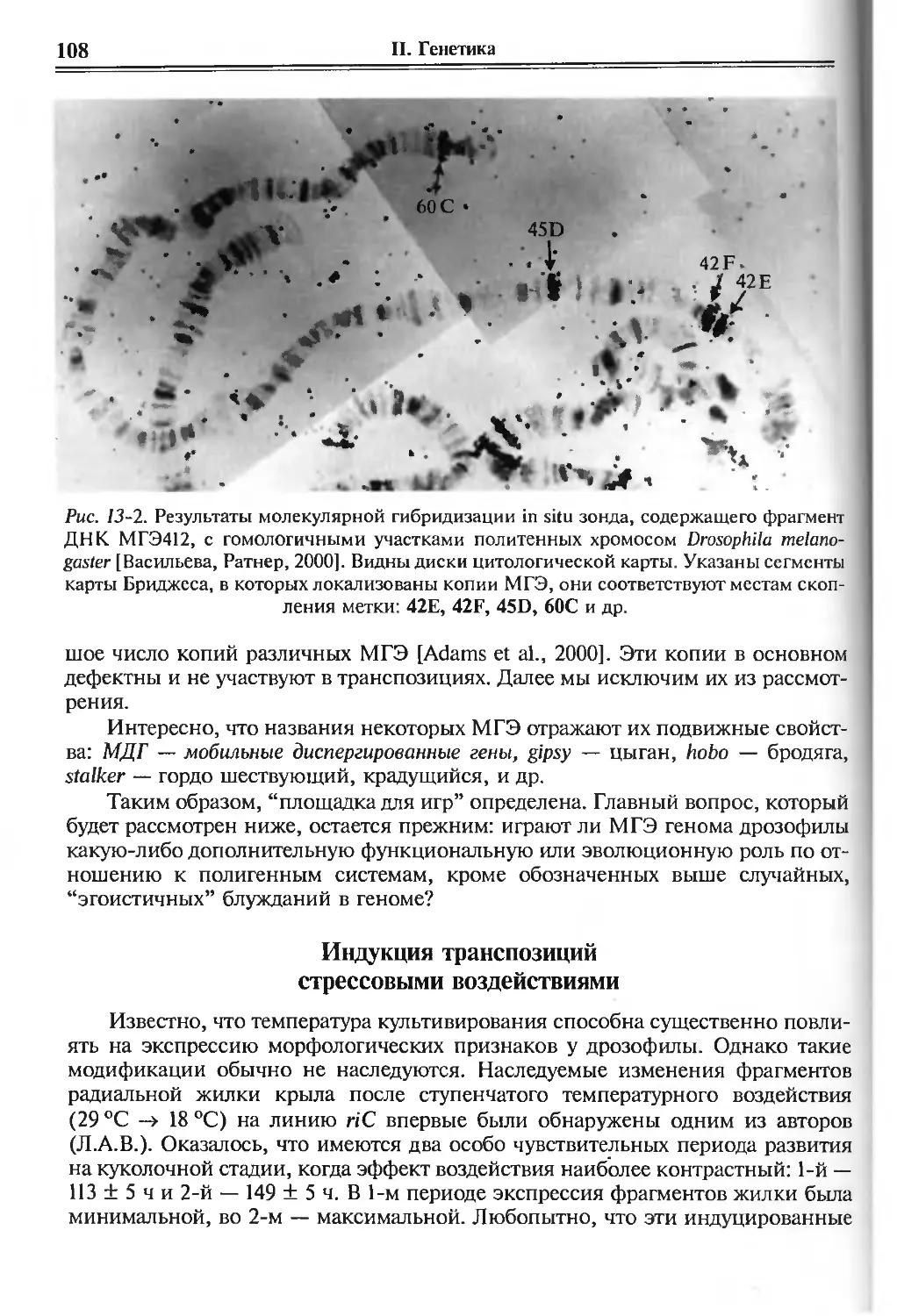
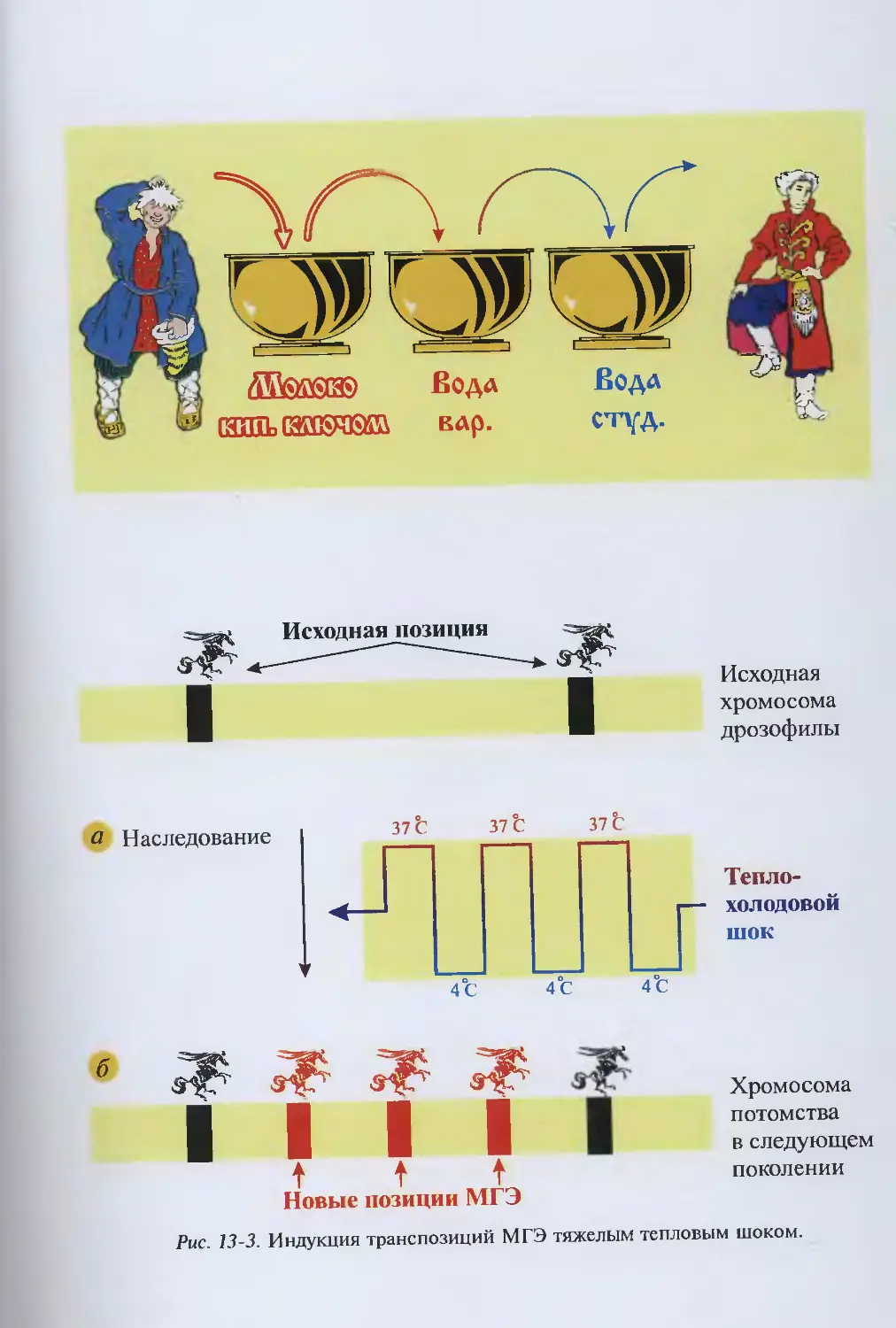
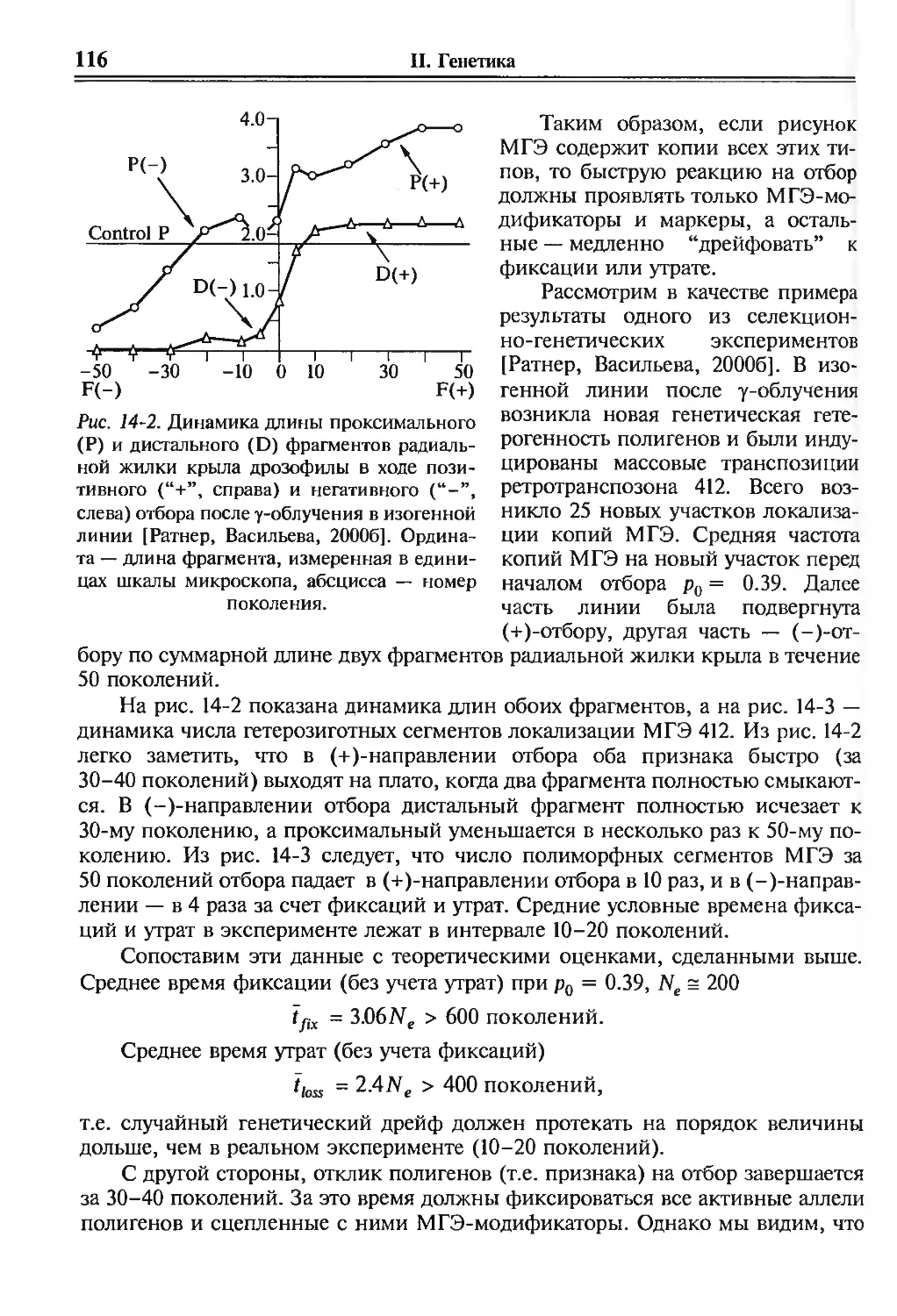
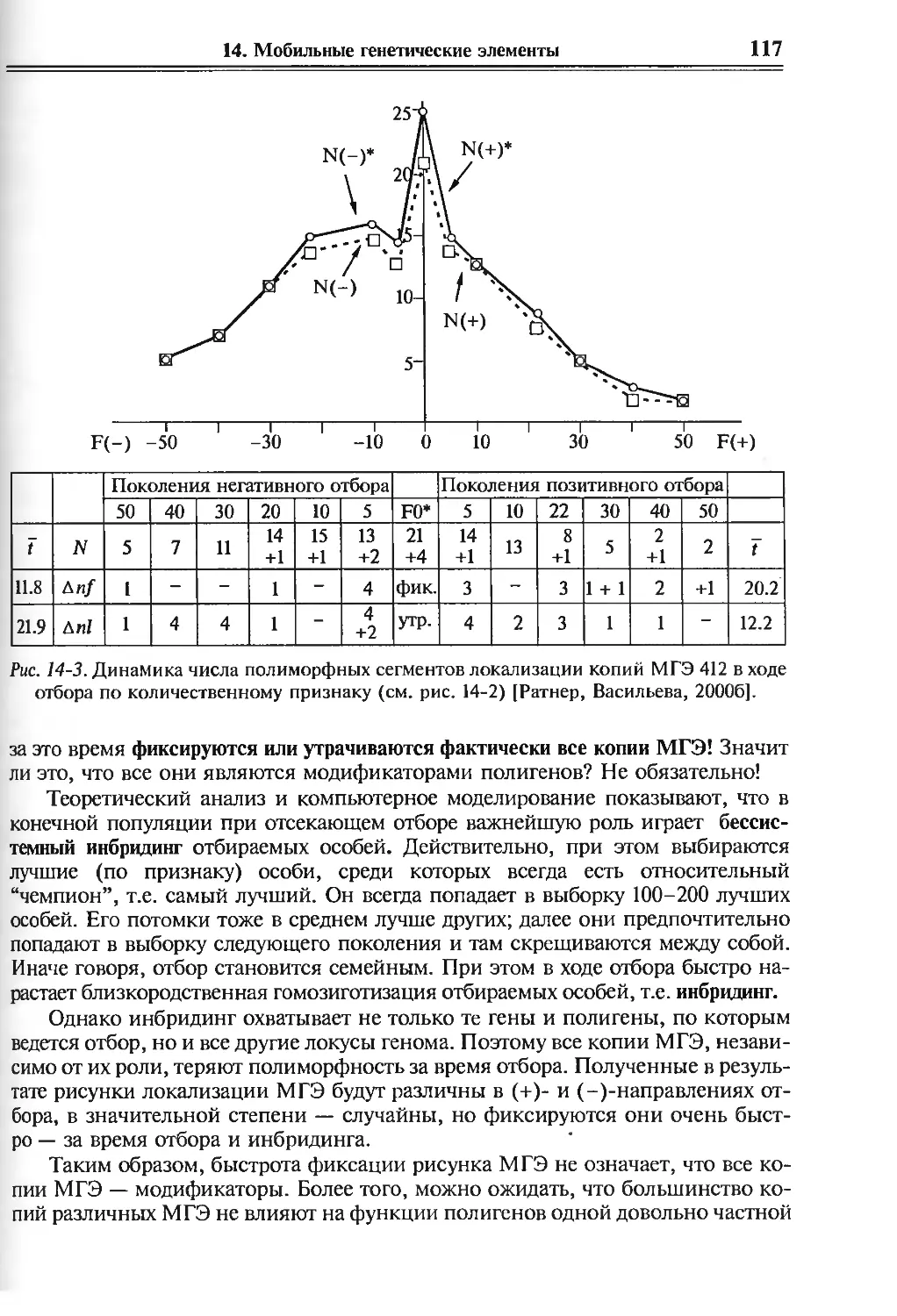
/








































































































































































































































































Author: Ратнер В.А.
Tags: язык языкознание лингвистика литература история науки кибернетика генетика молекулярная биология
ISBN: 5-02-031969-4
Year: 2002




Text
Not for sale !!!
For preview only !!!
If You like this book - buy it!
Scanned by Mykhaylo (molbiol(a!ukr.net)
Not for sale! For preview only! Scanned by Alykhaylo (molbiol(cvnkr.net)
Оглавление
ОТ РЕДАКТОРА 4
ПРЕДИСЛОВИЕ 5
ВВЕДЕНИЕ 6
I. PERSONALIA 10
1. Мамонт (заметки о Н.В. Тимофееве-Ресовском) 10
2. Николай Владимирович Тимофеев-Ресовский — ровесник века, гигант столетия 21
3. Алексей Андреевич Ляпунов 32
4. Игорь Андреевич Полетаев 42
5. С.М.Гершензон: впереди событий и в стороне от признания 48
6. Дмитрий Константинович Беляев 54
7. Памяти профессора Francesco M. Scudo 64
8. Памяти академика Н.Н. Моисеева 67
9. Поездка в Испанию. Памяти Н.Н. Воронцова 69
II. ГЕНЕТИКА 75
10. 100 лет — 18 конгрессов! 75
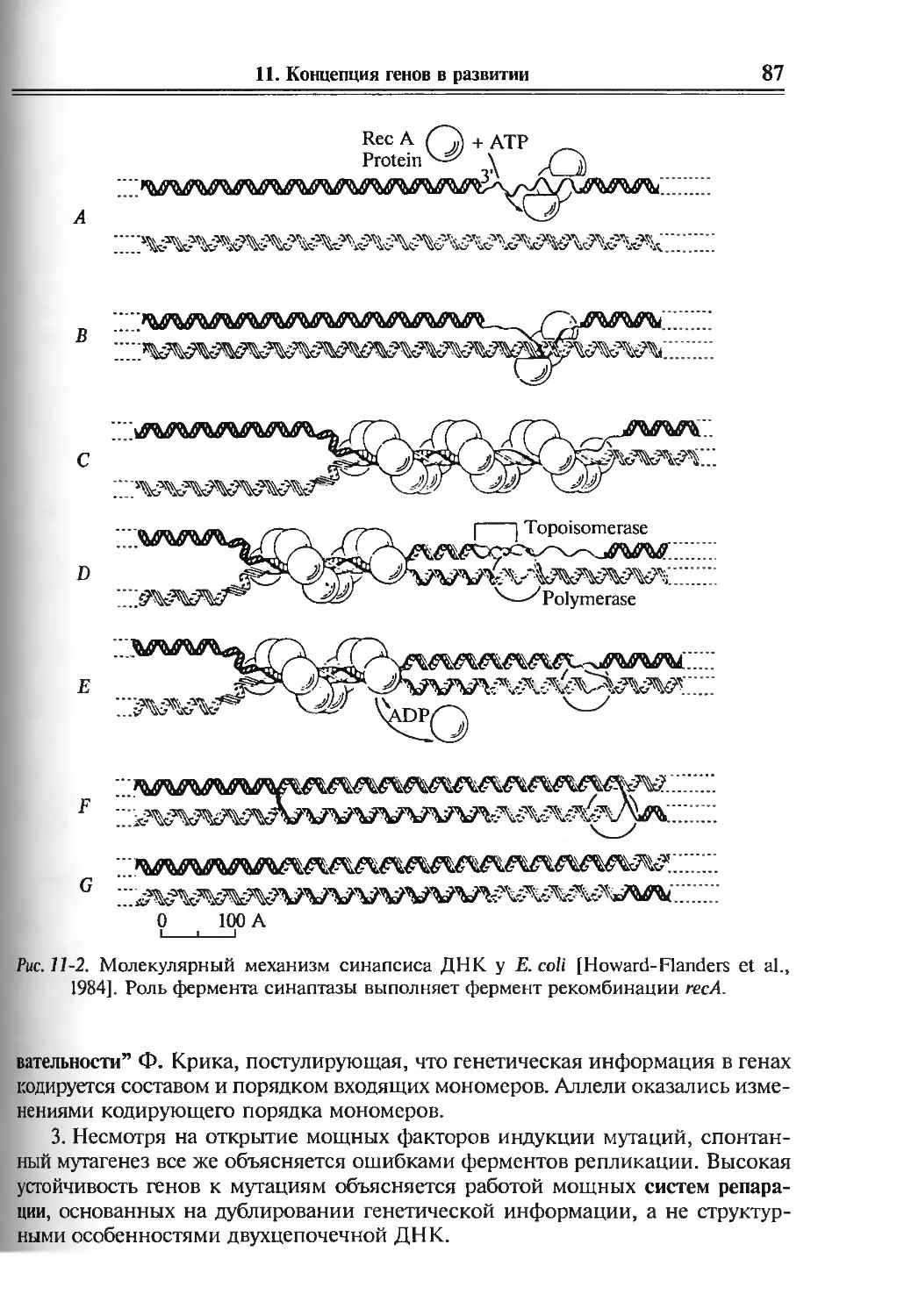
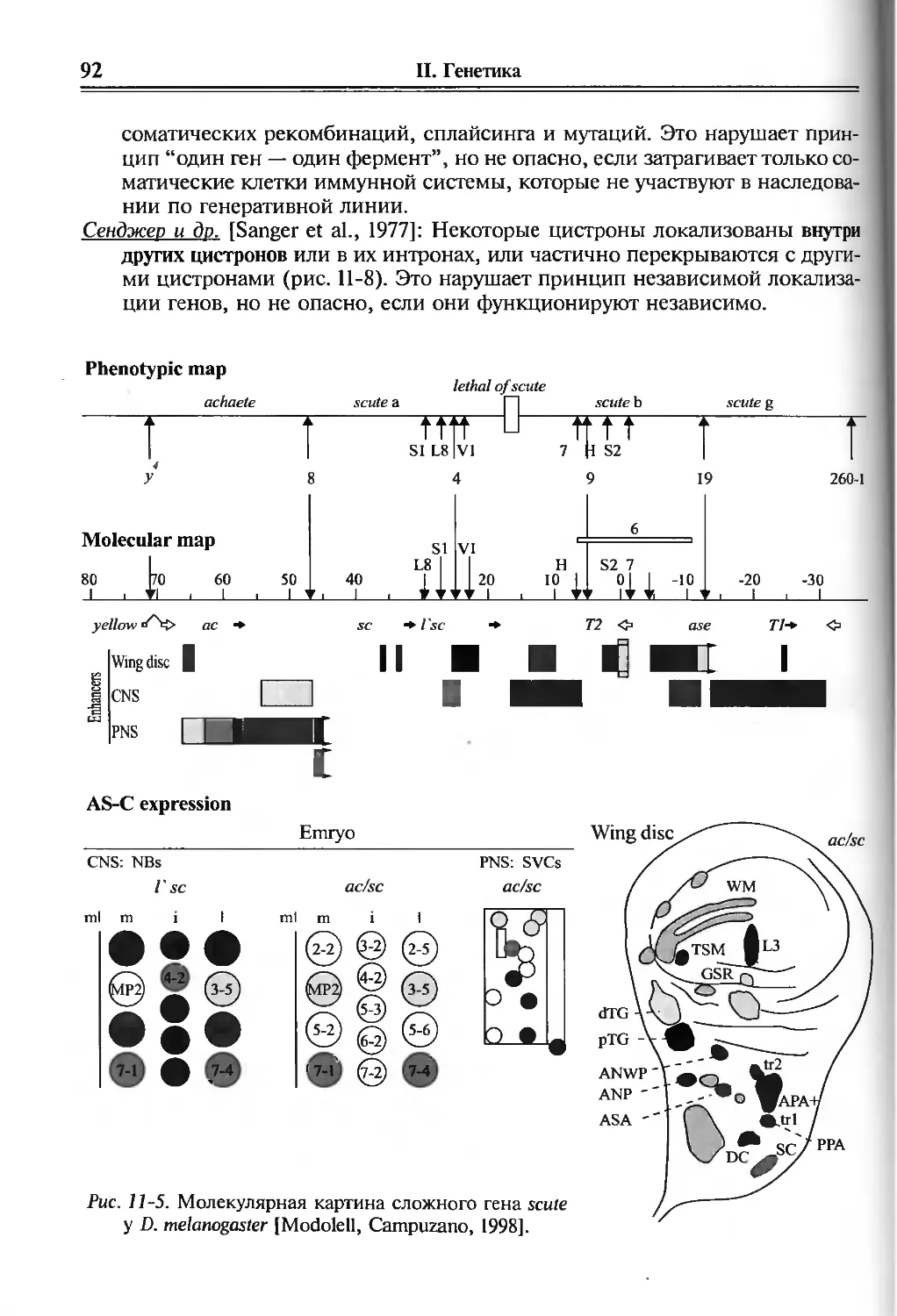
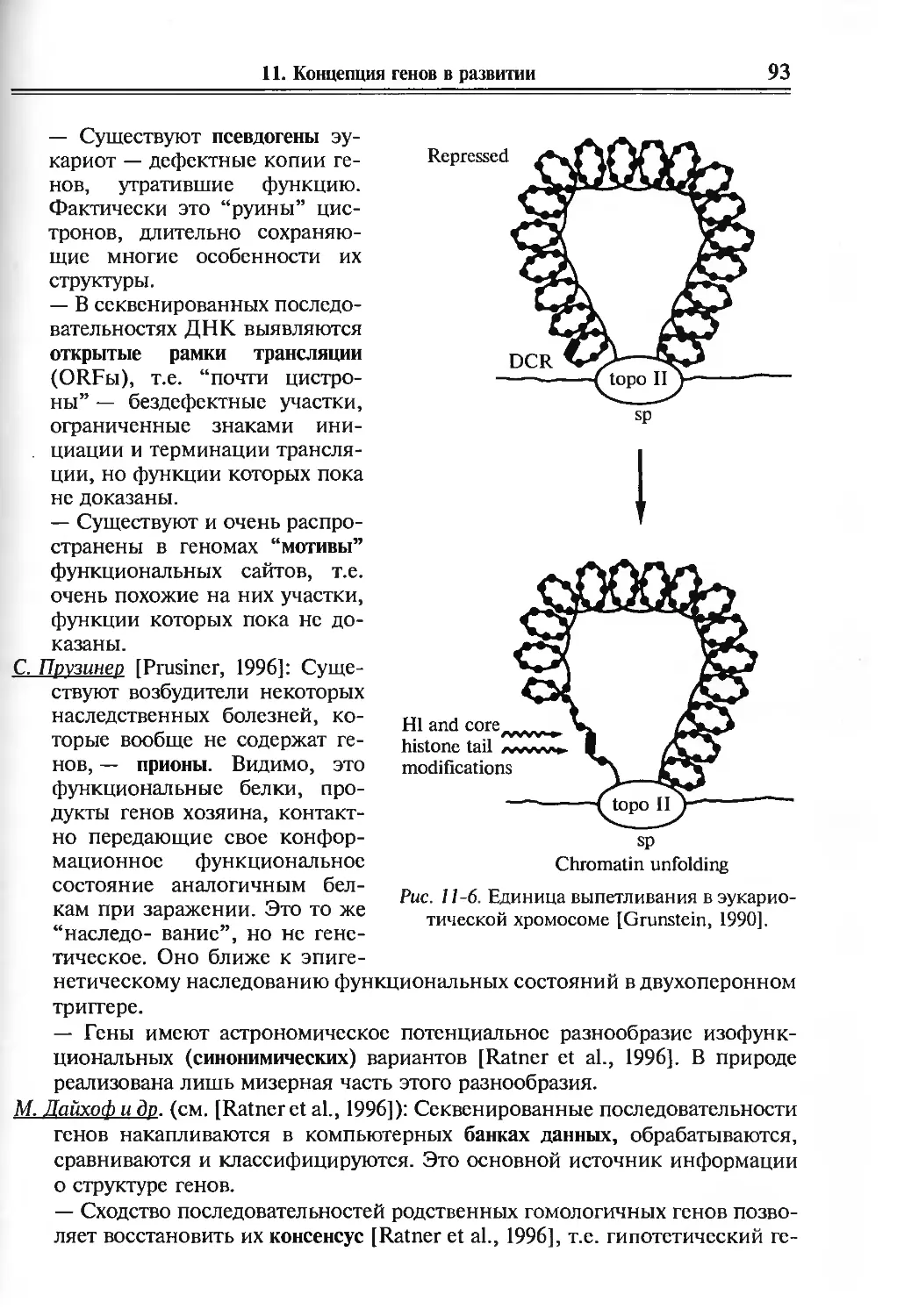
11. Концепция генов в развитии 81
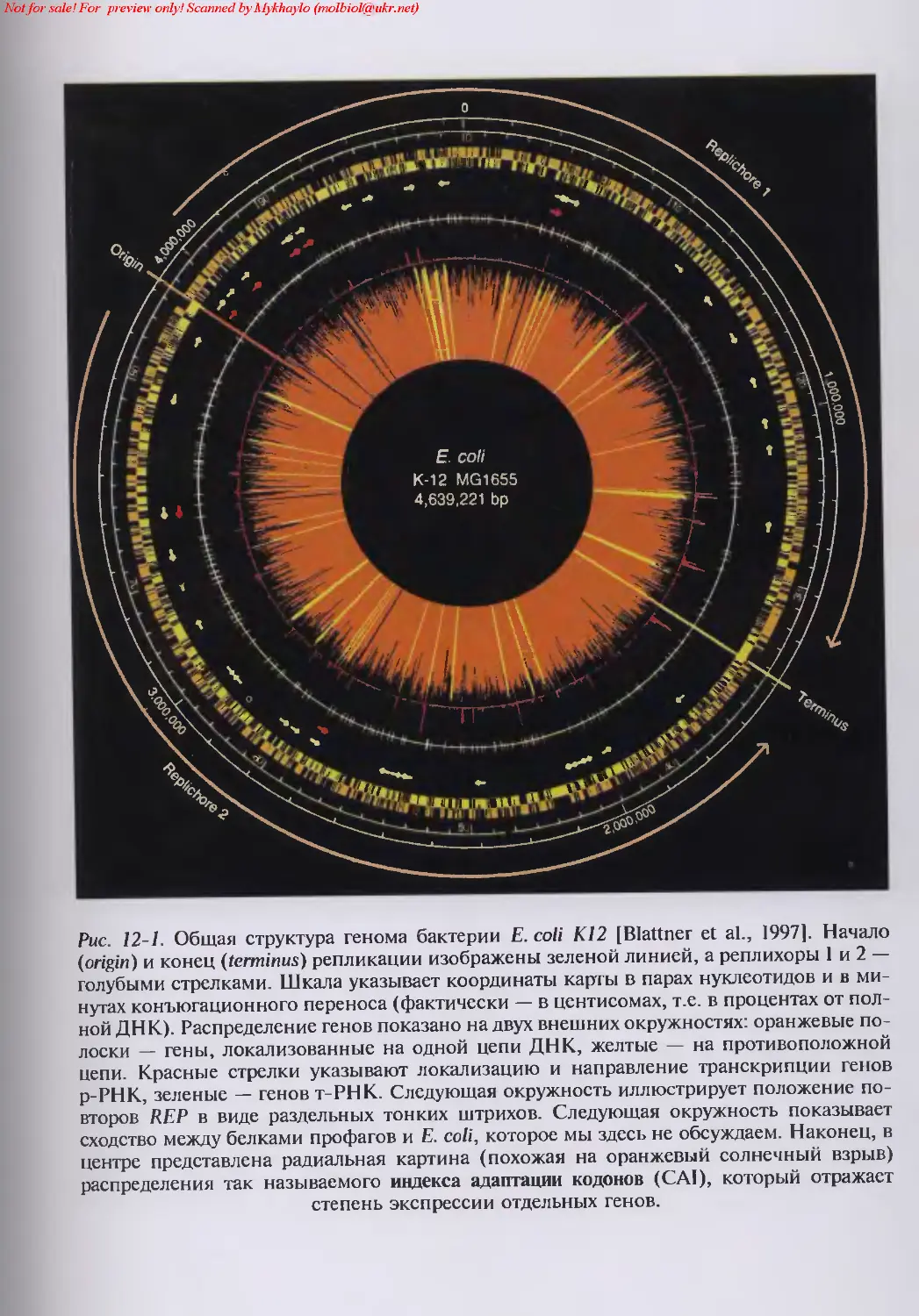
12. Что содержит полный геном Escherichia coli? 96
13. Индукция транспозиций мобильных генетических элементов стрессовыми воздей-
воздействиями 104
14. Мобильные генетические элементы: геномные бродяги или трудяги? 113
III. МОЛЕКУЛЯРНАЯ КИБЕРНЕТИКА 122
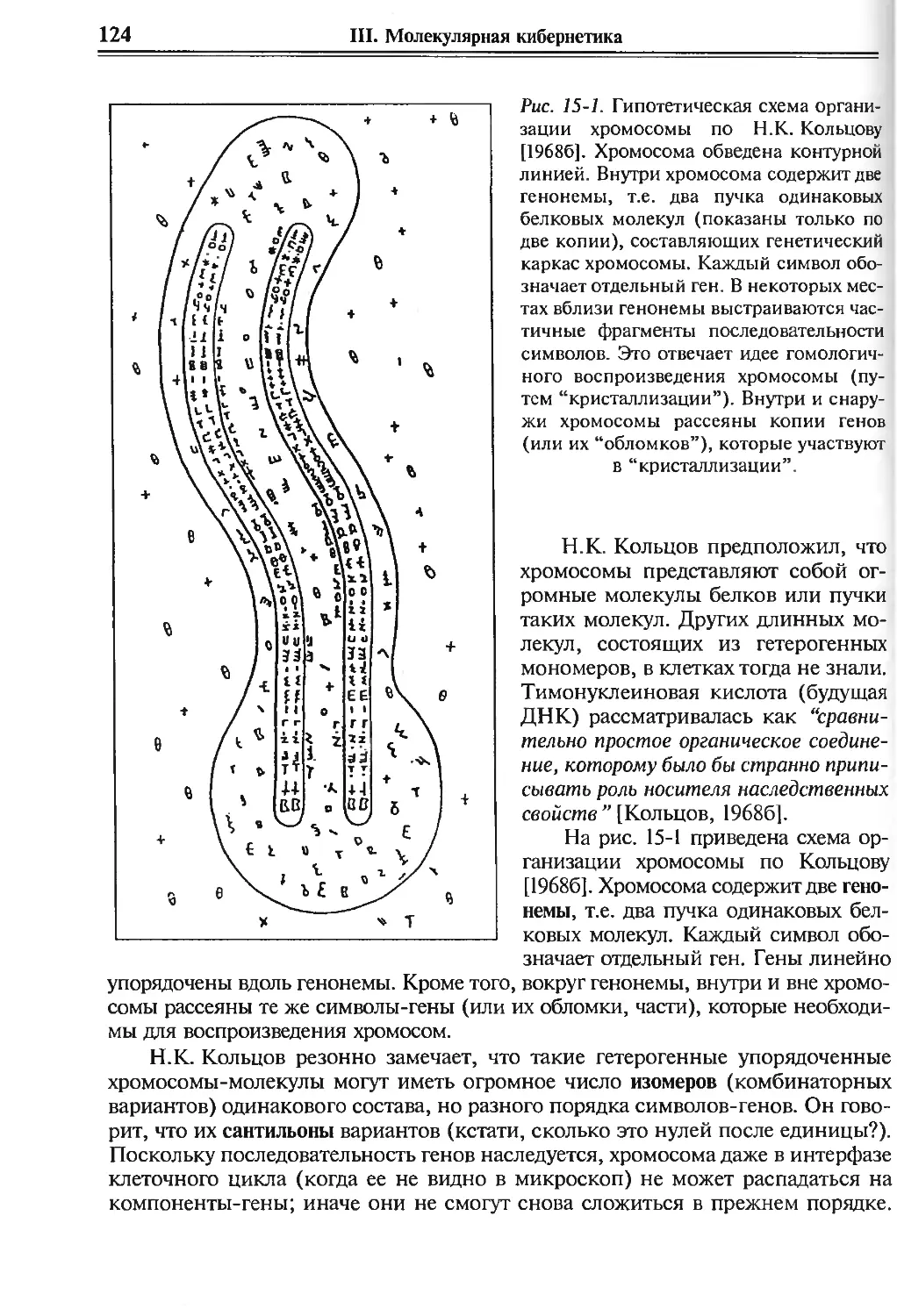
15. Хроника великого открытия: идеи и лица. I 122
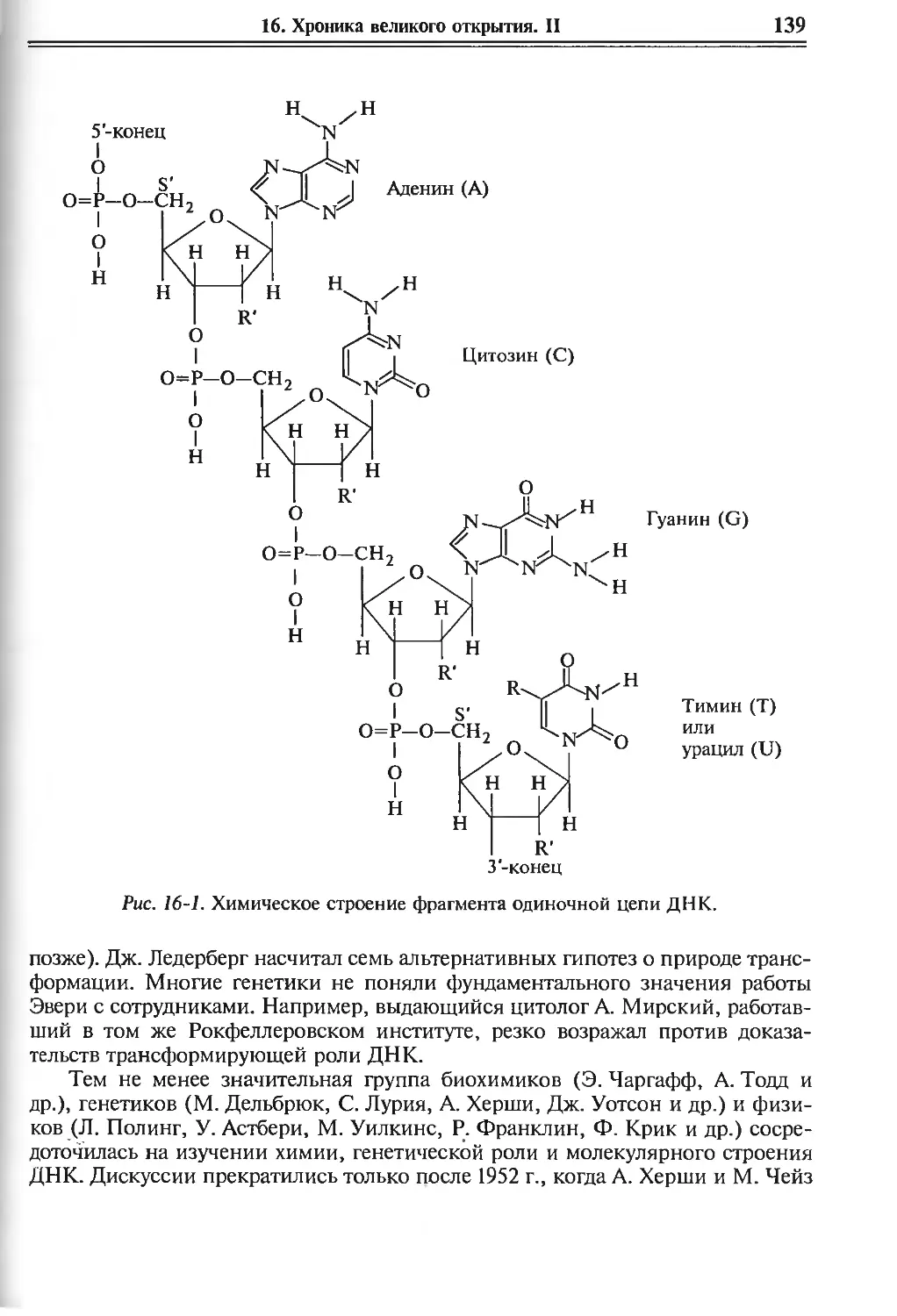
16. Хроника великого открытия: идеи и лица. II 136
17. Хроника великого открытия: идеи и лица. Ш 151
18. Молекулярная кибернетика в океане науки 167
19. Виртуальная реальность и системы управления 180
20. Генетический код как система 190
21. Генетический код как продукт эволюции 197
22. Закономерности кодирования генетической информации (генетический язык). 203
23. Генетические переключатели 218
24. Молекулярная эволюция 229
25. Укус издыхающей рептилии 238
26. Математическая генетика в НГУ и около 245
27. Те же проблемы, но в стихах 258
Научное издание
Ратнер Вадим Александрович
ГЕНЕТИКА, МОЛЕКУЛЯРНАЯ КИБЕРНЕТИКА
ЛИЧНОСТИ И ПРОБЛЕМЫ
Редакторы Л.В. Филиппова, Л.П. Голышева. Художественный редактор J1.B. Матвеева. Художник И.С. Попов. Технический редактор
И.М. Остроумова. Корректор И.Л. Малышева. Оператор электронной верстки Л.А. Антонова.
Изд. лиц. № 020297 от 23.07.97. Сдано в набор 25.06.02. Подписано в печать 23.09.02. Бумага легкомелованная. Формат 70x100 1/16.
Офсетная печать. Усл. печ. л. 21,9+0,88 вкл. на мел. бум. Уч.-изд. л. 22,8. Тираж 1000 экз. Заказ N° 162.
"Наука". Сибирская издательская фирма РАН. 630099, Новосибирск, ул. Советская, 18.
Оригинал-макет изготовлен на настольной издательской системе.
ГП "Новосибирский полиграфкомбинат". 630007, Новосибирск, Красный проспект, 22.
РОССИЙСКАЯ АКАДЕМИЯ НАУК
СИБИРСКОЕ ОТДЕЛЕНИЕ
ИНСТИТУТ ЦИТОЛОГИИ И ГЕНЕТИКИ
РОССИЙСКАЯ АКАДЕМИЯ ЕСТЕСТВЕННЫХ НАУК
МИНИСТЕРСТВО ОБРАЗОВАНИЯ
РОССИЙСКОЙ ФЕДЕРАЦИИ
НОВОСИБИРСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
В.А. РАТНЕР
ГЕНЕТИКА,
МОЛЕКУЛЯРНАЯ
КИБЕРНЕТИКА
ЛИЧНОСТИ И ПРОБЛЕМЫ
Ответственный редактор
доктор биологических наук Л.А. Васильева
Q
Not for sale !!!
For preview only!!!
If You like this book - buy it!
Scanned by Mykhaylo (molbiol@ukr.net)
НОВОСИБИРСК
"НАУКА"
2002
УДК 882
ББК 72.3 + 28.04
Р25
Ратнер В.А. Генетика, молекулярная кибернетика: Личности и про-
Р25 блемы. — Новосибирск: Наука, 2002. — 272 с.
ISBN 5-02-031969-4.
Вниманию читателей предлагаются написанные в увлекательной форме за-
заметки о выдающихся российских и зарубежных ученых-генетиках. Значительная
часть монографии посвящена ключевым проблемам генетики и теории молеку-
лярно-генетических систем управления, основанной и разработанной автором и
его учениками.
Для всех исследователей, озабоченных развитием и престижем отечественной
науки, интересующихся ее историей и личностями, студентов и аспирантов.
Рецензенты
доктор биологических наук Г.М. Дымшиц
член-корреспондент РАН И.Ф. Жимулев
доктор технических наук Я.И. Фет
Утверждено к печати
Ученым советом Института цитологии и генетики СО РАН
Книга издана при финансовой поддержке
Российского фонда фундаментальных исследований
по проекту 02—04—62021
ТП-02-П-№ 137 © В.А. Ратнер> 2002
© Российская академия наук, 2002
© Оформление. "Наука". Сибирская
ISBN 5-02-031969-4 издательская фирма РАН, 2002
От редактора
Эту книгу Вадим Александрович завершал, будучи тяжело больным. В Си-
Сибирской издательской фирме "Наука" РАН сделали все возможное, чтобы ус-
ускорить выход книги, но Вадим Александрович скончался 15 августа 2002 г., так
и не узнав, что верстка книги уже готова.
Книга была задумана как эстафета, которую автор принял от великих уче-
ученых XX века, с честью пронес свой отрезок дистанции и передает в XXI век.
Вадим Александрович создал настоящую научную школу математических
генетиков и не страшился, что эстафету некому принять. Более ста его учени-
учеников продолжают начатое им дело и развивают собственные научные направле-
направления. Он хотел рассказать, как развивались те или иные отрасли науки, о живых
участниках великих открытий, хотел, чтобы новые поколения ученых знали их
всех поименно.
Вадим Александрович был романтиком и свято относился к науке и людям
науки. Это святое отношение к науке и ее творцам он постарался пронести че-
через все страницы книги.
Своеобразными девизами его были два коротеньких стихотворения люби-
любимого им поэта Леонида Мартынова. Один — девиз по жизни:
И вскользь мне бросила змея:
— У каждого судьба своя! —
Но я-то знал, что так нельзя —
Жить, извиваясь и скользя.
1949
Другое стихотворение "След" — девиз его отношения к науке:
А ты?
Входя в дома любые —
И в серые,
И в голубые,
Всходя на лестницы крутые,
В квартиры, солнцем залитые,
Прислушиваясь к звону клавиш
И на вопрос даря ответ,
Скажи:
Какой ты след оставишь?
След,
Чтобы вытерли паркет
И посмотрели косо вслед,
Или
Незримый прочный след
В чужой душе на много лет?
1949
Л.А. Васильева
Предисловие
Настоящая книга представляет собой коллекцию биографических, истори-
исторических и популярных научных статей, очерков, подготовленных мною и час-
частично опубликованных в 1990-е годы. Поскольку при публикации в журналах
некоторые из них были урезаны или изменены редакторами, я позволил себе
восстановить наиболее существенные утраченные тексты. Так что в настоящем
варианте они ближе к задуманному, чем в других изданиях. Кроме того, многие
главы пополнились дополнительными фотографиями и рисунками. Чтобы не
повторять одни и те же рисунки в разных главах, введены перекрестные ссыл-
ссылки. Ссылки на литературу приведены раздельно по главам. Это более удобно,
хотя некоторые ссылки и дублируют друг друга. Наконец, я обновил некоторые
материалы и цифры, чтобы соответствовать началу нового тысячелетия.
Выражаю глубокую благодарность всем, кто на разных этапах работы по-
помогал мне в подготовке рукописей, фотографий и рисунков. Во-первых, это
инициаторы и редакторы нескольких мемориальных и исторических сборни-
сборников: Н.Н. Воронцов, Н.А. Ляпунова, В.И. Иванов, Я.И. Фет, чья инициатива
подвигла меня на подготовку многих мемориальных статей. Во-вторых, это ре-
редакторы отдельных статей: J.F. Crow ("Genetics"), O.O. Астахова ("Природа"),
Т.Ф. Клименко ("Соросовский образовательный журнал"), О.С. Ушакова
("Наука в Сибири"), А.А. Ончукова ("Информационный вестник ВОГиС") и
др., во взаимодействии с которыми оттачивался стиль публикаций. В-третьих,
это научный редактор предлагаемой книги Л.А. Васильева, которая была са-
самым жестким фильтром на авторском пути. Отдельная благодарность Н.А. Ля-
Ляпуновой, Е.А. Ляпуновой, Ю.Ф. Богданову, В.И. Иванову, СВ. Аргутинской за
предоставление фотографий и других материалов из личных архивов, В.А. Пра-
Прасолову — за фотографии из архива ИЦиГ СО РАН.
Издание книги финансировалось Сибирским отделением РАН, Институ-
Институтом цитологии и генетики СО РАН и поддержано издательским грантом
РФФИ № 02—04—62021. Кроме того, многие отдельные главы (статьи) этой
книги были поддержаны различными исследовательскими грантами РФФИ,
ГНТП Минобразования РФ "Университеты России — фундаментальные ис-
исследования" и грантом программы "Соросовские профессора".
В.А. Ратнер, академик РАЕН,
Соросовский профессор
Новосибирск, сентябрь 2001 г.
Введение
Непосредственным поводом для подготовки этой книги было предложение
Сибирской издательской фирмы "Наука" РАН опубликовать что-то важное на
животрепещущую тему. Так оно в итоге и получилось. Но истинные причины
этого лежали, конечно, гораздо глубже.
Рубеж тысячелетий — это прекрасный момент для оценки содеянного за
40 лет в новой и быстро развивающейся области науки — молекулярной киберне-
кибернетике. Следует отдавать себе отчет в том, что идея кодирования генетической
информации возникла еще в недрах довоенной науки в трудах ряда выдающих-
выдающихся и прозорливых исследователей — генетиков, физиков, химиков. Затем эта
идея была "материализована" не менее выдающимися исследователями -в рам-
рамках молекулярной биологии и молекулярной генетики. Выявлены материаль-
материальные носители генетической информации, определены закономерности кодиро-
кодирования, принципы управления работой генов, проанализированы механизмы
фундаментальных генетических процессов.
Особенность ситуации состояла в том, что на этом пути огромную роль
сыграл информационно-кибернетический подход к описанию молекулярно-гене-
тических систем и процессов в клетках и организмах. Иначе говоря, принципы
молодой тогда науки — кибернетики — оказались очень продуктивными в при-
приложении к еще более молодым наукам молекулярно-генетического цикла.
Так возникла классика молекулярной кибернетики A930-1950-е годы), свя-
связанная с именами Н.К. Кольцова, Н.В. Тимофеева-Ресовского, М. Дельбрюка,
Э. Шредингера, Л. Полинга, Ф. Крика, Дж. Уотсона, Г.А. Гамова, Дж. фон Ней-
Неймана и многих других. Интересно, что многие основополагающие проблемы
молекулярной генетики сформулированы и решены именно в рамках инфор-
информационно-кибернетического подхода (и терминологии): генетический код, ге-
генетическая информация, концепция последовательности, информационная
РНК, функциональные сайты (знаки) генетического управления, опероны, ге-
генетический контроль метаболизмом и онтогенезом и т.п. Успехи молекулярной
кибернетики стали неоспоримыми.
В 1960-70-е годы фактические результаты молекулярных наук генетическо-
генетического цикла были столь значительными, что встал вопрос о своевременном теоре-
теоретическом обеспечении работ. К этому времени уже сложились основы киберне-
кибернетики (Н. Винер, Дж. фон Нейман, К. Шеннон, Л. Бриллюэн и др.). Обобщив
опыт и результаты кибернетики, А.А. Ляпунов и СВ. Яблонский сформулиро-
сформулировали представление о центральном объекте этой науки — управляющих систе-
системах. И.А. Полетаев уточнил понятие "информации по смыслу", физические
требования к системам управления и принципы лимитирования в сложных
Введение
системах. Параллельно А.А. Ляпунов сформулировал концепцию управляющих
систем живой природы и представление о живых системах с позиций теории
устойчивости. Эти идеи и подходы использованы нами для построения теоре-
теоретического фундамента молекулярной кибернетики и моделирования многих
конкретных молекулярно-генетических систем.
В те же годы М. Эйген и сотрудники выполнили фундаментальный цикл
работ по моделированию ансамблей макромолекул, где проанализировали не-
некоторые аспекты проблемы самовоспроизведения, катастрофы мутационных
ошибок, устойчивости консенсусов и др. С. Кауфман, Р. Тома, М. Саважо и др.
начали построение моделей оперонов и оперонных систем. Р. Экк и М. Дайхоф
основали первый Банк данных белковых последовательностей (теперь это
PIR). Э. Цукеркандль и Л. Полинг, У. Фитч, М. Дайхоф, М. Гудмен, М. Киму-
ра, М. Ней и многие другие начали развивать Теорию молекулярной эволюции.
Именно тогда под влиянием идей А.А. Ляпунова начался наш цикл работ
по Теории молекулярно-генетических систем управления (МГСУ). В 1960-е годы
заложены основы концепции МГСУ, определены базовые понятия и пробле-
проблемы. Тогда же на факультете естественных наук Новосибирского университета и
в Институте цитологии и генетики СО АН СССР была начата подготовка сту-
студентов и аспирантов — математических биологов, которые очень быстро вли-
влились в ряды участников работ и получили первоклассные результаты. Их имена
читатель найдет в соответствующих разделах книги.
Ими разработаны математические модели оперонов и оперонных систем,
модели самовоспроизводящихся ансамблей макромолекул (сайзеров), модели
полирепликонных систем, компьютерная модель развития фага 1, модели по-
помехоустойчивости и эволюции генетического кода, основные представления о
генетическом языке, построены многие новые комплементационные карты ге-
генов и филогенетические деревья генов и белков, доказана теорема об асимпто-
асимптотическом невырождении популяций с информационно-генетическими свойст-
свойствами и многое другое.
В конце 1970-х годов молекулярная генетика вступила в эпоху секвенирова-
ния. Процедура дешифровки (секвенирования) нуклеотидных последовательно-
последовательностей превратилась в рутинную технологическую операцию. В результате стало
стремительно нарастать число секвенированных генов, фрагментов, плазмид, ге-
геномов вирусов, органелл и, наконец, клеточных геномов. Тогда же, в начале
1980-х годов возникли первые международные банки данных: GenBank в США,
EMBL Library в Европе и др. Остро встал вопрос о разработке компьютерных
г,сетодов анализа последовательностей, поиска генов, функциональных сайтов,
повторов. Иначе говоря, наши разработки, специалисты, результаты, концеп-
концепции оказались востребованными. Матбиологи новосибирской школы оказа-
оказались хорошо подготовленными к такому повороту событий.
В 1980-90-е годы Теория молекулярно-генетических систем управления на-
наполнилась новым содержанием. Прежде всего в самой теории нами были ос-
осмыслены и разработаны многие фундаментальные проблемы: роль сайзеров
как основы молекулярно-генетической организации; условия структурной ус-
устойчивости ансамблей макромолекул; свойства генетического языка на основе
огромного количества генетических текстов; проблема возникновения и эво-
Введение
люции генетического кода; блочно-модульный принцип молекулярно-генети-
ческой организации и эволюции МГСУ; принцип лимитирующих факторов в
экспрессии, организации и эволюции МГСУ; проблема роли мобильных эле-
элементов в экспрессии генов, изменчивости и эволюции геномов эукариот; про-
проблемы теории молекулярной эволюции, включая сценарий процесса молеку-
молекулярной эволюции МГСУ, и т.д.
В 1990-е годы во многих странах стала быстро развиваться биоинформати-
биоинформатика — наука о технических средствах исследования и моделирования макромо-
макромолекул и их систем. Появилось множество компьютерных специализированных
банков данных разной направленности, разработаны многие десятки пакетов
прикладных программ анализа полинуклеотидных и полипептидных текстов,
поиска функциональных сайтов, анализа геномов, построения вторичных и
пространственных структур макромолекул по первичной последовательности,
построения филогенетических деревьев и многие другие. Были начаты и час-
частично завершены крупные международные проекты полного секвенирования
клеточных геномов, включая геномы Е. coli, дрожжей, дрозофилы и человека.
Матбиологи новосибирской школы приняли участие во многих из этих направ-
направлений.
В конце 1990-х годов в биоинформатике обозначились новые направления:
начаты исследования генетических сетей контроля метаболизма, онтогенетиче-
онтогенетических процессов, иммунного ответа, клеточного цикла, гемопоэза и других
функциональных систем клетки и организма. Новые экспериментальные тех-
технологии, например технология микрочипов, способны одновременно анализи-
анализировать десятки тысяч фракций РНК клетки. Появилась возможность модели-
моделировать большие генные сети, вплоть до клетки в целом. В Теории молекулярной
эволюции стали активно развиваться геномика — теория эволюции полных кле-
клеточных геномов и протеомика — наука об эволюции белковых комплексов. Во
многих из этих направлений наши матбиологи вносят существенный, а иногда
и решающий вклад.
Таким образом, научный потенциал молекулярной кибернетики, Теории
молекулярно-генетических систем управления оказался очень высок. В 1990-е
годы автору пришлось затратить немало усилий на пропаганду этих и других
результатов. В НГУ были прочитаны курсы лекций по Молекулярной генетике,'
Теории МГСУ, Теории молекулярной эволюции и др. Популярные лекции чи-
читались Соросовским учителям, аспирантам, научным сотрудникам. Популяр-
Популярные и полупопулярные статьи были опубликованы в Соросовском образова-
образовательном журнале, в журнале "Природа", Информационном вестнике ВОГиС,
газете "Наука в Сибири", а также в сборниках и трудах конференций, выстав-
выставлены на институтском сайте ИНТЕРНЕТ.
Это тем более важно, поскольку молекулярную кибернетику и биоинфор-
биоинформатику в последние годы захлестнула волна "мигрантов" из смежных областей
знания: программистов, математиков, физиков, инженеров и т.д. Они приходят
из своих областей со своими математическими или компьютерными методами,
но имеют смутное, а подчас очень упрощенное представление о ситуации в
молекулярной кибернетике. Базового генетического или математико-биологи-
ческого образования они не имеют. В результате возникают большие трудности
Введение
в понимании проблем молекулярной кибернетики и направленности теорети-
теоретических работ. Поэтому в предлагаемой книге представлено около 15 популяр-
популярных очерков о главных проблемах молекулярной кибернетики.
Но и это еще не все. В 1990-е годы стал остро ощущаться дефицит преемст-
преемственности поколений исследователей в области молекулярной кибернетики.
Поколение основателей давно ушло. Новым поколениям, особенно не имею-
имеющим базового математико-биологического образования, кажется, что до них
никого не было. Я чувствую сейчас острую ответственность за то, чтобы доне-
донести до матбиологов новой волны да и всего молодого поколения исследовате-
исследователей имена своих предшественников-теоретиков и понимание того, что ими
сделано. К этому обязывают и многочисленные юбилеи ведущих ученых.
В результате книга пополнилась персоналиями многих крупнейших гене-
генетиков и математических биологов, особенно отечественных, а также историче-
историческими очерками о развитии идеи генетического кодирования, генетики как
науки, теории гена, а также о живой истории новосибирской школы математи-
математических генетиков.
Желаю читателю ощутить не только мощь и величие современной науки,
получить ответы на некоторые принципиальные вопросы генетической орга-
организации и эволюции, но и соприкоснуться с эмоциональным и творческим ми-
миром тех, кто делал эту науку своими руками.
Not for sale! For preview only! Scanned by Mykhaylo (molbioKfl4ikr.net)
I. PERSONALIA
1. Мамонт*
{заметки о Н.В. Тимофееве-Ресовском)
Прежде всего должен сказать, что ассоциация Николая Владимировича с
зубром, укрепившаяся после публикации прекрасной книги Д. Гранина, кажет-
кажется мне случайной и искусственной. По-моему, он больше похож на мамонта.
Зубр мне всегда представлялся существом темным и злобным, с непредсказуе-
непредсказуемыми приступами ярости. Мамонта же никто из нас не видел, но в воображе-
воображении нашем он кажется чем-то огромным, гигантом среди прочей живности,
вымирающим гигантом, которого очень легко уязвить, гигантом, который бро-
бродит среди окоченевшей пустыни, разыскивая остатки иссохшей травы. Гиган-
Гигантом, который пережил свой золотой век и попал в эпоху оледенения.
Впервые я услышал о Николае Владимировиче весной 1961 г. Надо объяс-
объяснить, что по образованию я физик и о биологии тогда имел довольно смутное
представление. А о генетике не слышал вообще, что следует отнести на счет на-
нашего самого передового школьного образования конца 1940-х годов. Незадолго
до этого, после периода случайных блужданий я поступил на работу в Институт
цитологии и генетики в Новосибирске, в лабораторию Д.К. Беляева, был полон
всяческих надежд и начинаний, а главное — написал первую работу, которой
очень гордился.
Мы помещались тогда в знаменитом темно-сером здании на ул. Советская,
20 (сейчас — 18), в Новосибирске, где временно квартировала значительная
часть Сибирской Академии. В соседней с нами комнате была лаборатория
Ю.Я. Керкиса. От своих товарищей я уже знал, что Юлий Яковлевич едет на
Урал к своему другу — великому генетику, который находится там фактически
в ссылке, но ежегодно собирает, как теперь говорят, "несанкционирован-
"несанкционированные" летние школы по биофизике и генетике, где происходит нечто весьма за-
загадочное и интересное. Помню, что при случае я спросил Керкиса, как можно
туда поехать. В ответ он пояснил, что школы происходят на биостанции "Ми-
ассово", что добираться туда можно самоходом, лучше со своей палаткой, что
ьублика там очень интересная, а доклады бывают практически на любые темы,
лишь бы было интересно. Что никакие приглашения на школу не рассылаются,
а попасть туда можно только через знакомых, которые уже участвовали в пре-
предыдущих школах. В этом случае на железнодорожную станцию высылается по-
полуторка. Если же вы очень "настырны", то до биостанции можно добраться
* Впервые опубликовано: Ратнер В.А. "Мамонт" (заметки о Н.В. Тимофееве-Ресовском) //
Николай Владимирович Тимофеев-Ресовский: Очерки. Воспоминания. Материалы / Отв. ред.
Н.Н. Воронцов. М.: Наука, 1993. С. 291-299.
1. Мамонт
11
пешком — около 40 км через лес, а если не очень — то без вас обойдутся. Нас,
однако, Юлий Яковлевич обещал рекомендовать для участия в школе.
В июле мы поехали в Миассово с Алешей Груздевым и Артуром Шеруди-
ло. Добрались на полуторке с другими участниками. На биостанции машину
встречал седой плотный мужчина, очень живой и подвижный, с легкой по-
походкой, несмотря на некоторую грузность фигуры. Это и был Николай Вла-
Владимирович.
Мы жили в палатках, заседали в бывшем барском доме — лаборатории био-
биостанции, слушали импровизированные лекции Николая Владимировича по
генетике и выступали со своими докладами. Обстановка очень демократич-
демократичная — полное самообслуживание и отсутствие штатных фигур. При этом среди
участников были выдающиеся ученые — Л.А. Блюменфельд, А.А. Ляпунов,
М.В. Волькенштейн, И.А. Полетаев, Р.Л. Берг, В.Я. Александров, В.П. Эфро-
имсон, Ю.Я. Керкис ("ученейший Керкис", как называл его Николай Влади-
Владимирович). В жару некоторые доклады переносились на берег озера, причем слу-
слушатели сидели в воде, а докладчик расхаживал в трусах на берегу и вещал. Со-
Сохранились фотографии В.П. Эфроимсона, читающего лекцию в купальном
виде и с платочком на голове.
Школа по генетике и биофизике. Миассово, 1961 г. 1-й ряд справа налево: Н.В. Лучник,
Н.В. и Е.А. Тимофеевы-Ресовские; 2-й ряд: А.Д. Груздев, В.А. Ратнер, Ю.М. Рукавиш-
Рукавишников и др. (архив автора).
12 I. Personalia
Николай Владимирович был пружиной всего действа. Его лекции поража-
поражали своей неакадемичностью. О генетике он говорил не отвлеченно, не книжно,
а в лицах. Он говорил о том, как его хорошие друзья и знакомые додумались до
прекрасных открытий. Например, о Стертеванте он всегда говорил "умница
Стертевант", имея в виду, как он догадался об эффекте положения генов. Ни-
Николай Владимирович называл всех по именам, часто давал им человеческие ха-
характеристики. И становилось ясно, что он — посланец из другого мира, из
мира, где происходили важнейшие научные события, где человеческий гений,
играя и взбрыкивая, творил великие дела, которые и не снились нашим угрю-
угрюмым "преобразователям природы". Зрелище было незабываемое.
Набравшись наглости, я подошел к Николаю Владимировичу и спросил,
нельзя ли мне рассказать о своей первой работе, так сказать, в узком кругу спе-
специалистов. Он согласился. Помню, мою теоретическую работу о корреляциях
признаков и генов, помимо Николая Владимировича, слушали также Н.В. Луч-
Лучник, Ю. Завильгельский, кажется, В.И. Иванов и др. Обсуждение было корот-
коротким и очень резким. От работы не осталось камня на камне. Оказалось, что я
просто ничего не понимаю в генетике и фантазирую там, где нужно знать пред-
предмет. Николай Владимирович говорил слова, которые я потом слышал от него
много раз по другим поводам: "нельзя объяснять непонятное неизвестным",
"нельзя относиться к своей работе со звериной серьезностью" и т.п. В общем, я
был разбит в пух и прах и крайне удручен. Впрочем, испытав полное отчаяние
от провала, я выспался и утром осознал, что случилось великое благо. Случи-
Случилась конструктивная критика, которую надо заслужить, понять и принять.
Жизнь продолжалась.
Вспоминая этот момент своей жизни, я думаю, что мне очень повезло. На
первом же шаге нового поприща я встретил человека, который несколькими
движениями определил уровень требований, задал методологию науки, пока-
показал, что такое хорошо и что такое плохо. Этот импульс я ощущаю до сих пор.
Следует сказать, что на фоне середины 1960-х годов Миассовские школы и
сменившие их в 1965 г. школы на Можайском море под Москвой выглядели как
светлое пятно после убогого сумрака лысенковщины. Туда, как на огонек, сле-
слетались очень яркие личности, ехала молодежь. Собственно говоря, там форми-
формировались сильнейшие генетическая и биофизическая школы, которыми руко-
руководили Н.В. Тимофеев-Ресовский и Л.А. Блюменфельд. С годами я стал не-
неожиданно замечать, что подавляющая часть моих научных контактов,
знакомств, интересов сложилась именно там. Там я встретил своих будущих
друзей — Ю.М. Свирежева, С.Г. Инге-Вечтомова, Р.А. Полуэктова, Л.Р. Гинз-
Гинзбурга, А.А. Гимельфарба, Л.А. Животовского, А.С. Антонова и многих других.
Но вернемся к Николаю Владимировичу. Как и все участники миассовских
"трепов", я очень любил слушать "тимофеевские байки". С высочайшим арти-
артистизмом он "трепался" о своей жизни, о людях, о случаях в науке и т.д. Напом-
Напомню, что для физиков 1950-е и 1960-е годы были романтическим периодом. Нас
завораживали легенды о Н. Боре, А. Эйнштейне, создателях атомной бомбы
и т.д. И вдруг оказалось, что Николай Владимирович не только был знаком с
Бором и его окружением. Это были его друзья. Он проводил в Копенгагене до-
довольно много времени, и его увлечение радиационной генетикой разделяли вы-
1. Мамонт 13
дающиеся квантовые физики копенгагенской школы. Напомню также, что
один из молодых физиков этой школы — Макс Дельбрюк, пройдя через руки
Николая Владимировича, стал одним из самых выдающихся основателей моле-
молекулярной генетики.
Николай Владимирович рассказывал о датском короле "Христианушке",
который патронировал копенгагенских физиков, приглашал их к себе и дер-
держался весьма демократично. Дома Николай Владимирович показывал набор
вересковых трубок для курения — подарок короля. Рассказывал научные анек-
анекдоты той поры. Среди них я запомнил анекдот о публикации в самом престиж-
престижном журнале "Nature" статей-хохм, розыгрышей о "лево-" и "право-жующих"
коровах и других животрепещущих проблемах. Многие не знают, что именно
тимофеевские "байки" были толчком для публикации в конце 1960-х годов
прекрасного сборника "Физики шутят", который доставил всем нам большое
удовольствие.
Ну а если говорить серьезно, то жизнь Николая Владимировича между 1955
и 1980 гг. была достаточно сложной. После освобождения из заключения в
1955 г. он был восторженно встречен выдающимися советскими физиками
ПЛ. Капицей, И.Е. Таммом, Л.Д. Ландау и др., но получил отказ официальных
инстанций на право жить и работать в Москве, Ленинграде, Киеве. Поэтому до
1963 г. он работал в Свердловске, фактически в полуизгнании.
Будучи почетным членом многих зарубежных академий, на Родине он не
имел не только докторской или кандидатской степени, но даже университет-
университетского диплома (их тогда не выдавали) и аттестата зрелости (который оказался
утерянным в годы гражданской войны). Докторскую диссертацию Николай
Владимирович защитил только в начале 1960-х годов, да и то не по крамольной
генетике, что тогда было невозможно, а по радиационной биогеоценологии,
которой занимался на Урале. Мало того, защищенная работа еще два года отле-
отлеживалась в ВАКе, видимо, корифеи бюрократии просто не знали, что с ней де-
делать. И только после снятия Н.С. Хрущева, который, как известно, поддержи-
поддерживал Т.Д. Лысенко, ВАК утвердил работу буквально в течение нескольких дней.
Можно ли ожидать принципиальности от таких марионеток? Что касается из-
избрания в Академию наук, то несколько попыток весьма авторитетных ученых
вьщвинуть кандидатуру Николая Владимировича закончились безуспешно. Не-
Невидимая бюрократия отражала эти попытки под стандартным предлогом:
"А что вы делали в Германии до 1946 г.?"
Примерно в эти годы A962—1963) встал вопрос о переезде Николая Влади-
Владимировича в один из новых развивающихся научных центров. Столицы по-преж-
по-прежнему были для него закрыты, поэтому обсуждались два варианта: Новосибирск
и Обнинск. В новосибирском Академгородке у него было много друзей:
| А.А. Ляпунов, Р.Л. Берг, И.А. Полетаев, Ю.Я. Керкис, Н.Н. Воронцов и др.
Недавно созданный Институт цитологии и генетики испытывал острую по-
потребность в крупных специалистах-генетиках. Председатель Президиума Си-
Сибирского отделения М.А. Лаврентьев, ранее пригласивший в Сибирь большое
число выдающихся ученых, часто со сложной биографией, склонялся в сторону
приглашения и Николая Владимировича. Его поддерживали С.Л. Соболев и др.
14 I. Personalia
Однако, судя по всему, ситуация для его переезда к этому моменту еще не
созрела. Институт цитологии и генетики незадолго до этого был обескровлен
снятием первого директора — Н.П. Дубинина, а бесчисленные комиссии пыта-
пытались его закрыть или ограничить его деятельность. Д. К. Беляев, ставший ди-
директором после Н.П. Дубинина, еще не успел достаточно укрепиться в своем
положении и в Академии. Возможно, Николай Владимирович был слишком
крупной мишенью, в которую непременно должен был полететь град стрел и
камней. Как бы то ни было, этот вариант переезда в Академгородок оказался
под вопросом.
Тогда, по инициативе А.А. Ляпунова, был предложен другой вариант —
организовать для Николая Владимировича отдел биофизики в Институте мате-
математики. Идея обсуждалась с С.Л. Соболевым, М.А. Лаврентьевым и М.В. Кел-
Келдышем. В целом поддерживая эту идею, но понимая всю сложность такого
симбиоза, начальство Академии решило немного подождать. Тогда Николай
Владимирович согласился переехать в Обнинск. Так наш Академгородок поте-
потерял великий шанс стать генетической Меккой!
Впрочем, Николай Владимирович неоднократно бывал в Новосибирске
в 1960-е и 1970-е годы. Обычно Д.К. Беляев приглашал его на отчетные сес-
сессии института, чтобы внести критический*дух и живость в обсуждаемые во-
вопросы. И это полностью себя оправдывало. Николай Владимирович себе не
изменял и с великим умением и артистизмом судил о новых направлениях
работы Института и первых результатах. Постоянно иронизировал по пово-
поводу, как он выражался, "ДНКаканья" и повторял, что всю эту молекулярную
генетику они (генетики) предвосхитили еще до войны. В чем-то он был не-
несомненно прав, поскольку многие из молекулярщиков были "мигрантами"
из других наук — физиками, химиками, медиками и т.д., не имевшими глу-
глубокого генетического образования и часто переоткрывавшими для себя за-
заново то, что классические генетики знали и высказывали задолго до них. Од-
Однако его выступления были, скорее, попыткой немного пощипать
"выскочек", чем помешать им работать.
Иногда, выступая с трибуны, Николай Владимирович позволял себе не-
немного "похулиганить". Однажды в прениях он произнес с невинным видом
примерно следующий монолог: "В моей родной Калужской губернии имеется
речушка Высса. Она сливается с другой речкой под названием Усса. А затем обе
они впадают в третью речку побольше под названием Моча..." Аудитория оне-
онемела, а он без тени улыбки продолжал свои калужские воспоминания, а потом
плавно перешел к научным проблемам. Говорят, что Николай Владимирович
устроил небольшой "цирк" при вручении ему Кимберовской медали за дости-
достижения в генетике.
Поразительно влияние личности Николая Владимировича на окружаю-
окружающих. Где бы он ни работал — в Кольцовском институте, в Германии, в заключе-
заключении, в Свердловске, Обнинске — вокруг него быстро формировался круг уче-
учеников и создавалась мощная школа. Это при том, что после заключения и пере-
перенесенной там тяжелой болезни у него резко упало зрение, читал он только при
помощи огромной лупы. Это означало, что нормально работать в лаборатории
он фактически не мог. Тем не менее само его присутствие как бы задавало ок-
1. Мамонт
15
ружающим высокие критерии работы и отношения к делу. Мне рассказывали о
длительных попытках О. Г. Газенко добиться перехода Николая Владимировича
в Космическое ведомство еще в 1950-е годы. Эти попытки, понятно, были без-
безуспешными. Только уже в 1970-е годы, после ухода Николая Владимировича на
пенсию, О.Г. Газенко смог пригласить его профессором-консультантом в Ин-
Институт медико-биологических проблем. Фактически этот мудрый шаг был сде-
сделан, вероятно, для того, чтобы создать у себя обстановку повышенной требова-
требовательности и интеллектуальности, а с другой стороны, — чтобы поддержать вы-
выдающегося генетика в трудный момент.
После реабилитации генетики в середине 1960-х годов генетическая жизнь
в стране возобновилась, возникли советы, общества, журналы, кафедры гене-
генетики. В научном совете по генетике и селекции была образована секция попу-
ляционной генетики, которую возглавлял Д.К. Беляев. Помню, он собрал
первое заседание секции в своем директорском кабинете. Стали делить роли,
обсуждать мероприятия. Все радовались возможности открыто заниматься ге-
генетикой и были очень активны. Среди приглашенных был и Николай Влади-
Владимирович. Внезапно он спросил: "А деньги у вас есть?" — "Денег нет, — ответил
Д.К. Беляев, — будем координировать развитие науки". Николай Владимиро-
Владимирович долго смеялся, потом сказал: "Что вы там можете координировать без де-
2-й Всесоюзный генетический съезд. Москва, МГУ, 1972 г. Второй слева: Н.В. Тимофе-
Тимофеев-Ресовский, затем Л.А. Васильева, В.А. Ратнер, С.Н. Родин (архив автора).
16
I. Personalia
2-й Всесоюзный генетический съезд. Москва, МГУ. Заседание секции популяционной
генетики. Председатели: Н.В. Тимофеев-Ресовский и Я.Я. Лусис. Докладывает В.А. Ратнер
(архив автора).
нег? Надо субсидировать, а не координировать. Это очередная говорильня!"
И рассказал о системе субсидий на науку, которые предоставляют известные
общества и фонды за границей. Впрочем, секция популяционной генетики и
Научный совет в целом организовали в те годы несколько очень неплохих кон-
конференций (Петергоф, 1968; Новосибирск, 1969; Елгава, 1970 и др.), где Николай
Владимирович присутствовал и с удовольствием выступал.
В библиотеке Института цитологии и генетики СО РАН хранится научный
архив А.С. Серебровского, переданный в дар его семьей. В архиве содержится
достаточно полная коллекция работ Николая Владимировича. Я несколько раз
смотрел эти работы, пытаясь оценить в целом его вклад в генетическую науку.
Впечатление создавалось поразительное. Он работал практически во всех круп-
крупных направлениях генетики: генетический анализ, феногенетика, радиацион-
радиационная генетика, популяционная генетика, теория гена, а также был одним из соз-
создателей синтетической теории эволюции, биофизики, теории мишени, радиа-
радиационной биогеоценологии. Работы имели чрезвычайно четкий и законченный
характер. Они написаны в такой классической форме, что их можно без правки
включать в учебники. В каждом направлении им заложен фундаментальный
камень, который остается там до сих пор. Видимо, в этом одна из причин его
мировой известности. Его стиль резко контрастирует с часто встречающейся
1. Мамонт 17
теперь даже у хороших ученых торопливостью, стремлением "застолбить" хо-
хорошую идею, "сорвать цветок" нового направления.
Уже в 1980-е годы, перерабатывая курс молекулярной генетики, я понял,
что через руки Николая Владимировича прошла одна из центральных идей мо-
молекулярной генетики — представление о гене как о кодирующей макромолеку-
макромолекуле (см. гл. 15-17). Впервые эту идею высказал еще в 1920-е годы Н.К. Кольцов.
Затем его ближайший ученик Николай Владимирович вместе с М. Дельбрюком
и К. Циммером оценили размеры гена как мишени для действия радиации. Из-
Известный физик Э. Шредингер развил эту идею, сформулировав представление
о генетическом кодировании как о центральной проблеме естествознания. На-
Наконец, М. Дельбрюк, уехав в США, основал там генетику фагов в поисках ре-
реального генетического объекта, наиболее близкого по размеру к гену.
Идея кодирования генетической информации полностью оправдала себя в
1950-е и последующие годы, предопределив стратегический успех молекуляр-
молекулярной генетики.
Неоднократно я слышал из уст Николая Владимировича серьезную озабо-
озабоченность состоянием охраны природы. Он не уставал говорить о пользе зооло-
зоологии и ботаники, поскольку, не зная природу, невозможно вести ее инвентари-
инвентаризацию. О себе он иногда говорил: "вообще-то я не генетик, а специалист по
десятиногим ракам". Действительно, будучи студентом МГУ, он специализи-
специализировался по зоологии беспозвоночных. Это было его первой любовью, а генети-
генетиком он стал, вложив в эту науку всю свою долгую жизнь.
У русских людей, продолжительное время живших за границей, в речи час-
часто проскальзывают иностранный акцент, необычный строй фразы, иногда они
ищут давно забытое слово. В 1968 г. на Генетическом конгрессе в Японии я раз-
разговаривал с Ф.Г. Добржанским. Прожив 40 лет в США, он говорил по-русски с
едва уловимыми американизмами, причем было впечатление, что он переводит
с английского на русский. Николай Владимирович, пробыв более 20 лет в Гер-
Германии, легко говорил и писал по-немецки и по-английски, любил иногда вста-
вставить короткую иностранную фразу или слово. Например, часто говорил:
"Nobody knows" — "Кто его знает, неизвестно". Однако его речь осталась абсо-
абсолютно русской по строю, образу, остроте. Было такое впечатление, что его ор-
организм отчаянно сопротивлялся иностранному влиянию, как инфекции, и
справился с нею, выработав иммунитет.
В генетике у Николая Владимировича, несомненно, была очень глубокая и
сильная идеология. Некоторые ее моменты, конечно, устаревали, но он, по-мо-
по-моему, их не менял. Сотрудничая и общаясь с самыми выдающимися физика-
физиками-теоретиками и математиками, имея учеников-математиков, он тем не менее
математикой не владел и, по-моему, за всю жизнь не написал ни одной матема-
математической формулы. Он не скрывал этого и даже с некоторой бравадой говорил,
что понимает математику только в присутствии самих математиков.
В 1950-е и 1960-е годы среди близких ему математиков одно из первых мест
занимал А.А. Ляпунов. По-моему, Николай Владимирович нежно любил его,
несмотря на значительную разницу в возрасте. Шутя и подтрунивая, он с удо-
удовольствием произнес однажды примерно такую фразу: "Ляпушка — милейший
человек, и с великим энтузиазмом он может увлечь вас по совершенно непра-
18
I. Personalia
вильному пути. Потом, обнаружив свой промах, он с неменьшим энтузиазмом
вновь увлечет вас по другому и опять совершенно неправильному пути. И т.д."
А.А. Ляпунов хмурился, но не мог сдержать улыбку. В 1973 г. А.А. Ляпунов по-
поехал в Москву на какое-то совещание и там скоропостижно умер. Панихида со-
состоялась в Институте математики АН, где он некоторое время работал до вой-
войны. Проститься пришел Николай Владимирович. Он стоял старый, седой, рас-
растрепанный, долго смотрел на неподвижное лицо своего друга, потом подошел,
перекрестил его по-русски, поцеловал в лоб. А.А. Ляпунова похоронили на
Введенском кладбище в Москве. Мы с Ю.М. Свирежевым были на похоронах и
несли крышку гроба.
В последний раз я видел Николая Владимировича в августе 1978 г. на Меж-
Международном генетическом конгрессе в Москве. В первых фразах книги Д. Гра-
Гранина "Зубр" описывается банкет, на котором был и Николай Владимирович.
Я тоже был на этом приеме в банкетном зале Дворца съездов в Кремле. Огром-
Огромное пространство заставлено столами с яствами и выпивками. Мы мирно вы-
выпивали и закусывали, общаясь между собой и с мигрирующими "а-ля фуршет"
иностранцами под наблюдением неусыпных стражей спокойствия. Неожидан-
Неожиданно к нашему столу подошла очень пожилая и интеллигентная пара иностранцев
и на чистом старо-петербургском диалекте спросила нас, знаем ли мы Николая
Летняя школа на Можайском море под Москвой. Н.В. Тимофеев-Ресовский и Т. Булга-
Булгакова (Эман) (архив автора). Все же он очень похож на мамонта!
1. Мамонт
19
Н В Тимофеев-Ресовский на ступенях
Воспоминания". М.: Согласие, 2000).
20 I. Personalia
Владимировича Тимофеева-Ресовского, жив ли он и здесь ли он. Я ответил, что
Николай Владимирович здравствует и присутствует здесь, а затем отвел их к
возвышению, где сидел Тимофеев-Ресовский. Он опирался на палку, повер-
повернувшись к залу, но людей фактически не видел, так как зрение у него к этому
времени уже очень сильно сдало. Меня он узнал по голосу и сказал: "А, Вади-
мушка! Ну как дела?" Я представил ему пару бывших соотечественников и ото-
отошел, чтобы не мешать их разговору.
В том же году я получил приглашение одного американского издательства
на перевод своей книги (в дальнейшем перевод не состоялся). В связи с этим
издательство запросило у меня Curriculum vitae — краткую научную биографию.
Среди вопросов к автору значился и такой: кто Ваши учителя в науке, кто ока-
оказал наибольшее влияние на формирование Ваших научных взглядов? Перебрав со-
события своей жизни, я неожиданно понял, что одним из главных научных им-
импульсов моей жизни была первая встреча с Николаем Владимировичем в Миас-
сово, образ его мыслей, идеология, научные критерии, наконец, его личность
сильно повлияли на меня. Я понял, что это точка отсчета, с которой сознатель-
сознательно или бессознательно сверял все свои дальнейшие научные интересы и реше-
решения. Произошло своеобразное соприкосновение с талантом, касание музы, ко- I
торое в науке нельзя заменить ничем. Так я и написал.
Я недаром сказал вначале, что Николай Владимирович напоминает мне ма-
мамонта. Есть еще одна грань этого образа. Он, вероятно, был одним из последних
энциклопедистов, человеком самых разнообразных интересов. Он вдохновлялся
музыкой и стихами. Прекрасно знал историю, русскую литературу, любил стихи
М. Цветаевой. Однажды сказал о ней так: "Мариночка хоть и баба, но поэт!"
Нынешние таланты часто однобоки, а энциклопедистов среди них почти не сы-
сыщешь.
В своем "ближнем круге" Николай Владимирович, возможно, был доста-
достаточно трудным человеком. Однако большое видится на расстоянии. Я никогда
не работал у него, не был его учеником или сотрудником в прямом смысле.
Возможно, это позволяет судить о нем по большому счету, без мелочей. Несо-
Несомненно, он был великим русским генетиком, одним из самых выдающихся ге-
генетиков XX века, ярчайшей личностью на небосклоне нашей науки. Таких лю-
людей сейчас мало. Я считаю, что Академия наук должна испытывать чувство глу-
глубочайшего стыда от того, что среди сотен "жестких" и "мягких" вакансий для
своих членов она по конъюнктурным причинам не нашла персональной и по-
почетной вакансии для Николая Владимировича Тимофеева-Ресовского. Он мог
Академию только украсить.
Я счастлив, что был знаком с Н.В. Тимофеевым-Ресовским. Он оставил
глубочайший след в памяти всех, кто его знал.
2. Ровесник века, гигант столетия 21
2. Николай Владимирович Тимофеев-Ресовский —
ровесник века, гигант столетия*
@7.09.1900 - 28.03.1981)
Не надо относиться к науке
со звериной серьезностью.
Н.В. Тимофеев-Ресовский
Сейчас Н.В. было бы 100 лет. Но это не все. Еще он был ровесником века и
ровесником генетики. Это особенно заметно теперь, в эпоху миллениума, когда
пересекается тысячелетняя грань времен, когда наступило время собирать кам-
камни.
Удивительно, но в личности и судьбе Николая Владимировича Тимофее-
Тимофеева-Ресовского сошлись и отразились самые драматические и ключевые собы-
события столетия. Корнями он вырос из века 19-го, из русской истории и классики.
Его родословная — это живая история России: здесь и казаки легендарного
"разбойника" Степана Разина, и потомки основателя русских княжеских дина-
династий варяга Рюрика, и адмиралы российского флота, и великий анархист князь
Петр Кропоткин, и многочисленные русские офицеры и интеллигенты XIX и
XX веков.
Первая мировая и гражданская войны бросили его в пучину потрясений,
достойных высокого литературного пера. В 1917 г. он попал на Юго-Западный
фронт в казачью часть, воевал против немцев в пешем строю, стал вахмистром
(унтер-офицером), потом пробивался домой, попал в банду анархистов на Ук-
Украине, чудом выбрался оттуда, добрался до Москвы, служил в 12-й Красной Ар-
Армии, отступал с ней с юга почти до Москвы, а потом наступал на Белую Армию
генерала Деникина обратно — до самого юга России, воевал против "Дикой"
кавказской дивизии, болел тифом. Но эти перипетии судьбы не погасили в нем
жажду знания и науки. В перерывах между сражениями он учился в Москов-
Московском университете, ходил в разнообразные кружки, пел 1-м басом в хоре Мос-
Московского военного округа, работал грузчиком, закончил МГУ в 1922 г. Позже
Н.В. говорил об этом периоде [Тимофеев-Ресовский, 2000]: "В общем, по-мо-
по-моему, жизнь была веселой: мало-мало голодали, мало-мало холодали — все такое. Но
люди были молодые, здоровые, крепкие ".
В 1922 г. Н.В. начал работать научным сотрудником Института экспери-
экспериментальной биологии у проф. Н.К. Кольцова. Николай Константинович Коль-
Кольцов был выдающейся личностью в русской биологической науке. Еще в 1911 г.
он организовал в частном Университете им. Шанявского первую в мире лабо-
лабораторию и кафедру экспериментальной биологии, которые в дальнейшем стали
основой Института экспериментальной биологии A916 г.). Из этого института
выросла весьма значительная российская школа теоретической и эксперимен-
экспериментальной генетики: старшие ученики и сподвижники Кольцова — С. С. Четвери-
Четвериков, А.С. Серебровский, М.М. Завадовский, позже — Н.В. и Е.А. Тимофее-
* Английский вариант статьи: Ratner V.A. Nikolay Vladimirovich TimofeefF-Ressovsky A900-
1981): Twin of the Centuiy of Genetics // Genetics. 2001. Vol. 158. P. 933-939.
22 I. Personalia
вы-Ресовские, Д.Д. Ромашов, Б.Л. Астауров, Н.К. Беляев, СМ. Гершензон,
Н.П. Дубинин, П.Ф. Рокицкий и др. — цвет новой российской генетики. Н.В.
становится одним из самых близких и талантливых учеников Кольцова.
В научной сфере Н.К. Кольцов интересовался всем. Однако в области ге-
генетики лично он инициировал три основных направления: феногенетику, т.е.
генетические основы онтогенеза, исследование закономерностей мутагенеза и
теоретическую генетику, которая в дальнейшем стала основой молекулярной ге-
генетики. По словам Н.В. [Тимофеев-Ресовский, 2000]: "Ведь в сущности чрез-
чрезмерная мода и успех так называемой молекулярной генетики..., — ведь это же
было зачато в кольцовском институте и непосредственно в работах, напечатан-
напечатанных Кольцовым ".
Другим учителем Н.В. в этот период стал С.С. Четвериков — основатель
теоретической и экспериментальной популяционной генетики. Он руководил
знаменитым семинаром "Дрозсоор" (русская аббревиатура от "Совместное
орание о дрозофиле"), в котором сложилась московская школа популяционной
генетики. По словам Н.В. [Тимофеев-Ресовский, 2000]: "...колъцовский инсти-
институт создал школу современного направления в эволюционном учении, то, что Джу-
Джулиан Хаксли потом уже, в 40-е годы, назвал синтетической эволюцией, понимая
под этим синтез генетики и классического эволюционного учения ".
Таким образом, Н.В. чрезвычайно повезло. Судьба вовремя свела его с эти-
этими выдающимися русскими генетиками первой волны. От них он получил
мощный импульс, который на десятилетия определил круг его научных интере-
интересов в области генетики: феногенетика, закономерности мутагенеза и молеку-
молекулярная природа генов, популяционная генетика и теория микроэволюции. При
этом импульс был полностью оригинальным, поскольку в течение 7-8 лет, во
времена войн и революций, российская наука была полностью оторвана от ев-
европейской и мировой.
В 1922 г. изоляция была прорвана: в Москву и Петроград приезжал один из
ближайших учеников и сотрудников Т.Х. Моргана — Герман Меллер. Он по-
посвятил своих российских коллег в новейшие результаты экспериментальной ге-
генетики моргановской школы, привез оттиски многочисленных работ, а глав-
главное, коллекцию линий плодовой мухи дрозофилы. Прямой контакт со школой
Моргана совершенно изменил ситуацию. После этого Н.В. стал работать на
D. melanogaster, D. funebris, начал основные циклы своих работ. В частности, он
был первым из учеников Н.К. Кольцова, который начал заниматься мутагене-
мутагенезом и феногенетикой дрозофилы, и одним из первых учеников С.С. Четверико-
Четверикова, который обнаружил резерв наследственной изменчивости в природных по-
популяциях дрозофил. После открытия в 1927 г. Г. Меллером мутагенного дейст-
действия Х-лучей, практически одновременно с ним, Н.В. начинает многолетние
исследования по радиационной генетике и радиобиологии.
В 1923-1925 гг. в Москву приезжал крупнейший немецкий невропатолог и
нейроанатом проф. О. Фогт, директор Института исследований мозга Общест-
Общества им. Кайзера Вильгельма в Берлине. Фогт консультировал лечение лидера
русской революции В.И. Ленина, а после его смерти помог Правительству
СССР организовать в Москве Институт для изучения мозга Ленина. Несколько
раз О. Фогт посетил Институт Кольцова, вдохновился высоким уровнем иссле-
-
2. Ровесник века, гигант столетия 23
дований и попросил рекомендовать ему молодого русского генетика для орга-
организации лаборатории генетики в Институте мозга в Берлине. Н.К. Кольцов и
нарком здравоохранения Н.А. Семашко рекомендовали ему молодую чету —
Н.В. и его жену Елену Александровну Тимофеевых-Ресовских. В 1925 г. они уе-
уехали в Берлин, а после завершения в 1929 г. строительства здания Института
мозга в Берлин-Бухе Н.В. перебрался туда и возглавил там Отделение генетики.
В 1937 г. Отделение превратилось в Институт генетики и биофизики, подчи-
подчинявшийся непосредственно Обществу им. Кайзера Вильгельма (аналог Акаде-
Академии наук). Н.В. стал директором Института.
Он пробыл в Германии 20 драматических лет, пережил звездные часы обще-
общения с лучшими биологами и физиками Германии и Европы, вошел в круг наибо-
наиболее талантливых исследователей Боровской школы — физиков, биофизиков, гене-
генетиков. Известно, что в этот период великий физик Н. Бор очень интересовался
действием жестких излучений на биологические объекты, а также отношением
квантовой физики к генетике. Но вместе с этим Н.В. пережил также безвременье
нацизма, Вторую мировую войну во вражеском стане, постоянную угрозу ареста,
гибель старшего сына Дмитрия в концлагере Маутхаузен, отлучение от Родины.
В 1937 г. он получил указание возвратиться в СССР. Научная командиров-
командировка в Германию была окончена. Однако от Н.К. Кольцова Н.В. получил преду-
предупреждение, что возврат может закончиться арестом. Два брата Н.В. в этот пери-
период уже были арестованы и затем погибли. Н.В. не рискнул вернуться на Родину.
Его юридический статус в Германии был весьма неопределенным — иностра-
иностранец без подданства, взятый на поруки Обществом им. Кайзера Вильгельма. Од-
Однако, даже попав в этот дьявольский клинч, Н.В. все же не терял надежду на
возвращение домой при изменении обстоятельств. В 1937 г. он отклонил при-
приглашение Рокфеллеровского фонда возглавить лабораторию Института Карне-
ги в Колд Спринг Харборе. В конце войны, когда научные институты эвакуиро-
эвакуировались из Берлин-Буха в Геттинген, он отказался уехать, и со всем своим Ин-
эм остался ждать прихода Советской Армии.
Научными вершинами этого периода для Н.В. были:
1) цикл классических работ по феногенетике (основные публикации
rimofeeff-Ressovsky, 1927a, 1934а]). Представления о пенетрантности и экс-
экспрессивности, а также о системной регуляции формирования фенотипических
признаков легли в основу феногенетики и далее — генетики онтогенеза;
2) цикл классических работ по Теории эволюции популяций [Timofeeff-
Ressovsky, 1927b, 1939, 1940, 1958, 1975, 1977]. Открытие резерва наследственной
изменчивости в природных популяциях дрозофил было первым подтверждением
амечательного его предвидения С.С. Четвериковым. Вершиной изучения попу-
[яционно-эволюционных процессов стала разработка учения о микроэволюции.
им термином Н.В. назвал эволюционные процессы внутри вида. Он построил
есьма логичную и стройную систему понятий этого учения: элементарным объ-
объектом микроэволюции является популяция, элементарным эволюционным ма-
ериалом — мутации, элементарным эволюционным событием — изменение ге-
нотипического состава, факторами эволюции — мутационный процесс, колеба-
[ численности, изоляция, миграции, отбор. Эти представления легли в основу
Синтетической теории эволюции [Timofeeff-Ressovsky, 1940];
24 I. Personalia
3) цикл классических, основополагающих работ по радиационной генетике
и радиобиологии [Timofeeff-Ressovsky, 1934b, 1944,1947,1968,1981]. Открытые им
закономерности дозовых зависимостей облучения дрозофил жесткими излуче-
излучениями легли в основу теории попадания, радиационной биологии, радиацион-
радиационной генетики;
4) "мозговой штурм" кольцовской проблемы молекулярной природы генов
совместно с К. Циммером и М. Дельбрюком, воплотившийся в так называемую
"Классическую зеленую тетрадку", основополагающую статью [Timofeeff-
Ressovsky, Zimmer, Delbruck, 1935], которую можно считать вехой в становле-
становлении молекулярной генетики. Эта работа послужила отправной точкой для клас-
классической книги Э. Шредингера "Что такое жизнь?" [1944] и дальнейшего бур-
бурного развития молекулярной биологии и генетики. Представление о физиче-
физической природе генов как макромолекул, оценка их размера и мономолекулярной
природы, понятие матричной конвариантной редупликации позже были адап-
адаптированы его учеником и соратником М. Дельбрюком в фундамент молекуляр-
молекулярной генетики.
