/



















































Author: Гвоздева В.А.
Tags: генетика




Text
К ст. В.А. Гвоздева
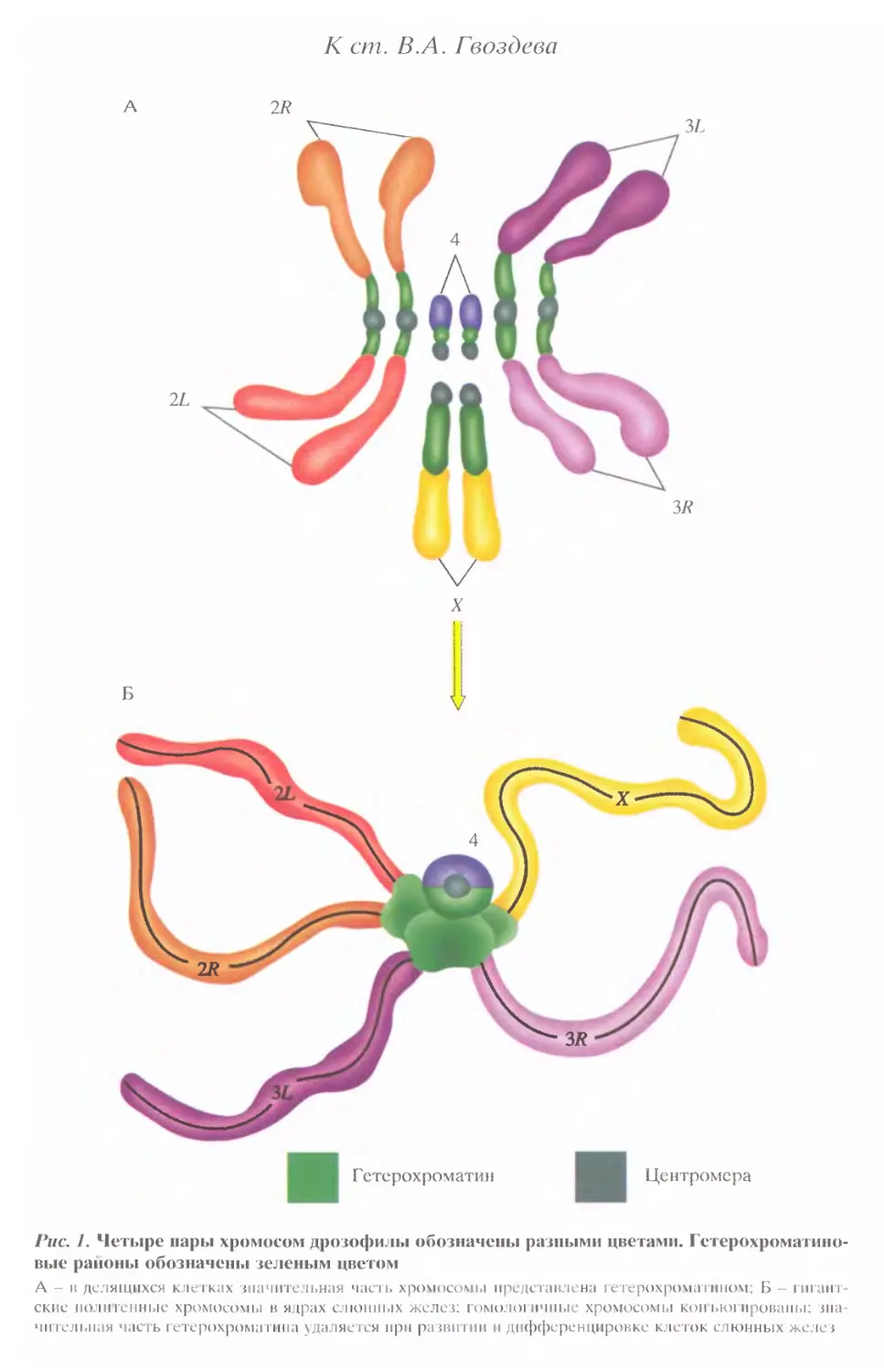
Рис. 1. Четыре нары хромосом дрозофилы обозначены разными цветами. Гетерохроматино-
вые районы обозначены зеленым цветом
А - в делящихся клетках значительная часть хромосомы представлена гетерохроматином; Б — гигант-
ские ноли генные хромосомы в ядрах слюнных желез; гомоло! ичпые хромосомы кон ькл провалы; зна-
чительная часть гетерохроматина удаляется при развитии и дифференцировке клеток слюнных желез
Рис. 3. Структура протяженного гетерохроматинового повтора (60 т. н. п.)
