона северной тайги. Северотаежные леса из даурской листвен-
ницы (Larix gmelinii) распространены на значительном пространстве от бас-
сейна Нижней Тунгуски на западе и до бассейна Колымы на востоке. Большая
часть территории, покрытой лиственничными лесами, представляет собой
плоскогорье, приподнятое на высоту более 250–300 м и расчлененное сравни-
тельно глубокими речными долинами. На значительно меньшей площади се-
веротаежные леса из даурской лиственницы располагаются на равнинах
(нижнее течение Лены, бассейн Омолоя, некоторые районы в бассейне Инди-
гирки и Колымы). Травяно-моховые, а также сфагновые болота и заболочен-
ные кустарники наблюдаются повсеместно, но небольшими участками, вкрап-
ленными в общий фон лесного ландшафта.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
168
Рис. 7.19. Карта растительности Якутии (по Атласу Якутской АССР)
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
169
Рис. 7.20. Аласы (контуры округлой формы) вблизи Дярыла, Якутия
Западнее р. Лены среди северотаежных лиственничников изредка встре-
чается ель, восточнее Верхоянского хребта она отсутствует совершенно. Неве-
лика роль в сложении северотаежных лиственничников и березы. Таким об-
разом, на всем пространстве северной тайги даурская лиственница является
всюду господствующей древесной породой.
Редкое стояние деревьев северотаежных лесов и сквозистые кроны лист-
венниц способствуют значительной освещенности под пологом леса (рис. 7.22).
Характерная особенность таких лесов – медленное восстановление гарей,
Рис. 7.21. Типичный алас на Центральноленской равнине (фото А. Исаева)
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
170
Рис. 7.22. Лиственничный лес с примесью березы (Betula platyphylla)
на цокольной террасе р. Лены (фото А. Галанина)
часто порастающих ерником (Betula exilis). В дальнейшем возобновляется лист-
венничный древостой. В более сырых местах на террасах и плоских водоразде-
лах в редкостойных лиственничных лесах хорошо выражен моховой покров с
участием сфагнов, а также кустарниковый ярус, в котором широко распро-
странен ерник. Местами, где близко сходятся истоки мелких речек, ерниковые
заросли сливаются в почти единый массив, прерываемый лишь небольшими
участками лиственничного леса. На склонах и на других более дренированных
местоположениях часто встречаются редкостойные лиственничные леса с по-
кровом из лишайников. Для кустарникового яруса лишайниковых листвен-
ничников характерны ерник (Betula exilis), кедровый стланик, голубика, при-
сутствуют некоторые травы и кустарнички (Carex pediformis, Empetrum nigrum,
Arctous alpina).
На плоских широких речных террасах встречаются зеленомошно-сфаг-
новые лиственничные ассоциации в кустарничковом ярусе которых господ-
ствует голубика. Плоские не дренированные водоразделы часто значительно
заболочены; среди сфагновых лиственничников на них встречаются бугри-
стые торфяники, а также низинные травяные болота, развитию которых бла-
гоприятствует минерализация грунтовых вод.
Своеобразная структура лиственничных лесов формируется на поло-
гоувалистых возвышенностях, сложенных палеозойскими карбонатными от-
ложениями. На аэро- и космоснимках они выделяются полосчато-струйчатым
рисунком. Для склонов характерна ступенчатость, связанная с различным ли-
тологическим составом палеозойских пород, в разной степени разрушенных
ложбинами стока. Этот микрорельеф подчеркивается распределением лист-
венничных лесов и изображается на космических снимках в виде чередую-
щихся полос (рис. 7.23).
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
171
Подзона средней тайги. На юго-западе Средне-Сибирской области в
строении среднетаежных лиственничников участвует лиственница сибирская,
а на всей остальной площади – лиственница даурская (Larix gmelinii). Эти
лиственницы, за исключением сравнительно узкой полосы их совместного
произрастания, оказываются лесообразующими породами в различных эко-
лого-географических условиях; сообщества их отличны друг от друга по своей
структуре.
Для подзоны характерны сухие парковые лиственничные леса из деревь-
ев, достигающих 150–200-летнего возраста. Они имеют хорошо развитую кро-
ну, средняя высота стволов 19–22 м. В кустарниковом ярусе – редкие экземпля-
ры спиреи средней (Spiraea media), ив (Salix bebbiana и S. brachypoda), ши-
повника (Rosa acicularis). В травяно-кустарничковом ярусе – брусника, майник
двулистный, линнея северная и лугово-степные травы: овсяница якутская
(Festuca jacutica), прострел (Pulsatilla multifida), полыни (Artemisia laciniata) и
др. Почвы палевые мерзлотно-таёжные. Пологие склоны северной экспози-
ции покрыты лиственничными с примесью сосны лесами. В подлеске – оль-
ховник (Duschekia fruticosa), береза тощая (Betula exilis), можжевельник
(Juniperus sibirica), багульник стелющийся (Ledum decumbens).
На бровках террас и супесчаных склонах южной экспозиции встречаются
толокнянковые и ягельниковые сосновые боры в сочетании с березовыми тра-
вянистыми ассоциациями, лугово-степными аласами с черноземовидными и
осолоделыми почвами, пятнами лугово-карбонатных солончаков.
Среднетаежные кустарничковые и травяно-брусничные лиственничники
покрывают большую площадь на Средне-Сибирском плоскогорье, в Централь-
ноякутской низменности и сочетаются с другими ассоциациями. В Централь-
ной Якутии на сухих местах среди лиственничных и сосново-лиственничных
Рис. 7.23. Полосчато-струйчатый рисунок ступенчатого рельефа
на космическом изображении (по Google Earth)
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
172
лесов встречаются сообщества остепненных лугов и луговых степей. В их со-
став входят мятлик кистевидный (Poa botryoides), житняк гребенчатый (Ag-
ropyron cristatum), овсяница ленская (Festuca lenensis), ковыль Крылова (Stipa
krylovii), полынь холодная (Artemisia frigida).
Общий облик среднетаежных травяно-кустарничковых и кустарничко-
вых лиственничников к востоку от Средне-Сибирского плоскогорья сохраня-
ется на большом протяжении с севера на юг, примерно от 65° с.ш. в верховьях
р. Вилюя и до 53° с.ш. в верховьях р. Лены. Этому способствует однообразие
климатического режима зимой и многолетнемерзлые грунты. С запада на во-
сток структура среднетаежных лиственничников изменяется в связи с более
существенными различиями в строении поверхности, а также в соответствии с
особенностями климата – более влажного на западе и более сухого на востоке.
На востоке на высоких террасах р. Алдана к травяно-кустарничковым лист-
венничным лесам близки и ассоциации лиственничников с хорошо развитым
кустарниковым ярусом из рододендрона (Rhododendron dauricum) или оль-
ховника. Обильный кустарник не препятствует хорошему развитию травяно-
кустарничкового покрова, в котором много брусники, злаков (Limnas stelleri,
Calamagrostis langsdorffii и др.) и разнотравья.
На юге средней тайги, на карбонатном плато в лиственничных лесах не-
редко присутствуют ель, сосна кедровая и пихта. Чаще встречаются сосновые
боры (Pinus sylvestris), даже на суглинистых почвах.
Современный облик лиственничных лесов на значительной площади
находится в зависимости от часто случающихся лесных пожаров. В листвен-
ничной тайге пожары легко распространяются на большую площадь. В извест-
ной мере пожары способствуют расселению лиственницы, так как на гарях она
возобновляется обычно успешно. Огромное влияние оказывают лесные пожа-
ры на кустарниковый ярус и напочвенный покров в лиственничных лесах. Из
числа кустарников и трав происходит отбор видов, меньше страдающих от ни-
зовых палов и развитию которых благоприятствуют изменения в почве, вы-
званные пожаром. Значительное распространение багульника в различных
ассоциациях лиственничного леса является следствием низовых пожаров.
Подзона южной тайги занимает Приангарское плато и Среднеан-
гарский кряж. Территория расчленена густой сетью притоков Ангары. Лето
сравнительно теплое (средняя температура июля 16–19 °С), а зима умеренно
холодная (температура января −19−24 °С). В течение года выпадает в среднем
350–450 мм осадков. Вечная мерзлота встречается лишь в виде островов и на
больших площадях отсутствует. Характерны дерново-подзолистые почвы.
В растительном покрове преобладают сосновые и сосново-лиственнич-
ные боры (Pinus sylvestris, Larix sibirica). Они занимают сложенные песчаным
аллювием поверхности террас Ангары и ее притоков, выходы диабазов и
участки распространения нижнепалеозойских песчаников. Леса обычно раз-
реженные, с хорошо развитым ярусом кустарников, преимущественно из ро-
додендрона даурского (Rhododendron dahuricum), и густыми зарослями луго-
во-лесных трав.
Среди сосновых лесов различают несколько типов. В сухих местообита-
ниях формируются высокобонитетные лишайниковые сосняки. Средние по
условиям увлажнения и дренажа участки заняты бруснично-разнотравными
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
173
борами, в которых заросли кустарничков – брусники, черники и голубики –
чередуются с полянами из вейника наземного (Calamagrostis epigeios), овся-
ницы приземистой (Festuca airiodes), прострела желтеющего (Pulsatilla orien-
tali-sibirica), касатика русского (Iris ruthenica). На более влажных местообита-
ниях обычны зеленомошные боры.
На наиболее возвышенных местах, на востоке, развита густая и влажная
темнохвойная тайга из пихты, ели и сосны сибирской (Abies sibirica, Picea
obovata, Pinus sibirica).
7.2.5. САХАЛИНСКАЯ ТАЕЖНАЯ ОБЛАСТЬ
Рельеф острова составлен средневысотными горами, низкогорьями и низ-
менными равнинами. Южная и центральная части острова характеризуются
горным рельефом и состоят из двух меридионально ориентированных горных
систем – Западно-Сахалинских и Восточно-Сахалинских гор, разделенных про-
дольной Тымь-Поронайской низменностью. Север острова (за исключением
полуострова Шмидта) представляет собой пологую холмистую равнину.
Климат Сахалина умеренно-муссонный морской с продолжительной хо-
лодной снежной зимой и умеренно теплым дождливым летом. Средняя тем-
пература января от −6 °С на юге до −24 °С на севере, августа – от 19 °С до 10 °С
соответственно. Наибольшее среднегодовое количество осадков 990 мм выпа-
дает на юге острова, наименьшее 476 мм – на севере. Зимой погода в значи-
тельной степени определяется Сибирским антициклоном: в это время преоб-
ладают северные и северо-западные ветры, могут стоять крепкие морозы, осо-
бенно в центральной части острова. В то же время с юга могут приходить
зимние циклоны, определяющие сильные и частые бураны. Летом контраст
между западным и восточным побережьями острова усиливается за счет соот-
ветственно теплого Цусимского течения Японского моря, которое достигает
юго-западной оконечности Сахалина, и холодного течения Охотского моря,
идущего вдоль восточного побережья с севера на юг.
Вытянутость острова Сахалин в меридиональном направлении создает
своеобразное сочетание бореальной охотской и южной маньчжурской флор. Во
флоре Сахалина насчитывается около 1100 видов, из них 35 эндемичных. От-
носительная многочисленность видов древесных растений является одной из
особенностей сахалинской флоры. На бореальный характер флоры указывает
ведущее положение в ее составе осок и ив, в то время как богатство представи-
телями семейств розоцветных, лютиковых, лилейных, орхидных и папоротнико-
видных свойственно флоре восточной умеренной Азии. По составу флоры север-
ный Сахалин имеет много общего с прилегающей материковой частью Татарско-
го пролива, флора южных частей Сахалина близка к флоре северной Японии.
Общее представление о растительности острова Сахалин дает карта
(рис. 7.24). На Северо-Сахалинской низменности распространены светлохвой-
ные леса из лиственницы даурской (рис. 7.25). Елово-пихтовые леса встречают-
ся только на хорошо дренированных склонах, на более богатых суглинистых
почвах. Долинные пойменные леса этой низменности состоят из ольхи воло-
систой (Alnus hirsuta), ивы, березы плосколистной (Betula platyphylla) и неко-
торых других пород.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
174
Рис. 7.24. Растительность о. Сахалин (Атлас Сахалинской области, 1967)
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
175
Рис. 7.24. (продолжение) Растительность о. Сахалин
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
176
Рис. 7.25. Лиственничное редколесье в сочетании с кустарниковой ивняковой
тундрой, Северный Сахалин (фото Н. Царенко)
Южнее средней части Восточно- и Западно-Сахалинских гор лиственнич-
ники сменяются пихтово-еловой тайгой из ели аянской (Picea ajanensis), ели
Глена (Picea glehnii) и пихты сахалинской (Abies sachalinensis), приурочен-
ной к нижним и средним частям горных склонов и к ровным незаболоченным
Рис. 7.26. Лес из пихты белокорой Abies nephrolepis на Среднем Сахалине
(фото Н. Царенко)
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
177
участкам (рис. 7.26). В древостое ель преобладает над пихтой. Напочвенный
покров состоит из ковра зеленых мхов. Выше по склону этот тип леса сменяет-
ся таким же пихтово-еловым, но с хорошо развитым кустарничковым ярусом
из черники овальнолистной (Vaccinium ovalifolium). Выше, в предгольцовом
поясе, пихтово-еловые леса сменяются каменноберезовыми лесами (Betula
ermanii) с зарослями курильского бамбука (Sasa kurilensis), а на еще большей
высоте – с зарослями кедрового стланика.
Господствующей формацией на большей части Южного Сахалина явля-
ется темнохвойная елово-пихтовая тайга. К югу от перешейка Поясок в древо-
стое пихта сахалинская и пихта Майра (Abies sachalinensis, A. mayriana) пре-
обладают над елью. Зеленомошники встречаются редко и к югу постепенно
вытесняются елово-пихтовым лесом с травяным покровом из папоротника
лепторуморы амурской (Leptorumohra amurensis).
На обращенных к морю склонах гор Южного Сахалина лес полностью
вырублен и замещен плотными зарослями курильского бамбука, препятствую-
щего восстановлению лесной растительности (рис. 7.27). В юго-западной части
острова, омываемой теплым морским течением, происходит переход к хвойно-
широколиственным лесам южно-маньчжурского типа, которые характеризу-
ются примесью широколиственных пород из бархата сахалинского (Phelloden-
dron sachalinense), дубов монгольского и курчавенького (Quercus mongolica,
Q. crispula), черемухи айнской (Padus ssiori), калопанакса (Kalopanax
septemlobus) и др. Особый колорит лесам придает множество лиан: актинидия
(Actinidia kolomikta, A. polygama), лимонник (Schisandra chinensis) и др., опле-
тающих деревья. Встречаются небольшие по площади дубравы с участием ти-
са (Taxus cuspidata).
Рис. 7.27. Заросли курильского бамбука (Sasa kurilensis) на месте лесных
вырубок
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
178
Рис. 7.28. Красника – Vaccinium
praestans (фото С. Никитина)
К плодово-ягодным растениям
относятся: два вида смородины (Ribes
sachalinensis, R. latifolium), жимолость
съедобная (Lonicera edulis), черемуха
айнская, рябина бузинолистная, три
вида шиповника, актинидия, лимон-
ник, два вида черники. Особо следует
сказать о краснике (рис. 7.28), назы-
ваемой местными жителями клопов-
кой (Vaccinium praestans). Ее собирают в больших количествах для изготовле-
ния соков, киселей и наливок. У человека хотя бы раз отведавшего клоповку,
образ Сахалина навсегда остается связанным с запахом этих ягод. В травяном
покрове господствуют различные виды папоротников. Леса зеленомошники
здесь не встречаются.
Луга на Сахалине располагаются по долинам рек, низменностям, речным
и морским террасам. Интересно и своеобразно сахалинское высокотравье
(рис. 7.29). Многие травы достигают 2–3 метров высоты, а некоторые превы-
шают 5 м. Таковы, прежде всего шеломайник (Filipendula camtschatica), бело-
копытник широкий (Petasites amplus), медвежий корень или дудник (Angelica
ursina), гречиха сахалинская (Reynoutria sachalinensis), крапива плосколист-
ная (камчатская) (Urtica platyphylla), крестовник (Senecio cannabifolius), бор-
щевик шерстистый (Heracleum lanatum).
7.2.6. ПРИМОРСКАЯ ОБЛАСТЬ ХВОЙНО-ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
Область находится между 43° и 52° с.ш.; на ее востоке расположены сред-
невысотные, выходящие к Японскому морю, хребты Сихотэ-Алиня. На таких
же широтах расположены субтропические области Крыма (Ялта 44°29′ с.ш.) и
Кавказа (Сочи 43°35′ с.ш.). Несмотря на южное расположение области, нижний
пояс гор, отвечающий ее зональной принадлежности, занят хвойно-широко-
лиственными лесами – подтайгой, что объясняется палеогеографическими и
современными климатическими особенностями. Климат умеренный муссон-
ный. Зима сухая и холодная, средняя температуря января составляет –20–24 °С.
Весна продолжительная, с частыми колебаниями температуры. Лето теплое
(температуры июля +20–24 °С) и влажное, на летние месяцы приходится мак-
симальное количество осадков, в некоторых районах до 90 % годовой сум-
мы. В Приханкайской низменности годовые осадки составляют 550–660 мм,
на побережье Японского моря 640–850 мм, в центральном Сихотэ-Алине 640–
920 мм. Осень теплая, солнечная и сухая. Летом преобладают юго-восточные
ветра с Тихого океана, а зимой северо-западные, приносящие холодную, но
ясную погоду с континента.
Весьма своеобразен почвенный покров хвойно-широколиственных лесов
Дальнего Востока. В отличие от дерново-подзолистых почв, преобладающих
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
179
Дудник медвежий (Angelica ursina)
Борщевик шерстистый (Heracleum lanatum)
Белокопытник широкий
(Petasites amplus)
Лабазник камчатский
(Filipendula camtschatica)
Гречиха сахалинская
(Reynoutria sachalinensis)
Бодяк камчатский
(Cirsium kamtschaticum)
Фото: P. Slichter, А. Чернышева, Б. Большакова, А. Барышенко
Рис. 7.29. Виды высокотравья
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
180
в хвойно-широколиственных лесах Русской равнины, здесь распространены
преимущественно бурые лесные почвы, формирующиеся на хорошо дрениро-
ванных холмистых и предгорных участках. Они сравнительно плодородны,
пригодны для распашки. Мощность этих почв различна, но часто невелика,
поэтому после уничтожения леса на склонах энергично идет процесс плос-
костного смыва почв.
В области произрастает около 2500 видов сосудистых растений, включая
около 250 видов деревьев, кустарников и деревянистых лиан, более трети –
российских папоротников, сотни – мхов и 600 видов лишайников. Регион от-
носится к восточно-азиатской флористической области.
Хвойно-широколиственные леса Дальнего Востока тяготеют к горным
склонам, где по сравнению с равнинами, наблюдается более ровный ход тем-
ператур, а также более мощный снежный покров и к узким горным долинам,
по которым поднимаются до высоты 500–600 (800) м над ур. м. на юге области
и до высоты 300 м на севере. Эти леса распространены на Сихотэ-Алине, Бу-
реинском и других, менее крупных хребтах. Общие черты растительного по-
крова Приморского края показаны на рис. 7.30.
Хвойно-широколиственные леса Дальнего Востока отличаются сложно-
стью их строения: древостой многоярусный с нечеткими границами между
подъярусами и обладает многовидовым составом. Обычно выделяется три
подъяруса в древесном ярусе, затем кустарники, лианы и до 3–4 подъярусов
травяного яруса. Обильны мхи-эпифиты на стволах деревьев. Количество ви-
дов деревьев и кустарников, формирующих лес, превышает 200. На 1 га мож-
но насчитать до 25 видов деревьев, до 30 видов кустарников, 5–6 видов лиан,
до 15 видов папоротников, 80–100 видов травянистых растений, более 50 ви-
дов мхов и лишайников.
Леса (рис. 7.31) включают, главным образом, сосну корейскую (Pinus
koraiensis), а также ели аянскую и корейскую (Picea ajanensis, P. koraiensis) и
пихты белокорую и цельнолистную (черную) (Abies nephrolepis, A. holophylla),
лиственницу даурскую (Larix gmelinii). Первое место среди лиственных
пород занимает дуб монгольский (Quercus mongolica). Обычны липы Та-
ке, амурская, маньчжурская (Tilia taquetii, T. amurensis, T. mandshurica),
береза ребристая (Betula costata), клен моно (Acer mono). Изредка встре-
чаются: тис остроконечный (Taxus cuspidata), вяз японский (Ulmus japo-
nica), ясени маньчжурский и носолистный (Fraxinus mandshurica, F. rhyn-
chophylla), орех маньчжурский (Juglans mandshurica), клен маньчжурский
(Acer mandshuricum), рябина ольхолистная (Sorbus alnifolia) (рис. 7 .32,
7.33).
В подлеске растут граб сердцелистный (Carpinus cordata), вяз разрезной
(Ulmus laciniata), груша уссурийская (Pyrus ussuriensis), клены (Acer ukurun-
duense, A. tegmentosum, A. pseudosiebolianum и др.), вишня Максимовича
(Cerasus maximowiczii), черемуха Маака (Padus Maackii) и целый ряд специ-
фичных видов кустарников: сирень амурская (Syringa amurensis), аралия вы-
сокая (Aralia elata), калина Саржента (Viburnum sargentii), барбарис амурский
(Berberis amurensis), чубушник тонколистный (Philadelphus tenuifolius), эле-
утерококк колючий (Eleutherococcus senticosus), малина боярышниколист-
ная (Rubus crataegifolius), калопанакс (Kalopanax septemlobus), различные виды
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
181
Рис. 7.30. Карта растительности Приморского края (http://www.fegi.ru)
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
182
Рис. 7.31. Кедрово-
широколиственный
лес Сихотэ-Алиня
(фото В. Галанина)
жимолостей (Lonicera maximoviczii, L. chrysantha и др.), лещины мань-
чжурская и разнолистная (Corylus mandshurica, C. heterophylla) и др.
(рис. 7 .34).
Многочисленны лианы: виноград амурский (Vitis amurensis), актинидии
острая и коломикта (Actinidia arguta, A. kolomicta), лимонник (Schisandra
chinensis). Очень разнообразны растения нижних ярусов; в их составе преоб-
ладают различные виды мезофитных папоротников и разнотравье (рис. 7.35).
В первом подъярусе представлены папоротники и высокотравье: щитовник
толстокорневищный (Dryopteris crassirhizoma), лунокучник густосорусовый
(Lunathyrium pycnosorum), борец Радде (Aconitum raddeanum), волжанка дву-
домная (Aruncus dioicus); во втором подъярусе (до 40 см): лепторумора амур-
ская (Leptirumohra amurensis), адиантум стоповидный (Adiantum pedatum),
лесной мак весенний (Hylomecon vernalis), осоки кривоносая, ржавопятни-
стая, уссурийская (Carex campylorhina, C. siderosticta, C. ussuriensis); в третьем
подъярусе – лесное мелкотравье: василистник нитчатый (Thalictrum filamen-
tosum), кислица обыкновенная (Oxalis acetosella), косоплодник сомнительный
(Plagiorhegma dubia), грушанка японская (Pyrola japonica) и др. Моховой по-
кров незначителен.
Характерной особенностью хвойно-широколиственных лесов Дальнего
Востока является широкое участие в них растений – реликтов: тиса остро-
конечного, можжевельника твердого (Juniperus rigida), бархата амурского
(Phellodendron amurense), калопанакса (Kalopanax septemlobus), аралии высо-
кой, маакии амурской (Maackia amurensis), сиреней амурской и Вольфа (Sy-
ringa amurensis, S. wolfii), вишни сахалинской (Cerasus sachalinensis), актини-
дии, винограда, лимонника, древогубца (Celastrus orbiculata), луносемянника
(Menispermum dauricum), чубушника, вейгелы (Weigela middendorffiana), эле-
утерококка и др. Они сами или их ближайшие предки входили в состав неоге-
новых тургайских лесов.
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
183
Сосна корейская (Pinus koraiensis)
Ель аянская (Picea ajanensis)
Пихта цельнолистная (Abies holophylla)
Липа амурская (Tilia amurensis)
Дуб монгольский (Quercus mongolica)
Бархат амурский (Phellodendron amurense)
Фото: П. Крестова, С. Никитина, Д. Кочеткова, С. Козлана, П. Шеенко
Рис. 7.32. Деревья хвойно-широколиственных лесов Дальнего Востока
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
184
Береза ребристая (Betula costata)
Клён маньчжурский (Acer mandshuricum)
осенью
Граб сердцелистный (Carpinus cordata)
Черёмуха Маака (Padus maackii)
Ясень носолистный (Fraxinus rhynchophylla) Орех маньчжурский (Juglans mandshurica)
Фото: А. Барышенко, С. Майорова, Ю. Семейкина, А. Чернышева, В. Папченкова
Рис. 7.33. Деревья хвойно-широколиственных лесов Дальнего Востока
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
185
Бересклет большекрылый
(Euonymus macroptera)
Жимолость Максимовича
(Lonicera maximowiczii)
Малина боярышниколистная
(Rubus crataegifolius)
Элеутерококк колючий
(Eleutherococcus senticosus)
Сирень амурская (Syringa amurensis)
Маакия амурская (Maackia amurensis)
Фото: А. Барышенко, А. Яковлева, Е. Волынец, М. Скотниковой
Рис. 7.34. Деревья и кустарники хвойно-широколиственных лесов Дальнего
Востока
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
186
Актинидия коломикта (Actinidia kolomikta) Лимонник китайский (Schisandra chinensis)
Женьшень обыкновенный
(Panax ginseng)
Василисник нитчатый
(Thaiictrum filamentosum)
Леспедеца двуцветная (Lespedeza bicolor)
Фрима азиатская (Phrуmа asiatica)
Фото: Н. Терехиной, В. Озерянко, Д. Кочеткова, А. Чернышева, Е. Волынец, В. Волкотруб
Рис. 7.35. Лианы и травы хвойно-широколиственных лесов Дальнего Востока
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
187
Хвойно-широколиственные леса Приморской области на севере грани-
чат с темнохвойными лесами южного побережья Охотского моря. Основной
лесообразующей породой здесь является ель аянская с примесью на севере
сибирской ели, в южной части охотского бассейна – белокорой пихты. Подле-
сок из видов рододендрона и кедрового стланика, образующего иногда густые
заросли. Кроме того, бывает развит ярус мелких кустарничков из багульника
(Ledum hypoleucum) и брусники. Тайга охотского типа выходит за пределы
зоны собственно бореальных таежных лесов, внедряясь за счет вертикальной
зональности в районы Приморья, где она утрачивает типично бореальный ха-
рактер благодаря обогащению широколиственными элементами.
В Приморье в горах господствуют елово-пихтовые, кедрово-еловые и
кедрово-широколиственные леса, переходящие на юго-западе в более богатые
и теплолюбивые чернопихтово-широколиственные леса. В речных долинах
растут увитые лианами широколиственные леса из ясеня, вяза и ореха маньч-
журского.
В высокогорьях преобладают типы растительности, близкие к камчат-
ским: каменноберезовые леса, заросли кедрового стланика и горные тундры c
золотистым и мелколистным рододендронами. На крутых южных склонах,
гребнях гор и каменистых почвах встречаются засухоустойчивые сообщества:
южные сосновые леса (сосна могильная – Pinus funebris), коренные дубняки,
дикие леса из абрикосов маньчжурского и сибирского (Armeniaca mandshurica,
A. sibirica). Вдоль речных русел тянутся ивняки, ольшаники, леса из различ-
ных тополей и чозении (Chosenia arbutifolia).
Из-за рубок и палов широкое распространение получили: леса из лист-
венницы даурской, берёзовые (из Betula platyphylla) и осиновые рощи, липо-
во-кленовые и особенно дубовые леса. В результате повторных пожаров часть
дубняков превратилась во влажные лесостепи. На месте сведенных долинных
лесов и заброшенных сельскохозяйственных угодий развились луга, переме-
жающиеся ивовыми редколесьями, березовыми колками и болотами.
Рассмотрим характерные особенности растительности Приморской обла-
сти на примере двух заповедников: Сихотэ-Алинского и Уссурийского.
Сихотэ-Алинский государственный природный биосферный
заповедник расположен в восточной и центральной водораздельной частях
хребта Сихотэ-Алинь, на территории Приморского края. Панораму Сихотэ-Али-
ня определяют сменяющие друг друга сложно разветвленные средневысотные
(500–800 м над ур. моря) горные гряды и отроги. Лишь отдельные вершины
превышают 1000 м над ур. моря. Высшая точка заповедника – г. Глухоманка
(1598 м). Климат Среднего Сихотэ-Алиня обусловлен взаимодействием мор-
ских и континентальных воздушных масс, проявляющимся в разные сезоны
резко различно. Летом территория заповедника находится под влиянием ти-
хоокеанского муссона, приносящего влажный морской воздух. Всего за теп-
лый период года выпадает 80–85 % годовой суммы осадков. Воздушные массы
арктического происхождения в теплый период года иногда вызывают резкие
похолодания. Главный фактор формирования погоды зимой – поступление
холодного и сухого арктического воздуха из области азиатского антициклона.
Отмечаются существенные различия температур на восточном и западном
макросклонах. На востоке сказывается отепляющее влияние Японского моря,
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
188
среднемесячная температура января колеблется от –9 до –16 °С. На западе
среднемесячные температуры января опускаются до –19...28 °С. Летом море
оказывает охлаждающее влияние – средняя температура июля в прибрежных
районах 14–15 °С; на западном же макросклоне на 2–4 °С выше.
В нижнем и среднем горных поясах под хвойно-широколиственными и
широколиственными лесами развиты бурые горнолесные почвы. Под кедров-
никами, рододендровыми и дубовыми лесами на узких скалистых водоразде-
лах и на крутых склонах при близком залегании, а местами и выходе на по-
верхность коренных пород развиты грубогумусовые бурые горнолесные поч-
вы. Под лесами верхнего горного пояса – пихтово-еловыми, лиственничными
и кедрово-еловыми – формируются буротаежные иллювиально-гумусовые
почвы (подбуры). Для низких речных пойм характерны примитивные аллю-
виальные, а для более высоких уровней долины – буроземно-аллювиальные
почвы. В приморской полосе под высокотравными древесно-кустарниковыми
зарослями развиты темноцветные дерново-глеевые почвы.
Заповедник по флористическому районированию А. Л. Тахтаджяна рас-
положен в Маньчжурской провинции. Особенности этой провинции – богатая
древесная флора, насыщенная большим числом эндемичных видов, состоя-
щая почти целиком из листопадных форм. Здесь много реликтовых растений,
входивших в состав так называемой тургайской флоры, господствовавшей на
материке Евразии в конце палеогена – начале неогена. Из видов, произраста-
ющих в Сихотэ-Алинском заповеднике, к характерным представителям этого
древнего комплекса относятся: тис остроконечный, орех маньчжурский, бар-
хат амурский, ясень маньчжурский, вязы японский и разрезной, клены, ара-
лия маньчжурская, заманиха высокая (Oplopanax elatus) и многие другие дре-
весно-кустарниковые растения. Среди травянистых растений, продолжаю-
щих тот же перечень, можно назвать чистоустник азиатский (Osmundastrum
asiaticum), оноклею чувствительную (Onoclea sensibilis), ряд видов семейства
орхидных.
К числу основных лесообразователей заповедника относятся ель аянская
и пихта белокорая – представители комплекса охотской флоры. Гораздо
меньшую роль в сообществах играют лиственница ольгинская (Larix olgensis)
и сопутствующие ей растения восточносибирской флоры. Из числа тихоокеан-
ских (берингийских) элементов флоры в заповеднике обычны береза шерсти-
стая (Betula lanata), кедровый стланик, рододендрон золотистый. Монголо-
даурское флористическое влияние проявляется в проникновении сюда от-
дельных степных видов – тонконого токийского (Koeleria tokiensis), овсяницы
овечьей, скабиозы шерстистолистной (Scabiosa lachnophylla).
Систематический состав флоры заповедника включает: лишайников –
214 видов (преобладают эпифитные формы), зеленых мхов – 100 видов, сосу-
дистых растений – более 1000 видов.
Растительность Сихотэ-Алинского заповедника относится к двум геобота-
ническим округам по схеме районирования Б. П. Колесникова (1961). Первый
из них Сихотэ-Алинский принадлежит Амурско-Сихотэ-Алинской провинции
Южно-Охотской темнохвойнолесной подобласти; второй – Тернейский отно-
сится к Дальневосточной провинции кедрово-широколиственных и дубовых
лесов Восточно-Азиатской хвойно-широколиственной области.
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
189
В Среднем Сихотэ-Алине преобладает лесной тип растительности (90 %
территории). В пестрой мозаике лесного покрова особое внимание привлека-
ют кедрово-широколиственные леса, а среди них – леса с резким преоблада-
нием в древостое сосны корейской (корейского кедра) – дальневосточные кед-
ровники. Большие площади в заповеднике занимают производные леса, воз-
никшие на месте гарей различной давности, – березняки (Betula platyphylla),
осинники, лиственничники, дубняки; кое-где пожары оставили за собой об-
наженные каменные россыпи.
Естественные закономерности размещения лесных формаций проявля-
ются весьма отчетливо, выражаясь прежде всего в высотной поясности. Гра-
ницы растительных поясов на западном макросклоне всегда расположены
выше, чем на приморском. Хорошо выделяются следующие семь высотных
поясов растительности: I. Прибрежная растительность. II. Приморские дубовые
леса. III. Кедрово-широколиственные леса. IV. Пихтово-еловые леса. V. Ка-
менноберезовые леса. VI. Заросли кедрового стланика. VII. Горно-тундровая
растительность. Из внепоясных лесных группировок в долинах рек растут
ивовые, ольховые, чозениевые и тополевые древостои в комплексе со смешан-
ными широколиственными и пихтово-еловыми лесами. Не привязаны к
определенному высотному поясу и лиственничники (как в долинах рек, так и
на горных склонах).
Узкую, прерывистую кайму вдоль берега моря образуют ассоциации
типично литоральных травянистых растений: осоки песколюбивой (Carex
arenicola), мертензии морской (Mertensia maritima), гонкении бутерлаковид-
ной (Honckenya peploides), волоснеца мягкого (Leymus mollis), чины японской
(Lathyrus japonicus) и др. Из кустарников для береговых валов наиболее ха-
рактерен шиповник морщинистый (Rosa rugosa).
На горных склонах приморской полосы, в высотном интервале до 500–
600 м, развит пояс лесов из дуба монгольского, которому обычно сопутствуют
липа амурская, клен моно, береза маньчжурская (Betula mandshurica) и то-
поль Давида (Populus davidiana). После пожаров дуб образует обильную по-
росль, со временем развивающуюся в почти чистый дубняк, сложенный низ-
корослыми корявыми экземплярами. Современные дубняки большей частью
производны, они возникли на месте своеобразных приморских кедровников с
дубом. Производные дубовые леса спорадически распространены и в поясах
кедрово-широколиственных, а также елово-кедрово-широколиственных лесов –
приблизительно до высоты 800 м над ур. моря и только на южных склонах.
Коренные дубняки изредка встречаются на скалистых гребнях водораз-
делов и на крутых склонах южной экспозиции. Однако и они в той или иной
степени повреждены лесными пожарами, особенно в прибрежной части запо-
ведника. На крутых горных склонах формируется труднопроходимое дубовое
криволесье высотой 1,5–2,5 м.
90 % всей площади лесов из дуба монгольского приходится на дубняки
горные, включающие особую группу высокогорных дубовых лесов, реликто-
вых по происхождению. В этих сообществах наряду с представителями тепло-
любивой маньчжурской флоры (клен моно, леспедеца двухцветная (Lespedeza
bicolor), лещина разнолистная, виноград амурский, актинидия коломикта)
встречаются в значительном количестве виды охотско-камчатской флоры (бе-
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
190
реза каменная, кедровый стланик, рододендрон золотистый), а также типич-
ные бореальные элементы (брусника, багульник, толокнянка).
На высоте 200–600 м над ур. моря самостоятельный пояс образуют кед-
рово-широколиственные леса – наиболее богатая и своеобразная во флорис-
тическом отношении лесная формация. Разнообразие лесорастительных усло-
вий и большое количество видов древесных и кустарниковых растений опре-
деляют сложный типологический состав кедровников. Значительная часть
видов растений, распространенных в кедровых лесах, относится к теплолюби-
вой маньчжурской флоре. В верхнем горном поясе сосна корейская и ее под-
рост встречаются в составе высокогорных пихтово-еловых лесов до высоты
1000–1200 м над ур. моря.
Кедрово-широколиственные леса делятся на два крупных эколого-типо-
логических комплекса: горные кедровники и долинные; последние к настоя-
щему моменту заменены производными лиственными лесами. Горные кед-
ровники наиболее сложны и разнообразны по типологическому составу. Они
включают следующие группы типов леса: сухие, периодически сухие, све-
жие и влажные леса. Первые приурочены к скалистым гребням невысоких
водоразделов и к крутым склонам южных экспозиций. На долю кедра (Pinus
koraiemsis) приходится 70–80 % состава. Постоянным спутником кедра явля-
ется дуб, иногда встречается береза даурская (Betula davurica). Фон в подлеске
образует рододендрон сихотинский (Rhododendron sichotense) с участием лес-
педецы двухцветной, бересклета малоцветкового (Euonymus pauciflora) и спи-
реи березолистной (Spiraea betulifolia). В травостое преобладают мезоксе-
рофитные растения – осоки возвратившаяся и уссурийская (Carex reventa,
C. ussuriensis), полыни Гмелина и побегоносная (Artemisia gmelinii, A. stoloni-
fera), мятлики дубравный и ложноболотный (Poa nemoralis, P. pseudopalus-
tris), патриния скальная (Patrinia rupestris).
Наиболее широко распространены свежие кедровники. Они приурочены
к среднекрутым склонам гор преимущественно южных экспозиций и высоким
террасам. Помимо кедра обычны липа амурская, береза ребристая (Betula
costata), вяз разрезной и дуб монгольский. Фон в подлеске образуют лещина
маньчжурская, чубушник тонколистный, элеутерококк и бересклет священ-
ный (Euonymus sacrosancta). Из лиан наиболее характерен виноград амур-
ский. В травостое преобладают кочедыжники, папоротник орляк, а также лан-
дыш Кейзке (Convallaria keiskei), марьянник щетинистый (Melampyrum
setaceum) и осока ланцетная (Carex lanceolata).
Для склонов северных экспозиций, гребней водоразделов и узких участ-
ков долин горных рек характерны кедровники с темнохвойными породами –
елью аянской и пихтой белокорой. В них встречаются такие древние релик-
товые растения, как тис остроконечный, вяз разрезной и – как крайняя ред-
кость – рододендрон короткоплодный (Rhododendron brachycarpum). По сво-
ему характеру эта группа кедровников является переходной к кедрово-еловым
лесам. В спелых древостоях на долю кедра приходится 60–70 % состава.
Обычны липа амурская, береза ребристая, клен моно, ель аянская, пихта бело-
корая, а в нижнем пологе – клены зеленокорый и желтый (Acer tegmentosum,
A. ukurunduense) и вишня Максимовича (Cerasus maximowiczii). В подлеске
преобладают чубушник, лещина маньчжурская, смородина Максимовича (Ribes
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
191
maximoviczianum), бересклет малоцветковый (Euonymus pauciflora) и абелия
корейская (Zabelia coreana). В травостое – причудливая смесь таежных и теп-
лолюбивых растений. Фон образуют крупные папоротники и осоки. Здесь же
растут самая маленькая из орхидей – калипсо луковичная (Calypso bulbosa),
крошечный касатик одноцветковый (Iris uniflora), энемион Радде (Enemion
raddeanum) и своеобразный небольшой папоротник гроздовник полулунный
(Botrychium lunaria). На стволах и в кронах деревьев хорошо развиты эпифит-
ные мхи.
Высотный интервал от 600–800 до 1100–1300 м над ур. моря определяет
положение пояса пихтово-еловых лесов. Ниже они проникают лишь по доли-
нам горных рек, где развиваются явления температурной инверсии. Помимо
основных лесообразующих пород для ельников заповедника характерно уча-
стие лиственницы ольгинской (Larix olgensis), берез ребристой и шерстистой,
кленов зеленокорого и желтого, кедра. Пихта белокорая наиболее обычна до
высоты 1000 м над ур. моря. Выше преобладают ельники с участием березы
шерстистой и лиственницы Комарова.
Еловые леса представлены двумя типологическими комплексами – гор-
ным и долинным. Горные ельники по особенностям флористического состава
и режима тепла и влаги разделяются на две категории – среднегорные и вы-
сокогорные. На первые приходится не менее 80 % площади еловых лесов. Ос-
новными группами типов в этих ельниках являются зеленомошные, папорот-
никовые и кустарниковые.
Высокогорные пихтово-еловые леса сформированы преимущественно
елью с участием пихты белокорой, березы шерстистой, рябин амурской и бу-
зинолистной (Sorbus amurensis, S. sambucifola). Наиболее широко распростра-
нены моховые и травяные ельники, а кустарниковые и бадановые (c Bergenia
pacifica) занимают меньшие площади. На склонах разной крутизны и на во-
доразделах близ верхней границы прямоствольного леса встречаются зелено-
мошные ельники. Пятнами в микропонижениях встречаются сфагновые мхи.
После пожаров зеленомошные ельники сменяются вейниковыми каменнобе-
резняками (с Calamagrostis purpurea).
Верхний высотный пояс лесной растительности – каменноберезовые ле-
са, непосредственно контактирующие с зарослями подгольцовых кустарников.
Образованы они березой шерстистой. На нижней границе распространения
для каменноберезняков характерно участие ели и пихты, а на верхней – кед-
рового стланика. Большей частью каменноберезняки не образуют сплошных
массивов и языками вклиниваются в пояс кедрового стланика, поднимаясь до
высоты 1400–1500 м над ур. моря и опускаются до высоты 1100–1000 м над
ур. моря в пояс высокогорных ельников.
На высоте 1200 м и более над ур. моря хорошо выражен пояс подгольцо-
вых кустарников. Он сформирован зарослями кедрового стланика. В этом поя-
се отмечены сообщества таких кустарников, как рододендроны золотистый и
сихотинский, вейгела Миддендорфа, можжевельник сибирский, багульник и
подбел. Рассеянно встречается высокогорная рябина бузинолистная. У ниж-
ней высотной границы своего распространения кедровый стланик образует
сплошные заросли, достигающие высоты 3–4 м и более. Наиболее широко
распространены кедровостланиковые заросли с покровом из зеленых мхов.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
192
У верхней границы своего распространения кедровый стланик не образует
сплошного полога; в этих условиях наиболее распространен кедровый стланик
лишайниковый.
Горные тундры встречаются лишь на отдельных вершинах, достигающих
высоты 1400 м и более над ур. моря. Чаще всего они представлены вересково-
лишайниковыми группировками. Местами встречаются густые заросли ба-
гульника приземистого и рододендрона мелколистного.
Из внепоясных лесных группировок в заповеднике наиболее распро-
странены лиственничники, которые встречаются как в долинах рек, так и на
склонах гор на разных высотах. В большинстве случаев они пришли на смену
коренным сообществам в результате пожаров. Обычны травянистые, кустар-
ничковые и моховые лиственничники.
В долинах горных рек распространены следующие лиственные лесные
формации: ивняки, ольшаники, чозенники, тополевники, а также смешанные
широколиственные группировки. Главные лесообразующие породы ивняков –
несколько видов ив (Шверина, росистая и удская – Salix schwerinii, S. rorida,
S. udensis). Кореянка земляничниколистная – чозения – формирует чистые
древостои в средней части бассейнов горных рек. Тополь Максимовича (Populus
maximowiczii) образует древостой на средних и высоких уровнях хорошо дре-
нированных пойменных террас. Долинные широколиственные леса, в основ-
ном представленные группировками из ясеня маньчжурского, встречаются
небольшими участками в нижней и средней частях бассейнов рек.
Уссурийский государственный природный заповедник имени
акад. В. Л. Комарова расположен на отрогах Южного Сихотэ-Алиня. Пре-
обладающая высота гребней водоразделов в заповеднике – 300–400 м над
ур. моря, отдельные вершины достигают 650–700 м. Климат района заповед-
ника можно характеризовать как муссонный летнетеплый и влажный с уме-
ренно суровой малоснежной зимой. Средняя температура января –17,9, авгу-
ста 19,7 °С. Годовая сумма осадков – 750–800 мм.
Под хвойно-широколиственными и широколиственными лесами на гор-
ных склонах развиты горнолесные бурые почвы. На крутых каменистых юж-
ных склонах под кедрово-дубовыми и дубовыми лесами формируются мало-
мощные, слабогумусированные почвы.
Для дренированных участков речных долин характерны дерново-аллюви-
альные и буроземно-аллювиальные почвы. В местах с застойным увлажнени-
ем и затрудненным дренажем выделяются почвы торфяно-глеевые, торфяни-
сто-перегнойно-глеевые и задернованные иловато-глеевые.
Уссурийский заповедник расположен в южной подзоне смешанных хвой-
но-широколиственных лесов. Безраздельное господство принадлежит здесь
маньчжурскому флористическому комплексу. Элементы охотского комплекса
переходят на второстепенные позиции. Флора Уссурийского заповедника сло-
жена почти исключительно лесными видами. Систематический состав флоры:
118 видов лишайников, 252 вида мохообразных, 825 видов сосудистых рас-
тений.
Основной формацией растительности заповедника являются леса, обра-
зованные главным образом кедровой сосной корейской (кедром корейским) в
сочетании с многочисленными широколиственными породами. Кедровники
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
193
характеризуются богатым флористическим составом, многоярусностью, высо-
кой сомкнутостью и разновозрастностью древостоев. Эти леса относятся к
южной ботанико-географической группе, о чем свидетельствует участие в их
составе пихты цельнолистной, граба сердцелистного, ряда теплолюбивых ли-
ан и представителей семейства аралиевых.
Часто встречаются участки кедровников, где на площади меньше гектара
можно обнаружить до 50–60 видов деревьев и кустарников, 4–5 лиан и более
сотни видов травянистых растений. Кедровники представлены большим чис-
лом типов; выделяются кедровники сухие, периодически сухие, свежие и
влажные.
Широко распространены влажные кедровники, на долю которых прихо-
дится более половины общей площади кедровой формации. Они приурочены
к пологим и среднекрутым склонам всех экспозиций. Древостои этих лесов
сложные по составу, сомкнутые, разновозрастные, с тремя-четырьмя полога-
ми. Кедр, достигающий высоты 35–40 м, преобладает. Из многочисленных
широколиственных пород обычны липа амурская и Таке, клены моно и мань-
чжурский, бархат амурский и орех маньчжурский. С высоты 400–500 м над ур.
моря в лесах возрастает участие ели аянской и пихты белокорой. Подлесок во
влажных кедровниках средней густоты, разнообразный по видовому составу.
Наиболее характерны чубушник, элеутерококк, жимолость Максимовича, бе-
ресклет большекрылый (Euonymus macroptera), лещина маньчжурская. Лиа-
ны – актинидии острая и коломикта, лимонник китайский. Подобные лиано-
вые кедровники внешне напоминают влажные леса субтропиков. В густом
травяном покрове фон создают щитовник толстокорневищный и кочедыжни-
ки с участием фримы (Phryma asiatica), василистника нитчатого (Thalictrum
filamentosum) и осок.
Влажные кедровники встречаются не только на горных склонах, но и в
долинах горных рек, где приурочены к дренированным участкам надпоймен-
ных террас. Здесь в составе древостоев появляются ясень маньчжурский, вяз
японский, ель аянская, пихта белокорая; возрастает участие ореха маньчжур-
ского и бархата амурского. Больших размеров достигают сирень амурская, че-
ремухи Маака и азиатская (Padus maackii, P. asiatica). Представители семей-
ства аралиевых – свободноягодник (элеутерококк) и калопанакс.
Леса из пихты цельнолистной, или черной, в Уссурийском заповеднике
находятся близ северной границы ареала. В их составе число видов древес-
ных растений очень велико. Выделяется две группы типов – свежие и влаж-
ные чернопихтарники. Влажные чернопихтарники – наиболее флористиче-
ски насыщенные леса заповедника. В их древостое помимо пихты цельно-
листной обычны липы амурская и Таке, ясень маньчжурский, вяз разрезной,
бархат амурский, орех маньчжурский и береза ребристая. Нижние полога
сложены кленами моно, маньчжурским, зеленокорым, грабом сердцелист-
ным, рябиной ольхолистной (Sorbus alnifolia) и маакией амурской (Maackia
amurensis). Из лиан наиболее мощно развита актинидия острая (Actinidia
arguta).
Кедрово-елово-широколиственные леса образуют переход от кедрово-
широколиственных к пихтово-еловым. В заповеднике они представлены груп-
пой формаций с елью аянской (преобладает) и елью корейской. В их соста-
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
194
ве обязательно встречаются сосна корейская и многочисленные лиственные
породы.
В верховьях рек в верхних частях крутых южных и западных склонов
встречаются своеобразные кедрово-елово-широколиственные леса с дубом мон-
гольским. Дуб в этих условиях достигает крупных размеров – высоты 18–22 м.
Леса из ели аянской, пихты белокорой характерны для склонов северных
экспозиций и к горным плато северной и северо-западной частей заповедни-
ка, где высоты превышают 500 м над ур. моря. В заповеднике пихтово-еловые
леса представлены тремя группами типов: папоротниковыми, зеленомошны-
ми и кустарниковыми ельниками.
Долинные лиственные леса в заповеднике представлены ивняками,
ольшаниками, тополевниками, чозенниками, вязовниками и ясеневниками,
образующими серийные сообщества. Отмечено несколько видов ив (сердце-
листная, росистая, тонкостолбиковая, скрытная, ниппонская, тарайкинская –
Salix cardiophylla, S. gracilistyla, S. abscontida, S. nipponica, S. taraikensis), но
древостой образует только ива Шверина (высота 10–12 м). Из тополевников
наиболее распространены тополевники папоротниковые, сложенные тополем
Максимовича, приуроченные к дренированным участкам средней и высокой
пойм горных рек. Чозенники занимают небольшие территории, приуроченые
к низким и средним уровням поймы со свежими песчано-галечниковыми
наносами. Кустарниковые и осоково-спиреевые ясеневники из ясеня мань-
чжурского приурочены к участкам высокой поймы и надпойменных террас, а
также встречаются на конусах выноса и на шлейфах горных склонов. Среди
вязовых лесов (вяз японский) наиболее распространены кустарниковые вя-
зовники, занимающие хорошо дренированные участки средней и высокой
поймы с плодородными почвами.
7.3. АНТРОПОГЕННЫЕ НАРУШЕНИЯ ЛАНДШАФТОВ ТАЙГИ
И ХВОЙНО-ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
Территория средней и особенно северной тайги стала интенсивно осваи-
ваться в России сравнительно недавно. Основу хозяйственной деятельности
населения долгое время составляли лесные промыслы: заготовка древесины,
сбор дикорастущих грибов и ягод, охота, рыболовство. Поначалу население
было немногочисленным и концентрировалось преимущественно в долинах
рек. Именно здесь стало развиваться животноводство, а затем и земледелие,
поскольку почвы пойм более плодородны. На месте вырубленных лесов по-
явились пастбища и пашни.
Очаговое земледельческое освоение среднетаежных регионов Европей-
ской России насчитывает не менее, а в южной тайге – более 1000 лет. В под-
зоне южной тайги, климат которой по сравнению с северными подзонами бо-
лее благоприятен для сельского хозяйства, площади освоенных земель стали
расширяться. Однако с середины ХХ века в связи с ликвидацией малых дере-
вень в тайге происходило постоянное сокращение площади сельскохозяй-
ственных угодий. За 1965–1998 гг. ее уменьшение в разных областях состави-
ло от 11 до 26 %. При этом наиболее существенное сокращение площади уго-
дий произошло в тайге Европейской территории России. Заброшенные земли
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
195
стали зарастать кустарниками и производными лесами. Заметим, что запусте-
ние сельскохозяйственных угодий наносит не только экономический ущерб,
но означает также утрату сельских пейзажей, с которыми ассоциируются кар-
тины родной русской природы.
Принципиальные изменения в структуре тайги произошли во второй
половине XX века. Объем заготовок древесины необычайно возрос. Расшири-
лось техническое вооружение лесозаготовителей, появилась сеть автомобиль-
ных и железных дорог, благодаря чему значительная часть территории тайги
стала сравнительно легкодоступной. Крупные районы освоения лесных ресур-
сов охватили практически всю европейскую часть таежных лесов, значитель-
ную часть Урала, большие площади в Западной, Средней и Восточной Сибири,
а также Дальнего Востока. В связи с расширением геологоразведочных работ,
добычей полезных ископаемых даже в труднодоступных местах таежные
ландшафты стали страдать от хозяйственной деятельности человека. Большой
урон тайге наносят разработка нефтяных месторождений, прокладка трубо-
проводов (рис. 7.36), лесозаготовки (рис. 7.37), а также пожары, выжигаю-
щие большие площади леса, ценные охотничьи, ягодные и грибные угодья
(рис. 7.38). Стабильность лесных экосистем нарушается, деревья начинают
страдать от болезней и массового размножения насекомых-вредителей.
Леса играют важную почвозащитную и водоохранную роль: там, где они
уничтожаются, увеличивается поверхностный сток и, как следствие, усилива-
ется смыв почвы, возникают овраги, уменьшается пополнение подземных вод,
малые реки мелеют в межень.
В XXI веке тенденции роста лесозаготовок, отчуждения таежных ланд-
шафтов для строительства городов и промышленных объектов будут сохра-
няться.
Рис. 7.36. Прокладка трубопровода в тайге
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ. Умеренный пояс
196
Рис. 7.37. Лесозаготовки
По данным ученых Мэрилэндского университета, работавших со снимка-
ми, сделанными со спутников серии Landsat, и проследивших изменения состо-
яния лесного покрова планеты с 2000 по 2012 год, Россия за этот период поте-
ряла больше лесов, чем любая другая страна мира: а именно, более 365 тысяч
из 8,5 миллиона кв. км леса. Из них около 162 тысяч квадратных километров
было восстановлено. Больше всего леса погибло в Якутии, в районе верхнего
течения реки Лены, в результате пожаров и хозяйственной деятельности.
Чтобы избежать неблагоприятных экологических последствий, человек
должен взять на себя затраты на восстановление экологического потенциала
таежных экосистем.
Рис. 7.38. Пожар в лесу
.
Глава 7. ЗОНА БОРЕАЛЬНЫХ ХВОЙНЫХ (ТАЕЖНЫХ) ЛЕСОВ
197
Вопросы к главе 7
1. Природные условия зоны тайги; экология растений, экологическая ординация со-
обществ.
2. Широтные особенности растительности зоны бореальных хвойных (таежных) лесов:
подзоны северной, средней, южной тайги и подтайги.
3. Региональные особенности растительности таежных лесов: Восточно-Скандинавская
(Карельская), Восточно-Европейская, Западно-Сибирская, Средне-Сибирская, Кам-
чатская, Сахалинская, Приморская области.
Рекомендуемая литература
Бобров Е. Г. Лесообразующие хвойные СССР. Л.: Наука, 1978. 187 с.
Болотные системы Западной Сибири и их природоохранное значение / Под ред. д.б.н.
проф. В. Б. Куваева. Тула: Гриф и К°, 2001. 584 с.
Боч М. С., Мазинг В. В. Экосистемы болот СССР. Л.: Наука, 1979. - 188 с.
Дальний Восток. М.: Изд-во АН СССР, 1961. 440 с.
Исаченко Г. А., Резников А. И. Динамика ландшафтов тайги Северо-Запада Европей-
ской России. СПб.: Изд-во Русск. Геогр. о-ва, 1996. 166 с.
Колесников Б. П. Растительность // Дальний Восток. М.: Изд-во АН СССР, 1961.
С. 183–245.
Лесные экосистемы Енисейского меридиана / Плешиков Ф. И ., Ваганов Е. А ., Вед-
рова Э. Ф. и др. Новосибирск: Изд-во СО РАН, 2002. 356 с.
Предбайкалье и Забайкалье. М.: Наука, 1965. 492 с.
Рысин Л. П. Кедровые леса России. М.: Т-во науч. изданий КМК, 2011. 240 с.
Рысин Л. П. Лиственничные леса России. М.: Т-во науч. изданий КМК, 2010. 343 с.
Рысин Л. П., Манько Ю. И., Бебия С. М. Пихтовые леса России. М.: Т-во науч. изданий
КМК, 2012. 200 с.
Рысин Л. П., Савельева Л. И. Еловые леса России. М.: Наука, 2002. 335 с.
Рысин Л. П., Савельева Л. И. Сосновые леса России. М.: Т-во науч. изданий КМК, 2008.
289 с.
Тиходеева М. Ю. Разнообразие лесов России: уч. пос. СПб.: Изд-во СПбГУ, 2011. 156 с.
Толмачев А. И. К истории возникновения и развития темнохвойной тайги. М.–Л.: Изд-
во АН СССР, 1954. 155 с.
Федорчук В. Н., Нешатаев В. Ю., Кузнецова М. Л. Лесные экосистемы северо-запад-
ных районов России: Типология, динамика, хозяйственные особенности. СПб.:
Хромис, 2005. 382с.
Шумилова Л. В. Ботаническая география Сибири. Томск: Изд-во Томского ун-та, 1962.
439 с.
Интернет-сайт «Леса России» http://lesa-rossii.ru/
Карты:
Атлас малонарушенных лесных территорий России / Ред. А . В. Щербаков. 2002.
(www.forest.ru)
Карта растительности Приморского края (http://www.fegi.ru)
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
198
нтразональная растительность – растительность, представленная со-
обществами, включенными в одну из основных зон растительности, и
не образующая самостоятельной зоны. Например, растительность материко-
вых (суходольных) лугов в таежной зоне, сфагновые болота в тундре и тайге.
8.1. МАТЕРИКОВЫЕ ЛУГА ЛЕСНОЙ ЗОНЫ
Луг как ботанико-географическое и геоботаническое понятие – это био-
геоценоз, образованный многолетними мезофильными травами, вегетирую-
щими без летнего перерыва. Растениями-эдификаторами в луговых фитоце-
нозах обычно являются злаки. Число видов травянистых растений, входящих
в состав конкретных луговых фитоценозов, сильно варьирует. Сообщества мо-
гут быть образованы 2–3 видами (на эталонной площади в 100 м2), а в сред-
нем насчитывают около 40 видов. Флористическое богатство луговых ценозов
зависит от нескольких причин: общего состава флоры района, условий произ-
растания, длительности существования сообщества, формы и степени влияния
деятельности человека и др.
Различают материковые и пойменные луга. Под материковыми лугами
понимают луга, расположенные на водораздельных междуречных равнинах
(плакорах), а также на холмах, гривах, в верхних частях склонов долин и овра-
гов. Предметом настоящей главы является описание интразональных матери-
ковых лугов, формирующихся в лесной зоне. Материковые луга разделяются
на суходольные и низинные. Суходольными материковыми лугами (или лу-
говыми суходолами) будем называть луга, увлажняемые главным образом
атмосферными осадками; грунтовые и почвенные воды залегают глубоко и в
увлажнении почвы значения не имеют или почти не имеют. Низинными лу-
гами называем материковые луга, увлажняемые не только атмосферными,
но также и грунтовыми или почвенными водами (проточными или застой-
ными).
Суходольные луга распространены в основном в подзонах средней, юж-
ной тайги и в подзоне хвойно-широколиственных лесов. В пределах Европей-
ской части России северная граница массового распространения суходольных
лугов проходит через южную Карелию и Вологодскую область в Пермское
Предуралье. Южная граница – Республика Беларусь, Московская и Горьков-
ская области, Южный Урал. Преобладание суходольных лугов в указанных
границах объясняется наибольшей населенностью этой части лесной зоны и
удовлетворительными эколого-географическими условиями. В северной по-
ловине лесной зоны, где население реже и приурочено больше к рекам с пой-
менными лугами, и где эколого-географические условия на суходолах ухуд-
шаются, суходольные луга встречаются, но небольшими массивами и редко.
Глава 8
ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
.
Глава 8. ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
199
Возникновение и развитие суходольных лугов. На всем протяже-
нии таежной зоны суходольные луга имеют вторичное происхождение, они
возникают на месте уничтоженной коренной лесной растительности в резуль-
тате пожаров, вырубок, создания пахотных угодий. Признаки лесного про-
шлого на суходольных лугах сохраняются долго, особенно на бедных почвах:
остатки лесной флоры, подзолистый горизонт в почве. Генетические взаимо-
связи лесной, луговой растительности и сельскохозяйственных угодий на их
месте показаны на рис. 8.1.
Лесные вырубки и гари на суходолах обычно зарастают иван-чаем
(Chamaenerion angustifolium), вейниками (Calamagrostis epigeios, а на более
плодородных почвах – С. arundinacea). Иногда на богатых почвах разрастаются
более сложные заросли высоких трав. Если деятельность человека не направле-
на на формирование луговых угодий на их месте, то происходит закустаривание
и, в конечном счете, – восстановление лесных сообществ. Наиболее глубокая
трансформация лесов происходит в результате создания на месте лесов пахот-
ных угодий. Если пашни выходят из севооборота, то на месте пашни (пара)
начинается процесс перехода ее в залежь, затем в перелог, который постепен-
но трансформируется в луг.