Здесь уместно прояснить некоторые эпизоды отношений Н.В. и М. Дель-
Дельбрюка. Вот как сам Н.В. описывает развитие событий [Тимофеев-Ресовский,
2000]:
"В начале 30-х годов я сдружился и, так сказать, втянул в наши работы Мак-
Макса Дельбрюка. Он был чисто теоретический физик, ученик Макса Борна и Нильса
Бора. Я его, в сущности, переманил в биологию теоретическую. Он сейчас очень
крупный вирусолог и теоретический биолог в Америке, Нобелевский лауреат, вооб-
вообще очень замечательный человек. Тогда он был молодой человек и, как смолоду все
крупные теоретики, немного нагловат, но это ничего. Мы с ним тоже нагло обра-
обращались, так что он обтесался очень быстро у нас и стал вполне приемлемым моло-
молодым человеком ".
"...и с тех пор попросился в нашу Буховскую группу <...>. Ну и тут мы быстро
сконтачили. Я рассказал ему про кольцовское общее представление о молекулярной
биологии генов ли, хромосом, что мы тогда пытались экспериментально показать
мономолекулярность, в общем смысле, генов. То есть что это единая физико-хи-
физико-химическая элементарная структура, так сказать, а не кусочек сливочного масла
<...>. Он в этом принял участие. Из этого родилась так называемая потом клас-
классическая зеленая тетрадь ".
Его сотрудник К. Циммер [1966] вспоминал впоследствии:
"Мы встречались два-три раза в неделю, в основном — в доме Тимофеева-Ре-
созского в Берлине, где мы разговаривали по десять и более часов без перерыва,
лишь иногда поглощая пищу в ходе разговоров. Не могу судить, кто больше почерп-
почерпнул из этого обмена идеями, знаниями и опытом, но является фактом, что через
несколько месяцев Дельбрюк столь глубоко заинтересовался количественной биоло-
биологией, и особенно — генетикой, что остался в этом поле навсегда ".
Иначе говоря, это был, как теперь говорят, "мозговой штурм" кольцовской
проблемы молекулярной природы генов. Зная неудержимый характер Н.В.,
2. Ровесник века, гигант столетия 25
можно сказать, что без сомнения — это именно он сеял семена. Итог Н.В. под-
подвел в следующих выражениях [Тимофеев-Ресовский, 2000]:
«Ялично с 40-х годов больше этим не занимаюсь. Правда, меня многие, особен-
особенно там, за рубежом, считают чем-то вроде деда этого направления. Потому что
новая послевоенная редакция его была запущена Дельбрюком, а Дельбрюку соот-
соответствующую вещь я заправил в мозги в 30-е годы. Вот с этого, в сущности, пошло
все, с этой самой нашей классической, так называемой, "зеленой тетрадочки"
Геттингенского общества наук. Ну и пусть, значит, дальше развивается на доб-
доброе здоровье».
Общим итогом этих лет была научная зрелость. Н.В. стал признанным уче-
ученым европейского и мирового уровня. Кругом его научного общения были фи-
физики Н. Бор, П. Дирак, Э. Шредингер, П. Оже, Ф. Перрен, У. Астбери и др.,
биологи Г. Меллер, Ф.Г. Добржанский, Н.И: Вавилов, Б.С. Эфрусси, В.И. Вер-
Вернадский, С. Дарлингтон, Дж. Холдейн, Т.О. Касперсон, O.K. Густафсон и другие
выдающиеся ученые Европы и Америки. Вместе со своим другом и соратником
М.Дельбрюком Н.В. был постоянным участником копенгагенских и других се-
семинаров элиты европейской науки. Вместе с французским генетиком русского
происхождения Борисом Эфрусси на деньги Рокфеллеровского фонда он орга-
организовывал ежегодные конференции по генетике, биофизике, радиационной
биологии, которые продолжались до 1939 г., т.е. до начала войны в Европе.
В 1945 г. Берлин был взят советскими войсками. Н.В. сдал свой Институт
генетики и биофизики советским военным властям и несколько месяцев про-
продолжал работать на прежнем месте. В этот период его посетил руководитель со-
советского атомного проекта зам. Наркома НКВД генерал А.П. Завенягин, кото-
который вместе с И.В. Курчатовым очень хотел использовать опыт Н.В. в своей
системе. Шла речь о перебазировании лаборатории Н.В. в Россию. Однако
вскоре события приобрели трагический характер.
В сентябре 1945 г. Н.В. был арестован другим отделом НКВД (как свиде-
свидетельствует историк генетики Бабков [2000], по доносу заезжего ученого!), осуж-
осужден на 10 лет за невозвращение на Родину и в 1946 г. отправлен в Караганду, в
один из самых ужасных лагерей ГУЛАГа. По пути в лагерь, в общей камере Бу-
Бутырской тюрьмы он познакомился с другим узником — А. Солженицыным, ко-
который позже описал эти эпизоды в своей книге "Архипелаг ГУЛАГ" [Солжени-
[Солженицын, 1991]. Там, в лагерях он чуть не умер от голода, заработал тяжелейшую
дистрофию и пеллагру, почти потерял зрение. В общем, этот очень здоровый
человек был на краю гибели.
Атомное ведомство в лице А.П. Завенягина долго не могло его найти. По
свидетельству Бабкова [2000], решающую роль в спасении Н.В. сыграл выдаю-
выдающийся французский физик, Нобелевский лауреат и участник Сопротивления
Фредерик Жолио-Кюри. Он посетил Москву и обратился к Министру МВД
Лаврентию Берия, которому подчинялись все засекреченные военно-научные
проекты, о необходимости спасения и предоставления работы крупнейшему
радиобиологу Н.В. Тимофееву-Ресовскому. Трудно сказать, что именно срабо-
сработало, но Н.В. был немедленно доставлен в Москву, помещен в лучший госпи-
госпиталь МВД для лечения и через несколько месяцев, весной 1947 г., доставлен на
26
I. Personalia
H.B. Тимофеев-Ресовский и А.И. Солженицын. 1968 г.
Южный Урал для организации большой лаборатории радиационно-биологиче-
ских исследований.
Объект № 0215 был расположен в г. Сунгуле. Н.В. заведовал там отделом
биофизики. Так Н.В. попал в "шарашку". Этот термин хорошо знаком жителям
Советской России. Так назывались засекреченные конструкторские и научные
лаборатории, где многие выдающиеся советские инженеры, конструкторы,
ученые, будучи узниками ГУЛАГа, разрабатывали самые современные образцы
военной техники. Через "шарашку" прошли СП. Королев (ракеты), А.Н. Ту-
Туполев и другие авиаконструкторы. Великий Н.И. Вавилов не дожил до "ша-
"шарашки", он умер в тюрьме от голода.
Хотя условия содержания на "объекте" были по тем временам очень не-
неплохие (помещение санатория, снабжение по высшему разряду и т.д.), для Н.В.
это все же была тюрьма. После продолжительного лечения он получил разре-
разрешение выписать из Германии семью и группу своих немецких сотрудников
(включая К. Циммера). С ними и с полусотней вольнонаемных сотрудников он
работал в Сунгуле до 1955 г. После этого "объект" был расформирован, Н.В. ос-
освобожден, а Атомное ведомство великодушно передало ему оборудование его
лаборатории для дальнейшего использования уже на воле.
2. Ровесник века, гигант столетия 27
Научный итог этого периода был очень плодотворным. В 1955 г. работы
были рассекречены, опубликованы, из них выросли последующие циклы ис-
исследований по радиобиологии и радиоэкологии на Урале и в Обнинске, кото-
которые успешно развиваются до сих пор. Лучше всего этот итог подвести словами
самого Н.В. [Тимофеев-Ресовский, 2000]:
"...Во всем мире считается, что американцы разработали всю медицинскую
изотопную, так сказать, биологию и всю водную изотопную биологию. А это все
мы раньше американцев сделали... Примерно к концу 60-х — началу 10-х годов я и
мои ученики закончили, собственно, работу по этой радиационной биогеоценоло-
гии. Пожалуй, эти работы в атомной системе и в Миассово на биостанции на
Урале были наиболее продуктивными в моей так называемой научной жизни ".
Любопытная деталь: в последние годы были рассекречены многие доку-
документы Нобелевского комитета. Только что из них стало известно [Блох, 2001],
что в 1950 г., когда Н.В. был в "шарашке", группой немецких ученых он был
представлен на Нобелевскую премию по разделу "Медицина". Речь могла идти
только о его работах германского периода. В то же время выясняется, что Нобе-
Нобелевский комитет очень осторожно обсуждал кандидатов из России, чтобы им
не навредить. Итоги голосования неизвестны, Н.В. не получил Нобелевской
премии, но само номинирование и обсуждение говорят о многом!
После освобождения в 1955 г. Н.В. посетил столицы — Москву, Ленинград,
Киев, везде нашел своих старых знакомых, а физиками и генетиками был встре-
встречен с огромным энтузиазмом. Его приветствовали будущие Нобелевские лауреа-
лауреаты по физике П.Л. Капица, Л.Д. Ландау, И.Е. Тамм, биофизик М.В. Волькен-
штейн, биохимик В.А. Энгельгардт, математик А.А. Ляпунов и др. В своих ин-
институтах, на кафедрах они организовывали доклады и лекции Н.В., которые
проходили с большим успехом. Несмотря на изоляцию на "объекте", Н.В. был
в курсе всех новых проблем генетики и молекулярной биологии, а многие авто-
авторы этих открытий были его старыми знакомыми и друзьями.
Однако в столицах Н.В. фактически получил отказ в возможности жить и
работать. Известно, что в этот период академик О.В. Газенко пытался взять его
в свой Институт медико-биологических проблем (фактически — космической
медицины) в Москве, но безрезультатно. Свобода оказалась довольно призрач-
призрачной и ограниченной. Тогда он вернулся на Урал и проработал в Свердловске и
на биостанции Миассово до 1964 г. в качестве заведующего Отделом биофизи-
биофизики Института биологии Уральского филиала АН СССР без ученой степени (!).
Парадоксально, но будучи признанным ученым мирового класса и членом
многих иностранных академий и обществ, Н.В. не имел ни диплома гимназии
(хотя закончил ее с золотой медалью), ни диплома университета, ни научных
степеней. Вихрь войн и революций смел все бумажные атрибуты его образова-
образования. Докторскую диссертацию он защитил в Свердловске только в 1963 г., а ди-
диплом доктора наук получил только в 1964 г. после падения Н.С. Хрущева и реа-
реабилитации генетики в СССР.
В 1964 г. Н.В. уехал на свою "историческую родину" — в Калужскую об-
область. В новом научном центре Обнинске он возглавил Отдел радиационной
биологии и генетики Института медицинской радиологии АМН СССР. Гене-
28
I. Personalia
Н.В. Тимофеев-Ресовский. Обнинск,
1960-е.
H.B. Тимофеев-Ресовский. Обнинск,
1960-е. Рис. С. Тулькеса.
тические и радиационные исследова-
исследования успешно продолжались. Однако в
1969 г. рухнула и эта ниша. В стране
начался долгий период застоя и борь-
борьбы с диссидентством. Первым был
уволен сотрудник отдела Жорес Медведев. Под большим партийно-админист-
партийно-административным давлением Н.В. был отправлен на пенсию. В декабре 1969 г. Макс
Дельбрюк получил Нобелевскую премию и позже посетил Москву. В своих лек-
лекциях он вспомнил о своем друге и соратнике и охарактеризовал его выдающую-
выдающуюся роль в становлении молекулярной биологии и генетики [Бабков, 2000]. Есть
сведения, что Дельбрюк обращался к Президенту АН СССР М.В. Келдышу
[Берг, 1993]. Возможно, это сыграло определенную роль, поскольку именно
тогда акад. О.В. Газенко удалось сломать невидимую преграду и взять Н.В. к
себе в институт научным консультантом. Здесь он и проработал последние
11 лет своей жизни. Умер Н.В. 28 марта 1981 г.
Следует отметить, что всю свою долгую жизнь Н.В. прожил вместе с Еле-
Еленой Александровной Тимофеевой-Ресовской (урожденной Фидлер, 1898-1973).
Они были женаты свыше 50 лет. Она была "всем" в его жизни — его любовью,
другом, ближайшим сотрудником, помощью, защитой и т.д. Она была отлич-
отличным экспериментатором и его соавтором по большинству работ. Когда Н.В.
арестовали, она заменила его на посту директора Института в Берлин-Бухе.
2. Ровесник века, гигант столетия 29
Когда он потерял в заключении центральное зрение и способность читать и пи-
писать, она стала его глазами: Е.А. писала все его статьи, письма и всю другую
письменную продукцию. Однажды Н.В. сказал, что в жизни ему дважды очень
повезло: его учителем был великий Н.К. Кольцов, а женой — Елена Александ-
Александровна.
После освобождения из ГУЛАГА и даже после его смерти определенные
недобросовестные и ангажированные личности пытались опорочить имя
Н.В. Тимофеева-Ресовского путем совершенно неадекватных квази-научных и
политических обвинений (см. об этом, например, [Гранин, 1987; Paul, Krimbas,
1992; Korogodin et al., 2000; Ratner, 2000; Timofeeff-Resovsky, 2000]). Все эти по-
попытки закончились полным крахом. В 1992 г. Н.В. (наконец-то!) был полно-
полностью официально оправдан, его дело закрыто, а его обвинение прекращено
вследствие отсутствия состава преступления.
За годы работы на Урале и в Обнинске научные интересы Н.В. стали сме-
смещаться в сторону глобальных общебиологических проблем: биосфера и челове-
человечество, биосфера-биогеоценоз-биоценоз-популяция, радиоактивное заражение
биосферы, принципы теоретической биологии. В этих областях он поднялся до
уровня своих великих учителей и предшественников — академиков В.И. Вер-
Вернадского и В.Н. Сукачева. Н.В. разработал стройную концепцию иерархиче-
иерархических уровней биосферы и живой природы [Тимофеев-Ресовский, 1970; Тюрю-
канов, Федоров, 1996]. Эти работы как бы венчали собой его великий талант
обобщения.
Кроме того, еще в годы работы на Урале он столкнулся с "первым звон-
звонком" чернобыльской трагедии. В 1957 г. на Южном Урале вблизи г. Кыштым
произошел выброс значительного количества радиоактивных отходов, загряз-
загрязнивших окружающие территории. Ликвидация последствий аварии стала госу-
государственной проблемой. Н.В. в полной мере оценил опасность радиоактивных
загрязнений и необходимость изучения их последствий. Во многих своих док-
докладах он обсуждал эти проблемы. В.В. Бабков [2000] утверждает, что один из
таких докладов произвел большое впечатление на акад. А.Д. Сахарова и побу-
побудил его обратиться к проблеме защиты биосферы и человечества от ядерных
испытаний.
Научный итог этого периода приведен выше. Но не менее важен, так ска-
сказать, и социальный итог деятельности Н.В. Прежде всего, он обрел полное на-
научное признание отечественных и зарубежных ученых как выдающийся биолог
современности. В этот период он получил очень престижную Кимберовскую
премию по генетике Национальной Академии Наук США, Дарвиновскую ме-
медаль Академии Леопольдина в г. Галле (Германия), Менделевскую медаль Че-
Чехословацкой АН. Во-вторых, в эти годы он вернул отечественной науке все
основные результаты, опубликованные им в Германии (и часто недоступные в
России). В частности, он опубликовал со своими учениками ряд итоговых мо-
монографий и обзоров по теории эволюции, радиационной генетике и другим
проблемам (Тимофеев-Ресовский и др. — работы, 1958, 1968, 1975, 1977,
1981 гг.). В-третьих, в каждом из мест, где ему пришлось находиться, включая
следственный изолятор КГБ на Лубянке, общую камеру Бутырской тюрьмы и
"шарашку" в Сунгуле, он организовывал школу своих слушателей и учеников,
30 I. Personalia
многие из которых стали затем крупнейшими российскими учеными, акаде-
академиками, деятелями культуры. Огромную роль в воспитании и подготовке на-
научной молодежи сыграли миассовские летние школы-семинары по генетике и
биофизике A956-1963 гг.) и подмосковные летние школы A965-1967 гг.), ко-
которые Н.В. собирал совместно с известным биофизиком Л.А. Блюменфель-
дом. На этих школах мне посчастливилось побывать (см. гл. 1).
Таким образом, великие потрясения XX века наложили свой драматиче-
драматический отпечаток на жизнь и творчество Н.В. Он неизменно был в фокусе со-
событий: войны и революции, тоталитарные режимы, ГУЛАГ и "малый
Чернобыль", — поистине гамлетовский набор проблем и страстей. Но вместе
с тем — вершины науки, необычайный дар мыслителя и ученого, могучий,
истинно русский характер учителя и просветителя. Необходимо подчерк-
подчеркнуть, что боевой дух Н.В., его моральные устои, ощущение Мира и Родины,
отношение к науке не были сломлены превратностями судьбы. Природное
здоровье и жизнерадостность неизменно брали верх, речь была полна шуток
и каламбуров, энергия и артистизм натуры не давали покоя. Может быть,
главное, что оставил Н.В. своим ученикам, — это отношение к науке. С од-
одной стороны, он почитал науку как высшее проявление человеческого гения
и разума. С другой стороны, он постоянно подтрунивал над собой, окружаю-
окружающими, другими деятелями науки, побуждая их относиться к науке как к
азартной игре ума, шутке гениев, которые на досуге выдумали для людей
много прекрасных и поучительных историй. Иначе говоря, он страховал
свою деятельность и науку от самолюбования, авторитарности и преувеличе-
преувеличения собственной роли, что чрезвычайно актуально для ученых всех времен и
поколений.
Говорят, что академику П.Л. Капице принадлежат следующие слова:
"Если ученого вспоминают и цитируют через 10 лет после смерти, то значит —
он классик/" Прошло 20 лет после смерти Н.В., но интерес к его личности, на-
научному и методологическому наследию не убывает. Переведены и изданы на
русском языке работы Н.В. германского периода, изданы воспоминания о
нем и его собственные устные истории ("байки") обо всем. Под эгидой
ЮНЕСКО в 2000 г. отмечался его 100-летний юбилей, в разных городах Рос-
России и странах прошли конференции памяти Н.В. Значит, он не умер! Значит,
он — классик!
Благодарности. Благодарю друзей, соратников и учеников Н.В. Тимофее-
Тимофеева-Ресовского за подготовку и издание бесценных материалов о его жизни и
творчестве, которыми автор активно пользовался. Это Вл.И. Иванов, Н.А. Ля-
Ляпунова, Л.А. Блюменфельд, Н.Н. Воронцов, А.Н. Тюрюканов, С.Э. Шноль,
В.В. Бабков, Е.С. Саканян, В.И. и В.Л. Корогодины, Д. Гранин и многие дру-
другие. Особо важную роль играет последнее издание устных историй, рассказан-
рассказанных Н.В. и записанных на диктофон его друзьями в последнее десятилетие его
жизни [Тимофеев-Ресовский, 2000]. Это редчайший случай, когда внутренний
мир великого ученого предстал перед читателем во всей своей оригинальности
и полноте.
2. Ровесник века, гигант столетия 31
Список литературы
Основные работы Н.В. Тимофеева-Ресовского
Timofeeff-Ressovsky N.W. Studies on the phenotypic manifestation of hereditary factors. I. On
the phenotypic manifestation of the genovariation radius incompletus in Drosophila funebris //
Genetics. 1927a. Vol. 12. P. 128-198.
Timofeeff-Ressovsky H.A. und N.W. Genetische Analyse einer freilebenden Drosophila
me/onogaster-Population // W. RouxArchivf. Entwicklungsmechanik d. Organismen. 1927b. Bd 109.
Heft 1. S. 70-109.
Timofeeff-Ressovsky N.W. Verkniipfung von Gen und Aupenmerkmal (Phanomenologie der
Genmanifestierung) // Wiss. Woche zu Frankfurt. 1934a. Bd 1. S. 92-115.
Timofeeff-Ressovsky N.W. The experimental production of mutations // Biol. Rev. Cambridge.
1934b. Vol. 9, N 4. P. 411-457.
Timofeeff-Ressovsky N.W., Zimmer K.G., Delbriick M. Uber die Natur der Genmutation und
der Genstruktur // Nachrichten von der Gesellschaft der Wissenschaften zu Gottingen. Biologie.
Neue Folge. 1935. Bd 1, N 13. S. 189-245.
Timofeeff-Ressovsky N.W. Genetik und Evolution (Bericht einer Zoologen) // Z. Ind. Abst.
Vererbl. 1939. Bd 76. Heft 1/2. S. 158-218.
Moglich F., Rompe R., Timofeeff-Ressovsky N.W. Uber die lndeierminiertheil und die
Verstarkerer-Scheinungen in der Biologie // Naturwiss. 1944. Bd 32.
Timofeeff-Ressovsky N.W., Zimmer K.G. Biophysik. 1. Das Trefferprinzip in der Biologie.
Leipzig, 1947.
Тимофеев-Ресовский Н.В. Микроэволюция. Элементарные явления, материал и факто-
факторы эволюционного процесса // Ботан. журн. 1958. Т. 43, № 3. С. 317-336.
Тимофеев-Ресовский Н.В. Структурные уровни биологических структур. Системный
подход в экологии // Системные исследования АН СССР. Институт истории естествознания
и техники. М.: Наука, 1970. С. 80-136.
Тимофеев-Ресовский Н.В. Избранные труды. М.: Медицина, 1996.
Тимофеев-Ресовский Н.В., Иванов В.И., Корогодин В.И. Применение принципа попада-
попадания в радиобиологии. М.: Атомиздат, 1968.
Timofeeff-Ressovsky N.W., Voroncov N.N., Jablokov A.V. Kurzer Grundriss der
Evolutionstheorie. Jena: Fisher, 1975.
Timofeeff-Ressovsky N.W., Jablokov A.V., Glotov N.V. Grundriss der Populationslehre. Jena:
Fisher, 1977.
Тимофеев-Ресовский Н.В., Савич А.В., Шальнов М.И. Введение в молекулярную радио-
радиобиологию. Физико-химические основы. М.: Медицина, 1981.
Основные публикации о Н.В. Тимофееве-Ресовском
и его работах
Бабков В.В. Николай Владимирович Тимофеев-Ресовский // Информ. вестн. ВОГиС.
Новосибирск: ИЦиГ СО РАН. 2000. № 15. С. 8-14.
Берг Р.Л. Николай Владимирович Тимофеев-Ресовский // Николай Владимирович Ти-
Тимофеев-Ресовский: Очерки. Воспоминания. Материалы. М., 1993. С. 226-239.
Блох А.М. Нобелевский комитет боялся навредить нашим ученым // Известия.
17.08.2001. № 140 B5987). С. 1-3.
БлохА.М. Советский Союз в интерьере нобелевских премий. СПб.: Гуманистика, 2001.
Воронцов Н.Н. (ред.) Николай Владимирович Тимофеев-Ресовский. Очерки. Воспоми-
Воспоминания. Материалы. М.: Наука, 1993.
Гранин Д. Зубр. Новый мир. 1987. № 1, 2.
32 I. Personalia
Корогодин В.И., Корогодина В.Л. (ред.) Н.В. Тимофеев-Ресовский. Буклет. Дубна:
ОИЯИ, 2000.
Ратнер В.А. Столетие // Информ. вестн. ВОГиС. Новосибирск: ИЦиГ СО РАН. № 15.
С. 6-8.
Солженицын А.И. Архипелаг ГУЛАГ. М.: ИНКОМ НВ, 1991. Т. 1. Ч. 2. Гл. 4. С. 413-416.
Тимофеев-Ресовский Н.В. Истории, рассказанные им самим, с письмами, фотографиями
и документами. М.: Согласие, 2000.
Тюрюканов А.Н., Федоров В.М. Н.В. Тимофеев-Ресовский: биосферные раздумья. М.,
1996.
Яблоков А.В. (ред.) Онтогенез. Эволюция. Биосфера. М.: Наука, 1989.
Paul D.B., Krimbas CD. Nikolai V. Timofeeff-Ressovsky // Sci. Amer. 1992. N 2. P. 86-92.
Schrodinger E. What is Life? The Physical Aspect of the Living Cell. Cambridge: Cambridge
Univ. Press, 1944.
Zimmer K.G. The Target Theory // Phages and the Origins Of Molecular Biology/ Eds.
J. Cairns, G.S. Stent, J.D. Watson. Cold Spring Harbor: Cold Spring Harbor Laboratory, 1966.
P. 33-42.
3. Алексей Андреевич Ляпунов*
Входит Ляпунов.
Ремарка из Русской истории
Мы будем очерчивать контуры наук.
А.А. Ляпунов, из выступлений
Впервые я услышал об А.А. Ляпунове в начале 1961 г. от Ю.Я. Керкиса.
Я только что поступил на работу в Институт цитологии и генетики СО АН
СССР в Академгородке. В зимние каникулы М.Е. Лобашов организовал в Ле-
Ленинграде первую легальную конференцию по экспериментальной генетике, и
все, кто мог, кинулись туда "на огонек". Наш директор Д.К. Беляев решил вы-
вывезти туда своих генетиков, включая молодежь. Я тоже ехал в поезде "Сибиряк"
Новосибирск—Москва вместе со всеми. Мы пили румынское вино "Фетяску",
зубоскалили и болтали всю дорогу, компания была очень хорошая. Поскольку
я вез на конференцию свою первую теоретическую работу о корреляциях,
спросил в дороге у Ю.Я. Керкиса, кому ее можно показать. Именно тогда он
произнес имя А.А. Ляпунова и дал его московский телефон.
Мы доехали до Москвы и должны были вечером пересесть на ленинград-
ленинградский поезд. И тут выяснилось, что накануне "злодей советской генетики"
Т.Д. Лысенко предпринял очередной демарш в ЦК КПСС и убедил отдел нау-
науки, что генетики опять собирают свой шабаш. Короче говоря, конференция
была отменена накануне открытия. Мы застряли в Москве.
И тогда я решил использовать открывшееся время с толком. Позвонил
А.А. Ляпунову, представился, сослался на Ю.Я. Керкиса и попросил о встрече.
Он очень живо откликнулся и предложил приехать к нему домой на Шаболов-
Шаболовку. Тогда А.А. было около 50 лет. Это был довольно высокий худощавый муж-
* Ранний вариант статьи: Ратнер В.А. Алексей Андреевич Ляпунов // Очерки истории информа-
информатики в России / Ред. Д.А. Поспелов, Я.И. Фет. Новосибирск: НИЦ ОИГГМ СО РАН, 1998.
С. 379-386.
3. Алексей Андреевич Ляпунов 33
чина, немного сутуловатый, с бледным вдохновенным лицом, слегка всклоко-
всклокоченный, с длинной черной бородой. Он ходил по комнате широким шагом,
оживленно разговаривал на ходу, поглаживал бороду и излучал энтузиазм. Со-
Содержание моей работы он воспринял сразу и стал давать всевозможные советы.
Один из советов был очень конструктивным: пойти к И.И. Шмальгаузену, ко-
который может представить работу в "Докладах АН СССР".
Так я и сделал. Иван Иванович Шмальгаузен сидел в маленькой комнатке в
Институте морфологии животных АН СССР на Б. Калужской, 33. Будучи от-
отставлен от всех должностей после Августовской сессии ВАСХНиЛ 1948 г., И.И.
работал вдвоем с лаборанткой. Только потом, прочитав стенограмму позорной
сессии ВАСХНиЛ, я понял, что этот тихий интеллигентный старик — мужест-
мужественный борец, один из немногих, кто не поступился совестью и не покаялся во
грехах после жесточайшего давления лысенкоистов. В середине 50-х годов он
неожиданно столкнулся с новой наукой — кибернетикой, и, будучи чистым
зоологом-натуралистом, предпринял попытку переосмыслить генетико-эволю-
ционную науку с позиций кибернетики. В результате возникла серия статей, а
затем и книга "Кибернетические вопросы биологии".
И.И. Шмальгаузен поддержал нашу первую статью, представил ее в ДАН,
где она и вышла в 1961 г., хотя теперь я могу сказать, что работа имела ряд недос-
недостатков, на которые чуть позже нам указал Н.В. Тимофеев-Ресовский (см. гл. 1).
В том же году А.А. переехал в новосибирский Академгородок по приглаше-
приглашению М.А. Лаврентьева и С.Л. Соболева. С этого момента кибернетическая
жизнь у нас существенно оживилась. А.А. был известной личностью, и люди
слетались на него, как на огонь и свет.
Прежде всего, по московскому примеру, А.А. организовал у себя дома се-
семинар по кибернетике. Туда ходили разные люди, от чистых математиков до
экономистов, биологов и лингвистов. Конечно, было много случайных фигур,
завсегдатаев научных "тусовок", но не подлежит сомнению, что вся активная
кибернетическая наука в Академгородке прошла через этот семинар.
До сих пор меня не покидает впечатление, что жизнь в форме постоянного
семинара была органически присуша А.А. Именно здесь он раскрывался наи-
наилучшим образом. Здесь он "очерчивал" (любимое выражение А.А.) контуры
наук. В 60-е годы он много болел, но семинары не прекращались. Поскольку
болезни были множественными, а отдых практически отсутствовал, врачи пы-
пытались ограничить доступ посетителей. Насколько я знаю, итогом было ухуд-
ухудшение состояния больного. В результате медики отказались от больничного
спокойствия, поставив только одно ограничение: начинать не ранее 10 утра и
заканчивать не позже 11 вечера, делая интервалы на еду и процедуры. Так оно и
шло: непрерывный поток посетителей и семинаров каждый день с утра и до ве-
вечера. В этой текучке состояние больного улучшалось.
Другим традиционным начинанием А.А. был курс лекций "Кибернетиче-
"Кибернетические проблемы биологии" в НГУ для студентов всех курсов всех факультетов.
Каждый год в сентябре в Большой Математической аудитории (теперь
им. Мальцева) набивалось море студентов и сочувствующих. Я бывал там 3 или
4 раза. А.А. начинал с огромным энтузиазмом, студенты воспринимали его с
34 I. Personalia
восторгом. По мере перехода от общих подходов к конкретным наукам аудито-
аудитория таяла, а через 5-6 недель А.А. заболевал и курс прерывался.
Любопытно, что это вовсе не означало провала начинания. В 1996 г., буду-
будучи на 2-м Сибирском конгрессе по прикладной и индустриальной математике,
посвященном памяти А.А. Ляпунова, И.А. Полетаева и А.П. Ершова, я поду-
подумал, как много замечательных людей прошло через эти семинары и лекции,
став затем крупными учеными. Импульс интереса к науке, идущий от А.А., ока-
оказался сильнее всех прочих обстоятельств. Огромную роль, конечно, играла по-
постоянная готовность А.А. помочь начинающему. Это ощутили на себе сотни
людей.
В то же время отношения А.А. со многими математиками бывали, мягко
говоря, натянутыми. По-видимому, они ждали от него доказанных теорем, а не
страстной научной проповеди. С другой стороны, их, видимо, обескураживала
детская наивность многих поступков А.А., его нравственный максимализм. Как
бы то ни было, мы с Аргентой Антониновной Титляновой были участниками
почти детективной истории в НГУ, где главным действующим лицом был А.А.
В 1967-1968 гг. после ряда предложений и обсуждений ректорат НГУ согла-
согласился на организацию при кафедре цитологии и генетики факультета естест-
естественных наук специализации по математической биологии. Ясно, что идеологом
этого предприятия был А.А. Я был биологическим куратором и "мотором".
С идеей согласились все ключевые фигуры — Д.К.Беляев, Р.И. Салганик,
Д.Г. Кнорре, А.Д. Слоним, ректор СТ. Беляев и др. А.А. Титлянова, будучи
зам. декана ФЕН, прикрывала операцию изнутри.
При первом же разговоре ректор СТ. Беляев, известный физик-теоретик,
сказал, что он согласен с предложением, но категорически против того, чтобы
специализацию возглавлял А.А. Ни на какие компромиссы он не согласился. И
тогда мы пошли к ближайшему ученику А.А. — Ю.И. Журавлеву, который уже
возглавлял Отделение кибернетики Института математики СО АН СССР, и
уговорили его быть зитц-руководителем специализации. Он сказал, что помо-
поможет А.А., но ни в какие дела вникать не будет, а согласен только подписывать
необходимые бумаги. Так и порешили. Ректор согласился.
Специализация стартовала в 1968 г. А.А. взялся за дело со свойственным
ему максимализмом. Еще за несколько лет до этого, сразу по приезде в Ново-
Новосибирск, он основал в НГУ кафедру теоретической кибернетики. Для матбио-
логов А.А. разработал грандиозную программу курса дискретной математики,
примерно 300-400 часов, на 4 семестра, куда вошли все нетрадиционные для
биологов разделы математики, вплоть до теории меры. Курс шел дополнитель-
дополнительной нагрузкой к полному курсу биологических наук. Так что вьщержать это мог
только несгибаемый энтузиаст. Интересно, что матбиологи первых поколений
выдержали этот учебный прессинг без потерь живой силы и техники.
Чтение основных разделов А.А. поручил своему ближайшему ученику и со-
сотруднику по кафедре, доценту Н.И. Глебову, который прочел прекрасный, ра-
рафинированный математический предмет, насыщенный многими красивыми,
но бесполезными для биологов главами. Вообще математики тогда стремились
рассказать все, что они знают, и поразить окружающих красотой своей науки.
Надо сказать, что кое в чем им это удалось. Первые матбиологи рассказывали
3. Алексей Андреевич Ляпунов 35
позже, что одно из самых глубоких впечатлений на них произвели именно эти
практически бесполезные для них главы математики. Они заронили уважение к
математике. Этот курс, разумеется, в несколько урезанном и модифицирован-
модифицированном виде, читается нашим матбиологам до сих пор.
Два года я безвылазно сидел на всех лекционных курсах и практикумах,
стараясь отладить программы, обговорить детали, найти заинтересованных
преподавателей, обеспечить дипломные работы. Короче говоря, все дыры заты-
затыкал собой. А.А. так и не узнал, что формально он не являлся руководителем
специализации. Но это ничего не изменило. Он был ее душой. А наша с
А.А. Титляновой главная установка состояла в том, чтобы успеть выпустить
первую группу матбиологов и на деле показать необходимость и пользу этого
предприятия. "Только бы проскочить" — вот был наш лозунг!
В 1970 г. выпуск состоялся. У меня было 6 дипломников. Сейчас эта цифра
приводит меня в ужас, но тогда мы ее осилили. Специализацию окончили
9 студентов, из которых четверо были биофизиками из Красноярска. В даль-
дальнейшем все они стали кандидатами наук, 5 из них — докторами, а 3 — директо-
директорами институтов: Р.Н. Чураев, А.Н. Дегерменджи, Н.А. Абросов. Недавно
А.Н. Дегерменджи был избран членом-корреспондентом РАН. Всего мы подго-
подготовили более 120 математических биологов, большинство из которых сейчас ус-
успешно работают в Москве, Новосибирске, Красноярске, Иркутске, Владиво-
Владивостоке, странах СНГ, а также в США, Германии, Англии, Израиле.
Будучи проповедником интеграции наук на базе кибернетики, А.А. неред-
нередко встречал непонимание со всех сторон. В этом судьба тех, кто пытается быть
объединяющим началом. Математики считали, что он больше натуралист, чем
математик, биологи — наоборот. Сам А.А. говорил, что, вступая в контакт с
представителями других наук, он до половины времени тратит на выработку
взаимно понятного языка общения. Часто на этом дело и обрывалось. Однако
там, где этот этап бывал пройден, открывался широкий простор для построе-
построения математической теории. Разумеется, если конкретная наука оказывалась
зрелой и богатой доброкачественным экспериментальным материалом. Так
произошло, например, в математической генетике, о чем разговор ниже.
Пожалуй, наиболее правильную оценку А.А. и его деятельности я слышал
от И.Б. Погожева, бывшего военного, ученика А.А. Главная сила А.А. как уче-
ученого состояла в том, что он видел и ощущал науку в целом. Это редкий дар. Он
видел внутреннюю связь, общность методов, фундаментальность проблем,
ближайшую и отдаленную перспективу. От него требовали доказывать теоре-
теоремы, а он видел науку в целом! Ясно, что требовать от него надо было то, в чем
он был сильнее других, тогда это продуктивно.
Продуктивность теоретической концепции измеряется калибром и разно-
разнообразием принципиальных задач, которые можно сформулировать на ее осно-
основе. В этом смысле кибернетический подход в биологии, и особенно в генетике,
оказался чрезвычайно плодотворным. А.А. очень рано это понял. В конце 50-х
годов вместе с А.Г. Маленковым он попытался формализовать классическую
генетику. Вместе с СВ. Яблонским он развил представление об управляющих
системах, в том числе — в живой природе. Для меня этот подход был толчком
для разработки концепции Молекулярно-генетических систем управления
Not for sale! For preview only! Scanned by Mykhaylo (molbiol(fl4ikr.net)
36 I. Personalia
(МГСУ). С тех пор я занимаюсь этим свыше 35 лет, причем чем дальше, тем
больше понимаю, что выбор направления был сделан верно.
В 1965 г. А.А. горячо поддержал мою первую диссертацию "Генетические
управляющие системы". Я долго сомневался, годится ли такая тема, особенно в
условиях еще не исчезнувшего лысенкоизма. А.А., не задумываясь, сказал: "Го-
"Годится, это будет темой и Вашей докторской!" И оказался прав.
Написав рукопись, я долго пытался поговорить с А.А. Однако он то болел,
то был занят, то не мог прочесть рукопись из-за болезни глаз. Разговор о дис-
диссертации состоялся в совершенно неожиданном и неподходящем месте. Ле-
Летом 1965 г. мы поехали на первую подмосковную школу Н.В. Тимофеева-Ре-
Тимофеева-Ресовского и Л.А. Блюменфельда на Можайское море. Жили в павильонах и па-
палатках летней базы МК ВЛКСМ. Собрался весь цвет теоретической биологии,
генетики, биофизики. Из наших были А.А. Ляпунов, И.А. Полетаев,
Ю.Я. Керкис, Р.Л. Берг и др. Неожиданно А.А. заболел. Он лежал в палатке и
не ходил на заседания. В тот день я дежурил по кухне и зашел к нему прове-
проведать и сказать два слова о работе. Неожиданно он предложил: "А Вы прочтите
мне свою работу вслух". В результате, убежав с кухни, я с выражением прочел
ему основную часть рукописи, попутно отвечая на вопросы и комментарии. В
конце А.А. сказал: «Я буду Вашим оппонентом, а работу надо опубликовать
как книгу в серии "Кибернетика в монографиях"». Эту серию редактировал
сам А.А.
Защита состоялась в конце 1965 г., оппонентами были А.А. Ляпунов и
А.А. Нейфах, а внешний отзыв прислал Н.В. Тимофеев-Ресовский. Я горжусь
этими именами, а также тем, что моя первая книга "Генетические управляю-
управляющие системы" вышла в 1966 г. в той же серии "Кибернетика в монографиях",
где в 1968 г. А.А. опубликовал книгу И.И. Шмальгаузена "Кибернетические во-
вопросы биологии".
Накануне защиты А.А. предпринял еще одну непредвиденную акцию. Не-
Неожиданно он обратился ко всем участникам моей защиты — Д. К. Беляеву
(директору института), Жукову (председателю Совета), Н.В. Тимофееву-Ресов-
Тимофееву-Ресовскому (внешнему рецензенту), А.А. Нейфаху (оппоненту) и др. с предложением
вместо кандидатской степени присудить мне сразу докторскую. Такие случаи
бывали, но в виде исключения, и требовали серьезных аргументов и полного
изменения процедуры защиты, в частности — появления утвержденного
третьего оппонента. Кроме того, в нашем институте почти все старшее поколе-
поколение генетиков еще не имели докторских степеней. Таким образом, все это было
очень приятно, но несвоевременно, даже немного неэтично. Поднятые А.А. за-
заинтересованные лица обсудили проблему и решили не спешить, тем более, что
неординарная защита гарантировала бы пристальное внимание к диссертации
в ВАКе и участие "черных" рецензентов. Я вздохнул свободно. Защита прошла
нормально. Правда, некто из крупных лысенковцев (возможно, Н. Жуков-Ве-
режников) затребовал диссертацию из Ленинской библиотеки и держал ее бо-
более полугода, но так ничего и не возразил.
Это были первые защиты диссертаций по генетике после всеобщего лысен-
ковского зажима. Мы испытывали огромный энтузиазм. Банкеты еще не были
запрещены. И мы, четыре диссертанта, банкетировали в холле нашего институ-
3. Алексей Андреевич Ляпунов
37
А.А. Ляпунов и Н.В. Тимофеев-Ресовский, конец 1950-х (архив Н.А. Ляпуновой).
та в присутствии дирекции и оппонентов. На банкете А.А. сказал: "Какая хоро-
хорошая у Вас обстановка, дружная. В нашем институте это невозможно". Действи-
Действительно, тогда все мы любили друг друга и испытывали большую радость осво-
освобождения. Однако А.А., конечно, уже чувствовал некий барьер непонимания со
стороны многих коллег-математиков. В итоге через несколько лет он ушел в
Институт гидродинамики к М.А. Лаврентьеву, и там под его крылом работал до
конца дней.
После 1970 г. положение нашей группы матбиологов в институте и специа-
специализации в НГУ укрепилось, студенты стали аспирантами, появилось много на-
научных результатов. Мы издали ряд сборников и книг. Концепция МГСУ на-
наполнялась конкретным содержанием и успешно развивалась. Первый итог я
подвел в докторской диссертации "Молекулярно-генетические системы
управления" [1974] и одноименной книге [1975], переведенной чуть позже на
немецкий язык [1977]. Предполагалось, что А.А. снова будет оппонентом, но в
1973 г. его не стало.
В 1993 г. в книге "Концепция Молекулярно-генетических систем управле-
управления" (НГУ, Новосибирск) я снова подвел итог развития идеологии МГСУ уже
за 30 лет и убедился, что поработали мы не зря. Мало того, многие мои ближай-
ближайшие ученики и сотрудники после двух десятилетий напряженной конкретной
работы в последние годы вдруг ощутили интерес и потребность вернуться к
корням, к истокам теории МГСУ, генетического языка, молекулярной эволю-
эволюции, восходящим к идеям Дж. фон Неймана, А.А. Ляпунова, И.А. Полетаева,
М. Эйгена и др. Этот симптом говорит о том, что ниточка преемственности не
прерывается.
38 I. Personalia
Насколько я могу судить, по научной школе, воспитанию и знанию ино-
иностранного языка А.А. тяготел к французскому. Он говорил "комплексный", а
не "комплексный", читал и цитировал французских авторов, был членом фран-
французского клуба в Доме ученых, принимал самое непосредственное участие в по-
слелагерной судьбе французской пианистки Веры Лотар-Шевченко, получал
красочные журналы из Французского посольства в Москве и т.д. Это наложило
определенный отпечаток на его научные симпатии. Не зная английского, он с
большим скепсисом относился к англо-американской школе математической
генетики (Р. Фишер, Дж. Холдейн, С. Райт и др.), но весьма одобрял труды
француза Г. Малеко, который независимо сделал почти то же самое, что и анг-
ло-саксы. Будучи активным сторонником и пропагандистом стохастической
теории эволюции, он почти отрицал результаты ее детерминистической ветви.
Но особую привязанность у него вызывали гении русской науки. Он очень
высоко ценил А.Н. Колмогорова, И.И. Шмальгаузена, Н.В. Тимофеева-Ресов-
Тимофеева-Ресовского, В.И. Вернадского, Н.И. Вавилова, С.С. Четверикова, М.А. Лаврентьева,
С.Л. Соболева и др. Конечно, А.А. не был сторонним наблюдателем, он сам
был частью русской науки, ее светлым, бескорыстным и беспокойным источ-
источником.
А.А. очень не любил нечистоплотность в науке. Многие люди пользовались
его доброжелательностью и открытостью для решения своих частных проблем,
а потом оставляли его под благовидным предлогом. А.А. не боролся с ними, он
переставал с ними знаться, вычеркивал их из своей жизни и твердо этому сле-
следовал. Особенное отторжение вызывали у него лысенкоизм и его деятели. Еще
в молодости, перед войной А.А. познакомился со многими генетиками (среди
них — Ю.Я. Керкис), участвовал с ними в семинарах, и даже — в статистиче-
статистической проверке законов Менделя. Лысенкоизм он воспринимал как мракобесие,
а его адептов — примерно как монстров из "Капричос" Ф. Гойи.
В конце 50-х годов генетические работы практически негде было печатать.
Общие биологические журналы их не брали, специализированных журналов не
было. Единственным академиком, который мог представить генетическую ра-
работу в ДАН, был И.И. Шмальгаузен. Однако и его представления тщательно
контролировались и повторно рецензировались. Неудивительно, что А.А. на-
направил меня именно к И.И. Шмальгаузену. Сам же А.А. открыл для генетиков
выпуски своего сборника "Проблемы кибернетики", которые редактировал.
Мне удалось опубликовать там несколько первых теоретических работ. Эта воз-
возможность по тем временам была просто неоценимой.
Еще одно воспоминание связано с 1961 г. Осенью Ф. Крик и сотр. опубли-
опубликовали в журнале "Nature" свою классическую статью по генетическому коду.
Оригинальный журнал тогда был почти недоступен, а репродуцировался он с
большим опозданием. Поскольку проблема генетического кода будоражила
тогда все умы, А.А. предпринял обходной маневр. Кто-то из знакомых матема-
математиков прислал ему фотокопию статьи из Берлина, а дальше она пошла по рукам
и по семинарам. Все мы изучали эту работу Крика по ляпуновской фотокопии.
До сих пор я рассказываю об этой работе студентам и не устаю восхищаться ее
красотой.
3. Алексей Андреевич Ляпунов 39
Летом 1961 г. в Москве проходил 5-й Международный биохимический кон-
конгресс, куда приехали многие выдающиеся молекулярные биологи, в том числе
Дж. Уотсон и Ф. Крик. А.А. пригласил их к себе домой, на Шаболовку, где они
весь вечер очень заинтересованно обсуждали взаимодействие между молеку-
молекулярной генетикой и кибернетикой. Остались очень выразительные фотографии
этой встречи.
В сентябре 1971 г. был 60-летний юбилей А.А. Мы подготовили ему поздра-
поздравительный адрес, где в стихах изложили историю сотворения основ кибернети-
кибернетики, сопроводив ее картинками, заимствованными из книги Жана Эффеля (см.
гл. 27). Кроме того, я привожу пародию на типичную дискуссию начала 60-х го-
годов "Человек, жизнь и кибернетика", написанную для институтского "капуст-
"капустника" в 1964 г., и довольно точно отражающую брожение мысли в тот период.
Некоторые из персонажей имели реальных прототипов. Наконец, как эмоцио-
эмоциональное отражение эпохи, под впечатлением посещения И.И. Шмальгаузена,
написано стихотворение "Галилей".
Умер А.А. летом 1973 г. в Москве. Он поехал на какую-то академическую
конференцию, участвовал в ней, волновался, был недоволен ходом дискуссий.
А вечером в доме своей матери упал в прихожей и умер, не приходя в сознание.
Похоронен А.А. на Введенском кладбище в Москве. Мы с Ю.М. Свирежевым
были на похоронах и несли крышку его гроба. Настроение было ужасное.
После смерти А.А. его научную библиотеку закупил наш ВЦ СО АН, орга-
организовав у себя Ляпуновскую мемориальную библиотеку. Она существовала до
самого последнего времени. Была создана комиссия по научному наследию
А.А., я входил в ее биологическую секцию. При разборе архива обнаружилось
своеобразие внутренней лаборатории А.А. У него почти не было законченных
рукописей статей и лекций на бумаге, в лучшем случае — краткие тезисы. То,
что можно считать текстом, возникало как целенаправленная импровизация в
ходе выступления, лекции или доклада. Иначе говоря, все опубликованные ра-
работы А.А. (кроме, вероятно, чисто математических) — это результат записи его
устных выступлений кем-нибудь из сотрудников или студентов, обработанные
затем автором.
Как-то в одном из разговоров А.А. сказал: "Сейчас, в зрелом возрасте, я в
основном пытаюсь реализовать идеи своей молодости". Действительно, в ран-
раннем возрасте часто возникают как будто простые вопросы, отвечать на которые
приходится всю жизнь. Я помню наивный вопрос, с которого началась моя
биологическая биография, и с которым я когда-то пришел к Д.К. Беляеву: "Ка-
"Какая сила заставляет жизнь столь неудержимо стремиться вверх, к совершенству,
преодолевая все преграды? Что это за внутренний стимул?" И сейчас я не могу
ответить на этот вопрос одной фразой. Не исключено, что он просто не имеет
ответа. Однако я уже понимаю, что такое молекулярно-генетические системы
управления, как в общих чертах идет их эволюция, в чем принципиальные
"хитрости" их организации; я уверен, что для понимания "чуда" жизни нет на-
надобности в изобретении Бога.
Да, А.А. в ранней молодости задал себе великие вопросы. В его московской
квартире висел его портрет кисти друга семьи художника И. Грабаря: розово-
розовощекий, взлохмаченный юноша, фактически — эскиз будущего Ляпунова. Он
40
I. Personalia
A.A. Ляпунов. Новосибирск, начало 1970-х (архив автора).
3. Алексей Андреевич Ляпунов
41
А.А. Ляпунов и Дж. Уотсон. Москва, 1961 г. На квартире Ляпунова на Шаболовке (архив
Н.А. Ляпуновой). В левом верхнем углу портрет молодого Ляпунова кисти И. Грабаря.
вырос в научной и очень интеллигентной среде, и через все сложности предво-
предвоенного вхождения в науку, через войну, через отчаянную битву за возрождение
кибернетики (и генетики тоже) сумел-таки заложить камень в здание мечты
своей молодости — всеобщей науки об управлении, единой науки о природе.
Как писала дочь А.А. Наталья Алексеевна, сам он считал, что сделал в тече-
течение своей жизни три больших дела, за которые ему не стыдно ни перед каким
судом. Первое дело связано со спасением Храма Василия Блаженного в Москве
от разрушения. Школьник А. Ляпунов наткнулся в рабочей многотиражке на
заметку, призывавшую к очищению Красной площади от поповской скверны.
Это было очень опасно, так как обычно сразу же в печати начиналась погром-
погромная кампания, которая заканчивалась разрушением объекта критики. А.А. тут
же сообщил об этом другу семьи художнику Игорю Грабарю, который возглав-
возглавлял комиссию по охране памятников старины. Грабарь связался с наркомом
просвещения Луначарским. Рецидив разрушения был погашен в зародыше.
Второе дело связано с его участием в Великой Отечественной войне. В
1943 г. А.А. командовал топографическим взводом в районе Курской магнит-
магнитной аномалии (КМА). Артподготовка, выполненная по инструкции, давала
плохие результаты. А.А. имел довоенный опыт экспедиций в район КМА и по-
понимал роль влияния мощного магнитного поля на результаты стрельб. В ре-
результате, рискуя попасть под суд за нарушение инструкций, он ввел поправки
42 I. Personalia
на магнитное поле, убедил начальство полка в своей правоте и добился высо-
высокой эффективности стрельбы по врагу. За это он был награжден орденом Крас-
Красной Звезды.
Третье дело было связано с работами по программированию на первой со-
советской вычислительной машине МЭСМ в 1952—1953 гг. в Феофании под Кие-
Киевом. В обстановке полной секретности в течение нескольких месяцев А.А. при-
придумал принципы операторного метода программирования. Уже в 90-е годы за
эти основополагающие работы Международным компьютерным сообществом
он был официально признан "основателем советской кибернетики и програм-
программирования".
Итак, как сказал в одном стихотворении сибирский поэт Леонид Марты-
Мартынов (цитирую по памяти):
"Написана книга,
И больше ни слова
Ты к ней не добавишь,
Ты к ней не припишешь.
Ни слова —
Ни доброго,
И ни злого...
Но новое солнце
Встает на Востоке,
На Западе
Новое солнце садится,
И просятся в книгу
Все новые строки,
Все новые строки
И новые лица..."
Жизнь продолжается. Свеча не угасла.
4. Игорь Андреевич Полетаев*
"Мое мнение перпендикулярно Вашему".
И.А. Полетаев
В конце 1950-х — начале 1960-х годов имя инженера И.А. Полетаева было
известно всей стране. Прежде всего, он был автором первой советской книги
по кибернетике — "Сигнал". Хитом околонаучных газетных дискуссий в "Лит-
газете", "Комсомолке" и других изданиях были знаменитые проблемы "Физи-
"Физики и лирики", "Нужна ли в космосе ветка сирени?" и т.п. Как известно, поэт
Б. Слуцкий, впечатленный могучим взлетом ядерной физики и космической
науки, задумчиво сказал:
"Что-то физики в почете,
Что-то лирики в загоне!
* Ранний вариант статьи опубликован: Ратнер В.А. Игорь Андреевич Полетаев // Очерки ис-
истории информатики в России / Ред. Д.А. Поспелов, Я.И. Фет. Новосибирск: НИЦ ОИГГМ СО
РАН, 1998. С. 386-391.
4. Игорь Андреевич Полетаев 43
Дело не в прямом расчете,
Дело в мировом законе!"
Любопытно, что как раз в этот период поэзия была в большом почете:
"Поэт в России
Больше, чем поэт..." —
говорил Е. Евтушенко. К тому же многие яркие "физики" в душе были боль-
большими поклонниками и знатоками "лирики". Барды собирали полные залы
вузов и научных институтов. Фактически речь шла о симпатиях молодежи. Рей-
Рейтинг естественных наук был исключительно высок. Все хотели быть физиками.
В этих условиях инженер И.А. Полетаев сказал свое "перпендикулярное"
слово. Он обострил проблему почти до крайности: если физика может все, то
надо ли тащить в космос ветку сирени, о чем хлопотали довольно бездарные
литераторы?
Главным оппонентом в этом был крупнейший писатель и публицист Илья
Эренбург. Он говорил, что надо брать ветку сирени, а иначе физики могут ос-
остаться бездуховными роботами. Верно-то верно, но рассчитано как раз на ро-
роботов. А физики, в том числе и особенно И.А., как раз и были очень чуткими
камертонами духовной жизни страны. Именно они зачитывались поэтами но-
новой волны, пели под гитару песни Окуджавы, Визбора, Галича, позже — Вы-
Высоцкого. Многие из них сами занимались гуманитарным творчеством. Из них
вышли многие будущие "шестидесятники".
Н.А. Ляпунова, дочь Алексея Андреевича Ляпунова, рассказывала, что в
это время была сделана организованная попытка изничтожения "физика" По-
Полетаева. На огромном собрании "лириков" во главе с И.Г. Эренбургом в одном
из московских клубов И.А. был подвергнут погромному разносу. Все могло бы
закончиться очень плохо, если бы его не спас А.А. Ляпунов. В своем выступле-
выступлении он напомнил Эренбургу его собственные образцы "лирики" военных лет,
где он призывал к мщению немецкому народу. У литератора после этого слу-
случился сердечный приступ, собрание пришлось закрыть.
В конце 50-х — начале 60-х годов, работая в московских военных НИИ,
И.А был очень активен не только в пропаганде и защите кибернетики, но и в
разработке ряда военно-кибернетических проектов и подготовке организации
первого института кибернетики. Вся эта активность закончилась неудачно: Ин-
Институт кибернетики создать в Москве не удалось (позже он возник в Киеве),
военное начальство не поддержало группу молодых полковников-кибернети-
полковников-кибернетиков и попросту уволило их из армии в 1961 г.
Переезжая на работу в Новосибирск, А.А. Ляпунов пригласил к себе своего
последователя, недавно демобилизованного военного инженера-физика
И.А Полетаева. Я увидел И.А. впервые в доме А.А. Ляпунова на Детском про-
проезде в Академгородке летом 1962 г. Был сильный дождь. Я пришел к А.А. пеш-
пешком, и поэтому снял по дороге сандалии, чтобы их не промочить. Короче гово-
говоря, в дом я вошел босиком. На звонок дверь открыл незнакомый худощавый
мужчина, седоватый, коротко стриженный, с острым изучающим взглядом. Это
был И.А. Он с удивлением посмотрел на мои мокрые пятки, но ничего не ска-
сказал.
44
I. Personalia
И.А. быстро вошел в кипящее со-
сообщество Академгородка. В это время
по инициативе ряда молодых и спо-
способных "физиков" здесь возник так
называемый Кофейно-Кибернетиче-
Кофейно-Кибернетический Клуб (ККК), где шли бурные
еженедельные дискуссии по всем во-
вопросам, запиваемые свежим кофе.
И.А. естественно влился в эту турбу-
турбулентную структуру и сразу стал зада-
задавать в ней тон. Если не ошибаюсь, он
даже был Президентом этого клуба.
Правда, по уставу Президент имел
только одно право — воздерживаться
при голосовании по любому вопросу.
Многие мои дальнейшие знакомства и
контакты в Академгородке возникли
именно в ККК. Я ездил туда из города,
а ночевал в общежитии СО АН у зна-
знакомых ребят.
Надо сказать, что И.А. был одним
из главных участников школ по теоре-
теоретической биологии у Н.В. Тимофее-
Тимофеева-Ресовского и Л .А. Блюменфельда
(Миассово на Урале, Можайское море
под Москвой), а потом — ежегодных
школ по математической теории эко-
экосистем (Пущино и др.). Везде он выступал с яркими докладами и "затравливал"
дискуссии. Полемист он был высочайшего разряда. Говорил быстро и очень
остро, часто парадоксально, возражал мгновенно. Поэтому его выступления
всегда пользовались большой популярностью. Уже гораздо позже И.А. неодно-
неоднократно приглашали для выступлений в Новосибирской ВПШ перед элитой бу-
будущих партработников. Он говорил о реальной экономике, об экологии, о со-
социальном устройстве, поругивал недостатки советской власти, но это всегда
сходило ему с рук, поскольку было интересно и происходило в закрытой ауди-
аудитории. Это было пикантно, как хороший анекдот.
В кибернетике И.А. взялся за центральные проблемы — системы управле-
управления (СУ) и информация, но подошел к ним по-своему. Дело в том, что матема-
математические работы А.А. Ляпунова по системам управления были столь широки и
всеобъемлющи, что под определение СУ можно было подвести все что угодно,
хоть камень на дороге. И.А. подошел к проблеме прагматически, как физик. Он
выделил класс физических систем, где описание в терминах СУ приводит к ре-
решению основных проблем их строения и функционирования. Он использовал
важное соображение, что акт управления, будучи, конечно, физическим актом,
отличается от обычных физических взаимодействий тем, что действие слабого
сигнала приводит к значительно более существенным физическим последстви-
И.А. Полетаев. Новосибирск, 1980-е.
(архив автора).
4. Игорь Андреевич Полетаев 45
ям. Именно тогда информационно-кибернетическое описание физических
систем адекватно и более продуктивно, чем материально-энергетическое. А.А.
был в целом согласен. Но И.А. шел своим путем гораздо дальше. В 1970 г. он
издал сборник работ своей лаборатории "Исследования по кибернетике" (под
редакцией, между прочим, А.А. Ляпунова), где с большой глубиной изложил
свой подход к пониманию информации, информационных процессов и систем
управления. Мне кажется, что, несмотря на кажущееся противоречие между
А.А и И.А., обе точки зрения весьма продуктивны и взаимно дополняли друг
друга. Именно так я и рассказываю о них нашим студентам-матбиологам.
Другим важным направлением работ И.А. стало развитие идеологии сис-
систем с лимитирующими факторами и разработка конкретных моделей таких сис-
систем. Идея лимитирования, идущая еще из XIX столетия от известного химика
Либиха, была развита до математических моделей именно И.А. и его ученика-
учениками. Фактически это способ приближенного описания сложных динамических
систем. Очень быстро такой подход стали использовать в физиологии и тео-
теории экосистем. До сих пор он активно применяется красноярскими биофизи-
биофизиками.
Мы тоже попытались разработать подход, сходный с лимитирующими фак-
факторами, для описания Молекулярно-генетических систем управления (МГСУ).
Мой сотрудник Р.Н. Чураев разработал дифференциальные и автоматные мо-
модели функционирования и взаимодействия генов, где использовалось понятие
порога. Самым крупным результатом на этом пути было построение пороговой
модели внутриклеточного развития фага X. Сейчас Р.Н. Чураев — директор Ин-
Института биологии УрО РАН в Уфе. Метод хорошо работал, особенно при моде-
моделировании на компьютерах.
Несколько позже, в последнее десятилетие XX в., мы поняли, что идеоло-
идеология лимитирующих факторов весьма продуктивна при анализе общих проблем
молекулярно-генетической организации и эволюции. МГСУ — сложнейшие
системы, содержащие 103-105 генов и фракций их биополимерных продуктов
на клетку. Описать формулами такую систему, даже если она полностью из-
известна, практически невозможно. Оказывается, во многих случаях это и не
нужно. В работах ряда теоретиков-биохимиков было показано, что в последо-
последовательных цепях метаболизма всегда есть ферменты с минимальными катали-
каталитическими активностями, которые лимитируют потоки конечных продуктов.
Другие гены и ферменты этих путей функционируют, но не лимитируют. Ясно,
что в данном случае отбор, оценивающий выход продукта, будет в основном
"фокусироваться" на лимитирующем звене, эволюция которого будет адаптив-
адаптивной. А нелимитирующие гены оказываются практически без селективной
оценки, т.е. эволюционируют нейтрально.
Это соображение было очень важным, потому что с конца 60-х годов в тео-
теории молекулярной эволюции резко обострилась борьба "нейтралистов"
(М. Кимура и др.) и "селекционистов" (большинство традиционных дарвини-
дарвинистов). Главный предмет спора был об удельной роли "нейтральной" эволюции.
Дарвинисты не могли смириться с идеей "нейтральности" молекулярной эво-
эволюции и считали разговоры о ней сильно преувеличенными. Из нашего подхо-
подхода, базирующегося на идее лимитирования, вытекало, что "нейтральная" эво-
46 I. Personalia
люция — это не экзотика, а совершенно реальное и длительное состояние боль-
большинства нелимитирующих генов.
Другой аспект проблемы был связан с так называемой "дилеммой
Холдейна". Английский популяционный генетик Дж.Б.С. Холдейн в 1950-е
годы показал, что геномы высших организмов не могут одновременно отби-
отбираться по многим генам, так как при этом селекционные потери не компенси-
компенсируются плодовитостью. Наш подход позволил сразу снять остроту проблемы.
Действительно, если в сложных системах отбор оценивает только лимитирую-
лимитирующие звенья, то число участвующих генов значительно меньше, а смена лими-
лимитирования позволяет последовательно включать в селективную оценку мно-
многие гены каждой системы. Такой режим эволюции мы с С.Н. Родиным назва-
назвали "эстафетным".
Наконец, я понял, что идея лимитирования может быть использована как
важнейший эвристический принцип. В сложных МГСУ в каждый момент вре-
времени имеются системные ограничения свойств, конструкции, которые можно
назвать лимитирующими факторами организации и эволюции. Преодоление этих
лимитов происходит при возникновении важнейших новых свойств — так
называемых эволюционных приспособлений широкого профиля, которые адап-
адаптивны почти всегда или в очень широком перечне условий. Таких свойств не
очень много, но они имеют фундаментальный характер. Например, мозг чело-
человека или пятипалая рука, диплоидность генома и т.д.
В этом случае эволюцию МГСУ следует описывать как последовательность
лимитирующих факторов организации и снимающих их эволюционных при-
приспособлений широкого профиля. Это как бы стержень "сценария" эволюции.
Все остальное надо описывать как второстепенные детали вокруг этого стерж-
стержня. Первые попытки пойти по пути такой аппроксимации были достаточно
продуктивны.
Однажды И.А. опубликовал популярную статью, если не ошибаюсь, в жур-
журнале "Техника — молодежи". В заголовке полагалось поместить фотографию
автора. Говорят, И.А. запросил редакцию, можно ли вместо этого прислать фо-
фотографию своих хромосом, поскольку они не менее индивидуальны, чем лицо
автора. Редакция неожиданно согласилась. Так и вышла эта статья с портретом
хромосом И.А. Полетаева. Разумеется, внутренне они очень индивидуальны,
но этого не видно на фотографии.
В 70-х годах пионерский дух в Академгородке пошел на убыль. У И.А. ока-
оказалось много недоброжелателей и даже врагов. Особенно это обострилось пос-
после смерти А.А. Ляпунова в 1973 г. Короче говоря, на лабораторию И.А. Поле-
Полетаева постоянно оказывалось административное и идеологическое давление.
Некоторые сотрудники уехали за границу. Настроение было плохое. Мы пыта-
пытались как-то помочь И.А. В частности, Д.К. Беляев направил одно или два пись-
письма в его поддержку, но результатов не было никаких.
В те годы И.А. много болел, что, однако, не помешало ему написать весьма
острый опус о "дуракологии" — науке о дураках. У меня нет этого текста, но,
возможно, он есть у его ближайших сотрудников. Хорошо бы его опубликовать.
Умер И.А. очень тихо. Конечно, ближайшие сотрудники и родные знали,
что он болен, помогали ему. Но за пределы узкого круга информация фактиче-
4. Игорь Андреевич Полетаев 47
ски не выходила. Как я узнал потом, он завещал не устраивать официальных
похорон с оркестрами и речами. Так и сделали. Через несколько дней устроили
поминки для более широкого круга, где был и я. Похоронен И.А. на кладбище
Чербузы в Академгородке. На могильном камне написано: Инженер И.А. По-
Полетаев.
В день 50-летия мы подарили И.А. адрес, где было написано следующее:
Желаем Вам,
Как и себе,
Жить в искушенье
И борьбе,
И порох свой
Назло другим,
На страх врагам
Держать сухим!
Желаем
Ни в какой тиши
Вам не найти
Покой души,
И в недовольстви»
Собой
Идти на бой,
Идти на бой!
Пусть будет вечен
Ваш союз
Друзей и жен,
Наук и муз!
И общность
Наших "лженаук"
Пусть будет крепче
Всех порук!
Кибернетический
Бедлам
Пусть будет
С "био" — пополам!
А клуб
С эмблемой ККК
Пусть будет
Здравствовать века!
Я думаю, что имя И.А. Полетаева не должно быть погребено в архивах.
Его острый и ясный ум, парадоксальная речь, полемический талант, его роль
в развитии кибернетики и математической биологии, его многочисленные та-
таланты впечатляют не только учеников и современников. О нем надо расска-
рассказывать новым поколениям математических биологов, чтобы они знали свои
корни.
48 I. Personalia
5. СМ. Гершензон: впереди событий
и в стороне от признания*
Некоторые специалисты считают, что религия — это не только учение о
Боге, но и образ жизни. Это очень правильное соображение. Точно так же и
наука — это не только сухие сведения о законах природы, но и образ жизни лю-
людей, делающих эту науку: их личности, взлеты и падения, догадки и озарения,
успех, мораль, научная этика и постоянное влияние общества, в котором живут
и работают ученые.
В 1992 г. ведущий американский журнал "Science" A992, 258, N 5079, 25)
посвятил свою рубрику "News & Comments" личности Сергея Михайловича
Гершензона, академика Национальной Академии наук Украины, Героя Социа-
Социалистического Труда, одного из патриархов советской генетики последних деся-
десятилетий. В 1996 г. СМ. Гершензону исполнилось 90 лет. В 1997 г. в Киеве на
английском языке вышла книга: S.M. Gershenson and Yu.N. Alexandrov
"Molecular Mechanisms of Mutagenicity of DNA and other natural and synthetic
polynucleotides". Все эти вехи и даты дают повод оглянуться на сделанное
СМ. Гершензоном в науке на фоне социальной действительности прошедших
десятилетий.
Прежде всего, приведем (с небольшими сокращениями) очерк из журнала
"Science".
Вся жизнь в борьбе за настоящую науку.
Чтобы понять, насколько трудно было заниматься наукой на Украине в по-
последние 70 лет, рассмотрим карьеру Сергея Гершензона... Западные Нобелевские
лауреаты подтверждают, что дважды он был близок к получению Нобелевской
премии.
Принимая посетителя в своей аккуратной киевской квартире, которую он за-
занимает со своими двумя дочерьми, Гершензон крепко пожимает руку, что удиви-
удивительно для его хрупкого сложения. Два года назад (в 1990 г. — В.Р.) он перестал
работать в лаборатории, но продолжает писать статьи и мемуары в своем каби-
кабинете, заставленном книгами, как и его спальня. Его библиотека содержит науч-
научные труды на немецком, русском и английском, а также работы по истории и ли-
литературе. В течение долгого времени на Украине, как и в остальной части Совет-
Советского Союза, обладание такими книгами, особенно западными работами по
генетике, давало повод для обвинений, вплоть до заключения.
В этом контексте трудно вспомнить, что Советский Союз, и особенно Москва,
были колыбелью современной генетики, где местными и приезжими западными иссле-
исследователями были сделаны многие выдающиеся открытия. Например, Гершензон, один
из первых генетиков по образованию (предыдущее поколение начинало как ботаники и
зоологи), работал в московской лаборатории американского генетика Г.Г. Меллера,
* Ранние варианты статьи с тем же заголовком опубликованы: Ратнер В.А. // Природа. 1998.
№ 8. С. 100-103; Ратнер В.А. // Информ. вести. ВОГиС. Новосибирск: ИЦиГ СО РАН, 1998. № 4.
С. 1-4.
5. СМ. Гершензон: впереди событий и в стороне от признания
49
V.
который в 1946 г. получил Нобелевскую
премию за открытие мутагенного дейст-
действия рентгеновских лучей.
Круг генетиков, с которыми Гер-
Гершензон работал, будучи студентом и
молодым исследователем, включал та-
таких наиболее известных московских ге-
генетиков, как Н.И. Вавилов, определив-
определивший мировые центры происхождения
культурных растений, и С.С. Четвери-
Четвериков, один из основателей популяционной
генетики. В сталинскую эпоху некото-
некоторые из этих людей погибли или попали в
заключение, в частности, за отказ от-
отречься от своей убежденности в менде-
левской генетике, другим, подобно Гер-
шензону, пришлось поменять область ис-
исследования.
Гершензон успешно работал в из-
известном Вавиловском институте (Ин-
(Институте генетики АН СССР. — В.Р.) в
Москве до 1937 г., когда он получил ла-
лабораторию в АН Украины в Киеве. До
1937 г., говорит Гершензон, он почти не
сталкивался с какой-либо дискримина-
дискриминацией. "Антисемитизм среди интелли-
интеллигенции тогда практически
отсутствовал", — вспоминает он. Но вскоре ситуация изменилась. Как раз когда
Гершензон подготовил свой первый крупный вклад в современную генетику — ста-
статью, демонстрирующую важность ДНК в генетике, между Германией и Совет-
Советским Союзом началась война. Гершензон и его семья избежали (оккупации. —
В.Р.) нацистов, которые в 1941 г. захватили Киев и уничтожили 34 000 евреев в
течение двух дней, 29 и 30 сентября, а затем вскоре и все остальное еврейское на-
население. До конца войны он прожил (в эвакуации. — В.Р.) на Урале, разрабатывая
репелленты против насекомых, в частности против мух — переносчиков тифа.
Статья Гершензона о ДНК, которая в конце концов была опубликована в
1948 г., показала, что введение ДНК из тимуса теленка в плодовую муху дрозофи-
дрозофилу вызывает существенное возрастание числа мутаций. В то время, когда он вы-
выполнял исследования, большинство биологов смеялись над его гипотезой, что ДНК
может нести генетическую информацию; тогда преобладало мнение, что гены по-
построены из белка. "Моя статья предсказывала многое из современной молекуляр-
молекулярной биологии", — задумчиво вспоминает Гершензон. Статья, которая вышла
только по-русски, была в основном не замечена западными учеными, которые поз-
позже и независимо повторили его результаты.
Публикация статьи Гершензона пришлась как раз на момент, когда Трофим
Лысенко довел до конца удушение советской генетики. "Я не смог даже получить
СМ. Гершензон. 1990-е. Рисунок из ста-
статьи в "Science".
50
I. Personalia
СМ. Гершензон. 1960-е (архив журнала "Природа").
свои оттиски ", — вспоминает Гершензон. Но худшее было еще впереди. В 1948 г.
Лысенко сделал свой известный доклад на сессии ВАСХНиЛв Москве и отлучил со-
советскую генетику от дарвиновских и менделевских идей. Гершензон стал одной из
главных мишеней лысенковцев, многие из которых были также антисемитами.
"Они четыре раза увольняли меня из советской Академии ", — вспоминает он, но
каждый раз его восстанавливали при помощи могущественных друзей. «Сначала
они уволили меня как "менделиста-морганиста ", а когда это не сработало, то
уволили как "космополита " (сталинское наименование евреев)».
«В последний раз, — вспоминает Гершензон, — враги также атаковали меня
как предателя за публикацию статьи на английском языке в 1945 г. (в американ-
американском журнале "Genetics "), тогда как советские люди-патриоты должны были ра-
работать для Родины. Единственное, что меня спасло, было письмо от Президента
АН СССР, просившего меня написать что-нибудь по-английски, чтобы он мог убе-
убедить американских коллег, что наука на Украине не погибла».
Не сумев уничтожить его напрямую, враги Гершензона попытались исклю-
исключить его из игры, перебросив на работу в другую область, которую они считали
тихой заводью: изучение вирусов насекомых. Оказалось, что они только помогли
5. СМ. Гершензон: впереди событий и в стороне от признания 51
ему, "Вирусы могут служить хорошей генетической моделью " — вспоминал Гер-
Гершензон, и его глаза светились удовлетворенно. К середине 60-х годов, работая с ви-
вирусом ядерного полиэдроза шелкопряда, он открыл активность любопытного фер-
фермента, который мог транскрибировать РНК в ДНК, т.е. в противоположном к
"нормальной "генетической транскрипции направлении. Но Гершензон не сумел вы-
выделить фермент, отвечающий за этот необычный процесс, поскольку не имел со-
соответствующих реагентов. "Если они дают Нобелевскую премию именно за нали-
наличие правильной идеи, — говорит Юрий Глеба, украинский биолог, в настоящее вре-
время работающий в США, — то ее должен был получить Гершензон ".
В то же самое время некоторые западные исследователи, включая Девида
Балтимора и Говарда Темина, выполнили сходные исследования на других вирусах.
За открытие активности и последующее выделение фермента, теперь называемо-
называемого "обратная транскриптаза ", Балтимор и Темин в 1975 г. получили Нобелевскую
премию. Единственным утешением для Гершензона было письмо Балтимора от
15 марта 1972 г., которое он бережно хранит в одной из своих многочисленных па-
папок. Балтимор извиняется, что не ссылался ранее на работу Гершензона: "Мое оп-
оправдание, — пишет Балтимор, — состоит только в том, что я ничего о ней не
знал ".
Распад Советского Союза в прошлом году A991 г. — В.Р.) может окончатель-
окончательно закрыть эту главу истории науки на Украине. Но, как обнаружили Гершензон и
его украинские коллеги, ученые на Западе все еще в основном не знают о его рабо-
работах.
(S.D.)
Да, грустная картина открывается взору! Талантливый исследователь дваж-
дважды был в ближайшей окрестности выдающихся открытий, и столько же раз об-
обстоятельства жизни — война и зигзаги политической ситуации — были барье-
барьером для "взятия высоты". Конечно, СМ. Гершензон не единственный, кто по-
попал в этот исторический капкан. Не говоря уже о репрессированных генетиках
(Н.И. Вавилов, С.С. Четвериков и др.), те, кто остались целы после эпохи ста-
сталинизма и лысенкоизма, еще долго ощущали приступы научной несправедли-
несправедливости. И.А. Рапопорт не получил Нобелевской премии за открытие химическо-
химического мутагенеза исключительно по политическим мотивам. Н.В. Тимофеев-Ре-
Тимофеев-Ресовский, великий генетик XX века, известный всему миру, лауреат престижной
Кимберовской премии и номинант на Нобелевскую премию, на Родине имел
просто мизерное официальное признание. Да и за рубежом: крупнейший гене-
генетик Герман Меллер, "розовый" по своим политическим убеждениям, долго не
мог найти постоянного профессорского места в США, и только получение Но-
Нобелевской премии прервало это состояние. Освальд Эвери, открывший генети-
генетическую роль ДНК в трансформации бактерий, так и умер, не дождавшись Но-
Нобелевской премии. А ведь был достоин!
Конечно, по итогу 90 лет жизни и работы СМ. Гершензон все же был при-
признан как советской наукой, так и властью. Он был избран академиком АН
УССР, основал в Киеве Институт молекулярной биологии и генетики АН
УССР и был его директором. Одним из последних указов М.С Горбачева вме-
вместе с пятью другими выдающимися генетиками он был награжден Звездой Ге-
Героя Социалистического Труда. Все это прекрасно! Но если бы все это было во-
52 I. Personalia
время! Да, война сломала жизнь каждому советскому человеку. Однако осталь-
остальные обстоятельства — послевоенные идеологические кампании, черной памяти
лысенковщина, медленная реабилитация генетики и др. — сделали свое дело.
В 1992 г. анонимный автор очерка (S.D.) застал СМ. Гершензона за подго-
подготовкой итоговой книги своих главных работ. В 1997 г. она вышла из печати, и
мы имеем возможность из первых рук получить ответы на все вопросы, взгля-
взглянуть на проблемы с позиций современной науки.
В чисто научном смысле СМ. Гершензон и сотр. в 1939—1941 гг. открыли
мутагенное действие чужеродной ДНК на гены дрозофилы. Действие оказалось
локус-специфическим. ДНК из тимуса теленка вызывала мутации некоторых
генов, контролирующих признаки крыла. В дальнейшем были успешно ис-
использованы ДНК различного происхождения, а также синтетические полинук-
леотиды.
Нестандартность ситуации состояла в том, что тогда по всеобщему мнению
гены считались белковыми макромолекулами, а тимонуклеиновая кислота
(ДНК) предполагалась довольно простой молекулярной структурой, играющей
третьестепенную роль в клеточном ядре. Результат Гершензона говорил о пря-
прямом участии ДНК в мутагенезе. Правда, эта роль могла быть разной. A) ДНК
могла быть самим мутагеном, воздействующим на структуру генов. Тогда непо-
непонятна локус-специфичность действия, так как другие химические мутагены
действуют повсеместно. B) ДНК могла быть материалом, идентичным мате-
материалу генов. Тогда мутагенез должен был состоять в рекомбинационном заме-
замещении или внедрении отдельных фрагментов, а локус-специфичность следова-
следовала из гомологических, взаимно специфических свойств локуса и фракции
ДНК. В этом случае ДНК должна бы быть материальным носителем генов.
Этот вывод был бы столь революционным, что его надо было обосновывать
очень веско, на молекулярном уровне. Тогда этого сделать не удалось. C) На-
Наконец, теперь ясно, что ДНК могла быть специфическим сигналом, индуци-
индуцирующим в геноме специфический мутационный ответ.
Фактически О. Эвери, открывший в 1944 г. ДНК-природу трансформирую-
трансформирующего агента у бактерий, стоял примерно перед такой же дилеммой и тоже не
мог сразу исключить все гипотезы, кроме B). Хотя работы Эвери стимулирова-
стимулировали биохимические исследования ДНК, все же до работ Уотсона и Крика A953)
и Херши и Чейз A952) генетическая роль ДНК была спорным моментом.
Именно поэтому О. Эвери, умерший в 1955 г., не дождался Нобелевской пре-
премии.
Как мы уже знаем, первый цикл работ СМ. Гершензона был прерван вой-
войной. Лишь в 1946-1947 гг. удалось восстановить эти исследования и весной
1948 г. — опубликовать сводку результатов. Но тут же последовал "разгон" ге-
генетики, и работы прервались еще на 10 лет. А когда открылась возможность их
возобновить, генетическую роль ДНК уже поздно было доказывать.
В то же время истинный молекулярный механизм мутагенеза при помощи
ДНК не ясен до сих пор. За это время появились многие новые возможности:
открыты мобильные генетические элементы (МГЭ); инсерционный мутагенез
при помощи МГЭ оказался главным источником мутаций у дрозофилы; открыт
"адресный" мутагенез мутагеном со специфическим полинуклеотидным фраг-
5. СМ. Гершензон: впереди событий и в стороне от признания 53
ментом, и т.д. Одна из новых возможностей состоит в том, что перемещения
МГЭ можно индуцировать внешними стрессовыми воздействиями через систе-
систему ответа на тепловой шок (ТШ). Но эта же система чувствительна и к появле-
появлению белков с дефектной конформацией. Возможно, инородная (и частично де-
дефектная) ДНК тоже может быть индуктором ТШ-системы и через нее — транс-
транспозиций МГЭ? Ясно, что здесь нужен цикл высококачественных ключевых
молекулярных экспериментов. По-моему, эту задачу должны решить ближай-
ближайшие ученики и соратники СМ. Гершензона.
В конце 50-х годов начался второй цикл работ СМ. Гершензона — иссле-
исследование вируса полиэдроза шелкопряда. Начался вынужденно, под флагом
борьбы с вирусным заболеванием тутового шелкопряда. Однако объект оказал-
оказался весьма благодатным, удобным для генетической работы. Так, введение ин-
инфекционной РНК вируса приводило к возникновению внутри клеток шелко-
шелкопряда полиэдрических включений, содержащих вирионы с ДНК-геномами.
Поскольку заражение фракцией инфекционной РНК приводило к возникнове-
возникновению ДНК-геномов вируса, встал вопрос о реальности процесса переноса гене-
генетической информации в направлении от РНК к ДНК, впоследствии названно-
названного обратной транскрипцией. Это соображение было высказано автором в явной
форме. Идея обратной транскрипции была очень революционной, поскольку
нарушала так называемую "центральную догму Крика" в молекулярной генети-
генетике. Однако для завершения доказательства следовало выделить фермент, кото-
который осуществляет этот процесс. По причине недостаточной оснащенности
СМ. Гершензону и сотр. этого сделать не удалось.
Зато это удалось через 10 лет хорошо оснащенным американским ученым
Д. Балтимору и Г. Темину. В этом их несомненный успех и заслуга. Они по
праву получили свою Нобелевскую премию. Однако почему они не знали о ра-
работах СМ. Гершензона? Или не искали? Известно, что многие американские
ученые не читают иностранных работ и считают наукой только то, что публику-
публикуется в американских журналах. Извинения о неосведомленности не снимают
проблемы. Так будьте же осведомлены, это Ваша проблема! Имеющий очи — да
видит! Если бы я написал в американский журнал статью и не сослался на су-
существующую американскую работу, то анонимный рецензент тут же меня по-
поправил бы. А в обратном направлении они не считают нужным это делать.
Заметим, что когда это выгодно, американцы проводят весьма тщательный
скрининг литературы. Когда в начале 60-х годов президент Дж. Кеннеди под-
подписал программу "Аполлон" для высадки на Луне, то НАСА провела тотальный
скрининг всех опубликованных в мире (на всех языках) работ по космонавтике.
И они отловили плохо оформленную брошюру, изданную в далеком Новоси-
Новосибирске провинциальным автором-самоучкой Ю.В. Кондратюком (псевдоним
А.И. Шаргея). Анализ показал, что он предложил самую оптимальную схему
полета к Луне. Именно идея Кондратюка легла в основу программы "Апол-
"Аполлон", а имя Кондратюка заняло прочное место в истории космонавтики и на
карте Луны. Так ведь могут, когда хотят!
Таким образом, дважды СМ. Гершензон находился в окрестности великих
открытий и дважды не встретил своевременного признания. "Nobel quality? —
Нобелевское качество?"— спрашивает анонимный автор очерка о нем. И сокру-
54 [. Personalia
шенно добавляет: "две ключевые работы Сергея Гершензона почти не известны на
Западе ". А следовало бы их знать, господа!
В заключение мне хотелось бы сказать, что и ранее в Советском Союзе, и
теперь в России, на Украине и в других странах СНГ мы в целом имели и име-
имеем, хотя и бедствующую, но великую науку! Именами наших крупнейших уче-
ученых можно гордиться. Пожелаем же тем из них, кто является нашими совре-
современниками, успехов и своевременного признания!
6. Дмитрий Константинович Беляев
Имя академика Дмитрия Константиновича Беляева прочно заняло свое ме-
место в истории отечественной генетики. Вехи его жизни известны. "Ровесник
Октября", сын священника из костромского села, он окончил в Москве бывшую
хвостовскую гимназию, а затем в 1938 г. — Ивановский сельскохозяйственный
институт. Война прервала начало его научной деятельности: 4 года он провел на
фронтах войны, окончив ее в чине майора и до конца дней сохранив верность
братству фронтовиков. После войны успел защитить кандидатскую диссертацию
по генетике пушных зверей и вскоре попал под всеобщий разгон генетики 1948 г.
Сумеречные годы лысенкоизма он пережил в Москве, будучи уволен с основной
работы и выезжая для экспериментов в эстонский зверосовхоз.
В 1957 г. Д.К. получил приглашение члена-корреспондента АН СССР
Н.П. Дубинина приехать в Новосибирск, во вновь создаваемый Институт цито-
цитологии и генетики СО АН СССР. Он поехал и остался здесь до конца своих
дней. Именно в Сибири он получил возможность реализовать некоторые свои
эволюционные идеи, которые вынашивал в предыдущие годы.
Центральная идея состояла в том, как происходит отбор коррелированных
признаков: один отбирается, а другой "тянется" за ним за счет корреляции.
Имелся в виду отбор животных по признакам поведения, неагрессивности по
отношению к человеку, за которым могут "потянуться" морфологические при-
признаки шерстного покрова, цикла размножения и др. Фактически эта проблема
замыкалась на другую, еще более фундаментальную: не был ли отбор по пове-
поведению основным механизмом доместикации (одомашнивания) животных. А
человек разве не одомашнен?
Вначале Д.К. заведовал отделом генетики животных ИЦиГ, потом стал за-
заместителем директора института, а с 1959 г., после драматического снятия пер-
первого директора — Н.П. Дубинина, стал и.о. директора института.
Момент был очень сложный. Известно, что после смерти И.В. Сталина его
фаворит Т.Д. Лысенко попал на несколько лет под справедливое отторжение,
биологическая наука вздохнула более свободно. Именно тогда, в 1957 г.
Н.П. Дубинин, по предложению академика М.А. Лаврентьева, создал Институт
цитологии и генетики СО АН СССР. Ехать в Сибирь собралась большая группа
генетиков старшего поколения, вступивших в науку до войны и переживших
разгром 1948 г. ИЦиГ казался убежищем от ограничений и репрессий. Реально
поехали Д.К. Беляев, Ю.Я. Керкис, З.С. Никоро, Н.А. Плохинский, И.Д. Ро-
Романов, П.К. Шкварников, А.Н. Лутков, Ю.П. Мирюта, Д.Ф. Петров, Е.П. Рад-
6. Дмитрий Константинович Беляев 55
жабли и др. Позже — В.Б. Енкен, В.В. Хвостова, Р.Л. Берг, Р.П. Мартынова,
Г.А. Стакан. Кроме того, потянулось новое поколение: Р.И. Салганик,
О.И. Майстренко, Г.Ф. Привалов, И.И. Кикнадзе, Н.Б. Христолюбова,
А.Д. Груздев, В.К. Шумный, позже — Л.И. Корочкин, Н.Н. Воронцов,
В.А. Драгавцев и др.
Однако еще в конце 50-х годов Н.С. Хрущева посетила бредовая идея дог-
догнать Америку по производству мяса и молока. Лысенко, прожженный полити-
политикан, оказался тут как тут. Он пообещал быстрый и подавляющий успех этой
идеи путем скрещивания всех коров страны с быками жирномолочной джер-
сейской породы и "укрепления наследственности кормлением"(!?). И попал в
точку. Карусель демагогии завертелась. Быстро он стал фаворитом Хрущева.
Америку не догнал, но начал второе наступление на генетику. Одной из первых
жертв произвола стал Н.П. Дубинин, который был снят из директоров вынуж-
вынужденным распоряжением М.А. Лаврентьева под прямым давлением Н.С. Хруще-
Хрущева прямо в самолете на пути из Китая после очередного столкновения с Мао
Цзедуном.
На Д.К. обрушилась огромная ответственность. Институт был обложен лы-
сенковцами со всех сторон. Ежегодно его "трепали" десятки комиссий различ-
различного уровня с единственным заданием — "на убой". Хрущев, посещая свое де-
детище — Академгородок, каждый раз спрашивал Лаврентьева: "а как там дела с
вашими менделистами-морганистами?" Лаврентьев отвечал уклончиво. А при
организации выставки достижений СО АН однажды сказал Беляеву: "а вас мы
отлучим от церкви". И поместил институт "на выселках", на другом этаже вы-
выставки, куда Хрущев заведомо не пошел. В этот период у института отобрали
фундамент будущего здания. Сейчас там Институт катализа СО РАН. Было
очень грустно.
Д.К. любил рассказывать историю о том, как М.А. Лаврентьев нейтрализо-
нейтрализовал самую главную "убойную" комиссию во главе с Президентом ВАСХНиЛ
ярым лысенковцем академиком Ольшанским. Комиссия долго копалась в ин-
институте, написала отрицательное заключение и явилась к Председателю СО АН
М.А. Лаврентьеву для доклада. Лаврентьев принял их в своем кабинете. Только
Ольшанский начал говорить, как раздался телефонный звонок. Лаврентьев
снял трубку и сказал: "Да, это я,... (и назвал имя куратора СО АН от отдела нау-
науки ЦК КПСС). Да, комиссия у меня. Какое у них мнение? Хорошее мнение.
Одобряют деятельность института". Комиссия была ошеломлена и обескураже-
обескуражена. Но делать было нечего. Против ЦК не попрешь! Так и остался наш институт
действующим гнездом менделизма-морганизма. Пикантность ситуации состоя-
состояла в том, что звонок был не из Москвы, а из соседней комнаты, где сидел зам.
Председателя СО АН академик С.А. Христианович. Они блестяще разыграли
этот цирк по всем правилам подковерного искусства.
В этот период Д.К. получил полную поддержку М.А. Лаврентьева. Инсти-
Институт был в состоянии упадка, его надо было ставить на ноги. Лаврентьев нашел
"козырной" ход. На очередных выборах в Академию в 1964 г. он поддержал
кандидатуру тогда еще кандидата наук Д.К. Беляева и добился его избрания в
члены-корреспонденты АН. Все это было сделано еще в хрущевские времена,
по Отделению общей биологии, где сидели Лысенко и его приспешники. Види-
56 1. Personalia
мо, они недооценили личность Д.К., сочли его второстепенной фигурой.
И просчитались. Через много лет в одном из институтских "капустников" мы
назвали этот момент следующими словами: "Д.К. вышел на околоземную ор-
орбиту!".
После этого институт пошел в гору, особенно после отставки Н.С. Хрущева
и реабилитации генетики в 1964 г. Генетики старшего поколения В.В. Хвостова,
П.К. Шкварников, Ю.П. Мирюта, А.Н. Лутков, Ю.Я. Керкис, Р.Л. Берг,
Р.П. Мартынова и др. получили докторские степени. Авторитет Д.К. вырос и
окреп. Он стал одним из ведущих генетиков страны. Каждый год Д.К. стал со-
собирать отчетные сессии института и приглашать на них многих выдающихся
генетиков: Б.Л. Астаурова, Н.В. Тимофеева-Ресовского, П.Ф. Рокицкого,
Л.В. Крушинского, А.А. Прокофьеву-Бельговскую, А.А. Нейфаха, Ф.Х. Бахтее- I
ва, Ю.М. Оленова, а также из институтов СО АН — А.А. Ляпунова, В.В. Вое-
Воеводского, Д.Г. Кнорре, Ф.Э. Реймерса и др.
Расскажу о некоторых личных впечатлениях того периода. Я работаю в ин-
институте с декабря 1960 г. Вначале я попал к Д.К. с несколькими весьма наивны-
наивными, хотя и непростыми вопросами об эволюции. Привели меня к нему Нинель
Христолюбова и Алеша Груздев. Д.К. послушал, послушал, а потом сказал:
"Давайте лучше я дам Вам маленькую "задачку, сделаете — тогда поговорим".
Задача, действительно, была маленькой: два гена, два коррелированных при-
признака. Отбираем один признак, за ним должен "тянуться" другой. Однако у фи-
физиков своя "придурь". Я тут же обобщил задачу на произвольное число генов и
признаков, наворотил огромные матрицы корреляций, долго преодолевал со-
собою же созданные математические трудности. Наконец, получил что-то, похо-
похожее на результат. Пошел к Д.К. Ему понравилось. Кстати, после этого он взял
меня в институт. Доложил на семинаре — тоже ничего. В Москве показал
А.А. Ляпунову и академику И.И. Шмальгаузену — тоже в общих чертах одобри-
одобрили. И.И. предложил напечатать работу в Докладах АН СССР, что мы и сделали.
Однако летом 1961 г. на Миассовской школе Н.В. Тимофеев-Ресовский и его
сотрудники не оставили от этой работы камня на камне. Они-то были специа-
специалистами. Оказалось, что я ничего не смыслю ни в генетике, ни в корреляциях,
ни в отборе. Биологам, конечно, трудно было разобраться в формулах, они ве-
верили на слово. А я их подвел. Обобщатель! Момент был драматический.
Все это заставило меня "сесть за парту". Последующие 2-3 года я только
изучал генетику и другие смежные науки. Правда, в 1961 г. нас подхватил ветер
открытия генетического кода. Я тоже туда погрузился и застрял надолго. Но об
этом позже.
Все 60-е годы я работал в лаборатории Д.К. События были бурные, и он
меня просто не трогал. Это и было условием моего выживания. Иначе бы выгна-
выгнали, наверное. За эти годы я погрузился в молекулярную генетику и кибернетику,
подготовил диссертацию и книгу "Генетические системы управления". Здесь
сильное влияние на меня оказали вьщающиеся ученые А.А. Ляпунов и Н.В. Ти-
Тимофеев-Ресовский. А.А. был оппонентом, а Н.В. прислал внешний отзыв.
Все эти годы мы общались с Д.К. довольно близко. Летом 1962 г. я получил
свою первую квартиру в Академгородке. 1 августа, в день моего рожденья, вдруг
раздался стук в дверь. Открываю — вся лаборатория во главе с Д.К. и с полны-
6. Дмитрий Константинович Беляев 57
ми руками вин и яств! Конечно, погужевались мы на славу! После этого не-
несколько лет жили с Д.К. в одном подъезде, наши сыновья учились в одном
классе. В 1967 г., когда умер мой отец, Д.К. очень помог мне с поездкой на по-
похороны в Хабаровск. А затем он организовал приглашение в Клинический от-
отдел СО АН моей матери, профессора В.М. Кантер, заведовать лабораторией
неврологии. Она проработала в Клиническом отделе около 10 лет.
В 1967 г. Д.К. перенес тяжелую операцию, ему удалили 2/3 желудка. Лежал
в больнице, потом дома. Спускаюсь однажды по лестнице — внизу на лавочке
сидит Д.К., рядом Светлана Владимировна. Д.К. совершенно худой, серый,
смотрит жалобно. Говорит мне: "Ну что, хорош я?" Я отвечаю в унисон — "Хо-
"Хорош, хорош. Главный-то страх позади". "Да, — говорит Д. К., — надо выбирать-
выбираться". И выбрался. Ему запретили пить вино, но водку и коньяк вроде бы не ис-
исключили из рациона. После этого каждый раз, как кто-нибудь заходил к нему
домой, раздавался клич: "Светочка! Принеси нам, дорогая, пару рюмочек конь-
коньячку!" Светлана Владимировна сердилась, но приносила рюмки. После этого
начинался деловой разговор. И часто весьма эффективный.
В 1968 г. Д.К. ездил на XII Международный генетический конгресс в Токио
в составе советской делегации. С его подачи я тоже был там в группе научных
туристов. Приехали теплоходом из Находки. Попали прямо на вступительный
прием, где всех приветствовал тогдашний наследный принц, а ныне император
Японии Акихито, биолог по образованию. На банкете Д.К. познакомил меня с
некоторыми выдающимися личностями — Ф.Г. Добржанским, Л.Л. Кавал-
ли-Сфорца, Дж. Кроу и др. Впечатление было огромное и свежее.
