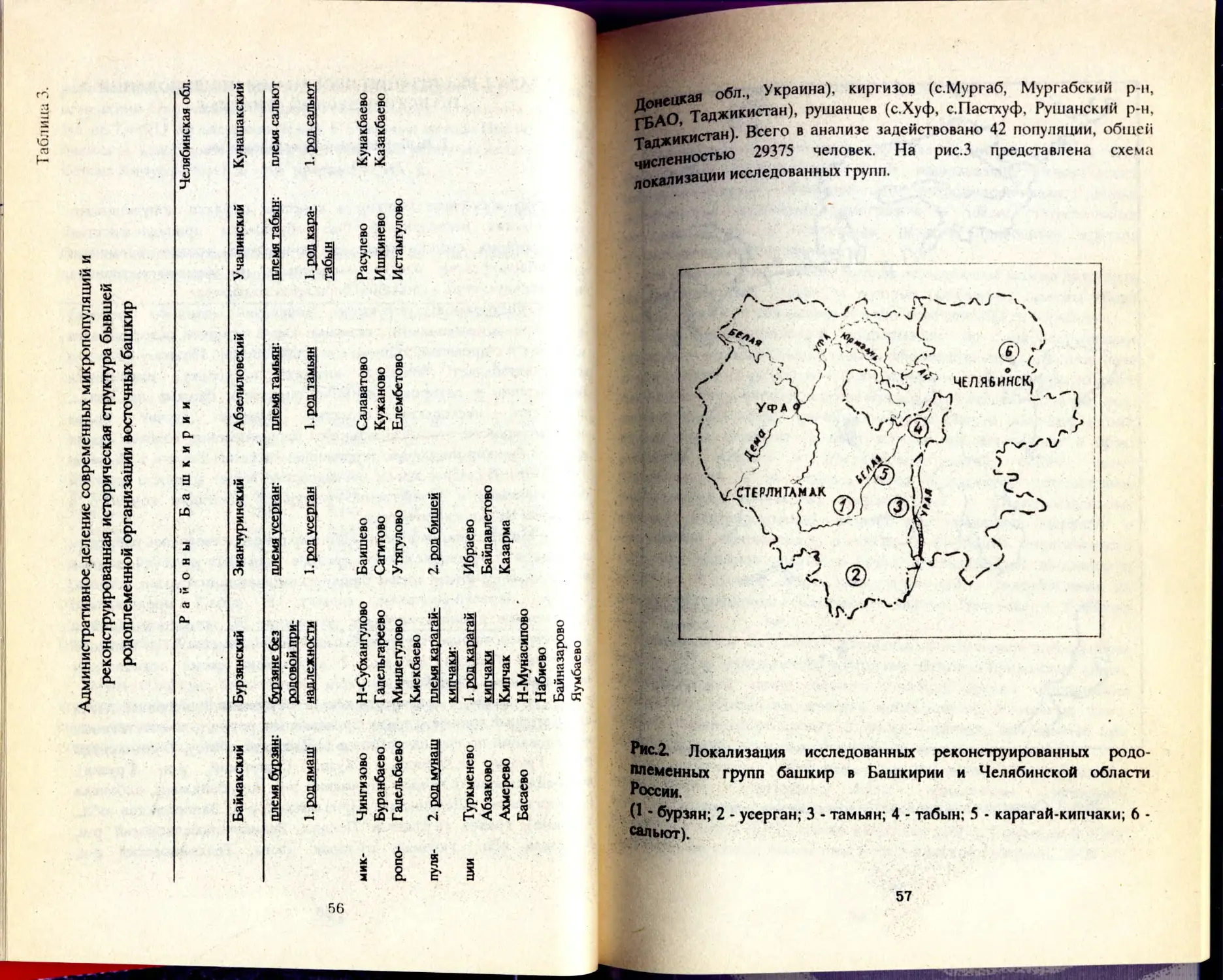
/










































































































Text
Н.Х. Спицына
ПРОБЛЕМЫ
ИСТОРИЧЕСКОЙ
ГЕНЕТИКИ
Москва
1993
/ЙО
РОССИЙСКАЯ АКАДЕМИЯ НАУК
институт этнологии И АНТРОПОЛОГИИ
им.Н.Н.Миклухо-Маклая
Н.X.Спицына
ПРОБЛЕМЫ ИСТОРИЧЕСКОЙ
ГЕНЕТИКИ
Москва
1993
fa 563993
+езо*44б.а ,оа\
+ mw
с.
Цевтраяьжа жулав
6i6/iortti
hi. В. L £«рзалсьхого
АН У к g at я
Ast J&19 &i~
Ответственный редактор: 13.11.Пасиков
ПРЕДИСЛОВИЕ.
Что из себя представляет историческая генетика?
Ее роль и эффективность в решении проблем
расо- и этногенеза.
С незапамятных времен людям было свойственно стремление
познать свою историю, их глубоко интересовали вопросы своего
происхождения, к числу которых принадлежат такие кардинальные
теоретические проблемы, как древность семейства гоминид,
прародина человечества, регионы, причина и динамика становления
расовых типов, хронологические аспекты дифференциации рас,
этнических групп.
Все эти проблемы решаются с помощью морфологических
методов, посредством детальных описаний и измерения различных
ископаемых находок древнего человека, путем сравнения и
сопоставления сотен человеческих популяций в разных регионах
земного шара.
Относительно недавно к этим классическим приемам прибавился
новый современный метод, находящийся сейчас в стадии своего
бурного развития. Его значение и влияние на разработку проблем
этно- и расогенеза ощущается в настоящее время особенно сильно.
Благодаря достижениям сначала иммуногенетики, затем
биохимической, и наконец, молекулярной генетики исследователи
получили возможность охарактеризовать популяции путем
установления в них относительного распространения сотен и тысяч
различных генов. Проведение подобных исследований стало
возможным и на групповом популяционном уровне по мере
"адаптации" этих методов к нуждам массовых исследований, в том
числе и в полевых исследованиях.
Современные достижения в области популяционной генетики,
биодемографии привели к становлению популяционной
антропологии, которая отличается не только более широким
биологическим подходом к анализу человека, но и к диалектическим
принципам, осознанием самых разнообразных связей человека с
э
исключительно разнообразной географической, биотической и
социальной средой его существования.
Таким образом идентифицировано огромное число
наследственных факторов, самих генов, специфических фрагментов
ДНК, которые в отличие от морфологических признаков, в течение
жизни индивида мало, а то и вовсе не испытывают на себе влияния
факторов окружающей среды... Этот новый путь - изучение
генетического состава групп населения, а тгцеже факторов,
способных модифицировать этот состав в рамках решения сложных и
многогранных вопросов расо- и этногенеза можно рассматривать как
историческую генетику.
Многие исследователи предлагают классифицировать население
исключительно по генетическим особенностям, оперируя доступным
числом достаточно просто определяемых генов, формируя
комбинации, при помощи которых расы дифференцируются лучшим
образом. И в то же время современные исследователи отдают себе
отчет в том, что нельзя изучать этническую антропологию в отрыве
от палеоантропологии. До сих пор в различных классификациях
значительное место отводилось особенностям скелета с тем, чтобы
мы могли обнаружить их в древних популяциях и сравнить древние
расы с современными. Более того, расовая классификация
традиционно построена с использованием приемов классической
антропологии, и она служит основой, базисом для последующих
разработок в этой области.
Успехи, достигнутые современной генетикой, столь ощутимы, что
изучение, например, на уровне антигенного состава или
полиморфизма ДНК стало возможным даже на древнем ископаемом
мумифицированном материале. Так начали определять не только
группы крови системы АВО в палеоантропологическом материале,
но и исследовать факторы системы тканевой совместомости (HLA) в
мумифицированных тканях древних обитателей Америки
доколумбовского периода (Hossaini, Allison, 1976). Это позволило
прийти к важным умозаключениям относительно характера
эволюционных изменений на протяжении относительно короткого
временного интервала. Совсем недавно стало возможным также
провести молекулярно-генетический анализ фрагментов ткани мозга
из мумифицированных останков человека, жившего 8000 лет назад;
этим впервые была показана возможность идентификации
4
«юлиморфи»ма ДНК на материале столь значительной древности,
что, безусловно, открывает большие перспективы для исследований в
области исторической генетики (Doran, Dickel, Ballinger and all.,
1986).
И все же современная генетика может привнести значительный
вклад в этническую антропологию, если пренебречь некоторыми
ограничениями, которые вводились рядом авторов, и принять во
внимание следующиее допущение: I) с точки зрения исторической
генетики существуют гены более или менее важные; 2)
обнаруживаются факторы, от которых зависит расовая
дифференциация.
К явным преимуществам генетико-исторического подхода следует
отнести, например, количественную оценку пропорций возможных
отдельных, расовых или этнических компонентов в исследуемой
популяции.
Кроме того, хорошо разработанная теория популяционной
генетики, при наличии достаточно полной информации о генах в
популяциях, во многих случаях позволяет оценить время
дифференциации этнических и расовых групп, реконструировать
становление той или иной популяции в хронологическом отношении.
Наконец, методы исторической генетики с успехом позволяют
решать проблемы динамики этнорасовых процессов при
установлении количественных характеристик таких эволюционных
факторов, как воздействие отбора, миграций, скорости мутационного
процесса и генетико-автоматических процессов.
На современном этапе историческая генетика наряду с другими
историческими и биологическими дисциплинами обладает
определенными возможностями для участия в решении целого ряда
спорных положений, полученных при сопоставлении классических
морфологических и генетических классификаций.
5
ГЛАВА I
ПРЕДЫСТОРИЯ ВОПРОСА. ДИНАМИКА И
ПРЕЕМСТВЕННОСТЬ РАЗВИТИЯ ЭТНОРАСОВЫХ
ГРУПП В РЕГИОНЕ ПРИУРАЛЬЯ.
ПРОБЛЕМА ЭТНОГЕНЕЗА БАШКИР.
Башкиры - один из крупных тюркоязычных народов Восточной
Европы. По данным переписи 1979 года численность башкир в
Советском Союзе достигает 1 млн. 371 тыс. человек, из них 1291 тыс.
человек расселены в РСФСР, при этом большая их часть - 75,5%
сосредоточена в Башкирской АССР (Население СССР по данным
Всесоюзной переписи населения 1979 г. 1980; Брук, 1981).
За пределами автономной республики башкиры расселены
относительно компактными группами в Челябинской, Курганской,
Пермской, Оренбургской, Куйбышевской и Саратовской областях,
таким образом, регионом основной их концентрации являете»
Южный Урал с прилегающими к нему районами.
На территории Башкирии основная часть башкир расселена в
долине реки Белой и ее притоков (Кузеев, Шитова, 1963). По своему
составу население республики многонационально, наиболее
многочисленными представителями других национальностей
являются русские (40,3%), татары (24,5%), чуваши, марийцы,
украинцы, мордва, немцы, удмурты и другие составляют около 11%.
Этногенез башкир принадлежит к числу малоизученных и
наиболее сложных в современной исторической науке (Кузеев, 1974).
Существует много гипотез и достаточно аргументированных
высказываний относительно происхождения башкирского народа,
которые в основном касаются древних этапов истории народа
(Рычков, 1762; Уйфальви, 1877; Татищев, 1962; и др.). В течение
долгого времени, пожалуй, наиболее дискуссионным оставался
вопрос об угорской или тюркской основе башкирского этноса. В
зависимости от подходов к решению этой проблемы и различаются
сторонники угро-мадьярской и тюркской гипотез происхождения
башкирского народа. Не было единого мнения и относительно
предков башкир, ими в разное время с известной долей уверенности
6
называли сарматов, финно-угров, гуннов и др. Историко-
фактологическая основа существующих гипотез о происхождении
башкир охватывает период с последних веков I тыс. н.э. - начала II
тыс. и базируется на дошедших до нас свидетельствах знаменитых
путешественников средневековья: Юлиана, Рубрука, Плано Карпини
(Аннинский, 1940; Эрдели, 1967; и др.). К этому же времени
относятся и дошедшие до нас первые письменные упоминания о
башкирах в Волго-Уральском регионе (Ковалевский, 1956;
Крачковский, 1957).
Идея угорско-мадьярского происхождения башкир впервые была
предложена Страленбергом (Stralenberg, 1730). А Д.А.Хвольсон
наиболее аргументировано рассмотрел эту гипотезу, основываясь на
филогенетическом анализе этнонимов башкорт и мадьяр, считая их
тождественными (Хвольсон, 1869). Данная точка зрения имеет своих
многочисленных сторонников, полагающих, что древнемадьярские
группы в составе тюркских племен Приуралья и Поволжья в целом
составили этническое ядро формирующейся башкирской народности
(Берс шн. 1852; Уйфальви, 1877; Толстов, 1948; Шастина, 1957;
Артамонов, 1962; и др.).
Другие исследователи отводили определяющую роль тюркским
племенам, ассимилировавшим в своем составе автохтонные, в том
числе и древнемадьярские группы (Флоринский, 1874; Малиев, 1876;
Назаров, 1890; Руденко, 1916, 1925, 1955). Свою концепцию они
основывали на данных в области антропологии и лингвистики.
Третьим направ,. _нием в изучении этногенеза башкир явилось
признание тюркской гипотезы происхождения данного народа,
однако с той или иной долей участия финно-угорского субстрата
(Yambery, 1885). К этому же заключению приводили, отмеченные
многими этнографами и антропологами, смешанность и
неоднородность этнического состава башкир (Аристов, 1895).
Значительный вклад в изучение этнической истории башкир
внесли археологические исследования, которые ввели в научный
оборот в ряде случаев достаточно хорошо датированный материал.
Однако, следует отметить, что до сих пор остаются недостаточно
познанными средневековые археологические памятники Башкирии.
Крупным событием в этнографии башкир явились работы
С.И.Руденко (Руденко, 1916, 1925, 1955). Собранные им
этнографические материалы составили основу Башкирской
7
коллекции музея (Авижанская, Кузеев, 1962). Не меньшую ценность
представляет также антропологическое изучение башкир, отличаясь
своей фундаментальностью, проведенное С.И.Руденко исследование
физического типа осуществлялось с обязательным учетом родовой
принадлежности и выявлением генеалогии лиц. Автор приходит к
выводу о сложности этнического и антропологического состава и о
ведущей роли пришлого тюркского компонента в формировании
башкирского этноса.
Весьма информативными для изучения родо-племенных структур
башкир и истории их формирования являются также материалы
декоративно-прикладного искусства, тамг, старинных родословных
(шежере) и т.д. (Кузеев, 1960; Шитова, 1971). Так, собранная
Д.Н.Соколовым коллекция, включающая свыше 800 башкирских
тамг, убедительно доказывает существование стабильных этнических
связей древних башкир с тюркскими народами Алтая и Средней Азии
(Соколов, 1898, 1904). Однако, при скудности сведений письменных
источников о происхождении башкир, а также недостаточности
археологических материалов средневековья, неразработанности
критериев выделения археологических культур и определения
этнической принадлежности их носителей, приблизиться к
сложнейшей задаче реконструкции этнической истории можно лишь
при условии соблюдения комплексного подхода к исследованию с
привлечением возможностей различных дисциплин.
Свидетельством несомненной значимости подобного подхода к
решению вопросов этногенеза башкир являются монографии
Р.Г.Кузеева (Кузеев, 1974, 1978 и др.). Обобщив богатый
предшествующий опыт исследований и дополнив его обширными
материалами, он выдвинул стройную концепцию происхождения
башкирского народа, начиная от времени возникновения первичных
этнических образований в глубине веков до формирования
народности с ее неповторимым этническим обликом. Была также
реконструирована родо-племенная организация башкир XVH-X1X
вв., определены особенности их расселения и контактов в регионе
Южного Урала. Совокупность всех данных позволила Р.Г.Кузееву
сделать вывод о том, что наблюдающаяся близость башкир в
элементах материальной культуры, фольклоре, этнонимии,
изобразительном искусстве с соседними народами объясняется тем,
что в iV-VIH вв. территорией пребывания древнебашкирских племен
8
являлся Восточный и Центральный Казахстан. В силу определенных
обстоятельств бурджане-баджгарды в IX веке были вынуждены
мигрировать в Нижнее Поволжье, Северный Кавказ, Приуралье,
часть их ушла на запад в Причерноморье. Именно с этого времени
начинается собственно уральский период истории древних башкир
(Кузеев, 1974).
Основу древнебашкирского этноса составили племена бурзян,
усерган, тангаур, тамьян, байлар. Однако очень существенно не
преуменьшить огромной роли происходивших метисационных
процессов, межэтнических контактов, смешения и ассимиляции
автохтонных племен, обитавших на Урале задолго до появления
пришельцев с юга (булгаро-мадьярская группа племен, бахмутинцы,
турбаслинцы и др.).
В последующие XI-X11I вв. происходит дальнейшее расселение
башкир по Южному Уралу, в Зауралье перекочевали кара-
тавлинская, тырнаклинская, сартовская и айлинская родовые группы.
По мнению исследователей, решающую роль в этнической
консолидации башкир сыграли XIII-X1V века (Кузеев, 1974; и др ).
1. Данные археологии.
Археологические исследования в Башкирии привели к
накоплению материалов, являющихся ценным источником по
изучению древнего населения края и позволяющими осветить
разные этапы истории заселения Южного Урала.
К одним из наиболее ранних археологических находок в
Восточной Башкирии относятся стоянки каменного века (Бибиков,
1950; Бадер, 1951; Сальников, 1952; Рюмин, 1960, 1961 и др.).
Археологические исследования в послевоенный период коснулись
прежде всего древнейшей истории Башкирии. Однако следует
отметить, что палеолитические находки на Южном Урале, в целом
довольно бедны остатками материальной культуры.
Предполагается, что заселение Урала человеком произошло с юга
в эпоху развитого ашеля (Бадер, 1971; Бибиков, 1959). Найденная в
верховьях р.Белой настенная живопись Каповой пещеры
подтверждает эту точку зрения.
9
Несколько полнее представлен мезолит Южного Урала.
Отмечаемое исследователями различие памятников
микролитических культур Предуралья от Зауральских в эпоху
мезолита указывает на близость предуральской культуры
(ильмурзинской) к Волго-Камью и зауральской (янгельской) - к
культурам Приобья, Прииртышья и Казахстана. Предположительно в
этот же период происходит заселение Южного Урала с юга
населением родственным ирано-среднеазиатскому. Отсутствие
антропологического материала не позволяет пока охарактеризовать
население периода мезолита (Матюшин, 1964; 1970).
Находки поздненеолитического и энеолитического времени с
территории Башкирии обнаруживают близость к так называемой
андроповской культуре, получившей к концу II и началу I тыс. н.э.
широкое распространение в Средней Азии и Южной Сибири
(Сальников, 1952).
Обобщающие работы по эпохам бронзы и железа позволили
констатировать существование этнических образований,
последовательно сменявших друг друга на территории Башкирии с
III тыс. до н.э. до рубежа нашей эры (Сальников, 1967; Стоколос,
1972; Збруев, 1952; Смирнов, 1952, 1957; Пшеничнюк, 1971). Также
установлена чрезвычайная сложность этнического состава населения
в I тыс. н.э., т.е. в эпоху формирования племенных образований,
сыгравших значительную роль в сложении древнебашкирского
этноса (Смирнов, 1957, 1971; Мажитов, 1968, 1977; Ахмеров, 1951,
1970; Генинг, 1964,1967 и др.).
Территория Башкирии с древнейших времен до конца I тыс. н.э.
была зоной активных и многочисленных контактов между
разноязычными (иранскими, финно-угорскими, позднее - тюркскими)
этническими группами, часть из которых впоследствии вошла в
состав племен, положивших начало башкирскому этносу. Следует
отметить, что в археологических работах ведущая роль в этногенезе
данного народа часто отводится племенам, обитавшим на Южном
Урале в VII-IX вв. и ранее (Мажитов, 1977; Халиков, 1972),
Например, многие археологи считают, что памятники Южного Урала
VII-IX вв. принадлежат ближайшим предкам башкир (Ахмеров, 1951,
1970; Сальников, 1952; Смирнов, 1957; и др). Напротив этрнографы
отдают приоритет тюркским мигрантам VII1-IX и последующих
веков, когда сюда из южных степей проникли древнебашкирские
10
племена, расселившиеся позже в северном направлении, в
горнолесные районы (Кузеев, 1974).
Периодизация средневековых памятников Прикамья была
проведена Спицыным, выделившим два периода: ломоватовский
(VIII-IX вв.) и поздний (Х-ХП вв.) (Спицын, 1902). Мнение
исследователей сходятся в том, что среди изученных археологических
памятников на территории Башкирии I тыс. н.э. не удалось выделить
культуру, носителей которой можно было бы с достаточной
определенностью считать прямыми предками башкир, хотя вопрос о
пребывании финно-угров, угров, ираноязычных сармато-аланов,
тюркизированных сармато-аланов, тюрков, тюрков-угров, племен
палеосибирского происхождения, самодийских племен достаточно
широко обсуждается в археологической литературе (Мажитов, 1956,
1977; Смирнов, 1957; Халиков, 1976, 1991, и др.). Заключение
большинства археологов также едино относительно того, что эти
разноязычные автохтонные племенные образования Южного Урала
V-VII1 вв. составили субстрат, на основе которого в дальнейшем и
шла консолидация башкир. Однако в разных работах неоднозначно
определение роли поздних кочевников-пришельцев в процессе
формирования этноса. Так, по мнению В.Ф.Генинга, собственно
башкирские племена находились в составе поздних групп
пришельцев в данный регион (Генинг, 1967).
По материалам археологических исследований восточная и
западная части Южного Урала в VII-VIII вв. были заселены
племенами, рода венными по происхождению. Об этом
свидетельствуют принадлежащие им находки из керамики
кушнаренковского, кара-якуповского и бакальского типов
(Сальников, 1956; Мажитов, 1977).
Качественно новым этапом истории Южного Урала являются IX-X
века. В это время получают повсеместное распространение
памятники типа Хусаиновских курганов: захоронения в гробах с
подстилкой и богато украшенными серебром предметами одежды, с
характерными женскими украшениями типа накосников и серег
салтовского вида (Мажитов, 1977). Исследователь отмечает также
идентичность поясных ремней, найденных в этих могильниках с
аналогичными предметами из Тувы, Южной Сибири и Алтая.
Сопоставление находок из курганов Хусаиновского типа с
памятниками VII-VIII вв. позволило выявить как сходство, так и
П
различия между ними, в свою очередь привело к выводу о том, что
ранние и поздние курганы связаны с поселениями подобными типу
Старо-Калмашевского городища. Тот факт, что поздние памятники
отличаются значительно более крупными размерами может служить
одним из указаний на оседлость образа жизни (Мажитов, 1977 и др.).
В 1Х-Х вв. на территории среднего течения р.Белой также как и в
последующие века обитали представители турбаслинских племен,
контакт которых с носителями кара-якуповской керамики явно
усилился, судя по изменению типа орнамента в сторону большего их
сходства.
Общение со степным миром кочевников, уходящее вглубь веков, с
конца 1 тысячелетия сыграло огромную роль в динамике
формирования башкир, так как отсюда в горно-лесные и
лесостепные районы проникали все новые волны носителей
тюркской культуры. Свидетельством этого являются существенные
изменения в характере материалов, связанных со временем
нашествия Золотой Орды, так заметно уменьшается количество
курганных типов захоронений, что в основном связано с переходом от
языческих обрядов к мусульманским: погребение без вещей,
ориентация головой на юг и т.д. (Акимова, 1974). Материалы
археологических раскопок, равно как и этнографические источники,
подтверждают высказывания Р.Г.Кузеева об особой важности XIII-
XIV вв., сыгравших основополагающую роль в этнической
консолидации башкир (Мажитов, 1977; Акимова, 1974 и др.).
Наибольший интерес в этом плане представляет изучение
краниологических находок того времени, проведенное
Р.М.Юсуповым, которое показало, что именно длительная метисация
являлась основной формой процесса расообразования в данном
регионе. Имеющиеся локальные различия, отмеченные
исследователем как в мужских, так и в женских сериях черепов были
не столь глубокими и сходство в целом по Башкирии проявилось в
полной мере (Юсупов, 1982).
2. Некоторые материалы ономастики,
лингвистики, фольклора.
Ономастика является молодой, но очень перспективной областью
знания, имеющей возможности решения целого ряда проблем
12
этногенетического, историко-географического и лингвистического
плана в широком диапазоне времени, начиная от глубокой древности
и до современности. Следует подчеркнуть, что при изучении
прошлого бесписьменных народов данные ономастики являются
одним из наиболее ценных источников, но ее значение не
утрачивается и в настоящее время, т.к. предметом исследования
служит продолжающийся непрерывно процесс, связанный с
появлением наименований новых различных предметов, новых имен
и др. Изучая закономерности возникновения этнонимов, топонимики,
антропонимики в различных этнических группах исследователи
выявляют связи с конкретными историческими и социально-
экономическими условиями среды (Никонов, 1973; Кузеев, 1973; и
др.).
Башкирский язык, относящийся к тюркским языкам, занимает
место в кипчакской группе вместе с киргизским, татарским,
казахским, каракалпакским, ногайским и другими языками
(Баскаков, 1960; Киекбаев, 1972). Башкирский язык очень самобытен
и анализ его, проведенный в советское время, позволил выявить
связи и параллели в развитии с якутским и туркменским языками, а
такж возможность монгольского и тюрко-угорского языкового
влияния (Баишев, 1955; Серебренников, 1963; и др.). Географические
особенности изоляции башкир от других тюркских племен
способствовали сохранению в языке наиболее архаичных черт,
сближающих их с представителями восточнохуннской ветви такими
как тувинцы, хакас..,, якуты и другими (Баскаков, 1962). С другой
стороны, этими же причинами объясняется существование у башкир
различных диалектов, характерных для отдельных племенных групп
(Киекбаев, 1958).
Башкирские диалектологи различают два диалекта - южный
(юрматинский) и восточный (куванакский), некоторые из них также
выделяют диалект северо-западных башкир (Юлдашбаев, 1968;
Гарипов, 1959). Последний в настоящее время очень близок к
татарскому языку, а в прошлом его отличало сходство с болгарским
(Ишбулатов, 1975; и др.).
Восточный диалект распространен в северо-восточной Башкирии,
а также Челябинской и Курганской областях. В нем, в свою очередь,
различают пять самостоятельных говоров (Максютова, 1976).
Башкиры, живущие в центральной, южной Башкирии,
13
Куйбышевской, Саратовской и Оренбургской областях говорят на
южном диалекте, в составе которого различают три говора
(Мирж.шова, 1979).
Возросший в Башкирии в течение последних десятилетий интерес
к изучению разных аспектов родоплеменной этнонимии привел к
накоплению, систематизации и анализу богатейшего материала, что
в итоге позволило выделить этнонимические пласты, с
установлением последовательности и хронологии включения их
носителей в этническое развитие башкирского народа (Гарипов,
Кузеев, 1971; Корнилов, 1971; Кузеев, 1973, 1974,1978,1987 и др.).
Всего в башкирской родоплеменной этнонимии исследователями
было выделено семь историко-стратиграфических пластов:
1. Ранний финно-угорский самодийский пласт, объединяющий три
группы названий, которые в свою очередь восходят к
древнесамодийским этнонимам. Отсутствие письменных источников,
относящихся к этому периоду затрудняет точность характеристик
этой группировки, т.к. основным материалом здесь служили
этногенетические предания - шежере. Более аргументированные
сведения получены в результате проведенных археологических
исследований, доказывающих наличие значительной миграции угро-
самодийских племен из Сибири в Волго-Уральский регион в 1II-VII
вв., а также анализ предметов материальной культуры и декоративно-
прикладного искусства, восходящих к этому периоду (Генинг, 1967;
Авижанская и др., 1964; Шитова, 1971; и др.).
2. Булгаро-мадьярский пласт, датируется VII1-IX вв., включает
племенные названия юрматы, юрми, гайна-тархан, буляр, танып и ряд
других более мелких групп (Куник, 1878; Эрдели, 1967; и др.).
3. Древнебашкирский пласт, объединяющий племенные названия
усерган или муйтен, бурзян, тамьян.тангаур, байлар, ун, бишул, кудей,
уран, сураш, ягалбай, унгар. В отличие от двух более ранних пластов,
большая часть перечисленных племенных образований уже была
объединена под общим названием башкорт (баджгард, башгард,
башкарт) (Гарькави, 1870). Датировка данного историко-
стратиграфического паста восходит к VIII-IX вв.
4. Огузо-кыпчакский пласт, датируется Х-ХП вв., объединяет
племенные названия ай, айле, тырнаклы, каратавлы, сарт, алтмаш-
колак, кумлы, абдел, кызылбаш, этимгэн, кусюк, истяк (Рашид-ад-
дин, 1952; Кононов, 1958).
5. Кыпчакский пласт, датируется XIII-XIV вв., включает целый ряд
собственно кыпчакской, катайской и табшнской родоплеменных
групп, включая также минскую. Влияние мощной кыпчакской
миграции этого периода во многом опередило направление развития
башкирского этноса и привело к формированию тех этнических
признаков, которые отличают его и сегодня.
6. Ногайский пласт, восходит к периоду связанному с возвышением
Ногайской орды и ее ролью в Волго-Уральском регионе в XV-XVI
вв. После отхода основной части ногайцев на юг остались
родоплеменные названия ногай-юрматы, ногай-кыпчак, ногай-
бурзян, ногайлар, кызылногай.
7. Седьмой историко-стратиграфический пласт объединяет целый
ряд мелких родовых подразделений и датируется более поздним
временем XVII-XVIII вв. В этот период особенно активными
становятся взаимодействия татар и башкир, особенно интенсивными
являются эти контакты в западной и северо-западной части
Башкирии.
На основании обширнейшего материала, собранного в последние
десятилетия, исследователи приходят к установлению картины
становления башкир и характера протекания этнических процессов в
Волго-Уральском регионе с древности и до настоящего времени:
Данные этнонимии позволяют проследить влияние самодийских и
угорских компонентов, пришлых из Сибири, при этом местный
финно-угорский субстрат проявляется несравненно слабее;
На ранних стадиях этногенеза башкир определенную роль
сыграло также участие булгаро-мадьярского компонента;
Основополагающую роль в формировании сыграли
древнебашкирские и кыпчакские волны миграций в VIII-XIV вв., и
особенно в XI1I-XIV вв. (Кузеев, 1973).
3. Антропологические данные
Палеантропология и антропология
современных башкир.
Изучение ранних стадий эволюции человека исследователями в
области палеоантропологии основано на данных физической
15
антропологии и археологии. Дошедшие до нас нт глубокой
древности первые палеолитические материалы, свидетельствуют о
далеком времени освоения человеком Южного Урала. Очевидно, это
был процесс, идущий с юга 40-30 тыс. лет назад, в эпоху развитого
ашеля и начала позднего палеолита (Бибиков, 1959; Бадер, 1971). К
сожалению, до сих пор не обнаружены палеоантропологические
материалы эпохи палеолита, а также мезолита.
К наиболее древним палеоантропологическим находкам с
территории современной Башкирии относятся несколько черепов
эпохи неолита (Бибиков, 1950 б). Судя по сохранившимся костным
остаткам, обитателями края были люди низкого роста, вполне
европеоидного вида, с незначительной монголоидной примесью.
Этот вариант, характерный для Урала, был назван Герасимовым
ранним вариантом субуральского типа (Герасимов, 1955). Однако
процесс метисации древнего европеоидного населения Урала был,
очевидно, не столь интенсивным, судя по черепам из Бурановской
пещеры (Бибиков, 1950). Так, единичный череп этого же периода,
обнаруженный в Давлеканово, характеризуется нешироким сильно
профилированным лицом с узким, резко выступающим носом, т.е.
имеет ярко выраженные черты южных европеоидов. Мозговая
коробка узкая и длинная, средневысокая, долихокранная, со
среднеразвитым переносьем, лицо ортогнатное, широкое (Акимова,
1974).
В эпоху бронзы наблюдается преемственность культур с
предшествующим периодом, а также усиление контактов населения
Приуралья с Зауральем. Последнее, по-видимому, объясняется
развитием и дальнейшим распространением скотоводства
(Матюшин, 1973).
Физический тип людей этого времени близок к европеоидному
энеолитаческому населению. Древние племена входили в огромную
Урало-Казахстанскую этнокультурную общность (Сальников, 1962). В
составе распространенных в эту эпоху абашевской, срубной,
горбуновской археологических культур выделяется доминирующая
роль элементов андроновской культуры Южного Зауралья и
Казахстана (Ахмеров, 1952).
Р.Б.Ахмеров также пишет о важной роли сармато-аланских и
оседлых финно-угорских племен, подчеркивая при этом длительное
почти тысячелетнее развитие ираноязычной сарматской культуры на
16
Южном Урале. Г.А.Федорова-Давыдова отмечает этнокультурное
влияние срубной культуры Поволжья, усиливающееся по мере
продвижения с востока на запад (Федорова-Давыдова, 1964).
Палеоантропологический материал этого периода
свидетельствует о неоднородности антропологического состава и
указывает на наличие пришлого компонента (Акимова, 1971). В
целом, население Южного Урала, как и в предыдущую эпоху,
европеоидное. Наметившиеся еще в неолите различия между
зауральскими и приуральскими племенами, привели в эпоху бронзы к
выраженной неоднородности населения, усиливающейся в
направлении с севера на юг. Предполагается, что абашевские
племена лесной полосы скорее всего относились к финно-угорский
языковой группе, а срубные и андроповские племена южных степных
районов - к ираноязычной (Сальников, 1961).
Памятники степного Приуралья А.П.Смирнов выделяет в
локальный самаро-уральский вариант савроматской культуры и
относит время формирования савроматской культуры на Южном
Урале к IV в. до н.э.; исследователь в то же время подчеркивает, что
восточный вариант савроматской культуры теснее связа" с северо-
иранским этническим миром. К рубежу нашей эры племена,
продвинувшиеся на север, были ассимилированы местными финно-
угорскими племенами, которые в свою очередь приобрели
некоторые сарматоидные черты (Смирнов, 1971). В последних веках
до н.э. заметно усиливаются торговые связи населения Приуралья и
Среднего Поволжья со Средней Азией и Кавказом (Ахмеров, 1952).
К числу доступных для краниологического изучения относятся
серии черепов из сарматских могильников, ископаемый материал в
которых датируется III в. до н.э. - I в. н.э. (Акимова, 1964).
Антропологические черты этого населения характеризовались
широко- и низколицим европеоидным обликом. Носителями этих
признаков являются сарматские племена, кочевавшие к югу от Уфы.
Однако, по мнению М.С.Акимовой, этот антропологический тип
возможно был не единственным в характеристике населения Южной
Башкирии рубежа I тыс. до н.э. и I тыс. н.э. (Акимова, 1971).
История сарматских племен развивалась на основе культуры
савроматов, являвшихся в свою очередь преемниками культур
срубных и андроновских племен. Исследователи по разному
оценивают вклад сарматов в формирование современного
17
антропологического типа башкир. К числу наиболее
последовательных сторонников значительности вклада сарматского
компонента в сложении физического типа башкир относится
С.И.Руденко, который связывал древних башкир с тиссагетами и
иирками Геродота, а также с савроматами, жившими в Приуралье в 1
тыс. до н.э. (Руденко, 1955). Мнение С.И.Руденко о том, что в основе
антропологического типа лежит тип тиссагетов и сармат, имевших
монголоидную примесь, был поддержан Н.А.Мажитовым, который в
решении проблемы этногенеза башкир доминирующую роль отводил
автохтонному населению, а не пришлым кочевым группам (Мажитов,
1969). В более поздних работах М.А.Мажитов несколько отошел от
этого взгляда. О большом вкладе сарматских племен в процессе
этногенеза башкир пишет А.П.Смирнов, подчеркивая возможность
сильного влияния их также и на население северных районов
Башкирии. В эпоху великого переселения народов сарматские
племена проникали далеко на север, вынуждая часть финно-угорских
племен отступить еще севернее и смешивались с оставшимися
группами. Но в отличие от других исследователей А.П.Смирнов
подчеркивает, что в башкирах следует признать пришлый народ,
смешавшийся с местными народами финно-угорской языковой
общности, которые ранее были тюркизированы пришельцами-
болгарами (Смирнов, 1957).
Несмотря на длительное пребывание сарматов на территории
современной Башкирии, в антропологическом отношении они
исследованы недостаточно. Поэтому весьма интересным
дополнением к имеющимся материалам сарматского периода
являются сравнительно недавно исследованные серии черепов с
территории Оренбургской области (могильники Мечет-сай и Новый
Кумак) и из Кармаскалинского района Башкирии (могильник
Старые Киишки), относящиеся к эпохе бронзы (Акимова, 1974).
Анализ показал близость савроматских черепов с Аландского и
Новокумакского могильников к черепам предшествующей срубной
культуры. Однако, на фоне преобладания европеоидных черт
имеются также выраженные монголоидные особенности. При
комплексном сравнении признаков отмечается близость к
савроматам. Более же поздние черепа, относящиеся к сарматам,
интересны четким проявлением различий между ними, что
позволяет, однако, объединить их в единый вариант большой
европеоидной расы. При сопоставлении с палеоантропологическими
материалами соседних территорий выявляется значительное сходство
с населением Западного Казахстана, Астраханской и Оренбургской
областей, а также прослеживается генетическая преемственность с
савроматами, т.е. налицо участие также и других компонентов,
помимо андроповского, в формировании автохтонного населения
(Акимова. 1974; Трофимов. 1952).
Краниологическая серия из Ново-Мурапталовских курганов
характеризует сармат как европеоидов с признаками,
обнаруживающими сходство с материалом из Кармаскалинского
района (Юсупов, 1979).
Обнаруженные в северных и центральных лесостепных районах
Башкирии памятники пьяноборской и караабызской культур
относятся к рубежу нашей эры. М.А.Акимовой исследован состав
населения караабызской культуры по материалам Охлебининского и
Биктпмировского могильников с IV в. до н.э. - II в. н.э. Серии черепов
в целом европеоидные, однако слабое выступание носа, некоторая
уплощенность лицевого скелета ука 1ывают на наличие в их составе
незначительной монголоидной примеси (Акимова, 1971).
Краниологическая серия, относящаяся к населению пьяноборской
культуры из Камышлы-Тамакского могильника Кушнаренковского
района, относится также к европеоидному типу южного
происхождения, однако в женской группе черепов отмечаются
некоторые монголоидные особенности, присущие племенам
западносибирского происхождения (Акимова, 1968; 1973; Toth Т,
1974).
Анализ палеоантропологических материалов эпохи железа
выявляет усиление различий в составе населения юга и севера
Башкирии, а также и в широтном направлении.
Первое тысячелетие является одним из наиболее сложных
периодов в формировании антропологических типов народов
Южного Урала, Приуралья и Среднего Поволжья, т.к. начиная с III в.
н.э. происходят значительные изменения в этно- и расогенетических
процессах, во многом активизированные волной великого
переселения народов, связанной с движением гуннских кочевых
племен на запад. В этом отношении формирование ананьинской
культурной общности представляет собой интерес своей
неоднородностью в антропологическом отношении, проявляющуюся
19
18
в дифференциации двух основных комплексов: низколицего
монголоидного и умеренно гиперморфного долихомезокранного
европеоидного (Ефимова, 1981, 1991).
Единичные ананьинские черепа Уфимского могильника, а также
немногочисленные находки пьяноборского периода отличаются
появлением на фоне выраженных европеоидных черт небольшой
примеси монголоидных.
Более значительными находками являются 29 черепов из
Бирского и Каратамакского могильников бахмугинского периода III-
VII вв. н.э. (Мажитов, 1959). Бирская серия отличается тем, что часть
черепов мезокранна, часть долихокранна, с невысоким и нешироким
лицом и умеренно выступающим носом. Все это является
свидетельством смешанности состава населения. Имеющаяся
монголоидная примесь характеризуется широким и сравнительно
высоким лицом, несильно выступающим носом и выраженной
уплощенностью (Акимова, 1968; Генинг, 1972; Мажитов, 1977; Toth
Т., 1974,1981).
Материалы V в. н.э. из Кушнаренковского могильника,
своеобразного археологического памятника, отличаются от
предыдущих серий резкой выраженностью европеоидных признаков,
не обнаруживающих аналогий в местном населении. То есть они
характеризуют пришлое население, при этом черепа в целом сходны
с черепами из Салтовского могильника (Алексеев, 1959).
Антропологический материал с Кушнаренковского могильника не
исключает влияния проникших с юга европеоидных групп (Акимова,
1964). Близка к этой серии и краниологическая выборка из Ново-
Турбаслинского могильника, в которой также отмечается комплекс
признаков пришлого населения, однако более смешанного по своему
составу. По мнению некоторых исследователей это могли быть уже
башкирские племена (Мажитов, 1977).
Следует особо подчеркнуть, что ни одну из племенных общностей
древности, существовавших на базе перечисленных выше культур и
культурных комплексов, невозможно считать собственно
башкирской. Эго подчеркивает и историко-этнографический анализ
этнического состава башкир, которые уже на раннем этапе
сформировались как родоплеменные образования (Кузеев, 1974).
К концу I тыс. н.э. в северной части территории современной
Башкирии появляются курганные погребения тюркских кочевников
20
(Мажитов, 1977). К этому периоду относятся и
палеоантропологические находки из Старо-Халиловских, Старо-
Мусинских, а также Мрясимовских курганов. Интересно, что будучи
морфологически близки между собой, они отличаются по
антропологическому облику как от автохтонного населения, так и от
групп с сопредельных с Уралом территорий, несколько сближаясь
только с материалами из Западной Сибири. Среди черепов имеются 4
искусственно деформированных. Менее представителен материал
Каратамакского могильника. По мнению М.С. Акимовой
монголоидность здесь еще более выражена (Акимова, 1968).
К сожалению, последующие этапы истории в антропологическом
отношении долгое время оставались малоизученными в связи с
фрагментарностью доступного краниологического материала, хотя
именно последние века I тысячелетия н.э. имели решающее значение
в формировании физического типа башкир, так как события этого
периода увязываются археологами и этнографами с началом
этнической истории башкирского народа (Кузеев, 1974; Мажитов,
1977; и др.). "В конце V1II-IX вв. в Волго-Уральской области
появляются древнебашкирские племена бурзян, усерган, тамьян,
байлар, бишул, ун, кудей, роды сураш, ягалбай" (Кузеев, 1974).
Формирование башкирского народа происходило в тесной
этнической и культурной связи с народами Средней Азии, Сибири,
Волго-Камья и Северного Кавказа, что, естественно, внесло
определенную мозаичность в антропологический материал
(Ефимова, 1987; Халиков, 1991). Поэтому краниологическая серия из
Бекешевских курганов, датируемая IX-X вв., представляет
несомненный интерес. Характеризуясь в целом как европеоидная, но
с некоторой монголоидной примесью, она по ряду признаков
занимает промежуточное положение между черепами кочевников
Восточно-Казахстанской области и Приаралья (Гинзбург, 1972;
Мажитов, 1977; Юсупов, 1982). Все это свидетельствует о том, что в
составе древнебашкирских племен Южного Урала в конце I тыс. н.э.
прослеживаются как признаки автохтонного европеоидного
населения, так и значительный пришлый монголоидный компонент.
Полученные Р.М.Юсуповым краниологические серии относятся к
более позднему периоду - XV1I-XIX вв. Этот материал с общим
объемом 380 черепов охватывает различные территориальные
группы башкир и позволяет дать их краниологическую
21
характеристику. Суммарно башкиры характеризуются средним
продольным и большим поперечным диаметром черепа
брахикранны. Лицевой скелет широкий, высокий,
среднепрофилирован на обоих уровнях горизонтальной плоскости и
несколько уплощен в вертикальной. Нос умеренно выступает над
линией профиля. Башкирское население того времени в
антропологическом отношении представляет собой смешанную
популяцию, в которой соотношение монголоидных и европеоидных
особенностей идет в сторону преобладания последних. По
межгрупповой вариации величин признаков и характеру их
распределения четко выделяются три локальных краниологических
комплекса, два из которых представляют собой различные варианты
европеоидного типа, а третий характеризуется наибольшей
сглаженностью европеоидных особенностей. Однако несмотря на
выявленные различия между территориальными группами
отмечается тенденция к антропологической консолидации, о чем
свидетельствуют низкие коэффициенты суммарных отличий между
сериями по способу 1 кнроуза (Юсупов, 198()).
История антропологического изучения современного населения
начинается с XIX в. Единичные работы этого периода носили в
основном описательный характер и были малы по числу наблюдений.
Однако они представляют определенную ценность, поскольку
описывают физический облик и антропологическую неоднородность
башкир (Рычков, 1762; Назаров, 1863; Казанцев, 1866; и др.).
Крупным первым антропологическим исследованием башкир
является работа профессора Н.М.Малиева, проведенная в Уфимском
и Белебеевском уездах Уфимской губернии (Малиев, 1876). Было
измерено 40 башкир по программе, включающей 32 признака.
Исследователь отметил наличие достаточно распространенных
монголоидных признаков, что позволило ему выделить в составе
башкир XIX века два антропологических типа - лесных и степных
Первый из которых характерен для башкир бассейна р.Сима и ее
притоков, второй - для групп, расселенных по рекам Дема и Уршак
(Малиев, 1876).
Антропометрические исследования в Оренбургской области также
выявили значительную неоднородность башкир с преобладанием
среди них тюркских признаков (Уйфальви, 1880; Сомье, 1891-1892).
Наиболее интересна, из проведенных в этой области, работа
сильной смешанности и
для всех башкир
не утратившим своего
ПС Назарова, который осуществил антропометрические
исследования с учетом родо-племенной принадлежности башкир
(Назаров, 1890).
Привлекает также внимание антропометрия 219 башкир в
Пермской, Уфимской и Оренбургской губерниях и изучение 49
черепов с Пермской области, в результате анализа которых автор
причислил башкир к "турецким народам" (Никольский, 1899).
Последующие изменения 122 башкир в Пермской и Уфимской
губерниях также подтвердили вывод о
отсутствии единого характерного
антропологического типа( Абрамов, 1907).
Наиболее фундаментальным трудом,
практического значения и в наши дни, является работа С.И.Руденко,
отличающаяся значительным числом обследованных из разных
родоплеменных групп башкир XX века. Различие в
антропологических особенностях их позволило выделить несколько
локальных вариантов, расположенных в восточной, юго-западной и
северо-западной областях Башкирии. Он отметил своеобразие
восточных башкир, отличающихся наибольшей пигментацией,
брахикранией, большой шириной лица и умеренным ростом. Северо-
западные башкиры имеют наименьшую пигментированность,
умеренную бранхикранию, сравнительно небольшие размеры черепа,
средний рост. Остальные группы занимают промежуточное
положение между этими крайними вариантами и характеризуются
средними антропологическими показателями в целом (Руденко,
1916).
Крупным антропологическим иммледованием современных
башкир, в достаточной мере полно охватившим все географические
группы, явилась работа, проведенная экспедицией Института и
кафедры антропологии МГУ в период с 1963 по 1967 гг. под
руководством М.С.Акимовой (Акимова, 1974). Исследовано 11
родоплеменных групп башкир: таныпцы, герейцы, канлынцы, минцы,
юрматинцы, кыпчаки, усергане, айлинцы, катайцы, табынцы - всего
1250 человек мужского пола. Последние три группы изучались в
Челябинской области. В географическом отношении это
представители северо-восточной, юго-западной и юго-восточной
*пгагеографических групп.
23
22
Проведенная антропометрия суммарно характеризует башкир
средней длиной тела, крупными размерами лица и головы, темными
прямыми волосами и промежуточными оттенками цвета глаз.
Однако, по целому ряду признаков наблюдается неоднородность
башкир, отмеченная многими исследователями ранее.
Анализ материала по современным башкирам позволил выделить
среди них четыре антропологических типа: субуральский, южно-
сибирский, светлый европеоидный и понтийский. Различия между
географическими группами определились различным соотношением
этих типов и были наиболее заметны при сравнении северо-западной
и северо-восточной групп. Это различие имеет большую давность,
так, данные палеоантропологии северо-запада характеризуют
местное население пьяноборской культуры как европеоидное со
средними размерами лица, при выраженной его профилировке и
несильном выступлении носа. На основе пьяноборских, позднее -
бахмутинских племен оформилась современная группа северо-
западных башкир с преобладанием субуральского и светлого
европеоидного типов. К южно-сибирскому типу, по мнению автора,
принадлежало местное население юга изучаемой территории эпохи
средневековья, с которыми вступили в контакт пришлые башкиры,
что наблюдается в отчетливом выявлении монголоидного
компонента в их современных потомках.
В решении вопроса о роли местных племен в сложении башкир
М.С.Акимовой высказывается мнение о значительной роли местного
компонента.
Сравнение антропологических особенностей западных башкир
(Верхнее Яркеево), юго-западных (Стерлибашево), восточных
(Архангельское) не показало каких-либо резких различий. Общим
антропологическим типом башкир является смешанный, при этом
монголоидный компонент в разных популяциях может
присутствовать в разных пропорциях. И.М.Золотаревой показано в
Илишевском районе ослабление монголоидных комплексов, что
интерпретируется исследователем как особенность
антропологического типа северо-западного региона республики.
Отмечается более светлая пигментация волос, радужной оболочки
глаз, более мягкие волосы, повышенный рост бороды, менее
развитая складка верхнего века, больше выражено выступление
спинки носа и др. Промежуточное положение по монголоидным
пизнакам заняли башкиры Стерлибашевского района, что особенно
сажается в степени пигментированности, особенностях строения
глаза, горизонтальной профилировке лица и меньшем выступлении
носа.
Представители коренного населения Архангельского района
отличаются наиболее выраженными признаками монголоидного
типа (I.M.Zolotareva, 1986; Kajanova, 1986).
Сопоставление с предыдущими антропологическими
характеристиками башкир по данным М.С.Акимовой (1963-1967 гг.)
показало, что за прошедшие 20 лет длина тела у башкир
увеличилась на 4 см. Так, у верхне-яркеевцев она возросла со 163,5 см
до 170 см, у стерлибашевцев от 163,5 см до 168 см. Головные индексы
были раньше равны 79,6 и 80,8, а в современном материале 79,3 и
80,6 соответственно.
Проведенное с помощью многомерного дискриминантного
анализа сравнение этих 3 групп с финноугорскими народами выявило
интересную картину. Башкиры в этом ряду оказались наиболее
пигментированы, а финны - наименее. И в целом широтные размеры
головы у башкир были наибольшими. Коми и марийцы
трансгрессируют между собой, но стоят ближе к башкирам, нежели к
финнам.
Сопоставление трех групп башкир между собой показало, что
мужчины-башкиры из Верхнее Яркеево оказались самыми близкими
к финнам (Kajanoja, 1986). Эти данные согласуются с основными
выводами по дерматоглифике о том, что в целом финноугорская
примесь у башкир незначительна.
Особенности морфологии зубо»з
Характер распределения одонтологических признаков в
башкирских микропопуляциях подробно изучен А.А.Зубовым и
С.Сегедой (Zoubov and Segeda, 1986). Исследователи подвергли
анализу группы башкир северо-западного (с. Верхней ркее во),
Центрального (с.Архангельское) и юго-западного регионов
республики. Одонтологические материалы были собраны и изучены
посредством методов разработанных ранее (Зубов, 1968,1973).
24
25
Для установления таксономического положения башкир в рамках
системы соседствующих этнических групп были использованы
сравнительные данные по одонтологии казахов, удмуртов, коми-
зырян и пермяков, хантов и манси (Зубов, 1972; Лксянова, 1979). На
основании информации о комплексе морфологических признаков
зубов были расчитаны средние таксономические расстояния (СТР)
между парами сопоставляемых групп (Зубов, Золотарева, 1980).
Анализ изменчивости основных признаков зубов среди
сравниваемых этно-территориальных групп свидетельствует, что
башкиры характеризуются смешанными монголоидно-
европеоидными особенностями с явным преобладанием "восточного
комплекса". В целом, полученные сведения показывают, что
интересующая нас группа отличается весьма своеобразным
сочетанием совокупности признаков зубов, который практически
полностью отсутствует у современных соседних популяций. При
сравнении с известным хорошо очерченным одонтологическим
комплексом башкиры занимают таксономическое положение между
европеоидными вариантами, локализованными в северо-восточной
части России (удмурты, коми-зыряне, коми-пермяки) и смешанными
группами Средней Азии. Однако некоторые особенности
башкирской популяции сближают ее с угорскими этническими
группами, особенно с хантами. Авторы идентифицировали
существенную межрайонную гетерогенность башкир в отношении
при знаков зубов. Так, было отмечено, что северо-западные башкиры
весьма существенно отличаются от других групп республики при
рассчитаном СГР=О,88. В целом это отличие ориентировано в
сторону усиления европеоидного компонента. В связи с наличием в
составе башкир кипчакского компонента, теоретически следовало бы
ожидать обнаружения сходства в строении признаков зубов у башкир
и казахов. Тем не менее, статистический анализ выявляет достоверно
значимые различия в плане морфологических характеристик между
этими двумя сопоставляемыми группами населения. Установленный
авторами эффект грацилизации зубов у башкир объясняется ими
возможными миграционными процессами финно-угорских племен.
На основании полученной информации делается вывод о
значительной роли финно-угорского субстрата в формировании
антропологического состава башкирского народа.
Таким образом, данные из области одонтологии позволяют
не только новую ценную информацию о расовом составе
П°й или иной группы населения, в данном случае башкирского
но и также оценить относительную долю вклада отдельных
народа» ч
исходных компонентов в историческом становлении изучаемой
пы и что особенно важно, соотнести формирование населения
на определенной территории и сопредельных областях во временных
масштабах.
Дерматоглифика
Первые материалы по этнической дерматоглифике башкир были
собраны экспедицией под руководством М.С.Акимовой в северо-
западной, северо-восточной, юго-западной, и юго-восточной
этногеографических группах у мужчин башкир, предки которых
являлись представителями 11 родопременных образований (Акимова,
1972).
Так, данные по дерматоглифике, полученные у 810 башкир,
характеризуют неоднородность групп, проявляющуюся в увеличении
монголоидных особенностей зауральских башкир. У них чаще
встречаются завитки на пальцах и соответственно наблюдается
увеличение дельтового индекса. На северо-западе, напротив, чаще
отмечается наличие центрального, двойных осевых и межпальцевых
трирадиусов.
Сравнение башкир с народами Восточной Европы и Скандинавии,
проведенное на уровне дифференциации этнических групп и расовых
типов в межгрупповом размахе вариации основных признаков,
выделило их в обширный субрегион, объединяющий Поволжье,
Прикамье, Башкирию и Калмыкию (Хить, 1983). Население данной
области отличается значительной разнородностью, с разной
степенью выраженности восточного комплекса, например, у мужчин
белорусов эта величина достигает 29%, а у башкир-айлинцев отмечен
максимум равный 75%. Наиболее отчетливо возрастание
концентраций средних и высоких величин восточного комплекса
признаков прослеживается в ряду популяций от чувашей, основной
массы татар, удмуртов, марийцев, с максимумом у башкир (Хитъ,
1983).
Z7
26
Сопоставление этнографических групп башкир между собой по
этому же принципу позволило авторам выявить различия у
зауральских групп в сравнении с приуральскими (Акимова, 1972;
Прокудина, 1971, 1972, 1974; Никольская, 1974; Долинова, 198(7; Хить
1974).
4. Информация по этнической геногеографии
и некоторые итоги.
Открытие L-Hirsfeki и H.Hirsfckl феномена существования
различий в распределении групп крови у разных народов явилось
началом широкого развертывания биологами и медиками работ в
этой области (1919 г.). В антропологии стали изучать полиморфизм
изоантигенов и ферментов крови для выяснения происхождения
народов, путей миграции, смешения, генохронологических
реконструкций и других задач эволюционного характера.
В Башкирии впервые данные серологи, как одна из существенных
характеристик антропологических особенностей, были применены
Л.Б.Вагнером в ЗО-х годах (Вагнер, 1925, 1926). Им были
исследованы две немногочисленные выборки Уфимского населения в
200 и 350 человек, которые выявили соответственно отличающиеся
распределения фенотипов системы АВО: О - 32,5%, А - 33,5%, В -
28,5%, АВ - 5,5% и О - 35,7%, А - 31,3%, В - 23,6%, АВ - 9,4%. В этих
работах характеристика башкир по системе групп крови АВО
представлена ограниченным числом обследованных в 135 человек и
имеет следующий вид: О - 29,5%, А - 36,3%, В - 25,2%, АВ - 8,9%.
