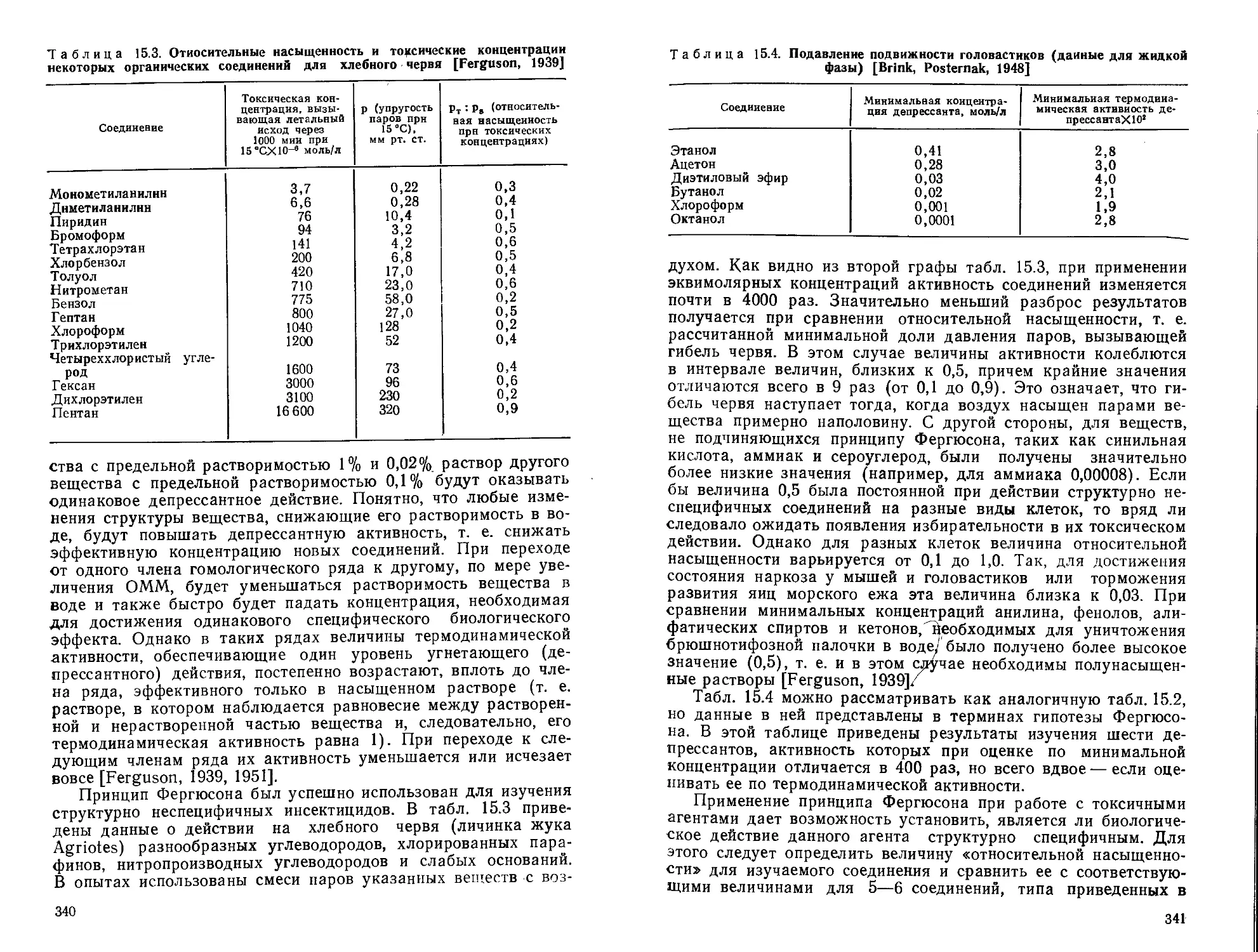
/





















































































































































































































Author: Альберт А.
Tags: лекарственные средства в соответствии с их активным веществом фармакология фармация токсикология химия основы терапии
ISBN: 5-225-01518-2
Year: 1989




Text
SELECTIVE
TOXICITY
The physico-chemical basis of therapy
SEVENTH EDITION
ADRIEN ALBERT
D.Sc. (bond.), Ph.D. Medicine (bond.), Fellow of the Australian Academy of Science Professor Emeritus, Department of Chemistry, Australian National University, Canberra;
Research Professor, Department of Pharmacological Sciences, School of Medicine, State University of New York, Stony Brook,
London New York
Chapman and Hall
А. АЛЬБЕРТ
ИЗБИРАТЕЛЬНАЯ ТОКСИЧНОСТЬ
ФИЗИКО-ХИМИЧЕСКИЕ
ОСНОВЫ ТЕРАПИИ
В 2 ТОМАХ
ТОМ 2
Перевод с английского канд. хим. наук М. А. ДУМПИС, М. Б. ГАНИНОЙ
Под редакцией проф. В. А. ФИЛОВА
МОСКВА«МЕДИЦИНА» 1989
ББК 52.8
А56
УДК 615.2/.3.065
Издание рекомендовано для перевода чл.-кор. АМН СССР |Лакиным К. м|, зав. каф. фармакологии ММСИ им. Н. А. Семашко
Альберт А.
А56 Избирательная токсичность. Физико-химические основы терапии. Пер. с англ. В 2 томах. Т. 2. — М.: Медицина, 1989, 432 с.: ил.
ISBN 5—225—01518—2.
ISBN 412—26010—7
В монографии рассмотрены современные вопросы избирательной токсичности различных веществ. Во 2-м томе описано токсическое действие антиметаболитов, ферментов, витаминов и др. Рассмотрены вопросы ионизации и ионизирующего действия отдельных веществ. Представлены известные и новые лекарственные вещества, влияющие на иммунные процессы и используемые в онкологии, психиатрии, кардиологии, анестезиологии и др. Освещено повреждающее и стабилизирующее действие биологически активных веществ.
Для практических врачей, фармакологов, токсикологов, биохимиков
и химиков.
4107030000—285
А----—----------75 — 89
039(01)—89
ISBN 5—225—01518—2
ISBN 412—26010—7
ББК 52.8
© Перевод на русский язык.
© Издательство «Медицина», Издательство «Медицина», Москва, 1989
ОГЛАВЛЕНИЕ
Глава 9. Антиметаболиты: аналоги коферментов и субстратов фермен-
тов, обладающие антагонистическим действием .... 8
9.0. Ферменты................................................ 8
9.L Антиметаболиты: определение, происхождение и механизм действия................................................ . 16
9.2. История изучения антиметаболитов до 1940 г. 24
9.3. Антагонисты фолиевой кислоты............................25
9.4. Другие аналоги метаболитов, играющие важную роль в профилактике и терапии.......................................45
9.5. Ингибиторы «переходного состояния»......................59
9.6. Последовательное блокирование...........................63
9.7. Аналоги метаболитов, образующие ковалентные связи 65
9.8. Особые случаи взаимосвязи между агонистами и антагонистами .......................................................70
9.9. Фармакогенетика.........................................72
Глава 10. Ионизация.................................................74
10.0. Природа ионизации.......................................74
10.1. Константа ионизации (Ка)................................76
10.2. Различия в ионизации, обеспечивающие избирательность 81
10.3. Вещества, обладающие большей биологической активностью в ионизированном состоянии ............................. 89
10.4. Вещества, менее активные в ионизированном состоянии 120
10.5. Вещества, проявляющие биологическое действие в виде ионов и неионизированных молекул........................122
10.6. Ионизация рецепторов...................................126
10.7. Заключение.............................................129
Глава 11. Вещества, связывающие металлы............................130
11.0. Металлы в живой клетке............................131
11.1. Биохимические различия, способствующие избирательности 144
11.2. Химизм хелатообразования..........................151
11.3. Количественные аспекты связывания металлов . . . 154
11.4. Химические различия, способствующие избирательности 160
11.5. Различные механизмы биологического действия хелатирующих агентов (введение) .................................... 166
11.6. Уменьшение токсического действия металла в результате хелатообразования, антидоты............................. . 169
11.7. Уменьшение токсического действия металлов хелатообразованием ....................................................173
11.8. Тетрациклины...........................................184
11.9. Вещества, биологическое действие которых отчасти обусловлено хелатообразованием ................................. 186
11.10. Особые случаи образования прочных комплексов . . 193
11.11. Основные принципы создания новых хелатирующих агентов, перспективы их применения...............................195
Глава 12. Стерические факторы......................................197
12.0. Некоторые основные принципы............................197
12.1. Оптическая изомерия....................................199
12.2. Геометрическая изомерия................................208
5
12.3. Конформация...........................................213
12.4. Рецепторы катехоламинов...............................218
12.5. Ацетилхолинэстераза...................................229
12.6. Холинорецепторы.......................................231
12.7. Рецепторы ГАМК и бензодиазепины.......................250
12.8. Морфин и рецепторы опиатов............................253
12.9. Психотерапевтические агенты...........................260
12.10. Заключение...........................................265
Глава 13. Ковалентная связь и избирательная токсичность 266
13.0. Препараты мышьяка, сурьмы и ртути...............266
13.1. Пенициллины...........................................272
13.2. Цефалоспорины и другие fJ-лактамные ингибиторы образования клеточных стенок.....................................279
13.3. Фосфорорганические соединения и карбаматы .... 284
13.4. Алкилирующие агенты...................................295
13.5. Летальный синтез......................................299
13.6. Различные примеры.....................................307
Глава 14. Химия поверхностных явлений, модификация мембран по-верхиостио-активиыми соединениями.................................310
14.0. Поверхностные явления in vitro........................310
14.1. Поверхностные явления и действие лекарств, диуретики, сердечные гликозиды...................................312
14.2. Ионофоры........................................319
14.3. Повреждение мембран биологически активными агентами 326
14.4. Защита мембран биологически активными агентами . . 332
Глава 15. Биологическая активность, не связанная со структурой 335
15.0. Общие биологические депрессанты (снотворные средства, общие анестетики, летучие инсектициды).....................335
15.1. Соединения, нарушающие митоз..........................350
Глава 16. Совершенствование путей поиска лекарственных веществ 351
16.0. Множественный регрессионный анализ....................352
16.1. Альтернативные методы.................................358
16.2. Стереохимический подход...............................360
Глава 17. Некоторые количественные данные........................370
17.0. Расчет степени ионизации по данным рКа и pH . . . 370
17.1. Фрагментарные гидрофобные константы и коэффициенты распределения ............................................. 370
17.2. Электронные эффекты (значения о-констант Гаммета и Тафта)......................................................373
17.3. Ядерный магнитный резонанс........................378
17.4. Поиск литературных данных.........................385
Список литературы.................................................388
Предметный указатель..............................................417
СПИСОК СОКРАЩЕНИЙ
АД — артериальное давление НАД-Н
АДФ — аденозиндифосфат
АМФ — аденозинмонофосфат НАДФН
АТФ — аденозинтрифосфат
АХ — ацетилхолин
АХЭ — ацетилхолинэстераза НИАЦФ
ВОЗ — Всемирная организа-
ция здравоохранения ОММ
ГАМК — гамма-аминомасля-
ная кислота
ГТФ — гуанозинтрифосфат ПАБ
ГЭБ — гематоэнцефаличе- пдп
ский барьер
дмдк — диметидилдитио- ПМР
карбаминовая кисло-
та рнк
ДНК — дезоксирибонуклеи-
новая кислота тгн
дгфк — дигидрофолиевая кис-
лота ттг
ДГФР — дигидрофолатредук- УТФ
таза ФАНИ
ИТФ — инозинтрифосфат
комт — катехоламино-О-ме-
тилтрансфераза ФАД
лг — лютеинезирующий
гормон ФАО
МАИР — Международное агент-
ство по изучению рака
МАО — моноаминоксидаза АхГ Г*
м. д. — миллионная доля ФС1
мдп — маниакально-депрес- цне
МКА сивный психоз
— минимальная концент-
рация, вызывающая ЦТФ
МСР анестезию — минимальные стери- ЭПР
МТР ческие различия — минимальные тополо- ЭДТА
гические различия ЭР
— никотинамидаденин-динуклеотид восстановленный
— никотинамидаденин-динуклеотидфосфат восстановленный
— необратимый ингибитор активного центра ферментов
— относительная молекулярная масса
— пара-аминобензойная кислота
— пептилдипептидаза
— протонный магнитный резонанс
— рибонуклеиновая кислота
— Д9-тетрагидроканна-бинол
— тиреотропный гормон
— уридинтрифосфат
— ферментативно-активируемый необратимый ингибитор
— флавинадениндинук-леотид
— Продовольственная и сельскохозяйственная организация Объединенных Наций
— фолликулостимулирующий гормон
— центральная нервная система
— цитидинтрифосфат
— электронный парамагнитный резонанс
— этилендиаминтетра-уксусная кислота
— эндоплазматический ретикулум
Глава 9
АНТИМЕТАБОЛИТЫ: АНАЛОГИ КОФЕРМЕНТОВ И СУБСТРАТОВ ФЕРМЕНТОВ, ОБЛАДАЮЩИЕ АНТАГОНИСТИЧЕСКИМ ДЕЙСТВИЕМ
В главе 2 (том 1) были рассмотрены примеры того, как незначительные изменения химической структуры избирательно действующих токсических агентов приводят к радикальным изменениям их биологической активности. В настоящей главе мы подробно рассмотрим один из случаев высокой специфичности, а именно соответствие между нормальным субстратом (или коферментом) фермента и агентом, ингибирующим этот фермент. В предыдущих изданиях мы называли эти агенты «аналогами метаболитов», но с тех пор слово «метаболит» приобрело более широкое значение.
С точки зрения биохимика «метаболиты» — это полупродукты метаболизма, такие как пируват-ион, жирные кислоты, кислород и нуклеотиды. Тем не менее многие фармакологи используют это слово для обозначения продуктов распада ксенобиотиков (инородных веществ). В настоящей книге под «антиметаболитами» мы будем иметь в виду аналоги коферментов и субстратов ферментов, обладающие антагонистическим действием. Такие вещества называют также «ингибиторами метаболитов».
9.0. Ферменты
Благодаря процессам роста и деления клетка находится в состоянии непрерывной химической активности. В большинстве случаев эта активность принимает форму химической реакции между ферментом и субстратом, при этом ферменты остаются неизменными, а субстраты превращаются в другие вещества в результате разрыва или образования ковалентных связей. Даже органические коферменты в процессе функционирования подвергаются подобным изменениям, например никотинамидаденинди-нуклеотид сначала присоединяет, а затем отщепляет атом водорода от атома углерода в положении 4. Аналогично дифосфотиамин сначала присоединяет ацетильную группу в положение 2, а затем теряет ее.
Некоторые из коферментов («кофакторов») представляют собой неорганические катионы, большинство же является простыми органическими молекулами. Однако многие ферменты не нуждаются в коферментах. Органические коферменты синтези-8
руются либо в данном организме, либо в другом, а затем попадают в первый с пищей. Белковая часть фермента, содержащая кофермент, называется апоферментом. К субстратам относится либо сама пища, либо простые молекулы, образующиеся из нее под действием разрушающих или синтезирующих ферментов. Известно более 2000 разных ферментов. Многие из них получены в чистом виде с помощью электрофореза или еще более эффективным методом — афинной хроматографией [Dixon, Webb, 1979]. Число ферментов значительно возрастает, если включить в их ряд еще и изоферменты (разд. 4.2). Некоторые ферменты представлены в клетках лишь несколькими молекулами, другие вообще отсутствуют, но в нужный момент их синтез начинается благодаря наличию соответствующей информации, заключенной в ДНК. В то же время известны и такие ферменты, которые составляют по крайней мере 50% сухого веса клетки, примером может служить миозин (аденозинтрифосфатаза) в мышечных клетках.
До недавних пор очистка ферментов обычными методами занимала много времени, а выход чистого фермента был невелик. Появление афинной хроматографии позволило резко ускорить этот процесс и повысить его эффективность. При этом специфический ингибитор данного фермента связывается ковалентно с полимерным адсорбентом с помощью подвижного мостика, например группы СН2СН2. Связанный фермент с ингибитором накапливается в колонке, с которой затем его легко вымывают, изменив pH элюента [Cuatrecasas, Wilcheck, Anfin-sen, 1968].
Первым синтетическим ферментом была рибонуклеаза. Ее синтезировали как в растворе, используя классические методы органического синтеза [Hirschmann et al., 1969], так и твердофазным методом Меррифилда [Gutte, Merrifield, 1969].
Для установления структуры ферментов наиболее успешно применяют метод рентгеноструктурного анализа. Самые точные результаты были получены с использованием аппаратуры с высоким разрешением (до ~0,2 нм), несмотря на то, что это достаточно дорого и занимает немало времени даже при наличии самой современной компьютерной техники. Этим методом удалось установить строение примерно 100 ферментов.
9.0.1. Конформационная адаптация фермента к субстрату
Для того чтобы подчеркнуть способность субстратов и коферментов изменить конформацию ферментов (и наоборот), Koshland (1964) ввел термин «наведенное соответствие». Результаты рентгеноструктурного анализа самих ферментов в сравнении с данными, полученными для комплекса фермента с медленно реагирующим субстратом или коферментом, подтвердили эту гипотезу. Так, например, было установлено, что апофермент лактатдегидрогеназы значительно деформируется в присутствии
9
своего кофермента НАД; кроме того, этот фермент, плотно скрученный, в нормальном состоянии был распрямлен с помощью апофермента [Adams et al., 1970]. Еще раньше отмечалось, что при образовании связей с активными центрами лизоцима конформация остатка ацетилглюкозамина переходит из формы «кресла» в форму «полукресла» [Blake et al., 1967]. В настоящее время известно, что активные центры фермента обычно состоят из двух или более аминокислотных остатков, расположенных в разных участках полипептидной цепи фермента. Например, для действия содержащегося в яичном белке фермента лизоцима на его субстрат в стенке бактериальных клеток (муреин, см. рис. 5.3, т. 1) необходимо, чтобы сблизились остатки 35 и 52 (глутаминовая и аспарагиновая кислоты соответственно) из разных ветвей фермента [Phillips, 1966]. Третичная структура лизоцима такова, что на его внутренней поверхности расположены преимущественно гидрофобные, а на внешней — гидрофильные группы. Такое строение типично для большинства ферментов.
С помощью рентгеноструктурного анализа было показано, что дрожжевая гексокиназа состоит из двух долей, образующих полость, в которую и попадает субстрат — глюкоза. При этом происходит смещение полипептидного остова фермента на 0,8 нм: две доли фермента смыкаются вокруг субстрата так, что снаружи остается лишь гидроксиметильная группа у атома С6 глюкозы [Steitz, Shoham, Bennett, 1981].
Уплотнение структуры фермента при взаимодействии с субстратом часто происходит и при связывании его с ингибиторами. Так, например, дигидрофолатредуктаза (разд. 9.3). — рецептор таких лекарственных веществ, как метотрексат (4.7), триметоприм (4.9) и хлоридин (4.8), легко гидролизуется протеолитическими ферментами в отсутствие этих препаратов [Burchall, 1966; Hakala, 1966]. Следующие примеры конформационной адаптации (наведенного соответствия), каждый из которых обладает совершенно индивидуальным механизмом, позволяют показать единую природу этого явления (рис. 12.2). Рибонуклеаза образует компактную структуру, гидрофильную снаружи и липофильную внутри, со щелью для захвата субстрата. В активном центре His-12 и His-119 располагаются рядом [Kartha, Bello, Harker, 1967]. Размеры сложенной молекулы рибонуклеазы примерно 3X3X3,8 нм (ОММ 15000). При насыщении субстратом, например цитидинфосфатом, один остаток гистидина образует связь с фосфатной группой, а другой — с сахаром. Аналогична третичная структура а-химотрипсина: в активном центре Ser-195 и His-57 из разных ветвей фермента расположены поблизости друг от друга [Matthews et al., 1967]. Гидроксильная группа серина участвует в алкоголизе пептидной связи, приводящем к образованию сложного эфира, который затем подвергается гидролизу, катализируемому имидазольным циклом гистидинового остатка.
10
Результаты рентгеноструктурного анализа с использованием ингибиторов фермента показали, что активный центр карбоангидразы человека расположен в полости глубиной 1,2 нм, на дне которой находится атом цинка, окруженный тремя гистидиновыми остатками (His-94, His-96 и His-119) и одной молекулой воды. В активном центре происходит связывание углерода и сульфаниламидного ингибитора [Kannan et al., 1975].
С помощью рентгеноструктурного анализа (разрешение 0,2 нм) было изучено строение карбоксипептидазы А, важного фермента поджелудочной железы, гидролизующего только пептиды с гидрофобными боковыми цепями. Было обнаружено, что атом цинка, являющийся необходимым кофактором, находится в небольшом углублении на поверхности молекулы рядом с глубокой липофильной полостью. Он связан с тремя аминокислотными остатками His-69, His-196 и Glu-72. Фермент состоит из 307 аминокислотных ферментов и имеет ОММ 34 500. Сравнение «контурных карт» фермента в присутствии и в отсутствие субстрата (глицилтирозина), припятствующего гидролизу, показало, что фенильная группа попадает в глубокую полость, вынуждая карбонильный атом кислорода пептида (субстрата) расположиться напротив атома цинка, теряющего при этом молекулу координационной воды. Свободная карбоксильная группа субстрата образует ионную связь с Arg-145, что вызывает смещение аргинина на 0,2 нм и соответственно разрыв близлежащих водородных связей. Эти изменения приводят к повороту свободной гидроксильной группы Tir-248 на 120 °C. При этом она входит в небольшое углубление, расположенное рядом с пептидной связью субстрата, и, вероятно, выступает в роли общего кислотного катализатора, участвующего в протонировании уходящей группы. Атакующим нуклеофилом в этом случае является, по-видимому, карбоксильная группа остатка Glu-270 [Lipscomb et al., 1969; Lipscomb, 1970; Breslow, Wernick, 1977]. Все эти изменения представлены на стереодиаграмме (рис. 9.1). Структуру гликопротеина —- нейраминидазы вируса гриппа — установили с помощью рентгеноструктурного анализа [Varghese, Laver, Colman, 1983].
Точная информация об активных центрах ферментов, получаемая с помощью рентгеноструктурного анализа, сделала бессмысленными умозрительные схемы, до недавнего времени часто встречавшиеся в литературе по энзимологии. С достаточной уверенностью можно полагать, что это же в скором времени произойдет и со схемами строения рецепторов лекарственных веществ (см., например, рис. 12.3).
9.0.2. Секвенирование
Последовательность аминокислот в молекуле фермента редко удается установить полностью с помощью рентгеноструктурного анализа. Быстрее и точнее это можно сделать путем сек-
11
Рис. 9.1. Стереодиаграмма активного участка карбоксипептидазы А. Субстрат (не показан) связан с цинком (Zn2+) и тремя аминокислотными остатками: His-69, Glu-72 и His-196. Остаток Glu-270, участвующий в последней стадии гидролиза, изображен в положении, которое он занимает, перед тем как остаток Arg-145 сдвигает его на 0,2 нм [Lipsomb, 1970].
венирования. Хотя оно и не позволяет получить информацию непосредственно об активных центрах ферментов, тем не менее это удается в тех случаях, когда реагент может образовывать ковалентную связь с одной из групп активного центра. В качестве субстратов для изучения протеолитических ферментов часто используют органические фосфаты (разд. 13.3). Так, диизо-пропилфторфосфат (13.26) взаимодействует с химотрипсином как псевдосубстрат и, фосфорилируя активный центр, делает невозможной реакцию с истинным субстратом. Было установлено, что в каждой молекуле фермента одна молекула ингибитора взаимодействует с одним активным центром. В результате гидролитического расщепления фермента, обработанного диизо-пропилфторфосфатом, образуется диизопропилфосфосерин. Это позволяет сделать вывод о том, что остаток серина играет важную роль в активном центре данного фермента [Shaffer, Мау, Summerson, 1954]. Подобный анализ продуктов расщепления показал, что в активных центрах химотрипсина (каждая его молекула содержит 230 аминокислот, 28 из которых являются остатками серина) и трипсина расположена последовательность Asp — Ser — Gly, в то время как в активных центрах АХЭ псевдохолинэстеразы и алиэстеразы печени присутствует последовательность Glu — Ser — Ala [Sanger, 1963].
Дополнительную информацию об активных центрах ферментов получают с помощью молекулярных зондов (разд. 2.4).
9.0.3. Кинетика ферментативных реакций
Жизненно важная роль ферментов в метаболизме заключается в том, что они ускоряют бесчисленное множество химических реакций, протекающих в клетке. Ферменты снижают энер
12
гетический барьер и тем самым повышают скорость протекания реакции. Например, с помощью фермента каталазы энергия активации реакции разложения перекиси водорода (Н2О2 —► —> Н2О + О) снижается с 18 до 2 ккал/моль, а скорость реакции увеличивается в 1,6Х 10й раза.
Высокая каталитическая активность ферментов основана на общих принципах физической и органической химии: а) молекула субстрата взаимодействует с ферментом с изменением конформации одного из них или обоих, при этом образуется переходное состояние, более энергетически выгодное по сравнению с субстратом; б) затем фермент может инициировать разрыв ковалентной связи вследствие близости реагирующих групп; в) этому способствует электрическое поле, поддерживающееся в очень малом объеме в необычно безводной среде, т. е. с низкой диэлектрической постоянной и г) боковые цепи аминокислотных остатков, расположенные в полости вокруг активного центра, могут образовывать короткоживущие связи, способствующие протеканию реакции. В бимолекулярных реакциях на ферменте создается значительно более высокая концентрация субстратов, чем в растворе, при этом они находятся ближе друг к другу и лучше ориентированы. Ферменты обладают двумя типами специфичности: 1) разной степенью сродства к субстратам и 2) разной скоростью реакции для различных субстратов после связывания. Эти типы называются специфичностью связывания и кинетической специфичностью и проявляются независимо друг от друга [Koshland, Neet, 1968].
Первым выделенным комплексом субстрата с ферментом был комплекс D-аланин, оксидаза D-аминокислоты и кофактор ФАД (флавинадениндинуклеотид). Этот комплекс представляет собой гексагональные кристаллы пурпурного цвета, стабильные в отсутствие воздуха. Исследования методом ЭПР показали, что в этих кристаллах кофактор присутствует в свободнорадикальной моногидроформе FADH [Yagi, 1965].
9.0.4. Простые модели ферментов
Все известные ферменты — это белки. Однако для изучения механизмов их действия и установления природы их высокой специфичности в отношении и продуктов и субстратов были созданы модельные соединения небелковой природы. Основой для них послужил гистидин, встречающийся в активных центрах многих ферментов. Наиболее важной функциональной группой гистидина является имидазольное кольцо (9.1), величина рКа которого близка 7. Это позволяет ему выступать в роли как донора протона, так и его акцептора. Имидазол и особенно 4'-(имидазол-4-ил) масляная кислота (9.2) значительно ускоряют гидролиз сложных эфиров по механизму обычного кислотно-основного катализа [Bruice, Benkovic, 1966].
13
н н
N N
—СН2СН2СН2СО2Н
Имидазол 4'-(имидазол-4-ил)масляная кислота
(9.1) (9.2)
По свойствам наиболее близки ферментам циклодекстрины (циклоамилазы), получаемые при расщеплении крахмала [Cramer, 1956]. Циклодекстрины действуют как эффективные эстеразы, субстратами которых служат фенилацетат и его производные. По данным рентгеноструктурного анализа молекулы циклодекстринов представляют собой тор из шести остатков a-D-глюкозы в обычной конформации. Гидроксильные группы образуют короны вокруг верхней и нижней части тора, внутренний диаметр которого составляет около 0,5 нм. При гидролизе карбонильная группа ацетата («гость») приближается к вторичным гидроксильным группам циклодекстрина («хозяин»): чем теснее этот контакт, тем быстрее проходит гидролиз. Сложный эфир («гость») ацилирует одну из гидроксильных групп «хозяина», высвобождая при этом фенол. Затем в результате реакции внутримолекулярного катализа происходит регенерация цнклодекстрина в неацилированную форму. Процесс при этом может повторяться неограниченно. Этот цикл реакций напоминает ферментативные превращения еще и тем, что он подчиняется кинетике Михаэлиса—Ментен (разд. 9.1) и легко ингибируется нормальными субстратами [Bender, Komiyama, 1978].
Ковалентным присоединением двух имидазольных групп к циклодекстрину можно получить искусственную рибонуклеазу, имеющую достаточно высокую специфичность в отношении как субстрата, так и продукта [Breslow, 1983]. Присоединяя ковалентно к циклодекстрину пиридоксамин, можно получить синтетическую трансаминазу. Эти модельные соединения вызывают ускорение соответствующих реакций в 200 раз, однако в ближайшем будущем можно надеяться на значительно большее увеличение скорости [Breslow, 1983].
Исследования этих модельных соединений и данные рентгеноструктурного анализа природных ферментов показали, что для гидролиза необходимы сближение нуклеофильной группы (аниона глутаминовой кислоты, аспарагиновой кислоты или имидазола) с электрофильным центром — атомом углерода карбонила амидной группы и одновременный подход протона к атому азота этой же группы. На циклодекстриновых моделях было установлено, что ферментативноподобная активность может быть свойственна веществам с ОММ несколько ниже 1000, имеющим простую структуру и симметрию. Более подробные сведения о моделях ферментов можно получить из работ Jencks (1969) и Bender, Komiyama (1978). В литературе есть данные
14
о синтезе микроцикла, ускоряющего ацетилирование аминокислот в 10" раз [Cram, 1983].
Действие ферментов в значительной степени зависит от мицелл амфифильных молекул, катализирующих многие химические реакции и нередко рассматриваемых как очень простые модели ферментов (определение понятий «амфифильный» и «мицелла» приведено в разд. 14.0). Расположение гидрофобных (внутри) и гидрофильных (снаружи) групп в мицелле напоминает расположение этих групп в молекуле фермента. Подобно ферментам мицеллы денатурируют нагреванием или действием мочевины; как и ферменты, они обладают избирательностью по отношению к субстратам [Jencks, 1969]. Реакции, сопровождающиеся высвобождением анионов, такие как гидролиз сложных эфиров, катализируются катионными мицеллами, например мицеллами цетилтриметиламмоний бромида (14, 16) [Menger, Portnoy, 1967]. Субстрат концентрируется в мицелле. При этом образуются ван-дер-ваальсовы связи, о чем свидетельствует повышение эффективности катализа при увеличении размеров молекул субстрата. Заряженные группы мицеллы создают заряд в среде, окружающей субстрат (это окружение может стабилизировать переходное состояние субстрата); предполагается, что фактически они и являются катализаторами. Низкая диэлектрическая постоянная остальной части мицеллы может приводить к дестабилизации любого заряда на субстрате, его переходном состоянии и продуктах.
Эффективность мицеллярного катализа иллюстрирует додецилсульфат натрия, ускоряющий связывание ионов Си2+ порфиринами в 500 000 раз [Lowe, Phillips, 1961]. О катализе мицеллами см. Cordes (1973), Fendler и Fendler (1975).
9.0.5. Биологическая регуляция ферментов
Ферменты, функционально связанные друг с другом в соответствии с последовательностью метаболических процессов, либо образуют частицы, либо встраиваются в мембраны. В этих системах ферменты соединяются химически, термодинамически или чисто физически. При этом продукт первой ферментативной реакции является субстратом второго фермента и т. д. Такие последовательности ферментов часто регулируются аллостерическим эффектом, суть которого заключается в следующем: у ферментов, катализирующих начальные реакции в цепи метаболических превращений, может быть второй рецептор (на некотором расстоянии от активного центра), способный реагировать с природным ингибитором (часто это продукт ферментативной реакции в одной из поздних стадий метаболических превращений). В результате этого взаимодействия молекула фермента претерпевает конформационные изменения, благодаря которым нормальный субстрат не может связаться с активным центром. Подобный механизм обратной связи играет важную
15
г" д Рис. 9.2. Регуляция син-
I____________ I теза катехоламинов.
Г * I
Тиро3ин Тирозин-- ДОФА Дофа* Дофамин д^фам*. Норадреналин гидроксилаза декарбоксилаза .^-гидроксилаза
роль в саморегуляции метаболических процессов в клетке [Mo-nod, Changeux, Jacob, 1963]. Показано, что присутствие природных пуринов в живой клетке может ингибировать их синтез до тех пор, пока их концентрация не упадет ниже нормального уровня. Подобный контроль с обратной связью был продемонстрирован также на примере синтетических и природных пиримидинов [Bresnick, Hitchings, 1961]. Схема регуляции синтеза катехоламинов по механизму обратной связи приведена на рис. 9.2. Подробнее об аллостерической регуляции см. Cohen (1980).
9.0.6. Ферменты и лекарственные вещества
Ценность многих лекарственных веществ избирательного действия заключается в их способности ингибировать определенный фермент. Однако в каждом конкретном случае необходимо убедиться в том, что: а) фермент в контактной клетке ингибируется также эффективно, как и в изолированном состоянии in vitro; б) концентрации агента, необходимые для ингибирования фермента, не превышают обычно используемые для фармакологического действия на клетку [Hunter, Lowry, 1956].
Основываясь на этих стандартных требованиях, удалось показать, что в основе биологического действия органических фосфатов, а также уретанов, лежит их способность ингибировать АХЭ, а действие оксимов, являющихся антидотами, заключается в том, что они снимают такое ингибирование (разд. 13.3). Большинство широко используемых и представляющих поэтому наибольший интерес агентов, способных подавлять действие ферментов, обнаруживают близкое сходство с субстратом или коферментом как по структуре, так и по характеру распределения электронов в молекуле. Такие вещества называются антиметаболитами.
О ферментах см. Williams (1969); Dixon и Webb (1979); Boyer (1970—1983). В разделе 4.2 обсуждаются изоферменты и изофункциональные ферменты. Классификацию ферментов см. в International Union of Biochemistry (1978).
9.1. Антиметаболиты: определение, происхождение и механизм действия
Биологические эффекты метаболитов (субстратов или коферментов), находящихся в незначительных количествах в клетке или ткани, могут быть подавлены действием их аналогов, называемых антиметаболитами. В молекуле каждого из
16
таких аналогов имеется участок, подобный участку метаболита, обеспечивающему взаимодействие аналога с ферментным белком. Для того чтобы аналог был эффективным, он должен иметь сходство с метаболитом не только в размерах, но и в распределении электронов, так как активные центры ферментов, как правило, высокополярны. Антагонизм между аналогом и метаболитом обусловлен тем, что антиметаболит занимает и блокирует активные участки фермента, обычно используемые метаболитом (разд. 9.2).
Хотя метаболит может быть превращен в аналог изменением химической структуры молекулы, это изменение не должно быть значительным, иначе метаболит потеряет биологическое действие, но не приобретет антагонистического. Так разнообразные модификации структуры тиамина путем присоединения дополнительной или отщепления имеющейся метильной группы снижают его активность как витамина, но не делают его антагонистом (глава 2). В целом, потеря или приобретение метильной группы молекулой метаболита — слишком большое изменение, чтобы он стал антагонистом, по крайней мере в случае небольших молекул, в которых измененный участок обычно находится внутри или очень близко к активному участку фермента. Целесообразнее сохранять стерические свойства метаболита (чтобы обеспечить связь с ферментом), но варьировать его электронные свойства (чтобы сделать новое вещество не подходящим для фермента в качестве субстрата).
Некоторые антагонисты имеют такую простую химическую природу, что их способность выступать в роли аналогов метаболитов часто остается незамеченной. Например, одни неорганические катионы конкурируют с другими (разд. 9.2 и 11.0). Даже ион водорода (по утверждению Митчелла1 один из наиболее важных метаболитов) участвует в конкуренции с органическими и неорганическими катионами (разд. 10.3.1) [Mitchell, 1979]. Этиловый спирт, входящий в состав алкогольных напитков, действует частично за счет конкуренции с водой (по крайней мере при распределении). Он может также снимать токсическое действие метилового спирта, вытесняя его с окисляющего фермента [Roe, 1955].
Известен ряд примеров конкуренции между простыми анионами. Так, перхлорат- и тиоцианат-анионы ингибируют накопление йодид-иона в щитовидной железе без сколько-нибудь заметного нарушения окислительного включения йодид-иона в тироксин [Stanbury, Wyngaarden, 1952]. Таким же образом способность бактерий Nitrobacter окислять нитриты в нитраты ингибируется цианат- или перхлорат-анионами: эффект последних легко снимается простой отмывкой [Lees, Simpson, 1957].
1 В соответствии с хемиосмотической концепцией Митчелла градиент протонов является единственной причиной перемещения электронов при синтезе АТФ.
2—734
17
Некоторые физиологические процессы регулируются парами метаболитов-аналогов. К последним, например, относятся полиеновые половые гормоны водорослей [Kuhn, 1940] и половые гормоны млекопитающих. Из двух простагландинов, содержащихся в легких человека, PGE2 расслабляет, a PGF2a сокращает мускулатуру бронхов. Таким же образом простагландин Di ингибирует увеличение проницаемости сосудов, вызываемое в коже крыс простагландинами Еь Е2 и D2 [Flower, Kingston, 1975]. Другая пара химически сходных регуляторов — это простациклин и тромбоксан у человека. Простациклин — наиболее сильный ингибитор агрегации тромбоцитов и мощный вазодилататор, в то время как тромбоксан обладает противоположными свойствами. Однако оба вещества образуются из одного и того же предшественника — простагландина эндопероксида (разд. 4.7).
Известно лишь несколько примеров антагонистов, созданных изменением расположения групп у асимметрического атома углерода. Одним из таких антагонистов является D-гистидин, ингибирующий фермент гистидазу, которая в норме размыкает имидазольное кольцо L-гистидина [Edlbacher, Baur, Becker, 1940]. Этот метод резко эффективен в применении к небольшим молекулам, вероятно, потому, что пространственное соответствие субстрата — метаболита, необходимое для адсорбции его ферментом, слишком резко нарушается при перемещении заместителей. Гораздо чаще обнаруживается, что если ( + )-изомер оказывает сильное физиологическое действие, то (—[-изомер обладает такого же рода действием, но значительно более слабым. Другими словами, он редко является антагонистом. По этой причине активность смеси двух оптических антиподов (или рацемата) обычно близка среднеарифметическому активности их обоих (например, в случае тироксина, атропина и DL-норадреналина). Применение неприродных оптических изомеров для создания лекарственных полипептидов оказалось полезным для защиты последних от действия желудочного сока пациента (см. аналоги энкефалина в разд. 12.8).
Надежный метод создания антагонистов для большой молекулы заключается в использовании малой молекулы, сходной с повторяющимся фрагментом структуры большой молекулы. Так, крахмал гидролизуется ферментом амилазой до мальтозы, представляющей собой ангидродимер глюкозы. Этот фермент активно ингибируется глюкозой, хотя и не образуется при гидролизе. Галактоза — стереоизомер глюкозы — менее сильный ингибитор амилазы, а фруктоза — изомер глюкозы, но не стереоизомер, вообще не оказывает ингибирующего действия [Wohl, Glimm, 1910].
Антагонистами небольших по размеру молекул могут быть их ближайшие гомологи, например малоновая кислота (9.3) — ингибитор окисления янтарной кислоты (9.4) сукцинатдегидрогеназой [Quastel, Wooldridge, 1927].
18
НО2С СО2Н
^С— н2 н2
Янтарная кислота
(9.4)
Пиридоксин (R=—СН:ОН)
(9-6)
НО2С СО2Н
'v
Н2
Малоновая кислота
Пиритиамин
(9-5)
Другой путь создания антагонистов-аналогов — это замена атомов в цикле, входящем в молекулу метаболита. Так, пиритиамин (9.5), получаемый заменой атома серы в молекуле тиамина (2.1) этиленовой группой, вызывает характерный симптом недостаточности тиамина у мышей [Woolley, 1950]. Замена тиазольного кольца пиридиновым кажется логичной вследствие их изостерности (по аналогии с тиофеном и бензолом) [Наг-tough, 1952]. Показано, что пиритиамин вытесняет тиамин с фермента, обычно фосфорилирующего его с образованием тиаминпирофосфата — кофермента пируватдегидрогеназы. К сожалению, пиритиамин не оправдал надежд в качестве лекарственного препарата, однако модификация его структуры привела к получению ампролиума (9.37), успешно используемого для лечения протозойных болезней птиц.
Целесообразен и противоположный путь — замена этиленового фрагмента серой, например превращения фенилаланина в тиенилаланин — сильный антагонист этой аминокислоты у микроорганизмов [Dittmer, 1949]. Подобная замена бензольного кольца тиофеновым эффективна и во многих других случаях. Например, а-тиенилалкиламины похожи на соответствующие фенилалкиламины по гипертензивной активности, а аминоалкиловые эфиры тиофен-2-карбоновой кислоты обладают сходным с соответствующими эфирами бензойных кислот местноанестезирующим действием [Hartough, 1952].
Один из лучших общих методов создания антагонистов — замещение одной электроноакцепторной группы другой. Так, карбоксильная группа (—СООН) может быть замещена группами —СОСНз, —SO2OH или —SO2NH2. Однако при заменах такого рода необходимо сохранять степень ионизации основной группы, если таковая имеется в молекуле. Так, например, аминогруппа в анионе пара-аминобензойной кислоты (ПАБ, 9.7) не ионизирована и, следовательно, замещение карбоксилатной группы сульфогруппой неприемлема, так как основная аминогруппа в присутствии последней превратилась бы в протонированную аммониевую группу и новое вещество отличалось бы от
2*
19
Таблица 9.1. Ваи-дер-ваальсовы радиусы некоторых заместителей в антагонистах метаболитов [Speakman, 1968]
Заместитель Радиус, им Заместитель Радиус, нм Заместитель Радиус, им
Н 0,12 N 0,155 СеН5 0,18
F 0,135 С1 0,18 СН3 0,20
С О 0,14 0,14 S 0,185 I 0,215
исходного слишком резко, чтобы быть эффективным антиметаболитом. Эксперименты свидетельствуют о правильности этого предположения. С другой стороны сульфаниламидный анион (9.8) оказался неплохим антагонистом аниона ПАБ (9.7) (разд. 9.3).
Эффективные антагонисты были получены заменой водорода фтором (присутствие которого в этих антагонистах приводит к обратимой блокаде метаболизирующих ферментов ЭР), например, синтетические антагонисты андрогенов и кортикостероидов [Gilman, Goodman, Gilman, 1980], противораковый препарат — фторурацил (4.0.2) и фунгицид — флуцитозин (4.23). Ряд подобных препаратов применяют в медицине, но замена лимонной кислоты на фторлимонную недопустима из-за высокой токсичности последней (разд. 13.5).
Нередко целесообразна замена метильной группы хлором. Примером могут служить антагонисты рибофлавина [Kuhn, Weygand, Moller, 1942]. Данные табл. 9.1 подтверждают правильность предположения о том, что такие изменения стериче-ски оправданы, и позволяют понять, почему замещение атома водорода на атом хлора или метильную группу обычно не приводит к получению эффективных антагонистов. Общим требованием при конструировании аналогов этого типа является их сходство с субстратом настолько, чтобы фермент, ошибаясь, использовал эти чужеродные молекулы вместо субстрата. В то же время антагонисты должны в достаточной степени отличаться от метаболита, чтобы, связавшись с ферментом, не вступать в химическую реакцию, которой в норме подвергается субстрат под действием фермента. Если же он в нее все-таки вступает, то образующийся продукт не должен взаимодействовать с ферментами, участвующими в дальнейшей цепи биохимических превращений нормального субстрата.
Антагонисты можно получить удалением из молекулы метаболита некоторых небольших групп, подвергающихся ковалентным превращениям в процессе нормального метаболизма. Так, дезоксипиридоксин [(9.6), R = CH3] вызывает у человека симптомы витаминной недостаточности пиридоксина, которые легко снимаются при введении последнего [(9.6) R = CH2OH] [Mueller,
20
Vilter, 1950]. Дезоксипиридоксин становится биологически активным в организме только после фосфорилирования с образованием производного, конкурирующего с пиридоксальфосфатом— коферментом декарбоксилазы аминокислоты.
Очевидно, антагонист может быть найден для молекулы любой величины. Например, ацетилирование тиреотропного гормона (ТТГ) — белка, содержащегося в гипофизе, приводит к образованию аналога, который накапливается в щитовидной железе и понижает гипертиреоидизм, блокируя действие ТТГ [Sonenberg, Money, 1957]. Небольшие химические изменения других полипептидных гормонов гипофиза, таких как окситоцин и вазопрессин, обусловливают образование антагонистов соответствующих гормонов [Dyckes et al., 1974; Manning et al., 1977]. Так, при замещении аминогруппы в положении 1 окситоцина пеницилламином получают сильный антагонист этого гормона.
В природе существует немало примеров антагонизма на по-липептидном уровне. Полипептид контрикал (ОММ 6512), обнаруживаемый в тканях млекопитающих в больших количествах, подавляет гипотензивный кинин (калликреин) поджелудочной железы и протеолитическое действие ферментов трипсина и плазмина. Он был с успехом использован при лечении панкреатитов [Trapnell, 1977]. Меланостатин и соматостатин — полипептиды, содержащиеся в гипоталамусе, ингибируют выделение передней долей гипофиза меланоцитстимулирующего гормона (полипептида) и соматотропина (гормона роста, белка) соответственно.
После того как Woods обнаружил антиметаболитную природу действия сульфаниламидов на бактерии (разд. 9.3.1), началось широкое исследование других антагонистов, которые могли бы стать полезными лекарственными препаратами. Это оказалось неожиданно трудной задачей, поскольку полезные и вредные клетки участвуют во многих общих биологических процессах. В течение некоторого периода времени успех ограничивался открытием лекарственных веществ, относящихся к группе антагонистов фолиевой кислоты. Затем были обнаружены (и случайно, и целенаправленно) многие другие антагонисты метаболитов, нашедшие применение в клинической практике (разд. 9.4).
Сегодня трудно рассчитывать на создание эффективных антагонистов тех или иных метаболитов, если нет строгих данных сравнительной биохимии, указывающих на участие этих метаболитов в биохимических процессах, протекающих только во вредной клетке. Из бесчисленного множества агентов, синтезированных за этот долгий период и не обладающих избирательным действием, многие были использованы в биохимии в качестве специфических блокаторов различных метаболических процессов in vitro. Об антагонистах метаболитов см. Hochster, Quastel [1963—1973].
21
Количественные аспекты. Взаимоотношения между метаболитами и антиметаболитами обычно имеют конкурентный характер, т. е. если х молекулам метаболита противодействует у молекул аналога, то для достижения того же биологического эффекта 10х молекулам метаболита потребуется 10у молекул аналога. Поскольку такие конкурентные реакции полностью-обратимы, антагонистическое действие, оказываемое у молекулами аналога на х молекул метаболита, может быть преодолена введением других х молекул метаболита и так далее. Конкурентные отношения этого типа наблюдаются между малоновой и янтарной кислотами, стрептоцидом (сульфаниламид) и пара-аминобензойной кислотой.
Для каждой пары веществ существует свой специфический индекс ингибирования, определяемый как соотношение числа молекул аналога к числу молекул метаболита, при котором достигается 50% ингибирование. Это соотношение имеет разные значения для различных биологических видов, но она всегда одинаково для каждого данного вида. Оно характеризует относительное сродство аналога и метаболита к рецептору и, кроме того, отражает различия в способности двух веществ проникать к месту действия, особенно тогда, когда это место труднодоступно. Поэтому уровень ингибирования, вызванного аналогом, определяется двумя факторами: во-первых, относительным сродством аналога и метаболита к рецептору и, во-вторых, относительными количествами аналога и метаболита в месте реакции.
Сродство субстрата к ферменту характеризуется уравнением: [Е] + [Sl=?=te[ESJ (I), где [Е—концентрация фермента, [S]—концентрация субстрата, a [ES]—концентрация комплекса, который они образуют. Естественно, сродство лекарственного препарата, являющегося ингибитором (I) фермента, может быть описано аналогичным выражением и тогда константа ингибирования (Ki), т. е. константа диссоциации комплекса фермент — ингибитор будет представлена уравнением:
к 1 [EI]
В отличие от ингибитора субстрат под действием фермента превращается в продукт (Р) и последовательность событий становится:
[Е] + [S] [ES] ----> IE] + [Р] (Ш)
Соотношение констант скоростей к'/к" есть ничто иное, как константа Михаэлиса — Ментен (Км). Она отражает диссоциацию комплекса фермент — субстрат [ES] на компоненты [Е] и [S] и может быть представлена равенством (IV), формально аналогичным приведенному выше для Ki (II):
[Е] [S] m [ES]
22
Индекс ингибирования, характерный для каждой пары ингибитор — метаболит, — это соотношение Ki Km — чем ниже его величина, тем эффективнее ингибитор.
Константа Михаэлиса, очевидно, обратно пропорциональна сродству фермента к субстрату и численно равна концентрации субстрата в тот момент, когда скорость реакции достигает половины максимальной. Размерность величин Ki и Кт выражается в г-моль/л. Однако это не истинные константы равновесия, а отношения констант скоростей прямой и обратной реакций фермента с субстратом или ингибитором. Наиболее удобно определять Кт графическим методом Lineweaver, Burk (1934), согласно которому строится график зависимости обратной величины начальной скорости образования комплекса [ES] от обратных величин концентрации субстрата. На графике должна получиться прямая линия, при этом точка пересечения полученной прямой с осью абсцисс соответствует величине Кт-
Поскольку интенсивные метаболические процессы в клетках гораздо чаще приводят к «стационарному» состоянию, чем к равновесному, целесообразно использовать уравнение Бриггса — Холдейна [Briggs — Haldane, 1925], которые показали, что совсем необязательно предполагать наличие равновесия между [Е] и [S]. Они вывели уравнение (V), формально аналогичное уравнению Михаэлиса — Ментен, но предполагающее условия стационарного состояния, а не равновесия.
Константа Кьь, имеющая более общий характер, чем предыдущая, является показателем степени одновременной диссоциации комплекса [ES], протекающей в двух противоположных направлениях. Если Кт заменить на Кьь, то индекс ингибирования становится равным соотношению Ki: Кьи- При проведении этих расчетов предполагается, что ни ингибитор, ни его комплекс с ферментом не разрушаются даже частично под действием других биологических веществ, которые могут присутствовать при реакции.
Молярный индекс ингибирования малоновой кислоты равен >/з [Thron, 1953]. Еще более эффективно ингибирует оксид углерода, когда замещает кислород в гемоглобине (1/210). Величины индекса ингибирования меньше единицы необычны, потому что очень редко аналог обладает большим сродством к природному рецептору, чем нормальный субстрат. Беспрецедентно высокий индекс ингибирования (1/10 000) для метотрексата (4.7) обусловлен, вероятно, тем, что самым основным атомом азота в молекуле этого ингибитора является N-1, в отличие от дигидрофолиевой кислоты, у которой таковым служит N-5. Очевидно, в этом заключена причина столь больших различий в соответствии метотрексата и дегидрофолиевой кислоты месту связывания их с рецептором (разд. 4.2 и 9.3.3). Такие индексы
23
ингибирования как у метотрексата (1/10 000)—предел обычно наблюдаемых для лекарственных веществ. Индексы ингибирования, величины которых лежат выше единицы, такие как, например, сульфаниламида по отношению к ПАБ (300/1) — другой предел, ниже которого не следует ожидать высокого терапевтического эффекта аналога.
Почему желательны минимальные величины индексов ингибирования для лекарственных препаратов? Потому что эволюция обусловила появление ферментов, использующих соответствующие природные субстраты с максимальной эффективностью. Другая причина относительно малой эффективности многих конкурентных ингибиторов — стационарное состояние живой клетки. Атакуемый фермент постоянно снабжается субстратом (от предыдущего фермента в цепи), а его продукт непрерывно удаляется последующим ферментом. Фермент, атакуемый ингибитором, обычно недостаточно насыщен субстратом и поэтому имеет большую резервную активность. При введении конкурентного ингибитора в стационарную систему целевой фермент сначала ингибируется, но по мере накопления нереагирующего нормального субстрата это ингибирование фермента прекращается. Таким образом, возникает новое стационарное состояние, в котором общий итог действия данного метаболического пути остается неизменным, поскольку концентрация субстрата для атакуемого фермента поддерживается на более высоком уровне с тем, чтобы уравновесить эффект ингибирования [Cleland, 1970]. В качестве примера можно привести различия между химией модельных систем, изучаемых в лаборатории, и более сложных систем, образовавшихся в живой клетке в процессе эволюции.
9.2. История изучения антиметаболитов до 1940 г.
Еще в 1883 г. Сидней Рингер обнаружил, что ионы натрия в растворе хлорида натрия не способны поддерживать сокращения изолированного сердца, если они не сбалансированы катионами кальция и калия. В результате этой работы были созданы сбалансированные физиологические растворы, названные в честь Рингера, Лока и Тирода.
В 1910 г. было установлено, что некоторые ферменты можно блокировать с помощью веществ, сходных по молекулярной структуре с нормальными субстратами этих ферментов. Так, амилаза, которая в обычных условиях гидролизует крахмал, ингибируется глюкозой (разд. 9.1). Малоновая кислота (9.3) конкурентно инактивирует фермент сукцинатдегидрогеназу, вытесняя с фермента нормальный субстрат — янтарную кислоту (9.4) [Quastel, Wooldridge, 1927]. Подобное явление в физиологии— это токсическое действие оксида углерода (С = О), обусловленное тем, что он вытесняет из комплекса с гемоглобином близкую по форме молекулу кислорода (О2) [Douglas, Haldane и Haldane, 1912].
24
Stedman показал (разд. 2.3), что алкалоид физостигмин (2.8) вызывает сокращение глазного зрачка, блокируя АХЭ и позволяя таким образом АХ (2.7) возбуждать мышцу [Stedman, 1926]. Активной частью молекулы физостигмина, ответственной за ингибирование АХЭ, является метилкарбамоилоксигруппа (2.9) [Stedman и Stedman, 1931]. В 1932 г. было обнаружено, что карбахолин (2.11), имеющий карбамоилоксигруппу, проявляет почти такое же биологическое действие, как и АХ (2.7), после чего стало ясно, что ингибирование АХЭ физостигмином объясняется наличием в его молекуле группы (2.9), обеспечивающей адсорбцию ферментом антагониста — физостигмина, вместо нормального субстрата — АХ.
В 1936 г. Ing опубликовал обзор, в котором указал, что алкалоид тубокурарин (2.6), блокирующий передачу импульса в нервно-мышечном соединении (разд. 2.1), вступает в конкурентные взаимодействия с АХ за рецепторы на мышцах и что оба вещества являются четвертичными аммониевыми основаниями. Ing предположил, что несколько более слабое кураре-подобное действие многочисленных аммониевых, фосфониевых, арсониевых, стибониевых и сульфониевых солей — результат их конкуренции с АХ [Ing, 1936]. (О физиологических функциях АХ и его антагонистов см. разд. 7.8, 7.6.3, 12.5 и 12.6.)
В 1938 г. Woolley и сотр. случайно открыли первый антивитамин. Они синтезировали два аналога никотиновой кислоты: 3-ацетилпиридин и пиридин-3-сульфокислоту. Полагая, что эти аналоги будут обладать, по крайней мере качественно, биологической активностью витамина, они дали их с пищей собакам, страдающим от недостатка никотиновой кислоты. К их удивлению состояние собак ухудшилось. Природа этого явления стала совершенно ясна, как только Woods (1940) показал, что антибактериальное действие сульфаниламида может снять ПАБ и объяснил это структурным подобием двух соединений.
9.3. Антагонисты фолиевой кислоты
Известны антагонисты и биосинтеза, и утилизации фолиевой кислоты. Об истории открытия антибактериальных сульфаниламидов— типичных представителей антагонистов ее биосинтеза, уже говорилось в разд. 2.1 и 6.3.1. В 1940 г. Woods показал, что антибактериальное действие стрептоцида определяется его конкуренцией с природным метаболитом — пара-аминобензой-ной кислотой (ПАБ) (9.7) [Woods, 1940]. Впоследствии было установлено, что этот процесс осуществляется на участке фермента дигидрофолатсинтетазы, которая использует ПАБ для построения молекулы дигидрофолиевой кислоты (2.14) [G. М. Brown, 1962].
Фермент ошибочно принимает стрептоцид за свой нормальный субстрат из-за большого сходства их электронной и пространственной структуры. ПАБ имеет рКа=4,9 и не является
25
амфотерным биполярным ионом, как глицин; по-видимому, биологически активная форма — ее анион (9.7). Стрептоцид — заметно более слабая кислота (рКа=10,3) и поэтому малоиони-зирован при физиологических значениях pH. Первичные аминогруппы обоих веществ малоосновны (рКа 2,5 и 2,6 соответственно) и неоионизированы при физиологически активных значениях pH. Размеры аниона ПАБ (2.12) и неионизированной молекулы стрептоцида (2.13) почти одинаковы. Обе молекулы плоские, в обеих первичная аминогруппа находится в параположении по отношению к электроноакцепторной группе. Таким образом, перечисленные факты говорят о высокой степени подобия двух молекул и, следовательно, о возможности проявления биологической активности молекулой аналога. Указанные размеры обсуждаемых веществ мало изменяются при ионизации [Bell, Roblin, 1942].
NH2
пара-Амииобензойная кислота (ПАБ) (9.7)
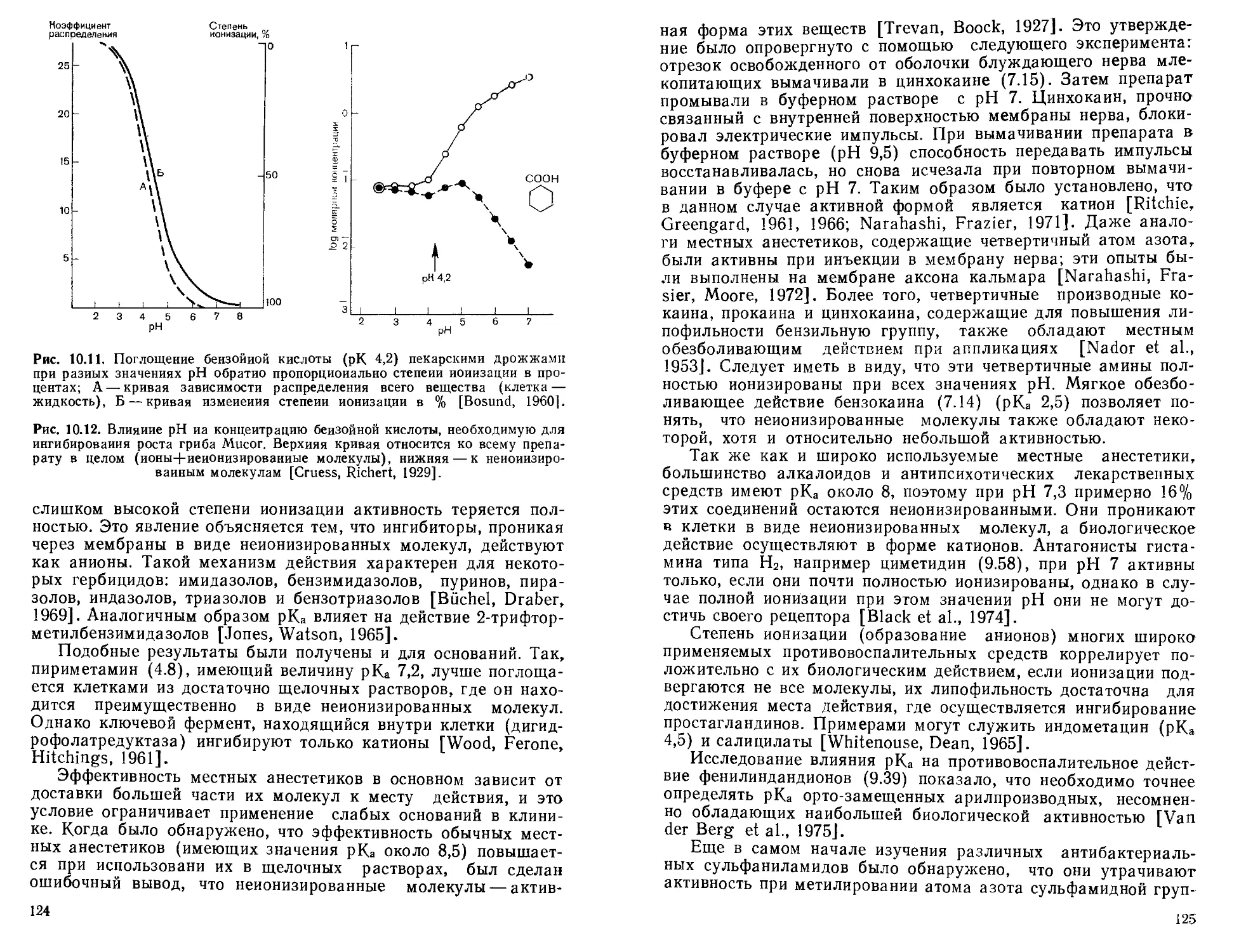
После введения стрептоцида (9.2) в клиническую практику были предприняты попытки модифицировать его молекулу с целью создания более активных аналогов. Было обнаружено, что наиболее пригодны для этого те сульфаниламиды, у которых радикал R в молекуле (9.8) представляет собой гетероцик-j лическое кольцо. Bell и Roblin (1942) показали, что это повышает степень кислотной ионизации и что сульфаниламиды, полностью ионизированные при pH 7, а следовательно, наиболее сходные с ПАБ, являются самыми сильными антибактериальными агентами (разд. 10.5). Сульфаниламиды, не способные к кислотной ионизации, также могут оказывать антибактериальное действие (например, дифенилсульфон, сульгин), однако оно всегда значительно слабее, чем у легко ионизирующихся сульфаниламидов. Так минимальная ингибирующая концентрация сульфазина [2-(4-аминофенил)сульфаниламидопиримидин] по отношению к Е. coli составляет 1,02 мкмоль/л, что примерно в. 100 раз ниже, чем стрептоцида [Kriiger-Thiemer, Biinger, 1965]. Это согласуется с большей легкостью ионизации сульфазина (рКа = 6,5), 75% которого превращается в анион при pH 7. Во всех этих N-замещенных сульфаниламидах радикал R, связанный с атомом азота, выведен из плоскости остальной молекулы и, следовательно, он не может служить препятствием для адсорбции ее на рецепторе, в норме занимаемом анионом ПАБ (9.7).
26
Избирательность антибактериального действия сульфаниламидов связана с тем, что млекопитающие неспособны синтезировать дигидрофолиевую кислоту и получают ее с пищей. В то же время патогенные бактерии не могут поглощать экзогенную дигидрофолиевую кислоту [Wood, Ferone, Hitchings, 1961] и, следовательно, уязвимы к действию сульфаниламидов, ингибирующих ее синтез.
Сульфапиридин [Ewins, Phillips, 1939], первый сульфаниламид с гетероциклическим заместителем, был вскоре вытеснен сульфатиазолом, который в свою очередь был заменен тремя более избирательными сульфопиримидинами, представленными в табл. 2.5 (т. 1). Эти пероральные препараты стали широко применяться при лечении большого числа бактериальных инфекций.
В настоящее время антибактериальные сульфаниламиды обычно используют в качестве уроантисептиков, например, при заболеваниях, вызываемых Е. coli и Proteus mirabilis. Их назначают также при нокардиозах легких или ступней, трахоме глаз, венерических лимфогранулемах, герпетических дерматитах. Велико их значение для профилактики стрептококковых инфекций у предрасположенных к ним больных, а также для предотвращения рецидивов ревматических воспалений.
Антибактериальные сульфаниламиды можно разделить на два основных класса: (а) быстро выводящиеся из организма и (б) длительно циркулирующие в кровотоке. Наиболее упот-требляемые соединения класса (а): 1) сульфазин, М'-(пирими-дин-2-ил) сульфаниламид (9.9), фактически является эталонным соединением, с которым сравнивают все остальные (сферу его применения расширяет его способность проникать в терапевтических концентрациях в спинномозговую жидкость); 2) сульфафуразол (9.10)—М'-(3,4-диметилизоксазол-5-ил)суль-фаниламидпрепарат широкого спектра действия, отличающийся более высокой по сравнению с сульфадиазином концентрацией в моче; 3) сульфаметоксазол (9.11), имеющий довольно большой для этого класса период полураспада, — один из лучших препаратов, благодаря его синергизму с триметопримом (разд. 9.6); 4) сульфацитин (9.12) и 5) сульфаметизол (9.13) наиболее предпочтительны в качестве уроантисептиков из-за короткого периода полураспада в кровотоке и отсутствия способности к специфическому накоплению.
NH2 6 Rt4J \о-
Стрептоцид (анион) (R=H)
(9.8)
27
в формуле (9.8):
О
Сульфафуразол
Me Me
Сульфаметоксазол
(9.И)
N ОМе
N N
Сульфацнтин Сульфаметизол
(9.12) (9.13)
(9.10)
Me N
Сульфапнрндазнн (9-14)
Сульфален
(9.15)
О и Me—С—
Сульфацил
(9.16)
Сульфаниламиды класса (а) так же, как и их ацетилпроиз-водные, в которые они всегда превращаются хотя бы частично, должны быстро выводиться из организма и соответственно иметь высокую растворимость в моче. Применение препаратов, не отвечающих этим требованиям, может создавать угрозу для жизни больных. Так, в 40-х годах было зарегистрировано много смертельных случаев вследствие блокады почек, вызванной приемом сульфатиазола. Проблемы подобного рода не возникают с сульфаниламидами класса (б), т. е. с теми, высокая концентрация которых в крови сохраняется так долго, что для достижения эффекта часто бывает достаточно однократного их приема. Основной недостаток этих препаратов — длительность вызываемых ими побочных реакций, иногда до нескольких дней. Наиболее опасными отрицательными реакциями на эти препараты являются синдром Стивенса — Джонса и многообразные эритремии, которые хотя и редко, но могут быть смертельными. Наиболее широко применяют следующие препараты этого класса: 1) сульфапиридазин (9.14) — N'-(6-метоксипиридазин-З-ил) сульфаниламид; 2) сульф а метокси ди азин, N'- (5-метоксипи-римидин-2-ил) сульфаниламид; 3) сульфаметопиразин, N'-(3-Me-токсипиразин-2-ил) сульфаниламид (9.15); 4) сульфадиметок-син, N'-(3,6-диметоксипиримидин-4-ил) сульфаниламид; 5) суль-фадоксин, N'- (5,6-диметоксипиримидин-4-ил) сульфаниламид — один из наименее токсичных сульфаниламидов, широко применяемый совместно с диаминопиримидином для достижения последовательного блокирования (разд. 9.6). Кроме того, в особых случаях применяют: сульфазин серебра (наружно при тяжелых ожогах), сульфацетамид натрия (9.16) (глазные инфекции), сульфапиридин (герпетические дерматиты), сульфазала-зин (колиты) и фталилсульфатиазол (перед операциями для подавления кишечной флоры).
Факторы, определяющие распределение сульфаниламидных препаратов, обсуждаются в разд. 10.5.
28
Известны многие аналоги ПАБ, не являющиеся сульфаниламидами. Из них наиболее широко применяют диафенилсуль-фон (9.17)—основной препарат для лечения проказы. Некоторые из препаратов этого типа не содержат атома серы, но обладают необходимым пространственным и электронным подобием ПАБ. Например, введение атома хлора в положение 2 или 3 ПАБ приводит к образованию активного антагониста ПАБ [Wyss, Rubin, Strandskov, 1943]. Диаминобензил (2.15) в несколько раз более активный антибактериальный препарат, чем стрептоцид, но его эффект обратим под действием ПАБ [Kuhn, Weygand, Moller, 1943]. К тому же пара-аминобензоларсоновая кислота — атоксил (6.2) обладает типичным сульфаниламидным действием [Albert, Falk, Rubbo, 1944]. Хотя вообще мышьяковые кислоты не являются антибактериальными препаратами, атоксил представляет исключение, поскольку он достаточно близок к ПАБ как по геометрическим, так и по электронным параметрам и может быть ее конкурентом.
nh2 nh2
Деафенилсульфон Конформационный аналог Мафенид
стрептоцида
(9.17) (9.18) (9.19)
Для того чтобы вещество могло взаимодействовать с дигид-рофолатсинтетазой вместо ПАБ, необходимы два условия. Первое и очень существенное—вещество должно содержать первичную ароматическую аминогруппу. В пара-положение вместо N-группы можно вводить только такие, которые в организме будут легко распадаться и высвобождать первичную аминогруппу. Очевидно, что азогруппы или азометиновые группировки в отличие от ациламино- или алкиламиногрупп расщепляются именно таким образом, например в сульфахризоидине (3.30) [Northey, 1948]. Второе условие — молекула должна содержать отрицательно заряженную группу, расположенную в пара-положении к аминогруппе и на том же расстоянии, что и в ПАБ. Значение расстояния между амино- и электроотрицательной группой для проявления антагонистических свойств можно проиллюстрировать на примере 4-амино-4'-сульфонамидодифенила (9.18), не обладающего этими свойствами.
Мафенид (4-аминометилбензолсульфонамид) (9.19), по структурной формуле напоминающий стрептоцид, — высокоосновное вещество, обладающее специфической активностью по
29
отношению к Clostridia (вызывающим газовую гангрену). Препарат не является антагонистом ПАБ [Jensen, Schmith, 1942] и, по-видимому, не играет никакой роли в метаболизме фолиевой кислоты.
Многие из широко применяемых препаратов, содержащих сульфаниламидные группы, не относятся к антибактериальным средствам, поскольку при их создании не стремились к аналогии с ПАБ; одни из них — диуретики (разд. 9.4.7), другие — антидиабетические агенты (разд. 12.4).
9.3.1. Механизмы ингибирования синтеза дигидрофолиевой кислоты сульфаниламидными препаратами
Открытие фолиевой кислоты позволило глубже понять механизм действия сульфаниламидных препаратов в качестве антагонистов ПАБ. Птероилглутаминовая кислота — ярко-желтое вещество, выделенное из печени, дрожжей, листьев и бактерий, предупреждающее малокровие у позвоночных, было названо фолиевой кислотой [Waller et al., 1948]. Структура этого соединения, подтвержденная синтезом, представлена формулой (9.20), но без двух атомов водорода в положениях 7 и 8, отщепляющихся от молекулы дигидрофолиевой кислоты при выделении. Молекула дигидрофилиевой кислоты (9.20) состоит из трех фрагментов: остатков глутаминовой кислоты, ПАБ и 2-амино-6-метил-4-оксоптеридина.
Бактерии используют ПАБ только для превращения ее в 7,8-дигидрофолиевую кислоту [Griffin, Brown, 1964]. Так бактерии Е. coli участвуют в конденсации ПАБ или пара- аминобензоилглутаминовой кислоты с 2-амино-4-гидроксиметил-7,8-оксоптеридином (9.21) (точнее его 6-пирофосфатом), что приводит к образованию дигидроптероиновой (или дигидрофолиевой) кислоты [Jaenicke, Chan, 1960]. Эту реакцию катализирует фермент дигидрофолатсинтетаза, которую конкурентно ингибируют сульфаниламидные препараты. Из бактерий Lactobacillus plantarum [Shiota et al., 1969a] были выделены кв а чистых фермента, ответственные за этот синтез. Первый из них катализирует этерификацию птеридина (9.21) в его 6-пирофосфорилпроизводное. Второй оказался дигидрофолатсинтетазой, обнаруженной ранее G. Brown (1962). Второй фермент также был выделен из нескольких штаммов Pneumococcus, причем было установлено, что его ОММ равна 90 000, а коферментами служат АТФ и Mg2+ [Ortiz, 1970].
СО2Н
CH—NH—СО -
СН2
I
СН2—СО2Н
2
Днгндрофолневая кислота
(9.20)
30
Таблица 9.2. Конкурентное и неконкурентное ингибирование ПАБ [Lam-pen, Jones, 1946]
Вещество Количество вещества, необходимое для 50% ингибирования действия сульфазнна на Streptococcus faecalis штамм Ральстона, мкг/мл
Сульфазин 1 10 100 1000
ПАБ 0,003 0,03 0,3 3,0
Птероилглутаминовая (фолиевая) кислота 0,0003 0,0003 0,0003
Тимин (4.16) 0,06 0,25 0,25 0,25
О
НОСН2Ч N || V^nh
Производное птеридина — промежуточное соединение в биосинтезе фолиевой кислоты (9.21)
Сульфаниламиды не ингибируют действие птероиновой и фолиевой кислот (а также их 7,8-дегидропроизводных). Наиболее четко это было показано на примере нескольких штаммод бактерий, способных поглощать фолиевую кислоту (например,, штаммы Streptococcus faecalis и некоторые представители Lactobacilli). Эти бактерии не патогенны для человека. В табл. 9.2' показана прямая корреляция между количеством сульфазина,, необходимым для угнетения роста бактерий Streptococcus faecalis (штамм Ральстона), и количеством ПАБ, требуемым для снятия ингибирующего действия. В отличие от этого количество-фолиевой кислоты, необходимое для снятия ингибирующего-эффекта, остается постоянным, независимо от количества использованного сульфазина. Это свидетельствует о том, что сульфазин нарушает синтез фолиевой кислоты из ПАБ, но не влияет на ее дальнейшие превращения в отличие, например, от метотрексата. Аналогично сульфаниламиды угнетают рост L. ага-binosus (которые нуждаются во внешнем источнике ПАБ),, и это угнетение конкурентно снимается ПАБ и неконкурентно — посредством фолиевой кислоты. Количество фолиевой кислоты (определяемое по разрушению бактерий и титрованием гомогенизированной бактериальной культуры бактериями L. casei, чувствительными к фолиевой кислоте, но не к ПАБ), которое вырабатывается этими микроорганизмами, обычно пропорционально количеству ПАБ в среде. Однако, если в среде присутствует и сульфаниламид, количество фолиевой кислоты уменьшается пропорционально его концентрации (в 10 000-кратном интервале ее измерения) [Nimmo-Smith, Lascalles, Woods, 1948].
31
Образование фолиевой кислоты в культуре Е. coli также ингибируется сульфаниламидами [Muller, 1944].
Подобное ингибирование ферментов сульфаниламидами, по-видимому, составляет основу терапевтического действия этих препаратов. Однако в некоторых экспериментах взаимодействие стрептоцида с птеридиновым интермедиатом (9.21) приводило к образованию устойчивого комплекса. Так, например, при выдерживании фермента Брауна в термостате в течение 2 ч в бесклеточной системе, содержащей стрептоцид, ингибирующее действие последнего не удалось снять с помощью ПАБ; в растущих клетках такой эффект не наблюдали [G. М. Brown, 1962]. Для изучения этого явления использовали фолатсинтезирующие ферменты из Е. coli: под их действием сульфаметоксазол (9.11) был превращен в аналог птероиновой кислоты — N'-З- (5-метилизок-сазолил) -N4- (7,8-дигидро-6-птероилметил) сульфаниламид, выделенный с помощью хроматографии. Полученное вещество оказалось идентичным синтетическому образцу [Bock et al., 1974].
Из всех реакций, катализируемых фолиевыми кислотами, синтез тимина наиболее чувствителен к недостатку этих кислот (разд. 9.3.2), хотя в некоторых микроорганизмах первым ингибируется синтез пуринов. Из данных, приведенных в табл. 9.2, можно видеть, что тимин способен подавлять противострепто-кокковую активность сульфазина, причем его действие является почти неконкурентным. Не удивительно, что при этом требуется большое количество тимина, так как тимин — не катализатор (подобно ПАБ и фолиевой кислоте), а компонент клетки и потребность в нем увеличивается по мере роста клетки.
Действие лекарственных препаратов — антагонистов ПАБ на большинство видов патогенных бактерий и простейших не снимается с помощью фолиевой кислоты и ее производных, так как они не способны проникать в эти микроорганизмы.
N
л СО—NH2
\ )-NH2
N
рибоза РО4
Амннонмндазол-карбоксамндриботнд
(9.22)
О
N || ХГГ
N N рибоза—РО4 Инозиновая кислота
(9.23)
9.3.2. Роль фолиевой кислоты и других птеридинов
Как было показано выше, производные фолиевой кислоты играют ключевую роль в биосинтезе пуринов и пиримидинов. Эти птеридины представляют собой коферменты, ответственные за включение атомов углерода в положения 2 и 8 в молекулах пуринов и за введение метильной группы в молекулу уридина при синтезе тимидина (4.1,6). При действии на бактерии суль-32
фаниламидных препаратов в низких концентрациях в культуральной жидкости накапливается рибонуклеотид 4-аминоими-дазол-5-карбоксамида (9.22). Это соединение является полупродуктом биосинтеза инозиновой кислоты (9.23), из которой образуются все пурины.
Коферменты образуются из дигидрофолиевой кислоты (9.20), которая превращается в тетрагидрофолиевую кислоту под действием фермента дигидрофолатредуктазы [Osborn, Freeman, Huennekens, 1958]. Тетрагидрофолиевая кислота затем модифицируется различными заместителями, содержащими один атом углерода. Роль кофермента, обеспечивающего введение атома С-2 в пурины, т. е. формилирования риботила (9.22) с образованием инозиновой кислоты (9.23), играет №°-формил-5,6,7,8-тетрагидрофолиевая кислота. На более ранней стадии введения атома С-8, т. е. формирования риботида, в результате которого (в конечном счете) образуется риботид соединения (9.22), участвует тот же фермент, а не №,М10-метенилтетрагидрофолиевая кислота (9.24), как полагали ранее. Коферментом, обеспечивающим введение метильной группы в молекулу уридиловой кислоты, т. е. образование тимидиловой кислоты, является N5,N10-Me-тилентетрагидрофолиевая кислота (9.25), которая ответственна также за взаимопревращения двух аминокислот — серина и глицина. Метилирование уридилата с образованием тимидилата происходит под действием тимидил атсинтетазы (ОММ67 000). Синтез метионина и катаболизм гистидина также катализируются птеридиновыми коферментами, каждый из которых действует совместно со специфичным апоферментом. Фолиевые кислоты, содержащие полипептиды, образованные из остатков глутаминовой кислоты, по-видимому, выполняют еще не выясненные функции коферментов. В плазме человека дигидрофолие-вая кислота содержит лишь один остаток глутаминовой кислоты, однако в природе число этих остатков часто увеличивается до 3—7; особенно это характерно для бактерий [Kisliuk, 1981]. Основное количество фолиевой кислоты человек получает с овощами в виде полиглутамилфолиевой кислоты. Богатыми источниками служат также печень и дрожжи. Биосинтез и функции фолиевых коферментов представлены на рис. 9.3.
RH+=CH RN—СН2
I I О I I О
н н
R= ] — СО—NH— СН(СО2Н)— СН2СН2СО2Н
Метенилтетр агндрофолневая Метнленте гр агндрофолневая
кислота кислота
(9.24)
(9.25)
3—734
33
।--------N10 -Формил-ТГфК-----------*-
Формиат
ДГФК—ТГФК n5 n 1q -Метинил-ТГФН
г
1----^5, N10 •Метилен-ТГФН----------
Серии
i N5. -Метил-ТГФК ------*“
Пурины
Тимидиловая нислота
Рис. 9.3. Биосинтез в функции коферментои фолиевой кислоты (ДГФК и ТГФК, т. е. ди- и тет-рагидрофолиевая кислоты соответственно).
Метионин
В природе биологически активные птеридины образуются при деградации гуанина (4.4) (в виде GTP), приводящей к получению 2-амино-4-оксо-6-(5-О-трифосфо-Е)-рибозиламино)-5-формиламинопиримидина (9.26) [Shiota et al., 1969b). Под действием фермента дигидронеоптеринтрифосфатсинтетазы (получаемой из Lactobacillus plantarum) он циклизуется за счет образования связи между экзоциклическим атомом азота и атомом С2 рибозы. Синтезированный таким образом дигидронеоптерин 2-амино-7,8-дигидро-4-оксо-6-(D-эритро-!',2',З'-тригидрок-сипропил)-птеридин под действием фермента гидронеоптериналь-долазы (получаемой из бактерий Е. coli) выделяет гликолевый альдегид, а остаток 2-амино-4-оксо-6-гидроксиметил-7,8-дигидро-птеридин легко встраивается в молекулу фолиевой кислоты (разд. 9.3.1).
(Т-трифосфатная группа)
(9.26)
Биоптерин (9.27)
Молибдоптерин (9.28)
Участие птеридинов в метаболизме не ограничивается производными фолиевой кислоты. Следует отметить, что наличие аминогруппы в положении 2 гетероцикла не имеет особого значения для проявления биологической активности. Так, биосинтез витамина рибофлавина осуществляется через 6,7-диметил-2,4-дезокси-8-рибитилптеридин. В этой реакции одна молекула птеридина предоставляет, а другая принимает четыре атома углерода для образования орто-ксилольного цикла рибофлавина
34
(Plaut, 1964]. Исходным соединением для данного птеридина служит GTP, рибозильная часть которого превращается в ри-битил-группу молекулы рибофлавина.
Биоптерин (9.27)— птеридин цвета слоновой кости, образуется в процессе биосинтеза у животных и широко распространен в природе; его химическое название—Ь-эритро-2-амино-6-(1,2-дигидроксипропил)-4-оксоптеридин. Исходным веществом для синтеза этого птеридина также является GTP. Восстановленная форма 5,6,7,8-тетрагидробиоптерина служит кофактором, необходимым для ряда ферментов, использующих молекулярный кислород для окисления фенилаланина в тирозин [Kaufman, 1964], тирозина в дофаминкарбоновую кислоту (3.43,6), триптофана в 5-гидрокситриптофан, а также, возможно, и при синтезе меланина. Этот кофактор использует молекулярный кислород. Его применяют в клинике при лечении одного из видов кетону-рии у детей [Cederbaum, 1979]. Тетрагидробиоптерин улучшает состояние больных при паркинсонизме [Narabayashi, 1982].
Птеридин считают основным рецептором электронов, высвобождающихся под действием света в процессе фотосинтеза [Fuller et al., 1971]; предполагают также, что птеридин, содержащийся в глазах млекопитающих, защищает их от ослепляющего действия света [Cremer-Bartels, 1975]. Раковые клетки расщепляют биоптерин до неоптерина, концентрация которого в моче может служить показателем при диагностике рака [Rokos et al., 1980].
Другим важным природным птеридином является молибдоптерин (9.28), кофермент ксантиндегидрогеназы, альдегидокси-лазы, нитратредуктазы, сульфитоксидазы и, видимо, других ферментов, для действия которых необходимо наличие молибдена и железа [Johnson, Rajagopolan, 1982]. Ксантоптерин — колим-фокин (разд. 5.2), ингибирует размножение лимфоцитов [Ziegler et al., 1983].
Химия и биология птеридинов см. Blair (1983).
Химия птеридинов во многих отношениях необычна, главным образом из-за наличия в их молекулах четырех двоесвязанных атомов азота, обладающих сильно выраженными электроноакцепторными свойствами, вследствие чего нарушается ароматический характер этих соединений.
Для птеридинового кольца характерна тенденция к ковалентному присоединению молекулы воды по двойной связи даже при комнатной температуре, например, ксантоптерин (разд. 2.5.1) [Albert, 1967, 1976]. Несколько легче происходит присоединение таких веществ, как ацетон, кетокислоты и меркаптаны. Еще одна особенность птеридинов и некоторых других азотсодержащих гетероциклов заключается в понижении их растворимости в воде за счет эффектов заместителей, способных к образованию внутри- или межмолекулярных водородных связей (например, —NHa или —NH—С = О группы) [Albert, Brown, Cheeseman, 1952]. У природных птеридинов обнаружена
3*
35
и количественно охарактеризована выраженная способность к хелатообразованию [Albert, 1953].
Биохимия фолиевой кислоты и родственных ей птеридинов см. Blakley (1969).
9.3.3. Лекарственные вещества, блокирующие действие дигидрофолатредуктазы
Фолиевая кислота — незаменимый провитамин для человека и других млекопитающих, недостаточность которого быстро приводит к макроцитарной анемии и заболеваниям желудочно-кишечного тракта. Поэтому химики первоначально с недоверием отнеслись к идее создания антагонистов метаболитов на основе птеридинового ядра. Тем не менее их работа оказалась успешной и в результате было создано много ценных лекарственных средств.
Птеридины в организме играют важную роль как катализаторы синтеза пуринов и пиримидинов, поэтому различные аналоги фолиевой кислоты были испытаны в качестве потенциальных противоопухолевых средств. Высокую активность проявил аминоптерин (производное фолиевой кислоты, в котором оксогруппа в положении 4 замещена аминогруппой). Клинические испытания, проводившиеся с 1958 г., показали, что большей избирательностью действия обладает метотрексат (9.29), поэтому в настоящее время именно его применяют в онкологии. Это соединение, созданное Seeger и сотр. в 1949 г., отличается от фолиевой кислоты всего лишь двумя заместителями: оксогруппа в положении 4 заменена аминогруппой, а атом водорода в положении 10 — метильной группой. В организме метотрексат частично превращается в производное, содержащее два дополнительных глутаминовых остатка [Covey, 1980].
Метотрексат вызывает заметную ремиссию симптомов острого лимфоцитарного лейкоза у больных молодого возраста [Farber, 1952] и в настоящее время он является ведущим лекарственным препаратом для лечения этого заболевания. К сожалению, лейкозные клетки постепенно вырабатывают резистентность к метотрексату. Это преодолевается с помощью последующей комплексной лекарственной терапии. Вначале проводится очень короткий курс сильнодействующего препарата — винкристина (5.18,а) (разд. 5.4.7). Затем начинают длительное лечение метотрексатом, часто в сочетании с каким-либо синергистом, обычно 6-меркаптопурином [Skipper, Schabel, Wilcox, 1964]. Таким образом, если несколько лет назад это заболевание быстро приводило к летальному исходу, то сейчас с помощью метотрексата удается добиться высокого процента излечения.
При двух других типах рака лечение метотрексатом приводит к полному излечению. Хориокарцинома (быстрорастущая злокачественная опухоль), появляющаяся во время беременности и обычно быстро приводящая к летальному исходу, полностью
36
излечивается метотрексатом [Ross et al., 1965]. До применения этого препарата из каждых шести женщин, пораженных этим типом рака, пять умирали в течение года. Злокачественная лимфома, обнаруженная Беркиттом у африканских детей, проживающих в областях с жарким и влажным климатом, часто начинается с подчелюстных желез, а затем быстро распространяется по всему организму и обычно приводит к смерти больного в течение шести месяцев. Метотрексат оказывает сильное-действие на лимфому Беркитта и во многих случаях приводит к полному выздоровлению [Bertino, Johns, 1967].
Метотрексат обладает умеренной избирательностью действия. Длительное применение его в больших дозах разрушает эритроциты, приводит к макроцитарной анемии; позже уменьшается образование лейкоцитов. Несмотря на это, терапевтическая широта и эффективность метотрексата были значительно повышены так называемой программой спасения.
При этом курсе терапии делают инъекции цитроворум-фак-тора (5-формилтетрагидрофолиевая кислота) (9.20) спустя день после временного прекращения введения метотрексата. Так, например, развитие остеогенной саркомы у больных молодого возраста можно контролировать с помощью высоких доз метотрексата, в сто раз превышающих обычную безопасную дозу, если в курс лечения ввести «периоды спасения» инъекциями цитроворум-фактора. Применение темидина с метотрексатом и цитроворум фактором
НО2С—СН2
Метотрексат (9.29)
может защитить организм от истощения тимина, при этом лечебный эффект достигается за счет недостаточности пуринов, что, по-видимому, обеспечивает большую избирательность действия препаратов [Frenkel, Hitchings, 1957; Tattersail, Jaffe, Frei, 1975].
Метотрексат используют также при лечении псориаза, распространенного и трудноизлечимого кожного заболевания [Weinstein, 1977]. При псориазе вследствие избыточного образования дигидрофолатредуктазы деление клеток эпидермиса происходит каждые 3 дня вместо 27 дней в норме. Лекарственный препарат необходимо принимать систематически; наличие побочных эффектов ограничивает продолжительность лечения.
А. Механизм действия метотрексата. Метотрексат ингибирует ДГФР, выделенную из клеток млекопитающих и бактерий,
37
несмотря на то, что обычно он не проникает в бактериальные клетки [Nichol, Welch, 1950; Werkheiser, 1953]. Действие метотрексата высокоспецифично: он вызывает 50% ингибирование ДГФР в концентрации 10~9 М, при которой не влияет на любой другой фермент. Этот ингибитор связывается с ДГФР примерно в 104 раз прочнее, чем субстрат (это очень необычное явление) (разд. 9.1), и, следовательно, его действие наименее обратимо по сравнению со всеми обратимыми ингибиторами. Отличия в ингибирующем действии метотрексата на ДГФР, выделенную из разных организмов, очень невелики (в отличие от диаминопиримидинов, см. ниже) [Werkheiser, 1963]. Отсутствие у метотрексата способности проникать в одноклеточные организмы обусловлено тем, что у них нет системы переноса, имеющейся у млекопитающих. Поскольку метотрексат имеет низкий log Р (—1,85), он не может свободно проникать через цитоплазматические мембраны.
Дигидрофолатредуктазы разных типов могут катализировать восстановление как фолата, так и дигидрофолата, хотя дигидрофолат реагирует с ферментами позвоночных в 4-—10 раз быстрее, чем фолат. Скорость восстановления фолата бактериальными ферментами очень незначительна [Blakley, 1969].
Рентгеноструктурный анализ дигидрофолатредуктазы (ДГФР), закристаллизованной с метотрексатом, позволил глубже понять механизм действия этого ингибитора и наглядно представить взаимодействие лекарственного вещества с рецептором [Matthews et al., 1977], позднее Bolin и сотр. (1982) использовали прибор с разрешением до 0,17 нм. Диаграмма фермента ДГФР, его кофермента (NADPH) и метотрексата представлена на рис. 9.4. Изображенный на нем фермент получен из бактерий Lactobacillus casei и те же авторы сообщили о ДГФР (закристаллизованной с метотрексатом), извлеченной из бактерий Е. coli. Однако им не удалось закристаллизовать метотрексат вместе с ДГФР, извлеченной из организма какого-либо позвоночного.
Метотрексат связывается с ферментом, полученным из L. casei, следующим образом. Анион Asp-26 фермента связан с катионом амидиния, образованным из атомов N-1 и 2-NH2 лекарственного вещества (значения рКа приведены в колонке N-1 табл. 9.3). Аминогруппы, связанные с атомом С-4 лекарственного вещества, образуют водородную связь с карбонильными группами Leu-4 и А1а-97. а-Дарбоксильная группа глутаминового фермента метотрексата образует ионную связь с основной боковой цепью Arg-57. Птеридиновое кольцо проникает в полость, ограниченную Leu-4, Ala-6, Leu-27, Phe-ЗО и А1а-97.
пара-Аминобензоильный фрагмент располагается в соседней полости, окруженной липофильными боковыми цепями Leu-27 и Phe-ЗО, с одной стороны, и Phe-49, Pro-50 и Leu-54 — с другой.
Молекула кофермента НАДФН полностью вытянута; никотинамидная часть располагается напротив птеридинового кольца 38
J А1а97Ц^ J
Leu4 /
TjMflPhe30j)
JA А Ф ^M.eul9
Thri16 L'®. JB
°=®с=й|Гтгр
SE «Я ж
Ala 97
Leu 4
Ц Aap26i
Thri16
Phe3<
.Leuig
Trp21
Рис. 9.4. Стереодиаграмма птеридинсвязывающего участка днгвдрофолатрс-дуктазы L. casei. Ингибитор метотрексат выделен жирными линиями, белок — светлыми, молекула НАДФ (показана только ее часть) — пунктиром; атомы углерода изображены небольшими, а атомы кислорода — большими светлыми кружками, атомы азота — черными кружками; цифры в кружках — связанные молекулы воды [Bolin et al., 1982].
в положении, облегчающем перенос гидрид-иона от пиридинового цикла к атому углерода в положении 6 птеридинового кольца [Filman et al., 1982].
Диаграмма для фермента, полученного из бактерий Е. coli, практически идентична представленной на рис. 9.4 [Bolin et al., 1982].
Отличительной особенностью метотрексата является то, что при связывании с ферментом птеридиновые ферменты антиметаболита и природного субстрата ориентированы противоположно. В отличие от метотрексата (см. выше) для дигидрофолата характерно следующее [Bolin et al., 1982]: атом N-1 не связан, 2-NH2, так же как 0-4, связана только с молекулами воды, атом N-3 образует водородную связь с Asp-26, атом N-5 не связан, а атом N-8 связан с Leu-4 силами ван-дер-ваальса. Таким образом, птеридиновый цикл нормального субстрата в отличие от метотрексата связан с ферментом значительно слабее. Связи, образуемые пара-аминобензоильным и глутаминовым фрагментом метаболита и его антагониста, идентичны. Именно в этом заключается причина необычно высокого индекса ингибирования (1:10 000) метотрексата по сравнению с дигидрофолиевой кислотой.
Полагают, что главная причина различной ориентации метотрексата и субстрата при связывании с ферментом заключается в разных положениях и основности наиболее основного атома азота в молекуле (см. табл. 9.3). Метотрексат почти в сто раз более сильное основание, чем дигидрофолиевая кислота, благо-
39
Таблица 9.3. Величины рКа фолиевой кислоты и ее производных (вводе), полученные: А — с помощью спектрофотометрии, В — с помощью ЯМР-образ-цов, обогащенных 13С по атому С-2 [Рое, 1977; Cocco et al., 1981]
Соединение N-1 (основание) N-5 (основание) CO-4/NH-3 (кислота)
Фолиевая кислота А 2,35 В 2,40 <—1,5 8,38 8,25
Дигидрофолиевая кислота А 1,38 3,84 9,54
Метотрексат А 5,71 В 5,73 <—1,5
даря замене циклической амидной группы (CO-4/NH-3) аминогруппой. Следует отметить, что наиболее основным центром метотрексата является амидиновая группа N1 —С2—NHa, находящаяся в сопряжении, в то время как наиболее основный центр в молекуле дигидрофолиевой кислоты расположен в другом цикле птеридинового ядра и не соприкасается с каким-либо противоположно заряженным центром молекулы фермента.
Наиболее основный центр молекулы метотрексата имеет рКа 5,71 и, следовательно, только 2% его будут существовать в виде катиона при pH 7,3 в воде (табл. 17.1). Поэтому если даже предположить, что молекула метотрексата образует лишь одну ион-дипольную связь с молекулой аспарагиновой кислоты, то и этого было бы достаточно для того, чтобы обеспечить большое конкурентное преимущество метотрексата по сравнению с дигидрофолиевой кислотой. Тем не менее следует ожидать, что величина рКа, определенная в воде, в действительности будет значительно выше внутри выстланной липидами полости, которой точно соответствует птеридиновое кольцо молекулы метотрексата, но не таковое дигидрофолата. Это повышение основности обусловлено понижением диэлектрической постоянной и отсутствием конкурирующих молекул воды. Предположение о повышении основности молекулы метотрексата при связывании с ферментом подтверждают также данные ЯМР-спектров, снятых для метотрексата, обогащенного изотопом 13С (до 90%) по атому С-2 [Cocco et al., 1981]. Установлено, что характеристичный сигнал атома С-2 в катионах сохраняется вплоть до pH 10, в то время как в аналогичных условиях основный рКа фолиевой кислоты, равный 2,4, не изменяется. Неизвестно, что именно происходит с дигидрофолиевой кислотой в этих условиях, так как ее недостаточно высокая стабильность не позволяет сделать обоснованных выводов. (Все исследования проводились на ДГФР, полученной из Streptococcus faecium.)
Б. Упрощение молекулы метотрексата. Этот противораковый препарат обладает низкой токсичностью по отношению к большинству бактерий и простейших [Wood, Ferone, Hitchings, 1961],
поскольку он не способен проникать в одноклеточные организмы. Вначале предполагали, что имеет смысл повысить его проникающую способность путем увеличения липофильности его молекулы. Однако вскоре стало очевидно, что антагонистическое действие дигидрофолата обусловлено в основном 2,4-диамино-птеридиновой частью молекулы. Примером попыток модификации структуры метотрексата могут служить следующие результаты.
Было показано, что 2,4-диаминоптеридин эффективен не менее, чем хинин, при малярии, возбудителем которой является Plasmodium gallinaceum. 2,4,7-Триамино-6-фенилптеридин (диуретик триамтерен) подавляет паразитемию у мышей и крыс, вызываемую Pl. berghei [Aviado, Brugler, Bellet, 1968]. 2,4-Ди-амино-6,7-диэтилптеридин, как оказалось, высоко активен против Vibro cholerai в лабораторных тестах [Collier, Waterhause, 1950], но не оказывает действия на многие другие виды бактерий. Ни один из этих результатов не привел к созданию новых эффективных лекарственных препаратов.
Наиболее успешными оказались попытки упрощения молекулы метотрексата, начатые Хитчингсом, который обнаружил, что удаление некоторых несущественных групп из молекулы (9.29) позволяет получить аналоги с большей проникающей способностью. Такие упрощенные производные метотрексата представляют собой 2,4-диамино-1,3,8-триазанафталины. Наиболее активен из них 6-бутил-2,4-диамино-5-метилпиридо[2,3-б]пи-римидин (9.30) [Robins, Hitchings, 1955]. Действие этого соединения на грамположительные и грамотрицательные бактерии облегчается наличием липофильных заместителей в положениях 5 и 6, а также за счет повышения основности вследствие удаления из цикла атома азота с двойной связью. Было обнаружено,, что ингибирование ДГФР, выделенной из разных бактерий, производными триазанафталина пропорционально их действию на интактные бактерии [Burchall, Hitchings, 1965; Hitchings,. Burchall, 1965].
К сожалению, применение этих триазанафталинов привело’ к появлению новой формы токсичности, поскольку они ингибируют действие гистамин-М-метилтрансферазы и приводят к накоплению у больных непереносимых концентраций гистамина. Таким образом, появилась необходимость модификации заместителя в положении 6, что привело к созданию 2,4-диамино-6-(2,5-диметоксибензил) -5-метилпиридо[2,3-б]пиримидина, аналога соединения (9.30). Это вещество оказалось столь же эффективным, как и метотрексат; оно ингибирует ДГФР лейкозных клеток человека и вызывает регрессию лейкоза и солидных опухолей у крыс и мышей [Grivsky et al., 1980; Duch et al., 1980]. Результаты клинических испытаний еще не получены. Известно, что некоторые производные хиназолина обладают антилейкозной активностью, однако о клиническом их применении до сих пор-не сообщалось.
41
Me—(СН2)
Me NH2
N N NH2
Диаминобутилтриазанафталии
(9.30)
(9.31)
Дальнейшее упрощение молекулы метотрексата с целью получения противораковых препаратов не принесло успеха, но показало, что его аналоги могут быть полезны в борьбе с микроорганизмами. Как уже указывалось в разд. 4.0, Хитчингс, продолжая изучение биологической активности фрагментов молекулы метотрексата, обнаружил, что простые производные 2,4-диаминопиримидина обладают мощной противомикробной активностью, выступая в роли антагонистов фолиевой кислоты. На основании этих данных он создал пириметамин — 2,4-диами-но-5- (4'-хлорфенил)-6-этилпиримидин (9.31) [Falco et al., 1951]— один из самых эффективных противомалярийных препаратов, в настоящее время широко применяемый в качестве профилактического средства. Пириметамин действует на преэритроцитные формы паразитов, особенно в печени. Его недельной дозы (25 мг) оказывается достаточно для высокоэффективного подавления малярии, вызванной Pl. falciparum. Однако для лечения острой формы малярии этот препарат не подходит. Циклогуанил (3.35), дигидротриазиновый аналог пириметамина, обсуждается в разд. 3.6. Большие дозы этого препарата иногда вводят в виде его нерастворимой соли с памоевой кислотой для пролонгирования профилактического действия. Комбинированное действие пириметамина и сульфаниламидов обсуждается в разд. 9.6 (последовательное блокирование).
Действие пириметамина на ДГФР, выделенную из разных организмов, представлено в табл. 4.1. Этот препарат не обладает высокой избирательностью действия по отношению к малярийным паразитам; фермент плазмодиев ингибируется пириметамином при концентрациях в 2000 раз более низких, чем необходимы для ингибирования аналогичных ферментов млекопитающих. Избирательность действия пириметамина при малярии обусловлена необыкновенно высокой чувствительностью фермента паразита по сравнению с ферментом организма хозяина. Но не только этим ограничивается отличие ДГФР паразита: фермент, выделенный из Pl. berghei, имеет ОММ около 200 000, что в 10 раз больше, чем у аналогичных ферментов млекопитающих и бактерий. Малярийный паразит, так же как бактерия, не способен поглощать экзогенную фолиевую кислоту.
Для позвоночных наиболее эффективными противомалярийными средствами являются 2,4-диаминопиримидины, содержащие заместитель с выраженными липофильными свойствами, например фенильную группу в положении 5 и алкильную труп*
42
пу в положении 6. Способность соединений ингибировать изолированную дегидрофолатгидрогеназу незначительно изменяется при замене фенильной группы в положении 5 пириметамина бутильной или 4-фенилбутильной группой, однако укорочение бутильной группы приводит к заметному снижению сродства к этому ферменту [Baker, Shapiro, 1966].
Для повышения антибактериальной активности 2,4-ди-аминопиримидинов у позвоночных необходимо, чтобы в объемный липофильный заместитель входила гидрофильная группа. Лучшим соединением такого типа оказался триметоприм (9.32) [Roth, Falco, Hitchings, 1962]. Триметоприм обладает такой же избирательностью действия по отношению к бактериальным ферментам, как пириметамин по отношению к ферментам малярийных паразитов. Что касается применения этих диаминопиримидинов в клинике, то их терапевтический индекс значительно выше, чем метотрексата. Триметоприм — сильнодействующий препарат: подавляя ДГФР грамположительных и грамотрица-тельных бактерий, он оставляет неповрежденными аналогичные ферменты млекопитающих. Индекс ингибирования триметоприма (для бактерий) в среднем равен 1 : 1000 [Roth, Cheng, 1982].
Высокая степень избирательности триметоприма по отношению к бактериальным ферментам (в сравнении с ферментами млекопитающих) не свойственна его аналогам; для повышения избирательности действия необходимо, чтобы все три положения в бензольном кольце (3-, 4-, 5-) таких соединений были заняты. Так, для 3,5-диметоксипроизводного (9.32) отношение способности ингибировать ДГФР бычьей печени и Е. coli (концентрации, необходимые для ингибирования чистой ДГФР) составляет 116 по сравнению с 1840 для триметоприма [Roth,. Cheng, 1982]. Аналог с незамещенным бензольным кольцом! избирательностью практически не обладает. Тем не менее 3,5-диалкоксипроизводные, содержащие в положении 4 изопро-пенильную, ацетильную или метоксикарбонильную группу, обладают высокой активностью и избирательностью действия. По-видимому, лучшими конкурентами триметоприма являются 3,5-диэтил-4-метокси- и 3,5-диметокси-4-метилтиопроизводные триметоприма. Однако эти соединения еще не прошли такой длительной клинической проверки, как триметоприм. Обычно в этой серии аналогов триметоприма повышение липофильности молекулы усиливает действие соединения на грамположитель-ные микроорганизмы и ослабляет на грамотрицательные. Помимо необходимости достижения нужного соотношения гидрофильных и липофильных свойств бензильного заместителя в пиримидиновом цикле, следует учитывать и строгие стерические требования [Roth, Cheng, 1982].
При кокцидиозе, протозойном заболевании домашней птицы,, сходном с малярией, применение ингибиторов ДГФР типа триметоприма предпочтительнее, чем аналогов пириметамина, но наиболее широко используют диаверидин (9.33).
43
Триметоприм (9.32)
Диаверндии
(9.33)
До сих пор отсутствуют данные рентгеноструктурного анализа ДГФР, связанной с каким-либо ингибитором диаминопиримидинового ряда. Результаты получены для 2,4-диамино-5,6-дигидро-6,6-диметил-5-(4'-метоксифенил)-1,3,5-триазина при разрешении 0,29 нм. Это соединение — близкий аналог антималя-рийного препарата циклогуанила (3.35) и отличается от него тем, что атом хлора замещен метоксигруппой [Volz et al., 1982]. Этот сильный ингибитор связывается с ДГФР малярийных паразитов и позвоночных (в данном случае — ферментом из печени цыпленка) и практически не реагирует с бактериальным ферментом.
Было обнаружено, что триазиновый и фенильный циклы занимают те же положения, что и соответственно пиримидиновый и пиразиновый циклы метотрексата в бактериальных ферментах. Важную роль в связывании этого ингибитора играют Clu-ЗО и Туг-31, каждый из которых сдвигается на 0,3 нм при связывании ингибитора с апоферментом. Кофермент NADPH полностью вытянут и располагается в длинной неглубокой впадине на поверхности наружной стороны апофермента [Volz et al., 1982].
Кроме незначительной степени связывания триметоприма с ДГФР печени цыпленка, рентгеноструктурный анализ показал, что при образовании связей с этим ферментом он находится в: конформации бабочки, энергетически менее выгодной, чем та, в которой он взаимодействует с ферментом Е. coli, когда два . цикла расположены почти перпендикулярно друг к другу [Matthews, 1981].
Открытие этих эффективных лекарственных веществ, производных 2,4-диаминопиримидина, явилось результатом научного предвидения. Установлено, что максимальную биологическую активность они проявляют при рКа между 7 и 8 [Roth, Strelitz, 1969]. Это указывает на их способность проникать через клеточ-. ные мембраны (разд. 10.5).
В медицинской практике триметоприм чаще всего применяют в комбинации со стрептоцидом с целью достижения последовательного блокирования (разд. 9.6). При беременности антаго-^ нисты фолиевой кислоты следует использовать осторожно, так как существует опасность повреждения плода. 1
Подробнее о биохимии фолиевой кислоты см. Blakley (1969);] обзоры Roth, Cheng (1982) и Roth, Bliss, Beddell (1983) посвя-i
44
щены рассмотрению связи структура — активность 2,4-диамин-пиримидинов, -птеридинов и -хиназолинов в качестве антибактериальных, противораковых и противомалярийных препаратов.
9.4. Другие аналоги метаболитов, играющие важную роль в профилактике и терапии
9.4.1. Введение
Создание аналогов метаболитов не представляет особых сложностей, однако найти среди них обладающий избирательным действием очень трудно, так как редко можно встретить метаболит, незаменимый для вредных видов и не имеющий особого значения для вида полезного. Если метаболит содержится во всех живых клетках (как, например, тиамин), то возможность получения аналога, обладающего избирательным действием, маловероятна. Тем не менее в нескольких случаях удалось обнаружить такие избирательно действующие вещества и применить их в качестве лекарственных средств. Успех нередко определяется тем, что один из видов обладает преимущественной способностью к поглощению аналога (по сравнению с другим). Даже относительно небольшие изменения молекулярной структуры соединений могут так повлиять на способность клетки к поглощению, что лекарственное вещество либо вообще нс будет попадать в клетки полезных видов, либо будет преимущественно поглощаться вредными или концентрироваться в той области, где сосредоточены вредные виды.
Следует отметить, что аналог метаболита может играть роль метаболита для одного вида и служить антагонистом для другого, родственного вида. Так, например, детиобиотин может заменять биотин (9.34) в качестве фактора роста дрожжей, но
Н (СН2)4—СО2Н
N |
Биотин Дикумарол
(9.34) (9.35)
Дихлон
(9.36)
45
в то же время является антагонистом биотина у Lactobacillus casei, у которых биотин ускоряет рост. Причина этого известна: дрожжи способны включать в молекулу биотина недостающий атом серы, а бактерии этой способностью не обладают [Ditmer, du Vigneaud, 1944]. Устойчивость гриба Endomyces к действию пиритиамина (9.5), приобретенная в процессе повторных пересевов на среду, содержащую это вещество, объясняется способностью устойчивого штамма расщеплять молекулу пиритиамина и синтезировать тиамин (2.1) из одного из образующихся больших фрагментов [Wooley, 1944].
Природный агонист может служить субстратом для нескольких ферментов, причем для блокирования этих ферментов могут быть необходимы разные ингибиторы. Так, например, дикума-рол (9.35) является избирательным антагонистом фактора свертывания крови у человека — витамина К (2.2), но не токсичен для микроорганизмов [Goth, 1945]. С другой стороны, высокая фунгицидная активность 2,5-дихлорнафтохинона (дихлон) (9.36) также связана с его ингибирующими действиями на витамин К у грибов, тогда как для млекопитающих он безвреден (Wooley, 1950,b).
Антиметаболиты обычно оказывают более эффективное разрушающее действие на клетки, нуждающиеся в притоке метаболитов извне; примером может служить пиритиамин [Wooley, White, 1943].
9.4.2. Аналоги витаминов и коферментов
Ампролиум (9.37), аналог тиамина (2.1), успешно применяют при лечении кокцидиоза у домашней птицы. Это лекарственное вещество, ингибирующее транспорт тиамина, в 50 раз сильнее действует на Eimeria, чем на клетки кишечника цыпленка [James, 1980].
2,2-Дихлорпропионат натрия (далапон) широко применяют для уничтожения однодольных сорняков на полях двудольных растений, например свеклы и люцерны. Исследование конкурентного действия 2,2-дихлорпропионата натрия на Е. coli показало, что он препятствует включению пантоевой кислоты [левая часть формулы (9.38)] в молекулу пантотеновой кислоты. Это основное направление его действия на однодольные сорняки; активность далапона снижается при добавлении пантотеновой кислоты [Hilton et al., 1959].
Me NH2
A,s/CH2y^n
H NT4,
M2N N CH2CH2Me Ампролиум (катион)
(9.37)
46
Me
HOCH2—С— СН(ОН)—СО I Me
NH—СН2—СН2—СО2Н
Пантотеновая кислота
(9.38)
ОН
^,СН(СвН5)—СН2СОМе ^О
Варфарин
(9.40)
Клинические испытания дикумарола (9.35), первого перорального антикоагулянта, были начаты в 1941 г. Было обнаружено, что это вещество [З.З'-метилен-бис (4-гидроксикумарин)], выделенное из подпорченного сена, вызывало кровотечения у крупного рогатого скота, поедавшего это сено. Длительный курс дикумарола показан больным гипертонией для профилактики тромбозов коронарных артерий и инсультов. Для проявления антикоагулянтного действия у кумаринов необходимо наличие 4-гидроксикумаринового цикла с атомом водорода или алкильным заместителем в положении 3. Механизм действия дикумарола и его более сильнодействующего аналога варфарина (9.40) заключается в ингибировании регенерации витамина К (2.2) из его 2,3-эпоксида [Zimmerman, Matschiner, 1974]. Эпоксид образуется при выполнении витамином К его биологической функции: введении дополнительной карбоксильной группы в десять остатков глутаминовой кислоты протромбина, что повышает способность этого белка хелатировать кальций [Stenflo et al., 1964].
Антикоагулянты кумаринового ряда неактивны in vitro, а для проявления их действия в организме необходимо 36 ч. Хотя механизм антагонизма между производными кумарина и витамином К очень прост (избыток одного из них нейтрализует другой), тем не менее для проявления их действия также необходимо много времени. Антикоагулянты индандионового типа, наиболее распространенным представителем которых является фениндион (9.39), действует несколько быстрее. Были предприняты попытки создания на их основе препарата, структура которого в большей мере соответствовала бы структуре витамина К (т. е. в положении 3 присутствовала бы липофильная группа, а фенольная группа была бы замещена кетонной).
9.4.3. Аналоги гормонов и нейромедиаторов
Agarwal в книге «Антигормоны» напоминает о том, что антиметаболиты способны ингибировать действие представителей каждого из пяти основных классов стероидных гормонов, а так
47
же тироксина, полипептидных мессенджеров и нейромедиаторов {Agarwal, 1979]. Некоторые из этих антагонистов применяют только в биохимических исследованиях, тогда как другие — в качестве эффективных лекарственных средств.
На первый взгляд может показаться парадоксальным, что аналоги, обладающие антагонистическим действием, иногда используют для достижения агонистического эффекга (по непрямому механизму). И наоборот, агонисты могут быть использованы для получения антагонистического эффекга (в зависимости от регулярности введения и биологических циклов организма). Хорошо известный и, пожалуй, уникальный пример — это применение эстрогена или прогестина, или (чаще всего) их смеси в одной «пилюле» в качестве контрацептивного средства. Эти препараты принимают около 50 млн женщин во всем мире. Чаще всего применяют такие эстрогены, как этинилэстрадиол и его 3-метиловый эфир, местранол. Эти соединения представляют собой агонистические аналоги женского гормона эстрадиола. Из прогестинов обычно применяют норгестрол и норэтиндрон — агонистические аналоги природного женского гормона прогестерона. Во всех этих контрацептивах структура природных гормонов изменена незначительно введением этинильной группы (—С^СН) для придания веществу активности при пероральном введении и для предохранения его от метаболического разрушения. Лекарственные препараты обоих типов предотвращают оплодотворение, подавляя овуляцию. Механизм действия эстрогена заключается в ингибировании выделения фолликулостимулирующего гормона (ФСТ), прогестин ингибирует выделение лютеинизирующего гормона (ЛГ); таким образом, искусствет-ным путем постоянно осуществляется то, что в природе происходит только периодически.
Известны также некоторые антиэстрогены, из которых чаще всего применяют тамоксифен (9.41), эффективный при лечении эстрогензависимых форм рака молочной железы. Тамоксифен связывается с белковыми рецепторами эстрадиола, понижая тем самым их концентрацию в цитоплазме. В процессе метаболизма он превращается в 4-гидроксипроизводное, которое считают активной формой тамоксифена [Borgna, Rochefort, 1981]. Аналоги тамоксифена — кломифен и нафоксидин. Тамоксифен напоминает по структуре диэтилстильбэстрол, но обладает (12.31) более сильным эстрогенным действием, чем природный гормон (12.32). Подробнее о стероидных гормонах см. разд. 2.4. и 12.2. Гормоны насекомых и их антиметаболиты см. разд. 4.7.
При лечении некоторых типов гормонозависимого рака молочной железы эффективным оказался аминоглутетимид (9.42)— предположительно ингибитор биосинтеза всех гормонов — производных холестерина [Henderson, Canellos, 1980].
К антиметаболитам, влияющим на действие нейромедиаторов, в первую очередь относятся аналоги АХ (2.7), блокирующие доступ этого нейромедиатора к рецепторам. В качестве
48
Аминоглутетимид
(9.42)
примера можно привести также миорелаксанты, применяемые при общей анестезии: тубокурарин (2.6), галламин (7.31), дити-лин (7.29) и простые аналоги атропина, такие как пропантелин (7.25). Лекарственные препараты типа физостигмина (7.33) усиливают действие АХ, блокируя его доступ к разрушающему ферменту (АХЭ), так же как и другие карбаматные ингибиторы этого фермента (разд. 13.3). Эти псевдоагонисты применяют при лечении глаукомы, миастении и атонии кишечника и мочевого пузыря. Более подробно об агонистах см. разд. 12.6, ингибиторы холиэстеразы — разд. 13.3 и 12.5. Модификации молекул агонистов с целью получения соединений, обладающих антагонистическим действием (в том числе на примере АХ), см. разд. 7.5.2 (том 1).
Адренергические антиметаболиты-—аналоги норадреналина (7.5) играют большую роль в терапии. Широту их применения можно проиллюстрировать следующими примерами. Метарами-нол (7.54), препятствующий захвату норадреналина пресинап-тическим окончанием, применяется для повышения низкого давления и выведения больного из шокового состояния. Эфедрин (9.43) накапливается в пресинаптическом окончании и способствует равномерному выделению норадреналина из депо в синаптическую щель; этот препарат эффективен при поллино-зах и других аллергических заболеваниях. Один из побочных эффектов эфедрина — возбуждающее действие на ЦНС. Фенамин (9.44) обладает сходными с эфедрином свойствами, однако значительно сильнее стимулирует ЦНС. Фенамин применяют для уменьшения аппетита и преодоления сонливости, однако его применение ограничено из-за возможности привыкания.
Три вышеперечисленных лекарственных препарата действуют за счет повышения эндогенного норадреналина больного, однако возможен и другой механизм. Октадин (7.55) накапливается в пресинаптических окончаниях и способствует равномерному выделению из них норадреналина, однако с такой низкой скоростью, что по мере высвобождения норадреналин успевает разрушаться. Применение этого лекарственного препарата дает
4-734
4 9*
альтернативный, хотя и недостаточно избирательный, метод лечения умеренных форм гипертонии. Орнид (12.4), дебрихозин и бетанидин действуют примерно таким же образом. Другие лекарственные вещества (например, мезатон, метилдофа), созданные на основе катехоламиновых нейромедиаторов, см. в разд. 12.4.
0[| ОН NHMe ^-СН—CH—Me
СН2—СН(Ме)—NH2
Эфедрин (9.43)
Фенамин
(9.44)
2
6-Гидроксидофамин (9.45)
Анаприлин (пропранолол) (12.56) и подобные ему соединения являются эффективными антагонистами норадреналина в постсинаптических рецепторах (разд. 12.4) и широко используются при лечении гипертонии.
6-Гидроксидофамин (9.45), избирательно разрушающийся в организме аналог норадреналина, широко применяют в экспериментальной физиологии для картирования рецепторов. При введении лабораторным животным он распределяется по всем симпатическим нервным окончаниям, где и окисляется в соответствующий орто-хинон. Последний ковалентно соединяется с этими окончаниями и, дезактивируя их, помогает картированию [Thoenen, Tranzer, 1968].
В 1940 г. было обнаружено, что гидразины с липофильными заместителями, например фенелзин (9.46), помогают при глубоких психических депрессиях. Известно, что механизм действия этих веществ связан с их взаимодействием с моноаминоксида-зой (МАО) в головном мозге. В результате этого ингибирования происходит накопление аминов — нейромедиаторов, таких как норадреналин, дофамин и 5-гидрокситриптамин. Однако из-за серьезных побочных эффектов, вызываемых производными гидразина, они были заменены липофильными первичными аминами, которые не соединяются так прочно с ферментом. Из первичных аминов в настоящее время наиболее широко применяют транилкипромин (9.47). В молекулах всех этих аминов содержатся стерические помехи, предотвращающие их дезаминирование под действием фермента.
Сх^\сн2—СН2—NH—NH2 CH—CH—NH2
сн2 Фенелзин Транилкипромин
(9.46) (9.47)
50
МАО человека представляет собой небольшую группу изоферментов, которые подразделяются на тип А, дезаминирующий преимущественно норадреналин и 5-гидрокситриптамин, и тип В, дезаминирующий дофамин и содержащие аминогруппу ксенобиотики, такие как бензиламин [Cawthon, Breakefield, 1979]. Побочные эффекты, вызываемые ингибиторами МАО из ряда первичных аминов, бывает трудно устранить из-за прочного связывания лекарственного вещества с ферментом. Поэтому при лечении психических депрессий иногда предпочтительнее применение трициклических антидепрессантов (разд. 12.9) (но не для кардиологических больных) или электрошоковой терапии, при которой происходит высвобождение нейромедиаторных аминов [Сгееп, Heale, Graham-Smith, 1977].
Подробнее о МАО и ее модификациях см. Singer, von Korf, Murphy (1979). Исследованию биохимических основ нейрофармакологии посвящена работа Cooper, Bloom (1982).
Широко применяемый инсектицид хлордимеформ (2.36) обладает мягким ингибирующим действием на МАО млекопитающих, а его инсектицидная активность связана, по-видимому, с его сходством с нейромедиатором членистоногих — октопами-ном (2.35) (разд. 2.6.1 и 6.4.1).
9.4.4. Аналоги аминокислот
Антибактериальный лекарственный препарат циклосерин (5.11) представляет собой структурный аналог D-аланина (5.12); биохимические основы его действия изложены в разд. 5.3. Циклосерин препятствует образованию D-аланина и его конденсации в О-аланил-О-аланин, в отсутствие которого прекращается синтез пентапептида, являющегося основой новых клеточных стенок [Strominger, Ito, Threnn, 1960]. Позднее был обнаружен более простой аналог — З-фтор-О-аланин, высоко эффективный in vivo при всех обычных грамположительных и грамотрицатель-ных бактериальных инфекциях. Антагонистом З-фтор-О-аланина может служить D-аланин (разд. 5.3). Более сильное действие оказывает L-алафосфалин (3.50), ингибирующий L-аланинраце-мазу — фермент, ответственный за образование D-аланина. D-Алафосфалин высоко эффективен in vivo по отношению к грамотрицательным бактериям (разд. 3.6). Эти аналоги аланинов до сих пор не нашли широкого клинического применения.
Антибиотик азасерин (9.49) — это серин, ацилированный диазоуксусной кислотой; он является структурным аналогом глутамина (9.49) и конкурирует с ним в синтезе пуринов (разд. 9.7.2) [Buchanan, 1957]. Азасерин применяют в химиотерапии рака в качестве агента, осуществляющего последовательное блокирование (разд. 9.6).
Антиметаболиты пептидов. Проблема, связанная с попытками создания новых полипептидных лекарственных препаратов, заключается в их быстром расщеплении в желудочно-кишечном
51
тракте и (даже если этого можно избежать заменой некоторых L-аминокислот D-энантиомерами) плохой всасываемости из желудка и кишечника. Преодолеть эти трудности удается заменой пептидной связи менее легко расщепляющимися полярными связями, обычно эфирной, или кетонной или даже неполярными звеньями, например циклогексановым кольцом. Такие продукты получили название «пептоидов» [Farms, Ariens, 1982].
Полагают, что вещество Р — ундекапептид, содержащийся в ЦНС, — служит нейромодулятором для устранения, а в более высоких дозах — для передачи болевых ощущений. Его антагонисты были получены путем удаления первых трех аминокислот и заменой на D-энантиомеры пролина-4 и триптофанов-7 и -9 [Caranikas et al., 1982]. Результаты клинических испытаний еще не получены.
Эффективные аналоги (агонисты и антагонисты) природных опиоидных олигопептидов (энкефалинов) см. разд. 12.8.
Пентапептид пентагастрин заменил природный гастрин (гептадекапептид) в качестве диагностического средства, стимулирующего желудочную секрецию. Пентагастрин представляет собой t-Boc-^-Ala-Trp-Met-Asp-Phe-NHa. Этот лекарственный препарат можно применять интраназально.
Ингибиторы ангиотензинтрансформирующего фермента, известного также как пептидилдипептидаза (ПДП), долгое время считались возможными источниками антигипертензивных лекарственных средств. По-видимому, существуют два механизма, лежащих в основе повышения давления крови этим ферментом. Во-первых, он удаляет концевой дипептид из ангиотензина I, в результате чего образуется октапептид ангиотензин II, вызывающий сильное сужение сосудов. Во-вторых, он дезактивирует брадикинин (нонапептид, находящийся в крови и обладающий сосудорасширяющим действием), отщепляя от него два дипептида.
Получаемый из яда змей батроп или терпротид (нонапептид— pyroClu-Trp-Pro-Arg-Pro-GIn-Ile-Pro-Pro) является ингибитором ПДП. Установлено, что концевой остаток пролина препятствует гидролизу пептида при действии фермента. Тем не менее терпротид оказался неэффективным при пероральном введении, так как пептидазы кишечника человека действуют менее избирательно. Поэтому было предпринято исследование пептоидов, включающих, например, сукциниламинокислоты, эквивалентные по длине дипептидному фрагменту. Каптоприл (9.50) был первым из этих соединений, нашедшим применение в клинике. Это сильно действующее средство для снижения повышенного давления крови имеет высокий терапевтический индекс и эффективно при пероральном введении [Ondetti, Rubin, Cushman, 1977; Ondetti, Cushman, 1981]. Эти исследования показали, что для проявления активности необходимы концевой пирролидиновый цикл, свободная карбоксильная группа (или ее изостера), еще одна карбонильная группа, меркапто- или фос
52
форильная группа для образования хелата с атомом цинка в ферменте, а также S,S-конфигурация молекулы (определение обозначения S дано в разд. 12.1).
В аналогичной работе был получен эналаприл [М-(3-фенил-пропил-I-этоксикарбонил)-L-аланин-Ь-пролин] и изучено его действие при гипертонии. Видимо, основные физиологические эффекты этих дипептидов связаны с их способностью непосредственно конкурировать с ангиотензином II [Unger, Ganten, Lang, 1983].
Ale _____
HSCH2CHCON I ^CO2H
Каптоприл (9.50)
9.4.5. Аналоги пуринов и пиримидинов
Различные аспекты использования аналогов природных пуринов и пиримидинов обсуждались в разд. 4.0. В химиотерапии рака применяют следующие соединения: 5-фторурацил (3.3), цитарабин (4.13), 6-меркаптопурин (3.14) и 8-азагуанин (4.40). К лекарственным средствам относятся такие противовирусные препараты, как ацикловир (4.19), являющийся производным гуанина, идоксуридин (4.14,а), трифлуридин (4.14,6) и видара-бин (4.16), аналог аденозина. Иммунодепрессант азатиоприн (3.41)—пролекарство 6-меркаптопурина, однако области их применения различны. Флуцитозин (4.23) и 6-азаурацил (4.41) — избирательно действующие фунгициды, первый назначают перорально при системных грибковых инфекциях у человека, второй используют в сельском хозяйстве. Азарибин, триацетильное производное 6-азауридина, назначают перорально при лечении псориаза.
Аллопуринол (9.51) — антиметаболит гипоксантина (9.52). В отличие от аналогов пуринов, рассмотренных в разд. 4.0, аллопуринол не встраивается в нуклеотид и его действие не связано с образованием ковалентных связей. Аллопуринол, 4-оксопиразоло-[3,4-<1]пиримидин, — одно из самых эффективных средств, позволяющих снижать содержание мочевой кислоты у больных, страдающих подагрой или заболеваниями, сопровождающимися избыточным образованием мочевой кислоты. Этот препарат при пероральном введении не имеет побочных эффектов. Механизм действия аллопуринола состоит в том, что он блокирует окисление гипоксантина в ксантин ферментом ксантинок-сидазой. Небольшая часть введенной в организм дозы препарата окисляется в соответствующий 2,4-диоксопиразолопиримидин (аллоксантин), блокирующий реакцию окисления ксантина в мочевую кислоту, катализируемую тем же ферментом [Elion et al., 1966].
53
Аллопуринол (9.51)
Гипоксантин
(9.52)
Антагонисты Hi-рецепторов. Антигистаминовые лекарственные препараты — это соединения, способные оккупировать гистаминовые рецепторы. Эффекты Hi-рецепторов в основном проявляются в аллергических реакциях кожи и слизистых оболочек. Наиболее точный тест на Hi-антагонистическое действие — проверка способности соединения уничтожать волдыри, возникающие у людей при подкожной инъекции гистамина (9.53) (существуют также точные тесты на морских свинках для ранних стадий скрининга), что считают проявлением обычной конкуренции между агонистом и антагонистом. Лекарственные препараты этого типа быстро помогают при насморке, вызванном пол-линозом, крапивнице, возникающей при укусах пчел, и зуде при чесотке. Поиск таких препаратов был начат Бове в середине 30-х годов в институте Пастера в Париже [Bovet, Staub, 1937], а 10 лет спустя был опубликован обзор, посвященный проблеме связи структура -— активность этого ряда веществ [Bovet, 1947].
Первым антигистаминовым препаратом, нашедшим применение в клинике, был антерган (9.54), открытый Halpern (1942). Значительно более сильно действующие вещества были получены при замене слабоосновного N-бензиланилинового фрагмента антергана более основной 2-бензиламинопиридиновой группой. Так был создан мепирамин (9.55,a) [Bovet, Horclois, Walthert, 1944], который используется до сих пор. В настоящее время известно, что многие производные П,П-диметилэтилендиамина являются Hi-антагонистами. В США был получен родственный лекарственный препарат трипеленамин (9.55,6). Значения рКа для менее основных групп этих двух соединений составляют 4,0 и 3,9 соответственно, тогда как более основная группа имеет рКа = 9, т. е. эти соединения менее основны, чем гистамин (рКа для менее основной группы 6,0), хотя более основная группа антигистаминов ионизируется так же легко, как и наиболее основная группа гистамина (рКа = 9,8).