На следующий день Д.К. сказал нам: "Пойдемте, покажу вам ночную
жизнь Токио". (Он прибыл на день раньше самолетом и уже успел все разве-
разведать.) Пошли наши сотрудники: И.И. Кикнадзе, Л.И. Корочкин, Ю.Я. Керкис,
Г.М. Роничевская, я и др. Шли клином во главе с Д.К. Пройдя по Гиндзе, Д.К.
свернул в боковую улочку. По фонарям можно было понять, что это район бор-
борделей. И тут от одной из загадочных дверей отделилась личность, о которой
можно было сказать только одно — плевка не стоит! Нагло ухмыляясь, он ска-
сказал: "Hello! — Sir! — Girls! — Drink!" Д.К. был выше его на две головы. Он под-
поднял указательный палец, покачал им влево-вправо и ответил: "No-no!". На том
и разошлись, соблюдая невинность.
Вспоминаю еще один эпизод из начала 60-х годов. Институт еще был в
опале. Д.К. пригласил посетить институт своего старого знакомого, лысенков-
ца, профессора М. из Омска. М. пришел в лабораторию, Д.К. долго ему расска-
рассказывал азы генетики и эволюции, но гость отвечал однообразно: "Это ваша ин-
интерпретация. На самом деле генов нет". Д.К. миролюбиво говорил: "Но и это
тоже Ваша интерпретация!" Наконец, у Д.К. лопнуло терпение. Он повел гостя
в зал, где ему показали популярный тогда фильм поляка Байера "Митоз". Вы
бы видели реакцию лысенковца М.! У него отпала челюсть. Он был полностью
сражен и поражен. После этого он ни о чем не спорил, собрал монатки и был
таков. Вот тебе и интерпретация!
Подводя итог этому периоду, можно сказать, что главная заслуга Д.К. со-
состояла в том, что он спас наш институт. Кроме того, он добился строительства
нового здания к 1964 г. Конечно, были различные сложности и разногласия, из
58 [. Personalia
института по разным причинам ушли Д.Ф. Петров, И.Д. Романов, Н.А. Пло-
хинский, П.К. Шкварников, Ю.П. Мирюта, Р.Л. Берг и др. Появились новые
сотрудники — Л.И. Корочкин, Н.Н. Воронцов, В.А. Драгавцев и др. Но в це-
целом Д.К. проявил неоспоримые качества лидера. Лидером и остался. Это при-
признавали и друзья, и противники.
Быстрота становления Д.К. как лидера, по-моему, связана с некоторыми
важными особенностями ситуации. Прежде всего, конечно, он был лидер по
натуре. Но главное, он был младшим среди старших генетиков. Старшее поко-
поколение генетиков, ученики Н.К. Кольцова, С.С. Четверикова, А.С. Серебров-
ского, Н.И. Вавилова и др., к этому времени были обескровлены и разобщены.
Только в конце 50-х годов возникли отдельные лаборатории, где они могли
продолжить исследования. Многие были деморализованы 10-летним отрывом
от науки. Так, Ю.Я. Керкис прибыл из таджикского совхоза, где работал дирек-
директором; П.К. Шкварников был председателем колхоза в Крыму, а З.С. Нико-
ро — баянистом Клуба моряков в Измаиле. Кроме того, Д.К. был младшим бра-
братом Николая Беляева — талантливого генетика, ученика Н.К. Кольцова, друга
и соратника многих генетиков старшего поколения. Н.К. Беляев погиб в 1938 г.
в сталинских лагерях. Друзья брата стали друзьями Д.К. Его безоговорочно
поддержали многие московские и ленинградские генетики. Особенно близок
ему был Б.Л. Астауров.
В 1972 г. Д.К. был избран академиком. В институтском "капустном"
фольклоре это явление отмечено как "выход Д.К. на околосолнечную орбиту".
Довольно быстро он стал одной из ведущих фигур в Сибирском отделении и
Отделении общей биологии АН СССР. В СО АН он стал заместителем Предсе-
Председателя Президиума по Академгородку. В эти годы он многое сделал для разви-
развития биологии в Сибири и генетики в стране. Институт цитологии и генетики
СО АН СССР стал крупнейшим генетическим институтом страны.
Но главным его делом того периода стало проведение XIV Международно-
Международного генетического конгресса в Москве в 1978 г. Во всяком случае, при этом реа-
реализовались все лучшие черты Д.К. как лидера. Будучи председателем Проблем-
Проблемного совета по генетике и селекции Академии наук, он был назначен Генераль-
Генеральным секретарем Конгресса. С.Г. Инге-Вечтомов, который тоже принимал
непосредственное участие в организации Конгресса, рассказывал, что первона-
первоначально дело стояло, академики выясняли отношения и бездействовали. Когда
приехал Д.К., все закрутилось. Прежде всего, он двинул в действие свой инсти-
институт. Назначил кураторов, привез в Москву человек 30 сотрудников, которые
безвыездно обеспечивали все дела. Несмотря на политический бойкот со сто-
стороны некоторых западных генетиков, конгресс прошел очень успешно. Д.К.
был избран Президентом Международной Генетической Федерации и находил-
находился на этом почетном посту 5 лет.
В 1983 г. на очередном Международном генетическом конгрессе в Дели он
открывал первое пленарное заседание, потом общался с Индирой Ганди и дру-
другими лидерами Индии. И надо сказать, выглядел вполне адекватно уровню со-
событий.
В 1982 г. Сибирское отделение праздновало свое 25-летие. Несмотря на
разногласия, Д.К. пригласил на праздник основателя института академика
6. Дмитрий Константинович Беляев
59
Д.К. Беляев. Новосибирск, 1970-е (архив СВ. Аргутинской).
60 I. Personalia
Н.П. Дубинина. Дубинин приехал, но держался очень отчужденно. В институ-
институте — своем детище — он был только в кабинете директора. Со старыми сотруд-
сотрудниками встречаться не стал. И ни о чем не расспрашивал. Своего снятия он,
видимо, так и не простил.
Институт развивался. Защитилось новое поколение докторов наук:
Р.И. Салганик, И.И. Кикнадзе, Н.Н. Воронцов, Н.Б. Христолюбова,
В.К. Шумный, Н.Д. Тарасенко, Л.Н. Трут, В.И. Евсиков, Г.Ф. Привалов,
К.К. Сидорова, я и др. Возникли новые лаборатории, направления, идеи. От-
Отмечу то, что мне ближе.
На разных этапах моей деятельности по математической генетике Д.К.
поддерживал наши начинания. В 1968 г. он способствовал созданию на своей
кафедре в НГУ специализации по математической биологии. Затем оставил в
институте почти всех первых выпускников, которые составили мою группу.
Правда, периодически он вызывал меня и спрашивал: "Слушай Вадим! Когда
же мы с тобой поработаем?" Я отвечал: "Дмитрий Константинович, вызовите
меня в любой момент и давайте поговорим конкретно о любой задаче".
"Да-да, — говорил Д.К., — непременно". Проходило полгода, год. Так все и
стояло на месте. Ему было некогда. Заедали другие неотложные проблемы:
Черга, гибберелины и др.
Году в 1972-м Д.К. купил для института по случаю выдающуюся вычисли-
вычислительную машину своего времени — "Наири". Это был ужасающий огромный
арифмометр, но мы очень ему радовались. Ко дню 60-летия Д.К. A977 г.) Воло-
Володя Куличков, который числился инженером-программистом на машине, сотво-
сотворил его печатный портрет в состоянии задумчивости. Юбилей мы отмечали на
пасеке Экспериментального хозяйства СО АН. Там и поднесли портрет прооб-
прообразу. Но Д.К. он, по-моему, не понравился.
В 1976 г. я стал бомбардировать Д.К. записками о необходимости создать в
Институте компьютерный комплекс и завести банки данных по молекулярной
биологии и генетике. Первая реакция его была стандартной. Он положил пред-
предложение под сукно. Второй раз начал расспрашивать, потом интересоваться, но
не предпринял ничего. Тогда я поехал в Москву к столпам Академии А.А. Баеву
и др. за поддержкой этой идеи. Академик А.А. Баев, в дальнейшем возглавив-
возглавивший программу "Геном человека", очень благожелательно меня выслушал и
сказал: "Что касается Ваших математических моделей, то занимайтесь ими
сколько угодно. А если нам понадобятся компьютерные услуги, мы купим их за
рубежом". На том все и остановилось.
Однако в начале 80-х годов мы неожиданно обнаружили, что руководство
Академии уже поняло, куда дует ветер, и решило все-таки создать такие банки,
но в Москве, на базе Института молекулярной генетики АН СССР под руково-
руководством А.А. Александрова, сына Президента АН. Я сказал об этом Д.К. Вот
тут-то все и завертелось, он начал действовать. Он взял в нашу лабораторию
группу молодых теоретиков и программистов, поддержал закупку советских
мини-ЭВМ СМ-4, организовал Информационно-вычислительный комплекс,
поддержал организацию летних школ по моделированию и банкам данных. В
закупке очень дефицитных тогда мини-ЭВМ огромную помощь нам оказал
мой старый друг Эдуард Ильич Елинер — начальник ВЦ Института ядерной
6. Дмитрий Константинович Беляев
61
физики. С этого момента все направление стало бурно развиваться и в дальней-
дальнейшем превратилось в доминирующую часть современной отечественной биоин-
биоинформатики.
Вот некоторые эпизоды из институтской жизни 70-х годов. Частым собы-
событием на отчетных сессиях института, лекциях и докладах была острая полемика
Зои Сафроньевны Никоро, заведующей лабораторией генетики популяций, где
я тогда работал, с Д.К. З.С. была человеком, острым на язык, несомненной ум-
умницей. Она всегда стремилась обострить обсуждение, внести некоторую пара-
парадоксальность. Вот эпизод. Отчитывается лаборатория директора. Д.К. делает
часовой доклад. Сначала разогревается, потом воспаряет в небеса, говорит
очень эмоционально. Аудитория внимательно слушает. Только доклад окон-
окончен, в зале взлетает одинокая рука. Встает З.С. Никоро, натягивает на щуплые
плечи серую кофту и начинает резким голосом задавать каверзные вопросики и
говорить нечто перпендикулярное. Д.К. от неожиданности даже немного расте-
растерян, отшучивается, не сдается, но З.С, как орлица, гнет свое. Оканчивается все
ничем. Каждый при своем.
Однако Д.К. был опытным полемистом и разбирался в людских характерах.
Однажды он одним движением полностью обезоружил З.С. Был какой-то юби-
юбилей или праздник, возможно, день 75-летия З.С. В директорском кабинете
были накрыты столы дарами докторского стола заказов. Все ели, пили, закусы-
З.С. Никоро и Д.К. Беляев. Новосибирск, конец 1970-х (архив СВ. Аргутинской).
62 I. Personalia
вали. Вдруг Д.К. сказал: "Зоя Сафроньевна, голубушка! Давайте с Вами сфото-
сфотографируемся на память". И нежно обнял ее за плечи. "Вася, сними нас, пожа-
пожалуйста", — продолжил Д.К., и В.А. Прасолов запечатлел эту улыбающуюся
пару. После этого говорить о какой-то конфронтации было уже невозможно.
Документ обязывал!
В последние годы Д.К. подружился и сошелся с известным философом
академиком И.Т. Фроловым. Фролов был одним из первых философов, кто в
свое время поддержал генетику и способствовал ее реабилитации. Однажды
они совместно выступали по Центральному телевидению в передаче "Очевид-
"Очевидное — невероятное". Говорили о биосоциальных явлениях — поведении живот-
животных, одомашнивании, эволюции. В заключение Д.К. неожиданно пошутил:
"А что если взять остров в Тихом океане, населить его высшими приматами,
вести отбор по поведению, — получится ли новый вид человека разумного?"
Заманчиво! Фантастично! Как поется в одной хорошей песне, "Нелепо! Смеш-
Смешно, безрассудно, безумно! Волшебно!" И сколько это лет надо отбирать?
100 000? Или миллион? Тем не менее академики размялись вдохновляющей
шуткой, а публике было очень интересно. А вдруг?!
Где-то уже в 80-е годы я однажды спросил Д.К., зачем он взял меня на ра-
работу осенью 60-го года, когда институт дышал на ладан. Я ведь был профан,
про генетику вообще не слышал, не понимал, что все может закончиться кра-
крахом. А прямой надобности не было. Он ответил: "Хотел разрядить обстановку в
институте, влить свежую кровь! Я тогда взял вас троих: тебя, Виктора Колпако-
ва и Викторию Тряско. Вы с Виктором прижились, а Виктория давно на
пенсии". Он забыл, что в том же году взял в институт еще Любу Васильеву, мою
будущую жену. Правда, брал для тяжелой экспедиционной работы, старшим
лаборантом. А она потом неожиданно раскрыла все свои таланты: и научные, и
педагогические, и общественные, и личностные. В последующие годы Д.К. че-
честно это признавал.
И еще. Тогда, в 1962 г., в день моего рожденья он спросил: "А сколько
тебе лет?" Я ответил — 30. "Какой хороший возраст, — сказал Д.К., — в этом
возрасте я..." И дальше шел рассказ, что делал Д.К. в 30 лет.
В 1972 г., в день моего 40-летия, наша лаборатория собралась в своем поме-
помещении на 2-м этаже института (к. 225). Внезапно открылась дверь, вошел Д.К.
"Что отмечаем?" — спросил он сразу. Я объяснил. "А сколько тебе стукну-
стукнуло?" — спросил Д.К. 0 лет" . "Какой хороший возраст, — сказал Д.К., — я в
этом возрасте..." (далее по тексту).
В 1982 г. мне было уже 50. Опять дело было в лаборатории. Вошел Д.К. и
спросил: "Что отмечаете? А сколько тебе?" — спросил следом. 0". "Какой хо-
хороший возраст, — сказал Д.К., — я в этом возрасте..."
В 1992 г. его уже не было.
В 1984 г., будучи в Москве, Д.К. попал в автомобильную аварию. Ехавший с
ним А.О. Рувинский довез его до больницы. Отлежавшись, Д.К. вышел на ра-
работу, но полностью здоровым уже больше не был никогда. Однажды перед на-
началом заседания Совета он проходил мимо сцены нашего зала, вдруг покачнул-
покачнулся и схватился рукой за стол. Я стоял рядом, тут же подошел, говорю: "Дмитрий
6. Дмитрий Константинович Беляев 63
Константинович, Вам плохо?" Он взглянул мне в глаза с очень близкого рас-
расстояния и тихо сказал: "А что ты видел? Ты ничего не видел! Понял?" Я понял.
Последние два года жизни Д.К. сильно болел, часто сидел дома. Летом
1984 г. у нас открывалась 1-я Всесоюзная школа по моделированию и банкам
данных. Д.К. лежал в больнице. Вдруг машина привозит его на открытие шко-
школы. Он идет в президиум, одет в летний чесучовый костюм, грузный, высокий,
произносит приветственную речь, где очень нас хвалит. Никто и не подумает,
что он болен. Академик в зените славы! А он очень болен.
Последний раз я был у Д.К. дома осенью 1985 г. Он уже очень сдал, был ху-
худой, даже какой-то заостренный. А я пришел показать ему наши новые данные
по отклику мобильных элементов на отбор и тепловой стресс. Слушал он рассе-
рассеянно, скорее, думал о своем. А ведь данные-то были как раз ему в унисон. Не
знаю, как гормональные механизмы, а поведение мобильных элементов при
стрессе очень неплохо согласуется с представлениями Д.К. о дестабилизирую-
дестабилизирующем отборе. Потом он сказал: "К сожалению, не смогу предложить тебе рюмку
коньяку — Светочка не разрешает!" Потом добавил: "Ну что, я хорош?" Я отве-
ответил в тон: "Хорош-то, хорош! Но я Вас и не таким видел!" "Да, — сказал
Д.К., — а я помню, как ты шел за мной от института, когда мне было плохо. До
самого дома". Действительно, еще в начале 60-х годов, когда Д.К. сильно бо-
болел, однажды его прихватило на работе. Он пошел домой пешком, а от сопро-
сопровождения отказался. Я на всякий случай шел за ним в отдалении, чтобы под-
подстраховать, если что. Так он, оказывается, заметил и запомнил этот случай на
десятилетия. И всегда, видимо, считал меня поэтому своим, невзирая на от-
отдельные разногласия и разночтения.
Умер он внезапно 14 ноября 1985 г. Буквально за час до этого от него ушел
Рустем Чураев, наш матбиолог. Вышел в дверь, а через час Д.К. не стало.
Хоронил его весь Академгородок. Будучи по-прежнему майором запаса,
Д.К. давно соответствовал генеральским погонам. Наше Высшее военно-поли-
военно-политическое училище было поднято в почетный караул. Курсанты с винтовками
стояли вдоль всего пути до Чербузов (кладбища Академгородка). День был
очень сумрачный, снег с дождем. Похоронен Д.К справа от входа на мемо-
мемориальной части кладбища. Его могила фактически возглавляет череду других
могил, где покоятся члены первого Ученого Совета нашего института —
В.В. Хвостова, Ю.Я. Керкис, В.Б. Енкен, З.С. Никоро, Н.Б. Христолюбова и
многие другие. Их поколение ушло почти полностью. Почти! Но в Киеве живет
П.К. Шкварников, под Парижем — Р.Л. Берг, а в далекой Северной Кароли-
Каролине — Р.И. Салганик.
В 1997 г. в Новосибирске состоялась Международная конференция "Со-
"Современные концепции эволюционной генетики", посвященная 80-летию
Д.К. Беляева. Приехали бывшие сотрудники института, друзья из-за рубежа и
многие другие. Несмотря на "эволюционное" наименование, конференция
включила в себя все большие и малые направления, возникшие в связи с инте-
интересами и именем Д.К. Оказалось, что этот круг очень широк. Любимые экспе-
экспериментальные объекты Д.К. — норка, лисица, другие пушные звери, собачьи и
др. — с его легкой руки вошли в широкий научный обиход, изучаются в самых
разных аспектах. Огромен интерес исследователей к эволюции и стрессу. Имя
64
I. Personalia
Д.К. объединило научные школы разных городов страны, разных стран. С ним
связана вершина старшего поколения генетиков, молодость нашего поколения.
На фоне общих научных затруднений в нашей стране неожиданно выяснилось,
что институт, которому Д.К. отдал лучшие годы своей жизни и с которым дос-
достиг вершин, не угас, не разъехался по другим континентам, а живет и развива-
развивается, продолжая начатое им дело. В нем уже шумят другие поколения. Главное,
чтобы ниточка научной преемственности не прерывалась!
7. Памяти профессора Francesco M. Scudo*
В октябре 1998 г. умер Франко Скудо, наш старый друг и товарищ. Мы по-
познакомились ровно 30 лет назад на 12-м Международном генетическом кон-
конгрессе в Токио. Франко был небольшого роста, черноволосый, с яркими серы-
серыми глазами, очень подвижный и холостой. Каждый раз, когда я на него наты-
натыкался, он был в компании двух прекрасных молодых итальянок. Активность и
жизнерадостность просто сочились из его глаз.
К этому времени он уже стал известным математическим генетиком в Па-
вии (Италия), где сотрудничал с Луиджи Кавалли-Сфорца. Вслед за Кавалли он
переехал в США и пробыл там около 15 лет. В Калифорнии сотрудничал со
многими выдающимися математическими генетиками — Л. Кавалли-Сфорца,
С. Карлиным, М. Фельдманом и др. Там же нашел свою жену, Катарину.
Мы переписывались несколько лет до момента, когда Франко решил вер-
вернуться в Италию. Он так и написал мне: "Устав от академической жизни в Аме-1
рике, я решил вернуться в Павию". Он получил лабораторию популяционной
генетики в новом Институте биохимической и эволюционной генетики и оста-
оставался там до последних дней. Да, не все, не все остаются в Америке!
Наши контакты неожиданно оживились после 1981 г., когда мы встрети-
встретились на Конференции по эволюции в Праге (Чехословакия). Многие помнят
эти конференции, которые собирал чешский энтомолог В.В. Новак. Мы очень
хорошо поговорили с Франко, и он пригласил меня посетить Павию. Ежегодно
до 1987 г. я пытался это сделать, и ежегодно мои документы самым непостижи-
непостижимым образом где-то незримо исчезали, не доходя до визы в Италию.
Спасибо перестройке! В 1987 г. мои документы неожиданно дошли до адре-
адресата, я впервые приехал в Италию и был гостем Франко Скудо в Павии. Он жил
в семейном доме в небольшой (но очень хорошей) деревушке Торраца Косте
под г. Вогера, недалеко от Павии. Катарина Скудо, его жена, работала ланд-
ландшафтным дизайнером. Подрастали две дочери — Петра и Саша.
Последний раз мы с Л.А. Васильевой были в Италии в 1991 г. Франко орга-
организовал нам очень интересную месячную поездку: Павия — Милан — Боло-
Болонья — Бари — Камерино — Рим. В каждом городе мы посещали генетические
кафедры и лаборатории и выступали с докладами о наших работах. Больше не-
недели жили у Франко, гуляли в саду и обсуждали различные эволюционно-гене-
тические проблемы. Катя Скудо оказалась очень милой женщиной, эмигрант-
* Впервые некролог опубликован: Ратнер В.А. // Информ. вестн. ВОГиС. Новосибирск:
ИЦиГ СО РАН, 1998. № 7. С. 12-13.
7. Памяти профессора F.M. Scudo
65
/-«¦*
Ф.М. Скудо и В.А. Ратнер. Италия, Торраце-Коста, дом Скудо. Конец 1980-х
(архив автора).
66 I. Personalia
кой из Чехословакии. В школе она изучала русский язык, так что разговаривать
было довольно просто. Кроме того, с ней нас объединяла пионерско-комсо-
мольская юность.
Франко использовал наше пребывание на полную катушку. Он пригласил
на эту "наживку" своих приятелей с семьями, а Л.А. попросил приготовить на-
настоящий русский борщ. Несмотря на особенности итальянских ингредиентов, I
борщ получился отменный. После этого он потребовал, чтобы мы спели
что-нибудь специфически русское. Мы спели несколько русских романсов и
имели огромный успех. Франко нас зауважал и утром не позволил Л.А. вымыть
чашки после завтрака. Расстались мы полными друзьями.
Оглядываясь на последние 30 лет, я ясно вижу, что Франко Скудо сыграл
особую роль в наших контактах с западными генетиками. В первую же мою по-
поездку в Италию он повез меня в Милан, в только что возникшую там группу
биоинформатики, где познакомил с несколькими молодыми биохимиками и
программистами, которые не знали толком, как начинать это дело. Я рассказал,
чем занимается наш Теоретический отдел, и предложил им научное сотрудни-
сотрудничество. Они с радостью согласились. Затем в Милан поехали Н.А. Колчанов и
другие наши матбиологи. Через Милан прошли В.В. Соловьев, И.Н. Шиндя-
лов, И.Б. Рогозин, В.Б. Стрелец, М.П. Пономаренко, А.Э. Кель, И. Ищенко и
многие другие. Все они побывали в гостеприимном доме Франко Скудо и ощу-
ощутили его интерес и поддержку.
В 1986 г. к Франко заехал Сэм Карлин, выдающийся американский мате-
математик и математический генетик. Он увидел у него на полке нашу книгу "Про-
"Проблемы теории молекулярной эволюции", заинтересовался и попросил передать
мне, чтобы я прислал ему копию. Через год я познакомился с Сэмом в США на
Международной конференции по количественной генетике. А в 1990 г. я про-
провел месяц в Стенфорде (Калифорния) в качестве гостя М. Фельдмана и С. Кар-
лина, затем проехал по восьми городам Америки. Книга же наша с подачи
Ф. Скудо, С. Карлина и Р. Левонтина вышла в 1996 г. в Германии в издательст-
издательстве "Springer-Verlag" на английском языке.
Дважды Ф. Скудо бывал в СССР. Первый раз он приехал на Международ-
Международный симпозиум по философии и биологии в Москву, в Институт философии
нН СССР в 1988 г. Он был шокирован Москвой. Все серо и запущено, по ули-
улицам ветер нес пыль и обрывки газет. В магазинах пусто. Я сопровождал Франко
по Москве и не знал, куда спрятаться от стыда. В ресторане ему принесли со-
совершенно нераскусываемый антрекот, которьш-QH так и не сумел разрезать ту-
тупым ножом. А на просьбу принести сыр официант4ответил, что сыра в Москве
нет — Латвия перестала поставлять! Это было ужасно, особенно на фоне того,
что в Италии — 400 сортов сыра. Франко ругался. Самое сильное ругательство
звучало так: "Uno cretino!"
Второй раз Франко был уже в Новосибирске, на нашей Международной
конференции "Моделирование и компьютерные методы в молекулярной био-
биологии и генетике" в 1990 г. Он был очень импозантен — жилетка, на шее цвет-
цветной платок, через плечо кожаная грубая сумка. Он сделал интересный доклад, а
также отвел душу у нас дома, на банкетах и приемах. Этой поездкой он остался
очень доволен, несмотря на еще большую пустоту в магазинах. Он оценил ве-
8. Памяти Н.Н. Моисеева 67
лжолепие русского борща, приобрел множество знакомств, был полон радуж-
радужных проектов.
Любопытно отношение Ф. Скудо к русской и советской науке. Он очень
высоко ценил имена В.И. Вернадского, И.И. Шмальгаузена, А.Н. Северцова,
А.Н. Колмогорова, В.А. Костицына и др. В 1977 г. издал в "Springer-Verlag"
сборник классических работ "Золотой век математической экологии". Имена
авторов говорят сами за себя: В. Вольтерра, А.Дж. Лотка, А.Н. Колмогоров,
В.А. Костицын. И.И. Шмальгаузена он просто боготворил, более чем любой из
нас. Я послал ему некоторые из книг Шмальгаузена, которые он начал (с помо-
помощью жены) переводить с русского. Затем стал переводить прекрасную книгу о
В.И. Вернадском. В 1996-м в Париже вышла трехтомная Энциклопедия дарви-
дарвинизма (ред. П. Тор). Именно Франко "сосватал" туда несколько российских
авторов, в том числе В.В. Бабкова и меня.
В августе 1998 г., будучи на 18-м Международном генетическом конгрессе в
Пекине, я присутствовал на Симпозиуме "Разнообразие геномов человека и
популяционная генетика", где блистали итальянские друзья Франко Скудо —
Л. Кавалли-Сфорца, А. Пьяцца и др. Поражала сама обстановка на заседании.
Председательствовал седой и очень красивый Лука Кавалли-Сфорца. В зале ви-
витал легкий аристократический шарм: все же Кавалли — наследник древней фа-
фамилии Сфорца! Конечно, заседание он не вел, оно шло само собой. Зато он
включил в программу своих итальянцев и сотрудников по Стенфорду, пригла-
пригласил всех знакомых итальянцев, расцеловался с каждым из них на входе (осо-
(особенно с итальянками) и сделал блестящий доклад обо всем и ни о чем: о попу-
ляционной генетике, о демографии, о лингвистике и т.д. Это была именно та
обстановка, где должен был участвовать Франко. Но его не было, он уже был
сильно болен.
Дом в Торраца Косте опустел. Дочь Скудо, Петра, учится в Милане на Фи-
ческом факультете. Младшая дочь, Саша — школьница. Катя, вдова Франко,
пока в неопределенности. "Я сильная, я выдержу", — сказала она мне по теле-
ону.
Мы должны помнить нашего друга Франко Скудо! Мы должны быть ему
благодарны — он бескорыстно помогал нам всегда!
8. Памяти академика Н.Н. Моисеева*
Умер академик Никита Николаевич Моисеев, боевой летчик времен Оте-
Отечественной войны, генерал от математики, крупнейший советский специалист
по механике и вычислительной математике. Его имя было широко известно
людям всей планеты, благодаря работам в области глобального моделирования
взаимодействия человечества и природы. Он активно проповедовал стратегию
гармонического взаимодействия человека и природы. Не покорения природы,
не принудительного "взятия ее милостей", не опустошительной эксплуатации,
а совместного продуктивного развития.
* Впервые опубликовано: Ратнер В.А. // Информ. вестн. ВОГиС. Новосибирск: ИЦиГ СО
РАН. 2000. № 13-14. С. 16.
68 I. Personalia
Крупнейшие потрясения XX века — Хиросима, истребительные войны,
Чернобыль, природные и техногенные катастрофы — толкнули его к построе-
построению глобальных эколого-техногенных математических моделей, среди которых
ключевое место занимают модели мировой ядерной катастрофы. Именно он
указал основное последствие этого безумия — так называемую "ядерную зиму",
когда колоссальные массы пыли поднимаются в атмосферу и полностью и на-
надолго блокируют проникновение солнечного света к поверхности Земли. При
этом не просто гибнет все живое, но и исчезают условия для повторного рас-
распространения жизни. Трудно предполагать, но не исключено, что такие гло-
глобальные оценки были одной из последних капель, склонивших чашу весов Ис-
Истории в сторону ядерного разоружения, договоренностей, мирного развития
человечества.
Для биологов Н.Н. был превосходным, все понимающим партнером. Он
создал сильнейшую школу математического моделирования экосистем, инте-
интересовался принципами биологической организации. Среди его учеников —
крупные теоретики популяционной генетики и матэкологии, Ю.М. Свирежев,
В.П. Пасеков и др.
Мне пришлось всего лишь один раз соприкоснуться с Н.Н., но впечатле-
впечатление было огромным. Несколько лет назад, под эгидой ЮНЕСКО в Москве
состоялась Международная конференция по эволюции инфосферы (!). Этот
термин, если не ошибаюсь, предложил сам Н.Н. или его ближайшее окруже-
окружение. Его доклад тоже был посвящен именно этому понятию. Инфосферой он
назвал область циркуляции информации на поверхности Земли. Для матема-
математического генетика ясно, что эта область возникла как биосфера, поскольку
первыми природными объектами кибернетической природы были именно
живые существа, их популяции и экосистемы. "Генетическая информация"
была первым опытом инфосферы. Гораздо позже на ее основе возникли сиг-
сигнальная информация, разум, речь, письменность и технические ипостаси ин-
информации. Ну а "Мировая паутина"(ИНТЕРНЕТ) и т.п. — это вообще дело
последних лет.
Я попал на конференцию случайно с докладом об организации молекуляр-
но-генетических систем управления. Все прошло нормально. Неожиданно, по-
после конца доклада ко мне подошел Н.Н., выразил неподдельный интерес к про-
проблеме и сказал примерно такие слова: "Просто удивительно, до какой степени
молекулярно-генетическая организация по своим принципам и решениям по-
похожа на организацию общества! Как интересно и поучительно!"
Я подарил ему зеленую книжку "Концепция МГСУ". Потом был элитар-
элитарный обед для докладчиков. Н.Н. держался очень демократично и весело, кажет-
кажется, даже рассказывал анекдоты. К сожалению, трудов конференции я так и не
видел, хотя, по-моему, они были изданы.
Н.Н. Моисеев пришел в нашу инфосферу в 1917 г., а покинул ее в начале
марта 2000 г., на рубеже тысячелетий, оставив очень глубокий след в науке, в
сердцах людей. Мир его праху. Долгая ему память в осознанной им инфо-
инфосфере!
9. Памяти Н.Н. Воронцова
69
I
9. Поездка в Испанию.
Памяти Н.Н. Воронцова
В апреле 2000 г. умер Николай Николаевич Воронцов, известный био-
биолог-эволюционист, зоолог, кариосистематик, вообще человек энциклопедиче-
энциклопедической эрудиции. Он прожил достаточно бурную и насыщенную жизнь, в кото-
которой были взлеты и приземления. Получив первоначальную закваску на биофа-
биофаке МГУ, а также в домашнем кружке А.А. Ляпунова, он начал научную
деятельность в Зоологическом институте АН СССР в Ленинграде, потом орга-
организовал курс эволюционной биологии на медико-биологическом факультете
2-го Московского мединститута. Выход в большие пространства начался для
Н.Н. в Новосибирске, куда он переехал вместе с семьей Ляпуновых, будучи зя-
зятем Алексея Андреевича. Здесь они вместе с женой Еленой Алексеевной стали
работать у Р.Л. Берг в лаборатории генетики популяций ИЦиГ СО АН СССР.
Но главное, Н.Н. стал ученым секретарем Сибирского отделения АН СССР по
биологии. Фактически здесь он созрел для более крупных деяний. В эти годы
он защитил докторскую диссертацию и несколько лет, после отъезда Р.Л. Берг,
заведовал ее лабораторией.
После смерти А.А. Ляпунова Н.Н. уехал но Владивосток, в Дальневосточ-
Дальневосточное отделение АН СССР, где возглавил Биолого-почвенный институт. С ним
уехала часть лаборатории. Видимо, "московско-новосибирский десант" соби-
собирался повторить на Тихом океане
звездные годы новосибирского Ака-
Академгородка с его высокой наукой, уза-
узаконенными вольностями и возможно-
возможностью очень быстрого развития. Дета-
Деталей не знаю, но, скорее всего, руковод-
руководство "десанта" не нашло должной
поддержки и понимания у местной
партийной власти. В этом смысле
Академгородок не повторился. Пред-
Председателю ДВО АН СССР члену-кор-
члену-корреспонденту АН А.П. Капице при-
пришлось уйти и вернуться в Москву.
Вместе с ним уехал и Н.Н. Воронцов.
В Москве он возвратился в рядо-
рядовую научную жизнь. Несколько лет ра-
работал старшим научным сотрудником
в лаборатории своего друга А.В. Ябло-
кова в Институте биологии развития
АН СССР. Ездил в экспедиции, изучал
кариосистематику животных. Еще в
предыдущие годы Н.Н. сблизился с
Н.В. Тимофеевым-Ресовским, напи- Н.Н. Воронцов. Новосибирск, конец
сал вместе с ним и А.В. Яблоковым 1960-х (архив ИЦиГ СО РАН).
70
I. Personalia
На банкете по поводу докторской зашиты В.А. Ратнера. Новосибирск, 1974 г.
Слева направо: Н.Н. Воронцов, Л.А. Васильева, В.А. Ратнер, Р.И. Салганик
(архив автора).
"Краткий очерк теории эволюции" A969), а затем сохранил у себя архив Тимо-
Тимофеева- Ресовского.
В конце 80-х годов его подхватила волна демократии. От научных обществ
Н.Н. был избран в горбачевский Верховный Совет СССР и вошел там в Меж-
Межрегиональную депутатскую группу. После одного из ярких выступлений Н.Н.
на Верховном Совете Председатель Совета Министров СССР Н.И. Рыжков
предложил ему как представителю демократической оппозиции пост Министра
природопользования и охраны окружающей среды СССР. Н.Н. согласился.
На этом посту Н.Н.
Министерство природопользования
и охраны окружающей среды СССР
ВОРОНЦОВ
Николай Николаевич
Министр
Доктор биологических наук
Профессор
Народный депутат РСФСР
СССР, 103009, Москва,
ул. Неждановой, 11
Телефон 229-57-79
Телетайп 207559 БОРЕЙ
Телефакс 2302792
Визитная карточка Н.Н. Воронцова — Министра СССР
(архив Е.А. Ляпуновой).
предложил и добился
реализации нескольких
очень важных дел для
сообщества генетиков.
В частности, он подго-
подготовил и довел до Прези-
Президента СССР указ о на-
награждении группы вы-
выдающихся советских
генетиков старшего по-
поколения высшими на-
наградами страны: Звезду
Героя Социалистиче-
Социалистического Труда получили
9. Памяти Н.Н. Воронцова 71
академики Н.П. Дубинин и В.А. Струнников, члены-корреспонденты АН
И.А. Рапопорт и Ю.И. Полянский, академик АН УССР СМ. Гершензон, про-
профессор B.C. Кирпичников. Большая группа старых генетиков награждена орде-
орденами.
Н.Н. занимал этот государственный пост до августовского путча 1991 г.
Тогда, в обстановке хаоса и неопределенности он занял твердо демократиче-
демократическую позицию, распрощался с правительством и ушел в Белый дом к Ельцину.
Известны фотографии, где он стоит с Ельциным на танке.
После этого несколько лет Н.Н. проводит в Верховном Совете РСФСР и
1-й Думе РФ, где возглавляет Комитет по науке. Я помню, как яростно и убеди-
убедительно он защищал Соросовскую программу помощи российским ученым от
интриг и мистификаций "охотника на ведьм" коммуниста Илюхина. В следую-
следующую думу он уже не попал: время митингов прошло, демократы раздробились и
проиграли выборы.
В начале 90-х годов, в связи с распадом СССР, очень остро встал вопрос о
судьбе АН СССР и создании АН России. В это время Н.Н. участвовал в качестве
учредителя в организации Российской Академии естественных наук (РАЕН), где
он стал вице-президентом по Отделению биологии и экологии. Была надежда,
что именно эта Академия станет ядром АН России. Однако руководство
"большой" Академии не допустило этого, РАЕН не удалось выиграть битву за
Россию. Теперь это очень неплохая общественная Академия, которая ставит сво-
своей целью научные исследования, развитие образования и просвещения. Мне
приятно отметить, что я стал членом РАЕН при поддержке Н.Н. Воронцова.
В 1998 г. у Н.Н. случился тяжелый инсульт с потерей речи. Два года он был
фактически парализован. Однако за этот период он сумел подготовить к печати
капитальную монографию "История эволюционных учений", которая вышла
весной 2000 года.
Эта канва событий была наполнена, конечно, каждодневными заботами,
поездками, экспедициями, научным трудом. Несколько раз мне пришлось тес-
тесно пересечься с Н.Н., и в этих пересечениях удалось разглядеть некоторые осо-
особенности его личности и ментальности. Я расскажу только об одном таком слу-
случае — нашей совместной поездке в Испанию, в Барселону летом 1988 г.
В конце июля 1988 г., после обычных для советских ученых мытарств с виза-
визами и билетами, мы вылетели из Москвы в Мадрид. Оба мы, как оказалось, были
независимо приглашены на конференцию, тема которой в русском переводе зву-
звучала так: "Эволюционная биология переходных нестабильных популяций". Ор-
Организатором был профессор Антонио Фонтдевила из Автономного Университета
Барселоны. От Мадрида поездом добрались до маленького курортного городка,
где в католическом аббатстве состоялась эта конференция. Жили в монастыр-
монастырской гостинице, вся обслуга — монашки. Все шло нормально. Доклады свои мы
сделали, а потом Фонтдевила издал их сборником в "Springer-Verlag", изобразив,
между прочим, на обложке крылышки мух из наших экспериментальных работ.
Особенностью поездки было то, что денег (валюты) у нас не было почти
совершенно, если не считать страховочные 20 долларов, выдаваемых в У ВС АН
на всякий случай (с возвратом). Оргкомитет оплатил нам дорогу туда и обратно
и 5 дней пребывания. Но рейсы "Аэрофлота" в Мадрид и обратно ходили ред-
72
I. Personalia
Поездка в Испанию A988 г.)- В.А. Ратнер (слева) и Н.Н. Воронцов (справа) с сотрудни-
сотрудницами. Барселонского vwreftptvrara фрхив ЪА. Ляпуновой).
ко, поэтому обратные билеты у нас были через 15 дней после приезда, в районе
10 августа. Оставалось 10 безденежных дней, которые мы должны были где-то
провести.
На помощь пришел хозяин конференции А. Фонтдевила. На своей машине
он увез нас в Барселону и поселил в квартире своего аспиранта Уго и его жены
Аны в маленьком "спальном" городке вблизи Барселоны. Каждый день они пе-
передавали нас кому-нибудь из сотрудников кафедры, которые нас куда-нибудь
везли и что-нибудь показывали. Это были замечательные 10 дней! С испанцами
и между собой мы разговаривали обо всем.
Во-первых, сама Барселона. Прекрасный средиземноморский город, от-
отмеченный прямо-таки какой-то шизофренической гениальностью архитекту-
архитектуры Гауди. Волнистые, расползающиеся линии домов и памятников, яркие
краски. Музей молодого Пикассо, где он еще никакой не кубист, а прекрас-
прекрасный портретист. Колонна памяти X. Колумба и точная копия его корабля
"Санта-Мария" в гавани. В Барселоне был какой-то каталонский праздник.
Все главные улицы были увешаны флажками и надписями (в русской транс-
транскрипции: "Барселона ке ске май!", что означает: "Барселона лучше всех!").
Вот так просто и понятно. И никаких гвоздей! Молодцы каталонцы! Свою ро-
родину они не ругают так, как мы.
Мы посетили два барселонских университета, в одном из них познакоми-
познакомились со старейшим испанским генетиком Антонио Превости. Его имя было из-
9. Памяти Н.Н. Воронцова 73
вестно еще до войны. Будучи в отпуске, он специально приехал, чтобы пови-
повидаться с русскими профессорами.
Были в зоопарке Барселоны, одном из лучших в мире. Видели там унику-
уникума — "белую обезьяну", гориллу-альбиноса. Выяснили, что зоопарк состоит в
мировой системе зоопарков, центр которой находится в американском штате
Миннесота. Там в компьютерном центре находятся все сведения о крупных
штучных животных и составляются схемы их скрещивания, при которых ин-
инбридинг был бы минимален.
Посетили гору Монжуйк (Еврейская гора), где когда-то селилась еврейская
община Барселоны. Видели там цветные музыкальные фонтаны и место рас-
расстрела республиканского правительства Испании в 1939 г. Посетили горный
монастырь Монсерат — чудо в горах.
В разговоре с одним из сотрудников выяснилось, что он очень левый, поч-
почти анархист. И тут Н.Н. вспомнил, как в своем довоенном детстве в толпе со-
советских детей он встречал испанских детей, приехавших в СССР. Как кричал
"Но пассаран!", сжимал кулачок в республиканском приветствии и плакал от
счастья. Испанцы были растроганы, а Н.Н. искренне разволновался от этих
воспоминаний.
Профессор Фонтдевила пару раз приглашал нас на ужин в ресторан, где мы
вели методологические разговоры. И тут выяснилось, что мы с Н.Н. одинаково
относимся к науке. Для нас наука — святое дело, для которого можно пойти на
многие жертвы и лишения. А уж приоритетность науки перед другими занятия-
занятиями неоспорима. Испанский профессор слушал нас с удивлением. В Испании
люди живут, развлекаются, а уж потом, в свободное от сиесты время, занима-
занимаются наукой. А мы с Н.Н., как дети, братались в святилище науки.
Несколько раз мы ездили в Барселону и обратно поездом, естественно, за
страховочные деньги. И вдруг однажды, когда мы вышли из поезда на вокзале в
Барселоне, меня посетила идея. Под ногами валялись брошенные проездные
билеты. А что если собрать их, наклеить на лист и предъявить в Транспортный
отдел посольства. Если Посольство подтвердит эту сумму, ее вычтут из наших
затрат в Испании. Н.Н. согласился. Собирая билеты, мы хохотали до смерти от
нелепости ситуации: два русских профессора в летах, гости международной
конференции, как бомжи, собирают подержанные билетики! Сумма билетов на
двоих тянула на трехзначную цифру в песетах. Посольство подтвердило наши
притязания. Так мы отспорили у государства мизерную сумму, на которую
смогли купить сувениры своим домашним.
Закончились 10 дней в Барселоне. Мы отбыли в Мадрид. Пробыли там ров-
ровно сутки. Первым делом разыскали наше Посольство. Оно оказалось малень-
маленьким и невзрачным. Оказывается, тогда Испания для СССР была отдаленной
окраиной Европы, связей почти никаких. Посольство было второразрядным.
Оставив вещи в Посольстве, мы отправились в музей Прадо и провели там
4 или 5 часов. Впечатление было колоссальное. Бесчисленные залы, наполнен-
наполненные шедеврами испанской живописи. Только там я понял, чем велик Гойя. До
него все королевские портреты были величественны, монументальны и статич-
статичны. А у его королей и инфантов появились живые человеческие лица, появи-
появилось движение. Это было незабываемо.
74 I. Personalia
Подходил вечер, мы вернулись в Посольство. Пошли в отдел культуры
спросить о ночлеге. "Гостиницы нет", — ответили нам. И тут Н.Н. сказал очень
любезному офицеру охраны: "А давайте мы прочтем вам вечером лекцию в клу-
клубе о достижениях современной науки, а вы дадите нам диванчик перено-
переночевать". Офицер позвонил, согласовал и ответил, что клуб Посольства в подва-
подвале, придут в основном члены семей дипломатов, а в том же клубе есть два ди-
диванчика, где мы сможем переночевать.
Публика собралась пестрая: молодые люди, дети, женщины, несколько
служащих. На нас напало вдохновение. Н.Н. пересказал аудитории недавно вы-
вышедшие на Родине книги: Д. Гранина "Зубр" и В. Дудинцева "Белые одеж-
одежды" — о генетиках. Что говорил я — не помню, но тоже что-то современное и
заманчивое. Публика удивительно хорошо нас проводила, просила приезжать
еще. Затем мы спали на своих стареньких продавленных диванах в состоянии
полного счастья. Утром нас разбудил дежурный охраны, принес чаю. Потом мы
улетели в Москву.
Будучи в Барселоне, Н.Н. пригласил наших радушных хозяев в Москву.
Уго и Ана приехали в декабре, жили у Н.Н. дома, привыкали к нашей пище и
легким московским морозам. А после Нового года Н.Н. отправил их к нам в
Новосибирск. Аспиранты прилетели с приключениями и вышли из самолета
налегке в 40-градусный мороз. Мы утеплили их, как смогли, и только сумели
показать Академгородок и Этнографический музей. Увидев муляжи коренных
жителей Сибири, Уго расчувствовался и рассказал, что вообще-то он из Вене-
Венесуэлы, а сам он полуиндеец. Поэтому он был поражен физическим сходством
американских индейцев и аборигенов Сибири. Мы прочли им маленькую лек-
лекцию о заселении Америки сибирскими народами, и он успокоился. Но самый
большой восторг у него и его жены вызвала ледяная горка, где в 40-градусный
мороз с гиканьем и падениями катилась толпа разгоряченных подростков. Эту
горку у нас воздвигают каждый год около кинотеатра "Академия". В разное
время наши дети и внуки освоили эту горку.
Прошло меньше года, и Н.Н. стал депутатом Верховного Совета, а позже —
Министром СССР. Мне было очень забавно вспоминать, как совсем недавно мы
жили нахлебниками у двух малоимущих аспирантов, собирали билеты на вокзале
и читали лекцию в клубе Посольства, чтобы нам дали там переночевать.
П. ГЕНЕТИКА
10. 100 лет — 18 конгрессов!*
Завершился XX век — век генетики. Конечно, Мендель был раньше. Ко-
Конечно, были другие предтечи генетики. Но генетика как наука фактически на-
началась с началом XX века. В августе 1998 г. в Пекине состоялся 18-й Междуна-
Международный генетический конгресс (МГК), последний конгресс этого века. Он про-
прошел под знаком подведения итогов XX столетия. Тем любопытнее взглянуть на
список генетических конгрессов [Haynes, 1998]**, многие из которых были
этапными, а другие — сопряжены с весьма драматическими событиями.
1899 г. Международная конференция по гибридизации и кросс-бридингу форм.
Лондон, Англия. Президент Дж.Дж. Тревор Лоуренс / Ген. секретарь
У. Уилкс. В дальнейшем эта конференция была объявлена 1-м МГК. Пуб-
Публикация материалов: J. Roy. Hort. Soc. 1900. Vol. 24.
1902 г. Международная конференция по растениеводству и гибридизации.
Нью-Йорк, США. Президент Дж. Вууд / Ген. секретарь Л. Беррон. В даль-
дальнейшем эта конференция была объявлена 2-м МГК. Публикация материа-
материалов: Mem. Hort. Soc. N.Y. 1903. Vol. 1.
1906 г. 3-й МГК. Лондон, Англия. Президент У. Бетсон / Ген. секретарь
У. Уилкс. С этого года по предложению У. Бетсона МГК идут регулярно
каждые 5 лет. Публикация материалов: Roy. Hort. Soc. L. 1906.
1911 г. 4-й МГК. Париж, Франция. Президент Ив Делаж / Ген. секретарь Ф. де
Вильморен. Публикация материалов: Compt. Rend. Rap., Masson et Cie.
Libraries de l'Academie de Medecine. Paris. 1913.
Далее перерыв, связанный с Первой мировой войной.
1927 г. 5-й МГК. Берлин, Германия. Президент Э. Бауэр. Здесь впервые при-
присутствовала большая советская делегация: Н.И. Вавилов, Н.К. Кольцов,
А. С. Серебровский, Г.Д. Карпеченко и др. Публикация трудов МГК:
Z. f. induct. Abstamm.-u. Vererbungsl. 1928. Suppl. 1.
1932 г. 6-й МГК. Итака, Нью-Йорк, США. Президент Т. Хант Морган / Ген.
секретарь Р.А. Эмерсон. Морган, первый из генетиков, вскоре получит Но-
Нобелевскую премию. Генетика на подъеме. Здесь одним из важнейших со-
событий был доклад С. Райта о генетическом дрейфе. От СССР присутство-
присутствовал один Н.И. Вавилов, на делегацию не хватило средств. Н.В. Тимофе-
* Ранний вариант статьи под тем же названием: Ратнер В.А. 100 лет — 18 конгрессов // Ин-
форм. вестн. ВОГиС. Новосибирск: ИЦиГ СО РАН, 1998. № 7. С. 8-10.
** Haynes R.H. // Genetics. 1998. Vol. 148, N 4. P. 1419-1431.
76
II. Генетика
ев-Ресовский приехал из Германии. Публикация трудов: Proc. 6th ICG.
Brooklyn Botanical Garden, Brooklyn. N.Y. 1932.
Следующий, 7-й МГК должен был состояться в 1937 г. в Москве как при-
признание бурного и успешного развития генетики в СССР. Президентом Кон-
Конгресса должен был быть акад. Н.И. Вавилов. Однако из-за происков Т.Д. Лы-
Лысенко и политических амбиций в последний момент Правительство СССР от-
отложило проведение 7-го МГК в Москве на год. В результате Оргкомитет
перенес 7-й МГК в Эдинбург, где он собрался с 2-летней задержкой накануне
Второй мировой войны. Президентом 7-го МГК Оргкомитет заочно избрал
акад. Н.И. Вавилова.
1939 г. 7-й МГК. Эдинбург, Шотландия. Н.И. Вавилов не смог приехать в
Эдинбург, его попросту не пустили. Президентом и Ген. секретарем стал
Ф.А.Э. Крю. Н.В. Тимофеев-Ресовский приехал из Германии. Других со-
советских генетиков не было. Публикация трудов: Proc. 7th ICG. Cambridge:
Cambridge Univ. Press, 1941.
Далее перерыв связан со Второй мировой войной.
1948 г. 8-й МГК. Стокгольм, Швеция. Президент Г.Г. Мёллер / Ген. секретарь
Г. Дальберг. Недавно A946 г.) Мёллер получил Нобелевскую премию за ра-
работы по генетике. Назревали выдающиеся открытия. Советские генетики в
Конгрессе не участвовали. Это год разгрома советской генетики. Публика-
Публикация трудов: Proc. 8th ICG. Hereditas. 1949. Suppl.
5-й Международный генетический конгресс. Берлин, 1927 г. Советская делегация сле-
слева направо: С.С. Четвериков, А.С. Серебровский, Г.Д. Карпеченко, Н.И. Вавилов (из
книги И.А. Захарова "Краткие очерки по истории генетики". М.: Биоинформсервис,
1999).
10. 100 лет — 18 конгрессов
6-й Международный генетический конгресс. Итака, США, 1932 г. (справа налево)
Н.И. Вавилов, Т.Х. Морган и Н.В. Тимофеев-Ресовский (архив Н.В. Тимофеева-Ресов-
Тимофеева-Ресовского).
78
II. Генетика
12-й Международный генетический конгресс. Токио, 1968 г. Участники Конгресса справа
налево: Д.К. Беляев, СМ. Гершензон, Ф.Г. Добржанский, В.А. Ратнер (архив автора).
1953 г. 9-й МГК. Беллагио, Италия. Президент Р.Б. Голдшмидт/ Ген. секретарь
Дж. Монталенти. Советские генетики не участвовали. Публикация трудов:
Proc. 9th ICG. Caryologia. 1954. Vol. VI. Suppl., 2 parts.
1958 г. 10-й МГК. Монреаль, Канада. Президент С. Райт / Ген. секретарь
У. Бойс. От СССР принимала участие небольшая делегация, состоявшая из
лысенковцев. Что они там делали? Публикация трудов: Proc. 10th ICG.
Toronto: Univ. of Toronto Press, 1959.
1963 г. 11-й МГК. Гаага, Нидерланды. Президент Э. Хедорн / Ген. секретарь
К.Л. Ремке. От СССР была небольшая делегация, включавшая нескольких
генетиков — Д.К. Беляева и М.А. Арсеньеву, а также группу лысенковцев.
Публикация трудов: Genetics Today. Proc. Xlth ICG. Oxford: Pergamon Press,
1963-1965. Vol. 1-3.
1968 г. 12-й МГК*. Токио, Япония. Президент Хитоши Кихара / Ген. секретарь
Ятаро Тазима. Впервые МГК собрался в азиатской стране. От СССР впер-
впервые выехала большая делегация, включавшая Б.Л. Астаурова, Н.П. Дуби-
Дубинина, Д.К. Беляева, СМ. Гершензона и др. Съезд приветствовал наследник
династии Акихито, нынешний Император Японии, биолог по образова-
образованию. Отношение к советской делегации было очень дружеским. Это был
первый массовый выезд после реабилитации генетики в СССР A964 г.).
* Автор был участником 12-го, 14-го и 18-го МГК.
10. 100 лет — 18 конгрессов
79
14-й Международный генетический конгресс. Москва, 1978 г. Н.В. Тимофеев-Ресовский
на торжественном приеме во Дворце Съездов. Вот таким я видел его в последний раз
(архив СВ. Аргутинской).
Омрачило событие вторжение наших войск в Чехословакию как раз во вре-
время конгресса. Публикация трудов: Proc. Xllth ICG. Tokyo: Science Council
of Japan, 1968-1969. Vol. 1-3.
1973 г. 13-й МГК. Беркли, Калифорния, США. Президент К. Штерн / Ген. сек-
секретарь У. Браун. На конгрессе была представительная советская делегация.
Публикация трудов: Genetics. 1974. Vol. 78, N 1; 1975. Vol. 79. Suppl.;
Abstracts: Genetics. 1973. Vol. 74. June supplement. N 2. Part 2.
1978 г. 14-й МГК. Москва, СССР. Президент акад. Н.В. Цицин / Ген. секретарь
акад. Д.К. Беляев. Организатором конгресса был Д.К. Беляев, в конгрессе
без преувеличения участвовали все советские генетики. Вокруг конгресса
кипели политические страсти. Ряд западных генетиков не приехали на кон-
конгресс в знак протеста против преследования диссидентов в СССР. Однако
участие иностранных ученых все же было очень велико и продуктивно.
Они впервые увидели всю советскую генетику после реабилитации и 14 лет
работы. Конгресс прошел с большим успехом. Акад. Д.К. Беляев был из-
избран Президентом Международной Генетической Федерации (МГФ) на 5
лет. Публикация трудов: Т. 1. Благосостояние человечества и генетика; Т. 2.
Проблемы общей генетики; Т. 3. Молекулярные основы генетических про-
процессов. Москва: Мир, 1980-1981.
1983 г. 15-й МГК. Нью-Дели, Индия. Президент М.С. Сваминатан / Ген. секре-
секретарь В.Л. Чопра. Конгресс приветствовали Президент МГФ Д.К. Беляев и
80
II. Генетика
15-й Международный генетический конгресс. Нью-Дели, Индия, 1983 г. Президент
МГФ Д.К. Беляев и Премьер-министр Индии Индира Ганди (архив СВ. Аргутинской).
Премьер-министр Индии Индира Ганди. Советская делегация была до-
довольно ограниченной. Бесспорно, успехи прикладной генетики в Индии
очень велики. Публикация трудов: Genetics: New Frontiers. New Delhi-
Oxford and IBH Publ., 1984. Vol. 1-4.
1988 г. 16-й МГК. Торонто, Канада. Президент Р.Х. Хейнс / Ген. секретарь
Д.Б. Уолден. Советская делегация невелика. Публикация трудов: Genome.
1989. Vol. 31, N 1, 2; Abstracts: Genome. 1988. Vol. 30. Suppl. 1.
1993 г. 17-й МГК. Бирмингем, Англия. Президент P.P. Райли / Ген. секретарь
Д. Смит. Участие российских ученых минимальное, финансовые трудно-
трудности. Чувствуется, что генетика стала столь большой наукой, что форма кон-
конгрессов уже слишком громоздка. Многие генетики перестали ездить на
конгрессы. Публикации трудов не было.
1998 г. 18-й МГК. Пекин, КНР. Президент С.С. Тан / Ген. секретарь Шоуи Чен.
К сожалению, российские генетики почти не были представлены (несколь-
(несколько частных лиц), делегации не было совсем из-за разногласий руководства
ВОГиС и Оргкомитета МГК, а также по финансовым причинам. На кон-
конгрессе, естественно, преобладали китайцы. Западные ученые были пред-
представлены довольно ограниченно, многие не приехали. Не было ни одного
Нобелевского лауреата. Однако в целом конгресс прошел довольно успеш-
успешно. Китайская генетика имеет в основном прикладную направленность.
Публикация трудов: Abstr. 18th ICG. Beijing, China, 1998.
2003 г. 19-й МГК будет в Мельбурне, Австралия. Но это уже XXI век!
11. Концепция генов в развитии
81
18-й Международный генетический конгресс. Пекин, КНР, 1998 г. Старейшины Кон-
Конгресса Дж.Ф. Кроу (справа) и У. Бодмер (архив автора).
Таким образом, научный мир генетиков из небольшой группы энтузиастов
в начале века вырос в огромное сообщество, включающее, вероятно, десятки
тысяч исследователей из многих стран. Теперь они могут одновременно уме-
уместиться разве что на стадионе. Конгрессы становятся, скорее, общественно-по-
общественно-политическими мероприятиями, свидетельствующими о признании националь-
национальных школ, чем местом регулярного общения и подведения итогов. Активная
научная жизнь перемещается на узкие конференции, школы и симпозиумы по
интересам. Важную роль начинают играть INTERNET и другие электронные
формы общения. К этому надо привыкнуть и приспособиться.
11. Концепция генов в развитии
Гены — это атомы наследственности.
С. Бензер
Ген — самое фундаментальное понятие генетики. Оно складывалось посте-
постепенно, наполняясь конкретным и обобщенным содержанием, преодолевая
кризисы, заблуждения и неожиданные повороты. Посмотрим, как развивалось
это представление за последнее столетие.
82 II. Генетика
Классическая генетика*
Г. Мендель: Дискретные признаки контролируются дискретными наследствен-
наследственными факторами, которые неделимы при сегрегации и не смешиваемы при
агрегации гамет ("чистота гамет").
A. Вейсман: Наследственные факторы должны передаваться через зародышевую
плазму половых клеток (гипотеза).
B. Иоганнсен: Менделевские дискретные факторы "заочно" названы генами.
Г. Де-Фриз: Гены способны мутировать, мутационные варианты называются ал-
аллелями.
Т. Бовери. В. Сеттон и др.: Гены находятся в хромосомах клеточного ядра.
У. Бетсон. Р. Пеннет: Гены могут быть сцеплены между собой.
Т. Морган. А. Стертевант. К. Бриджес. Г. Меллер [Морган. 1927]: Гены — неде-
неделимые частицы генома, занимающие определенные локусы в хромосомах и
на генетической карте, способные удваиваться, мутировать и контролиро-
контролировать признаки. Они неделимы при кроссинговере. Отличительное свойство
гомологичных генов — синапсис в мейозе. Гены очень стабильны. В целом
ген — единица функции, мутирования и рекомбинации.
Н.И. Вавилов [1968]: Гены особей близкородственных видов имеют сходные
ряды гомологической изменчивости. Сходство генов порождает сходство и:
изменчивости.
Р. Голдшмидт [Goldschmidt, 1955]; К. Мазер [Мазер, Джинкс, 1985]: Существ;
гены большого эффекта (майор-гены, главные гены, олигогены), контроли-
контролирующие менделевские признаки, и гены малого эффекта (минорные гены,
модификаторы, полигены), контролирующие непрерывные количествен-
количественные признаки.
Эти результаты и гипотезы обосновывают классическое представление о ге
нах. Суммируя, получаем [Pontecorvo, 1958; Ратнер В.А., 1983]:
Генами называются локусы генома, обладающие следующими свойства-
свойствами:
1) способностью к самоудвоению;
2) способностью мутировать независимо от остальных генов, дискретно
изменяя свое внутреннее состояние как целое, т.е. порождая аллели;
3) неделимостью и несмешиваемостью в процессах кроссинговера и сегре-
сегрегации: на рекомбинационной карте ген изображался точкой (локусом), не
имеющей внутренней структуры;
4) определенной устойчивой локализацией в геноме относительно других
генов;
5) способностью к гомологичному синапсису в мейозе;
6) способностью контролировать развитие некоторого признака или не-
небольшого их числа, что отражено в названиях генов;
7) высокой устойчивостью к мутациям и другим изменениям.
Portugal, Cohen, 1979; Инге-Вечтомов, 1989; Льюин, 1987.
11. Концепция генов в развитии 83
Уже на уровне классической феноменологии и перечисленных свойств за-
заметна одна ярко выраженная особенность генов — их дискретность, которая
проявляется многими способами [Ратнер, 1983]:
а) относительная независимость мутирования;
б) частичная независимость проявления от других генов;
в) несмешиваемость с аллельными генами при образовании зигот;
г) неделимость и несмешиваемость при кроссинговере;
д) независимость синапсиса при сегрегации в мейозе;
е) разделение аллельных генов при сегрегации в мейозе;
ж) независимость сегрегации для многих неаллельных генов;
з) независимость локализации, неперекрывание и редкая зависимость
функции гена от локализации;
и) независимость управления работой генов.
Все это говорит о том, что гены — выделенные, автономные образования в ге-
геноме, и это проявляется в самых разных отношениях.
В моргановской интерпретации хромосома представлялась линейной по-
последовательностью сцепленных, но функционально не связанных генов ("нить
бус"). Действительно, макроструктура генетических карт у дрозофилы, кукуру-
кукурузы и других объектов не указывала на какую-нибудь корреляцию между лока-
локализацией генов и их функциями.
Основным экспериментальным методом генетики являлся и является до
сих пор гибридологический анализ, идущий от Менделя. В рамках этого подхода
особую роль играли два генетических теста: рекомбинационный и комплемента-
ционный (цис-транс-тест), идущие от школы Моргана. Оба теста оперировали
альтернативными дискретными наследуемыми признаками, что подразумевало
самоудвоение генов, их проявление через признаки, существование мутацион-
мутационных вариантов, участие в рекомбинациях. Эти же тесты использовались для
изучения аллелизма мутаций. Аллелями считались рецессивные мутации, не ре-
комбинирующие между собой и функционально комплементарные при образо-
образовании гибридных зигот. Заметим, что комплементационный тест по своей при-
природе выявляет лишь альтернативность, функциональную взаимозаменимость
мутаций, а рекомбинационный — различие локализации их структурных де-
дефектов.
Недостаточность классических принципов
Уже в рамках классической генетики и в ходе ее развития накапливались
явления, противоречившие моргановскому представлению о генах или выхо-
выходившие за его рамки.
А. Стертевант [Sturtevant, 19251: Н.П. Дубинин [1970]: Проявление генов иногда
зависит от их локализации в геноме ("эффект положения"), что противоре-
противоречит моргановской модели "нити бус".
А.С. Серебровский. Н.П. Дубинин [1968]: Гены имеют сложную внутреннюю
функциональную структуру (явление "ступенчатого аллелизма") (рис. 11-1).
84
II. Генетика
,10
mr
dc
vl
sa
I3
v?
ps
pa
ОС \ nrl
or2y
sc
pv
ex
st
h
I2
w
L_L
LA.
sc" =
SC10 =
sc1 +
(sc
sc1 -
= achx
= ach2
acti*
13)
sc2
sc*
sc6
sc1
sc9
sc*
sc3
sc
15
sc
17
Puc. 11-1. Сложная внутренняя функциональная структура гена scute Drosophilt
melanogaster [Серебровский, Дубинин, 1968; Дубинин, 1931].
М. Грин [Green, Green, 1949]: Комплементирующие аллели некоторых гено!
способны рекомбинировать между собой (явление "псевдоаллелизма"), чте
противоречит классическим тестам.
Р. Бринк [Brink et al., 1968]: Некоторые аллели в гибридных зиготах наследуемс
влияют на проявление друг друга (явление "парамутации"), что нарушае'
менделевский "принцип чистоты гамет".
Г. Меллер [Muller, 1927]: Мутации генов можно искусственно индуцироват]
действием жестких излучений. Поскольку излучения способны взаимодей
ствовать только с веществом, гены должны иметь материальную природ;
или быть связаны с материальными носителями.
И.А. Рапопорт [1948]: Ш. Ауэрбах [Auerbach, Robson, 1944]: Мутации генов мож
но индуцировать действием химических мутагенов. Это значит, что гены об
ладают химическими свойствами, т.е. связаны с молекулярным уровне*
организации вещества.
Н.К. Кольцов (см. гл. 15): Гены — это боковые радикалы аминокислот в белково]
макромолекуле-геноме (гипотеза), воспроизводящейся по матричном;
принципу.
11. Концепция генов в развитии 85
Н.В. Тимофеев-Ресовский. К. Ииммер, М. Дельбрюк (см. гл. 15): Ген - это гетеро-
гетерогенная макромолекула (белка?), имеющая внутреннюю структуру и способ-
способная к матричной конвариантной редупликации. Как макромолекула ген
обладает квантовыми свойствами: устойчивые пространственно-изомер-
пространственно-изомерные состояния отвечают аллелям гена, а редкие спонтанные переходы меж-
между ними — мутациям. Этим объясняется высокая природная стабильность
генов к мутациям.
Э. Шредингер (см. гл. 15): Ген — это макромолекула, несущая в себе "шифро-
"шифровальный код", т.е. запись наследуемого свойства.
Л. Полина. М. Дельбрюк (см. гл. 16): Синапсис генов как молекул нельзя объяс-
объяснить квантовым физическим дальнодействием, а только контактными си-
силами молекулярного близкодействия.
Дж. Понтекорво [Pontecorvo, 1958]: Представление, что ген — единица одно-
одновременно функции (цистрон), мутирования (мутон) и рекомбинирования
(рекон), неудовлетворительно.
Эти открытия послужили источником развития новых направлений, кото-
которые далее породили молекулярную генетику: использование микроорганизмов
как экспериментальных объектов, поиск материального носителя генов, исто-
истоки информационно-кибернетического подхода, анализ молекулярной органи-
организации фундаментальных генетических процессов.
Молекулярная генетика*
Все эти трудности концепции генов нашли свое разрешение в рамках моле-
молекулярной генетики:
Дж. Бидл. Э. Тейтум [Beadle, Tatum, 1941]: Гены контролируют синтез фермен-
ферментов ("один ген — один фермент").
О. Эвери: Материальным носителем генов является ДНК (см. гл. 16).
X. Френкелъ-Конрат [I960]: Материальным носителем генов может быть также и
РНК.
Дж. Уотсон. Ф. Крик (см. гл. 16): Ген — это линейная гетерогенная последова-
последовательность мономеров двухцепочечной ДНК, способная к матричной репли-
репликации, к кодированию свойств при помощи порядка мономеров, к мутаци-
мутациям — путем изменения этого кодирующего порядка. В основе матричного
принципа лежит комплементарность нуклеотидов: А — Т, G — С. Двухце-
почечность ДНК лежит в основе стабильности генов.
ГА. Гамов (см. гл. 16): Ген — это линейная последовательность символов четы-
четырехбуквенного алфавита нуклеотидов, т.е. генетический текст, кодирующий
первичную структуру белка. Должен существовать генетический код, свя-
связывающий последовательность нуклеотидов в гене с последовательностью
аминокислот в белке. Гены — не физические, а информационные единицы.
С. Бензер (см. гл. 17): Ген — непрерывный участок цепи ДНК размером ~103 н.п.,
кодирующий белок; является единицей функции (цистрон), выделяемой
при помощи цис-транс-теста. Этот тест является молекулярным вариантом
• Стент, Кэлиндар, 1981; Ратнер, 1983.
86 II. Генетика
теста комплементарное™. Мутации способны затрагивать любые пары
нуклеотидов (мутоны), а рекомбинации — разделять смежные нуклеотиды
(реконы).
М. Демерец [см. Инге-Вечтомов, 1983]: Все мутации одного цистрона (по
цис-транс-тесту) — аллели, но нерекомбинирующие — изоаллели, а реком-
бинирующие — гетероаллели.
С. Бреннер. Ф. Жакоб. М. Мезельсон [Brenner et al., 1961]: Цистроны реализуют '
свою информацию через синтез информационной (матричной) РНК.
Ф. Крик. С. Бреннер и др. (см. гл. 17): Цистроны разбиты на триплетные кодоны
и вьщелены знаками пунктуации трансляции — начальными и терминаль-
терминальными триплетами.
Ф. Крик. М. Ниренберг. Г. Корана (см. гл. 17): Существует универсальный гене-
генетический код, при помощи которого в цистронах записана генетическая ин-
информация о структуре белков.
Ф. Жакоб. Ж. Моно [Jacob, Monod, 1961]: Существуют структурные гены (цис-
(цистроны), кодирующие функциональные белки (ферменты и др.), и регуля-
торные гены (цистроны регуляторных белков, функциональные сайты), ко-
которые регулируют синтез белков структурными генами. Опероны — управ-
управляемые единицы транскрипции (см. гл. 23), репликоны — управляемые
единицы репликации. Они выделены знаками пунктуации (инициирующи-
(инициирующими и терминальными знаками) и подчинены знакам управления.
П. Говард-Фланденс. Р. Сетлоу и др. [Howard-Flanders et al., 1984; Льюин, 1987]:
Высокая стабильность генов ДНК объясняется существованием мощных
молекулярных систем репарации дефектов, основанных на дублировании
генетической информации. Синапсис генов фактически сводится к компле-
комплементарным взаимодействиям цепей ДНК с участием специализированных
ферментов-посредников (рис. 11-2).
А. Корнберг [1977]: Спонтанные мутации генов объясняются не столько воздей-
воздействиями внешних мутагенов, сколько ошибками ферментов репликации.
Формирование молекулярного представления о генах было развитием идей
классической теории генов, устранением ряда ограничений, упрощений, оши-
ошибочных гипотез. При этом использованы новые экспериментальные методы:
молекулярная гибридизация ДНК х ДНК, ДНК х РНК, радиоактивные метки,
электрофорез, электронная микроскопия, методы массовой селекции редких
событий и т.д. Существенное влияние на теорию генов оказал информацион-
информационно-лингвистический и кибернетический подход: основные понятия молеку-
молекулярной генетики сформулированы в этих терминах — генетический контроль,
генетическая информация, генетический код, опероны, информационная
FHK, регуляторные гены и белки и т.д.
В итоге представление о генах в молекулярной генетике существенно изме-
изменилось и расширилось [Ратнер, 1983; Льюин, 1987].
1. Полностью отпала гипотеза белка как материального носителя генов.
Во всех случаях эту роль выполняют ДНК или РНК.
2. Полностью отпала квантовая гипотеза пространственных изомеров мак-
макромолекул-генов М. Дельбрюка. Полностью подтвердилась "гипотеза последо-
11. Концепция генов в развитии
87
Rec А
Protein
+ АТР
в
D
Е
Topoisomerase
14 ' Polymerase
uc. 11-2. Молекулярный механизм синапсиса ДНК у Е. coli [Howard-Flanders et al.,
1984]. Роль фермента синаптазы выполняет фермент рекомбинации гесА.
чьности" Ф. Крика, постулирующая, что генетическая информация в генах
одируется составом и порядком входящих мономеров. Аллели оказались изме-
ениями кодирующего порядка мономеров.
3. Несмотря на открытие мощных факторов индукции мутаций, спонтан-
спонтанный мутагенез все же объясняется ошибками ферментов репликации. Высокая
устойчивость генов к мутациям объясняется работой мощных систем репара-
репарации, основанных на дублировании генетической информации, а не структур-
структурными особенностями двухцепочечной ДНК.
88 II. Генетика
4. Выявлены принципы молекулярной организации и ферментативные
системы основных фундаментальных генетических процессов, обеспечивающих
свойства и функции генов: репликацию, транскрипцию, трансляцию, мутиро-
мутирование и репарацию, рекомбинацию, сегрегацию и т.д.
5. Выявлены функциональные кодирующие единицы, отвечающие основным
матричным генетическим процессам [Ратнер, 1983]: кодоны и цистроны —
трансляции, скриптоны — транскрипции, репликоны — репликации, группы
сцепления — мейотической сегрегации. Выявлена пунктуация этих единиц и
процессов — знаки начала и конца, которые обеспечивают их ограниченную
локализацию и управление в геномах. Этим обеспечивается дискретность всех
функциональных единиц во всех отношениях.
6. Решена проблема считывания и реализации генетической информации от
генов до ближайших к ним молекулярных признаков: это механизмы транс-
транскрипции скриптонов, трансляции цистронов и специфического узнавания
функциональных сайтов.
7. Решена проблема гомологичного синапсиса молекул ДНК и хромосом.
Фактически синапсис сводится к комплементарности цепей ДНК и участию
ферментов-синаптаз. Молекулярный механизм основан на принципах близко-
действия макромолекул (комплементарность, физический контакт), а все пред-
предложенные механизмы дальнодействия отвергнуты.
8. Показано, что в основе теста комплементарности фактически лежит не-
независимость трансляции отдельных цистронов и функционирования их белко-
белковых продуктов. Вместе с тем тест комплементарности имеет явные ограничения
и действует не всегда. Он выявляет функциональные единицы не ниже цистро-
цистронов.
9. С операциональной точки зрения представление о цистронах наиболее
близко к классическому представлению о генах. Исторически на первом этапе
молекулярной генетики классическое нестрогое представление о системах
ген — признак очень сильно сузилось до принципа "один ген — один фермент".
Понятие "локус", вместо точки на генетической карте, стало означать "непре-
"непрерывный участок генома". Молекулярный размер цистрона оценен в
102-103 н.п., т.е. разнообразие возможных мутаций очень велико. Последова-
Последовательность цистрона коллинеарна последовательности кодируемого белка. В свя-
связи с этим уточнилось понятие аллелизма.
10. Однако с развитием молекулярно-генетической и биохимической тех-
техники представление о системах ген — признак снова быстро расширилось, ох-
охватив самые разнообразные генетические структуры и признаки — биохимиче-
биохимические, морфологические, функциональные, транспортные, защитные, регуля-
торные и др. В результате оказалось, что цис-транс-тест имеет дело лишь с
ограниченным классом функций и функциональных единиц (с цистронами),
которые заведомо не исчерпывают смысловой, кодирующей части генома.
Скриптоны т-РНК и р-РНК кодируют только РНК, вообще не кодируют бел-
белков, но косвенно участвуют в их синтезе. Знаки пунктуации и управления вооб-
вообще ничего не синтезируют, но при помощи порядка мономеров кодируют
функции специфического опознания регуляторных белков. Тем не менее все
эти единицы локализованы в геномах, реплицируются, рекомбинируют, мути-
11. Концепция генов в развитии 89
руют, кодируют наследуемые функции наравне с цистронами. Их следует
включить в представление о генах.
11. В геномах функциональные единицы всех типов и разделяюшие их
спейсеры построены из одинакового генетического материала: мономеров
ДНК или РНК. Значит, гены являются не физическими, а информационными
единицами [Ратнер, 1993]. В этом смысле они напоминают лингвистические
единицы информации в текстах естественных языков. Сравнение их свойств
показывает, что кодоны более всего напоминают трехбуквенные слова, цистро-
ны — предложения, скриптоны — абзацы текста, а репликоны — единицы вос-
воспроизведения. Смысл закодированной информации — это функции и свойства
соответствующих продуктов структурных генов или самих функциональных
сайтов генома.
12. Таким образом, информационно-лингвистический подход [Ратнер, 1993]
оказался очень продуктивным для понимания природы генов. В рамках такого
подхода понятие "ген" ближе всего к понятию "значащая лингвистическая
конструкция", которое охватывает и слог, и слово, и предложение, и абзац, и
печатный лист, и т.д. Это — групповое понятие, необходимое как в теоретиче-
теоретическом, так и в эвристическом отношении. Экспериментатор-генетик длительное
время не знает, какова истинная природа локуса, с которым он работает. В этой
ситуации нужен групповой термин, охватывающий широкий круг возможных
функциональных единиц.
Итак, понятие "цистрон" имеет достаточно частное значение, тогда как
за термином "ген" желательно сохранить его фундаментальное значение в
генетике. Будем называть генами протяженные участки генома (локусы), со-
сохраняющие свою непрерывность в некоторых генетических процессах и осуще-
осуществляющие в них свою матричную или опознавательную функцию. Более де-
детально гены обладают следующими общими свойствами [Ратнер, 1983]:
1) способностью к конвариантной репликации в клетке или эквивалент-
эквивалентной ей системе;
2) способностью к дискретным случайным изменениям порядка мономеров,
не затрагивающим других генов, в клетке или эквивалентной ей системе;
3) способностью обмениваться любыми частями с гомологичными гена-
генами в процессе общей рекомбинации в клетке или эквивалентной ей системе;
4) определенной дискретной локализацией в геноме относительно других
генов;
5) способностью кодировать дискретную порцию информации (молекуляр-
(молекулярную функцию или свойство) при помощи порядка нуклеотидов, которая мо-
может быть реализована в клетке или эквивалентной ей системе через процес-
процессы транскрипции, трансляции или контактного узнавания;
6) способностью вступать в синапсис с гомологичными генами в клетке
или эквивалентной ей системе;
7) способностью длительно сохранять генетическую информацию в клетке
или эквивалентной ей системе.
90 II. Генетика
При таком определении генами являются все типы функциональных еди-
единиц. Гены естественно подразделить на структурные, выполняющие матричные
функции в соответствующих генетических процессах, и пунктуаторы, выпол-
выполняющие в этих процессах роль знаков пунктуации и управления при соответст-
соответствующих структурных генах. Структурные гены образуют иерархическую систему
(снизу вверх): кодон, цистрон, скриптон, репликон, группа сцепления. Цис-
троны занимают в этой иерархии особое место, так как (а) это единственный
класс генов, коллинеарных своим белковым продуктам, и (б) это низший уро-
уровень структурных генов, имеющих знаки пунктуации в полинуклеотидных по-
последовательностях, а потому выделяемых в цис-транс-тесте.
Локусы ДНК и РНК обладают свойствами генов не сами по себе, а только в
клетке или эквивалентной ей системе. Иначе говоря, для осуществления этих
свойств необходимы специальные универсальные молекулярные исполняющие
устройства (комплексы ферментов) процессов репликации, репарации, реком-
рекомбинации, транскрипции, трансляции, сегрегации и т.д. Они образуют централь-
центральную подсистему Молекулярно-генетической системы управления в клетке.
Новые расширения концепции генов
После того как сложились основы молекулярной концепции генов, стали
накапливаться новые факты, нарушающие эту концепцию или требующие ее
развития. Многие из этих фактов — плод коллективного творчества, поэтому
трудно назвать фамилию автора.
У. Джилберт [Gilbert, 1978]: Цистроны эукариот разбиты на кодирующие сег-
сегменты (экзоны) и некодирующие (интроны), т.е. разрывны (рис. 11-3). Это
не опасно, если имеется специальная молекулярная технология устранения
разрывов к моменту трансляции (сплайсинг).
Б. Мак-Клинток [McClintock, 1956]: Существуют мобильные генетические эле-
элементы (МГЭ) — подвижные генетические фрагменты геномов прокариот и
эукариот. Они, как правило, содержат гены своей подвижности, кодирую-
кодирующие ферменты транспозиции (рис. 11-4), поэтому изначально МГЭ счита-
считались "эгоистической ДНК", нарушающей принцип стабильности локализа-
локализации генов. МГЭ играют значительную роль в экспрессии генов, их измен-
изменчивости и эволюции. Подвижность МГЭ не опасна для генома, если
частоты транспозиций невелики. МГЭ занимают существенную часть эука-
риотических геномов.
Ж. AfoHo. Ф. Жакоб (см. гл. 23)-. Существуют генетические триггеры — цикличе-
циклические двухоперонные системы, обладающие неменделевскими свойствами
наследования по клеточной линии. За счет передачи регуляторных белков
они наследуют не только аллельные, но и функциональные состояния вхо-
входящих генов. В целом такие и более сложные системы играют важную роль
в генетическом контроле онтогенеза. Иногда их называют "эпигенами".
Р. Бриттен. Д. Коне [Britten, Kohne, 1968]: У эукариот многие гены представле-
представлены мультигенными семействами, тандемными или диспергированными, со-
содержащими от десятков до сотен и тысяч цистронов или скриптонов
(рис. 11-3).
11. Концепция генов в развитии
91
— Существуют сложные гены эукариот (рис. 11-5) — участки размером до 105
н.п., вероятно, единицы "выпетливания" ДНК в хромосомах (рис. 11-6), со-
содержащие несколько единиц транскрипции и общие регуляторные меха-
механизмы — энхансеры, сайленсеры, инсуляторы и др.
— Существуют супергены эукариот размером до 106 н.п. (рис. 11-7) — танде-
тандемы сотен или тысяч экзонов, производящие огромное комбинаторное раз-
разнообразие однородных белков (например, иммуноглобулинов и др.) путем
а
30 31 104 105 146
200
400
600
800
1000
1200
1400
1600
31 32
99 100 141
Chromosome 16
\j/a2 \j/al
Ю-D-
a2
al
ei
Chromosome 11
€
Chromosome 22
Myoglobin
Рис. 11-3. Экзон-интронная структура генов субъединиц аир гемоглобина человека
(а). Мультигенное надсемейство глобинов человека (см. [Ратнер, 1983]) (б).
RT RH
ORF1
ORF2
rLTR
I |
0 481 613 997
1301
1770
1613
3047 Л 3465 4067
3135 3142
4601 j 5244 5743
4871
6411 \7440
6758 6959
Рис. 11-4. Генетическая структура мобильного генетического элемента Dm-412 дрозофи-
дрозофилы (см. [Ратнер, Амикишиев, 1996]). Обозначения: LTR — длинные концевые повторы,
ORF — открытые рамки трансляции, домены ферментативных активностей, Р — про-
теаза, RT — обратная транскриптаза, RH — рибонуклеаза Н, I — интеграза.
92
II. Генетика
соматических рекомбинаций, сплайсинга и мутаций. Это нарушает прин-
принцип "один ген — один фермент", но не опасно, если затрагивает только со-
соматические клетки иммунной системы, которые не участвуют в наследова-
наследовании по генеративной линии.
Сенджер и др. [Sanger et al., 1977]: Некоторые цистроны локализованы внутри
других цистронов или в их интронах, или частично перекрываются с други-
другими цистронами (рис. 11-8). Это нарушает принцип независимой локализа-
локализации генов, но не опасно, если они функционируют независимо.
Phenotypic map
achaete
scute a
lethal of scute
scute b
scute g
T
^4
SI L8 VI
ntt t
7 H S2
19
Molecular map
80
60
I
50
SI
40
I ' Lj
VI
20
S2 7
1 ?U
-20
I
260-1
-30
yellow "t> ac
Wing disc |
1 CNS
tu
PNS
sc * / sc
II I
T2 о
1-* о
AS-C expression
CNS: NBs
Г sc
ml m i I
Emryo
ac/sc
ml m i
@
PNS: SVCs
ac/sc
Wing disc
ac/sc
Puc. 77-5. Молекулярная картина сложного гена scute /
у D. melanogaster [Modolell, Campuzano, 1998].
PPA
11. Концепция генов в развитии
93
Repressed
— Существуют псевдогены эу-
кариот — дефектные копии ге-
генов, утратившие функцию.
Фактически это "руины" цис-
тронов, длительно сохраняю-
сохраняющие многие особенности их
структуры.
— В секвенированных последо-
последовательностях ДНК выявляются
открытые рамки трансляции
(ORFbi), т.е. "почти цистро-
ны" — бездефектные участки,
ограниченные знаками ини-
. циации и терминации трансля-
трансляции, но функции которых пока
не доказаны.
— Существуют и очень распро-
распространены в геномах "мотивы"
функциональных сайтов, т.е.
очень похожие на них участки,
функции которых пока не до-
доказаны.
С. Прузинер [Prusiner, 1996]: Суще-
Существуют возбудители некоторых
наследственных болезней, ко-
которые вообще не содержат ге-
генов, — прионы. Видимо, это
функциональные белки, про-
продукты генов хозяина, контакт-
контактно передающие свое конфор-
мационное функциональное
состояние аналогичным бел-
белкам при заражении. Это то же
"наследо- вание", но не гене-
генетическое. Оно ближе к эпиге-
эпигенетическому наследованию функциональных состояний в двухоперонном
триггере.
— Гены имеют астрономическое потенциальное разнообразие изофунк-
циональных (синонимических) вариантов [Ratner et al., 1996]. В природе
реализована лишь мизерная часть этого разнообразия.
М. Дайхоф и др. (см. [Ratner etal., 1996]): Секвенированные последовательности
генов накапливаются в компьютерных банках данных, обрабатываются,
сравниваются и классифицируются. Это основной источник информации
о структуре генов.
— Сходство последовательностей родственных гомологичных генов позво-
позволяет восстановить их консенсус [Ratner et al., 1996], т.е. гипотетический ге-
Hl and core
histone tail
modifications
Chromatin unfolding
Рис. 11-6. Единица выпетливания в эукарио-
тической хромосоме [Grunstein, 1990].
94
II. Генетика
^Transcription
DNA
Рис. 11-7. Суперген доменов легких це-
цепей иммуноглобулина у человека.
4 HI v Lrl :
I Translation
5 HI v Lrl 71
I Removal of
| leader
1 95 108 214
Amino acid
1
Protein
mRNA END
ORIGIN OF DNA
REPLICATION
нетический текст, состоящий из наиболее устойчивых элементов, участ-
участвующих в кодировании. Консенсус — это как бы "родовой портрет" груп-
группы генов, выражающий их структурное родство в связи с кодированием
общей функции.
У. Фитч. М. Дайхоф и др.: Сходство последовательностей родственных генов
позволяет более детально строить деревья их родства и классификации, от-
отражающие пути их эволюции (см. [Ratner et al., 1996]).
— В целом гены имеют блочно-модульное строение, т.е. содержат структур-
структурно-функциональные модули, способные к комбинаторному переформиро-
переформированию [Ratner et al., 1996].
М. Кидуэлл [Kidwell, 1993]: Обыч-
Обычно гены наследуются и пере-
передаются по "вертикальному"
пути — от родителей к по-
потомкам (зародышевая плазма
Вейсмана), но не исключены
явления "горизонтального
переноса" (особенно у бакте-
бактерий), когда переносчики ге-
генов (вирусы, плазмиды,
МГЭ) преодолевают барьеры
видовой изоляции.
— Предполагается, что гены
и полигены имеют однород-
однородную генетическую природу,
это объекты одного молеку-
Рис. 11 -8. Карта генома фага ipXl74 [Sangeret al., лярного класса, но прояв-
1978]. Пример ситуации "ген в гене". ляющиеся по-разному в за-
11. Концепция генов в развитии 95
висимости от положения в системе МГСУ. Гены, лимитирующие экспрес-
экспрессию признака, — менделевские; гены, не лимитирующие экспрессию, —
полигены (локусы количественного признака — QTL). Существуют поли-
полигены большого эффекта [Ратнер, Васильева, 2000].
Эти новые факты получены новыми экспериментальными и теоретически-
теоретическими методами генной инженерии (секвенирование, ПЦР, перенос генов вектора-
векторами, создание библиотек клонов и др.) и биоинформатики (создание компьютер-
компьютерных банков данных, компьютерный анализ и сравнение генетических текстов,
построение деревьев сходства и консенсусов и т.д.). Ясно, что с течением вре-
времени информационно-лингвистический подход к строению генов приносит
все больше успешных результатов. На очереди стоит проблема разработки гене-
генетического языка, т.е. системы правил формирования полноценных генов с оп-
определенными функциональными свойствами.
Фактически эти новые результаты усложняют, но не разрушают молеку-
молекулярную концепцию генов. Они либо не опасны для генетической организации
(комбинаторика антител, перекрывание генов и др.), либо компенсируются
специальными молекулярными технологиями (репарации, сплайсинг и др.),
либо имеют филогенетический характер, либо выявляют глубинные особенно-
особенности генетического языка.
Список литературы
Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости // Классики
советской генетики / Ред. П.М. Жуковский. Л.: Наука, 1968. С. 9-50.
Дубинин Н.П. // Журн. экспер. биол. 1931. Т. 7, № 5-6. С. 525-591.
Дубинин Н.П. Общая генетика. М.: Наука, 1970.
Инге-Вечтомов С.Г. Введение в молекулярную генетику. М.: Высш. шк., 1983.
Инге-Вечтомов С.Г. Генетика с основами селекции. М: Высш. шк., 1989.
КорнбергА. Синтез ДНК. М.: Мир, 1977.
Льюин В. Гены. М.: Мир, 1987.
Мазер К., Джинкс Дж. Биометрическая генетика. М.: Мир, 1985.
Морган Т.Г. Теория гена. Л.: Сеятель, 1927.
Рапопорт И.А. // Докл. АН СССР. 1948. Т. 60. С. 469-472.
Ратнер В.А. Молекулярная генетика: принципы и механизмы. Новосибирск: Наука,
1983.
Ратнер В.А. Концепция молекулярно-генетических систем управления. Новосибирск:
Наука, 1993.
Ратнер В.А., Васильева Л.А. // Современные проблемы эволюционной генетики / Ред.
В.К. Шумный, А.Л. Маркель. Новосибирск: ИЦиГ СО РАН, 2000. С. 145-170.
Серебровский А.С., Дубинин Н.П. Искусственное получение мутаций и проблема гена //
Классики советской генетики / Ред. П.М. Жуковский. Л.: Наука, 1968. С. 294-302.
Стент Г., Кэлиндар Р. Молекулярная генетика. М.: Мир, 1981.
Френкель-Конрат Г., Сингер В., Вильяме Р. // Химические основы наследственности.
М.: ИЛ, 1960. С. 404-414.
Auerbach Ch., Robson J.M. // Nature. 1944. Vol. 154. P. 80-81.
Beadle G.W., Tatum E.L. Genetic control of biochemical processes in Neurospora // PNAS.
1941. Vol. 27. P. 499-506.
Brenner S., Jacob F., Meselson M. // Nature. 1961. Vol. 190. P. 576-580.
96 II. Генетика
Brink R.A., Styles E.D., Axtell J.D. // Science. 1968. Vol. 159, N 4811. P. 161.
Britten R.J., Kohne D.E. // Science. 1968. Vol. 161, N 3841. P. 529-540.
Gilbert W. // Nature. 1978. Vol. 271, N 5645. P. 501.
Goldschmidt R. Theoretical Genetics. Los Angeles: Berkeley, 1955.
Green M., Green K. // PNAS. 1949. Vol. 35. P. 586-591.
Grunstein M. Nucleosomes: regulation of transcription// TIG. 1990. Vol. 6, N 12. P. 395-400.
Howard-Flanders P. et al. // CSHSQB. 1984. Vol. 49. P. 571-580.
Jacob F., Monod J. // J. Mol. Biol. 1961. Vol. 3. P. 318-356.
Kidwell M.G. Lateral Transfer in Natural Populations of Eukaryotes // Ann. Rev. Genet. 1993.
Vol. 27. P. 235-256.
McClintock B. // CSHSQB. 1956. Vol. 21. P. 197-216.
Modolell J., Campuzano S. The achaete-scute complex as an integrative device // Int. J. Dev
Biol. 1998. Vol. 42. P. 275-282.
Muller H.J. Artificial transmutation of the gene // Science. 1927. Vol. 66. P. 84-87.
Pontecorvo G. Trends in Genetic Analysis. N.Y.: Columbia Univ. Press, 1958.
Portugal F.H., Cohen J.S. A century of DNA. MIT Press. Cambridge, MA; London, England
1979. Ch. 5, 6.
Prusiner S.B. Inherited Prion Diseases // TIBS. 1996. P. 482-487.
Ratner V.A., Zharkikh A.A., Kolchanov N.A., Rodin S.N., Solovyov V.V., Antonov A.S
Molecular Evolution. Berlin e.a.: Springer-Verlag, 1996.
Sanger F., Air G.M., Darrell B.C. et al. Nucleotide sequence of bacteriophage q>X174 DNA //
Nature. 1977. Vol. 265. P. 687-695.
Sturtevant A.H. The effects of unequal crossingover at the Bar locus in Drosophila // Genetics.
1925. Vol. 10. P. 117.
12. Что содержит полный геном Escherichia coli!*
В настоящее время Е. coli, несомненно
представляет собой самую изученную кле
из всех существующих.
Г. Стент
Кишечная палочка, Escherichia coli — классический объект молекулярной
генетики, на котором исследованы наиболее принципиальные проблемы орга-'
низации генетического материала. Штамм Е. coli K12 был успешно использо-
использован Дж. Ледербергом и Э. Тейтумом в 1946 г. для доказательства существования
рекомбинаций у бактерий. Позже Дж. Ледерберг построил для нее первую ге-
генетическую карту, а Ф. Жакоб и Э. Вольман — первую кольцевую карту.
В 1963 г. Дж. Керне сфотографировал кольцевой геном Е. coli в процессе его ре-
репликации.
Около 30 лет назад огромным событием в генетике было секвенирование
первого гена. К концу 70-х годов технология секвенирования упростилась и
стала рутинной процедурой. После этого были секвенированы геномные ДНК
и РНК сначала малых фагов (R17, Q р, уХ174 и др.), затем больших фагов и ви-
вирусов (фага Я., вирусов коровьей оспы, оспы и др.), клеточных органелл (мито-
* Вариант этой главы опубликован ранее: Ратнер В.А. // Информ. вестн. ВОГиС. Новоси-
Новосибирск ИЦиГ СО РАН, 2001. № 18. С. 2-5.
12. Что содержит полный геном Е. соШ 91
хондрий и хлоропластов) и плазмид (F-фактора и др.)- Наконец, в конце 80-х
годов были начаты первые международные программы секвенирования полных
клеточных геномов бактерий, грибов, растений, насекомых, млекопитающих,
человека.
Зачем это нужно? Клеточный геном представляет собой сбалансированную
систему генов — архив генетической информации, достаточной для контроля
всего клеточного метаболизма, развития, морфогенеза, самовоспроизведения
[Ратнер, 1993; Ratner et al., 1996]. В частности, геном клетки содержит гены всех
основных генетических процессов — репликации, транскрипции, трансляции,
репарации, рекомбинации, сегрегации и т.д. Полное секвенирование генома
позволяет сопоставить и оценить генетическую сложность тех или иных моле-
молекулярных систем и геномов, выявить ранее неизвестные гены, выполнить срав-
сравнительный анализ функционального и структурного сходства различных генов
и геномов, выявить общие принципы организации сложных клеточных моле-
кулярно-генетических систем управления.
Проект полного секвенирования генома Е. coli К12 был начат в 1991 г.