При сравнительно-антропологическом изучении народностей,
населяющих территорию Башкирской АССР, предпринятом
Антропологическим отрядом Академии наук СССР с 1928 г.,
исследования групп крови системы АВО были включены в
программу работ и проведены М.Ш.Асфаган в первый же год ее
осуществления (Асфаган, Кузьмина, 1929). По этим материалам
картина распределения фенотипов группы крови АВО в Аргаяшском
кантоне, представленная выборкой 2168 человек, была следующей: О
- 30,1%, А - 32,3%, В - 30,3%, АВ - 7,3%. Частоты генов: г - 0,5490, р
• 0,2230, q - 0,2100. Иное соотношение по этой же системе
далось среди населения Бирского кантона в выборке из 1 ЮО
НЯ Л*°к* О - 29,8%, А - 41%, В - 22,2%, АВ - 7%. Частоты генов: г -
Чв$46О р • 0,2790, q - 0,1590. Отмечались также возрастные
° • ичйя в характере распределения групп крови системы АВО при
полном отсутствии различий зависящих от пола исследуемого (см.
табл. !)•
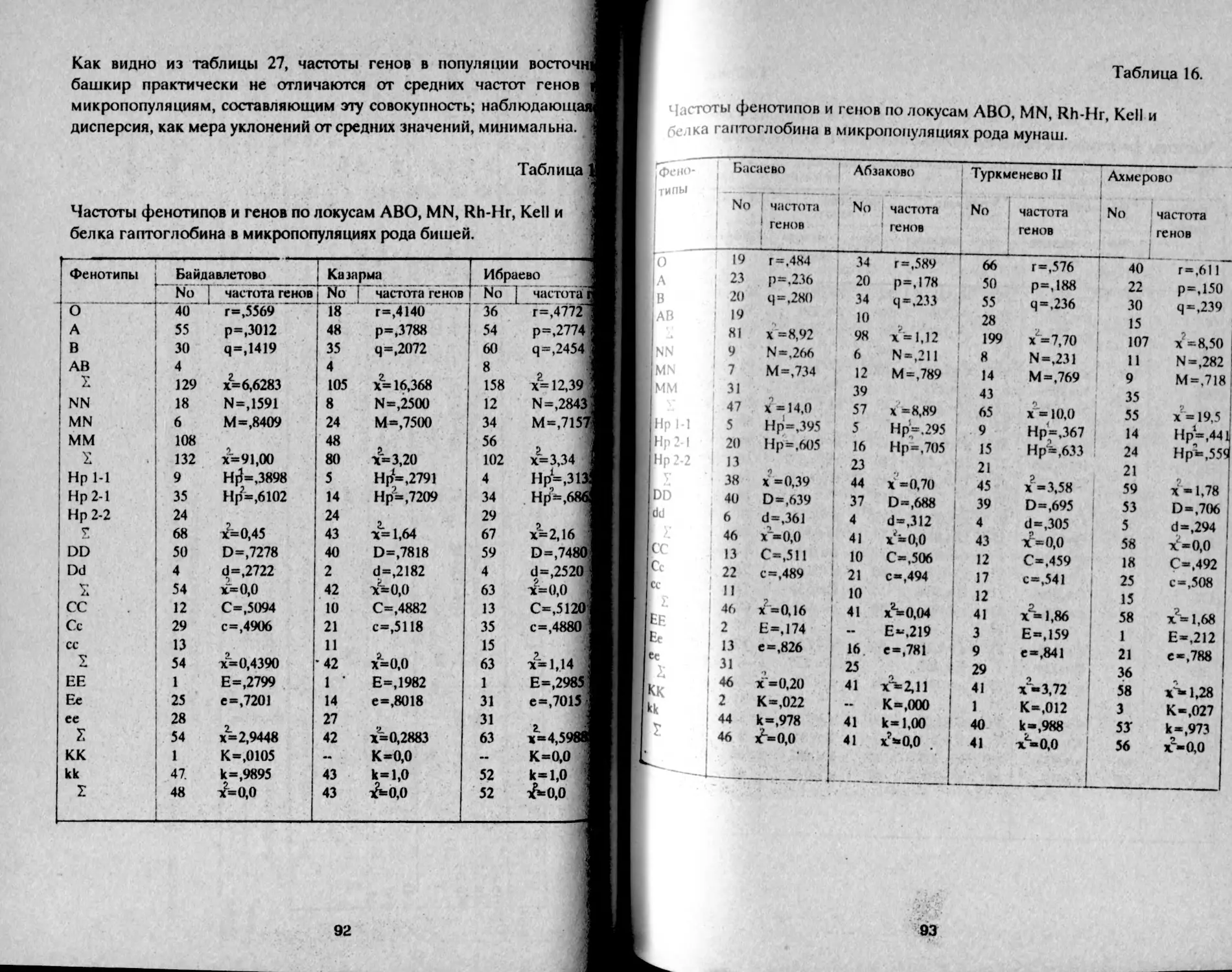
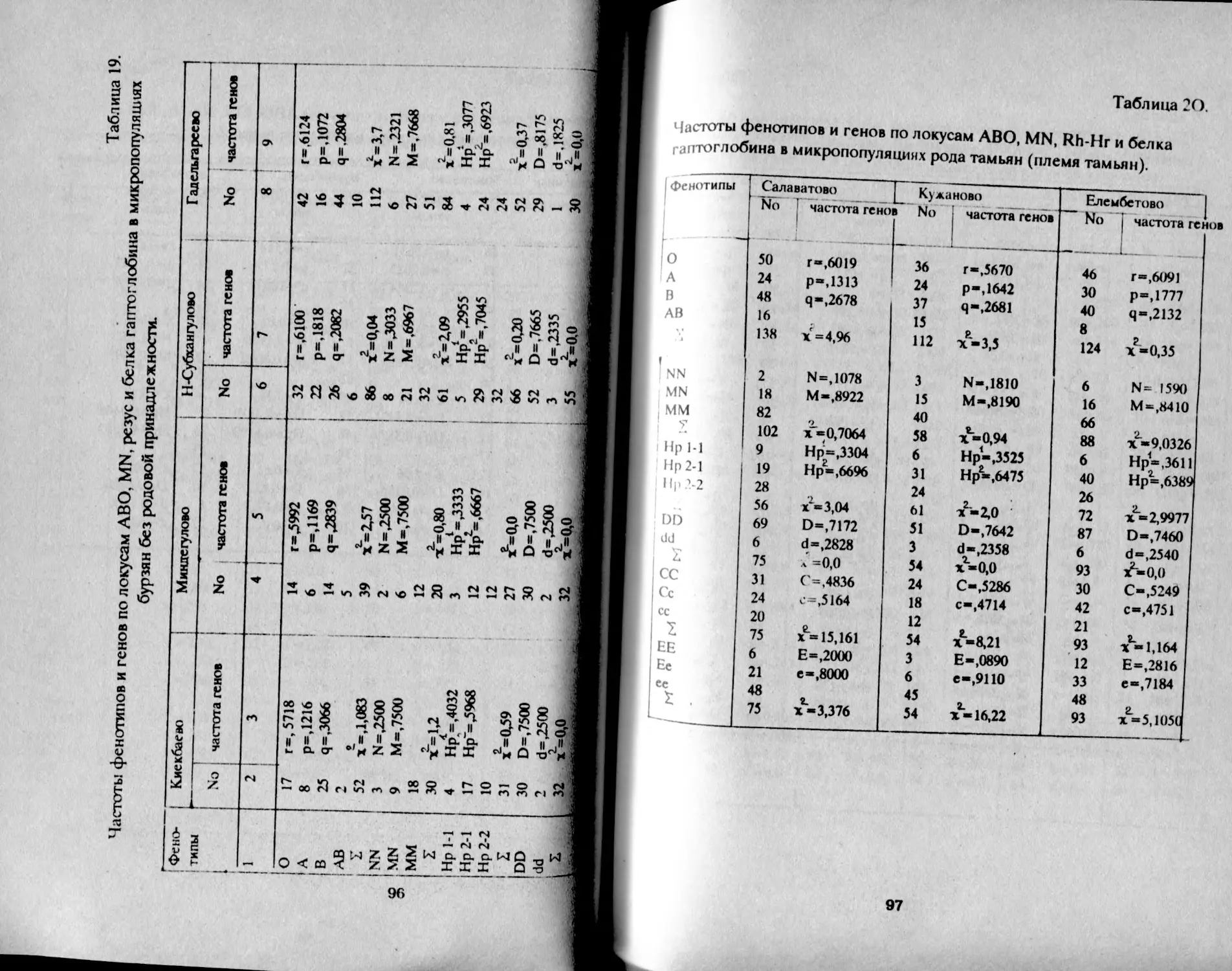
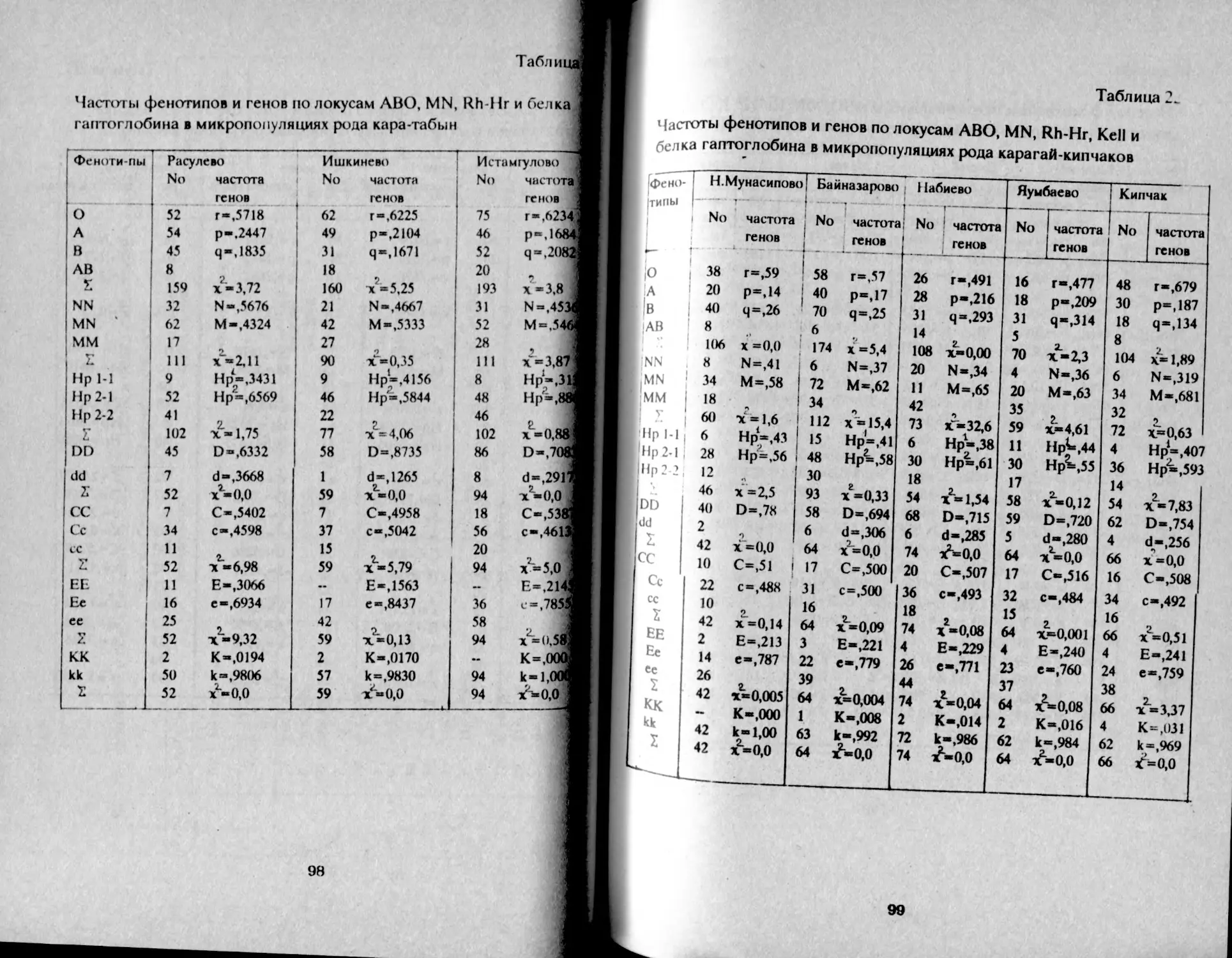
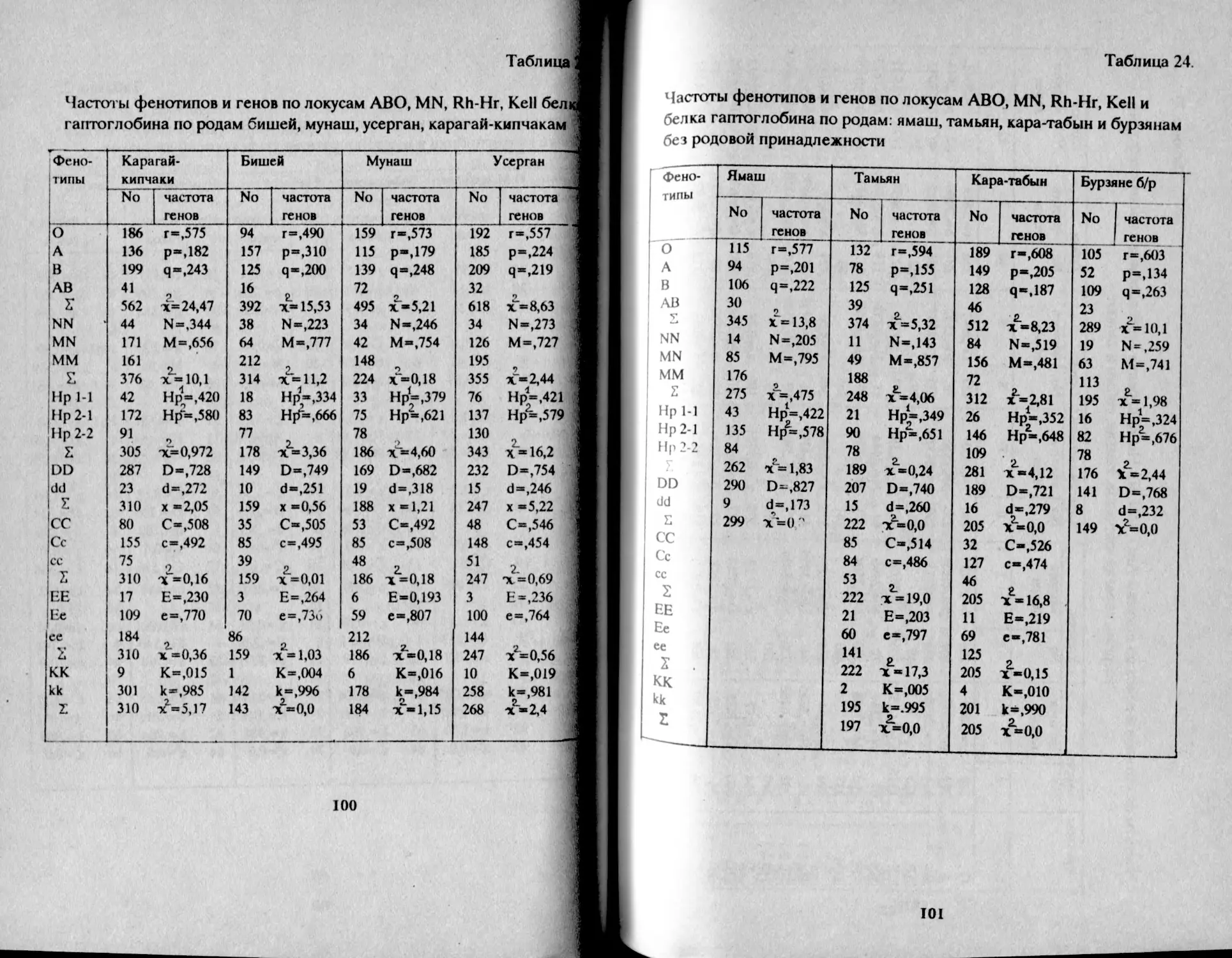
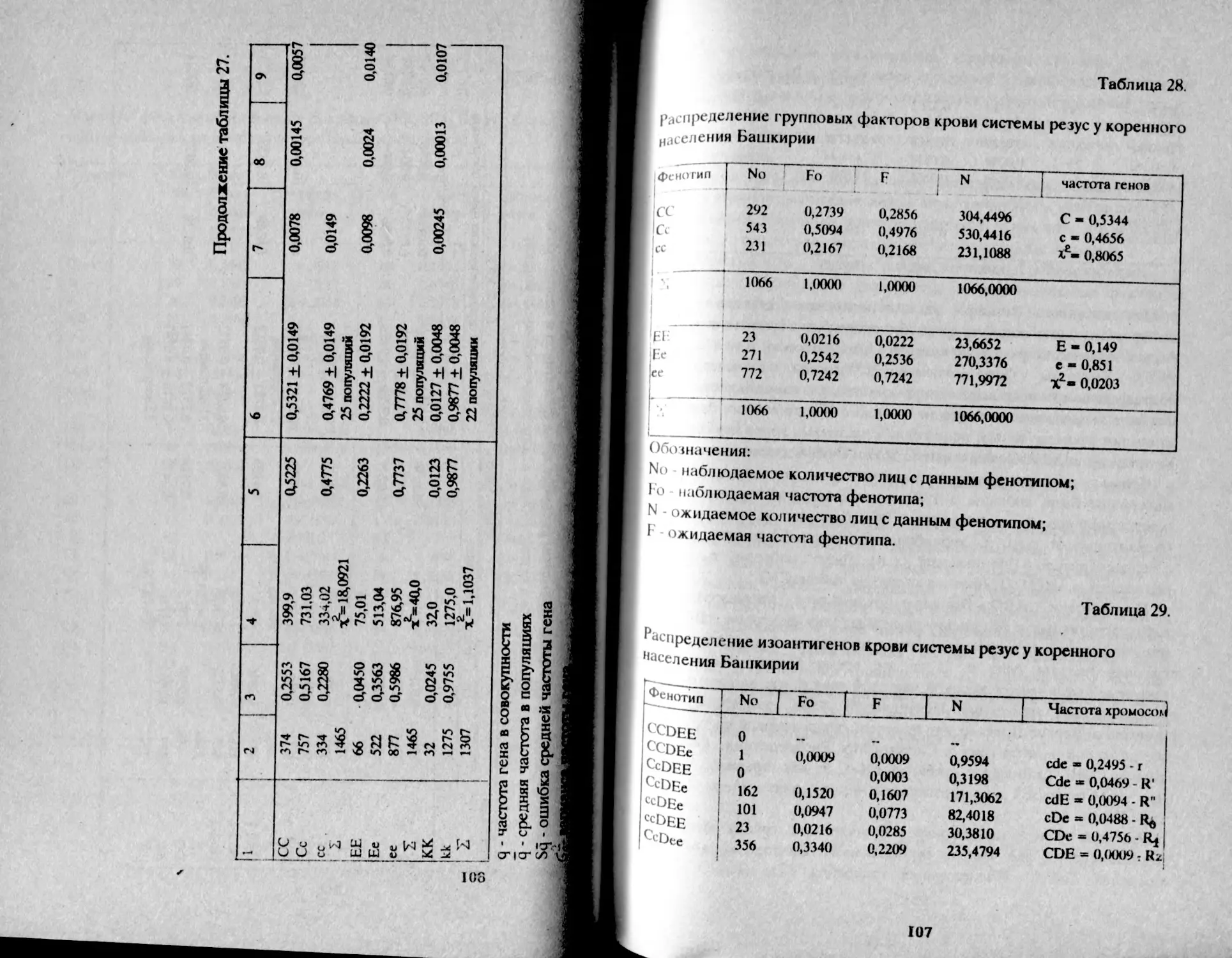
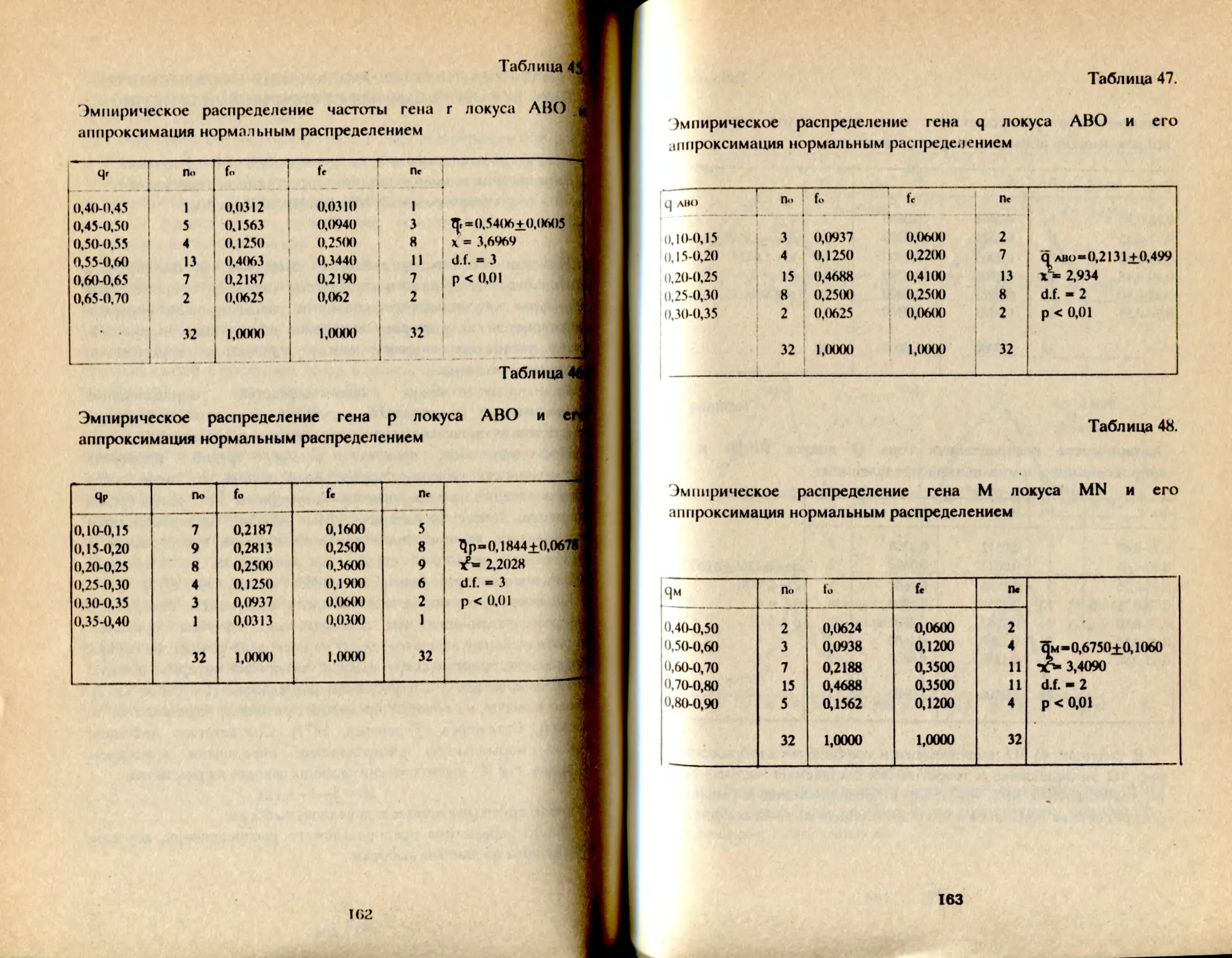
Таблица 1.
Распределение фенотипов и частот генов групп крови АВО
у башкир Аргаяшского и Бирского кантонов
Народность К-во о 1 А В АВ Г Р Ч
Башкиры Агаяшские мужчины К189 30,2 32,6 29.8 7,4 55,0 22,5 20,7 98,2
женщины 1079 30,1 32,2 30,5 7,2 54,9 22,2 21,0 98,1
Всего 2168 30,1 32,3 30.3 7,3 54,9 22,3 21,0 98,2
Башкиры 'Бирские мужчины 577 29,9 41,5 22.2 6,4 54,7 27,8 15,5 98,0
женщины 523 29,8 39,8 22,8 7,6 54,7 27,5 16,5 98,6
всего i 1М» • 29,8 41,0 22,2 7,0 54,6 27,9 15,9 98.4 1
В целом, башкиры аргаяшские отличаются более высокой
частотой фенотипа В - 30% и частотой гена q = 21% в сравнении с
бирскими: В - 22% и q = 16%, т.е. восточная группа отличается так
называемым монголоидным распределением групп крови (Асфаган,
Кузьмина, 1929).
В эти же годы предприняты попытки обобщения распределений
групп крови у разных народов и составлены первые
геногеографические карты (Steffan, 1932; Steffan, Wellisch, 1936;
Mourant et al., 1958). Однако некоторые неточности границ
геногеографических зон Восточной Европы, представленные на
карте Моуранта, позволили В.В.Бунаку составить новую карту
распределения фенотипов и частот генов с учетом появившихся
Данных тех лет. Согласно этой карте башкиры суммарно в выборке
28
29
4051 человек имеют следующие соотношения фенотипов состемы
групп крови АВО: О - 29,6%, А - 34,9%, В - 26%, АВ - 7,5%в
соответственно частоты генов: г - 0,5580, р - 0,2430, q - 0,1990
(Бунак, 1969). Следует отметить, что данная картина распределения
частот фенотипов и генов была представлена без учета родо.
племенной принадлежности башкир.
Планированное, целенаправленное исследование башкирского
населения было предпринято в 7О-х годах антропологической
экспедицией МГУ под руководством М.С.Акимовой. Были
обследованы башкиры, проживающие в северных, северо-западных,
юго-западных и южных районах Башкирии, предки которых
восходили к 11 существовавшим в прошлом родовым группам:
таныпцы, герейцы, кыргызы, канлынцы, минцы, юрматинцы,
кыпчаки, усергане, айлинцы, китайцы и табынцы. Последние три
группы изучались в Челябинской области (Акимова, 1973, 1974).
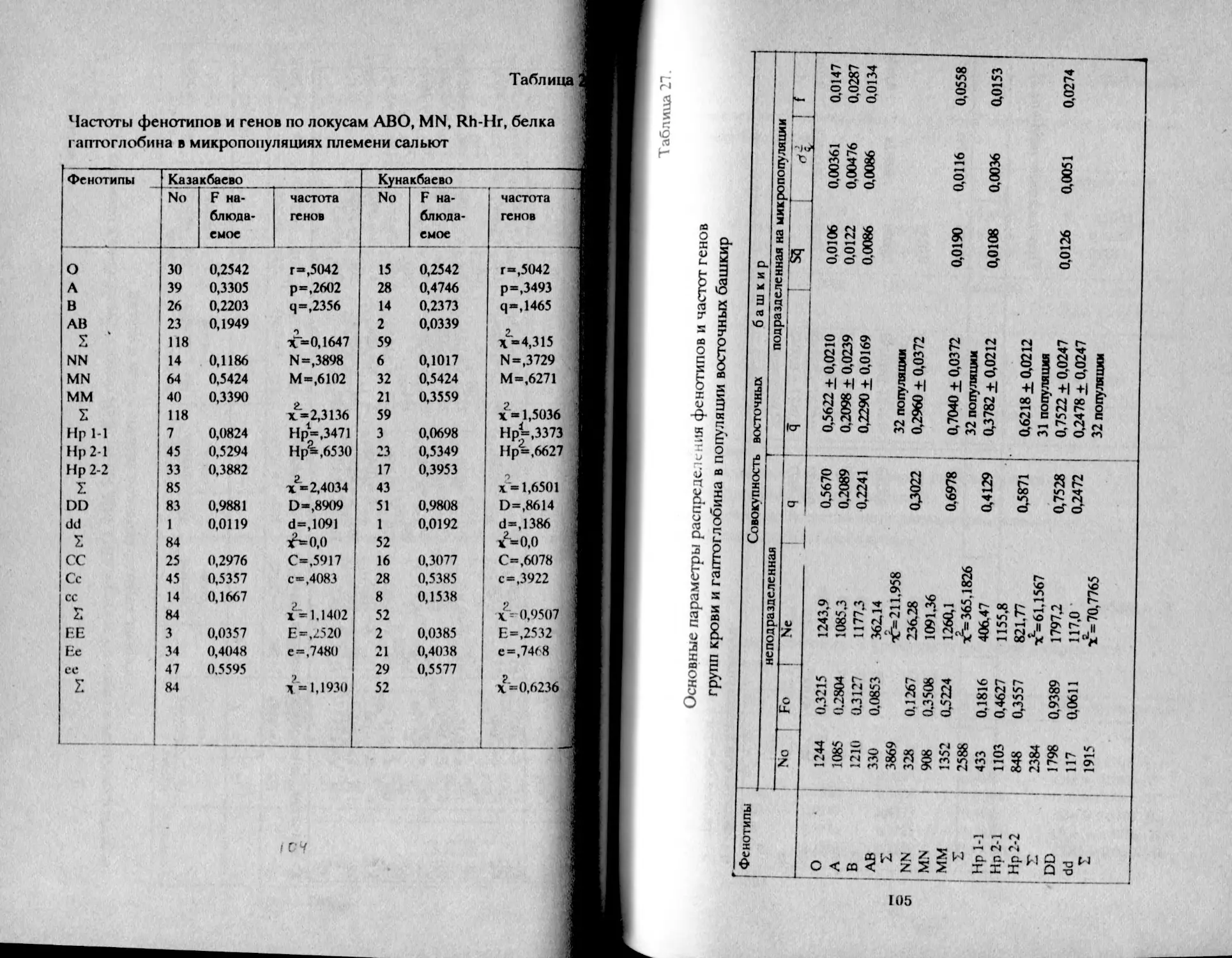
Данные по системам групп крови ABO, MN характеризуют
выборку башкир в количестве 1476 человек. Сумарно по башкирам
получено следующее распределение фенотипов по локусу АВО: О -
27,44%, А - 32,05%, В - 31,23%, АВ - 9,28% и соотношение частот
генов: г- 0,5340, р - 0,2360, q - 0,2300. В северо-западной и
зауральской группах соответственно: О - 24,63%, А - 37,79%, В -
27,98%, АВ - 9,70% и О - 24,45%, А - 31,27%, В - 29,82%, АВ - 9,46%.
Соотношение частот генов: г - 0,5100, р - 0,2790, q - 0,2120; и г -
0,5480, р - 0,2310, q - 0,2210. Распределение фенотипов по системе
MN суммарно следующее: ММ - 39,16%, NN - 14,16%, MN - 46,68%;
пропорции частот генов: М - 0,6250, N - 0,3750. В северо-западной
и зауральской группах соответственно: ММ - 39,16%, NN - 14,16% и
MN - 46,63%, ММ - 46,55%, NN - 17,09%, MN - 36,36%. Соотношение
частот генов в северо-западной группе: М - 0,5730, N - 0,4270; в
зауральской: М - 0,6470, N - 0,3530.
Отмечая наблюдающуюся некоторую тенденцию увеличения
частоты встречаемости группы В у зауральских башкир, автор все же
оставляет приоритет за антропологическими и
дерматоглифичвскими исследованиями в решении задач
внутригруппового анализа смешанных групп населения (Акимова,
1973). Однако следует заметить, что серологические исследования по
системам групп крови АВО и MN были проведены в выборке,
состоящей из мужчин башкир в возрасте от 20 до 50 лет. Такое
. рмирование выборки, в которой отсутствует элемент-
™ ,)айности, и не все индивиды имеют равные шансы попасть в нее,
Сстественно, могло отразиться на результатах исследования и, в
целом привести к суждению, снижающему значимость
серологических материалов для антропологии.
Обзор литературы по разным дисциплинам (этнографии,
археологии, антропологии, ономастики, лингвистики, фольклора)
унимающимися проблемами этногенеза башкир, позволяет
констатировать сложность и неоднозначность этнической истории,
отдельные этапы которой изучены довольно фрагментарно.
Страницы древнейшей истории Волго-Уральского региона
освещены несколько полнее. Охарактеризованы также основные
процессы формирования физического типа автохтонных племен,
расселявшихся еще с эпохи неолита, на территории древней
фашкирии. Южный Урал, отделявший Европу от Азии, издавна
являлся областью этнических контактов различных народов. Так,
европеоидное население в прошлом неоднократно испытывало
влияние пришельцев - южных европеоидов и монголоидов с
усилением в дальнейшем роли последних. Неравномерность
процессов ассимиляции и смешения, очевидно, и обусловили отличия
населения Приуралья от Зауралья, проявившиеся еще в мезолите
(Халиков, 1991).
Однако, ни одно из племенных общностей древности не является
собственно башкирской - это основной вывод по исследовательским
данным палеоантропологических и археологических работ.
Сложение башкирского народа произошло значительно позднее. По
мнению С.И.Руденко, впервые достаточно убедительно
сформулировавшего тюркскую концепцию происхождения башкир -
это относится к началу I тыс. н.э. (Руденко, 1955).
Согласно аргументированным высказываниям в исследованиях
последних лет, вероятнее всего в консолидации народа были
решающие VIII-IX вв. н.э. (Кузеев, 1974; Акимова, 1974 и др.).
Подобное расхождение взглядов о времени прихода башкир на Урал
объясняется отсутствием палеоантропологических данных начиная с
VII до XVII вв. н.э.
Основные племена, образующие древнебашкирский пласт, и, по-
видимому, составившие ядро в формировании башкирской
народности, генетически связаны с народами Центральной Азии и
31
30
Алтая. Об этом свидетельствуют многочисленные этнонимические
параллели башкир с тюркскими народами Алтая, а также сходство в
материальной и духовной культуре (Потапов, 1969).
Археологи, этнографы, антропологи на основе анализа
различного материала, охватывающего более позднее время
приходят к единому выводу о смешанности и неоднородности
отдельных групп башкир по своему составу и происхождению
(Смирнов, 1952; Сальников, 1966; Мажитов, 1977; Руденко, 1916;
Кузеев, 1974, 1978, 1987; Акимова, 1974).
Чрезвычайная сложность этногенетических процессов в эпоху
широкой миграции тюркских образований в Среднюю Азию и
Восточную Европу не позволяет посредством традиционных методов
исторических дисциплин достоверно установить те основные
компоненты, которые явились основой формирования башкирского
этноса.
По этнической антропологии башкирского народа в последние
десятилетия появились значительные исследования. Однако именно
это обстоятельство с очевидностью показало неясность или
дискуссионность целого ряда вопросов этнической истории башкир.
Применяемые методы антропологического анализа, опирающиеся
на расоводиагностические признаки: цвет кожи, волос, глаз,
различные размеры головы, лица не позволяют отделить проявление
генетической компоненты изменчивости в картине соотношения
антропологических типов башкирского населения от средовой
компоненты: т.к. основным приемом исследования в этнической
антропологии при сравнении антропологических характеристик
выборок из этнических групп является статистическое сравнение и
значительно реже используется изучение обобщающих оценок
различий или сходства. Назрела необходимость генетического
освещения динамики происходящих процессов в плане этнической
антропологии. Большое значение приобретает последовательная
реконструкция генетической структуры популяции башкир на разных
этапах ее истории с выяснением характера взаимодейсвия
автохтонного и пришлого компонентов, а также выявлении вклада
каждого из них в формировании башкирского народа.
Поэтому методы исторической генетики привлекают внимание
антропологов и этнографов возможностями анализа именно
генетических различий между популяциями человека, тем более,
32
истории - это поиск происхождения, родства, путей
контактов разных народов. Историко-генетические
будучи эволюционными по своей природе, могут
, т.к.
популяции принадлежат к разным этносам, доступны изучению,
еСЛИ в них наследственные признаки стабильны и не
а исследуе
яются в течение жизни индивидов. 11опуляционно-генетическии
ализ дает возможность воспроизведения исторических звеньев
ней цепи популяций, связанных с проникновением на ту или иную
^гшторию, естественно-исторический опыт которых запечатлен в
генетической структуре современных популяций. Кроме того, в
общем плане ................... “..........
расселения и
исследования,
существенным образом способствовать решению этих проблем
установление этапов истории народа в приложении биологических
принципов есть ни что иное, как исследование эволюции внутри вида.
В плане биологии - это проблема адаптации популяции к природным
условиям окружающей среды и выработка различных хозяйственно-
культурных типов ведения хозяйства.
В нашей стране популяционно-генетический материал для
освещения проблем этнической антропологии впервые был
использован Ю.Г.Рычковым с сотрудниками при изучении популяций
коренного населения Северной Азии (Рычков, 1965, 1969, 1973,
1978,1980 и др.). Изучение генетики популяций разных видов
проводилось Ю.П.Алтуховым (1983), Е.К.Гинтером(198О, 1982 и др.),
А.А.Ревазовым (1984, 1991 и дрДА. Г. Гаджиевым (1971),
Г.М.Афанасьевой (1990), Яблоковым (1987) и другими.
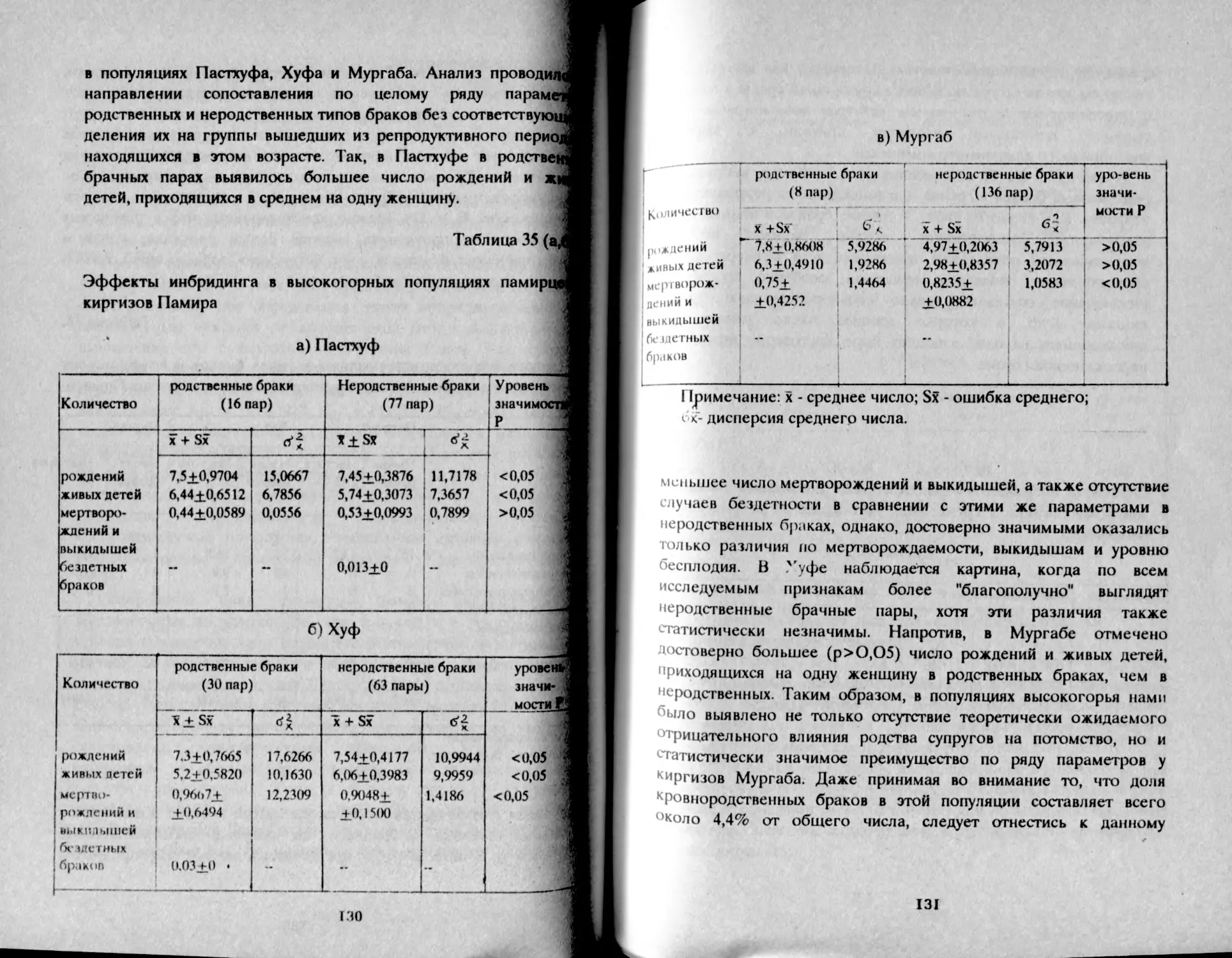
Результаты популяционно-генетических исследований в Башкирии
могут внести определенный вклад в решение этногенетических
вопросов, касающихся последовательности хронологии отдельных
этапов истории народа. Автором настоящей работы на конкретном
материале были применены методы исторической генетики,
позволившие определить основные параметры динамики
формирования башкирской популяции степень их генного
разнообразия, роль факторов микроэволюции, вклад отдельных
расовых компонент и др. Осуществлена также реконструкция
генетической структуры популяций древнебашкирского пласта на
основе исследования восточных башкир, являющихся современными
потомками бурзянцев, усерганцев, тамьянцев, карагай-кипчаков,
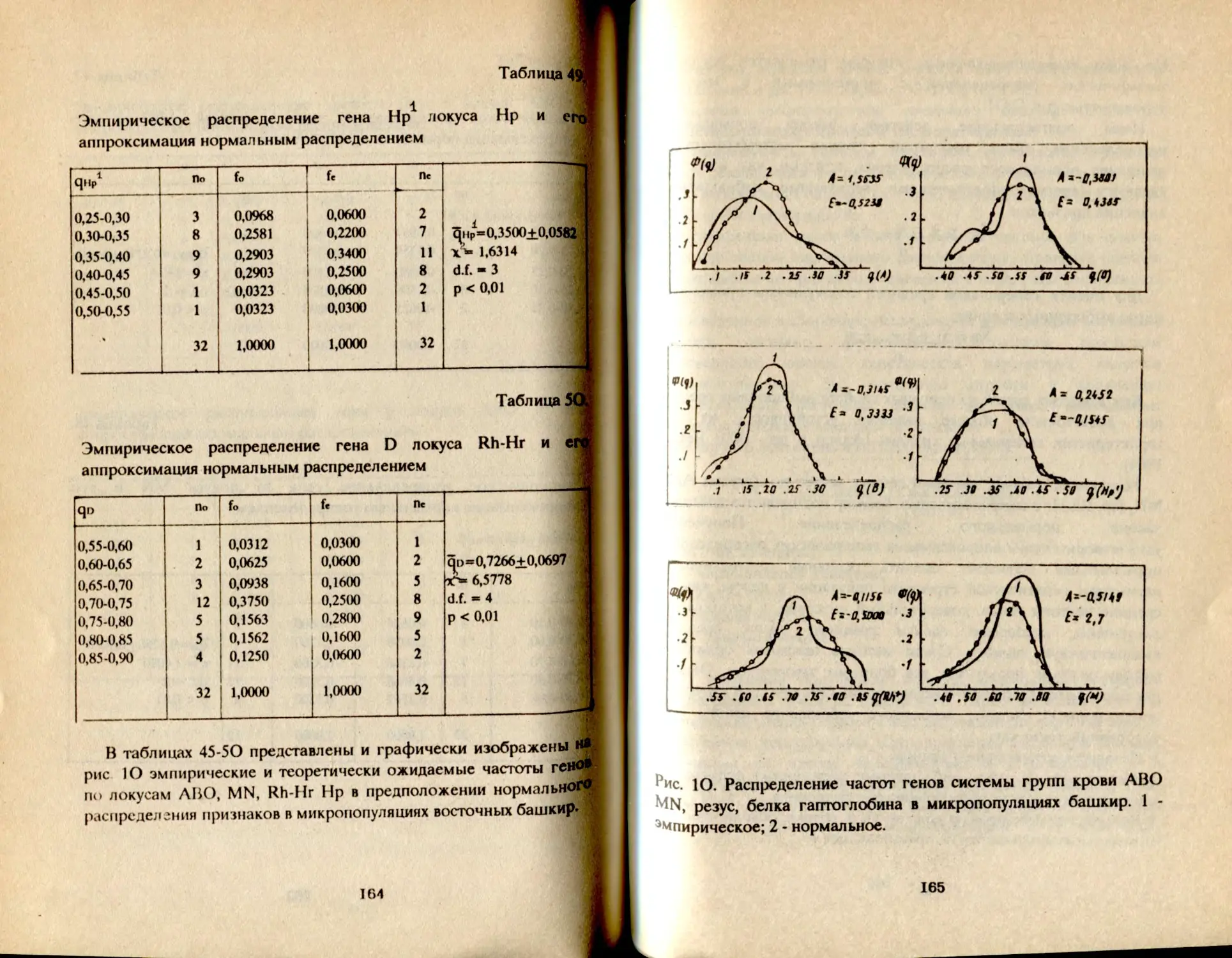
табынцев, сальютцев. Все это сделало возможным ретроспективно
оценить время дифференциации популяции башкир,
Центральна меукоах *
б1бл1отека
„ 1м. В. I. Вернадского 1
АН У к pai ни
Дат №19^3"
—------------— ' -30
реконструировать родословное древо, и тем самым ввести в научный
оборот новые интересные данные для познания этногенеза башкир.
В основу книги положены материалы, собранные автором лично
либо при его непосредственном участии, в ходе экспедиций 1973-
1990 гг. на Южном Урале и Приуралье, Кавказе, Западном Памире и
Приазовье.
Пользуясь случаем, выражаю глубокую признательность
сотрудникам Отдела биохимии и цитохимии БФ АН СССР, на базе
которого было развернуто популяционно-генетическое изучение
башкир, а также Института этнологии и антропологии Российской
Академии наук, где завершалась обработка материалов.
ГЛАВА 2. ПОПУЛЯЦИОННАЯ ГЕНЕТИКА КАК
ИСТОЧНИК ИНФОРМАЦИИ О РАСО- И ЭТНОГЕНЕТИЧЕСКИХ
ПРОЦЕССАХ.
1. Генетические маркеры. Частоты генов.
Наследственный полиморфизм популяций.
Начало XX столетия ознаменовалось важнейшим открытием
наследственного полиморфизма у человека. В 1900 г.
К.Ландшейнер, производя перекрестные реакции между
эритроцитами и сывороткой обнаружил, что одни эритроциты
агглютинируются некоторыми сыворотками, а другие - нет. Именно
это наблюдение и позволило ему описать первую генетическую
систему АВО. Исследование групп крови АВО оказалось жизненно
важным с точки зрения гемотрансфузионной терапии, а также
профилактики и лечения гемолитической болезни новорожденных.
Вскоре после этого было открыто множество других серологических
систем, полиморфизмов, определяемых двумя или большим числом
генов, встречающихся с полиморфными частотами, которые
обнаруживают существенную вариабельность между различными
этническими группами. В дополнение к этому следует заметить, что
было также идентифицировано большое количество серологических
пределах которых все или почти вес люди обладают
систем, гомозиготным фенотипом и только некоторые
единств обяадакУГ отличающимся наследственным типом. Техника
' этих иммунологических маркеров в высшей степени
ИЗУЧеНотизована и реагенты для идентификации соответствующих
СТаН--1«» повсеместно распространены, так что группы крови и
антиген*-’0
остаются широко используемыми маркерными системами в
П<зсемейном анализе, в случаях установления спорного отцовства в
бной медицине, в популяционно-генетических исследованиях. К
сожалению, за исключением точно установленной причины
возникновения гемолитической болезни новорожденных, вследствие
несовместимости между организмом матери и плода, очень мало
известно о нормальной функциональной нагрузке антигенов групп
крови, равно как и об их взаимодействиях с внешней средой.
В дальнейшем последовател ыю были обнаружены другие
эритроцитарные группы крови. Так, работами того же Ландштейнера
были открыты системы MN, Р, а в дальнейшем и резус.
Значение исследования генетического полиморфизма в изучении
вариаций человеческих рас и других иерархических
антропологических объединений получило оценку еще в работе
Hizszfeld L., Hizszfekl Н. (1919). Фундаментальные исследования
Мурента с соавтором (1976) по целому ряду наследственных систем
при сопоставлении с данными классической антропологии
подтвердили значение определения генетических маркеров для
решения задач этнической антропологии.
С той поры благодаря использованию новых методов изучения
анти-эритроцитарных антител (в частности, пробы Кумбса, Мурента,
Рейса) антигенная мозаика эритроцитов стала быстро обогащаться;
были одна за другой открыты антигенные системы Льюис, Лютеран,
Келл, Даффи, Кидд, Джей, Диего и другие.
В зависимости от методических приемов анализа, успехов
иммуногематологии, биохимической генетики, гистохимии,
энзимологии, молекулярной генетики, в хронологическом порядке
изучение маркеров генов, самого наследственного полиморфизма
человека можно дифференцировать на несколько четко очерченных
стадий. Это изучение: 1) групп крови или эритроцитарных
агглютиногенов; 2) белков плазмы крови; 3) эритроцитарных
ферментов и гемоглобинов; 4) антигенов гистосовместимости HLA и
34
35
аллотипов иммуноглобулинов; 5) маркеров ДНК (или, Та*
называемый полиморфизм длины рестрикционный фрагменте^
ДНК).
К настоящему времени установлено большое число белко^
плазмы с их детально определенными функциями и которые
охарактеризованы в отношении их физических и химических свойст»,
Обнаружение данных полиморфизмов целиком зависело от
успешной разработки электрофоретических приемов анализа в
биохимии. Функция большинства этих белков в организме хорощ0
известны и в некоторых случаях идентифицированы различные
физиологические эффекты разных аллелей. Например, в случае
аллея PiZ и PiS, когда они отвечают за продукцию недостаточного
количества белка плазмы '/j-антитрипсина, что, как правило,
ассоциируется с целым рядом тяжелых патологических состояний.
Известно большое число полиморфных белков сыворотки крови,
характеризующихся множественным полиморфизмом. Таким
образом, эта вторая стадия изучения генетических маркеров связана с
именем О.Смитиса (O.Smithies, 1955), который разработал весьма
эффективный метод электрофореза в крахмальном геле. Этот
надежный аналитический прием позволил дифференцировать
отдельные белковые компоненты из сложной смеси в зависимости от
заряда, формы и размеров их молекул. Использование таких тонких
приемов анализа привело первоначально к обнаружению
множественного наследственного полиморфизма по белкам,
несущим преимущественно транспортную функцию в организме,
таким как гаптоглобин (Нр), трансферрины (Tf), церулоплазмины
(Ср), альбумины (Al)/*i-антитрипсин или ингибитор протеаз (о^ АТ,
Pi), множество систем компонентов комплемента (Ci.........С9),
орозомукоид (ORM). Комплексные электрофоретические,
иммуноэлектрофоретические и иммунологические исследования
позволили вскоре идентифицировать полиморфизм новых локусов,
таких как группо-специфический компонент Gc, липопротеины (Ag,
Ах и др.); весьма важные с точки зрения медицинской генетики и
этнической антропологии иммунологически определяемые HLA, и
ряд других (см. обзоры в монографиях В.В.Томилина и А.С.Гладких,
1981; В.А.Спицына, 1985). ,
Основным белком эритроцитов, как известно, является
гемоглобин, который содержит очень большое число различных
Наследственная изменчивость гемоглобина имеет
ое значение в некоторых регионах земного шара, например.
°ГР°рритории Закавказья, Средней Азии, а также в Африке, Южной
^Юго-Восточной Азии. Все большее и большее число ферментов
обнаруживают состояние полиморфизма, которые определяются в
основном методом установления скорости миграции их различных
изоформ в процессе электрофореза в различных гелевых носителях с
последующей их идентификацией посредством гистохимических
приемов анализа. Эта стадия изучения генетического полиморфизма
связана с именами таких исследователей как Гопкинсон, Харрис.
Спенсер* и ее можно датировать началом 60х годов, когда Гопкинсон
в 1963 г. открыл систему кислой эритроцитарной фосфатазы Ас1\:
N.Spencer с соавт. в 1964 г. обнаружил полиморфизм
фосфоглюкомутазы PGM. Вслед за этим интенсивные исследования
привели к обнаружению таких эритроцитарных ферментных систем
как аденилаткиназа (АК), аденозин дезаминаза (ADA), глюкозо-6-
фосфат-дегидрогеназа (G-6-PD), 6-фосфоглюконат дегидрогеназа
(6-PGD), глиоксалаза I (GLO-1), эстераза (EsD), карбоангидраза
(CAI, САП), супепроксиддисмутаза (SOD-A), глутамат-пируват
трансаминаза (GPT) и целого ряда других Эритроцитарных
полиморфизмов (см. обзоры В.В.Томилина, Ф.С.Гладких, 1981;
В.АСпицына, 1985, в которых эти системы представлены более
подробно).
Так же, как и в случае различных белков плазмы физико-
химические свойства и функциональное значение рассматриваемых
ферментов широко известны. Изменение активности какого-либо
фермента в результате генетического дефекта, связанного с
мутантным их вариантом, приводит подчас к тяжелым
наследственным нарушениям. Более того, различные генные
продукты одних и тех же ферментов различаются в отношении их
химической активности, и это, как было показано в работах Н. Walter
(1976), A. Mourant (1976), В.А.Спицына (1985) приводит к различиям
в ответе разных фенотипов на воздействие внешнесредовых
факторов. Полученные факты привели к обнаружению адаптивной
значимости ферментных полиморфизмов.
Следует подчеркнуть реальность существования еще одного этапа
в изучении полиморфных наследственных систем. Этот этап связан с
новым весьма эффективным приемом анализа - изоэлектрическим
36
37
фокусированием - тонким и весьма эффективным аналитически^
методом, позволившим идентифицировать множественный широкий
полиморфизм тех систем, которые до сих пор рассматривались к;1(,
мономорфные или почти нс обладавшие генетической
изменчивостью. С помощью изоэлсктрофокусирования стад0
возможным выявлять аллели, которые не обнаруживаются обычным
электрофоретическим методом исследования. Так, стало возможным
идентифицировать новые дополнительные аллеи в системах If, Gc
PGM, Pi. Многие из них оказались весьма эффективными ддя
изучения с позиций этнической антропологии, в частности для
дифференциации по этим маркерам американских индейцев от
других антропологических групп.
5. Маркеры ДНК. В обсуждаемом контексте представляет
несомненный интерес так называемый полиморфизм длины
рестрикционных фрагментов ДНК (ПДРФ). ПДРФ представляет
собой нормальную изменчивость в последовательности оснований
ДНК, которая не связана с какими-либо выраженными эффектами у
индивида, и наследуется как кодоминантный признак. ПДРФ
идентифицируется с помощью применения особых ферментов
рестрикционных эндонуклеаз (или рестриктирующих эндонуклеаз),
Эти ферменты режут молекулы ДНК в специфических сайтах (то
есть в строго определенных местах) последовательности оснований.
В случае проявления такого полиморфизма, характеризующегося в
различиях длины образующихся в результате разрезания фрагментов
ДНК, можно сравнивать индивидов между собой по различиям в
отношении ПДРФ. Эти фрагменты определяются в
электрофоретическом геле посредством так называемого блоттинга
по Саузерну при гибридизации с соответствующим каким-либо
зондом ДНК, комплементарным к данному специфическому району
исследуемой ДНК. Этот процесс можно проилюстрировать на
рисунке.
В данном случае имеется в виду полифоризм в сайте рестрикции В
(рис. 1); отсутствие сайта при этом являет собой аллель 1 , а
присутствие сайта предполагает наличие аллеля 2. Когда
рестрикционный фермент режет ДНК в одной хромосоме в сайтах А
и С, образуется единственный фрагмент размером IO kb (I kb я
ЮОО пар оснований), который соответствует аллелю 1. Ехли же
фермент режет ДНК не только в сайтах А и С, но и в сайтах В, то
У1ОТСя два фрагмента размерами 1 kb и 3 kb, что соответствует
°°" 1 2 Поэтому полиморфные генотипы могут быть выведены из
аЛЛ педеления особенностей электрофоретических фракций,
видимых на агаровом геле после соответствующей обработки. В
водимом примере один из пораженных болезнью родителей
является гетерозиготным (1-2) в отношении этого полиморфизма,
также как и два других пораженных ребенка (П> и Пг), тогда как
самый младший потомок оказывается здоровым (Пз) и является
гомозиготным по аллелю -2.