Вскоре было обнаружено, что для получения эффективных антигистаминных препаратов можно использовать этаноламин (вместо этилендиамина). Так был получен димедрол (9.56). Представители этого класса наряду с антигистаминными обла
54
дают также седативными свойствами. Молекулы этих соединений содержат только одну ионизирующую группу (рКа 9,0).
Вг
i=j—СН2—СН2—NH2
hn'^'n
—N—СН2—С Н2—NMe2
Гистамин
(9.53)
Антерган
(9.54)
а) Мепирамии (R = OMe) б) Трипеленамин (R = H)
(9.55)
Седативное действие, ярко выраженное у димедрола, присуще и другим антигистаминным препаратам. Поскольку иногда оно может быть нежелательным, были предприняты попытки уменьшить седативное действие антигистаминов. Так был создан хлорфенамин (9.57), в котором второй полярный атом (N или О) удален из алифатической цепочки. Величины рКа хлорфенамина (4,0 и 9,2) близки к соответствующим значениям рКа предыдущих соединений. Другой тип соединений с пониженным снотворным действием, примером которого может служить хлорцикли-зин (9.56а), содержит пиперазиновый цикл, образованный путем соединения атомов азота этилендиаминовой части цепочкой из двух насыщенных атомов углерода. Поиск Hi-антагонистов, не способных преодолевать ГЭБ и, следовательно, не обладающих седативными свойствами, привел к созданию стерически затрудненного астемизола, 2-[1-[1-(4-метоксифенил)этил]пипе-рид-4-иламино]-1-(4-фторбензил) бензимидазола, применяемого при поллинозах [Laduron et al., 1982].
О роли стерического фактора в механизме действия Hi-анти-гистаминных препаратов см. Casy, Ison (1970) и Rocha, Silva (1978).
СН—О—СН2—СН2—NMe2
Димедрол
(9.56)
55
Mex /2H2SCH2CH2NH—C—NHMe
\ / и
Метиамид
(9.61)
Me2N—СН2—
—CH2SCH,CH2NH—С—NHMe
II
CH
I no2
Ранитидин
(9.62)
9.4.7. Другие аналоги
Гербицид амитрол (4.67) (3-амино-1,2,4-триазол), по-видимому, является антиметаболитом некоторых природных имидазолов. Он вызывает накопление имидазолглицерофосфата в дрожжевых клетках [Weyter, Broquist, I960]. Ингибирующее действие амитрола на рост дрожжей и водорослей Prototheca снимается гистидином [Casselton, 1964].
Теперь рассмотрим аналог очень простой химической структуры— бикарбонат-иона. Было обнаружено, что под действием стрептоцида (9.8) куры несут яйца без скорлупы. Кроме того, оказалось, что стрептоцид вызывает щелочной диурез. Оба эти наблюдения привели к установлению того факта, что сульфаниламид-ион (9.63) конкурентно блокирует доступ бикарбонат-иона (9.64) к ферменту карбоангидразе, что препятствует его дегидратированию и превращению в диоксид углерода. Расстояние между атомами кислорода в анионе сульфаниламида 0,24 нм, в угольной кислоте 0,232 нм. Накопление бикарбонат-иона в высоких концентрациях приводит к его выделению вместе с эквивалентным количеством ионов натрия, что вызывает сильный диурез. При наличии заместителя в группе C=NH блокирования рецептора не происходит и поэтому большинство антибактериальных сульфаниламидов подобным действием не обладают. Результаты этих исследований доказали целесообразность введения в состав аниона гетероциклических групп, таких как R в (9.63). Это позволяет получить соединения в большей степени ионизированные, чем стрептоцид, и в 2000 раз более активные, причем с возрастанием степени ионизации активность увеличивается [Miller, Dessert, Roblin, 1950]. Лучший из этих препаратов диакарб (9.85)—сильное мочегонное пероральное средство, оказался тахифилаксическим (с самопрекращающимся действием) и поэтому вскоре был вытеснен другими, более новыми диуретиками (разд. 14.1). Однако при острой глаукоме внутривенные инъекции диакарба могут спасти зрение, уменьшая выделение гуморальной жидкости.
58
NH
R-S
OH
Сульфаниламид-ион Бикарбонат-ион
(9.63) (9.64)
/NHCOMe
HN:SO2"
Диакарб-ион
(9.65)
Тиоглюкоза, в которой атом кислорода цикла заменен атомом серы (с образованием 5-тио-О-глюкозы), избирательно действует на гипоксические раковые клетки. Действие этого препарата изучали в сочетании с лучевой терапией. Тиоглюкоза также защищает оксические клетки от повреждения радиацией. Этот препарат способен избирательно ингибировать транспорт и поглощение глюкозы клетками. Тиоглюкоза проходит испытания в качестве мужского контрацептивного средства с обратимым действием [Zysk et al., 1975].
9.5. Ингибиторы «переходного состояния»
Существует много аналогов, препятствующих действию ферментов, занимая место нормального субстрата или кофермента и лишая их способности взаимодействовать с этим ферментом. Таким образом они блокируют нормальные биохимические реакции этого фермента. Если, согласно Э. Фишеру, описывать соответствие субстрата и фермента как «ключ к замку», то в данном случае можно сказать, что ингибитор будет несоответствующим ключом, не способным открыть замок и в то же время препятствующим введению нужного ключа. Диссоциация комплекса ингибитора с ферментом обычно протекает быстро, поэтому трудно установить, произошло ли только образование первичного комплекса ингибитор — фермент или этот комплекс перешел в нестабильное промежуточное соединение путем образования ковалентной связи между субстратом и ферментом1. Рентгеноструктурный анализ позволяет разрешить эту проблему при условии, что имеются источники чистых ферментов. Для
1 Под переходным состоянием понимают наиболее нестабильное состояние на всем пути реакции. На диаграмме изменения свободных энергий реагентов в зависимости от времени переходное состояние располагается в максимуме. В переходном состоянии старые химические связи находятся в процессе разрыва, а новые — в процессе образования. В ферментативных реакциях, протекающих через образование нестабильного интермедиата, он содержит вновь образовавшиеся связи и обладает энергией, средней между энергиями двух переходных состояний. Hammond (1955) постулировал, что переходное состояние должно быть близким по структуре нестабильному интермедиату, строение которого часто можно установить спектроскопически. Следовательно, при необходимости создания аналога переходного состояния можно руководствоваться структурой нестабильного интермедиата.
59
метотрексата, который прочно связывается с дигидрофолатре-дуктазой, установлено, что сокристаллизация фермента, кофермента и субстрата не приводит к образованию ковалентной связи (разд. 9.33). Высокий индекс ингибирования, характерный для метотрексата, объясняется тем, что он более эффективно, чем субстрат, взаимодействует с активным центром фермента.
С этим же явлением связана способность некоторых ингибиторов вызывать более сильные конформационные изменения фермента, чем те, которые происходят при действии субстрата. Так кинетика образования связи между аллопуринолом (9.51)—антиметаболитом гипоксантина (9.52), и ксантиноксида-зой указывает на то, что этот процесс происходит медленно, но в результате образуется прочная связь; устойчивое промежуточное соединение не зафиксировано [Cha, Agarwal, Parks, 1975J. Авторы полагают, что подобное течение реакции свидетельствует о сильных конформационных изменениях.
Рассмотрим ингибиторы, которые создаются по аналогии не с самим субстратом, а с его переходным состоянием. Цель такой работы — получение ингибиторов, способных образовывать значительно более прочные связи с ферментом. Интерес к ним возник в связи с широко распространенным мнением о том, что активный центр свободного фермента комплементарен переходному состоянию субстрата, а не свободному субстрату [Pauling, 1948].
Во многих случаях неизвестно, какое именно переходное состояние образуется при соединении субстрата с ферментом. Поэтому обычно предполагают, что это состояние эквивалентно переходному состоянию, установленному для аналогичной реакции, проводимой в небелковом окружении в присутствии неорганического катализатора, такого как кислота или щелочь. Рассмотрим некоторые общие случаи. При гидролизе амидной (или пептидной) связи на первой стади происходит присоединение молекулы воды по двойной связи С=О с образованием тетраэдрического аддукта. Так, при взаимодействии амида (9.66) с водой происходит образование интермедиата (9.67), который быстро диссоциирует до соответствующей карбоксильной кислоты (9.68). Другим примером может служить конденсация первичного амина с соединением, содержащим альдегидную группу (9.69) (как в пиридоксале) с образованием нестабильного интермедиата (9.70) и последующей его диссоциацией до азометина (основания Шиффа) (9.71).
О НО ОН О
II \ / II
R—С—NH2 + Н2О R—С —NH3 R—С—ОН
----» । ----------->
nh2
Амид Тетраэдрический Карбоновая
интермедиат кислота
(9.66) (9.67) (9.68)
60
О НО NH—R N—R
II \ / II
—CH + R—NH2 — С —Н20—СН
------ N --------------’
Альдегид Тетраэдрический Азометин
интермедиат
(9.69) (9.70) (9.71)
Пентостатин (4.18) (дезоксикоформицин) и его рибозидный аналог коформицин обладают сильным ингибирующим действием по отношению к аденозиндеаминазе, ферменту, превращающему аденозин (9.72) в инозин (9.74) предположительно через интермедиат (9.73). Эту реакцию, подобную гидролизу амидина с образованием амида, необходимо ингибировать при лечении больных препаратами, содержащими адениновый фрагмент, например видарабином (4.16). Молекула пентостатина (4.18) по структуре напоминает молекулу интермедиата (9.73) в области протекания реакции.
Пентостатин и коформицин связаны с ферментом очень прочно, практически необратимо [Cha, Agarwal, Parks 1975].
NH2
HO NH2
N \/
N N
R
Тетраэдрический интермедиат (R-рибоза)
(9.73)
О
N ||
N N
R
Цитидин
(9.75)
Инозин
(9.74)
OH
I
H CH2
N \/
N N
3,4,5,6-Тетрагидроуридин 1,6-Дигидро-б-гидроксиметилпури»
(9.76)
(9.77)
Точно так же 3,4,5,6-тетрагидроуридин (9.76) — сильный и специфичный ингибитор реакции дезаминирования цитидина (9.75) цитидиндеаминазой [Carmenier, 1968]. Прочная связь, образуемая этим ингибитором с ферментом, объясняется его структурным сходством с тетраэдрическим интермедиатом, подобным (9.75). Аналогично 6-гидроксиметил-1,6-дигидропурин> (9.77)—сильный ингибитор аденозиндезаминазы [Evans, Wol-fenden, 1970].
Борные кислоты обладают сильным ингибирующим действием на пептидазы за счет стабильности тетраэдрической структуры, образованной гидроксильными группами, что делает их
61?
аналогами нестабильных интермедиатов, образующихся при гидролизе пептидной связи (9.67). Соединение (9.78) специфично действует на химотрипсин, так как необходимым условием для субстрата химотрипсина служит наличие ароматического ядра в боковой цепи [Koehler, Lienhard, 1971].
Альдегиды также представляют собой сильнодействующие ингибиторы пептидазы и находятся в состоянии равновесия с тетраэдрической гидратированной структурой, например (9.79). При этом, чем больше электроноакцепторных заместителей содержит молекула, тем сильнее равновесие смещено в сторону образования метиленгликоля. Некоторые организмы вырабатывают альдегиды в качестве специфичных ингибиторов протеолитических ферментов. Так, например, лейпептины (типичный представитель — ацетил-Ь-лейцил-Ь-лейцилангиналь), вырабатываемые по меньшей мере 17 видами Streptomyces, эффективно ингибируют плазмин, трипсин и папаин, но не химотрипсин. При пероральном введении мышам лейпептины оказывают противовоспалительное действие, кроме того, они обладают радиозащитными свойствами, ингибируют свертывание крови и возникновение опухолей [Umezava, 1972].
Фенилэтан-2-борная кислота (гидрат аниона)
(9.78)
ОН
I н—с-он
I
н
Енолят-анион Нарбоксилат-анион
(9.80) (9.81)
Формальдегид (гидратированный) (9.79)
Q- Q
Н О’
Переходное состояние гексозы Лактон (каноническая формула) (9.82) (9.83)
Механизм действия ингибиторов «переходного состояния» .другого типа связан с карбанионами и карбениевыми катионами. Установлено, что аналоги карбоксилат-иона (9.81) способны эффективно ингибировать некоторые ферментативные реакции, в которых переходное состояние, по-видимому, напоминает енолят-анион (9.80). Таким же образом лактоны, аналогичные по структуре субстрату гексозе, являются сильно действующими ингибиторами большинства гликозилтрансфераз. Переходное состояние в этих реакциях напоминает алкоксикарбениевый
«2
ион (9.82), который, так же как и лактон (9.83), имеет конформацию полукресла и положительный заряд на атоме кислорода в цикле.
Было высказано предположение, хотя и недостаточно обоснованное, о том, что при взаимодействии пенициллина с пептидогликантранспептидазой (разд. 13.1) сначала происходит ингибирование по типу «переходного состояния», а затем уже ацилирование антибиотика ферментом [Lee, 1971]. Некоторые 6-дигидро-8-азапурины показали высокие терапевтические индексы при действии на опухоли мышей [Albert, 1980]. Эти экспериментальные препараты разрабатывались как ингибиторы «переходного состояния» биосинтеза пуринов. Вероятно, такие ингибиторы «переходного состояния» имеют большое будущее, однако достижения в этой области пока невелики.
Подробнее об ингибиторах «переходного состояния» см. Lindquist (1975) и Lienhard (1972). В некоторых случаях существует второе место связывания необратимо действующих ингибиторов ферментом (разд. 9.7).
9.6. Последовательное блокирование
Терапевтическую эффективность антиметаболитов можно повысить, создавая пары таких агентов, которые могли бы поодиночке блокировать две последовательные стадии процесса метаболизма клеток [Hitchings, 1952]. Природные агонисты в-клетке образуются в результате ступенчатого синтеза при участии ряда ферментов, подобно тому, как собираются детали на заводском конвейере. На каждой стадии синтеза функционирует свой особый фермент. Сущность последовательного блокирования заключается в следующем: если первый из ферментов-цепи блокирован на 90%, то только 10% синтезированного промежуточного продукта достигает второго фермента. Если удается найти способ блокировать второй фермент тоже на 90%, то на этой стадии образуется только 1 % следующего промежуточного продукта и этого количества может оказаться слишком мало для поддержания жизнедеятельности паразита. Может возникнуть вопрос, нельзя ли, повысив концентрацию первого лекарственного вещества, блокировать первый фермент сразу на 100%? Ответ на этот вопрос можно получить на основании анализа кривой, выражающей зависимость эффекта от дозы: увеличение концентрации вещества сверх определенного значения лишь в редких случаях приводит к заметному усилению-эффекта. Это происходит потому, что кривая адсорбции из раствора обычно имеет вид гиперболы (изотерма Ленгмюра, разд. 8.1). Поэтому для достижения 100% ингибирования фермента, блокированного ранее на 90%, необходим такой избыток лекарственного вещества, который может оказаться опасным для жизни больного.
Примером использования метода последовательного блокирования может служить совместное применение сульфазина
63-
с хлоридином (9.31) при лечении токсоплазмоза [Wettingfeld, Rowe, Eyler, 1956]. В этой комбинации сульфазин блокирует включение ПАБ в молекулу дигидрофолиевой кислоты, а хло-ридин тормозит реакцию восстановления дигидрофолиевой кислоты в тетрагидрофолиевую (разд. 9.3.2 и 9.3.3). Еще в 1959 г. Hurly обнаружил, что сульфазин и хлоридин настолько усиливают действие друг друга, что применение их комбинации при малярии, вызванной Pl. falciparum, позволяет добиться полного излечения. Так, менее 0,1 м. э. д. (минимальная эффективная доза) хлоридина и 0,25 м. э. д. сульфазина вместе так же эффективны, как 1 м. э. д. каждого препарата в отдельности. В настоящее время при лечении малярии, вызываемой устойчивыми к применявшемуся ранее хингамину Pl. falciparum, успешно используют малоприм — комбинацию хлоридина и дифенилсульфата (9.17) (у последнего, как и у хлоридина, низкая скорость выведения).Такую замену можно считать большой удачей, так как эта разновидность паразитов вызывает наибольшее число заболеваний с летальным исходом. Другая комбинация для лечения этого вида малярии методом последовательного блокирования, фансидар, представляет собой смесь хлоридина и суль-фадоксина (9.84) ЬГ-б.б-диметоксипиримидин^-илсульфанил-амид). Хлоридин имеет большой период полураспада в организме— 4 дня (что значительно дольше, чем у циклогуанила), поэтому во всех сочетаниях серосодержащий компонент должен также обладать низкой скоростью выведения. Подробнее об эффективности применения подобных комбинаций при лечении устойчивой к лекарственным препаратам малярии см. WHO (1973а). К сожалению, в настоящее время появились виды, устойчивые как к действию хингамина, так и к последовательным комбинациям. В таких случаях применяют хинин (довольно токсичный для организма-хозяина) или, в крайнем случае, мефлокин.
nh2
I
МеО
%=/ д N Z \ О O’
Сульфадоксин (анион)
(9.84)
Аналогичные комбинации весьма успешно применяют при лечении болезней домашней птицы, вызванных простейшими Eimeria.
Применение метода последовательного блокирования позволило получить практические результаты и в противобактери-альной химиотерапии. Синергический эффект совместного при-
«64
Рис. 9.5. Синергизм действия триметоприма и сульфазина при лечении мышей, инфицированных Proteus vulgaris. Указаны дозы препаратов, обеспечивающие выживание максимум 50% мышей; каждая точка иа графике представляет собой комбинацию дробных частей этих доз (для сульфазина эта доза 0,14 мг, для триметоприма — 4 мг на мышь) [Hit-chings, Burchall, 1965].
менения триметоприма (9.32) и сульфазина наглядно иллюстрирует рис. 9.5. Наиболее интересные и важные клинические результаты были получены в результате отбора сульфаниламида с таким же характером распределения и устойчивостью, как у диаминопиримидина, а именно триметоприма. Таким образом, был выбран сульфаметоксазол (9.11), 3-(4-аминофенилсульфа-мидо)-5-метилизоксазол, и создана комбинация его с триметопримом — бактрим. Препараты в таком сочетании последовательно блокируют сначала синтез, а затем восстановление дигидрофолиевой кислоты. Бактрим широко применяют в клинике при лечении бактериальной дизентерии, бронхитов, инфекционных заболеваний мочевых путей, вызванных Е. coli, Proteus mi-rabilis, Neisseria gonorrhoeae, Klebsiella, Streptococcus и Staphylococcus [Cattell et al., 1971].
Подробнее о химиотерапии, основанной на применении метода последовательного блокирования процессов метаболизма фолиевой кислоты, см. Hitchings (1983).
9.7. Аналоги метаболитов, образующие ковалентные связи
9.7.1. Необратимые ингибиторы активного центра ферментов (НИАЦФ)
Известны случаи, когда аналог метаболита сначала притягивается ферментом, с которым обычно взаимодействует природный метаболит, а затем образует с этим ферментом ковалентную связь. Так, пенициллин (разд. 13.1), который, по-видимому, является антагонистом П-алаиил-П-аланина, хотя и не конкурирует с ним при взаимодействии с транспептидазой, но необратимо связывается с этим ферментом ковалентной связью. Кроме того, метйодид пиридин-2-альдоксима (13.37) благодаря наличию в молекуле четвертичной аммониевой группы взаимодействует с тем участком АХЭ, который обычно предназнача-,ется для четвертичной аммониевой группы АХ. Если же этот фермент инактивирован фосфорилированием, то при действии
5-731
65
антидота (13.27) его активность восстанавливается в результате переноса фосфорильного остатка на гидроксильную группу антидота (разд. 13.3).
Одним из путей создания других ценных лекарственных средств является синтез аналогов метаболитов, обладающих способностью адсорбироваться на ферменте и содержащих в молекуле такой заместитель, который может образовывать ковалентную связь с ферментом и тем самым необратимо инактивировать его. Б. Бейкер, используя в качестве модели лактатдегидрогеназу, обнаружил, что алкилирующие ингибиторы можно разделить на два типа: ингибиторы первого типа алкилируют в ферменте активный центр, который обычно связывает нормальный субстрат, а ингибиторы второго типа ингибируют смежные с ним центры. Эти два типа ингибиторов получили
название эндо- и экзоалкилирующих агентов соответственно. Исследуя ингибиторы экзотипа, Бейкер обнаружил, что салициловая кислота (9.85), подобная по структуре молочной кислоте (9.86), является ингибитором лактатдегидрогеназы (хотя и обратимым), и синтезировал 4-йодацетамидсалициловую кислоту (9.87). Это соединение необратимо ингибирует не только лактатдегидро-, но и L-глутаматдегидрогеназу. В противоположность незамещенному йодацетамиду его производное (9.87) ингибирует эти ферменты, поскольку благодаря структурному сходству с салициловой кислотой этот ингибитор концентрируется на ферменте [Baker et al., 1962]. Это ингибирование необратимо, так как происходит отщепление йодистого водорода и образование ковалентной связи между ферментом и ингибитором. Этот ингибитор способен различать даже изоферменты. Так, он ингибирует лактатдегидрогеназу скелетной машины, но не действует на тот же фермент сердца. Родственное соединение, 5-(феноксикарбониламино)салициловая кислота, необратимо ингибирует изофермент сердца, но не действиет на фермент скелетной мышцы, а его 4-изомер оказывает прямо противоположное действие [Baker, Patel, 1964; Baker, 1967].
Более подробно о необратимых ингибиторах активных центров фермента см. Baker (1967).
а ОН Н ^ОН
МеС—ОН ICH2CONHj|
СО2Н СО2Н \=#^со2н
Салициловая кислота Молочная кислота 4-Йодацетамидсалициловая
(9.85) (9.86) кислота (9.87)
9.7.2. Ферментативно-активируемые необратимые ингибиторы (ФАНИ)
Этот новейший тип ингибиторов отличается от других тем, что прежде, чем образуется ковалентная связь с ферментом (как это происходит с НИАЦФ), они под действием этого фер
66
мента должны превращаться в истинно активный агент, т. е. все ингибиторы этого типа являются пролекарствами (разд. 3.6). Их называют «ферментативно-активируемыми необратимыми ингибиторами» (ФАНИ) или «Irreversible Mechanism — Based Inhibitors» (IMBI). Иногда в шутку, но не совсем правильно их называют «ингибиторами-самоубийцами». Широкий интерес к исследованию подобных ингибиторов возник после появления обзора Abeles, Maycock (1976); впервые действие ФАНИ случайно наблюдал К. Блох [Morisaki, Bloch, 1972]. Для создания лекарственных препаратов такого типа необходимо, основываясь на результатах изучения кинетики, установить механизм действия выбранного фермента. Этот механизм, например, может заключаться: а) в отщеплении атомов водорода и образовании сопряженной двойной связи в молекуле субстрата или б) в превращении субстрата в электрофильное соединение протонированием или окислением.
До настоящего времени наиболее распространенным из известных типов реакции между ферментом и ФАНИ является конденсация Михаэля. Поэтому пролекарство должно под действием фермента превращаться в акцептор Михаэля. Такой акцептор [Michael, 1887] обязательно содержит двойную связь, поляризованную (а следовательно, активированную) наличием электроноакцепторного заместителя [Bergmann, Ginsburg, Рар-ро, 1959].
Как правило, молекула ФАНИ содержит активирующую группу, например, фторметильную, кетонную, сложноэфир-ную или нитрильную, а фермент выполняет свою обычную функцию — создание двойной связи. Конечно, молекула ФАНИ должна иметь характеристическую группу нормального субстрата, так как в противном случае она не будет взаимодействовать с ферментом, который должна блокировать.
Удачным примером ФАНИ такого типа может служить 2-дифторметилорнитин (9.88) —лекарственное вещество, необратимо ингибирующее орнитиндекарбоксилазу. Этот фермент с помощью кофермента миридоксаля декарбоксилирует орнитин, превращая его в путресцин (эта реакция имеет большое значение для некоторых простейших, а также для раковых клеток). Пролекарство, очень близкое по структуре к орнитину, ковалентно связывается с коферментом-пиридоксалем. Таким образом, аминогруппа в положении 2 молекулы ФАНИ и альдегидная группа пиридоксаля (9.90) образуют азометин (основание Шиффа) аналогично тому, как это происходит с орнитином. На этой стадии фермент обычно отщепляет атом водорода от молекулы орнитина или атом фтора, близкий по размеру атому водорода (см. табл. 9.1), но обладающий значительно большей электроотрицательностью. Отщепляя атом фтора от молекулы ФАНИ, фермент превращает его в акцептор Михаэля (9.89). Таким образом, ФАНИ становится истинным лекарственным веществом, способным атаковать донор Михаэля в молекуле
5* 67
фермента (предполагают, что это SH-группа) и необратимо его' ингибировать [Prakash et al., 1980].
Дифторметилорнитин (9.88) имеет высокий терапевтический индекс и показал хорошие результаты при клинических испытаниях (лечение лейкоза и малярии). К сожалению, короткий период полупревращения этого препарата в организме человека создает трудности в поддержании терапевтических доз.
HCF2
H2N—с—со2н 1 (СН2)з
nh2
Акцептор Михаэля (9.89) H2CF
h2nAh—СО2Н
СНО
Пиридоксаль
(9.90)
2-Дифторметилорнитин
(9.88)
З-Фтораланин
(9.91)
Аналогичный механизм действия характерен для антибактериального ФАНИ, D-3-фтораланина (9.91), который проходит клинические испытания. Это соединение необратимо ингибирует аланинрацемазу — фермент, который поставляет D-аланин, необходимый для синтеза клеточных стенок бактерий [Wang, Walshj 1978].
Классическими донорами Михаэля являются карбанионы, полученные из активированной метиленовой группы (например, в диэтилмалонате). Однако в молекулах ферментов донорами Михаэля чаще всего являются анионы либо NH-группы имидазола (как в оригинальной работе К. Блоха), либо гидроксигруппы серина или меркаптогруппы цистеина. Для активации можно использовать также винильный или этинильный заме^ ститель. Такие соединения были получены для повышения концентрации ГАМК (12.91) в мозге. ГАМК — это тормозной нейромедиатор. Однако при инъекционном введении она не обладает противосудорожным действием, так как не способна проникать через ГЭБ. При периферическом введении мышам 4-амино-5-гексеновой кислоты (4-винилпроизводного ГАМК) содержание ГАМК в мозге повышается втрое. Это происходит вследствие ингибирования трансаминазы 4-аминомасляной кислоты (ГАМКазы), механизм которого, по-видимому, состоит в следующем: ФАНИ (9.92), соединяясь с коферментом-пири-доксалем, образует азометин, от которого фермент отрывает протон, что приводит к образованию двойной связи, т. е. к получению акцептора Михаэля, способного необратимо дезактивировать ГАМК-разрушающий фермент [Lippert et al., 1977].
Р8
Этот ФАНИ не взаимодействует с коферментом, находящимся в форме пиридоксамина, и обладает значительной специфичностью, так как не действует на ГАМКазу бактерий. Кроме того, он не взаимодействует с некоторыми другими, требующими пиридоксаль ферментами: глутаматдекарбоксилазой и аспартатаминотрансферазой млекопитающих. Последняя тем не менее ингибируется винилглицином (2-амино-З-бутеновая кислота) [Rando, 1974].
НС=СН2
H2N—СН—СН2СН2СО2Н 4-Амиио-5-гексеновая кислота (9.92)
С=СН I H2N—СН—СН2СН2СО2Н
4-Амино-5-гексиновая кислота
(9.93)
Аналог 4-амино-5-гексиновая кислота (9.93) также повышает уровень ГАМК в мозге мышей [Jung, Metcalf, 1975]. Более того, при внутрибрюшинных инъекциях 4-амино-5-фторвале-риановой кислоты мышам уровень ГАМК в мозге понижается в 16 раз [Silverman et al., 1981]. Установлено, что механизм действия этого вещества аналогичен описываемому выше, за исключением того, что кофермент находится в форме пиридоксамина [Silverman, Levy, 1981]. В этом случае отщепление фтора происходит при алкилировании аминогруппы кофермента с помощью ФАНИ, а затем фермент отрывает протон, что приводит к образованию акцептора Михаэля.
Образование акцептора Михаэля может происходить и другим путем — при окислении ФАНИ ферментом. Так, например, паргилен (3.45) под действием МАО, первого фермента биосинтеза катехоламинов, окисляется в имин (PI1CH2. N+Me=CH— С==СН). Таким же образом осуществляется превращение гидроксиацетилена в кетоацетилен. Продукт превращения паргиле-на присоединяется к донору Михаэля, которым, как известно, является атом азота центрального цикла кофермента рибофлавина [Abeles, Maycock, 1970]. Транилципромин (9.47) N-(l-метилциклопропил) бензиламин, ФАНИ, относящийся к наиболее распространенным антидепрессантам, окисляется под действием МАО с образованием реакционноспособного циклопропиминие-вого ион-радикала, который, по-видимому, атакует кофермент рибофлавин [Silverman, Hoffman, 1981].
Другие подходы применяются при создании ФАНИ для ферментов, не способных инициировать реакции Михаэля, но имеющих тенденцию образовывать электрофильные соединения. Так, например, диазокетон под действием фермента может про-тонироваться и превращаться в ион диазония, который затем алкилирует активный центр фермента. Антибиотик азасерин (9.48), подобно глутамину (9.49), связывается с ферментом формилглицинамидаминотрансферазой и инактивирует его. Это препятствует превращению амидной группы субстрата (формил-
69
глицинамидрибоиуклеотида) в амидиновую группу и, таким образом, ингибирует биосинтез пуринов. Глутамин в этой реакции является кофактором (источником аммония (Buchanan, 1978]. Азасерин, вероятно, наиболее известный ФАНИ, так как он применяется в исследованиях и лечении рака с 1954 г.
Существуют ферменты, способные превращать пролекарство в электрофильное соединение введением атома кислорода. Так, оксидазы со смешанными функциями из ЭР (разд. 3.5) окисляют дифторметилпроизводные нормального субстрата с образованием ацилфторида, ацилирующего активный центр фермента. Ароматаза, например, представляет собой фермент, ответственный за синтез эстрона из андрост-4-ен-3,17-диона. 19,19-Диф-торпроизводное этого субстрата действует в качестве ФАНИ и, как уже описывалось выше, необратимо связывается с ферментом в положении 19 [Marcotte, Robinson, 1982]. Возможно, этот лекарственный препарат окажется полезным при лечении рака молочной железы.
Подобным же образом действуют ферменты, способные активировать а-цианоаналог субстрата и окислять его до гидрок-сикетенамина: NC—CH2R—>-HN=C = C(OH)R. Примером может служить 4-гидроксибензилцианид—ФАНИ, ингибирующий дофамин ^-гидроксилазу. Блокирование этого фермента снижает количество синтезируемого адреналина, поэтому предполагают, что 4-гидроксибензилцианид может оказаться полезным для понижения симпатического тонуса [Baldoni, Villafran-са, Mallette, 1980].
Подробнее о ФАНИ см. Penning (1983) и Seiler, Jung, Koch-Weser (1978).
9.8. Особые случаи взаимосвязи между агонистами и антагонистами
Антиметаболиты—это не обязательно аналоги метаболитов. Существуют соединения, не обнаруживающие никакого сходства с метаболитами, но тем не менее способные взаимодействовать с апоферментом, ферментом, субстратом или разрушать их [Fildes, 1940]. Единственное, что для этого необходимо, — это наличие выраженного сродства, что часто приводит к образованию ковалентной связи. Эти антагонисты не вытесняются субстратом. Примером такого антагониста может служить порфирин, гемин, тиамин, расщепленный на две части, из которых одна включает пиримидиновый, а другая — тиазольный цикл. Поэтому у животных, употребляющих в пищу рыбу, в избытке содержащую это вещество, развиваются признаки недостаточности тиамина [Woolley, 1952].
Наличие конкурентного антагонизма между двумя веществами еще не означает, что одно из них непременно является метаболитом. Например, сильные сокращения кишечника, вызываемые хлоридом бария, конкурентно ингибируются сульфатом
70
натрия, так как между этими двумя веществами происходит химическая реакция, в результате которой токсический ион бария осаждается в виде нерастворимого сульфата. Очевидно, прежде чем отнести какое-либо вещество к группе метаболитов, нужно доказать, что оно встречается в природе, а также выяснить его происхождение и функции. Примером пары соединений, конкурирующих друг с другом, хотя ни один из них природным метаболитом не является, могут служить морфин и аллилнорморфин [Unna, 1943] (разд. 7.3). Точно так же антагонистами можно назвать 2,4-дихлорфеноксиуксусную кислоту, искусственный регулятор роста растений, и ее искусственный аналог — 2,4-дихлорофенилтиоизомасляную кислоту [Griffits et al., 1966]. Это примеры классического эффекта «терапевтической интерференции» (разд. 6.6).
Исследованиям действия лекарственных веществ, вступающих в антагонистические отношения друг с другом, способствовало введение Schild (1947) в употребление величины рАх. Это отрицательный логарифм молярной концентрации антагониста, при которой эффект, полученный от х-кратной дозы лекарственного вещества, уменьшается до эффекта, получаемого от одной дозы. Удобно при этом пользоваться значениями х, равными 2 и 10. Смысл этой величины заключается в следующем: предполагается, что агонисты, действующие на одни и те же рецепторы, будут иметь одинаковые величины рАх по отношению ко всем конкурентным антагонистам, что является следствием закона действующих масс и применимо к агонистам любой эффективности и сродства.
Значения рАх могут быть использованы для классификации лекарственных веществ, поскольку только вещества, действующие на одни и те же рецепторы, будут иметь одинаковые значения рАх по отношению к данному конкурентному антагонисту. Так, например, был изменен антагонистический эффект 2,4-ди-фенилгидрамина (9.56) по отношению к двум агонистам, вызывающим сокращения подвздошной кишки у морских свинок,— гистамину и 2-(2-пиридил)этиламину (9.94). Для них были получены одинаковые величины рАх, что доказывает, что оба агониста действуют на один и тот же рецептор.
п
^n"^ch2ch8nh8
2- (2-пир идил)этиламии
(9.94)
Более того, одинаковые значения величин рАх для рецепторов в разных тканях, полученные для данной пары лекарственное вещество — антагонист, с большой вероятностью указывают на сходство этих рецепторов. Так, одинаковые величины рАх были получены для антагонистического эффекта АХ в таких
71
разных образцах, как сердце лягушки, амнион зародыша цыпленка, кишечник крысы и легкие морских свинок [Arunlakshana, Schild, 1959].
9.9. Фармакогенетика
При исследовании ингибиторов ферментов всегда следует учитывать, что наборы ферментов у отдельных индивидуумов могут отличаться от того, что характерно для популяции в целом. Так, у многих людей, особенно в странах Африки и Среднего Востока, отмечается дефицит глюкозо-6-фосфатдегидроге-назы. Такой биохимический дефект не проявляется до тех пор, пока в организм не попадает противомалярийный препарат примахин (3.36). В этом случае у больных очень быстро развивается гемолитическая анемия, вызванная биохимическим дефектом, наследуемым вместе с неполностью доминантным геном [Beutler, 1959; Fraser, Vessel!, 1968]. Причина анемии — гемолиз, вызванный хиноном, образующимся на первой стадии метаболита; фермент, который обычно разрушает это вещество, отсутствует.
Вариабельность реакций ацетилирования — хорошо изученное фармакогенетикое явление. Оно влияет на ацетилирование N-ацетилтрансферазой как в печени, так и в тонкой кишке. У человека различают два фенотипа — быстрые и медленные ацетиляторы, причем соотношение этих фенотипов для разных стран неодинаково [Karim, Elfellah, Evans, 1981]. Около 90% японцев и китайцев относятся к генетически быстрым N-ацети-ляторам изониазида (6.13), в то время как для населения США черной или белой расы эта цифра составляет только 40% [Ка-low, 1962]. Медленные ацетиляторы должны применять большие дозы, а следовательно, накапливать более высокий уровень лекарственного вещества в крови, чем быстрые ацетиляторы. Однако при применении изониазида ситуация осложняется тем, что у быстрых ацетиляторов происходит образование ацетил-гидразина, вызывающего поражение печени [Mitchell, Thorgeirs-sen, Black, 1975].
Таким же образом генетически контролируются процессы ацетилирования (дезактивации) антибактериальных сульфаниламидов, антиаритмических лекарственных препаратов, ново-каинамида (7.56) [Woosley et al., 1978], гипотензивного препарата апрессина (11.47) и амина, образующегося при метаболизме седативного средства нитразепама (12.98). В каждом из этих случаев быстрое ацетилирование приводит к снижению эффекта лекарственного препарата. Однако у медленных ацетиляторов апрессин может вызывать системную красную волчанку с артритоподобными симптомами [Batchelor et al., 1980].
Фермент алкогольдегидрогеназа также может быть двух типов— быстрый и медленный, что наследуется генетически. Быстрый тип характерен для жителей Востока, у которых 72
вследствие быстрого высвобождения ацетальдегида отмечается покалывание и прилив крови к лицу [Propping, 1978]. Дебризо-хин (9.95) — гипотензивный лекарственный препарат, действующий подобно октадину (7.55), превращается в неактивное 4-гидроксипроизводное, причем это более характерно для жителей Кавказа, чем Востока [Kalow et al., 1980].
^Х/Х NH
N—C—NH2
Дебризохии
(9.95)
Другим нежелательным фармакогенетическим эффектом является повышение внутриглазного давления при введении в глаз глюкокортикоидов. Такую реакцию наблюдали у 5% жителей Кавказа. Установлено, что этот эффект наследуется, так же как и наличие атипичной малоактивной холинэстеразы в плазме, что приводит к пролонгированию действия дитилина (7.29) на таких больных во время операций [Kalow, 1962].
Подробнее о фармакогенетике см. Kalow (1962, 1980) и World Health Organization (19736).
Г л а в a 10
ИОНИЗАЦИЯ
Алифатические четвертичные аммониевые основания, такие как АХ, полностью ионизированы при всех значениях pH. Это обусловлено их структурой: у иих иет протона, способного отщепляться, а атом азота может образовывать не более четырех ковалентных связей. Однако большинство биологически активных веществ — это слабые кислоты или основания и поэтому степень их ионизации зависит от pH. Константы ионизации этих веществ (а следовательно, и степень ионизации при определенном pH) можно варьировать в широких пределах, внося незначительные изменения в их химическую структуру. Степень ионизации можно менять двумя путями: во-первых, можно использовать одно и то же вещество при различных pH и, во-вторых, брать несколько родственных веществ (с разными константами ионизации) при фиксированном pH. Эти два способа очень важны для исследования, поскольку ионы и незаряженные молекулы весьма различны по свойствам. Так, например, ионы и незаряженные молекулы вступают в разные химические реакции, по-разному проникают через мембраны и адсорбируются на различных типах веществ.
10.0. Природа ионизации
Многие вещества при растворении в воде не повышают ее электропроводности. Это так называемые неэлектролиты (примерами могут служить хлороформ и сахароза), они понижают температуру замерзания воды пропорционально их молярной концентрации. С другой стороны, кислоты, основания и соли повышают электропроводность воды. Большинство биологически активных веществ представляют собой кислоты, основания и соли, а следовательно, являются электролитами. Все электролиты понижают температуру замерзания воды в значительно большей степени, чем можно было бы ожидать, исходя из их молярной концентрации. В разбавленных растворах соляной кислоты, гидроксида натрия и хлорида натрия это понижение оказалось в два раза больше ожидаемого. Это послужило шведскому химику Аррениусу основанием для создания теории ионизации электролитов (1884—1887 гг.). В водном растворе хлористый водород (соляная кислота) полностью диссоциирован на катионы водорода и анионы хлора (Н+ и С1~), гидроокись
74
натрия — иа катионы натрия и анионы гидроксила (Na+ и ОН-), хлорид натрия — на катионы натрия и анионы хлора (Na+ и С1-). В случае сульфата натрия понижение температуры замерзания воды оказалось в три раза больше ожидаемого, и это объясняется тем, что вместо одной молекулы Na2SO4 в растворе присутствуют три иона, а именно два катиона натрия и один анион сульфата (SO42-).
Соли. Как правило, соли в разбавленных растворах полностью ионизированы. Исключения из этого правила немногочисленны: наиболее известные из них — галогениды ртути, кадмия и свинца. Вследствие полной ионизации солей их биологические свойства целиком определяются свойствами составляющих их ионов. Так, физиологическое действие хлорида кальция определяется действием, свойственным ионам кальция и ионам хлора. Однако эта простая концепция оказывается неверной в тех случаях, когда соль образована слабой кислотой или слабым основанием, так как в результате гидролиза происходит высвобождение некоторого количества незаряженных частиц, биологическое действие которых суммируется с эффектами, вызванными ионами.
Вообще физиологическое действие полностью ионизирующейся соли не может быть больше или меньше суммы действия ее ионов. Так, например, Hata, исследуя на мышах токсичность 3,6-диамино-10-метилакридинийхлорида (6.5) и соответствующего йодида, установил, что йодид в два раза менее активен, чем хлорид (при одинаковых по весу дозах. Сравнение действия этих веществ на мышах, инфицированных стрептококками, показало, что йодид и в этом случае вдвое менее активен. Так как оба аниона биологически инертны при использованных концентрациях, то активность должна быть пропорциональна количеству катионов в исследуемых соединениях, которые в нейтральных растворах полностью ионизированы [Albert, 1966]. ОММ этих соединений 260 и 351 соответственно, поэтому активность йодида должна составлять 74% активности хлорида. Результаты биологических исследований при различных разбавлениях соответствуют приведенному расчету. Поэтому такие эксперименты позволяют только сравнить биологическое действие катиона диаминометилакридиния с его незаряженной молекулой. Изучение антибактериальных свойств некоторых акридиниевых солей (сульфатов, нитратов, гидрохлоридов и гидройодидов) показало, что, как и предполагалось, анион не вносит практически никакой специфики в биологическое действие [Browning et al., 1922].
Кислоты и основания. В отличие от солей кислоты и основания не обязательно должны быть полностью ионизированы в растворе. Сильные кислоты (например, соляная) и сильные основания (например, гидроксид натрия) полностью ионизированы при значениях pH от 0 до 14, в то время как слабые кислоты и основания в этих пределах pH имеют разную степень
75
ионизации. Даже небольшие отклонения pH в любую сторону от нейтрального значения (pH 7 ) могут существенно повлиять на ионизацию таких лекарственных средств, как барбитураты, алкалоиды, местные анестетики и антигистаминные препараты. Рассмотрим несколько примеров.
Соли слабых кислот (или слабых оснований) в растворе частично гидролизованы до исходных, малодиссоциированных кислот (иди оснований). Реально степень диссоциации в растворе определяется только двумя факторами: pH раствора и рКа кислоты (или основания). Последняя из этих величин (она будет охарактеризована ниже) является константой для любой кислоты и основания. Поэтому при определенной величине pH степень ионизации зависит только от природы кислоты (или основания), при этом не важно, были они предварительно нейтрализованы или нет. Так, отношение концентрации ионов атропина к концентрации его неионизированных молекул остается неизменным независимо от того, добавляется ли к буферному раствору при pH 7 атропин или его гидрохлорид или сульфат. При повышении pH раствора количество ионов атропина уменьшится, но новое соотношение также не будет зависеть от того, в какой форме находится добавляемый атропин. Во избежание путаницы при употреблении терминов «свободные» или «неиони-зированные» кислоты и основания обычно применяют термин «молекула» или «нейтральная частица» для обозначения всех незаряженных форм.
В 1944 г. американские бактериологи опубликовали работу, в которой было показано, что бактерицидное действие 9-амино-акридина (10.8) (по отношению к Pneumococcus типа III, выращенных на бульоне с глюкозой) в 64 раза сильнее, чем действие его гидрохлорида. Авторы не указывают, обладала ли использованная ими среда буферными свойствами. Если был использован буфер, то в обоих случаях должен получиться один и тот же результат и, следовательно, их методика была неверна. Если же среда не была буферной, то количественные данные не имеют смысла вследствие большой разницы в pH растворов сильного основания (рКа 10,0) и его солей. Значительное повышение антибактериальной активности акридинов при увеличении pH известно уже давно [Graham-Smith, 1919; Browning, Gulbransen, Kennaway, 1919].
Подробнее о кислотах, основаниях и ионизации см. Albert, Sergeant (1984) и Bell (1973).
10.1 Константа ионизации (Ка)
Существенной частью теории ионизации Аррениуса явилось применение закона действия масс для описания состояния ионного равновесия. Так, уксусная кислота (СНзСООН) — это слабая кислота, ионизирующаяся в воде с образованием некоторого количества ионов водорода (Н+) и ацетат-ионов 76
(СНзСОО-). Отношение произведения концентраций этих ионов ([Н+][СН3СОО-]) к концентрации неионизированных молекул [СНзСООН] всегда является постоянной величиной. Это соотношение и определяет константу кислотности (Ка) или просто константу ионизации данной кислоты.
_ [Н-] [СН3СОО-]
Ка [СНзСООН] ' W
Величина Ка уксусной кислоты была определена экспериментально, она равна 1,75х10-5 (при 25°C).
Иногда выражение «константа диссоциации» используют вместо константы ионизации. Последнее название точнее, так как многие комплексы, например системы ферментов, «диссоциируют» на составляющие, а мицеллы — на мономеры. Устанавливающееся при этом равновесие можно выразить через константы диссоциации, также выведенные из закона действия
масс, но эти константы не являются константами ионизации.
Состояние ионизации слабых оснований также может быть охарактеризовано константами кислотности. Например, аммиак— слабое основание, которое может присоединять ионы водорода с образованием ионов аммония. В таком случае ион NH4+ можно рассматривать как слабую кислоту, ионизированную в воде на ионы водорода (Н+) и молекулы аммиака (NH3). Тогда константа ионизации может быть соотношением:
[Н+] [NH3]
а [NH4+] ’
(И)
а экспериментальное ее значение равно 5.5Х10-10 (при 25°C).
В 1929 г. Бренстед предложил использовать константы кислотности для описания ионизации оснований, так как это позво
ляет применить одну и ту же шкалу для ионизации и кислот, и оснований, так же как единая шкала pH используется для измерения кислотности и щелочности.
До этого для оснований применяли отдельную шкалу и константу основности выражали соотношением:
_ [ОН-] [NH4+]
[NH4OH] '
(П1)
Экспериментальное значение такой константы 1,8хЮ-5 (при 25°C). Уравнение III нереально, так как соединения, содержащие азот с пятью ковалентными связями, например NH4OH, существовать не могут. Уравнения I и II показывают, что кислота отщепляет ионы водорода, а основание их присоединяет. Таким образом, и кислоту, и основание можно характеризовать количественно одной величиной — сродством к иону водорода. Это позволяет применять константы кислотной ионизации как для кислот, так и для оснований.
А. Определение рКа- Величины констант ионизации слишком малы, поэтому значительно удобнее пользоваться их отрица
77
тельными логарифмами (обозначенными как рКа). Так, рКа уксусной кислоты 4,76, а аммиака 9,26. В более ранней литературе для оснований часто встречаются только значения рКь (например, 4,74 для аммиака). Зная эту величину, можно перейти к значениям рКа; для этого из отрицательного логарифма ионного произведения воды (pKw) при данной температуре надо вычесть значение рКь (величина pKw равна 14,16 при 20 °C, 14,00 при 25°C и 13,58 при 37°C).
Очевидно, что значения рКа дают возможность легко сравнивать силу разных кислот (или оснований). Чем сильнее кислота, тем ниже значение ее рКа (чем сильнее основание, тем выше его рКа). Значение pH, при котором кислота или основание ионизированы наполовину, эквивалентно их рКа. Если pH на одну единицу ниже, чем рКа, то кислота ионизирована на 9%, а основание — на 91% (разд. 17.0).
Любая частично нейтрализованная кислота или основание представляет собой эффективный буфер в пределах от рН = = рКа — 1 до рКа + 1. Поэтому 6ИОЛОГИ ИМвЮТ ВОЗМОЖНОСТЬ ПОД-бирать буфер для каждого конкретного эксперимента. Если для исследуемой системы необходимы катионы металлов, то анионные буферы (например, цитратный и фосфатный) обычно не используются, так как они способны образовывать комплексы с катионами. В качестве примера можно привести фосфатный буфер, ингибирующий действие изоцитратдегидрогеназы (в сердце свиньи) за счет удаления марганца [Lotspeich, Peters, 1951]. Вместо анионных буферов часто применяют катионные, такие как N-этилморфолин (pH от 7,0 до 8,2), «Трис» (трисгид-роксиметиламинометан) (pH от 7,5 до 8,7) и цвиттерионы («Ге-пес» и «Тэс», pH от 6,8 до 8,1) [Good et al., 1966]. Исследованию буферов посвящена книга Perrin; Dempsey (1974).
Б. Установление значений рКа. Значения рКа более чем для 500 соединений приведены в справочнике Albert, Serjeant (1984). Более полные данные можно найти в сборниках ИЮПАК: для органических оснований Perrin (19656), дополнение (1972); для органических кислот Kortiim, Vogel, Andrussow (1961), дополнение Serjeant, Dempsey (1979); для неорганических кислот и оснований Perrin (1983). В табл. 10.1 приведены величины рКа некоторых наиболее распространенных кислот и оснований. Данные таблицы следует запомнить; это позволит сравнивать те новые величины рКа, с которыми можно столкнуться при чтении, с приведенными в ней значениями. Кислоты и основания равной силы расположены в таблице друг против друга.
Величина рКа около 5 (ср. с рКа уксусной кислоты) типична для большинства монокарбоновых кислот, как алифатических, так и ароматических. Величина рКа 10 типична для фенолов. Кислоты, рКа которых выше 7, практически не изменяют цвета нейтральной индикаторной бумаги, а выше 10 — не имеют даже кислотного вкуса.
78
Таблица 10.1. Относительная сила наиболее распространенных кислот и оснований
Кислоты рКа Основания Р«а
Соляная кислота
Фосфорная кислота (отрыв первого протона)
Уксусная кислота
Угольная кислота
Фосфорная кислота (отрыв второго протона)
Синильная кислота, борная кислота
Феиол
Фосфорная кислота (отрыв третьего протона)
Глюкоза
<0
2
3
4
5
6
7
8
9
10
11
12
13
Гидроксид натрия
Этиламин
Аммиак
Хинин, стрихнин
Анилин, пиридин
п-Нитроанилии
>14
12
11
10
9
8
7
6
5
4
3
2 1
Величина рКа 11 (как, например, у этиламина) типична для алифатических оснований, а значение рКа 5 — для ароматических, которые гораздо слабее алифатических. Величины рКа представляют собой десятичные логарифмы, следовательно, основность этиламина и анилина различается в миллион раз (т. е. антилогарифм 6). Многие алкалоиды и другие биологически активные основания имеют значения рКа около 8. Основания, рКа которых ниже 7, практически не изменяют цвета нейтральной индикаторной бумаги.
Введение в молекулу электронодонорных групп (например —СН3) увеличивает ее основность и уменьшает кислотность, а электроноакцепторных (например, —NO2) — ослабляет основность и усиливает кислотность. Сведения об электронных свойствах различных заместителей приведены в разд. 17.2. Влияние заместителей на кислотность и основность в настоящее время настолько хорошо изучено, даже количественно, что соответствующие константы ионизации могут быть рассчитаны еще до синтеза соединения [Perrin, Dempsey, Serjeant, 1981]. Это позволяет экономить большое количество времени, которое могло бы быть затрачено на синтез соединений с нежелательными кислотно-основными свойствами. Нет необходимости говорить о том, что после того, как вещество получено, следует определять истинную константу ионизации.
Подобные практические указания по определению констант ионизации экспериментальным путем даны в справочнике Albert, Serjeant (1984).
В. Расчет степени ионизации. Степень ионизации любого основания в водном растворе можно рассчитать с помощью урав-
79
Таблица 10.2.’ Расчет степени ионизации по заданным величинам рКа и pH
рКа-рН % ионизации (по анионному типу) % ионизации (по катионному типу)
—4 99,99 0,01
-3 99,94 0,10
—2 99,01 0,99
—1 90,91 9,09
0 50,00 50,00
1 9,09 90,91
2 0,99 99,01
3 0,10 99,94
4 0,01 99,99
В развернутом виде эта таблица приведена в разд. 17.0.
нения (IV) при условии, что известны pH раствора и рКа вещества:
100
Степень ионизации (%) =----------------------. (IVI
1 + антилогарифм (pH — рКа) ' ?
Это уравнение показывает, что степень ионизации данной кислоты (или основания) зависит от pH. Эта зависимость не является линейной, а выражается сигмоидной кривой, как, например, представленной на рис. 10.1.
Анализ этой кривой показывает, что небольшое изменение pH может вызвать сильное изменение ионизации, особенно если значение pH раствора близко к величине рКа исследуемого вещества. Это хорошо видно и из данных табл. 10.2. Например, при работе с нитрофенолом, имеющим рКа 7, при значении pH, также равном 7, половина вещества будет находиться в ионизированном состоянии. При повышении pH до 8 фенол будет ионизирован почти полностью, а при понижении его до 6 почти весь фенол перейдет в неионизированное состояние.
Г. Полиэлектролиты. Полиэлектролиты — это вещества, в молекулах которых имеется много групп, способных к ионизации, причем все они могут иметь одинаковые, как, например, в полиакриловой кислоте, или разные, как в белках, заряды. Кривые титрования полиэлектролитов имеют довольно неопределенную «размытую» форму. В отличие от моно- и диионов полиион окружен большим количеством противоионов. Подробнее о противоионах см. Albert, Serjeant (1984).
Д. Цвиттерионы (биполярные ионы). До сих пор ничего не говорилось о цвиттерионах — внутренних солях, имеющих одновременно положительный и отрицательный заряды. Выясняя, является ли данное соединение цвиттерионом или нет, необходимо уточнить pH среды, так как в щелочном растворе цвиттерион будет вести себя как анион, а в кислом — как катион.
Например, кислотная группа глицина имеет рКа 2,2, а основная — рКа 9,9. Из данных табл. 10.2 следует, что при pH 3,2
89
Рис. 10.1. Типичная кривая, полученная при потенциометрическом титровании кислоты (борная кислота, рКа9,21 при 20 °C).
ионизированными окажутся только 90 %) кислотных групп (и все основные); следовательно, при этом pH 90% вещества будет представлять собой цвиттерион, а 10% — катион. При pH от 3,2 до 8,9 глицин почти полностью будет в форме цвиттериона.
Не все вещества, обладаю
Степень ионизации
щие и кислотными, и основными
группами, являются цвиттерионами (внутренними солями), так как возможно, что ни при одном значении pH обе группы не будут ионизированы одновременно. Другой важный фактор — это pH раствора. Так, вещество будет на 90% или более в форме цвиттериона, если pH по меньшей мере на одну единицу выше рКа кислотной группы и на одну единицу ниже рКа основной группы. Если применить это правило к ПАБ (рКа кислотной группы 4,8, рКа основной 2,7, то легко заметить, что это вещество в отличие от глицина и в нейтральных, и в щелочных растворах находится только в анионной форме. Другие тесты, позволяющие отличить цвиттерионы от просто амфотерных соединений (например, глицин и ПАБ соответственно), приведены в книге Albert, Serjeant (1984).
В цвиттерионе каждая ионизирующаяся группа имеет две константы — для молекулы со второй ионизированной группой, и со второй неионизированной группой. Эти константы называются микроскопическими, а макроскопические константы (определяемые спектрометрией или потенциометрическим титрованием) представляют собой сумму их составляющих. Тирозин был одним из первых цвиттерионов, микроскопические константы которого были определены [Edsall, Martin, Hollingworth 1958] с помощью методов, применяющихся и до сих пор. Микроскопические константы серии катехоламинов приведены в разд. 12.4.
10.2. Различия в ионизации, обеспечивающие избирательность
Какое значение имеет степень ионизации при оценке биологического действия? Действительно ли слабоионизированные вещества образуют хотя бы несколько ионов и по мере их удале
6-734
81
ния неионизированные молекулы поставляют новые ионы в соответствии с законом действия масс (уравнение I)?
Однако следует помнить, что ионы не образуют с рецепторами ковалентных связей, а следовательно, могут легко отрываться. Поэтому для поддержания рецептора в насыщенном состоянии необходимо, чтобы в растворе, окружающем рецептор, постоянно находился избыток данных ионов. Например, кристаллический фиолетовый [N, N', N''-гексаметилпроизводное соединения (10.5)] оказывает бактериостатическое действие на Е. coli при разбавлении 1 : 10 000, но не 1 :20 000, хотя оба раствора окрашены в интенсивный фиолетовый цвет. Фиолетовая окраска указывает на присутствие катионов, являющихся активной формой этого антибактериального препарата (неионизиро-ранные молекулы бесцветны).
Объяснить это явление помогают уравнения (V) и (VI). Из уравнения (V) видно, что величина Ка (константа ионизации) определяет соотношение числа катионов (ВН+) к числу неиони-зированных молекул (В) при любой заданной концентрации ионов водорода [Н+]. Аналогичным образом уравнение (VI) определяет величину Ks, представляющую собой константу устойчивости ионной пары (АВН), образующейся путем присоединения катиона лекарственного вещества (ВН+) к уязвимому анионному участку бактерии (А-). Этот комплекс существует за счет ионного связывания, обычно усиленного водородными дли ван-дер-ваальсовыми связями:
_ [В][Н+] [ВН+]
(константа ионизации),
(V)
[АВН] [А-][ВН+]
(константа устойчивости).
(VI)
Если величины Ка и Ks сравнимы, то недостаток катионов в растворе будет пополняться не только за счет неионизирован-ного лекарственного препарата (В), но и за счет комплекса лекарственное вещество — бактерия (АВН). В этих условиях константа ионизации лекарственного вещества становится лимитирующим фактором, определяющим возможности использования этого препарата: ионы должны не просто присутствовать, а находиться в избытке. Из этого вовсе не следует, что кислоты, содержащиеся в рецепторах, должны быть слабыми. Дело в том, что время существования комплекса адсорбированный катион — рецептор (если ионная связь не приобретает дополнительную прочность за счет водородных связей или ван-дер-ваальсовых сил) очень коротко вследствие обмена с ионами натрия или кадия, присутствующими в окружающем солевом растворе.
Многие бактерии значительно более чувствительны к кристаллическому фиолетовому, чем Е. coli. Так, например, Streptococcus pyogenes ингибируется раствором с разбавлением 1:320 000 (при разбавлении 1:640 000 ингибирующее действие отсутствует), а раствор с разбавлением 1:2000 000 убивает
82
Staphylococcus aureus. Эти отличия могут быть обусловлены различиями в местах потерь, однако известно, что константы стабильности различаются для разных организмов и для разных тканей одного организма-хозяина. Это и создает основу для избирательного действия катионных лекарственных средств.
Ионизация может способствовать проявлению избирательности действия и в тех случаях, когда место действия лекарственного вещества имеет необычное значение pH, как, например, в желудочном соке или моче. В разд. 4.2 уже отмечалось, что опухолевые клетки имеют аномально низкие величины pH. Bisker (1974) предсказывал, что глюкурониды цитостатических фенолов и амидов должны обладать избирательным действием на раковые клетки, так как для действия ^-глюкуронидазы оптимальная величина pH 5.2. Тем не менее использование различий в pH нормальных и опухолевых клеток не должно зависеть от процесса обнаружения и гидролиза пролекарства ферментом. 4-Додецилпиридин (10.1) (рКа 5,5) циркулирует в токе крови мыши и не является токсичным для этого организма-хозяина при внутрибрюшинных инъекциях в дозе 0,5 г/кг-1. Тем не менее патогенные клетки вследствие повышенной кислотности поглощают катион этого соединения, который обладает выраженными поверхностно-активными свойствами и может воздействовать на эти вредные клетки [Firestone, Pisano, Bonney, 1979J.
(CH2)n—Me
ОН
0Н
Т_Ь'С—СН2ОН
4-Додецилпиридин
Аскорбиновая кислота
(ЮЛ)
(Ю.2)
Другие типы избирательного действия зависят от свойств, отличающих ионы от неионизированных молекул. Эти различия, рассматриваемые ниже, могут быть подразделены на три группы: а) ковалентная реакционная способность (образование и разрыв ковалентных связей); б) адсорбция на поверхности и в) проникновение через мембраны.
А. Ковалентная реакционная способность. Нитрование незаряженной молекулы анилина происходит в орто- и пара-положениях, в то время как его катион нитруется в основном в мета-положении. Таким образом, изменяя кислотность нитрующей смеси, можно контролировать соотношение образующихся изомеров. С точки зрения биохимии большой интерес представляет аскорбиновая кислота (10.2), дианион которой легко окисляется на воздухе, тогда как моноанион и неионизированиая молекула (любая из этих форм может стать основной при соответствующем значении pH) к окислению устойчивы [Weisberger, Lu Valle, 1944]. Из данных, представленных в верхней части
6* 83
Таблица 10.3. Расчет степени ионизации для случаев, когда величины рКа и pH близки
рка-рн % ионизации (по апнойному типу) % ионизации (по катионному типу)
—0,5 75,97 24,03
—0,4 71,53 28,47
—0,3 66,61 33,39
—0,2 61,32 38,68
—0,1 55,73 44,27
0 50,00 50,00
+0,1 44,27 55,73
+0,2 38,68 61,32
+0,3 33,39 66,61
+0,4 28,47 71,53
+0,5 24,03 75,97
табл. 10.4, видно, что при уменьшении pH скорость окисления снижается пропорционально падению концентрации дианиона и мало зависит от концентрации двух других форм.
В присутствии ионов меди аутоокисление аскорбиновой кислоты происходит значительно быстрее, но в этом случае меняется механизм реакции, так как скорость реакции пропорциональна концентрации моноаниона, а не концентрации дианиона или неионизированной молекулы (табл. 10.4, нижняя часть).
Разрыв ковалентных связей ферментами сильно влияет на действие избирательно токсичных агентов, так как при этом они могут превращаться в более активные или, наоборот, инертные вещества. Однако, поскольку скорости реакции с ионами и молекулами различаются очень сильно, изменения рКа внутри серии веществ могут привести к существенным различиям.
Б. Адсорбция на поверхности. Существуют два вида адсорбции — неспецифическая и специфическая. Неспецифическая адсорбция характерна для амфифильных веществ, имеющих концевую гидрофильную группу, связанную с относительно большим гидрофобным остатком. Примером подобных соединений может служить обыкновенное мыло. Следует напомнить, что молекулы воды связаны друг с другом водородными связями и вещества растворяются в воде только благодаря их способности разрывать некоторые из них и образовывать новые связи с молекулами воды. Амфифильные молекулы, растворенные в воде, находятся в состоянии подвижного равновесия, так как их углеводородная часть непрерывно вытесняется молекулами воды, стремящимися соединиться друг с другом.
84
Таблица 10.4. Самоокисление аскорбиновой кислоты [Weissberger, Lu Valle, 1944]
pH Концентрация диаииоиа, % Концентрация моиоаиио-иа, % Концентрация иеиоии-зироваииых молекул, % Коистанта скорости, мии”1Х 103
9,21 0,5 99,5 22 В отсутствие ионов ме-
8,71 0,2 99,8 — 11 талла
7,61 0,01 99,99 0,7
5,80 — 97,8 2,2 0,1
4,70 — 79,4 20,6 0,03
9,31 0,6 99,4 101 В присутствии медного
7,49 0,01 99,99 — 170 катализатора
6,12 — 99,0 1,0 134
5,08 — 90,1 9,9 91
3,87 — 36,0 64,0 26
2,59 — 2,9 97,1 2
Эти свойства воды лежат в основе неспецифической адсорбции, поскольку такие вещества занимают любую доступную им поверхность, независимо от ее химической природы. Например, в сосуде, содержащем мыльный раствор, мыло накапливается не только на поверхности раздела воздух — вода, но и на поверхности раздела стекло — вода. Кроме того, если в мыльный раствор погружать различные предметы, то мыло адсорбируется также и на их поверхности. Это пример типичной неспецифической адсорбции, при которой нейтральные молекулы адсорбируются сильнее, чем ионы. Так, олеиновая кислота адсорбируется сильнее, чем олеат натрия. Это происходит потому, что ион гидратируется сильнее, чем соответствующие неионизиро-ванные молекулы, и поэтому последние легче вытесняются из воды.
Специфическая адсорбция играет более важную роль при действии лекарственных веществ. Она свойственна гидрофильным веществам, которые стремятся выйти из воды и разместиться на поверхности, имеющей химически комплементарный характер. Простейшим примером такой комплементар-ности может служить притяжение аниона к положительно заряженному участку поверхности, а катиона — к отрицательно заряженному. В таких случаях ион будет адсорбироваться сильнее, чем неионизированная молекула. Наибольший интерес представляют фосфорилированные витамины группы В, накапливающиеся на комплементарных поверхностях ферментных систем с поразительной быстротой и эффективностью, несмотря на то что они хорошо растворяются в воде и содержатся в пище в малых количествах.
В. Проникновение через мембраны. Каждая живая клетка окружена полупроницаемой мембраной, толщина которой со-
85
ставляет две-четыре молекулы. Такая мембрана (обычно липопротеидной природы) имеет высокий заряд и практически непроницаема для ионов. Трудность прохождения ионов через мембрану обусловлена следующими причинами: 1) ионы имеют относительно большую величину вследствие гидратации и 2) заряд ионов либо аналогичен по знаку той части белковой поверхности, к которой он приближается (что приводит к отталкиванию), либо противоположен (что приводит к его фиксации).
Незаряженные молекулы обычно легко проникают через мембраны при условии, что они имеют не более трех гидрофильных групп и их ОММ не превышает 150 (разд. 3.2). Пример ионизационного контроля проницаемости через мембраны представлен на рис. 10.11. Синтезированы новые амины, содержащие 1231, с такими величинами рКа, что они могут использовать градиент pH, существующий между кровью (pH 7,4) и мозгом (pH 7,0). Эти соединения проникают в мозг в виде неиони-зированных молекул и попадают в цитоплазму с более низким pH, вследствие чего доля катионов, не способных к диффузии, значительно повышается [Tramposch, Kung, Blau, 1983J.
Однако не следует считать, что ионы вообще не способны проникать через нейтральные мембраны. Во-первых, существуют особые механизмы поглощения для каждого из тех ионов, которые необходимы клетке. Так, поглощение ионов холиния происходит по механизму «Тип 2» (см. разд. 3.2.2), а ионов Na+ и К+ — по механизму «Тип 3», требующему затрат энергии (разд. 3.2.3). Кишечник человека легко пропускает ионы натрия и хлора, несколько хуже — цитрат- и ацетат-ионы; что касается ионов Са2+, Mg2+, Fe2+ и фосфат-иона, то для них существует специфический механизм захвата, характеризующийся быстрым насыщением, по которому поглощаются только следы этих ионов, необходимых организму; сульфат- и тартрат-анионы практически не поглощаются. Во-вторых (это особенно относится к синтетическим лекарственным препаратам), ионы, не обладающие способностью проникать через мембрану, могут приобретать эту способность при введении липофильной группы. Атом хлора и другие липофильные заместители, присутствующие в молекулах противомалярийных препаратов мепакрина (атебрин) и хлорохина, способствуют их проникновению в эритроциты.
Теперь перейдем к рассмотрению другого аспекта влияния ионизации на проникновение препаратов через мембраны. Эффективность лекарственного средства безусловно зависит от способности его проникновения к рецептору. Если значение рКа данного агента между 6 и 8, то при физиологическом значении pH 7 его молекулы всегда находятся в равновесии, по крайней мере с 10% самой минорной ионной формы (см. табл. 10.2). Такой агент может войти в соприкосновение с мембраной, через которую смогут проникнуть только незаряженные молекулы. Но пройдя через мембрану, они, вероятно, попадут в среду с тем 86
же значением pH, где будут вновь образовывать ионы, пока по обе стороны мембраны не установится равная степень ионизации для данного агента. Величины рКа многих лекарственных средств находятся в пределах 6—8, хотя известны агенты с рКа, сильно отличающимися от этих величин, и поэтому они имеют другой характер распределения и иной тип действия.
Г. Псевдооснования. Время, необходимое для создания ионного равновесия в растворе, настолько мало (например, 10~7 с), что ионные реакции можно считать мгновенными. Однако существуют немногочисленные виды катионов, которые медленно образуют ковалентную связь с гидроксилионами, давая неионизи-рованные вещества, известные под названием псевдооснований. При действии кислоты из псевдооснования может быть снова выделен исходный катион. В разных случаях для установления равновесия может потребоваться различное время, от минуты до недели. Две наиболее известные группы псевдооснований — это гетероароматические четвертичные соединения и трифенилметановые красители. Методы расчета равновесных констант ионизации (обозначаемых как pKaEq) и констант скорости установления этого равновесия были предложены Goldacre, Phillips (1949) для четвертичных соединений и Cigen (1958) —для производных трифенилметана.
Me Me
Катион 5-метилфенантридиння Карбинольное (или псевдо-) основание
Парафуксин
Карбинольное (нли псевдо-) основание'
(10.5)
(10.6)
Образование псевдооснований может способствовать проникновению вещества через мембраны, поэтому следует рассмотреть химизм этого процесса. Катион 5-метилфенантридиния (10.3), такой как содержащийся в хлориде N-метилфенантриди-ния, может служить примером четвертичных аммониевых оснований, у которых атом азота' гетероцикла имеет двойную связь
87
Таблица 10.5. Равновесие между четвертичными катионами и соответствующими псевдооснованиям и [Bunting, Meathrel, 1972]
Четвертичные катионы Р^кон
1 -Метилхино лнинн 2-Метнлизохннолнннн 10-Метилакридиний 5-Метилфеиаитрндиний (10.3) 1 -Метнл-З-нитрохинолиний 2-Метнл-4- ннтроизохинолиний 1 -Метилхнноксалиний З-Метнлхиназолиний 16 15 9,9 10,4 6,7 5,0 8,6 <7
(катионы соответствующих третичных аминов не образуют псевдооснований). Азот не может иметь более четырех ковалентных связей и, следовательно, в соединении (10.3) он обладает максимальной валентностью; кроме того, метильная группа не так подвижна, как ион водорода, поэтому неионизированной молекулы, соответствующей иону (10.3), не существует. Таким образом, можно было бы предположить, что это соединение остается ионизированным при любом pH. Однако этот ион медленно' реагирует с ионом гидроксила (реакция обратима), в результате чего образуется ковалентная связь и возникает неионизиро-ванная молекула (10.4) [Magrath, Phillips, 1949]. Этот вторичный спирт представляет собой псевдооснование соединения (10.3). Псевдооснования обладают гораздо большей растворимостью в липидах, чем соответствующие катионы, из которых они образованы, возможно, эти ионы проникают в клетки именно в форме псевдооснований. Трипаноцидное действие четвертичных гетероциклических соединений (разд. 10.3.5) можно, объяснить только с учетом равновесия, такого как, например, (10.3) (10.4), и соответствующего времени полупревраще-
ния (to.e)-
Тенденция к образованию псевдооснований возрастает с усложнением гетероциклического ядра. Так, если для катиона 1-метилпиридиния образование псевдооснований совершенно не характерно, а ионы 1-метилхинолиния и 2-метилизохинолиния превращаются в псевдооснования только при очень высоких значениях pH, то при дальнейшем аннелировании (например, как в 10-метилакридинии) реакция протекает значительно легче. Такое же действие оказывает и введение в цикл электроноакцепторного заместителя (такого, как группа NO2) или второ-ю атома азота с двойной связью, как в катионах хиноксалиния и хиназолиния. Некоторые примеры приведены в табл. 10.5, где рКцон обозначает то значение pH, при котором концентрации псевдооснования и четвертичного катиона равны. Формально' величина рКцон аналогична величине рКа, с той лишь разницей,
88
что равновесие в этом случае устанавливается значительно медленнее.
Формулой (10.5) изображена структура катиона парафуксина, типичного трифенилметанового красителя, медленно реагирующего с гидроксил-ионами с образованием псевдооснования (10.6). Псевдооснования обоих типов (10.4) и (10.6) быстро реагируют со спиртами, образуя бесцветные эфиры. Поэтому при количественном определении красителя (колориметрическим методом), поглощаемого биологическими объектами, следует избегать экстракции спиртом.
Цвиттерионы обычно довольно инертны с точки зрения фармаколога. Так, например, гистидин, представляющий собой цвиттерион, не обладает теми важными физиологическими свойствами, которые характерны для родственного ему катионного соединения гистамина. При окислении винильной группы хинина в молекуле появляются карбоксильная группа, придающая свойства цвиттериона, при этом соединение теряет противомалярийную активность. При этерификации этой кислоты (ките-нин) соединение теряет цвиттерионную природу, становится положительно заряженным и вновь приобретает противомалярийную активность [Goodson, Henry, MacFie, 1930]. Аналогичные примеры встречаются и в серии производных акридина (разд. 10.3.1).
10.3. Вещества, обладающие большей биологической активностью в ионизированном состоянии
Только во второй четверти нашего столетия было обнаружено, что многие органические катионы обладают антибактериальной активностью. Ранее было известно, что алифатические амины, существующие при pH 7 главным образом в виде катионов, проявляют бактерицидное действие в отличие от ароматических аминов, которые существуют при pH 7 в основном в виде неио-низированных молекул и этого действия не проявляют. Однако тогда не связывали антисептическое действие с наличием органических катионов; предполагалось, что антибактериальная активность зависит от «наличия гидроксильных ионов, высвобождаемых при ионизации гидроксидов алкиламмония, образующихся при взаимодействии аминов с водой» [Morgan, Cooper, 1912]. Иными словами, наличие бактерицидного действия объяснялось щелочностью антисептических растворов, при этом не учитывалось, что те же основания, находящиеся в форме солей или в нейтральном буферном растворе, также обладают бактерицидными свойствами. Существенный вклад в решение этой проблемы был внесен в 1924 г. Stearn и Stearn, высказавшими предположение о том, что антибактериальная активность трифенилметановых красителей объясняется взаимодействием катиона красителя с определенными анионными группами бактерий с образованием слабо диссоциированных комплексов того
89
же типа, который обсуждался в разд. 10.2 (ср. уравнение VI). Хотя эти исследователи не знали степени диссоциации исследуемых красителей1, они предсказали, что соли многих сильных оснований могут обладать антибактериальным действием, так как они способны образовывать достаточное количество катионов при физиологических значениях pH. Они показали также, что при повышении pH среды антибактериальное действие усиливается за счет повышения степени ионизации (анионной) рецепторов бактерий. Однако это увеличение щелочности среды не следует доводить до такой степени, чтобы начиналось подавление ионизации самого антисептика.
Первое строгое доказательство взаимосвязи между ионизацией и биологическим действием было получено лишь через 17 лет [Albert, Rubbo, Goldacre, 1941]. Исследование серии аминоакридинов показало, что существует количественная зависимость между их антибактериальным действием и степенью ионизации по катионному типу. Позднее эта корреляция была подтверждена и расширена на примере других соединений [Albert et al., 1945].
NH2
Профлавин Аминакрин
(3,6-диаминоакридии) (9-аминоакридин)
(10.7) (10.8)
10.3.1. Аминоакридины, обладающие антибактериальным действием
Акридин (2.16) имеет плоскую молекулу и обладает слабыми основными свойствами. Тем не менее два из пяти возможных изомеров моноаминоакридина, а именно 3- и 9-аминоакридины, являются сильными основаниями, благодаря сопряжению в их катионах (разд. 2.2). В молекулах 2- и 4-амино-акридинов дополнительное сопряжение существовать не может вообще, а в 1-изомере оно проявляется лишь в небольшой степени, так как орто-хиноидное расположение связей, необходимое для возникновения сопряжения, энергетически невыгодно [Albert, Goldacre, 1946; Albert, Goldacre, Phillip, 1948].
В 1913 г. шотландский патолог К. Browning предложил использовать аминоакридины в качестве антибактериальных средств при ранениях. Выяснилось, что один из них, профлавин (10.7) (3,6-диаминоакридин), токсичен по отношению к широкому ряду грамположительных и грамотрицательных бактерий,
1 В то время еще нельзя было оценить степень диссоциации псевдоосно-ваиий по чисто техническим причинам.
90
Таблица 10.6. Зависимость бактериостатического действия некоторых производных акридина от ионизации [Albert et al., 1945]
Производные акридина Минимальная бактериостатическая концентрация для 48-часовой инкубации при 37 “С в бульоне с 10% сыворотки при 7,3 Степень ноавмщнн по катионному типу (pH 7,3, 37 °C)
Акридин 1 5000 1
4-Амнно- 1 5000
2-Амино- 1 10000 2
1-Амино- 1 10 000 2
4,5-Диамино- 1 <5000
2,7-Диамино- 1 20 000 4
З-Амино- 1 80000 72
9-Амино-(аминакрин), (10.8) 1 160 000 99
3,9-Диамино- 1 160 000 100
3,7-Диамино- 1 160 000 76
3,6-Диамино-(профлавин) (10.7) 1 160000 99
2-Амино-9-метил- 1 20 000 3
1 -Амино-4-метил- 1 20 000 1
4-Амино-5-метил- 1 <5000 <1
2-Амино-6-хлоро- 1 <5000
З-Амино-9-хлоро- 1 <5000 11
З-Амино-7-хлоро- 1 40 000 20
З-Амино-6-хлоро- 1 40000 24
9-Амнно-1 -метил- 1 160000 99
9-Амино-2-метил- 1 160 000 99
9-Амино-З-метил- 1 160 000 99
9-Амино-4-метил- 1 320 000 99
9-Амино-1 -хлоро- 1 160 000 86
9-Амино-2-хлоро- 1 160 000 94
9-Амино-З-хлоро- 1 160 000 98
9-Амино-4-хлоро- 1 80 000 83
но безвреден для тканей человека [Browning, Gilmour, 1913; Browning et al., War Office, 1922]. Акридины, содержащие четвертичный атом азота, например эуфлавин, более токсичны для млекопитающих, но в то же время являются и более эффективными антибактериальными средствами, чем некватернизирован-ные, например профлавин. В серии акридинов псевдооснования могут образовываться только из кватернизированных соединений, поэтому удалось избежать трудностей, встречающихся при работе с трифенилметановыми красителями.
Предварительные эксперименты, проведенные Albert и сотр. в 1939 г., показали, что антибактериальная активность акридинов возрастает при повышении степени их катионной ионизации [Albert, Rubbo, Goldacre, 1941]. Эти результаты были подтверждены при испытаниях 101 производного акридина на 22 видах бактерий [Albert et al., 1945; Albert, Goldacre, 1948]. Некоторые наиболее типичные результаты представлены в табл.10.6, расширяющей данные табл. 2.2 об изученных производных ак
91
ридина. Результаты для 100 производных акридина опубликованы в книге Albert (1966).
Из пяти возможных моноаминоакридинов два сильно ионизированы при pH 7, остальные — слабо (объяснение причины этих различий приведено в разд. 2.2). Из табл. 10.6 видно, что два изомера с высокой степенью ионизации обладают сильным антибактериальным действием, а эффективность слабоионизи-рованных аминоакридинов очень незначительна. Такая же корреляция наблюдается и для остальных исследованных соединений (см. табл. 10.6, ср. активность сильно- и слабоионизиро-ванных метиламиноакридинов и активность сильноионизирован-ных хлораминоакридинов с их слабоионизированными изомера-ми). Во всех случаях ионизация сопровождается резким повышением активности, как правило, в 8—16 раз. Таким образом,, в этой серии соединений изменения структуры незначительно изменяют активность, но только в том случае, если они не влияют на ионизацию. Эти выводы подтверждаются испытаниями на многих видах бактерий (см. табл. 10.7).
Подробные данные для серии 100 производных акридина й многих разновидностей бактерий (анаэробных и аэробных, грамположительных и грамотрицательных), подтверждающие наличие корреляции между бактериостатическим действием и степенью ионизации, см. Albert (1966).
Ионизация, определяющая наличие антибактериального действия производных акридина, должна быть катионной по своему характеру. Вводя кислотные группы в циклы сильноосновного акридина, можно легко получить соединение, представляющее собой цвиттерион и, следовательно, уже не катион, как, например, 9-аминоакридин-2-карбоновая кислота (табл. 10.8). Эта кислота не обладает антибактериальными свойствами, но вновь приобретает их при этерификации, приводящей к образованию катионного соединения. Вводя кислотные группы в цикл слабоосновных производных акридина, можно получить соединение анионного характера, такое как акридин-9-карбоновая кислота (см. табл. 10.8). Это соединение, его эфир, а также сам акридин (все эти вещества — не катионы) не проявляют сколько-нибудь заметной антибактериальной активности.
Механизм действия акридинов был изучен более подробно на организме Е. coli, способном существовать как в кислой так и в щелочной среде. На рис. 10.2 показана зависимость минимальных бактериостатических концентраций (выраженных в логарифмах) четырех производных акридина от pH среды в диапазоне от 5,5 до 8,5. Только один из этих акридинов достаточно сильное основание, чтобы ионизироваться при указанных значениях pH. Таким образом, не удалось выявить какой-либо закономерности. Однако, если на графике по оси ординат отложить концентрацию катионов (вместо общей концентрации вещества), то получаются параллельные прямые, что позволяет сделать определенные выводы.
92
Таблица 10.7. Сравнительная чувствительность 22 видов бактерий к действию некоторых производных акридина
Микроорганизм 9-Амино-акридин (10.8) (амиио-акрин) 4-Метил-9-амияо-акридин 3,6-Диамиио-акридин (10.7) (профлавин) 3» 7-Диамино-4,5 диметил-акридии 3,7-Диа-мииоак-ридин
Грамположнтельные бактерии Cl. welchii 1в: 320 000 640 000 160 000 1 280 000 320 000
Cl. sporogenes 160 000 320 000 160 000 640 000 160 000
В. subtilis 80 000 160 000 80 000 320 000 80 000
В. anthracis 160 000 320 000 80 000 640000 80 000
Strept. pneumoniae 320 000 320 000 160 000 640 000 160 000
Strept. pyogenes A 160 000 320 000 160 000 640 000 160 000-
Strept. viridans 160 000 640 000 160 000 640 000 160 000
Staph, aureus 40 000 80 000 40 000 160000 40 000
Lactobacillus 320 000 320 000 160 000 640 000 160 000
C. diphtheriae 160 000 320 000 40 000 320 000 160 000
Mycobact. phlei 160 000 320 000 160 000 640 000 160 000
Среднее разбавление, необходимое для ингибирования 160 000 300 000 110 000 540 000 140 000
Грамотрицательные бактерии Е. coli 40 000 40 000 20 000 40 000 40 000
Proteus vulgaris 40 000 40 000 10000 20 000 40 ОСО
Ps. pyocyanea В. Friedlanderi 80 000 АСсолк 80 000 )тно не актш 20 000 1НЫ 160 000 80 000
Eberthella typhosum 80 000 160 000 80 000 80 000 80 000
Shigella dysenteriae 320 000 320 000 160 000 320 000 320 000
Vibrio cholerae 160 000 160 000 160 000 320 000 160 000
Pastuerella 160 000 160 000 40 000 160 000 160 000
Brucella 640 000 640 000 320 000 280 000 320 000
Haem, influenzae 320 000 640 000 320 000 640 000 160 000
Neiss. meningitidis 320 000 640 000 320 000 02 560 000 320 000
Среднее разбавление, необходимое для ингибирования 100 000 130 000 60 000 150 000 100 000
Примечание. Сильное разбавление препятствует заметному росту бактерий в-течение 48 ч при 37 °C (среда: бульон с 10% сыворотки, pH 7,3). При данных условиях все вещества практически полностью ионизированы [Albert et al., 1945].
Во-первых, основным фактором, определяющим бактериостатическое действие производных акридина при любом pH является концентрация катиона (это зависит от pH среды и рКа. соединения, см. табл. 10.2), а не общее количество вещества (катионы и неионизированные молекулы). Во-вторых, наклон прямой показывает, что существует конкуренция между катионами акридина и ионами водорода. Простейшее объяснение механизма действия катионов акридина заключается в том, что
93
Таблица 10.8. Зависимость бактериостатического действия производных акридина от степени катионной ионизации [Albert et al., 1945]
Соединение Минимальная бактериостатическая концентрация после 48-часовой инкубации при 37 *С (среда: бульон с 10% сыворотки, pH 7,3) Степень ионизации (pH 7,3; 37 °C)
катион анион цвиттерион нейтральная молекула
9-Аминоакрнднн (10.8) 9-Амнноакрнднн-2- 1в:160 000 99 0 0 0
карбоновая кислота Метиловый эфир 9-амн-ноакрндин-2-карбоно- <5000 0 0,2 99,8 0
БОЙ КИСЛОТЫ 160 000 89 0 0 11
Акридин Акрнднн-9-карбоновая 5000 0,3 0 0 99,7
кислота Метиловый эфир акри-дин-9-карбоновой кисло- <5000 0 99,3 0,7 0
ТЫ <5000 0 0 0 100
они конкурируют с ионами водорода за жизненно важную анионную группу бактерии. О местонахождении этой группы речь пойдет ниже. Из данных табл. 10.9 видно, что катионы того же размера и такие же плоские, как и рассматриваемые нами, способны вытеснять ионы водорода из бактерий.
Графики, приведенные на рис. 10.3, свидетельствуют о том, что количество аминоакридина, применяемого при ранениях, можно уменьшить, если предупредить развитие в ране кислой реакции. Исследования в этом направлении проводились в годы второй мировой войны в австралийской армии, где наиболее широко применяли гидрохлорид 9-аминоакридина, наиболее эффективного из этого ряда соединений. В результате было рекомендовано перед обработкой ран гидрохлоридом 9-аминоакридина (10.8) промывать их раствором бикарбоната натрия. Это прекрасный метод для профилактики и лечения сепсиса даже при глубоких гнойных и запущенных ранах. После лабораторных испытаний (см. табл. 10.6 и 10.7) и успешного применения в клинике для лечения тяжелых ранений [Poate, 1944; Turnbull, 1944] 9-аминоакридин был включен в Британскую фармакопею под названием «аминакрин».
Точное место действия аминоакридинов было установлено в результате нескольких открытий: во-первых, стало ясно, что активны именно плоские молекулы [Albert, Rubbo, Burvill, 1949]; во-вторых, было обнаружено, что аминоакридины окрашивают только ДНК и РНК, содержащиеся в клетках позвоночных [Strugger, 1940]; в-третьих, установлено, что аминоакридины интеркалируют в ДНК; в-четвертых, выяснилось, что аминоакридины способны ингибировать ДНК-полимеразу бакте-
94
Рнс. 10.2. Конкуренция между нонами водорода н акридинами (смесь катионов с неионизированными молекулами, организм Е. coli).
Минимальная бактериостатическая концентрация лекарственного вещества в целом
9-Аминоакридин
2-Аминоакридин
рий путем связывания с матричной ДНК [Hurwitz et al., 1962]. Доказательства второго и четвертого положений рассмотрены ниже, первого и третьего — в разд. 10.3.2.
А. Окрашивание
живых систем. Доказано, что живые клетки млекопитающих, в том числе и мозга [Manifold, 1941], практически не повреждаются при обработке аминоакридинами [Russel, Falconer, 1941, 1943; Selbie, McIntosh, 1943]. В 1940 г. две группы исследователей независимо друг от друга установили, что аминоакридины способны накапливаться в нуклеиновых кислотах, содержащихся в живых клетках позвоночных, не причиняя им никакого вреда. Было обнаружено, что при этом ядерная и цитоплазматическая РНК флуоресцирует огненно-красным цветом, а ядерная ДНК — зеленым (в поле флуоресцентного микроскопа) [Strugger, 1940; Bukatsch, Haitinger, 1940]. Эти красители не повреждают даже чувствительные половые клетки и, следовательно, не влияют на процессы размножения. Таким образом, была усовершенствована методика окрашивания живых систем, предложенная П. Эрлихом, и получено наглядное доказательство того, что нуклеиновые кислоты способны избирательно накапливать аминоакридины в живых клетках.
Способность к поглощению и концентрированию аминоакридинов в ядре была показана на клетках легких, сердца, печени, селезенки, языка, почек и тонкого кишечника крыс, мышей и кроликов [De Bruyn, Robertson, Farr, 1950]. Тот факт, что белки клеток не способны накапливать аминоакридины, не вызывает удивления, так как известно, что при проведении экспериментов in vitro белок, содержащийся в сыворотке, не влияет на антибактериальное действие этих соединений [Browning, Gilmour, 1913]. Нуклеиновые кислоты, напротив, эффективно ингибируют бактериостатическое действие профлавина и акрифлавина [McIlwain, 1941]. Максимальное количество моноаминоакридина, которое может связать растворенная ДНК, варьирует в соответствии с изменением антибактериальной активности соединений в ряду 9->3->l->2->4-NH2 [Jackson, Mason, 1971].
Б. Ингибирование ДНК- и РНК-полимер азы. Два факта ука-
95
Таблица 10.9. Выделение ионов водорода из бактерий Bacillus bellus, вызванное адсорбцией катионов кристаллического фиолетового1 (pH измеря-________________ли стеклянным электродом) [McCalla, 1941]_________________
pH бактериальной суспензии (1) pH раствора кристаллического фиолетового (2) pH смеси (I) и (2) Снижение pH, вызываемое адсорбцией
8,5 8,5 6,6 1,9
6,5 6,3 4,8 1.7
5,5 5,5 3,8 1,7
5,0 5,1 3,5 1,6
4,1 4,2 3,0 1,1
‘ №,Ы\М''-гексаметплпронзводное парафукснна (10.5).
зывают на то, что местом действия аминоакридинов является внешняя поверхность плазматической мембраны бактерии. Во-первых, антибактериальная активность этих соединений не снижается при повышении степени ионизации от 70 до 100%, несмотря на то, что при этом исчезают нейтральные молекулы, способные легче катиона проникать через мембрану. А во-вторых, при увеличении липофильности аминоакридинов происходит резкое уменьшение их антибактериального действия [Albert et al., 1945].
В бактериях большая часть ДНК клетки содержится в единственной кольцеобразной хромосоме, присоединенной к цитоплазматической мембране в одной точке [Ryter, Jacob, 1964]. РНК содержится в цитоплазматической мембране [Yudkin, Davis, 1965] и в рибосомах.
Профлавин (30 мкМ) in vitro ингибирует бактериальную ДНК-полимеразу на 85%, а РНК-полимеразу — на 30% [Hurwitz et al., 1962]. Некоторые наиболее типичные данные для РНК-систем представлены на рис. 10.4. На вставке показан график зависимости изменения величины обратной скорости полимеризации от обратной концентрации ДНК-матрицы, указывающий на то, что именно матрица является местом действия профлавина, ингибирующего таким образом синтез ДНК и РНК в живых бактериях. Механизм интеркаляции, с помощью которого аминоакридины соединяются со спиралью ДНК и препятствуют разделению ее цепей, будет рассмотрен в разд. 10.3.2.
Более подробно был изучен механизм действия аминоакридинов на ДНК-зависимый синтез РНК на примере Е. coli [Nicholson, Peacocke 1966], бактериофага [Sarris, Niles, Canellakis, 1977], Azotobacter [Canellakis et al., 1976]. Хотя ингибирование ферментов у эукариот нетипично, тем не менее 9-аминоак-ридин (500 мкМ) полностью ингибирует включение уридинмонофосфата (УМФ) при попытках проведения транскрипции ДНК вилочковой железы теленка РНК-полимеразой печени крысы [Zoncheddu et al., 1980].
Рассмотрим вкратце некоторые выдвигавшиеся ранее гипотезы о механизме действия аминоакридинов (помимо идеи об S6
Рис. 10.3. Конкуренция между ионами водорода и катионами акридинов (организм Е. coli).
Минимальная бактериостатическая
интеркаляции этих соединений в ДНК), представляющие интерес хотя бы с исторической точки зрения. Предполагалось, что антибактериальное действие акридинов обусловлено их способностью ингибировать различные ферменты. Однако для этого обычно требуются очень высокие концентрации аминоакридинов, в то время как даже в низких концентрациях эти соединения подавляют синтез и нормальное функционирование нуклеиновых кислот. [Albert, 1966].
Были сделаны попытки связать антибактериальное действие аминоакридинов с их окислительно-восстановительными потенциалами [Breyer, Buchanan, Duewell, 1944]. Так, для веществ с сильным антибактериальным действием (например, 3-амино-9-амино- и 3,6-диаминоакридин) Ео' на первой стадии восстановления при pH 7,3 лежит между —0,47 и —0,92 В, тогда как соответствующее значение Ео' для соединений со слабым антибактериальным действием— между —0,31 и —0,39 В1. Таким образом, очень небольшое различие окислительно-восстановительных потенциалов (между —0,47 и —0,39 В) должно определить, обладает акридин антибактериальным действием или нет. Однако из рис. 10.3 видно, что в действительности не это различие определяет их активность. Так, при pH 7,3 одни акридины (например, 2-аминоакридин), слабо ионизированные при данном значении pH, обладают слабым, а другие — сильным антибактериальным действием, но активность их становится одинаковой при понижении pH до такой величины (например, 5,4), при которой обе группы акридинов сильно ионизированы. Поскольку снижение pH не уменьшает разности окислительно-восстановительных потенциалов 2- и 3-аминоакридинов,
1 Значения потенциалов ионизации приведены относительно нормального водородного электрода, Ео охарактеризовано в разд. 11.4, а значения Ео, указанные в тексте, определены при pH 7,3.
7-734
97
Рнс. 10.4. Ингибирование РНК-полимеразы Е. coli профлавином, измеренное по уменьшению потребления УМФ в присутствии избытка ДНК, АТФ, ГДФ и ЦТФ. На вставке представлен график зависимости обратной скорости полимеризации от обратной концентрации ДНК-матрицы в присутствии (верхняя линия) и в отсутствие (нижняя линия) профлавина [Hurwitz et al., 19621.
следует заключить, что окислительно-восстановительные потенциалы не влияют на антибактериальную активность производных акридина.
Гипотеза о том, что антибактериальное действие аминоакридинов определяется наличием свободных радикалов, образующихся на первой стадии (1Н) восстановления, также неверна, так как на этой стадии все акридины образуют свободные радикалы [Кауе, 1950]. Более того, вещества, не поддающиеся восстановлению, такие как 2-антранилгуанидин (табл. 10.10), проявляют акридиноподобное антибактериальное действие. Примечательно, что антрацены не восстанавливаются даже при —2,0 В [Zanker, Schnith, 1959].
В. Фунгицидное действие. Хотя аминоакридины подобно большинству антибактериальных препаратов почти не действуют на грибы, некоторые из них обладают сильным трансформирующим действием на дрожжи. Они способны встраиваться в ДНК дрожжей, находящуюся в митохондриях, вызывая тем самым наследственные изменения (утрату способности к окислительному фосфорилированию), но не затрагивая при этом ядра. Наиболее активным в этом отношении является эуфлавин (хлорид 3,6-диамино-10-метилакридиния) (6,5). [Ephrussi, Hot-tinguer, 1950; Marcovich, 1953]. Два акридина, оказавшиеся наименее токсичными для тканей человека и наиболее полезными в хирургии (9-аминоакридин и 3,6-диаминоакридин), не оказывают действия на дрожжи.
Г. Применение акридинов в современной медицине. Амина-крин и профлавин, обладающие антибактериальным действием, наиболее широко используют в Англии, континентальной Европе и Австралии для оказания первой помощи при ранениях и
98
ожогах в полевых госпиталях и больницах, а также при перевязках в домашних условиях. Этакридин (риванол) (3,9-диами-но-7-этоксиакридин) применяют перорально при инфекциях тонкой и толстой кишки, в основном в странах Западной Европы. Мепакрин все еще широко используют при лечении малярии и лямблиоза в Азии и Африке (разд. 10.3.5). Два соединения акридинового ряда, амсакрин и ледакрин, представляют собой противораковые препараты и в настоящее время проходят клинические испытания (разд. 10.3.4).
10.3.2. Катионные антибактериальные средства, действующие по типу акридинов.
Выше была рассмотрена зависимость между степенью катионной ионизации и антибактериальной активностью производных акридина. Подобная зависимость существует также по крайней мере для семи других серий соединений: бензоакридинов (10.9) — (10.11) (всего исследовано 16 представителей), фе-нантридинов (10.12) (исследовано 6 представителей) и трех изомерных бензохинолинов (10.13) — (10.15) (исследовано 21 соединение) [Albert, Rubbo, Burvill, 1949]. Некоторые из этих примеров приведены в табл. 10.10.
Бензо(с)акридин Бензо(Ь)акридин Бензо(а)акридин
(10.9) (10.10) (10.11)
9
Фенатридин Бензо(Г)хинолин Бензо(£)хинолнн
(нумерация по IUPAC)
(10.12) (10.13) (10.14)
4
Бензо(Ь)хинолии
(10.15)
А. Необходимость плоской структуры молекул. Albert и сотр. установили, что соединения, образованные удалением одного
свн5х
PCI—СвН4/
СН—N
N—Me
Хлорфенамин
(9.57)
Хлорцнклизнн (9.56a)
Антагонисты Н2-рецепторов. Н2-рецепторы гистамина ответственны за стимуляцию матки, предсердия и секрецию кислоты в желудке. Поиски антагонистов Н2-рецепторов гистамина были начаты Дж. Блэком в надежде, что эти лекарственные препараты смогут принести облегчение или даже полечить язву желудка. Так был создан циметидин (9.58), нашедший широкое применение в клинике [Gray et al., 1977].
Ключом к решению многих проблем стало открытие того, что 5-метилгистамин (9.59) представляет собой агонист только Н2-рецепторов (но не Hi-рецепторов). Подобное явление уже отмечали в случае метахолина ^-метилпроизводного АХ, проявляющего такую же, как у АХ, активность по отношению к мускариновым рецепторам, но не действующего на никотиновые (разд. 12.6). Было обнаружено, что сильное агонистическое действие производных гистамина на Н2-рецепторы непосредственно определяется возможностью таутомерии (перемещение атома водорода от одного атома азота к другому). Отсутствие такой возможности приводит к появлению агонистического действия на Hi-рецепторы, как, например, в случае 2-(2-аминоэтил) пиридина [Durant, Ganellin, Parsons, 1975].
Me----- • CH2—S—CH2—CH2—NH—С—NHMe т
hn^Jn N—СН
Циметиднн
(9.58)
Me-.
\. ------ch2ch2nh2
I I N NH
4-Метнлгистамнн
(9.59) .CH2CH2CH2CH2NH—C—NHMe hn^'n
Буринамнд
(9.60)
В разд. 7.5.2 обсуждался вопрос о том, что молекула антагониста должна иметь значительно более сложную структуру, чем молекула агониста. Примером этого могут служить антаго-
56
цисты Hi-рецепторов. Агонистическое действие исчезает при наличии в молекуле лекарственного вещества неионизирующей-ся боковой цепи; антагонистическая активность повышается при удлинении боковой цепи. Подходящим концевым фрагментом боковой цепи считают метилмочевину и некоторые другие плоские структуры с двойной связью, также способствующие адсорбции. На основе вышеизложенного был создан первый Н2-аго-нист—буринамид (9.60). Было показано, что он действует как конкурентный и специфический антагонист гистамина на предсердие морских свинок и матку крыс, на рецепторы которых Н.-антагонисты не действуют. К сожалению, действие буринами-да оказалось слишком слабым для применения его в клинике [Black et al., 1972], кроме того, при пероральном введении он плохо всасывается.
Те же исследователи заменили в молекуле буринамида одну группу—СН2 — боковой цепи атомом серы (—S—).
Это не только облегчило процесс синтеза, но и привело к снижению основности цикла, которая была слишком высокой (рКа = 7,3) по сравнению с гистамином (для имидазольного кольца рКа = 6). Введение метильной группы в положение 5 имидазольного кольца позволило получить новый лекарственный препарат метиамид (9.61) с рКа = 6,8, хорошо всасывающийся при пероральном применении. Так, при pH 7,3 процент ионизированной формы (катиона) буринамида, метиамида и гистамина составляет 50, 24 и 5% соответственно. С помощью метиамида было вылечено несколько сотен больных с язвой желудка. Однако после того, как было обнаружено, что этот препарат обладает способностью снижать количество гранулоцитов в крови у небольшой группы больных, он был изъят из применения [Durant et al., 1977].
Этот побочный эффект был вызван наличием в молекуле буринамида тиомочевинного фрагмента. Поиск неионизирую-щейся изостерной группы привел к замене ее цианогуанидиновым фрагментом. Полученный в результате циметидин (9.58), М-метил-М'-циано-КГ' - [(5-метилимидазол-4-ил) метилтиоэтил] гуанидин, имеет такую же величину рКа = 6,8, как и метиамид [Ganellin, 1981]. Аналогичными клиническими свойствами обладает препарат ранитидин (9.62), созданный несколько позднее. Его молекула отличается от молекулы циметидина тем, что ее боковая цепь вместо группы С—N—CN содержит изостерную ей группу =СН—NO2. Кроме того, имидазольный цикл молекулы циметидина заменен в этой молекуле фурановым, не обладающим основными свойствами и снижающим основность диметиламиногруппы боковой цепи. Способность ранитидина понижать секрецию кислоты в желудке обсуждается в работе Bohman (1980). Заменой имидазольного цикла мочевиной был получен препарат димаприт (7.45) (разд. 7.3).
Об антигистаминных лекарственных препаратах см. Rocha, Silva (1978).
57
Таблица 10.10. Зависимость бактериостатического действия от а) степей» ионизации и б) площади плоской поверхности молекулы в ряду катионных лекарственных средств1 [Albert et al., 1949]
Соединение Минимальная бактериостатическая концентрация для Streptococcus pyogenes после 48-часовой инкубации при 37 °C (среда: бульон с 10% сыворотки, pH 7,3) Степень ионизации (pH 7,3, 37 °C), % Площадь плоской поверхности, рассчитанная, как указано на рис. 9.5, им2
Акридин 1в:5000 1 0,385
9-Амнноакридин (10.16) 160 000 100 0,385
Бензо (с) акридин <5000 1 0,487
11-Аминобензо(с) акридин 160 000 97 0,487
Бензо (Ь) акридин (10.10) 10 000 1 0,489
11-Аминобензо (в) акридин 320 000 100 0,489
Бензо(а)акридин <5000 <^1 0,487
11-Амннобензо (а) акридин 160 000 98 0,487
Фенантрндин (10.12) 10 000 <1 0,383
6-Аминофенантридин 40 000 50 0,383
3,6,8-Трнамннофенантридин 80 000 98 0,383
Бензо (f) хннолин (10.13) 5000 <1 0,383
1-Ам ннобензо (f) хннолин 20 000 98 0,383
Бензо(g)хинолин 10 000
4-Ам ннобензо (g) хинолин 40 000 100 1 0,383
Бензо(h)хинолин <5000
4-Аминобензо (Ь) хннолин 80 000 96 >
Хинолин <5000 <1 0,279
4-Амннохннолнн (10.17) 4-Амнио-2-стнрнлхннолнн <5000 98 0,279
(10.20) 80 000 99 0,499
Пнрнднн <5000 <^1 0,174
4-Амннопирнднн (10.18) 4-Амнно-2,6-днстнрнлпнрн- <5000 98 0,174
днн 9-Амнно-5,6,7,8-тетрагндро- 80 000 95 0,616
акридин (10.-9) 5000 100 0,279
Феннлгуаннднн <5000 100 0,17
2-Антрнлгуаннднн (10.21) 40 000 100 0,385
Феннл-N 1-днгуаннд <5000 100 0,385
2-AHTpH.n-N i-днгу аннд 40 000 100 0,385
Данные для других соединений и бактерий см. Albert, Rubbo, Burvill (1949).
или двух колец из молекулы 9-аминоакридина (10.16), не обладают антибактериальной активностью, даже несмотря на то, что их молекулы по-прежнему полностью ионизированы. Это подтверждается данными табл. 10.10 для 9-аминоакридина (10.16) и его структурных аналогов 4-аминохинолина (10.17) и 4-аминопиридина (10.18). Отсутствие антибактериальных свойств у последних двух соединений связано с недостаточной величиной плоской поверхности их молекул [Albert et al., 1949J. Если очертить контуры их колец (как показано на рис. 10.5), то полученная площадь составит 2,8 нм2 для хинолина и 1,7 нм2 для пиридина, тогда как для акридина она равна 3,8 нм2.
100
Рис. 10.5. Минимальный прямоугольный контур, равноудаленный от атомов углерода и азота молекулы на 0,08 нм.
4-Амниохинолнн (<1 : 5000)
(10.17)
4-Аминопнридин «1 : 5000)
(10.18)
4-Амиио-2'Стирвлхниолин (1 : 80 000)
(10.20)
NH2 I
9-Амииоакрндии (1 : 160 000)
(10.16)
nh2
9-Амниотетрагндроакрндин (1 : 5000)
(10.19)
(1:160 000 и др. — минимальная бактериостатическая концентрация, как в табл. 10.10).
Все гетероциклы, о которых до сих пор шла речь, совершенно плоские, поскольку они представляют собой полностью сопряженные системы (т. е. в молекуле каждая вторая связь — двойная). Однако при гидрировании одного из колец 9-аминоакридина (10.16) в молекуле образовавшегося 9-аминотетрагидро-акридина (10.19) останется только 2,8 нм2 плоской поверхности, так как гидрированные кольца трехмерны. Такое уменьшение плоской поверхности сопровождается почти полной утратой антибактериальной активности.
Исследование связи между плоской структурой молекулы и антибактериальной активностью позволили сделать выводы о возможности создания высокоэффективных антибактериальных средств путем введения в молекулы хинолина и пиридина заместителей, увеличивающих площадь их плоской поверхности. По данным рентгеноструктурного анализа стильбен, в отличие
101
Рис. 10.6. Шаровые модели катионов 9-амииоакридииа (две проекции слева) и его 1,2,3,4-тетрагидропроизводиого (справа).
от дифенила [Karie, Brockway, 1944] имеет совершенно плоскую молекулу, поэтому возникло предположение, что введение стирильной группы в 4-аминохинолин с образованием соединения (10.20) может привести к получению чрезвычайно активного антибактериального препарата. Это было подтверждено экспериментально (см. табл. 10.10). Введение двух стирильных групп в молекулу 4-аминопиридина также привело к хорошим результатам [Albert et al., 1949].
Следует отметить, что все соединения от (10.16) до (10.20) полностью ионизированы при pH 7. Слабоионизированные аналоги соединения (10.20) не обладают антибактериальной активностью. В конденсированных системах, имеющих достаточно большую плоскую поверхность (например, в молекуле акридина), важным лимитирующим фактором является катионная ионизация; в менее крупных системах (таких как хинолин и пиридин) лимитирующим фактором служит минимальная величина плоской поверхности молекулы.
На рис. 10.6 с помощью пространственных моделей, точно воспроизводящих межатомные и ван-дер-ваальсовы расстояния, изображены катионы 9-аминоакридина (10.16) и его тетрагидропроизводного (10.19). Некопланарность тетрагидропроизводного особенно хорошо видна сбоку.
Почему так важен именно размер плоской поверхности этих молекул? Очевидно, что увеличение числа атомов в молекуле повышает ее способность адсорбироваться на рецепторе. Это 102
объясняется тем, что слабые ван-дер-ваальсовы силы, связывающие каждый атом агента с каждым атомом рецептора, в совокупности становятся действительно мощной силой, если во взаимодействии с обеих сторон участвует большое число атомов. Этой силе противостоит кинетическая энергия движения молекул, которая не увеличивается при возрастании размеров молекул. Ионные пары, образованные катионом (лекарственное вещество) и анионом рецептора, существовали бы очень недолго в отсутствие этих дополнительных связей (разд. 8.0). Для рассматриваемой серии веществ особенно важно копланарное расположение атомов лекарственного вещества. Это означает, что атомы биологической поверхности, с которыми они вступают во взаимодействие, также располагаются в одной плоскости.
Можно ли получить эффективные антибактериальные средства акридинового типа (т. е. воздействующие и на грамполо-жительные и на грамотрицательные бактерии при большом разбавлении без ослабления эффекта в присутствии белка) из всех молекул с достаточной степенью катионной ионизации и плоскостью поверхности? К подобным веществам относится метиленовый синий, имеющий плоскую молекулу и высокую степень ионизации. Однако его окислительно-восстановительный потенциал так высок, что катионы метиленового синего восстанавливаются продуктами метаболизма большинства бактерий, например Е. coli; метиленовый синий токсичен для родственного штамма Aerobacter aerogenes, не способного восстанавливать его катионы (продукт восстановления не ионизируется при pH 7). Однако число плоских гетероароматических соединений, способных образовывать сильно ионизированные аминопроизводные, очень ограничено, поскольку введение добавочного гетероатома приводит к резкому снижению основности [Albert, Goldacre, Phillips, 1948; Albert, 1968].
Это можно преодолеть, введя короткую высокоосновную боковую цепь в плоское ароматическое ядро с площадью поверхности 3,8 нм2 таким образом, чтобы боковая цепь и ароматический цикл находились в сопряжении. Такой боковой цепью может быть остаток гуанидина или дигуанида. В этом случае необязательно, чтобы цикл был гетероароматическим: пригоден ароматический углеводород подходящих размеров (например, антрацен). Осуществляя эту гипотезу на практике, мы обнаружили соединения, обладающие истинной антибактериальной активностью акридинового типа, не содержащие гетероцикла; примерами могут служить два производных антрацена, представленные в табл. 10.10 [Albert, Rubbo, Burvill, 1949].
Удивительно сходство биохимического действия аналогов акридина с действием самих акридинов. Так, например, этидиум и профлавин, ингибируют одни и те же ДНК-зависимые полимеразы, а именно синтезирующие как ДНК, так и (менее эффективно) РНК (в обоих случаях они соединяются с ДНК-матрицей) [Elliott, 1963; Waring, 1965].
103
Б. Механизм связывания акридинов и их аналогов с нуклеиновыми кислотами. В экспериментах in vitro было продемонстрировано, что связывание профлавина (10.7) (3,6-диаминоакри-дин) с ДНК идет по двум механизмам: во-первых, по реакции первого порядка, в которой равновесие достигается при соотношении 4 или 5 нуклеотидов на одну молекулу профлавина, и, во-вторых, в результате более медленной реакции высокого порядка, в которой одна молекула профлавина связывается с одной молекулой нуклеотида [Peacocke, Skerrett, 1956]. Второй процесс заключается в неупорядоченной адсорбции дополнительных молекул акридина на внешней стороне спирали ДНК.
2-Аитрилгуаиидин
(10.21)
2-Антрилдигуанид
(10.22)
Этидий (10.23)
В 1961 г. Lerman предположил, что молекулы 3,6-аминоак-ридина более прочно присоединяются к ДНК, благодаря интеркаляции между двумя слоями пар азотистых оснований, при этом первичные аминогруппы связываются ионной связью с двумя остатками фосфорной кислоты в спирали Уотсона — Крика, а плоский скелет акридинового цикла удерживается на молекулах пурина и пиримидина ван-дер-ваальсовыми силами [Lerman, 1964а]. На рис. 10.7 представлена эта структура (вид сбоку). Необходимость большой плоскости поверхности и высокой степени ионизации у молекул, обладающих антибактериальным действием, очевидно, и объясняется образованием этой структуры.
По данным рентгеноструктурного анализа слои азотистых оснований ДНК обычно связаны между собой сверху и снизу
104
Рис. 10.7. Схема вторичной структуры нормальной ДНК (слева) и ДНК, включающей интеркалироваиные молекулы профлавина (справа). Спираль рассматриваетси с отдаленной точки под таким углом, что видны лишь боковые проекции пар азотистых оснований и интеркалированиых молекул профлавина, а фосфорибозильный остов выглидит как регулярная спираль.
ван-дер-ваальсовыми силами. Расстояние между центрами атомов соседних пар оснований в цепи составляет 0,336 нм. Это означает, что молекулы аминоакридинов, имеющие точно такую же толщину, что и пуриновые и пиримидиновые основания, могут проникать в оставшиеся 0,336 нм (от общего расстояния 0,672 нм). Это пространство может образоваться за счет небольшого раскручивания двойной спирали (угол поворота варьируется для каждого вещества; так для этидиума он составляет 26° [Wang, 1974], для профлавина и алкалоида эллип-тицина он несколько меньше [Kohn et al., 1975]).
Теперь рассмотрим, как Lerman пришел к теории интеркаляции. При взаимодействии профлавина с ДНК происходит трехкратное увеличение вязкости. Это объясняли тем, что внедряющиеся молекулы не только вытягивают спираль, но и делают ее жесткой и спрямленной. Было установлено, что комплекс ДНК —профлавин имеет более низкий коэффициент седиментации, чем свободная ДНК- Это происходит за счет потери массы на единицу длины (ОММ профлавина составляет менее половины массы равного объема ДНК). Эти результаты были получены в разбавленном водном растворе. Было также обнаружено, что нити, вытягиваемые из комплекса, дают значительно более простые рентгенограммы, чем получаемые для чистой ДНК. Меридианальный рефлекс, соответствующий расстоянию 0,34 нм между соседними слоями, сохраняется, но новые положения первых экваториальных рефлексов свидетельствуют о том, что каждая молекула ДНК теперь имеет более плотную упаковку, а следовательно, и меньший диаметр, чем молекула чистой ДНК [Lerman, 1961]. В 1963 г. Lerman показал, что соотношение интенсивностей флуоресценции движущегося и неподвижного раствора согласуется с представлением о перпендикулярном расположении молекул акридина по отношению к оси спирали. Затем он обнаружил резкое падение скорости диазотирования первичных аминогрупп профлавина в присутствии ДНК- Это означало, что ДНК защищает аминогруппы от действия азотистой кислоты [Lerman, 1964]. Было замечено, что для денатурации ДНК после образования комплекса с 9-амино-акридином требуется более высокая температура [Lerman, 1964b] (ср. Chambron, ниже).
Теория интеркаляции была подтверждена данными, полученными другими методами. Так, радиоавтография ДНК (содер-
105
жащих [3Н]тимин), выделенной из Т2-колифага, проводилась до и после погружения в разбавленный раствор профлавина. При этом было показано, что аминоакридин удлиняет молекулу ДНК от 45 до 75 мкм [Cairns, 1962]. Столь же убедителен был и тот факт, что благодаря интеркаляции профлавина температура плавления (Тт) ДНК повышалась на 20°, а при «расплавлении» комплекса происходило внезапное выделение большей части связанного профлавина (т. е. нити двойной спирали разделялись) [Chambron, Daune, Sadron, 1966]. Впоследствии это явление было подтверждено Kleinwachter, Bakarova, Bohacek (1969).
Прямое доказательство правильности теории интеркаляции было получено при рентгеноструктурном изучении комплекса профлавин — ДНК., показавшем, что молекула аминоакридина располагается параллельно парам оснований ДНК в соотношении 1:3 [Neville, Davies, 1966]. Исследование растворов комплексов пяти моноаминоакридинов с ДНК методами линейного и кругового дихроизма подтвердило, что катионы акридина лежат в плоскостях, параллельных тем, в которых находятся пары оснований [Jackson, Mason, 1971]. И наконец, было обнаружено, что свободная энергия связывания аминоакридинов с ДНК близка энергии, характерной для процессов интеркаляции, но слишком высока для любого другого вида связывания на внешней стороне цепей ДНК [Jordan, 1968].
С точки зрения электростатических взаимодействий область ДНК, доступная для интеркаляции, несет высокий отрицательный заряд, образующийся не только за счет фосфат-анионов, пуриновых и пиримидиновых оснований (за исключением аденина) и частичных отрицательных зарядов атомов кислорода дезоксирибозы. Именно в это отрицательно заряженное пространство и втягиваются катионы лекарственного вещества, почти полностью нейтрализуя избыточный отрицательный заряд. Последующее образование ван-дер-ваальсовых связей с основаниями ограничивает дальнейшее продвижение молекулы и обеспечивает относительную устойчивость комплекса. На рис. 10.8 показана интеркаляция профлавина в молекулу ДНК (изображение построено по данным рентгеноструктурного анализа). Согласно расчетам, средняя площадь пары оснований в спирали ДНК или РНК составляет около 0,5 нм2, что превышает площадь молекулы акридина (0,385 нм2), рассчитанную методом, показанным на рис. 10.5. Однако в любой одноцепочечной модели небольшая часть каждой молекулы акридина будет выпячиваться наружу. Липофильные аналоги, такие как акридиновый оранжевый (3,6-бисдиметиламиноакридин), не только интеркалируют, но и прочно прикрепляются к внешней стороне ДНК.
В. Другие соединения, интеркалирующие в ДНК. Как это часто случается с новыми идеями, концепция интеркаляция поначалу не получила должного признания. Однако вскоре, и это 106
Рис. 10.8. Интеркаляция 3,6-диамииоакридииа между слоями пар оснований ДНК. Левое кольцо акридина противостоит почти точно над цитозином, а правое кольцо — почти точно над пиримидиновым циклом гуанина; область ван-дер-ваальсовых связей молекулы акридина обозначена сплошной линией [Ler-man, 1964b].
тоже не редкость, после ее признания очень многим лекарственным веществам самых разных типов стали приписывать именно этот механизм действия. Для проверки подобных утверждений был введен специальный тест на интеркаляцию, показывающий, вызывает ли исследуемое вещество локальное раскручивание двойной спирали суперспирализованной кольцевой ДНК, например ДНК колифага [Waring, 1970]. При первичном добавлении аминоакридина количество правозакрученных суперспиралей равномерно уменьшается; при достижении критического соотношения добавленных молекул суперспирали исчезают и ДНК превращается в раскрученное открытое кольцо, имеющее благодаря интеркаляции большой диаметр. По мере дальнейшего добавления лекарственного вещества нарастание напряжений в кольце ДНК снова приводит к появлению суперспиралей, но теперь уже левозакрученных. Так как суперспирали более компактны, чем раскрытые кольца, они быстрее осаждаются, что позволяет судить о ходе процесса по изменению коэффициента седиментации, проходящего через минимум [Waring, 1970].
Применение этого метода позволило установить, что механизм интеркаляции характерен для некоторых аминофенантри-динов, включая (10.23), для антималярийного препарата хин-гамин (10.31), трех карциностатических антибиотиков: ногало-мицина, даунорубицина и актиномицина D (4.38) (разд. 4.0). Однако для хлорпромазина и диэтиламида лизергиновой кислоты (ЛСД) такого подтверждения получено не было, хотя ранее им приписывали интеркаляционный механизм действия. Было обнаружено, что спермин, стрептомицин, диминазен и митрами-цин активно взаимодействуют с ДНК, но не образуют интерка-лятов [Waring, 1970].
107
Рассмотрим другие, не менее важные примеры интеркаляции. Парафуксин (10.5), краситель трифенилметанового ряда, интеркалирует в ДНК, однако не так плотно, как профлавин [Armstrong, Panzer, 1972]. Антибиотик эхиномицин содержит полипептидную цепь, с которой связаны два хиноксалиновых цикла, далеко отстоящие друг от друга. Эти циклы одновременно интеркалируют в разные участки ДНК. Происходящее вследствие этого раскручивание и удлинение спирали в два раза превышает то, которое вызывается действием аминоакридинов [Waring, Wakelin, 1974].
Путем введения акридиновых циклов в молекулы путресцина (11.6) и спермина (11.4) были получены синтетические аналоги, способные ингибировать ДНК-зависимую РНК-полимера-зу и повышать температуру плавления ДНК сильнее, чем сами аминоакридины [Canellakis et al., 1976]. Противораковое действие соединений, образующих двойные интеркаляты, рассматривается в разд. 10.3.4.
Биологические свойства лекарственных веществ, действие которых основано на интеркаляции, обсуждаются в обзоре [Schwartz, 1979].
Почему же аминоакридины и соединения с подобным механизмом действия обладают такой высокой избирательностью действия по отношению к бактериям и практически не действуют на клетки млекопитающих? Установлено, что для серии аминоакридинов характерно наличие антибактериальных свойств, высокая степень ионизации и интеркаляционный механизм действия [Jackson, Mason, 1971]. Избирательность их действия обусловлена, по-видимому, двумя факторами: доступностью одиночных хромосом бактерий (разд. 5.3), а также тем, что эти лекарственные вещества действуют преимущественно на кольцевые ДНК [Waring, 1970]. Кроме избирательного действия на бактерии, аминоакридины действуют избирательно и на другие кольцевые ДНК: они способны отщеплять бактериальные плазмиды [Bouanchaud et al., 1968], а также вызывать наследственные изменения у дрожжей путем подавления репликации ДНК в митохондриях, не затрагивая при этом ядра (разд. 10.3.1) [Hollenberg, Borst, van Bruggen, 1970].
Далее, аминоакридины и аминофенантридины могут или подавлять кинетопласт в трипаносомах или (при более низких концентрациях) разрушать его кольцевую ДНК [Riou, Delain, 1969].
Рассчитано, что, поскольку эти лекарственные вещества уменьшают суперспирализацию кольцевой ДНК, выделение свободной энергии повышает сродство к агенту, в то время как аналогичная однонитевая ДНК имеет пониженное сродство [Bauer, Vinograd, 1970]. По-видимому, этот случай можно отнести к проявлению избирательности за счет накопления (гл. 3).
108
10.3.3. Катионные и анионные антибактериальные средства с другим типом действия
Многие антибактериальные средства действуют также в форме катионов, но механизм их действия отличается от акридинового, для которого характерно следующее: а) бактериостатическое действие на многие разновидности и грамположительных, и грамотрицательных бактерий (чи при большом разбавлении) и б) неизменность антибактериального действия в присутствии сывороточных белков.
Трифенилметановые красители, например, парафуксин (10.5), кристаллический фиолетовый, бриллиантовый зеленый и малахитовый зеленый представляет собой катионные антибактериальные средства особого химического типа (разд. 10.2). Равновесие между катионом и его основанием (в действительности — псевдооснованием) устанавливается очень медленно. Поэтому следует учитывать и константу равновесия, и константу ионизации, которые были определены Goldacre и Phillips (1949). Из табл. 2.6 (том 1) видно, что антибактериальное действие соединений этого ряда непосредственно связано со степенью ионизации. Однако в отличие от аминоакридинов они малоактивны по отношению к грамотрицательным бактериям, а их действие на грамполжительные бактерии сильно подавляется сывороточным белком [Goldacre, Phillips, 1949]. Трифенилметановые псевдооснования жирорастворимы и легко проникают в клетки. Молекулы этих псевдооснований не имеют жесткой структуры, тогда как их катионы жесткие и совершенно плоские. Эти катионы легко образуют ковалентные связи с нуклеофильными реагентами любого типа.
Алифатические амины (включая и четвертичные аммониевые основания) также обладают антибактериальным действием, совершенно отличным от действия акридинов. Они вызывают лизис клеточной мембраны (разд. 14.3) и поэтому бактерицидное действие достигается быстро, тогда как акридины действуют медленно и применяются в основном как бактериостатические средства. Алифатические амины, молекулы которых содержат боковые цепи с двенадцатью и более атомами углерода, обладают сильным литическим действием. Молекулы таких соединений нежесткие и легко адсорбируются кислотными группами сывороточных белков, эту адсорбцию можно легко снизить, превращая амины в четвертичные производные, содержащие по меньшей мере один объемный заместитель, например бензильную группу, в бензалконийхлорид. Хорошо известен такой представитель класса четвертичных аминов, как бромид цетилтриметиламмония. В этой серии соединений также отмечается повышение активности при возрастании pH, хотя лекарственное вещество остается постоянно ионизированным [Blu-baugh, Botts, Gerwe, 1940; Gershenfeld, Molanick, 1941].
Соли и эфиры некоторых алифатических сульфокислот, со
юз
держащие цепи с двенадцатью и более атомами углерода (например тетрадецилсульфат), обладают умеренной антибактериальной активностью. Они действуют в виде анионов [Gershen-feld, Milanick, 1941; Putnam, Neurath, 1944].
Глюконат хлоргексидина (10.24) [1,6-бис (N-n-хлорферил-дигуанидогексан] применяют для дезинфекции кожи и слизистых. Этот препарат высокоэффективен при больших разведениях по отношению к грамполбжительным и грамотрицатель-ным организмам и сохраняет'бактерицидные свойства в крови. Подробнее см. разд. 14.3.
NH NH
/=\ II II /=\
С1—(. NH—С—NH—С—NH—(CH2)e—NH— С—NH—С—NH—< С1
и и
NH NH
Хлорогексидин
(10.24)
10.3.4. Противораковые препараты
Аминакрин (9-аминоакридин) обладает, хотя и слабым, избирательным действием на опухолевые клетки. У 9-анилиноак-ридина это действие усиливается, хотя катионная ионизация при pH 7 сохраняется. Исходя из этого, группа новозеландских исследователей во главе с В. Cain в результате долгой работы синтезировала широко применяемый в клинике препарат амса-крин (10.25), 4-(9-акридиниламино)метансульфон-мета-анизи-дид. Он эффективен при лейкозе и обладает высоким терапевтическим индексом [Legho et al., 1979].
Вместо присутствующих в молекуле амсакрина заместителей можно вводить и целый ряд других групп. Варьирование заместителей не приводит к потере антилейкозной активности до тех пор, пока сохраняется катионная ионизация и определенная невысокая липофильность, что подтверждается при измерении коэффициента распределения [Denny, Atwell, Cain, 1979]. Установление количественной зависимости между силой связывания с ДНК и эффективной дозой лекарственного вещества (т. е. дозой, необходимой для удлинения на 40% времени жизни мышей с лейкозом L1210) показало, что местом действия соединений этого ряда является ДНК [Baguley et al., 1981]. Силу связывания с ДНК определяли флуориметрически по количеству этидиума (10.23), вытесняемого из вилочковой железы теленка.
Амсакрин действует на ДНК довольно быстро (разрывая ее цепи, что приводит к изменениям в расщеплении хромосом при митозе и последующей гибели клетки [Baguley et al., 1981]. Сильное воздействие амсакрина на костный мозг препятствует его использованию для лечения солидных опухолей. Baguley и сотр. обнаружили, что введение двух заместителей в поло-110
жения 5 и 6 акридинового цикла подавляет этот эффект, не вызывая уменьшения цитотоксического действия.
Наличие сопряжения между акридиновым и анилиновым циклами амсакрина впоследствии было подтверждено данными рентгеноструктурного анализа, свидетельствующими о том, что атом углерода С-9 связан с анилиновым атомом азота частично двойной связью. Акридиновый цикл интеркалирует между слоями оснований ДНК, а анилиновое кольцо располагается в малой бороздке [Baguley et al., 1981; Waring, 1976].
Амсакрия
(10.25)
NO2 NH(CH2)s—NMe2
Нитракрин
(10.26)
Нитракрин (10.26), созданный группой польских исследователей во главе с Ledochowski, не оказывает действия на костный мозг. Применение этого препарата в клинике при послеоперационном лечении рака толстой кишки показало, что он обладает сильным карциностатическим действием. У тридцати больных, принимавших нитракрин, в течение пяти лет не наблюдали рецидивов [Bratcowska-Seniow et al., 1976]. Этот лекарственный препарат имеет рКа 6,2 (ядро) и 9,7 (боковая цепь), ср. 7,9 и 10,5 для мепакрина. Полагают, что действие нитракрина осуществляется посредством восстановления нитрогруппы в гидроксиламинную, которая сшивает ДНК [Konopa, Koldej, Pawlak, 1976; Ledochowski, 1976]. Единственным недостатком этого препарата, предположительно имеющего высокий терапевтический индекс, является раздражение, появляющееся на месте инъекции.
Подробные сведения об использовании акридинов для лечения рака можно найти в работе Denny и сотр. (1983).