Треугольники, выделенные бледно-розовым, красным и оранжевым цветами, обозначают внедрение
представителей разных семейств рстротранспозонов со сходным механизмом перемещений (ретротранс-
позоны с длинными концевыми повторами). Зеленым и синим цветами обозначена локализация других
ретротранспозонов, не содержащих длинных концевых повторов. Черные прямоугольники показывают
расположение поврежденных повторов Stellate, желтым цветом обозначен фрагмент кластера генов ри-
босомной РНК
РНК-интерференция Косупрессия Подавление экспрессии генов
Stellate
Рис. 4. Механизм РНК-интерференции
Транскрипция обеих цепей Y-повторов обеспечивает образование двухнитевой РНК. Компоненты бел-
ковых комплексов, участвующих в “интерференции” и деградации мРНК, обозначены разноцветными
шариками. Старты транскрипции показаны сломанными стрелками
ген
винкулина
(разорван)
Сателлит
Распространение инактивации
600 т.н.п.
Г етерохроматин
нет инактивации
Рис. 5. Инактивация генов эухроматина Х-хромосомы в случае эу-гетерохроматиновых пере-
строек
Разнокачественные блоки гетерохроматина изображены разными цветами. Ц-центромера. Эухромати-
новый ген, кодирующий винкулин (см. статью В.Е. Алаторцева), разорван в результате эу-гстсрохрома-
тиновой перестройки. Длина горизонтальных стрелок отражает степень инактивации генов. При умень-
шении размеров блока гетерохроматина наблюдается постепенное снижение степени инактивации, пока-
занное вертикальной стрелкой. Инверсия фрагмента гетерохроматинового блока полностью снимает
инактивацию (нижняя строка)
Рис. 6. Нестабильная инактивация гена-репортера w, вызванная прилегающими повторами
Stellate
Участки ткани с резко ослабленной окраской свидетельствуют об инактивации гена. Обнаруживаемая
мозаичная окраска иллюстрирует нестабильность инактивации гена и,+. Рядом изображен глаз мухи с
нормальной равномерной окраской в отсутствие эффекта положения гена
К cm. Е.Г. Пасюковой
U3 RU5
U3 R U5
U3 RU5
U3 RU5
ДКП
Рис. 7. Строение ретровирусов (А) и ретротранспозонов (Б)
U5. К U3 - регуляторные районы длинных концевых повторов (ДКП), стрелка - старт транскрипции,
красная полоса - сайт связывания транспортной РНК, зеленая полоса - полипуриновый тракт, gag, pal,
ellv - гены
Рис. 8. Взаимодействие ретротранспозон - хозяин
Р-ретротранспозон; 1 - встройка ретротранспозона в геном влияет на признаки хозяина, на которые мо-
жет действовать естественный отбор; 2 - ретротранспозон сам регулирует свои жизненный цикл: 3 - ю
ны хозяина регулируют жизненный цикл ретротранспозона
Рис. 9. Локализация ретротранспозона copia на политенных хромосомах дрозофилы
Места локализации указаны стрелками и видны как черные диски на фоне фиолетовой окраски хромосом
К ст. Ю.Я. Шевелева
Чувствительность к ДНКазе I
Рис. 13. Примеры кластерной организации генов млекопитающих
Родственные гены имеют одинаковый цвет, овалы - энхансеры, квадраты или прямоугольни
торы, синие полосы - инсуляторы. В некоторых случаях над кластером указана кривая чув<
сти хроматина к ДНКазе I
Yu
CG13581
Crtp
Инсулятор?
Энхансер? и Инсулятор?
II *
CG13590
CK2$tes Pros28.1B
anon60Da
Рис. 14. Структурная организация кластера семенник-специфичных генов
Гены, входящие в кластер, закрашены зеленым цветом, гены вне кластера нс закрашены. Красные ова-
лы - предсказанные компьютерной программой места связывания ДНК с ядерным матриксом. Указаны
возможные области расположения энхансера и инсуляторов
10Е 14В
СК2$ Pros28.1
Рис. 15. Эволюционная схема возникновения кластера
60D Энхансер |
14В
Pros28.1
□□
CG13581
CK2$tes Pros28.1B
К ст. В.Е. Алаторцева
87В13
Дупликации генов
Повторы сатДНК 1.688
152АЗ.7 Сур4е4 CG17777
tw _ Pf>d 152A33Cytfd2pnI7E2.1dMetI рех К10 Or2F 25Е83 Vml
ph-d ph-p bcn92waplCyp4dlCyp4d8dNnid3103B4.2mstamstsVbic kz crn 10330 256725E8210146 11715
Igrl CG14053 30В8330В8.6 25E8.1 25E8.4
I-----1----1-----1----1-----1-----1--—I-----1-----1-----1-----1----1-----1-----1----I-----1----1-----1-----1----1
0 10 20 30 40 50 60 70 80 90 100 110 120 130 140 150 160 170 180 190 200 т.п.н
Рис. 17. Элементы генома в районе 2DF Х-хромосомы дрозофилы
Участки синтеза РНК, определенные в результате исследований отдельных генов района и анализа кло-
нов кДНК, обозначены прямоугольниками. Оранжевым цветом показаны транскрипты, синтезируемые
в направлении слева направо, желтым - по комплементарной цепи. Описания соответствующих генов да-
ны в базе данных FlyBasc (http://flybase.bio.indiana.edu:82/).