Зонд
Рис. 1. Графическое представление аутосомного ПДРФ (наверху);
наследование ПДРФ в пределах семьи с тремя ее членами,
пораженными заболеванием, наследуемым по аутосомно-
кодоминантному типу (посредине); внешний вид рестрикционных
фрагментов после проведения блоттинга по Саузерну (внизу) (цит
по Эмери, 1986).
К настоящему времени интерес, проявляемый к ПДРФ, огромен.
Используя множество различных рестриктаз (эндонуклеаз), а также
применяя для исследования различные зоны ДНК, можно получить
поистине неисчерпаемое число генотипов, контролируемых весьма
большим числом генов, что открывает совершенно новые
Уникальные возможности использования генетического
38
39
полиморфизма для изучения в области этнической антропологи^
Более того, в принципе, при этом возможно дифференцировать
селективно-нейтральные локусы, от локусов, подвержеши.ц
давлению естественного отбора. Применение первых особенно
эффективно для решения этно-расогенетических задач. 8О-е годы
ознаменовались поистине удивительными открытиями в области
этнической антропологии, когда этнорасовые группы стали
исследоваться при помощи метода ПДРФ, с использованием в
качестве материала митохондриальной формы ДНК (M.JJohnson,
D.S.Wallace et al., 1983).
Еще в 1908 г. Харди и Вайнберг указали на частоту гена, как на
важную характеристику в популяционной генетике. Они
сформулировали основной принцип, позволяющий анализировать
факторы, изменяющие частоты генов.
Если из N диплоидных особей D несут только доминантные
аллели (АА), Н гетерозиготы (Аа), a R обладают только
рецессивными аллелями (аа), то D + H+R = N. Для простоты можно
обозначить такую группу индивидов как (D, Н, R). Эти три символа
соответствуют АА, Аа, аа. Наличие индивидов трех типов в данной
группе обусловлено существованием только двух аллельных генов А
и а. В сумме N индивидов несут 2 N генов. Поскольку каждый
индивид АА имеет два гена А, а каждый индивид Аа - лишь один ген
этого типа, то полное число генов А в группе составляет 2 D + H.
Следовательно, доля генов А равна:
Р = 2D + H = D + 1/2 Н
2N N
Это соотношение называется частотой гена А. Аналогично
определяется частота гена а : q=(H + 2R) / 2N=(1/2H + R) / N; так
что р + q = 1 (Ли, 1978).
Принципы расчета генных частот в зависимости от характера
наследования, а именно, можно ли идентифицировать гетерозиготы,
будут даны в соответствующей главе III.
Наследственный полиморфизм. Полиморфизм можно определить
как закую ситуацию, при которой в той или иной популяции два или
более аллелей данного генного локуса встречаются с частотой не
менее 0,01 (Crawford. 1973). Поданным Харриса (1973), в каждой из
обследованных расовых групп полиморфизм, по-видимому,
----1 всех локусов изученных
несомненно, на самом деле
ястречается примерно для четверти
Ферментных систем. Однако, несо:----------, ----- ------
Ф орфизм должен встречаться гораздо чаще уже по одному
полимор что обследование проводилось им только по
Т°МУ-грофоретическим различиям. Более того, разрешающая
^особность этого метода для разных ферментов оказывается весьма
зличной и вполне вероятно, что некоторые случаи полиморфной
изменчивости остались незамеченными (Харрис, 1973). Следует
целиком и полностью согласиться с прозорливым предсказанием
этого автора, когда в настоящее время при использовании более
совершенных приемов анализа - изоэлектрофокусирования удалось
значительно расширить рамки определения генетических различий
Так для систем, считавшихся в ряде случаев мономорфными,
установлен весьма высокий полиморфизм, например, для
трансферринового локуса. Таким образом, результаты современных
исследований прямо указывают на то, что для значительной части
всех генных локусов, кодирующих структуру полипептидов,
свойственен полиморфизм.
Информативность отдельных генетических систем в изучении
происхождения, становления человеческих рас, отдельных
антропологических типов и этнических групп оказывается
неравноценной. В этой связи для эффективного решения задач,
касающихся этнической антропологии нельзя отбирать гены
произвольно или руководствуясь удобством в идентификации, а надо
искать среди них только те, которые представляют наибольший
научный интерес (Оливье, 1964). Так Саттер (Sutter, 1956, цит. по
Ж.Оливье, 1964) весьма образно сравнивает всю массу генов в
организме человека с жидкостью, находящейся в сосуде, на дне
которого располагаются наиболее часто встречающиеся, постоянные
гены, на поверхности - наиболее редкие, их можно привести в
движение волнами, тогда как в глубине все остается спокойным
Можно использовать этот образ, расположив у дна гены, присущие
данному виду, на поверхности - гены индивидуальные и летальные, а
ниже поместить гены, определяющие расовые характеристики.
Можно даже представить себе и особый слой генов,
соответствующих морфологическому типу индивида, например,
брахиморфному или долихоморфному. Эта схема, на наш взгляд.
41
40
выразительно иллюстрирует мысль о том, что не все гены одинаковы
но значению в отношении расовой классификации.
Генетическим маркером называется дискретный, сегрегирующий
признак, при помощи которого можно охарактеризовать
исследуемую популяцию в терминах "присутствия - отсутствия"
признака, более высокой или более низкой его частоты (Crawford,
1973). Требования, выдвигаемые к генетическим маркерам, сводятся
в основном к точно определенному независимому типу наследования
признака и стабильности его проявления в течение всей жизни
индивида. Важным моментом является также различная, но
достаточно высокая частота встречаемости его в популяциях, что
дает возможность считать репрезентативным выборочное
исследование в разных популяциях. Существенно необходимым
является также простота и доступность для широкого популяционно-
генетического исследования.
Накопленный в литературе материал по распределению большого
числа иммунологических и биохимических маркеров в популяциях
позволяет провести классификацию типов распространения генных
частот в зависимости от степени полиморфизма, и тем самым
оценить таксономическую ценность наследственных систем в
антропологическом отношении. В наиболее подробной степени такая
классификация проведена в работе В.А.Спицына (1985), где было
выделено шесть специфических категорий генетической
изменчивости в популяциях человека. Наибольшее внимание
привлекает категория так называемых эндемичных полиморфизмов,
характерной особенностью которых является наличие особых
аллелей, или гаплотипов, ограниченных распространением в
определенных крупных регионах. Такие аллели обнаруживаются с
высокой частотой в отдельных расовых группировках, тогда как
среди прочего населения земного шара они отсутствуют. Типичной
иллюстрацией их могут служить гаплотипы cDe, Gm1’5’14,17,
маркирующие негроидные популяции Африки; Gm1,3,5,13
характерный фактор для представителей южноазиатских
монголоидов; Gm3’5’10,13,14 , достигающий у европейцев частоты
почти 80%; Gm1’13'15’16 и рецессивный аллель d ушной серы сухой
консистенции, распространенные с исключительно высокой
концентрацией у центрально- и североазиатских монголоидов. В
отдельную категорию выделяется также локальная изменчивость
42
частот, при которой имеется не только обычный ген, но и
генных спецИфические аллели данного локуса, распространенные с
другие с ПОЛИморфной или близкой к этому состоянию
меныпеи, и
протон Распространение их по большей части ограничивается
пеленными этнорасовыми и сопряженными с ними группами: НЬ
°П поостранен среди южноазиатских монголоидов; аллель RDS2
гЬеомента роданезы и фактор альбумина "наскапи" (А)*"’) отмечаются
V североамериканских индейцев; Gm2-17*21 характерен для айнов;
дт1,5,17 и АсР свойственны бушменам; GcAb,!'встречается
у аборигенов Австралии и папуасов Новой Гвинеи; АсР присущ почти
исключительно европеоидам. Наконец, третий, интересующий нас
тип полиморфизма,
изменчивостью. Он
свойственного только одной этнической группе. Эти факторы
назван "приватной" генетической
отличается наличием особого аллеля,
обычно наблюдаются с весьма низкими концентрациями, хотя в ряде
случаев их плотность может достигать высоких полиморфных частот.
Например, аллель СА> карбоангидразы в группе маманва
о.Минданао (Филиппины) с частотой 0,21, а эндемичный для
негритосов аэта аллель GcrN обнаружен на о.Лузон (Филиппины) с
высокой полиморфной концентрацией 0,11 (Омото, 1982). Следует
заметить, что приватные аллели могут иметь место как в поли- так и
в мономорфных локусах.
Подчеркиваем, что далеко не все из вышеназванных систем
являются доступными для массового популяционного исследования.
Поэтому другим важнейшим принципом исторической генетики
является принцип комплексности, когда при наличии большого числа
изученных генов независимых локусов можно с успехом проводить
сопоставления этнических и расовых групп для решения тех или иных
задач этнической антропологии. К числу таких признаков относятся
эритроцитарные группы крови, многие сывороточные белки,
липопротеины и ферменты крови, слюны, типы ушной серы и т.д
(Рогинский, 1947; Wiener, 1967; Рычков, 1969; Хить, 1961; Glass, 1954,
1956; Mourant et al., 1976; Tibera-Dumitru, 1968; Waissblut et al., 1971;
Keiichi Ornoto, 1973; Adachi 1937; Norakmal, 1979; Matsunaga, 1962;
Mc.Cullongh et al., 1970; Chakravartti et al., 1978; et al., 1987).
Весьма заманчива для исследователя возможность проследить
историческую динамику признака во времени. Однако,
применительно к популяциям башкир, также как и ко многим другим
43
народам - это возможно лишь в интервале последних десятилетии
Связано это с тем, что собственно накопление данных По
распределению маркеров берет свое начало начиная с 1901 года ц
интенсивно развивается в последние годы.
В дальнейшем изложении мы стремились применить оба
принципа исторической генетики в практических конкретных целях
изучения этногенеза башкирского народа и сопоставления
полученной информации о башкирской популяции с имеющимися в
литературе сведениями по сопредельным антропологическим
группам.
2. Роль микроэволюционных факторов в
дифференциации и интеграции популяций.
Значение популяционной генетики в познании сложнейшего
явления живой природы - эволюции - огромно. Основой современной
теории исторического развития является изучение преобразования
генетической структуры популяций в поколениях.
Согласно синтетической теории эволюции основными факторами,
определяющими формирование генофонда популяций разных видов,
являются давление отбора, миграций, мутаций и случайные события.
В результате их суммарного воздействия в конкретных условиях
среды происходит интеграция уникальных особенностей структуры
элементарных групп (Fisher, 1930; Wright, 1931; Mayr, 1965; Левонтин,
1978; и др.). Условия окружающей среды, в которых существует
популяция, во многом определяют вклад каждого из эволюционных
факторов. Соответственно этому интенсивность последних способна
измениться при смене местообитания, (связанного с кочевым
образом жизни), либо с изменениями климатогеографических
параметров среды (в отношении оседлых групп). В совокупности это
во многом определяет наблюдающуюся дифференциацию популяций
в пространстве признаков. Таким образом, эволюцию популяций в
подобном случае можно представить в виде траектории в
многомерном пространстве "время-признаки" (Cavalli-Sforza, 1964;
1965а; 1967 и др.). Описание эволюции популяций в виде дивергенции,
конвергенции, параллелизма и истории происхождения отражает
44
в системе популяций взаимопротивоположные
протека^ диффереициации и интеграции во времени и в
ПР° нстве. Именно в единстве и многообразии форм организации
ПР°й природы и проявляется ее динамизм. При этом познание
ожного механизма микроэволюционных преобразований является
основой познания самой эволюции популяций. Скорость
эволюционных изменений за поколение для любой генетической
системы является пропорциональным дисперсии концентрации генов
следующего поколения. В свою очередь, величина дисперсии зависит
от концентраций генов в кагорте родителей.
Большая часть истории человечества связана с популяциями
малой численности. Генетическая изоляция являлась основным
фактором расообразования (Алексеев, 1968; Рычков, 1969 и др.). В
популяциях с малым эффективно-репродуктивным объемом генные
частоты способны претерпевать изменения, которые совершенно
независимы от влияния мутаций, миграций и отбора, то есть в них
велик вклад стохастических процессов. Известно также, что в
прошлом эволюция во многом определялась действием отбора так,
смертность от инфекционных заболеваний была очень высокой
(Бочков, 1984; и др.). Изоляция, приводя к росту степени родства
членов группы, неизбежно в поколениях способствовала росту
инбредности потомков. Напротив, смешение и давление мутаций не
могло играть существенной роли в происходящих процессах
дифференциации, сущность которой сводилась, в терминах
популяционной генетики, в постепенном повышении доли гомозигот.
Именно поэтому в изолированных группах, связанных общностью
происхождения, могут наблюдаться концентрации генов,
достигающие мирового размаха изменчивости (Хить, 1961; Рычков,
1969; Спицын, 1985; и др.; Wright, 1931).
Динамика современных процессов микроэволюционных
преобразований достаточно сложна и неоднозначна, в целом,
значительно отличается от такового в малых изолированных
популяциях прошлого. Происходящая повсеместно ломка границ
изолятов, небывалый рост урбанизации в мире, существенно
изменило картину воздействия эволюционных факторов. Так,
проявление их в основном связано с большими подразделенными
популяциями. Степень аутбридинга резко возросла в последние
Десятилетия, что привело к увеличению эффективной величины
45
популяций и значительному росту в них уровня генетическое
изменчивости. (Майр, 1974). Высокая степень панмиксии, смешения
уровня миграций, тенденция роста давления мутаций, а также
происходящее одновременно снижение давления отбора и рОди
генетико-автоматических процессов - вот далеко не полный перечень
факторов, формирующих генетические особенности современных
популяций.
Дрейф генов.
Если размер популяции ограничен, то генные частоты способны
претерпевать в поколениях изменения, которые совершенно
независимы от влияния мутаций, миграций, отбора. Возникают
различия между частотами генов потомков и когортой родителей.
Случайные колебания частот генов в популяциях ограниченной
численности очень интересны с точки зрения познания эволюции
(Алтухов, 1983 и др.).
Увеличение дисперсии генных концентраций за поколение при
случайном генетическом дрейфе служит мерой скорости эволюции.
И чем меньше группа, тем менее вероятно полное воспроизведение
генетической информации родительского поколения. Именно этим
объясняются происходящие колебания частот генов во времени. При
этом эволюция в результате дрейфа генов-ненаправленная. И если
частота какого-либо гена мала, то чисто случайно, со временем она
может упасть до нуля, и наоборот, ген, встречающийся в популяции с
достаточно высокой частотой имеет шанс к случайному закреплению
(Wright, 1931). Как в случае утраты гена, так и при его закреплении
дальнейшего колебания по данному аллелю не будет. Значит, для
возобновления дрейфа необходима либо мутация, либо занос генов
извне. Теоретически подобная ситуация допустима, но на практике
существование популяций, формирование генетической структуры
которых происходит без определяющего влияния систематических
факторов эволюции, и полностью обусловлено случайными
событиями, является нереальным. С полной очевидностью встает
вопрос в отношении каждой конкретной исследуемой популяции о
совместном действии систематических и случайных факторов
46
ппиводящих в итоге к уникальности каждой популяции в
эволюиии>
вида. При этом дрейф генов выступает в роли активатора
Хлюпни (Рычков, 1968, 1973; и др.). Изменчивость является
свойством, присушим любой природной популяции, вследствие того,
что отбор часто благоприятствует гетерозиготам в разнообразных
условиях среды. (Майр. 1974).
7 Имеется прямая связь между инбридингом и дрейфом. При
панмиксии в бесконечно большой популяции, два индивида,
вступающие в брак, не являются родственниками. Однако в малых
популяциях, прошедших длительный исторический путь развития, все
индивиды в какой-то степени оказываются связанными узами
родства. И в них даже при отсутсвии мутаций коэффициент родства
достигает при неограниченном времени своего максимума. При
совместном действии инбридинга и дрейфа эффект роста
гомозиготности проявляется наиболее полно. В присутствии мутаций
или других линейных систематических давлений (b), f достигает
значения (1 + 4bN)"‘ (Malecot, 1950).
Динамика современных популяционно-генетических процессов в
мире, связанная во многом с растущей урбанизацией, разрушением
системы изолятов, а также неудержимым ростом миграций, приводит
к постепенному снижению значения генетического дрейфа, как
эволюционного фактора.
Миграции генов.
В формировании генетической структуры популяций миграции
играют очень важную роль. Миграции являются одной из основных
эволюционных сил, действующих на популяцию. Они выступают в
роли основного фактора, противодействующего изоляции и
определяют степень генетической дифференциации. Сам процесс
миграции населения является одновременно миграцией генов, так
как появление мигрантов в популяции связано с внесением в нее
новых генов. Ясно, что люди, прибывшие из отдаленных групп,
являются потенциальными носителями иных генетических свойств.
Обогащение генного пула новой генетической информацией
математически эквивалентно мутациям (Рычков, 1982).
47
Если рассматривать миграции отдельно от влияния других
эволюционных факторов на популяцию и проследить судьбу
единичного "мигрантного" гена во времени, то она, очевидно, будет
аналогичной судьбе единичного мутантного гена, у котового
вероятность закрепления в поколениях ничтожно мала. Влияние же
давления повторяющихся миграций (генного потока) на генофонд
популяции достаточно велико. И чем выше интенсивность и
информативность миграций, тем сильнее изменение динамики
генетического процесса в рассматриваемой группе. Немаловажную
роль при этом имеют также и размеры популяции (особенно
величина репродуктивной группы), в которую прибывают
пришельцы. Так, для изолятов и малых популяций миграции
являются одним из основных источников поддержания генетического
разнообразия и при совместном действии с другими эволюционными
силами (дрейф генов, отбор) имеется шанс закрепления даже
единичных привнесенных генов. В больших популяциях изменения в
генетической структуре происходят, как правило, при миграциях
большей интенсивности. (Рычков, 1982; Алтухов, и др., 1984; Бочков и
др., 1984).
В результате давления систематически повторяющихся миграций в
поколениях формируется сходство с генофондом популяции,
являющейся источником миграций, что ведет к группированию
частот генов около средних значений для системы популяций в
целом, непосредственно влияя на дисперсию средней величины гена
в сторону ее уменьшения (Ли, 1978).
Таким образом, в эволюционном плане миграции служат для
одних популяций источником генетического разнообразия, в других,
напротив, нивелируют их генетическое своеобразие, приводя в итоге
к гетерозиготации населения. •
Направление, интенсивность и характер миграций могут меняться
во времени. Последние десятилетия характеризуются ростом
масштабов миграций во всем мире. Растущая урбанизация в России,
совершенствование средств передвижения, освоение новых регионов
и другие факторы способствуют резкому увеличению миграционной
активности населения. Так, повсеместно происходит полная или
частичная ломка старых границ брачных ареалов и расширение круга
брачных связей. Как правило, миграции перестают зависеть от
расстояния. В качестве одного из примеров можно привести рост
48
межнациональных, а также гетеролокальных браков. Миграции
Я°ЛИ и связанная с ней рекомбинация генов создает предпосылки
геН° образования новых генотипов, к изменению адаптивных
комплексов, выработанных в поколениях. (Соловенчук, 1984).
Происходящий процесс аутбридинга и наблюдающаяся тенденция
к его росту требуют систематического изучения в плане выработки
адаптивных свойств к конкретным средовым факторам. Особенно это
становится актуальным для регионов с экстремальными условиями
среды.
Естественный отбор.
Естественный отбор сыграл решающую роль в эволюции всех
видов, в том числе и человека, путем преобразования генетической
структуры популяций в поколениях. Под влиянием этого
систематического фактора эволюции осуществляется отбор гамет
или диплоидных организмов, которые оказываются менее
приспособленными в определенных условиях среды. Причем
направление отбора, его характер и интенсивность меняется с
изменением условий окружающей среды.
Схема, в которой эволюцию можно представить в виде эволюции
отдельных локусов, слишком упрощена. В действительности
объектом отбора всегда является особь — поскольку
приспособленность отдельных локусов сконцентрирована в общей
приспособленности организма. Так, изменение условий окружающей
среды приводит к изменению приспособленности отдельных особей,
следствием чего является формирование иных частот генотипов и
связанное с этим изменение средней величины приспособленности
популяции. Согласно фундаментальной теории Фишера, скорость
отбора прямо пропорциональна генетической вариансе
приспособленности (Fisher, 1930; Левонтин, 1978; и др.). Именно
отбор привел к существованию балансированного полиморфизма в
человеческих популяциях, обеспечивая тем самым уникальную
генотипическую индивидуальность каждого человека.
В современный период в промышленно развитых странах
благодаря успехам здравоохранения и социальных преобразований
49
давление естественного отбора значительно снизилось. Однако этот
систематический фактор в человеческих популяциях продолжает
действовать, хотя его проявления в основном относятся к
внутриутробному периоду. Известно, например, что более половину
зачатий не заканчивается родами, вследствие гибели эмбриона или
плода на разных стадиях развития. Спонтанные аборты составляют
около 15% всех зачатий, мертворождения - 2%, также достаточно
часто отмечается бесплодие (Кулешов, 1979; Бочков и др., 1984).
Вследствие сложности селективных процессов естественный отбор
у человека весьма труден для изучения. Так как каждый организм
находится по влиянием процесса в течение всей своей жизни и в
результате одни генотипы оставляют больше потомков, чем другие,
то его интенсивность может быть адекватно оценена только у
индивидов предшествующего поколения.
Существующие разные подходы к определению давления
естественного отбора на человеческую популяцию можно условно
подразделить на две группы: собственно генетические и генетико-
демографические методы.
В свою очередь, в группе собственно генетических методов
выделяются два направления. Так, в одном из них делается акцент на
физиологических сторонах проявления действия естественного
отбора. Схема их сводится к сбору обширного материала по разным
типам браков и выявлению систематических отклонений пропорций
генотипов в браках от ожидаемых Менделевских распределений и
Харди-Вейнберга. Анализ отклонений от ожидаемых пропорций
генотипов и позволяет судить об интенсивности давления
естественного отбора. (Morton. 1950; Brues, 1954; и др.). Другое
направление генетических методов основано на принципе
популяционно-генетического исследования. Предполагается, что
если естественный отбор "ответственен" за формирование
генетической структуры популяций, то изучение ее позволит судить о
вкладе этого важного систематического фактора эволюции
(Livingstone, 1960).
Генетико-демографические методы наиболее широко
применяются при исследовании давления естественного отбора и
позволяют вычленить его компоненты, связанные с
дифференциальной плодовитостью (If) и дифференциальной
смертностью (Im) (Crow, 1958). Так, согласно Кроу для измерения
50
тотального отбора (индекс максимально возможного
КСЗ индекс Кроу и т.д.) нужно определить дифференциальные
ОТ°^ сти выживания и плодовитости в ряде последующих поколений.
Следовательно, это есть изменение скорости эволюции популяции
п и предположении, что все различия в приспособленности являются
генетически детерминированными. В действительности же скорость
эволюции будет значительно ниже, так как только часть различий в
приспособленности будет иметь генетическую природу. При этом за
приспособленность брачной пары принимается ее эффективная
плодовитость, а приспособленность популяции оценивается
отношением межсемейной дисперсии по эффективной плодовитости
к квадрату среднего значения данного параметра в изучаемой
популяции_2
I = V / W , где
V-дисперсия числа потомков;
W - средняя биологическая приспособленность (среднее число
детей).
Индекс потенциального отбора, согласно Кроу, равен:
1= Im + И ; Im = Pd ; Ps = 1 - Pd ; If = Vf
Ps Ps xe
где Pd - доля потомков, недоживших до репродуктивного
возраста;
Ps - доля потомков, доживших до репродуктивного возраста;
Vf и х - дисперсия и среднее число рождений на одну женщину.
Все эти величины сравнительно легко могут быть подсчитаны из
материалов демографии. Так, если Pd уменьшается, то
соответственно снижается и компонента дифференциальной
смертности (1m), тем самым остается большой простор для отбора,
благоприятствующего различиям в плодовитости. В настоящее время
такое явление наблюдается в большинстве развитых стран, когда
растет вклад компоненты дифференциальной плодовитости. В свою
очередь в них отмечается также тенденция к стиранию различий в
плодовитости (регулируемая рождаемость), связанная с социальными
изменениями в обществе, а также достижениями современной
медицины. В развивающихся странах, напротив, наблюдается высокая
рождаемость, как правило, коррелирующая с высокой детской
смертностью. (Cavalli-Sforza, Bodmer, 1971).
51
Однако, применяя для анализа естественного отбора в
человеческих популяциях метод Кроу, следует помнить, что
получаемые при этом значения являются максимально возможными
поскольку с привлечением данных демографии (плодовитость,
смертность) мы анализируем итог действия целого ряда факторов
как биологической, так и небиологической природы, так или иначе
оказывающих непосредственное влияние на воспроизводство
народонаселения.
Мутации генов.
Формирование популяций человека на протяжении всей истории
развития происходило при постоянном влиянии давления мутаций.
Мутации вызывают изменение последовательности нуклеотидов
ДНК путем замены, добавления или утраты части генетического
материала. С точки зрения генетика под мутацией следует понимать
превращение состояния гена. Эффект при этом зависит от того,
насколько велико произошедшее изменение.
Мутации являются довольно редкими событиями и в отношении
конкретного локуса частота их достигает порядка 1 • 1О — 110.
Судьба единичного мутантного гена, прослеженная во времени,
свидетельствует о том, что шанс утраты его возрастает
пропорционально времени, прошедшему с его возникновения, и
достигает максимума равного 1,0 после 127 поколений (Fisher,
1930). Значение повторяющихся мутаций в эволюции популяций,
напротив, очень велико. При этом весьма важным генетическим
параметром является доля лиц, находящихся в репродуктивном
возрасте, так как именно с ней связан процесс передачи
наследственной информации новому поколению. Скорость
мутационного процесса Qu) может быть оценена на основании
использования данных о частотах редких вариантов белков:
1) ._м = I , где
2Nto
,м - мутационный темп на ген -/ поколение;
I - среднее число мутантных аллелей на локус;
N - репродуктивный размер популяции;
Ь - среднее время, необходимое для элиминации мутантного
аллеля;
52
(Kimura. Ohta, 1969).
2) v> = Is / 4 In / (2 nq), где
- - скорость мутации на один локус;
Is - ожидаемое число редких аллелей;
N - эффективный размер популяции;
q . средняя частота гена в рассматриваемой популяции.
(Nei. 1977).
Скорость мутационных изменений (м) способна измениться под
влиянием радиации, воздействия температур, а также разнообразных
химических веществ. Так, примерно четвертая часть всех мутаций
индуцирована энергией естественного фона радиации, а остальная
часть обусловлена влиянием других средовых факторов. При этом
скорость непосредственно зависит от состояния окружающей среды
(Алтухов, 1984; Бочков, 1984; Спицын, 1985; и др ). Поэтому с полной
очевидностью встает вопрос о необходимости решения проблем
охраны биосферы от загрязнений мутагенными веществами.
Возникновение целого ряда научных дисциплин, таких как
радиационная биология, радиационная генетика, экс ’огическая
генетика и другие, связана с актуальностью изучения влияния
индуцированного мутагенеза, а также накопления мутационного
груза в популяциях разных видов, в том числе и человека.
Так, например, успехи здравоохранения способствовали выработке
определенных тенденций к ослаблению интенсивности естественного
отбора в человеческих коллективах. Подобная ситуация, даже при
неизменной скорости мутационного давления, приводит к
неизбежному росту отягощенности мутационным грузом. В реальной
жизни современных популяций данное явление выражается в
повышении частоты наследственных аномалий, генетически
обусловленной предрасположенности к целому ряду заболеваний, а
также снижением приспособленности и роста нега™вных эффектов в
воспроизводительной функции отдельных индивидов.
В этой связи представляются весьма интересными имеющиеся
сведения о различиях в скорости мутационного процесса в разных
этнических группах. Так, у южно-американских индейцев данная
величина на один, иногда два порядка выше, чем в европейских
популяциях (Neel, 1973, 1974). В таблице 2 приводятся оценки
скоростей мутационного процесса в популяциях России (цит. по
53
работе Спицына, 1985). Как видно из табл. 2 скорость мутационных
изменений в популяциях Северной Азии варьирует в пределах от 3,0-
1О до 7,4 • 10 на локус/поколение. С помощью метода Нея получена
оценка 9, вдвое превышающая величину, рассчитанную на основании
метода Кимуры-Оты (2,6'10' против 1,1 • 10 ).
Таблица 2.
Оценки скорости мутационного процесса на локус/поколение в
некоторых группах населения СССР.
Группа ЧИСЛО локу- сов средний объем выборки редкие аллели локус Оценка мутационного темпа методами
Нея(9) Кимуры-Огы
на
Чукчи 9 305,4 0,222 4,6.10* 6 3,4. 10* 6
Коряки 10 77,6 0,100 1,6.10* 2,9.10*с
Нивхи 10 72,5 0,200 6,3.10-'’ 1,1.10-'
Якуты 13 411,6 0,538 1,6. КГ6 1,1. 10* ь
Эвенки 9 206,1 0,556 8,1. JO"6 3,0.10’7
Тувинцы 9 139,0 0,333 1,2. 10- 3,0. 10*?
Тофалары 10 64,2 0,1 (Ю 7,4. 10’5 5,0. IO'5
Шорцы 6 79,7 0,333 2,3. 10-_ 4,0. 10
Ненцы 10 202,3 0,200 4,1. Ю-5 2,6. 10’5
Евреи 7 115,0 0,142 1,1. 10'3 3,7. 10’ь
бухарские
(цит. по работе Спицына, 1985).
Таким образом, способность мутаций изменять генетическую
структуру популяций позволяет считать их одной из наиболее важных
систематических сил эволюции, определяющих динамику
популяционно-генетических процессов.
54
ГЛАВА?. РЕАЛИЗАЦИЯ ПРОГРАММЫ ИССЛЕДОВАНИЙ
ПОИСТОРИЧЕ( КОЙ ГЕНЕТИКЕ.
I. Выбор объектов исследования.
Выбор восточных башкир в качестве объекта популяционно-
генетических исследований был обусловлен принадлежностью
составляющих субнопуляций на этническом уровне племен к
древнебашкирскому пласту, сыгравшему, по мнению историков,
значительную роль в сложении башкирского народа.
В генетической структуре популяций данного региона
запечатлены особенности, связанные как с историей освоения этих
мест, так и с древними этапами миграции племен. Поэтому изучение
их представляет большой интерес, поскольку, имеющиеся
расхождения в датировке времени появления башкир на Урале,
достигают нескольких столетий. Однако звучит более
аргументировано гипоте за позднего проникновения кочевых групп
древних башкир на данную территорию - в конце I тыс н э. (Кузеев,
1974, 1978). В связи с этим с очевидностью встает вопрос о времени
биологической и хозяйственно-культурной адаптации кочевого в
прошлом башкирского народа.
При выборе микропопуляций башкир руководствовались работами
историков о принадлежности предков коренных жителей данных
населенных пунктов к древне-башкирским племенам: бурзян, усерган,
тамьян, карагай-кипчакам, сальют. В табл.З представлено
современное административное положение 30 микропопуляций и
реконструированная структура бывшей родо-племенной организации
восточных башкир. На рис. 2 изображена схема локализации
исследованных групп башкир.
В качестве сравнительного материала использовались
литературные данные, а также привлекались результаты собственных
исследований по группам абхазов (с.Джгерда, с.Члоу, Очамчирский
Р-н, Грузия; с.Дурипш, с.Хуап, Гудаутский р-н, Грузия),
азербайджанцев (с.Абдал, Агдамский р-н, Азербайджан), албанцев
(о.Георгиевка, с.Девнинское, Приазовский р-н, Запорожская обл.,
Украина), греков (с.Красная Поляна, Великоновоселковский р-н,
Донецкая обл., Украина; с.Старая Ласпа, Тельмановский р-н,
55
Таблица 3.
Административное деление современных микропопуляций и
реконструированная историческая структура бывшей
родоплеменной организации восточных башкир
Рай о н ы Баш к и р И И Челябинская обл.
Баймакский Бурзянский Зианчуринский Абзелиловский Учалинский Кунашакский
племя бурзян: 1. род ямаш бурзяне без родовой при- надлежносги племя усерган: 1. род усерган племя тамьян: 1. род тамьян племя табын: 1. род кара- табын племя сальют 1. род салькгг
МИК- ропо- пу ЛИ- НИИ Чингизово Буранбаево Гадельбаево 2. род мунаш Туркменево Абзаково Ахмерево Басаеве Н-Субхангулово Гадельгареево Миндегулово Киекбаево племя карагай- кипчаки: 1. род карагай кипчаки Кипчак Н-Мунасипово ’ Набиево Байназарово Яумбаево Баишево Сагитово Утягулово 2- род бишей Ибраево Байдавлетово Казарма Салаватово Кужаново Елембетово Расулево Ишкинево Истамгулово Кунакбаево Казакбаево
СП
обл., Украина), киргизов (с.Мургаб, Мургабский р-н
Рис. 3. Схема локализации исследованных популяций.
58
2. Организация сбора материала.
Популяционно-генетическое изучение групп включает в себя
енение совокупности методов исследования: генетических,
ПР унологических , биохимических, антропологических, данных
исторически* дисциплин, демографии, а также статистических
методов и ДР- В настоящем разделе приводится краткая
характеристика методов анализа.
Все применяемые в работе методы исследования можно разделить
на две категории, первая из которых объединяет приемы сбора
информации, а вторая включает различные способы ее обработки.
Сбор демографической информации по микропопуляциям
проводится по специальным схемам. Индивидуальная карта анамнеза
семьи состоит из паспортной части (фамилия, имя, отчество, возраст,
национальность, место рождения мужа, жены), сведений о детях (пол,
возраст живых детей, причина и возраст смерти умерших детей),
акушерского анамнеза (возраст матери при вступлении в брак,
рождении первого и последующих детей, общее число
беременностей и их исходы, число и характер отягощенных
беременостей, количество искусственных и самопроизвольных
абортов, мертворождений, возраст при появлении menarche и
наступлении climacterium), а также информация генетического
характера (кровное родство в браке, наследственные аномалии и
дефекты). В данной работе применялась схема, разработанная на
основе популяционно-генетической анкеты Пасекова и Ревазова
(1975).
Используются материалы официальных источников информации
(записи актов гражданского состояния, церковно-приходские книги,
похозяйственные книги сельских Советов), архивы медицинских
учреждений (больницы, женские консультации, родильные дома,
станции переливания крови), а также сведения, получаемые при
работе с информаторами в обследуемых населенных пунктах.
Для изучения динамики формирования популяционно-
генетической структуры были привлечены историко-
Демографические материалы: "Уфимская губерния. Список
населенных мест. 1870", статьи Уйфальви (1877), Соколова (1904),
фавитный список населенных мест Уфимской губернии, 1906 г.",
59
"Сведения о количестве населения Башкирии. 1920 г", "Списщ
населенных мест Башреспублики. 1926 г.", ежемесячный журн^
"Хозяйство Башкирии" (1928, 1929), "Материалы по исторИ|1
Башкирской АССР" (1936, 1949, 1956, 1960), "Итоги Всесоюзной
переписи населения 1970 г." (1980), "Население мир»
этнодемографический справочник" (Брук, 1981), "Демографический
энциклопедический словарь (1985), 'Таджики долины Хуф (BepxoBbJ|
Аму-Дарьи)" (Андреев, 1958), и некоторые другие сведения.
3. Идентификация систем групп крови и биохимических
маркеров генов в башкирских популяциях.
Генетическую структуру микропопуляций исследовали
выборочным способом путем анализа систем эритроцитарных групп
крови и сывороточных белков. Стандартными серологическими
методами с использованием реакции гемагглютинации проводили
определение групп крови АВО, MN в полевых лабораториях в день
взятия образцов крови. Систему резус (D) определяли с помощью
стандартного универсального реагента антирезус в пробирках без
подогрева (Башлай, 1960). В работе использовались специфические
сыворотки анти-М, анти-N производства Ленинградского НИИ
вакцин и сывороток, универсальный реагент антирезус и
изосыворотки АВО производства Московской городской станции
переливания крови. Изоантигены системы Rh - Hr и Келл-Челлано
выявляли методом конглютинации в желатине (Башлай, 1960).
Биохимическими методами определялись фенотипы
сывороточных белков гаптоглобина, трансферрина,
группоспецифического компонента. Типирование выполнено в
лаборатории генетики Отдела биохимии и цитохимии БФАН СССР
электрофоретическим методом (Giblett, 1969; Hoppe et al., 1972;
Hopkinson, 1973). Методом изоэлектрофокусирования определялись
фенотипы Ri -антитрипсина (Laurell, Erikson, 1963).
фосфоглюкомутазы PGM-1 (Spencer et al., 1964) в лабораторий
экологической генетики Медико-генетического центра АМН России.
60
4. Статистический анализ данных.
каждой генетической системы
и генов. С помощью критерия
фенотипов « _ ______ _________г г
ожидаемыми частотами (Лакин, 1969).
вычислялись частоты
соответствия х
' делились различия между эмпирическими и теоретическими
Частоты генов системы АВО рассчитывали общепринятыми
способами (Race, Sanger, 1962; Bernstein, 1930; Stevens, 1938):
р' = 1- Jb + О ; q' = 1- Га + О ; г'-4сГ
с последующим введением поправок, равных половине разности
между единицей и суммой:
р = (1 +J_ ). (I-Jd + В);
2Д ____________,
q = (l +_L_).(1-[O + А);
2Д г-,
r = (l +J_).(MO+_L)
2Д 2Д
Двуаллельные системы групп крови, сывороточных белков и
эритроцитарных ферментов с кодоминантным наследованием
представлены в популяциях тремя фенотипами - гетерозиготным и
двумя гомозиготными. Расчеты частот системы MN упрощаются, т.к.
частота фенотипа равна частоте генотипа:
М = М + MN ; N = N + MN
2 2
Аналогичны вычисления частот генов сывороточного белка
гаптоглобина, фенотопирование которого проводили после
Дискэлектрофореза в 7.5% полиакриламидном геле с применением
трисглицинового буфера pH = 8,3. Продолжительность
электрофореза 1,5 часа при силе тока 3 mA на один образец
сыворотки (Рафиков, 1977).
61
Нр = Нр 1-1 4- Нр 2-1; Нр2= Нр 2-2 + Нр 2-1
2 2
Для систем с полным доминированием (Rh-Hr и Kell) частоту
рецессивных генов (d*, с*, е*, к*) определялись путем извлечения
корня квадратного из частоты соответствующего фенотипа. Таким
образом, частота рецессивного гена будет равна:
q = \|d*, с*, е*... к*,
а пропорция соответствующего доминантного аллеля: Q= 1- q ,
При этом стандартную ошибку можно определить как: Sq=j(l- q )/4N?
где N является выборочным объемом (Дж.Ниль, У.Шэлл, 1958).
Дисперсии и квадратические ошибки для системы АВО вычислены
по формулам, предложенным в работе Нила и Шэлла (1958).
Дисперсия для системы MN и других кодоминантных систем
определены по формуле:
= q • (1 - q) , где
2N
q - частота гена т,
(1 - q) - частота гена п,
N - общее число наблюдений.
Дисперсия для систем Р, резус определена по формуле:
= l >гДе
2N
q - частота рецессивного гена,
N - число наблюдений.
Подобным же способом вычисляются дисперсии для систем
Льюис, ABH-секреции, ощущения вкуса фснилтиокарбамида и типов
ушной серы.
При оценке генетических параметров соблюдали формально-
статистические критерии достоверности, предъявляемые к
выборочным данным из совокупности. Репрезентативность
выборочного объема проверяли по формуле (Гмурман, 1972):
n = •g'2 ; О = t • , где
Q
62
. минимальный объем выборки;
" находится по таблице функций Лапласа;
- точность оценки,
>2. дисперсия частоты гена.
Анализ генетической дифференциации осуществлялся с помощью
F статистик Райта, эмпирические значения их определяются через
вариансы генных частот (Nei, 1965).
Пропорции генотипов панмиксной популяции, находящейся в
равновесии Харди-Вайнберга, определяются разложением бинома
Ньютона: 2 2 2
(pi + рг) = Pi + 2pi . pi + pi
и, таким образом, доступны исследованию благодаря четко
выраженной алгебраической природе Менделевской теории
наследственности, относящейся к области формальной теории
(Харди, 1908: Вайнберг, 1908). Количественная оценка отклонения
от равновесия Харди-Вайнберга анализировалась методом X2
(Рокицкий, 1978):
Г = .где
Е
Е - теоретическая численность генотипов;
О - наблюдаемая численность,
с последующей проверкой значения по таблице распределения при
вероятности р = 0,05 (Fischer, 1950).
Проверку популяционной системы на однородность по 2-х
аллельным локусам проводили по общепринятой формуле:
х2 » 2рх-рГх , где
М
х - удвоенная частота одного гена, а р » 2/2 N.
При наличии в локусе более чем 2-х аллелей применяли формулу:
х2 =.Spp. Ipp + Sqq, Iqq t ZSpq, Ipq, где
Zn
SPP, Sqq, Srr - сумма квадратов p, q, r.
IPP> Iqq, Ipq - количество информации в выборках (Stevens, 1938,
1950,1952).
63
По данным выборочных исследований определена средняя частота
гена по микропопуляциям, дисперсия и ошибка средней частоты, й
случае 2-х аллельных локусов (гаптоглобин, MN, Келл-Челлапо, J
также система резус, контролируемая тремя парами аллельных генов
Dd, Сс, Ее), определяется простая биномиальная димперсия:
V . (р1) = р (1 - р), где
2G
G = 2No, т.е. удвоенному объему выборки.
Имеются специальные таблицы оценок частоты генов системы
АВО, полученных по методу максимального правдоподобия (Ли, 197,
1978).
Каждая микропопуляция, являющаяся структурной единицей в
популяционной системе, реально существует благодаря
преемственности поколений и устойчивости наследственных
признаков, т.е. имеет длительный период исторического
формирования элементов ее структуры (Тимофеев-Ресовский, 1973;
Li, 1976: Рычков, 1976, 1982, 1985 и др.). Поэтому способы
определения тотального размера популяции (Nt), доли
репродуктивного (Nr) и эффективно-репродуктивного (Ne) объемов
в нем приобретают особое значение. Статистически наиболее просто
определяется общий объем популяции. Критериями для определения
границ репродуктивного возраста у женщин являются данные
генеалогии и анамнеза, позволяющие определить возраст при
появлении menarche и наступлении climacterium, а также при
рождении первого и последнего ребенка. Для мужчин к таким
характеристикам отностися возраст при рождении первого и
последнего ребенка. Интерпретация генеалогических данных в
дальнейшем связана с эффективно-репродуктивным объемом
популяции. Все эти особенности подчеркивают значимость данной
величины среди других демографических параметров.
Существует ряд формул для расчета реальной величины
эффективно-репродуктивного размера популяции (Wright, 1939; Ли,
1955,1976):
1. По неравной численности лиц репродуктивного возраста разного
пола:
64
Ne = 4No . Ni , где
No + N.
No и Ni лица репродуктивного возраста разного пола;
Г четом различия лиц по репродуктивной способности:
W Ne = NK.j(NK-n _ , где
(N -1) с к + NK(K-l)
jj.среднее число потомков, приходящееся на одного индивида;
бк - дисперсия этого числа;
3 С учетом периодического уменьшения численности популяции:
_1_ = _1_ . _1_ + -. + _1_ ,
Ne t Ni Nt
где Ni и Nt -численность популяции во времени;
4. Сучетом влияния инбридинга:
Ne = N , где F - коэффициент инбридинга.
Кг. 1 + F
В дальнейших расчетах использовалась гармоническая средняя
величина HNe, определяющая эффективную численность популяции
в различных поколениях (согласно формуле 3).
Подробное описание популяционно-генетических моделей и
методов определения миграций, инбридинга, отбора,
гетерозиготности, метисации, реконструкции родословных древ,
генетических расстояний и т.д. приводятся в соответствующих главах
книги.
65
ГЛАВА 4. ВРЕМЕННАЯ И
ПРОСТРАНСТВЕННАЯ ОРГАНИЗАЦОД
ПОПУЛЯЦИЙ
башкирской популяции претерпела наиболее
: изменения в последние десятилетия в связи с
социально-экономическими преобразованиями
Структура
существенные
крупными
произошедшими в стране. На смену единоличному способу ведения
хозяйства, сохранившему во многом элементы патриархально-
родовых отношений, пришло коллективное. Многоотраслевое
колхозное производство в современных башкирских селениях
объединило существовавшее ранее скотоводство, бортничество,
земледелие, охоту и другие формы хозяйствования. Новые
экономические отношения в сочетании с тенденцией укрупнения
хозяйств, централизацией руководства и административного деления
районов Башкирии значительно изменили характер и интенсивность
связей между бывшими родоплеменными образованиями.
Последствия произошедших воздействий, естественно, нашли свое
отражение в генетической структуре микропопуляций.
По оценочным данным второй половины XIX в. население
усерганских волостей составляло 42 тысячи человек (Соколов, 1904).
Совокупность бурзянских башкир насчитывала около 50 тыс.
(Уйфальви, 1877). Численность тамьянцев в конце XVIII в.
приближалась к 21 тыс. человек (МИБ, 1949). По противоречивым
данным демографических источников XVIII в. невозможно сколь-
нибудь точно оценить численность кыпчаков и сальютов. Население
табынских волостей в начале XVIII в. составляло примерно 60 тыс.
человек, а восточных табынцев - 26,5 тыс. человек (МИБ, 1949).
Численность башкир в СССР по данным переписи 1970 г. составляла
1240 тыс. человек, переписи 1978 г. - 1371 тыс. человек, в России по
переписи 1989 г. - 1345 тыс. человек.
Характеристика демографической структуры популяции башкир
проводилась на основе имеющихся архивных материалов переписей
населения, а также данных, собранных в ходе экспедиций. В таблице 4
представлена динамика роста численности башкир и их хозяйств в
селениях за период с 1870 по 1974 гг. Как видно из сведений табл. 4,
отражающих демографическую картину некоторых населенных
пунктов восточных башкир, численность их возросла в течении
последних десятилетий и оставалась достаточно стабильной в
Динамика численности восточных башкир за столетие (1870 -1974)
66
S!
I
00
о
— оли- ество жителе 577 820 585 505 403
ы: г *
5 | » ® О> О Г- ”Т ГI Ч- г*-, —' тг
Т п — —. ~ ~ о
)Х
Коли- чество жителе 567 708 525
5 |
|| 119 145 94
Коли- чество жителей = 8 8 38 й in 00 гл m v-j
5 £
х о \О
7 S 8 2 3 £ =
>х
1оли- ество жителе МП Я
ж У R >Л 40
«ело оров
5“ « II н>
3
trXuoti Баишево Ибраево Идияшево Утягулово Байлавлетов
Микро
67
прошлом. Отсутствие волнообразных колебаний в ДинамцЛ
численности населения и устойчивость среднего размера семЗ
позволяет сохранить преемственность генетической стР}ктурЭ
популяционной системы башкир. Аналогичную ситуацию, когм
численность мало меняется в течение последних полутора столетийJ
проявляется тенденция к росту ее лишь в последние десятилетия,
отметили исследователи в популяциях манси (вогулов) Паткано»
1911; Давыдова, 1989).
1. Возрастная структура.
Временная организация системы микропопуляций восточн^Н
башкир исследована по возрастной структуре и структуре поколений,
включающей характеристики продолжительности и скорости смены?
поколений. В табл. 5 и на рис. 4 представлены данные о возрастной
структуре северо-восточной субпопуляций. В них выявляется^
удовлетворительное совпадение особенностей возрастной структуры,
с некоторым уменьшением доли женщин в возрасте 20-40 лет у
северо-восточной группы. Однако при сравнении микропопуляций
обнаруживаются более значительные вариации возрастных пирамид I
Аналогичный дефицит возрастной группы 20-30 лет наблюдал’
Ю.Г.Рычков в популяции эвенков Средней Сибири. Отметив, что
время рождений данной возрастной группы приходится на период с
1942 по 1951 гг., он сделал вывод о снижении рождаемости в это
время, а также о возможном влиянии урбанизации населения и
связанной с ней миграцией молодежи в города (Рычков и др., 1972,
1974).
68
Таблица 5.
пл оасгная структура субпопуляций северо-восточных
^° го-восточных башкир и их средний возраст
рю
jojpacTHbie группы Восточные башкиры
Северо- восточные (7044 чел.) Юго-восточные (4565 чел.) Суммарно (11609 чел.)
мужч. женщ. мужч. женщ. мужч. женщ.
0-19 48,4 56,9 37,2 48,4 42,8 53,6
<20-54 38,1 35,8 52,4 44,5 45,3 39,2
55- 13,5 7,3 10,4 7,1 11,9 7,2
Средний возраст 26,8 25,9 26,3
Рис. 4. Распределение восточных башкир по возрасту и полу
(женщины в левой части пирамид)
А - северо-восточная группа (табын, карагай-кипчаки, сальют)
Б* юго-восточная группа (бурзян, усерган, тамьян)
69
Таблицаб
Соотношения возрастных групп в популяциях греков и албанцев
Приазовья.
греки албанцы '—
Возрастные группы Красная Поляна (4619 че- ловек). Старо- ласпа (955 че- ловек). Девнинс- кое (677 человек). Георгиев-ка (539 человек).
дорепродуктивная
репродуктивная 19,9 21.7 21,6 23,0
пострепродуктивная 1 41,09 39,0 39,0 39,3 40,3 38,1 39,0 38,0
Таблица 7.
Соотношения возрастных групп в высокогорных популяциях
памирцев и киргизов.
Возрастные группы памирцы киргизы
Пастхуф (2000 м Хуф (3000 м над Мургаб (3800-
над уровнем моря уровнем моря (673 4200 м над
(700 человек; человека) уровнем моря (892 человека)
59,0 52,3 45,0
доре продуктивная 24,0 28,5 41,0
репродуктивная пострепродуктивная 17,0 19,2 — 14,0 -
На рис. 5 графически изображены половозрастные
характеристики исследованных групп греков, албанцев, памирцев и
киргизов. Как видно из таблицы 6 и 7 в греческих и албанских
70
Рис. 5. Распределение возрастных групп в популяциях Приазовья и Памира (А - греки; Б - албанцы;
памирцы; Г - киргизы).
71
группах отмечается превалирование числа женщин, тогда как среди
башкирок это наблюдается только в дорепродуктивной когорт
Обращает на себя внимание резкая деформация возрастные
пирамид, в популяциях Приазовья, в значительной стененц
обусловленная снижением дорепродуктивной возрастной группу
Другим существенным отличием является также значительно более
высокий удельный вес лиц пострепродуктивного возраста.
Демографическая ситуация в высокогорных популяциях наиболее
оптимальна с большой долей молодых возрастов, составляют^
более половины от общей численности у памирцев и немного
меньше этого значения у киргизов. Подобная тенденция отмечена
рядом исследователей среди популяций алтайцев, манси и других
народов Сибири и Дальнего Востока (Лузина, 1987; Давыдова, 1989;
Гольцова, Сукерник, 1979; и др.).
Важным параметром возрастной структуры популяции является
также средний возраст последней. В табл. 5 представлен анализ
возрастной структуры субпопуляций башкир. Как видно из таблицы 5,
средний возраст в северо-восточной группе (26,8 лет), несколько
выше, чем в юго-восточной (25,9 лет), в целом средний возраст
популяции восточных башкир составляет 26,3 лет. Сравнительно
невысокое значениме данной характеристики можно объяснить
вкладом возрастной категории лиц до 19 лет (Шереметьева,
Горшков, 1982; Лузина, 1987 и др.).
2. Поколение, его длительность
и темп смены в популяциях.
Длительность поколения в микропопуляциях восточных башкир
оценивалась на основании фактических данных анализа родословных
За реальную характеристику поколения принимается средний возраст
составляющих лиц и дисперсия данной величины. В табл. 8 и на рис. 6
представлены результаты оценки продолжительности поколений и
скорость смены в популяции башкир. Характерной особенностью
является наблюдающееся перекрывание поколений, что, по
видимому, обеспечивает генетическую преемственность мсжДУ
поколениями в популяциях, за счет нивелирования возможных резких
изменений в передаче информации (Рычков, 1982;).
72
образом, в системе популяций башкир адаптивный
Таким *
эм обеспечивает достижение среднего возраста,
“рентного длительности одного поколения, равного 25 годам,
эквИ п(ПВОЛЯет осуществить передачу и реализацию генетической
ЧТфОрмации в поколениях потомков
Таблица 8.
Характеристика поколений в популяции восточных башкир
X - средний год рождения с 99% доверительными интервалами,
§х - стандартное отклонение, F - число индивидуумов,
генеалогически отнесенных в данное поколение.
Точка отчета - 1977 год
Поколения Nn Nn/Nt x + tm Sx
Fo 4503 0,3886 1963 + 0,33 7,49
Fl 5248 0,4529 1950 + 0,48 11,62
F2 1836 0,1584 1925 +0,90 12,84
Nt 11587
Рис. 6. Структура популяции восточных башкир по поколениям и
протяженность поколений во времени.
По оси ординат - процентная доля от общего объема популяции и
номер поколения в 99%-ных доверительных интервалах.
73
Темп передачи генетической информации на Урале, равный 26,3 г|
очень близок с алтайцами и несколько выше, чем у некоторЬ|’
народов Сибири и Дальнего Востока (Рычков, 1978; Лузина, 19^-у.
Бахолдина, 1981; и др.).
3. Характеристика общего, репродуктивного и
эффективно-репродуктивного размеров.
Объем популяций человека, является характеристикой, сильно
варьирующей у разных народов (Saugstad L, 1972; Ferak V., Kroiipova
Z., 1976; Schull W.I., 1977; Брук, 1986; Фогель, Мотульски, 1990; и
др.). Размер популяций может колебаться от гигантских городских
агломераций с численностью от десятков, сотен тысяч человек до
малых изолированных популяций, состоящих из десятков и сотен
индивидов. В истории человека города возникли сравнительно
поздно, основная часть исторического процесса связана с
популяциями малой численности. Так, основываясь на анализе
современных групп охотников и собирателей Австралии и Африки,
можно предположить, что численность человеческих сообществ в
палеолите не превышала нескольких сотен индивидумоз,
представляющими несколько перекрывающихся поколений, с
естественным характером репродукции и размером семей,
подверженным случайным колебаниям (Li, 1955; Harrison et. al., 1972;
Харрисон, 1979). Несмотря на наблюдающиеся различия между
народами по размеру популяций величина их в каждом конкретном
случае является оптимальной, выработанной в процессе адаптации и
способствующей устойчивости популяции к неблагоприятным
условиям окружающей среды (Рычков, Шереметьева, 1972).
Очевидно, небольшие коллективы в прошлом являлись адаптивными
комплексами, свойственными определенному типу хозяйственно*
культурного уклада.
Другой важнейшей демографической характеристикой популяции,
тесно связанной с генетической структурой, является ее
репродуктивный объем, или объем размножающейся группы (Nr).
Его нельзя рассматривать в отрыве от возрастной структуры и от
общего размера популяции, так как формирование генофонда
74
его поколения связано лишь с той частью генов, носители
слеДУ10 находятся в репродуктивном возрасте. Данный период
^°^яет У индивидов меньше половины продолжительности
Объем размножающейся группы в популяциях человека
*ИЗчительно ниже тотального, как правило, не достигает 2/3 от
зна го а в ряде случаев отмечен ниже чем 1/3 (Morton N., 1969; De
Oliveira a. Salsano, 1969; и др.). Например, в популяциях береговых
чекчей и азиатских эскимосов п-ва Чукотка он достигает 27-30% от
тотального объема, несколько выше (37%) у илимпийских тунгусов
(Рычков, и др, 1972, 1974, 1978). Социально обусловленное
уменьшение доли лиц репродуктивного возраста, связанное с
оттоком молодежи из популяций присуще многим современным
группам. Так, у хакасов отмечено снижение отношения Nr/Nt с 38% в
1940 году до 31% в 1980 (Казаченко, 1986).