Этот процесс проходит через ряд смен видов-доминантов, относящихся
к разным биоморфам. Первая стадия перехода пара в залежь – бурьянистая –
Восста-
новив-
шиеся
корен-
ные
леса
Мелко-
лист-
венные
леса
Лесные
гари
Корен-
ные
леса
Лесные
выруб-
ки
Произ-
водные
мелко-
лист-
венные
леса
Восста-
новив-
шиеся
корен-
ные
леса
Пашня
Перелог
Луговые
болота
Луговые
ценозы
Болота
Кустарниково-
луговые ценозы
Производные
мелколиственные
леса
Восстановившиеся
коренные леса
Пашня
Перелог
Луговые
ценозы
Кустарниково-
луговые ценозы
Производные
мелколиственные
леса
Восстановившиеся
коренные леса
Рис. 8.1. Генетические взаимосвязи лесной, луговой растительности
и сельскохозяйственных угодий на их месте
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
200
представлена стержнекорневыми растениями: одуванчик лекарственный (Ta-
raxacum officinale), полынь горькая (Artemisia absinthium), бодяк полевой
(Cirsium arvense), бедренец камнеломка (Pimpinella saxifraga), купырь лесной
(Anthriscus sylvestris), цикорий обыкновенный (Cichorium intybus), клевер лу-
говой (Trifolium pratense), люцерна серповидная (Medicago falcata); в первые
годы здесь также сохраняются остатки сорно-полевой флоры: щавелек обык-
новенный (Rumex acetosella), василек синий (Centaurea cyanus), трехреберник
непахучий (Tripleurospermum inodorum), пастушья сумка обыкновенная
(Capsella bursa-pastoris), ярутка полевая (Thlaspi arvense), сурепка обыкно-
венная (Barbarea vulgaris), осот полевой (Sonchus arvensis).
Следующий этап – дерновый процесс, который сопровождается обога-
щением сообществ луговыми видами. Стадии дернового процесса заключают-
ся в последовательной смене среди эдификаторов: длиннокорневищных (Ely-
trigia repens, Bromopsis inermis), рыхлокустовых (Phleum pratense, Alopecurus
pratensis, Dactylis glomerata) и плотнокустовых (Deschampsia cespitosa, Nar-
dus stricta, Festuca ovina) злаков (рис. 8.2). Формирование плотного дернового
горизонта ухудшает аэрацию, разложение органических остатков, условия пи-
тания; нередко дерновое уплотнение почвы вызывает слабое заболачивание.
Если антропогенная деятельность на лугах не производится, то происходит
закустаривание и лесное сообщество вновь восстанавливается.
Рис. 8.2. Дерновинные злаки и типы их кущения (по А. П. Шенникову):
1 – длиннокорневищный (Elytrigia repens); 2 – рыхлокустовой (Phleum pratense);
3 – плотнокустовой (Deschampsia cespitosa)
.
Глава 8. ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
201
Фитоценотическая структура луговых сообществ. Фитоценоти-
ческая структура луга образована ярусом травянистых растений. По мнению
Т. А. Работнова, подразделение травяного яруса на подъярусы нецелесообраз-
но, что связано с образованием его растениями одной жизненной формы –
многолетними травами; относительно небольшой высотой травостоев; при-
сутствием в пределах популяций отдельных видов особей различной высоты; с
отсутствием у высоких растений приверженности листьев к какому-то опреде-
ленному горизонту; и в целом, постепенным кулисообразным заполнением
пространства растениями, имеющими разную высоту. В травяных фитоцено-
зах ярусность выражена лишь там, где развит моховой покров и можно разли-
чать два яруса: травяной и моховой. Там, где ярусность не выражена, нет смыс-
ла искусственно выделять подъярусы, важнее изучать разновысотное разме-
щение листьев отдельных видов. В целом, по характеру сложения луговые
травостои обычно делят на высокорослые, средне- и низкорослые.
Видовой состав и сложение сообщества формируются таким образом,
чтобы эффективно использовать различные надземные и подземные горизон-
ты в разные периоды вегетации. В результате адаптаций растений к совмест-
ному произрастанию вырабатывались структура ценозов и специфика ритмов
их сезонного развития (рис. 8.3). Высотное распределение органов меняется в
течение сезона в связи с динамикой роста и увеличением числа побегов на
единицу площади.
Подземные органы растений на лугах также распределены на разных
глубинах, но основная масса их сосредоточена в дернине, т. е. примерно в
20–30 -сантиметровом верхнем горизонте почвы. Обычно растения на лугах
представлены не единичными экземплярами, относящимися к какому-
либо виду, а совокупностью особей этого вида – ценотической популяцией.
Рис. 8.3. Вертикальное сложение лугового сообщества
(по H. Walter, S.-W. Breckle, с изменениями)
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
202
Каждая ценотическая популяция характеризуется числом растений, ее со-
ставляющих, и соотношением их возрастных групп. Помимо вертикальной
структуры, луговые ценозы обладают горизонтальным расчленением травяно-
го покрова, что объясняется небольшими различиями в условиях местообита-
ния и проявляется в чередовании участков, или пятен, различающихся по ви-
довому составу. Такие участки получили название микрогруппировок, а явле-
ние – мозаичностью.
Сезонная изменчивость луговых сообществ. Луговые ценозы ха-
рактеризуются хорошо выраженной сезонной изменчивостью. В течение ве-
гетационного периода происходит смена фенологического состояния компо-
нентов сообществ, особенно ярко проявляющаяся в динамике их цветения:
постепенном увеличении числа цветущих видов от весны к лету, вплоть до
«апогея фенологического развития», а затем в довольно быстром уменьше-
нии их. Массовое цветение отдельных видов или групп видов, последова-
тельно сменяющих друг друга в течение вегетационного периода, преобра-
жают на каждом этапе вид луга, создавая особое цветовое пятно – аспект
(рис. 8.4). Во многих фитоценозах основное значение в выделении аспектов
имеют сменяющиеся за вегетационный сезон фазы цветения некоторых ви-
дов. В ряде случаев не меньшую роль играет фаза плодоношения, например,
в сообществах с большим участием одуванчика различают аспекты как в фа-
зе его цветения, так и в фазе его плодоношения. Наиболее отчетливо смена
аспектов выражена в многовидовых травяных фитоценозах с обильным уча-
стием видов разнотравья, имеющих крупные цветки или соцветия, нередко
яркоокрашенных.
Отмечаются значительные изменения в количественном соотношении
видов по вегетационному сезону, вплоть до смены доминантов, в результате
чего один и тот же участок по описаниям, проведенным в разные сроки, мож-
но отнести к различным сообществам.
Эколого-топологическая ординация луговых сообществ. Эко-
логические соотношения между сообществами луговой растительности и их
переходы в соседние нелуговые сообщества изображаются в виде схемы эко-
логических рядов (рис. 8.5). Эта схема одновременно выражает и эколого-то-
пологические закономерности их размещения и фитоценобиотические раз-
личия.
Ряд от настоящих лугов к остепненным наблюдается при переходе от лу-
гов лесных областей к лугам степных областей (географический ряд) и в обеих
областях – при переходе от средневлажных к сухим и достаточно теплым и
богатым местообитаниям (топологический ряд). Настоящие луга сменяются
пустошами в северных и высокогорных (холодных) областях (географический
ряд) и в условиях бедных, но не заболоченных почв (топологический ряд). Как
остепнение, так и переход в пустоши сопровождаются уменьшением эдифика-
торного значения настоящих мезофитов.
Болотистые и торфянистые луга встречаются в гумидных и даже суб-
аридных, в широтных и высотных ботанико-географических зонах, в каждой
из них имея свои особенности, обусловленные климатом и эколого-топологи-
ческими условиями.
.
Глава 8. ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
203
Рис. 8.4. Фенологический спектр
мелкозлаково-разнотравного
краткопоемного луга около
р. Вологда (Шенников, 1964,
с изменениями)
Сезонные стадии луга: 1 – предвесен-
няя; 2 – ранневесенняя; 3 – середины
весны; 4 – конца весны – начала лета;
5–разгарлета;6–концалета;7–
осенняя.
Фенологические фазы: а – вегетатив-
ная; б – бутонизации; в – цветения;
г – созревания семян; д – обсемене-
ния; е – отмирания.
Виды: 1 – Viola arenaria; 2 –
Equisetum pratense; 3 – Glechoma
hederacea; 4 – Geum rivale; 5 –
Taraxacum officinale; 6 – Polygala
amarella; 7 – Alchemilla xanthochlora;
8 – Carum carvi; 9 – Trollius europaeus;
10 – Fragaria vesca; 11 – Ranunculus
acris; 12 – Plantago media; 13 – Luzu-
la campestris; 14 – Trifolium pratense;
15 – Cerastium holosteoides; 16 – Bis-
torta major; 17 – Veronica chamae-
drys; 18 – Anthriscus sylvestris; 19 –
Rumex thyrsiflorus; 20 – Ranunculus
polyanthemos; 21 – Polygala comosa;
22 – Stellaria graminea; 23 – Amoria
repens; 24 – Vicia cracca; 25 –
Tragopogon pratensis; 26 – Euphrasia
hirtella; 27 – Leucanthemum vul-
gare;28 – Poa pratensis; 29 – Festuca
rubra; 30 – Lathyrus pratensis; 31 –
Geranium pratense; 32 – Oberna litto-
ralis; 33 – Heracleum sibiricum; 34 –
Rhinanthus serotinus; 35 – Leontodon
autumnalis; 36 – Galium mollugo;
37 – Galium boreale; 38 – Campanula
glomerata; 39 – Deschampsia cespi-
tosa; 40 – Prunella vulgaris; 41 –
Achillea millefolium; 42 – Brisa media;
43 – Festuca pratensis; 45 – Dactylis
glomerata; 46 – Pimpinella saxifraga;
47 – Agrostis gigantea; 48 – Delphini-
um elatum; 49 – Phleum pratense;
50 – Elytrigia repens; 51 – Hieracium
umbellatum; 52 – Tanacetum vulgare;
53 – Antennaria dioica.
Ширина полос видовых спектров со-
ответствует проценту встречаемости
вида, ширина полос генеративной
фазы – проценту встречаемости гене-
ративных пробегов
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
204
Рис. 8.5. Схема экологических рядов луговой растительности и смежных типов
растительности (по А. П. Шенникову)
Настоящие луга приурочены к средним условиям умеренно и устойчи-
во увлажняемой почвы, достаточно теплой, достаточно богатой элементами
минерального питания в легко усвояемой форме, без заболачивания, без засо-
ления. Эти условия могут быть обеспечены и географическим и топологиче-
ским положением. Стрелки в схеме (см. рис. 8.5) указывают направления смен,
наблюдаемых между сообществами. Между крайними положениями имеются
промежуточные и границы между ними условны. Многие ассоциации зани-
мают место между основными рядами, как это показано в схеме.
Конкретизируя схему экологических рядов А. П. Шенникова, приведем
схему эдафо-фитоценотической ординации суходольных лугов средней и юж-
ной тайги Восточно-Европейской равнины (рис. 8.6), отражающей смены рас-
тительности в связи с изменением богатства и режима увлажнения почв.
По богатству различаются бедные, средние и богатые почвы, по режиму
увлажнения – устойчивый, переменный, обильно застойный и обильно про-
точный режимы. При составлении этой схемы использована методология
В. Н. Сукачева для ординации растительных сообществ в виде эдафо-фитоце-
нотического креста.
.
Глава 8. ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
205
В центре креста расположены местообитания с оптимальными условия-
ми для формирования настоящих лугов: устойчивым увлажнением и относи-
тельно богатыми дерново-подзолистыми супесчаными и суглинистыми поч-
вами.
В растительном покрове господствуют мезофиты. Доминантами луговых
сообществ являются полевица тонкая (Agrostis tenuis) и душистый колосок
(Anthoxanthum odoratum); сопутствующие виды – овсяница луговая (Festuca
pratensis), тимофеевка луговая (Phleum pratense), мятлик луговой (Poa pratensis);
разнотравную основу составляют – нивяник (Leucanthemum vulgare), коло-
кольчик раскидистый (Campanula patula), манжетка желто-зеленая (Alchemilla
xanthochlora), клевер луговой (Trifolium pratense), черноголовка (Prunella
vulgaris) и др. (рис. 8.7).
При переходе к условиям переменного режима увлажнения, обусловлен-
ного низкой влагоемкостью почв, и снижения их плодородия последовательно
формируются элементы ряда А. Местообитания сухие; в естественной расти-
тельности господствуют ксерофиты и психрофиты. Мезофиты, если они име-
ются, представлены особенно выносливыми видами и всегда сильно угнетены.
Переменному режиму увлажнения соответствуют сухоразнотравные сообще-
ства, представленные мелкотравными лугами – малопродуктивными сообще-
ствами, в которых доминируют овсяница овечья (Festuca ovina) и луговик или
овсик извилистый (Avenella flexuosa). Из разнотравья к ним примешиваются
смолка клейкая (Viscaria vulgaris), кульбаба щетинистая (Leontodon hispidus),
ястребиночка обыкновенная (Pilosella officinarum), кошачья лапка (Antenna-
ria dioica), колокольчик круглолистный (Campanula rotundifolia), лапчатка
серебристая (Potentilla argentea) и др. (рис. 8.8). Завершающим элементом
ряда являются абсолютные суходолы, обычно с бедными песчаными почвами,
Рис. 8.6. Эдафо-фитоценотическая схема для суходольных лугов средней
и южной тайги Восточно-Европейской равнины
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
206
Полевица тонкая (Agrostis tenuis)
Овсяница луговая (Festuca pratensis)
Тимофеевка луговая
(Phleum pratense)
Душистый колосок
(Anthoxanthum odoratum)
Нивяник обыкновенный
(Leucanthemum vulgare)
Герань лесная
(Geranium sylvaticum)
Фото: G. Wessels, В. Гуменюка, А. Цветковой, А. Любченко, M. Новиковой, Е. Антоновой
Рис. 8.7. Доминанты настоящих лугов
.
Глава 8. ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
207
Овсяница овечья (Festuca ovina)
Овсик извилистый (Avenella flexuosa)
Кошачья лапка двудомная
(Antennaria dioica)
Ястребиночка обыкновенная
(Pilosella officinarum)
Белоус торчащий (Nardus stricta)
Смолка клейкая (Viscaria vulgaris)
Фото: J. C . Schow, P. Filippov, Д. Орешкина, Н. Гамовой, В. Гуменкжа
Рис. 8.8. Виды растений луговых сообществ ряда А
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
208
характеризующиеся резко переменным режимом увлажнения. Здесь наблю-
дается доминирование овсяницы овечьей, белоуса и разрастание ястребинки
волосистой. Луга переходят в категорию пустошного типа растительности,
особенно при развитии на них мохово-лишайникового покрова. Такие сооб-
щества образуются после вырубки сухого зеленомошно-лишайникового сос-
нового леса.
Особый эколого-топологический ряд Б формируется в бессточных ни-
зинах с обильно застойным режимом увлажнения, слабой аэрацией, повы-
шенной кислотностью почв. Настоящие луга уступают место сырым лугам с
господством щучки (Deschampsia cespitosa), ситников (Juncus effusus), осок.
Преобладают сивцово-мелкоосоковые, мелкоосоково-щучковые долгомошные
и сфагновые луга. Участие в сообществе мхов и прогрессирующий процесс
торфонакопления ведут к формированию верхового сфагнового болота.
Ряд В направлен в сторону увеличения богатства почв при сохранении
устойчивого режима увлажнения. В этих условиях формируются высокопро-
дуктивные крупнотравные луга с широким участием неморальных элемен-
тов – так называемые «листвяги». Травостой лугов очень густой, видовая
насыщенность не менее 45–50 видов в сообществе; в особо благоприятных
условиях многие представители разнотравья достигают здесь высоты до 1 м,
например, герань лесная, гравилат речной (Geum rivale), лютики золотистый,
едкий, многоцветковый (Ranunculus auricomus, R. acris, R. polyanthemos).
В сложении лугового травостоя ведущая роль принадлежит представителям
эумезофитного разнотравья; доминантами обычно являются: манжетка, ге-
рань лесная, василек фригийский (Centaurea phrygia); в качестве содоминан-
тов могут выступать лютики едкий и золотистый, нивяник обыкновенный, ты-
сячелистник (Achillea millefolium) и различные представители семейства зон-
тичных: сныть, дудник лесной (Angelica sylvestris), купырь лесной и др. Злаки
играют второстепенную роль, чаще всего встречаются душистый колосок, ежа
сборная (Dactylis glomerata), тимофеевка и щучка дернистая. Бобовые пред-
ставлены единичными видами клеверов лугового и среднего (Trifolium pratense,
T. medium), горошков мышиного и заборного (Vicia cracca, V. sepium). Луг такого
типа представлен на рис. 8.9.
Ряд Г направлен в сторо-
ну увеличения обильнопроточ-
ного увлажнения и несколько
более богатого минерального
питания. Основной тип лугов –
щучково-разнотравные гид-
ромезофитные луга, где на
фоне щучки и овсяницы крас-
ной (Festuca rubra) ярко вы-
деляется красочный травостой
Рис. 8.9. Злаково-разнотрав-
ный луг ряда В (фото Н. Тере-
хиной)
.
Глава 8. ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
209
Сныть обыкновенная
(Aegopodium podagraria)
Василёк фригийский
(Centaurea phrygia)
Гравилат речной (Geum rivale)
Купырь лесной (Anthriscus sylvestris)
Змеевик большой (Bistorta major)
Вербейник обыкновенный (Lysimachia vulgaris)
Фото: В. Гуменюка, В. Покатайкина, Э. Измайлова, Н. Демченко, Е. Журба
Рис. 8.10. Виды растений луговых сообществ рядов В и Г
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
210
из сивца лугового (Succisa pratensis), кукушкина цвета (Coccyganthe flos-cuculi),
буквицы лекарственной (Betonica officinalis), змеевика лекарственного (Bistorta
officinalis) купальницы (Trollius europaeus), василька фригийского, вербейника
обыкновенного (Lysimachia vulgaris), некоторых видов орхидей и пр. (рис. 8.10).
При увеличении обильно проточного увлажнения в ложбинах рельефа
или у подножья возвышенностей, где выклиниваются грунтовые воды, форми-
руются приручьевые гигрофитные луга, переходящие в высокотравные низин-
ные болота. Доминантами растительного покрова здесь выступают лабазник
вязолистный (Filipendula ulmaria), осоки дернистая и острая (Carex cespitosa,
C. acuta), камыш озерный (Scirpus lacustris), гравилат речной, герань лесная.
В заключение отметим, что суходольные луга лесной зоны, будучи про-
изводными от коренной растительности, находятся в динамичном состоянии,
образуя короткие сукцессионные ряды, никогда не завершающиеся состояни-
ем устойчивого (климаксового) лугового сообщества. Их состав и структура во
многом определяются эколого-топологическими условиями, в которых суще-
ствовала коренная растительность.
Луговые сообщества представляют характерную черту окультуренных
средне- и южно-таежных ландшафтов, сохраняя видимость устойчивых сооб-
ществ. Этот облик поддерживается в течение длительного времени благодаря
постоянному сенокосу и выпасу. При полном прекращении хозяйственной дея-
тельности происходят последовательные процессы восстановления в плакор-
ных условиях квазикоренных лесов, формирование верховых, а при обильно-
проточном увлажнении – низинных болот.
Вопросы к параграфу 8.1
1. Природные условия материковых лугов лесной зоны.
2. Экология растений и экологическая ординация сообществ.
3. Растительность материковых лугов лесной зоны и ее динамика.
Рекомендуемая литература
Андреев Н. Г. Луговедение. М.: Агропромиздат, 1985. 255 с.
Горчаковский П. Л. Антропогенная трансформация и восстановление продуктивности
луговых фитоценозов. Изд-во «Екатеринбург», 1999. 155 с.
Павлова Г. Г. Суходольные луга юга Средней Сибири. – Новосибирск: Наука. Сиб. отд-
ние, 1980. 216 с.
Работнов Т. А. Луговедение. М.: Изд-во МГУ, 1974. 384 с.
Шенников А. П. Луговедение. Изд-во ЛГУ, 1941. 511 с.
8.2. БОЛОТА
Болота – особые природные комплексы, характеризующиеся обильным
застойным увлажнением, своеобразными сообществами растений и накопле-
нием полуразложившихся растительных остатков (торфа). Это – интразо-
нальный тип растительности, распространенный в России преимущественно в
тундре и в тайге. Необходимым условием для формирования болот является
.
Глава 8. ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
211
влажный климат, когда количество атмосферных осадков превышает испаря-
емость.
Болота выполняют важные экологические функции:
–
гидрологическая функция заключается в регулировании стока рек и
уровня грунтовых вод;
–
газовая функция заключается в способности растительности сокращать
содержание СО2 в атмосфере, изымая из планетарного цикла углерод и кон-
сервируя его в торфяниках на тысячи лет;
–
геохимическая функция заключается в способности торфяных болот
выступать естественными фильтрами, поглощающими токсичные элементы;
–
природоохранная функция выражается в том, что болота представляют
собой местообитания специфической флоры и фауны и способствуют сохра-
нению их генофонда;
–
ресурсная функция заключается в том, что болота сохраняют значи-
тельные объемы чистой пресной воды, содержат запасы торфа, являются ис-
точником ягод и лекарственных растений.
В зависимости от гидрологического режима, определяющего характер
питания, болота можно разделить на низинные, переходные и верховые. Ни-
зинные болота, связанные в своем развитии с грунтовыми водами, обогащен-
ными элементами минерального питания, являются эвтрофными (богатого
питания). Это моховые (гипновые и сфагновые), осоковые и лесные или ку-
старниковые болота. Мощность торфяной толщи низинных болот обычно не-
большая – до 1,5–2 м, редко более 3 м. В связи с постоянным торфонакопле-
нием поверхностный слой болота повышается, и роль грунтовых вод, питаю-
щих растительный покров, постепенно уменьшается: уже не все корни
достигают минерального грунта. В снабжении растений водой все большую
роль начинают играть атмосферные осадки. Низинные болота вступают в ста-
дию переходных мезотрофных (средних по богатству минерального питания)
болот. Это преимущественно сфагновые болота с мощностью торфа более 1 м.
При значительном повышении поверхности болота, приводящем к полному
отрыву корневых систем растений от грунтовых вод, растения начинают пи-
таться преимущественно атмосферными осадками бедными минеральными
солями, и болото переходит в стадию верхового сфагнового олиготрофного
(бедного питания) болота. Мощность торфа сфагновых болот в среднем 3–6 м.
8.2.1. ЭКОЛОГО-ТОПОЛОГИЧЕСКИЕ УСЛОВИЯ ОБРАЗОВАНИЯ
ВЕРХОВЫХ СФАГНОВЫХ БОЛОТ
Сфагновые мхи (рис. 8.11) являются растениями-эдификаторами, пол-
ностью контролирующими весь комплекс экологических условий: гидрологи-
ческий и газовый режимы, температуру корнеобитаемого горизонта, образо-
вание форм мезо- и микрорельефа, процессы почвообразования, точнее фор-
мирования торфяной залежи. Растительности крупных массивов верховых
болот присуща ярко выраженная комплексность, обусловленная хорошо раз-
витым болотным микрорельефом (рис. 8.12). В образовании микрорельефа
болота (а также в значительной части мезорельефа) большую роль играет са-
ма растительность.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
212
Рис. 8.11. Сфагновый мох и
болотная кочка – начальная
стадия формирования
суходольного верхового
сфагнового болота
Сфагнум впитывает атмосферные осадки, как губка. Среди клеток, со-
ставляющих его листья и стебель, имеются крупные мертвые клетки с отвер-
стиями в оболочке, которые легко наполняются водой. В сухой сезон года го-
ловки сфагнума могут высыхать до воздушно-сухого состояния. Однако если
вырвать пучок мха из болотной кочки, можно увидеть, что в своей нижней ча-
сти он напитан водой, которая легко выжимается. Раздвинув куртину мха, ви-
дим, что стебельки мха внизу теряют свою индивидуальность, все сливается в
сплошную коричневую массу полуразложившихся частей болотных растений –
это торф. Почвы в обычном понимании здесь нет. Все растения укореняются
или в еще живом сфагнуме или в торфе. Ощущается быстрое понижение тем-
пературы с глубиной. Если температура воздуха 25 °С, то на глубине 20 см она
может уменьшиться вдвое, а на глубине 50 см опуститься до 5–6 °С. Резкое
снижение температуры с глубиной объясняется плохой теплопроводностью
как живого сфагнума, так и торфа.
Растения холодных и влажных местообитаний образуют особую эколо-
гическую группу – психрофиты. Они характерны для растительного покрова
тундровых биомов. Таким образом, облик верховых болот таежной зоны как бы
переносит нас в расположенный в арктических широтах тундровый ландшафт.
Как для верхового болота, так и для тундры, кроме ковра из мхов и лишайников,
Рис. 8.12. Грядово-мочажин-
ный комплекс в Полистов-
ском заповеднике, Псковская
область (фото А. Колотилина)
.
Глава 8. ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
213
Подбел обыкновенный
(Andromeda polifolia)
Хамедафне прицветничковая
(Болотный мирт) (Chamaedaphne calyculata)
Багульник болотный (Ledum palustre)
Вереск обыкновенный (Calluna vulgaris)
Клюква болотная (Oxycoccus palustris)
Пушица узколистная
(Eriophorum angustifolium)
Фото: Л. Онищенко, М. Новиковой, Е. Глазуновой, Е. Ивановой, Д. Кочеткова, Г. Окатова,
В. Гелюты, Д. Павлова
Рис. 8.13. Растения верховых болот
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
214
характерны мелкие кустарнички: подбел (Andromeda polifolia), вереск обыкно-
венный (Calluna vulgaris), багульник болотный (Ledum palustre), два вида клюк-
вы (Oxycoccus), голубика (Vaccinium uliginosum), болотный мирт (Chamaedaphne
calyculata) и др. Это все вечнозеленые растения, относящиеся к одному семей-
ству вересковых (Ericaceae). Характерны также виды пушицы (Eriophorum),
осок (Carex) и шейхцерия (Scheuchzeria palustris) (рис. 8.13).
Для болотных растений характерны признаки ксероморфизма: кожи-
стые, иногда свернутые листья с волосками или восковым налетом, глубоко
расположенные устьица, вечнозеленость, мелколиственность. Это объясня-
ется целым комплексом факторов: в период летней засухи увлажненность
сфагнового ковра и верхнего слоя торфа резко сокращается, и в этих услови-
ях растениям приходится ограничивать расход воды; корнеобитаемый гори-
зонт имеет низкую температуру и плохую аэрацию; почвенная влага харак-
теризуется высокой кислотностью; сильная освещенность на фоне понижен-
ных температур; недостаточность азотного питания на фоне общей бедности
минеральными солями.
На поверхности мохового покрова растет росянка (Drosera rotundifolia,
D. anglica) – небольшое травянистое растение с розеткой прикорневых листь-
ев. Поверхность листа росянки покрыта волосками, которые имеют на кончи-
ке каплю жидкости, блестящей на солнце, как роса. Под действием сока, вы-
деляемого волосками, росянка переваривает попавших на лист мелких на-
секомых (рис. 8.14). Питаясь таким образом, росянка получает с белками
недостающий ей азот. Содержащиеся в росянке химические элементы посте-
пенно переходят в торф, что частично повышает его минеральный состав.
Из древесных пород на верховых болотах растет преимущественно сосна
(Pinus sylvestris), образующая здесь ряд экологических форм, резко отличаю-
щихся от высокоствольных деревьев, растущих в лесу на минеральном грунте.
Болотная сосна никогда не достигает больших размеров: мутовки ветвей
близко расположены одна к другой, что указывает на малый годичный при-
рост. Возраст сосенки высотой около метра может достигать 50–70 лет. На
болотах с быстрым приростом сфагнума (3–4 см в год) встречается своеоб-
разная кустарниковая форма сосны, у которой на поверхности видны только
молодые побеги, покрытые хвоей (рис. 8.15).
Тундровый облик растительно-
сти верховых болот хранит следы
послеледниковья. Сфагновые болота
начали формироваться по перифе-
рии отступавшего ледника в голо-
цене, около 10 тыс. лет назад. Спе-
цифические условия существования
растительности на верховых болотах
позволили им сохранить свой тунд-
ровый облик в таежной зоне до на-
ших дней.
Рис. 8.14. Росянка (Drosera rotundifo-
lia), поймавшая насекомое
.
Глава 8. ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
215
Рис. 8.15. Экологические формы сосны (Pinus silvestris), произрастающей
на олиготрофных болотах (по С. Н. Тюремнову):
1 – f. pumila; 2 – f. willkommii; 3 – f. litwinowii; 4 – f. uliginosa
8.2.2. СТАДИИ РАЗВИТИЯ ВЕРХОВЫХ БОЛОТ
При благоприятных условиях в процессе формирования массивов верхо-
вых болот выделяется несколько стадий (рис. 8.16). В разрезе торфяной зале-
жи сохраняется память о стадиях ее развития в виде слоев торфа разного бо-
танического состава (рис. 8.17). В обильно увлажненных, плоходренируемых
местообитаниях обычно образуются травяные низинные болота, чаще всего
осоковые, разнотравно-осоковые, гипново-осоковые, камышовые. Сначала
поверхность болота слегка вогнутая, затем, по мере отложения торфа, она ста-
новится ровной, торф уплотняется, и на болоте поселяются деревья, чаще все-
го береза. Дальнейшее развитие болот определяется, прежде всего, постоянно
накапливающимися отложениями торфа, что приводит к изменению условий
существования: повышается поверхность, увеличивается площадь, меняются
растительный покров, рельеф, условия стока, формируется своя гидрографи-
ческая сеть. Иными словами, по мере увеличения толщи торфяной залежи
болото все в меньшей степени зависит от условий окружающей местности.
С увеличением толщины торфяного слоя обедняется водно-минеральное
питание, так как корневые системы растений изолируются от минерального
грунта. По мере отложения торфа более требовательные виды сменяются ме-
нее требовательными, и болото из низинного эволюционирует в переходное, а
затем в верховое.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
216
Рис. 8.16. Развитие болотных массивов (по В. П. Денисенкову)
Условные обозначения: 1 – сосна; 2 – вахта; 3 – кустарнички; 4 – осоки; 5 – пушица; 6 –
тростник; 7 – сфагновые мхи; 8 – низинный торф и сапропель; 9 – переходный торф; 10 –
верховой торф; 11 – гряды, мочажины, озерки; 12 – направление стока. Объяснение в тексте
Рис. 8.17. Схема строения торфяной залежи верхового сфагнового болота
(по В. Н. Сукачеву)
.
Глава 8. ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
217
После того как поверхность болота становится плоской, приток поверх-
ностно-сточной воды с окружающих суходолов уменьшается и до центральных
частей болота, как правило, не доходит. Поэтому центральные части болот по-
лучают питание только с атмосферными осадками. Здесь появляются наи-
менее требовательные к питанию сфагновые олиготрофные мхи Sphagnum
fuscum, S. magellanicum, S. angustifolium. Их остатки в условиях сильной кис-
лотности слабо разлагаются; торф быстро накапливается, вследствие этого в
центре болота постепенно появляется выпуклость. На окраинах болот еще
долго остаются более благоприятные условия водно-минерального питания
(поверхностно-сточные воды с суходолов и близкий к поверхности уровень
грунтовых вод). В этих условиях деятельность микроорганизмов, участвующих
в разложении растительных остатков, повышена, и прирост торфа здесь
меньше, чем в центральной части болота. Поэтому окраины болот отстают в
росте от центральной части. С появлением выпуклости на болоте его сток пе-
рераспределяется, он направляется от центра к окраинам. По мере накопле-
ния торфа выпуклость болота продолжает еще некоторое время расти, дости-
гая относительной высоты 3–6 м и более.
Последняя стадия развития выпуклых олиготрофных болот представле-
на плосковыпуклыми болотами. Характерным признаком является плоская,
почти без уклона, сильно переувлажненная поверхность. Максимального раз-
вития получают грядово-мочажинные и грядово-озерковые комплексы. В бо-
лотных ландшафтах Широтного Приобья и Васюганья (Западная Сибирь) за-
озеренность достигает 80 % и более (рис. 8.18). Торфяная залежь наиболее
глубокая, ее мощность от центральных частей к окраинам меняется незначи-
тельно.
Рис. 8.18. Ландшафт слившихся массивов олиготрофных сфагновых болот
(Широтное Приобье, Западная Сибирь)
Элементы ландшафта: грядово-озерковый комплекс, осложненный термокарстовыми
озерами округлой формы
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
218
Рис. 8.19. Аапа-болото в Карелии
Особый тип представляют аапа-болота – широкие равнинные или слабо
наклоненные заболоченные территории. Они получают питательные веще-
ства от окружающих территорий. Как правило, лишенные деревьев аапа-бо-
лота окружены топями с низкорослыми соснами. Характерная особенность
аапа-болот – залегание в сильно обводненных понижениях с вогнутым попе-
речным профилем, часто достаточно выраженный грядово-мочажинный мик-
рорельеф с расположением гряд и их фрагментов перпендикулярно направле-
нию стока (рис. 8.19).
8.3. АНТРОПОГЕННЫЕ НАРУШЕНИЯ БОЛОТНЫХ ЭКОСИСТЕМ
Верховые (сфагновые) болота таежной зоны – уникальный природный
комплекс – долгое время рассматривался с хозяйственной точки зрения как
источник торфа на топливо или, после осушения, как сельскохозяйственные
угодья. Во второй половине XX века на болота шло массированное наступле-
ние. Однако оказалось, что экономическая польза от осушения болот очень
небольшая, а отрицательные последствия огромны.
С целью добычи торфа, повышения продуктивности лесов, сельскохозяй-
ственного освоения территорий проводилось планомерное осушение болот
путем прокладки специальной сети осушительных канав (рис. 8.20). При ис-
пользовании осушенных торфяников под пахотные сельхозугодья слой торфа
быстро срабатывается. Этот процесс усиливается в южной тайге. За год тол-
щина торфа уменьшается в среднем на 1–2 см; максимальная скорость дости-
гает 12 см в год. Двух-, трехметровый торфяной слой разрушается до мине-
рального грунта через 10–20 лет после осушения.
Пересушенный торф обладает гидрофобными свойствами: он с трудом
смачивается водой и поэтому очень пожароопасен. Важнейшей особенностью
.
Глава 8. ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
219
Рис. 8.20. Торфоразработки
торфяных пожаров является то, что во время горения торфяной залежи пожа-
ры распространяются медленно и продолжаются долго – в течение многих
месяцев, а иногда даже в течение нескольких лет. Торф не горит открытым
огнем, а тлеет, выделяя большое количество дыма (рис. 8.21). Тление торфа
может продолжаться даже зимой.
Рис. 8.21. Тушение пожара торфяной залежи
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
220
Осушение болот ведет к нарушению режима питания рек: мелеют и ис-
чезают вытекающие из болот ручьи и малые реки – истоки крупных рек. Сни-
жается уровень грунтовых вод на прилегающих к осушенным болотам терри-
ториях. Усыхают леса, сокращается разнообразие болотных растений, живот-
ных, количество перелетных птиц. Прекращаются традиционные промыслы –
сбор ягод, лекарственных трав, охота.
Вопросы к параграфу 8.2
1. Природные условия болот и их экологические функции.
2. Экология растений болот.
3. Стадии формирования верховых болот.
4. Антропогенные нарушения болот.
Рекомендуемая литература
Боч М. С., Мазинг В. В. Экосистемы болот СССР. Л.: Наука, 1979. 188 с.
Денисенков В. П. Основы болотоведения: Учеб. пособие. СПб.: Изд-во С.-Петерб. ун-та,
2000. 224 с.
Елина Г. А. Многоликие болота / Отв. ред. В. Д. Лопатин. Л.: Наука, 1987. 192 с.
Инишева Л. И. Болотоведение. Учебник. Томск: Изд-во ТГПУ, 2009. 210 с.
Михалев С. С. Основы болотоведения. М.: Изд-во МСХА, 2005. 257 с.
Ниценко А. А. Краткий курс болотоведения. Москва, 1967. 148 с.
.
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
221
Луг как ботанико-географическое и геоботаническое понятие – это био
она неморальных широколиственных лесов подразделяется на подзоны
широколиственных лесов и лесостепи.
Подзона широколиственных лесов в России, как и подзона хвойно-
широколиственных, представлена в двух разобщенных частях континента: в
Европейской и Дальневосточной (см. рис. 7.10), что позволяет выделять Во-
сточноевропейскую и Дальневосточную области широколиственных лесов.
В составе дендрофлоры этих территорий много общих родов и есть некоторые
близкие виды (например, Tilia cordata, Fraxinus excelsior, Acer tataricum в ев-
ропейской части России, Tilia аmurensis, Fraxinus mandshurica, Acer ginnala –
в Приморье). Травы широколиственных лесов на Русской равнине и на Даль-
нем Востоке в большем количестве, чем деревья и кустарники, представлены
близкими видами, по существу подвидами, или географическими расами,
обособление которых произошло, видимо, недавно. Особенности флористиче-
ского состава европейских и дальневосточных широколиственных лесов в ос-
новных чертах сложились в неогене, в значительной мере еще тогда, когда
широколиственные формации в северном полушарии образовывали сплош-
ную зону (тургайские леса). Разрыв ареалов широколиственных лесов был
обусловлен эпохой оледенений в плейстоцене. В Западной Сибири широко-
лиственные леса заменены коренными березовыми лесами, входящими в со-
став подтайги и лесостепи.
Современные широколиственные леса относятся к группе летнезеленых
лесов умеренного пояса. На Восточноевропейской равнине они находятся под
влиянием западного переноса, на Дальнем Востоке на их формирование влия-
ет муссонный климат.
Климатические условия неморальной зоны определяют общие для всех
листопадных широколиственных древесных пород черты строения и разви-
тия. Стволы и ветви защищены от зимних холодов достаточно толстой коркой,
а почки у большинства пород – смолистыми чешуями. Еще задолго до листо-
пада в основании листового черешка появляется разъединяющий слой клеток.
Связь листа с ветвью становится очень непрочной, и осенью даже легкий по-
рыв ветра срывает лист. Опавшая осенью листва образует мощный слой под-
стилки. Умеренно-теплый и влажный климат, активная деятельность почвен-
ных организмов способствуют быстрому разложению опавших листьев и
накоплению гумуса. Наиболее типичны для широколиственных лесов серые,
темно-серые и бурые лесные почвы. Широколиственные леса занимают места
с тяжелыми почвами, уступая песчаные и супесчаные разности соснякам и
смешанным лесам.
Состав и структура широколиственных лесов намного богаче и сложнее,
чем у лесов бореальной зоны и подтайги. В их сложении, как правило, прини-
мают участие три яруса: древесный, в котором различается до трех подъяру-
сов, кустарниковый (до двух подъярусов) и травяной (с двумя-тремя подъяру-
Глава 9
ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
222
сами). Особый облик обретают широколиственные леса ранней весной, когда
еще не сформировалась густая листва древесного полога, – в наземном ярусе
расцветают многочисленные «подснежники» – эфемеры и эфемероиды.
На Восточноевропейской равнине подзона представлена дубовыми, ясе-
нево-дубовыми, кленово-липово-дубовыми, сосново-широколиственными
(Quercus robur, Tilia cordata, Acer platanoides, Fraxinus excelsior, Pinus sylvestris)
кустарниковыми (Euonymus europaea, E. verrucosa, Corylus avellana, Acer
campestre) лесами; черноольховыми (Alnus glutinosa) топями; в Заволжье − ду-
бово-липовыми, липовыми, кленово-липово-дубовыми (Tilia cordata, Quercus
robur, Acer platanoides, Ulmus glabra) кустарниковыми (Euonymus europaea,
E. verrucosa, Corylus avellana, Lonicera xylosteum) лесами; в Маньчжурии −
липово-дубовыми (Quercus mongolica, Tilia amurensis, Betula davurica) ку-
старниковыми (Corylus mandshurica, C. heterophylla, Spiraea ussuriensis) па-
поротниково-травяными лесами.
Подзона лесостепи является переходом от широколиственных лесов
к степной зоне и представляет собой мозаичный экотон, в котором массивы
лесов чередуются с пространствами остепненных лугов и луговых степей. В на-
стоящее время до 80 % территории заняты пашней, а участки луговых степей
сохранились только на крутых склонах и на охраняемых территориях. Лесо-
степная подзона протягивается непрерывной полосой от границы с Украиной
на западе до предгорий Алтая на востоке. На Русской равнине ее южную гра-
ницу проводят по границе распространения древесной растительности на
плакорах, которая, по мере продвижения к Уралу, смещается к северу; на За-
падно-Сибирской равнине подзона имеет широтное простирание.
Рельеф лесостепной подзоны в пределах Восточноевропейской равнины
определяется Среднерусской и Приволжской возвышенностями, а также Вы-
соким Заволжьем и Донецким кряжем. Возвышенности сильно расчленены
овражно-балочной сетью. Местами в оврагах обнажаются меловые горные по-
роды, к которым приурочена специфичная меловая флора. Лесостепные
ландшафты Западной Сибири характеризуются плоскоравнинным рельефом,
что обусловливает слабую дренированность территории. Близкое к поверхно-
сти расположение соленосных палеогеновых и неогеновых отложений и зна-
чительная испаряемость на юге подзоны вызывают засоленность грунтов и
формирование солонцов и солончаков. Со слабой расчлененностью и дрени-
рованностью территории связано обилие озер.
Для лесостепной подзоны характерен умеренный континентальный
климат, переходный от влажного к сухому. Средняя температура июля в пре-
делах подзоны колеблется от +19 до 25 °С. Январские температуры понижают-
ся с запада на восток от –6 °С (абсолютный минимум –34,5 °С) в районе Белго-
рода до –15 °С (абсолютный минимум –47,9 °С) в Кургане. В европейской части
лесостепной подзоны за год выпадает 550–600 мм осадков, в азиатской – 300–
450 мм. Лесостепные ландшафты формируются в той части умеренного пояса,
где годовое количество влаги, поступающей с атмосферными осадками, при-
близительно равно испаряемости (ГТК около 1). При переходе к степной зоне
испаряемость увеличивается, и сухой сезон проявляется все более отчетливо.
На севере лесостепной подзоны европейской части России развиты глав-
ным образом серые лесные почвы, на юге – выщелоченные и мощные черно-
.
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
223
земы, встречаются и обыкновенные черноземы. В Западной Сибири на севере
подзоны преобладают выщелоченные и оподзоленные черноземы, сильно
измененные под древесной растительностью; в средней части подзоны они
сменяются плодородными тучными черноземами, которые вблизи южной
границы лесостепи постепенно переходят в черноземы обыкновенные. На се-
вере подзоны распространены также лугово-черноземные, темно-серые лес-
ные и болотные почвы, на юге – солоди, солонцы и солончаки.
Растительность подзоны лесостепи представлена: на Восточноевропей-
ской равнине − липово-дубовой лесостепью (Quercus robur; Tilia cordata, Acer
platanoides; Stipa pulcherrima, S. tirsa, S. dasyphylla, S. pennata, Bromopsis ri-
paria, B. inermis, Poa angustifolia, Phleum phleoides, Helictotrichon schellianum,
Carex humilis, Filipendula vulgaris, Salvia stepposa, Galium verum, Trifolium
montanum, T. alpestre); в Заволжье − кленово-липово-дубовой лесостепью
(Quercus robur; Tilia cordata, Acer platanoides; Stipa pulcherrima, S. dasyphylla,
S. tirsa, S. zalesskii, S. pennata, Bromopsis riparia, B. inermis, Poa angustifolia,
Calamagrostis epigeios, Phleum phleoides, Helictotrichon schellianum,
H. desertorum, Salvia pratensis, Filipendula vulgaris, Trifolium alpestre); в За-
падной Сибири − колочной березовой и осиновой лесостепью (Betula pendula,
Populus tremula; Stipa tirsa, S. zalesskii, Bromopsis inermis, Poa angustifolia,
Calamagrostis epigeios, Phleum phleoides, Helictotrichon schellianum, Carex pedi-
formis, Filipendula vulgaris, Salvia pratensis, Peucedanum morisonii, Vicia uniju-
ga, Trifolium lupinaster); на Дальнем Востоке − дубовой лесостепью (Quercus
mongolica, Betula davurica; Arundinella anomala, Filifolium sibiricum, Clematis
hexapetala) и вязово-дубовой лесостепью (Quercus mongolica, Ulmus pumila,
Pinus funebris; Corylus heterophylla, Lespedeza bicolor, L. juncea).
9.1. БОТАНИКО-ГЕОГРАФИЧЕСКИЕ ОБЛАСТИ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
Зона широколиственных лесов на территории России подразделяется на
две области: Восточно-Европейскую и Дальневосточную. Западно-Сибирская
область представлена лесостепью.
9.1.1. ВОСТОЧНО-ЕВРОПЕЙСКАЯ ОБЛАСТЬ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
Подзона широколиственных лесов господствует в Западной Евро-
пе. В Европейской части России, где заметно усиливается континентальность
климата, леса меняют свой состав и постепенно выклиниваются. В общих чер-
тах, область распространения широколиственных лесов на Восточно-Европей-
ской равнине напоминает по форме треугольник, вершиной обращенный к
Уралу. Его северная граница проходит приблизительно по линии Брянск –
Москва – Уфа, а южная – по линии Курск – Тула – Пенза – Уфа. К востоку от
Волги и южнее 53° с.ш. леса встречаются отдельными массивами, чередуясь с
луговыми степями. На Урале широколиственные леса становится компонен-
том высотной поясности. В северной части своего ареала восточноевропейские
широколиственные леса занимают водоразделы, в лесостепной зоне – при-
урочены к возвышенным плато и высоким берегам рек.
Соотношение тепла и влаги в подзоне широколиственных лесов близко
к оптимальному (ГТК около 1). Продолжительность вегетации удлиняется до
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
224
6–7 месяцев. Зимы мягкие. Средняя температура января –8...0 °С, июля +20–
24 °С. Среднее годовое количество атмосферных осадков 600–800 мм.
Характерной особенностью восточноевропейских широколиственных ле-
сов является постепенное обеднение флоры в направлении с запада на восток
в результате удаления от основных юго-западных рефугиумов древней немо-
ральной флоры и усиления континентальности климата. Важным ботанико-
географическим рубежом является Волга, по направлению к которой посте-
пенно выпадают многие западноевропейские неморальные виды. В направ-
лении с севера на юг обедняется собственно неморальная флора, поскольку
выпадают наиболее мезофильные виды этой группы. Отмечается заметное
участие во флоре подзоны на востоке южносибирских светолюбивых и холо-
достойких видов. Роль бореальных видов наиболее существенна в северной
части подзоны и на Урале.
На крайнем западе России в Калининградской области в виде отдельных
фрагментов встречаются широколиственные леса с участием бука (Fagus
sylvatica) по составу похожие на среднеевропейские буковые леса. Грабовые и
грабово-дубовые (из Carpinus betulus, Quercus robur) леса отмечены в Брян-
ской области. В центральной части Восточно-Европейской равнины широко-
лиственные леса представлены дубравами. В них обычно присутствуют липа
сердцевидная (Tilia cordata), клен остролистный (Acer platanoides), вяз, ясень.
Основной эдификатор – дуб черешчатый – сравнительно зимостойкий и од-
новременно достаточно засухоустойчивый вид. Его ареал охватывает подзону
южной тайги и зону степей, где он образует островные лески. Существенными
факторами естественного возобновления являются освещенность, водный ре-
жим почвы, мощность подстилки и развитие травяного покрова. Под пологом
старовозрастных дубрав самосева дуба бывает значительно меньше, чем са-
мосева клена остролистного и ясеня; желуди нередко растаскиваются мыше-
видными грызунами, все это является факторами, затрудняющими естест-
венное возобновление дуба. На месте, в прошлом занятом дубравами, встре-
чаются производные смешанные насаждения ясеня, липы, кленов и других
пород. В подлеске присутствуют лещина (Corylus avellana), виды боярышника
(Crataegus), бересклеты бородавчатый и европейский (Euonymus verrucosa,
E. europaea), волчеягодник (Daphne mezereum) и другие кустарники.
Своеобразный аспект широколиственные леса приобретают весной, ко-
гда господствуют эфемеры и эфемероиды (рис. 9.1): пролеска (Scilla siberica),
хохлатка полая (Corydalis cava), гусиный лук (Gagea lutea), ветреница дубрав-
ная (Anemone nemorosa), чина весенняя (Lathyrus vernus), печеночница бла-
городная (Hepatica nobilis).
Травянистые растения широколиственных лесов, имеющие широкие
нежные листовые пластинки, относят к экологической группе дубравного ши-
рокотравья. Некоторые из них проникают в таежную зону, где их называют
неморальными элементами. В частности, они представляют характерную чер-
ту сложных (неморальных) ельников. Это копытень европейский (Asarum
europaeum), осока волосистая (Carex pilosa), зеленчук желтый (Galeobdolon
luteum), сныть обыкновенная (Aegopodium podagraria), медуница неясная
(Pulmonaria obscura), пролесник многолетний (Mercurialis perennis), купена
душистая (Polygonatum odoratum), лютик кашубский (Ranunculus cassubicus)
.
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
225
Пролеска сибирская (Scilla siberica)
Ветреница дубравная (Апeтопе nemorosa)
Печеночница благородная (Hepatica nobilis)
Хохлатка полая (Corydalis cava)
Гусиный лук желтый (Gagea lutea)
Чина весенняя (Lathyrus vernus)
Фото: В. Чалых, С. Черепушкина, В. Прохорова, Р. Лежоева, С. Майорова, Н. Демченко
Рис. 9.1. Эфемеры широколиственных лесов
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
226
Рис. 9.2. Липняк щитовниковый, Костромская область
и др. Напочвенный моховой покров широколиственным лесам не свойстве-
нен, хотя на гниющей древесине и в основании стволов мхи постоянно присут-
ствуют.
В широколиственных лесах сравнительно обильно представлена липа
сердцевидная, являющаяся наиболее типичным элементом широколиствен-
ных лесов на востоке Восточно-Европейской области (рис. 9.2). Роль доминан-
та или субдоминанта липа играет преимущественно в приволжских, а особен-
но в приуральских широколиственных лесах.
В настоящее время широколиственные леса сохранились в виде островов
среди безлесных окультуренных территорий, которые, будучи предоставлены
сами себе, часто сменяются растительностью лугово-степного типа.
Ввиду ограниченного числа заповедников в подзоне широколиственных
лесов уместно привести описание широколиственных лесов в заповеднике
«Лес на Ворскле», хотя он расположен несколько южнее зоны широколист-
венных лесов. Леса здесь представляют собой уникальный набор сообществ,
дающий наиболее полное представление о составе и структуре растительно-
сти. Особенно ценным является то, что в основу описания растительности
заповедника «Лес на Ворскле» положены результаты полувекового мони-
торинга, осуществленного Ю. Н. Нешатаевым и В. Н. Ухачевой.
Заповедник «Лес на Ворскле» расположен в Белгородской области
около с. Борисовка. Он занимает высокий правый берег р. Ворсклы (перепад
абсолютных высот 135–217 м), расчлененный несколькими ярами (логами).
Площадь массива около 1100 га, из которых 55 % заняты естественными ле-
сами, а 41 % приходится на лесные культуры. Естественные леса в основном
.
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
227
представлены производными сообществами, в которых наряду с дубом че-
решчатым принимают участие: клен остролистный, липа мелколистная и
ясень обыкновенный. Первостепенный интерес представляет высокостволь-
ная дубрава как наименее нарушенный и наиболее близкий к коренному типу.
В настоящее время высокоствольная дубрава на 20 % заменена производными
сообществами (липняками и кленовниками) (рис. 9.3). Отличительной чертой
высокоствольных дубняков является их обедненный видовой состав, отмечено
всего 92 вида (деревьев – 14, кустарников – 8, трав – 70).
Древесный ярус имеет сложную структуру и расчленен на 2–4 полога.
Его составляют три основные породы: дуб, липа и клен. Древесный подрост
достаточно развит и представлен липой, вязом и особенно кленом остролист-
ным. Подрост дуба единичен.
Кустарниковый ярус как структурно-ценотическая единица не выражен.
Основной вид – бересклет европейский – обычно встречается в куртинно-сте-
лющейся форме и не выходит за пределы травостоя. Травяной покров развит
достаточно хорошо, особенно в снытевой серии ассоциаций, где его проектив-
ное покрытие обычно составляет 80–100 %. Доминирующими видами явля-
ются: будра плющевидная (Glechoma hederacea), звездчатка ланцетовидная
(Stellaria holostea), крапива двудомная (Urtica dioica), недотрога обыкновен-
ная (Impatiens noli-tangere), осока волосистая (Carex pilosa), пролесник мно-
голетний (Mercurialis perennis), чесночница черешковая (Alliaria petiolata) и
яснотка пятнистая (Lamium maculatum); часто встречаются также: герань Ро-
берта (Geranium robertianum), копытень европейский, купена многоцветковая
(Polygonatum multiflorum), медуница неясная, подмаренник цепкий (Galium
aparine).
Рис. 9.3. Кленово-дубовый снытевый лес, заповедник «Лес на Ворскле»
(фото Д. Мирина)
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
228
Во второй половине XX века произошли изменения в составе и структуре
высокоствольной дубравы, свидетельствующие о ее мезофилизации – увели-
чение фитоценотической роли более влаголюбивых видов, в древостое – липы
и клена, в травяном покрове – экспансия сныти. Соответственно уменьшилась
фитоценотическая роль более засухоустойчивых видов (дуб, груша, тёрн, осока
волосистая, звездчатка ланцетовидная, мятлик дубравный).
Лесостепная подзона. В прошлом естественная растительность обла-
сти представляла собой сочетания широколиственных лесов с открытыми
пространствами луговых степей. В настоящее время древесно-кустарниковая
растительность представлена преимущественно байрачными лесами. Они рас-
тут по балкам и высоким правым берегам рек, где формируется свой микро-
климат. Байрачные леса, в связи с разнообразием условий существования, об-
разуют большое количество вариантов, различающихся составом древесных
пород, а также составом кустарникового и травяного ярусов. Основные по-
роды, образующие подобные леса – дуб, клен, вяз, ясень, липа, груша, яблоня.
В некоторых районах также встречаются черешня (Cerasus avium), граб
(Carpinus betulus). В подлеске чаще всего произрастают клен татарский (Acer
tataricum), боярышник (Crataegus sp.), калина гордовина (Viburnum lantana),
лещина, бересклеты, местами скумпия (Cotinus coggygria), на опушках – тёрн
(Prunus stepposa), шиповник, вишня степная (Cerasus fruticosa), бобовник
(Laburnum anagyroides). Байрачные леса Ростовской области представлены
дубравами, смешанными насаждениями с кленом, липой и ясенем. Хотя в неко-
торых лесах Верхнедонского района недалеко от границы с Воронежской обла-
стью преобладает сосна. Байрачные леса Донецкого кряжа – это чистые дубра-
вы. Постепенно, с выравниванием рельефа, исчезают и сами байрачные леса.
Луговые степи сложены мезофитным разнотравьем (Filipendula vulgaris,
Salvia pratensis, Leucanthemum vulgare, Galium verum, Myosotis sylvatica,
Trifolium montanum и мн. др.), мезофитными и ксеромезофитными преиму-
щественно корневищными злаками (Poa angustifolia, Calamagrostis epigeios,
Agrostis vinealis) и осоками (Carex praecox), в меньшем количестве рыхлоку-
стовыми луговыми злаками (Festuca pratensis). Обычна примесь более или
менее ксерофитных дерновинных злаков (типчак, ковыли).
В открытой степи встречаются плодовые деревья и кустарники: яблони,
груши, боярышник, шиповник и тёрн. В заповеднике «Белогорье», в Ямской
степи, помимо одиночных деревьев, произрастают куртины, в которых яблоню
и грушу сопровождают клен татарский, клен американский (Acer negundo) –
интродуцированный вид, широко внедрившийся в естественную растительность.
Остановимся на более детальной характеристике растительности лесо-
степной подзоны на примере двух заповедников: Центрально-Черноземный
им. В. В. Алехина и «Приволжская лесостепь».
Центрально-Черноземный заповедник. Степи под Курском про-
фессор В.В. Алехин в начале XX века назвал «Курской растительной аномали-
ей». Но эта «аномальность» связана не с тем, что растительность нетипична, а
с тем, что здесь сохранились в условиях сплошной распашки одни из немно-
гих нетронутых целинных участков луговой степи. На этих землях в пределах
Курской области организован Центрально-Черноземный государственный
заповедник им. В. В. Алехина (Гусев, Оликова, Гусева, 1989).
.
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
229
Рис. 9.4. Луговая степь, Центрально-Черноземный заповедник
(фото http://www.zapovedniki-mira.com)
Заповедная территория лежит на Среднерусской возвышенности, по-
верхность которой прорезана глубокими оврагами и балками. Часть террито-
рии заповедника занята лесной растительностью, преимущественно дубрава-
ми (в основном порослевого происхождения), образованными дубом череш-
чатым с небольшой примесью осины, ивы козьей (Salix caprea), яблони,
груши, рябины, черемухи и др. Степной травостой густой и многоярусный
(рис. 9.4).
Флора заповедника насчитывает 1287 видов сосудистых растений, вклю-
чая адвентивные (заносные) травянистые растения и древесные интродуценты.
В их числе 86 редких видов. Реликтами считаются такие виды, как: проломник
Козо-Полянского (Androsace koso-poljanskii), володушка многожилковая
(Bupleurum multinerve), волчеягодник боровой (Daphne cneorum), шиверекия
подольская (Schivereckia podolica), хризантема Завадского (Chrysanthemum
zawadskii). Они имеют разорванные ареалы, части которых лежат в горах
Урала, Сибири или Западной Европы.
Однодольные представлены 24 семействами, крупнейшим из которых
являются семейства: мятликовые (108 видов), осоковые (47 видов), орхидные
(12 видов – все они внесены в региональную Красную книгу, а 2 вида из них –
венерин башмачок настоящий, глянцелистник Лёзеля (Liparis loeselii) – в Крас-
ную книгу России), лилейные (8 видов, включая 2 вида из Красной книги России
(рябчики русский и шахматный – Fritillaria ruthenica, F. meleagris) и 1 вид из
Красной книги Курской области (лилия царские кудри, саранка – Lilium
martagon). Двудольные представлены 93 семействами, крупнейшим являются:
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
230
Рис. 9.5. Весенне-летние аспекты
в луговой степи:
а – начало апреля: бурый аспект с ли-
ловыми пятнами Pulsatilla patens; жел-
тые – мужские колоски Саrех humilis,
первые цветки Adonis vernalis;
б – вторая половина апреля: желтый
аспект от Adonis vernalis. Кроме того,
цветет
нежно-голубой Hyacinthus
leucophaeus. Степь еще не зазеленела;
в – конец мая: голубой аспект от
Myosotis sylvatica, отдельные цветущие
экземпляры Anemone sylvestris и жел-
тые пятна Tephroseris integrifolia. За-
цветает ковыль;
г – июнь, слева направо: Trifolium re-
pens, Salvia pratensis, Festuca valesiaca,
Viola rupestris, Trifolium montanum, Sti-
pa pennata, Carex caryophyllea, Ranun-
culus polyanthemos, Leucanthemum
vulgare, Trommsdorffia maculata, An-
thoxanthum odoratum, Eremogone sax-
atilis, Filipendula vulgaris, Carex mon-
tana, Agrostis vinealis, Viola canina,
Scorzonera purpurea, Euphorbia subtilis,
Potentilla heptaphylla, Festuca rubra,
Astragalus danicus, Phlomoides tuberosa,
Koeleria delavignei, Galium boreale,
Echium russicum, Carex montana.
Вертикальная проекция, квадраты де-
циметровые (по Г. Вальтеру)
астровые (165 видов, из них Красную книгу региона внесено 12 видов), розо-
цветные (114 видов, из них в Красную книгу России внесен 1 вид – кизиль-
ник алаунский, а в Красную книгу Курской области – еще 5 видов), бобовые
(67 видов, из них 6 видов внесены в Красную книгу Курской области), яснот-
ковые (61 вид, из них 4 вида внесены в Красную книгу региона), капустные
(60 видов, из них 5 видов представлены в Красной книге Курской области).
Луговые степи необыкновенно красочны. В течение вегетационного пе-
риода меняется видовой состав цветущих видов, в результате чего изменяется
облик растительного покрова (рис. 9.5).
.
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
231
Приведем характеристику основных аспектов луговой степи заповедника
(рис. 9.6).
Послезимний аспект. Снег в степи только что сошел и почва покрыта
остатками прошлогодних растений.
I. Предвесенний аспект. Достаточно нескольких теплых дней в апреле,
чтобы появились фиолетовые цветки прострела раскрытого (Pulsatilla patens);
одновременно начинает «пылить» осока низкая (Carex humilis).
II. Ранневесенний аспект. Уже в первой половине мая повсюду появля-
ются крупные золотистые звезды адониса весеннего (Adonis vernalis), красиво
выделяющиеся на фоне еще цветущего прострела, между ними проглядывают
нежные светло-голубые соцветия гиацинтика пепельно-серого (Hyacinthella
leucophaea).
III. Средневесенний аспект. Степь уже зеленеет, раннецветущие виды
исчезают. Во второй половине мая их сменяют белые соцветия чины (Lathyrus
pannonicus), лиловые цветки ириса безлистного (Iris aphylla) и крупные белые
звезды ветреницы лесной (Anemone sylvestris).
IV. Поздневесенний аспект. В конце мая – начале июня степь снова пре-
ображается. Это ее самая красочная фаза: на зеленом фоне живописно разбро-
саны бесчисленные нежно-голубые незабудки (Myosotis sylvatica), желтый
пепельник цельнолистный (Tephroseris integrifolia) и лютик (Ranunculus poly-
anthemos), появляются первые легкие перья ковыля (Stipa pennata).
V. Раннелетний аспект. Весенний период закончился, начинается фе-
нологическое лето. Степь отливает фиолетово-голубым цветом от изобилия
цветков лугового шалфея (Salvia pratensis). Между ними колышутся на ветру
длинные перистые ости видов Stipa, поднимаются метелки высокого костра
берегового (Bromopsis riparia). Ранним утром повсюду светятся золотистые
звезды козлобородника (Tragopogon pratensis), закрывающего свои цветки
уже в полдень.