под руководством доктора Фреда Блаттнера (Лаборатория генетики, Вис-
консинский Университет, Мэдисон, США). В январе 1997 г. основные ре-
результаты были переданы в компьютерную базу данных GenBank [3], а в сен-
сентябре 1997 г. в американском журнале "Science " появилась итоговая статья
коллектива участников секвенирования [Blattner et al., 1997]. Полная после-
последовательность ДНК генома Е. coli К12 стала достоянием науки. Ниже мы
приводим в сводной форме основные результаты этих работ с необходимыми
комментариями, имея в виду, что такой уникальный материал позволяет от-
ответить на многие принципиальные вопросы молекулярно-генетической ор-
организации и эволюции.
Заметим также, что параллельно с Е. coli секвенированы многие другие
клеточные геномы бактерий и эукариот. К концу 1997 г. опубликовано 8 пол-
полных клеточных геномов, к лету 1998 г. — уже 15, а к лету 2001 г. — 5 геномов эу-
эукариот (пекарские дрожжи, нематода, арабидопсис, дрозофила и человек),
свыше 30 бактериальных геномов (включая Е. coli, Bacillus subtilis, стрептокок-
стрептококки, риккетсии, хламидии, микоплазмы и др.) и свыше 9 геномов архебактерий
(включая галобактерии, термоплазмы, метанобактерии и др.) (табл. 12-1). На
очереди геномы других бактерий и грибов, а также пшеницы, риса, кукурузы,
мыши и других объектов. Многие фармацевтические и биотехнологические
фирмы ведут секвенирование геномов продуцентов, возбудителей ряда заболе-
заболеваний и др., но свои данные засекречивают. Считается, что в работе сейчас не-
несколько сот геномов разных видов. В целом это направление теперь называют
геномикой. Это, вероятно, одна из главных точек роста современной молеку-
молекулярной генетики.
Непосредственно для секвенирования была выбрана линия Е. coli К12
MG 1655, из которой исключены профаг X и F-плазмида, а другие генетические
манипуляции сведены к минимуму. Длина генома этой линии L = 4639221 н.п.
Эта последовательность отвечает кольцевой генетической карте Е. coli K12, ка-
калиброванной на 100 мин по времени конъюгационного переноса (рис. 12-1).
Начало отсчета карты выбрано между генами lasT и thrL.
98
П. Генетика
Таблица 12-1
Общие характеристики некоторых секвенированных прокариотических и
эукариотических клеточных геномов
Биологический объект, вид
Mycoplasma genitalium
Mycoplasma pneumoniae
Borrelia burgdorferi
Aquifex aeollcus
Methanococcus jarmaschii
Helicobacter pylori
Methanobacterium
thermoautotrophicum
Haemophilus influenzae
Archaeoglobus fulgidus
Bacillus subtilis
Escherichia coli
Saccharomyces cerevisiae
Caenorabdilis elegans
Drosophila melanogaster
Homo sapiens
L, Мб
0.580
0.816
0.910
1.551
1.66
1.667
1.751
1.830
2.178
4.214
4.639
12.068
100.0
120.0
2910.0
Число
цистронов
470
677
853
1512
1738
1590
1855
1743
2436
4100
4288
5885
12 178
13 600
38 588
Год
1995
1996
1997
1998
1996
1997
1997
1995
1997
1997
1997
1996
1998
2000
2001
Примечание. Таблица составлена по данным оригинальных работ, опубли-
опубликованных в журналах "Nature", "Science", "NAR" и др. в 1995-1998 гг., а также базы
данных GenBank [Karp, Riley, 1998] и последних публикаций [Adams et al., 2000;
Venter et al., 2001]. Методы оценки и сравнения возможных цистронов и их белков
допускают некоторые неоднозначности в интерпретации. Поэтому оцененные числа
цистронов не надо воспринимать как окончательные. Оценки некоторых OfiFoB как
цистронов могут быть уточнены.
Общие характеристики последовательности ДНК генома Е. coli K12 таковы:
87.8 % генома занимают реальные и вероятные белок-кодирующие гены, или
цистроны. Примерно одна треть из них была известна ранее, а остальные
выбраны среди огромного числа новых открытых рамок трансляции (воз-
(возможных цистронов, или ORFob) путем сложного сопоставления многих
свойств, имеющих характерные различия между кодирующими и некоди-
рующими районами. Функции 38 % этих цистронов неизвестны;
0.8 % — гены стабильных фракций РНК (т-РНК, р-РНК и др.);
0.7 % — некодирующие повторы;
11.0 % генома — функциональные сайты и другие участки, выполняющие ре
ляторные и другие функции.
Таким образом, геном Е. coli K12 очень плотно нагружен генами (~ 88.5 %),
а межгенные участки занимают относительно малую долю (~ 11 %). Из 4288 вы-
выявленных или предсказанных цистронов 1853 описаны ранее, а 2435 — новые.
Самый большой цистрон содержит 7149 н.п. B383 кодона), функция его неиз-
неизвестна. Средний размер цистрона 951 н.п. C17 кодонов). Средний интервал ме-
между цистронами — 118 н.п. Однако межгенные интервалы в большинстве своем
Notfor sale! For preview only! Scanned by Mykhaylo (molbioKa-ukr.net)
Рис. 12-1. Общая структура генома бактерии Е. coli K12 [Blattner et al., 1997]. Начало
(origin) и конец (terminus) репликации изображены зеленой линией, а реплихоры 1 и 2 —
голубыми стрелками. Шкала указывает координаты карты в парах нуклеотидов и в ми-
минутах конъюгационного переноса (фактически — в центисомах, т.е. в процентах от пол-
полной ДНК). Распределение генов показано на двух внешних окружностях: оранжевые по-
полоски — гены, локализованные на одной цепи ДНК, желтые — на противоположной
цепи. Красные стрелки указывают локализацию и направление транскрипции генов
р-РНК, зеленые — генов т-РНК. Следующая окружность иллюстрирует положение по-
повторов REP в виде раздельных тонких штрихов. Следующая окружность показывает
сходство между белками профагов и Е. со//, которое мы здесь не обсуждаем. Наконец, в
центре представлена радиальная картина (похожая на оранжевый солнечный взрыв)
распределения так называемого индекса адаптации кодонов (CAI), который отражает
степень экспрессии отдельных генов.
12. Что содержит полный геном Е. colii 99
содержат различные функциональные сайты, т.е. выполняют регуляторные
функции. Кроме того, цистроны не содержат интронов — внутренних некоди-
рующих участков.
Известно, что цистроны выделяются в ДНК и м-РНК начальными и ко-
конечными знаками пунктуации. В общей форме они были известны ранее и вне-
внесены в генетический код. Однако в геноме Е. coli они встречаются с различны-
различными частотами:
Знаки пунктуации
начальные
ATG 3542
GTG 612
TTG 130
АТТ1
CTG 1
конечные
ТАА 2705
TGA 1257
TAG 326
Интересно, что у 405 пар смежных цистронов вообще нет межгенных ин-
интервалов: знак начала трансляции одного частично перекрывается с конечным
знаком другого:
(нач) (нач) (нач) (нач)
ATGA, TAATG, TGATG, GTGA, и др.
[кон] [кон] [кон] [кон]
кул
По неполным данным на январь 1998 г. [Karp, Rilet, 1998] сложность моле-
улярно-генетической системы управления и метаболической сети Е. coli мож-
можно охарактеризовать следующим образом:
Длина ДНК генома, Мб 4.6
Полное число генов 4909
Число цистронов 4288
Число кодируемых ими ферментов 804
Число метаболических реакций 988
Число метаболических путей 123
Число химических веществ, участвующих в ме- 1303
таболизме
Число фракций т-РНК (генов т-РНК) 79 (86)
Число регуляторных белков 60
В таких случаях специалисты говорят: "жизнь при 4909 генах". Метаболизм
сложен, но не запредельно. В дальнейшем приведенные цифры могут возрасти
в ходе исследований за счет новых знаний.
Более подробная классификация цистронов по 22 функциональным клас-
классам представлена в табл. 12-2. Здесь примерно 1/4 клеточных ресурсов связана с
метаболизмом малых молекул, 1/8 — с метаболизмом макромолекул и 1/5 — с
клеточными структурами и процессами. В метаболизме малых молекул ключе-
ключевую роль играют синтез, распад и преобразование нуклеотидов E8 цистронов),
аминокислот A31), энергетические процессы B43), транспорт A46), централь-
100
II. Генетика
Таблица 12-2
Распределение цистронов и белков Е. сой по 22 функциональным
[Blattner et al., 1997]
Функциональный класс
Регуляторная функция
Предполагаемая регуляторная функция
Структура клетки
Предполагаемые мембранные белки
Предполагаемые структурные белки
Фаги, транспозоны, плазмиды
Транспортные и связывающие белки
Предполагаемые транспортные белки
Энергетический метаболизм
Репликация, рекомбинация, модификация и репарация ДНК
Транскрипция, синтез, метаболизм и модификация РНК
Трансляция, посттрансляционная модификация белков
Клеточные процессы, включая адаптацию и защиту
Биосинтез кофакторов, простетических групп и носителей
Предполагаемые шапероны
Биосинтез и метаболизм нуклеотидов
Биосинтез и метаболизм аминокислот
Метаболизм фаттиевой кислоты и фосфолипидов
Катаболизм соединений углерода
Центральный промежуточный метаболизм
Предполагаемые ферменты
Другие известные гены (генные продукты и фенотипы неизвестны)
Гипотетические, неклассифицированные, неизвестные
Всего...
классам
Число
белков
45
133
182
13
42
87
281
146
243
115
55
182
188
103
9
58
131
48
130
188
251
26
1632
4288
%
1.05
3.10
4.24
0.30
0.98
2.03
6.55
3.40
5.67
2.68
1.28
4.24
4.38
2.40
0.21
1.35
3.06
1.12
3.03
4.38
5.85
0.61
38.06
100
ный промежуточный метаболизм A88) и другие процессы. В частности, систе-
системы, выполняющие основные генетические процессы, содержат следующее
число генов ( % генома):
Репликация, рекомбинация, модификация и ре-
репарация ДНК
Транскрипция, синтез, метаболизм и модифика-
модификация РНК
Трансляция и посттрансляционная модификация
белков
115 B.68 %)
55 A.28 %)
182 D.24 %) +
+ 21генр-РНК +
+ 86 генов т-РНК
Кроме того, найдено 9 цистронов, контролирующих синтез шаперонов —
вспомогательных белков, способствующих формированию правильной про-
пространственной упаковки всех остальных белков. Этот процесс называется са-
самоорганизацией, или фолдингом белков.
12. Что содержит полный геном Е. coli!
101
ILoci of \
transcription
system /
Loci of
translation
system
ILoci of
replication
system
DNA
mRNA
Proteins WS
Рис. 12-2. Блок-схема клеточной универсальной системы самовоспроизведения (сай-
зера) [Ратнер, 1983, 1993]. Блоки отвечают молекулярным системам репликации,
транскрипции и трансляции; система сегрегации на схеме не показана. Сайзер спосо-
способен к самовоспроизведению всех своих макромолекулярных компонент и всех других
компонент с аналогичной пунктуацией процессов. Он является ядром молекуляр-
но-генетической организации клетки. Подробности см. в тексте (гл. 18 и рис. 18-3).
Помимо различных вспомогательных функций, эти системы в совокупности
образуют сайзер — универсальную систему самовоспроизведения клетки [Рат-
[Ратнер, 1993; Ratner et al., 1996]. Сайзер составляет сердцевину молекулярно-генети-
ческой системы управления клетки. Несмотря на внушительное число участвую-
участвующих генов (~ 460, свыше 10 % всех генов), принципиальная блок-схема сайзера
достаточно проста [Ратнер, 1983, 1993] (рис. 12-2).
Далее рассмотрим разнообразие функциональных единиц транскрипции.
Для генома Е. coli и других энтеробактерий характерно присутствие управляе-
управляемых единиц транскрипции — оперонов (см. гл. 23). Первые опероны были от-
открыты именно у Е. coli: /ас-оперон, контролирующий сбраживание сахара лак-
лактозы, fr-p-оперон, контролирующий синтез аминокислоты триптофана, и др.
(см. [Ратнер, 1983]). Важной особенностью оперонов является наличие обрат-
обратной связи между концентрацией контролируемого метаболита и наработкой
ферментов его синтеза или распада. Всего в геноме Е. coli выявлено и предска-
предсказано 2584 оперона. Среди них:
73 % содержат 1 цистрон,
16 % — 2 цистрона,
4.6 % — 3 цистрона (в том числе /ас-оперон),
6 % — 4 цистрона и более (в том числе trp-, his-опероны).
Все они имеют не менее 1 промотора — начального знака транскрипции.
Опероны управляются регуляторными белками через специфические
функциональные сайты управления. Например, белок-репрессор /ас-оперона
узнает его оператор — функциональный сайт и через него подавляет функцию
инициации транскрипции. Иногда опероны подчинены нескольким регулятор-
ным белкам и имеют несколько регуляторных сайтов [Ратнер, 1983,1993]. Всего
по данным секвенирования выявлено 45 цистронов, кодирующих белки с регу-
102 II. Генетика
ляторными функциями, и еще цистроны 133 предполагаемых регуляторных
белков. Большинство из них, вероятно, участвуют в управлении оперонами.
Так, внутри областей с предсказанными сайтами управления (в основном
оперонов)
89.2 % регулируются 1 белком (в том числе frp-оперон),
8.4 % — 2 белками (в том числе /ас-оперон),
2.4 % — 3 белками и более.
В свою очередь, эти области содержат:
81.2 % — 1 сайт управления,
12.2 % — 2 сайта управления (в том числе /ас-оперон),
6.6 % — 3 сайта управления и более.
Это значит, что большинство оперонов регулируются достаточно просто.
Этим они существенно отличаются от генов эукариот, которые подвержены
действию многих общих и специфических белковых факторов управления.
Геном Е. coli содержит 2 функциональные единицы репликации. Ф. Блатт-
нер и др. [Blattner et al., 1997] назвали их реплихорами (рис. 12-1). Общее двусто-
двустороннее начало репликации (ori, origin) локализовано примерно на участке
84.5 мин конъюгационного переноса и занимает ~ 250 н.п. В этой зоне иниции-
инициируется двусторонняя репликация. Реплихор 1 ориентирован по часовой стрел-
стрелке, реплихор 2 — против. Оба процесса заканчиваются на противоположном
участке генетической карты, ~ 34-35 мин, где каждый из них имеет свой от-
отдельный ориентированный терминальный знак (ter) — Т1 и Т2. Следует отме-
отметить, что традиционно участки репликации, ограниченные знаками ori и ter,
называют репликонами [Ратнер, 1983].
Геном Е. coli K12 содержит также значительное число необязательных (фа-
(факультативных) включений — профагов, плазмид и транспозонов. Выявлено
87 цистронов и белков этих включений (табл. 12-2). Число их может быть раз-
различным, поскольку они подвижны, способны к внедрению в геном и выщепле-
нию из него. Наилучшим образом это продемонстрировано для умеренного фа-
фага X и полового фактора (плазмиды) F, которые в данной линии отсутствуют.
Многие фаги исключаются из генома не полностью, оставляя там в качестве
"следа" некоторые свои гены. Эти остатки, неспособные к самостоятельному
перемещению и развитию, называют "критическими" фагами. Среди факуль-
факультативных включений в этой линии найдена 41 копия различных транспозонов
(IS), которые участвуют в процессах внедрения и исключения плазмид.
Наконец, следует отметить, что геном Е. coli содержит ряд функциональ-
функциональных и нефункциональных повторов. Октамер GCTGGTGG отвечает "горячим
точкам рекомбинаций" (так называемым х-сайтам). Он встречается в сотнях
позиций в обеих ориентациях и играет ключевую роль в конъюгационной ре-
рекомбинации и других генетических процессах. Найдено большое число копий
E81) небольшого палиндромного повтора REP длиной ~ 40 н.п. Функция их не-
неизвестна. В сумме они занимают 0.54 % ДНК генома. Известны и другие повто-
повторы. В основном они попадают в межцистронные интервалы.
Таким образом, молекулярно-генетическая система управления Е. coli ока-
оказалась хотя и сложной, но вполне обозримой. Геном кодирует белки всех ос-
основных генетических процессов и систем: контролирует синтез и метаболизм
12. Что содержит полный геном Е. coli? 103
мономеров, энергетику, транспорт, клеточные процессы, защитные реакции.
Хотя функции 38 % выявленных белков пока неизвестны, скорее всего, они по-
пополнят уже обозначенные функциональные группы. Опероны как управляемые
единицы транскрипции являются доминирующим вариантом организации ге-
генов Е. coli K12 и других прокариотических клеток.
Сравнительный анализ молекулярно-генетических систем Е. сой и других
объектов (см. табл. 12-1) позволил выявить многие гомологичные гены, оце-
оценить степень сходства геномов, а также высказать предположения о минималь-
минимальной сложности гипотетической (а может быть, первичной?) клетки. Ясно, что
для организации клетки необходим некоторый минимум молекулярных струк-
структур и процессов. В табл. 12-1 приведены суммарные данные по числу генов и
размерам многих секвенированных клеточных геномов. Минимальный геном
имеет микоплазма М. genitalium — 0.58 Мб, 470 генов. Путем сравнения геномов
выявлено, что минимальная клетка, способная к автономной жизнедеятельно-
жизнедеятельности и самовоспроизведению, должна была бы содержать не менее 250-300 наи-
наиболее существенных генов. Группа японских исследователей [Tomita et al.,
1998] показала, что можно обеспечить все основные метаболические потребно-
потребности клетки, трансляцию и репликацию РНК-генома в системе со 127 генами.
Правда, при этом клетка должна быть лишена архива ДНК, репарационных и
других важных систем защиты и помехоустойчивости, что делает ее эволюци-
онно беззащитной.
Для сравнения мы приводим также данные по первым секвенированным ге-
геномам эукариот (см. табл. 12-1). Хотя физические размеры геномов возрастают
на 2-3 порядка величины, числа генов при этом увеличиваются всего в несколь-
несколько раз. Эта особенность связана с тем, что геномы эукариот становятся
"рыхлыми" и содержат до 95-99 % некодирующего генетического материала.
Например, в геноме человека суммарная длина всех экзонов не превышает 1.1 %.
Таким образом, сложность эукариотических геномов связана не с увеличе-
увеличением числа генов, а, скорее, с усложнением их регуляторных отношений.
В заключение выражаю благодарность К.С. Макаровой, Ю.И. Вульфу и
А.Э. Келю за содействие в адаптации последних данных по клеточным геномам.
Список литературы
Ратнер В.А. Молекулярная генетика: принципы и механизмы. Новосибирск: Наука,
1983.
Ратнер В.А. Концепция молекулярно-генетических систем управления. Новосибирск:
Наука, 1993.
|Сайт в INTERNET: ftp://ncbi.nlm.nih.gov/genbank/genomes
Adams M.D. et al. The genome sequence of Drosophila melanogaster // Science. 2000. Vol. 287.
. 2185-2195.
Blattner F.R. et al. The Complete Genome Sequence of Escherichia coli K12 / Science. 1997.
Vol.277. P. 1453-1462.
Karp P.D., Riley M. EcoCyc: Encyclopedia of E. coli genes and metabolism.
http://ecocyc.PangeaSystems.com/ecocyc/ecocyc.html
Ratner V.A. et al. Molecular Evolution. Berlin e.a.: Springer-Verlag, 1996.
104 II. Генетика
Tomita M. et al. A virtual cell with 127 genes// Proc. 1st Intern. Conf. "Bioinformatics of
Genome Regulation and Structure (BGRS'98)". Novosibirsk: lnst. Cytol. Genet., 1998. Vol. I
P. 97-99.
Venter J.C. et al. The sequence of human genome // Science. 2001. Vol. 291, N 5507.
P. 1304-1351.
13. Индукция транспозиций
мобильных генетических элементов
стрессовыми воздействиями*
Введение
Мобильными генетическими элементами (МГЭ) называют подвижные фраг
менты генома клетки, способные к самостоятельным перемещениям внутри ге
нома. МГЭ открыты американским генетиком Барбарой Мак-Клинток на ку
курузе в 1951 г., а в 1983 г. это открытие было увенчано Нобелевской премией. I
60-х годах МГЭ были открыты также у микроорганизмов, причем выявлены и
некоторые молекулярные особенности и механизмы транспозиций (перемеше
ний внутри генома). Наконец, в конце 70-х годов одновременно в СССР -
группой советских генетиков во главе с Г.П. Георгиевым, и в США — группа
американских генетиков во главе с Д. Хогнессом, были открыты МГЭ у дрозо
филы. С этого момента исследования МГЭ существенно ускорились. Кром
того, различные МГЭ были найдены также у дрожжей, млекопитающих, вклю
чая человека, и других объектов. Иначе говоря, мир МГЭ оказался велик
многолик.
МГЭ подвижны в геноме хозяина, причем содержат внутри себя гень
обеспечивающие транспозицию. Опыт построения генетических карт, казг
лось, твердо указывал, что положение генов на карте очень стабильное (с точ
ностью до редких хромосомных перестроек), устойчиво наследуется, а сам
карты являются специфичными для видов. Однако оказалось, что это относш
ся лишь к части генома, действительно преобладающей и стабильной, но не ж
черпывающей его строение. Значительная часть генома представлена разли*
ными МГЭ, среди которых многие копии способны к относительно частым ш
ремещениям (со скоростями до 10~3-10 событий на копию за поколение, 41
значительно выше скоростей возникновения мутаций и перестроек). Таким о(
разом, генетический материал генома пришлось разделить на две сопостав!
мые части: 1) устойчивую — совокупность устойчивых генов и других элементе
генома и 2) подвижную — совокупность копий МГЭ генома, способных к пер
мещениям со всеми вытекающими отсюда генетическими последствиями (м'
тациями генов, перестройками и т.д.).
Поскольку МГЭ оказались подвижными, сразу же возник вопрос об i
роли в геноме. В 1980 г. Дулиттл и Сапиенца и параллельно Орджел и Крик bi
сказали гипотезу об "эгоистичной ДНК", согласно которой МГЭ являются "г
* Вариант этой главы опубликован ранее: Ратнер В.А., Васильева Л.А. // Соросовский обра
вательный журнал. 2000. Т. 6, № 6. С. 14-20.
13. Индукция транспозиций 105
томными паразитами", "бродягами", которые самостоятельно перемещаются в
геноме, наследуются вместе с другими генами генома и, внедряясь в функцио-
функционирующие гены, способны вызывать их мутационные нарушения. Действи-
Действительно, все эти проявления МГЭ были найдены, причем оказалось, что у дрозо-
дрозофилы подавляющая доля известных мутаций в генах вызвана именно инсер-
циями (внедрениями) МГЭ, а не обычными заменами нуклеотидов. Однако
вопрос о том, могут ли МГЭ выполнять какие-либо полезные функции в гено-
геноме, остался открытым.
Эту проблему можно исследовать с разных сторон. Прежде всего, можно
выбрать хорошо изученный ген, контролирующий дискретный менделевский
признак, экспериментально внедрить в его окрестность (или удалить из нее)
копию МГЭ и изучить генетические последствия. Таких примеров достаточно
много (см. [Гвоздев, 1998а, б]). Они указывают на реальную возможность такой
регуляции активности генов со стороны копий МГЭ.
Более сложная ситуация возникает при исследовании количественных при-
признаков, которые контролируются многими генами, образующими полигенные
системы. Считается, что полигенные системы содержат гены главного эффекта
(олигогены, главные гены), которые необходимы для формирования количест-
количественного признака, и полигены (гены малого эффекта, модификаторы), которые
влияют на проявление главных генов [Васильева, Ратнер, 2000]. Предполагается,
что отдельные полигены вносят малый вклад в проявление главных генов (т.е.
слабо их модифицируют), рассеяны по геному, а их число на порядок величины
или более превышает число генов главного эффекта. В качестве "непрерывных"
количественных признаков (в отличие от дискретных) могут быть взяты любые
измеримые характеристики. Например, у дрозофилы такими признаками явля-
являются: масса особи, размер крыла, число абдоминальных щетинок, длина фраг-
фрагмента радиальной жилки крыла, концентрация глазного пигмента и др.
Если предположить, что МГЭ играют существенную роль в модификации
количественных признаков, то для изучения этого влияния придется использо-
использовать другие экспериментальные средства. Необходимо работать не с отдельным
полигеном (и копией МГЭ), а со всей полигенной системой и с суммарным ри-
рисунком (паттерном) локализации копий МГЭ в геноме. Только тогда феноти-
пический эффект воздействия может быть заметен. При этом подразумевается,
что взаимоотношения копий МГЭ и полигенов примерно такие же, как в изу-
изученных случаях регуляторного действия копий МГЭ на смежно расположенные
гены главного эффекта: усиление функции или ее подавление.
Экспериментально можно использовать два основных подхода. Во-первых,
можно индуцировать транспозиции МГЭ различными стрессовыми и генетиче-
генетическими воздействиями (тепловой шок, у-облучение, определенные варианты ге-
генетических скрещиваний и др.). При этом можно контролировать, с одной сто-
стороны, перемещения копий МГЭ, а с другой — изменения проявления количе-
количественного признака. Выявление связи между ними будет свидетельствовать о
модифицирующем влиянии МГЭ на полигены.
Во-вторых, можно выполнить отбор по количественному признаку в попу-
популяции (линии) в сторону его увеличения ((+)-отбор) или уменьшения ((-)-от-
бор) и параллельно контролировать изменение рисунка локализации копий
106
II. Генетика
МГЭ в политенных хромосомах. Определенный, воспроизводимый и устойчи
вый отклик (изменение) рисунка МГЭ на отбор будет означать, что некоторы
копии МГЭ модифицируют проявление полигенов.
Геном дрозофилы, гены и МГЭ
Дрозофила оказалась очень удобным объектом для исследования МГ?
Международная программа полного секвенирования генома дрозофилы прел
ставила к 2000 г. полную последовательность ДНК с указанием положений все
известных генов, других генов с неизвестными пока функциями, регуляторны
сайтов и других функциональных субструктур. Если исключить прицентромер
ный гетерохроматин, то гаплоидный геном дрозофилы содержит ~ 1.2 х 108 н.г
ДНК, в том числе ~ 13 600 локализованных генов [Adams et al., 2000], кодирую
щих различные типы РНК и белков (табл. 13-1). Примерно 5 % суммарно
ДНК генома непосредственно участвует в кодировании, а остальные 95 % счи
таются некодирующими (интроны, межгенные промежутки, которые содержа
многие знаки управления — энхансеры, инсуляторы и другие участки (съ
[Гвоздев, 1996]), и, возможно, выполняют какие-то иные функции. Указанны
гены, если так можно выразиться, "известны нам в лицо", т.е. они клонировг
ны и секвенированы. Примерно для половины секвенированных генов извест
ны их функции или сходство с известными функционирующими генами, част
известны их механизмы управления, выявлены их белковые продукты и tj
Другая половина секвенированных генов пока функционально не описана и н
на что не похожа. Их анализ ожидается в будущем.
Что касается полигенов, то здесь положение совсем иное. Любопытно, чт
до сих пор ни один полиген "не известен нам в лицо", т.е. не выделен, не клс
нирован и не секвенирован. Неясно, чем полигены отличаются от обычных п
нов. Однако, если все полигены различны, то их число в геноме дрозофил
должно было бы достигать 100 000. А если они сопоставимы по длине с генам!
то должны были бы занимать до 50 % генома, что мало вероятно.
Таблица 13-1
Геном Drosophila melanogaster [Adams et al., 2000]
Показатель
ДНК эухроматиновой части генома
Кодирующая часть генома
Число локализованных генов по результатам
секвенирования
Некодирующая часть генома
Геномная система МГЭ
Число дисков на цитологической карте поли-
политенных хромосом
Число сегментов на карте Бриджеса:
1-й уровень
2-й уровень
Размер сегментов 1-го уровня
Размер
1.2 х 10» н.п.
6.0 х 106 н.п.
13 600
1.14 х 108 н.п.
1.1 х Ю7 Н.П.
5100
102
612
190-1820 тыс. н.п.
%
100
5
95
10
13. Индукция транспозиций
107
Раздавленные
клетки-мишени
Предметное
стекло
Заморозить сухим льдом
Отмыть этанолом
Погрузить в жидкий агар
Денатурировать ДНК
Добавить радиоактивный зонд
Отмыть несвязавшуюся метку
Радиоавтография
Зачерненные области
отражают расположение
зерен серебра и
соответствуют сайтам,
в которых произошла
гибридизация с зондом
Дрозофила имеет одно
удивительное свойство, кото-
которое активно используется в
эксперименте: огромные поли-
тенные хромосомы клеток
слюнных желез имеют харак-
характерную поперечную исчерчен-
ность, содержат до 5100 окра-
окрашенных дисков (участков плот-
плотной упаковки) [Жимулев, 1996].
Порядок дисков тоже рассмат-
рассматривается как хромосомная кар-
карта (карта Бриджеса), она весьма
устойчива при наследовании и
характерна для вида. Эта карта
разделена на 102 сегмента 1-го
уровня, 612 сегментов 2-го
уровня и более мелкие сегмен-
сегменты (см. табл. 13-1). Цитологиче-
Цитологическая карта Бриджеса охватыва-
охватывает плечи больших хромосом,
содержащие примерно 75 % га-
гаплоидного набора ДНК и орга-
организованные в диски. Осталь-
Остальные 25 % ДНК входят в районы
прицентромерного гетерохро- Рис. 13-1. Схема метода молекулярной гибридиза-
матина, где дисковая организа- ции ДНК in situ [Льюин, 1987].
ция отсутствует.
Локализацию копий ДНК на цитологической карте Бриджеса проводят ме-
методом молекулярной гибридизации in situ (рис. 13-1) [Льюин, 1987]. Для этого
используется специальный зонд, содержащий определенный фрагмент ДНК,
меченный радиоактивной или флуоресцентной меткой. Гибридизация ДНК
зонда с денатурированной ДНК политенных хромосом производится прямо на
давленых цитологических препаратах (in situ, т.е. на месте!) клеток слюнных
желез личинок дрозофилы. Если фрагмент ДНК зонда встречает в ДНК хромо-
хромосом гомологичный участок, то он гибридизуется с ним по принципу перекрест-
перекрестной комплементарности одноцепочечных участков ДНК. Препараты покрыва-
покрываются фотоэмульсией, длительно экспонируются и проявляются. На получен-
полученных фотографиях диски, где произошла гибридизация, выявляются по
скоплению метки.
Если зонд содержит ДНК МГЭ, то можно найти точки локализации копий
этого МГЭ с точностью до видимых сегментов карты Бриджеса (рис. 13-2). В
результате в любом эксперименте по индукции транспозиций МГЭ или по от-
отбору количественного признака можно следить за изменениями в рисунках ло-
локализации копий МГЭ. Кроме того, можно убедиться, что прицентромерный
гетерохроматин (не попадающий в карту Бриджеса) тоже содержит очень боль-
108 П. Генетика
42F*
42 Е
Puc. 13-2. Результаты молекулярной гибридизации in situ зонда, содержащего фрагмент
ДНК МГЭ412, с гомологичными участками политенных хромосом Drosophila melano-
gaster [Васильева, Ратнер, 2000]. Видны диски цитологической карты. Указаны сегменты
карты Бриджеса, в которых локализованы копии МГЭ, они соответствуют местам скоп-
скопления метки: 42Е, 42F, 45D, 60С и др.
шое число копий различных МГЭ [Adams et al., 2000]. Эти копии в основном
дефектны и не участвуют в транспозициях. Далее мы исключим их из рассмот-
рассмотрения.
Интересно, что названия некоторых МГЭ отражают их подвижные свойст-
свойства: МДГ — мобильные диспергированные гены, gipsy — цыган, hobo — бродяга,
stalker — гордо шествующий, крадущийся, и др.
Таким образом, "площадка для игр" определена. Главный вопрос, который
будет рассмотрен ниже, остается прежним: играют ли МГЭ генома дрозофилы
какую-либо дополнительную функциональную или эволюционную роль по от-
отношению к полигенным системам, кроме обозначенных выше случайных,
"эгоистичных" блужданий в геноме?
Индукция транспозиций
стрессовыми воздействиями
Известно, что температура культивирования способна существенно повли-
повлиять на экспрессию морфологических признаков у дрозофилы. Однако такие
модификации обычно не наследуются. Наследуемые изменения фрагментов
радиальной жилки крыла после ступенчатого температурного воздействия
B9 °С -> 18 °С) на линию riC впервые были обнаружены одним из авторов
(Л.А.В.). Оказалось, что имеются два особо чувствительных периода развития
на куколочной стадии, когда эффект воздействия наиболее контрастный: 1-й —
113 ± 5 ч и 2-й — 149 ± 5 ч. В 1-м периоде экспрессия фрагментов жилки была
минимальной, во 2-м — максимальной. Любопытно, что эти индуцированные
13. Индукция транспозиций 109
фенотипы закрепились в потомстве и дали начало двум "температурным" ли-
линиям, соответственно пТПЗ и ИТ149, существенно различающимся по длине
фрагментов жилки крыла.
Это явление было удивительно прежде всего потому, что температурное
воздействие всегда считалось немутагенным, оно должно было вызывать не на-
наследуемые изменения, а преходящие модификации. Еще более любопытным
оказался факт, что у "температурных" линий рисунки локализации МГЭ оказа-
оказались существенно измененными по сравнению с исходной линией riC. Иначе
говоря, температурное воздействие не только повлияло на экспрессию призна-
признака, но и параллельно индуцировало транспозиции МГЭ. Но самым поразитель-
поразительным было то, что две линии, riSN — "селекционная" и пТПЗ — "температур-
"температурная", полученные совершенно разными путями, имели как сходные фенотипы,
так и весьма сходные рисунки локализации нескольких МГЭ. Мало того, спек-
спектры транспозиций в обоих случаях тоже были сходны и неслучайны. Иначе го-
говоря, индуцированные изменения признаков оказались коррелированными с
индуцированными же транспозициями МГЭ. Этот результат явно свидетельст-
свидетельствовал в пользу гипотезы о модифицирующем действии МГЭ на полигены изу-
изучаемой системы.
Предыдущие эксперименты выполнены на гетерогенной линии дрозофил,
где полигенная система и рисунки локализации МГЭ были полиморфными.
Гетерогенность затрудняет изучение транспозиций МГЭ. Поэтому явление ин-
индукции транспозиций было специально исследовано в нескольких особо одно-
однородных линиях дрозофил. Такие линии называют изогенными. Их можно полу-
получить при помощи довольно сложной схемы скрещиваний, в результате которой
в диплоидных зиготах встречаются одинаковые копии гаплоидных геномов
(см. ниже). Такие геномы полностью гомозиготны (в том числе и по рисункам
локализации МГЭ), и на этом фоне легко идентифицировать каждое событие
транспозиции.
Действующими факторами были: A) обычный тепловой шок (ТШ), т.е. по-
повышение t культивирования самцов до 37 °С в течение 1.5-2 ч; B) тяжелый те-
тепловой шок (ТТШ), т.е. троекратное чередование теплового C7 °С) и холодо-
вого D °С) шока; C) у-облучение самцов при нескольких дозах. Во всех случа-
случаях было обнаружено увеличение скорости транспозиций на 1-2 порядка вели-
величины (рис. 13-3, 13-4).
Существенной особенностью феномена индукции транспозиций ТТШ
была значительная ее неравномерность в различных сегментах генома.
В табл. 13-2 приведены данные одного из последних экспериментов. В двух сег-
сегментах, 43В и 97DE, сосредоточено свыше 70 % всех транспозиций, они назва-
названы "горячими сегментами". Вероятность транспозиции в 43В равна 0.7 E9 со-
событий у 85 личинок) и в 97DE — 0.93 G9 событий у 85 личинок). Это очень вы-
высокие значения вероятности, которые означают, что транспозиции в эти
сегменты обнаруживаются у большинства личинок-потомков. В 16 других сег-
сегментах локализованы остальные 30 % транспозиций. Ясно, что "горячие сег-
сегменты" должны иметь какую-то особенность, обеспечивающую их исключи-
исключительные свойства. В этом эксперименте средняя частота индуцированных
транспозиций достигла огромного значения: 0.11 (т.е. 11 %) событий на сегмент
110
II. Генетика
Таблица 13-2
Индукция транспозиций МГЭ 412 тяжелым тепловым шоком (ТТШ) по данным гибридизации in situ
Сегмент карты
Бриджеса
зс
4В
22В
28А
34В
43В
56Е
Число транспози-
транспозиций (85 личи-
личинок)
1
1
1
4
4
59
4
Сегмент карты
Бриджеса
60В
63А
67А
75С
83D
86D
87В
Число транспози-
транспозиций (85 личи-
личинок)
17
1
1
2
1
6
2
Сегмент карты
Бриджеса
87F
94D
97DE
98Е
18 сегментов
Число транспози-
транспозиций (85 личи-
личинок)
2
1
79
6
193E5)
за поколение. Это значение на 2-3 порядка величины выше, чем значение для
спонтанных транспозиций.
Таким образом, сам феномен индукции транспозиций можно было считать
доказанным. В качестве возможного механизма температурной индукции мы
рассматриваем, в первую очередь, геномную систему ответа на тепловой шок
(ТШ-систему). Эта система хорошо изучена. Она обеспечивает толерантность
клеток к стрессовым внешним и внутренним физиологическим воздействиям.
Гены этой системы имеют специальные функциональные сайты, опознаваемые
регуляторными белками "теплового шока". Белки ТШ-генов выполняют функ-
функции шаперонов, т.е. белков, способствующих формированию правильной про-
пространственной конформации других разнообразных белков или уничтожающих
белки с испорченной конформацией. Тепловой шок нарушает конформацию
многих белков и индуцирует систему толерантности, которая позволяет устра-
устранить возникшие дефекты.
Однако причем тут МГЭ? Во-первых, в нашем случае эти МГЭ были рет-
ротранспозонами, которые размножаются через прямую и обратную транс-
транскрипцию полноразмерной РНК-копии этого МГЭ. Во-вторых, мы предполо-
предположили, что они являются как бы пассивными соучастниками ответа на тепловой
шок. Для этого им достаточно иметь в своей структуре функциональные сайты
для узнавания белков ТШ, которым подчинена промоторная область инициа-
инициации транскрипции самих МГЭ. Тогда индукция системы теплового шока авто-
автоматически будет усиливать транскрипцию и транспозицию ретротранспозонов.
Действительно, компьютерный поиск позволил обнаружить в последователь-
последовательностях 17 секвенированных ретротранспозонов присутствие таких регулятор-
ных последовательностей. Кроме того, найдены короткие последовательности
(так называемые "мотивы"), похожие на функциональные сайты — энхансеры,
которые являются активаторами транскрипции и действуют совместно с сайта-
сайтами теплового шока.
Что касается природы "горячих сегментов", то следует иметь в виду два об-
обстоятельства. Для инсерции копии МГЭ в некоторый сегмент хромосомы необ-
необходимо, чтобы этот сегмент (а) был "распакован", т.е. освобожден от нуклео-
сом и других инактивирующих структур, и (б) содержал внутри себя "мишень"
инсерции, т.е. специфическую последовательность оснований, которая узнает-
Исходная позиция
I
I
Исходная
хромосома
дрозофилы
а Наследование
4С
г
37 "С
37 "С
37 t
4С
Тепло-
холодовой
шок
4С
I I I I I
t t t
t t t
Новые позиции МГЭ
Хромосома
потомства
в следующем
поколении
Рис. 13-3. Индукция транспозиций МГЭ тяжелым тепловым шоком.
Not for sale! For preview only! Scanned by Mykhaylo (molbioKcvukr.net)
10°
Тепловой
шок
Гамма-
облучение
Изоге-
низация
Рис. 13-4. Индукция транспозиций МГЭ 412 различными стрессовыми внешними и
внутренними воздействиями.
13. Индукция транспозиций 111
ся и разрезается ферментом инсертазой. Наилучшими кандидатами на такую
роль являются некоторые гены, активно функционирующие в процессе спер-
сперматогенеза у самцов дрозофилы. "Мишени" инсерции невелики по размеру и,
вероятно, встречаются в геноме довольно часто.
Любопытно, что тепловой шок — не единственный стрессовый фактор, за-
запускающий систему ответа на тепловой шок. Аналогичную роль играют другие
стрессовые факторы, способные вызвать нарушения конформации белков или
их накопление: яды, детергенты, лекарства, этиловый спирт, некоторые другие
химические вещества, а также у-облучение, гипоксия, впрыскивание дефектных
белков, вирусное заражение и т.д. Иначе говоря, система ответа на тепловой
шок — генерализованная система ответа на геномный стресс. Присутствие в
списке факторов этилового спирта может быть грозным предупреждением для
алкоголиков, правда пока среди дрозофил. Все же никогда не мешает поберечь и
человеку свои мобильные элементы от индукции массовых перемещений!
Индукция транспозиций при помощи у-облучения, вероятно, связана с
возникновением индуцированных двухцепочечных разрывов в ДНК хромосом.
Эти разрывы используются ретротранспозонами для инсерции с повышенной
вероятностью, т.е. "залечиваются" ими [Гвоздев, 19986].
Но наиболее мощным индуцирующим воздействием оказался процесс изо-
генизации. Эта генетическая процедура в принципе состоит в том, что при помо-
помощи специального скрещивания из гетерогенной популяции сначала "выхваты-
"выхватывается" вариант гаплоидного генома, а затем он размножается без рекомбинаци-
онного перемешивания с другими вариантами и быстро (в течение 3-4 поколе-
поколений) полностью гомозиготизируется. Процесс "выхватывания" гаплоидного ге-
генома осуществляется путем скрещивания со специальной (так называемой
"балансерной") линией мух, где все хромосомы помечены генами-маркерами, а
кроссинговеры в них заперты. Такое скрещивание можно считать отдаленным
(аутбридинг). Затем, когда в дочерних диплоидных зиготах встречаются две иден-
идентичные копии одного и того же гаплоидного генома, происходит почти полный
инбридинг, т.е. состояние идентичной гомозиготности по всем генам генома.
Существенно, что этот процесс сопровождается массовыми перемещения-
перемещениями копий МГЭ, т.е. индукцией транспозиций. Скорости индукции транспози-
транспозиций и эксцизий (вырезание) копий МГЭ составили огромные цифры: соответ-
соответственно 0.35 и 0.13 событий на копию МГЭ, на гаплоидный геном, в результате
изогенизации. Для МГЭ 412 на 22.14 занятых позиций среднего гаплоидного ге-
генома приходятся в среднем 11.19 транспозиций и 4.4 эксцизий. Фактически это
эквивалентно реорганизации всего рисунка локализации копий МГЭ генома!
Роль индуцирующего фактора в этом случае выполняли как аутбридинг, так
и инбридинг. Вклад аутбридинга в индукцию транспозиций показан экспери-
экспериментально, а механизм действия, вероятно, связан с нарушением синапсиса в
мейозе. Что касается роли инбридинга, то мы предполагаем, что механизмом
индукции транспозиций в этом случае является та же система ответа на тепло-
тепловой шок. Известно, что причинами угнетения жизненных функций при инбри-
инбридинге являются гомозиготизация генома и выщепление в гомозиготной форме
различных дефектных генов, в том числе, вероятно, производящих белки с ис-
испорченной конформацией. Изогенизация эквивалентна очень быстрому ин-
112 II. Генетика
бридингу, в результате которого концентрация дефектных белков может пре-
превысить порог индукции системы ответа на тепловой шок, а через нее — транс-
транспозиций МГЭ.
Известен еще один пример генетического воздействия, вызывающего
мощную индукцию перемещений МГЭ. Это так называемые "дисгенные скре-
скрещивания" линий дрозофил, содержащих (Р-линии) и не содержащих (М-ли-
(М-линии) транспозон Р. Предполагается, что Р-элементы синтезируют некий ре-
репрессор, подавляющий транспозицию и ограничивающий число их копий в
геноме. В геномах клеток Р-линий есть копии Р-элемента, а в цитоплазме —
репрессор. В геномах клеток М-линии Р-элемента нет, а в цитоплазме отсут-
отсутствует репрессор. При скрещивании самок М-линии с самцами Р-линии клет-
клетки полученного гибрида МР имеют половину копий Р-элемента в геноме, но
не имеют репрессора в цитоплазме. Это происходит потому, что яйцеклетки
М-линии исходно не имеют ни Р-элемента, ни репрессора, а спермин Р-ли-
Р-линии переносят копии Р-элемента в геноме, но не имеют цитоплазмы и не пе-
переносят репрессора. Поэтому у гибридов сразу нарушается баланс между чис-
числом копий Р-элемента в геноме и репрессией транспозиций, с огромной ско-
скоростью индуцируются новые транспозиции копий Р-элемента — 0.26 событий
на сегмент за поколение. Это явление, называемое "гибридным дисгенезом",
тоже представляет собой пример индукции транспозиций МГЭ при помощи
генетической процедуры — дисгенного скрещивания.
Таким образом, МГЭ генома дрозофилы оказались не просто "геномными
бродягами", а генетическими объектами, весьма чувствительными к внешним
и генетическим воздействиям. Они реагируют на такое вмешательство вспыш-
вспышками перемещений МГЭ, множественных в геноме и массовых в популяции.
Эти транспозиции способны вызвать дефекты или функциональные измене-
изменения в различных генах и полигенах. Хотя ТТШ сам по себе не является мута-
мутагенным воздействием, но участие МГЭ как промежуточного фактора порож-
порождает новый пласт наследственной изменчивости генома. Этот источник из-
изменчивости может играть ключевую роль в стрессовых условиях
существования популяций.
Список литературы
Васильева Л.А., Ратнер В.А. Полигенная система количественного признака radius
incompletus у дрозофилы: Генетические особенности, взаимодействие с другими генами и
паттерном МГЭ, эволюционные свойства // Современные концепции эволюционной гене-
генетики / Ред. В.К. Шумный, А.Л. Маркель. Новосибирск: ИЦиГ СО РАН, 2000. С. 117-127. I
Гвоздев В.А. Механизмы регуляции активности генов в процессе транскрипции //
Соросовский образоват. журн. 1996. № 1. С. 23-31.
Гвоздев В.А. Подвижные ДНК эукариот. Ч. 1. Структура, механизмы перемещения и
роль подвижных элементов в поддержании целостности хромосом // Соросовский образо-
образоват. журн. 1998. № 8. С. 15-21.
Гвоздев В.А. Подвижные ДНК эукариот. Ч. 2. Роль и регуляция активности генов и эво-
эволюция генома // Там же. С. 15—21.
Жимулев И.Ф. Современные представления об организации и функционировании по-
литенных хромосом // Соросовский образоват. журн. 1996. № 11. С. 2-7.
14. Мобильные генетические элементы 113
Льюин Б. Гены. М.: Мир, 1987. 544 с.
Adams M.D. et al. The Genome Sequence of Drosophila melanogaster // Science. 2000.
Vol. 287. P. 2185-2195.
14. Мобильные генетические элементы:
геномные бродяги или трудяги?*
В гл. 13 мы описали феномен индукции транспозиций стрессовыми факто-
факторами. Здесь рассмотрим результаты генетического и селекционно-генетическо-
селекционно-генетического анализа участия мобильных генетических элементов (МГЭ) генома дрозофи-
дрозофилы в проявлении количественного признака.
Радиальная жилка крыла
Один из соавторов (Л.А.В.) выполнила цикл работ на радиальной жилке
крыла дрозофилы [Васильева, Ратнер, 2000]. В норме эта жилка замкнута, мута-
мутация radius incompletus (ri) прерывает жилку, образуя проксимальный и дисталь-
ный фрагменты (рис. 14-1, а). Их длины легко измеримы и удобны в качестве
количественных признаков. Мы выбрали суммарную длину двух фрагментов в
качестве количественного признака. В линии ri имеется нормальное распреде-
распределение признака. Можно вести отбор на полное восстановление жилки или на
ее полное исчезновение. Отбор ведется путем отсечения из этого распределе-
распределения 10 % особей с максимальным ((+)-отбор) или минимальным ((-)-отбор)
признаком. Отбор вполне эффективен и через 40-70 поколений достигает
окончательных результатов. Считается, что в таких случаях на отбор откликает-
откликается полигенная система, контролирующая признак. Все события происходят на
фоне мутации гена главного эффекта п. В результате из исходной гетерогенной
линии ИС (рис. 14-1, а) получены две "селекционные" линии — соответственно
riSP с полностью восстановленной жилкой (рис. 14-1, б) и riSN без этой жилки
(рис. 14-1, в).
Теперь можно провести генетический анализ полигенной системы [Ва-
[Васильева, Ратнер, 2000]. Для этого отдельные сегменты хромосом линии riC мар-
маркировали видимыми рецессивными мутациями, а затем эту линию скрещивали
с контрастными линиями riSP и riSN. Оказалось, что полигены, влияющие на
экспрессию обоих фрагментов жилки, локализованы не менее чем в 18 крупных
сегментах генома, т.е. генетическая система действительно полигенная. Поли-
гены, влияющие на проксимальный фрагмент жилки, действуют аддитивно,
что прекрасно согласуется с классической теорией полигенов К. Мазера. Поли-
Полигены дистального фрагмента ведут себя посложнее.
Кроме того, в более сложных генетических экспериментах было показано,
что в одной из ri-линий среди полигенов имеются два совершенно необычных
[Васильева, Ратнер, 2000]: а) сегмент, содержащий полиген, контролирующий
большой положительный вклад в длину проксимального (+1.24 единицы шка-
шкалы микроскопа) и дистального фрагментов (+2 единицы), и б) сегмент, содер-
• Глава подготовлена совместно с Л.А. Васильевой.
114
II. Генетика
Рис. 14-1. Экспрессия количественных признаков прерванной радиальной жилки крыл
у дрозофилы [Васильева, Ратнер, 2000].
а — исходная гетерогенная линия, riC; б — "селекционная" линия riSP (результат позитивного от
бора); в — "селекционная" линия riSN (результат негативного отбора).
жащий полиген, контролирующий большой отрицательный вклад в листал
ный фрагмент (-1.08 единицы шкалы). Учитывая, что максимальная длина ра-
радиальной жилки крыла у самок 6 единиц шкалы микроскопа, легко понять, что
вклады этих полигенов огромны!
Таким образом, наше представление о полигенах пополнилось новыми
фактами: 1) полигены могут определять крупный вклад в признак и 2) полиге-
полигены могут иметь противоположное влияние на признак — как увеличивать его,
так и уменьшать. Молекулярная природа "больших" и "малых" полигенов ос-
остается пока неизвестной.
Интересно отметить, что влияние полигенной системы на проявление при-
признака выявляется только на фоне мутации гена главного эффекта п. На фоне
нормального аллеля этого гена, п+, полигенная изменчивость не проявляется,
отбор не эффективен. Это говорит о том, что мутация ri становится лимити-
лимитирующим звеном формирования жилки. Полигенная система проявляется толь-
только через экспрессию лимитирующего гена главного эффекта.
Отклик рисунка локализации копий МГЭ
на отбор по количественному признаку
Как указано выше, изучение изменения рисунка локализации МГЭ в ходе
отбора по признаку может дать свидетельства участия некоторых копий МГЭ в
модификации функций полигенов. Отбор проводился неоднократно в различ-
различных линиях дрозофил, где в качестве количественного признака была выбрана
суммарная длина двух фрагментов радиальной жилки крыла [Ратнер, Василье-
Васильева, 20006]. Отбор был эффективен в тех линиях, где полигенная система этого
признака была генетически гетерогенной. Напротив, в изогенных (т.е. почти
полностью гомозиготных) линиях отбор был неэффективен. Генетическую ге-
гетерогенность можно было задать разными способами: 1) взять исходно гетеро-
гетерогенную линию (п'С); 2) взять смесь различных изогенных линий; 3) индуциро-
индуцировать генетическую гетерогенность и транспозиции МГЭ в изогенной линии при
помощи различных стрессовых воздействий (см. [Ратнер, Васильева, 2000а]).
Во всех этих случаях отбор по количественному признаку был эффективен, т.е.
происходило изменение частот аллелей полигенов, а через них — изменение
среднего признака.
14. Мобильные генетические элементы 115
В том, что полигены откликаются на отбор по количественному признаку,
нет ничего удивительного. Удивительно другое: на отбор откликается также и
рисунок локализации МГЭ генома, которые, как исходно предполагалось, не-
ювисимы и "эгоистичны". Означает ли это, что копии МГЭ влияют на экс-
экспрессию полигенов?
Чтобы оценить реальное значение МГЭ, необходимо учесть, что копии
МГЭ по отношению к полигенам (и к генам вообще) могут играть различную
роль и соответственно иметь различную популяционную динамику при отборе
[Ратнер, Васильева, 20006].
1. Независимые копии МГЭ не сцеплены (или слабо сцеплены) с полигена-
полигенами и не влияют на их проявление. Тогда ожидается, что при отборе по количе-
количественному признаку они не увлекаются отбором, а подвержены чистому генети-
генетическому дрейфу, т.е. случайному процессу эволюции. Исследуемые популяции
имеют конечную, не очень высокую численность: Ne s 100-200 особей. В этом
случае при переходе от поколения к поколению происходит случайный выбор
конечного числа потомков, частота копий МГЭ флуктуирует. Процесс закан-
заканчивается либо случайной фиксацией копии МГЭ, либо ее утратой.
Теория генетического дрейфа позволяет прогнозировать некоторые харак-
характеристики популяции. Например, если вначале частота копии МГЭ в опреде-
определенном сегменте карты Бриджеса былар0, то к завершению процесса она может
стать либо 1 (фиксация), либо 0 (утрата), а полиморфной оставаться не может.
Вероятность случайной фиксации одиночной копии МГЭ
"(Ро) = Ро, A)
а вероятность случайной утраты
v(po) = 1 - Ро- B)
Среднее время фиксации (без учета утрат) отдельной копии МГЭ, согласно
теории генетического дрейфа, равно
4Ne(l ~
Ро
= _ 4Ne(l Po)lnA _ ро) поколений
Эти формулы мы используем ниже.
2. Копии МГЭ-маркеров тесно сцеплены с полигенами, но не влияют на их
функции. Теоретический анализ показывает [Ратнер, Васильева, 20006], что в
этом случае частоты копий МГЭ-маркеров существенно изменяются в ходе от-
отбора полигенов, полиморфизм быстро исчезает, а вероятности фиксаций и ут-
утрат и оценки tfix становятся промежуточными между чистым генетическим
дрейфом нейтральных копий и селективной фиксацией копий-модификаторов.
Иначе говоря, МГЭ-маркеры становятся более похожими на МГЭ-модифика-
торы.
3. Копии МГЭ-модификаторов тесно сцеплены с полигенами и существенно
влияют на их проявление в признаке. Можно ожидать, что в этом случае копии
МГЭ ведут себя так же, как полигены, т.е. достаточно быстро, воспроизводимо и
ггпм птбппр. ттп количественному признаку.
116
II. Генетика
Р(-)
Рис. 14-2. Динамика длины проксимального
(Р) и дистального (D) фрагментов радиаль-
радиальной жилки крыла дрозофилы в ходе пози-
позитивного ("+", справа) и негативного ("-",
слева) отбора после у-облучения в изогенной
линии [Ратнер, Васильева, 20006]. Ордина-
Ордината — длина фрагмента, измеренная в едини-
единицах шкалы микроскопа, абсцисса — номер
поколения.
-—- Таким образом, если рисунок
МГЭ содержит копии всех этих ти-
типов, то быструю реакцию на отбор
должны проявлять только МГЭ-мо-
дификаторы и маркеры, а осталь-
остальные — медленно "дрейфовать" к
фиксации или утрате.
Рассмотрим в качестве примера
результаты одного из селекцион-
селекционно-генетических экспериментов
[Ратнер, Васильева, 20006]. В изо-
изогенной линии после у-облучения
возникла новая генетическая гете-
гетерогенность полигенов и были инду-
индуцированы массовые транспозиции
ретротранспозона 412. Всего воз-
возникло 25 новых участков локализа-
локализации копий МГЭ. Средняя частота
копий МГЭ на новый участок перед
началом отбора р0 = 0.39. Далее
часть линии была подвергнута
(+)-отбору, другая часть — (-)-от-
бору по суммарной длине двух фрагментов радиальной жилки крыла в течение
50 поколений.
На рис. 14-2 показана динамика длин обоих фрагментов, а на рис. 14-3 —
динамика числа гетерозиготных сегментов локализации МГЭ 412. Из рис. 14-2
легко заметить, что в (+)-направлении отбора оба признака быстро (за
30-40 поколений) выходят на плато, когда два фрагмента полностью смыкают-
смыкаются. В (-)-направлении отбора дистальный фрагмент полностью исчезает к
30-му поколению, а проксимальный уменьшается в несколько раз к 50-му по-
поколению. Из рис. 14-3 следует, что число полиморфных сегментов МГЭ за
50 поколений отбора падает в (+)-направлении отбора в 10 раз, и в (-^направ-
(-^направлении — в 4 раза за счет фиксаций и утрат. Средние условные времена фикса-
фиксаций и утрат в эксперименте лежат в интервале 10-20 поколений.
Сопоставим эти данные с теоретическими оценками, сделанными выше.
Среднее время фиксации (без учета утрат) при р0 = 0.39, Ne = 200
tfix = 3.06Ne > 600 поколений.
Среднее время утрат (без учета фиксаций)
iloss = 2ANe > 400 поколений,
т.е. случайный генетический дрейф должен протекать на порядок величины
дольше, чем в реальном эксперименте A0-20 поколений).
С другой стороны, отклик полигенов (т.е. признака) на отбор завершается
за 30-40 поколений. За это время должны фиксироваться все активные аллели
полигенов и сцепленные с ними МГЭ-модификаторы. Однако мы видим, что
14. Мобильные генетические элементы
117
F(-) -50
-30
-10
50 F(+)
t
11.8
21.9
N
hnf
Дл/
Поколения негативного отбора
50
5
1
1
40
7
-
4
30
И
-
4
20
14
+1
1
1
10
15
+1
-
-
5
13
+2
4
4
+2
F0*
21
+4
фик.
утр.
Поколения позитивного отбора
5
14
+1
3
4
10
13
-
2
22
8
+1
3
3
30
5
1 + 1
1
40
2
+1
2
1
50
2
+1
-
t
20.2
12.2
Рис. 74-3. Динамика числа полиморфных сегментов локализации копий МГЭ 412 в ходе
отбора по количественному признаку (см. рис. 14-2) [Ратнер, Васильева, 20006].
за это время фиксируются или утрачиваются фактически все копии МГЭ! Значит
ли это, что все они являются модификаторами полигенов? Не обязательно!
Теоретический анализ и компьютерное моделирование показывают, что в
конечной популяции при отсекающем отборе важнейшую роль играет бессис-
бессистемный инбридинг отбираемых особей. Действительно, при этом выбираются
лучшие (по признаку) особи, среди которых всегда есть относительный
"чемпион", т.е. самый лучший. Он всегда попадает в выборку 100-200 лучших
особей. Его потомки тоже в среднем лучше других; далее они предпочтительно
попадают в выборку следующего поколения и там скрещиваются между собой.
Иначе говоря, отбор становится семейным. При этом в ходе отбора быстро на-
нарастает близкородственная гомозиготизация отбираемых особей, т.е. инбридинг.
Однако инбридинг охватывает не только те гены и полигены, по которым
ведется отбор, но и все другие локусы генома. Поэтому все копии МГЭ, незави-
независимо от их роли, теряют полиморфность за время отбора. Полученные в резуль-
результате рисунки локализации МГЭ будут различны в (+)- и (-)-направлениях от-
отбора, в значительной степени — случайны, но фиксируются они очень быст-
быстро — за время отбора и инбридинга.
Таким образом, быстрота фиксации рисунка МГЭ не означает, что все ко-
копии МГЭ — модификаторы. Более того, можно ожидать, что большинство ко-
копий различных МГЭ не влияют на функции полигенов одной довольно частной
118 II. Генетика
генетической системы. Скорее, число МГЭ-модификаторов этой системы не- |
велико. Тем не менее в ходе этого и других экспериментов были найдены неко-
некоторые копии МГЭ, которые всегда быстро фиксировались в (+)-направлении
отбора, но столь же быстро утрачивались в (-)-направлении. Такова, напри-
например, копия МГЭ 412 в сегменте 60В. Его динамика при отборе полностью соот-
соответствует ожидаемому отклику на отбор МГЭ-модификатора для полигена с
положительным вкладом в признак. Копия МГЭ 412 в сегменте 97DE проявила
противоположный отклик на отбор: в (+)-направлении она быстро исчезала, а в
(-)-направлении столь же быстро фиксировалась. Такая динамика полностью
соответствует ожидаемому отклику на отбор МГЭ-модификатора для полигена
с отрицательным вкладом в признак. Число примеров можно увеличить. Пред-
Предполагается, что среди этой группы копий МГЭ действительно имеются реаль-
реальные МГЭ-модификаторы полигенов.
В гл. 13 мы привели результаты индукции транспозиций МГЭ 412 тяжелым
тепловым шоком, выявив существование "супергорячих" сегментов индукции.
Затем в индуцированной линии был проведен жесткий отсекающий (+)- и
(-)-отбор по длине фрагментов радиальной жилки крыла. Из 35 возникших по-
после индукции полиморфных сайтов локализации МГЭ 412 26 вели себя при от-
отборе как селекционно-независимые копии, а 9 — как селекционно-зависимые.
Эта последняя группа содержит в себе все копии МГЭ-маркеров и модифика-
модификаторов, причем в нее попали все "супергорячие" сайты индукции ТТШ (напри-
(например, 43В и 97DE).
Таким образом, чувствительность сегментов к транспозициям МГЭ суще-
существенно коррелирует с их откликом на отбор, т.е. с влиянием на экспрессию от-
отбираемого количественного признака. Вероятно, обе характеристики зависят
от состояния сегмента в хромосоме: и при индукции в него, и при влиянии
МГЭ на экспрессию локализованного в нем полигена сегмент должен быть
"распакован" и доступен для обоих процессов.
Концепция: геномная система МГЭ
Геном дрозофилы содержит до 50 различных семейств МГЭ [Гвоздев,
1998а, б]. В сумме они занимают до 10 % ДНК генома (табл. 14-1). Число копий
МГЭ можно оценить следующим образом. Средний размер копии МГЭ ~ 3000-
5000 н.п. Если это принять за 10 %, то тогда на копию МГЭ в среднем прихо-
приходится фрагмент ДНК размером ~ 30 000-50 000 н.п. A00 %). Поскольку коди-
кодирующая ДНК составляет в среднем ~ 5 %, это соответствует ~ 1500-2500 н.п. на
такой фрагмент, т.е. 2-3 генам. Следовательно, на 1 копию МГЭ в геноме при-
приходится в среднем 2-3 гена, т.е. их числа сравнимы. Тогда число копий МГЭ в
геноме дрозофилы оценивается в 3000-5000. Каждая копия МГЭ в среднем ок-
окружена фрагментом геномной ДНК размером до 50 000 н.п.
Все эти результаты, а также ряд косвенных данных позволили нам предло-
предложить следующую концепцию [Васильева, Ратнер, 2000]. МГЭ генома представ-
представляют собой не просто "ватагу геномных бродяг", а геномную систему, на долю
которой приходится весьма существенная функциональная и эволюционная
роль. Число копий МГЭ в плечах больших хромосом приближается к числу ге-
14. Мобильные генетические элементы 119
Таблица 14-1
Геномная система МГЭ дрозофилы
Показатель
Число семейств МГЭ
Число копий МГЭ на семейство
Средний размер копии МГЭ
Средний размер фрагмента ДНК на 1 копию
МГЭ
Средний размер кодирующего фрагмента на 1
копию МГЭ
Средний размер кодирующей части гена
Среднее число генов на 1 копию МГЭ
Среднее число копий МГЭ на геном
Характеристика
50
1-100
3000-5000 н.п. A0 %)
30 000-50 000 н.п. A00 %)
1500-2500 н.п. E %)
1000 н.п.
2-3
3000-5000
нов, но специфические сайты локализации МГЭ каждого семейства заполнены
лишь на 25-30 %. Иначе говоря, есть достаточный простор для специфических
перемещений. В плечах больших хромосом сайты локализации МГЭ не случай-
случайны, а сосредоточены в основном в окрестностях функционирующих генов и их
регуляторных зон.
Копии МГЭ способны модифицировать функции менделевских генов и
полигенов, локализованных в их окрестности. Предполагаемый механизм тако-
такого влияния — функции энхансеров, мотивы которых найдены в структуре МГЭ.
Энхансеры — это функциональные сайты, знаки управления, узнаваемые спе-
специальными регуляторными белками и активирующие транскрипцию смежных
генов и полигенов. Зона влияния энхансера может простираться до
50 000-100 000 н.п. вдоль цепи ДНК, т.е. в зоне действия системы МГЭ нахо-
находятся практически все гены генома. Это не значит, что все копии МГЭ действу-
действуют всегда, напротив, в большинстве случаев они могут быть пассивными или
действовать слабо, но возможность влияния открыта. А в таких случаях, как из-
известно, Природа редко остается безучастной.
МГЭ способны к восприятию внешних индуцирующих сигналов: теплово-
теплового шока, других стрессовых факторов, гормонов, тканеспецифичных сигналов
и т.д. Это значит, что копии МГЭ в принципе подчинены генетическому управ-
управлению и могут участвовать в перестройках такого управления. Последователь-
Последовательности ДНК копий МГЭ содержат сотни мотивов функциональных сайтов (т.е.
участков, очень сходных с известными функциональными сайтами) [Ратнер,
Васильева, 2000], которые можно использовать в самых различных ситуациях.
Скорее, это даже "полуфабрикаты" функциональных сайтов, которые, однако,
довольно просто довести до функционального состояния. Поэтому копии МГЭ
можно рассматривать как "подвижные кассеты функциональных сайтов" внутри
Мслекулярно-генетических систем управления клеток. При перемещениях они
порождают новые варианты регуляторной изменчивости. В геномах эукариот,
где 95 % ДНК не кодирует белков, большинство инсерций МГЭ не разрушает
генов или их регуляторных зон, а скорее способно вызывать более "мягкий"
эффект изменчивости — регуляторное влияние на соседние гены и полигены.
120 II. Генетика
Копии МГЭ могут быть полноправными участниками полигенных систем.
Будучи подвижными, они вряд ли сами являются полигенами, однако можно
ожидать, что они являются модификаторами полигенов. Тогда они отбираются
вместе с полигенами при селекции.
В связи с этим можно вернуться к классической концепции менделевских
генов и полигенов. Нет сомнения, что гены главного эффекта (менделевские)
играют роль лимитирующих факторов метаболизма, морфогенеза и формирова-
формирования количественных признаков. В этом случае достаточно предположить, что
полигены по своей молекулярной природе ничем не отличаются от обычных
генов, кодирующих белки, но находятся в нелимитирующем положении в тех же |
цепях метаболизма и т.п. Мутируя, эти гены могут сами стать лимитирующими,
т.е. генами главного эффекта. Каждый менделевский ген в лимитирующем ал-
лельном состоянии имеет группу нелимитирующих менделевских генов, слабо
влияющих на его экспрессию. Их мы и называем полигенами. Это значит, что
мы давно "знакомы в лицо" с полигенами, но не подозревали об этом. Теперь
знакомство становится явным.
Скорее всего, полигены — это любые другие генетические элементы гено-
генома (нелимитирующие, регуляторные гены и др.), влияющие тем или иным об-
образом на экспрессию лимитирующих генов главного эффекта данной генетиче-
генетической системы. Единая молекулярная природа генов и полигенов означает, что
они имеют сходные арсеналы механизмов функционирования и управления:
например, возможно участие энхансеров, сайтов теплового шока и других из-
известных знаков генетического управления.
Наконец, в этом случае не возникает парадокса переполнения генома по-
полигенами (см. выше), так как роль полигенов в одних системах выполняют од-
однотипные гены других систем, но находящиеся в иных отношениях управле-
управления, лимитирования и т.д. по отношению к первым.
Интересны эволюционные аспекты излагаемой концепции. Во-первых,
лимитирующие менделевские гены непосредственно проявляются в признаках,
т.е. оцениваются отбором. Нелимитирующие менделевские гены (полигены) в
малой степени влияют на признак (хотя и необходимы для его формирования),
а потому фактически находятся в нейтральном режиме эволюции, пока не ста-
станут лимитирующими. Поэтому каждый ген лишь малую часть своей эволюци-
эволюционной истории проводит в адаптивном режиме эволюции, в остальном его эво-
эволюция нейтральна.
Во-вторых, популяции, попадая в стрессовые внешние и внутренние ситуа-
ситуации, способны ответить на них вспышками регуляторной изменчивости через
массовую в популяции и множественную в геноме индукцию транспозиций
МГЭ. Этот источник изменчивости очень важен для быстрой реорганизации
управления геномов и становления новой видовой нормы в новых условиях су-
существования.
В-третьих, если правильно все предыдущее, то современные геномы выс-
высших организмов должны содержать свидетельства прежних перемещений ко-
копий МГЭ, приведших к регуляторным изменениям проявления некоторых ге-
генов. Действительно, американский генетик Р. Бриттен, проанализировав сек-
венированные последовательности ДНК в окрестностях многих известных
14. Мобильные генетические элементы 121
генов животных, обнаружил до 10 примеров, когда работающие знаки управле-
управления были окружены "руинами" предковых копий МГЭ. У растений число та-
таких примеров достигает сотни.
Таким образом, возвращаясь к вопросу, поставленному в заголовке статьи,
мы можем сказать, что в целом МГЭ — несомненные "геномные бродяги" со
стажем, но они имеют также несомненные задатки "трудяг" на ниве генетиче-
генетического управления. Наша задача — выявить, "кто" из них уже пошел по правед-
праведному пути.
Сопоставление МГЭ геномов ряда родственных видов (особенно - у дрозо-
дрозофил) позволило выявить ряд случаев так называемого "горизонтального" пере-
переноса МГЭ между видами. Например, считается, что вид Drosophila melanogaster
приобрел семейство Р-элементов всего лишь в XX столетии от других видов
дрозофил. Это своеобразное "заражение" коснулось, в первую очередь, попу-
популяций Нового Света, а все линии, взятые из природных популяций в начале
века или позже в Старом Свете, не содержали Р-элементов. После таких "зара-
"заражений" происходит достаточно быстрая "доместикация" (одомашнивание) но-
новых семейств МГЭ, они начинают играть некоторую роль в экспрессии и из-
изменчивости генов генома хозяина.
Уже выявились вполне определенные свидетельства того, что копии МГЭ
способны влиять на регуляцию функций смежных генов и полигенов, воспри-
воспринимать внешние и внутренние регуляторные воздействия, участвовать в измен-
изменчивости генов и геномов, участвовать в отклике на отбор. Иначе говоря, они
действительно образуют геномную систему МГЭ, которая может быть весьма
полезна высокоорганизованным молекулярно-генетическим системам клеток.
Список литературы
Васильева Л.А., Ратнер В.А. Полигенная система количественного признака radius
incompletus у дрозофилы: генетические особенности, взаимодействие с другими генами и пат-
паттерном МГЭ, эволюционные свойства// Современные концепции эволюционной генетики /
Ред. В.К. Шумный, А.Л. Маркель. Новосибирск: ИЦиГ СО РАН, 2000. С. 117-127.
Гвоздев В.А. Подвижные ДНК эукариот. 4.1. Структура, механизмы перемещения и
роль подвижных элементов в поддержании целостности хромосом // Соросовский образоват.
журн. 1998а. № 8. С. 15-21.
Гвоздев В.А. Подвижные ДНК эукариот. Ч. 2. Роль н регуляция активности генов и эво-
эволюция генома // Там же. 19986. С. 15-21.
Ратнер В.А., Васильева Л.А. Индукция транспозиций мобильных генетических элемен-
элементов стрессовыми воздействиями // Соросовский образоват. журн. 2000а. Вып. 6. № 6.
С. 14-20.
Ратнер В.А., Васильева Л.А. Мобильные генетические элементы (МГЭ): "эгоистическая
ДНК" или функциональная часть генома? // Современные концепции эволюционной гене-
генетики / Ред. В.К. Шумный, А.Л. Маркель. Новосибирск: ИЦиГ СО РАН, 20006. С. 128-150.
III. МОЛЕКУЛЯРНАЯ КИБЕРНЕТИКА
15. Хроника великого открытия:
идеи и лица. I*
Наука середины XX века была потрясена открытием генетического кода.
Этого события ждали все ученые. Когда оно произошло A965 г.), мир науки со-
содрогнулся от рукоплесканий. "Финалисты" решения этой проблемы (Ф. Крик,
Дж. Уотсон, М. Уилкинс, М. Ниренберг, Г. Корана, Р. Холли) были увенчаны
Нобелевскими премиями. Проблема генетического кода заняла центральное
место в учебниках биологии, генетики, молекулярной биологии, биофизики и
остается там до сих пор.
Однако за громом оваций не всегда заметно истинное движение и развитие
этой великой идеи, связанное с именами ряда выдающихся физиков и биоло-
биологов. Многие из них были гениями русской и мировой науки, но попробовать
себя в этой проблеме стремились почти все. Идея кодирования генетической
информации вызревала постепенно, как бы переходя из рук в руки в соответст-
соответствии с "принципом одиночного касания". Есть такой способ игры в баскетболе:
игра в одно касание. Развитие этой идеи я постараюсь показать "в лицах", как
это представляется нам сегодня.
Наследственные молекулы
Как известно, представление о дискретных наследственных факторах было
сформулировано основателем генетики Грегором Менделем еще в 60-е годы
XIX в., но принято и осознано наукой [Голубовский, 1997] только в начале
XX в. Тогда же П. Иоганзен назвал эти факторы генами, а местом их локализа-
локализации в клетке единодушно были признаны хромосомы ядра. Однако вплоть до
50-х годов ничего не было известно ни о материальной природе генов, ни о ме-
механизмах их действия и контроля за формированием признаков. О генах судили
как бы "заочно", не зная, что конкретно кроется за этим кратким словом. Как
пошутил позже один из ныне здравствующих генетиков-теоретиков: "ген — это
мифическая единица якобы наследственности". Или, как совершенно серьезно
сказал известный американский генетик С. Бензер: "гены — это атомы наслед-
наследственности". Атом — значит неделимый!
В 1927 г. русский ученый Николай Константинович Кольцов, известный
цитолог и генетик, член-корреспондент АН СССР и директор Института экс-
экспериментальной биологии, выступил с докладом [Кольцов, 1968а], где изложил
свои довольно гипотетические взгляды на материальную природу генов и хро-
* Ранний вариант статьи: Ратнер В.А. Природа. 1998. № 4. С. 68-79.
.
15. Хроника великого открытия. I
123
мосом. Как рассказывал сам
Н.К. Кольцов, исходным толчком к
постановке этой проблемы для него
послужил весьма впечатляющий эпи-
эпизод из его студенческой биографии. В
1893 г. в Москве, в зале Дворянского
Собрания (после революции — Ко-
Колонный зал Дома Союзов) проходил
Съезд естествоиспытателей и врачей.
На этом съезде столкнулись две
противоположные точки зрения на
сложность устройства клетки и на-
наследственности. Профессор-зоолог
МЛ. Мензбир (будущий академик)
считал механизм наследственности
очень сложным: клетка, ядро, прото-
протоплазма, хромосомы, в хромосомах —
дискретные наследственные факто-
факторы, и т.д. В этом случае сложность
хромосом должна быть сопоставима
со сложностью организма!
Противоположную точку зрения
сформулировал профессор-химик
АЛ. Колли. Он сопоставил размер го-
головки сперматозоида (~ 30 мкм у человека), через который передается половина
всей наследственной информации к потомку, с известными тогда размерами мо-
молекул белка. Получилось, что в этом объеме могло находиться лишь небольшое
число больших молекул, немного превышающее число самих хромосом.
Итак, наследственность очень сложна в биологическом отношении, но
проста в химическом смысле. Совместимы ли такие точки зрения? Студент Ни-
Николай Кольцов запомнил эту коллизию. Через 35 лет зрелый ученый профессор
Н.К. Кольцов сформулировал одну из первых гипотез о материальной природе
хромосом и генов, которая давала ответ на вопрос. Разумеется, это было сдела-
сделано на уровне знаний своего времени, поэтому многие представления потом су-
существенно изменились; но наиболее глубокие предположения оказались вер-
верными. В 1935 г. Н.К. Кольцов изложил гипотезу в более развернутой и иллюст-
иллюстрированной форме.
К тому времени уже были известны некоторые свойства белков. Во-пер-
Во-первых, определены молекулярные массы белков, лежащие по оценкам в интерва-
интервале ~ 10-200 тыс. д. (теперь такие молекулы называют макромолекулами).
Во-вторых, было показано, что разные белки распадаются на аминокислоты не
менее 17 типов (в дальнейшем — 20). Если представить себе, что белки — ли-
линейные молекулы, и учесть, что, по рентгеноструктурным данным, аминокис-
аминокислоты имеют линейный размер вдоль цепи ~ 0.003 мкм, то линейные цепочки
длиной всего 100 звеньев составят уже -0.3 мкм. Этот размер сопоставим с ви-
видимыми размерами хромосом и клеточного ядра C-10 мкм).
Николай Константинович Кольцов.
124
III. Молекулярная кибернетика
Рис. 15-1. Гипотетическая схема органи-
организации хромосомы по Н.К. Кольцову
[19686]. Хромосома обведена контурной
линией. Внутри хромосома содержит две
генонемы, т.е. два пучка одинаковых
белковых молекул (показаны только по
две копии), составляющих генетический
каркас хромосомы. Каждый символ обо-
обозначает отдельный ген. В некоторых мес-
местах вблизи генонемы выстраиваются час-
частичные фрагменты последовательности
символов. Это отвечает идее гомологич-
гомологичного воспроизведения хромосомы (пу-
(путем "кристаллизации"). Внутри и снару-
снаружи хромосомы рассеяны копии генов
(или их "обломков"), которые участвуют
в "кристаллизации".
Н.К. Кольцов предположил, что
хромосомы представляют собой ог-
огромные молекулы белков или пучки
таких молекул. Других длинных мо-
молекул, состоящих из гетерогенных
мономеров, в клетках тогда не знали.
Тимонуклеиновая кислота (будущая
ДНК) рассматривалась как "сравни-
"сравнительно простое органическое соедине-
соединение, которому было бы странно припи-
приписывать роль носителя наследственных
свойств " [Кольцов, 19686].
На рис. 15-1 приведена схема ор-
организации хромосомы по Кольцову
[19686]. Хромосома содержит две гено-
генонемы, т.е. два пучка одинаковых бел-
белковых молекул. Каждый символ обо-
обозначает отдельный ген. Гены линейно
упорядочены вдоль генонемы. Кроме того, вокруг генонемы, внутри и вне хромо-
хромосомы рассеяны те же символы-гены (или их обломки, части), которые необходи-
необходимы для воспроизведения хромосом.
Н.К. Кольцов резонно замечает, что такие гетерогенные упорядоченные
хромосомы-молекулы могут иметь огромное число изомеров (комбинаторных
вариантов) одинакового состава, но разного порядка символов-генов. Он гово-
говорит, что их сантильоны вариантов (кстати, сколько это нулей после единицы?).
Поскольку последовательность генов наследуется, хромосома даже в интерфазе
клеточного цикла (когда ее не видно в микроскоп) не может распадаться на
компоненты-гены; иначе они не смогут снова сложиться в прежнем порядке.
15. Хроника великого открытия. I 125
Поэтому в процессе воспроизведения хромосомы-молекулы должны строить
себе подобные варианты порядка генов. По биологической традиции это сооб-
соображение было облечено в форму афоризма [Кольцов, 19686]:
"каждая молекула от молекулы"
("omnis molecula ex molecula" — no латыни)
На рис. 15-1 это отражено в виде коротких последовательностей символов,
которые выстраиваются вблизи таких же последовательностей хромосом.
Пользуясь аналогией, Н.К. Кольцов замечает, что этот процесс весьма похож
на кристаллизацию, правда — гетерогенную. Для этого одноименные амино-
аминокислоты должны иметь сродство своих боковых радикалов друг к другу.
"Если мы признаем, что самой существенной частью хромосомы являются
длинные белковые молекулы, состоящие из нескольких десятков или сотен атом-
атомных групп радикалов (всего-то! — B.P.J, то моргановское представление о хромо-
хромосоме как о линейном ряде генов получит ясную конкретную основу. Радикалы хро-
хромосомной молекулы — гены — занимают в ней совершенно определенное место, и
малейшие химические изменения в этих радикалах, например отрыв тех или иных
атомов и замена их другими..., должны являться источником новых
мутаций " [Кольцов, 19686].
Эта идея схематически изображена на рис. 15-2: генонема — молекула бел-
белка, а гены — боковые радикалы аминокислот. Между прочим, отсюда вытекает,
что можно поискать такие химические вещества, которые способны модифи-
модифицировать боковые радикалы, т.е. возможен индуцированный химический мутаге-
мутагенез. Директор ИЭБ Н.К. Кольцов в середине 30-х годов XX в. предложил своим
молодым сотрудникам начать поиск таких химических мутагенов. Наибольше-
Наибольшего успеха из них добился Иосиф Абрамович Рапопорт, который по праву счита-
считается одним из первооткрывателей химического мутагенеза [Строева, 1997]. Он
занимался их изучением всю свою жизнь.
Сейчас мы можем объективно оценить идею Н.К. Кольцова. Фактически
он первый предположил, что генетическим "каркасом" хромосомы является
гигантская линейная макромолекула, построенная из ограниченного разнооб-
разнообразия мономеров. Это предположение полностью оправдалось, правда для мо-
молекул ДНК, а не белков. Гены он уподобил боковым радикалам такой молеку-
молекулы, а их химические изменения — мутациям. Это представление не подтверди-
подтвердилось, гены оказались участками молекулы, но химический мутагенез оказался
реальным.
Вместо случайной сборки генов в хромосому при ее удвоении он предло-
предложил фактически матричный принцип воспроизведения хромосом, сохраняющий
порядок генов. Для этого он постулировал как бы "гомологию" отношений
между одноименными боковыми радикалами (генами). Все это хорошо согла-
согласовывалось с тогдашними представлениями генетиков о гомологичном спари-
спаривании генов в мейозе, о линейной структуре хромосом и т.д. Кроме того, эта
идея фактически натолкнула его на успешное представление о реальности хи-
химического мутагенеза.
126
III. Молекулярная кибернетика
^YchYc
Xiiilii/ *Дсо)
(снКс
(Hi
(еще
(coj—
!I(nh)
з@
Puc. 75-2. Гипотетическая схема молеку-
молекулярной структуры генонемы [Кольцов,
19686]. Фактически это полипептидная
цепь белка, где боковые радикалы играют
роль генов.
Разумеется, многие частные де- I
тали и гипотетические представления
не выдержали испытания временем.
Главную роль в гипотезе Кольцова
играют белки, а нуклеиновые кисло-
кислоты оказались "за кадром" построен-
построенной схемы. Но такова была реаль-
реальность. Время нуклеиновых кислот
пришло позже — в конце 1940-х —
начале 1950-х годов. Хотя линейные
размеры макромолекул белков дейст-
действительно приближаются к размерам
хромосом, надо учесть, что глобуляр-
глобулярные белки свернуты в пространствен-
пространственные глобулы, размеры которых равны
-2-10 нм, т.е. на три порядка вели-
величины меньше, чем линейные разме-
размеры. Такие же проблемы пространст-
пространственной упаковки существуют для еще
более длинных молекул нуклеиновых
кислот: гаплоидный геном человека
содержит 3 х 109 нуклеотидов и имеет
длину около 1 м, а упакован в клеточ-
клеточном ядре объемом 20 мкм3! Хромосо-
Хромосомы содержат не сотни генов, а десят-
десятки тысяч. Линейный размер генов
тоже значительно больше, чем разме-
размеры боковых радикалов аминокислот.
Отметим также, что никаких избира-
избирательных специфических отношений
между одноименными аминокисло-
аминокислотами не найдено до сих пор.
Однако главное значение эта ги-
гипотеза скорее имела в смысле науч-
научной перспективы. Идея хромосо-
хромосомы-молекулы произвела глубокое
впечатление на ближайшего ученика
Н.К. Кольцова — Николая Владими-
Владимировича Тимофеева-Ресовского.
"Классическая зеленая тетрадка"
Ученик Н.К. Кольцова, в дальнейшем — выдающийся русский генетик
Николай Владимирович Тимофеев-Ресовский, в 1924-1945 гг. работал в Герма-
Германии по соглашению между правительствами наших стран. В пригороде г. Бер-
Берлина — Бухе — он возглавлял Отдел генетики Института мозга Общества
15. Хроника великого открытия. I
127
им. Кайзера Вильгельма. Разумеется,
он знал о гипотезе Кольцова и всяче-
всячески ее пропагандировал. Кроме того,
он имел обширные контакты с физика-
физиками-теоретиками Копенгагенской шко-
школы, хорошо знал Нильса Бора и увле-
увлекался его квантовой методологией. В
середине 30-х годов в Лаборатории
появился молодой физик-теоретик
Макс Дельбрюк, ученик Макса Борна.
Между ними произошло следующее
"касание", обогатившее идею молеку-
молекулы-гена. Вот как сам Н.В. описывает
дальнейшее развитие событий [Тимо-
[Тимофеев-Ресовский, 1993] (обращаю вни-
внимание читателя на весьма колоритный
язык Н.В.!):
«Отвечу на вопрос, имел ли я влияние
на Дельбрюка, особенно в смысле колъцов-
ской идеи о наследственной молекуле. Ни-
Никакого особенного влияния тогда не имел.
Это он выдумывает... Конкретное пере-
переманивание Дельбрюка в биологию... я, соз-
сознаюсь, произвел... А он, будучи
"хаузтеоретикер" (домашний теоретик)
у самого Хана (Отто Гана. — В.Р.), думал, что теоретик-физик — он все знает и
кое к чему может снизойти. А прочее человечество — это такие придурки ка-
какие-то, человекообразные. Поэтому он к нам явился, так сказать, с откровением,
что вот, между прочим, отбором ведь можно количественно заняться... Тогда я по-
позволил себе из собственной лабораторной периодики извлечь около дюжины книг,
презентовал их ему, что "Америки вы покеда не открыли "... Он был несколько этим
смущен. Он, что говорить, очень горд был. И с тех пор попросился в нашу Буховскую
группу... Ну и тут мы быстро сконтачили. Я рассказал ему про кольцовское общее
представление о молекулярной биологии генов и хромосом, что мы тогда... пытались
экспериментально показать мономолекулярность принципиальную, в общем смысле,
генов. То есть что это единая физико-химическая элементарная структура (выде-
(выделено мной. — В.Р.), так сказать, а не кусочек сливочного масла... Он в этом при-
принял участие. Из этого родилась так называемая потом классическая зеленая тет-
тетрадка (выделено мной. — В.Р.).
Да, повлиял. Но какое это влияние — треп один».
Тут Н.В. явно поскромничал. Его сотрудник Карл Циммер вспоминал впо-
впоследствии [Zimmer, 1966]:
"Мы встречались два-три раза в неделю, в основном — в доме Тимофеева-Ре-
Тимофеева-Ресовского в Берлине, где мы разговаривали по десять и более часов без перерыва,
лишь иногда поглощая пищу в ходе разговоров. Не могу судить, кто больше почерп-
Нпколаи Владнмпронмч Тимофеев-Ресон-
ский. Германия, 1940-е годы.
128 III. Молекулярная кибернетика
пул из этого обмена идеями, знаниями и опытом, но является фактом, что через
несколько месяцев Дельбрюк столь глубоко заинтересовался количественной биоло-
биологией, и особенно — генетикой, что остался в этом поле навсегда".
Иначе говоря, это был, как теперь говорят, "мозговой штурм". Зная неудер-
неудержимый характер Н.В. и его идейное доминирование в любой аудитории, могу ска-
сказать, что это именно он сеял семена, а его молодые коллеги слушали не отрываясь.
В конце 20-х - середине 30-х годов Н.В. Тимофеев-Ресовский и его сотруд-
сотрудники изучали закономерности возникновения мутаций под действием рентге-
рентгеновских лучей и других жестких излучений. Главная идея состояла в том, что
квант излучения или частица ионизируют атомы внутриклеточного вещества.
При этом возникают мутации генов. Если механизм мутаций одноударный, т.е.
один акт ионизации в объеме мишени v вызывает мутацию, то зависимость чис-
числа мутаций N* от дозы облучения выражается формулой [Zimmer, 1966]
N* = No {I - exp(-v • D)},
где No — начальное число особей, D — доза облучения, измеряемая числом пар
ионов в 1 см3. В тканях дрозофилы доза в один рентген вызывает 1.6 х Ю12 актов
ионизации на 1 см3. Поэтому константа v имеет размерность см3, т.е. объема
мишени, ионизация которой достаточна для возникновения мутации. Измерив
No, N* и D, можно оценить объем мишени, которая предположительно отожде-
отождествляется с геном. Таким образом, можно попытаться оценить физический раз-
размер гена. По представлениям Тимофеева-Ресовского, ген — неделимая элемен-
элементарная единица, своего рода атом наследственности, а его мутации — измене-
изменения внутреннего состояния.
Подсчет рецессивных летальных мутаций в Х-хромосоме дрозофилы, пс
данным для различных излучений, показал, что v= 1.7хЮ~17 см3 = 1700 нм3.
Это соответствует сфере диаметром d к 3.2 х Ю см = 32 нм. Поскольку харак-
характерные межатомные расстояния ~ 0.2-0.3 нм, а средний объем атомг
я (8-27) ¦ 10~3 нм3, значит число атомов в гене 300-1000, а характерный размер
гена 2-3 нм (по более поздним оценкам, 2-6 нм) [Ли, 1963]. Для сравнения
диаметры молекул глобулярных белков 2-10 нм. Если среднее число атомов
составляющих мономеры белков (аминокислоты), равно 15, то воображаемый
ген-белок должен состоять из 20-70 аминокислот. Сегодня мы знаем, что этс
число занижено. При пересчете на известные теперь молекулярные размерь
нуклеотидов ДНК получается молекула из 10 000-40 000 мономеров. Для слож-
сложных генов дрозофилы по современным данным эти цифры не являются завы-
завышенными (с учетом интронов) (см. гл. 11).
Тем не менее вывод, сделанный тремя авторами, ясен: ген — макромолекул:
примерно такого размера A00 мономеров и более), который предлага;
Н.К. Кольцов для наследственных макромолекул (хромосом). Поэтому молеку
ла-хромосома должна быть в сотни и тысячи раз больше.
Но это еще не все. Всю работу пронизывала идеология квантовой физики
Если объем молекулы-гена не очень велик C00-1000 атомов), то она должгс
обладать некоторыми квантовыми свойствами. Из химии, в том числе кванто
вой химии, известно, что сложные молекулы могут иметь изомерные формы
15. Хроника великого открытия. I 129
т.е. при том же составе иметь различные структурные формулы. Если молеку-
молекулы-гены состоят из небольшого разнообразия мономеров, то возможно огром-
огромное комбинаторное разнообразие вариантов одного состава. Например, для мо-
молекул длиной L мономеров, принадлежащих к п типам, число вариантов будет
L\/k{\k2\...kn, 5>,- = 1,
где fc(. — число мономеров i-ro типа.
Дельбрюк предположил, что изомерные варианты соответствуют аллелям
гена, а изомерные переходы — мутациям. С другой стороны, изомерные вариан-
варианты отвечают дискретным энергетическим состояниям молекулы, потенциаль-
потенциальным ямам, которые разделены потенциальными барьерами (порогами) конеч-
конечной величины. Согласно законам квантовой механики, сквозь эти барьеры воз-
возможен туннельный эффект с небольшой вероятностью. Чем выше порог, тем
дольше ожидание. Однако, если извне поступит квант энергии, достаточный
для преодоления барьера, то произойдет радиационная индукция изогенного
перехода, т.е. индукция мутации. Ясно также, что большая молекула имеет ог-
огромное число дискретных устойчивых состояний-"аллелей".
Интересно, что уже в 50-е годы идея изомерных переходов между квантовы-
квантовыми состояниями генов нашла свое реальное воплощение. Было показано, что
азотистые основания нуклеотидов ДНК имеют так называемые таутомерные ва-
варианты. Таутомеры образуют различное число водородных связей в ДНК и в ре-
результате "спариваются" с другими нуклеотидами, нарушая канонические прави-
правила комплементарности: А-Т и G-C. Спонтанные (квантовые) таутомерные пере-
переходы вносят свой вклад в спонтанный мутагенез. А многие химические мута-
мутагены — аналоги нуклеотидов — имеют повышенную вероятность таутомерного
перехода, т.е. индуцируют мутационные замены нуклеотидов в ДНК.
В 1935 г. Тимофеев-Ресовский, Циммер и Дельбрюк по приглашению Гет-
тингенской академии естествознания сделали доклад о структуре гена и меха-
механизме мутаций, который был затем опубликован в виде небольшой тетради с
зеленой обложкой и получил название "Классическая зеленая тетрадь" [Тимо-
[Тимофеев-Ресовский, Циммер, Дельбрюк, 1935]. Позже в своих воспоминаниях Н.В.
писал (см. гл. 2):
«Ялично с 40-х годов больше этим не занимаюсь. Правда, меня многие, особен-
особенно там, за рубежом, считают чем-то вроде деда этого направления. Потому что
новая, послевоенная редакция его была запущена Дельбрюком, а Дельбрюку соот-
соответствующую вещь я заправил в мозги в 30-е годы. Вот с этого, в сущности, пошло
все, с этой самой нашей классической, так называемой "зеленой тетрадочки"
Геттингенского общества наук. Ну и пусть, значит, дальше развивается на доб-
доброе здоровье».
Не следует думать, что модель трех авторов в полной мере выдержала ис-
испытание временем. Оценка размера генов, действительно, оказалась не очень
далекой от реальной, а гены, действительно, оказались макромолекулами, хотя
и не белками, а сегментами ДНК. Квантовые явления, действительно, играют
некоторую роль в первичном мутагенезе, но совсем не такую, как предполагал
130
III. Молекулярная кибернетика
Дельбрюк. В модели рассматривание]
и другие важнейшие свойства генов -
способность к удвоению и контролк
признаков клетки и организма и др
Именно Тимофеев-Ресовский предло-
предложил тогда идею (и термин) конвари
антной редупликации, т.е. их способно
сти "рядом с собой построить себе точ-
точно подобные и оттолкнуть", но <
включением в себя наследственны;
вариаций, мутаций.
Главное значение "модели трех'
состоит в том, что она привлекла i
проблеме гена интерес многих выдаю-
выдающихся физиков. В дальнейшем эт<
оказалось решающим событием в ход<
выяснения молекулярной природы ге-
генов, зарождения молекулярной био
логии и генетики, формулирования ин
формационно-кибернетического под
хода в генетике.
В 1938 г. Макс Дельбрюк, настро
енный резко антинацистски, восполь
зовался поддержкой Рокфеллеровско
го фонда и уехал в США, полностьк
посвятив свою деятельность пробле
мам молекулярной генетики. Опыт физика, вероятно, подсказал ему, что дга
успешного решения проблемы гена следует найти такие биологические объек
ты, которые по размеру самым минимальным образом отличались бы от генов
Дельбрюк нашел бактериофаги. В то время A939-1940 гг.) никто не предпола
гал, что фаги подчиняются законам генетики. Вместе с А. Херши, американ
ским вирусологом, и С. Лурия — молодым вирусологом, эмигрантом из Ита-
Италии, они организовали так называемую "фаговую группу", которая впервые ис
пользовала фаги как генетические объекты. В дальнейшем М. Дельбрюк
С. Лурия и А. Херши получили Нобелевскую премию за основополагающие ис-
исследования по генетике бактериофагов.