Однако ситуации в реальной жизни отличаются от предложенной
модели и не все лица репродуктивного возраста вносят свой вклад в
генофонд нового поколения, так как существует целый ряд причин
социальной и биологической природы, способствующих сокращению
репродуктивного объема популяции. Одним из приближений
яв.'ин гея эффективно-репродуктивный объем. Он отличается от
репродуктивного в той мере, в какой существует неравный вклад лиц
родительского поколения в генофонд последующего. Таким
образом, число индивидов, способных оставить потомство, не
единственный критерий, определяющий стабильность популяции по
отношению к случайным колебаниям.
Величина общего (Nt), репродуктивного (Nr) и эффективно-
репродуктивного (Ne) объемов в микропопуляциях башкир, а также
гармонические средние этих основных параметров приведены в табл.
9. Сопоставление микропопуляций в таблице произведено с учетом
восстановленной исследователями родо-племенной принадлежности
предков современных жителей данных населенных пунктов. Как
видно из табл. 9 колебания численности в микропопуляциях
наблюдаются в пределах от 372 до 810 человек, значительно меньше
Различаются гармонические средние величины тотальных объемов
племен (HNt) составляющие от 420 до 498 человек.
Демографические сведения, необходимые для определения
границ репродуктивного возраста, представлены в табл. Ю, где
отражены средние величины по родам: ямаш, тамьян, бурзян без
75
Таблица 9.
Оценка репродуктивного (Nr) и эффективно-репродуктивного (Ne)
размеров и их долей в общем объеме микропопуляций башкир (Nt).
Популяции 1 Nt 2 Nr I Nr/Nt 4 К 5 1 N л© 1 Nel 7 Ne2 8 I Ne/Nt 1 9
Чингизово 549 131-116 0,24-0,21 3,93 4,2230 128-114 123 0,23-0,21
Буранбаево 417 99-97 0,24-0ДЗ 307 7,9700 69-68 98 0,100,16
Гадельбаево 403 113-111 0,28-0,27 3,71 4,2840 109-107 112 0,27-0,27
Н-Субхангулово 425 99-95 0,23-0,22 2,94 3,8550 9086 97 0,21-0,27
Гальдергареево 468 113-107 0,2441,23 2,43 3,9843 90-85 НО 0,19-0,18
Туркменево II 410 100-98 0,24-0,24 3,28 7,9800 70-68 99 0,17-0,16
Абзаково 403 98-92 0,24-0,23 3,69 4,373 91-86 95 0,23-0,21
Ахмерово 479 115-109 0,24-0,23 2,49 3,9650 92-84 105 0,19-0,18
Басаеве 409 Н 436 99-91 119 0.24-0,22 308 5,8111 80-71 95 104 0,200,17
Баишево 577 131-127 0.23-0,22 2,30 3,9284 100-97 129 0.17-0,17
Сагитове 407 91-97 0,22-004 2,86 4,5963 75-80 94 0.18-0.20
Утягулово 505 119-109 0,24-0,22 4,14 6,7749 103-95 114 0,20-0,19
Идияшево 585 141-137 0,24-0,23 4,84 6,6340 131-127 139 0,22-0,22
Байдаалетоьо 403 97-91 004-0,23 307 5,7763 79-74 94 0.20-0.18 1
Казарма 406 91-89 0,22-0,22 3,11 4,6164 79-77 90 0.19-0,19
Ибраево 810 Н 498 192-169 125 0,24-0,21 4,13 6,1632 172-151 180 113 0,21-0.19 ,L
Продолжение таблицы 9.
Г ” ' ' i П 2 3 | ‘ 1 . 1 5 ч 7 ’ 8 9 \
Салаватово 453 117-102 0,26-0,23 2,76 4,6343 94-82 109 0,21-0,18 1
Куханово 407 98-91 0,24-0,22 3.10 4.2452 88-81 95 0,22-0,20 1
Елембетово 403 101-99 0.25-0,25 2,84 4,4598 84-82 100 0.21-0,20
Н 420 103 101
И шкиве во 532 122*106 0,23-0,20 3.30 10,877 72-63 113 0,14-0,12
Истамгулово 457 118-101 0,26-0,22 2,80 10,726 59-50 109 0,13-0,11
Расулево 416 104-96 0,25-0,23 2,60 7,730 59-55 100 0,14-0,13
Н 434 109 107
Капчах 557 129-121 0,23-0,22 2,29 3,8971 99-90 124 0,18-0.16
Н-Мунасипово 407 92-88 0,23-0,22 3,16 4,6091 78-70 84 0,19-0.17
Набиево 430 115-107 0,27-0,25 2.81 4.6551 95-81 108 0.22-0,19
Байна за рово 583 137-129 0.23-0,22 2,41 3,9841 102-91 123 0.17-0,16
Яумбаево 392 95-89 0,24-0,23 ЗД1 5,7169 77-69 88 0,20-0,18
Н 461 108 103
К - среднее число гамет, передаваемых родителями репродуктивного возраста следующему поколению.
- дисперсия среднего числа гамет
Nei - эффективно-репродуктивный размер с учетом дифференциальной плодовитости.
Nez - эффективно-репродуктивный размер с учетом неравного числа особей разного пола
репродуктивного возраста.
Н - гармоническая средняя.
3
Данные к определению границ репродуктивного периода.
S5
min/ггк 12-20 20-53 37-55 5
X ОС t—
3 м 4/1 я 3 Я 00
Л
род с 342 я гч 5© V) гч 3
бурзяне б/р К 15 12-18 5 я Т—« S?
с 241 JO •—• я
min/max 14-23 4- 20-61 05
3
яма и 5© я ГЧ 3
род с 311 237 211
о з р а с т Начала menarche При рождении 1 ребенка При рождении последнего ребенка Начала climacterium Мужчины при рождении 1 ребенка Мужчины при рождении последнего гбенка
род кара-табын 1 i |К с 331 15 12-28 ! 348 22 16-41 309 33 18-51
род б и ш е й п х min/max —1 352 15 12-21 262 22 16-35 258 37 20-48
род усерган п х min/max 474 15 11-24 340 23 17-39 332 34 20-46
Карагай-кипчаки п * х min/max 1. 379 15 12-20 2. 295 22 17-38 3. 251 38 18-43
40-56 I 153 48 35-56 I 98 46 35-60 67 46 32-55
5
78
вОЛ принадлежности, карагай-кипчаки, усерган, бишей, кара-
' Г а также приведены максимальные и минимальные значения,
^блюдаемые в микропопуляциях.
«ризиологическая граница начала репродуктивного периода у
ок в среднем колеблется от 15 до 16 лет и различия между
максимальным и минимальным значениями лежат в интервале от 11
28 лет. Соответственно этому меняется и разрыв между социально-
условленной границей и максимальным физиологическим
значением. По литературным данным в эвенкийской популяции
средняя величина начала репродуктивного возраста составила 14 лет
(Рычков и др., 1974). У кызыльских и сагайских женщин-хакасок
физиологические границы репродуктивного периода по данным
акушерского анамнеза находятся в пределах 15,4 - 45,7 и 15,6 - 48,3
года соответственно. (Казаченко, 1986), что сопоставимо с
аналогичными параметрами у алтаек (Лузина, 1987),
Средний возраст башкирок при рождении 1 ребенка составил 22-
23 года, а мужчин - 25 лет. Несколько отличается этот возраст у
эвенкийских женщин - 21-22 года, у мужчин - также равен 25 годам.
Для хакасов характерно сближение данных значений и в целом оно
колеблется от 25,6 для кызыльцев и 25,5 - сагайцев.
Средний возраст начала climacterium для башкирок составил 45-48
лет. Таким образом, физиологическая продолжительность
репродуктивного периода у женщин находится в интервале от 23 до
35 лет. Мы провели оценку репродуктивного объема популяции
башкир в границах 18-45 лет для женщин и 18-50 лет для мужчин.
Относительность этих данных, а также существующий разрыв между
социально-обусловленными границами и физиологической
продолжительностью репродуктивного возраста затрудняет
получение точной оценки величины размножающейся группы в
популяции. Репродуктивная часть (Nr) в популяциях, составляет 21-
28% от тотального объема, соответственно гермоническая средняя
величина колеблется между племенами в пределах 103-125 человек.
Величина эффективно-репродуктивного объема (Ne) по
микропопуляциям, определенная с учетом дифференциальной
плодовитости, оказалась значительно меньше репродуктивного. По-
видимому, одной из причин снижения эффективного объема в
МИкропопуляциях башкир является нерегулируемая рождаемость, т.к.
Дисперсия ( д к) среднего числа детей, приходящихся на одного
79
индивида (К), существенно превышает среднее число. Среди других
теоретически возможных факторов, снижающих эффективную
численность, следует назвать инбридинг, случаи безбрачия,
бесплодия, дифференциальную смертность, а также колебание
численности популяции в поколениях (Ли, 1976). Поэтому в
дальнейших расчетах использовали гармонические средние значения
HNe, вычисленные с учетом неравной численности особей разного
пола: предел, в котором находятся эти значения, несколько выше - от
1О1 до 107 человек в племенах-субпопуляциях и HNe (средняя
гармоническая) является наиболее реальным значением Ne, при
колеблющихся оценках по отдельным популяциям и этническим
группам. Теоретическое обоснование применения HNe, как
величины, отражающей эффективную численность в поколениях,
дано Райтом (Wright, 1939).
Сравнительный материал по оценке основных параметров
популяционной структуры высокогорных групп Памира (памирцы
к.Пастхуфа, Хуфа, киргизы к.Мургаба), абхазов (с.Джгерда, Члоу,
Хуап, Дурипш), греков Приазовья (с.Красная Поляна, Староласпа),
албанцев Приазовья (с.Девнинское, Георгиевка) представлен в табл.
11. Данный материал вызывает большой интерес в связи с
различиями в протекании микроэволюционных процессов,
особенностями динамики формирования элементов популяционно-
генетической структуры, связей с окружающими популяциями,
исторически обусловленным своеобразием адаптивных процессов и
пр.
Как видно из данных табл. 11 наиболее благоприятная
демографическая ситуация наблюдается в высокогорных популяциях
Памира. Отсутствие в них искусственного контроля рождаемости с
сохранением естественного характера репродукции в популяциях,
относящихся к разным этносам й отличающихся по типу ведения
хозяйства, тесно связано с особенностями популяционной структуры,
для которой характерен высокий удельный вес младшей возрастной
группы, затем в порядке уменьшения располагаются средняя и
старшая. Обращает на себя внимание также высокое значение
среднего числа детей в семьях от 5,9-6,4 на среднегорье до 3,08 на
высоте 38000-42000 м. над уровнем моря. Как и у башкир
дисперсия среднего значения очень высока (от 4,7420 в Мургабе, до
10,5310 в Хуфе).
80
Оценка общего, репродуктивного и эффективно-репродуктивного объемов в популяциях памирцев
(к.Пастхуф, к.Хуф), киргизов (к.Мургаб) Памира, абхазов (с.Джгерда, Члоу, Хуап, Дурипш), греков
(с.Красная Поляна, Староласпа), албанцев (с.Девнинское, Георгиевка).
Другой группой популяций достаточно хорошо сопоставимых
репродуктивной структурой башкирских популяций являют
абхазские. Однако в них дисперсия среднего числа детей в семь
превышает это значение незначительно, а в с.Дурипш отмечй|
снижение рождаемости (менее 2 детей на брачную пару). Из данш
табл. 11 видно, что у абхазов объем репродуктивной группы Л1
составляет от 33 до 51% от общей численности. Эта величиг
минимальна в популяции Дурипш и максимальна • в Члоу и Хуан
Также как и у башкир эффективно-репродуктивный объем сниже
что объясняется уменьшением доли младших возрастных труп
дефицитом лиц в возрасте 20 39 лет, связанный с некоторы
снижением рождаемости за последние десятилетия, а также росте
миграции сельской молодежи в город. К важной особенное
абхазских семей относится сравнительное постоянство в них чиа
детей с весьма незначительными вариациями, что служ
благоприятным фактором устойчивости воспроизводства популяш
в поколениях.
Примером популяций, в которых действует система планирован!
размеров семей и применяется практика искусственного контрол
рождаемости являются греческие и албанские в Приазовы
Приведенные выше в главе 4.1 возрастные пирамиды резк
деформированы за счет снижения дорепродуктивной возрастно
группы. Об этом же свидетельствует низкое значение среднего числ
детей, приходящееся на одну женщину - от 1,78 (пос.Староласпа), д
2,36 (пос.Георгиевка). Почти во всех группах дисперсия средней ниж
самого значения среднего числа детей. Столь малое отклонени
также является подтверждением стабильности средней, но в данно!
случае это скорее показатель устойчивости неблагоприятно
демографической ситуации, когда популяция не полность»
воспроизводит себя в новом поколении, либо (п.Красная Полян<
Георгиевка) приближается к простому воспроизводству. Поскольк
различия в дифференциальной плодовитости индивидов оказание
минимальными, то рассчет эффективно-репродуктивного объем
был произведен нами с учетом возросшего в данном случае вклал
неравенства численности лиц разного пола (Wright, 1938).
сравнении с башкирскими популяциями пострепродуктивный объе
велик, колеблется от 38-39% и практически равен репродуктивном;
приводя к эффекту "старения" популяции.
82
| ц целом характеризуя структуру популяционной системы
I -.сточных башкир, выделить следующие основные моменты:
Микропопуляции, образующие систему, конечны, величина их
колеблется от 372 до 810 человек; гармонические средние общих
объемов (HNe) в субпопуляциях достигают 420-498 человек.
Репродуктивный объем (Nr) составляет 21-28% от общего размера
I микропопуляций; средние величины его между субпопуляциями
равны 103-125 человек.
Эффективная численность по популяции в целом невелика, HNe =
J 105 человек.
Дифференциальная плодовитость, как одно из проявлений
нерегулируемой рождаемости, является фактором, существенно
снижающим эффективно-репродуктивный объем популяции.
Возрастная структура популяции представлена индивидуумами
трех поколений. которые значительно хронологически
перекрываются. Средняя продолжительность поколения несколько
выше 25 лет, обычно принимаемых по данным литературы за средний
В возраст популяции.
Анализ динамики численности микропопуляций за последние 1ОО
I лет выявил устойчивость популяционной системы в пространстве и во
I времени.
ГЛАВА 5. ГЕНЕТИЧЕСКАЯ СТРУКТУРА
БАШКИРСКОЙ ПОПУЛЯЦИИ
Наследственный полиморфизм популяции по
Данным о группах крови. Биохимические маркеры генов и
другие формы генетической изменчивости.
I l .
"етическая структура башкирской популяции исследована на
Новании собранного нами обширного нового материала по
Ропределению генетических маркеров среди основных
^Гн°гРафических групп башкир (Рафиков, Юмагужина, 1974;
>МагУ*ина, 1977; Рафиков, Юмагужина, Кузеев, 1977; Юмагужина,
83
Рафиков, Кузеев, Чекмин, 1978; Рафиков, Кузеев, Юмагужина, 1<1
Рафиков, Белова, Юмагужина, Кузеев, 1981). Положенный в осн
работы принцип исторической преемственности развития популя)
выразился в комплексном историко-генетическом подходе к airvii
структуры изучаемых групп.
В таблицах 12-29 представлены распределения частот генов
локусам АВО, MN, Rh-Hr, Kell, сывороточному белку гаптоглобин
микропопуляциях башкир, расположенных в соответствии
восстановленной работами историков бывшей родо-племенн
организацией предков коренных жителей данных селений.
Как видно из табл. 12 выборка из башкирского рода яик-суби-м
характеризуется следующим распределением частот генов гр
локусов: группы крови системы АВО - r>p>q; система MN-M>
гаптоглобин - Нр > Нр‘. В табл. 13 (род миркат-мин) распределен
частот генов в селах Казбуруново и Турсугали имеет вид r>q>p
r>p>q в Мурадымово и Турумбетово. По системе MN пропори
аллелей во всех микропопуляциях весьма сходные: М = 0,6692-0,68
и N = O,3102-0,3300. У жителей села Турумбетово обнаруж!
относительно низкий уровень аллеля rh (11,4%) и СС (0,4%) i
системе Rh-Hr. Частота аллеля К варьирует от 1,87 до 3, 98%.
В Аскинском районе, расположенном в северо-западной час
Башкирии, было обследовано население сел Мутабаш, Мута Ел г
Казанчи и в Татышлинском районе - Старый Кайпан и Буль-Кайпь
(принадлежавшие в прошлом к родовым группам казанчи и кайпан
Как видно из табл. 14 при относительном сопоставлении с другиь
группами башкир у кайпан по системе АВО преобладают частот
аллелей г и q и высокая частота гена Нр. В микропопуляциях ро;
казанчи по системе АВО распределение аллелей имеет вид г>р>
Здесь отмечена также относительно высокая частота аллеля Нр и г!
В таблицах с 15 по 22 представлены распределения часп
фенотипов и генов по локусам групп крови АВО, MN, Rh-Hr, Кг
белка гаптоглобина в микропопуляциях башкирских родов: бише:
мунаш, усерган, ямаш, бурзян без родовой принадлежности, а такж
тамьян, кара-табын, карагай-кипчаков. Суммарные сведения и
вышеперечисленным родам даны в табл. 23, 24.
В таблице 25 приведены суммарные частоты фенотипов и генов п
локусам АВО, MN, Rh-Hr, Kell и белка гаптоглобина и рассчитан)
84
критерии однородностей внутри башкирских племен бурзян, усерган,
к чрагай-кипчаков, тамьян, табын. сальют.
Как видно из таблицы 25 в субпопуляциях частота гена г локуса
дВ() отличается значительной гомогенностью и колеблется в
пределах от 0,5402 до 0,5941. Напротив, наблюдается значительное
колебание частот фенотипов Л и В и генов р и q по отдельным
группам; распределение r>p>q отмечено внутри племен усерган,
табын, сальют и r>q>p - внутри племен бурзян, карагай-кипчаков и
тамьян, таким образом, отмечается большая "монголоидность"
второй совокупности племен.
Сравнение по локусу MN выявляет наиболее характерное для
субпопуляций распределение типа M>N, и лишь у табын
наблюдается N>M.
Распределение частот фенотипов и генов белка гаптоглобина
сходно для всех сравниваемых субпопуляций и частота гена Нр
соответствует виду Нр > Нр‘.
Сравнение субпопуляций по системе Rh-Hr выявило
приблизительно одинаковую для них картину распределения
фенотипов и генов:
Наибольшая частота резус отрицательных фенотипов и
соответственно частота гена d = O,3333 наблюдается в племени
табын, а наименьшая величина - в племени сальют, где d = O,1218.
Легко видеть, что племя сальют характеризуется весьма малой
пропорцией резус отрицательных фенотипов, что в целом
свойственно коренному азиатскому населению, однако, по системе
АВО представители этого племени сближаются с европейскими
группами.
По системе Келл-Челлано, как и следовало ожидать, отмечается
Распределение к>К; наибольшая частота антигена К=О,О164
наблюдается в субпопуляции бурзян и наименьшая К =0,0051 у
1:1мьян. К сожалению, мы не располагаем данными по этой системе
Аля племени сальют.
В таблицах 23-24 представлены результаты сопоставления на
Однородность микропопуляций в пределах субпопуляций. Нулевая
гипотеза об однородности составляющих микропопуляций
пР°верялась сравнением значений тг с табличными при р=О,О5;
'исло степеней свободы при этом равно произведению числа
85
независимых параметров в локусе на уменьшенное на единицу чиа
серий наблюдений (Ниль и Шэлл, 1958).
Изучение однородности микропопуляций на разных иерархическ
уровнях популяционной системы восточных башкир выяви]
значительное колебание ее по генетическим системам и увеличен)
гетерогенности по мере усложнения структуры. Так, внутри род
наблюдается наибольшая однородность микропопуляций.
Как видно из таблиц 23-24 наблюдается однородное
микропопуляций у бурзян без родового наименования и муна.
Выявляется гетерогенность по системе групп крови АВО у карага
кипчаков (хг= 24,37, d.f. = 8) и рода ямаш (х‘= 13,87, d.f. = 8). Д
микропопуляций рода бишей гетерогенность обнаружена по локу
АВО (х’ = 15,53, d.f. = 4) и MN (х2 = 11,24, d.f. = 2); групп;
включающиеся в родтамьян, неоднородны по антигенам Е-е систем
Rh-Hr (х; = 14,83, d.f. = 2). Наибольшая гетерогенность установле
между микропопуляциями рода кара-табын по локусу АВО
антигенам системы резус.
Анализ однородности на уровне племен выявил значительную
гетерогенность. Как видно из таблицы 25, в племени бурзг
включающего роды ямаш, мунаш и бурзян без родов,
принадлежности отмечается неоднородность микропопуляций 1
локусу АВО (х = 14,55, d.f. = 4), Rh-Hr системы резус (xz= 21,87). Д
усерган, включающих роды бишей и усерган, также характер
гетерогенность по системе АВО (х = 20,78, d.f. = 2); в отличие
бурзян между ними наблюдается однородность по системе резус,
отсутствует по MN (хГ= 17,53, d.f. = 7) и гаптоглобину (х~= 27,19, <
= 7). Для Зауральских башкир Челябинской области, представленн
племенем сальют (табл. 26), характерна неоднородность по систе
групп крови АВО (х^= 15,11, d.f. = 2) и антигена (Е-е) системы ре:
(х"= 9,28, d.f. = 1). Однородность микропопуляций внутри плем
тамьян (род тамьян), табын (род кара-табын) и карагай-кипчак;
(род карагай-кипчаки) обсуждена выше.
В системе микропопуляций восточных башкир наблюдаез
гетерогенность по всем локусам, кроме локуса гаптоглобина
системы Келл-Челлано.
В таблице 27 приведены основные параметры распределен
фенотипов и частот генов групп крови системы АВО, MN, Rh-I
Келл-Челлано и белка гаптоглобина в популяции восточных башк.
86
3
Л
Фенотипы, распределение частот генов и гетерогенность популяций по
локусам групп крови АВО, MN, белку гаптоглобину (род яик-суби-мин).
к
$
2 2
52
X X
ан zj
О < со •? z 5 2
и
87
Таблица 13.
Фенотипы, распределение частот генов и гетерогенность микропопуляций по локусам систем групп
крови АВО, MN, Rh-Hr, Kell и белку гаптоглобину (род миркат-мин).
Фенотипы Казбуруново Мурадымово Турсугали Турумбетово
No частота генов No частота генов No частота генов No частота генов
1 2 3 4 5 6 7 8 9
О 27 г=0,4932 36 г=0,5071 30 52 г=0,5774
А 33 р=0,2419 52 р=0,2857 17 г=0,6411 34 р=0,2651
В 39 q=0,2649 40 Ч=0,2072 21 р=0,1613 52 4 = 0,2575
АВ 12 12 5 4=0,1976 18
X 111 140 73 156
NN 12 11 8 8
MN 43 N=0,3102 39 N=0,3112 29 N=0,3215 31 N =0,3308
ММ 53 М=0,6898 48 М=0,6888 33 М=0,6785 32 М=0,6692
I 108 98 70 71
Нр 1-1 6 8 12 5 1
Нр2-1 46 Нр1=0,3223 42 Нр1=0,3671 47 Нр =0,4329 40 Нр =0,3472
Нр2-2 38 Нр^=0,6777 29 Нр2^ 0,6329 23 Нр2=0,5671 27 Нрг=0,6523
1 К...._ 90 79 82 72
Продолжение таблицы 13.
1 । ~ j 3 4 5 . 6 7 : 8 i 9 i
DD Dd i D=0,6939 63 0,7289 70 D=0,7417 76 i D-0 886
dd X 6 64 d=0,3061 ; 5 68 0,2711 5 75 d=0,2583 1 77 d = 0,1140
сс Сс 14 28 C=0,4138 23 25 C=0,4577 21 27 C=0,4000 17 50 C=0 6396
СС 5 22 64 c=0,5862 20 68 c=0,5423 27 75 c=0,6000 10 77 c=0,3604 1
ЕЕ Ее 2 14 E=0,1340 2 18 E=0,1598 8 18 E=0,1917 1 21 E=0,1548 e = 0,8452
ее Z 48 64 e=0.8660 48 68 e=0.8402 49 ' 75 e = 0.8083 55 77
кк 5 4 7
Кк кк Е । L. 59 64 K=0,0398 к=0,9602 60 64 K=0,0317 к=0,9683 52 54 K=0,0187 k = 0,9813 74 79 К=0,0322 к=0,9678
Таблица 14.
Фенотипы, распределение частот генов и гетерогенность микропопуляций по локусам групп крови АВО,
MN, Rh-Нг, Kell и белку гаптоглобину (племя танып).
Фено- типы Род кайпан Род казанчи нов
Кайпаново Буль-Кайпаново Мутабаш Мута-Елга Казанчи
No частота генов No частота генов No частота генов No частота генов No ’ частота ге
1 2 3 4 5 6 7 8 9 10 11
о 75 37 30 31 52 г=0,5435
А 82 г=0,5213 40 г=О,51О5 25 г=5296 34 г=0,554 56 р=0,2398
В 86 р=0,2329 44 р-0,2259 40 р=1873 26 р=0,2483 64 q=0,2167
АВ 33 q=0,2458 21 q=0,2636 12 q=0,2831 10 q=0,1977 4
£ 276 142 107 101 176
NN 43 14 38 25 51
MN 88 N=0,3937 35 N =0,2532 . 46 N=0,5596 42 N=0,4646 79 N=0,5143
ММ 90 М =0,6063 73 М=0,7418 25 М =0.4404 32 М=0,5353 46 М=0,4857
£ 221 122 109 99 176
Как видно из таблицы 27, частоты генов в популяции восточг
башкир практически не отличаются от средних частот генов
микропопуляциям, составляющим эту совокупность; наблюдаюша
дисперсия, как мера уклонений от средних значений, минимальна.
Таблица
Частоты фенотипов и генов по локусам АВО, MN, Rh-Hr, Kell и
белка гаптоглобина в микропопуляциях рода бишей.
Фенотипы Байдавлетово Казарма
No частота генов No I частота генов
О А В АВ NN MN ММ X Нр 1-1 Нр2-1 Нр 2-2 S DD ! Dd СС Сс СС V ЕЕ Ее ее X КК kk 40 г=,5569 55 р=,3012 30 q=,1419 4 129 х=6,6283 18 N=,1591 6 М=,8409 108 132 £=91,00 9 Н[3=,3898 35 Нр=,6102 24 г 68 х=0,45 50 D=,7278 4 d=,2722 54 г-о,о 12 С=,5094 29 с=,4906 13 54 х= 0,4390 1 Е=,2799 25 е-,7201 28 г 54 х=2,9448 1 К=,0105 47. к=,9895 48 х=0,0 18 г—,4140 48 р=,3788 35 q=,2072 4 105 х= 16,368 8 N=,2500 24 М=,7500 48 80 4=3,20 5 Нр‘=,2791 14 Нр=,7209 24 г 43 Х=1,64 40 D=,7818 2 d=,2!82 42 £=0,0 10 С=,4882 21 с=,5118 11 - 42 х-0,0 1 ' Е=,1982 14 е=,8018 27 42 х=0,2883 К-0,0 43 к-1,0 43 £о,О
Ибраево
No [ частота
36
54
60
8
158
12
34
56
102
4
34
29
67
59
4
63
13
35
15
63
1
31
31
63
52
52
г=,4772
р=,2774
q=,2454
2
х= 12,39
N=,2843
М-,715'
х=3,34
Нр=,31;
Нр=,6Ж
х=2,16
D=,7480
d=,2520
£=0,0
С=,5120
с=,4880
£•1,14
Е=,2985
е=,7015
х=4,5981
К=0,0
к=1,0
£=0,0
92
Таблица 16.
Частоты фенотипов и генов по локусам ЛВО, MN, Rh-Hr, Kell и
ч 1ка гаптоглобина в микропопуляциях рода мунаш.
Г
А
В
АВ
Inn
Imn
мм
Нр 1-1
Нр2-1
Нр 2-2
pb
с.
сс
г
сс
I Е
Ее
с
Г*
Кк
к
Басаеве Абзаково Туркменево И Ахмерове
No Г । частота | генов No частота генов No частота генов No частота генов
19 г = ,484 34 г = ,589 66 г»,576 40 г=,611
23 р=,236 20 р = ,178 50 р = ,188 22 р=,150
2(1 q = ,280 34 q-,233 55 q = ,236 30 q = ,239
19 10 28 15
81 х =8,92 98 х = 1,12 199 x'-7,70 107 х?=8,50
9 N=,266 6 N-,211 8 N-,231 11 N=,282
7 М=,734 12 М = ,789 14 М=,769 9 М = ,718
31 39 43 35
47 х =14,0 57 х =8,89 65 х> 10,0 55 х = 19,5
5 Нр=,395 5 Нр = ,295 9 Нр-,367 14 Нр»,441|
20 Нр=,6О5 16 Нр-,705 15 Нр=,633 24 Нр:=,555
13 23 21 21
38 X =0,39 44 X =0,70 45 2 х-Х5« 59 9 х-1,78
40 D = ,639 37 D = ,688 39 D=,695 53 D = ,7O6
6 d=,361 4 d=,312 4 d=,3O5 5 d=,294
46 хг=0,0 4) х=0,0 43 >С=0,0 58 хг=0,0
13 С»,511 10 С-,506 12 С-,459 18 С=,492
22 с=,489 21 с-,494 17 с=,541 25 с-,508
11 10 12 15
46 х?=0,16 41 хг=0,04 41 хг»1,86 58 хг=1,68
2 Е=,174 — Е-,219 3 Е=,159 1 Е-,212
13 е=,826 16 е-,781 9 е»,841 21 е-,788
1 3> 25 29 36
1 46 х=0,20 41 х^2,11 41 х-3,72 58 х^-1,28
2 К=,022 — К=,000 1 К=,012 3 К-,027
44 к=,978 41 к= 1,00 40 к = ,988 53- к-,973
46 1 Л xt=0,0 41 х’*0,0 . 41 х^-0,0 56 хЗ-о,о
93
Таблица 17.’
Частоты фенотипов и генов по локусам АВО, MN, Rh-Hr, Kell и
белка гаптоглобина в микропопуляциях рода усерган.
Фено- типы Идияшево Утягулово Байте во Сагитово
No частота генов No частота генов No частота генов No
часто генов
о 44 г=,553 26 г-,532 92 г=,568 30 г=,559
А 36 р=,193 26 р=,22О 90 р=,230 33 р=,25;
В 52 q=,254 34 q=,248 94 q=,202 29 q=,19C
АВ 12 6 10 4
X 144 х~=0,41 92 XZ=2,18 286 хг=13,9 96 х=4,0
NN 9 N=,286 8 N=,293 9 N=,289 8 N=,22
MN 18 М=,714 32 М=,707 48 М=,711 28 М=,77
ММ 36 42 57 60
£ 63 тс =5,61 82 хМ.16 114 х^0,015 96 2- А Л/ x=2,9J
Нр 1-1 13, Нр=,365 27 Н{/=,516 18 Нр=,352 18 Нр>,5
Нр 2-1 36 Нр=,635 42 Нр=,484 45 Нр=,648 14 Нр=,3
Нр 2-2 36 24 52 18
£ 85 х-,63 93 ТС=,&1 115 х-=1,20 50 х=9,6
DD . 56 D=,814 55 D=,813 73 D=,704 48 D=,72i
dd 2 d=,l«6 2 d«,187 7 d=,296 4 d=,271
£ 58 х=0,0 57 Х'=0,0 80 х=0,0 52 х=0,0
СС 12 С=,606 11 С=,5О4 14 С=,553 11 С=,53
Сс 37 с=,394 32 с=,496 50 с=,447 29 *с =, 48G
СС 9 14 16 12
Е 58 %г«7,18 57 хг=1,27 80 х=1,22 52 хЛ= 1,0
ЕЕ 1 Е=,234 1 Е=,262 1 Е-,225 — E-.2J
Ее 23 е=,766 25 е=,738 31 е-,775 21 е=,77^
ее 34 31 48 31
£ 58 <х'-7,18 57 х-1,27 80 х=7,22 52 x-=l,0i
кк 4 К=,030 1 К=,011 3 К=,017 2 к=о,а
kk 64 к=,970 47 к=,989 85 к=,983 62 к=,90|
с — 68 х=0,0 48 х^«0,0 88 xZ=0,0 64 х:=0Д
94
Таблица 18.
Частоты фенотипов и генов по локусам АВО, MN, Rh-Hr, Kell и
белка гаптоглобина в микропопуляциях рода ямаш.
'фенотипы ‘ Чингизово Буранбаево Гадельбаево
No частота генов No частота генов No частота генов
1° 46 г-0,5631 27 г-0,5145 42 г-0,6547
А 39 р-0,2025 37 р-0,2776 18 р-0,1277
|в 45 q-0,2344 27 q-0,2079 м q-0,2176
। АВ 15 11 4
2k 145 х-0,144 102 Х--0.05 98 Х-0,44
NN 9 N-0,2178 1 N-0,1950 4 N-0,2027
MN 26 М-0,7822 37 М-0,8050 22 М-0,797’
ММ 66 62 48
101 х-6,04 100 х-3,18 74 X2—0,47
Нр 1-1 23 Нр-0,4421 12 Нр-0,3830 8 Нр-0,4468
Нр2-1 61 Нр-0,5579 48 Нр-0,6170 26 Нр-0,5532
Ир 2-2 37 34 13
121 х-0,06 94 к*-0,61 47 Х^-О.б?
D1? 142 D-0,8156 98 D-0,8277 50 D-0,8600
dd 5 d=0,1844 3 d-0,1723 1 d-0,1400
147 <-0,0 101 хг-о,о 51 Х1-0,0
95
§
Таблица 19.
Частоты фенотипов и генов по локусам АВО, MN, резус и белка гаптоглобина в микропопуляциях
бурзян без родовой принадлежности.
1 Фено I типы 1 Кмекбаеэо Миндегулово Н-Субхангулово Гадельгареево
No частота генов No частота генов No частота генов No 1 частота генов :
1 1 2 3 4 5 6 7 8 9
|о 1А !в : ав I 2 INN 1 MN ММ X Нр 1-1 |Нр2-1 |Нр2-2 DD dd 1 Z 17 r=,5718 8 p=,1216 25 q=,3066 *» 52 x'=,1,083 3 N=,2500 9 M=,75OO 18 30 х“=1Д 4 Hp*=,4O32 17 Hp"=^968 10 31 хг=0Л9 30 D=,7500 2 d=,2500 32 хг=0,0 14 г=,5992 6 р=,1169 14 q=0839 5 39 Х=2,57 2 N = ,2500 6 М-,7500 12 20 х=0,80 3 Нр1=,3333 12 Hpi=,6667 12 27 х~=0,0 30 D=,7500 2 d=,2500 32 xZ=0,0 32 г=,6100 22 р=,1818 26 q=,2O82 6 86 хг=0,04 8 N=3033 21 М = ,6967 32 61 хг«2,09 5 Нр‘=,2955 29 Нр2=,7045 I 32 66 хг=0^0 52 D=,7665 3 d=,2335 i 55 х^О.О 42 г=.6124 16 р=,1072 44 q = .28O4 10 112 Хг=3,7 6 N=,2321 27 М=,7668 51 84 хг=0.81 4 Нр1=,3077 24 Нрг=,6923 24 52 х‘=0,37 29 D=,8175 1 d = ,1825 30 х=00
2 2 z
о
2 z
я
з: £
II
£
г
г
к
2
_ ос о ое
2 §
m *
< а
I?
2 z
» ।
8 ©
8
Й О' О 8о £ О'
о s &
Е
X I *
3
г
о
S i
S
г
2
£
CD
О
к
Табл и
Частоты фенотипов и генов по локусам АВО, MN, Rh-Hr и белка
гаптоглобина в микропопуляциях рода кара-табын
Фенотипы Расулево Ишкинево
No частота No частота
генов генов
О 52 г-,5718 62 г-.6225
А 54 р-,2447 49 р-,2104
В 45 q-,1835 31 q-,1671
АВ 8 18
Л 159 Х-3,72 160 х?-5,25
NN 32 N-,5676 21 N-,4667
MN 62 М-,4324 42 М-,5333
ММ 17 27
Е 111 х’-2,11 90 х-0,35
Нр 1-1 9 Нр=,3431 9 Нр'-,4156
Нр2-1 52 Нр-,6569 46 Нр=,5844
Нр 2-2 41 22
Е 102 X -1,75 77 х=4,06
DD 45 D-,6332 58 D-.8735
dd 7 d = ,3668 1 d-,1265
X 52 хг-о,о 59 хг-о,о
СС 7 С-,5402 7 С»,4958
Сс 34 с-,4598 37 с-,5042
СС И 15
г 52 хг-6,98 59 хг-5,79
ЕЕ 11 Е-,3066 — Е-,1563
Ее 16 е-,6934 17 е-,8437
ее 25 42
2 52 хг-9,32 59 хг-о,13
К.К 2 К-,0194 2 К»,0170
кк 50 к=,9806 57 к=,9830
52 Х^-О.О 59 хг-0,0
Истамгулово
No часто
75
46
52
20
193
31
52
28
111
8
48
46
102
86
генов
г -.623
р-,16«
q = ,208
2
х-3,8
N-,45'
М=,54
8 d=,291
94 а1-0,0
18 С-,531
56 с-,461
20
94 х^=5,0
Е-,214
36 с -,785
58
94 хг-(>,51
К=,0
94 к-1,(
94 xS-0,1
98
Таблица 2_
Частоты фенотипов и генов по локусам АВО, MN, Rh-Hr, Kell и
белка гаптоглобина в микропопуляциях рода карагай-кипчаков
фено- ТИПЫ О А В *? NN MN ММ Нр 1-1 Нр 2-1 Нр2 2 2. DD dd X сс Сс сс X ее Ее ее I КК kk Н.Мунасипово Байнаэарово 1 (абиево Яуибаево Кипчак
No частота генов No частота генов No 1 частота генов No частота генов No частота генов
38 г=,59 20 р=,14 40 q=,26 8 106 х =0,0 8 N=,41 34 М=,58 18 60 тс = 1,6 6 Нр‘=,43 28 Нр=,56 12 „ 46 х=2,5 40 D-.78 2 42 х=0,0 10 С-,51 22 с=,488 10 2. 42 х=0,14 2 Е-,213 14 е-,787 26 42 х=0,005 К-,000 42 к =1,00 42 /-0,0 58 г-,57 40 р=,17 70 q-,25 6 174 7=5,4 6 N=,37 72 М-,62 34 112 х”=15,4 15 Нр'=,41 48 Нр=,58 30 93 хг=О,33 58 D=,694 6 d=,306 64 xL=0,0 17 С=,500 31 с=,500 16 64 /-0,09 3 Е-,221 22 е-,779 39 64 7=0,004 1 К=,008 63 к-,992 64 /-0,0 26 г-,491 28 р—,216 31 q=,293 14 108 х-0,00 20 N-,34 11 М=,65 42 73 7=32,6 6 Нр-,38 30 Нр=,61 18 54 Х2-1,54 68 D-,715 6 d=,285 74 7=0,0 20 С-,507 36 с-,493 18 74 хг«0,08 4 Е-,229 26 е-,771 44 74 /-0,04 2 К-,014 72 к-,986 74 /-0,0 16 г-,477 18 р-,209 31 q—,314 5 70 7-2,3 4 N-,36 20 М-,63 35 z 59 х-4,61 И Нр^-,44 30 Нр-,55 17 58 х-0,12 59 D-.720 5 d-,280 64 xL-0,0 17 С-,516 32 с-,484 15 z 64 х-0,001 4 Е-,240 23 е-,760 37 64 7=0,08 2 К-,016 62 к=,984 64 7-0,0 48 г-.679 30 р=,187 18 q = ,134 8 104 7-1,89 6 N = ,319 34 М-,681 32 , 72 х=0,63 4 Нр‘=,407 36 Нр-,593 14 54 х2-7,83 62 D-,754 4 d-,256 66 х”-0,0 16 С-,508 34 с—,492 16 66 х2=0,51 4 Е-,241 24 е-,759 38 66 xZ=3,37 4 К=,031 62 к=,969 66 7=0,0
99
Таблица
Частоты фенотипов и генов по локусам АВО, MN, Rh-Hr, Kell бел
гаптоглобина по родам бишей, мунаш, усерган, карагай-кипчакам
Фено- типы Карагай- кипчаки Бишей Мунаш Усерган
No частота генов No частота генов No частота генов No частота генов
о 186 г-,575 94 г=,490 159 г-,573 192 г-,557
А 136 р-,182 157 р=,зю 115 р-,179 185 р=,224
В 199 q = ,243 125 q-,200 139 q=,248 209 q = ,219
АВ К 41 562 2. *=24,47 16 392 X-15,53 72 495 х-5,21 32 618 х-8,63
NN 44 N=,344 38 N=,223 34 N-.246 34 N=,273
MN 171 М=,656 64 М=,777 42 М=,754 126 М=,727
ММ £ 161 376 x’-lO.l 212 314 те=Н,2 148 224 х>0,18 195 355 х-2,44
Нр 1-1 42 Нр=,420 18 Нр = ,334 33 Нр=,379 76 Нр=,421
Нр2-1 172 Нр=,580 83 HfT=,666 75 Нр=,621 137 Нр=,579
Нр 2-2 £ 91 305 тЗ» 0,972 77 178 4= 3,36 78 186 Х=4,60 130 343 <=16,2
DD 287 D-,728 149 D=,749 169 D-.682 232 D«,754
dd 23 d-,272 10 d=,251 19 d=,318 15 d=,246
Е 310 х -2,05 159 х -0,56 188 х =1,21 247 х =5,22
СС 80 С»,508 35 С-,505 53 С-,492 48 С = ,546
Сс 155 с»,492 85 с=,495 85 с=,508 148 с=,454
СС 75 39 48 51
Z 310 % -0,16 159 х=0,01 186 х=0,18 247 Х = 0,69
ЕЕ 17 Е=,23О 3 Е=,264 6 Е-0,193 3 Е=,236
Ее 109 е=,770 70 е=,73<> 59 е=,807 100 е = ,764
ее 184 86 212 144
2 310 х-0,36 159 х = 1,03 186 х=0,18 247 х=0,56
кк 9 К=,015 1 К = ,004 6 К=,016 10 К=,019
kk Z 301 310 к «,985 э?»5,17 142 143 к=,996 Л =0,0 178 184 к=,984 х-1,15 258 268 к=,981 х=2,4
100
Таблица 24.
Частоты фенотипов и генов по локусам АВО, MN, Rh-Hr, Kell и
белка гаптоглобина по родам: ямаш, тамьян, кара-табын и бурзянам
без родовой принадлежности
Фено- типы Ямаш Тамьян Кара-табын Бурзяне б/р
No частота генов No частота генов No частота генов No частота генов
О А В АВ NN MN ММ 2 Нр 1-1 Нр2-1 Нр2-2 DD dd Е СС Сс сс S ЕЕ Ее ее КК kk 115 г-,577 94 р=,201 106 q=,222 30 345 к = 13,8 14 N = ,205 85 М=,795 176 275 х’=,475 43 Нр=,422 135 Нрг=,578 84 262 -хЫ,83 290 D=,827 9 d=,173 299 х=0 ” 132 г=,594 78 р-,155 125 q»,251 39 374 х=»5,32 11 N-,143 49 М=,857 188 248 х^=4,06 21 Нр=,349 90 Нр=,651 78 189 х'=0,24 207 D=,74O 15 d=,260 222 х“=0,0 85 С=,514 84 с=,486 53 г 222 х = 19,0 21 Е=,203 60 е-,797 141 г 222 х-17,3 2 К=,005 195 к=.995 197 х=0,0 189 г=,608 149 р-,205 128 q-,187 46 512 х=8,23 84 N-.519 156 М=,481 72 312 х=2,81 26 Нр=,352 146 Нр=,648 109 281 х=4,12 189 D-.721 16 d«,279 205 Х^-О.О 32 С-,526 127 с-,474 46 г 205 Х-16,8 11 Е=,219 69 е-,781 125 г 205 х-0,15 4 К-,010 201 к=,990 205 xZ=0,0 105 г=,603 52 р=,134 109 q=,263 23 289 хг=Ю,1 19 N=,259 63 М-,741 113 г 195 х-1,98 16 Нр=,324 82 Нр=,676 78 г 176 *=2,44 141 D=,768 8 d=,232 149 Хг=0,0
101
Таблица 25.
Частоты фено i ипов и генов по локусам АВО, MN, Rh-Нг, Kell, белка гаптоглобина в племенах бурзян,
усерган, карагай-кипчаков, тамьян, табын и сальют
J 02
‘ Фенотипы ... ... Бурзян Усерган Карагай-кипчаки Тамьян Табын Сальют
No частота генов No частота генов No частота генов No частота генов No частота генов No частота генов
1 1 1 2 3 4 5 6 7 8 9 10 11 12 13
' О 379 г-.5585 286 г-3322 186 г-,5753 132 г-3941 216 г=3869 45 г = 3042
А 261 р=,1977 342 р=,2563 136 р—,1816 78 р—,1552 201 р=,2286 67 р=,2913
В 354 ц=Д438 334 q=,2115 199 q-,2431 125 q=,2507 158 q—,1845 40 q = ,2045 1
хв 125 48 41 39 52 25 -
I 1119 Х~= 14,55 1010 х== 20,78 562 X-24Д8 374 х=5,32 627 х = 28,38 177 X» 15.11
1 NN 67 N-.2334 72 N-.2496 44 N=,3444 11 N-,1432 114 N=,5189 20 N-.3823
1 MN 190 М=,7666 190 М-,7504 171 М-,6556 49 М—,8568 212 М-,4811 96 М-,6158
ММ 437 407 161 х’-10,09 188 98 61
д 694 х“-3,94 669 £•1733 376 248 хг=4,06 424 хг=2,91 177 X =0.126
Нр 1-1 92 НРу3813 94 Нр=,3916 42 Нр-,4197 21 Нр=,3492 38 Нр=,3690 10 Нр>,3437
Нр2-1 292 Нр“=,б186 220 Нр—,6084 172 Нр=,58ОЗ 90 Нр=,6508 203 Нр=.631О 68 Нр=,6563
Нр2-2 240 z 207 z 91 •> 78 137 50
Е 624 х-8,47 521 X =27,19 305 Х~=0.9721 189 X =0,24 378 х“=7.1638 128 х =0.015
DD 600 D-,7621 381 D-.7518 287 D—0,7276 207 D=,7400 280 D=,6667 134 D = ,8788
Jd 36 d=,2379 25 d=,2482 23 d=3724 15 d=,2600 35 d=3333 7 d=,1218
1 - 636 хг= ,21,87 406 хг= 0,0 310 х=2,05 222 Х-0,63 315 х= 20,92 136 х= 0,241
Продолжение таблицы 25.
1 2 3 41 5 6 7 8 9 10 11 12 i 13
СС 53 С=,4920 83 С=3292 80 С=,5082 85 С=,5114 59 С-3353 41 С=3979
Сс 85 с=,5080 233 с=,4708 155 с=,4918 84 с=,4886 188 с=.4647 73 с=,4021
сс 48 90 75 53 68 22
Т 186 Х~=0,18 406 £—0,108 310 х*=0.16 222 £-1,13 315 х—2,78 136 5 X =0,054
ЕЕ 6 Е—,1934 6 Е=,2473 17 Е-,2296 21 Е-,2031 32 Е-,1973 5 Е-Д525
Ее 59 е=,8066 170 е=,7527 109 е = ,77О4 60 е=,7969 81 е=,8027 55 е=,7475
ее 121 230 184 141 203 76
X 186 Х-0,18 406 Х-0.461 310 £=0356 222 х = 14,83 315 2. х = 17.64 136 х=9,28
кк 6 К=,0164 И К-,0135 9 К-.0146 2 К=,0051 4 К-,0098
kk 178 к=.9836 400 к=,9865 301 к = ,9854 195 к=,9949 201 к=,9902
Z 184 тд=1,15 — 411 — £=2,42 310 £-5,17 197 Х=0 205 x*=6,51
- значения однородности внутри племени при р = 0,05.
Основные параметры распределения фенотипов и частот генов
групп крови и гаптоглобина в популяции восточных башкир
Таблица 21
г— Фенотипы No Fo «подразделенная Ne Совокупность ч восточных башкир г “ - подразделенная на микропопуляции q I | ~Т—
!О 1 А 1244 0.3215 1243.9 0.5670
i А 1085 0.2804 1085.3 0.2089
В 1210 0.3127 11773 03241
АВ 330 0.0853 362.14
А 3869 <=211,958
сл NN 328 0.1267 236.28 03022
M.N 908 0.3508 1091.36
ММ 1352 03224 1260,1 0.6978
2588 xS 365.1826
Нр 1-1 433 0.1816 406,47 0,4129
1 Нр 2-1 1103 0,4627 1155.8
Нр2-2 848 03557 821.77 03871
1 2384 <=61.1567
DD 1798 0.9389 17973 0,7528
1 i dd 117 0,0611 117,0 0,2472
— 1915 70,7765
0,5622 + 0,0210 0,2098 + 0,0239 03290 + 0,0169 0,0106 0.0122 0.0086 0.00361 0,00476 0,0086 0.0147 0.0287 0.0134
32 популяции 1 0,2960 + 0,0372
0,7040 + 0,0372 32 популяции 0,0190 0,0116 0,0558
0,3782 + 0,0212 0,0108 0,0036 0,0153
0,6218 + 0,0212 31 популяция 0,7522 + 0,0247 0,2478 + 0,0247 32 популяции 0.0126 0,0051 0,0274
108
Таблица 28.
1>.|Спределение групповых факторов крови системы резус у коренного
населения Башкирии
фенотип No Fo F N частота генов
сс 292 0,2739 0,2856 304,4496 С - 0,5344
Сс 543 0,5094 0,4976 530,4416 с - 0,4656
сс 231 0,2167 0,2168 231,1088 Х£- 0,8065
1066 1,0000 1,0000 1066,0000
Е1 23 0,0216 0,0222 23,6652 Е - 0,149
|Ее 271 0,2542 0,2536 270,3376 е - 0,851
ее 772 0,7242 0,7242 771,9972 X2- 0,0203
1066 1,0000 1,0000 1066,0000
Обозначения:
No наблюдаемое количество лиц с данным фенотипом;
1 о наблюдаемая частота фенотипа;
N ожидаемое количество лице данным фенотипом;
I ожидаемая частота фенотипа.
Таблица 29.
Распределение изоантигенов крови системы резус у коренного
Населения Башкирии
Фенотип No Fo F N Частота хромосом
CCDEE 0
CCDEc 1 0,0009 0,0009 0,9594 cde - 0,2495 - r
c'cDEE 0 0,0003 0,3198 Cde - 0,0469 R'
CcDEe 162 0,1520 0,1607 171,3062 cdE » 0,0094 - R"
ссЬце 101 0,0947 0,0773 82,4018 cDe » 0,0488 - R^
ccDEE 23 0,0216 0,0285 30,3810 CDe - 0,4756 - Rj
ccDee 356 0,3340 0,2209 235,4794 CDE = 0,0009 - Rz
107
Продолжение таблицы 29
CCDee 291 0,2730 0,2262 241,1292 cDE = 0,161
ccDee .14 0,0319 0,0244 26,0104
ccddee 66 0,0619 0,0623 66,4118 -X-256.395
Ccddee 25 0,0234 0,0234 24,9444
ccddEe 7 0,(Х)66 0,0165 17,589
Все (К-
тальные
возможные 0 — 0,1586 169,076
1066 1,0000 1,0000 1066,00
Исследование башкир по генетически обусловлена
диморфизму ушной серы по ее консистенции было проведено в се
Лесы (Белорецкий р-н) и Узунларово (Архангельский р-н) (Спин
1985). Так, на фоне имеющихся данных в литерат
подтверждающих ярко выраженную клинальную изменчивость ге
d и w с запада и юго-запада на восток и северо-восток, башк!
занимают промежуточное положение с частотами генов w=O,43O
Узунларово и 0,3406 в Ассы и d = O,5697 и 0,6594 соответственно.
Новые популяционно-генетические и антропологичеа
материалы были собраны в 1983 г. во время работы совмесп
советско-финнской экспедиции. У башкир с.АрхангельО
(Архангельский р-н), с.Стерлибашево (Стерлибашевский р-н)
с.Верхне-Яркеево (Илишевский р-н) было собрано в об|
сложности около 300 образцов крови, см. таблицу 30.
По системе АВО башкиры отличаются промежуточн
характеристиками в отношении распределения факторов АВО.
них свойственна относительно низкая частота гена г(о), достатс
высокая частота гена р. В то же время башкиры некотс
популяций отличаются высокой частотой гена q, что характерно
монголоидных популяций Центральной Азии, за исключе
популяции Верхне-Яркеево, где отмечена очень высокая частота
р, при малой частоте гена q. Система MN. Распределение факт
системы MN отличается стабильностью, и, как правило, час
аллеля М почти в 2 раза превышает концентрацию альтернатив
гена (N).
Система резус, характеризует башкир как группу метис
происхождения, где частота европеоидного фактора колеблез
пределах 22-23%. Концентрация гаплотипа CDe (Ri) сбли
I он
башкир с южными европеоидами, например, турками. Частота
гЛПлотипа cDE (R?) сближается с таковой у финноязычных групп
(финны, лопари и др.), и занимают промежуточное положение между
европеоидами и монголоидами. Анализ остальных гаплотипов
системы резус не представляет особого интереса, поскольку частота
их крайне мала.
Система Kell. По этой системе башкиры также обнаруживают
промежуточное положение между европеоидными и монголоидными
популяциями, поскольку частота фактора К у них хотя и в меньшей
степени, но существенно превышает "монголоидные значения".
Система Daffy. Частоты генов системы Daffy стабильны, и
изученные популяции не отличаются по распределению факторов
этой системы. При этом частота Fy немного превышает частоту
аллеля Fy.’ Ген Fy встречен у башкир с более высокой частотой
(0,55), чем у русских (0,50) и существенно выше, чем у северо-
азиатских монголоидов (0,23), но ниже, чем у марийцев (0,60).
Чистота Р. Концентрация гена р+ у башкир очень велика, и в
среднем составляет 82%, однако межпопуляционных различий не
установлено.
И ”/чена также система Lewis. Наиболее характерным фенотипом
которой является I.e (а- Ь-), частоты ее колеблются от 0,74 до 0,88 в
разных группах башкир.
Система супероксиддимутазы (SOD). Для большинства групп
населения земного шара эта система оказывается мономорфной.
। Мировой максимум редкого аллеля SOD 2 наблюдается у
финноязычных групп Скандинавии. Представляет интерес то
°бст()ятельство, что у башкир также был обнаружен редкий аллель
SOI) 2, что в какой-то мере может свидетельствовать, по мнению
I Финнских коллег, принимавших участие в исследовании этой
I п°иуляции, о включении финноугорского компонента в составе
ба,пкир. Для монголоидной расы является типичным наличие гена Di
СИстемы Диего (0,007), что является ниже, чем частота этого
а ' 'еля, ранее установленного для марийцев
L
109
Iаблица
Частоты генов в популяциях башкир Архангельское, Стерлибашев
Верхне-Яркеево (цит. по Raphikov et al., 1986)
Аллели Стерлибашево Архангельские Верхне-Яркеев<
Г 0,5693 0,6148 0,5693
р 0,2299 0,1919 0,2777
q 0,2008 0,1938 0,1340
m 0,6435 0,6603 0,6443
n 0,3564 0,3396 0,3557
D 0,7640 0,7621 0,7809
d 0,2356 0,2379 0,2191
c • 0,4814 0,4528 0,4856
c 0,5186 0,5471 0,5144
E 0,2731 0,2170 0,2644
e 0,7269 0,7830 0,7356
к 0,0377 0,000 0,0048
k 0,9623 1,000 0,9952
Fyu 0,5324 0,5425 0,5529
Fy° 0,4676 0,4575 0,4471
P+ 0,8149 0,8348 0,8302
P- 0,1851 0,1652 0,1698
Le (a + b-) 0,2653 0,1407 0,1118
Le (a* b-) 0,7447 0,8593 0,8882
SOD 1 0,9950 0,9854 1,000
SOD 2 0,0050 0,0146 O,(XX)
(у них 0,013), Так монголоидный маркер ген Di с наибольшей част
всгречен в с.Архангельском, что вдвое выше, чем в Верхне-Ярк<
и Стерлибашеве.
Башкиры имеют среднюю частоту гена Jk = 50%, что о
сближает их с русскими, финнами и другими европейс!
популяциями. Популяция Стерлибашево имеет значительно б
высокую частоту гена Jlc, чем марийцы и коми. Для сравнения еле
заметить, что ген ,1k встречается с высокой частотой (около 70'
всех африканских популяций, с низкой частотой (около 30?