VI. Среднелетний аспект. Во второй половине июня степь становится
белой от бесчисленных соцветий клевера горного (Trifolium montanum) и ни-
вяника (Leucanthemum margaritae), лабазника обыкновенного (Filipendula
vulgaris). В общий фон вкраплены колокольчики (Campanula sibirica и С. per-
sicifolia), соцветия короставника (Knautia arvensis) и др.
VII. Аспект разгара лета. В начале июля начинают доминировать розо-
вые соцветия эспарцета (Onobrychis arenaria). Однако пестрые краски уже
блекнут, число цветущих видов уменьшается, степь приобретает монотонную
окраску. В это время цветет желтый подмаренник настоящий (Galium verum).
Растительность плакорных луговых степей заповедника в настоящее
время представлена в основном разнотравно-широколиственнозлаковыми
сообществами со значительным участием дерновинных злаков и бобовых. Из
мезофильных злаков довольно обильны мятлик узколистный (Poa angustifolia),
овсец пушистый (Helictotrichon pubescens), полевица виноградниковая (Agrostis
vinealis), ежа сборная (Dactylis glomerata), тимофеевки степная и луговая
(Phleum phleoides, Ph. pratense). Из крупнодерновинных злаков наиболее ха-
рактерен и обилен ковыль перистый (Stipa pennata), реже встречаются ковы-
ли узколистный и опушеннолистный (Stipa tirsa, S. dasyphylla); из мелкодерно-
винных – типчак (Festuca valesiaca), тонконог гребенчатый (Koeleria cristata).
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
232
I
II
III
Прострел раскрытый
(Pulsatilla patens)
Адонис весенний
(Adonis vernalis)
Ирис безлистный
(Iris aphylla)
IV
V
V
Незабудка Попова
(Myosotis popovii)
Шалфей луговой
(Salvia pratensis)
Ковыль Лессинга
(Stipa lessingiana)
VI
VI
VII
Колокольчик персиколистный
(Campanula persicifolia)
Клевер горный
(Trifolium montanum)
Подмаренник настоящий
(Galium verum)
Фото: В Куропаткина, Т. Никулиной, Ю. Пирогова, Г. Гуль, И. Полостюк, В. Гелюты,
А. Иванова, М. Скотниковой, Н. Апполоновой
Рис. 9.6. Виды, определяющие смену сезонных аспектов растительности
в Центрально-Черноземном заповеднике: I–VII – пояснения в тексте
.
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
233
Среди разнотравья особенно обильны такие виды, как: адонис весенний
(Adonis vernalis), первоцвет весенний (Primula veris), лютик многоцветковый
(Ranunculus polyanthemos), земляника зеленая (Fragaria viridis), лабазник
обыкновенный (Filipendula vulgaris), шалфей луговой (Salvia pratensis), мытник
Кауфмана (Pedicularis kaufmannii), василек шероховатый (Centaurea scabiosa),
подмаренник настоящий (Galium verum), резак обыкновенный (Falcaria
vulgaris), горичник горный (Peucedanum oreoselinum) и др. Из бобовых значи-
мую роль играют: клевера горный и альпийский (Trifolium montanum, T. alpes-
tre), горошек тонколистный (Vicia tenuifolia), эспарцет песчаный (Onobrychis
arenaria) и др. Степные кустарники представлены вишней степной (Cerasus
fruticosa), ракитничком русским (Chamaecytisus ruthenicus), тёрном (Prunus
spinosa); полукустарничек – чабрец Маршалла (Thymus marschallianus).
Леса заповедника имеют островной характер и представлены, главным
образом, дубравами. Они занимают плакорные местообитания (водораздель-
ные дубравы), островами разбросаны по пойме (пойменные дубравы), по скло-
нам и днищам балок (байрачные дубравы). Лесные насаждения представлены
множеством производных сообществ от коренного ясене-дубняка лещиново-
снытевого, среди которых наиболее часто встречается дубняк снытево-крапив-
ный. Современный этап развития лесных сообществ заповедника характери-
зуется усложнением строения древостоев в связи с расширением их видового
разнообразия. Первый подъярус древесного яруса образован обычно дубом,
реже осиной, кленом остролистным, вязом. Второй подъярус выражен слабо,
максимальную площадь проекций крон здесь имеют груша, ива козья, яблоня.
В подлеске преобладают: черемуха, лещина, бересклет европейский, жостер
слабительный (Rhamnus cathartica) и тёрн. Меньшее распространение имеют
бересклет бородавчатый, крушина ломкая (Frangula alnus) и рябина. Доми-
нантами травостоя, наряду с типично лесными видами, такими как сныть
обыкновенная, копытень европейский, ландыш майский и др., часто являются
и рудеральные виды – крапива двудомная, чистотел большой (Chelidonium
majus) и др.
Заповедник «Приволжская лесостепь», созданный в 1989 г., рас-
положен в Среднем Поволжье в Пензенской области. Он находится на юго-
западе Приволжской возвышенности, в пределах главного водораздела между
Волгой и Доном.
Флора заповедника чрезвычайно разнообразна. В настоящее время на
его территории выявлено 825 видов сосудистых растений, 108 видов лишай-
ников, 72 − мохообразных и 119 − грибов. В Красную Книгу России включены
встречающиеся в заповеднике 8 видов сосудистых растений: рябчик русский
(Fritillaria ruthenica), гнездоцветка клобучковая (Neottianthe cucullata), пыль-
цеголовник красный (Cephalanthera rubra), надбородник безлистный
(Epipogium aphyllum) и 4 вида ковылей: опушеннолистный (Stipa dasyphylla),
перистый (S. pennata), красивейший (S. pulcherrima) и ковыль залесского (S.
zalesskii) (рис. 9.7).
Среди луговых степей заповедника выделяется несколько основных рас-
тительных сообществ: луговые степи, остепненные луга, кустарниковые луго-
вые степи, кустарниковые остепненные луга, настоящие луга, болотистые луга,
степные кустарники . Неповторимый облик луговым степям придают ковыли.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
234
Рябчик русский (Fritillaria ruthenica)
Гнездоцветка клобучковая
(Neottianthe cucullata)
Надбородник безлистный
(Epipogium aphyllum)
Пыльцеголовник красный
(Cephalanthera rubra)
Фото: В. Гелюты, А. Эбсля, В. Савчука, А. Поповича
Рис. 9.7. Растения заповедника «Приволжская лесостепь», внесенные
в «Красную книгу России»
Всего в заповеднике выявлено 9 видов ковылей. Кроме ковылей большую
роль в формировании луговых степей и остепненных лугов играют такие зла-
ки как типчак, кострецы безостый и береговой (Bromopsis inermis, B. riparia),
вейник наземный (Calamagrostis epigeios), овсец пустынный (Helictotrichon
desertorum). Вместе с разнотравьем они создают пестрый ковер, поражающий
обилием красок в июне и июле, во время цветения большинства видов трав.
.
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
235
Из степных кустарников многочисленны: тёрн обыкновенный (Prunus
spinosa), миндаль низкий (Amygdalus nana), вишня кустарниковая (Cerasus
fruitcosa), спирея городчатая (Spiraea crenata), ракитничек русский
(Chamaecytisus ruthenicus), шиповник майский (Rosa majalis) (рис. 9.8).
В качестве иллюстрации приведем карту растительности одного из участ-
ков заповедника «Попереченская степь» (рис. 9.9).
Среди луговых степей на территории заповедника встречаются также не-
большие участки леса – так называемые, лесные колки. Они образованы оси-
ной, черемухой, кленом татарским, или дубом и березой.
Лесная растительность занимает около 80 % территории заповедника,
отличаясь исключительным разнообразием. Дендрофлора заповедника вклю-
чает 26 видов деревьев и 34 вида кустарников. Главными лесообразующими
породами являются сосна обыкновенная, дуб черешчатый, березы повислая и
пушистая, осина обыкновенная. Широко распространена липа мелколистная,
несколько меньше – клен остролистный, вяз голый и вяз гладкий (Ulmus
glabra, U. laevis). В поймах рек и ручьев по сырым и заболоченным местам
растут ольха черная (Alnus glutinosa), ивы белая и понтийская (Salix alba,
S. euxina), изредка тополя белый и черный (Populus alba, P. nigra). Перечис-
ленные виды деревьев образуют верхние подъярусы древостоя, которые ока-
зывают значительное влияние на всю структуру лесных сообществ. Второй
подъярус составляют низкорослые деревья: рябина, черемуха, клен татарский,
ива козья, яблоня ранняя (Malus praecox) и груша обыкновенная. В состав
кустарникового яруса входят: бересклет бородавчатый, жимолость татарская
и обыкновенная (Lonicera tatarica, L. xylosteum), калина обыкновенная, круши-
на ломкая, шиповник, волчеягодник (Daphne mezereum), ракитничек русский
(Chamaecytisus ruthenicus) и дрок красильный (Genista tinctoria). Последние два
вида растут как в степях, так и в подлеске сухих сосновых боров. Редко встреча-
ются боярышник кроваво-красный (Crataegus sanguinea), бузина кистевидная
(Sambucus racemosa) и барбарис обыкновенный (Berberis vulgaris).
Довольно четко проявляется зависимость распределения групп типов ле-
са от рельефа. Так, сосновые леса приурочены, главным образом, к водораз-
дельным пространствам и верхним частям склонов, а также к надпойменным
песчаным террасам. Для водораздела Верховьев Суры характерно наличие
среди сосняков значительного количества болот, образующихся в понижениях
между буграми и грядами. Группа дубовых лесов занимает выровненные
средние части склонов от водораздела к поймам рек. Коренные березовые ле-
са распределены небольшими участками по понижениям на водоразделе и на
нижних частях склонов, переходных к поймам. В поймах рек и ручьев растут
черноольшаники, местами ивняки (ветляники).
Группа сосновых коренных лесов отличается наибольшим типологиче-
ским разнообразием. В ней преобладают сосняки орляково-разнотравные, за-
тем следуют сосняки травяно-мшистые. Меньшие площади занимают злако-
во-разнотравные и лещинно-липовые сосняки. Редкими типами являются
сосняки черничники, лишайниковые и майниково-черничные. Среди свежих
сосняков простой и сложной субори (орляково-разнотравные и лещинно-
липовые) по вершинам песчаных холмов вкраплены небольшие участки сухих
местообитаний – субори и сложной субори. Сложные сосняки имеют в своем
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
236
Миндаль низкий (Anmygdatus nana)
Вишня кустарниковая (Cerasus fruticosa)
Тёрн колючий (Prunus spinosa)
Ракитничек русский (Chamaecytisus rutheniсus)
Шиповник майский (Rosa majalis)
Спирея городчатая (Spiraea crenata)
Фото: А. Иванова, P.C. Ушаковой, Д. Мирина, В. Беспалова, Е. Тихоновой, Э. Измайлова
Рис. 9.8. Степные кустарники заповедника «Приволжская лесостепь»
.
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
237
Рис. 9.9. Карта растительности участка «Попереченская степь» заповедника
«Приволжская лесостепь» (по Л. А. Новиковой)
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
238
Рис. 9.10. Обнажения меловых пород.
Заповедник «Дивногорье»
составе широколиственные породы:
липу и дуб, которые здесь долгое
время находятся в угнетенном состо-
янии, сначала в виде стелющейся
формы, а в осветленных участках – в
виде полноценного подроста. В тра-
вяно-кустарниковом ярусе сухих сос-
няков на песчаных дюнах преобла-
дают светолюбивые боровые виды
(кошачья лапка (Antennaria dioica),
вейник наземный (Calamagrostis
epigeios), ястребинка зонтичная
(Hieracium umbellatum), купена ду-
шистая (Polygonatum odoratum), жаб-
рица порезниковая (Seseli libanotis) и
др.), а также некоторые лесостепные
виды (ракитничек русский, дрок кра-
сильный). Во влажных сосняках обы-
чны такие таежные виды, как майник
двулистный, костяника, седмичник
европейский, вечнозеленые гру-
шанковые: зимолюбка зонтичная
(Chimaphila umbellata) и ортилия однобокая (Orthilia secunda).
Вторая по величине группа типов леса – дубравы, они приурочены к бо-
лее богатым суглинкам. Эта группа представлена, в основном, лещинно-липо-
выми, снытьево-разнотравными и злаково-разнотравными дубняками. До-
вольно широко распространены смешанные дубняки. Это – молодняки порос-
левого происхождения, развивающиеся на месте сведенных коренных дубрав.
В их одноярусных древостоях кроме дуба всегда присутствуют береза, осина и
липа. Местами они преобладают над дубом в различных соотношениях или
полностью его заменяют.
Меловая флора. Выходы мела служат убежищем для своеобразной
кальцефильной флоры. Приведем описание меловой флоры на примере при-
родного, архитектурно-археологического музея-заповедника «Див-
ногорье» и южной части Сенгилеевской возвышенности в Самарской обла-
сти. Заповедник «Дивногорье» расположен в Воронежской области на правом
берегу реки Дон. Свое название заповедник получил благодаря меловым
столбам, которые местное население называет «дивами» (рис. 9.10).
Особые почвы появляются там, где в качестве почвообразующей породы
выступает элювий мела, подстилаемый пластами писчего мела. Продукты вы-
ветривания мела отличаются очень высоким содержанием карбоната кальция.
Эти породы бедны элементами питания растений.
На меловых обнажениях безраздельно господствуют представители ме-
ловой флоры (кальцефиты). На меловом субстрате в ряде мест хорошо сохра-
.
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
239
нились своеобразные тимьянниковые степи и кальцефитно-петрофитные лу-
говые степи («сниженные альпы»).
Флора «Дивногорья» включает ряд редких и эндемичных видов: осока
низкая – «мамонтова трава» (Carex humilis), оносма простейшая (Onosma
simplicissima), проломник Козо-Полянского (Androsace koso-poljanskii), живуч-
ка хиосская (Ajuga chia), хвойник (эфедра) двухколосковый (Ephedra distachya),
бурачок Гмелина (Alyssum gmelinii), тимьян меловой (Thymus calcareus). Все
эти виды, как правило, облигатные кальцефиты (рис. 9.11), которые приуро-
чены исключительно к меловым обнажениям.
Южная часть Сингелеевской возвышенности протягивается вдоль пра-
вого берега реки Волги на северо-западе Самарской области. Для нее харак-
терны белокаменные пирамиды меловых обнажений (рис. 9.12). Флора со-
судистых растений карбонатных субстратов здесь насчитывает 111 видов,
принадлежащих к 79 родам, 32 семействам. Ведущими по числу видов семей-
ствами являются: Asteraceae – 16 видов; Fabaceae – 14; Poaceae – 11; Brassi-
caceae – 10; Lamiaceae – 8; Scrophulariaceae – 5. Преобладание данных се-
мейств является характерной чертой средиземноморской флоры.
К облигатным кальцефилам, растущим почти исключительно на мело-
вых обнажениях, лишенных почв, относятся 57 видов: Diplotaxis cretacea,
Matthiola fragrans, Linum ucranicum, Polygala cretacea, Helianthemum cretaceum,
Thymus dubjanskyi, Asperula exasperata, Anthemis trotzkiana, Ephedra distachya,
Hedysarum grandiflorum, Gypsophila altissima, Bupleurum falcatum и др. Груп-
па факультативных кальцефилов представлена 12 видами, которые произ-
растают необязательно на карбонатных субстратах, но на них более обычны
или более обильны. К ним относятся: Reseda lutea, Astragalus rupifragus и др.
Рис. 9.11. Сенгилеевские горы
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
240
Оносма простейшая (Onosma simplicissima)
Живучка хиосская (Ajuga chia)
Проломник Козо-Полянского
(Androsace koso-poljanskii)
Бурачок Гмелина
(Alyssum gmelinii)
Эфедра двухколосковая (Ephedra distachya)
Тимьян меловой (Thymus calcareus)
Фото: Ю. Пирогова, M. Скотниковой, М. Зайцева, П. Евсеенкова, И. Турбанова, Ю. Ковыгина
Рис. 9.12. Эндемичные виды «Дивногорья», приуроченные к меловым
обнажениям (преимущественно облигатные кальцефиты)
.
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
241
В состав меловой флоры Сенгилеевской возвышенности входит также
ряд третичных реликтов: сосна меловая (форма сосны обыкновенной), ла-
зурник трехлопастный (Laser trilobum) и шаровница точечная (Globularia
punctata).
9.1.2. ЗАПАДНО-СИБИРСКАЯ ЛЕСОСТЕПНАЯ ОБЛАСТЬ
Подзона широколиственных лесов на юге Западной Сибири отсутствует,
здесь она замещается березовыми лесами (см. 7.2.3). Подзона лесостепи охва-
тывает Ишимскую и Барабинскую равнины, центральную часть Приобского и
Зауральского плато и представлена луговыми степями и остепненными луга-
ми в сочетании с осиново-березовыми лесами (колками).
Леса связаны с оподзоленным черноземным почвам, выщелоченным
черноземам и солодям. Узор лесостепной подзоны подобен кружеву, образо-
ванному чередованием леса, луговых полян и небольших озер (рис. 9.13).
Ландшафты лесостепи носят вторичный характер, березовые леса в процессе
освоения территории интенсивно вырубались, они сохранились лишь в запа-
динах или по склонам и днищам логов и балок (рис. 9.14). Наиболее распро-
странены осиново-березовые и березово-осиновые остепненные злаково-
разнотравные леса местами в сочетании с разнотравно-злаковыми лугами.
В северной лесостепи остепнение выражено лишь на повышенных фор-
мах рельефа, колки встречаются часто и на разных элементах рельефа. Зо-
нальными для этой полосы считаются остепненные разнотравно-злаковые
луга, в прошлом занимавшие гривы и высокие водоразделы с выщелоченны-
ми черноземами и лугово-черноземными почвами, богаторазнотравно-злако-
вые луговые степи и осиново-березовые остепненные леса. Разнотравно-зла-
ковые остепненные луга, составляющие сочетания с массивами лесов, сильно
Рис. 9.13. Космическое изображение ландшафта лесостепи Западно-Сибирской
равнины
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
242
Рис. 9.14. Березовая роща в западине в ландшафте луговой степи
изменены антропогенными воздействиями – сенокошением, палами, пасть-
бой, часто они имеют вторичный характер, развиваются на месте сведенных
лесов. Древостой представлен березой повислой (Betula pendula) и осиной
(Populus tremula). Кустарниковый ярус выражен слабо, состоит из Rosa majalis,
Lonicera tatarica, Crataegus sanguinea. Травостой лугов полидоминантный,
высокий и густой. Ведущее положение занимают злаки – Elytrigia repens,
Bromopsis inermis, Calamagrostis epigeios, Poa angustifolia, Brachypodium
pinnatum, Phleum phleoides, Festuca ovina. Характерно присутствие небольшо-
го количества мезофилов – Rubus saxatilis, Heracleum sibiricum, Crepis sibirica,
Galium boreale, Achillea millefolium, Solidago virgaurea, Sanguisorba officinalis,
Angelica silvestris, Heracleum dissectum, Pleurospermum uralense, Heiracium
umbellatum, Pulmonaria mollis. Обычны ксеромезофильные виды: Filipendula
vulgaris, Seseli libanotis, Phlomoides tuberosa, Fragaria viridis, Veronica spuria,
Campanula sibirica, Ranunculus polyanthemos, Dianthus versicolor, Artemisia
sericea, Galium verum. Хорошо развита группа бобовых трав, представленная
ксеромезофильными Astragalus danicus и Medicago falcata и мезофильными
Trifolium lupinaster, Lathyrus pratensis, Vicia cracca, V. sepium видами. Степ-
няки Peucedanum morisonii, Veronica spicata, Plantago urvillei, Allium lineare,
Stipa pennata – на таких лугах немногочисленны.
Галофитно-разнотравные и галофитно-разнотравно-злаковые луговые
(солонцеватые) степи развиты на сильно солонцеватых черноземах, средне- и
глубокостолбчатых солонцах по склонам и шлейфам грив, межгривным при-
поднятым участкам. Для этих степей характерно многообразие экологических
и жизненных форм. Среди доминирующих видов дерновинные, рыхлодерно-
винные и корневищные злаки – Festuca pseudovina, Koeleria cristata, Poa
.
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
243
transbaicalica, Elytrigia repens, Calamagrostis epigeios. Значительное обилие
имеют корневищные осоки – Саrех рrаесох, С. supina. Среди многочисленного
разнотравья наряду с обычными видами луговых степей постоянно присутству-
ют галофильные растения (лугово-степные и лугово-солончаковые) – Galatella
biflora, Glycyrrhiza uralensis, Limonium gmelini, Plantago salsa, Artemisia pontica,
A. nitrosa, A. laciniata, A. rupestris. С корковыми солонцами связаны фитоце-
нозы бескильницевых и бескильвицево-полынных ксерогалофитных лугов,
отличающихся бедностью видового состава и изреженностью травяного по-
крова. В составе травяного покрова преобладают Puccinellia tenuissima, Artemisia
nitrosa. Из постоянно присутствующих видов следует назвать Limonium gmelinii,
Plantago salsa, Festuca pseudovina, Artemisia rupestris, Saussurea salsa, Salicornia
herbacea, Suaeda corniculata, Halimione verrucifera.
Комплексная растительность солонцеватых степей сохранилась благода-
ря приуроченности к почвам, не пригодным для распашки: территории, заня-
тые галофитными вариантами остепненных лугов издавна используются под
выпас сельскохозяйственных животных. При интенсивном постоянном выпасе
в составе травяного покрова увеличивается обилие пастбищеустойчивых расте-
ний Festuca pseudovina, Роа angustifolia, Plantago media, Taraxacum officinale,
Trifolium repens, формирующих пастбищные модификации остепненных лу-
гов – разнотравно-осочково-мятликовые (Саrех рrаесох), типчаково-вейнико-
во-осочковые, разнотравно-вейниково-типчаковые.
На слабодренированных водоразделах формируются многочисленные
блюдцеобразные бессточные впадины. Во впадинах за счет притока поверх-
ностных или грунтовых вод развиваются болота, среди которых наиболее ха-
рактерны крупнотравные, преимущественно тростниковые, осоково-тростни-
ковые и крупноосоковые болота.
Для растительного покрова лесостепи типично обилие европейских ви-
дов (70–90 %). При сравнении целинных плакорных луговых степей и остеп-
ненных лугов Западной Сибири с таковыми в Восточно-Европейской области,
отмечается большая мезофильность и флористическая обедненность первых,
отсутствие в их составе ряда европейских степных видов, участие восточных
видов (Peucedanum morisonii, Iris ruthenica, Geranium asiaticum, Artemisia
glauca, A. laciniata и др.), бедность эфемероидами, представленными лишь
единичными видами гусиного лука (Gagea granulosa, G. minima).
9.1.3. ДАЛЬНЕВОСТОЧНАЯ ОБЛАСТЬ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
Широколиственные леса Дальнего Востока как коренные сообщества
особо характерны для равнин (Уссурийской, Приханкайской и Среднеамур-
ской) и низких предгорий, расположенных на юге Приамурья, Сихотэ-Алиня
и Приморья (см. рис. 7.10 и 7.36). В горах они часто выступают как производ-
ные на месте хвойно-широколиственных лесов. Так же, как в хвойно-широ-
колиственных лесах Дальнего Востока, подавляющее большинство листвен-
ных пород, формирующих широколиственные леса, здесь являются породами
реликтовыми, входившими в состав неогеновых тургайских лесов.
В восточной части неморальной зоны в соответствии с муссонным типом
климата зима сухая и холодная, лето теплое и дождливое. Средняя температу-
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
244
ра самого холодного месяца –16 °С; самого теплого – около 20 °С. За год
выпадает примерно столько же осадков, сколько и в европейских широко-
лиственных лесах (500–700 мм), но местами их количество может достигать
1600 мм. По сезонам они распределяются крайне неравномерно. Большую
часть осадков приносит летний муссон; всю зиму – сухой сезон. В январе-
феврале выпадает всего 5–10 мм, снежный покров – маломощный. Почвы
преимущественно бурые оподзоленные.
Господствующими лесообразующими породами служат дубы и липы.
Дуб монгольский (Quercus mongolica) – относительно засухоустойчивый и мо-
розостойкий; дуб зубчатый (Quercus dentata) – более теплолюбивый и тяготе-
ющий к районам с морским климатом. В некоторых ассоциациях в качестве
характерного содоминанта встречаются липа амурская (Tilia amurensis), липа
таке (Tilia taquetii); в долинных лесах Дальнего Востока – липа маньчжурская
(Tilia mandshurica). В составе дубняков с липой амурской часто принимают
участие клён моно (Acer mono), березы ребристая и даурская (Betula costata,
B. davurica), а также осина обыкновенная. Местами дубняки формируют дре-
востой с небольшим участием березы даурской (Betula davurica) в комплексе с
лещиной маньчжурской (Corylus mandshurica) и ярусом из рододендрона ост-
роконечного (Rhododendron mucronulatum). Граб сердцелистный (Carpinus
cordata), растущий на юге Приморья, – обычно не превышающий 15 м высо-
ты – располагается во втором или третьем подъярусе, в местоположениях,
наиболее защищенных от низких зимних температур. На юге Приморья по
сухим склонам, преимущественно в дубняках, произрастает ясень носолист-
ный (Fraxinus rhynchophylla).
Леса многоярусные, увитые лианами. В подлеске встречаются жимо-
лость, сирень, рододендрон, аралия высокая и др. Большинство этих родов
представлено многочисленными видами, отсутствующими в Европе. Распро-
странены лианы: виноград амурский (Vitis amurensis), актинидия (Actinidia
kolomikta), лимонник (Schisandra chinensis). На юге Дальнего Востока в лесах
растет легендарный женьшень (Panax ginseng).
В травяном ярусе обычны горошек амурский (Vicia amurensis), герань
Максимовича (Geranium maximowiczii), вероника лекарственная (Veronica
officinalis), косоплодник сомнительный (Plagiorhegma dubia), кислица обрат-
нотреугольная (Oxalis obtriangulata), стеблелист мощный (Caulophyllum ro-
bustum), веретенник яйцевидный (Atractylodes ovata), деллингерия шершавая
(Doellingeria scabra) и многие другие; папоротники – адиантум стоповид-
ный (Adiantum pedatum), кочедыжник китайский (Athyrium sinense), луно-
кучник густосорусовый (Lunathyrium pycnosorum), щитовник толстокорне-
вищный (Dryopteris crassirhizoma), чистоустник азиатский (Osmundastrum
asiaticum).
На южных отрогах Буреинского хребта встречаются дубняки со сложным
подлеском из рододендрона даурского (Rhododendron dauricum), лещины
разнолистной (Corylus heterophylla), спиреи (Spiraea ussuriensis), бересклета
(Euonymus pauciflora), леспедецы двуцветной (Lespedeza bicolor). Травяни-
стый покров чрезвычайно богат и разнообразен, часто с полынью Гмелина
(Artemisia gmelinii), ландышем и осоками. Почти всюду в приамурских дубра-
вах встречается примесь лиственницы даурской, поэтому переходы от хвойно-
.
Глава 9. ЗОНА НЕМОРАЛЬНЫХ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
245
широколиственных лесов к широколиственным прослеживаются с трудом.
Бывают и остепненные дубравы, в которых травяной ярус слагают полынь
Гмелина, касатик одноцветковый, тонконог гребенчатый, пижма, типчак и
другие травянистые ксерофиты.
Долинные, приречные леса высокоствольны, в них преобладают вяз япон-
ский (Ulmus japonica), орех маньчжурский (Juglans mandshurica), ясень мань-
чжурский (Fraxinus mandshurica), бархат амурский (Phellodendron amurense) и
более низкие маакия (Maakia amurensis), груша уссурийская (Pyrus ussuriensis),
яблоня маньчжурская (Malus mandshurica), сирень амурская (Syringa
amurensis), боярышник перистонадрезанный (Crataegus pinnatifida) и др.
По осоково-тростниковым болотам в низовьях рек, редины обычно образу-
ет ольха японская (Alnus japonica), реже она формирует чистые насаждения
на местообитаниях с проточным увлажнением. Ее высота редко превышает
10–15 м.
Современные чистые дубняки часто растут на месте хвойно-широко-
лиственных лесов. Возникли они в результате пожаров и вырубки хвойных
пород. В прошлом чистые дубняки были широко распространены на низ-
менностях, которые ныне частично распаханы. Вместо девственных лесов на
низменностях и в предгорьях рассеяны низкорослые древесно-кустарнико-
вые заросли, представляющие характерный элемент ландшафта многих рай-
онов юга Дальнего Востока. Дубняки также занимают местообитания, не
подходящие для хвойных деревьев: сухие южные склоны с маломощными
скелетными почвами.
9.2. АНТРОПОГЕНННЫЕ НАРУШЕНИЯ ШИРОКОЛИСТВЕННЫХ ЛЕСОВ
И ЛЕСОСТЕПИ
Широколиственные леса произрастают на плодородных почвах и в мяг-
ком климате. Не случайно многие дворянские усадьбы, в том числе родовые
имения великих русских писателей, поэтов и композиторов, располагались на
территории хвойно-широколиственных и широколиственных лесов. Они бы-
ли заселены раньше и с большей плотностью, чем другие зоны России.
В конце голоцена, в результате интенсивного антропогенного воздей-
ствия, произошло значительное сокращение площади лесов, их граница от-
ступила на север. Ко времени образования Киевской Руси девственные лесо-
степные ландшафты уже отсутствовали. В то же время леса служили местами
укрытия от набегов кочевников и играли огромную оборонительную роль. По-
этому в пограничных районах лесные массивы всячески оберегались. Кочев-
ники, наоборот, выжигали леса, чтобы подорвать оборону славян и расширить
участки с травянистой растительностью для скота. В результате противопо-
ложного отношения русских племен и кочевников к растительности, леса и
луга (степи) в лесостепи неоднократно сменяли друг друга.
С конца XIX и в течение XX века разрушение естественных ландшафтов
происходило все возрастающими темпами. В настоящее время большая часть
территории широколиственных лесов распахана, занята населенными пунк-
тами, объектами транспортной инфраструктуры и др. (рис. 9.15). Участки
естественных экосистем сохранились лишь в немногих заповедниках.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
246
Рис. 9.15. Космическое изображение территории Тульской области
(зона широколиственных лесов). На рисунке доминируют прямоугольные
участки пахотных угодий (по Google Earth)
Вопросы к главе 9
1. Природные условия зоны неморальных широколиственных лесов; экология расте-
ний.
2. Широтные особенности растительности зоны неморальных широколиственных ле-
сов: подзоны широколиственных лесов и лесостепи.
3. Региональные особенности растительности неморальных широколиственных лесов:
Восточно-Европейская и Дальневосточная области широколиственных лесов, За-
падно-Сибирская лесостепная область.
Рекомендуемая литература
Восточноевропейские широколиственные леса / Под ред. О. В. Смирновой. М.: Наука,
1994. 364 с.
Горчаковский П. Л. Широколиственные леса и их место в растительном покрове Юж-
ного Урала. М.: Наука, 1972. 146 с.
Клеопов Ю. Д. Анализ флоры широколиственных лесов Европейской части СССР. Ки-
ев: Изд-во «Наукова думка», 1990. 352 с.
Курнаев С. Ф. Теневые широколиственные леса Русской равнины и Урала. М.: Наука,
1980. 315 с.
Мильков Ф. Н. Лесостепь Русской равнины: Опыт ландшафтной характеристики. М.:
Изд-во АН СССР, 1950. 296 с.
Рысин Л. П. Липовые леса Русской равнины. М.: Т-во науч. изд. КМК, 2014. 194 с.
.
Глава 10. ЗОНА СТЕПЕЙ
247
Луг как ботанико-географическое и геоботаническое понятие – это био
о мере продвижения на юг, где климат становится более теплым, все
отчетливее проявляется роль увлажнения как лимитирующего эколо-
гического фактора. Широколиственные леса и лесостепи сменяются зоной
степей, которая простирается от венгерских пушт через всю центральную
часть Евразии до Алтая и далее через Забайкалье в Монголию и Северо-
Восточный Китай.
Степную растительность образуют виды, сформировавшиеся в основном
в теплых и сухих условиях средиземноморского климата. В доледниковое вре-
мя степные формации уже отмечались в пределах древнего Средиземноморья
(в Передней, Средней Азии и в Иране), а также в горах Карпат, на Балканском
полуострове, Кавказе. Все эти центры дали основное ядро ксерофитной флоры
современных степей Евразии. С наступлением похолоданий в плейстоцене эта
ксеротермофитная флора обогатилась новыми видами и трансформировалась
в криоксерофитную перигляциальную растительность. В пределах современ-
ного ареала степи стали формироваться в основном в начале голоцена. Север-
ные более влаголюбивые виды (широкотравье, корневищные луговые мезо-
фитные злаки и осоки) проникли в луговые степи из лесной зоны.
Луга и степи относятся к типам растительности, в которых ценозообра-
зующая роль принадлежит травянистым растениям; первые мезофитными
видами, вторые – в основном ксерофитными. Настоящими степями называют
травянистые сообщества с более или менее сомкнутым травостоем, состоящим
главным образом из ксерофитных дерновинных злаков.
Природные условия. Рельеф степной зоны Восточно-Европейской и
Западно-Сибирской равнин определяется теми же геоморфологическими осо-
бенностями, которые характеризуют рельеф лесостепной подзоны. На Восточ-
но-Европейской равнине они связаны с возвышенностями, сильно расчленен-
ными овражно-балочной сетью. В Западной Сибири, отличающейся равнин-
ным рельефом, в зоне степей распространены блюдцеобразные западины и
озера, наполняющиеся водой только в весеннее время. Особые природные
условия характеризуют Прикаспийскую низменность, расположенную ниже
уровня моря. Она представляет собой молодую первичную морскую равнину с
очень плоским рельефом.
Для климата степной зоны характерно возрастание континентальности с
севера на юг и по мере удаления от океанов. Степная зона на западе Восточно-
Европейской равнины доступна теплым и влажным воздушным массам, при-
ходящим с Атлантики. Средние температуры января в Ростове-на-Дону –3 °С
(абсолютный минимум –31,9 °С); в Кызыле (Тува), находящейся во власти
зимнего антициклона, средняя температура января –28,6 °С (абсолютный ми-
нимум –52,6 °С). Значения температур июля мало различаются по мере дви-
жения по широте: в Ростове-на-Дону 23,4 °С, на Южном Урале 22 °С, в Кы-
зыле 20,4 °С.
Глава 10
ЗОНА СТЕПЕЙ
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
248
Рис. 10.1. Климатограмма Нижнего
Поволжья
----
количество выпадающих осадков;
–––
среднемесячные температуры воз-
духа
Важным фактором формиро-
вания степной растительности яв-
ляется количество и ритмика вы-
падения атмосферных осадков. Го-
довое количество осадков сущест-
венно меняется с запада на восток.
В Ростове-на-Дону за год выпадает до 614 мм осадков, испаряемость меньше
этого показателя (ГТК > 1), сухой сезон полностью отсутствует. К юго-востоку
количество осадков снижается. В Нижнем Поволжье среднегодовое количество
осадков составляет 406 мм, испаряемость значительно выше (ГТК < 1), засуш-
ливый период устанавливается с июня до середины сентября (рис. 10.1).
Если в европейской части России и в Западной Сибири сезонный ход вы-
падения осадков определяется западным переносом, то в Забайкалье – муссо-
нами (рис. 10.2). Здесь наблюдается летний пик увлажнения на фоне засушли-
вых и холодных весны и осени, что хорошо видно на климатограмме (рис. 10.3).
Под действием летнего муссона дожди выпадают в течение 1–1,5 месяцев
(в июле–августе). Поскольку в этот же период наблюдается максимум тепла,
интенсивность жизнедеятельности растений высокая.
Рис. 10.2. Сезонная ритмика выпадения атмосферных осадков в европейской
части России и в Забайкалье (по В. Г. Мордковичу)
.
Глава 10. ЗОНА СТЕПЕЙ
249
Рис. 10.3. Климатограмма
Забайкалья:
----
количество выпадающих осадков;
–––
кривая среднемесячных температур
воздуха
В растительных сообществах
степей мощно развиты как надзем-
ная, так и подземная части расте-
ний (рис. 10.4). Степные травы еже-
годно продуцируют несколько тонн
зеленой массы на гектар. Эта масса
потребляется травоядными живот-
ными; значительная ее часть разла-
гается редуцентами. Накопление гумуса в почве является следствием непол-
ного цикла деятельности микроорганизмов, которые из-за летней засухи и
зимних морозов успевают осуществить лишь первые стадии трансформации
органического вещества. Это приводит к доминированию процессов гумифи-
кации органических остатков над процессами их минерализации. Известьсо-
держащие материнские породы способствуют связыванию гуминовых кислот
в труднорастворимые гуматы кальция. Таким образом осуществляется главный
процесс формирования почв в степной
зоне – гумусонакопление. Почвы степной
зоны представлены различными подти-
пами черноземов; по мере продвижения
на юго-восток условия для образования
черноземов в связи с усилением засуш-
ливости климата становятся все менее
благоприятными и здесь формируются
подтипы каштановых почв.
Адаптация степных растений.
Степные растения во время вегетации
испытывают влияние высоких темпера-
тур, сочетающихся с недостаточной ув-
лажненностью. Эти факторы определяют
приспособительные особенности степ-
ных растений. Основными растениями,
слагающими растительный покров сте-
пей, являются ксероморфные злаки. К та-
ким злакам относятся: виды ковыля
(Stipa), овсянница валлисская или типчак
(Festuca valesiaca), житняк гребенчатый
Рис. 10.4. Вертикальная структура
степного сообщества (по М. С. Шалыту):
1 – Euphorbia seguieriana; 2 – Festuca valesiaca;
3 – Salvia nutans; 4 – Stipa lessingiana
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
250
Ковыль Лессинга (Stipa lessingiana)
Житник гребенчатый (Agropyron cristatum)
Овсяница овечья (Festuca ovina)
Тонконог жестколистный
(Koeleria sclerophylla)
Рис. 10.5. Ксероморфные злаки с узкими и шиловидными листьями
.
Глава 10. ЗОНА СТЕПЕЙ
251
Рис. 10.6. Поперечный разрез листа ковыля:
1 – пластинка листа свернутая в трубку; 2 – пластинка листа
развернутая. Сильное развитие механической ткани (заштри-
ховано), бороздчатость, устьица в бороздках, транспирация в
замкнутую полость свернутого листа.
Рисунок В. К. Василевской (по А. П. Шенникову)
(Agropyron cristatum), житняк гребневидный (A. pectinatum), тонконог сизый
(Koeleria glauca), тонконог гребенчатый (K. cristata). Общей особенностью
этих растений являются плотные дерновины, содержащие десятки, а иногда и
сотни надземных побегов. После окончания вегетационного периода надзем-
ные части растений отмирают, образуя массу ветоши, которая часто служит
причиной пожаров. Однако почки возобновления злаков защищены дерни-
ной, а пепел пожарищ благоприятствует росту растений в новый вегетацион-
ный период. Вода после снеготаяния и выпадающие осадки частично задер-
живаются в дерновине, что благоприятствует началу весенней вегетации.
Основная масса корней степных злаков расположена в гумусовом гори-
зонте, часть корней проникает значительно глубже: почти до 2 м у ковыля
украинского (Stipa ucrainica), на 180 см у ковыля волосатика (S. capillata), бо-
лее чем на 120 см у типчака и до 80 см у тонконога. Однако корни не достига-
ют глубоко залегающих грунтовых вод. По своему экологическому типу степ-
ные растения относятся к омброфитам, жизнедеятельность которых обеспечи-
вается влагой атмосферных осадков.
Листовые пластинки степных злаков узкие и даже шиловидные (рис. 10.5).
В период летней засухи листья свертываются устьичной стороной внутрь
(рис. 10.6). Свернутость бывает полная, когда края листа смыкаются, образуя
полость. Нередко свернутость бывает неполная, и загнутые края листа лишь
отчасти прикрывают нижнюю сторону листа. В сырую погоду свернутость
уменьшается, в сухую – увеличивается. Свернутый лист ковыля, типчака и
других степных злаков защищен от чрезмерной потери воды расположением
устьиц в узких бороздках, открывающихся в почти замкнутую полость. Водя-
ной пар, выделяемый устьицами в полость, увлажняет в ней воздух, и транс-
пирация замедляется или прекращается.
Степное разнотравье в отличие от единообразной группы злаков, имеет
различные особенности строения, обеспечивающие существование в засуш-
ливых условиях. Растения обладают приспособлениями, способствующими
уменьшению расхода воды. Так у одних растений уменьшению транспирации
способствует густое опушение стеблей, листьев и цветков. У других приспособ-
лением служит жестколистность (склерофильность). Эти растения имеют жест-
кие листья с толстой кутикулой и с сильно развитыми механическими тканя-
ми, что придает им высокую засухоустойчивость.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
252
В степном разнотравье есть еще одна очень характерная для степей груп-
па растений, получившая название перекати-поле. К ней принадлежат виды
из различных семейств. Надземные части этих растений перед окончанием
вегетации принимают форму шара. Перекати-поле после созревания плодов
отрываются от почвы и, гонимые ветром, долго перекатываются по степи, по-
степенно рассыпая семена. Одни из них являются однолетниками – верблюдка
отклоненная (Corispermum declinataum), другие – двулетниками – солянка
холмовая (Salsola collina), к многолетникам относится качим метельчатый (Gyp-
sophila paniculata).
Своеобразную группу жизненных форм образуют эфемеры (однолетние
растения) и эфемероиды (многолетние растения), которые избегают летней
засухи благодаря развитию в весенний и раннелетний период, когда влаги в
почве еще достаточно. Эфемеры и эфемероиды образуют несколько красоч-
ных аспектов, которые были рассмотрены нами при описании луговых степей
Центрально-Черноземного заповедника (см. раздел 9.1.1).
Степи характеризуются весьма высокими показателями видового разно-
образия, которые изменяются с запада на восток и с севера на юг. Так, если в
европейских степях насчитывается свыше 800 видов, то на востоке в сухих
степях Даурии всего 400–450 видов.
Существенные различия природных условий определяют разнообразие
формаций, ассоциаций, экологических вариантов степной растительности как
с севера на юг, так и с запада на восток, что позволяет различать подзоны и
ботанико-географические области степей.
10.1. ПОДЗОНЫ СТЕПЕЙ
В степной зоне различают три подзоны: северную, среднюю и южную,
которые сменяют друг друга широтными полосами. Причинами их выделения
является различное соотношение тепла и влаги, нарастание континенталь-
ности климата, что отражается на распространении почвенно-растительного
покрова. Природе и растительности южных опустыненных степей посвящен
раздел 10.3.
Северостепная подзона занимает бóльшую часть зоны. В условиях
более влажного климата здесь сформировались черноземы обыкновенные, к
которым приурочены разнотравно-дерновиннозлаковые степи. На Восточноев-
ропейской равнине они представлены разнотравно-типчаково-ковыльными
(Stipa ucrainica, S. tirsa, S. dasyphylla, S. anomala, S. capillata, S. lessingiana,
Helictotrichon schellianum, Festuca valesiaca, Koeleria cristata, Bromopsis riparia,
Filipendula vulgaris, Trifolium montanum, Medicago romanica, Salvia nutans,
Euphorbia stepposa, Carex humilis) степями, местами с кустарниками (Cerasus
fruticosa, Prunus spinosa); в Заволжье − разнотравно-ковыльными, разнотрав-
но-тырсовыми (Stipa zalesskii, S. capillata, S. tirsa, S. lessingiana, S. pennata,
S. korshinskyi, Helictotrichon desertorum, Festuca valesiaca, Koeleria cristata,
Salvia stepposa, Filipendula vulgaris, Medicago romanica, Pulsatilla multifida,
Carex humilis) степями, местами с кустарниками (Rosa majalis, Chamaecytisus
ruthenicus, Amygdalus nana, Spiraea crenata, S. hypericifolia); в Западной Си-
бири − разнотравно-залесскоковыльными, разнотравно-овсецовыми, разно-
.
Глава 10. ЗОНА СТЕПЕЙ
253
травно-тырсовыми (Stipa zalesskii, S. capillata, S. lessingiana, S. pennata,
S. korshinskyi, Helictotrichon desertorum, Festuca valesiaca, Koeleria cristata,
Peucedanum morisonii, P. ruthenicum, Pulsatilla orientali-sibirica, Salvia stepposa,
Filipendula vulgaris, Seseli krylovii, Artemisia frigida, Carex supina) степями,
местами с кустарниками (Spiraea hypericifolia, S. crenata, Caragana frutex); в
Даурии − разнотравно-типчаково-ковыльными, разнотравно-вострецово-ко-
выльными, нителистниково-ковыльными (Stipa baicalensis, Leymus chinensis,
Festuca lenensis, Clematis hexapetala, Hemerocallis minor, Phlojodicarpus
sibiricus, Filifolium sibiricum) степями.
Среднестепная подзона. К менее плодородными южными чернозе-
мами приурочены дерновиннозлаковые степи, которые по видовому составу
значительно беднее и имеют более разреженный растительный покров. Рас-
тительность представлена: в Причерноморье − типчаково-ковыльными, тип-
чаково-тырсовыми (Stipa ucrainica, S. lessingiana, S. brauneri, S. capillata,
Festuca valesiaca, Leymus ramosus, Tanacetum odessanum, Linaria biebersteinii,
Iris pumila, Crinitaria villosa, Artemisia austriaca, Carex supina) степями; на
Восточноевропейской равнине − типчаково-ковыльными, типчаково-тырсо-
выми (Stipa ucrainica, S. lessingiana, S. capillata, Festuca valesiaca, Leymus
ramosus, Tanacetum achilleifolium, Iris pumila, Galatella villosa, G. tatarica,
Artemisia austriaca, Carex supina) степями; в Заволжье − типчаково-ковылко-
выми, типчаково-тырсовыми, типчаково-ковыльными, типчаковыми (наряду с
перечисленными для предыдущих типов степей видами, по мере продвижения
на восток существенную роль начинают играть Stipa pennata, S. korshinskyi,
S. kirghisorum, а S. ucrainica выходит из сообществ) степями, местами с ку-
старниками (Spiraea hypericifolia, Caragana frutex, C. pumila); в Даурии − мелко-
дерновиннозлаковыми, вострецовыми, типчаково-ковыльными (Stipa krylovii,
Cleistogenes squarrosa, Koeleria cristata, Festuca lenensis, Agropyron cristatum,
Poa botryoides, Leymus chinensis, Artemisia frigida, Carex duriuscula) степями.
Подзона южных опустыненных степей выделяется на северо-
западе Прикаспия. Коренной зональный тип растительности здесь представлен
полукустарничково-дерновиннозлаковыми сообществами на незасоленных
светло-каштановых супесчаных и бурых песчаных почвах. Доминантами явля-
ются злаки (Stipa capillata, S. lessingiana, Agropyron desertorum, A. pectinatum,
Festuca valesiaca) и полукустарнички: прутняк (Kochia prostrata), полыни ав-
стрийская и белая (Artemisia austriaca, A. lercheana). На солонцах формиру-
ются интразональные группировки полукустарничков ксерогалофитов – по-
лыни черной и полыни сантонинной (Artemisia pauciflora, A. santonica).
Большая комплексность почвенно-растительного покрова характерна для
ландшафтов Сарпинской низменности, где высокоминерализованные мор-
ские отложения хвалынских террас с интразональными группировками ксеро-
галофитов местами перекрываются незасоленными флювиальными отложе-
ниями легкого гранулометрического состава с зональными сообществами
дерновинных злаков. В ландшафтах Черных земель, почво-грунты которых
формируются на однородном покрове флювиальных отложений, комплекс-
ность не выражена, здесь господствует зональный тип растительности.
В Север-Западном Прикаспии в естественных условиях не происходит
характерная для растительности подзоны северных пустынь вытеснение дер-
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
254
новинных злаков полынью. Полынь белая, обладающая широкой экологиче-
ской толерантностью и высокой жизненностью, вытесняет дерновинные злаки
в результате интенсивного выпаса.
10.2. БОТАНИКО-ГЕОГРАФИЧЕСКИЕ ОБЛАСТИ СТЕПЕЙ
Подзоны северных и средних степей на территории России подразделя-
ются на области: Восточно-Европейскую (Донско-Волжскую), Заволжско-
Уральскую, Западно-Сибирскую, Дауро-Монгольскую.
10.2.1. ВОСТОЧНО-ЕВРОПЕЙСКАЯ (ДОНСКО-ВОЛЖСКАЯ) ОБЛАСТЬ
Область занимает территорию к югу от лесостепи, простираясь с запада
на восток от излучины р. Дона до Нижнего Поволжья. С северо-запада на юго-
восток происходят следующие эколого-фитоценотические изменения в струк-
туре Восточно-Европейских степей:
–
уменьшается густота травостоя степных сообществ, нарастает их есте-
ственная изреженность;
–
уменьшается биологическая продуктивность травостоя степей;
–
увеличивается ксероморфность длительновегетирующих компонентов
степного травостоя, в том числе дерновинных злаков;
–
в связи с увеличением в южных степях продолжительности жаркого и
сухого летнего периода увеличивается количество видов эфемероидов и эфе-
меров, использующих почвенную влагу в первую, менее засушливую половину
вегетационного периода.
Северные разнотравно-типчаково-ковыльные степи ранее, до
распашки, покрывали плакоры (водоразделы) к югу от лесостепи. От нижнего
течения Дона эти степи доходят до предгорий Кавказа. Они связаны на плако-
рах с типичными (обыкновенными) и отчасти южными черноземами.
Основу травостоя на водоразделах и пологих склонах на суглинистых
черноземах образуют эвксерофильные и некоторые мезоксерофильные плот-
нодерновинные злаки, преимущественно перистые ковыли (Stipa lessingiana,
S. zalesskii, S. ucrainica, S. tirsa).
На более легких почвах или в
условиях усиленного выпаса до-
минирует ковыль волосатик –
тырса (S. capillata), относящий-
ся к волосовидным ковылям.
Из мелкодерновинных злаков
обычно обилен типчак или ов-
сяница валлийсская (Festuca
valesiaca). Там, где доминанта-
ми становятся ковыли, их ости
в период цветения придают
степи серебристый оттенок.
Рис. 10.7. Ковыльная степь
.
Глава 10. ЗОНА СТЕПЕЙ
255
Степь напоминает седое волнующееся море, когда травостой колышется от
легкого ветра (рис. 10.7). Для многих сообществ плакоров характерен корот-
кокорневищный кострец береговой (Bromopsis riparia); обычно присутствует
в небольшом количестве мятлик (Poa angustifolia).
Характерно присутствие эфемероидов, заканчивающих вегетацию в нача-
ле или середине лета и отсутствующих в северных луговых степях. Видовое
разнообразие эфемероидов весьма значительно: Adonis volgensis, Bellevalia
sarmatica, Serratula erucifolia, Poa bulbosa (разрастается на сильно выбитых
участках степей), Gagea pusilla, G. bulbifera, Tulipa gesneriana, T. biebersteiniana,
Hyacinthella leucophaea, H. pallasiana, Ornithogalum navaschinii, Crocus reticu-
latus и др. Эфемеры обычно присутствуют, но в небольшом числе особей, чаще
встречаются: Holosteum umbellatum, Arenaria serpyllifolia, Alyssum turkestanicum
var. desertorum, Erophila verna, Draba nemorosa, Viola kitaibeliana, Androsace
elongata, Veronica verna.
В разные времена года ковыльные степи, подобно луговым степям, дают
ряд аспектов. Приведем в качестве примера смену аспектов разнотравной ко-
выльной степи (рис. 10.8).
I. Предвесенний аспект. Тотчас после схода снега, во второй половине
марта, между остатками прошлогодних растений начинают зеленеть мхи
(Syntrichia ruralis) и развивается густой покров сине-зеленой водоросли
(Nostoc commune).
II. Ранневесенний аспект. Во второй половине апреля появляются цве-
тущие растения: разноцветные пятна низких Iris pumila, ярко-желтые круп-
ные звезды Adonis volgensis и желтовато-зелёные звёздочки Gagea pusilla и
G. bulbifera. Весьма многочисленны маленькие озимые однолетники (Erophila
verna и Holosteum umbellatum); на слабозасоленных почвах цветут тюльпаны
(Tulipa gesneriana и Т. biebersteiniana).
III. Аспект разгара весны. Вскоре степь зеленеет, и на общем зеленом
фоне «вспыхивают» многочисленные светлые соцветия Lathyrus pannonicus и
L. pallescens, Anemone sylvestris, Valeriana tuberosa, желтые цветки Potentilla
humifusa и P. patula. Но лучшим украшением степей служат пионы (Paeonia
tenuifolia), предпочитающие северные склоны. В это же время появляются
первые ковыли (Stipa pennata). Начинают цвести кустарники, образующие
сплошные заросли на склонах и каменистых почвах: тёрн (Prunus spinosa),
миндаль степной (Amygdalus nana) и карагана кустарниковая (Caragana
frutex).
IV. Раннелетний аспект. Во второй половине мая и до середины июня
степь достигает наибольшей красочности. Цветут и плодоносят ковыли
(особенно Stipa lessingiana), различные другие злаки (Festuca rupicola,
Koeleria cristata, Bromopsis riparia). Между белыми метелками ковылей про-
ступают синие соцветия шалфея (Salvia nutans), белые соцветия Filipendula
vulgaris, Trifolium montanum и Eremogone saxatilis, светло-голубые незабудки
(Myosotis sylvatica), голубые колокольчики (Campanula stevenii), красные го-
ловки наголоватки паутинистой (Jurinea arachnoidea) и высокие свечки синя-
ка русского (Echium russicum).
V. Аспект разгара лета. Постепенно красочность степей тускнеет. Уже
во второй половине июня степь начинает приобретать бурый оттенок. Цветут
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
256
I
II
Тюльпан Биберштейна
(Tulipa biebersteiniana)
Пион тонколистный (Paeonia tenuifolia)
III
III
Ковыль Лессинга (Stipa lessingiana)
Колокольчик Стевена (Campanula stevenii)
IV
V
Углостебельник татарский (Goniolimon tataricum) Жабрица извилистая (Seseli tortuosum)
Фото: В. Гелюты, Д. Ганжи, В. Коломийчука, А. Цветковой, С. Банкетовой, Т. Гореловой
Рис. 10.8. Виды, определяющие смену сезонных аспектов растительности
разнотравной ковыльной степи: I–VI – пояснения в тексте
.
Глава 10. ЗОНА СТЕПЕЙ
257
лишь немногие виды: желтые Galium verum и Centaurea ruthenica, подорож-
ник средний Plantago media, фиолетово-красные Serratula radiata, Jurinea
multiflora и местами Salvia nemorosa.
В это время года цветет и большинство растений, образующих перекати-
поле (Serratula erucifolia, Phlomis pungens и др.). Преобладают желтый и голу-
бой с красноватым оттенком цвета.
VI. Позднелетний аспект. Со второй половины июля до начала сентября
в облике степи доминируют жесткие, с металлическим блеском ости ковыля
(Stipa capillata) и многие зонтичные (Peucedanum ruthenicum, Ferula feru-
laeoides, Seseli tortuosum, Falcaria vulgaris и др.). В это время цветет также
грудница мохнатая (Galatella villosa).
VII. Осенний аспект. С середины октября до снегопада степь безжизнен-
на. Осенняя влага пробуждает к жизни только водоросли и мхи.
Иногда в травостое попадаются отдельные побеги караганы кустарнико-
вой – дерезы Caragana frutex, образующей более или менее компактные за-
росли по склонам балок или северным склонам водоразделов. В зарослях
степных кустарников принимают также участие Amygdalus nana и Spiraea
crenata.
Средние типчаково-ковыльные степи с господством Stipa lessingi-
ana связаны с южными черноземами и темно-каштановыми почвами. Они
расположены на равнине Крымского полуострова, в бассейне нижнего и сред-
него течения Дона; в долине р. Медведицы степи по правобережью Волги до-
вольно далеко продвигаются на север.
Типчаково-ковыльные степи отличаются от разнотравно-типчаково-ко-
выльных более ксерофитным составом господствующих дерновинных степных
злаков и более бедным разнотравьем. Северное степное разнотравье или со-
вершенно отсутствует, или, если встречается, то приурочено к лугово-степным
и остепненно-луговым сообществам сухих глубоких ложбин стока или нижних
частей склонов балок.
Травостой в этих степях изреженный, летний период полупокоя выражен
отчетливо. Господствуют эвксерофильные плотнодерновинные злаки, пре-
имущественно ковыли (Stipa lessingiana, S. ucrainica, S. capillata) и типчак
(Festuca valesiaca); обычен, но в меньшем количестве тонконог (Koeleria cris-
tata). Местами большую роль, часто образуя более или менее густые заросли,
играет волоснец (Leymus ramosus). Степное разнотравье представлено ксеро-
фильными видами (Dianthus guttatus, Ferula orientalis, Goniolimon tataricum,
Limonium sareptanum, L. bungei, Galatella villosa, Tanacetum millefolium и др.);
часто довольно обильно развиваются Medicago romanica, Falcaria vulgaris,
Artemisia austriaca. Увеличивается число видов эфемероидов (Astragalus hen-
ningii, A. dolichophyllus, Ferula caspica, Carduus uncinatus, Prangos odontalgica,
Adonis volgensis, Serratula erucifolia).
Начиная с середины XIX века плодородные земли степной зоны все ак-
тивнее вовлекались в сельскохозяйственное производство. В настоящее время
они почти полностью распаханы. Небольшие фрагменты степной раститель-
ности сохранились в заповедниках.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
258
10.2.2. ЗАВОЛЖСКО-УРАЛЬСКАЯ СТЕПНАЯ ОБЛАСТЬ
Заволжско-Уральская область простирается к востоку от Нижней Волги
до Южного Урала. В пределах области выражены подзоны северных разно-
травно-типчаково-ковыльных и средних типчаково-ковыльных степей на су-
глинистых черноземах и каштановых почвах. В районах распространения
южных черноземов и разнотравно-типчаково-ковыльных степей увеличивает-
ся засоление почв, а южнее, в полосе темно-каштановых и особенно каштано-
вых, светло-каштановых почв и типчаково-ковыльных степей засоленность
становится почти повсеместной, большие площади заняты солонцами.
Северные разнотравно-типчаково-ковыльные степи. В степной
части башкирского Приуралья на тучных черноземах, южнее переходящих в
обыкновенные и нередко карбонатные, в доагрикультурное время господство-
вали разнотравно-тырсовые степи с доминированием Stipa capillata, местами
с участием S. lessingiana и S. zalesskii, а также S. korshinskyi, S. sareptana, ино-
гда Helictotrichon desertorum, с большим участием Festuca valesiaca и Koeleria
cristata. Из разнотравья в этих степях встречались: из бобовых – Astragalus
austriacus, A. onobrychis, Medicago falcata s. I, Onobrychis sibirica, Oxytropis
pilosa, из других семейств – Silene chlorantha, Sisymbrium polymorphum,
Filipendula vulgaris, Euphorbia seguieriana, Seseli libanotis, Xanthoselinum
alsaticum, Onosma simplicissima, Phlomoides tuberosa, Salvia stepposa, Thymus
marschallianus, Scabiosa ochroleuca, Veronica incana, Artemisia austriaca,
A. dracunculus, A. glauca, A. latifolia, A. marschalliana, Aster alpinus, Galatella
angustissima, Pilosella echioides, Jurinea arachnoidea, J. multiflora, Tanacetum
millefolium, Galatella villosa, Plantago urvillei и др.
В юго-западной части башкирского Предуралья на маломощных карбо-
натных черноземах были развиты более ксерофитные варианты разнотравно-
типчаково-ковыльных степей с господством Stipa sareptana, S. zalesskii, Helic-
totrichon desertorum, с относительно богато представленным ксерофильным и
со слабо развитым мезофильным разнотравьем. Здесь же на карбонатных
черноземах были распространены «разнотравно-ковыльные» степи с боль-
шим количеством Stipa pennata, S. pulcherrima, местами также Нelictotrichon
desertorum и Stipa zalesskii. На выщелоченных черноземах южных районов
Башкирии, например, вдоль р. Белой, существовали степи с мощным мезо-
фитным разнотравьем и с большим количеством S. tirsa.
Для данной подзоны степей характерны также заросли степных кустар-
ников (Spiraea crenata, S. hypericifоlia, Cerasus fruticosa, Amygdalus nana,
Caragana frutex), встречающиеся по степным склонам и другим местообита-
ниям.
Средние типчаково-ковыльные степи формируются на темно-
каштановых почвах, несолонцеватых или слабосолонцеватых. Доминирую-
щими видами являются Stipa lessingiana, S. capillata и Festuca valesiaca. На
легких разностях темно-каштановых почв растительный покров представлен
тырсовыми степями, в случае тяжелого механического состава роль тырсы
(Stipa capillata) уменьшается, в степном травостое господствует ковыль (Stipa
lessingiana); повсюду значительно участие типчака. Для тырсовых и ковылково-
тырсовых степей характерны полукустарнички Artemisia austriaca, A. lerchiana;
корневищная осока Carex supina; разнотравье – Galium verum, Galatella villosa,
.
Глава 10. ЗОНА СТЕПЕЙ
259
Tanacetum achilleifolium, Galatella villosa, G. tatarica; только в тырсовой степи –
Veronica incana и только в ковылково-тырсовой – Taraxacum serotinum; места-
ми встречаются кустарники – Spiraea hypericifolia, Caragana frutex, C. pumila.