Эллиптицин — алкалоид, выделяемый из тропического вечнозеленого дерева, представляет собой 5,11-диметилпир идо-14, 3-Ь]-карбазол [Woodward, lacobucci, Hochstein, 1959]. Это соединение имеет планарную циклическую структуру и способно интеркалировать в ДНК- Эллиптицин проходит клинические испытания на больных лейкозом. Эхиномицин, химическая структура которого обсуждалась в разд. 10.3.2, проявляет высокую эффективность при лимфоцитарном лейкозе и меланоме у мышей, но не оказывает заметного действия при других формах рака [Carter, Sakurai, Umezawa, 1981]. Молекула антибиотика карцинофилина А содержит сложную бициклическую систему, образованную с помощью эфирной и сложноэфирной
111
связей, основного атома азота и двух нафталиновых циклов. Последние интеркалируют в ДНК, а остальная часть молекулы занимает малую бороздку ДНК- Кроме того, содержащиеся в молекуле карцинофилина А азиридиновые циклы алкилируют гуаниновые группы ДНК после того, как антибиотик занимает окончательное положение. Этот лекарственный препарат применяют для лечения рака в Японии с начала 60-х годов, однако его использование ограничено сильным токсическим действием на нормальные клетки'ДЬохуп, Hanstock, 1982].
М,Ь1'-(9,9'-диакридинил)-1,6-диаминогексан относится к акридинам, способным интеркалировать в ДНК двумя фрагментами (разд. 10.3.2); в настоящее время он проходит клинические испытания. Соединения этого типа так прочно интеркалируют в ДНК, что свободное вещество не удается обнаружить [Kuhlmann, Mosher, 1981].
Доксорубицин (4.27) и родственный ему даунорубицин широко применяют в клинике для лечения солидных опухолей и лейкозов соответственно. Они являются сильными интеркалято-рами и действуют в форме катионов благодаря наличию аминогруппы, связанной с углеводородной частью молекулы. Однако механизм действия этих соединений до конца не изучен (разд. 4.0.4). Обзор Schwartz (1979) посвящен противоопухолевому действию веществ, способных интеркалировать в ДНК-
10.3.5. Препараты для лечения протозойных инфекций
Многие эффективные биоцидные средства, обладающие избирательным действием на простейших, представляют собой соли сильных органических оснований, а один из трипаноцид-ных препаратов является солью сильной органической кислоты.
А. Жгутиковые. Потребление огромного количества пищи трипаносомами, необходимое для их постоянного движения, всегда обращало на себя внимание исследователей (разд. 4.9). Именно на эту «ахиллесову пяту» и воздействуют трипаноцид-ные препараты, содержащие мышьяк, особенно меласопрол (13.3). Однако эти препараты применяют в основном на поздних стадиях болезни. Другой путь для избирательного воздействия рассматривался в разд. 10.3.2, а именно способность аминоакридинов и аминофенантридинов изменять и, в конечном счете, подавлять кинетопласт этих жгутиковых путем соединения с их кольцевой ДНК. Кинетопласт несет информацию для создания митохондрий в альтернативной форме паразита, существующей в насекомых-переносчиках (мухах). Эти лекарственные препараты, воздействуя на кинетопласт, эффективно препятствуют передаче его вторичному организму-хозяину, прекращая таким образом распространение эпидемии, но не оказывают помощи зараженному больному. Лечебное действие катионных лекарственных средств определяется их способностью либо воздействовать на ядрышки, рибосомы и лизосомы,
112
либо подавлять избыточный углеводородный метаболизм паразитов [Williamson, Macadam, Dixon, 1975].
При американском трипаносомозе эффективно воздействующее на Т. cruzi соединение нифуртимокс (6.20), которое не ионизируется. Для профилактики и лечения африканской сонной болезни используют сурамин (6.8), анионное лекарственное вещество и пентамидин (10.27), являющийся катионом. Ароматические амидины — это сильные. основания (рКа 11); хинапи-рамин и этидиум — четвертичные аммониевые основания, находящиеся в равновесии с псевдооенованиями (разд. 10.2). Крупный рогатый скот лечат диминазеном (10.28), этидиумом (10.23) и хинапирамином (10.29); эти лекарственные препараты действуют также на разновидности простейших, заражающих лошадей и верблюдов. Биологическая природа трипаносомоза рассматривалась в разд. 1.1. Все известные трипаноцид-ные лекарственные средства вызывают у больных токсические побочные эффекты.
нж __ ,nh2
.С—" ,,—О(СН2)6О—" ,,—с
Hn/ V_7 Ч-NH
Пентамидин
(10.27)
H2N. -- ,NH
)-NH-N=N-( )-cf
NH^ Y/ \nh2
Хииапирамии (дикатиои)
(10.29)
Поскольку патогенные трипаносомы трудно культивировать in vitro, большинство опытов было поставлено на родственном им простейшем Crithidia oncopelti (известном также под названием Strigomonas), который легко выращивать. Впоследствии эти трудности удалось преодолеть, и в настоящее время опыты проводятся на высокопатогенных трипаносомах (Тг. rhodesiense). Исследовали in vitro действие лекарственного вещества на: а) потерю подвижности трипаносом как показатель нарушения энергетического метаболизма и б) заражаемость мышей как показатель нарушения процесса деления клеток. Третий тест — лечение зараженных штаммов Тг. rhodesiense мышей исследуемыми лекарственными препаратами. Было уста
8—734
113
новлено, что сурамин (6.8) сначала повреждает структуру рибосом, а затем подавляет деление клетки. Этидиум (10.23) и хинапирамин (10.29) поражают кинетопласт и подавляют деление клеток. Пентамидин (10.27) в основном подавляет гликолиз, но также повреждает кинетопласт (ДНК) и ядрышки (РНК) [Williamson, Macadam, Dixon, 1975].
Пентамидин быстро поглощается трипаносомами и сразу же проявляет трипаностатическое действие как in vitro, так и in vivo [Hawking, 1944]. Через несколько дней паразиты погибают под действием защитных сил организма. Данные о распределении этих и других аминов в трипаносомах можно найти в работе Williamson, Macadam (1965). Излечению способствует еще один косвенный фактор. Паразиты, выжившие после непосредственного воздействия лекарственного вещества, сохраняют инфек-ционность, но при этом они содержат множество гранул с лекарственным веществом. Когда некоторые из них погибают, высвобождается лекарственное вещество, а также антиген, стимулирующий выработку адекватного количества антител организмом-хозяином. Такой опосредованный процесс объясняет замедленное развитие лечебного действия диамидиновых лекарственных средств и их пролонгированный профилактический эффект JFulton, Grant, 1965]. Если блокировать иммунную реакцию у крысы удалением селезенки или инъекцией меди, то трипаносомоз поддается лечению стильбамидином, сурамином и хинапи-рамином [Ormerod, 1961]. В настоящее время жителям тропических областей Африки с целью профилактики трипаносомоза делают инъекции пентамидина или сурамина через каждые 3 мес.
Ни пентамидин, ни диминазен не интеркалируют [Waring, 1970], но оба они способны связываться в малой бороздке кольцевых спиралей ДНК кинетопластов, причем сшивка осуществляется путем присоединения каждой амидиновой группы к фосфатному аниону [Festy, Sturm, Daune, 1975]. Еще более ражную роль при лечении играет их способность избирательно тормозить метаболизм углеводов посредством ингибирования а-глицерофосфатдегидрогеназы, даже при разбавлении 2 мкМ fFairlamb, Bowman, 1975], благодаря противодействию коферментному эффекту спермидина (11.5) [Bacchi, 1981].
Алифатические полиамины, так же как и диамидины, относятся к соединениям, действующим на ДНК- Как показали результаты рентгеноструктурного анализа, спермин, широко распространенный в природе [Cohen, 1971], соединяется с ДНК, стягивая двойную спираль по малой бороздке и образуя ионные связи с фосфатными анионами [Suwalsky et al., 1969]. Расстояние между фосфатными группами составляет 1,0 нм в малой бороздке и 2,0 нм в большой. Исключительная прочность ронных связей, образуемых амидинами, обсуждалась в разд. 8.4.
Кроме трипаносомоза, диамидины применяют для лечения других протозойных болезней крупного рогатого скота, особенно группы «пироплазмозов», вызываемых простейшими видами 114
Babesia и Theileria. Помимо пентамидина и диминазена хорошие результаты получены при использовании амикарбалида [Ь1,М'-ди-(3-амидинофенил) мочевина] и родственного ему ими-докарба. Диамидины также успешно применяют для лечения лейшманиозов человека, которые не поддаются действию препаратов сурьмы.
Возвращаясь к трипаносомозу и широко используемому анионному трипаноцидному препарату сурамину (6.8), следует отметить, что особенность структуры этого лекарственного вещества заключается в том, что удаление двух метильных групп приводит к полной потере его активности. Тем не менее, полианионы совершенно иной природы, например декстрансульфат, обладают сильным трипаноцидным действием, хотя и недостаточно широко используются в медицине. Уже давно известно, что сурамин подавляет инфекционность трипаносом, не влияя при этом на их подвижность [Roehl, 1926Ь]. С помощью электронного микроскопа удалось установить, что это лекарственное вещество избирательно поражает рибосомы паразитов, утрачивающие вследствие этого полисомальный характер и образующие «цитоплазматические гранулы», строение которых было описано ранее. По-видимому, атака в основном направлена на РНК-полимеразу; ядра, ядрышки и кинетопласт остаются незатронутыми, и синтез ДНК продолжается [Macadam, Williamson, 1974].
При онхоцеркозе после ослабления диэтилкарбамазином (6.35, б) микрофилярий макрофилярии легко поражаются су-рамином.
Б. Споровики. В сравнении со жгутиковыми эти простейшие ведут более спокойное существование, однако имеют более сложный жизненный цикл. Род Plasmodium вызывает различные виды малярии.
Акридины приобретают высокую антималярийную активность при введении в их молекулу в положение 9 боковой цепи основного характера. Акридины, содержащие четвертичный атом азота, неактивны. Акрихин (10.30) был первым акридиновым противомалярийным средством, получившим широкое распространение. Более подробно о зависимости между строением и действием акридиновых противомалярийных препаратов см. Albert (1966).
Акрихин
(10.30)
Me—СН—(CH2)3NEt2
NH
Cl N
Хингамин
(10.31)
8*
115
Акрихин, применявшийся с 1933 г., не получил такого широкого распространения, как значительно менее активный хинин, из-за побочного эффекта — способности придавать коже желтую окраску. В начале 1940-х годов был получен более простой бесцветный аналог акрихина — хингамин (10.31) (7-хлоро-4-о-диэтиламино-а-метилбутиламинохинолин). В настоящее время его широко применяют при лечении малярии, хотя и в более высоких дозах, чем акрихин, который до сих пор выпускается промышленностью. Эти противомалярийные препараты не обладают трипаноцидным действием, а трипаноциды не проявляют противомалярийной активности.
В то время как противомалярийные препараты, нарушающие метаболизм фолиевой кислоты (пириметамин, сульфадиазин, бигумаль см. разд. 9.3.3) поражают шизонтов только на стадии их деления, лекарственные вещества, соединяющиеся с нуклеиновыми кислотами (особенно акрихин, хингамин), и хинин уничтожают шизонтов на любой стадии [Josephson et al., 1953] при условии, что эти препараты попадают в эритроциты. Благодаря высокой избирательности, акрихин и хингамин в дозах, эффективно поражающих шизонтов, находящихся в эритроцитах (трофозоитов), не приносят вреда организмам-хозяевам (млекопитающим и птицам). К сожалению, они не действуют на шизонтов, находящихся вне эритроцитов или на спорогенных стадиях. Причина этих различий, по-видимому, заключается в том, что эритроциты, содержащие паразитов, концентрируют эти препараты в тысячу раз больше. Так при лечении хингамином в крови больного находится только 10~6 этого препарата, но в 10~3 М больше внутри эритроцитов [ср. Macomber, O’Brien, Hahn, 1966].
Принято считать, что противомалярийное действие акрихина и хингамина в основном обусловлено их способностью соединяться с ДНК паразита. Плоский ароматический цикл (в обоих случаях) интеркалирует, и основная группа образует ионную связь с фосфатной группой ДНК [O’Brien, Olenick, Hahn, 1966; Waring, 1970]. Lerman (1963) показал, что интеркаляция акрихина происходит в ДНК так же легко, как и у простых акридинов (разд. 10.3.2). Об аналогичной интеркаляции хингамина свидетельствует понижение скорости седиментации, увеличение вязкости и повышение температуры плавления ДНК- Хингамин плохо реагирует с одноцепочечной ДНК [Allison, O’Brien, Hahn, 1965]. Он ингибирует бактериальную ДНК-полимеразу, соединяясь с ДНК-стартером [Cohen, Yielding, 1965]. С ДНК Plasmodia эксперименты не проводились. Эти три противомалярийных средства ингибируют включение фосфата (32Р) в ДНК и РНК Pl. gallinaceum у цыплят при концентрации 10~5 М и в Pl. berghei, паразитирующих в крови [Schellenberg, Coatney, 1961]. Следует отметить, что 8-амино-хинолин, обладающий гаметоцидным действием, не ингибирует включение фосфора.
116
Боковая цепь акрихина и хингамина, содержащая два атома азота, разделенных четырьмя атомами углерода, напоминает путресцин (1,4-диаминобутан), который вырабатывается орнитиндекарбоксилазой малярийных паразитов.
CF3
Мефлокин (10.34)
Хингамин обладает сильным ингибирующим действием на этот фермент, декарбоксилирующий орнитин (10.32). Это свойство может быть важным для терапевтического действия [Кб-nigk, Putfarken, 1983J. Вопросам химии полиаминов посвящены работы Cohen (1971) и Taber, Taber (1983).
Боковая цепь, являющаяся важной частью молекул акрихина и хингамина, связывается с двумя смежными (по вертикали) фосфатными анионами одной и той же цепи ДНК. Более простые диамины, например этилендиамин, прочно связываются (и придают жесткость) с одноцепочечными полинуклеотидами, а также со спиралью ДНК. Длины дикатиона этилендиамина достаточно для связывания соседних анионов (расстояние между ними 0,7 нм), но мало для связывания двух цепей двойной спирали ДНК через малую бороздку (1,0 нм) [Gabbay, 1968J. Расстояние между атомами азота боковой цепи хингамина по данным рентгеноструктурного анализа 0,554 нм [Preston, Steward, 1970]; расстояние в молекуле акрихина лишь слегка отличается.
Ионизационные свойства боковой цепи акрихина также связаны с его активностью. рКа атома азота на свободном конце боковой цепи составляет 10,48, а на фиксированном конце — только 7,9 (при 20°, ср. 10,1 и 7,0 для этилендиамина). Оба конца боковой цепи ионизированы при pH 7 (см. табл. 10.3). При любом укорачивании боковой цепи более низкое значение рКа значительно понижается, в результате чего и степень ионизации и эффективность противомалярийного действия резко уменьшаются [Albert, 1966J. Такое понижение обусловлено ку
117
лоновским эффектом (сила отталкивания между одноименными зарядами изменяется обратно пропорционально квадрату расстояния).
Хинин (10.33) до сих пор применяют для лечения разных видов малярии, устойчивых к действию хингамина. Хинин очень, незначительно интеркалирует в ДНК, механизм его действия на паразитов до сих пор не установлен, но он отличен от механизма действия хингамина [Estensen, Кгеу, Hahn, 1969J. Хинин необходимо принимать большими дозами, так как большая его часть окисляется (в положении 2) печенью больного, а продукт реакции не обладает противомалярийным действием. Этот дефект удалось устранить путем введения в положение 2 заместителей, не поддающихся окислению. Особенно удачным препаратом такого типа оказался мефлохин (10.34); однократный прием этого препарата (в дозе 0,5 г) может привести к клиническому излечению больного, жизни которого угрожала заболевание, вызванное Plasmodium falciparum, устойчивыми к действию хингамина. Это лекарственное вещество не взаимодействует с ДНК [Davilson et al., 1977].
Период полураспада мефлохина в человеческом организме в среднем 15 дней. В противоположность хинину мефлохин имеет высокий терапевтический индекс. Интенсивные клинические исследования показали, что мефлохин безопасен и эффективен при лечении малярии, вызванной и falciparum и vivax, несмотря на то, что они устойчивы к действию хингамина. Поскольку слишком беспорядочное употребление мефлохина может привести к вырабатыванию устойчивости к этому препарату, мефлохин успешно применяют для лечения видов малярии, вызванных устойчивыми к хингамину штаммами Plasmodium. В профилактических целях его назначают в комбинации с парой лекарственных препаратов пириметамин — сульфадоксин для того, чтобы понизить вероятность развития устойчивости к мефло-хину (в соответствии с принципами, изложенными в разд. 6.5). Подробнее о клиническом использовании мефлохина см. Schmidt и сотр. (1978); полные сведения о мефлохине можно найти в материалах WHO (1983).
При переходе от хинина (10.33) к мефлохину (10.34) происходит заметное упрощение фармакологически активной группы —О—С—С—N—, свойства которой уже обсуждались в разд. 7.2; однако возможны и дальнейшие упрощения в соответствии с принципами, изложенными в разд. 7.3. Ученые, создавшие препарат мефлохин, провели исследования с целью упрощения боковой цепи и ядра молекулы препарата с сохранением его противомалярийной активности. Размыкание пиперидинового цикла с образованием диалкиламинометильной группы привело к созданию такого препарата, как WR 30090, представляющего собой хинолиновый аналог соединения (10.36) и обладающего высокой противомалярийной активностью (что подтвердили клинические испытания).
118
WR 33063
(10.35)
CH(OH)CH2NBu2
Нафталиновый аналог хинина
(10.36)
Наличие подобных боковых цепей придает и негетероциклическим соединениям противомалярийные свойства. Хорошие результаты были получены при лабораторных и полевых испытаниях против инфекций, вызванных Pl. falciparum, таких фенантреновых препаратов, как WR 33063 (10.35) [Canfield, Rozman, 1974] и WR 122455, 3,6-бис (трифторметил-а, 2-пиперидил-9-фе-нантренметанол) [Rinehart, Arnold, Canfield, 1976]. Хинолин также можно заменить нафталином, так как 3-(дибутиламино (-5,7-дихлор-1 -нафтил-3-(пара-хлорфенил)пропан-1-ол (10.36) высокоактивен по отношению к Pl. berghei у мышей и терапевтически эквивалентен хинолиновому препарату с аналогичными заместителями [Shamblee, Gillespie, 1979].
10.3.6. Фармакодинамические средства
АХ (7.4), важнейший медиатор, передающий нервный импульс, представляет собой алифатический четвертичный амин и поэтому он полностью ионизирован при любых условиях. Активная форма никотина (7.26), имитирующая действие АХ на некоторых участках, является катионом, так как его йодмети-лат (кватернизированный атом азота пирролидинового цикла) не менее активен [Barlow, Hamilton, 1962]. Катион ареколина (12.80) также обладает сильными АХ-подобными свойствами (разд. 12.6). В экспериментах на подвздошной кишке морских свинок (pH 6,0—9,36) активность нейтральных молекул ареколина составляла 2% активности катионов [Burgen, 1965].
Подавляющее большинство алкалоидов, местных анестетиков, барбитуратов и нейролептиков имеет величины рКа в пределах от 6 до 8, поэтому при физиологических значениях pH ионы и нейтральные молекулы находятся в равновесии. Эти вещества будут рассмотрены в разд. 10.4 и 10.5. Из всех алкалоидов выделяются высокой степенью ионизации атропин (7.16) (рКа Ю) и тубокурарин (2.6). Тубокурарин — четвертичный амин и поэтому полностью ионизирован при всех значениях pH. В медицине применяют и другие четвертичные амины: карбахол (2.11), неостигмин (7.34), соли декаметония (7.28) и суксаметония (7.29) и галламин (7.31).
119
Антигистаминные препараты и антидепрессанты обычно имеют величину рКа около 9. Ионизация норадреналина и его N-алкильных гомологов рассматривается в разд. 12.4.
Хотя многие химиотерапевтические лекарственные средства действуют в виде анионов, среди фармакодинамических средств-таких немного. Однако бутадион (10.37, а), применяемый при лечении подагры (он резко снижает количество мочевой кислоты в крови), действует в форме аниона. Величина рКа бутадиена составляет 4,4, а pH мочи — 4,8 или выше, поэтому концентрация аниона в мочевом тракте и в моче ниже оптимальной. Этот недостаток был устранен в препарате сульфинпира-зоне (10.37,6), имеющем более низкое значение рКа (2,8), что обусловливает повышение концентрации аниона в моче и, следовательно, более высокий терапевтический индекс [Burns et al., 1960] (изменение рКа обусловлено образованием водородных связей). Эти лекарственные вещества блокируют всасывание мочевой кислоты в цилиндрических клетках почечного эпителия.
свн5
ВДл N ОН
О к
а) Бутадион, R —— (СН2)з—Me
б) Сульфинпиразон, R=—(СН2)2—SO—СбНв
(10.37)
10.4 . Вещества, менее активные в ионизированном состоянии
Неионизированные вещества могут обладать очень сильным физиологическим действием. Это относится, например, к таким соединениям, как эфир, хлороформ, и к мощным лакриматорам— бромацетону и а-хлорацетофенону. В 1921 г. Vermast обнаружил, что многие слабые кислоты наиболее полно проявляют биологическую активность при тех значениях pH, при которых они наименее ионизированы. В этом можно убедиться на примере ингибирования деления клеток яиц иглокожих салициловой кислотой [Н. Smith, 1925]. Из рис. 10.9 можно видеть, что эта кислота наиболее активна при pH 5. В этом случае количество недиссоциированной салициловой кислоты (рКа 3,0) составляет 0,99%, что значительно больше, чем при любом более высоком pH (разд. 17.0). Самое простое объяснение заключается в том, что нейтральные молекулы, а не анионы, салициловой кислоты ингибируют деление клеток. Можно ожидать, что при более низких значениях pH салициловая кислота будет более сильным ингибитором, так как в этих услови-120
Рис. 10.9. Влияние pH иа суммарную концентрацию иоиов и иеионизированиых молекул салициловой кислоты, необходимой для роста Echinarachnius parva [Smith, 1925].
ях отношение концентрации неионизирован-ных молекул к концентрации ионов повышается. Однако при попытках снизить pH воз- * никает ряд трудностей: во-первых, организм | может проявлять чувствительность к измене- | нию pH, во-вторых, снижение pH может при- § вести к изменению ионизации рецептора и со-ответственно изменению сродства к этому | агенту (разд. 10.6). I
Все представители ряда 30 барбитуратов g проникают в яйца и личинки морского ежа Arbacia только в форме неионизированных молекул [Clowes, Keltch, Krahl, 1940]. Более того, оказалось, что ингибирование деления клеток и дыхания обусловлено именно ими. Известно также, что неионизированный теофиллин (в отличие от аниона) стимулирует деятельность сердца черепахи [Hardman, 1962]. -I
В настоящее время вызывает беспокойство производные азотистой кислоты, попадающие в
2
1_1__I___I___I
5 6 7 8
PH
тот факт, что организм чело-
3
века с табачным дымом и консервированным мясом, возможно, реагируют в желудке с вторичными аминами, образуя нитрозоамины, обладающие канцерогенными свойствами (см. разд. 13.5 о канцерогенезе). В связи с этим следует отметить, что с азотистой кислотой быстрее реагируют неионизированные вторичные амины. Поэтому амины, слабо ионизированные при pH 2, должны быть наиболее чувствительны к действию азотистой кислоты [Sander, Schweinsberg, Menz, 1968]. По-видимо-
му, такие малоосновные амины редко встречаются в содержимом желудка.
Изучая действие слабых кислот на биологические объекты, нетрудно заметить, что количество вещества, необходимое для достижения эффекта, остается постоянным независимо от pH среды при условии, что pH по меньшей мере на одну единицу ниже, чем рКа, что обеспечивает отсутствие ионизации токсичного агента. Этот вывод подтверждается графиками (рис. 10.10), наглядно свидетельствующими о влиянии pH на концентрацию фенола и уксусной кислоты, необходимой для прекращения роста различных плесневых грибов. В изученных пределах pH (от 2 до 6) количество фенола остается постоянным, а количество уксусной кислоты уменьшается при снижении pH. Это объясняется тем, что рКа фенола 9,9 и, следовательно, в указанных пределах pH он не ионизирован, тогда как рКа уксус-
121
1931] (табл. 10.11). При
Рис. 10.10. Влияние pH иа концентрацию фенола (рКаЭ.9) и уксусной кислоты (рКа4,8), необходимые для ингибирования роста плесневых грибов [Hoffman et al., 1940, 19411.
ной кислоты 4,8 и поэтому при pH 5,8 она ионизирована на 90%, а при pH 3,8 — только на 9% и т. д.
Все подобные эксперименты не дадут удовлетворительного результата, если не будет подобран контроль, позволяющий выяснить, не влияют ли изменения pH на подопытный организм. Это было сделано при исследовании наркотизирующего действия разных веществ на червя Arenicola [Clowes, Keltch, этом отмечалось, что действие неэлект
ролитов, например хлороформа, не зависит от pH. Опыты с этими
неионизирующимися веществами показывают, что изменение pH не влияет на организм червей. Слабые основания, такие как кокаин, действуют иначе. Их эффективность возрастает с повышением pH, т. е. пропорционально подавлению их ионизации. Аналогичным образом эффективность слабых кислот (четыре изомерных барбитурата) повышается при уменьшении pH, что также связано с подавлением ионизации. В любом из этих случаев наркотическое действие снимается отмыванием червей морской водой. Проведение исследований с червем Arenicola значительно упрощается потому, что изменение pH не влияет на этот организм. Менее удачными оказались опыты по изучению действия производных хинина на бактерии. Авторы,
проводившие эти эксперименты, полагали, что им удалось по-
казать, что неионизированные молекулы активнее, чем ионы, так как эффективность этих лекарственных веществ возрастала при повышении щелочности среды [Z. Immunitats, 1922, 34, 194]. К сожалению они не учли ионизирующего действия щелочи на рецепторы бактерий.
10.5 . Вещества, проявляющие биологическое действие в виде ионов и неионизированных молекул
Вполне вероятно, что многие вещества, особенно со значениями рКа в пределах от 6 до 8, проникнув в клетки в виде неионизированных молекул, проявляют свое биологическое действие, как ионы (разд. 10.2). Так, на рис. 10.11 показано, что поглощение бензойной кислоты дрожжами обратно пропорционально степени ионизации.
Известно много веществ, у которых биологическая активность определяется в основном долей присутствующих неиони-122
Таблица 10.11. Зависимость между степенью ионизации различных наркотических средств и их действием на червя Arenicola (минимальные дозы в г/100 мл морской воды, вызывающие обездвиживание червя через 5 мин) [Clowes, Keltch, 1931]
pH 7,0 pH 8,0 pH 9,0
Неэлектролиты
Изопропиловый спирт 2,5 2,5 2,5
Изоамиловый спирт 0,1 0,1 0,1
Хлороформ 0,012 0,012 0,025
Хлорбутол 0,025 0,025 0,025
Слабые основания (рКа около 8,5)
Кокаин 0,01 0,005 0,0025
Прокаин 0,002 0,001 0,0005
Бутакаин 0,001 0,0002 0,0002
Барбитуровые кислоты (рКа око-
ло 8,0)
Изоамил, этил- 0,006 0,025 0,05
Пропилметилкарбинил, этил- 0,003 0,006 0,012
Диэтилкарбинил, этил- 0,006 0,012 0,05
н-Амил, этил- 0,006 0,012 0,05
зированных молекул, однако и ионизированная часть вещества также вносит свой вклад в биологическую активность.
При изучении биологической активности слабой кислоты обычно оказывается, что для получения стандартной реакции необходимо одинаковое количество вещества при любых значениях pH, которые на одну единицу или более ниже значения рКа. При этих условиях кислота слабо ионизирована (разд. 10.1), а следовательно, ее биологическая активность обусловлена прежде всего неионизированными молекулами. Это иллюстрирует левая часть рис. 10.12. Однако если значения pH начинают превышать значения рКа, то для получения постоянной биологической реакции потребуется все возрастающее количество вещества, т. е. для получения постоянного биологического ответа необходимо либо постоянное число молекул (ср. разд. 10.4), либо постоянно убывающее в связи с тем, что анион обладает тем же биологическим действием, что и неионизирован-ные молекулы, но менее выраженным. На рис. 10.12 показано действие бензойной кислоты на плесневой гриб Mucor.
Метод построения подобных кривых был разработан Simon (1950), который обнаружил, что у большинства веществ, наиболее активных в наименее ионизированном состоянии, ионы тем не менее обладают некоторой активностью. Примером может служить фунгицидное действие 2,4-динитрофенола (рКа 4,0) на грибы Trichoderma viride при различных значениях pH [Simon, Beevers, 1952J.
Многие азотсодержащие гетероциклы, ионизирующиеся с образованием аниона, сильно ингибируют реакцию Хилла при фотосинтезе (разд. 4.6). Однако было обнаружено, что при
123
Коэффициент распределения
Степень ионизации, %
Рис. 10.11. Поглощение бензойной кислоты (рК 4,2) пекарскими дрожжами при разных значениях pH обратно пропорционально степени ионизации в процентах; А — кривая зависимости распределения всего вещества (клетка — жидкость), Б — кривая изменения степени ионизации в % [Bosund, I960).
Рис. 10.12. Влияние pH на концентрацию бензойной кислоты, необходимую для ингибирования роста гриба Мисог. Верхняя кривая относится ко всему препарату в целом (ионы+иеионизированиые молекулы), нижняя — к неиоиизиро-ваиным молекулам [Cruess, Richert, 1929].
слишком высокой степени ионизации активность теряется полностью. Это явление объясняется тем, что ингибиторы, проникая через мембраны в виде неионизированных молекул, действуют как анионы. Такой механизм действия характерен для некоторых гербицидов: имидазолов, бензимидазолов, пуринов, пира-золов, индазолов, триазолов и бензотриазолов [Btichel, Draber, 1969]. Аналогичным образом рКа влияет на действие 2-трифтор-метилбензимидазолов [Jones, Watson, 1965].
Подобные результаты были получены и для оснований. Так, пириметамин (4.8), имеющий величину рКа 7,2, лучше поглощается клетками из достаточно щелочных растворов, где он находится преимущественно в виде неионизированных молекул. Однако ключевой фермент, находящийся внутри клетки (дигидрофо латредуктаза) ингибируют только катионы [Wood, Ferone, Hitchings, 1961].
Эффективность местных анестетиков в основном зависит от доставки большей части их молекул к месту действия, и это условие ограничивает применение слабых оснований в клинике. Когда было обнаружено, что эффективность обычных местных анестетиков (имеющих значения рКа около 8,5) повышается при использовани их в щелочных растворах, был сделан ошибочный вывод, что неионизированные молекулы — актив
124
ная форма этих веществ [Trevan, Вооск, 1927]. Это утверждение было опровергнуто с помощью следующего эксперимента, отрезок освобожденного от оболочки блуждающего нерва млекопитающих вымачивали в цинхокаине (7.15). Затем препарат промывали в буферном растворе с pH 7. Цинхокаин, прочно связанный с внутренней поверхностью мембраны нерва, блокировал электрические импульсы. При вымачивании препарата в буферном растворе (pH 9,5) способность передавать импульсы восстанавливалась, но снова исчезала при повторном вымачивании в буфере с pH 7. Таким образом было установлено, что в данном случае активной формой является катион [Ritchie, Greengard, 1961, 1966; Narahashi, Frazier, 1971]. Даже аналоги местных анестетиков, содержащие четвертичный атом азота, были активны при инъекции в мембрану нерва; эти опыты были выполнены на мембране аксона кальмара [Narahashi, Frasier, Moore, 1972]. Более того, четвертичные производные кокаина, прокаина и цинхокаина, содержащие для повышения липофильности бензильную группу, также обладают местным обезболивающим действием при аппликациях [Nador et al., 1953]. Следует иметь в виду, что эти четвертичные амины полностью ионизированы при всех значениях pH. Мягкое обезболивающее действие бензокаина (7.14) (рКа 2,5) позволяет понять, что неионизированные молекулы также обладают некоторой, хотя и относительно небольшой активностью.
Так же как и широко используемые местные анестетики, большинство алкалоидов и антипсихотических лекарственных средств имеют рКа около 8, поэтому при pH 7,3 примерно 16% этих соединений остаются неионизированными. Они проникают в клетки в виде неионизированных молекул, а биологическое действие осуществляют в форме катионов. Антагонисты гистамина типа Н2, например циметидин (9.58), при pH 7 активны только, если они почти полностью ионизированы, однако в случае полной ионизации при этом значении pH они не могут достичь своего рецептора [Black et al., 1974].
Степень ионизации (образование анионов) многих широко применяемых противовоспалительных средств коррелирует положительно с их биологическим действием, если ионизации подвергаются не все молекулы, их липофильность достаточна для достижения места действия, где осуществляется ингибирование простагландинов. Примерами могут служить индометацин (рКа 4,5) и салицилаты [Whitenouse, Dean, 1965].
Исследование влияния рКа на противовоспалительное действие фенилиндандионов (9.39) показало, что необходимо точнее определять рКа орто-замещенных арилпроизводных, несомненно обладающих наибольшей биологической активностью [Van der Berg et al., 1975].
Еще в самом начале изучения различных антибактериальных сульфаниламидов было обнаружено, что они утрачивают активность при метилировании атома азота сульфамидной груп-
125
Рис. 10.13. Влияние изменения величины рКа на бактериостатическое действие сульфаниламидов (организм Е. coli, pH 7, синтетическая питательная среда). Вещества, значения рКа которых ниже 6 (точки в левой части рисунка) в этих условиях, сильно ионизированы (по анионному типу) [Bell, Robin, 19421.
пы, что препятствует образованию анионов. Впервые это было установлено на примере сульфапиридина и сульфатиазола [Shepherd, Bratton, Blanchard, 1942]. Впоследствии соотношение между ионизацией и антибактериальной активностью было исследовано Bell, Roblin (1942) (рис. 10.13). Оптимальное значение рКа сульфонамидной группы находится в пределах от 6 до 8. Это указывает на то, что в клетки проникают неионизи-рованные молекулы сульфаниламидов, но их биологическое действие обусловлено ионами. Подобная работа была проведена для широкой серии более близких по структуре сульфаниламидов и оптимальные значения рКа подтвердились, хотя полученная кривая была более пологой [Seydel, 1981].
Тот факт, что сульфаниламиды действуют как анионы, но проникают в бактериальные клетки в виде неионизированных молекул, был позднее подтвержден сравнением поведения сульфаниламидов в бесклеточном фолатсинтезирующем препарате Е. coli и на интактных клетках Е. coli. Оказалось, что антибактериальное действие прямо пропорционально проценту ионизации в первом случае, а во втором — зависит еще и от липофильности молекул [Miller, Doukos, Seydel, 1972].
10.6 . Ионизация рецепторов
Значения рКа рецепторов нельзя предсказать заранее, так как их химическая структура еще мало изучена. Очевидно, катионные лекарственные препараты связываются с анионными рецепторами, которые могут иметь значения рКа от 2 до 7 (при наличии фосфатных групп), от 2 до 6 (карбоксильных групп) или 10 (в присутствии остатков тирозина, пиримидина, цистеина). Катионные рецепторы должны иметь значения рКа 4 (аденин), 7 (гистидин), 10 (лизин), 13 (аргинин).
Наружные клеточные рецепторы. Далеко не все рецепторы находятся внутри клеток. Например, ацетилхолин и метиленовый синий оказывают противоположное действие на сердце ля-126
гушки, не проникая в клетки [Clark, 1933], так как на наружной поверхности мембраны находится множество ферментов и пермеаз. Так, на наружной поверхности дрожжей находятся аденозинтрифосфатаза и некоторые гидролазы.
Величину рКа наружного рецептора клетки во многих случаях можно измерить, определяя реакцию на лекарственный препарат при различных pH, если только известно, что клетка не повреждается при изменениях pH, а степень ионизации лекарственного вещества в этих пределах pH остается постоянной. О применении этого подхода к изучению места действия антибактериальных аминоакридинов на Е. coli см. разд. 10.3.1.
Не всегда исследователи обращают внимание на то, что степень ионизации рецептора может изменяться при изменении pH. Например, было обнаружено, что дыхание (цельных или гемолизованных) эритроцитов птиц при pH 10 в 2,5 раза сильнее, чем при pH 5. Поскольку рКа хинина составляет 8,4, было сделано заключение, что ингибирование вызывается действием неионизированных молекул, а не ионов. Возможность увеличения степени ионизации кислотного рецептора при повышении pH (после чего он может связывать дополнительное количество катионов хинина) не была принята во внимание [Biochem. Zeitschr., 1922, 128, 169].
ОХх, NH
II О
5,5'Диметил оксазол идин-2,4-дион
(10.38)
Б. Внутриклеточные рецепторы. Рецептор можно считать внутриклеточным, если вещества с липофильными группами более эффективны, чем не имеющие их, и если вещества, образующие (при pH среды эксперимента) только 70% ионной формы, более активны, чем ионизированные полностью. В этом случае изучение влияния ионизации становится более трудным, так как особенно серьезное значение приобретает pH среды, окружающей рецептор.
Согласно методу Waddell, Butler (1959), для определения pH внутри клетки необходимо определить три параметра: pH наружной среды и концентрация иона-индикатора внутри и вне клетки (клеточная мембрана при этом должна быть непроницаема для иона и полностью проницаема для неионизированных молекул). Авторы метода использовали бесцветный флуоресцентный индикатор 5,5-диметил-2,4-диоксооксазолидин (ДМО, 10.38), являющийся слабой кислотой (рКа 6,13 при 37 °C). С помощью этого метода было обнаружено, что pH внутри мышцы в состоянии покоя (у собаки) 7,04, вдыхание двуокиси углерода
127
понижает его до 6,6. Применяя радиоактивную форму индикатора, Addanki, Cahill, Sotos (1967) установили, что внутри митохондрии в состоянии покоя pH 7,74. Считают, что этот индикатор для пациентов безвреден.
Другой метод определения pH внутри клетки заключается в измерении сдвига 31Р фосфатной группы методом ЯМР; точность определения составляет ±0,02 единицы pH [Navon et al., 1977]. Применяя другой вариант этого метода, Roberts и сотр. (1980) установили, что в клетках растений pH составляет 7,1 в цитоплазме и 5,5 в вакуолях. Использованный ими метод заключался в определении химического сдвига пика 31Р в глюко-зо-6-фосфате (изменение этой величины в зависимости от значений pH описывается обычной сигмоидной кривой типа кривой титрования, приведенной на рис. 10.1). Глюкозо-6-фосфат был выбран как наиболее распространенный фосфорилированный компонент клетки растений.
Существуют специальные микроэлектроды, которые вводят в живую клетку и с их помощью регистрируют изменения pH, происходящие в течение нескольких часов [Thomas, 1974].
С применением индикаторов было установлено, что pH цитоплазмы животных и растительных клеток составляет 6,8±0,2, а эти жидкости — хорошие буферы. pH нуклеоплазмы 7,6. Внешняя поверхность органелл в том случае, если она несет отрицательный заряд (например, белковые поверхности), может содержать больше водородных ионов, чем цитоплазма вследствие эффекта дзета-потенциала. У амеб pH цитоплазмы 6,7; pH растительных вакуолей в среднем составляет 5,2, хотя у некоторых видов вакуолей эта величина значительно ниже; pH злокачественных тканей млекопитающих часто ниже, чем окружающих органов, что может быть использовано в терапии (разд. 4.2). Столь низкое значение pH злокачественных тканей, связанное с высокой скоростью анаэробного дыхания, требует более тщательного подбора значений рКа при создании карциностатических средств типа азотистых ипритов [Ross, 1961].
Вирусы животных могут заражать организм хозяина, проникая в его внутриклеточные пузырьки по путям, где возможно временное понижение pH. Такие величины pH необходимы, вероятно, и для соответствующих конформационных изменений молекул белка [Helenius, Marsh, White, 1980].
Значения pH внутри бактериальных клеток может меняться с изменением pH буферных сред, окружающих эти клетки, что подтверждается следующим экспериментом. Тирозиндекарбоксилаза Strept. faecalis обладает максимальной активностью in vitro при pH 5,5. В интактной бактериальной клетке активность этого фермента невелика, если окружающая среда нейтральная или щелочная, однако, если поместить бактерии в буфер с pH 5,0—5,5, активность фермента сильно возрастает [Gale, 1946]. Была обнаружена линейная зависимость между pH вне- и внут-128
риклеточной жидкости для ткани сердца черепахи в пределах 6,5—9,5 (рНвнутр = рНвнешн+2,98) [Waddell, Hardman, 1960].
При проведении фармакологических исследований желательно работать с буферными растворами, даже если известно, что рецептор находится внутри клетки. Наличие внешнего буфера создает однородные условия благодаря стандартизации степени ионизации лекарственного препарата, поступающего в клетку, независимо от формы его введения и состояния химических групп, ответственных за адсорбцию лекарственного вещества на поверхности клетки и за его проникновение в клетку.
Результативность применения буферов определяется количеством клеток, с которыми буфер вступает в контакт. В массе культуры тканей лекарственное вещество может достичь рецепторов только после прохождения через другие клетки и внутриклеточную жидкость. Результаты подобных экспериментов нельзя сравнивать с результатами опытов, в которых все клетки находятся в непосредственном контакте с лекарственным веществом при известном pH [Simon, Beevers, 1951].
10.7 . Заключение
Рассмотрение основных принципов, изложенных в этой главе, позволяет понять, насколько важно знать константы ионизации веществ при изучении их биологического действия.
Далее необходимо установить, в какой форме эти вещества более эффективны — в виде ионов или неионизированных молекул. Это можно сделать двумя способами, которые не исключают, а дополняют друг друга. Первый заключается в том, что введением различных заместителей в молекулу лекарственного вещества меняется величина его рКа и, таким образом, синтезируется соединение с заданной величиной рКа в любых рядах соединений (разд. 17.2). Преимущество этого метода заключается в том, что ионизация рецептора остается неизменной, а живые клетки постоянно находятся при оптимальном для них pH. Недостатком является то, что химические изменения в молекуле могут быть ответственны за изменение биологической реакции; этих затруднений можно избежать, если проводить эксперимент с двумя аналогами, имеющими одно и то же необходимое значение рКа, и считать результат верным в том случае, если он совпадает для обоих опытов. Второй метод заключается в том, что структура лекарственного вещества остается постоянной, но варьируется pH среды. Он имеет то преимущество, что в данном опыте живые клетки подвергаются воздействию только одного лекарственного вещества; влияние изменений pH на рецептор и на жизнеспособность самой клетки следует проверять отдельно в контрольных опытах.
Поскольку оба метода дают возможность контролировать степень ионизации (один из наиболее изменчивых параметров), то их применение может помочь при планировании дальнейших более сложных экспериментов.
9—734
129
Глава 11
ВЕЩЕСТВА, СВЯЗЫВАЮЩИЕ МЕТАЛЛЫ
Тяжелые металлы в следовых количествах необходимы для всех форм жизни. Они проникают в живую клетку в виде катионов, но поглощение их строго регулируется, поскольку большинство (если не все) тяжелых металлов в больших количествах токсичны. Следует отметить одну особенность, заключающуюся в том, что только в очень редких случаях избыток одного жизненно важного металла может восполнить ущерб, нанесенный недостатком другого. В действительности избыток одного металла чаще всего усугубляет вред, обусловленный недостаточностью другого.
Вещества, способные связываться с металлами, часто образуют с ними хелатные соединения1. К последним принадлежат многие эффективные лекарственные средства и другие вещества, обладающие избирательной токсичностью. Они производятся промышленностью в огромных количествах, особенно фунгициды. Их действие состоит в нарушении весьма чувствительного баланса между катионами тяжелых металлов в природе. Некоторые из них способны вытеснять металлы из живых тканей, но большинство усиливает присущую тяжелым металлам токсичность.
Помимо использования в сельском хозяйстве в качестве фунгицидов, связывающие металлы вещества применяют в медицине и ветеринарии с учетом основных принципов их действия. Те из них, которые дифференцированно действуют на позвоночных и их паразитов, используются для борьбы с грибами, бактериями и вирусами (разд. 11.7—11.9). Другие вещества применяют как антидоты, так как они по-разному связывают жизненно важные или токсичные для организма металлы (разд. 11.6). Соединения, действующие различно при нормальных и патологических процессах, применяют для лечения ревматизма, сердечно-сосудистых заболеваний и рака (разд. 11.9— 11.11).
1 Вещества, связывающие металлы, называются лигандами. Если металл при связывании с лигандом оказывается заключенным в лиганде между любыми двумя элементами, такими как N, О нлн S, то образуется хелатное кольцо. В таком соединении металл более прочно связан с лигандом, чем в нехелатированном, где он не является частью кольца. Термин «хелат», предложенный Morgan, Drew (1920), происходит от греческого слова khele — клешня (краба).
130
11.0. Металлы в живой клетке
Человек, так же как и другие позвоночные, нуждается в катионах нижеперечисленных металлов, обеспечивающих протекание многих жизненно важных процессов. Более того, многие из этих металлов необходимы для всех других форм жизни. К ним относятся: а) тяжелые металлы (кобальт, медь, железо, марганец, молибден, цинк и в меньших количествах хром, ванадий, никель и свинец); б) легкие металлы, обычно встречающиеся в большем количестве (кальций, магний, калий и натрий).
Медь, железо, молибден, кобальт и иногда марганец принимают участие в окислительно-восстановительных процессах; действие цинка, магния и марганца связано с процессами гидролиза и переноса групп; кальций играет наиболее важную роль при создании гибких или жестких структур, а также может инициировать реакцию, по-видимому вызывая структурные изменения (подобное действие иногда проявляет и магний). Натрий и калий, благодаря их распространенности, служат переносчиками заряда; они очень слабо связываются и поэтому могут быстро обмениваться. Рассмотрим некоторые примеры биологического действия тяжелых металлов в той последовательности, которая указана выше. Положение и валентность металлов в ферментах определяют с помощью электронного парамагнитного резонанса (ЭПР) и другими современными методами.
Кобальт является необходимым кофактором некоторых ферментов, например карбокситрансфосфорилазы (ответственной за перенос фосфатных групп), гликольдегидразы (вызывающей перегруппировку) и карбоксипептидазы, разрушающей различные кинины (пептиды, циркулирующие в крови млекопитающих и вызывающие воспалительные процессы) и играющей важную роль в усвоении полипептидов [Davies, Lowe, 1966]. Кобальт прочно связывается большим тетрапиррольным циклом коферментов группы кобаламидов. Для заместительной терапии при пернициозной анемии используют два вещества — гидроксикобаламин и цианкобаламин (витамин В12), образующиеся при разрушении коферментов во время их выделения из дрожжей или печени. Эти коферменты катализируют восстановление рибонуклеотидов в дезоксирибонуклеотиды, взаимопревращения анионов p-метиласпаргиновой и глутаминовой кислот, анионов метилмалоната и сукцината, а также метилирование гомоцистеина в метионин.
В 1935 г. две группы австралийских ученых обнаружили, что сильная анемия у овец, пасущихся на обширных обильных пастбищах Австралии и Новой Зеландии, обусловлена недостатком кобальта — элемента, потребность в котором у растений незначительна. При содержании кобальта в траве в соотношении, меньшем чем 0,2: 1 000 000, овцы начинают слабеть и
9*
131
погибать, что объясняется неспособностью флоры рубца овец синтезировать содержащие кобальт коферменты [Marston, Allen, Smith, 1961]. В настоящее время для предупреждения этого заболевания в рацион овец добавляют кобальт.
Для лечения пернициозной анемии у человека достаточно суточной дозы ( 1 мкг) витамина В12 (внутримышечно). Для человека с массой тела 70 кг это составляет 2-10~ 10% массы тела.
Медь_ существенная составная часть многих ферментов, в том числе фенолазы (тирозиназы), играющей важную роль в процессе метаболизма растений, грибов и пигментных клеток млекопитающих, а также поставляющей дубильные белковые вещества, придающие твердость оболочке насекомых. С помощью ЭПР было показано, что медь в родственном ферменте лакказе в исходном состоянии двухвалентна, но переходит в одновалентное состояние (Си+), когда фермент вступает в реакцию. В молекуле цитохромоксидазы содержатся два атома железа и два атома меди; железо в отличие от меди в живой природе всегда связано с порфиринами. Медь также необходима для действия допамингидроксилазы (ключевой фермент в синтезе катехоламиновых гормонов), МАО и оксидазы аскорбиновой кислоты.
Супероксиддисмутаза — фермент зеленого цвета, присутствующий во всех тканях эукариот, содержит медь и железо, связанные с одним и тем же имидазольным циклом (His—61) апофермента. Медь проходит цикл окисления и восстановления по мере того, как она разрушает токсичные перекисные радикалы (О2~), появляющиеся в реакциях с использованием атмосферного кислорода.
В супероксиддисмутазе бычьих эритроцитов (а возможно, и в других клетках) цинк и медь связаны с гистидиновым остатком. Аналогичные ферменты, содержащие марганец вместо меди, встречаются в организмах млекопитающих и бактерий [Oberley, Buettner, 1979]. Подробнее об этих необычных металлосодержащих ферментах см. Oberley (1982) и Rodgers, Powers (1981).
Исследования влияния следовых количеств меди на рост овса показали, что как недостаток, так и избыток ее в среде одинаково вредны для растений. Недостаток меди оказался причиной многочисленных неурожаев на окультуренных участках земли в Голландии и Дании. Недостаток меди у домашних животных приводит к развитию анемии, демиелинизации спинного мозга и потере пигментации. Избыточное накопление меди в печени овец приводит к гемолизу и смерти. У овец, в пище которых недостает меди, шерсть теряет волнистость, а так как именно это качество особенно ценится при продаже, многие фермеры несут большие убытки.
Достоверно установлена и широко используется способность меди избирательно действовать на беспозвоночных животных, 132
грибы и водоросли. Такая избирательность определяется выработанным в процессе эволюции более тонким устройством позвоночных, позволяющим им изолировать, переносить и выделять этот высокоактивный с химической точки зрения элемент. У здоровых млекопитающих более 90% всей меди связано с белком церулоплазмином, синтезируемым печенью и циркулирующим в кровотоке. Его физиологические функции заключаются в уменьшении концентрации ионов двухвалентной меди в плазме ниже токсического уровня, снабжении медью медьсодержащих ферментов и обеспечении постепенного высвобождения железа из депо. Любой избыток комплекса медь — церулоплазмин выделяется с желчью в экскременты. Не связанная с церулоплазмином медь циркулирует в сыворотке крови в виде легко распадающегося комплекса с альбумином (разд. 11.1).
С помощью ЭПР удалось показать, что в церулоплазмине медь частично одновалентная, частично двухвалентная. Отсутствие церулоплазмина в организме является причиной наследственной гепатолентикулярной дегенерации (болезнь Вильсона), которая у детей в отсутствие лечения приводит к смерти вследствие разрушающего действия ионов двухвалентной меди на нервную систему. О лечении этого заболевания хелатообразующими лекарственными препаратами см. разд. 11.6.
Медьсодержащий пигмент пластоцианин (ярко-синего цвета) принимает участие в процессе переноса электронов при фотосинтезе у растений. Входя в состав дыхательного пигмента гемостатина (синего цвета) определенных моллюсков и ракообразных, медь играет такую же роль, как железо в дыхательных пигментах высших организмов. Медь необходима также для образования гемоглобина у человека.
Известно, что медь накапливается (в несколько раз больше по сравнению с другими металлами) в симпатических нервных окончаниях, в синаптических пузырьках мозга и других нервных тканях. Это способствует накоплению норадреналина в тройном комплексе с АТФ [Colburn, Maas, 1965], а также 5-гидрокситриптамина [Roberts, 1966]. Ионы двухвалентной меди, АТФ и норадреналина образуют прочно связанный тройной комплекс в соотношении 1:1:1, имеющий log К 12,15 (комплекс ион меди — норадреналин (1:1) имеет logK 10,25). Норадреналин образует несколько более прочный комплекс, дофамин — менее прочный. Применяемые в медицине аналоги, такие как фенамин и фенилэфрин, связываются слабее. Очевидно, что оба атома кислорода катехольного фрагмента играют в связывании важную роль. Прочность связывания этих адренергических медиаторов и лекарственных средств с гранулами си-наптосом изменяется в том же порядке, что и константы стабильности их комплексов [Rajan, Davis, Colburn, 1974].
Более подробно о роли меди в биохимических процессах см. Sigel (1981).
133
Железо — важнейший компонент порфириновых ферментов^ каталазы, пероксидаз и различных цитохромов, выполняющих’ роль переносчиков электронов и необходимых для всех живых клеток. Гемоглобин — дыхательный пигмент, содержащийся в эритроцитах млекопитающих, птиц, рептилий, земноводных й рыб. Гемоглобин обнаружен также в плазме нескольких видов моллюсков и кольчатых червей (некоторые виды кольчатых червей имеют железосодержащий дыхательный пигмент зеленого цвета — хлорокруорин). Миоглобин — пигмент мышц, являющийся переносчиком кислорода, по структуре очень напоминает гемоглобин.
Гликопептид трансферрин (ОММ 86 000) прочно связывает ионы трехвалентного железа между имидазольным и тирозиновым остатками и снижает их содержание в сыворотке крови; в свободном состоянии ионы железа — вредны для сердца. Для образования гемоглобина ретикулоцитами может быть использовано только железо, связанное с трансферрином. Трансферрин, так же как и аналогичный ему ^-глобулин, содержащийся в куриных яйцах (кональбумин), способен настолько понижать концентрацию ионов трехвалентного железа в питательной среде, что в ней прекращается рост бактерий. Аналогичный полипептид— лактоферрин, был обнаружен в поте, слезах, молоке, а также носовых и бронхиальных выделениях. Полагают, что благодаря способности этих соединений препятствовать поглощению железа бактериями они служат эффективными средствами защиты от бактериальных инфекций, так как железо повышает болезнетворное действие многих бактерий в 100 000 раз [Bullen, Rogers, 1969].
У позвоночных основными соединениями, накапливающими железо, являются ферритин, содержащийся главным образом в печени, и гемосидерин — в селезенке и мышцах.
Железо также важнейшая составляющая некоторых непорфириновых ферментов, например аконитазы, альдолазы и сукцинатдегидрогеназы. Ингибирование синтеза глюкозы триптофаном в клетках животных зависит от образования хелатных связей. В процессе метаболизма триптофан превращается в пиридин-2,3-дикарбоновую кислоту, которая с двухвалентным железом образует комплекс, необходимый для действия фосфо-енолпируваткарбоксикиназы (ключевого фермента глюконеогенеза) [Veneziale et al., 1967]. Для захвата необходимого им для питания железа бактерии и грибы используют сидерохромы — набор фенолов и гидроксамовых кислот (разд. 11.1).
Ферредоксины представляют собой белки, содержащие равное количество атомов железа и серы в каждом активном центре. Некоторые из этих соединений переносят электроны при потенциалах, более низких, чем потенциал водородного электрода (ферредоксин Fes 1—при —305 мВ, а ферредоксин Fes 2 — при —20 мВ. Другие (например FeS s-З) действуют при значительно более высоких значениях потенциала: -[-60 мВ (эти 134
цифры относятся к Ео по стандартному водородному потенциалу при pH 7,2) [Sweeney, Rabinowitz, 1980; Spiro, 1982]. ферредоксины 8Fe-8S встречаются у наиболее примитивных организмов (анаэробных бродильных грибов и фотосинтезирующих бактерий) и служат переносчиками электронов в пиру-ватфосфорокластической системе; ферредоксины типа 4Fe-4S находятся, видимо, на следующей ступени эволюции, так как они обнаружены у сульфат- и нитратвосстанавливающих бактерий. 2Fe-2S-ферредоксины найдены у растений и животных, которым они необходимы для окислительного фосфорилирования в митохондриях, для фосфорилирования, происходящего под действием фотосинтеза в хлоропластах, и для синтеза катехоламиновых гормонов. Структура ферредоксинов различных типов была установлена с помощью спектроскопии ЭПР и мессбауэровской спектроскопии.
Хотя в почве железо содержится в больших количествах, оно часто недоступно для растений, особенно в щелочных почвах. Однако если в почве находятся соответствующие бактерии, то железо становится доступным для растений, так как микроорганизмы растворяют его с помощью гидроксаматсидерофо-ров (разд. 11.1). Такие железо(III)-органические комплексы накапливаются в почвах в количествах, достаточных для питания растений [Powell et al., 1980]. Отсутствие полезных бактерий можно компенсировать опрыскиванием почвы этилендиаминтет-рауксусной кислотой (ЭДТА), экстрагирующей железо путем образования комплекса железо(Ш)—ЭДТА, который всасывается корнями растений. Эксперименты с растениями томатов, выращенных на среде, к которой был добавлен комплекс железо— ЭДТА, содержащий меченые атомы 59Fe и С14 (в положении 2 ацетатной группы), показали, что растение всасывает и транспортирует интактный комплекс. В процессе метаболизма органическая часть комплекса разрушается, а неорганическая (железо)—остается. Если почва бедна железом, то аналогичные результаты можно получить при опрыскивании ее раствором комплекса ЭДТА — железо (III).
Марганец, хотя и представляет собой металл с переменной валентностью, редко участвует в биологических процессах окисления. Он необходим для действия многих катаболических ферментов — оксалоацетатдекарбоксилазы, аргиназы, пируват-оксидазы и пролидазы, а также для ферментов, под действием которых происходит обмен таких объемных групп, как остатки сахаров.
Марганец необходим в процессе фотосинтеза у растений, так как он способствует протеканию той части реакции Хилла, в которой происходит отщепление гидроксид-иона, сопровождающееся выделением молекулярного кислорода.
Молибден выполняет важнейшие функции в составе таких ферментов, как ксантиноксидаза, альдегидоксидаза, нитратре-Дуктаза и нитрогеназа. С помощью спектроскопии ЭПР уста-
135
ъовлено, что во всех этих ферментах он связан с серой и изменяет свою валентность Мо5+^Мо6+; обычно в этих ферментах •содержится также железо. Молибден необходим для фиксации азота различными видами Rhizobium и Azotobacter, обитающими в клубеньках корней растений.
Формиатдегидрогеназа — фермент плазматической мембраны бактерий Е. coli, содержит функциональные атомы молибдена и железа и прочно связанный селен [Enoch, Lester, 1975].
Известно, что в некоторых бактериях вольфрам занимает место молибдена [Andreesen, Ljungdahl, 1974].
Цинк — важнейший компонент, необходимый для действия по меньшей мере двадцати ферментов с весьма различными функциями. К ним относятся алкогольдегидрогеназы дрожжей и печени млекопитающих, глицеральдегидфосфатдегидрогеназа, фосфогликомутаза дрожжей, щелочная фосфатаза бактерий, карбоксипептидаза млекопитающих, карбоангидраза, АМФ-гид-ролаза, пируваткарбоксилаза (дрожжи) и альдолаза (дрожжи и бактерии). Каждая молекула щелочной фосфатазы бактерий Е. coli содержит четыре атома цинка, два из которых определяют четвертичную структуру фермента и могут быть заменены ионами Мп2+, Со2+ или Си2+, тогда как два других играют важную роль в функционировании фермента [Trotman, Greenwood, 1971]. Кроме того, цинк необходим для действия ДНК- и РНК-полимераз в бактериях и фагах, что было установлено с помощью диализа против хелатирующих агентов [Coleman, 1974; Poiesz, Seal, Loeb, 1974].
Обратная транскриптаза, выделенная из некоторых вирусов, содержит по одному атому цинка в каждой молекуле фермента [Vallee, 1975]. РНК-полимераза вируса гриппа В дезактивируется in vitro под действием орто-фенантролинов, селеноциста-мина и других хелатирующих агентов [Oxford, Perrin, 1975].
Употребление высокоочищенных пищевых продуктов в высокоразвитых странах приводит к появлению в пожилом возрасте цинковой недостаточности, которая характеризуется потерей остроты вкусовых ощущений и медленным заживлением ран и ссадин. В почках млекопитающих содержится белок металло-тионеин, связывающий избыток цинка, кадмия и ртути [Rupp, Voelter, Weser, 1974].
Недостаток цинка вызывает тяжелые заболевания яблонь, цитрусовых и виноградной лозы. Отсутствие цинка в почве служит причиной неурожая зерновых. В 1930-х годах в Южной Австралии одна из равнин, поросшая редким кустарником, путем регулярного опрыскивания с воздуха раствором цинковых солей была превращена в пышные пастбища, на которых теперь пасутся большие стада овец.
Хром (Сг3+). Хром, содержащийся в дрожжах в виде комплекса с никотиновой кислотой и алифатическими аминокислотами, считают «фактором толерантности к глюкозе», необходимым для нормального углеводного обмена в организме челове-136
ка. Его действие заключается в усилении гипогликемического действия инсулина [Mertz, 1975J. В высокоразвитых странах, где пища слишком рафинирована, а также в развивающихся странах, где ее недостаточно, нехватка хрома — обычное явление [Gurson, Saner, 1971; Levine, Streeten, Doisy, 1968].
Ванадий необходим для питания позвоночных [Lambert et al., 1970] и, по-видимому, является природным ингибитором На+,К+-АТфазы [Cantley et al., 1978]. В организмах низших хордовых-оболочников (асцидии), занимающих предыдущую перед позвоночными ступень в филогенетическом древе, ванадий изменяет железо в пигментах крови, переносящих кислород.
Никель. В экспериментах на свиньях и крысах было установлено, что никель жизненно важен для их питания [Nielsen, 1975]. Необходимые количества никеля настолько малы по» сравнению с другими металлами, рассмотренными ранее, что для его обнаружения потребовалась разработка нового аналитического метода — нейтронного активационного анализа. Фермент уреаза, содержащийся в бобах, является никельзависимым [Dixon et al., 1975]. Некоторые бактерии, например Methano-bacterium и Desulphovibrio, содержат редуктазы, для действия которых необходим никель [Thauer, 1980], как и для бактериальных дегидрогеназ оксида углерода (II) [Deake, Hu, Ward, 1980].
Полагают, что олово необходимо для питания животных в следовых количествах [Schwarz, Milne, Vinard, 1970].
Алюминий, считающийся биологически инертным, после растворения в вакуолях растений, имеющих кислую среду, проявляет некоторую токсичность.
Кальций широко распространен во многих живых организмах (у бактерий его значительно меньше). В больших количествах он содержится в зубах, костях, раковинах, а также в растениях в виде кристаллических оксалатов. Основная роль кальция в природе — обеспечение твердости тканей, испытывающих большие нагрузки; например, сухожилия, которые при удалении кальция путем растворения в ЭД ТА (11.27) разделяются на тонкие коллагеновые волокна. Возмещение потерь кальция в значительной степени их восстанавливает. Хрящ превращается в кость при включении в него ионов кальция и фосфата; этот процесс конкурентно обратим при помощи ионов магния.
У млекопитающих уровень циркулирующего в крови кальция регулируется тиреоидными и паратиреоидными гормонами. Концентрации кальция внутри и вне клеток составляют примерно 10~6 и 10~3 М соответственно. Высвобождение кальция из вне- или внутриклеточных депо может вызвать множество разнообразных биологических реакций. Уровень концентрации кальция контролируется плазматическими мембранами, мембранами ЭР (в мышце — мембранами саркоплазматического ре
137
тикулума) и внутренними мембранами митохондрий. Одна из наиболее важных функций кальция —регуляция проницаемости полупроницаемых мембран, которые при наличии кальция сохраняют целостность, а в его отсутствие становятся пористыми и легко проницаемыми. Такая регуляция определяется большой распространенностью кальция, его двухвалентностью и наличием в мембранах фосфат-анионов [Amer, Tsukada, Nesbett, 1967].
Другая важная функция ионов кальция — роль «вторичного мессенджера». При участии кальция происходит сопряжение химического стимула с секрецией, как, например, при высвобождении нейромедиаторов под действием нервного импульса в синапсах или нервно-мышечных соединениях [Miledi, Slater, 1966] или выделении адреналина из мозгового слоя надпочечников под действием ацетилхолина [Douglas, 1968]. Точно так же кальций необходим для сопряжения высвобождения нейромедиатора при сокращении мышц. По-видимому, кальций инициирует пролиферацию различных типов клеток, таких как мышечные, лимфоциты и фибробласты. Он также играет важную роль в фагоцитозе. Многие из перечисленных эффектов могут быть вызваны ионофорами (разд. 14.2), осуществляющими транспорт кальция в клетку. Полагают, что на молекулярном уровне действие кальция заключается в облегчении доступа АТФазы к ее субстрату (АТФ) и обеспечении названных процессов энергией.
Кальций играет жизненно важную роль по меньшей мере в трех стадиях цикла сокращения — расслабления мышц позвоночных [Taylor, Lymn, Moll, 1970]. В саркоплазматическом ретикулуме кальций находится в кальсеквестрине, белке, обладающем высоким сродством к кальцию — каждая его молекула содержит 43 атома кальция. Под действием нервного импульса происходит высвобождение кальция, инициирующего цепь реакций, вызывающих сокращение мышцы, после чего кальций поступает обратно в ретикулум с помощью кальций-магнийза-висимой АТФазы.
То, что кальций способствует свертыванию крови, известно давно. В некоторых тканях, особенно у низших животных, кальций может частично или полностью заменять натрий во входящем токе в нервной проводимости.
Только митохондрии млекопитающих способны накапливать кальций и при необходимости выделять его; возможно, таким образом осуществляется контроль за обызвествлением тканей. Митохондрии сердца накапливают кальций в обмен на натрий, а в митохондриях печени он заменяется ионами водорода, однако при этом сохраняется независимый канал для медленного проникновения ионов кальция. Сокращение сердечной мышцы вызывается высвобождением кальция из митохондрий, которое в свою очередь происходит под действием ионов натрия или АМФ.
138
Кальмодулин — внутриклеточный, связанный с кальцием белок, участвует во многих процессах, происходящих в организме, например, в секреции, активации миозинкиназы и метаболизма циклических нуклеотидов. Подобный белок — тропо-нин-с, регулирует конформационные изменения скелетной мышцы. Контроль за сокращением скелетной мышцы полностью осуществляется внутриклеточным кальцием. Именно поэтому такие вещества, как нифедипин (разд. 14.2), блокирующие каналы, не оказывают на них никакого действия. С другой стороны, уровень кальция во внеклеточной жидкости сильно влияет на гладкую и сердечную мышцы.
Калпайн — это кальцийзависимая цистеинпротеаза, действующая в качестве необратимого регулятора почти во всех клетках млекопитающих [Murachi, 1983]. Протеины, содержащие у-карбоксиглутаматные остатки (у-карбоксигруппы образованы с помощью витамина К), являются важными факторами свертываемости крови. Другие протеины этого типа находятся в дентине зубов и в матрице кости.
Для определения следовых количеств кальция в биохимии широко используют аэкворин. Флуоресценция этого белка, ответственного за голубое свечение медуз Aequorea, обусловлена в основном наличием кальция в среде [Shimomura, Johnson, 1969].
Роль кальция в механизмах действия лекарственных веществ см. Weiss (1978), биологические аспекты — Angirilieri. (1982) и Cheung (1980).
Магний. Катион магния занимает второе место по распространенности внутри клетки подобно тому, как кальций — вне ее. Магний является незаменимым звеном, обусловливающим целостность рибосом и соединяющим их с шРНК. Он служит кофактором всех ферментов, использующих АТФ при переносе фосфатных групп, и для многих других ферментов, действие которых связано с переносом групп или гидролизом. Магний ингибирует высвобождение АХ в концевой пластине двигательного нерва и во многих случаях действует как обратимый антагонист кальция. У бактерий магний — наиболее распространенный двухвалентный металл. Магний — ключевой компонент хлорофилла, занимающий в его молекуле центральное положение. Повышенная частота смертных случаев от сердечных заболеваний в местностях с мягкой водой обусловлена, по-видимому, отсутствием Mg2+.
Калий и натрий, ионы которых похожи с точки зрения физики, различаются по способности проникать сквозь клеточные' мембраны в состоянии покоя и возбуждения, по сродству к механизму активного транспорта и возможности активировать-ферменты. Так, например, мембрана нервного волокна в состоянии покоя в 30 раз более проницаема для К+, чем для Na+, а в-активном состоянии — в 10 раз более проницаема для Na+, чем для К+ (ср. разд. 7.5.1). Размер катионов в гидратированном
139’
или негидратированном состоянии не играет существенной роли в избирательности, различия ее определяются разницей в значениях свободных энергий: '
АРкисл. центра — AFOCH. центра АРкисл, ьоды + AFqch. воды»
где АРцентра —свободная энергия притяжения между катионом и отрицательно заряженным центром; ДРВОды — свободная энергия гидратации катиона. Основной переменной величиной, влияющей на соотношение избирательности Na+: К+, является величина отрицательного заряда на мембране, отражающаяся в величине рКа; для Na+ предпочтительнее сильные кислоты, а точка перехода находится при рКа 3 [Diamond, Wright, 1969].
Калий — важнейший внутриклеточный ион всех живых организмов. Натрия в большинстве клеток содержится мало, он появляется внутри клетки только в момент возбуждения. Наземные растения всегда содержат очень мало натрия.
Даже одновалентные ионы могут существовать изолированно и часть ионов натрия в живых тканях животных находится в осмотически неактивной форме. Катионы натрия удерживаются полимерами, повторяющиеся звенья которых имеют по меньшей мере одну кислотную группу (примеры: нуклеиновые кислоты, мукополисахариды). Изолированные ионы натрия могут быть мгновенно вытеснены ионами металлов более высокой валентности; концентрация ионов натрия измеряется с помощью натрийселективного электрода [Palaty, 1966].
В последнее время экспериментально найдено много агентов, изменяющих проницаемость мембран для этих ионов. Тет-родоксин (разд. 7.5.1) снижает проницаемость мембран для ионов натрия, а валиномицин (разд. 4.2) способствует поглощению калия [Moore, Pressman, 1964]. Образуя комплекс с ионами калия, тетрафенилборат натрия (NaBPtu) может диспергировать ткани на отдельные клетки [Rappaport, Howze, 1966].
Почки в норме выделяют натрий, но задерживают каЛий. Чем меньше лекарственные средства нарушают этот механизм, тем более хорошими диуретиками они являются (см. разд. 14.1).
Литий обычно не является компонентом клетки, но в отличие от всех остальных известных ионов может выполнять, хотя и с меньшей эффективностью, функцию натрия при проведении нервного импульса. Литий также проходит через изолированную кожу лягушки, используя механизм, предназначенный для переноса натрия [Schou, 1957]. Успешное применение лития для лечения хронических психозов было начато в Австралии [Cade, 1949]; благодаря исследованиям Schou этот метод стал основным при лечении подобных заболеваний. Использование лития не сопровождается летаргией. К сожалению, его токсическая доза только в три раза превышает терапевтическую.
Антагонизм катионов. Многие болезни растений и животных могут быть вызваны нарушением баланса металлов. У растений 140
сои, обработанных избытком марганца, быстро развиваются признаку недостаточности железа, которые могут быть легко устранен^ добавлением соответствующих его количеств. Если же растения сои выращивать на почве с чрезмерно высоким содержанием \келеза, то у них появляются симптомы недостаточности марганца [Somers, Shive, 1942]. В Англии у выращиваемых на разлйцных пастбищах ягнят нередка болезнь «изогнутая спина» (энзоотическая атаксия), приносящая большие экономические убытки. Эта болезнь связана с повышением содержания в траве цинка или свинца и излечивается добавлением в корм меди. На других пастбищах, богатых молибденом, у овец появляются симптомы недостаточности меди, которая излечивается добавлением ее в корм. Избыток цинка в пище вызывает у крыс анемию, исчезающую при добавлении в рацион меди [Smith, Larson, 1946]. Для предотвращения заболевания, вызываемого недостатком меди, в Голландии ею обогащали почву, в результате чего в выращенных плодах и травах нередко отмечали недостаток марганца, хотя содержание его в почве было нормальным. Эти виды антагонизма напоминают простой антагонизм ионов, ответственный, как показал Ringer в 1883 г., за регуляцию частоты сердечных сокращений (кальций против калия). Аналогичный антагонизм между магнием и кальцией связан с процессом сокращения мышц.
Многие примеры подобного антагонизма представляют собой как бы неорганический эквивалент взаимоотношений метаболитов и антиметаболитов (гл. 9).
Токсическое действие чужеродных металлов часто обусловлено антагонизмом катионов. Так, например, свинец — известный нейротоксин, вытесняет кальций из некоторых отделов нервной системы и тем самым препятствует выделению нейромедиаторов [Kober, Cooper, 1976]. Около 1960 г. широко распространившееся загрязнение Восточного побережья Японии кадмием в сочетании с низким уровнем поглощения кальция привело к развитию мучительной болезни «итаи-итаи» — одной из разновидностей остеомиелита. Выяснилось, что причина этого заболевания — антагонизм кадмия и кальция [Friberg, 1974].
Двухфазная ответная реакция. Известно, что реакция организма на тяжелые металлы является двухфазной. Если организм получает слишком мало металлов, ему наносится тяжелый ущерб. Это объясняется тем, что в организме содержится множество ферментов, которые могут функционировать только в присутствии тяжелых металлов хотя бы в следовых количествах. Однако если организм получает слишком много металла, то наступает вторая фаза, связанная с токсическим действием избыточного количества.
Примером такой двухфазной реакции может служить действие меди на овес [Piper, 1942]: как избыточное, так и недостаточное количество этого металла наносит вред его росткам
141
Рис. 11.1. Влияние меди иа рост овса. Содержание меди в питательной среде (слева направо): 3, 6, 10, 20, 100, 500, 2000 и 3000 мкг/л [Piper, 1942].
(рис. 11.1). В эксперименте овес выращивали в сосудах, концентрацию меди в которых изменяли от 0 до 3000 мкг/л, причем максимальная длина ростков овса была зафиксирована при 500 мкг меди на литр. Рост микроорганизмов часто зависит от концентрации одного или нескольких катионов металлов в питательной среде: следует избегать как избыточных, так и недостаточных концентраций, так как в любом случае рост будет заторможен.
Обедненные металлами среды. Тем, кто только начинает изучать микроэлементы, часто кажется странным, что перед началом эксперимента необходимо очищать среды от металлов. К сожалению, «химически чистые» и «чистые для анализа» реагенты содержат очень много примесей тяжелых металлов. Это неизбежно, так как вещество, очищенное на 99,99%, содержит бХЮ17 молекул посторонних примесей на каждый грамм. Это значение рассчитано с помощью числа Авогадро (бХЮ23 молекул в 1 моле любого вещества), причем средняя величина ОММ принималась равной 100. Фактически реактивы марки «чда» обычно содержат менее 99,99% основного вещества. Более того, содержание металлов в бактериологических средах, 142
как4правило, велико. Типичный случай загрязнения реактивов примесями металлов был выявлен при проведении эксперимента с жгутиковой водорослью Euglenia gracilis, клетке которой необходимо только 4800 атомов (5Х10-19 г) кобальта для образования нового организма. В предварительном опыте было обнаружено,\что «аналитически чистая» соль железа, применявшаяся для Приготовления культуральной среды, содержит кобальта в тридцать три раза больше нужного количества [Hutner, 1949]. Л
Существует'следующий удобный метод приготовления сред с незначительным содержанием металла. Питательные соли растворяют в воде. Этот раствор многократно встряхивают с раствором дитизона в хлороформе, затем экстрагируют хлороформом для удаления дитизона и пропускают воздух (для удаления хлороформа) [Piper, 1942]. Аналогичным образом используют и другие вещества, обладающие высоким сродством к металлам, например 8-гидроксихинолин применяют для удаления металлов из бактериологических сред [Rubbo, Albert, Gibson, 1950]. В работе Donald, Passey, Swaby (1952) рассмотрены 38 способов удаления металлов из питательных сред.
При проведении экспериментов часто возникает необходимость поддержать определенную концентрацию иона металла таким образом, чтобы она не поднималась выше и не опускалась ниже нужного значения. Эта проблема решается с помощью буферов, содержащих ион металла и сохраняющих постоянное значение рМ, подобно тому, как буферы, содержащие ион водорода, имеют неизменное значение pH. В этих системах происходит постоянное восполнение удаляемых в процессе реакции свободных ионов металла за счет металла, находящегося в виде комплекса. Вначале для этих целей использовали такие комплексообразующие агенты, как цитрат- и тарт-рат-ионы, однако более широкое применение получили ЭДТА (11.27), диэтилентриаминпентауксусная кислота (ДТПА) и нитрилотриуксусная кислота (НТА). Количественные аспекты этого метода рассмотрены в работе Perrin, Dempsey (1974).
К неметаллическим питательным микроэлементам относятся бор (для растений), йод и фтор (для позвоночных), селен и кремний. Селен — необходимая составная часть важного фермента млекопитающих, глутатионпероксидазы, которая защищает мембраны от окисления и является одним из микробио-цидных ферментов фагоцитов [Stadtman, 1980]. Ферменты, содержащие селен, железо и молибден, уже рассматривались при обсуждении свойств молибдена. Полагают, что кремний также вносит вклад в построение и придание упругости соединительным тканям позвоночных. Кремний необходим для роста и развития организма крыс. По-видимому, кремний содержится в виде силанолята с мостиками Si—О—R в таких полисахаридах, как гепарин и гиалуроновая кислота [Schwarz, 1973].
143
Подробнее о роли микроэлементов в питании см. Uijder-wood (1977); неорганические аспекты биологических процессов рассматриваются в работе Williams (1976). /
11.1. Биохимические различия, способствующие избирательности /
Любую биологическую систему можно рассматривать как арену постоянной борьбы за следовые количества/катионов тяжелых металлов. При поражении позвоночных/микроорганиз-мами в организме-хозяине быстро понижается /уровень железа и цинка в плазме. Из всех биологически значимых тяжелых металлов наиболее сильным действием на бактерии обладает железо, а цинк воздействует на дрожжи и другие грибы. Уровень железа в плазме млекопитающих понижается за счет переноса его в печень; в то же время уменьшается его всасывание в кишечнике [Weinberg, 1974]. При тяжелых хронических инфекциях уровень меди в плазме повышается на 50—90% [Weinberg, 1972]; это повышенное содержание меди, как полагают, является антипаразитическим.
Аналогичные явления отмечают у онкологических больных, у которых понижается уровень содержания железа в плазме [Konaka, Matsuoka, 1967] и повышается концентрация меди [Mortazani et al., 1972]. Иногда из-за наличия злокачественной опухоли понижается концентрация цинка в плазме [Davies, Musa, Dormandy, 1968], что, по-видимому, является реакцией организма, так как у многих животных наблюдали ингибирование роста опухоли при исключении из рациона цинка [De Wys, Pories, 1972].
С помощью каких же веществ происходит связывание катионов тяжелых металлов в живых клетках? Следует отметить, что таких веществ великое множество, но в первую очередь внимания заслуживают аминокислоты. Большинство аминокислот, входящих в состав белков, имеет примерно такие же константы связывания, как глицин (11.1). Необычно высокой способностью связывать металлы обладают две аминокислоты: гистидин (11.2) благодаря наличию в его молекуле имидазольного кольца и цистеин (11.3) за счет тиольной группы [Albert, 1952]. К этим двум примерам можно добавить цистин — эффективный, специфически действующий агент, способный связывать медь [Hawkins, Perrin, 1963]. В настоящее время очевидно, что белки связывают металлы в основном с помощью гистидиновых и цистеиновых остатков. Так рентгеноструктурный анализ миоглобина показывает, что между атомом железа и атомом азота имидазольного кольца существует прочная связь.
Компьютерный анализ результатов потенциометрического изучения 160 равновесных взаимодействий между Cu2+, Zn2+ и 17 аминокислотами (все присутствовали одновременно) показал, что 85% меди образует комплекс гистидин—медь—цис-144
теинв соотношении 1:1:1 и 67% цинка связано с цистеином л гистидином [Hallman, Perrin, Watt, 1971].
\ /СН2—CH—nh2
H2N—СН2—СО2Н | I CO2H HS—CH2—CH(NH2)CO2H
" v HN N
Глицин ' Гистидин Цистеин
(11.1) (11.2) (11.3)
В кровотоке аминокислоты связаны с очень незначительным количеством меди и цинка. Около 90% всего количества меди находится в связанном состоянии, образуя с белком церулоплазмином комплекс ярко-синего цвета, причем связь эта настолько прочна, что никакого обмена не наблюдается. Концентрации обменивающихся ионов меди и цинка в плазме крови составляют 1,1 мкМ и 15 мкМ соответственно [Agarwal, Perrin, 1976]. Почти все обменивающиеся ионы меди связаны с концевыми остатками альбумина NH2-Asp-Thr-His (в плазме быка) и NH2-Asp-Ala-His (в плазме человека). В связывании участвуют МН2-группы, два пептидных атома азота и имидазольный атом азота гистидинового остатка. Условная константа связывания (выражаемая как log К1) составляет 12,1 при pH 7,4 для комплекса медь — альбумин (1:1) и 7,6 для комплекса цинк — альбумин (1 : 1), хотя характер связывания цинка и меди различается. Цинк в отличие от меди легко удаляется из; плазмы крови при инъекции комплекса кальций—ЭДТА. (11.27).
Металл в качестве кофактора необходим примерно 12% всех известных ферментов. Какую же роль он играет в этих, случаях? По-видимому, металл часто служит мостиком между субстратом и белком, причем последний способен активировать металл, акцептируя его свободные электроны. Образование избыточного положительного заряда на атоме металла приводит к тому, что электронная плотность смещается от субстрата к металлу. При этом уменьшение свободной энергии активации субстрата происходит за счет изменения его электронной-структуры под действием металла.
Подобная картина наблюдалась и при исследовании двух содержащих марганец ферментов — креатинкиназы и мышечной енолазы методом ЭПР. Было показано, что у первого из них марганец образует связь с коферментом (АДФ) и с субстратом, но не с белком, а у второго — атом марганца, по-видимому, служит мостиком между субстратом и белком [Cohn, Leigh, 1962]. В других случаях роль металла, по-видимому, сводится исключительно к обеспечению необходимой третичной структуры белка (по аналогии с дисульфидным мостиком S—S). При образовании требуемой третичной структуры часто происходит сближение двух или трех остатков аминокислот, образующих активный центр, тогда как в вытянутой полипептидной цепи
10—734
145
эти остатки располагаются далеко друг от друга. Примером •может служить атом цинка в молекуле щелочной фосфатазы •бактерий Е. coli: при удалении атома цинка спектр кругового дихроизма этого фермента резко изменяется под действием 6М мочевины, так как природная конформация превращается в беспорядочную спираль [Trotman, Greenwood, 1971J. Подробнее об этом ферменте см. разд. 11.0.
Многие ферменты, например трипсин, функционируют без участия металла. Однако те ферменты, которым необходимы тяжелые металлы, обычно удерживают их очень прочно. Поэтому во многих случаях металл не удается удалить диализом и даже сильные хелатирующие агенты проникают в клетки микроорганизмов, не причиняя им вреда (разд. 11.7.1). Вероятнее всего, это обусловлено спецификой третичной структуры, а не высокими значениями констант устойчивости (см. выше).
Кроме аминокислот, белков и пептидов существует много других металлосвязывающих соединений, играющих важную роль во всех живых клетках. К ним прежде всего относятся птеридины (в том числе фолиевая кислота) и пурины, для комплексов которых с металлами были определены константы устойчивости [Albert, 1953; Albert, Serjeant, I960]; рибофлавин, обладающий наибольшим сродством к металлу в частично восстановленном состоянии [Hemmerich, Veeger, Wood, 1965], спермин (11.4) и диамины спермидин (11.5) и путресцин (11.6).
H2N(CH2)3NH(CH2)4NH(CH2)3NH2 H2N(CH2)3NH(CH£)4NH2
Спермин Спермидин
(11.4) (11.5)
H2NCH2CH2CH2CH2NH2
Путресцин (11.6)
Все виды фосфатов способны связывать металлы в живых клетках. Говоря о неорганических фосфатах, следует отметить, что для орто-фосфатов (Са2+, Mg2+, Zn2+, Cu2+) значения констант устойчивости были заново определены и выяснилось, что комплексы, образованные в соотношении 1 : 1, например СаНРО4, имеют тенденцию к образованию димеров типа Са2Н2(РО4)2 [Childs, 1970]. Данные для аналогичных пирофосфатов можно найти в работе Wolhoff, Overbeck (1959). Определены также константы устойчивости комплексов АТФ с Mg2+, Са2+, Mn2+, Со2+, Ni2+, Zn2+ и Cu2+ [Perrin, Sharma, 1966] и аналогичные величины для АМФ, АДФ, УТФ, ГТФ, ЦТФ, ИТФ [Phillips, 1966]. Как показывают потенциометрическое титрование и метод ЯМР 31Р, ионы магния прочно связаны с фосфатными группами АТФ [Tuck, Baker, 1973]. Константы устойчивости комплексов Си2+, Са2+ и Fe3+ с ДНК имеют тот же порядок, что и комплексы этих металлов с АМФ, имею-146
щим 'значительно меньшее сродство к катионам металлов, чем:-АТФ [\Bryan, Frieden, 1967]. Некоторые катионы более тяжелых металлов в природе связаны с молекулами нуклеиновых кислот. Предполагают, что ионы цинка образуют координационную связь с пуриновыми основаниями при обратимом скручивании и раскручивании спирали ДНК [Shin, Eichhorn, 1968]. Жирные кислоты цикла лимонной кислоты также связывают катионы металлов.
Насколько известно, константы устойчивости большинства перечисленных веществ близки по величине к константам устойчивости комплексов аминокислот (табл. 11.1). Только у порфиринов константы имеют более высокие значения; порфирины настолько прочно удерживают железо, что не удалось обнаружить никаких признаков обмена с его радиоактивным изотопом [Hahn et al., 1940].
Из вышесказанного ясно, что опыты с микроэлементами нельзя проводить в присутствии фосфатного или цитратного буфера. Для pH в пределах 7,0—8,2 удобным буфером, не образующим хелатных связей, служит N-этилморфин (температура кипения 138—139 °C). Подробнее о других буферах см. Perrin, Dempsey (1974).
Хелатообразующие соединения, избирательно действующие на организмы разных классов. Исследования в области сравнительной биохимии постоянно выявляют новые видовые различия в связывании и использовании металлов (разд. 4.2 и 11.0).
Аэробным бактериальным клеткам необходимо пополнять запас железа из окружающей их среды, где оно содержится, в основном в виде почти нерастворимого оксида железа (III) (количество растворяющегося вещества составляет 10~39 М). Однако существуют соединения, способные растворять и переносить железо, а также превращать его в биологически доступную форму. Эти так называемые сидерохромы содержатся в бактериях, дрожжах и грибах. Из них наиболее распространены феноляты, например энтерохелин (энтеробактин) (11.7) и гидрок-саматы, например дефероксамин В (11.9). Сидерохромы обоих типов представляют собой слабые кислоты (рКа около 9) и способны очень прочно связывать железо (III) (log К 45).
Наиболее важный сидерохром типа енолятов — энтерохелин, циклический триэфир N-(2,3-дигидроксибензоил)-Ь-серина. Это-соединение выделено из бактерий Е. coli [O’Brien, Gibson, 1970]. Gibson и Nielands (1974) установили механизм действия энтерохелина, использовав серию мутантов, каждый из которых был лишен способности осуществлять одну из биохимических стадий. Рентгеноструктурный анализ показал, что в молекуле энтерохелина каждый катион железа (III) связан с шестью атомами кислорода, предоставляемыми тремя фрагментами катехолдианионов (11.8), что и придает комплексу пурпурную окраску [Anderson et al., 1976]. Бактерии выделяют энтерохе-10* 147
лин, который образует комплекс с железом из окружающей среды; затем этот комплекс поглощается бактериями И/Тидро-лизуется специальными ферментами с образованием железа (O’Brien, Сох, Gibson, 1971J.
Фрагмент соединения (11.7).энтерохелин, образующий комплекс с железом (11.8)
H2N(CH2)5—N—С(СН2)2—CO-NH(CH2)5-N-C(CH2)2-CO-NH(CH2)5-N-G-Me
НО О
но о
но о
Деферонсамин (11-9)
1 R-алкильная цепочка(С^д)
Микобактин Т (11.10)
Дефероксамины, подобные соединению (11.9), представляют собой гексапептиды, выделяемые из Streptomyces. Рентгеноструктурный анализ показал, что шесть атомов кислорода, принадлежащие трем группам гидроксамовой кислоты, образуют тетраэдрическую структуру, связывающую ион железа (III). Для этого соединения log К, равный 31, значительно выше, чем для ЭДТА (11.27) (log К 24), но ниже, чем log К энтерохелина [Schwarzenbach, Schwarzenbach, 1963].
148
Сидерохром микобактин Т (11.10) встречается у возбудителей туберкулеза человека. Восемь других микобактинов были получены из других разновидностей рода Mycobacteria. Эти соединения имеют только две гидроксаматных группы, однако наличие фенольной группы, участвующей в трехцентровом связывании иона железа, делает комплекс микобактинов с железом таким же прочным, как и комплексы с другими сидерохро-мами [Snow, 1970J. Это единственные природные соединения железа, имеющие хорошую липофильность.
Значительные различия в использовании железа бактериями и млекопитающими открывают большие возможности для химиотерапии. О применении сидерохромов в качестве антидотов при отравлениях железом см. разд. 11.6.
Многие грибы, например Aspergillus aerogenes, выделяют энтерохелин, тогда как другие (в том числе Aspergillus niger и Penicillium reticulosum) удерживают железо с помощью феррихрома, циклического гексапептида, в молекуле которого ион железа прочно связан тремя гидроксаматными группами. Грибы содержат множество хелатирующих пиронов, таких, как койевая кислота (3-окси-7-пирон) (11.11), суммарное значение логарифма константы устойчивости (£)3) которой для железа (III) равно 25. Биологическая роль этих соединений неизвестна, так же как и роль полигидроксиантрахинонов, содержащихся в грибах. Все дрожжи выделяют производное гидроксамовой кислоты — родоторуловую кислоту (11.12) для доставки железа в клетки.
Микробный метаболизм железа см. обзор Nielands (1974).
О
ОН МеС—Nk ,СН2 о СН2 С°2Н СО2Н СО2Н Мугинеивая кислота (11.13) OH^JI Q-^CHaOH Койевая кислота (11.11) N ОН /СН2\ /СН2Ч | Y ЧСН/ \N—СМе х А А к СН^ N 0 0 Н Родоторуловая кислота (Н.12) 1 1 НО—О—СН2— СН—СО21 1 Г7 nh2 Тироксин (Н.14)
149
Таблица 11.1. Логарифмы констант устойчивости некоторых обозначены
Лигаид Р*а Fea+ Си2*
Глицин (11.1) 9,9; 2,4 10 8,5; 215
Цистеин (11.3) 10,3; 8,4; 2,0 см.1 см.1
Гистидин (11.2) 9,2; 6,0; 1,8 ? 10,5; 219
Гистамин (7.6) Этилендиамин 9,8; 6,0 4 10; 216
(рис. 11.2) 10,1; 7,0 р 11; 2202
ЭДТА (11.27) Птероилглутаминовая 11,0; 6,3; 2,7; 2,0 24 19
кислота (фолиевая кислота (см. 9.20) 8,3 ? 28
Гипоксантин (9.52) 8,9; 2,0 ? 6
Гуанозин 9,3 ? 6
Аденин (4.3) 8-Гидроксихннолин (ок- 9,8; 4,3 ? 214
син) (11.30) орто-Фенантролин 11,2; 3,8 12; 224; 336 12; 223
(11.18) 4,9 314 220
Бипиридил (11.19) 4,3 р 317
Щавелевая кислота 4,2; 1,2 10 6
Салициловая кислота 13,4; 3,0 16; 228 11; 219
Тетрациклин (11.36) Гидразид изоникотнно- 9,6; 7,8; 3,4 10; 325 8; 213
вон кислоты (изониазид) (11.37) 10,8; 3,5; 1,9 ? 8
Диметилдитиокарбами-
новая кислота (3.57) 3,2 р 11; 222
1 Цистеин окисляется этим катионом.
2 Ср. Ч1 для Си+.
4 Для Са2+ также ~3.
Комплекс марганца с бнпиридилом 2 : 1 (log р 6) обозначен как 26, а ЭДТА 1 : I
Данные для тетрациклина см. [Albert, Rees, 1956], изониазида [Albert, 1956], сали днметнлдитнокарбаминовой кислоты [Janssen, 1958], аминокислот [Albert, 1950, 1952],
Растения также содержат специальные химические соединения, способные связывать железо. Многие злаковые культуры выделяют в почву мугинеивую кислоту (11.13), способную образовывать с ионом железа (Fe3+) комплекс, который затем поглощается и используется растениями. Мугинеивая кислота представляет собой производное азотсодержащего циклического соединения азетидина. В отсутствие необходимого количества железа у растений наступает хлоротическое состояние и они увядают [Mino et al., 1981].
Некоторые природные агонисты млекопитающих, а именно тироксин (11.14), норадреналин (7.5) и гистамин (7.6) способны связывать металлы. Только у млекопитающих в митохондриях накапливается кальций.
150
комплексообразующих агентов (в воде при 20 °C; насыщенные константы как Jog р)
Ni2+ Zn!+ Со2+ Fe2+ Мп2* | Mg2*
6; 211 5; 29 5; 29 4; 28 3; 25,5 2; 24
10; 219 10; 218 316 6 4 <4
9; 216 7; 212 7; 213 5; 29 3,5 <4
7; 211 5; 29 5; 29 4 3 ?
8; 318 6; 312 6; 314 4; 39,5 3; 25 0,4
18 16 16 14 13 9
29 27,5 28 28 26 ?
5 7 7 4 ?
4 4,5 3 4 3 3
4 213 4; 28 ? 3 3
10; 218 8,5; 216 9; 217 8; 215 7; 212 4,5
324 317 320 321 310 1
320 313 316 317,5 26 0,5
5,5 5 4,5 4,5 4 З3
7; 212 7 7; 211 6; 211 6; 210 ?
6; 211 5; 29 5; 210 5; 29 4; 28 4
5,5 5 5 ? ? ?
? 29 ? ? ? ?
(log р 13) — как 13.
цнловон кислоты [Perrin, 1958], бнпириднла и орто-фенантролнна [Irving, Mellor, 1962], остальные значения [Sillen, Martell, 1964, и дополнения].
11.2. Химизм хелатообразования
По сравнению с ферментами, обладающими высокой специфичностью в отношении определенного металла, среди синтетических металлосвязывающих агентов подобная избирательность встречается значительно реже. Металлы по своему сродству к большинству хелатирующих агентов располагаются примерно в следующем порядке (от наибольшего сродства к наименьшему):
Fe3+, Hg2+, Cu2+, Al3+, Ni2+, Pb2+, Co2+, Zn2+, Fe2+, Cd2+, Mn2+, Mg2+, Ca2+, Li+, Na+, K+.
Впервые общую тенденцию такого рода обнаружили Mellor и Maley (1947). Некоторые из приведенных двухвалентных металлов расположены друг за другом в периодической системе
151
следующим образом (в скобках указаны атомные номера): М.п (25), Fe (26), Со (27), Ni (28), Си (29), Zn (30). В этом ряду, называемом первым рядом переходных элементов, сродство к хелатирующим агентам последовательно увеличивается, достигая максимума у меди (Си2+). Повышение сродства к хелатирующим агентам является следствием уменьшения ионного радиуса [Irving, Williams, 1953]. Эта зависимость позволяет объяснить последовательность расположения металлов в приведенном выше ряду 11.1. С увеличением валентности металла происходит уменьшение его радиуса, поэтому неудивительно, что Fe3+ имеет большее сродство, чем Fe2+, а ионы трехвалентных металлов располагаются в начале ряда, тогда как одновалентные в конце (ионные радиусы приведены в. табл. 11.3). Тем не менее выполнению этого простого правила иногда препятствуют некоторые вторичные факторы, хотя приведенный порядок увеличения сродства металлов к хелатирующим агентам сохраняется для большинства лигандов.
Особого внимания заслуживают те немногочисленные хелатирующие агенты, которые не подчиняются общему правилу. Прежде чем перейти к обсуждению таких исключений, следует рассмотреть принципы нормального хелатообразования. Значение тяжелых металлов в биологии обусловлено их способностью образовывать связи более прочные, чем обычные ионные, поскольку они отчасти координационные (разд. 8.0).
Некоторые немногочисленные лиганды (так называемые мультидентатные, например, ЭДТА) образуют с ионом металла более чем одно хелатное кольцо (комплекс 1:1). Рассмотрим три основные вида бидентатных лигандов, т. е. образующих одно кольцо в комплексе 1:1. Некоторые бидентатные лиганды (например, этилендиамин и 2.2/-бипиридил) содержат две электронодонорные группы, в этом случае заряд катиона металла при образовании хелатного соединения не изменяется. Лиганды могут содержать также одну электронодонорную и одну анионную группу, как в глицине. В этом случае заряд металла уменьшается на единицу. И наконец, лиганд может содержать две анионные группы (например, щавелевая кислота), в этом случае заряд металла уменьшается на две единицы. Образование хелатных связей атомами кислорода и азота происходит обычно лишь в тех случаях, когда при этом получаются пяти- и шестичленные циклы. Пятичленные циклы значительно стабильнее. Однако при образовании хелатных связей через серу возникают стойчивые четырехчленные циклы [Peyronel, 1940; Deskin, 1958]. На рис. 11.2 представлены три основных типа хелатных соединений. Стрелки в кольце показывают направление смещения в норме неподеленной пары электронов от атомов О, N или S к металлу. В присутствии избытка лигандов могут образовываться комплексы в соотношении 2:1. Лиганды типа щавелевой ислоты используют заряд для образования комплексов 1 : 1, эднако последние могут соединяться в дальнейшем с комплек-152
сами этилендиаминового типа, образуя комплексы смешанного типа [например, Watters, DeWitt, 1960]. Комплексы 1:1 глицинового типа способны соединяться с другими лигандами этого типа. Комплекс 1 : 1, образованный лигандами, подобными этилендиамину, может соединяться с любым из трех типов ли-
гандов.
Координационное число двухвалентной меди обычно равно четырем и полное насыщение наступает при соединении ее с двумя молекулами лигандов (одинаковыми или разными). Это справедливо также для двухвалентных металлов — кальция, магния и марганца. Однако координационное число двухвалентных ионов железа, кобальта и цинка равно шести для лигандов типа этилендиамина, а у трехвалентных ионов оно равно шести также и для лигандов типа глицина. Располагаясь вокруг металла, лиганды образуют разные конфигурации в зависимости от направления валентностей катионов металла. Большинство перечисленных выше металлов обычно образуют тетраэдрические комплексы с основными типами лигандов, изображенными на рис. 11.2, но для Си2+ предпочтительнее образование планарных комплексов, а для Fe2+ и Fe3+— октаэдрических комп-
лексов.
Следует подчеркнуть, что понятие «лиганд» относится не ко всем присутствующим в системе молекулам органического соединения, а только к той их части, которая находится в соответствующей ионной форме, т. е. может связывать катион металла. В случае этилендиамина, глицина и щавелевой кислоты
лигандами могут служить неионизированные молекулы, моно-и дианионы соответственно. Поэтому, если при сравнении относительной реакционной способности лигандов при физиологических условиях пользуются константами устойчивости, необходимо учитывать и значения рКа лигандов (разд. 11.3).
Сера может входить в простые четырехчленные циклы, такие как в диметилдитиокарбамате меди (II) (11.15), образующемся из ионов двухвалентной меди и диметилдитиокарбами-
(Me)2N—C=S
। -L
S—Cu+
Комплекс
цинк—меркаптоуксусная кислота
(11.16)
Комплекс
медь — диметилдитиокарбамат
(11.15)
новой кислоты (3.57). Однако серосодержащие лиганды способны образовывать также равновесные смеси из многих комп-
153
Рис. 11.2. Три основных типа комплексов 1:1.
лексов. Так, цинк с меркаптоуксусной кислотой (L2-) образуют следующий ряд комплексов:
ZnL, ZnL22-, ZnL34“, Zn2L32-, Zn3L42-.
Последний член этого ряда (2пзЬ42~), изображенный формулой (11.16), наиболее устойчив. Сведения об этих соединениях были получены при обработке данных титрования на электронно-вычислительной машине. Таким же образом для цистеина (L2-) был рассчитан следующий ряд комплексов с цинком [Perrin, Sayce, 1968]:
ZnHL2~, ZnH2L2, ZnL22~, ZnL42-.
В более кислых растворах присутствуют также протонированные формы комплекса цинк — цистеин [Shindo, Braun, 1965]. Очевидно, что лиганды, содержащие серу, образуют более сложные комплексы, чем кислород- и азотсодержащие лиганды; которые, однако, более широко распространены.
О конформациях и абсолютной конфигурации комплексов с хелатными связями см. Hawkins (1971). Основные закономерности образования и применения хелатных соединений с металлами см. Bell (1977) и Perrin (1964) и в серии книг Sigel (после 1973).
11.3. Количественные аспекты связывания металлов
Для оценки прочности связей (варьирующейся в широких пределах для разных комплексов) применяют константы устойчивости, характеризующие равновесие между одним или несколькими лигандами и одним ионом металла, подчиняющееся закону действия масс. Так, для комплекса глицина с двухвалентной медью в соотношении 1 : 1 константа устойчивости (Ki) рассчитывается следующим образом:
[H2N—СН2—СО2—Си+]
1 [Cu24 [H2N-CH2-CO2-] ’
а для комплекса 2: 1 она может быть выражена так:
к = [H2N—СН2—СО2СиО2С—СН2—NH2]
2 [H2N—СН2—CO2Cu+] [H2N—СН2—СО2"] ‘
154
В каждом случае в числителе находится концентрация комплекса, а в знаменателе — концентрация образующих его компонентов.
Часто необходимо знать общую константу устойчивости (0), представляющую собой произведение частных констант. Произведение двух частных констант обозначают как 02, трех — 03.
Константы устойчивости обычно определяют потенциометрическим титрованием лигандов (со стеклянным электродом) в присутствии и в отсутствие металла и обработкой результатов посредством довольно сложных вычислений [подробнее см. Albert, Serjeant, 1984].
Вкратце метод заключается в следующем: кислотная группа возможного хелатообразующего агента титруется щелочью, причем значение pH регистрируется после добавления каждой десятой доли эквивалента. Затем титруется смесь (1:1) исследуемого вещества и соли (например, перхлората или нитрата меди). Если образования комплекса не происходит, то новая кривая последовательно воспроизводит индивидуальные кривые обоих компонентов (рис. 11.3). Если же комплекс образовался, то катионы водорода, выделившиеся в процессе хелатообразования, сдвигают кривую в сторону более низких значений pH (рис. 11.4). Вещества в форме солей с кислотами, не имеющие кислотного характера, можно титровать щелочью.
Потенциометрический метод можно с некоторыми предосторожностями использовать даже для разбавленных растворов слаборастворимых веществ, например, титрование 0,0001 М раствора аденина в присутствии ионов двухвалентной меди (8 мкМ) в воде позволяет получить достаточно точное значение константы устойчивости [Albert, Serjeant, 1960]. При определении констант веществ, имеющих биологическое значение, нельзя использовать никакие другие растворители, кроме во-
155
Рис. 11.4. Кривые потенциометрического титрования, свидетельствующие о том, что глицин является сильным хелатообразующим агентом (А — глицин, 0,01 моль; Б — нитрат меди, 0,005 моль; В — глицин, 0,01 моль+нитрат меди, 0,005 моль).
ды. Применение смесей воды и органических растворителей приводит к неверным результатам [Albert, Serjeant, 1984].
При потенциометрическом титровании стеклянный электрод иногда с успехом можно заменить медным, если объектом исследования служит именно этот металл. Для особенно труднорастворимых комплексов вместо потенциометрического титрования применяют обменные методы, например, два лиганда конкурируют за один металл или, наоборот, два металла — за
один лиганд. Измерения удобно производить, если один из компонентов содержит изотопную метку [Schubert, 1956]. Распределительные методы — методы, основанные на различиях в распределении между двумя несмешивающимися жидкостями, пригодны в тех случаях, когда значения констант настолько велики, что кажущаяся концентрация свободного [Н+] выше, чем общая концентрация [Н+] (свободного и связанного); в этом случае потенциометрию использовать нельзя. Вкратце рекомендуется следующее: раствор лиганда известной концентрации в несмешивающемся с водой растворителе встряхивают с водным раствором катиона металла (известной концентрации в водном буфере). Затем концентрацию лиганда в неводном растворе измеряют спектрофотометрически [McBryde, 1967]. Только УФ-спектроскопия четких результатов не дает. В особых случаях используют другие методы, такие как потенциометрия с катионселективными электродами, инфракрасная спектроскопия в D2O, методы ЭПР и ЯМР дисперсию, оптическое вращение, кондуктометрия, метод термической релаксации (температурного скачка) и полярография. Однако для общих целей предпочтительнее всего метод прямой потенциометрии.
Рассчитать значения констант устойчивости по данным потенциометрического титрования не представлялось возможным до тех пор, пока Bjerrum (1941) не показал, что они связаны с двумя переменными (п) и [L] следующим уравнением:
(1-п) [L]’
156
где [L] — концентрация лиганда, точнее той формы комплексообразующего агента, в которой он участвует в образовании хелатного соединения. Эту величину нельзя рассчитать, зная только значение [А] — концентрацию комплексообразующего агента в момент начала титрования. Так, в случае глицина в-значение [А] входят концентрации трех ионных форм глицина и неионизированной формы, т. е. полная концентрация глицина. А значение [L] относится только к концентрации аниона глицина, так как только он участвует в образовании хелата. Величину й можно определить как долю лиганда, связанную с одним атомом металла. Для комплекса состава 1 : 1 эта величина находится в пределах от 0 до 1.
Величины [L] и й изменяются в процессе титрования и рассчитываются следующим образом:
[L] = ([А] - [Н+] + [ОН-] + [НС1] - [КОН]) :Р n=([A]-Q[L]):[M0],
где [Мо]—начальная концентрация металла, а [L] и [А] были определены выше; [Н+] и [ОН-] варьируются в зависимости от pH, a [HC1J и [КОН] также варьируются в зависимости от используемого титранта. Остается определить только величины Р 11 Q:
Р= [Н+]:Ка + 2[Н+р:КаК'а
Q = [Н+] :Ка + [Н+]2:КаК'а + 1,
где Ка — первая константа ионизации лиганда.
Простое объяснение процесса, происходящего во время титрования, заключается в том, что по мере добавления щелочи все большее число ионов водорода отрывается от хелатирующего агента, замещаясь катионами металла. Однако, как показывают приведенные выше уравнения, их соотношение не является линейным.
Величина константы устойчивости не зависит от pH, поэтому применение уравнения (1) дает один и тот же результат в пределах значения й от 0 до I. При й>1 следует использовать соответствующее уравнение для Кг:
_
(2-п) [L]
Так как величина f) может быть представлена произведением индивидуальных констант устойчивости, то для комплекса-меди с глицином и во всех случаях, в которых происходит насыщение валентности катиона металла после соединения с двумя молекулами лиганда, ее можно выразить как f) = Ki Кг-В случае комплекса цинка с этилендиамином насыщение происходит при соединении цинка с тремя молекулами лиганда, поэтому Р = К1КгКз, а для комплекса с металлом, валентности которого насыщаются при соединении с одним лигандом (например, цинк и ЭДТА), & = Ki-
157
Для многих комплексов, в которых металл связан с двумя змолекулами лиганда, по данным титрования можно построить график, где по одной оси откладываются величины^: (n J)[L], .а по другой — соответствующие величины (2—n)[L]:(n—1), .в результате чего получается прямая линия, угол наклона которой равен а отрезок, отсекаемый на оси ординат, равен —Кь Эту процедуру удобнее выполнять с использованием микрокалькулятора, снабженного программой обработки данных по методу наименьших квадратов [Albert, Serjeant, 1984].
Поскольку некоторые соединения металлов с лигандами могут образовываться не столь обычным путем, какой мы рассматривали до сих пор, каждое титрование следует повторить минимум два раза для того, чтобы изучить два разных соотношения металла с лигандом. Если получаются одни и те же значения log Ki и р, то любыми конкурентными равновесиями можно пренебречь.
Типичными случаями отклонения от нормы являются: а) образование хелатных соединений цвиттерионами аминокислот и получение в результате насыщенного положительно заряженного комплекса («протонированный комплекс») и б) комплексы, в которых металл образует связи не только с лигандами, но и с гидроксил-ионом («гидратированные комплексы») [Perrin, Sharma, 1967]. В таких случаях следует прибегать к более тщательным расчетам с помощью компьютера [Leggett, 1983].
Интерпретация результатов титрования растворов, в которых одновременно присутствуют один металл и два различных лиганда, не представляется сложной [Perrin, Sayce, Sharma, 1967; Perrin, Sharma, 1968, 1969]. Если два лиганда А и В «обладают одинаковым сродством к металлу М, то вероятность образования смешанного комплекса АМВ в два раза выше, чем простого комплекса типа АМА. Аналогично, если в растворе присутствуют три подобных лиганда (А, В и С), вероятность образования комплекса МАВС выше в 6 раз [Watters, De Witt, 1960].
Константы устойчивости для некоторых веществ, имеющих биологическое значение, приведены в табл. 11.1. Значения констант устойчивости других комплексных соединений см. Sillen, Martell (1964, 1971) и Perrin (1979).
Константу устойчивости соединения невозможно предсказать точно до завершения его синтеза и выполнения соответствующих измерений, однако можно предсказать приблизительные значения первой, руководствуясь следующим положением: в любом ряду соединений чем прочнее лиганд связан с ионами водорода (что определяется значениями рК), тем прочнее он будет связывать металл. Если константы ионизации еще не описаны в литературе, их можно рассчитать приблизительно [Perrin, Dempsey, Serjeant, 1981]. Однако следует подчеркнуть, что это правило подходит только для серии близких по строению веществ и перестает действовать в тех случаях, когда по-(158
являются стерические помехи вследствие введения объемных: заместителей (см. ниже).
При сравнении соединений, принадлежащих к различным сериям органических соединений, следует учитывать, что наиболее общим фактором, влияющим на соотношение рКа: log Ki,-является сопряжение в хелатном цикле. В таких случаях очень важно правильно установить положение двойных связей в цикле, так как факторы, уменьшающие кратность связей, обычно уменьшают и устойчивость [Calvin, Wilson, 1945]. Например, в комплексе меди с ацетилацетоном хелатный цикл содержит две целочисленные двойные связи и поэтому связи с металлом более прочны, чем в комплексе меди с салицилальдегидом, в котором из-за сопряжения с бензольным кольцом есть «полуторные» связи.
Помочь предсказывать константы устойчивости сможет развитие теории поля лигандов. В основу этого подхода положен постулат теории кристаллического поля, смысл которого заключается в том, что пять d-орбиталей тяжелых металлов, в норме-вырожденные, дифференцируются, попадая в электростатическое поле лигандов. В частности, энергия d-орбиталей, направленных в сторону лигандов, возрастает, а направленных в противоположную — снижается. Донорные электроны лигандов отталкивают d-электроны металла; это отталкивание уменьшается за счет перемещения d-электронов на d-орбитали, наиболее удаленные от лигандов. Для ароматических лигандов и некоторых металлов (особенно железа, никеля и кобальта) необходимо сделать поправку на наличие л-связей. С учетом этой поправки теория кристаллического поля в сочетании с теорией молекулярных орбиталей образует так называемую теорию поля лигандов [Basolo, Pearson, 1967].
Ранжирование при фиксированных значениях pH. Если члены данного ряда хелатообразующих агентов имеют почти одинаковые константы ионизации, то по относительным величинам констант устойчивости можно определить их способность к связыванию с различными катионами металлов при любом данном значении pH. Однако для хелатообразующих агентов с сильно различающимися константами ионизации константы устойчивости не могут служить критерием оценки сродства к катионам металлов. Это объясняется способностью таких веществ ионизироваться в разной степени при данном значении pH. Вещества, обладающие меньшим сродством к металлам (на что указывает более низкая константа устойчивости), за счет различий в значениях рКа могут образовываться значительно больше анионов, чем другие агенты. В этом случае вещество, обладающее меньшим сродством к металлу, может присоединять большее количество катионов металла, чем вещества, у которых это сродство больше. Это объясняется тем, что для хелатообразования необходимо не только наличие сродства между лигандом и металлом, но также и быстрое образование
159л
Т а б л и ц а 11.2. Распределение катионов между некоторыми лигандами в нейтральных растворах
Лиганд Cu2+ Fe2+ Fea+
Глицин Салициловая кислота Оксин 45(8,5) 1(10,5) 200 000(12,2) 30(4,3) 1(6,6) 150 000(8,0) 1(10,0) 400(16,4) 200(12,3)
Первые цифры — пропорциональные числа, в скобках приведены логарифмы констант устойчивости комплексов состава 1 : 1; столбцы читать по вертикали [Perrin, 1958].
анионов лиганда из агента (или молекул лиганда, если агент — основание). Такого рода конкуренция между константами устойчивости и константами ионизации иллюстрирует табл. 11.2. В случае двупротонной кислоты, например щавелевой, хелатообразующим является дианион; в случае амфолита, например глицина, хелатное соединение образует моноанион, а если агент представляет собой двукислотное основание, например этилендиамин, то хелат образуется нейтральной молекулой.
Для того чтобы рассчитать количество металла (С), связанного при данном pH, можно использовать следующие формулы (I — для комплексов 1:1 и II — для комплексов 2:1):
с = lOOKjWA
1 + KiWA + KiK2W2A2 W
= 100KjK2W2A2
1 + К^А + KiK2W2A2 ’
Здесь А — концентрация лиганда, не участвующего в образовании комплекса. Если имеется большой избыток лиганда, то величину А можно заменить общей концентрацией лиганда. Можно также рассчитать значение А методом последовательных приближений. При этом:
W= 1:Ц + 10<РКа1-рН) + io<PKal+PKM-2PH)lb
Значения констант устойчивости см. Sillen, Martell (1964, 1971), Perrin (1979), Martell, Smith (1974—1982).
11. 4. Химические различия, способствующие избирательности
Наряду с закономерностями, определяющими сродство к металлу, существует ряд явлений, не подчиняющихся этим закономерностям. Избирательность биологического действия некоторых ионов часто зависит именно от таких исключений из правила.
В большинстве случаев образующиеся хелатные комплексы представляют собой тетраэдры, но железо (и двух- и трехва-160
Таблица 11.3. Кристаллические радиусы катионов (нм) [Pauling, 1960]
Катион металла Радиус Катион металла Радиус Катион металла Радиус
Li+ 0,060 Be2+ 0,031 AP+ 0,050
Na+ 0,095 Mg2+ 0,065 Fe3+ 0,064
К+ 0,133 Са2+ 0,099 Ca3+ 0,062
Rb+ 0,149 Sr2+ 0,113 Tl3+ 0,095
Cs+ 0,169 Ва2+ 0,135 Co»+ 0,063
Cu+ T1+ Ag+ NH<+ 0,096 0,140 0,126 0,148 Mn2+ Fe2+ Co2+ Ni2+ Cu2+ Zn2+ Cd2+ Hg2+ Pb2+ 0,080 0,076 0,074 0,072 0,072 0,074 0,097 0,110 0,120 (J4+ Pb4+ 0,097 0,084
лентное) образует октаэдрические комплексы. Однако наибольшей избирательностью обладают ионы меди, образующие плоские комплексы. Это означает, что медь чрезвычайно чувствительна к стерическим изменениям, обусловленным присутствием объемных заместителей. Так, в табл. 11.1 константы устойчивости комплексов меди (II) с бипиридилом (11.19), фенантролином (11.18) и фолиевой кислотой расположены ниже констант для соответствующих комплексов никеля, что противоречит последовательности, обсуждавшейся в разд. 11.2.
Стерический эффект другого типа возникает, когда объемные заместители располагаются вблизи хелатообразующих групп. Это препятствует сближению двух лигандов настолько, чтобы стало возможным образование прочного комплекса в соотношении 2: 1, если же катион металла имеет небольшой диаметр, то он вообще не может быть захвачен. Так, 2-метилоксин (11.17) связывает все катионы значительно менее прочно, чем оксин, причем эта разница максимальна для иона алюминия [Irving, Rossotti, 1956], имеющего наименьший кристаллический радиус (0,05 нм) по сравнению со всеми обычными металлами (табл. 11.3). Две молекулы 2,9-диметил-орто-фенантролина (11.18, R—СНз) стерически мешают друг другу настолько, что не могут удержать ион железа (II). В этом случае взаимное отталкивание двух молекул лиганда приводит к тому, что два атома азота не могут участвовать в образовании октаэдрического комплекса с ионом железа (II); даже плоский комплекс с медью (Си2+) образуется с трудом. Тем не менее такое расположение молекул лиганда благоприятно для связывания катиона Сп+ (образующего тетраэдрические комплексы). Такие комплексы одновалентной меди отличаются большой устойчивостью, что позволяет использовать их для анализа.
11—734
161
он
2-Метилоксин (11.17)
орто-Фенантролин (R = H)
(11.18)
2,2'-Бипиридил (11.19)
О О
II II
СО" "ОС
HgC^ \н2
HgC drigj
\:сг -ос^
Два днаниоиа янтарной кислоты
(11.20)
Рассмотрим теперь лиганды, объем которых слишком велик для того, чтобы они могли сблизиться и образовать достаточно устойчивый комплекс в соотношении 2:1с катионом небольшого диаметра. Радиус иона магния на 0,034 нм меньше радиуса иона кальция (см. табл. 11.3). Обычно Mg2+ связывается прочнее, чем кальций, но для лигандов, молекулы которых слишком велики для того, чтобы их пара могла удержать металл, эта последовательность нарушается. Такой эффект характерен для всех дикарбоновых кислот, например, для янтарной кислоты в (11.20) (см. также щавелевую кислоту на рис. 11.5). Однако винная кислота отличается тем, что связывает Са2+ значительно прочнее, чем Mg2+ [Williams, 1952]. Радиусы некоторых ионов приведены в табл. 11.3. Следует отметить, что катионы Fe2+, Со2+, Ni2+ и Zn2+ имеют настолько сходные диаметры, что их нельзя разделить, пользуясь этим стерическим эффектом.
Величины радиусов негидратированных катионов постоянно пересматриваются [Ladd, 1968]. Все используемые методы основаны на заключениях и предположениях. Наиболее точными и широко используемыми остаются данные, приведенные в табл. 11.3.
Следует помнить, что в химии хелатных соединений применяют именно радиусы негидратированных ионов (радиусы гидратированных ионов приведены в табл. 12.2). Реакции, обсуждавшиеся до сих пор, контролируются равновесным стерическим эффектом. Подробнее о кинетическом стерическом эффекте см. Wilkins (1962).
Избирательное усиление прочности связи с железом было обнаружено у некоторых ароматических лигандов. Оно обуслов
162
лено образованием «обратной двойной связи», т. е. л-связи за счет передачи железом неиспользованных электронов Зб-орби-тали (при благоприятном ее расположении) на свободные молекулярные орбитали лиганда. При этом смещение электронной плотности происходит в направлении, показанном в формуле (11.22), вместо обычного смещения, как в (11.21). Вследствие этого повышается прочность связи между металлом и лигандом и возрастает устойчивость. Ион двухвалентного железа обычно образует парамагнитные почти бесцветные комплексы, однако с гетероароматическими лигандами, содержащими азот, соединенный двойной связью, Fe2+ образует интенсивно окрашенные комплексы (обычно красного цвета), не обладающие парамагнитными свойствами. Этот эффект можно наблюдать на примере бипиридила, фолиевой кислоты и орто-фенантроли-на (табл. 11.1), у которых константы устойчивости комплексов с двухвалентным железом значительно выше, чем у комплексов с цинком. Никель и кобальт также способны образовывать «обратные двойные связи», но в меньшей степени.
Сравнение значений log К для орто-фенантролина и 2,2'-би-пиридила (табл. 11.1) показывает, что они подчиняются двум закономерностям, рассмотренным выше. Ион Си+ обычно образует комплексы с планарной структурой, третья молекула ли-
RT2
Fe2+ Ч <7
->N 1 I -Fe»+
Обычное смещение «Обратная двойная связь»
электронной плотности
(11.21) (11.22)
ганда включается в комплекс с большим трудом. Поэтому двухвалентная медь занимает в таблице необычно низкое место — ниже Ni2+. Необычно высокие значения имеет ион двухвалентного железа. Это обусловлено очень высоким значением третьей частной константы (Кз), так как в этом случае возможно образование «обратной двойной связи» в комплексе со спаренными спинами. Именно поэтому Fe2+ занимает в таблице необычно высокое место — выше Zn2+ и Со2+. Вследствие комбинации указанных принципов Fe2+ образует такие же прочные связи, как и Cu2+ [Irving, Mellor, 1962].
Другой характер имеют кривые зависимости констант устойчивости от атомного номера в ряду: кальций, магний, марганец, железо, кобальт, никель, медь и цинк (рис. 11.5). Наиболее резкий подъем кривых отмечается для тех лигандов, у которых в образовании хелатов участвуют два атома азота (например, для этилендиамина). Меньше крутизна кривых для тех лигандов, у которых в образовании хелатов участвуют один атом азота и один атом кислорода (например, для глицина). Наиболее 11* 163
Рис. 11.5. Относительное сродство двухвалентных катионов к кнслород-и азотсодержащим хелатирующим агентам (на осн координат отложены логарифмы первых констант устойчивости комплексов, по осн абсцисс — катионы металлов с возрастающими атомными номерами).
пологие кривые наблюдаются для лигандов, у которых в образовании комплексов участвуют два атома кислорода (например, для щавелевой кислоты) [Irving, Williams, 1953]. Кривые для соединений, принадлежащих к этим трем классам, пересекаются между Мп2+
и Fe2+ (см. рис. 11.5). Не только Mg2+ и Са2+, но и Fe3+, Т13+, Мо5+ и V5+ проявляют большее сродство к кислороду, чем к азоту.
Большинство металлов легче соединяется с лигандами, содержащими кислород, чем с серосодержащими лигандами. Однако Cu+, Ag+, Au+, Hg2+, As3+ и Sb3+ отдают предпочтение сере; именно этим объясняется успешное применение димерка-прола (11.23) в качестве антидота при отравлениях некоторыми из этих катионов. У трех других катионов Cu2+, Ni2+ и Со2+ сродство к сере несколько выше, чем к кислороду, если сера находится в неионизированном состоянии, как, например, в органических сульфидах.
Другим фактором, влияющим на относительное сродство ряда металлов, служит изменение окислительно-восстановителъ: ного потенциала металлов1, вызванное образованием хелатных соединений с металлами, имеющими переменную валентность (например, Си, Fe, Со, Мп, Mo, V). Вследствие хелатообразования такие металлы могут даже изменить первоначальную валентность на более высокую или низкую (количественные дан-
’ Окислительно-восстановительный потенциал (Ео) —это константа, рассчитанная при pH 0 по следующему уравнению:
_ _ RT , [окисл. форма]
Ео = Ен ~ 37 1п "7 Т 7“, пг [восст. форма]
где Е„—потенциал, измеренный экспериментально; R — универсальная газовая постоянная; Т—абсолютная температура; F — число Фарадея; п—число участвующих электронов, а In — натуральный логарифм. Прн любом pH, отличном от 0, используют символ Е' н указывают pH. Потенциалы, определенные с помощью полярографии, часто измеряют относительно «насыщенного каломельного электрода»; для того чтобы перевести их в упоминавшуюся выше шкалу (относительно нормального водородного электрода), необходимо прибавить 0,246 В.
164
Таблица 11.4. Влияние лигандов на окислительно-восстановительный потенциал (в основном при 20 °C)
Ион металла Ео, в Лиганд* Ео, в
Fe(H2O)62+ = Fe(H2O)63+ +0,77 ЭДТА ’Салициловая кислота 6Цианид-нон Глицин 1'28-Гидроксихннолин 2Окситетрацнклин 3Бипириднл ’орто-Фенантролин +0,14 +0,20—0,22 +0,36 +0,38 +0,52+0,27 +0,57+0,43 +1,06 +1,06
Со(Н2О)62+= —0,83 —0,22 +0,14
= Со(Н20)6’+ +1,84 ®Цианид-ион 3Этнлендиамин 6Аммиак
Си(Н2О)х+ = —0,38 —0,16 +0,12 +0,19+0,24 +0,20+0,27 +0,34 +0,26+0,35
= Си(Н2О)42+ +0,71 2Этилендиамин 2Глицин 2Бипиридил 1 2Метилтиоэтиламин ‘•Пиридин 2Аммиак 2Имидазол
1 Индексы перед названиями лигандов имеют то же значение, что и в табл. 11.1 (например, 8Бипирндил означает комплекс бнпнрнднла 3; 1). Большинство данных взято из работ Perrin (1959), Hawkins, Perrin (1962).
ные см. в табл. 11.4). Следует помнить, что величины потенциалов колеблются от +2 В для наиболее сильных окислителей до —2 В для самых сильных восстановителей. В качестве примера можно привести соединения кобальта. Соли двухвалентного кобальта обычно устойчивы в водных растворах, тогда как соли трехвалентного кобальта мгновенно разлагаются водой с выделением кислорода. Тем не менее после образования хелатного соединения с этилендиамином потенциал падает настолько резко, что комплекс с двухвалентным кобальтом легко окисляется до более стабильного соединения с кобальтом (III).
Потенциал комплекса определяется: 1) ионным зарядом на лиганде; 2) «обратной связью» металла с лигандом и 3) эффектом кристаллического поля [Perrin, 1959]. Например, относительно пункта 1) известно, что при анионном характере лиганда предпочтительнее высшая валентность металла; относительно пункта 2)—что образованию «обратной связи» железа с лигандами способствует низшая валентность металла, и пункта 3)—что для эффекта кристаллического поля предпочтительнее оказываются Си и Fe в двухвалентном состоянии. К сожалению, на основе этих фактов пока еще нельзя предсказывать потенциал комплекса без проведения необходимых измерений.
165
Из данных табл. 11.4 видно, что некоторые комплексы являются значительно более сильными окислителями, чем свободные катионы металлов. Такие комплексы способны вызывать разрушение природных метаболитов, например, хелаты меди и диэтилдитиокарбаматов разлагают тиоктовую кислоту (2.28) [Sijpesteijn, Janssen, 1959].
Рассматривая другие аспекты взаимодействия металл — лиганд, следует отметить, что металл может изменять избирательность органического лиганда: а) влияя на распределение электронов в лиганде; б) повышая реакционную способность активного центра лиганда; в) вызывая изменение конформации лиганда; г) обеспечивая возможность присоединения или отрыва электрона; д) увеличивая липофильность лиганда и, следовательно, его способность проникать в живую клетку.
В разд. 14.2 рассматриваются ионофоры, способные без образования хелатных соединений осуществлять транспорт одновалентных ионов (например, Na+) через мембраны.
11.5. Различные механизмы биологического действия хелатирующих агентов (введение)
Наличие двухфазной реакции организма на металлы свидетельствует о существовании двух разных механизмов действия хелатирующего агента в биологических системах: I — удаление металлов из клетки или «маскировка» их в клетке (в виде комплексов) и II — накопление в клетке металлов в большем количестве (или при большем окислительно-восстановительном потенциале), чем в обычных условиях. Дальнейшее подразделение зависит от того, являются ли исследуемые металлы жизненно важными или токсичными для организма.
А. Механизм I. Большинство хелатирующих агентов, биологическое действие которых осуществляется по механизму I, получили распространение в качестве антидотов, предназначенных для «маскировки» или удаления из организма токсичных металлов, случайно попавших в организм высших млекопитающих. Эти антидоты рассматриваются в разд. 11.6.
Лишь в редких случаях сам агент, связывающий металл, оказывается токсичным для организма. Так, оксин (11.30) проникает в клетки бактерий и грибов, не нанося им заметного вреда (разд. 11.7.1). Это объясняется тем, что обычные стерические факторы и сродство к металлам позволяют активным участкам хелатирующего агента накапливать и удерживать металл.
Наиболее изученное соединение, чье повреждающее действие связано с явлением «маскировки», — синильная кислота. Она связывает свободные валентности железа в цитохромоксидазе, не затрагивая при этом четырех его связей с порфириновым ядром. Таким образом, фермент лишается возможности соединяться со своим субстратом, и дыхание прекращается. У многих видов это приводит к немедленной гибели организма.