кит <йцев в целом (Mourant et al., 1976).
ПО
Гаптоглобин. Частота Нр'варьирует от 0,43 до 0,47, т.е. является
ВЬ1СОКОЙ частотой даже для смешанных групп в масштабе Европы.
Эритроцитарная кислая фосфатаза. Европеоидный ген С
эритроцитарной кислой фосфатазы колеблется от 0,025 в
Стерлибашево, до 0,038 в с.Архангельское.
Группоспецифический компонент Gc. В среднем по 3 районам
частота генов колеблется следующим образом: Gc'^ = 0,25; Gc1'5 =
0,49; Gc 2 - 0,26.
I г
Для сравнения в Европе это распределение выглядит: Gc = 0,14;
Gc = 0.55; Gc 2 = 0,27; в Японии: Gc = 0,49; Gc ‘ = 0,23; Gc 2 =
0,26 соответственно.
Как видим, башкиры занимают промежуточное положение,
наиболее существенные различия проявляются лишь по гену Gc .
Трансферрин. В суммарной выборке башкир из трех районов
имеется следующее распределение субтипов трансферрина: TfC1 =
0,77; Tf 2 = 0,16; Tf<c = 0,035; тЛм = 0,003. Популяция
Архангельское резко отличается от Верхнего Яркеева и
(терлибашева в сторону большей монголоидности по системе
трансферрина. Например, здесь Tf ген = 0,22, в Верхнем Яркеево
частота его достигает 0,13; Стерлибашево 0,14. Так же отличается и
распределение гена ТГчастота его в Архангельском равна 0,06;
Илишевском р-не 0,016 и в Стерлибашево - 0,010. (Raphikov H.S.
et al., 1986; R.Frants et al., 1986; Рафиков и др., 1985, 1987).
ГЛАВА 6. ОСОБЕННОСТИ ГЕНЕТИЧЕСКОЙ
ДИФФЕРЕНЦИАЦИИ ПОПУЛЯЦИЙ; ФАКТОРЫ,
СПОСОБСТВУЮЩИЕ ЕЕ ПОДРАЗДЕЛЕННОСТИ.
1. Степень генного разнообразия на различных
уровнях иерархической структуры.
Каждому из нас не составляет большого труда распознать друг
J'Pyra по внешнему облику, за исключением монозиготных
близнецов. Однако, как поступить в том случае, когда мы
нРедполагасм оценить степень сходства индивидов в пределах одной
III
расы, этнической группы, точнее тотальное население каждой из них,
между собой? Имеют ли микропопуляции свои характерные,
особенности, подобно тому, как они видны при физиономическом
сравнении людей в них? Далее, у исследователя антрополога или
этнографа возникают вопросы о величинах генетических различий
между родами в пределах расы и т.д.
Несомненный научный интерес представляют ответы на такие
вопросы. Продолжая эти рассуждения, остановимся на объективном,
хотя и тривиальном факте, который свидетельствует о том, чтх
каждый субъект отличает представителя, например, монголоидно!
расы от европейца или африканца. Однако, африканцу во
европейцы физиономически представляются очень сходными, ил|
почти сходными. Кажущаяся гомогенность каждой расы при
сопоставлении с бросающимися в глаза различиями между крупным!
расовыми группами, обусловлена историческими причинами, а такж
тем обстоятельством, что люди фиксируют в своем воображени!
наиболее яркие, "броские" признаки: цвет кожи, форма глазно:
щели, особенности волосяного покрова и ряд других. То есть, реч
идет о таксономически значимых признаках, которым придаете
особое и основное значение со стороны специалистов, имеющих дел*
с классической антропологией.
Нее эти вопросы имеют прямое отношение к проблеме оцено!
внутри- и межгрупповой изменчивости в популяциях человека
Решение этих вопросов на основании точных научных фактов и:
области популяционной генетики имеет большое социально*
значение в борьбе с наблюдающимися и поныне в некоторы]
регионах мира расовыми предубеждениями и предрассудками.
Все эти вопросы были поставлены и успешно решены таким!
известными эволюционистами-генетиками, как Ней (Nei, 1973)
Левонтин (Ixvontm, 1978).
Известно, что общую генетическую изменчивость можи
расчленить на компоненты, относящиеся к разным уровня
агрегации популяций. Гак, согласно Nei (1973), тотальное генн<
разнообразие (Нт) дифференцируется на составляющие Нс и Ds
где первая величина определяет внутрипопуляционное разнообрази
вторая - характеризует межпопуляционные различия. В общем случ<
генное разнообразие в тотальной популяции определяет
выражением Ну = 1 — 1у. Если в локусе существует п-аллелей
112
частота к-го аллеля в данной популяции суть Хк, то вероятность
идентичности двух случайно отобранных генов определяется как
1 ='^хгск иН = 1-1 соответственно. Вероятность идентичности-
ц служит мерилом генной изменчивости в популяции и носит
название гетерозиготности. Таким образом, генное разнообразие в
тотальной популяции можно выразить через Ну = Hg + DgT, где Hg
генное разнообразие внутри популяций, a Dgy - среднее генное
различие между субпопуляциями. Если абсолютная величина генной
дифференциации среди субпопуляций выражается через величину
|)ST, то генная дифференциация по отношению к тотальной
популяции может быть получена через Gjy = D$T/Hy . При
пониженных величинах Нс значения могут быть весьма
значительными. Параметр Ggy оказывается эквивалентным
райтовским FST и назван Неем коэффициентом генной
дифференциации. При подразделенности каждой субпопуляции на
некоторое число локальных групп Hj может быть
дифференцирована на генные различия внутри и между локальными
группами (Нс и Dy соответственно). Поэтому Hy=Hg+ D^y + Dgy.
•тот тип анализа может быть продолжен до любой степени
иерархической подразделенности.
Итак, генное разнообразие тотальной популяции (НуСможет быть
подразделено на генное разнообразие внутри субпопуляций (Нс) и
генное различие между субпопуляциями (DjT) и Ну = Hf + D^.. В
общем случае Ну = Up + X Di, где i соответствует уровню
иерархии. ZDi удобнее представить в виде Di + Dz + Dn. В каждом
случае Di представляет вклад генной дифференциации между
субпопуляциями на соответствующем уровне иерархии в общее
значение Ну .
Первоначально для изучения генетических процессов в
" ’родонаселении СССР с позиций исторической генетики такой
1 >лиз был применен Ю.Г.Рычковым и Е.Ящук (1980) в рамках
НЬ|полнения серии работ "Генетика и этногенез". Важность подхода к
11 ’Учению наследственных характеристик подразделенных популяций
‘"означается в установлении преобладания генетического вклада
носов в разнообразие популяций, а также выявлении содержания
скватной информации о всех этапах сложения этнической
^'ЧНОСТИ как популяционной системы в современном генетическом
Р‘: "’ообразии популяций. В этой же работе Рычков, Ящук, 1980)
ИЗ
было показано, что соотношение внутри- и межпопуляционньп
компонент общего разнообразия остается постоянным S
народонаселении других регионов мира.
Этот метод для исследования подразделенности популяци!
Северной Азии был применен В.А.Спицыным с соавторами (1ч8О)
Авторы стремились с помощью этого анализа идентифицировав
наиболее адекватные структурные композиционные единицы дл;
исследования этно-расовой систематики. Предварительный анали
тотальной гетерозиготности диктуется тем обстоятельством, ЧТ1
между популяциями дифференциация может оказаться стол
незначительной, что возникает вопрос - правомочно ли сравнены
отдельно взятых популяций? Получим ли мы реальнуи
классификацию, если будем сопоставлять единичные популяций
между собой? И, если нецелесообразность сопоставления их 1
систематическом плане окажется реальностью, то вправе задаться
целью - с каких более крупных таксономических единиц
(совокупностей) нужно проводить сравнение, чтобы приблизиться к
тому уровню, начиная с которого вычленяются характерные типы,
лежащие в основе антропологической классификации (Спицын с
соавт., 1980). В работах Левонтина (levontin, 1974), Рычкова, Ящук,
(1980), Спицына с соавт. (1980) принимались во внимание
основном крупные таксономические уровни организации популяций,
начиная с этнического и выше. Известно немного работ, касающихся
иерархической структурированности небольших агрегаций
популяций, среди которых следует упомянуть результаты, полученные
Мальхотрой с сотрудниками (Malhotra et al., 1978) для кастово-
религиозных групп штата Махараштра (Индия). В настоящем
исследовании была поставлена задача установить степень генного
разнообразия, наблюдающуюся на субэтническом уровне, включая
генное разнообразие внутри самих микропопуляций, генные
различия между любыми популяциями башкир в пределах среднего
рода, между родами в пределах среднего объединения на уровне
племени. 31 микропопуляция башкир исследовалась в отношении
распределения i рупп крови систем АВО, MN, Rh(D), а также
гетерогенности сывороточного белка гаптоглобина (Нр). Таким
образом, для анализа генной дифференциации использовались
аллелей 4-х независимых локусов.
114
Для дальнейшего анализа была построена иерархическая схема
звеньев цепи постоянно усложняющихся систем структуры коренного
населения Башкортостана и на каждом ее уровне (звене) проведен
анализ тотального генного разнообразия (Нт) и его составляющих
НС и SDg-p .Таким образом, 31 изученная локальная группа башкир
состоит из 9 соответствующих, согласно историческим источникам,
племен, объединяющихся в тотальную башкирскую общность
(табл. 28).
Проведенный анализ генного разнообразия представлен в
таблице 31 .
Полученные результаты свидетельствуют о том, что генетическое
разнообразие среди тотальной популяции башкир, измеренное с
помощью средней гетерозиготности на локус, составляет 0,460505.
Сходные величины этого параметра были получены для этнических
групп Сибири, варьирующие в пределах 0,3923-0,5013 при среднем
значении 0,4316 (по данным Спицына с сотр., 1980), а также 0,415
(Рычков, Ящук, 1983). В среднем по 11 генетическим локусам
характеристика общего генетического разнообразия для популяции
Зарубежной Европы по данным Рычкова и Яшук оказалась также
весьма близкой (0,439); для этих же популяций с учетом их
этнической принадлежности величина Hqp оказалась равной 0,445,
те. весьма сходной с аналогичной оценкой, полученной для
Башкирии.
II?
Таблица
Дифференциация тотального генного разнообразия для башкирок
популяции
Генное разнообразие на разных уровнях иерархии Показатели разнообразия Нт= Нс + Di + D2 + D3
Абсолютные Относительные
Тотальная популяция башкир (1Ьр) 0,460505 100%
Внутри популяций (H(j) Между популяциями в пределах рода 0,446828 97,03%
(D1) 0,003592 0,78%
Между родами в пределах племени (1_>2) 0,000081 0,21%
Между племенами (D3) 0,009104 1,98%
Коэффициент генной дифференциации («ST) 0,02970
Полученные нами результаты также достаточно Хорог
согласуются с данными Мальхотры с соавт. (Malhotra et al., 1971
который получил величины генетической гетерогенности для кажд
из изученных им касговых групп порядка 39-47%, а также с данньп
Даса и др. (Das et al., 1975) для пяти религиозных групп Северн'
Индии (46-51%). В вышеуказанных работах, как и в нашем случае
расчет принимались только полиморфные локусы. Наши данш
находятся в противоречии с результатами Ройчудхури (Roychoudhu
1974), который включил кроме полиморфных, достаточно больш
число мономорфных локусов и получил весьма низкие значен
генетической гетерогенности порядка 8-11%. Следователь*
полученные значения тотальной гетерозиготности во многом завис
от соотношения охарактеризованных поли- и мономорфных локусе
а также от степени их полиморфизма. Приводимые в работе Рычко
и Яшук (1983) данные понижений общей гетерозиготности Л
Америки и Африки (0,386 и 0,324 соответственно) име»
биологический смысл и не являются следствием использования д
расчетов локусов с пониженным полиморфизмом.
116
0з рассмотрения данных, приведенных в таблице 31 для
башкирской популяции в целом, следует, что при расчленении генной
рариабельности на внутри- и межпопуляционные компоненты,
оСНовная доля генного разнообразия приходится на
вНутрипопуляционный уровень, что в количественном выражении
составляет 0,446828 или 97,03%. Как тут не вспомнить аналогию с
физиономическим разнообразием, которое мы наблюдали в любой
локальной популяции. Доля межпопуляционных различий, напротив,
оказывается составляет весьма незначительный компонент, а именно
всего 0,78% от общего генного разнообразия. Еще меньше
генетические различия (примерно в 3 раза) от предыдущей величины
установлены между родами в пределах любого племени башкир.
Объяснение этого явления лежит в самой истории народа. Анализ
имеющихся данных о формировании башкирского этноса привел
ученых к заключению, что отдельные его части различаются между
собой по происхождению и неоднородны по составу (Кузеев и др.,
1974). Не углубляясь детально в сложные этнические процессы,
имевшие многовековую имторию, хотелось бы только подчеркнуть те
особенности, которые, на наш взгляд, послужили причи' ой весьма
незначительных различий между родами, включающимися в одно
племя или объединение племен. Так известно, что сложившаяся
система браков у башкир характеризуется отчетливо выраженным
стремлением к экзогамности брачных отношений внутри родов. При
кочевом образе жизни - это было общение близких родов и племен,
1,1 как смена местообитания в процессе миграций у них совпадала.
Переход к полукочевому образу жизни, а потом и к оседлости не
изменил существующих издревле традиций экзогамности браков
11 денко, 1955; Кузеев, 1974 и др.). В конечном счете это приводило к
Осокой эндогамности племени. В проведенных популяционно-
Генетических исследованиях современных башкир была также
'1’еделена значительная эндогамность браков внутри племени, со
Дним значением индекса эндогамии равным 0,9306 (Рафиков и
- 1980; Кузеев и др., 1982).
1аким образом, если бы мы смогли представить каждую из 31
^Икропопуляции в виде группы из трех десятков лиц, то их сходство
казалось бы впечатляющим. Это же сравнение можно провести и в
"’ошении сопоставления группировок на уровне родовой структуры.
117
Данные табл. 3! повествуют еще о весьма интересном свойст
иерархической структуры башкирской популяции. Оказывается, ч
генетическое различие между племенами достаточно существенно
оказывается более чем в два раза выше, чем различия меж,
популяциями. Это, в свою очередь, во-первых, свидетельствует
сложности и многокомпонентное™ состава башкирских племен,
формировании которых приняли участие в различных соотношени
европеоидные и монголоидные элементы. Во-вторых дает указание
пользу эффективности сопоставления башкирских племен nj
решении ряда вопросов этнической истории, этногенеза башкирско!
народа (Кузеев, 1974).
С помощью коэффициента генной дифференциации мож»
оценить степень дифференциации (изолированности групп
пределах тотальной популяции; в данном случае башкир).
При структурированности сходного класса популяционнс
системы в количественном выражении, для одной из кастой
религиозных групп Махараштры Сев.Индия, Мальхотра установи
значения Gg-p, колеблющиеся от 0,049 до 0,078, что существен»
выше, чем в изученной нами популяции.
Материалы анализа генетического разнообразия на различи!
уровнях агрегации популяций в системе имеет принципиаль»
значение для изучения меры изменчивости у человека, так как п|
исследовании проблемы этно-расогенеза для решения различи!
вопросов расовой систематики специалисты прежде всег
акцентируют внимание на биологических различиях межд
человеческими популяциями. И это вполне естественно, ибо о
установления характера и степени таких различий зависят ответы И
поставленные вопросы. Именно в этом заключается основной подх<
исследователя,
занимающегося
этническо
проблемами
антропологии независимо от особенностей примененных методо!
будь то антропологические, антропоскопически
дерматоглифические, одонтологические или серологическ.
приемы. Анализ по степени сходства, специальный поис
направленный на максимальный учет сходных черт, напротив, буд<
как бы нивелировать различия между крупными популяционным
объединениями, затрудняя решение задач расовой систематик;
Именно поэтому вполне оправдан поиск новых таксоном и чесн
важных разграничительных антропологических признаков иЛ
118
генетических факторов, характеристик, маркирующих ту или иную
расовую группировку. В этом отношении достигнуты за последние
годы большие успехи: достижения в области биохимической и
молекулярной, популяционной генетики позволили
идентифицировать маркеры или гены, присущие с характерными
существенными концентрациями в определенных этно-расовых
группах, что способствует решению многих вопросов происхождения,
а также истории становления тех или иных групп.
Однако задумаемся над вопросом - насколько велики различия
между человеческими расами. Убедительный ответ на этот вопрос
дают оценки, полученные Левонтиным (Levontin, 1974) с помощью
метода, сходного с методом, примененным Неем. На основании
анализа совокупностей большого числа локусов оказалось, что 85%
всего генетического разнообразия у человека наблюдается в
пределах популяций, а различия между разными нациями в пределах
одной расы и между расами составляют только лишь 7,5%.
Конкретно, основная доля аг тотального генного разнообразия
находится в пределах отдельных популяций (0,849), различия между
популяциями в пределах расы и между расами '•оставляют
одинаковые величины, а именно по 0,075 (Левонтин, 1978, цит. по
Levontin, 1972). Проводя дифференциацию человеческих
группировок на основании морфологических или лингвистических
критериев мы получим ту же 85%-ную внутригрупповую
изменчивость (Левонтин, 1978).
Следует подчеркнуть, что недостаточно полная информация о
генетических маркерах, равно как и неравномерность исследования
различных популяций по Земному шару, приводят, по-видимому, к
заниженной оценке величины внутрипопуляционного генного
разнообразия; истинная величина внутрипопуляционной
Изменчивости у человека по мере пополнения экспериментального
Материала скорее составляет оценку, близкую к 95%. Таким образом,
на долю межрасовых различий приходится лишь несколько
процентов от тотальной величины генного разнообразия
Человечества. Этот вывод, полученный советскими и зарубежными
Исследователями весьма важен с позиций борьбы с расистской
Идеологией, с пережитками, базирующимися на родоплеменной
дифференцировке. Данные современной популяционной генетики
'идетельствуют о весьма зна'нггельной близости рас,
119
антропологических типов, этносов между собой с точки зрен
биологического сходства, а та, существующая незначительная до
межгрупповых различий, с успехом используется исследователя!
для познания сложных и интересных проблем истории человечест
(Спицын и др., 1990).
2. Оценка генетической дифференциации
микропопуляций.
Анализ генетической дифференциации на уровне локальн!
микропопуляций представляет весьма важный материал для оцен:
роли факторов микроэволюции в динамике системы популяций к
единого целого.
Поскольку для изучения эволюции популяций необходимо име
данные по генетической структуре как во времени, так и
пространстве, а сбор генетических сведений в поколени
практически является трудно выполнимым в полном объеме,
исследователям зачастую приходится ограничиваться информацие
по ряду популяций в "поперечном сечении".
Исследования популяций в одном временном срезе мож!
проводить в направлении анализа изменчивости в различных аллел
или локусах, либо на основании демографических сведен,
прогнозировать уровень генетической вариации (Cavalli-Sfon
Bodmer, 1971).
Так, если популяции имеют ограниченную численност
представлены в системе подразделенной на микропопуляции
существуют длительный период времени, то предельную величи,
ожидаемой генетической дифференциации (fe) можно вырази
следующей формулой (Wright, 1965):
1)Те = (4Ne me + 1)1
где fe - прогнозируемый уровень генетической дифференциации;
Ne - эффективно-репродуктивная численность;
те - коэффициент миграции при ступенчатой модели.
Данное выражение показывает зависимость уровня различий в
популяциях от величины эффективной численности, интенсивности и
120
характера генных миграций. Она справедлива для популяций, в
которых миграции зависят от расстояния, поэтому при ступенчатой
м0Дели структур учитывается величина эффективной миграции
(Wright, 1965; Рычков, 1976, 1977, 1984).
Степень генетических различий также связана со значением
вариансы частоты гена. Чем выше изоляция, тем больше варианса
частот генов в микропопуляциях:
2) f = Ур , где Vp варианса частоты гена;
pq p,q - средние частоты генов.
Отсюда для островной модели: _
3) Vp = f pq = pq I (1 +4 Neme)
Для ступенчатой модели:
4) Vp = f pq = pq / (1 + 4 Neme)
Таким образом, с применением данных о демографической
структуре, а именно эффективной числености и типа миграционной
структуры, можно прогнозировать теоретически ожидаемую
величину генетической изменчивости локальных поп ляций. И
наоборот, используя данные по частотам генов в популяциях, при
условии, что они связаны единством происхождения, можно
определить замкнутость популяции.
Так в популяции башкир, подразделенной на микропопуляции с
эффективным объемом Ne = 105 и эффективной величиной
коэффициента миграции те = 0,0407 (см. гл. 6.5.) ожидаемая
величина генетической дифференциации составляет fe = 0,0553.
В табл. 32 представлены результаты наблюдения реального уровня
генетической изменчивости по разным локусам (fo). '
Следует отметить, что проведенная нами стандартизация генных
Дисперсий, позволяет сравнивать их вне зависимости от исходных
концентраций генов.
121
Таблица 3
Распределение наблюдаемых стандартизованных варианс по
отдельным локусам в популяционно-генетической системе башкир.
Г' _ Г- Локус Аллель Ч fo
АВО Г 0,5622 0,0147
р (1,2098 0,0287
ч 0,2290 0,0134
MN m 0,7040 0,0556
Нр Нр- 0,3782 0,0153
' Rh(D) d 0,2478 0,0274
Rh (с) С 0.4769 0,0057
Rh (е) е 0,7778 0,0140
Kell (к) к 0,9877 0,0107
Gc Gc 0,5020 0,0657
to=0,0251
Примечание:
q - средняя частота гена
fo - стандартизованная варианса (наблюдаемая)
Как видно из табл. 32 истинная величина генетически
изменчивости варьирует у башкир по разным локусам
минимального значения fo = 0,0057 по локусу Rh (с), ,
максимального fo = 0,0657 по Gc, составляя в среднем их 1
оценок по 8 локусам в популяции башкир fo = 0,0251. Сравнен;
наблюдаемого уровня замкнутости, полученного по генетическ
структуре популяций, с теоретически ожидаемым значением I
данным демографии (fe = 0,0553). По всем остальнь
генетическим системам выявляются различия. И средняя_ I
совокупности локусов наблюдаемая генная дифференциация fo
0,0251 более чем в два раза ниже ожидаемой равновесн<
величины. Другими словами, соотношение fo<<fe показывае
что достигнутый в популяции башкир уровень фактическ
дифференциации значительно ниже предельно возможного
Имеющиеся в литературе сведения по генетичес
дифференциации в популяциях литовцев, уйгур, дунган, алеу"
тофаларов, нивхов, коряков, эвенков, алтайцев и других наро
122
СССР в основном свидетельствуют о близости в этих группах
значений fo и fe (Рычков. 1982, 1986; Бахолдина, 1982; Воронина,
1983; Кучинскас, 1987; Лузина, 1987 и др.).
1 |риступая к анализу обособленного положения башкир среди
других народов, следует, очевидно, остановиться в первую очередь
на самой модели определения fe, которая строится в
предположении решающей роли эффективной численности,
особенностей генных миграций, а также допущения селективной
нейтральности генов. Так, отдавая должное малой эффективной
численности в популяциях башкир, как одной из основных причин
возникновения изменчивости вследствие генного дрейфа, нельзя
бъяснить разброс данных по стандартизованным вариансам по
локусам селективно-нейтральным типом развития. Различия в
значениях стандартизованных варианс по локусам возникают под
давлением отбора, а сами различия во вкладе отбора в отношении
конкретных локусов можно определить по разнице между
наблюдаемой и ожидаемой величине стандартного отклонения
(Рычков, 1976; Nei. 1976).
Рассматривая под таким углом зрения динамику генетической
изменчивости в исследуемых микропопуляциях, следует отметить,
что лишь по локусу MN наблюдается селективная нейтральность
процесса. В отношении остальных - налицо существование
давления отбора различной силы. Наблюдаемая очень низкая
величина стандартного отклонения в микропопуляциях по локусу
Rh (с) позволяет предположить действие по данной системе
тбора стабилизирующего типа (табл. 32) (подробно анализ
интенсивности естественного отбора в популяциях башкир
приводится в гл. 6.4.).
Истинную оценку роли каждого из микроэволюционных
факторов в формировании генетической изменчивости локальных
популяций башкир невозможно провести без рассмотрения
’’атериалов по этногенезу данного народа, поскольку
обнаруженное нами соотношение fe<<fe сразу выдвигает вопрос
0 малом времени существования популяции. Однако имеющиеся
многочисленные работы историков, этнографов, археологов,
'’нтропологов и других, подтверждают длительность
Многовековой истории развития, глубоко уходящую корнями на
в°сток нашей страны (Руденко, 1955; Кузеев, 1971; и др ). Поэтому
123
обнаруженная "молодость" древних родоплеменных образован
башкир, очевидно, связана лишь со временем появления башк
на Урале. С другой стороны, сама степень генетическ
дифференциации свидетельствует о сравнительно неболыщ
интервале времени в историческом масштабе, прошедшем в hobi
для пришлых тюркоязычных групп экологических условиях Волг
Уральского региона, времени, которого оказалось яв|
недостаточно для выработки в поколениях устойчиво
динамического равновесия генетической структуры популяции.
Все это может служить одним из аргументов в ноль
сторонников позднего прихода башкир на Урал.
6.3. Оценки уровня инбридинга на основании
анализа генных частот и родословных.
Одним из вариантов отклонения от панмиксии в человеческ:
группах является образование кровнородственных браков
частотой, превышающей таковую при чисто случайном подбо)
пар.
Основной эффект инбридинга заключается в уменьшенй
гетерозигот и преобладании гомозигот в генофонде популяцж
При этом, в оценке величины коэффициента кровного родств
интересна не вероятность того, что предки являются кровным
родственниками, а вероятность встреч в поколениях гено
идентичных по происхождению. Именно в этом и заключаете
основное отличие от генов идентичных по свойствам (Xappucof
1979; Alan E.H.Emery, 1986).-
Изучение обособленных групп человека выявило значигельну»
вариабельность в уровнях инбридинга, как правило, связанную С
традиционными нормами брачных отношений, с существующей
них системой избегания, или, наоборот, предпочтени
кровнородственных браков. Теоретически в небольших п
численности изолятах степень инбридинга заведомо должна быт
достаточно высокой, так как при конечном размере популяции
практически все ее члены оказываются в какой-то ме|
124
-вязанными родственными отношениями (Харрисон, 1979;
дфанасьева, 1990; и др.).
Эффект гомозиготации оказывает большое влияние на
^стояние здоровья потомства в этих супружеских парах, так как
основная часть наследственных заболеваний проявляется именно в
гомозиготном соотношении рецессивных генов. И, если в брак
вСтупают родственники, являющиеся скрытыми носителями генов
наследственных заболеваний, то очень велик риск рождения детей,
отягощенных разными формами наследственной патологии.
Такова сущность инбредной депрессии, оказывающей негативное
воздействие на общий статус здоровья популяции (Sutter et al.,
1951, 1952; Гинтер и др., 1980, 1982, 1983; Бочков и др., 1984;
Парадеева, 1987; Куандыков и др., 1988; Dinesh Parmar et al., 1990;
и др.). Классическими методами оценки коэффициента
инбридинга является определение его из родословных.
Определение генеалогического инбридинга дано Малеко, а его
меру охарактеризовал Райт. Биологический смысл данного
коэффициента заключен в вероятности того, что у данного
индивида пара аллелей данного локуса идентичны, то-есть
происходят от общего предка, или доля локусов его генотипа
состоит из аллелей идентичных по происхождению. При переходе
от индивидуального уровня определения инбридинга к
популяционному используют среднее арифметическое значение
коэффициентов всех ее индивидов.
К одному из существенных недостатков данного метода
относится трудная уловимость отдаленной степени родства, как
правило, информация ограничивается охватом 4-х, максимум 5-ти
поколений.
Предложенный Кроу и Мандж метод определения
коэффициента инбридинга на основании анализа частоты
11 юнимных браков может быть применен в популяциях, в которых
Фамильное родство соответствует биологическому на протяжении
многих поколений. (Crow, Mange, 1965). Этому условию не
отвечают памирские популяции и башкирские, так у последних
фамилии существуют менее одного века.
Согласно методу изонимии:
Fgp . где
125
F$t является случайной компонентой инбридинга, возникающе
результате конечности популяции. Тотальный инбридинг |
определяется делением доли изонимных браков на четыре; Fi
компонента, обусловленная брачной ассортативност,
получается из следующего выражения:
FIS- Fit-Fst
1-Fst
Оценка коэффициента инбридинга на основании анали
дисперсии генных частот в субпопуляциях и отклонения часз
гетерозигот от ож" щемых по закону Харди-Вайнбер|
предложенная Райтом, полностью приложима к популяц
башкир, так как здесь соблюдается единственное требуем
условие о подразделенности популяций. Применение F - статист
Райта основано на свойстве инбридинга выступать в качестве ме|
интенсивности генетического дрейфа, при этом возрасти
количества гомозигот может быть обусловлено за сч
подразделения (случайный инбридинг), а также в результа
особой структуры браков внутри субпопуляций в СИСТС1
(неслучайный инбридинг). В свою очередь тотальш
коэффициент инбридинга Ijy складывается из случайно
компонента F^ и неслучайного компонента F. П|
использовании селективно-нейтральных или близких к это:
состоянию локусов получаются оценки инбридинга, близкие
значению к таковым при использовании метода родословн!
(Morton et al., 1971).
В популяциях человека величина инбридинга подверже
большой изменчивости. Так, в группах, которые максималы
панмиксны, значение соответствующего коэффициента мены
0,00001 (0,001%). Напротив, в популяциях, практикующ
кровнородственные браки, уровень инбридинга может достигну
5-ти % уровня и выше (см. сводку Crawford et al., 1973; в табл. 33).
Таблица 3
Коэффициенты инбридинга в разных популяциях человека (:
по Crawford et al, 1973).
популяции с F>O,OOOO1
126
Продолжение таблицы 33
I Нидерланды
Панама
популяции с F между 0,00001 и 0,0001
Чехословакия (Брно) после 1950 г.
CII1A (католики)
популяции с F между 0,0001 и 0,001
Бельгия
Чехословакия (Брно) до 1950 г.
Италия (Север)
Швейцария
Канада (французы)
Канада(католики)
Чили (католики)
Уругвай (католики)
Аргентина (католики)
Куба (католики)
Мексика(католики)
Боливия(католики)
Бразилия(католики)
популяции с F между 0,001 и 0,01
Япония (различные группы)
[ Франция
Венгрия (Ивад)
Италия (Сардиния и Элинские о-ва)
Испания
( 111 А (Сев.Каролина)
США (Мормоны)
США (Индейцы рама)
Индия (Бомбей)
Бразилия (Северо-восточная)
популяции с F между 0,01 и 0,05
Япония (Хосошима)
Индия (Побережье штата Андхра Прадеш)
Индия (Сельские р-ны штата Андхра Прадеш)
"Дия (Каванур, Мадрас)
"Дия (Каста Телага штата Андхра Прадеш)
*°Рдания и Израиль (Самаритане)
11,пет (Нубия)
127
Продолжение таблицы j
Гвинея (Фоута Джалон)
США (Дункеры, Пенсильвания)
США (Гуттериты, Южная Дакота и Миннесота)
Тристан да Кунья
Бразилия (Шаванте)
Бразилия (Северо-восток)
^Другие популяции с близкими кровнородственными браками/
которых отсутствуют точные оценки коэффициентов инбридинг^
Авсгралия (Аборигены), Цейлон, Бирма, Бедуины.
Анализ структуры браков в микропопуляциях башкир выла
систему тщательного избегания кровного родства (см. миграци
гл.6.5.), поэтому для определения уровня инбридинга наибо/ц
оптимальным оказался не классический метод родословных,!
вычисление его с помощью F-статистик Райта. Так, средн!
значения F-статистик, для популяций башкир, определенные 1
совокупности локусов (АВО, MN и гаптоглобину) оказали!
следующими: F1T = 0,00927; Fjj = 0,0251; Fj3 =-0,0162. К*
видно из этих значений, тотальный инбридинг в башкирсы
популяциях складывается в основном только за счет случайн(|
составляющей компонента инбридинга FST, при этом неслучайн!
компонента имеет отрицательное значение, т.е. исследуем!
популяция характеризуется отрицательной брачнс
ассортативностью по родству. Аналогичная ситуация отмече^
исследователями в адыгейской популяции (Кадошникова и др
1991).
Совершенно иная структура браков наблюдается ]
высокогорных популяциях Памира, а также и в Азербайджан!
причем выявляются ярко выраженные этнические различия. Та
из табл. 34 видно, что значения коэффициентов инбридинга очей
высоки у памирцев кишлаков Пастхуф и Хуф, составляя 0,0594
0,0545 соответственно, напротив, у киргизов кишлака Мургаб F 1
0,0056. Подобная демографическая ситуация во много!
обусловлена экономическими причинами, когда крайн
ограничено количество земли, пригодной для земледелия. СаМ
история происхождения Хуфа, а затем и Пастхуфа полностН
подтверждает это (Андреев, 1958). В подобных условий
кровнородственные браки во многих случаях могут оказаться даЖ
предпочтительными, т.к. препятствуют дальнейшему дроблени!
128
земельных участков. Традиции нерегулируемой рождаемости,
наблюдаемые в исследуемых памирских группах, обладают
аналогичным действием, приводя к быстрому росту численности
популяции. С другой стороны, высокая рождаемость сама по себе
способствует поддержанию высокого уровня инбридинга, так как
понятие инбридинг целиком относится к потомству от
кровнородственного брака (в случае бездетности такого
супружества F = О). Можно констатировать, что в памирских
высокогорных популяциях имеется целый ряд социальных и
биологических факторов, способствующих поддержанию такого
значительного уровня инбридинга.
Таблица 34.
Частота встречаемости различных типов браков и коэффициент
инбридинга в высокогорных группах памирцев и киргизов Памира.
Тип брака Пастхуф Хуф Мургаб
Число браков Частота в% Число браков Частота в% Число браков Част %
Двоюродные сибсы 15 14,7 17 16,8 2 U
1 роюродные сибсы 0 0 10 9,9 4 2,2
Четвероюродные сибсы 0 0 3 2,9 0 0
Пятиюродные сибсы 0 0 0 0 2 1,1
Дядя-двоюродная 1 0,98 0 0 0 0
ллемянниа
Неинбредные браки 86 84,32 71 70,4 92 95,6
Всего 102 1,000 101 1,000 179 1,000
J
Средний коэффициент
ИиГ|ридинга в популяции _ _ _
F - 0,0594 F - 0,0545 F = 0,0056
В связи с этим представляет несомненный интерес возможность
оценки влияния инбридинга на популяционно-генетическую
груктуру, В табл. 35 (а,б,в) представлены данные по репродукции
129
в популяциях Пастхуфа, Хуфа и Мургаба. Анализ проводи!
направлении сопоставления по целому ряду парам»
родственных и неродственных типов браков без соответствую!
деления их на группы вышедших из репродуктивного перио
находящихся в этом возрасте. Так, в Пастхуфе в родстве!
брачных парах выявилось большее число рождений и ж
детей, приходящихся в среднем на одну женщину.
Таблица 35 (а
Эффекты инбридинга в высокогорных популяциях памир|
киргизов Памира
а) Пастхуф
Количество родственные браки (16 пар) Неродственные браки (77 пар) Уровень значимое Р
х + Sx rf* T±SX Л
рождений 7,5+0,9704 15,0667 7,45+0,3876 11,7178 <0,05
живых детей 6,44±0,6512 6,7856 5,74±О,ЗО73 7,3657 <0,05
мертворо- ждений и выкидышей бездетных браков 0,44 + 0,0589 0,0556 0,53+0,0993 0,013±0 0,7899 >0,05
б) Хуф
Количество родственные браки (30 пар) неродственные браки (63 пары) уровен! значи- мости J
1 + Sx 4 х + Sx
рождений живых ветей мерз по- рождений и кыкпдышей бездетных браков 7,3 + 0,7665 5,2+0,5820 0,96б7± ±0,6494 0,03+0 • 17,6266 10,1630 12,2309 7,54+0,4177 6,06 + 0,3983 0.9048+ + 0,1500 10,9944 9,9959 1.4186 <0,05 ' <0,05 | <0,05
130
в) Мургаб
Ivi |ичеспю родственные браки (8 пар) неродственные браки (136 пар) уро-вень значи- мости Р
х +Sx 6 А х + Sx 6;
рождений 7.8 ±((,86(18 5,9286 4,97 + 0,2063 5,7913 >0,05
живых детей 6,3+0,4910 1,9286 2,98+0,8357 3,2072 >0,05
мертворож- дений и выкидышей бе шетных браков 0,75 + ±0,4252 1,4464 0,8235 + +0,0882 1,0583 <0,05
1фимечание: х - среднее число; Sx - ошибка среднего;
i к- дисперсия среднего числа.
меньшее число мертворождений и выкидышей, а также отсутствие
случаев бездетности в сравнении с этими же параметрами в
неродственных браках, однако, достоверно значимыми оказались
только различия по мертворождаемости, выкидышам и уровню
бесплодия. В .'’уфе наблюдается картина, когда по всем
исследуемым признакам более "благополучно" выглядят
неродственные брачные пары, хотя эти различил также
статистически незначимы. Напротив, в Мургабе отмечено
к>стоверно большее (р>О,О5) число рождений и живых детей,
приходящихся на одну женщину в родственных браках, чем в
неродственных. Таким образом, в популяциях высокогорья нами
было выявлено не только отсутствие теоретически ожидаемого
1,трицательного влияния родства супругов на потомство, но и
статистически значимое преимущество по ряду параметров у
киргизов Мургаба. Даже принимая во внимание то, что доля
кровнородственных браков в этой популяции составляет всего
«коло 4,4% от общего числа, следует отнестись к данному
131
феномену со всей серьезностью. Очевидно, мы имеем дед
явлением, когда экстремальность окружающей среды в сочета
с определенным соотношением действия эволюционных си
условиях длительной изоляции привели к вырабс
своеобразного адаптивного комплекса.
В табл. 36 (а, б, в) представлены эффекты инбридинг
родственных браках в группе лиц вышедших из репродуктив
возраста. Как видно из табл. 36 анализ браков в полных cei
закончивших репродукцию, выявил сходную по
исследованным популяциям тенденцию, хотя размеры выб
соответственно оказались меньшими, особенно по Мур
Исключение составили только кровнородственные брак
кишлаке Хуф, в которых среднее число живых д
приходящихся на одну женщину было достоверно ниже, ч
неродственной группе.
Табли!
Эффекты инбридинга в высокогорных популяциях памир!
киргизов Памира в группе лиц, вышедших из репродуктив
возраста.
а) Пастхуф
Количество родственные браки (6 пар) неродственные браки (37 пар)
х + Sx х + Sx
рождений живых детей мертворо- ждений и выкидышей бездетных браков 9,67+4,8700 6,00+1,5490 0,33±0,1730 21,0667 14,4000 0,1790 8,49+1,1264 5.70+1,0860 0,87+0,0864 0,03±0,0 10,9517 8,5856 0,1581
<0,05
<0,05
>0,05
! уровень
| значи-
мости Р
132
б) Хуф
Продолжение таблицы 36
Количество родственные браки (13 пар) неродственные браки (39 пар) уровень значи- мости Р
х + Sx Г; х +. Sx <>2
рождении 8,46+1,1580 17,4358 7,44+1,0082 11,4020 <0,05
дивых детей 4,46 + 0,5848 4,665(1 5,77+0,8511 10,9879 >0,05
мертворо- ждений и выкидышей бездетных браков 1,08 + 0,3962 2,0403 0,87 + 0,1413 (),03±0 1,9877 <0,05 к.
в) Мургаб
Количество родственные браки (4 пары) неродственные браки (46 пар) уровень значи- мости Р
х + Sx x±Sx
рождений 9,00+1,4714 8,6600 8,06+ 0,9881 7,8794 >0,05
живых детей 5,0+0,8155 2,6700 3,6 + 0,7067 5,2490 >0,05
мертворо- ждений и выкидышей бездетных браков 1,5+0,6100 1,5000 0,78 + 0,0083 0,02+0 0,0856 >0,05
Примечание: "X - среднее число; SX - ошибка среднего;
"л- дисперсия среднего числа.
Примером популяций с исторически сложившейся системой
браков, выражающейся в предпочтении браков между
родственниками, являются азербайджанцы. Ниже приведены
Результаты полного генеалогического изучения, проведенного в
1986 г. , жителей села Абдал, расположенного в Агдамском р-не
Азербайджана (Павленко, Спицына, 1987). Были учтены все типы
браков, проанализировано их потомство и произведено сравнение
Различных аспектов плодовитости в инбредной и неинбредной
частях популяции.
133
В популяции Абдал выявлено 109 кровнородственных 6pi
что составляет 27,5% от общего числа. Представляет интерес
структура родства, так, преобладающее большинство cocraej
браки между двоюродными сибсами - 68,8%, доля троюродн
четвероюродных соответственно равна 17,4% и 13,7%. Полно
отсутствуют браки типа дядя-племянница, тетя-племянник,
одноперекрестные, также не выявлены случаи множествен
родства. В целом по популяции средний коэффициент инбрид
F = 0,0642, в инбредной его части F=0,2468. Столь выс
значение коэффициента обуславливается предпочтением бли
степени родства в браках, а также достаточно вьи
плодовитостью в этих семьях. Следует, однако, учесть, <
действительности уровень инбридинга в данной г
азербайджанцев может быть еще выше и не улавлис
примененным нами генеалогическим методом, коз
ограничивается учетом не очень далекой степени родства,
таком подходе почти полностью выпадает из поля 3|
исследователя доля отдаленного кровного родства супруго
если принять во внимание такие демографические параметры
сравнительно небольшая численность этой популяции, выс
степень эндогамности, а также устойчивость системы бра'
традиций в историческом аспекте, то можно предположить,
вклад отдаленного кровного родства в коэфициент инбрид
мог бы быть еще более значительным. К этому же еле,
добавить выявленные нами единичные случаи нежел]
отдельных индивидов сообщить действительную информаци
характере брака.
К сожалению, мы не располагаем соответствующими дани!
по предшествующим поколениям, хотя по аналогии с друг
группами населения нашей страны можно предположить, Ч
прошлом величина коэффициента инбридинга была неско;
выше (Гольцова, 1981; Афанасьева, 1990).
В табл 37 представлены данные по репродукции у женщин I
Абдал.
134
Таблица 37.
Эффекты инбридинга в кровнородственных браках популяции
Абдал
Количество родственные браки (104 пар) неродственные браки (222 пары) уро-вень значи- мости Р
х + S« б; f + SX «X
рождений 6,08 + 0,6864 11,7787 6,19 + 0,3888 11,0391 0,05
живых детей 4,88 + 0,5280 6,9703 5,05 + 0,3221 7,5779 0,05
мертворо- ждений и 0,24+0,0513 0,6518 0,1233+0,053 0,2063 0,05
выкидышей бездетных браков 0,039+0 — .. 0,0130+0 -
Примечание: х - среднее число; Sx - ошибка среднего;
с дисперсия среднего числа.
Как видно из таблицы 37, среднее число рождений,
приходящихся на одну женщину в группе кровнородственных
браков равнялось 6,08, в неродственной 6,19. Среднее число
живых детей соответственно было 4,88 и 5,05. Однако
статистически значимыми оказались лишь различия по
прентальной смертности, которая была в инбредных браках в 2
раза выше, а также в величинах бесплодия, в 3 раза
превышающую эту характеристику в сравнении с неродственной
частью популяции.
Так как эффекты инбридинга трудно дифференцировать в
группе женщин всех возрастов, сравнение между которыми
проводилось только по принципу деления их на родственные и
неродственные браки, то возникла необходимость исследования
полных семей, т.е. таких брачных пар, которые уже завершили
Репродукцию. В табл. 38 представлен анализ различных аспектов
плодовитости в полных кровнородственных браках в сравнении с
полными неродственными семьями.
135
Таблица
Эффекты инбридинга в азербайджанской популяции Абдад
группе лиц, вышедших из репродуктивного возраста
родственные браки (63 пары) неродственные браки (144 пары) уровень значи-
Количество X+.SX А if±sr 6 к м(нти Р
рождений живых детей мертво|м>- жлений и выкидышей бездетных браков 7,86+0,7389 6,27 + 0,0072 0,20±0,1360 8,1904 0,6767 0.2770 6,75±0,4819 5,4683 + 0,43 0,06±0,0450 11,1489 8,8203 0,0985 0,05 0,05 0,05
Примечание: х - среднее число; Sx - ошибка среднего; дисперсия среднего числа.
Как видно из табл, 38 по всем сравниваемым критерг
среднему числу рождений, живых детей, а также мертворожде
и выкидышей, приходящихся на одну женщ
кровнородственные браки имеют достоверно высокие значе
чем неродственная группа семей. В популяции Лбдал в инбред
браках выявлена картина повышенного темпа рождаемо
соответственно среднего числа детей и сравнительно б<
высокий уровень мертворож,1аемости и выкидышей. Дан
ситуацию можно объяснить, очевидно, с одной стороны - луч
совместимостью гамет в кровнородственных браках, а с др]
стороны - неблагоприятным влиянием роста гомозиготности,
следствия инбридинга. Сходный результат был получен
популяциях Японии (Schull et al., 1970) и при исследова!
популяции в Южной Индии (Katherine Напп, 1985), где из 1
женщин 37,74% находились в кровнородственных браках, 60,7
в неродственных браках и 1,46 - неизвестно были ли эти бр
кровнородственными. Данные обнаружили, что женщины
родственных браках, завершившие свой репродуктивный nepi
136
показывают оо
а,ербайджанок
поенатальная
ее высокую плодовитость. Однако, в отличие от
Абдала, в Южной Индии наблюдается низкая
смертность, более высокая постнатальная
смертность, низкие уровни бесплодия у женщин в родственных
фраках, что весьма сходно с результатами, полученными нами в
п0Г1уляциях памирцев Пастхуфа и киргизов Мургаба. При этом
сЛедует отметить, что в высокогорных группах Памира эти
отличия статистически достоверны Данные об отсутствии
инбредной депрессии в Пастхуфе, Мургабе и отчасти в Абдале
перекликаются также с немногочисленными сообщениями Нил и
ЦЬлл (1472) в Японии, Rao и Bonne у самаритане (1473), Rao по
К) Индии (1474) и др. Представляют интерес результаты работы
по изучению связи родственных браков с долгожительством в
Азербайджане. Так, генеалогия 316 человек (138 мужчин и 178
женщин) в возрасте 80-100 лет и старше показала, что 22,46%
мужчин и 12,24% женщин являлись потомками от
кровнородственных браков (Асадов, 1990).
Таким образом, инбридинг исследованный методом F-
статистик Райта в микропопуляциях башкир, и генеалогическим
мег мм в высокогорных группах памирцев и киргизов Памира, а
также азербайджанцев популяции Абдал выявил разные его
уровни от F=O,00927 у башкир, F=O,OO56 у киргизов,
1 =0,0545 и 0,0594 у памирцев до F=O,O642 у азербайджанцев.
На конкретном материале анализа видно, что суждение о
характере влияния родства супругов на состояние здоровья
потомства не может быть однозначным.
Инбридинг - это сложное биологическое явление, с которым
человеческие группы сталкиваются в той или иной степени на
протяжении всей истории своего развития и поэтому влияние его
На формирование структуры популяций довольно значизельно.
В качестве объяснения различного влияния инбридинга на
Г|||пуляционно-генетическую структуру предлагаем следующую
гипотезу о существовании двоякого эффекта инбридинга - с одной
^'роны инбридинг ведет к росту гомозиготности, который может
"Рй носительстве патологических генов быть причиной
8ь,,иепления рецессивных генов наследственных заболеваний, что
"Р°является в снижении фертильности, росте пре- и
137
постнатальной смертности, бесплодии, все это в конечном с’
снижает эффективный размер популяции;
- с другой стороны индивиды в кровнородственном браке им
лучшую совместимость гамет, что ведет к повыше
фертильности через снижение токсикозов беременности.
Считаем, что в популяциях действуют одновременно оба
противоположные процессы и от соотношения их между соб<
также особенностей структуры популяции и зависит итон
действие инбридинга на репродуктивную стр;
выражающееся в наличии либо отсутствии инбредной депре<
4. Соотношение вкладов дифференциальной смертности и
дифференциальной плодовитости, как компонентов
приспособленности, в структуру естественного
отбора в популяциях человека.
В условиях социально-экономических преобразова
происходящих в мире, большое значение в популяцио
генетическом плане приобретает анализ вклада средо
компонентов в формирование структуры популяций раз.
этносов. Изучение адаптации их к среде обитания явля*
важным в аспектах определения роли факторов микроэволющ
биологии человека в целом, для контроля за проводим!
санитарно-гигиеническими мероприятиями и уст
профилактического направления медицины, в плане эгничес
антропологии с точки зрения познания и выявления этничес
особенностей популяций. Динамика современных этничес
процессов также нуждается в исследованиях этничес
общностей в историко-генетическом отношении.
Изучение интенсивности давления естественного отб
представляет несомненный интерес в связи с вопросами адапта!
населения в конкретных условиях среды, поскольку вырабо
адаптивных комплексов является длительным процесс
охватывающим десятки и сотни поколений, то она тесно связа»
историей формирования народа. Исходя из этого, a priori моЖ
предположить, что периодическая смена местообитаН!
' ЛН
,анная с кочевым образом жизни башкир, выдвигала отбор на
р.цвый план среди других эволюционных факторов,
«Армирующих популяционно-генетическую структуру.
табл. 39 представлены индексы потенциального отбора и
значения составляющих их компонент в 12 микропопуляциях
gall)KHp из северо-восточной и юго-восточной этно-
геограФических ГРУПП' По методу Кроу исследовалась когорта
лип вышедших из репродуктивного возраста. Как видно из табл.39
индекс потенциального отбора (Itot) варьирует по
микропопуляциям от 0,30 (Утягулово), до 1,64 (Байдавлетово),
равняясь в общебашкирской совокупности 0,82. Однако
наибольший интерес, на наш взгляд, представляют соотношения
компонент, составляющих данный индекс. Так, компонента
[дифференциальной смертности (1m) колеблется от 0,01
(Истамгулово) до 0,64 (Расулево), а дифференциальной
плодовитости (If) от 0,19 (Утягулово), до 0,61 (Байдавлетово).
Обращает на себя внимание относительно высокая величина If,
которая в восьми случаях из двенадцати микропопуляций равна
или даже превышает Im.
В '.|бл,4О приведены параметры репродуктивной способности
женщин гречанок и албанок Приазовья, закончивших период
репродукции. Одинокие женщины в анализ не были включены.
Учитывая значител! чый объем пострепродуктивной возрастной
группы в исследуемых популяциях, составляющий до 39,3% от
общей численности, оказалось возможным вычленить в ней две
выборки семей: раннюю возрастную когорту составили женщины,
завершившие репродукцию к 1960 году (I) и позднюю (II),
закончившие репродукцию в 80-е годы. По такому важному
показателю, как среднее число беременностей, приходящееся на
°-1ну женщину, завершившую репродукцию, в пос.Красная Поляна
Различий между ранней (I) и поздней (II) когортами не
°бнаружено. А в Староласпе, Девнинском и Георгиевке отмечен
Рост этой величины у лиц группы II. В группе 1 во всех популяциях
сРеднее число живорождений выше. Напротив, среднее число
абортов в несколько раз больше у лиц когорты П. Следует
°тМетить, что
139
Таблица 39.
Индексы потенциального отбора и составляющие их
компоненты в популяциях Башкирии (по методу Crow)
' Популяции ' Vf Ps 1m и If / Ps Itot
Среднее число Дисперсия Доля детей i Диффе- Диффе- Индекс
детей на 1 среднего доживших рении- рении- потен-
женщину числа детей до ре про- альная альная циального
дуктивнос- смерт- гиюдовм- отбора
I 1 1 тм ность гость
i Истамгулово 3333 4,3341 0,99 0,01 0,39 0,39 0,4042 !
| Расулево 4,100 5,46 0,61 0,64 0,32 033 1,17
Абзаково 3,83 6.27 0,81 0.23 0.43 033 0,76
Чингизово 4,08 5,36 0,98 0,02 032 0,33 0,35
I Буранбаево 4,77 1037 0,91 0,10 0,45 0,50 0,60
Ибраево 4,18 10,06 0,74 035 037 0,78 1,13
i Казарма 4,62 5,72 0,79 0,27 0,27 0,34 0,60
| Баишево 3,84 4,47 0,93 0,07 0,30 0,33 0,40
| У тягу лево 5,20 i 5,08 0,91 0,10 0,19 0,21 0,30
| Сагитово 5,05 5.71 0,64 0,56 0,22 035 0.91
> Илияшсво 4,92 5,30 0,76 0,31 0,22 0,29 0.60
j Байдавлетово 4,00 9,75 0,61 0,64 0,61 0.99 1,64
j Башкирия 4Д7 6,58 0,74 0,35 0,34 0,47 0,82
i суммарно 1 ' , i
Г
141
!
и
OI
Б
О
11
ill
2.12
2,37
Продолжение таблицы 40
2,36 I 2,19 ;
2,41 i 2.204
2.41
2.62
Примечание: I-ранняя возрастная когорта (женщины,
завершившие репродукцию к 1960 г.); П-поздняя (женщины,
завершившие репродукцию в 8О-е годы); Ш-суммарн,
показатели репродуктивной способности.
особенно велико различие у албанок- в Ю и более раз, хс
суммарные характеристики данного параметра значительнее
гречанок. Во всех четырех популяциях наблюдается резк
снижение детской смертности в поздней группе лиц
соответственно, в суммарной характеристике; особенно вели
различия в Староласпе и Георгиевке. Сокращение чис
рождений, отмечаемое выше в поздней когорте, привело
снижению в них среднего числа детей, доживших
репродуктивного возраста, а также отразилось в уменьшен,
суммарного значения.
Представляет интерес, произошедший в популяциях резв
переход к мерам по планированию размеров семьи. Так, есл
раньше у албанок только 0,03-0,05% от общего чис
беременностей завершалось абортом, а у гречанок эта величи
составляла 16-31%, то в группе II она составляет соответствен
45-46% и 57-66%. И в итоге, по суммарным характеристикам от
до 36% у албанок и от 45 до 56% беременностей у гречанок
заканчивается родами.
В табл.41 представлены индексы потенциального отбора I
методу Кроу и составляющие его компоненты в популяци
Приазовья. Беременности, завершившиеся абортом, в анализ
были включены. Также приведен сравнительный материал I
популяциям Памира и литературным источником. Данные табл?
показывают, что у женщин пос.Красная Поляна различия П
индексам дифференциальной смертности (1Ш
дифференциальной плодовитости (If) и индексу потенциально,
отбора (hot) незначительны. Напротив, в пос.Староласг
Девнинское и Г'еоргиевка ранняя когорта (1) имеет более ч<
двукратное превышение индексов потенциального отбора
М2
составляющих его компонентов в сравнении с поздней (II). В
популяциях памирцев и киргизов Памира существует совершенно
иной характер репродукции и механизм действия естественного
(Отбора. В них полностью отсутствуют меры по искусственному
регулированию размеров семьи (так, отмечено всего 2 случая
прерывания беременности). Соответственно среднее число детей,
приходящееся на одну женщину, существенно выше аналогичного
показателя женщин Приазовья, но имеет тенденцию к понижению
с увеличением высоты местообитания групп над уровнем моря - от
5,97 в Пастхуфе (2000 м) до 4,43 в Мургабе (3800-4200 м).
Также с подъемом в высоту закономерно снижается доля лиц,
доживших до репродукции и, соответственно, возрастает
компонента дифференциальной смертности от 0,54 до 1,44.
(Угмечается четко выраженный градиент роста давления отбора и
экстремальностью условий высокогорья от hot « 0,92 (Пастхуф),
Itot = 1,12 (Хуф) до Itot = 2,29 (Мургаб). Следует отметить, что
экологические условия в данном примере выступают в качестве
основного фактора естественного отбора, действующего в
высокогорных популяциях, нивелируя различия, обусловленные
этническими, хозяйственно-культурными и другими особенностями
адаптивных процессов групп.