10.2.3. ЗАПАДНО-СИБИРСКАЯ СТЕПНАЯ ОБЛАСТЬ
Степная растительность на юге Западно-Сибирской равнины протягива-
ется широкой полосой от предгорий Урала до Оби, занимая плоские равнины
(Ишимскую, Барабинскую, Кулундинскую), а также холмисто-увалистые пла-
то – Зауральское и Приобское. Большая часть ареала степей области находит-
ся на территории Республики Казахстан. Ишимская степь располагается на
водораздельных пространствах рек Тобола и Иртыша. В пределах Обь-Иртыш-
ского междуречья расположена Барабинская степь. Южнее Барабы на Обь-
Иртышском водоразделе находится Кулундинская степь. В центральной части
она представляет собой слабовогнутую чашу. В строении рельефа Кулунды
большая роль принадлежит системе древних ложбин стока. Для всех назван-
ных степных ландшафтов характерно наличие крупных и мелких бессточных
впадин с многочисленными озерами разных размеров.
Степная растительность связана с выщелоченными, обыкновенными и
южными черноземами, а также каштановыми почвами различного механиче-
ского состава. Существенное влияние на развитие почвенного и растительного
покрова степей оказывает засоленность грунтов и грунтовых вод. Развитию
процессов засоления содействовали равнинность рельефа и отсутствие по-
верхностного стока. На водораздельных понижениях равнин широко распро-
странены комплексы и сочетания обыкновенных, солонцеватых черноземов,
луговых солонцеватых и солончаковых почв, солонцов и солончаков. В низких
недренируемых западинах лугово-черноземные почвы сочетаются с солонца-
ми, солончаками и солодями. Приозерные понижения и древние ложбины
стока занимают солончаки. В северных районах распространения степной рас-
тительности преобладает содовое засоление, в южных – хлоридами и сульфа-
тами с участием соды.
Степные сообщества состоят преимущественно из дерновинных злаков.
Корневищные злаки в настоящих степях принимают небольшое участие.
Большинство степных сообществ имеет полидоминантный состав с тремя или
большим числом доминирующих видов. Доминантными и эдификаторными
видами являются дерновинные злаки Stipa pennata, S. zalesskii, S. capillata,
S. lessingiana, S. korshinskyi, Festuca pseudovina, F. valesiaca, реже Koeleria
cristata, Helictotrichon desertorum. Кроме дерновинных злаков в качестве по-
стоянных доминантов выступают представители лугово-степного и степного
разнотравья Peucedanum morisonii, Seseli libanotis, S. ledebourii, Filipendula
vulgaris, Pulsatilla orientali-sibirica, Galatella biflora, Artemisia glauca, A. latifolia,
A. sericea и полукустарнички Artemisia austriaca, A. frigida, A. nitrosa, Thymus
marschallianus. Нередко в степных сообществах доминируют мезоксерофиль-
ные корневищные осоки Саrех supina, С. duriuscula. В составе степных сооб-
ществ также присутствуют степные кустарники, встречающиеся единично или
образующие заросли – Spiraea crenata, S. hypericifolia, Caragana frutex, а в бас-
сейне Тобола европейские виды – Cerasus fruticosa и Chamaecytisus ruthenicus.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
260
Коротковегетирующие эфемероиды и эфемеры для степей Сибири не типич-
ны и представлены лишь единичными видами Androsace septentrionalis,
A. maxima, Gagea granulosa, G. minima, Alyssum turkestanicum var. desertorum,
Erophila verna.
Подзона северных степей представлена разнотравно-типчаково-ко-
выльными степями, простирающимися по южной окраине Западно-Сибир-
ской равнины. Широкое распространение эти степи имели на Прииртышской
и в особенности на Кулундинской равнинах, где занимали обширные площа-
ди. При движении с севера на юг ксерофитизация степей все более усилива-
лась. Основные признаки плакорных степных сообществ этой подзоны − до-
минирование дерновинных ксерофитных и мезоксерофитных злаков – Stipa
zalesskii (рис. 10.9), S. capillata, S. pennata, Festuca pseudovina, F. valesiaca, по-
стоянное участие богатой видами группы разнотравья (Peucedanum morisonii,
Seseli ledebourii, Artemisia latifolia, A. dracunculus, A. glauca), а для некоторых
эдафических вариантов этих степей характерно обильное развитие полуку-
старничковых полыней (Artemisia marschalliana, A. austriaca, A. frigida).
Значительное разнообразие экологических условий в пределах подзоны
обусловливает широкое распространение экологических вариантов разно-
травно-типчаково-ковыльных степей, связанных с условиями местообитания,
отличными от плакорных.
Большое разнообразие растительных сообществ встречается на южных
разностях черноземов и каштановых почв различной степени солонцеватости
и на солонцах. Наиболее распространены галофитные варианты разнотравно-
типчаково-ковыльных степей. В полосе распространения зональных типчако-
во-ковыльных степей на Кулундинской равнине галофитно-разнотравно-
типчаковые и ковыльно-типчаковые степи с галофитно-степными сообще-
ствами сохранились лишь в депрессионных формах мезорельефа с солонцева-
тыми разностями темно-каштановых почв или южных черноземов и солон-
цов. На средне- и особенно высокостолбчатых солонцах, которые постоянно
входят в комплекс с солонцеватыми темно-каштановыми почвами, развива-
ются типчаково-полынные сообщества с участием пустынно-степных полуку-
старничков и южных ксерогалофитных растений (Косhia prostrata, Artemisia
schrenkiana, A. gracilescens, A. nitrosa, Goniolimon speciosum, Limonium gmelinii,
L. coralloides).
Псаммофитные разнотравно-
песчаноковыльные и гемипсаммо-
фитные разнотравно-типчаково-
тырсовые степи довольно широко
распространены в полосе северных
разнотравно-типчаково-ковыльных
степей. Приурочены они к легкосу-
глинистым и супесчаным разнос-
тям черноземов и песчаным почвам,
Рис. 10.9. Настоящие степи со Stipa
zalesskii (фото И. Э. Смелянского)
.
Глава 10. ЗОНА СТЕПЕЙ
261
распространенным на современных и древних речных террасах Тобола и Ир-
тыша, а также на аллювиальных равнинах Обь-Иртышского междуречья.
Здесь растительный покров имеет более мезофильный характер по сравнению
с плакорными участками. На легкосуглинистых почвах возрастает значение
ковыля Залесского (S. zalesskii), и псаммофитно-разнотравно-песчаноковыль-
ные сообщества замещаются разнотравно-ковыльными. Основа их состоит из
S. zalesskii, S. pennata, Festuca valesiaca, в небольшом количестве примеши-
ваются Phleum phleoides и Bromopsis inermis. В травостое много полупсам-
мофитов и псаммофитов (Silene borystenica, Gypsophila paniculata, Pulsatilla
orientali-sibirica, Thymus marschallianus, Jurinea cyanoides). Хорошо развиты
полыни (Artemisia austriaca, A. latifolia, A. glauca, A. dracunculus, A. sericea,
A. marschalliana).
Кальцефитные разнотравно-ковыльные и разнотравно-овсецово-ковыль-
ные степи встречаются на Ишим-Иртышском водоразделе на склонах увалов
и высоких террасах озер, на плоских равнинных пространствах между ними.
Индикаторное значение в сообществах кальцефитных степей принадлежит
ковылю коржинского (Stipa korshinskyi) и овсецу пустынному (Helictotrichon
desertorum), являющимися содоминантами и соэдификаторами. Степи с гос-
подством ковыля коржинского связаны только с карбонатными черноземами.
В разнотравно-овсецово-ковыльных кальцефитных степях основу травостоя со-
ставляют ксерофитные и мезоксерофитные злаки: Stipa zalesskii, S. korshinskyi,
Festuca valesiaca, Helictotrichon desertorum, к ним примешиваются Koeleria
cristata, Stipa lessingiana, Phleum phleoides и корневищные мезофитные Elytrigia
repens, Bromopsis inermis. Группа разнотравья состоит в основном из мезоксе-
рофитов и ксерофитов, наиболее обычны Veronica incana, Potentilla humifusa,
Artemisia dracunculus, Seseli ledebourii, Peucedanum morisonii, Pulsatilla orientali-
sibirica, Galium verum, Phlomoides tuberosa, Artemisia latifolia, Eryngium planum,
Achillea nobilis, Salvia stepposa. Характерно участие в травостое растений бо-
лее южных степей Dianthus leptopetalus, Phlomoides agraria, Galatella villosa.
Бобовые, встречающиеся в небольшом количестве, представлены Astragalus
austriacus, A. onobrychis, Hedysarum gmelinii, Medicago falcata.
Средние полынно-типчаково-тырсовые степи со Stipa capillata
и S. pennata на юге Западной Сибири имеют распространение на плакорах Ку-
лундинской равнины и на Иртыш-Омском междуречье. Почвы под ними юж-
ные и обыкновенные черноземы от тяжелого до легкого гранулометрического
состава. Эдификаторная роль в этих степях принадлежит плотнодерновинным
ксерофитам Stipa capillata и Festuca valesiaca с участием Calamagrostis epigeios,
в качестве постоянных доминантов выступают Koeleria cristata и мезоксе-
рофитный Stipa pennata. Характерно присутствие разнообразных полыней
(Artemisia glauca, A. dracunculus, A. scoparia, A. pontica, A. latifolia, A. sericea,
A. marschalliana, A. austriaca), степных кустарников (Spiraea hypericifolia,
S. crenata), единично встречаются Filipendula vulgaris. Inula salicina, Phlomoides
tuberosa, Seseli libanotis, Peucedanum morisonii.
На легкосуглинистых и супесчаных разностях южных черноземов среди
злаков в большом количестве появляется Helictotrichon desertorum. Из корне-
вищных злаков в таких степях присутствуют Bromopsis inermis и Calamagrostis
epigeios, в некоторых сообществах довольно хорошо развиваются рыхлодер-
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
262
новинные злаки Phleum phleoides и Cleistogenes squarrosa показатели легких
почв. Доминирующая роль принадлежит Artemisia austriaca, меньшее разви-
тие имеют Artemisia glauca и A. marschalliana, единично встречаются Potentilla
bifurca, Seseli ledebourii, Veronica spicata, Onosma simplicissima, Thymus
marschallianus, Hieracium virosum, H. echioides, Helichrysum arenarium,
Orostachys spinosa. Из бобовых постоянно участие Medicago falcata, Astragalus
onobrychis.
10.1.4. ДАУРО-МОНГОЛЬСКАЯ СТЕПНАЯ ОБЛАСТЬ
В пределах России данная область занимает Забайкалье и Даурию. В про-
тивоположность огромным пространствам степных равнин Восточной Европы
и Западной Сибири степи здесь известны под названием «островных». Свое
название они получили благодаря приуроченности к межгорным котловинам,
характерным для гор Алтая и Забайкалья (рис. 10.10). Рассмотрим особенно-
сти островных степей на примере межгорных котловин Бурятии. Они разме-
щаются в Селенгинском среднегорье и Баргузинской котловине. Степные
ландшафты Бурятии возникли в процессе естественноисторического развития
территории в межгорных понижениях. Их природное саморазвитие транс-
формировалось вследствие возрастающего антропогенного воздействия.
К настоящим степям относятся: Боргойская, Болдокская, Селенгинская,
Баргузинская и Кяхтинская степи. Несмотря на такую территориальную раз-
общенность, они имеют однотипный флористический состав. Настоящие
степи образованы из типичных ксерофитов, преимущественно дерновинных
злаков. Примесь мезофильных форм невелика. Отличительные черты насто-
ящих степей здесь: низкий несомкнутый травяной покров; полидоминант-
ность сообществ при невысоком видовом разнообразии (на 1 м2 встречается
до 12 видов растений); ряд видов образуют подушки и куртинки: пустинница
волосовидная (Eremogone capillaris), звездчатка вильчатая (Stellaria dicho-
toma), лапчатка бесстебельная (Potentilla acaulis), – так называемое распла-
станное разнотравье (такие формы роста связаны с процессом адаптации к
холодным и сухим местообитаниям в условиях резко континентального кли-
мата); мощная корневая система, масса которой превосходит надземную в
десятки раз.
Степи Бурятии образованы сообществами мелкодерновинных и крупно-
дерновинных злаков. Из мелко-
дерновинных степей в Забайкалье
преобладают мятликовые, типча-
ковые, житняковые, тонконоговые
сообщества. Основные массивы
этих степей используются в каче-
стве естественных кормовых уго-
дий для выпаса овец.
Рис. 10.10. Чуйская степь
(фото Н. Белозеровой)
.
Глава 10. ЗОНА СТЕПЕЙ
263
Типчаковые степи – мятликово-типчаковые, ковыльно-типчаковые, мел-
козлаково-типчаковые – встречаются в развитых долинах рек и узкой полосой
окаймляют южные крутые склоны хребта Хамар-Дабан. Почвы типчаковых
степей горно-каштановые, бескарбонатные, маломощные, сильно щебнистые.
Типчаковые сообщества – низкотравные, основу травостоя образует овсяница
ленская (Festuca lenensis). Из содоминантных видов пятнистость создают си-
нузии Artemisia frigida и Potentilla acaulis. Кроме вышеперечисленных видов
постоянно присутствуют тонконог, житняк, змеёвка (Cleistogenes squarrosa),
ковыль Крылова (Stipa krylovii). Из видов с меньшим обилием характерны арк-
тогерон злаковидный (Arctogeron gramineum), пустынница волосовидная
(Eremogone capillaris), хамеродос трехнадрезный (Chamaerhodos trifida), смо-
левка енисейская (Silene jeniseensis), полынь пижмолистная (Artemisia
tanacetifolia). Бобовые представлены единичными растениями: астрагал зая-
чий (Astragalus laguroides), остролодочник остролистный (Oxytropis oxyphylla).
Из кустарников выделяется карагана карликовая (Caragana pygmaea), расту-
щая отдельными экземплярами.
Мятликовые степи с доминированием ксерофитного, многолетнего рых-
лодерновинного злака мятлика кистевидного (Poa botryoides), широко распро-
страненным по всей степной области Евразии. Они встречаются в долинах и
на террасах среднего течения рек. В их состав входят сообщества: прострелово-
мятликовая, осочково-мятликовая, чабрецово-мятликовая и полынно-мятлико-
вая. Разнотравье представлено: прострелом Турчанинова (Pulsatilla turczani-
novii), лапчаткой бесстебельной (Potentilla acaulis), подмаренником настоя-
щим (Galium verum), полынью пижмолистной (A. tanacetifolia), тимьяном
монгольским (Thymus mongolicus), рассечённокотовником многонадрезан-
ным (Schizonepeta multifida), проломником седым (Androsace incana). На серо-
зеленом фоне степи из кустарников часто встречаются крупные, с золотистой
корой, кусты караганы карликовой (Caragana pygmaea).
Житняковые степи – житняково-ковыльные, осоково-житняково-ко-
выльные, мятликово-житняковые – распространены в основном небольшими
участками среди других степных массивов. Доминирующую роль в травостое
житняковых степей играет житняк гребенчатый (Agropyron cristatum) засухо-
устойчивый, морозостойкий, неприхотливый к почвам злак. Обычны ковыль
Крылова (Stipa krylovii), реже тонконог (Koeleria), мятлики кистевидный (Poa
botryoides) и оттянутый (P. attenuata), а также отмечается змеевка растопы-
ренная (Cleistogenes squarrosa). Из разнотравья рассеянно встречаются: хамеро-
дос алтайский (Chamaerhodos altaica), бурачок ленский (Alyssum lenense), зоп-
ник клубненосный (Phlomoides tuberosa), пустынница волосовидная (Eremogone
capillaris), серебристый фон образует полукустарничек Artemisia frigida. Плот-
но прижавшись к почве, растут зеленые розетки суккулентного монокарпика
горноколосника колючего (Orostachys spinosa).
Крупнодерновинные ковыльные степи – ковыльно-полынные, осоково-
ковыльные, лапчатково-ковыльные – широко распространены на подгорных
шлейфах и нижних склонах гор Бурятии. Почвы темно-каштановые и кашта-
новые, легкого гранулометрического состава. В этих степях травостой образо-
ван преимущественно злаками, доминирующую роль играют два вида ковыля –
байкальский (Stipa baicalensis) и Крылова. Кроме ковылей встречаются тип-
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
264
чак, тонконог, змеевка, колосняк (вострец) китайский (Leymus chinensis). Ин-
дикаторами пасторальной дигрессии выступают полынь холодная (Artemisia
frigida), лапчатка бесстебельная (Potentilla acaulis), осока твердоватая (Carex
duriuscula).
Сухие дерновинно- и корневищнозлаковые степи характерны для меж-
горных впадин юга Бурятии по р. Селенге, где они занимают самые понижен-
ные части, а также в центре островной Баргузинской степи. По составу преоб-
ладают сообщества ковыля Крылова и колосняка китайского, в качестве со-
доминантов выступают Koeleria cristata и Cleistogenes squarrosa, a Agropyron
cristatum, Poa botryoides, хотя и встречаются постоянно, но необильны. Ха-
рактерные виды разнотравья – лапчатка бесстебельная (Potentilla acaulis), лук
неравноногий (Allium anisopodium), лапчатка вильчатая (P. bifurca), володушка
козелецелистная (Bupleurum scorzonerifolium), цимбария даурская (Cymbaria
daurica), осока твердоватая, полынь холодная и др.
По всей Бурятии распространены мелкодерновиннозлаковые вострецо-
вые и галофитно-сазово-вострецовые сообщества, которые могут рассматри-
ваться как типичные сухостепные. В галофитно-сазово-вострецовниках наибо-
лее резко выражена господствующая роль востреца китайского. Это чистые
вострецовники, где нет согосподствующих видов; сопутствующих видов мало:
полынь холодная, полынь веничная (A. scoparia), прутняк (Kochia prostrata),
осока твердоватая, термопсис ланцетный (Thermopsis lanceolata), володушка
козелецелистная, змеевка растопыренная и др. Состав мелкодерновиннозла-
ковых вострецовников более разнообразен; для них характерна хорошо выра-
женная синузия мелкодерновинных ксерофильных злаков: мятлик кистевид-
ный, тонконог гребенчатый, житняк гребенчатый, змеевка растопыренная.
В большом обилии встречаются осока твердоватая и довольно разнообраз-
ное по составу ксерофильное разнотравье – лапчатка вильчатая (Potentilla
bifurca), вероника седая (Veronica incana) и др.
10.2.5. ПРИКАСПИЙСКАЯ ОБЛАСТЬ ЮЖНЫХ ОПУСТЫНЕННЫХ СТЕПЕЙ
Опустыненные степи, занимающие территорию Северо-Западного При-
каспия, представляют собой западную окраину пояса Великих азиатских пу-
стынь. Здесь, на равнинах юго-востока европейской части России, происходит
переход растительности степной зоны к растительности пустынь. Важнейши-
ми факторами, определяющими своеобразие растительного покрова, служат
геоморфологическое строение, климат и почвы. Основную территорию Севе-
ро-Западного Прикаспия занимают равнины Сарпинской низменности и Чер-
ных Земель, ограниченные на северо-западе и западе Ергенинской возвышен-
ностью, на северо-востоке и востоке – Волго-Ахтубинской поймой, ново-
каспийской террасой и побережьем Каспийского моря, на юге – Манычской
ложбиной. Эти равнины, расположенные ниже уровня Мирового океана, ха-
рактеризуются плоским низменным рельефом хвалынских морских террас и
бугристым эоловым рельефом, возникшим на песчаных отложениях Палео-
волги, веерные дельты которой следовали вслед за отступавшим морем.
Основные черты климата – континентальность и сухость. Средняя тем-
пература июля 35 °С, абсолютный максимум 45 °С; средняя температура янва-
.
Глава 10. ЗОНА СТЕПЕЙ
265
ря от –9 до –4 °С, абсолютный минимум –35 °С. Анализ многолетнего хода
выпадения атмосферных осадков показывает чередование периодов относи-
тельно сухих и влажных. Из десяти лет приблизительно шесть лет бывают
благоприятными для вегетации растений и четыре – засушливыми. Количе-
ство атмосферных осадков может колебаться от 132 до 436 мм. Увлажнение
определяется не только суммой выпавших осадков, но и количеством испаря-
емой влаги. В Северо-Западном Прикаспии испаряемость колеблется от 850
до 1100 мм. Интегральным экологическим показателем условий формирова-
ния зональной растительности служит гидротермический коэффициент Селя-
нинова (ГТК). В среднестепной подзоне Восточно-Европейской (Донско-Вол-
жской) области ГТК 1. В Северо-Западном Прикаспии, где ГТК 0,5, условия
увлажнения достаточны для формирования степных сообществ дерновинных
злаков.
Почвенный покров сформировался при взаимодействии геоморфолого-
литологического и климатического факторов. Основными типами почв явля-
ются: светло-каштановые суглинистые и супесчаные, а также бурые песчаные
на слабоминерализованных флювиальных отложениях; лугово-каштановые
суглинистые почвы в суффозионных западинах; солонцы и солончаки на за-
соленных отложениях хвалынских террас.
Растительный покров Северо-Западного Прикаспия характеризуется вы-
сокой пространственно-временной изменчивостью, связанной с разнообразием
эколого-топологических условий и интенсивностью хозяйственной деятельно-
сти. Коренная растительность плакоров является зональной, она образована
дерновинными ксерофитными злаками и полукустарничками, омброфитами –
растениями с поверхностной корневой системой, жизнедеятельность которых
обеспечивается влагой атмосферных осадков. В суффозионных западинах на
лугово-каштановых почвах формируется особый вариант разнотравно-злако-
вых сообществ. На солонцах – интразональные сообщества полукустарничков
ксерогалофитов омброфитов, представленные в основном чернополынными
группировками (Artemisia pauciflora + A. santonica). Трудности определения зо-
нального статуса растительности Северо-Западного Прикаспия обусловлены
комплексностью почвенно-растительного покрова и разнообразием произ-
водных сообществ, вызванных хозяйственной деятельностью человека.
В качестве типичной на территории Северо-Западного Прикаспия вы-
ступает растительность ландшафтов Сарпинской низменности и Черных зе-
мель.
Ландшафты и растительность Сарпинской низменности.
Сарпинская низменность расположена по правобережью Волги, к югу от
г. Волгограда; с запада ограничена возвышенностью Ергеней, на юге – при-
близительно 47° с.ш. Для нее характерен плоский рельеф хвалынских террас,
сложенных высокоминерализованными «шоколадными» глинами, широко
распространены суффузионные западины. Поверхность террас местами пере-
крыта флювиальными песчаными отложениями. Общие черты геоморфоло-
гического строения Сарпинской низменности показаны на карте (рис. 10.11).
Почвенно-растительный покров характеризуется комплексностью, свя-
занной с пестрой мозаикой почво-грунтов; большое влияние на него оказыва-
ет также выпас скота, земледелие и орошение.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
266
Рис. 10.11. Ландшафтно-геоморфологическая карта Сарпинской низменности
(по В. Г. Можаевой)
Условные обозначения: I – ландшафт плоской морской аккумулятивной равнины средне-
хвалынского возраста; II – ландшафт пологоволнистой равнины на месте древней дельты
сарпинского рукава Волги; 1 – береговая зона стояния хвалынского бассейна, 2 – древние
морские лиманы, 3 –древние береговые валы с позднейшей эоловой обработкой, 4 – запа-
дины суффозионного происхождения, 5 – русла временных современных водотоков, 6 –
ключевые участки, место заложения эколого-топологических профилей в 1961 и 2008 гг.
Ландшафт Волго-Сарпинского района тянется широкой полосой вдоль
Волги. Это пологоволнистая равнина на месте древней дельты сарпинского
рукава Волги, расчлененная сложноветвящимися руслами. К сухим руслам и
озерным впадинам приурочены заросли тростника. В первой половине ХХ
века, когда растительный покров был еще слабо изменен в результате хозяй-
ственной деятельности, на водораздельных участках на светло-каштановых
почвах господствовали зональные белополынно-дерновиннозлаковые сооб-
щества, а на солонцах – интразональные сообщества полукустарничков ксеро-
галофитов Artemisia pauciflora + A. santonica.
.
Глава 10. ЗОНА СТЕПЕЙ
267
Для анализа пространственно-временной динамики растительности рас-
смотрим детальные описания выполненные в 1961 г. и в 2008 г. вдоль эколо-
го-топологического профиля, заложенного на ключевом участке у пос. Ики-
Бухус (см. рис. 10.11, А; рис. 10.12).
Характерные черты растительности вдоль профиля в 1961 г. опреде-
лялись сочетанием интразональных сообществ полыней черной и сантонин-
ной (Artemisia pauciflora + A. santonica) с участием камфоросмы (Camphorosma
monspeliaca) на солонцах и сообществ белополынно-бекманиевых (Beckmannia
eruciformis + Artemisia lercheana) с участием житняков (Agropyron desertorum,
A. pectinatum, A. fragile) на светло-каштановых почвах. В 70-е годы эти земли
были распаханы, затем заброшены и в начале XXI в. представляли собой за-
лежь, используемую под пастбище. К 2008 г. здесь сформировались сообщества
лугово-степного типа длиннокорневищного злака мезофита пырея (Elytrigia
repens) с участием полыней Лерха (белой) и австрийской (Artemisia lercheana,
A. austriaca) и степные сообщества дерновинного злака житняка гребневид-
ного (Agropyron pectinatum) с участием полыней Лерха, австрийской и лечеб-
ной (Artemisia abrotanum) . Эти сообщества стали играть господствующую роль.
Рис. 10.12 . Эколого-топологический профиль через пологоволнистую
равнину на месте древней дельты сарпинского рукава Волги у пос. Ики-Бухус
(см. рис. 10.11, А), составленный по материалам Е. Н. Кудрявцевой,
В. Г. Можаевой, К. Е. Нефедова, Н. Н. Семеновой (по В. Г. Можаевой)
Условные обозначения: Р а с т и т е л ь н о с т ь: 1 – полынь черная (Artemisia pauciflora); 2 – по-
лынь Лерха (Artemisia lercheana); 3 – полынь сантонинная (Artemisia santonica); 4 – полынь ле-
чебная (Artemisia abrotanum); 5 – полынь песчаная (Artemisia arenaria); 6 – прутняк (Kochia pros-
trata); 7 – камфоросма монпелийская (Camphorosma monspeliaca); 8 – пижма (Tanacetum achillei-
folium); 9 – типчак (Festuca valesiaca); 10 – ковыль сарептский (Stipa sareptana); 11 – житняк
пустынный (Agropyron desertorum); 12 – житняк гребневидный (Agropyron pectinatum); 13 –
житняк сибирский (Agropyron sibiricum); 14 – пырей (Elytrigia repens); 15 – бекмания (Beckman-
nia eruciformis); 16 – осока черноколосая (Carex melanostachya); 17 – девясил британский (Inula
britannica); 18 – солодка голая (Glycyrrhiza glabra); 19 – ксерофильное разнотравье; 20 – сорное
разнотравье; П о ч в ы: Клсп – лугово-каштановые супесчаные; Кл – лугово-каштановые; Кп
1 – свет-
ло-каштановые песчаные; К1
сн
–
светло-каштановые солонцеватые; Сн2 – солонцы глубокие; Снср –
солонцы средние; Снк – солонцы корковые; Снл – солонцы луговые; Снл
сч
–
солонцы луговые со-
лончаковатые; Лл
ос
–
лугово-лиманные осолоделые; Лл
сч
–
лугово-лиманные солончаковатые; Лб –
лугово-болотные; Л и т о л о г и я: 21 – глина серого цвета тонкослоистая с линзами суглинков и
песков; 22 – глина шоколадного цвета тонкослоистая, у кровли сильно засоленная, местами тре-
щиноватая, пластичная, с линзами песка сильно ожелезненного разнозернистого; 23 – суглинок
тяжелый серо-бурый, коричнево-бурый, бурый; 24 – суглинок средний бурого цвета; 25 – сугли-
нок легкий желто-бурого цвета; 26 – супесь желто-бурого цвета; 27 – песок желтый и желто-бурый
разнозернистый, несортированный, местами глинистый
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
268
Представители ксеро-галофитного комплекса полынь малоцветковая (Artemisia
pauciflora) и кермек (Limonium platyphyllum) в сложении сообществ заняли
второстепенное место. Несмотря на смену растительных сообществ во време-
ни, во флористическом составе разногодичных описаний общими видами яв-
ляются представители зональной флоры Agropyron pectinatum, Tanacetum
achilleifolium и Artemisia lercheana, а также интразональный вид ксерогало-
фит Artemisia pauciflora.
Ландшафт Большецарынского района приурочен к плоской хвалын-
ской морской равнине, характеризующейся сочетанием невысоких водоразде-
лов (микроплакоров) и суффозионных западин (рис. 10.13). На микроплако-
рах на светло-каштановых почвах преобладают типчаково-ромашниковые и
ковыльно-типчаковые сообщества с доминированием злаков: Festuca valesi-
aca, Stipa sareptana, Agropyron pectinatum, A. sibiricum, Beckmannia eruci-
formis и пижмы (Tanacetum achilleifolium) с участием прутняка (Kochia pros-
trata), полыни Лерха (Artemisia lercheana) и др. К солонцам приурочены чер-
нополынные сообщества (Artemisia pauciflora + Bassia sedoides). В западинах
на лугово-каштановых суглинистых почвах формируются сообщества пырея
(Elytrigia repens) с участием разнотравья (Polygonum aviculare, Chenopodium
album, Eremopyrum orientale).
Описания растительности были выполнены на ключевом участке, при-
уроченном к лиману Большой Царын (см. рис. 10.11, Б). В 1961 г. характер-
ные черты растительности вдоль профиля определялись господством сооб-
ществ с доминированием степных видов (Beckmannia eruciformis, Agropyron
desertorum, A. pectinatum, A. fragile, Festuca valesiaca, местами Stipa lessingiana,
S. sareptana, S. capillata) и пижмы (Tanacetum achilleifolium) на светло-каш-
тановых почвах; на солонцах господствовала полынь черная и сантонинная
(Artemisia pauciflora, A. santonica). В 80-е годы XX века исследуемый участок
был занят рисовыми чеками, в начале XXI века орошение прекратилось, и ри-
совые чеки трансформировались в залежь. К 2008 г. в результате восстанови-
тельной сукцессии на светло-каштановых почвах сформировалась раститель-
ность, которая стала использоваться под пастбища, в результате чего домини-
рующую роль приобрел мортук (Eremopyrum triticeum), злак, индицирующий
сильную пасторальную дигрессию.
Группировки галофита бассии
(Bassia sedoides) связаны со вто-
ричным засолением почв. Микро-
западины с характерными для них
сообществами в 2008 г. не были
отмечены. Несмотря на смену рас-
тительных сообществ во времени,
во флористическом составе разно-
Рис. 10.13. Аэрофотоизображение
ключевого участка:
1 – суффозионные западины, 2 – плос-
кие микроплакоры
.
Глава 10. ЗОНА СТЕПЕЙ
269
Рис. 10.14. Эколого-топологический профиль через плоскую морскую равнину,
осложненную песчаными буграми у пос. Зергент, составленный по материалам
Е. К. Вавилова, Б. В. Виноградова, В. Г. Можаевой, К. Е. Нефедова
(по В. Г. Можаевой). Условные обозначения см. на рис. 10.12
годичных описаний общими видами являются представители зональной фло-
ры Agropyron fragile и Artemisia lercheana.
Ландшафт Сарпинского района представляет собой плоскую морскую
равнину, осложненную песчаными буграми, образованными на месте переве-
янных древних береговых валов. Растительность на буграх представлена бе-
лополынно-дерновиннозлаковыми сообществами на светло-каштановых и
бурых песчаных почвах. На равнине распространены сообщества полыни ма-
лоцветковой (Artemisia pauciflora) с участием солянок на солонцах; к много-
численным западинам приурочены сообщества колосняково-разнотравные
(Leymus ramosus + разнотравье) и пырея (Elytrigia repens) на лугово-кашта-
новых почвах.
Описания растительности эколого-топологического профиля выпол-
нены в 1961 г. и в 2008 г. на ключевом участке у пос. Зергент (см. рис. 10.11, В;
рис. 10.14).
В 1961 г. на плоской морской равнине в начале и в конце профиля осо-
бенности растительного покрова определялись сочетанием сообществ
Beckmannia eruciformis + Tanacetum achilleifolium с участием полыни белой на
старопахотных светло-каштановых солонцеватых почвах и Artemisia pauciflora +
A. santonica с участием солодки (Glycyrrhiza glabra) на солонцах. Раститель-
ность песчаных бугров была представлена сообществом дерновинных злаков
Agropyron fragile + Festuca valesiaca с участием злака мезофита Beckmannia
eruciformi, а также прутняка (Kochia prostrata) и полыни белой на бурых пес-
чаных почвах.
Сравнение геоботанических описаний 1961 и 2008 гг. показывает, что
на песчаных буграх сохранялась степная растительность. На древней морской
равнине к 2008 г. проявилась комплексность, представленная микроценозами
с участием злаков и галофитов. Несмотря на смену растительных сообществ во
времени, во флористическом составе разногодичных описаний общими вида-
ми являются представители зональной флоры Agropyron fragile, Tanacetum
achilleifolium и Artemisia lercheana.
Ландшафты и растительность Черных земель. К югу от Сар-
пинской низменности расположены Черные земли, на востоке они ограничены
ландшафтом бэровских бугров и бровкой новокаспийской террасы, на юге –
долиной р. Кума. Их облик определяется ландшафтами пологоволнистой
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
270
Рис. 10.15. Ландшафты Черных земель и прилежащих территорий (по К. М. Петрову)
Условные обозначения: 1 – морские мелководья с зарослями прибрежно-водной растительности и пес-
чано-ракушечными отмелями; 2 – современное низменное побережье Каспийского моря с тростнико-
выми плавнями. кермеково-тростниково-бескильницевыми и сантониннополынными сообществами;
3 – новокаспийская равнина с сантониннополынными сообществами. песчаными массивами. солонча-
ками и сухими руслами; 4 – верхнехвалынская супесчаная пологоволнистая равнина с дерновиннозла-
ково-прутняково-белополынными сообществами и производными сообществами пасторальных серий;
5 – верхнехвалынская песчаная бугристая равнина с дерновиннозлаково-прутняково-белополынными
сообществами на закрепленных песках и зарослями псаммофитов на эоловых массивах; 6 – дельта Вол-
ги. внутренняя дельта Кумы и северного рукава Терека; 7 – ландшафт бэровских бугров и подстепных
ильменей; 8–15 – урочища доминанты: 8 – бэровские бугры; 9 – песчаные массивы; 10 – границы
устьевой части Палеоволги; 11 – границы долины Кумы; 12 – протоки; «ерики»; 13 – ильменные озера
и лиманы; 14 – сухие русла; 15 – песчано-ракушечные отмели на мелководье Каспийского моря
.
Глава 10. ЗОНА СТЕПЕЙ
271
супесчаной и бугристой песчаной равнин, сформированных на месте веерных
дельт Палеоволги (рис. 10.15). Покров флювиальных отложений на хвалын-
ских террасах определяет относительную однородность почв и растительно-
сти. На Черных землях не выражены солонцы, солончаки и суффозионные
западины, определяющие комплексность почвенно-растительного покрова,
свойственную морским отложениям хвалынских террас Сарпинской низмен-
ности.
Для ландшафтов Черных земель характерны два экологических вари-
анта зональной растительности. Первый вариант представлен кустарничково-
дерновиннозлаковыми сообществами на светло-каштановых супесчаных поч-
вах (рис. 10.16). Главные ценозообразователи – дерновинные злаки: ковыль
(Stipa capillata, S. lessingiana), житняк (Agropyron desertorum, A. pectinatum),
типчак (Festuca valesiaca) и полукустарнички: прутняк (Kochia prostrata), по-
лыни (Artemisia austriaca, A. lercheana). Участие полыни Лерха возрастает по
мере усиления пастбищной нагрузки. Сильная пасторальная дигрессия ведет к
замене зональной растительности пасторально-бурьянным комплексом, для
которого характерны полынь белая, рогач песчаный или эбелек (Ceratocarpus
arenarius), дурнишник (Xanthium strumarium), увеличивается участие гало-
фитов (Bassia sedoides, Atriplex tatarica, Salsola australis).
Второй вариант – кустарничково-дерновиннозлаковые сообщества на
мелкобугристых закрепленных песках со светло-каштановыми супесчаными и
бурыми песчаными почвами. Основные ценозообразователи здесь те же, что в
первом варианте, отличие состоит в более широком участии псаммофитов.
Для слабо закрепленных песков, возникающих в результате усиленного выпа-
са и уничтожения коренной растительности, характерны жузгун (Calligonum
aphyllum), полынь песчаная (Artemisia arenaria), волоснец кистистый (Leymus
racemosus), вейник (Calamagrostis epigeios), кумарчик (Agriophyllum squar-
rosum), верблюдка (Corispermum aralo-caspicum).
В весенний период во влажные годы в обоих вариантах сообществ
обильно развиваются эфемеры и эфемероиды Роа bulbosa, Carex stenophylla,
Tulipa schrenkii, Т. biebersteiniana, Т. biflora, Alyssum turkestanicum и др.
Процессы восстановления нарушенной растительности четко просле-
живаются в биосферном заповеднике «Черные Земли», имеющем статус био-
сферного резервата мирового значения. На начальных этапах восстановления
формируется покров пионеров за-
растания эоловых песков: нерав-
ноцветник кровельный (Anisantha
tectorum), кумарчик (Agriophyllum
squarrosum), волоснец кистистый
(Leymus racemosus), ковыль во-
лосатик (Stipa capillata) и др.
(рис. 10.17).
Рис. 10.16. Белополынно-прут-
няково-житняковое сообщество
на верхнехвалынской супесчаной
пологоволнистой равнине
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
272
Рис. 10.17. Пионеры зарастания:
кумарчик (Agriophyllum squar-
rosum), полынь песчаная (Arte-
misia arenaria), житняк пустын-
ный (Agropyron desertorum) и др.
на эоловых песках
На следующем этапе умень-
шается роль однолетних и сорных
видов, возрастает значимость ко-
выля и степного разнотравья.
В течение 6–8 лет после прекра-
щения выпаса простые и пио-
нерные группировки эфемеров
и однолетников вытесняются
ковыльными сообществами. На заключительной стадии восстановительной
сукцессии характерные черты растительного покрова определяются сообщест-
вом ковыля (Stipa lessingiana + S. capillata) с участием Agropyron fragile, Kochia
prostrata, Artemisia lercheana, степного разнотравья, эфемеров и однолетни-
ков (рис. 10.18).
10.3. АНТРОПОГЕННЫЕ НАРУШЕНИЯ ЛАНДШАФТОВ СТЕПЕЙ
Вся послеледниковая история степей – это в то же время и история от-
ношений между человеком и природой. Начало земледелия в России было
положено в лесостепной зоне. Основу сельского хозяйства составляло подсеч-
но-огневое земледелие. Конец XVIII века стал поворотным этапом в освоении
южных окраин России. В результате присоединения Крыма страна получила
свободный выход к Черному морю и начала осваивать богатства вновь обре-
тенного края. Быстрая колонизация южнорусских степей сопровождалась пе-
реселением в новые районы крестьян и городского населения из северных
губерний, постройкой городов и селений, распашкой бескрайних степных
просторов. Естественных местообитаний становилось все меньше. Исчезали
многие характерные представители фауны степных ландшафтов.
В советское время интенсивное освоение степей продолжалось. В годы
Великой Отечественной войны юг
Европейской территории России
стал ареной боевых действий. По-
сле войны в 1946–47 гг. эти зем-
ли охватила засуха. Для восста-
новления природного потенциа-
ла степных ландшафтов в 1948
году было принято постановление
Рис. 10.18. Сообщество Stipa
lessingiana в заповеднике
«Черные Земли»
.
Глава 10. ЗОНА СТЕПЕЙ
273
«О плане полезащитных лесонасаждений, внедрения травопольных севообо-
ротов, строительства прудов и водоемов для обеспечения высоких устойчивых
урожаев в степных и лесостепных районах Европейской части СССР». В печати
указанный документ назвали «Сталинским планом преобразования приро-
ды». Идея охраны почв с помощью создания полезащитных насаждений была
предложена В.В. Докучаевым еще в начале XX века. В ходе реализации плана
были созданы государственные полезащитные полосы общей протяженно-
стью более 5300 км, площадью 2,3 млн. га. Одновременно было создано около
4 тыс. водохранилищ, которые позволили резко улучшить окружающую среду.
Осуществленные мероприятия привели к росту урожайности зерновых, ово-
щей, трав; созданию прочной кормовой базы для развития животноводства.
Однако после смерти Сталина в 1950-е годы выполнение плана было сверну-
то. Многие лесополосы были вырублены, несколько тысяч прудов и водоемов
были заброшены. Роковым для степных ландшафтов явилось принятое в 1954
году решение советского правительства, направленное на освоение целинных
земель в Зауралье – на юге Западной Сибири и в Северном Казахстане. Нача-
лась беспощадная охота за степной целиной не только в Сибири, но и на юге
Европейской части России. Во время этой эпопеи не было создано ни одного
заповедника. Наоборот, уже существующие были закрыты, а их территории
распаханы. Были обработаны клочки девственных степей даже на научных
стационарах. В результате осуществления этого проекта степные ландшафты
были почти полностью уничтожены и заменены на пахотные угодья, что при-
вело к серьезным негативным экологическим последствиям. В результате рас-
пашки и уничтожения естественного растительного покрова почва стала уяз-
вимой для всех видов эрозии (рис. 10.19). Тонкие частицы гумуса и мелкозема
легко уносились потоками воды или ветром, оставляющими только грубый
материал.
К неблагоприятным явлениям в степной зоне относятся засухи, суховеи
и пыльные бури. Плодородный гумусный слой может быть унесен с пашни
одной сильной бурей. Поистине, планетарный характер имели пыльные бу-
ри середины 1960-х годов, пронесшиеся над степями СССР. В зоне опусты-
ненных степей в Северо-Западном Прикаспии ведущим фактором деграда-
ции растительности (опустынивания) является чрезмерная пастбищная на-
грузка.
В настоящее время степная зона стала почти полностью земледельче-
ской. Интенсификация сельскохозяйственных работ повышается благодаря
использованию современной техники. В некоторых районах пашни занимают
70 % территории, а местами и более.
Главным методом сохранения степей является организация особо охра-
няемых природных территорий (ООПТ). Основную роль в этой системе игра-
ют заповедники и заказники, в меньшей степени национальные парки и па-
мятники природы. В начале XXI века площадь всех ООПТ в степной зоне со-
ставляла лишь 0,4 % ее территории. Большинство степных заповедников
расположено в Европейской России. В 1989 году создан «Оренбургский» госу-
дарственный природный заповедник. На востоке в межгорных котловинах
функционируют два международных биосферных заповедника, расположен-
ных по границе с Монголией.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
274
Рис. 10.19. Роль степной растительности в защите почвенного слоя
Вопросы к главе 10
1. Природные условия зоны степей; экология растений.
2. Широтные особенности растительности зоны степей: подзоны северных, средних и
южных степей.
3. Региональные особенности растительности зоны степей: Восточно-Европейская
(Донско-Волжская), Заволжско-Уральская, Прикаспийская, Западно-Сибирская, Дау-
ро-Монгольская области степей.
Рекомендуемая литература
Дамбиев Э. Ц., Намзалов Б. Б., Холбоева С. А. Ландшафтная экология степей Бурятии.
Улан-Удэ: Изд-во Бурятского гос. ун-та, 2006. 185 с.
Ковыли и ковыльные степи Белгородской, Курской, Орловской областей: кадастр сведе-
ний, вопросы охраны / Н. И. Золотухин, А. В. Полуянов, Л. Л. Киселева, И. Б. Золоту-
хина, О. М. Пригоряну, О. В. Рыжков, Т. Д. Филатова, П. А. Дорофеева, О. И. Фандее-
ва, О. П. Власова, Н. В. Вышегородских. Курск, 2015. 487 с.
Лавренко Е. М. Степи Евразиатской степной области, их география, динамика и исто-
рия // Вопросы ботаники. 1954. Вып. 1. М.-Л.: Изд-во АН СССР, 1954. С. 609–625.
.
Глава 10. ЗОНА СТЕПЕЙ
275
Лавренко Е. М. Степи СССР // Е. М. Лавренко. Избранные труды. СПб.: Изд-во С.-Пе-
тербург. ун-та, 2000. С. 11–222.
Мордкович В. Г. Степные экосистемы / Отв. ред. И. Э. Смелянский. 2-е изд. испр. и доп.
Новосибирск: Академическое издательство «Гео», 2014. 170 с.
Мордкович В. Г., Гиляров А. М., Тишков А. А., Баландин С. В. Судьба степей. Новоси-
бирск: Мангазея, 1997. 300 с.
Намзалов Б. Б. Степи Тувы и Юго-Восточного Алтая. Новосибирск: Академическое
издательство «Гео», 2015. 294 с.
Чибилев А. А. Степной мир Евразии от Венгрии до Монголии. – Институт степи УрО
РАН, Оренбургское региональное отделение Всероссийской общественной органи-
зации «Русское географическое общество». Оренбург, 2013. 118 с.
Чибилев А. А. Степная Евразия: региональный обзор природного разнообразия М.;
Оренбург: Институт степи РАН; РГО, 2016. 324 с.
Шифферс Е. В. Растительность Северного Кавказа и его природные кормовые угодья.
М.–Л.: Изд-во АН СССР, 1953. 400 с.
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
276
Субтропический пояс является переходным от умеренного к тропическому.
В Евразии его северная граница проходит на широте около 40°. Климатиче-
ские особенности субтропиков в Европе летом формируются под влиянием
Азорского максимума, определяющего сухую и жаркую погоду. Зимой, когда
область высокого давления смещается к югу, с Атлантического океана прихо-
дят циклоны, несущие дожди; температура воздуха редко опускается ниже
0 °С. Горы, выступающие в виде барьеров, вызывают увеличение атмосфер-
ных осадков.
Глава 11
ЗОНА СУБТРОПИКОВ
России субтропическая зона представляет собой экстразональный вы-
ступ, формирование которого определяется влиянием теплого Черного
моря и барьерной ролью гор Крыма и Кавказа, усиливающих выпадение осад-
ков при вхождениях влажных воздушных течений с юга и препятствующих
продвижению на юг холодных воздушных масс с севера. Отличительные чер-
ты климата определяют своеобразие субтропиков средиземноморского типа и
влажных субтропиков. Первые характеризуются жарким и сухим летом, про-
хладной и влажной зимой (температуры редко опускаются ниже 0 °С); они
представлены на южном берегу Крыма и черноморском побережье в районе
Геленджика. Вторые – более или менее теплой зимой и большим количеством
атмосферных осадков, выпадающих равномерно в течение года; на террито-
рии России они расположены на побережье Черного моря в районе Сочи. Рас-
тительность здесь выступает основанием высотного ряда поясности гор и яв-
ляется переходной от умеренной к субтропической. Она отличается большой
древностью, представляя собой мало измененную неогеновую (эвксинскую)
растительность. Рассмотрим особенности растительности южного берега Крыма
и западного Кавказа.
11.1. СУБТРОПИКИ ЮЖНОГО БЕРЕГА КРЫМА
Над Южным берегом Крыма (ЮБК) возвышается крутой уступ Главной
Крымской гряды (рис. 11.1). ЮБК, закрытый с севера горами и омываемый
теплыми водами Черного моря, обладает необычно теплой для этих широт
зимой. Средняя температура января в Ялте +3,7 °С. Зимой на юге Крыма часто
идут дожди или выпадает снег, который тут же тает. Самый влажный месяц в
Ялте – январь. В разгар зимы на ЮБК цветут лещина (Corylus avellana) и фи-
алка душистая (Viola odorata). Лето жаркое: средняя температура июля в Ялте
24 °С. Наиболее сухой месяц – август.
Субтропический пояс
.
Глава 11. ЗОНА СУБТРОПИКОВ
277
Рис. 11.1. Растительность Крыма
(по Н. И. Рубцову, М. Н. Котовой, Л. В. Махаевой, 1964)
Условные обозначения: 1 – низкоствольные (шибляковые) леса из древовидного можже-
вельника, 2 – леса из крымской сосны и сосны Коха, 3 – буковые леса с примесью клена
Стевена, 4 – скальнодубовые леса с примесью грабовых и ясеневых лесов, 5 – пушистодубо-
вые леса и грабинники на их месте, 6 – редколесья из дуба пушистого в комплексе с тимь-
янниками, 7 – разнотравно-ковыльные настоящие степи предгорья, 8 – луговые степи из
ковыля тонколистного, тимьянники и производные группировки на их месте, 9 – горные
луговые степи из осоки низкой, типчака скального, бромопсиса каппадокийского, луга и
тимьянники, 10 – солончаки, 11 – засоленные степи и луга, 12 – сады и виноградники на
месте пушистодубовых лесов и разнотравно-ковыльных настоящих степей, 13 –сады и ви-
ноградники на месте ковыльно-типчаковых степей, 14 – сельскохозяйственные угодья зер-
новых, пропашных культур на месте разнотравно-ковыльных степей и пушистодубовых
лесов, 15 – сельскохозяйственные угодья зерновых, пропашных культур на месте типчако-
во-ковыльных степей, 16 – населенные пункты, 17 – растительность обнажений мела, 18 –
растительность обнажений кислых кристаллических пород
(http://krymology.info/index.php?curid=29042)
Растительность ЮБК имеет ясно выраженные средиземноморские чер-
ты. Из 2300 видов высших растений Горного Крыма около половины – среди-
земноморские. До 75 % видов растений Горного Крыма являются общими с
субтропиками черноморского побережья Кавказа.
В приморском поясе до высоты 350–400 м преобладают засухоустойчи-
вые леса из низкоствольных дуба пушистого (Quercus pubescens) (рис. 11.2)
и можжевельника высокого (Juniperus excelsa), а также кустарниковые за-
росли. К видам древней средиземноморской флоры относятся вечнозеленые
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
278
Рис. 11 .2. Заросли дуба пушистого – Quercus pubescens
(фото В. Гелюты)
кустарники: иглица подъязычная и колючая (Ruscus hypoglossum, R. aculeatus)
и ладанник крымский (Cistus tauricus), а из деревьев – земляничник мелко-
плодный (Arbutus andrachne) (рис. 11.3). В составе лесов выделяют три основ-
ные группы: 1) пушистого дуба; 2) дуба и древовидного можжевельника и
3) дуба и фисташки туполистной (Pistacia mutica).
Кустарниковые заросли (шибляк)
из низкорослого (2–4 м высотой) порос-
левого, большей частью кустообразного
пушистого дуба наиболее широко рас-
пространены в пределах приморского
пояса от Севастополя до Феодосии. Кро-
ме дуба в зарослях много грабинника
(Carpinus orientalis) и колючих кустар-
ников: держи-дерева (Paliurus spina-
christi), различных видов шиповника
(Rosa). В травяном покрове преобла-
дают степные злаки: типчак, бородач
(Bothriochloa), эгилопс (Aegilops), яч-
мень луковичный (Hordeun bulbosum)
и полукустарнички: виды чабреца и
солнцецвета (Helianthemum), фумана
(Fumana).
Рис. 11.3. Земляничное дерево – Arbutus
andrachne (фото И. Турбанова)
.
Глава 11. ЗОНА СУБТРОПИКОВ
279
Рис. 11.4. Растительность на склонах Карадага
Можжевелово-дубовые заросли наибольшие площади занимают между
Симеизом и Алуштой. К основным древесным породам примешиваются виды
ясеня – высокий и остроплодный (Fraxinus excelsior, F. oxycarpa) а также сос-
на крымская (Pinus pallasiana). В кустарниковом ярусе много грабинника, ме-
стами кизила (Cornus mas), видов иглицы (Ruscus), ладанника (Cistus tauricus),
можжевельника красного (Juniperus oxycedrus), жасмина кустарникового
(Jasminum fruticans), пираканты ярко-красной (Pyracantha coccinea). В за-
падной части ЮБК в приморском поясе на хорошо обогреваемых солнцем
склонах встречается земляничник мелкоплодный, который вместе с вечнозе-
леными иглицей и ладанником кое-где образует самостоятельные микросо-
общества, представляющие собой обедненные варианты средиземноморского
маквиса.
Дубово-фисташковые заросли распространены небольшими массивами
вдоль морского побережья до высоты 200–250 м. Заросли из кустообразных
дуба пушистого и фисташки туполистной не превышают 3–4 м. Полагают, что
такие формы пород возникли под влиянием длительных рубок или из-за небла-
гоприятных условий обитания. В Крыму плоды у фисташки не созревают, по-
этому она здесь размножается вегетативно. Кроме дуба и фисташки, в составе
сообществ имеются низкорослый можжевельник высокий, миндаль обыкно-
венный (Amygdalus communis), ясень, груша лохолистная (Pyrus elaeagrifolia),
а из кустарников – держи-дерево, реже сумах (Rhus coriaria), скумпия
кожевенная (Cotinus coggygria), виды боярышников (Crataegus orientalis,
C. karadaghensis, С. sphaenophylla, C. pojarkovae), ладанник (рис. 11.4).
Наряду с этим, на крайнем западе и на востоке ЮБК, в районах Батили-
мана, Судака и Нового Света, распространено своеобразное редколесье из эн-
демичной сосны пицундской Pinus pityusa (известной также как сосна судак-
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
280
ская, или сосна Станкевича) (рис. 11.5) и можжевельника вонючего (Juniperus
foetidissima). В травяном покрове преобладают степные виды: ковыль, тонко-
ног, кострец береговой (Bromopsis riparia), асфоделина (Asphodeline). Изуче-
ние растительности ЮБК показывает, что по мере возрастания хозяйственного
воздействия происходит замена лесов из древесных пород дуба, можжевельни-
ка, фисташки кустарниковыми зарослями. В свою очередь, они уступают место
степным или полукустарничковым сообществам, чередующимся с голыми
скалами. В настоящее время в связи с природоохранными мероприятиями
наблюдаются медленные восстановительные смены растительности. Проис-
ходит постепенное смыкание полога кустарников, главным образом, в резуль-
тате вегетативного разрастания грабинников.
Пояс лесов из сосны крымской (Pinus pallasiana) (рис. 11.6) простирается
в полосе от 350–400 до 900–1000 м над ур.м. и наиболее полно выражен от
Симеиза до Гурзуфа. Леса из сосны крымской, или сосны Палласа, покрывают
все склоны алупкинского, ялтинского и гурзуфского амфитеатров, а отдель-
ные ее экземпляры ютятся на карнизах обрывов яйлинских массивов, прида-
вая им эффектный неприступный вид.
К категории редких следует отнести ксерофильные трагакантники, ос-
новной биоморфой которых является астраканта (Astracantha arnacantha) в
сообществе с ксероморфным разнотравьем (Artemisia caucasica, Alyssum obtu-
sifolium, Poa bulbosa, Eryngium campestre, Phlomis pungens, Bromopsis riparia
и др.), встречающиеся в виде небольших зарослей в нижних частях юго-
западного и южного склонов в окрестностях г. Судака и пгт. Планерского.
Уникальными являются дроковые кустарниковые сообщества из до-
минанта-эндемика Genista verae с участием Dorycnium graecum, Dianthus
humilis, Rumex acetosella, Inula aspera, Asplenium septentrionale, A. trichomanes,
Cerastium schmalhausenii, C. brachypetalum, Milium vernale, Erophila praecox,
Рис. 11.5. Сосна пицундская или Станкевича – Pinus pityusa (фото В. Шатко)
.
Глава 11. ЗОНА СУБТРОПИКОВ
281
Рис. 11.6. Сосна крымская (Pinus pallasiana)
Arabis sagittata и др. Эти сообщества развиты только на восточном склоне
горы Аю-Даг. Более распространенным является дрок крымский – Genista
taurica (рис. 11.7), часто встречаю-
щийся на сухих известковых скло-
нах.
Редким и оригинальным сооб-
ществом является скальнодубовый
(Quercus petraea) лес на вершине
горы Аю-Даг с синузией весенних
эфемероидов (Viola odorata, Galan-
thus plicatus, Arum elongatum, Scilla
bifolia, Ornithogalum fimbriatum,
Corydalis paczoskii, Allium decipiens,
Dentaria quinquefolia, Crocus an-
gustifolius, Veronica umbrosa, Lathy-
rus laxiflorus, Potentilla micrantha,
Dactylorhiza romana и др.), весьма
обильных, местами достигающих
100 % проективного покрытия.
Рис. 11.7. Дрок крымский (Genista
taurica)
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
282
11.2. СУБТРОПИКИ ЧЕРНОМОРСКОГО ПОБЕРЕЖЬЯ КАВКАЗА
На Черноморском побережье Кавказа выделяется две подзоны: расти-
тельность средиземноморского типа и растительность влажных субтропиков.
Растительность средиземноморского типа занимает нижний
пояс гор на черноморском побережье Кавказа от Новороссийска до Геленджи-
ка. Лето здесь жаркое и сухое, осадки выпадают в основном в холодное время
года. Для июля средняя температура воздуха составляет 23,9 °С. Средняя тем-
пература воздуха самого холодного месяца 4,3 °С. Среднее годовое количество
атмосферных осадков 630 мм.
Особенно ярко облик средиземноморской растительности выражен на
склонах гор, обрамляющих Цемесскую (г. Новороссийск) и Геленджикскую
бухты (рис. 11.8). Прибрежная терраса до высоты 150–200 м покрыта редко-
лесьем из дуба пушистого (Quercus pubescens) с примесью грабинника
(Carpinus orientalis) и шибляком на коричневых почвах. Шибляк представля-
ет собой вторичные густые и непроходимые ксероморфные сообщества с пре-
обладанием кустарников с опадающей листвой, формирующиеся на месте вы-
рубок леса из дуба пушистого. В составе шибляка доминирует колючий ку-
старник держи-дерево (Paliurus spina-christi) (рис. 11.9). Изредка в составе
шибляка встречается фисташка туполистная (Pistacia mutica), свойственная и
южному берегу Крыма. В непосредственной близости к морю и местами на
приморском обрыве произрастают леса из сосны пицундской (Pinus pityusa).
На высоте от 150 до 400–450 м развито можжевеловое редколесье, об-
разованное двумя видами древовидного можжевельника (Juniperus excelsa,
J. foetidissima) (рис. 11.10). На известковых склонах в составе можжевелового
Рис. 11.8. Панорама побережья Новороссийск – Геленджик
.
Глава 11. ЗОНА СУБТРОПИКОВ
283
Рис. 11.9. Держи-дерево
(Paliurus spina-christi)
в составе шибляка
(фото М. Лучкина)
редколесья обычны: жостер Палласа (Rhamnus pallasii), спирея (Spiraea
hypericifolia), эфедра (Ephedra procera), кизил (Cornus mas), скумпия (Cotinus
coggygria), ирга (Amelanchier ovalis) и др.; на неизвестковых склонах встреча-
ются как некоторые из перечисленных выше кустарников, так и жимолость
(Lonicera iberica), можжевельник (Juniperus oblonga), шиповник (Rosa iberica),
барбарис (Berberis vulgaris).
Растительность влажных субтропиков распространена на юго-
восточном побережье Черного моря. На территории России она представлена
у подножья гор в районе г. Сочи и далее в Абхазии и Грузии. В г. Сочи средняя
температура самого теплого ме-
сяца (августа) 25,2 °С, средняя
температура января 6,1 °С, сред-
негодовое количество осадков
1550 мм. Во влажных субтропи-
ках почвы лесные бурые.
В неогене в нижней части у
склонов гор существовали влаж-
ные субтропические (эвксинские)
леса, которые затем распростра-
нились на сформировавшиеся в
четвертичный период молодые
прибрежные равнины. Совре-
менные влажные субтропические
леса насыщены реликтовыми
видами и отличаются сложной
фитоценотической структурой:
Рис. 11.10. Древовидный можже-
вельник – Juniperus excelsa
(фото М. Лучкина)
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
284
Рис. 11.11. Лавровишня – Laurocerasus
officinalis (фото И. Гайворонской)
кроме древесного яруса широколиствен-
ных пород, ярко выражен подлесок с уча-
стием вечнозеленых растений, синузии
эпифитов и лиан. Господствующие фор-
мации эвксинского типа представлены,
главным образом, дубовыми из дуба име-
ретинского (Quercus imeretina), дуба гру-
зинского (Q. iberica), буковыми из Fagus
orientalis и буково-каштановыми лесами с
Castanea sativa. В качестве примеси по-
стоянно встречаются клен ложноплатано-
вый (Acer pseudoplatanus), вязы (Ulmus
minor, U. glabra), липа бегониелистная
(Tilia begoniefolia), ясень высокий (Fraxi-
nus excelsior). Характерно наличие вечно-
зеленого подлеска: рододендрон понтий-
ский (Rhododendron ponticum), самшит
(Buxus colchica), лавровишня (Laurocerasus officinalis) (рис. 11.11), а также
листопадного кустарника – черники кавказской (Vaccinium arctostaphylos).
Самый нижний вечнозеленый ярус подлеска создает иглица (Ruscus aculeatus),
реже встречается волчеягодник (Daphne pontica) и падуб (Ilex aquifolium).
Из лиан наиболее характерны плющи
(Hedera helix, H. colchica), сассапариль
высокий (Smilax excelsa), ломоносы
(Clematis vitalba, С. viticella), хмель (Hu-
mulus lupulus), обвойник (Periploca grae-
ca), тамус (Tamus communis) (рис. 11.12).
В первичных лесах, вероятно, встречался
виноград (Vitis sylvestris); в настоящее
время в лесах как первичных, так и вто-
ричных, широко распространен виноград
Изабелла (Vitis labrusca) – лиана, искус-
ственно насаженная человеком. Эпифи-
ты представлены разнообразными папо-
ротниками и мхами; например, папорот-
ник многоножка южная (Polypodium aust-
rale). Травяной ярус включает большое
разнообразие папоротников, например,
Рис. 11.12. Лианы:
А – сассапариль высокий Smilax excelsa (фото
В. Прохорова); Б – ломонос Clematis vitalba
(фото О. Щедровой)
.
Глава 11. ЗОНА СУБТРОПИКОВ
285
Рис. 11.13. Тисо-самшитовая
роща с Taxus baccata и Buxus
colchica
папоротник листовик (Phyllitis
scolopendrium), папоротник пте-
рис критский (Pteris cretica).