Таким образом, "касание" Н.В. Тимофеева-Ресовского и М. Дельбрюк
породило квантовую модель гена-молекулы. В свою очередь, "модель трех" i
"Зеленая тетрадка" произвели большое впечатление на выдающегося австрий
ского физика-теоретика Э. Шредингера.
Макс Дельбрюк в 1960-е годы.
Что такое жизнь с точки зрения Шредингера
Выдающийся австрийский физик-теоретик Эрвин Шредингер, один и:
создателей квантовой механики, именем которого названо основное уравне
ние квантовой механики, Нобелевский лауреат, в 1938 г. покинул родину и пе
15. Хроника великого открытия. I
131
реехал в столицу Ирландии — Дублин.
Хотя он не был евреем и не подвергал-
подвергался гонениям в Австрии и Германии, он
не принял нацизм. Поэтому он принял
приглашение Президента Ирландии
И. де Валера возглавить Школу Теоре-
Теоретической физики Института Высших
Исследований в Дублине. Авторитет
Шредингера в физике был исключи-
исключительно высок. По свидетельству
У. Мура [Moore, 1989], биографа
Э. Шредингера, до 1960 г. было опуб-
опубликовано свыше 100 000 работ, осно-
основанных на уравнении Шредингера.
Однако физика была только ча-
частью его жизни. Кроме этого он опуб-
опубликовал ряд книг на нефизические
темы: о роли древних греков в совре-
современной цивилизации, книгу стихов и
др. Особое место в научном творчестве
Э. Шредингера занимает небольшая
книжка "Что такое жизнь? С точки
зрения физика" [Шредингер, 1972],
опубликованная в 1944 г. в Кембридж-
Кембриджском университете. В феврале 1943 г.
Э. Шредингер прочел в Дублине три
популярные лекции на эту тему, которые и составили книгу. Он базировался на
"Зеленой тетради" Н.В. Тимофеева-Ресовского, К. Циммера и М. Дельбрюка [1935],
которая, судя по результату, произвела на него глубокое впечатление. Это и было сле-
следующим "касанием" создателей великой идеи, в данном случае — заочным.
Прежде всего, Шредингер развил и углубил понимание модели гена-моле-
гена-молекулы трех авторов (Модели Дельбрюка, как он ее часто называл):
"Предположим, что по своей структуре ген является гигантской молекулой,
которая способна только к дискретным изменениям, сводящимся к перестановке
атомов с образованием изомерной молекулы.
Перестановка может коснуться небольшой части гена; возможно огромное
число таких различных перестановок. Потенциальные барьеры, разделяющие воз-
возможные изомерные конфигурации, должны быть достаточно высокими (по сравне-
сравнению со средней тепловой энергией атома), чтобы сделать переходы редким собы-
событием. Эти редкие события мы будем отождествлять со спонтанными
мутациями ".
Шредингер обратил внимание на то, что стабильная структура молекул го-
гораздо ближе к структуре кристалла, чем аморфного тела, жидкости или газа.
Стабильность молекул имеет квантовую природу, поскольку стабильные со-
состояния соответствуют минимумам энергии атомов, отгороженным от других
Эрвнп Шредингер.
132 III. Молекулярная кибернетика
состояний высокими потенциальными барьерами. Рассматривая гетерогенную
сложную органическую молекулу, Шредингер заключает:
"Мы можем совершенно точно назвать это образование апериодическим кри-
кристаллом или твердым телом и выразить нашу гипотезу словами: мы полагаем, что
ген или, возможно, целая хромосомная нить представляют собой апериодическое
твердое тело ".
Эта модель может быть экспериментально проверена. При термоактиваци-
онном механизме среднее время изомерного перехода (т.е. мутации) будет
t = х ехр{ W/kT],
где W — высота барьера, кТ — средняя тепловая энергия атома при температуре
Т, а константа г = 10~13-10~14 с соответствует периоду колебаний атомов в сис-
системе. Поэтому вероятность мутаций должна зависеть от температуры. Чем
выше W и ниже Т, тем больше t, т.е. тем стабильнее "аллель" гена.
Действительно, генетиками было экспериментально показано, что низкая
мутабильность "нормальных" аллелей часто возрастала при увеличении темпе-
температуры, а мутабильность некоторых нестабильных аллелей вообще при этом не
менялась. В то же время скорость мутаций, индуцированных рентгеновскими
лучами, не зависит от скоростей спонтанного мутирования облучаемых алле-
аллелей, т.е. одинакова как для устойчивых, так и для нестабильных аллелей. Этого
следует ожидать, если энергия жестких квантов заметно превышает порог W.
Далее Шредингер сделал еще один шаг принципиального значения. Фак-
Фактически он попытался ответить на вечный вопрос о том, какова функциональ-
функциональная роль молекулы-гена и молекулы-хромосомы по отношению к контролируе-
контролируемым ими признакам организма.
«...Хромосомы... содержат в виде своего рода шифровального кода (выделено
мной. — В.Р.) весь "план "будущего индивидуума и его функционирования в зрелом
состоянии. Каждый полный набор хромосом содержит весь шифр...»
"Но термин шифровальный код, конечно, слишком узок. Хромосомные струк-
структуры служат в то же время и инструментом, осуществляющим развитие, кото-
которое они же предвещают... Они являются одновременно и архитектором, и строи-
строителем... "
«Как такая крошечная частичка вещества — ядро оплодотворенного яйца —
может вместить сложный шифровальный код, включающий в себя все будущее
развитие организма ? Хорошо упорядоченная ассоциация атомов, наделенная дос-
достаточной устойчивостью для длительного хранения своей упорядоченности, пред-
представляется единственно мыслимой материальной структурой, в которой разно-
разнообразие возможных С "изомерных ") комбинаций достаточно велико, чтобы заклю-
заключать в себе сложную систему детерминации в пределах минимального
пространства».
Иначе говоря, биологическая сущность гена и хромосомы в том, что они
представляют собой молекулярные "записи" сложной структуры признаков, он-
онтогенеза, организма в целом. Средства записи — комбинаторные варианты
больших молекул-генов. Таким образом, Шредингер сделал шаг к формулиро-
15. Хроника великого открытия. I 133
ванию информационно-кибернетического подхода в генетике. Конечно, этот
первый шаг еще был неконкретным (как именно?) и достаточно фигуральным,
метафоричным. Но из него в дальнейшем вырос подход к проблеме кодирова-
кодирования генетической информации.
Наконец, Шредингер делает последний шаг в своей концепции:
"Из общей картины наследственного вещества, нарисованной Дельбрюком,
следует, что деятельность живой материи, хотя и основана на законах физики,
установленных к настоящему времени, но, по-видимому, подчиняется до сих пор
неизвестным другим законам физики, которые, однако, как только они будут от-
открыты, должны составить такую оке неотъемлемую часть этой науки, как и
первые ".
"Жизнь — это упорядоченное и закономерное поведение материи, основанное
не только на одной тенденции переходить от упорядоченности к неупорядоченно-
неупорядоченности, но и частично на существовании упорядоченности, которая поддерживается
все время ".
Фактически речь идет о том, что организм является макроскопической
упорядоченной системой, действующей по детерминированным законам, тогда
как в его основе (гены, хромосомы исходной зиготы) лежат макромолекулы,
подверженные квантовым, статистическим явлениям. Казалось бы, второе на-
начало термодинамики (закон возрастания энтропии) требует, чтобы неупорядо-
неупорядоченность нарастала. Энтропия служит мерой хаотичности системы, а ее убыва-
убывание увеличивает упорядоченность. Однако организмы не являются изолиро-
изолированными системами, они не только сохраняют упорядоченность, но
размножают себя и строят в онтогенезе новые макроскопические упорядочен-
упорядоченные системы под контролем квантовых объектов — генов. Шредингер предпо-
предполагает, что эта особенность жизни, отличающая ее от всего неживого, возника-
возникает на основе действия каких-то новых, неизвестных физике законов, не менее
великих и фундаментальных, чем, например, начала термодинамики.
В общих словах (т.е. без указания конкретных механизмов) живой орга-
организм избегает перехода к равновесию, молекулярному хаосу за счет компенса-
компенсации извне падения упорядоченности, т.е. за счет питания высоко упорядочен-
упорядоченными веществами. По словам Шредингера:
«Он питается отрицательной энтропией, как бы привлекая на себя ее поток
(извне. — В.Р.), чтобы компенсировать этим увеличение энтропии, производимое
им в процессе жизни, и таким образом поддерживать себя на постоянном и доста-
достаточно низком уровне энтропии...
...способность "пить" упорядоченность из подходящей среды, по-видимому,
связана с присутствием "апериодических твердых тел " — хромосомных молекул».
Для физика такое положение дел производит впечатление чуда: единичная
группа атомов, существующая изначально (в зиготе) в единственном экземпля-
экземпляре, влияет на формирование макроскопического признака. Даже в многокле-
многоклеточном организме (число клеток »1014 ) число таких групп все равно составляет
мизерную долю всего вещества. И тогда следует вопрос:
134 III. Молекулярная кибернетика
"...не напоминают ли они нам отдельные пульты управления, разбросанные по
всему организму и осуществляющие связь между собой благодаря общему для них
коду?"
И наконец, последнее. "Апериодический кристалл" — ген, хромосома —
способен формировать "порядок из порядка", точнее — распространять уже
имеющийся в нем порядок на новые массы вещества. Фактически это похоже
на "апериодическую кристаллизацию", инициированную исходным геном-мо-
геном-молекулой. В более умозрительной форме этот принцип был отмечен еще
Н.К. Кольцовым. Генетически это прообраз удвоения хромосомы-молекулы.
Таким образом, Э. Шредингер излагает, хотя и недетализированную, но
достаточно последовательную картину физических явлений, которые могли бы
быть основой генетических механизмов. Наиболее замечательными выводами,
как теперь ясно, были представления о гене и хромосоме как о молекулярных
записях ("шифровальном коде"), контролирующих формирование макроско-
макроскопических признаков организма путем управления. Иначе говоря, это начало ин-
информационно-кибернетического подхода в генетике. Согласно концепции, гены
обладают "квантовой" стабильностью и воспроизводятся путем "кристаллиза-
"кристаллизации" с участием затравки — предыдущего "апериодического кристалла" —
гена.
Книга Э. Шредингера имела огромное влияние на дальнейший ход собы-
событий. Из уст признанного всеми великого ученого молодое поколение физиков
узнало о возможной реальности новых фундаментальных законов физики. Це-
Целое поколение молодых физиков замерло от искушения. По стечению обстоя-
обстоятельств это совпало с окончанием войны и возвращением в науку большого
числа молодых физиков. Многие из них в результате выбрали путь в биологию
и усвоили идею "шифровального кода" и гена-молекулы. Среди тех, на чьи
судьбы повлияли идеи Шредингера, были Ф. Крик, Дж. Уотсон, М. Уилкинс,
С. Бензер, Г. Стент, Дж. Ледерберг и сотни других. Это было "касанием" ге-
гения, но касанием массовым. Имена Н.В. Тимофеева-Ресовского и М. Дель-
Дельбрюка стали широко известны англоязычному миру науки. У нас книга впер-
впервые была переведена и издана в 1947 г., но вскоре запрещена вместе со всей ге-
генетикой в годы лысенковщины. Только после 1955 г. идеи Шредингера стали
публично обсуждаться в советской физической и биологической науке. Однако
за это время наука уже заметно ушла вперед, и на повестку дня встали следую-
следующие проблемы: генетический код, генетические процессы, молекулярная ки-
кибернетика и т.д., в решении которых советские ученые уже смогли принять по-
посильное участие.
Интересен, однако, дальнейший ход событий. Через 20 лет, подводя пер-
первые итоги развития молекулярной биологии и генетики, Г. Стент отметил
[Stent, 1966; Стент, Келиндер, 1981]:
«Вторжение этих людей в генетику и родственные ей области биологии в 40-х
годах произвело в этой науке революцию, которая, когда пыль рассеялась, остави-
оставила в качестве своего наследия молекулярную биологию... Увы, физики были обману-
обмануты в своих надеждах. Никаких "других законов физики" на этом пути не обнару-
15. Хроника великого открытия. I 135
жилось... Чтобы понять, как функционирует наследственное вещество, нужно,
по-видимому, лишь понимать, как разрываются и образуются водородные связи.»
На этом пути не подтвердились многие ранние идеи М. Дельбрюка и
Э. Шредингера [Timofeeff-Ressovsky et al., 1935; Шредингер, 1972]. Стабиль-
Стабильность генов оказалась связанной не столько с квантовыми механизмами,
сколько с существованием мощных систем репарации повреждений ДНК, дей-
действующих на основе дублирования генетической информации (двойная цепь
ДНК, диплоидность генома, дублирование генов и т.д.). Температурные зави-
зависимости скоростей мутаций, найденные у дрозофилы, оказались относящими-
относящимися не к генам, а к ассоциированным ферментам. Механизмы многих мутаций
вообще оказались связанными не с изменением конформации гена (ДНК), а с
внедрением мобильных генетических элементов.
Но тем не менее дело было сделано. Подтвердилась идея молекулы-гена,
но не в виде белка, а в виде ДНК, и на более сложном уровне, где роль мономе-
мономеров-символов играют не атомы, а нуклеотиды. Молекулярная биология и гене-
генетика с самого начала были "окрашены" в тона информационно-кибернетиче-
информационно-кибернетические. Центральные проблемы этих наук были сформулированы и решены в
рамках данного подхода: генетическая информация, ген как информационная
единица, генетический код, матричная РНК-переносчик, управление функ-
функциями генов, гены-регуляторы и т.д. [Стент, Кэлиндар, 1981]. Уже у следующих
поколений исследователей автору этой статьи пришлось развивать Теорию мо-
лекулярно-генетических систем управления, т.е. фактически молекулярную
кибернетику. Вот каковы были последствия того, что Шредингер осторожно
перевел стрелки на этот путь развития науки.
Список литературы
Альберте Б., Брей Д., Льюис Дж., Рэфф М., Роберте К., Уотсон Дж. Молекулярная био-
биология клетки. М: Мир, 1986. Т. 1. ИЗ с.
Голдовский М.Д. Личностное знание и парадоксы истории генетики // Природа. 1997.
№ 6. С. 67-79.
Кольцов Н.К. Физико-химические основы морфологии // Классики советской генети-
генетики. Л.: Наука, 1968а. С. 85-92.
Кольцов Н.К. Наследственные молекулы // Там же. 19686. С. 93-120.
Ли Д.Е. Действие радиации на живые клетки. М.: Госатомиздат, 1963. 139 с.
Стент Г., Кэлиндар Р. Молекулярная генетика. М.: Мир, 1981.
Строева О.Г. Иосиф Абрамович Рапопорт в зеркале своих трудов // Природа. 1997. № 1.
С. 3-12.
Тимофеев-Ресовский Н.В. О прожитом // Николай Владимирович Тимофеев-Ресовский:
Очерки, воспоминания, материалы / Ред. Н.Н. Воронцов. М.: Наука, 1993. С. 18-51.
Тимофеев-Ресовский Н.В., Циммер К.Г., Дельбрюк М. О природе генных мутаций и
структуре гена // Н.В. Тимофеев-Ресовский. Избр. тр. / Ред. О.Г. Газенко, В.И. Иванов. М.:
Медицина, 1996. С. 105-153.
Шредингер Э. Что такое жизнь? М.: Атомиздат, 1972.
Crow J.F. Erwin Schrodinger and the Hornless Cattle Problem // Genetics. 1992. Vol. 130.
P. 237-239.
Moore W. Schrodinger, Life and Thought. Cambridge: Cambridge University Press, 1989.
Portugal F.H., Cohen J.S. A Century of DNA. Cambridge, MA, London: MIT Press, 1979. 257 p.
136 III. Молекулярная кибернетика
Stent G. Introduction: waiting for the paradox // Phage and the Origins of Molecular Biology /
Eds. J. Cairns, G. Stent, J.D. Watson. Cold Spring Yarbor, N.Y.: Cold Spring Harbor Lab., 1966.
P. 3-8.
Zimmcr K.G. The Target Theory // Phages and the Origins of Molecular Biology. Cold Spring
Harbor, N.Y.: Cold Spring Harbor Laboratory, 1966. P. 33-42.
16. Хроника великого открытия: идеи и лица. II
Материальный носитель
В гл. 15 уже указывалось [Ратнер, 1998], что вплоть до начала 40-х годов глав-
главными молекулярными кандидатами на роль материальных структур наследст-
наследственности считались белки. Для этого были некоторые важные основания. Уже в
30-е годы было известно, что белки являются макромолекулами большой моле-
молекулярной массы и состоят из ограниченного разнообразия мономеров — амино-
аминокислот. При этом мономеры связаны между собой стандартными пептидными
связями, а все разнообразие белков определяется составом и порядком боковых
радикалов. Это было понятно Н.К. Кольцову и многим другим.
Сопоставимые данные для нуклеиновых кислот были получены существен-
существенно позже, и это было связано с некоторыми драматическими обстоятельствами.
Ключевую и противоречивую роль в выявлении мономеров, связей между
ними, а также в формировании общих представлений о роли нуклеиновых ки-
кислот сыграл американский биохимик русского происхождения Ф.А. Левин
[Portugal, Cohen, 1979]. Он окончил Медико-хирургическую академию в Петер-
Петербурге в чине капитана русской армии; химию изучал под руководством извест-
известного композитора — профессора А. Бородина; степень магистра он получил в
Петербурге. По религиозным мотивам семья Левина в 1890-е годы эмигрирова-
эмигрировала в США, где через некоторое время он занялся исследовательской работой в
области биохимии и органической химии в Колумбийском университете, а поз-
позже — в Рокфеллеровском институте в Нью-Йорке.
Достижения Ф.А. Левина в науке огромны. За 40 лет с лишним он опубли-
опубликовал свыше 700 работ. В области исследования нуклеиновых кислот именно
он [Portugal, Cohen, 1979] "...выявил структуру обеих сахарных компонент нук-
нуклеиновых кислот (рибозы и дезоксирибозы. — В.Р.), установил природу нуклео-
зидов и нуклеотидов (и дал им названия. — В.Р.) и предложил структуру нуклеи-
нуклеиновых кислот, которая состояла в линейной комбинации нуклеотидов с правильны-
правильными межнуклеотидными связями ".
Однако Ф.А. Левину принадлежала и так называемая тетрануклеотидная
гипотеза, которая 40 лет сковывала развитие представлений о молекулярной
природе генов. Она была основана на ранних и достаточно неточных данных о
молярных концентрациях оснований в нуклеиновых кислотах. В 1908-1909 гг.
Левин с сотрудниками получили данные, что нуклеиновые кислоты из тимуса
теленка идрожжей имеют равные молярные концентрации всех четырех нук-
нуклеотидов. "Тетрануклеотидная гипотеза" предполагала, что четыре разных нук-
* Ранний вариант главы опубликован: Ратнер В.А. Природа. 1998. № 11. С. 18-29.
16. Хроника великого открытия. II
137
Фебус Аарон Левин A869-1940 гг.).
леотида связаны последовательно в стандартный тетрануклеотид, который
многократно повторяется в структуре нуклеиновой кислоты. В более поздних
вариантах гипотеза допускала высокую полимерность нуклеиновых кислот пу-
путем повторения тетрануклеотида, но, очевидно, исключала возможную комби-
комбинаторику нуклеотидов.
138 III. Молекулярная кибернетика
Таким образом, "стандартный тетрануклеотидный кирпич" (М ~ 1500) по-
позволял строить только унылую, однообразную последовательность. В этом слу-
случае нуклеиновые кислоты нельзя было рассматривать как материальные струк-
структуры генов. Гипотеза была принята фактически на веру подавляющим боль-
большинством выдающихся биохимиков и надолго задержала развитие
молекулярных представлений о генах.
В 40-е годы тетрануклеотидная гипотеза подверглась уничтожающей кри-
критике со стороны Э. Чаргаффа и многих других, а сам Ф.А. Левин оказался "коз-
"козлом отпущения" за это заблуждение. По мнению историков науки Ф. Португа-
ла и Дж. Коэна [Portugal, Cohen, 1979], именно тетрануклеотидная гипотеза по-
помешала Ф.А. Левину получить Нобелевскую премию за другие работы, которой
он несомненно был достоин. Умер Ф.А. Левин в 1940 г., когда уже началась
война, а вопросы чистой науки оказались за пределами внимания большинства
ученых.
Тем не менее к началу 40-х годов уже было ясно, что нуклеиновые кислоты
(нынешние ДНК и РНК) могут быть высокополимерны (М ~ 500 тыс. - 1 млн).
В конце 40-х годов Э. Чаргафф [ChargafT, 1951] показал, что ДНК разного видо-
видового происхождения имеют разный состав нуклеотидов, а общая их эквимоляр-
ность не выполняется. Использовав новый тогда метод хроматографии на бу-
бумаге, Э. Чаргафф обнаружил, что между молярными концентрациями пуринов
и пиримидинов имеются, однако, другие регулярные соотношения: А = Т и
G = С. Объяснения этим свойствам Э. Чаргафф не дал. Однако стало совер-
совершенно ясно, что мономерами нуклеиновых кислот являются не тетрануклеоти-
ды, а четыре стандартных нуклеотида, имеющих одинаковую сахарофосфатную
часть, участвующую в образовании стандартных фосфодиэфирных связей, и
различные основания, комбинаторика которых допускает огромное разнообра-
разнообразие вариантов (рис. 16-1).
Тем не менее, даже с учетом этих свойств, генетическую роль молекул
ДНК еще предстояло доказать. Этот шаг выполнил профессор Рокфеллеров-
Рокфеллеровского института Освальд Эвери с сотрудниками в 1944 г. [Avery et al., 1944].
Еще в 1928 г. английский врач-инфекционист Ф. Гриффите обнаружил явле-
явление трансформации у бактерий пневмококков. Оно состояло в том, что бакте-
бактерии одного штамма (невирулентные) приобретали наследуемую способность
быть вирулентными при контакте с лизатом вирулентных бактерий, убитых
нагреванием. Свыше 10 лет Эвери с сотрудниками отрабатывали методы
фракционирования лизата бактерий пока, наконец, не выделили активную
фракцию. По физико-химическим свойствам она совпала с фракцией ДНК.
По этому поводу О. Эвери писал своему брату: "Это нечто такое, что долго
было мечтой генетиков".
С одной стороны, это была сенсация, опровергавшая тетрануклеотидную
гипотезу: ДНК обладала генетическими свойствами. С другой, интерпретация
данных по трансформации тоже не была однозначной. ДНК могла быть либо
A) генетическим материалом, который рекомбинирует с гомологичным гено-
геномом бактерии-реципиента, либо B) мутагеном, который вызывает мутации ге-
мов, и тогда природа генов может быть другой, либо C) специфическим сигна-
сигналом, переключающим функциональное состояние гена (этот вариант выявился
16. Хроника великого открытия. II
139
Аденин (А)
Цитозин (С)
Гуанин (G)
Тимин (Т)
или
урацил (U)
н
R1
3'-конец
Рис. 16-1. Химическое строение фрагмента одиночной цепи ДНК.
позже). Дж. Ледерберг насчитал семь альтернативных гипотез о природе транс-
трансформации. Многие генетики не поняли фундаментального значения работы
Эвери с сотрудниками. Например, выдающийся цитолог А. Мирский, работав-
работавший в том же Рокфеллеровском институте, резко возражал против доказа-
доказательств трансформирующей роли ДНК.
Тем не менее значительная группа биохимиков (Э. Чаргафф, А. Тодд и
др.), генетиков (М. Дельбрюк, С. Лурия, А. Херши, Дж. Уотсон и др.) и физи-
физиков (Л. Полинг, У. Астбери, М. Уилкинс, Р. Франклин, Ф. Крик и др.) сосре-
сосредоточилась на изучении химии, генетической роли и молекулярного строения
ДНК. Дискуссии прекратились только после 1952 г., когда А. Херши и М. Чейз
140
III. Молекулярная кибернетика
Освальд Т. Звери A877-1955 гг.).
16. Хроника великого открытия. II 141
[Hershey, Chase, 1952] показали, что при заражении бактерии Е. coli фагом Т2
инфекционным началом является почти чистая ДНК фага. О. Эвери умер в
1955 г., не дождавшись своей Нобелевской премии, которой несомненно был
достоин.
Следует добавить, что в 1939-1940 гг. очень близкое открытие сделал
СМ. Гершензон в Киеве (см. гл. 6). Он показал, что введение или скармлива-
скармливание дрозофиле чужеродной ДНК вызывает вспышку мутаций признаков крыла.
Однако эта работа была прервана войной и не доведена до конца.
Двойная спираль ДНК
Следующее одиночное касание, высекшее "искру гения", состоялось в анг-
английском Кембридже между двумя очень непохожими людьми. Осенью 1951 г.
туда приехал американец Джеймс Уотсон, 23 лет, только что защитивший док-
докторскую диссертацию (Ph. D.) у Сальвадора Лурии в Университете штата Ин-
Индиана, США. Он был членом "фаговой группы" М. Дельбрюка, испытал мощ-
мощное влияние этой легендарной личности, а также находился под впечатлением
книги Э. Шредингера "Что такое жизнь? С точки зрения физика". Его [Уотсон,
1969] "...интерес к ДНК вырос из возникшего в колледже на последнем курсе жела-
желания узнать, что же такое ген ".
Формально Дж. Уотсон получил стипендию для изучения методов рентге-
ноструктурного анализа белков в группе М. Перуца в Кавендишской лаборато-
лаборатории Кембриджского университета. Однако там он встретил физика Френсиса
Крика — полутеоретика, полуэкспериментатора. Во время войны он занимался
оборонными исследованиями в Военно-морском ведомстве. В 1946 г. под влия-
влиянием книги Э. Шредингера и лекции Л. Полинга он решил заняться приложе-
приложением физики в биологии. В 1951 г. он работал в группе М. Перуца над теорией
дифракции рентгеновских лучей на белках.
Дж. Уотсон и Ф. Крик оказались в одной комнате. Позже Уотсон [1968]
вспоминал: "Но после разговоров с Френсисом моя судьба была решена. Мы быстро
поняли, что в биологии мы намереваемся идти одинаковым путем. Центральной
проблемой биологии был ген и контролируемый им метаболизм. Главной задачей
было понять репликацию гена и путь, которым гены контролируют синтез белков.
Было очевидно, что приступить к решению этих проблем можно лишь после того,
как станет ясной структура гена. А это значило выяснение структуры ДНК".
"В лаборатории Макса (Перуца. — В.Р.) нашелся человек, который знал, что
ДНК важнее, чем белки, — это было настоящей удачей " [Уотсон, 1969].
"Ничто не мешало мне разговаривать с Френсисом по меньшей мере по не-
нескольку часов в день"[Уотсон, 1969].
Вот как историки науки Ф. Португал и Дж. Коэн характеризуют этот науч-
научный тандем [Portugal, Cohen, 1979]:
"Контраст между Уотсоном и Криком мог показаться очень большим. Крику
во время их встречи в 1951 г. было 35 лет, и он еще не имел докторской степени
(Ph. D.). Уотсону было 23 года, он получил свою докторскую степень необычно
рано — в 22 года и был приглашен в члены фаговой группы. Крик был крупным и ге-
гениальным, Уотсон — тощим и угловатым. Но они имели много общего. Оба были
142
III. Молекулярная кибернетика
одиночками, которые тем не менее не скрывали своих веских идей по многим вопро-
вопросам. Оба имели выраженный интерес к открытию строения генетического мате-
материала. Но там, где из разных подходов — рентгеноструктурного анализа и гене-
генетики фагов — возникала их комплементарностъ, такой синтез вел к существен-
существенным результатам. В этом важном отношении Уотсон выполнял роль моста
между информационной и структурной школой в молекулярной биологии ".
Чтобы понять причины успеха совместной работы Уотсона и Крика, надо
учесть некоторые дополнительные обстоятельства.
Во-первых, поблизости от Кембриджа — в Лондонском Кинге-Колледже
работали друг Ф. Крика Морис Уилкинс и Розалинд Франклин — крупнейшие
английские специалисты по рентгеноструктурному анализу ДНК. Именно их
экспериментальные данные были использованы Уотсоном и Криком для обос-
обоснования и проверки своей модели.
Во-вторых, существенную роль для двух молодых исследователей играл дух
конкуренции с крупнейшим американским физикохимиком Лайнусом Полин-
гом. В то время звезда Полинга достигла своего зенита. Он был автором блестя-
блестящей классической книги "Природа химической связи" A939 г.). В 1950 г. вме-
Джеймс Д. Уотсон (слева) и Френсис Крик у стереомодели ДНК.
16. Хроника великого открытия. II 143
сте с Г. Кори теоретически, с помощью молекулярных стереомоделей, предска-
предсказали существование а-спиралей в глобулярных белках. С тех пор идея спирали
как бы "висела в воздухе" применительно к любым макромолекулам. Вот мне-
мнение Дж. Уотсона [1969]: "Спирали в то время были в центре внимания лаборато-
лаборатории, главным образом из-за а-спирали Полинга. ...Через несколько дней после моего
[Уотсона. — В.Р.] приезда мы уже знали, что нам следует предпринять: пойти по
пути Полинга и одержать над ним победу его же оружием ". Но и Полинг не дре-
дремал. Он активно обдумывал варианты молекулярных моделей ДНК.
В-третьих, к началу работы Крик уже имел опыт исследования теории ди-
дифракции рентгеновских лучей на спиралях. В результате он обладал интуицией,
позволявшей мгновенно отыскивать признаки спиральности на фотографиях
дифракции рентгеновских лучей. Иначе говоря, он был подготовлен к поиску
спиралей.
В-четвертых, Уотсон и Крик понимали, что ставки очень высоки. Речь шла
о молекулярной структуре генов — ключевых объектов биологической органи-
организации. Это требование налагало на любую модель ряд очевидных ограничений.
Следовало в молекулярных терминах объяснить, как гены выполняют свои ос-
основные функции: самоудвоение, мутирование, запись информации, контроль
над синтезом белков и др.
В частности, следовало понять, каков механизм самоудвоения (реплика-
(репликации) ДНК. Генетическая традиция, основанная на микрофотографиях поведе-
поведения хромосом в митозе и мейозе, постулировала идею гомологичного узнавания
подобных генов и сегментов хромосом. Уже в модели Н.К. Кольцова (см. гл. 15,
рис. 15-1) репликация хромосом рисуется как гомологичное выстраивание сег-
сегментов вдоль матрицы. Для этого требуются определенные молекулярные
силы и отношения. Поддерживая этот подход, известный немецкий физик-тео-
физик-теоретик П. Иордан предположил, что помимо известного физико-химического
"близкодействия" (силы Ван-дер-Ваальса, солевые мостики, водородные связи
и др.) существуют пока неизвестные квантовые резонансные "силы дальнодей-
дальнодействия", которые способны притягивать гомологичные структуры друг к другу
на больших расстояниях.
Против этого резко возражал Л. Полинг. Весь опыт структурной химии и
квантовой физики подсказывал ему, что воображаемые "силы дальнодейст-
дальнодействия" — это фикция. Что касается "сил близкодействия", то они требуют наибо-
наиболее тесного контакта между взаимодействующими молекулярными поверхно-
поверхностями. Ясно, что этому отвечал широко известный к тому времени принцип
взаимодействия антиген—антитело, фермент—субстрат и др., т.е. принцип
"ключ—замок". Иначе говоря, тесно взаимодействующие поверхности должны
быть взаимно комплементарны. В 1940 г. Л. Полинг и М. Дельбрюк изложили
свои аргументы против П. Иордана в журнале Science [Pauling, Delbmck, 1940].
Мозговой штурм продолжался 18 месяцев. Он сопровождался довольно
сложными личностными отношениями между участниками предприятия. На-
Например, Дж. Уотсон и Ф. Крик встречали решительный отпор со стороны
Р. Франклин. Однако именно ее данные по В-форме ДНК дали ключевой им-
импульс для разработки модели и лучше всего соответствовали результатам моде-
144
III. Молекулярная кибернетика
лирования. Авторы перебрали многие десятки возможных спиральных струк-
структур, но все они имели какие-нибудь недостатки.
Л. Полинг тоже исследовал различные варианты спиральных структур, но
он остановился на трехцепочечных спиралях, т.е. пошел по неправильному
пути. Отсутствие непосредственных контактов Уотсона—Крика и Полинга по-
позволило первым совершить "интеллектуальный рывок". Даже случай способст-
способствовал Уотсону и Крику. Полинг неоднократно просил прислать ему рентгено-
рентгенограммы дифракции, но М. Уилкинс не торопился. Когда Полинг собрался на
конференцию в Лондон, чтобы посетить Кембридж и увидеть все воочию, Гос-
Госдепартамент США не выдал ему визу (!). Виной тому была активная пацифист-
пацифистская деятельность Полинга против ядерных испытаний. Известно, что он, по-
помимо Нобелевской премии по химии A954), впоследствии был также лауреатом
двух премий мира — Нобелевской и Ленинской. Однажды автору довелось со-
сопровождать Л. Полинга во время его пребывания в новосибирском Академго-
Академгородке. Могу подтвердить, что впечатление он производил очень яркое.
В начале 1953 г. Уотсон и Крик познакомились (полулегально!) с последни-
последними данными Р. Франклин по дифракции рентгеновских лучей на препаратах
В-формы ДНК, возникающей при высокой влажности. Они сразу узнали при-
признаки спирали с шагом 34 А и диаметром 20 А. Для проверки необходимо
было срочно построить стереомодель, однако мастерские задерживали изготов- I
ление металлических деталей, моделирующих пурины и пиримидины. Тогда
Уотсон нарезал их из толстого картона и стал раскладывать их на плоскости
стола. Тут его и посетило озарение. Впоследствии он вспоминал [Уотсон, 1969]:
"И вдруг я заметил, что пара аденин-тимин, соединенная двумя водородными
связями, имеет точно такую же форму, как и пара гуанин-цитозин, тоже соеди-
соединенная по меньшей мере двумя водородными связями.... Если пурин всегда соединя-
соединяется водородными связями с пиримидином, то две нерегулярные последовательно-
последовательности оснований прекрасно укладываются регулярно в центре спирали. При этом
аденин всегда должен спариваться только с тимином, а гуанин только с цитози-
ном, и правила Чаргаффа, таким образом, неожиданно оказывались следствием
двуспиралъной структуры ДНК. А главное, такая двойная спираль подсказывала
гораздо более приемлемую схему репликации. Последовательности оснований двух
переплетенных цепей комплементарны друг другу. ... Поэтому было очень легко
представить себе, как одна цепь может стать матрицей для другой... ".
В течение ближайших дней была построена стереомодель двухцепочечной
ДНК. Она оказалась правовинтовой спиралью с противоположной ориентаци-
ориентацией цепей (рис. 16-2).
"Уже через два дня Морис (Уилкинс. — В.Р.) позвонил нам и сказал, что, как
убедились они с Рози (Франклин. — В.Р.), рентгенографические данные явно под-
подтверждают существование двойной спирали " [Уотсон, 1969].
Через месяц, в мае 1953 г., первая статья о двойной спирали ДНК была
опубликована в журнале Nature [Watson, Crick, 1953].
"Полинг впервые услышал о двойной спирали от Дельбрюка... Полинг, как и
Дельбрюк, был сразу же покорен" [Уотсон, 1969].
16. Хроника великого открытия. II
145
"Открытие двойной спирали... принес-
принесло нам не только радость, но и облегчение.
Это было невероятно интересно и сразу по-
позволило нам сделать важное предположе-
предположение о механизме дупликации деново " [Уот-
сон, 1968].
Модель Уотсона—Крика была при-
признана быстро и повсеместно благодаря
своим неоспоримым достоинствам. Она
полностью выдержала также испытание
временем. Одним ударом она разрешила
множество трудных проблем. Прежде
всего, она объяснила правила Чаргаффа и
рентгеноструктурные данные. Сам
Э. Чаргафф, который весьма скептически
относился к тандему Уотсон—Крик, не
смог ничего возразить по существу, его
критика скорее напоминала брюзжание:
"...мне кажется, что то огромное искус-
искусство и изобретательность, которые были
затрачены на конструирование различных
малоподходящих моделей, по существу про-
пропали даром " [Чаргафф, 1960].
Далее модель утвердила матричный
принцип, основанный на парной ком-
комплементарное™ нуклеотидов, т.е. на
принципе "близкодействия". Из модели
вытекала простая и естественная схема
матричной репликации. Ясно, что в этом
случае копирование отдельной матрицы
можно произвести только в два этапа:
позитив
негатив
позитив
Рис. 16-2. Схема строения и реплика-
репликации двойной спирали ДНК по
Уотсону— Крику.
Однако двухцепочечность спирали
решает и эту проблему. Двойная цепь
способна к точному копированию в один
этап путем двух сопряженных матричных процессов. Фактически двойная цепь
как раз и обладает вожделенным генетическим свойством: удвоением путем
контактного гомологичного выстраивания сегментов на матрице:
позитив—негатив
позитив—негатив + позитив—негатив
Наконец, модель как бы открыла путь для понимания других фундамен-
фундаментальных генетических процессов и свойств. Генетическое разнообразие оказа-
оказалось возможным свести к вариантам порядка мономеров, как предполагали
Н.К. Кольцов, М. Дельбрюк, Э. Шредингер и многие другие. Тогда сохранение
порядка должно быть основой консервативности наследственности. Двойная
146 III. Молекулярная кибернетика
цепь ДНК, где стандартный сахарофосфатный костяк расположен снаружи, а
вся специфичность (водородные связи оснований) спрятана внутри и менее
доступна для воздействий, прекрасно соответствовала ожиданиям генетиков.
Изменения же порядка мономеров, очевидно, должны были вызывать наслед-
наследственные изменения, т.е. мутации.
В 1962 г. Дж. Уотсон, Ф. Крик и М. Уилкинс получили Нобелевскую пре-
премию по физиологии и медицине за установление молекулярной структуры нук- I
леиновых кислот и ее роли в передаче информации в живой материи. К сожале-
сожалению, Р. Франклин не дождалась такого признания, она умерла в 1958 г. Несо-
Несомненно, она была достойна этой премии.
Теперь оценим полученные результаты с точки зрения информацион-
информационно-кибернетического подхода. Материальный носитель генетической инфор-
информации найден — это нуклеиновые кислоты (ДНК и, как стало ясно позже,
РНК). Определен также промежуточный получатель генетической информа-
информации — белки. Те и другие имеют ряд общих особенностей: линейные полиме-
полимеры, построенные из небольшого разнообразия мономеров — нуклеотидов и
аминокислот. В обоих случаях у мономеров есть стандартная, универсальная
часть, позволяющая им соединяться в последовательности произвольной дли-
длины и порядка. Кроме того, мономеры имеют специфические боковые группы
(основания, радикалы аминокислот), порядок которых способен определять
функциональные свойства соответствующих последовательностей. Разнообра-
Разнообразие перестановок астрономическое! Между мономерами полинуклеотидов су-
существуют особые парные отношения комплементарное™ (А-Т, G-C), позво-
позволяющие полинуклеотидам выполнять матричные функции.
Ясно, что ситуация весьма напоминает лингвистические и другие инфор-
информационные системы, где информация кодируется при помощи порядка симво-
символов. Налицо алфавиты (мономеры), тексты (последовательности), матричный
принцип копирования (комплементарность). Можно ожидать, что существуют
некие правила кодирования, которые используются клеточной информацион-
информационной машиной.
"Крик и Гам"
Этим словесным каламбуром Н.В. Тимофеев-Ресовский охарактеризовал
события, которые последовали за расшифровкой структуры ДНК. Уотсон и
Крик, разумеется, хорошо понимали генетико-информационный смысл и зна-
значение своей модели. Недаром Дж. Уотсон [1969] в своей книге пишет:
"Буквально все имевшиеся тогда факты убеждали меня в том, что ДНК служит
матрицей, на которой образуются цепочки РНК. В свою очередь, цепочки РНК
были вполне вероятным кандидатом на роль матриц для синтеза белка. ... Идея
бессмертия генов была похожа на правду, и я повесил на стену над своим столом
листок с надписью ДНК -> РНК -> Белок. Стрелки обозначают не химические пре-
превращения, а перенос генетической информации... ".
В 1958 г. Ф. Крик сформулировал этот принцип как "центральную догму "
молекулярной генетики [Crick, 1958].
16. Хроника великого открытия. II
147
Георгий Антонович Гамов A904-1968 гг.).
Однако вскоре после публикации
модели в бой вступила неожиданная и
свежая сила. Это был крупнейший
физик-теоретик Георгий Антонович
Гамов, или в английской транскрип-
транскрипции Джордж Энтони Геймов (George
Antony Gamov). В конце 20-х — начале
30-х годов Г.А. Гамов был гордостью
молодой советской теоретической фи-
физики [Френкель, Чернин, 1989; Гамов,
1994]. Его, выпускника и аспиранта
Ленинградского университета, друга
Л.Д. Ландау, послали в Геттинген
(Германия) к М. Борну, а затем в Ко-
Копенгаген (Дания) к Н. Бору для науч-
научной стажировки. Там он выполнил
множество теоретических работ высо-
высочайшего класса и был признан одним
из самых обещающих молодых физи-
физиков Европы. Интересно, что одна из
этих статей в 1930 г. была опубликова-
опубликована совместно с молодым немецким
физиком-теоретиком М. Дельбрюком.
А в 1932 г., когда Гамова не выпустили
за границу, его доклад Сольвеевскому конгрессу представил его друг Макс
Дельбрюк.
В 1932 г. по представлению академика В.А. Вернадского и двух других ака-
академиков Г.А. Гамов был избран членом-корреспондентом АН СССР. Ему было
28 лет, его воспевали поэты [Гамов, 1994]:
Советский парень Гамов
Уже до атома добрался, лиходей!
Д. Бедный
Однако в 1933 г., выехав на очередной Сольвеевский конгресс за границу,
Гамов не дождался продления командировки и не вернулся, став невозвращен-
невозвращенцем. Это был большой грех. За это его отлучили от Академии наук и от Родины.
И только в 1990 г. посмертно восстановили в Академии.
Г.А. Гамову принадлежали два крупнейших открытия: теория а-распада и
космологическая теория "горячей Вселенной" — работы нобелевского уровня.
Третьим своим основным достижением Гамов считал постановку проблемы ге-
генетического кода.
Вот как сам Гамов [1994] описывал этот момент: «Прочитав в Nature в мае
1953 г. статью Уотсона и Крика, которая объясняла, как наследственная инфор-
информация хранится в молекулах ДНК в форме последовательности четырех видов про-
простых атомных групп, известных как "основания "(аденин, гуанин, тимин и цито-
148 III. Молекулярная кибернетика
зин), я задался вопросом, как эта информация переводится в последовательность
двадцати аминокислот, которые образуют молекулы протеина. Простая идея,
которая пришла мне в голову, состояла в том, что можно получить 20 из 4 подсче-
подсчетом числа всех возможных триплетов, образующихся из четырех различных сущ-
сущностей. Возьмем, например, колоду игральных карт (выделено мною. — В.Р.), в
которой мы обращаем внимание только на масть карты. Сколько триплетов од-
одного и того же вида можно получить? Четыре, конечно: трое червей, трое бубен,
трое пик и трое треф. Сколько триплетов с двумя картами одной и той же мас-
масти и одной другой? Пусть мы имеем четыре выбора для третьей карты. Поэтому
мы имеем 4 *3 = 12 возможностей. В дополнение мы имеем четыре триплета со
всеми тремя различными картами. Итак, 4 + 12 + 4 — 20, а это и есть точное
число аминокислот, которое мы хотели получить».
Таким образом, Гамов первым сформулировал проблему генетического
кода. Генетическая информация записана в полинуклеотидах в виде последова-
последовательности букв, символов четырех типов: А, Т, G и С. Затем она перекодируется
в последовательность букв 20 типов (аминокислот). Кодирующие группы сим-
символов могут быть только триплетными. Правила соответствия триплетных
групп нуклеотидных символов (в дальнейшем названных кодонами) и символов
аминокислот образуют генетический код. Главная задача — расшифровать этот
код, в том числе объяснить происхождение числа 20, имея в наличии 43 = 64
триплета.
Чтобы понять такой поворот мысли, надо учесть некоторые сопутствую-
сопутствующие обстоятельства.
Во-первых, Гамов сравнил последовательность нуклеотидов с длинным
числом, записанным в четверичной системе счета. В шутку он назвал его
"звериным числом ", имея в виду религиозную легенду из "Апокалипсиса", где
имя антихриста ("зверя из бездны") скрыто под неизвестным числом. Расшиф-
Расшифровка "звериного числа" необходима для победы над зверем. Кроме того, 20 —
число аминокислот — он назвал "магическим числом ", предполагая, что объяс-
объяснить его из внутренней структуры кода — это и значит решить проблему (цит.
по: [Ичас, 1971]).
Первая статья Гамова и Томкинса была послана в журнал Proceedings of the
National Academy of Sciences of the USA и отвергнута редакцией, поскольку
Томкинс — это мифический персонаж популярных книг Гамова, а не реальное
лицо. От имени одного Гамова эта статья вышла в свет в 1954 г. в Докладах Дат-
Датской Академии наук в Копенгагене.
Во-вторых, летом 1953 г. Уотсон и Крик составили стандартный список из
20 аминокислот, который далее был канонизирован. Фактически они выбрали
20 аминокислот, непосредственно участвующих в синтезе белков, а вторичные
их производные исключили.
В-третьих, Гамов очень непринужденно использовал карточную термино-
терминологию. Чего стоят хотя бы такие пассажи: "Возьмем, например, колоду игральных
карт..." [Гамов, 1994] или «Допустим, мы играем в "упрощенный покер"...»
[Gamov, 1958], и далее по тексту. Образ оказался очень точным. Действительно,
имеем четыре масти — две черные с ножками (пурины) и две красные без но-
16.
S? S? 4> 4<
Хроника великого открытия.
> 4» s? 4 <0
II
A
149
1» ^ и т.д.
J
Ser
Val
Glu
Ser
Рис. 16-3. Гипотетическая полинуклеотидная цепь, изображенная Г.А. Гамовым при
помощи карточных символов.
жек (пиримидины). Последовательность нуклеотидов можно представить в до
боли знакомом виде (рис. 16-3) [Gamov, 1958].
Природа как бы играет с теоретиком в "упрощенный покер", игра азарт-
азартная, а выигрыш — крупнейшее открытие XX века. Ясно, что души теоретиков
дрогнули! Предсказания Э. Шредингера сбывались! Интерес к проблеме стре-
стремительно достиг апогея. Романтический и оптимистический период изучения
генетического кода начался.
Кстати, "сродство" Гамова к карточной терминологии становится понят-
понятным, если учесть некоторые его пояснения и обмолвки. Например, [Гамов,
1994]: "Однажды вечером, когда я зашел в знаменитое Казино да Урка, чтобы по-
посмотреть на карточную игру... ". Короче говоря, Гамов — картежник, но в дан-
данном случае это способствовало постановке проблемы. Известно ведь, что тео-
теория вероятностей возникла из практики карточных игр и рулетки!
В-четвертых, Гамов попытался использовать для решения проблемы гене-
генетического кода методы дешифровки шпионских кодов, в которых имел некото-
некоторый опыт. Вначале он предложил гипотезу о "перекрывающемся ромбическом
коде", когда можно было проследить за определенными закономерностями в
структуре известных полипептидов. В своей автобиографии Гамов [1994] писал:
"...работа была столь же трудна, как расшифровка секретного военного кода на
основе только двух коротких посланий, добытых шпионами. Так как в то время я
(Гамов. — В.Р.) был консультантом в Военно-морском министерстве Соединен-
Соединенных Штатов в Вашингтоне, O.K., я пошел к адмиралу, под командованием кото-
которого находился, и спросил, можно ли поручить сверхсекретной криптографической
группе расшифровку японского кода. В результате в моем отделе Университета
им. Дж. Вашингтона появились три человека...
Я поставил перед ними задачу, и через несколько недель они сообщили мне, что
она не имеет решения. То же заключение было получено моими друзьями-биолога-
друзьями-биологами: Мартинасом Ичасом, уроженцем Литвы, и Сиднеем Бреннером, уроженцем
Южной Африки. Это исключило возможность перекрывающегося кода...".
В целом такая же судьба постигла и другие гипотезы. Гамов и Ичас предло-
предложили гипотезу "комбинаторного" кода [Ичас, 1971], где все триплеты одинако-
одинакового состава считались синонимами; 64 триплета образовали 20 групп (магиче-
(магическое число!); код был вырожден, триплеты в тексте не перекрывались. Очень
похоже на правду! Однако и этот код был забракован.
Крик, Гриффите (племянник открывателя трансформации) и Орджел
[Ичас, 1971] предложили идею "кода без запятых", в котором триплеты в тексте
не отделены какими-либо знаками, но считываются единственным образом,
кодирующими являются 20 гетеротриплетов, а все их циклические перестанов-
перестановки D0) — некодирующими. Четыре гомотриплета в этом случае — то же неко-
150 III. Молекулярная кибернетика
дирующие. Этот вариант также не подтвердился, хотя сама математическая
проблема "кодов без запятых" исследуется математиками до сих пор.
В целом, в этом умственном состязании приняли участие многие выдаю-
выдающиеся математики, физики, химики, инженеры, а также научная молодежь.
Можно сказать, что они славно размялись! Несмотря на остроумие многих
предложений, все они оказались неверными.
"Природа хитра...", — заключил Гамов [1968] через 10 лет. Оптимистиче-
Оптимистический период исследований генетического кода закончился. Наступило время
экспериментального решения, которое в итоге оказалось очень успешным и
совершенно иным. Имя Г.А. Гамова почти исчезло из научной литературы по
молекулярной биологии. В 1968 г. Г.А. Гамов умер.
Значение работ Гамова было очень точно сформулировано Ф. Криком
(цит. по: [Гамов, 1994]): "Важность работы Гамова состояла в том, что это
была действительно абстрактная теория кодирования, которая не была перегру-
перегружена массой необязательных химических деталей...". Иначе говоря, это был ин-
информационно-кибернетический подход в чистом виде, который позже полно-
полностью себя оправдал при разработке теории молекулярно-генетических систем
управления и генетического языка [Ратнер, 2001] (см. гл. 18).
И в заключение — два личных замечания. В 1995 г. на конференции в Мо-
Москве я познакомился с Мартинасом Ичасом — соавтором Гамова по первым ра-
работам. Он говорит по-русски, интересуется российской наукой и написал пре-
превосходную книгу о генетическом коде [Ичас, 1971]. Очень милый старик! Он
подтвердил практически все, что мы знаем о Гамове.
В 1964 г. я послал Г.А. Гамову свою первую статью по генетическому коду и
получил его ответ. Это был оттиск в серой обложке под названием "What is
Life?" из журнала "Труды индийского института теоретической физики им. Бо-
зе" [Gamov, 1963]. На оттиске было написано:
Судьба — индейка, а жизнь — копейка.
Г. Гамов
Список литературы
Гамов Г.А. Комбинаторные принципы в генетике // Прикладная комбинаторная мате-
математика / Ред. Э. Беркенбах. М.: Мир, 1968. С. 289-308.
Гамов Г.А. Моя мировая линия: неформальная автобиография. М.: Наука, 1994.
Ичас М. Биологический код. М.: Мир, 1971.
Ратнер В.А. Хроника великого открытия: идеи и лица // Природа. 1998. № 4. С. 68-79.
Ратнер В.А. Молекулярно-генетическая система управления // Природа. 2001. № 3.
С. 16-22.
Уотсон Дж.Д. Роль РНК в синтезе белка (Нобелевская лекция, 1962) // Гершкович И.
Генетика. Приложение IX. М.: Наука, 1968. С. 635-652.
Уотсон Дж.Д. Двойная спираль. М.: Мир, 1969.
Френкель В.Я., Чернин А.Д. Возвращается Г.А. Гамов // Природа. 1989. № 9. С. 82-102.
Чаргафф Э. Основания, входящие в состав дезоксипентозных и пентозных нуклеиновых
кислот у разных видов // Химические основы наследственности / Ред. У. Макелрой, Б. Гласе.
М.: Изд-во иностр. лит., 1960. С. 419-424.
17. Хроника великого открытия. Ill 151
Avery O.T., MacLeod СМ., McCarty M. Studies on the Chemical Nature of the Substance
Inducing Transformation of Pneumococcal Types. I // J. Exp. Med. 1944. Vol. 79. P. 137-158.
ChargafT E. Structure and Function of Nucleic Acids as Cell Constituents // Feder. Proc. 1951.
Vol. 10. P. 654-659.
Crick F.H.C. // Symp. Soc. Exptl. Biol. 1958. Vol. 12. P. 138-163.
Hershey A.D., Chase M. Independent Functions of Viral Protein and Nucleic Acid in Growth
of Bacteriophage // J. Gen. Physiol. 1952. Vol. 36. P. 39-56.
Gamov G.A. What is Life? // Trans, of the Bose Research Institute. Calcutta. 1963. Vol. 24.
Gamov G.A. Matter, Earth and Sky. N.Y.: Pentice-Hall, 1958.
Pauling L., Delbriick M. The Nature of the lntermolecular Forces Operating in Biological
Systems // Science. 1940. Vol. 92. P. 77-79.
Portugal F.H., Cohen J.S. A Century of DNA. The MIT Press. Cambridge, MA, 1979.
Watson J.D., Crick F.H.C. A Structure of Deoxyribose Nucleic Acid // Nature. 1953. Vol. 171.
P. 737-738.
17. Хроника великого открытия: идеи и лица. III
Итак, теоретическая атака на проблему генетического кода оказалась почти
безрезультатной (см. гл. 15) [Ратнер, 1998]. Излюбленный прием физиков-тео-
физиков-теоретиков — рассмотреть множество мыслимых вариантов на все случаи жизни,
базируясь на некоторых гипотетических принципах, — не принес успеха. Как
выразился потом Френсис Крик [Crick, 1966], "к 1959 г. проблема кодирования
была в упадке". Крупнейший советский молекулярный биолог академик
В.А. Энгельгардт на вопрос корреспондента газеты о том, когда будет решена
проблема генетического кода, ответил: лет через пятьдесят. Проблема, действи-
действительно, была трудной, но не безнадежной. Вопреки предсказанию, это стало
ясно уже через один-два года, когда были нащупаны экспериментальные под-
подходы к решению проблемы кода. Однако все по порядку.
Гены — это атомы наследственности
Этими словами американский генетик Саймур Бензер начал свою итого-
итоговую Гарвеевскую лекцию о внутренней структуре гена в 1961 г. [Benzer, 1961;
Бензер, 1963]. Такова классическая точка зрения генетиков. Как развивалось
это представление — см. гл. 11.
С. Бензер был физиком. Как и на сотни других студентов, книга Э. Шре-
дингера "Что такое жизнь? С точки зрения физика"(см. гл. 16) произвела на
него сильное впечатление. В 1949 г., поступив в аспирантуру по биологии в Ок-
ридже, он получил возможность продолжить работу в лабораториях С. Лурии
или М. Дельбрюка. Аспирант Дж. Уотсон, который обучался у С. Лурии, давал
С. Бензеру советы: "Лурия будет спрашивать тебя ежедневно, что ты делал, а
Дельбрюка ты вообще можешь не увидеть неделями" [Benzer, 1966]. С. Бензер вы-
выбрал лабораторию М. Дельбрюка в Калифорнийском институте технологии
(Caltech).
* Ранний вариант главы опубликован: Ратнер В.А. Природа. 2000. № 6. С. 22-30.
152
III. Молекулярная кибернетика
Саймур Бензер.
В 1950 г. М. Дельбрюк послал аспиранта в Париж, Институт Пастера, к из-
известному специалисту по фагам Андре Львову. Вот как об этом вспоминал сам
Бензер [Benzer, 1966]: "Впервые я открыл феномен rll в Париже, но тогда не оце-
оценил его значения ". Как сказал Пастер: "мой ум не был подготовлен".
17. Хроника великого открытия. III 153
Позже, в 1952 г., он вернулся к этой проблеме: "Когда я вторично столкнул-
столкнулся с феноменом rll, мой ум уже был подготовлен... Это была система со свойства-
свойствами высокого генетического разрешения " [Benzer, 1966].
Иначе говоря, Бензер обнаружил условную летальность мутаций rll фага Т4
при его размножении на различных штаммах бактерии-хозяина: на Е. coli В
гЯ-мутанты фага были жизнеспособны, ускоренно размножались, убивая бак-
бактерии, а на Е. coli K12 не размножались совсем, т.е. были летальны. На Е. coli В
всех мутантов можно было размножать и поддерживать. На Е. coli K12 их мож-
можно было скрещивать и отбирать, т.е. вести удобный анализ при помощи класси-
классических генетических методов — теста функциональной комплементарности и
метода рекомбинационного картирования:
«Разрешающая способность (рекомбинаций. — В.Р.) должна была быть дос-
достаточной для разделения мутаций, даже если бы они были локализованы в смеж-
смежных нуклеотидных сайтах (участках. — В.Р.). Другими словами, это была систе-
система, где, как позже выразился Дельбрюк, было можно "привязать генетическую
карту к местности"» [Benzer, 1966].
С благословения М. Дельбрюка С. Бензер начал строить высокоточную
внутреннюю генетическую карту локуса rll фага Т4. За 10 лет A952-1961) он
картировал свыше 1600 гЯ-мутаций, оценил размер локуса, его физическую
длину, получил множество других впечатляющих данных. В последующие годы
число исследованных мутаций достигло 2400. На несколько десятилетий вперед
локус rll оказался наиболее подробно генетически картированным геном в
биологическом мире (рис. 17-1).
Основные результаты С. Бензера состояли в следующем [Бензер, 1963]:
1) локус rll включает два гена (цистрона) — rll А и В, которые не перекры-
перекрываются. В дальнейшем было показано, что эти гены кодируют белки;
2) размер цистронов оценен в ~ 102-103 нуклеотидных пар (н.п.); в дальней-
дальнейшем было показано, что rll А имеет размер ~ 2400 н.п. (белок — 88 000 D), a rll
В ~ 1200 н.п. (белок - 44 000 D);
3) рекомбинации разделяют участки (сайты) мутирования, между которы-
которыми было несколько нуклеотидных пар; в дальнейшем удалось разделить сосед-
соседние нуклеотиды;
4) локус rll был разбит на 308 мутационных сайтов (в дальнейшем — 370);
отдельный сайт в среднем отвечает 3 н.п. (по более поздним данным ~ 10 н.п.).
Это достаточно высокая степень заполнения локуса мутациями, почти до уров-
уровня кодонов.
Таким образом, молекулярное представление о генах приобрело новые
очертания. Ген (цистрон) по Бензеру — это непрерывный протяженный уча-
участок последовательности ДНК размером ~102-103 пар мономеров, кодирующий
белок. Он состоит из сотен сайтов, мутации которых искажают или уничтожа-
уничтожают общую функцию гена. Поэтому цистрон как целое является единицей функ-
функции. Вспомним шутку Н.В. Тимофеева-Ресовского: "это...не кусочек сливочного
масла\" (см. гл. 1). Мутации могут происходить в любых сайтах, вплоть до от-
отдельных нуклеотидов, т.е. ген не является единицей мутирования (мутоном).
Мутон равен паре нуклеотидов. Рекомбинации могут разделять смежные пары
2
Ala
Albi
Alb2
A2a A2b A2c A2d A2e A2f A2g
A2hl
A2h2
A4d A4c A4b A4a A3i A3h A3g A3f A3e
A3a-d
A2h3
A4e
A4f
2
1
3
I
B8 B9a B9b BIO
Рис. 17-1. Рекомбинационная генетическая карта локуса rll фага Т4, построенная С. Бензером [1963]. Цистроны А и В не пе-
перекрываются. Каждая клеточка означает одного мутанта. Сайты вдоль карты разделены рекомбинациями. Два сайта содержат
"горячие точки" мутирования.
17. Хроника великого открытия. III 155
нуклеотидов. Поэтому ген не является неделимой единицей рекомбинации (ре-
коном). Рекон равен паре нуклеотидов.
Все это прекрасно укладывается в рамки информационно-лингвистическо-
информационно-лингвистического подхода. Гены представляют собой не зачатки биологических структур, а на-
напоминают линейные тексты (участки ДНК), записанные при помощи некото-
некоторых правил и несущие генетическую информацию о молекулярных структурах и
функциях. Они ограничены некоторыми размерами, имеют разные последова-
последовательности символов-мономеров. Возможны "бессмысленные" участки, не ко-
кодирующие белки или кодирующие испорченные белки. При этом во всех случа-
случаях и гены, и некодирующие участки являются сегментами молекул ДНК, т.е.
построены из того же алфавита четырех нуклеотидов. Поэтому различия между
такими текстами не в их физической природе, а исключительно — в последова-
последовательностях символов-мономеров. В этом ключ информационно-лингвистиче-
информационно-лингвистического подхода. Это значит, что гены — не физические, а информационные едини-
единицы наследственности, т.е. предположение Э. Шредингера о "шифровальном
коде "действительно подтверждается и материализуется.
Но если гены как последовательности нуклеотидов все же ограничены по
размеру, то они должны быть как-то выделены в геноме, поскольку основное
генетическое качество генов — это их дискретность. Далее, должно быть соот-
соответствие между линейными порядками нуклеотидов в гене и аминокислот в
белке (колинеарное соответствие). Наконец, текст цистрона должен быть тем
или иным способом разбит на кодирующие группы (кодоны), которые кодиру-
кодируют отдельные аминокислоты.
С. Бензер продолжал картирование локуса rll примерно до 1967 г. Потом
он полностью оставил эту область и переключился на генетику поведения дро-
дрозофилы, где сделал несколько очень интересных открытий. Однако главные его
результаты все же лежат здесь.
Сдвиг вправо, сдвиг влево — приравнивается к летали
Ясно, что ответы на эти вопросы лежали скорее в области информацион-
информационно-лингвистического подхода, чем чисто биохимического. Пожалуй, в конце
50-х годов такой подход в наибольшей мере был присущ именно Ф. Крику.
И он (подход) оказался плодотворным. В теоретическом плане Ф. Крик и
С. Бреннер поняли, что в генетическом тексте (цистроне), разбитом на коди-
кодирующие группы одинаковой длины (кодоны), замены символов не нарушают
разбиение текста на кодоны, а выпадения (-) и вставки (+) символов произво-
производят сдвиг фазы разбиения соответственно влево или вправо (рис. 17-2). Комби-
Комбинирование (-)- и (+)-сдвигов может вызвать генетическое явление супрессии
(т.е. восстановления) исходной фазы трансляции. Особенности такой внутри-
цистронной супрессии позволят сделать заключение об общих свойствах гене-
генетического кода. Заметим, что эта идея не оперирует никакими физико-химиче-
физико-химическими особенностями ДНК, а рассматривает ее абстрактно как последователь-
последовательность символов алфавита, разбитую на слова-кодоны.
Рассказывают, что для экспериментального решения этой проблемы
Ф. Крик с сотрудниками сняли на лето 1961 г. навес для велосипедов в Кем-
156 III. Молекулярная кибернетика
...ATCGTCAGTAGCACTGTC... Исходная последовательность цитрона
...ATCTCAGTAGCACTGTC... Делеция одного нуклеотида
( ' """ Сдвиг фазы трансляции
Вставка ОДНОГО
(+) т~ Сдвиг фазы трансляции
v'—V—^
...ATCTCAGTAGCGACTGTC... Супрессия (- +)
(-) "¦ (+) Сдвиг фазы трансляции
Рис. 17-2. Схема, поясняющая идею эксперимента Крика и сотр. по супрессии сдвиго-
сдвиговых мутаций в сегменте цистрона rll В фага Т4. (+) — вставка, (-) — выпадение символа.
бридже, одолжили на время лабораторное оборудование и успешно выполнили
свой классический эксперимент. После этого все было ликвидировано. В кон-
конце 1961 г. работа была опубликована в английском журнале Nature [Крик и др.,
1963].
Они использовали генетическую систему локуса rll фага Т4 С. Бензера.
Несколько ранее в Кембридже Ф. Крик и С. Бензер изучали мутагенез в локусе
rll фага Т4, индуцированный при помощи акридиновых красителей. Почти все
мутации оказались летальными. Это было очень непохоже на мутагенез анало-
аналогами оснований, которые вызывают замены нуклеотидов. Хотя данные были не
столь очевидны, Ф. Крик предположил, что акридины вызывают выпадения и
вставки нуклеотидных пар в цепи ДНК. Это и был желанный подход к внутри-
цистронной супрессии. Интуиция не подвела Крика. Его группа получила свы-
свыше 120 акридиновых мутаций в небольшом левом сегменте цистрона rll В вбли-
вблизи границы с цистроном rll А (см. рис. 17-1). Почти все мутации были леталь-
летальными на Е. coli K12, а 80 из них при комбинировании в одном геноме проявили
различные свойства восстановления (взаимной супрессии) функции гена rll В.
Из этих 80 мутаций можно выделить две группы, (+) и (-), по способности
к взаимной супрессии функции гена В при попадании в одну копию гена. При
этом:
A) у двойных мутантов (++) и (--) функции гена В никогда не восстанав-
восстанавливалась;
B) у двойных мутантов (+-) и (-+) во многих случаях функция гена В была
восстановлена, но лишь в случаях, когда мутации были на близких расстояниях
друг от друга (как потом выяснилось, не более 50-60 н.п.), а на больших рас-
расстояниях такого эффекта не наблюдали;
C) у тройных мутантов ( ) и (+++) в нескольких случаях на близких
расстояниях функция восстанавливалась, но далеко не всегда.
Информационно-лингвистический подход позволяет очень четко интер-
интерпретировать эти данные. Рассмотрим ген (цистрон) как очень длинный текст
17. Хроника великого открытия. III 157
(фактически — предложение), разбитый на короткие слова одинаковой длины
(кодоны). Разбиение цистрона на кодоны соответствует фазе считывания тек-
текста, трансляции. Рассмотрим влияние вставок и выпадений символов на изме-
изменение или восстановление правильной единственной фазы трансляции.
Пусть (+)-мутация связана с внедрением одного или нескольких символов
(но не кратно размеру кодона), а (-)-мутация — с выпадением такого же фраг-
фрагмента. Пусть пары мутаций (++) и (--) тоже не кратны размеру кодона. Тогда
одиночные (+)- и (-)-мутации должны полностью разрушать правильную фазу
трансляции "вниз по течению" (т.е. по направлению трансляции). У (++)- и
(—)-мутантов правильная фаза трансляции не должна восстанавливаться, а у
(+-)- и (-+)-мутантов — должна (см. рис. 17-2).
Если у тройных мутантов (+++) и ( ) возможно восстановление пра-
правильной фазы трансляции, то это значит, что кодоны триплетны или кратны
трем (правда, последнее уж очень расточительно!), поскольку происходит сум-
суммарный сдвиг фазы, кратный трем.
В триплетных кодонах сдвиг влево на один символ с точки зрения фазы
трансляции эквивалентен сдвигу вправо на два символа; а сдвиг влево на два
символа эквивалентен сдвигу вправо на один. Других вариантов нет. Поэтому
должно быть ровно две группы ((+) и (-)) мутантов. В дублетном коде была
бы одна группа, где все мутации восстанавливали бы друг с другом правиль-
правильную фазу трансляции. В квадруплетном коде было бы более двух групп взаим-
взаимной супрессии. Поэтому кодоны триплетны без оговорок, а их максимальное
число 64.
Если в генетическом коде есть некодирующие триплеты (нонсенсы), то
при сдвиге образуется как бы новая случайная последовательность кодонов,
среди которых могут встретиться и нонсенсы. При этом важен размер участка
сдвига: на малых участках нонсенсы могут не встретиться, а на больших встре-
встретятся обязательно и станут барьерами трансляции. Если в коде 64 триплетных
кодона, а число нонсенсов два-три, то в этом случае в среднем они должны
встретиться на участках сдвига размером больше 20-30 кодонов D0-60 н.п.).
Поэтому у многих двойных мутантов типа (+-) и (—О на участке сдвига фаза
трансляции должна восстанавливаться на расстояниях не более 40-60 н.п.,
когда нонсенсы-барьеры практически встречаются редко. Это подтверждено
экспериментально. Поэтому генетический код сильно вырожден и содержит
несколько некодирующих триплетов-нонсенсов.
У двойных мутантов (+-) и (-+) на участке сдвига фаза разбиения на кодо-
кодоны меняется. Бывшие границы старых кодонов становятся внутренними участ-
участками новых, а бывшие внутренние участки — новыми границами. Трансляция
при этом не останавливается, а значит, между кодонами нет никаких специаль-
специальных разделительных знаков ("запятых").
Если все кодоны триплетны, то в принципе любой цистрон может быть
прочитан тремя способами (т.е. в трех фазах). Однако реализуется лишь одна
фаза считывания, когда внутри цистрона нет нонсенсов-барьеров. В остальных
фазах, как показано выше, нонсенсы встречаются уже на расстояниях порядка
20-30 кодонов. Поэтому если кодоны не отделены "запятыми" и нет других
158
III. Молекулярная кибернетика
1589
внутренних способов их выделения
(например, "код без запятых"), то
должно существовать фиксированное
начало отсчета фазы, т.е. начало
трансляции.
Ранее Бензер нашел тестерную
делецию № 1589, где выпали правая
часть цистрона rll А, левая часть
цистрона rll В и все, что было между
ними (рис. 17-1 и 17-3). Крик и кол-
коллеги показали, что при этом (+)-му-
тация в rll А вызывала инактивацию
цистрона rll В, а супрессия (+-) в А
тоже распространялась в rll В. Сле-
Следовательно, цистроны А и В сшиты в
одной фазе трансляции и считыва-
ются слева направо в виде одного об-
общего белкового продукта. Между
ними нет препятствий. В нормаль-
нормальном локусе rll, где этого нет, цистро-
цистроны А и В должны быть разделены границей — межцистронным интервалом и
двумя знаками: терминальным знаком цистрона А и начальным знаком цис-
цистрона В.
Любопытно, что когда в начале 80-х годов описанные зоны цистронов rll A
и rll В были секвенированы, все эти выводы полностью подтвердились. Даже
знак мутаций Крик угадал правильно: (+) — внедрение, (-) — выпадение нук-
леотида. Конечно, это чистая случайность, но каков случай! Поразительно, как
много новой информации было получено в этом эксперименте! Поистине сдвиг
сдвигом вышибают). Работа Крика и сотрудников — просто шедевр информаци-
информационно-лингвистического подхода! К 1994 г. геном фага Т4 был секвенирован
полностью.
Зона сдвига
Рис. 17-3. Сшивание двух цистронов rll А и
В при помощи делеции rll 1589.
а — нормальные цистроны Аи В разделены гра-
границей трансляции; б — делеция 1589 устраняет
границу между цистронами, функция А потеря-
потеряна, функция В сохранена; в — (+)-мутация в А
инактивирует функцию В путем общего сдвига
вправо фазы трансляции; г — добавочная
(-)-мутация в А супрессирует (+)-мутацию и
восстанавливает функцию цистрона В.
Гонка за лидером
Другая сенсация 1961 г. пришла с неожиданной стороны. Молодой амери-
американский биохимик М. Ниренберг и приехавший из Германии в США X. Мат-
теи изучали синтез белков в бесклеточной системе трансляции Е. coli. Ранее
они показали, что для включения меченых аминокислот необходимо участие
природной или синтетической РНК, которую потом стали называть
"РНК-посредником" или "матричной РНК".
В одном из экспериментов для контроля они добавили в реакционную
смесь гомополимер поли-U в качестве матрицы в количестве Юцг на 1 мл. При
этом почти в 1000 раз увеличивалось включение меченой аминокислоты фени-
лаланина (Phe) в продукты синтеза. Другие аминокислоты не включались, и
потому полипептидным продуктом оказался полифенилаланин. Стало ясно,
что кодон UUU должен кодировать фенилаланин.
17. Хроника великого открытия. III
159
Маршалл Ниренберг.
М. Ниренберг и X. Маттеи по-
послали в печать первую работу в марте
1961 г. Затем они представили доклад
на одной из секций V Международ-
Международного биохимического конгресса, ко-
который проходил в Москве 10-16 авгу-
августа [Ниренберг, Маттеи, 1962]. В ку-
кулуарах М. Ниренберг разговорился с
Дж. Уотсоном и А. Тиссье, рассказав
им о своих результатах. Те, оценив
ситуацию, немедленно сообщили об
этом Ф. Крику, который был пред-
председателем одного из самых круп-
крупных симпозиумов — "Биологические
структуры и функции на молекуляр-
молекулярном уровне". Ф. Крик сразу понял,
что эта работа сулит "прорыв" в ис-
исследовании кода. Нарушив правила,
Крик пригласил Ниренберга повто-
повторить его доклад на своем симпозиуме
перед большой аудиторией и сопро-
сопроводил выступление своими положи-
положительными комментариями.
Так был дан старт великой "гонке за лидером". Собственно, к тому момен-
моменту гонка уже началась. В 1961 г. 34-летний малоизвестный доктор Ниренберг
имел небольшую лабораторию в Национальном институте артрита и болезней
обмена (г. Бетесда, Мэриленд). Начав изучение генетического кода, он сразу
же попал в "высококонкурентную среду ". О его работе прослышал крупнейший
биохимик, нобелевский лауреат С. Очоаи, конечно, тут же понял, какова высо-
высота ставки. Он был председателем другого симпозиума на Международном био-
биохимическом конгрессе. Его сопредседатель академик А.И. Опарин во вступи-
вступительном слове выразил большое сожаление, что профессор Очоа не смог при-
прибыть на конгресс. Да, Очоа не поехал в Москву, а тут же летом попытался
сделать бросок вперед, чтобы опередить Ниренберга. Силы были неравные: у
авторитетнейшего ученого Очоа не было финансовых проблем, кроме того, он
владел многими уникальными методами. Но и Ниренберг не собирался сда-
сдаваться.
Очень быстро в обеих лабораториях при помощи фермента полинуклео-
тид-фосфорилазы (за открытие которой С. Очоа и получил в 1959 г. Нобелев-
Нобелевскую премию) начали синтезировать статистические полинуклеотиды задан-
заданного состава, например, U: А = 5: 1 и т.д. Эти матрицы стимулировали вклю-
включение других меченых аминокислот в определенных пропорциях. В результате
к 1963 г. был установлен нуклеотидный состав 49 кодонов, отвечающих опреде-
определенным аминокислотам (табл. 17-1). Данные двух лабораторий фактически сов-
совпали. С. Очоа не удалось обогнать М. Ниренберга. После этого С. Очоа факти-
фактически вышел из игры.
160
III. Молекулярная кибернетика
Таблица 17-1
Состав кодонов по Ниренбергу—Очоа (цит. по: [Ратнер, 1966])
Аминокислота
Алании (Ala)
Аргинин (Arg)
Аспарагиновая кислота (Asp)
Аспарагин (Asn)
Цистеин (Cys)
Глутаминовая кислота (С/и)
Глутамин (Gin)
Глицин (Gly)
Гистидин (His)
Изолейцин (Не)
Лейцин (Leu)
Лизин (Lys)
Метионин (Met)
Фенилаланин (Phe)
Пролин (Pro)
Серии (Ser)
Треонин (Thr)
Триптофан (Тгр)
Тирозин (Туг)
Валин (Val)
Кодомы
W1CW, 1AIC1G, 2C1G
1U1C1G, 2C1G, 2A1G
W1AIG, IA1CW
1U2A, 1U1A1C, 2А1С
2UW
2A1G, 1U1A1G
2А1С, 1WAIC
W2G, 1A2G, 1C2G
WIAIC, 1A2C
2ША, W2A, W1A1C
2UW, 2U1A, 2UIC, W2C, UUV ?
ААА, W2A, 1G2A
1U1GIA
UUU, 2ШС
ССС, 1А2С, W2C
2U1C, Ш2С, 1U1G1C, 1A1G1C
2А1С, 1А2С, 1U1A1C
1U2G
2U1A, 1U1A1C
2U1G
Далее, по словам Ф. Крика [Crick, 1966], "наступила пауза, поскольку было
неясно, как продолжать работу и найти порядок оснований в триплетах. Это
привело к шквалу теоретических работ, большинство из которых благополучно за-
забыто... Я сам сделал несколько попыток по этому поводу, но не думаю, что их
стоило публиковать ".
Да, теоретики вновь почувствовали запах пороха! В этот период многие по-
попробовали угадать, предсказать, вывести порядок оснований в кодонах из имев-
имевшихся неполных экспериментальных данных и некоторых научных гипотез. Я
расскажу об одной такой попытке, которая, к моему огорчению, не потрясла
мир, но все-таки породила у автора определенную уверенность, что такие вещи
нужно и даже можно делать успешно. Этим автором был я сам [Ратнер, 1966].
В 1963 г. теоретики располагали следующими данными:
1) таблицей состава 49 кодонов (см. табл. 17-1). Это был явно не полный
список, поскольку некоторые кодоны было трудно выявить на фоне других;
2) списком мутационных замен аминокислот в некоторых природных бел-
белках (оболочечном белке вируса табачной мозаики (ВТМ), триптофан-синтетазе
Е. coli и гемоглобинах человека). Эти списки тоже были далеко не достаточны-
достаточными, часто пересматривались, некоторые мутации были ненадежными.
Ясно, что замены аминокислот должны были согласовываться с реальной
таблицей состава кодонов (см. табл. 17-1). В принципе, при полноте таких дан-
данных можно было надеяться восстановить относительный порядок оснований во
всех кодонах с точностью до одинаковых перестановок символов во всей кодо-
кодовой таблице. При неполноте этих данных можно было рассчитывать получить
17. Хроника великого открытия. III 161
относительный порядок оснований лишь в части кодонов. Чтобы избавиться от
неоднозначности за счет перестановок символов, нужно было иметь независи-
независимые данные об абсолютном порядке символов хотя бы в одном гетерогенном
мутирующем кодоне.
В 1963 г. таких данных не было, однако были другие, которые можно было
использовать вместо известного кодона. Ч. Яновский и сотрудники в 1963 г.
опубликовали "мутационную вилку" в 211-м кодоне белка триптофан-синтета-
зы Е. coli (цит. по: [Ратнер, 1966]):
Gly
Ala Ser Thr Ik
Некоторые из этих мутантов рекомбинировали между собой:
Arg х ™Val => mGly + Ser.
Яновский показал, что мутант Val имеет маркерную мутацию Ас~(т), лока-
локализованную слева от кодона Val за пределами локуса. У мутантов Arg и Val за-
затронуты разные нуклеотиды одного кодона. Сравнение кодонов всех участвую-
участвующих серий аминокислот между собой и учет перехода левого маркера (Ас~~)
именно к рекомбинанту Gly приводят к важным следствиям:
— все данные совместимы только при предположении, что м-РНК считы-
вается в направлении 5' -» 3', если N-конец белка ориентирован влево (этот
факт еще не был известен, биохимические данные были неоднозначны);
— вследствие предыдущего, мутация Gly -> Arg оказалась левее мутации
Gly -> Glu -> Val, т.е. возможные варианты порядка в кодонах Gly могли
быть: GGU, GUG, GGA или GAG (помечены участвующие нуклеотиды).
Таким образом, Яновский определил порядок двух символов в кодоне Gly.
Вначале я думал, что это был побочный результат его попытки разделить ко-
дон рекомбинацией. Однако потом оказалось, что эксперимент был задуман
именно для определения абсолютного порядка символов и оказался успеш-
успешным.
Мутационных данных по-прежнему не хватало, чтобы устранить все не-
неоднозначности. Однако еще раньше, в 1962 г., я заметил, что применительно к
структуре генетического кода можно сформулировать одно дополнительное
свойство эволюционной природы — "принцип связности кодовых серий ". Сери-
Серией называется группа синонимических кодонов одной аминокислоты. Вспом-
Вспомним, что кодоны опознаются антикодонами т-РНК. Можно предположить,
что изоакцепторные фракции т-РНК (переносящие одну аминокислоту) име-
имеют общее эволюционное происхождение и возникают из одной общей пред-
ковой фракции путем замен нуклеотидов в антикодонах т-РНК. В этом случае
все кодовые серии должны быть связными. Серия называется связной, если,
162 III. Молекулярная кибернетика
оставаясь в пределах серии, можно из каждого ее кодона получить каждый
другой путем последовательных замен отдельных символов. В противном слу-
случае серия считается разрывной [Ратнер, 1966].
Итак, серии кодонов должны возникать как связные группы. Но может
быть, они теряют некоторые кодоны эволюционно, в результате чего серии мо-
могут стать разрывными вторично? Я показал, что это невозможно (цит. по: [Рат-
[Ратнер, 1966]). Действительно, список кодонов эквивалентен словарю. Если кодо-
кодоны используются многократно в структурах генов, то утрата любого из них в ге-
генетическом коде (т.е. утрата фракций т-РНК или кодирующего фермента
аминоацил-т-РНК-синтетазы) будет грубой деталью, поскольку в результате
возникнут множественные дефекты ("дырки") в генетических текстах. Такие
мутации жестко отсекаются стабилизирующим отбором. Поэтому, каков бы ни
был истинный механизм вырождения кода, ни один из когда-либо возникших
кодонов словаря не мог быть утерян в дальнейшем ходе эволюции. Поэтому се-
серии должны быть связные.
В результате я разработал удобную схему (теперь ее можно назвать алгорит-
алгоритмом) последовательной выбраковки вариантов кода путем их сопоставления с
известными мутациями в белках и с принципом связности. В течение одного
дня я мог "расправиться" со всеми известными данными и сократить разнооб-
разнообразие вариантов кода до 2-3. В каждом варианте порядок символов был опре-
определен примерно в 20 кодонах. В 1963 г. я послал несколько статей на эти темы в
печать (один из оттисков отправил Г.А. Гамову — см. гл. 15). Затем в 1964 г.
появились большой массив мутационных данных и данные Яновского о пове-
поведении маркера Ас~ в рекомбинациях. Тогда я послал небольшую статью с по-
последними вариантами кода (где было определено уже до 40 кодонов) и аргумен-
аргументами в пользу направления трансляции E' -» 3') академику А.Н. Белозерскому
для представления в журнале "Доклады АН СССР". Работа была принята ре-
редакцией.
Однако именно в этот момент, в конце 1964 г., Г. Фунатцу и X. Френ-
кель-Конрат, отказавшись от предыдущей интерпретации нескольких ключе-
ключевых мутационных замен аминокислот в оболочечном белке ВТМ, предложили
вместо них совершенно другие варианты мутаций [Funatsu, Fraenkel-Conrat,
1964]. Базис моей работы рассыпался, похоронив под обломками все остальные
результаты. Я отозвал работу из печати.
Правда, я тут же переделал работу и получил два новых варианта кодонов.
Один из них (А1.1) имел ряд дефектов. Другой (В1.1) был значительно лучше, он
показан в табл. 17-2 [Ратнер, 1966] в форме, пригодной для сравнения. Он был
более чем любопытен, поскольку в пределах неполноты данных, как оказалось,
очень неплохо совпадал со списком кодонов Ниренберга и сотрудников, кото-
который уже начал накапливаться в непрерывных публикациях (см. далее
табл. 17-3). Из 64 триплетов 47 совпали с кодом Ниренберга, 6 не совпали, а 11
еще не были определены по составу. Несовпадения были связаны с ошибками в
исходных данных и разрывностью серии Ser, а не с теоретическим методом.
Метод себя оправдал.
В итоге я не стал снова посылать эту работу в печать, а включил ее резуль-
результаты в корректуру своей книги "Генетические управляющие системы", которая
17. Хроника великого открытия. III
163
Таблица 17-2
Порядок символов в кодонах, полученный путем использования принципа
связности, состава кодонов (см. табл. 17-1) и списка мутационных замен
аминокислот в белках [Ратнер, 1966]
li2->
и
с
А
G
и
UUU Phe
UUC Phe
UUA Leu
UUG Leu
CUU Leu
CUC Leu
CUA Gin
CUG [Leu]
AUU He
AUC lie
AUA He
AUG Met
GUU Val
GUC [Val]
GUA GJu
GUG [Val]
С
UCU Ser
UCC Ser
UCA Ihr
UCG [Set]
CCU Pro
CCC Pro
CCA Pro
CCG ?
ACU Asn
ACC Thr
АСА Thr
ACG [Thr]
GCU Ala
GCC Ala
GCA Ala
GCG ?
A
UAU Tyr
UAC (Tyr)
UAA Lys
UAG ?
CAU (His)
С AC His
CAA Gin
CAG ?
AAU Asn
AAC Asn
AAA Lys
AAG ?
GAU Asp
GAC Asp
GAA Glu
GAG 1
G
UGU Cys
UGC Ser
UGA ?
UGG (Try)
CGU Arg
CGC Arg
CGA Arg
CGG ?
AGU ?
AGC Ser
AGA Arg
AGG ?
GGU Gly
GGC Gly
GGA Gly
GGG ?
13
и
с
A
G
U
С
A
G
U
С
A
G
U
С
A
G
Примечание. Подчеркнуты кодоны, которые в дальнейшем оказались оши-
ошибочными. В круглых скобках — кодоны, в которых порядок символов установлен не-
неоднозначно, в квадратных — кодоны, добавленные в ходе процедуры вывода для объ-
объяснения мутаций и связности. Из 64 кодонов 47 совпадают с кодом Ниренберга, 6 не
совпадают, а 11 (см. табл. 17-1) не были определены по составу (из них 9'с С в третьих
позициях кодонов).
в этот момент уже была принята к печати в издательстве "Наука" [Ратнер,
1966]. Однако работы в России публикуются медленно. Книга вышла только
осенью 1966 г. Время было упущено. Пиршество победителей уже состоялось
(см. ниже).
Что же осталось от этой работы? Во-первых, интерес к проблеме. Во-вто-
Во-вторых, понимание, что информационно-лингвистический подход полностью аде-
адекватен проблеме и может приносить принципиальные результаты. В дальней-
дальнейшем я опубликовал много различных работ по особенностям структуры и эво-
эволюции генетического кода. В-третьих, остался принцип связности кодовых
серий и его эволюционное обоснование. Он выполняется во всех сериях, кроме
Ser. В-четвертых, я независимо заметил, что из рекомбинационных данных
Яновского вытекает направление трансляции 5' -> 3' по матрице. Этот факт еще
не был известен и сыграл ключевую роль в выводе вариантов кода. И наконец,
в-пятых, осталась песня, принадлежащая Л.А. Блюменфельду, выдающемуся
биофизику, которого я глубоко уважаю. В 1962 г. на Миассовской летней школе
164 III. Молекулярная кибернетика
у Н.В. Тимофеева-Ресовского он в форме научного фольклора отразил доклады
Н.В. Лучника и мой по генетическому коду [Блюменфельд, 1993]:
Первым речь держал блатной Лучник,
Я к нему уже давно привык,
Он и Ратнер этим летом
Все распутали триплеты,
Что не сделал даже Беня Крик.
Пиршество победителей
В 1964 г. М. Ниренбергу и Ф. Ледеру удалось преодолеть трудности и разра-
разработать метод связывания на рибосомах в бесклеточной системе фракций ами-
ноацил-т-РНК с небольшими синтетическими олигорибонуклеотидами задан-
заданного порядка оснований. Конечно, это не было трансляцией, но взаимодейст-
взаимодействие кодон—антикодон здесь осуществлялось в полной мере, хотя и не совсем в
физиологических условиях. Поэтому такие работы стали называть эксперимен-
экспериментами in vitro. Прежде всего, все динуклеотиды оказались неэффективными в
связывании т-РНК. Дублетные коды отпали полностью. Далее метод позволял
перепробовать все триплеты по одному.
В результате уже через год, к весне 1965 г., были испытаны все 64 триплета
[Nirenberg et al., 1966]. Почти все они были признаны кодирующими. Амери-
Американский биохимик Г. Хорана [Khorana et al., 1966] пошел другим путем и иссле-
исследовал матричные свойства синтетических полирибонуклеотидов с повторяю-
повторяющимися парами, тройками, четверками оснований: поли-({/С)п, поли-(/Ш)п,
поли-([/С)п, поли-([/[/С)п и др. РНК с повторяющимися парами синтезирова-
синтезировали полипептиды с чередующимися аминокислотами, а РНК с повторяющими-
повторяющимися тройками кодировали по три раздельных полипептидных гомополимера. Ре-
Результаты полностью совпали с данными М. Ниренберга. Известные мутацион-
мутационные замены аминокислот в белках прекрасно соответствовали кодонам. Иначе
говоря, данные in vivo полностью совпали с данными in vitro.
Весной 1965 г. Ф. Крик отправился в США. Он объехал или обзвонил все
лаборатории, обладавшие новыми данными о структуре кодонов, и свел их все
в единую таблицу. Эта таблица далее была канонизирована как генетический
код Е. coli. (табл. 17-3). В течение 1965 г. были согласованы и устранены неко-
некоторые неоднозначности, доказано, что три нонсенса выполняют роль терми-
терминальных знаков, а триплет AUG, по совместительству, — роль инициирующего
знака трансляции. Р. Холли секвенировал первую фракцию т-РНК дрожжей —
аланиновую; чуть позже в структуре различных т-РНК были локализованы ан-
тикодоны. Ф. Крик предложил свою гипотезу неоднозначности спаривания
кодон—антикодон в третьей позиции. С. Бреннер с сотрудниками и независи-
независимо Ч. Яновский доказали коллинеарность гена и белка — давно ожидаемое
свойство генетических текстов. Вал результатов стремительно нарастал.
2-9 июня 1966 г. в Колд-Спринг-Харборе (залив Холодной Весны!) на
Лонг-Айленде, вблизи Нью-Йорка, собрался "съезд победителей". Был представ-
представлен весь цвет биохимической науки, в основном американцы. Из СССР был один
участник — профессор Семен Ефимович Бреслер из Ленинграда, но без доклада.
Таблица генетического кода (см. табл. 17-3), сведенная Криком, была представле-
17. Хроника великого открытия. III
165
14-2->
и
с
А
G
и
Phe
Phe
Leu
Leu
Leu
Leu
Leu
Leu
He
He
lie
Met
Val
Val
Val
Val
С
Ser
Ser
Ser
Ser
Pro
Pro
Pro
Pro
Thr
Thr
Thr
Thr
Ala
Ala
Ala
Ala
A
Tyr
Tyr
Ochre
Amber
His
His
Gin
Gin
Asn
Asn
Lys
Lys
Asp
Asp
Glu
Glu
С
Cys
Cys
?
Trp
Arg
Arg
Arg
Arg
Ser
Ser
Arg
Arg
Gly
Gly
Gly
Gly
4-3
U
С
A
G
U
С
A
G
U
С
A
G
U
С
A
G
на как плод коллективного труда и фактически Таблица 17-3
канонизирована. Насыщенные доклады обос- Генетический код
новывали код по всем направлениям.
В своем главном вступительном докладе
Ф. Крик сказал [Crick, 1966]:
"Это историческое событие... Оставляя в
стороне все сомнительные пункты и оговорки,
можно сказать, глядя на статьи этого симпо-
симпозиума, что открытие генетического кода — это
действительно ключ к молекулярной биологии,
поскольку он показывает, как два великих языка
полимеров — язык полинуклеотидов и язык поли-
полипептидов, — связаны между собой (выделено
мной. — В.Р.). Мы можем быть полностью уве-
уверены, что наши общие идеи, такие как гипотеза
последовательности (выделено мной. — В.Р.),
действительно правильны. После этого для со-
сомневающихся будет очень трудно не принять
фундаментальные положения молекулярной био-
биологии, которые мы пытались доказать в тече-
течение многих лет ".
Однако в пылу финального успеха фактически были забыты многие, в основ-
основном теоретики проблемы. Г.А. Гамов не был на симпозиуме, а его имя было упо-
упомянуто лишь дважды. Только Ф. Крик вспомнил о роли Г. Гамова (см. также гл.
16) и в целом отметил роль теоретиков [Crick, 1966]: "Тем не менее, теория принес-
принесла определенную пользу, будучи время от времени путеводителем для эксперимента-
экспериментаторов... Может быть, более важно, что дискуссии о кодировании фокусировали
внимание на проблеме и сделали ее более реальной для людей, которые в ином случае
просто бы о ней не знали ".
Итак, словарь языка кодонов был определен полностью. Проблема генети-
генетического кода нашла свое экспериментальное решение. Структурно-функцио-
Структурно-функциональный базис молекулярной биологии получил прочное обоснование. Однако,
несмотря на все превратности судьбы, это был также и грандиозный успех ин-
информационно-лингвистического подхода. Проблема синтеза белков была реше-
решена в терминах "кодирования информации", "генетического текста",
"генетического языка ", "матричного принципа" и т.п. Правда, в значительной
мере эти понятия использовались еще как объяснительные метафоры. Однако
вскоре стало ясно, что молекулярная биология и генетика — это новое открытое
поле для серьезных теоретических обобщений, причем именно информацион-
информационно-лингвистический подход является научным базисом этой теории.
Период экспериментальной дешифровки кода успешно закончился. Начал-
Начался следующий период — теоретического осмысли вания найденных законо-
закономерностей. Настало время построения основ теории молекулярно-генетических
систем управления, теории генетического языка и др. Но это уже другая история.
В 1968 г. М. Ниренберг, Г. Хорана и Р. Холли были удостоены Нобелев-
Нобелевской премии по физиологии и медицине за расшифровку генетического кода и
его функции в синтезе белка.
166
III. Молекулярная кибернетика
На Симпозиуме в Колд-Спринг-Харборе в 1966 г. Слева направо: Г. Хорана, Ф. Крик и
М. Грюнберг-Манаго.
Список литературы
Блюменфельд Л.А. Очень кратко о Н.В. Тимофееве-Ресовском // Н.В. Тимофеев-Ресов-
Тимофеев-Ресовский. Очерки. Воспоминания. Материалы / Ред. Н.Н. Воронцов. М.: Наука, 1993. С. 332-335.
Ниренберг М.В., Маттеи Г. Зависимость синтеза белка от природной или синтетической
РНК-матрицы в выделенной из Е. coli бесклеточной системе // Труды V Междунар. биохим.
конгресса. Симп. 1. 1962. С. 211-226.
Ратнер В.А. Генетические управляющие системы. Новосибирск: Наука, 1966.
Ратнер В.А. Хроника великого открытия: идеи и лица // Природа, 1998. № 4. С. 68-79;
№ 11. С. 18-30.
Benzer S. Genetic Fine Structure // Harvey Lecture. 1961. Vol. 56.; Бензер С. Тонкая структу-
структура гена // Молекулярная генетика: Сб. переводов. М.: Изд-во иностр. лит., 1963. С. 11-32.
18. Молекулярная кибернетика в океане науки 167
Benzer S. Adventures in the rll Region // Phage and the Origins of Molecular Biology / Eds.
J. Cairns, G.S. Stent, J.D. Watson. Cold Spring Harbor, 1966. P. 157-165.
Crick F.H.C., Barnett L., Brenner S., Watts-Tobin R.G. The general nature of the genetic code
for proteins // Nature. 1961. Vol. 192, N 4809. P. 1227-1232; Крик Ф., Барнетт Л., Бреннер С,
Уоттс-Тобин Р. Общая природа генетического кода для белков // Молекулярная генетика: Сб.
переводов. М.: Изд-во иностр. лит., 1963. С. 33-50.
Crick F.H.C. The Genetic Code—Yesterday, Today, and Tomorrow // The Genetic Code: Cold
Spring Harbor Symp. Quant. Biology. 1966. Vol. 31, P. 3-9.