Показаны локальные дупликации генов и расположение повторов сатДНК 1.688
Транскрипты
Распространение инактивации генов на ~1 м.п.н.
50% -50%
Г етрохроматин
।-------1-------1-------1------1-------1-------1-------1-------1-------1-------1-------1------1
0 10 20 30 40 50 60 70 80 90 100 110 Ш т.п.н
Рис. 20. Инактивация генов района 2DF при приближении к гетерохроматину
Показана область эу-гетерохроматической границы в перестройке Х-хромосомы In(lLR)pn2a. Гены, для
которых были проведены количественные измерения активности, отмечены красным цветом
К ст. И.П. Арман, А.Б. Девина
Рис. 21. Клетки Saccharomyces cerevisiae
Вверху: дрожжевые клетки, активно размножающиеся почкованием; внизу: отдельная пара материнског
(более крупная) и дочерней клеток; на поверхности материнской клетки заметны “шрамы” от предыду-
щих почкований
Аск со спорами
Рис. 22. Жизненный цикл дрожжей Saccharomyces cerevisiae
Гаплоидная клетка содержит набор из п различных хромосом, у диплоидов каждая хромосома гаплоидно! о набора
представлена двумя гомологами (2л). Индивидуальные гаплоидные клетки обладают типом спаривания, или полом;
полов у них два - а и а. Зиготы образуются при слиянии клеток противоположного пола
ПРИРОДНАЯ ХРОМОСОМА ДРОЖЖЕЙ
TEL Ml CEN М2 ARS TEL
YAC
Рис. 24. Рекомбинантные векторы и искусственная хромосома дрожжей
А - природная хромосома и ее основные элементы: TEL - теломера, Ml и М2 - гены-маркеры, ARS - ав-
тономно реплицирующаяся последовательности, CEN - центромера; Б - Yip - кольцевой вектор бакте-
рии, содержащий ген-маркер дрожжей - Ml; В - YRp - в состав Yip включена ARS-последовательность;
Г - YCp - в состав YRp включена CEN-последовательность; Д - YLp - линейный вектор: YRp фланкиро-
ван TEL последовательностями; Е — YAC - в состав YLp включена CEN-последовательность, гены Ml и
М2 расположены по обе стороны CEN и маркируют правое и левое плечи искусственной хромосомы (см.
табл. 1.1)
ДНК
Трансформация сферопластов
Рис. 25. Схема TAR-клонирования: конструирование YAC in vivo
ДНК человека вводят в клетки дрожжей одновременно с двумя линеаризованными векторами, в составе
которых гены дрожжей (Ml и М2) и CEN, а на флангах каждого вектора - TEL- и Alu-последовательно-
сти. YAC образуется за счет процессов гомологичной рекомбинации между Alu-послсдоватслыюстями
(черные блоки) ДНК человека и векторов (Larionov et aL, 1996а)
Рис. 26. Этапы отбора колоний трансформантов с YAC, содержащими ДНК человека, при
TAR-клонировании (Larionov et al., 1996а)
1 - колонии первичных трансформантов; 2 - генетическая селекция; 3 - изоляция YAC методом PFGE;
4 - окраска бромистым этидием и гибридизация с зондом Д1IK человека
LicB
LTRnp-LicB
Рис. 30. Качественное определение активности лихеназы в клетках дрожжей, содержащих
контрольную плазмиду HERV-K LicB и плазмиды HERV-K LTR-LicB
Колонии трех контрольных (плазмида LicB) и трех независимых трансформантов, содержащих плазми-
ду LTR-LicB, инкубировали в течение 3-10 ч на среде с лихенаном. В зависимости от времени инкубации
усиливается ореол свечения вокруг растущих колоний. Контрольные колонии ореола нс образуют
Рис. 27. Гибридизация продуктов Alu PCR ДНК YAC, содержащих HERV-K LTR, с метафаз-
ными хромосомами человека (метод FISH)
Стрелки указывают на сигнал гибридизации
Доза, Гр
Рис. 34. Кривые выживания после 7-облучения диплоидов, гомо-гетерозиготных по мутаци-
ям rad9A и rad53
Кривые соответствуют результатам усреднения экспериментальных значений радиочувствительности
клеток, полученных в каждом случае для 3 штаммов одинакового генотипа; показаны среднеквадратич-
ные ошибки. Для сравнения дана кривая, ожидаемая при аддитивности влияния двух мутаций на радио-
чувствительность клеток
К ст. В.Г. Никифорова
Рис. 36. Модель активного центра (А) и трехмерная структура минимальной РНК-полимера-
зы Thermus aquaticus (Б)
К ст. С.З. Миндлина и др.