Также произрастают подснежни-
ки (Galanthus caucasicus, G. kras-
novii, G. woronowii), цикламены
(Cyclamen coum, C. adsharicum).
Особый тип реликтового ле-
са представляет тисо-самшитовая
(Taxus baccata, Buxus colchica)
роща в долине р. Хосты, неповторимый облик которой придают космы эпи-
фитных мхов – мха неккера (Neckera complanata, N. crispa) – свисающие с
ветвей и стволов самшита (рис. 11.13). Травяной покров в связи с затене-
нием почвы не развит. С 2014 года роща находится под угрозой уничтожения
в связи с инвазией насекомого-вредителя – самшитовой огневки (Cydalima
perspectalis), гусеницы которой поедают вечнозеленые листья самшита.
На Пицундском мысу сохранился реликтовый лес из сосны пицундской
(Pinus pityusa) (рис. 11.14).
Интродукция субтропических растений. Известный русский бота-
нико-географ А. Н. Краснов, совершив-
ший длительные экспедиции по изуче-
нию тропической растительности Юго-
Восточной Азии, считал возможным,
учитывая мягкий климат Черноморско-
го побережья Кавказа, интродуцировать
сюда разнообразные субтропические
растения. С этой целью по его инициа-
тиве в Батуми в 1912 г. был создан бота-
нический сад. В наши дни в ботаниче-
ских садах и в зеленых насаждениях
Южного берега Крыма и Черноморского
побережья Кавказа широко представле-
ны интродуцированные виды субтропи-
ческих плодовых и декоративных расте-
ний. Богатые коллекции субтропических
культур собраны в Никитском ботани-
ческом саду и в Сочинском дендрарии
(рис. 11.15, 11.16).
Рис. 11.14. Сосновый лес
(Pinus pithyusa) в Пицунде
(Автономная Республика Абхазия)
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
286
Веерные пальмы на набережной
им. Ленина в Ялте (фото: shkolnie-turi.ru)
Кипарисовая аллея в Симеизе
(фото: arhiveuatravels.in.ua)
Павловния войлочная (Paulownia tomentosa)
(фото Людмилы Саплицкой)
Смоковница обыкновенная (Ficus carica)
(фото Сергея Одинца)
Кактусовая горка в Никитском
ботаническом саду (фото: mybookit.ru)
Альбиция шелковая (Albizia julibrissin)
(фото: azon-liliya.ru)
Рис. 11.15. Растения-интродуценты из Восточной и Передней Азии,
Средиземноморья и кактусы из Центральной Америки на Южном берегу
Крыма
.
Глава 11. ЗОНА СУБТРОПИКОВ
287
Магнолия Суланжа (Magnolia soulangeana)
(фото daily.ru)
Акация серебристая (Acacia dealbata)
(фото yuga.ru)
Юкка алоэлистная (Yucca aloifolia)
(фото Владимира Иванова, www.plantarium.ru)
Финик канарский (Phoenix canariensis)
в парке Дендрарий в Сочи (фото: priroda.su)
Банан (Musa sp.) в парке Дендрарий в Сочи
(фото: http://rajskij-sad.livejournal.com)
Кортадерия Селло (Cortaderia selloana)
(фото Николая Голубева)
Рис. 11.16. Интродуценты из Южной Америки, Австралии, Юго-Восточной
Азии, Средиземноморья на Черноморском побережье Кавказа
.
Ч а с т ь II. РАСТИТЕЛЬНОСТЬ ПРИРОДНЫХ ЗОН РОССИИ
288
Вопросы к главе 11
1. Природные условия зоны субтропиков.
2. Особенности растительности субтропиков Южного берега Крыма и Черноморского
побережья Кавказа
Рекомендуемая литература
География Крыма / Под ред. канд. геогр. наук. проф. П. Д. Подгорельского, канд. геогр.
наук., доц. В.Б. Кудрявцева. Симферополь: Крымское учебно-педагогическое гос.
изд-во, 1995. 224 с.
Долуханов А. Г. Растительный покров // Кавказ. М.: Наука, 1966. С. 223–255.
Рубцов Н. И. Растительный мир Крыма. Научно-популярный очерк. Симферополь:
Таврия, 1978. 129 с.
Солодько А. С. К геоботаническому районированию Сочинского Причерноморья //
Бот. журн. 1999. Т. 84, No 1. С. 45–56
Солодько А. С., Нагалевский М. В., Кирий П. В. Атлас флоры Сочинского Причерно-
морья. Дикорастущие сосудистые растения. Сочи, 2006. 287 с.
Национальный парк Сочинский http://oopt.info/index.php?oopt=984
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
289
Часть III
РАСТИТЕЛЬНОСТЬ
ГОРНЫХ СТРАН
Растительность гор характеризуется высотной поясностью, которая ме-
няется в зависимости от положения горной страны внутри той или иной при-
родной зоны. Кроме того, положение гор вблизи океана или на удалении от
него обусловливает формирование поясности океанического или континен-
тального типа. Немаловажна ориентировка горных хребтов по отношению к
направлению движения воздушных масс, несущих осадки, – количество
осадков на наветренных склонах может превышать осадки на подветренных
склонах более чем в десять раз. Этим объясняется асимметрия поясности
горных систем.
Флора гор намного богаче флор прилегающих равнин, что связано с вы-
сотной поясностью, обилием горных изолятов, в которых протекают видооб-
разовательные процессы, и с сохранением в горных убежищах реликтов.
В горных флорах, как правило, большая доля эндемиков, особенно молодых.
При подъеме в горы снижается температура, на больших высотах в юж-
ных широтах даже в летние месяцы температура не поднимается выше 0 °С.
Соответственно снижается испаряемость, усиливаются ультрафиолетовая ра-
диация и освещенность местообитаний, сокращается вегетационный период.
В зависимости от увлажненности и температуры в высокогорьях проявляются
криофитизм и психрофитизм, т. е. формы приспособления растений к сухой и
холодной или влажной и холодной среде. Для криофитов типичны преобла-
дание подушечных жизненных форм и многолетников, значительная (до 400
лет) индивидуальная продолжительность жизни, адаптация к сильной уль-
трафиолетовой радиации, снижение транспирации, высокое (до 25 атм.) ос-
мотическое давление клеточного сока, повышенный водный дефицит, за-
медленное поглощение азота, усиление процессов дыхания, рост интенсив-
ности фотосинтеза. Для психрофитов характерно низкое осмотическое
давление клеточного сока, слабая активность обмена веществ, малое коли-
чество хлорофилла, склерофильность тканей, слабая водоудерживающая си-
ла цитоплазмы.
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
290
ысотная поясность в горах, будучи функцией рельефа, связана с пони-
жением температуры и, как правило, с повышением увлажненности с
высотой. Высотная поясность начинается с той широтной зоны, в которой
находится горная страна. Там, где высота гор позволяет, поясность замыкается
вечными снегами и льдами. При этом в арктическом поясе снеговая граница
опускается до уровня моря; в умеренном поясе она поднимается. В каждой
природной зоне формируется свой набор высотных поясов. На склонах гор
разной экспозиции наблюдается асимметрия высотной поясности.
Горные массивы России расположены в основном на юге (Кавказ), юго-
востоке (Алтай, Прибайкалье) и востоке (Северо-Восток Сибири); горный мас-
сив Камчатки вытянут вдоль Тихоокеанского побережья. Уральский хребет
является естественной границей между европейской и азиатской частями
страны.
Приведем описания высотной поясности, наиболее полно раскрываю-
щие особенности растительности основных горных стран России.
12.1. РАСТИТЕЛЬНОСТЬ УРАЛА
Древний хребет Уральских гор протянулся с севера на юг, примерно, от
70° до 52° с.ш., общей длиной свыше 2500 км, пересекая, таким образом, не-
сколько широтных растительных зон. Урал делится на пять частей: Поляр-
ный, Приполярный, Северный, Средний и Южный.
Полярный Урал своей крайней точкой на севере имеет вершину Кон-
стантинов камень, высотой 483 м, его самая высокая гора – гора Пайер 1499 м.
Высочайшей вершиной Приполярного Урала и всего Уральского хребта явля-
ется гора Народная высотой 1895 м. Средний Урал простирается от горы
Ишерим, 1331 м высоты, до горы Юрма, достигающей высоты 1003 м. Наивыс-
шая вершина Среднего Урала – Конжаковский камень, высотой 1569 м. В Юж-
ном Урале, который тянется от горы Юрма до среднего течения р. Урал, горы
повышаются. Высочайшей точкой Южного Урала служит гора Яман-тау, до-
стигающая высоты 1640 м. Вершины Среднего и Южного Урала не достигают
снеговой линии. Чрезвычайно велики различия западного и восточного скло-
нов Урала: западный склон пологий, имеющий предгорья, богаче осадками по
сравнению с восточным сибирским, круто обрывающимся в Западно-Сибир-
скую равнину. На западных склонах Северного и Приполярного Урала за год
выпадает более 800 мм атмосферных осадков. В средней части хребта выпада-
ет 600 мм и даже больше осадков, причем на западном склоне – осадков на
150 мм больше, чем на восточном. На Южном Урале количество осадков
уменьшается до 500–300 мм.
Флора Урала насчитывает около 1600 видов растений, из них на долю
эндемиков приходится лишь 5 % – качим уральский (Gypsophila uralensis),
Глава 12
РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
291
астрагал Гельма (Astragalus helmii), гвоздика иглолистная (Dianthus acicularis),
минуарция Крашенинникова (Minuartia krascheninnikovii), чина Литвинова
(Lathyrus litvinovii) и др. Бедность Урала эндемичными видами объясняется
его срединным положением на материке, доступностью для расселения и
смешения различных флор, которые преодолевали горы, не образуя изолиро-
ванных ареалов. Так, многие сибирские древесные хвойные породы перевали-
ли через Урал, и западная граница их ареала ныне проходит по Русской рав-
нине.
На обобщенном профиле показаны особенности вертикальной поясно-
сти западного и восточного склонов Урала (рис. 12.1). На крайнем севере от
предгорных равнин до горных вершин распространены тундры (рис. 12.2).
Равнинные тундры на склонах сменяются горными. Почвы – тундрово-гле-
евые суглинистые и тундровые подбуры на каменисто-щебнистом элювии и
делювии коренных пород. Близ полярного круга тундры становятся высотным
поясом, занимающим склоны и вершины гор, а к их подножиям подходят
редкостойные леса.
Леса являются самым распространенным типом растительности. Они
сплошной полосой тянутся по горным склонам Урала от полярного круга до
субширотного отрезка реки Сакмара (южнее 52° c.ш.), а по предгорьям до
Уфимского плато и южнее Екатеринбурга. Леса Урала разнообразны по соста-
ву: хвойные, широколиственные, мелколиственные. Наиболее широко рас-
пространены пихтово-еловые леса. В состав темнохвойных лесов (рис. 12.3),
характерных для Предуралья и западных склонов гор, входят ель сибирская
(Picea obovata), пихта сибирская (Abies sibirica), сосна обыкновенная (Pinus
sylvestris) и сосна сибирская (кедр) (Pinus sibirica). Для восточных склонов
Урала более типичны сосновые леса. На их долю приходится около трети всех
хвойных лесов. Лиственница сибирская (Larix sibirica) встречается в северных
районах, а по восточным склонам гор доходит до южных районов Урала, но
чистых лиственничных лесов на Урале практически нет. Таежные почвы
представлены глееподзолистыми, подзолистыми и дерново-подзолистыми в
сочетании с болотными.
В южной части тайги Предуралья (южнее 58° c.ш.) в составе хвойных ле-
сов появляется примесь широколиственных пород: липы, клена остролистно-
го, вяза. К югу их роль возрастает, однако они чаще не выходят в древесный
ярус, оставаясь в подлеске, и лишь изредка образуют второй полог древостоя.
Настоящие хвойно-широколиственные и широколиственные леса распро-
странены лишь на западных склонах гор Южного Урала. Широко известны
липовые леса Башкирии (рис. 12.4). Здесь же распространены дубовые леса.
Однако широколиственные леса занимают на Урале не более 4–5 % лесопо-
крытой площади. На восточном склоне этих лесов нет. Из широколиственных
пород одна липа заходит за Урал.
Значительно шире представлены на Урале мелколиственные березовые
и березово-осиновые леса. Они распространены по всему Уралу, но особенно
много их на Южном и Среднем. Есть коренные березовые леса, чаще вторич-
ные, возникшие на месте вырубленных хвойных лесов.
Верхняя граница леса на Северном Урале проходит на высоте 500–800 м,
вершины Среднего Урала практически не выходят за пределы лесного пояса
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
292
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
293
Рис.12.2. Горная тундра, на
переднем плане дриада
восьмилепестковая (Dryas
octopetala), остролодочник
грязноватый (Oxytropis
sordida), Полярный Урал
(фото С. Корневой)
Рис. 12.3. Темнохвойный
лес на Урале
Рис. 12.4. Липовый лес
в Башкирии
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
294
(800–900 м), а на Южном Урале граница леса поднимается до 1200 м. Выше
ее находится неширокий пояс, основу растительности которого образуют низ-
корослые редкостойные леса в сочетании с лугами. Он сменяется горными
тундрами, которые на Северном и Полярном Урале сменяются гольцами.
Как в западных, так и в восточных предгорьях Среднего Урала появляются
острова лесостепи. На Южном Урале лесостепи подходят к подножию гор сна-
чала на восточном, а затем на западном склоне. В Предуралье разнотравные
степи сочетаются с небольшими дубовыми и березовыми островами, в Заура-
лье – с березовыми и осиново-березовыми перелесками. Под широколиствен-
ными лесами формируются серые лесные почвы с постепенно увеличивающи-
мися к югу пятнами черноземов оподзоленных, выщелоченных и типичных.
Юго-восток Зауралья и крайний юг гор заняты степями разнотравно-
дерновинно-злаковыми и дерновинно-злаковыми. Среди них встречаются
заросли степных кустарников: вишни кустарниковой, спиреи, караганы.
12.2. РАСТИТЕЛЬНОСТЬ КАВКАЗА
Кавказ – обширная горная страна, расположенная к югу от Русской рав-
нины, представляющая сложную систему высоких горных хребтов альпийской
складчатости, нагорий, плато. На юго-западе ее омывает Черное море, на во-
стоке – Каспийское. Северную часть Кавказа образует Предкавказье, прости-
рающееся до подножия Большого Кавказа. Для Большого Кавказа, вытянутого
с северо-запада на юго-восток на 1100 км, характерны высокогорья, альпий-
ский тип рельефа, встречаются крупные ледники длиной до 12 км и более.
Несколько вершин относится к категории высочайших, т.е. имеют высоту бо-
лее 5000 м, в том числе Эльбрус (5642 м).
Флора Кавказа насчитывает около 6500 видов. Кавказ располагается на
стыке Циркумбореальной, Средиземноморской и Ирано-Туранской областей
Голарктического флористического царства, и отдельные его территории отно-
сятся к разным флористическим провинциям – Восточноевропейской, Крым-
ско-Новороссийской, Эвксинской, Кавказской, Армено-Иранской, Гирканской
и Туранской (Тахтаджян, 1978).
На Кавказе сочленяется растительность умеренного и субтропического
поясов. Растительный покров Кавказа необычайно разнообразен (рис. 12.5).
Это объясняется пестротой орографических и климатических условий, а также
расположением Кавказского перешейка на стыке многих растительных сооб-
ществ различного генезиса – европейских лесов, растительности евразиатских
высокогорий, восточноевропейских степей, переднеазиатских нагорных ксе-
рофитов и сложных ценозов Средиземноморья.
Типы высотной поясности Кавказа показаны на рисунке 12.6, где в пре-
делах групп поясности даны типы поясности (колонки), которые представля-
ют спектры поясности растительности. Условные знаки в легенде содержат
название каждого пояса. Наиболее наглядно особенности растительности
Кавказа раскрываются при описании северного склона Большого Кавказа.
Изменение климата и орографической структуры обусловливает выделение
трех районов: Западного, Центрального и Восточного. Границы между ними
проходят по меридиану г. Эльбрус и долине р. Терек соответственно.
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
295
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
296
Рис. 12.5 (продолжение). Легенда к карте растительности Кавказа
Номера условных обозначений соответствуют общей нумерации условных знаков в легенде
«Карта растительности Европейской части СССР» (1974)
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
297
Рис. 12.5 (продолжение). Легенда к карте растительности Кавказа
Номера условных обозначений соответствуют общей нумерации условных знаков в легенде
«Карта растительности Европейской части СССР» (1974)
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
298
Рис. 12.6. Типы высотной поясности Кавказа (Зоны и типы..., 1999):
54 – Нивально-альпийско-субальпийско-широколиственнолесо-лесостепной (Кубанский);
55 – Нивально-альпийско-субальпийско-широколиственнолесо-лесостепной (Эльбрусский);
56 – Нивально-альпийско-субальпийско-широколиственнолесо-лесостепной (Терский); 57 –
Альпийско-субальпийско-лесо-аридноредколесно-степной (Дагестанский); 58 – Нивально-
альпийско-субальпийско-субтропическолесной (Колхидский); 59 – Нивально-альпийско-
субальпийско-широколиственнолесо-лесостепной (Южноосетинский); 60 – Горностепно-
субтропическостепной (Талышский); 73 – Нивально-альпийско-субальпийско-лесо-аридно-
редколесный (Лагодехско-Закатальский); 74 – Альпийско-субальпийско-широколиствен-
нолесо-аридноредколесный (Центрально-Малокавказский); 75 – Альпийско-субальпийско-
лесо-аридноредколесный (Карабах-Зангезурский); 76 – Альпийско-субальпийско-лесо-арид-
норедколесный-нагорноксерофитно-степно-пустынынй (Армянский)
Западный район. Его восточной границей является меридиан, прохо-
дящей по западной окраине Ставропольской возвышенности, южной – водо-
раздел Главного Кавказского хребта.
Лесостепной пояс начинается в предгорьях на высоте 200−250 м и за-
нимает полосу до 500−600 м (рис. 12.7). Среднее годовое количество осадков
800 мм. Растительность сильно изменена в результате вырубок и выпаса скота.
Леса заменены зарослями кустарников и встречаются только выше, где они че-
редуются с лугово-степными участками. Древесный ярус составлен дубом че-
решчатым (Quercus robur) и дубом Гартвиса (Q. hartwissiana), реже встречают-
ся граб (Carpinus betulus), ясень (Fraxinus excelsior) и вяз малый (Ulmus minor).
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
299
Рис. 12.7. Лесостепь, предгорья
Западного Кавказа
Во втором подъярусе: клен поле-
вой (Acer campestre), ближе к
опушкам груша (Pyrus communis)
и яблоня восточная (Malus orien-
talis). В подлеске: лещина (Cory-
lus avellana), слива вишненосная
(Prunus cerasifera), кизил (Cor-
nus mas), свидина (Swida san-
guinea), боярышник (Crataegus
monogyna), крушина (Frangula
alnus), калина гордовина (Viburnum lantana), рододендрон желтый (Rhodo-
dendron luteum), ежевика (Rubus discolor) и жимолость козья (Lonicera capri-
folium).
Пояс широколиственных лесов занимает склоны гор до высоты 1400–
1500 м. До высоты 900–1000 м он образован дубом скальным (Quercus
petraea) с примесью дуба черешчатого (рис. 12.8). В их состав входят: граб, вяз
малый, клен полевой, ясень, липа, каштан (Castanea sativa) (рис. 12.9), яблоня
восточная, груша кавказская (Pyrus caucasica), вишня птичья или черешня
(Cerasus avium) и др. В кустарниковом ярусе: лещина, бересклет, кизил, бо-
ярышник и др. Выше, до высоты примерно 1400–1500 м, располагаются бу-
ковые леса (рис. 12.10) из бука восточного (Fagus orientalis) с примесью граба
(Carpinus betulus), явора (Acer pseudoplatanus), липы, клена остролистного
(Acer platanoides), вяза горного (Ulmus laciniata) и др.; в подлеске – рододенд-
рон желтый (Rhododendron luteum), черника кавказская (Vaccinium
arctostaphylos), бузина черная (Sambucus nigra), падуб (Ilex colchica) и др.
Почвы – темно-серые и серые лесные. Встречаются монодоминантные мерт-
вопокровые буковые леса.
Пояс темнохвойных пихтово-еловых лесов располагается на высоте
от 1400–1500 м до 2200–2300 м над ур. м. Преобладают насаждения пихты
кавказской (Abies nordmanniana)
(рис. 12.11) и ели восточной (Picea
orientalis), к которым примеши-
ваются осина, береза, сосна крюч-
коватая (Pinus uncinata) и др.; в
подлеске – рододендрон понтий-
ский (Rhododendron ponticum),
падуб, лавровишня, черника кав-
казская; в травяно-кустарнич-
ковом ярусе – папоротники. На
верхней границе распространения
Рис. 12.8. Дуб черешчатый
(Quercus robur)
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
300
Рис. 12.9. Каштан – Castanea sativa
(фото Н. Дегтярева)
лес сильно изрежен, высота деревьев умень-
шается, ветви и стволы густо покрыты ли-
шайниками. Почвенный покров представ-
лен ненасыщенными выщелоченными и
оподзоленными бурыми лесными почвами.
Субальпийский пояс расположен выше
пояса темнохвойных лесов. Чаще всего он
представлен криволесьем (рис. 12.12), ред-
колесьем (Betula litwinowii, B. raddeana,
Acer trautvetteri, Fagus orientalis), высоко-
горными стланиками (Juniperus hemi-
sphaerica, J. sabina) и субальпийским вы-
сокотравьем в сочетании с различными
лугами. Очень характерно участие мелких
кустарничков из семейства вересковых:
черники (Vaccinium myrtillus), брусники
(Vaccinium vitis-idaea), голубики (Vaccinium uliginosum) и толокнянки (Arcto-
staphylos uva-ursi). На хорошо увлаженных склонах северной экспозиции ши-
роко распространены заросли рододендрона кавказского (Rhododendron
caucasicum) (рис. 12.13). Кусты рододендрона образуют густую, почти непро-
ходимую заросль, создающую сильное затенение. Под ними формируются
торфянистые почвы.
Субальпийское высокотравье в западном районе Северного Кавказа ха-
рактеризуется пышными разнотравными лугами с богатым красочным траво-
стоем, достигающим 70–80 см высоты и значительной густоты. Чаще всего
здесь отмечаются: ветреник нарциссоцветковый (Anemonastrum narcissiflorum),
звездовка большая (Astrantia maxima), буквица крупноцветковая (Betonica
macrantha), головчатка (Cephalaria sp.), колокольчик холмовой (Campanula
collina) , девясил (Inula sp.) , василек (Centaurea sp.) , лён зверобоелистный
Рис. 12.10. Буковый лес весной
Рис. 12.11. Пихтовый лес
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
301
Рис. 12.12. Криволесье
в субальпийском поясе
Рис. 12.13. Рододендрон кавказский
(Rhododendron caucasicum)
(Linum hypericifolium), незабудка альпийская (Myosotis alpestris), бедренец ро-
зовоцветный (Pimpinella rhodantha), змеевик мясо-красный (Bistorta carnea),
поповник красный (Pyrethrum coccineum), лютик кавказский (Ranunculus
caucasicus), скабиоза кавказская (Scabiosa caucasica). В этом высокотравье
обычно один подъярус, ибо создаваемое им затенение мешает развиться низ-
корослым растениям.
Злаковые луга представлены, главным образом, сообществами с корот-
коножкой (Brachypodium rupestre) и вейником (Calamagrostis arundinacea).
Из других злаков особо характерны такие виды, как тимофеевка (Phleum
nodosum, P. alpinum), трясунка (Briza media), мятлики (Poa iberica, P. meyeri)
и др. Злаково-разнотравные луга характериуются высоким видовым разнооб-
разием. Кроме злаков в сложении сообществ принимают участие: клевер (Tri-
folium canescens), лютик (Ranunculus caucasicus), вероника (Veronica gentia-
noides) и др. В западной половине Большого Кавказа типичным представите-
лем разнотравных лугов являются луга с купальницей (Trollius ranunculinus) c
участием герани (Geranium gymnocaulon), змеевика (Bistorta carnea), полеви-
цы (Agrostis planifolia) и тимофеевки (Phleum alpinum).
Высокогорья и характерные для них альпийские луга в Западном районе
занимают небольшую площадь.
Центральный район. С востока граница района проходит по Даге-
станскому нагорью, с юга – по водоразделу Главного Кавказского хребта. Об-
щие условия формирования климата Центрального Кавказа те же, что и для
Западного Кавказа, за исключением того, что количество атмосферных осад-
ков уменьшается. В результате в Центральном районе почти полностью выпа-
дают мезофильные древесные породы: каштан, ель, пихта, лавровишня и др.
Лесостепной пояс сильно видоизменен хозяйственной деятельностью
человека, и поэтому его верхняя и нижняя границы трудно определимы.
Лишь по остаткам лесной растительности предгорий можно предположить
нижнюю границу пояса. Она проходит приблизительно на высоте 250–300 м
над ур. м. Островки лесной растительности образованы дубом скальным, гра-
бом, грабинником, ясенем, кленом полевым и др. Куртины этих лесов в ос-
новном порослевого происхождения, возобновившиеся после сплошной руб-
ки. Кустарниковый ярус представлен кизилом, свидиной, боярышником, ро-
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
302
додендроном желтым, лещиной и др. Почвы – черноземы на участках, заня-
тых первичной степной растительностью; под лесами формируются серые
лесные почвы.
Пояс широколиственных лесов занимает предгорья и нижние части
склонов хребтов до высоты 1200–1500 м над ур. м. Растительность представ-
лена лесами из тех же древесных пород, что и в предыдущем поясе: дуба
скального, ясеня, граба, клена полевого, липы и др. В подлеске – боярышник,
кизил, бересклет, свидина, лещина, рододендрон желтый и др. На склонах
Главного Кавказского хребта благодаря увеличению влажности климата дубо-
вые леса сменяются буковыми. К буку восточному примешивается граб, липа,
клен остролистный, яблоня восточная, груша кавказская. Подлесок состоит из
бузины черной, бересклета и др.
Пояс сосново-березовых лесов замещает здесь темнохвойные леса харак-
терные для Западного района. Леса представлены как смешанными, так и чи-
стыми сосновыми и березовыми насаждениями. К сосне (Pinus kochiana) и бе-
резе (Betula litwinowii, B. raddeana) примешиваются осина, черемуха, ива ко-
зья и др. В подлеске – жимолость восточная (Lonicera orientalis), ракитничек
русский (Chamaecytisus ruthenicus) и др.
Высокогорные пояса в пределах Центрального района выражены наибо-
лее полно ввиду того, что Большой Кавказский хребет достигает здесь макси-
мальной высоты.
Субальпийский пояс располагается на высоте 2000–2200 м над ур. м.
Растительность представлена криволесьем и редколесьем, субальпийским
высокотравьем и субальпийскими лугами. В состав криволесья и редколесья
входят: березы, кустарниковая форма бука (Fagus orientalis), клен Траутфет-
тера (Acer trautvetteri), ива козья, рябина и заросли рододендрона кавказского
(Rhododendron caucasicum). В субальпийском поясе Центрального и Восточ-
ного районов Большого Кавказа характерно зонтичное высокотравье – виды
борщевика (Heracleum sp.) (рис. 12.14). В сочетании с ним произрастают и дру-
гие виды: аконит (Aconitum orientale), живокость (Delphinium flexuosum), аде-
ностилес (Adenostyles platyphylloides и др.), цицербита (Cicerbita macrophylla),
дороникум (Doronicum macrophyllum), окопник (Symphytum asperum), коз-
лятник (Galega orientalis) и др. Благодаря своим крупным размерам борщевик
придает особый облик всей группировке. Почвенный покров образован лугово-
лесными, горнолуговыми дерновыми и торфянисто-подзолистыми почвами.
Альпийский пояс. В альпийском
поясе (на высоте более 2800 м) выде-
ляются два типа сообществ: плотно-
дерновинные злаковые и осоковые
луга и альпийские ковры (рис. 12.15),
где задернение почвы производится
элементами разнотравья и где злаки и
осоки играют второстепенную роль.
Рис. 12.14. Борщевик сосновского
(Heracleum sosnowskyi)
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
303
Рис. 12.15. Альпийские луга,
Приэльбрусье
Дерновинные альпийские луга под-
разделяются на чистые и смешан-
ные. К смешанным лугам относятся
типчаково-осоковые луга, образо-
ванные Festuca airoides, F. ruprechtii
и Carex meinshauseniana. Среди дер-
на, образованного этими двумя ком-
понентами, находят себе приют
овсец (Helictotrichon adzharicum),
душистый колосок (Anthoxanthum
odoratum), клевер (Trifolium ambiguum), скабиоза (Scabiosa caucasica) и др.
В центральной и восточной половине Большого Кавказа распространены
смешанные осоково-типчаковые альпийские луга с таким же богатым флори-
стическим составом. Особым типом альпийского луга являются белоусники,
образованные жестколистным злаком Nardus stricta.
Альпийские ковры, состоящие из разнотравного мелкотравья, развива-
ются на мелкоземах и являются в достаточной степени плотно сомкнутыми.
Альпийские ковры на склонах образованы высокогорным одуванчиком (Ta-
raxacum stevenii), лапчаткой (Potentilla crantzii), осокой (Carex micropodioides)
и др. Распространенным типом ковров является колокольчиковое мелкотра-
вье (Campanula tridentata и С. biebersteiniana). Часто встречаются манжетко-
вые ковры, образованные Alchemilla caucasica. На склонах с более мощной
почвой и лучшими условиями увлажнения развиваются верониковые и
незабудковые ковры; местами, в условиях среднего увлажнения – клеверные
типы ковров, в которых наибольшее участие принимает Trifolium ambiguum.
Непосредственно вокруг снежных пятен формируется особый тип ковров,
для которых характерно обилие альпийских лютиков (Ranunculus buhsei,
R. oreophilus).
Нивальный пояс – область распространения вечных снегов и ледников.
Начиная с Эльбруса и восточнее до Казбекского массива включительно, оле-
денение Главного Кавказского хребта достигает своего максимума.
Восточный район простирается до восточного окончания Главного
Кавказского хребта. В него входят: Внутренний Дагестан, для которого харак-
терны складчатые известняковые хребты и плато, внешние склоны окаймля-
ющих его хребтов, восточная часть Главного Кавказского хребта и полоса
низменности вдоль Каспийского моря (равнинный Дагестан).
Горные хребты отгораживают Внутренний Дагестан от воздушных масс,
приносящих влагу. Осадков выпадает в среднем за год 200–300 мм на низ-
менности и в предгорьях, 400–600 мм – во Внутригорном и 600 и более
1000 мм – в Высокогорном Дагестане. Самые теплые месяцы на низменности
и в предгорьях – июль, август (17–24 °С). Средняя температура самого холод-
ного месяца января в низменной части около 1–2 °С, в предгорьях от 2 до 3 °С
и в горной части от –5 до –11 °С. На низменности Дагестана осадки выпадают
в основном в холодное время года, в теплое время года испарение превышает
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
304
Рис. 12.16. Климатограмма
г. Дербента:
---
количество выпадающих осадков;
–––
кривая среднемесячных температур
воздуха
количество выпадающих осадков, с
мая по сентябрь – засушливый пе-
риод (рис. 12.16). Общие черты
растительности Восточного района
отображены на карте (рис. 12.17).
Естественная растительность
равнинного Дагестана, вытянутого узкой полосой вдоль побережья Каспий-
ского моря, была представлена полынно-злаковыми степями (Stipa tirsa,
S. dagestanica, Bothriochloa ischaemum) на светло-каштановых почвах. В насто-
ящее время этот район в основном занят сельскохозяйственными угодьями.
Уникальной среди аридных ландшафтов нижнего пояса гор является
растительность федерального заказника «Самурский лес». Территория заказ-
ника представляет собой широкую равнину, изрезанную рукавами рек Самур
и Гюльгеричай, режим которых поддерживает снеговое, дождевое и грунтовое
питание. Приречные тугайные леса сформированы ольхой бородатой (Alnus
barbata) и тополями черным и сереющим (Populus nigra, P. canescens). Среди
низинных лесов преобладает коренная формация дубовых насаждений на лу-
гово-лесных почвах. В качестве доминанта выступает дуб черешчатый; в при-
меси дубы ножкоцветный и пушистый (Quercus pedunculiflora, Q. pubescens).
В связи с чрезмерными рубками площади дубовых лесов сокращаются, значи-
тельная часть древостоев низкоствольна, имеет порослевое происхождение.
Сопутствующими древесными породами являются: орех грецкий (Juglans
regia), вяз малый (Ulmus minor), тополь белый (Populus alba), лапина крыло-
плодная (Pterocarya pterocarpa), ясень обыкновенный (Fraxinus excelsior),
клен полевой (Acer campestre), груша кавказская (Pyrus caucasica), шелковица
белая (Morus alba), граб обыкновенный (Carpinus betulus), яблоня восточная
(Malus orientalis); в подлеске боярышник пятипестичный и вееролистный
(Crataegus pentagyna, С. rhipidophylla), свидина южная (Swida australis), муш-
мула (Mespilus germanica), лещина обыкновенная (Corylus avellana), бересклет
бородавчатый и европейский (Euonymus verrucosa, E. europaea), тёрн (Prunus
spinosa), шиповник собачий и щитконосный (Rosa canina, R. corymbifera),
калина обыкновенная (Viburnum opulus), жостер слабительный (Rhamnus
cathartica). Деревья обвиты лианами (рис. 12.18): сассапариль высокий (Smilax
excelsa), плющ Пастухова (Hedera pastuchowii), обвойник греческий (Periploca
graeca), виноград лесной (Vitis sylvestris), ломонос восточный и горный (Clematis
orientalis, C. montana). Травяной покров образован: осокой лесной (Carex
sylvatica), коротконожкой лесной (Brachypodium sylvaticum), овсяницей лес-
ной (Festuca drymeja), ежой сборной (Dactylis glomerata), фиалкой белой (Viola
alba), эгонихоном пурпурно-синим (Aegonychon purpureocoeruleum), подлесни-
ком европейским (Sanicula europaea), гравилатом горным (Orogeum montanum),
физалисом обыкновенным (Physalis alkekengi), молочаем миндалевидным
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
305
ВЫСОКОГОРНАЯ ПЕТРОФИЛЬНАЯ (КАМЕНИСТАЯ)
Скальная, осыпная, каменисто-щебнистая (минуария Биберштейна, камнеломка му-
скатная, крупка моховидная, лжепузырка пальчатая, низкозонтичник бесстебельный,
осока горная)
ЛУГОВАЯ
Альпийские луга
Мезофильные рыхлодернинные разнотравные и плотнодернинные разнотравно-осо-
ково-злаковые (лисохвост ледниковый, тимофеевка альпийская, мятлик альпийский,
первоцвет холодный)
Субальпийские луга
Мезофильные разнотравно-вейниковые, остепненные и костровые и пестроовсянице-
вые (вейник тростниковидный клевер волосистоголовый, звездовка Биберштейна, голо-
стебельный, костер пестрый, овсяница пестрая)
Рис. 12.17. Карта растительности нагорного Дагестана
(Атлас Республики Дагестан, 1999. Карта растительности, фрагмент), номера
условных обозначений соответствуют общей нумерации условных знаков
в легенде к карте растительности Дагестана
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
306
Послелесные луга среднегорного лесного пояса
Мезофильные полевицевые и остепненные низкоосоковь (полевица белая, тимофеевка
луговая, овсяница луговая осока низкая, манжетка шелковистая, кульбаба щетинистая
Послелесные предгорные луга
Мезофильные бобово-разнотравные, остепненные разнотравно-злаковые (мятлик лес-
ной, мятлик луговой, буквица крупноцветковая, пиретрум розовый, ежа сборная, лабаз-
ник шестилепестный, зверобой продырявленный, пырей ползучий, лапчатка прямая,
трясунка средняя)
ШИБЛЯКОВАЯ (СУХАЯ КУСТАРНИКОВАЯ)
Палиурусовая, боярышниковая, пушисто-дубовая (палиурус (держи-дерево), боярыш-
ник восточный, дуб пушистый)
НАГОРНО-КСЕРОФИЛЬНАЯ
Ксерофильные редколесья можжевельника многоплодного
Ксерофильные редколесья можжевельника казацкого
Ксерофильные спирейные кустарниковые сообщества (спирея городчатая, спирея зве-
робоелистная)
Ксерофильные трагакантные кустарниковые сообщества (астрагал Маршалла (трагакан-
товый астрагал)
ЛЕСНАЯ
Хвойные, смешанные леса
Сосновые, смешанные (сосна Коха, береза бородавчатая, береза Литвинова, береза
поникшая, бук восточный, граб обыкновенный, клен Траутфеттера, рябина кавказород-
ная, дуб восточный, лещина древовидная, чуб ушник кавказский)
Мелколиственные леса
Березовые мелколесья, криволесья (береза бородавчатая, береза повислая, береза
Литвинова, береза Радде)
Широколиственные леса
Влажные предгорные буковые (бук восточный, бук лесной, граб кавказский, черемуха
обыкновенная, черешня обыкновенная, клен остролистный, вяз шершавый, липа обык-
новенная, липа кавказская, бузина черная. лещина обыкновенная, бересклет широко-
листный)
Сухие предгорные дубовые (дуб скальный, дуб пушистый, боярышник согнутостолбико-
вый, клен полевой, клен гирканский, вяз пробковый, ясень обыкновенный, алыча рас-
топыренная, шиповник собачий, бересклет бородавчатый, жостер слабительный, айва
обыкновенная, груша кавказская)
Влажные равнинные приречные леса
Тополевые, ивовые, черешчато-дубовые, ольховые, грабовые (тополь гибридный, то-
поль белый, тополь черный, ива белая, яблоня восточная, айва обыкновенная облепиха
крушиновая, пираканта ярко-красная, алыча растопыренная алыча каспийская, крушина
ломкая, калина обыкновенная, ольха бородатая, ольха клейкая, ольха серая, ломонос
виноградолистный, ломонос восточный, виноград лесной, плющ Пастухова, обвойник
греческий, сассапариль высокий)
Рис. 12.17 (продолжение). Легенда к карте растительности Дагестана
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
307
Рис. 12.18. Самурский лес
(фото Г. С. Джамирзоева)
(Euphorbia amygdaloides), луком па-
радоксальным (Allium paradoxum),
кирказоном
ломоносовидным
(Aristolochia clematitis).
Пояс аридного редколесья за-
нимает предгорья и представляет
собой лесостепь южного типа на
коричневых почвах. В Восточном
районе аридное редколесье явля-
ется переходным от лесостепи ши-
роколиственных лесов (дуб, груша,
яблоня) к аридному редколесью, образованному несколькими видами дре-
вовидного можжевельника (Juniperus polycarpos, J. foetidissima, J. oxycedrus,
J. excelsa). Заросли кустарников состоят из держи-дерева, жимолости восточ-
ной, жостера Палласа (Rhamnus pallasii), барбариса, спиреи и др.
Пояс широколиственных лесов начинается с высоты 300–400 м и дости-
гает 1000 м над ур. м. Леса образованы дубом грузинским (Quercus iberica),
грабом, ясенем, кленом полевым, кленом светлым (Acer laetum), вязом ма-
лым, грушей кавказской, яблоней восточной; в подлеске – мушмула, кизил,
бересклет и др. Почвенный покров образован бурыми лесными и перегнойно-
карбонатными почвами.
На склонах гор от 900–1000 до 1500 м над ур. м. растут буковые леса,
включающие: бук восточный, граб кавказский, дуб крупнопыльниковый
(Quercus macranthera), клен полевой, черешня и др. В подлеске – бересклет,
бузина черная, жимолость восточная, смородина и др.
Пояс березово-сосновых лесов хорошо выражен на Кавказском хребте на
высоте от 1500 до 2100–2200 м над ур.м. В лесах вместе с сосной Коха (Pinus
kochiana), березой повислой, березой Литвинова встречаются рябина, липа,
ива козья, крушина и др. Почвенный покров в основном состоит из оподзо-
ленных бурых лесных почв разной мощности.
Высокогорные пояса выражены на Главном Кавказском хребте на высоте
от 2200–2300 до 2400–2500 м над ур. м. –
субальпийский и на высоте от
2400–2500 м и выше – альпийский. Характер растительности высокогорий
Восточного района аналогичен растительности субальпийского и альпийского
поясов Центрального района.
Климат Внутреннего Дагестана сухой континентальный, годовое количе-
ство осадков не превышает 500–550 мм. На горных склонах, на скелетных
почвах развиваются формации нагорных ксерофитов средиземноморско-
иранского типа, в том числе фригана и трагакантники (рис. 12.19), которые
занимают пояс от 1200–1300 до 1800–2000 м над ур. моря.
Фригана – группировка невысоких ксероморфных полукустарников или
полукустарничков с участием эфемерного и эфемероидного разнотравья;
обычно с большей или меньшей примесью терофитов и геофитов. Она приуро-
чена к крутым и сухим склонам. В составе фриганы на известковых склонах
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
308
Астраканта обнаженная
(Astracantha denudata)
Астрагал волосистоязычковый
(Astragalus lasioglottis)
Каперсы травянистые (Capparis herbacea)
Эспарцет рогатый (Onobrychis cornuta)
Шалфей седеющий (Salvia canescens)
Скабиоза гумбетская (Scabiosa gumbetica)
Фото: С. Банкетова, А. Иванова
Рис. 12.19. Представители формаций нагорных ксерофитов
средиземноморско-иранского типа
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
309
самый характерный представитель – шалфей (Salvia canescens), весьма мно-
гочисленны: астраканта обнаженная (Astracantha denudata), астрагал волоси-
стоязычковый (Astragalus lasioglottis), каперсы (Capparis herbacea), скабиоза
(Scabiosa gumbetica), эспарцет рогатый (Onobrychis cornuta), вьюнок (Convol-
vulus ruprechtii), наголоватка (Jurinea ruprechtii) и др. Для фриганы на неиз-
вестковых склонах характерен другой вид шалфея (Salvia beckeri), а также ряд
травянистых растений: гелиотроп (Heliotropium styligerum), ковыль (Stipa da-
ghestanica), кермековидка (Limoniopsis owerinii) и др.
Трагакантники представляют собой сообщества, образованные колю-
чими кустарниками характерной подушкообразной формы; доминантами
в большинстве случаев в них являются трагакантовые астраканты: астра-
канта кавказская, золотистая, обнаженная (Astracantha caucasica, A. aurea,
A. denudata), реже колючий эспарцет рогатый. В качестве субдоминантов ча-
сто выступают полукустарничковые виды: тимьяны дагестанский и Елизаве-
ты (Thymus daghestanicus и Т. elisabethae), шлемник многозубый (Scutellaria
polyodon) или степные злаки – типчак (Festuca valesiaca), тырса (Stipa capillata),
пырей стройный (Elytrigia gracillima), кострец береговой (Bromopsis riparia),
а также бородач (Bothriochloa ischaemum); нередко попадается осока низкая
(Carex humilis).
12.3. РАСТИТЕЛЬНОСТЬ АЛТАЯ
Алтайские горы представляют сложную систему самых высоких в Сибири
хребтов, разделенных глубокими долинами рек и обширными внутригорными
и межгорными котловинами (рис. 12.20). Наибольших высот алтайские горы
достигают в области Катунского хребта и Чуйских белков (3500–4500 м), яв-
ляющихся мощным центром современного оледенения. Самая высокая точка
горного Алтая – гора Белуха (4506 м) (рис. 12.21). Алтай делится на Южный
(Юго-Западный), Юго-Восточный и Восточный, Центральный, Северный и
Северо-Восточный, Северо-Западный. От северных и западных предгорий Ал-
тая к юго-востоку наблюдается постепенное повышение. Горный характер ре-
льефа нарушается целым рядом котловин разного типа. Периферийные хреб-
ты в значительной мере перехватывают влагу западных воздушных потоков.
Поэтому в юго-восточном направлении происходит значительное увеличение
сухости и нарастание континентальности климата, что является следствием
экранирующей роли хребтов. Для большей части Горного Алтая характерен
континентальный климат с суровой зимой и жарким летом. Повышение кон-
тинентальности климата сказывается в усилении ксерофитности раститель-
ных сообществ всех высотных поясов. Важным моментом в климатической
обстановке Алтая является перераспределение тепла и влаги на горных скло-
нах разных экспозиций. Солярная и ветровая экспозиция склонов определяет
асимметрию растительности макросклонов хребтов.
Флора Алтая насчитывает около 2000 видов высших споровых, голосе-
менных и цветковых растений. Количество эндемичных видов составляет
11,5 %. Поэтому Алтай в свою очередь является мощным центром распростра-
нения видов для большей части Сибири, прилегающих районов Казахстана и
Монголии.
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
310
Рис. 12.20. Орографическая схема Алтая (Огуреева, 1980):
а – горные хребты главные; б – прочие крупные хребты; в – отметки высот, м; г – межгор-
ные котловины: 1 – Абайская; 2 – Уймонская; 3 – Катандинская; 4 – Ануйская; 5 – Курай-
ская; 6 – Улаганская; 7 – Чуйская; 8 – Джулукуйская; 9 – Бертекская; 10 – Самахинская
Общие закономерности распределения растительности Алтая показаны
на карте (рис. 12.22). Отдельными контурами на этой карте выделены круп-
ные подразделения растительного покрова. В легенде для каждого типа кон-
туров перечислены только самые распространенные в этом контуре сообще-
ства.
Основную структурную единицу растительного покрова на Алтае, как и
везде в горах, представляет пояс растительности. Он, как определенная высот-
ная ступень, состоит из климатически обусловленных сообществ одного или
нескольких типов растительно-
сти. Внутри поясов растительно-
сти выделяются подпояса. Они в
свою очередь могут подразде-
ляться на высотно-климатичес-
кие полосы. Выделяются шесть
основных растительных поясов
Алтая: степной, лесостепной, лес-
ной, субальпийский, альпийско-
тундровый, нивальный.
Рис. 12.21. Вид на гору Белуха
с юга
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
311
Рис. 12.22. Карта растительности Алтая (Копыл, Огуреева, 1978)
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
312
Рис. 12.23. Типы высотной поясности для Алтае-Саянской горной страны
(Зоны и типы..., 1999):
26 – альпийско-субальпийско-таежный (Североалтайский); 26.1 – Северо-Восточноалтай-
ский; 26.2 – Западно-Восточносаянский; 26а – горнотаежный (Салаирский кряж и Кузнец-
кий Алатау); 26б – горнотаежно-лесостепной (Кузнецкая котловина); 27 – нивально-
альпийско-субальпийско-таежно-лесостепной (Центральноазиатский); 27б – таежно-лесо-
степной (Северный Алтай); 28 – альпийско-субальпийско-таежно-кустарниково-лесостеп-
но-степной (Западноалтайский)
Варианты высотной поясности для Алтае-Саянской горной страны изоб-
ражены на рис. 12.23. В пределах групп поясности даны типы и другие града-
ции поясности (колонки), которые представляют спектры поясности расти-
тельности. Условные знаки в легенде содержат название каждого пояса.
Степной пояс на Алтае расположен в широком диапазоне высот (400–
2400 м над ур.м.). Он представлен относительно широкой полосой в северных
и юго-восточных районах Алтая, более узкой и в виде разрозненных пятен – в
центральной части горной страны. В западной части Алтая степной пояс под-
разделяется на два подпояса. В нижнем подпоясе (400–500 м над ур.м.) рас-
пространены разнотравно-типчаково-ковыльные (Stipa zalesskii) степи; харак-
терны дерновинные злаки широкого распространения (Festuca valesiaca,
Koeleria cristata). В верхнем подпоясе предгорий и низкогорий Западного Ал-
тая (500–800 м над ур.м.) широко распространены сообщества кустарников с
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
313
лугово-степным травостоем. Основными эдификаторами кустарниковых
степных сообществ являются Lonicera tatarica, Spiraea hypericifolia, S. crenata,
Caragana frutex, а также Cotoneaster melanocarpus, Rosa hugonis.
С увеличением континентальности климата во внутригорных районах
Алтая основная роль в степном поясе переходит к сообществам ксерофитно-
разнотравно-мелкодерновиннозлаковых степей на каштановых почвах.
Своеобразным феноменом структуры растительного покрова Алтая
являются «островные степи», сформированные в межгорных котловинах
(рис. 12.24). Растительный покров теплых внутригорных котловин (Абайской,
Уймонской и Катандинской) имеет лесостепной характер. В котловинах Канн-
ской, Улаганской и Теньгинской, где количество атмосферных осадков
уменьшается до 350 мм и менее, растительный покров приобретает более ксе-
рофитный облик, здесь характерно развитие настоящих степей с господством
дерновинных злаков, преимущественно типчаковых (Festuca valesiaca), тип-
чаково-ковыльных (Stipa capillata), полынно-мелкодерновиннозлаковых
(Artemisia frigida, Koeleria cristata, Poa attenuata, Agropyron cristatum) на
каштановых почвах. Подпояс центральноазиатских опустыненных степей хо-
рошо выражен в пределах Чуйской высокогорной котловины на высотах
1700–2000 м над ур.м. Опустыненные степи на светло-каштановых почвах
носят явно выраженный петрофитный характер.
Лесостепной пояс Алтая формируется в условиях среднегорного рельефа
и наиболее широко представлен в диапазоне высот 500–1700 м над ур.м. в
долинах крупных рек, склоны которых отличаются значительной расчленен-
ностью. Он характерен также для останцов, отрогов горных хребтов, обрамля-
ющих межгорные котловины.
В целом лесостепной пояс горного Алтая существует при амплитуде
среднегодовых осадков от 250 до 750–800 мм и при средних годовых темпера-
турах воздуха от –4 до 2 °С. Пояс включает неоднородные, часто контрастные,
растительные сообщества. Для склонов южной экспозиции характерно разви-
тие сложных степных сообществ, которые включают многие ксеропетрофит-
ные виды, представляющие характерный элемент скальной флоры Алтая.
Среди них нужно отметить спирею трехлопастную (Spiraea trilobata), являю-
щуюся постоянным видом на каменистых склонах. Среди других видов можно
назвать эндемичные для Алтая хризантема выемчатолистная (Chrysanthemum
sinuatum), патриния скальная (Patrinia rupestris), качим Патрэна (Gypsophila
patrinii), змееголовник поникаю-
щий и иноземный (Dracocephalum
nutans, D. peregrinum) и др.
На склонах северной экспози-
ции развиваются леса из листвен-
ницы сибирской (Larix sibirica).
Находясь в непосредственном кон-
такте со степной растительностью
южных склонов , они претерпели
Рис. 12.24. Степь в котловине,
Восточный Алтай
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
314
Рис. 12.25. Остепненные листвен-
ничники, Алтай (фото А. Иванова)
существенные изменения и во
многих районах Алтая представ-
лены типами, не встречающимися
за пределами лесостепного пояса.
Прежде всего, это касается остеп-
нения травяного покрова лист-
венничных лесов и развития под
ними серых лесных и черноземо-
видных почв (рис. 12.25).
На северо-западе и западе Алтая, где увлажнение больше, лесостепной
пояс занимает полосу в пределах высот порядка 500–700 м. Нижняя и верх-
няя его границы здесь обусловлены годовым количеством осадков 600–800 мм.
В этих районах наблюдается преобладание увлажнения над испаряемостью.
Для северных периферийных частей предгорий характерно сочетание раз-
личных вариантов луговых степей и остепненных лугов, сообществ степных
кустарников с березовыми и осиново-березовыми перелесками по склонам
северных экспозиций. Доля лесов колеблется от 10–20 до 30–40 %. Во внут-
ренних районах северных предгорий лесной компонент представлен преиму-
щественно лиственничными и березово-лиственничными травяными лесами.
Для лесостепных сообществ Центрального Алтая характерно гораздо
меньшее разнообразие лесов по сравнению с районами северных низкогорий.
Преобладают южносибирские лиственничные и березово-лиственничные ле-
са. На юго-востоке Алтая лесостепной пояс практически выклинивается, и от-
дельные изолированные фрагменты лесов входят в контакт со своеобразными
сообществами тундрово-степного облика, занимая высоты 2000–2200 м над
ур.м. Таким образом, пояс горной, преимущественно лиственничной лесосте-
пи, выступает как самобытная особенность растительности гор Южной Сиби-
ри, в которой лесостепные сообщества занимают одно из ведущих мест.
Лесной пояс делится на два подпояса. Первый из них, черневой подпо-
яс, расположен в пределах высот 400–1200 м на ур.м., получающих до 800–
1000 мм осадков в год. Он включает горные темнохвойные леса из кедро-
вой сосны, пихты и ели сибирских (рис. 12.26).
Флороценотический комплекс
черневой тайги складывается из
видов неморальной группы (Galium
odoratum, Asarum europaeum,
Stachys silvatica, Paris quadrifolia,
Festuca altissima, F. gigantea,
Brachypodium sylvaticum) и видов
темнохвойнотаежного комплекса
Рис. 12.26. Темнохвойный лес,
Телецкое озеро
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
315
Рис. 12.27. Лиственничный лес
на склоне у Телецкого озера
(Aconitum septentrionale, Saussurea
latifolia, Crepis lyrata, Alfredia cernua,
Lathyrus gmelinii, Viola biflora).
Нижняя полоса черневого подпояса
занимает высоты 400–800 м. Она
образована из осиново-пихтовых
высокотравных лесов и производ-
ных осиново-березовых на их месте.
В пределах нижней полосы черне-
вого подпояса северных и западных предгорий распространены сосновые ле-
са. Они связаны с песчаными и супесчаными почвами речных террас. Верхняя
полоса черневого подпояса (800–1200 м) образована кедрово-пихтовыми вы-
сокотравными лесами.
Второй – горнотаежный подпояс занимает широкий диапазон абсолют-
ных высот от 400 до 2465 м. Он состоит из светлохвойных лиственничных и
темнохвойных формаций (рис. 12.27). Березово-лиственничные леса разно-
травных групп характерны для предгорных и центральных районов Алтая.
Резко континентальный климат внутригорных районов Центрального Алтая
способствует широкому распространению лиственничных лесов из Larix
sibirica. Среднегорные лиственничники отличаются богатством и разнообра-
зием видового состава подлеска и травяного покрова: Poa sibirica, Potentilla
chrysantha, Geranium pseudosibiricum, Primula pallasii, Bupleurum longifolium
subsp. aureum.
В растительности высокогорий Алтая выделяются субальпийский, аль-
пийско-тундровый и нивальный пояса. Субальпийский пояс хорошо просле-
живается на всем протяжении высокогорий. Альпийско-тундровый, включая
альпийские луга, встречается во всех высокогорных районах Алтая. Ниваль-
ный пояс занимает высоты свыше 3000–3200 м и выражен только на наибо-
лее высоких хребтах, где имеется современное оледенение. Он характеризует-
ся обилием скал, каменистых россыпей, наличием ледников и практически
лишен растительности.
Субальпийский пояс выражен везде, где горные склоны поднимаются
выше границы леса, на абсолютных высотах 1800–2600 м. Он отличается са-
мобытным набором растительных формаций, их разнообразием и специфи-
кой флористического состава. В пределах субальпийского пояса выражены
три подпояса (субальпийские луга, ерники, тундро-степь), по-разному пред-
ставленных в различных районах Алтая.
Подпояс субальпийских лугов характерен для относительно увлажнен-
ных с мощным снежным покровом западных районов Центрального Алтая.
Субальпийские луга встречаются обычно в нижней части субальпийского поя-
са и представлены высокотравными и низкотравными формациями. Высоко-
травные луга хорошо развиты непосредственно у верхней границы леса на
горно-луговых почвах. Хорошо выражен переход кедровых и лиственничных
редколесий к субальпийским разнотравьям (рис. 12.28).
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
316
Рис. 12.28. Переход кедрово-
пихтового леса к высокотравью
Среди высокотравных лугов на-
иболее характерны луга формации
Stemmacantha carthamoides. В их
составе участвуют до 64 % лесных
(Saussurea latifolia, Aconitum septen-
trionale, Trollius asiaticus, Bupleurum
longifolium subsp. aureum, Poa sibiri-
ca, Delphinium elatum, Pleurosper-
mum uralense) и до 53 % альпийских
и аркто-альпийских (Dracocephalum
grandiflorum, Phlomoides oreophila,
Viola altaica, Aquilegia glandulosa) видов. Переход от высокотравных лугов к
низкотравным постепенный.
Среди низкотравных субальпийских лугов широко распространены ман-
жетково-гераниевые луга на горно-луговых бурых, слабо скелетных почвах,
для которых характерны Geranium albiflorum, Alchemilla sibirica, Sanguisorba
alpina, Phleum alpinum, Seseli condensatum и др. В юго-восточных районах Ал-
тая, в условиях резко континентального климата, субальпийские луга посте-
пенно выпадают из растительного покрова высокогорий.
Ерниковый подпояс образован Betula rotundifolia (рис. 12.29). Опти-
мальные условия ерник имеет на нижней ступени субальпийского пояса, где
образует кустарниковые сообщества высокой сомкнутости, высотой 50–70 см.
Ерники образуют почти чистые одновидовые сообщества и лишь иногда в них
отмечаются другие кустарники, чаще всего ивы (Salix arbuscula, S. krylovii,
S. berberifolia, S. glauca, S. reticulata, S. vestita) или Juniperus sibirica, Spiraea
alpina, Cotoneaster uniflorus. В некоторых районах в ерниковом подпоясе ивы
образуют самостоятельные сообщества. Высота кустарникового яруса обычно
соответствует мощности снежного покрова. В пределах подпояса преобладают
ерниковые сообщества с мохово-лишайниковым покровом. Наибольшие пло-
щади занимают ерники моховой группы. Среди них выделяются зеленомошные
с доминированием Aulacomnium palustre, Hylocomium splendens, Camptothecium
nitens и политриховые с преоблада-
нием Polytrichum strictum. В соста-
ве лишайниковых ерников щебни-
стых местообитаний принимает
участие Dryas oxyodonta. Встреча-
ются ерники с осоково-сфагновым
покровом по пониженным элемен-
там рельефа.
Рис. 12.29. Ерник из березы кругло-
листной (Betula rotundifolia)
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
317
Рис. 12.30. Злаковые остепненные
высокогорные тундры
Тундрово-степной подпояс
развит только в системе высотной
поясности Юго-Восточного Алтая
на высотах 2200–2800 м. С этим
подпоясом связано распростра-
нение криофитно-пустошной рас-
тительности, представляющей
собой остепненные варианты зла-
ковых и осоково-кобрезиевых лу-
говых тундр (рис. 12.30). Тундрово-степные сообщества наиболее широко раз-
виты по южным склонам моренных холмов, по выположенным участкам
древних поверхностей выравнивания. Криофитно-пустошные сообщества от-
личаются плотным задернением; они нередко прерываются фрагментами
других тундровых или степных сообществ, образуя с ними различные типы
комплексов.
Альпийско-тундровый пояс. Горные тундры Алтая представлены в ос-
новном несколькими группами луговых, мохово-лишайниковых формаций и
распространены на абсолютных высотах 2000–3500 м. Выделяют осоково-
дерновиннозлаковые и осоково-кобрезиевые тундры на горно-тундровых дер-
новых почвах. Сухие дриадово-лишайниковые тундры развиваются в условиях
дренированных и открытых местообитаний. Для напочвенного покрова ха-
рактерно преобладание мхов и лишайников. Значительная часть альпийско-
тундрового пояса занята открытыми каменистыми россыпями, где цветковых
растений крайне мало, и они образуют сильно разреженные сообщества. Кам-
ни покрыты лишайниками и мхами (рис. 12.31). Каменистые тундры, харак-
терные для верхней полосы альпийско-тундрового пояса, богаты эндемичны-
ми видами. С ними связана верхняя граница распространения цветковых рас-
тений на Алтае. Они приурочены к снежникам, понижениям, защищенным от
ветра и увлажненным местообитаниям.
Для охраны уникальной
природы Алтая создан ряд особо
охраняемых территорий миро-
вого значения, среди которых
Алтайский, Катунский заповед-
ники и плоскогорье Укок обра-
зуют объект Всемирного насле-
дия ЮНЕСКО, именуемый «Ал-
тай – Золотые горы».
Рис. 12.31. Каменистые тундры
Алтая
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
318
12.4. РАСТИТЕЛЬНОСТЬ ГОР ВОСТОЧНОГО ПРИБАЙКАЛЬЯ
Восточное Прибайкалье – горная страна, омываемая на западе водами
Байкала. Она характеризуется мощными хребтами и обширными межгорны-
ми котловинами. Площадь гор более чем в 4 раза превышает площадь, зани-
маемую низменностями. К озеру Байкал примыкают высокие хребты Прибай-
калья. Они имеют форму кряжей, вытянутых преимущественно с юго-запада
на северо-восток. Южную часть Восточного Прибайкалья окаймляет плоско-
вершинный хребет Улан-Бургасы. От берегов Байкала его отделяют более
низкий Морской хребет (высота до 1500 м) и 35-ти километровая полоса хол-
мистой поверхности. Хребет имеет сравнительно мягкие очертания и плоские,
выровненные процессами длительной денудации вершины, которые отделены
друг от друга на значительном протяжении низкогорными залесенными
участками; его склоны, обращенные к Байкалу, относительно пологи и скали-
стых выступов не образуют. Склоны хребта сильно изрезаны долинами рек,
часто встречаются сухие безводные долины, или распадки, как на склонах
хребта, так и в предгорьях.
Климат Восточного Прибайкалья – резко континентальный, сочетаю-
щийся с муссонным режимом выпадения атмосферных осадков в теплое вре-
мя года. Высокое положение хребта Улан-Бургасы над уровнем моря и его се-
веро-восточное простирание обеспечивают перехват влаги из западного пере-
носа воздушных масс. На западном макросклоне хребта проявляется заметное
смягчающее влияние Байкала. Это сказывается как на температурном режи-
ме, так и на перераспределении осадков. Среднее годовое количество осадков
в верхней части склонов и на гребнях составляет 800–1000 мм, у подножья
250 мм.