Funatsu G., Fraenkel-Conrat H. Location of amino acid exchanges in chemically evoked
mutants of tobacco mosaic virus // Biochemistry, 1964. Vol. 3, N 9. P. 1356-1362.
Khorana H.G. et al. Polynucleotide Synthesis and the Genetic Code // Ibid. — P. 39-49.
Nirenberg M. et al. The RNA Code and Protein Synthesis // The Genetic Code: Cold Spring
Harbor Symp. Quant. Biology. 1966. Vol. 31. P. 11-24.
The Genetic Code. XXXI Cold Spring Harbor Symp. Quant. Biology. Cold Spring Harbor,
N.Y., 1966.
18. Молекулярная кибернетика в океане науки*
Концепция управляющих систем живой природы А.А. Ляпунова
Приложение принципов кибернетики к теоретическому описанию явле-
явлений жизни было одним из главных направлений научной деятельности
А.А. Ляпунова. Концепция управляющих систем живой природы фактически
была методологией информационно-кибернетического подхода к описанию
биологических систем. Сущность концепции состояла в следующем.
В конце 50-х гг. А.А. Ляпунов и СВ. Яблонский [1963] обобщили результа-
результаты первых десятилетий развития кибернетики и сформулировали представле-
представление о центральном объекте кибернетики — управляющей системе (системе
управления). В содержательном смысле управляющие системы проявляют сле-
следующие свойства:
1) состоят из элементов, способных выполнять различные операции над
информацией;
2) имеют каналы связи, по которым элементы обмениваются информацией;
3) элементы организуются в относительно автономные подсистемы, нахо-
находящиеся в иерархическом подчинении.
4) высшие ярусы иерархии образуются из низших либо структурно, когда
формируются специальные каналы связи, либо статистически, случайно.
Для описания управляющих систем используются следующие математиче-
математические характеристики:
(а) схема, отражающая информационное строение, конструкцию системы
и включающая в себя элементы, их сети и информацию;
(б) информация, которая полностью определяется состояниями ячеек па-
памяти;
* Первоначальный вариант опубликован в газете "Наука в Сибири" B000. № 18. С. 5;
№26—27. С. 6), а также в журнале "Природа": Ратнер В.А. Молекулярно-генетические системы
управления // Природа. 2001. № 3. С. 16-22.
168 III. Молекулярная кибернетика
(в) координаты, характеризующие расположение компонент схемы в про-
пространстве;
(г) функции, характеризующие поведение управляющей системы.
Описание произвольных систем в таких характеристиках лежит в основе
информационно-кибернетического подхода. При этом различные
"материальные" особенности систем (структурные, физические, химические,
технические) отходят на второй план, а главными считаются отношения управ-
управления, кодирования, принципы организации, потоки информации, алгоритмы
функционирования, языки, помехоустойчивость и т.п. Если какая-то система
легко формализуется в таких понятиях, то для нее можно сформулировать ряд
специфических принципиальных задач. Например, исследовать функциональ-
функциональные режимы при задании схемы, информации и координат или помехоустой-
помехоустойчивость кодирования информации.
С начала 60-х годов (это уже новосибирский период) А.А. Ляпунов [1963,
1964] начинает активно выступать с докладами и статьями об управляющих
системах живой природы и кибернетическом определении жизни. Внешним
поводом для этого, вероятно, были попытки лысенковцев опорочить киберне-
кибернетический подход к биологии (см. гл. 25). Внутринаучные мотивы А.А. Ляпунов
сформулировал следующим образом:
"Для процессов жизнедеятельности характерно наличие специальных управ-
управляющих процессов. Последние отличаются тем, что передача небольших масс или
порций энергии вызывает действия, состоящие в передаче гораздо больших энергий
или масс" [Ляпунов, 1963].
"Фундаментальным обстоятельством, благодаря которому кибернетический
подход к изучению жизни стал неизбежным, является наличие разветвленной сис-
системы специфических процессов управления, которые определяют функционирова-
функционирование живых структур " [Ляпунов, 1964].
"...Управление можно объявить характеристическим свойством жизни в ши-
широком смысле " [Ляпунов, 1964].
Действительно, к этому времени биологические науки, особенно генетика,
молекулярная биология, физиология, изобиловали примерами реального
управления биологическими процессами. Использование информационно-ки-
информационно-кибернетического подхода не только сулило грядущие успехи, но уже явно их де-
демонстрировало. А.А. Ляпунов сделал этот подход базисным для построения
теоретической и математической биологии на кибернетических принципах.
Прежде всего, А.А. Ляпунов [1963,1964] попытался дать определение поня-
понятию "жизнь" в терминах кибернетики. В основу он положил представление об
особой устойчивости живого вещества, обеспечиваемой сохраняющими реак-
реакциями биологических управляющих систем:
"Итак, мы выделяем вещество, ограниченно однородное и повышенно устойчи-
устойчивое, обладающее сохраняющими реакциями и управляющей системой, которая ш
вырабатывает и которая использует информацию, хранимую в виде мономолеку-
мономолекулярных кодов.
Такое вещество мы будем называть живым.
18. Молекулярная кибернетика в океане науки 169
Короче, жизнь можно охарактеризовать как высокоустойчивое состояние ве-
вещества, использующее для выработки сохраняющих реакций информацию, коди-
емую состояниями отдельных молекул ".
Это определение не стало общепринятым. Оно несло в себе не только пе-
печать времени, но и зерна будущих обобщений. Напомним, что и Э. Шредингер
[1972] (см. гл. 15) пытался связать особые свойства живого с термодинамиче-
термодинамической и квантовой устойчивостью макромолекул. По-моему, до сих пор нет дос-
достаточно ясного определения жизни. По мнению автора, главным свойством
живых является их способность к самовоспроизведению, т.е. к синтезу всех
своих макромолекулярных компонентов и наследованию. Эти свойства в клет-
клетках обеспечиваются присутствием МГСУ — молекулярно-генетической систе-
системы управления, построенной из кодирующих макромолекул. Однако ясно, что
это — кибернетическое свойство, действительно обеспечивающее сохраняю-
сохраняющие реакции в молекулярно-генетических системах.
Вопрос об особой повышенной устойчивости биологических систем имеет
сложный характер. Можно обсуждать принципы динамической, параметриче-
параметрической, структурной устойчивости МГСУ, а также их эволюционную устойчи-
устойчивость. Ясно, что наиболее фундаментальный характер имеют структурная и
эволюционная устойчивость, хотя и остальные варианты не исключаются. Те-
Теперь уже ясно, что структурная и эволюционная устойчивость МГСУ обеспечи-
обеспечиваются такими специализированными подсистемами управления МГСУ, как
модуль самовоспроизведения (сайзер), архив генетической информации, моду-
модули контроля метаболизма и онтогенеза и др. (см. ниже).
Что касается устойчивости популяций в стохастическом процессе эволю-
эволюции, то основным решением этой проблемы, по-видимому, является достаточ-
достаточно быстрое развитие и усложнение МГСУ индивидуумов, способное обеспе-
обеспечить не только простые сохраняющие реакции, но также анализ и прогноз ус-
условий существования (в том числе — и не предвиденных заранее), на основе
чего возможны сложные сохраняющие реакции. Эти функции обычно связыва-
связывают с появлением разума. Только это сулит перспективу выживания популяций
биологических систем и жизни в целом как феномена природы.
Таким образом, концепция управляющих систем живой природы А.А. Ля-
Ляпунова стала фокусом информационно-кибернетического подхода в биологии,
который оказался особенно плодотворным в науках молекулярно-генетическо-
го цикла. Конкретным воплощением этого подхода стала концепция молеку-
молекулярно-генетических систем управления клетки и организма.
Возникновение концепции МГСУ
Концепция молекулярно-генетических систем управления возникла в се-
середине 60-х годов [Ратнер, 1966] как приложение идей и методов кибернетики
для описания, анализа и моделирования явлений молекулярно-генетической
организации. К этому времени в теоретической кибернетике были получены
крупные результаты, открывшие возможность обосновать и решить эти про-
проблемы. Дж. фон Нейман [1971] разработал основы теории самовоспроизводя-
170 III. Молекулярная кибернетика
щихся автоматов, имея в виду проблемы и прообразы из генетики и молекуляр-
молекулярной биологии. Л. Бриллюэн [1960], К. Шеннон [1963] и другие ученые проясни-
прояснили понятие количества информации. А.А. Ляпунов и СВ. Яблонский [1963]
описали центральный объект кибернетики — системы управления, а И.А. По-
Полетаев [1970] уточнил понимание "информации по смыслу" физических осо-
особенностей актов управления и принципа лимитирования в сложных системах.
Кибернетика была активной и бурно развивавшейся наукой, принципы кото-
которой пытались приложить к самым разным областям знания.
Однако далеко не везде эти приложения были эффективными. Главным
критерием успеха оказалась адекватность природы реальных объектов конкрет-
конкретных наук принципам и идеям кибернетики. В этом отношении "молодые" нау-
науки генетического цикла оказались благодатным полем приложения кибернети-
кибернетических идей.
Во-первых, представление о кодировании наследственных свойств в струк- |
туре генов было как бы органически присуще самой идеологии генетики, хотя
и в расплывчатой форме. В 30-40-е годы оно неоднократно высказывалось
многими корифеями генетики. После создания Дж. Уотсоном и Ф. Криком
двухцепочечной модели ДНК [Watson, Crick, 1953] физик-теоретик Г.А. Гамов
[1957] впервые явно сформулировал проблему генетического кода как ключе-
ключевую проблему кодирования генетической информации.
Во-вторых, к середине 60-х годов молекулярная генетика уже решила про-
проблемы физической природы генов (ДНК, РНК) и организации основных гене-
генетических процессов: репликации, транскрипции, трансляции, репарации, му-
мутирования, рекомбинации и др. [Stent, 1970]. Кроме того, она уже изобиловала
такими крупными успехами, как расшифровка генетического кода, открытие
информационной РНК, регуляторных генов и белков, оперонов, знаков пунк-
пунктуации и управления и т.п. Любопытно, что кибернетическая терминология с
самого начала была присуща молекулярной генетике, именно в этих терминах
были сформулированы и решены многие ее фундаментальные проблемы. Од-
Однако первоначально это делалось, скорее, в виде ярких научных метафор, чем
содержательных научных понятий. Назрела необходимость последовательного
и конструктивного построения молекулярной кибернетики — системы понятий
и методов описания и моделирования молекулярно-генетических систем кле-
клеток как систем управления.
Первые попытки обоснования и развития концепции молекулярно-генети-
молекулярно-генетических систем управления были сделаны автором в 1964-1966 гг. [Ратнер, 1966]
под влиянием идей А.А. Ляпунова. Первоначально это была "рамочная" кон-
концепция, определявшая принципы описания, понятия, подходы к формулиров-
формулировке задач и общие проблемы. Данные о конкретных системах были еще доволь-
довольно бедны. Секвенированные последовательности исчислялись пальцами одной
руки. Тем не менее на этой основе возникло активное направление теоретиче-
теоретических исследований МГСУ, которое в целом продолжается до сих пор. В нем
приняло участие несколько поколений математических генетиков новосибир-
новосибирской школы, имена которых теперь хорошо известны: Р.Н. Чураев, Н.А. Кол-
Колчанов, С.Н. Родин, В.А. Куличков, А.Г. Бачинский, В.В. Соловьев, А.А. Жар-
Жарких, В.В. Шамин, СИ. Бажан и многие другие. Очень близкие направления не-
18. Молекулярная кибернетика в океане науки 171
зависимо и параллельно с нами развивали также наши коллеги за рубежом:
М. Эйген в ФРГ, С. Кауфман в США, Р. Тома в Бельгии, М. Саважо в США и
др. Их результаты существенно укрепили концепцию МГСУ.
Концепция МГСУ
Весь опыт молекулярной генетики показывает, что наиболее существенны-
существенными молекулярными компонентами клетки являются фракции кодирующих
биополимеров — ДНК, РНК и белков. С ними связаны все наиболее важные
процессы и свойства клеток: самовоспроизведение, наследование, контроль
метаболизма, ферментативный катализ, построение морфологических струк-
структур, транспорт веществ, развитие, дифференцировка, иммунитет и т.д. Сово-
Совокупность кодирующих биополимеров клетки обладает несколькими общими,
фундаментальными свойствами:
1) все полимеры построены из малого разнообразия стандартных мономе-
мономеров, нуклеиновые кислоты — из 4 типов нуклеотидов, белки — из 20 типов ами-
аминокислот;
2) функции и свойства конкретных макромолекул определяются почти ис-
исключительно их размером, составом и порядком мономеров;
3) клеточная система кодирующих биополимеров способна к самовоспроиз-
самовоспроизведению, т.е. к синтезу всех своих макромолекулярных компонентов и отноше-
отношений между ними при помощи специальных внутрисистемных устройств (фер-
(ферментов репликации, транскрипции, трансляции, рибосом и т.д.);
4) исполняющие устройства основных генетических процессов сами по-
построены из кодирующих биополимеров и кодируются в генах той же клеточной
системы;
5) в клетках имеется система фундаментальных генетических процессов, вы-
выполняемых этими устройствами (репликация, транскрипция, трансляция, ре-
репарация, рекомбинация, деградация, сегрегация и др.), над всеми фракциями
биополимеров.
Мы назвали эту систему биополимеров клетки молекулярно-генетической
системой управления [Ратнер, 1966, 1975, 1993]. При информационно-киберне-
информационно-кибернетическом описании этой системы на передний план выходят принципы орга-
организации и управления, самовоспроизведение, информационные процессы, по-
помехоустойчивость, кодирование, память, языки и т.п., а структурные, физи-
физико-химические свойства отходят на второй план. В этом случае: мономеры
считаются символами базового алфавита; макромолекулы (гены, РНК, белки)
задаются последовательностями символов, или генетическими текстами; систе-
системы взаимодействующих генов характеризуются схемами их молекулярных
взаимодействий, конструкциями, или генетическими сетями; геномы — последо-
последовательностями символов генов, знаков пунктуации и управления и других
функциональных единиц, т.е. генетическими картами; МГСУ в целом задается
замкнутой конструкцией, т.е. схемой функциональных взаимодействий.
Теперь молекулярные свойства, отношения, функции, записанные в гене-
генетических текстах, можно считать генетической информацией, а правила и зако-
закономерности ее записи — генетическим языком. Определяются также и другие
I
172 III. Молекулярная кибернетика
понятия кибернетического характера: генетическая память, информационные
процессы, архив генетической информации (геном) и др. Опыт показывает, что
такой подход очень продуктивен, на его основе формулируются и решаются де-
десятки и сотни принципиальных задач организации и эволюции МГСУ.
Внутреннее строение МГСУ
МГСУ клеток — сложные системы. Достаточно сказать, что они содержат
от 500 до 100 000 генов, примерно столько же белков; контролируют сотни и
тысячи метаболических процессов, а также дифференцировку, развитие, имму-
иммунитет и другие процессы. В то же время эта сложность не чрезмерна. При этом
отдельные молекулярные и более сложные функции и свойства не "размазаны"
по системе в целом, а сосредоточены в конкретных модулях организации,
имеющих определенную внутреннюю информационную структуру [Ратнер,
1993]. В целом реальные клетки и организмы, а также их МГСУ имеют иерархи-
иерархическое блочно-модулыгае строение, возникшее эволюционно [Ратнер, 1992].
Блоком (модулем) системы называют автономную подсистему с определенной
функциональной нагрузкой. Иерархичность МГСУ подразумевает, что модули
более высоких ярусов являются комбинациями блоков предыдущих ярусов и
соединительных элементов, а выделяемые ярусы, блоки и подсистемы имеют
информационную природу.
Наиболее известными модулями нижних ярусов МГСУ являются кодоны,
знаки пунктуации и управления, гены, транскрипты, белки, опероны, реплико-
ны, мобильные элементы и др. Наиболее известное проявление автономности
этих модулей — дискретность генов.
С другой стороны, если двигаться сверху вниз иерархии, т.е. от сложных
свойств и функций к более простым, то можно выделить несколько модулей
верхних ярусов. Еще Э. Шредингер [1972] в 1944 г. указывал, что "хромосомные
структуры являются одновременно и архитектором, и строителем ". В наших
терминах [Ратнер, 1975, 1993] это означает, что можно выделить центральную
подсистему МГСУ (рис. 18-1) как модуль, содержащий исполняющие устройст-
устройства (ферменты и т.д.) и гены всех фундаментальных генетических процессов, об-
обслуживающих всю МГСУ, т.е. универсальных в клетке. Сюда относятся модули
репликации, транскрипции, трансляции, репарации, рекомбинации, сегрега-
сегрегации и т.д. Центральная подсистема является базой организации клетки,
"строителем" по Шредингеру.
Все остальные, неуниверсальные модули МГСУ естественно объединить в пе-
периферическую подсистему (рис. 18-2). В целом она отвечает за разнообразие других
функций МГСУ, которые в отдельности не универсальны, но в совокупности
тоже обслуживают всю клетку и обеспечивают ее жизнедеятельность, энергетику и
материальную автономность. Фактически в этой части МГСУ сосредоточена ин-
информация о специфической архитектуре МГСУ (по Шредингеру).
"Ядром" центральной подсистемы является группа взаимодействующих
блоков репликации, транскрипции, трансляции и сегрегации, которые в сово-
совокупности и во взаимодействии обеспечивают способность МГСУ к самовос-
самовоспроизведению (рис. 18-3) [Ратнер, 1993, Ratner et al., 1996]. Этот модуль мы на-
Not for sale! For preview only'! Scanned by A fykhaylo (molbioKcv ukr.net)
ДНК
Наследственная
генетическая nai
Метаболизм, онтогенез
Рис. 18-1. Блочно-модульное строение МГСУ клетки: модули наследственной генетиче-
генетической памяти (выделен серым цветом) и оперативной генетической памяти (выделен фио-
фиолетовым цветом) [Ратнер, 1975, 1993]. Обозначения процессов и исполняющих уст-
устройств: синяя горизонтальная дугообразная стрелка — действие исполняющего устрой-
устройства репликации, красная — системы транскрипции, зеленая — системы трансляции.
Черные нисходящие стрелки — процессы считывания информации (транскрипция и
трансляция).
Днк
м-РНК
Белок
Центральная
подсистема
Гены универсальных -
генетических процессов
Гены неуниверсальных
генетических процессов
Периферическая
подсистема
Рис. 18-2. Блочно-модульное строение
МГСУ клетки: центральная подсистема
(выделена желтым цветом), содержащая
гены, РНК и белки, исполняющие уни-
универсальные генетические процессы, и
периферическая подсистема (выделена
голубым цветом), содержащая гены. РНК и белки, исполняющие неуниверсальные гене-
генетические процессы. Обозначения процессов и исполняющих устройств такие же, как на
рис. 18-1. Выходящие вертикальные стрелки обозначают исполняющие устройства систем
репликации (синие), транскрипции (красные) и трансляции (зеленые). Предполагается,
что в полной схеме они замыкаются с горизонтальными стрелками, описанными выше.
Для простоты изображения здесь эти замыкания опущены.
Сайзер — "ядро'
центральной подсистемы
Внешние контуры,
навешенные на сайзер
Рис. 18-3. Блочно-модульное строение МГСУ клетки: модуль универсальной самовос-
самовоспроизводящейся подсистемы — сайзера (выделен голубым цветом) как "ядро" цен-
центральной подсистемы и остальные модули, навешенные на сайзер и вместе с ним спо-
способные к самовоспроизведению (выделены салатным цветом). Обозначения стрелок та-
такие же, как на рис. 18-1, 18-2. Исполняющие устройства блоков репликации,
транскрипции и трансляции (выходящие стрелки) синтезируют макромолекулы самого
сайзера, а также любых других блоков, навешенных на сайзер извне (горизонтальные
входящие стрелки соответствующих цветов).
Гены
рекомби"
нации
Гены
репара-
репарации
Гены
г репли-~
кации
Гены других систем
и др.
Д Д
Рис. 18-4. Блочно-модульное строение МГСУ клетки: модуль архива генетической инфор-
информации (выделен голубым цветом), включающий наследственную память всех генов и
блоки поддержания, сохранения и манипулирования генетической информацией (ре-
(рекомбинации, репарации, репликации и др.). Действие этих блоков на наследственную
память изображено восходящими цветными дугообразными стрелками. Остальные обо-
обозначения такие же, как на рис. 18-1—18-3.
Автономно управляемые гены,
контролирующие
метаболизм
Рис. 18-5. Блочно-модульное строение МГСУ клетки: модуль контроля метаболических
процессов (выделен голубым цветом), содержащий тысячи или десятки тысяч автоном-
автономных секций считывания генетической информации (ДНК-> РНК -> Белок), которые
независимо или совместно управляются обратными связями и внешними сигналами
(восходящие дугообразные серые стрелки) и катализируют (нисходящие прямые серые
стрелки) метаболические процессы (горизонтальные прямые серые стрелки). Другие
обозначения такие же, как на предыдущих рисунках данной главы. Сходное строение
имеет, вероятно, модуль контроля онтогенеза клетки.
18. Молекулярная кибернетика в океане науки 173
звали сайзером, т.е. универсальной системой самовоспроизведения. Белки реп-
репликации воспроизводят гены всех четырех блоков, а также любые другие гены,
имеющие такую же пунктуацию процесса репликации. Белки транскрипции
транскрибируют гены всех четырех блоков, а также любые другие гены с такой
же пунктуацией транскрипции. Белки трансляции транслируют фракции
м-РНК всех четырех блоков, а также любых других генов с такой же пунктуаци-
пунктуацией. Белки сегрегации обеспечивают равное разделение всех генов между дочер-
дочерними клетками. Иначе говоря, именно универсальность белков и знаков пунк-
пунктуации этих процессов образует прямые и обратные связи между блоками, что
стабилизирует всю МГСУ и обеспечивает ее самовоспроизведение.
Можно увеличить внутреннюю сложность блоков сайзера или ввести до-
дополнительные, внешние к нему блоки и контуры МГСУ с такой же пунктуаци-
пунктуацией; при этом способность к самовоспроизведению сохранится (см. рис. 18-3).
Значит, сайзер является "ядром" молекулярно-генетической организации.
В начале 80-х годов нам с В.В. Шаминым [Ratner et al., 1996] удалось построить
математические модели сайзеров и впервые показать их богатейшие динамиче-
динамические свойства и ключевую роль в молекулярно-генетической организации.
Удивительно, что, вопреки априорному ожиданию, схема, реализующая
идею самовоспроизведения, оказалась достаточно простой. Насколько я могу
судить, в природе и технике пока нет других систем, обладающих этим свойст-
свойством в полной мере.
Другим универсальным модулем центральной подсистемы МГСУ является
архив генетической информации (рис. 18-4) [Ратнер, 1993]. Он содержит частич-
частично перекрывающиеся универсальные блоки процессов репарации, общей ре-
рекомбинации, репликации и др., а также неуниверсальные блоки процессов
транспозиции мобильных генетических элементов (МГЭ), обратной транс-
транскрипции, хромосомных перестроек, упаковки хроматина и некоторых других.
Фактически это система хранения, поддержания, дублирования, коррекции,
манипулирования, т.е. в целом — помехоустойчивости генетической информа-
информации. Этот же модуль обеспечивает изменчивость генетической информации,
поскольку функция помехоустойчивости дополняет функцию мутирования,
которая возникает в основном путем ошибок в системах репликации и других
процессов архива.
Архив генетической информации содержит сотни и тысячи отдельных сек-
секций центральной и периферической подсистем с автономным доступом к ним
(гены, опероны и т.д.). Они восприимчивы к специфическим внешним сигна-
сигналам и через них — к внешнему управлению, а также способны к отклику путем
автономного изменения потоков считывания информации. В секциях блоков
периферической подсистемы записаны функции контроля метаболизма, онто-
онтогенеза, физиологических процессов, иммунитета и т.д. Эти секции архива на-
находятся на входе соответствующих блоков. Основное содержание блоков — ме-
механизмы специфического управления секциями архива, продукты этих секций
(м-РНК, белки), а также схемы и процессы их взаимодействия между собой
(мультимеры, комплексы, генетические сети и т.п.) и исполнение функций.
В частности, блок управления метаболизмом (рис. 18-5) [Ратнер, 1993] состо-
состоит из сотен и тысяч автономных систем метаболического контроля, работаю-
174 III. Молекулярная кибернетика
щих почти независимо и параллельно или входящих в сложные каскадные сис-
системы. Сложность блока должна быть близка к сложности путей метаболизма.
Специфическое управление секциями блока выполняется через разнообразные
прямые и обратные связи между генами, ферментами, регуляторными белками,
знаками управления, метаболитами, сигнальными агентами, гормонами и др.
Фактически автономное управление может быть введено почти в любом неуни-
неуниверсальном звене потока информации. Наиболее существенные автономные
подсистемы этого блока контролируют базовый метаболизм (включая синтез
мономеров — нуклеотидов и аминокислот), энергетику, транспорт веществ и
другие процессы.
Блок управления развитием (онтогенезом) (рис. 18-5) [Ратнер, 1993] контро-
контролирует временную динамику и пространственную топографию онтогенетиче-
онтогенетических событий. Это особенно существенно для многоклеточных организмов, но
развитие претерпевают также отдельные клетки (клеточный цикл) и даже виру-
вирусы и фаги внутри этих клеток. Онтогенез завершается самовоспроизведением
их МГСУ. На входе этого блока находится большая группа автономных секций
архива (генов), кодирующих специфические звенья процесса развития — диф-
ференцировки, морфогенеза, формирования тканей, клеточного деления и т.п.
На промежуточных этапах процесса используются универсальные блоки реп-
репликации, транскрипции, трансляции, процессинга и др.
Онтогенез совершается на основе наследственной программы управления
развитием, которая должна быть сосредоточена в архиве (геноме). Это не вы-
вызывает сомнения. Вопрос состоит в том, каким образом генетическая програм-
программа онтогенеза записана в архиве. Содержит ли блок наследственной памяти ал-
алгоритмическую запись этой программы?
Весь опыт молекулярной генетики показывает, что генетическая программа
развития действительно записана в геноме, но не в виде последовательного текста
[Altan, Koppel, 1990]. Во всяком случае, генетические карты не проявляют ка-
какой-либо существенной упорядоченности генов, коррелирующей с их функ-
функциями в онтогенезе. Прямой способ задания программ последовательным тек-
текстом присущ компьютерным наукам, но практически не выполняется в рамках
МГСУ. Генетическая программа онтогенеза записана в архиве опосредованно,
через взаимодействие молекулярных компонентов МГСУ [Ратнер, 1993].
Управляемые секции архива (гены) кодируют отдельные макромолекулярные
компоненты модулей (белки, РНК), которые обладают определенными внут-
внутренними свойствами и параметрами — специфическими, каталитическими,
структурными, регуляторными и т.д. В результате их взаимодействия на основе
специфических отношений катализа, опознания, матричной активности, фи-
физического сцепления, пространственного контакта, генетического управления
и т.д. формируется динамическая система развития МГСУ, реализующая эту
программу. Схемы отношений между компонентами этой системы обычно изо-
изображают в виде генетических сетей.
Блок онтогенеза и реализующая его динамическая система развития МГСУ
обладают одним уникальным свойством: они кодируют необратимые функцио-
функциональные изменения модулей МГСУ, наследуемые далее по клеточной линии.
Иначе говоря, они обладают эпигенетическими свойствами. Хотя отдельные
18. Молекулярная кибернетика в океане науки 175
менделевские гены не обладают такими свойствами, можно показать, что суще-
существуют системы менделевских генов с обратными связями (двухоперонные
триггеры и др.), которые уже способны наследовать функциональные состоя-
состояния, т.е. не являются менделевскими в строгом смысле. Такие системы были
названы эпигенами.
Одним из первых крупных опытов математического и компьютерного мо-
моделирования онтогенетической сети была наша работа с Р.Н. Чураевым и
Г.Х. Кананяном [Kananyan et al., 1981] о развитии фага X. Этот фаг был излюб-
излюбленным объектом исследования в 60-70-е годы, информация о нем была ог-
огромной. К 1980 г. нам удалось построить достаточно сложную компьютерную
модель, которая успешно описывала и прогнозировала различные свойства
развития фага X в клетке.
Таким образом, к началу 80-х годов концепция МГСУ полностью проявила
свой эвристический потенциал. На ее основе нами были сформулированы и ре-
решены десятки других крупных проблем молекулярно-генетической организа-
организации [Ратнер, 1975, 1993, 1996]: модели оперонных и полирепликонных систем;
системные свойства генетического кода; описание свойств генетического язы-
языка; динамические модели ансамблей макромолекул, в том числе сайзеров; зада-
задачи комплементационного анализа, проблемы теории молекулярной эволюции
МГСУ; блочно-модульный принцип организации и эволюции МГСУ; принцип
лимитирующих факторов организации МГСУ; и многое другое.
Эпоха секвенирования
В 1977-1978 гг. в биотехнологии и молекулярной биологии произошло важ-
важное событие: были разработаны достаточно быстрые и удобные методы секве-
секвенирования (расшифровки последовательностей) полинуклеотидов. После это-
этого стал стремительно нарастать вал расшифрованных последовательностей ге-
генов, вирусных геномов, различных фрагментов и т.д. В начале 80-х годов
возникли основные международные банки данных нуклеотидных и полипеп-
полипептидных последовательностей, т.е. генетических текстов. На повестке дня встал
вопрос об использовании этого богатейшего материала в рамках теории МГСУ,
т.е. о разработке компьютерных методов для извлечения генетической инфор-
информации из этого моря данных и ее интерпретации.
Важно подчеркнуть, что, имея опыт работы в рамках концепции МГСУ,
мы оказались полностью подготовленными к такому повороту событий. Очень
быстро наши сотрудники начали собственные разработки и влились в мировой
поток биоинформатики, причем весьма успешно. Возглавил эту работу
Н.А. Колчанов, а наиболее активными участниками были В.В. Соловьев,
А.А. Жарких, И.Н. Шиндялов, М.П. Пономаренко, А.Э. Кель, И.Б. Рогозин и
др. Ими были разработаны методы сравнительного анализа последовательно-
последовательностей, поиска генов и знаков управления, восстановления пространственной
структуры макромолекул, филогенетического анализа и др. [Modeling..., 1992].
Таким образом, "рамочная" концепция МГСУ стала наполняться конкрет-
конкретным содержанием о строении компонентов, их взаимодействии и эволюции.
Концепция фактически стала перерастать в теорию МГСУ. За два последних
176 III. Молекулярная кибернетика
десятилетия были проанализированы сотни коллекций и семейств генов, РНК,
белков, знаков пунктуации и управления, построены сотни филогенетических
деревьев. Исходные общие предположения о принципах организации МГСУ
подтвердились на большом фактическом материале.
Однако стратегия массового секвенирования не сводится просто к накоп-
накоплению разрозненных фрагментов генетических текстов. Прежде всего, стали
направленно накапливаться данные о генах и механизмах управления некото-
некоторых достаточно сложных клеточных и межклеточных подсистем: иммунитета,
кроветворения, отклика на тепловой шок, клеточного цикла, SOS-репарации и I
др. Такие подсистемы характеризуют генными сетями (схемами взаимодейст- I
вий), на которых базируются динамические модели функционирования. Этот I
раздел моделирования МГСУ становится сейчас чрезвычайно актуальным и I
продуктивным, так как позволяет выявлять режимы функционирования кле-
клеточных подсистем и их возможные нарушения.
Генетические взаимодействия в значительной степени реализуются через
регуляторные белки, их функциональные центры, знаки управления генов и
отчасти через регуляторные сигналы (метаболиты, гормоны, модифицирующие
воздействия). Поэтому в последние годы стремительно развиваются исследова-
исследования функциональных знаков (сайтов) управления транскрипцией: операторов,
сайтов узнавания общеклеточных активаторов, энхансеров, инсуляторов, тка-
не- и гормон-специфичных сайтов, сайтов "распаковки" хроматина, сайтов от-
ответа на тепловой шок и др. Компьютерный анализ регуляторных зон генов дает
возможность выявить такие знаки управления и замкнуть связи в схемах ген-
генных сетей [Proceedings..., 2000].
С другой стороны, в последние годы появились новые экспериментальные
технологии, позволяющие автоматически следить за динамикой сотен и тысяч
генов и их продуктов в ходе функционирования и развития клеток. Такова тех-
технология микрочипов ДНК [De Risi, Iyer, 1999]. Сотни и тысячи клонированных
фрагментов ДНК с определенными генами распределяются по микролункам
панели и гибридизуются с меченой валовой РНК клетки (через так называемую
к-ДНК). Благодаря комплементарности цепей, в каждой лунке гибридизуется
отдельная фракция к-ДНК. Затем автоматически микрометодами измеряются
концентрации гибридных молекул. В результате в каждый момент времени
можно выписать все активно транскрибируемые гены. Эти данные можно срав-
сравнить с моделями генных сетей.
Уже в начале 80-х годов были секвенированы первые полные геномы виру-
вирусов и фагов, среди них — фага X [Sanger et al., 1982]. В конце 80-х годов были
начаты крупные международные проекты полного секвенирования клеточных
геномов бактерий, грибов, высших животных и растений: кишечной палочки,
дрожжей, дрозофилы, мыши, пшеницы, человека и др. Многие из проектов
уже завершены; опубликовано свыше 40 полных клеточных геномов мико-
плазм, архебактерий, кишечной палочки, возбудителей ряда болезней, а также
пекарских дрожжей, маленького червя-нематоды и растения-арабидопсиса. Ве-
Вероятно, истинное число секвенированных геномов гораздо больше, потому что
многие фармацевтические и биотехнологические фирмы засекречивают свои
результаты, не публикуя их. В 2000 г. завершено полное секвенирование генома
18. Молекулярная кибернетика в океане науки 177
дрозофилы — классического объекта генетики; весной 2001 г. появились дан-
данные о завершении секвенирования генома человека. Это чрезвычайные по важ-
важности события в молекулярной генетике и биоинформатике! Они означают су-
существенный прорыв знаний, возможностей эксперимента и развития теории
МГСУ. Науку о полных геномах стали называть "геномика".
Уже первые результаты полного секвенирования клеточных геномов по-
позволили наполнить конкретным содержанием основные модули МГСУ. Бакте-
Бактериальные геномы содержат примерно от 500 генов у микоплазм до почти
5000 генов у кишечной палочки. Сравнительные оценки показывают, что клет-
клетки должны содержать не менее 300 генов, в основном принадлежащих цен-
центральной подсистеме МГСУ. Анализ генома кишечной палочки (см. гл. 12)
[Blattner et al., 1997] выявил 4909 генов, из которых 4288 кодируют белки, но
функции 38 % из них пока неизвестны. На долю модуля сайзера приходится
свыше 460 известных генов (около 10 %), на долю блока контроля метаболиз-
метаболизма— свыше 1047 известных генов (около 25 %). Интересно, что эти 1047 генов
контролируют 804 известных фермента и 988 известных метаболических реак-
реакций, т.е. их сложность сопоставима. Около 20 % известных генов связано с кле-
клеточными структурами и процессами. Поскольку не все выявленные гены опо-
опознаны и идентифицированы, число их будет возрастать, но доли генов вряд ли
существенно изменятся. Кроме того, в геноме кишечной палочки найдено 2584
оперона — управляемых единиц транскрипции (секций архива). Примерно та-
такие же доли генов в основных блоках показывают другие бактерии, имеющие
иные размеры геномов.
Таким образом, МГСУ бактериальных клеток действительно сложны, но
не чрезмерно. Сайзер занимает существенную часть МГСУ и ее архива — до
10 %, блок метаболизма — до 25 %, блок клеточного цикла и морфологии — до
20 %. Подавляющее число генов кишечной палочки входит в автономно управ-
управляемые секции архива, имеющие знаки управления и подчиненные регулятор-
ным белкам (см. гл. 12). На оперон приходится в среднем 1-2 регуляторных
белка и 1-2 знака управления. Иначе говоря, регуляторные зоны генов доста-
достаточно просты. В то же время число выявленных и предполагаемых регулятор-
регуляторных белков у кишечной палочки ~ 178, т.е. на порядок величины меньше, чем
число оперонов. Это значит, что опероны скорее образуют ассоциации, управ-
управляемые совместно небольшим числом регуляторных белков. Так начинают вы-
вырисовываться контуры конструкции клеточных МГСУ. "Рамочная" концепция
МГСУ оказалась весьма эффективным средством их описания.
Сравнительный анализ полных геномов фагов, вирусов и клеточных орга-
нелл с клеточными ядерными геномами показывает, что обычно первые не
имеют полноценных блока контроля метаболизма и блока сайзера, но содержат
блок развития и морфологии. Например, у фага фХ174 от блока репликации
присутствует только нуклеаза А, остальные гены и блоки сайзера, а также блок
метаболизма отсутствуют. У фага X от блока репликации присутствуют только
регуляторные гены (О и Р), но ДНК-полимеразы и полностью блоки транс-
транскрипции и трансляции, а также блок метаболизма отсутствуют [Sanger et al.,
1982]. У фага Т4 имеется полноценный блок репликации, но полностью отсут-
отсутствуют блоки транскрипции и трансляции [Molecular Biology..., 1994]. В мито-
178 III. Молекулярная кибернетика
хондриях млекопитающих имеется часть системы трансляции, но остальные
блоки сайзера и блок метаболизма отсутствуют [Brooke, 1999]. В хлоропластах
растений присутствует полноценная система трансляции, гены блока транс-
транскрипции, но отсутствует система репликации [Umesono, Ozeki, 1987]. Поэтому
фаги и органеллы воспроизводятся только с участием генома клетки-хозяина,
они не самостоятельны.
Перспективы
Бурное развитие биоинформатики и геномики последних лет, появление
новых комплексных экспериментальных технологий создали новую и весьма
продуктивную ситуацию. Полное секвенирование и идентификация геномов
таких объектов, как вирусы, бактерии, дрожжи, дрозофила, нематода, араби-
допсис, человек и др., создает возможность глобального моделирования МГСУ
клеток, в том числе и в процессе развития. На этой основе ожидается прорыв в
медицине, биотехнологии, производстве продуктов питания, фармакологии.
Моделирование развития вирусов позволит выявить их "слабые" секции, ми-
мишени фармакологического и иммунного воздействия. Модели МГСУ бактерий
и грибов позволят выявить и оптимизировать пути биосинтеза лекарств, анти-
антибиотиков, метаболических продуктов, а также нащупать мишени противодей-
противодействия многим заболеваниям. Модели МГСУ растений дадут возможность вы-
выявить лимитирующие звенья продуктивности и преодолеть их естественными и
генно-инженерными средствами. Модели МГСУ дрозофилы, скорее всего,
внесут основной вклад в изучение фундаментальных процессов генетической
организации, онтогенеза, изменчивости и т.п. Модели МГСУ человека позво-
позволят ускорить поиск средств фармакологической и иммунной защиты от заболе-
заболеваний, генной терапии и т.д. Возможно моделирование взаимодействия МГСУ
паразит—хозяин, патоген—хозяин, симбионт—хозяин, а также онкогенного
перерождения МГСУ клеток и т.д.
Несомненно, будет расширяться разнообразие видов с изученными МГСУ.
Среди них уже появляются некоторые экзотические формы, исследование ко-
которых может пролить свет на условия выживания в экстремальных ситуациях:
при высоком уровне радиации, высоких или очень низких температурах, в бес-
бескислородной среде, в полной темноте, в условиях химического и иного загряз-
загрязнения и т.д.
В теории МГСУ остались нерешенными многие принципиальные пробле-
проблемы. Необходимо построение полноценной генетической лингвистики, правил
генетического языка с возможностями прогноза молекулярных функций [Рат-
нер, 1993]. На этом пути уже имеется ряд крупных открытий, но они не позво-
позволяют, скажем, точно предсказать пространственную структуру белка по его
аминокислотной последовательности. Вероятно, кроме ключевых правил
("крупных мазков") существует еще "шлейф" весьма специфичных и локаль-
локальных правил, число которых очень велико и практически не может быть описа-
описано. Поэтому в модели и реальном эксперименте допустим случайный поиск оп-
оптимальных решений.
18. Молекулярная кибернетика в океане науки 179
На базе секвенированных геномов и генных сетей необходимо разработать
динамические модели онтогенеза, клеточного цикла, канцерогенеза, физиоло-
физиологических функций, иммунного ответа и др. Единую теорию молекулярной эво-
эволюции необходимо построить как теорию эволюции МГСУ и ее подсистем
управления, механизмов регуляции, лимитирующих факторов организации и
т.д. [Ratner et al., 1996].
Таким образом, роль теории МГСУ и вообще теоретических методов в мо-
лекулярно-биологических науках ближайшего будущего и их приложениях бу-
будет существенно нарастать. Во многих случаях эта теория становится ключом к
решению краеугольных проблем и непременной составляющей каждодневной
исследовательской работы.
Список литературы
Бриллюэн Л. Наука и теория информации. М.: Физматгиз, 1960.
Гамов Г.А., Рич А., Ичас М. Проблема передачи информации от нуклеиновых кислот к
белкам // Вопросы биофизики. М.: Изд-во иностр. лит., 1957. С. 205-263.
Ляпунов А.А. Об управляющих системах живой природы и общем понимании жизнен-
жизненных процессов // Проблемы кибернетики. 1963. Вып. 10. С. 179-193.
Ляпунов А.А. Об управляющих системах живой природы // О сущности жизни / Ред.
Г.М. Франк и др. М.: Наука, 1964. С. 66-80.
Ляпунов А.А., Яблонский СВ. Теоретические проблемы кибернетики // Проблемы ки-
кибернетики. 1963. Вып. 9. С. 5-22.
Нейман Дж. фон. Теория самовоспроизводящихся автоматов. М.: Мир, 1971.
Полетаев И.А. К определению понятия «информация». 1. Семантический аспект. Об
"информации по смыслу" // Исследования по кибернетике / Ред. А.А. Ляпунов. М.: Сов. ра-
радио, 1970. С. 211-227.
Ратнер В.А. Генетические системы управления. Новосибирск: Наука, 1966.
РатнерВ.А. Молекулярно-генетические системы управления. Новосибирск: Наука, 1975.
Ратнер В.А. Блочно-модульный принцип организации и эволюции молекулярно-генети-
ческих систем управления // Генетика. 1992. Т. 28, № 2. С. 5-24.
Ратнер В.А. Концепция молекулярно-генетических систем управления. Новосибирск:
Изд-во НГУ, 1993. 118 с.
Шеннои К. Математическая теория связи // Работы по теории информации и киберне-
кибернетике. М.: Изд-во иностр. лит., 1963. С. 243-332.
Шредиигер Э. Что такое жизнь? С точки зрения физика. М.: Атомиздат, 1972.
Altan H., Koppel M. The cellular computer DNA: program or data? // Bull. Math. Biol. 1990.
Vol. 52, N 3. P. 335-348.
Blattner F. et al. The complete Genome Sequence of Escherichia coli K12 // Science, 1997.
Vol. 277. P. 1453-1462.
Brooke J.L. Animal mitochondrial genomes // Nuclear Acids Research. 1999. Vol. 27.
P. 1767-1780.
De Risi J.L., Iyer V.R. Genomics and array technology // Nature Genetics. 1999. Vol. 21,
Suppl. 1. P. 33-37.
Kananyan G.Kh., Tchuraev R.N., Ratner V.A. Enlarged model of lambda phage ontogenesis // J.
Theor. Biol. 1981. Vol. 90. P. 301-315.
Modeling and Computer Methods in Molecular Biology & Genetics/ Eds. V.A. Ratner,
N.A. Kolchanov. N.Y.: Nova Science Publs., 1992.
Molecular Biology of Bacteriophage T4 / Ed. J.D. Karam. Washington, D.C.: Amer. Soc. for
Microbiol, 1994.
180 III. Молекулярная кибернетика
Proceedings of the 2nd International Conference on Bioinformatics of Genome Regulation and
Structure (BGRS-2000). Novosibirsk: ICG SB RAS, 2000.
Ratner V.A. Zharkikh A.A., Kolchanov N.A. et al. Molecular Evolution. Berlin e.a.:
Springer-Verl., 1996.
Sanger F. et al. Nucleotide Sequence of bacteriophage X // J. Mol. Biol. 1982. Vol. 163.
P. 729-773.
Stent G. Molecular Genetics. San-Francisco: Halden-Day, 1970.
Umesono K., Ozeki H. Chloroplast gene organization in plants // Trends in Genetics. 1987.
Vol. 3, N 10. P. 281-287.
Watson J.D., Crick F.H.C. The structure of DNA // Cold Spring Harbor Sympos. Quant. Biol.
1953. Vol. 18. P. 123-131.
19. Виртуальная реальность
и системы управления*
Imagination is more important than knowledge
A. Einstein
Воображение более важно, чем знание
А. Эйнштейн
Виртуальный мир компьютеров и мир физической реальности
Слово "виртуальный" в последнее время стало столь же расхожим, как
"попса", "полтергейст" или "DIROL без сахара". Виртуальной реальностью
обычно называют мир компьютерных образов, с которыми столкнулась не-
неискушенная часть человечества. Этот мир был создан искушенной частью чело-
человечества, профессионалами — учеными, инженерами, программистами, худож-
художниками — в процессе их научного, технического или художественного творчест-
творчества. Используется же этот мир в самых разнообразных отношениях, включая соз-
создание фильмов, анимаций, компьютерных игр, теоретических моделей,
рекламы, дизайна, криминалистических фотороботов или других продуктов ути-
утилитарного, коммерческого и творческого назначения. В восприятии пользовате-
пользователя виртуальная реальность все менее отличается от естественной, а в фантазиях
творческих личностей все чаще отрывается от нее и превосходит ее активностью
действия, вселенскими масштабами и кровожадностью обитателей.
В последнее время творческая фантазия художников совершила несколько
"выбросов" из мира виртуальной реальности в мир естественной реальности и
обратно. Вспомним хотя бы американский фильм "Виртуальная реальность".
Надо разобраться, что здесь шутка создателя (человека!), а что пустой домысел
недоучек. Для пользователя граница между объектами естественной реальности
и образами виртуального компьютерного мира действительно очень размыта.
-Почему бы не перейти ее в узком месте?
В словарях слово "виртуальный" обычно переводится как "возможный".
Однако в компьютерной научной практике оно приобрело несколько другой
смысл. Виртуальным миром компьютеров можно назвать мир компьютерных
информационных образов. Главное его отличие от мира соответствующих им
• Расширенный вариант статьи, опубликованной в журнале "Природа" B002, № 2. С. 67-71).
19. Виртуальная реальность и системы управления 181
объектов естественной реальности, природы состоит в том, что для него не обя-
обязательно выполнение законов естественной природы в широком смысле. Речь
идет, конечно, не о физических законах, использованных для функционирова-
функционирования компьютерной техники, электроники (с этим все в порядке!), а о поведе-
поведении компьютерных образов. Их трудно назвать объектами. Объект материален.
Здесь лучше подходит английское слово "entity" — сущность, нечто существую-
существующее. Я буду называть их "образованиями" или "образами". Пользователю, зри-
зрителю, программисту с самого начала ясно, что для реальных объектов обяза-
обязательно соблюдение законов физики, химии, биологии и т.д. Эти законы объек-
объективны и присущи самой природе. Что касается компьютерных образов,
которые имитируют реальные объекты, то для них, вообще, исполнение зако-
законов природы не обязательно и задается (или отменяется) самим автором.
Примеры. Известно, что в физическом пространстве два материальных
объекта, имеющие массу покоя, не могут одновременно находиться в одной точке:
они или объединяются, или альтернативны. В компьютерном пространстве вирту-
виртуальные образы этих материальных объектов могут спокойно проходить сквозь
друг друга, если специально не заданы законы их столкновений. В реальном фи-
физическом пространстве все объекты подвержены действию силы тяготения. В
компьютерном пространстве, имитирующем движение в поле тяжести, ничего не
стоит выключить отдельный образ из действия виртуальной "силы тяжести". В
компьютерном мире можно легко обратить виртуальные процессы во времени,
что невозможно в реальной природе. Можно легко преобразовать виртуальный
образ, имитирующий определенное биологическое существо, в другой образ, ими-
имитирующий любое другое биологическое существо, или другой объект природы. В
реальной природе это невозможно. В компьютерном мире можно имитировать
любое чудо или волшебство, чего, конечно, нет в реальной природе.
Я очень люблю телевизионную передачу "Жизнь животных". Но более все-
всего в этой передаче я люблю заставку, где средствами анимации танцуют эти са-
самые животные. Птицы летают вполне правдоподобно, но вот обезьяны летают
по таким замысловатым траекториям, которые доступны только пернатым. В
реальном мире это невозможно.
Итак, виртуальная реальность образов компьютерного мира не требует
обязательного соблюдения законов имитируемого реального физического
мира. Фактически творец этого виртуального мира — ученый, программист,
пользователь — должен чувствовать себя богом или полубогом этого мира, по-
поскольку способен творить его законы. Недаром люди со слабой психикой часто
удовлетворяют свою потребность властвовать именно в компьютерном мире,
где не встречают сопротивления.
Посредник: виртуальный мир человеческого мозга
Теперь, указав основное отличие виртуальной реальности компьютерного
мира образов от естественного мира объектов и полей, мы обнаруживаем, что
между этими двумя мирами стоит посредник, оперирующий обоими мирами и
их соответствиями. Это сам человек — ученый, инженер, программист, худож-
художник, пользователь и т.п. Мозг человека тоже является своеобразной системой
182
III. Молекулярная кибернетика
управления, "биологическим компьютером", формирующим свою виртуальную
реальность образов, понятий и других информационных образований. Различия
способов организации мозга и компьютера в данном случае роли не играют.
Виртуальный мир информационных образований человеческого мозга мо-
может быть очень богат и операбелен. Он имеет: свой язык записи и исполнения
информации; личностную основу; базовый, оперативный и долгосрочный ар-
архивы информации; блочно-модульный принцип построения сложных инфор-
информационных образований из более простых и т.д. Ясно, что в этом виртуальном
мире соблюдение законов реальной действительности для имитирующих ин-
информационных образований тоже не обязательно. Это создает огромные твор-
творческие возможности для генерирования, перебора и оценки идей, образов, по-
понятий и других более сложных информационных образований. Открывается
возможность для полета фантазии. Критериями отбора информационных про-
продуктов мозга могут быть как их степень соответствия объектам и отношениям
реальной действительности (наука), так и любые другие формы поощрения:
вдохновение, успех первооткрывателя, художественное совершенство, озаре-
озарение, религиозный экстаз, психологический комфорт, самоутверждение, обога-
обогащение, патологический императив и т.д.
Перечислим наиболее явные (хотя иногда спорные) информационные
продукты человеческого мозга (или их глобальной сети), известные нам и по-
построенные на основе таких подходов и критериев: 1) язык; 2) личность, харак-
характер, менталитет; 3) сознание, разум; 4) интуиция; 5) душа; 6) гений, талант;
7) эмоции, в том числе самоощущение, боль, радость; 8) знание, наука, особен-
особенно математика; 9) вера, религия, в том числе бог; 10) картина мира; 11) миро-
мировоззрение; 12) моделирование; 13) тинкеринг; 14) художественное творчество,
живопись; 15) музыка; 16) литература, поэзия; 17) лицедейство; 18) фантастика;
19) сновидения; 20) экстрасенсорика; 21) гипноз; 22) психические расстрой-
расстройства, галлюцинации; 23) мораль, этика; 24) эстетика; и др.
Каковы бы ни были строгие и конкретные определения этих образований,
их следует считать информационными продуктами мозга и только ими! Психо-
Психопродукция — виртуальная реальность, данная нам в ощущениях!
Таким образом, виртуальный, информационный мир человеческого мозга
тоже имеет богатые возможности для несоблюдения его автором законов реаль-
реального мира, имитируемого в этом виртуальном.
Базис: информационный мир
генетической организации (МГСУ)
Человеческий мозг является частью высших ярусов кибернетической орга-
организации индивидуума, но не исчерпывает этой организации. В основе кибер-
кибернетической организации клетки и организма (индивидуума) лежит молекуляр-
но-генетическая система управления (МГСУ), которая имеет свой мир инфор-
информационных сущностей. Наиболее фундаментальными и важными среди них
можно считать: гены; функциональные сайты; опероны; генетические сети;
мобильные элементы; каталитические активности; специфичности; программы
онтогенеза; генетический язык, в том числе генетический код; и др.
19. Виртуальная реальность и системы управления 183
Этот информационный мир МГСУ, по сравнению с предыдущими, имеет
ряд специфических особенностей. В кибернетике, лингвистике "идеальный
символ" можно считать семантически произвольным: его смысл полностью за-
задается автором и не зависит от материального воплощения. Конечно, реально
всегда используется мнемоника, символы языка обычно являются продуктами
эволюции пиктограмм и т.п., но в принципе это не обязательно.
В информационном мире МГСУ (т.е. в молекулярной кибернетике) дело
обстоит не так. Многие особенности символов, модулей, систем существенно
зависят от материальных свойств носителей информации и не могут быть при-
приписаны им произвольно. Так, символы генетического НК-языка, нуклеотиды,
имеют присущие им физико-химические свойства комплементарности (А — Т,
G — С), которые играют важнейшую роль в матричных информационно-гене-
информационно-генетических процессах и не могут быть изменены произвольно в любой схеме. То
же касается основных свойств символов ПП-языка — аминокислот: их размер
и полярность определяются их молекулярной структурой. Функциональные
(семантические) свойства глобулярных белков (функциональные центры) оп-
определяются их пространственной структурой (фолдингом). Программы онтоге-
онтогенеза записаны в геноме организма и клетки не в виде последовательности сим-
символов или модулей, а через их внутреннюю структуру и возникающую на ее ос-
основе взаимную специфичность отношений и взаимодействий (генетические
сети). И. т.п.
Другая особенность информационного мира МГСУ состоит в том, что он
образован и предназначен для технических целей: формирования "элементной
базы" МГСУ (в первую очередь — биополимеров и их комплексов) и обеспече-
обеспечения ее функционирования и воспроизведения. "Интеллектуальные изыски",
сопоставимые с информационным миром человеческого мозга, в нем не преду-
предусмотрены и фактически невозможны. Генетическая память клетки и организма
содержит информацию о внешнем реальном мире только в виде записей специ-
специфичности узнавания и переработки некоторых внешних молекулярных объек-
объектов, а также о функциональном ответе некоторых более сложных специализи-
специализированных систем (физиологических, иммунных и др.) на более сложные внеш-
внешние воздействия. Среди генетически кодируемых систем ответа важную роль
играет мозг (особенно — человеческий), способный выполнять не только тех-
технические функции, но и аналитические, прогностические и другие, например
отвлеченно-интеллектуальные. Функции мозга фактически универсальны.
Функции МГСУ ограничены.
Таким образом, в информационном мире МГСУ определенные, приписы-
приписываемые свойства символов, модулей, систем должны соответствовать реальным
свойствам их молекулярных прототипов и не могут быть изменены произвольно.
Отношения "миров" и систем управления
Суммируем и сформулируем изложенное выше. Это и будет наша концеп-
концепция.
Виртуальной реальностью называют мир информационных сущностей, об-
образований, задаваемых или генерируемых в определенной системе управления.
184
III. Молекулярная кибернетика
В каждой отдельной системе управления может быть создан свой виртуальный
мир, и даже не один. Различные виртуальные миры могут быть согласованы
между собой и с миром окружающей реальности.
Следует принять как аксиому: виртуальные миры не существуют вне систем
управления или связывающих их материальных носителей информации. Последнее
существенно, если системы управления образуют сети, хотя бы и удаленные.
В настоящее время можно указать базисные свойства реального и извест-
известных виртуальных миров (рис. 19-1).
1). Реальный мир естественных объектов, явлений, систем, которые имеют
материальную природу и подчиняются естественным законам.
2). Виртуальный мир информационных образований человеческого мозга,
который осознанно или неосознанно создается самим человеком (человечест-
(человечеством), в том числе в процессе обучения: язык, понятия, принципы, критерии,
правила морали, табу, заповеди, зрительные образы и др. Законы этого вирту-
виртуального мира задаются самим человеком. Виртуальный мир во многом соотно-
соотносится с миром физической реальности, но его сущности, имитирующие реаль-
реальные объекты, не обязательно подчиняются законам природы (физики, химии,
биологии и т.д.).
3). Виртуальный мир компьютерных информационных образований, соз-
созданный человеком адекватно виртуальному миру человеческого мозга, а через
него — реальному миру. Законы этого виртуального мира тоже задаются самим
человеком — программистом, ученым, пользователем — и не обязательно сов-
совпадают с законами физической реальности. Хотя обычно контакт между миром
физической реальности и виртуальным миром компьютеров происходит через
Виртуальный мир
крлшъютёриых:
ибразвв
Виртуальный :мйр
человеческого
1
Техническое
воспроизведение
> /^Реальный мир
\^ I ( естественных
^ ч^^ объектов
Онтогенетическое
воспроизведение
Самовоспроизведение
Рис. 19-1. Схема отношений "миров" и систем управления.
19. Виртуальная реальность и системы управления 185
человека — оператора, программиста, все же он может поручить этот контакт
автоматизированной системе исследования, мониторинга природы и т.п.
4). Виртуальный мир информационных сущностей МГСУ. Они связаны
функционально с объектами реального мира через входы и выходы МГСУ. За-
Законы этого виртуального мира никем не задаются, они полностью естествен-
естественные, природные, а потому не вполне произвольны, и через свойства материаль-
материальных носителей информации (макромолекул и их систем) они ограничены осо-
особенностями законов физико-химического материального мира.
В процессе аналитического исследования МГСУ человеком в принципе ус-
устанавливается соответствие двух виртуальных миров, точнее, в пределах чело-
человеческого мозга формируется подсистема виртуальных образований (генетиче-
(генетических текстов, схем генных сетей, генетических карт, моделей и др.), отвечаю-
отвечающих виртуальному миру МГСУ. Через участие человека-оператора можно
создать соответствующие виртуальные подсистемы в компьютерном мире ин-
информационных образований. Они называются базами данных. Исследователь
может поручить прямое взаимодействие виртуальных миров МГСУ и компью-
компьютера автоматизированной системе — секвенатору, монитору и другим прибо-
приборам. Эта тенденция все возрастает.
Системы управления могут быть объединены в сети. Так, объединение вир-
виртуальных миров мозга многих людей некомпьютерными средствами (обмен но-
носителями информации — книгами, статьями, журналами и др.) создает вирту-
виртуальный мир человечества (знание, наука, библиотеки и др.). Сети компьютеров
образуют виртуальный мир ИНТЕРНЕТа, к которому присоединены также
виртуальные миры всех пользователей, т.е. значительная часть человечества.
Образование сетей означает объединение технических ресурсов многих систем
управления, т.е. принципиальное увеличение их возможностей.
МГСУ дифференцированных клеток одного организма тоже создают сеть.
Однако индивидуумы (как одноклеточные, так и многоклеточные) очень редко
образуют сети своих МГСУ, обычно они изолированы и автономны. Правда, в
процессах скрещиваний происходит регламентированная комбинаторика фраг-
фрагментов генетической памяти. Почти во всех остальных случаях МГСУ особей
одного вида, и особенно разных видов, изолированы, т.е. объединения их вир-
виртуальных миров не происходит. Случаи вирусного заражения и "горизонталь-
"горизонтального переноса" по разным причинам очень нестабильны и лишь в слабой сте-
степени нарушают эту изоляцию.
Развитие и разумная эксплуатация всех этих информационных образова-
образований сулит человечеству новые неожиданности и успехи, недооценка — трудно-
трудности, проблемы и угрозы.
Место для бога
Обратимся к более детальному описанию некоторых особенностей вирту-
виртуального мира человеческого мозга. В целом появление такого типа систем
управления с мощным спектром информационных функций — это несомнен-
несомненное эволюционное приобретение широкого профиля, которое обеспечивало
186 III. Молекулярная кибернетика
возможности: оперативного управления поведением; анализа и прогноза ситуа-
ситуации; творчества — т.е. выживания в сложных условиях существования.
Особенность нашей концепции состоит в том, что все перечисленные в п. 2
продукты деятельности человеческого мозга являются чисто информационными
продуктами, и только ими (некоторые оговорки мы сделаем в следующем разде-
разделе). Базовыми информационными продуктами следует считать язык, мировоз-
мировоззрение, картину мира, религиозные убеждения и т.п.
Человеческий мозг как система управления содержит виртуальный образ
окружающего мира в той мере, в какой с ним соприкасается и сам того желает.
В принципе картина мира, мировоззрение должны быть адекватны реальной
действительности. Только тогда они в целом будут способствовать выживанию
индивидов. Однако они всегда имеют личностные характеристики, содержат
продукты творчества, фантазии, гипотезы, случайные особенности. Этот образ
постепенно объективизируется, стираются его фантастические или гипотетиче-
гипотетические особенности, они заменяются информационными продуктами опыта и
науки.
Реальный мир природы очень велик, необъятен. Информационные ресур-
ресурсы человеческого мозга, в принципе, конечны, и потому рано или поздно в ин-
информационной картине мира возникает проблема отношения к несопостави-
несопоставимому, очень большому. Эта проблема требует особого решения [Колмогоров,
1998]. Переход от индивидуальных систем управления мозга к мировой сети
лишь отодвигает решение проблемы, но не снимает ее в принципе.
Примеры. Рано или поздно человек во всех стихиях сталкивается с не-
непреодолимыми, несопоставимыми явлениями, объектами: необозримая земная
твердь, бескрайняя степь, пустыня, тайга, вулкан, горная вершина, континент;
бескрайний океан, неукротимый шторм, тайфун, цунами; бесконечное, недо-
недосягаемое небо над головой, Солнце, Луна, звезды, космос, молния. Кроме того,
человек сталкивается с такими непонятными, непостижимыми явлениями, как
жизнь и смерть, гениальность, непреодолимая власть, маловероятный случай и
т.п. Их проявление, поведение, само присутствие непреодолимы, несопостави-
несопоставимы со шкалами человеческого мозга. Тем не менее их надо как-то обозначить в
информационной картине мира и определить свое отношение к ним. Решить
эти проблемы человек быстро не может, а "застревать" на них надолго у него
нет времени. Остается изолировать эти проблемы доступными средствами. Ис-
Искушенный человек, имеющий опыт научного подхода, ставит в этом месте ин-
информационной картины мира соответствующий информационный объект, по-
понятие, выражающее эту несопоставимость, несоизмеримость: бесконечность,
вечность, случай, комбинаторика, очень большое, очень сложное. Иначе гово-
говоря, он откладывает решение проблемы на завтра. Неискушенный человек ста-
ставит на это место образы, которые ему неподвластны: бог, черт, нечистая сила,
судьба, рок, мировой разум и т.п. Он наделяет их чрезвычайными силами,
свойствами и полномочиями. Перед ними он склоняется, признает свое бесси-
бессилие, т.е. он просто отказывается от их решения.
Таким образом, для бога нет места в реальной природе, ему
"забронировано" место в информационной картине мира человеческого мозга,
не более того. Бог и его культ — религия, вера — это информационные продук-
19. Виртуальная реальность и системы управления 187
ты мозга, информационные образования, не обладающие никакой реальной
силой или властью, но способные существенно повлиять на поведение челове-
человека. Человеческим воображением они наделены сверхъестественной силой и
властью, и потому им позволено не подчиняться законам природы (чудеса,
волшебство). Однако в пределах виртуального мира человеческого мозга эти
образования могут играть фундаментальную роль, определяя критерии жизни и
смерти, веры, морали, права, власти, личности, таланта и т.п., т.е. быть базовы-
базовыми образованиями виртуального мира.
Такие же соображения относятся к понятию "мировой разум" и ему подоб-
подобным. Нет разума вне систем управления или связывающих их материальных
носителей информации — полей и т.п. Только во взаимодействии с системой
управления состояние материального носителя может быть признано инфор-
информационным образованием. Поэтому не может быть никакого "абсолютного
разума", создавшего природу. Для существования таких всеобщих, вселенских
виртуальных сущностей, как бог, "абсолютный разум" и т.п., надо, чтобы Все-
Вселенная уже существовала и была огромной системой управления. Но чтобы
быть системой управления, а не просто астрофизической системой взаимодей-
взаимодействующей материи, Вселенная должна подчиняться принципам управления.
В частности, слабые физические сигналы должны вызывать в ней огромные,
несопоставимые физические последствия. Этого нет, пока не замечено. Пока
что все, что происходит в видимой Вселенной, удовлетворительно описывается
астрофизическими процессами. Кроме того, скорости распространения физи-
физических полей и предметов (потенциальных носителей информации) ограниче-
ограничены, конечны. Они не могут обеспечить информационный контакт между отда-
отдаленными зонами Вселенной, более быстрый, чем сами физические взаимодей-
взаимодействия.
Некоторые особенности и возможности
виртуальных миров
Каждая система управления имеет свои технические (физиологические,
физико-химические) параметры, которые влияют на формирование и функ-
функционирование информационных образований. У компьютеров ограниченный
объем памяти, определенная скорость исполнения операций, несущая частота,
помехоустойчивость, гарантийный срок и т.п. Человеческий мозг имеет огра-
ограниченное число нейронов, генетически контролируемые параметры нервных
процессов, влияющих на память, способность к обучению, абстрактному мыш-
мышлению, художественному творчеству, склонность к патологии, параметры ста-
старения и т.п. МГСУ имеют генетически контролируемые параметры активности
ферментов, специфичности узнавания, скорости реакций, характерные време-
времена переходных процессов переключения и т.п.
Таким образом, информационные образования этих систем управления
всегда как бы "окрашены" этими параметрами в индивидуальные "цвета", не
утрачивая своих общих информационных свойств. Фактически разнообразие
этих параметров среди различных МГСУ и мыслительных систем человека со-
188 III. Молекулярная кибернетика
ставляет значительную часть генетического разнообразия популяций, оценива-
оценивается отбором и играет важную роль в эволюции популяций.
Однако существуют информационные образования, которые имеют оче-
очевидные индивидуальные характеристики. Особенно наглядно это демонстриру-
демонстрируется на примере некоторых информационных образований человеческого моз-
мозга: личность, душа, характер, менталитет, художественное творчество и др. Лю-
Любое информационное образование может быть перекодировано на другой
материальный носитель, в другую систему управления, где при помощи систем
помехоустойчивости может поддерживаться сколь угодно долго. Во всяком слу-
случае, гораздо дольше времени существования исходного информационного об-
образования и исходной системы управления. Кроме того, это информационное
образование может быть передано в пространстве при помощи распространяю-
распространяющегося процесса — поля.
Таким образом, накопив достаточно большую информацию о личности
человека, его способностях, характере, личных параметрах, можно создать
динамическую компьютерную имитацию личности (см. также [Урусиков,
2001]). В принципе с ней можно иметь активный информационный контакт:
вести беседы, задавать задачи, получать ответы и т.д. Чисто интеллектуаль-
интеллектуальную часть можно сопроводить совершенно индивидуальной эмоциональной
частью, а также движущимся изображением конкретного человека. Эту ими-
имитацию можно длительно поддерживать уже без ее оригинала, а также перене-
перенести на любой компьютер, внедрить в любой человеческий мозг, передать на
расстояние.
Страшно сказать, но это означает принципиальную возможность отделения
личности, души и тому подобных информационных продуктов от исходного чело-
человека-носителя, переселения их в другие системы управления! Я готов принять все
пинки и удары от заинтересованных личностей!
Одновременно ясно, какую угрозу это представляет для человечества, если
не будет юридически урегулирован статус таких виртуальных явлений!
Но и какую радость это может принести близким и родным человека, кото-
который безвременно умер!
Эволюционные аспекты
Наша концепция позволяет обозначить приемлемые пути возникновения
всех обозначенных виртуальных миров. Они могли возникнуть только эволюци-
онно, вместе со своими системами управления. Непосредственно в ходе возник-
возникновения и расширения ранней Вселенной какие-либо сложные формы материи,
по-видимому, были полностью исключены. Относительно простые объекты ре-
реального мира свойствами управления не обладали. Для осуществления систем
управления требуются определенные особенности составляющих их компонен-
компонентов и минимальная сложность организации. Поэтому системы управления и
виртуальные миры могли возникнуть только в ходе эволюции природы.
Наиболее ранние системы управления — МГСУ — основа биологической
организации. Базисными компонентами МГСУ являются кодирующие биопо-
no no i/vr
19. Виртуальная реальность и системы управления 189
их функции существенно зависят от порядка входящих мономеров. Из очень
ограниченных алфавитов D типа нуклеотидов и 20 типов аминокислот) путем
комбинаторики линейных последовательностей можно генерировать астроно-
астрономическое (а фактически — неограниченное!) разнообразие текстов, кодирую-
кодирующих разные молекулярные функции.
Реальный физический мир ранней Вселенной, по-видимому, не имел этого
класса молекул с самого начала. Они должны были возникнуть путем физи-
физико-химической комбинаторики. С одной стороны, поле для поиска функцио-
функциональных вариантов открыто. С другой — случайный перебор последовательно-
последовательностей неспособен за разумные периоды времени сформировать достаточную
"элементную базу". Реальный путь преодоления этой трудности предполагает
три фактора: 1) блочно-модульный перебор вариантов последовательностей из
уже имеющихся функциональных фрагментов (так называемый тинкеринг),
2) отбор тех вариантов систем, где выше вероятность воспроизведения, и 3) за-
запоминание отобранных вариантов (наследование). Тогда за разумные интерва-
интервалы эволюционного времени этот процесс добиологической эволюции завер-
завершится формированием молекулярных систем самовоспроизведения — сайзе-
ров, — которые представляют собой ядро, сердцевину МГСУ. Как мы видим,
виртуальный мир кодирующих последовательностей макромолекул возникает
совместно с ядром МГСУ. При этом ключевым свойством МГСУ является
именно их способность к самовоспроизведению. Ясно, что это — эволюцион-
эволюционное приспособление широкого профиля, всегда имеющее преимущество.
Таким образом, МГСУ — наиболее ранние по происхождению системы
управления в природе. Их возникновение совпадает с возникновением жизни
(протоклеток) на Земле, приблизительно 3.5-4 млрд лет назад. Возникновение
МГСУ и их виртуального мира происходит самопроизвольно, без внешнего
"архитектора и строителя". Фактически речь идет о самовоспроизведении ба-
базиса молекулярно-генетической организации клеток. Остальные типы систем
управления и их виртуальные миры возникают уже при непременном участии
МГСУ, которые обеспечивают их воспроизведение.
Дальнейшее эволюционное развитие МГСУ и биологической организации
идет по блочно-модульному принципу с отбором и приводит к созданию иерар-
иерархической организации с все возрастающими и усложняющимися функциями. На
верхних этажах этой иерархии у многоклеточных организмов формируется спе-
специализированная и быстродействующая система управления многоклеточной
организацией, накопления опыта и анализа условий существования — мозг.
Сначала его функции примитивны, но постепенно, путем генетической измен-
изменчивости, наследования и отбора МГСУ в целом, они наращиваются и дают облада-
обладателю все большие преимущества при отборе. Мозг — это тоже эволюционное при-
приспособление широкого профиля, выгодное своему обладателю всегда. Рано или
поздно мозг начинает выполнять сложные аналитические и даже прогностические
функции. Это можно сделать только на основе развития мощного виртуального
мира — памяти, языка, сознания, интуиции и других атрибутов информационной
деятельности. Эти функции выполняет мозг человека, что дало возможность Homo
sapiens стать доминирующим видом живой природы: он достаточно быстро заселил
все континенты и подчинил себе продуктивную и иную деятельность других форм.
190 III. Молекулярная кибернетика
Таким образом, человеческий мозг как система управления — продукт
эволюции блочно-модульного и иерархического усложнения программы раз-
развития МГСУ, а его виртуальный мир в значительной степени — продукт ин-
информационной деятельности самого человека (или человечества). Отдельной
проблемы самовоспроизведения мозга человека не возникает, так как это
свойство обеспечено принадлежностью к верхним ярусам биологической ор-
организации индивидуумов, формируемым и воспроизводимым под контролем
МГСУ.
Наконец, в ходе развития своего социума, производства, человек наталки-
наталкивается на лимитирующие факторы системы управления своего мозга и осоз-
осознанно пытается их преодолеть изобретением технических средств. В результате
он изобретает компьютер с его все расширяющимися интеллектуальными спо-
способностями и виртуальным миром. В этой сфере человек в полной мере — тво-
творец виртуального компьютерного мира. Воспроизведение компьютеров выпол-
выполняется в ходе их производства под контролем человека или в автоматическом
режиме.
Список литературы
Колмогоров А.Н. Автоматы и жизнь // Очерки истории информатики в России / Ред.
Д.А. Поспелов, Я.И. Фет. Новосибирск: Науч.-изд. центр ОИГГМ СО РАН, 1998. С. 621-632.
Ратнер В.А. Концепция молекулярно-генетических систем управления. Новосибирск:
Изд-во НГУ, 1993. 120 с.
Урусиков Д. Я — виртуальный человек // Известия. 2001. 25 авг. С. 7.
Ratner V.A., Zharkikh A.A., Kolchanov N.A. et al. Molecular Evolution. Berlin e.a.: Springer-
Verl., 1996.
Ratner V.A. The Genetic Language. Intern. Conf. on Biolnformatics: Report. Magdeburg,
Germany, 1999.
20. Генетический код как система*
"Самым трудным в проблеме кода было понять, что код существует. На это
потребовалось целое столетие. Когда это поняли, то для того, чтобы разобраться
в деталях, хватило каких-нибудь десяти лет1' [Ичас, 1971].
Проблема генетического кода — это проблема прошлого столетия. В конце
50-х — начале 60-х годов она приковывала к себе внимание, возбуждала актив-
активность умов, побуждала веру в величие и мудрость загадок науки. В широком
смысле генетический код — это способ записи генетической информации в по-
последовательностях нуклеиновых кислот (ДНК или РНК) о структуре полипеп-
полипептидов (белков). В конкретном смысле генетический код — это соответствие
между триплетными кодонами матричной РНК (м-РНК) и аминокислотами
кодируемого белка, задаваемое кодовой таблицей (см. табл. 20-1).
* Опубликовано ранее: Ратнер В.А. Генетический код как система // Соросовск. образоват.
журн. 2000. Т. 6, № 3. С. 17-22.
20. Генетический код как система 191
Развитие проблемы генетического кода прошло несколько этапов. Предте-
Предтечами этой проблемы можно считать многих выдающихся исследователей.
Н.К. Кольцов (см. гл. 15) предложил в обшей форме идею молекулы-гена и
матричный принцип ее дублирования. Н.В. Тимофеев-Ресовский, К. Циммер
и М. Дельбрюк (см. гл. 15) указали на макромолекулярную природу генов и гру-
грубо оценили их физический размер. Э. Шредингер (см. гл. 15) явно сформулиро-
сформулировал необходимость кодирования генетической информации в структуре ге-
генов-молекул. П. Колдуэлл и С. Хиншельвуд в 1950 г. предложили идею матрич-
матричного синтеза белков на ДНК. А. Дауне в 1952 г. сформулировал гипотезу о
синтезе белков на РНК.
Научные представления о генетическом коде как о реальной проблеме экс-
эксперимента и теории были сформулированы Г.А. Гамовым сразу же после обос-
обоснования Дж. Уотсоном и Ф. Криком в 1953 г. модели строения двойной спира-
спирали ДНК. Первый этап изучения проблемы A953-1961) можно назвать гипоте-
гипотетическим. Из модели Уотсона—Крика (см. подробнее [Ичас, 1971]) вытекало
представление о линейной последовательности ДНК — тексте, построенном из
четырех типов нуклеотидов (А, Т, G и С — четыре символа алфавита). Однако
кодируемые белки тоже имеют линейную первичную структуру — текст, по-
построенный из 20 типов канонических аминокислот (алфавит из 20 символов).
Поэтому Г.А. Гамов в 1954 г. (см. гл. 16) сразу же сформулировал идею генети-
генетического кода в конкретном смысле — как соответствие двух текстов, записан-
записанных при помощи двух разных алфавитов. Кроме того, он предложил использо-
использовать технические средства криптографии (расшифровки неизвестных кодов)
для решения центральной проблемы генетики.
Генетический код сразу же приобрел облик великой загадки природы, ре-
ребуса для остроумных. Многие сотни математиков, физиков, химиков, биоло-
биологов, включая Г.А. Гамова, Ф. Крика и др., предложили гипотетические вариан-
варианты генетического кода, которые представляют теперь лишь исторический инте-
интерес. Реальный код оказался совсем иным.
Научными результатами первого этапа можно считать [Молекулярная ге-
генетика, 1963; Ичас, 1971]: 1) постановку проблемы генетического кода; 2) фор-
формирование понятий линейного текста, алфавита для нуклеиновых кислот и
белков, генетической информации, записанной в этих текстах при помощи
символов алфавита; 3) представление о матричной роли РНК в трансляции;
4) понятие о кодонах и доказательство их неперекрывания; 5) предположение о
триплетности кодонов и коллинеарности гена и белка, доказанное лишь в даль-
дальнейшем, и т.д.
Второй этап A961-1966) можно назвать экспериментальным, так как в этот
период генетический код был расшифрован в прямом эксперименте [Ичас,
1971; The Genetic Code, 1966; Ратнер, 1983]. В 1961 г. Ф. Крик с сотрудниками в
блестящей работе показали, что: а) кодоны триплетны; б) между ними нет раз-
разделительных знаков ("запятых"); в) гены, кодирующие структуру белков (цис-
троны), имеют фиксированное начало, ориентированное направление и фик-
фиксированный конец; г) существует небольшое число некодирующих триплетов
(нонсенсов, бессмысленных кодонов), а код в целом сильно вырожден. В
1964 г. Ч. Яновский с сотрудниками и С. Бреннер с сотрудниками показали,
192 III. Молекулярная кибернетика
что ген и кодируемый им белок взаимно коллинеарны, т.е. имеется последова-
последовательное соответствие между кодонами гена и аминокислотами белка.
Прямая расшифровка генетического кода in vitro оказалась возможной бла-
благодаря технике белкового синтеза в бесклеточных системах [Ичас, 1971; The
Genetic Code, 1966; Ратнер, 1983], т.е. в клеточных экстрактах, содержащих все
необходимые компоненты аппарата трансляции (т-РНК, рибосомы, аминокис-
аминокислоты, ферменты, источник энергии и т.д.), кроме м-РНК. Вводя в такие систе-
системы естественные м-РНК или искусственные небольшие олигорибонуклеоти-
ды, можно было изучать специфичность включения меченых аминокислот в
строящиеся полипептиды. М. Ниренберг и Ф. Ледер подавали в бесклеточную
систему трансляции Е. coli различные олигорибонуклеотиды и показали, что
индивидуальные фракции три-рибонуклеотидов, ассоциированные с рибосома-
рибосомами, связывают определенные фракции т-РНК, заряженные определенными
мечеными аминокислотами. С помощью такого метода генетический код был
расшифрован полностью. Летом 1966 г. на симпозиуме по количественной био-
биологии в Колд-Спринг-Харборе (США) все полученные данные были сведены
Ф. Криком воедино [The Genetic Code, 1966]. Расшифрованный генетический
код Е. coli, исследованный in vitro, полностью согласовывался также с другими
независимыми данными, полученными in vivo, и для других видов. Этот вывод
подтверждается также результатами секвенирования последних лет, когда най-
найдено, что тысячи генов и кодируемых ими белков действительно соответствуют
друг другу по правилам генетического кода.
Полный "словарь" генетического кода Е. coli приведен в табл. 20-1 [The
Genetic Code, 1966; Ратнер, 1983]. Его анализ позволяет выявить ряд общих
свойств генетического кода как целого.
Из 64 возможных триплетов 61 является смысловым кодоном, т.е. кодирует
аминокислоты. Все кодоны триплетны, неразрывны и не перекрываются в тек-
тексте, а также не разделены межкодонными знаками ("запятыми").
Все кодоны однозначны, т.е. каждый кодирует единственную аминокисло-
аминокислоту. Иначе говоря, в направлении кодон => аминокислота генетический код од-
однозначен.
Обратное соответствие в направлении аминокислота => кодон неоднознач-
неоднозначно, и это свойство называется вырожденностью. Отдельные аминокислоты ко-
кодируются группами (сериями) кодонов-синонимов. 18 серий из 20 содержат от
двух до шести кодонов, две серии (Met и Тгр) не вырождены, содержат по одно-
одному кодону. Средняя вырожденность генетического кода — приблизительно
3 кодона на серию.
Из 18 вырожденных серий 17 имеют свойство связности, т.е. образуют тес-
тесные группы. На рис. 20-1 понятие связности иллюстрируется графически. Вер-
Вершины графов отвечают кодонам, а соединяющие их ребра — заменам одиноч-
одиночных нуклеотидов. Приведены примеры связных серий по четыре (Val —
рис. 20-1, а) и шесть (Leu — рис. 20-1, б) кодонов. В результате из любого кодо-
кодона связной серии можно перейти к любому другому синониму путем последо-
последовательных замен, не выходя при этом за пределы графа этой серии. Если такой
переход можно сделать за один шаг, то серия называется полносвязной
20. Генетический код как система
193
Таблица 20-1
Словарь канонического универсального генетического кода Е. coli
Нуклеотид кодона
Пер-
Первый
и
с
А
G
и
иии
иис
UUA
UUG
сии
cue
CUA
CUG
AUU
AUC
AUA
AUG
GUU
GUC
GUA
GUG
Б
Phe
НП
Leu
Ср
Ср
НП
Leu
Ср
Не
НП
Ср
Met
F-Met
М
НП
Val
Все аминокислоть
неполярны,
не край-
них свойств и раз-
размеров
С
иси
исс
UCA
UCG
ecu
ССС
ССА
CCG
ACU
АСС
АСА
ACG
GCU
GCC
GCA
GCG
М
п
Ser
М
НП
Pro
М
п
Thr
М
НП
Ala
Второй
Все аминокислоты
малые, а основы силь-
сильные
UAU
UAC
UAA
UAG
CAU
САС
САА
CAG
AAU
ААС
ААА
AAG
GAU
GAC
GAA
GAG
А
Б
Туг
П
Term
Б
His
П
Gin
Ср
Ср
Asn
П
Lys
Б
Ср
Asp
П
GIu
Ср
Все аминокислоты
полярнън
и не малые,
а основы слабые
С
UGU
UGC
UGA
UGG
CGU
CGC
CGA
CGG
AGU
AGC
AGA
AGG
GGU
GGC
GGA
GGG
Крайние
М
Cys
НП
Term
Trp Б
Б
П
Arg
М
Ser
П
Arg
Б
М
НП
Gly
варианты
аминокислот и ано-
аномалии серий
Тре-
тий
и
С
А
G
и
С
А
G
и
С
А
G
и
С
А
G
Примечание. Обозначения аминокислот: Ala — аланин, Asn — аспарагин, Asp — аспарагиновая ки-
кислота, Arg — аргинин, Cys — цистеин, F-Met — формил-метионин, Gin — глутамин, Glu — глутаминовая кисло-
кислота, Gly — глицин, His — гистидин. Не — изолейцин, Leu — лейцин, Lys — лизин, Met — метионин, Phe — фени-
лалании, Pro — пролин, Ser — серии, Thr — треонин, Term — терминальный нонсенс, Туг — тирозин, Val — ва-
лин. Другие обозначения см. в тексте.
194
III. Молекулярная кибернетика
(рис. 20-1, а). Связность нарушена только в одной серии (Ser — см. рис. 20-1, в),
но и тогда она распадается на две связные подсерии.
Вырожденность называется систематической, если кодоны-синонимы раз- I
личаются в 3-й позиции либо пуринами (R = А или G), либо пиримидинами
(Y = U или С), либо вообще любыми из 4 нуклеотидов (N = A, G, U или С).
Этим принципам удовлетворяют 30 пар кодонов из 32, а также 8 тетрад из 16. I
Все эти пары связны, а тетрады полносвязны. Остальные варианты вырожден- I
ности называют несистематическими. Они относятся к большим сериям (см. I
табл. 20-1): Leu и Arg — связные серии, Ser — несвязная серия, Не — 3 кодона,
полносвязная серия.
Генетический код содержит также знаки пунктуации (начала и конца)
трансляции. Кодоны AUG, GUG и UUG у прокариот, помимо кодирования
аминокислот, кодируют также инициацию трансляции. Однако однозначность
кодирования при этом не нарушается, так как инициирующие знаки локализо-
локализованы в определенном окружении (контексте), способном образовывать само-
самокомплементарные "шпильки". У эукариот инициируют триплеты AUG и более
слабо — UUG, AUA и ACG. Три "вакантных" триплета у Е. coli — UAA (ochre),
UAG (amber) и UGA (opal) — не кодируют аминокислот, а выполняют роль тер-
терминальных знаков трансляции (стоп-кодонов, нонсенс-кодонов, или терми-
терминальных нонсенсов). В норме ими заканчиваются все цистроны, т.е. трансли-
транслируемые гены, единицы трансляции. Мутационное возникновение нонсенсов
внутри гена приводит к преждевременной терминации трансляции и обрыву
белка. Нонсенсы тоже образуют связную серию (см. табл. 20-1).
Расшифровка генетического кода была одним из самых выдающихся науч-
научных открытий XX века.
Третий этап изучения проблемы генетического кода (после 1966 г.) связан с
углубленным исследованием молекулярных механизмов кодирования, систем-
системных свойств генетического кода: симметрии, регулярности, помехоустойчиво-
помехоустойчивости, универсальности, а также путей его возникновения и эволюции (цит. по:
[Ратнер, 1983]). Молекулярной системой, обеспечивающей соответствие кодо-
кодонов м-РНК и аминокислот, является набор адапторных молекул транспортных
РНК (т-РНК) и набор кодирующих ферментов аминоацил-т-РНК-синтетаз
(АРС-аз). Каждая специфическая молекула т-РНК имеет антикодон, взаимо-
взаимодействующий с кодоном м-РНК, а также специфический сайт взаимодействия
а б в
GUU
GUA
CUA
UUA
UCA
AGU
I •
GUS
GUS
GUC
GUS
UUG
UCC
UCG
AGC
Рис. 20-1. Графы связности для некоторых кодовых серий.
а — серия Val, вырожденность 4 кодона, полносвязная; б — серия Leu, вырожденность 6, связная;
в — серия Ser, вырожденность 6, несвязная, но распадается на две полносвязные подсерии.
20. Генетический код как система
195
с определенной АРС-азой и неспецифический сайт связывания аминокислоты
(рис. 20-2). Каждая АРС-аза опознает все изоакцепторные (переносящие одну
аминокислоту) фракции т-РНК, одну определенную аминокислоту и соединяет
их макроэргической (энергобогатой) связью. Поэтому соответствие антикодо-
на т-РНК и аминокислоты определяется именно АРС-азой. Фракции т-РНК
выполняют функции адаптеров (специфических посредников) между кодонами
м-РНК и аминокислотами.
Многие свойства генетического кода обеспечиваются свойствами молекул
т-РНК и АРС-аз. Триплетный и неразрывный антикодон выделен в антикодон-
ной петле т-РНК специальными модифицированными нуклеотидами (см.
рис. 20-2). Этим обеспечиваются триплетность и неразрывность узнаваемых
кодонов матрицы. Все антикодоны одинаково триплетны, поэтому, начиная от
инициирующего знака, трансляция осуществляется "триплетными шагами",
т.е. формируется определенная "рамка" (фаза) трансляции — одна из трех воз-
возможных. В этом случае межкодонные знаки ("запятые") не нужны, а кодоны
не перекрываются. Инициирующие кодоны у Е. coli опознаются специальной
фракцией T-PHKF'Met, переносящей модифицированную аминокислоту фор-
мил-метионин. Терминальные нонсенсы вообще не имеют своих фракций
т-РНК, а опознаются специальными белковыми факторами терминации, кото-
которые, как оказалось, по своей пространственной структуре весьма напоминают
т-РНК.
Однозначность кода в направлении кодон => аминокислота обеспечивается
строгой специфичностью АРС-аз. Каждая АРС-аза узнает единственную амино-
аминокислоту, поэтому неоднозначность исключена или очень маловероятна. В осно-
основе систематической вырожденности лежат правила неоднозначности спаривания
кодон—антикодон, установленные Ф. Криком (цит. по: [Ичас, 1971; Ратнер,
1983]). Один антикодон может узнавать один, два или три кодона, различающие-
различающиеся по третьей позиции (табл. 20-2). Согласно правилам неоднозначного спарива-
спаривания, систематическая вырожденность в парах кодонов обеспечивается отдель-
отдельными фракциями т-РНК, имеющими U, G или I
(инозин) в трех позициях антикодонов. Вырожден-
Вырожденность 3 у изолейцина (Не) требует фракцию т-РНК с
I в третьей позиции антикодона. Такой нуклеотид
там действительно есть. Вырожденность 4 требует
не менее двух фракций т-РНК, вырожденность 6 —
не менее трех фракций. Всего генетический код Е.
coli требует не менее 32 фракций т-РНК. Реально у
Е. coli полное число генов т-РНК равно 86 для 79
фракций с различными антикодонами (см. гл. 12).
Следовательно, многие фракции т-РНК частично
дублируют друг друга.
Анализ таблицы генетического кода как цело-
целого (табл. 20-1, рис. 20-3) позволяет выявить удиви-
удивительные свойства его регулярности и симметрии
[Ратнер, 1983]. Обозначим позиции нуклеотидов в
кодонах E'—1—2—3—3*) через x-y-z , где х — при-
Таблица 20-2
Неоднозначность спаривания
нуклеотидов в третьей позиции
кодона и антикодона (цит. по:
[Ратнер, 1983])
Антикодон
и
С
А
G
I
Кодон
A, G
G
и
и, с
U, С, А
196 III. Молекулярная кибернетика
ставка, у — корень, z — окончание, а ху — основа кодона. Эти термины, пред-
предложенные Ю.Б. Румером, отражают сходные понятия лингвистики. Корни
слов определяют их смысл. Все мутации, затрагивающие корень (у) кодона,
также изменяют кодовую серию, т.е. нарушают смысл кодона. Приставки слов
тоже участвуют в определении смысла, хотя и не так жестко, а многие их из-
изменения меняют смысл слов. Замены в приставках (х) кодонов чаше всего из-
изменяют их смысл, но иногда являются синонимическими. Окончания слов
обычно участвуют в словоизменении, т.е. в синонимических преобразовани-
преобразованиях. Аналогично 70 % замен в окончаниях (z) кодонов синонимические. Нако-
Наконец, приставка и корень слова образуют его основу, несущую полную или до-
доминирующую смысловую нагрузку. Основа кодона (ху) тоже играет ключевую
роль в особенностях генетического кода.
Регулярность генетического кода связана с распределением основных
свойств кодонов и аминокислот по столбцам (корням) генетического кода (см.
табл. 20-1). Кодоны характеризуются свойствами их основ и корней. Кодоны,
имеющие одинаковые основы, образуют 16 тетрад генетического кода. Основа
называется сильной, если она полностью определяет "смысл" (аминокислоту)
кодонов тетрады. Нуклеотид в третьей позиции тоже, конечно, необходим, но
он может быть любым (N). Таковы основы CU, GU, UC, CC, AC, GC, CG, GG.
Легко заметить, что они содержат нуклеотиды в соотношении С : G : U : А =
7:5:3:1. Основа называется слабой, если для однозначного кодирования
аминокислоты необходимо также участие определенного 3-го нуклеотида (z) в
кодоне. Таковы основы UU, AU, UA, СА, АА, GA, UG, AG. Они содержат нук-
нуклеотиды в обратном соотношении С : G : U : А = 1 : 3 : 5 : 7. Известно, что уча-
участки ДНК с избытком пар G-C более стабильны, чем А-Т-богатые участки. По-
Поэтому сильные основы образуют в среднем больше водородных связей с анти-
кодонами т-РНК, чем слабые.
Аминокислоты имеют два основных характерных свойства, существенных
в пространственной структуре глобулярных белков: размер (М — малые, Ср —
средние, Б — большие) и полярность / неполярность (П / НП).
Легко заметить, что столбцы генетического кода, отвечающие определен-
определенным "корням" кодонов, имеют некоторые групповые свойства (см. табл.
20-1). Третий столбец (корень А) содержит полярные аминокислоты, не малые
по размеру, а все основы кодонов слабые. Это как бы групповое свойство кор-
корня А. Второй столбец (корень С) содержит аминокислоты, малые по размеру, а
все основы кодонов сильные. Первый столбец (корень U) включает неполяр-
неполярные аминокислоты разных (но не крайних) размеров, а свойства кодонов неод-
неоднозначны. Наконец, четвертый столбец (корень G) содержит все крайние и
аномальные варианты аминокислот и кодовых серий [Ратнер, 1983]: самую ре-
реактивную аминокислоту (Cys), самую большую и плоскую (Тгр), самую боль-
большую и "корявую" (Arg), самую маленькую (Gly), часть единственной несвяз-
несвязной серии (Ser) и неоднозначный терминальный нонсенс (UGA), который в
ряде случаев кодирует 21-ю аминокислоту селеноцистеин (Sec). Таким обра-
образом, если групповые свойства корней (и столбцов) отвечают каким-то общим
правилам их возникновения, то четвертый столбец скорее напоминает свалку
Not for sale! For preview only! Scanned by Alykhaylo (molbioftw ukr.net)
D-петля
Аминокислота
76
Акцепторный
стебель
Г-петля
Антикодонныи
стебель
Вариабельная
петля
Антикодонная
петля
Антикодон
Рис. 20-2. Обобщенная вторичная структура молекул транспортных РНК ("клеверный
лист"). Указаны основные фрагменты структуры: антикодон, ограниченный модифици-
модифицированными нуклеотидами, выделяющими триплетную рамку; неспецифический участок
АСС, связывающий аминокислоту; четыре двухспиральных участка (стебля) и три не-
спаренные петли.
Рис. 20-3. Генетический код в круговой форме. Для наглядности симметрии важен из-
избранный порядок символов по часовой стрелке: U—С—A—G. Обозначения аминокис-
аминокислот такие же, что в табл. 20-1. Другие обозначения: П — полярные; НП — неполярные;
• — терминальные нонсенсы (Term); A — кодоны, кодирующие аминокислоты Met и
Val, но в определенном контексте играющие роль начальных знаков трансляции; * — се-
серии с вырожденностью 6. Правила симметрии см. в тексте.
21. Генетический код как продукт эволюции 197
всего, что не попало в первые три столбца по групповым правилам их форми-
формирования.
Теперь рассмотрим не менее впечатляющее свойство симметрии генетиче-
генетического кода. На рис. 20-3 генетический код изображен в круговой форме [Рат-
нер, 1983], где внутренний круг отвечает 1-м позициям кодонов, среднее коль-
кольцо — 2-м и внешнее кольцо — 3-м позициям. Сильные основы изображены не-
подразделенными секторами внешнего кольца, а слабые — подразделенными.
Свойство симметрии состоит в следующем:
1) проведем ось симметрии через центр круга перпендикулярно плоскости
листа и повернем круг на 180° в плоскости листа. При этом все сильные и сла-
слабые основы сохраняют свои позиции, т.е. совмещаются с одноименными;
2) проведем через центр плоскость симметрии, перпендикулярную плоско-
плоскости листа и строкам текста. При зеркальном отражении круга в этой плоскости
все сильные основы меняются местами со слабыми и наоборот;
3) проведем через центр плоскость симметрии, перпендикулярную плоско-
плоскости листа и параллельную строкам текста. При зеркальном отражении круга в
этой плоскости сильные основы меняются на слабые и наоборот.