гидролиз
р-лактамного
кольца
О „
II ZS\ /СН3
I III Хсн3
nh2 о=с-------N СН—соон
но н
Рис, 37. Инактивация антибиотика ампициллина ферментом р-лактамазой
Ген Ыа на плазмиде - ген, кодирующий р-лактамазу. Красным обозначено интактное
цо ампициллина, синим - гидролизованное кольцо неактивной формы
плазмиды
Рис, 38. Перемещение плазмид из одной бактерии в другую
Розовым кольцом обозначена конъюгативная плазмида
Hg2+ Hg°
Рис. 39. Механизм устойчивости бактерий к неорганическим соединениям ртути (по: Osborn
et al., 1997)
Описание дано в тексте
Рис. 40. Генетическая организация транспозона Tn501 (Grinsted et al., 1990)
Условные обозначения: неправильные прямоугольники соответствуют генам, заостренные концы ука-
зывают направление транскрипции. Гены mer-оперона закрашены красным: R - ген merR, регулирует ин-
дукцию оперона ионами Hg2+; Т, Р - гены тегТ, тегР, участвуют в транспорте ионов Hg2+ внутрь клетки;
ген тег А, кодирует фермент ртуть-редуктазу, который восстанавливает токсичные ионы Hg2+ до метал-
лической ртути; D - ген merD, участвует в дополнительной регуляции оперона; 1 (orfl), 2 (prf2) - гены с
неизвестной функцией. Гены, необходимые для транспозиции, закрашены зеленым: ген tnpP кодирует
резольвазу и ген tnp/\ — транспозаз'7; res — / ^-область; треугольники на концах транспозона - инвертиро-
ванные повторы
Tn501
•о
Тп5041
Тп5044
Тп5053
Тп5070
и
1 kb
I------1
Рис. 41. Транспозоны устойчивости к ртути различных типов (Kholodii et al., 1995,1997,2000)
Обозначения mer-генов как на рис. 40. Дополнительные гены тег-оперона: С, ген тегС, участвует в
транспорте ионов Hg2+ внутрь клетки; F, ген тегЕ, функция неизвестна; гены orfE и W//-858 - гены с не-
известной функцией; sigY - сигмаподобный фактор, входящий в состав шег-оперона Тп5044. Обозначе-
ния генов, необходимых для транспозиции, Тп501, Тп5041, Тп5044 и Тп5070 как на рис. 40. Гены tniA, tniB,
tniO контролируют транспозицию, tniB - резолюцию Tn5053; tnpC, 408 (m/408), 237 (prf23T), orf[, orfQ,
orfP, oX, R — гены с неизвестной функцией, входящие в состав транспозонов
pKLH2
pKLH272
pKLH247
1 kb
Рис. 42. Мозаичный игег-оперон “рКЬН2”-типа
Вверху представлена генетическая организация шег-оперона плазмиды pKLH2 (Kholodii et aL, 1993a).
Внизу - схематически изображена структура мозаичного шег-оперона плазмиды pKLH272 Enterobacter
aqglomerans КН72 и плазмиды pKLH247 Acinetobacter sp. LS47 (Yurieva et al., 1997). Обозначения генов,
как на рис. 40 и 41; вертикальными стрелками отмечено место инвертированного повтора (IR) и res-сай-
та {res). Участки прямоугольников, соответствующие последовательностям pKLH2, заштрихованы, сх и
Р - ранее неизвестные последовательности; у - последовательность, гомологичная Тп501
Tn5041
merA
orfY tnpC
Tn5046
Тп5085
Тп5084
Тп5083
Tn5044
Рис. 43. Генетическая организация рекомбинантного транспозона Tn5046 (Mindlin et al., 2001)
Обозначения, как на рис. 41. Участки Тп5046, гомологичные Тп5041, выделены оранжевым; участки
Тп5046, гомологичные Тп5044, - синим
комплекс генов
Рис. 44. Схематическое изображение мозаичной структуры \w5084 и Tn50&3 (Bogdanova et
al., 1998, 2001)
R1 и R2 обозначают гены mctRI и merR2, кодирующие разные белки-регуляторы. ORFs обозначает гены
ог/2, orf3 и ог/4, функции которых не вполне ясны (Wang et al., 1989; Gupta et al., 1999; ilobman. Brown,
1997; Huang et aL, 1999b).