На территории Восточного Прибайкалья сочетаются светлохвойные леса
из сосны и лиственницы, темнохвойная тайга из ели, пихты и сосны кедровой,
занимающие склоны гор и степи в межгорных котловинах. Обобщенные чер-
ты растительности Восточного Прибайкалья отображены на геоботанической
карте (рис. 12.32). Легенда к карте представлена в виде матрицы, где строки
соответствуют типам растительных сообществ, а столбцы – ступеням высот и
формам рельефа. В ячейках таблицы отмечено наличие или отсутствие опре-
деленного выдела на данной ступени высот.
Горный рельеф Восточного Прибайкалья определяет характер высот-
ной поясности растительности, элементы которой хорошо видны на блоке
(рис. 12.33), вырезанном из карты растительности. Для западных склонов
хребта Улан-Бургасы, обращенных к Байкалу, от подножия к вершине хребта
наблюдается последовательная смена светлохвойных (сосновых и листвен-
ничных), темнохвойных (еловых, кедровых, пихтовых) лесов, субальпийских
лугов и высокогорной растительности (рис. 12.34). Приведем описание основ-
ных сообществ высотных поясов западного склона хребта Улан-Бургасы.
Лесостепной пояс представлен на высотах от 500 до 800 м на южных и
юго-западных склонах. Наиболее распространена сосновая, лиственнично-
сосновая и сосново-лиственничная лесостепь с примесью березы и осины.
Главная лесообразующая порода – сосна обыкновенная (Pinus sylvestris). Ха-
рактерны для лесостепи заросли различных кустарников: Spiraea aquilegifolia,
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
319
Растительность
450–850 850–1250 1250–1650 1650–2050 Выше 2050
1. Горные тундры на склонах гор разных экспозиций
каменисто-щебенчатые и мохово-лишайниковые в соче-
таниях с пустошами, нивальными луговинами, ерниками
и сообществами кедрового стланика
–
–
–
+
+
2. Горные злаково-разнотравные луга, заросли папорот-
ников в сочетаниях с ассоциациями кедрового стланика,
ольховников, ерников, ивняков, с рединами пихты и
березы
–
–
+
+
–
3. Кедровый стланик в верхнем поясе гор
–
+
+
+
–
4. Листгенничние и кедрово-лиственничные на горных
склонах: кустарниково-моховые и мохово-лишайнико-
вые нередко с ярусом ерников, кедрового стланика, ду-
шекии. Леса с преобладанием лиственницы сибирской
–
+
+
+
–
Рис. 12.32. Карта растительности Восточного Прибайкалья, м-б 1:500 000
(составлена А. Унагаевым на основе классификации элементов космического изображения)
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
320
Растительность
450–850 850–1250 1250–1650 1650–2050 Выше 2050
5. Лиственничные и кедрово-лиственничные на приво-
дораздельных склонах: зеленомошные, бадановые,
каменистые, бруснично-багульниковые, иногда с ярусом
из ерников, рододендрона даурского и душекии
–
–
+
+
–
6. Лиственничные на на приводораздельных склонах:
травяно-брусничные, травяно-моховые, каменистые с
ярусом из душекии
+
+
+
+
–
7. Лиственничные и сосново-лиственничные на склонах
гор: травяно-брусничные, брусничные с ярусом родо-
дендрона даурского
+
+
–
–
–
8. Сосновые и лиственнично-сосновые на горных скло-
нах и в предгорьях: травяно-брусничные, остепкенные,
брусничные, иногда с ярус ста рододендрона даурского
и ольхи кустарниковой
+
+
+
–
–
9. Сосновые и лиственнично-сосновые на герньк скло-
нах: травяно-брусничньв, брусничные, бруснично-
рододендроновые
+
+
–
–
–
10. Сосновые на песчаных отложениях в днищах котло-
вин: кустарничково-мохово-лишайниковые и травяно-
брусничные, бруснично-толокнянковые с разреженным
ярусом рододендрона даурского и ольхи кустарниковой
+
–
–
–
–
11. Сосновые и лиственнично-сосновые в сочетаниях с
мелколиственными лесами по склонам гор и днищам
котловин
+
+
–
–
–
12. Кедрово-пихтовые леса на склонах гор и в долинах:
кустарничково-зеленомошные, травяно-зеленомошные,
бадановые
–
+
+
+
+
13. Кедровые и лиственнично-кедровые леса на горных
склонах: кустарничково-зеленомошные, лишайниково-
зеленомошные, каменистые, бадановые
–
+
+
+
–
14. Березовые и березово-осиновые: разнотравно-
брусничные, злаково-разнотравные, остепненные-
восстановительные серии на месте светлохвойных лесов
+
+
–
–
–
15. Березовые кустарничково-зеленомошные, травя-
но-зеленомошные, вейниковые, разнотравные вос-
становительные серии на месте темнохвойных лесов
+
–
–
–
–
16. Тонконогово-типчаковые и кистевидно-мятлико-
вые степи
+
+
+
–
–
17. Луга долинные и пойменные: злаковые, осоково-
злаковые, злаково-разнотравные в сочетаниях с за-
рослями ив, зарослей боярышника, черемухи, ябло-
ни, других деревьев и кустарников
+
–
–
–
–
18. Луга заболоченные и травяные болота: осоковые,
осоково-злаковые, разнотравно-злаковые
+
+
–
–
–
19. Болота приозерных низменностей осоково-сфаг-
новые, злаково-сфагновые с участием ив и ерников
+
–
–
–
–
20. Участки с разреженным растительным покровом
(редколесья, горелые и вьрубленные леса)
+
+
+
+
+
Рис. 12.32. (продолжение) Легенда к карте растительности Восточного
Прибайкалья
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
321
Рис. 12.33 . Высотная поясность на западном и восточном склонах хребта
Улан-Бургасы
Условные обозначения см. на рис. 12.32.
S. dahurica, S. media, Salix bebbiana, Cotoneaster melanocarpus, Rosa acicularis,
Ulmus pumila, Caragana pygmaea и др. Травяной покров в лесостепи доста-
точно разнообразен по видовому составу. В его распределении отмечается
следующая закономерность: при поднятии вверх по склону наблюдается уве-
личение влаголюбивого разнотравья и уменьшение злаковых компонентов.
Сообщества многовидовые.
Лесной пояс. Его нижнюю часть занимают светлохвойные леса. Они часто
образованы однопородным древостоем из сосны или лиственницей сибирской.
Рис. 12.34. Высотная поясность западного склона хребта Улан-Бургасы
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
322
Рис. 12.35. Светлохвойный сосново-
лиственничный лес
Преобладают сосняки: спирейно-раз-
нотравные,
разнотравно-злаковые,
рододендроновые, бруснично-зеле-
номошные, можжевельниково-раз-
нотравные (рис. 12.35).
Сосняки спирейно-разнотравные
приурочены к юго-западным и юго-
восточным склонам. Древостой высо-
кий, разреженный. Проективное по-
крытие травяного покрова около 25–
35 %, преобладают виды: Calamagrostis arundinacea, С. lapponica, Poa attenuata,
P. pratensis, Astragalus adsurgens, A. propinquus, Lathyrus humilis, Veronica incana.
Сосняки рододендроновые развиты на северных склонах. В древостое
единично участвует лиственница сибирская и сосна кедровая. Подлесок из
Rhododendron dauricum (рис. 12.36), встречается шиповник (Rosa acicularis).
Проективное покрытие травяно-кустарничкового яруса 40–70 %. Доминируют
Vaccinium vitis-idaea, Pyrola incarnata, P. minor, Pulsatilla orientali-sibirica,
Maianthemum bifolium, Lathyrus humilis. Мохово-лишайниковый ярус не вы-
ражен. Мхи встречаются в приствольной части деревьев и на валежинах.
Сосняки бруснично-зеленомошные с подлеском из можжевельника обык-
новенного распространены на юго-западных склонах. В подлеске, кроме доми-
нирующего Juniperus communis, присутствуют Pinus pumila, Lonicera pallasii,
Salix caprea, Spiraea media. В травяно-кустарничковом покрове преобладают
Vaccinium vitis-idaea, V. myrtillus, Maianthemum bifolium, Melica nutans, Viola
uniflora, Linnaea borealis, Pyrola incarnata. Моховой покров (Pleurozium
schreberi, Dicranum polysetum, Ptilium crista-castrensis) мозаичный, покрытие
в понижениях до 80 %; на выпуклых участках присутствуют лишайники.
Лиственничники разнотравные приурочены к склонам восточной экспо-
зиции. Травяной ярус хорошо развит, проективное покрытие травостоя до-
стигает 90–95 %. Доминируют виды: Calamagrostis epigeios, С. lapponica,
Thalictrum appendiculatum, Т. simplex, Vicia unijuga, V. cracca. Мхи встречаются
лишь у основания стволов.
Темнохвойные леса встречаются
в местах с устойчивым увлажнением
и развиты в верхней половине лесно-
го пояса. В восточной части хребта
верхняя граница леса образована
сосной кедровой и проходит на высо-
тах 1450–1700 м, в западной – пихтой
сибирской на высотах 1200–1450 м.
В составе темнохвойных лесов прини-
Рис. 12.36. Весна, цветет рододендрон
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
323
Рис. 12.37. Темнохвойный лес
Рис. 12.38. Субальпийский
высокотравный луг
мают участие лиственница сибирская, сосна обыкновенная и береза плоско-
листная (рис. 12.37). Кустарниковый ярус образован можжевельником, ро-
додендроном, ивами, местами ольховником. В травяном покрове доминиру-
ют черемша (Allium microdictyon), чемерица (Veratrum lobelianum), бадан
(Bergenia crassifolia), купальница, грушанка, черника, брусника, голубика, ба-
гульник. В мохово-лишайниковом ярусе преобладают зеленые мхи.
Пихтовые леса из Abies sibirica представлены пихтарниками папоротни-
ковыми, чернично-зеленомошными, рододендрово-зеленомошными, разно-
травно-хвощовыми, хвощовыми. Пихтарник рододендрово-зеленомошный
(с Rhododendron aureum) распространен в верховьях рек на склонах северной
экспозиции. В травяно-кустарничковом покрове распространены Bergenia
crassifolia, Vaccinium myrtillus, Maianthemum bifolium, Linnaea borealis. Мохо-
вой покров мощный, сплошной, с покрытием 95 % и образован из Ptilium cris-
ta-castrensis, Hylocomium splendens, Dicranum polysetum.
Кедровые леса из Pinus sibirica развиты на склонах северной и восточной
экспозиций. Выделены ассоциации: кедровник бадановый, брусничный, осо-
ково-моховый, кедровник с кедровым стлаником в подлеске.
Вторичные сообщества формируются на местах вырубок и пожаров,
представленные березняками (из Betula platyphylla, B. pendula) злаковыми,
злаково-разнотравными, осинниками разнотравно-злаковыми. В березняках
единично встречаются сосна кедровая, сосна обыкновенная. Травостой густой,
равномерный, доминируют: Calamagrostis epigeios, C. lapponica, Maianthemum
bifolium, Linnaea borealis, Lathyrus humilis. Моховой покров не выражен.
Субальпийский пояс представлен высокотравными лугами в сочетании с
пихтовым редколесьем по склонам различной экспозиции на высоте от 1500
до 1650 м (рис. 12.38). Единично встречаются кустарники: Lonicera pallasii,
Pinus pumila, Juniperus communis, Sambucus sibirica. Проективное покрытие
травостоя 85–100 %, высота до 1,5 м, преобладают: Geranium krylovii, Solidago
dahurica, Cirsium helenioides, Anthriscus sylvestris, Pedicularis incarnata, Gali-
um boreale, Calamagrostis purpurea, Milium effusum, Trollius ledebourii. По-
крытие мохового покрова до 40 %, он представлен Polytrichum commune,
P. strictum, Dicranum scoparium.
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
324
Рис. 12.39. Кедровый стланик
Рис. 12.40 Горно-тундровый
гольцовый пояс
Выше расположен пояс кедрового стланика (Pinus pumila) с Salix krylovii,
Betula divaricata, B. exilis, Rhododendron aureum с участием субальпийских
лугов (рис. 12.39). Травяно-кустарничковый ярус образован Vaccinium vitis-
idaea, Sorbaria pallassii, Silene chamarensis, Patrinia sibirica, Minuartia arctica.
Проективное покрытие мхов и лишайников составляет 10–15 %.
Начиная с высоты 1800–2000 м, простирается пояс горных тундр
(рис. 12.40). Между камнями единично присутствуют цветковые растения
Hierochloe alpina, Silene chamarensis, Luzula sibirica. В ложбинах, связанных с
водотоками, встречаются пятнами Rhodiola rosea, Saxifraga nelsoniana. Скали-
стые вершины гор представлены гольцами.
12.5. РАСТИТЕЛЬНОСТЬ ГОР СЕВЕРО-ВОСТОКА СИБИРИ
Обширная территория, лежащая к востоку от нижнего течения Лены, се-
вернее низовьев Алдана и ограниченная на востоке горными массивами Тихо-
океанского водораздела, образует горную страну Северо-Восточной Сибири.
Это – страна молодых и древних структур, выраженных горными системами,
кряжами, нагорьями, плоскогорьями, приморскими и межгорными равнина-
ми (рис. 12.41). Низменности здесь занимают немногим более 20 % площади.
Важнейшие орографические элементы – горные системы окраинных хребтов
Верхоянского и Колымского нагорья – образуют выпуклую к югу дугу длиной
4000 км. Наиболее высокие горы расположены на юге. Средняя высота их –
1500–2000 м, однако многие вершины поднимаются выше 2300–2800 м, а
самая высокая из них – гора Победа – достигает 3003 м. Характерны альпий-
ские формы рельефа: горные пики, крутые скалистые склоны, глубокие реч-
ные долины, к верховьям которых приурочены фирновые поля и ледники.
Снеговая линия лежит здесь высоко – на отметках от 2100 до 2600 м, что объ-
ясняется преобладанием даже на этих высотах континентального климата.
Большинство ледников занимает склоны северной, северо-западной и северо-
восточной экспозиции. В северной половине страны горные хребты ниже и
многие из них протягиваются в близком к меридианальному направлению.
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
325
Рис. 12.41. Орографическая схема Северо-Восточной Сибири
(по Н. А. Гвоздецкому, Н. И. Михайлову)
Климат страны суровый, резко континентальный, зимой формирующий-
ся под влиянием Азиатского антициклона. Летом влияние Тихоокеанского
муссона ослаблено горными барьерами. Субмеридианальное простирание
горных хребтов определяет свободное проникновение холодных арктических
масс воздуха с севера. Средняя температура января варьирует от –32 до
– 48 °С; бывают морозы до –60÷–68 °С. Средняя температура июля на севере
составляет 3,8 °С, на юге 14 °С, местами 19 °С; иногда жара достигает 30–
36 °С. Годовая сумма осадков составляет от 200 мм на севере до 400 мм на юге,
увеличиваясь в горах до 600 мм. Осадки выпадают в основном летом и осенью.
Зима малоснежная. Страна Северо-Восточной Сибири расположена в аркти-
ческом, субарктическом и умеренном климатических поясах.
Суровый климат страны обусловливает интенсивное промерзание гор-
ных пород и сплошное распространение многолетней мерзлоты, оказываю-
щей существенное влияние на формирование ландшафтов. Северо-Восточная
Сибирь выделяется большой мощностью многолетней мерзлоты, которая в
большинстве горных областей составляет от 200 до 400 м.
Растительности гор свойственна высотная поясность, характер которой
меняется с севера на юг (рис. 12.42). В пределах Верхоянской горной системы
нижние части склонов, на севере – до высоты 200–300 м, на юге – до 1000–
1100 м, заняты редкостойными лесами из лиственницы Каяндера (Larix
cajanderi). Под разреженным покровом невысоких (до 10–15 м) лиственниц
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
326
Рис. 12.42. Высотная поясность гор Северо-Востока Сибири
(по Ю. П. Пармузину)
располагаются заросли низкорослых кустарников: березы (тощей – Betula
exilis, приземистой – B. humilis и растопыренной – B. divaricata), ольховника
(Duschekia fruticosa), можжевельника (Juniperus sibirica), рододендронов
(Rhododendron parvifolium и R. adamsii), различных ив (Salix bebbiana,
S. glauca, S. lanata), или почва покрыта почти сплошным ковром зеленых мхов
и кустистых лишайников – кладоний и цетрарий. Под лесами преобладают
своеобразные горные таежно-мерзлотные почвы с кислой реакцией и без ясно
выраженных генетических горизонтов (за исключением гумусового). Особенно-
сти этих почв связаны с неглубоким залеганием мерзлоты, низкими темпера-
турами, слабым испарением. Летом такие почвы испытывают временное пере-
увлажнение, что обусловливает слабую их аэрацию и появление признаков
оглеения. В нижней части горно-таежного пояса на западном и южном скло-
нах окраинных хребтов изредка встречаются ель (Picea obovata) и сосна (Pinus
sylvestris); в долинах рек есть леса из тополя душистого (Populus suaveolens) и
кореянки (Chosenia arbutifolia); из кустарников – ива удская, аляскинская,
Шверина, красивая (Salix udensis, S. alaxensis, S. schwerinii, S. pulchra).
В верховьях Яны и Индигирки, на склонах южной экспозиции и на дне
некоторых долин встречаются участки лугово-степной и степной растительно-
сти на горных мерзлотно-таежно-степных почвах легкого механического со-
става. В травостое здесь преобладают злаки – типчак (Festuca lenensis), тонко-
ног (Koeleria gracilis), овсец (Helictotrichon krylovii), мятлик (Poa attenuata),
ковыли, осоки (Carex duriuscula, C. korshinskyi), кобрезия (Kobresia filifolia) –
и довольно богатое разнотравье из полыни (Artemisia commutata), вероники
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
327
(Veronica incana), прострела (Pulsatilla orientali-sibirica), лапчатки (Potentilla
nivea), зонтичных и т. д.
Выше верхней границы древесной растительности располагается неши-
рокий пояс низкорослых кустарников (Betula exilis. B. divaricata, Duschekia
fruticosa, Rhododendron parvifolium, Ledum decumbens); на западе преиму-
щественно из ольховника и на востоке – из кедрового стланика. Еще выше
лежит зона горных тундр, представленная лишайниковыми, кустарничко-
выми тундрами и кобрезиевниками (Dryas punctata, Cassiope tetragona, Carex
arctisibirica, C. concolor, Kobresia myosuroides). Наиболее высокие вершины
входят в зону гольцов, где преобладают накипные лишайники, изредка встре-
чаются цветковые растения (Cassiope ericoides, Luzula confusa, Artemisia
lagocephala, Salix berberifolia).
На юге страны, в пределах Юдомо-Майского нагорья значительная пло-
щадь занята тайгой (Larix gmelinii, L. sibirica, Pinus sibirica, Picea obovata) на
горно-таежных карбонатных почвах. Пониженные приречные участки покры-
ты среднетаежными кустарничковыми и травяно-кустарничковыми листвен-
ничными лесами борового типа, багульниково-моховыми лиственничниками
и сосновыми борами. Выше 450–500 м эти леса сменяются среднегорной тай-
гой, доходящей до 900–1000 м. Здесь наиболее обычны зеленомошные и
сфагновые лиственничники с подлеском из ерников. Местами встречается ель
аянская (Picea ajanensis) и береза шерстистая (Betula lanata). Наиболее вы-
сокие участки пояса заняты подгольцовыми редкостойными лиственничны-
ми лесами с почти сплошным мохово-лишайниковым покровом. Выше рас-
положен пояс кустарников (Pinus pumila, Betula divaricata, Rhododendron
aureum), сменяющийся гольцово-тундровым поясом с преобладанием лишай-
никовых и кустарничковых сообществ (Dryas punctata, Loiseleuria procumbens,
Cassiope tetragona, Diapensia obovata, Phyllodoce caerulea, Rhododendron
redowskianum).
12.6. РАСТИТЕЛЬНОСТЬ КАМЧАТКИ
Полуостров Камчатка на западе омывается водами Охотского моря, на
востоке – водами Тихого океана и Берингова моря. По строению поверхности
Камчатка характеризуется преобладанием гор. По северо-восточной оси по-
средине полуострова протягивается широкой полосой система Срединного
хребта, наивысшие вершины которой имеют высокогорный характер и дости-
гают высоты более 3500 м (наивысшая точка – вулкан Ичишский, 3621 м). По
западному побережью Камчатки простирается наклонная Западно-Камчат-
ская равнина с высотами до 300 м. Значительно меньшая по площади Цен-
трально-Камчатская межгорная равнина расположена восточнее Срединного
хребта. Восточный борт равнины образован средневысотным Восточным
хребтом и высокогорной Восточно-Камчатской вулканической горной груп-
пой. Всего на Камчатке насчитывается более 300 вулканов, среди них к актив-
ным и потенциально активным вулканам относят от 28 до 36.
Характерной особенностью климата района является сравнительно рав-
номерное распределение осадков по сезонам и многоснежная зима, что объ-
ясняется близостью этой территории к зимним путям циклонов. Благодаря
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
328
частому проникновению океанического воздуха в пределы района зима здесь
относительно теплая, а лето прохладное.
Почвы Камчатки весьма специфичны, что обусловлено сочетанием ряда
факторов почвообразования, не встречающихся в других частях материка: хо-
лодным, избыточно влажным в приморских районах климатом, разнообрази-
ем рельефа, особенностями растительности. Своеобразие почвенного покрова
Камчатки тесно связано с современным вулканизмом: на территории полу-
острова преобладают вулканические почвы (андосоли). Отдел вулканических
почв представлен несколькими типами: охристые, перегнойно-охристые,
охристо-подзолистые, агроохристые. Их строение отличается своеобразной
«этажностью» – залеганием под современным серии горизонтов, погребенных
вулканическим пеплом.
Значительные площади на Западно-Камчатской равнине заняты болот-
ными почвами. Особо следует отметить наличие прослоев вулканических пеп-
лов в торфяных почвах и смены сфагновых верховых торфов, перекрытых
пеплом, вышележащими осоковыми и гипновыми торфами.
Характер растительного покрова по-
луострова Камчатка определяется его гео-
графическим положением и гористым ре-
льефом. В зональном отношении расти-
тельность равнин относится к северной и
средней тайге. В горах выражена высот-
ная поясность, характер которой меняет-
ся в зависимости от экспозиции склонов.
Характер растительного покрова отобра-
жен на карте (рис. 12.43).
Растительный мир Камчатки насчи-
тывает около 1200 видов сосудистых рас-
тений. Дендрофлора региона насчитывает
более 100 видов деревьев, кустарников, по-
лукустарников, кустарничков, лиан и про-
чих растений с деревенеющими стеблями,
что составляет примерно 8 % от флоры со-
судистых растений региона. Из деревьев
самая распространенная порода – береза
Эрмана или каменная (Betula ermanii), об-
разующая редкостойные леса (рис. 12.44)
по всему краю и доходящая на севере до юга
Корякского нагорья. В оптимальных усло-
виях береза каменная – довольно крупное
дерево до 15–20 м высотой и 90 см
Рис. 12.43. Карта растительности
п-ова Камчатки (по И. А. Соколову):
1 – хвойные леса; 2 – белоберезовые леса (из
Betula platyphylla); 3 – каменноберезовые леса; 4
– стланики; 5 – тундры; 6 – болота
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
329
Рис. 12.44. Береза каменная – Betula
ermanii (фото Н. Терехиной)
Рис. 12.45. Кедровый стланик,
иван-чай и ольховник на склоне
вулкана Шивелуч (фото Н. Терехиной)
в диаметре. Однако на океанском побережье, у верхней границы леса и на се-
вере полуострова вследствие неблагоприятных климатических факторов ее
ствол сильно искривлен и редко достигает даже 10 м высоты.
В южных и восточных районах Камчатки наиболее широко представле-
ны сообщества высокотравных и кустарниково-разнотравных каменноберез-
няков. В северо-восточных районах полуострова распространены вейниковые
и низкотравные (папоротниково-дереновые) каменноберезняки, а также ка-
менноберезняки с подлеском из кедрового стланика. В северо-западных райо-
нах и на верхнем пределе распространения каменноберезовых лесов в горах
преобладают низкотравные и вейниково-папоротниковые каменноберезняки.
Крупные хвойные деревья: лиственница камчатская (Larix kamtschatica)
и ель аянская (Picea ajanensis) образуют леса преимущественно в долине реки
Камчатки. Эндемик полуострова – пихта грациозная (Abies gracilis) встречается
только в приустьевой части реки Семячик на восточном побережье Камчатки.
Тополь душистый (Populus suaveolens), осина, ольха волосистая (Alnus
hirsuta), кореянка (Chosenia arbutifolia), ива удская (Salix udensis) растут в ос-
новном в поймах рек. В центральных и южных районах Камчатки в подлеске
встречаются черемуха азиатская (Padus asiatica), боярышник зеленомясый
(Crataegus chlorosarca), бузина камчатская (Sambucus kamtschatica), рябина
сибирская (Sorbus sibirica), ива козья и др. По склонам гор и на плато широко
распространены кедровый стланик (Pinus pumila) и ольховник камчатский
(Duschekia kamtschatica), образующие на огромных площадях своеобразные
стланиковые заросли (рис. 12.45).
Для Камчатки, особенно для прибрежных территорий, характерно высо-
котравье, достигающее высоты 3–4 м, в состав которых входят: борщевик шер-
стистый (Heracleum lanatum), лабазник камчатский (Filipendula camtschatica),
дудник медвежий (Angelica ursina), дудник преломленный (Angelica genuflexa),
дудник Гмелина (Angelica gmelinii), ребросемянник уральский (Pleurospermum
uralense),крестовник коноплеволистный (Senecio cannabifolius), какалия кам-
чатская (Cacalia kamtschatica) и др. Черемша или медвежий чеснок (Allium
ochotense) – типичное камчатское растение.
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
330
Западно-Камчатская лугово-лесистая заболоченная равнина в южной
части представляет совершенно плоскую, сплошь заболоченную приморскую
низменность. Соседство с Охотским морем, хотя и замерзающим, обусловли-
вает сравнительно теплую зиму. Только во второй половине января и в первой
половине февраля, да и то на севере, суточные температуры воздуха держатся
ниже –15 °C. Лето прохладное и дождливое; лишь в июле и августе суточные
температуры воздуха превышают 10 °C.
На низкой равнине преобладают сфагновые плащеобразные болота, ко-
торые практически не зависят от форм рельефа благодаря интенсивным про-
цессам торфонакопления. Эти болота безлесные, с хорошо развитыми грядо-
во-мочажинными комплексами. На грядах, кочках, буграх растут сфагновые
мхи (Sphagnum fuscum, Sph. magellanicum), болотные кустарнички (багуль-
ник, водяника), береза растопыренная (Betula divaricata); в мочажинах –
Sphagnum lindbergii, Sph. papillosum, осока топяная (Carex limosa). На сухих
грядах обычны лишайники (виды рода Cladonia)
На полого-увалистой и холмисто-увалистой равнине произрастают пар-
ковые леса из березы каменной со злаково-папоротниковым и высокотрав-
ным покровом. По долинам многочисленных рек распространены чозениево-
тополевые леса, высокотравные и разнотравные луга. Участки низких гор за-
няты стелющимися лесами кедрового стланика и ольховника. В северной ча-
сти равнины у побережья развиты кедровниково-лишайниковые лесотундры
и лишайниковые тундры; плоские низменности по долинам рек покрыты осо-
ково-сфагновыми и бугристыми болотами. Почвы дерновые, болотные и
тундровые глееватые.
Горно-тундрово-лесистый средневысотный Срединный хребет занимает
центральную часть Камчатки, простираясь в северо-восточном направлении.
Горный рельеф обусловливает относительно более низкие зимние температуры
по сравнению с побережьем: весь январь суточные температуры держатся здесь
ниже –20 °C. Лето, однако, не теплее, чем на побережье, что связано с высотой
местности. В июле и августе суточные температуры держатся выше 10 °C. Здесь
несколько суше, чем на побережье. Почвы горно-дерновые и дерновые; по севе-
ро-восточному склону – горно-тундровые торфянисто-перегнойные.
В южной половине Срединного хребта ниже гольцового пояса верхний
предел растительности образуют горные тундры и альпийские луга с рододенд-
роном. Ниже господствуют стелющиеся леса из ольховника и особенно кедро-
вого стланика с отдельными участками высокотравных лугов. Еще ниже по
склонам распространены парковые каменноберезовые леса в комплексе с за-
рослями лабазника и ивово-тополевыми лесами с чозенией по долинам рек,
поднимающимися высоко в горы. В северной половине Срединного хребта вы-
сотная поясность растительности иная. В верхнем поясе растительность пред-
ставлена лишайниковыми и мохово-лишайниковыми тундрами с участками
каменных россыпей. Пояс стелющихся лесов кедрового стланика и ольховника
значительно шире, чем на юге; каменноберезовые леса сохраняются лишь по
долинам.
Срединный хребет отделен от восточных гор Центрально-Камчатской
межгорной депрессией, вытянутой также в северо-восточном направлении.
Она представляет собой лесистую равнину. По дну долины протекает самая
крупная река полуострова – Камчатка, текущая на север, южная часть долины
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
331
дренируется р. Быстрой, текущей на юг. Депрессия относится к средней тайге,
в растительном покрове которой преобладают хвойные леса из лиственницы и
ели. Ельники зеленомошники в наибольшей степени соответствуют понятию
зонального сообщества для лесного пояса Центральной Камчатки. Сопутствую-
щими видами древесных пород являются береза каменная и рябина сибирская
(Sorbus sibirica); встречаются отдельные экземпляры лиственницы Каяндера
(Larix cajanderi). В подлеске обычны жимолости (Lonicera edulis, L. chamissoi),
рябина бузинолистная (Sorbus sambucifola), кедровый стланик (Pinus pumila),
иногда можжевельник (Juniperus sibirica), единично встречаются спирея Бо-
вера (Spiraea beauverdiana) и шиповник иглистый (Rosa acicularis). Харак-
терно присутствие лианы – княжика охотского (Atragene ochotensis). Травяно-
кустарничковый ярус представлен двумя подъярусами: для первого характерны
хвощи (Equisetum pratense, E. arvense, E. hyemale, E. sylvaticum), вейник
Лангсдорфа (Calamagrostis langsdorffii), иван-чай узколистный (Chamaenerion
angustifolium); для второго подъяруса – осока бледная (Carex pallida) и группа
бореальных видов таежного низкотравья (Linnaea borealis, Orthilia secunda,
Lycopodium annotinum, Diphasiastrum complanatum, Moneses uniflora, Pyrola
incarnata, P. minor, Gymnocarpium dryopteris, Trientalis eurpaea, Maianthemum
bifolium, Goodyera repens). В мохово-лишайниковом ярусе, проективное покры-
тие которого местами достигает 70–80 %, доминируют Pleurozium schreberi,
Polytrichum commune, Dicranum majus. Кроме того, часто встречаются ельни-
ки низкотравные, хвощевые, разнотравно-зеленомошные; на склонах гор –
ельники кустарниково-разнотравные.
В Центральной долине Камчатки широко распространены группы ассоци-
аций из Larix cajanderi: лиственничники кустарниково-разнотравные, листвен-
ничники багульниковые и лиственничники кедровостланиковые в сочетании с
кустарничково-сфагновыми листвен-
ничными марями. Встречаются также
лиственничники лишайниковые, хво-
щевые, зеленомошные. В образовании
высотного ряда принимают участие со-
общества субальпийских лиственнич-
ных редколесий, которые местами под-
нимаются до высоты 1150–1200 м.
На месте лиственничников и ель-
ников довольно обширные площади
занимают белоберезняки (рис. 12.46).
Особенно много их в южной части Цен-
тральной долины Камчатки. Белобе-
резняки и каменноберезняки имеют
здесь сходный подлесок из ив, боя-
рышника, шиповника, жимолости и
густой травяной покров. В почвенном
покрове преобладают охристые почвы.
Рис. 12.46. Белоберезняк из Betula
platyphylla (фото Н. Терехиной)
.
Часть III. РАСТИТЕЛЬНОСТЬ ГОРНЫХ СТРАН
332
На низких террасах р. Камчатки и на озерных равнинах встречаются сфагно-
вые лиственничные редколесья и открытые сфагновые болота.
На Восточном хребте и Восточно-Камчатской вулканической горной груп-
пе четко выражена высотная поясность. Высокогорные территории покрыты
скудной растительностью. Здесь преобладают накипные лишайники, встречают-
ся участки лавы и пепла, лишенные растительности. Горные тундры представ-
лены лишайниками и верещатниками (рис. 12.47) из голубики (Vaccinium uligi-
nosum), брусники (Vaccinium vitis-idaea var. minus), водяники (Empetrum
nigrum), арктоуса (Arctous alpina), филлодоци (Phyllodoce caerulea, Ph. aleutica),
кассиопеи плауновидной (Cassiope lycopodioides), мохоцветника гмелини
(Bryanthus gmelinii), дриады точечной (Dryas punctata), диапенсии обратно-
яйцевидной (Diapensia obovata), рододендронов (Rhododendron aureum, Rh.
camtschaticum) и жимолости (Lonicera caerulea). Плоские вершины заняты
яркими альпийскими лугами. Средние части склонов вулканов и высоких гор
покрыты зарослями кедрового стланика и ольховника, которые образуют ком-
плекс с лугами субальпийского типа, обычными видами для которых являются
иван-чай (Chamaenerion angustifolium), соссюрея ложно-тилезиева (Saussurea
pseudotilesii), змеевик живородящий (Bistorta vivipara), хвощи лесной и луговой
(Equisetum sylvaticum, E. pratense), герань пушистоцветковая (Geranium erian-
thum), лук охотский (Allium ochotense), мытник перевернутый (Pedicularis resupi-
nata), чемерица остродольная (Veratrum oxysepalum) и др. Ширина этого пояса
составляет 300–400 м. На склонах складчатых хребтов и в краевой зоне вулка-
нических плато развиты леса из березы каменной (верхняя граница березняков
обычно не превышает 900–960 м), в широких долинах на высоких террасах –
парковые березняки, на более низких террасах – высокотравные луга, а на пой-
мах – ивово-тополевые леса с зарослями гигантского лабазника камчатского
(Filipendula camtschatica). В предгорьях вулкана Шивелуч сохранились участки
старовозрастных ельников из Picea ajanensis (рис. 12.48). Вулканогенные нару-
шения в районах активного вулканизма выражаются в уничтожении горно-
таежного пояса хвойных лесов (на восточном макросклоне Ключевской группы
вулканов). Здесь вместо типичного горно-таежного пояса выражен производный
пояс, образованный длительнопроизводными белоберезняками (Betula platy-
phylla), возникшими на месте уничтоженных еловых и лиственничных лесов.
Рис. 12.47. Верещатник на склоне
вулкана Шивелуч (фото Н. Терехиной)
Рис. 12.48. Старовозрастный еловый
лес у подножия вулкана Шивелуч
(фото Н. Терехиной)
.
Глава 12. РАСТИТЕЛЬНОСТЬ ОСНОВНЫХ ГОРНЫХ СТРАН РОССИИ
333
Западное и Восточное побережья полуострова Камчатка относятся к се-
веротаежной подзоне. Здесь преобладают каменноберезняки (Betula ermanii),
большие площади заняты кедровыми (Pinus pumila) и ольховыми (Duschekia
fruticosa) стланиками, образующими группировки, распространенные от бере-
га моря до горных тундр и альпийских лугов в горах. Обычны болота грядово-
мочажинные осоково-кустарничково-гипново-сфагновые (аапа), и своеобраз-
ные «верещатники», в составе которых преобладают стелющиеся по земле ку-
старнички, особенно водяника.
Вопросы к главе 12
1. Природные условия и особенности высотной поясности Урала.
2. Природные условия и особенности высотной поясности Кавказа.
3. Природные условия и особенности высотной поясности Алтая.
4. Природные условия и особенности высотной поясности гор Восточного Прибайкалья.
5. Природные условия и особенности высотной поясности гор Северо-Востока Сибири.
6. Природные условия и особенности высотной поясности гор Камчатки.
Рекомендуемая литература
Алтайский край: Атлас. М.: Барнаул, 1978. Т. 1.
Байкал. Атлас. М.: РАН Сиб. отд. Федеральная служба геодезии и картографии России, 1993.
Гулисашвили В. З. и др. Растительность Кавказа. М., 1975. 233 с.
Заповедники России. Заповедники Сибири. Т. 1. М.: Логата, 1999. 304 с.
Заповедники СССР. Заповедники Дальнего Востока. М.: Мысль, 1985. 319 с.
Заповедники СССР. Заповедники европейской части РСФСР. Ч. I. М.: Мысль, 1988. 288 с.
Заповедники СССР. Заповедники Кавказа. М., 1990. 365 с.
Карпачевский Л. О., Алабина И. О., Захарихинa Л. В., Макеев Л. О., Маречек М. С., Ра-
дюкин Л. О., Шоба С. А. Почвы Камчатки. М.: ГЕОС, 2009. 224 с.
Конспект флоры Кавказа. Т. 1. СПб., 2003.
Конспект флоры Кавказа. Т. 2. СПб., 2006.
Коропачинский И. Ю. Дендрофлора Алтайско-Саянской горной области. Новосибирск:
Наука, 1975. 290 с.
Нешатаева В. Ю. Растительность полуострова Камчатка. М.: Товарищество научных изда-
ний КМК, 2009. 537 с.
Огуреева Г. Н. Ботаническая география Алтая. М.: Наука, 1980. 192 с.
Флора и растительность Южно-Уральского государственного природного заповедника //
Кол. авторов. Под ред. Б. М. Миркина. Уфа: Гилем, 2008. 528 с.
Шумилова Л. В. Ботаническая география Сибири. Томск: изд-во Томского ун-та, 1962. 439 с.
Байкальский биосферный резерват http://oopt.info/index.php?oopt=204
Баргузинский биосферный резерват http://oopt.info/index.php?oopt=212
Висимский биосферный резерват http://oopt.info/index.php?oopt=142
Джергинский заповедник http://oopt.info/index.php?oopt=244
Забайкальский национальный парк http://oopt.info/index.php?oopt=349
Заповедник Денежкин камень http://oopt.info/index.php?oopt=452
Кабардино-Балкарский заповедник http://oopt.info/index.php?oopt=924
Кавказский биосферный резерват http://oopt.info/index.php?oopt=933
Магаданский заповедник http://oopt.info/index.php?oopt=1292
Национальный парк Зюраткуль http://oopt.info/index.php?oopt=500
Национальный парк Приэльбрусье http://oopt.info/index.php?oopt=975
Олекминский заповедник http://oopt.info/index.php?oopt=1301
Северо-Осетинский заповедник http://oopt.info/index.php?oopt=944
Тебердинский биосферный резерват http://oopt.info/index.php?oopt=952
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
334
УКАЗАТЕЛЬ РУССКИХ НАИМЕНОВАНИЙ
Абелия корейская (Zabelia coreana) 191*
Абрикос маньчжурский (Armeniaca mands-
hurica) 187
– сибирский (Armeniaca sibirica) 187
Аденостилес (Adenostyles) 302
Адиантум стоповидный (Adiantum pedatum)
182, 244
Адонис весенний (Adonis vernalis) 231–233
Акация серебристая (Acacia dealbata) 287
Акомастилис
ледяной,
(Acomastylis
glacialis) 102
Аконит (Aconitum) 302
Актинидия (Actinidia) 177, 244
– коломикта (Actinidia kolomikta) 182, 186,
189, 193
– многодомная (Actinidia polygama) 177
– острая (Actinidia arguta) 182, 193
Альбиция шелковая (Albizia julibrissin) 286
Аралия высокая (Aralia elata) 180, 182, 244
Арктогерон злаковый (Arctogeron gramine-
um) 263
Арктополевица тростниковидная (Arctagro-
stis arundinacea) 109
Арктоус (Arctous) 86, 91, 95, 332
– альпийский (Arctous alpina) 109, 145
Арктофила рыжеватая (Arctophila fulva) 104
Арника Ильина (Arnica iljinii) 103
– фенноскандинавская (Arnica fennoscandi-
ca) 140
Астрагал (Astragalus) 81
– альпийский (Astragalus alpinus) 103
– волосистоязычковый (Astragalus lasioglot-
tis) 307, 308
– Гельма (Astragalus helmii) 291
– заячий (Astragalus laguroides) 263
– зонтичный (Astragalus umbellatus) 103
– холодный (Astragalus frigidus) 103
Астраканта (Astracantha) 280
– золотистая (Astracantha aurea) 309
– кавказская (Astracantha caucasica) 309
– обнаженная (Astracantha denudate) 308,
309
Асфоделина (Asphodeline) 280
* Латинские и русские наименования рас-
тений приводятся в соответствии с названиями,
принятыми в интернет-атласе видов «Планта-
риум» (http://www.plantarium.ru/).
Багульник (Ledum) 81, 89, 142, 187, 330
– болотный (Ledum palustre) 81, 213, 214
– подбел (Ledum hypoleucum) 191
– стелющийся (Ledum decumbens) 81, 109,
171
Бадан толстолистный (Bergenia crassifolia)
322
Банан (Musa) 287
Барбарис (Berberis) 283, 306
– амурский (Berberis amurensis) 180
– обыкновенный (Berberis vulgaris) 235
Бартсия альпийская (Bartsia alpine) 145
Бархат (Phellodendron) 193
– амурский (Phellodendron amurense) 182,
183, 188, 193, 245
– сахалинский (Phellodendron sachalinense)
177
Бассия (Bassia) 268
Бедренец (Pimpinella) 200
– розовоцветный (Pimpinella rhodantha) 301
– камнеломка (Pimpinella saxifraga) 200
Бекмания (Beckmannia) 267
Белокопытник широкий (Petasites amplus)
178, 179
Белоус (Nardus) 28, 208
– торчащий (Nardus stricta) 207
Берёза (Betula) 9, 90, 97, 98, 144, 145, 147,
215, 238, 299, 302
– даурская (Betula davurica) 190, 244
– жёлтая (Betula costata) 190, 193
– каменная (Betula ermanii) 190, 328–332
– карликовая (Betula nana) 78, 80, 89, 98
– круглолистная (Betula rotundifolia) 316
– Литвинова (Betula litwinowii) 303, 307
– маньчжурская (Betula mandshurica) 189
– плосколистная (Betula platyphylla) 173,
323, 324
– повислая (Betula pendula) 140, 141, 158,
163, 235, 242, 307
– приземистая (Betula humilis) 325
– пушистая (Betula pubescens) 140, 141, 144,
158, 163, 165, 235
– Радде (Betula raddeana) 306
– растопыренная (Betula divaricata) 80, 326,
330
– ребристая (Betula costata) 180, 184, 190,
244
– тощая (Betula exilis) 80, 81, 109, 171, 326
– Черепанова (Betula czerepanovii) 86, 144
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ*
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
335
– шерстистая (Betula lanata) 188, 191, 327
– Эрмана (Betula ermanii) 328, 332
Бересклет (Euonymus) 228, 233, 244, 299,
302, 306, 307
– большекрылый (Euonymus macroptera)
185, 193
– бородавчатый (Euonymus verrucosa) 151,
224, 233, 235, 304, 306
– европейский (Euonymus europaea) 224,
227, 233, 304
– малоцветковый (Euonymus pauciflora)
190, 191
– священный (Euonymus sacrosancta) 190
Бобовник (Laburnum) 228
Бодяк камчатский (Cirsium kamtschaticum)
179
– полевой (Cirsium arvense) 200
Болотный мирт (Chamaedaphne calyculata)
157, 214
Борец Радде (Aconitum raddeanum) 182
Бородач (Bothriochloa) 278, 309
Бородник шароносный (Jovibarba globifera)
153
Борщевик (Heracleum) 302
– Сосновского (Heracleum sosnowskyi) 302
– шерстистый (Heracleum lanatum) 115,
178, 179, 330
Боярышник (Crataegus) 163, 224, 228, 279,
299, 301, 304, 320, 321, 331
– вееролистный (Crataegus rhipidophylla)
304
– восточный (Crataegus orientalis) 306
– зеленомясый (Crataegus chlorosarca) 329
– кроваво-красный (Crataegus sanguinea) 235
– перистонадрезанный (Crataegus pinnati-
fida) 245
– пятипестичный (Crataegus pentagyna)
304
– согнутостолбиковый (Crataegus rhipido-
phylla) 306
Брусника (Vaccinium) 77, 81, 86, 89 95, 99,
101, 142, 144 155, 157–161, 171, 173,
190, 300, 322, 323
– малая (Vaccinium vitis-idaea var. minus)
109
Будра плющевидная (Glechoma hederacea)
227
Бузина камчатская (Sambucus kamtschatica)
329
– кистевидная (Sambucus racemosa) 235
– черная (Sambucus nigra) 299, 302, 306, 307
Бузульник сибирский (Ligularia sibirica) 153
Бук (Fagus) 9, 125, 128, 138, 224, 302
– восточный (Fagus orientalis) 299, 302, 306,
307
– лесной (Fagus sylvatica) 306
Буквица крупноцветковая (Betonica macran-
tha) 300, 306
– лекарственная (Betonica officinalis) 210
Бурачок Гмелина (Alyssum gmelinii) 239,
240
– ленский (Alyssum lenense) 263
Василек (Centaurea) 300
– синий (Centaurea cyanus) 200
– фригийский (Centaurea phrygia) 208, 209,
210
– шероховатый (Centaurea scabiosa) 233
Василистник альпийский (Thalictrum al-
pinum) 89
– нитчатый (Thalictrum filamentosum) 182,
193
Вейгела (Weigela) 182
– Миддендорфа (Weigela middendorffiana)
191
Вейник (Calamagrostis) 163, 164, 199, 271,
3010
– Лангсдорфа (Calamagrostis langsdorffii)
160, 330
– наземный (Calamagrostis epigeios) 173,
234, 238
– притупленный (Calamagrostis obtusata)
160
– пурпурный ( Calamagrostis purpurea) 115
– тростниковидный (Calamagrostis phrag-
mitoides) 162
– тростниковый (Calamagrostis arundina-
cea) 143
Венерин башмачок крупноцветковый (Cyp-
ripedium macranthon) 114
– башмачок настоящий (Cypripedium cal-
ceolus) 145, 153, 229
Вербейник обыкновенный (Lysimachia vul-
garis) 209, 210
Верблюдка (Corispermum) 271
– повислая (Corispermum declinatum) 252
Вереск (Calluna) 86, 142
– обыкновенный (Calluna vulgaris) 213, 214
Веретенник
яйцевидный (Atractylodes
ovata) 244
Вероника (Veronica) 301, 326
– дубравная (Veronica chamaedrys) 143
– лекарственная (Veronica officinalis) 244
– седая (Veronica incana) 264
Ветреник нарциссоцветковый (Anemonas-
trum narcissiflorum) 117, 300
Ветреница дубравная (Anemone nemorosa)
224, 225
– лесная (Anemone sylvestris) 145, 232
Виноград (Vitis) 182, 284
– амурский (Vitis amurensis) 182, 189, 190,
244
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
336
– Изабелла (Vitis labrusca) 284
– лесной (Vitis sylvestris) 304, 306
Вишня кустарниковая (Cerasus fruticosa)
235, 236, 294
– Максимовича (Cerasus maximowiczii) 180,
190
– птичья (Cerasus avium) 299
– сахалинская (Cerasus sachalinensis) 182
– степная (Cerasus fruticosa) 228, 233
Водяника (Empetrum) 77, 81, 86, 89, 118,
140, 144, 157, 330, 332, 333
– сибирская (Empetrum sibiricum) 117
– черная (Empetrum nigrum) 109
Волжанка двудомная (Aruncus dioicus) 182
Володушка козелецелистная (Bupleurum
scorzonerifolium) 264
– многожилковая (Bupleurum multinerve) 229
Волоснец (Leymus) 257
– кистистый (Leymus racemosus) 271
Волчеягодник (Daphne) 151, 224, 235, 284
– боровой (Daphne cneorum) 229
Воронец красноплодный (Actaea erythro-
carpa) 144, 151
Вороний глаз четырехлистный (Paris quad-
rifolia) 144
Вьюнок (Convolvulus) 309
Вяз (Ulmus) 17, 147, 150, 151, 187, 233, 284
– гладкий (Ulmus laevis) 151, 155, 235
– голый (Ulmus glabra) 235
– малый (Ulmus minor) 297, 299, 304, 307
– разрезной (Ulmus laciniata) 180, 188, 190,
193
– японский (Ulmus japonica) 180, 188, 193,
245
Гариманелла моховидная (Harrimanella
hypnoides) 141
Гвоздика иглолистная (Dianthus acicularis)
291
Гелиотроп (Heliotropium) 309
Герань лесная (Geranium sylvaticum) 206,
208, 210
– Максимовича (Geranium maximowiczii)
244
– пушистоцветковая (Geranium erianthum)
332
– Роберта (Geranium robertianum) 227
–холмовая (Geranium collinum) 28, 29
Гиацинтик пепельно-серый (Hyacinthella
leucophaea) 232
Гилокомиум блестящий (Hylocomium splen-
dens) 159
Гирчовник татарский (Conioselinum tatari-
cum) 145
Глянцелистник Лёзеля (Liparis loeselii) 153,
229
Гнездовка настоящая (Neottia nidus-avis)
153
Гнездоцветка клобучковая (Neottianthe cu-
cullata) 233, 234
Головчатка (Cephalaria) 300
Голокучник обыкновенный (Gymnocarpium
dryopteris) 157
Голубика (Vaccinium) 77, 81, 86, 89, 101,
112, 140, 142, 144, 157, 170, 173, 214,
300, 323, 332
– обыкновенная мелколистная (Vaccinium
uliginosum ssp. microphyllum) 109
Гонкения бутерлаковидная (Honckenya pep-
loides) 189
Горицвет (Coccyganthe flos-cuculi) 210
Горичник горный (Peucedanum oreoselinum)
233
Горноколосник колючий (Orostachys spi-
nosa) 263
Горошек амурский (Vicia amurensis) 244
– заборный (Vicia sepium) 208
– мышиный (Vicia cracca) 208
– тонколистный (Vicia tenuifolia) 233
Граб (Carpinus) 138, 228, 297, 299, 307
– обыкновенный (Carpinus betulus) 304
– сердцелистный (Carpinus cordata) 180,
184, 193, 244
Грабинник (Carpinus orientalis) 278, 282
Гравилат (Geum) 28
– горный (Orogeum montanum) 304
– речной (Geum rivale) 208, 209, 210
Гречиха сахалинская (Reynoutria sacha-
linensis) 178, 179
Гроздовник многораздельный (Botrychium
multifidum) 152
– полулунный (Botrychium lunaria) 191
Грудница мохнатая (Galatella villosa) 257
Груша (Pyrus) 228, 233, 299, 307
– кавказская (Pyrus caucasica) 299, 302, 304,
306, 307
– лохолистная (Pyrus elaeagrifolia) 279
– обыкновенная (Pyrus communis) 235
– уссурийская (Pyrus ussuriensis) 180, 245
Грушанка (Pyrola) 323
– японская (Pyrola japonica) 182
Гудайера ползучая (Goodyera repens) 150,
160
Гусиный лук (Gagea) 28, 224
– желтый (Gagea lutea) 225
Двурядник сплюснутый (Diphasiastrum
complanatum) 144
Девясил (Inula) 300
– британский (Inula britannica) 267
Деллингерия
шершавая
(Doellingeria
scabra) 244
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
337
Дереза (Caragana frutex) 257
Держи-дерево (Paliurus) 278, 282, 283, 306,
307
Дёрен (Chamaepericlymenum) 86
– шведский (Chamaepericlymenum sueci-
cum) 19, 21, 118
Диапенсия лапландская (Diapensia lapponi-
ca) 141
– обратнояйцевидная (Diapensia obovata)
109, 332
Дороникум (Doronicum) 302
Древогубец (Celastrus) 182
Дремлик лесной (Epipactis helleborine) 150
– темно-красный (Epipactis atrorubens) 145
Дриада (Dryas) 68, 81, 88, 91, 109
– восьмилепестковая (Dryas octopetala) 141,
145, 293
– точечная (Dryas punctata) 76, 101, 332
Дриада (Dryas) 68, 88
Дрок красильный (Genista tinctoria) 235,
238
– крымский (Genista taurica) 281
Дуб (Quercus) 8, 128, 147, 150, 151, 153, 154,
189, 190,194, 224, 227, 228, 233, 235,
238, 244, 278–280, 307
– Гартвиса (Quercus hartwissiana) 297
– грузинский (Quercus iberica) 284, 306
– зубчатый (Quercus dentata) 244
– имеретинский (Quercus imeretina) 284
– крупнопыльниковый
(Quercus
macranthera) 307
– курчавенький (Quercus crispula) 177
– монгольский (Quercus mongolica) 177,
180, 183, 189, 190, 194, 244
– ножкоцветный (Quercus pedunculiflora)
304
– пушистый (Quercus pubescens) 277–279,
282, 304, 306
– скальный (Quercus petraea) 299, 301, 302,
306
– черешчатый (Quercus robur) 224, 227,
229, 235, 297, 299, 304
Дудник (Angelica) 178
– Гмелина (Angelica gmelinii) 115, 329
– преломленный (Angelica genuflexa) 329
– лесной (Angelica sylvestris) 208
– медвежий (Angelica ursine) 179, 329
Дурнишник (Xanthium) 271
Душистый колосок (Anthoxanthum odora-
tum) 205, 206, 208, 302
Дюпонция Фишера (Dupontia fisheri) 99
Ежа (Dactylis) 164
– сборная (Dactylis glomerata) 208, 232, 304,
306
Ежевика (Rubus) 299
Ель (Picea) 130, 141, 144, 146, 158, 173, 177,
301, 326
– аянская (Picea ajanensis) 136, 176, 180,
183, 187, 188, 190, 193, 194, 327, 329
– восточная (Picea orientalis) 299
– Глена (Picea glehnii) 177
– европейская (Picea abies) 140, 147
– корейская (Picea koraiensis) 180, 193
– сибирская (Picea obovata) 86, 89, 96, 140,
143, 147, 157, 291, 314
– финская (Picea x fennica) 136
Жабрица извилистая (Seseli tortuosum) 256
– порезниковая (Seseli libanotis) 238
Жасмин кустарниковый (Jasminum fruti-
cans) 279
Женьшень (Panax) 244
– обыкновенный (Panax ginseng) 186
Живокость (Delphinium) 302
Живучка хиосская (Ajuga chia) 239, 240
Жимолость (Lonicera) 160, 161, 163, 182,
283, 331, 332
– восточная (Lonicera orientalis) 302, 307
– козья (Lonicera caprifolium) 299
– Максимовича (Lonicera maximoviczii) 185,
193
– обыкновенная (Lonicera xylosteum) 151
– съедобная (Lonicera edulis) 178
– татарская (Lonicera tatarica) 235
Жирянка лопатчатая (Pinguicula spathulata)
111
Житняк (Agropyron) 267, 271
– гребенчатый (Agropyron cristatum) 172,
249, 263, 313
– гребневидный (Agropyron pectinatum) 267
– пустынный (Agropyron desertorum) 267,
272
– сибирский (Agropyron sibiricum) 267
Жостер Палласа (Rhamnus pallasii) 289, 307
– слабительный (Rhamnus cathartica) 233,
304, 306
Жузгун (Calligonum) 271
Заманиха высокая (Oplopanax elatus) 188
Звездовка большая (Astrantia maxima) 300
Звездчатка вильчатая (Stellaria dichotoma)
262
– ланцетолистная (Stellaria holostea) 147, 227
Зеленчук желтый (Galeobdolon luteum) 147,
224
Земляника (Fragaria) 28
– зеленая (Fragaria viridis) 233
Земляничник мелкоплодный (Arbutus andr-
achne) 278, 279
Зимолюбка зонтичная (Chimaphila umbella-
ta) 153, 238
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
338
Змеевик (Bistorta) 301
– большой (Bistorta officinalis) 89, 209, 210
– живородящий (Bistorta vivipara) 88, 117,
332
– мясо-красный (Bistorta carnea) 300
Змеёвка (Cleistogenes) 264
– растопыренная (Cleistogenes squarrosa)
263
Змееголовник иноземный (Dracocephalum
peregrinum) 313
– поникающий (Dracocephalum nutans) 313
– Рюйша (Dracocephalum ruyschiana) 145
Золотарник (Solidago) 322
– обыкновенный (Solidago virgaurea) 144,
160
Золотой корень (Rhodiola rosea) 88
Зопник клубненосный (Phlomoides tuberosa)
263
Ива (Salix) 66, 67, 82, 86, 88, 95, 109, 129,