Применение подсчета индекса Кроу и составляющих его
компонентов для исследования величины максимально
возможного отбора является корректным в памирских популяциях,
отвечающих требованиям, необходимым для проведения данного
анализа. Цифры же, полученные нами для групп Приазовья,
значительно отличаются от реальных, поскольку небиологические
факторы (социальные меры регуляции численности) сводят на нет
'диетические различия в плодовитости, смертности и искажают
истинную силу давления отбора. (Crow, 1958).
Поэтому особый интерес представляет подсчет индексов
"отенциального отбора с учетом эмбриональных потерь по
Джонстону и Кесингеру (Johnston F.E., Kesinger К.М., 1971).
143
Таблица 4
Индексы Кроу в популяциях греков и албанцев Приазовья,
памирцев и киргизов Памира.
популяции X Vf Ps Im If Itot
Красная Поляна 1 3,00 2,8500 0,82 0,22 0,3167 0,6060
II 2,125 1,5424 9,78 0,28 0,3416 0,7179
111 2,372 1,8940 0,80 0,25 0,3365 0,6706
Староласпа 2,609 2,8600 0,69 0,45 0,4305 1,0587
2,36 1,64(М> 0,89 0,12 0,2927 0,4525
III 2,415 2,0300 0,79 0,26 0,3483 0,7067
j Девнинское 2,222 2,4180 0,61 0,64 0,4897 1,4420
II 2,1940 1,8180 0,75 • 0,33 0,3776 0,8368
III 2,204 1,9760 0,71 0,41 0,4070 0,9816
Георгиевка I 2,905 3,6640 f 0,61 0,64 0,4343 1,3512
11 2,407 1,7950 0,82 0,22 0,3097 0,5972
III 2,625 2,5520 0,71 0,41 0,3704 0,9301
Пастхуф 5,970 8,9700 0,65 0,54 0,2500 0,9200
ХуФ 5,320 10,690 0,65 0,54 0,3800 1,1200
Мургаб 4,4.30 6,2600 0,41 1,44 0,3500 2,2900
Примечание: I-ранняя возрастная когорта (женщины,
завершившие репродукцию к 1960 г.), И-поздняя (женщины,
завершившие репродукцию в 8О-е годы), 111-показатели по
популяции в целом;
Xs - среднее число детей, приходящееся на одну женщину; Vf -
дисперсия среднего числа детей; Ps - доля потомков, доживших до
репродуктивного возраста; Im - компонента дифференциальной
плодовитости; Itot - индекс потенциального отбора.
В табл.42 представлены индексы потенциального отбора и его
компоненты, связанные с дифференциальной смертностью и
дифференциальной плодовитостью в различных популяциях
земного шара. Как видно из табл, отмечается широкая вариация
значений индекса Кроу в анализируемых группах. С наибольшей
144
интенсивностью действие отбора проявляется в человеческих
коллективах, находящихся на низших ступенях социально-
экономического развития, так. в примитивных племенах,
например, у яномама оно достигает максимума hot = 4,24 (Neel,
1975). Также давление отбора достигает довольно высокой силы в
группах, живущих в экстремальных условиях среды. Отмечается
достоверная корреляция значений индексов потенциального
отбора с высотой местообитания популяций над уровнем моря
(рис. 7). Например, у памирцев кишлака Пастхуф (среднегорье,
высота 2 тыс. м) Itot = 0,92; памирцев кишлака Хуф (высота 3 тыс.
м) ltot= 1,12, киргизов села Мургаб (высота от 3800 м до 4200 м)
Itot = 2,29 (Спицына и др., в печати). Жизнь на высокогорье
выдвигает необходимость выработки механизма адаптации к
тдким жестким природным условиям как: резкие перепады
суточных и сезонных температур, пониженное парциальное
давление кислорода, сухость, повышенный естественный фон
радиации, инсоляции и др. А свыше 4 тыс. м над уровнем моря
подъем вверх на каждые сто метров связан со значительным
ростом экстремальности среды.
Так, в популяции индейцев кечуа (Перу, высота 4 тыс. м)
Itot=0,83, а у индейцев аймара (Чили, высота 4300 м)
интенсивность отборпа почти в два раза выше Itot = 1,58 (Крус-Кок,
1981).
Как видно из табл.42 башкиры характеризуются одной из
минимальных величин индекса Кроу Itot=0,82. При этом
соотношения компонент дифференциальной смертности и
дифференциальной плодовитости почти равны и составляют 0,35
и 0,34 соответственно. Факт этот, с одной стороны
свидетельствует об ослаблении
145
Таблица 42
Индексы потенциального отбора (Itot) и его компоненты,
связанные с дифференциальной смертностью (Im) и
дифференциальной плодовитостью (If) в различных популяциях
земного шара.
Популяция । Im » If/Ps 1«)| Источник
1.Население США 0,03 0,89 0,92 0,95 Neel, Chagron, 1968
европейск. происх. 2.Население США (за 0,05 1,34 1.41 1,46
исключением европеоидов) З.Кашнначуа (Перу) . 0,79 0,11 0,98 Jonston, Kesinder, 1971
4.Снн-Пабло (Мексика) 1,63 0.31 2,46 Neel and Chagnon, 1968
| 5.Скот оводы Чили 1,38 0,17 1,78 Crow. 1966
1 б.Тлакеала (Мексика) 0,59 0,35 1,14 • Halhcrstein and Crawfopn 1972
7.Населенне Франции,1Ч)Н 0,26 0,84 1,32 Jacquard, 1974
({.Население Австралии, 0,25 0,40 0,75 Cavalli-Storza, Bodmer, I
1 ЧОО 1971
9. Горожане Чили 0,15 0,45 0,67 Crow, 1966
Ю.Население Юты США) 0,07 0,47 0,57 Tyzzer, 1974
1 Ыаселеиие Аш лни и 0,04 1,21 1,29 Sphuler, 1963
12.с. Марагатерья У >л1и ‘ 0,22 0,47 0,80 Bernis, 1974
13.с Кампродон' Испания i (Испания। 0,04 0,28 0,33 Torrejon and Bertranpetit, 1987
14. с. Ногалес (Испания) 0,14 0,47 0,67 Fuster, 1982
15.Сельское население. 0.33 0,22 0,62 Crow, 1966
,, .. Чили .. , _ 16.с. ААЫ1уджар|)а,Нс11;п1ия(1,13 0,30 0,47 Luna F., and Moral, 1990
17.Индейцы кечуа .Перу ! (4000м) 0,52 0,20 0,83 Крус-Кок, 1981
| 18,Навахи, США 0,37 0,52 0,71 1,07 Morgan, 1973
| 19.Индейцы аймара, Чили 1,18 0,18 1,58 Крус-Кок, 1981
1(4300м) ' 20. И идей цы шаванте, 0,49 0,41 0,90 Neel, Chagnon, 1968
Ю.Америка 21. И золят в Бразилии 0,28 0,20 0,53 Magalhaes, 1987
22.Яномама, Ю.Америка 1.22 1,36 4,24 Neel, 1975
23.Банту (Уганда) 1,70 0,54 2,46 3,16 Neel, Chagnon, 1968
24.Сельская группа, найка, Гуджарат, Индия 0,31 0,13 0,48 Padmanabham, 1985
146
Продолжение таблицы 42
25.Урбанизированная 0,46 0,26 1 0,83
группа, найка, Гуджарат, Индия Зб.Сельское население 0,45 0,22 0,32 0,77 Neel, Chagnon, 1968
Восточный Пакистан 27.Москва, 1975 0,05 0,37 0,39 0,44 Курбатова, 1977
28. Русские, г.Вологда 0,19 0,35 0,60 Ревазов, 1984
29.Русские, г.Галич 0,10 0,37 0,52
ЗО.Русские, г.Шарья 0,11 0,36 0,51 м
31.Русские, г.Егорьевск 0,30 0,15 0,19 0,49 Спицын, (в печати)
32 Русские, Пинега 0,69 0,50 0,85 1,54 Ревазов, 1979
33.Русские, Архангельская, 0,45 0,54 1.23 Большакова
Вологодская обл. 34.Греки, с.Красная 0,25 0,34 0,42 0,67 Спицына, данное сообш.
Поляна Украина 35 Г реки, с.Староласпа, 0,26 1 0,35 0,44 0,71
Украина 16.Албанцы, сДевнинское, 0.41 0,41 0,57 0,98
Украина '37.Албанцы, с.Георгиевка, 0,41 0,37 0,52 0,93
Украина 38. Башкиры 0,35 0,34 0,47 0,82 Спицына, данное сообщ.
ЗЧ.Памирны, к.Пастхуф 0,54 0,25 0,38 0,92 Спицына и др. (в печати]
(Памир, 2000 м) 4О.Памирцы, Хуф (Памир, 0,54 0,38 0,58 1,12 *«
2000 м) 41 Киргизы, с.Мургаб, 1,44 0,35 0,85 2,29 и
(Памир, 3800-4200 м) 42.Горцы Зап. Памира 0,26 0,45 0,57 0,83 Дементьва, 1974
43.Популяция г.Алма-Аты 0,13 0,38 0,56 Святова, 1989
44.Туркмены, к.Нохур 0,33 0,45 0,93 Ревазов и др., 1984
45.Узбеки сельские 0,38 0,41 0,95 Большакова, 1986.
46.Нганасаны, п-ов Таймыр 1,17 0,18 0,39 1,56 Гольцова, 1979
47. Якуты, Якутская АССР 0,82 0,23 0,41 1.23 Боева (в печати)
48Десные ненцы 0,79 0,20 1.15 Абанина, 1983
49 Ханты 0,45 0,24 0,80 Пуэырсв, 1988
,50.Эскимосы Азии 0,82 0,22 1,22 Соловенчук, 1989
|-<’ Эскимосы Чукотского 1,04 0,15 1,35 Вибс, 1990
п-аа *2.Береговые чукчи 0,96 0,20 1,36 Соловенчук, 1989
Э-Оленные чукчи 0,60 0,21 0,94
1 Коряки 0.54 0,31 1,01 II
Эвены Чукотки 0,59 0,22 0,94
56 Эвены Якутии 0,30 0,25 0,32 0,62 Посух и др., 1990
Т47
Продолжение таблицы 42
57.Эвенки, Ср.Сибирь 0,82 0,46 0,84 1,66 Рычков и др.. 1974
58Хакасы (кызыльцы) 0,23 0,35 0,44 0,67 Казаченко. 1986
59Хакасы (сагайцы) 0,22 0,39 0,48 0,70
бО.Буряты, о.Ольхон 0,20 0,32 0,59 СПИЦЫН И Др. (В IH '1.1 ги]
61.Буряты, с.Гоханы 0,33 0,29 0,72
62. Буряты, Агинский бурятский АО 0,24 0,56 0,94
63.Буряты, Иркутская обл. 0,26 0,30 0,64 .. ..
64.Нивхи, о.Сахалин 0,28 0,11 0,14 0,42 Казаченко и др., 1983 d
65.Русские, Краснодарский край 0,19 0,23 0,40 Голубцов, 1991
66. Адыгейцы, Краснодарский край 0,22 0,27 0,43 - _J
вклада дифференциальной смертности в поддержание,
стабильности возрастной структуры и численности башкир. С
другой стороны - о значительном влиянии социальных
преобразований, происходящих в Башкирии, а также в снижении
детской смертности. Аналогичный эффект был получен
Тимаковым В.В. с соавт. (1991), показавшими, что компонента 1m
за период с 1926- 19Н7 гг. для СССР в целом уменьшилась более;
чем в 1О раз и в настоящее время по союзным республикам
варьирует от 0,020 до 0,094 для городского населения и от;
0,030 до 0,121 для сельского. Сходная тенденция отмечается
также в популяциях греков и албанцев Приазовья (Спицына, в|
печати).
Полученные нами значения интенсивности максимально
возможного отбора являются одним из доказательств^
формирования благоприятной демографической ситуации, также
выработки адаптивных комплексов в исследуемой популяции
башкир.
148
Рис.7. Возрастание значения индекса максимального возможного
отбора у коренного населения Памира в зависимости от высоты
над уровнем моря.
На рис.Я представлено графическое распределение популяций в
зависимости от соотношения величин компонентов отбора; на оси
ординат отложены значения компоненты дифференциальной
плодовитости, на оси абцисс - значения компоненты
лифференциальной смертности. В соответствии с величинами этих
компонентов весь имеющийся массив мировых данных (табл. 42)
Распался на три комплекса, которые взаимно-перекрываются.
входная картина была получена исследователями при анализе
меньшего числа популяций (Luna at al., 1990).
149
1.5
a 0,5
Рис.8. Распределение популяций
(компонентов отбора).
1.0 1,5
в пространстве признаков
Комплекс I образован популяциями США, Англии, Франции,
Австралии, Москвы, Алма-Аты, т.е. урбанизированными,
индустриально высокоразвитыми группами. Популяции
характеризуются большой численностью, а также практикой
планирования размеров семьи. В них отмечаются низкие значения
компоненты дифференциальной смертности Im и высокие -
дифференциальной плодовитости If. Комплекс И наиболее
представителен с наивысшей плотностью популяций. Образован в
основном сельскими популяциями городского типа и городами
малой численности: Галич, Шарья, Вологда, Кострома, Егорьевск,
Пинега, Испания (село), греки Приазовья, башкиры, узбеки
(кишлак I loxyp), туркмены, хакасы, нивхи (Сахалин) и др. Помимо
свойственной им средней численности наблюдается также
отсутствие или не жесткая детерминация контроля над
рождаемостью. Характерны также низкие и средние (в мировом
масштабе различий) значения компонент дифференциальной
смертности и плодовитости, причем их вклад в значени»
150
тотального индекса отбора hot приблизительно одинаков.
Комплекс III образован популяциями, находящимися на более
низкой ступени экономического развития (охотники, рыболовы,
собиратели, примитивные племена), или расположенные в
экстремальных экологических условиях среды: индейцы кечуа,
аймара. Уганда, Чили, памирцы и киргизы Памира, нганасаны,
чукчи, якуты, коряки, эвенки Средней Сибири, эскимосы Чукотки,
лесные ненцы, яномама и т.д. Этому массиву свойственны высокие
и низкие и средние значения дифференциальной плодовитости.
Представлен популяциями средней, малой численности и
изолятами с естественным характером репродукции и традицией
нерегулируемой рождаемости. Если на полученную картину
распределения популяций на рис. нанести значения индексов
максимально возможного отбора, то получим U-образный тип
распределения индексов, где комплексы популяций I и III имеют
наивысшие значения, а средний (II) - наименьший.
Таким образом, в урбанизированных, высокоразвитых в
индустриальном отношении популяциях уровень максимально
возможного отбора выше, чем в городских с небольшой
численностью и сельских городского типа популяциях. Вклад
компоненты дифференциальной смертности в них минимален, и, в
основном, величина коэффициента Кроу определяется
различиями в эффективной плодовитости. Можно сделать
предположение о существовании "экологического оптимума" для
популяций, непосредственно связанного с историей
происхождения, численностью оптимальной в конкретных
условиях, а также характером и скоростью воспроизводства.
Гигантские городские агломерации - явление в эволюционном
плане возникшее относительно недавно, и, естественно,
интенсивность отбора в них выше в основном за счет различий в
'Ффективной плодовитости, т.к. в них велик вклад социальных
факторов (более высокие в сравнении с сельскими популяциями
Уровни сердечно-сосудистой, онкологической, профессиональной
патологии, нервных заболеваний и др.). Следует также упомянуть
° возможном влиянии аутбридинга - явления в большей мере
свойственного городскому населению, итоговое действие
которого приводит к росту гетерозиготности в популяции (Бочков,
1у«4; Святова и др., 1988; Куандыков и др., 1988 и др ). В качестве
151
другого механизма, способствующего поддержанию
генетического разнообразия, можно назвать существование
репродуктивной компенсации, что особенно ярко проявляется в
случаях неблагоприятных исходов беременностей. В популяциях с
естественным характером репродукции компенсирование
происходит за счет укорочения интергенетического интервала, в
группах практикующих планирование размеров семьи
наблюдается и увеличивается контингент женщин рожающих в
более старшем возрасте (Большакова, 1989).
Изучение нами распределения фенотипических и генных частот
системы -антитрипсина (ингибитора протеаз) среди коренных
жителей Памира, проживающих на высокогорье, с
одновременным биодемографическим обследованием показало,
что в браках, где один из супругов является носителем редкого
варианта Р1, отмечено статистически значимое увеличение
благоприятных исходов беременности (снижение выкидышей и
мертворождений), а также низкое число умерших детей.
Результаты свидетельствуют о репродуктивной компенсации,
поддерживающей уровень генетического разнообразия по генам
PI в популяции (Спицын и др., 1989).
Таким образом, изучение разных сторон сложнейшего
процесса адаптации популяций к конкретным экологическим
условиям среды представляет не только огромный теоретический
интерес, но необходимо при планировании и организации медико-
генетической помощи населению.
5. Ступенчатый характер 'енных миграций - как наиболее
адекватная модель для популяции башкир.
Одной из наиболее значительных систематических сил
эволюции, противодействующих изоляции, и, как следствию
подразделенности популяции - инбридингу, являются миграции!
История развития человечества неразрывно связана с миграцией *<{
процессом, идущим из глубины веков, с освоения человеком
ойкумены, переселения людей, групп и целых народов на новые
152
территории и до настоящего времени. Естественно, что менялся
характер, направление и интенсивность этого процесса.
Миграции, как фактор популяционной динамики, влияют на
генетические процессы, определяя степень генетической
дифференциации и эволюционной пластичности системы.
В реальных популяциях совершаются сложные процессы
взаимодействия, смешения, итогом которого в генетическом
смысле является обмен генами, приводящий к возникновению
совершенно новых комбинаций генов. Изменение частоты того
или иного гена под влиянием миграции во многом зависит от
числа иммигрантов и от частоты гена среди них (Харрисон и др.,
1979). Поэтому определение величины миграционного потока
представляется необходимым при установлении генетической
структуры популяционной системы. Кроме того, анализ брачно-
миграционной структуры позволяет оценить истинные границы
исследуемых популяций, степень их замкнутости и интенсивность
связей с окружающими популяциями (Кадошникова и др., 1991;
Кучер и др., 1991).
Вклад генов, полученный популяцией от мигрантов может
быть различным, т.к. зависит от структуры присущей самим генам
и степень информативности их может колебаться даже при равных
абсолютных числах, выражающих отношение мигрантов к общему
числу индивидуумов (от аутбредных браков до смешения в
пределах одного дема). При одинаковой интенсивности
генетическая эффективность миграции тем значительнее, чем
больше генетическое различие между популяциями,
обменивающимися мигрантами.
Пропорция генов, полученная популяцией извне, заключена в
коэффициенте миграции m (Kimura, Weis, 1964; Malecot, 1950) и
иногда 2m (Cavalli-Sforza, Bodmer, 1971), тогда как (1 - m) - это
пропорция генов, воспроизведенная в популяции основной
группой.
Существуют разные математические модели миграции, выбор
которых зависит от структуры генных миграций в объектах
исследования. Так, к популяционным системам, занимающим
сравнительно большие площади, более приемлема модель
Дифференциации островного типа, включающая 3 возможных
п°дтипа взаимодействия, которые объединены предельным
153
упрощением, допускающим полную независимость миграций ой
расстояния. Эффективность миграции определяется только!
числом источников миграции, независимо от того, близки или
далеки популяции друг от друга (Wright, 1943). Области
применения модели миграций островного типа различны - от
одновременного изучения ряда сходных изолятов, до популяции,
распространенной на обширной материковой территории.
качестве примера анализа структуры сходных изолятов можно
привести работу по изучению 12 популяций в Японии - из них 7
расположены на островах, а 5 - на горных пастбищах страны.
Островная модель миграции пригодна в этом случае потому, что
изоляты не обмениваются мигрантами с материка и не зависят
друг от друга. По данным о распределении групп крови системы
АВО получена оценка коэффициента сходства или F$T
подразделенности равная 0,0024:
.2, -1
fe = '>/ pq = (1 + 4Nm)
отсюда, m = O,O6 при среднем эффективно-репродуктивном
объеме Ne=196O (Nei, Imaizumi, 1966).
При исследовании совокупностей внутри дискретных и
распространенных на большой территории популяций более
корректным является применение линейных моделей (Ли, 1978;
Fix-Alan G., 1978; Morton et al., 1973, 1976; Gace T„ 1979).
Ленточный тип миграций (одномерный, двухмерный и
теоретически изучаемый трехмерный варианты в приложении к
популяциям непрерывного и прерывистого типов) более
приближен к реальности, в сравнении с островным. (Wright, 1943,
1946, 1951; Malecot 1966, 1967; Kitnura, Weis, 1964). В то же время ।
отмечается односторонность детерминистических работ Райта и
Малеко по влиянию миграций (Morton, 1972).
Популяционная структура башкир близка к модели
пространственной подразделенной популяции, предложенной
Малеко; микропопуляции, составляющие систему конечны И
имеют различную плотность в пространстве, интенсивно
обмениваются мигрантами. С увеличением расстояния обмен
генами уменьшается - об этом свидетельствует величина
коэффициента корреляции г=О,366. Географической
154
особенностью территории, занимаемой популяцией, является
срединное положение Уральских гор, рассекающих ее с севера
почти до южных границ. Все это определило выбор одномерной
ступенчатой модели миграций прерывистого типа (близкий к
крайнему варианту - кругу) представляющий в реальности
полукруг, огибающий горный массив (рис. 9).
Миграцию предполагаем изотропической, т.е. величина
коэффициента миграции m складывается из т/2 прямых и т/2
обратных миграций, причем, скорости их равны между собой.
Анализ круга брачных связей у башкир выявил одну из
основных особенностей брачного института - строгую родовую
экзогамию, которая для них традиционно характерна. Так, не
допускались браки между членами рода по отцу. Патрилинейный
характер генеалогического родства свойственен многим
тюркоязычным народам (Исмагулов, 1977; Лузина, 1987; и др.).
Анализ материалов по бракам у башкир, позволил
констатировать, что брачные ареалы их жителей в прошлом в
основном замыкались на определенных ближайших селениях в
пределах одного племени. В итоге это приводило к высокой
степени эндогамности внутри племен при отчетливо выраженной
экзогамности составляющих их родов. Поэтому изучение
миграции на этническом уровне племен позволило нам отказаться
от учета расстояния, как меры реальности шагов в матрице
миграций. Соотнесение мигрантов с тем или иным башкирским
родом, производили исходя из населенного пункта, где родились
их предки; этническая принадлежность коренного населения
поселков определена работами этнографов. При этом следует
отметить, что подвижность и многоступенчатость родо-племенной
организации башкир приводит к неустойчивости традиционных
показателей, поэтому для определения границ родоплеменных
образований служит целый комплекс признаков. "Следовательно,
когда мы пользуемся общепринятыми в исторической литературе
терминами "племя”, "род", "родовое подразделение", то мы делаем
тто условно, как бы "подводя" родо-племенную структуру под
классическую схему племенной структуры древних народов"
(Кузеев, 1974). К сожалению метод изонимии для определения
Родовой структуры, предложенный Ю.Г.Рычковым, неприменим к
155
популяции башкир, т.к. фамилии существуют в ней менее столетия ?
(Рычков и др., 1976).
Время внесло свои коррективы в сложившуюся в веках *
структуру браков. Происходящее повсеместно изменение границ
брачных ареалов наблюдается также и в сельских популяциях I
Башкирии. Растущая в стране урбанизация неизбежно связана с
оттоком молодежи в города на учебу и на работу. Так,
существенно растет число смешанных семей, а в одноэтнических ?
браках наблюдаются представители отдаленных районов
республики. В терминах демографической генетики в последние
десятилетия в популяции башкир наблюдается усиление
генетической эффективности миграции (Юмагужина, 1982).
Рис. 9. Схема одномерной ступенчатой модели миграций
популяции башкир
Восточные башкиры
.............Западные башкиры
Однако, учитывая, что вышеназванные изменения структу|
миграций касаются в основном представителей двух последи
поколений, при построении матрицы миграций на этничесК’
уровне мы сочли рациональным выделить миграцию меЖ
племенами - субпопуляциями в первый шаг, а миграции из друг
156
популяций расположенных вокруг — во второй. В таблице 43
приведена матрица миграций в субпопуляциях башкир.
Эффективная оценка миграций зависит от частот генов, т.е. от
степени родства: чем ближе между собой популяции, тем ниже
эффективность миграции и наоборот:
пъ = mi »(1-г) где
т, = i k - mt = 0,06006
। i
-1
(1- r) =77/ 2 . (loge. 4_) = 0,6785
'F2m <v‘
mi
Таблица 43.
Матрица миграции на этническом уровне субпопуляций восточных
башкир
Субпопуляции Еп Ех Ki К.2
1. Габын 0,9699 0,0301 0,0267 0,0033
2. Бурзян 0,9407 0,0593 0,0575 0,0018
3. Усерган 0,9302 0,0698 0,0662 0,0033
4.Тамьян 0,9408 0,0592 0,0535 0.0057
5. Кара гай-кипчаки 0,8945 0,1055 0,0964 0,0091
X 0,9306 0,0648 0,0601 0,0047
_ * 0,0003 0,0003 0,0002 0,000004
Еп - индекс эндогамии
Ех - индекс экзогамии
К1 - миграция между племенами
Кз • миграция из других популяций
х среднее значение величины, Sx - ее ошибка.
157
Таблица 44,1
Значение коэффициентов миграции по локусам групп крови АВО,
MN, Rhesus и белку гаптоглобину в популяции восточных башкир
Локусы 1 АВО MN Гены р- р q N 0,1600 0,0805 0,1757 0,0404 Нр Rh-Hr
Нр1 Rh(-)
0,1540 0,0845
отсюда, пъ = 0,0407, где
К - число шагов между популяциями;
ль - интенсивность миграции на К-шагов;
г» - генетическая корреляция популяций, разделенных на К-шагов. I
Как видно из расчетов, поправка на корреляцию частот генов
равная 0,6785, значительно ниже единицы и является основной
причиной уменьшения эффективной величины происходящей
миграции. С другой стороны, это также свидетельствует о
существовании положительной брачной ассортативности по
этнической принадлежности супругов приводя к обмену брачными
партнерами между разными селениями башкир.
В формировании структуры популяционной системы давление
миграций определяет группировку генных концентраций около
средних значений в системе, те. непосредственно влияет на
дисперсию средней частоты гена в сторону ее уменьшения (Ли,
1978). Поэтому можно вычислить значение коэффициента
миграции непосредственно через концентрации генных частот ]в
предположении стационарности их распределения:
= q-(l-q) ; отсюда m = яЛЫ) _ V4N
4 4Nm + 1
В таблице 44 представлены значения коэффициентов миграции
определенные через концентрации генов групп крови АВО, MN и
белка гаптоглобина. Как видно из таблицы 44 оценки показателей
миграции в популяции восточных башкир сильно варьируют л°
разным локусам. Аналогичную картину наблюдал Рычков •
популяции Сибири и Ливингстон - в Западной Африке (Рычков,
158
|Ц69; Livingstone, I960). Отмечается сходство оценки
коэффициента миграции пъ = 0,0407, полученной из
демографических данных, с величиной ш = О,О4О4 для локуса MN
С с]п. = 0,7040 и
=лп; - пц'’ =0,01162
°- •" п - I
Применение в качестве последнего этапа проверки полученной
величины nir в формуле определения вариансы частоты гена
локуса MN с введением гармонической средней величины
эффективного объема HNe=105 показало полное совпадение с
вариансой рассчитаной выше, что свидетельствует о правильности
построения миграционной структуры популяции восточных
башкир и реальности ее эффективного размера (Cavalli-Sfor/a,
Bodmer, 1971; Livingstone, I960; Cavalli-Sforza, 1986).
Таким образом, по своей величине популяция восточных
башкир относится к большой подразделенной на частично
изолированные субпопуляции с эффективной величиной
коэффициента миграции между ними пъ = 0,0407, т е. 4 человека
из р< продуктивного объема HNe=lO5 мигрирукхг из популяции
или в нее за поколение (25 лет). Такая же величина получена
исследователями у адыгов пъ=О,О4О7 (Кадошникова и др., 1991).
Интенсивность миграции у алтайцев несколько выше, там
происходит замена 5.5% генов за поколение (Лузина, 1987). В пять
раз ниже средняя интенсивность миграции между народами
Северной Азии (Рычков, 1973; Рычков и др., 1974; Воронина, 1983
и др.).
Полученная величина коэффициента миграции при анализе
одного поколения башкир, имеет длительную историю.
Имеющиеся этнографические, археологические, исторические
Данные подтверждают длительный миграционный путь
Древнебашкирских племен: Алтай, Западная Сибирь, Северный
Кавказ, Поволжье и неоднократная смена мест обитания уже на
территории Башкирии - весь этот длительный этап кочевого пути
сопровождается, как и в любой человеческой популяции,
"Роцессами миграции и смешения. Но все это не привело к
Достижению полного отождествления с окружающими народами,
очевидно, из-за отсутствия длительного периода соседства и
159
обмена мигрантами. С другой стороны можно предположить, что,
это было общение близкородственных групп племен- "плавающих,
островов" в людском океане.
Совершенно очевидно, что степень информативности ступеней,
менявшаяся в процессе истории, а также структура браков с
отчетливо выраженным стремлением к экзогамности внутри родов
и приводящая к эндогамности в племени - сформировали во
многом существующее своеобразие генетической струк гуры
популяции восточных башкир.
Учитывая возникшую в последние десятилетия тенденцию
изменения демографической ситуации у башкир, можно было бы
предположить высокое значение коэффициента миграции.
Однако, как видно на конкретном материале, сложившаяся
система браков, несмотря на значительное расширение круга
брачных связей в последние годы, в целом, оказывается довольно
устойчивой, и эта стабильность существенно снижает
эффективность миграций. Налицо существование
сформировавшегося динамического равновесия устойчивости и
изменчивости в поколениях. И при условии сохранения генного
потока данной интенсивности, а также при неизменности размеров
популяций, потребовалось бы длительное время для того, чтобц
совершились значительные изменения в генетической структур®
сельских популяций башкир. В действительности же оба ЭТИ
условия трудно выполнимы, так как происходящая повсеместна
интенсификация сельского хозяйства в Башкортостане, связана »
значительной мере с укрупнением хозяйств (равно и популяций),»
это, в свою очередь, неизбежно усиливает миграционную
активность населения.
И в будущем в популяции башкир можно ожидать дальнейший
рост эффективной величины миграций, с возрастанием доли
гетеролокальных и межнациональных браков.
160
ГЛАВА 7. ИСТОРИЯ ФОРМИРОВАНИЯ БАШКИРСКОГО
НАРОДА В СВЕТЕ ДАННЫХ ИСТОРИЧЕСКОЙ ГЕНЕТИКИ.
1. Таксономический анализ групп, выделение структурных
единиц и их взаимоположение в пространстве 'енетических
факторов АВО, MN, Rh, Нр, Kell систем.
Из этнической истории башкир известно, что они прошли
длительный путь развития. Поэтому изучение генетической
структуры популяционной системы на разных уровнях иерархии
направляется как в сторону выявления общих закономерностей,
так и установления связей между отдельными структурными
единицами системы.
К числу общих закономерностей, определяющих
формирование генетической структуры относится подчинение
генетических признаков закону нормального распределения.
Поскольку на практике весьма трудно проверить
многомерность действия нормального закона, то чаще всего
исследователи ограничиваются двумерным и одномерным
анализом его проявления. Мы сочли целесообразным
остановиться на последнем варианте анализа гистограммы с
дальнейшей проверкой степени их совпадения по известным
статистическим критериям (Cattel, 1965; Рычков, 1969, 1975).
Коротко, сущность метода состоит в следующем: интервал, в
котором заключены все наблюдаемые значения признака,
(первоначальные варианты) делят на несколько равных частичных
интервалов, практически, в каждый частичный интервал должно
попасть не менее 5-8 первоначальных вариант (Гмурман, 1972).
Число классов, на которые производят разбивку, определяется по
правилу Стаджерса (Терентьев, 1977). Соответствие действию
Закона нормального распределения определяли критерием
1 'ирсона, где К - число степени свободы находят из равенства:
К = S-1-r, где
" число групп (частичных интервалов) выборки;
" число параметров предполагаемого распределения, которые
’"Ределены поданным выборки.
161
Таблица 4
Эмпирическое распределение частоты гена г локуса АВО
аппроксимация нормальным распределением
Ч' По (<. fe Пе
0,40-0,45 1 0,0312 0,0310 1
0,45-0,50 5 0.1563 0,0940 3 0,5406 + 0,0605
0,50-0,55 4 0,1250 0,2500 « X. = 3,6969
0,55-0,60 13 0,4063 0,3440 11 d.f. - 3
0,60-0,65 7 0,2187 0,2190 7 р < 0,01
0,65-0,70 2 0,0625 0,062 2
32 1.0000 1,0000 32
Таблица
Эмпирическое распределение гена р локуса АВО и el
аппроксимация нормальным распределением
Чр По fo fc Пе
0,10-0,15 7 0,2187 0,1600 5
0,15-0,20 9 0,2813 0,2500 8 Чр- 0,1844 + 0,0671
0,20-0,25 8 0,2500 0,3600 9 2,2028
0,25-0,30 4 0,1250 0,1900 6 d.f. - 3
0,30-0,35 3 0,0937 0,0600 2 р < 0,01
0,35-0,40 1 0,0313 0,0300 1
32 1,1X100 l.(XXX) 32
162
Таблица 47.
Эмпирическое распределение гена q локуса АВО и его
аппроксимация нормальным распределением
' “ТЕнИТ q ано По fo ft Пе —
0,10-0,15 3 0,0937 0,0600 2
0.15-0,20 4 0,1250 0,2200 7 а лво-0,2131+0,499
0.20-0,25 15 0,4688 0,4100 13 х> 2,934
0,25-0,30 8 0,2500 0,2500 8 d.f. - 2
0.30-0,35 2 0,0625 0,0600 2 р < 0,01
32 1,(КММ) 1,0000 32
Таблица 48.
Эмпирическое распределение гена М локуса MN и его
аппроксимация нормальным распределением
qM По to fe Пе
0,40-0,50 2 0,0624 0,0600 2
10,50-0,60 3 0,0938 0,1200 4 Ям-0,6750±0,1060
0,60-0,70 7 0,2188 0,3500 11 тб- 3,4090
0,70-0,80 15 0,4688 0,3500 11 d.f. - 2
0,80-0,90 5 0,1562 0,1200 4 р < 0,01
32 1,0000 1,0000 32
163
Таблица
Эмпирическое распределение гена Нр локуса Нр и ei
аппроксимация нормальным распределением
qiip1 По fo Пе
0,25-0,30 3 0,0968 0,0600 2
0,30-0,35 8 0,2581 0,2200 7 ф1р-0,3500±0,0582
0,35-0,40 9 0,2903 0.3400 11 х’ь- 1,6314
0,40-0,45 9 0,2903 0,2500 8 d.f. - 3
0,45-0,50 1 0,0323 0,0600 2 р < 0,01
0,50-0,55 1 0,0323 0,0300 1
32 1,0000 1,0000 — 32
Таблица
Эмпирическое распределение гена D локуса Rh-Hr и е
аппроксимация нормальным распределением
qo По fo fe Пс
0,55-0,60 1 0,0312 0,0300 1
0,60-0,65 2 0,0625 0,0600 2 3d=0,7266+ 0,0697
0,65-0,70 3 0,0938 0,1600 5 х> 6,5778
0,70-0,75 12 0,3750 0,2500 8 d.f. = 4
0,75-0,80 5 0,1563 0,2800 9 р < 0,01
0,80-0,85 5 0,1562 0,1600 5
0,85-0,90 4 0,1250 0,0600 2
32 1,0000 1,0000 32
В таблицах 45-50 представлены и графически изображены
рис IO эмпирические и теоретически ожидаемые частоты rei
по локусам ЛВО, MN, Rh-Hr Нр в предположении нормал ьн<
распределения признаков в микропопуляциях восточных башкир
164
Рис. 1О. Распределение частот генов системы групп крови АВО
MN, резус, белка гаптоглобина в микропопуляциях башкир. 1 -
Эмпирическое; 2 - нормальное.
165
По всем вышеперечисленным локусам отмечается согласие
эмпирических распределений с нормальными с высокой
вероятностью р < 0,01.
Имея подтверждение действия закона нормального
распределения, были рассчитаны оценки математического
ожидания нормального распределения, которые, как и честно
являются среднеарифметическими величинами наблюдаемых
значений признаков:
X = XI 4- Xi 4- - 4- Хл
п
Для оценки генеральной среднгей совокупности применялся
метод выборочных средних:
Хв = XI 4-Х2 4-.,,4-Хп
п
Как известно, одной из основных свойств выборочных средних
при достаточном объеме является устойчивость их как
характеристик генеральных средних (Адлер и др., 1971; Бейли,
1964).
Результаты средних значений генных частот по локусам АВО,
MN, Нр, Rh-Hr в микропопуляциях башкир подчиняются действию
закона нормального распределения. Полученная
удовлетворительная аппроксимация эмпирических распределений
нормальными позволяет оценить основные статистические
параметры генетической структуры популяции в целом, как то:
средние частоты генов, доверительный интервал, в козором они
заключены, дисперсию оценки среднего числа, среднюю
квадратическую ошибку. Среди методов непрямой ординацИИ
методы анализа пользуются все большим признанием. Однако,
для проведения факторного исследования генетической структура
популяционной системы необходимо соблюдение целого ряДД
требований,таких как:
1. Независимость исходных данных.
2. Используемый набор наблюдаемых переменных подчиняется
многомерному закону нормального распределения.
3. Количество факторов не должно быть велико, обычно считай*
оптимальным количество, не превышающее 15.
166
4. Выборка исходных данных должна быть представительной
(Дубровский и др., 1974; Харман, 1972; Cattel, 1965).
Система микропопуляций восточных башкир полностью
удовлетворяет перечисленным выше требованиям:
применяемые в качестве факторов генетические признаки
имеют независимый тип наследования; общее число их равно 15, а
число независимых аллелей (n- 1) по локусам АВО, MN, Нр, Rh-
jlr, Келл-Челлано равно 8;
распределение генов в популяционной системе подчиняется
действию закона нормального распределения., проверка степени
их совпадения по критерию ^приведена в параграфе 2 настоящей
главы;
проведенное выборочное исследование в популяции восточных
башкир, согласно формально-статистическим критериям
достоверности оценок генетических параметров является
репрезентативным. Это позволило перейти к сравнению
микропопуляций восточных башкир по сумме мономерных
признаков методами числовой таксономии:
1. Метод М.Б.Малютова и В.П.Пасекова (1971):
/7 * г ~~
R =! i . larccos t (£,1 \ xik, yikjjг
/ £ .(Ni-1)
где M - число систем;
N - число аллелей а системе,
1 - каждая система,
к - каждый аллель,
х, у - генные частоты.
Па основании полученной мазрицы расстояний, построена
Дендрограмма с помощью взвешенного парно-группового метода
•9 микропопуляций образуют 406 сравниваемых пар). На рис. 11
Представлена дендрограмма микропопуляций восточных башкир,
Полученная по методу М.Б.Малютова и В.П.Пасекова и с
кластеризацией по Лансу и Виллиямсу (Малютов, Пасеков, 1971;
*-ance G.N., Williams W.T., 1966). Как видно из рис. 11 отчетливо
выявляется группирование микропопуляций в 6 кластеров,
167
которые объединяются в систему на разных этапах итерации,
соответственно которым произведено их обозначение.
На первой ступени итерации объединяются 4 кластера: 1.
Яумбаево, Набиево, Истамгулово, Байназарово, Н-Мунасипово; 2.
Туркменево II, Абзаково, Баишево, Ибраево, Басаево, Идияшево,
Н-Субхангулово, Сагитово, Ахмерово, Кипчак; 3. Елембетово,$
Кужаново, Салаватово, Миндегулово, Киекбаево, Гадельгареевор:
4. Бурунбаево, Чингизово, Утягулово, Гадельбаево.
На второй итерации микропопуляции Казарма и Байдавлетово;
образуют кластер 5, который вместе с кластером 6 объединяется
основной кластер 8, который вместе с седьмым завершаем
объединение популяционной системы восточных башкир.
Pi.с. 11. Дендрограмма микропопуляций восточных башкир
методу М.Б.Малютова и B.IТПасекова)
1-9 - образовавшиеся кластеры.
168
Рис. 12. Дендрограмма микропопуляций восточных башкир (по
методу Г.С.Розенберга)
1-10 - образовавшиеся кластеры
МП - уровни итерации
И. Метод Г.С. Розенберга (1976):
rij
где
rik - генетическое расстояние между объектами,
xij - коэффициенты сопряженности объектов,
N - количество объектов, включенных в анализ.
На рис. 12 представлена построенная дендрограмма
Микропопуляций восточных башкир по программе для БЭСМ-4
169
(Розенберг, 1976). Как видно из рис. 12 образуется 9 сгущений1!
(кластеров), из которых 7 объединяются на первом этапе итерации|
1: Чингизово, Гадельбаево, Буранбаево;
Миндегулово.Гадельгареево, Киекбаево, Ахмерово, Байназарове
Кипчак, Н-Мунасипово; 3: Набиево, Яумбаево, Идияшево; 4;
Туркменево II, Елембетово, Кужаново, Абзаково, Салаватово I
Басаеве, Байназарово; 5: Н-Субхангулево, Баишево; 6: Ишкинево
Казарма, Ибраево.
На второй итерации отмечается сближение микропопуляций
Утягулово и Сагитово.
На третьей - отмечается сближение Расулево и Истамгулово. '
На четвертом этапе построения дендрограммы завершается
объединение всех кластеров, образованных 29 микропопуляциями.
Полученная дендрограмма в целом соответствует предыдущей,
однако, классический метод построения дендрограммы выявляет
более ранние ступени дифференциации популяционной системы в
сравнении с методом Г.С.Розенберга и лучшую сопоставимость с
историко-этнологическими данными (Кузеев, 1974, 1978). Эго
подтверждает имеющиеся в литературе сведения о хорошей
сопоставимости дендрограмм, построенных по методам
Малютова-Пасекова и Кнуссмана на примере популяций Чукотки
и Камчатки (Алексеева и др., 1978), а также других популяций
(Sangvi, 1972, 1973; Leuscher D., 1974), история изучения которых
методами числовой таксономии началась с работ Р.Сокала И
Р.Снита (Socal, Sneath, 1963).
Возможная коррекция генных частот с расстоянием •
популяции восточных башкир была проверена анализом •
соответствии с методом, предложенным Маруяма, позволяющим
определить взаимосвязь между генной идентичностью W
географическим расстоянием (Maruyama. Т., 1973):
у = Ao. Ai . х , где
у - генетическое расстояние,
г - географическое расстояние,
Ao, А: - коэффициенты регрессионного уравнения (в анализ®
микропопуляций башкир Ао = 0,08055, А| = 0,000196). .д
На рис. 13 приведена полученная картина положительней
прямой зависимости генетических расстояний меЖДУ
170
микропопуляциями восточных башкир от географических.
Проверка криволинейности проводилась по формуле:
г х/у - г 0,01 , где
г х/у - прямое корреляционное отношение,
г - коэффициент корреляции,
г у/х = 'п \ (у/х Му)
\ х(у - Му)
где у/х - значения эмпирической регрессии,
М - среднеарифметическая ряда,
у - варианта ряда,
п - число вариант в частной группе (Зайцев, 1973).
Ijhc.13. Регрессионный анализ зависимости генетических
Расстояний между микропопуляциями башкир от географических.
х - географическое расстояние в км.
у - генетическое расстояние.
171
Как видно из рис. 13 значения, соответствующие 406 пара^;
возможных сочетаний, образованных 29 микропопуляциями
расположены в системе координат, где на оси х - отложено
расстояние в км, а на оси у - генетическое расстояние. Зависимость
в образованном полиноме, несомненно, прямолинейна, т.к.
кривизна равна 0,0000000283.
Однако существует большой разброс вокруг линии регрессии
что указывает на слабость связи между географическим и
генетическим расстоянием.
В литературе имеются данные о взаимосвязи между генной
идентичностью и географическим расстоянием в II кастовых
группах (микропопуляциях) Индии. Отметив существующее
отклонение 55 точек, образованных попарным сравнением II
групп, от линии регрессии (А = 0,015), авторы делают вывод о
том, что рассматриваемые касты отражают раннюю стадию
дифференциации (Malhotra К.С. et al., 1978). Сходные результаты
были получены и другими исследователями (Blanco R., et al.,1975;
Chakraborty R., 1976; Chakraborty R., et al., 1976).
Таким образом, проведенный многомерный статистический
анализ микропопуляций позволил вычленить популяцию
восточных башкир как сложную систему, иерархический тип
структуры которой сформировался в результате преемственности
поколений. Полученная величина коэффициента корреляции
генных частот с расстоянием г = 0,366, указывает на то, что дрейф
генов является не единственным фактором, определившим
произошедшую дифференциацию популяционной системы
восточных башкир.
Наблюдающаяся сложность генетической структуры
популяционной системы восточных башкир, представляющих
наиболее древний пласт в этническом составе башкир,
преемственность развития которого в поколениях анализируете^
нами в реально существующем множестве современных
башкирских микропопуляций, является свидетельством
длительной генетической дифференциации в процесс®
4» рмирования системы.
Для сравнения генетической сгруктуры различных популяций
по целому ряду независимых признаков удобно применение
простого и наглядного метода векторов или метода полигоне®
172
(Серебровский, 1970; Дебец, 1948). Этот метод применен для
анализа генетической структуры субпопуляций на уровне племен
восточных башкир. Сопоставляется материал по 7-ми локусам,
включающим 15 генов, из которых 8 независимы, т.е. могут
и пленяться независимо друг от друга в 8-ми направлениях.
Полученное качественное сравнение представлено на рис. 14. Как
видно из рис. 14 в форме полигонов приведена характеристика
субпопуляций бурзян, усерган, тамьян, табын, карагай-кипчаков и
всей совокупности восточных башкир по 15 генам (О, А, В, N, М,
Нр, 1 Ipr D, d, С, с, Е, е, К, к). Подобный анализ не проводился по
племени сальют, ввиду отсутствия данных о распределении в нем
антигенов системы Келл-Челлано. На полигонах по радиусам
отложены частоты генов по локусам АВО, MN, Нр, Rh-Hr, Kell;
частота генов, равная 1, помещена в центре круга. Частота
каждого отдельного гена в таком полигоне является средней
частотой в субпопуляции. При сопоставлении полигонов
выявляется устойчивая картина распределения генов, хотя они
могли изменяться в 8-ми независимых направлениях. Это служит
указанием устойчивости средних частот генов и возможную
знач1 тельную роль систематических факторов в формировании
генетической структуры популяции восточных башкир.
восточных башкир по локусам АВО, MN, Rh-Hr, Келл-Челлано и
1 лптоглобину.
173
Частота О - по периметру;
Частота 1 - в центре круга.
Я
Подобный вывод был сделан при сравнении генетическоЦ
структуры популяции коренного населения Америки с популяцией
Северной Азии (Шереметьева, 1976; Шереметьева, Рычков. 1978).-
Для определения сходства или различий в генетической
структуре популяций используются различные методы числовой
таксономии (Sangvi, 1953; Edwards, Cavalli-Sforza, 1964;
Balakrishnan, Sangvi, 1968; Kurczynski, 1970; Sangvi, Balakrishnan,
1972). Применение обобщенного генетического расстояния
одновременно по совокупности генетических маркеров позволяет
получить средний квадрат различий во всех независимых генных
частотах, подвергнутых угловой трансформации (Cavalli-Sforz^,
Edwards, 1967).
Ok = arccos Z \l xi. xi'
2_ g -где
e = z ok/n
к
xi и xi' - концентрации i-того аллеля в двух рассматриваемый
популяциях, а суммирование проводится по всем аллелям к-тоЙ
системы.
Этот метод применен нами для анализа различий Л
генетической структуре популяции башкир на уровне родов, как
наиболее подвижном звене родоплеменной организации*
свойственном кочевым народам (Кузеев, 1974). В таблице Я
приведена построенная матрица генетических расстояний при 8
степенях свободы, между субпопуляциями башкир
соответствующих родам. Как видно из таблицы 51, величин^
генетических расстояний находятся в интервале от 0,0379 Д°
0,1652 радиана, что позволяет предположить наличие
существенных различий в генетической структуре родовых
подразделений без формальной оценки величины самого
ра стояния, считая близкое к 0,0379 - малым, а к 0,1652 Д
большим.
2.
Наименьшее расстояние €) = 0,0379 наблюдается в пар® ;
усерган-карагай-кипчаки, наибольшее - между кара-табын и ямаДО-
174
175
0“ = 0,1652. Характеризуя величины обобщенных генетических
расстояний при рассмотрении каждого рода в отдельности следует
отметить, что карагай-кипчаки оказываются близкими к
усерганам (О = 0,0379), а также мунашам (0 = 0,1010).
Бурзяне без родовой принадлежности близки к мунаш (О =
0,0577) и существенно разнятся от табын и сальют. Род ямащ
одинаково близок как к роду усерган (О" = 0,0562), так и к
бурзянам без родовой принадлежности (О = 0,0743) и тамьянам
(0г = 0,0746). Такая же зависимость наблюдается и по роду
мунаш, где генетичес- ое расстояние с бурзянами без родовой
принадлежности равно 0,0577 радиан. Род усерган оказывается
ближе к карагай-кипчакам (0 = 0,0379) и мунашам, ямашам (О2
= 0,04X1), бишей (0 = 0,0615). Род бишей характеризует его
близость к усерганам (0 = 0,0615), сравнительно дальше он
отстоит от других родов. Внутри племени бурзян, объединяющего
три рода (ямаш, мунаш и бурзян без родовой принадлежности),
близки бурзяне без родовой принадлежности и мунаш (9 =
0,0577); роды ямаш и мунаш отличаются близостью как к
бурзянам, так и к роду усерган из племени усерган,
представленном парой родов бишей-усерган, генетическое
расстояние О = 0,0615. Это расстояние является наименьшим для
бишей, но усерган соотносится значительно ближе к родам из
других племен: карагай-кипчакам (0 = 0,0379), манашам (0 =
0,04X1). Характеризуя роды, являющиеся для нас единственными
представителями племен тамьян, сальют, табын и карагай-
кипчаков, следует отметить близость тамьян-мунаш (0 = 0,0581),
значительную удаленность кара-табын и сальют как между собой,
так и от других сравниваемых родоплеменных подразделений.
2. Сравнительный вклад европеоидного и монголоидного
компонентов в формирование популяции.
Вклад европеоидного и монголоидного компонентов •
формирование антропологического типа башкир был доказан на
основании всестороннего исследования палеоантропологического
материала, с помощью анализа антропометрически*»
176
антропоскопических, одонтологических и дерматоглифических
данных. Комплекс всей этой информации свидетельствует о том,
что башкиры занимают промежуточное положение между
европеоидными группами и центрально- и североазиатскими
монголоидами. Однако, при интерпретации этих данных возникает
целый ряд нерешенных вопросов. Во-первых, встает проблема
самой возможности количественной оценки соотношения
исходных расовых компонентов, принявших участие в этногенезе
башкир. Во-вторых, возможность непосредственной оценки
монголоидное™ и европеоидности в башкирской популяции. Как
справедливо замечено выше процесс формирования башкирского
народа был значительно более сложный и многокомпонентный,
чем проводимая рядом авторов констатация факта присутствия
европеоидного и монголоидного субстратов.
Развитие теории популяционной генетики, и в особенности
получение новой генетической информации на базе достижений
биохимической и молекулярной генетики - привело к новому этапу
в изучении этно- и расогенеза. Использование материалов по
геногеографии не только способствовало подтверждению ранее
полученных результатов в рамках проблемы этнической
антропологии, но и что самое важное, молекулярно-генетические
и генетико-биохимические данные содействовали получению
новых, важных, подчас неожиданных сведений, которые позволяют
по-новому читать страницы увлекательнейшей истории
формирования человека современного вида.
Рассмотрим отличие собственно генетического подхода от
традиционного антропологического,
С увеличением знаний о полиморфных генетических системах у
человека, становится ясно, что многие из них обладают особой
таксономической ценностью" для прослеживания исторических
взаимоотношений между различными антропологическими
группами. Накопленный в литературе материал о распределении
большого числа. серологических, иммунологических,
биохимических, а также молекулярных маркеров в популяциях
человека позволяет провести классификацию типов
распространения генных частот в зависимости от степени
полиморфизма и особенностей его пространственного
Распространения (Спицын, 1985). Согласно этой классификации,
177
устанавливающей 6 типов генетической изменчивости в
популяциях человека, для прослеживания и анализа этно-расовых
процессов представляют особый интерес следующие из них:
1) Эндемичные полиморфизмы, характерной особенностью
которых является наличие особых аллелей, или гаплотипов,
ограниченных распространением в определенных крупных
регионах. Такие аллели обнаруживаются с высокой частотой в
отдельных этнорасовых группировках, а среди прочего населения
Земного шара встречаются весьма редко или полностью
отсутствуют. Типичной иллюстрацией их могут служить гаплотип
cDe системы Rh, Gml’^ll^.l? , маркирующие негроидные
популяции Африки характерный фактор для
представителей южно-азиатской расы; Gn?'^найденный
только у европеоидов и достигающий среди них 8О%-ной
концентрации; Gm^: 13; 15,16 t и аллель d ушной серы сухой
консистенции, распространенные у центрально- и северо-азиатских
монголоидов. Именно такой тип изменчивости позволил
Шенфильду, Алексеевой и Кроуфорду установить в составе
северо-азиатских монголоидов наличие определенной, хотя и
незначительной пропорции южно-монголоидной примеси
(Schanfield, Alexeeva, Crawford, 1980).
2) Локальная изменчивость генных частот, при которой имеется не
только обычный аллель, но и другие аллели данного локуса,
распространенные с полиморфной или близкой к этому уровню
частотой. Распространение их также ограничивается
определенными этно-расовыми группами: 11Ь' встречается
преимущественно у негров, 1 lb1 распространен среди южно-
азиатских монголоидов;
RDS ген системы роданезы и тип альбумина At (Наскапи)
отмечается у североамериканских индейцев; Gm‘ ' I'"’, 21 харак-
терен только для айнов; Gml' 5, 1? , свойственен бушменам;
Gc^borC8 встречается у аборигенов Австралии и папуасов
Новой Гвинеи; АсР, присущ европеоидам (Спицын, 1985).
Использование именно этих наследственных систем, не говоря
уже о PDRF (полиморфизма длины рестрикционных фрагментов
ДНК) позволяет дать ответ на многие интересные,
противоречивые и важные вопросы этногенеза.
178
Немного теории. Если известно, что в сравнительно недавнее
чремя произошло смешение двух групп, то в ряде случаев
концепции генетики популяций дают возможность достаточно
точно оценить, в какой мере каждая из первоначальных групп
представлена в третьей, произошедшей от них популяции, или,
если известны первоначальные частоты генов и численность
смешивающихся групп, можно рассчитать ожидаемую частоту в
возникшей популяции:
qn - cq< + dqu
с + d , где
qn - ожидаемая частота гена х;
qc - частота гена х в популяции С;
qa - частота гена х в популяции D; с - число лиц в популяции С,
включенных в новую популяцию; d - число лиц в популяции D,
включенных в новую популяцию.
Таким образом, ожидаемая частота представляет собой
взвешенную среднюю из частот исходных популяций.
В работах по антропологии чаше возникает вопрос не о составе
некоторой гипотической производной популяции, а о данной
частоте гена qn, возникшей при смешении двух популяций с
частотами qc и qa. При этом необходимо узнать в каком
соотношении произошло смешение этих популяций. Выведенная
из предыдущей формула имеет вид:
С _ qn - Qd
с + d qc - qa (Ниль, Шелл, 1958).
Применив подобный подход можно решить вопрос, например, о
соотношении двух исходных расовых компонент в
рассматриваемой популяции.
Более сложные задачи, однако, требуют применения других
методов теоретической популяционной генетики. Так, метод
аппроксимации, предложенный Банке (A.Wanke, 1962) позволяет
определить относительный вес отдельных расовых компонент (в
процентном соотношении):
рС ° 1OQ , d
Z • dk ,где
179
v . (ci - xi) к=1 .где
ist
m - число независимых признаков;
I - исследуемая популяция;
г - число расовых типов, с которыми сравнивается популяция;
ci - частоты соответствующих аллелей в предполагаемых
исходных популяциях;
xi - частоты соответствующих аллелей в исследуемой популяции.