Bl, B2, ВЗ обозначают гены /??е/В1, /??с/В£ и ше/ВЗ, кодирующие различные органомеркурпат-лпазы:
tnpR, tnpK и tnpA.' обозначают гены, кодирующие резольвазу, транспозазу и фрагмент транспозазы. IRr,
IRt и res обозначают терминальные повторы и предполагаемый гел-сайт 1. Участки, закрашенные синим
и черным на схемах Тп5084 и Тп5083, отличаются от соответствующих участков Тп5085 на 6-10 и 40% со-
ответственно. Участки гена тег А. на схеме Тп5083, закрашенные желтым, отличаются от соответствую-
щих участков Тп5085 на 5-8%
рекомбинация
тег
res}/2
Рис. 45. Гипотетические схемы происхождения рекомбинантных транспозонов (Kholodii et
al., 1993b; Mindlin et al., 2001)
A - возникновение рекомбинантного транспозона в результате транспозиции одного мобильного элемента в
другой, неродственный ему элемент, и последующей внутренней делеции; Б - возникновение рекомбинант-
ного транспозозна в результате специфической рекомбинации по /^-сайтам между родственными транспо-
зонами. тег обозначает тле/'-оперон; tnp\ и tnp2 (tni) - транспозиционные модули взаимодействующих транс-
позонов; res 1 и res2 - соответствующие res-сайты. Концевые повторы обозначены треугольниками
К ст. В.З. Тарантула
Рис. 49. Структура генома вируса иммунодефицита человека типа 1 (HIV-1)
Рис. 46. Предполагаемый механизм перераспределения плазмидных генов с помощью новой
системы резолюции CinH/RS2 (Kholodii, 2001)
1 - слияние плазмидных репликонов с образованием коинтеграта (рекомбинация между /ел-саитами X и
X'); 2 - разрешение коинтеграта с образованием рекомбинантных плазмид (рекомбинация между /тл-сай-
тами Y и Y')
Рис. 50. Динамика роста трансформированных генов tat HIV-1 поликлональных культур кле-
ток PC 12 и Rat-2 в стандартных условиях культивирования
Фиолетовые линии и значки - контрольные культуры клеток, трансфицированные плазмидой с геном
пест, зеленые линии и значки - культуры клеток, трансфицированные геном tat HIV-1, красные линии и
значки - культуры клеток, трансфицированные геном tat HIV-1 и культивируемые в присутствии индук-
тора металлотионеинового промотора (ионов цинка)
Рис. 52. Динамика роста трансформированной геном nef HIV-1 поликлональной культуры
клеток Rat-2 в стандартных условиях культивирования
мРНК лимфомы
(трейсер)
мРНК нормальных клеток
(драйвер)
Обратная транскрипция
с праймером Т1
Поли(А)
Поли(А)
Обратная транскрипция
с праймером Т2
кДНК
Праймер Т1
кДНК
Праймер Т2
Пришивка олиго(dА)
Олиго(йА)
ПЦР с праймером Т2
ПЦР с праймером Т1
Один цикл ПЦР с
праймерами L1 и L2
Праймер L1
Праймер L2
Биотинилирование
Заполнение концов
Лигирование с вектором
Рис. 54. Схема метода вычитающей гибридизации, использованная для выявления лим<|
специфических генов в HIV/SIV-ассоциированных лимфомах
К ст. Л.Е. Андреевой, В.З. Тарантула
Рис. 60. Микроскоп с оптикой интерференционного контраста и микроманипулятор, исполь-
зуемые для микроинъекций экзогенной ДНК в яйцеклетки млекопитающих: общий план
(А), камера с яйцеклетками (Б)
Рис. 61. Микроинъекция экзогенной ДНК в зиготы мыши: зиготы мыши (А), зигота мыши
перед микроинъекцией раствора с экзогенной ДНК (Б)
Рис. 63. Зиготы свиньи до центрифугирования (А), микроинъекция экзогенной ДНК в зиго
ту свиньи после центрифугирования (Б)
Рис. 65. Экспрессия репортерного гена CMV-lacZ в 3-суточных предличипках вьюна после
оплодотворения икринок сперматозоидами, которые предварительно инкубировались в рас-
творе с плазмидой, содержащей этот ген