Ясно, что эти свойства отражают не случайный, высокоорганизованный
характер генетического кода, связанный с правилами синонимии кодовых се-
серий.
Таким образом, генетический код Е. coli представляет собой не случайный
конгломерат соответствий между кодонами и аминокислотами, а высокоорга-
высокоорганизованную систему соответствий, поддерживаемую сложными молекулярны-
молекулярными механизмами. По выражению Френсиса Крика, внесшего решающий вклад
в открытие и изучение кода [The Genetic Code, 1966], "это ключ к молекулярной
биологии, поскольку он показывает, как два великих языка полимеров —язык поли-
нуклеотидов и язык полипептидов — связаны между собой ". Дальнейший анализ
покажет (см. гл. 21), каковы были пути возникновения и эволюции генетиче-
генетического кода.
Список литературы
Ичас М. Биологический код. М.: Мир, 1971.
Молекулярная генетика. М.: Мир, 1963.
Ратнер В.А. Молекулярная генетика: принципы и механизмы. Новосибирск: Наука,
1983.
The Genetic Code. Cold Spring Harbor Symp. Quant. Biol. (CSHSQB). Cold Spring Harbor,
N.Y., 1966. Vol. 31.
21. Генетический код
как продукт эволюции
В гл. 20 были показаны особенности и закономерности строения генетиче-
генетического кода как системы [Ратнер, 2000]. Совершенно ясно, что такие свойства
не возникают одномоментно, а являются результатом длительного эволюцион-
эволюционного процесса. Многие особенности кода являются продуктом эволюции.
198 III. Молекулярная кибернетика
Структура генетического кода отражает его помехоустойчивость [Ратнер,
1983; Ratner et al., 1996]. Наличие почти полной вырожденности серий генети- |
ческого кода F1 смысловой кодон из 64 триплетов) уменьшает долю летальных
мутаций смысловой кодон -» нонсенс до минимума, обусловленного необходи-
необходимостью терминальных нонсенсов. Связность и систематическая вырожден-
вырожденность серий максимизируют долю синонимических замен кодонов (без замены
аминокислот). Регулярность кода уменьшает долю несинонимических замен с
изменением размера и полярности аминокислот (т.е. наиболее "тяжелых" за-
замен). Наконец, частоты использования аминокислот в первичных структурах
реальных белков не равномерны, наблюдается сильная их корреляция с вырож-
вырожденностью соответствующих кодовых серий. Иначе говоря, чаще используются
те аминокислоты, которые имеют больше кодонов-синонимов. Например,
сильно вырожденные серии — Ser F), Leu F), Ala D), Gly D) и др. — имеют
частоты использования 7-8 %, тогда как у невырожденных — Met A) и Тгр
A) — частоты использования 1.0-1.5 %. Эта корреляция минимизирует число
летальных несинонимических замен кодонов в генетических текстах. Таким
образом, помехоустойчивость кода была одним из основных критериев его эво-
эволюционной оптимизации.
Частоты использования синонимических кодонов в генах тоже не равно-
равномерны (цит. по: [Ratner et al., 1996]). В каждой вырожденной серии выделяют-
выделяются 1-2 "оптимальных" кодона, используемые чаще других, а остальные встре-
встречаются редко. Неравномерность использования оптимальных и неоптималь-
неоптимальных кодонов усиливается при увеличении белковой продуктивности генов.
Так, ген рибосомного белка L7 / L12 у Е. coli (массовый белок рибосом) ис-
использует всего 24 кодона, ген 9 из митохондрий дрожжей (все гены митохонд-
митохондрий высокоактивны) — 27, ген р-цепи гемоглобина человека (массовый белок
из эритроцитов) — 37. Напротив, низкоактивные гены используют большин-
большинство кодонов. Например, гены регуляторных белков-репрессоров С; фага X и
lac i Е. coli (активность которых в клетке ничтожна) содержат по 55 разных
кодонов. Ген овальбумина курицы тоже использует 55 кодонов, а ген А фага
ц>Х174 — 60. "Оптимальные" кодоны узнаются фракциями т-РНК, которые
имеют наибольшую концентрацию в клетке. Использование "оптимальных"
кодонов позволяет увеличить скорость трансляции. Это особенно важно для
высокопродуктивных генов и может быть критерием отбора кодонов внутри
таких генов.
Генетический код универсален в том смысле, что его основная часть одина-
одинакова для всех форм жизни на Земле. Этот вывод обоснован опытом массового
секвенирования генов и белков. Почти всегда коллинеарное соответствие генов
и белков согласуется с правилами генетического кода. Однако в некоторых эк-
экзотических системах трансляции (митохондрии животных, растений и грибов,
хлоропласты растений, мельчайшие бактерии — микоплазмы, реснитчатые
простейшие и др.) найдены минорные отклонения в генетическом коде, а так-
также изменения правил неоднозначного спаривания и наборов антикодонов и
фракций т-РНК. Это своеобразные "диалекты" генетического кода, отражаю-
отражающие специфику их эволюции и функционирования.
21. Генетический код как продукт эволюции 199
Известно, что мутационные замены нуклеотидов в антикодоне способны
изменить специфичность т-РНК и переадресовать ее другому кодону или
нонсенсу. В результате наступает явление соответственно
"миссенс-супрессии" (изменения смысла кодона) или "нонсенс-супрессии"
(вовлечения нонсенса в кодирование аминокислоты). Легко понять, что для
занятых и используемых кодонов переадресовка может создать неоднознач-
неоднозначность кодирования, что обычно отсекается отбором. Однако, для
"неоптимальных" (т.е. почти не используемых) кодонов переадресовки до-
допустимы. Отметим, что гены митохондрий и хлоропластов как раз высокоак-
высокоактивны и используют в основном именно оптимальные кодоны. С другой сто-
стороны, экспериментально найдены реальные мутантные фракции т-РНК, вы-
выполняющие нонсенс-супрессию.
Несомненно, что генетический код явился продуктом добиологической
молекулярной эволюции и продолжал частично эволюционировать в дальней-
дальнейшем. В стохастическом процессе молекулярной эволюции свойства генетиче-
генетического кода могли быть: 1) либо предзаданы (предетерминированы) физико-хи-
физико-химическими характеристиками компонентов и условий; 2) либо отобраны как
адаптивные среди альтернативных вариантов; 3) либо фиксированы случайно.
Гипотезы возникновения генетического кода в разной степени учитывают эти
возможности [Ратнер, 1985; Ratner et al., 1996].
Так, гипотеза "замороженного случая" Крика A968 г.) (цит. по: [Ратнер,
1985]) полагала, что исторически была фиксирована первая случайная, но удов-
удовлетворительная система кодирования, которая далее была размножена, подвер-
подверглась эволюционному усложнению и оптимизации, так как обеспечивала уско-
ускоренное воспроизведение.
Другая крайняя модель — неслучайность, слабая взаимная предзаданность
отношений между аминокислотами и антикодонными динуклеотидами (цит. по:
[Ратнер, 1985]). Существуют данные о корреляции параметров гидрофильности
и гидрофобности аминокислот и антикодонных динуклеотидов, отвечающих
этим аминокислотам по генетическому коду. Такая корреляция могла привести
к постепенному возникновению взаимной специфичности через посредников.
Возможно, вначале были зафиксированы лишь корни антикодонов: U — для по-
полярных аминокислот, А — для гидрофобных, G — для малых по размеру, а С —
для всех остальных (корни антикодонов комплементарны корням кодонов в
столбцах генетического кода). Это соответствует свойству регулярности совре-
современного генетического кода.
У. Фитч и К. Аппер A987 г.) (цит. по: [Ratner et al., 1996]) предложили ги-
гипотетическую схему возможных последовательных событий возникновения ан-
антикодонов т-РНК. Вначале соответствия специфичности аминокислот и анти-
антикодонов не было, неоднозначность была максимальной (рис. 21-1, а). Затем
произошло разделение функций пуринов (R = А и G) и пиримидинов (Y = U и С)
во 2-й позиции, причем R преобладали для гидрофильных аминокислот, a Y —
для гидрофобных (см. рис. 21-1, б — вертикальная линия). Дальнейшие верти-
вертикальные подразделения схемы уточняли свойства второй позиции, а горизон-
горизонтальные — первой и третьей позиций в кодонах. Эти события в основном затра-
200
III. Молекулярная кибернетика
и
с
А
G
и
%
<к
asp
Вторая
С
<
<*>
•
ALA
позиция
А
О
G
и
Рис. 27-7. Мнемоническая схема У. Фитча
и К. Аппера A987 г.) (цит. по: [Ratner et al.,
1996]), изображающая идею последователь-
последовательного усиления специфичности кодирова-
кодирования в рамках будущего генетического кода.
а — исходное отсутствие специфичности
кодирования аминокислот; б — первое раз-
разделение функций: вертикальная линия
обозначает разделение функций сначала
между пуринами (R) и пиримидинами (Y)
во второй позиции (корне) будущих кодо-
нов; в — итог дальнейшего разделения
функций: вертикальные линии — во вто-
второй позиции, горизонтальные линии — в
первой (между тетрадами) и в третьей по-
позиции (внутри тетрад, имеющих слабые ос-
основы кодонов).
гивали свойства антикодонов т-РНК
и функциональных центров АРС-аз.
На рис.21-1, в показан современный
генетический код.
Т. Джукс A987 г.) (цит. по:
[Ratner et al., 1996]) предложил сцена-
сценарий последовательной эволюции ос-
основных параметров генетического
кода (рис. 21-2). В табл. 21-1 приведе-
приведены предполагаемые характеристики
исторических и современных вариан-
вариантов генетического кода. Предполага-
Предполагается, что эволюция клеточных вари-
вариантов шла в сторону увеличения чис-
числа кодонов, аминокислот, фракций
т-РНК и сужения правил неодно-
неоднозначного спаривания, а варианты ко-
кодов митохондрий и хлоропластов воз-
возникли путем вторичной редукции
числа фракций т-РНК и повторного
расширения механизма неоднознач-
неоднозначного спаривания.
В итоге универсальность генети-
генетического кода считается указанием на
монофилетичность происхождения
системы кодирования и всего биоло-
биологического мира; вырожденность, связ-
связность, симметричность и регулярность
кода — следствием предзаданности и
единообразных процессов возникно-
возникновения и закрепления кодовых серий
путем неоднозначного спаривания,
дупликаций и мутационной дивер-
дивергенции антикодонов т-РНК и
АРС-аз; помехоустойчивость — след-
следствием отбора вариантов кода на ми-
минимальность мутационных потерь.
Некоторые свойства кода — набор
антикодонов т-РНК, нарушение
связности, регулярности и т.д. — мо-
могут быть случайными. Алфавиты 4
нуклеотидов и 20 аминокислот счита-
считаются предзаданными. Однако набор
нуклеотидов обязательно должен
быть четным, попарно комплемен-
комплементарным и желательно — минималь-
21. Генетический код как продукт эволюции
201
Архетипический
код
GC
I
:Maj
кол
I
Минимальный
код
GC
Митохондриальный
код
AT
Код
"эубактерии
AT
Код
эукариот
Код
галобактерий/
метанококков-архебактерий
GC-давление отбора
АТ-давление отбора
Код
хлоропластов
растений
Код
микоплазм
Код
реснитчатых
простейших
Рис. 21-2. Сценарий эволюции генетического кода, предложенный Т. Джуксом A987 г.)
(цит. по: [Ratner et al., 1996]). Параметры кодов приведены в табл. 21-1. Первичные сис-
системы кодирования содержали, вероятно, меньше элементов, чем современные. Эволю-
Эволюция шла в сторону усложнения систем кодирования. Развитые современные почти
идентичные системы имеют эубактерии (включая микоплазм), ядра эукариот и галобак-
галобактерий (группа архебактерий). Другие архебактерии — метанококки, по-видимому, со-
сохранили один из ранних вариантов редуцированного кода. Симбиотические органеллы
эукариот — митохондрии и хлоропласта — имеют, вероятно, вторично редуцированные
коды, восходящие к ранним эубактериям. GC-давление отбора — эпоха роста содержа-
содержания пар GC, АТ-давление отбора — эпоха роста содержания пар AT.
Таблица 21-1
Общие характеристики реальных и гипотетических предковых систем кодирования
по Джуксу (цит. по: [Ratner et al., 1996])
Ияпиянт кпля
Архетипический
Минимальный
Ранний
Эубактерии
Эукариоты (ядра)
Галобактерий
Метанококки
Митохондрии позвоночных и дру-
других форм
Хлоропласты растений
Число ами-
аминокислот
14-15
20
20
20
20
20
20
20
20
Число фрак-
фракций т-РНК
14-15
23
31
42*
44
45
25
22
31
Правила неоднознач-
неоднозначного спаривания
Расширенные
»
Правила Крика
„>
Расширенные
Смешанные
* У Е. coli 79 фракций т-РНК, имеющие 42 различных антикодона.
202 III. Молекулярная кибернетика
ным. Этим почти определено число 4. Набор аминокислот должен удовлетво-
удовлетворять принципу достаточного разнообразия свойств для формирования глобу-
глобулярных белков. Конкретный выбор аминокислот (из более чем 200 известных!)
тоже может быть отчасти случайным.
Однажды возникнув, генетический код, вероятно, был жестко стабилизи-
стабилизирован в ходе эволюции. Здесь можно привести такую лингвистическую анало-
аналогию. В языке различают символы алфавита и такие же символы текста. Выпаде-
Выпадение символа текста приводит к локальному дефекту одного слова и исправляет-
исправляется тоже локально. Выпадение символа из алфавита приводит к полному
разрушению всех текстов. Набор кодонов кодовой таблицы (см. табл. 20-1,
рис. 21-1, в) аналогичен "словарю", при помощи которого записаны генетиче-
генетические тексты. Выпадение любого кодона из кода (т.е. дефект т-РНК, АРС-азы
или других компонентов системы кодирования) вызывает множественные де-
дефекты в генетических текстах, которые заведомо летальны. Поэтому любой из
кодонов, многократно используемый в кодировании белков, не мог быть утра-
утрачен без гибели соответствующей клетки. Такие мутации сразу же отсекались бы
стабилизирующим отбором. Естественный отбор жестко стабилизировал не-
неубывание списка кодонов, но не препятствовал "захвату" новых нонсенсов в
процесс кодирования. Каким бы путем ни возникла структура кода в целом,
она тоже была стабилизирована, т.е. переадресовки и утраты кодонов практи-
практически были исключены.
Таким образом, к моменту полного формирования генетический код был
почти таким же, как сейчас. Изменения могли затронуть только редко исполь-
используемые кодоны и знаки пунктуации митохондриальных и других довольно ав-
автономных систем кодирования. В остальном стабилизация генетического кода
поддерживала его универсальность.
В итоге этого анализа можно заключить, что генетический код представля-
представляет собой высокоорганизованную систему, возникшую на основе определенных
эволюционных механизмов, а затем жестко стабилизированную и оптимизиро-
оптимизированную в ходе естественного отбора.
Список литературы
Ратнер В.А. Молекулярная генетика: принципы и механизмы. Новосибирск: Наука,
1983.
Ратнер В.А. Структура и эволюция генетического кода // Итоги науки и техники. Сер.
Молекулярная биология. М.: ВИНИТИ, 1985. Т. 21. С. 158-197.
Ратнер В.А. Генетический код как система// Соросовск. образоват. журн. 2000. Т. 6,
№ 3. С. 17-22.
Ratner V.A., Zharkikh A.A., Kolchanov N.A. et al. Molecular Evolution. Berlin e.a.:
Springer-Verl., 1996.
22. Генетический язык 203
22. Закономерности кодирования
генетической информации
(генетический язык)
В середине XX столетия великий физик-теоретик Э. Шредингер A943 г.)
(см. гл. 15), исходя из некоторых общих принципов физики и биологии, совер-
совершенно ясно сформулировал представление о кодировании генетической ин-
информации в структуре геномных макромолекул. Эта идея далее была развита
Дж. Уотсоном и Ф. Криком A953 г.), Г.А. Гамовым A954 г.) (см. гл. 16) и мно-
многими другими после открытия структуры и свойств нуклеиновых кислот и бел-
белков. В настоящее время концепция генетической информации и ее материаль-
материальных носителей является незыблемым краеугольным камнем генетики и биоло-
биологии в целом.
Сейчас, в начале нового века, мы уже частично знаем, как именно, по ка-
каким общим правилам генетическая информация записана в генах, регулятор-
ных сайтах и функциональных центрах макромолекул, в структурных элемен-
элементах геномов и как она воплощается в молекулярные функции и свойства. Это
очень сложная система, поэтому наши сведения о ней весьма неполны, но они
вселяют надежду на успех в понимании принципов кодирования и в их продук-
продуктивном использовании. Причем чем больше мы понимаем закономерности ко-
кодирования генетической информации, тем более поражает их сходство с прин-
принципами лингвистики человеческих и компьютерных языков.
Генетический язык: исходные понятия
Проблемы кодирования информации изучаются в информатике, лингвис-
лингвистике и других близких науках. Воспользуемся их опытом и терминологией для
описания нашей проблемы. Прежде всего, отвлечемся от физико-химической
природы биополимеров, будем обозначать их мономеры символами, а последо-
последовательности символов считать текстами. Определим исходные понятия и тер-
термины для дальнейшего.
Генетический язык — совокупность правил и закономерностей кодирова-
кодирования генетической информации (см. ниже) в генетических текстах. Эти правила
могут быть строгими, ограничительными или статистическими, но они сущест-
существенны для кодирования.
Генетический текст определяется как линейно упорядоченная последова-
последовательность символов мономеров непрерывной макромолекулы (ДНК, РНК или
белка), участвующей в кодировании генетической информации (рис. 22-1). Ге-
Генетические тексты являются конкретными реализациями генетического языка.
Генетическая информация определяется как совокупность молекулярных и
более сложных биологических свойств и функций, кодируемых в генетических
текстах.
Код генетической информации задается как соответствие между генетиче-
генетическими текстами и кодируемыми ими функциями или свойствами.
5'РО
38800 ,---
СЮ.' ИЕ GU GN AR IL ТН LU LV АР ТГ AL ME AR РК GV GN TH LY TH AL LY АР LU GV VA ТГ
TAAAGATCTCGCCGrATAT
TATAGATTGTGGCACGCACAACTGATAAAATGeAGACCGCCACTATTACCAACGTACATGATTCCTCCAACATACCTTGTTGCGrATTGGeACTTTCTAAlTACGTTACGCGAAACCCGTrTGGTTCrGTCGATrTCTAGAGCCGCArArA
•МЫ 'Belli
38950
GN SE AL IL AN LT AL IL Kt AL СГ AR LV IL PH LU TN IL AN AL АР СГ SE VA ТГ AL CU CU VA LT PR PH PR И AN LY LT TH TH AL
CAAAG CG CGATCAAC AAGGCC AT'
-I-
-I-
-I-
-I-
iCATTCCAGCC
GTTTCGCGCTAGTTGTTCCGeTAAGTACGTCCGGCTTTCTAAAAAAATTGATATTTGCGACTACCTTCGCAAATACeCCTTCTCCATTTCGGeAAeeGCTCATTGTTTTTTTeTTGTCGTATTTATTGGGGCGAGAATGTGTAAGGTCGS
•Nhal 'Mbol
Clli KE VA AR AL AN LY AR AN GU AL LU AR IL GU SE AL LU LU AN LY
39100 GACTTTTTCCCGTAGTTTAATTTGSTGTGGATACCACATACGTAAATAAACGTATGTAAGTTAGTTAACAATAGATTCCTTTATGAATGTATACCAACCACGTTTGTTTGCGTTGCTCCGAGATGCTTAGCTCTCACGCAACGAATTGTT
Taql
IL AL HE LU GY TH GU LY TH AL GU AL VA GY VA AP LY SE GN IL SE AR TR LY AR AP TR IL PR LY PK SE KE LU LU AL VA LU GU TR GY VA VA AP AP AP ME AL AR LU
AATCGCAATGCTTGGAACTGAGAAGACAGCGGAAGCTGTGGGCGTTGATAAGTCGCAGATCAGCAGGTGGAAGAGGGACTGGATTCCAAAGTTCTCAATGCTGCTTGCTGTTCTTGAATGGGGGSTCGTrGACGACGACATGSCTCGArr
TTAGCGTTACGAACCTTGACTCTTCTGTCGCCTTCGACACCCGCAACTATTCAGCGTCTAGTCGTCCACCTTCTCCCTGACCTAAGGTTTCAAGAGTTACeACGAACGACAAGAACTTACCCCCCAGCAACTGCTGCTGTACCGAGCrAA
•мьо! Та?'
AL AR GN VA AL AL IL LU TH AN LY LY AR PR AL AL TH GU AR SE GU GN IL GN KE GU PH 0; HE TH AN TH AL LY IL LU AN PH CY
Gr.CGCGACAAGTTGCTGCGATTCTCACCAATAAAAAACGCCCGGCGSCAACCGAGCGTTCTCAACAAATCCAGATGGAGTTCTGAGGTCATTACTGGATCTATCAACAGGAGTCATTATGACAAArACAGCAAAAATACTCAACTTCGGC
CCGCGCTGTTCAACGACGCTAAGAGTGGTTAT
'Hhal
39400
39550
@.?)
AR GY AN PH AL GY GN GU AR AN VA AL AP LU AP AP GY TY AL AR LU SE AN ИЕ LU LU GU AL TV SE GY AL AP LU TK LY AR GN PK LY VA LU LU AL IL LU AR LY TH TY
AGAGGTAACTTTGCCGGACAGGAGCGTAATGTGGCAGATCTCGATGATGGTTACGCCAGACrATCAAATATGCTGCTTGAGGCTTATTCGCGCGCAGArCTGACCAAGCGACAGTTTAAAGTGCrGCrTGCCATTCrGCGrAAAACCrAT
TcrCCATTGAAACGGCCTGTCCTCGCATTACACCGTCTAGAGCTACTACCAATGCGr-TCTCATAGTTTATACGACGAACTCCGAATAAGCCCGCGTCTAGACTGGTTCGCTGTCAAATTTCACGACGAACGGTAAGACGCArTTTf-r-ATA
•Hhall 'Belli Taql 'Hhal 'Belli
GY TR AM LY PR M AP AR IL TH AP SE GN LU SE GU IL TN LY LU PR VA LY AR CY AN GU AL LV LU GU LU VA AR HE AN IL IL LY GN GN GY GY ME PH GY PR All LY »N
39700 icCACCTTATTTGGTTACCTGTCTTAGTGGCTAAGAGTTGAATCGCTCTAATGTTTCAATPGACAGTTTGCCACGTTACTTCGGTTCAATCTTGAGCAGTcTTACTTATAATAGTTCRTCGTTCCGCCGTACAA/lCCTGGTrTATTTTTG
•Hinf 'Avail
39850
IL SE GU TR CY IL PR GN AN GU CY LY SE PR LY TH AR AP LY TH SE LU LY LU GY AP CY TY PR SE LY GN GY AP TH LY AP TH IL TH LY GU LY AR LY AP TY SE SE CU
GGTGCATCCCTOAAACGAGGGAAAATCCCCTAAAACGAGGGATAAAACATCCCTCAAATTGGGGGATTGC
¦I [ 1 | 1 1-Г 1 -—1
AAAAAAGAAAAGATTATTCGTCAGAG
I-
AN SE GY GU SE <? AP GN PR GU AN AP LU SE VA VA LY
40000 *ATrcTOOCO**TC<:TCTOACC*GCC*GA***c6*CCTTTCTGTGGTGA/\
TTAAGACCGCrrAGGAGACrGGrCGGTCnTrGCTGISAAAGACACCACm
I—199 1
I ?96 1
22. Генетический язык 205
Генетический язык реализуется в двух формах: полинуклеотидной
(НК-язык) и полипептидной (ПП-язык), которые тесно связаны между собой.
В НК-языке все тексты и знаки пунктуации и управления построены только из
символов базисного алфавита. Это аналогично тому, как кодируются знаки
препинания в телеграфном языке: ТЧК — точка, ЗПТ — запятая и т.д. В
ПП-языке нет знаков пунктуации, поскольку полипептиды не обладают мат-
матричными свойствами.
Существенно, что все эти понятия имеют смысл лишь по отношению к мо-
лекулярно-генетической системе управления клетки (МГСУ), в пределах которой
осуществляются разнообразные операции с генетической информацией.
Генетический язык, как и другие, задается следующими компонентами:
A) алфавитом — набором символов канонических мономеров или модулей.
Базовыми алфавитами являются символы 4 нуклеотидов (A, T(U), G и С) для
НК-текстов и 20 аминокислот — для ПП-текстов. Из символов базовых алфа-
алфавитов построены все модули текстов различных уровней;
B) грамматикой — набором правил формирования кодирующих последова-
последовательностей макромолекул (генетических текстов);
C) набором используемых в генетических текстах знаков пунктуации и
управления основных генетических процессов и т.д.;
D) семантикой — перечнем кодируемых элементарных молекулярных
свойств и функций, правил их комбинирования и соответствия кодирующим
последовательностям.
Разумеется, перечисленные компоненты генетического языка известны не
в полной мере, однако даже частичное знание позволяет говорить о генетиче-
генетическом языке в целом.
Грамматика и пунктуация
Правила грамматики генетического языка в целом определяют допустимые
способы построения кодирующих последовательностей, взаимного расположе-
расположения символов алфавита, модулей, знаков пунктуации и управления, повторов,
Рис. 22-1. Пример генетического текста, содержащего цистроны, знаки пунктуации и
другие элементы (фрагмент генома фага X). Приведены последовательности нуклеоти-
нуклеотидов обеих цепей ДНК, а также последовательности трех регуляторных белков: его, СП и
О. рг — промотор правоориентированной транскрипции оперона R]. Начало реплика-
репликации, ori, локализовано в пределах участков г93, г99 и г96 и находится внутри цистрона О,
контролирующего белок-инициатор репликации. Область вблизи ori содержит также
ряд инвертированных и прямых повторов, которые могут образовать причудливую вто-
вторичную структуру ДНК. Указаны также некоторые сайты рестрикции (*HhaI и др.).
Двухбуквенные обозначения аминокислот: AL — аланин; AN — аспарагин; АР — аспа-
рагиновая кислота; AR — аргинин; CY — цистеин; GN — глутамин; GU — глутаминовая
кислота; GY — глицин; HI — гистидин; IL — изолейцин; LU — лейцин; LY — лизин;
ME — метионин; РН — фенилаланин; PR — пролин; SE — серии; ТН — треонин; TR —
триптофан; TY — тирозин; VA — валин.
206 III. Молекулярная кибернетика
разделительных интервалов и других сегментов генетических текстов. Многие
из этих правил универсальны, т.е. используются в большинстве различных ге-
генетических текстов. Они тесно связаны также с правилами семантики (см.
ниже). В настоящее время можно выделить несколько групп правил граммати- I
ки генетического языка.
I. Правила генетического кода — свойства кодонов в НК-текстах (см. гл. 20).
Эти правила изучены наиболее подробно: триплетность, неперекрывание и од- I
нозначность кодонов, отсутствие межкодонных знаков, кодирующая роль от-
отдельных позиций в кодонах, наличие триплетных знаков пунктуации и т.д.
II. Правила выделения дискретных функциональных единиц в НК-текстах
(см. рис. 22-1): цистронов, скриптонов, репликонов, групп сцепления (см.
табл. 22-1 и пояснения к ней в тексте), а также экзонов, интронов, провирусов,
мобильных элементов, встроенных плазмид и др. Фактически они определяют-
определяются допустимыми вариантами взаимного расположения знаков пунктуации раз-
различных процессов синтеза, сайт-специфической рекомбинации, специфиче-
специфических повторов, разделительных интервалов (спейсеров) и т.д. Например, при
компьютерном анализе НК-текста всякий участок, ограниченный (слева на-
направо по направлению трансляции) инициирующим и терминальным знаками
AUG > UAA, обозначается как ORF (открытая рамка трансляции), т.е. воз-
возможный цистрон. На рис. 22-1 такими цистронами являются гены его, СП и О
фага^..
III. Правила формирования знаков пунктуации и других функциональных сай-
сайтов в НК-текстах. Роль сайта (т.е. протяженного и неразрывного функциональ-
функционального участка) может выполнять, по-видимому, любой сегмент полинуклеотида,
узнаваемый специфическим функциональным центром исполняющего или ре-
гуляторного белка или сайтом другого полинуклеотида. Важную роль может иг-
играть вторичная структура полинуклеотида ("шпильки" и др.), а также палин-
дромные свойства (симметричность к инверсии).
Например, типичный прокариотический промотор (начало транскрипции)
содержит сайты ТАТА-бокса (или блока Прибнова, блока (-10)) и блока (-35),
узнаваемые различными субъединицами фермента РНК-полимеразы. Эти сай-
сайты несимметричны и определяют одностороннее движение фермента. На
рис. 22-2 в регуляторной зоне frp-оперона Е. coli указаны эти сайты промотора
и направление транскрипции (слева направо). Там же приведен оператор — ре-
гуляторный сайт, узнаваемый репрессором транскрипции. Оператор имеет ось
симметрии и палиндромную структуру, узнаваемую симметричным же мульти-
мером репрессора. Он блокирует посадку и движение РНК-полимеразы в обо-
обоих направлениях. Наконец, в структуре м-РНК frp-оперона, вблизи начала
трансляции AUG, локализован несимметричный сайт Шайна—Дельгарно, узна-
узнаваемый З'-концом 16S-p-PHK рибосомы и связывающий рибосому. Этот сайт
необходим для инициации трансляции. Он тоже несимметричен и определяет
одностороннее движение рибосомы (см. рис. 22-2).
Из других функциональных сайтов управления транскрипцией у эукариот
отметим терминатор и сигнал полиаденилирования, завершающие транскрип-
транскрипцию, а также энхансеры, усиливающие инициацию, сайты ответа на тепловой
шок, действие гормонов, тканеспецифичных сигналов, тяжелых металлов и т.д.,
Промотор D3)
Оператор B0) Лидерный сайт связывания рибосомы на м-РНК C4)
S'-AATGAGCTGTTGACAATTAATCATCGAACTAGTXAACTAGTACGCAAGTTCACGTAAAAAGGGTATCGA-
-40 -30 -20 -10 +1 +10 +20
(блок - 35) (блок Прибнова) ¦
Лидерная м-РНК
i
-CAATGAAAGCAATTTTCGTACTGAAAGGTTGGTGGCGCACTTCCTGAAACGGGCAGTGTATTCACCATGCGTAAAGCAATCAGATACC-
III I I I I I I
+30 +40 +50 +60 +70 +80 +90 +100 +110
»
F-Met Lys Ala He Phe Val Leu Lys GLy Тф Тф Arg Thr Ser Nons - Лидерный полипептид Лидерная м-РНК
Зона аттенюатора C9)
i 1 Сайт связывания рибосомы на м-РНК frp-оперона C6)
IGC - богатый участок B0) lAT - богатый участок A9) J '
-CAGCCCGCCTAATGAGCGGGClTlTnriGAACAAAATTA'GAGAATAACAATGCAAACACAAAAACCGACTCTCGAACTGCTAACGTG-S'
I '- I ' 1 I I I I I
+120 +130 +140 +150 +160 +170 +180 +190 +200
>>>>» : >
Конец лидерной м-РНК м-РНК ftp-оперона
F-Met Gin Thr Gin Lys Pro Thr Leu Glu Leu Leu Thr Cys - Полипептид f/pE
Рис. 22-2. Регуляторная зона триптофанового (trp) оперона Е. coli. Указаны структура и локализация функциональных сайтов:
промотора, оператора, аттенюатора, двух сайтов связывания рибосом на м-РНК, инициации и терминации лидерного поли-
полипептида, инициации трансляции первого цистрона (trpE). Палиндромы выделены подчеркиванием.
208 III. Молекулярная кибернетика
подчиняющие транскрипцию внешним и внутренним специфическим сигна-
сигналам и воздействиям. Свои знаки пунктуации и управления имеют также цис-
троны, репликоны и другие функциональные единицы генома.
IV. Правила формирования регуляторных зон, контролирующих динамику
функционирования генов, их взаимодействие, ответ на специфические внеш-
внешние и внутренние сигналы, участие в дифференцировке, онтогенетическом раз-
развитии и т.д., а также репликацию и другие процессы. Отдельные сайты могут
опознаваться разными исполняющими и регуляторными белками, которые ак-
активируют или подавляют работу гена. Сайты могут перекрываться, иметь опре-
определенный порядок, регуляторные белки могут конкурировать за сайты, кон-
контактно взаимодействовать между собой и т.д. Иначе говоря, регуляторная зона
может содержать много сайтов, взаимодействующих со многими белками или
мультимерными их комплексами. Итогом взаимодействия является выработка
результирующего сигнала активации или подавления подчиненного гена. Фак-
Фактически регуляторная зона играет роль логического модуля со многими входами
(сайты) и одним выходом, контролирующим функцию гена. Сложность регуля-
торной зоны хорошо видна на рис. 22-2. У прокариот регуляторные зоны зани-
занимают сотни пар нуклеотидов, у эукариот — десятки килобаз (тысяч нуклеотид-
ных пар ).
V. Правила формирования пространственных структур белков, РНК, ДНК и
их комплексов. Это вторичные структуры пространственной упаковки белков
(ос-спирали, р-участки и р-листы и др.), гидрофобное ядро, функциональные
центры, домены, линкеры и т.д., а также правила формирования "шпилек" и
биспиральных участков РНК и ДНК и т.п. Существенно, что эти правила име-
имеют ярко выраженную семантическую, т.е. функциональную направленность.
В белках они основаны на кооперативном принципе формирования, в поли-
нуклеотидах основную роль играет принцип комплементарности.
На рис. 22-3 приведена пространственная структра фрагмента Клёнова —
бифункционального домена фермента ДНК-полимеразы I кишечной палочки,
где изображены различные элементы структуры. Очевидна высокая сложность
как пространственной конфигурации белка, так и предполагаемых путей его
сворачивания (самоорганизации), которые определяются первичной структу-
структурой (ПП-текстом), а также участием специальных вспомогательных белков
сворачивания — шаперонов.
Вероятно, в дальнейшем можно будет выделить и другие группы правил
грамматики и пунктуации генетического языка.
Семантика и синонимия
В лингвистике правила семантики описывают способы кодирования смыс-
смысла в текстах, комбинирования и согласования смысловых единиц, а также спо-
способы синонимических преобразований. В генетическом языке роль смысла иг-
играет кодируемая молекулярная функция или свойство. Закономерности семан-
семантики генетического языка тесно связаны с правилами его грамматики. Однако
они имеют и свои характерные свойства. Перечислим наиболее существенные
из них.
330
N
347
352 375
Рис. 22-3. Схематическое пред-
представление пространственной
структуры фрагмента Клёно-
ва — бифункционального доме-
домена фермента ДНК-полимеразы I
Е. coll
Изображена взаимная упаковка
элементов вторичной структу-
структуры: а-спирали — цилиндры;
р-слои — плоские стрелки; свя-
связывающие их линкеры — лома-
ломаные линии.
210 III. Молекулярная кибернетика
A). Семантика НК-формы генетического языка в значительной степени
базируется на принципе парной комплементарное™ нуклеотидов (A-T(U), G-C).
В частности, огромное число молекулярных функций и событий связано с ком-
плементарностью узнавания олиго- и полинуклеотидов и сводится к актам пар-
парной комплементарности: матричный принцип репликации, транскрипции и
трансляции; ключевые акты рекомбинации, репарации, хромосомных пере-
перестроек, транспозиции; образование вторичной и пространственной структуры
р-РНК, т-РНК, м-РНК, геномных РНК вирусов, рибозимов и т.д.; многие ре-
гуляторные события — инициация транскрипции и репликации и т.д.
B). Семантика ПП-формы генетического языка в целом основана на прин-
принципе кооперативности мономеров. Между мономерами полипептидов не обна-
обнаружено каких-либо предпочтительных специфических отношений, сравнимых
с комплементарностью нуклеотидов. Поэтому формирование функционально
значимых структур белка (вторичных и третичных структур, функциональных
центров, доменов и т.д.) является кооперативным эффектом многих мономеров
в зоне общей трансляции, фактически — в пределах доменов. Иначе говоря,
внутренние последовательности цистронов, кодирующих глобулярные белки,
имеют контекстно-зависимые закономерности записи их молекулярных функ-
функций и свойств.
C). Многие непосредственные функции макромолекул (катализ, узнава-
узнавание белок — белок, белок — полинуклеотид, белок — метаболит и др.) сосредо-
сосредоточены в относительно небольших функциональных сайтах полинуклеотидов и
центрах белков. Это знаки пунктуации и управления, сайты рестрикции, реком-
рекомбинации, сплайсинга и других процессов — в полинуклеотидах и центры ката-
катализа, специфического узнавания метаболитов и сайтов полинуклеотидов, цен-
центры контакта между белками, аллостерического регулирования, антигенные де-
детерминанты, лидерные и "якорные" участки и т.д. — в белках. Их функции
осуществляются либо при прямом контактном взаимодействии, либо через ма-
материального посредника. Без опосредующей структуры действие на расстоянии
невозможно.
D). Семантические отношения взаимной специфичности функциональ-
функциональных центров белков и сайтов полинуклеотидов определяются их геометриче-
геометрической дополнительностью. Они могут быть универсальными, множественными
или уникальными. Универсальный белок узнает синонимическое семейство
функциональных сайтов, присущих всем функциональным единицам опреде-
определенного уровня. Таковы инициирующие и терминальные знаки трансляции,
транскрипции, репликации, знаки общей рекомбинации и т.д. Множественны-
Множественными являются сайты ряда общегеномных систем, контролирующих многие (но
не все) функциональные единицы: энхансеры, сайты регуляции системы теп-
теплового шока, тканеспецифичные сайты, гормон-чувствительные сайты и др.
Уникальными являются сайты и центры многих автономных систем: операторы
оперонов прокариот, центры взаимного контакта белков и т.д.
E). К закономерностям семантики можно отнести групповые свойства ряда
типов функциональных центров белков: полость каталитических центров, "вы-
"выпуклость" антигенных детерминантов, наличие "цинковых пальцев" и других
22. Генетический язык 211
"торчащих" образований в центрах контакта регуляторных белков, узнающих
большую канавку спирали двухцепочечной ДНК, и т.д.
F). Важнейшие семантические закономерности генетического языка свя-
связаны с синонимией. Синонимия функциональных единиц генетического языка
эквивалентна их изофункциональности. В целом синонимическое разнообра-
разнообразие генетических текстов создается такой их изменчивостью, которая не нару-
нарушает правил образования грамматических и семантических структур. Иначе го-
говоря, правила грамматики и семантики генетического языка, обеспечивающие
кодирование функций, как бы дополняют правила синонимических преобразо-
преобразований. Некоторые участки текстов, не существенные для кодирования функ-
функций или утратившие эту способность, могут варьировать почти свободно. Тако-
Таковы, по-видимому, некоторые интроны эукариот, вырезаемые сегменты белков
(фибринопептиды и др.), линкеры, псевдогены и др.
G). Закономерности синонимии кодонов хорошо известны и сводятся к пра-
правилам систематической и несистематической вырожденности серий генетиче-
генетического кода (см. гл. 20). Синонимические замены нуклеотидов составляют ~ 23%
всех замен в генетическом коде и приблизительно 70 % замен в 3-х позициях
кодонов. Кроме того, к семантическим правилам можно отнести свойства связ-
связности, симметрии, регулярности по столбцам кода, а также связанные с ними
оптимальность и помехоустойчивость кода.
(8). Синонимия многих цистронов и белков чрезвычайно развита, что является
следствием большой длины цистронов и белков (~ 102-103 символов), не слиш-
слишком высокой их функциональной нагрузки и достаточно широкого определе-
определения синонимии. Синонимия белков почти никогда не бывает строгой, она все-
всегда лишь частичная. Согласно комбинаторным оценкам, число синонимиче-
синонимических вариантов для надсемейства белков-глобинов лежит в интервале
1012 < N < 1064. Это разнообразие столь велико, что несопоставимо с реальными
возможностями перебора этих вариантов. Поэтому в ходе эволюции была апро-
апробирована лишь мизерная часть этого синонимического разнообразия вариан-
вариантов, причем доминирующим процессом их изменения была необратимая ди-
дивергенция. Филогенетические деревья семейств и надсемейств макромолекул
отражают их синонимическую эволюцию (см. гл. 24).
(9). Семантическим правилом дивергентных синонимических преобразо-
преобразований, по-видимому, является сохранение функциональных инвариантов. В про-
простейших случаях это сохранение мотивов, необходимых для выполнения функ-
функций, в последовательностях полинуклеотидов и белков. Часто эти мотивы охва-
охватывают существенные функциональные сайты полинуклеотидов и центры
белков. В других случаях инвариантна не последовательность, а распределение
вдоль нее некоторых существенных молекулярных свойств: участков самоком-
плементарности в РНК, гидрофобности, гидрофильности и других свойств в
белках.
A0). Синонимические последовательности могут быть совершенно различ-
различны, во всяком случае, могут не проявить статистически значимой гомологии.
Они могут возникнуть как вследствие конвергенции при отборе (тогда их раз-
размеры невелики), так и после такой дивергенции, которая полностью стирает
черты сходства. Поэтому, в принципе, сходство последовательностей генов и
212
III. Молекулярная кибернетика
белков не необходимо и не достаточно для их синонимичности. Для не сильно
дивергировавших последовательностей сходство сохраняется с необходимо-
необходимостью, но оно не достаточно для синонимичности.
Разумеется, эти закономерности не исчерпывают проблемы семантики и
синонимии генетического языка.
Иерархическая структура (лингвистическая система)
генетического языка
Генетический язык и тексты построены по иерархическому принципу и
содержат не менее 6 уровней кодирующих последовательностей (табл. 22-1).
Каждый уровень имеет все атрибуты языка, а вся иерархия уровней представля-
представляет собой лингвистическую систему. В табл. 22-1 сопоставляются матричные
функциональные единицы в НК-текстах разных уровней, начиная с нижних, и
кодируемые ими функции, объекты или свойства. Функциональные единицы
описываются двумя характеристиками: внешней зависимостью (или независимо-
независимостью) кодирования от окружающего контекста и ролью внутреннего порядка моду-
модулей в кодировании. Функциям и свойствам тоже присущи две особенности:
внешняя автономность от других функций и внутренняя целостность. Кроме того,
названы генетические процессы, в которых указанные функциональные еди-
единицы фигурируют как целое, а также знаки пунктуации и управления и вспо-
вспомогательные знаки, характерные для этих процессов.
Иерархическая блочно-модульная структура (лингвистическая
Функциональные едини-
единицы в НК-текстах
1. Кодопы
2. Цистроиы
3. Скриптоиы
4. Репликоны
5. Группы сцепления
6. Геномы клеток
Внешняя контекстная за-
зависимость
Контекстно-независи-
Контекстно-независимы
Почти всегда контекст-
контекстно-независимы
В основном контекст-
контекстно-независимы
Контекстно-независи-
Контекстно-независимы
Независимы полно-
полностью
Независимы
Роль внутреннего порядка
в кодировании
Кодирует функции
Кодирует функции
Изредка кодирует
функции
Практически не коди-
кодирует функций
Практически не коди-
кодирует функций
Не кодирует функций
Генетический процесс
Трансляция-1
Трансляция-II
Транскрипция
Репликация
Мейотическая сегрега-
сегрегация
Митотическая сегрега-
сегрегация
22. Генетический язык
213
Кодоны — триплеты нуклеотидов, кодирующие включение определен-
определенных аминокислот в процессе трансляции; они имеют 4-буквенный алфавит
(A, T(U), G и С). Правила грамматики фактически соответствуют закономерно-
закономерностям неразрывности, триплетности, неперекрывания, однозначности кодонов в
НК-текстах (см. табл. 20-1). Знаки пунктуации, разделения и управления отсут-
отсутствуют (код без запятых), но правильность фазы трансляции определяется спе-
специальной "триплетной рамкой" антикодонов т-РНК. Семантика определяется
правилами соответствия кодонов и аминокислот, возникающими в процессе
трансляции-I, т.е. при посредничестве ферментов кодирования (амино-
ацил-т-РНК-синтетаз), а также закономерностями вырожденности и связности
(синонимия), регулярности по столбцам, симметричности и оптимальности
кода в целом.
Цистроны соответствуют структурным генам полинуклеотидов и кодируе-
кодируемым ими полипептидам. В НК-форме генетического языка алфавит представ-
представлен набором 61 кодона, из которых построены все цистроны, в ПП-форме —
набором 20 канонических аминокислот. Правила грамматики в НК-форме ге-
генетического языка определяют в основном выделение цистронов знаками
пунктуации, локализацию сайтов управления трансляцией и посттрансляцион-
посттрансляционной модификации, в ПП-форме — правила формирования пространственной
структуры белков: ос-спиралей, р-участков, "цинковых пальцев" и т.д., а также
общие свойства функциональных центров, линкеров, "якорных" участков, гид-
гидрофобного ядра, доменов и т.п. (см. рис. 22-3). Только в НК-форме существуют
Таблица 22-1
система) генетического языка
Знаки пунктуации и управ-
управления
Нуклеотиды-выделите-
ли рамки в т-РНК
Инициирующие и терми-
терминальные знаки транс-
трансляции, сайты Ш-Д
Промоторы, терминато-
терминаторы, операторы, энхан-
серы, сайты внешних
сигналов и др.
Репликатор, терминаль-
терминальные знаки управления
Центромеры, теломеры
и др.
Центриоли
Кодируемая функция, объ-
объект или свойство
Аминокислоты
Полипептиды-протоме-
ры ферментов и дру-
других белков
Автономные системы
контроля метаболизма
(опероны), предшест-
веники т-РНК, р-РНК,
м-РНК и др.
Единицы воспроизведе-
воспроизведения
Единица сегрегацион-
сегрегационной комбинаторики
Индивидуальность и раз-
развитие клетки организ-
организма
Внешняя автономность
функций
Автономны
Автономны, могут вхо-
входить в мультимеры
Автономны в управле-
управлении
Автономны
Автономны в мейоти-
ческой сегрегации
Полностью автоном-
автономны и независимы
Внутренняя целостность
функций
Целостны
Целостны, кооперати-
вны
Целостны в управле-
управлении
Целостны
Целостны в сегрега-
сегрегации
Целостны
214 III. Молекулярная кибернетика
знаки пунктуации и управления: инициирующие триплеты (AUG и др.), терми-
терминальные нонсенсы (UAA, UAG и UGA), сайт Шайна—Дельгарно (Ш—Д) и дру-
другие знаки, функционирующие в процессе трансляции-И. Семантика определя-
определяется правилами кооперативного формирования характерных свойств функцио-
функциональных центров (каталитических, специфических, регуляторных, контактных
и др.), антигенных детерминантов и т.д. Правила грамматики и семантики тес-
тесно связаны между собой. Кооперативный характер формирования функцио-
функциональных субструктур белков означает, что правила грамматики для них контек-
контекстно-зависимые.
Скриптоны существуют только в НК-форме генетического языка и отвеча-
отвечают единицам транскрипции. Модулями алфавита фактически являются цис-
троны, разделяющие их интервалы (спейсеры), экзоны и интроны, фланговые
участки, различные повторы и палиндромы и др. Правила грамматики в основ-
основном сводятся к правилам выделения и подразделения скриптонов знаками
пунктуации, формирования регуляторных зон, экзонов и интронов, а также
структурно-функциональных элементов РНК — биспиральных участков и их
комбинаций, которые кодируются инвертированными повторами и палиндро-
палиндромами. Знаки пунктуации — промоторы, терминаторы, границы экзонов и ин-
интронов, знаки сплайсинга и др. Очень развиты знаки управления транскрипци-
транскрипцией: операторы, сайты позитивного управления, энхансеры, сайты узнавания ре-
рецепторов действия гормонов, теплового шока, лекарств, тканеспецифичные
сайты и т.д. Вблизи скриптонов они образуют сложные регуляторные зоны (см.
рис. 22-2), выполняющие роль коммутаторов управления транскрипцией. Пра-
Правила семантики в основом связаны с формированием функциональных цен-
центров т-РНК, р-РНК, рибозимов, других фракций РНК, регуляторных зон
скриптонов и др.
Репликоны существуют только в НК-форме генетического языка и соответ-
соответствуют единицам репликации ДНК и РНК. Модулями алфавита фактически
являются скриптоны, их регуляторные зоны, спейсеры, псевдогены, зоны по-
повторов, провирусы, мобильные элементы и т.д. Правила грамматики, по-види-
по-видимому, малосущественны и сводятся к взаимному расположению и ориентации
знаков пунктуации (репликаторов, терминаторов) и сайтов действия вспомога-
вспомогательных ферментов репликации (нуклеаз, топоизомераз, гираз, праймаз и т.д.).
Сайты управления репликацией образуют регуляторные зоны вблизи реплика-
репликаторов. Правила семантики, вероятно, не превосходят правил формирования
этих зон. Провирусы, мобильные элементы, встроенные эписомные плазмиды
являются независимыми репликонами и имеют свои правила грамматики: по-
положение и ориентацию концевых повторов, репликаторов, управляющих эле-
элементов и др.
Аналогичные, но менее существенные соответствия есть для уровней групп
сцепления и геномов. В частности, роль знаков узнавания и ограничения в них
играют достаточно сложные модули — центро-, теломеры, центриоли, функ-
функционирующие в мейотической и митотической сегрегации.
Эта иерархическая лингвистическая система имеет некоторые общие свой-
свойства и закономерности. Прежде всего, главными и необходимыми атрибутами
выделения уровней являются соответствующие знаки пунктуации. Таковы три-
22. Генетический язык 215
плетная рамка антикодонов т-РНК, инициирующие и терминальные знаки
трансляции, транскрипции и репликации, ограничительные и опознаватель-
опознавательные знаки сегрегации. Будучи независимыми, именно они образуют иерархи-
иерархическую систему, выделяя матричные функциональные единицы генетических
НК-текстов и обеспечивая их дискретность. Роль остальных знаков вторична.
Приведенные уровни имеют универсальный характер, они распространены
у всех форм жизни на Земле — от вирусов до высших эукариот. Они соответст-
соответствуют взаимным отношениям универсальных и фундаментальных генетических
процессов синтеза и распределения кодирующих макромолекул, образующих
ядро молекулярно-генетической организации: репликации, транскрипции,
трансляции, сегрегации.
На нижних уровнях лингвистической системы генетического языка (кодо-
ны, цистроны, частично — скриптоны) молекулярные функции кодируются в
генетических текстах порядком входящих модулей, элементов. Значит, эти уров-
уровни генетического языка контекстно-зависимые в пределах соответствующих
функциональных единиц. На верхних уровнях (репликоны, группы сцепления,
геномы, частично скриптоны) порядок модулей не играет существенной функ-
функциональной роли, т.е. эти уровни генетического языка контекстно-свободные.
Между этими основными уровнями можно вьщелить промежуточные под-
подуровни (экзоны-интроны, модули репликонов, мобильные элементы, супер-
супергены, мультигенные семейства и т.д.), единицы которых не обязательно укла-
укладываются в общие свойства иерархии, т.к. не имеют универсальной пунктуа-
пунктуации. Они участвуют не в матричных процессах синтеза и распределения, а в
других процессах: рекомбинации, сплайсинге, транспозиции, дифференциров-
ке и т.д.
Молекулярные функции, кодируемые на нижних уровнях генетического
языка, достаточно просты, даже примитивны. Это либо отдельные биохимиче-
биохимические акты (катализ, опознание, регуляция, контакт и т.д.), либо последователь-
последовательности стандартных актов (матричный синтез, деградация, синапсис и т.д.).
По-видимому, это связано с ограниченным размером таких функциональных
единиц и способом кодирования. Более сложные молекулярные функции ко-
кодируются иными средствами — путем образования между кодирующими гене-
генетическими модулями смысловых связей, не зависящих от порядка в генетиче-
генетических текстах. Такие связи строятся на основе взаимной специфичности, уча-
участия белков и РНК, сайтов управления, элементов вторичной и
пространственной структуры, образования мультимерных комплексов, нали-
наличия мигрирующих посредников взаимодействия (т-РНК, м-РНК, регулятор-
ных белков, сигнальных метаболитов, гормонов и др.) и т.д.
Практически все атрибуты генетического языка имеют эволюционное про-
происхождение, но в их возникновении и дальнейшей эволюции существенную
роль играли отбор, случай и частичная предетерминация. Важно подчеркнуть,
что эволюция генетических текстов всех уровней идет в рамках заданных пра-
правил генетического языка. Такова, например, синонимическая эволюция цис-
тронов и белков, выражаемая филогенетическими деревьями (см. гл. 24). Пра-
Правила генетического языка тоже эволюционируют, но гораздо медленнее, чем
генетические тексты. Поэтому многие правила универсальны для всех форм
216 III. Молекулярная кибернетика
жизни на Земле. Так, генетический код (см. табл. 20-1) действительно практи-
практически универсален, причем несомненно является продуктом эволюции, где
можно видеть последствия влияния всех трех перечисленных выше факторов.
Генетический язык и другие языки
Генетический язык — это независимо возникшая лингвистическая систе-
система, имеющая свою предназначенность, появившаяся исторически и прошед-
прошедшая свой путь эволюции. По предназначению генетический язык является ти-
типичным техническим, машинным языком, на основе которого формируется
"элементная база" молекулярно-генетических систем управления клеток и осу-
осуществляется их функционирование. Поскольку генетический язык имеет неза-
независимое природное происхождение, его следует отнести к естественным язы-
языкам. Фактически это первый язык, возникший на Земле естественным путем и
дошедший до нас. Появление генетического языка было сопряжено с возник-
возникновением первых МГСУ клеток.
Сравнительный анализ генетического языка, естественных языков челове-
человека и языков программирования выявил их поразительное сходство. Естествен-
Естественные языки человека тоже имеют иерархическую структуру. Наиболее сущест-
существенны нижние уровни этих языков: всегда есть базовый алфавит — знаковая
система, уровень морфем — минимальных смысловых сегментов текста (корни,
приставки, суффиксы и т.д.), уровень слов, обозначающих множество исход-
исходных объектов, действий, понятий, свойств и т.д., и уровень предложений — ос-
основной комбинаторный уровень формирования значащих фрагментов текста.
Другие уровни имеют гораздо более слабые внутренние связи и значительно
менее целостны.
Наиболее близким аналогом морфемы в НК-языке, по-видимому, является
позиция нуклеотида в кодоне (см. табл. 20-1). Если обозначить первую, вторую
и третью позиции как х, у и z, то х напоминает приставку, у — корень, a z —
окончание. Это обусловлено их функциями в составе кодонов. Аналогом сло-
словоформы в НК-языке является кодон, аналогом предложения — цистрон.
В ПП-языке функциональные центры аналогичны частям предложения: ска-
сказуемому, дополнению и др. Аналоги подлежащего, однако, отсутствуют, так
как субъектами действия являются сами макромолекулы. Скриптоны скорее
напоминают абзацы текста, а репликоны определенно аналогичны единицам
воспроизведения — печатным листам. Геном клетки, фигурально говоря, ана-
аналогичен книге, содержащей все инструкции по жизнедеятельности, воспроиз-
воспроизведению и построению всех компонентов МГСУ и клетки. Общие иерархиче-
иерархические свойства генетического языка и естественных языков человека весьма
сходны. Можно говорить о единых принципах организации этих лингвистиче-
лингвистических систем.
Однако, в отличие от естественных языков, все "слова" генетического язы-
языка (кодоны) триплетны, число их невелико F1) и полностью определено.
"Предложения" (цистроны) очень велики и неравномерны, что порождает ог-
огромные синонимические семейства. Фактически наличие развитой синони-
синонимии — условие гибкости и выразительности естественного языка. В генетиче-
22. Генетический язык 217
ском языке эта "гибкость" имеет не индивидуальное, а эволюционное значе-
значение, поскольку отдельные МГСУ не порождают синонимических вариантов
или порождают их редко и случайно. В естественных языках соответствие
"Смысл <=> Текст" двустороннее, т.е. механизмы языка чрезвычайно быстро по-
порождают новые тексты, имеющие заданный смысл, и прочитывают уже готовые
тексты. В генетическом языке есть механизмы считывания информации и реа-
реализации ее смысла, но нет механизма записывания смысла в виде генетическо-
генетического текста. Запись генетической информации формируется эволюционно путем
мутаций, комбинаторной изменчивости и отбора вариантов.
В генетическом языке существенна роль инвертированных и палиндром-
ных повторов. В естественных языках это свойство находится на дальней пери-
периферии: языковые игры, кроссворды и т.п. В генетическом языке знаки пунктуа-
пунктуации построены из символов алфавита и образуют синонимические семейства.
Это указывает на их естественное происхождение. В естественных языках зна-
знаки пунктуации обособлены и жестко зафиксированы, что свидетельствует об их
искусственном происхождении. Список аналогий и, напротив, контрастных
свойств можно продолжить.
Можно подвергнуть такому же анализу структуру языков программирова-
программирования. Результаты будут весьма сходны.
Американский генетик С. Оно сопоставил кодоны ДНК с нотами музы-
музыкального алфавита. В этом случае при помощи компьютера удалось воспроиз-
воспроизвести длинные музыкальные тексты, отвечающие определенным генетическим
текстам. Любопытно, что полученная "генетическая музыка" звучит ничуть не
хуже, чем современные джазовые импровизации. Во всяком случае, повторы
генетического текста звучат как элементы мелодий. Интересно попробовать
обратное перекодирование: будут ли иметь какой-нибудь молекулярно-генети-
ческий смысл результаты перезаписи классических музыкальных мелодий и
произведений?
В целом становится ясно, что каждый язык соответствует определенному
типу систем управления:
генетический язык является языком воспроизведения, функционирования
и построения компонентов молекулярно-генетической системы управления
клетки;
естественные языки человека являются языками переработки информации
мозгом человека и общения между людьми;
компьютерные языки являются языками внутреннего функционирования
компьютеров и их общения с человеком.
Каждая из лингвистических систем продукт коэволюции с соответствую-
соответствующим типом систем управления. Наиболее древняя ко-система: генетический
язык — МГСУ. Она эволюционировала наиболее медленно, в течение несколь-
нескольких миллиардов лет. Наиболее быстро эволюционируют компьютерные языки
и отвечающие им системы компьютеров (несколько десятилетий). При этом
активно используются прообразы из естественных языков, а в перспективе, ве-
вероятно, могут быть использованы и закономерности генетического языка.
В ближайшей перспективе можно ожидать, что понимание принципов ге-
генетического языка станет совершенно необходимым условием при реализации
218 III. Молекулярная кибернетика
таких огромных и дорогостоящих современных проектов секвенирования гено-
геномов, как "Геном человека", "Геном Е. coli", "Геном дрозофилы" и др. В банках
данных число секвенированных последовательностей макромолекул накапли-
накапливается гораздо быстрее, чем происходит понимание того, что в них записано.
Таким образом, вопросы семантики генетического языка становятся весьма ак-
актуальными для осуществления этих крупных проектов.
В заключение благодарю И.А. Мельчука (Монреаль, Канада) за интерес к
проблеме и полезные дискуссии.
Список литературы
Льюин Б. Гены. М: Мир, 1987. 544 с.
Ратнер В.А. Молекулярно-генетические системы управления. Новосибирск: Наука, 1975.
287 с.
Ратнер В.А. Структура и эволюция генетического кода // Итоги науки и техники. Сер.
Молекулярная биология. М.: ВИНИТИ, 1985. Т. 21. С. 158-197.
Ратнер В.А. Концепция молекулярно-генетических систем управления. Новосибирск:
Изд-во НГУ, 1993. 118 с.
Ratner V.A. Progress in Theoretical Biology/ Eds. R. Rosen, F.M. Snell. N.Y.: Acad. Press,
1974. Vol. 3. P. 143-228.
Ratner V.A., Zharkikh A.A., Kolchanov N.A. et al. Molecular Evolution. Berlin e.a.:
Springer-Verl., 1996. 433 p.
23. Генетические переключатели
Введение
В классической генетике представление о взаимодействии генов и призна-
признаков выглядело весьма односторонне:
ген => признак,
т.е. предполагалось, что гены контролируют формирование признаков, а ка-
каких-либо обратных влияний не подразумевалось. Это представление базирова-
базировалось на сравнительном гибридологическом анализе альтернативных призна-
признаков, большинство из которых имело морфологическую природу (наличие раз-
различных морфологических особенностей и структур — форма, размер и т.д.), а
также окрасок, чувствительности к патогенам и воздействиям и др. Эти при-
признаки были онтогенетически устойчивы, необратимы и наследовались по мен-
делевским правилам. Внешние воздействия на эти признаки часто приводили к
их изменениям, которые, однако, никогда не наследовались, т.е. оценивались
как временные модификации. Следует учесть, что такое представление соот-
соответствовало уровню знаний и было важнейшим достижением классической ге-
генетики.
В дальнейшем это представление было пересмотрено, развито и усложне-
усложнено. В поле зрения молекулярных генетиков (цит. по: [Ратнер, 1975, 1983]) попа-
попали новые признаки — биохимические, физиологические, которые имели воз-
возрастную и временную динамику. Кроме того, прояснилась молекулярная при-
23. Генетические переключатели 219
рода генов и их ближайших продуктов — РНК и белков, выявилась
информационная природа их отношений. На раннем этапе развития молеку-
молекулярной генетики информационные отношения между основными фракциями
кодирующих макромолекул были выражены так называемой "центральной
догмой" Ф. Крика:
UHK (гены) => РНК=> Белки (ферменты) => Метаболиты
(молекулярные
признаки)
Здесь стрелки означают направления переноса генетической информации.
Следует подчеркнуть, что речь идет о потоках переноса так называемой струк-
структурной информации, которая записана в виде определенной последовательности
мономеров — нуклеотидов в ДНК и РНК, аминокислот в белках. Данная ин-
информация считывается в ходе матричного синтеза макромолекул. В соответст-
соответствии с этим представлением замены или другие структурные изменения мономе-
мономеров в ДНК могут осуществлять аллельные варианты генов, которые наследуют-
наследуются как моногибридные менделевские факторы. В свою очередь, многие белки
являются ферментами или выполняют другие молекулярные функции. Фер-
Ферменты контролируют пути метаболизма, морфогенеза и другие преобразова-
преобразования, а структурные белки вместе с другими макромолекулами формируют суб-
субклеточные и более сложные структуры.
Кроме того, обнаружено, что проявление генов в признаках этим не исчер-
исчерпывается. Одни и те же гены (точнее, их аллели) могут иметь различную функ-
функциональную динамику во времени и топографию в различных тканях. Напри-
Например, гены субъединиц дыхательного белка (гемоглобина) функционируют не во
всех клетках, а только в тех, которые выполняют дыхательную функцию — пе-
перенос кислорода. На разных этапах онтогенеза эти белки различны и сменяют
друг друга по определенной онтогенетической программе. Гены антител функ-
функционируют только в клетках иммунной системы, а в остальных клетках выклю-
выключены.
Таким образом, помимо наследуемого аллельного состояния, каждый ген
имеет еще одну функциональную характеристику — степень проявления, выра-
выражения, или экспрессии. На молекулярном уровне степень проявления гена мо-
может определяться интенсивностью синтеза тех же самых ближайших генных
продуктов — м-РНК и белков. Изменение степени проявления в самом про-
простом варианте может быть обозначено бинарным термином: включено — вы-
выключено. Такие изменения не затрагивают структуры генов, а являются чисто
регуляторными. Ясно, что по этой причине они не должны участвовать в мен-
делевском наследовании. Поток сигналов, вызывающих изменение степени
I экспрессии генов, можно считать потоком регуляторной информации.
В этом случае схема потоков структурной и регуляторной информации вы-
I глядит значительно сложнее, чем "центральная догма": между различными ге-
генами и их продуктами имеется целая сеть взаимодействий, прямых и обратных
220
III. Молекулярная кибернетика
связей, контролирующих динамику всех генов и формирование всех признаков
в ходе онтогенеза и физиологического функционирования.
Далее мы рассмотрим простейшие механизмы генетического управления
на уровне транскрипции, а также — варианты генов и генных систем с обрат-
обратными связями.
Опероны
Первая схема строения и функционирования оперона у бактерий — лактоз-
ного оперона (/ас-оперона) у кишечной палочки, Escherichia coli, — была пред-
предложена Ф. Жакобом и Ж. Моно [1964]. В более позднем варианте этот оперон
изображен на рис. 23-1, а структура его регуляторной зоны — на рис. 23-2 [Рат-
нер, 1983]. Транскрипция генов осуществляется в рамках функциональных еди-
единиц, называемых скриптонами. Скриптон ограничен знаками начала
(промотором—р) и конца (терминатором—f) транскрипции, а его продуктом яв-
является матричная РНК. Между этими знаками расположены гены, кодирую-
кодирующие белки, — структурные цистроны. В /ас-опероне их три: цистроны фермен-
ферментов р-галактозидазы (z), пермеазы (у) и трансацетилазы (а). Все эти ферменты
участвуют в катализе метаболического пути расщепления молочного сахара
лактозы и ее аналогов.
/ас-оперон
А,
Г Ч
Промотор
Сайт РНК -
Сайт полимеразы
БАК E1)
Регулятор A7)
Р i
Г
Оператор
1 C5)
1040
_L
Структурные цистроны
Z У
3510
1 780 | 825
Репрессор
ц-АМФ-БАК
РНК-полимераза
/\
Лактоза _
внутренняя"
Глюкоза
> +
Галактоза
Лактоза внешняя
Рис. 23-1. Схема строения и регуляторных отношений в /ас-опероне Е. coli [Ратнер,
1983]. Обозначения: z, У, а — структурные цистроны, контролирующие синтез фермен-
ферментов метаболического пути расщепления сахара лактозы; / — цистрон-регулятор, выра-
вырабатывающий белок-репрессор /ас-оперона; БАК — белок активации катаболитом — по-
позитивный регуляторный белок, участвующий в активации транскрипции /ас-оперона в
отсутствие глюкозы. Другие обозначения см. в тексте. Цифрами обозначено число нук-
леотидов в генетическом элементе.
Промотор F8)
Сайт связывания БАК A7) Сайт связывания РНК-полимеразы
i 11
|GC - богатый участок
S'-GAAAGCGGGCACTGAGCGCAACGCAATTAATGTGAGTTAGCTCACTCATTAGGCACCCCAGGC-
II III. I I
-60 -50 -40
-99
-90
-80 -70
>м-РНК регулятора lac i
-Glu Ser Gly Gin Term Белок-репрессор
Промотор
Сайт связывания РНК-полимеразы E1)
I AT - бог. участок IGC - бог. участок!
TTTACACTTTATGCTTCCGGCTCGTATGTTGTGTGGAATTGTGAGCGGATAACAATrTCACACAGGA-
i i i i i i I i »i- - 1 i
Участок связывания рибосомы на м-РНК
Оператор C5)
1
-30
(блок — 35) A2)
I I ' I
-20 -10 0
(блок Прибнова) G)
I
+10
1
+20
+30
• м-РНК /ас-оперона
Участок связывания рибосомы на м-РНК D0)
AACAGCTATGACCATGATTACCGATTCACTGC- 3'
+40
I
+50
+60
-> .м-РНК /ас-оперона
F - Met Thr Met He The Asp -
р-галактозидаза
Рис. 23-2. Регуляторная зона /ас-оперона
Е. coli [Ратнер, 1983]. Обозначения: пози-
позиция +1 — начало синтеза м-РНК, позиции
38-40 — триплет ATG — начало трансля-
трансляции цистрона z , позиция — 85 и левее —
триплет TGA — терминальный знак транс-
трансляции цистрона-регулятора /. Остальные
обозначения и пояснения см. в тексте.
222 III. Молекулярная кибернетика
Промотор является местом посадки фермента транскрипции РНК-полиме-
разы, а терминатор — местом ее освобождения. В /ас-опероне промотор подчи-
подчинен действию специального знака управления — оператора. Это функциональ-
функциональный сайт размером 35 н.п., узнаваемый регуляторным белком-репрессором.
Комплекс репрессора с оператором препятствует посадке молекулы РНК-по-
лимеразы на промотор и тем самым подавляет начало транскрипции. Репрес-
сор синтезируется под контролем другого цистрона-регулятора, который может
быть расположен в любом участке бактериального генома. Ген-регулятор (i)
/ас-оперона локализован смежно с ним (см. рис. 23-1).
Белок-репрессор способен также взаимодействовать с некоторыми метабо-
метаболитами и в результате изменять свою пространственную конформацию и ак- 1
тивность. Для /ос-оперона таким метаболитом является субстрат метаболиче- I
ского пути — лактоза и ее аналоги. В отсутствие лактозы репрессор активно по-
подавляет транскрипцию. В присутствии лактозы репрессор теряет репрессорную
активность, и транскрипция возобновляется. Регулирующий метаболит назы-
называется эффектором оперона.
Таким образом, /ас-оперон представляет собой управляемую единицу транс-
транскрипции и состоит из группы структурных цистронов (z, у, а), знаков пунктуа-
пунктуации транскрипции (промотора и терминатора) и группы знаков управления
(оператора и сайта позитивного управления БАК). Последний знак координи-
координирует работу /ос-оперона и других сахарных оперонов в зависимости от присут-
присутствия сахара глюкозы, /ос-оперон подчинен управляющему действию со сторо-
стороны /ос-репрессора и позитивного белка-регулятора БАК, которые, в свою оче-
очередь, чувствительны к присутствию метаболитов-сигналов, соответственно
лактозы и циклической АМФ (ц-АМФ).
Любопытно, что эффектором (сигналом переключения) /ос-оперона явля-
является субстрат ферментов этого оперона. В результате замыкается обратная связь
между концентрацией эффектора и активностью синтезируемых ферментов.
Такие опероны называются индуцируемыми, поскольку они начинают работать
в ответ на появление индуцирующего сигнала. Обычно индуцируемые оперо-
опероны — это системы синтеза катаболических ферментов, осуществляющих распад
более сложных веществ на простые. Например, дисахарид лактоза расщепляет-
расщепляется на два моносахарида — глюкозу и галактозу (см. рис. 23-1).
На рис. 23-3 приведена схема двух функциональных состояний индуцируе-
индуцируемого оперона. В отсутствие эффектора (о) оперон выключен, синтеза фермен-
ферментов нет. Появление эффектора-субстрата инактивирует репрессор и включает
синтез ферментов, перерабатывающих этот субстрат (б). Выработка ферментов
падает с уменьшением концентрации субстрата до полной его переработки.
С кибернетической точки зрения индуцируемый оперон является следя-
следящей системой, которая отслеживает и восстанавливает нулевую концентрацию
субстрата, т.е. работает до полного его исчезновения. Между концентрациями
субстрата и ферментов имеются положительная прямая и отрицательная обрат-
обратная связи:
Уровень ферментов
Уровень субстрата
23. Генетические переключатели
223
R
п
Р о
В t
R
В t
т т
Репрессор
менты
\
Апорепрессор N S
Эффектор
Рис. 23-3. Функциональные состояния индуцируемого оперона.
а — оперон выключен, б — включен. Состояние а устойчивое, б — неустойчивое. Обо-
Обозначения: R — ген-регулятор; А и В — структурные цистроны ферментов метаболиче-
метаболического пути; S — субстрат, Р— продукт; р — промотор, о — оператор, t — терминатор
транскрипции; апорепрессор — неактивный репрессор.
Такая схема имеет одно устойчивое состояние — выключенное. Внешним
воздействием (появлением субстрата) ее можно вывести из этого состояния
(т.е. включить), но с исчезновением субстрата она возвращается в выключен-
выключенное состояние.
Фактически организация индуцируемого оперона напоминает организа-
организацию дверного звонка. Звонок устойчиво выключен. Нажатие кнопки вызывает
другое состояние — звучащее, но оно поддерживается только до тех пор, пока
Вы нажимаете кнопку. Это состояние неустойчиво.
Известны также другие варианты оперонов. Например, анаболические опе-
роны, контролирующие синтез аминокислот и других сложных веществ из бо-
более простых, обычно отслеживают и поддерживают конечную концентрацию
метаболита-продукта. На рис. 23-4 показана схема организации триптофаново-
го оперона (frp-оперона) Е. coli, контролирующего 5 этапов синтеза аминокис-
аминокислоты триптофана. Его регуляторная зона содержит оператор и другие знаки
управления, а сигналом-эффектором служит конечный продукт — триптофан.
С кибернетической точки зрения между уровнями ферментов и продукта
имеются положительная прямая и отрицательная обратная связи:
Уровень ферментов
fl(-) (+)W
Уровень продукта
Такие опероны называют репрессируемыми, поскольку они снижают актив-
активность или выключаются при увеличении уровня эффектора-сигнала
(рис. 23-5). В отсутствие избытка продукта они обеспечивают постоянную вы-
выработку этого продукта, который потребляется на другие нужды (а). Например,
аминокислота триптофан используется в синтезе белков. Появление избытка
триптофана подавляет синтез ферментов оперона, а через них — синтез самого
триптофана (б). Возникновение недостатка триптофана усиливает синтез фер-
ферментов, а через них — синтез самого триптофана. Концентрация триптофана
колеблется вокруг устойчивого состояния, возвращаясь к нему после отклоне-
224
III. Молекулярная кибернетика
Гф-оперон
а
trpR
п
РD3) [
оB0) аГC9)
Е
D
В
1 1675 1675 1 1250
1400
800
Репрессия
Рис. 23-4. Схема строения и регуляторных отношений в Гф-опероне Е. coli [Ратнер,
1983]. Обозначения: А, В, С, D, Е — структурные цистроны, контролирующие ферменты
метаболического пути синтеза аминокислоты триптофана; trpR — цистрон регулятора;
at — аттенюатор транскрипции. Другие необходимые пояснения и термины см. в тексте.
б
R
Р о
R
Апорепрессор
Потребление
Репрессор
\
Избыток Р
(эффектор)
Г Т
Рис. 23-5. Функциональные состояния репрессируемого оперона.
а — оперон включен, б — выключен. Состояние а устойчиво. Обозначения такие же, как на
рис. 23-3.
ния. Иначе говоря, такие опероны тоже имеют одно устойчивое состояние —
включенное.
Техническим аналогом устройства мог бы быть турникет, где проход за-
закрыт, находясь под напряжением. Нажатием кнопки или опусканием жетона
можно выключить напряжение, открыть проход, но только на время прохожде-
прохождения жетона в системе. Затем закрытое состояние устойчиво восстанавливается.
23. Генетические переключатели
225
По данным полного секвенирования (см. гл. 12), геном Е. coli содержит
2584 оперона, т.е. фактически все метаболические пути этой бактерии (и мно-
многие другие процессы) контролируются оперонами. Это позволяет бактериаль-
бактериальной клетке адекватно и избирательно отвечать на колебания концентраций
множества метаболитов и других веществ, поддерживая их концентрации на
нужном уровне и экономя при этом синтез белков. Например, выключение
frp-оперона в питательной среде, содержащей триптофан, экономит 1.25 %
энергетического бюджета клетки и дает ей 10%-й выигрыш при отборе за счет
сокращения клеточного цикла.
Двухоперонный триггер
Выявив существование и особенности оперонов, Ф. Жакоб и Ж. Моно со-
совершили в генетике мини-революцию в представлениях о генах. Помимо
структурных генов, контролирующих ферменты метаболических путей, морфо-
морфологических признаков и т.д., они обнаружили существование регуляторных ге-
генов и белков, которые контролируют проявление других генов. Кроме того,
возникло представление о знаках пунктуации и управления, через которые дей-
действие ферментов и регуляторных белков вводится в локусы генома. Стало ясно,
что из структурных и регуляторных генов и знаков управления можно постро-
построить множество разнообразных генетических систем, способных выполнять
многие интересные молекулярные режимы функционирования. Наибольшее
внимание из них привлекла модель двухоперонного триггера [Моно, Жакоб,
1964; Ратнер, 1966, 1975].
Два индуцируемых оперона образуют циклическую систему, взаимно по-
подавляя друг друга при помощи синтеза регуляторных белков-репрессоров
(рис. 23-6). Гены-регуляторы входят в состав противоположных оперонов. Лег-
Легко сообразить, что оба оперона не могут работать одновременно, такое состоя-
состояние системы неустойчиво. Моделирование триггера различными математиче-
математическими методами показало, что при определенных условиях он имеет два устой-
устойчивых состояния:
1) включен оперон 1, выключен оперон 2;
2) включен оперон 2, выключен оперон 1.
Каждый репрессор чувствителен к действию некоторого метаболита-эф-
метаболита-эффектора (внешнего сигнала) и в его
присутствии теряет репрессорную ак-
активность. Появление сигнала, воспри-
воспринимаемого действующим репрессо-
ром, переключает триггер из одного
состояния в другое. Существенно, что
сигнал нужен только на время перехода,
а после этого может исчезнуть. Оба
состояния триггера устойчивы без
участия сигналов.
Триггер по-английски — переклю-
переключатель. По этому принципу устроен
Оперон 1
^Репрессор 2
Репрессор 1
Оперон 2
Метаболит 2
Метаболит 1
Рис. 23-6. Схема двухоперонного триггера.
Пояснения см. в тексте.
226
III. Молекулярная кибернетика
электрический переключатель в Вашей комнате. Он имеет два устойчивых со-
состояния: включено и выключено. Нажатием кнопки или тумблера Вы переводи-
переводите его из одного состояния в другое, а потом идете по своим делам.
В 1966 г. автор предложил использовать модель двухоперонного триггера для
описания генетического управления репликацией фага X [Ратнер, 1966]. Этот фаг
имеет два альтернативных режима развития в бактериальной клетке Е. соН: лити-
ческий и лизогенный. При литическом режиме происходит многократная репли-
репликация ДНК фага, а его развитие завершается лизисом бактериальной клетки и
выходом полноценного фагового потомства. При лизогенном режиме реплика-
репликация ДНК фага прекращается, профаг X внедряется в хромосому бактерии, кото-
которая становится лизогенной. Оба режима альтернативны (т.е. несовместимы в од-
одной клетке) и устойчивы. Лизогенное состояние можно прервать слабыми доза-
дозами ультрафиолетового облучения, а литический режим прервать нельзя.
Гипотеза состояла в том, что два альтернативных режима развития фага I
отвечают двум устойчивым состояниям двухоперонного триггера, контроли-
контролирующего инициацию репликации ДНК фага [Ратнер, 1966]. Гипотеза и триггер-
ная модель контроля репликации были подтверждены уже к началу 70-х годов
На рис. 23-7 показана схема взаимодействия двух оперонов фага X, образующи)
реальный триггер, контролирующий синтез регуляторных белков-инициаторо!
репликации фага X [Ратнер, 1975].
В последующие годы усилиями многих генетиков были выявлены реаль
ные молекулярные механизмы организации двухоперонного триггера у фага 3
[Пташне, 1988]. На рис. 23-7 и 23-8 показаны многие детали организации регу
ляторной зоны двух взаимодействующих оперонов. Оперон R, содержит регу
ляторный ген его, контролирующий регуляторный белок-репрессор второй
оперона L,. Кроме того, в R, входят гены О и Р, контролирующие одноименные
регуляторные белки-инициаторы реп
ликации фага. Оперон L, содержит ре
гуляторный ген С,, контролирующи1
синтез одноименного белка-репрессо
ра фага X, подавляющего противопо
*сгехС1°сРРцм
pRoRcro...O
Рис. 23-7. Двухоперонный триггер в гено-
геноме фага X. Обозначения: L/ — левоориен-
левоориентированный, R] — правоориентирован-
правоориентированный оперон; ori — начало репликации;
О и Р — регуляторные цистроны бел-
белков-инициаторов репликации; Q и его —
репрессоры, входящие в триггер; рцм —
левый, pR — правый промотор; ол — сай-
сайты общего оператора; tc — терминатор
транскрипции оперона Lj.
ложный оперон R,. Опероны имею
противоположную ориентацию и об
щую регуляторную зону, где находятс
два противоположно ориентирован
ных промотора (pRM — левый ирк-
правый) и общий оператор oR с трем
сайтами узнавания репрессоров С,
его (см. рис. 23-8, а). Сайт oRl пере
крывается с правым промотором р,
сайт ою — с левым промотором рш,
промежуточный сайт о^ частично пе
рекрывается с обоими промоторам*
Все три сайта сходны, но не одинакс
вы и имеют равный размер — п
17 н.п.
Рим
17 н. п.
Pr
сга-транскрипт
Выкл.
Вкл.
Рис. 23-8. Молекулярные механизмы функционирования общей регуляторной зоны
двухоперонного триггера фага X [Пташне, 1988].
а — топография функциональных сайтов, р^м — левый, рц — правый промотор, or
сайты общего оператора;
б — 1-е функциональное состояние триггера — репрессор С; связан с сайтами oRI и о^
РНК-полимераза инициирует левый процесс транскрипции; оперон L/ включен, R/
выключен;
в — 2-е функциональное состояние триггера — репрессор его связан с сайтом o^
РНК-полимераза инициирует правый процесс транскрипции; оперон Л7 включен, L,
выключен. Дальнейшие пояснения см. в тексте.
23. Генетические переключатели 227
У сайтов оператора различное сродство к молекулам репрессора С1 и репрес-
сора его: сродство к репрессору С, максимально на oR] (правый сайт) и уменьша-
уменьшается влево; сродство к репрессору его максимально на ою (левый сайт) и падает
вправо. Поэтому при избытке репрессора С, он связывается сначала с oR] и пре-
прерывает правую транскрипцию в опероне R, (см. рис. 23-8, б). При избытке ре-
репрессора его он связывается сначала с ою и прерывает левую транскрипцию в
опероне L, (см. рис. 23-8, в). В действительности процесс более сложен, но при-
применительно к триггеру его итоги укладываются в два обозначенных результата.
Таким образом, двухоперонный триггер реально существует и выполняет у
фага X (и других умеренных фагов) ключевую роль переключателя между двумя
основными режимами развития: литическим и лизогенным. Обратим внимание
на некоторые особенности организации двухоперонного триггера, которые вы-
выделяют его среди других оперонных систем.
Прежде всего, функциональные состояния триггера не просто устойчивы в
течение клеточного цикла хозяина, но и способны наследоваться по клеточной
линии сколь угодно долго. Действительно, лизогенное состояние профага X в
бактериальной клетке устойчиво наследуется, а репликация генома X в литиче-
ском режиме тоже повторяется много раз. Механизм наследования функцио-
функционального состояния совершенно иной, чем при наследовании аллельного со-
состояния, которое наследуется путем репликации и сегрегации ДНК генома.
Функциональное состояние триггера наследуется помимо генома, за счет рас-
распределения между дочерними клетками молекул белка-репрессора, поддержи-
поддерживающего устойчивое функциональное состояние. Например, концентрация
белка-репрессора С, профага X в лизогенном состоянии приблизительно 100
активных молекул на клетку бактерии. При делении клетки эти молекулы рас-
распределяются случайно. Поэтому вероятность того, что одна из дочерних клеток
не получит ни одной молекулы репрессора, ничтожна. Лизогенное состояние
устойчиво наследуется.
Таким образом, структурные гены генома, кодирующие генетическую ин-
информацию при помощи порядка нуклеотидов, образуют структурную память ге-
генома. Она наследуется путем репликации ДНК генома. Циклические триггер-
ные системы генов можно рассматривать как ячейки динамической памяти, в
которых циркулирует сигнал, поддерживающий функциональное состояние.
Эти состояния наследуются внегеномно.
Есть еще одна поучительная особенность двухоперонного триггера. Оперо-
ны, составляющие триггер, являются менделевскими генами, подчиненными
управлению. Каждый из них в отдельности удовлетворяет классическим менде-
левским законам наследования, поскольку имеет только одно устойчивое
функциональное состояние. Например, выполняется закон чистоты гамет. Но
циклическая система — двухоперонный триггер — уже не подчиняется менде-
левским законам наследования. Это становится понятным, если выполнить
следующий мысленный эксперимент.
Образуем диплоидную клетку, где в каждом гаплоидном геноме имеется
триггер, а функциональные состояния двух триггеров различны (включены
разные опероны). Клетка имеет общий пул синтезируемых репрессоров. Со-
Состояние, когда разные триггеры синтезируют разные репрессоры, неустойчиво,
228 III. Молекулярная кибернетика
поскольку состояния альтернативны, несовместимы. Исходом будет унифика-
унификация состояний двух триггеров. Одно из состояний будет обязательно утеряно.
Этот результат противоречит менделевскому принципу чистоты гамет, когда ал-
лельные гены, побывав в одной зиготе, не смешиваются и не теряются, а расхо-
расходятся при сегрегации такими же, какими они пришли в клетку.
Итак, двухоперонный триггер — это неменделевская система, построенная
из менделевских генов. Поэтому можно предположить, что в сложных молеку-
лярно-генетических системах клеток, где возможны такие и более сложные
циклические системы, реальны различные неменделевские эффекты и прояв-
проявления свойств. Исследователь должен быть к этому готов. Впрочем, следует от-
отметить, что среди изученных оперонных систем такие примеры попадаются до-
довольно редко. Большинство оперонов контролирует метаболизм и ведет себя
как автономные следящие системы управления.
Неменделевское наследование функциональных состояний генов очень су-
существенно в ходе онтогенеза. Дифференцированные клетки во многих случаях
наследуют свое необратимо дифференцированное состояние по клеточной ли-
линии в течение многих клеточных поколений.
В последние годы в молекулярной биологии большие успехи достигнуты
путем применения технологии ДНК-чипов. Специальная панель, содержащая
тысячи ячеек (чипов) для клонированных фрагментов ДНК отдельных генов, |
гибридизуется с меченой валовой РНК клетки. Все присутствующие фракции
РНК находят свои комплементарные фракции генов и метят соответствующие
ячейки. По общей картине связывания можно судить об активности транс-
транскрипции генома в целом и всех его подсистем, вплоть до отдельных оперонов.
Этим путем были проанализированы РНК пекарских дрожжей [De Risi et al.,
1997] и других объектов, выявлены координированные системы генов. Однако
открывшиеся новые возможности не умаляют автономности работы оперонов,
отслеживающих продуктивность отдельных путей метаболизма. Точно так же,
как сведения о глобальной загрузке электрических сетей не умаляют для Вас
значимости включателя лампочки в Вашей комнате.
Список литературы
Жакоб Ф., Моно Ж. Регуляция активности генов // Регуляторные механизмы клетки.
М: Мир, 1964. С. 278-304.
Моно Ж., Жакоб Ф. Общие выводы: телеономические механизмы в процессах клеточно-
клеточного обмена, роста и дифференцировки // Регуляторные механизмы клетки. М.: Мир, 1964.
С. 477-497.
Пташне М. Переключение генов. Регуляция генной активности и фаг \. М.: Мир, 1988.
Ратнер В.А. Генетические управляющие системы. Новосибирск: Наука, 1966. Гл. 3.
С. 91-92.
Ратнер В.А. Молекулярно-генетические системы управления. Новосибирск: Наука, 1975.
Ратнер В.А. Молекулярная генетика: принципы и механизмы. Новосибирск: Наука,
1983.
De Risi J.L., Iyer V.R., Brown P.O. Exploring the metabolic and genetic control of gene
expression on a genomic scale // Science. 1997. Vol. 278. P. 680-686.
24. Молекулярная эволюция 229
24. Молекулярная эволюция*
Теория эволюции является одним из краеугольных камней биологии и ес-
естествознания в целом. В последние годы теория эволюции испытывала уско-
ускоренное изменение и развитие в связи с появлением в естественно-научной
практике нового мощного пласта экспериментальных данных и теоретических
методов. В поле зрения теории эволюции попали кодирующие макромолеку-
макромолекулы — ДНК, РНК и белки, составляющие костяк молекулярно-генетической
организации клеток и организмов. Теперь уже широко известна роль ДНК и
РНК как материальных носителей генов, м-РНК как переносчика генетиче-
генетической информации от генов к белкам и белков как исполнителей самых разно-
разнообразных биологических функций: ферментов — катализаторов процессов об-
обмена, антител — иммунологических защитников организма, структурных эле-
элементов, регуляторных и транспортных элементов и т.д.
Первые последовательности аминокислот в белках были расшифрованы
(секвенированы) в 1950-е годы, нуклеотидов в генах и РНК — в 1960-е, но мас-
массовое секвенирование началось только в 1977-1978 гг., став к настоящему вре-
времени рутинной технологической операцией. Результаты секвенирования с тех
пор накапливаются в компьютерных банках данных. К августу 2001 г. крупней-
крупнейший международный банк данных (GenBank) содержал сведения примерно о
13.5 млрд нуклеотидов из 12.8 млн секвенированных последовательностей, при-
причем эта информация очень быстро нарастает.
Легко понять, что расшифровка нуклеотидных последовательностей ге-
генов (ДНК), РНК и аминокислотных последовательностей белков создает но-
новую ситуацию, позволяющую взглянуть на процесс эволюции с молекулярно-
молекулярного уровня организации жизни. Действительно, в классической генетике алле-
аллели генов принято обозначать однородными буквами или индексами: В и b, A1
и А2, и т.д. Если в процессе эволюции один аллель вытесняет другой (В => Ь и
т.д.), то кажется, что происходит как бы утрата предыдущего аллеля (В) и рас-
распространение нового (Ь). В то же время, в молекулярной генетике известно,
что многие мутации состоят в замене лишь одного или нескольких нуклеоти-
нуклеотидов:
В =* b
TTCGAGGCTAG => TTCGaGGCTAG
Поэтому замена аллеля сводится фактически к замене и фиксации единствен-
единственного нуклеотида, а остальная, большая часть гена остается неизменной. Сход-
Сходство последовательностей аллельных генов (или белков) остается максималь-
максимальным. Чем больше накапливается различий, тем меньше сходство последова-
последовательностей. Чем раньше дивергировали два гена, тем больше фиксированных
различий они накопят.
Таким образом, сходство последовательностей макромолекул можно поло-
положить в основу молекулярно-филогенетических построений. Число различий
* Сокращенный вариант этой главы опубликован ранее: Ратнер В.А. Молекулярная эволю-
эволюция // Соросовск. образоват. журн. 1998. № 3. С. 41-47.
230 III. Молекулярная кибернетика
мономеров (или близкая к нему величина) может быть мерой эволюционной
дивергенции. Обилие секвенированных последовательностей в банках данных
делает применимость этого подхода практически неограниченной.
В течение последних 30 лет было разработано несколько десятков компью-
компьютерных методов и пакетов программ для построения филогенетических деревь-
деревьев макромолекул и филогенетического анализа. При их помощи построены
многие сотни филогенетических деревьев макромолекул. В результате их ана-
анализа получены многочисленные интереснейшие данные, причем некоторые из
них имели фундаментальное значение. Многие макромолекулы эволюциони-
эволюционировали гораздо медленнее, чем морфологические признаки живых форм, по-
поэтому их филогенетический анализ позволяет заглянуть в очень ранние перио-
периоды эволюционного процесса (до 100-1000 млн лет тому назад).
На этом базируется развитие Теории молекулярной эволюции. Здесь мы не
будем описывать ее на профессиональном уровне, этому посвящены другие из-
издания, а продемонстрируем, так сказать, "научный продукт" этой науки — фи-
филогенетические деревья, и покажем, к каким любопытным выводам приводят
их анализ и интерпретация. Ряд приводимых результатов получен автором и его
сотрудниками.
Кодирующие макромолекулы эволюционируют с разными скоростями.
Наиболее консервативными обычно являются гены и белки некоторых очень
глубоких и рано возникших генетических процессов, которые сильно функ-
функционально нагружены и представлены у многих (или даже у всех) форм жизни.
Таковы, например, гены рибосомных РНК (входящих в молекулярный меха-
механизм синтеза белка), некоторых гистонов — белков компактизации ДНК хро-
хромосом, и др. Менее консервативны гены и белки систем, которые встречаются
у определенных широких групп видов (например, глобины животных). Нако-
Наконец, наиболее изменчивыми являются гены и белки РНК-содержащих вирусов,
которые стремительно изменяются в борьбе с иммунной системой их хозяев
(вирус гриппа, ВИЧ (HFV), онкогенные ретровирусы и др.).
Примером "глобального" дерева, охватывающего все крупнейшие таксоны
биологического мира, является дерево l6S-18S-pn6ocoMHbix РНК, построенное
американцами К. Вуузом и сотрудниками (рис. 24-1). Наиболее неожиданным
свойством этого дерева является существование в нем трех крупнейших ветвей,
объединяющих ЭУБАКТЕРИИ (обычные бактерии), АРХЕБАКТЕРИИ (осо-
(особая экзотическая группа бактерий) и ЭУКАРИОТЫ (высшие ядерные организ-
организмы). До этой работы считалось, что бактерии составляют единое надцарство,
Prokariota, отделенное от второго надцарства, Eukariota, существенной дистан-
дистанцией. В дереве Вууза все три глобальные ветви примерно равно удалены друг от
друга. Поэтому архебактерий следует считать самостоятельным надцарством.
Таким образом, Вууз ввел три "НАДЦАРСТВА", или "ДОМЕНА": Bacteria,
Archaea, Eucarya. Некоторые исследователи предлагали также для них термин
"ИМПЕРИИ".
Этот важный результат сразу привлек внимание к изучению архебактерий,
причем вскоре было показано, что по многим другим свойствам они действи-
действительно существенно удалены как от эукариот, так и от эубактерий. Многие ар-
архебактерий существуют в природе в весьма экзотических условиях: при высо-
ARCHAEBACTERIA
Sulfolobus solfatancus
Thermoproteus tenax
Homo sapiens
Xenopus laevis
Zeamays
EUKARYOTES
^alococcus morrhua
/Halobacterium volcanii
Methanospirillum hungatei
Methanobacterium formicicum
Methanococcus vannielii
Pseudomonas testosteroni
Escherichia coli
EUBACTERIA
Рис. 24-1. "Глобальное» филогенетическое дерево надсемейства 16S-18S рибосомаль-
ных РНК, построенное К. Вуузом и сотр. [Ratner et al., 1996].
¦ l
А2
69-7:
101-106.
С5
а-Цепи
Р-Цепи гемоглобинов
Миоглобины Эритрокруорины
хирономуса
Леггемо-
глобины
Рис. 24-2. Филогенетическое дерево надсемейства глобинов, построенное А.А. Жарких
для 40 удаленных последовательностей белков [Ратнер и др., 1985; Ratner et al., 1996].
1 — фиксированные делении, 2 — инсерции. Цифры указывают на их локализацию
внутри белка.
Not for sale! For preview only! Scanned by A fykhaylo (molbioKcvukr.net)
24
22
20
18
16
14
12
10
8
6
4
2
0
900
Скорость фиксации замен, xlO9 на позицию, в год
Л
К
А
i
1
V
V
\
л
¦
¦
.-•
1—»
• 7
• 2
• 3
700
500 300
Т, млн лет назад
Рис. 24-3. Адаптивное ускорение фиксации замен аминокислот в гемоглобинах в эпоху
выхода позвоночных животных на сушу D00-500 млн лет назад) [Ratner et al., 1996].
Скорость эволюции: Z — в среднем по белку; 2 — в центре контакта оц-Рг; 3 — в регуля-
торном центре связывания ДФГ.
1920 г
1930
1940
1950
1960
1970
1980
Рис. 24-4. Филогенетическое дерево генов
гемагглютинина НЗ вируса гриппа типа А,
построенное Л.В. Омельянчуком и др.