166
Б. Механизм II. В разд. 11.0 было рассмотрено использование хелатообразования для подкормки деревьев железом. Другой пример — инъекции больным, страдающим недостаточностью кальция, комплекса кальция с глюконовой кислотой, медленно разрушающегося в организме с выделением ионов кальция. Наиболее изученные примеры действия хелатирующих агентов по механизму II можно найти среди бактерицидов и фунгицидов (разд. 11.7). Сведения, необходимые для понимания механизма II («кооперативный эффект» и эффект распределения), излагаются ниже.
«Кооперативный эффект». Явление возрастания химической активности вследствие хелатообразования подтверждается на примере гемоглобина, способного связывать кислород, и гемсодержащих ферментов, являющихся сильными окислителями. Неорганические соли железа обладают некоторой каталазной и пероксидазной активностью, возрастающей во много раз при включении железа в порфириновое ядро, связанное со специфичным белком. Аналогично ионы меди катализируют окисление аскорбиновой кислоты на воздухе, но этот эффект многократно увеличивается при включении меди в аскорбин-оксидазу (см. табл. 10.4).
Подобный «кооперативный эффект» может иметь место и в отсутствие белков. Нередко при добавлении комплексообразующего соединения с целью дезактивации металла наблюдается обратное явление: образующийся комплекс оказывается более активным катализатором. Токсическим действием обычно обладают металлы, способные изменять валентность, особенно, медь и железо. «Кооперативный эффект» чаще всего проявляется в тех случаях, когда прибавляется недостаточное количество комплексообразующего агента, т. е. образуется ненасыщенный комплекс. Приведем несколько таких примеров.
Комплекс орто-фенантролина (11.18) с ионом одновалентной меди в соотношении 2 : 1 в присутствии перекиси водорода способен мгновенно расщеплять двойную спираль ДНК- Эта реакция происходит за счет рециклизации с ионами меди, имеющими две свободные валентности. Это подтверждается отсутствием расщепления под действием 2,9-диметил-орто-фе-нантролина, который вследствие стереохимических ограничений способен соединяться только с одновалентной медью [Pope et al., 1982]. Потемнение дигидроксифенилаланина в результате окисления в присутствии сульфата двухвалентной меди значительно ускоряется при добавлении орто-фенантролина [Isaka, 1957]. Аутоокисление глутатиона в экстракте ткани хрусталика глаза (катализируемое следами железа, содержащегося в экстракте) ускоряется в присутствии ЭДТА [Pirie, Van Неу-ninger, 1954]. Как орто-фенантролин, так и бипиридил в 100 раз повышают скорость разложения перекиси водорода, катализируемой железом: эти соединения можно рассматривать как модели гемопротеиновых ферментов.
167
Гидролиз диизопропилфторфосфата, катализируемый медью,*; резко ускоряется аминокислотами, этилендиамином, орто-фе4 нантролином и бипиридилом. Наиболее удачным считают соота ношение компонентов, при котором образуется комплекс 1: Р [Wagner-Jauregg et al., 1955]. ЭДТА не влияет на реакции-гидролиза, так как это соединение не проявляет «кооператива ного эффекта» с медью и может предотвращать действие оксина, как такового [Byrde, Woodcock, 1957].
Механизм действия многих избирательно токсичных агентов основан на «кооперативном эффекте» (разд. 11.7). Обычно хелатные соединения металлов обладают большей избирательностью, чем их неорганические соли.
Эффект распределения. Известно, что клеточные мембраны точно регулируют поглощение катионов тяжелых металлов, поскольку даже жизненно важные катионы, необходимые в следовых количествах, в избытке оказываются токсичными. Так, железо токсичнее, чем обычно полагают; пероральное введение больших доз сульфата двухвалентного железа может привести к некрозу печени человека, наступающему через 48 ч [Luongo, Bjornson, 1954]. Токсическое действие вируса энцефаломиелита, содержащего много железа, объясняют переносом этого металла вирусами через гемато-энцефалический барьер (ГЭБ), который в норме для него непроницаем [Racket, Krims-ky, 1947]. Однако комплексы, не имеющие заряда, жирорастворимы и поэтому способны проникать через клеточные мембраны, которые не могут регулировать их проникновение. Как показано на рис. 11.2, комплексы двухвалентных металлов с щавелевой кислотой, их комплексы с глицином в соотношении 2: 1 и им подобные не имеют заряда, а поэтому легко проникают в клетки.
(Комплексы полидентатных соединений типа ЭДТА (11.27) часто содержат избыток гидрофильных групп, способных соединяться с металлом, и поэтому не могут проникать через обычные клеточные мембраны.) За счет образования таких жирорастворимых комплексов хелатирующий агент способен осуществлять транспорт металла в клетку в количествах, превышающих обычные. Некоторые хелатные соединения металлов оказывают действие, находясь вне микроорганизма, чей характерный отрицательный заряд вызывает притяжение и накопление таких положительно заряженных комплексов, как а) хелатные комплексы с агентами типа глицина, обладающие положительным зарядом при неполном насыщении (например, комплексы с двухвалентными металлами 1:1), и б) хелатные комплексы с агентами типа этилендиамина, положительно заряженные при любой степени насыщения (см. рис. 11.2).
Существуют предположения о том, что хелатообразование подавляет токсическое действие лекарственных препаратов на организм, «маскируя» токсикогенные группы за счет связывания их с металлом [Gosalver et al., 1978; Perrin, 1970].
168
11.6. Уменьшение токсического действия металла в результате хелатообразования, антидоты
Некоторые хелатирующие агенты широко используются в клинике в качестве антидотов при профессиональных и бытовых отравлениях металлами, хронических интоксикациях металлами, вызванных передозировками лекарственных препаратов, а также для ускорения выведения из организма радиоактивных элементов. Для этих целей хелатирующие вещества начали применять только с 1945 г. Такие антидоты циркулируют в крови, не вызывая уменьшения концентрации жизненно важных тяжелых металлов. При этом необходимо строго контролировать их дозу. Для того чтобы антидот смог проникать в клетки в небольших количествах и быстро выводиться из организма, его молекулы должны содержать полярные (желательно легко ионизирующиеся) группы, например ОН, СО2Н, SH, NH2. Эти группы должны присутствовать в избытке, чтобы по меньшей мере одна из них оставалась свободной после насыщения антидота металлом. Кроме того, антидот обычно создают такой, чтобы его хелатные комплексы не могли проникать в клетки из кровотока и легко выводились почками.
Первый антидот — димеркапрол (11.23) был синтезирован в 1940 г. в качестве антидота при отравлениях мышьяксодержащим боевым отравляющим веществом [Peters, Stocken, Thompson, 1945] и получил название «Британский антилюизит» (БАЛ). В настоящее время его часто применяют для лечения отравлений соединениями золота, ртути (в виде неорганических и органических соединений), сурьмы и мышьяка. Связывание металла происходит, как показано в формуле (11.24). Димеркапрол вводят внутримышечно через каждые 4 ч в течение первого дня, а затем — в зависимости от состояния больного, Димеркапрол не только снимает токсическое действие элементов, перечисленных выше, но и выводит их из организма в виде комплексов.
СН2—SH I
СН2—SH
СН2—он
СН2—ОН
Димеркапрол (11.23)
Связывание иоиа ртути димеркапролом
(11.24)
СО2Н
НС—SH
НС—SH
СО2Н
Димеркапто-яитариая кислота
(11.25)
Me
Me—С—SH
CH—NH2
I
COaH
Пеницилламин
(11.26)
Позднее в употребление была введена 2,3-димеркаптоянтар-ная кислота (11.25), имеющая преимущества по сравнению с димеркапролом: она активна при пероральном введении, проникает через ГЭБ и фактически выводит ион метилртути из мозга [Aaseth, Friedheim, 1978]. Этот ион часто попадает в орга
169
низм людей при употреблении в пищу продуктов — зерновых или рыбы, подвергшихся воздействию промышленных отходов, содержащих ртуть. Другой аналог димеркапрола, 2,3-димеркап-топропан-1-сульфоновая кислота, обладает активностью при пероральном введении.
Пеницилламин (11.26), применяемый аналогично при отравлении медью, не обладает токсическим действием. Этот препарат используется при лечении болезни Вильсона — наследственное нарушение процесса метаболизма, при котором содержащаяся в пище медь не выводится, а накапливается в мозге, почках и печени в таких количествах, что это приводит к смерти уже в раннем возрасте. Применение пеницилламина (перорально) предотвращает подобный исход и может устранить патологические изменения, вызываемые ионами меди. Комплекс медь — пеницилламин легко выделяется с мочой [Walshe, 1968]. Пеницилламин, эффективно действующий при пероральном введении, целесообразно назначать на поздних стадиях хронических отравлений свинцом, которые лечат внутривенными введениями ЭДТА.
Этилендиаминтетраацетат кальция (11.28) — наиболее эффективное лекарственное средство при отравлениях свинцом: его вводят внутривенно в виде натриевой соли [Bessman, Rubin, Leikin, 1954]. Комплекс ЭДТА с натрием, т. е. то же вещество, но не содержащее кальция, применяли для удаления кальция при стенозе клапанов сердца. Несмотря на то, что уровень кальция в крови падал, состояние больных не ухудшалось даже после длительных курсов инъекций: по 3 г (в виде 0,5% раствора) каждые 24 ч 5 дней в неделю и три недели в месяц [Seven, 1969].
Этилендиаминтетраацетат натрия успешно применяют для лечения сердечной аритмии, вызываемой препаратами на-
СО2Н I сн,
I
НО2С— Н2С—N
На
СО2Н I сн2
N—СН2—СО2Н
Н2
ЭДТА (Этилеидиамиитетрауксусиая кислота)
(11.27)
О2С СО2
Hz \ /сн2 но2с—ch2n nch2—со2н
^42С—СН2
Этилеидиамиитетраацетат кальция
(11.28)
170
CH2—SH ch2-nh2 Цистеамин (11.29) перстянки, так как он способен восстанавливать баланс К+/Са2+ [Szekely, Wynne, 1963].
При отравлении плутонием лиц, работающих в атомной энергетике, лечение необходимо начинать немедленно, так как радиоактивность может разрушить капилляры, по которым антидот проникает в ткани. В этих случаях для удаления плутония можно использовать те же препараты, что и для выведения железа, например, дефероксамин (11.9). Удачным дополнением к нему является диэтилентриаминпентауксусная кислота (ДТПА) [Smith, Chapman, Marlow, 1969]. Оба этих препарата вводятся парентерально.
Дефероксамин представляет собой стандартный препарат для лечения отравлений железом (разд. 11.1), которые могут появляться после нескольких повторных переливаний крови при двух генетически обусловленных болезнях: талассемии большой (анемия Кули) и серповидно-клеточной анемии. При талассемии бета-цепь молекулы гемоглобина синтезируется в недостаточном количестве, что приводит к ухудшению переноса кислорода кровью. Больные этой болезнью умирают в детстве, если не проводить повторяющиеся переливания крови, однако смерть нередко наступает и в подростковом возрасте или около двадцати лет, так как вследствие этих переливаний происходит накопление неорганических соединений железа в почках, печени и сердце. Это заболевание наиболее широко распространено среди населения Средиземноморья и Юго-Восточной Азии. В некоторых районах Нигерии до 30% населения являются носителями гена серповидно-клеточной анемии, болезни, при которой нет недостатка гемоглобина, но большая его часть несколько отличается по химической структуре от нормального. Около 0,15% негритянских детей в США страдают этой болезнью, но, несмотря на то что смертность среди них в детстве высока, часть из них выживают и имеют детей. При серповидно-клеточной анемии переливания крови необходимы только во время мучительных гипоксических кризов.
Другие соединения, образующие хелатные комплексы с железом, проходят клинические испытания; к ним относятся родо-торуловая кислота (11.12), получаемая из дрожжей (инъекционное введение), 2,3-дигидроксибензойная кислота, являющаяся частью молекулы энтерохелина (11.7), активная при пероральном применении, и этилендиамин-бис(2-гидроксифенилук-сусная кислота), соединение, родственное ЭДТА.
При отравлении бериллием применяют большие дозы салицилата натрия или более низкие дозы ауринтрикарбоновой кислоты, содержащей три остатка салициловой кислоты, связан
171
ные с центральным атомом углерода. Ауринтрикарбоновая кислота в отличие от салициловой не способствует выведению бериллия из организма, а удерживает его в тканях в виде нерастворимого комплекса.
При отравлениях таллием нередко назначают диэтилдитиокарбамат натрия, ср. (3.57).
2-Меркаптоэтиламин (11.29) (цистеамин) успешно применяют для защиты от лучевого поражения. Было показано, что у мыши, получившей 750 рад (разовая доза при общем облучении), значительно возрастает концентрация железа и меди в костном мозге, надпочечниках, селезенке, печени, легких и вилочковой железе, а также содержание железа в мышцах и почках [Yendell, Tupper, Wills, 1967]. Некоторые исследователи считают, что радиозащитные свойства цистеамина обусловлены его способностью захватывать катионы тяжелых металлов, выделяющиеся из клеток при облучении [Jones, 1960]. Другие гипотезы можно найти в работе Р. Brown (1967).
Под воздействием высоких энергий в воде происходит образование гидроксильного анион-радикала (ОН-), который затем атакует ДНК. Было высказано предположение о том, что этот процесс приводит к образованию гидроперекисей пиримидиновых оснований ДНК, например, 5,6-дигидро-6-гидроперокси-6-гидрокситимина. Существует гипотеза, согласно которой эти гидроперекиси взаимодействуют с ионами переходных металлов с образованием свободных радикалов, вызывающих химические изменения структуры соседних оснований, что приводит к нарушениям в образовании пар при репликации [Thomas et al., 1976].
Очевидно, что эффективное связывание ионов меди и железа приводит к блокированию любого деструктивного (окислительного) цепного процесса, катализируемого этими металлами. Многие вещества, сходные по структуре с цистеамином, могут предотвращать лучевую болезнь. Примером может служить 2-меркаптоэтилгуанидин, образующийся в организме из S-(2-аминоэтил)изотиурониевых солей [Doherty, Shapira, Burnett, 1957].
В некоторых случаях радиоактивные катионы могут быть удалены из организма посредством «обменного разведения», т. е. введения того же катиона, но не радиоактивного, а в виде комплекса со средней устойчивостью. Так, для удаления радиоактивного циркония применяют цитрат циркония [Schubert, 1957].
Противораковое действие гидроксимочевины заключается в уменьшении биологического (но не токсического) действия металла во вредных клетках. В этих клетках гидроксимочевина связывает железо, необходимое дегидрогеназе, превращающей рибозиды в дезоксирибозиды (разд. 4.0).
О применении хелатообразующих агентов при отравлениях тяжелыми металлами см. Levine (1978).
172
11.7. Уменьшение токсического действия металлов хелатообразованием
Спектр антибактериального действия 8-гидроксихинолина (11.30) аналогичен спектру действия пенициллина. Хотя эти препараты и не отличаются такой избирательностью действия, как пенициллин, они начиная с 1895 г. постоянно применялись для обработки и лечении ран. Преимущество 8-гидроксихинолина и его производных перед другими антибактериальными препаратами заключается в том, что они действуют быстро и обладают также фунгицидным действием, однако их активность по отношению к грамотрицательным бактериям не слишком высока. Мази, содержащие 8-гидроксихинолин, используют в дерматологии, например, при сыпи, вызванной устойчивыми стафилококками, при вторичных инфекционных экземах, а также в качестве фунгицидов. Комплекс 8-гидроксихинолина с медью применяют для обработки материалов различных конструкций с целью придания им устойчивости, например для защиты брезента палаток от поражения грибами.
После того как в 1944 г. было обнаружено, что бактерицидное действие 8-гидроксихинолина обусловлено хелатообразованием [Albert, 1944; Albert et al., 1947], возникло предположение, что многие другие антибактериальные и противогрибковые средства действуют аналогично. Вскоре было установлено, что оксин (одно из названий 8-гидроксихинолина) малоактивен в средах, не содержащих тяжелые металлы [Rubbo, Albert, Gibson, 1950; Albert, Gibson, Rubbo, 1953]. Следовательно, его действие осуществляется не за счет удаления жизненно важных металлов, а благодаря образованию высокотоксичного комплекса с любым металлом, присутствующим в среде (разд. 11.7.1). Впоследствии было показано, что в основе антибактериального действия некоторых веществ, близких по структуре к оксину (разд. 11.7.2), а также соединений, химически от него отличных, лежит один и тот же механизм, сущность которого составляет «кооперативный эффект», обсуждавшийся в разд. 11.5. По-видимому, только металлы с переменной валентностью способны активизировать эти агенты.
11.7.1. Механизм действия 8-гидроксихинолина (оксин)
Антибактериальное действие оксина (11.30) обусловлено хелатообразованием, что было доказано следующим образом [Albert et al., 1947]. Оксин долгое время применяли в аналитической химии в качестве комплексообразующего агента. Он обладает ярко выраженными хелатообразующими свойствами (см. табл. 11.1). Все остальные шесть гидроксихинолинов, изомерных оксину, не способны образовывать хелаты и не обладают антибактериальной активностью. Сам же оксин останавливает рост стафилококков даже в концентрации две части на миллион
173
Таблица 11.5. Влияние увеличения концентрации оксина на его бактерицидное действие, Staph, aureus в мясном бульоне при pH 7,0—7,3 (20 °C)
Концентрация оксина, 1/М Рост, ч
0 1 3 24
800 +++ 4-4-4- 4—F4- 4-
1600 +++ 4-4-4- 4-4-4- 4--
3200 4—1—ь 4-4-4- 4- 4-
6400 4—1—(- 4-4—1- 4-
12 000 4-4-4- 4- 4- —
25000 4-4—F 4- —
50 000 4-4-4- 4- — —
100 000 4-4-4- — .— —
200 000 4-4-4- 4-4-4- 4-4-4- 4-4—F
Обозначения: (—) — роста нет; ( + ) — до 50 колоний; ( + + ) — 50—100 колоний; ( + + + ) — обильный рост [Albert, Gibson, Rubbo, 1953].
Примечание. В этой и следующих таблицах определение бактерицидного действия основано на тесте Miles, Misra (1938). Через определенные промежутки времени отбирают пробы, которые затем разбавляют и высевают на чашки с кровяным агаром. Чашки просматривают через 48 ч при 37 °C.
(М/100 000). Позднее было показано, что О- и N-метилпроиз-водные (11. 31) и (11.32) оксина также не способны образовывать хелаты, так как СН3-группа, в отличие от протона, не может замещаться металлом и не обладает антибактериальной активностью. Таким образом было установлено существование зависимости между способностью оксина к хелатообразованию и его антибактериальным действием.
Необходимо было выяснить, обусловлено ли токсическое действие оксина удалением жизненно важных металлов, как предполагалось ранее [Zentmyer, 1944], или же он усиливает токсическое действие металлов, обычно присутствующих в питательной среде. Справедливым оказалось предположение, причем как для агентов с бактериостатическим, так и бактерицидным действием (разд. 2.3) [Rubbo et al., 1950; Albert et al., 1953]. Первым доказательством этой гипотезы послужило явление, известное под названием «обращение эффекта концентрации».
8-Гидроксихинолии (окснн) 8-Метоксихинолнн
(11.30) (11.31)
Оксин метохлорид
(11.32)
Обращение эффекта концентрации. Трудно представить себе, что эффективность биологически активного вещества может уменьшаться с возрастанием его концентрации. Однако именно такая зависимость наблюдается в случае оксина. Как видно из табл. 11.5, стафилококки, погибающие в течение часа
174
при концентрации оксина М/100 000, при концентрации M/I600 не погибают даже через 3 ч (фактически они остаются живыми даже в насыщенном растворе оксина, т. е. при концентрации М/200). Некоторое токсическое действие, однако, наблюдается через 24 ч [Albert et al., 1953]. Аналогичным образом оксин действует и на стрептококки. Сущность этого явления стала понятной только после того, как было обнаружено, что оно происходит только в бульоне, но не в дистиллированной воде.
Эксперименты с оксином в дистиллированной воде. Жизнеспособность стафилококков в дистиллированной воде в течение по меньшей мере 24 ч позволяет сделать определенные выводы. Из данных, приведенных в табл. 2.3. (том 1), видно, что в дистиллированной воде оксин (0,01 мМ) биологически инертен, но в присутствии эквивалентного количества железа приобретает бактерицидное действие. Очевидно, токсическим агентом служит не сам оксин и не железо, а их комплекс. При замене воды бульоном исчезает необходимость в добавлении железа, так как в этой среде оно присутствует в достаточном количестве. При увеличении концентрации оксина до 1,25 мМ бактерицидное действие исчезает. Это обусловлено образованием комплекса (2:1) оксин — железо (11.23), не обладающего антибактериальным действием. При добавлении к бульону железа в количествах, необходимых для образования комплекса в соотношении 1 : 1 (2.25), бактерицидные свойства восстанавливаются [Albert et al., 1953].
Комплекс оксина с двухвалентным железом в соотиошеиии 2 ; 1 (насыщенный)
(11.33)
«Кооперативный эффект комплексов металлов с оксином. В отсутствие тяжелых металлов оксин проникает в бактериальные клетки Staph, aureus, не причиняя им никакого вреда [Beckett, Vahora, Robinson, 1958]. Аналогичным образом он проникает в клетки грибов Aspergillus niger, не вызывая никаких повреждений [Greathouse et al., 1954]. Последнее исследование было проведено с двумя образцами радиоактивного оксина, один из которых был получен из [14С]-анилина, другой — из [14С]-глицерина (с радиоактивными метками в бензольном и пиридиновом циклах соответственно). Однако в присутствии соответствующего металла оксин оказывается высоко токсичным для этих микроорганизмов.
175
Оксин оказывает повреждающее действие на грамположи-тельные бактерии только в том случае, если в питательной среде присутствует один из следующих катионов: Cu2+, Fe2+ или Fe3+, особенно, видимо, важны для действия ионы железа. В аэробных культурах быстро устанавливается равновесие между комплексами оксина с двух- и трехвалентным железом. На большинство грамотрицательных бактерий оксин действует очень слабо, причем действие не зависит от наличия или отсутствия металлов [Rubbo et al., 1950]. Оксин повреждает мицелий грибов только в присутствии ионов меди в питательной среде, в данном случае железо не может заменить медь [Anderson, Swaby, 1951; Block, 1956], это же характерно и для дрожжей [Nordbring-Hertz, 1955].
Антагонизм кобальта к токсическому действию комплекса оксина с железом. Очевидно, что добавление большого избытка (200 эквивалентов) инертного металла могло бы предотвратить токсическое действие комплекса оксин — железо, если бы константа устойчивости нового комплекса была выше или лишь немногим ниже, чем таковая комплекса оксин — железо. При этом согласно закону действия масс оксин должен почти полностью соединяться с инертным металлом. Кадмий, кобальт, цинк и никель в этих условиях оказывают защитное действие, а марганец, магний и кальций не эффективны (соответствующие константы устойчивости см. в табл. 11.1).
Особое положение занимает кобальт. Он оказывает защитное действие не только в больших, но и в следовых количествах. Сульфат двухвалентного кобальта даже в очень низкой концентрации (М/25 000) полностью снимает бактериостатическое действие оксина в концентрации М/100 000 (рис. 11.6) [Rubbo et al., 1950]. Данные, приведенные в табл. 11.6, характеризуют эффективность действия кобальта в качестве антагониста бактерицидного действия комплекса оксин — железо. Эффективность комплекса оксин — медь лишь ненамного ниже.
Кобальт защищает также дрожжи от действия комплекса оксин — медь [Nordbring-Hertz, 1955], но не мицелий грибов, очевидно, вследствие их медленного роста [Anderson, Swaby, 1951]. Тем не менее кобальт оказывает защитное действие на ДМДК (диметилдитиокарбаминовая кислота) (разд. 11.7.3). Данные, приведенные в табл. 11.7, показывают, что кобальт защищает трипаносомы от летального действия оксина [Williamson, 1959].
Каково же объяснение подобного защитного действия кобальта? На первый взгляд может показаться, что кобальт, соединяясь с оксином, препятствует образованию его комплекса с железом. Но тогда никель должен бы быть еще более эффективным, так как константа устойчивости комплекса никель — оксин значительно выше, чем таковая комплекса оксин — кобальт (см. табл. 11.1) [Albert et al., 1953]. Однако никель в низких концентрациях защитным действием не обладает.
176
Рис. 11.6. Антагонизм между оксином н следами кобальта (Staphylococcus aureus в питательном бульоне, pH 7,2).
К более правильным выводам можно прийти на основании того, что некоторые жизненно важные компоненты клеток, в частности меркаптосоединения, например дигидролипоевая кислота (2.28) (разд. 11.7.3), легко окисляются кислородом воздуха в присутствии следов железа или меди. В результате этих реакций образуются перекись водорода и супероксидный радикал (О2_), которые в присутствии катионов металлов вызывают взрывоподобную цепную реакцию, так что очень небольшое количество металла может катализировать широко распространяющийся процесс разрушения. На примере некоторых модельных реакций такого типа было показано, что следы кобальта прерывают эти цепные реакции и тормозят процесс разрушения (рис. 11.7) [Dainton, 1966].
Оксин представляет собой «кооперативный» хелатирующий агент (разд. 11.5). Так, смесь неорганической соли железа с оксином катализирует окисление воздухом SH-групп в нуклеопротеидах печени крыс и икры рыб, тогда как соли железа без оксина неэффективны [Bernheim, Bernheim, 1939]. Исключительно высокая каталитическая активность комплекса оксин — железо, вероятно, обусловлена перестройкой орбиталей катиона трехвалентного железа в процессе хелатообразования (что подтверждается необычно глубокой окраской этих комплексов оксина: красной — с двухвалентным железом, зеленой — с трехвалентным).
Логично предположить, что токсическая форма оксина — это его комплекс с трехвалентным железом 1: 1, так как он
12—734
177
Таблица 11.6. Защитный эффект кобальта при бактерицидном действии комплексов оксин — железо и оксин — медь на Staph, aureus (бульон, очищенный от металлов; pH 7,3, 20 °C; концентрация оксина в каждой пробирке M/25 000 Albert et al., 1953]
Пробирка, № Концентрация добавленного металла (1/М) Рост, ч
FeSO4 C11SO4 C0S04 0 2 3 24
1 0 0 0 Н—1—н +++ —L +++
2 50 000 0 0 +++ — —- —
3 50 000 0 50 000 ++ ++
4 0 50000 0 +++ —
5 0 50000 50000 +++ — — —
6 0 50 000 10 000 Ч—1—h +++ +++ 4—+
Таблица 11.7. Защитный эффект кобальта при трипаиоцидном действии оксина на Т. rhodesiense в физиологическом растворе, содержащем лошадиную сыворотку и глюкозу; инкубация прн 37 °C в течение 4 ч [Williamson, 1959]
Пробирка, Ns Концентрация 1/М. Относительное’ число выживших трипаносом по сравнению с контролем, %
Оксин Со2* Любой из следующих катионов: Си^+, Ni*+, Zn2+, Fe2+, МпМ-, Mg2+
1 0 0 0 100
2 800 000 0 0
3 800 000 400 000 0 117
4—9 800 000 0 400 000 <3
ненасыщен, т. е. обладает необходимой для катализаторов способностью к связыванию. С другой стороны, комплекс 2: 1 (11.33) полностью насыщен и, по-видимому, не способен быть катализатором. (В аэробных системах комплексы двух- и трехвалентного железа находятся в равновесии, а, следовательно, комплексы трехвалентного железа с соотношением 1:1 и 2: 1 — (Ненасыщены и обладают каталитической активностью, тогда как комплекс состава 3: 1 насыщен и инертен.) Эти выводы подтверждают аналогичные заключения, сделанные на основе концепций «обращенного эффекта концентрации» (разд. 11.7.1).
Место действия комплекса оксин-железо в бактериях неизвестно, однако ключом к его обнаружению могут служить данные о месте действия его в грибах; в этом случае он способствует окислительной деструкции дигидролипоевой кислоты (разд. 11.7.3). Информацию о месте действия можно получить И из данных по липофильности. Производные оксина с низким коэффициентом распределения в системе масло — вода не обладают антибактериальным действием. Например, комплексы ок-син-5-сульфокислоты имеют такие же константы устойчивости, как и комплексы оксина [Albert, 1953], но в отличие от него 178
Рис. 11.7. Действие кобальта, предотвращающее окисление цистеина (М/40, 20 °C) кислородом, катализируемое медью (А — сульфат меди, М/10 000; Б — сульфат меди, М/100 000; В — аналогично Б, но с добавлением сульфата кобальта, M/500) tBaur, Preis, 1936].
они не переходят из воды в липиды и совершенно лишены антибактериальных свойств. Чтобы подтвердить важность высокого коэффициента распределения для антибактериального действия оксина, был синтезирован [Albert, Hampton, 1952, 1954] и исследован [Albert et al., 1954] ряд производных оксина, не имеющих электрического заряда и отличающихся низким коэффициентом распределения. Антибактериальное действие этих азаоксинов (разд. 11.7.2) ослабляется или усиливается пропорционально уменьшению или увеличению коэффициентов распределения, вызванных незначительными изменениями их молекулярной структуры. Таким образом, высокий коэффициент распределения играет важную роль в проявлении активности оксина и родственных ему соединений. Это доказывает, что действие оксина осуществляется внутри клетки или, по крайней мере, в цитоплазматической мембране.
Производные оксина. Галогенирование оксина увеличивает эффективность его действия на грамотрицательные бактерии, многие из этих производных широко применяют на практике. Халхинол (смесь 5-хлоро- и 5,7-дихлороксина) и хлорхинал-дол (5,7-дихлоро-2-метилоксин) входят в состав мазей, используемых в дерматологии. 5,7-ДийоДооксин (дийодогидроксихин) и 5-хлоро-7-йодооксин (клиохинол) применяют при бактериальной дизентерии. В период между 1955 и 1970 г. около 10 000 японцев страдали подострым миелоглазным нервным заболеванием, мучительной, калечащей людей болезнью, иногда приводящей к смерти. Полагают, что эта болезнь была вызвана хлорхинолом. Из-за этого в Японии была приостановлена продажа хлорхинола. Однако в 1975 г. Meade обнаружил, что полученные ранее данные плохо коррелируют между собой. В 1977 г. эти данные были пересмотрены и был сделан вывод, что при применении доз хлорхинола, необходимых для лечения диареи, опасность заболевания незначительна. Подробнее о потенциальной опасности галогенизированных оксинов см. Gilman, Goodman, Gilman (1980).
12* 179
Таблица 11.8. Ослабление и усиление бактериостатического действия в зависимости от изменений коэффициентов распределения Staph, pyogenes, мясной бульон при pH 7,3 (37°) [Albert et al., 1954]
Соединение Коэффициент распределения в системе олеиновый спирт — вода Минимальное разбавление, при котором наблюдается ингибирующий эффект, 1/М Логарифм первой константы устойчивости (с NiJ+)
Оксин 67 200 000 9,8
5- Азаоксин 0,02 800 5,8
7-Азаоксин 0,1 800 6,7
6-Азаоксин 1 800 5,9
З-Азаоксин 5 13000 7,6
2-Азаокснн 6 13 000 7,8
4-Азаокснн 8 6 400 7,6
4-Метнл-2-азаоксин 16 25000 8,1
4-Метил-З-азаоксин 17 50000 7,9
4-Пропнл-З-азаоксии 135 100 ОСО 7,9
7-Аллил-З-азаокснн 310 100000 7,9
5-Метилоксин успешно применяли во Франции при эпидемиях холеры [Zribi, Ben-Rachid, 1973]. Этот препарат высокоактивен in vitro по отношению к Plasmodium falciparum и действует даже на штаммы, устойчивые к хлорохину [Scheibel, Adler,
11.7.2. Вещества, близкие к оксину по химическому строению
Как было обнаружено, конденсация молекулы оксина с бензольным или пиримидиновым кольцом не вызывает потери активности; при этом, однако, не должны замещаться положения 2 и 3, поскольку в этих случаях возникают стерические препятствия к хелатообразованию [Albert et al., 1947].
Увеличение коэффициента распределения оксина в четыре раза путем введения липофильных заместителей не сопровождается заметным повышением его активности in vitro: он и так достаточно высок [Albert, Rees, Tomlinson, 1956]. Поэтому более интересно выяснить, что происходит при понижении его коэффициента распределения. В случае оксина этого легко достигнуть, синтезировав ряд аналогов, у которых одна или несколько = СН-групп замещены значительно более гидрофильным двоесвязанным атомом азота (=N—) [Albert, Hampton, 1952, 1954]. Все эти вещества используются под тривиальными названиями, например 8-гидроксициннолин, 8-гидроксихиназо-лин, 8-гидроксихиноксалин, однако правильнее и удобнее для наших целей пользоваться названиями 2-азаоксин, 3-азаоксин (11.34) и т. д.
180
Введение атомов азота и кислорода приводит к понижению коэффициента распределения, а введение атомов галогенов и алкильных групп — к его повышению. Данные, приведенные в табл. 11.8, показывают, что при замене =СН-групп азотом ( = N—) коэффициент распределения понижается. При этом антибактериальная активность уменьшается, часто почти до нуля. Не удаляя дополнительный атом азота, можно повысить коэффициент распределения введением алкильной группы небольшого размера (содержащей не более трех атомов углерода) до величины, характерной для оксина и даже превышающей ее. При этом антибактериальная активность также возрастает до максимальной величины (см. табл. 11.8) [Albert et al., 1954].
оа а | N NS
ОН 4 |
О---Fe++
З-Азаоксин Комплекс пиритиона
(11.34) (11.35)
Вследствие индукционного эффекта второго атома азота не удается достигнуть ни высокой константы устойчивости, ни столь же эффективного антибактериального действия, как у оксина (антибактериальная активность уменьшается уже при двукратном последовательном разбавлении). Тем не менее последние семь соединений в табл. 11.8 имеют константы устойчивости того же порядка, что и у оксина. У этих соединений антибактериальное действие изменяется параллельно изменению коэффициента распределения, что подтверждено соответствующей корреляцией.
11.7.3. Вещества аналогичного действия, отличающиеся от оксина по химической структуре
N-Оксиды пиридина, хинолина и бензохинолина обладают антибактериальной активностью при условии, что в положении 2 находится легкоионизирующаяся группа, необходимая для образования хелатных соединений. Наиболее подходящей является меркаптогруппа. Так, например, 2-меркаптопиридин-Н-ок-сид (1-гидроксипиридин-2-тион) (2.26) обладает такой же антибактериальной активностью, как и оксин. Несмотря на то что строение хелатных комплексов этих веществ, например (11.35) и оксина (2.24), совершенно различно, можно утверждать, что механизм их действия одинаков. Об этом можно судить потрем следующим критериям: оба эти вещества проявляют бактерицидное действие только в присутствии железа, а ингибитором служит кобальт или избыток самого вещества [Albert et al., 1956].
181
Таблица 11.9. Безопасность N-оксида 2-меркаптопиридина и отсутствие железа (тест на бактерицидность и иоде, дистиллироианной и стекле; Staph, aureus, pH 6—7, 20 °C) [Albert et al., 1956]
Концентрация N-окснда 2-меркаптопнридина (пн-рнтнон) (2.26) Fe3+ Рост, 1 ч
80 000 0 +++
0 80 000 +++
80 000 80 000
Примечание. + + + означает обильный рост с большим числом колоний, не поддающихся подсчету; — заметного роста нет.
Безвредность этого хелатного соединения в отсутствие железа показана в табл. 11.9. Его широко применяют в моющих средствах. В качестве компонента шампуней этот комплекс оказался весьма эффективен против Pityrosporum ovale, вызывающих образование перхоти. Он также широко используется в промышленности в качестве фунгицида (для обработки тканей, охладительных систем и добавления в смазочные масла).
Диметилдитиокарбаминовая кислота, или ДМДК (3.57),— активный фунгицид, применяющийся в сельском хозяйстве в виде натриевой соли и комплексов с железом и цинком. В формуле (11.15) изображен комплекс с медью в соотношении 1:1.
Изучение показало, что это соединение действует как антифунгицид только в виде комплекса с медью [Goksoyr, 1955; Sijpesteijn, Janssen, van der Kerk, 1957]. Некоторые результаты этих работ приведены в табл. 11.10.
Из данных табл. 11.10, указывающих на наличие «обращенного эффекта концентрации», следует необходимость ионов металла (медь) для действия ДМДК на Aspergillus niger. Железо в исследованном случае оказывается неэффективным. К нему нечувствительны и другие плесневые грибы [Sijpesteijn, Janssen, 1959]. Из табл. 11.10 видно, что на активность решающим образом влияет соотношение металла и хелатирующего агента, так же как и в случае оксина [табл. 2.3; Albert et al., 1956]. Голландские ученые считают, что существование зоны отсутствия ингибирования (находящейся между 10 и 50 частями на миллион ДМДК) объясняется крайне малой растворимостью комплекса ДМДК с медью (в соотношении 2: 1) в воде. Растворимость комплекса в воде 0,01 части на миллион, что ограничивает поступление агента в клетку. Третья зона, токсическая (появляющаяся при концентрациях 50 частей на миллион), была приписана токсическому действию ДМДК, не связанному с Комплексообразованием [Sijpesteijn et al., 1957].
Кобальт является антагонистом действия диметилдитиокарбамата натрия на Aspergillus niger, так же как и оксина (катализатор — медь) на дрожжи (разд. 11.7). Бактерицидное действие ДМДК, активируемое медью, см. Liebermeister (1950). 182
Таблица 11.10. Влияние меди на ингибирование роста Aspergillus niger диметилдитиокарбаматом натрия (и течение 3 дней при 25 °C; среда: глюкоза, соли и витамины и иоде, дистиллированной в стеклинной аппаратуре, pH 7) [Sijpesteijn, Janssen, 1959]
Си2+ 1/М Концентрация ДМДК-Na 1/М
0 70 000 280 000 140 000 70 000 280 000 140 000 7000
0 + + + + + + + +
83 000 + + + + + + + +
25 000 + + — — — + + +
Накопление пировиноградной кислоты в опытах с Aspergillus указывает на молекулярное место действия ДМДК, оксина и пиритиона: они катализируют окислительное разрушение ди-гидролипоевой (тиоктовой кислоты) (2.28) [Sijpesteijn, Janssen, 1959]. Дигидролипоевая кислота представляет собой важный кофермент окислительного декарбоксилирования пировиноградной кислоты дигидролипоилацетилтрансферазой, компонентом мулътиферментного комплекса, называемого пируватдегидрогеназой.
Подробнее о диметилдитиокарбаматах см. Thorn, Ludwig (1962). В сельском хозяйстве комплексы ДМДК с цинком и железом используют в основном из-за их адгезивных свойств, позволяющих им удерживаться на растениях даже во время длительных дождей. Применяют также тетраметилтиурамдисуль-фид, полученный окислением ДМДК- Этот дисульфид в полевых условиях медленно превращается в ДМДК-
Испытания in vitro на Plasmodium falciparum хелатирующих агентов, действующих подобно оксину (оксин, 5-метилок-син, N-оксид 2-меркаптопиридин, N-оксид 2-меркаптохинолин и диэтилдитиокарбамат натрия), показали, что эти соединения являются по меньшей мере такими же сильными ингибиторами, как хинин (использовавшийся в качестве контрольного образца) [Scheibel, Adler, 1982]. До сих пор неизвестно, аналогичен ли механизм биологической активности 2,2'-бипиридила таковому оксина. Однако для его производных характерен «обращенный эффект концентрации», что справедливо и по отношению к тем производным, которые обладают активностью на культуру Mycoplasma gallisepticum [Linscoten et al., 1984]. Микоплазмы, мельчайшие из самовоспроизводящихся прокариот, лишены клеточных стенок; для их роста необходим холестерин (этим они отличаются от бактерий). М. gallisepticum часто вызывают респираторные заболевания у цыплят, выращиваемых в закрытых помещениях. Этот организм также ингибируется орто-фенантролином (11.18), причем особенно сильно если метильные группы находятся в орто-положениях к атомам азота. Для биологического действия этих соединений необходимо присутствие стехиометрических количеств меди. Это по-
183
Таблица 11.11. Физические свойства гидразидов и их противотуберкулезное действие [Albert, 1956]
Гндразнд Р*а Гидра-зндной группы log константы устойчивости комплекса (1 : I)1 со следующими металлами Сравнительная активность против М. tuberculosis H3?Rv
Си2+ Ni2+ Со2+ Zn2+ in vitro in vivo
Изоникотиновой кислоты2 10,77 8,0 5,5 4,8 5,4 1 1
Никотиновой кислоты2 11,47 8,7 6,0 5,4 ? 0,001
Пиколиновой кислоты2 12,27 12,4 10,7 9,6 8,4 0,017
Бензойной кислоты 12,45 9,0 6,3 ? ? 0,002
Цианоуксусной КИСЛОТЫ 11,17 8,5 6,0 5,3 ? 0,008 0,2
1 Константы устойчивости некоторых комплексов состава 2 : 1 см. Albert (1956).
2 Эти три вещества представляют собой изомеры. Они содержат гидразидную труп-пу в положениях 4, 3, 2 соответственно.
зволило сделать вывод о том, что активная форма агента — комплекс с медью [Antic et al., 1977].
1,10-Фенантролин и его С-метилпроизводные обладают близкими антибактериальными свойствами. Их комплексы с металлами также обладают антибактериальными свойствами, однако при понижении способности металла к комплексообразованию скорость действия этих соединений увеличивается (табл. 11.1). Бактерицидное действие резко возрастает при введении в молекулу шести метильных групп [Shulman, Dwyer, 1964; Butler et al., 1969].
11.8. Тетрациклины
Тетрациклин (11.36) и его производные представляют собой октагидронафтацены. Они широко используются для лечения системных бактериальных инфекций. В отличие от оксина действие тетрациклинов на бактерии развивается медленно и не ускоряется в присутствии железа [Albert, Rees, 1956]. Тетра-
Тетрациклин (1136)
184
циклимы обладают очень слабым фунгицидным действием (о тетрациклинах см. разд. 4.1).
Albert и Rees (1956) впервые продемонстрировали эффективные хелатирующие свойства тетрациклинов и обнаружили, что константы устойчивости этих соединений близки к таковым глицина (см. табл. 11.1).
Тетрациклин представляет собой сильную кислоту, в которой гидроксильная группа в положении 3 имеет рКа 3,3 (она является частью трикарбонилметановой системы, образованной атомами кислорода в положениях 1, 2 и 3). Остальные значения рКа (7,8 и 9,6) отражают одновременную ионизацию диметиламиногруппы и «фенольной» {}-дикетонной системы (последняя включает атомы в положениях 10, 11 и 12) [Leeson, Krueger, Nash, 1963; Rigler et al., 1965]. Такая одновременная ионизация двух групп, значения рКа которых отличаются не менее чем на две единицы, характерна для цистеина (Edsall et al., 1958].
Изучение связи структура — активность для тетрациклинов показало, что различные изменения в положениях 5, 6 и 7, не влияют на антибактериальную активность [McCormick et al., 1960; Blackwood, 1970]. Значительно более важны уменьшения в других областях молекулы. Трикарбонилметановая часть молекулы, как уже говорилось выше, играет существенную роль в процессе связывания металлов, а также в антибактериальном действии. При добавлении к раствору тетрациклина ионов Са2+, Си2+, Мп2+ и Со2+ в спектрах ПМР наблюдают уширение сигналов в соответствующей области, однако при добавлении появляется аномальный сигнал, свидетельствующий о том, что только этот металл вызывает изменение конформации тетрациклина перед образованием связи [Williamson, Everett, 1975]. Известно, что наличие и доступность «фенолдикетонной» области (положения 10, 11, 12) необходимы для связывания Mg2+ и Са2+ [Mitscher, Bonacci, Sokoloski, 1968; Doluizio, Martin, 1963]. Известно также, что определяющее значение имеет стереохимия фрагмента 4а—12а, т. е. соединения третьего и четвертого кольца, если считать слева направо в молекуле (11.36). Рассмотрение последних трех зависимостей позволяет определить расположение катионов магния в хелатном комплексе. И наконец, для антибактериального действия in vivo (но не in vitro) большое значение имеет диметиламиногруппа.
Как уже отмечалось в гл. 3 (том 1), избирательность действия тетрациклинов основана на их способности накапливаться в бактериях в значительно большей степени, чем в клетках млекопитающих. Существуют доказательства того, что эти лекарственные препараты образуют липофильный комплекс с магнием в плазматической мембране бактерий, что позволяет им проникать в цитоплазму [Franklin, 1971; Dockter, Magnuson, 1973; Franklin, Snow, 1981]. Основным местом действия тетрациклинов в бактерии являются рибосомы, отличающиеся повы
185
шенным содержанием магния и прекращающие синтез белка при действии этих антибиотиков. Ионы никеля, кобальта и цинка, образующие с тетрациклином значительно более прочные связи, чем магний, в отличие от последнего не снимают действия этих лекарственных веществ [Weinberg, 1954]; с помощью флуоресцентных методов удалось показать, что образование хелатных соединений с магнием в рибосомах бактерий предшествует ингибированию синтеза белков тетрациклинами [White, Cantor, 1971].
Благодаря способности к хелатообразованию, лекарственные препараты тетрациклинового ряда инактивируются во внутренних органах больного ионами кальция или магния, поступающими с пищей, например с молоком, или с антацидными лекарственными средствами. Подобная инактивация приводит к тому, что для многих больных эффект применения этих антибиотиков, значительно снижается. Препараты тетрациклинового ряда обычно принимают перорально. Сам тетрациклин назначают довольно часто, однако, кроме него, существует ряд тетрациклиновых препаратов, эффективных в более низких дозах, — демек-лоциклин, метациклин и их подкласс — доксициклин и миноциклин, отличающиеся пролонгированным действием.
11.9. Вещества, биологическое действие которых отчасти обусловлено хелатообразованием
Кроме перечисленных выше, в клинике широко применяют и многие другие хелатирующие агенты. Строение этих веществ таково, что они не могут не соединяться с тяжелыми металлами в тканях, однако механизм их действия не настолько изучен, чтобы можно было утверждать, что их действие обусловлено только хелатообразованием. Независимо от того, в какой степени механизм действия этих веществ определяется их способностью к хелатообразованию, это свойство заслуживает дальнейшего изучения в связи с его ролью в процессах адсорбции и возникновения побочных эффектов.
А. Изониазид (11.37). Об открытии этого вещества см. разд. 6.3.1. В 1956 г. A. Albert установил, что этот лекарственный препарат — гидразид изоникотиновой кислоты, представляет собой эффективный хелатирующий агент, константы устойчивости которого имеют тот же порядок, что и у глицина [Albert, 1956]. Изониазид (11.37) входит в состав комплекса в виде аниона (11.38). Метильное производное изониазида М-метил-ЬТизоникотиноилгидразин (11.40) не образует аниона и фактически не обладает противотуберкулезной активностью [Cymerman-Craig, Willis, 1955; Cymerman-Craig et al., 1955]. Так, изониазид ингибирует Mycobacterium tuberculosis H37RV (in vitro в присутствии 10% сыворотки) в концентрации М/5000 000, а вещество (11.40) не активно.
В присутствии ионов меди М. tuberculosis H37Rv (в культу-
186
Изониазид Аннон Комплекс с металлом
(11.37) (11.38) (11.39)
О NH2
II I
С—N—Me
N
Метнлпронзводное
(11.40)
ре) значительно быстрее поглощают изониазид [Youatt, 1962]. Содержание меди в сыворотке крови поддерживается на низком уровне (см. разд. 11.0), но, учитывая, что инфекция вызывается небольшим количеством бактерий, а лечение довольно продолжительно, меди оказывается достаточно для того, чтобы превратить терапевтические концентрации лекарственного вещества в хелатное соединение. Однако действие изониазида нельзя отнести полностью за счет хелатообразования, так как пи один из двух его изомеров не оказывает сколько-нибудь заметного действия на М. tuberculosis даже in vitro хотя их константы устойчивости несколько выше [Albert, 1956], что показано в табл. 11.11. Известно, что изониазид проникает только в клетки нерезистентных бактерий туберкулеза, но не в клетки резистентных штаммов; его не поглощают также такие распространенные бактерии, как Staph, aureus и Е. coli, на которые изониазид вообще не действует [Youatt, 1958].
Очевидно образование хелатного соединения со следовыми количествами меди придает лекарственному веществу дополнительную липофильность, необходимую для проникновения через наиболее гидрофобную из бактериальных оболочек. Kruger-Thiemer (1957) и Seydel и сотр. (1976а) установили, что изониазид представляет собой транспортную форму (пролекарство) истинного лекарства изоникотинамида (в который он окисляется в организме). Этот амид превращается далее в аналог никотинамидадениндинуклеотида (НАД). Замещение НАД этим аналогом приводит к нарушению метаболизма липидов. Несомненно то, что терапевтические концентрации изониазида вызывают понижение содержания НАД в микобактериях, что приводит к прекращению синтеза миколевой кислоты — типичного воскообразного материала, образующего защитную пленку вокруг бактерий штамма Н37 [Winder, Collins, 1970].
Seydel и сотр. (1976b) синтезировали изоникотиновый аналог изониазида и обнаружили, что он идентичен соединению, выделенному из штамма М. tuberculosis Н37. Изониазид действует только на те штаммы М. tuberculosis, которые содержат пероксидазу (высокопатогенные штаммы, не резистентные к лекарственным препаратам, содержат избыток пероксидазы).
187
Пероксидаза превращает изониазид в изоникотиновую кислоту in vitro (в бесклеточной системе), поэтому при лечении изониазидом изоникотиновая кислота накапливается в бактериальных клетках. Никотиновая кислота и ее гидразид могут служить антагонистами бактерицидного действия изониазида in vitro.
Ранее полагали, что изониазид действует на Mycobacteria, соединяясь с пиридоксалем. Однако вскоре это утверждение было опровергнуто, хотя некоторые побочные эффекты, наблюдаемые у больных, можно отнести и за счет этой реакции.
Б. Другие хелатообразующие противотуберкулезные препараты. Тиоацетазон (тиосемикарбазон-пара-ацетамидобензальде-гида) (11.41), известный также под названием амитиозон, обладает специфическим действием на туберкулезные бактерии, которое, однако, значительно слабее, чем у изониазида. Между этими двумя лекарственными веществами не возникает перекрестной резистентности. Тиосемикарбазоновая группа образует хелатную связь с тяжелыми металлами; антибактериальное действие тиоацетазона усиливается в присутствии меди, которую он восстанавливает до одновалентного состояния (связь, образуемая с цинком прочнее, чем с медью) [Stiinzi, 1982].
Противотуберкулезные свойства тиосемикарбазон-2-ацетил-пиридина усиливаются в результате комплексообразования с железом или медью[Scovill, Klayman, Franchino, 1982].
S
MeCO—HN
CH=N— NH—C—NH2
Тиоацетазон J
(Н.41) OH—сн4 CH2OH
CH—NH—CH2—CH2—NH— CH
I I
C2H5 C2H5
Этамбутол 1
(11.42)
Днанноя салициловой кислоты
(11.43)
При лечении туберкулеза для предупреждения развития резистентности изониазид обычно применяют в сочетании с рифампицином (4.37) [Fox, 1977]. Однако из-за высокой стоимости рифампицина его обычно заменяют высокоэффективным этам-
188
бутолом (11.42), который, однако, совершенно неактивен в отношении других видов бактерий. Этамбутол представляет собой простое гидроксиалкилпроизводное этилендиамина (см. табл. 11.1), образующего хелатные соединения с металлами подобно незамещенному соединению. Избирательность этамбу-тола обусловлена стерическими препятствиями, возникающими из-за наличия а-ветвей в боковых цепях [Shepherd, Wilkinson, 1962]. В процессе метаболизма это лекарственное вещество превращается в соответствующую дикарбоновую кислоту, обладающую значительно более сильной способностью связывать медь и цинк [Cole, May, Williams, 1981].
Пара-аминосалициловая кислота и стрептомицин (первые адъюванты, применявшиеся вместе с изониазидом) почти вышли из употребления. Салициловая кислота — сильный хелатирующий агент при условии достаточной щелочности среды для образования лиганда дианиона (11.43) [Perrin, 1958]. Салициловая кислота обладает мягким антибактериальным и фунгицидным действием; она широко применяется для размягчения огрубевшей кожи, действуя как мягкая форма фенола. Производные салициловой кислоты, такие как салициланилид, салициламид и салициловый альдегид, не способны образовывать карбокси-лат-анионы и являются менее сильными хелатирующими агентами [Perrin, 1958]. Ацетилсалициловая кислота, в молекуле которой блокирована только фенольная группа, не образует хелатов, однако при лечении ревматических болезней она легко гидролизуется в организме до салициловой кислоты, которая, как полагают Sih и Takeguchi (1973), образует хелатный комплекс с медью в простагландинсинтетазе. Антибиотик антимицин имеет структуру 3-формамидосалициламида и способен прочно связывать ион трехвалентного железа [Farley, Strong, Bydalek, 1965].
Налидиксовая кислота (4.33) связывает тяжелые металлы с карбоксильной и оксо-группами [Ctumplin, Midgley, Smith, 1980]. Неизвестно, играет ли это какую-либо роль в ее биологическом действии, заключающемся в ингибировании синтеза ДНК (разд. 4.0.5). Антибактериальное действие койевой кислоты (11.11), пирона, выделяемого из некоторых грибов, усиливается катионами металлов [Weinberg, 1957]. Полипептидный антибиотик бацитрацин (разд. 13.2) в присутствии ЭДТА утрачивает антибактериальное действие на Staph, aureus; однако в присутствии катионов двухвалентных металлов оно восстанавливается [Adler, Snoke, 1962].
В. Противовирусные лекарственные средства. Феноксиуксус-ная кислота (разд. 4.0.2, том 1) образует с цинком и другими двухвалентными металлами шестичленный цикл [Stiinzi, Perrin, 1979]. Полагают, что противовирусная активность метиса-зона (6.19) и его аналогов (разд. 6.3.1, том 1) обусловлена образованием хелатного соединения, в котором металл удерживается между атомом кислорода и вторым атомом азота боковой
189
цепи [O’Sullivan, Sadler, 1961]. Это лекарственное вещество восстанавливает медь до одновалентного состояния, а с цинком образует более прочную связь, чем с медью [Stiinzi, 1982]. Ме-тисазон способен дезактивировать в культуре ткани многие РНК-содержащие вирусы, вызывающие рак [Levinson, Wood-son, Jackson, 1971]. Тиосемикарбазон 2-ацетил-пиридина сильно ингибирует in vivo репликацию вируса герпеса симплекс типа 1 и 2, практически не затрагивая клеточную ДНК или синтез белков [Shipman et al., 1981].
Г. Фунгицидные препараты. Кроме сильнодействующих фунгицидов (разд. 11.7) и салициловой кислоты (см. выше), особое значение имеют простые производные трополона (11.44), содержащиеся в сердцевине некоторых пород хвойных деревьев. Эти соединения связывают медь, появляющуюся случайно в незначительных количествах, и тем самым обеспечивают защиту древесины от грибов на несколько десятилетий (что обусловливает сохранность неокрашенных деревянных зданий). Мо-ноизопропилпроизводные соединения трополона, туйяплицины распространены довольно широко. Полагают, что в основе механизма их действия лежит ингибирование образования ацетил-КоА из ацетат-ионов [Raa, Goksoyr, 1966].
Д. Противораковые препараты. Тиосемикарбазоны обладают противовирусными и противотуберкулезными свойствами, а также противораковой активностью, например тиосемикарбазон 5-гидроксипиколинового альдегида (4.12) (разд. 4.0.1, том I). Эти соединения эффективно ингибируют рибонуклеозид-дифосфатредуктазу, фермент, превращающий рибо- в дезоксирибонуклеотиды, что приводит к подавлению синтеза ДНК. Предполагают, что в основе действия тиосемикарбазонов лежит взаимодействие с железом, необходимым для ферментов [Agra-wal et aL, 1972]. Аналогично действует и гидроксимочевина (4.10), применяемая в клинике для лечения меланомы (разд. 4.0.1).
Тиосемикарбазон 5-гидроксипиколинового альдегида, так же как и тиоацетазон (11.41), является бидентатным лигандом (т. е. может образовывать только две связи с каждым атомом металла), поэтому были предприняты попытки отыскать агенты, более прочно связывающие металл, среди тридентатных производных этого альдегида [Ankel, Petering, 1980] и среди квадрадентатных бис-тиосемикарбазонов 1,2-дикарбонильных соединений, таких как З-этокси-2-оксобутиральдегид [Petering, 1972]. Ионы двухвалентной меди могут переносить это соединение в живые клетки [Sartorelli, Creasey, 1969].
Блеомицин (4.28)—наиболее широко применяемый в клинике противораковый металлосвязывающий препарат, использует железо для активации кислорода воздуха и затем разрушает ДНК (разд. 4.0.4).
Сильнее всего D-пеницилламин (11.26) действует на аденокарциномы, имплантированные крысам. Это происходит за счет 190
дезактивации фермента супероксиддисмутазы, контролируемого медью и цинком, нормальные клетки компенсируют его недостаточность за счет аналогичного фермента, контролируемого-магнием, которого в раковых клетках мало [Okuyama, Hitoshi». 1981; Oberley, 1982].
Диметилглиоксим (11.45) действует на карциному Эрлиха и саркому Крокера на мышах, однако он проявляет противоопухолевую активность при добавлении в среду незначительных нетоксичных количеств ионов двухвалентной меди [Takamiya, 1960].
/Ч /ОН
/ MeC=NOH
।
\MeC=NOH
Троп ол он Днметнлглнокснм
(11.44) (11.45)
Конканавалин А — белок, выделяемый из канавалии мечевидной, способен связывать углеводороды и обладает свойствами, характерными для «лектина». Этот белок агглютинирует злокачественные клетки и подавляет их рост [Sharon, Lis, 1972], однако при этом агглютинируются также и эритроциты. Каждый мономер конканавалина А содержит один участок, связывающий ионы кальция, и другой, связывающий катионы Zn2+, Со2+ или Мп2+; для проявления биологической активности оба этих участка должны быть заняты соответствующим металлом.
Е. Препараты для лечения протозойных инфекций. Биала-микол (11.46), появившийся в 1950 г., был первым синтетическим препаратом, непосредственно атакующим паразитов при амебиазе. Он обладает хелатирующими свойствами глицинового типа [Dill et al., 1957]. Тиосемикарбазон 2-ацетилпиридина в низких дозах (20 мг/кг) излечивает инфекции, вызванные Pl. berghei у мышей (20 мг/кг) [Klaymann et al., 1979].
Такие хелатирующие агенты, как кофейная кислота (3,4-ди-гидроксикоричная кислота), неокупреин (2,9-диметил-ортофе-нантролин) и 2-пиридилгидразон пиридин-2-аля, при внутривенном введении излечивали всех мышей, которым вводили летальную дозу Trypanosoma brucei [Shapiro et al., 1982]. Считается, что эти агенты лишают паразитов жизненно важного железа, содержащегося в глицерофосфатоксидазе, которая (только в этих организмах) осуществляет окисление в конце метаболического пути.
Ж. Антигельминтные препараты. Даже простые Ь1,ЬГ-хела-тирующие агенты обладают антигельминтными свойствами, особенно 2,2/-бипиридил (11.19) и орто-фенантролин (11.18) [Baldwin, 1948b; Dwyer et al., 1952]. Широко используют также вермицид, тиабендазол (6.36), который прочно связыва-
191
ет катионы переходных металлов двумя атомами азота [Kowa-la et aL, 1971].
NH—NHS
%/4^N
Бналамнкол
(11.46)
Апрессин
(И-47)
3. Фармакодинамические средства. Металлосвязывающие свойства адреналина, тироксина и гистамина уже обсуждались в разд. 11.1. Действие полипептидных гормонов, содержащих дисульфидный мостик (—S—S—), усиливается катионами двухвалентных металлов. Так, сокращение матки под действием окситоцина усиливается катионами Со2+, Мп2+ и Ni2+, но ослабляется Са2+ и Си2+. Полагают, что металл образует тройной комплекс с гормоном и его рецептором [Schild, 1969].
Хелатирующее действие приписывают иногда и кортизону, однако доказательства этого факта до сих пор не получены. Мнение о том, что индолил-3-уксусная кислота является регулятором роста растений вследствие хелатирующего действия, было опровергнуто после того, как удалось доказать, что это вещество не обладает хелатообразующими свойствами [Perrin, 1961].
Многие металлосвязывающие агенты способны снижать повышенное давление крови у человека, действуя непосредственно на мускулатуру кровеносных сосудов [Schroeder, Perry, Menhard, 1955], например апрессин (11.47), димеркапрол (11.23) и тиоцианат натрия, высвобождающийся при внутривенном введении нитропруссида натрия при гипертонических кризах. Апрессин — одно из наиболее распространенных лекарственных средств, назначаемых перорально при умеренных формах гипертонии. Он образует хелатное соединение с катионами переходных металлов, связывая их подобно этилендиамину между двумя атомами азота. Аналогичные гидразиновые производные других азотсодержащих гетероциклов обладают такими же терапевтическими свойствами.
Амринон (5-амино-3,4'-бипиридил-6-он) в настоящее время применяют перорально при застойной сердечной недостаточности [Benotti et al., 1978]. Этот препарат способен связывать катионы металлов между заместителями в положениях 5 и 6. Об ионофоретических сердечных средствах см. разд. 14.2.
Антидиабетическое лекарственное средство фенформин (fj-фенил-этилбигуанид) образует хелатные соединения с ионами двухвалентной меди и обладает сродством, близким к таковому глицина. Среди родственных ему соединений способность 192
к хелатообразованию с медью изменяется параллельно антидиабетической активности, выражающейся в ингибировании глюконеогенеза в печени [Foye, O’Laughlin, Duvall, 1961].
11.10. Особые случаи образования прочных комплексов
В водных растворах комплексы большинства катионов металлов находятся в динамическом равновесии с их компонентами. При нарушении этого равновесия мгновенно возникает новое. Однако особопрочные комплексы могут существовать в течение нескольких часов (или даже дней); их биологические свойства значительно отличаются от свойств их компонентов. Эти комплексы образуются ионами металлов, имеющих 3, 4 (низкоспиновые), 5 или 6 d-электронов, что обусловливает высокие затраты энергии для стабилизации поля лигандов при образовании комплекса. Особопрочные комплексы наиболее часто образуются переходными металлами: платиной, иридием, осмием, палладием, родием, рутением, а также (несколько реже) никелем, кобальтом и железом. Наиболее легко особопрочные комплексы с этими переходными металлами образуют галоид- и особенно цианид-анионы, затем следуют другие азот-и кислородсодержащие анионы, обладающие подобными свойствами в меньшей степени.
В 1938 г. Beccari обнаружил, что некоторые особопрочные хелатные соединения обладают курареподобными свойствами, например, координационный комплекс, образованный ионом двухвалентного железа и тремя молекулами 2,2'-бипиридила и имеющий заряд, равный двум, вызывает слабую форму паралича и остановку дыхания у кроликов. Позднее ему удалось показать, что кролики, так же как и лягушки, выделяют этот комплекс в неизмененном виде [Beccari, 1941]. Продолжая эти исследования, Dwyer, Gyarfas, O’Dwyer (1951) изучали комплексы железа, никеля, кобальта, рутения и осмия и обнаружили, что комплекс рутения с 2,2',2/л-терпиридилом обладает наибольшей активностью, равной одной десятой активности тубо-курарина. Активными были только его положительно заряженные комплексы. В экспериментах на мышах или на изолированной диафрагме крыс было обнаружено, что парализованные мышцы отвечают на прямую стимуляцию электрическим током, а паралич, так же как и вызванный тубокурарином, снимается действием физостигмина. Эти комплексы не содержат незащищенных химически активных групп и их окислительно-восстановительные потенциалы находятся вне биологических пределов. Таким образом, геометрия и заряд особопрочных комплексов определяют их биологическое действие, а роль металла сводится только к удерживанию остальных атомов в конфигурации, дополняющей таковую рецептора [Shulman, Dwyer, 1964]. Позднее были синтезированы соединения, содержащие два положительно заряженных центра на таком же расстоянии друг
13—734
193
от друга, как и в молекуле тубокурарина, и обладающие большей, чем у тубокурарина, активностью [Taylor, Callahan, Shaikh, 1975].
Препарат цис-дихлордиаминплатина (11.48) способен вызывать регрессию больших солидных опухолей саркомы-180 [Rosenberg, Van Camp, 1970]. Это лекарственное вещество, известное под названием цисплатин, оказалось лучшим для лечения опухолей семенников и яичников, нередко полностью их излечивая. Цисплатин часто применяют и при лечении рака мочевого пузыря и др. [Rosencweig et aL, 1977; Einhorn, Williams, 1979]. Основные токсические эффекты цисплатина — рвота, миелосуппрессия, ототоксичность и (наиболее тяжелое) повреждение почек. Препарат вводят внутривенно; нефротоксичность удается предотвратить, повышая диурез.
Цисплатин представляет собой пролекарство. Он становится биологически активным только после того, как оба атома хлора замещаются гидроксигруппами. Соответствующий транс-изомер неактивен. Механизм действия этого препарата заключается, по-видимому, в связывании двух цепочек ДНК, а также в усилении фосфорилирования хроматина в ядрах. Аналоги цисплатина, имеющие более высокий терапевтический индекс, были получены заменой двух молекул аммиака двумя молекулами циклогексиламина [Connors et al., 1972]. В настоящее время в клинике прошел испытания карбоплатин (11.49), который обеспечивает быструю ремиссию при раке яичников, не обладает ни нефро-, ни ототоксичностью и в меньшей степени вызывает тошноту [Calvert et al., 1982].
Подробнее о противораковых препаратах, содержащих платину, см. Roberts, Pera (1983).
Спирогерманий (11.50)—особопрочный комплекс, применяемый при раке толстой кишки и яичников, один из немногих противоопухолевых препаратов, не оказывающий повреждающего действия на костный мозг. Большие дозы спирогермания токсичны для ЦНС [Slavik et al., 1982].
Все комплексы золота, применяемые для лечения тяжелейших форм артрита, содержат этот металл, связанный в одновалентном состоянии атомом серы. Эти соединения можно отнести к комплексам только на основании того, что они медленно образуются, но быстро обмениваются. До недавнего времени все эти препараты вводили только инъекционным путем, однако теперь существует пероральный препарат ауранофин (11.51)
О
H3N\ /С1
H3N4 /О-С,
Цнсплатнн
(11.48)
О Карбоплатин (11.49)
194
[Berglof, 1977]. Его химическая формула — 1-тио-рД)-глюкопи-ранозато (триэтил фосфин) золото-2,3,4,6-тетраацетат.
Ауранофин (11.51)
11.11. Основные принципы создания новых хелатирующих агентов, перспективы их применения
При изыскании новых хелатирующих агентов следует прежде всего иметь в виду, что большинство этих соединений лишено биологической активности. Например, среди металлосвязы-вающих агентов, обычно применяемых для аналитических целей, лишь немногие обладают антибактериальным действием [Albert et al., 1947; Schraufstatter, 1950].
Ни один хелатирующий агент не активен в биологической среде, если константы устойчивости его компонентов по меньшей мере не столь же высоки, как у комплексов обычных аминокислот, например глицина (см. табл. 11.1). Однако и у веществ с очень высокими константами устойчивости трудно ожидать наличия биологической активности, так как потеря таких веществ в результате их насыщения металлом происходит еще до того, как они достигнут места действия. В некоторых случаях для проявления этими соединениями активности должен происходить хотя бы умеренный обмен катионов [Schubert, 1957].
При поиске новых хелатирующих агентов необходимо знать место их биологического действия: внутри или снаружи клетки. Если молекула должна проникать в клетку, то в нее необходимо ввести липофильные группы. Для этой цели используют атомы углерода, галогенов, водорода и серы, а атомы азота и кислорода придают молекулам гидрофильные свойства; контроль можно проводить по величине коэффициентов распределения в системе масло — вода. Результаты введения гидрофильных и липофильных групп в молекулу оксина см. в табл. 11.8. В комплексы, подобные образуемым этилендиамином (см. рис. 11.2), в которых металл сохраняет первоначальный заряд, необходимо вводить липофильные группы для повышения их способности проникать в клетку, причем количество таких групп должно быть больше, чем для незаряженных комплексов.
Очень небольшие изменения в химической структуре молекулы могут вызвать значительные сдвиги в величинах коэффициентов распределения. Так, введение в цикл оксина дополнительного атома азота с образованием 3-азаоксина (8-гидрокси-
13*
195
хиназолина) резко повышает гидрофильность молекулы — коэффициент распределения в системе масло — вода уменьшается от 67 до 5 (см. табл. 11.8). Введение в боковую цепь заместителя, состоящего всего из трех атомов углерода (пропильная группа), восстанавливает коэффициент распределения до прежнего значения.
Увеличить способность вещества проникать в клетки можно, не только руководствуясь коэффициентами распределения, но и используя лиганды, сходные с природными субстратами, например аминокислоты, углеводороды, холин, пурины и пиримидины.
Стерические и другие химические различия, которые могут способствовать избирательности, рассматривались в разд. 11.4.
Многие биологически активные хелатирующие агенты непригодны в качестве пероральных средств для человека просто потому, что они недостаточно избирательны. Необходимо всегда соблюдать осторожность с металлосвязывающими веществами, поскольку некоторые из них, такие как диметилдитиокарбамат натрия [Kadota, Midorikawa, 1951], повреждают островки Лангерганса и вызывают диабет у подопытных животных. Аналогичным образом действуют и 5-амино- и 2-метилпроизводные оксина [Kadota, Abe, 1954].
Известны некоторые случаи, в которых хелатирующие агенты способны преодолевать резистентность по отношению к лекарственным препаратам (разд. 6.5, том 1).
Современные проблемы поиска хелатирующих средств охватывают все вопросы, обсуждавшиеся в разд. 11.9, а также современные методы детоксикации и обеззараживания тканей, поврежденных химически токсичными или радиоактивными металлами, попытки предупреждения хелатообразования как причины кариеса зубов, а также предотвращение потерь кальция из костной ткани в пожилом возрасте, которые являются причиной преждевременного старения.
Глава 12 СТЕРИЧЕСКИЕ ФАКТОРЫ
Биологическое действие химического соединения в большой степени зависит от формы и размера его молекул. Для проявления некоторых типов биологической активности необходимым требованием является планарность молекулы, для других — объемность, трехмерность структуры. Например, бензольное кольцо (12.1)—плоское, и все шесть связей, направленные от кольца к атомам и группам, также лежат в этой плоскости. Плоское строение имеют и все другие сопряженные системы (т. е. системы, у которых каждая вторая связь двойная).
В отличие от них молекулы алифатических и алициклических соединений непланарны. Такова, например, молекула циклогексана (12.2), отличающаяся от бензола наличием шести дополнительных атомов водорода; только четыре из шести атомов углерода лежат в одной плоскости, а все связи направлены к ней под разными углами. Атропин (7.16)—лекарственное вещество с неплоским кольцом, тогда как акридин (10.8)—типичный пример лекарственного вещества с планарной молекулой. И плоские, и трехмерные структуры часто встречаются в лекарственных веществах, иногда даже входят в одну молекулу, как, например, в акрихине (10.30) и никотине (7.26).
Ме
1
Ме—С-—
I ХОН
Ме
Бензол
(12.1)
Циклогексан Триметилуксусная кислота Орнид
(12 2) (12.3) (12.4)
12.0. Некоторые основные принципы
Зависимость биологической активности от площади планарной части молекулы в ряду химиотерапевтических лекарственных средств была впервые показана для производных аминоакридина, в молекулах которых площадь, необходимая для проявления противобактериальных свойств, равна 0,38 нм2 [Albert,
197
Rubbo, Burvill, 1949], что обусловливает взаимодействие такой молекулы с пуриновыми и пиримидиновыми основаниями ДНК (разд. 10.3.2). Если бензольное или пиридиновое кольцо молекулы взаимодействует с плоским участком, окружающим рецептор, то дополнительный выигрыш в энергии адгезии, обусловленный ван-дер-ваальсовыми связями, составляет2—Зккал/моль (8—12 кДж/моль).
При замене ароматического цикла в молекуле лекарственного вещества на более гидрированный необходимо учитывать геометрическую форму нового цикла. Молекула циклопентана вследствие тесного расположения заместителей (даже если это атомы водорода), почти плоская, так же как и пятичленный цикл рибозы. Циклопентеновый цикл тоже имеет плоскую структуру, тогда как молекулы циклогексена и циклобутана и особенно циклогексана (12.2) деформированы. Даже если бы циклобутановое кольцо имело плоскую структуру, энергия его ван-дер-ваальсова взаимодействия составила бы всего 2/3 от таковой бензольного кольца. К снижению энергии ван-дер-ваальсова взаимодействия приводит и то, что каждый заместитель, непосредственно связанный с небензоидным циклом, выведен из его плоскости.
Замена фенильной группы в молекуле лекарственного вещества другими циклами позволяет установить, насколько необходима плоская структура для биологической активности данного соединения. Так, при замене бензольного кольца в молекуле фенамина и других лекарственных веществ, производных фенетиламина или фенилэтаноламина, циклобутановым, цикло-пентановым, циклогексеновым и циклогексановым кольцами, всегда наблюдается ослабление фармакодинамического действия [Burger et al., 1961, 1963].
В органическом синтезе хорошо известно влияние стериче-ских затруднений на протекание химических реакций (разд.2.5 и 2.5.1). Например, реакция этерификации триметилуксусной кислоты (12.3) протекает в более жестких условиях, чем уксусной кислоты.
Стерические препятствия, возникающие между двумя группами, находящимися по одну сторону двойной связи, могут приводить к нарушению планарности этой связи и изменению физических и химических свойств молекулы. В разд. 13.1 описано, каким образом можно создать стерические препятствия ферментативному гидролизу в молекулах полусинтетических пенициллинов. Точно так же при синтезе лидокаина (7.13) учитывали, что две орто-метильные группы в его молекуле будут затруднять гидролиз амидной связи, поэтому его действие будет длительнее по сравнению с другими местными анестетиками.
В некоторых случаях стерические затруднения оказываются необходимыми для проявления фармакологического эффекта. Примером такого соединения может служить гипотензивный препарат орнид (12.4). Биологическая активность сохраняется
198
при замене в его молекуле атома брома на хлор, йод, метильную или нитрогруппу, но исчезает при замене атома брома на атом водорода. Следовательно, роль орто-заместителя заключается в выведении группировки, обладающей основным характером, из плоскости бензольного кольца [Biura, Copp, Green, 1959]. Стерическое взаимодействие двух этильных групп обусловливает эстрогенные свойства стильбэстрола (разд. 12.2).
Для решения стереохимических проблем следует использовать молекулярные модели. Существуют два типа таких моделей; первый — это скелетные или рамочные модели, предложенные Дрейдингом или Кендрью. В них обозначены центры связей, соединяющих атомы, и они очень удобны для поиска конформаций, обеспечивающих взаимодействие двух молекул. Другой тип — шариковые модели, позволяет оценить не только форму молекулы, но и занимаемый ею объем. Поэтому они очень удобны для рассмотрения общей формы, поверхности и объема молекулы. На практике химики пользуются всеми тремя видами моделей. Существуют еще и CCS-модели, которые, являясь в принципе скелетными, могут быть легко преобразованы в шариковые и наоборот [Clarke, 1977].
Построение молекулярных моделей см. Walton (1978).
12.1. Оптическая изомерия
Атом углерода более всех других атомов способен к образованию различных стереоизомеров, о которых пойдет речь в этом и двух последующих разделах. Большинство оптических изомеров возникает из-за того, что в молекуле вещества есть атом углерода, связанный простыми связями с четырьмя разными атомами или группами атомов. Атом углерода с четырьмя разными заместителями называется асимметрическим.
Четыре связи атома углерода, имеющего сферическую форму, направлены к вершинам тетраэдра, в центре которого расположен сам атом углерода. Углы между связями равны 109°. У простой молекулы, образованной атомом углерода, соединенным с четырьмя заместителями А, В, С и D, при сохранении порядка расположения заместителей все проекции равноценны. Так, формулой (12.5) представлен вариант, когда заместители В, С и D лежат в одной плоскости, а заместитель А находится над ней. Этот же тетраэдр можно повернуть так, что любой из заместителей (В, С или D) окажется над плоскостью, в которой расположены три остальные. В формулах (12.5—12.7) представлены различные расположения тетраэдрической молекулы относительно одной плоскости. Однако если поменять местами два заместителя [ср. формулы (12.5) и (12.8)], то получается другое соединение, т. е. другой представитель хиральной пары. Рассмотрим этот вопрос подробнее на примере молочной кислоты (12.9).
199
н I Me—С—CO2H
ОН
Молочная кислота
(12.9)
Если к центральному атому углерода присоединены четыре разных атома или группы, как в молекуле молочной кислоты (12.9), то возможны два различных расположения этих заместителей в пространстве, а именно (12.10) и (12.11). Эти две структуры являются зеркальным отображением друг друга, как правая и левая рука. От греческого слова «kneir» произошло слово «хиральный», поэтому такая пара соединений называется хиральной или энантиомер (раньше применяли и такие названия, как энантиоморфы или оптические антиподы).
Оба энантиомера обладают идентичными химическими свойствами. У них одинаковы и физические свойства, за исключением одного: каждый вращает плоскость поляризации света на один и тот же угол в противоположные стороны. У всех хи-ральных молекул есть оптические изомеры. Оптически активные вещества избирательно абсорбируются на таких оптически активных поверхностях, как поверхность белковых молекул, и поэтому один и тот же биологический эффект двух оптических изомеров может быть выражен в разной степени.
в в
С—-D D-----------1---С
А А
(12.10) (12.11)
При написании структурной формулы значительно проще изображать молекулу в плоскости, располагая в центре атом углерода и отходящие от него четыре связи к заместителям — в четырех углах тетраэдра. При такой записи изображением одного энантиомера молочной кислоты служит формула (12.10), а другого — формула (12.11). Клинообразное изображение связи обозначает, что атом на утолщенном конце клина находится над плоскостью бумаги, а атомы на конце пунктирных линий — под плоскостью бумаги, на которой расположен центральный атом углерода. Плоское изображение (12.10) соответствует объемному изображению (12.6). В сложных молекулах или для ускорения записи клинышки в изображении связей можно заменить утолщенными линиями, как, например в (7.11), но пунктирные линии нельзя заменять тремя точками, так как последними обозначают водородные связи.
Часто при нагревании, а также под действием кислоты или щелочи один чистый энантиомер может превращаться в смесь, 200
содержащую равное количество обоих энантиомеров; при этом оптическая активность пропадает. Такое явление называют рацемизацией. Разделение рацемической смеси оптических изомеров возможно с помощью оптически активных ионов с противоположным (относительно энантиомеров) зарядом или ферментов, разрушающих лишь один энантиомер [Wilen, 1971; Wilen, Collet, Jacques, 1977]. Иногда удается разделить энантиомеры хроматографией на асимметрических носителях [Albert, Serjeant, 1964].
По данным 1980 г. из 2000 наиболее широко применяемых лекарственных веществ около половины составляют хиральные соединения. Из синтетических препаратов в виде оптически активных энантиомеров применяют только 88. Из 550 пестицидов лишь 103 имеют хиральный центр.
Молекула с двумя различными асимметрическими атомами углерода существует в виде четырех энантиомеров, образующих две различные рацемические смеси. Энантиомеры из двух рацемических смесей не являются зеркальными отображениями друг друга и называются диастереомерами, например, четыре стереоизомера эфедрина (12.12) — (12.15) [Witcop, Foltz, 1957]. Хорошо видно, что, для того чтобы молекулы (12.12) и (12.13) стали зеркальным отражением друг друга, необходима более чем одна перестановка заместителей. В конце -раздела 12.3 приведен другой способ изображения диастереомеров (12.42).
ОН NHMe Н Н
II II
Ph—С—С—Me Ph—С—С—Ме
II II
НН ОН NHMe
(—)-Эфедрин (протнвоастмати- (4-)-Эфедрин (в медицине
ческнй препарат) не применяют)
(12.12) (12.13)
Н NHMe ОН Н
II II
Ph—С—С—Me Ph—С—С—Ме
I I II
ОН Н Н NHMe
(Ч-)’Псевдоэфедрин (обладает местным сосудорасширяющим действием)
(—)-Псевдоэфедрин (в медицине не применяют)
(12.14) (12.15)
Четыре диастереомерные формы эфедрина
12.1.1. Определение абсолютной конфигурации
Со времен Био и Пастера любое соединение, вращающее плоскость поляризации света влево, обозначили знаком (—), а его энантиомер соответственно ( + ). Эту классификацию, отражающую только свойства, но не строение соединения, применяют и сейчас в тех случаях, когда неизвестна абсолютная конфигурация (строчные буквы lad вышли из употреб
201
ления). Однако уже к концу XIX в. стало ясно, что эти обозначения не несут информации об истинном строении молекул. Поляриметрические данные свидетельствуют лишь о том, что в молекуле есть асимметрический центр, но не указывают, какова его конфигурация, так как направление и величина угла вращения зависят от наличия в молекуле других заместителей, растворителя, температуры и длины волны поляризованного света. Поэтому многие соединения в разных условиях могут давать как отрицательный (—), так и положительный ( + ) углы вращения. Даже при измерении оптической активности при 25 °C в воде на длине волны D-линии натриевой лампы наблюдают много аномалий. Поэтому уже в конце XIX в. предпринимались неоднократные попытки ввести какой-либо стандарт, с помощью которого можно было бы определить истинное строение асимметрического центра независимо от показаний поляриметра.
В 1906 г. М. Rosanoff предложил в качестве стандарта «абсолютной конфигурации» ( + )-глицериновый альдегид, конфигурация асимметрического центра которого обозначалась маленькой буквой D [Hudson, 1948]. Глицериновый альдегид был выбран как простейшее вещество со свойствами сахара. В то время он еще не был получен, его синтез и разделение на оптические изомеры были осуществлены позднее Wohl и Momber (1917). Поскольку это вещество [НОСНзСН(ОН)СНО] образуется в организме при действии ферментов на глюкозу и фруктозу, оно стало «ключевым соединением» в химии углеводов. Все соединения, для которых путем химических переходов, не затрагивающих хиральный атом, может быть установлена генетическая связь с этим «ключом», относятся к D-ряду.
Именно так были установлены абсолютные конфигурации D-( + )-глюкозы в крови, D-(—)-фруктозы во фруктах и D-(—)-дезоксирибозы в ДНК.
Пространственная конфигурация, обозначенная буквой D, была приписана глицеральдегиду случайно, истинное же его строение оставалось неизвестным. Поэтому предпринимались неоднократные попытки его установления с помощью различных физических методов. Не удалось решить этот вопрос и методом рентгенографии. С помощью последнего можно точно измерить длины всех связей и углы между ними, но оказалось, что для хиральных пар все эти величины одинаковы. И только в 1951 г. Bijvoet разработал новый метод для определения пространственной конфигурации, впервые рентгенографически обнаружив разницу между двумя стереоизомерами.
Bijvoet применил метод задержки по фазе, возбуждая флуоресценцию одного атома около хирального центра пучком света с определенной длиной волны. Используя натриево-рубидие-вую соль винной кислоты, он показал, что приписываемая правовращающей винной кислоте проекционная формула правильно отражает реальную конфигурацию. Так как химическими ме-
202
годами уже была доказана ее принадлежность к D-ряду, то «абсолютная конфигурация», обозначаемая буквой D, действительно оказалась абсолютной [Bijvoet, Peerdemar, Van Bommel, 1951].
Предложенный Bijvoet метод слишком трудоемок для рутинных исследований [Klyne, Buckingham, 1978; периодическое издание «Molecular Structure and Dimensions»]. Обычный рентгеноструктурный анализ помогает установить конфигурацию соединений с несколькими хиральными центрами при условии, что конфигурация первого центра уже известна.
Особенно важно было установить абсолютную конфигурацию аминокислот белка. В 1950 г. D-( + )-глицериновый альдегид в пять стадий был превращен в D-( + )-серин. Так было показано, что (—)-серин, выделенный из белков, является L-(—)-серином. После этого было уже нетрудно установить, что все аминокислоты, содержащиеся в белках, относятся к L-ряду, хотя некоторые из них (как аланин) вращают плоскость поляризации вправо [Brewster et al., 1950].
В действительности в природе обычно встречаются обе энантиоморфные формы одного вещества. Так, например, одинаково распространены ( + )- и (—)-вращающие кварцы, ( + )- и (—)-молочные кислоты. Лишение бактерий как L, так и D-аланина приводит к их гибели (разд. 5.3), и хотя белок не может состоять из смеси L- и D-аминокислот, они часто присутствуют в полипептидах (разд. 14.3), облегчая их способность к циклизации. В коре хинного дерева образуются два энантиомера, известные под названиями ( + )-хинидин и (—)-хинин (10.33), в которых вторичная гидроксильная группа присоединена к асимметрическому атому углерода. Оба изомера применяют в медицине: хинидин —при сердечных аритмиях, хинин — при малярии.
Для установления абсолютной конфигурации значительно менее надежен (чем химический или рентгеноструктурный) метод дисперсии оптического вращения, в котором измеряется оптическое вращение вещества при разных длинах волн [Djerassi, 1960]. Он дает надежные результаты лишь для простых молекул.
Для упрощения номенклатуры оптически активных молекул Cahn, Ongold, Prelog (1956) ввели «правило последовательности». Использование символов (R) и (S) позволяет избежать неоднозначностей, возникающих при использовании системы D, L, для соединений, не родственных углеводам и аминокислотам, и помогает систематизации стереохимической информации и построению молекулярных моделей. Эта система наиболее удобна, если в одной молекуле есть два или три хи-ральных атома. Упрощенный вариант этих правил см. в работе Cahn (1964).
Применяя правило последовательности к описанию конфигурации молекулы, следует начинать с хирального центра с наи-
203
меньшим номером, определенным согласно правилам ИЮПАК. Его хиральность может быть взята из «Стереохимического атласа» или определена одним из вышеописанных методов. Затем четыре атома, соединенные с асимметрическим атомом углерода, обозначаются символами а, Ь, с и d в порядке уменьшения их относительной атомной массы. Если два из них — атомы углерода, то очередность устанавливается в зависимости от атомов, с которыми они связаны. Затем на хиральный центр смотрят со стороны, удаленной от младшего заместителя (d): если старшинство остальных заместителей уменьшается по часовой стрелке а->-Ь->-с, то такая конфигурация обозначается R, (лат. rectus), а если против часовой стрелки, то конфигурация обозначается символом S (лат. sinister). Примеры такого обозначения приведены рядом с формулами эфедрина (12.12) и псевдоэфедрина (12.13). Природный треонин имеет (2S, 3R)-конфигурацию.
Правило последовательности в настоящее время общепринято и широко используется. Однако в некоторых случаях оно не удобно, например, когда в серии родственных соединений близкие по строению вещества получают различные индексы [Cahn, Dermer, 1979]. Например, большинство аминокислот, входящих в состав белков, в том числе и серин, имеют S-конфигурацию, однако цистеин, отличающийся от серина лишь заменой атома О на атом S, имеет R-конфигурацию. Это отклонение возникает из-за того, что предпочтение отдается атому серы, имеющему большую массу. Поэтому Kahn и Dermer рекомендуют не пользоваться правилом последовательностей для углеводов аминокислот, стероидов, циклитолов и липидов, для которых уже существуют вполне приемлемые системы обозначения.
Вернемся к структурным формулам. Для изображения углеводов используют три различных вида формул, из них самый старый — проекционные формулы Фишера для глюкозы (12.16). При таком изображении углеродная цепь располагается вертикально, а альдегидная (полуацетальная) группа наверху. Предполагается, Что заместители в углеродной цепи лежат в плоскости бумаги. D-Конфигурацию приписывают энантиомеру, имеющему у асимметрических атомов углерода большее число старших заместителей справа.
Позднее Haworth предложил более соответствующий действительности способ изображения, например формула (12.17). В этом случае ацетальная группа расположена справа, а часть кольца, расположенная ближе к наблюдателю, изображена утолщенными линиями. Отчетливо видно, какие заместители располагаются над плоскостью кольца, а какие — под плоскостью. Например, группа —СНгОН расположена над плоскостью, а атом водорода, присоединенный к тому же атому углерода,— под плоскостью (атомы углерода кольца не обозначаются). Однако из данных рентгеноструктурного анализа 204
известно, что кольцо у гексоз не плоское, что изображается так, как это указано в формуле (12.18). Каждый способ изображения используют в зависимости от обстоятельств.
1 неон
2 НСОН
3 носн
4 НСОН
5 НСО---'
6 СН2ОН
Формула D-глюнозы
(12.17)
Изображение нонформации D-глюнозы
(12.18)
D-глюноза
(12.16)
В «Стереохимическом атласе» приведены абсолютные конфигурации (со ссылками на источники) примерно для 3000 органических соединений. Кроме оптических изомеров, туда включены вещества, хиральность которых связана с изотопным замещением, хиральными осями или плоскостями, а также соединения, у которых хиральным атомом является не углерод [Klyne, Buckingham, 1978].
12.1.2. Фармакологические аспекты хиральности
Cushny (1926) был первым, кто установил корреляцию между биологической активностью энантиомеров и способностью хиральных форм к взаимодействию с рецептором. Если же разницы в биологической активности у энантиомеров нет, то следует предположить, что асимметрический атом не контактирует с рецептором или же между рецептором и молекулой осуществляется только двухточечный контакт. Так, (+)- и (—)-формы барбитуратов (2.34), оптическая активность которых обусловлена наличием асимметрического атома углерода алкильной группы в положении 5, обладают одинаковым фармакологическим действием [Kleiderer, Shonle, 1934]. Как ( + )-, так и (—)-формы кокаина (7.11) являются одинаково активными местными анестетиками [Gottlieb, 1923]. Среди синтетических лекарственных веществ можно найти много подобных примеров. Так, одинаковым противомалярийным действием обладают ( + )- и (—)-хингамины [Riegel, Sherwood, 1949].
Антагонизм между оптическими изомерами, из которых один активнее другого, встречается редко. Это объясняется тем, что при переходе от D- к L-форме (или наоборот) резко изменяется конфигурация, которая и определяет адсорбцию на рецепторе. Поэтому биологическая активность смеси двух оптических ан-
205
Контакт в двух точках, отсутствует реакция
2- (2,4,5-трихлорфенокси) пропионовой ляется в присутствии L-энантиомера man, 1952]. В последнем случае это,
Рис. 12.1. Диаграмма, иллюстрирующая комплемен-тарность поверхности одному из двух оптических изомеров.
типодов (рацемата) обычно равна средней величине активностей обеих форм, и при этом нет антагонизма. Однако известны и некоторые исключения из этого правила: D-гистидин ингибирует размыкание имидазольного цикла гистидазой в L-гисти-дине [Edlbacher, Bauer, Becker, 1940], а гормональное действие на растения D-формы кислоты резко ослаб-[Smith, Wain, Wight-видимо, связано с тем,
что ауксины взаимодействуют с двумя последовательно расположенными рецепторами, из которых первый не обладает стереоспецифичностью (разд. 2.4). Известно огромное число примеров значительных различий в фармакологическом действии энантиомеров. Так, D-(—)-изопропилнорадреналин (изад-рин) обладает в 800 раз более сильным бронхорасширяющим действием, чем L-(+)-изомер [Luduena et al., 1957]. Аналогично, природный D-(—)-изомер адреналина в различных тестах в 12—20 раз активнее своего энантиомера [Taintcr, 1930; Blacsh-ko, 1950]. Ь-( + )-форма ацетилф-метилхолина оказывает на кишечник в 200 раз более сильное действие, чем D-(—)-форма. Необычным представляется то, что у никотина разница в активности природной L-(—)-формы и D-изомера сильно варьируется в различных тест-системах (от 1 : 1 до 1:40). Но если активность одной из форм выше (а так обычно и бывает), то всегда более активна L-форма [Barlow, Hamilton, 1965].
Исследовали влияние оптической изомерии на прессорную активность многих адренергических аминов. Предполагают, что контакт молекулы D-адреналина с рецептором осуществляется в трех точках — по аминогруппе, бензольному кольцу с его двумя фенольными гидроксильными группами и по спиртовой гидроксильной группе боковой цепи. Биологически менее активный L-адреналин может контактировать с рецептором только двумя группами (рис. 12.1). Отсюда следует, что дезоксиадреналин (эпинин) должен иметь такую же активность, как и L-адреналин, что и соответствует действительности [Easson,
206
Stedmann, 1933]. Эта гипотеза была подтверждена Badger (1947) и Stedman (1947).
Многие агенты с высокой биологической активностью не обладают оптической изомерией. Но если у родственных им соединений в молекулу вводят асимметрический центр, то появляется и разница между биологическим действием изомеров. Молекулы ауксинов (регуляторов роста растений) обычно не содержат асимметрического атома углерода и, следовательно, не образуют пары оптических изомеров. Однако, если вводят такой атом в молекулу, как, например в 2-(2,4,5-трихлорфенокси)-пропионовую кислоту (12.19), то оказывается, что активность связана преимущественно с D-( + )-формой. Поэтому была выдвинута гипотеза о том, что ауксиноподобное действие требует трехточечного контакта вещества с рецептором, как в случае адреналина (см. рис. 12.1).
Ме
О—С—СО2Н
L 1| н
2- (2,4,5-трихлорфенокси)про-пионовая кислота
(12.19)
Лабеталол
(12.20)
У пропранолола (12.56), первого вошедшего в практику ^-адреноблокатора (разд. 12.4), левовращающий (S) изомер обладает значительно более сильной гипотензивной активностью, однако мембраностабилизирующие и местноанестези-рующис свойства изомеров одинаковы [Barrett, Cullum, 1968].
В молекуле лабеталола (12.20) два асимметрических центра (отмечены звездочками), поэтому он может существовать в виде четырех стереоизомеров. (R,S)- и (S,S)-изомеры практически неактивны, (S,R)-изомер обладает а-адреноблокирующей активностью, a (R,R)-изомер — p-адреноблокирующей активностью, при этом это единственный из изомеров со сравнимой с пропранололом гипотензивной активностью [Gold et al., 1982].
Оптическая изомерия связана не только с наличием в молекуле асимметрического атома углерода; она возникает и тогда, когда затрудненное вращение вокруг ординарной связи создает в молекуле центр асимметрии. Наиболее известным примером таких молекул служат бифенилы, которые можно разделить на ( + )- и (—)-изомеры в том случае, если оба бензольных кольца
207
имеют в орто-положениях объемные заместители,/ Эти орто-группы создают стерические препятствия свободному вращению вокруг простой связи, внося, тем самым, в молекулу необходимый элемент асимметрии. При этом оба ортогзаместителя могут быть одинаковыми. Наиболее простым соединением такого типа является бифенил-2,2'-дисульфокислота (12.21).
В алленах две двойные связи перпендикулярны друг другу и поэтому такие производные, как (12.22), могут быть разделены на оптические изомеры (X может быть, например, бензольным циклом, a Y —нафталиновым).
Подробные сведения по стереохимии см. Mislow (1965); молекулярная асимметрия в биологии — Bentley (1969).
Хиральный бифенил
(12.21)
/Х с=с=с;
Хиральный аллен
(12.22)
НС—СО2Н НС—СОйН
II II
нс—со2н но2с—сн
цис- транс-
(12.23) (12.24)
12.2. Геометрическая изомерия
Геометрические изомеры возникают, если свободное вращение атомов в молекуле ограничено вследствие наличия двойной связи. Примером такой пары изомеров могут служить малеиновая (12.23) и фумаровая (12.24) кислоты (цис- и транс- соответственно). По химическому строению геометрические изомеры очень похожи, но они не являются зеркальными отображениями друг друга и не вращают плоскость поляризации света. Как правило, цис- и транс-изомеры значительно отличаются по физическим свойствам. Например, малеиновая кислота (12.23) плавится при 130 °C, величина ее рКа 1,9, она очень хорошо растворима в холодной воде (79 г на 100 мл); константы ее геометрического изомера — фумаровой кислоты (12.24) соответственно 287 °C, 3,0 и 0,7 г на 100 мл. Неудивительно, что геометрические изомеры обладают разными биологическими свойствами [Butler, 1944] и поэтому очень важно при изучении химической формулы нового соединения учитывать все возможности существования изомерии такого типа.
Цис- и транс-изомеры можно легко разделить кристаллизацией или хроматографически. Общего метода для превращения одного изомера в другой не существует, однако при нагревании, как правило, образуется наиболее стабильный изомер, а под действием света — менее стабильный. Зрение человека зависит от превращения 11-цис-изомера ретиналя в 11-транс-форму под действием света. Как только возбуждающий луч света исчезает,
208
этот каротиноидный пигмент снова переходит в цис-форму, прерывая тем сй^ым идущий к мозгу импульс [Gilardi et al., 1971].
Цис- и транс-изомеры существуют и у плоского циклопентанового кольца, представляющего собой как бы большую двойную связь. Хотя циклогексановое кольцо вообще не плоское, оно тем не менее достаточно плоское для образования цис- и трансизомеров. Так, существуют и доступны как цис- (12.25), так и транс- (12.26) формы диаминоциклогексана. Одна и та же молекула может образовывать и геометрические, и оптические изомеры. Например, транс-изомер (12.26) может быть разделен на (S,S) (12.27) и (R,R) (12.28) хиральные изомеры. Однако цис-изомер на хиральные формы разделен быть не может, так как в нем существует плоскость симметрии. У бензольного кольца нет геометрических изомеров, так как у каждого атома углерода кольца только один заместитель.
Цис- Гране- Транс-(5 S) Tpanc-(R,R)
(12.25) (12.26) (12.27) (12.28)
Стереоизомеры 1,2-диаминоцинлогенсана
Иногда бывает трудно выбрать два из четырех заместителей при двойной связи для определения цис- или транс-конфигурации. Правило последовательности предписывает выбирать заместителей с наиболее тяжелыми атомами, при этом цис-форма обозначается буквой Z (от немецкого слова zusammen), а трансформа— буквой Е (entgegen). Иногда в названиях соединений, в которых геометрическая изомерия может проявиться многократно, заместитель, имеющий самый маленький номер (по правилу нумерации), обозначают буквой г, а обозначения с-(цис) и t-(транс) перед другими заместителями показывают их положение по отношению к г-заместителю.
Аналогично индол-3-илуксусной кислоте (4.82), стимулирующей рост растительных клеток, могут действовать и другие карбоновые кислоты, карбоксильная группа которых находится под углом к плоскости ароматического кольца. Геометрическая изомерия ограничивает возможность такого расположения двух заместителей, поэтому из коричных кислот активен только цисизомер [Haagen-Smit, Went, 1935]. У 2-фенилциклопропан-1-карбоновой и 1,2,3,4-тетрагидронафталиден-1-уксусной кислот также активны только цис-изомеры [Veldstra, Van der Weste-ringh, 1951]. На молекулярных моделях видно, что кольцо и карбоксильная группа в транс-изомере (неактивном) этих веществ лежат в одной плоскости, в то время как в цис-форме (активной) они некопланарны. Впервые указал на эту связь между
14—734 209
некопланарностью и стимулирующей рост активностью Veldstra. Некопланарность может возникнуть и вследствие стерических препятствий. Так, бензойная кислота имеет плоскую форму и не активна, а 2,6-дихлорбензойная и 8-метил-1-нафтойная кислоты непланарны и биологически активны [Veldstra, 1963].
В аналогах ауксинов карбоксильная группа может быть заменена и на другие электроноакцепторные группы (—CN, —NO2, —SO3H), при этом биологическая активность лишь незначительно уменьшается. О связи между структурой и действием в этой серии см. Koepfli, Thimann, Went (1938) и Veldstra (1963).
Геометрическая изомерия стероидов заслуживает специального рассмотрения. На формуле (12.29) приведена общая структура этой группы природных насыщенных соединений (показана нумерация атомов углерода и буквенные обозначения четырех циклов). В природных стероидах кольца В и С находятся в транс-сочленении, причем оба они закреплены в конформации кресла. В сердечных гликозидах сочленение циклов С и D имеет цис-конфигурацию, но в гормонах животных, стеринах и желчных кислотах — транс-сочленение. У большинства биологически активных стероидов кольца А и В находятся в транс-сочленении («5а»-ряд, называвшийся ранее «алло»). Каждое из колец в молекуле стероидов образует складки, что хорошо видно на боковой проекции формулы (12.30).
Обозначение «5а» говорит о том, что атом водорода в положении 5 находится ниже общей плоскости колец. Все заместители, расположенные ниже этой плоскости, обозначаются символом «а», а выше — символом «р». а-Заместители обозначают пунктирными линиями, а ^-заместители — сплошными. Символы а- и ^-используют и для других полициклических соединений, например, тритерпенов и алкалоидов. Сложность строения этих соединений затрудняет использование R- и S-номенклатуры.
Как правило, у млекопитающих высокая биологическая активность стероидных соединений связана с отсутствием а-заме-стителей в положениях 1, 9, 11—13, 17 и отсутствием р-замести-телей в положениях 4—8, 14, 15. Боковая проекция молекулы
210
гидрокортизона иллюстрирует это правило [Sarett, Patchett, Steelman, 1963]. Первой стадией в биологическом действии стероидных гормонов является их влияние на специфический транспорт белков (разд. 2.4). Предполагают, что с белками стероиды взаимодействуют плоской нижней стороной (a-поверхность) молекулы.
Различные стероиды отличаются друг от друга в основном заместителями R1, R2 и R3 (12.29), но иногда и степенью ненасыщенности или наличием других заместителей вне колец. Для того чтобы стероид обладал прогестиновой, андрогенной и кортикоидной активностью, как правило, необходимо наличие циклогексеноновой структуры кольца А. Для проявления активности кортизонового типа необходимы атомы кислорода в положениях 3, 11 и 17 и характеристичная группа —СО—СН2ОН в положении 17. Андрогенная и кортикоидная активность в большей степени зависит от этих деталей строения молекулы, однако прогестиновая активность сохраняется, если ацетильная группа в положении 17 находится в a-конфигурации, не встречающейся в природных соединениях, а замена метильной группы в положении 18 на этильную приводит даже к усилению этой активности (пероральный контрацептив норгестрел).
Из всех стероидных гормонов наименее жесткие требования к структуре предъявляют соединения, обладающие эстрогенной активностью. При условии ароматизации кольца А и наличии кислой гидроксильной группы в положении 3 строение остальной части молекулы имеет второстепенное значение. В 1938 г. появились простые высокоэффективные бензольные аналоги стероидных эстрогенов. Хотя считалось, что их молекулы по форме похожи на молекулы стероидов, в действительности между ними не так уж много общего. По данным рентгеноструктурного анализа молекула диэтилстильбэстрола (12.31) имеет транс-конфигурацию, искаженную из-за стерических затруднений, создаваемых метиленовыми фрагментами двух этильных групп. Поэтому два бензольных кольца образуют с этиленовым фрагментом двугранный угол 63°, что делает форму молекулы совершенно не похожей на стероидную. Однако в этих молекулах примерно одинаково расстояние между атомами кислорода: 1,21 нм в диэтилстильбэстроле и от 1,07 до 1,11 нм в стероидных эстрогенах, но все эти молекулы слишком жесткие и поэтому не способны взаимодействовать с одним и тем же рецептором, в котором фиксировано расстояние между точками связывания. Известно, что для проявления эстрогенной активности необходимо образование двух водородных связей рецептора с атомами кислорода лиганда, следовательно, рецептор должен обладать определенной гибкостью [Weeks, Cooper, Norton, 1970]. Oki, Urushibara (1952) впервые отметили, что действие диэтилстильбэстрола связано и с толщиной его молекулы, которая составляет 0,45 нм и равна толщине молекул стероидных эстрогенов [Weeks et al., 1970].
14*
211
Цпэтплстилъбэстрсл
(12.31)
Me ОН
а) Синэстрол (R = Me) б) Норгексэстрол (R=H)
(12.33)
Диэтилстильбэстрол 3,4-ди (4-гидроксифенил) гекс-3-ен (12.31), эффективный и недорогой заменитель основного женского гормона эстрадиола (12.32), появился в 1938 г. [Dodds et al., 1938]. Этот синтетический лекарственный препарат, отличающийся от природного гормона высокой эффективностью при пероральном применении и большей длительностью действия, стал основным препаратом в эндокринной терапии. Некоторое недоверие к этому препарату возникло на основании двух факторов: во-первых, его неограниченно применяли для увеличения веса сельскохозяйственных животных, а во-вторых, были отмечены случаи заболевания раком у женщин, матери которых принимали его во время беременности. Несмотря на это, диэтилстильбэстрол продолжают широко применять и считают безопасным средством; его не назначают только в первые три месяца беременности (что, кстати, в равной степени относится и к природному гормону). О применении его фосфата для лечения рака предстательной железы см. разд. 4.2. Сходными эстрогенными свойствами обладает и дигидропроизводное диэтилстильбэстрола—синэстрол (12.33, а) (конфигурация 3R, 4S). С эстрогенсвязывающим белком (разд. 2.4) он соединяется сильнее, чем сам диэтилстильбэстрол, а еще сильнее взаимодействует с этим белком его низший гомолог—норгексэстрол (12.33, б) [Landvatter, Katzenellenbogen, 1982].
Сердечные гликозиды рассматриваются в разд. 14.1. Подробное изложение химии и стереохимии стероидов см. Shoppee (1964), биохимии и фармокологии — Briggs, Christie (1977).
Геометрическая изомерия 4-аминокротоновой кислоты, которая помогла определить активную конформацию нейромедиатора гамма-аминомасляной кислоты, рассматривается в разд. 12.7.
212
12.3. Конформация
Несмотря на то что вокруг одинарной связи возможно свободное вращение, данные инфракрасных спектров часто показывают, что атомы в молекуле занимают различные «предпочтительные» положения. В общих чертах правила конформационного анализа были установлены Hassel (1943), a Barton (1950) систематизировал их. Кроме инфракрасных спектров, для конформационного анализа используют рентгеноструктурный анализ, дифракцию электронов, микроволновые спектры, данные о дипольных моментах, изучение реакционной способности молекул (см. ниже) и спектры ПМР, часто с применением уравнения Karplus (разд. 17.3). Понятие конформации противостоит понятию конфигурации. «Конформационными изомерами» (или «конформерами», «ротамерами») называются стереоизомеры, превращающиеся друг в друга при вращении атомов или групп вокруг одинарной связи. «Конфигурационные изомеры» для своего взаимопревращения требуют разрыва связи (примеры такого типа рассмотрены в разд. 12.1 и 12.2). Хотя физические методы изучения структуры веществ и указывают на существование пары конформеров, каждый из которых обладает своими собственными физическими характеристиками, выделить каждый из них в чистом виде невозможно из-за высокой скорости взаимопревращения. Однако в случае достаточно сложных молекул, имеющих как конформационные, так и оптические изомеры, иногда можно разделить пару изомеров, обладающих различными физическими свойствами (см. ниже).
12.3.1. Конформационные изомеры, образуемые двумя заместителями
Когда два заместителя (тяжелее водорода) располагаются возможно дальше друг от друга, то такая конформация называется «вытянутой» (ранее ее называли транс). Если эти две группы расположены друг против друга, то такая конформация называется «заслоненной» («гош»-конформация представляет собой промежуточную между «вытянутой» и «заслоненной»). Эти конформации (на примере 1-хлорпропана) показаны на формулах (12.34) и (12.35).
Me хН Me Cl
/-с--—с^н
н A ci нхА хн н н
1-Хлорпропан ГХлорпропан
(вытянутая) (заслоненная)
(12.34) (12.35)
Конформационный анализ широко применяется при изучении алициклических соединений как для определения положе
213
ния заместителей в пространстве, так и формы цикла. Например, циклогексан может существовать в трех конформациях: кресла (12.36), ванны (12.37) и скрученной (или полукресла) (12.38).
Форма кресла — наименее напряженная и поэтому для молекулы циклогексана наиболее предпочтительна, при этом каждый аксиальный атом водорода удален на 0,25 нм от других двух аксиальных атомов водорода, расположенных по эту же сторону цикла. Скрученная (или твист-) форма занимает промежуточное положение (между формой кресла и лодки), а самая напряженная — форма ванны. Последняя, однако, может быть стабилизирована двумя или более конденсированными циклами, содержащими соответствующие заместители. Молекула декагидронафталина (декалина) может существовать в виде двух устойчивых форм, структура которых была определена методом дифракции электронов, показавшим, что транс-форма (температура плавления —30 °C, температура кипения 117 °C при 100 мм рт. ст.) состоит из двух транс-конденсированных циклов в конформации кресла, а цис-форма (температура плавления —43 °C, температура кипения 124 °C при 100 мм рт. ст.) состоит из двух цис-конденсированных циклов в конформации кресла. Цис-форма переходит в транс-форму при повышенной температуре и в присутствии катализатора. Молекула декалина представляет собой пример геометрической изомерии относительно мостиковых атомов углерода, но каждый цикл остается конформационно мобильным.
Интерес представляет конформационный анализ связи —СО—Н—, определяющей структуру пептидов. В белках ее конформация всегда «вытянутая» (транс), хотя в пептидах, содержащих пролин, т. е. третичную амидную группу, достаточно велика доля равновесной «заслоненной» (цис) конформации (до 40%) [Rabenstein, Anvarhuesein, 1982]. Во вторичных формамидах существует равновесная смесь «вытянутого» и «заслоненного» конформеров в соотношении 8 : 92 в случае N-метилформамида (12.39, 12.40). Их существование установлено по наличию двух раздельных сигналов в спектрах ПМР. С увеличением объема заместителя при атоме азота доля «вытянутого» конформера возрастает, достигая 18% при замене метильной группы на трет-бутильную [La Planche, Robers, 1964].
214
Н. М HL /Ме
Си -Ме О7 'Н
«Заслоненный» «Вытянутый»
(12.39) (12.40)
Два конформера М-метилформамида
12.3.2. Конформационные положения одного заместителя
Две основные конформации, в которых может находиться заместитель относительно цикла, называются экваториальная (в плоскости цикла) и аксиальная (перпендикулярно этой плоскости). В монозамещенных циклогексане преобладает экваториальный изомер, поскольку в этой форме наименьшее перекрывание атомов водорода. Оба конформера имеют форму кресла, и хотя наличие смеси можно установить по данным физических методов, разделить ее невозможно, так как конформеры быстро превращаются друг в друга. В полизамещенных циклах некоторые группы обязательно занимают аксиальное положение. Аксиальные группы испытывают наибольшие сте-рические затруднения, поэтому экваториальные гидрокси- и карбоксигруппы легче образуют эфиры, которые в свою очередь легко гидролизуются. Часто конформационная изомерия накладывается на геометрическую, как, например, в паре кокаин (7.11) и ф-кокаин— у первого метоксикарбоксильная группа занимает аксиальное, у второго — экваториальное положение.
В тироксине (11.14) и более активном трийодтиронине атомы йода в положениях 3 и 4 заставляют два кольца расположиться перпендикулярно друг другу. Дальнейшее изучение аналогов этих соединений показало, что такое расположение необходимо для проявления их тиреоидного действия [Dietrich et al., 1977]. О стереохимии взаимодействия тиреоидных гормонов с рецептором см. разд. 2.4.
На первый взгляд молекулы эметина и циклогексимида совершенно не похожи, однако фрагменты этих молекул имеют сходную конформацию, и поэтому оба эти соединения ингибируют рибосомный синтез белка в большинстве живых клеток (разд. 4.1).
В растворе и кристаллическом состоянии АХ имеет одинаковую конформацию (по данным спектров ЯМР в D2O и рентгеноструктурного анализа) (разд. 12.6). Но до какой степени распространяется эта корреляция? Byrn, Graber, Midland (1976) просмотрели всю литературу по этой теме и вместо трудоемкого рентгеноструктурного анализа предложили простой тест: сравнение инфракрасных спектров данного соединения в твердом состоянии и в хлороформе. Хотя этот тест требует дальнейшего изучения, следует отметить, что эти авторы получили очень похожие инфракрасные спектры в твердом со
215
стоянии и в растворе для холинхлорида и антигистаминового лекарственного вещества метапирилена, тогда как спектры гистамина и другого антигистаминового лекарственного вещества димедрола значительно отличались друг от друга.
12.3.3. Взаимная адаптация лекарственного вещества и рецептора
Хотя в предыдущем разделе указывалось, что можно установить конформацию вещества в растворе и твердом состоянии (представляющем собой специфический случай адсорбции вещества на себе самом), до сих пор ничего не известно о конформации вещества на рецепторе. Некоторые рецепторы представляют собой ферменты, и известны случаи изменения конформации субстрата при взаимодействии с ферментом. Например, методом ЯМР с Фурье-преобразованием было показано сильное искажение тетрауглеводного фрагмента муреина (разд. 5.3) при связывании его с ферментом лизоцимом [Sykes, Patt, Dolphin, 1971]. С другой стороны, из сравнения данных спектров 13С-ЯМР и рентгеноструктурного анализа следует, что, при взаимодействии с химотрипсином конформация N-ацетил-Ь-триптофана не меняется [Rodgers, Roberts, 1973].
Вопрос, может ли лекарственное вещество вызвать конформационные изменения рецептора, обсуждается в разделе 7.5.2 («Аллостерическая гипотеза»). Из данных рентгеноструктурного анализа четко видно, что некоторые субстраты вызывают конформационные изменения ферментов (см., например, в разд. 9.0 о карбоксипептидазе). С помощью этого же метода было показано изменение конформации гемоглобина при связывании кислорода (превращении в оксигемоглобин). При этом изменении ближайший к порфириновому циклу остаток гистидина удаляется от плоскости цикла на расстояние около 0,085 нм (рис. 12.2). Поскольку атом железа жестко связан с этим остатком гистидина, этот сдвиг приводит и к другим изменениям третичной структуры: в центре молекулы спираль F глобина приближается к спирали Н и выталкивает остаток тирозина (140) из полости, в которой он располагался между этими двумя спиралями. В свою очередь тирозин тянет за собой аргинин (141), разрушая его ионные связи. Такое изменение конформации разрыхляет плотную структуру четырех гемов в молекуле гемоглобина и облегчает подход второго атома кислорода.
Можно привести еще один пример: по данным ЯМР-спект-ров конформационное равновесие панкреатической РНКазы быка изменяется в различной степени под действием ингибиторов — цитидин-2'-, 3'- и 5'-монофосфата, причем изменения пропорциональны ингибированию [Meadows, Roberts, Jardetz-ky, 1969].
216
Конформационная специфичность была показана для НАДН (12.41). Некоторые апоферменты, использующие этот кофермент, способны различать аксиальный и экваториальный атомы водорода в положении 4. Это удалось легко показать, замещая по очереди каждый из этих атомов водорода на атомы дейтерия. Абсолютная конформация лабильных атомов водорода в молекуле NADH была определена Cornforth и сотр. (1962).
NADH (R-рибозилдифосфоаденозин) (12.41)
NHMe
Эфедрин (формула Ньюмена) (12.42)
NHMe
С6Н3(ОН)2
Адреналин
(12.43)
Следует учитывать, что изменение конформации может изменить все физические свойства молекулы и, следовательно, биологическую активность. Это особенно важно в тех случаях, когда пару конформеров можно разделить и подвергнуть раздельно биологическим испытаниям. Это удается редко, в основном для молекул, содержащих еще и хиральный центр. При этом стерические затруднения, возникающие в энантиомерах, искажают остов молекулы и препятствуют переходу одного конформера в другой. Эта же цель может быть достигнута направленным созданием стерических затруднений в насыщенных бициклических молекулах (примеры см. в разд. 12.8). Один из таких изомеров может быть липофильнее другого, или же более сильным основанием. Поэтому биологическая активность может зависеть не от изменения формы молекулы, а от новых физических свойств, связанных с этим изменением. Если для создания лекарственных веществ эти новые физические свойства желательны, то следует проверить их важность другими путями, не связанными с изменением конформации молекул.
Достаточно полную информацию об оптической изомерии и (или) конформации дают формулы Ньюмена, например (12.42), ср. с обычным способом изображения (12.12). Эти формулы особенно удобны, когда оба из двух последовательных атомов углерода остова молекулы хиральны. Эти два атома совмещены таким образом, что шесть заместителей лежат на радиусах одной окружности. Для сравнения такая формула может быть нарисована и для родственного соединения, имеющего лишь один хиральный центр, например адреналина (12.43).
Для описания конформаций часто требуется знание торсионного угла. Для группы атомов ХАВ —это угол, образованный плоскостью ХАВ в плоскостью АВ. Четыре торсионных угла АХ обсуждаются в разд. 12.6.
217
Рис. 12.2. Диаграмма конформационных изменений третичной структуры субъединицы гемоглобина при взаимодействии с кислородом [Perutz, 1970].
Подробно вопросы конформационной изомерии изложены Barton, Cookson (1956) и Eliel и сотр. (1965). Конформации макромолекул рассмотрены Hopfinger (1973), гетероциклов — Riddell (1980).
Обзор общих проблем стереохимии приведен Eliel, Basolo (1968) и Bentley (1969), уделившему особое внимание биологи
ческим аспектам. О лекарственных веществах, содержащих гетероциклы, см. Armarego (1977).
В дальнейшем в этой главе будут приведены данные о природе некоторых рецепторов, представляющих интерес для фар-
макодинамики, так как при создании лекарственных веществ конечной целью является их действие на рецепторы.
12.4. Рецепторы катехоламинов
Первые работы, посвященные вопросам связи структура — активность в ряду симпатомиметических средств — производных фенетиламина, были проведены Barger, Dale (1910). Было установлено, что наиболее сильное прямое действие оказывают гормоны и медиаторы, имеющие гидроксильные группы в положениях 3 и 4 бензольного кольца, т. е. «катехоламины», например норадреналин, адреналин и дофамин. Непрямое действие их аналогов, не имеющих гидроксильных групп, рассмотрено в разд. 9.4.3 и 7.6.3. Биологическая активность D-катехоламинов выше, чем их L-энантиомеров (см. рис. 12.1).
Предположение о существовании двух различных адренергических рецепторов было впервые выдвинуто Ahlquist (1948). Эффекты, легко вызываемые норадреналином, труднее адреналином (7.43), еще труднее изадрином (12.44) и труднее всего N-трет-бутилнорадреналином, связаны со стимуляцией а-ре-цепторов, а эффекты, легче всего вызываемые N-трет-бутилнорадреналином и труднее всего норадреналином, — fj-рецепто-ров. Типичными a-эффектами считают сужение кровеносных сосудов, стимуляцию матки, расслабление кишечника, а типичными ^-эффектами — расширение кровеносных сосудов, расслабление мускулатуры матки, стимуляцию гликогенолиза в мышцах, тахикардию [Levy, Ahlquist, 1961].
Для полной характеристики рецепторов катехоламинов идеальным было бы их выделение. Хотя этого сделать еще не уда-218
лось, определенный успех в изучении этих рецепторов достигнут применением избирательной метки. Для того чтобы обратимо пометить рецепторы гладкой мышцы матки, применяют алкалоид [3Н]-дигидроэргокриптин, сильный а-адреноблокатор. С его помощью было установлено, что специфичность и эффективность взаимодействия а-адренергических агонистов и антагонистов с рецептором хорошо коррелируют с данными их активности, полученными на целом органе [Williams, Mullikin, Lefkowitz, 1976].
НО
НО
Me н н / С—С—N—СН
L В I Н Н \ /ЧЧ ОН Me
сн2—сн=сн2
н /Ме
ОСН2—CH—СН2—N—CH
ОН
^Ме
Изадрнн Алпренолол
(12.44) (12.45)
Пнндолол
(12.46)
Для аналогичного изучения ^-рецепторов был использован (—)-[3Н]-дигидроалпренолол, меченое производное лекарственного вещества алпренолола (12.45), сильного ^-адреноблока-тора. Он избирательно и обратимо связывается с ^-адреноре-цепторами в адипоцитах крысы. С помощью этого маркера удалось показать, что ^-адренергические агонисты и антагонисты конкурируют за связывающие места рецептора стереоспецифично и с той же сравнительной эффективностью, как и на целом организме [Williams, Jarett, Lefkowitz, 1976]. В качестве обратимосвязывающего маркера использовали и производное другого p-адреноблокатора пиндолола (12.46), а именно С-[1251]-йодгидроксибензилпиндолол. С его помощью из мембран эритроцитов индюка удалось выделить (в микроколичествах) меченый рецептор [Brown Е. et al., 1976]. Этот же маркер использовали на фибробластах человека и клетках глиомы крыс. Он стереоспецифично и примерно с той же эффективностью, что и в опытах на лабораторных животных, вытесняется другими p-адренергическими агонистами и антагонистами [Maguire et al., 1976]. Связывание этих меченых агонистов насыщаемо и может быть ингибировано предварительным инкубированием с немеченым соединением. Измерение констант
219
Таблица 12.1. Процентное содержание ионизированных форм катехоламинов и их агонистов в воде прн pH 7,4 (25 °C)
Агонист Катион Цвиттерион Нейтральная молекула Анион
Норадреналин 94,4 3,9 1,6 0,03
Адреналин 95,5 4,5 — 0,01
Изадрин 94,8 5,2 — 0,01
Салбутамол 97,9 1,5 0,6 —
Тербуталин 95,6 4,4 — 0,01
Орцнпреиалин 95,5 4,5 — 0,01
диссоциации показало, что эритроциты птиц содержат около 1000 р-адренорецепторов.
Адреналин — это (R)-(—)-1-(3,4-дигидроксифенил)-2-ме-тиламиноэтанол. В твердом состоянии [Carlstrom, Bergin, 1967] и в растворе [Ison, Partington, Roberts, 1973] он существует преимущественно в полностью вытянутой конформации. Рассмотрим ионизацию трех катехоламинов и трех их агонистов. Хотя норадреналин и его N-алкильные гомологи существуют в основном в виде катионов в интервале pH 6,0—8,0, наличие в одной молекуле кислотных и основных групп, константы ионизации которых различаются на 2 единицы рКа и противоположны по знаку, приводит к существованию в растворе сложного набора минорных ионов. Из двух видимых констант по методу, использованному ранее для тирозина [Edsall, Martin, Hollingworth, 1958], были рассчитаны четыре микроскопические константы. Наличие четырех констант связано с тем, что аминогруппа имеет две константы (при неионизированной и ионизированной фенольной группе). По этой же причине одна из фенольных групп также имеет две константы. (Ионизация первой фенольной группы по закону Кулона полностью подавляет ионизацию второй группы.)
Для норадреналина макроскопические константы равны 8,63 (фенольная) и 9,73 (основная). С помощью УФ-спектроскопии были рассчитаны следующие микроскопические константы: pHiz 8,78, рКгг 9,58, pKm 9,16 и рК2ы 9,20, где Z и N относятся к цвиттериону и нейтральной молекуле соответственно. С использованием этих четырех констант рассчитаны данные для норадреналина, приведенные в табл. 12.1. Из них видно, что и норадреналин, и пять других адреномиметиков в водных растворах при нейтральных pH присутствуют преимущественно в виде катионов [Ijzerman et al., 1984]. Это же указывалось и в более ранней работе Sinistri, Villa (1962). Поэтому наиболее вероятно, что биологической активностью обладает катион. Более того, между а- и ^-агонистами нет различий в степени ионизации.
До тех пор пока не удастся выделить адренорецептор, дальнейшие попытки понимания его функций должны сконцен-220
трироваться на изучении связи структура — активность агонистов и антагонистов катехоламинов в различных биологических системах. Предполагают, что основной вклад во взаимодействие катехоламинов с рецептором вносит ионная связь. В усилении связывания, по-видимому, определенную роль играет и водородная связь, образуемая с участием р-гидроксильной группы, так как; ее удаление из молекулы, по данным Barger, Dale (1919), резко снижает активность соединений (см. обсуждение рис. 12.1).
А. а-Рецеторы. Воздействие на а-рецепторы в гладких мышцах приводит к увеличению проницаемости клеточной мембраны для неорганических ионов [Bulbring, Tomita, 1969]. В мышцах кишечника увеличивается проницаемость для ионов К+, что приводит к гиперполяризации и последующей релаксации мышц. В большинстве других типов гладких мышц увеличивается проницаемость для ионов Na+ и Са2+, при этом мембранный потенциал падает и вслед за возбуждением наступает контрактура. .
В настоящее время а-адренорецепторы подразделяются на два типа: постсинаптические ai-рецепторы и о^-рецепторы, чаще всего пресинаптические, но иногда и постсинаптические. Активация постсинаптических рецепторов приводит к сужению сосудов, а пресинаптические рецепторы участвуют в обратном захвате норадреналина. Лекарственные вещества, а-адреномиме-тики, применяют для повышения АД при гипотонии, а также как препараты общего сосудосуживающего действия при ринитах, конъюнктивитах глаз и для анестезии в стоматологии. Природным агонистом а-адренорецепторов является норадреналин. Однако чаще применяют синтетический препарат мезатон (12.47), отличающийся от норадреналина отсутствием одного из фенольных гидроксилов. По сравнению с норадреналином мезатон действует немного слабее, но его эффект более длителен, он активен при пероральном применении и не стимулирует ЦНС. Впервые этот препарат исследовали Barger, Dale (1910). Теперь известно, что мезатон проявляет чисто агонистическое действие на ai-адренорецепторы, т. е. как истинный агонист действует на постсинаптический рецептор; он не является псевдомедиатором (см. определение в разд. 7.6.3).
СН2 I
Фентоламин
(12.48)
221
Me I PhOCH2—CH ^NCHsCHsCl PhCH2/
Феноксибензамин
(12.49)
Вначале а-адреноблокаторы не разделяли на ai- и а2-типы. В терапии их применяли редко, например при обморожениях или в других случаях длительного нарушения циркуляции. Типичный пример имидазолины, такие как фентоламин (12.48), и хлорэтиламины, такие как феноксибензамин (12.49), который активируется самокватернизацией (определение см. в разд. 13.4) и затем необратимо алкилирует оц-рецептор.
Позднее для лечения гипертонии стали применять чистые ai-адреноблокаторы, особенно празоцин (7.57) и похожий на него индорамин [Docherty, MacDonald, McCrath, 1979]. Эти лекарственные препараты применяют для лечения гипертонии у больных астмой, регулярно принимающих fj-адреномиметики, или же у плохо переносящих обычно используемую комбинацию диуретиков с р-адреноблокаторами. Поскольку ои-адрено-блокаторы могут вызывать ортостатический коллапс, в легких случаях их не применяют. Интересен новый лекарственный препарат лабеталол (12.20), сочетающий в себе ai- и ^-блокирующие свойства. Его потенциальные возможности заключаются
в следующем:
cci-Адреноблокаторы понижают АД, ингибируя стимулирующий эффект норадреналина на а-адренорецепторы сосудов. Поэтому они вызывают расширение периферических сосудов и таким образом уменьшают периферическое сопротивление, в результате чего у больных с повышенным давлением оно значительно понижается. Однако ai-адреноблокаторы имеют два неприятных побочных эффекта: ортостатический коллапс (поэтому больные после приема препарата должны находиться некоторое время в положении лежа) и рефлекторная тахикардия. Что касается ^-блокаторов, их антигипертензивный эффект сопровождается урежением сердечных сокращений с одновременным небольшим рефлекторным возрастанием периферического сопротивления. Лекарственные препараты, блокирующие и сх|- и ^-адренорецепторы, оказывают благоприятное действие на больных, так как при сохранении положительных качеств каждого блокатора их побочные эффекты взаимно компенсируются [Richards, Prichard, 1978].
Me
СН2—С—NH2
Метилдофа
(12.50)
I СО2Н
Метилдофамин
(12.51)
222
но
НО
ОН Me
с—с—nh2 н н
а-Метилнорадреналив
(12.52)
Для лечения тяжелых форм гипертонии применяют два аг-адреномиметика — клонидин и метилдофа. Оба вещества избирательно накапливаются и действуют в мозге.
Метилдофа (12.50), 3,4-дигидрокси-2-метилфенилаланин, применяют перорально [Gillespie et al., 1962]. В ЦНС она декарбоксилируется до метилдофамина (12.51), который затем гидроксилируется (см. рис. 9.2) до а-метилнорадреналина (12.52). Предполагают, что оба этих метаболита понижают АД, стимулируя центральные «2-адренорецепторы [Iversen, 1967; Freed, Quintero, Murphy, 1978]. Эти рецепторы, по-видимому, расположены на ингибиторных нейронах, активация которых приводит к торможению периферической симпатической системы.