SD SA
Рис. 66. Общая схема строения ретровирусного вектора, используемого для трансгеноза
LTR - длинные концевые повторы ретровируса, SA - акцепторный сайт сплайсинга, SD - донорный сайт
сплайсинга, - участок, отвечающий за упаковку вируса
Рис. 67. Гистологический анализ костного мозга контрольных и трансгенных мышей Fj-no-
коления с геном ЭПО (бордовые столбики - контрольные мыши, синие - трансгенные)
ПЭ - проэритробласты; БЭ — базофильные эритробласты; ПХЭ - полихроматофильные эритробласты;
НБ-нормобласты; РЦ-ретикулоциты; МК - миелоидные клетки; ЭК - эритроидные клетки; ЛЦ-лим-
фоциты
Рис. 68. Трансгенные эмбрионы вьюна с геном CMV-lacZ через 19 ч после оплодотворения
Рис. 69. Клоны клеток, экспрессирующих ген CMV-lacZ в мышцах 4-суточной личинки вьюна
70. Локализация экспрессии гена CMV-lacZ в 14-суточном эмбрионе мыши
Рис. 71. Поздняя бластоциста козы на 4-е сутки культивирования в среде для ЭСК
Рис. 72. Окраска на щелочную фосфатазу островков клеток козы, подобных эмбриональным
стволовым клеткам
•. 73. Эмбриоидное тело, образованное клетками козы, подобными эмбриональным ство-
клеткам
Рис, 74. Инъекция ЭСК в полость бластоцисты мыши: бластоциста перед инъекцией бласто-
циста с ЭСК внутри бластоцеля (Б)
Б а неоплодотворенная яйцеклетка
зигота перед энуклеацией
проноклеусы
в перенос ядра
перенос ядра
б энуклеация
б энуклеация
микропипетка
блестящая
оболочка
пронуклеусы
в кариопласте
Рис. 77. Схема получения клонированных животных с помощью пересадки ядер
А - реципиент ядра - оплодотворенная яйцеклетка (зигота) на стадии пронуклеусов, а - мыкропипетка
для удаления ядра находится напротив одного из пронуклсусов: б - оба пронуклеуса, окруженные не-
большим количеством цитоплазмы и плазматической мембраной, втянуты в энуклеационную микропп-
петку. в - кариопласт или целая клетка с донорским ядром помещается под блестящую оболочку и сли-
вается с цитопластом путем электрослияния или. например, с помощью инактивированного вируса Сен-
дай Б - реципиент ядра - овулировавший нсоплодотвореннып ооцит, а - ооцит на стадии метафазы вто-
рого мейоч ичсского деления с метафазной пластинкой и веретеном деления на анимальном полюсе: б -
плазматическая мембрана и небольшое количество цитоплазмы, содержащие веретено деления и мета-
фазные хромосомы втягиваются в энуклеационную микропипетку; в - кариопласт или целая клетка с до-
норским ядром помещается под блестящую оболочку и сливается с цитопластом путем электрослияния
или инактивированного вируса Сендай, либо “голое” донорское ядро инъецируется непосредственно в
цитоплазму (Seller, 2000)
К ст. А.И. Николаева
Рис. 78. Альтернативные судьбы молекул экзогенной ДНК после ее переноса в клетки-ми-
шени
А - интеграция с хромосомой клетки-мишени; Б - автономное существование в виде экстрахромосомной
кольцевой ДНК
Копииность
100
50
Время после
трансфекции, (дни)
Рис. 80. Локализация хромосомного ori в локусе гена с-тус и конструкция автономного век-
тора, содержащего ARS-фрагмент этого ori (McWhinney, Leffak, 1990)
А - экзонно-интронная схема гена с-тус с прилегающей 5-областью, содержащей ARS-элсмснт и про-
моторы этого гена. Е - EcoRI, Н - Hindlll. X - Xhol. Красным цветом выделен ARS-элемент размером
2,4 т. п.н. Б - структура рекомбинантной автономной плазмиды pNeo.Myc - 2.4, содержащей ARS-эле-
мент, размером 2,4 т. п.н.. ген пео под контролем ММТ-промо гора и сигнала полиадснилирования SV40.