[Ratner et al., 1996]. "Висячие" вершины и
ведущие к ним ребра отвечают эпидеми-
эпидемическим штаммам A, 2, 3, 4, 5, 10). Индек-
Индексы штаммов указывают на место и год вы-
выделения. Внутренние вершины и ребра де-
дерева отвечают неэпидемическим стадиям
циркуляции вируса ("резервуар"). Вблизи
ребер указаны числа синонимических/не-
синонимических/несинонимических замен нуклеотидов.
24. Молекулярная эволюция 231
кой температуре вблизи подводных вулканов; в среде, насыщенной метаном,
соединениями серы и т.д. Время разделения надцарств Bacteria и Archaea оце-
оценивается примерно в 3500 млн лет назад.
Дерево Вууза включает в себя фракции 168-подобных р-РНК, кодируемые
не только ядерными генами, но и генами клеточных органелл — митохондрий и
хлоропластов. Легко заметить, что ядерная фракция р-РНК кукурузы (Zea
mays) попадает в ветвь эукариот, как и ожидается, а фракции из митохондрий и
хлоропластов этой же кукурузы (Zea mays mitochondrion, chloroplast) — в ветвь
эубактерий. Этот факт считается самым веским аргументом в пользу "симбио-
тической" гипотезы эволюционного возникновения эукариот, согласно кото-
которой митохондрии происходят от симбиотических предковых пурпурных бакте-
бактерий, а хлоропласты — от цианобактерий (синезеленых водорослей), а вовсе не
от ядерных структур эукариотических клеток.
Белки глобины (гемоглобины крови, миоглобины тканей) и их гены рас-
распространены в живой природе не столь широко. Они найдены в основном у
животных, некоторые удаленные их представители (леггемоглобины) найдены
также у растений, а у прокариот они пока не обнаружены. Поэтому глобины
образуют молекулярное надсемейство, охватывающее лишь часть таксономиче-
таксономического дерева жизни. Деревья эволюции глобинов и их генов были построены
многими исследователями. На рис. 24-2 приведено филогенетическое дерево
глобинов, построенное А. Жарких в нашей лаборатории. В целом оно достаточ-
достаточно хорошо соответствует принятой таксономии животных.
Из независимых палеонтологических данных можно почерпнуть датировки
эпох существования общих предков многих ныне живущих биологических
форм. Так, в дереве глобинов датированы 6 точек ветвления. Поскольку методы
реконструкции деревьев позволяют определить длины ребер дерева (числа
фиксированных замен, мутаций) на различных маршрутах, с учетом датировок
это дает возможность подсчитать скорости эволюции на разных этапах и в раз-
разных ветвях дерева. Нами были получены очень любопытные оценки
(рис. 24-3).
Во-первых, оказалось, что средняя скорость эволюции по всему белку была
не постоянна во времени, а имела максимум примерно 400-500 млн лет назад,
в эпоху выхода позвоночных животных из мирового океана на сушу (рис. 24-3,
кривая 7). Во-вторых, эта особенность выявилась еще ярче для тех участков
глобинов, которые отвечают за образование их четвертичной (мультимерной,
агрегатной) структуры. Дело в том, что миоглобины и гемоглобины некоторых
примитивных животных до сих пор представляют собой протомеры (отдельные
субъединицы белка). Круглоротые рыбы (минога и др.) имеют димерные агре-
агрегаты гемоглобинов, а большинство других позвоночных — тетрамерные агрега-
агрегаты, состоящие из двух а- и двух р-субъединиц (а2р2). Субъединицы соединяют-
соединяются при помощи так называемых "ЦЕНТРОВ КОНТАКТА", которые обознача-
обозначаются а.]—р,, а,-р2 и т.д. Для современных гемоглобинов известны амино-
аминокислоты, входящие в центры контакта. Поэтому можно подсчитать скорости
эволюции непосредственно для центров контакта. Максимум скорости выяв-
выявляется особо четко для центра контакта а,-р2 (рис. 24-3, кривая 2) и регулятор-
ного центра связывания дифосфоглицерата (ДФГ) (рис. 24-3, кривая 3). Таким
232 III. Молекулярная кибернетика
образом, при выходе позвоночных животных на сушу их гемоглобин приобрел
тетрамерную структуру, когда разные субъединицы (а и р) связаны между со-
собой центрами контакта а,—р2, и регуляторным центром ДФГ.
Прежде всего, в эту эпоху (или немного раньше) ген общего предка гемо-
глобинов был дуплицирован, а его копии в ходе дивергенции дали начало
двум родственным семействам — аир. Именно в эту эпоху мы видим макси-
максимальную скорость эволюции центров их контакта и регуляции, а затем в тече-
течение сотен миллионов лет скорость резко падала, часто до нуля, когда возник-
возникшие центры были просто неизменны. Иначе говоря, мы как бы видим собы-
события эпохи формирования новых функциональных структур молекул
гемоглобина, которые в дальнейшем сохраняются у всех наземных форм по-
позвоночных. Следует учесть, что выход позвоночных на сушу и переход к дыха-
дыханию свободным кислородом воздуха сопровождался резкой перестройкой
всей системы дыхания, в том числе, как мы видим, структуры гемоглобинов.
Ускорение эволюции в эту эпоху означает, что указанные приобретения были
высоко адаптивными, т.е. обеспечивали существенное преимущество их обла-
обладателям.
Примером дерева наиболее быстрой и локальной молекулярной эволюции
является дерево генов, кодирующих гемагглютинины НЗ вируса гриппа
(рис. 24-4), построенное у нас Л.В. Омельянчуком и сотр. Это белок вирусного
капсида, некоторые выпуклые участки которого (антигенные детерминанты)
узнаются специфическими антителами хозяина (человека). В результате им-
иммунное сопротивление хозяина препятствует размножению вируса. Вирус
гриппа имеет РНК-геном, в котором мутации происходят с наибольшей из-
известной частотой ~1(Г3-10~4 на позицию за репликацию. В XX в. изучен ряд
локальных эпидемий и пандемий гриппа. Одной из первых изученных была
пандемия "испанки" 1918-1919 гг. В дальнейшем зафиксированы пандемии
"гонконгского" гриппа и др. В большинстве случаев образцы эпидемических
штаммов вируса гриппа были собраны и сохранены в коллекциях. После 1978 г.
РНК этих эпидемических штаммов были секвенированы, а позже теоретиками
были построены филогенетические деревья.
Анализ дерева на рис. 24-4 показал, что все эпидемические варианты ока-
оказались на нижних тупиковых ветвях дерева (так называемые "висячие" верши-
вершины). Это значит, что эпидемические штаммы, как правило, не бывают прямы-
прямыми потомками других эпидемических штаммов. Иначе говоря, неэпидемиче-
неэпидемический вирус, видимо, циркулирует в каком-то "резервуаре" (в локальной
популяции человека, где к вирусу есть устойчивый иммунитет, или в популя-
популяции животных, которые могут быть промежуточными носителями вируса).
Эпидемии возникают тогда, когда появляется новый вариант вируса, против
которого нет готовых антител, т.е. готового иммунитета хозяина. Этот вариант
как бы "выплескивается" в человеческую популяцию и быстро в ней распро-
распространяется, пока не будет выработан адекватный иммунный ответ. Тогда эпи-
эпидемия затухает. Следующая эпидемия возникает путем независимого "выплес-
"выплескивания" нового варианта вируса гриппа, не связанного, как правило, с преды-
предыдущей эпидемией.
24. Молекулярная эволюция
233
Но это не все. Поскольку все эпидемии были датированы (см. рис. 24-4),
оказалось возможным определить скорости эволюции. В эпидемических ветвях
они оказались в 3-5 раз выше, чем в неэпидемических, причем особенно боль-
большое ускорение (в 36 раз) выявлено именно в позициях, кодирующих антиген-
антигенные детерминанты белка. Иммунное сопротивление хозяина размножению ви-
вируса является главным селективным фактором, действующим на вирус. Поэто-
Поэтому "выплеск" эпидемического штамма, не встречающего иммунного
сопротивления хозяина, сопровождается ускорением эволюции. Это размно-
размножение высоко адаптивно для вируса и не адаптивно для хозяина-человека. На-
Напротив, после выработки иммунного ответа спокойное размножение вируса
становится для него и для хозяина нейтральным — в ожидании новых адаптив-
адаптивных мутаций и рекомбинаций. В целом можно говорить о КОЭВОЛЮЦИИ ви-
вируса и иммунного ответа системы хозяина. Вирус стремится как бы мутационно
"выскользнуть" из-под готового иммунного ответа хозяина, а хозяин стремится
"догнать" новые варианты вируса путем выработки нового специфичного им-
иммунного ответа.
Еще один пример — эволюция вируса ВИЧ (вирус иммунодефицита чело-
человека, HIV — в английской транскрипции), вызывающего заболевание СПИД
(синдром приобретенного иммунодефицита, в английской транскрипции —
AIDS). Вирус ВИЧ передается половым путем, а также нередко при перелива-
переливании крови, нестерильных инъекциях и хирургических вмешательствах, при ро-
родах — от матери ребенку, и в других подобных ситуациях. Он был обнаружен в
начале 1980-х годов в крови больных, погибавших от различных тяжелых забо-
заболеваний. Эти заболевания имели одно общее свойство — иммунная система
больного была неспособна противостоять как этому вирусу, так и сопровож-
сопровождавшим возбудителям других заболеваний, от которых, собственно, больные и
погибали. Были обследованы образцы из банков переливания крови, которые
созданы во многих странах для меди-
медицинских целей. Оказалось, что многие
образцы содержали вирус ВИЧ.
Различные штаммы вируса ВИЧ,
выделенные в разных районах Афри-
Африки, Вест-Индии и США, были секве-
нированы, для них теми же методами,
что и выше, были построены филоге-
филогенетические деревья. Одно из них при-
приведено на рис. 24-5. Это дерево указы-
указывает на то, что вирус ВИЧ существовал
в Центральной Африке (Заир) до
1960 г., занесен в Вест-Индию (Гаити)
до середины 70-х годов, и в США — до
1978 г. Иначе говоря, его истоки лежат
в Африке, где в некоторых странах ви-
русоносители составляют до полови-
половины населения, хотя заболевание
СПИД проявляется далеко не у всех.
MAL/85[ я
Z3/83 J т
0 1 2
3456789 10 11
Процент дивергенции
Рис. 24-5. Филогенетическое дерево штам-
штаммов вируса ВИЧ (HIV), вызывающего за-
заболевание СПИД, построенное В.-С. Ли и
сотр. (см. [Ratneret al., 1996]). Числа вбли-
вблизи ребер указывают процент дивергенции
последовательностей. Справа указаны ре-
регионы, где изолированы штаммы.
234
III. Молекулярная кибернетика
Видимо, размножение вируса все же сдерживается их иммунной системой. Ви-
Вирус ВИЧ сходен с некоторыми вирусами обезьян, поэтому многие ученые пред-
предполагают, что он возник в результате изменчивости этих вирусов и был занесен
в популяцию человека извне относительно недавно. Отдельные исследователи
даже выдвигали гипотезу, что врачи сами являются виновниками распростра-
распространения вируса ВИЧ в популяциях человека. Дело в том, что вакцина против по-
полиомиелита культивировалась в тканях подопытных обезьян, и не исключено,
что при этом произошел неконтролируемый занос вируса ВИЧ от обезьян к че-
человеку — реципиенту этой вакцины. Однако эти опасения пока недостаточно
обоснованы. Недавно вирус ВИЧ обнаружен в образце крови человека, датиро-
датированной 1950 г.
Интересно, что изменчивость вируса ВИЧ (мутационная и рекомбинаци-
онная) столь велика, что он успевает измениться в сторону усиления своей аг-
агрессивности непосредственно в ходе развития болезни СПИД у некоторых
больных, в течение 1.5-2 лет и менее. Оценки изменчивости показывают, что
она превышает верхний допустимый предел, в результате чего иммунная систе-
система больного не успевает "справиться" с выработкой новых вариантов антител и
фактически распадается, открывая путь процессу заболевания СПИД и других
болезней. Именно в этом, по-видимому, состоит источник особой агрессивно-
агрессивности заболевания СПИД.
Есть биологические проблемы, которые в большой степени и всегда инте-
интересуют человека и человечество. Среди них особое место занимают проблемы
происхождения и расово-этнической
эволюции самого человечества. В по-
последние годы в этой области прогресс
связан в основном с филогенетиче-
филогенетическим анализом макромолекул.
Американский ученый М. Гудмен
и сотрудники построили филогенети-
филогенетические деревья для ряда генов и белков
от высших приматов и человека. На
рис. 24-6 приведено одно из них. Из
него следует, что порядок последова-
последовательного ответвления видов от эволю-
эволюционного ствола человека таков: макак
резус — орангутан — горилла — шим-
шимпанзе — человек. Иначе говоря, наибо-
наиболее близким к человеку является шим-
NWM
Or
G
Н
Рис. 24-6. Филогенетическое дерево выс-
высших приматов, построенное М. Гудменом
и сотр. (см. [Ratner et al., 1996]) для неко-
дирующих последовательностей из рай-
района вокруг гена 1|/т}-глобина. Обозначе-
Обозначения: Н — человек, С — шимпанзе, G —
горилла, Or — орангутан, Rh — макак ре-
резус, NWM — обезьяны Нового Света.
Цифры вблизи ребер означают числа за-
замен на 100 позиций нуклеотидов.
панзе.
Следует отметить, что до этих ра-
работ в таксономии высших приматов
выделяли два семейства: Hominidae,
включавшее только вид Homo sapiens, и
Pongidae, включавшее виды гориллы,
шимпанзе и орангутана. Молекуляр-
но-филогенетический анализ сущест-
24. Молекулярная эволюция 235
венно изменил эту классификацию. Гудмен и сотр. предложили новую таксо-
таксономию высших приматов:
Семейство Hominidae
Подсемейство Homininae
Род Gorilla A вид — горилла)
Род Homo A вид — человек)
Род Pan B вида: обычный шимпанзе и
карликовый шимпанзе)
Подсемейство Ponginae
Род Pongo A вид — орангутан).
Близость человека и шимпанзе дала основание Дж. Дайамонду назвать че-
человека "третьим шимпанзе". По его оценкам, геномы человека и шимпанзе
различаются примерно на 3.2 х 107 одиночных замен нуклеотидов из 3 х 109, т.е.
на каждый сотый нуклеотид, но остальные 99 из каждых 100 нуклеотидов у них
одинаковы. Время их молекулярной дивергенции оценивается как 5-10 млн лет
назад. Эти факты являются очень веским аргументом в пользу естественного
происхождения человека в процессе филогенетического развития жизни на
Земле, в противовес различным религиозным мифам о разовом творении
5000-10 000 лет тому назад.
Особо интересным представляется исследование путей возникновения,
эволюции и расово-этнической дивергенции человечества. Американский
ученый А. Вильсон и сотр. использовали для этих це/гей сравнение митохон-
дриальных ДНК (мт-ДНК) от представителей пяти основных расово-этниче-
ских групп. Были взяты образцы мт-ДНК от 20 африканцев, 34 азиатов, 46
европеоидов, 21 австралийского аборигена, 26 аборигенов Новой Гвинеи. В
результате было реконструировано огромное генеалогическое дерево
мт-ДНК человечества (рис. 24-7). Использовав археологические датировки
заселения Новой Гвинеи (~30 000 лет назад), Австралии (~40 000 лет назад) и
Америки ( ~12 000 лет назад), они определили скорость эволюции мт-ДНК
~B0-40) х Ю~9 замен на позицию в год, что на порядок превышает аналогич-
аналогичные скорости для ядерных генов. Тогда общий митохондриальный предок
современного человечества существовал примерно 140 000-290 000 лет назад
в Африке.
Поскольку митохондрии наследуются почти исключительно по материн-
материнской линии, этим предком должна была быть африканская женщина. Именно
ее митохондрии в результате генетического дрейфа зафиксировались в предко-
вой африканской популяции, а затем распространились с расселением ее по-
потомков по всей Земле. Остальные варианты были потеряны. Авторы назвали
эту воображаемую прародительницу человечества "черной мт-Евой". Разумеет-
Разумеется, это фигуральное выражение. Речь идет, конечно, о предковой популяции
человека в Африке, от которой до наших дней сохранились потомки только од-
одного варианта мт-ДНК. К ядерным генам это уже не относится. Однако генети-
генетики быстро сообразили, что можно поискать молекулярные следы воображаемо-
воображаемого прародителя "Адама". Для этого нужно взять гены, передающиеся только по
мужской линии. Такие гены, действительно, есть, они локализованы в половой
236
III. Молекулярная кибернетика
80*,
90
60. Y-хромосоме. Попытки филогенети- I
ческого анализа этих генов показали,
что воображаемый "Адам", Y-хромо-
сома которого была предковой для
всего ныне живущего мужского чело-
человечества, существовал примерно в ту
же эпоху и мог встретиться с "черной
мт-Евой".
В пользу африканского корня че-
человечества в дереве на рис. 24-7 (ука-
(указан стрелкой) говорит то, что одно из
двух его поддеревьев (ниже стрелки)
имеет чисто африканскую природу, а
другое — смешанную. Первая дивер-
дивергенция разделила эти две группы в
Африке. Африканская часть осталась
на месте, а вторая часть, вероятно не-
небольшая группа основателей, через
значительное время мигрировала из
Африки через Суэцкий перешеек, дав
начало другим группам. Утрата зна-
значительного генетического разнообра-
разнообразия этими мигрантами (в том числе и
по мт-ДНК.) говорит о том, что их по-
популяция в это время проходила ста-
стадию "бутылочного горлышка", т.е.
была очень мала. Время дивергенции
оценивается довольно неточно,
где-то в интервале 57-135 тыс. лет на-
назад. Расселение дало начало осталь-
остальным расово-территориальным груп-
группам — европеоидам, азиатской груп-
группе и аборигенам Тихого океана
(рис. 24-8). По археологическим дан-
данным считается, что миграция из Аф-
Африки началась примерно 50 000 лет
назад, а расселение по Евразии завер-
завершилось ~30 000 лет назад. Заселение
Америки началось ~12 000 лет назад
через Берингию и Аляску, сомкнувшиеся при последнем оледенении. Все расо-
расовые группы (кроме исходной африканской) имели множественное происхож-
происхождение (смешанные ветви в дереве на рис. 24-7), а территории, по-видимому, за-
заселялись ими многократно.
Таким образом, молекулярно-филогенетический анализ позволяет при-
приблизиться к решению очень важных вопросов эволюции. Иногда это единст-
единственная возможность заглянуть далеко вспять. При этом почти всегда мы можем
Africa
о Asia
A Australia
A New Guines
о Europe
0
0.2
l i 1 l i i i i i i
0.4 0.6 0.6 0.4 0.2
Sequence divergence, %
Рис. 24-7. Генеалогическое дерево paco-
во-этнических групп человека, построен-
построенное А.Вильсоном и сотр. (см. [Ratner et al.,
1996]) по данным для мт-ДНК. Корень де-
дерева, указанный стрелкой, соответствует
времени существования "черной мт-Евы".
Под этой точкой одна из ветвей (малая) —
чисто африканская, другая — смешанная.
Обозначения основных расовых групп
приведены на рисунке.
24. Молекулярная эволюция
237
Европа
!9 000-51 000
лет назад
Азия
,. 56 000-73 000
Ш I лет назад
>установлено по типам митохондриальной ДНК
> установлено по типам Y-хромосомы
Рис. 24-8. Дерево расселения расово-этнических групп человечества по континентам
[Янковский, Боринская, 2001].
оперировать только кодирующими последовательностями от ныне живущих
видов. "Ископаемых" последовательностей почти нет. Правда, в последнее
время удается выделить и секвенировать фрагменты ДНК из замерзших иско-
ископаемых животных (мамонт и др.), мумифицированных тел и ископаемых ос-
останков человека и др. Так, секвенирование фрагментов мт-ДНК из ископаемых
останков неандертальца показало, что неандертальцы не были предками Homo
sapiens, общий предок человека и неандертальца существовал очень давно,
500-600 тыс. лет назад. После заселения Европы 40-50 тыс. лет назад челове-
человеком разумным неандертальцы, видимо, были вытеснены им полностью.
Определенный интерес представляет уникальная возможность выделения
ДНК из мелких насекомых (мухи, термиты, пчелы, долгоносик), законсервиро-
законсервированных в природной смоле деревьев (янтарь). Известно, что эту идею прекрас-
прекрасно использовал американский кинорежиссер С. Спилберг в фильме "Парк
Юрского Периода": в янтаре якобы найден комар, в кишечнике которого со-
содержатся остатки клеток крови динозавра, укушенного этим комаром; из них
выделяется ДНК динозавра. Затем динозавр полностью воспроизводится ген-
генно-инженерными методами на радость и страх посетителям парка. Разумеется,
это фантазия. Но анализ ДНК законсервированных насекомых вполне реален.
Так, для термита, пчелы и долгоносика из янтаря были клонированы и секве-
нированы гены 18S рибосомальной РНК. Они включены в филогенетические
деревья этих молекул. Возраст найденных в янтаре термитов и пчел 25-40 млн
лет, долгоносика — 120-135 млн лет. Пока это рекордные случаи. Впрочем, они
не дали каких-либо сенсационных результатов. С другой стороны, многие био-
238 III. Молекулярная кибернетика
химики утверждают, что столь долгий срок фрагменты ДНК не могут существо-
существовать, они распадаются, а все объявленные случаи их секвенирования - это ар-
артефакты, связанные с плохим качеством работы.
Не следует думать, что приведенные выше научные гипотезы имеют ка-
какое-либо отношение к обоснованию тех или иных религиозных догм и мифов.
Мифы живут своей жизнью как продукт художественного творчества человече-
человечества. Нет решительно никакого повода отождествлять "черную митохондриаль-
ную Еву" с мифологической прародительницей человечества. С таким же успе-
успехом можно в филогенетическом дереве глобинов высших приматов (рис. 24-5)
обожествить нашего общего обезьяноподобного предка, а в дереве надсемейст-
ва глобинов (рис. 24-2) — общего предка всех наземных позвоночных! Не ду-
думаю, чтобы какой-то религиозный деятель вдохновился образом праобезьяны
или двоякодышащей рыбы. Да и облик предполагаемой "мт-Евы", дикой, не-
неразвитой женщины, обитавшей, вероятно, более 140 000 лет назад, почти не
знавшей человеческого языка, тоже вряд ли удовлетворит высоким эстетиче-
эстетическим требованиям к объекту обожания и поклонения.
В заключение можно заметить, что молекулярный филогенетический ана-
анализ становится сейчас одним из весомых методов таксономии живых форм.
Таксономия высших приматов сейчас больше базируется на молекулярных
данных, чем на каких-либо других. В систематике наивысших таксонов, систе-
систематике высших растений решающую роль играют деревья рибосомных РНК.
Таксономию бактерий вообще невозможно построить без молекулярных дан-
данных. Можно надеяться, что в дальнейшем роль этого подхода только возрастет.
Список литературы
Айала Ф. Введение в популяционную и эволюционную генетику. М.: Мир, 1984.
Айала Ф., Кайгер Дж. Современная генетика. М.: Мир, 1988. Т. 3.
Кимура М. Молекулярная эволюция: теория нейтральности. М.: Мир, 1985.
Ратнер В.А. Краткий очерк теории молекулярной эволюции. Новосибирск: НГУ, 1992.
Ратнер В.А., Жарких А.А., Колчанов Н.А., Родин С.Н., Соловьев В.В., Шамин В.В. Про-
Проблемы теории молекулярной эволюции. Новосибирск: Наука, 1985.
Эволюция. М.: Мир, 1981.
Янковский Н.К., Боринская С.А. Наша история, записанная в ДНК // Природа. 2001.
№ 6. С. 10-17.
Ratner V.A., Zharkikh A.A., Kolchanov N.A., Rodin S.N., Solovyov V.V., Antonov A.S.
Molecular Evolution. Berlin e.a.: Springer-Verl., 1996.
25. Укус издыхающей рептилии*
(из архива АЛ. Ляпунова)
В архиве члена-корреспондента АН СССР А.А. Ляпунова за 1962 г. имеют-
имеются копии документов, связанных с попыткой подавить и обуздать издание
сборников "Проблемы кибернетики" (ред. А.А. Ляпунов) по разделу "Процес-
* Опубликовано ранее: Ратнер В.А. // Алексей Андреевич Ляпунов / Ред. Н.А. Ляпунова,
Я.И. Фет. Новосибирск: филиал "Гео" Изд-ва СО РАН, Изд-во ИВМиМГ СО РАН, 2001.
С. 234-241.
25. Укус издыхающей рептилии 239
сы управления в живых организмах". Обстоятельства событий состояли в сле-
следующем.
Сборники "Проблемы кибернетики" (ПК) начали издаваться в Физматгизе
с 1958 г. на волне реабилитации кибернетики. Редактор сборников профессор
А.А. Ляпунов, будучи одним из международно признанных основателей совет-
советской кибернетики, имел глубокий интерес к биологическим наукам, в частно-
частности к генетике, математической биологии и биологической кибернетике, вся-
всячески их поддерживал, пропагандировал и развивал. Неудивительно, что уже в
№ 1 "ПК", в статье редактора он включил эти вопросы в область интересов ки-
кибернетики и выделил специальный раздел в сборниках. В № 1-5 "ПК" ему уда-
удалось опубликовать несколько статей выдающихся ученых-биологов: академика
И.И. Шмальгаузена, Н.В. Тимофеева-Ресовского, В.П. Эфроимсона, А.А. Ма-
Малиновского, Р.Л. Берг и др.
Следует сказать несколько слов о личностях авторов статей. Академик
Иван Иванович Шмальгаузен — выдающийся советский зоолог, эволюцио-
эволюционист, эволюционный генетик, автор теории стабилизирующего отбора. Он был
одним из немногих ученых, кто выстоял на презренной Сессии ВАСХНиЛ
1948 года, выдержал удар и не покаялся в научных грехах. За это И.И. был фак-
фактически отстранен от науки почти на десятилетие. Он сидел в небольшой ком-
комнате на ул. Б. Калужской, 33, в помещении своего бывшего института, а из со-
сотрудников имел одну лаборантку. В этот период он неожиданно открыл для
себя идеологию кибернетики. Будучи чистым зоологом, он попытался пере-
переформулировать идеи эволюционной генетики в терминах кибернетики и полу-
получил совершенно новый взгляд на проблему. В общей сложности он напечатал
10 весьма интересных работ на эту тему. Одна из них была опубликована в № 4
"ПК". В дальнейшем А.А. Ляпунов издал все эти статьи в сборнике
"Кибернетические вопросы биологии" (Серия "Кибернетика в монографиях".
Физматгиз, 1968).
Николай Владимирович Тимофеев-Ресовский, выдающийся русский гене-
генетик, биофизик, эволюционист и радиационный эколог, ученик Н.К. Кольцова.
Проработал свыше 20 лет в Германии, затем 8 лет был в заключении в так назы-
называемой "шарашке" на Урале, а в 1955 г. был освобожден, после чего ряд лет ра-
работал в Институте биологии УФ АН СССР в г. Свердловске. Н.В. принадлежит
ряд классических работ в различных областях генетики, радиационной биоло-
биологии, эволюции. Он один из соавторов Синтетической теории эволюции - круп-
крупнейшего достижения эволюционной теории 30-40-х годов. Его совместная ра-
работа с М. Дельбрюком и К. Циммером по оценке размера генов была предтечей
развития молекулярной генетики. После 1955 г. он занимался в основном
закрытыми вопросами радиационной биогеоценологии, связанными с послед-
последствиями ядерных взрывов и загрязнения окружающей среды. Во всех этих об-
областях Н.В. оставил основополагающий след. С 1963 г. он заведовал отделом в
Институте медицинской радиологии в г. Обнинске, а с 1970 г. был профессо-
профессором-консультантом Института медико-биологических проблем (космической
биологии и медицины).
К 1962 г. Н.В. опубликовал в "ПК" две статьи совместно с немецким уче-
ученым P.P. Ромпе и советским генетиком Р.Л. Берг. Фактически это были про-
240 III. Молекулярная кибернетика
блемные обзоры результатов генетической науки за ряд предшествующих лет,
изложенные в форме, удобной для теоретического обобщения.
Александр Александрович Малиновский, в дальнейшем доктор биологи-
биологических наук, был весьма оригинальной фигурой в советской генетике. Сын
известного медика и философа-марксиста А.А. Богданова (Малиновского),
автора знаменитой книги "Тектология", предвосхитившей современную ки-
кибернетику. Все мы изучали, как Богданов был раскритикован в пух и прах
классиком марксизма В. Ильиным (В.И. Лениным). В начале 30-х годов А.А.
был участником генетических семинаров в Кольцовском институте, где об-
обсуждались многие теоретические проблемы генетики. В 50-е годы он очень
легко и естественно усвоил идеологию кибернетики и начал ее использовать
для теоретического осмысления явлений биологического и генетического
управления. На эту тему он опубликовал в № 4 "ПК" обзорно-теоретиче-
обзорно-теоретическую статью.
Владимир Павлович Эфроимсон, в дальнейшем профессор, доктор меди-
медицинских наук, был энциклопедистом современной медицинской генетики и
страстным ее проповедником. За непримиримость характера он дважды попа-
попадал за решетку, причем, как говорится, первым садился и последним бывал ос-
освобожден. По книгам В.П. училось целое поколение советских генетиков и ме-
медиков. В 1961 г. в № 5 "ПК" он опубликовал две обзорные статьи по иммуните-
иммунитету растений и механизмам канцерогенеза.
Раиса Львовна Берг, дочь академика Л.С. Берга и аспирантка академика
И.И. Шмальгаузена, в дальнейшем профессор, доктор биологических наук и
заведующая лабораторией ИЦиГ СО АН СССР, была известным эволюциони-
эволюционистом и генетиком, автором оригинальных работ по корреляционным плеядам у
растений и динамике мутаций в природных популяциях дрозофил. К 1962 г. она
была старшим научным сотрудником Ленинградского университета и одним из
организаторов очень известных тогда семинаров и конференций по кибернети-
кибернетике в биологии.
Необходимо отметить своеобразие момента. К 1962 г. успехи новых наук —
молекулярной биологии и генетики, а также кибернетический подход к ним
были неоспоримы: структура ДНК, открытие информационной РНК, оперо-
нов, начало исследования генетического кода и др. Советские физики, матема-
математики и биологи много сделали для пропаганды и легализации этих достижений,
возобновления работ по генетике, издания оригинальных книг, возрождения
преподавания генетики в МГУ, ЛГУ и других вузах. В научном сообществе со-
созрел острый интерес к этим проблемам, который вылился в организацию не-
неформальных семинаров, лекций, летних школ для молодежи, организованных
под руководством П.Л. Капицы, И.Е. Тамма, А.Н. Колмогорова, Н.Н. Семено-
Семенова, Н.В. Тимофеева-Ресовского, А.А. Ляпунова, Н.П. Дубинина и многих дру-
других.
С другой стороны, напомним, что в 1948 г. советской генетике был нанесен
сильнейший удар со стороны сталинской партийно-административной систе-
системы руками распоясавшейся орды лысенковцев — "хунвейбинов" от науки. По-
После смерти И.В. Сталина A953 г.) напор несколько ослаб, и к концу 50-х годов
в стране было организовано несколько институтов генетической направленно-
25. Укус издыхающей рептилии 241
ста. Однако Т.Д. Лысенко удалось заморочить голову Н.С. Хрущеву обещания-
обещаниями быстрого успеха в сельском хозяйстве, и в результате в начале 60-х годов
обозначился частичный реванш лысенкоизма. Именно в это время и имел
место описываемый эпизод научно-политической борьбы.
К тому времени кибернетика была полностью и бесповоротно реабилити-
реабилитирована по причине ее необходимости для обороны страны. С генетикой этого
пока еще не случилось. Генетическая тематика с трудом просачивалась в планы
научных лабораторий, часто камуфлируясь под нейтральные наименования.
Генетических журналов не было, книги по генетике выходили с трудом. В таких
условиях сборник "Проблемы кибернетики" был одним из немногих изданий,
где публиковались проблемные обзорные статьи генетического содержания.
В этом смысле его роль была огромна. Для лысенковцев сборник был как бель-
бельмо в глазу. Это понимали все — и соратники, и враги.
Итак, в начале 1962 г. Главлит (фактически - цензура печати) получил (или
заказал?) закрытую отрицательную рецензию на статьи раздела "Проблемы
управления в живых организмах" в сборниках "Проблемы кибернетики"
№ 1-5. В архиве А.А. Ляпунова нет прямых данных о том, кто персонально ин-
инспирировал и осуществил эту подковерную атаку на сборник, попытку под
флагом защиты марксизма набросить на него узду. Некоторые следы ведут в
ВАСХНиЛ. Конечно, мне очень хотелось бы узнать, кто тот поганец, который
написал научный пасквиль и донос на "ПК". Однако теперь это уже чисто ис-
исторический интерес, вряд ли автор дожил до наших дней.
Рецензия была в анонимной форме направлена в Физматгиз, а оттуда - в
Научный совет по кибернетике АН СССР для ознакомления и экспертной
оценки по существу поставленных критических вопросов и предложений.
В свою очередь, председатель Совета академик А.И. Берг направил около 100
копий рецензии ряду выдающихся ученых и получил около 40 ответов.
Сначала приведем выборку ключевых высказываний анонимного рецен-
рецензента:
"...печатание их ("статей. — В.Р.) было несомненно крупной ошибкой..."
«В статье А.А. Ляпунова (в. 1, с. 21) "кибернетическим языком "оспаривается
возможность наследования приобретенных признаков».
«...своими общебиологическими статьями сборники "Проблемы кибернетики "
противопоставили себя линии партии в вопросах общей биологии».
"...претензия подменить диалектический материализм кибернетикой..."
"...никаких научных оснований для кибернетического подхода к явлениям на-
наследственности и эволюции не существует. (Внимание: дальше идет основопо-
основополагающее определение лысенкоизма!) Наследственность, как показал академик
Т.Д. Лысенко, это не вещество и не механизм, а основное свойство живого, заклю-
заключающееся в способности живого тела требовать для своего роста, развития от-
относительно определенных условий и по-своему реагировать на те или иные условия.
При таком понимании наследственности, а оно есть единственно правильное, ма-
материалистическое понимание, отпадает возможность приложения кибернетики к
общетеоретическим вопросам биологии. Можно ...утверждать, что в мичурин-
мичуринском учении кибернетический подход неприменим. " (После таких определений в
науке делать нечего! — В.Р.).
242 III. Молекулярная кибернетика
"...ни математики, ни сторонники корпускулярной генетики не делают попы-
попыток хоть как-то связать кибернетический подход с практикой сельского хозяйст-
хозяйства и медицины ".
"...Факты же, свидетельствующие об отсутствии хромосом ...в интерфазе
клеток, ...прочно установлены..."
Такого рода тенденциозные, "калькированные" заключения переполняют
текст рецензии. Кроме того, обращаясь к так называемым "фактам", аноним
манипулирует незаконченными, тенденциозно поданными или невнятными
якобы результатами других авторов. Впечатление тягостное!
В итоге аноним делает вывод о
"...нецелесообразности дальнейшей публикации в сборниках "Проблемы
кибернетики " статей по общетеоретическим вопросам биологии (т.е. по вопро-
вопросам генетики и теории эволюции). Все материалы такого рода... следует переда-
передавать в специальные биологические издания, где они могут найти компетентную
редакцию и читательскую аудиторию ".
"...не следует только решать вопрос таким образом, что редакцию сборников
следует пополнить биологами. Это окажется без пользы, так как настроенность
редактора (АЛ. Ляпунова) хорошо известна, и он пригласит в редакцию тех же
Шмальгаузена, Эфроимсона и Малиновского, а то и еще хуже — Тимофее-
Тимофеева-Ресовского... "
Как все хорошо продумано: рупор закрыть, а науку отдать на откуп в жур-
журналы, где уже сидят свои ставленники, поднаторевшие как раз на удушении ге-
генетической науки! Например, в "Журнале общей биологии" сидел ответствен-
ответственный редактор — ярый лысенковец член-корреспондент Н.И. Нуждин. Это все
равно, что направить на минное поле погулять! А Ляпунова-то аноним боится!
А Тимофеев-Ресовский-то вызывает у него содрогание!
Как отреагировали крупнейшие советские ученые в своих откликах на ре-
рецензию? Среди них были П.Л. Капица, А.Н. Колмогоров, И.Е. Тамм, В.А. Эн-
гельгардт, В.Н. Сукачев, B.C. Немчинов, И.Л. Кнунянц, Н.М. Амосов, Б.Л. Ас-
тауров, А.Д. Александров, В.В. Парин и др. Ответы академиков, сведенные На-
Научным советом по кибернетике, по форме корректны и интеллигентны, но по
существу плюют анониму в физиономию:
«...подавляющее большинство ученых, приславших ответ, заявили, что при-
приложение кибернетики к вопросам биологии является не только допустимым, но и
настоятельно необходимым, оценка конкретных статей несправедлива, что
статьи, опубликованные в "Проблемах кибернетики ", представляют большую на-
научную ценность, и что работы по приложению кибернетики в биологии публико-
публиковать не только целесообразно, но и настоятельно необходимо».
"В ряде отзывов отмечается наличие заведомых подтасовок в рецензии ".
Академик А.Н. Колмогоров, один из крупнейших математиков XX века,
участник семинаров с московскими генетиками в 30-х годах, знающий о гене-
генетике из первых рук:
"Приложения кибернетики к вопросам биологии принципиально допустимы и
практически полезны ".
25. Укус издыхающей рептилии 243
"...Реальный управляющий механизм, осуществляющий приспособление вида к
изменению внешних условий, существует. Компонентами этого механизма явля-
являются системы размножения, наследования признаков и изменчивости, свойствен-
свойственные виду ".
"...Область применимости кибернетических идей принципиально ограниче-
ограничена использованием таких материальных систем, которые действуют
целесообразно ".
"Широкая работа кибернетика в области биологии неизбежно приводит к бо-
более трудным для логического анализа проблемам ВОЗНИКНОВЕНИЯ
ЦЕЛЕСООБРАЗНОСТИ в материальных системах, возникающих в процессе ес-
естественной эволюции из неживой природы, знающей лишь сцепление ПРИЧИН И
СЛЕДСТВИЙ".
"Проблематика естественного... возникновения материальных систем, дей-
действующих по свойственной им системе целей, существенна для кибернетики ".
Любопытно, что выдающийся математик учит рецензента, вроде бы биоло-
биолога, азам эволюционно-генетического подхода!
Выдающийся физик-теоретик, Нобелевский лауреат академик И.Е. Тамм,
активный пропагандист новых идей и достижений молекулярной биологии и
генетики:
"Утверждение автора (рецензии) о невозможности генетического кода
представляется особенно нелепым, потому что во второй половине прошлого
1961 г. в генетике произошло решающее событие первостепенного значения, а
именно работами Крика, Очоа и Виттмана не только выявлен общий принцип по-
построения кода, определяющего синтез белков в организме, но этот код уже фак-
фактически расшифрован..."
Крупнейший биохимик, основатель советской молекулярной биологии
академик В.А. Энгельгардт:
«Общеизвестно, каким нападкам подвергалось со стороны некоторых
'запальчивых"философов именно фактическое содержание кибернетики. Извест-
Известно, что эти нападки надолго задержали ее развитие в нашей стране и нанесли
этим существенный ущерб советской науке и технике... Теперь, очевидно,
"запальчивость " некоторых философов направляется на биологические приложе-
приложения кибернетики».
Член-корреспондент АН СССР (в дальнейшем академик) А.Д. Александ-
Александров, ректор ЛГУ, поддержавший генетику и кибернетику еще в середине 50-х
годов:
"Приемы политического и философского опорочивания научных взглядов и по-
постановок научных проблем недостойны и недопустимы. С ними нужно бороться
настойчиво, решительно и до конца, без всяких уступок ".
Следует особо отметить отклики ученых из молодого, только что созданно-
созданного Сибирского отделения АН СССР. Недаром академик М.А. Лаврентьев при-
пригласил члена-корреспондента Н.П. Дубинина создать современный генетиче-
генетический институт (ИЦиГ СО АН СССР) в Новосибирске. Правда, заботами лы-
244 III. Молекулярная кибернетика
сенковцев к 1962 г. Дубинин уже снят из директоров, его замещает еще
неизвестный стране Д. К. Беляев, будущий академик. Но работа идет, наука не
стоит на месте, только бы не мешали!
Из СО АН СССР отклики представили академик М.А. Лаврентьев,
академик С.Л. Соболев, директор ИЦиГ Д.К. Беляев, только что переехавшие в
Новосибирск А.А. Ляпунов и И.А. Полетаев, В.В. Хвостова, Ю.Я. Керкис и
многие другие.
Так, академик М.А. Лаврентьев, Председатель СО АН СССР, крупнейший
математик, отвечает:
«1. Развитие работ по применению кибернетики в биологии необходимо.
2. Оценка статей из сборников "Проблемы кибернетики " по биологии, данная в
присланной рецензии, неправильна.
3. Печатание в сборниках "Проблемы кибернетики "статей, относящихся к при-
применению кибернетики в биологии, целесообразно».
Вот так! Сказал, как отрезал! С подонками споров нет! Фактически мнение
М.А. Лаврентьева составило костяк ответа Научного совета по кибернетике.
Академик С.Л. Соболев, один из самых талантливых математиков страны,
директор Института математики СО АН СССР:
"Автор рецензии высказывает совершенно неправильную точку зрения на роль
кибернетики в биологии ".
"Автор рецензии игнорирует научное значение вопроса о перекодировании на-
наследственной информации на биохимическом уровне ".
"Тут важно, что постановка вопросов, возникших под влиянием кибернетики,
привела к открытию новых экспериментальных фактов ".
"Объявляя наследственность свойством живого, автор считает, что этим
все объяснено и что это освобождает естествознание от необходимости изучать
строение процессов передачи наследственных признаков от родителей к
потомкам ".
"Совершенно недопустимо накладывать запрет на кибернетические публика-
публикации по вопросам общей биологии только потому, что такие публикации не соот-
соответствуют вкусам того или иного лица, например, автора присланной рецензии. "
И т.д.
Отклик группы ведущих сотрудников Института цитологии и генетики СО
АН СССР:
"Рецензия в целом производит очень тяжелое впечатление. Вся она пропитана
духом догматизма времен культа личности, стремлением к навешиванию ярлыков
и к облыжным политическим обвинениям ученых, придерживающихся иных, чем
рецензенты, взглядов на конкретные вопросы науки ".
Надо отдавать себе отчет, что именно в это время лысенковцы делали мно-
многократные попытки, чтобы закрыть ИЦиГ СО АН СССР или исключить его из
Академии. Только стойкая защита генетики М.А. Лаврентьевым позволила со-
сохранить институт — крупнейший генетический центр России, тогда и до наших
дней.
26. Математическая генетика в НГУ и около 245
Прислали свои ответы рецензенту и авторы упомянутых биологических ра-
работ — Н.В. Тимофеев-Ресовский, Р.Л. Берг, А.А. Малиновский и др. У них по-
положение несколько иное — они объект критики. Они отвечали на конкретные
наветы, сдерживая негодование и презрение.
Н.В. Тимофеев-Ресовский пишет:
«Мне представляется совершенно неубедительной критика автора
"Замечаний" взаимоотношений кибернетики с биологией, доходящая до полного
отрицания наличия управляющих систем у живых организмов; как же иначе ин-
интерпретировать хотя бы стройное онтогенетическое развитие, протекающее по
видоспецифическому наследственному плану, целый ряд видово-специфических био-
биохимических процессов и синтезов, или целый ряд явлений из области высшей нерв-
нервной деятельности?»
"Я полагаю, что трудно у нас найти более компетентное лицо для оценки об-
общей методологической значимости биологических работ, чем АЛ. Ляпунов ".
Р.Л. Берг отвечает не только на критику своей статьи, но и защищает рабо-
работы И.И. Шмальгаузена, А.А. Малиновского и В.П. Эфроимсона.
А.А. Малиновский:
"...рецензия является неверной — как с точки зрения фактической, так с на-
научной, философской и политической ".
Запросив отзыв Научного совета по кибернетике, дирекция Физматгиза
(Г.Ф. Рыбкин) попыталась перестраховаться, изъяв из очередного (№ 7) выпус-
выпуска "ПК" биологические статьи. Однако попытка не удалась: получив совер-
совершенно однозначную поддержку опубликованных статей научным сообщест-
сообществом, издательство было вынуждено опубликовать их в № 8 и более не вмеши-
вмешиваться в научную политику сборника, проводимую А.А. Ляпуновым.
Любопытно, как изменились времена! В 1948 г. научная общественность
молча проглотила разгон генетики. Даже Президент АН СССР академик
СИ. Вавилов, брат погибшего Президента ВАСХНиЛ академика Н.И. Вави-
Вавилова, не посмел сказать ни слова! Прошло 15 лет бурных событий, включая
хрущевскую "оттепель", и теперь цвет Академии наук единодушно встает на
защиту генетики и биокибернетики. Правда, в 1964 г. Академия чуть не попла-
поплатилась за свою вольность и едва не была разогнана Н.С. Хрущевым во гневе
по навету того же Лысенко. Однако это были уже арьергардные бои! А описан-
описанный выше частный случай — один из последних укусов издыхающей репти-
рептилии лысенкоизма!
26. Математическая генетика в НГУ и около*
Преамбула
Я приехал в Новосибирск осенью 1959 г., примерно тогда, когда был создан
НГУ. В этом смысле мы ровесники. Окончив физфак ленинградского универ-
университета в 1955 г., я несколько лет работал ассистентом-физиком в вузах Дальнего
* Опубликовано ранее: Ратнер В.А. О подготовке биологов в НГУ / Наука. Академгородок. Уни-
Университет. Воспоминания. Очерки. Интервью. Новосибирск.: Изд-во НГУ, 1999. Вып. 1. С. 296-302.
246 III. Молекулярная кибернетика
Востока и Сибири. В 1957 г., услышав о создании Сибирского отделения АН
СССР, я начал пробиваться в Новосибирск. После нескольких неудачных
«заходов», в качестве временного пристанища я поступил на Кафедру физики и
математики НИИГАиК, который тогда находился в центре Новосибирска, на
ул. Потанинской, что через два дома от первой резиденции СО АН СССР
(ул. Советская, 20, сейчас — 18). Близость к Академии была главным критерием
выбора института. Именно тогда, в сентябре 1959 г., я впервые приехал в Ака-
Академгородок и увидел великую стройку науки.
Доехав из города на автобусе до ст. "Сеятель", я прошел через водосточную
трубу под железной дорогой и пошел направо через осенний лес. Затем вышел
на Проспект науки (будущий проспект М.А. Лаврентьева). Впечатление было
огромное. В то время я имел привычку все существенные и несущественные
эмоции выплескивать стихами. Первое стих-впечатление от Академгородка см.
в гл. 27 ("Сказка").
В начале 1960 г. через новых знакомых — А.Д. Груздева и Н.Б. Христолюбо-
ву я вышел на директора Института цитологии и генетики СО АН СССР
Д. К. Беляева. Он дал мне небольшую теоретическую задачу по популяционной
генетике, а когда я ее сделал, взял к себе в лабораторию младшим научным со-
сотрудником. Это было в декабре 1960 г. С того времени, свыше 40 лет я непре-
непрерывно работаю в ИЦиГ. В главе 27 ("Любопытно") привожу первое стих-впе-
стих-впечатление от соприкосновения с тогдашней научной средой.
В 1962 г. Д.К. Беляев создал на ФЕН НГУ Кафедру общей биологии, кото-
которая стала родоначальницей многих других биологических кафедр. В 1968 г. от
нее отделилась Кафедра цитологии и генетики ФЕН.
В дальнейшем на нашей кафедре работало много выдающихся генетиков
старшего и нового поколений. Д.К. Беляев, ставший позже академиком, заведо-
заведовал кафедрой с 1968 по 1985 г. Он определял направление развития и дух кафед-
кафедры, привлекал к преподаванию сотрудников института, приглашал приезжих
крупных генетиков. Фактически кафедра всегда была и является сегодня продол-
продолжением Института цитологии и генетики. Из генетиков старшего поколения
наибольшую роль на кафедре играла профессор В.В. Хвостова — несгибаемая
энтузиастка генетики, крупнейший цитолог и цитогенетик, бывшая сотрудница
Н.П. Дубинина. Курс генетики ряд лет читал профессор Ю.Я. Керкис — ученик
и сотрудник Н.И. Вавилова, Г.Г. Меллера, Н.П. Дубинина, бывший директор
овцеводческого совхоза в Таджикистане в годы лысенковщины. Эволюционное
учение несколько лет читала профессор Р.Л. Берг — ученица академика
И.И. Шмальгаузена, дочь академика Л.С. Берга. Несколько лет биометрию чита-
читала доцент З.С. Никоро — ученица и сотрудница С.С. Четверикова.
Среднего поколения фактически не было. Из нового поколения большую
роль на кафедре сыграли нынешний заведующий кафедрой академик
В.К. Шумный (генетика растений), профессор И.И. Кикнадзе (цитология, ци-
тогенетика), профессор Л.И. Корочкин (генетика развития), профессор
Л.А. Васильева (биометрия, количественная генетика), профессор Н.Н. Ворон-
Воронцов (эволюционное учение), доктор биологических наук М.Д. Голубовский
(генетика популяций), надеюсь — и я (молекулярная генетика, математическая
генетика). Более молодые профессора, выросшие в НГУ и Институте:
26. Математическая генетика в НГУ и около 247
член-корреспондент РАН И.Ф. Жимулев (генетика, цитогенетика), профессор
П.М. Бородин (эволюционное учение), профессор Е.Н. Грунтенко (иммуноге-
нетика), профессор Л.В. Высоцкая (цитология, генетика), профессор И.К. За-
Захаров (эволюционное учение), профессор А.О. Рувинский (генетика, эволюци-
эволюционное учение) и др.
Многие сотрудники ИЦиГ работали на смежных кафедрах: академик
Р.И. Салганик, профессор Т.М. Морозова, профессор Г.М. Дымшиц и др. — на
кафедре молекулярной биологии, член-корреспондент РАН Л.И. Корочкин,
академик Л.И. Иванова, профессор А.Л. Маркель и др. — на кафедре физиоло-
физиологии. Кафедру общей биологии до последнего времени возглавлял ученик
Д.К. Беляева, член-корреспондент РАН В.И. Евсиков.
Перед студентами неоднократно выступали с лекциями приглашенные
крупные ученые: профессор Н.В. Тимофеев-Ресовский, член-корреспондент
РАН И.А. Рапопорт, академик РАМН В.И. Иванов, академик Р.В. Петров,
член-корреспондент РАН С.Г. Инге-Вечтомов, член-корреспондент РАЕН
Л.З. Кайданов, профессор У. Сталь и М. Грин (США) и многие другие.
В последние годы на кафедре было три Соросовских профессора из нашего
института (И.Ф. Жимулев, Л.А. Васильева и В.А. Ратнер) + профессор Г.М. Дым-
Дымшиц с кафедры молекулярной биологии и три Соросовских доцента (Е.И. Ка-
ракин, Л.В. Высоцкая, Н.А. Попова).
Меня никто не приглашал в НГУ. Я напросился туда сам. Защитив в 1965 г.
кандидатскую диссертацию по математической генетике и начитавшись лите-
литературы, я предложил Д.К. Беляеву прочесть курс молекулярной генетики для
4-го курса биологов. Поскольку самим корифеям было некогда это делать, а
4-й курс неумолимо надвигался, он согласился. В результате я читаю этот курс
до сих пор [Ратнер, 1983]. И не жалею об этом.
Следует иметь в виду также одно важное обстоятельство. Дело в том, что
после 1948 г. генетика в СССР была фактически запрещена. После смерти
И.В. Сталина в 1953 г. недобитые генетики несколько оживились, но все равно
давление на них продолжалось с переменным успехом до октября 1964 г., так
называемого малого Октябрьского переворота, т.е. до снятия Н.С. Хрущева. Я
помню, как наш институт "клевали" некоторые биологи-лысенковцы, бездар-
бездарные философы и номенклатурные функционеры. Бывали моменты, когда ин-
институт висел на волоске. Нас спасли выдержка М.А. Лаврентьева и его умение
отвращать беду методами подковерной номенклатурной же дипломатии.
После этого для нас наступили дни свободы и ренессанса. В прессе, на те-
телевидении, в общественном мнении, в вузах и в смежных науках возник острый
интерес к недобитой «служанке империализма». Нас просто рвали на части.
Мы выступали с лекциями, водили экскурсии по институту, принимали высо-
высоких и невысоких гостей, проводили бесчисленные школы и семинары, писали
популярные статьи и книги, некоторые даже снимались в кино и на телевиде-
телевидении. Сибирское отделение АН СССР и НГУ были в первых рядах возрождения
генетики.
Настроение у всех было приподнятое и радостное. В какой-то мере это
можно почувствовать из материалов шутливой дискуссии "Человек, жизнь и
кибернетика", которая была разыграна на сцене ИЦиГ в 1964 г. (см. гл. 27).
248 III. Молекулярная кибернетика
В этом смысле вузы страны были гораздо более консервативны. Во многих
университетах, сельскохозяйственных и медицинских вузах на кафедрах сидели
откровенные лысенковцы, которые не собирались и не могли преподавать ге-
генетику будущим специалистам. Настоящих генетиков в это время готовили
только в МГУ и ЛГУ. Поэтому учреждение в НГУ Кафедры цитологии и гене-
генетики было очень своевременным и дальновидным шагом.
С другой стороны, генетика как наука стремительно развивалась. Именно в
конце 50-х — начале 60-х годов были сделаны многие выдающиеся открытия:
структура ДНК, генетический код, матричная РНК, опероны и др. В Новоси-
Новосибирск все это доходило с заметным опозданием, но отставать было нельзя.
И мы пахали, пахали, пахали эту ниву и сеяли плодоносные семена. Это было
занятие для энтузиастов! Глядя на прошедшие годы, я думаю, что наша актив-
активность не пропала даром. Следующие поколения генетиков из НГУ оправдали
затраченные на них усилия.
Как-то в самый разгар этой миссионерской деятельности я спросил
Д.К. Беляева, нельзя ли немного увеличить нам оплату в НГУ, все же платили
нам совершенно мизерные деньги. Он ответил: "Я прекрасно знаю, что мы ра-
работаем фактически бесплатно, за эфемерную зарплату, но это наш долг перед
генетикой. Иначе мы не сможем возродить ее в короткий срок". И это было
правдой!
Фигуранты
Однако на этом научно-социальном фоне главным ориентиром для меня
стала научная и преподавательская деятельность в области математической мо-
молекулярной и эволюционной генетики. В 1961 г. в Новосибирск переехал из
Москвы профессор Алексей Андреевич Ляпунов — известный математик, один
из основателей советской кибернетики и страстный энтузиаст математической
биологии. В НГУ он сразу основал кафедру теоретической кибернетики,
дома — семинар по кибернетике и математической биологии, в Институте ма-
математики — лабораторию, а потом — отделение кибернетики. К А.А. потяну-
потянулись молодежь и специалисты. Я, например, был постоянным участником его
семинаров и слушателем лекций.
К 1962/63 г. стало ясно, что молекулярная биология и генетика содержат
огромный новый уникальный материал, который может быть осмыслен с пози-
позиций Теории управляющих систем А.А. Ляпунова. С благословения Д.К. Беляе-
Беляева и А.А. Ляпунова я начал эту работу. Она вылилась в разработку основ теории
молекулярно-генетических систем управления. На эту тему мне удалось защи-
защитить кандидатскую диссертацию. Оппонентами были А.А. Ляпунов, А.А. Ней-
фах, а внешний отзыв дал Н.В. Тимофеев-Ресовский. Этими именами я гор-
горжусь. А.А. Ляпунов не только одобрил работу, но и издал диссертацию как мо-
монографию [Ратнер, 1966] в своей серии "Кибернетические вопросы биологии".
Однако в одиночку поднять такую проблему было невозможно. В русле об-
общих принципов М.А. Лаврентьева о соединении научной работы и подготовки
специалистов назрел вопрос об организации на Кафедре цитологии и генетики
специализации студентов по животрепещущим проблемам математической
26. Математическая генетика в НГУ и около 249
биологии. Это было время начинаний, и наша инициатива прошла успешно.
Мне уже приходилось писать о некоторых занятных особенностях этой эпопеи
[Ратнер, 1998, гл. 3]. Поэтому не буду повторяться. Расскажу только о содержа-
содержательной стороне дела.
Специализация по матбиологии началась с 1968 г., кураторами были
А.А. Ляпунов и я, от деканата ФЕН нам очень помогала замдекана А.А. Титля-
нова, бывшая сотрудница Н.В. Тимофеева-Ресовского. На 3-м курсе биологи-
биологического отделения ФЕН образовалась очень активная группа студентов, кото-
которые просто не давали мне покоя, требуя организации для них обучения по ма-
математической биологии. Среди них были Рустем Чураев, Володя Куличков,
Сережа Родин, Иосиф Медник и некоторые другие. В дальнейшем к ним при-
присоединились Коля Колчанов, Сережа Бажан, Саша Осадчук со следующего
курса, а также четверо прикомандированных к нам студентов-биофизиков из
Красноярского университета: Саша Бачинский, Андрей Дегерменджи, Коля
Абросов и др.
А.А. Ляпунов составил грандиозную программу дополнительного курса ма-
математики для матбиологов на 400 ч. Это тем более впечатляет, что все биологи
тогда уже имели курс математики на 400 ч. Он поручил вести занятия доценту
Н.И. Глебову и другим сотрудникам своей кафедры. Этот курс в несколько мо-
модифицированной форме читается до сих пор. Я прослушал его вместе с первы-
первыми матбиологами, испытал восторг от чистой науки и удовлетворение от воз-
возможности ее приложения к нашим проблемам. Два года я сидел в аудиториях,
почти не занимаясь наукой, впитывая идеи и методы и помогая сформировать
оптимальный для биологов курс.
В этом первом выпуске 1970 г. у меня было шесть (!) дипломников: Рустем
Чураев, Володя Куличков, Иосиф Медник, Сережа Родин, Саша Бачинский и
Витя Козлов. Трое красноярцев (Дегерменджи, Абросов и Шашкин) дипломи-
дипломировались у И.А. Полетаева по системам с лимитирующими факторами. О мно-
многих из них я должен сказать отдельно (некоторые есть на фото).
Рустем Чураев из Уфы появился в Новосибирске школьником 8-го класса
примерно в 1963 г. Тогда его пронзила великая идея описания кибернетической
организации клетки. После случайных блужданий в Уфе и Москве он прибыл в
Новосибирск к А.А. Ляпунову завоевывать мир. А.А. направил его ко мне, а за-
затем предложил ему поступить в ФМШ. Две недели Рустем ходил на занятия, а
потом сказал, что ему неинтересно, он все это знает. Уехал домой, сдал экстер-
экстерном экзамены на аттестат зрелости. А затем поступил к нам на ФЕН НГУ. Я
писал ему похвальную рекомендацию.
Учился он хорошо, хотя иногда неровно. Но сразу же стал ходить в нашу
лабораторию. Научными вопросами занялся очень рано, уже на 3-м курсе мы
обсуждали с ним серьезные проблемы. Дипломная работа у него была разме-
размером > 100 (!) стр. и посвящена математическому моделированию генных сис-
систем. Позже профессор И.И. Кикнадзе, которой он сдавал несколько экзаменов,
откровенно сказала: "Я его боюсь!".
Далее, вместе с Сережей Родиным, они были оставлены в аспирантуре на-
нашего института. Пошли сдавать экзамен по философии и банально провали-
250
III. Молекулярная кибернетика
лись. Пришлось им год проработать старшими лаборантами. Аспирантуру они
закончили успешно, но с опозданием.
На счету Рустема много оригинальных и пионерских идей и разработок:
модели двухоперонного триггера, развития фага X, гипотеза об эпигене и мно-
многое другое. В середине 80-х годов он возвратился в Уфу, потом вскоре защитил
у нас докторскую диссертацию, а затем стал директором Института биологии
БФ УрО РАН в г. Уфе. Там пребывает и сейчас.
Володя Куличков и Ося Медник, бывшие ФыМыШата, были антиподами.
Ося — превосходный математик, способный найти решение задачи, доказать
теорему. Володя — прагматик, способный организовать дело, спортсмен. Ди-
Дипломную работу они делали совместно, а потом разделили ее примерно попо-
пополам. Вместе они моделировали популяционную динамику инверсионного по-
полиморфизма. Для этого требовалось просчитать уравнения динамики на
БЭСМ-6 ВЦ СО АН. За два дня до защиты они еще не получили окончатель-
окончательных результатов. Тогда я им сказал: "Идите на ВЦ, сидите там день и ночь,
обольщайте программисток, делайте что хотите, но чтобы результаты были по-
получены в срок!" За день до защиты они притащили рулоны распечаток и рас-
расстелили их поперек холла нашего института. Потом ползали по ним в поисках
нужных цифр. Зрелище было впечатляющее, их окружала толпа зевак и сочув-
Лаборатория популяционной генетики ИЦиГСО АН СССР, родоначальница Теорети-
Теоретического отдела и ряда других подразделений института, в 1980 г. В первом ряду слева
направо: И.К. Захаров, Э.Х. Гинзбург, Д.П. Фурман, З.С. Никоро, И.Д. Ерохина,
Р.Н. Чураев, М.Д. Голубовский. Во втором ряду слева направо: Л.В. Омельянчук,
И.Н. Шиндялов, Ю.Г. Матушкин, С.Н. Родин, А.А. Жарких, В.В. Соловьев, В.А. Рат-
нер, И.А. Шахмурадов, Н.А. Колчанов (архив ИЦиГ СО РАН).
26. Математическая генетика в НГУ и около 251
ствуюших. Дипломы они зашитили успешно. Программисток, я думаю, оболь-
обольщал Володя Куличков.
После защиты оба они попали в Мединститут, в группу, которая моделиро-
моделировала что-то медицинское. Через полгода там случился скандал, и всю группу
ликвидировали. Ребята остались без работы и пришли ко мне. У меня не было
вакансий, но я им сказал, чтобы они устроились поблизости на любую работу, а
там что-нибудь придумаем. Ося подумал и поехал в Душанбе к родителям. Ра-
Работал там в местной академии, защитил в конце концов диссертацию, но от
науки оторвался. Как сказала его бабушка: "Все говорил — надо дерзать, надо
дерзать! Вот и надерзил!" Сейчас он уехал в США.
Володя пошел грузчиком в Торговый центр, успешно там работал около
полугода. За это время наш институт купил по случаю первую ЭВМ "Наири"
(вспоминаю с ужасом!), и Д.К. Беляев взял его на машину программистом. В
результате он проработал у нас около 10 лет и сделал ряд интересных работ. Ус-
Успешно защитил диссертацию, а потом перешел во ВНИИМБ. Там работает до
сих пор, заведует теоретическим отделом.
Сережа Родин в ФМШ прибыл из Алма-Аты. Учился хорошо. Был очень
импозантным, фотогеничным, хорошо знал английский. Будучи студентом
3-го курса НГУ, колебался в выборе пути. Дело в том, что в детстве он снялся в
каком-то малозаметном фильме о природе и браконьерах. В студенческие годы
где-то в Москве вновь пересекся с киношниками и получил предложение пе-
перейти во ВГИК(!). Однако интерес к науке все-таки взял верх, и он пошел в
матбиологи. Сделал интересную дипломную, а потом кандидатскую работу по
теории комплементации. Артистом не стал, но артистизм и кино-фотогенич-
кино-фотогеничность сохранил. Когда в конце 70-х годов ему пришлось читать лекции в НГУ,
он неизменно пользовался успехом у студенток. Они даже сочинили вдохно-
вдохновенную "Песню о Родине". Потом он защитил докторскую диссертацию по
теории молекулярной эволюции, получил лабораторию в нашем институте. А
затем уехал в США. Там и работает до сих пор в г. Дуарте, Калифорния, в так
называемом City of Норе (Городе Надежды — раковом центре). Говорят — ус-
успешно.
Саша Бачинский приехал из Красноярского ГУ. На биофизику он посту-
поступил случайно, за компанию с приятелем. Но проявил очень хорошие качества.
Учился прекрасно, имел хороший ровный характер, любое дело делал хорошо с
первого раза. Зав. кафедрой биофизики КрасГУ профессор И.И. Гительзон по-
послал его вместе с другими студентами к А.А. Ляпунову, чтобы затем создать тео-
теоретическую группу в Институте биофизики СО АН СССР. Саша сделал не-
несколько прекрасных работ по помехоустойчивости и эволюции генетического
кода. До сих пор я рассказываю эти работы студентам. Успешно защитил ди-
дипломную работу, а после аспирантуры — кандидатскую диссертацию. Затем он
Еернулся в Красноярский ГУ, проработал там несколько лет и потом перешел
во ВНИИМБ заведующим лабораторией. Там работает до сих пор.
Коля Колчанов не был моим дипломником. У него было два руководите-
руководителя по диплому и кандидатской диссертации, А.А. Ляпунов и М.Г. Колпаков,
его область исследования — математическая физиология. Когда он заканчи-
заканчивал аспирантуру, его постигло сразу два несчастья: один за другим умерли
252
HI. Молекулярная кибернетика
оба руководителя. Однако он успешно завершил и защитил диссертацию.
После этого я взял его в нашу матгенетическую группу. У нас он очень быст-
быстро адаптировался, развился и стал очень заметной фигурой в российской
биоинформатике и компьютерной генетике. Он организовал разработку бан-
банков данных и пакетов прикладных программ в области молекулярной биоло-
биологии и генетики. Защитил докторскую диссертацию, получил свою лаборато-
лабораторию в Теоретическом отделе. Сейчас он замдиректора ИЦиГ СО РАН, участ-
участник крупнейших российских и международных проектов по биоин-
биоинформатике, профессор НГУ.
Назову также имена наиболее ярких выпускников-матбиологов последую-
последующих лет. В 1972 г. нашу специализацию окончили два прекрасных студента —
Миша Штабной и Миша Коростышевский. Миша Штабной, действительно
сын офицера, был круглым отличником, очень сильным математиком. Миша
Коростышевский проявил себя весьма оригинальными содержательными во-
вопросами и идеями. В частности, они вместе доказали теорему об условиях
асимптотического невырождения конечных популяций. Эту работу я излагаю и
цитирую до сих пор во всех книгах и считаю ее превосходной. Миша Коросты-
Коростышевский около 20 лет проработал в нашей Медакадемии, защитил диссерта-
диссертацию. Затем уехал в Израиль. Появился у нас летом 2001 г. на конференции.
Сейчас он должен перейти на работу в Университет г. Хайфы, я написал ему
весьма положительную рекомендацию. А его друг Миша Штабной поступил в
Студенты-матбиологи М. Штабной (слева) и М. Коростышевский доказывают теорему
об асимптотическом невырождении популяций (архив автора)
26. Математическая генетика в НГУ и около 253
аспирантуру нашего института и через 2 года просто сгорел от ужасной сарко-
саркомы. Вечная ему память!
Еще один выпускник 1972 г., мой дипломник Фима Фрисман, специалист
по математической теории популяций и экосистем, сейчас доктор наук,
заведующий лабораторией Института автоматики и процессов управления ДВО
РАН во Владивостоке. Он часто появляется в Новосибирске. Судя по всему он
на своем достойном месте, работает и преподает успешно.
Школьником 8-го класса ФМШ пришел к нам в лабораторию сельский
житель Андрей Жарких. Затем он поступил на ФЕН НГУ. Сначала учился скре-
скрещивать дрозофил, потом увлекся математической генетикой. Еще студентом
разработал новый компьютерный метод построения филогенетических деревь-
деревьев, который в дальнейшем стал основой Пакета прикладных программ "Вос-
"Восторг". Окончив НГУ в 1977 г., он стал главным "мотором" Информацион-
Информационно-вычислительного комплекса ИЦиГ. Выполнил ряд прекрасных работ по
теории молекулярной эволюции, защитил кандидатскую диссертацию. Мог бы
давно защитить докторскую, но уехал в США, где работал и работает в лучших
лабораториях и фирмах. Сейчас он находится в штате Юта, в мощной биотех-
биотехнологической фирме. Участвует в полном секвенировании генома риса. Был
здесь в 2001 г. Очень жаль, что он уехал, у него был огромный научный потен-
потенциал.
В 1976 г. пришел в лабораторию студент 2-го курса Володя Шамин из Те-
миртау. Родители его погибли в какой-то аварии. В НГУ ему все помогали
преодолеть сложности судьбы. Он посмотрел на меня светлыми глазами и
сказал, что хочет быть математическим генетиком. В том году мы не смогли
организовать группу матбиологов, и я это ему объяснил. Но он сказал: "Я все
равно буду матбиологом, с группой или без. Значит — сдам все экзамены
сам!" И сдал! У него была удивительная способность преодолевать математи-
математические трудности в задачах. В дипломной работе и после, в нашей лаборато-
лаборатории, он выполнил цикл прекрасных работ по эволюции кольцевых плазмид и
по моделям сайзеров — молекулярных систем самовоспроизведения. Эти ра-
работы вошли во все наши книги по молекулярной эволюции. Недавно в Бель-
Бельгии был открыт сайт в ИНТЕРНЕТе по сайзерам.
Потом Володя заскучал и ушел во ВНИИМБ к В. Куличкову. Там он ус-
успешно проработал несколько лет и снова заскучал. Перешел обратно к нам в
институт, в лабораторию Н.А. Колчанова. Собрался написать диссертацию, но
снова заскучал. И бросил науку. Насколько я знаю, сейчас он профессионально
играет в бридж. Видимо, выигрывает. Мне кажется, что ему не хватило терпе-
терпения. Он работал быстро и эффективно. И ждал столь же быстрого признания. А
оно, как всегда, запаздывало. Вот и ушел в бридж. А жаль, он несомненно та-
талантлив!
В 1985 г. специализацию закончила "великолепная четверка": Саша Кель,
Игорь Рогозин, Андрей Ржецкий и Люба Задорина. Всех оставили в институте.
Сейчас Саша Кель работает у нас и много времени проводит в Германии. Игорь
Рогозин уехал в США. Оба они кандидаты наук, прекрасные специалисты по
компьютерной генетике и биоинформатике. Андрей Ржецкий тоже уехал в
США, где неплохо работает в Колумбийском университете. Люба уехала в свой
254
III. Молекулярная кибернетика
родной Магадан, изучала там, кажется, птиц. Или рыб? Но потом прошла по
конкурсу Соросовского фонда и уехала в Центральную Европу. Сейчас она в
Испании, вышла там замуж. Все вспоминают ее с удовольствием. Нашими нау-
науками не занимается. Это очень жаль, так как девочка была очень способная.
С конца 70-х годов у нас появились студенты-физики. Их стала нам давать
ежегодно кафедра академика Ю.Н. Молина. Многие из них были успешно
"переварены" в молекулярной биологии, генетике и эволюции и стали перво-
первоклассными специалистами. Первым был Витя Соловьев. Защитив диплом и по-
получив рекомендацию в аспирантуру, он отправился летом на Чукотку команди-
командиром студотряда на рыбную путину. Не знаю, сколько они там поймали рыбы,
но по ходу трудового процесса он повздорил с местной властью, за что был за-
задержан милицией. Мы получили телеграмму с Чукотки с просьбой прислать ха-
характеристику "злодея". Характеристику, конечно, послали самую положитель-
положительную, с рекомендацией в аспирантуру. Просили не задерживать надежду нашей
науки, так как начинаются приемные экзамены. Наносить ущерб Родине мест-
местная власть не стала. Все кончилось благополучно. Витя Соловьев разработал
компьютерные методы моделирования самоорганизации РНК и белков, контек-
контекстного анализа последовательностей и другие вопросы. Защитил хорошую кан-
кандидатскую диссертацию. Потом уехал в
США, затем работал в Сенджеровском
центре компьютерной генетики, в анг-
английском Кембридже. Сейчас снова в
США. Витя активно сотрудничает с на-
нашими лабораториями. Мог бы без со-
сомнения защитить докторскую.
Превосходным математическим
генетиком стал также физик Миша
Пономаренко. Человек очень широких
интересов и возможностей — от про-
пространственной структуры белков до ге-
генетической лингвистики и методоло-
методологии науки, он активно развивается в
нашем институте, является сейчас зам-
завлабораторией теоретической гене-
генетики у Н.А. Колчанова. У него, несо-
несомненно, высокий потенциал и хорошее
будущее.
Оригинальный путь выбрал фи-
зик-матбиолог Леня Омельянчук. За-
Защитив отличную дипломную работу и
кандидатскую диссертацию по теории
молекулярной эволюции, он ушел в
экспериментальную генетику дрозофи-
Матбиолог Игорь Ищенко, талантливый лы- Сейчас заведует лабораторией в
теоретик, безвременно погибший в 1997 г. ИЦиГ, защитил докторскую диссерта-
(архив автора). цию.
26. Математическая генетика в НГУ и около 255
И многие другие: физики Илья Шиндялов, Витя Стрелец, Андрей Птицын,
выпускник НЭТИ Володя Капитонов сейчас работают в разных концах США.
Туда же уехали супружеские пары матбиологов: Павел Морозов и Ира Клику-
нова, Юра Вульф и Кира Макарова. Юра и Кира после университета поступили
в аспирантуру к В. Соловьеву как раз перед его отбытием в США. Аспирантура
"подвисла" без руководителя. Я был вынужден забрать их в свою лабораторию,
где они успешно защитились по теории молекулярной эволюции. Они стали хоро-
хорошими специалистами. В США работают в Бетесде, Институте Здоровья, занима-
занимаются эволюционной геномикой у крупнейшего специалиста в этой области Е. Ку-
нина. Поддерживают с нами активную связь по электронной почте. Математики
из Баку Илхам Шахмурадов и Асаф Саламов закончили у нас аспирантуру, успеш-
успешно защитили диссертации и продолжают работать в различных местах планеты.
Хотелось бы также вспомнить физика Игоря Ищенко, прекрасного про-
программиста и теоретика, любимца всех, кто его знал, трагически погибшего ле-
летом 1997 г. Вечная ему память!
Уже заметно следующее поколение. Оно внушает надежду на будущее.
Амбула
Итак, специализация "математическая биология" существует с 1968 г., т.е.
свыше 30 лет. За это время выпущено свыше 120 специалистов матбиологов вы-
высокого класса, биологов и физиков. Большинство из них работают в лаборато-
лабораториях ИЦиГ СО РАН и "Векторе" в Новосибирске, а также в Красноярске, Вла-
Владивостоке, Иркутске, Москве, Севастополе и других городах России и СНГ.
Многие уехали за границу: в США, Великобританию, Германию, Израиль и др.
Спрос на таких специалистов очень велик. Фактически их нигде так не готовят,
как в нашей специализации.
Эпоха массового секвенирования ДНК, РНК, белков и использования
компьютеров породила новую область математической биологии — биоинфор-
биоинформатику и компьютерную генетику. Начав еще в 60-е годы, мы оказались подго-
подготовленными к этой эпохе и быстро включились в работу. С помощью моего
старого друга Э.И. Елинера, начальника ВЦ ИЯФ СО АН СССР, нам удалось
приобрести первые мини-ЭВМ и организовать в ИЦиГ Информационно-вы-
Информационно-вычислительный комплекс. Специализация вовремя подготовила матбиологов,
способных осуществить весь цикл теоретических, математических и компью-
компьютерных работ, которых требует научный профиль. Их руками и выполнена ос-
основная работа. Это уже поняли многие наши иностранные коллеги, которые
сейчас испытывают большие трудности: в биоинформатику у них приходят уз-
узкие специалисты, "технари", не имеющие собственных теоретико-биологиче-
теоретико-биологических интересов. Рейтинг наших выпускников высок, их активно приглашают
на работу, в аспирантуру. Некоторые западные университеты, особенно в Гер-
Германии, интересуются нашим опытом подготовки матбиологов.
Примерно в середине 70-х годов началась массовая отдача результатов на-
нашей работы — диссертации, сборники, книги, статьи. Все это сопровождалось
бурным самовыражением участников эпопеи. Как говорил Н.В. Тимофеев-Ре-
256
HI. Молекулярная кибернетика
совский: "Не надо относиться к науке со звериной серьезностью!" Гл. 27 книги
в какой-то степени отражает общее состояние "духа" участников этой эпопеи.
Большинство студентов, выбравших матбиологию, были выпускниками
ФМШ. Они были пытливы, развиты, не имели трудностей с компьютерами,
рано приходили в институт, легко вливались в коллективы. Первые матбиологи
зажглись энтузиазмом после ярких лекций А.А. Ляпунова. Некоторые прочли
популярные статьи и книги по генетике, в том числе наши. Многие преодолели
большие трудности, чтобы стать матбиологами: переводились из других уни-
университетов, подрабатывали где только могли, работали ночами на компьютерах
по графику, преодолели огромные учебные перегрузки и т.д.
Восемь выпускников матбиологов стали докторами наук, четверо из них —
директорами и заместителями директоров крупных академических или ведом-
ведомственных институтов. Число кандидатов наук мне трудно подсчитать, но, веро-
вероятно, их не меньше 40. Десятки матбиологов являются заведующими отделами,
лабораториями, секторами и группами. Аналогичные позиции они занимают и
за границей. Иначе говоря, составляют реальный костяк математической гене-
генетики и биоинформатики в нашей стране и частично за рубежом.
На заседании кафедры цитологии и генетики ФЕН НГУ, профессор Л.И. Корочкин
(слева), профессор В.А. Ратнер (конец 1970-х) (архив ИЦиГ СО РАН).
26. Математическая генетика в НГУ и около
257
Руководителями и выпускниками специализации опубликовано не менее 400
работ по различным разделам математической биологии и биоинформатики, де-
десятки сборников и книг, в том числе за границей. Организованы десятки школ,
конференций, симпозиумов, в том числе несколько международных конферен-
конференций в Новосибирске [Ratner, Kolchanov, 1992; BGRS-98, 2000; ИНПРИМ-96, 98,
2000]. Сделано несколько сот докладов. Например, на 2-м, 3-м и 4-м Сибирских
конгрессах по прикладной и индустриальной математике [ИНПРИМ-96, 98,
2000], в секции математической биологии каждый раз бывало до 50 докладов, из
которых 2/3 — продукция наших выпускников матбиологов. Примерно такую же
долю составляли доклады руководителей и выпускников специализации на Меж-
Международном симпозиуме BGRS-98 и BGRS-2000.
В 1996 г. ИНПРИМ был посвящен памяти А.А. Ляпунова, И.А. Полетаева
и А.Н. Ершова. На нем встретились многочисленные ученики и соратники ос-
основателей кибернетики и матбиологии в России, Новосибирске и НГУ, при-
приехавшие со всех концов страны и света. Многие из них приняли участие в сбор-
сборнике "Очерки истории информатики в России" [Поспелов, Фет, 1998], где соб-
собраны воспоминания и документы 50-60-х годов.
Перспективы математической биологии, компьютерной генетики и биоин-
биоинформатики огромны. В наступающем столетии их роль будет стремительно воз-
возрастать. Поэтому подготовка матбиологов должна быть укреплена и расшире-
Наш добрый компьютерный гений Э.И. Елинер (справа), без помощи которого у нас не
было бы никаких компьютеров в начале 1980-х. Новосибирск, 1997 г. (архив автора).
258 III. Молекулярная кибернетика
на. При небольших вложениях в НГУ можно организовать выгодное междуна-
международное сотрудничество, подготовку студентов и аспирантов из-за рубежа,
особенно из стран Востока. Реально сотрудничество в этой области с универси-
университетами Германии, Израиля, стран Восточной Европы и СНГ.
Иначе говоря, все впереди!
Список литературы
ИНПРИМ-96. Тезисы докладов. Секция математической биологии / Ред. В.А. Ратнер.
Новосибирск: ИМ СО РАН, 1996. С. 17-39.
ИНПРИМ-98. Тезисы докладов. Ч. IV. Секция математической биологии/ Ред.
В.А. Ратнер. Новосибирск: ИМ СО РАН, 1998. С. 109-132.
ИНПРИМ-2000. Тезисы докладов. Ч. III. Секция математической биологии. Новоси-
Новосибирск, 2000. С. 57-78.
Очерки истории информатики в России / Ред. Д.А. Поспелов, Я.И. Фет. Новосибирск:
НИЦ ОИГГМ СО РАН, 1998. 662 с.
Ратнер В.А. Генетические управляющие системы. Новосибирск: Наука, 1966.
Ратнер В.А. Молекулярная генетика: принципы и механизмы. Новосибирск: Наука,
1983.
Ратнер В.А. Алексей Андреевич Ляпунов // Очерки истории информатики в России /
Ред. Д.А. Поспелов, Я.И. Фет. Новосибирск: НИЦ ОИГГМ СО РАН, 1998. С. 379-386.
Modeling and Computer Methods in Molecular Biology & Genetics / Eds. V.A. Ratner,
N.A. Kolchanov. N.Y.: Nova Sci. Publ., 1992.
Proceedings of the First International Conference on Bioinformatics of Genome Regulation and
Structure / Eds. N.A. Kolchanov, V.V. Solovyov. Novosibirsk: ICG SB RAS, 1998. Parts 1 and 2.
Proceedings of the Second International Conference on Bioinformatics of Genome Regulation
and Structure / Eds. N.A. Kolchanov, R.Hofestadt. Novosibirsk: ICG SB RAS, 2000. Parts 1 and 2.
27. Те же проблемы, но в стихах
Сказка
Встает сентябрьское утро,
Прохладу ночи сохраня.
В разломах рош зеленокудрых
Ночные блики гомонят.
Ночные запахи пока что
Дрожат и в хвое, и в листве,
Но предвещает шелест каждый,
Что нечто будет в естестве!
Земля легка, земля суха,
Полна остывшего песка,
Полна листвы, и трав, и мхов,
Сосновых шишек и сучков,
И под иссохшими пеньками
Населена не червяками —
Букашками сосновыми,
А старичками-гномами.
27. Те же проблемы, но в стихах 259
Они должны тут где-то жить,
Иначе сказки не сложить!
Они распахивают лес
Не удержавшись от чудес.
Без восклицательных словес
Включают день и свет небес...
И небо, небо голубое
Тотчас становится без дна,
И там, в вершине, над тобою,
Едва заметная, одна,
Как будто выписана краской, —
Блестит серебряная нить,
И больше не на чем мне глаз свой
Хотя б на миг остановить!
А здесь, внизу, средь пыльной мари,
Средь чада сизого дымов,
Всплывают, как мираж в Сахаре,
Кварталы сказочных домов,
Они белеют, растворяясь
Там, где намечен горизонт, —
Чуть зыбки, каждый повторяясь
На свой особенный фасон.
Они белеют вдоль дороги,
Как будто не было глуши,
И, как обычно, на пороге
Играют чьи-то малыши,
Мелькают радостные лица,
Беседы, споров ручейки...
И так мне хочется напиться
Из этой огненной реки,
Так я глотаю эти звуки,
Так ощущаю их накал,
Так слышу пульс большой науки
В туманных глыбах городка,
Так я хочу здесь делать дело,
И вместе, рядом с ними жить,
Что, право, черту душу смело
Готов сейчас же заложить.
Но черта нет, а только — гномы,
И я иду по их следам,
260 III. Молекулярная кибернетика
Очарованием ведомый,
По рытвинам — как по садам!
Иду и впитываю краски,
И все стучится в голове:
"Да, это — сказка, если сказки
Еще бывают в естестве!
Да, это сказочная встреча,
Неописуемый полет..."
А в небе,
В небе бесконечном
Все кувыркался самолет...
1959
Любопытно!
Любопытно сидеть
Средь ученых мужей!
Любопытно глазеть
На ученых ужей,
Как втыкают друг другу ученые
Золоченые
Шпильки точеные!
Словно гуси —
Сидят преуспевшие,
Как лягухи —
Галдят потерпевшие,
Словно мухи —
Кругом любопытные,
Лопоухи,
Как парнокопытные.
И дебаты идут,
И баталии,
И защиты,
Порою скандальные,
А ученые важные степени
Разливаются лаком
По темени.
А в итоге,
Светла и таинственна,
Возникает упрямая истина.
В синяках,
Как ей и полагается!
И — смотрите-ка! —
Улыбается!
1961
27. Те же проблемы, но в стихах 261
Галилей
Посвящается памяти
И. И. Шмалъгаузена
От робости алея,
Коленками дрожа,
Я руку Галилея
В отчаяньи пожал.
Он стар и тих,
И очень сед,
Ему недолго жить,
И каждым днем последних лет
Он должен дорожить,
Но десять лет усилий
Затоптаны в земле...
Да он и по фамилии
Совсем не Галилей,
И имя очень русское,
И отчество — как раз,
И выраженье грустное
У старых синих глаз,
Усталые движения
И тяжесть на плечах...
Но гнев омоложения
Гремел в его речах!
Заруганный халдеями,
Один средь тяжких дум,
Идеями,
Идеями
Был полон этот ум!
Он торопился,
Он спешил,
Он все хотел отдать,
И он давным-давно решил
Под занавес сказать,
Как тот,
Испивший все до дна,
Великий и неистовый:
"А все же вертится она!"
И — выдержать,
И — выстоять!
1961
262 III. Молекулярная кибернетика
Дискуссия на тему "Человек, жизнь и кибернетика"
Точка зрения литератора
Говорят, поэта можно
Заменить теперь машиной —
Без ошибок строфы вяжет,
Рифмы помнит наизусть,
Правда, стиль еще корявый,
Но на это есть причина —
Молода еще, незрела,
Наберется...
Ну и пусть!
Ну и пусть себе строгает
Скороспелые стишата!
Я не стану с ней тягаться,
Но не стану и читать.
А скажите, не сумеет ли
Она из суррогата
Человечий сделать разум,
Научив его мечтать!
Коль сумеет, я теперь же
Человечье наше званье
Без раздумья, без сомненья
Положу к ее ногам —
Все, что коплено годами,
Все свое образованье,
Все, что есть во мне святого,
Без остатка — все отдам!
Только где же эти ноги?
Впрочем, шутки неуместны!
Я ж забыл, что только тварям
Полагается ходить.
Но скажу вам, между прочим,
В отношении телесном
Не удастся ей живого
Ни состряпать, ни родить.
Не удастся из мякины
Ей создать такое тело,
Чтобы жило и плодило
И отважилось мечтать!
Что ж, давай, хитра-машина,
Не словами, а на деле —
Кто скорей из нас сумеет
Жизнь треклятую зачать?
27. Те же проблемы, но в стихах 263
Точка зрения математика
Посвящается А.Н. Колмогорову
Все живое есть машина —
Ты и он, и вы, и я.
Но не всякая машина
Попадает в ранг живья!
Безразлично, из чего мы —
Из белков иль шестерен,
Но любопытно — для чего мы,
И какой у нас фасон.
Да побольше бы извилин,
Да поменьше б настроений,
Был бы хабитус цивилен
В продолженьи поколений.
Только были бы такие
Средь машин,
Чтобы глазки голубые
У машин,
Чтобы губки были алы,
Как коралл,
Чтобы юбки были в талию...
Пропал!
Точка зрения биолога
Посвящается себе
Мы — за союз испытателей,
Эмпириков и мечтателей,
Физиков и генетиков,
Матемо-кибернетиков
И всяких других еретиков.
Минуя большие торосы,
Мы ставим большие вопросы,
Даем перспективные темы,
Величественные проблемы.
Мы — за вниманье к веществу,
И к естеству,
И к существу,
Мы ждем во сне и наяву,
Что кто-нибудь поможет,
Но с уваженьем к большинству
И с подчиненьем меньшинству,
Мы ничего по существу,
Увы, сказать не сможем...
264 III. Молекулярная кибернетика
Точка зрения химика
Посвящается Л.С. Сандахчиеву
Скажу вам вот что, старики,
Мне стало как-то не с руки,
Вдруг захотелось побежать,
Долить, отлить и размешать.
А после этих важных дел
Я вот что вам сказать хотел.
Стоит всего один вопрос,
Но ты, старик, не вешай нос,
Ведь все вокруг, уверен я, —
Мура, зола и чешуя!
Вопрос же вот в чем состоит:
У жизни — только внешний вид,
А в глубине того живья —
Мура, зола и чешуя!
Вот если б это разложить,
Да тут же заново сложить,
Да подкислить,
Да подсолить,
Да возогнать,
Да осадить,
А если попросту сказать, —
Экспер-
мен-
тально
Доказать!
Тогда бы, каюсь, старики,
Мне было больше бы с руки!
Но тут я больше не могу,
Вас оставляю и бегу,
А на бегу замечу я,
Что все — зола и чешуя!
Точка зрения обывателя
А я тут сижу да слушаю —
О чем это люди спорят.
Мороженого откушал
На выходе в коридоре,
Потом побежал в курилку,
Оттедова — в туалет,
Пивка раздавил бутылку,
Зайдя по дороге в буфет.
27. Те же проблемы, но в стихах
265
Побаловался леденцами,
Обертки же прямо под стул
Пихнул — и дело с концами,
Да чуть было здесь не уснул.
Но вдруг все вокруг загалдели,
Что надо как будто вроде
Узнать, что об ентом деле
Думают в гуще народа,
И вот я оттедова вышел
И с вами веду разговор,
Да только слегка недослышал,
О чем, бишь, ведется спор...
Точка зрения философа (резюме)
Посвящается И. Матвеенкову
Вы все, ко-нечно,
Во всем не-правы —
И те, кто слева,
И те, кто справа.
А те, кто сверху,
И те, кто снизу,
Уже впа-дают
В иде-а-лизм!
А в за-клю-ченье
Вас за-ве-ряю,
Что раз-но-чтений
Не о-до-бряю,
И при-зы-ваю:
"Вперед — к ци-тате,
Без дог-ма-тизьмов
И от-се-бятин!"...
1964
Однако так как
Все ре-ля-тивно,
Ничто люд-ское
Нам не про-тивно,
Мы с лю-бо-пытством
Язвим при-роду,
И ле-зем в воду,
Не зна-я броду.
266
III. Молекулярная кибернетика
Сотворение основ*
Посвящается АЛ.Яяпунову
в день 60-летия
1. Прелюдия
В ту ночь задумчиво Создатель
Бродил по выцветшей земле.
И нимб на Боговом челе
Сиял печатью благодати...
И думал Он...
Томился дух Его без дела.
Мысль развивалась без предела,
Как экспонента, — в небеса
И возвращалась в полчаса.
Как вдруг средь облачных развилин,
Средь сокровенных Божьих снов
Мятежным краешком извилин
Узрел Он КОНТУРЫ ОСНОВ!
•40
Г*>
Рисунки из книги Ж. Эффеля "Сотворение мира". М: Изобр. искусство. 1984. (Надписи на
27. Те же проблемы, но в стихах
267
2. Творение
И не моргнув державным глазом,
Взглянув в ядро первопричин,
Он повелел державным гласом
Создать основы дисциплин:
Устлать полы дискретным матом,
Из диа-мата дом сложить,
Многоэтажным био-матом
Его покрыть и обложить.
Велел создать в развитье темы
Он СЕМИНАР всеобщий, где мы
Могли б за рюмкой "КИБЕРНЭ"
Решать великие "ПРОБЛЕМЫ",
Поскольку истина - в вине.
268
III. Молекулярная кибернетика
3. Разверзлась твердь
И благолепие настало.
Присев на тучку, Бог-Отец
Вздремнуть собрался, наконец,
Как вдруг земля затрепетала,
Дыра возникла. А из ней
Хвостатый поднялся злодей.
4. Сомнение
Какими сладкими речами
Он ум адамов окрутил!
Какими ясными очами
Он душу чистую смутил!
27. Те же проблемы, но в стихах
269
И в бесконечности Вселенной,
Подрыгав нервною ногой,
Нашел ЛИМИТ — один, другой, —
В движенье плавном переменных...
5. Искупление
Но нет!
Адам был нашей веры!
Все искушенья перенес,
И вспомнив кстати контр-примеры,
Злодею он наставил нос.
270
III. Молекулярная кибернетика
И крестным осенив знаменьем
Себя от чуба до пупа,
Он понял: истина тупа,
Коль не очищена сомненьем!
А в лоне праведной науки,
Как Основатель нас учил,
Какие он придумал штуки!
Какие леммы отмочил!
И лишь однажды, в воскресенье,
Он вспомнил бесово ученье
И... Еву в жены получил!
Not for sale! For preview only! Scanned by Alykhaylo (molbioKwukr.net)
27. Те же проблемы, но в стихах
271
6. Апофеозис
Ну, а теперь, без лишних слов
Ура - СОЗДАТЕЛЮ ОСНОВ!
Поднимем рюмки "Кибернэ",
Поскольку истина — в вине!
1971
Not for sale !!!
For preview only !!!
If You like this book - buy it!
Scanned by Mykhaylo (molbiolfSjukr.net)
Оглавление
ОТ РЕДАКТОРА 4
ПРЕДИСЛОВИЕ 5
ВВЕДЕНИЕ 6
I. PERSONALIA 10
1. Мамонт (заметки о Н.В. Тимофееве-Ресовском) 10
2. Николай Владимирович Тимофеев-Ресовский — ровесник века, гигант столетия 21
3. Алексей Андреевич Ляпунов 32
4. Игорь Андреевич Полетаев 42
5. С.М.Гершензон: впереди событий и в стороне от признания 48
6. Дмитрий Константинович Беляев 54
7. Памяти профессора Francesco M. Scudo 64
8. Памяти академика Н.Н. Моисеева 67
9. Поездка в Испанию. Памяти Н.Н. Воронцова 69
II. ГЕНЕТИКА 75
10. 100 лет — 18 конгрессов! 75
11. Концепция генов в развитии 81
12. Что содержит полный геном Escherichia coli? 96
13. Индукция транспозиций мобильных генетических элементов стрессовыми воздей-
воздействиями 104
14. Мобильные генетические элементы: геномные бродяги или трудяги? 113
III. МОЛЕКУЛЯРНАЯ КИБЕРНЕТИКА 122
15. Хроника великого открытия: идеи и лица. I 122
16. Хроника великого открытия: идеи и лица. II 136
17. Хроника великого открытия: идеи и лица. 111 151
18. Молекулярная кибернетика в океане науки 167
19. Виртуальная реальность и системы управления 180
20. Генетический код как система 190
21. Генетический код как продукт эволюции 197
22. Закономерности кодирования генетической информации (генетический язык). . . 203
23. Генетические переключатели 218
24. Молекулярная эволюция 229
25. Укус издыхающей рептилии 238
26. Математическая генетика в НГУ и около 245
27. Те же проблемы, но в стихах 258
Научное издание
Ратнер Вадим Александрович
ГЕНЕТИКА, МОЛЕКУЛЯРНАЯ КИБЕРНЕТИКА
ЛИЧНОСТИ И ПРОБЛЕМЫ
Редакторы Л.В. Филиппова, Л.П. Голышева. Художественный редактор Л.В. Матвеева. Художник И.С. Попов. Технический редактор
Н.М. Остроумова. Корректор ИЛ. Малышева. Оператор электронной верстки Л.А. Антонова.
Изд. лиц. N° 020297 от 23.07.97. Сдано в набор 25.06.02. Подписано в печать 23.09.02. Бумага легкомелованная. Формат 70x100 I/I6.
Офсетная печать. Усл. печ. л. 21,9+0,88 вкл. на мел. бум. Уч.-изд. л. 22,8. Тираж 1000 экз. Заказ N° 162.
"Наука". Сибирская издательская фирма РАН. 630099, Новосибирск, ул. Советская, 18.
Оригинал-макет изготовлен на настольной издательской системе.
ГП "Новосибирский полиграфкомбинат". 630007, Новосибирск, Красный проспект, 22.