Таков же и механизм действия клонидина (12.53), 2-(2,6-дихлоранилино)-2-имидазолина [Kobinger, 1978; Timmermans, Van Zwieten, 1977]. Здесь приведена формула наиболее предпочтительного в кристаллах таутомера (по данным рентгеноструктурного анализа). Любое заметное изменение структуры клонидина приводит к значительному уменьшению эффективности действия. Так, при замене азотного мостика на —СН2— активность снижается в 25 раз. Два атома хлора, находящиеся в положениях 2 и 6, придают молекуле липофильность, достаточную для проникновения через ГЭБ. Наличие этих заместителей приводит к тому, что молекула становится непланарнойг торсионный угол между плоскостями двух циклов равен 75°. В результате этого поворота нарушается резонанс гуанидинового остатка с бензольным кольцом и уменьшается величина рКа- Поэтому в равновесной смеси всегда есть определенная доля нейтральных молекул, легче проникающих через барьер [Stable, 1982].
СН(ОН)—CH2NHtBu
HN х/ NH
Клонидии (12.53)
Салбутамол
(12.54)
223
он
он
Тербуталин
(12.55)
В клинической практике «2-блокаторы назначают редко. Изучается возможность применения их в качестве антидепрессантов, повышающих уровень норадреналина в мозге [Сагооп et al., 1982].
Б. ^-Рецепторы. Установлено существование двух типов р-рецепторов [Lands et al., 1967]. pi-Рецепторы увеличивают скорость и силу сердечных сокращений, расширяют коронарные сосуды и расслабляют гладкую мускулатуру кишечника, тогда как ^-рецепторы расслабляют гладкую мускулатуру бронхов, матки и артерий, идущих к скелетным мышцам. Окружение Pi-рецепторов менее липофильно, чем р2-рецепторов [Basil et al., 1976].
За последние годы были найдены эффективные р-адреноми-метики, применяемые в виде аэрозолей для расширения бронхов у больных астмой. В качестве основы при создании этих препаратов была выбрана молекула изадрина (12.44), в которую вносились изменения для усиления взаимодействия с р2-рецепторами. Прежде всего было необходимо устранить тахикардию, вызываемую действием вещества на ргрецепторы. Затем следовало изменить расположение гидроксильных групп для повышения устойчивости содинений к действию фермента КОМТ, разрушающего катехоламины (для взаимодействия с Рг-рецепторами достаточно одного фенольного гидроксила). Типичными примерами таких лекарственных веществ являются салбутамол (12.54) и тербуталин (12.55). В общем, активность р2-адреномиметиков возрастает с увеличением размера N-ал-кильной группы вплоть до трет-бутильной.
В волокнах полосатых мышц (например, мышц диафрагмы) и мышце матки отсутствует симпатическая иннервация; эти мышцы содержат только р2-рецепторы. Поэтому на них сильно действует гормон (адреналин) и не действует нейромедиатор (норадреналин). В то же время ткани с симпатической иннервацией (например, мышцы сердца и двенадцатиперстной кишки) содержат больше ^-рецепторов, чем р2-рецепторов.
Многие ингибиторы адренергической передачи, сами по себе или в комбинации с диуретиками, назначают для снижения небольших подъемов АД. Они известны под названием р-блокаторов и представляют собой наиболее широко применяемый класс лекарственных веществ. При первых попытках синтеза ^-блокаторов были получены лишь частичные агонисты. При этом стало ясно, что для достижения цели необходимо зна
224
чительное изменение молекулы адреналина. Успех пришел тогда, когда Дж. Блэк и его коллеги обнаружили, что боковая цепь — ОСН2—СН(ОН)—CH2NHR (где R— изопропил или трет-бутил) придает ароматическим углеводородам свойства чистых ^-антагонистов [Black et al., 1964, Le Count, 1982].
Наиболее широко применяют ^-блокатор анаприлин (пропранолол) (12.56) [1-изопропиламино-3-(нафт-1-ил)пропан-2-ол], исключительные клинические свойства которого описали Prichard, Gillam (1964). Предполагают, что он действует одновременно по трем механизмам. Благодаря высокой липофильности [Р (октанол — вода) 3,6] пропранолол проникает в мозг и оказывает слабое действие на ЦНС. Значительно более вы-раженно он действует на ^-рецепторы сердца, уменьшая частоту сердечных сокращений, и на [}2-рецепторы мышц сосудов, вызывая их расслабление [Lewis, Haeusler, 1975]. Пропранолол широко применяют и для лечения стенокардии. Исторический обзор открытия [J-блокаторов см. Le Count (1982).
ОН I
О—CHj—CH—CH2NHCHMe2
Анаприлин
(12.56)
нН
Н\
но/\/у [Н
О—СН2—CH—CH2NH—СМе3
Надолол
(12.57)
ОН
МеО—СН2СН2—
I
—ОСН2СН—СН2—NHiPr
Метопролол
(12.58)
ОН I О—СН2СН—CH2NHtBu
Тимолол
(12.59)
Невозможно перечислить все существующие лекарственные вещества, молекулы которых являются модификациями молекулы пропранолола. Надолол (12.57) имеет сходное фармаколо-
15-734
225
гическое действие, но более продолжительное. Атенолол и метопролол (12.58) действуют более избирательно на ^-рецепторы сердца, поэтому некоторые врачи предпочитают назначать больным с заболеваниями бронхов именно эти препараты. Малое изменение в (12.58) может уменьшать действие на ^-рецепторы [Machin et al., 1983]. Пиндолол (12.46) и окспренолол классифицируются как псевдоагонисты (определение см. разд. 2.1.1); они стимулируют ^-рецепторы сердца. Алпренолол (12.45) и соталол по действию напоминают пропранолол. Тимолол (12.59) также похож на пропранолол, но его действие значительно сильнее. В составе глазных капель его применяют для снижения внутриглазного давления при глаукоме. Исследования показали, что при регулярном приеме тимолол снижает смертность от сердечных приступов [Pedersen, 1981].
В. Природа симпатомиметических рецепторов. Стимуляция £1- и [^-рецепторов приводит к активации аденилатциклазы, и этот фермент начинает синтезировать аденозин-3',5'-цикломо-нофосфат (цАМФ) (12.60). Установлено, что ^-рецепторы и аденилатциклаза расположены недалеко друг от друга: рецептор на внешней, а аденилатциклаза на внутренней поверхности мембраны. При связывании нейромедиатора с рецептором циклаза (с участием гуанозинтрифосфата) начинает синтезировать цАМФ [Percins, 1973; Levitzki, 1977; Citri, Schramm, 1980]. При блокировании аденилатциклазы гуанил-5'-ил имидодифосфатом в ней происходят конформационные изменения, ингибирующие связывание агонистов с ^-рецептором, однако не влияющие на связывание антагонистов [Lefkovitz, Mullikin, Caron, 1976]. Следует отметить, что антагонисты, включая пропранолол (12.56), ингибируют связывание медиатора с рецептором, но не блокируют аденилатциклазу; в то же время этот фермент может быть инактивирован нагреванием, однако при этом связывание с рецептором не нарушается [Schramm et al., 1977]. Инъекция цАМФ может вызывать примерно такие же физиологические эффекты, как и катехоламины.
цАМФ ; (12.60)
Стимуляция «2-рецепторов ингибирует аденилатциклазу (для этого требуется ГТФ), и одновременно уровень цАМФ
226
падает. Стимуляция ои-рецепторов не влияет на адеиилатцик-лазу, а приводит к возникновению нервного импульса в результате высвобождения ионов кальция.
Г. Циклический аденозинмонофосфат. Физиологическая роль цАМФ, высвобождение которого косвенно вызывается катехоламинами, заключается в активации фермента протеинкиназы, который, как считают, может фосфорилировать специфический постсинаптический мембранный белок. В результате этого в мембране открывается канал для неорганических катионов, закрывающийся при быстром ферментативном гидролизе фосфо-эфирной группы [Greengard, 1976]. Много работ было посвящено выяснению молекулярного механизма возникновения постсинаптического потенциала. Такие метилированные ксантины, как кофеин (7.52, а) и теофиллин, специфично, но слабо препятствуют гидролизу цАМФ фосфодиэстеразой.
Предполагают, что стимулирующий эффект кофеина на ЦНС обусловлен его сильным антагонистическим действием на аденозиновые рецепторы [Daly, Brons, Snyder, 1981].
цАМФ и родственный ему цГМФ являются вторичными мессенджерами не только в эффектах катехоламинов, но и гормонов. Например, они играют главную роль в регуляции выделения инсулина из поджелудочной железы in vivo. Бутамид (12.61) и подобные ему сульфонилмочевины (такие, как хлор-пропамид, толазамид, глибенкламид), применяемые для лечения диабета у пожилых людей, достаточно избирательно ингибируют панкреатическую фосфодиэстеразу и тем самым уменьшают гидролиз цАМФ [Goldfine, Perlman, Roth, 1971]. В ряду сульфонил мочевин активность соединений хорошо коррелирует с липофильностью и со способностью этих соединений связываться с сывороточным альбумином [Seydel, Ahrens, Losert, 1975]. Эти циклические фосфаты являются вторыми мессенджерами в диуретическом действии вазопрессина АКТГ и многих других гормонов [Sutherland, Qye, Butcher, 1965].
Me—SO2NH—CO—NHBu
Бутамид (12.61)
Д. Дофамин. Этот нейромедиатор (7.48) менее изучен, чем норадреналин, поэтому в настоящее время широко исследуется природа его рецепторов. Как недостаток, так и избыток дофамина может быть причиной различных заболеваний. Болезнь Паркинсона возникает от недостатка этого медиатора в полосатом теле и черном веществе головного мозга; введение леводофы восполняет этот недостаток. Считают, что шизофрения связана с нарушением метаболизма дофамина, приводящим к увеличению содержания в мозге [Haracz, 1982]. Такие нейролептики, как аминазин, применяемые для лечения шизофрении, 15* 227
блокируют в полосатом теле рецепторы дофамина. Ведутся исследования по выделению дофаминовых рецепторов.
Сульпирид
(12.62)
Домперидон
(12.63)
Существует несколько классификаций дофаминовых рецепторов. Широко применяют классификацию, предложенную Kebabian, Caine (1979), в основу которой положено различие в активации аденилатциклазы: D-1 дофаминовые рецепторы ее активируют, a D-2 не активируют. Для облегчения пользования этой классификацией см. Leff, Creese (1983). В альтернативной классификации периферических дофаминовых рецепторов, предложенной Goldberg, Kohli (1983), выделяют два типа рецепторов — DAi и DA2. Воздействие на первые вызывает релаксацию мышц, приводящую к расширению сосудов. Для этих рецепторов апоморфин—слабый агонист, (К)-сульпирид (12.62)—сильный антагонист, а домперидон (12.63) не активен. Возбуждение ПА2-рецепторов ингибирует действие норадреналина, высвобождающегося из симпатических нервных окончаний. Для этих рецепторов сильным агонистом служит апоморфин, а (5)-сульпирид и домперидон—сильными антагонистами. Дофаминовые рецепторы ЦНС относятся, по-видимому, к ПА2-типу.
2-Амино-6,7-дигидрокси-1,2,3,4-тетрагидронафталин представляет собой жесткий циклический аналог дофамина. При введении в мозг интактным крысам это соединение захватывается синаптосомами мозга и действует подобно дофамину [Elkhawad, Woodruff, 1975]. Olsen и сотр. (1981) предложили гипотетическую трехмерную модель D-2-рецептора.
228
О лечении паркинсонизма предшественниками дофамина см. разд. 3.6.1; об использовании антагонистов дофамина при лечении шизофрении см. разд. 12.9.
Е. 5-Гидрокситриптамин (серотонин). Этот аминофенол (7.49) является лишь отдаленным родственником катехоламинов. Предполагают, что существует два типа его рецепторов, но они практически не изучены. Из них 5-HTi-рецептор определяют как место связывания с 5-гидрокситриптамином и другими агонистами, что вызывает сужение периферических кровяных сосудов. Рецептор 5-НТг определяют по его способности связывать такие специфические ингибиторы, как метизергид (сложное производное эрготалкалоидов) и ципрогептадин (12.64), препятствующие захвату изотопномеченого 5-гидрокси-триптамина [Fozard, 1983].
Ни 5-гидрокситриптамин, ни его агонисты в клинике не применяют. Предполагают, что трициклические антидепрессанты (разд. 12.9) ингибируют захват как 5-гидрокситриптамина, так и катехоламинов. Для профилактики мигрени и снятия демпинг-синдрома назначают перорально два его антагониста: ципрогептадин (12.64) и метизергид (бутаноламид-1 -метил-лизергиновой кислоты).
Более подробно об адренорецепторах, их агонистах и антагонистах см. Kunos (1981—1983), Cooper, Bloom (1982) и Yamamura, Enna, Kuhar (1978) *.
12.5. Ацетилхолинэстераза
Хотя АХЭ можно выделить в чистом виде, ее высокая ОММ (320000) сильно затрудняет изучение структуры с помощью дифракции рентгеновских лучей. По данным С-концевого аминокислотного анализа в состав фермента входят два различных полипептида с одинаковой ОММ. Молекула АХЭ состоит из четырех пептидных цепей, по две каждого типа [Leuzinger, 1971]. В высокоочищенном состоянии АХЭ можно выделить экстракцией из электрического органа угря Electroforus electri-cus. Она представляет собой фермент, гидролизующий эфирные связи в различных соединениях, однако отличается от других эстераз значительно большей эффективностью действия на АХ.
АХ (12.65) всегда существует в виде катиона: степень его ионизации не зависит от pH среды. Исследования молекулярного механизма гидролиза АХ начались с работы Wilson, Bergmann (1950), определивших константу диссоциации комплекса АХ — фермент, как равную 2.6Х10-4. Изучая зависимость ско-
1 Использование теоретического конформационного анализа для изучения строения связывающего центра а-адренорецепторов см. в работе: В. А. Говы-рин, Б. С. Жоров. Физиол. журн. СССР им. Сеченова, 1984, т. 70, с. 529— 551. — Примеч. ред.
229
Анионный центр
Эстератичесиий центр
Рис. 12.3. Активный участок ацетилхолинэстеразы [Wilson, Bergmann 1950; Wilson, 1962] (символ GH обозначает имидазольный цикл гистидинового остатка).
рости гидролиза от pH среды1, они установили, что этот фермент имеет основную группу с рКа 7,2 (вероятнее всего, имидазольный цикл остатка гистидина) и кислотную группу с рКа 9,3 (по-видимому, остаток тирозина). Они предположили, что фермент-субстратный комплекс образуется ионной связью между четвертичной аммониевой группой АХ и анионной группой с рКа 9,3 фермента, а также диполь-дипольным взаимодействием атома азота при двойной связи имидазольного цикла (в ферменте) с дробным положительным зарядом атома углерода С=О группы эфира. Таким образом, у фермента предполагается наличие двух связывающих мест, а именно анионного, взаимодействующего с катионной группой субстрата, и «эстер-атического», сначала связывающего, а потом гидролизующего сложноэфирную группу [термин «эстератический» введен Wilson, Bergmann (1950)].
Схематическое изображение активного участка АХЭ показано на рис. 12.3 [Wilson, 1962]. С помощью ингибиторов с жесткими молекулами типа цис- и трансизомеров йодида 2-три-метиламмонийциклопентанола было установлено, что расстояние между связывающими местами фермента составляет около 0,25 нм [Friess, Baldridge, 1956].
Позднее было высказано предположение об образовании ковалентной связи между атомом кислорода карбонильной группы ацетилхолина и гидроксильной группы остатка серина. Основанием для этого послужила способность органических фосфатов, например диизопропилфторфосфата (13.26), фосфорилировать остаток серина в ферменте на участке с аминокислотной последовательностью (Glu-Ser-Ala) [Schaffer, May, Summerson, 1954; см. разд. 9.0]. Был сделан вывод, что ацетильная группа АХ притягивается к гидроксильной группе серинового остатка и ацетилирует ее. Функция имидазольной группы, расположенной на другой складке ферментативного белка, видимо, заключается в облегчении гидролиза ацетилированной группы серина [Wilson, Cabib, 1956].
О О
+ II II
Me3N+—СН2—СН2—О—СМе Ме3С—СН2—СН2—О—С—Ме
Ацетилхолин (катион) 3,3-Диметилбутилацетат
(12.65) (12.66)
1 Этот метод определения величин рКа для ферментов может приводить к ошибкам [Kosower, 1962], так как при изменении ионизации различных групп в ферменте возможны изменения конформаций самого фермента,
230
После того как было обнаружено, что 3,3-диметилбутилаце-тат (12.66) и АХ одинаково легко гидролизуются АХЭ, стало ясно, что ван-дер-ваальсовы взаимодействия метильных групп (присутствующих в катионной головке АХ) могут играть более важную роль в связывании природного медиатора с ферментом, чем положительный заряд [Whittaker, 1951]. В настоящее время считают, что «анионное место связывания» в действительности является «липофильным местом связывания», взаимодействующим с высоколипофильной частью молекулы АХ, и не зависит от ее заряда [O’Brien, 1971].
В отсутствие фермента АХ в холодном водном растворе устойчив в кислой среде, но неустойчив в щелочной при pH более 10,0. Величины констант скорости его гидролиза указывают на то, что заряженный атом азота притягивает атакующие гидроксильные ионы к соседней сложноэфирной группе; поэтому АХ значительно легче гидролизуется в щелочной среде, чем обычные эфиры, например этилацетат [Butterworth, Eley, Stone, 1953].
На поверхности концевой пластинки мышцы содержится достаточное количество АХЭ для расщепления 1 млн молекул АХ в 1 мсек, что в тысячу раз превышает число молекул АХ, необходимое для деполяризации концевой пластинки [Nach-mansohn, 1940].
Кроме ингибирования органическими фосфатами и другими ацилирующими агентами (разд. 13.3), АХЭ может обратимо блокироваться и простыми четвертичными аммонийными соединениями. Установлено, что при увеличении размера алкильных групп в бисчетвертичных соединениях повышение сродства (измеряемое как —AF) на каждую метиленовую группу (СН2) составляет более 300 кал/мол. При данной длине цепи моночетвертичные ингибиторы связываются сильнее, чем бисчетвертичные [Bergmann, Segal, 1954]. Об изучении гидрофобной области холинэстераз см. Kabachnik и сотр. (1970).
Прекращение действия АХ в сердце бивалентных моллюсков, не имеющих АХЭ, осуществляется по иному механизму. Высвобождение АХ из нервных окончаний вызывает выделение из мышцы АТФ-подобного вещества, снижающего чувствительность холинорецептора по аллостерическому механизму. Такой же механизм существует и в сердце высших животных, однако там его перекрывает более эффективное действие АХЭ [Turpa-ev, Sakharov, 1973].
12.6. Холинорецепторы
АХ служит природным медиатором в нескольких различных видах синапсов, например: а) соединения произвольный нерв -* произвольная мышца, б) ганглионарные синапсы, представляющие собой соединения нерв-»-нерв в автономной системе, в) парасимпатические постганглионарные синапсы. АХ
231
выполняет роль медиатора и в ЦНС, например между волокнами спинальных корешков и клетками Реншоу.
Никотин имитирует действие АХ в синапсах первых двух видов (а) и (б), а мускарин — в синапсе третьего вида (в) (см. табл. 7.1, том 1). Впервые деление холинорецепторов на Никотиновые и мускариновые было предложено Dale (1914). Мускариновые и никотиновые рецепторы работают в совершенно различных временных режимах. Единичный нервный импульс в мускариновом рецепторе длится 500 мсек с латентным периодом около 100 мсек. В противоположность этому время стимуляции никотинового рецептора в произвольном нервно-мышечном соединении составляет около 0,2 мсек и даже в медленных ганглионарных синапсах — только 60 мсек. Считают, что в возникновении ответов мускариновых рецепторов принимает участие второй мессенджер цГМФ. Гладкие мышцы реагируют на действие АХ значительно медленнее, чем поперечнополосатые. АХ уменьшает поляризацию во всех мышцах, кроме сердечной, где под его действием поляризация усиливается, что приводит к замедлению частоты сердечных сокращений.
Поскольку рецептор в отличие от АХЭ не гидролизует АХ, то, следовательно, он не обладает эстератическим центром. Другие их существенные различия заключаются в следующем: а) диметилбутилацетат (12.66)—хороший субстрат для фермента, но он не активирует рецептор (для этого требуется основная группа; б) мускарин не является субстратом для фермента, но проявляет свойства сильного агониста мускариновых рецепторов; в) диизопропилфторфосфат (13.26) связывается с активным центром АХЭ, но не холинорецептора; г) ацетил-бунгаротоксин специфично связывается с никотиновым холино-рецептором, но не с ферментом.
Создается впечатление, что структурные требования к агонистам никотиновых рецепторов менее жесткие, чем для мускариновых агонистов. К тому же никотиновый рецептор, который можно выделять из рыб, более доступен. Однако наличие в никотиновом рецепторе нескольких мест связывания (реагирующих с разными скоростями) осложняет его изучение.
12.6.1. Мускариновый холинорецептор
Среди отличительных особенностей мускаринового рецептора следует отметить не только длительность его ответа на АХ, но и быстроту возникновения этого ответа [Greengard, 1976]. Этот рецептор выделяют экстракцией из мозга быка, при этом берут только каудатные ядра, содержащие основную часть АХ мозга. Эти ядра являются частью полосатого тела, структуры лимбической системы. Ткань этих ядер экстрагируют водным раствором холата натрия, и после центрифугирования разделяют на фракции электрофорезом. Затем добавляют [3Н]-хинук-
232
лидин-3-иловый эфир бензиловой кислоты (12.67), специфично связывающийся с мускариновыми рецепторами. (Это соединение появилось в результате поиска специфичных для мускаринового рецептора эфиров холина.) Вначале был найден холиновый эфир бензиловой кислоты (метацин), а соединение (12.67) можно рассматривать как его производное, так как структура холина входит в структуру хинуклидинола. После очистки комплекса реагент отмывают диализом и выделяют чистый мускариновый рецептор. Мускариновые антагонисты (атропин, гиосцин, изопропамид) блокируют связывание соединения (12.67) с рецептором, тогда как никотиновые антагонисты (тубокурарин, гексаметоний) и агонист (никотин) на связывание не влияют [Carson, 1982; Yamamura, Snyder, 1974].
Мускариновый рецептор выделен также из головок фруктовой мушки Drosophila [Dudai, Ben-Barak, 1977], но в головках обычной мухи — совершенно другой тип холинорецепторов (разд. 12.6.2).
Ph
। Г7>-0-СО-С-О Н М к
X '-СН2-Ы+Ме3.
Хинуклидин-З-иловый эфир бензиловой кислоты
(12.67)
1_-(+)-Ацетил-0 -метилхолик Метахолин
(12.68)
Me О
- 1 11
Me3N+CH2—СН—О—CNH2
Бетанехол (натион) (12.69)
Большинство сведений о структуре мускаринового рецептов ра получено при изучении связи структура — активность агонистов, имитирующих действие АХ в постганглионарных синапсах. Эти рецепторы традиционно называются мускариновыми, однако следует помнить, что это название чисто историческое, так как мускарин в медицине для лечения больных не применяется. Антагонисты мускариновых рецепторов очень сильно различаются по структуре и поэтому среди них трудно установить связь структуры с активностью. Однако из данных о действии антагониста пирензипина (бензодиазепинона) можно сделать вывод, что существуют два подтипа мускаринового рецептора.
Наиболее типичный мускариновый агонист — метахолин (12.68), а-ацетил-^-метилхолин. Эта простая молекула, отличающаяся от АХ наличием дополнительной метильной группы, обладает такой же мускариновой активностью, что и природный медиатор, но совершенно не проявляет никотиноподобноё
233
действие. Следует отметить, что L-изомер в 200 раз активнее D-изомера [Ellenbroek, Van Rossum, 1960]. В медицинской практике метахолин в основном применяют в послеоперационном периоде для понижения тонуса кишечника. Чистым мускариновым агонистом является и бетанехол (12.69), гибрид метахолина и карбахолина (2.11), применяющийся в клинике по тем же показаниям, что и метахолин. Эти лекарственные вещества обладают важным преимуществом перед АХ: они значительно медленнее гидролизуются АХЭ.
Равновесное расстояние между четвертичным атомом азота в молекуле АХ и отрицательно заряженной группой рецептора рассчитано по разности свободных энергий взаимодействия рецептора с ацетилхолином (12.65) и с диметилбутилацетатом (12.66): оно равно 0,329 нм. Диметилбутилацетат имеет не обладающую основными свойствами головку, изостеричную катионной головке АХ. Это расстояние практически равно расстоянию максимального сближения, найденному на молекулярных моделях [Burgen, 1965].
В последние годы структура многих жестких аналогов АХ была определена рентгеноструктурным анализом для предсказания: а) конформации молекулы АХ (из множества возможных для такой гибкой молекулы), взаимодействующей с данным рецептором, и на основании этого б) стереохимии самого рецептора. Такой подход к установлению структуры рецептора достаточно дискуссионен, так как известна способность рецепторов деформироваться при взаимодействии с агонистами (разд. 12.3).
Формулой (12.70) представлена двумерная проекция молекулы ацетилхолинбромида по данным рентгеноструктурного анализа [Сапера, Pauling, Sorum, 1966]. Межатомное расстояние между N-метильной группой и эфирным атомом кислорода (т. е. атом кислорода, связанный с двумя атомами углерода) равно 0,302 нм, а между атомом азота и эфирным атомом кислорода 0,329 нм (оба эти расстояния короче, чем обычно). Оба атома кислорода находятся в одной плоскости с ближайшими тремя атомами углерода, расположенными в правой части формулы (12.70). Таким образом, геометрии молекул ацетилхолина и мускарина (12.72) [рентгеноструктурные данные Jellinek, 1957] в твердом состоянии похожи. Однако в водном растворе конформация молекулы АХ может быть совершенно иной, что обусловлено отсутствием влияния соседних молекул того же самого вещества. Поэтому конформации АХ изучали по анализу вицинальных констант в спектрах ЯМР в растворе D2O [Culvenor, Ham, 1966]. Полученные результаты подтверждают данные рентгеноструктурного анализа, за исключением того, что сложноэфирная группа находится в конформации, более свойственной сложным эфирам. Гош-конформация цепи +NCCO, существующая во всех трех структурах (12.70), (12.71) и (12.72), является предпочтительной для мно-
234
гих 1,2-дизамещенных этанов в растворе. Таким образом, предпочтительная конформация АХ весьма обычна; однако взаимодействие с рецептором может изменить ее полностью.
Ме
Me
//2125
О
О 0J32 Ме
Ацетилхолин
(по данным рентгёнострунтуриого анализа;длины связей в нм)
(12.70)
Ме^
Ме N —Ме
Ацетилхолин (поданным ЯМР) (12.71)
Меч+/Ма
0,148 0147 Q 0,147 Ме
Me ХСХ
\ А
С-—'°'.
''О
Мускарин (поданнымрентгеноструктурного анализа) (12.72)
Жесткая циклическая структура молекулы мускарина позволяет получить более полную информацию о структуре мускаринового холинорецептора, чем гибкая молекула АХ [Waser, 1961]. Из семи стереоизомеров мускарина только природный L-( + )-изомер обладает высокой ацетилхолиноподобной активностью. Атом азота должен быть четвертичным, а атом кислорода в цикле нельзя заменить даже на серу, иначе активность соединения пропадает. На основе этих данных Waser постулировал связывание мускарина с рецептором посредством атомов азота и кислорода, входящего в цикл. Конформационные возможности молекулы мускарина значительно меньше, чем у молекулы АХ, в нем подвижна лишь боковая цепь с триметиламмонийной головкой [Waser, 1961].
Величины отношений активности L-мускарина к активности АХ для разных постганглионарных холинорецепторов изменяются от 0,1 до 5,4. Из возможных изомеров и аналогов мускарина активностью обладают лишь те, в которых метильная, гидроксильная и ониевая группы (боковой цепи) расположены так же, как и в молекуле мускарина.
Какие еще структурные модификации молекул АХ и мускарина можно проводить, сохраняя при этом сильную ацетилхолиноподобную активность? Все эти данные являются лишь по-луколичественными, и для их уточнения требуется проведение
235
дополнительных исследований. Дело в том, что «активность» каждого соединения складывается из двух составляющих: эффективности и сродства (разд. 7.5.2). Так, катион диметилами-ноэтилацетата (12.73), нечетвертичного аналога АХ (12.65), практически не обнаруживает мускариновой активности. В действительности же его внутренняя активность выше, чем у АХ, а низкая активность обусловлена малым сродством — в 1000 раз меньшим, чем у АХ [Gloge, Liillmann, Mutschler, 1966].
Особый интерес представляет 2-ацетоксициклопропилтри-метиламмоний-йодид (12.74), так как в этом соединении структура N—С—С—О АХ жестко закреплена ковалентными связями. Это соединение было получено в виде смеси четырех изомеров, конфигурация которых была определена рентгеноструктурным анализом [Chothia, Pauling, 1970].
I . /|Н‘ о
MeaN—СН2—СНа—О—С—Me Me3N+—СИ ~
\С— о—С—Ме
Н
Диметиламиноэтилацетат 2-Ацетоксициклопропилтриметиламмоиий
(катион)
(12.73) (12.74)
Ме^^О^4СНа—N+Me3
5-Метилфурметид (12.75)
Полной мускариноподобной активностью обладает (+)-транс-изомер, остальные три малоактивны. (Активный изомер обладает лишь 1 % активности ацетилхолина, остальные изомеры еще менее активны.) Поэтому был сделан вывод, что по крайней мере мускариновая активность АХ связана с конформацией, похожей на таковую активного (IS, 2S)-изомера соединения (12.74). 5-Метил-2-триметиламмониометилфуран (12.75) (5-метилфурметид) обладает такой же мускариновой активностью, как и АХ, и почти не обладает никотиновой активностью [Armitage, Ing, 1954]. Подобно мускарину это соединение имеет в молекуле жесткий плоский цикл, но (благодаря двойным связям)—СНг-группа фрагмента —СНа—+NMe3 жестко закреплена в плоскости цикла, и поэтому форма молекулы четко определена. Следует отметить, что в отличие от мускарина в соединении (12.75) отсутствует гидроксильная группа и стереоизомерия здесь невозможна. Из данных кристаллографических исследований молекулы 5-метилфурметида [Baker et al., 1971] видно, что, хотя строение некоторых фрагментов этой молекулы совпадает со строением молекул мускарина и АХ, в другом она сильно от них отличается, поэтому эти
236
данные ничего не дали для выяснения структуры мускарин" вых холинорецепторов. Удаление метильной группы из соединения (12.75) резко снижает мускариновую активность, так как приводит «к понижению электронной плотности на эфирном атоме кислорода, но не меняет конформацию. Из этого можно сделать вывод, что для проявления мускариновой активности значительно важнее распределение электронов в молекуле, чем ее конформация. Как фурметид, так и 5-метилфурметид служат примерами, подтверждающими правило «пяти атомов боковой цепи» (см. ниже).
Синтез жестких агонистов АХ — задача довольно сложная. Обширные конформационные возможности молекулы АХ (12.76) зависят от четырех торсионных углов: С5—С4—N—СЗ, 01—С5—С4—N, С6—01—С5—С4 и 02—С6—01—С5. По аналогии со всеми похожими структурами первый из этих углов, вероятнее всего, постоянен — это анти-планарная вытянутая цепочка с торсионным углом 180°. Причиной этой жесткости являются стерические препятствия, создаваемые N-метильными группами, так как их ван-дер-ваальсовы радиусы мешают более плотной упаковке (разд. 8.0). Из-за планарности эфирных групп (связь С6—01) фиксирован и последний из этих углов (02—С6—01—С5). Рентгеноструктурные исследования кристаллов различных агонистов АХ показали, что два других торсионных угла могут изменяться в широких интервалах. Одна из групп исследователей пришла к выводу, что для проявления мускариновой активности необходимо только расположение атомов С1 и С7 АХ по одну сторону молекулы [Baker et al., 1971]. Другая группа на основании похожих доказательств (в сочетании с данными для антагонистов, менее значимыми) постулировала, что АХ взаимодействует с мускариновыми рецепторами эфирным атомом кислорода и аммонийным катионом [Beers, Reich, 1970].
? Z7
сз—N—С4—С5—Ol—С6 R—N+Me3
С2 4x02
Нумерация атомов в молекуле ацетилхолина Четвертичные амины
(12.76) (12.77)
А. Катионная головка ацетилхолина. Целесообразно сравнить действие АХ и неорганических катионов, находящихся, по-видимому, на поверхности рецептора. В то время как калий оказывает стимулирующее действие на всю мышцу, действие АХ обычно ограничивается незначительной областью мышцы — концевой пластинкой. Экранирующее влияние алкильных групп (при атоме азота четвертичного амина) приводит к тому, что в водных растворах ион фактически не гидратируется [Robinson, Stokes, 1959]. Поэтому можно считать, что эффективный ионный радиус в растворах равен ионному радиусу, получен-
237
Таблица 12.2. Размер некоторых катионов
Радиусы негидратированиых ионов | Предполагаемые радиусы гидра-
Катион (в кристаллическом состоянии)1, | тированых ионов (по/подвижио-
нм | сти в воде при 25 °C)2, нм
L1+ 0,060 0,37'
Na+ 0,095 0,33
К+ 0,133 4
NH4+ 0,148
NMe4+ 0,2413
Mg2+ 0,065 0,44
Ca2+ 0,099 0,42
1 Pauling (I960).
2 По закону Дж. Г. Стокса с исправлениями R. Robinson, R. Stokes (1959).
3 Johnson (1960).
4 Примерное значение 0,13 им, наружный слой воды связан слабо, как у ионов С&+ в Rb+.
ному из данных рентгеноструктурного анализа. Ионный радиус тетраметиламмония равен 0,241 нм (табл. 12.2) и таким же должен быть радиус катионной головки АХ.
Радиусы всех неорганических ионов в безводном состоянии хорошо известны, однако ионы легких металлов сильно гидратированы в водных растворах и их эффективный радиус значительно больше. Насколько больше — сказать определенно нельзя, так как точных методов для таких измерений пока не существует. Все же величины, приведенные для ионов Li+ и Na+ в последнем столбце табл. 12.2, показательны. Катионная головка— самая большая часть молекулы АХ. Что произойдет, если она станет еще больше? Но прежде чем это обсуждать, несколько слов об ацетилхолиноподобном действии простых алифатических четвертичных аминов. Они оказывают лишь слабое действие на мускариновые рецепторы и сильное — на никотиновые (см. ниже). Например, действие солей тетраметиламмония на кишечник и сердце примерно в тысячу раз слабее, чем действие АХ [Clark, Raventos, 1937]. Соли тетраэтиламмо-ния и его высших гомологов являются только антагонистами. Тем не менее гомологический ряд солей алкилтриметиламмония (12.77) представляет большой интерес. Сродство к рецептору в этом ряду увеличивается с возрастанием длины алкильной цепи, а эффективность снижается от тетраметиламмония к три-метилэтиламмонию, а затем возрастает, достигая максимума у солей н-пентилтриметиламмония; исследовали их действие на кишечник млекопитающих Stephenson (1956), Van Rossum, Ariens (1959), на сердце лягушки Raventos (1937), на давление крови у собак Alles, Knoefel (1939). Эти и другие факты дали возможность сформулировать «правило пяти атомов» (см. ниже) . Однако активность даже самых эффективных соединений этого ряда не превышает 1 % активности АХ.
Б. Изменение катионной головки ацетилхолина. Последовательная замена метильных групп в молекуле АХ (12.65) на
238
Оксотреморин (12.79)
атомйх водорода или этильные группы приводит к резкому снижению хвсех видов парасимпатомиметической активности [Ing, 1949]. Выше упоминалось, что замещение на атомы водорода уменыпаё^ сродство и увеличивает эффективность. Этильная группа, погвидимому, увеличивает сродство, но снижает эффективность [Barlow, Scott, Stephenson, 1963]. Третичные амины типа (12.73) при pH 7,3 ионизированы по меньшей мере на 99%. Следовательно, падение основности не является причиной снижения сродства. Холинорецептор, вероятно, связан сильным ван-дер-ваальсовым взаимодействием с головкой АХ [Holton, Ing, 1949]. На основании этих данных можно предположить наличие в рецепторе чашеобразного углубления, форма которого обеспечивает максимальное ван-дер-ваальсово взаимодействие с четырьмя атомами углерода, расположенными в вершинах тетраэдра относительно четвертичного атома азота [Ве1-leau, Ригапеп, 1963].
Снижение эффективности при замещении трех метильных групп тремя этильными наблюдали и у многих других веществ с ацетилхолиноподобными свойствами [Barlow, 1968].
О С"
11 1 +
R—С—Y R—C=Y+
Резонансные гибриды (12.78)
Хотя простые третичные амины не проявляют заметной мускариновой активности, она может возникнуть, если остальная часть молекулы этих аминов приобретет способность прочно связываться с рецептором. Такое увеличение способности к связыванию карбонильной группы может быть достигнуто, если неподеленная пара электронов на атоме кислорода будет делокализована сильнее, чем это имеет место в сложных эфирах [Y = OR в резонансных гибридах на формуле (12.78)[. Такой делокализации еще более способствует амидная группа (Y = NH2 или NR2), так как атом азота легче приобретает положительный заряд, чем атом кислорода. О величине этого эффекта можно судить по положению полосы карбонильной группы в ИК-спектре. Смещение полосы поглощения от частоты, типичной для эфиров (1735 см-1), до частоты, типичной для амидов (1690 см-1 для свободных и 1650 см-1 для ассоциированных амидов), сопровождается резким повышением мускариновой активности третичных оснований, вплоть до активности четвертичных оснований, как, например, у оксотреморина (12.79) [Bebbington, Brimblecombe, Shakeshaft, 1966]. Оксо-треморин — это М-холиномиметик, применяемый в экспериментальной фармакологии; амидная группа включена в пирролидоновый цикл [Cho, Haslett, Jenden, 1962]. Необходимо упомянуть и два мускариновых антагониста, имеющих третичную
239
о
II ЧЧ—С—ОМе
N
I
Ме
Ареколин (12.80)
аминогруппу и сложноэфирную группу. Жители Восточной Индии для достижения легкой эйфории жуют листья/бетеля, содержащие ареколин (12.80). Высокая мускаринова/ актив/ ность ареколина становится очевидной, если рассматривать его как «обращенный ацетилхолин», прототипом которого являете^ (12.85). Ареколин обладает и слабым никотиновым/действием, однако оно в медицине не используется. В пилокарпине (12.81), слабом мускариновом агонисте, сложноэфирная группа вклю? чена в цикл (лактон). Он используется преимущественно для снижения внутриглазного давления при глаукоме.
MeN-------СН2—СН--CH—СН2—Ме
k JI н,с с-о
N V
Пилокарпин
(12.81)
Активность аналогов ацетилхолина, в которых атом азота замещен атомом фосфора или мышьяка, составляет всего лишь от 1 до 10% активности АХ (на различных участках). В таких соединениях замена атома азота не приводит к изменению углов связей, но увеличивается на 27—35% среднее расстояние между метильными группами, поскольку связи Р—С и As—С длиннее, чем связь N—С [Holton, Ing, 1949].
В. Правило «пяти атомов в боковой цепи». Необходимость наличия пяти атомов в боковой цепи для высокой эффективности ацетилхолиноподобных соединений впервые была отмечена Alles, Knoefel (1939), а затем подтверждена Ing (1949). У АХ и его аналогов типа R—+М(СНз)з самым активным членом в любом гомологическом ряду обычно оказывается тот, у которого заместитель R представлен цепью из пяти атомов (не считая атомов водорода). Эта закономерность уже обсуждалась выше для соединений, у которых R-алкильная группа, однако это справедливо и для случаев замены атомов углерода другими атомами, например кислородом. Так, АХ (12.65) значительно активнее формил- или пропионилхолинов, а бутирил-и валероилхолины имеют едва выраженную мускариновую активность. И азотистый, и азотнокислый эфиры холина, содержащие боковую цепь из 5 атомов, обладают значительной мускариновой активностью [Dale, 1914]. Уксусные эфиры спиртов НО—СН2—+NMe3 и НО—(СН2)з—+NMe3 менее активны, чем соответствующий эфир холина НО—(СН2)3—+NMe3 [Hunt, Та-veau, 1911].
Точно так же этоксихолин, этиловый эфир холина (12.82), более активен, чем метиловый или пропиловый эфиры [Dale, 1914], а н-пропиловый эфир — самый активный в ряду эфиров НО—СН2—+NMe3. В ряду производных диоксолана (или «аце
240
талей»), типа (12.83), обладающих мускариновым действием., более активны соединения с R=Me, чем таковые с R = H или Et Tourneau et al., 1944]. Изображенный на формуле (12.83)е Ь( + )-цйр]изомер в шесть раз активнее АХ [Belleau, Lacasse,. 964]. Факторы, лежащие в основе правила пяти атомов, обсуждаются циже в разделе «Антагонисты ацетилхолина» (ни одно из упомянутых в этом параграфе соединений не применяется в медицинской практике).
Н О----СНг
Ме—СН2—О—СН2—СН«—N+Me. Ч | |
4 3 2 1 R—С CH—СН2—N+Mes
Этоксихолин (катион)
(12.82)
Диоксолановый агоннст
(12.83)
Г. Оптимальное положение атома кислорода в боковой цепи. Активность эфира (12.82) составляет примерно от 1 до 10% активности АХ, причем у него более выражено мускариновое, а не никотиновое действие. Эта активность снижается,, если атом кислорода переместить из положения 3 в положение' 2 или 4. Кетон (12.84) обладает слабой мускариновой активностью, а величина его никотиновой активности на разных препаратах составляет от 0,2 до 100% активности АХ. Его активность снижается, если карбонильную группу переместить из положения 4 в положение 3 или 2 [Ing, Kordik, Williams, 1952].. Таким образом, активность этих соединений максимальна, если? эфирный и карбонильный атомы кислорода расположены в молекуле так же, как и в АХ (12.65).
Это согласуется с измерениями, проведенными на моделях Beers, Reich (1970), сделавших вывод, что мускариновый и никотиновый рецепторы связываются ацетилхолином через атомы? азота и кислорода, эфирный в соединении (12.82) и кетонный-в соединении (12.84). Эти рецепторы перекрываются соответствующими фрагментами, содержащимися во всех мускариновых и никотиновых агонистах и антагонистах.
Расстояния между атомом азота и эфирным атомом кислорода (0,44 нм) и между атомом азота и кето-атомом кислорода (0,59 нм) на моделях несколько длиннее, чем по данным» рентгеноструктурного анализа, вероятно, из-за того, что на моделях не учитывается частично двоесвязанный характер эфирной связи. Однако относительные величины этих двух расстояний установлены. Отсюда следует, что между рецептором ив атомами кислорода образуются водородные связи.
О о
II II
Me—С—СН2—СН2—CH2N+Me3 Me—О—С—СН2—СН2—N+Me3
4 3 2 1
Кетон — аналог ацетилхолина «Обращенный ацетилхолин»
(12.84) (12.85)
16—734
246
Метиловый эфир p-триметиламмониопропионовой кислоты (12.85) можно рассматривать как АХ, в котором эф^ный и карбонильный атомы кислорода поменяли местами. Это соединение является плохим субстратом для АХЭ и не способно ингибировать этот фермент. Однако такой эфир оказцйает сильное мускариновое и умеренное никотиновое действие [Bass et al., 1950].
Д. Антагонисты мускариновых рецепторов. Изучение строения антагонистов дает значительно меньше информации о строении рецепторов, чем изучение агонистов, так как антагонисты ло своей природе не обладают точным структурным соответствием рецептору, а лишь перекрывают его. Наиболее изучен атропин (7.16) (влияние структурных изменений в молекуле атропина на активность см. разд. 7.3.3).
Гомологический ряд производных алкилтриметиламмония (12.77) может служить иллюстрацией широко известного принципа превращения стимуляторов в антагонисты путем увеличения ОММ (табл. 7.2, том 1). Кинетические исследования показали, что скорость образования комплекса вещества с рецептором для всех членов этого ряда одинакова, но с увеличением числа метиленовых групп скорость диссоциации этого комплекса снижается (разд. 10.3.2). Поэтому низшие члены этого ряда являются чистыми мускариновыми агонистами, но при Р=СеН1з появляется атропиноподобная блокирующая активность, а при R=Ci2H25 действие соединения чисто атропиноподобное [Paton, 1961]. Появление антагонистического действия при R>5 может объяснить, почему максимум мускариновой .активности во многих сериях соединений приходится на вещества, содержащие в боковой цепи пять атомов (см. выше). Как предложил Chothia (1970), уменьшение активности при удалении концевой метильной группы вызвано ослаблением взаимодействия с соответствующей липофильной областью в рецепторе.
Мускарон, получаемый окислением вторичной спиртовой труппы мускарина (12.72) до карбонильной, обладает не только более выраженным мускариновым, но и никотиновым действием. Молекула мускарона содержит и эфирный, и карбонильный атомы кислорода, но расстояние между ними больше, чем в молекуле АХ. О применении мускариновых антагонистов в терапии см. разд. 7.3 (при обсуждении атропина).
12.6.2. Никотиновые холинорецепторы
Никотиновый холинорецептор в отличие от мускаринового значительно быстрее реагирует на появление стимула. Он был выделен из электрических органов рыб (разд. 2.1, рис. 2.1, том 1). Этот гликопротеин можно пометить змеиным токсином и а-бунгаротоксином, обычно используемым в виде [3Н]-ацетил-производного. Реагент специфично связывается с ацетилхолин-242
связыйающим местом рецептора. Такой комплекс токсин — рецептор можно использовать для очистки рецептора, отделяя токсин в, конце очистки длительным диализом. В 1980 г. очищенный рецептор был встроен в искусственную плоскую липидную мембрану с целью демонстрации его пермеазной природы. С помощью дифракции рентгеновских лучей было установлено,, что рецептор пронизывает мембрану, выступая понемногу с каждой стороны. Диаметр рецептора равен приблизительно» 8,5 нм, и он включает в себя ионный канал диаметром 0,65 нм. Такой реконструированный рецептор под действием АХ увеличивал проницаемость мембраны для катионов, причем эффект-АХ блокировался тубокурарином [Nelson et al., 1980].
Каждая рецепторная розетка (см. рис. 2.1) включает в себя пять параллельных цепочек, образующих ионный канал. На двух цепочках, называемых а-субъединицами, расположены места связывания ацетилхолина. Остальные три играют чисто структурную роль1 *.
На выделенной из Torpedo californica иРНК, кодирующей а-субъединицу, была синтезирована и проклонирована комплементарная ДНК- Так была получена иРНК в количествах, достаточных для установления нуклеотидной последовательности, по которой в свою очередь была определена первичная структура а-субъединицы с ОММ 50116, содержащей 437 аминокислотных остатков.
Дисульфидный мостик около ацетилхолинсвязывающего места, по-видимому, образован двумя остатками Cys-128 и 142. Что касается самого ацетилхолинсвязывающего места, то предполагается, что в его состав входят остатки Asp-138 (или Clu-129) и His-134. Наиболее вероятное место присоединения глюкозы — атом азота остатка Asp-141 [Noda et al., 1982].
Для возникновения физиологического ответа как минимум две молекулы АХ должны связаться с рецептором. Выброс АХ, длящийся 1 мкс, вызывает в скелетной мышце ответ продолжительностью не менее 10 мкс. Судя по этим цифрам, открывание ионного канала контролируется непосредственно рецептором и для этого не требуется предварительного фосфорилирования. Таким образом, механизм никотинового ответа является более простым и более прямым, чем постулированный Greenyard механизм мускаринового ответа (разд. 12.6.1).
Все приведенные выше данные получены для рецептора, выделенного из электрического органа рыб, однако сходный белок был выделен из скелетных мышц млекопитающих [Dolly, Barnard, 1977; Froehner, Reiness, Hall, 1977] и из коры головного мозга морских свинок. Последний имеет ОММ около» 86 000, а связывание с ним [3Н]-бунгаротоксина игибируется тубокурарином и галламином [Bosman, 1972].
1 О структуре никотинового холинорецептора см. также обзор В. И. Скок,
Биол. мембраны, 1985, т. 2, с. 245—255. — Примеч. ред.
16* 243
Более подробно о никотиновых рецепторах см. Karlin /(1980) я Conty-Tronconi, Raferty (1982).
Задолго до выделения никотинового рецептора было начато изучение связи структура — активность в ряду агонистов, действующих на активируемые АХ места, традиционно называемые никотиновыми. Этот термин, так же как и термин «мускариновый», имеет чисто историческое значение, так как никотин клинического применения не имеет. Известно, что никотин проявляет холиномиметические свойства у позвоночных в трех структурах: а) на моторной концевой пластинке произвольной мышцы в нервно-мышечном соединении; б) в ганглиях симпатических и парасимпатических нервов и в) в коре головного мозга. Наиболее доступна для действия никотина структура t(a) (это следует хотя бы из того, что никотин является ядом для млекопитающих). Однако у насекомых, в нервно-мышечных соединениях которых не содержится АХ, основное действие никотина сосредоточено в ганглиях ЦНС [Yaeger, Munson, 1945]. В настоящее время никотин мало используется в качестве инсектицида. Синтезированы никотиноподобные инсектициды {Kamiura et al., 1963]: для повышения их активности необходимо наличие основного заместителя в положении 3 пиридинового цикла, причем заместитель может быть и алифатический, как, например в (7.27), но не должен содержать четвертичную труппу, так как это затрудняет проникновение инсектицида в «организм насекомого. При кватернизации атома азота пирролидинового цикла в молекуле никотина у соединения сохраняется никотиноподобное действие на млекопитающих. Сложность использования никотина и его аналогов для изучения природы никотинового рецептора заключается в том, что на ганглии избыток никотина оказывает антиацетилхолиновое (блокирующее) действие [Barlow, Hamilton, 1962]. Возникновение эффекта эйфории при курении связано, вероятно, с действием никотина на структуры (в) (см. выше).
Простейшее соединение, обладающее сильным никотиновым (но не мускариновым) действием, основной агонист — это тет-раметиламмоний-хлорид (12.77, R=Me). В ряду Н-холиноми-метиков это такое же стандартное соединение, как и метахо-дин (12.68) в ряду М-холиномиметиков.
По силе действия на автономные ганглии соли тетраметил-аммония не отличаются от АХ [Burns, Dale, 1915], однако на нервно-мышечное соединение скелетных мышц (никотиновая активность) они действуют в сто раз слабее, чем АХ, а на соединение с гладкими мышцами (мускариновая активность) — в тысячу раз слабее. Взаимодействие медиатора с автономными ганглиями кратковременно, а в нервно-мышечном соединении этот процесс более длителен, и для него предпочтительнее молекулы, способные образовывать водородные связи, например, как АХ — за счет кето-атома кислорода. Очевидно, катион тетраметиламмония определяет биологическую активность АХ;
244
при необходимости длительного взаимодействия важную роль играет и остальная часть молекулы, обеспечивая дополнительное связывание с рецептором.
Из соединений ряда катионов алкилтриметиламмония (родоначальником которого является тетраметиламмоний) максимальной активностью обладают соли н-пентилтриметиламмо-пия, их действие на никотиновые рецепторы такое же, как у никотина, и в восемь раз сильнее, чем у АХ [Willey, 1955]. Выраженной никотиновой, но слабой мускариновой активностью обладают D-лактоилхолин [Sastry, Lasslo, Pfeiffer, 1960], а также фениловые эфиры холина типа (12.86) [Неу, 1952]. Однако соотношение этих активностей можно изменить введением метильных групп в оба орто-положения бензольного кольца, это свидетельствует о том, что стерические затруднения в молекуле в большей степени влияют на явление никотинового, чем мускаринового, действия (о влиянии С-метильной группы в метахолине см. разд. 12.6.1).
О а+ «
Me3N+—СН2—СН2—S—С—Ме
О—СН2—СН2—N+Me3
Фениловый эфир холина Ацетилтиохолнн
(12.86) (12.87)
Молекулы никотиновых агонистов, вероятно, должны находиться в гош-конформации (определение см. разд. 12.3). Установлено, что в кристаллах и растворах большинство этих агонистов действительно имеют гош-конформацию (синклинальную) фрагмента —+N—С—С—О [Baker et al., 1971; Calvenor, Ham, 1966; Сапера et al., 1966]. Исключение составляет карбахолин (2.11), существующий в кристаллах в вытянутой (антипланар-ной) конформации, а в растворе — в гош-конформации [Ваг-rans, Clastre, 1970; Baker et al., 1971]. Однако данные по конформации агонистов, содержащих фрагменты —+N—С—С—S— и —+N—С—С—Se—, для которых вытянутая (антипланарная) конформация является преимущественной как в кристаллах [Shefter, Mautner, 1969], так и (по данным ЯМР) в растворах [Cashley, Mautner, 1970], ставят под сомнение правомерность гош-гипотезы. Например, тиохолин и S-метилтиохолин (оба в вытянутой конформации) — сильные агонисты, тогда как их кислородные аналоги холин и О-метилхолин (оба в гош-конформации) не активны [Mautner, Bartels, Webb, 1966]. Более того, замена карбонильного атома кислорода в молекуле АХ атомом серы не приводит к изменению гош-конформации [Mautner, Dexter, Low, 1972] и как сам АХ, так и ацетилтиохолин (12.87) высокоактивны в различных биологических тестах [Scott, Mautner, 1967; Mautner et al., 1966; Mautner, 1969]. Эти данные свидетельствуют о том, что, вероятно, для проявления агонистической активности исходная конформация не менее
245
важна, чем распределение электронов. Было подсчитано, что для изменения преимущественной гош-конформации молекулы АХ в растянутую рецептору необходимо затратить лишь 1 ккал (4,18 кдж) [Liquori, Damiani, de Coen, 1968].
При изучении рецепторного связывающего места был применен иммунологический подход, для чего были получены кроличьи антитела к фениловому эфиру холина (12.86). К сожалению, при попытке использования этих антител в качестве модели никотинового рецептора выяснилось, что они не способны различать ни вещества мускаринового и никотинового типа действия, ни агонисты и антагонисты [Marlow, Metcalf, Burgen, 1969].
Антагонисты никотиновых рецепторов. Ни один чистый никотиновый агонист не используется в медицине. Карбахолин (2.11) обладает не только никотиновыми, но и выраженными мускариновыми свойствами. Антихолинэстеразные лекарственные вещества усиливают действие эндогенного АХ, что означает одновременную активацию мускариновых и никотиновых структур. В качестве примера можно привести использование физостигмина (2.8) для лечения острой глаукомы, фосфорорганических соединений (разд. 13.3)—для лечения хронических случаев этого заболевания, часто встречающегося у пожилых людей, и прозерина (2.10) —для снятия нервно-мышечного блока при миастении гравис. Последний лекарственный препарат обладает слабым никотиновым действием и на произвольные мышцы.
Хотя стимуляцию никотиновых рецепторов мало используют в терапии, в хирургии широко применяют никотиновые антагонисты, позволяющие добиваться глубокого мышечного расслабления и снижения доз общих анестетиков. В 1942 г. для этой цели был применен тубокурарин (2.6), затем в дополнение к нему появились другие четвертичные аммониевые катионы. В настоящее время известны два основных типа таких лекарственных веществ. Представители первого типа, например тубокурарин, препятствуют деполяризации мембран ацетилхолином, но сами не оказывают на них никакого ацетилхолиноподобного действия. Они являются, конкурентами АХ при его взаимодействии с рецептором, но не препятствуют его синтезу, высвобождению и распаду. Блокирование взаимодействия АХ с рецептором приводит к полному расслаблению мышцы. Основываясь на том, что катионные головки тубокурарина (2.6} (у которого только одна четвертичная аминогруппа) и галла-мина (7.31), по-видимому, стерически не соответствуют рецептору АХ, Waser (1960) предположил, что молекулы этих соединений заслоняют собой углубление, в котором находится холи-норецептор. Второй класс антагонистов, действующих на нервно-мышечное соединение, представляют вещества, например, декаметоний (7.28) и дитилин (7.29), препятствующие деполяризации постсинаптических мембран под действием АХ, но об-
246
Таблица 12.3. Зависимость специфичности действия от длины цепи бис-триметиламмоиийных катионов на два типа холииорецепторов (антиникотиновое действие)
Тип холинергического синапса Число метиленовых групп
4 5 6 7 8 9 10 11 12_
Нерв — нерв Нерв — мышца 0,01 0,8 1,0 0,1 0,02 0,07 0,7 0,1 0,55 0,2
Примечание. Приведены относительные активности, для каждого типа макси-мальная величина активности 1,0 [Paton, Zaimis, см. текст].
ладающие достаточно высокой ацетилхолиноподобной активностью для того, чтобы вызвать деполяризацию до того, как наступит нервно-мышечный блок. Триметиламмонийные группы этих соединений не испытывают стерических затруднений, поэтому возможно взаимодействие по меньшей мере одного конца молекулы с анионным местом холинорецептора, хотя вся молекула в целом достаточно велика, чтобы закрыть полость.
Сродство этих бис-ониевых солей к трем основным типам холииорецепторов отличается высокой специфичностью. Все они практически не действуют на постганглионарные рецепторы, однако их активность в ганглиях и нервно-мышечных соединениях может быть исключительно высокой при условии, что для каждого из этих двух видов рецепторов выбран надлежащий гомолог. Декаметоний в дозах, в сто раз превышающих дозы, полностью блокирующие нервно-мышечное соединение, совершенно не действует на ганглии. Гексаметоний, наоборот, в стократной дозе по сравнению с дозой, блокирующей ганглии, не действует на нервно-мышечное соединение [Paton, Zaimis, 1949, 1952]. Эта специфичность проиллюстрирована данными в табл. 12.3.
Ганглиоблокаторы типа гексаметония (7.30) вошли в медицинскую практику около 1950 г. в качестве препаратов для лечения гипертонии. Однако их действие на организм оказалось настолько неспецифичным, с множеством побочных эффектов, что в последствии эти препараты были вытеснены ^-адрено-блокаторами и лекарственными веществами, расслабляющими мышцы непосредственно в малых кровеносных сосудах.
Лекарственное вещество, способное связываться одновременно с двумя местами связывания на рецепторе, должно отличаться очень высоким сродством к последнему, так как: а) если один конец молекулы в результате диссоциации удаляется от поверхности, то другой конец будет ее удерживать на близком расстоянии от поверхности и тем самым способствовать рекомбинации и б) молекулы лекарственного вещества будут соединены более прочно за счет ван-дер-ваальсовых сил с частью биологической поверхности, лежащей между двумя местами связывания. Введение эфирной группы в молекулы бис-ониевых
247
блокаторов нервно-мышечной передачи резко повышает их способность ингибировать деполяризацию, как, например, холиновые эфиры дикарбоновых кислот типа (12.88). Из этих соединений в хирургии наиболее широко применяют дитилин (7.29) ' из-за его способности медленно гидролизоваться холинэстера- 1 зой плазмы, что способствует прекращению действия препарата на больного после операции. (В разд. 3.5.4 упоминалось новое лекарственное вещество атракурий, гидролизующийся под действием воды.)
Усиление действия соединений типа (12.88) при введении эфирной группы указывает на наличие в рецепторе групп, способных к образованию водородных связей. В ряду подобных соединений вторая триметиламмонийная группа может быть без ущерба для действия заменена атомом водорода [Danilov et al., 1974]. Замена одной триметиламмонийной группы в молекуле декаметония на другую гидрофильную (например, метокси) группу не нарушает действие вещества и, следовательно, для связывания никотиновых антагонистов необходима лишь одна кислотная группа в рецепторе.
О О
х II II
Me3N+—СН2СН2—О—С—(CH2)n—С—О—СН2СН2—N+Me3
Холиновые эфиры дикарбоновых кислот
(12.88)
Паннуроний (динатион) . (12.89)
О о
х II II
Me3+N—СН2—СН2—О—С—NH— (СН2)6—NH—С—О—СН2—СН2+—NMe3
Карболоиий (дикатион)
(12.90)
О расстоянии между двумя местами связывания в холино-рецепторе можно судить по размерам молекулы панкуроний-бромида (12.89), сильного неполяризующего миорелаксанта, применяющегося в клинической практике [Baird, Reid, 1967]. Жесткость его молекулы была подтверждена спектрами ЯМР в растворе. По данным рентгеноструктурного анализа расстоя-248
ние между двумя атомами азота в молекуле панкурония равно 1,108 нм, что сравнимо с таковым у М,О,О'-триметил-6-тубоку-рарин-дийодида в кристаллах, равным 1,07 нм (вероятно, таково же расстояние между атомами азота и в самом тубокура-рине). Следует отметить, что панкуроний структурно ближе АХ, чем соединения бис-ониевого типа, так как в его молекуле дважды повторяется фрагмент Me—С (О)—О—CHR—CHR— —N+R3 [Savage et al., 1970].
Молекула декаметония (7.28) в кристаллическом состоянии полностью вытянута и атомы азота удалены друг от друга на расстояние 1,37 нм [Lonsdale, Milledge, Pant, 1965], однако в растворе это расстояние, видимо, меньше. А у молекулы дити-лина, гибкой, как и у всех деполяризующих агентов, расстояние между атомами азота в кристаллическом состоянии зависит от аниона, образующего данную соль, и варьируется от 0,78 до 1,19 нм.
Бис-ониевые соединения, содержащие между атомами азота цепочку из 16 атомов (не обязательно углерода), обозначают N-16-N, или «С-16». Примеры таких соединений — суберилди-холин (субехолин), себацинилдихолин и карболоний-бромид (12.90), изредка применяемый в хирургии. Холинорецептор, чувствительный к соединениям «С-16», появился на ранней стадии эволюции и широко распространен в животном царстве. Нервно-мышечный синапс моллюсков, иглокожих и первичнохордовых ингибируется соединениями «С-16», но не «С-10»; некоторые аннелиды чувствительны к соединениям «С-10», однако все черви — к соединениям «С-16». Амфибии значительно более чувствительны к «С-16»-, чем к «С-10»-ингибиторам, а высшие позвоночные примерно одинаково чувствительны к обоим типам. Из того факта, что невозможно блокировать рецепторы для соединений структуры «С-16», не заблокировав рецепторы для соединений структуры «С-10» (и наоборот), следует, что эти рецепторы должны иметь общие группы [Khromov-Borisov, Michelson, 1966; Михельсон, Земаль, 1970]. На основании этих данных была создана тетрамерная модель мышечного холинорецептора, согласно которой четыре анионных связывающих места расположены в вершинах квадрата. Соединения «С-10» взаимодействуют с двумя центрами, расположенными на одной стороне квадрата (например, дитилин), а соединения «С-16» — с центрами на диагонали [Khromov-Borisov, Michelson, 1966] *.
Об истории создания и применении никотиновых антагонистов см. разд. 7.3.
1 Более подробно об исследовании структуры холинорецептора химикофармакологическими методами см. Н. В. Хромов-Борисов в ки.: Биохимическая фармакологии (под ред. П. В. Сергеева). М., Медицина, 1982. — Примеч. ред.
249
Третий тип холинорецепторов. Из головок домашних мух экстракцией удалось выделить два холинорецептивных белка, обладающих гибридным никотино-мускариновым характером (с ОММ 94 000 и 64 000 соответственно) и способных взаимодействовать с некоторыми типичными никотиновыми и муска- I риновыми агонистами и антагонистами. Однако ни один из них не блокируется а-бунгаротоксином и хинуклидинилбензилатом, типичными блокаторами никотиновых и мускариновых рецепторов соответственно [Tripathi, Tripathi, O’Brien, 1969]. Подробнее о связывании агонистов и антагонистов АХ с рецепторами см. Triggle, Triggle (1976), Cooper, Bloom (1982) и Yamamura, Enna, Kuhar (1978).
12.7. Рецепторы ГАМК и бензодиазепины
Из всех существующих в ЦНС нейромедиаторов у-амино-масляная кислота (ГАМК) наиболее доступна для исследования. Она служит передатчиком нервных импульсов в системе тормозных нейронов в непосредственной близости от спинного мозга и участвует в пресинаптическом интраспинальном торможении. По меньшей мере в 12 образованиях мозга, в том числе в основных эфферентах клеток коры мозжечка, ГАМК служит тормозным нейромедиатором. Для изучения ГАМКергической передачи обычно используют алкалоиды судорожного действия пикротоксин бикукулин, в медицине не применяющиеся [Curtis et al., 1971].
Для этой же цели используют более простые соединения, например мусцимол, воспроизводящий эффекты ГАМК, и окси-аминоуксусную кислоту, тормозящую обратный захват ГАМК [Johnston, 1978; Iversen, 1978]. Нипекотовая кислота (12.92) (пиперидин-3-карбоновая кислота) также тормозит захват ГАМК в пресинаптических нервных окончаниях, не влияя на соответствующие постсинаптические рецепторы. Однако изомерная пиперидин-4-карбоновая кислота (изонипекотовая кислота) представляет собой специфический высокоактивный агонист постсинаптического рецептора ГАМК. По-видимому, ГАМК существует в двух различных конформациях, взаимодействующих с пре- и постсинаптическими рецепторными мишенями [Krogsgaard-Larsen, Scheel-Kriiger, Kofod, 1979].
“О2С—СН2 /\ТХ'С°2’
I I
CH2 О
I + N+
СН2—NHS pj/ \pj
ГАМК Нипекотовая кислота
(12.91) (12.92)
Аналог ГАМК, цис-4-аминокротоновая кислота, существует в сложенной конформации (12.94) и биологически неактивна, 250
тогда как транс-изомер (12.93) влияет на центральную нервную систему млекопитающих так же активно, как и ГАМК. Это указывает на необходимость растянутой, а не сложенной конформации ГАМК [Johnston et al., 1975].
Обычное физиологическое действие ГАМК связано с влиянием на рецепторный комплекс, имеющий более сложную структуру, чем никотиновый холинорецептор. В нем расположены рецептор для ГАМК, рецептор для бензодиазепинов и третий участок, связывающий пикротоксин: все три рецептора расположены вокруг входа в канал для ионов хлора (С1~) [Olsen, 1982]. Вначале предполагалось, что существует несколько типов ГАМК-рецепторов [Hill, Bowery, 1981] и бензодиазепиновый рецептор. Однако тщательные кинетические исследования показали наличие отдельной популяции бензодиазепинового рецептора, существующего в двух конформациях. Агонисты бензодиазепиновых рецепторов сибазон (12.95), а также их антагонисты, известный под названием RO 15-1788 (12.96), индуцируют взаимный переход этих двух конформационных состояний [Chiu, Dryden, Rosenberg, 1982]. В конформации, обладающей высоким сродством к ГАМК, бензодиазепиновый рецептор облегчает ГАМКергическую передачу и повышает проводимость для ионов хлора. Бензодиазепиновый антагонист (12.96) неактивен, видимо, из-за затрудненной пространственной конфигурации молекулы. Метиловый эфир 6,7-диметокси-4-этил-[5-кар-болин-3-карбоновой кислоты (12.99), вызывающий связывается с бензодиазепиновым каналом закрывает ионный 1983].
судороги, таким образом, что [Chiu, Rosenberg,
канал для ионов хлора
HSN
со2-
СО2-
\nh
Снбазон (диазепам)
(12.95)
транс-4-Аминокротоновая кислота
(12.93)
цис-4-Аминокротоновая кислота
(12.94)
RO 15-1788
(12.96)
Баклофен, ^-(пара-хлорфенил) производное ГАМК, — единственный аналог ГАМК, применяющийся в клинической меди
251
цине; используется для ослабления спастических состояний после повреждений ЦНС и при рассеянном склерозе [Davidoff, 1978].
Бензодиазепины. История этих широко известных веществ; началась с открытия Sternbach в 1959 г. хлозепида (12.97), который был применен в клинике в качестве транквилизатора [Tobin, Lewis, 1960]. Впоследствии был синтезирован и испытан в клинике его более активный аналог—сибазон (12.95) [Randall, 1961]. Бензодиазепины используют в качестве седативных, снотворных, противосудорожных, антифобических средств и миорелаксантов, пригодных для премедикации. У разных веществ то или иное свойство превалирует над другими. В качестве снотворных препаратов, применяемых в высоких дозах, специально отбирались вещества с относительно коротким сроком действия: из них следует отметить нитразепам (12.98), флунитразепам, флуразепам и темазепам. Для лечения эпилепсии предпочтительнее клоназепам. В разделе 3.6 отмечалось, что хлозепид, сибазон и темазепам метаболизируют в организме до нордазепама (3.47), период полусуществования которого в плазме крови 90 ч и который, видимо, является активным началом всех вышеуказанных соединений.
Хлозенид
(12.97)
Нитразепам (12.98)
Этиловый эфир карболин-3-карбоновой кислоты
(12.99)
В умеренных дозах действие бензодиазепинов сходно с действием снотворных веществ типа барбитуратов, однако, в отличие от них первые более избирательны, так как с повышением дозы не становятся токсичными для больных. Для общего наркоза вместо тиопентала можно применять диазепам внутривенно. Установлено, что барбитураты влияют на чувствительные к пикротоксину рецепторы, входящие в состав ГАМК-ре-цепторного комплекса [Olsen, 1982], усиливая действие ГАМК. Этот эффект стереоспецифичен и характерен для (S)-изомера этаминала [Huang, Barker, 1980]. Вклад рецепторов ГАМК в эффекты барбитуратов, по-видимому, незначителен, учитывая,
252
что последние считаются структурно-независимыми липофильными веществами (разд. 15.0).
Бензодиазепины вначале относили к транквилизаторам, затем к малым транквилизаторам, однако скорее всего они являются седативными веществами. Для лечения психозов их не-применяют.
Бензодиазепины связываются с рецептором в непосредственной близости от участка связывания ГАМК. Тем самым они: могут увеличивать проницаемость нейрональных мембран для; ионов хлора и усиливать индуцированную ГАМК стабилизацию^ электрического потенциала клетки — фактора, снижающего ее-возбудимость [Simmonds, 1980]. Некоторые исследователи считают, что природным лигандом бензодиазепинового участка связывания в рецепторе ГАМК служит белок, названный моду-лином [Costa, Di Chiara, Gessa, 1981], или непентин [Wolf,. Nixon, 1981], другие называют выделенный из мочи человека этиловый эфир карболин-3-карбоновой кислоты (12.99), обладающий сродством к бензодиазепиновому рецептору [Braestrup,. Nielsen, Olsen, 1980].
Макроциклический лактон ивермектин, все чаще используемый в качестве высокоизбирательного инсектицида и глистогонного средства, представляет собой агонист ГАМК в периферической нервной системе насекомых и нематод. Он не действует на гельминтов и ленточных червей, содержащих другие нейромедиаторы. У млекопитающих ГАМКергическая передача ограничена ЦНС и поэтому от ивермектина, применяемого для: лечения сельскохозяйственных животных, их защищает ГЭБ,. Вещество липофильно, и после однократного введения внутрь, или подкожно его эффекты сохраняются в течение нескольких: недель.
Подробнее о бензодиазепинах см. Garattini, Mussini, Randall (1973) и Usdin и сотр. (1983).
12.8. Морфин и рецепторы опиатов
Термином опиаты (синоним ненаркотические анальгетики), обозначают природные и синтетические вещества с морфиноподобными аналгезирующими свойствами, влияющие на ЦНС и гладкие мышцы. Морфин, основной алкалоид опия, служит-стандартом для оценки других опиатов. Его используют для-облегчения послеоперационного периода в качестве болеутоляющего, противокашлевого и успокаивающего средства. В разд.. 7.3 рассматривались конфигурация молекулы морфина и поиск опиатов с менее сложной химической структурой, обладающих аналгезирующими свойствами и не вызывающих подобно морфину эйфории. Там же обсуждался и первый из антагонистов морфина — налорфин (7.35). В настоящее время ясно, что первоначальное предположение о важности аксиального расположения ароматического кольца в молекулах синтетических
25&
анальгетиков [Beckett, 1959; Beckett, Casy, 1962] несущественно (по данным рентгеноструктурного анализа, ЯМР-спектроско-пии и спектров кругового дихроизма, особенно при исследованиях с использованием более жестких молекул).
Наиболее отчетливые доказательства независимости аналге-тического действия от конформации молекулы получены на примере транс-1-метил-4-пропионилокси-4-фенил-декагидрохиноли-на. Это соединение можно хроматографически разделить на два конформера (12.100) и (12.101) с фенильной группой в экваториальном и аксиальном положении соответственно. При обычных условиях из-за сильных стерических затруднений они не переходят друг в друга. Сами эти соединения оптически не активны (но каждый из них может быть разделен на два энантиомера). Оба конформера (12.100) и (12.101) проявляют одинаковую анальгетическую активность, в десять раз меньшую, чем морфин (у мышей).
(’2.102) (12.103)
Две пары стереоизомерных опиатов
Исследования другой пары жестких взаимонепревращаю-'.щихся изомеров еще более способствовали решению данной проблемы. Из двух эпимерных 5-карбоэтокси-2-метил-5-фенил-;2-аэобицикло [2.2.1] гептанов [(12.102) экзофенил и (12.103) эндофенил], которые можно рассматривать как жесткие аналоги лидола (7.37), эндоэпимер в 2 и 4 раза соответственно превышал по анальгетической активности петидин и экзоэпимер (на мышах). Установлены также различия в степени их проникновения через ГЭБ, коэффициентах распределения и рКа [Portoghese, Mikhail, Kupferberg, 1968]. Эти данные не только показывают, что изменения пространственного расположения
‘.254
ароматического кольца могут не влиять на анальгетическую активность вещества, но и указывают на то, что изменения конформации приводят к изменению физических свойств молекулы, именно этим, может определяться физиологический ответ.
Из данных для указанной выше пары соединений следует, что конформационные требования к веществам, обладающим анальгетической активностью, не очень строги: расстояния от центра бензольного кольца до атома азота значительно отличаются около 0,6 нм в (12.102) и только 0,4 нм в (12.103).
Молекула морфина жесткая и поэтому дает возможность оценивать расстояние между различными группами в морфиновом рецепторе. Три структурные особенности определяют анальгетическую активность морфина: наличие в его молекуле-бензольного кольца, атома азота и, видимо, «^-ненасыщенной вторичной спиртовой группы. На основании рентгеноструктурного анализа были рассчитаны расстояния между этими центрами в молекулах морфина и кодеина, а также альфапродина и метадона [Jung, Lami, 1970; Jung, Koffel, Lami, 1971], показавшие существование нескольких популяций опиатных рецепторов.
Леворфанол Налонсон
(12.104) (12.105)
А. Опиатные рецепторы. Впервые существование опиатного рецептора в мозге мышей было показано при исследовании стереоспецифического взаимодействия леворфанола (аналога морфина) (12.104) и его антагониста налоксона с субклеточными фракциями мозга [Goldstein, Lowney, Pal, 1971]. Был выделен липопротеин с ОММ 60 000, и авторы предположили, что взаимодействие рецептора с агонистом (но не антагонистом) индуцирует в нем конформационные перестройки и биологический ответ, приводящий к аналгезии. Pert, Snyder (1973) подтвердили присутствие опиатных рецепторов в нервной ткани и установили различное влияние ионов натрия на связывание специфических агонистов и антагонистов с рецепторами. Они считали, что ионы натрия, увеличивая количество доступных для антагонистов участков, вызывают конформационные изменения, обратные происходящим при взаимодействии с агонистами.
Изучение физиологической значимости опиатного рецептора затруднялось отсутствием в арсенале исследователей его эндо-
255-
генного агониста. Hughes (1975) экстрагировал из мозга пентапептид (H-Tyr-Gly-Gly-Phe-Met-OH), названный мет-эндоке-«фалином, и небольшое количество его менее активного аналога, лей-энкефалина. Оба энкефалина воспроизводят спазмолитический эффект морфина в гладкомышечных препаратах кишечника морской свинки и семявыносящего протока мыши и ингибируют стереоспецифическое связывание опиатного рецептора с [3Н]-налоксоном (12.105) в гомогенатах мозга. Однако при введении через канюлю энкефалинов в мозг мышей (энкефалины не проходят через ГЭБ и за 10 мин гидролизуются ферментами мозга) анальгетический эффект более активного мет-эн-кефалина в 20 раз слабее, чем у морфина. Повторные инъекции опиатов вызывают развитие толерантности; при этом возникает и перекрестная толерантность к морфину [Hughes et al., 1975].
С помощью структурных аналогов энкефалинов установлено, что проявление их активности связано с наличием ОН- и NH-групп остатка тирозина и липофильной части остатка фенилаланина, которую можно заменить и другими липофильными группами [Gorin et al., 1980]. Швейцарские исследователи, поэтапно изменяя структуру мет-энкефалина, синтезировали -соединение «FK 33-824» при введении в желудочки мозга лабораторных животных в 1000 раз более активное, чем морфин [Roemer et al., 1977], но вызывающее анафилаксию у человека. Затем группа французских ученых синтезировала тиорфан [В,Ь-3-меркапто-2-бензилпропанолглицин (12.106)], дипептид, ингибирующий энкефалиназу (этот фермент гидролизует связь между Gly и Phe)n повышающий вследствие этого уровень эндогенных энкефалинов [Roques et al., 1980]. Предположение, что некоторые анальгетики влияют не на рецепторы энкефалинов, а на энкефалиназу, подкрепляется данными о блокировании налоксоном действия тиорфана.
Другой ряд эндогенных анальгетических полипептидов образуется при гидролизе липотропина в мозге [Guillemin, 1978]. Наиболее активный из них p-эндорфин содержит аминокислотную последовательность 61—91 молекулы липотропина. У крыс •он в 2 раза активнее морфина и вызывает более длительную аналгезию, очень быстро уменьшаемую налоксоном (12.105). Аминокислотная последовательность 61—65 в молекуле р-эн-дорфина повторяет последовательность мет-энкефалина, однако известно, что в организме энкефалины из эндорфинов не синтезируются.
Из гипофиза был выделен гептадека пептид динорфин — в 200 раз более активный анальгетик, чем морфин [Goldstein -et al., 1981]. Роль динорфинов, эндорфинов и энкефалинов в модуляции боли освещена в обзоре Terenius (1982). В настоящее время наметилась тенденция рассматривать энкефалины в качестве нейромедиаторов, а р-эндорфин— как гормон, подавляющий высвобождение других нейромедиаторов. Следует от-;256
метить, что в организме человека анальгетики начинают выделяться только при возникновении боли.
У млекопитающих различают несколько типов опиатных рецепторов. Их классификация построена главным образом на данных фармакологического анализа поведенческих эффектов опиатов; она может быть принята, по крайней мере, до установления структуры рецепторов. Вполне возможно, что в действительности типов рецепторов значительно меньше, так как разные типы могут представлять собой конформеры [Lord et al., 1977; Iwamoti, Martin, 1981].
Наиболее важны мю-рецепторы, через которые опосредуется анальгетическое действие морфина. Эндогенные энкефали-ны значительно в большей степени связываются с дельта-, а не мю-рецепторами. Тем не менее при замещении трех аминокислот в молекуле эндогенного энкефалина был синтезирован высокоактивный лиганд мю-рецепторов D-Ala2, Phe4, С1у-ол5-метэнкефалин. Ранее мю-рецепторы были идентифицированы с помощью меченного тритием дигидроморфина. Налоксон-—антагонист мю-рецепторов, а также сигма- и каппа-рецепторов. Пентазоцин, используемый в клинике в качестве анальгетика,— антагонист мю-рецепторов, но агонист каппа- и сигма-рецепторов. Спинальная анестезия и седативный эффект морфина обусловлены его связыванием с каппа-рецепторами.
Активация налорфином и пентазоцином сигма-рецепторов усиливает дыхание и вызывает галлюцинации. Специфическим агонистом данного типа рецепторов является N-аллилнормета-зоцин, однако их специфический антагонист неизвестен.
Дельта-рецептор, очевидно, расположен в непосредственной близости от мю-рецептора [Smith, Lee, Loh, 1983]. Бета-эндорфин в равной степени связывается с мю- и дельта-рецепторами, а энкефалины — в основном с дельта-рецепторами [Chang et al., 1981; Snyder, 1980], в большей степени опосредующими спазмолитические эффекты энкефалинов, а не их анальгетическую активность. Эторфин и D-Pen2, ПС-Реп5-энкефалин (где Реп — диметилцистеин)—специфические агонисты (последний используется и как специфическая метка для рецептора), а М,М-диаллил-Туг-О1у-С1у-(СН25)-РЬе-Реи(ОН) — специфический антагонист дельта-рецепторов [Shaw et al., 1982]. Для сравнительного анализа влияния опиатов на эти типы опиатных рецепторов целесообразно использовать препараты семявыво-дящего протока мышей (более сильная дельта- и слабая мю-активность) и тонкого кишечника морской свинки [Chang et al., 1981].
При гидролизе казеина молока высвобождается тетрапептид H2N-Tyr-Pro-Phe-Pro(OH), чья анальгетическая активность связана с влиянием на мю-рецепторы, а не на дельта-рецепторы [Chang et al., 1981]. Динорфин действует на каппа-рецепторы.
Действие частичного агониста и антагониста налорфина (7.35, б) описано в разд. 7.3.7.
17—734
257
Б. Опиаты, содержащиеся в пище. При расщеплении казеина образуется опиатный тетрапептид. С помощью точных современных аналитических методов доказано, что человек постоянно получает следовые количества морфина (до 500 нг/кг сырого веса), который содержится в салате-латуке, а также в материнском и коровьем молоке [Hazum et al., 1981]. В кофе присутствует в значительных количествах вещество непептидной природы с ОММ менее 3500, активно связывающееся с мю-рецепторами и являющееся их частичным агонистом или антагонистом [Boublik et al., 1983].
В. Толерантность и лекарственная зависимость. Интерес к этим явлениям появился после того, как было установлено, что
возникновение толерантности к опиатам у животных не со
провождается увеличением скорости их разрушения в организме [Klee, Strety, 1974]. Точно так же количество опиатных рецепторов и их сродство к опиатам не различаются у интактных и зависимых от морфина крыс. Исследование действия опиатов на культуру нервных клеток позволило предположить, что морфин ингибирует аденилатциклазу, причем после удаления морфина возникает гиперактивация фермента. В результате увеличивается содержание цАМФ, что хорошо видно в мозге крыс, проявляющих симптомы морфиновой абстиненции. Более того, инъекция цАМФ вследствие ингибирования соответствующей фосфодиэстеразы (проверено на гомогенатах мозга) вызывает симптомы морфиновой абстиненции у крыс, никогда не подвергавшихся воздействию морфина [Sharma, Klee, Nirenberg, 1975; Collier, Francis, 1975].
Субъективные и объективные симптомы отмены опиатов у животных с зависимостью от них быстро устраняются клониди-ном. Предполагают, что его эффекты связаны с блокадой альфа-адренорецепторов, количество которых в мозге крыс увеличивалось при хроническом введении морфина [Redmond, Kleber, 1980]. В некоторых клиниках клонидин используют как составной элемент общей программы лечения абстиненции.
HS—CH2-CH(CH2Ph)CO—NH-CH2CO2H
Ингибитор энкефалиназы
(12.106)
Налтрексон (12.107)
Впервые лечение привыкания к героину метадоном (7.38)» (перорально) было начато в США [Dole, Hyswander, Kreek,. 1966]. Оно эффективно устраняет эмоциональную неустойчивость, вызванную героином. Лечение метадоном можно прово
258
дить в амбулаторных условиях, ежедневно уменьшая дозу метадона (1 мг метадона на каждые 2 мг героина) на 20%. Лишь у небольшой части больных лечение метадоном неэффективно и требует применения налоксона (12.105) с последующим назначением более длительно действующего налтрексона (12.107). У многих больных может возникнуть рецидив длительностью до нескольких недель из-за таких побочных явлений, как бессонница, недомогание и усиление моторики кишечника. В этом случае следует вернуться к лечению метадоном и попытаться замедлить развитие абстиненции.
Лечение продолжают, применяя метадилацетат, имеющий период полусуществования 72 ч, тогда как у метадона он равен 20 ч [Ling, Klett, Gillis, 1978].
Хотя поиск сильных анальгетиков без эйфорических свойств (т. е. потенциально не вызывающих привыкания) для лечения описанной абстиненции развивается успешно, больным после операции требуются и такие анальгетики эйфорического типа, как морфин. Ключ к синтезу «чистого» анальгетика был найден несколько позже. Были синтезированы несколько чрезвычайно активных анальгетиков, не взаимодействующих, по-видимому, с опиатными рецепторами. Например, сильный анальгетик мептазинол (12.108) не вызывает эйфории и нарушает дыхание слабее, чем морфин и пентазоцин [Pearce, Robson, 1980]. Сильное анальгетическое действие нантрадола (12.109) не блокируется налоксоном. Этот препарат, производное фенантреди-на, проходил клинические испытания в качестве противорвот-ного средства при химиотерапии рака [Johnson, Milne, 1980], но широкого применения в этой области не нашел.
Мептазинол - (12.108)
Г. Разное. Вызванная акупунктурой аналгезия у мышей, предположительно связанная с высвобождением эндорфина, исчезает после инъекции налоксона [Mayer et al., 1977; Pomeranz, Ghiu, 1976]. У больных под влиянием внутримозговой электростимуляции, примененной для ослабления хронического болевого синдрома, отмечено повышение уровня циркулирующих опиатов [Terenius, Wahlstrom, 1978].
Хотя индукция сна не является основным эффектом опиатов, следует отметить выделенный из мозга кролика вызывающий сон нонапептид «dsip». Сходный эффект данного пептида, Trp-Ala-Gly-Gly-Asp-Ala-Ser-Gly-Glu-, подтвержден в исследо-
17*
259
ваниях на здоровых добровольцах на пяти различных моделях сна [Schoenenberger, Schneider-Helment, 1983].
О зависимости биологической активности опиатов от их химического строения см. Szekely (1982) и Kosterlitz, Collier, Villareal (1972).
12.9. Психотерапевтические агенты
К середине 1950-х годов стало ясно, что даже самые тяжелые психические заболевания значительно легче поддаются медикаментозному лечению, чем психотерапии. Благодаря лекарственным веществам многие больные смогли вернуться к полноценной жизни. Но понимание биохимической природы заболеваний не должно заслонять и важной роли в возникновении этих расстройств различных вредных факторов, действующих на человека в быту и на работе [Baldessarini, 1977].
Большинство психозов («сумасшествие» или «безумие») делятся на два типа: шизофрению и маниакально-депрессивный психоз (МДП). Шизофрения характеризуется нарушениями мышления, снижением эмоциональности и нередко слуховыми и зрительными галлюцинациями. Во время делирия больной впадает в буйство. При МДП сохраняются память и ориентация, но сильно страдают эмоциональная сфера, мышление и поведение. Это заболевание может быть монополярным (т. е. только депрессивным) или биполярным (с чередованием двух типов).
12.9.1. Нейролептики
Хотя эти препараты используют в качестве средств первой помощи при маниакальных приступах ( МДП), основной областью их применения является лечение шизофрении, считавшейся раньше наименее поддающимся лечению видом умопомешательства. До 1952 г. две трети госпитализированных больных шизофренией проводили в больницах более двух лет и лишь менее 10% возвращались к нормальной жизни. Это положение резко изменилось после того, как Delay, Deniker, Harl (1952) ввели в клиническую практику производное фенотиазина—аминазин (12.110). С его помощью можно купировать галлюцинации у больных, в связи с чем отпадает необходимость ограничения их самостоятельности. Это привело к резкому изменению атмосферы в психиатрических клиниках, так как стало возможным занимать больных разными несложными работами, например в саду. Но все-таки излечение не было полным, так как больные должны были постоянно принимать препарат и во многих случаях не могли вернуться к полноценной жизни.
Хотя аминазин обладает сильным успокаивающим действием, в отличие от депрессантов общего действия (гл. 15) при 260
его применении сознание остается ясным. Молекула аминазина неоднократно модифицировалась, в частности и для увеличения продолжительности его действия. Так, при инъекции 25 мг флуфеназина энантата, растворенного в масле, эффект продолжается от 2 до 3 нед. Флуфеназин 10-{3-[4-(2-гидроксиэтил) пиперазин-1-ил] пропил}-2-трифторметилфенотиазин является производным аминазина 10- (3-диметиламинопропил)-2-хлор-фенотиазин (12.110).
Аминазин
(12.110)
Хлорпротиксен
(12.111)
О
Г алоперидол
(12.112)
Тиоридазин
(12.113)
Такое нейролептическое (раньше называвшееся «транквилизирующим») действие позднее было найдено и у других классов химических соединений, особенно у тиоксантенов, например хлорпротиксен (12.111), и бутирофенонов, например галоперидол (12.112). Вначале из всех трех классов препаратов в наименьших дозах применяли производные бутирофенона, однако сейчас известны высокоактивные препараты и в двух классах: этаперазин, трифтазин, фторфеназин и тиотиксен. (Уменьшению дозы способствует замена легко метаболизирующей диметиламиногруппы пиперазином.)
В экспериментах на мозге мышей [Carlsson, Lundqvist, 1963; Seeman, 1977] и при обследованиях больных шизофренией [Horn, Snyder, 1971] было показано, что механизм действия нейролептиков заключается в блокаде дофаминовых рецепторов мозга. Дополнительные доказательства были получены с помощью флупентиксола, сходного по строению с хлорпротиксе-ном (12.111). Наличие двойной связи в положении 9 делает
261
возможным существование двух геометрических изомеров, из которых клинической эффективностью обладает лишь цис-изомер. Было показано, что in vitro дофаминовые рецепторы мозга тоже блокирует только цис-изомер [Crow, Johnstone, 1977]. Болезнь Паркинсона вызвана недостатком в мозге дофамина (о ее лечении см. разд. 3.6.5). При шизофрении, наоборот, нарушение деятельности мозга вызвано накоплением дофамина в переднем мозге, а возможно, и в других структурах. Поэтому неудивительно, что одним из побочных эффектов при лечении шизофрении нейролептиками является легкая форма паркинсонизма. Экстрапирамидные симптомы, включающие тремор и ригидность, исчезают при уменьшении дозы и назначении ан-тихолинергических препаратов, например, циклодола (7.24). При длительном применении нейролептиков примерно у 20% больных возникает «запоздалая дискинезия», часто проявляющаяся в непроизвольных повторяющихся подергиваниях лицевых мышц, учащающихся с возрастом. Это побочное действие реже проявляется при применении препаратов, используемых в малых дозах (см. выше); из них одними из лучших являются: из производных фенотиазина — тиоридазин (12.113), а из производных бутирофенона — тимиперон, проходящий в настоящее время клинические испытания. Некоторые надежды внушает и находящийся на клинических испытаниях сульпирид, 5 - аминосульфонил-2-метокси - N - [ (1-этилпирролидин-2-ил) метил] бензамид (12.62), по химическому строению сильно отличающийся от большинства нейролептиков [Bratfos, Haug, 1979].
Предполагают, что при шизофрении нарушаются и норадренергические системы [Mason, 1983], что подсказывает возможность других терапевтических подходов. Фенотиазины легко окисляются до свободных радикалов (красного цвета) [Borg, Cotzias, 1962], однако для нейролептического действия это не имеет значения.
Имизин Амитриптилин
(12.114) (12.115)
12.9.2. Антидепрессанты
В 1949 г. Cade показал, что с помощью солей лития можно контролировать маниакальную фазу МДП. Таким образом, эти соли оказались первыми эффективными терапевтическими агентами, даже более ранними, чем аминазин. В настоящее время применение солей лития является стандартным методом лечения (см. подробнее разд. 11.0).
262
Впервые контроль над депрессивной фазой установил Kuhn (1958), обнаруживший, что трициклическое соединение имизин (имипрамин) (12.114) эффективно при лечении больных с депрессивными состояниями. Эффект наступает через 1—2 нед. после начала лечения, продолжающегося обычно до 6 мес. Высокие дозы препарата противопоказаны больным с сердечной недостаточностью, которая может усилиться под действием препарата.
Высокоэффективными антидепрессантами и нейролептиками служат соединения, содержащие три конденсированных цикла, однако эти два свойства никогда не встречаются вместе у одного соединения. Исследования связи структура — активность [Wilhelm, Kuhn, 1970] показали, что тип действия зависит от топологии трициклического скелета, а детали его проявления — от боковых цепей.
Форма скелета (циклической системы) задается четырьмя стерическими параметрами: углом изгиба а, углом аннелиро-вания р, торсионным углом у и межатомным расстоянием б. Фармакологическое действие определяет в первую очередь угол изгиба. Относительно плоские молекулы типа фенотиазина и тиоксантена (а=25°) проявляют преимущественно свойства нейролептиков, тогда как соединения с такими относительно изогнутыми скелетами, как, например, дибензоциклогептадиен (55°), имидодибензил (55°) или дибензодиазепин (65°)—антидепрессанты (тимолептики). Так, например, имизин — это производное имидодибензила, а амитриптилин — дибензоциклогептадиена. Угол изгиба а — это угол, образованный плоскостями двух боковых ароматических циклов, спроецированных друг на друга. Угол аннелирования [3 представляет собой меру отклонения двух боковых ароматических циклов от прямой линии. Он равен 0° для дигидроантрацена, 60° — для дигидрофенантрена и 40° — для имидодибензила. Торсионный угол 7 — угол поворота плоскости одного бокового ароматического цикла относительно плоскости другого, возникающий из-за наличия центрального семичленного цикла. Для вышеупомянутых лекарственных соединений он изменяется в интервале от 0° до 20°.
Для того чтобы соединение обладало выраженными нейролептическими свойствами, его боковая цепь должна быть не-разветвленной и содержать три атома углерода. В тиоридазине (12.113) один из этих атомов углерода входит в состав пиперидинового цикла. Фенотиазиновое ядро входит и в молекулы некоторых антигистаминовых лекарственных веществ, например дипразина (пипольфен), однако для препаратов этого типа действия достаточно наличия боковой цепи лишь из двух атомов углерода. Антидепрессанты могут содержать боковую цепь как из двух, так и из трех атомов углерода. По данным рентгеноструктурного анализа боковая цепь должна быть такой, чтобы концевая основная группа могла располагаться рядом
263
с бензольным циклом (у нейролептиков) или перекрывать его (у антидепрессантов). Предполагают, что полости для этих веществ на рецепторах невелики по размеру.
Если больные плохо переносят такие побочные эффекты имизина и амитриптилина, как сухость во рту, вызванную их антимускариновым действием, то можно применять вторичные амины, имеющие в боковой цепи метиламиногруппу (например, дезипрамин и нортриптилин).
Ценным свойством трициклических антидепрессантов является, по-видимому, их способность повышать уровень норадреналина и 5-гидрокситриптамина. Тщательное изучение как их про- и анти-5-гидрокситриптаминовых эффектов, так и про- и анти-норадреналиновых эффектов проводилось на разных стадиях лечения и в различных структурах мозга [Кое, 1983; Fillion, Rillion, 1981].
Величина рКа трициклических антидепрессантов обычно лежит в интервале 9,5—10, а коэффициент распределения в системе октанол — вода равен 4. Улучшение их терапевтических свойств с целью получения соединений, не действующих на сердце, шло по двум направлениям. Сначала появились такие тетрациклические антидепрессанты, как мапротилин (12.116) и миансерин (12.117), затем антидепрессанты с более простым ядром типа номифензина (12.118). Возможно, что эти новые препараты по биохимическому механизму действия отличаются от имизина и амитриптилина и обладают преимуществами, за исключением, пожалуй, лишь их применения для лечения кардиологических больных.
Мапротилин (12J16)
Миансерин
Номифензин
(12.117) (12.118)
К больным, на которых не оказывают действия трициклические антидепрессанты, возможно применение двух альтернативных терапевтических подходов, связанных с повышением уровня нейромедиаторных аминов в мозге. Высокую эффективность проявляют ингибиторы МАО (разд. 9.4), однако их применение требует ограничений в диете и, кроме того, они могут вызывать понижение АД. Стойкого улучшения состояния больного можно достичь, применяя электрошоковую терапию, практически безболезненную для больных [Green, Heale, Grahame-Smith, 1977].
264
12.10. Заключение
Изучение пространственного строения молекул биологически активных веществ очень важно для понимания связи структура — активность, а также дает возможность исследовать структуру различных рецепторов (некоторые из них обсуждались в этой главе). Аналогичные методы были использованы и при изучении других рецепторов, например рецепторов инсулина и тироксина1. Безусловно, интерес к таким исследованиям будет возрастать, поскольку будут возрастать возможности установления механизма действия агонистов.
Изучение природы рецепторов часто начинают с изучения связывания меченых соединений, сходных по химической структуре с соответствующим лекарственным веществом. Такие исследования часто дают неверные результаты из-за наличия «мест потерь», также эффективно связывающих лиганд. Тщательная очистка фракций, связывающих лиганд, еще не гарантирует действительного выделения рецептора. Чтобы быть уверенным в том, что мы имеем дело с рецептором, необходимо подтвердить и сохранность других, характеристичных для него свойств. Необходимо показать, что с ним связываются и другие соответствующие агонисты и антагонисты (причем так же, как и с нативным рецептором), продемонстрировать влияние различных ионов на связывание и определить другие кинетические характеристики данного рецептора.
Пока не получены новые точные данные о строении рецептора (вероятно, это возможно только после выделения чистого рецептора), можно использовать имеющиеся в литературе старые предположительные схемы строения рецептора (типа приведенной на рис. 12.3), подборку таких данных см. Ehrenpreis, Fleisch, Mittag (1969).
Более подробно о влиянии стерических факторов на биологическую активность см. Bentley (1969).
1 Автор рассматривает далеко не все рецепторы, известные в настоящее время. В частности, совершенно не упоминаются привлекающие большое внимание рецепторы аминокислот. Нейрофармакологическим и нейрохимическим аспектам действия этих соединений посвящена монография: К. С. Раевский, В. П. Георгиев. Медиаторные аминокислоты. М„ Медицина, 1986, 239 с.; связь структура — активность в ряду возбуждающих аминокислот рассмотрена в обзоре: Л. Б. Пиотровский. Хим.-фарм. журя, 1987, № 7, с. 773—782. — При-меч. ред.
Глава 13
КОВАЛЕНТНАЯ СВЯЗЬ И ИЗБИРАТЕЛЬНАЯ ТОКСИЧНОСТЬ
Большинство биологически активных соединений образуют с рецепторами непрочные связи, легко разрывающиеся при отмывании. Однако существует немногочисленная группа агентов, связывающихся с рецепторами более прочно ковалентными связями (разд. 8.0). Эти связи между атомами углерода разрываются только под действием химических соединений с высокой реакционной способностью или под воздействием высоких температур но, как правило, устойчивы к воздействию мягких реагентов в физиологическом интервале температур.
Значительно легче разрываются ковалентные связи между неуглеродными атомами, хотя для этого необходим большой избыток реагента (разд. 12.0, 12.3).
Разрыв ковалентных связей уже рассматривался в разд. 3.6 при обсуждении превращений пролекарств в истинные активные агенты. Кроме того, образование и разрыв множества ковалентных связей происходят в клетках при участии ферментов в ходе нормальных метаболических процессов или при метаболизме ксенобиотиков (разд. 3.5). В этой главе речь пойдет только об образовании ковалентных связей между агентом и комплементарной ему группой атомов рецептора.
13.0. Препараты мышьяка, сурьмы и ртути
Из всех лекарственных средств именно для препаратов мышьяка впервые было установлено, что их биологическое действие обусловлено образованием ковалентных связей. Еще в 1909 г. Эрлих высказал предположение о том, что некоторые производные мышьяка взаимодействуют с важными меркаптогруппами (сульфгидрильными, тиольными, SH-группами) в организме паразита. Он пришел к этому выводу на основании того факта, что арсеноксиды чрезвычайно легко взаимодействуют с сульфгидрильными группами, даже в присутствии большого избытка воды (см. формулу 13.1). Более того, в биологических системах производные мышьяка могут вступать только в эту реакцию. Для мышьяка возможны три уровня окисления: (13.2), (6.4) и (6.3), валентность его при этом равна пяти (13.2) и трем (6.4) и (6.3) (арсеноксид и арсенобензол соответственно) . Несмотря на то что мышьяк трехвалентен в двух последних
266
Рис. 13.1. Паразитоцидное действие (А) арсфенамииа (сальварсана), (Б) оксофенарсина (мафарзида) и (В) смеси оксофенарсина с восстановленным глютатионом на крыс, инфицированных трипаносомами [Voegtlin, 19251.
соединениях, уровень его окисления в арсенобензоле ниже, чем в арсеноксиде. По данным рентгеноструктурного анализа арсенобензолы в твердом состоянии существуют в виде тримеров [Hedberg, Huges, Waser, 1961], а эбулиоскопические измерения указывают на то, что бензоларсеноксиды ассоциированы в тетрамеры [Blicke, Smith, 1930].
Эрлих установил, что соединения пятивалентного мышьяка приобретают биологическую активность только после восстановления до арсеноксида, но то, что и арсенобензол для проявления биологической активности должен окислиться до арсеноксида, было показано значительно позже [Voegtlin, 1925]. Voegtlin
HS—СН2—
HS—СН2—
.S—СН2— —
^S—СН2—
он
НО I NH—СОМе
Ацетарсол (13.2)
Соединение препаратов мышьяка с меркаптанами
(13.1)
показал, что терапевтической активностью обладают только производные арсеноксидов. Ему удалось показать наличие меркаптогрупп в трипаносомах. На рис. 13.1 видно, что в присутствии оксофенарсина (6.4) содержание трипаносом в крови крыс резко снижается через 0,5 ч после введения препарата, тогда как при введении соответствующего ему арсенобензола арсфе-намина (6.3) подобный эффект достигается более чем через 5 ч. Это свидетельствует о том, что арсфенамин приобретает активность только после превращения в активное соединение. Если арсфенамин перед введением окислить в арсеноксид (оксофенарсин), пропуская воздух через его раствор, то обнаруживается характерное для арсеноксида быстрое действие. То же самое происходит при инкубации арсфенамииа в присутствии тканей, способных его окислять, например с тканью печени.
Из рис. 13.1 также видно, что соединения, содержащие сульфгидрильные группы, например глутатион, цистеин или натриевую соль тиогликолевой кислоты, действуют как антагони-
267
сты арсеноксидов. Этот эффект кратковремен, если количество тиола невелико, так как продукт взаимодействия арсеноксида с меркаптопроизводными легко гидролизуется с образованием исходных компонентов. В присутствии избытка тиольного производного эффект более продолжителен (в соответствии с законом действия масс).
Связывание арсеноксидов с тиольными группами паразитов обратимо, поэтому если на паразитов подействовать несколькими летальными дозами препаратов мышьяка, а затем избытком соединений, содержащих тиольные группы, то они могут выжить.
В настоящее время считают, что быстрая гибель паразита (в течение 30 мин после введения препарата) наступает из-за нарушения процессов дыхания или гликолиза, а не вследствие нарушения синтеза нуклеиновых кислот или белков [Macadam, Williamson, 1974]. В состав каких соединений в организме паразита входят жизненно важные SH-группы? Предполагалось, что это тиольные группы глутатиона или белков [Rosenthal, Voegtlin, 1930]. В настоящее время считают, что блокируются SH-группы ферментов, причем показано, что арсеноксиды могут ингибировать очищенные ферменты, причем в этом случае различные тиолы проявляют защитный эффект.
Наиболее чувствительны к действию мышьяксодержащих препаратов следующие ферменты: 1) дегидрогеназа липоевой кислоты и 2) а-оксидазы (например, пируватоксидаза), для которых дигидролипоевая кислота (2.28) (тиоктовая кислота) служит коферментом. В разных цепях молекулы дегидрогеназы липоевой кислоты присутствуют два остатка цистеина, сближающиеся при образовании третичной структуры [Massay, Hofmann, Palmer, 1962]. Мышьяксодержащие соединения могут блокировать SH-группы этих остатков цистеина (см. формулу 13.1), а молекулы липоевой (6,8-димеркаптокаприловой) кислоты легко сшиваются мостиком из атомов мышьяка, как это показано на формуле (13.1) [Peters, 1963].
Чем объясняется избирательность поражающего действия препаратов мышьяка на паразита? Во введении к разд. 3 уже упоминалось о том, что клетки паразитов содержат относительно больше свободных SH-групп, чем клетки организма-хозяина. Кроме того, было обнаружено, что некоторые ферменты трипаносом, например фосфопируваткиназа, крайне чувствительны к действию препаратов мышьяка, тогда как на аналогичные ферменты млекопитающих эти соединения действуют значительно слабее [Grant, Sargent, 1960]. Избирательность действия в большой степени определяется наличием ароматической структуры в молекуле лекарственного соединения и повышается при введении заместителей в ароматическое кольцо (6.4). Неорганические и алифатические соединеиня мышьяка не менее токсичны для трипаносом, чем ароматические, но избирательностью действия обладают только последние. Этот вывод был подтвержден
268
исследованиями устойчивости паразитов к лекарственным веществам (см. разд. 6.5), связанной с избирательной адсорбцией.
А. Механизм действия препаратов мышьяка при лечении сифилиса. Работали Rosenthal (1932) и Eagle (1938) было показано, что препараты мышьяка действуют на спирохеты по тому же механизму, как и на трипаносомы. Независимо от степени окисления мышьяка, входящего в молекулу лекарственного препарата, связывание вещества с сульфгидрильными группами паразита происходит только после превращения в соответствующий арсеноксид. О зависимости между строением и избирательностью действия препаратов мышьяка, применяемых для лечения сифилиса, см. обзор Eagle (1951).
Эрлих считал, что арсеноксиды слишком токсичны для человека и поэтому в течение четверти века для лечения сифилиса применяли необоснованно высокие дозы таких препаратов мышьяка, как арсфенамин и неоарсфенамин. При введении этих огромных доз лишь небольшая часть препаратов в организме превращается в истинно действующий арсеноксид, а основное количество вступает в побочные реакции, ведущие к образованию токсичных соединений. Применение для лечения сифилиса оксофенарсина — арсеноксида, структурного аналога арсфенами-на, неизмеримо более токсичного для паразита, чем для хозяина, дало возможность снизить используемые дозы [Tatum, Cooper, 1934]. В настоящее время для лечения сифилйса вместо препаратов мышьяка применяют пенициллин.
Б. Химическое действие антидотов, содержащих SH-группы. Связь атомов As—S так легко разрывается под действием меркаптосоединений, так как атом мышьяка в отличие от атомов азота, углерода и кислорода имеет вакантную d-орбиталь, на которую может переходить неподеленная пара электронов от атома серы цистеина. Это приводит к образованию переходного комплекса более высокой валентности, где три атома серы связаны с одним атомом мышьяка (два атома принадлежат рецепторному белку, а третий — цистеину). Такой переходный комплекс нестабилен и вероятность разрыва всех связей As—S одинакова, следовательно, в одном случае из трех вновь образованная связь сохранится. Если цистеин находится в избытке, то количество мышьяксодержащего препарата, связанного с рецептором, в соответствии с законом действия масс окажется незначительным. Аналогичная ситуация создается при гидролизе тетрахлорида кремния, быстро разрушающегося под действием воды, что объясняется наличием свободной d-орбитали у атома кремния. У атома углерода вакантной d-орбитали нет и в противоположность тетрахлориду кремния четыреххлористый углерод представляет собой очень устойчивое соединение.
В некоторых соединениях существуют еще более прочные связи As—S. Люизит (С1СН = СН—AsCU)—самое сильное боевое отравляющее вещество нарывного действия из всех соедине-
269
Таблица 13.1. Действие препаратов мышьяка на бактерии (Предельные разбавления, при которых еще наблюдается полное ингибирование роста в течение 48 ч при 37 °C). Среда: бульон с пептоном, содержащий 10% сыворотки, pH 7,2 [Albert, Falko, Rubbo, 1944}
Соединения Микрооргани эмы
С1. welch И Strept. haem. А. Staph, aureus E. coli Proteus
Ацетарсол (З-ацетамидо-4-гидроксифениларсеновая кислота) (13.2) * * sis * *
Оксофенарсин (З-амино-4-гидроксифениларсено-ксид) (6.4) 1:160 000 1 :80 000 1 :160 000 1:10 000 1:10 000
Неоарсфенамин (З-амино-4-гидроксиарсенобензол-N-метиленсульфоксилат) 1:10 000 1:10 000 1:10 000 * ♦
Оксофенарсин в бульоне, содержащем 0,1% тио-гликолята 1:5000 1 :10 000 * *
Хлористая ртуть (контроль) 1:40 000 1:160 000 1 :40 000 1:80 000 1:80 000
* — отсутствие ингибирующей активности даже при концентрации 1 : 5000.
ний мышьяка, вызывает в организме человека необратимые повреждения, не снимающиеся ни одним монотиолом. Установлено, что поражающее действие люизита на кожу и быстро развивающееся токсическое действие обусловлены его способностью блокировать SH-группы пируватоксидазы [Peters, 1963]. Поражения, вызванные люизитом, можно полностью снять с помощью дитиольных соединений, у которых SH-группы расположены в непосредственной близости друг от друга [Peters, Stocken, Thompson, 1945], т. е. даже очень прочная связь мышьяка с SH-группами может быть разрушена с помощью высокоактивного антидота. Преимущество дитиолов состоит в том, что если в молекуле временно разрывается одна из связей, например под действием теплоты, то другая сохраняется. Лучший антидот из ряда дитиолов 2,3-димеркаптопропанол (11.23) (уни-тиол) служит прекрасным противоядием при отравлениях мышьяком (разд. 11.6).
В. Препараты мышьяка, обладающие антибактериальной активностью. До 1944 г. считалось, что препараты мышьяка не действуют на обычные патогенные бактерии. Это неверное представление возникло из-за того, что в биологических тестах использовали соединения мышьяка с несоответствующей степенью окисления. Проверка соединений, содержащих мышьяк нужной степени окисления, позволила установить, что они проявляют выраженную антибактериальную активность [Albert, Falk, Rubbo, 1944]. Данные, приведенные в табл. 13.1, свидетельствуют о том, что бактериостатической активностью обла
270
дают производные арсеноксидов. Микроорганизмы способны в незначительной степени превращать производные арсенобензола (но не соединения пятивалентного мышьяка) в соответствующие арсенокси'ды, но к производным пятивалентного мышьяка это не относится. Показано также, что антибактериальная активность арсеноксидов подавляется тиогликолятом натрия.
Г. Препараты мышьяка в медицине. Вскоре после создания Эрлихом в 1910 г. арсфенамина применение препаратов мышьяка достигло максимума. Создание этого лекарственного средства явилось большим достижением химиотерапии не только потому, что было найдено средство для лечения сифилиса, ранее считавшегося неизлечимым. Арсфенамин был практически первым лекарственным средством для лечения бактериальных инфекций и оставался единственным вплоть до 1935 г., когда Домагк открыл сульфаниламиды. Однако гораздо важнее то, что это дало основание Эрлиху говорить о химической природе взаимодействия лекарственного вещества с рецептором. Правда, он основывался лишь на одном факте — образовании связи между атомами серы и мышьяка, и в течение многих лет эта общепринятая в настоящее время концепция опиралась только на этот единственный пример.
После того как в 1943 г. для лечения сифилиса стали применять пенициллин, препараты мышьяка использовали все реже, и в настоящее время их применяют только для лечения африканского трипаносомоза, при котором поражается мозг. Для лечения этого заболевания наиболее эффективен меларсопрол (13.3), «замаскированный арсеноксид», в отличие от обычных трипаноцидных лекарственных средств легко проникающий в ЦНС [Friedheim, 1951]. В молекуле этого лекарственного препарата соединены структуры арсеноксида и 1,2-димеркаптоэта-на; он легко проходит через ГЭБ и проникает в мозг, где расщепляется с образованием арсеноксида. Меларсопрол оказывает быстрое лечебное действие и не обладает опасным токсическим действием на глаза, что отличает его от применявшегося ранее трипарсамида (6.7).
Меларсопрол
(13.3)
Д. Препараты сурьмы. Предполагают, что в основе биологического действия препаратов сурьмы лежит взаимодействие с жизненно важными SH-группами. (Реакционноспособным является трехвалентное состояние, до которого сурьма может восстанавливаться в организме.) Однако твердых доказательств этой гипотезы пока нет. Установлено, что взаимодействие препаратов трехвалетной сурьмы с фосфофруктокиназой шистосом
271
не блокируется меркаптоэтанолом [Mansour, Bending, 1954]» но действие более сильного реагента унитиола не исследовалось. О влиянии препаратов сурьмы на метаболизм углеводов см. табл. 4.6 (разд. 4.4, том 1), о лечении ими лейцшаниоза и шистосомоза— разд. 6.3.3 и 6.3.5 соответственно. /
Г. Препараты ртути. Противобактериальнрб действие таких соединений ртути, как хлористая ртуть (Сулема) и нитрат фенилртути, обусловлено их способностью связываться с жизненно важными SH-группами бактерий [Fildes, 1940]. Однако действие этих препаратов обращается тиолами типа тиоглико-левой кислоты и даже сероводородом [Chick, 1908].
В течение долгих лет соединения ртути служили наиболее сильнодействующими диуретиками, но после создания в 1958 г. хлортиазида, а вслед за ним целого ряда «чисто органических» диуретиков, гораздо менее токсичных, чем препараты ртути» последние более не применяют в медицине. Все органические препараты ртути, обладающие мочегонным действием, расщепляются под действием кислой среды в почках с образованием небольшого количества ионов ртути. Эти ионы блокируют основные меркаптогруппы ферментов почек, ответственных за резорбцию хлористого натрия.
13.1. Пенициллины
Бактерии резко отличаются от всех живых организмов необычным химическим составом клеточных стенок (разд. 5.3). Самый характерный компонент бактериальной стенки ацетил-мурамовая кислота (5.9) связана с полипептидами типа (5.10). Пенициллин действует на бактерии, блокируя включение ацетилмурамовых пептидов в строящиеся клеточные стенки [Rogers, Mandelstam, 1962]. Для подавления этого синтеза у бактерий требуются концентрации лекарственных веществ, в 2—3 раза меньшие, чем для ингибирования роста этих же бактерий, причем как грамотрицательных, так и грамположитель-ных.
RCO-HN-НС---------Iх \/Ме
СО2Н HN-----[ Ме
СО2Н
а) 6-Аминопенициллановая нислота (R=H) Пеницилл овые нислоты
б)Бензилпенициллин (RsCeHsCHjCO —) (13.5)
(13.4)
Для бактерий характерно необычайно высокое внутреннее давление. Пенициллин поражает бактерии в фазе роста, ослабляя их клеточные стенки, которые не выдерживают высокого давления и разрываются. Однако разрыва клеточных стенок бактерий не происходит, если их поместить в среду с высоким осмотическим давлением (например, в 0,3 М раствор сахаро
272
зы). Механизм лизиса бактерий детально изучен на Е.соК [Lederberg, 1957] и подтвержден на других микроорганизмах. Разрушению^ клеточных стенок бактерий под действием пенициллина поможет, по-видимому, и фермент мукопептидаза, имеющийся почти у всех видов бактерий. Если биосинтез этого фермента подавлен левомицетином, эффективность пенициллина оказывается заметно сниженной [Rogers, 1967].
Циклосерин (5.11) (разд. 9.4.3), ванкомицин и бацитрацин^ а также многие другие химические соединения, структурно близкие пенициллину и вмешивающиеся в процессы синтеза компонентов клеточных оболочек, рассматриваются в разд. 13.2.
Целый ряд пенициллинов можно выделить из культуральной среды, в которой выращивается активный штамм Penicillum notatum (разд. 6.3.1). Все они имеют общую структуру (13.4, а) и отличаются только строением боковой цепи. Наиболее распространенный из пенициллинов — «бензилпенициллин», под названием пенициллин подразумевают обычно именно его (5.13; 13.4,6). По стерическому строению пенициллин (3S), (5R), (6R) —изомер [Pitt, 1952; McGregor, 1974].
Первые работы с пенициллином были осложнены легкостью его гидролиза под действием щелочей, двухвалентных ионов меди или ^-лактамазы (обычно называемой пенициллазой) с образованием пеницилловой (13.5), пенициллановой кислот и пеницилламина (11.26). Биосинтез пенициллина в природе осуществляется конденсацией 6-пеницилламина, D-валина и фенилуксусной кислоты. Благодаря наличию в молекуле четырехчленного лактамного кольца пенициллин представляет собой активный ацилирующий агент специфического действия. Лактамный цикл легко размыкается по связи С7-—Ni [Johnstone, Woodward, Robinson, 1949]. По данным рентгеноструктурного анализа установлено, что два цикла в молекуле пенициллина расположены почти под прямым углом друг к другу. Тем не менее в настоящее время известно, что напряженность скелета и отсутствие сопряжения в молекулах пенициллина и цефалоспорина ранее переоценивались [Proctor, Gensmamtel, Page, 1982].
Изучение инактивации бензилпенициллина показало, что он преимущественно ацилирует [3-аминомеркаптаны, в меньшей степени другие амины и не ацилирует совсем другие меркаптаны. На формуле (13.6) представлен продукт ацилирования ^-меркаптоэтиламина по аминогруппе. Роль меркаптогруппы, по-видимому, состоит в образовании водородной связи с атомом кислорода лактамного кольца и соответственным увеличением поляризации С=О связи в пенициллине.
Эксперименты с меченым 35Б-пенициллином показали, что устойчивые штаммы стафилококков (даже не продуцирующие пенициллазу), не поглощают пенициллин из растворов, тогда как чувствительные штаммы связывают от 200 до 750 молекул пенициллина на каждую клетку. Эта связь очень прочна: пеницил
18—734
273
лин не отмывается и не обменивается с немеченным пенициллином [Rowley et al., 1950; Maass, Johnson, 1949]. Первые молекулы пенициллина связываются инертными веществами, «о при насыщении образуется ковалентная связь пенициллина с рецептором, играющим ключевую роль в биосинтезе клеточной стенки бактерий. Клетки млекопитающих не поглощают пенициллин, тогда как бактериальные клетки связываются с ним менее чем за 2 мин. Компонент бактериальной клетки, связывающий пенициллин, находится, по-видимому, на цитоплазматической мембране в месте синтеза клеточной оболочки, и обнаруживается во фракции, содержащей фосфолипиды [Cooper, 1956].
S Me Н/ \ I /Ме R_c—N—сн—С С
II Н о
HS—(СНФ,—\н—с
II о
Продукт реакции Р-меркаптоэтамина с пенициллином
(13.6)
Пенициллин действует главным образом на мембранно-связанную транспептидазу цитоплазматической мембраны бактерий. Сначала фермент связывается с терминальной D-аланил-D-аланиновой группировкой мономера, при полимеризации которого образуется новая клеточная оболочка. Этот мономер представляет собой пептидогликан, состоящий из субъединиц муропептида (разд. 5.3), связанных углеводными компонентами. D-Аланин играет роль опознавательного сигнала для фермента. Следующий шаг — ацилирование фермента пептидогликаном, сопровождающееся потерей одной молекулы D-аланина:
Фермент R-D-Ala-D-Ala -----> R-D-Ala-фермент J- D-Ala
Некоторые факты свидетельствуют о том, что реакция ацилирования происходит по —ОН — группе остатка серина.
Затем транспептидаза регенерируется, связывая концевой остаток D-аланина молекулы пептидогликана с цепочкой из четырех молекул глицина (соединенных с остатком лизина), второй молекулы пептидогликана. В разных видах бактерий встречаются различные варианты образования этой связи, но в общем виде этот процесс может быть представлен следующим образом: [Franklin, Snow, 1981; Gale et al., 1982].
R-D-Ala-фермент J-Gly ---->- R-D-Ala-Gly + фермент
Гликопептидтранспептидаза, выделенная в чистом виде из Е. coli, необратимо ингибируется пенициллином в концентрациях, летальных для интактных бактериальных клеток [Izaki, Matsuhashi, Strominger, 1966]. Предполагают, что сначала пенициллин связывается с транспептидазным сайтом фермента и необратимо ингибирует его, используя в качестве ацилирующего
274
агента рХцактамный цикл антибиотика [Izaki, Matsuhashi, Strominger, 19^8]. Такой механизм действия пенициллина представляется наиболее вероятным и подтверждается данными рентгеноструктурного анализа В-аланил-В-аланинкарбокситранспеп-тидазы, выделёцной из Streptomyces-R 61 в присутствии или без-орто-йодофенилпенициллина, и в опытах на модельном субстрате а-трет-бутоксикарбонил-е-трифторацетил-Ь-лизил-В-глута-мил-В-аланине. Оба субстрата связываются с одним активным участком на ферменте, при этом их карбоксильные группы вступают в реакцию с концевыми аминогруппами двух спиралей фермента [Kelly et al., 1982].
Сравнение молекулярных моделей пенициллина и В-аланил-В-аланина свидетельствуют о том, что пенициллин можно считать антиметаболитом этого дипептида [Tipper, Strominger, 1965]. Так, расстояние между N' и N" в обеих молекулах [см. проекции молекулярных моделей (13.7) и (13.8)] одинаково и равно 0,33 нм. Одинаково у них и расстояние между N" и С1, а именно 0,25 нм. Расстояние от N' до С1 составляет 0,54 нм для пенициллина и 0,57 нм для дипептида. В-конфигурация пенициллина обеспечивается наличием атома серы, связанного с атомом углерода, общим для обоих циклов в молекуле. Эпимеризация по положению 6 (см. 13.4) приводит к полной потере антибактериальной активности. Наиболее реакционноспособная ([1-лактамная) связь в молекуле пенициллина расположена соответственно пептидной связи, соединяющей два В-аланиновых остатка. Однако между этими связями существуют и различия: связь между остатками аланина на 25% длиннее. Угол C-CO-N в В-А1а-В-А1а равен 117°, а в пенициллине только 91°. Более того, двугранный угол, образованный ^-лактамным циклом и лактамной связью, почти равен 135°, что значительно меньше угла пептидной связи (180°) и резко отличается от тетраэдрического переходного комплекса (разд. 9.5), который В-аланин предположительно образует с ферментом [Lee, 1971]. Еще более серьезным аргументом против вышеописанной структурной аналогии служит отсутствие противобактериальной активности у 6-метилбензилпенициллина, по химическому строению еще более близкого В-А1а-В-А1а [Boehme et al., 1971].
Ме2
I I I
О О Me
Пенициллины (проекция молекулярных моделей) (Ацил)-В-алании-Р-аланин
(13.7) (13.8)
18*
275
А. Модифицированные пенициллины. Бензилпенициллин до настоящего времени является антибиотиком, наиболее широко используемым в медицинской практике, особенно для инъекций, тогда как пенициллин V, т. е. феноксиметилпешфиллин (13.4; R = C6H5OCH2CO—), чаще применяют перорально. В 1957 г. из культуральной жидкости Penicillum chrysogendm [Batchelor et al., 1961] была выделена 6-аминопенициллановая кислота (6-АРА), послужившая исходным соединением для создания целой серии полусинтетических аналогов пенициллина. Многие из них представляют собой ацилированные производные 6-АРА (ср. с бензилпенициллином 13.4,6), содержащие стерические затрудненные заместители в метиленовой (СН2) группе. Эти соединения, менее устойчивые, чем бензилпенициллин, к кислому гидролизу, которому они подвергаются в желудке, явились предшественниками современных пенициллинов. Целью модификаций молекулы пенициллина было не только создание соединений, устойчивых к гидролизу под действием ^-лактамазы, присутствующей в резистентных типах бактерий, но и расширение области терапевтического действия пенициллинов созданием соединений, действующих не только на грамположительные, но п на грамотрицательные бактерии, а затем создание веществ, обладающих одновременно хотя бы двумя из вышеуказанных свойств. Ниже будут приведены примеры модифицированных пенициллинов, наиболее часто употребляющихся в медицинской практике.
Устойчивые к кислому гидролизу и легко инактивируемые ^-лактамазой амоксициллин (13.9, а) и ампициллин (13.9,6) подобно пенициллину прекрасно всасываются при пероральном введении, особенно ампициллин. Введение основной группы в молекулу обеспечивает ряду амоксициллинов широкий спектр активности, и они значительно превосходят традиционные бензилпенициллин и пенициллин V, действуя на многие виды грам-отрицательных микроорганизмов. Однако в связи с отрицательным побочным действием на флору кишечника в некоторых случаях (когда заражение вызвано не грамотрицательными бактериями) их применять не следует.
/X
CH(NHa)CO- у —-J-CO—
у о
а) Амоксициллин (Х=НО—) в) Оксациллин (X = Y = H)
б) Ампициллин (Х-=Н—) б) Клоксациллин (X = H; Y = C1)
в) Диклоксациллин (X = Y=C1)
Включены в качестве заместителя Включены в качестве заместителя (R—) в (13.4, a) (R—) в (13.4, а)
(13.9) (13.10)
В противоположность этому ряду пенициллинов группа ме-тациллина обладает узким спектром действия и эффективна 276
при инфёкциях, вызванных Staphylococcus aureus, содержащих р-лактамайу и, следовательно, устойчивых к действию бензилпенициллина. Необходимость в таких антибиотиках связана с появлением в больницах резистентных микроорганизмов, являю-
О-СН-С0- ,-----—СН—СО—
со2н со2-<^
Карбенициллин Тикарциллин
(13.11) (13.12)
EIn/ СО—NH—CH(Ph)— СО—
УЧ о о
Пиперациллин
(13.13)
Включены в качестве заместителя (R—) в (13.4, а)
щихся причиной контагиозных нозокомиальных инфекционных заболеваний. Первый представитель этого ряда метициллин, 6-(2,6-диметоксибензамидо)пенициллановая кислота, легко гидролизуется в желудке. Поэтому его заменили оксациллины (13.10), проявившие высокую эффективность при пероральном введении. Наиболее сильным и эффективным антибиотиком из группы оксациллинов является диклоксациллин (13.10,в), однако по действию на нерезистентные виды стафилококков даже он не может сравниться с бензпенициллином. Низкая всасываемость нафциллина, еще одного представителя этой группы, ограничивает его применение.
Существуют типы Staphylococcus aureus, резистентность которых обусловлена иными факторами, возможно, изменениями в составе клеточной стенки. Эти линии стафилококков, часто встречающиеся в больницах, устойчивы также и к действию цефалоспоринов, эритромицина, аминогликозидов и тетрациклинов. Эффективен против них лишь довольно неизбирательный антибиотик ванкомицин (разд. 13.2), но обычно можно применять и такие синтетические противобактериальные препараты, как нитрофурантоин и триметоприм.
Методами рентгеноструктурного анализа, ЯМР- и ИК-спект-роскопии установлено [Blanpain et al., 1980], что устойчивость антибиотиков группы метициллина к биодеградации связана в первую очередь с ригидностью боковой цепи, обусловленной силами ван-дер-ваальсовых взаимодействий, и не дающей возможности сближения ^-лактамного цикла лекарственного вещества и активного центра фермента. В молекулах оксациллинов существует дополнительное сопряжение между амидным, изоксазольным и фенильным заместителями.
277
Антибиотики группы карбенициллина чрезвычайно эффективны против грамотрицательных микроорганизмов, цключая нечувствительные к действию антибиотиков группы амоксициллина, а именно Pseudomonas, Enterobacter, Proteus и Klebsiella, широко распространенные относительно безвредные микроорганизмы, опасные для больных, чья иммунная система частично подавлена (например, после прививок). Препараты группы карбенициллина не активны при пероральном применении, легко разрушаются ^-лактамазой и не действуют на некоторые распространенные грамположительные бактерии, такие как Staphylococcus aureus.
Однако для больных они зачастую спасительны. Этерификацией карбоксильной группы карбенициллина (13.11) получают лекарственные вещества, эффективные при пероральном применении. Наиболее активные представители этой группы антибиотиков— тикарциллин (13.12) и пиперациллин (13.13).
Таким образом, создание полусинтетических антибиотиков значительно расширило область терапевтического применения пенициллинов, ограниченное, однако, их высокой стоимостью. Для уменьшения выведения пенициллинов через почки препараты группы амоксициллина назначают перорально вместе с пробенецидом (3.16).