163, 165, 171, 192, 194, 316, 323, 325,
331
– аляскинская (Salix alaxensis) 326
– белая (Salix alba) 235, 306
– козья (Salix caprea) 160, 229, 233, 235,
302, 307, 329
– красивая (Salix pulchra) 99, 101, 326
– монетолистная (Salix nummularia) 76
– мохнатая (Salix lanata) 78, 96
– ниппонская (Salix nipponica) 194
– пепельная (Salix cinerea) 142
– ползучая (Salix reptans) 89, 99, 101
– полярная (Salix polaris) 67, 76, 102
– понтийская (Salix euxina) 235
– росистая (Salix rorida) 194
– сердцелистная (Salix cardiophylla) 194
– сетчатая (Salix reticulata) 141, 145
– сизая (Salix glauca) 96, 101
– скрытная (Salix abscontida) 194
– тарайкинская (Salix taraikensis) 194
– тонкостолбиковая (Salix gracilistyla) 194
– удская (Salix udensis) 192, 326, 328
– филиколистная (Salix phylicifolia) 96
– Шверина (Salix schwerinii) 192, 194, 326
Иван-чай (Chamaenerion) 68, 199, 329, 332
– узколистный (Chamaenerion angustifo-
lium) 330
Иглица (Ruscus) 279, 284
– колючая (Ruscus aculeatus) 278
– подъязычная (Ruscus hypoglossum) 278
Ильм горный (Ulmus lacinata) 299
Ирга (Amelanchier) 283
Ирис безлистный (Iris aphylla) 231, 232
Калина (Viburnum) 163
– городовина (Viburnum lantana) 228, 299
– обыкновенная (Viburnum opulus) 235, 304,
306
– Саржента (Viburnum Sargentii) 180
Калипсо луковичная (Calypso bulbosa) 145,
191
Калопанакс (Kalopanax) 180, 182
Калужница (Caltha) 68
Камнеломка (Saxifraga) 67, 68, 72, 99
– снежная (Saxifraga nivalis) 70
Камфоросма монпелийская (Camphorosma
monspeliaca) 267
Камыш озерный (Scirpus lacustris) 210
Каперсы (Capparis) 309
– травянистые (Capparis herbacea) 308
Карагана (Caragana) 161, 294
– карликовая (Caragana pygmaea) 263
– кустарниковая (Caragana frutex) 255, 257
Касатик одноцветковый (Iris uniflora) 191, 245
– русский (Iris ruthenica) 173
Кассиопея плауновидная (Cassiope lycopo-
dioides) 117, 331
– четырехгранная (Cassiope tetragona) 76,
101, 109
Качим метельчатый (Gypsophila paniculata)
252
– Патрэна (Gypsophila patrinii) 313
– уральский (Gypsophila uralensis) 145, 290
Каштан (Castanea) 299, 300
Кедровый стланик (Pinus pumila) 78, 80,
112, 136, 170, 182, 188, 190–192, 319,
321–323, 326, 329, 331, 332
Кермек (Limonium) 268
Кермековидка (Limoniopsis) 309
Кизил (Cornus) 279, 283, 299, 301
Кизильник алаунский (Cotoneaster melano-
carpus) 230
– одноцветковый (Cotoneaster uniflorus) 145
Кирказон ломоносовидный (Aristolochia
clematitis) 307
Кислица (Oxalis) 18, 154, 160, 161
– обратнотреугольная (Oxalis obtriangulata)
244
– обыкновенная (Oxalis acetosella) 182
Кисличник (Oxyria) 67
Кладония звездчатая (Cladonia stellaris) 158
– лесная (Cladonia arbuscula) 158
– оленья (Cladonia rangiferina) 158
Клевер (Trifolium) 301, 303
– волосистоголовый (Trifolium trichocep-
halum) 305
– горный (Trifolium montanum) 231–233
– луговой (Trifolium pratense) 28, 29, 200,
205, 208
– средний (Trifolium medium) 208
Клён (Acer) 17, 124, 147, 151, 154, 180, 188,
224, 227, 228
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
339
– американский (Acer negundo) 228
– гирканский (Acer hyrcanum) 306
– желтый (Acer ukurunduense) 190, 191
– зеленокорый (Acer tegmentosum) 190, 191,
193
– ложноплатановый (Acer pseudoplatanus)
284, 285, 299
– маньчжурский (Acer mandshuricum) 180,
184, 193
– мелколистный (Acer mono) 193, 244
– моно (Acer mono) 139, 180, 189, 190, 193,
244
– остролистный (Acer platanoides) 151, 152,
224, 227, 233, 235, 291, 299, 302, 306
– полевой (Acer campestre) 299, 301, 302,
304, 306, 307
– светлый (Acer laetum) 307
– Стевена (Acer stevenii) 277
– татарский (Acer tataricum) 228, 235
– Траутфеттера (Acer trautvetteri) 306
Клюква (Oxycoccus) 18, 77, 214
– болотная (Oxycoccus palustris) 213
– мелкоплодная (Oxycoccus microcarpus)
111
Княжик (Atragene) 150
– охотский (Atragene ochotensis) 330
– сибирский (Atragene sibirica) 145, 151
Кобрезия (Kobresia) 326
Ковыль (Stipa) 228, 230–233, 249, 254, 255,
257, 258, 271, 272, 280, 309
– байкальский (Stipa baicalensis) 263
– волосатик (Stipa capillata) 251, 254, 271
– Залесского (Stipa zalesskii) 233, 261
– Коржинского (Stipa korshinskyi) 261
– красивейший (Stipa pulcherrima) 233
– Крылова (Stipa krylovii) 172, 263, 264
– Лессинга (Stipa lessingiana) 250, 256
– опушеннолистный (Stipa dasyphylla) 232,
233
– перистый (Stipa pennata) 232, 233
– сарептский (Stipa sareptana) 267
– узколистный (Stipa tirsa) 232
– украинский (Stipa ucrainica) 251
Козлобородник (Tragopogon) 232
– коротконосиковый
(Tragopogon
brevirostris) 29
Козлятник (Galega) 302
Колокольчик круглолистный (Campanula
rotundifolia) 205
– персиколистный (Campanula persicifolia)
231
– раскидистый (Campanula patula) 205
– Стевена (Campanula stevenii) 256
– холмовой (Campanula collina) 300
Колосняк (Leymus) 113, 115
– китайский, вострец (Leymus chinensis) 264
– мягкий (Leymus mollis) 189
Копеечник
альпийский (Hedysarum
alpinum) 89
– арктический (Hedysarum arcticum) 103
Копытень европейский (Asarum europaeum)
147, 224, 227, 233
Кореянка (Chosenia) 326, 329
Кореянка земляничниколистная (Chosenia
arbutifolia) 192
Коровяк (Verbascum) 18, 28
Короставник (Knautia) 232
– полевой (Knautia arvensis) 29
Коротконожка (Brachypodium) 164, 301
– лесная (Brachypodium sylvaticum) 304
Кортадерия Селло (Cortaderia selloana) 287
Косоплодник сомнительный (Plagiorhegma
dubia) 182, 244
Кострец безостый (Bromopsis inermis) 234
– береговой (Bromopsis riparia) 232, 234,
255, 280, 309
Костяника (Rubus saxatilis) 28, 143, 161, 238
Кохия простертая (Kochia prostrate) 264
Кочедыжник (Athyrium) 193
– китайский (Athyrium sinense) 244
Кошачья лапка двудомная (Antennaria
dioica) 205, 207, 238
Крапива двудомная (Urtica dioica) 227, 233
– плосколистная (Urtica platyphylla) 178
Красника (Vaccinium praestans) 178
Крестовник (Senecio) 178
– коноплеволистный (Senecio cannabifolius)
115, 329
Криптограмма курчавая (Cryptogramma cri-
spa) 140
Крупка (Draba) 19, 20, 68, 81, 99
– волосистая (Draba pilosa) 76
– моховидная (Draba bryoides) 305
Крушина (Frangula) 142, 151, 298, 307
– ломкая (Frangula alnus) 166, 233, 235, 306
Кубышка малая (Nuphar pumila) 145
Кукушкин лён (Polytrichum commune) 89,
144
Кульбаба щетинистая (Leontodon hispidus)
205, 306
Кумарчик (Agriophyllum) 271, 272
Купальница (Trollius) 28, 210, 330, 332
Купена душистая (Polygonatum odoratum)
142, 224, 238
– многоцветковая (Polygonatum multiflorum)
227
Купырь лесной (Anthriscus sylvestris) 200,
209
Курильский бамбук (Sasa kurilensis) 177
Куропаточья трава (Dryas) 67, 68, 88
Лабазник (Filipendula) 279, 330
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
340
– вязолистный (Filipendula ulmaria) 210
– камчатский (Filipendula camtschatica) 179,
329, 332
– обыкновенный (Filipendula vulgaris) 232,
233
Лавровишня (Laurocerasus) 284, 299, 301
Ладанник (Cistus) 279
– крымский (Cistus tauricus) 278
Ладьян
трехраздельный (Corallorrhiza
trifida) 150, 153
Лазурник трехлопастный (Laser trilobum)
241
Ландыш (Convallaria) 244
– Кейзке (Convallaria keiskei) 190
– майский (Convallaria majalis) 233
Лапина крылоплодная (Pterocarya ptero-
carpa) 303
Лапчатка (Potentilla) 67, 81, 303, 327
– бесстебельная (Potentilla acaulis) 262–264
– вильчатая (Potentilla bifurca) 264
– прямая (Potentilla recta) 306
– серебристая (Potentilla argentea) 205
– снежная (Potentilla nivea) 103
Латук сибирский (Lactuca sibirica) 145
Лён зверобоелистный (Linum hypericifolium)
300
Лепторумора
амурская
(Leptirumohra
amurensis) 177, 182
Лесной мак весенний (Hylomecon vernalis)
182
Леспедеца двуцветная (Lespedeza bicolor)
186, 189, 190, 244
Лещина (Corylus) 151, 224, 228, 233, 276,
299, 302
– маньчжурская (Corylus mandshurica) 182,
190, 193, 244
– обыкновенная (Corylus avellana) 304, 306
– разнолистная (Corylus heterophylla) 182,
189, 244
Лжегравилат калужницелистный (Parageum
calthifolium) 117, 118
Лилия царские кудри (Lilium martagon) 229
– саранка (Lilium martagon) 153, 229
Лимонник (Schisandra) 177, 178, 182, 244
– китайский (Schisandra chinensis) 186, 193
Линнея (Linnaea) 157, 161
– северная (Linnaea borealis) 19, 160, 171
Липа (Tilia) 147, 151, 155, 161, 226–228,
238, 291, 299, 302, 307
– амурская (Tilia amurensis) 180, 183, 189,
190, 193, 244
– бегониелистная (Tilia begoniifolia) 284
– кавказская (Tilia begoniifolia) 306
– маньчжурская (Tilia mandshurica) 180,
244
– мелколистная (Tilia cordata) 227, 235
– обыкновенная (Tilia vulgaris) 305
– сердцевидная (Tilia cordata) 166, 224, 226
– Таке (Tilia taquetii) 180, 193, 244
Лисохвост (Alopecurus) 72
– альпийский (Alopecurus alpinus) 70, 99
Лиственница (Larix) 140, 150, 158, 171, 321
– даурская (Larix gmelinii) 81, 101, 135, 167,
171, 173, 180, 187, 244
– камчатская (Larix kamtschatica) 328
– Каяндера (Larix cajanderi) 105, 330
– ольгинская (Larix olgensis) 188, 191
– сибирская (Larix sibirica) 96, 135, 143,
155, 157, 171, 291, 313, 319, 321–323
Листовик (Phyllitis) 284
Ллойдия (Lloydia) 99
Лобария легочная (Lobaria pulmonaria) 150
Лобелия Дортмана (Lobelia dortmanna) 153
Ложечница арктическая (Cochlearia arctica)
67
Ломонос (Clematis) 284
– виноградолистный (Clematis vitalba) 306
– восточный (Clematis orientalis) 304, 306
– горный (Clematis montana) 304
Луговик извилистый (Avenella flexuosa) 142,
143, 205
Лук неравноногий (Allium anisopodium) 264
– охотский (Allium ochotense) 332
– парадоксальный (Allium paradoxum) 307
Лунокучник густосорусовый (Lunathyrium
pycnosorum) 182, 244
Луносемянник (Menispermum) 182
Любка двулистная (Platanthera bifolia) 150
– камчатская (Neolindleya camtschatica) 114
Лютик (Ranunculus) 66, 68, 232, 301, 303
– едкий (Ranunculus acris) 28, 208
– золотистый (Ranunculus auricomus) 208
– кавказский (Ranunculus caucasicus) 301
– кашубский (Ranunculus cassubicus) 224
– многоцветковый (Ranunculus polyanthe-
mos) 208, 233
– серно-желтый (Ranunculus sulphureus) 70
Люцерна серповидная (Medicago falcata)
200
Маакия амурская (Maackia amurensis) 182,
185, 193
Магнолия Суланжа (Magnolia soulangeana)
287
Майник (Maianthemum) 154, 161
– двулистный (Maianthemum bifolium) 160,
171, 238
Мак (Papaver) 66, 72
– подушковидный (Papaver pulvinatum) 103
– полярный (Papaver polare) 70
– югорский (Papaver lapponicum ssp. jugo-
ricum) 88
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
341
Малина (Rubus) 151, 160, 163
– боярышниколистная (Rubus crataegi-
folius) 180, 185
Манжетка (Alchemilla) 208
– альпийская (Alchemilla alpina) 140
– желто-зеленая (Alchemilla xanthochlora)
205
Марьянник луговой (Melampyrum pretense)
143, 160
– щетинистый (Melampyrum setaceum) 190
Медвежий чеснок (Allium ochotense) 329
Медуница (Pulmonaria) 150, 161
– неясная (Pulmonaria obscura) 145, 147,
224, 227
Мертензия морская (Mertensia maritima)
189
Меч-трава обыкновенная (Cladium mariscus)
153
Миндаль низкий (Amygdalus nana) 235
– обыкновенный (Amygdalus communis)
279
– степной (Amygdalus nana) 255
Минуарция красноватая (Minuartia rubella)
88
– Крашенинникова (Minuartia krascheninni-
kovii) 291
– крупноплодная (Minuartia macrocarpa) 99
Многоножка южная (Polypodium australe)
284
Многорядник копьевидный (Polystichum
lonchitis) 140
Можжевельник (Juniperus) 141, 159, 166,
171, 278, 282, 283, 307, 326, 331
– вонючий (Juniperus foetidissima) 278, 280
– высокий (Juniperus excelsa) 277, 279
– казацкий (Juniperus sabina) 307
– красный (Juniperus oxycedrus) 279
– обыкновенный (Juniperus communis) 321
– сибирский (Juniperus sibirica) 191
– твердый (Juniperus rigida) 182
Молочай
миндалевидный (Euphorbia
amygdaloides) 303
Морошка (Rubus chamaemorus) 77, 81, 86,
89, 112, 140, 144, 157
Мортук (Eremopyrum) 268
Мохоцветник Гмелина (Bryanthus gmelinii)
117, 332
Мушмула (Mespilus) 304, 307
Мытник (Pedicularis) 66, 67, 99
– Кауфмана (Pedicularis kaufmannii) 233
– перевернутый (Pedicularis resupinata) 332
– приятный (Pedicularis amoena) 103
– царский скипетр (Pedicularis sceptrum-
carolinum) 153
Мякотница однолистная (Malaxis mono-
phyllos) 150, 153
Мятлик (Poa) 72, 99, 255, 301, 326
– альпигенный (Poa alpigena) 99
– альпийский (Poa alpina) 305
– арктический (Poa arctica) 88
– дубравный (Poa nemoralis) 28, 190
– кистевидный (Poa botryoides) 172, 263,
264
– крупночашуйный (Poa macrocalyx) 118
– лесной (Poa nemoralis) 228, 306
– ложноболотный (Poa pseudopalustris) 190
– ложноукороченный (Poa pseudoabbrevi-
ata) 103
– луговой (Poa pratensis) 28, 29, 205, 306
– оттянутый (Poa attenuata) 263
– расставленный (Poa remota) 145
– сизый (Poa glauca) 103
– Танфильева (Poa tanfiljewii) 145
– узколистный (Poa angustifolia) 232
Наголоватка (Jurinea) 308
– паутинистая (Jurinea arachnoidea) 255
Надбородник безлистный (Epipogium
aphyllum) 150, 233, 234
Недоспелка
камчатская
(Cacalia
kamtschatica) 329
– копьевидная (Cacalia hastata) 145
Недотрога обыкновенная (Impatiens noli-
tangere) 227
Незабудка (Myosotis) 71, 233, 255
–альпийская (Myosotis alpestris) 301
– Попова (Myosotis popovii) 231
Незабудочник волосистый (Eritrichium vil-
losum) 71
Неравноцветник кровельный (Anisantha
tectorum) 271
Нефрома арктическая (Nephroma arcticum)
159
Нивяник (Leucanthemum) 205, 232
– обыкновенный (Leucanthemum vulgare)
206, 208
Обвойник (Periploca) 284
– греческий (Periploca graeca) 304, 306
Овсец (Helictotrichon) 303, 326
– пустынный (Helictotrichon desertorum)
234, 261
– пушистый (Helictotrichon pubescens) 232
Овсик извилистый (Avenella flexuosa) 205,
207
Овсяница (Festuca) 99
– валлисская (Festuca valesiaca) 249, 254
– красная (Festuca rubra) 208
– ленская (Festuca lenensis) 172, 263
– лесная (Festuca drymeja) 304
– луговая (Festuca pratensis) 28, 205, 206,
306
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
342
– овечья (Festuca ovina) 142, 144, 188, 205,
207, 208, 250
– приземистая (Festuca airiodes) 88, 173
– Ричардсона (Festuca richardsonii) 103
– ушковатая (Festuca auriculata) 103
– якутская (Festuca jacutica) 171
Одуванчик лекарственный (Taraxacum offic-
inale) 200
Ожика (Luzula) 72, 99
– волосистая (Luzula pilosa) 144
Окопник (Symphytum) 302
Оксиграфис ледяной (Oxygraphis glacialis)
103
Ольха (Alnus) 9
– бородатая (Alnus barbata) 304, 306
– волосистая (Alnus hirsuta) 173, 328
– клейкая (Alnus glutinosa) 305
– серая (Alnus incana) 129, 140, 143, 305
– черная (Alnus glutinosa) 235
– японская (Alnus japonica) 245
Ольховник (Duschekia) 96, 171, 323, 326,
327, 329, 330, 332
– камчатский (Duschekia kamtschatica) 329
– кустарниковый (Duschekia fruitcosa) 78,
112, 114
Оноклея чувствительная (Onoclea sensibilis)
188
Оносма простейшая (Onosma simplicissima)
239, 240
Орех грецкий (Juglans regia) 304
– маньчжурский (Juglans mandshurica) 180,
184, 187, 188, 193, 245
Орляк (Pteridium) 19, 20
Ортилия однобокая (Orthilia secunda) 160,
238
Осина (Populus) 128, 129, 140, 143, 147, 163,
233, 238, 242, 302
– обыкновенная (Populus tremula) 235
Осока (Carex) 71, 95, 99, 214, 228, 258, 267,
303, 326
– Бигелоу (Carex bigelowii) 88, 140
– бледная (Carex pallida) 331
– Буксбаума (Carex buxbaumii) 164
– возвратившаяся (Carex reventa) 190
– волосистая (Carex pilosa) 224, 227, 228
– двурядная (Carex disticha) 164
– дернистая (Carex cespitosa) 164, 210
– длинноостистая (Carex macrochaeta) 115
– кривоносая (Carex campylorhina) 182
– ланцетная (Carex lanceolata) 190
– лесная (Carex sylvatica) 304
– низкая (Carex humilis) 232, 239, 306, 309
– одноцветная (Carex concolor) 99, 102, 104
– острая (Carex acuta) 210
– островная (Carex nesophila) 112
– песколюбивая (Carex arenicola) 189
– приземистая (Carex supina) 309
– редкоцветковая (Carex rariflora) 102
– ржавопятнистая (Carex siderosticta) 182
– твердоватая (Carex duriuscula) 264
– топяная (Carex limosa) 330
– траурная (Carex lugens) 109
– уссурийская (Carex ussuriensis) 182, 190
– шаровидная (Carex globularis) 144, 157
Осот полевой (Sonchus arvensis) 200
Остролодочник грязноватый (Oxytropis
sordida) 89, 293
– каргинский (Oxytropis karga) 103
– Миддендорфа (Oxytropis middendorffii) 103
– остролистный (Oxytropis oxyphylla) 263
Павловния войлочная (Paulownia tomen-
tosa) 286
Падуб (Ilex) 284, 299
Пальчатокоренник
длиннолистный
(Dactylorhiza baltica) 153
– мясо-красный (Dactylorhiza incarnata)
150
– пятнистый (Dactylorhiza maculata) 150
– Траунштейнера (Dactylorhiza traunstei-
neri) 150
– Фукса (Dactylorhiza fuchsii) 150
Пастушья сумка обыкновенная (Capsella
bursa-pastoris) 200
Патриния скальная (Patrinia rupestris) 190,
313
Пельтигера пупырчатая (Peltigera aphthosa)
159
– собачья (Peltigera canina) 159
Пепельник тундровый (Tephroseris tundri-
cola) 103
– цельнолистный (Tephroseris integrifolia)
89, 232
Первоцвет весенний (Primula veris) 233
– холодный (Primula algida) 305
Перловник (Melica) 164
Песчанка ложнохолодная (Arenaria pseudo-
frigida) 88
Петров крест чешуйчатый (Lathraea squa-
maria) 153
Печеночница благородная (Hepatica nobilis)
21, 22, 142, 151, 224, 225
Пижма (Tanacetum) 245, 267, 268
Пион (Paeonia) 255
– Марьин корень (Paeonia anomala) 145
– тонколистный (Paeonia tenuifolia) 256
Пираканта ярко-красная (Pyracantha cocci-
nea) 279, 306
Пихта (Abies) 140, 147, 150, 151, 158, 159,
161, 166, 172, 301
– белокорая (Abies nephrolepis) 176, 180,
187, 188, 190, 191, 193, 197
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
343
– грациозная (Abies gracilis) 329
– Майра (Abies mayriana) 177
– кавказская (Abies nordmanniana) 299
– сахалинская (Abies sachalinensis) 136, 176,
177
– сибирская (Abies sibirica) 143, 147, 155,
173, 291, 314
– цельнолистная (Abies holophylla) 183
Плаун (Lycopodium) 160
– булавовидный (Lycopodium clavatum) 144
– годичный (Lycopodium annotinum) 157
Плевроциум Шребера (Pleurozium schreberi)
159
Плющ (Hedera) 284
– Пастухова (Hedera pastuchowii) 303, 305
Подбел (Andromeda) 27, 214
– обыкновенный (Andromeda polifolia) 213
Подлесник европейский (Sanicula europaea)
304
Подмаренник душистый (Galium odoratum)
147
– настоящий (Galium verum) 28, 29, 231–
233, 263
– цепкий (Galium aparine) 17, 227
Подорожник средний (Plantago media) 257
Подснежник (Galanthus) 285
Полевица (Agrostis) 301
– виноградниковая (Agrostis vinealis) 232
– тонкая (Agrostis tenuis) 205, 206
Полушник азиатский (Isoetes asiatica) 114
– щетинистый (Isoetes setacea) 153
Полынь (Artemisia) 171, 260, 261, 271
– австрийская (Artemisia austriaca) 253, 267
– арктическая (Artemisia arctica) 117
– белая (Artemisia lercheana) 253, 267, 271
– веничная (Artemisia scoparia) 264
– Гмелина (Artemisia gmelinii) 190, 244, 245
– горькая (Artemisia absinthium) 200
– Лерха (Artemisia lercheana) 267, 268, 271
– лечебная (Artemisia abrotanum) 267
– малоцветковая (Artemisia pauciflora) 268,
269
– песчаная (Artemisia arenaria) 267, 271,
272
– пижмолистная (Artemisia tanacetifolia)
263
– побегоносная (Artemisia stolonifera) 190
– пышная (Artemisia opulenta) 115
– сантонинная (Artemisia santonica) 253,
267, 268
– холодная (Artemisia frigida) 172, 264
– чёрная (Artemisia pauciflora) 253, 267, 268
– шелковистая (Artemisia sericea) 259, 261
Поповник красный (Pyrethrum coccineum)
301
Пролеска (Scilla) 224
– сибирская (Scilla siberica) 225
Пролесник многолетний (Mercurialis peren-
nis) 224, 227
Проломник арктосибирский (Androsace
arctisibirica) 88
– Козо-Полянского (Androsace koso-poljan-
skii) 229, 239, 240
– седой (Androsace incana) 263
Прострел (Pulsatilla) 151, 171, 232, 327
– весенний (Pulsatilla vernalis) 140
– желтеющий (Pulsatilla orientali-sibirica)
173
– раскрытый (Pulsatilla patens) 231, 232
– Турчанинова (Pulsatilla turczaninovii) 263
Прутняк (Kochia prostrata) 253, 267–269,
271
Птерис критский (Pteris cretica) 285
Пустынница волосовидная (Eremogone
capillaris) 262, 263
Пухонос альпийский (Trichophorum al-
pinum) 153
Пушица (Eriophorum) 18, 67, 71, 81, 89, 95,
99, 105, 214, 216
– влагалищная (Eriophorum vaginatum)
213, 214
– узколистная (Eriophorum angustifolium)
99, 102
– Шейхцера (Eriophorum scheuchzeri) 99,
104
Пыльцеголовник красный (Cephalanthera
rubra) 233, 234
Пырей (Elytrigia) 267–269
– ползучий (Elytrigia repens) 28, 164, 306
– стройный (Elytrigia gracillima) 309
Ракитничек русский (Chamaecytisus ru-
thenicus) 166, 233, 235, 236, 238, 302
Ребросемянник уральский (Pleurospermum
uralense) 329
Резак обыкновенный (Falcaria vulgaris) 233
Рогач песчаный (Ceratocarpus arenarius) 271
Рододендрон (Rhododendron) 172, 184, 244,
300, 322, 323, 326, 330, 332
– даурский (Rhododendron dauricum) 172,
244, 320, 321
– желтый (Rhododendron luteum) 299, 302
– золотистый (Rhododendron aureum) 116,
187, 188, 190, 191
– кавказский (Rhododendron caucasicum)
300–302
– камчатский (Rhododendron camtschati-
cum) 112
– короткоплодный (Rhododendron brachy-
carpum) 190
– мелколистный (Rhododendron parvifoli-
um) 109, 187, 192
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
344
– остроконечный (Rhododendron mucronu-
latum) 244
– понтийский (Rhododendron ponticum)
284, 299
– сихотинский (Rhododendron sichotense)
190, 191
Росянка (Drosera) 214
Рябина (Sorbus) 129, 142, 151, 159, 161, 163,
233, 235, 302, 307
– амурская (Sorbus amurensis) 191
– бузинолистная (Sorbus sambucifola) 178,
191, 330
– ольхолистная (Sorbus alnifolia) 180, 193
– сибирская (Sorbus sibirica) 160, 329, 331
Рябчик русский (Fritillaria ruthenica) 229,
233, 234
– шахматный (Fritillaria meleagris) 229
Самшит (Buxus) 284, 285
Сассапариль высокий (Smilax excelsa) 284,
303, 305
Свидина (Swida) 299, 301
– южная (Swida australis) 304
Седмичник (Trientalis) 142, 157
– европейский (Trientalis europaea) 160, 238
Сиверсия ледяная (Acomastylis glacialis) 102
Сивец луговой (Succisa pratensis) 210
Синяк русский (Echium russicum) 255
Сирень (Syringa) 244
– амурская (Syringa amurensis) 180, 182,
185, 193, 245
– Вольфа (Syringa wolfii) 182
Ситник (Juncus) 208
– трехраздельный (Juncus trifidus) 88
Скабиоза (Scabiosa) 303, 309
– гумбетская (Scabiosa gumbetica) 308
– кавказская (Scabiosa caucasica) 301
– шерстистолистная (Scabiosa lachnophylla)
188
Скумпия (Cotinus) 228, 283
– кожевенная (Cotinus coggygria) 279
Слива вишненосная (Prunus cerasifera) 299
Смоковница обыкновенная (Ficus carica) 286
Смолевка (Silene) 81
– бесстебельная (Silene acaulis) 71, 87
– енисейская (Silene jeniseensis) 263
– малолистная (Silene paucifolia) 103
– скальная (Silene rupestris) 140
Смолка клейкая (Viscaria vulgaris) 205, 207
Смородина (Ribes) 101, 178, 307
– Максимовича (Ribes maximoviczianum) 190
– черная (Ribes nigrum) 163
Сныть (Aegopodium) 142–144, 161, 208
– обыкновенная (Aegopodium podagraria)
165, 209, 224, 233
Солнцецвет (Helianthemum) 278
Солодка (Glycyrrhiza) 269
– голая (Glycyrrhiza glabra) 267
Солянка холмовая (Salsola collina) 252
Сосна (Pinus) 86, 128, 140, 143, 144, 147,
157, 214, 216, 218, 228, 302, 322, 326
– кедровая (Pinus sibirica) 128, 144, 145,
155, 166, 172, 313, 317, 321–323
– корейская (Pinus koraiensis) 138, 180, 183,
189, 192, 194
– Коха (Pinus kochiana) 277, 306, 307
– крымская (Pinus pallasiana) 277, 279–281
– крючковатая (Pinus uncinata) 299
– могильная (Pinus funebris) 187
– обыкновенная (Pinus sylvestris) 129, 140,
155, 235, 241, 291, 318, 323, 324
– пицундская (Pinus pityusa) 279, 280, 282,
285
– сибирская (Pinus sibirica) 158, 159, 160,
173, 291
Соссюрея альпийская (Saussurea alpina) 89
– ложно-тилезиева (Saussurea pseudotilesii)
332
Спирея (Spiraea) 244, 283, 294, 306
– берёзолистная (Spiraea betulifolia) 190
– Бовера (Spiraea beauverdiana) 331
– городчатая (Spiraea crenata) 235, 236, 306
– зверобоелистная (Spiraea hypericifolia)
306
– иволистная (Spiraea salicifolia) 163
– средняя (Spiraea media) 145, 163, 171
– трехлопастная (Spiraea trilobata) 313
Стеблелист мощный (Caulophyllum ro-
bustum) 244
Сумах (Rhus) 279
Сфагнум (Sphagnum) 18, 89, 91, 214, 221
– бурый (Sphagnum fuscum) 89
Схизонепета многонадрезанная (Schizone-
peta multifida) 263
Таволга обыкновенная (Filipendula vulgaris)
28, 29
Таймырская роза (Acomastylis glacialis) 102
Тайник сердцевидный (Listera cordata) 145,
153
– яйцевидный (Listera ovata) 150
Тамус (Tamus) 284
Термопсис ланцетный (Thermopsis lanceola-
ta) 264
Тёрн (Prunus) 228, 233, 255, 304
– обыкновенный (Prunus spinosa) 235, 236
Тимофеевка (Phleum) 28, 208, 301
– альпийская (Phleum alpinum) 305
– луговая (Phleum pratense) 205, 206, 232, 306
– степная (Phleum phleoides) 232
Тимьян дагестанский (Thymus daghestani-
cus) 309
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
345
– Елизаветы (Thymus elisabethae) 309
– крайний (Thymus extremus) 103
– меловой (Thymus calcareus) 239, 240
– монгольский (Thymus mongolicus) 263
– ползучий (Thymus serpyllum) 142
– Талиева (Thymus talijevii) 145
Типчак (Festuca) 228, 232, 234, 245, 249,
257, 258, 263, 267, 271, 278, 309, 326
Тис (Taxus) 177
– остроконечный (Taxus cuspidata) 180, 182,
188, 190
Толокнянка (Arctostaphylos) 77, 101, 155,
190, 300, 331
– обыкновенная (Arctostaphylos uva-ursi) 81
Тонконог (Koeleria) 251, 257, 263, 280, 326
– азиатский (Koeleria asiatica) 103
– гребенчатый (Koeleria cristata) 232, 245,
251, 264
– жестколистный (Koeleria scleraphylla) 250
– сизый (Koeleria glauca) 251
– токийский (Koeleria tokiensis) 188
Тополь белый (Populus alba) 235, 304, 306
– Давида (Populus davidiana) 189
– душистый (Populus suaveolens) 326, 329
– Максимовича (Populus maximowiczii) 192,
194
– сереющий (Populus canescens) 304
– черный (Populus nigra) 235, 304, 306
Трехреберник непахучий (Tripleurosper-
mum inodorum) 200
Тризетокелерия таймырская (Trisetokoeleria
taimyrica) 99
Трищетинник колосистый (Trisetum spi-
catum) 103
Трясунка (Briza) 301
Тырса (Stipa capillata) 254, 258, 309
Тысячелистник (Achillea) 208
– обыкновенный (Achillea millefolium) 28, 29
Тюльпан (Tulipa) 28, 255
– Биберштейна (Tulipa biebersteiniana) 256
Углостебельник татарский (Goniolimon ta-
taricum) 256
Ужовник обыкновенный (Ophioglossum vul-
gatum) 151
Фиалка белая (Viola alba) 304
– душистая (Viola odorata) 276
Физалис обыкновенный (Physalis alkekengi)
304
Филлодоце (Phyllodoce) 332
Финик канарский (Phoenix canariensis) 287
Фиппсия холодная (Phippsia algida) 103
Фисташка туполистная (Pistacia mutica)
278, 279, 280, 282
Фрима (Phryma) 193
– азиатская (Phryma asiatica) 186
Фумана (Fumana) 278
Хамедафне
прицветничковая
(Chamaedaphne calyculata) 89, 213
Хамеродос алтайский (Chamaerhodos al-
taica) 263
– трехнадрезной (Chamaerhodos trifida) 263
Хаммарбия болотная (Hammarbya palu-
dosa) 153
Хвойник двухколосковый (Ephedra dis-
tachya) 239
Хвощ (Equisetum) 157, 160
– лесной (Equisetum sylvaticum) 161, 332
– луговой (Equisetum pratense) 332
Хмель (Humulus) 284
Хохлатка
дымянкообразная
(Corydalis
capnoides) 145
– полая (Corydalis cava) 224, 225
Хризантема выемчатолистная (Chrysanthe-
mum sinuatum) 312
– Завадского (Chrysanthemum zawadskii)
229
Цкламен (Cyclamen) 28, 285
Цикорий обыкновенный (Cichorium intybus)
200
Цимбария даурская (Cymbaria daurica) 264
Цицербита (Cicerbita) 302
Чабрец (Thymus) 278
– Маршалла (Thymus marschallianus) 233
Чемерица Лобеля (Veratrum lobelianum)
323
– остродольная (Veratrum oxysepalum) 332
Черёмуха (Padus) 154, 161, 233, 235, 302
– айнская (Padus ssiori) 177, 178
– азиатская (Padus asiatica) 193, 329
– Маака (Padus maackii) 180, 184, 193
– обыкновенная (Padus avium) 306
Черемша (Allium microdictyon) 323
Черемша (Allium ochotense) 329
Черешня (Cerasus avium) 299
Черника (Vaccinium) 18, 142, 144, 159, 160,
161, 173, 299, 322, 323
– кавказская (Vaccinium arctostaphylos) 284,
299
– овальнолистная (Vaccinium ovalifolium) 177
Черноголовка (Prunella) 205
Чертополох (Carduus) 17
Чесночница черешковая (Alliaria petiolata)
227
Чина (Lathyrus) 232
– весенняя (Lathyrus vernus) 224, 225
– Литвинова (Lathyrus litvinovii) 291
– японская (Lathyrus japonicas) 189
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
346
Чистотел большой (Chelidonium majus) 233
Чистоустник азиатский (Osmundastrum
asiaticum) 188, 244
Чозения (Chosenia) 187, 192, 330
Чубушник (Philadelphus) 182, 193
– тонколистный (Philadelphus tenuifolius)
180, 190
Шалфей (Salvia) 255, 309
– луговой (Salvia pratensis) 231–233
– мутовчатый (Salvia verticillata) 153
– седеющий (Salvia canescens) 308
Шаровница точечная (Globularia punctata)
241
Шейхцерия (Scheuchzeria) 214
Шеломайник (Filipendula camtschatica) 178
Шиверекия
подольская
(Schivereckia
podolica) 229
Шиповник (Rosa) 101, 142, 159, 171, 178,
228, 235, 278, 283, 322, 331
– иглистый (Rosa acicularis) 160, 166, 330
– майский (Rosa majalis) 163, 235, 236
– морщинистый (Rosa rugosa) 189
– собачий (Rosa canina) 304, 306
– щитконосный (Rosa corymbifera) 304
Шлемник многозубый (Scutellaria polyodon)
309
Щавелёк обыкновенный (Rumex acetosella)
200
Щитовник мужской (Dryopteris filix-mas)
147
Щитовник толстокорневищный (Dryopteris
crassirhizoma) 182, 193, 244
Щучка (Deschampsia) 208
– берингийская (Deschampsia beringensis)
118
– дернистая (Deschampsia cespitosa) 28, 208
– коротколистная (Deschampsia brevifolia)
70, 102
Эгилопс (Aegilops) 278
Эгонихон пурпурно-голубой (Aegonychon
purpureocoeruleum) 304
Элеутерокок (Eleutherococcus) 182, 190, 193
– колючий (Eleutherococcus senticosus) 180,
185
Энемион Радде (Enemion raddeanum) 191
Эспарцет (Onobrychis) 232
– песчаный (Onobrychis arenaria) 233
– рогатый (Onobrychis cornuta) 308, 309
Эфедра (Ephedra) 239, 283
– двухколосковая (Ephedra distachya) 240
Юкка алоэлистная (Yucca aloifolia) 287
Яблоня (Malus) 228, 233, 307
– восточная (Malus orientalis) 298, 302, 304,
306
– маньчжурская (Malus mandshurica) 245
– ранняя (Malus praecox) 235
Явор (Acer pseudoplatanus) 299
Ярутка полевая (Thlaspi arvense) 200
Ясень (Fraxinus) 187, 224, 228, 279, 297,
299, 307
– высокий (Fraxinus excelsior) 279, 284
– маньчжурский (Fraxinus mandshurica)
180, 188, 192–194, 245
– носолистный (Fraxinus rhynchophylla)
180, 184, 244
– обыкновенный (Fraxinus excelsior) 227,
303, 305
– остроплодный (Fraxinus oxycarpa) 279
Ясколка (Cerastium) 99
Яснотка пятнистая (Lamium maculatum)
227
Ястребинка
зонтичная
(Hieracium
umbellatum) 238
Ястребиночка
обыкновенная
(Pilosella
officinarum) 205, 207
Ятрышник шлемоносный (Orchis militaris)
153
Ячмень луковичный (Hordeun bulbosum) 278
УКАЗАТЕЛЬ ЛАТИНСКИХ НАИМЕНОВАНИЙ
Abies gracilis (Пихта грациозная) 329
– holophylla (Пихта цельнолистная) 180, 183
– mayriana (Пихта Майра) 177
– nephrolepis (Пихта белокорая) 136, 176,
180
– nordmanniana (Пихта Нордманна, п. кав-
казская) 299
– sachalinensis (Пихта сахалинская) 136,
176, 177
– sibirica (Пихта сибирская) 136–140, 143,
147, 153, 173, 291, 323
Acacia dealbata (Акация подбелённая, а. се-
ребристая) 287
Acer campestre (Клён полевой) 222, 299,
304
– ginnala (Клён гиннала) 221
– laetum (Клён светлый) 307
– mandshuricum (Клён маньчжурский) 180,
184
– mono (Клён моно, к. мелколистный) 139,
180, 244
– negundo (Клён американский) 228
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
347
– platanoides (Клён остролистный, к. плата-
новидный) 138, 139, 147, 153, 222–224,
299
– pseudoplatanus (Клён ложноплатановый,
явор) 153, 284, 299
– pseudosiebolianum (Клён ложнозиболь-
дов) 180
– tataricum (Клён татарский) 221, 228
– tegmentosum (Клён зеленокорый) 180, 190
– trautvetteri (Клён Траутфеттера) 299, 300
– ukurunduense (Клён укурунду, к. желтый)
180, 190
Achillea millefolium (Тысячелистник обык-
новенный) 163, 164,165, 203, 208, 242
– nobilis (Тысячелистник благородный) 261
Acomastylis glacialis (Акомастилис ледяной,
таймырская роза) 102, 109
Aconitum maximum (Борец большой) 115
– orientale (Борец восточный, аконит во-
сточный) 302
– raddeanum (Борец Радде) 182
– septentrionale (Борец северный) 89, 162,
163, 315, 316
– volubile (Борец вьющийся) 164
Aconogonon ochreatum var. laxmannii (Таран
широкораструбовый Лаксманна) 111
– tripterocarpum (Таран трехкрылоплод-
ный) 111
Actaea erythrocarpa (Воронец красноплод-
ный) 144, 151
Actinidia arguta (Актинидия острая) 182,
193
– kolomikta (Актинидия коломикта) 177,
182, 186, 244
– polygama (Актинидия многодомная) 177
Adenostyles platyphylloides (Аденостилес
плосколистный) 302
Adiantum pedatum (Адиантум стоповидный)
182, 244
Adonis vernalis (Адонис весенний) 230–233
– volgensis (Адонис волжский) 255, 257
Aegilops (Эгилопс) 278
Aegonychon purpureocoeruleum (Эгонихон
пурпурно-голубой) 303
Aegopodium podagraria (Сныть обыкновен-
ная) 139, 142, 144, 161–165, 209, 224
Agapetes (Агапетес) 66
Agrimonia pilosa (Репешок волосистый) 164
Agriophyllum squarrosum (Кумарчик расто-
пыренный) 271, 272
Agropyron cristatum (Житняк гребенчатый)
172, 250, 251, 253, 263, 264, 312
– desertorum (Житняк пустынный) 253, 267,
268, 271, 272
– fragile (Житняк ломкий) 267, 268, 269,
272
– pectinatum (Житняк гребневидный) 251,
253, 267, 268, 271
– sibiricum (Житняк сибирский) 109, 268
Agrostis gigantea (Полевица гигантская)
164, 203
– planifolia (Полевица плосколистная) 301
– tenuis (Полевица тонкая) 205, 206
– vinealis (Полевица виноградниковая) 228,
230, 232
Ajuga chia (Живучка хиосская) 239, 240
Albizia julibrissin (Альбиция шелковая) 286
Alchemilla alpina (Манжетка альпийская)
140
– caucasica (Манжетка кавказская) 302
– sibirica (Манжетка сибирская) 315
– xanthochlora (Манжетка желто-зеленая)
203, 205
Alectoria (Алектория) 71
– nigricans (Алектория черноватая) 97
– ochroleuca (Алектория бледно-охряная)
75, 105
Alfredia cernua (Альфредия поникшая) 315
Alliaria petiolata (Чесночница черешковая)
227
Allium anisopodium (Лук неравноногий) 264
– decipiens (Лук обманчивый) 281
– lineare (Лук линейный) 242
– microdictyon (Лук мелкосетчатый, черем-
ша) 323
– ochotense (Лук охотский, черемша) 329,
332
– paradoxum (Лук парадоксальный) 307
Alnus barbata (Ольха бородатая) 304
-
glutinosa (Ольха черная, о. клейкая) 137,
138, 222, 235
- hirsuta (Ольха волосистая) 173, 329
- incana (Ольха серая) 140
- japonica (Ольха японская) 245
Alopecurus alpinus (Лисохвост альпийский)
70–72, 82, 99
– pratensis (Лисохвост луговой) 200
Alyssum gmelinii (Бурачок Гмелина) 239,
240
– lenense (Бурачок ленский) 263
– obtusifolium (Бурачок туполистный) 280
– turkestanicum (Бурачок туркестанский)
271
– turkestanicum var. desertorum (Бурачок
туркестанский пустынный) 255, 260
Amelanchier ovalis (Ирга овальная) 283
Amygdalus communis (Миндаль обыкновен-
ный) 279
– nana (Миндаль низкий, м. степной) 235,
252, 255, 257, 258
Andromeda polifolia (Подбел обыкновен-
ный) 90, 91, 113, 156, 213, 214
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
348
Androsace arctisibirica (Проломник аркто-
сибирский) 88
– elongata (Проломник удлиненный) 255
– incana (Проломник седой) 263
– koso-poljanskii (Проломник Козо-Полян-
ского) 229, 239, 240
– maxima (Проломник большой) 260
–
septentrionalis (Проломник северный)
260
Anemonastrum narcissiflorum (Ветреник
нарциссоцветковый) 115–117, 300
– sibiricum (Ветреник сибирский) 109
Anemone nemorosa (Ветреница дубравная)
224, 225
– sylvestris (Ветреница лесная) 145, 230,
232, 255
Angelica genuflexa (Дудник коленчатосогну-
тый, д. преломленный) 115, 329
– gmelinii (Дудник Гмелина) 115, 329
– sylvestris (Дудник лесной) 163, 164, 208,
242
– ursina (Дудник медвежий) 178, 179, 329
Anisantha tectorum (Неравноцветник кро-
вельный) 271
Antennaria dioica (Кошачья лапка двудом-
ная) 166, 203, 205, 207, 238
Anthemis trotzkiana (Пупавка Корнух-Троц-
кого) 239
Anthoxanthum odoratum (Душистый коло-
сок обыкновенный) 205, 206, 230, 303
Anthriscus sylvestris (Купырь лесной) 200,
203, 209, 323
Aquilegia glandulosa (Водосбор железистый)
316
Arabis sagittata (Резуха стреловидная) 281
Aralia elata (Аралия высокая) 180
Arbutus andrachne (Земляничник мелко-
плодный) 278
Arctagrostis (Арктополевица) 71
– arundinacea (Арктополевица тростнико-
видная) 109
Arctogeron gramineum (Арктогерон злако-
вый) 263
Arctophila fulva (Северолюбка арктофила
рыжеватая) 104
Arctopoa eminens (Арктомятлик выделяю-
щийся) 115
Arctostaphylos (Толокнянка) 67
– uva-ursi (Толокнянка обыкновенная) 81,
136, 155, 300
Arctous alpina (Арктоус альпийский) 86, 88,
90, 91, 95, 97, 109, 145, 170, 331
Arenaria pseudofrigida (Песчанка ложнохо-
лодная) 88
– serpyllifolia (Песчанка тимьянолистная)
255
Aristolochia clematitis (Кирказон ломоносо-
видный) 307
Armeniaca mandshurica (Абрикос мань-
чжурский) 187
– sibirica (Абрикос сибирский) 187
Arnica fennoscandica (Арника фенноскан-
динавская) 140
– iljinii (Арника Ильина) 103
Artemisia abrotanum (Полынь лечебная)
267
– absinthium (Полынь горькая) 200
– arctica (Полынь арктическая) 116, 117
–arenaria (Полынь песчаная) 271, 272
– austriaca (Полынь австрийская) 253, 257–
262, 271
– caucasica (Полынь кавказская) 280
– commutata (Полынь замещающая) 326
– dracunculus (Полынь эстрагон) 260, 261
– frigida (Полынь холодная) 172, 253, 259,
260, 263, 264, 313
– glauca (Полынь сизая) 243, 259, 260, 261,
262
– globularia (Полынь шаровидная) 109
– gmelinii (Полынь Гмелина) 190, 244
– gracilescens (Полынь тонковатая) 260
– laciniata (Полынь рассеченная) 171, 243
–
lagocephala (Полынь куроголовчатая)
326
– latifolia (Полынь широколистная) 258–
261
– lercheana (Полынь Лерха, п. белая) 253,
258, 267–269, 271, 272
– marschalliana (Полынь Маршалла) 258,
260–262
– nitrosa (Полынь селитряная) 243, 259,
260
– opulenta (Полынь пышная) 115
– pauciflora (Полынь малоцветковая, п.
чёрная) 253, 265–269
– pontica (Полынь понтийская) 243, 261
– rupestris (Полынь каменная) 243
– santonica (Полынь сантонинная) 253,
265–269
– schrenkiana (Полынь Шренка) 260
– scoparia (Полынь веничная) 261, 264
– sericea (Полынь шелковистая) 242, 259,
261
– stolonifera (Полынь побегоносная) 190
– tanacetifolia (Полынь пижмолистная) 263
– tilesii (Полынь Тилезиуса) 111
Arum elongatum (Аронник удлиненный) 281
Aruncus dioicus (Волжанка двудомная) 182
Arundinella anomala (Тростничок уклоня-
ющийся) 223
Asarum europaeum (Копытень европейский)
147, 224, 313
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
349
Asperula exasperata (Ясменник шерохова-
тый) 239
Asphodeline (Асфоделина) 280
Asplenium septentrionale (Костенец север-
ный) 280
– trichomanes (Костенец волосовидный) 280
Aster alpinus (Астра альпийская) 111, 258
Astracantha arnacantha (Астраканта колюч-
ковая) 280
– aurea (Астраканта золотистая) 309
– caucasica (Астраканта кавказская) 309
– denudata (Астраканта обнаженная) 308,
309
Astragalus (Астрагал) 81
– adsurgens (Астрагал приподнимающийся)
322
– alpinus (Астрагал альпийский) 103
– austriacus (Астрагал австрийский) 258,
261
– danicus (Астрагал датский) 230, 242
– dolichophyllus (Астрагал длиннолистный)
257
– frigidus (Астрагал холодный) 103
– helmii (Астрагал Гельма) 291
– henningii (Астрагал Хеннинга) 257
– laguroides (Астрагал заячий) 263
– lasioglottis (Астрагал волосистоязычко-
вый) 308, 309
– onobrychis (Астрагал эспарцетовый) 258,
261, 262
– propinquus (Астрагал сходный) 321
– rupifragus (Астрагал камнеломный) 239
– umbellatus (Астрагал зонтичный) 103, 109
Astrantia maxima (Звездовка большая) 300
Athyrium filix-femina (Кочедыжник жен-
ский) 161, 163
– sinense (Кочедыжник китайский) 244
Atractylodes ovata (Веретенник яйцевид-
ный) 244
Atragene ochotensis (Княжик охотский)
331
– sibirica (Княжик сибирский) 145, 151
Atriplex tatarica (Лебеда татарская) 271
Aulacomnium palustre (Аулакомниум болот-
ный) 88, 316
– turgidum (Аулакомниум вздутый) 74, 88,
91, 95, 97, 98, 101
Avenella flexuosa (Овсик извилистый, луго-
вик извилистый) 89, 90, 115, 142, 144,
205, 207
Barbarea vulgaris (Сурепка обыкновенная)
200
Bartsia alpine (Бартсия альпийская) 145
Bassia sedoides (Бассия очитковидная) 268,
271
Beckmannia eruciformis (Бекмания обыкно-
венная) 267–269
Bellevalia sarmatica (Бельвалия сарматская)
255
Berberis amurensis (Барбарис амурский) 180
– vulgaris (Барбарис обыкновенный) 235,
283
Bergenia crassifolia (Бадан толстолистный)
323
– pacifica (Бадан тихоокеанский) 191
Betonica macrantha (Буквица крупноцвет-
ковая) 300
– officinalis (Буквица лекарственная) 210
Betula cajanderi (Берёза Каяндера) 136
– costata (Берёза ребристая, б. жёлтая) 180,
184, 190, 244
– czerepanovii (Берёза Черепанова) 82, 86,
90, 140, 144, 145
– davurica (Берёза даурская) 190, 222, 223,
244
– divaricata (Берёза растопыренная) 80,
111, 136, 324, 326, 327, 330
– ermanii (Берёза Эрмана, б. каменная) 137,
177, 328, 329, 333
– exilis (Берёза тощая) 73, 80, 109, 111, 113,
116, 136, 170, 171, 324, 326, 327
– fruticosa (Берёза кустарниковая) 136, 137
– humilis (Берёза приземистая) 325
– lanata (Берёза шерстистая) 188, 327
– litwinowii (Берёза Литвинова) 300, 302
– mandshurica (Берёза маньчжурская) 189
– nana (Берёза карликовая) 73, 80, 82, 89,
90, 96, 97, 136, 144, 145, 156, 157
– ovalifolia (Берёза овальнолистная) 137
– pendula (Берёза повислая) 136, 137, 139,
140, 143, 163, 164, 223, 242, 323
– platyphylla (Берёза плосколистная) 136,
137, 170, 173, 323, 328, 331, 332
– pubescens (Берёза пушистая) 90, 97, 98,
136, 137, 140, 143–145, 157, 158, 163,
164
– raddeana (Берёза Радде) 300, 302
– rotundifolia (Берёза круглолистная) 316
– tortuosa (Берёза извилистая) 97
Bistorta carnea (Змеевик мясо-красный)
301
– major (Змеевик большой) 209
– officinalis (Змеевик лекарственный) 89, 90,
109, 210
– vivipara (Змеевик живородящий) 88, 115,
117, 332
Bothriochloa (Бородач) 278
– ischaemum (Бородач обыкновенный) 304,
309
Botrychium lunaria (Гроздовник полулун-
ный) 191
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
350
– multifidum (Гроздовник многораздель-
ный) 152
Brachypodium pinnatum (Коротконожка
перистая) 139, 163–165, 242
– rupestre (Коротконожка скальная) 301
– sylvaticum (Коротконожка лесная) 304,
314
Braya (Брайя) 66
Briza media (Трясунка средняя) 301
Bromopsis inermis (Кострец безостый) 200,
223, 234, 242, 261
– riparia (Кострец береговой) 223, 232, 234,
252, 255, 280, 309
Bryanthus gmelinii (Мохоцветник Гмелина)
117, 331
Bryum (Бриум) 70
Bupleurum falcatum (Володушка серповид-
ная) 239
– longifolium subsp. aureum (Володушка
золотистая) 315, 316
– multinerve (Володушка многожилковая)
229
– scorzonerifolium (Володушка козелецели-
стная) 264
– triradiatum (Володушка трехлучевая) 111
Buxus colchica (Самшит колхидский) 284,
285
Cacalia hastata (Недоспелка копьевидная)
145, 164
– kamtschatica (Недоспелка камчатская)
115, 329
Calamagrostis arundinacea (Вейник трост-
никовый) 139, 143, 163, 164, 166, 301,
321
– canescens (Вейник седеющий) 163, 164,
165
– epigeios (Вейник наземный) 163–165, 173,
199, 223, 228, 234, 238, 242, 243, 261,
271, 322, 323
– langsdorffii (Вейник Лангсдорфа) 111,
136, 137, 160, 163, 172, 331
– lapponica (Вейник лапландский) 322, 323
– neglecta (Вейник незамечаемый) 164
– obtusata (Вейник притупленный) 160,
161, 163
– phragmitoides (Вейник тростниковидный)
162
– purpurascens (Вейник краснеющий) 111
– purpurea (Вейник пурпурный) 115, 191,
323
Calliergon (Каллиергон) 157
Calligonum aphyllum (Жузгун безлистный)
271
Calluna vulgaris (Вереск обыкновенный) 73,
86, 135, 136, 142, 213, 214
Calypso bulbosa (Калипсо луковичная) 145,
191
Campanula biebersteiniana (Колокольчик
Биберштейна) 302
– collina (Колокольчик холмовой) 300
– glomerata (Колокольчик скученный) 203
– patula (Колокольчик раскидистый) 205
– persicifolia (Колокольчик персиколист-
ный) 231, 232
– rotundifolia (Колокольчик круглолист-
ный) 205
– sibirica (Колокольчик сибирский) 232, 242
– stevenii (Колокольчик Стевена) 255, 256
– tridentata (Колокольчик трехзубый) 303
Camphorosma monspeliaca (Камфоросма
монпелийская) 267
Camptothecium nitens (Камптотеций) 316
Capparis herbacea (Каперсы травянистые)
308, 309
Capsella bursa-pastoris (Пастушья сумка
обыкновенная) 200
Caragana arborescens (Карагана древовид-
ная) 161, 166
– frutex (Карагана кустарниковая) 253, 255,
257–259, 313
– pumila (Карагана низкорослая) 253
– pygmaea (Карагана карликовая) 263, 320
Carduus uncinatus (Чертополох крючкова-
тый) 257
Carex (Осока) 214
– acuta (Осока острая) 210
– arctisibirica (Осока арктосибирская) 72,
73, 95, 99, 327
– arenicola (Осока песколюбивая) 189
– bigelowii (Осока Бигелоу) 71, 73, 88, 89,
97, 140
– brunnescens (Осока буроватая) 164
– buxbaumii (Осока Буксбаума) 164
– campylorhina (Осока кривоносая) 182
– caryophyllea (Осока гвоздичная) 230
– cespitosa (Осока дернистая) 164, 210
– chordorrhiza (Осока плетевидная) 86
– cinerea (Осока седеющая) 90
– concolor (Осока одноцветная) 89, 95, 99,
102, 104, 113, 327
– cryptocarpa (Осока скрытоплодная) 115
– disticha (Осока двурядная) 164, 165
– duriuscula (Осока твердоватая) 111, 253,
264, 327
– globularis (Осока шаровидная) 89, 90, 91,
97, 144, 157
– gmelinii (Осока Гмелина) 115
– humilis (Осока низкая) 223, 232, 239, 252,
309
– juncella (Осока ситничковая) 136
– korshinskyi (Осока Коржинского) 327
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
351
– lanceolata (Осока ланцетная) 190
– lasiocarpa (Осока волосистоплодная) 142
– limosa (Осока топяная) 113, 330
– lugens (Осока траурная) 72, 73, 109
– macrochaeta (Осока длинноостистая) 115,
116
– macroura (Осока большехвостая) 137, 163
– meinshauseniana (Осока Мейнсгаузена)
303
Carex membranacea (Осока перепончатая)
109
– micropoda (Осока коротконожковая) 116
– micropodioides (Осока ложнокоротконож-
ковая) 303
– montana (Осока горная) 230
– nesophila (Осока островная) 109, 112
– obtusata (Осока притупленная) 111
– pallida (Осока бледная) 331
– pediformis (Осока стоповидная) 170, 223
– pilosa (Осока волосистая) 224, 227
– praecox (Осока ранняя) 228
– rariflora (Осока редкоцветковая) 86, 102
– reventa (Осока возвратившаяся) 190
– riishirensis (Осока риширинская) 116
– rostrata (Осока носатая) 142, 164
– rotundata (Осока кругловатая) 86
– rupestris (Осока наскальная) 71, 109, 112
– scirpoidea (Осока камышевидная) 109
– siderosticta (Осока ржавопятнистая) 182
– spaniocarpa (Осока малоплодная) 111
– stenophylla (Осока узколистная) 271
– supina (Осока приземистая) 253, 258
– sylvatica (Осока лесная) 304
– ussuriensis (Осока уссурийская) 182, 190
– vaginata (Осока влагалищная) 109, 164
Carpinus betulus (Граб обыкновенный) 138,
153, 224, 228, 297, 298, 304
– cordata (Граб сердцелистный) 138, 180,
184, 244
– orientalis (Граб восточный, грабинник)
278, 282
Carum carvi (Тмин обыкновенный) 203
Cassiope
ericoides
(Кассиопея
вересковидная) 327
– lycopodioides (Кассиопея плауновидная)
117, 332
– tetragona (Кассиопея четырехгранная) 72,
73, 76, 101, 109, 327
Castanea sativa (Каштан посевной) 284, 299,
300
Caulophyllum robustum (Стеблелист мощ-
ный) 244
Celastrus orbiculata (Древогубец кругло-
листный) 182
Centaurea (Василек) 300
– cyanus (Василек синий) 200
– phrygia (Василек фригийский) 208, 209
– ruthenica (Василек русский) 257
– scabiosa (Василек шероховатый) 164, 165,
233
Cephalanthera rubra (Пыльцеголовник
красный) 233, 234
Cephalaria (Головчатка) 300
Cerastium arcticum (Ясколка арктическая)
81
– bialynickii (Ясколка
Бялыницкого-
Бирули) 82
– brachypetalum (Ясколка коротколепестко-
вая) 280
– holosteoides (Ясколка костенцовая) 203
– maximum (Ясколка крупная) 99
– regelii (Ясколка Регеля) 82
– regelii ssp. caespitosum (Ясколка Регеля
дернистая) 81
– schmalhausenii (Ясколка Шмальгаузена)
280
Cerasus avium (Вишня птичья, черешня)
299
– fruticosa (Вишня кустарниковая) 235, 236,
252, 258, 259
– maximowiczii (Вишня Максимовича) 180,
190
–sachalinensis (Вишня сахалинская) 182
Ceratocarpus arenarius (Рогач песчаный,
эбелек) 271
Cetraria islandica (Цетрария исландская) 70,
75, 140
Cetrariella delisei (Цетрариелла Делиса) 70
Chamaecytisus ruthenicus (Ракитничек
русский) 166, 233, 235, 236, 252, 259,
302
Chamaedaphne calyculata (Хамедафне при-
цветничковая) 89, 113, 142, 156, 157,
213, 214
Chamaenerion angustifolium (Иван-чай уз-
колистный) 164, 199, 330, 332
Chamaepericlymenum suecicum (Дёрен
шведский) 19, 21, 86, 117
Chamaerhodos altaica (Хамеродос алтай-
ский) 263
– trifida (Хамеродос трехнадрезной) 263
Chelidonium majus (Чистотел большой) 233
Chenopodium album (Марь белая) 268
Chimaphila umbellata (Зимолюбка зонтич-
ная) 153, 166, 238
Chosenia arbutifolia (Кореянка землянични-
колистная) 187, 327, 329
Chrysanthemum sinuatum (Хризантема вы-
емчатолистная) 313
– zawadskii (Хризантема Завадского) 229
Cicerbita macrophylla (Цицербита крупно-
листная) 302
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
352
Cichorium intybus (Цикорий обыкновенный)
200
Circae alpina (Двулепестник альпийский)
137
Cirsium arvense (Бодяк полевой) 200
– helenioides (Бодяк девясиловидный) 89,
323
– heterophyllum (Бодяк разнолистный) 164
– kamtschaticum (Бодяк камчатский) 115,
179
Cistus tauricus (Ладанник крымский) 278,
279
Cladium mariscus (Меч-трава обыкновен-
ная) 153
Cladonia (Кладония) 71, 86, 155, 329
– amaurocraea (Кладония темно-мясная)
101
– arbuscula (Кладония лесная) 140, 144,
156, 158, 166
– cornuta (Кладония рогатая) 98
– gracilis ssp. elongata (Кладония удлинен-
ная) 89
– macroceras (Кладония крупнорогая) 95,
96
– mitis (Кладония мягкая) 75, 88, 89, 95, 96,
144
– rangiferina (Кладония оленья) 75, 88, 95,
96, 98, 101, 140, 144, 157, 158, 166
– stellaris (Кладония звездчатая) 86, 98, 101,
140, 141, 156–158, 166
– uncialis (Кладония дюймовая) 98
Claytonia sibirica (Клайтония сибирская)
115
– tuberosa (Клайтония клубненосная) 109
Cleistogenes squarrosa (Змеёвка растопы-
ренная) 253, 262–264
Clematis hexapetala (Ломонос шестилепест-
ковый) 223, 253
– montana (Ломонос горный) 304
– orientalis (Ломонос восточный) 304
– vitalba (Ломонос виноградолистный) 284
– viticella (Ломонос фиолетовый) 284
Climacium dendroides (Климациум древо-
видный) 161
Cnidium cnidiifolium (Жгун-корень книдие-
листный) 111
Coccyganthe flos-cuculi (Кукушкин цвет
обыкновенный, горицвет) 210
Cochlearia arctica (Ложечница арктическая)
67
Comarum palustre (Сабельник болотный)
115, 164
Conioselinum chinense (Гирчовник китай-
ский) 115
– tataricum (Гирчовник татарский) 145
Convallaria keiskei (Ландыш Кейзке) 190
– majalis (Ландыш майский) 137
Convolvulus ruprechtii (Вьюнок Рупрехта)
308
Coptis trifolia (Коптис трехлистный) 115
Corallorrhiza trifida (Ладьян трехраздель-
ный) 150, 153
Corispermum aralo-caspicum (Верблюдка
арало-каспийская) 271
– declinatum (Верблюдка отклонённая, в.