Нами были применены оба метода для исследования
антропологического состава башкирской популяции. Принимая во
внимание упрощенную модель, оценивающую доли
монголоидного и европеоидного компонентов в количественном
отношении у башкир, были использованы следующие
информативные локусы: 1) система Резус; 2) система Cerumen
(ушная сера); 3) система кислой эритроцитарной фосфатазы АсР. |
Теоретически возможно также использование для подобных
целей локуса эритроцитарной эстеразы Д (EsD), однако, изученная
выборка мужчин башкир показала, по-видимому за счет
недостаточной репрезентативности, весьма отклоняющиеся
частоты от теоретически ожидаемых значений. Поэтому данный
локус исключен из дальнейших расчетов.
Зная средние величины частот этих факторов в исходных
европеоидных популяциях можно оценить относительные доли
этих компонент в смешанной башкирской популяции.
Исходные величины частот аллелей для башкир, сравниваемые
с данными по европеоидам и монголоидам, представлены в
таблице 52. В анализ включены из мировых сводок о
распределении генных частот системы Rh в популяциях (Mourant
et al., 1976; Беневоленская, 1974). Подробная информация о
группах, включенных в анализ, приводится в пояснении к табл. 43.
Используя формулу:
- 9?
qi - q2 ,где
qt>- частота гена у башкир;
qi - соответствующая частота у европеоидов;
qj. —" — у монголоидов,
180
получаем следующие соотношения европеоидных и монголоидных
компонент в башкирских группах, приведенные в таблице 53.
Таблица 52.
Исходные частоты генов, включенных в анализ соотношения
европеоидного и монголоидного компонентов в башкирской
популяции.
1 Локус Ген евро- пеоиды МОНГО- ЛОИДЫ башкиры
Rh d cde (r) 0,36 0,037 0,2356 (Стерлибашево) 0,2379 (Архангельское) 0,2191 (Илишевский р-н) 0,25
Cerumen d* 0,217 0,910 0,5697 (Узунларово) 0,6594 (Ассы)
Ac Pi P r 0,058 ОДНИ 0,038
Gc Gc1T 0,13 0,49 0,25
Gcis 0,60 0,25 0,48
Duffy Fy° 0,4 0,9 0,55
Kell К 0,04 0,00 0,026
Tf 0,84 0,74 0,77
Принимая во внимание весь комплекс данных о
таксономической значимости генетических полиморфизмов,
изученных в башкирских группах, можно сформулировать вывод о
том, что соотношения европеоидного и монголоидного
компонентов в целом по популяции почти одинаковы с
несущественным преобладанием пропорции европеоидности
Некоторые отличия результатов, полученных на основании
анализа системы Cerumen можно объяснить двояким образом: а) в
отношении этой системы изучены группы, в состав которых
исторически входили более монголоидные родо-племенные
объединения; б) система Cerumen обладает существенно большей
таксономической емкостью и позволяет идентифицировать даже
самую незначительную долю монголоидной примеси в таких
Популяциях как эстонцы, марийцы и других.
181
Таблица 53
Распределение соотношений расовых компонент в башкирских
популяциях по отдельным локусам и в целом по совокупности
локусов.
Популяции башкир Локус Аллель или галлоти Пропорция П репорт I европеоид- монголом
п ности в % ности в %
1.Стерлибашево Rh d 61 39
2. Архангельское Rh d 62 38
З.Илишево Rh d 56 44
Популяция 1-3 Rh d 66 34
Башкирия (случайная AcPi P 63 37
выборка) У зунларово Cerumen d* 49 51
Ассы Cerumen d* 36 64
Башкирия Gc Gt e 66 34
Башкирия Gc Gc 15 66 34
Башкирия Duffy Fy1 70 30
Башкирия Kell К 65 35
Башкирия Tf Tfe< 30 70
Башкирия в целом по всем локусам (генам) X £ 57,5 42,5
Результаты по анализу локусов Cerumen и Tf в плане оценки
соотношения расовых компонент обнаруживают завышение
монголоидное™ в башкирской популяции в целом. В рамках
дискуссии можно предположитьб следующие объяснения этому
феномену:
1) хорошо известно, что локус Tf в существенной мере
подвержен в человеческих популяциях действию отбора, •
результате чего в экологических условиях Южного Урала •
большей степени благоприятным является тип TfC1;
2) локусы Cerumen и Tf в сильнейшей степени ассоциируют с
морфологичепскими признаками, эта связь также, возможно,
182
повлияла на эксцесс степени монголоидности в башкирской
популяции.
Тем не менее, только дальнейшие исследования в этом плане
по всему Башкортостану позволят с большей корректностью
ответить на поставленные вопросы.
Однако, простая констатация соотношения долей
европеоидности и монголоидности в данном случае вряд ли внесет
дополнительную информацию в исследуемую проблему (за
исключением точных количественных оценок этих компонентов).
Другой подход, частичное теоретическое обоснование которого
приводилось несколько выше, предусматривает анализ всех
возможных антропологических элементов, исторически
включившихся в состав башкирской популяции. Действительно, на
протяжении многих веков Южный Урал являлся областью
смешения и дифференциации множества племен, этнических
групп, отличавшихся в антропологическом отношении. Динамика
этого процесса прослеживается и в настоящее время, учитывая все
возрастающую миграционную активность населения, следствием
которого является усиление генного потока в восточном
направлении.
Историко-этнографические, археологические, палеоант-
ропологические, лингвистические материалы свидетельствуют о
наличии вполне определенных этнических компонентов,
диффундировавших в состав башкирской популяции. В процессе
изложения эти вопросы уже подробно рассматривались выше.
Нижним наиболее ранним предковым субстратом в процессе
этногенеза в данном регионе является финно-угорский, который в
последующие века воспринял как элементы, ассоциирущиеся с
"южной европеоидностью", так и центрально- и северо-азиатские
монголоидные компоненты. Более того, как было подчеркнуто
ранее, современная популяционная генетика в рамках ее
геногеографического аспекта, позволяет идентифицировать
наличие пространственно весьма отдаленных антропологических
черт (характерных генов) в составе аборигенного населения,
например. Северной Азии.
Поэтому воспользуемся методом аппроксимации Ванке для
Установления комплекса гипотетически возможных расовых
Компонентов в составе башкирского народа. К сожалению.
183
следует констатировать, что в Башкортостане до сих пор Не
проводилось изучение таких таксономически важны**
наследственных систем, как иммуноглобулины (Gm), система
тканевой совместимости (HLA), анализ которых пролил бы свет на
многие неясные моменты этнической истории башкирского
народа. Наряду с этим достаточной таксономической ценностью
обладает также гаплотипическая система резус, с помощью
которой в определенной мере охарактеризована башкирская
популяция.
Важную информацию в расо-этногенетическом плане может
также принести система ингибитора протеаз Pi (или 74-
антитрипсин ^-АТ). Частоты обычных субаллелей,
контролирующих синтез субтипов Mi, М2 и Мз оказываются
расоспецифическими, т.е. каждая из крупных расовых единиц
отличается своим особым сочетанием субаллелей Pi*Mi, Pi*M2,
Р1*М.з. Кроме того, эта система хорошо дифференцирует
популяции Европы от азиатских европеоидов, что имеет,
например, особое значение при установлении соотношения двух
различных европеоидных компонентов в составе исследуемой
группы населения.
В таблице 54 представлены особенности частот хромосом
системы Резус для 4 крупных расовых единиц, которые в той или
иной мере в разные исторические периоды могли оказать влияние
на становление башкир.
Таблица 54.
Характеристика особенностей распределения гаплотипов системы
резус для 4-х крупных расовых единиц Евразии в сравнении с
башкирами.
расовые группы Частоты гаплотипов системы Резус
CDe(R') cDE(R”) cde(r) cDe(Ro)
1.европеоиды Европы 0,43 0,16 0,36 0,022
2.европеоиды Азии 0,61 0,08 0,17 0,09
З.северные и центрально- 0,51 0,45 0,06 0,036
; азиатские монголоиды
4. южно-азиатские 0,78 0,18 0,00 0,056
МОНГОЛОИДЫ
З.башкиры (суммарно) 0,50 0,21 0,25 0,04
Примечание:
1 - шведы, финны, датчане, австрийцы, французы, итальянцы,
болгары, словаки, поляки, русские, украинцы, белорусы; (113
групп).
2 - туркмены, азербайджанцы, турки, таджики, персы, афганцы,
курды (76 групп).
3 - якуты, монголы, буряты, тувинцы, эвенки (38 групп).
4 - бирманцы, таиландцы, кхмеры, вьетнамцы, малайцы, южные
китайцы (59 групп).
Посредством метода Ванке оценим вклад этих четырех
компонентов в составе современных башкир. В таблице 55
приведены результаты проведенного анализа.
Из данных, представленных в таблице 55, явствует, что
пропорция европеоидности, определенная с использованием
системы Резус, также составляет 50%, как и по отдельным другим
системам (см. выше табл. 54). Однако доля монголоидности в
последнем случае (табл. 55) существенно снижается за счет ее
замещения" азиатско-европеоидным компонентом.
1Я'
Iаолица
Соотношения исходных компонентов в составе башкирской
популяции.
Исходные антропологические компоненты Пропорции компонентов в составе башкир (в %)
1.европеоиды Европы 49,86
2.европеоиды Азии 26,(М
3.северные и центрально- 17,00
азиатские монголоиды
4.к>жно-азиасткие монголоиды 7,10
Графическое представление
включенных компонентов
Примечание: перечень популяций,
включенных в 1,2,3,4 - приведен в табл. 54
I □ -европеоиды Европы
[•—европеоиды Азии
северные и цент-
рально-азиатские
монголоиды
L южно-азиатские
монголоиды
Однако здесь нет противоречий между результатами,
полученными двумя различными способами. Прежде всего не
вызывает сомнений наличие в башкирской популяции пласта,
связанного с большой индо-средиземноморской расой.
В аспекте рассматриваемой проблемы считаем необходимым
подчеркнуть, что исследования генетико-биохимического
полиморфизма (наследственной гетерогенности большого ряд* 8
ферментных и других белков), а также полиморфизма длины
рестрикционных фрагментов ДНК (ПДРФ), ядерной
митохондриальной ДНК - все эти материалы свидетельствуют 0
выраженной близости между азиатским европеоидным •*
монголоидным стволами человека современного вИД8,
186
( 1сдовательно, генетические различия популяций, принадлежащих к
91им расовым вариантам, настолько незначительны, что выявляются
иппь с помощью специальных приемов анализа и во многих случаях
,jrn два компонента генетически объединяются в один кластер.
Объединив данные компоненты у башкир получим величину 43%, что
и среднем соответствует монголоидности в башкирской популяции,
как и следует и»табл. 55 (42,5%).
Итак, популяционно-генетический анализ вносит принципиально
новую информацию о процессе метисации, сыгравшем, пожалуй,
основную роль в сложной истории формирования башкирского
народа.
3. Оценки времени генетической
дифференциации популяций.
Наиболее сложной проблемой исторической генетики является
получение временных оценок дифференциации человеческих
популяций. Это обусловлено целым рядом причин и, прежде всего,
сложностью миграционных процессов во всей истории становления
Homo sapiens.
Исследователями были предприняты работы как по определению
времени дифференциации огромных в пространственно-временных
координатах групп, так и для определения времени дифференциации
больших рас. Принимая во внимание пространственное
распространение расо-специфических гаплотипов Gm и
археологическую информацию была осуществлена попытка
определения динамики распространения носителей определенных
генных комплексов Gm во времени (Спицын, 1985). Наибольшие
перспективы в этом отношении открывают возможности
0('наружения огромной генетической изменчивости в локусах на
Уровне анализа ДНК, что позволит корректнее определить время
Дивергенции популяций разного иерархического уровня.
Чрезвычайная сложность этногенетических процессов в эпоху
Миграции тюркских образований в Среднюю Азию и Восточную
‘ нроиу не позволяет посредством традиционных методов
Исторических дисциплин достоверно установить те основные
187
компоненты, которые явились этнической основой формирования
башкирского этноса. В этом свете требуют подтверждения
интересные выводы об определяющем значении этнических связей
ранних башкир с тюркскими народами Алтая и Южной Сибири
(Киекбаев, 1958: Кузеев, 1974).
По мнению СИ.Руденко и некоторых других исследователей
сложение древнебашкирского этноса на Южном Урале произошло
еще до начала I тысячелетия н.э. (Руденко, 1955). Однако,
осуществленные в последние годы исследования в области
этнографии, археологии, антропологии, исторической лингвистике и
их сопоставление позволяют считать, что этноним "башкорт" в
Среднем Поволжье появился не ранее последней четверти I тыс. н.э.,
когда основная масса "великих болгар" отошла на запад, а другие
группы мигрировали на Среднюю Волгу, где позднее большей
частью были ассимилированы племенами, пришлыми из 11риаралья и
Западной Сибири (Акимова, 1968; Генинг, 1972; Кузеев, 1971). В то
же время этот тезис, касающийся этнической истории не только
башкир, но и других народов Волго-Уральского региона, нуждается в
дальнейшей аргументации с привлечением новых датированных
материалов.
Средневековые этапы этнической истории башкир, казалось бы,
известны значительно лучше, благодаря шежере и совокупности
источников, названных С.М.Абрамзоном и Л.П.Потаповым
"народной этнотенией" (Кузеев, 1960; Абрамзон, I ктгапов, 1975).
Однако, в связи с сильным акцентом, который многие
исследователи, особенно языковеды и археологи, придают
древнейшим и древним этапам этнической истории башкир (равно и
татар), роль печенежско-кипчакских племен в окончательном
сложении этнокультурных характеристик оказалась заметно
преуменьшеной (Ефимова, 1991; Халиков, 1991). Все это приводит к
мысли о необходимости генетического освещения динамики
происходивших процессов в плане этнической антропологии.
Большое значение приобретает в этом свете последовательная
реконструкция генетической структуры на разных этапах истории с
выяснением характера взаимодействия автохтонного и пришлого
компонентов, а также выявлением вклада каждого из них •
формирование башкирского народа.
188
Основными моментами, необходимыми для реконструкции
родословного древа восточных башкир, является уточнение
параметров эволюционного древа, согласно задачам
филогенетической таксономии: установление типа древа, характера
ветвления и временных характеристик реконструированных точек
ветвления (Майр, 1965).
Объем воспроизводящей части (Ne) определялся с учетом
дифференциальной плодовитости и неравного соотношения лиц
разного пола, находящихся в репродуктивном возрасте. При расчете
времени дивергенции использовали гармоническую среднюю
величину (HNe = 105).
При определении концентраций генов реконструированных
вершин родословного древа применяли гармонические средние
величины эффективно-репродуктивных объемов субпопуляций, даже
если в сравниваемой паре известно только одно значение (таблица
S6).
Анализ генетической структуры современных популяций
позволяет определить родословное древо, т.е. реконструировать
прошлую генетическую историю этнических групп: бурзян, усерган,
тамьян, табын и карагай-кипчаков. Мы опустили в данном анализе
сальютцев, учитывая массовый уход их в XVI-XV1I вв. в Зауралье и
ослабление этнических связей с вышеперечисленными племенами
(Кузеев, 1974). Реконструкция возможна потому, что колебания
частот генов в ряду популяций математически равнозначны
колебаниям их в поколениях (Рычков и др., 1973). Модель
генеалогического древа признает решающую роль генетического
Дрейфа в микроэволюционных преобразованиях, т.е. диффузный тип
Динамики изменения концентраций 7-ми локусов (15 аллелей, из
которых 8 независимы). При этом не учитывается влияние
систематических факторов, как то: отбора, миграций, мутаций. Таким
'словиям больше всего отвечают малые популяции, в структуре
которых отражена большая часть истории башкирского народа.
Сияние факторов генетической изоляции в них несравненно выше,
Чем в городских популяциях, что благоприятствует накоплению
*’икроэволюционных изменений и, в конечном счете, эволюции,
становление же этапов истории народа в приложении
^«логических принципов, есть не что иное, как исследование
^8°люции внутри вида. Так как исходными данными являются
189
концентрации генов, а в них фиксируется влияние систематически^
факторов эволюции, то систематическими ошибками можц^
пренебречь и считать модель полностью приложимой к конкретному
материалу. Существуют различные методы генеалогической
реконструкции (Cavalli-Sforza, Edwards, 1965; Edwards, 1967; Малюгов
Пасеков, 1971: Пасеков, Коростелев, 1971 и др ).
Имеющиеся в литературе сведения о проведенных
генохронологических реконструкциях популяции Аландского
архипелага, Восточных Саян в Сибири и Командорских островов
подтверждают соответствие модели для изучения малых
изолированных популяций (Ма.потов, Пасеков, 1971; Malyutov
Passekov, Rychkov, 1972; Рычков, Шереметьева, 1972).
Нами применен метод Малютова и Пасекова к популяциям
башкир с неравными эффективно-репродуктивными объемами и
данными по частотам генов 6 локусов групп крови, а также
сывороточного белка гаптоглобина.
В таблице 57 приведены исходные концентрации генов по локусам
ABO, MN, Up, Rh-Hr (1), С, Е) и Келл-Челлано в субпопуляциях
башкир. На рис. 15 изображено реконструированное родословное
древо восточных башкир.
Коротко изложим основные этапы реконструкции:
1. ('начала между каждыми двумя популяциями вычисляем
обобщенное генетическое расстояние, являющееся своеобразно#
мерой генетических ра зличий между ними (угловая трансформация я
радианах). В таблице 58 приведена построенная матрице
генетических расстояний.
Таблица 56.
Эффективно-репродуктивные объемы субпопуляций восточных
башкир (Н - гармоническая средняя)
1 Бурзян I Усерган 1 Тамьян Табын Кара гай- Восточные
HNe 1 кипчаки башкиры
104 113 101 107 103 105
190
Таблица 57.
[ енетическая характеристика субпопуляций восточных башкир (q -
средняя концентрация генов)
Гены Бурзян Ч Усерган q Тамьян q Табын q Карагай- кипчаки q
Г 0,5585 0,5322 0,5941 0,6076 0,5753
р 0,1977 0,2563 0,1552 0,2049 0,1816
q 0,2438 0,2115 0,2507 0,1875 0,2531
N 0,2334 0,2496 0,1432 0,5192 0,3333
М 0,7666 0,7504 0,8568 0,4808 0,6556
Нр1 0,3813 0,3916 0,3492 0,3523 0,4197
Нрй 0,6186 0,6084 0,6508 0,6477 0,5803
D 0,7621 0,7518 0,7400 0,7207 0,7276
d 0,2379 0,2482 0,2600 0,2793 0,2724
С 0,4920 0,5292 0,5114 0,5263 0,5082
с 0,5080 0,4708 0,4886 0,4735 0,4918
Е 0,1934 0,2473 0,2031 0,2192 0,2296
Iе 0,8066 0,7527 0,7969 0,7808 0,7704
К 0,0164 0,0135 0,0051 0,0098 0,0146
к 0,9836 0,9865 0,9949 0,9902 0,9854
Рис. 15. Реконструированное время дивергенции субпопуляций
‘«сточных башкир (в поколениях)
191
Таблица 58.
Матрица генетических расстояний (О ij) между субпопуляциями
башкир
1 .. _ 2 3 L. 4 _ .. _ . .. |_ 5
1.Бурзяне 0,0469 0,0538 0,1120 0,0510
2.Усерган — 0,0728 0,1068 0,0538
З.Тамьян — 0,1517 0,0922
4.Та6ын — 0,0755
5.Карагай- —
кипчаки
2. В предположении дихотомического характера ветвления, для
соотнесения 5 популяций с двумя родоначальными ветвями,
применили разделение на кластеры по методу Пасекова и
Коростелева (Пасеков, Коростелев, 1971).
р =р • ij = max
кол-во слагаемых
где Ni и Nj - эффективно-репродуктивные объемы исследуемых
популяций.
3. Определяем время расхождения популяций 1 и 2 по формуле:
02= t/4Ne
В таблице 59 приведены временные характеристики
реконструированных точек ветвления.
4. Вычисляем концентрации генов в реконструированной точке би
показатель ошибки концентраций генов.
d = t ВС_________(В, С - разошедшиеся популяции)
8. (Nb + Nc)
В таблице 60 представлены концентрации генов ®
реконструированных точках ветвления. 2
5. I (аходим генетическое расстояние О з-б = 0,06.
6 Определяем время расхождения популяций 3 и 6 из точки 8.
T9?
7. Уточняем общее время расхождения популяций 1,2,3 из точки 8.
'I' = Ь + max (t <, to)
8. Определяем концентрации генов и показатель ошибки вер-
шины 8.
9. Устанавливаем время расхождения популяций 4 и 5 из точки 7
аналогично (3).
1О. Определяем концентрации генов вершины 7 и показатель
•шибки полученных концентраций аналогично (4).
11. Находим генетическое расстояние 0'?» = 0,1086.
12. Вычисляем время расхождения двух родоначальных ветвей из
очки 9 и общее время дифференциации исследуемых популяций:
Т = (ч + max (t») = 88,1 поколений.
Гаким образом, по схемам, предложенным Малютовым и
I (асековым, Рычковым и Шереметьевой, проведен
филогенетический анализ субпопуляций башкир усерган, бурзян,
гамьян (древнебашкирский пласт), а также карагай-кипчаков и
табыи, имеющих сложную структуру. Реконструируемый по
генетическим и демографическим данным интервал времени
дивергенции прапопуляции башкир датируется III в. до н.э., что по
историческим сведениям является временем нахождения восточно-
поркских предков башкир в Центральной Азии. Однако на наш
взгляд наиболее интересна та часть древа, которая отражает время
формирования современного этнического облика подразделений
башкир с характерной для них генетической структурой: X век для
гамьян, XII в. для табын и карагай-кипчаков, XVI в. для бурзян и
усерган, что соответствует времени нахождения башкир на Урале.
Подводя основные итоги проведенной работы по изучению
истории формирования башкирского народа в свете данных
исторической генетики, следует подчеркнуть, что анализ
дифференциации древнебашкирского пласта является первой
попыткой приложения популяционно-генетических методов и данных
к проблеме этногенеза и этнической антропологии башкир. "В
отечественной литературе при фундаментальной разработке
проблем этнической антропологии и этногенеза популяционная
иенетика человека в связи с вопросами истории популяции
представлена лишь единично" (Шереметьева, Рычков, 1978, Вибе,
193
1аблица 59
Реконструированное время (t) в точках ветвления и общая оценка
времени (Т) в поколениях и годах родословного древа восточных
башкир
Точки ветвления Поколения II Число лет
U 17,3 433
t? 32,0 800
to 25,2 630
t« 45,6 1140
Ti 42,5 1060
T 45,6+42,5=88,1 2203 1
Таблица 60.
Генохронологическая характеристика прапопуляции восточных
башкир
[Гены точки 6 7 8
ветвления
древа
г 0,5458 0,5911 0,5694
р 0,2248 0,1929 0,1889
4 0,2281 0,2147 0,2390
1 N 0,2411 0,4263 0,1897
1 м 0,7588 0,5658 0,8062
1пр 0,3861 0,3856 0,3677
.Нр2 0,6137 0,6131 0,6317
lD 0.7571 0,7242 0,7486
Id 0,2428 0,2757 0,2512
Iе 0,5096 0,5171 0,5105
Iе 0,4900 0,4829 0,4893
|В 0,2184 0,2245 0,2108
е 0,7806 0,7755 0,7885
к 0,0150 0,0121 0,0095
к 0,9849 0,9878 0,9896
144
,991; Карафет, 1992). Подобное же положение наблюдается и в
(арубежной литературе, где также редки работы, сочетающие
популяционную генетику с проблемами этногенеза.
Популяция башкир занимает .особое место в Волго-Уральском
регионе , т.к. многовековой исторический путь народа таит в себе
много неясного и дискуссионного. В аспекте биологии - это
ложнейшая проблема адаптации кочевого в прошлом народа к
меняющимся условиям среды. И в конечном счете, механизму
адаптации подчиняются все элементы популяционной структуры, как
енетическая, пространственная, временные аспекты, так и
вырабатываются специфические комплексы хозяйственно-
культурного уклада, оптимальные для предгорий Урала.
Изучение динамики роста численности башкир, а также хозяйств в
поселках за период с 1870 до 1974 гг. выявило более менее
равномерное увеличение численности дворов в селениях, количества
жителей и сохранение соотношения полов (1:1). Устойчивым
оказался средний размер семьи (от 5 до 6,3 человека). Следует
отметить еще одну особенность популяции восточных башкир - какой
является средний размер микропопуляций 428-497 человек,
пред.тавляющий исторически преемственную величину, по-видимому
определявшуюся в прошлом хозяйственно-культурным укладом
кочевников (Уйфальви, 1877; Соколов, 1904; Асфаган, 1929 и др.).
Соответственно размеру популяции объем эффективно-
репродуктивной группы составляет в среднем 101-107 человек, а
гармоническая средняя этих объемов по микропопуляциям
HNe=lO5.
Чем же объяснить устойчивость этих основных характеристик в
популяциях башкир? Как известно, из теории популяционной
генетики, значительное возрастание численности групп мало влияет
на ее эффективную численность, если за данным возрастание следует
'ответствующее снижение. Учитывая динамику численности башки):
XIX-XX вв. следовало бы ожидать количественно более
‘начительный эффективный объем, нежели HNe=lO5.
Следовательно, необходимо проанализировать действие всех
возможных факторов, могущих привести к уменьшению
эффективной численности башкир: неравное соотношение полов,
Дифференциальная плодовитость, колебания численности в
поколениях, инбридинг, безбрачие, бесплодие, детская смертность и
195
др. Но, как подчеркивалось выше, среди причин снижающих Ne
популяции башкир нельзя привести неравенство лиц разного пола
остается рассмотреть возможность влияния инбридинга и различий в
плодовитости, как наиболее сильно влияющих факторов на данную
величину. Тщательное избегание кровного родства и поколенная
экзогамия - являются характерной особенностью структуры браков в
популяции башкир, поэтому отделенным кровным родством
определяющим истинную величину коэффициента инбридинга F, как
функции подразделенности, F=O,O148, можно пренебречь, считая ее
достаточно низкой (хотя и обладающей способностью накапливаться
в поколениях). В микропопуляциях башкир сложилась благоприятная
демографическая ситуация с индексом максимально возможного
отбора по Кроу 1=0,84 и соотношением компонент плодовитости и
смертности 0,34 и 0,35 соответственно. Это свидетельствует об
ослаблении вклада дифференциальной смертности в поддержание
устойчивости возрастной структуры, а также численности в
популяциях. В процессе воспроизводства нового поколения
определена картина, при которой дисперсия среднего числа
существенно превышает само среднее число детей, приходящихся на
одного индивида б"- > к, что свидетельствует о явлении
нерегулируемой рождаемости, которая, очевидно, и является
основным фактором, снижающим эффективно-репродуктивнуй
объем популяции башкир. Таким образом, размер размножающейся
группы популяции восточных башкир невелик, но достаточен для
устойчивого воспроизводства популяции и сохранения равенства
соотношения полов в репродуктивной группе.
Генетическая структура неразрывно связана с основными
параметрами демографической структуры популяции, которые во
многом определяют особенности ее формирования, т.к. генная
дифференциация тесно связана с самой историей происхождения.
Прямая корреляция их проявляется в том, что воспроизведение
нового поколения потомков поколением родителей в конечной
популяции неизбежно зависит от подбора родительских гамет и чем
меньше данная популяция, тем ограниченнее выбор. Формирование в
пределах ареала уникальной генетической структуры популяции
восточных башкир не являющихся исторически аборигенными в этом
крае, выработка характерных типов распределений частот генов
является и значительной мерс адаптивным процессом *
196
континентальным условиям Урала. Удовлетворительная
аппроксимация эмпирических распределений нормальными
позволила оценить основные статистические параметры
генетической структуры популяции: средние частоты генов,
доверительный интервал в котором они заключены, дисперсию
среднего числа, среднюю квадратическую ошибку.
Генетическая дифференциация, изученная на индивидуальном и
популяционном уровнях иерархии совокупности восточных башкир
I микропопуляции - поселки, субпопуляции - роды, популяция -
совокупность восточных башкир), наиболее выражена на низших
дупенях и снижается по мере усложнения структуры.
Рис. 16. Уровни иерархии в популяциях восточных башкир.
Частоты генов в совокупности восточных башкир практически не
отличаются от средних частот генов по микропопуляциям;
наблюдающаяся дисперсия, как мера уклонений от средних значений,
минимальна. Это может явиться указанием на устойчивость их во
времени, т.к. известно, что колебания частот генов в ряду популяций
математически разнозначны колебаниям их в поколениях.
Аналогичный вывод сделан Рычковым при исследовании Сибирского
изолята: "В совокупности, дифференцированной на субпопуляции,
чаибольшей устойчивостью во времени обладают средние значения
Г|о сумме составляющих, а не средние отдельных составляющих.
197
независимо от сложности
(Рычков, 1969).
генетической детерминации признаков
выделение
структуры
Таким образом, устойчивость параметров генетической структуру
популяции восточных башкир свидетельствует о древней истории
формирования, что подтверждает историческое
дроевнебашкирского пласта (Кузеев, 1974).
Наблюдающаяся сложность генетической
популяционной системы восточных башкир, преемственность
развития которых в поколениях проанализирована в реально
существующем множестве современных башкирских
микропопуляций, является свидетельством длительной генетической
дифференциации в процессе формирования системы. А выявленная
гетерогенность внутри субпопуляций по многим аллелям является
следствием происходящих процессов дифференциации.
Проведенный факторный анализ микропопуляций восточных
башкир позволил вычленить популяцию как сложную систему,
иерархический тип структуры которой сформировался в результате
преемственности поколений. Обнаруженная с помощью БЭСМ-4
корреляция генетических расстояний между микропопуляциями
может явиться указанием на местонахождение "эффекта
родоначальника" в пространстве в той мере, в которой можно
применить это суждение к кочевым в прошлом народам (Cavalli-
Sforza et al., 1964, 1967). Кимура и Вейс показали в своей работе
уменьшение корреляции частот генов групп крови системы ЛВО с
расстоянием между парами сравниваемых колоний или пунктов. Они
пришли к выводу, что если дифференциация происходит случайно за
счет генетического дрейфа, генные частоты близких областей имеют
тенденцию быть более похожими, чем частоты отдаленных областей
(Kimura, Weis 1964). Ней и др., изучая популяцию японцев, также
получил уменьшение корреляции частот генов системы групп крови
ЛВО, MN между префектурами с расстоянием. Полученные клины в
распределении частот генов он объяснил совместным проявлением
действия диффузного эффекта миграции внутри популяции с
географическим градиентом интенсивности отбора. Степень генных
различий изучалась Неем с помощью коэффициентов инбридинга,
полученных из ковариации генных частот, а также сравнением
наГ подасмых и ожидаемых корреляций между частотами генов (Net,
lm.<i?umi |Чбг.
Именно влиянием случайного генного дрейфа можно объяснить
наблюдающуюся неоднородность внутри субпопуляций башкир на
гНическом уровне родов и племен. Оптимальным для этого
процесса, видимо, является малый эффективный репродуктивный
обьем популяции (HNe=lO5). Анализ дрейфа генов, как функции
малого размера популяции, хорошо продемонстрирован в работах
«авалли-Сфорца и др., Рычкова Ю.Г., Гаджиева А.Г., и др. (Cavalli-
Sforza, Bodmer, 1971; Рычков, 1972; Гаджиев 1971, и др.). К факторам,
противодействующим дрейфу генов в подразделенной популяции,
относится в первую очередь систематическое давление миграций.
Для сравнения интенсивности генных миграций в Восточной
Башкирии приведем имеющиеся литературные данные по популяции
Сибири, которую можно рассматривать как рассеянный на огромной
территории малочисленный изолят, микропопуляции которого
разделены большими расстояниями (Рычков, 1973; Рычков и др.,
1974). Изучение структуры миграций общесибирской совокупности на
угническом уровне в предположении островного типа миграций,
выявило иерархическое строение этой системы со средней
интенсивностью миграции т=О,О1О7. На полную независимость
мигрций от расстояния указывает возможность сетевого характера
миграции, когда отдельные народы могли пройти большие
расстояния не обменявшись мигрантами с соседними популяциями.
Структура же самих миграций достаточно сложна, источник
генетической информации помещен внутри системы и функцию ядра
выполняют популяции тунгусов-эвенков и эвенов.
Анализ миграции в популяции эвенков Средней Сибири с
применением островной модели позволил Ю.Г.Рычкову с
сотрудниками определить величину коэффициента миграции
m=0,0607. Отмечая некоторую корреляцию величины ее с
расстоянием, была впоследствии применена ступенчатая модель
Кимуры и Вейса, где в качестве миграций извне был приняз
общесибирский коэффициент миграции т=О,О1О7, отсюда
91=0,0607 ± 0,0107.
Таким образом, отмечается своеобразие популяции эвенков, где
малочисленность связана с высокой степенью кровного родства и
Миграцией значительной интенсивности. Предложенная Райтом
теоретическая оценка меры генетической дифференциации была
также применена к изучению циркумполярных популяций в
199
предположении разных типов структур миграции в них. Как
контрольный способ проверки применяемых моделей Ю.Г.Рычковыц
предложен метод определения уровня генетической
дифференциации, независимый от генетических параметров, а также
от данных о размере популяции и характера миграций генов. Таким
источником могут служить сведения о родовой структуре
эвенкийской популяции, определенные по частоте изонимных браков.
Одним из примеров применения подобного подхода на практике
является работа Ф.А.Лузиной, исследовавшей популяции алтайцев
(1987).
Сравнение полученных стандартизованных варианс при
островной, ступенчатой моделях миграции со средней
стандартизированной вариансой по генным частотам, а также по
родовой структуре и удовлетворительная аппроксимация
эмпирических и теоретических распределений, привели к выводу о
том, что пространственная структура популяции эвенков Средней
Сибири соответствует промежуточному типу между островной и
ступенчатой моделями (Рычков, и др., 1976).
Определение интенсивности генных миграций в Восточной
Башкирии между субпопуляциями на уровне племен проводили
независимым от концентраций генов способом через
демографическую структуру популяции с применением ступенчатой
модели миграции, как наиболее приближенной к реальности в
популяции с ленточным ареалом и с учетом обнаруженной
корреляции генных частот с расстоянием (г=О,366). Следует
отметить, однако, что в популяции башкир только генетические
расстояния можно считать величиной связанной с историей
происхождения, поскольку географические расстояния для кочевого
в прошлом и полукочевого в XV’I1I-X1X вв. являлись непостоянными.
Современное расположение на территории, где максимальное
расстояние в сравниваемых парах равно 370 км, а также
относительно невысокое значение коэффициента корреляции генных
частот с расстоянием, позволяет придавать дрейфу генов
существенное значение в произошедшей дифференциации
популяционной системы.
Полученная небольшая величина эффективной оценки
коэффициента миграции тс=О,О4О7, означает, что 4 человека в
популяции с репродуктивным объемом HNe=1O5 мигрируют из
200
популяции или в нее за поколение (25 лет). Среди основных причин,
снижающих эффективную оценку миграции в популяции башкир,
следует назвать сложившуюся систему браков с ярко выраженной
поколеной экзогамией внутри родов, что в конечном счете
приводило к высокой эндогамности племени. Таким образом,
пределенная величина миграции с эффективным коэффициентом
миграции гпс = О,О4О7, естественно, не могла выступать в роли
(хзновной эволюционной силы, сформировавшей генетическую
структуру популяции восточных башкир и не могла
противодействовать дифференцирующему влиянию случайных сил
эволюции.
Популяция восточных башкир представляет собой большую
подразделенную на множество частично изолированных групп с
HNt=46O, и миграцией между ними mc=O,O4O7. С точки зрения
эволюции - это наиболее благоприятное состояние, так как изоляция
приводит к разным формам дифференциации, а миграции между
ними предотвращают полную фиксацию аллелей. Отбор и
мутационное давление в сравнении с большой популяцией сглажены
и не могут быть ведущими факторами эволюционных изменений.
Таким образом, все эволюционные силы действуют одновременно и
популяция развивается непрерывно (Ли, 1978). Однако, если по
!отельному объему популяцию восточных башкир можно отнести к
популяции средней численности, то по величине эффективно-
репродуктивного обз.ема, составляющей менее 1/3 от общего
HNe=105), она стоит значительно ближе к малым популяциям,
которые при миграции с коэффициентом те=О,О4О7, являются\
полуизолированными. И таким образом, анализ факторов
микроэволюции показал, что при данной репродуктивной структуре
именно случайные факторы эволюции являются определяющими, а
выработка адаптивных комплексов: средний размер микропопуляций,
шуизолированность, относительно невысокая скорость миграции и
'беспечивают генетическую пластачность системы популяций
меточных башкир.
К выводу о решающей роли генетического дрейфа мы пришли
также в результате анализа степени генетической дифференциации
'етодом обобщенного генетического расстояния по всем
‘’сследованным локусам между субпопуляциями башкир на
‘'бэтническом уровне родов и племен. Табл. 51. Происходящая
201
дифференциация обнаруживается на уровне субпопуляций системы
гетерогенность которых по многим локусам является мерой
дивергенции микропопуляций имеюших единое происхождение.
Полученные генетические расстояния между субпопуляциями
башкир достаточно велики и находятся в пределе от 0,0379 до
0,1652 радиан, соответственно время реконструкции - 88 поколений
т.е. восходит к IV в. до н.э., а временные характеристики
реконструированных вершин - X, XII, XVI вв. н.э.
Подобную картину больших генетических расстояний в заведомо
ограниченном и этнически однородном районе Восточных Саян
между тюркскими популяциями тофаларов и тувинцев получили
Рычков с сотрудниками и Малютов и др. (Рычков и др., 1969;
Malyutov et al., 1972). При этом величины расстояний находятся в
интервале значений от 0,0180 до 0,1302 и отмечается хорошее
соответствие временных характеристик историческим сведениям.
Напротив, предположение о решающей роли генетического дрейфа
в популяции Берингии не подтвердилось, т.к. полученные
генетические расстояния существенно снизили характеристику
времени дивергенции этой древней, порядка 2-13 тысячелетий по
историко-археологическим данным, популяции (Шереметьева,
Рычков, 1978).
Известное положение о необходимости сопоставление временных
характеристик реконструкции с историческим древом
подтверждается на примере двух популяций Командорских островов
и пяти популяций азиатских эскимосов и береговых чукчей п-ва
Чукотка. Полученное общее время дифференциации эскимосских и
чукотских популяций равно 9,7 поколений, хотя времй
преемственного существования одних только эскимосов на этой
территории должно оцениваться не менее чем 80 поколений.
Изучение произошедшей лингвистической дифференциации
эскимосов и чукчей подтверждает длительность процесса во времени
порядка нескольких тысячелетий. Метод также не оправдал себя при
определении типа ветвления древа. Это позволяет сделать вывод °
том. что дрейф генов является нс единственным фактором,
онре (еляющим генетическую структуру популяций северной части
Гихого океана. При имеющемся минимальном давлении миграций
m 0,00236 + 0,00432, которое достоверно не отличается от нуля’
,i i.ikac малом эффективном размере популяции с HNe = 72, силой.
способной противостоять дрейфу генов, является стабилизирующий
тип отбора, приводящий к эффекту сокращения времени
(Шереметьева, 1975; Шереметьева, Рычков, 1978).
Сопоставление полученных временных характеристик
реконструированного родословного древа восточных башкир с
историческим древом выявляет соответствие основным выводом
сторонников позднего прихода башкир на Урал (Кузеев, 1974, 1978).
Учитывая появление тюрок на мировой арене в VII в. н.э. эта
гипотеза, подтверждаемая нашими исследованиями, исторически
более реальна (Гумилев, 1967). Полученные при
генохронологической реконструкции временные точки: X в. для
тамьян, XII в. для табын и карагай-кинчаков, XVI в. для усерган и
бурзян, свидетельствуют о том, что окончательная дифференциация
произошла на территории современной Башкирии. Отмеченную при
реконструкции "молодость" вышеперечисленных племен, являющихся
весьма древними по историческим сведениям, можно объяснить
двумя причинами.
Первой, является возможность сильного смешения между
племенами и, как результат, высокая степень близости. Для бурзян,
усерган, тамьян наше предположение вполне логично, если учесть
единое центральноазиатское происхождение, кочевой образ жизни и
совместные пути миграции в Приаралье, долину Сырдарьи, на
Бугульминскую возвышенность и Южный Урал через
< еверокавказские степи и Поволжье. Наше допущение не
противоречит также этнической истории карагай-кинчаков и табын,
разработанной на основе историко-этнографических данных. Обе
популяции являются сложными конгломератами, характерными для
предмонгольского и монгольского времени в степях Поволжья и
Казахстана. Однако, полученная величина коэффициента миграции
m=0,0407 не подтверждает наше предположение об интенсивных
процессах смешения между двумя этими древними племенами, но п
не отрицает их полностью, т.к. скорость миграций, равная обмену
четырьмя мигрантами за поколение (25 лет), может привести к
'пределенным изменениям генегической структуры популяции с
малым эффективно-репродуктивным объемом.
Другой причиной относительно близкого к современности
времени формирования групп может быть сильное влияние на
пришлый генофонд тюркоязычных кочевников местных сармато-
203
аланских, булгаро-мадьярских, финно-угорских и других племен
ассимилированных в ходе исторического развития (Акимова, 1974).
11очти подное отсутствие следов угорского языкового компонента в
башкирском языке объяснима быстротой смены событий, т.е.
отсутствием периода длительного соседства с ними и обмена
мигрантами. Судя по форме древа, генетическая структура
популяции вплоть до конца 1 тыс. н.э. не испытывала значительных
изменений, способных привести к отдаленным дивергенциям.
Другими словами, этапы большого и длительного по времени
миграционного пути на запад через Приаралье и долину Сырдарьи
не отразились в форме древа, что свидетельствует о тесном общении
близкородственных кочевых групп, населявших эти области.
Полученные при реконструкции родословного древа восточных
башкир предварительные данные, интересные в плане этнической
антропологии, требуют подтверждения и дальнейшей разработки с
привлечением материала по западным группам башкир, а также
другим популяциям Волго-Уральского региона. Средней Азии,
Южной Сибири и Северного Кавказа
204
ЗАКЛЮЧЕНИЕ
Каким представляется будущее популяции с позиций исторической
генетики?
Сразу же позволим себе заметить, что наши рассуждения, хотя и
«атрагивают в большей степени одну из конкрез'ных популяций
уральского региона, могут быть со всей определеностью
распространены на большинство представленных в настоящее время
типичных" популяций этносов, народов.
Вопрос, с которого начата заключительная часть изложения
настоящей работы, в принципе не нов. В сущности будущее - это есть
глобальная основная проблема, волнующая все человечество,
прогнозы в отношении которого пытаются сформулировать люди
самых разных специальностей.
К настоящему времени достижения современной генетики, успехи
в области биологии и медицины позволяют с позиций этих дисциплин
объективно оценить состояние тех или иных популяций в отношении
общей приспособленности, статуса здоровья, степени
адашированности к окружающей среде.
В этом контексте правомерно возникновение вопроса: существует
ли реальная основа для динамики микроэволюционных процессов в
современных человеческих популяциях? Если ответ утвердителен, то
какова эта динамика и как меняются отдельные соотношения во
времени; в частности, продолжает ли естественный отбор интенсивно
действовать на человека? По-видимому, нет оснований для сомнения
в том, что эффективность генного дрейфа, как функции
ограниченной численности популяций, в качестве одного из
микроэволюционных факторов, в прошлом игравших одну из
основных ролей в дифференциации человеческих подразделений,
неуклонно продолжает падать. Напротив, в связи с ростом
интенсивности миграционных процессов, перемещением массы
людей в различных направлениях в глобальном масштабе возросла
^ответственно и частота смешанных браков (в том числе и
межнациональных), а следовательно и метисация. Резкое увеличение
'утбридинга у современного человека привело к существенному
човышению эффективно-репродуктивного объема популяций и
значительному росту генетической изменчивости. Все это
205
совершенно справедливо считают развитием в благоприятном
направлении, поскольку при этом уменьшается опасность
вышепления гомозигот по вредным генам. Можно, однако, думать
что благоприятный эффект в данном случае не абсолютен, так как
подобный аутбридинг лишь "отдаляет день расплаты" и генетические
последствия зависят от частоты мутаций безотносительно к размеру
популяции (Майр, 1974). К тому же при этом исключено образование
рекомбинационного груза, что может привести к разрушению
хорошо интегрированных генных комбинаций. В этой же связи, как
при исследовании нами памирских изолятов, заметим, что инбридинг
далеко не всегда может приводить только лишь к отрицательным
последствиям. Среди некоторых племен Австралии, Америки,
Африки браки заключаются между индивидами одного и того же
племени, или одной и той же группы, что с течением длительного
времени приводит к тому, что все члены этих популяций оказываются
связанными между собой различной степенью кровного родства. В
качестве примера положительной брачной ассортативности по
родству в нашей стране можно привести структуру браков в ряде
республик Средней Азии, Азербайджане, где несмотря на все усилия
медико-генетической пропаганды о возможных негативных
последствиях этого явления для потомства, инбридинг остается
достаточно высоким (Дементьева, 1974; Гарькавцева, 1984; Ревазов,
1985, 1986; Павленко, 1989; и др.).
Принимая во внимание аспекты взаимодействия человека и
окружающей среды, мы вправе поставить вопрос о том, продолжает
ли естественный отбор в качестве систематического
микроэволюционного фактора воздействовать с достаточной
интенсивностью на человека. И далее, какова динамика этого
процесса, насколько мы становимся независимыми от него?
Материалы антропологии со всей определенностью
свидетельствуют, что ЗО-тысячный период становления
современного человека от кроманьонской его фазы, проходил без
видимой биологической эволюции. Поэтому ряд авторов считает, чт°
человек совершенно или почти полностью освободился от давления
естестве иного отбора. Однако факты, в частности, генетико-
цемографичсского характера свидетельствуют об обратном-
Существенная гибель зигот (прослеживаемая в онтогенезе &
зачатия до достижения репродуктивного возраста), а так*®
206
значительные различия между индивидами в отношении
плодовитости подтверждают реальность существования отбора в
человеческих популяциях и в настоящее время. Даже в
высокоразвитых индустриальных странах, где удельный вес
дорепродуктивной смертности как компоненты отбора сведен к
минимуму за счет прогресса медицины и здравоохранения, со всей
очевидностью продолжают действовать другие факторы, как то:
неравный вклад индивидов в процесс воспроизводства. Таким
образом, дифференциальная плодовитость сейчас становится одним
из основных структурных элементов отбора по сравнению с той же
ситуацией, которая имела место несколько десятилетий назад. Также
не менее важным представляется вопрос, который непосредственно
связан с предыдущим, о возможностях и степени адаптации человека
к меняющимся условиям среды. Для нашей огромной по масштабам
страны по-прежнему остаются актуальными экотипические
адаптации, позволяющие людям жить и трудиться в суровых и
специфичных условиях, например, в аридных или циркумполярных
зонах. Так, по мнению Майра (1974), по мере роста независимости
цивилизованного человека от селективных факторов физической
среды, значение экотипической адаптации неуклонно уменьшается, и
его постепенная утрата не скажется на удовлетворительной
адаптации к новой среде обитания. Однако, при всей авторитетности
подобного высказывания этого исследователя с последним тезисом
трудно согласиться, в чем можно убедиться из дальнейшего
изложения.
Резко возросшая миграционная активность населения, связанная с
планированием народонаселения, размещения трудовых ресурсов в
связи с освоением новых промышленных и сельскохозяйственных
районов в России, приводит к перемещению десятков тысяч людей на
новые места обитания с иными подчас экстремально суровыми
условиями внешней среды: регионы Крайнего Севера, аридные зоны,
высокогорные районы и др. Однако, далеко не все из них, даже
несмотря на удовлетворительные условия жизни и труда, физически
оказываются в состоянии адаптироваться к несвойственным им
экологическим условиям. Результаты современных генетико-
биохимических и популяционных исследований убедительно
свидетельствуют о существовании индивидуальных генетических
Различий в ответ на воздействия однотипных факторов среды.
207
Поэтому уже давно назрела проблема дифференцированного
подхода к сложному процессу планирования народонаселения с
учетом генетического статуса отдельных индивидов, моральные и
экономические преимущества подобного решения очевидны.
Именно в связи с вопросами адаптации следует рассматривать еще
одно явление, весьма важное в жизни аборигенного населения этих
регионов, издревле приспособленных к специфическим условиям
своей среды, а именно смену, зачастую и ломку зрадиционного
уклада (Симченко, 196Я). Так, эндокринная и ферментная адаптация к
новым физическим условиям и изменению статуса питания может
сыграть особую роль в здоровье коренных жителей Крайнего Севера.
Например, при пероральном тесте с нагрузкой глюкозой
североамериканские индейцы и эскимосы дают показатели,
свойственные тяжелым формам диабета. Это явление объясняют
недостаточностью у аборигенов некоторых биологически активных
соединений, стимулирующих выделение инсулина в ответ на
повышение в крови концентрации глюкозы. Коренное население
циркумполярной зоны на протяжении десятков тысяч лет выработало
механизм биологической адаптации к особой диете, поэтому рост
потребления ферментируемых углеводов приводит у них к
специфическим особенностям метаболизма сахаров, что зачастую
имеет печальные последствия. Игнорирование наследственных
различий между лицами, а также нарушения сбалансированных
адаптивных комплексов может привести, а в ряде случаев уже
приводит к резко отрицательной экогенетической ситуации как среди
мигрантов, так и среди коренных жителей этих регионов.
Теперь мы ясно представляем себе, по мере накопления фактов,
что естественная физическая среда избирательно оказывает влияние
на здоровье людей в зависимости от их индивидуальных
наследственных характеристик. Что же происходит, когда человек,
воруженный современной технологией и техникой, наукой атомного
века, вторгается в извечные тайны природы, нарушая экологическое
равновесие? Мы начинаем призывать к разуму, к общественому
мнению, и средствам массовой информации, большей частью тогда,
когда непоправимое уже свершилось. Так, например, после выбросов
химических соединений, приводящих к массовой гибели
растительного покрова, представителей животного мира, наносящих
непоправимый ущерб здоровью людей, осуществляются попытки
208
исправить положение вещей. Однако только знание экологических
взаимосвязей, строгое научное обоснование предлагаемых
мероприятий по использованию продуктов производственной
деятельности позволяют избежать отрицательного влияния человека
как на природную среду, так и на самого человека.
В природно-экологическом отношении Урал является
жемчужиной нашей страны; климат в данном регионе отличается
целым рядом особенностей, благоприятных для здоровья людей.
Гигантский скачок в направлении индустриализации, произошедшей
в Башкирии, особенно с начала 4О-х годов, превратил ее в один из
наиболее развитых промышленных центров нашей страны. Мощный
натиск цивилизации привел к тому, что в настоящее время в этом,
некогда аграрном крае России, с полной очевидностью встали
проблемы взаимоотношений человека и среды, сохранения
экологического равновесия, и другие. Продолжающееся расширение
горнодобывающей, перерабатывающей промышленностей,
нефтехимии остро поставило вопрос об охране здоровья людей.
Наличие в городах Башкортостана концентрации крупных
промышленных центров диктует необходимость скорейшего решения
аспектов профессионального отбора и профориентации в
зависимости от индивидуальных наследственных особенностей
людей. Игнорирование этого может привести в будущем к
непредсказуемым сдвигам в генетической структуре популяции. При
этом врачам, медицинским генетикам, профпатологам,
сангигиенистам следует обратить особое внимание на такие важные
в экогенетическом отношении системы, как, например,
наследственный дефицит -антитрипсина (неблагоприятные
эффекты у индивидов проявляются при загрязнении среды),
варианты трансферрина (Сг), определенные факторы фермента
глутатион - S - трансферазы (GST). Эти и другие биохимические
истемы, отличающиеся множественным наследственным
полиморфизмом, необходимо учитывать при наличии
неблагоприятных искусственных факторов среды: смог,
‘апыленность, воздействие различных ксенобиотиков и пр. в
основном касающиеся жителей города. Интенсивное применение в
льском хозяйстве пестицидов, таких как параоксон (Е 600),
Попадание которого в организм вследствие наследственно
обусловленной аномалии фермента параоксоназы может приводить
209
к поражению 30-50% людей (ориентировочная частота для региона
Урала). Это реальность современного состояния региона, которая в
обозримом или отдаленном будущем определенным образом может
сказаться на состоянии генетической структуры населения.
Вторая особенность заключается в следующем. Регион Урала
являлся и является исторической зоной контактов европеоидного и
монголоидного населения страны. Последние десятилетия
сопровождаются интенсивной миграцией из самых разных уголков
России, подчас самых отдаленных. Все это не может не оказывать
влияния на генетическую и социальную структуру популяции. Столь
выраженный аутбридинг диктует необходимость своего дальнейшего
изучения. Его последствия, в целом, несомненно, благоприятные,
возможно в незначительном отношении могут привести к
накоплению рекомбинационного груза, что также следует учитывать
в медико-генетической практике.
Этот феномен чрезвычайно интересен с позиций антропологии,
поскольку динамика популяционно-генетических изменений,
происходящих сейчас в Поволжье и на Урале может служить
моделью для познавания этих сложных явлений в мировом
народонаселении.
Наконец, экологическая ситуация здесь во многом зависит от
воздействия ряда биотических факторов, в частности от коренным
образом изменившегося и продолжающего меняться статуса
питания. Адаптация ферментных и эндокринных систем к новым
условиям диеты может играть роль в становлении многих
специфических особенностей здоровья населения рассматриваемого
региона. Так, коренное население Южного Урала - кочевники и
скотоводы в недалеком прошлом, отличались на протяжении многих
и многих поколений выработанным специфическим статусом
питания, характерным для данного образа жизни. Биологически они
не были "подготовлены" к переходу на новый режим питания с иным
комплексом пищевых продуктов, богатых ферментируемыми
углеводами. Поэтому, надо полагать, существует определенное
противоречие между эволюционно сложившейся системой усвоения,
ориентированной на традиционные пищевые продукты, и
поступлением новых продуктов питания. Ярко выраженный
наследственный полиморфизм по таким фермента как лактаза,
способствующему усвоению молочного сахара, липаза, пепсиноген,
210
отражающие межрасовые и межнациональные различия,
свидетельствуют о том, что каждая группа населения в
специфических условиях ведения определенных типов хозяйств, была
приспособлена к ним на уровне основных метаболических путей и
процессов. Поэтому с большей долей вероятности можно полагать,
что значительная распространенность печеночной и желудочно-
кишечной патологии среди коренного населения в настоящее время
во многом обусловлена этим дисбалансом. Как показал в главе 6.4
анализ компонентов максимально возможного отбора, то в целом,
генетико-демографическая ситуация в Башкортостане остается
достаточно благоприятной.
Таким образом, в заключительной части работы мы остановились
на наиболее актуальных задачах исторической генетики, тесно
смыкающихся с жизненно важными проблемами экологии, от
скорейшего решения которых зависит судьба населения не только
региона, на описании которого было акцентировано
преимущественное внимание, но и судьба многих групп мирового
народонаселения.
211
ЛИТЕРАТУРА
Абанина Т.А. Генетико-демографическое изучение лесных немцев
Западной Сибири. Брачная структура. Репродукция: Автореф.
дис.... канд. мед. наук. М. 1983. с. 23.
Абрамов А.Н. Башкиры (антропологический очерк) Ц Русский
антропологический журнал. М. 1907. кн. 27-28. N 3-4, с. 1-55.
Авижанская С.А., Кузеев Р.Г. Этнографические коллекции по
башкирам Государсп.'иного музея этнографии народов СССР//
АЭБ, Уфа, 1962, т.1, с. 344-360.
Авижанская С.А., Бикбулатов Н.В., Кузеев Р.Г. Декоративно-
прикладное искусство башкир. Уфа, 1964, с. 259.
Адлер Ю.П., Маркова Е.В., Грановский Ю.В. Планирование
эксперимента при поиске оптимальных условий. М.: Наука, 1971,
с.282.
Акимова М.С. Антропологический состав населения пьяноборской
культуры. - Вопросы антропологии, 1961, в. 8, с. 130-140.
Акимова М.С. Антропологические материалы бахмутинской
культуры. - В кн.: Археология и этнография Башкирии, Уфа, изд-
во БФ АН СССР, 1961, т.1, с. 361-365.
Акимова М.С. Древнее население Башкирии по антропологическим
данным. - В кн.: Археология и этнография Башкирии, Уфа, изд-
во БФ АН СССР, 1964, т.1, с.158-163.