В структуре молекул всех применяющихся в клинике пенициллинов рядом с ацильной группой в положении 7 находится ароматический цикл. Наличие аналогичного фрагмента в цефалоспоринах позволяет предположить существование плоского участка в структуре рецептора, с которым они взаимодействуют.
В последнее время практикуется совместное применение пенициллина с клавулановой кислотой (13.14), выделенной из Streptomyces clavuligerus. В структуру этого соединения, обладающего слабой противобактериальной активностью, входит [3-лактамный цикл, что обусловливает ее способность сильно ингибировать ^-лактамазу. Совместное введение ампициллина или амоксициллина и клавулановой кислоты дает ярко выраженный синергический эффект.
Пролекарства, превращающиеся в пенициллины, рассмотрены в разд. 3.6.
Более подробно о ^-лактамных антибиотиках см. Morin, Corman (1982).
Нлавулановая кислота
(13.14)
278
13.2. Цефалоспорины и другие р-лактамные ингибиторы образования клеточных стенок
В 1948 г. Brotzu обнаружил антибиотические свойства плесени Cephalosporium acremonium, активным компонентом которой оказался цефалоспорин С [структура представлена формулой (13.15), где X — аминоаципиновая кислота и Y—ацетоксигруппа (—ОСОМе)] [Newton, Abraham, 1956]. Основу молекулы представляет собой ^-лактамный цикл, сконденсированный с дигидротиазиновым циклом, и напоминает молекулу пенициллина. Однако цефалоспорин С недостаточно эффективен, чтобы его применять в качестве противобактериального агента. Аналогично пенициллину, биологическая активность этого антибиотика усиливается при замене алифатической цепочки в положении 7 ароматическим циклом. Для этого цефалоспорин гидролизуют в 7-аминоцефалоспорановую кислоту (13.15) (Х=Н и Y = —ОСОМе), похожую на 6-аминопенициллановую кислоту (13.4, а), из которой получают полусинтетические цефалоспорины.
со2н со2н
С“елет гголенулы цефалоспорина Цефокситин
(13.15) (13.16)
Цефалоспорины, так же как пенициллин, блокируют D-ала-нинузнающую полимеразу, ответственную за синтез новой клеточной оболочки бактерий [Abraham, 1962]. Более дорогостоящие, чем пенициллин, и менее активные цефалоспорины обычно не применяют при инфекциях, вызванных грамположительнымн бактериями. Однако они чрезвычайно нужны сравнительно не большому числу больных, чувствительных к пенициллину, хотя следует помнить, о том, что у некоторых людей обнаруживается и чувствительность к цефалоспорину. Гораздо чаще цефалоспорины применяют против штаммов Staphylococcus aureus, не чувствительных к бензилпенициллину, содержащих действующую только на пенициллины [i-лактамазу (так называемую пеницил-лазу). Учитывая тот факт, что существуют штаммы, содержащие цефалоспориназы, прежде чем начинать лечение, следует проводить первичные проверки в патолабораториях. Цефаман-дол и цефокситин (см. ниже) действуют против грамотрицатель-ных микроорганизмов подобно группе карбенициллина.
В табл. 13.2 представлены химическая структура и международные названия основных типов цефалоспоринов.
Цефалоти н — первый полусинтетический цефалоспорин, более других устойчив к действию ^-лактамазы стафилококков
279
Таблица 13.2. Антибиотики группы цефалоспорина
Название
Включены в качестве заместителя в (13.15)
положение 7
положение 3
Цефалотин1
Цефалоридин
Цефалексин2
J-СН2СО— S
аналогично цефалотину
—О—СОМе
Цефамандол
Цефотаксим
—СН—СО—
NH2
СН—СО—
ОН
—ОМе
N-
С—СО—
Цефуроксим
S I ОМе
,-----С-СО—
О | ОМе
—О—CONH2
1 Цефарипин действует аналогично.
2 Аналогичным действием при пероральном введении обладают цефаклор, пефадрок-сил, цефрадин.
и наиболее широко применяется в медицинской практике. Вводится внутривенно, так как плохо всасывается при пероральном введении, а внутримышечные инъекции болезненны.
Цефалоридин — также один из первых синтетических цефалоспоринов, его биологическое действие сходно с действием цефалотина, но изменения структуры молекулы привели к уменьшению болезненности внутримышечных инъекций. Он обладает токсическим действием на почки. Цефалексин — первый цефалоспорин для перорального применения, содержит, подобно ампициллину (13.9) в боковой цепи глицин. Он эффективнее всего действует против грамположительных микроорганизмов и похож на цефалотин, но слабее последнего. Для уменьшения выведения всех этих антибиотиков с мочой их вводят вместе с пробенецидом (3.16).
Цефамандол — первый цефалоспорин, уничтожающий грамотрицательные микроорганизмы, — действие, еще более сильно выраженное у цефотаксима и цефуроксима. Эти лекарственные препараты вводятся в инъекционной форме, они очень
280
эффективны при нозокомиальных инфекциях и являются альтернативой для группы карбенициллина. Применяют их и в качестве уроантисептиков.
Цефокситин (13.16) выделен не из грибка, а из бактерий (Streptomyces sp.), содержащих цефомицин, в молекуле которого, как и у цефалоспорина С, в боковой цепи находится амино-адипиновая кислота. При замене этой боковой цепи получается цефокситин, высокоэффективный против грамотрицательных (но не грамположительных) микроорганизмов (вводят внутримышечно). Цефокситин — самый эффективный из цефалоспоринов при лечении тяжелых кишечных инфекционных заболеваний, вызываемых анаэробными Bacterioides fragilis. Устойчивость цефокситина к действию ^-лактамаз, секретируемых грамотрицательными микроорганизмами, связана со стерически-ми затруднениями, возникающими в результате «двойного замещения» в положении 7 (метоксигруппа присутствует и в молекуле природного соединения, выделяемого бактериями).
О цефалоспоринах см. Neu (1982) и Morin, Gorman (1982).
Механизм действия р-лактамных антибиотиков. Двойная связь в шестичленном цикле молекул цефалоспоринов, по-видимому, активирует р-лактамную группу аналогично торзионному эффекту, который оказывает пятичленный цикл пенициллинов [Sweet, Dahl, 1970]. Перемещение двойной связи в положение 2, 3 лишает соединение биологической активности. Предполагают, что на ацилирующую способность цефалоспоринов, усиливающуюся двойной связью в положениях 3 и 4, влияет делокализация электронов, вызванная енаминовым сопряжением вне лактамного цикла [Sweet, Dahl, 1970]. Однако после создания других типов ^-лактамных противобактериальных агентов стало очевидно, что не только эта структура определяет взаимодействие лекарственного вещества и полимеразой клеточной стенки. Прежде всего вызывает удивление тот факт, что наличие в соединении даже такого простого цикла, как азетидин-2-он, может обеспечивать ему пенициллиноподобную активность, как, например, у азтреонама (13.17) [Kamiya et al., 1981; Cohen, 1983].
Молекула синтетического соединения 2-пентилпенема (13.18, б) отличается от пенициллина наличием двойной связи, расположенной, так же как в цефалоспорине. Это вещество довольно стабильно и его антибактериальная активность сравнима с таковой пенициллина. Как и у пенициллина, активен только (5R) энантиомер [Lang et al., 1979]. Структура этого соединения удивительно проста, более того, антибактериальная активность сохраняется даже у соединения (13.18, а), в котором отсутствует пентильный заместитель. Длины связей 3, 4 и 4, 7, определенные с помощью рентгеноструктурного анализа, указывают на то, что это соединение содержит пирамидальный атом азота (как в пенициллине), т. е. атом азота лежит вне плоскости связи —С—О, тогда как в обычной амидной группе вследствие сопряжения он находится в одной плоскости с этими
281
атомами. Атом азота в двух изомерных карбапенемах (13.19} и (13.20) также пирамидален. Все три соединения обладают умеренной устойчивостью к гидролизу, как и пенициллин. Изучение этих соединений показало, что два из них (13.18, а) и (13.19) обладают антибактериальным действием, тогда как соединение (13.20) его не имеет [Pfaendler et al., 1981]. На основании этих данных был сделан вывод, что ни умеренная активность fJ-лактамной группы, ни пирамидальная геометрия атома азота не могут объяснить или предсказать антибактериальную активность пенициллиноподобных соединений. Cohen (1983) предполагает, что искомый определяющей фактор — это конформация карбоксильной группы, которая, видимо, должна быть в экваториальном положении в соединениях, обладающих антибактериальным действием, как в (13.18) и (13.19), а не в аксиальном, как в (13.20). Однако в молекуле пенициллина (в растворе) конформация этой группы не фиксирована.
Азтреонам
(13.17)
со2н
Пенемы a) R=H 6)R = C5H„ (13.18)
Д2-Карбапенем
(13.19)
а) Тиенамицин (R=H)
б) Производное амидина (R = -CH-’NH)
(13.22)
В настоящее время ряд новейших ^-лактамных препаратов проходит клинические испытания (см. клавулановая кислота в разд. 13.1). Латамоксеф (13.21)—полусинтетический аналог цефалоспоринов, в молекуле которого атом серы заменен атомом кислорода (ср. со структурой цефамандола, табл. 13.2). Так как он лучше других цефалоспоринов проникает в СМЖ, его применяют для лечения менингита [Schaad, 1981; Neu, 1982]. Тиенамицин (13.22,а)—карбапенем, выделенный из 282
Streptomyces cattleya, обладает широким спектром действия и высокоэффективен против Pseudomonas. Поскольку он ацилирует собственную аминогруппу, он нестабилен в концентрированных растворах, но его амидиновое производное (13.22,6) более стабильно. Оно обладает двумя полезными свойствами: ингибирует [J-лактамазы и синтез бактериальной клеточной стенки [Tally, Jacobus, Gorbach, 1978].
Более подробно о новых 0-лактамных антибактериальных агентах см. Neu (1982).
Ванкомицин (ОММ около 1500)—хлорированный полифениловый эфир, в состав молекулы которого входят углеводный остаток и аминокислотная боковая цепь, выделен из Streptomyces orientalis. О строении его молекулы см. Sheldrick и сотр. (1978). Ванкомицин соединяется с D-аланил-П-аланиновым участком мономера, служащего исходным соединением для построения клеточной стенки бактерий, поэтому растущие клетки погибают вследствие разрыва плазматической мембраны [Anderson et al., 1965]. Этот лекарственный препарат вводят внутривенно, но никогда не сочетают с другими препаратами, усиливающими его нефро-и ототоксичность. Ванкомицин, как правило, назначают только больным, инфицированным резистентными стафилококками, если эта резистентность вызвана не ^-лактамазой.
Бацитрацин, выделенный из В. subtilis, представляет собой декапептид, содержащий четыре остатка D-аминокислот и тиазолиновый цикл. Механизм его действия заключается в образовании комплекса с переносчиком углеводов — ундекапренилпирофосфатом, в плазматической мембране, т. е. вмешательстве, в раннюю стадию синтеза клеточной стенки {Stone, Strominger, 1971] (разд. 5.3). Для осуществления этого процесса требуется присутствие ионов двухвалентного металла (разд. 11.9). Из-за высокой нефротоксичности бацитрацин применяют только в качестве основного компонента антисептических мазей.
Фосфономицин (13.23), 1,3-эпоксипропилфосфоновая кислота, выделен из вида Streptomyces. Подобно фосфоенолпи-рувату он ковалентно связывается с остатком цистеина в УДФ-ацетилглюкозаминпирувилтрансферазе. В результате этого ингибируется ранняя стадия биосинтеза пептидогликана и предотвращается образование новой клеточной стенки. В Италии его применяют в педиатрии, например для лечения менингита, вызванного грамотрицательными бактериями [Kahan et al., 1974].
н Н
Фосфономицин
(13.23)
283
13.3. Фосфорорганические соединения и карбаматы
Соединения, рассматриваемые в этом разделе, обладают способностью ацилировать эстеразы, особенно АХЭ. Существует корреляция между способностью этих соединений вызывать гибель насекомых и ингибировать изоферменты АХЭ грудной клетки (но не мозга) [Bigley, 1966; Farnham, Gregory, Sawicki, 1966]. Точно так же токсическое действие органических фосфатов на млекопитающих прямо пропорционально их ингибирующему действию на АХЭ [Aldridge, Barnes, 1952].
Одним из первых инсектицидов группы фосфорорганических соединений был тетраэтилпирофосфат, обладавший не очень высокой избирательностью [Schrader, 1936, 1944]. В 1945 г. был создан паратион (13.24), получивший сразу же широкое применение [Schrader, 1963]; его избирательность была несколько выше, но все еще далека от современных требований, предъявляемых к инсектицидам.
ЕЮ\ ,0—C6H4NO2(p) XPZ
Eto/
Паратион
(13.24)
ЕЮ. ,0—C6H4NO2(p)
XPZ
Et»/
Параоксон
(13.25)
Me2CHO
Me2CHO
Днфлос
(13.26)
А. Фосфорорганические соединения: химизм действия. Еще в самом начале изучения этих соединений было замечено, что у лиц, работающих с соединениями типа дифлоса (13.26), возможны одышка, повышенная чувствительность к свету, временная потеря сознания и обратимая потеря зрения [Lange, Kreuger, 1932; Schrader, 1963]. Указанное действие фосфорорганических соединений предположительно связывали с ингибированием АХЭ1. Изучение фосфорилирования фермента с применением радиоактивного изотопа 32Р, интенсивного источника fl-частиц с периодом полураспада 14 дней, показало, что дифлос (0,0-дии-зопропилфторфосфат) (13.26) ацилирует гидроксигруппу остатка серина в ферменте, являющуюся каталитическим сайтом для АХ. При этом происходит отщепление фтористого водорода [Schaffer, May, Summerson, 1954]. При гидролизе фосфорилированного фермента выделяется О,О-диизопропилфосфорилсерин.
1 Во время второй мировой войны в Германии были созданы некоторые фосфорорганические соединения с целью применения их в качестве боевых отравляющих веществ («нервные газы»). Из трех таких соединений — зомаиа, табуиа и зарина — наиболее активен последний [Schrader, 1963; Windholz, 1976].
284
Дифлос использовали для снижения внутриглазного давления при глаукоме, но в настоящее время для этой цели созданы новые, менее токсичные лекарственные препараты.
Действие органических фосфатов изучали также на примере параоксона (13.25), представляющего собой активную форму препарата паратиона (13.24) [Gage, 1953]. Параоксон подобно дифлосу фосфорилирует гидроксигруппу серина, при этом выделяется пара-нитрофенол [Hartley, Kilby, 1952]. На основании этих исследований был сделан вывод, что в АХЭ существует только один каталитический активный участок—• гидроксигруппа серина. Однако субстрату АХЭ равно необходим второй активный участок, связывание субстрата с которым вызывает конформационные изменения фермента, способствующие каталитическому действию активного центра [O’Brien, 1971]. Предполагают, что есть три типа таких «вторичных» активных центров: первый — для АХЭ и тетраэтиламмониевых солей, второй — для фосфорорганических соединений и третий —для карбаматов. Таким образом, «вторичный» активный центр (см. модель рецептора на рис. 12.3) является не анионным, а липофильным.
Кинетические исследования показали, что фосфорорганические соединения и фермент сначала образуют переходный комплекс, характеризующийся константой аффинности К [Aldridge, 1950; Main, 1964]. В результате фосфорилирования фермента этот комплекс распадается и выталкивает уходящую группу. Характеристикой этого практически необратимого процесса служит кр — константа фосфорилирования (или инактивации). Значения К и кр определены для многих фосфорорганических соединений и АХЭ, выделенных из разных источников.
Эти кинетические исследования послужили основой для изучения связи структура — активность. Например, для ряда аналогов параоксона (13.25) при изменении алкоксигрупп (от метокси до бутокси) и варьировании электроноакцепторных заместителей в бензольном кольце было установлено, что ни алкоксигруппы, ни природа уходящей группы не влияют на величину кр. Это справедливо и в случае изменения четырех алкильных заместителей в молекуле малаоксона (13.27). Однако значения К для производных параоксона в 2—10 раз выше, чем для малаоксона. По-видимому, вышеуказанные изменения химической структуры могут изменять сродство к ферменту, но не влиять на фосфорилирующую активность [Chiu, Main, Dauterman, 1969]. Присутствие в молекуле электроноакцепторных заместителей в уходящей группе способствует повышению анти-холинэстеразной активности до определенного максимума, однако значительное уменьшение прочности связи уходящей группы с остальной частью молекулы приводит к образованию соединений, легко гидролизующихся под действием воды [Aldridge, Barnes, 1952; Aldridge, Davison, 1952]. Ниже этого максимума существует линейная зависимость между логарифмом константы гидролиза и логарифмом константы скорости бимолекуляр-
285
о о
(MeO)2P-F -г НО-В (MeO)2P-OR + HF
Фермент Ингибированный фермент
। ;
Х2с =NO'(H+)
Реактивирующий агент
!
i о
HCR + (МеО)2Р-ОЫ:СХ2
Реактивированный фермент
Выдержка во времени
i
I
i
о
О\МеО'?Р-ОВ
Необратимо ингибированный фермент
Рис. 13.2. Инактивация фосфорилированием и реактивация фермента при условии отсутствия его «старения».
ной реакции взаимодействия фосфорорганического соединения с ферментом.
Эффективные инсектициды найдены среди эфиров фосфорной, фосфиновой и фосфоновой кислот. В эфирах фосфиновой кислоты обе алкильные группы присоединены непосредственно к атому фосфора, в эфирах фосфорной кислоты обе присоединены через кислород; эфиры фосфоновой кислоты занимают промежуточное положение.
На рис. 13.2 изображены основные процессы, происходящие при фосфорилировании фермента. Если еще не произошло «старения» ингибируемого фермента, то он может быть реактивирован в результате взаимодействия с анионной группой гидроксиламина. Явление «старения» было открыто Hobbiger (1955), обнаружившим, что сложные эфиры вторичных спиртов способствуют «старению» в большей степени, чем соответствующие эфиры первичных спиртов. Правильное представление о «старении», как о процессе, скорость которого пропорциональна скорости гидролиза алкоксигруппы, превращающейся при этом в анион, появилось позднее [Berends et al., 1959]. Нечувствительность инактивированного, «постаревшего» фермента к действию антидота обусловлена силами кулоновского отталкивания.
В опытах на насекомых было показано, что под действием фосфорорганических соединений в первый час сильно ингибируется АХЭ и резко возрастает уровень свободного АХ [Small-man, Fiske, 1958]. О действии этих агентов на нервную систему насекомых см. разд. 6.4.1.
Ацетилхолинэстеразы, выделенные из различных видов насекомых, значительно отличаются по чувствительности к фосфорорганическим инсектицидам [R. O’Brien, 1967]. АХЭ у насекомых менее доступна для экзогенных соединений, чем у млекопитающих, поэтому ионизированные аналоги не могут быть использованы в качестве инсектицидов, так как чужеродные ионы не могут проникать через мембрану (разд. 3.2).
Б. Фосфорорганические соединения: повышение их избирательности. Инсектициды повышенной избирательности, относящиеся к классу фосфорорганических соединений, следует искать
286
среди новых соединений, по-разному метаболизирующих в организмах насекомых и позвоночных, т. е. создавать либо инертные соединения, превращающиеся в активные вещества только в организме насекомых, либо активные соединения, детоксицирующиеся только в организме млекопитающих. Одно из таких соединений — наратион (О,О-диэтил-О-пара-нитрофенилтиофос-фат) (13.24); в организме насекомых он превращается в активное соединение параоксан (13.25), что обусловлено наличием у насекомых фермента, ответственного за превращение связи P = S в связь Р = О. В организме позвоночных этот процесс идет в минимальной степени, чем достигается избирательность действия инсектицида. При замене двух этоксигрупп метоксигруппами был получен препарат «паратион-метил», обладающий большей избирательностью и менее токсичный для позвоночных.
Фенитротион, 0,0 - диметил - О-(З-метил-4-нитрофенил) тиофосфат, отличается от паратион-метила только наличием метильной группы в положении 3 фенильного кольца, однако он неизмеримо более избирателен, его ЛД50 равна 500 мг/кг, тогда как для паратиона и паратион-метила — 8 и 20 мг/кг соответственно. Аналог фенитротиона фенитрооксон, в котором сера заменена кислородом, сильно ингибирует АХЭ комнатных мух в таких низких концентрациях, в которых оп почти не действует на ферменты-аналоги бычьих эритроцитов и мозга мышей [Но1-lingworth, Fukoti, Metcalf, 1967]. Кроме того, существуют инсектициды, избирательно действующие на вредных насекомых. Так, О,О-диизопропильный аналог параоксона (13.25) избирательно1 ингибирует АХЭ мух, но не медоносных пчел (преимущество, приписываемое стерическому эффекту) [Camp, Fukuto, Metcalf, 1969].
Пока велись работы по изучению аналогов паратиона, были созданы вещества с «двойным механизмом» избирательности. Первым из таких соединений был малатион — диметил-5-1,2-бисэтоксикарбонилэтилдитиофосфат (13.27). В организме насекомых он превращается в истинно токсичный агент малаоксон, в котором группа P = S заменена на Р = О. В организме млекопитающих обе эфирные связи малатиона быстро гидролизуются карбоксиэстеразами, а образующаяся при этом кислота тотчас же выводится из организма. Хотя в организме насекомых происходит замена P = S на Р = О, гидролиз протекает лишь в незначительной степени. Таким образом, один и тот же субстрат подвергается разным метаболическим превращениям, что и обусловливает его избирательное действие [Krueger, O’Brien, 1959]. В организме насекомых превращение связи P = S в связь Р = О осуществляется микросомами кишечника, нервных волокон и жирных клеток. Существует возможность выделения микросом насекомых для использования их в экспериментах [Nakatsugawa, Dahm, 1965]. Карбоксиэстеразы насекомых относительно малоактивны; кроме того, доза, получаемая насе-
287
Рис. 13.3. Уровень малаок-соиа после инъекции 30 м. д. малатнона-32Р [Krueger, O’Brien, 1959].
комыми по сравнению с позвоночными (на единицу массы тела) значительно выше.
В организме мыши, например, лишь незначительная часть малатиона превращается в токсичный агент малаоксон, тогда как в организме насекомых не только выше процент превращения малатиона, но и гораздо дольше время токсического воздействия (рис. 13.3). Приведенные данные (табл. 13.3 и рис. 13.3)
Таблица 13.3. Токсичность фосфорорганических инсектицидов для млекопитающих и насекомых, ЛДйо
Вид Малатион Малаоксон
Мышь (внутрибрюшинное введение) 1590 75
Домашняя муха 30 15
Таракан 120 15
свидетельствуют о том, что мышам можно довольно безболезненно вводить огромные дозы малатиона, а в организме тараканов и особенно домашних мух этот препарат чрезвычайно легко превращается в высокотоксичный малаоксон. Аналогично этому карбоксиамидазы млекопитающих способны гидролизовать
МеО. yS—СН—СО.2Е1
Хр/ |
Met)/ 'Ч? CH2CO2Et
а) Малаоксон (R=O) б) Малатион (R=S)
(13.27)
амидные группы, в то время как насекомые такой способностью не обладают (см. ниже о диметоате).
Широко используемый малатион представляет собой исключительно безвредное соединение. Еще один инсектицид, обладающий «двойным механизмом» избирательности, — это диазинон, О,О-диэтил-О- (2-изопропил-6-метилпиримидин-4-ил) тиофосфат (13.28), созданный в 1952 г. В организме млекопитающих происходит быстрое С-гидроксилирование изопропильной груп-288
пы с образованием соединения, легко выводящегося с мочой, а в насекомых связь P=S превращается в Р=О, в результате чего образуется исключительно токсичный агент [Miyazaki et al., 1970; Krueger, O’Brien, Dauterman, I960]. Диазинон успешно применяют для защиты овец от падальных мух.
Диазинон
(13.28)
МеО^р/О—СН=СС18
МеО/
Дихлофос
(13.29)
Хлорфенвинфос (R=Et, Х=Н) Тетрахлорвиифос (R=Me, Х=С1>
(13.30)
МеО^ /СН(ОН)—СС13
MecZ
а) б)
Трихлорфон
(13.31)
В 1950 г. были найдены токсичные агенты, обладающие иным механизмом детоксикации в организме млекопитающих, чем замена связи P = S связью Р = О: алифатические эфиры фосфорной кислоты, содержащие короткую боковую цепь с двумя или более электроноакцепторными группами (такими, как двойная связь и атом галогена). Первым соединением этого типа был дихлофос, О,О-диметил-О-(2,2-дихлорвинил) фосфат (13.29). В настоящее время его широко применяют для уничтожения домашних мух. Попадая в организм млекопитающих через кожу или легкие, он быстро гидролизуется (в плазме или печени) до диметилфосфата и О-метил-О-(2,2-дихлорвинил) фосфата, а последний— в О-(2,2-дихлорвинил) фосфорную кислоту и далее до неорганического фосфата [Casida, McBryde, Niedermeir, 1962]. Однако для насекомых его токсичность кумулятивна [Hodgson, Casida, 1962]. Допустимое для человека содержание дихлофоса в воздухе 1 мг/л [Ashe, 1964].
Изменение молекулы дихлофоса привело к созданию хлорфенвинфоса [О- [ 1 - (2,4-дихлорфенил) -2-хлорвинил] -О-О-диэтил-фосфат] (13.30, а), применяемого для борьбы с целым рядом насекомых-вредителей. При распылении этот агент задерживается
19—734
289
Таблица 13.4. Сравнительная токсичность фосфорорганических инсектицидов [Edson et al., 1964, 1966]
Вещество ЛД&о для крыс, мг/кг ЛДбо для насекомых
пероральное введение (острая токсичность) нанесение на кожу Megoura viciae (тля внковая), % Tetranychus telarius (клещ), % Pieris brassicae (бабочка белянка; личинки), кг/га , Musca domestica (половозрелая ком-1 натная муха, сам-| ка) мг/м3
половозрелые особи яйца
Тетраэтилпирофосфат Паратион Шрадан Дихлофос Диметоат Диазинон Трихлорфон Меназон Малатион Галоксон 0,5 3 5 25 200 300 650 1200 1400 (900—2000) 20 50 50 75 750 >1200 >2800 >500 >4000 ? 2 2 30 3 12 4 18 75 65 35 2 14 90 125 550 60 175 50 175 <0,018 0,77 0,044 >и 0,484 «а л 1 1 1life!
на листьях в течение 3 нед и в почве — 2 мес. Однократная обработка раствором хлорфенвинфоса овец и рогатого скота на длительное время защищает их от эктопаразитов (мух, вшей и разных видов клещей). Гораздо менее токсичный для млекопитающих тетрахлорвинфос (13.30, б) используют в сельском хозяйстве и животноводстве для борьбы с эктопаразитами (хотя действие его менее продолжительно).
Сравнительная токсичность некоторых инсектицидов приведена в табл. 13.4, из данных которой видно, что достигнутое за последние годы уменьшение токсичности для млекопитающих не сопровождалось ее снижением для насекомых. Подробнее см. Tolkmith (1966).
В 1982 г. для уничтожения комаров анофелес в тропиках при проведении противомалярийной кампании ВОЗ рекомендовала опрыскивание фенитротионом (разд. 13.3). Против черных мух-переносчиков и распространителей онхоцеркоза в Западной Африке распыляют ларвициды, темефос, ди [4-(О,О-диметилтио-фосфорил) фенил] сульфид, и хлорофоксим, О,О-диэтил-2-хлор-а-цианобензилиденаминотиофосфат. Из фосфорных инсектицидов ФАО разрешены к применению трихлорфон (13.31), малатион (13.27, б), диазинон (13.28), хлорпирифос (13.32), фосфа-мидон (13.33) и диметоат (13.35).
Некоторые фосфорорганические соединения настолько избирательны, что могут применяться перорально. Так, одна пероральная доза трихлорфона (13.31) защищает крупный рогатый
290
скот от оводов. В медицине его применяют под названием мет-рифонат, О,О-диметил-О- (1-гидрокси-2,2,2-трихлорэтил) фосфо-нат. Метрифонат представляет собой пролекарство, так как в организме он дегидратируется и изомеризуется в дихлофос (13.29).
Me С1
С1
Хлорпирифос
(13.32)
I I
МеОч /О—С=С—С—NEtg XPZ II
Met)/ ХО 0
Фосфамидои
(13.33)
В. Системные фосфорорганические инсектициды. В 1950 г. Schrader открыл первый инсектицид, который при внесении в почву захватывается корнями и распределяется по всему растению. Это обеспечивает защиту растений от тли и других сосущих насекомых в течение 6 нед; поскольку у растений отсутствует нервная система, препарат для них совершенно безвреден. При этом погибают только насекомые, поедающие растения, а полезные насекомые остаются невредимыми. Препарат, созданный Schrader (систокс), представлял собой смесь изомеров: О,О-диэтил-О-(2-этилтиоэтил) тиофосфата (13.34, а) и О,О-ди-этил-S-(2-этилтиоэтил) тиофосфата (13.34,6). Эти изомеры в дальнейшем были выделены в чистом виде и один из них модифицирован: обе этоксигруппы заменили метоксигруппами и получили препарат метил-деметон. Затем были созданы диметоат (13.35) и фосфамидон (13.33), рекомендованные к применению ФАО.
О
a) (EtO)2P(=S)— OCH2CH2SEt
б) (EtO)2P(=O)— SCH2CH2SEt
MeOv ,Р.
S—СН2—С—NHMe
S
Деметон (13.34)
Диметоат (13.35)
Диметоат обладает внутренней избирательностью и неизмеримо более токсичен для насекомых, чем для млекопитающих. Примечательно, что его избирательность обусловлена главным образом преимущественным действием на него амидазы млекопитающих и очень мало зависит от конверсии S------> О (как,
например, в случае малатиона или диазинона) [Krueger, O’Brien, Dauterman, 1960]. Это открытие значительно расширило возможности поиска избирательных фосфорорганических инсектицидов.
Г. Фосфорорганические антигельминтные средства. Кумафос (13.36, а)—первый высокоизбирательный препарат из класса фосфорорганических соединений, эффективный в качестве гли
19*
291
стогонного перорального средства для крупного рогатого скота. В молекулу кумафоса была введена кумариновая компонента с учетом того факта, что в организме крупного рогатого скота содержащийся в клевере кумарин метаболизирует чрезвычайно легко. При изменении структуры молекулы кумафоса был получен малотоксичный для млекопитающих галоксон (13.36,6): его ЛДэо для крыс 900 мг/кг, а ягнята легко переносят пятикратную лечебную дозу этого препарата [Brown N. et al., 1962]. Избирательность галоксона в большей степени определяется заменой атома водорода атомом хлора, чем заменой атома кислорода атомом серы. Галоксон применяют перорально против нематод у крупного рогатого скота и овец; остриц и аскарид у лошадей; нескольких разновидностей глистов у свиней и домашней птицы. Высокоизбирательное действие галоксона связано с существованием ферментов-аналогов (разд. 4.2). Этот токсический агент необратимо ингибирует АХЭ Haemonchus contortus, тогда как соответствующий фермент в эритроцитах овец блокируется только временно и быстро восстанавливается [Lee, Hodsden, 1963]; АХЭ телят восстанавливается на 50% в течение 30 мин [Pickering, Malone, 1967]'.
Дихлофос (13.29) применяют перорально в качестве антигельминтного препарата широкого спектра действия для свиней, лошадей и собак. Об использовании метрифоната при шистосомозе у людей см. разд. 6.3.5.
а) Кумафос (Х = О, Y=H) б) Галоксон (X—S, Y=CI)
(13.36)
CH=NOH CH=NOH
6 6
N+ N+
\СН2ОСН2/
CXchZ-0H
N+
Me
Пралидоксим
(13.37)
Обидоксим
(13.38)
Д. Антидоты. Многие фосфорорганические инсектициды высокотоксичны для человека, поэтому в арсенале лекарственных средств необходимо иметь их антидоты. В случае отравлений фосфорорганическими соединениями следует прежде всего ввести атропин (действующий преимущественно на мускарино-
292
вне рецепторы), далее лечение проводят с помощью оксимов, избирательно действующих на никотиновые рецепторы. Оксимы обеспечивают реактивацию фосфорилированной АХЭ в организме больных [Holmes, Robins, 1955], точно так же как при действии оксимов на изолированный фермент [Wilson, Meislich, 1953; Childs et al., 1955]. В результате нуклеофильной атаки антидотом атома фосфора происходит разрыв ковалентной связи между этим атомом и атомом кислорода в остатке серина и образуется новая ковалентная связь с остатком гидроксиламина (см. рис. 13.2). Один из эффективных антидотов— пралидоксим (2-ПАМ), (13.37)—антиизомер метилпири-диний-2-альдоксима. Этот препарат можно принимать перорально, но его эффективность при парентеральном введении гораздо выше. Позднее появился более сильный антидот обидоксим (13.38), обычно применяемый в виде внутримышечных инъекций [Hobbiger, Vojvodic, 1966]. При отравлениях карбаматами оксимы неэффективны.
Триарилфосфаты (например, трикрезилфосфат) и некоторые фторсодержащие фосфорорганические соединения обладают сильным нейротоксическим действием на человека, не влияя при этом на АХЭ. Это действие проявляется медленно и продолжительно (до 2 лет) и характеризуется набуханием нейронов. Применение оксимов не дает эффекта.
Подробнее о химии фосфорорганических инсектицидов см. Fest, Schmidt (1982).
Е. Карбаматы (уретаны). Карбаматы применяют с 1947 г.; они менее токсичны для людей и действуют более избирательно на насекомых, чем инсектициды других классов соединений. Так же как фосфорорганические соединения, они сначала образуют соединения включения с ферментом, а затем его карбамоилиру-ют [O’Brien, 1968]. Карбаматы ацилируют АХЭ, перенося группу R2NCO на гидроксильную группу серина и образуя карбамоильные производные фермента [Hobbiger, 1954; Wilson, Harrison, Ginsburg, 1961]. Спонтанный процесс реактивации карбамоильных производных АХЭ идет гораздо быстрее, чем фосфорилированных, что объясняется неустойчивостью N-алкильной группы [Davies, Campbell, Kearns, 1970]. Восстановление после отравления карбаматами проходит быстрее, чем в случае фосфорорганических инсектицидов. Период полураспада большинства инсектицидов группы N-метилкарбамата в организме человека около 15 мин в противоположность большинству фосфорсодержащих аналогов, имеющих период полураспада 8 ч. При изучении различных карбаматов было установлено, что оптимальный заместитель при атоме азота — метильная группа. Введение электроноакцепторных заместителей, усиливающее действие фосфорорганических инсектицидов, не влияет на эффективность карбаматов; большое значение имеют и стерические факторы [O’Brien, 1971].
Инсектициды класса карбаматов в среднем более гидрофиль
293
ны, чем фосфорорганические, поэтому их используют в качестве системных. Повышенная гидрофильность способствует тому, что эти соединения хорошо проникают в ткани растений и нервные ткани насекомых. В то же время следует отметить, что ионизированные карбаматы, аналогичные применяемым в медицине, в нервные ткани насекомых не проникают.
Первым инсектицидом класса карбаматов, получившим применение в сельском хозяйстве и ветеринарии, был карбарил, М-метил-М-(нафт-1-ил)карбамат, (13.39), малотоксичный для млекопитающих (ЛД50 для крыс при пероральном введении 540 мг/кг). В организме млекопитающих карбарил сначала гидроксилируется по кольцу, а затем фенольный гидроксил образует конъюгат [Fukuto, 1972]. Используется в качестве системного инсектицида, например, для защиты растений от листогрызущих гусениц. Позднее в практику вошел инсектицид с более простой молекулой — пропоксур, О- (2-изопропоксифенил) -N-метилкарбамат, (13.40) и алифатический карбамат метомил (13.41). Карбаматы часто применяют в сочетании с синергистами, усиливающими действие пиретринов (разд. 3.5).
Из карбаматов ФАО рекомендованы к применению только карбарил и метомил. Для борьбы с эктопаразитами домашних животных часто используют карбарил и пропоксур. Монокрото-фос (13.42), в молекуле которого винилкарбаматная группа объединена с фосфатной, применяют в сельском хозяйстве, распыляя с воздуха над большими площадями, при этом явного вреда для людей не отмечалось.
Подробнее о карбаматных инсектицидах см. O’Brien (1971), Kuhr (1976). О карбаматных фунгицидах см. разд. 6.4.
О
О—С—NHMe
i—РгО
Пропоксур (13.40) Ме
Карбарил
(13.39)
MeS—С=N—О—С—NHMe
I Н
Me О
Метомил
(13.41)
МеОк ZOC=CH—CONHMe
МеО/
Монокротофос
(13.42)
В медицине используют изостеричный АХ (2.7) карбахол (карбамоилхолинхлорид) (2.11). Он блокирует АХЭ и довольно медленно гидролизуется ею. Проявляя и мускариновую, и никотиновую активность, свойственные АХ, он обладает более интенсивным и длительным действием. Для лечения миастении и глаукомы применяют также такие карбаматы, как физостигмин
294
(2.8) и прозерин (2.10) (разд. 7.6.3). Карбарил весьма эффективен при педикулезе.
13.4. Алкилирующие агенты
Все алкилирующие агенты токсичны для млекопитающих, например йодистый метил и диазометан. Однако существуют вещества, содержащие две алкилирующие группы и избирательно действующие на клетки опухолей млекопитающих. Эти соединения, так называемые азотистые иприты, были первыми противоопухолевыми препаратами (историю их создания см. в разд. 6.3.6).
Алкилирующие агенты способны этерифицировать анионы фосфорных и карбоновых кислот, присутствующих в живых клетках, а также анионы фенолов и меркаптанов. Наиболее активные из них алкилируют и нейтральные аминогруппы (первичные, вторичные и даже третичные). Реакция обычно протекает по Sn 1-механизму (реакция нуклеофильного замещения первого порядка) [Ross, 1962]. По этому механизму катион лекарственного вещества, например (13.43), образующийся с относительно низкой скоростью, быстро атакует анион биополимера, и получается нейтральный (незаряженный) продукт. Альтернативой является быстрая атака катионом нейтрального амина с высвобождением иона Н+. Скорость реакции определяется наличием мест возможного алкилирования, а не концентрацией алкилирующего агента. Первым лекарственным веществом из класса азотистых ипритов, применявшимся в клинике, был эмбихин. Его до сих пор используют для лечения болезни Ходжкина и других видов лимфом. Эмбихин вводят внутривенно и применяют либо самостоятельно, либо чаще в системе комплексной терапии (вместе с винкристином, прокарбазином, преднизоном). В присутствии воды эмбихин (4.25) медленно циклизуется в азиридиниевый катион (13.43), который и является истинным лекарственным веществом. Эмбихин оказывает мощное антипролиферирующее действие на опухолевые клетки, хотя в организме большая часть введенного препарата гидролизуется или задерживается в «местах потерь». Токсический эффект азотистых ипритов проявляется в поражении костного мозга и влиянии на гемо- и лимфопоэз [Ross, 1962]. Для лечения солидных опухолей эти лекарственные вещества применяют редко, в основном после иссечения первичной опухоли. Слабая избирательность азотистых ипритон усиливается введением их непосредственно в опухоль или использованием наложения жгута или градиента температур.
СН2
Me—N—СН2 НО2С—(СН2) 3—/ N (СН2С Н2С1) 2
|
СН2—СН2С1
Промежуточное соединение, Хлорбутин
образуемое эмбихииом
(13.43) (13.44)
295
HO2C-CH(NH2)CH2—N(CHsCHsCl)s
Сарколизии
(13.45)
Хлорбутин, [пара-бис-(2-хлорэтил) аминофенилмасляная кислота] (13.44), полученный в 1953 г. Everett, Roberts, Ross — один из «ароматических азотистых ипритов». Эти соединения — слабые основания (подобно анилину), вследствие чего образуют азиридиниевые соли крайне медленно. Это позволяет им достигать места действия прежде, чем они успевают вступить в какие-либо побочные реакции. Из всех азотистых ипритов, используемых в клинике, хлорбутин наименее токсичен и действует медленнее всех остальных. Его успешно применяют перорально для продолжения лечения хронического лимфоцитарного лейкоза амбулаторно после курса циклофосфана или эмбихина, проведенного в клинике.
При создании родственного лекарственного вещества L-cap-колизина (13.45) (производного фенилаланина) предполагали, что благодаря аминокислотной природе он будет проникать через мембраны с помощью систем активного транспорта (разд. 3.2). О достижении поставленной цели свидетельствует неактивность D-изомера [Be'rgel, Stock, 1954]. По своей активности L-сарколизин занимает среднее место между эмбихином и хлорбутином. Аналогичный подход использования систем активного транспорта природных веществ, в данном случае глюкозы, был применен при создании дегранола [ 1,6-бис-(2-хлорэтил) амино-1,6-дидезок-симаннитол] [Vargha et al., 1957]. Препарат эффективен, однако особых преимуществ перед другими не имеет.
В клинике применяется также алкилирующий агент совершенно иного типа — миелосан (13.46), (бис.-метилсульфоновый эфир бутандиола-1,4), созданный G. Timmis. Он высокоэффективен и оказывает избирательное действие при лечении хронического миелолейкоза, но обладает побочным действием [Haddow, Timmis, 1953]. Его действие заключается в алкилировании цистеиновых остатков в белках и пептидах с последующим отщеплением серы [Roberts, Warwick (1961)]. Реакция протекает без предварительной ионизации, по Sn 2-механизму; скорость процесса зависит от концентраций алкилирующего агента и мест нуклеофильной атаки на молекуле биополимера.
MeSO2—О—(СН2)4—О—SO2Me
Миелосан
(13.46)
Если азотистые иприты в основном можно отнести к пролекарствам, то циклофосфан (3.38) представляет собой «пролекарство пролекарства», потому что он способен образовывать
296
азиридиниевый катион (подобно соединению (13.43) только после раскрытия фосфонолактонового цикла, происходящего в опухолевых клетках (см. разд. 3.6). Изучение этой реакции с применением 31Р показало, что алкилирующая способность азиридиниевого иона возрастает при падении pH от 7 до 5. Таким образом, избирательность направлена против злокачественных клеток с пониженным значением pH [Eagle, Zon, Egan, 1979].
Циклофосфан — пероральный препарат с относительно широким противоопухолевым спектром действия; его применяют при болезни Ходжкина, лимфосаркоме, лимфоме Беркитта, остром лимфоцитарном лейкозе у детей, а также после хирургического лечения рака молочной железы в период пременопаузы [Fairley, Simistdr, 1965]. В химиотерапии рака этот препарат используется чаще, чем все другие алкилирующие агенты.
В терапевтических дозах эмбихин и циклофосфан ингибируют синтез ДНК [Wheeler, Alexander, 1969]. Предполагают, что действие бифункциональных алкилирующих агентов состоит в образовании поперечных сшивок между цепями ДНК, что препятствует ее репликации. По-видимому, этими поперечными сшивками соединяются атомы азота двух остатков гуанина в положении 7, о чем свидетельствует наличие большого количества ди (гуанин-7-илэтил) сульфида в гидролизате после взаимодействия ди (2-хлорэтил) сульфида с ДНК. На РНК эти препараты влияют значительно меньше [Lawley, Brookes, 1967]. Эмбихин взаимодействует с ДНК следующим образом:
О
,СН2— CHSC1
\СН2-СН2С1
Долгое время считали, что, для того чтобы лекарственный препарат обладал противоопухолевым действием, в его молеку-
297
лу должны входить две алкилирующие группы (например, —С1), разделенные примерно пятью атомами, как в соединениях (4.25) и (13.45) [Haddow et al., 1948]. Необходимо, чтобы реакционная способность этих алкилирующих групп была не настолько велика, чтобы соединение легко гидролизовалось в присутствии воды, однако они должны сохранять способность к взаимодействию с биологическими рецепторами. Предположение об образовании бифункциональными агентами поперечных сшивок в хромосомах подтверждается существованием критического расстояния между функциональными группами в молекулах таких соединений и аномалиями митоза, часто наблюдаемыми в микроскоп [Goldaere, Loveless, Ross, 1949]. Такой механизм действия не может обеспечить высокую избирательность, и у азотистых ипритов она не велика.
Митомицин С (4.26)—антибиотик фиолетового цвета, который метаболизирует в организме млекопитающих с образованием бифункционального алкилирующего агента, необратимо связывающего две цепочки ДНК [Iyer, Szybalski, 1964]. Его применение (при раке желудка и кишечника) довольно ограничено из-за высокой токсичности.
Действие бифункциональных алкилирующих агентов подобно радиации, так как оно неспецифично для отдельных стадий клеточного цикла и проявляется сильнее при использовании больших однократных доз.
Поиски новых алкилирующих агентов привели к открытию нитрозомочевин [Johnston, McCaleb, Montgomery, 1963], эффективных при лечении целого ряда опухолей. Известно, что эти соединения действуют на некоторые виды опухолей мозга, так как благодаря способности растворяться в липидах нитрозомочевины легко проникают через ГЭБ. К сожалению, нельзя применять высокие дозы нитрозомочевин из-за выраженной способности этих препаратов подавлять миелопоэз. Из соединений этого ряда наиболее широко применяют пероральный препарат ломустин (13.47), 1-нитрозо-1-(2-хлорэтил)-3-циклогексилмоче-вина, и его метилированный аналог семустин. В некоторых случаях для лечения опухолей поджелудочной железы применяют антибиотик стрептозоцин, также относящийся к классу нитрозомочевины.
Вероятно, действие нитрозомочевин направлено преимущественно на ДНК, которая алкилируется продуктом неэнзиматического расщепления 2-хлорэтилкарбениевым ионом (С1СН2СН2+) [Cheng et al., 1972; Connors, Hare, 1974; Carter, Sakurai, Umezawa, 1981]. При спонтанной деградации образуется второй продукт циклогексилизоцианат (СбНц = С = О), способный карбамоилировать белки (т. е. образовывать производные мочевины). Однако он обладает слабым терапевтическим действием.
Другим пролекарством, высвобождающим алкилирующий агент (образующийся под влиянием ЭР печени), является де-
298
NO О
I II H
Cl—CH2—CH2—N—C—N—
Ломустин
(13.47)
N
—СО—NH2
—N=N—NMe2 N
H
Декарбазин
(13.48)
карбазин (13.48), 5-(3-диметил-1-триазено)имидазол-4-карбокса-мид [Shealy, Krauth, 1966]. С помощью масс-спектрометрии было показано, что при внутривенном введении он в организме дважды деметилируется с образованием 5-аминоимидазол-4-кар-боксамида (природного метаболита) и метильного радикала. Этот радикал — истинное активное начало; он метилирует остаток гуанина РНК (в положении 7) и белки, но почти не действует на ДНК [Bono, 1976]. Его применение в 20% случаев приводит к регрессии злокачественной меланомы — заболевания, при лечении которого не удавалось получить таких результатов с помощью других препаратов. Действие дакарбазина на миелопоэз не позволяет увеличивать дозу [Carter, Friedman, 1972].
В разд. 6.4.2 обсуждаются некоторые фунгициды, например каптан (6.58) и хлорталонил (6.59), алкилирующие меркаптогруппы в жизненно важных ферментах.
Более подробно об алкилирующих агентах см. Wilman, Connors, (1983). Канцерогенное действие алкилирующих агентов обсуждается в разд. 13.5.
13.5. Летальный синтез
Термин «летальный синтез» был введен Peters для описания биохимических превращений фторуксусной кислоты. Сама по1 себе она не токсична, но метаболизирует в организме млекопитающих с образованием токсичного соединения — фторлимонной кислоты.
Фторуксусная кислота стерически очень похожа на уксусную и поэтому может служить субстратом многих ферментов цикла трикарбоновых кислот. Она подвергается тем же метаболическим превращениям, что и уксусная, до тех пор, пока не превращается во фторлимонную кислоту (13.49) [Peters et al., 1953; Liebecq, Peters, 1949]. Токсический эффект этой кислоты связан с ингибированием фермента, дегидратирующего эту7 кислоту в цис-аконитиновую. Таким образом, фторлимонная кислота служит метаболитом-аналогом лимонной кислоты.
«Деградация до действия» лекарственного вещества уже обсуждалась в разд. 3.6. Отличие летального синтеза состоит в том, что это действительно синтез, а не деградация: соединение только с двумя атомами углерода превращается в соединение с шестью атомами. Более того, в синтезе принимают участие по
299
меньшей мере три ферментные системы, постадийно превращающие исходное соединение в токсичный продукт.
Фторуксусная кислота содержится в южноафриканском растении Dichapetalum, из-за чего в этой стране погибло много скота. Предполагают, что ее токсическое действие связано не с реакционной способностью атома фтора, а с его малой величиной (см. табл. 9.1) [Bartlett, Barron, 1947]: на первых стадиях превращений субстратная специфичность ферментов не очень высока и поэтому возможна замена в субстрате атома водорода атомом фтора, тогда как на поздних стадиях, где действуют более специфичные ферменты, эта замена уже существенна. Фторуксусная кислота, применяемая для уничтожения кроликов в норах, токсична для человека, так как избирательность ее действия не очень высока.
F-CH-CO2H но-с-со2н
СН2'СО2Н
Фторлимонная кислота
(13.49)
(13.50) и, А
Форбол
(13.51)
Канцерогены — это частично избирательные химические соединения, зачастую сами по себе безвредные, но способные метаболизировать в организме, образуя действительно канцерогенные агенты. Впервые связь между возникновением рака и химическими соединениями, присутствующими в окружающей среде, была установлена Potts в 1775 г. Он связал частоту возникновения рака мошонки у молодых трубочистов с действием смолы и сажи. Через полтора века из каменноугольной смолы было выделено химическое соединение, являющееся причиной возникновения болезни — им оказался бензо[а]пирен (13.50) [Cook, Hewett, Hieger, 1933].
До этого считали, что причина рака — старение организма. В настоящее время, на основании данных по сравнительному изучению различных популяций стало очевидно, что причиной возникновения рака во многих случаях служит загрязнение окружающей среды. Для разных популяций в различных частях света наблюдаются значительные вариации частоты возникновения и типов рака. У людей, меняющих место жительства, возникновение рака связано также с их способностью адаптироваться к укладу жизни, существующему в новом месте. Международное агентство по изучению рака (МАИР) (1976) сообщило, что более 80% случаев возникновения рака связано с воздействием солнечных лучей, потреблением табака, алкоголя
300
или загрязнением воздуха ,[ср. Cairns, 1975]. Имеет значение и генетическая предрасположенность.
В понимании процесса канцерогенеза важную роль сыграло открытие Berenblum (1941) его двустадийности. Он обнаружил, что после, активации соединениями на первый взгляд химически инертными (промоторами) малая доза канцерогенного углеводорода, безвредная в других случаях, инициирует рост опухоли. Б настоящее время наиболее часто используют такие промоторы, как кротоновое масло или его активное начало — эфиры тетрациклического многоатомного спирта форбола (13.51). Очень часто применяют одно из его производных 12-О-тетраде-каноилфорбол-13-ацетат. Предполагают, что эти промоторы активируют в плазме протеинкиназу С [Castagna et al., 1982]. Berenblum (1974) отмечал, что если действие инициатора предшествует действию промотора, то рак не возникает.
В настоящее время известно много агентов, ускоряющих рост клеток и уменьшающих их способность к агрегации (разд. 5.1). Эта так называемая трансформация может вызываться некоторыми вирусами, лектинами овощей и различными химическими веществами. Трансформация обратима, но если она не обратима, то она наследуется и, следовательно, представляет собой мутацию (генетическое изменение, передающееся по наследству). Для превращения трансформированных клеток в злокачественные требуется обязательно вмешательство канцерогена.
Недавно был открыт другой двустадийный процесс канцерогенеза. При действии таких известных канцерогенов, как диметилсульфат или бенз [а] пирен (13.50), здоровые клетки приобретают способность к неограниченной пролиферации. Эти клетки сохраняют все свойства здоровых и в отличие от трансформированных клеток обладают нормальными характеристиками роста, но способны к неограниченному числу делений. Однако они могут быть превращены в злокачественные в одну стадию — инкубированием с онкогеном (например, с онкогеном, известным под названием EJ). После такой обработки клетки становятся истинно раковыми и не воспринимают никаких сигналов ограничения роста от здоровых соседних клеток [Newbold, Overell, 1983]. (В таком превращении здоровых клеток в раковые канцерогенное химическое соединение может быть заменено другим онкогеном.) В настоящее время предполагают, что возникновение некоторых видов рака у человека связано с мутациями, превращающими нормальные гены в онкогены [Fox, 1983].
В отдельных случаях различия в химическом строении онкогена и соответствующего ему нормального гена минимальны. Так, рак мочевого пузыря может развиться вследствие замены всего лишь одного нуклеотида, что вызывает в свою очередь замену одной аминокислоты, а именно валина на глицин в положении 12 от N-конца белка [Tabin et al., 1982]. Маловероятно, что в большинстве случаев превращение нормального гена в онкоген происходит в результате замены лишь одного нуклеотида,
301
однако следует напомнить, что замена только одной ^минoкиc-лоты (валина на глутаминовую кислоту) в гемоглобине приводит к серповидно-клеточной анемии [Pauling et ah, 1949].
Однако лишь об очень немногих соединениях можно с уверенностью сказать, что они канцерогенные для человека. Это положение обусловлено двумя факторами: во-первых, большими трудностями в получении соответствующей информации и, во-вторых, наличием у человека высокоэффективных механизмов репарации, функционирование которых препятствует получению точных данных. Большую работу по проверке канцерогенности различных веществ проводит МАИР (одно из подразделений ВОЗ). С 1982 г. было проверено 585 соединений, подозреваемых в способности вызывать рак у человека. Из них 147 оказались, канцерогенными в опытах на лабораторных животных, и только для 44 были получены доказательства их канцерогенности для человека в основном при анализе историй болезни рабочих определенного производства. Например, у рабочих, занятых на производстве бензидина (4,4'-диаминобифенил, полупродукт для получения красителей) после 15—20-летней работы частота возникновения рака мочевого пузыря возрастала от 25 до 75%. В большинстве развитых стран производство бензидина в настоящее время запрещено. Однако в некоторых странах оно все еще существует, причем на основе бензидина получают различные красители, например Прямой черный 38, способный в организме человека разрушаться с выделением бензидина.
Другой пример: МАИР установило, что из 1000 человек, работающих в помещениях с концентрацией бензола в воздухе 100 м. д., 140—170 погибают от лейкоза. В настоящее время использование бензола в качестве растворителя практически запрещено, но так как он служит исходным соединением в производстве большинства лекарственных веществ и других ценных продуктов, его продолжают производить и применять под строгим контролем.
В список других химических соединений, канцерогенных для человека, входят: а) некоторые неорганические вещества — мышьяк, асбест (особенно голубой асбест), кадмий, хроматы,, гематит (один из видов железной руды), никель (только очищенный); б) некоторые органические полупродукты в производстве красителей и пластмасс —2-нафтиламин (в настоящее время применение запрещено), 4-аминобифенил, винилхлорид (активен его метаболит: оксид хлорэтилена); в) лекарственные вещества — азатиоприн, хлорбутин, циклофосфан, диэтилстильбэстрол, мелфалан, смесь эмбихина, винкристина, прокарбазина и преднизона. Опасность возникновения рака под действием химических соединений существует для работающих на производстве (и ремонте) обуви, производстве резины и мебели, синтезе бензальдегида из хлорированного толуола и при любых видах деятельности, где возможен контакт с сажей, смолой или неочищенными маслами.
302
Риск заболевания раком возрастает при курении и потреблении пищи, зараженной продуктом жизнедеятельности грибов — афлатоксином. Некоторый риск возникает и при применении лекарственный веществ, перечисленных выше, однако он невелик, если их применяют под тщательным контролем. Предполагают, что кофеин, синтетические сладкие вещества, формальдегид и хлорированные углеводороды также обладают канцерогенными свойствами, однако доказательств их канцерогенности не было найдено [The International Agency for Research on Cancer (JARC), 1982].
Канцерогены вызывают у человека рак лишь через 10—20 лет, даже после постоянного контакта с ними дома или на работе. Проверка канцерогенных свойств соединений на мышах, продолжительность жизни которых около двух лет, требует многих месяцев. Что же происходит за это время? По двухстадийной теории возникновения злокачественных клеток Berenblum, у канцерогенов существует неопределенная фаза активации, но в настоящее время установлено, что большинство этих агентов должны вступить в несколько химических реакций, прежде чем превратиться из проканцерогена в истинный (называемый также проксимальным) канцероген, который и вызывает злокачественные изменения. Этот процесс протекает с увеличением ОММ и приводит к положительно заряженному продукту [Miller, 1970]. Такие канцерогены являются электрофильными реагентами и (подобно азотистым ипритам) способны образовывать ковалентные связи с основаниями в ДНК- Однако они не могут сшивать две цепи ДНК, в отличие от азотистых ипритов, и поэтому лишены полезных свойств в последних [Brookes, 1966].
Поскольку лабораторные крысы и мыши значительно более подвержены раковым заболеваниям, чем человек, не существует однозначных критериев для экстраполяции данных, полученных в экспериментах на грызунах, на человека, хотя делаются попытки найти формулу для пересчета этих данных (МАИР, 1982). Существуют достаточно обоснованные данные, указывающие на то, что процессы репарации ДНК у человека более эффективны, чем у грызунов. Во всяком случае, наблюдения за людьми, подвергшимися радиационному облучению, свидетельствуют о том, что поврежденная генетическая информация восстанавливается у человека в значительной степени. Постоянные тщательные наблюдения, проводившиеся за жителями Хиросимы и Нагасаки в течение 34 лет, показали, что через 40 лет после интенсивного облучения в этой популяции не наблюдается н а-следуемых дефектов, т. е. облучение родителей не привело к статистически достоверному повышению уровня мутаций у потомства (изучались следующие характеристики: неблагоприятное протекание беременности, смертность в детском возрасте, частота хромосомных аберраций у детей, в качестве контроля была выбрана популяция в областях, не подвергавшихся воздействию радиации) [Schul, Otake, Neel, 1981].
303
Рассмотрим теперь химические изменения, происходящие с канцерогенными ароматическими аминами при их метаболическом превращении из проканцерогена в истинный .канцероген в организме млекопитающих. В общем случае снача^ гидроксилируется аминогруппа (с образованием гидрокс^йамина). Затем гидроксильная группа переносится на соседний (орто-) атом углерода и превращается там в эфир серной кислоты, который в свою очередь гидролизуется с образованием карбениевого иона типа (13.52), получающегося из 2-ацетамидофлуорена [Cramer, Miller, Miller, 1960]. По-видимому, именно эти карбение-вые ионы и являются истинными канцерогенами. Инъекция неорганического сульфата крысам, получавшим 2-ацетамидо-М-гидро-ксифлуорен, резко повышает токсичность этого промежуточного продукта [De Batin et al., 1970]. Аналогичные результаты получены и для таких канцерогенов, как бензидин и 4-ацетамидоби-фенил [Miller, Miller, Hartmann, 1961], 2-аминонафталин [BoylancC Dukes, Grover, 1963] и 4-гидроксиаминохолин-1-оксид (полученного биологическим восстановлением 4-нитрохинолин-1-оксида) [Endo, Ono, Sugimu'ra, 1970]. 4-Диметиламиноазобен-зол (называемый масляный желтый, так как раньше он применялся для придания цвета маргарину) метаболизирует до монометильного аналога, подвергающегося в свою очередь таким же превращениям, что и 2-ацетамидофлуорен [Poirier et al.» 1967].
—N(CH2CH2C1)
Пример истинного канцерогена (катион) (13.52)
2
Анилиновый нпрнт
(13.53)
Как было впервые показано Brookes, Lawley (1964), канцерогенность полициклических углеводородов зависит от их способности после серии метаболических изменений связываться с нуклеиновыми кислотами. Видимо, ДНК митохондрий значительно более чувствительна к их действию, чем ядерная [Nudd, Wilkie, 1983]. Бенз[а]пирен (13.50), загрязняющий воздух в больших городах, в клетках млекопитающих метаболизирует до соответствующего 7, 8-дигидрокси-7,8,9,10-тетрагидро-9,10-эпок-сида, прочно связывающегося с ДНК [Sims et al., 1974]. В результате нуклеофильной атаки эпоксидный цикл размыкается и образуется связь между атомом углерода в положении 10 углеводорода и экзоциклическими аминогруппами гуанина и аденина в ДНК [Straub et al., 1977].
Boyland (1950) первым предположил, что метаболическое превращение в эпоксид может быть решающим для проявления углеводородом канцерогенных свойств. В то время общепринятой была другая гипотеза: связь углеводорода с мишенью образуется в «К-области», при этом канцерогенность возрастает с увеличением электронной плотности в этой области [Robinson, 304
1964; Pullman, Pullman, 1955]. Примерами «К-областей» в полициклических молекулах могут служить двойная связь в положениях 9, Ю фенантрена и связь 4, 5 в соединении (13.50). Эта гипотеза осйовывалась на известных данных об отсутствии канцерогенных свойств у таких полициклических углеводородов, как 1,2-бензантрацен, приобретающих их, однако, при введении в соответствующее положение в молекуле метильной группы. В настоящее время гипотеза «К-области» потеряла свое значение, тогда как предположение о роли эпоксидов в проявлении канцерогенных свойств оправдалось для многих канцерогенных полициклических углеводородов.
Часть молекулы углеводорода, образующая связь с нуклеиновыми кислотами, называется «областью ниши»; примером может служить область между положениями 10 и 11 в молекуле (13.50). Чаще всего эпоксидирование протекает именно в этой области, однако ему могут препятствовать объемные заместители, расположенные рядом с ней. Хотя антрацены сходны с канцерогенными фенантренами, в них «область ниши» отсутствует и поэтому они не канцерогенны.
Предполагают, что в основе мутагенного действия некоторых лекарственных веществ, большинство из которых являются антигельминтными препаратами (гикантон, амосканат, метрифонат, оксамнихин, мебендазол, метронидазол и диазепам), лежат химические изменения, происходящие под действием бактериальной флоры кишечника. Их можно предотвратить, вводя экспериментальным животным такие пищевые антиоксиданты, как 2-третбутил-4-гидроксианизол и 1,2-дигидро-кси-2,2,4-триметил-6-этоксихинолин (этоксихин). Применение этих соединений может предупредить действие даже таких канцерогенов, как бенз [а] пирен, нитрозамины и уретан. Предполагают, что эти эффекты антиоксидантов связаны с активацией глутатион-Э-трансферазы и эпоксидгидратазы [Bueding et al., 1979].
Алкилирующие агенты, применяемые в медицине (см. предыдущий раздел), обычно алкилируют гуанин в положении 7. В противоположность этим препаратам, высококанцерогенные алкилирующие агенты алкилируют атом кислорода по Sn 2-механизму. Так, этилнитрозомочевина и М'-нитро-К-нитро-зоэтилгуанидин атакуют в РНК атом 0-2 цитозина и урацила, а в двунитчатой ДНК—атом О-2 тимина и атом 0-6 гуанина [Singer, 1976].
Об общих вопросах канцерогенеза см. Freudenthal, Jones, (1976) и Arcos, Argus (1968); о канцерогенезе, обусловленном действием радикалов на липиды и последующим перекисным окислением см. McBrien, Slater (1982); вопросы регистрации лекарственных веществ и пищевых добавок см. Coulston (1979).
Проверка на канцерогенность. Эти дорогостоящие исследования, длящиеся от 2 до 3 лет и требующие большого количества лабораторных животных для получения статистически достовер-
20—734
305
ных данных, обычно проводят на крысах. Предпочтительнее тесты, требующие меньших затрат времени, но их результаты менее надежны. В настоящее время большое внимание привлекают тесты на мутагенность, основанные на предположении, что канцерогенез напоминает мутации, так как различные химические соединения вызывают разные виды р^ка, при этом каждый вид в дальнейшем репродуцируется без изменений. Один из таких тестов основан на использовании гистидинзависи-мых мутантов бактерии Salmonella typhimurium, погибающих в средах, не содержащих гистидин. Однако при наличии в среде подходящего мутагена может произойти мутация со сдвигом рамки, в результате которой бактерии вновь приобретают способность синтезировать гистидин [Ames, Sims, Grover, 1972; Ames et al., 1973].
Химические соединения, вызывающие обратную мутацию, встречаются достаточно часто, например, таким действием обладают многие соединения с жесткой структурой. В известном тесте Ames используют мутанты безоболочечного типа бактерий, так как углеводная оболочка может мешать свободной диффузии лекарственного вещества. Обычно проводят двойной тест с использованием S9 микросомальной фракции печени крысы с тем, чтобы проверить возможность образования канцерогенных метаболитов при действии на вещество оксидаз со смешанными функциями (разд. 3.5.1).
Однако этот тест иногда дает и неверные результаты (как положительные, так и отрицательные). По этому тесту канцерогенны два природных вещества, встречающиеся в животных клетках — цистеин и глутатион, хотя известно, что оба эти соединения играют определенную роль в предотвращении канцерогенеза [Glatt, Protic-Sabljic, Oesch, 1983]. Диоксин, чья ярко выраженная биологическая активность носит мутагенный характер (по крайней мере, на чувствительных линиях мышей), по этому тесту не классифицируется как мутаген. Такие отклонения указывают на то, что тестом Ames следует пользоваться лишь для первичного отбора, несмотря на высокую степень совпадений между мутагенностью, установленной по этому тесту, и канцерогенностью, определенной в опытах на грызунах.
В настоящее время предпринимаются попытки совместить тест Амеса со вторым экспресс-тестом, таким как количественный анализ повреждения хромосом человека с использованием лимфоцитов, культивированных в крови. При параллельном проведении обоих тестов точный ответ можно получить через 6 нед. Широко используется альтернативный тест трансформации клеток Styles [Purchase et al., 1976], основанный на биологической особенности клеток тканей млекопитающих: они не могут расти на полутвердом агаре, так как эта среда мешает им контактировать друг с другом, а в таких условиях клетки погибают. Однако если химическое соединение может вызывать трансформацию клеток (разд. 13.5), то его добавление к среде приводит к росту .306
изолированных клеток. Для этого теста Syres использовал клетки печени человека и почек хомяка. В некоторых случаях результаты этого теста противоположены получаемым по тесту Ames. Например, по тесту Ames 2-аминонафталин канцерогенен, а масляный желтый — нет, тогда как по тесту Стайлса — наоборот (хотя известно, что оба эти соединения канцерогенны для человека).
Существует широко распространенное мнение, что каждый мутаген — это потенциальный канцероген. Однако следует помнить, что мутагенность, определяемая по тесту Ames (способность вызывать эффект сдвига рамки), отличается от мутагенности, которой обладают раковые клетки (неограниченный рост и ускорение клеточного цикла). Известны серии соединений, в которых при усилении канцерогенных свойств ослабляются мутагенные и наоборот. Такова, например, серия из 15 производных анилиновых ипритов (13.53) [Leo et al., 1981]. Мутагенные свойства этих соединений определяли по тесту Ames (с инкубацией и без инкубации в S9 микросомальной фракции печени), а канцерогенные-—по способности увеличивать число опухолей в легких у линии мышей, склонных к таким опухолям (шестимесячный тест). Самым сильным мутагеном и самым слабым канцерогеном оказалось 4-феноксипроизводное, тогда как для 3,5-диуреидо-аналога (—NH—CO—NH2)2 было установлено противоположное.
Другие примеры летального синтеза. Среди аналогов пуриновых и пиримидиновых оснований нуклеиновых кислот (разд. 4.0) наиболее эффективны те, которые вызывают в клетке летальный синтез путем включения в аналоги дезоксирибозидов и -риботидов, конкурирующих с нормальными нуклеозидами и нуклеотидами. К таким соединениям относятся противораковые препараты 5-фторурацил (3.3), цитарабин (4.13) и 6-меркапто-пурин (3.14), противовирусные препараты видарабин (4.16) и ацикловир (4.19), иммунодепрессант азатиоприн (3.41) и пероральный противогрибковый препарат флуцитозин (4.23). Первый эффективный противовирусный препарат 5-йоддезоксиури-дин (4.14) встраивается в ДНК организма и поэтому как потенциальный мутаген применяется только наружно. На синтез РНК влияют: противораковый препарат 8-азагуанин (4.40) и сельскохозяйственный фунгицид 6-азаурацил (4.41) (разд. 4.0).
Сенситизация больных к пенициллину возникает обычно в результате ацилирования е-аминогруппы лизина в у-глобулине fj-лактамным циклом антибиотика [Hamilton-Miller, Abraham, 1971].
13.6. Различные примеры
Фармакологические эффекты на ЦНС резерпина и гидразинов, ингибирующих МАО (разд. 9.4.3), необратимы, что позволяет предполагать образование ковалентной связи при их проявлении.
20*
307
Гормон, синтезирующий тироксин, по-видимому, представляет собой белок с йодсульфенильной группой (—CSI). Сцйтают, что действие таких серосодержащих гетероциклов, как 2-тиоурацил (13.54), и его 6-пропил- и 6-метилпроизводных, применяемых в клинической практике для лечения тиреотоксикоза, заключается в образовании ковалентной связи —S—S— с этой группой и высвобождении HI [Cunningham, 1964; Jirousek, Pritchard, 1971].
О
2-Тиоурацил (13.54)
Четыреххлористый углерод, применявшийся ранее против кишечного паразита (анкилостомы) и используемый до сих пор как растворитель и как гаситель пламени, а также как глистогонное средство у крупного рогатого скота, овец и птиц, вызывает у человека обширные поражения печени. Флавопротеиновый фермент превращает этот агент в свободный радикал СС1з [Slater, 1966]. Реакция сильно подавляется такими ловушками свободных радикалов, как пропиловый эфир галловой кислоты, прометазин и М,М'-дифенил-пара-фенилендиамин [Slater, Sawyer, 1971]. Известно, что для инициирования и протекания реакции перекисного окисления липидов необходимы свободные радикалы. Даже низкие концентрации четыреххлористого углерода могут стимулировать перекисное окисление в гомогенате печени при 37 °C. Четыреххлористый углерод поражает печень, вызывая перекисное окисление двойной связи в ^-алифатической цепи фосфолипидов мембран ЭР, приводящее к необратимому разрушению этой структуры [Мау, МсСау, 1968]. Более подробно см. McBrien, Slater (1982).
Хлор в элементарном виде может существовать в водных растворах только при величине pH меньше 2,0. В менее кислых растворах он превращается в хлорноватистую кислоту:
С12 + Н2О = НОС1 + Н+ + Cl-
Величина рК хлорноватистой кислоты равна 7,2, а ее дезинфицирующая способность пропорциональна концентрации нейтральной молекулы. Высокая активность хлора, проявляющаяся даже при его содержании в воде 0,2 м. д., указывает на специфичность его действия. Предполагают, что после превращения в хлорноватистую кислоту, проникающую в клетку в виде нейтральной молекулы, атаке подвергаются сульфгидрильные SH-группы соответствующего фермента. Альдолаза Е. coli — один из существенных ферментов гликолиза, чувствительна к хлорно-308
ватистой кислоте, чем и объясняется бактерицидное действие этого вещества [Knox et al., 1948]. Некоторые организмы можно защитить от действия хлорноватистой кислоты тиосульфатом натрия [Mudge, Smith, 1935].
При реакции хлорноватистой кислоты с аммиаком образуется менее активное соединение хлорамин (Н2С1), часто используемое, когда необходимо длительное действие, например, для дезинфекции воды в плавательных бассейнах. Точно так же хлорноватистая кислота образует замещенные хлорамины с аминокислотами и белками как внутри, так и вне клетки. Эти соединения могут действовать и как депо-формы, так как находятся в равновесии с хлорноватистой кислотой. Наконец, синтетическим хлорамином, пара-толуолсульфонилхлорамином (хлорамин-Т), пользуются туристы в качестве дезинфицирующего средства (источника хлорноватистой кислоты).
Йод в водных растворах существует в элементарном виде, и именно элементарный йод обладает спороцидным действием. При pH 8,5 соотношение йодноватистая кислота — йод составляет 1 : 1, при этом спороцидная активность падает (йодноватистая кислота имеет рК 10 и относится к слабым кислотам). Йод не образует депо-формы при реакции с аммиаком, но такими свойствами обладает его комплекс с калием (К1з). Для связывания йода в этот комплекс в водных растворах требуется большой избыток йодида калия [Wyss, Strandskov, 1945].
Считают, что бактерицидное действие формальдегида вызвано его способностью связываться с аминогруппами белков, что приводит к их денатурации. Такие сильные окислители, как перманганат калия и перекись водорода, разрушают любые органические молекулы. Перекись водорода в присутствии легко-окисляемых субстратов (например, тиола или аскорбиновой кислоты) и следов катионов тяжелых металлов (например, железа) может инициировать самоподдерживающуюся цепную реакцию.
Дифторметилорнитин (9.88) и другие необратимые ингибиторы ферментов (как НИАЦФ и ФАНИ) обсуждаются в разд. 9.7.
Глава 14
ХИМИЯ ПОВЕРХНОСТНЫХ ЯВЛЕНИЙ, МОДИФИКАЦИЯ МЕМБРАН
ПОВЕРХНОСТНО-АКТИВНЫМИ СОЕДИНЕНИЯМИ
Интенсивные цитологические исследования последних двух десятилетий свидетельствуют о том, что клетки, как и большинство содержащихся в них органелл, покрыты липопротеидными мембранами (разд. 5.4). Совершенно очевидно, что жизнь невозможна в отсутствие двухфазных (липофильных внутри и гидрофильных с обеих поверхностей) мембран, обеспечивающих разделение реагирующих веществ. Эти мембраны определяют последовательность протекания реакций в том порядке, который было бы невозможно обеспечить иным путем.
Ферменты, находящиеся обычно внутри мембран (но иногда и на поверхности), обеспечивают последним очень высокую биологическую избирательность. Макромолекулы, в том числе ферменты и другие крупные белковые молекулы, либо в составе частиц, либо растворенные в основной фазе, представляют собой поверхность, на которой протекают реакции. Сыворотка крови человека, поверхность белка в 1 см3 которой достигает 100 м2, является весьма наглядным примером того, сколь велика может быть такая поверхность в «растворах» белков.
14.0. Поверхностные явления in vitro
Короткодействующие ван-дер-ваальсовы силы (разд. 8.0) обеспечивают взаимное притяжение всех молекул, находящихся в контакте друг с другом. Наличие этих сил в жидкостях становится особенно очевидным у поверхности. В объеме жидкости они действуют во всех направлениях с одинаковой интенсивностью, тогда как на границе раздела воздух — вода молекулы испытывают лишь ничтожное воздействие газовой фазы и, следовательно, притяжение их жидкостью (водой) почти не имеет противодействия. В результате расположенные на поверхности молекулы втягиваются внутрь жидкой фазы и поверхность приобретает конфигурацию с минимальной возможной площадью — именно этим и объясняется сферическая форма капель жидкости и пузырьков газа (рис. 14.1). Между молекулами растворителя, находящимися в поверхностном слое и внутри основного объема раствора, происходит постоянный обмен молекулами растворителя.
310
Рис. 14.1. Схема, иллюстрирующая притяжение, испытываемое молекулами иа границе раздела воздух — вода, и ориентацию молекул амфифильного вещества на поверхности раздела масло — вода.
Граница раздела жидкость — жидкость (т. е. поверхность между двумя несмешивающимися жидкостями) по свойствам подобна границе раздела воздух — вода, за тем лишь существенным исключением, что разница в силах притяжения каждой из жидкостей, действующих на молекулы в поверхностном слое, в этом случае значительно меньше. Во многих случаях поверхностное натяжение у границы раздела жидкость — жидкость почти не отличается от разности величин поверхностного натяжения каждой из жидкостей на ее границе с воздухом.
Амфифильные вещества стремятся сконцентрироваться на границах раздела. Молекулы этих веществ обычно состоят из длинных углеводородных цепей, связанных с короткой полярной «головкой». В большинстве случаев полярность «головки» обусловлена наличием атомов азота или кислорода, неподеленные пары электронов которых образуют водородные связи с молекулами воды. С другой стороны, для попадания в воду углеводородная цепь должна разорвать водородные связи между молекулами воды, которые энергетически препятствуют этому разрыву. Поэтому амфифильные вещества, используя минимум энергии, располагаются на границе раздела масло — вода таким образом, что гидрофильная «головка» находится в воде, а липофильные углеводородные цепи размещаются в масле, взаимодействуя с подобными себе цепями растворителя (см. рис. 14.1,6). Накопление амфифильного вещества на границе раздела прекращается сразу же после того, как на ней образуется мономо-лекулярный слой этого вещества. Однако такой монослой всегда будет местом постоянного турбулентного обмена с другими молекулами амфифильного вещества, стремящимися занять место в пограничном слое; последний имеет пониженное поверхностное натяжение и легко деформируется. На границе раздела
311
могут накапливаться как растворимые, так и нерастворимые вещества.
Упорядоченное, ориентированное расположение молекул в поверхностном слое и повышенная концентрация вещества в нем обусловливают отличие химической реакционной способности молекул в поверхностном слое от таковых, находящихся внутри фаз по обе стороны от него. Например, в поверхностном слое молекулы транс-изомера ненасыщенной алифатической кислоты могут быть настолько сближены, что двойные связи становятся недоступны для перманганат-иона из прилежащего водного слоя. Поэтому в этих условиях окисляться будет только цис-изомер, тогда как внутри фазы оба изомера окисляются с одинаковой скоростью [Pideal, 1945].
Мицеллы. Разбавленные водные растворы поверхностно-активных веществ имеют обычные физические свойства. Однако при определенной высокой концентрации (характерной для каждого вещества) наступает резкое изменение поверхностного натяжения, осмотического давления и электропроводности, обусловленное появлением новой диспергированной фазы, образованной агрегатами, называемыми мицеллами. Обычно они имеют почти сферическую форму, так как вследствие взаимодействия с окружающей их водой гидрофильные группы вещества располагаются на поверхности сферы, а липофильные углеводородные цепи — внутри нее. Минимальная концентрация вещества, при которой возможно образование мицелл, называется критической мицеллярной концентрацией.
Более подробно о коллоидах и мицеллах см. Fisher, Oaken-full (1977) и Tanford (1980).
14.1. Поверхностные явления и действие лекарств, диуретики, сердечные гликозиды
Большинство лекарственных веществ действуют на поверхности, поэтому физико-химические параметры этих соединений, определяемые после адсорбции вещества на мономолекулярной пленке, имели бы большую биологическую значимость. С применением этого метода были получены интересные результаты, но пока лишь немногие из них пригодны для создания единой стройной картины изучаемых явлений. Низкая интенсивность развития этих исследований обусловлена не только острым дефицитом специалистов, широко эрудированных как в физике, так и в биологии, но и тем, что не все фундаментальные законы химии поверхностей изучены досконально.
14.1.1. Мицеллярные равновесия
Один из наиболее важных разделов химии поверхностно-активных веществ, с помощью которого могут быть решены многие неясные вопросы избирательной токсичности, связан с проб-
312
Рис. 14.2. Бактерицидное действие 4-бензилфеиола (0,0016 моль) при 20°C иа Е. coli (2Х109 клеток/мл) в водных растворах лаурата калия разной концентрации [Berry, Cook, Wills, 1956[.
2
х 1.8
£
g 1,6 X
0,02 0,04 0,06 0,08 0,10 0,12 0,14 Молярная концентрация лаурата калия
лемами равновесия между мономерами и мицеллами в водных растворах. Например, ионы агентов с ОММ выше 150 в концентрированных растворах обычно обратимо полимеризуются, образуя мицеллы. Сравнительное изучение влияния заместителей на критическую мицеллярную концентрацию (КМК) и биологическую активность лекарственных веществ — перспективная область исследований. Методами изучения КМК следует считать прецезионную оптическую интерферометрию, исследование отклонений от закона Бугера — Ламберта — Бэра (пропорциональность между оптической плотностью и концентрацией вещества) с помощью УФ-спектроскопии, изучение специфической электропроводности (мицеллы обладают большей электропроводностью, чем составляющие их ионы), определение внезапного относительного снижения концентрации противоиона (например, С1~ или Na+) при повышении концентрации лекарственного вещества.
Применение мыла для повышения растворимости в воде фенолов, используемых для дезинфекции, основано на образовании смешанных мицелл фенола и мыла. Оказалось, что при изменении их относительной концентрации (или соотношения между ними) отмечается несколько зон бактерицидной активности (рис. 14.2) [Berry, Cook, Wills, 1956]. Первая из них, со слабой активностью, существует при концентрации лаурата калия ниже 0,03 М. Максимальный бактерицидный эффект достигается при величине КМК 0,03 М для мыла. Следовательно, бактерицидное действие — это совместное действие фенола (в основном) и мыла на протоплазматическую мембрану (о повреждении мембран см. разд. 14.3). При повышении концентрации мыла до 0,045 М происходит резкое снижение бактерицидного эффекта, что объясняется включением все большего числа молекул фенола в
313
мицеллы, и, следовательно, все меньшая часть из них способна оказывать дезинфицирующее действие. Дальнейшее повышение концентрации мыла приводит к усилению дезинфицирующего действия (третья зона), вызванного токсичностью самого мыла. Все фенолы, используемые обычно как дезинфицирующие средства, в том числе и 4-хлор-мета-крозол, образуют подобные зоны.
Антигельминтное действие фенолов аналогично усиливается мылами, но и в этом случае необходимо избегать избытка мыла, поскольку мицеллы могут удерживать большую часть фенола и тем самым препятствовать его действию на глистов [Alexander, Trim, 1946]. При концентрациях мыла, не превышающих его КМК, оно повышает антигельминтный эффект фенола. При этом само мыло в организм глистов не проникает.
Способность к образованию смешанных мицелл хорошо известна в физической химии. Например, концентрация миристата калия, необходимая для образования мицелл, составляет четверть от той, которая нужна в случае лаурата калия, однако всего 15% миристата калия, добавленных к лаурату калия, снижают его КМК вдвое благодаря образованию смешанных мицелл [Klevens, 1948].
Родственным явлением можно считать связывание сывороточным альбумином лекарственных веществ, обладающих достаточно высоким коэффициентом распределения масло/вода [Brodie, Hogben, 1957] (разд. 3.4). Рассмотрим следующий пример: для роста некоторых видов бактерий необходимо ничтожное количество олеиновой кислоты, незначительное увеличение которого сразу же подавляет их развитие. Тем не менее в присутствии сывороточного альбумина эти же бактерии оказываются устойчивыми к действию таких количеств кислоты, в сотни раз превышающих бактериостатическую концентрацию. Это объясняется тем, что каждая молекула сывороточного альбумина способна связывать девять молекул жирной кислоты настолько прочно, что остающейся кислоты недостаточно даже для проявления гемолитического действия. Однако смешанные мицеллы при этом остаются в равновесии со свободной кислотой так, что ее оказывается достаточно для питания бактерий [Davis, 1947].
Разрушающее действие фенола на кожу человека, как впервые было показано Bechhold и Ehrlich (1906), ослабляется при введении в бензольное кольцо липофильных заместителей, например алкильных групп или атомов хлора. Однако получаемые при этом фенолы вследствие более высоких значений коэффициентов распределения между маслом и водой, значительно легче, чем незамещенный фенол, инактивируются сывороткой за счет связывания с альбумином. Поэтому растворы такого типичного представителя хлорированных фенолов, как 4-хлор-мета-крезол, используются для быстрой дезинфекции неповрежденных кожи и слизистых оболочек, но оказываются вредными при несоблюдении этого условия.
314
Мицеллообразование между фосфолипидами и солями желчных кислот, способствующее их растворению, играет существенную роль в транспорте жиров в кровоток и тонкий кишечник соответственно.
Более подробно о биологическом значении мицелл см. Elworthy, Florence, Macfarlane (1968).
14.1.2. Плазматические мембраны
Каждая клетка, как и каждая органелла, окружена тонкой полупроницаемой мембраной, состоящей из ламеллярной, а частично и мицеллярной мозаики липидов и белков (разд. 5.1). Проницаемость этих природных мембран обсуждалась в разд. 3.2. Взаимодействие плазматических мембран с диуретиками, сердечными гликозидами и другие ионофорные эффекты будут рассмотрены ниже.
Подробнее о мембранах см. Robertson (1983).
14.1.3. Диуретики
В основе действия всех диуретиков лежит изменение проницаемости мембран почечных канальцев, приводящее к активизации нормального процесса перекачивания ионов натрия из мочи в почечные канальцы. До 1957 г. из сильных диуретиков были известны только ртутьорганические препараты (как мер-салил), применяемые в форме инъекций. В почках эти препараты выделяют ионы ртути, связывающиеся с меркаптогруппой пермеазы, осуществляющей транспорт катионов [Clarkson, Rothstein, Sutherland, 1965]. Широко используемые ныне диуретики не содержат ионов ртути. Наиболее известный из них — гидрохлортиазид (14.1, a) [Stevens et al., 1958] обладает более сильным диуретическим действием, чем обнаруженный несколькими годами ранее хлортиазид (14.1,6) [Novello, Sprague, 1957]. Это типичные представители большой группы диуретиков, производных 1,2,4-бензотиадиазин-1,1-диоксида (обычно называемых «тиазидами»). Они оказывают действие на дистальный сегмент канальцев почек, препятствуя всасыванию мембранами ионов натрия (а следовательно, пассивно и ионов хлора). Хотя исходно они являются сульфаниламидами, однако в отличие от ацетазоламида (9.65) они лишь очень слабо влияют на активность карбоангидразы. Тиазиды обладают побочным эффектом, вызывая у больных утомляемость вследствие повышенного выведения ионов калия. Для предупреждения этого одновременно принимают триамтерен (см. ниже). Основное назначение пероральных тиазидов — снижение повышенного давления крови за счет потери воды (часто единственное лечение, необходимое в случаях легкой гипертонии).
Несколькими годами позже были открыты «суперактивные» диуретики. Наиболее широко применяемый из них — фуросемид
315
(14.2), повышающий уровень диуреза до величины практически недостижимой для тиазидов. Его назначают при отеках (водянке) [Muschaweck, Hajdu, 1964]. Действие фуросемида обусловлено подавлением всасывания хлорид-ионов (и, следовательно, пассивно ионов натрия) в восходящих почечных канальцах ниже места действия тиазидов. Он совершенно не влияет на карбоангидразу. Феноксиуксусная кислота действует подобно фуросемиду, но при ее применении необходим более строгий контроль.
о/ ^0
а) Гидрохлортиазид (точно соот- Фуросемид
встствует формуле)
б) Хлортиазид (двойная связь
в положении 3.4)
(14.1) (14.2)
Триамтерен
(14.3)
Амилорид
(14.4)
Триамтерен (14.3)—калийсберегающий диуретик, интенсивно подавляющий выведение калия и всасывание натрия в верхней части восходящих канальцев почек. Его эффекты не связаны с действием альдостерона. Триамтерен — эффективный диуретик, но часто применяется как агент, корректирующий побочное действие тиазидов (выведение калия). Несмотря на то что он является производным птеридина, у него совершенно отсутствует антифолатная активность [Wiebelhaus et al., 1965]. Другой диуретик этого типа — амилорид, относящийся к производным пиразина, содержит основную боковую цепь.
Подробнее о диуретиках см. Cragoe (1983).
14.1.4. Сердечные гликозиды
Эти природные вещества широко используют для замедления ритма сердца с одновременным усилением полноты сердечных сокращений, что делает их уникальными среди всех сердечно-сосудистых средств. Основными природными источниками сердечных гликозидов являются два вида наперстянки (Digitalis) и различные виды Strophanthus (источник одаболи-на); ландыш майский и пролеска (сцилла) больше не используются. Молекулы всех сердечных гликозидов содержат 19-уг-
316
леродное стероидное ядро (о химии этих соединений см. разд. 12.2). ^-Гидроксильная группа, расположенная у атома углерода С14, обеспечивает необычное цис-связывание колец С и D, что, по-видимому, определяет их положительное инотропное действие на сердечную мышцу больных с сердечной недостаточностью. Наиболее часто используют дигитоксин — гликозид, содержащий стероидный агликон дигитоксигенин (14.5), связанный с одной молекулой глюкозы и тремя молекулами диги-токсозы в положении 3. Агликон не используют в лечебной практике, несмотря на то, что он всасывается и распределяется в организме быстрее гликозидов, так как он слишком быстро метаболизирует, образуя сульфаты и глюкурониды по гидроксильной группе в положении 3. Помимо защитного эффекта, остаток сахара около стероидного ядра играет определенную роль в связывании лекарственного вещества с рецептором. Замена этого остатка на аминоалкильную боковую цепь приводит к увеличению активности [Thomas, 1981].
При пероральном введении некоторые молекулы гликозида достигают сердца, где они сильно ингибируют Na.K-АТФазу [Bonting, 1970]. Этот фермент либо является рецептором гликозида, либо подвергается конформационным изменениям вслед за рецептором. При прибавлении меченых гликозидов, экстрагированных из сердец различных млекопитающих, к фракциям, содержащим этот фермент, у таких высокочувствительных к сердечным гликозидам видов, как человек и собака, наблюдается высокая степень ассоциации гликозида с ферментом. И наоборот, у низкочувствительных к гликозидам млекопитающих, таких как крысы, сродство гликозидов к аналогичному ферменту оказалось низким [Schwartz, Matsui, Laughter, 1968]. Те же авторы установили, что химические аналоги гликозидов, не проявляющие инотропной активности, не связываются с ферментами человека.
После ингибирования упомянутого фермента повышается концентрация ионов натрия в клетке, что вызывает высвобождение избыточных ионов кальция [Klaus, Lee, 1969]. В свою очередь ионы Са2+, ингибируя тропонин, вызывают сокращение
317
сердечной мышцы. (Нервный импульс, инициирующий каждое биение сердца, сопровождается выделением примерно 50 микромолей ионов кальция на килограмм сердечной мышцы.)
Наличие в молекуле сердечных гликозидов лактонного кольца, связанного со стероидным циклом, побудило исследовать инотропную активность простейших лактонов. Вслед за предварительными исследованиями Chen в 1942 г. Giarmann (1949) изучил действие 28 простых нестероидных лактонов и нашел один, активность которого составляла примерно 5% активности лучших сердечных гликозидов. Однако последующая работа показала, что лактонное кольцо гликозидов может быть заменено ненасыщенной эфирной или нитрильной группами без потери биологической активности [Thomas, Boutagy, Gelbart, 1974].
Лактонное кольцо может быть замещено разными группировками, но обязательным требованием к последним является необходимость их расположения в одной плоскости. Так, лактонное кольцо можно заменить группой СН = СН—CR=X, где X — гетероатом (О или N), а радикал R может быть водородом или алкилом, но не большим, чем метокси-группа. Активность таких аналогов полностью исчезает, как при восстановлении этиленовой группировки, так и при увеличении цепи сопряжения. Вероятно, взаимодействие сердечных гликозидов с рецептором начинается с притяжения атома С20 (связанного с атомом С17 стероидного ядра) к гетероатому, несущему эффективный положительный заряд [Thomas, 1981].
Другие вещества, действующие подобно гликозидам наперстянки, такие как бисгуанилгидразон преднизолона и кассаин — один из алкалоидов Erythrophleum [Thorp, Cobbin, 1967], по-видимому, не являются их химическими аналогами. Однако боковые цепи этих соединений соответствуют предложенной выше модели рецептора. В каждом случае наличие объемного ядра может играть существенную роль, сдерживая конформационные изменения рецептора, связанные с работой катионного насоса. У стероидов такой эффект связывают с липофильностью поверхности а-кольца [Thomas, 1981].
Нерешенным остается важный вопрос: почему столь высока специфичность сердечных гликозидов к сердечной мышце, если в организме они распределяются кровотоком и в почки, и в печень, и в другие органы, богатые АТФазой?
Уровень безопасности применения сердечных гликозидов очень низок, и вполне возможно, что если бы они были вновь открыты в наше время, не получили бы рекомендации к клиническому применению. Тем не менее множество людей, в том числе пожилых и болезненных, продлили свою жизнь на много лет, благодаря разумному применению этих лекарств, и трагические случаи редки. И все же открытие новых более избирательных типов лекарств, предпочтительно не содержащих стероидного ядра, было бы чрезвычайно желательным.
.318
14.2. Ионофоры
Липидный бислой природных мембран представляет собой непроницаемый барьер для свободной диффузии ионов, обладающих низкой липофильностью из-за сильной гидратации. Ионофоры предназначены для того, чтобы облегчить транспорт ионов через эти мембраны. Существуют два типа ионофоров: подвижные и стационарные. Последние («ионные насосы») представляют собой пронизывающие бислой каналы, наполненные водой. Многое из того, что известно о ионофорах, установлено при изучении подвижных типов ионофоров, выделенных из микробов (например, валиномицин, грамицидин). Такие экзогенные ионофоры, если они эффективны, высокотоксичны для клеток млекопитающих, но не в низких дозах.
Хотя диуретики и сердечные гликозиды так или иначе связаны с переносом ионов, их не следует рассматривать как ионофоры. Даже На,К-АТФаза осуществляет лишь вспомогательное действие. Так, в натриевом насосе этот фермент, связанный с мембраной, используется для выведения натрия и введения калия в клетку. Энергия, необходимая для протекания этого процесса, выделяется при гидролизе АТФ. Фермент связывает и высвобождает катионы на разных стадиях цикла работы ионного насоса. Конформационные изменения во время цикла приводят к тому, что катион, войдя с одной стороны мембраны, выделяется на противоположной. Частично механизм заключается в следующем: гидролиз АТФ вызывает натрийзависимое фосфорилирование аспартильной группы фермента, сопровождающееся калийзависимым переносом этой фосфорильной группы фосфатазой от аспартата в воду. Существует мнение о том, что по крайней мере у некоторых видов ванадат-анион, конкурирующий с АТФ за места связывания, принимает участие в физиологической регуляции этого фермента. Родственный фермент Са,М§-АТФаза выступает в роли совершенно аналогичного насоса, нагнетающего катионы магния в клетку и выводящего кальций. Природа ионофоров подвижного типа у млекопитающих изучена слабо.
В противоположность процессу хелатирования (разд. 11),. при котором атом водорода лиганда замещается неорганическим катионом, транспорт последних ионофорами нуждается лишь в постадийном замещении ионофором молекул воды, включенных в координационную сферу катиона. Энергия десольватации компенсируется энергией связывания ионофора. В процессе теплового движения сольватированный катион сначала сталкивается с полярными группами лиганда-ионофора, который затем обволакивает катион, и образующийся комплекс, имеющий липофильную внешнюю сторону, поглощается цитоплазматической мембраной. Конформационные изменения, сопровождающие захват катиона, протекают очень быстро, но могут быть обнаружены (для валиномицина) с помощью релак
31»
сационных методов (например, быстрых температурных скачков или поглощения звуковых волн) [Grell, Eggers, Funck, 1972].
Некоторые метаболиты грибов повышают пассивный захват калия. Одни из них, например ионактин и монактин (14.6), относятся к классу макротетролидов (многозвенных циклических сложных эфиров), другие (валиномицин)—к классу депсипеп-тидов. Валиномицин (14.7) (из Streptomyces fulvissimus) представляет собой 36-членный макроцикл, включающий по три остатка L-валина, D-валина, L-молочной кислоты и остаток a-D-гидроксиизовалериановой кислоты, связанных сложноэфир-ными и амидными связями [Shemyakin et al., 1963]. Он растворим в липидах и нерастворим в воде. Периметр этой плоской молекулы липофилен, тогда как внутренняя часть кольца выстлана незаряженными гидрофильными группами. В результате конформационных изменений, вызванных захватом катиона калия, молекула приобретает браслетообразную форму.
Валиномицин способствует проникновению ионов калия в эритроциты, митохондрии [Moore, Pressman, 1964] и через бактериальные плазматические мембраны [Harold, Baarda, 1967],
но почти не влияет на проникновение ионов натрия, лития и водорода. Нонактин обладает подобным действием, а моноактин, кроме того, слегка повышает проницаемость для ионов натрия [Henderson, McGivan, Chappell, 1969]. Валиномицин проявляет противоопухолевые свойства (на мышах) [Carter, Sakurai, Umezawa, 1981].
Me
Me
а) Ноиактии (R*H> б) Монаитин (R>Me)
04.6)
Рентгеноструктурный анализ показал, что ионы калия координируют с четырьмя эфирными и четырьмя карбонильными атомами кислорода нонактина [Dobler, Dunitz, Kilbourn, 1969] и с шестью атомами кислорода карбонильных групп валиноми-цина. Таким образом, ранее гидратированный ион калия приобретает внешнюю липофильную оболочку и поэтому способен
320
Л X1
Me Vn'CH'C~<’
X L
.СН
/° L 0=0 уСН / Me "-CH D
NH
yMe CH—Ma
Me I CH—Me
CH—CH D
Ml \q
О \
L CH—Me
<5=0
I
/СН Me—CH x
I
Me
L
NH, L
/^ch
О / Me
NH
L иг /Me nU\ / / чсн n /°So Хме DO
D JO X л --NH
O-C-CH NH \>
11 X Me
о ;сн
Me Me
Валиномицин
(14.7)
достаточно легко проникать в клеточные мембраны. При столкновении этого комплекса с наружной границей между мембраной и водой часть ионов калия проходит через нее, а другая часть обменивает молекулу нонактина на воду (медленная стадия пенетрации). В этом движении анион (например, С1~) должен сопровождать К+, но на небольшом расстоянии и постоянно подвергаясь обмену. Предпочтительность захвата но-нактином ионов калия, а не натрия, определяется скорее более низкой энергией дегидратации первого, чем размерами внутренней полости циклической молекулы [Prestegard, Chan, 1970].
С помощью аламетицина — линейного пептида, выделенного из грибов Trichoderma viride, можно установить, как образуются калиевые каналы. Этот пептид, кроме терминального L-фе-нилаланинола, содержит 20 аминокислотных остатков, в том числе восемь остатков а-аминомасляной кислоты, а другие — обычные аминокислоты, входящие в состав белков. Данные рентгеноструктурного анализа кристаллов этого соединения свидетельствуют о том, что гидрофильные фрагменты направлены в одну сторону, а липофильные — в другую.
21—734
321
С помощью молекулярных моделей удалось показать, что это соединение может образовывать агрегаты, причем каждые восемь молекул образуют гидрофильные поры диаметром около 0,45 нм, которые могут быть блокированы глутаминовым остатком (Gln-7) в ответ на изменения дипольного момента. С уверенностью можно сказать, что аламетицин сам спонтанно включается в липидные мембраны и сквозь его поры, открытые воротным потенциалом, начинают проходить неорганические катионы [Fox, Richards, 1982].
Простые синтетические и недорогие циклические полиэфиры, называемые «краун-эфирами», способны переносить катионы щелочных и щелочно-земельных металлов из водного слоя в липофильный с очень высокой избирательностью. Одно из наиболее активных соединений — дициклогексил- 18-краун-6 (14.8), представляет собой кольцо, в котором две молекулы 1,2-дигид -роксициклогексана связаны двумя молекулами диэтиленгликоля. Комплексообразующая способность краун-эфиров относительно невелика, несмотря на то, что катион взаимодействует с несколькими атомами кислорода одновременно, например с шестью в эфире (14.8) [Izatt et al., 1971]. Этот краун-эфир связывает преимущественно калий, а не натрий; эфир с меньшим числом атомов кислорода — натрий, а еще с меньшим — литий. При захвате солей эти молекулы часто изменяют свою конформацию [Pedersen, 1970].
Вслед за открытием Pedersen краун-эфиров в США, во Франции начали работать с криптандами. Эти смешанные макромолекулярные аминоэфиры не просто включают катион в цикл подобно краун-эфирам, но прочно связывают его по всем направлениям [Lehn, Sauvage, Dietrich, 1970]. Так, соединение (14.9) является мощным комплексообразователем катионов калия. И краун-эфиры, и криптанды не нашли широкого применения в биологии из-за высокой токсичности. Соединение (14.9), например, обладает раздражающим действием на глаза и кожу, а его высшие гомологи поражают ЦНС [Pedersen, 1972]. И все-таки эти соединения очень полезны — их можно рассматривать как модели многочисленных и очень дорогих природных ионофоров. Большинство наиболее липофильных краун-эфиров или криптандов связывают преимущественно одновалентные, а не двухвалентные катионы, т. е. натрий имеет преимущество перед кальцием, несмотря на то, что размеры этих катионов в негид-ратированном состоянии почти одинаковы.
Краун-эфиры были использованы в качестве модели транспорта анионов сквозь мембраны против градиента концентрации (ср. с транспортом типа 2, разд. 3.2.2). Транспортируемыми анионами были N-бензоилированные аминокислоты и короткие пептиды; мембрана представляла собой перемешиваемый слой хлороформа, с обеих сторон соприкасающийся с водой. Так как краун-эфиры малорастворимы в воде, они накапливались в хлороформе. К первой границе раздела вода — хлороформ были 322
подведены ионы калия. Это привело к экстракции ионов в хлороформенный слой с образованием тройного комплекса (анион,
Дициклогексил-18-краун-6
(14.8)
CH2CH2OEt
Mev .CH2OCH2CONC7Hi5 4 С
MeZ '4CH2OCH2CONC7H15
CH2CH2OEt
Литиевый ионофор
(14.10)
K+, ионофор), из которого оба иона выделялись во вторую водную фазу (за мембраной). Освобожденный ионофор снова захватывал анион и катион из первой водной фазы, т. е. цикл повторялся [Tsukube, 1982]. О комплексообразовании анионов криптандами см. Dietrich и сотр. (1978).
Ациклический ионофор (14.10) для катионов лития образует вокруг иона липофильную гексадентатную оболочку иона, имеющую октаэдрическую структуру. Предпочтительность захвата катиона лития в 40 раз выше по сравнению с катионом натрия. Это соединение при внутривенном введении повышает захват лития мозгом крыс [Shanzer, Samuel, Korenstein, 1983].
Анионные ионофоры. Нейтральные ионофоры, связав неорганический катион, приобретают положительный заряд и способность повреждать природные мембраны (подобно катионным антибактериальным препаратам, см. разд. 14.3). В то же время известно уже около ста природных ионофоров, содержащих карбоксильную группу.
Аватек (14.11)—производное салициловой кислоты, в котором бензольное кольцо цепочкой из семи метиленовых групп связано с тетрагидрофурановым циклом, в свою очередь соединенным с тетрагидропиранильным радикалом, был выделен из бактерий Streptomyces. Рентгеноструктурный анализ показал, что каждый атом кислорода, за исключением фенольного гид-
21*
323
роксила, связан с катионом (в данном случае барий). Синтезирован большой ряд производных этого соединения, изучены их коэффициенты распределения и константы ионизации [Westley et al., 1973]. Это соединение легко высвобождает ионы кальция в биологических мембранах, так же как и катехоламины [Pressman, Guzman, 1974]. Изотропное действие этого вещества на сердце напоминает эффекты препаратов наперстянки [Osborne, Wenger, Zanko, 1977], однако селективность, по-видимому, близка к границе допустимой. Под названием «ласалоцид» это соединение широко используется для лечения протозойного заболевания — кокцидиоза у кур.
Монензин (14.12), выделенный из Streptomyces cinnamonen-sis, полностью гидрированное производное диоксаспиродецилмасляной кислоты, содержащее тетрагидрофурановые и тетрагидропирановое кольца, с ОММ 670 [Lutz, Winkler, Dunitz, 1971]. Натриевая соль этого соединения лучше растворима в углеводородах, чем в воде. Монензин используется для ускорения роста крупного рогатого скота, влияя на активный транспорт у анаэробных бактерий в рубце, что благоприятствует жизнедеятельности других штаммов, ответственных за синтез необходимых для питания алифатических кислот. Этот препарат применяют также для лечения кокцидиоза у цыплят. Грамицидин D может служить примером подобных соединений (разд. 14.3).
Ионофоры микробного происхождения, рассмотренные в этом разделе, удобно использовать для изучения транспорта ионов, однако наивысшая избирательность в биологических процессах может быть достигнута с помощью ионофоров-белков. К ним относится порин, выделенный из Е. coli. В липидном бислое 524
этих микроорганизмов он образует каналы диаметром около 0,9 нм [Chen et al., 1982].
Блокаторы кальциевых каналов. В 1964 г. Ffeckenstein обнаружил, что верапамил (14.13) имитирует эффект выведения кальция в мышце [Fleckenstein, 1977]. За прошедшйй с тех пор период времени было найдено много других соединений самой разной структуры, способных блокировать функции кальциевых каналов. Их можно разделить на две группы. Представители первой из них, например верапамил, нифедипин (14.14) и другие дигидропиридины, а также дилтиазем, очень избирательны по отношению к Са2+. Члены второго ряда соединений, такие как прексиламин, фендилин и теродилин (все они являются производными дифенилпропиламина), блокируют, кроме того, и натриевые токи. Верапамил и дилтиазем обладают примерно одинаковой активностью на гладких мышцах сердца и сосудов, тогда как инфедипин (и другие дигидропиридины) действует избирательно на мышцы сосудов [Towart, Schramm, 1984].
I I MeN— (CH2)3—CH—CN
Верапамил
(14.13)
Нифедипин
(14.14)
Нифедипин часто используют вместе с пропранололом (разд. 12.4) для снижения высокого давления крови. Более токсичный верапамил применяют для лечения стенокардии и сердечной аритмии. Об ингибиторах захвата натрия и калия см. в разд. 14.1, о блокаторах кальциевых каналов — Janis, Triggle (1983) и Rahwan, Witiak (1982). Аналоги нифедипина, не содержащие заместителей в орто-положениях бензольного кольца, оказались неактивными. Это свидетельствует о том, что пиридиновое и бензольное кольца не должны располагаться в одной плоскости, а образовывать громоздкую пробкоподобную молекулу; никакого хелатирования при этом не происходит. Замещение нитрогруппы нифедипина трифторметильной группой превращает его из блокатора в агонист, известный под индексом BAY-K 8644 [Towart, Schramm, 1984]. Подробнее о блокаторах кальциевых каналов см. Flaim, Zelis (1982) и Fleckenstein (1983); о транспорте через биологические мембраны см. Antoloni, Gliozzi, Gorio (1982) и Martonosi (1982).
325
Рис. 14.3. Разрыв клеточных оболочек Е. coli (3X10? клеток/мл) под действием гекснлрезорцинола при 25 °C [Beckett, Patki, Robinson, 1959].
14.3. Повреждение мембран биологически активными агентами
Холестерин в разных количествах входит в состав плазматических мембран млекопитающих. При этом увеличение его количества обычно приводит к ослаблению функций мембраны. Так, АТФаза мембран саркоплазматического ретикулума постепенно теряет способность к транспорту кальция по мере замещения фосфолипидов холестерином. Подобно этому активность АТФазы, сукцинатдегидрогеназы и ^-гидроксибутиратде-гидрогеназы митохондрий сердца быка падает после выдерживания их с холестерином [Warren et al., 1975].
Алкилфенолы, например (14.15), четвертичные аммониевые основания с длинной цепью, например (14.16), и полипептидные антибиотики расщепляют стенки бактериальных клеток [Gale, Taylor, 1947]. В экспериментах на протопластах в гипертонической среде было показано, что этот эффект является вторичным. Это не что иное, как результат спонтанного разрушения цитоплазматических мембран под влиянием соединений любого из трех перечисленных классов. Разрушающее действие этих веществ было изучено также путем измерения времени, затрачиваемого на вытеснение содержимого клетки. Для этого удобно использовать график зависимости изменения оптической плотности при 260 нм от времени, как это показано на рис. 14.3 на примере действия гекснлрезорцинола на Е. coli, которое, как видно из рисунка, прекращается примерно через 2 мин. Подобные результаты были получены и для четвертичных аммониевых оснований, таких как ЦТАБ (14.16) [Salton, 1951]. Природа этого явления заключается, по-видимому, в образовании пор, 326
достаточно больших для того, чтобы даже такие крупные молекулы, как цитохром С, могли выходить из клетки.
ОН сН2 СН2 Ме
но
Гексилрезорцниол
(14.15)
Ме
Нг Н2 Н2 Н2 Н2 Нг Нг Нг Нг |
С С С С С С С С с—N—Ме
Ме^X 'ХС< "'(Г '"с" '’'С ХС С I
Н2 Н2 Н2 Н2 Н2 Н2 Н2 Н2 Ме
В Г
Гидрофобная цепочка
Натион
Анион
Бромид цетилтриметиламмония (14,16)
Полимиксин В (14.17)—одно из лекарственных веществ, наиболее мягко повреждающих цитоплазматические мембраны. Этот препарат обычно применяют (внутримышечно) для лечения больных, инфицированных грамотрицательными бактериями, особенно Pseudomonas, при которых неэффективны другие антибактериальные средства. Он выделен из штамма Bacillus polymyxa и представляет собой циклический гептапептид с три-пептидной боковой цепью, N-ацилированной липофильной жирной кислотой [Suzuki et al., 1964]. Другие полимиксины, включающие две D-аминокислоты, обладают очень высокой почечной токсичностью [Wilkinson, Lowe, 1966]. Наличие в их молекуле шести остатков 2,4-диаминомасляной кислоты обусловливает их высокую основность. Это свойство и присутствие длинной углеводородной цепи ацилирующей группы придает этим антибиотикам амфифильный характер и наряду со структурным подобием определяет родство этих антибиотиков с ЦТАБ. Алкилфенолы, четвертичные аммониевые основания и полипептидные антибиотики применяют в основном как антибактериальные препараты местного действия.
/Dab—D—Phe—Thr Моа—Dab—Thr—Dab—Dab |
"''''•Thr—Dab-Dab
Полимиксин Bi Moa — 6-метилоктановая кислота, Dab — диамииомасляиая кислота, Thr — треонин, D-Phe—D-феинлаланин
(14.17)
327
• L=Orn-
L=Leu----D=Phe
L=Val
L=Pro
L=Pro
L=Val
D=Phe----L=Leu---L=Orn
Грамицидине
(14.18)
Выделенный из палочковидных бактерий, циклический де-капептидный антибиотик грамицидин S (14.18) содержит два остатка необычной аминокислоты — D-фенилаланина. По данным рентгеноструктурного анализа и спектров ЯМР молекула грамицидина S имеет структуру, подобную лестнице, ступеньками которой служат четыре связи N—Н’"О [Conti, 1959]. Этот антибиотик разрушает мембраны, не не участвует в транспорте катионов. Тироцидин В имеет очень похожую структуру. Грамицидин D — линейный полипептид, состоящий из 15 аминокислотных остатков, в том числе нескольких с D-конфигурацией. Он повреждает бактериальные клетки, удаляя из них калий (подобно валиномицину, но значительно сильнее), без разрушения плазматических мембран. Подобен ему и грамицидин А — основной компонент коммерческого грамицидина [Haydon, Hladky, 1972]; его используют в виде антибактериальных мазей. Две молекулы грамицидина D образуют спиральный ионный канал, диаметр которого 0,5 нм, а длина 3,2 нм.
Алкилфенолы, четвертичные аммониевые основания, и поли-пептидные антибиотики являются поверхностно-активными веществами, однако этого их свойства недостаточно для реализации биологического действия [Luduena et al., 1955]. Обычные поверхностно-активные вещества, различающиеся по типу заряда на три группы — катионные, анионные и нейтральные, при равном поверхностно-активном действии могут обладать как сильным, так и слабым антибактериальным действием или быть просто инертными, т. е. поверхностная активность вещества сама по себе не определяет его антибактериальное действие, а лишь способствует концентрации вещества на поверхности бактерий. Любые повреждения бактерий связаны со структурноспецифичным высвобождением компонентов клеточной мембраны.
Повреждение мембран, о котором идет речь в этом разделе, по сути дела состоящее в разрыве пленок смешанного состава, можно изучать на модельных системах. Если адсорбированное поверхностно-активное вещество образует новый монослой поверх пленки, то происходит лишь простое слипание отдельных 328
участков пленки друг с другом (явление агглютинации). Для того чтобы произошел разрыв пленки, поверхностно-активное вещество должно либо нейтрализовать ее заряд (например, при действии катиона на анионную пленку), либо проникнуть внутрь пленки, как это происходит, если агент обладает большей поверхностной активностью, чем компоненты пленки, или если он может образовывать прочные связи с одним из компонентов пленки и тем самым замещать другой компонент. Именно этот механизм лежит в основе гемолиза эритроцитов сапонинами или фосфолипидами [Schulman, Rideal, 1937]. Иногда в результате внедрения поверхностно-активного агента пленка становится более прочной и, следовательно, разрыва не происходит. Но и в этом случае ее нормальные биологические функции будут утрачены. Рассмотрим другие примеры агентов, повреждающих мембраны.
Поверхностно-активными веществами катионного типа являются два широко применяемых фунгицида — додин (додецил-гуанидин) и глиодин (2-гептадецил-2-имидазолин). Они разрывают плазматические мембраны некоторых грибов, а других убивают тем, что резко повышают проницаемость этих мембран, после чего внедряются в клетку и разрушают структуру внутриклеточных мембран [Somers, Pring, 1966].
Большинство клеток Е. coli погибает, если их поместить на 5 мин при 20 °C в водный раствор (1:10 000) хлоргексидина (10.24), широко применяемого местного анестетика [Rye, Wiseman, 1966]. Как анилид тетрахлорсалициловой кислоты, так и гексахлорофан вызывают выделение из бактериальных клеток веществ с максимумом поглощения УФ-света 260 нм (критерий разрушения плазматической мембраны); бактерии при этом погибают. Оба этих вещества используют в качестве жирорастворимых средств для дезинфекции кожи [Woodroffe, Wilkinson, 1966; Joswick, 1961].
Стрептомицин, гентамицин и другие аминогликозидные антибиотики подавляют синтез белка на рибосомах. Однако из-за высокой полярности они не способны пассивно диффундировать через клеточные мембраны. Эффективным средством доставки этих гликозидов в бактериальные клетки служит нормальный транспорт электронов, связанный с окислительным фосфорилированием. Через некоторый небольшой промежуток времени скорость резко увеличивается, вероятно, вследствие повреждения плазматической мембраны лекарственным веществом [Anand, Davis, Armitage, 1960; Bryan, Van Den Elzen, 1977].
Полиеновые антибиотики, выделенные из некоторых видов Streptomyces, найденных в почве, эффективны против системных грибковых заболеваний [Dutcher, Boyak, Fox, 1954].
Эти полиеновые противогрибковые препараты — макролиды, одна из частей лактонного цикла которых содержит много гидроксильных групп, а другая — от четырех до семи сопряженных кратных связей. По числу этих связей их называют тетраенами,
329
пентаенами и т. д. Сопряженная часть не содержит заместителей и все кратные связи имеют транс-конфигурацию. Как видно из формулы молекулы нистатина (14.19), кольцо имеет относительно плоскую липофильную часть и менее жесткую гидрофильную область. К лактонному циклу могут быть присоединены разные заместители например аминосахара, карбоксильные и эпоксидные группы, алифатические и ароматические боковые цепи. Полиеновые антибиотики губительны для большинства грибов, простейших и водорослей, практически не действуют на бактерии, а чувствительность высших животных и растений к действию этих антибиотиков средняя между таковой грибов и бактерий. Клиническая избирательность этих фунгицидов связана с различиями между холестерином (во всех мембранах животных) и растительными стеринами.
Эти полиены повреждают грибы, связываясь с эргостерином и ситостерином плазматических мембран [Hamilton-Miller, 1973]. Так, филипин не действует на Mycoplasma laidlawii, если их рост идет в отсутствие стеринов, в присутствии последних происходит лизис бактериальных клеток [Weber, Kinsky, 1965]. Если ввести полиеновые антибиотики под монослой смешанных липидов, находящийся в поверхностной впадине, они
Нистатин
(14.19)
4 - Д одецилпнрнднн
(14.20)
внедряются в пленку и увеличивают ее поверхность. В результате переориентации стериновых компонентов, происходящей при взаимодействии с полиеном, слой, по-видимому, становится текучим [Demel, Van Deenen, Kinsky, 1955]. Мембраны митохондрий и ядер, а также клеточные стенки не подвержены действию полиенов [Kinsky, 1962].
Весь спектр повреждений плазматических мембран можно получить, используя разные полиеновые антибиотики от фили-пина, вызывающего огромные повреждения и разрывы, до N-ацетилкандидина, приводящего только к утечке ионов калия и сорбозы, и N-сукцинилперимицина, высвобождающего только ионы калия [Borowski, Cybulska, 1967]. В целом, низкомолеку
330
лярные члены ряда вызывают наибольшие повреждения [Cirillo, Harsch, Lampen, 1964]. При действии нистатина и амфотерицина В (5.14) в мембранах грибов образуются ионопроницаемые гидрофильные поры диаметром около 0,4 нм [Dennis, Stead, Andreoli, 1970].
В клинике наиболее широко используют два полиеновых антибиотика—нистатин (14.19), структура которого была установлена несколькими авторами [Birch et al., 1964; Chong, Rickards, 1970] и амфотерицин (5.14). Последний по данным рентгеноструктурного анализа [Ganis et al., 1971] содержит 35-членный гептаеновый лактонный цикл. В его длинной тонкой молекуле, включающей десять гидроксильных групп, способных к образованию водородных связей, гидрофильная и липофильна области сильно разделены.
Нистатин при пероральном введении (часто в виде суспензий, из-за низкой растворимости в воде) широко используют для лечения кандидозов гортани, влагалища и прямой кишки. Он не всасывается из кишечника. Амфотерицин В вводят внутривенно в виде коллоидной суспензии при лечении глубоких грибковых инфекций легких, костного мозга и головного мозга. Его избирательность не очень высока, но в настоящее время заменить его нечем.
Фунгициды — производные миконазола, обсуждавшиеся в разд. 6.3.4, вмешиваются в процесс образования плазматических мембран грибов.
Известны попытки найти агенты, способные селективно разрушать мембраны раковых клеток. Например, амины, такие как 4-додецилпиридин (14.20), с величиной рКа в пределах 5—9 избирательно накапливаются в лизосомах, поскольку подобно раковым клеткам они имеют достаточно высокую кислотность плазмы (разд. 4.2). Эти не ионизированные и липофильные амины свободно проникают в клетку, но, превратившись в ней в катионы, они фактически попадают в ловушку, из которой уже не могут выбраться. Поскольку катионы в отличие от неионизированных аминов являются поверхностно-активными агентами, под их действием лизосомные мембраны разрываются. Можно надеяться на то, что это явление будет использовано в химиотерапии опухолей. 4-Додецилпиридин не проявляет токсического действия при внутрибрюшинном введении мышам в дозе 0,5 г/кг [Firestone, Pisano, Bonney, 1979].
Сильные боли и воспаления, возникающие у больных подагрой, вызываются содержимым лизосом, которые разрушаются, когда кристаллы натриевой соли мочевой кислоты, фагоцитируемые из кровотока полиморфно-ядерными лейкоцитами, ослабляют плазматические мембраны этих клеток.
331
14.4. Защита мембран биологически активными агентами
Биологические мембраны находятся под постоянной угрозой разрыва под действием вредных веществ, присутствующих в той же самой или в соседних клетках. Эти соединения, обычно связываемые другими мембранами, могут высвобождаться в результате патологических процессов (инфекции, аллергии или аутоиммунные процессы). Одно из таких веществ — гистамин, вызывает сильную, хотя и кратковременную, реакцию, однако известны и длительно действующие аналоги гистамина.
Прежде чем обсудить, что следует делать для защиты мембраны живых клеток, полезно отметить некоторые детали механизма, лежащего в основе повреждения одной клетки другой. Наиболее агрессивны так называемые тучные клетки и их циркулирующие аналоги базофильные лейкоциты. (Тучные клетки располагаются у внешней части ткани, их функции мало изучены.) Каждая тучная клетка имеет более 5 млн рецепторов антител, известных под индексом IgE. Простое связывание IgE с клеткой само по себе не приводит к высвобождению опасных веществ, необходимо еще, чтобы циркулирующий антиген мог первым связаться с IgE. Образовавшаяся частица активирует бездействующие каналы, по которым в клетку поступают ионы Са2+, — процесс, приводящий в том числе к высвобождению связанного гистамина. Присутствующий в клеточной мембране липид фосфатидилхолин способствует образованию этих каналов.
Астматический бронхоспазм у человека вызывают вещества с более длительным действием, чем гистамин. Раньше их называли МРВ (медленно реагирующие вещества). Однако в настоящее время известно, что причина возникновения бронхоспазма — четыре лейкотриена — выделенные из легких человека соединения, родственные простагландинам. Из них самый типичный лейкотриен D (14.21) [Morris et al., 1980].
Мощный и избирательный ингибитор биосинтеза лейкотриенов — 5,6-метано-производное лейкотриена А4 (приставка метано- в данном случае означает наличие трехчленного цикла, образованного двухвалентным заместителем группой СН2) [Nico-laou, Petasis, Seitz, 1981]. Другой ингибитор — препарат пира-золинового ряда (14.22), известный под индексом BW 755С, введенный в клиническую практику в 1980 г. Он блокирует фермент, ответственный за биосинтез лейкотриенов [Higgs, Flower, Vane, 1979].
В отсутствие эффективных антилейкотриеновых препаратов в клинике астматические явления в большинстве случаев успешно снимают ингаляциями интала (14.23) (хромолин натрия) [Сох, 1967; Howell, Altounyan, 1967]. Этот препарат предотвращает разрыв тучных клеток легких, по-видимому, за счет влияния на транспорт кальция [Johnson, 1978]. Это соединение лишь прекращает приступ, но не препятствует его повторению.
332
Действие интала строго направлено, он не защищает тучные клетки кожи и не всасывается при пероральном приеме. Весьма о
Лейкотриен
(14.21)
3-амино-1-(3-трнфторметилфенил)-2-лнразолив
(14.22)
успешно его применяют для предупреждения сенной лихорадки и в качестве профилактического средства (при пероральном введении) против пищевых аллергенов в кишечнике.
Впоследствии было найдено много аналогов интала (14.23), представляющих собой как бы половину его молекулы. Все они содержат кислотные группы, имеют плоскую структуру и атом азота или кислорода в кольце, расположенном диаметрально противоположно карбонильной группе (С=О). Кроме действия на тучные клетки, некоторые из этих препаратов защищают плазматические мембраны лимфоцитов. В большинстве случаев результаты их клинического применения оказались неожиданными. Так, 2-(орто-пропоксифенил)-6-оксо-8-азапурин (14.24) из серии 8-азапуринов в 40 раз более эффективен, чем интал [Broughton et al., 1975]. Существенные стерические затруднения, создаваемые изопропилоксигруппой в этой молекуле, выводят бензольное ядро из плоскости пуринового цикла, увеличивая тем самым поверхность молекулы.
О О—СН2—СН—СН2—О О
(Na+)-OaC
Хромогликат натрия (хромолни натрия)
(14.23)
2-орто-пропилоксифенил-6-оксо-8-азапурнн (14.24)
333
При различных патологических состояниях выделяются и многие другие повреждающие агенты, например брадикинин (и иные кинины), некоторые из простагландинов (хотя известны и такие, которые обладают лечебным действием) и даже ионы калия в определенных ситуациях. Важную роль в защите мембраны играет кальций. В дерматологии для восстановления поврежденных тканей и ускорения их заживления широко применяют препараты цинка и алюминия. Достаточно известно и использование в дерматологии кортикоидных препаратов для стабилизации мембран [Weissman, Dingle, 1961].
Уже утвердилось мнение о том, что стабилизация мембран ЦНС лежит в основе антиэпилептического действия фенобарбитала и фенитоина. Ингибирование агрегации склеротических бляшек вальпроатом натрия и антиаритмическое действие фенитоина на сердце являются проявлением их мембранозащитных свойств. О роли стабилизации мембран в реализации седативного и общеанестетического действия лекарственных веществ уже говорилось в разд. 15.
Весьма перспективным можно считать и лечение артритов и других воспалительных процессов путем стабилизации мембран.
Глава 15
БИОЛОГИЧЕСКАЯ АКТИВНОСТЬ, НЕ СВЯЗАННАЯ СО СТРУКТУРОЙ
Большинство биологически активных соединений чрезвычайно чувствительно к малейшим изменениям структуры. Однако существует класс веществ, не подчиняющихся этому правилу, а именно биодепрессанты. К ним относятся соединения, подавляющие многие функции клеток у разных видов живых организмов. В медицине их применяют в качестве снотворных средств и местных анестетиков. Если благодаря высокой летучести соединения можно создавать его высокие концентрации в воздухе, то такие соединения можно использовать как инсектициды, особенно для окуривания зернохранилищ. Антимитоти-ческие эффекты (упомянутые в разд. 15.1) обнаруживаются при низких концентрациях и, по-видимому, не зависят от структуры.
15.0. Общие биологические депрессанты (снотворные средства, общие анестетики, летучие инсектициды)
Большинство соединений, относящихся к классу биодепрессантов, не способно к ионизации. Это могут быть углеводороды (алифатические и ароматические), хлорированные углеводороды, спирты, эфиры, кетоны, сульфоны, слабые кислоты, слабые основания, алифатические нитросоединения. Действие альдегидов, сложных эфиров, сильных кислот и сильных оснований обычно отличается от такового биодепрессантов. Для объяснения биологического действия последних было выдвинуто множество гипотез.
Первая наиболее связная гипотеза [Ernest Overton, 1901; Hans Meyer, 1899] основывалась на трех следующих принципах: а) все химически инертные вещества, растворимые в липидах, обладают свойствами депрессантов; б) наиболее быстро и сильно эффекты депрессантов проявляются в клетках, богатых липидами; в) действие депрессантов, даже в рядах структурно не связанных соединений, усиливается с увеличением коэффициента распределения веществ в системе липид/вода. Более липофильные вещества имеют высокие коэффициенты распределения и наиболее выраженное депрессантное действие1.
1 В немецком оригинале — наркотическое действие.
335
Эта гипотеза представляет собой основу современных представлений о биологическом действии депрессантов, однако теперь известно, что принцип (в) может бь1ть описан уравнением параболы, а не прямой линии [Hansch et al., 1968; Hansch, 1971, с. 297, 300]. Это следует из того факта, что если вещество настолько липофильно, что практически нерастворимо в воде, то оно может не обладать и свойствами депрессанта, так как будет необратимо накапливаться в первом же липофильном «месте потерь», с которым столкнется в организме.
Наиболее важная область использования депрессантов в медицине— применение их в качестве снотворных средств, например, тиопентал (15.1,6). При внутривенном введении в больших дозах он, кроме того, вызывает и мышечную релаксацию. Внедрение специфичных миорелаксантов, таких как короткодействующий дитилин (7.29) или длительно действующий тубокура-рин (2.6), позволило резко снизить дозы общих анестетиков по сравнению с используемыми до 1940 г. В развитых странах широко применяют анестетик галотан (2-бром-1,1,1-трифтор-2-хлорэтан). Почти так же интенсивно используют появившийся значительно позже энфлуран (1,1,2,-трифтор-2-хлор-этилдифтор-метиловый эфир). Другой галогенированный эфир — метокси-флуран, применяют несколько реже из-за токсического действия на почки. У этих анестетиков значительно сильнее выражено снотворное действие, чем обезболивающее, поэтому обычно их применяют вместе с 20% закисью азота, обезболивающее действие которого значительно сильнее снотворного. Диэтиловый эфир, дивиниловый эфир, хлористый этил, циклопропан, этилен, трихлорэтилен, хлороформ в развитых странах вышли из употребления; их применяют только в случае крайней необходимости в полевых условиях или в тех слаборазвитых странах, где нет современной (сложной и дорогой) анестезиологической аппаратуры, как впрочем и квалифицированных специалистов, способных ею пользоваться. Таким образом, те анестетики, к которым еще 25 лет назад относились скептически, благодаря миорелаксантам теперь доминируют в клинике. Успех, достигнутый в создании столь избирательных агентов, во многом способствовал улучшению состояния больных и резкому сокращению времени их реабилитации.
Для индукции обезболивающего действия галотана часто применяют барбитураты, в частности тиопентал (15.1,6); после внутривенной инъекции в предплечье больной обычно засыпает в течение 15 сек.