повислая) 252
Cornicularia (Корникулярия) 71
Cornus mas (Кизил мужской) 279, 283, 299
Cortaderia selloana (Кортадерия Селло)
287
Corydalis capnoides (Хохлатка дымянкооб-
разная) 145
– cava (Хохлатка полая) 224, 225
– paczoskii (Хохлатка Пачоского) 281
Corylus avellana (Лещина обыкновенная)
151, 222, 224, 276, 299, 304
–heterophylla (Лещина разнолистная) 182,
222, 223, 244
– mandshurica (Лещина маньчжурская) 182,
222, 244
Cotinus coggygria (Скумпия кожевенная)
228, 279, 283
Cotoneaster
melanocarpus (Кизильник
алаунский) 164, 313, 321
– uniflorus (Кизильник одноцветковый) 145,
316
Crataegus (Боярышник) 224, 228, 301
– chlorosarca (Боярышник зеленомясый)
330
– karadaghensis (Боярышник карадагский)
279
– monogyna (Боярышник однопестичный)
299
– orientalis (Боярышник восточный) 279
– pentagyna (Боярышник пятипестичный)
304
–pinnatifida (Боярышник перистонадрезан-
ный) 245
– pojarkovae (Боярышник Поярковой) 279
– rhipidophylla (Боярышник вееролистный,
б. согнутостолбиковый) 304
– sanguinea (Боярышник кроваво-красный)
163, 235, 242
– sphaenophylla (Боярышник клинолист-
ный) 279
Crepis lyrata (Скерда лировидная) 315
– sibirica (Скерда сибирская) 163, 164, 242
Crocus angustifolius (Шафран узколистный)
281
– reticulatus (Шафран сетчатый) 255
Cryptogramma crispa (Криптограмма кур-
чавая) 140
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
353
Cyclamen adsharicum (Дряква аджарская)
285
– coum (Дряква косская) 285
Cymbaria daurica (Цимбария даурская) 264
Cypripedium calceolus (Венерин башмачок
настоящий) 145, 150, 153
– macranthon (Венерин башмачок крупно-
цветковый) 114
– yatabeanum (Венерин башмачок Ятабе)
116
Dactylina (Дактилина) 71
Dactylis glomerata (Ежа сборная) 164, 165,
200, 203, 208, 232, 304
Dactylorhiza aristata (Пальчатокоренник
остистый) 115
– baltica (Пальчатокоренник длиннолист-
ный) 153
– fuchsii (Пальчатокоренник Фукса) 150
– incarnata (Пальчатокоренник
мясо-
красный) 150
– maculata (Пальчатокоренник пятнистый)
150
– romana (Пальчатокоренник римский) 281
– traunsteineri (Пальчатокоренник Траун-
штейнера) 150
Daphne cneorum (Волчеягодник боровой) 229
– mezereum (Волчеягодник обыкновенный)
151, 224, 235
– pontica (Волчеягодник понтийский) 284
Delphinium elatum (Живокость высокая)
163, 203, 316
– flexuosum (Живокость извилистая) 302
Dentaria quinquefolia (Зубянка пятилист-
ная) 281
Deschampsia alpina (Щучка альпийская) 81
– beringensis (Щучка берингийская) 118
– brevifolia (Щучка коротколистная) 70, 71,
82, 102
– cespitosa (Щучка дернистая) 200, 203, 208
Dianthus acicularis (Гвоздика иглолистная)
291
– guttatus (Гвоздика пятнистая) 257
– humilis (Гвоздика низкая) 280
– leptopetalus (Гвоздика узколепестная) 261
– versicolor (Гвоздика разноцветная) 242
Diapensia lapponica (Диапенсия лапланд-
ская) 141
– obovata (Диапенсия обратнояйцевидная)
109, 327, 331
Dicranum angustum (Дикранум узкий) 95,
96
– elongatum (Дикранум удлиненный) 86, 88,
91, 157
– flexcaule (Дикранум извилистостебель-
ный) 86, 88, 157
– fuscescens (Дикранум буроватый) 88
– majus (Дикранум большой) 90, 330
– polysetum (Дикранум многоножковый)
90, 156, 160, 166, 322, 323
– scoparium (Дикранум метловидный) 90,
324
Diphasiastrum complanatum (Двурядник
сплюснутый) 144, 160, 330
Diplazium sibiricum (Диплазиум сибирский)
161
Diplotaxis cretacea (Двурядка меловая) 239
Ditrichum flexicaule (Дитрихум кривосте-
бельный) 70
Doellingeria scabra (Деллингерия шерша-
вая) 244
Doronicum macrophyllum (Дороникум круп-
нолистный) 302
Dorycnium graecum (Птилистник гречес-
кий) 280
Draba macrocarpa (Крупка крупноплодная)
99
– nemorosa (Крупка дубравная) 81, 255
– pauciflora (Крупка малоцветковая) 99
– pilosa (Крупка волосистая) 76
– pohlei (Крупка Поле) 99
– subcapitata (Крупка почти-головчатая) 19,
20
Dracocephalum grandiflorum (Змееголовник
крупноцветковый) 316
– nutans (Змееголовник поникающий) 313
– peregrinum (Змееголовник иноземный) 313
– ruyschiana (Змееголовник Рюйша) 145
Drepanocladus (Дрепанокладус) 157
Drosera anglica (Росянка английская) 214
– rotundifolia (Росянка круглолистная) 156,
214
Dryas (Дриада, куропаточья трава) 67, 71
– integrifolia (Дриада цельнолистная) 72,
73, 81, 109
– octopetala (Дриада восьмилепестковая) 72,
81, 88, 109, 141, 145, 293
– oxyodonta (Дриада острозубчатая) 316
– punctata (Дриада точечная) 72, 81, 91, 95,
97, 109, 327, 332
Dryopteris carthusiana (Щитовник картузи-
анский) 161, 163
– crassirhizoma (Щитовник толстокорне-
вищный) 182, 244
– filix-mas (Щитовник мужской) 147
Dupontia fisheri (Дюпонция Фишера) 71, 99
Duschekia fruticosa (Ольховник кустарнико-
вый) 73, 78, 96, 111, 171, 326, 327, 333
– kamtschatica (Ольховник камчатский)
111, 329
– manshurica (Ольховник маньчжурский)
137
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
354
Echium russicum (Синяк русский) 230, 255
Eleutherococcus senticosus (Свободноягод-
ник колючий, элеутерокок колючий)
185
Elymus confusus (Пырейник смешиваемый)
111
Elytrigia gracillima (Пырей стройный) 309
– repens (Пырей ползучий) 28, 164, 200,
203, 242, 243, 261, 267–269
Empetrum (Водяника) 81
– hermaphroditum (Водяника гермафродит-
ная) 73, 86, 88–91, 135, 140
– nigrum (Водяника черная) 77, 98, 109,
137, 144, 170, 332
– sibiricum (Водяника сибирская) 117
– subholarcticum
(Водяника
почти-
голарктическая) 73, 136
Endocellion glaciale (Эндоцеллион ледяной)
109
Enemion raddeanum (Энемион Радде) 191
Ephedra distachya (Хвойник двухколоско-
вый эфедра) 239, 240
– procera (Хвойник рослый) 283
Epilobium hornemannii (Кипрей Горнемана)
115
Epipactis atrorubens (Дремлик темно-крас-
ный) 145
– helleborine (Дремлик лесной) 150
Epipogium aphyllum (Надбородник без-
листный) 150, 233, 234
Equisetum arvense (Хвощ полевой) 115, 331
– hyemale (Хвощ зимующий) 331
– pratense (Хвощ луговой) 157, 203, 331,
332
– scirpoides (Хвощ камышовый) 157
– sylvaticum (Хвощ лесной) 90, 91, 157, 160,
164, 331, 332
Eremogone capillaris (Пустынница волосо-
видная) 111, 262, 263
– saxatilis (Пустынница скальная) 230, 255
Eremopyrum orientale (Мортук восточный)
268
– triticeum (Мортук пшеничный) 268
Eriophorum (Пушица) 67, 81, 89
– angustifolium (Пушица узколистная) 71,
95, 96, 99, 102, 213
– medium (Пушица средняя) 102
– russeolum (Пушица рыжеватая) 86
– scheuchzeri (Пушица Шейхцера) 71, 99,
104
– vaginatum (Пушица влагалищная) 73, 91,
96, 97, 105, 113, 156, 214
Eritrichium sericeum (Незабудочник шелко-
вистый) 111
– villosum (Незабудочник волосистый) 71
Erophila praecox (Веснянка ранняя) 280
– verna (Веснянка весенняя) 255, 260
Eryngium campestre (Синеголовник поле-
вой) 280
– planum (Синеголовник плоский) 261
Euonymus europaea (Бересклет европей-
ский) 222, 224, 304
– macroptera (Бересклет большекрылый)
185, 193
– pauciflora (Бересклет малоцветковый)
190, 191, 244
– sacrosancta (Бересклет священный) 190
– verrucosa (Бересклет бородавчатый) 151,
222, 224, 304
Euphorbia amygdaloides (Молочай минда-
левидный) 307
– seguieriana (Молочай Сегье) 249, 258
–
stepposa (Молочай степной) 252
– subtilis (Молочай тонкий) 230
Euphrasia hirtella (Очанка мохнатая) 203
Eutrema edwardsii (Эвтрема Эдвардса) 109
Fagus orientalis (Бук восточный) 284, 298,
300, 302
– sylvatica (Бук лесной) 138, 153, 224
Falcaria vulgaris (Резак обыкновенный) 233,
257
Ferula caspica (Ферула каспийская) 257
– ferulaeoides (Ферула феруловидная) 257
– orientalis (Ферула восточная) 257
Festuca airiodes (Овсяница приземистая) 88,
89, 173, 303
– altaica (Овсяница алтайская) 109
– altissima (Овсяница высочайшая) 314
– auriculata (Овсяница ушковатая) 103
– brachyphylla (Овсяница коротколистная)
99
– drymeja (Овсяница лесная) 304
– gigantea (Овсяница гигантская) 314
– jacutica (Овсяница якутская) 171
– lenensis (Овсяница ленская) 111, 172, 253,
263, 326
– ovina (Овсяница овечья) 89, 142, 144, 200,
205, 207, 242, 250
– pratensis (Овсяница луговая) 164, 165,
203, 205, 206, 228
– pseudovina (Овсяница ложноовечья) 242,
243, 259, 260
– richardsonii (Овсяница Ричардсона) 99,
103
– rubra (Овсяница красная) 165, 203, 208,
230
– rupicola (Овсяница каменистая) 255
– ruprechtii (Овсяница Рупрехта) 303
– valesiaca (Овсяница валлисская, типчак)
165, 230, 232, 249, 252–254, 257–261,
268–271, 309, 312, 313
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
355
Ficus carica (Смоковница обыкновенная)
286
Filifolium sibiricum (Нителистник сибир-
ский) 223
Filipendula camtschatica (Лабазник камчат-
ский, шеломайник) 115, 178, 179, 329,
332
– ulmaria (Лабазник вязолистный, таволга)
154, 164, 165, 210
– vulgaris (Лабазник обыкновенный) 163,
223, 228, 230, 232, 233, 242, 252, 253,
255, 258, 259, 261
Flavocetraria (Флавоцетрария) 71
– cucullata (Флавоцетрария клобучковая)
70, 75, 88, 95, 96, 105, 155
– nivalis (Флавоцетрария снежная) 86, 141
Fragaria vesca (Земляника лесная) 203
– viridis (Земляника зеленая) 233, 242
Frangula alnus (Крушина ольховидная) 142,
151, 233, 299
Fraxinus excelsior (Ясень высокий, я. обык-
новенный) 138, 153, 221, 222, 279, 284,
298, 304
– mandshurica (Ясень маньчжурский) 180,
221, 245
– oxycarpa (Ясень остроплодный) 279
– rhynchophylla (Ясень носолистный) 139,
180, 184, 244, 180
Fritillaria meleagris (Рябчик шахматный)
229
–
ruthenica (Рябчик русский) 229, 233,
234
Fumana (Фумана) 278
Gagea bulbifera (Гусиный лук луковиценос-
ный) 255
– granulosa (Гусиный лук зернистый) 243,
260
– lutea (Гусиный лук желтый) 224, 225
– minima (Гусиный лук маленький) 243,
260
– pusilla (Гусиный лук низкий) 255
Galanthus caucasicus (Подснежник кавказ-
ский) 285
– krasnovii (Подснежник Краснова) 285
– plicatus (Подснежник складчатый) 281
– woronowii (Подснежник Воронова) 285
Galatella angustissima (Солонечник узко-
листный) 258
–
biflora (Солонечник двуцветковый) 243,
259
– tatarica (Солонечник татарский) 253, 259
– villosa (Солонечник мохнатый, грудница
мохнатая) 253, 257–259, 261
Galega orientalis (Козлятник восточный)
301
Galeobdolon luteum (Зеленчук желтый) 137,
147, 224
Galium aparine (Подмаренник цепкий) 227
– boreale (Подмаренник северный) 163,
165, 166, 203, 230, 242, 323
– mollugo (Подмаренник мягкий) 203
– odoratum (Подмаренник душистый) 147,
314
– verum (Подмаренник настоящий) 223,
228, 231–233, 242, 257, 258, 261, 263
Genista taurica (Дрок крымский) 281
– tinctoria (Дрок красильный) 235
– verae (Дрок Веры) 280
Gentiana glauca (Горечавка сизая) 116
– pneumonanthe (Горечавка легочная) 163
Geranium albiflorum (Герань белоцветковая)
316
– asiaticum (Герань азиатская) 243
– erianthum (Герань пушистоцветковая)
115, 332
– gymnocaulon (Герань голостебельная) 301
– krylovii (Герань Крылова) 323
– maximowiczii (Герань Максимовича) 244
– pretense (Герань луговая) 203
– pseudosibiricum (Герань ложносибирская)
163, 315
– robertianum (Герань Роберта) 227
– sylvaticum (Герань лесная) 165, 166, 206
Geum rivale (Гравилат речной) 203, 208,
209
Glechoma hederacea (Будра плющевидная)
203, 227
Globularia punctata (Шаровница точечная)
241
Glyceria alnasteretum (Манник ольховнико-
вый) 115
Glycyrrhiza glabra (Солодка голая) 269
– uralensis (Солодка уральская) 243
Goniolimon speciosum (Углостебельник кра-
сивый) 260
– tataricum (Углостебельник татарский)
256, 257
Goodyera repens (Гудайера ползучая) 150,
160, 331
Gymnocarpium dryopteris (Голокучник
обыкновенный) 157, 160, 163, 331
Gypsophila altissima (Качим высочайший)
239
– paniculata (Качим метельчатый) 252, 261
– patrinii (Качим Патрэна) 313
– uralensis (Качим уральский) 145, 290
Halimione verrucifera (Халимион бородав-
чатый) 243
Hammarbya paludosa (Хаммарбия болот-
ная) 153
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
356
Harrimanella hypnoides (Гариманелла мо-
ховидная) 141
Hedera colchica (Плющ колхидский) 284
– helix (Плющ обыкновенный) 284
– pastuchowii (Плющ Пастухова) 304
Hedysarum alpinum (Копеечник альпий-
ский) 89
– arcticum (Копеечник арктический) 103,
109
– gmelinii (Копеечник Гмелина) 261
– grandiflorum (Копеечник крупноцветко-
вый) 239
Helianthemum cretaceum (Солнцецвет мело-
вой) 239
Helichrysum arenarium (Цмин песчаный)
262
Helictotrichon adzharicum (Овсец аджар-
ский) 303
– desertorum (Овсец пустынный) 223, 234,
252, 253, 258, 259, 261
– krylovii (Овсец Крылова) 111, 326
– pubescens (Овсец пушистый) 232
– schellianum (Овсец Шелля) 223, 252
Heliotropium styligerum (Гелиотроп столби-
коносный) 309
Hemerocallis minor (Краснодев малый)
253
Hepatica nobilis (Печеночница благородная)
21, 22, 137, 142, 151, 224, 225
Heracleum (Борщевик) 302
– dissectum (Борщевик рассеченный) 164,
242
– lanatum (Борщевик шерстистый) 115, 178,
179, 329
– sibiricum (Борщевик сибирский) 164, 203,
242
– sosnowskyi (Борщевик Сосновского) 302
Hieracium umbellatum (Ястребинка зонтич-
ная) 163–166, 203, 238
– virosum (Ястребинка ядовитая) 262
Hierochloe alpine (Зубровка альпийская)
109, 324
Holosteum umbellatum (Костенец зонтич-
ный) 255
Honckenya peploides (Гонкения бутерлако-
видная) 189
Hordeun bulbosum (Ячмень луковичный)
278
Humulus lupulus (Хмель обыкновенный)
284
Hyacinthella leucophaea (Гиацинтик пепель-
но-серый) 232, 255
– pallasiana (Гиацинтик Палласа) 255
Hylocomium splendens (Гилокомиум бле-
стящий) 74, 88, 96, 140, 143, 157, 160,
161, 316, 323
– splendens var. obtusifolium (Гилокомиум
блестящий круглолистный) 101
Hylomecon vernalis (Лесной мак весенний)
182
Hylotelephium triphyllum (Очитник трех-
листный) 165
Ilex aquifolium (Падуб остролистный) 284
– colchica (Падуб колхидский) 299
Impatiens noli-tangere (Недотрога обыкно-
венная) 227
Inula (Девясил) 300
– aspera (Девясил шероховатый) 280
– salicina (Девясил иволистный) 261
Iris aphylla (Ирис безлистный) 231, 232
– pumila (Ирис карликовый) 253, 255
– ruthenica (Касатик русский) 173, 243
– setosa (Ирис щетинистый) 115, 116
– uniflora (Касатик одноцветковый) 191
Isoetes asiatica (Полушник азиатский) 114
– setacea (Полушник щетинистый) 153
Jasminum fruticans (Жасмин кустарнико-
вый) 279
Jovibarba sobolifera (Бородник шаронос-
ный) 153
Juglans mandshurica (Орех маньчжурский)
180, 184, 245
– regia (Орех грецкий) 304
Juncus beringensis (Ситник беринговский)
116
– effusus (Ситник развесистый) 208
– trifidus (Ситник трехраздельный) 88
Juniperus communis (Можжевельник обык-
новенный) 97, 141, 144, 166, 322, 323
– excelsa (Можжевельник высокий) 277,
282, 283, 307
– foetidissima (Можжевельник вонючий)
278, 280, 282, 307
– hemisphaerica (Можжевельник полушаро-
видный) 300
– oblonga (Можжевельник длиннолистный)
283
– oxycedrus (Можжевельник красный) 279,
307
– polycarpos (Можжевельник многоплод-
ный) 307
– rigida (Можжевельник твердый) 182
– sabina (Можжевельник казацкий) 300
– sibirica (Можжевельник сибирский) 171,
316, 326, 331
Jurinea arachnoidea (Наголоватка паутини-
стая) 255, 258
– cyanoides (Наголоватка васильковая) 261
– multiflora (Наголоватка многоцветковая)
257, 258
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
357
– ruprechtii (Наголоватка Рупрехта) 309
Kadenia dubia (Кадения сомнительная)
163–165
Kalopanax septemlobus (Калопанакс семи-
лопастный) 177, 180, 182
Knautia arvensis (Короставник полевой) 232
Kobresia filifolia (Кобрезия нителистная)
326
– myosuroides (Кобрезия мышехвостнико-
вая) 327
Kochia prostrata (Кохия простертая, прут-
няк) 253, 264, 268, 269, 271, 272
Koeleria (Тонконог) 263
– asiatica (Тонконог азиатский) 103
– cristata (Тонконог гребенчатый) 232, 242,
251, 252, 253, 255, 257–259, 261, 264,
312, 313
– delavignei (Тонконог Делявиня) 230
– glauca (Тонконог сизый) 251
– gracilis (Тонконог гребенчатый) 326
– sclerophylla (Тонконог жестколистный)
250
– tokiensis (Тонконог токийский) 188
Laburnum anagyroides (Бобовник анагиру-
совидный) 228
Lactuca sibirica (Латук сибирский) 145
Lagotis glauca (Лаготис сизый) 116
Lamium maculatum (Яснотка пятнистая)
227
Larix cajanderi (Лиственница Каяндера)
105, 136, 325, 330, 331
– gmelinii (Лиственница даурская) 101, 136,
137, 167, 171, 180, 327
– kamtschatica (Лиственница камчатская)
136, 329
– olgensis (Лиственница ольгинская) 188,
191
– sibirica (Лиственница сибирская) 90, 96,
135–137, 140, 143, 157, 158, 172, 291,
313, 315, 326
Laser trilobum (Лазурник трехлопастный)
241
Lathraea squamaria (Петров крест чешуйча-
тый) 153
Lathyrus gmelinii (Чина Гмелина) 315
– humilis (Чина приземистая) 321, 322, 323
– japonicus (Чина японская) 189
– laxiflorus (Чина рыхлоцветковая) 281
– litvinovii (Чина Литвинова) 291
– pallescens (Чина бледнеющая) 255
– pannonicus (Чина венгерская) 232, 255
– pilosus (Чина волосистая) 115
– pisiformis (Чина гороховидная) 164, 165
– pratensis (Чина луговая) 163, 203, 242
– vernus (Чина весенняя) 224, 225
Laurocerasus officinalis (Лавровишня лекар-
ственная) 284
Ledum (Багульник) 81
– decumbens (Багульник стелющийся) 81,
96–98, 109, 111, 171, 326
– hypoleucum (Багульник подбел) 136, 187
– palustre (Багульник болотный) 81, 86, 89,
90, 96, 113, 135, 136, 140, 142, 144, 156,
213, 214
Leontodon autumnalis (Кульбаба осенняя)
203
– hispidus (Кульбаба щетинистая) 205
Leptirumohra
amurensis (Лепторумора
амурская) 177, 182
Lespedeza bicolor (Леспедеца двуцветная)
186, 189, 223, 244
– juncea (Леспедеца ситниковая) 223
Leucanthemum margaritae (Нивяник Мар-
гариты) 232
– vulgare (Нивяник обыкновенный) 203,
205, 206, 228, 230
Leymus chinensis (Колосняк китайский,
вострец) 253, 264
– mollis (Колосняк мягкий) 115, 189
– racemosus (Волоснец кистистый) 271
– ramosus (Колосняк ветвистый) 253, 257,
269
Ligularia sibirica (Бузульник сибирский) 153
Ligusticum scoticum (Лигустикум шотланд-
ский) 115
Lilium martagon (Лилия царские кудри,
лилия саранка) 153, 164, 229
Limnas stelleri (Болотник Стеллера) 136, 172
Limoniopsis owerinii (Кермековидка Овери-
на) 308
Limonium bungei (Кермек Бунге) 257
– coralloides (Кермек коралловидный) 260
– gmelini (Кермек Гмелина) 243, 260
– platyphyllum (Кермек плосколистный)
268
– sareptanum (Кермек сарептский) 257
Linaria biebersteinii (Льнянка Биберштейна)
253
Linnaea borealis (Линнея северная) 19, 21,
90, 136, 157, 161, 322, 323, 331
Linum hypericifolium (Лён зверобоелист-
ный) 301
– ucranicum (Лён украинский) 239
Liparis loeselii (Глянцелистник Лёзеля) 153,
229
Listera cordata (Тайник сердцевидный) 145,
153
– ovata (Тайник яйцевидный) 150
Lloydia serotina (Ллойдия поздняя) 99
Lobaria pulmonaria (Лобария легочная) 150
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
358
Lobelia dortmanna (Лобелия Дортмана) 153
Loiseleuria procumbens (Луазелерия лежа-
чая) 88, 327
Lonicera altaica (Жимолость алтайская) 160
– caerulea (Жимолость голубая) 332
– caprifolium (Жимолость козья) 299
– chamissoi (Жимолость Шамиссо) 331
– chrysantha (Жимолость золотистая) 182
– edulis (Жимолость съедобная) 178, 331
– iberica (Жимолость грузинская) 283
– maximoviczii (Жимолость Максимовича)
182, 185
– orientalis (Жимолость восточная) 302
– pallasii (Жимолость Палласа) 322, 323
– tatarica (Жимолость татарская) 235, 242,
314
– xylosteum (Жимолость обыкновенная)
151, 161, 163, 222, 235
Lunathyrium pycnosorum (Лунокучник гу-
стосорусовый) 182, 244
Luzula campestris (Ожика равнинная) 203
– confusa (Ожика спутанная) 72, 327
– nivalis (Ожика снежная) 72, 95, 96, 109
– pilosa (Ожика волосистая) 136, 144
– sibirica (Ожика сибирская) 324
– tundricola (Ожика тундровая) 109
Lychnis samojedorum (Зорька самоедов) 111
Lycopodium annotinum (Плаун годичный)
111, 157, 160, 331
– clavatum (Плаун булавовидный) 144
– dubium (Плаун сомнительный) 111
Lysimachia vulgaris (Вербейник обыкновен-
ный) 209, 210
Maackia amurensis (Маакия амурская) 182,
185, 193
Magnolia soulangeana (Магнолия Суланжа)
287
Maianthemum bifolium (Майник двулист-
ный) 136, 154, 161–163, 166, 322–324,
331
Malaxis monophyllos (Мякотница однолист-
ная) 150, 153
Malus mandshurica (Яблоня маньчжурская)
245
– orientalis (Яблоня восточная) 298, 304
– praecox (Яблоня ранняя) 235
Matteuccia struthiopteris (Страусник обык-
новенный) 163
Matthiola fragrans (Левкой пахучий) 239
Medicago falcata (Люцерна серповидная)
200, 242, 258, 261, 262
– romanica (Люцерна румынская) 252, 257
Melampyrum pretense (Марьянник луговой)
90, 143, 160, 166
– setaceum (Марьянник щетинистый) 190
Melica nutans (Перловник поникающий)
161, 164, 322
Menispermum dauricum (Луносемянник
даурский) 182
Mercurialis perennis (Пролесник многолет-
ний) 224, 227
Mertensia maritima (Мертензия морская)
189
Mespilus germanica (Мушмула германская)
304
Milium effusum (Бор развесистый) 161, 323
– vernale (Бор весенний) 280
Minuartia arctica (Минуарция арктическая)
323
– krascheninnikovii (Минуарция Крашенин-
никова) 291
– macrocarpa (Минуарция крупноплодная)
99
– rubella (Минуарция красноватая) 88
Moehringia lateriflora (Мерингия бокоцвет-
ковая) 115
Moneses uniflora (Одноцветка обыкновен-
ная) 331
Morus alba (Шелковица белая) 304
Musa (Банан) 287
Myosotis alpestris (Незабудка альпийская)
71, 301
– popovii (Незабудка Попова) 231
– sylvatica (Незабудка лесная) 228, 230, 232,
255
Myrica gale (Восковник болотный) 138
– tomentosa (Восковник войлочный) 137
Nardus stricta (Белоус торчащий) 200, 207,
303
Neckera complanata (Неккера сплюснутая)
285
– crispa (Неккера курчавая) 285
Neolindleya camtschatica (Любка камчат-
ская) 114
Neottia nidus-avis (Гнездовка настоящая)
150, 153
Neottianthe cucullata (Гнездоцветка клобуч-
ковая) 233, 234
Nephroma arcticum (Нефрома арктическая)
89, 98, 155
Nostoc commune (Носток обыкновенный)
255
Nuphar pumila (Кубышка малая) 145
Oberna littoralis (Хлопушка прибрежная)
203
Oncophorus wahlenbergii (Онкофорус Ва-
ленберга) 70
Onobrychis arenaria (Эспарцет песчаный)
232, 233
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
359
– cornuta (Эспарцет рогатый) 308, 309
– sibirica (Эспарцет сибирский) 258
Onoclea sensibilis (Оноклея чувствительная)
188
Onosma simplicissima (Оносма простейшая)
239, 240, 258, 262
Ophioglossum vulgatum (Ужовник обыкно-
венный) 152
Oplopanax elatus (Заманиха высокая) 188
Orchis militaris (Ятрышник шлемоносный)
153
Origanum vulgare (Душица обыкновенная)
164
Ornithogalum fimbriatum (Птицемлечник
бахромчатый) 281
– navaschinii (Птицемлечник Навашина)
255
Orogeum montanum (Гравилат горный) 304
Orostachys spinosa (Горноколосник колю-
чий) 262, 263
Orthilia secunda (Ортилия однобокая) 136,
160, 166, 238, 331
Orthothecium chryseon (Ортотециум золоти-
стый) 70
Osmundastrum asiaticum (Чистоустник ази-
атский) 188, 244
Oxalis acetosella (Кислица обыкновенная)
137, 154, 162, 182
– obtriangulata (Кислица обратнотреуголь-
ная) 244
Oxycoccus (Клюква) 214
– microcarpus (Клюква мелкоплодная) 91,
111
– palustris (Клюква болотная) 77, 213
Oxygraphis glacialis (Оксиграфис ледяной)
103
Oxyria digyna (Кисличник двустолбиковый)
67
Oxytropis campestris (Остролодочник поле-
вой) 111
– karga (Остролодочник каргинский) 103
– middendorffii (Остролодочник Мидден-
дорфа) 103, 111
– oxyphylla (Остролодочник остролистный)
263
– pilosa (Остролодочник волосистый) 258
– sordida (Остролодочник грязноватый) 89,
293
– vassilczenkoi var. substepposa (Остроло-
дочник Васильченко вар. остепненный)
111
Padus asiatica (Черёмуха азиатская) 193,
329
– avium (Черёмуха обыкновенная) 154, 161,
164
– maackii (Черёмуха Маака) 180, 184, 193
– ssiori (Черёмуха Сьори, ч. айнская) 177
Paeonia anomala (Пион уклоняющийся,
Марьин корень) 145
– tenuifolia (Пион тонколистный) 255, 256
Paliurus spina-christi (Держи-дерево колю-
чее) 278, 282, 283
Panax ginseng (Женьшень обыкновенный)
186, 244
Papaver (Мак) 72
– lapponicum ssp. jugoricum (Мак югорский)
88
– polare (Мак полярный) 66, 81
– pulvinatum (Мак подушковидный) 103
– radicatum (Мак корневой) 71
Parageum calthifolium (Лжегравилат калуж-
ницелистный) 115–117
Paris quadrifolia (Вороний глаз четырех-
листный) 144, 162, 163, 314
Parnassia palustris (Белозор болотный) 115
Patrinia rupestris (Патриния скальная) 190,
313
– sibirica (Патриния сибирская) 324
Paulownia tomentosa (Павловния войлоч-
ная) 286
Pedicularis (Мытник) 66, 67, 99
– amoena (Мытник приятный) 99, 103
– capitata (Мытник головчатый) 109
– incarnata (Мытник мясо-красный) 323
– kaufmannii (Мытник Кауфмана) 233
– lapponica (Мытник лапландский) 113
– resupinata (Мытник перевернутый) 332
– sceptrum-carolinum (Мытник царский
скипетр) 153
– verticillata (Мытник мутовчатый) 99
Peltigera aphthosa (Пельтигера пупырчатая)
89, 98, 155
– malacea (Пельтигера мягкая) 155
Periploca graeca (Обвойник греческий) 284,
304
Petasites amplus (Белокопытник широкий)
178, 179
– frigidus (Белокопытник холодный) 89
Peucedanum morisonii (Горичник Морисона)
223, 242, 243, 253, 259–261
– oreoselinum (Горичник горный) 233
– ruthenicum (Горичник русский) 253, 257
Phegopteris connectilis (Буковник обыкно-
венный) 163
Phellodendron amurense (Бархат амурский)
182, 183, 245
– sachalinense (Бархат сахалинский) 177
Philadelphus tenuifolius (Чубушник тонко-
листный) 180, 182
Phippsia algida (Фиппсия холодная) 71, 82,
103
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
360
Phleum alpinum (Тимофеевка альпийская)
301, 316
– nodosum (Тимофеевка узловатая) 301
– phleoides (Тимофеевка степная) 163–165,
223, 232, 242, 261, 262
– pratense (Тимофеевка луговая) 165, 200,
203, 205, 206, 232
Phlojodicarpus sibiricus (Вздутоплодник си-
бирский) 253
Phlomis pungens (Зопник колючий) 257, 280
Phlomoides agrarian (Зопничек полевой)
261
– oreophila (Зопничек горолюбивый) 316
– tuberosa (Зопник клубненосный) 165, 230,
242, 258, 261, 263
Phoenix canariensis (Финик канарский) 287
Phryma asiatica (Фрима азиатская) 186, 193
Phyllitis scolopendrium (Листовик обыкно-
венный) 285
Phyllodoce aleutica (Филлодоце алеутская)
332
– caerulea (Филлодоце голубая) 88, 327, 332
Physalis alkekengi (Физалис обыкновенный)
304
Picea abies (Ель европейская) 130, 135–141,
144, 146, 147, 153
– ajanensis (Ель аянская) 136, 176, 180, 183,
327, 329, 332
– fennica (Ель финская) 136
– glehnii (Ель Глена) 176
– koraiensis (Ель корейская) 180
– obovata (Ель сибирская) 86, 89, 96, 130,
135–137, 139–141, 144–147, 153, 157,
158, 173, 291, 326, 327
– orientalis (Ель восточная) 298
Pilosella echioides (Ястребинка румянковид-
ная) 258
– officinarum (Ястребиночка обыкновенная)
205, 207
Pimpinella rhodantha (Бедренец розово-
цветный) 300
– saxifraga (Бедренец камнеломка) 200, 203
Pinguicula spathulata (Жирянка лопатчатая)
111
Pinus friesiana (Сосна Фриза) 135
– funebris (Сосна могильная) 187, 223
– kochiana (Сосна Коха) 302, 307
– koraiensis (Сосна корейская) 129, 138, 139,
180, 183
– pallasiana (Сосна Палласова, с. крымская)
279–281
– pityusa (Сосна пицундская) 279, 280, 282,
285
– pumila (Сосна низкая, кедровый стланик)
78, 80, 136, 322, 324, 327, 329, 331,
333
– sibirica (Сосна сибирская, с. кедровая) 129,
135–137, 144, 158, 173, 291, 323, 327
– sylvestris (Сосна обыкновенная) 86, 135–
139, 143, 157, 161, 165, 172, 214, 222,
291, 318, 327
– sylvestris f. litwinowii (Сосна обыкновен-
ная форма Ливинова) 215
– sylvestris f. pumila (Сосна обыкновенная
форма низкая) 215
– sylvestris f. uliginosa (Сосна обыкновенная
форма топяная) 215
– sylvestris f. willkommii (Сосна обыкновен-
ная форма Вилькома) 215
– uncinata (Сосна крючковатая) 299
Pistacia mutica (Фисташка туполистная)
278, 282
Plagiorhegma dubia (Косоплодник сомни-
тельный) 182, 244
Plantago media (Подорожник средний) 203,
243, 257
– salsa (Подорожник солончаковый) 243
– urvillei (Подорожник Урвилла) 242, 258
Platanthera bifolia (Любка двулистная) 150
Pleurospermum uralense (Ребросемянник
уральский) 162–164, 242, 316, 329
Pleurozium schreberi (Плевроциум Шребера)
74, 88, 90, 97, 98, 140, 143, 156, 157,
160, 166, 322, 331
Poa alpigena (Мятлик альпигенный) 99
– alpina (Мятлик альпийский) 72
– angustifolia (Мятлик узколистный) 223,
228, 232, 242, 243, 255
– arctica (Мятлик арктический) 71, 88, 109
– attenuata (Мятлик оттянутый) 263, 313,
322, 326
– botryoides (Мятлик кистевидный) 172,
253, 263, 264
– bulbosa (Мятлик луковичный) 255, 280
– glauca (Мятлик сизый) 99, 109
– iberica (Мятлик грузинский) 301
– lanata (Мятлик шерстистый) 116
– macrocalyx (Мятлик крупночашуйный)
115, 118
– malacantha (Мятлик мягкоцветковый)
115
– meyeri (Мятлик Мейера) 301
– nemoralis (Мятлик дубравный) 190
– pratensis (Мятлик луговой) 164, 203, 205,
321
– pseudoabbreviata (Мятлик ложноукоро-
ченный) 103
– pseudopalustris (Мятлик ложноболотный)
190
– remota (Мятлик расставленный) 145
– sibirica (Мятлик сибирский) 315, 316
– tanfiljewii (Мятлик Танфильева) 145
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
361
– transbaicalica (Мятлик забайкальский)
242
– trivialis (Мятлик обыкновенный) 164
Polemonium acutiflorum (Синюха остроцвет-
ковая) 115, 116
– caeruleum (Синюха голубая) 164
Polygala amarella (Истод горьковатый) 203
– comosa (Истод хохлатый) 203
– cretacea (Истод меловой) 239
Polygonatum multiflorum (Купена много-
цветковая) 227
– odoratum (Купена душистая) 142, 164,
224, 238
Polygonum aviculare (Спорыш птичий) 268
Polypodium australe (Многоножка южная)
284
Polystichum lonchitis (Многорядник копье-
видный) 140
Polytrichum commune (Политрихум обык-
новенный, кукушкин лён) 89,–91, 98,
143, 160, 323, 331
– strictum (Политрихум сжатый) 74, 86, 88,
96, 98, 316, 323
Populus alba (Тополь белый) 235, 304
– canescens (Тополь сереющий) 304
– davidiana (Тополь Давида) 189
– maximowiczii (Тополь Максимовича) 192
– nigra (Тополь черный) 235, 304
– suaveolens (Тополь душистый) 326, 329
– tremula (Осина обыкновенная) 137, 139,
140, 223, 242
Potentilla (Лапчатка) 67, 81
– acaulis (Лапчатка бесстебельная) 262–264
– argentea (Лапчатка серебристая) 205
– biflora (Лапчатка двухцветковая) 109
– bifurca (Лапчатка вильчатая) 262, 264
– chrysantha (Лапчатка золотистоцветко-
вая) 315
– crantzii (Лапчатка Кранца) 303
– heptaphylla (Лапчатка семилисточковая)
230
– humifusa (Лапчатка распростертая) 255,
261
– micrantha (Лапчатка мелкоцветковая) 281
– nivea (Лапчатка снежная) 103, 111, 326
– patula (Лапчатка оттопыренно-пушистая)
255
Prangos odontalgica (Прангос противозуб-
ной) 257
Primula pallasii (Первоцвет Палласа) 314
– veris (Первоцвет весенний) 233
Prunella vulgaris (Черноголовка обыкно-
венная) 203, 205
Prunus cerasifera (Слива вишненосная) 298
– spinosa (Слива колючая, тёрн обыкновен-
ный) 233, 235, 236, 252, 255, 304
– stepposa (Слива степная, тёрн степной)
228
Pseudobryum cinclidioides (Псевдобриум
цинклидиевидный) 161
Psoroma hypnorum (Псорома гипновая) 70
Pteridium aquilinum (Орляк обыкновенный)
19, 20, 163
Pteris cretica (Птерис критский) 285
Pterocarya pterocarpa (Лапина крылоплод-
ная) 304
Ptilidium ciliare (Птеридиум реснитчатый)
88, 98, 101
Ptilium crista-castrensis (Птилиум гребенча-
тый) 143, 157, 160, 166, 322, 323
Puccinellia angustatа (Бескильница сужен-
ная) 66
– tenuissima (Бескильница тончайшая) 243
– vahliana (Бескильница Валя) 66
– wrightii (Бескильница Райта) 66
Pulmonaria mollis (Медуница мягкая) 161,
163–165, 242
– obscura (Медуница неясная) 137, 145, 147,
166, 224
Pulsatilla multifida (Прострел многораз-
дельный) 111, 171, 252
– orientali-sibirica (Прострел желтеющий)
173, 253, 259, 261, 322, 327
– patens (Прострел раскрытый) 151, 230,
231, 232
–
turczaninovii (Прострел Турчанинова)
263
– vernalis (Прострел весенний) 140
Pyracantha coccinea (Пираканта ярко-
красная) 279
Pyrethrum coccineum (Поповник красный)
301
Pyrola (Грушанка) 136
– incarnata (Грушанка мясо-красная) 322,
331
– japonica (Грушанка японская) 182
– minor (Грушанка малая) 163, 166, 322,
331
– rotundifolia (Грушанка круглолистная)
163, 166
Pyrus caucasica (Груша кавказская) 299, 304
– communis (Груша обыкновенная) 299
– elaeagrifolia (Груша лохолистная) 279
– ussuriensis (Груша уссурийская) 180, 245
Quercus crispula (Дуб курчавенький) 177
– dentata (Дуб зубчатый) 244
– hartwissiana (Дуб Гартвиса) 297
– iberica (Дуб грузинский) 284, 307
– imeretina (Дуб имеретинский) 284
– macranthera (Дуб крупнопыльниковый)
307
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
362
– mongolica (Дуб монгольский) 139, 177,
180, 183, 222, 223, 244
– pedunculiflora (Дуб ножкоцветный) 304
– petraea (Дуб скальный) 281, 299
– pubescens (Дуб пушистый) 277, 278, 282,
304
– robur (Дуб черешчатый) 138, 139, 147,
222–224, 297, 299
Racomitrium (Ракомитрум) 70
– lanuginosum (Ракомитрум шерстистый)
91, 95
Ranunculus acris (Лютик едкий) 203, 208
– auricomus (Лютик золотистый) 208
– buhsei (Лютик Бузе) 303
– cassubicus (Лютик кашубский) 224
– caucasicus (Лютик кавказский) 301
– oreophilus (Лютик горолюбивый) 303
– polyanthemos (Лютик многоцветковый)
203, 208, 230, 232, 233, 242
– propinquus (Лютик близкий) 95
– repens (Лютик ползучий) 115
– sabinei (Лютик Сабина) 66
– sulphureus (Лютик серно-желтый) 70
Reseda lutea (Резеда желтая) 239
Reynoutria sachalinensis (Рейнутрия саха-
линская, гречиха сахалинская) 178,
179
Rhamnus cathartica (Жостер слабительный)
233, 304
– pallasii (Жостер Палласа) 283, 307
Rhinanthus serotinus (Погремок поздний)
203
Rhizomatopteris sudetica (Корневищник су-
детский) 161
Rhodiola rosea (Родиола розовая, золотой
корень) 88, 324
Rhododendron adamsii (Рододендрон Адам-
са) 326
– aureum (Рододендрон золотистый) 111,
116, 117, 323, 324, 327, 332
– brachycarpum (Рододендрон коротко-
плодный) 190
– camtschaticum (Рододендрон камчатский)
73, 112, 332
– caucasicum (Рододендрон кавказский)
300–302
– dauricum (Рододендрон даурский) 137,
172, 244, 321
– luteum (Рододендрон желтый) 299
– mucronulatum (Рододендрон остроконеч-
ный) 244
– parvifolium (Рододендрон мелколистный)
109, 326, 327
– ponticum (Рододендрон понтийский) 284,
299
– redowskianum (Рододендрон Редовского)
327
– sichotense (Рододендрон сихотинский) 190
Rhus coriaria (Сумах дубильный) 279
Rhytidiadelphus triquetrus (Ритидиадельфус
трехгранный) 143, 161
Rhytidium rugosum (Ритидиум морщини-
стый) 88
Ribes latifolium (Смородина широколист-
ная) 178
– maximoviczianum (Смородина Максимо-
вича) 190
– nigrum (Смородина черная) 163, 164
– sachalinensis (Смородина сахалинская)
178
Rosa (Шиповник) 278
– acicularis (Шиповник иглистый) 90, 142,
144, 160, 166, 171, 321, 322, 331
– canina (Шиповник собачий) 304
– corymbifera (Шиповник щитконосный)
304
– iberica (Шиповник грузинский) 283
– majalis (Шиповник майский) 163, 164,
235, 236, 242, 252
– rugosa (Шиповник морщинистый) 189
– hugonis (Роза Хугона) 313
Rubus arcticus (Княженика обыкновенная)
163
– chamaemorus (Морошка) 77, 81, 86, 89–
91, 96, 111, 113, 144, 156, 157
– crataegifolius (Малина боярышниколист-
ная) 180, 185
– discolor (Ежевика гигантская) 298
– idaeus (Малина обыкновенная) 151, 163
– saxatilis (Костяника) 143, 161, 164, 166,
242
– stellatus (Княженика звездчатая) 115
Rumex acetosella (Щавелёк обыкновенный)
200, 280
– crispus (Щавель курчавый) 164
– thyrsiflorus (Щавель пирамидальный) 203
Ruscus (Иглица) 279, 284
– aculeatus (Иглица колючая) 278, 284
– hypoglossum (Иглица подъязычная) 278
Salicornia herbacea (Солерос европейский)
243
Salix (Ива) 71
– abscondita (Ива скрытная) 194
– alaxensis (Ива аляскинская) 73, 111, 326
– alba (Ива белая) 235
– arbuscula (Ива деревцовидная) 316
– arctica (Ива арктическая) 66, 115
– bebbiana (Ива Бебба) 171, 321, 326
– berberifolia (Ива барбарисолистная) 316,
327
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
363
– boganidensis (Ива боганидская) 73
–
brachypoda (Ива коротконожковая) 171
– caprea (Ива козья) 160, 163, 229, 322
– cardiophylla (Ива сердцелистная) 194
– cinerea (Ива пепельная) 142, 163
– crassijulis (Ива толстосережковая) 116
– euxina (Ива понтийская) 235
– fuscescens (Ива буреющая) 73
– glauca (Ива сизая) 82, 88–90, 96, 101, 109,
111, 316, 326
– gmelinii (Ива Гмелина) 89
– gracilistyla (Ива тонкостолбиковая) 194
– herbacea (Ива травянистая) 86
– krylovii (Ива Крылова) 73, 111, 316, 324
– lanata (Ива мохнатая) 78, 82, 88, 89, 95,
96, 116, 326
– lapponum (Ива лапландская) 82, 89, 90,
144, 145
– nipponica (Ива ниппонская) 194
– nummularia (Ива монетолистная) 71, 76,
95
–
phlebophylla (Ива жилколистная) 72,
109
– phylicifolia (Ива филиколистная) 82, 89,
90, 96, 144, 145
– polaris (Ива полярная) 67, 72, 76, 95, 102
– pulchra (Ива красивая) 99, 101, 109, 111,
116, 326
– reptans (Ива ползучая) 72, 89, 99, 101
– reticulata (Ива сетчатая) 141, 145, 316
– richardsonii (Ива Ричардсона) 73, 111
– rorida (Ива росистая) 192, 194
– rotundifolia (Ива круглолистная) 72, 109
– schwerinii (Ива Шверина) 111, 192, 326
– sphenophylla (Ива узколистная) 109
– taraikensis (Ива тарайкинская) 194
– udensis (Ива удская) 192, 326, 329
– vestita (Ива нарядная) 316
Salsola australis (Солянка южная) 271
– collina (Солянка холмовая) 252
– beckeri (Шалфей Беккера) 309
Salvia canescens (Шалфей седеющий) 308,
309
– nemorosa (Шалфей дубравный) 257
– nutans (Шалфей поникающий) 249, 252,
255
– pratensis (Шалфей луговой) 223, 228, 230,
231–233
– stepposa (Шалфей степной) 223, 252, 253,
258, 261
– verticillata (Шалфей мутовчатый) 153
Sambucus kamtschatica (Бузина камчатская)
329
– nigra (Бузина черная) 299
– racemosa (Бузина кистевидная) 235
– sibirica (Бузина сибирская) 323
Sanguisorba alpina (Кровохлебка альпий-
ская) 316
– officinalis (Кровохлебка лекарственная)
164, 242
Sanicula europaea (Подлесник европейский)
304
Sasa kurilensis (Саза курильская, куриль-
ский бамбук) 177
Saussurea alpina (Соссюрея альпийская)
89
– latifolia (Соссюрея широколистная) 315,
316
– pseudotilesii (Соссюрея ложно-тилезиева)
332
– salsa (Соссюрея солончаковая) 243
– tilesii (Соссюрея Тилезиева) 109
Saxifraga (Камнеломка) 67
– cernua (Камнеломка поникающая) 82
– cespitosa (Камнеломка дернистая) 71, 72,
82
– nelsoniana (Камнеломка Нельсона) 109,
324
– nivalis (Камнеломка снежная) 70, 72, 82
– oppositifolia (Камнеломка супротивно-
листная) 71, 82
– platysepala (Камнеломка плоскочашеч-
ная) 72
Scabiosa caucasica (Скабиоза кавказская)
301, 303
– gumbetica (Скабиоза гумбетская) 308, 309
– lachnophylla (Скабиоза шерстистолист-
ная) 188
– ochroleuca (Скабиоза бледно-желтая) 258
Scheuchzeria palustris (Шейхцерия болот-
ная) 142, 214
Schisandra chinensis (Лимонник китайский)
177, 182, 186, 244
Schivereckia podolica (Шиверекия подоль-
ская) 229
Schizonepeta multifida (Схизонепета много-
надрезанная) 263
Scilla bifolia (Пролеска двулистная) 281
– siberica (Пролеска сибирская) 224
Scirpus lacustris (Камыш озерный) 210
Scorpidium revolvens (Скорпидиум отверну-
тый) 95
Scorzonera purpurea (Козелец пурпурный)
230
Scutellaria polyodon (Шлемник многозубый)
309
Selaginella rupestris (Плаунок наскальный)
111
Senecio cannabifolius (Крестовник конопле-
волистный) 115, 178, 329
– pseudoarnica (Крестовник ложноарнико-
вый) 115
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
364
Serratula coronata (Серпуха венценосная)
164
– erucifolia (Серпуха эруколистная) 255, 257
– radiata (Серпуха лучистая) 257
Seseli condensatum (Жабрица густоцветко-
вая) 316
– krylovii (Жабрица Крылова) 253
– ledebourii (Жабрица Ледебура) 260–262
– libanotis (Жабрица порезниковая) 163,
165, 238, 242, 258, 259, 261
– tortuosum (Жабрица извилистая) 256, 257
Silene acaulis (Смолевка бесстебельная) 71,
81, 88, 109
– borystenica (Смолевка днепровская) 261
– chamarensis (Смолевка хамарская) 324
– chlorantha (Смолевка зеленоцветковая)
258
– jeniseensis (Смолевка енисейская) 263
– paucifolia (Смолевка малолистная) 103
– repens (Смолевка ползучая) 111
– rupestris (Смолевка скальная) 140
Sisymbrium polymorphum (Гулявник из-
менчивый) 258
Smelowskia calycina var. porsildii (Смелов-
ския Порсилда) 111
Smilax excelsa (Сассапариль высокий) 284,
304
Solidago dahurica (Золотарник даурский)
323
– virgaurea (Золотарник обыкновенный)
90, 144, 160, 162, 165, 242
Solorina crocea (Солорина шафрановая) 70
Sonchus arvensis (Осот полевой) 200
Sorbaria pallassii (Рябинник Палласа) 324
Sorbus alnifolia (Рябина ольхолистная) 180,
193
– amurensis (Рябина амурская) 191
– aucuparia (Рябина обыкновенная) 142,
144, 151
– sambucifola (Рябина бузинолистная) 191,
331
– sibirica (Рябина сибирская) 160, 161, 163,
164, 330, 331
Sphaerophorus globosus (Сферофорус шаро-
видный) 70, 88
Sphagnum angustifolium (Сфагнум узко-
листный) 91, 142, 217
– balticum (Сфагнум балтийский) 96, 156
– capillifolium (Сфагнум волосолистный)
142
– fallax (Сфагнум обманчивый) 142, 160
– fimbriatum (Сфагнум бахромчатый) 95
– fuscum (Сфагнум бурый) 89, 142, 156, 217,
330
– girgensohnii (Сфагнум Гиргензона) 89, 91,
98
– lenense (Сфагнум ленский) 96
– lindbergii (Сфагнум Линдберга) 86, 89, 96,
330
– magellanicum (Сфагнум магелланский)
217, 330
– majus (Сфагнум большой) 86
– obtusum (Сфагнум тупой) 142
– papillosum (Сфагнум папиллозный) 142,
330
– riparium (Сфагнум береговой) 86
– russowii (Сфагнум Руссова) 89, 160
Spiraea alpina (Спирея альпийская) 316
– aquilegifolia (Спирея водосборолистная)
317
– beauverdiana (Спирея Бовера) 111, 331
– betulifolia (Спирея берёзолистная) 190
– crenata (Спирея городчатая) 235, 236, 252,
253, 257–259, 261, 313
– dahurica (Спирея даурская) 321
– hypericifolia (Спирея зверобоелистная)
252–254, 258, 259, 261, 283, 313
– media (Спирея средняя) 145, 163, 166,
171, 321, 322
– salicifolia (Спирея иволистная) 163
– trilobata (Спирея трехлопастная) 313
– ussuriensis (Спирея уссурийская) 222, 244
Stellaria bungeana (Звездчатка Бунге) 137
– calycantha (Звездчатка чашечкоцветко-
вая) 115
– dichotoma (Звездчатка вильчатая) 262
– graminea (Звездчатка злаковидная) 163,
203
– holostea (Звездчатка ланцетолистная) 137,
147, 166, 227
Stemmacantha carthamoides (Большеголов-
ник сафлоровидный) 316
Stereocaulon (Стереокаулон) 70, 71
– paschale (Стереокаулон голый) 86, 98, 155
Stipa (Ковыль) 231, 232, 249
– anomala (Ковыль уклоняющийся) 252
– baicalensis (Ковыль байкальский) 253, 263
– brauneri (Ковыль Браунера) 253
– capillata (Ковыль волосатик, тырса) 251–
254, 257–261, 268, 271, 272, 309, 313
– daghestanica (Ковыль дагестанский) 304,
309
– dasyphylla (Ковыль опушеннолистный)
223, 232, 233, 252
– kirghisorum (Ковыль киргизский) 253
– korshinskyi (Ковыль Коржинского) 252,
253, 258, 259, 261
– krylovii (Ковыль Крылова) 172, 253, 263
– lessingiana (Ковыль Лессинга, ковылок)
249, 250, 252–259, 261, 268, 271, 272
– pennata (Ковыль перистый) 223, 230, 232,
233, 242, 252, 253, 255, 258–261
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
365
– pulcherrima (Ковыль красивейший) 223,
233, 258
– sareptana (Ковыль сарептский) 258, 268
– tirsa (Ковыль узколистный) 223, 232, 252,
254, 258, 304
– ucrainica (Ковыль украинский) 251, 252,
253, 254, 257
– zalesskii (Ковыль Залесского) 223, 233,
252–254, 258–261, 312
Suaeda corniculata (Сведа рожконосная) 243
Succisa pratensis (Сивец луговой) 210
Swida australis (Свида южная, свидина юж-
ная) 304
– sanguinea (Свидина кроваво-красная) 299
Symphytum asperum (Окопник шершавый)
302
Syntrichia ruralis (Синтрихия сельская) 255
Syringa amurensis (Сирень амурская) 180,
182, 185, 245
– wolfii (Сирень Вольфа) 182
Tamus communis (Тамус обыкновенный)
284
Tanacetum achilleifolium (Пижма тысяче-
листниколистная) 253, 259, 268, 269
– millefolium (Пижма тысячелистная) 257,
258
– odessanum (Пижма одесская) 253
– vulgare (Пижма обыкновенная) 164, 165,
203
Taraxacum officinale (Одуванчик лекар-
ственный) 200, 203, 243
– serotinum (Одуванчик поздний) 259
– stevenii (Одуванчик Стевена) 303
Taxus baccata (Тис ягодный) 285
– cuspidata (Тис остроконечный) 177, 180
Tephroseris integrifolia (Пепельник цельно-
листный) 89, 230, 232
– tundricola (Пепельник тундровый) 103
Thalictrum alpinum (Василистник альпий-
ский) 89
– appendiculatum (Василистник придатко-
вый) 323
– filamentosum (Василистник нитчатый)
182, 193
– minus (Василистник малый) 162
– simplex (Василистник простой) 164, 165
Thamnolia vermicularis (Тамнолия червеоб-
разная) 70
Thermopsis lanceolata (Термопсис ланцет-
ный) 264
Thlaspi arvense (Ярутка полевая) 200
Thymus calcareus (Тимьян меловой) 239,
240
– daghestanicus (Тимьян дагестанский) 309
– dubjanskyi (Тимьян Дубянского) 239
– elisabethae (Тимьян Елизаветы) 309
– extremus (Тимьян крайний) 103
– marschallianus (Тимьян Маршалла, ча-
брец Маршалла) 233, 258, 259, 261, 262
– mongolicus (Тимьян монгольский) 263
– serpyllum (Тимьян ползучий) 111, 142
– talijevii (Тимьян Талиева) 145
Tilia amurensis (Липа амурская) 139, 180,
183, 221, 222, 244
– begoniifolia (Липа бегониелистная, л. кав-
казская) 284
– cordata (Липа сердцевидная, л. мелко-
листная) 137–139, 147, 153, 221–224
– mandshurica (Липа маньчжурская) 180,
244
– sibirica (Липа сибирская) 161
– taquetii (Липа Таке) 180, 244
Tomentypnum nitens (Томентгипнум бле-
стящий) 88, 96, 101
Tragopogon brevirostris (Козлобородник
коротконосиковый) 29
– pratensis (Козлобородник луговой) 203,
232
Trichophorum alpinum (Пухонос альпий-
ский) 153
Trientalis europaea (Седмичник европей-
ский) 115, 136, 142, 157, 162, 163, 331
Trifolium alpestre (Клевер альпийский) 223,
233
– ambiguum (Клевер сомнительный) 303
– canescens (Клевер седоватый) 301
– lupinaster (Клевер люпиновидный) 223,
242
– medium (Клевер средний) 208
– montanum (Клевер горный) 223, 228,
230–233, 252, 255
– pratense (Клевер луговой) 165, 200, 203,
205, 208
– repens (Клевер ползучий) 165, 230, 243
Tripleurospermum inodorum (Трехреберник
непахучий) 200
Trisetokoeleria taimyrica (Тризетокелерия
таймырская) 99
Trisetum spicatum (Трищетинник колоси-
стый) 103
Trollius asiaticus (Купальница азиатская)
316
– europaeus (Купальница европейская) 89,
203, 210
– ledebourii (Купальница Ледебура) 323
– ranunculinus (Купальница лютиковая) 301
– riederianus (Купальница Ридера) 115
Trommsdorffia maculata (Троммсдорфия
крапчатая) 166, 230
Tulipa biebersteiniana (Тюльпан Бибер-
штейна) 255, 256, 271
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
366
– biflora (Тюльпан двуцветковый) 271
– gesneriana (Тюльпан Геснера) 255
– schrenkii (Тюльпан Геснера, т. Шренка)
271
Ulmus glabra (Вяз голый) 138, 139, 153,
222, 235, 284
– japonica (Вяз японский) 180, 245
– laciniata (Вяз разрезной, ильм горный)
180, 299
– laevis (Вяз гладкий) 138, 151, 155, 235
– minor (Вяз малый) 284, 297, 304
– pumila (Вяз низкий) 223, 321
Urtica dioica (Крапива двудомная) 227
– platyphylla (Крапива плосколистная) 178
Vaccinium (Вакциниум) 66, 67
– arctostaphylos (Черника кавказская) 284,
299
– myrtillus (Черника обыкновенная) 89–91,
135, 136, 140, 142, 166, 299, 322, 323
– ovalifolium (Черника овальнолистная) 177
– praestans (Красника) 178
– uliginosum (Голубика обыкновенная) 77,
81, 86, 88–91, 98, 111, 113, 135–137,
140, 142, 144, 214, 300, 332
– uliginosum ssp. microphyllum (Голубика
обыкновенная мелколистная) 96, 109
– vitis-idaea (Брусника обыкновенная) 77,
81, 86, 88–90, 135–137, 140, 142, 155,
157, 166, 300, 322, 324
– vitis-idaea var. minus (Брусника малая) 95,
96, 109, 332
Valeriana tuberosa (Валериана клубненос-
ная) 255
Veratrum lobelianum (Чемерица Лобеля)
323
– oxysepalum (Чемерица остродольная) 115,
332
Verbascum (Коровяк) 18
Veronica chamaedrys (Вероника дубравная)
143, 203
– gentianoides (Вероника горечавковая) 302
– incana (Вероника седая) 258, 259, 261,
264, 322, 327
– officinalis (Вероника лекарственная) 244
– spicata (Вероника колосистая) 242, 262
– spuria (Вероника настоящая) 242
– umbrosa (Вероника теневая) 281
– verna (Вероника весенняя) 255
Viburnum lantana (Калина городовина) 228,
299
– opulus (Калина обыкновенная) 163, 164,
304
– Sargentii (Калина Саржента) 180
Vicia amurensis (Горошек амурский) 244
– cracca (Горошек мышиный) 163, 164, 203,
208, 242, 322
– megalotropis (Горошек крупнолодочный)
164
– sepium (Горошек заборный) 163–165, 208,
242
– sylvatica (Горошек лесной) 164
– tenuifolia (Горошек тонколистный) 233
– unijuga (Горошек однопарный) 223, 322
Viola alba (Фиалка белая) 304
– altaica (Фиалка алтайская) 316
– biflora (Фиалка двуцветковая) 315
– canina (Фиалка собачья) 230
– epipsiloides (Фиалка лысоватая) 115
– kitaibeliana (Фиалка Китайбеля) 255
– langsdorfii (Фиалка Лангсдорфа) 115
– mirabilis (Фиалка удивительная) 164
– odorata (Фиалка душистая) 276, 281
– rupestris (Фиалка скальная) 230
– uniflora (Фиалка одноцветковая) 322
Viscaria vulgaris (Смолка клейкая) 205, 207
Vitis amurensis (Виноград амурский) 182,
244
– labrusca (Виноград лабруска, в. Изабелла)
284
– sylvestris (Виноград лесной) 284, 304
Warnstorfia exannulatus (Варнсторфия бес-
колечковая) 88
– fluitans (Варнсторфия плавающая) 86
– sarmentosa (Варнсторфия лозовидная) 95
Weigela middendorffiana (Вейгела Мидден-
дорфа) 182
Xanthium strumarium (Дурнишник зобо-
видный) 271
Xanthoselinum alsaticum (Златогоричник
эльзасский) 258
Yucca aloifolia (Юкка алоэлистная) 287
Zabelia coreana (Абелия корейская) 191
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
367
СВЕДЕНИЯ ОБ АВТОРАХ
ПЕТРОВ КИРИЛЛ МИХАЙЛОВИЧ, доктор географических наук, профессор ка-
федры биогеографии и охраны природы Института наук о Земле Санкт-
Петербургского государственного университета (СПбГУ), заслуженный деятель
науки РФ, почётный профессор СПбГУ. Окончил кафедру ботанической геогра-
фии Ленинградского госуниверситета (ныне кафедра биогеографии и охраны
природы СПбГУ) в 1956 г. Область научных интересов: изучение растительности
и ландшафтов природных зон России, подводных ландшафтов морских мелко-
водий. Участник и организатор многочисленных экспедиций в различные регио-
ны России, морских и подводных исследований в Южных и Дальневосточных
морях России, на коралловых рифах Индийского океана. Автор более 200 науч-
ных публикаций, 19 монографий и учебников, в том числе: Подводные ланд-
шафты: теория, методы исследования (1989), Общая экология: взаимодействие
общества и природы (1998), Экология человека и культура (2000), Геоэкология
(2004), Философские проблемы географии: натурфилософская парадигма (2005),
Биогеография (2006), Биогеография океана (2008).
ТЕРЕХИНА НАТАЛИЯ ВЛАДИМИРОВНА, кандидат географических наук, доцент
кафедры биогеографии и охраны природы Института наук о Земле СПбГУ. Окон-
чила кафедру биогеографии (ныне кафедра биогеографии и охраны природы)
СПбГУ в 1994 г. Область научных интересов: ботаническая география, фитоинди-
кация, биогеохимия естественных и антропогенно нарушенных биогеоценозов.
Автор и соавтор более 70 научных работ, в том числе 3 монографий и 3 учебных
и методических пособий.
.
УКАЗАТЕЛЬ НАИМЕНОВАНИЙ РАСТЕНИЙ
368
УЧЕБНИК ДЛЯ ВУЗОВ
ПЕТРОВ Кирилл Михайлович
ТЕРЕХИНА Наталия Владимировна
РАСТИТЕЛЬНЫЙ ПОКРОВ
РОССИИ
Компьютерная верстка Т. М. Лебедевой
Подписано в печать 10.07.17. Формат бумаги 70 × 1001/16.
Бумага офсетная No 1. Печать офсетная. Усл. печ. л. 00.
Тираж 350 экз. Зак. No
. С.34.
ХИМИЗДАТ
191023, Санкт-Петербург, Апраксин пер., 4
Тел. коммерческой группы для оптовых покупателей
(812) 570-39-86
Е-mail: chimizdat@yandex.ru
айт: http://www.chimizdat.ucoz.ru
Отпечатано с PDF-файла
.