В - пролонгированное поддержание автономно-реплицирующейся конструкции pNeo.Myc - 2.4 в культу-
ре клеток человека HcLa по сравнению с простой плазмидной конструкцией
Ori P
Белок
EBNA-1
Трансформирующее
действие на клетки-
мишени
Терапевтическое
де не тг.ие на клетки-
МИ1ПСН11
Рис. 81. Структура разработанных автономных векторов, основанных на использовании
ori Р вируса EBV, активируемого вирусным белком EBNA-1 (Krysan et al., 1989)
Family of repeats - модуль ori, взаимодействующий с ядерным матриксом. Dyad - модуль ori, необходимый
для инициации репликации. EBNA-1 - ген EBV, кодирующий транс-действующий фактор, активирующий
ori. обладающий онкопотенциалом. Гетерологичная кДНК - последовательность вектора, кодирующая
белок, обладающий терапевтическим действием в клетках-мишенях, выбранных для генной терапии
г
Время, дни
Рис. 82. Подход к созданию и характеристики экстрахромосомпого поддержания безопасно-
го автономного вирусного вектора, основанного на модифицированном трансдействующем
вирусном факторе (Cooper et al., 1997)
А - схема полипептидной цепи T-ag с генетическими детерминантами, определяющими его взаимодейст-
вие с белками-супрессорами злокачественной трансформации р53, РВ, а также с ori SV40. Б - модифика-
ция T-ag с помощью замены двух аминокислотных остатков (107 и 402), приводящая к снижению онко-
потенциала за счет потери взаимодействия модифицированного белка с р53 и РВ. В - поддержание ре-
комбинантных автономно-реплицирующихся конструкций, основанных на функции ori SV40 и содержа-
щих гены дикого и модифицированного T-ag (транс-действующего фактора ori) в культуре клеток Рад-
жи. Г - уровень репортсрного продукта гена LacZ, клонированного в модифицированной автономно-ре-
плицирующейся конструкции, регулируемой модифицированным T-ag, по сравнению с вариантом про-
стой плазмидной конструкции
1 Т. II. н.
Рис. 84. Формирование стабильного экстрахромосомного трансгена рг8а тутового шелкопря-
да в результате серии перестроек трансгенов (Николаев и др., 1991, 1993а, б)
А - схема формирования стабильного экстрахромосомного трансгсна в результате серии перестроек
трансгенов при переносе экзогенной ДНК н при передаче по наследству.
Красная линия - ARS-содсржащая область трансгенов и эндогенной экстрахромосомнои ДНК. Желтая
линия - фра! мент клеточной ДНК шелкопряда Б()-ноколсния (желтая линия). Зеленая линия - фрагмент
клеточной ДНК шелкопряда Р^поколения (зеленая линия). Черная линия - фрагмент плазмиды pBR322
(черная линия).
Б - физическая карта рг8а с указанием рестрикционных сайтов. Красная линия - ARS-элсмснт рг8а. Чер-
ная линия - фрагмент последовательности pBR322
A
Рис. 86. Схема перестроек стабильного трансгена шелкопряда после его переноса и переда-
чи по наследству в трансгенных мышах (Николаев и др., 1998)
А - схема последовательных делеций рг8а после переноса ее в зиготу мыши. Del 1 - обнаружена после пе-
реноса рг8а в пронуклеус мышей. Del2 - обнаружена после передачи трансгена по наследству от основа-
телей к мышам Fj-поколения. Del3 - обнаружена после передачи трансгена по наследству от мыши Fr
поколения к мышам Р2-поколения. Красная линия - ARS мыши. Лиловая линия - фрагмент ARS рг8а, де-
легированный на стадии dell, и не требующийся для поддержания автономного статуса трансгенов в мы-
шах. Б - стабильный трансген, обнаруживаемый в мышах Р2-поколения, являющийся образованной in
vivo производной рг8а. Выявление этого трансгена в составе суммарной ДНК трансгенных мышей F2-no-
коления представлено выше (рис. 85). Размеры плазмид и делеций даны в т. п.н.
Sequence
Рис. 88. Обнаружение в автономных трансгенах участков изогнутой ДНК (Nikolaev et al.,
неопубликовано)
А - стабильный экстрахромосомный трансгсн мышей pmyc(H-P) (Sudo et al., 1990), в ARS-элементс ко-
торого ранее было экспериментально показано присутствие участка изогнутой ДНК (Trivedi et al., 1998).