Исследование мембран, покрывающих нервные волокна, показало необычно высокое соотношение холестерина и фосфолипидов (1:3) [Chacko et al., 1976]. Поскольку спинной и головной мозг покрыт мембранами, богатыми липидами, депрессанты действуют на ЦНС в значительно меньших дозах, чем на мускулатуру. Так, коэффициент распределения галотана (между газом и тканью) для мозга (серое вещество) составляет 6,80,
336
Таблица 15.1. Коэффициенты распределения анестетиков н снотворных средств, используемых в медицине (см. ,также табл. 17.3) [Hansch et aL 1968; Glave, Hansch, 1972]
Препарат log Р (октанол — вода)
Г алотан 1,81
Метоксифлуран Хлороформ Трихлорэтилен Диэтиловый эфир1 Дипропиловый эфир Этхлорвинил Барбитураты 2,21 1,97 2,29 0,77 2,03 2,00 1,95
Изоамилэтил (амобарбитал)
1-Метилбутилэтил (пентобарбитал, иембутал) 1,95
Аллил, i-метилбутил (секобарбитал) 2,15
Фенилэтил (фенобарбитал) 1,42
1-Метилбутил-2-тиоэтил (тиопентал)2 Хлордиазепоксид (либриум) 3,00 2,44
Диазепам3 2,82
* Как анестетик в пять раз слабее, чем дипропнловый эфир.
2 Считают, что препарат имеет дополнительный источник активности.
3 О месте действия см. разд. 12.7.
а для мышцы — всего 2,92 (при 37 °C) [Lowe, 1968]. Место действия общих анестетиков в ЦНС точно не установлено. Одни считают, что это полисинаптические области, другие — цитоплазматические мембраны.
Независимо от строения анестетики и снотворные обладают высокой липофильностью, приближающейся к logP = 2, причем биологическая активность исчезает (или по крайней мере ослабляется) как при снижении, так и повышении этой величины [Hansch, 1971, разд. 7.6]. Наиболее типичные данные представлены в табл. 15.1. Следует добавить, что коэффициенты распределения многих анестезирующих газов определены методом газовой хроматографии [Hansch et al., 1975; Leo et al., 1975].
Общие сведения о коэффициентах распределения см. в разд. 3.3 и 17.1.
В табл. 15.2 приведены выборочные данные корреляции биологической активности депрессантов с коэффициентами распределения [Meyer, Hemmi, 1935]. Поскольку в качестве тест-объ-ектов использовали немлекопитающих позвоночных, а большинство изученных веществ не применяется в медицине, эти результаты позволяют оценить границы применения данного подхода. В табл. 15.2 приведены результаты для веществ, значительно отличающихся друг от друга величинами коэффициентов распределения, что дает возможность оценить биологически эффективную концентрацию. Для оценки концентрации агента в липидах клетки необходимо перемножить величины первых двух колонок. Величины в последней колонке округлены до двух значащих цифр. Результаты, полученные ранее при использова-22—734 337
Таблица 15.2. Корреляция минимальной дозы снотворного и коэффициента распределения /
Соединение Коэффициент распределения (олеиновый спирт — вода) Концентрация, вызывающая обездвиживание головастиков, моль/л Рассчитан^ ная угнетающая концентрация, моль/л
Этанол 0,10 0,33 0,333
Бутанол 0,65 0,03 0,020
Валерамид 0,30 0,07 0,021
Бензамид 2,50 0,013 0,033
Салициламид 5,90 0,0033 0,021
«Фенобарбитал 5,90 0,008 0,048
о-Нитроанилин 14,0 0,0025 0,035
Тимол 950,0 4,7Х10-5 0,045
нии в качестве липидной фазы оливкового масла, имели больший разброс, но позволили сделать определенные выводы (см. табл. 2.1, т. 1). Коэффицент корреляции (г) по данным для всех 17 веществ, изученных Meyer и Hemmi (из них лишь 8 приведены в табл. 15.2), равен 0,99, что свидетельствует о статистической значимости этих результатов [Hansch, 1971]. В разд. 3.3 уже говорилось о том, как данные, полученные для одного неводного растворителя, могут быть использованы для сравнения с данными для другого растворителя, тот же порядок ранжирования был сохранен и здесь.
А. Принцип Фергюсона. В 1939 г. корреляция биологического действия депрессантов с их липофильностью была рассмотрена Фергюсоном в рамках более широкого термодинамического подхода. Он сформулировал эту проблему в терминах изменения свободной энергии (AG), наблюдающегося при переходе молекулы из основного в равновесное состояние. Производное G, использованное Фергюсоном, — это парциальная молярная свободная энергия, определяемая как химический потенциал (ц). Lewis и Bandall (1923) показали, что
р = Це + PTInA, (I)
где цо — химический потенциал стандартного состояния, А — (термодинамическая) активность (но не концентрация). Из этого уравнения следует, что изменение химического потенциала пропорционально термодинамической активности. Отсюда химические потенциалы всех снотворных веществ, обладающих одинаковым биологическим действием, в растворах должны быть одинаковы.
Во времена Фергюсона трудно определяемые величины термодинамической активности были известны только для некоторых соединений. Поэтому Фергюсон ввел следующее допущение (только для летучих веществ): если термодинамическую активность чистого вещества (т. е. вещества, находящегося под дав
338
лением своих собственных паров) принять за единицу, тогда термодинамическая активность паров этого вещества в воздухе при любой концентрации будет равна относительной насыщенности его паров, следовательно:
A = pt:ps, (И)
где рт — парциальное давление паров вещества в воздухе, a ps — давление насыщенных паров этого вещества при данной температуре.
Иначе говоря, любой компонент, распределяющийся равномерно между двумя фазами, должен иметь одно и то же парциальное давление в каждой фазе.
Одной из слабых сторон этого подхода является предположение о биологическом равновесии. Более того, возникает вопрос: как быстро достигается равновесие при введении общих анестетиков в организм? Так, при прекращении подачи анестезирующего газа к больному сразу же в какой-то мере возвращается сознание; при возобновлении подачи анестетика у больного быстро наступает более глубокий уровень анестезии. Фактически из-за сложности рассматриваемых систем следует скорее говорить о достижении не равновесного, а стационарного состояния, но для того, чтобы иметь возможность использовать простейшие принципы термодинамики. Фергюсон счел возможным в первом приближении считать подобные состояния как равновесные.
В соответствии с этим допущением можно предполагать, что термодинамический потенциал депрессанта одинаков во всех фазах. Тогда для определения термодинамической активности достаточно измерить ее только во внешней фазе, так как термодинамическая активность в биофазе будет такой же. Это верна и в тех случаях, когда ничего не известно о месте расположения и химической природе биофазы (под этим термином подразумевают область, в которой реализуется действие депрессанта, например, мембрана нерва и т. п.). Природа биофазы обсуждается ниже. \
Уравнение (II) позволяет определить термодинамическую активность летучих веществ, применяемых в виАе^г^за. В водных растворах ее можно принять равной относительной насыщенности раствора при условии, что вещество не очень хорошо растворимо.
Следовательно, для корреляции действия депрессантов важнейшим параметром является относительная насыщенность среды (воздуха или воды) этими веществами, а не их концентрация. Этот принцип выдержал проверку на разных биологических тест-объектах [Ferguson, 1939; 1951; Ferguson, Pirie, 1948; Burtt, 1945].
Установлено, что многие структурно неспецифические агенты вызывают одинаковый биологический эффект при примерно равной относительной насыщенности. Так, 0,2% раствор веще-
22*
339
Таблица 15.3. Относительные насыщенность и токсические концентрации некоторых органических соединений для хлебного червя [Ferguson, 1939]
Соединение Токсическая концентрация, вызывающая летальный исход через 1000 мии при 15°СХ10-в моль/л Р (упругость паров при 15 °C), мм рт. ст. Рт • Р» (относительная насыщенность прн токсических концентрациях)
Монометиланилнн 3,7 0,22 0,3
Днметиланилнн 6,6 0,28 0,4
Пиридин 76 10,4 0,1
Бромоформ 94 3,2 0,5
Тетрахлорэтан 141 4,2 0,6
Хлорбензол 200 6,8 0,5
Толуол 420 17,0 0,4
Нитрометан 710 23,0 0,6
Бензол 775 58,0 0,2
Гептан 800 27,0 0,5
Хлороформ 1040 128 0,2
Трихлорэтилен Четыреххлористый угле- 1200 52 0,4
род 1600 73 0,4
Гексан 3000 96 0,6
Дихлорэтилен 3100 230 0,2
Пентан 16 600 320 0,9
ства с предельной растворимостью 1% и 0,02% раствор другого вещества с предельной растворимостью 0,1% будут оказывать одинаковое депрессантное действие. Понятно, что любые изменения структуры вещества, снижающие его растворимость в воде, будут повышать депрессантную активность, т. е. снижать эффективную концентрацию новых соединений. При переходе от одного члена гомологического ряда к другому, по мере увеличения ОММ, будет уменьшаться растворимость вещества в воде и также быстро будет падать концентрация, необходимая для достижения одинакового специфического биологического эффекта. Однако в таких рядах величины термодинамической активности, обеспечивающие один уровень угнетающего (де-прессантного) действия, постепенно возрастают, вплоть до члена ряда, эффективного только в насыщенном растворе (т. е. растворе, в котором наблюдается равновесие между растворенной и нерастворенной частью вещества и, следовательно, его термодинамическая активность равна 1). При переходе к следующим членам ряда их активность уменьшается или исчезает вовсе [Ferguson, 1939, 1951].
Принцип Фергюсона был успешно использован для изучения структурно неспецифичных инсектицидов. В табл. 15.3 приведены данные о действии на хлебного червя (личинка жука Agriotes) разнообразных углеводородов, хлорированных парафинов, нитропроизводных углеводородов и слабых оснований. В опытах использованы смеси паров указанных веществ с воз-
340
Таблица 15.4. Подавление подвижности головастиков (данные для жидкой фазы) [Brink, Posternak, 1948]
Соединение Минимальная концентрация депрессанта, моль/л Минимальная термодинамическая активность де-прессантаХЮ*
Этанол 0,41 2,8
Ацетон 0,28 3,0
Диэтиловый эфир 0,03 4,0
Бутанол 0,02 2,1
Хлороформ 0,001 1,9
Октанол 0,0001 2,8
духом. Как видно из второй графы табл. 15.3, при применении эквимолярных концентраций активность соединений изменяется почти в 4000 раз. Значительно менывий разброс результатов получается при сравнении относительной насыщенности, т. е. рассчитанной минимальной доли давления паров, вызывающей гибель червя. В этом случае величины активности колеблются в интервале величин, близких к 0,5, причем крайние значения отличаются всего в 9 раз (от 0,1 до 0,9). Это означает, что гибель червя наступает тогда, когда воздух насыщен парами вещества примерно наполовину. С другой стороны, для веществ, не подчиняющихся принципу Фергюсона, таких как синильная кислота, аммиак и сероуглерод, были получены значительно более низкие значения (например, для аммиака 0,00008). Если бы величина 0,5 была постоянной при действии структурно неспецифичных соединений на разные виды клеток, то вряд ли следовало ожидать появления избирательности в их токсическом действии. Однако для разных клеток величина относительной насыщенности варьируется от 0,1 до 1,0. Так, для достижения состояния наркоза у мышей и головастиков или торможения развития яиц морского ежа эта величина близка к 0,03. При сравнении минимальных концентраций анилина, фенолов, алифатических спиртов и кетонов,^Необходимых для уничтожения брюшнотифозной палочки в воде,' было получено более высокое значение (0,5), т. е. и в этом случае необходимы полунасыщен-ные растворы [Ferguson, 1939]/
Табл. 15.4 можно рассматривать как аналогичную табл. 15.2, но данные в ней представлены в терминах гипотезы Фергюсона. В этой таблице приведены результаты изучения шести депрессантов, активность которых при оценке по минимальной концентрации отличается в 400 раз, но всего вдвое — если оценивать ее по термодинамической активности.
Применение принципа Фергюсона при работе с токсичными агентами дает возможность установить, является ли биологическое действие данного агента структурно специфичным. Для этого следует определить величину «относительной насыщенности» для изучаемого соединения и сравнить ее с соответствующими величинами для 5—6 соединений, типа приведенных в
341
табл. 15.1—15.3, о которых известно, что их биологическое действие структурно неспецифично. Если полученные результаты сильно различаются по выбранному биологическому тесту, то можно с высокой степенью вероятности утверждать, что испытуемый агент действует по специфическому механизму, и для его установления требуется выполнить большую программу исследований. Если же этот тест не покажет существенной разницы в величинах относительной насыщенности, то это будет означать наличие структурно неспецифического типа действия* для уточнения этого вывода достаточно более короткой программы исследований.
За много лет, прошедших с момента появления принципа Фергюсона, несмотря на интенсивную дискуссию, он не был ни расширен, ни уточнен. Предположение Фергюсона о том, что парциальное давление может быть использовано в качестве термодинамической функции, не было опровергнуто, однако* с другой стороны, Фергюсон не сумел предсказать параболического изменения биологического ответа при возрастании коэффициентов распределения [Hansch, 1971]. В настоящее время коэффициенты распределения широко применяют для количественных исследований связи структура — активность, как один из методов оптимизации поиска новых биологически активных соединений [Hansch, Fujita, 1964; Hansch, 1971] (см. также разд. 3.3 и гл. 16). Однако оба этих метода, основанные на термодинамическом подходе, исключают друг друга.
Б. Анестезирующее действие инертных газов. При исследовании действия анестетиков было обнаружено, что инертные газы могут быть такими же биологическими депрессантами, как и любые другие вещества, в том числе и перечисленные в таблицах 15.1—15.4 (сравнение проводили по термодинамической активности). В качестве примера можно привести наличие анестезирующих свойств у азота. Еще более ста лет назад водолазы при работе под водой отметили появление головокружений и потерю ориентации — симптомы, которые связывались с вдыханием под давлением азотной компоненты сжатого воздуха [Unsworth, 1861]. Обследования нескольких водолазов в лабораторных условиях показали, что при дыхании смесями кислорода (20%) с азотом или аргоном или с обоими газами при атмосферном давлении не наблюдалось никаких нарушений жизнедеятельности, тогда как при повышенном давлении, соответствующем погружению на глубину 30—100 м, проявлялся выраженный гипнотический эффект [Behnke, Yarbrough, 1939]. Эти два инертных газа липофильны, причем коэффициент распределения аргона в системе масло — вода выше коэффициента распределения эфира.
Наркотическое действие инертных газов возрастает с увеличением их ОММ. Гелий — самый легкий из инертных газов, этим действием не обладает. Эффект аргона (ОММ 40) выше, чем азота (ОММ 28). Находясь под водой на глубине примерно
342
70 м, водолаз работает под давлением, в 7 раз превышающим то, которое он испытывает на суше. Если при этом используемый для дыхания воздух обогащен азотом, то резко снижается способность к ясному мышлению и принятию разумных решений. Для предотвращения «азотного наркоза» в настоящее время используют для дыхания смесь кислорода с гелием (1:4), позволяющую работать на глубинах, превышающих 300 м.
Ксенон (ОММ 131) вызывает временную потерю рефлексов у мышей [Lawrence et al., 1946]. В этом эксперименте термодинамическая активность газа составляла примерно 0,01, что свидетельствует о весьма сильном депрессантном действии. Для ответа на вопрос, вызывает ли ксенон анестезию у человека, не получавшего ранее никаких депрессантов (ни барбитуратов, ни миорелаксантов) больным при операции давали дышать ксеноном, разбавленным воздухом (20%) [Cullen, Gross, 1951]. При этом наблюдали быструю потерю сознания, хорошую релаксацию мышц и быстрое пробуждение. Анестезия была столь же глубокой, как и в случае этилена, но слабее, чем при применении эфира. Под таким наркозом была осуществлена орхиэктомия и перевязывание фаллопиевых труб. Ксенон был оценен как удовлетворительный легкий анестетик. На основании этих результатов можно сделать вывод: анестезия инертными веществами (из которых наиболее предпочтителен ксенон из-за его высокой эффективности и низкой химической реакционной способности) наступает после того, как определенная часть некоторой фазы нервной клетки заполняется чужеродными молекулами.
В. Место действия снотворных средств. Структурно неспецифичные агенты, по-видимому, накапливаются в каких-то жизненно важных частях клетки, вызывая тем самым дезорганизацию нормальных метаболических процессов, т. е. они действуют просто как чужеродные тела. Эти агенты могут аккумулироваться клетками, благодаря одному физическому свойству, обусловленному их химическим строением и зависящему только от баланса двух переменных молекулярной структуры, а именно числа гидрофильных и липофильных атомов. Необходимое соотношение этих атомов всегда может быть достигнуто в пределах бесчисленных гомологических рядов различных типов химических соединений.
Какова же природа этой биофазы, столь чувствительной к аггестетикам? Часто в качестве наиболее простой из возможных модмей нервной клетки используют простейшее одноклеточное амебу. \Hiller (1927) показал, что амебу можно анестезировать, поместив ее в разбавленный раствор депрессанта, но не вводя его внутрь клетки. При удалении депрессанта от клетки чувствительность амебы восстанавливалась до нормальной.
У млекопитающих наиболее чувствительной фазой являются липопротеидные мембраны (митохондриальные, синаптосомаль-ные или плазматические), расположенные в ЦНС. Накопление
343
депрессанта в такой мембране может вызвать ее набухание. Физика этого явления проста: липофильные вещества способны встраиваться между углеводородными боковыми цепями белков и, возможно, липидов в мембране (разд. 5.4.1). Хотя и было показано, что липопротеидные мембраны несколько растягиваются под действием инертных газов [Clements, Wilson, 1962], так же как и мембраны эритроцитов человека под действием галотана и эфира [Seeman, Roth, 1972], эти изменения сравнительно незначительны (лишь несколько частей на 1000 при использовании концентраций депрессантов, применяющихся в клинике) и, по мнению Franks, Lieb (1981), не могут служить основой для объяснения общей анестезии. Некоторые данные указывают на то, что депрессанты просто сорбируются чувствительными областями и тем самым предотвращают открытие ионных каналов [Franks, Lieb, 1982].
Г. Влияние температуры и давления. При повышении температуры активность газообразных анестетиков понижается из-за усиления перехода молекул анестетика с места действия в газовую фазу. Этот факт свидетельствует о справедливости гипотезы, связывающей механизм действия общих анестетиков с простой адсорбцией, и не согласуется с гипотезами: 1) критического объема [Miller et al., 1973]; 2) фазового перехода [Trudell, Hubbell, 1976] и 3) текучести липидного бислоя [Metcalfe, Seeman, Burgen, 1968], согласно каждой из которых активность анестетиков должна увеличиваться при повышении температуры. Гипотеза критического объема (1) утверждает, что анестезия наступает, когда депрессант вызывает расширение места действия сверх определенного объема, что и приводит к закрыванию ионных каналов. Гипотеза фазового перехода (2) оказалась несостоятельной, когда было показано, что два анестетика, имеющие равную активность, сдвигают равновесие между твердой и жидкой фазой в противоположные стороны [Pringle, Brown, Miller, 1981]. Опыты, в которых использовали концентрации анестетиков, применяемые в клинике, не подтвердили гипотезу липидного бислоя (3), основанную на предположении об увеличении текучести под влиянием анестетиков.
Хотя повышение давления может вызвать состояние наркоза, избыточное давление может снять его. Так, анестезия тритонов азотом под давлением 34 атм снималась повышением давления до 140 атм. Аналогично анестезию тритонов эфиром, галотаном или пентабарбиталом (без давления) снимали повышением гидростатического давления окружающей юг-воды [Lever et al., 1971]. Этот эффект избыточного давления поначалу пытались объяснить уменьшением критического объема [Miller et al., [1973]. Однако предпочтительнее более простое объяснение, заключающееся в том, что молекула анестетика, попавшая в результате адсорбции в место действия, выдавливается оттуда при повышении давления [Franks, Lieb, 1982].
344
Д. Опровергнутые гипотезы. Проблема механизма действия депрессантов до конца не решена. Traube (1904) полагал, что эффекты депрессантов пропорциональны снижению поверхностного натяжения на границе раздела воздух — вода. Warburg (1921) считал, что снотворные средства предохраняют окислительные ферменты подобно тому, как тонкая пленка смазки защищает железо от ржавления. Обе эти гипотезы были опровергнуты Меуег и Hemmi (1935).
Установлено, что в ряду депрессантов нет никакой связи между анестезирующими дозами и способностью ингибировать активность ряда ферментов дыхательной цепи [Butler, 1950]. Концентрации депрессантов, необходимые для ингибирования дыхания в тканях in vitro, всегда существенно превышали используемые для анестезии в хирургической практике. Таким образом, вызываемое депрессантами падение клеточного дыхания определяется, вероятнее всего, не непосредственным их воздействием, а снижением активности нервной деятельности [McIlwain, 1962].
Л. Полинг и С. Миллер независимо друг от друга высказали мнение о том, что каждая молекула анестетика выступает в роли стабилизирующего центра, вокруг которого молекулы воды концентрируются в более компактную структуру, что и вызывает анестезию. Такая структурированная вода может образовывать «футляр» вокруг мембраны нервной клетки, затрудняя прохождение ионов, что в свою очередь препятствует деполяризации. Одним из существенных доказательств правильности выдвинутой гипотезы авторы считали наличие строгой корреляции между парциальным давлением, необходимым для анестезии различными газами, и парциальным давлением их гидратов, фактически отношение этих двух величин равно константе [Pauling, 1961; Miller, 1961] (о структуре воды см. разд. 3.1).
Хотя углеводороды и инертные газы образуют истинные гидраты, последние устойчивы лишь при давлении в несколько атмосфер [ср. Stackelberg, 1954]. Согласно гипотезе Полинга, анестетики подобно пропану образуют клатратные, льдоподобные структуры. Поскольку такие клатраты не стабильны при 37 °C и атмосферном давлении, было сделано еще одно допущение — стабильность клатратов обусловлена их взаимодействием с заряженными боковыми цепями молекул белков, что и обеспечивает снижение нервной проводимости. Следует отметить, что все изложенные гипотезы имеют общее положение, а именно: все они связывают действие депрессантов с процессом обратимой дезорганизации. Однако гидратная гипотеза Полинга и Миллера имеет ряд слабых мест.
Для того чтобы определить, вызвано ли структурирование воды воздействием неполярных групп, Grunwald и Ralph (1967) измерили константы скорости разрыва водородных связей между молекулами третичных аминов и воды как R3N—НОН. Здесь R — углеводородная группа, размер которой постоянно
345
увеличивается. Однако полученные результаты не дали никаких доказательств образования льдоподобной структуры под влиянием гидрофобных групп. Противоречит гидратной гипотезе и тот факт, что в полости клатратов льда могут встраиваться лишь сферические молекулы и, следовательно, по мере увеличения углеводородного радикала способность к образованию гидратов должна падать, тогда как активность анестетиков при этом возрастает. К тому же самый типичный анестетик диэтиловый эфир, имеющий довольно плоскую молекулу, напоминающую по форме бабочку, не образует кристаллических гидратов.
Критерием правильности каждой гипотезы является ее применимость к фторированным углеводородам — наиболее широко применяемым анестетикам. Эти соединения из-за слабости межмолекулярных связей могут служить оптимальным тестом для проверки гидратной гипотезы. Способность различных анестетиков образовывать гидраты была оценена графически, по зависимости давления диссоциации гидрата при О °C от эффективности анестезирующего действия. Оказалось, что в концентрациях, вызывающих одинаковую степень анестезии, фторсодержащие соединения значительно менее гидратированы, чем другие анестетики [Miller, Paton, Smith, 1965].
Для сравнения гидратной и липидной гипотез изучали зависимость минимальной концентрации, вызывающей анестезию (МКА), от соответствующих физических свойств для следующих веществ: тетрафторметана, гексафторида серы, закиси азота, ксенона, циклопропана, фторексена (трифторэтилвинилового эфира), диэтилового эфира, енфлурана (см. выше), галотана, хлороформа и метоксифлурана (см. выше) [Eger et al., 1969] (ряд составлен в соответствии с увеличением липофильности). В результате была получена прекрасная корреляция активности анестетиков с их липофильностью и слабая — со способностью к образованию гидратов. Так, МКА приведенных анестетиков изменялась в пределах 0,0023—26 атм (т. е. в И ООО раз), тогда как коэффициент распределения варьировался лишь от 1,44 до 2,86, т. е. увеличивался всего в 2 раза. С другой стороны, соотношение МКА к давлению, необходимому для диссоциации гидратов, варьировало от 0,12 до 6,4 атм( в 53 раза) (при этом надо учитывать, что галотан, эфир, метоксифлуран, фторексен и енфлуран вообще не образуют гидратов).
Смысл сравнения рассматриваемых гипотез механизмов действия общих анестетиков заключается в том, что даже наиболее современная из них, например гипотеза Franks и Lieb (1982), адекватно объясняющая действие снотворных средств, может оказаться слишком упрощенной для общих анестетиков. Последние должны обладать и снотворным и обезболивающим действием (как диэтиловый эфир). Однако в современной практике используют смесь галотана (обладающего в основном снотворным действием) с закисью азота (проявляющей главным образом обезболивающий эффект).
346
Е. Барбитураты. Эти соединения были предложены в 1903 г. в качестве снотворных средств для замены простых алифатических амидов и сульфонов. Терапевтический индекс барбитуратов со временем был улучшен введением в молекулу боковой цепи, способной к биодеградации в организме (по обычным механизмам метаболизма ксенобиотиков) в течение нескольких часов после введения (разд. 3.5). Первый снотворный препарат ряда барбитуратов барбитал (веронал) не подвержен метаболическому окислению и выводится из организма очень медленно, вследствие чего в организме больного создается повышенная концентрация препарата. Типичным представителем группы легкоокисляемых барбитуратов является пентобарбитал или нембутал (15.1,а), относительная безопасность которого обусловлена разветвленностью боковой цепи, облегчающей окисление. Подобный эффект обеспечивает и наличие двойной связи в боковой цепи.
Для увеличения скорости проявления эффекта препарата коэффициенты распределения барбитуратов могут быть повышены одним из трех следующих способов: 1) увеличением числа атомов углерода насыщенной углеводородной боковой цепи; 2) замещением атома кислорода серой, как в тиопентале (15.1,6)—анестетике для внутривенного введения; 3) введением метильной группы к одному из атомов азота цикла. Если липофильность барбитурата становится слишком высокой, например в результате метилирования обоих атомов азота или при увеличении длины боковой цепи сверх шести атомов углерода, он приобретает способность вызывать судороги. Еще более липофильные молекулы, например дибензилбарбитуровая кислота, неактивны.
В настоящее время барбитураты, как снотворные применяют все меньше из-за того, что они вызывают привыкание и даже через 12 ч после однократного приема обычной дозы препарата нередко проявляются расстройства мышления и рассеивание внимания. В настоящее время барбитураты в основном заменяют бенздиазепинами (разд. 12.7), которые хотя и вызывают расстройства, подобные перечисленным, но значительно менее токсичны (имеют более высокие значения ЛД50). Хроническая бессонница лучше излечивается психотерапевтическими методами, чем снотворными средствами.
СаН6
—СН—СН2—СН2—Me
О Ме
HN
Фенобарбитал
(15.2)
а) Пентобарбитал (Х = О) б) Тиопентал (X»=S)
(15.1)
347
реакций). Этанол усиливает это ненормальное состояние. Более высокие дозы марихуаны вызывают спутанность сознания, состояния беспокойства и галлюцинации [Thacore, Shulka, 1976].
15.1. Соединения, нарушающие митоз
Многие простые биологические депрессанты неблагоприятно влияют на митоз. Они не прекращают деления хромосом, но дочерние хромосомы не расходятся к двум противоположным полюсам веретена, как обычно, а остаются друг около друга. Это приводит к остановке митоза в метафазе. Из большого числа инертных веществ, действующих подобным образом, можно упомянуть следующие: гексан и другие алифатические углеводороды, спирты, эфир, хлороформ, ацетон, паральдегид, ацетамид, мочевина, ацетофенон, бензофенон, ацетанилид, бензол, хлорбензол, сульфонал [Ostergren, 1951], а также азот и аргон под давлением [Ferguson, Hawkins, Doxey, 1950].
Этот эффект, являющийся характерным признаком биологической депрессии, исчезает при уменьшении концентрации агента. Колхицин относится к другому типу агентов, нарушающих митоз (разд. 5.4.7). Он эффективен при очень низких значениях термодинамической активности, причем даже небольшие изменения молекулярной структуры существенно сказываются на •его биологическом действии. В заключение следует отметить, что вещества, вызывающие такие грубые и необратимые нарушения, как разрывы хромосом (фенолы, кумарины, иприты), резко отличаются по своим физическим свойствам от тех депрессантов, биологическое действие которых не связано со структурой.
Глава 16
СОВЕРШЕНСТВОВАНИЕ ПУТЕЙ ПОИСКА ЛЕКАРСТВЕННЫХ ВЕЩЕСТВ
Биологически активные вещества с оптимальным набором свойств можно найти, либо используя принцип избирательной токсичности, либо случайно, либо в результате утомительного скрининга. Обычно активность и избирательность действия таких соединений недостаточно высоки, чтобы считать их перспективными для применения в клинической практике. Однако при модификации такого соединения можно получить лекарственный препарат. Каким же образом следует искать более избирательно действующие аналоги среди многочисленных производных? Как можно решить эту задачу, синтезируя и изучая минимальное число соединений?
Некоторые подходы к этому описаны выше. Например,, в разд. 10.7 идет речь о тесте, позволяющем определить, что является действующим началом — ион или неионизированная молекула. Способность соединения образовывать хелаты с катионами металлов можно определить по его структуре (разд. 11.2) в сочетании с быстрым pH-тестом (разд. 11.3). Является ли действие исследуемого соединения структурно неспецифичным, можно выяснить, сравнив минимальную снотворную дозу с соответствующими дозами изученных соединений,, таких как хлороформ, или сопоставив их коэффициенты распределения (разд. 15.0).
Далее пути исследования расходятся. Один из них, в том виде, в котором он использовался в этой книге до сих пор, упорное проведение цитологических и биохимических исследований и изучение распределения препарата по органам и тканям. Другой путь — построение статистически достоверных корреляций между несколькими физическими свойствами базовой молекулы и ее биологическими эффектами. Следует подчеркнуть, что статистика скрадывает неточности экспериментальных данных и может, в лучшем случае, свидетельствовать о правдоподобности этих корреляций. Несмотря на это, статистические методы применяют очень широко, особенно в промышленности,, и, по-видимому, целесообразно дать здесь краткое описание приложения их к обсуждаемой проблеме количественных соотношений «структура — активность» (КССА) химических соединений.
351
16.0. Множественный регрессионный анализ
Наиболее распространенным из этих подходов является предложенный Хэншем в 1968 г. множественный регрессионный анализ, основным уравнением для которого является следующее:
log 1/C = — k(logP)2 + k2(logP) -f- k3crp + k4Es -f- k6.
По существу это уравнение регрессии, приведенное в разд. 3.3, по в него введены два дополнительных члена кзор и k4Es. Наиболее важными переменными в данном уравнении являются: 1) коэффициенты распределения (Р) (в системе этанол — вода); 2) о- и р-константы Гаммета, используемые в аналогичном упомянутом уравнении, основанном на принципе линейности свободных энергий (разд. 17.2), и 3) стерические константы Тафта (Es), позволяющие оценить свободный объем, остающийся между молекулой и ее рецептором при их взаимодействии, а также размер заместителей, введение которых в исходную молекулу не приведет к нарушению взаимодействия из-за сте-рических затруднений (разд. 16.2). Обсудим теперь подробнее каждый из этих типов дескрипторов.
С — наименьшая концентрация, при которой вещество проявляет биологическое действие, и Р — коэффициент распределения см. в разд. 3.3 (т. 1). Коэффициенты распределения большого числа соединений определены экспериментально, величины Р (log Р) несколько тысяч соединений приведены в обзоре Leo и сотр. (1971). Если значения Р изучаемого вещества не известны, лучше определить их экспериментально, например методом, описанным в разд. 17.1. Если результаты нужны срочно или нет экспериментальных источников, величину Р можно рассчитать (приблизительно) как сумму констант ее фрагментов, перечисленных в табл. 17.2. Перед тем как приступить к расчетам подобного типа, следует точно уяснить границы их применимости и случаи, в которых необходимо использовать некоторые модификации [Rekker, 1977].
Физический смысл о- и р-констант Гаммета (как и о-констант Тафта для алифатических соединений) обсуждается в разд. 17.2. Величины, определенные экспериментально, наиболее часто используемые, приведены в табл. 17.4 и в книге Perrin, Dempsey, Serjeant (1981); о- и р-константы см. также Hansch, Leo (1979).
При проведении op-анализа полизамещенных соединений расчеты следует начинать с использования о-констант одного из заместителей или любой комбинации заместителей (суммированием их о-констант) до тех пор, пока не будет получен максимально высокий коэффициент корреляции (г) для коэффициента к3 у этих переменных.
Один из основных недостатков о-констант заключается в том, что они относительно мало отличаются друг от друга, по-352
скольку варьируются в интервале 1,8 логарифмических единиц. Более того, не следует особенно доверять второму знаку после запятой в значениях о-констант, поскольку все они являются результатом вычитания двух логарифмических чисел, сотые доли каждого из которых или их обоих определены недостаточно надежно [Albert, Serjeant, 1984]. Несмотря на эти ограничения, о-константы наиболее широко используют в регрессионных уравнениях для учета электронных эффектов заместителей — эффектов, во многом определяющих изменения биологической активности соединений.
Альтернативный подход, по-видимому еще редко используемый, заключается во введении непосредственно в регрессионное уравнение констант ионизации. Это не только позволяет избежать ошибки, возможной при вычитании двух логарифмических величин (процесс Гаммета), но и дает более точное представление о распределении электронной плотности во всей молекуле в целом. Для веществ, частично ионизированных при физиологических значениях pH, степень ионизации может быть использована в качестве наиболее точного дескриптора. В случае неионизирующихся веществ можно использовать величины дипольных моментов, рассчитанные, в частности, как сумма локальных диполей, вычисленных по диаграмме электронной плотности на основании расчетов методом Паризера — Парра [Kier, 1971].
В качестве дескриптора, учитывающего стерические факторы, до сих пор наиболее широко использовали так называемые стерические константы Es. Эти величины предназначены для введения в регрессионное уравнение параметров, характеризующих: 1) объем заместителей и 2) особенности пространственного строения, которые обеспечивают наличие сродства (для наиболее важных участков молекулы) и возможность доступа лекарственного вещества к рецептору. Константа Es представляет собой стерический параметр, введенный Тафтом и полученный им на основании большого числа разных экспериментов. Тафт исследовал состояния равновесия в реакциях соединений, молекулы которых содержат объемные заместители, так или иначе связанные химическим (но не биологическим) реакционным центром. Хэнш ввел этот дескриптор в свое уравнение и часто его использовал. В тех случаях, когда этот параметр не действует, в качестве альтернативного варианта можно применять величины молекулярной рефракции. Константа Тафта относится к одному заместителю [Taft, 1956], в то время как молекулярная рефракция может быть отнесена как к одному заместителю, так (при суммировании) и ко всей молекуле [Vogel, 1948; Hansch et al., 1973].
Применение какого-либо дескриптора позволяет иногда получить удовлетворительную корреляцию, однако в большинстве случаев одного дескриптора недостаточно. Таким образом, для определения свойств молекулы, обусловливающих ее взаимодей
23—734
353
ствие с рецептором, как правило, необходим набор параметров. Обычно химики (сознательно или бессознательно) пренебрегают последним (стерическим) членом в уравнении Хэнша, сохраняя его постоянным для серии соединений, пространственные характеристики которых, по их мнению, изменяются незначительно. Однако уже из огромного различия в биологическом действии R- и S-стереоизомеров отчетливо видно, что вклад стерического фактора в уравнение Хэнша может быть наиболее важным. Следует отметить, что определить величины констант Es очень трудно. В настоящее время предпринимаются попытки разработать менее сложныё способы количественной оценки стерических эффектов заместителей: их обсуждению посвящен разд. 16.2.
В качестве дополнительного члена в регрессионном уравнении было предложено использовать степень образования водородных связей [Fujita, Nishoika, Nakajima, 1977]; этот и другие потенциально возможные члены уравнения обсуждаются в работе Hansch и Leo (1979). Однако регулярно ни один из них не применяется.
А. Планирование статистической значимости. В связи с высокой стоимостью компьютерного времени, необходимого для корреляционного анализа, очень важно, чтобы необработанных данных было заведомо достаточно для получения результатов, позволяющих сделать значимые выводы как по качеству, так и по количеству. Для этого необходимо, во-первых, иметь статистически достаточное число соединений в обучающей и экзаменуемой выборках. Обе эти выборки должны состоять, по крайней мере, из пяти членов для каждого дескриптора, включенного в уравнение регрессии. Так, если пространственные характеристики молекул в выборках остаются практически неизменными, то каждая выборка должна содержать как минимум десять соединений: пять для дескриптора — коэффициента распределения и пять — для констант Гаммета. Таким образом, в общей сложности необходимо иметь двадцать разных соединений, хотя и родственных химически и структурно, причем эти соединения должны содержать заместители, охватывающие широкий ряд значений двух выбранных дескрипторов. Численные значения констант Гаммета и логарифмов коэффициентов распределения для десяти соединений математически обрабатывают на компьютере по методу наименьших квадратов для определения величин коэффициентов (к).
Результаты затем выражают через n, s и г, где п — число соединений, равное 10; s представляет собой стандартное отклонение регрессии, аг — коэффициент корреляции, который в идеальном случае должен быть равен 1,00, но удовлетворительным считают значения 0,95 и выше. Полученные таким образом коэффициенты (к) используют для второй выборки из 10 соединений (для этой половины компьютер для расчета не применяют). Если в результате получают такие же высокие значения г
354
SO NH
• 2 2 •MeSQ, oonh2
г
no2 ””0,75
•GN
CH>’’cO*Me
cf3so2
CF3 SFs
.OCF3
co2H-'0'25
-0.8 -0.4 f 0,4
—(—i—°—b MeCONH
Cl 0,8
SMe
a—0,25 Me OMe’
1,6
-H
t-Butyl
OH
•NMe2
-0,75.
Рис. 16.1. Взаимосвязь о-констант Гаммета и л-констант Хэнша некоторых наиболее распространенных заместителей [Craig, 1971; Redl, Cramer, Berkoff, 1974].
и низкие значения s, то значит задача решена, так как исследователю ясно, какие именно дескрипторы и в каком соотношении имеют существенное значение в корреляции структура — активность данной выборки веществ [Topliss, Costello, 1972; Hansch, Unger, Forsythe, 1973]. После этого можно переходить к изучению широкой серии соединений, члены которой значительно больше отличаются друг от друга, в том числе стерически. Однако, если для экзаменуемой выборки значения величин s и г будут не столь высоки, как для обучающей выборки, то необходимо пересмотреть постановку задачи и начать решать ее сначала.
При первичном отборе соединений для проведения множественного регрессионного анализа большую помощь может оказать о — л-диаграмма Крэйта (рис. 16.1). Для определения о/л-области максимальной активности заместители в соединениях представленного ряда должны быть выбраны в каждом из четырех квадратов.
При выборе дескрипторов регрессионного уравнения обычно проводят некоторые предварительные исследования. Если в выборке присутствуют два соединения, имеющие резко отличающиеся о-величины, но примерно одинаковую степень биологической активности (1/С), представляется очень удобным исключить значения о-констант и, следовательно, иметь дело с меньшей серией соединений. Однако этого не следует делать. Так, например, при изучении загрязнения озер было установлено токсическое действие на рыб ряда фенолов, значения рКа которых изменяются от И (слабые кислоты) до 5 (сильные кислоты). Оказалось, что токсичность соединений примерно пропорциональна log Р, а включение о-констант не улучшает результата и не позволяет получить удовлетворительные величины
23*
355
s и г. Более тщательное исследование показало, что по мере снижения величины рКа степень ионизации фенола при биологических значениях pH возрастает, и это приводит к увеличению его токсичности в опытах, где фенолу не требуется преодолевать биологическую мембрану. Так как ион практически не способен переходить в неводную фазу, в опытах на интактных рыбах два эффекта ионизации практически гасили друг друга [Saarikoski, Viluksela, 1982].
Б. Обработка результатов. Для получения стандартного отклонения (s) необходимо возвести в квадрат все величины, соответствующие разности между данным значением и средним арифметическим всех полученных значений. Сумма этих квадратов делится на число исследуемых соединений. Квадратный корень из этой величины представляет собой стандартное отклонение. В физико-химических экспериментах обычный предел для стандартного отклонения ±0,02; эта величина так мала потому, что все выпадающие значения взаимно уничтожаются. Однако биологические результаты варьируются в значительно более широких пределах, поэтому в регрессионных уравнениях, подобных уравнению Хэнша, допускаются значительно большие стандартные отклонения (обычно ±0,15). Значение стандартного отклонения заключается в том, что оно показывает, в каких пределах варьируется полученный результат по отношению к среднему значению. Стантартное отклонение используется также при расчете коэффициента корреляции (г). Если г = 0,9, это означает, что 81% всех колебаний х можно отности за счет изменений у; аналогично, если г —0,95, то эта величина возрастает до 90%. Подробнее о статистике такого рода см. Crossland (1981).
В. Исследования без применения компьютера. Исследователям, предпочитающим работать без компьютеров и статистической обработки, большую помощь может оказать «древо решений» Топлисса [Topliss, 1972]. Он начинает с рассмотрения серии соединений, в которой основу родоначальной молекулы составляет бензольное кольцо. В этом случае следует рассмотреть сначала пара-хлорпроизводпое и сравнить его активность с таковой родоначального соединения. Если активность возрастает, этот рост может быть отнесен за счет констант + л, +о, или их обеих, вместе взятых (знаки констант заместителей можно найти в табл. 17.2 и 17.3). Если активность соединений одинакова, то это можно объяснить наличием небольшого положительного л-эффекта (почти все лекарственные вещества обладают небольшим биологически благоприятным л-эффектом), которому противопоставлено неблагоприятное о-взаимодействие. Для проверки этого промежуточного вывода необходимо получить 4-метилпроизводное, которое должно обладать большей активностью вследствие +л- и —о-эффектов. Пониженная активность 4-метилпроизводного по сравнению с 4-хлорзамещен-ным может быть отнесена за счет неблагоприятных стерических
356
эффектов пара-заместителя. Поэтому далее следует получить аналоги, содержащие атом хлора и метильную группу в положении 3. И, наконец, если активность 4-хлорпроизводного значительно ниже активности родоначального вещества, то причина, вероятнее всего, заключается в большем влиянии —о-эф-фекта, так как влияние —л-эффекта проявляется значительно реже. Это следует проверить, получив 4-диметил а минопроизводное, в котором проявление —о-эффектов будет более явным. Топлисс предложил аналогичную процедуру и для боковой цепи: в этом случае следует начинать с монотонного повышения величины л-константы, сохраняя постоянными стерические эффекты, например, отдавая предпочтение циклобутильным заместителям, а не группам с открытой цепью. Дальнейшее развитие эти положения получили в книге Topliss (1977).
Г. Некоторые размышления по поводу регрессионного анализа. Статистика — это часть математики, и хотя ее роль в медицине огромна, она не может заменить метод «проб и ошибок». Множественный регрессионный анализ наиболее полезен тогда, когда он используется не обособленно, а для того, чтобы направить исследования по новым направлениям. Поэтому наиболее ценным результатом применения регрессионного анализа может быть выявление этих ранее недооцениваемых направлений. Например, в ряду хинолина с заместителями в гетероцикле химикам было предложено отобрать 20 соединений с альтернативными заместителями. По окончании работы, по их мнению, был получен представительный ряд. Однако сравнение величин о-констант еще до начала регрессионного анализа показало, что одна часть шкалы о-констант Гаммета в этом ряду не представлена вовсе. Это произошло потому, что любой метод синтеза, якобы позволяющий получить бесчисленное множество продуктов, в действительности всегда приводит к образованию преимущественно соединений, о-константы заместителей которых располагаются в одной части шкалы Гаммета (причина этого достаточно очевидна — от природы заместителя, т. е. от его о-константы, существенно зависит направление реакции). Поэтому для законченного решения поставленной задачи необходимо применять разные методы синтеза, а если подходящей реакции не существует, то искать обходные пути.
Безусловно полезна взаимная критическая оценка работ исследователями, занимающимися множественным регрессионным анализом [например, Cammarata et al., 1970; Martin, 1970]. Всегда следует помнить следующую цитату группы ученых из Pomona School: «Ограниченное пространство не позволяет обсуждать множество примеров, в которых никакая комбинация констант не позволяет получить даже умеренной корреляции» [Hansch, 1971]. После предостережений по поводу переоценки статистических критериев в ущерб здравому смыслу Unger и Hansch (1973) добавляют: «В отсутствие такой качественной перспективы исследователь рискует получить не-
357
Бемегрид (15.3)
Фенитоин
(15.4)
Отличаются ли механизмы действия барбитуратов от других снотворных средств? Действие барбитуратов, по-видимому, качественно отличается от действия бензодиазепинов (к тому же оно слабее), несмотря на то, что барбитураты способны адсорбироваться на рецепторах ГАМК (разд. 12.7). Фактически по своему действию они настолько напоминают структурно неспецифичные депрессанты, что при клинически эффективных дозах им можно приписать механизм действия, предложенный Franks и Lieb (1982) (разд. 15.0). Совершенно очевидно, что зависимость от барбитуратов и симптомы их отмены связаны с системой ГАМК [Но, Harris, 1981].
Барбитураты, применяемые в качестве снотворных, слабо ионизируются в организме, и небольшие количества заряженной формы практически не оказывают влияния на их действие [Mautner, Clemson, 1970; Hansch, Anderson, 1967].
Сразу же после ограничения свободной продажи барбитуратов на фармацевтическом рынке появились аналогичные барбитуратам производные пиперидина (в циклах которых содержится один атом азота), такие как метиприлон (нолудар) и глутетимид (дориден), избирательность которых не выше таковой барбитуратов [Mautner, Clemson, 1970; Hansch et al., 1968].
Бемегрид (р,р-метилэтилглутаримид) (15.3)—мощный антагонист барбитуратов, оказался очень полезным при лечении отравлений ими. Структурное сходство этого соединения с барбитуратами заметить нетрудно, но его аналептическое действие не специфично по отношению к барбитуратам. В разд. 7.5.2 (т. 1) было указано, что увеличение размеров алкильных групп превращает такие стимуляторы в вещества, обладающие снотворным действием [Laycock, Shulman, 1967], тогда как для барбитуратов справедливо противоположное. Как было показано с помощью компьютерного графического анализа, пространственная структура барбитуратов ответственна за проявление ими способности вызывать судороги [Andrews et al., 1983].
Ж. Противоэпилептические препараты. Обладая снотворным действием, фенобарбитал (15.2) к тому же и сильное противо-эпилептическое средство. Этот эффект обусловлен наличием в молекуле фенильного цикла. Фенильные заместители содержатся и в молекулах таких противоэпилептических препаратов, как дифенин (15.4) (5,5-дифенилгидантоин) и мефенитоин (3.32), не обладающих в отличие от фенобарбитала снотворным дейст
348
вием. Противоэпилептические препараты специфически подавляют высоковольтные, высокочастотные разряды, возникающие в мозге во время большого эпилептического припадка [Green-gard, Mclllwain, 1955]. Дифенин способен специфично ингибировать изменение натриевой проницаемости мембран в ЦНС, что, по-видимому, и лежит в основе механизма действия препаратов подобного типа [Perry, McKinney, De Weer, 1978]. Механизм действия вальпроата (дипропилацетата) натрия иной — он ингибирует фермент, разрушающий ГАМК [Bruni, Wilder, 1979].
3. Cannabis indica. Д9-Тетрагидроканнабинол (ТГК) (15.5) — действующее начало марихуаны, одновременно и похож и отличается от биологических депрессантов. Его высокая липофильность (коэффициент распределения в системе октанол — вода примерно 8000), возможно, определяет специфические фармакокинетические характеристики, в частности его медленное распределение по органам и тканям и длительность выведения из организма. Этот препарат приводит к состоянию мягкого наркоза, продолжающегося несколько часов, но не вызывает глубокой анестезии, что напоминает эффект отключения, обнаруженный при изучении зависимости действия алифатических спиртов от постоянного увеличения ОММ (разд. 2.5.1, рис. 2.3) [Paton, 1975].
З'-Гидрокси- и 3',11 -дигидроксипроизводные ТГК, образующиеся в процессе его метаболизма, сохраняют большинство-фармакологических свойств исходного соединения [Handrick et al., 1982]. Поскольку существует несколько альтернативных способов нумерации положений в цикле, следует отметить, что 3' означает замещение в боковой цепи, all — в метильной группе соединения (15.5).
Ate I
Тетрагндроканнабннол
(15.5)
При низких дозах седативный эффект, получаемый при курении марихуаны, подобен тому, который вызывают барбитураты, но с более ярко выраженной эйфорией, сопровождающейся неконтролируемым потоком мыслей, нарушением восприятия времени и места. Действие одной сигареты продолжается в течение 2—3 ч, во время которых у курильщика настолько снижается координация, что он неспособен управлять автомобилем (понижена способность к принятию решений и замедлено время
34»
ких статистических единорогов — животных, существующих только на бумаге, а не в действительности. Так, например, недавно стало очевидно, что можно коррелировать серию зависимых переменных, используя случайные числа в качестве независимых переменных». См. также Topliss и Costello (1972).
Подробнее о соотношениях свободных энергий, которые обсуждались выше, см. Chapman, Shorter (1972), а также перечень работ в конце этой главы.
16.1. Альтернативные методы
А. Аддитивный метод Фри — Вильсона. Этот метод представляет собой усовершенствованную старую идею о том, что биологическая активность — это аддитивное свойство, в которое каждый заместитель вносит определенный математический вклад [Free, Wilson, 1964]. Как отмечал Хэнш [Hansch (1971)], этот подход совершенно не обременен попытками приложения термодинамики и физической химии. Вместо этого для определения констант заместителей методом наименьших квадратов используют компьютер, что позволяет по данным биологической активности, полученным в стандартных лабораторных тестах, найти «наилучшие положения» каждого заместителя в молекуле лекарственного вещества. На основании этих констант можно оценить степень биологической активности при комбинации разных заместителей в одной молекуле. Преимущество метода Фри — Вильсона заключается в том, что его можно использовать, когда нет никаких физических данных. Однако в этих случаях данные для уравнения множественного регрессионного анализа могут быть взяты из таблиц. Недостаток метода Фри — Вильсона заключается в том, что необходимо иметь много примеров соединений, содержащих данный заместитель, чтобы получить решение уравнения со множеством неизвестных. Поэтому этот метод обычно применяют для усовершенствования тех лекарственных веществ или пестицидов, для которых можно получить большое число очень близких по структуре аналогов. Подробнее об этом методе см. James (1974) и Purcell, Bass, Clayton (1973).
Б. Квантово-химические методы. В начале 50-х годов Pullman и Pullman применили квантовую механику для изучения связи канцерогенности полициклических углеводородов с их структурой. С тех пор было предпринято немало попыток расширить метод КССА, используя квантово-химические расчеты. Основные препятствия для достижения успехов в этой области это: 1) высокая стоимость компьютерного времени и 2) абстрактная природа получаемых дескрипторов, которые трудно представить химикам-практикам и биологам. Кроме того, в связи с большим количеством параметров, получаемых для каждого соединения, необходимо, чтобы множество аналогов давали хорошую корреляцию. По-видимому, наиболее успешным можно считать применение этого метода для корреляции конформаци
358
онных параметров с биологической активностью, как, например, при создании новых антагонистов и агонистов, родственных природным нейромедиаторам [Weintraub, Hopfinger, 1973]. Информация, получаемая с помощью метода молекулярных орбиталей, относится в основном к электронным свойствам, однако в следующем разделе (16.2) мы рассмотрим некоторые попытки использования таких расчетов для изучения пространственного строения лекарственных веществ. Несомненно, что в дальнейшем квантовая химия будет играть важную роль в объяснении и прогнозировании биологических эффектов лекарственных веществ, однако в настоящее время для расчетов используют такое множество разных методов, что очень трудно провести сравнение результатов, полученных разными авторами.
Подробнее об этом методе см. Rihards (1983), Kier, (1971), Purcell, Bass, Clayton (1973).
В. Молекулярная связность. В химии нефти давно используются расчетные методы, учитывающие разветвленность молекул, для предсказания таких физических свойств, как температура кипения, вязкость, теплота образования. Было обнаружено, что эти свойства сильно отличаются у разных изомеров и зависят от степени компактности молекулы. Со временем подобный подход был выражен в количественной форме в виде матриц [Randic, 1974], a Kier нашел ему применение в биологии [Kier, Hall, 1976]. Преимущество этого метода заключается в том, что расчеты могут быть сделаны быстро с помощью портативного калькулятора и при этом нет необходимости измерять физические свойства исследуемых соединений. Этот метод можно применить отдельно для липофильных соединений, причем получаемые с его помощью результаты совпадают с результатами, получаемыми по методу Хэнша (logP), что было показано при изучении общих анестетиков и действия наркотических веществ на головастиков. Для амфифильных соединений этот метод может быть использован с целью приблизительной оценки объема молекулы; в этом случае его можно комбинировать с экспериментально определенными значениями logP и константами Гаммета в уравнении множественного регрессионного анализа. Однако данный метод обладает серьезным недостатком, так как он не отражает тонких стереохимических различий, например, между цис- и транс-изомерами, R и S оптическими изомерами или между конформерами, т. е. свойств, от которых в значительной степени зависит биологическая активность вещества. Другой недостаток этого метода состоит в том, что для его применения из-за множества вводимых индикаторных переменных1 (по одной на каждый атом в молекуле, кроме водорода) необходимы серии, включающие большое число соединений.
1 В отличие от дескриптора, который может иметь полный набор значений, индикаторная переменная обозначает свойство, которое либо присутствует, либо отсутствует.
359
Расчеты методом молекулярной связности начинают с изображения структурной формулы в виде связей, не включая в нее атомы водорода. Атом, находящийся в конце каждой связи или в месте соединения связей, называется вершиной (вертекс) и ему приписывают значение, равное числу связанных с ним атомов, отличных от водорода. Следовательно, для атома углерода возможны цифры 1, 2, 3 или 4, называемые «вершинные валентности». Затем валентности, располагающиеся на концах каждой связи, перемножают и получают величину, называемую «произведением вершин» («вертекс-произведением») или «краевым членом» и обозначаемую буквой «V». Затем рассчитывают величины, обратные корню квадратному из каждого «краевого члена». Сумма этих величин представляет собой «индекс связности» для данного соединения и обозначается как «х». Так, например, индекс связности х для изомера гептана триметил-бутана составляет 2,943 или (2x0,577 + 3X0,5 + 0,289), а для нормального гептана 3,414. Различие между этими двумя величинами служит мерой различий в разветвленности двух молекул.
Чтобы применить свой метод к большинству лекарственных веществ, Kier и Hall вынуждены были рассчитать «вершинные валентности» для гетероатомов азота, кислорода, серы, которые в предыдущих работах не рассматривались. Эти величины иногда удавалось получить эмпирическим путем, а в некоторых случаях с помощью регрессионной программы расчета. Поскольку валентные углы у гетероатомов больше, чем у тетраэдрического атома углерода, величины их «вершинных валентностей» выше. Сложности возрастают в случае гетероаромати-ческих молекул, содержащих больше одного гетероатома.
16.2. Стереохимический подход
Количественно описать объем и конформацию молекулы в целом или отдельных ее фрагментов — это пока недостижимая цель для исследователей, занимающихся корреляционным анализом. С помощью молекулярных моделей легко представить себе, как именно агонист взаимодействует с рецептором, понять таким образом их пространственное строение. Этот подход, но в значительно более сложном виде, применен в системе цветных рисунков, созданной Evans и Sutherland (разд. 12.0). С помощью таких наглядных методов можно с достаточной правдоподобностью моделировать взаимодействие комплементарных участков молекул лекарственного вещества и рецептора, дополняющих друг друга по липофильным свойствам, электрическим зарядам и (или) пространственному строению. Так осуществляют визуальный контроль пространственного соответствия, причем в рассматриваемую молекулу или ее фрагмент может входить множество разных атомов. Точно так же можно изучать конформационные изменения молекулы и лекарственного вещества, и рецептора.
360
Эти представления значительно сложнее выразить математически и не менее трудно представить зрительно, если они выражены численно в виде коэффициентов у неопределенных дескрипторов. В таком случае химик может синтезировать аналоги наиболее активного лекарственного вещества, содержащие объемные заместители в различных положениях. Затем с помощью биологических тестов можно получить информацию о пространстве, окружающем каждое положение в молекуле. Поскольку эта работа слишком трудоемка, усилия часто направляются на быстрое достижение цели с помощью расчетов.
Некоторые методы направлены на изучение конфигурации целой молекулы, определяющей, по-видимому, некоторые типы биологических ответов. Так, Holan установил, что высокой инсектицидной активностью обладают лишь те аналоги ДДТ и пи-ретрина, кеторые сохраняют клиновидную конфигурацию (и размер) исходной молекулы (клинья подобно мельчайшим шипам вскрывают натриевые каналы насекомых) (разд. 7.6.5), хотя, возможно, что действие большинства агентов определяется главным образом конфигурацией некоторых определенных участков молекулы. Для таких соединений с помощью описываемых методов были получены данные для отдельных фрагментов молекулы и предприняты попытки определить, какой именно участок (и почему) определяет биологическую активность. Применение этих методов объясняется большим желанием исследователей оптимизировать активность уже известных лекарственных соединений.
16.2.1. Подходы, основанные на учете всего объема молекулы
Биологическое действие хлорированных циклодиеновых инсектицидов резко отличается от действия инсектицидов группы ДДТ — пиретрина, стерические требования для этих соединений также совершенно иные. В 1965 г. Soloway исследовал 106 циклодиенов и пришел к выводу, что для проявления биологической, активности эти соединения должны иметь конфигурацию, близкую к сферической. Он спроецировал модель молекулы альдрина на плоскость и получил круг с очень неровными краями;, эффективными инсектицидами являются те вещества, молекулы которых могут проникнуть в отверстие таких размеров [см. формулу (6.47)]. Кроме того, в молекуле обязательно должны присутствовать два электроноакцепторных заместителя (обычно-атомы хлора).
Полезнее этих общих представлений о конфигурации молекул был бы метод расчета их ван-дер-ваальсовых объемов, т. е. объема, занимаемого молекулой с учетом ван-дер-ваальсовых радиусов краевых атомов (разд. 8.0.4). Обычно его рассчитывают косвенным путем с помощью молекулярной рефракции (см. ниже). Были предприняты попытки определить этот объем*.
36 k
^покрывая молекулярные модели целлюлозой, обладающей адгезионными свойствами, с последующим погружением их в воду и измерением объема вытесненной воды [Leo, Hansch, Jow, 1976]. Однако при использовании этого способа легко допустить просчеты [некоторые из них указаны Pearlman (1980)]. Компьютерный расчет ван-дер-ваальсовых объемов на основе геометрических данных связан с определенными трудностями (например, окклюзия частей атомов, изображаемых в виде сфер), даже, если при этом применяют рекомендуемый метод суммирования долей1. Pearlman (1980) предлагал использовать в корреляционном анализе величины площадей поверхности молекул, которые линейно связаны с их объемами, но рассчитываются .значительно легче.
В методе «коэффициентов подобия», созданном Amoore (1964), для корреляции запаха вещества со структурой его молекулы использован другой подход к определению объема молекулы, которым незаслуженно пренебрегают. Вкратце метод заключается в том, что Amoore фотографировал молекулярные модели с трех точек, расположенных под прямым углом друг к другу. На этих фотографиях он вычерчивал радиусы с интервалом 10°, а затем суммировал 108 измерений. Это позволяло получить достаточно хорошую корреляцию, несмотря на низкую точность биологического теста, использованного Amoore (оценка интенсивности запахов людьми). Простота метода Amoore позволяет рекомендовать его для решения некоторых фармакологических задач, однако он нуждается в системе более объективных биологических тестов.
Группа румынских исследователей под руководством Bala-Ьап разработала три метода, предназначенных для установления соответствия между лекарственным веществом и его рецептором на основе объемных соотношений. Эти методы получили название МСР, ,'МТР и РМК (расположены в порядке увеличения сложности).
Для использования метода МСР (минимальные стерические различия) необходимо выбрать вещество, обладающее наиболее высокой биологической активностью данного типа. Затем следует нарисовать ето структурную формулу в виде схемы связей, пропуская связи с атомами водорода. Аналогичным образом все исследуемые вещества со сходной химической структурой изображают на прозрачном материале и накладывают на изображение молекулы исходного соединения. Затем несовпадающие .атомы отмечают звездочкой, не обращая при этом внимания на объемность структуры молекул (например, не делают различий между бензолом и циклогексаном). Каждому атому, отмечен
1 В этом случае.под долями подразумевают сегменты сферы, напоминающие приблизительно одну из четвертей апельсина. Pearlman рассчитал большое количество таких сегментов для каждого атома, уменьшая объем в случаях взаимодействия соседних атомов (ван-дер-ваальсовы силы отталкивания частично перекрываются внутри молекулы).
362
ному звездочкой, приписывают «штраф» (или «число несовпадения»), равный 1 для С, N или О, но больший для тяжелых атомов, т. е. 1,5 для S и С1 и 2 для I. Хотя различиями в длинах связей и величинах углов между ними пренебрегают, следует быть осторожным и не совмещать атомы, соединенные связью, с несвязанными атомами, так как ковалентная связь (обычно от 0,1 до 0,2 нм) значительно короче тех расстояний, на которых проявляются ван-дер-ваальсовы взаимодействия (0,3—0,4 нм).
По мнению Balaban, сродство лекарственного вещества к рецептору является линейно уменьшающейся функцией суммы пеперекрывающихся объемов потенциальных лекарственных веществ и полости рецептора. Предполагают, что наиболее активное лекарственное вещество отражает общую формулу полости рецептора. Однако, если заместитель в самом активном лекарственном веществе не всегда перекрывается заместителями в других соединениях, его следует удалить, так как (по расчетам Balaban) на изменение конформации рецептора, обеспечивающее его подстраивание под молекулу лекарственного вещества, требуется 0,85 ккал. Поэтому удаление этого заместителя должно привести к образованию более активного лекарственного вещества. Этот и два других метода более подробно изложены в книге Balaban и сотр. (1980). Метод МСР больше всего подходит для больших молекул, в которых необходимо заменить только один небольшой фрагмент. Декапептид, известный как лютеинизирующий рилизинг-гормон, становится в 2,48 раза более активным после замены концевого остатка глицинамида метиламином. Метиламин не получает «штрафа» по методу МСР, тогда как глицинамид получает три «штрафа», и это их отличие открывает новые возможности для совершенствования биологического действия природного гормона [Fujino et al., 1973; Balaban et al., 1980]. Для молекул меньшего размера метод МСР дает не такие хорошие результаты, как метод молекулярной рефракции и метод Verloop (см. ниже). Подробнее о МСР см. Simon и сотр. (1984).
При использовании второго метода Balaban — МТР (минимальные топологические различия), наиболее подходящего для циклических и других молекул, имеющих жесткую структуру, также следует выбрать наиболее активное лекарственное вещество и нарисовать его структурную формулу в виде схемы связей, пропуская связи с атомами водорода. Однако в этом методе используют также углы между связями и другие пространственные характеристики. Затем совмещают формулы потенциальных лекарственных веществ и отмечают уровни расположения атомов над и под средней плоскостью. Это делают для того, чтобы определить сложную «гипермолекулу», предназначенную для взаимодействия с трехмерной полостью рецептора. При совмещении формул полярную группу помещают над полярной, а атом, образующий водородные связи, над подобным ему атомом. Счет ведется следующим образом: 1 присваивается атому,
363
касающемуся стенки полости рецептора; 0 — атомам, попадающим внутрь этой полости, и —1—тем, которые находятся вне полости. Это распределение проводится экспериментально или. с помощью компьютерной программы, опубликованной в книге Balaban. Сумма величин +1 хорошо коррелирует с биологиче-, ской активностью. Этот метод был применен к серии веществ,, обладающих эстрогенной активностью [Popoviciu, 1978]. Кор-, реляция оказалась очень удачной (г = 0,971) для серии из-25 стероидов, содержащих кислород в положениях 3 и 17, но коэффициент корреляции уменьшился до 0,741 при исследовании 25 других стероидов, еще хуже корреляция для нестероидных эстрогенов. Преимуществом этого метода является то, что он позволяет обнаружить заместители или участки молекул, лежащие вне предполагаемой полости рецептора и которые можно удалить. Кроме того, он может дать информацию о незанятых областях рецептора и, следовательно, указать новые потенциальные лекарственные вещества, которые могли бы занять эти области. МТР не подходит для гибких (алифатических) молекул, в этих случаях лучше использовать МСР.
Третий метод Balaban — РМК (расчеты по методу Монте-Карло), создан для расчета ван-дер-ваальсовых объемов молекулы в нм3. Этот расчет начинают с составления программы для: получения большого набора случайных величин, которые используются в совокупности с формулой молекулы лекарственного вещества для расчета огромного числа их радиусов, а по радиусам — объемов. Хотя этот метод достаточно сложен, с его помощью были получены результаты, подтверждающие возможность использования величин молекулярной рефракции в качестве меры ван-дер-ваальсовых объемов (см. ниже).
16.2.2. Подходы, учитывающие специфические участки молекул
При изучении равновесия в процессах гидролиза сложных эфиров, а также стерических препятствий, создаваемых соседними группами в этих соединениях, Taft (1953) ввел теперь уже хорошо известные стерические параметры (Es). И хотя он не предвидел возможности использования их в биологии, Хэнш в 1965 г. предложил включить эти параметры в уравнения множественного регрессионного анализа в качестве меры стерических препятствий, оказываемых заместителем при адсорбции наиболее активной группы лекарственного вещества на рецепторе. Величины этих параметров можно найти в работах Taft (1953) и Hansch, Leo (1979). Величины Es могут составлять О для метильной группы, —0,9 — для фенила, —1,5 — для трет-бутильного заместителя и —2,1—для трихлорметильной группы; возможны и более высокие значения для других замести-, телей.
364
Некоторые константы Тафта можно использовать в одном регрессионном уравнении, как это сделали Хэнш и сотр., изучавшие предупреждающее действие замещенных бензойных кислот на взаимодействие антигена яичного белка с его антителом, при котором каждая молекула бензойной кислоты включается в антиген в качестве гаптена [Kutter, Hansch, 1969]. Они получили следующее уравнение множественного регрессионного анализа:
log К = 0,863E°s + 0,081 Esm + 0,446Es₽ - 0,695. и =22 s=0,177 г = 0,974
В этом уравнении имеется член, характеризующий каждое из трех положений замещения, причем очевидно, что наибольший вклад вносят заместители в орто- и пара-положениях. Дополнительное введение logP или константы Гаммета приводит .лишь к небольшому улучшению потому, что коэффициент корреляции (г) и так очень высок.
Максимальная рефракция, несмотря на название, определяется отдельными заместителями, хотя их величины можно использовать в совокупности для характеристики всей молекулы в целом. Наиболее удачным методом расчета величин молекулярной рефракции (MR) является применение уравнения Лоренц-Лоренца, позволяющее получить величину M.R в см3/моль:
MR = MW(n2 — 1): d(n2 + 2), где d — плотность жидкости, ап — показатель преломления. Многие значения MR, полученные разными способами, приведены в работе Vogel (1948), а также в книге Hansch, Leo (1979). В 1973 г. Хэнш ввел величину MR в уравнение множественного регрессионного анализа в качестве стерического параметра как альтернативу Е5-константам (он предупреждал, что MR имеет также электронную компоненту, однако стерическая компонента является преобладающей) [Hansch et al., 1973].
При использовании «стеримолекулярного» метода, предложенного Verloop, структурные формулы каждой группы в молекуле лекарственного вещества (включая водород) обозначают одиночными линиями, и эту информацию вводят в компьютер последовательно, описывая атом за атомом. Дополнительно вводят также информацию о длинах и углах связей и ван-дер-ваальсовых радиусах атомов. Это позволяет рассчитать ван-дер-ваальсову огибающую вокруг рассматриваемой группы. Затем программа по команде выдает численное значение (L)’ для длины этой группы, начиная с точки присоединения, и измеряет под прямыми углами к длинным осям величины минимальной ширины (EL) и максимальной ширины (В4). Голландские авторы также опубликовали таблицы величин (L) и (В) для всех обычных групп и многих их последовательностей. Например, для группы —CH2CN стеримолекулярные величины
365
составляют 3,99 (L), 1,52, 1,90 и 4,12 (В1-4) [Verloop, Hoogen-straaten, Tipker, 1976].
В этом методе не используется информация о конформации изучаемых соединений, за исключением того, что авторы всегда рассматривают форму молекулы с самой низкой энергией (наиболее вытянутую), считая, что дальнейшие превращения молекулы происходят на рецепторе. Возвратившись к анализу предупреждающего действия бензойных кислот на соединение антигена с антителом, выполненному Kutter, Hansch (см. выше), Verloop и сотр. (1976) заменили использованное ими уравнение другим:
[ogK= — 1,46В!0 + 0,50В4° 4-0,84.
и = 36 8 =0,302 г = 0,955
Следует отметить, что величина (L) при таком анализе значения не имеет, поэтому Verloop и сотр. могли включить в корреляцию большой ряд соединений, которые Kutter и Hansch вынуждены были отбросить как «выпадающие».
При изучении серии ларвицидов голландские ученые получили прекрасную корреляцию, сочетая величины коэффициентов распределения и ц-констант Гаммета с двумя «стеримоле-кулярными» параметрами: (L)2 и (В4). В последующих работах метод Verloop был усовершенствован для повышения его прогнозирующей способности; была выпущена новая таблица значений параметров, используемых в этом методе для разных заместителей, а также выведено арифметическое соотношение между величинами (BJ и (Es) [Verloop, 1981, 1983].
При использовании метода Verloop желательно синтезировать ряд аналогов, в котором максимальная (или минимальная} ширина молекул разных соединений располагается в разных точках длинной оси молекулы.
В методе анализа молекулярной конфигурации Hopfinger предложен новый подход для нахождения стерического члена уравнения регрессионного анализа. Его основным дескриптором является объем стерического перекрывания (Vo), представляющий собой долю молекулярного объема, общую для потенциального лекарственного вещества и его конформера, обладающего наибольшей биологической активностью [Hopfinger, 1980]. Величины Vo получают путем сложных расчетов, в которых преимущественно применяют методы молекулярной механики (с использованием как вращательных, так и трансляционных компонент), а не методы молекулярных орбиталей.
Для молекул с гибкой структурой, с помощью программы CHEM.LAB (доступной благодаря CIS, см. разд. 17.4) можно изменять торзионный угол (углы) с интервалом в 30°, до получения одной или нескольких конформаций с минимальными энергиями. Затем строится график зависимости величин энергий (выраженных в ккал/мол) от величины угла, для того чтобы найти минимумы потенциальной энергии. Модели таких графи
366
ков для веществ с наиболее высокой биологической активностью-’ могут служить стандартами для классификации других соединений (по разнице в энергии). Если какой-либо член ряда имеет более одного стабильного конформера, каждый из них рассматривается отдельно. Данные для каждого члена ряда объединяют (в виде матриц) с величинами длин ван-дер-ваальсовых связей и углов связей и рассчитывают величины объемов стерического перекрывания. Хотя этот метод позволяет получить сведения о молекуле в целом, в нем используются данные о конфигурациях отдельных участков.
H2Nz N NH2
2,4-Диамиио-5-бензилпиримидии
(16.1)
Примером использования метода Hopfinger может служить обработка серии 2,4-диамино-5-бензилпиримидинов (16.1), самым активным из которых является антибактериальный препарат триметоприм (9.32) (разд. 9.3.3). По результатам теста (ингибирования ДГФР печени быка)1 было составлено следующее уравнение:
logl/C = -21,3V0+ 1,39(V0)2 + 0,44л3>4 + 52,2.
п = 23 s=0,l ’ r= 0,93
В молекулах соединений этой серии, обладающих высокой активностью против ДГФР Е. coli, угол между бензольным кольцом и метиленовым мостиком составляет 30°, а последний расположен под углом 90° к пиримидиновому циклу (что установлено с помощью конформационного анализа, описанного выше) . Следовательно, бензольное кольцо в этих молекулах расположено примерно перпендикулярно пиримидиновому циклу и почти покрывает его. Для членов этого ряда, наиболее активных по отношению к ферменту млекопитающих, более выгодной конформации не найдено.
В приведенном выше уравнении константа отражает липофильность (вычислена из величин log Р, см. разд. 3.3, т. 1),. а индексы 3 и 4 указывают на то, что в данном случае использована сумма л-констант заместителей в положениях 3 и 4. Попытки ввести в это уравнение а-контакты Гаммета не привели к улучшению результатов. Данные, полученные Hopfinger,
1 Это тест на токсичность для больных, так как триметоприм малоэффек-
тивен по отношению к ДГФР млекопитающих (см-..табл; 4.2, том 1).
367
оказались почти точной копией результатов Blaney и сотр,, которые вывели следующие уравнения (1979):
log 1/С = 0,62л3 + 0,32о31 4> 5 + 4,99. n=23 s=0,15 г = 0,93
Таким образом, последние два уравнения отчетливо показали уровень, на котором находится современный регрессионный анализ в приложении к биологии: отличная корреляция была получена как с учетом всей доступной информации о стериче-«ских параметрах, так и без нее.
Hopfinger исследовал также ряд производных 2,4-диамино-6,6-диметил-фенил-1,3,5-триазина, включающий в том числе известный противомалярийный препарат циклогуанил (3.35) — триазин, содержащий атом хлора в положении 4 бензольного кольца. В приведенном ниже уравнении С обозначает дозу вещества, вызывающую 50% ингибирование ДГФР опухоли молочной железы. При этом были рассмотрены 35 соединений, отличающихся по ингибирующей активности в 10 000 раз. При применении So была получена лучшая корреляция, чем при использовании Vo; параметр So есть не что иное, как Vo2/3 с той же размерностью, что и площадь поверхности, но, как предупреждали авторы, резко от нее отличающийся и поэтому трудно представляемый наглядно [Hopfinger, 1980]. Тем не менее было получено уравнение с хорошими корреляционными параметрами:
logl/C = - 1,47S3 + 0,02(S3)2+0,38л3< 4 — 17,1. n = 25 s=0,40 г = 0,96
Это уравнение следует сравнить с уравнением, полученным группой американских ученых для более широкой серии соединений, включающей и все те, которые изучал Hopfinger:
log 1/C =0,89it3 — 10,13(«s)s + 0,15MR4 + 6,62. n = 65 sc=0,33 r = 0,91
Изучение другой серии ингибиторов ДГФР — 2,4-диамино-хиназолинов, методами анализа молекулярной формы [Batters-hell, Malhotra, Hopfinger, 1981] дало следующие уравнения:
log 1/C = 0,35So — 0,002(S0)2 + 0,49it5j e —0,090 — 6,95, n=>35 st=0,36 r=0,97
где 6 — двугранный угол, образуемый фенильными циклами в в положениях 5 и 6.
Позже Hopfinger описал новый метод анализа молекулярной формы, основанный на использовании полей молекулярной потенциальной энергии [Hopfinger, 1983]. В этом случае рассчитывают сумму энергий ван-дер-ваальсова отталкивания и притяжения зарядов с фиксированным положением. Как и прежде, никакого внимания не уделяют рецептору, за исключением предположения о том, что наиболее активное вещество является зеркальным отражением структуры рецептора. Де-368
скрипторы были определены вычитанием полей попарных наложений молекул лекарственных веществ друг на друга. Применение этого метода для обработки данных об активности обсуждавшейся выше серии 2,4-Диамино-5-бензилпиримидинов привело к тем же выводам, при условии одновременного включения в уравнение константы лз,4.
16.2.3. Заключение
В этой важной области в настоящее время ведется значительно больше работ, чем было упомянуто здесь, но многое еще предстоит сделать. Поскольку очищенные рецепторы становятся доступными в достаточных количествах, могут быть достигнуты большие успехи в экспериментальных исследованиях, особенно в рентгеноструктурном анализе, подобно тому, как это было сделано с рецепторами АХ (разд. 12.6) и лекарственных веществ, ингибирующих ДГФР (разд. 9.3).
О стратегии и тактике исследований в области КССА (корреляционный анализ) см. Topliss (1983), Hahn (1975) и Martin (1978, 1979); о физико-химических свойствах лекарственных веществ и методах их определения см. Hansch, Leo (1979), Yalkowsky, Sinkula, Valvoni (1980), Lyman, Rechl, Rosenblatt (1982).
24—734
Глава 17
НЕКОТОРЫЕ КОЛИЧЕСТВЕННЫЕ ДАННЫЕ
17.0. Расчет степени ионизации по данным рКа и pH
Табл. 17.1, в которой приведены значения степени ионизации (в %), вычисленной из величины рКа и pH, появилась еще в первом издании этой книги (1951).
Для кислот этот расчет осуществляется по формуле (1), а для оснований — по уравнению (2):
„ юо
Степень ионизации (%) =-------- . (!') для кислот
1 + 10(рКа-рН) ,100 Степень ионизации (%) =-------- . (2) для основании
1 + 10(рН-рКа>
17.1. Фрагментарные гидрофобные константы и коэффициенты распределения
Эта глава дополняет разд. 3.3 и посвящена обсуждению вопросов использования коэффициентов распределения.
Коэффициенты распределения твердых и жидких веществ удобно измерять в плотно закрытых центрифужных стаканах объемом 250 мл, куда помещают навеску изучаемого вещества и оба растворителя в соотношении, установленном ранее для других подобных соединений или определенном в предварительных опытах. Стакан закрывают и интенсивно встряхивают 100 раз в течение 5 мин (хотя равновесие вполне может установиться уже в первые 2 мин). Смесь центрифугируют, дают отстояться и проверяют, не образовались ли хотя бы следы эмульсии (если она образуется, опыт проводят заново). После этого в обеих фазах определяют количество растворенного вещества [Leo, Hansch, Elkins, 1971].
Для определения коэффициента распределения газа его пропускают через смесь двух растворителей, затем берут пробу из органического слоя и анализируют ее с помощью газожидкостной хроматографии [Leo et al., 1975; Hansch et al., 1975].
Таблица, в которой приведены значения 5800 коэффициентов распределения, опубликована в работе Leo, Hansch, Elkins (1971), некоторые из них даны в табл. 17.3.
В табл. 17.2 представлены фрагментарные константы (f), рассчитанные из значений коэффициентов распределения в сис-370
Таблица 17.1. Степень ноннзацни, вычисленная по данным рКа и pH
рКа — pH Концентрация аннона, % Концентрация катиона, % рКа-рН Концентрация аниона, % Концентрация катиона, 1%'
—6,0 99,99990 0,0000999 0 50,00 50,00
—5,0 99,99900 0,0009999 +0,1 44,27 55,73
—4,0 99,9900 0,0009990 +0,2 36,68 61,32
—3,5 99,968 0,0316 +0,3 33,39 66,61
—3,4 99,960 0,0398 +0,4 28,47 71,53
—3,3 99,950 0,0501 +0,5 24,03 75,97
—3,2 99,937 0,0630 +0,6 20,07 79,93
—3,1 99,921 0,0794 +0,7 16,63 83,37
—3,0 99,90 0,09991 +0,8 13,70 86,30
—2,9 99,87 0,1257 +0,9 11,19 88,81
—2,8 99,84 0,1582 +1,0 9,09 90,91
—2,7 99,80 0,1991 +1.1 7,36 92,64
—2,6 99,75 0,2505 + 1,2 5,93 94,07
—2,5 99,68 0,3152 + 1,3 4,77 95,23
—2,4 99,60 0,3966 + 1,4 3,83 96,17
—2,3 99,50 0,4987 + 1,5 3,07 96,93
—2,2 99,37 0,6270 + 1,6 2,450 97,55
—2,1 99,21 0,7879 + U 1,956 98,04
—2,0 99,01 0,990 + 1,8 1,560 94,44
—1,9 98,76 1,243 +1.9 1,243 98,76
—1,8 98,44 1,560 +2,0 0,990 99,01
—1,7 98,04 1,956 +2,1 0,7879 99,21
—1,6 97,55 2,450 +2,2 0,6270 99,37
—1,5 96,93 3,07 +2,3 0,4987 99,50
—1,4 96,17 3,83 +2,4 0,3966 99,60
—1,3 95,23 4,77 +2,5 0,3152 99,68
—1,2 94,07 5,93 +2,6 0,2505 99,75
—1,1 92,64 7,36 +2,7 0,1991 99,80
—1,0 90,91 9,09 +2,8 0,1582 99,84
—0,9 88,81 11,19 +2,9 0,1257 99,87
—0,8 86,30 13,70 +3,0 0,09991 99,90
—0,7 83,37 16,63 +3,1 0,0794 99,921
—0,6 79,93 20,07 -4-3,2 0,0630 99,937
—0,5 75,97 24,03 +3,3 0,0501 99,950
—0,4 71,53 28,47 +3,4 0,0398 99,960
—0,3 66,61 33,39 +3,5 0,0316 99,968
—0,2 61,32 38,68 +4,0 0,0099990 99,9900
—0,1 55,73 44,27 +5,0 0,0009999 99,99900
+6,0 0,0000999 99,99990
теме октанол — вода. Константы для сложных фрагментов могут быть вычислены суммированием приведенных в таблице значений для более простых фрагментов. Константы f, заменяющие л-константы, в большинстве случае превышают их на 0,2 единицы. Они выбраны из 100 значений, рассчитанных по результатам 1 000 измерений logP.
Из данных табл. 17.2 видно, что введение углеводородного радикала в молекулу повышает ее липофильность. Наличие двойной или тройной связи повышает гидрофильность, но вклад последней существенно выше (—0,55 и —1,42 соответственно). Для алифатических соединений величина вклада фенильной
24*
371
Таблица 17.2. Некоторые фрагментарные константы для органических молекул [Rekker, de Kort, 1979]
А. Фрагменты, не содержащие углерода и водорода В. Фрагменты с атомом углерода, ие содержащие водород
Алифатические Ароматические Алифатические Ароматические
—I —Вг —CI —F —N< —no2 —О— —so2n —so— —SO2— —s— 0,56 0,26 0,06 0,49 —2,07 —0,93 —1,59 —2,75 —0,51 1,43 1,21 0,93 0,38 —0,91 —0,07 —0,43 —2,39 —2,05 —1,87 0,11 —с— —CN —CON< —СО— —СО2— —СО2— —SCN -ссь —CFS -О—С (=О) — 0,13 -1,5 —2,88 —1,67 —1,27 —5,00 —0,34 2,09 0,75-—0,41 0,19 —2,02 —0,81 —0,41 —4,14 1,33 —0,99
Б. Фрагменты, имеющие атомы водорода и не содержащие углерода
—н —NH— —ОН— —NH2 —SH —SO2NH2 0,17 — 1,82 —1,47 —1,42 0,00 —1,48 0,17 —0,95 —0,31 —0,85 0,62 — 1,94
Г. Фрагменты, содержащие атомы углерода н водорода Д. Гетероциклические фрагменты
Алифатические Ароматические Имидазолил= Пирролил= Пиридил= Хинолил= Акридинил= Ин долил= Бензимидазолил= Урацилил= Барбитурил= —0,10 0,60 0,53 1,82 3,11 1,89 1,19 —1,31 — 1,55
—СНз1 —СН2— —СН —СН = СН2 -С6н5 —с6н4 —CONH —О—CONH- —СО2Н —CONH» —NHSO2CF3 0,69 0,53 0,33 0,91 —2,43 —1,92 —0,94 —1,97 1,84 1,66 — 1,56 —1,34 —0,08 —1,10 1,22
1 Углеводородным заместителям в бензольном кольце давы значения для алифатических фрагментов.
372
Таблица 17.3. Коэффициенты распределении лекарственных веществ между собой и октанолом1
Вещество Структурная формула К» log Р
Тетрациклин 11.36 —1,47
Аденозин 9.72 —1,10
Сульфазин 9.9 —0,13
Кофеин 7.52 —0,07
Сульфаметазол 9.13 0,54
Морфин 7.35 0,76
Эфедрин 12.12 0,93
Преднизолон 1,42
Атропин 7.16 1,79
Бензилпенициллин 5.13 1,83а
Новокаин 7.8 1,87
Дифенилгидантоин 15.4 2,47
Эритромицин 4.47 2,48
Диазепам 12.95 2,82
Нафталин 3,37
Прогестерон 3,87
Совкаин 7.15 4,40
Имипрамин 12.114 4,62
Аминазин 12.110 5,16
1 Коэффициенты распределения наркотических веществ и общих анестетиков приведены в табл, 15.1, в среднем для них log Р—2,0 [Leo, Hansch, Elkins, 1971].
2 Это значение откорректировано с учетом ионизации, но поскольку данное соединение является достаточно сильной кислотой (рКа«'2,8), значение эффективного log Р будет иметь отрицательную величину.
группы лежит между величинами вкладов этильной и пропиль-ной групп, тогда как вклады нафтильной и метильной групп примерно одинаковы. Наличие неподеленной пары электронов обычно придает соединению гидрофильный характер, что отчетливо видно из данных для азот- и кислородсодержащих групп.
17.2. Электронные эффекты (значения о-констант Гаммета и Тафта)
Зная, как тот или иной заместитель влияет на любые ионизирующиеся группы, можно легко отнести каждый из них к электронодонорным или электроноакцепторным. Количественную меру этих эффектов ввел Гаммет (примерно в 1940 г.), использовав уравнение, основанное на принципе линейности свободных энергий:
log К — log Ко = р<т,
где Ко — константа ионизации бензойной кислоты, используемой в качестве стандарта, а К — константа ионизации замещенной бензойной кислоты; о — константа заместителя — мера его электронного влияния; р — реакционная константа — мера чувствительности исследуемой реакции к эффекту заместителей,
373
В обсуждаемом случае — ионизации бензойных кислот. По определению р=1,00 для процесса ионизации бензойных кислот в воде при 20 °C. Проблема ро-анализа подробно освещена в книге Hammett (1970). Список о-констант был составлен Ехпег '(1972), а также Perrin, Dempsey, Serjeant (1981).
А. а-Константы Гаммета заместителей в ароматических соединениях. Подробный перечень этих констант, составленный непосредственно для проведения множественного регрессионного анализа, см. в книге Hansch и Leo (1979). Небольшая выборка из него, приведенная в табл. 17.4, вполне достаточна для того, чтобы продемонстрировать последовательность, в которой, располагаются заместители в соответствии с их электронными; эффектами. ам-Константы мета-заместителей, расположенные в первой колонке, характеризуют в основном величину индукционного эффекта. При определении этих констант для разных серий соединений они воспроизводятся лучше, чем оп-константы пара-заместителей, очевидно, потому, что последние отражают главным образом эффект сопряжения заместителя и в значительно меньшей — индукционное влияние. Величина эффекта сопряжения иногда может изменяться за счет возникновения так называемого прямого полярного сопряжения [Clark, Perrin, 1964]. Это явление можно иллюстрировать следующим примером:
1 + 11
ОН +он
Прямое полярное сопряжение в молекуле пара-нитрофенола
(17.1)
В табл. 17.4 заместители, имеющие отрицательные значения а-констант, являются электроноакцепторными, а положительные— электронодонорными. Полный интервал изменений а-кон-стант не превышает двух единиц, поэтому небольшое изменение о-константы соответствует весьма существенному изменению электронного эффекта. Не следует обращать особого внимания на различия в сотых долях этих констант, поскольку каждая из них представляет собой разницу между двумя значениями рКа, а погрешность определения последних обычно составляет 0,02—0,05 логарифмической единицы. Следует отметить, что если значения о-констант вычислены по данным других реакционных серий (по среднему значению р), а не из величин рКа бензойных кислот, значения их будут отличаться от приведенных в табл. 17.4, но порядок расположения заместителей в ряду останется неизменным. Используя цвиттер-ионные молекулы (например, мета-аминобензойную кислоту, содержащую в от-
374
Таблица 17.4. ст-Константы заместителей [Perrin, Dempsey, Serjeant, 1981]
Заместитель Для бензойных кислот Для уксусных кислот
мета-ам пара-ап 0*
—NHMe —0,30 —0,84 —0,81
—CH2SiMe3 —0,17 —0,27 —0,31
—NMe2 —0,15 —0,83 0,32
—t-Bu —0,09 —0,15 —0,30
—Me —0,06 —0,14 0,00
-NH—OH —0,04 —0,34 0,30
—NH—CO—NH2 —0,03 —0,24 1,31
—NH—NH2 —0,02 —0,55 0,40
—H 0,00 0,00
—NH2 0,00 —0,57 0,62
—C6H5 0,05 —0,01 0,75
—OMe 0,11 —0,28 1,81
—NH—COMe 0,12 —0,09 1,40
—OH 0,13 —0,38 1,34
—SMe 0,14 0,00 1,56
-SH 0,25 0,15 0,68
—M3 0,27 0,15 2,62
—co—nh2 0,28 0,31 1,68
—CO—OMe 0,32 0,39 2,00
—CO—OH 0,35 0,44 2,08
—CHO 0,36 0,44 2,15
—COMe 0,36 0,47 1,81
—OCFj 0,36 0,33
—SCFj 0,38 0,50 2,75
—F 0,34 0,06 3,21
—Cl 0,37 0,24 2,96
—Br 0,39 0,22 2,84
—I 0,35 0,21 2,46
—О—COMe 0,39 0,31 2,56
ccis 0,40 0,46 2,65
—CN 0,41 0,52 3,43
—CF3 0,46 0,53 , 2,61
—SO2NH2 0,46 0,57 : 2,61
—CN 0,62 0,70 | 3,30
—SO2Me 0,64 0,73 | 3,68
—no2 0,74 0,78 ! 4,25
—so2cf3 0,76 0,95 : 4,50
Ионы —s--co2--NH3+ —0,36 0,09 0,67 —0,05 0,53 —1,06 3,76
—-NMe3+ 0,99 0,96 4,55
* См. инже в тексте»
375
Личие от пара-аминобензойной кислоты большое количество цвиттер-ионной формы), можно определить о-константы ионных заместителей. Некоторые из них приведены в конце табл. 17.4. В этом случае в уравнение Гаммета следует включить дополнительный член, учитывающий электростатическое взаимодействие. Обычно его обозначают буквой г, поскольку он представ-' Ляет собой расстояние между взаимодействующими зарядами-’ (в нм) [Hoefnagel, Hoefnagel, Wepster, 1978]. Таким образом,-уравнение примет следующий вид: ;
Эффект заместителя = ро-3,1/ч.
Этот эффект может быть особенно важен в случае алифатических соединений, так как в их молекулах расстояние между зарядами больше.
Влияние эффекта прямого полярного сопряжения на величину ор особенно заметно в соединениях, содержащих нитро-, циано-, формильную или ацильную группы в пара-положении-к гидроксильной или аминогруппам. Так, обычное значение" оп = 0,78 для нитрогруппы под влиянием прямого полярного сопряжения увеличивается до 1,24, а в случае формильной группы изменяется от 0,44 до 1,03 [Perrin, Dempsey, Serjeant, 1981].
Выше уже говорилось о том, что ом-константы в небольшой мере отражают вклад эффекта сопряжения заместителя. Taft, Lewis (1959) первыми попыталить выделить два компонента ом-константы, сравнив их со значениями о-констант, полученными Тафтом для алифатических карбоновых кислот (см. ниже) и, следовательно, свободными от вклада эффекта сопряжения. Позже Swain, Lupton (1968), впервые показавшие, что эффекты поля (т. е. почти те же, что и индукционные) одной и той же группы, находящейся в мета- и пара-положении, заметно отличаются, разделили о-константу на две составляющих: F-полевую и R-резонансную [см. также Reynolds, Topson, 1984]. Hansch и сотр. (1973), обнаружив в работе Swain, Lupton (1968) систематическую арифметическую ошибку, исправили ее и опубликовали сводку примерно 200 таких констант. Порядок расположения заместителей в соответствии с изменением эффекта сопряжения от отрицательного к положительному: —NH2, —ОМе, F, —NHAc, Cl, Me, tBu, Ph, H (нулевой), —NMe+ (нулевой), —CO2-, —NO?. Swain, Lupton вывели также ряд заместителей для ароматических соединений в соответствии с увеличением полевого (индукционного) от отрицательного эффекта к положительному: —СОО~, tBu, Me, Н (нулевой), —NH2, Ph, —ОМе, —NHAc, —СООН, Cl, F, —NO2, —NMe+3; естественно, этот же ряд сохраняется и для алифатических соединений. Подробнее о разделении вкладов эффектов поля и сопряжения см. Taft, Grob (1974).
а-Константы орто-заместителей представляют собой столь же сложный, составной параметр, как и оп-константы, но в них 376
добавляются еще два вклада: 1) взаимное пространственное влияние заместителей, находящихся в орто-положении и 2) возможность образования между ними водородных связей. Решение этой сложной проблемы предложили Fujita и Nishioka (1975), предположив, что <т0 равна сумме оп-константы, полевой константы Swain, Lupton (поскольку заместители максимально сближены друг с другом) и стерической Es-константы Тафта (разд. 16.0). Если между орто-заместителями образуется водородная связь, как, например, в орто-нитрофеноле, необходимо ввести дополнительные параметры. Таблицу оо-коцстант 35 заместителей, расположенных в орто-положении к амино-, гидрокси- или пиридиногруппам, см. [Perrin, Dempsey, Serjeant (1981), приложение 5].
Б. о-Константы Тафта для алифатических заместителей. После того как Гаммет успешно решил проблему количественной оценки электронного влияния заместителя в ароматических соединениях, Тафт попытался оценить индукционный эффект заместителей в алифатическом ряду. Из нескольких альтернативных вариантов предложенных им констант [Taft, 1956], наиболее полезной при расчетах рКа и в множественном регрессионном анализе оказалась константа о*. Изначально Тафт рассчитал их величины, сравнивая константы скоростей кислотного и щелочного гидролиза а-замещенных метил ацетатов. Более приближенно, но значительно проще их можно вычислять по уравнению:
а = ( — ДрКа — 0,06)/0,63,
где А рКа получена вычитанием из величины рКа уксусной кислоты величины рКа замещенной уксусной кислоты, содержащей заместитель, значение о* которого нужно определить. При вычислении коэффициентов для метильной группы было принято о* = о.
По мере удаления заместителя от реакционного центра его эффект заметно снижается: до 40% при сдвиге на одну группу СН2, до 16 и 6% — на две и три соответственно. Введение группы СН=СН снижает эффект заместителя точно так же, как и введение метиленовой группы. Значения о*-констант заместителей обычно больше, чем цм, вследствие большего расстояния между заместителем и реакционным центром в последнем случае. В табл. 17.4 значения о* приведены в третьей графе. Более полные таблицы этих констант можно найти в книгах Perrin Dempsey, Serjeant (1981) /и Hansch, Leo (1979). При работе’ с гетероциклическими заместителями следует помнить, что двое-связанный атом азота в цикле (как в пиридине)—сильный электроноакцептор, а связанный простыми связями (как в пирроле)— донор электронов.
* См. табл. 17.4.
377
В. Константа р в уравнении Гаммета. Первоначально Гам-’ мет использовал величину р как коэффициент пересчета констант заместителей, определенных в реакциях с разрывом ко-1 валентных связей, в о-константы, рассчитанные из величин рК*г замещенных бензойных кислот (для ионизации бензойной кис-1 лоты принято р=1). Затем р-константы были использованы для] пересчета величин, полученных вычитанием из значения рКа] фенола (9,99) величин рКа его производных (р = 2,23). И наоборот, с ее помощью можно рассчитать величину рКа: например для 3-нитрофенола в рКа = 9,99— 2,23X0,74 = 8,34, а экс-, перимент дает величину 8,36 (значение 0,74 взято из табл. 17.4).< Для анилина р = 2,81. Таблицу величин р-констант, рассчитан-; ных по данным для фенола и анилина, см. Hansch, Leo (1979).
Другой пример использования констант р — расчет о-кон-стант для других ароматических циклов по даным для бензойных кислот. Впервые это было сделано для нафталина и хинолина, а затем для ряда других гетероциклов [Perrin, 1965; Perrin, Dempsey, Serjeant, 1981].
17.3. Ядерный магнитный резонанс
А. Введение. Атомы с нечетными величинами атомных масс или атомных номеров обладают магнитными моментами, обусловливающими их прецессию при наложении сильного магнитного поля. Частота колебаний, прямо пропорциональная напряженности магнитного поля, является характеристикой данного типа ядра и попадает в диапазон радиочастот электромагнитного спектра. Из элементов, наиболее распространенных в органической химии, этот эффект проявляют 44, 13С, I5N, I9F и 31Р, а 12С и 1ВО подобным эффектом не обладают.
Б. Протонный магнитный резонанс (ПМР). Раствор исследуемого соединения помещают в сильное магнитное поле с частотой электромагнитного излучения 2,35 Т (Т — тэсла, 2,35 Т эквивалентны 100 МГц или 23 500 циклов в секунду) или более. Под действием электромагнитного поля происходит переход ядер атомов водорода на более высокий энергетический уровень, который можно измерить по поглощению образцом энергии радиочастотного поля, проявляющегося в снижении радиочастотного напряжения на катушке (в случае простейшего датчика). Это напряжение сравнивается у напряжением на эталонной катушке без образца. Разностное Напряжение подается в приемник, затем усиливается, детектируется и регистрируется.
Напряженность магнитного поля йри каждом ядре водорода обычно меньше, чем приложенного внешнего магнитного поля вследствие экранирующих диамагнитных эффектов электронных облаков, окружающих ядра. В перзых моделях ЯМР-спектро-метров изменили напряженность магнитного поля, а частоту поддерживали постоянной, т. е. применяли так называемый стационарный метод ЯМР. В настоящее время используют им-378
Таблица 17.5. Приблизительные величины химических сдвигов протонов в различном окружении
Тип протоиа 6, м. д. Тип протоиа в, м. д.
сн3—с 0,9 NH2—Аг 3,5х
С—CHj—C СНз—О—СО 3,8
C-CH(R)-C 1,2—1,5 СНз—О—Аг 3,8
HaN—CR2 1,6х СН2=С 4,7
СНз c = c 1,8 HR(R) = C 5,3
СНз—c=o 2,0 НО—С 5,3х
СНз-S 2,0 H2N—СО 7,01
СНз—N 2,2 Н—Аг 7—8
СНз—Аг 2,4 НО—Аг 7,7х
СНз—N(R)—СО 2,8 R—СНО 9,8 ,
СНз—о 3,3 R—СООН 10,8
1 Значение этих химических сдвигов может изменяться при образовании водородных связей.
пульсную технику ЯМР с преобразователем Фурье, обладающую принципиально важными достоинствами. Возбуждая одновременно все ядра образца с помощью короткого импульса мощного радиоизлучения и «прослушивая» излучаемые ими частоты по мере возвращения ядер к равновесному распределению по энергии, можно получить интерференционную картину, содержащую всю информацию о нормальном спектре образца. Эта процедура занимает примерно 1 сек. При математической обработке (преобразованием Фурье) этой интерференционной картины можно получить обычный спектр с разверткой по частоте. Это преобразование осуществляется с помощью компьютера и требует небольшого дополнительного времени после накопления. Спектр поглощения, получаемый таким образом, подобен тем, которые получали на спектрометрах ранней конструкции, однако разрешение в этом случае значительно выше. Разность резонансных частот разных ядер водорода называется «химическим сдвигом», который обычно измеряется в миллионных долях. Чаще всего стандартом, которому приписывают величину 6, равную 0,00 м. д., являются протоны тетраметилсила-на; это дает возможность составить таблицу величин химических сдвигов (например, табл. 17.5). Большая величина 6 указывает на понижение напряженности приложенного внешнего магнитного поля, обусловленное сильным экранированием ядра в данной молекуле. Ранее существовала иная шкала, в которой тетраметилсилану приписывалась величина химического сдвига, равная 10; величины всех остальных химических сдвигов записывались в так называемой шкале т (т=10—6).
Обычно в химии метод ЯМР используют для того, чтобы определить, присутствуют ли в молекуле те или иные определенные группы; его широко применяют для установления строения вновь синтезированных или выделенных соединений. Метод
379
ДМР играет важную роль в исследовании потенциально таутомерных соединений. В случаях, когда таутомерное равновесие устанавливается медленно (как это часто происходит, если подвижный протон соединен с атомом углерода в одной из таутомерных форм), сигналы различных таутомерных форм регистрируются независимо и появляется возможность провеет^ кинетическое исследование. Если же подвижный протон связан! с атомами азота или кислорода, равновесие устанавливается почти мгновенно.
Образование водородных связей изменяет окружение атомов водорода и, следовательно, может оказывать существенное влияние на величину химических сдвигов. Так, например, 6 протона, ОН-группы в концентрированных растворах спиртов в хлоро-? форме обычно равна 5,3, но при достаточном разбавлении (или. нагревании) сигнал смещается в область слабого поля и б становится равной 0,5 м. д. вследствие разрыва межмолекулярных связей. Однако химические сдвиги протонов веществ, имеющих внутримолекулярные водородные связи, не изменяются при разбавлении, что позволяет различить меж- и внутримолекулярные водородные связи. При съемке спектров при различных температурах это различие становится еще более явным даже для таких сложных молекул, как полипептиды [Kessler, 1982].
Кроме эффектов экранирования, о которых говорилось выше, магнитное поле прибора может вызывать электронные токи, приводящие к сильному экранированию в случае протонов ацетилена (6 = 2,3), в то время как ароматические протоны сильно деэкранированы (6 = 7—8).
Из-за спин-спинового взаимодействия между протонами с разными химическими сдвигами сигналы в спектре ПМР часто представляют собой дублеты, триплеты или квартеты. Это рас- ? щепление выражается в виде частоты, обозначаемой буквой J, и называется константой спин-спинового взаимодействия. Вза- • имодействие протонов через сигма-связи обычно бывает сильным, но быстро ослабляется с расстоянием, в то время как взаимодействие через л-электронные системы в молекулах ароматических соединений слабо и медленно уменьшается с увеличением расстояния.
Для съемки спектров ПМР чаще всего используют или апротонные растворители (например, четыреххлористый углерод), или такие, в которых сигналы протонов лежат в слабом поле (например, трифторуксусная кислота), или же полностью дейтерированные растворители (например, дейтерированные диметилсульфоксид, ацетон, хлороформ). Для биологически важных соединений наиболее подходящий растворитель вода, однако, сигнал ее протонов может перекрываться с сигналами исследуемых веществ. Этот сигнал можно устранить с помощью импульсного метода. В качестве растворителя применяют оксид дейтерия. Он сам не дает сигналов в спектре, но приводит к исчезновению сигналов протонов, связанных с атомами азота, 380
кислорода или серы, способных быстро обмениваться на дейтерий.
Каталог интерпретированных спектров см. Simons и Zanger (1972), введение в ПМР-спектроскопию — Jackman, Sternhell (1969), обзор по ПМР-спектроскопии азотсодержащих гетероциклов— Batterham (1973).
В. Другие виды ядерного магнитного резонанса. Метод ЯМР на ядрах 13С по частоте использования занимает второе место после метода ПМР и позволяет получить информацию, служащую дополнением к данным протонных спектров. Естественное содержание изотопа 13С составляет 1,1%, поэтому для съемки спектра необходимо использовать ампулы большого объема. Получаемые слабые сигналы усиливаются посредством быстрого усреднения во времени, предшествующего преобразованию Фурье. Применение соответствующих радиочастот позволяет получить спектры с полным подавлением С—Н-взаимодействий (С—С-взаимодействие не представляет сложной проблемы). В спектрах ЯМР 13С по сравнению со спектрами ПМР реже проявляются локальные эффекты, кольцевые токи и анизотропия также выражена значительно слабее, поэтому сигналы в спектрах 13С хорошо разрешены и легче интерпретируются.
Обычно в спектрах ЯМР 13С используется область 0— 220 м. д., причем в качестве внутреннего стандарта, химический сдвиг которого равен 0, используется тетраметилсилан (ТМС). Сигналы атомов углерода метильных групп обычно лежат в области 10—30 м. д.; при 50 м. д. появляются сигналы атомов углерода СН- и МСНг-групп, затем сигналы ароматических и гетероароматических атомов углерода (около 125 м. д.) и, наконец, сигналы атомов углерода карбоксильных групп и амидов (около 170 м. д.). При работе с биологически важными веществами идеальным растворителем является вода, так как при этом можно использовать буферные растворы. Однако можно применять и углеродсодержащие растворители, если их сигнал не лежит в исследуемой области спектра.
Вопросам интерпретации спектров ЯМР 13С посвящена книга Wehrli, Wirthlin (1976).
Естественное содержание l5N составляет 0,37%, однако применение преобразования Фурье позволяет широко использовать ЯМР 15N. Стандартом при этом служит ион аммония, а спектр снимают в области от 8 до 800 м. д. Исследуемый образец помещают в большие ампулы. Сигналы алкиламинов лежат около 0 м. д., амидов — около 100 м. д., пуринов и пиримидинов — около 200 м. д., нитрогрупп — около 350 м. д., а нитрозогрупп— около 750 м. д. Таким образом, сигналы в спектре распределяются неравномерно, большинство из них располагается в области сильного поля.
Наличие неподеленной пары электронов определяет многие свойства атома азота, отсутствующие у углерода. Протонирование изменяет резонансные вклады по величине и направле
381
нию, что является характерным для каждого типа азотсодержащих групп. Положение сигналов в спектрах ЯМР 15N часто значительно более чувствительно к изменению растворителя (по сравнению с сигналами атомов углерода), занимающего аналогичное положение, что имеет большое значение при анализе спектра. Спектры часто снимают в воде, диметилсульфокси-де или трифторэтаноле. Альтернативными вариантами внутреннего стандарта являются нитрометан и нитрат-анион. С помощью ЯМР 15N была уточнена геометрия алкалоидов и синтетических лекарственных веществ; установлено, что атом азота цис-изомеров больше экранирован по сравнению с трансизомерами. Образование N ... Н водородных связей играет важную роль в связывании лекарственных веществ с рецептором.
Интерпретацию спектров ЯМР 15N см. Levy, Lichter (1979),.
Природное содержание изотопа 31Р составляет 100%, поэтому спектроскопию ЯМР на ядрах 31Р широко применяют при исследовании биологических объектов, в том числе и организма человека. Интенсивность сигналов в этих спектрах несколько ниже, чем в спектрах ЯМР ’Н и 19F, но сигналы значительно более четкие. В качестве внутреннего стандарта обычно используют фосфорную кислоту, а спектр лежит в области от —5 до 4-25 м. д. Сигналы фосфатов и фосфолипидов располагаются около 0, нуклеотидов — в области 5—22 м. д., а пирофосфатов— около 12 м. д., т. е. сигналы расположены довольно близко друг к другу. Спин-спиновое взаимодействие с атомами водорода очень интенсивно и разнообразно. В качестве растворителя обычно применяют воду. Метод ЯМР 31Р используют во многих случаях, в том числе и для изучения фосфорсодержащих инсектицидов, АТФ и нуклеиновых кислот.
Содержание 19F также составляет 100%, что позволяет широко менять этот тип ЯМР-спектроскопии и пользоваться при съемке спектра ампулами небольшого диаметра. Положение сигналов варьируется от 150 до —150 м. д.; за 0 принимают сигнал трифторуксусной кислоты. Разрешение в этих спектрах очень высокое, однако интерпретация сигналов затруднена вследствие разных эффектов окружения. Метод ЯМР 19F используют для анализа фторсодержащих соединений, таких как фторотан, или для анализа «меченных» фтором (например, в виде трифторацетильной группы) лекарственных веществ, обычно его не содержащих.
Г. Релаксационные методы. Эти методы позволяют получить данные о стереохимии ключевого атома в молекуле посредством временного ограничения влияния соседних атомов. С помощью одного из широко используемых методов этого типа определяют величину Т]—время спин-решеточной релаксации. Для создания неравновесного намагничивания (обычно это относится к СН-группам при съемке спектра ЯМР 13С) используют последовательность радиочастотных импульсов. Время, необходимое для восстановления равновесия, может служить мерой динами
382
ческих свойств лекарственного вещества в растворе, таких как вращательное движение молекулярного остова или внутреннее вращение боковой цепи. Применение этого метода подробно рассмотрено в работе Allerhand, Komorowski (1973) на примере грамицидина S.
При исследовании СН-групп методом 13С широко применяют ядерный эффект Оверхаузера. Благодаря этому эффекту интенсивность одного сигнала обычно повышается, в то время как сигналы соседних атомов остаются неизменными. Более точные выводы получаются при воздействии на два соседних атома. С помощью этого метода можно получить информацию о молекулярном движении, конформациях больших молекул, например пептидов, и приблизительно оценить межъядерные расстояния.
Д. Применение ЯМР в биологической и медицинской химии. В этих областях ЯМР используют, во-первых, для установления конфигурации или даже конформации небольших молекул биологически активных веществ в растворе. Много таких задач было решено с помощью уравнения Карплюса, позволяющего коррелировать константы расщепления между вицинальными протонами и величины торзионных углов (определение см. в разд. 12.3) между различными углерод-водородными плоскостями [Roberts, 1968]. Так была установлена конформация четырех изомеров молекулы кокаина (7.11) [Sinnema et al., 1968]. Другой пример — определение с помощью ЯМР конформации ацетилхолина в водном растворе (разд. 12.6.1). Изучение спектров ЯМР ’Н и 13С гипотензивного лекарственного средства каптоприла (9.50) в воде при различных значениях pH показало, что это вещество в растворе находится в виде равновесной смеси двух изомеров, представляющих собой разные конформеры по амидной связи. Преобладающей является транс-конформация, причем ее доля определяется ионизацией как карбокси-, так и меркаптогрупп. Карбоксильная группа транс-изомера менее кислая, чем цис-изомера (разница 0,66 единиц рК). По данным спектров было установлено наличие внутримолекулярной водородной связи между амидной карбонильной группой и карбоксилом транс-конформера [Rabenstein, Anvarhusein, 1982]. Другие примеры конформационных исследований методом ЯМР можно найти в работе Casy (1971).
Метод ЯМР применяют также для изучения взаимодействия низкомолекулярных веществ с биополимерами, причем он наиболее эффективен среди используемых в этой области других физико-химических методов. В спектре биополимера многие резонансные сигналы перекрываются, поэтому для получения четкого спектра приходится использовать особые методы. Так, можно изменять резонансные частоты, вводя в молекулу или в среду парамагнитные ионы, например лантаниды (лантан, европий), или спиновую метку, представляющую собой стабильный сводный радикал (например, нитроксил). Применяют также и релаксационный метод (например, с помощью Т], как
383
описано выше). Кроме того, можно изменять молекулу лекарственного вещества, вводя в нее группы, содержащие 13С или 19F. Подробнее об этих методах см. Jardetzky, Roberts (1981).
Спин-спиновая релаксация. Адсорбция лекарственного вещества макромолекулой вызывает уширение сигналов в определенной части спектра ЯМР агента. Метод спин-спино-вой релаксации основан на том, что скорость релаксации (1/Т2)» а следовательно и ширина ЯМР-сигналов определяется движением молекулы относительно одной оси. При ограничении движения молекулы время релаксации ядра уменьшается, а шири-» на его сигнала в спектре ЯМР увеличивается. Таким образом, при связывании молекулы лекарственного вещества с макромолекулой происходит уширение линий в ее спектре ЯМР в 100—1000 раз, что в первом приближении пропорционально увеличению эффективной относительной молекулярной массы. Если в связывании участвует только одна группа молекулы агента, то происходит избирательное уширение сигнала только этой группы, а сигналы несвязанных групп остаются сравнительно узкими. Это позволяет определить часть молекулы, связанную с макромолекулой [Jardetzky, Jardetzky, 1962]. Например, при связывании атропина (7.16) с ацетилхолинэстеразой происходит уширение сигналов фенильной и N-метильной групп, т. е. оба конца молекулы прочно связываются с ферментом. Аналогично было показано, что молекула физостигмина образует связи за счет обеих N-метильных групп [Kato, Yung, Ihnat, 1970; Kato, 1975].
Установлено, что существуют два типа комплексов: 1) ван-дер-ваальсовы, в которых прочность связывания (а также скорость релаксации для связанной молекулы) увеличивается с возрастанием полярности растворителя и 2) электростатические, для которых справедливо обратное. В качестве примера образования ван-дер-ваальсовых комплексов можно привести связывание боковых цепей молекулы пенициллина или сульфаниламидного остатка с сывороточным альбумином [Fischer, Jardetzky, 1965]; примером электростатического комплекса может служить соединение производных холина с антихолиновым антителом. Такое антитело было получено для феноксихолина при создании иммунохимической модели холинергических рецепторов. К сожалению, избирательность этого антитела оказалась слишком низкой для выявления различий между агонистами и антагонистами или между мускариновыми и никотиновыми хо-линомиметиками [Marlow, Metcalf, Burgen, 1969].
С помощью рассматриваемого метода удобно изучать комплексы обычного типа, для которых константы связывания Ks лежат в пределах 103—105. Такие комплексы находятся в состоянии быстрого равновесия с исходными компонентами, скорость их образования kR имеет обычно порядок 108—1010 лХ Хмоль-1Хз-1, а скорость диссоциации ко 103—10s s-1 (величину последней получают из соотношения Ks=kR/kD).
384
С помощью методов ЯМР 'Н, 13С и 31Р была обнаружена интеркаляция актиномицина (4.38) между пурин-пиримидиновыми парами дезоксиолигонуклеозидов по изменению химических сдвигов сигналов пурин-пиримидиновых фрагментов, вызываемому полями кольцевых токов ядра феноксазона. Это-позволило яснее определить структуру интеркалирующих компонентов и подтвердило, что интеркаляция происходит в местецентральных пар Г—Ц [Patel, 1974]. Дополнительные данные-были получены при изучении актиномицина, обогащенного 15М [Brown, Shafer, Mirau, 1982].
С помощью спектроскопии 31Р ЯМР буферных водных растворов дихлордиэтиламида амидофосфорной кислоты (3.39), активного продукта метаболизма циклофосфамида (3.38), были, определены константы скорости первого порядка превращения соединения (3.39) в азиридиниевый катион (разд. 3.6, т. 1). Изменение скорости в зависимости от pH указывает на то, что при pH от 7,4 до 6,0 распад соединения (3.39) происходит к десять раз быстрее. Это позволяет предположить, что повышенная кислотность раковых клеток может обеспечить избирательность действия препарата [Engle, Zon, Egan, 1979]. Циклофос-фан в настоящее время выпускают в виде рацемата, однако его можно разделить на ( + )- и (—)-энантиомеры, хорошо различаемые в спектрах ЯМР *Н или 31Р в присутствии соединений европия. Этот метод обладает некоторыми преимуществами по сравнению с традиционными измерениями оптического вращения [Ludeman, Bartlett, Zon, 1979].
При исследовании систем, в которых происходят быстрые обменные процессы для установления различий между возможными структурами образующегося комплекса данных ЯМР-спектроскопии недостаточно. Однако на основании этих данных можно предположить, какие из ассоциатов предпочтительнее, а затем по данным рентгеноструктурного анализа выбрать подходящую [см. обзор Williams, 1977].
Более подробно см. Cohen (1980), Campbell, Dobson (1979),. Jardetzky, Roberts (1981) и Dweck (1977).
17.4. Поиск литературных данных
Нередко у исследователей возникает вопрос: как отыскать все, что уже написано о каком-либо лекарственном веществе, если известно только его название? Начинать следует с тривиального названия препарата (например, ацикловир), а не с торгового (зовиракс). Оба эти наименования указаны в Martindale’s Extra Pharmacopoeia [Reynolds, 1982], где приводите» также полное химическое наименование, но не дается структурная формула1. Эта фармакопея содержит сведения об облает»
1 В СССР таким официальным изданием является справочник М. Д. Машковского «Лекарственные средства».—М.: «Медицина», 1985. — Примеч. ред‘.
25—734
385
применения препарата, его побочных эффектах (со ссылками на самые последние источники), дозах, лекарственных формах и синонимах (включая торговые названия). Структурную формулу можно отыскать в Merck Index (1983). В этом справочнике материал расположен не строго в алфавитном порядке, юднако работу облегчает указатель. Дополнительную информацию о химических свойствах вещества можно получить в Heil-bron’s Dictionary of Organic Compounds. Словарь состоит из семи томов, в которых приведены данные о химической структуре, химических свойствах и спектральных характеристиках большого числа органических соединений. Существует также Chemical Abstracts Service Registry Numbers (CAS). Тривиальные и торговые названия препаратов, используемых в сельском хозяйстве, можно найти в книге Worthing (1983).
В Pharmacological Basis of Therapeutics — широко известной книге Gilman, Goodman, Gilman с исчерпывающей полнотой приведены характеристики практически всех препаратов, применяемых в настоящее время в клинике, особое внимание при. этом уделено их биологической активности. В книге приведенье структурные формулы соединений и рассмотрены физиологические, а часто и биохимические аспекты их механизма действия; каждому препарату дана относительная оценка с точки зрения его эффективности при лечении того или иного заболевания. В Burger’s Medicini Chemistry в центре внимания находится ; проблема изменения биологического действия при вариации заместителей в рядах соединений [Wolff, 1981]. Для ознакомления с новыми препаратами, предложенными для клинического применения, особенно в США, очень полезен ежегодник Drugs in Current Use and New Drugs.
Многие количественные характеристики биологически активных соединений можно найти в различных справочниках, либб в компьютерных банках данных. Константы ионизации этик веществ приведены в книге Albert, Serjeant (1984) (некоторые из них помещены в гл. 10); новые данные о константах ионизации легко отыскать в Chemical Abstracts с помощью общего предметного указателя (под рубрикой «Ионизация в жидкостях») или указателя химических соединений (по названию или эмпирической формуле конкретного вещества). При необходимости предсказания констант ионизации по структурным параметрам следует обратиться к книге Perrin, Dempsey, Serjeant (1981). Некоторые сведения о константах устойчивости комп-* лексов металлов помещены в конце разд. 11.3.
В работе Hansch, Leo, Elkins (1971) приведены 5800 коэффициентов распределения для системы октанол — вода. В книгё Hansch и Leo (1979) «Константы заместителей для корреляционного анализа» особенно полезны две таблицы. Первая —-«Заместители в ароматических соединениях» содержит следующую информацию о 166 группировках: 1) гидрофобные л-кон-станты заместителей; 2) способность заместителя к образова-386
нию водородных связей; 3) молекулярная рефракция; 4) значения полевых и резонансных констант Swain и Lupton^ 5) Ом- и Оп-константы Гаммета, а также стерические Es-константы заместителей для алифатических и ароматических соединений. Во второй таблице приведены следующие данные для ПО заместителей в алифатических соединениях: 1) фрагментарные гидрофобные константы; 2) способность заместителя к образованию водородных связей; 3) молекулярные рефракции; 4) значения полевых констант Swain-Lupton. Значения о* помещены в приложении 1.
US National Institute for Occupational Safety and Health ежегодно публикует Registry of Toxic Effects of Chemical Substances (RTECS). Компьютеризованным вариантом этих данных можно воспользоваться с помощью системы TOXLINE Национальной Медицинской библиотеки и Службы химической информации (CIS) под эгидой Национального института здоровья и Агентства по защите окружающей среды. CIS обладает также мощной поисковой системой SANSS (Структурно-номенклатурная поисковая система), позволяющей очень быстро получить сведения о любом из включенных в нее 200 000 соединений. С ее помощью можно отобрать группу веществ, имеющих такой же или такие же структурные параметры, как изучаемое вещество. Поиск ведут по структуре, номеру по регистру Chemical Abstracts (CAS) или названию (полному, тривиальному^ торговому и синонимам).
Исчерпывающий поиск родственных субструктур можно привести и по CAS Online System, в которую из Chemical Abstracts включены данные о 600 000 соединениях. Во Франции существует подобная система DARC. С помощью этих систем' можно получить и некоторые биологические данные, но, зная регистрационный номер вещества в CAS, удобнее пользоваться системой MEDLARS Национального института здоровья США. Известны банки данных по секвенированию нуклеиновых кислот как в Европе [Walgate, 1982], так и в США [Lewin, 1982]. Банк данных по кристаллической структуре белков создан Национальной лабораторией Брукхейвен (Нью-Йорк, США).
Многие поисковые системы подобного рода с помощью терминалов можно использовать в разных странах мира. Пря разумном подходе они экономят время и повышают продуктивность исследований. Их недостаток — тенденция к развитию^ поиска на основе подобия структуры, очевидно, что биологические свойства веществ существенно зависят от их физических свойств, а последние могут оказаться идентичными у веществ, с совершенно разной структурой. В разработанной Службой; химической информации (CIS) системе LOGP, содержащей данные для большого числа соединений, можно вести поиск коэффициентов распределения для конкретных веществ или же искать все соединения с заданной величиной log Р. В систему включены также ссылки на литературные источники.
25*
СПИСОК ЛИТЕРАТУРЫ
Aaseth J., Friedheim E. — Acta Pharmacol. Toxicol., 1978, vol. 42, p. 248.
Abeles R., Maycock A. — Acc. Chem. Res., 1976, vol. 9, p. 313.
Abraham E.— Pharmacol. Rev., 1962, vol. 14, p. 473.
Adams M., McPherson A., Rossmann M. et al. — J. Molec. Biol., 1970, vol. 51, p. 31.
Addanki S„ Chaill F., Sotos J. — Nature, 1976, vol. 214, p. 400.
Adler R., Snoke J. —J. Bact., 1962, vol. 83, p. 1315.
Agarwal M. (ed.) Antihormones. — Amsterdam: Elsevier, 1979.
Agarwal R., Perrin D. — Agents and Actions, 1976, vol. 6, p. 667.
Agrawal K., Booth B., Moore E. et al. — J. Med. Chem., 1972, vol. 15, p. 1184.
Ahlquist R. — Amer. J. Physiol., 1948, vol. 153, p. 586.
Albert A. — Med. J. Austral., 1944, i, p. 245.
Albert A. — Biochem. J., 1950, vol. 47, p. 531.
Albert A. — Biochem. J., 1952, vol. 50, p. 690.
Albert A. — Biochem. J., 1953, vol. 54, p. 646.
Albert A. — Nature, bond., 1956, vol. 177, p. 525.
Albert A. The Acridines, their Preparation, Properties and Uses, 2nd edn. — London: Edward Arnold, 1966.
Albert A. — Angew. Chem., Internat. edn, 1967, vol. 6, p. 919.
Albert A. Heterocyclic Chemistry, an Introduction, 2nd edn. — London: Athlone Press, 1968.
Albert A. —Adv. Heterocycl. Chem., 1976, vol. 20, p. 117.
Albert A. —J. Chem. Soc., Perkin trans. I, 1980, p. 2918.
Albert A., Brown D., Cheeseman G. — J. Chem. Soc., 1952, (a), p. 1620; (b) p. 4219.
Albert A., Falk J., Rubbo S. — Nature, Lond., 1944, vol. 153, p. 712.
Albert A., Gibson M., Rubbo S. — Brit. J. Exper. Path., 1953, vol. 34, p. 119.
Albert A., Goldacre R. — J. Chem. Soc., 1946, p. 706.
Albert A., Goldacre R. — Nature, Lond., 1948, vol. 161, p. 95.
Albert A., Goldacre R„ Phillips J.— J. Chem. Soc., 1948, p. 2240.
Albert A., Hampton A. — J. Chem. Soc., 1952, p. 4985.
Albert A., Hampton A. — J. Chem. Soc., 1954, p. 505.
Albert A., Hampton A., Selbie F. et al. — Brit. J. Exper. Path., 1954, vol. 35, p. 75.
Albert A., Rees C. — Nature, Lond., 1956, vol. 177, p. 433.
Albert A., Rees C„ Tomlinson A. — Brit. J. Exper. Path., 1956, vol. 37, p. 500.
Albert A., Rubbo S., Burvill M. — Brit. J. Exper. Path., 1949, vol. 30, p. 159.
Albert A., Rubbo S„ Goldacre R. — Nature, Lond,, 1941, vol. 147, p. 332.
Albert A., Rubbo S., Coldacre R. et al. — Brit. J. Exper. Path., 1947, vol. 28, p. 69.
Albert A., Rubbo S„ Goldacre R. et al. — Brit. J. Exper. Path., 1945, vol. 26, p. 160.
Albert A., Serjeant E. — Biochem. J., 1960, vol. 76, p. 621.
Albert A., Serjeant E. — J. Chem. Soc., 1964, p. 3357.
Albert A., Serjeant E. The Determination of Ionization Constants. 3rd edn.— London and New York: Chapman and Hall, 1984.
Aldridge W. — Biochem. J., 1950, vol. 46, p. 451.
Aldridge W., Barnes J. — Nature, Lond., 1952, vol. 169, p. 345.
Aldridge W., Davison A. — Biochem. J., 1952, vol. 52, p. 663.
Alexander A., Trim T. — Proc. Roy. Soc. B., 1946, vol. 113, p. 220.
Allerhand A., Komorowskl R. — J. Amer. Chem. Soc., 1973, vol. 95, p. 8228.
:388
Alles G., Knoefel P.— Univ. Calif. Publ. Pharmacol., 1939, vol. 1, p. 187.
Allison J„ O’Brien, R., Hahn F. — Science, 1965, vol. 149, p. 1111; Antimicrob.
Agents. Chemother., 1965, p. 310.
Ames A., Tsukada T., Nesbett F.— J. Neurochem., 1967, vol. 14, p. 145.
Ames B„ Durston W., Yamasaki E. et al. — Proc. Natl. Acad. Sci., U.S.A., 1973, vol. 70, p. 2281.
Ames. B., Sims P.. Grover P. — Science, 1972, vol. 176, p. 47.
Amoore J. — Ann. N.Y. Acad. Sci., 1964, vol. 116, p. 457.
Anand N., Davis B., Armitage A. — Nature, Lond., 1960, vol. 185, p. 23.
Anderson B., Buckingham D., Robertson G. et al. — Nature, Lond., 1976, vol. 262, p. 722.
Anderson D„ Swaby R. — Austral. J. Sci. Res. B., 1951, vol. 4, p. 275.
Anderson J., Matsubashi M., Hoskin M. et al. — Proc. Natl. Acad. Sci., U.S.A., 1965, vol. 53, p. 881.
Andreesen J., Ljungdahl L. — J. Bact., 1974, vol. 120, p. 6.
Andrews P., Mark L.. Winkler D. et al.—J. Med. Chem., 1983, vol. 26, p. 1223.
Anghilieri L. (ed). Role of Calcium in Biological Systems. 3 vols. — Boca Raton, Florida: CRC Press, 1982.
Ankel E„ Petering D. — Biochem. Pharmacol., 1980, vol. 29, p. 1833.
Antic B., Van der Goot H„ Nauta W. et al. — Eur. J. Med. Chem., 1977, vol. 12, p. 573.
Antoloni R., Gliozzi A., Gorio A. (eds). Transport in Biomembranes: Model Systems and Reconstitution. — New York: Raven Press, 1982.
Arcos J., Argus M. Chemical Induction of Cancer. — New York: Academic Press, 1968.
Armarego W. Stereochemistry of Heterocyclic Compounds. 2 vols. — New York: Wiley-Interscience, 1977.
Armitage A., Ing H. — Brit. J. Pharmacol., 1954, vol. 9, p. 376.
Armstrong R., Panzer ЛГ. — J. Amer. Chem. Soc., 1972, vol. 94, p. 7650.
Arunlakshana O., Schild H. — Brit. J. Pharmacol., 1959, vol. 14, p. 48.
Ashe H. — Arch. Environ. Health, 1964, vol. 9, p. 545.
Aviado D„ Brugler B., Bellet J. — Exper. Path., 1968, vol. 23, p. 294.
Bacchi G. — J. Protozool., 1981, vol. 28, p. 20.
Bacon F. Novum Organum. — Lugd. Batavorum, Leyden, 1620.
Badger G. — Nature, Lond., 1947, vol. 159, p. 194.
Baguley B., Denny W., Atwell G. et al. — J. Med. Chem., 1981, vol. 24, p. 520.
Baird W., Reid A. — Brit. J. Anaesth., 1967, vol. 39, p. 755.
Baker B. Design of Active-Site Directed Irreversible Enzyme Inhibitors. — New York: Wiley, 1967.