Акимова М.С. Материалы к антропологии древнего населения
Южного Урала // АЭБ, Уфа, 1968, т.З, с. 391-426.
Акимова М.С. Этногенез башкир по данным антропологии. - В кн.:
Археология и этнография Башкирии, Уфа, изд-во БФ АН СССР,
1971, т.4, с. 38-43.
Акимова М.С. Значение данных антропологии, дерматоглифики и
серологии для изучения смешанных групп (на башкирском
материале). - Вопросы антропологии, 1973, в. 44, с. 85-91.
Акимова М.С. Антропологические исследования в Башкирии //
Антропология и геногеография. М. 1974, с. 85-96.
Аксянова Г.А. Население бассейна Печеры и нижней Оби И
Этническая одонтология СССР. М. 1979.
Алексеева Т.Н. Географическая среда и биология человека. М, 1977,
с. 301.
212
Алексеева Т.Н., Алексеев В.П., Спицын В.А., Круковская О.Б. и др.
Ферментные и другие белки крови и дифференциация популяций
Северо-Восточной Азии // Вопросы антропологии, 1978, в. 68, с.
23-41.
Алексеев В.II. География человеческих рас. М. 1974, с. 351.
Алексеев В.II. Историческая антропология и этногенез. М. 1989.
Алтухов Ю.П. Генетические процессы в популяциях. М. 1983, с. 277.
Алфавит волостей и селений Уфимской губернии. Уфа, 1902, с.
435.
Андреев М.С. Таджики долины Хуф (верховье Аму-Дарьи). 1958.
Аннинский С.А. Известия венгерских миссионеров XI1I-X1V вв. о
татарах и Восточной Европе 33 Исторический архив. М.-Л. 1940,
с. 71-112.
Аргун Ю.Г., Павленко А.П., Спицына Н.Х. Фамильно-родовая
система и брачные ареалы // Абхазское долгожительство. М., с.
67-80.
Артамонов М.И. История хазар. М.-Л. 1962, с. 523.
Аристов Н.А. Опыт выяснения этнического состава киргиз-казаков
Большой орды и каракиргизов на основании ро .основных
сказаний и сведений о существующих родовых делениях и о
родовых тамгах, а также исторических данных о начинающихся
антропологических исследованиях. СПб, 1895, с. 157.
Хсадов 111.А. Связь родственных браков с долгожительством в
Азербайджане// Цитология и генетика, 1990, -24, N 5, с. 67-70.
Асфаган М.Ш., Кузьмина Т.Я. Результаты исследования групп крови
у народностей Б АССР // Хозяйство Башкирии. Уфа, 1929, т.6-7, с.
172-182.
Афанасьева Г.М. Традиционная система воспроизводства нганасан
(проблемы репродукции обособленных популяций). М. 1990, с.
871.
Ххмеров Р.Б. Уфимские погребения VI-VI1 вв. н.э. Ц Краткие
сообщения. Ин-т истории материальной культуры, М. 1951, т. 40,
с. 125-137.
Ххмсров Р.Б. Некоторые вопросы этногенеза башкир по
археологическим данным // Сов. этнография, 1952, N 3, с. 36-49.
Ахмеров Р.Б. Уфимские погребения IV-VII вв. н.э. и их место в
древней истории Башкирии // Древности Башкирии, М. 197 , с. 69-
93.
213
Бадер О.Н. Палеолит Башкирии // АЭБ, Уфа, 1971, т.4, с. 64-68.
Баскаков Н.А. Введение в изучение тюркских языков. М. 1962, с. 332.
Бахолдина В.Ю. Антропология и популяционная генетика нивхов:
Автореф. дис.... канд. биол. наук. М. 1982, с. 23.
Бейли Н. Статистические методы в биологии. М. 1964, с. 271.
Беневоленская Ю.Д. О характере полиморфизма популяций по
системе Резус// Антропология и геногеография. М. 1974, с. 103-
135.
Березин И.Н. Булгар на Волге // Ученые записки Казанского
университета. Казань 1852, N 3, с. 251-291.
Бибиков С.Н. Пещерные палеолитические местонахождения в
нагорной полосе Южного Урала // Сов. археология. 1950, т. 12, с.
66-105.
Большакова Л.П. Механизмы репродуктивной компенсации в
популяциях с разным характером репродукции // Генетика, 1989,
т.25, N 9, с. 1652-1657.
Бочков Н.П., Захаров А.Ф., Иванов В.И. Медицинская генетика. М.
1984, с. 365.
Брук С.И. Население СССР по данным Всесоюзной переписи 1979 г.
М. 1981.
Брук С.И. Население мира. Этнодемографический справочник. М.
1986, с. 830.
Бунак В.В. Геногеографические зоны Восточной Европы,
выделяемые по факторам крови АВО // Вопросы антропологии,
1965, в. 32, с. 6-27.
Бунак В.В. Род Номо, его возникновение и последующая эволюция.
М. 1980, с. 328.
Вагнер Л.Б. Кровяные группы с точки зрения антропологии //
Врачебное дело. М. N 3, 1925, с. 31-59.
Вагнер Л.Б. Сводка распределения кровяных групп среди населения
земного шара к 1926 г. // Русский антропологический журнал. М-
1926, т. 15, в. 1-2.
Воронина В.Г. Генетико-антропологическая характеристика
коренного населения Приморского края. Сообщение !•
Популяционно-генетические данные о народах Приморья И
Вопросы антропологии. М. 1875, в. 5 , с. 46-67.
Гаджиев А.Г. Антропология малых популяций Дагестана.
Махачкала, 1971, с. 367.
214
Гарькавцева РФ., Гинтер Е.К., Ревазов А. А. Региональные
особенности распространения наследственной патологии в
У збекистане // Вестник Акад. мед. наук СССР. 1984. N 7, с. 69-75.
I еннинг В.Ф. Мазунинская культура в Среднем Прикамье // Вопросы
археологии Урала. Свердловск, 1967, в. 7, с. 114-137.
Геннинг В.Ф. Южное Приуралье III-VII вв. н.э. (проблема этноса и
его происхождение) // Проблемы археологии и древней истории
угров. М. 1972, с. 247-25 .
I ерасимов М.М. Восстановление лица по черепу // Труды Ин-та
этнографии АН СССР, Новая серия. М. 1955, т. XXVIII, с. 413-
414.
I инзбург В.В., Трофимова Т.А. Палеоантропология Средней Азии.
М. 1972, с. 372.
I интер Е.К., Ревазов А.А., Краснопольская К.Д., Ахмина Н.И.,
Южина Г.П., Нечволодова О.А., Гринио Л.П., и др. Медико-
генетическое изучение населения Туркмении. Сообщение 1.
Наследственная патология в пяти районах Ашхабадской области
//Генетика, 1980, т. 16, N8, с 1487-1494.
Гинтер Е.К., Ревазов А.А., Краснопольская К.Д., Тур? ва Ш.М.,
Инсарова Н.Г. и др. Медико-генетическое изучение населения
Туркмении. Сообщение 2. Изучение распространения
наследственной патологии в пяти районах Ташаузской области //
Генетика, 1982, т.18, N 6, с. 1018-1023.
интер Е.К., Тураева U1.M., Ревазов А.А., Пантелеева О.А. и др.
Медико-генетическое изучение населения Туркмении.
Сообщение 3. Наследственная патология у туркмен-нохурли //
Генетика, 1983, т. 19, с. 1344-1352.
интер Е.К., Будагова К.А., Ревазов А.А. и др. Медико-генетическое
изучение населения Узбекистана. Сообщение 4:
Территориальное распределение наследственных болезней в
населении четырех районов Хорезмской области и . его
отягощенность наследственной патологией // Генетика, 1986, N ,
с. 519-52.
мурман В.Е. Теория вероятности и математическая статистика. М.
1972, с. 368.
। олубцов В.И. Влияние факторов популяционной динамики на
отягощенность наследственными болезнями населения
Краснодарского края: Дисс. ... докт. мед. наук. Киев, 1991.
215
Гольцова Т.В., Сукерник Р.И. Генетическая структура обособленной
группы коренного населения Северной Сибири нганасан
(тавгийцев) Таймыра. Сообщение 4. Изучение популяционной
динамики // Генетика, 1979, т. 15, N 4, с. 734-744.
Гольцова Т.В. Родственные браки, инбридинг и его эффекты у
нганасан Таймыра // Генетика, 1981, т. 17, N 5, с. 896-905.
Гольцова Т.В., Сукерник Р.И. Популяционная структура лесных
ненцев. Демографические характеристики: структура браков,
миграции, анализ смешения // Генетика, 1982, т. 18, N 11, с. 1884-
1893.
Гумилев Л.Н. Древние тюрки. М. 1967, с. 509.
Давыдова Г.М. Антропология манси. М. 1989, с. 128.
Дебец Г.Ф. Палеоантропология СССР. М.-Л., 1948, с. 333.
Дементьева Е.С. Медико-генетическое изучение населения
Западного Памира. Сообщение 4. Локальная генетическая
дифференциация в популяциях Западного Памира // Генетика, N,
1974. Демографический энциклопедический словарь. М. 1985, с.
607.
Долинова Н.А. Материалы по дерматоглифике коми // Полевые
исследования Ин-та этнографии в 1978 г. М. 1980, с. 199-206.
Дубровский С.А., Зейгер Е.М., Френкель А.А. Факторный анализ.
Методы и приложения // Многомерный статистический анализ в
социально-экономических исследованиях. М. 1974, с. 229-294.
Ефимова С.Г. К краниологии Волго-Камья эпохи раннего железа //
Вопр. антропологии, 1981, в. 67.
Ефимова С.Г. Палеоантропология Поволжья и Приуралья. М., 1991,
с. 93.
Ефимова С.Г. Новые краниологические материалы к проблеме
формирования антропологического типа башкир // Докл.
МОИП. Общая биология, М., 1987.
Зайцев Г.Н. Методика биометрических расчетов. Мат. статистика в
эксперим. ботанике. М. 1973, с. 256.
Збруева А.В. История населения Прикамья в ананьинскую эпоху И
Материалы и исследования по археологии СССР. М., 1952, N 3 , с-
323.
Зубов А.А. Материалы по одонтологии казахов // Сов. этнограф*0*’
1972, N 3.
Зубов А.А. Этническая одонтология. М. 1973.
216
Зубов А.А., Золотарева И М. Монголы в мировой систематике
одонтологических типов// Вопр. антропологии, 1980, в. 64.
Зубов А.А. О расово-диагностическом значении некоторых
одонтологических признаков//Сов. этнография, 1986, N 3.
Исмагулов О. Этническая геногеография Казахстана Алма-Ата,
1977, с. 160.
Итоги Всесоюзной переписи населения 1970 года (в 7-ми т.) М.,
1973, т. 1, с. 176.
Кадошникова М.Ю., Голубцов В.И., Ельчинова Г.И., Почешхова
Э.А., Ревазов А.А. Брачно-миграционная структура и
коэффициент инбридинга в адыгейской популяции // Генетика, т.
27, N 2, 1991, с. 327-334.
Казанцев Н. Описание башкирцев. СПб., 1866, с. 97.
Казаченко Б.Н. Генетико-демографический подход в
антропологических исследованиях. Половозрастная и семейная
структура хакасов// Вопр. антропологии 1986, в. 76, с. 78-91.
Киекбаев Д.Г. О звуках с, з и п и их развитие в башкирском,
туркменском и якутском языках // Ученые записки Башкирского
Государственного университета. Уфа, 1958, в. 6, сер.
филологическая, N 5, с. 11-96.
Ковалевский А.П. Книга Ахмеда ибн-Фадлана о его путешествии на
Волгу в 921-922 гг. Харьков, 1956, с. 347.
Козлов В.И. Введение. Об изучении долгожительства в
Азербайджане Ц Долгожительство в Азербайджане. М., 1989, с.
5-15.
Козлов В.И, Максименко Т.В. О психологических факторах
активного долголетия среди азербайджанцев. Там же.
Крачковский И.К). Арабская географическая литература. М.-Л.,
1957, соч. т. 4, с. 919.
Крус-Кок Р. Генетическое описание высокогорных популяций //
Биология жителей высокогорья. М. 1981, с. 58-78.
Куандыков Е.У., Святова Г.С., Михайличенко Н.П. Исход
беременности и оценка состояния здоровья новорожденных в
различных типах браков // Факторы риска акушерской и
гинекологической патологии. Алма-Ата, 1988, с. 69-74.
Кузеев Р.Г. Башкирские шежере. Уфа, 1960, с. 298.
узеев Р.Г., Шитова С.Н. Башкиры // Очерки общей этнографии
Европейской части СССР. М. 1968, с. 410-423.
217
Кузеев Р.Г. Урало-Аральские этнические связи в конце I тысячелетия
н.э. и история формирования башкирской народности //
Археология и этнография Башкирии. Уфа, 1971, т. 4, с. 11-17.
Кузеев Р.Г. Происхождение башкирского народа. Этнический
состав, история расселения. М., 1974, с. 574.
Кузеев Р.Г. Историческая этнография башкирского народа. Уфа,
1978, с. 262.
Кузеев Р.Г., Рафиков Х.С., Юмагужина Н.Х. (Спицына Н.Х.).
Этногенез и генетическая дивергенция восточных башкир // Сов.
этнография, 1982, N 4 с. 26-34.
Кузеев Р.Г. Этнические процессы в Башкирии в новое и новейшее
время. Уфа, 1991.
Курбатова О.Л. Генетические процессы в городском населении
(опыт генодемографического исследования популяции
г.Москвы): Автореф. дис.... канд. биол. наук. М. 1977, с. 22.
Кучер А.Н., Курбатова О.Л. Популяционно-генетическое
исследование дифференциальной плодовитости в городском
населении // Генетика, 1986, т. 22, N 2, с. 304-311.
Кучер А.Н., Пузырев В.П., Назаренко Л.П. Генетические процессы в
сельских популяциях Томской области (характеристика
миграций)//Генетика, 1991, т. 27, N 6, с. 1084-1094.
Кучинскас В.К. Результаты изучения динамики генетических
процессов в сельских популяциях Литовской ССР и отражение
этих процессов в распространении наследственных болезней //
Вопр. проф. наследств, болезней у детей. Вильнюс, 1987, с. 64-78.
Лакин Г.Ф. Биометрия. М., 1980, с. 292.
Левонтин Р. Генетические основы эволюции. М., 1978, с. 350.
Ли Ч. Введение в популяционную генетику. М., 1978, с 555.
Лузина Ф.А. Наследственный полиморфизм и генетические
процессы в коренном населении горного Алтая: Автореф. дисс. -
канд. биол. наук, М., 1978, с. 22.
Мажитов НА. Курганный могильник в д.Ново-Турбаслы И
Башкирский археологический сборник. Уфа, СССР, 1959, с. 143.
Мажитов Н.А. Бахмутинская культура. Этническая история
населения Северной Башкирии середины I тысячелетия н.э. ***•
1968, с. 118. “Л S
Мажитов Н А., Пшеничнюк А.Х. Камышлы-Тамакский могильник
Археология и этнография Башкирии. Уфа, 1969, т. 3, с. 38-58.
218
Мажитов Н.А. Южный Урал в VII-XIV вв. М. 1977, с. 24 .
Майр Э. Популяции, виды и эволюция. М. 1974, с. 46 .
Максютова Н.Х. Восточный диалект башкирского языка в
сравнительно-историческом освещении. М., 1976, с. 292.
Малиев Н.М. Антропологический очерк башкир // Труды о-ва
естествоиспытателей при Казанском университете). Казань, 1876,
т.4, в. 5, с. 32.
Малютов М.Б., Пасеков В.П. Реконструкция родословных деревьев
изолированных популяций. М. 1971, МГУ, Препринт N 19, с. 52.
Материалы по истории Башкирской АССР. Часть 1, М.-Л., 1936, с.
631; Том 3, М.-Л., 1949, с. 691; Том 4, М.-Л., 1956, с. 494.
Матюшин Т.Н. К изучению мезолита и неолита в Башкирии /
Археология и этнография Башкирии. Уфа, 1964, т, 3, с. 32-42.
Матюшин Т.Н. Пути развития Южного Урала в эпоху мезолита и
неолита // Археология и этнография Башкирии, Уфа, 1979, т. 4, с.
85-88.
Матюшин Т.Н. Энеолит Башкирии // Археология и этнография
Башкирии, Уфа, 1973, т. 4, с. 159-161.
Микулич А.И. Наша генетическая память. Минск, 1987, с. 72.
Миржанова С.Ф. Южный диалект башкирского языка. М., 1979, с.
272.
Могильников В.А. К вопросу о связях населения Башкирии и
Зауралья в конце I тысячелетия до н.э. - I тысячелетия н.э. Ц
Археология и этнография Башкирии. Уфа, 1972, т. 4. с. 151-157.
1азаров П.С. Башкиры о башкирцах Ц Современник, СПб., 1863, т. 2,
с. 57-84.
11азаров П.С. К антропологии башкир // Известия об-ва любителей
естествознания. СПб., 1889, т. 18.
Назаров П.С. К этнографии башкир // Этнографическое обозрение.
М. 1890, N 1, с. 169.
икольский Д.П. Башкиры. Этнографическое и санитарно-
антропологическое исследование. СПб, 1899, с. 377.
икольская Н.А. Новые материалы по дерматоглифике
финноязычных народов // Этногенез финно-угорских народов по
данным антропологии. М. 1974, с. 119-137.
иконов В.А. Актуальные процессы в антропонимии татар и башкир
И Ономастика Поволжья, Уфа, 1973, с. 29-40.
^иль Дж., Шэлл У. Наследственность человека. М., 1958 с 3X8
219
Оливье Ж. Генетика и человеческие расы Ц Современная
антропология, М., 1964, с. 284-287.
Павленко А.П., Спицына Н.Х. Традиционная брачная система
азербайджанцев и долгожительство // Долгожительство в
Азербайджане, М. 1989, с. 51-73.
Пасеков В.П., Ревазов А.А. К популяционной генетике населения
Европейского Севера СССР. Сообщение 1. Данные по структуре
шести деревень Архангельской области // Генетика, 1975, т. 2, N
7, с. 145-155.
Пасеков В.П., Корооелев АН. Об оценке параметров
эволюционного процесса дивергенции популяций в результате
случайного генного дрейфа // Успехи соврем, биологии. М., 1971,
т. 72, в. 3(6), с. 410-430.
Патканов С.К. О приросте инородческого населения Сибири. СПб,
1911.
Посух О.Л., Вибе В.П., Сукерник Р.И. Генетическое и экологическое
изучение коренных жителей северо-востока Сибири. Сообщение
3. Демографическая структура трех современных популяций
эвенов Якутии // Генетика, 1990, т. 26, N 9, с. 1628-1636.
Прокудина Н.А. Дерматоглифика русских Восточной Европы //
Вопр. антропологии, 1971, в. 39, с. 138-152.
Прокудина Н.А. Дерматоглифические исследования русских и
народов Поволжья // Итоги полевых работ Ин-та этнографии в
1971 г. М„ 1072,4. 2, с. 321-324.
Прокудина Н.А. Дерматоглифические исследования некоторых
народов европейской части СССР // Новое в этнографических и
антропологических исследованиях, М., 1974.
I (узырев В.П. Народности Севера: генетические аспекты здоровья (к
концепции развития). Препринт. Томск, 1988, с. 27.
Пузырев В.П. Медико-генетическое исследование население
приполярных регионов. Томск, 1991, с. 196.
Пшеничнюк А.Х. Племена железного века в Башкирии И
Археология и этнография Башкирии. Уфа, 1971, т.4, с. 210.
Рафиков X С., Юмагужина Н.Х. (Спицына Н.Х.). Распределение
некоторых генетических систем (групп крови АВО, резус и
гаптоглобина) среди коренного населения Башкирии // Вопр-
биохимии и иммунологии человека и животных. Уфа, 1974, с. Н
119.
220
Рафиков Х.С., Чекмин И.Ф., Кузеев Р.Г., Башлай А.Г., Юмагужина
Н.Х. (Спицына Н.Х.) и др. Исследование изоантигенов крови
системы резус у коренного населения Башкирии // Вопр.
антропологии, 1979, в. 62, с. 7 -75.
Рафиков Х.С., Юмагужина Н.Х. (Спицына Н.Х ), Кузеев Р.Г.
Генетическая дивергенция восточных башкир в аспектах
этногенеза // Популяционно-генетические исследования народов
Южного Урала. Уфа, 1981, с. 37-54.
Рафиков Х.С., Белова И.Ю., Юмагужина Н.Х. (Спицына Н.Х.),
Кузеев Р.Г. Структура популяции башкир в регионе Среднего
Поволжья и Урала. Уфа, 1981, с. 3-36.
Рафиков Х.С., Хуснутдинова Э.К., Долматова И.Ю., Франте Р. и др.
Структура и генетическая близость башкир к народам Волго-
Уральского региона и Сибири // Антропология и популяционная
генетика башкир. Уфа, 1987, с. 6 -76.
Ревазов А.А. Динамика параметров популяционной структуры как
результат социально-демографических преобразований //
Вестник АМН СССР, 1984, N 7, с. 21-26.
Рычков П.И. Топография Оренбургская или полное географическое
описание Оренбургской губернии, ч. 1-2. СПб., 1762.
Рычков ЮГ. Некоторые аспекты серологических исследований в
антропологии// Вопр. антропологии, 1965, в. 19, с. 95-105.
Рычков ЮГ. Популяционная генетика алеутов Командорских
островов (в связи с проблемами истории народов и адаптации
населения Древней Берингии)// Вопр. антропологии, 1972, в. 40,
с. 45-70.
Рычков Ю.Г., Шереметьева В.А. Факторы генетической
дифференциации популяционной системы коренного населения
Северной Азии // Генетика, М. 1974, N 8, с. 137-149.
Рычков Ю.Г., Ящук Е.В. Генетика и этногенез// Вопр. антропологии,
1980, в. 64, с.23-39.
Рычков Ю.Г., Ящук Е.В., Веселовская Е.В. Генетика и этногенез (о
генетической прапамяти систем коренного населения Северной
Азии и Америки // Вопр. антропологии, 1982, в. 69, с. 3-18.
^ычков Ю.Г. Генетика демографических процессов в
народонаселении И Вопр. антропологии, 1982, в. 70, с. 3-13.
'|Чков Ю.Г., Ящук Е.В. Генетика и этногенез. Историческая
упорядоченность генетической дифференциации популяций
22!
человека (модель и реальность) // Вопр. антропологии, 1985, в
75, с. 97-116.
Розенберг Г.С. Семь программ количественных методов анализа
растительности на БЭСМ-4. Препринт. Уфа, 1976, с. 30.
Рогинский Я.Я. Закономерности пространственного распределения
групп крови у человека. (К проблеме антропологии "окраинных
народов") // Труды Ин-та этнографии АН СССР. М., 1947, т.1, с.
216-231.
Рокицкий П.Ф. Введение в статистическую генетику. Минск. 1978, с.
466.
Руденко С.И. Башкиры. Опыт этнологической монографии. Часть 1.
Физический тип башкир // Записки Русского географического
общества. 1916, Пгд, т.43, в. 1, с. 260.
Руденко С.И. Башкиры. М.-Л., 1955, с. 386.
Рюмин А.В. Тайна древнего Урала И Вокруг света, 1960, N 4, с. 44.
Сальников К.В. Об этническом составе населения лесостепного
Зауралья в сарматское время // Сов. этнография, 1966, N 5, с. 118-
124.
Сальников К.В. Очерки древней истории Южного Урала. М., 1967, с.
47. Сведения о количестве населения Башкирии 1920 г. Уфа,
1927, с. 293.
Святова Г.С. Генетико-демографическая характеристика большого
многонационального города: Автореф. дисс. ... канд. мед. наук,
М., 1989, с. 24.
Серебровский А.С. Генетический анализ. М., 1970, с. 342.
Симченко Ю.Б. Некоторые данные о древнем этническом субстрате
в составе народов Северной Евразии Ц Проблемы антропологии
и исторической этнографии Азии. М., 1968.
Смирнов А.П. Очерки древней и средневековой истории народов
Среднего Поволжья и Прикамья. М., 1952, с. 276.
Смирнов К.В. Савроматы. М., 1964, с. 278.
Соколов Д.Н. О башкирских тамгах (с приложением таблицы
башкирских тамг). - Труды Оренбургской ученой архивн
комиссии. Оренбург, 1904, т.12, с. 93.
Соловчук Л.Л. Корреляция генетических и демографически*
различий между популяциями коренного населения Север0
Востока СССР // Генетика, 1989, т.25, N 4, с. 744-752.
222
Сомье. О башкирах // Записки Общества любителей естествознания
Екатеринбург, 1891-1892, т. 13, с. 22-34.
Список населенных мест Башреспублики 1926 г., Уфа.
Спицын В.А., Казаченко Б.Н., Шебан Г.В. Генная дифференциация
среди коренного населения Северной Азии: степень генного
разнообразия на различных уровнях иерархической структуры //
Популяционно-генетические исследования народов Южного
Урала. Уфа, 198 , с. 72-82.
Спицын В.А. Биохимический полиморфизм человека. М., 1985, с.
214.
Спицын В.А., Новорадовский А.Г., Спицына Н.Х., Парик Ю.Я.
Полиморфизм -антитрипсина в популяциях Памира.
Репродуктивная компенсация-возможный механизм поддержания
генетического разнообразия по PI генам И Генетика, 1989, т. 25,
N 12, с. 2218-2224.
Спицын В.А., Новорадовский А.Г., Спицына Н.Х., Парик Ю.Я.
Экогенетическое исследование население высокогорья. I.
антитрипсин. Эффект репродуктивной компенсации - возможный
механизм поддержания генетического разнообразия по генам PI
// Сборник научн. трудов Ин-та мед. генетики АМН СССР, ДСП,
N 44, с. 26-43.
Спицын В.А., Спицына Н.Х. Генетическая дифференциация башкир
на разных уровнях иерархической структуры // Сравнительная
антропология башкирского народа. Уфа, 1990, с. 78-84.
Спицын В.А., Новорадовский А.Г., Исполатов А.Д., Спицына Н.Х.,
Игнатенко Н.Р. Генетические исследования субтипов
фосфоглюкомутазы (PGM1): популяционный аспекты //
Генетика, 1991, т. 27, N 4, с. 709-718.
Стоколос В.С. Культура населения бронзового века Южного
Зауралья (хронология и периодизация). М., 1972, с. 168.
Татищев В.Н. История Российская. М.-Л., 1962, с. 500.
ерентьев П.В., Ростова Н.С. Практикум по биометрии. Л., 1977, с.
151.
I имаков В.В., Курбатова О.Л. Значение индексов потенциального
отбора для населения СССР Ц Генетика, т. 27, N 5, с. 928-937.
1 имофеев-Ресовский Н.В., Яблоков А.В., Глотов Н.В. Очерк учения
о популяции. М., 1973, с. 277.
223
К
Тишков В.А. Советская этнография: преодоление кризиса //
Этнографическое обозрение, 1991, N 1, с. 5-20.
Толстов СИ. Древний Хорезм. М., 1948, с. 352.
Трофимова Т.А. Антропологические материалы из аланского
могильника возле Стерлитамака в Башкирии // Краткие
сообщения Ин-та этнографии АН СССР. 1962, в. 17, с. 56-65.
Томилин В.В., Гладких А.С. Судебно-медицинское исследование
крови. М., 1981, с. 239.
Тураева Ш.М., Гинтер Е.К., Ревазов А.А., Гарькавцева Р.Ф. и др.
Медико-генетическое изучение населения Туркмении.
Сообщение 6. Внутрипопуляционная изменчивость при анализе
брачных миграций и маркерных систем АВО и Нр // Генетика,
1985, т. 21,N6,c. 1039-1046.
Тураева Ш.М. Геногеография наследственных болезней в
Туркменской ССР и разработка региональных принципов
медико-генетического консультирования: Автореф. ... докт. дисс.
М. 1991.
Уйфальви Г. Башкиры, мещеряки, тептяри // Известия Русского
географического об-ва, СПб, 1877, т. 13, в. 2, с. 127.
Уфимская губерния // Волости и важнейшие селения Европейской
России. СПб., 1885, в. 6, с. 103.
Федорова-Давыдова Э.А. К вопросу о периодизации памятников
эпохи бронзы в Южном Приуралье Ц Археология и этнография
Башкирии. Уфа, 1964, с. 84-92.
Флоринский В.М. Башкирия и башкиры // Вестник Европы. СПб.,
1874, кн. 12, с. 721-765.
Фогель Ф., Мотульски А. Генетика человека. М., т. 2, 1990, с.
378.
Хайду Петер. Уральские языки и народы. М., 1985, с. 430.
Халиков А.Х. К вопросу о начале тюркизации населения Поволжья
и Приуралья И Сов. этнография, 1972, N 1, с. 40-53.
Халиков А.Х. Татарский народ и его предки. Казань, 1989.
Халиков А.Х. Основы этногенеза народов Среднего Поволжья
Приуралья. Казань. 1991, с. 106. '
Хар| неон Дж., Уайнер Дж., Теннер Дж. и др. Биология человека,
1979, с. 611.
Харман Г. Современный факторный анализ. М., 1972, с. 486.
Хвольсон Д.А. Известия о хазарах, буртасах, болгарах, мадьярах,
славянах и руссах Абу-Али Ахмеда бен Омара ибн-Даста,
неизвестного доселе арабского писателя X века, по рукописи
Британского музея в первый раз издал, перевел и объяснил
Д А.Хвольсон, СПб., 1868, с. 114.
Хить Г.Л., Халдеева Н И. Дерматоглифические и одонтологические
исследования в Хакасии // Новое в этнографических и
антропологических исследованиях, М. 1974, с. 144-147.
Хить Г.Л. Дерматоглифика народов СССР. М. 1983. Хозяйство
Башкирии. Уфа, 1928, 1929 гг., вып. 1-7.
Шастина Н.П. Путешествие на восток Плано Карпини и Гильома
Рубрука, М., 1967, с. 113.
Шереметьева В.А., Рычков Ю.Г. Популяционная генетика народов
Северо-Восточной Азии. М., 1978, с. 151.
Шереметьева В. А., Горшков В. А. Коряки Камча тки.
Пространственная структура. Эффективный размер и миграции
генов в популяции // Генетика, 1982, т. 18, N 8, с. 1363-1369.
Шитова С.Н. Этнокультурные связи башкир по данным
материальной культуры и декоративно-прикладного искусства Ц
Археология и этнография Башкирии. Уфа, 1971, т.4, с. 161-174.
Эрдели И. Извлечения из хроники вегерского Аниома Ц Вопр.
археологии Урала. Свердловск, 1967, в. 7, с. 71-89.
Юмагужина Н.Х. (Спицына Н.Х.). Структура генных миграций в
популяции восточных башкир // Тез. докл. Четвертый съезд
Всесоюзного об-ва генетиков и селекционеров им. Н.И.Вавилова,
Кишинев, 1982, с. 151.
Юсупов Р.М. Краниология башкир. Ленинград, 1989, с. 198.
Яблоков А.В. Популяционная биология. М., 1987.
225
Adachi В. Das Ohrenschmals als Rassenmerkmal und der Rassengeruch
("Achselgeruch") nebst dem Rassenunterschied der Scheveissdrusen //
Z.Rassenk, 1937, Bd.6,s.273.
Alan E.H.Emery. Metodology in Medical Genetics/ An introduction to
statistical methods. Edinburgh London Melbourne and New York,
1986, p. 197.
Balakrishnan V., Sanghvi L. Distance between Populations on the Basis of
Atribute Date // Biometrics, 1968, v. 24, N 4, p. 517-523.
Basu S., Kshatrija G., Jindal A. Fertility and mortality differentials among
the tribal population groups of Bactar district, Madhya Pradesh, India
// Hum. Biol., 1988. - v. 60, N 3, p. 407-416.
Bernis C. Estudio biodemografico de la poblacion maragata. - Doctoral
Thesis. 1974//Univ. Complutense, Madrid.
Bernstein F. Uber die Erblichkeit der Blutgruppen // Ztschr.f.Abstamm. -
u.Vererbungslehre, 1930, Bd. 54, s. 400-426.
Bernstein F. Fortgesetzte Untersuchungen aus der Theorie der
Blutgruppen // Ztschr.f.Abstamm.u.Vererbungslehre, 1930, Bd. 56, s.
233-273.
Blanco R., Chakraborty R. Genetic Distance Analysis of Twenty-Two
South American Indian Tribes // Hum. Hered., 1975, v. 25, p. 177-193.
Brues A.M. Models of race and cline // Amer.J.Phys. Anthrop. - 1972. V.
37, N 3, p. 289-399.
Cattell R.B. Factor Analysis: an Introduction to Essentials // Biometrica,
1965, v. 21, N l,p. 117-121.
Cattell R.B. Factor Analysis: an Introduction to Essentials // Biometrica,
1965, v. 21, N 2, p. 57-62.
120. Cavalli-Sforsa L, Barrai J., Edwards A. Analysis of Human
Evolution under Random Genetic Drift. "Gold Spring Harbor
Symposia on Quantitative Bioligy", 1964, v. 29, p. 9.
Cavalli-Sforza L., Edwards A. Analysis of Human Evolution H Genetics
Today, 1965a, v. 3, p. 932. I
Cavalli-Sforza L, Edwards A. Phylogenetic Analysis Models an
Estimation Procedures// Evolution, 1967, v. 21, N 3, p. 550.
Cavalli-Sforza L.L., Bodmer W.F. The Genetics of Human population
I iceman a.Co., San Francisco, 1971, p. 782. Jл
Cavalli-Sforza L.L. African Pigmies. Academic Press, INC., 1986. P. 46 •
Chakraborty R., Blanco R., Rothhammer F., Llop E. Genetic Variability
in Chilean, Indian Populations and its Association with Geography,
language and Culture // Soc. Boil., 1976, v. 23, p. 73-82.
Chakraborty R. Cultural, Language and Geographical Correlates of
Genetic Variability in Andean Highland Indians // Nature, 1976, v.
264, p. 350-352.
Chakravartti MR., Chakravartti R. Cerumen Types in Some Population
Groups// Z.Morphol. und Anthropo!., 1978, Bd. 68, N 1, S. 75-85.
Crauford M.N., Dykes D.D., Skradski K., Polesky H.F. Gene Flow and
Genetic Microdifferentation of a Transplanted Teaxcaltecan Indian
Population: Saltillo // Amer.J.Phys.Anthropol., 1979, v. 50, N 3, p.
401-412.
Crawford M.N., Workman P.L Methods and Theories of Anthropological
Genetics. 1973. University of New Mexico Publ.
Crow J.F. Some possibilities for measuring selection intensities in man //
Human Biol., 1958,30. P. 1-13.
( row .I P., Mange A.P. Measurement of inbreeding from the frequency of
marriges between persons of the same surname // Eug. Quarterly.
1965. V. 12, N 4. P. 199-203.
Das * K„ Mukherjee B.N., Malhotra K.S., Majumder P.P. Serological and
biochemical investigations among five endogamous groups of Delhi,
India// Proc. 2-nd Annu.Conf, ISHG. Calcutta, 1975.
De Oliveira A.F., Sal. ano F.M. Genetic Implications of the Demography
of Brasilian Juruna Indians // Sonal Biology, 1969, v. 16, N 3, p. 27.
Dinesh Parmar, L.K.Scncupta. Inbreeding and its clinical conhsequences:
a brief review with special reference to India // Bionature, IO (1,2),
1990, p. 61-64.
1 dwards A., Cavalli-Sforza L Reconstraction of Evolutionary Trees //
Phenetic and Phylogenetic Classification. (Ed. by 1964, v. 6, p. 67).
1 erak V., Kroupova Z. Changes of the Population Structure in Slovakia //
Symp. Med. Genetics, Debrecen-Haiduszoloszio, 1976, Abstracts, p.
76-89.
Hscher R.A. Statistical Methods for Research Workers, 11th ed., New
York, Hafner Publishing Co., 1950, p. 73.
fisher R.A. The genetical theory of natural selection. Oxford: Clarendon
Press, 1930. P. 272.
f'x A.C. The Role of Kin-Structured Migration in Genetic
Microdifferentation // Ann.Hum.Genet., 1978, v. 41, N 3, p. 329-339.
227
I rants PR., Kirjarinta M., Khan P.M., Raphikov U.S., Eriksson A.W.
Genetic polymorphisms in serum and erythrocytes of the Bashkirs //
Somatology and Population Genetics of the Bashkirs. Helsinki, I486 //
Annales Acad. Sci. Series A, V.Medica, 175. Fennicae. P. 17-24.
Gage T.B. The Effect of Out Migration on the Genetic Structure of
American Asamoa // Amer.J.Phys.Anthropol., 1979, v. 50, N 3, p.
440-451.
Gajdusek D.C., I^eyshon W.C., Kirk R.L., Blake N.M., Bronya Keats,
McDermid E.M. Genetic Differentiation among Populations in
Western New Guinea // Amer.J.Phys.Anthropol., 1978, v. 48, N 1, p.
47-64.
Glass B. Genetic Changes in Human Populations, Especially Those Due
to Gene Flow and Genetic Drift Ц Advances in Genetics, 1954, v. 6, p.
95-139.
Glass B. On the Evidence of Random Genetic Drift in Human
Populations//Amer.J.Phys.Anthropol., 1956, v. 14, N 14.
Guz K., Dedeoglu N. and Luleci G. The frequency and medical effects of
consanquineous marriages in Antalja, Turkey // Hereditas III: 79-83,
1989.
Halberstein R.A., and Crawford M.N. Human biology in Tlaxcala,
Mexico: demography// Amer.J. of Phys. Anthrop. 1972. 36. P. 199-212.
Hann Katherine L. Inbreeding and fertility in South Indian Population //
Annals of Human Biology, 1985. V. 12, N 3. P. 267-274.
Hardy G.H. Mendelian Proportions in a Mixed Population // Science,
1908, v. 28, p. 49-50.
Harrison G.A., Boyce A.J. The Structure of Human Populations. 1972.
Oxford University Press, London, p. 175.
Hed H.M.E. Opportunity for selection in the Barish of Tuna, Sweden //
I lum.I lercd. 1987. T. 37, N 1. P. 30-55.
Hirszfeld L., Hirszfeld Hanna. Serological differences between the blood
of different races. The result of researches in Makedonian front //
Lancet. 1919, II. P. 675-9.
Horkinson D.A., Spenser N., Harris H. Red cell acid phosphatase
variants: a new human polimorphism // Nature (L.). 1963. V 199.
Jacquard A. Gcnctiquc des populations humaines// Presses Universitaries
de France. 1974.
lacquard A., Ward R H. The Genetic Consequences of Changing
Reproductive Behavior//.!, of Hum. ['vol. 1976, N 5. P. 139-154.
Johnson М.1., Wallace D.C., Ferris S.D., Rattarri M.C. and Cavalli-Sforza
L.L. Radiation of human mitochondria DNA types analyzes by
restriction endonuclease cleavage patterns // J/MoI.Evol. 1983. V. 19,
N 3/4. P. 225-271.
Jonston F.E., Kesinger K.M. Fertility and mortality differentials and their
implications for microevolutionary change among the Cashinahua Ц
Hum.Biol. 1971. V. 43, N 3. P. 356-364.
Keiichi (). Polymorphic Traits in Peoples of Eastern Asia and Pacific //
Genet. Polymorphisms and Diseases Man.Sheba Int. Syrnp. Tel Aviv,
1973. New York - 1-ondon, 1974, p. 69-89.
Kimura M., Weiss G.H. The Stepping-Stone Model of Population
Structure and Decrease of Genetic Correlation with Distance //
Genetics, 1964, v. 49, p. 561-567.
Kurczynski T. Generalized Distance and Discrete Variables // Biometrics,
197, v. 26, N3, p. 186-199.
Ixince G.N., Williams W.T. A Generalized Sorting Strategy for Computer
Classifications // Nature, 1966, v. 219, p. 218.
I xturell C.B., Eriksson S. The electrophoretic alpha, globulin pattern of
serum antitrypsin deficiency // Scand.J.Clin. Lab. Invest., 1963. V. 15.
P.132.
Ixturenson S.F., Benton M.A., Bishop A.J. and Mascie-Taylor C.G.N.
Fertility at low and high altitude in Central Nepal Ц Social Biol., 1985.
V. 32, N 1-2. P. 65 "'О.
Ix:uschner D. Einfuhrung in die numerische Taxonomie. Jena, 1974, S.
192.
Li Ch.Ch. Population Genetics. The University of Chicago Press. Chikago
- London, 1955, p. 322-327.
Li Ch.Ch. Table of Variance of ABO Gene Frequency Estimates //
Ann.Hum.Genet., 1970, v.34, p.189-194.
Little-Wood R.A., Lajeunesse R.M. Founder's Effect and Inbreeding in
the Cape Barren Island Isolate, 1815-1974 // Amer.J.Phys.Anthropol.,
1979, v. 50, N 3, p. 459.
ivingstone F.M. Natural Selection, Disease and Ongoing Human
Evolution as Illustrated by the ABO Blood Groups Ц Hum.Biol., 1960,
v. 1, p. 17.
una Francisco and Moral Pedro. Mechanisms of natural selection in
Human rural populations, survey of a Mediterranen region (1л
229
Alpujarra, SE Spain) // Annals of Human Biology. 1990. V. 17, N 2. p
153-158.
Malhotra K.C., Chakraborty R., Chakraborty A. Gene Differentiation
among the Dhangar Caste-Cluster of Maharashtra, India //
Hum.Hered., 1978, v. 28, p. 26-36.
Malecot G. Decrease of Relationship with Distance // Cold Spring Harbor
Symp.Quant.Biol., 1950, p. 52-53.
Malecot G. Identical loci and Relationship. Proc. 5th Berkeley
Symp.Math.Stat.Prob., 1966, p. 317-332.
Malyutov M.B., Passekov V.P., Rychkov Yu.G. On the Reconstruction of
Evolutionary Trees of Human Populations Resulting from Random
Genetic Drift // In* Proceeding of the International Symposium 'The
Assement of Population Affinities in Man", ed. J.S. Weiner, J.Huizinga.
The Claredom Press, Oxford, 1972, p.56.
Mayr E. Numerical Phenetics and Taxonomic Theory // Systematic
Zoology, 1965, v. 14, p. 73.
Maruyama T. Isolation by Distance, Genetic Variability the Time
Required for a Gene Substitution, and Local Differentiation in Finite
Geographically Structured Populations Morton, Genetic Structure of
Population Univ, of Hawaii Press, Honolulu, 1973, p. 80-81.
Matsunaga E. The Dimorphism in Human Normal Cerumen //
Ann.Hum.Genet., 1962, v. 25, p. 273.
Morton N. Human Population Structure // Annual Rev.Genet., 1969, v.3,
p.241.
Morton N.E. Population Genetics // Human Genet., Amsterdam, 1972, p
131-132.
Morton N.E., Lalouel J.M. Bioassay of Kinship in Micronesia /.
Amer.J.Phys. Anthropo!., 1973, v. 38, N 3, p. 709-720.
Morton N.E., Smith C., Hill R., Frackiewicz A., Law P., Yee S. Population
Structure of Barra (Outer Hebrides) // Ann.Hum.Genet., 1976, v. 39,
N 3, p. 339-352.
McCullough T.M., Giles E. Human Cerumen Types in Mexico and Ne*
Guinea: a I lumidity-Related Polymorphism in "Mongoloid" Peoples
Nature, 1970, v. 226, N 2, p. 348-361. :
Moui.mt A.E., Kohek A.C., Domanewska K. ABO Blood Groups, Oxfor
1953, p. 119.
230
Mourant Л.Е., Kapcs Л.С., Domaniewska-Sobczak K. The Distribution of
the Human Blood Groups and Other Polymorphisms Oxford Univ.
Press. London, 1978, p. 1055.
Neel J.V. and Chagnon N.A. The demography of tow tribes of primitive
relatively unacculturated American Indians // Proceedings of the
National Academy of Sciences. 1968. V. 59. P. 680-689.
Neel J.V., and K.M.Weiss. The genetyic structure of a tribal population,
the Yanomama Indians. XII. Biodemographic studies // Amer.J. of
Phys. Anthropology. 1975. V. 42. P. 25-52.
Neel J.V. The Circumstance of Human Evolution // J.Hopkins Med.J.,
1976, v. 138, N 6. p. 233-244.
Nei M. Variations and covariation of gene frequencies in subdivided
populations // Evolution. 1965. V. 19, N 2. P. 256-258.
Nei M., Imaizumi Y. Genetic Structure of Human Populations. V. Local
Differentiation of Blood Group Gene Frequences in Japan //
Heredity, 1966, v. 21, N 3, p. 461-472.
Nei M. Analysis of gene diversity in subdivided population //
Proc.Nay.Acad.Sci.USA. 1973. V. 3, N 70. P. 3321-3323.
Norakmal J., Tan S.G. Cerumen Polymorphism in the Three M jor Ethnic
Groups of Malaysia Ц Jap.J.Hum.Genet., 1979, v. 24, N. 2, p. 119-121.
)moto K. Allelic diversity of human populations in the Asian-Pacific
Area. - In: Molecular evolution protein polimorphism and the neutral
theory (ed. M.Kimura)// Jap.Sci.Soc.Press. Tokyo; Springer-Verlag.
Padmanabham P.B.S.V. Selection intensity among the rural and urban
Naika of Gujarat, India//Anthropologic, XXIII/2,1985. P. 101-103.
Prokop O., Uhlenbruck G. Lehrbuch der menschlischen Blut und
Serumgruppen. Leipzig, 1966, S. 239.
Часе R., Sanger Blood Groups in Man. 4th ed., Blackwell Scientific
Publications. Oxford, 1962, p. 193.
anghvi L. Comparison of Genetical and Morphological Differences //
Amer.J.Phys.Anthropol. (n.s.), 1953, v.l 1, N 4, p. 210.
Sanghvi L., Balakrishnan V. Comparison of Different Measures of
Genetic Distance between Human Populations // In: The Assesment
of Population Affinities in Man. Oxford, 1972, p. 131.
1 ao P.S., Inbaraj S.G. Inbreeding effects on fertility and sterility in
Southern India//J.Med.Genet. 1979. V. 16. P. 24.
oberts D.F., Bonne B. Reproduction and inbreeding among the
Samaritance//Soc.Biol. 1973. V. 20. P.64.
231
Rajanikumari I., Srikumari C.R., Rao T.V. Variability of selection
opportunities with changing socio-cultural environments ц
Hum.Hered. 1985. T. 35, N 4. P. 218-222.
Roychondhury A.K. Gene differentiation among caste and linguistic
populations of India Ц Hum.Hered. 1974. N 24. P. 217-233.
Sanghvi L., Balakrishnan V. Comparison of Different Measures of
Genetic Distance between Human Population // In: The Assesment of
Population Affinities in Man. Oxford, 1972, p. 171.
Sanghvi L. Genetic Distance: a Review Bull, of the Decean College
Rcs.Inst., 1973, v. XXX XXXII, Poona, p. 331-339.
172. Saugstad L.F. Inbreeding in Norway//Ann.Hum.Genet., 1977, v. 40,
N4, p. 481-492.
Schanfied M.S., Alexeyeva T.U., Crawford M.N. Studies on the
immunoglobulin allotypes of asiatic population. VIII. Immunoglobulin
allotypes among the tuvinians of the USSR // Hum.Hered. 1980. V.
30. P. 343-349.
Schull W.J., Furusho T., Yamamoto M. I'he effects of parental inbreeding
in Hirada Japan. IV. Fertility and reproductive compensation. Human
Genet. 197. N 9. P. 294-315.
Schull WJ. Primitive Populations. Some Contributions to the
Understanding of Human Population Genetics // Human Genet.,
Amsterdam, 1972, p. 112-123.
Schwidetzki J. Die neue Rassenkunde. Stuttgart, 1962.
Sokal R.E., Sneath P.H.A. Principles of Numerical Taxogony. San
Francisco, W.H.Freeman and Co., 1963, p. 186.
Spencer N., Hopkinson D.A., Harris H. Phosphoglucomutase
polimorphism in man // Nature (L), 1964. V. 204.
Spuhler J.N. Anthropological genetics: an overview /
theories of anthropological genetics I Eds. Crawford M.N., Workman
P.L. New Mexico: Univ. New Mexico Press, 1973. P. 423.
Spuhler I.M. The maximum opportunity for natural selection in some
human populations // Demographic anthropology: quantitative
approaches. Albuquerque, 1978. P. 185-226.
Steffan P. Handbuch der Blutgruppenkunde. Munchen, 1932.
Steffan P., Wellisch P. Die geographische Verteilung der Blutgruppen fur
Rassenphysiologie. 1928-1936, Bd. 2-7.
Stevens W.L. Estimation of Blood Group Gene Frequences. Ann.Eugen'’
1938, v.8, p. 362-375.
Strahlenberg J. Das Nord- und Ostlische Thiel von Europa und Asia.
Stockholm, 1930.
Stevcncc W.L. Statistical Analysis of the A-B-O Blood Groups // Human
Biol., 1950, v. 22. p. 191-217.
Stevens W.L. A-B-O System in Mixed Populations // Human Biol., 1952, v.
24, p. 12-24.
Tibera-Dumitru. La frequence des groupes sanguins du facteur gustatif
(PTC) du systems d’haptoglobine dans la region Oltenia de Poumanie,
1968, p. 552-557.
forrejon I., and Bertranpetit J. Estructura biodemografica de la
problacion del Valle de Camprodon // Trabajons de Antropologia.
1987. V. 20. P. 393-557.
lyzzer R.N. An investigation of the demographic and genetic structure of
a south-western American Indian population, the southern Ute tribe
of Colorado// Doctiral Dissertation, University of Colorado, 1974.
l oth T. Somatologiya i paleoantropologiya naseleniya Vengrii. - Doktori
disszcrtacio, l-II. (Somatology and paleoanthropology of the
Hungarians. Doct. dissertation, 1-11). - Budapest, 1974. P. 649.
I loth T. Some anthropological aspects in the ethnogenesis of I 'ungarians.
Ц Cong.Quintus Intern. Fenno-Ugristarum (Turku, 1980). 1981. V. 8.
P. 427-437.
Waissbluth J.G. Langman M.J.S. ABO Blood Groups, Secretor Status,
Salivary Protein, and Serum and Salivary Immunoglobulin
Concentrations. - Gut, 1971, v. 12, N 8, p. 646-649.
I 'V.dter H., Steegmuller H. Studies of the geographical and racial
distribution of the Hp and Gc polimorphisms // Hum.Hered. 1969. V.
19, N 2.
// Methods and V liter H. On the geographical variability of the red cell PGMi and acid
u Kt u/nrkman phosphatase gene frequencies// Hum.Hered. 1976. V. 26, N 1.
issenberg S. Ein Beitrag zur Anthropologic dcr Turkvolkel Baschkiren
und Metschezjaken // Zeitschrift fur Ethnologic, Bd. XXIV, Berlin,
1892.
< mberg W. Uber den Nachweis der Vererbung beim Menschen, Jahresh
Verein f.Vaterl.Naturk.in Wruttemberg, 1908, Bd. 64, S. 368-382.
ner A S. History of Blood Group Nomenclature with a Questionaire
of Rh-Hr Nomenclature//J.Forensic. Med., 1967, v. 14, N 1, p. 3-12.
rkman P.L. Population Studies in the Aland Islands, Finland: 1750-
1950// AmerJ.Phys.Anthropo!., 1979, v. 50, N 3, p. 494.
233
232
ОГЛАВЛЕНИЕ
Wright S. Evolution in Mendelian Populations // Genetics, 1931, v. 16, ft
2, p. 281-312.
Wright S. Statistical Genetics in Relation to Evolution. Paris, Hermann
and Cie, 1939, p. 152.
Wright S. Isolation by Distance I I Genetics, 1943, v. 28. p. 114.
Wright S. Isolation by Distance under Divers System of Mating //
Genetics, 1946, v. 31, p. 39-59.
Wright S. The Genetical Structure of Populations // Ann.Eugenics, 1951,
v. 15, p. 323-354.
Yambery H. Das Tirkenvolk in seinen ethnologischen und
ethnographischen Beziechungen. Baschkirischen. Leipzig, 1885.
Zubov A.A., Segeda S.P. Dental morphology of the Bashkirs //
Somatology and population genetics of the Bashkirs I Ed. P.Kajanqja,
A.A.Zubov. Helsinki, 1986.
1редисловие. Что из себя представляет историческая
I генетика? Ее роль и эффективность в решении проблем расо-
II этногенеза-........................................... 3
I лава 1. Предыстория вопроса. Динамика и преемственность
I развития этнорасовых групп в регионе Приуралья. Проблема
гногенеза башкир......................................... 6
1. Археологические источники.
2. Некоторые материалы ономастики, лингвистики,
фольклора.
3. Антропологические данные (палеоантропология,
антропология, одонтология, дерматоглифика).
4. Информация по этнической геногеографии и
некоторые итоги.
,ава 2- Популяционная генетика как источник информации о
асо- и этногенетических процессах....................... 34
1. Генетические маркеры. Частоты генов. Наследственный
полиморфизм популяций.
2. Роль микроэволюционных факторов в дифференциации
и интеграции популяций. Дрейф генов. Миграции генов.
Систематические события (отбор). Мутационный процесс.
ава 3. Реализация программы исследований по
дорической генетике................................... 55
1. Выбор объектов исследования.
2. Организация сбора материала.
3. Идентификация систем групп крови и биохимических
маркеров генов в популяциях.
4. Статистический анализ данных.
ава__4. Временная и пространственная организация
Ьпуляции................................................ 66
1. Возрастная структура.
2. Поколение, его длительность и темпы смены в
популяциях.
3. Характеристика общего, репродуктивного и
эффективно-репродуктивного размеров.
234
235
Глава 5- Генетическая структура башкирской популяции...
1. Наследственный полиморфизм популяций по данным о
группах крови.
2. Биохимические маркеры генов и другие формы
генетической изменчивости.
Глава 6. Особенности генетической дифференциации
популяций; факторы, способствовавшие ее подразделенности... щ
1. Степень генного разнообразия на различных уровнях
иерархической структуры.
2. Теоретическая оценка уровня генетической
дифференциации популяций по демографическим и
генетическим данным.
3. Оценки уровня инбридинга на основании анализа
генных частот и родословных.
4. Соотношение вкладов дифференциальной смертности и
дифференциальной плодовитости, как компонентов
приспособленности, в структуру естественного отбора в
популяциях человека.
5. Ступенчатый характер генных миграций - как наиболее
адекватная модель для популяции башкир. Зависимость
генетических расстояний между группами от
географических дистанций - еще одно свидетельство о
направленном генном потоке.
Глава 7- История формирования башкирского народа в свете
данных исторической генетики..........................161
1. Таксономический анализ групп, выделение структурных
единиц и их взаимоположение в пространстве
генетических факторов АВО, MN, Rh-Hr, Нр, Kell систем.
2. Сравнительный вклад европеоидной и монголоидной
компонент в формирование популяции.
3. Оценки времени генетической дифференциации
популяций.
Заключение............................................205
Художественное оформление: А.И.САПЛИН
Подготовила к изданию: С.Н.КВАШНИНА
Утверждено к печати Институтом этнологии и антропологии
Российской Академии Наук
Пвя з-го1 *оо»4о-г
Тодписано к печати 14.12.92
0,6 уч.-изд л. Тираж 300 экз.
Формат Ы1Х84/16 Объем: 14 пл.
Цена договорная Заказ
часток оперативной полиграфии Института этнологии и антропологии
17334 Москва Ленинский проспект 32 А
236
Автор монографии предлагает новыми S.3TL
рико-генетический подход для решения слож-
нейших вопросов истории и этногенеза разных
народов, представляющий собой синтез етодов
исторических и генетических дисциплин. Работа
построена на оринальном собственном материале
по популяциям бывшего Советского Союза с прив
лечением обширных мировых данных.
Монография рассчитана на специалистов
антропологов, этнологов, археологов, историков, а
также на широкий круг читателей, интересую-
щихся проблемами истории и генетических попу-
ляций.
The author has offered a new historico-genetic
approach to resolve the most complicated problems of
history and ethnogenesis of different peoples.
This approach is based on the synthesis of
historical and genetic method of science.
The author has examined population of the
former USSR. The study is based on her own original
materials and on the general world date as well.
The book may be recommended for the speci-
alists in anthropology, ethnology, archaeology, history
and for those who take interest in the matters in
question as well.