Ь - стабильный трансгсн шелкопряда рг8а (представлена гистограмма части EcoRI-RvuII фрагмента
трансгена). Детектирование участков изогнутой ДНК проведено с помощью программы Bent-lt, разме-
щенной на сайте: www2,icgcb.trieste.it/dna/bend_it.html, согласно которой участок предположительно изо-
гнутой ДНК должен характеризоваться уменьшенным ГЦ-содержанисм и увеличенным свыше 5° углом
поворота двойной спирали в расчете на один виток. Зеленая линия - изогнутая ДНК
Д 1 ctgtttttctcaccacatttttttcattattgtttccttcctcttttaccccagagcctt
61 tttagaaacttttttctgattgattctcccattatattttaatcacatatacactgtgta
121 t r*t я 111 яеяГ at er-t яГ gt яГ яГ <~t гГ.яГ. t rt ata rat, t.aa att aaa aa gtata3 30t t
181 aatqttttattattqcaaaqaatqataaaacaattaaaaattaaattaaaaqqctattga
241 cattttactt aactt gat aactat att ctt agagcaacttttttgaaaat agatttttgt
301 gggtacataagaggcgtctgtgtttatggaataagtgaaatatttttatataggcagaca
361 attcataataatcacagcagggtgaatggagtatccaatgcttcaaacatttatcatttc
421 t ctctgctacatacatcccatttat act attttagttattttataatatataataaatta
481 tcattaacagtagtcatcctattgagctatcgaatactaaat cat att cattttttctaa
841 ctatattttttgtgcccgttagccatctccattttcccccatatgatccagcaatcccac
601 t actgggtatttatccaaaggaaatgaaatcagtatgtcaacaaaatatttacactccca
661 tgtttattgcagcactattcacaatagctaagattt ggaagcaacttaggtgtctaacag
721 atgaatggataaagaaaatatggcacttatacacaatggagtactacttagccataaaaa
781 agaatgagagatcctgtcatttgcaagaacatggatggagetggaggacattatgtteat
8 41 t ggacaacatt aaaccca11gatetgcacatacctaaccacactt gaatetet aggасаa
901 attccacttg gtcat gat gaatgattgttttaatgt gtt ctt
Б
AKS "
OCT 1 сайт
REE- подобный элемент
Изогнутая
ДНК
5’flank ocINF (94%) А / Т MER 27 (77%) LINE 1 (69%)
---------------------------------------------------------------►
О 100 200 300 400 500 600 700 800 900 п.н.
Рис. 89. Генетическая и физическая карта EcoRI-Pvull фрагмента ARS рг8а
А - последовательность нуклеотидов этого фрагмента, на которой выделены: синяя линия - участок изо-
гнутой ДНК, красная линия — ACS-элементы. Б - карта фрагмента с указанием генетических детерми-
нант: MER — повторяющаяся последовательность со ‘средней ' частотой встречаемости в геноме челове-
ка; LINE - повторяющаяся транспозонподобная последовательность генома человека; АТ - участок
ДНК, обогащенный А и Т нуклеотидами; INF (интерферон) — ген человека; Octi — сайт связывания фак-
тора транскрипции/репликации Octi; REE - энхансер репликации
К ст. И.А. Гривенникова и др.
BPS 1
Рис. 92. Схема топологии белка пресенилина-1 человека (Li, Greenwald, 1998)
Трансмембранные домены обозначены римскими цифрами. Серые прямоугольники - дополнительные
гидрофобные домены. Красные горизонтальные стрелки показывают локализацию мутаций M146V и
CHOY, использованных в работе. Вертикальный треугольник обозначает место протеолитическою рас-
щепления белка
К ст. И.А. Гривенникова и др.
Рис. 93. Трансфекция псевдонормальных клеток крысы Rat-2
А - монослой клеток в контрольных вариантах трансфекции плазмидами pBR322, pSV2-neo и pEJdl.
Б - многослойные фокусы морфологической трансформации при трансфекции плазмидой pEJ6.6
К ст. И.А. Гривенникова, Е.С. Мануйловой
Рис. 104. Колонии ЭС клеток мыши на фидерном слое эмбриональных фибробластов, х 40
а - колония клеток линии R1, б - клетки фидерного слоя
Рис, 106, Выявление активности щелочной фосфатазы в культуре ЭС клеток мыши, X 20
а - окрашены колонии, сформированные ЭС клетками; б - неокрашенные клетки фидерного слоя эмбриональных фибробластов. Окраска с
помощью нафтола AS-BI-фосфата и ВВ-красного
Рис. 107. Эмбриоидные тела, образовавшиеся из ЭС клеток на 3-4-е сутки после прикрепле-
ния их к желатиновой дорожке, х 5
а - эмбриоидное тело; б - миграция клеток из эмбриоидного тела
4
Рис. 109. Влияние регуляторных генов nef и tat вируса иммунодефицита человека на проли-
ферацию ЭС клеток мыши in vitro
По оси абсцисс дни культивирования, по оси ординат количество клеток. Зеленые столбики - контроль-
ные клетки. Синие столбики - клетки трансфицированные геном tat. Малиновые столбики - клетки
трансфицированные геном nef
Рис. ПО. Флуоресцентная микрофотография эмбриональных стволовых клеток в культуре,
трансфицированных плазмидой с геном “зеленого белка”. X 100
К ст. С.А. Лимборской, П.А. Сломинского
0,4
0,35
0,3
0,25
0,2
0,15
0,1
0,05
0
Больные ПМС
Случайная выборка
Больные диабетом (II)
Больные ИБС
Больные ПМС
Больные ИБС
Z Больные диабетом (II)
Случайная выборка
Рис. 115. Распределение аллельных вариантов (СА) микросателлитного повтора в 3' нетранс-
лируемом экзоне гена ангиотензиногена в московской популяции в случайной выборке и у
больных различными заболеваниями
Число повторов