Baker B., Lee W., Martinez A. et al. — J. Org. Chem., 1962, vol. 27, p. 3283.
Baker B., Patel R. — J. Pharm. Sci., 1964, vol. 53, p. 717.
Baker B., Shapiro H. — J. Pharm. Sci., 1966, vol. 55, p. 308.
Baker R., Chotia C., Pauling P. et al. — Nature, Lond., 1971, vol. 230, p. 439.
Balaban A., Chiriac A., Motoc J. et al. Steric Fit in Quantitative Structure — Activity Relations. — Berlin: Springer, 1980.
Baldessarini R. Chemotherapy in Psychiatry. — Cambridge, Mass: Harvard University Press, 1977.
Baldoni J., Villafranca J., Mallettee M. — Fed. Proc., 1980, vol. 39, p. 2568.
Baldwin E. — Brit. Pharmacol. Chemother., 1948b, vol. 4, p. ’9L
Barger G., Dale H. —J. Physiol., 1910, vol. 41, p. 19.
Barlow R. Chemical Pharmacology, 2nd edn — London: Methuen, 1968.
Barlow R., Hamilton J. — Brit. J. Pharmacol., 1962, vol. 18, pp. 510, 543.
Barlow R., Hamilton J. — Brit. J. Pharmacol., 1965, vol. 25, p. 206.
Barlow R., Scott K„ Stephenson R. — Brit. J. Pharmacol., 1963, vol. 21, p. 509.
Barrett A., Cullum V. — Brit. J. Pharmacol., 1968, vol. 34, p. 43.
Barrnas Y„ Clastre I, — Compt. rend. Acad. Sci., Paris, 1970 vol 270 C p. 306.
Bartlett G., Barron E. — J. Biol. Chem., 1947, vol. 170, p. 67.
Barton D„ Cookson R. — Quart. Rev. Chem. Soc., 1956, vol. 10, p. 44.
Basil B., Clark J., Coffe E. et al.— J. Med. Chem,., 1976, vol. 19, p. 399.
389
Basolo F., Pearson R. Mechanisms of Inorganic Reactions, 2nd edn. — New York: Wiley, 1967.
Bass IF., Schueler F., Featherstone R. et al. — J. Pharmacol., 1950, vol. 100, p. 465.
Batchelor F., Chain E., Richards M. et al. — Proc. Roy. Soc. B, 1961, vol. 154, p. 522.
Batchelor }., Welsh K., Tinoco P. et al. — Lancet, i, 1980, p. 1107.
Batterham T. NMR Spectra of Simple Heterocycles. — New York: Wiley, 1973, Battershell C., Malhotra D„ Hopfinger A.—J. Med. Chem., 1981, vol. 24, p. 812; Bauer W„ Vinograd J. — J. Molec. Biol., 1970, vol. 47, p. 419.
Baur E„ Preis H. — Z. phys. Chem., 1936, Bd 32B, S. 65.
Bebbington A., Brimblecom.be R„ Shakeshaft D. — Brit. J. Pharmacol. Chemother., 1966, vol. 26, p. 56.
Beccari E. — Boll. Soc. ital. Biol, sper., 1938, vol. 13, p. 6.
Beccari E. — Boll. Soc. ital. Biol, sper., 1941, vol. 16, p. 214; Arch. Sci. biol., Bologna, 1941, vol. 27, p. 204.
Bechhold H„ Ehrlich P. — Z. physiol. Chem., 1906, Bd 47, S. 173.
Beckett A. — Arzneim. Forsch., 1959, Bd 1, S. 455.
Beckett A., Casy A. — Progr. Med. Chem., 1962, vol. 2, p. 43.
Beckett A., Patki S., Robinson A. — J. Pharm. Pharmacol., 1959, vol. 11, p. 360;
Beckett A., Vahora A., Robinson A.—J. Pharm. Pharmacol., 1958, vol. 10*. p. 160 T.
Beers W., Reich E. — Nature, Lond., 1970, vol. 228, p. 917.
Behnke A., Yarbrough O. — Amer. J. Physiol., 1939, vol. 126, p. 4091.
Bell C. Principles and Applications of Metal Chelation. — Oxford: University Press 1977.
Bell P., Roblin R. — J. Amer. Chem. Soc., 1942, vol. 64, p. 2905.
Bell R. The Proton in Chemistry, 2nd edn. — London and New York: Chapman" and Hall, 1973.
Belleau B„ Lacasse G. —J. Med. Chem., 1964, vol. 7, p. 768.
Belleau B., Puranen J. — J. Med. Chem., 1963, vol. 6, p. 325.
Bender M., Ko mi у am a M. Cyclodextrin Chemistry. — Berlin: Springer, 1978.
Benotti J., Grossman W., Braunwald E. et a/.— New Engl. J. Med., 1978, vol. 299, p. 1373.
Bentley R. Molecular Asymmetry in Biology. 2 vols. — London: Academic Press, 1969.
Berenblum I. — Cancer Res., 1941, vol. 1, p. 807.
Berenblum I. — In: The Physiopathology of Cancer. 3rd edn./Ed. W. Homburger, Basel: Karger, 1974, p. 393.
Berends F., Posthumus C., Van der Sluys I. et al. — Biochim. Biophys. Acta, 1959, vol. 34, p. 576.
Bergel F„ Stock J. — J. Chem. Soc., 1954, p. 2409.
Berglof F.-E. — Arthritis Rheum., 1977, vol. 20, p. 108.
Bergmann E., Ginsburg D., Pappo R. — Organic Reactions, 1959 vol. 10*. p. 179.
Bergmann F., Segal R. — Biochem. J., 1954, vol. 58, p. 692.
Bernheim F., Bernheim M. — Cold Spring Harbor Symp. Quant. Biol., 19391*. vol. 7, p. 174.
Berry H., Cook A., Wills B. —J. Pharm. Pharmacol., 1956, vol. 8, p. 425.
Bertino L, Johns D. — Ann. Rev. Med., 1967, vol. 18, p. 27.
Bessman S., Rubin M., Leikin S. — Pediatrics, 1954, vol. 14, p. 201.
Beutler E. — Blood, 1959, vol. 14, p. 103.
Bigley W. — J. Econ. Entomol., 1966, vol. 59, p. 60.
Bijvoet J., Peerdeman A., Van Bommel A. — Nature, Lond., 1951 vol 168 p. 271.
Birch A., Holzapfel C., Rickards R. et al. — Tetr. Lett., 1964, p. 1491.
Bjerrum J. Metal Ammine Formation in Aqueous Solution. — Copenhagen; Haase, 1941.
Black Crowther A., Shanks R. et al. — Lancet, i, 1964, p. 1080.
Black J., Duncan W., Durant G. et al. — Nature, Lond., 1972, vol. 236, p. 385.
390
BlackJ., Durant G., Emmet J., GanellinR.— Nature, Lond,, 1974, vol. 248, p. 65. Blackwood R. — Adv. Appl. Microbiol., 1970, vol. 13, p. 237.
Blair J. (ed). Pteridines and Folic Acid Derivatives. — Berlin: de Gruyter, 1983. ,
Blake C„ Johnson L„ Main G. et aZ. —Proc. Roy. Soc. B, 1967, vol. 167, p. 378.
Blakeley R. The Biochemistry of Folic Acid and Related Pteridines. — Amsterdam: North-Holland, 1969.
Blaney J., Dietrich S., Reynolds M., Hahsch C. — J. Med. Chem., 1979, vol. 22, p. 614.
Blaney J., Jorgensen E., Connolly M. et al. — J. Med. Chem., 1982, vol. 25, p. 785.
Blanpain P., Nagy J., Laurent G., Durant F. — J. Med. Chem., 1980, vol. 23, p. 1283.
Blaschka ff. — Ргос. Roy. Soc. B, 1950, vol. 137, p. 307.
Blicke F., Smith F. — J. Amer. Chem. Soc., 1930, vol. 52, p. 2946.
Block S. — J. Agric. Food. Chem., 1956, vol. 4, p. 1042.
Blubaugh L., Botts C„ Gerwe A. — J. Bact., 1940, vol. 39, p. 51.
Bock L., Miller G., Schaper K-, Seydel J. — J. Med. Chem., 1974, vol. 17, p. 23.
Boehme E., Applegate H., Toeplitz B. et al. — J. Amer. Chem. Soc., 1971, vol. 93, p. 4324.
Bohman T. — Scand. J. Gastroenterol., 1980, vol. 15, p. 183.
Bolin J., Filman D., Matthews D. et al.—J. Biol. Chem., 1982, vol. 257, p. 13650.
Bono V. — Cancer Treat. Rep., 1976, vol. 60, p. 141.
Bonting S. — In: Membranes and Ion Transport/Ed. E. Bittar, New York: Wiley-Interscience, 1970, vol. 1, ch. 8.
Borge D., Cotzias G. — Proc. Natl. Acad. Sci., U.S.A., 1962, vol. 48, pp. 623, 643.
Borgna J.-L., Rochefort H.— J. Biol. Chem., 1981, vol. 256, p. 859.
Borowski E„ Cybulska B. — Nature, Lond., 1967, vol. 213, p. 1034.
Bosman H. —J. Biol. Chem., 1972, vol. 247, p. 130.
Bosund I. — Physiol. Planta., 1960, vol. 13, p. 793.
Bouanchaud D,, Scavizzi M., Chabbert Y.—J. Gen. Microbiol., 1968, vol. 54, p. 417.
Boublik J., Quinn M., Clements J. et al. — Nature, Lond., 1983, vol. 301, p. 246.
Boura A., Copp F., Green A. — Nature, Lond., 1959, vol. 184, p. 70.
Bovet D. — Rendiconti Istituto super. Sanita, Rome, 1947, vol. 10, p. 1161.
Bovet D., Horclois R., Walthert F. — Comptes rend Biol., Paris, 1944, vol. 138, p. 99.
Bovet D., Staub A. — Comptes rend. Soc. Biol., Paris, 1937, vol. 124, p. 547.
Boyer P. (ed.). The Enzymes, 3rd edn. — New York: Academic Press (in 16 vols), 1970—1983.
Boyland E. — Biochem. Soc. Symp., 1950, vol. 5, p. 40.
Boyland E., Dukes C., Grover P. — Brit. J. Cancer, 1963, vol. 17, p. 79.
Braestrup C., Nielsen M., Olsen C. — Proc. Natl. Acad. Sci., U.S.A., 1980, vol. 77, p. 2288.
Bratfos O.,Haug I. — Acta Psychiat. Scand.— 1979, vol. 60, p. 1.
Bratkowska-Seniow B., Kaniak T., Stumpf A. et al. — Materia Medica Polona, Warsaw, 1976, vol. 8, p. 316.
Breslow R. — Chem. Brit., 1983, vol. 19, p. 126.
Breslow R., Wernick D. — Proc, Natl. Acad. Sci., U.S.A., 1977, vol. 74, p. 1303.
Bresnick E., Hitchings G. — Cancer Res., 1961, vol. 21, p. 105.
Brewster P„ Hughes E., Ingold C., Rao P. — Nature, Lond., 1950, vol. 166, p. 178.
Breyer B„ Buchanan G., Duewell H. — J. Chem. Soc., 1944, p. 360.
Briggs G„ Haldane J. — Biochem. J., 1925, vol. 19, p. 338.
Briggs M„ Christie G. (eds). Advances in Steroid Biochemistry and Pharmacology. — New York: Academic Press, 1977.
Brink F., Pasternak J. — J. Cell. Compar. Physiol., 1948, vol. 32, p. 211.
391
Brodie В., Hogben С. — J. Pharm. Pharmacol., 1957, vol. 9, p. 345.
Brookes P. — Cancer Res., 1966, vol. 26, p. 1994.
Brookes P., Lawley P. — Nature, Lond., 1964, vol. 202, p. 781.
Brotzu G. Lav. 1st. Igiene Cagliari. — Sardinia, 1948.
Broughton G., Chaplen P., Knowles P. et al. — J. Med. Chem., 1975, vol. 18, p. 1117.
Brown E., Aurbach G., Hauser D., Troxler F. — J. Biol. Chem., 1976, vol. 251, p. 1232.
Brown G. — J. Biol. Chem., 1962, vol. 237, p. 536.
Brown N., Hollinshead D., Kingbury P., Malone J. — Nature, Lond., 1962, vol. 194, p. 379.
Brown P. — Nature, Lond., 1967, vol. 213, p. 363.
Brown S., Shafer R., Mirau P. — J. Amer. Chem. Soc., 1982, vol. 104, p. 5504.
Browning C., Cohen J., Gaunt R., Gulbransen R. — Proc. Roy. Soc. B, 1922, vol. 93, p. 329.
Browning C„ Gilmour W.— J. Path. Bact., 1913, vol. 18, p. 144.
Browning C., Gulbransen R., Kennaway E.— J. Path. Bact., 1919, vol. 23, p. 106.
Browning C., Gulbransen R., Kennaway E., Thornton L.— Brit. Med. J., 1917, i„ p. 73.
Bruice T., Benkovic S. Bioorganic Mechanisms (2 vols). — New York, Benjamin, 1966.
Bruni J., Wilder B. — Arch. Neurol., 1979, vol. 36, p. 393.
Bryan L„ Van den Elzen H.— Antimicrob. Agents Chemother., 1977, vol. 12, p. 163.
Bryan S., Frieden E. — Biochemistry, 1967, vol. 6, p. 2728.
Buchanan J. — In: Chemistry and Biology of Purines/Eds. G. Wolstenholme, C. O’Connor, London: Churchill, 1957, p. 233.
Buchanan J. — In: Enzyme-Activated Irreversible Inhibitors/Eds. N. Seiler, M. Jung, J. Koch-Weser, Amsterdam: North Holland, 1978.
Bilchel K, Draber W.— Progr. Photosyn. Res., 1969, vol. 3, p. 1777.
Bueding E., Batzinger R„ Cha Y. et al. — Pharmacol. Rev., 1979, vol. 3<K p. 547.
Bukatsch F., Haitinger M. — Protoplasma, 1940, vol. 34, p. 515.
BUlbring E„ Tomita T. — Proc. Roy. Soc. B, 1969, vol. 172, p. 103.
Bullen J., Rogers H. — Nature, Lond., 1969, vol. 224, p. 380.
Bunting J., Meathrel W. — Canad. J. Chem., 1972, vol. 50, p. 917.
Burchall J. — Fed. Proc., 1966, vol. 25, p. 277.
Burchall J., Hitchings G. — Molec. Pharmacol., 1965, vol. 1, p. 126.
Burgen A. — Brit. J. Pharmacol. Chemother., 1965, vol. 25, p. 4.
Burgen A., Standridge R„ Arlens E. — J. Med. Chem., 1963, vol. 6, p. 221.
Burger A., Standridge R., Stjernstrom N., Marchini P. — J. Med. Pharm. Chem., 1961, vol. 4, p. 517.
Burn J., Dale H. — J. Pharmacol., 1915, vol. 6, p. 417.
Burns J., Yu T., Dayton P. et al. — Ann. N.Y. Acad. Sci., 1960, vol. 86, p. 253.
Burtt E. —Ann. Appl. Biol., 1945, vol. 32, p. 247.
Butler H., Hurse A., Thursky E., Shulman A. — Austral. J. Exper. Biol. Med.
Sci., 1969, vol. 47, p. 541.
Butler T. — J. Pharmacol., 1944, vol. 81, p. 72.
Butler T. — Pharmacol. Rev., 1950, vol. 2, p. 121.
Buttlerworth J., Eley D., Stone G. — Biochem. J., 1953, vol. 53, p. 30.
Byrde R., Woodcock D. — Nature, Lond., 1957, vol. 179, p. 539.
Byrn S., Graber C., Midland S. — J. Org. Chem., 1976, vol. 41, p. 2283.
Cade J. — Med. J. Austral., 1949, vol. 36, p. 349.
Cahn R.—J. Chem. Educ., 1964, vol. 41, pp. 116, 503.
Cahn R., Dermer O. Introduction to Chemical Nomenclature, 5th edn. — London: Butterworths, 1979.
Cahn R., Ingold C., Prelog V. — Experientia, 1956, vol. 12, p. 81.
Cairns J. — Cold Spring Harbor Symp. Quant. Biol., 1962, vol. 27, p. 311.
Cairns J. — Sci. Amer., 1975, vol. 233, p. 64.
392
Calvert A., Harland S., Newell D. et al. — Cancer Chemother. Pharmacol., 1982, vol. 9, p. 140.
Calvin M., Wilson К— J. Amer. Chem. Soc., 1945, vol. 67, p. 2003.
Cammarata A., Yau S., Collett J., Martin A. — Molec. Pharmacol., 1970, vol. 6, p. 61.
Camp H„ Fukoto T., Metcalf R.— J. Agric. Food Chem., 1969, vol. 17, p. 243. Campbell I., Dobson D. — Methods Biochem. Anal., 1979, vol. 25, p. 1.
Canellakis E., Bono V., Bellantone A. et al. — Biochim. Biophys. Acta, 1976, vol. 418, p. 300.
Сапера F., Pauling P., Sorum H. — Nature, Lond., 1966, vol. 210, p. 907. Canfield C„ Rozman R. — Bull. World Health Org., 1974, vol. 50, p. 203. Cantley L., Resh M„ Guidotti G.— Nature, Lond., 1978, vol. 272, p. 552. Caranikas S., Mizrahi J., Escher E., Regoli D. — J. Med. Chem., 1982, vol. 25, p. 1313.
Carlsson A., Lundqvist M. — Acta Pharmacol. Toxicol., 1963, vol. 20, p. 140.
Carlstrom D., Bergin R. —Acta Crystallogr., 1967, vol. 23, p. 313.
Carmenier G. — Biochem. Pharmacol., 1968, vol. 17, p. 1981.
Caroon J., Clark R., Kluge A. et al. — J. Med. Chem., 1982, vol. 25, p. 666.
Carson S. — Biochem. Pharmacol., 1982, vol. 31, p. 1806.
Carter S., Friedman M. — Eur. J. Cancer, 1972, vol. 8, p. 85.
Carter S., Sakurai Y„ Umezawa H. (eds.). New Drugs in Cancer Chemotherapy. — New York: Springer, 1981.
Casida J., McBryde L., Niedermeir R. — J. Agric. Food Chem., 1962, vol. 10, p. 370.
Casselton P. — Nature, Lond., 1964, vol. 204, p. 93.
Castagna M., Takai Y., Kaibuchi K. et al. — J. Biol. Chem., 1982, vol. 257, p. 7847.
Casy A. PMR Spectroscopy in Medicinal and Biological Chemistry. — London: Academic Press, 1971.
Casy A., Ison R. — J. Pharm. Pharmacol., 1970, vol. 22, p. 270.
Cattell W., Chamberlain D.t Fry 1. et al. — Brit. Med. J., 1971, i, p. 377. Cawthon R., Breakefield X. — Nature, Lond., 1979, vol. 281, p. 692.
Cederbaum D. — New Engl. J. Med., 1979, p. 301.
Cha S., Agarwal R„ Parks R. — Biochem. Pharmacol., 1975, vol. 24, p. 2187.
Chacko G„ Villegas G., Barnola F. et al. — Biochim. Biophys. Acta, 1976, vol. 443, p. 19.
Chambron J., Daune M., Sadron C. — Biochim. Biophys. Acta, 1966, vol. 123, pp. 306, 319.
Chang К, Hazum E., Kilian A., Cuatrecasas P. — Molec. Pharmacol., 1981, vol. 1, p. 20.
Chapman N., Shorter J. Advances in Free Energy Relationships. — London: Plenum Press, 1972.
Chen R., Kraemer C., Schmidmayr W. et al. — Biochem. J., 1982, vol. 203, p. 33.
Cheng C„ Fugimura S., Grunberger D., Weinstein I. — Cancer Res., 1972, vol. 32, p. 22.
Cheung W. Calcium and Cell Function (3 vols). — New York: Academic Press, 1980—82.
Chick H. — J. Hyg., 1908, vol. 8, p. 92.
Childs A., Davies D„ Green A., Rutland J.—Brit. J. Pharmacol., 1955, vol. 10, p. 462.
Childs C. — Inorg. Chem., 1970, vol. 9, p. 2465.
Chiu T., Dryden D., Rosenberg H.— Molec. Pharmacol., 1982, vol. 21, p. 57.
Chiu T., Rosenberg H.— Trends Pharmacol. Sci., 1983, vol. 4, p. 348.
Chiu Y., Main A., Dauterman W. — Biochem. Pharmacol., 1969, vol. 18, p. 2171.
Cho A., Haslett W, Jenden D. — J. Pharmacol., 1962, vol. 138, p. 249.
Chong C., Rickards R. — Tetr. Lett., 1970, p. 5145.
Chothia C. — Nature, Lond., 1970, vol. 225, p. 36.
Chothia C., Pauling P. — Nature, Lond., 1970, vol. 226, p. 541.
Cigen R. — Acta Chem. Scand., 1958, vol. 12, p. 1456.
Cirillo V., Harsch M., Lampen J.— J. Gen. Microbiol., 1964, vol. 35, p. 249.
393
Citri Y„ Schramm M. — Nature, Lond., 1980, vol. 287, p. 297.
Clark A. The Mode of Action of Drugs on Cells. — London: Edvard Arnold, 1933. Clark A., Raventos J. — J. Exper. Physiol., 1937, vol. 26, p. 375.
Clark J., Perrin D. — Quart. Rev. Chem. Soc., London, 1964, vol. 18, p. 295.
Clarke F. — J. Chem. Educ., 1977, vol. 54, p. 230.
Clarkson T., Rothstein A., Sutherland R. — Brit. J. Pharmacol., Chemother., 1965, vol. 24, p. 1.
Cleland W.— In: The Enzymes. 3rd edn./Ed. P. Boyer. —New York: Academic Press, 1970.
Clements J., Wilson К— Proc. Natl. Acad. Sci., U.S.A., 1962, vol. 48, p. 1008.
Clowes G„ Keltch A.— Proc. Soc. Exper. Biol. Med., 1931, vol. 29, p. 312:
Clowes G., Keltch A., Krahl M. — J. Pharmacol., 1940, vol. 68, p. 312.
Cocco L., Groff J., Temple C. et al. — Biochemistry, 1981, vol. 20, p. 3972.
Cohen J. (ed). Magnetic Resonance in Biology, vol. I. — New York: Wiley, 1980.
Cohen N. — J. Med. Chem., 1983, vol. 26, p. 259.
Cohen P. Control of Enzyme Activity. — London, New York: Chapman and Hall, 1980.
Cohen S. Polyamines. — New Jersey: Prentice-Hall, 1971.
Cohen S., Yielding К— Proc. Natl. Acad. Sci., U.S.A., 1965, vol. 54, p. 521.
Cohn M., Leigh J. — Nature, Lond., 1962, vol. 193, p. 1037.
Colburn R., Maas J. — Nature, Lond., 1965, vol. 208, p. 37.
Cole A., May P., Williams D. — Agents and Actions, 1981, vol. 11, p. 296.
Colebrook L., Kenny M. — Lancet, 1936, i, p. 1279.
Coleman J. — Biochem. Biophys. Res. Commun., 1974, vol. 60, p. 641.
Collier H., Francis D. — Nature, Lond., 1975, vol. 255, p. 159.
Collier H„ Waterhouse P.—Ann. Trop. Med. Parasit., 1950, vol. 44, p. 156.
Connors T., Hare J. — Brit. J. Cancer, 1974, vol. 30, p. 477.
Connors T., Jones M., Ross W. et al.—Chem. Biol. Interactions, 1972, vol. 5, p. 415.
Conti F. — Nature, Lond., 1969, vol. 221, p. 777.
Conti-Tronconi B., Raferty M. — Ann. Rev. Biochem., 1982, vol. 51, p. 491.
Cook J., Hewett C., Hieger I. — J. Chem. Soc., 1933, p. 396.
Cooper J., Bloom F. Biochemical Basis of Neuropharmacology. — Oxford: University Press, 1982.
Cooper P. — Bact. Rev., 1956, vol. 20, p. 28.
Cordes E. Reaction Kinetics in Micelles. — New York: Plenum Press, 1973.
Cornforth J., Ryback G„ Popjak G. et al. — Biochem. Biophys. Res. Commun., 1962, vol. 9, p. 371.
Costa E., Di Chiara G., Gessa G. (eds). GABA and Benzodiazepine Receptors.— In: Adv. Biochem. Pharmacol., New York: Raven Press, 1981, vol. 26.
Couston F. (ed.) Regulatory Aspects of Carcinogenesis and Food Additives; the Delaney Clause, New York: Academic Press, 1979.
Covey J. — Life Sci., 1980, vol. 26, p. 665.
Cox J. — Nature, Lond., 1967, vol. 216, p. 1328.
Cragoe E. (ed.) Diuretics. — New York: Wiley, 1983.
Craig P. —J. Med. Chem., 1971, vol. 14, p. 680.
Cram D. — J. Amer. Chem. Soc., 1983, vol. 105, p. 135. !
Cramer F. — Angew. Chem., 1956, Bd 68, S. 115.
Cramer J., Miller J., Miller E. — J. Biol. Chem., 1960, vol. 235, p. 885.
Cremer-Bartels G. — In: Chemistry and Biology of Pteridines/Ed. W. Pfleide-rer, Berlin: de Gruyter, 1975.
Crossland J. (ed.) Lewis’s Pharmacology, 5th edn. — London: Churchill, 1981. Crow T., Johnstone E. —Brit. J. Pharmacol., 1977, vol. 59, p. 466 P.
Cruess W., Richert P.—J. Bact., 1929, vol. 17, p. 363.
Crumplin G., Midgley J., Smith J. — Top. Antibiot. Chem., 1980, vol. 3, p. 1.
Cuatrecasas P., Wilcheck M., Anfinsen C. — Proc. Natl. Acad. Sci., U.S.A., 1968, vol. 61, p. 636.
Cullen S., Gross E. — Science, 1951, vol. 113, p. 580.
Culvenor C„ Ham N. — Chem. Commun., Chem., Soc., London, 1966, p. 537.
394
Cunningham L. — Biochemistry, 1964, vol. 3, p. 1629.
Curtis D., Duggan A., Felix D. et al. — Brain Res., 1971, vol. 33, p. 57. Cushley R., Mautner H. — Tetrahedron, 1970, vol. 26, p. 2151.
Cushny A. Biological Relations of Optically Isomeric Substances. — Baltimore: Williams and Wilkins, 1926.
Cymerman-Craig J., Rubbo S., Willis D.t Edgar J. — Nature, Lond., 1955, vol. 176, p. 35.
Cymerman-Craig J., Willis D. — J. Chem. Soc., 1955, p. 4315.
Dalnton F. Chain Reactions. — London and New York: Methuen, 1966.
Dale H.—J. Pharmacol., 1914, vol. 6, p. 147.
Daly. J., Brons R., Snyder S. — Life Sci., 1981, vol. 28, p. 2083.
Danilov A., Guli-Kevkhyan R„ Laverentieva V. et al. — Arch, internal. Pharma-codyn. Then, 1974, vol. 208, p. 35.
Davidoff R. — Neurology, 1978, vol. 28, p. 46.
Davidson M., Griggs B., Boykin D., Wilson W. — J. Med. Chem., 1977, vol. 20, p. 1117.
Davies F., Musa M., Dormandy T. — J. Clin. Path., 1968, vol. 21, p. 363.
Davies G., Lowe J. — Brit. J. Pharmacol. Chemother., 1966, vol. 27, p. 107.
Davies J., Campbell W., Kearns C. — Biochem. J., 1970, vol. 117, p. 221.
Davis B., Dumos R. —J. Exper. Med., 1947, vol. 86, p. 215.
Deake H„ Hu S.-I., Ward H. —J. Biol. Chem., 1980, vol. 255, p. 7174.
De Baun J., Smith J„ Miller E., Miller J. — Science, 1970, vol. 167, p. 184.
De Bruyn P., Robertson R., Farr F. — Anat. Rec., 1950, vol. 108, p. 279.
Delay J., Deniker P., Harl J. — Ann. M6d-psychol., 1952, vol. 110, p. 112. Demel R., Van Deenen L., Kinsky S. — J. Biol. Chem., 1965, vol. 240, p. 2749. Dennis V., Stead N., Andreoli T. — J. Gen. Physiol., 1970, vol. 55, p. 375.
Denny W., Atwell G„ Cain B.—J. Med. Chem., 1979, vol. 22, p. 1453.
Denny W„ Baguley B., Cain B., Waring M. — In: Molecular Aspects of Anticancer Drug Action/Eds. S. Neidle, M. Waring. — London: Macmillan, 1983.
Deskin W. — J. Amer. Chem. Soc., 1958, vol. 80, p. 5680.
De Wys W., Paries W. — J. Natl. Cancer Inst., 1972, vol. 48, p. 375.
Diamond J., Wright E. — Ann. Rev. Physiol., 1969, vol. 31, p. 581.
Dietrich B., Fyles T., Lehn J. et al. — Chem. Commun., Chem. Soc., Lond., 1978, p. 934.
Dietrich S., Bolger M., Kollman P., Jorgensen E. — J. Med. Chem., 1977, vol. 20, p. 863.
Dill W., Fisken R., Reutner T. et al. — Antibiot. Chemother., 1957, vol. 7, p. 99.
Dittmer К — J. Amer. Chem. Soc., 1949, vol. 71, p. 1205.
Dittmer K., du Vigneaud V. — Science, 1944, vol. 100, p. 129.
Dixon M., Webb E. Enzymes. 3rd edn. — London: Longmans; New York: Academic Press, 1979.
Dixon N., Gazzola C., Blakeley R., Zerner B.—J. Amer. Chem. Soc., 1975, vol. 97, p. 4131.
Djerassi C. Optical Rotatory Dispersion. — New York: McGraw-Hill, 1960. Dobler M., Dunitz J., Kilbourn B.—Helv. Chim. Acta, 1969, vol. 52, p. 2573.
Docherty J., MacDonald A., McGrath J. — Brit. J. Pharmacol., 1979, vol. 67, p. 421.
Dockter M., Magnuson J. — Biochem. Biophys. Res. Commun., 1973, vol. 54, p. 790.
Dodds E., Goldberg L., Lawson W., Robinson R. — Nature, Lond., 1938, vol. 142, p. 34.
Doherty D., Shapira R., Burnett W. — J. Amer. Chem. Soc., 1957, vol. 79, p. 5667.
Dole V., Nyswander M„ Kreek M. — Arch. Intern. Med., 1966, vol. 118, p. 304. Dolly J., Barnard E. — Biochemistry, 1977, vol. 16, p. 5053.
Doluizio J., Martin A. —Med. Chem., 1963, vol. 6, pp. 16, 20.
Domagk G. — Dtsch. med. Woch., 1935, Bd 61, S. 250.
Donald C., Passey B., Swaby R.—J. Gen. Microbiol., 1952, vol. 7, p. 211.
Douglas C., Haldane J. S., Haldane J. B. S. — J. Physiol., 1912, vol. 44, p. 275,
395
Douglas W.— Brit. J. Pharmacol., 1968, vol. 34, p. 451.
Duch D., Sigel C„ Bowers S. et al. — Curr. Chemother. Infect. Dis., Proc. Int.
Congr., 1980, vol. 2, p. 1597.
Dudai У., Ben-Barak J. — FEBS Lett., 1977, vol. 81, p. 134.
Durant G., Ganellin C., Parsons M.— J. Med. Chem., 1975, vol. 18, p. 905.
Durant G„ Ganellin C., Parsons M. — Agents and Actions, 1977, vol. 7, p. 39.
Dutcher J., Boyak G., Fox S. Antibiotic Annual. — New York: Medical Encyclopedia Inc., 1954.
Dweck R. (ed.) Nuclear Magnetic Resonance in Biology. — New York: Academic Press, 1977.
Dwyer F., Gyarfas E., Koch J., Rogers W.— Nature, Lond., 1952, vol. 170, p. 190.
Dwyer F„ Gyarfas E., O'Dwyer M.— Nature, Lond., 1951, vol. 167, p. 1036.
Dyckes D., Nestor J., Ferger N., du Vigneaud V. —J. Med. Chem., 1974, vol. 17,
p. 969.
Eagle H. — J. Pharmacol., 1939, vol. 66, pp. 10, 423, 436.
Eagle H.— Pharmacol. Rev., 1951, vol. 3, p. 107.
Eagle T., Zon G., Egan W. — J. Med. Chem., 1979, vol. 22, p. 897.
Easson L„ Stedman E. —Biochem. J., 1933, vol. 27, p. 1257.
Edlbacher S„ Baur H., Becker M.— Z. physiol. Chem., 1940, bd 265, S. 61.
Edsall J., Martin R„ Hollingworth B. — Proc. Natl. Acad. Sci., U.S.A., 1958).
vol. 44, p. 505.
Edson E., Sanderson D., Noakes D. — World Rev. Pest Control, 1964, vol. 4, p. 36.
Edson E., Sanderson D., Noakes D. — World Rev. Pest Control., 1966, vol. 5, p. 143.
Eger E., Lundgren C., Miller S., Stevens W.— Anaesthesiology, 1969, vol. 30, p. 129.
Ehrenpreis S., Fleisch J., Mittag T. — Pharmacol., 1969, vol. 21, p. 131.
Ehrlich P. — Ber. dtsch. chem. Ges., 1909, Bd 42, S. 17.
Einhorn L., Williams S. — New Engl. J. Med., 1979, vol. 300, p. 289.
Eliel E., Allinger N., Angyal S., Morrison G. Conformational Analysis. — New York: Wiley-Interscience, 1965.
Eliel E., Basolo F. Elements of Stereochemistry. — New York: Wiley, 1969.
Elion G., Rovensky A., Hitchings G. et al. — Biochem. Pharmacol., 1966, vol. 15fc p. 863.
Elkhawad A., Woodruff G. — Brit. J. Pharmacol., 1975, vol. 54, p. 107.
Ellenbroek B., Van Rossum J. — Arch. Internet. Pharmacodyn. Ther., 1960; vol. 125, p. 216.
Elliott IF.— Biochem. J., 1963, vol. 86, p. 562.
Elworthy P., Florence A., Macfarlane C. Solubilization by Surface-Active Agents. — London and New York: Chapman and Hall, 1968.
Endo H., Ono T., Sugimura T. Chemistry and Biological Actions of 4-Nitroquino-line-l-oxide. — Berlin: Springer, 1970.
Engle T., Zon G., Egan IF. — J. Med. Chem., 1979, vol. 22, p. 897.
Enoch H., Lester R. —J. Biol. Chem., 1975, vol. 250, p. 6693.
Ephrussi B., Hottinguer H. — Nature, Lond:, 1950, vol. 166, p. 956.
Estensen R., Krey A., Hahn F. — Molec. Pharmacol., 1969, vol. 5, p. 532.
Evans B., Wolfenden R. — J. Amer. Chem. Soc., 1970, vol. 92, p. 4751.
Everett J., Roberts J., Ross IF. — J. Chem. Soc., 1953, p. 2386.
Ewins A., Phillips M. — Brit. Pat., 1939, vol. 512, p. 145.
Exner O. — In: Advances in Free Energy Relationships/Eds. N. Chapman and J. Shorter, London: Plenum Press, 1972.
Fairlamb A., Bowman 1. — Trans. Roy. Soc. Trop Med. Hyg. 1975, vol. 69, p. 268.
Fairley G., Simister J. (eds). Cyclophosphamide. — Baltimore: Williams and Wilkins, 1965.
Falco E., Goodwin L., Hitchings G. et al. — Brit. J. Pharmacol, 1951, vol. 6, p. 185.
Farber S. — Blood, 1952, vol. 7, p. 107.
Farley T., Strong F., Bydalek T. — J. Amer. Chem. Soc., 1965, vol. 87, p. 3501,
396
Farmer О., Ariens E. — Trends Pharmacol. Sci., 1682, vol. 3, p. 362.
Farnham A., Gregory G., Sawicki R.— Bull. Entomol. Res., 1966, vol. 57; p. 107.
Fendler J., Fendler E. Catalysis in Micellar and Macromolecular Systems. — New York: Academic Press, 1975.
Ferguson J. — Proc. Roy. Soc. B, 1939, vol. 127, p. 387.
Ferguson J. Colloques intrnatioaux du Centre national de la Recherche scienti-fique. No. 26: Mecanisme de la Narcose. — Paris, 1951, p. 25.
Ferguson J., Hawkins S., Doxey D. — Nature, Lond., 1950, vol. 165, p. 1021.
Ferguson J., Pirie H. — Ann. Appl. Biol., 1948, vol. 35, p. 532.
Fest C., Schmidt K-J. The Chemistry of Organophosphorus Pesticides. 2nd? edn. — Berlin: Springer, 1982.
Festy B., Sturm J., Daune M.— Biochim. Biophys. Acta, 1975, vol. 407, p. 24l
Fildes P. — Lancet, 1940a, i, p. 955.
Fildes P. — Brit. J. Exper. Path., 1940b, vol. 21, p. 67.
Fillion G„ Fillion M.— Nature, Lond., 1981, vol. 292, p. 349.
Filman D., Bolin J., Matthews D., Kraut J. — J. Biol. Chem., 1982, vol. 257,. p. 13663.
Firestone R., Pisano J., Bonney R. — J. Med. Chem., 1979, vol. 22, p. 1130.
Fischer J., Jardetzky O. — J. Amer. Chem. Soc., 1965, vol. 87, p. 3237.
Fisher L., Oakenfull D. — Chem. Soc. Rev., 1977, vol. 6, p. 25.
Flaim S., Zelis R. (eds). Calcium Blockers. Mechanisms of Action and ClinicaJ Applications. — Baltimore: Urban and Schwarzenberg, 1982.
Fleckenstein A. — Annu. Rev. Pharmacol. Toxicol., 1977, vol. 17, p. 149.
Fleckenstein A, Calcium Antagonism in Heart and Smooth Muscle. — New York г Wiley-Interscience, 1983.
Flower R., Kingston W. — Brit. J. Pharmacol., 1975, vol. 55, p. 239 P.
Fourneau E., Bovet D., Bovet F., Montezin G. — Bull. Soc. Chim. Biol., 1944; vol. 26, pp. 134, 516.
Fox J. — Chem. Eng. News, 1983, vol. 61 (March 14th), p. 8.
Fox R„ Richards F. — Nature, Lond., 1982, vol. 300, p. 325.
Fox W. — Proc. Roy, Soc. Med., 1977, vol. 70, p. 4.
Foye W., O’Laughlin R., Duvall R. — J. Pharm. Sci„ 1961, vol. 50, p. 641.
Fozard J. — Trends Pharmacol. Sci., 1983, vol. 4, p. 288.
Franklin T. — Biochem. J., 1971, vol. 123, p. 267.
Franklin T., Snow G. Biochemistry of Antimicrobial Action, 3rd edn. — London* and New York: Chapman and Hall, 1981.
Franks N., Lieb W.— Nature, Lond., 1981, vol. 292, p. 248.
Franks N., Lieb W.— Nature, Lond., 1982, vol. 300, p. 487, but cf. ibid. 1984;. vol. 310, p. 599.
Fraser I., Vessell E. — Ann. N.Y. Acad. Sci., 1968, vol. 151, p. 777.
Free S., Wilson J. — J. Med. Chem., 1964, vol. 7, p. 395.
Freed C., Quintero E., Murphy C. — Life Sci., 1978, vol. 23, p. 313.
Frenkel J., Hitchings G. —Antibiot. Chemother., 1957, vol. 7, p. 630.
Freudenthal R.t Jones P. (eds) Carcinogenesis. — New York: Raven Press, 1976; vol. 1.
Friberg L. Cadmium in the Environment (2nd edn). — Boca Raton, Florida: CRCi Press, 1974.
Friedheim E. — Amer. J. Trop. Med., 1951, vol. 31, p. 218.
Friess S., Baldridge H. — J. Amer. Chem. Soc., 1956, vol. 78, p. 2482.
Froehner S., Reiness C„ Hall Z.— J. Biol. Chem,., 1977, vol. 252, p. 8589.
Fujino M., Shinagawa S., Obayashi M. et al. — J. Med. Chem., 1973, vol. 16;
p. 1144.
Fujita T., Nishioka T. — Progr. Phys. Org. Chem., 1975, vol. 12, p. 49.
Fujita T., Nishioka T., Nakajima M. — J. Med. Chem., 1977, vol. 20, p. 1071.
Fukuto T. — Drug Metab. Rev., 1972, vol. 1, p. 117.
Fuller R., Kidder G., Nugent N. et al. — Photochem. Photobiol., 1971, vol. 14„ p. 359.
Fulton J., Grant P. — Ann. Trop. Med. Parasit., 1956, vol. 50, p. 381.
Gabbay E. — J. Amer. Chem. Soc., 1968, vol. 90, p. 5257.
Gage J. — Biochem. J., 1953, vol. 54, p. 426.
397?
Gale E. — Adv. Enzymol., 1946, vol. 6, p. 1.
Gale E., Cundliffe E., Reynolds P. et al. The Molecular Basis of Antibiotic Action.
2nd edn.—London: Wiley, 1981.
Gale E., Taylor E. —J. Gen. Microbiol., 1947, vol. 1, p. 77.
Ganellin C. — J. Med. Chem., 1981, vol. 24, p. 913.
Ganis P., Avitable G., Mechlinskl IF., Schaffner C. — J. Amer. Chem. Soc., 1971, vol. 93, p. 4560.
Garattini S., Mussini E., Randall L. (eds) The Benzodiazepines. — New York: Raven Press, 1973.
Gershenfield, Milanick C. — Amer. J. Pharm., 1941, vol. 113, p. 306.
Giarmann N.—J. Pharmacol., 1949, vol. 96, p. 119.
Gilardi R., Karie L, Karie J. Sperling W.—Nature, Lond., 1971, vol. 232, p. 187.
Gillespie L., Oates J., Croat J., Sjoerdsma A.—Circulation, 1962, vol. 25, p. 281.
Gilman A. G., Goodman L., Gilman A. (eds) Goodman and Gilman’s Pharmacological Basis of Therapeutics. — New York: Macmillan, 1980.
Glatt H., Protic-Sablfic M., Oesch F. — Science, 1983, vol. 220, p. 961.
Glave W., Hansch C. — J. Pharm. Sci., 1972, vol. 61, p. 589.
'Gloge H., Luttmann H., Mutschler E. — Brit. J. Pharmacol. Chemother., 1966, vol. 27, p. 185.
Goksoyr J. — Nature, Lond., 1955, vol. 175, p.. 820; Physiol. Plant., 1955, vol. 8, p. 719.
Gold E., Chang W., Cohen M. et al. — J. Med. Chem., 1982, vol. 25, p. 1363.
Goldacre R., Loveless A., Ross W.— Nature, Lond., 1949, vol. 163, p. 667.
Goldacre R., Phillips J. —J. Chem. Soc., 1949, p. 1724.
Goldberg L., Kohli J. —Trends Pharmacol. Sci., 1983, vol. 4, p. 64.
Goldfine L, Perlman R., Roth J. — Nature, Lond., 1971, vol. 234, p. 295.
•Goldstein A., Fischli W., Lowney C. et al. — Proc. Natl. Acad. Sci., U.S.A., 1981, vol. 78, p. 7219.
Goldstein A., Lowney L., Pal B.— Proc. Natl. Acad. Sci., U.S.A,, 1971, vol. 68, p. 1742.
Goodson J., Henry T., MacFie J. — Biochem. J., 1930, vol. 24, p. 874.
Gorin F., Balasubramanian T., Cicero T. et al.—J. Med. Chem., 1980, vol. 23, p. 1113.
Gosalvez M., Blanco M., Vivero C., Valles F. — Eur. J. Cancer, 1978, vol. 14, p. 1185.
Goth A. — Science, 1945, vol. 101, p. 383.
Gottlieb R. — Arch, exper. Path. Pharmacol., 1923, vol. 97, p. 113.
Grant P., Sargent J. — Biochem. J., 1960, vol. 76, p. 229.
Gray G., Smith L, McKenzie I. et al. — Lancet, 1977, i, p. 4.
Greathouse Gr, Block S., Kovak E. et al. Research on Chemical Compounds for Inhibition of Fungi. US Corps of Engineers, Fort Belvoir, Virginia, 1954.
Green A., Heale D., Grahame-Smith D. — Psychopharmacology, 1977, vol. 52, p. 195.
Greengard O., McIlwain H. — Biochem. J., 1955, vol. 61, p. 61.
Greengard P. — Nature, Lond., 1976, vol. 260, p. 101.
Grell E„ Eggers F., Funck T. — Chimia, 1972, vol. 26, p. 632.
Griffin M., Brown G. —J. Biol. Chem., 1964, vol. 239, p. 310.
Griffiths L, Intarakoslt P., Taylor H., Wain R. — Ann. Appl. Biol., 1966, vol. 58, p. 183.
Grivsky E„ Lee S., Sigel C. et al. — J. Med. Chem., 1980, vol. 23, p. 327.
Grunwald E., Ralph E. — J. Amer. Chem. Soc., 1967, vol. 89, p. 4405.
Guillemin R. — Science, 1978, vol. 202, p. 390.
Gurson C., Saner G. — Amer. J. Clin. Nutr., 1971, vol. 24, p. 1313.
Gutte B„ Merrifield R. — J. Amer. Chem. Soc., 1969, vol. 91, p. 501.
Haagen-Smit A., Went F. — Proc. k. ned. Akad. Wetenschap, 1935, vol. 38, p. 852.
Haddow A., Harris R., Kon G., Roe E. — Proc. Roy. Soc. B, 1948, vol. 241, p. 147.
Haddow A., Timmis G. — Lancet, 1953, vol. 1, p. 207.
Hahn F. — Naturwiss., 1975, Bd 62, S. 449.
5398
Hahn P., Bale W„ Ross I. et al. — Science, 1940, vol. 92, p. 131.
Hakala M.— Fed. Proc., 1966, vol. 25, p. 750.
Haldane J. Enzymes. — London: Longmans, 1930.
Hallman P„ Perrin D„ Watt A. — Biochem. J., 1971, vol. 121, p. 549.
Halpern B. — Arch. Internal. Pharmacodyn. Ther., 1942, vol. 68, p. 3391 Hamilton-Miller J. — Bact. Rev., 1973, vol. 37, p. 166.
Hamilton-Miller J., Abraham E. — Biochem. J., 1971, vol. 123, p. 183.
Hammett L. Physical Organic Chemistry. 2nd edn. — New York: McGraw-Hill, 1970.
Hammond G. — J. Amer. Chem. Soc., 1955, vol. 77, p. 334.
Handrick G., Duffley R., Lambert G. et al. — J. Med. Chem., 1982, vol. 25^ p. 1447.
Hansch C.—J. Med. Chem., 1968, vol. 11, p. 920.
Hansch C. — Drug Design, 1971, vol. 1, p. 271.
Hansch C., Anderson S. — J. Med. Chem., 1967, vol. 10, p. 745.
Hansch C., Fujita T. —J. Amer. Chem. Soc., 1964, vol. 86, p. 1610.
Hansch C., Rim K., Sarrna R. — J. Amer. Chem. Soc., 1973, vol. 95, p. 6447.
Hansch C., Leo A. Sustituent Constants for Correlation Analysis in Chemistry and Biology. — New York: Wiley, 1979.
Hansch C., Leo A., Unger S. et al.—J. Med. Chem., 1973, vol. 16, p. 1207.
Hansch C., Li R., Blaney J., Langridge R.—J. Med. Chem., 1982, vol. 25, p. 777.
Hansch C., Quinlan J., Lawrence G. — J. Org. Chem., 1968, vol. 33, p. 347.
Hansch C., Steward A., Anderson S., Bentley D. — J. Med. Chem., 1968, vol. 11, p. 1.
Hansch C„ Unger S„ Forsythe A. — J. Med. Chem., 1973, vol. 16, p. 1217.
Hansch C„ Vittoria A., Silipo C., Low P. — J. Med. Chem., 1975, vol. 18, p. 546.
Haracz J. — Schizophrenia Bull., 1982, vol. 8, p. 438.
Hardman H. — Circ. Res., 1962, vol. 10, p. 598.
Harold F„ Baarda J. — J. Bact., 1967, vol. 94, p. 53.
Hartley B., Kilby B. — Biochem. J., 1952, vol. 50, p. 672.
Hartough H. Thiophene and its Derivatives.—New York: Interscience, chap. 2, 1952.
Hata A. — Kitasato Arch. Exper. Med,, 1932, vol. 9, p. 1.
Hawking F. — J. Pharmacol., 1944, vol. 82, p. 31.
Hawkins C. Absolute Configuration of Metal Complexes. — London: Wiley>. 1971.
Hawkins C., Perrin D. — J. Chem. Soc., 1962, p. 1351.
Hawkins C., Perrin D. — Inorg. Chem., 1963, vol. 2, p. 843.
Haydon D., Hiadky S. — Quart. Rev. Biophys., 1972, vol. 5, p. 187.
Нагит E., Sabatka J., Chang L.-J. et al. — Science, 1981, vol. 213, p. 1010.
Hedberg K-, Hughes E., Waser J. — Acta Crystallogr., 1961, vol. 14, p. 369.
Helenius A., Marsh M., White J. — Trands Biochem. Sci., 1980, vol. 5, p. 104-Hemmerich P., Veeger C., Wood H. — Angew, Chem., 1965, Bd 77, S. 1.
Henderson I., Canellos G. — New Engl. J. Med., 1980, vol. 302, p. 78.
Henderson P., McGivan J., Chappell J.—Biochem. J., 1969, vol. Ill, p. 521.
Hey P.—Brit. J. Pharmacol., 1952, vol. 7, p. 117.
Higgs G., Flower R., Vane J. — Biochem. Pharmacol., 1979, vol. 28, p. 1959 Hill D., Bowery N. —Nature, Lond., 1981, vol. 290, p. 149.
Hiller S. — Proc. Soc. Exper. Biol. Med., 1927, vol. 24, p. 427.
Hilton J., Ard J., Jansen L., Gentner W. — Weeds, 1959, vol, 7, p. 381.
Hirschmann R., Nutt R„ Veher D. et al.—J. Amer. Chem. Soc., 1969, vol. 9I'> p. 507.
Hitchings G. — Trans. Roy. Soc. Trop. Med. Hyg., 1952, vol. 46, p. 467.
Hitchings G. (ed.). Inhibition of Folate Metabolism in Chemotherapy. — Berlin*.
Springer, 1983 (in English).
Hitchings G., Burchall J. — Adv. Enzymol., 1965, vol. 27, p. 417.
Ho L, Harris R. — Ann. Rev. Pharmacol., 1981, vol. 21, p. 83.
Hobbiger F.—Chem. Indust., 1954, p. 415.
Hobbiger F. — Brit. J. Pharmacol., 1955, vol. 10, p. 356.
399*
Hobbiger F., Vojvodic V. — Biochem. Pharmacol., 1966, vol. 15, p. 1677.
Hochster R., Quastel J. (eds). Metabolic Inhibitors. (4 vols). — New York: Academic Press, 1963—73.
Hodgson E., Casida J. —J. Agric. Food Chem., 1962, vol. 10, p. 208.
Hoefnagel A., Hoefnagel M., Wepster B. —J. Org. Chem., 1978, vol. 43, p. 4720.
Hoffman C., Schweitzer T„ Dalby G. — J. Amer. Chem. Soc., 1940, vol. 62, p. 988.
Hoffman C., Schweitzer T., Dalby G. — Indust. Eng. Chem., 1941, vol. 33, p. 749.
Hollenberg C., Borst P., Van Bruggen E.— Biochim. Biophys. Acta 1970, vol. 209, p. 1.
Hollingworth R., Fukoto T., Metcalf R.—J, Agric. Food Chem., 1967, vol. 15, p. 235.
Holmes R., Robins E. — Brit. J. Pharmacol., 1955, vol. 10, p. 490.
Holton P., Ing H. — Brit. J. Pharmacol., 1949, vol. 4, p. 190.
Hopfinger A. Conformational Properties of Macromolecules. — New York: Academic Press, 1973.
Hopfinger A. —J. Amer. Chem. Soc., 1980, vol. 102, p. 7196.
Hopfinger A. — J. Med. Chem., 1983, vol. 26, p. 990.
Horn A., Snyder S. — Proc. Natl. Acad. Sci., U.S.A., 1971, vol. 68, p. 2325.
Howell J., Altounyan R. — Lancet, 1967, ii, p. 539.
Huang L., Barker J. — Science, 1980, vol. 207, p. 195.
Hudson C. — Adv. Carbohyd. Chem., 1948, vol. 3, p. 1.
Hughes J. — Brain Res., 1975, vol. 88, p. 295.
Hughes I., Smith T„ Kosterlitz H. et al. — Nature, Lond., 1975, vol. 258, p. 577.
Hunt R., Taveau R.— Bull. Hyg. Lab., U.S. Treasury, 1911, No. 73.
Hunter F., Lowry O. —Pharmacol. Rev., 1956, vol. 8, p. 89.
Hurly M.— Trans. Roy. Soc. Trop. Med. Hyg., 1959, vol. 53, pp. 410, 412.
Hurwitz J., Furth J., Malamy M., Alexander M. — Proc. Natl. Acad. Sci., U.S.A., 1962, vol. 48, p. 1222.
Hutner S. — Science, 1949, vol. 110, p. 548.
Jjzerman A„ Bultsma T., Timmerman H., Zaagsma J. — J. Pharm. Pharmacol., 1984, vol. 36, p. 11—15.
Ing H. — Physiol. Rev., 1936, vol. 16, p. 527.
Ing H. — Science, 1949, vol. 109, p. 264.
Ing H„ Kordik P., Williams T. — Brit. J. Pharmacol., 1952, vol. 7, p. 103.
International Agency for Research on Cancer (WHO). Monographs on Evaluation of Carcinogenic Risk of Chemicals to Humans. — Lyon (France), 1982, vol. 29.
International Union of Biochemistry. Enzyme Nomenclature. — New York: Academic Press, 1978.
Irving H., Mellor D. —J. Chem. Soc., 1962, pp. 5222, 5237.
Irving H., Rossotti H. — Acta. Chem. Scand., 1956, vol. 10, p. 72.
Irving H., Williams R.—J. Chem. Soc., 1953, p. 3192.
Isaka S. — Nature, Lond., 1957, vol. 179, p. 578.
Ison R., Partington P., Roberts G. — Molec. Pharmacol., 1973, vol. 9, p. 756.
Iversen L. The Uptake and Storage of Noradrenaline in Sympathetic Nerves. — Cambridge: University Press, 1967.
Iversen L. — In; Psychopnarmacology— A Generation of Progress/Eds. M. Lipton, A. Di Mascio, K. Killam, New York: Raven Press, 1978.
Iwamoto E., Martin W. — Medicinal Res. Rev., 1981, vol. 1, p. 411.
Iyer V., Szybalski W.— Schience, 1964, vol. 145, p. 55.
Izaki R., Matsuhashi M., Strominger J. — Proc. Natl. Acad. Sci. U.S.A., 1966, vol. 55, p. 656.
Izaki K-, Matsuhashi M„ Strominger J.—J. Biol. Chem., 1968, vol. 243, p. 3180.
Izatt R., Nelson D., Rylting J. et al. — J. Amer. Chem. Soc., 1971, vol. 93, p. 1619.
Hackman L„ Sternhell S. Applications of N.M.R. Spectroscopy in Organic Chemistry. 2nd edn. — Oxford; Pergamon Press, 1969.
.Jackson K„ Mason S. — Trans. Farad. Soc., 1971, vol. 67, p. 966.
Jaenicke L., Chan P. — Angew. Chem., 1/960, Bd 72, S. 753.
400
James К. —Progr. Med. Chem., 1974, vol. 10, p. 205.
James S. — Parasitology, 1980, vol. 80, p. 313.
Janis R., Triggle D. —J. Med. Chem., 1983, vol. 26, p. 795.
Janssen M. — J. Inorg. Nuclear Chem., 1958, vol. 8, p. 340.
Jardetzky O., Jardetzky C.— Methods Biochem. Anal., 1962, vol. 9, p. 235.
Jardetzky O., Roberts G. NMR in Molecular Biology. — New York: Academic Press, 1981.
Jellinek F. — Acta Crystallogr., 1957, vol. 10, p. 277.
Jencks W. Catalysis in Chemistry and Enzymology. — New York: McGraw-Hill, 1969.
Jensen R„ Schmith R.— Z. Immunitats., 1942, Bd 102, S. 261.
Jirousek L., Pritchard E.—Biochim. Biophys. Acta, 1971, vol. 243, p. 230.
Johnson H.— In: Immediate Hypersensitivity/Ed. M. Bach. New York: Marcel Dekker, 1978, p. 533.
Johnson J., Rajagopolan R. — Biochemistry, 1982, vol. 79, p. 6856.
Johnson J., Woodward R., Robinson R. (eds) The Chemistry of Penicillin.— Princeton, N.J.: University Press, 1949.
Johnson M., Milne G. —J. Het. Chem., 1980, vol. 17, p. 1817.
Johnson Q. — U.S. Atom. Energy Commiss., 1960, UCRL 9350.
Johnston G, — Ann Rev. Pharmacol., 1978, vol. 18, p. 269.
Johnston G„ Curtis D., Beart P. et al.—J. Neurochem., 1975, vol. 24, p. 433.
Johnston T., McCaleb G., Montgomery J.—J. Med. Chem., 1963, vol. 6, p. 669.
Jones M. — Nature, Lond., 1960, vol. 185, p. 96.
Jones O., Watson W.— Nature, Lond., 1965, vol. 208, p. 1169.
Jordan D. — In: Molecular Associations in Biology./Ed. B. Pullman, New York: Academic Press, 1968, p. 221.
Josephson E., Taylor D., Greenberg J., Coatney G. —J. Infect. Dis., 1953, vol. 93, p. 257.
Joswick H. Ph. D. thesis: Mode of Action of Hexachlorophene. — Ann Arbor: University of Michigan, 1961.
Jung L., Roffel J.-C„ Lami H.— Bull. Chim. then, 1971, vol. 6, p. 341.
Jung L„ Lami H. — Bull. Chim. ther., 1970, vol. 5, p. 391.
Jung M., Metcalf B.—Biochem. Piophys. Res. Commun., 1975, vol. 67, p. 301.
Rabachnik M., Brestkin A., Godovikov N. et al. — Pharmacol. Rev., 1970, vol. 22, p. 355.
Radota L, Abe T. — J. Lab. Clin. Med., 1954, vol. 43, p. 375.
Radota L, Midorikawa O. —J. Lab. Clin. Med., 1951, vol. 38, p. 671.
Rahan F., Rahan J., Cassidy P., Rroph H.—Ann. N.Y. Acad. Sci, 1974, vol. 235, p. 364.
Ralow W. Pharmacogenetics, Heredity and Response to Drugs. — Philadelphia: Saunders, 1962.
Ralow W.—Trends Pharmacol. Sci., 1980, vol. 1, p. 403.
Ralow W., Otton S., Rador D. et al.—Canad. J. Physiol. Pharmacol., 1980, vol. 58, p. 1142.
Ramiura H., Matsumoto A., Miyazake Y_, Yamamoto I. — Agric. Biol. Chem., Tokyo, 1963, vol. 27, p. 684.
Ramiya R., Takamoto M., Wada Y., Asai M.—Acta Crystallogr. B, 1981, vol. 37, p. 1626.
Rannan R., Notstrand B., Fridborg R. et al. — Proc. Natl. Acad. Sci., U.S.A., 1975, vol. 72, p. 51.
Rarim A., Elfellah M., Evans D. — J. Med. Genet., 1981, vol. 19, p. 325.
Rarle I., Brockway L. — J. Amer. Chem. Soc., 1944, vol. 66, p. 1974.’
Rarlin A. —Cell Surface Rev., 1980, vol. 6, p. 191.
Rartha G., Bello J., Harker D. — Nature, Lond., 1967, vol. 213, p. 862.
Rato G. — J. Pharm. Sci., 1975, vol. 64, p. 488.
Rato G„ Yung J., Ihnat M. — Molec. Pharmacol., 1970, vol. 6, p. 588.
Raufman S. — In: Pteridine Chemistry/Eds. W. Pfleiderer, E. Taylor, Oxford: Pergamon Press, 1964, p. 307:
Raye R. —J. Pharm. Pharmacol., 1950, vol. 2, p. 902.
Rebabian J„ Caine D. — Nature, Lond., 1979, vol. 277, p. 93.
Relly J., Moews P., Rnox J. et al. — Science,’ 1982, vol. 218, p. 479.
26—734
401
Kessler H.— Angew. Chem. Internal. Edn, 1982, vol. 21, p. 512.
Khromov-Borisov N„ Michelson M. — Pharmacol. Rev., 1966, vol. 18, p. 1051.
Kier д. _ Molecular Orbital Theory in Drug Research. — New York: Academic Press, 1971.
Kier L„ Hall L. — Molecular Connectivity in Chemistry and Drug Research. — New York: Academic Press, 1976.
Kinsky S.— Proc. Natl. Acad. Sci., U.S.A., 1962, vol. 48, p. 1049; J. Bact, 1962, vol. 83, p. 351.
Kisliuk R.— Molec. Cell Biochem., 1981, vol. 39, p. 331.
Klaus W., Lee K. — J. Pharmacol., 1969, vol. 166, p. 68.
Klayman D., Scovill !., Bartosevich J., Mason C.—J. Med. Chem., 1979, vol. 22, p. 1367.
Klee W„ Streaty R. — Nature, Lond., 1974, vol. 248, p. 63.
Kleiderer E., Shonle H. — J. Amer. Chem. Soc., 1934, vol. 56, p. 1772.
Kleinwachter V., Bakarova Z., Bohacek J. — Biochim. Biophys. Acta, 1969, vol. 174, p. 188.
Klevens H. — J. Phys. Colloid Chem., 1948, vol. 52, p. 130.
Klyne W„ Buckingham J. Atlas of Stereochemistry. 2nd edn. (2 vols). — London and New York: Champan and Hall, 1978.
Knox W., Stampf P., Green D., Auerbach V. — J. Bact., 1948, vol. 55, p. 451.
Kober T., Cooper G. — Nature, Lond., 1976, vol. 262, p. 704.
Kobinger W.— Rev. Physiol. Biochem. Pharmacol., 1978, vol. 81, p. 39.
Кое B. — Trends Pharmacol. Sci., 1983, vol. 4, p. 110.
Koehler K, Lienhard G. — Biochemistry, 1971, vol. 10, p. 2477.
Koepfli J., Thimann K, Went F. — J. Biol. Chem., 1938, vol. 122, p. 763.
Kohn K., Waring M., Glaubiger D., Friedman C. — Cancer Res., 1975, vol. 35, p. 71.
Kanaka K„ Matsuoka T. — Kumamoto Med. J., 1967, vol. 20, p. 196.
Konigk E., Putfarken B. — Tropenmed. Parasit., 1983, vol. 34. p. 1.
Konopa J., Koldej K, Pawlak J. — Chem. Biol. Interact., 1976, vol. 13, p. 99.
Kortilm G., Vogel W., Andrussow K. Dissoziationskonstanten organischer Sauren in wasseriger Losung, compiled for International Union of Pure and Applied Chemistry. — London: Butterworths, 1961.
Koshland D. — Fed. Proc., 1964, vol. 23, p. 719.
Koshland D., Neet K. — Ann. Rev. Biochem., 1968, vol. 37, p. 359.
Kosower E. Molecular Biochemistry. — New York: McGraw-Hill, 1962.
Kosterlitz H., Collier H., Villareal J. (eds) Agonist and Antagonist Actions of Narcotic Analgesic Drugs. — London: Macmillan; Baltimore: University Park Press, 1972.
Kowala C., Murray K-, Swan J., West B. — Austral. J. Chem., 1971, vol. 24, p. 1369.
Krogsgaard-Larsen P., Scheel-Kruger N., Kofod H. (eds) GABA-Neurotransmit-ters, Pharmacochemical, Biochemical, and Pharmacological Aspects. — Copenhagen: Mundsgaard, 1979.
Krueger H., O'Brien R. — J. Econ. Entomol., 1959, vol. 52, p. 1063.
Krueger H., O'Brien R., Dauterman IF.— J. Econ. Entomol., 1960, vol. 53, p. 25.
Krilger-Thiemer E. — Berichte Borstel, Germany, 1957, Bd 4, S. 299.
KrUger-Thiemer E., Bilnger P. — Chemotherapie, 1965, vol. 10, pp. 61, 129.
Kuhlmann K., Mosher C. — J. Med. Chem., 1981, vol. 24, p. 1333.
Kuhn R. — Angew. Chem., 1930, Bd 53, S. 1.
Kuhn R. — Amer. J. Psychiat., 1958, vol. 115, p. 459.
Kuhn R., Weygand F., Moller E. — Ber. dtsch. chem. Ges, 1943, Bd 76 S. 1044.
Kuhr R. Carbamate Insecticides. — Boca Raton, Florida: CRC-press, 1976.
Kunos G. Adrenoceptors and Catecholamine Action (2 vols). — New York: Wiley 1981—83.
Kutter E., Hansch C. — J. Med. Chem., 1969, vol. 12, p. 647.
Ladd M. — Theoret. Chim. Acta, Berlin, 4968, Bd 12, S. 333.
Laduron P., Janssen P., Gommeren W., Leysen J. — Molec. Pharmacol., 1982, vol. 21, p. 294.
402
Lambert J., Simpson R., Mohr H., Hopkins L.—J. Assoc. Offic. Anal. Chem., 1970, vol. 53, p. 1145.
Lampen J., Jones M.— J. Biol. Chem., 1946, vol. 166, p. 435.
Lands A., Arnold A., McAuliff J. et al. — Nature, Lond., 1967, vol. 214, p. 597. Landvatter II7., Katzenellenbogen J. — J. Med. Chem., 1982, vol. 25, p. 1300. Lang M., Prasad K, Holick U7. et al.—J. Amer. Chem. Soc., 1979, vol. 101, p. 6296.
Lange W., von Kreuger G.— Ber. dtsch. chem. Ges., 1932, Bd 65, S. 1598.
La Planche L., Rogers M. — J. Amer. Chem. Soc., 1964, vol. 86, p. 337.
Lawley P., Brookes P. — J. Molec. Biol., 1967, vol. 25, p. 143.
Lawrence J., Loomis U7., Tobias C„ Turpin F. — J. Physiol., 1946, vol. 105, p. 197.
Laycock G., Shutman A. —Nature, Lond., 1967, vol. 213, p. 995.
Le Count D. — In: Chronicles of Drug Discovery/Eds. J. Bindra, D. Lednicer, New York: Wiley, 1982.
Lederberg J. — J. Bact., 1957, vol. 73, p. 144.
Ledochowski A. — Materia Medica Polona, Warsaw, 1976, vol. 8, p. 237.
Lee B. — J. Molec. Biol., 1971, vol. 61, p. 463.
Lee R„ Hodsden M. — Biochem. Pharmacol., 1963, vol. 12, p. 1241.
Lees H., Simpson J. — Biochem. J„ 1957, vol. 65, p. 297.
Leeson L„ Krueger J., Nash A. — Tetrahed. Lett., 1963, p. 1155.
Leff S., Creese I. — Trends Pharmacol. Sci., 1983, vol. 4, p. 463.
Lefkovitz R., Mullikin D., Caron Л1.— J. Biol. Chem., 1976, vol. 251, p. 4686.
Leggett D. (ed.) Computational Methods for Determination of Stability Constants.— New York: Plenum Press, 1983.
Legho S., Blumenschein G., Buzdar A. et al. — Cancer Treat. Rep., 1979, vol. 63, p. 1961.
Lehn J., Sauvage J., Dietrich B. — J. Amer. Chem. Soc., 1970, vol. 92, p. 2916.
Leo A., Hansch C. — J. Org. Chem., 1971, vol. 36, p. 1539.
Leo A., Hansch C„ Elkins D. — Chem. Rev., 1971, vol. 71, p. 525.
Leo A.t Hansch C., Jow P. —J. Med. Chem., 1976, vol. 19, p. 611.
Leo A., Jow P., Silipo C., Hansch C. — J. Med. Chem., 1975, vol. 18, p. 865.
Leo A., Panthananickal A., Hansch C. et al. — J. Med. Chem., 1981, vol. 24, p. 859.
Lerman L. —J. Molec. Biol., 1961, vol. 3, p. 18.
Lerman L. —Proc. Natl. Acad. Sci., U.S.A., 1963, vol. 49, p. 94.
Lerman L.—J. Molec. Biol., 1964a, vol. 10, p. 367.
Lerman L.—J. Cell. Comp. Physiol., 1964b, vol. 64, Suppl. 1, p. 1.
Leuzinger W.— Biochem. J., 1971, vol. 123, p. 139.
Lever M., Miller K, Paton W„ Smith E. — Nature, Lond., 1971, vol. 231, p. 368.
Levine R., Streeten D„ Doisy R. — Metabolism, 1968, vol. 17, p. 114.
Levine W. (ed.) Chelation of Heavy Metals. — Oxford: Pergamon Press, 1978.
Levinson W., Woodson B., Jackson J. — Nature New Biol., 1971 vol. 232, p. 116.
Levitzki A. — Biochem. Biophys. Res. Commun., 1977, vol. 74, p. 1154.
Levy B., Ahlquist R. — J. Pharmacol., 1961, vol. 133, p. 202.
Levy G., Lichter R. Nitrogen-15 Nuclear Resonance Spectroscopy. — New York: Wiley-Interscience, 1979.
Lewin R. — Science, 1982, vol. 218, p. 817.
Lewis G., Randall M. Thermodynamics and Free Energy of Chemical Substances.— New York: McGraw-Hill, 1923.
Lewis P., Haeusler G. — Nature, Lond., 1975, vol. 256, p. 440.
Liebecq C., Peters R. — Biochim. Biophys. Acta., 1949,’vol. 3, p. 215.
Liebermeister K. — Z. Naturforsch., 1950, Bd 5b, S. 79.
Lienhard G. — Ann. Rep. Med. Chem., 1972, vol. 7, p. 249.
Lindquist R. — Drug Design, 1975, vol. 5, p. 23.
Lineweaver H., Burk D.—J. Amer. Chem. Soc., 1934, vol. 56, p. 658.
Ling W., Klett C„ Gillis R. — Arch. Gen. Psychiat., 1978, vol. 35, p. 345.
Linschoten M„ Gaisser H.-D., Van der Goot H.t Timmerman H. — Eur. J. Med. Chem., 1984, vol. 19, p. 137.
26*
403
Lippert B„ Metcalf B„ Jung M., Casara P. — Eur. J. Biochem., 1977, vol. 74, p. 441.
Lipscomb W.— Acc. Chem. Res., Amer. Chem. Soc., 1970, vol. 3, p. 81.
Lipscomb W., Hartsuck J., Quiocho F., Reeke G. — Proc. Natl. Acad. Sci., U.S.A., 1969, vol. 64, p. 28.
Liquori A., Damiani A., de Coen J.—J. Molec. Biol., 1968, vol. 33, p. 445.
Lonsdale K„ Milledge H„ Pant L. — Acta Crystallogr., 1965, vol. 19, p. 827.
Lord J., Waterfield A., Hughes J., Kosterlitz H. — Nature, Lond., 1977, vol. 267, p. 495.
Lotspeich IV., Peters R. — Biochem. J., 1951, vol. 49, p. 704.
Lowe H. — In: Theory and application of Gas Chromatography/Eds. H. Kro-man, S. Bender, New York: Grune and Stratton, 1968.
Lowe M., Phillips J. — Nature, Lond., 1961, vol. 190, p. 262.
Lown W., Hanstock C. — J. Amer. Chem. Soc., 1982, vol. 104, p. 3213.
Lowney L., Schulz K„ Lowrey P., Goldstein A. — Science, 1974, vol. 183, p. 749.
Ludeman S., Bartlett D., Zon G. — J. Org. Chem., 1979, vol. 44, p. 1163.
Luduena F„ von Euler L„ Tullar B„ Lands A. —Arch, internal. Pharmacodyn, Ther., 1957, vol. Ill, p. 392.
Luduena F„ Hoppe J., Nachod F. et al. — Arch, internat. Pharmacodyn. Ther., 1955, vol. 101, p. 17.
Luongo M., Bjornson S. — New Engl. J. Med., 1954, vol. 251, p. 995.
Lutz W., Winkler F., Dunitz J. — Helv. Chim. Acta, 1971, vol. 54, p. 1103.
Lyman W., Rechl W„ Rosenblatt D. (eds) Handbook of Chemical Property Estimation Methods. — New York: McGraw-Hill, 1982.
Maass E„ Johnson M. — J. Bact., 1949, vol. 57, p. 415.
Macadam R„ Williamson J.—Ann. Trop. Med. Parasit., 1974, vol. 68, p. 291.
McBrien D„ Slater T. (eds) Free Radicals, Lipid Peroxidation, and Cancer.— London: Academic Press, 1982.
McBryde W. — Canad. J. Chem., 1967, vol. 45, p. 2093.
McCalla T. — Proc. Soil. Sci. Soc. Amer., 1941, vol. 6, p. 165.
McCormick J., Jensen E., Miller P., Doerschuk A. — J. Amer., Chem. Soc., 1960, vol. 82, p. 3381.
McGregor D. — Froschr. Chem. Org. Natur., 1974, vol. 31, p. 1.
Machin P., Hurst D., Bradshaw R. et al. — J. Med. Chem., 1983, vol. 26, p. 1570.
McIlwain H. — Biochem. J., 1941, vol. 35, p. 1311.
McIlwain H. — In: Enzymes and Drug Action/Eds. J. Mongar, A. de Reuck, London: Churchill, 1962.
Macomber P., O’Brien R., Hahn F. — Science, 1966, vol. 152, p. 1374.
Magrath D., Phillips J. — J. Chem. Soc., 1949, p. 1940.
Maguire M., Wiklund R., Anderson H., Gilman A. — J. Biol. Chem., 1976, vol. 251, p. 1221.
Main A. — Science, 1964, vol. 144, p. 992.
Manifold M. — Brit. J. Exper. Path., 1941, vol. 22, p. 111.
Manning M., Lowbridge J., Holdon J., Sawyer W. — Fed. Proc., 1977, vol. 36, p. 1848.
Mansour T„ Bueding E. — Brit. J. Pharmacol. Chemother., 1954, vol. 9, p. 459.
Marcotte P., Robinson C. —Biochemistry, 1982, vol. 21, p. 2733.
Marcovich H. —Ann. Inst. Pasteur, 1953, vol. 85, p. 199, 443.
Marlow H., Metcalf J„ Burgen A. — Molec. Pharmacol., 1969, vol. 5, p. 156.
Marston H. — Proc. Roy. Soc. B, 1949, vol. 199, p. 273.
Marston H., Allen S., Smith R. — Nature, Lond., 1961, vol. 190, p. 1085.
Martell A., Smith R. Critical Stability Constants. — New York: Plenum Press, 1974— 1982.
Martin Y.— J. Med. Chem., 1970, vol. 13, p. 145.
Martin Y. — Quantitative Drug Design. — New York: Marcel Dekker, 1978.
Martin Y. Drug Design, 1979, vol. 8, p. 1.
Martonosi A. Membranes and Transport. (2 vols). — New York: Plenum Press, 1982.
404
Mason S. —Trends Pharmacol. Sci., 1983, vol. 4, p. 353.
Massey V., Hofmann T., Palmer G.— J. Biol. Chem., 1962, vol. 237, p. 3820.
Matthews B., Sigler P., Henderson R., Blow D. — Nature, Lond., 1967, vol. 214, p. 652.
Matthews D. Proc. 12th Congress Internet. Union Crystallogr., Associated Meeting on Molecular Structure and Biological Activity. — Buffalo, New York: Elsevier-North Holland, 1981.
Matthews D., Alden R„ Bolin J. et al. — Sciense, 1977, vol. 197, p. 452.
Mautner H.— J. Gen. Physiol., 1969, vol. 54, p. 271S.
Mautner H., Bartels E., Webb G.— Biochem. Pharmacol., 1966, vol. 15, p. 187.
Mautner H., Clemson H. — In: Medicinal Chemistry. 3rd edn./Ed. A. Burger, New York: Wiley, 1970.
Mautner H., Dexter B., Low D. — Nature, New Biol., 1972, vol. 238, p. 87.
May H„ McCay P. — J. Biol. Chem., 1968, vol. 243, p. 2288.
Mayer D„ Price D., Rafii A. — Brain Res., 1977, vol. 121, p. 368.
Meadows D., Roberts G„ Jardetzky O. — J. Molec. Biol., 1969, vol. 45, p. 491.
Mellor D„ Maley L. — Nature, Lond., 1947, vol. 159, p. 370.
Menger F„ Portnoy C.— J. Amer. Chem. Soc., 1967, vol. 89, p. 4698.
Mertz W.— Nutr. Rev., 1975, vol. 33, p. 129.
Metcalfe J., Seeman P., Burgen A.—Molec. Pharmacol., 1968, vol. 4, p. 87.
Meyer H. — Arch. Exper. Path. Pharmacol., 1899, vol. 42, p. 109.
Meyer K-, Hemmi H. — Biochem. Z., 1935, Bd 277, S. 39.
Michael A. — J. prakt. Chem., 1887, Bd [21 35, S. 349.
Michaelis L., Menten M. — Biochem. Z., 1913, Bd 13, S. 333.
Михельсон M. Я., Зеймаль Э. В. Ацетилхолин. О молекулярном механизме действия. — Л., «Наука», 1970.
Miledi R., Slater С. — J. Physiol., 1966, vol. 184, р. 473.
Miles A., Misra S. — J. Hyg., Camb., 1938, vol. 38, p. 732.
Miller A. — Proc. Soc. Exper Biol. Med., 1944, vol. 57, p. 151.
Miller E., Miller J., Hartmann H. — Cancer Res., 1961, vol. 21, p. 815.
Miller G., Doukos P., Seydel J. — J. Med. Chem., 1972, vol. 15, p. 700.
Miller J. — Cancer Res., 1970, vol. 30, p. 559.
Miller К.-, Paton W., Smith E. — Nature, Lond., 1965, vol. 206, p. 574.
Miller K-, Paton W„ Smith E., Smith R. — Molec. Pharmacol., 1973, vol. 9, p. 131.
Miller S. — Proc. Natl. Acad. Sci., U.S.A., 1961, vol. 47, p. 1515.
Miller W., Dessert A., Roblin R.—J. Amer. Chem. Soc., 1950, vol. 72, p. 4893.
Mino Y., Ishida T., Ota N. et al. — J. Amer. Chem. Soc., 1981, vol. 103, p. 6979.
Mislow K. Interoduction to Stereochemistry. — New York: Benjamin, 1965.
Mitchell I., Thorgeirssen U., Black M. — Clin. Pharmacol. Ther., 1975, vol. 18, p. 70.
Mitchell P.— Science, 1979, vol. 206, p. 1148 (Nobel Lecture).
Mitscher L., Bonacci A., Sokoloski T. — Antimicrob. Agents Chemother., 1968, p. 78.
Miyazaki H., Tojinbara L, Watanabe Y. et al. — Proc. 1st Symp. Drug. Metab. Action, Chiba, Japan, 1970, p. 135.
Monod J., Changeux J.-P., Jacob F.—J. Molec. Biol., 1963, vol. 6, p. 306.
Moore C., Pressman B. — Biochem. Biophys. Res. Commun., 1964, vol. 15, p. 562.
Morgan G., Cooper E. — Rep. 8th Internet Congr. Appl, Chem., 1912, vol. 19, p. 243.
Morin R., Gorman M. (eds) Chemistry and Biology of fl-Lactam Antibiotics. — New York: Academic Press, 1982.
Morisaki M., Bloch C. — Biochemistry, 1972, vol. 11, p. 309.
Morris H„ Taylor G., Piper P., Tippins J. — Nature, Lond., 1980, vol. 285, p. 104.
Mortazani S., Bari-Hashemi A., Mozafari M., Raffl A. — Cancer 1972, vol. 29, p. 1193.
Mudge C., Smith F. — Amer. J. Publ. Health, 1935, vol. 25, p. 442.
Mueller J., Vilter R. — J. Clin. Invest., 1950, vol. 29, p. 193.
405
Murachi T. — Trends Biochem. Sci., 1983, vol. 8, p. 167.
Muschaweck R„ Hajdu P.—Arzneim. Forsch., 1964, Bd 14, S. 44.
Nachmansohn D.— Yale J. Biol. Med., 1940, vol. 12, p. 565.
Nador K„ Herr K., Pataky G„ Borsy J. — Arch. Exper. Path. Pharmakoi., 1953, vol. 217, p. 447.
Nakatsugawa T., Dahm P. —J. Econ. Entomol., 1965, vol. 58, p. 500.
Narabayashi H., Kondo T., Nagatsu T. et al. — Proc. Japan Acad., B, 1982, vol. 58, p. 283.
Narahashi T„ Frazier D. — Neurosci. Res., 1971, vol. 4, p. 65, (Review).
Narahashi T., Frazier D., Moore J. — J. Neurobiol., 1972, vol. 3, p. 267.
Navon G., Ogawa S„ Shulman R., Tamane T. — Proc. Natl. Acad. Sci., U.S.A., 1977, vol. 74, p. 888.
Nelson N., Anholt R., Lindstrom J., Montal M. — Proc. Natl. Acad. Sci., U.S.A., 1980, vol. 77, p. 3057.
Neu H. — Ann. Rev. Pharmacol. Toxicol., 1982, vol. 22, p. 599.
Neville D., Davies D.— J. Molec. Biol., 1966, vol. 17, p. 57.
Newbold R., Overell R. — Nature, Lond., 1983, vol. 304, p. 648.
Newton G., Abraham E.-—Biochem. J., 1956, vol. 62, p. 651.
Nichol C., Welch A. — Proc. Soc. Exper. Biol. Med., 1950, vol. 74, p. 403.
Nicholson B., Peacocke A. — Biochem. J., 1966, vol. 100, p. 50.
Nicolaou K„ Petasis N., Seitz S. — Chem. Commun., Chem. Soc., London, 1981, p. 1195.
Nielands J. (ed.) Microbial Iron Metabolism. — New York: Academic Press, 1974.
Nielsen F. — J. Nutr., 1975, vol. 105, p. 1620.
Nimmo-Smith R„ Lascelles J., Woods D. — Brit. J. Exper. Path., 1948 vol. 29, p. 264.
Noda M., Takahashi H„ Tanabe T. et al. — Nature Lond., 1982, vol. 299, p. 793.
Nordbring-Hertz B. — Physiol. Plantarum, 1955, vol. 8, p. 691.
Northey E. The Sulfonamides and Allied Compouds. — New York: Reinhold, 1948.
Novella F„ Sprague J. — J. Amer. Chem. Soc., 1957, vol. 79, p. 2028.
Nudd R„ Wilkie D. — Chem. Brit., 1983, vol. 19, p. 911.
Oberley L. (ed.) Superoxide Dismutase (2 vols). — Boca Raton, Florida: CRC Press, 1982.
Oberley L., Buettner G.— Cancer Res., 1979, vol. 39, p. 1141.
O’Brien I., Cox G., Gibson F. — Biochim. Biophys. Acta, 1971, vol. 237, p. 537.
O’Brien L, Gibson F. — Biochim. Biophys. Acta., 1970, vol. 215, p. 393.
O’Brien R. Insecticides, Action and Metabolism. — New York; Academic Press, 1967.
O’Brien R. — Molec. Pharmacol., 1968, vol. 4, p. 121.
O’Brien R. — Drug Design, 1971, vol. 2, p. 161.
O'Brien R., Olenick J., Hahn F. — Proc Natl. Acad. Sci., U.S.A., 1966, vol. 55, p. 1511.
Oki M., Urushibara Y. — Bull. Chem. Soc. Japan., 1952, vol. 25, p. 109.
Okuyama S„ Hitoshi M — Tohoku J. Exper. Med,, 1981, vol. 135, p. 215.
Olsen R. — Ann. Rev. Pharmacol. Toxicol., 1982, vol. 22, p. 245.
Olsen G., Cheung H.-С., Morgan K. et al. — J. Med. Chem. 1981, vol. 24, p. 1026.
Ondetti M„ Cushman D.—J. Med. Chem., 1981, vol. 24, p. 355 (Review).
Ondeiti M„ Rubin B„ Cushman D. — Science, 1977, vol. 196, p. 441.
Ormerod W. — Proc. Roy. Soc. Trop. Med. Hyg., 1961, vol. 55, p. 313.
Ortiz P. — Biochemistry, 1970, vol. 9, p. 355.
Osborn M., Freeman M„ Huennekens F. — Proc. Soc. Exper. Biol. Med., 1958, vol. 97, p. 429.
Osborne M., Wenger L, Zanko M. — J. Pharmacol., 1977, vol. 200, p. 195.
Ostergren G. Colloques internationaux du Centre national de la Recherche scien-tifique.— Paris, 1951, No. 26, Mecanisme de la Narcose, p. 77.
O’Sullivan D„ Sadler P. — Nature, Lond., 1961, vol. 192, p. 341.
Overton E. Studien fiber die Narkose. — Jena: Gustav Fischer, 1901, 195 S.
Oxford J., Perrin D. — J, Gen. Virol., 1974, vol. 23, p. 59.
406
Palaty V. — Nature, Lond., 1966, vol. 211, p. 1177.
Patel D. — Biochemistry, 1974, vol. 13, pp. 1476, 2396.
Paton II7.— Proc. Roy. Soc. B., 1961, vol. 154, p. 21.
Paton U7.— Ann Rev. Pharmacol., 1975, vol. 15, p. 191.
Paton U7., Zaimis E. — Pharmacol. Rev., 1952, vol. 4, p. 219.
Paton W., Zaimis E. — Pharmacol. Rev., 1952, vol. 4, p. 219.
Pauling L. — Amer. Sci., 1948, vol. 36, p. 51.
Pauling L. Nature of the Chemical Bond. 3rd edn. — Ithaca, N.Y.: Cornell University Press, 1960 (1st edn., 1939).
Pauling L, — Science, 1961, vol. 134, p. 15.
Pauling L„ Jtano H., Singer S., Wells I.— Science, 1949, vol. 110, p. 543.
Peacocke A., Skerrett J. — Trans. Farad. Soc., 1956, vol. 52, p. 261.
Pearce V„ Robson P.— Postgrad. Med. J., 1980, vol. 56, p. 474.
Pearlman R. — In: Physical Chemical Properties of Drugs./Eds. S. Yalkowsky, A. Sinkula, S. Valvoni, New York: Marcel Dekker, 1980.
Pedersen C. — J. Amer. Chem. Soc., 1970, vol. 92, pp. 386, 391.
Pedersen C. — Organ. Synth., 1972, vol. 52, p. 66.
Pedersen T. and the Norwegian Multicentre Study Group. — New Engl. J. Med., 1981, vol. 304, p. 801.
Penning T. — Trends Pharmacol. Sci., 1983, vol. 4, p. 212.
Perkins J. — Adv. Cyclic Nucleotide Res., 1973, vol. 3, p. 1.
Perrin D. — Nature, Lond., 1958, vol. 182, p. 741.
Perrin D. — Rev. Appl. Chem. (Austral.), 1959, vol. 9, p. 257.
Perrin D. — Nature, Lond., 1961, vol. 191, p. 253.
Perrin D, Organic Complexing Reagents. — New York: Wiley-Interscience, 1964,
Perrin D. Dissociation Constants of Organic Bases in Aqueous Solution, compiled for International Union of Pure and Applied Chemistry. — London: Butterworths, 1965b.
Perrin D. — J. Chem. Soc., 1965c, p. 5590.
Perrin D. Masking and Demasking of Chemical Reactions. — New York: Wiley-Interscience, 1970.
Perrin D. Dissociation Constants of Organic Bases in Aqueous Solution, First Supplement. — Oxford: Pergamon Press, 1972.
Perrin D. Stability of Metal-Ion Complexes, Second Supplement (see Sillen and Martell for 1st Suppl.), compiled for IUPAC. — Oxford: Pergamon Press, 1979.
Perrin D. Dissociation Constants of Inorganic Acids and Bases (2nd edn) compiled for IUPAC. — London: Butterworths, 1983.
Perrin D., Dempsey B. Buffers for pH and Metal Ion Control. — London and New York: Chapman and Hall, 1974.
Perrin D., Dempsey B., Serjeant E. pKa Prediction for Organic Acids and Bases. — London and New York: Chapman and Hall, 1981.
Perrin D„ Sayce J. — J. Chem. Soc. A., 1968, p. 82.
Perrin D., Sayce J., Sharma V. — J. Chem. Soc. A, 1967, p. 1755.
Perrin D., Sharma V. — Biochim. Biophys. Acta, 1966, vol. 127, p. 35.
Perrin D., Sharma V. — J. Chem. Soc. A, 1967, p. 724.
Perrin D., Sharma V. —J. Chem. Soc. A, 1968, p. 446.
Perrin D., Sharma V. —J. Chem. Soc. A, 1969, p. 2060.
Perry A, McKinney L., De Weer P. — Nature, Lond., 1978, vol. 272, p. 271.
Pert C„ Snyder S. — Science, 1973, vol. 179, p. 1011.
Pert C„ Snyder S. — Molec. Pharmacol., 1974, vol. 10, p. 868.
Perutz M. — Nature, Lond., 1970, vol. 228, p. 726.
Petering D. — Bioinorg. Chem., 1972, vol. 1, p. 255.
Peters R. Biochemical Lesions and Lethal Synthesis. — Oxford: Pergamon Press, 1963.
Peters R„ Stocken L., Thompson R. — Nature, Lond., 1945, vol. 156, p. 616.
Peters R., Wakelin R., Rivett S„ Thomas L. — Nature, Lond., 1953, vol. 171.
Peyronel G. — Z. Krist., 1940, vol. 103, pp. 139, 157.
Pfaendler H_, Gosteli J,, Woodward R., Riis G. — J. Amer. Chem. Soc., 1981, vol. 103, p. 4526.
407
Phillips D. — Sci. Amer., 1966, vol. 215, p. 78.
Phillips R.—Chem. Rev., 1966, vol. 66, p. 501.
Pickering W., Malone J. —Biochem. Pharmacol., 1967, vol. 16, p. 1183.
Piper C. —J. Agric. Sci., 1942, vol. 32, p. 143.
Pirie A., Van Heyningen R. — Nature, Lond., 1954, vol. 173, p. 873.
Pitt G. —Acta Crystallogr., 1952, vol. 5, p. 770.
Plant G. — In: Pteridine Chemistry/Eds. W. Pfleiderer, E. Taylor, Oxford: Per-gamon Press, 1964, p. 443.
Poate H. — Lancet, 1944, ii, p. 238. Med. J. Austral., 1944, i, p. 242.
Poiesz B, Seal G., Loeb L. — Proc. Natl. Acad. Sci., U.S.A., 1974, vol. 71, p. 4892.
Poirier L., Miller J., Miller E„ Sato K.— Cancer Res., 1967, vol. 27, p. 1600.
Pomeranz B„ Chiu D. — Life Sci., 1976, vol. 19, p. 1757.
Pope L., Reich K„ Graham D., Sigman D. — J. Biol. Chem., 1982, vol. 257, p. 12121.
Popoviciu V. — Studia Biophys. (Berlin), 1978, Bd 69, S. 75.
Portoghese P., Mikhail A., Kapferberg H. —J. Med. Chem., 1968, vol. 11, p. 219.
Powell P., Cline G., Reid C„ Szaniszlo P. — Nature, Lond., 1980, vol. 287, p. 833.
Prakash N„ Schechter P„ Mamont P. et al. — Life Sci., 1980, vol. 26, p. 181.
Pressman B., Guzman N. — Ann. N.Y. Acad. Sci., 1974, vol. 227, p. 380.
Prestegard J., Chan S. — J. Amer. Chem. Soc., 1970, vol. 92, p. 4440.
Preston H., Steward J. — Chem. Commun. Chem. Soc., Lond., 1970, p. 1142.
Prichard B„ Gillam P. — Brit. Med. J., 1964, ii, p. 725.
Pringle M., Brown K., Miller K.—Molec. Pharmacol., 1981, vol. 19, p. 49.
Proctor P., Gensmantel N., Page M. — J. Chem. Soc., Perkin 11, 1982, p. 1185.
Propping P. — Rev. Physiol. Biochem. Pharmacol., 1978, vol. 82, p. 123.
Pullman A., Pullman B. Cancerisation par les Substances chimiques et Structures moleculaires. — Paris: Masson, 1955.
Purcell W., Bass G., Glayton J. Strategy of Drug Design. — New York: Wiley-Interscience, 1973.
Purchase I., Longstaff E.t Ashby J. et al. — Nature, Lond., 1976, vol. 264, p. 624.
Putnam F., Neurath H.—J. Amer. Chem. Soc., 1944, vol. 66, p. 1992.
Quastel J., Wooldridge W. —Biochem. J., 1927, vol. 21, p. 1224.
Raa J., Goksoyr J. — Physiol. Plantarum, 1966, vol. 19, p. 840. '
Rabenstein D„ Anvarhusein A. — Anal. Chem., 1982, vol. 54, p. 526.
Racker E„ Krimsky I. — J. Exper Med., 1947, vol. 85, p. 715.
Rahwan R., Witiak D. (eds) Calcium Regulation oy Calcium Antagonists. — Washington: American Chemical Society, 1982.
Rajan K, Davis J., Colburn R. — J. Neurochem., 1974, vol. 22, p. 137.
Randall L. — Dis. Nerv. Syst., 1961, vol. 22, p. 7.
Randic M. — J. Chem. Phys., 1974, vol. 60, p. 3920.
Rando R. — Biochemistry, 1974, vol. 13, p. 3859.
Rappaport C., Howze G. — Proc. Soc. Exper. Biol. Med., 1966, vol. 121, pp 1010, 1016.
Raventos J. — Quart. J. Exper. Physiol., 1937, vol. 26, p. 361.
Redl G., Cramer R„ Berkoff C. — Chem. Soc. Rev. Lond., 1974, vol. 3, p. 273.
Redmond D., Kleber H. — Psychiat. Res., 1980, vol. 2, p. 37.
Rekker R. The Hydrophobic Fragmental Constant. — Amsterdam: Elsevier, 1977. Rekker R., De Kort H. — Eur. J. Med. Chem., 1979, vol. 14, p. 479.
Reynolds J. (ed.) Martindale, the Extra Pharmacopoeia. — London: Pharmaceutical Press, 1982.
Reynolds W„ Topsom R. —J. Org. Chem., 1984, vol. 49, p. 1989.
Richards D„ Pritchard B. — Clin. Pharmacol. Ther., 1978, vol. 23, p. 253.
Richards G„ Sackwild V. — Chem. Brit., 1982, vol. 18, p. 635.
Richards W. Quantum Pharmacology. — London: Butterworths, 1983.
Riddell F. Conformational Analysis of Heterocyclic Compounds. — London: Academic Press, 1980.
Rideal E. — Endeavour, 1945, vol. 4, p. 83.
Riegel B„ Sherwood L. —J. Amer. Chem. Soc., 1949, vol. 71, p. 1129.
Rigler N„ Bag S., Leyden D. et el. —Anal. Chem., 1965, vol. 37, p. 872.
408
Rinehart J., Arnold J., Canfield C. — Amer. J. Trop. Med. Hyg., 1976, vol. 25, p. 769.
Ringer S. —J. Physiol., 1883, vol. 4, p. 29.
Riou G., Delain E.— Proc. Natl. Acad. Sci., U.S.A., 1969, vol. 64, p. 618.
Ritchie J., Greengard P. — J. Pharmacol., 1961, vol. 133, p. 401.
Ritchie J., Greengard P. — Ann. Rev. Pharmacol., 1966, vol. 6, p. 405.
Roberts G.— Biochem. J., 1966, vol. 100, 30P.
Roberts G., Feeney J., Birdsall B. et al. — In: Nuclear Magnestic Resonance in Biology/Ed. R. Dweck, New York: Academic Press, 1977, p. 95.
Roberts J. — In: Structural Chemistry and Molecular Biology./Eds. A. Rich, N. Davidson, San Francisco: Freeman, 1968.
Roberts J., Pera M. — In: Molecular Aspects of Anit-cancer Drug Action./Eds S. Neidle, M. Waring, London: Macmillan Press, 1983.
Roberts J., Ray P., Ward-Jardetzky N., Jardetzky O. — Nature, Lond., 1980, vol. 283, p. 870. ,
Roberts J., Warwick G. — Biochem. Pharmacol., 1961, vol. 6, p. 217. :
Robertson R. The Lifely Membrane. — Cambridge: University Press, 1983.
Robins R„ Hitchings G. — J. Amer. Chem. Soc., 1955, vol. 77, p. 2256.
Robinson R. — Brit. Med. J., 1946, i, p. 945.
Robinson R., Stokes R. Electrolyte Solutions. 2nd edn. — London: Butterworths, 1959.
Rocha e Silva M. (ed.) Histamine and Anti-Histaminics, Handbuch exper. Pharmacol.— Berlin: Springer, 1978, vol. 18.
Rodgers M., Powers E. Oxygen and Oxy-Radicals in Chemistry and Biology. — New York: Academic Press, 1981.
Rodgers P„ Roberts G. — FEBS Lett., 1973, vol. 36, p. 330.
Roe O. — Pharmacol. Rev., 1955, vol. 7, p. 399.
Roehl W.— Archiv. Schiffs Tropenhyg., 1926b, Bd 30, Beiheft 1, S. 103.
Roemer D., Buescher H., Hill R. et al. — Nature, Lond., 1977, vol. 268, p. 547.
Rogers H. — Nature, Lond., 1967, vol. 213, p. 31.
Rogers H„ Mandelstam J. — Biochem. J., 1962, vol. 84, p. 299.
Rokos J., Rokos K., Frisius H. et al. — Clin. Chim. Acta, 1980, vol. 105, p. 275.
Roques B., Forunie-Zaluski M„ Soroca E. et al. — Nature, Lond., 1980, vol. 288, p. 286.
Rosanoff M. — J. Amer. Chem. Soc., 1906, vol. 28, p. 114.
Rosenberg B., Van Camp L. — Cancer Res., 1970, vol. 30, p. 1799.
Rosencweig M., Van Hoff D., Slavik M., Muggia F. — Ann. Intern. Med., 1977, vol. 86, p. 803.
Rosenthal S. — U.S. Pub], Health Repts, 1932, vol. 47, p. 933.
Rosenthal S„ Voegtlin C. — J. Pharmacol., 1930, vol. 39, p. 347.
Ross G., Goldstein D., Hertz R. et al. — Amer. J. Obstet. Gynecol., 1965, vol. 93, p. 223.
Ross W. — Biochem. Pharmacol., 1961, vol. 8, p. 235.
Ross W. Biological Alkylating Agents. — London: Butterworths, 1962.
Roth B., Bliss E., Beddell C. — In: Molecular Aspects of Anit-cancer Drug Action./Eds S. Neidle, M. Waring, London: Macmillan Press, 1983.
Roth B., Cheng C. — Progr. Med. Chem., 1982, vol. 19, p. 270.
Roth B., Falco E., Hitchings G. — J. Med. Pharm. Chem., 1962, vol. 5, p. 1103.
Roth B., Strelitz J. — J. Org. Chem., 1969, vol. 34, p. 821.
Rowley D., Cooper R., Roberts P. et al. — Biochem. J.,-1950, vol. 46, p. 157. Rubbo S., Albert A., Gibson M.—Brit. J. Exper. Path., 1950, vol. 31, p. 425. Rupp H„ Voelter W., Weser U. — Fed. Eur. Biochem. Soc. Lett., 1974, vol. 40, p. 176.
Russell D., Falconer M. — Brit. J. Surg., 1941, vol. 28, p. 472.
Russell D„ Falconer M. — Lancet, 1943, i, p. 580.
Rye R,, Wiseman D. —J. Pharm. Pharmacol., 1966, vol. 18, p. 114 S.
Ryter A., Jacob F. — Ann. Inst. Pasteur, 1964, vol. 107, p. 384.
Saarikoski J., Viluksela M — Ecotoxicol. Environ. Safety, 1982, vol. 6, p. 501.
Salton M. —J. Gen. Microbiol., 1951, vol. 5, p. 391.
Sander J., Schweinsberg F., Menz H. — Z. physiol. Chem., 1968, Bd 349, S. 1691.
Sanger F. — Proc. Chem. Soc., Lond., 1963, p. 76.
409
Sarett L„ Patchett A., Steelman S. — Progr. Drug Res., 1963, vol. 5, p. 11.
Sarris A., Niles E., Cannellakis E.—Biochim. Biophys. Acta, 1977, vol. 474, p. 268.
Sartorelli A., Creasey W.— Ann. Rev. Pharmacol., 1969, vol. 9, p. 51.
Sastry B., Lasslo A., Pfeifer C. — J. Pharmacol., 1960, vol. 130, p. 346.
Savage D„ Cameron A., Ferguson G. et al.—J. Chem. Soc. B, 1970, p. 410.
Schaad U.—J. Pediatr., 1981, vol. 98, p. 129.
Schaffer N., May S„ Summerson W.— J. Biol. Chem., 1954, vol. 206, p. 201.
Scheibet L„ Adler A. — Molec. Pharmacol., 1982, vol. 22, p. 140.
Schellenberg K-, Coatney G. — Biochem. Pharmacol., 1961, vol. 6, p. 143.
Schild H.— Brit. J. Pharmacol., 1947, vol. 2, p. 189.
Schild H. — Brit. J. Pharmacol., 1969, vol. 36, p. 329.
Schoenenberger G., Schneider-Helment D. — Trends Pharmacol. Sci., 1983, vol. 4, p. 307.
Schou M.— Pharmacol. Rev., 1957, vol. 9, p. 17.
Schrader G. Die Entwicklung neuer Insektizider Phosphorsaure. 3rd edn.— Weinheim: Verlag Chemie, 1963.
Schramm M., Orly J., Eimerl S., Korner M.— Nature, Lond., 1977, vol. 268, p. 310.
Schraufstdtter E. — Z. Naturforsch., 1950, Bd 5b, S. 190.
Schroeder H., Perry H., Menhard E.— J. Lab. Clin. Med., 1955, vol. 46, p. 416.
Schubert J. — Methods Biochem. Anal., 1956, vol. 3, p. 247.
Schubert J. — Chimia, 1957, vol. 11, p. 113.
Schul W., Oiake M., Neel J. — Science, 1981, vol. 213, p. 1220.
Schulman J., Rideal E. — Proc. Roy. Soc. B, 1937, vol. 122, p. 46.
Schwarcz R., Creese I., Coyle J., Snyder S. — Nature, Lond., 1978, vol. 271, p. 766.
Schwartz A., Maisui H„ Laughter A. — Science, 1968, vol. 160, p. 323.
Schwartz H.— Adv. Cancer Chemother., 1979, vol. 1, p. 1.
Schwarz K.— Proc. Natl. Acad. Sci., U.S.A., 1973, vol. 70, p. 1608.
Schwarz K., Milne D., Vinard E.—Biochem. Biophys. Res. Commun., 1970, vol. 40, p. 22.
Schwarzenbach G., Schwarzenbach K.— Helv. Chim. Acta, 1963, vol. 46, p. 1390.
Scott K, Mautner H. — Biochem. Pharmacol., 1967, vol. 16, p. 1903.
Scovill J., Klayman D„ Franchino C.—J. Med. Chem., 1982, vol. 25, p. 1261.
Seeger D., Cosulich D., Smith J., Hultquist M — J. Amer. Chem. Soc., 1949, vol. 71, p. 1753.
Seeman P. — Biochem. Pharmacol., 1977, vol. 26, p. 1741.
Seeman P„ Roth S. — Biochim. Biophys. Acta, 1972, vol. 255, p. 171.
Seiler N., Jung N.t Koch-Weser E. (eds)—Enzyme-Activated Irreversible Inhibitors.— Amsterdam: Elsevier, 1978.
Selbie F., McIntosh J. — J. Path. Bact., 1943, vol. 55, p. 477.
Serjeant E., Dempsey B. — Ionization Constants of Organic Acids in Aqueous Solution (supplement to the volume listed under Kortum G. et al.).— Oxford: Pergamon Press, 1979.
Seven M. — In: Metal-Binding in Medicine./Eds. M. Seven, L. Johnson, Philadelphia: Lippincott, 1960, p. 95.
Seydel J. —J. Quant. Chem., 1981, vol. 20, p. 131.
Seydel J., Ahrens H., Losert W. — J. Med. Chem., 1975, vol. 18, p. 234.
Seydel J., Schaper K-, Wempe E., Cordes H. — J. Med. Chem., 1976a vol. 19 p. 483.
Seydel J„ Топо-Oka S., Schaper K. et al. — Arzneim. Forsch., 1976b Bd 26 S. 477.
Shamblee D., Gillespie J. — J. Med. Chem., 1979, vol. 22, p. 86.
Shanzer A., Samuel D., Korenstein R. — J. Amer Chem. Soc, 1983, vol 105 p. 3815.
Shapiro A., Nathan H„ Hutner S. et al. — J. Protozool., 1982, vol. 29, p. 85.
Sharma S., Klee W., Nirenberg M. — Proc. Natl. Acad. Sci,, U.S.A., 1975>, vol 72 p. 3097.
410
Sharon N„ Lis H. — Science, 1972, vol. 177, p. 946.
Shealy Y„ Krauth C. —J. Med. Chem., 1966, vol. 9, p. 34.
Shefter E„ Mautner H. — Proc. Natl. Acad. Sci., U.S.A., 1969, vol. 63, p. 1253.
Shelrick G., Jones P., Kennard O. et at. — Nature, Lond., 1978, vol. 271, p. 223.
Shemyakin M„ Aldanova E., Vinogradova E., Feigina M. — Tetr. Lett., 1963, vol. 28, p. 1921.
Shepherd R., Bratton A., Blanchard K. — J- Amer. Chem. Soc., 1942, vol. 64, p. 2532.
Shepherd R., Wilkinson R. — J. Med. Pharm. Chem., 1962, vol. 5, p. 823.
Shimomura 0., Johnson F.— Biochemistry, 1969, vol. 8, p. 3991.
Shin Y., Eichhorn G. — Biochemistry, 1968, vol. 7, p. 1026.
Shindo H„ Brown T. — J. Amer. Chem. Soc., 1965, vol. 87, p. 1904.
Shiota T., Baugh C., Jackson R„ Dillard R. — Biochemistry, 1969a, vol. 8, p. 5022.
Shiota T., Baugh C., Myrick J. — Biochim. Biophys. Acta, 1969b, vol. 192, p. 205.
Shipman C., Smith S., Drack J., Klayman D. — Antimicrob. Agents Chemother., 1981, vol. 19, p. 682.
Shoppee C. Chemistry of the Steroids. 2nd edn. — London: Butterworths, 1964..
Shulman A., Dwyer F. — In: Chelating Agents and Metal Chelates./Eds F. Dryer, D. Mellor, New York: Academic Press, 1964, chap. 9.
Sigel H. (ed.) Metal Ions in Biological Systems. — New York: Marcel Dekker; 1973.
Sih C., Takeguchi C. — In: The Prostaglandins./Ed. P. Ramwell, New York: Plenum Press, 1973, vol. 1, p. 83.
Sijpesteijn A., Janssen M. — Antonie Van Leeuwenhoek, 1959, vol. 25, p. 422.
Sijpesteijn A., Janssen M„ Van der Kerk G. — Biochim. Biophys. Acta, 1957, vol. 23, p. 550.
Sillen L., Martell A. Stability Constants of Metal-Ion Complexes, compiled for IUPAC. — London: The Chemical Society, 1964.
Sillen L., Martell A. First supplement to previous entry, 1971 (for second supplement, see Perrin, 1979).
Silverman R., Hofjman S. — Biochem. Biophys. Res. Common, 1981, vol. 101, p. 1396.
Silverman R., Levy M — Biochemistry, 1981, vol. 20, p. 1197.
Silverman R., Levy M., Muztar A., Hirsch J. — Biochem. Biophys. Res. Common, 1981, vol. 102, p. 520.
Simmonds M. — Nature, Lond, 1980, vol. 284, p. 558.
Simon E. — Nature, Lond, 1950, vol. 166, p. 343.
Simon E„ Beevers H.— Science, 1951, vol. 114, p. 124.
Simon E., Beevers H. — New Phytol, 1952, vol. 51, p. 163.
Simon Z., Chiriac A, Holban S. et al. — New York: Wiley, 1984.
Simons W., Zanger M. The Sadtler Guide to NMR Spectra. — Philadelphia: Sadtler Research Laboratories, 1972.
Sims P., Grover P., Swaisland A. et al. — Nature, Lond, 1974, vol. 252, p. 326.
Singer B. — Nature, Lond, 1976, vol. 264, p. 33.
Singer T., von Korf R., Murphy D. (eds) Monoamine Oxidase: Structure, Function, and Altered Funktions. — New York: Academic Press, 1979.
Sinistri C„ Villa L. — Il Farmaco, Ed. sci, 1962, vol. 17, p. 949.
Sinnema A., Maat L., Van der Gugten A., Beyerman H. — Rec. Trav. chim. Pays-Bas, 1968, vol. 87, p. 1027.
Skipper H., Schabel F., Wilcox W. — Cancer Chempther. Rpts, 1964, vol. 35, p. 1.
Slater T. — Nature, Lond, 1966, vol. 209, p. 36.
Slater T., Sawyer B. — Biochem. J, 1971, vol. 123, pp. 815, 823.
Slavik M., Elias L., Mrema J., Saiers J. — Drugs Exper. Clin. Res. (Switzerland), 1982, vol. 8, p. 379.
Smallman B., Fiske R. — Canad. J. Biochem. Physiol, 1958, vol. 36, p. 575.
Smissman E., Steinman M — J. Med. Chem, 1966, vol. 9, p. 455.
Smith A., Lee N., Loh H. — Trends Pharmacol. Sci, 1983, vol. 4, p. 163.
Smith H. — Amer. J. Physiol, 1925, vol. 72, p. 347.
411
Smith H., Chapman J., Marlow С. — Nature, Lond., 1969, vol. 222, p. 676.
Smith M., Wain R., Wightman F. — Ann. Appl. Biol., 1952, vol. 39, p. 295.
Smith S., Larson E. — J. Biol. Chem., 1946, vol. 163, p. 29.
Snow G. — Bact. Rev., 1970, vol. 34, p. 99.
Snyder S. — Science, 1980, vol. 209, p. 9761.
Soloway S. — Adv. Pest Control, 1965, vol. 6, p. 85.
Somers E., Pring R. — Ann. Appl. Biol., 1966, vol. 58, p. 457.
Somers L, Shive J. — Plant Physiol., 1942, vol. 17, p. 582.
Sonenberg M„ Money W. — Endocrinology, 1957, vol. 61, p. 12.
Speakman J. A Valency Primer. — London: Edward Arnold, 1968.
Spiro T. (ed.) Iron-Sulfur Proteins. — New York: Wiley, 1982.
Stackelberg M. — Z. Elektrochem., 1954, vol. 58, pp. 25, 162.
Stadtman T. — Ann. Rev. Biochem., 1980, vol. 49, p. 93.
Stale H. — In; Chronicles of Drug Discovery./Eds. J. Bindra, D. Lednicer, New York: Wiley, 1982.
Stanbury Wyngaarden 1. — Metabolism, 1952, vol. 1, p. 533.
Stearn A„ Stearn E. — J. Bact., 1924, vol. 9, p. 491.
Stedman E. — Biochem. J., 1926, vol. 20, p. 719.
Stedman E. — Nature, Lond., 1947, vol. 159, p. 194.
Stedman E., Stedman E. — Biochem. J., 1931, vol. 25, p. 1147.
Steitz T„ Shoham M., Bennett W. — Philos. Trans. Roy. Soc., London, B, 1981, vol. 293, p. 43.
Stenflo J., Fernlund P., Egan W., Roepstorff P. — Proc. Natl. Acad. Sci., U.S.A., 1964, vol. 71, p. 2730.
Stephenson R. —Brit. J. Pharmacol., 1956, vol. 11, p. 379.
de Stevens G., Werner L„ Halamandaris A„ Ricca S. — Experientia, 1958, vol. 14, p. 463.
Stone K„ Strominger J. — Proc. Natl. Acad. Sci., U.S.A., 1971, vol. 68, p. 3223. Straub K„ Meehan T., Burlingame A., Calvin M. — Proc. Natl, Acad. Sci., U.S.A., 1977, vol. 74, p. 5285.
Strominger J., Ito E., Threnn R. — J. Amer. Chem. Soc„ 1960, vol. 82, p. 998.
Strugger S. — Jena. Z. Med. Naturwiss., 1940, Bd 73, S. 97.
Stiinzi H. —Austral. J. Chem., 1982, vol. 35, p. 1145.
Stilnzi H., Perrin D. — J. Inorg. Biochem., 1979, vol. 10, p. 309.
Sunderman F. — Amer. J. Med. Sci., 1967, vol. 253, p. 209.
Sutherland E„ 0ye I., Butcher R. — Recent Progr. Horm. Res., 1965, vol. 21, p. 623.
Suwalsky M., Traub W„ Shmueli U., Subirana J.—J. Molec. Biol., 1969, vol. 42, p. 363.
Suzuki T., Hayashi K„ Fujikawa K.., Tsukamoto K. — J. Biochem., Tokyo, 1964, vol. 56, p. 335.
Swain C., Lupton E. — J. Amer. Chem. Soc., 1968, vol. 90, p. 4328.
Sweeney W„ Rabinowitz J. — Ann. Rev. Biochem., 1980, vol. 49, p. 139.
Sweet R„ Dahl L. — J. Amer. Chem. Soc., 1970, vol. 92, p. 5489.
Sykes B., Patt S., Dolphin D. — Gold. Spring Harbor Symp. Quant. Biol., 1971, vol. 36, p. 29.
Szekely J. (ed.) Opioid Peptides, (3 vols). — Boca Raton, Florida: CRC Press, 1982
Szekely P„ Wynne N. — Brit. Heart J., 1963, vol. 25, p. 589.
Tabin C„ Bradley S„ Bargmann C. et al. — Nature, Lond., 1982 vol. 300, p. 143.
Tabor H., Tabor C. Polyamines. — New York: Academic Press, 1983.
Taft R. — J. Amer. Chem. Soc., 1953, vol. 75, p. 4231.
Taft R. Steric Effects in Organic Chemistry./Ed. M. Newman, New York: Wiley, 1956.
Taft R., Grob C. — J. Amer. Chem. Soc., 1974, vol. 96, p. 1236.
Taft R„ Lewis I. — J. Amer. Chem. Soc., 1959, vol. 81, p. 5343.
Tainter M. — J. Pharmacol., 1930, vol. 40, p. 43.
Takamiya K. — Nature, Lond., 1960, vol. 185, p. 190-
Tally F., Jacobus N., Gorbach S. — Antimicrob. Agents Chemother, 1978 vol 14, p. 436.
412
Tanford C. The Hydrophobic Effect: Formation of Micelle and Biological Membranes. 2nd edn. — New York: Wiley, 1980.
Tattersail M., Jaffe N., Frei E. — In: Pharmacological Basis of Cancer Chemo-therapy./Eds R. Cumley, J. McCay, Baltimore: Williams and Wilkins, 1975.
Tatum A., Cooper G.— J. Pharmacol., 1934, vol. 50, p. 198.
Taylor D., Callahan K„ Shaikh 1.—J. Med. Chem., 1975, vol. 18, p. 1088.
Taylor E., Lymn R., Moll G.— Biochemistry, 1970, vol. 9, p. 2984.
Terenius L. — Adv. Neurol., 1982, vol. 33, p. 59.
Terenius L„ Wahlstrom A. — In: Centrally-Acting Peptides./Ed. J. Hughes, London: Macmillan, 1978.
Thacore V., Shukla S. — Arch. Gen. Psychiat., 1976, vol. 33, p. 383.
Thauer R.— Trends Biochem. Sci., 1980, vol. 11, p. 304.
Thoenen H., Tranzer J. — Arch, exper. Path. Pharmakoi., 1968, Bd 261, S. 271.
Thomas H., Herriott R„ Hahn B„ Wang S. — Nature, Lond., 1976, vol. 259, p. 342.
Thomas R. — J. Physiol., 1974, vol. 238, p. 159.
Thomas R. — In: Burger’s Medicinal Chemistry./Ed. M. Wolff, New York; Wiley, 1981, 4th edn., p. 47.
Thomas R„ Boutagy J., Gelbart A. — J. Pharmacol., 1974(a), vol. 191, p. 219;
(b) J. Pharm. Sci., 1974, vol. 63, p. 1649.
Thorn M — Biochem. J., 1953, vol. 54, p. 540.
Thorn G., Ludwig R. The Dithiocarbamates and Related Compounds.—Amsterdam: Elsevier, 1962.
Thorp R„ Cobbin L. — In: Cardiac Stimulant Substances./Ed. G. de Stevens, New York: Academic Press, 1967.
Timmermans P„ Van Zweiten P. — J. Med. Chem., 1977, vol. 20, p. 1636.
Tipper D., Strominger J. — Proc. Natl. Acad. Sci., U.S.A., 1965, vol. 54, p. 1133.
Tobin J., Lewis N. — J. Amer. Med. Assoc., 1960, vol. 174, p. 1242.
Tolkmith H. — Ann. N.Y. Acad, Sci., 1966, vol. 136 (art. 3), p. 59.
Topliss J. —J. Med. Chem., 1972, vol. 15, p. 1006.
Topliss J —J. Med. Chem., 1977, vol. 20, p. 463.
Topliss J. (ed.) Quantitative Structure-Activity Relationships of Drugs. — New York: Academic Press, 1983.
Topliess J., Costello R.—J. Med. Chem., 1972, vol. 15, p. 1066.
Towart R., Schramm M. — Trends Pharmacol. Sci., 1984, vol. 5, p. 111.
Tramposch K., Kung H.t Blau M. —J. Med. Chem., 1983, vol. 26, p. 121.
Trapnell J. — In: Chemistry and Biology of the Kallikrein-Kinin System in Health and Disease./Eds J. Pisano and K. Austen, Washington: U.S. Government Printing Office, 1977.
Traube J. —Arch. ges. Physiol., 1904, Bd 105, S. 541.
Trevan J., Boock E. —Brit. J. Exper. Path., 1927, vol. 8, p. 307.
Triggle D., Triggle C. Chemical Pharmacology of the Synapse. — London: Academic Press, 1976.
Tripathi R., Tripathi H., O’Brien R. — Biochim. Biophys. Acta, 1979, vol. 586, p. 624.
Trotman C., Greenwood C. — Biochem. J., 1971, vol. 124, p. 25.
Trudell J., Hubbell W. — Anesthesiology, 1976, vol. 44, p. 202.
Tsukube H. —J. Chem. Soc., Perkin Trans., 1982, vol. 1, p. 2359.
Tuck L., Baker J. — Chem. Biol. Interact., 1973, vol. 7, p. 355.
Turnbull H. — Austral. N. Z. J. Surg., 1944, vol. 14, p. 3.
Turpaev T., Sakharov D. — In: Comparative Pharmacology./Ed. M. Michelson, Oxford: Pergamon Press, 1973, vol. 1, p. 251.
Umezawa H. Enzyme Inhibitors of Natural Origin. — Baltimore: University Park Press, 1972.
Umezawa H. — Biomedicine, 1973, vol. 18, p. 459.
Underwood E. Trace Elements in Human and Animal Nutrition. 4th edn. — New York: Academic Press, 1977.
Unger S., Hansch C. —J. Med. Chem., 1973, vol. 16, p. 745.
Unger T., Ganten D., Lang R. — Trends Pharmacol. Sci., 1983, vol. 4, p. 514.
Unna K- —J. Pharmacol., 1943, vol. 79, p. 27.
Unsworth 1. — St Mary's Hospital Gazette, London, 1861, vol. 66, p. 272.
413
Usdin E., Skolnick P., Tallman J. et al. Biochemistry of S-Adenylmethionine and Related Compounds. — London: Macmillan Press, 1983.
Vallee B. — Biochem. Biophys. Res. Commun., 1975, vol. 62, p. 296.
Van der Berg G., Bultsma T., Rekker R., Nauia W.— Eur. J. Med. Chem., 1975, vol. 10, p. 242.
Van Rossum J., Arlens E. — Arch, internal. Pharmacodyn. Ther., 1959, vol. 118, p. 418.
Vargha L.. Toldy L„ Feher 0., Lendval S. — J. Chem. Soc., 1957, p. 805.
Varghese J., Laver W., Colman P. — Nature, Lond., 1983, vol. 303, pp. 35, 41.
Veldstra H.— In: Comprehensive Biochemistry./Eds. M. Florkin, E. Stotz, Amsterdam: Elsevier, 1963.
Veldstra H., Van der Westeringh C. — Rev. Trav. chim. Rays-Bas, 1951, vol. 70, p. 1127.
Veneziale C„ Walter P., Kneer N., Lardy H.—Biochemistry, 1967, vol. 6, p. 2129.
Verloop A.— Philos. Trans. Roy. Soc. B, 1981, vol. 295, p. 45.
Verloop A. — Pesticide Chemistry./Eds. J. Miyamoto, P. Kearney, Oxford: Per-gamon Press, 1983.
Verloop A„ Hoogenstraaten W., Tipker J. — Drug Design, 1976, vol. 7, p. 165.
Vermast P.—Biochem. Z., 1921, Bd 125, S. 106.
Voegtlin C, — Physiol. Rev., 1925, vol. 5, p. 63.
Vogel A. — J. Chem. Soc., 1948, p. 1833.
Volz K., Matthews D.t Alden R. et al.—J. Biol. Chem., 1982, vol. 257, p. 2528.
Waddell W., Butler T. — J. Clin. Invest., 1959, vol. 38, p. 720.
Waddell W., Hardman H.—Amer. J. Physiol., 1960, vol. 199, p. 1112.
Wagner-Jauregg T., Hackley B., Lies T. et al. — J. Amer. Chem. Soc., 1955, vol. 77, p. 922.
Walgate R. — Nature, Lond., 1982, vol. 296, p. 596.
Waller C., Hutchings B., Mowat J. et al. — J. Amer. Chem. Soc., 1948, vol. 70, p. 19.
Walshe J. — Lancet, 1968, i, p. 775.
Walton A. Molecular and Crystal Structure Models. — Chichester: Ellis Horwood; New York: 'Wiley, 1978.
Wang E„ Walsh C. — Biochemistry, 1978, vol. 17, p. 1313.
Wang J. — J. Molec. Biol., 1974, vol. 89, p. 783.
Warburg O. — Biochem. Z., 1921, Bd 119, S. 134.
Waring M. — Molec. Pharmacol., 1965, vol. 1, p. 1.
Waring M. — J. Molec. Biol., 1970, vol. 54, p. 247.
Waring M. — Eur. J. Cancer, 1976, vol. 12, p. 995.
Waring M„ Wakelin L. — Nature, Lond., 1974, vol. 252, p. 653.
War Office, Great Britain. The Official History of the War. — London: H. M. Stationery, Office, 1922, vol. 1.
Warren G., Housley M., Metcalfe J„ Birdsall N. — Nature, Lond., 1975, vol. 255, p. 684.
Waser P. — J. Pharm. Pharmacol., 1960, vol. 12, p. 577.
Waser P. — Pharmacol. Rev., 1961, vol. 13, p. 465.
Watters J., De Witt R. — J. Amer. Chem. Soc., 1960, vol. 82, p. 1333.
Weber M., Kinsky S. — J. Bact., 1965, vol. 89, p. 306.
Weeks C„ Cooper A., Norton D.—Acta Crystallogr. B, 1970, vol. 26, p. 429.
Wehrli F., Wirihlin T. Interpretation of Carbon-13 Spectra. — London; Heyden, 1976,
Weinberg E.—Antibiot. Chemother., 1954, vol. 4, p. 35.
Weinberg E. — Ann. N.Y. Acad. Sci., 1972, vol. 199, p. 274.
Weinberg E. — Science, 1974, vol. 184, p. 952.
Weinstein G. — Ann. Intern. Med., 1977, vol. 86, p. 199.
Weintraub H., Hopfinger A. — S. Theor. Biol., 1973, vol. 41, p. 53.
Weiss G. (ed.) Calcium in Drug Action.— New York: Plenum Press, 1978.
Weissberger A., Lu Valle S. — J. Amer. Chem. Soc., 1944, vol. 66, p. 700.
Weissman G., Dingle J. — Exper. Cell. Res., 1961, vol. 25, p. 207.
Werkheiser IF.— Cancer Res., 1963, vol. 23, p. 1277.
Westley J., Oliveto E., Berger J. et al. —J. Med. Chem., 1973, vol. 16, p. 397.
414
Weltingfeld R., Rowe J., Eyles D.—Ann. Intern. Med., 1956, vol. 44, p. 557.
Weyter F„ Broquist H.— Biochim. Biophys. Acta, 1960, vol. 40, p. 567.
Wheeler G„ Alexander J. — Cancer Res., I960, vol. 29, p. 98.
White J., Cantor C. — J. Molec. Biol., 1971, vol. 58, p. 397.
Whitehouse M, Dean P. — Biochem. Pharmacol., 1965, vol. 14, p. 557.
Whittaker V.— Physiol. Rev., 1951, vol. 31, p. 312.
WHO, see World Health Organization.
Wiebelhaus V., Weinstock Л, Maass A. et al.—J. Pharmacol., 1965, vol. 149, p. 397.
Wilen S. — Top. Stereochem., 1971, vol. 6, p. 107.
Wilen S., Collet A., Jacques J. — Tetrahedron, 1977, vol. 33, p. 2725.
Wilhelm W., Kuhn R. — Pharmakopsych. Neuro-psychopharmakol., 1970, Bd 3, S. 317.
Wilkins R. — J. Chem. Soc., 1962, p. 4475.
Wilkinson S., Lowe L.— Nature, Lond., 1966, vol. 212, p. 311.
Willey G. — Brit. J. Pharmacol. Chemother., 1955, vol. 10, p. 466.
Williams A. Chemistry of Enzyme Action. — London: McGraw-Hill, 1969.
Williams D. (ed.) Bio-inorganic Chemistry. — Springfield, Illinois: Charles C.
Thomas, 1976.
Williams L., Jarett L., Lefkowitz R. — J. Biol. Chem., 1976, vol. 251, p. 3096.
Williams L., Mullikin D., Lefkowitz R. — J. Biol. Chem., 1976, vol. 251, p. 6915.
Williams R. — J. Chem. Soc., 1952, p. 3770.
Williams R. —Angew. Chem. Internal. Edn., Engl., 1977, vol. 16, p. 766.
Williamson D., Everett G. — J. Amer. Chem. Soc., 1975, vol. 97, p. 2397.
Williamson J.-—Brit. J. Pharmacol. Chemother., 1959, vol. 14, p. 443.
Williamson J., Macadam R. — Trans. Roy. Soc. Trop. Med. Hyg., 1965, vol. 59, p. 367.
Williamson J„ Macadam R., Dixon H. — Biochem. Pharmacol., 1975, vol. 24, p. 147.
Wilman D., Connors T. — In: Molecular Aspects of Anti-Cancer Drug Action./ Eds S. Neidle, M. Waring, London: Macmillan Press, 1983.
Wilson 1. — In: Enzymes and Drug Action./Eds J. Mongar, A. deReuck, London: Churchill, 1962.
Wilson 1., Bergmann F. — J. Biol. Chem., 1950, vol. 186, p. 682.
Wilson Cabib E. — J. Amer. Chem. Soc., 1956, vol. 78, p. 202.
Wilson L, Harrison M„ Ginsburg S.—J. Biol. Chem., 1961, vol. 236, p. 1498.
Wilson L, Meislich E. — J. Amer. Chem. Soc., 1953, vol. 75, p. 4628.
Winder F., Collins P. — J. Gen. Microbiol., 1970, vol. 63, p. 41.
Witkop B., Foltz C. — J. Amer. Chern. Soc., 1957, vol. 79, p. 197.
Wohl A., Glimm E. — Biochem. Z., 1910, Bd 27, S. 349.
Wohl A., Momber F. — Ber. dtsch. chem. Ges., 1917, Bd 50, S. 455.
Wolff M. (ed.) Burger s Medicinal Chemistry (3 vols). 4th edn. — New York: Wiley, 1981.
Wolhoff J., Overbeck 7. — Rec. Trav. chim. Pays-Bas, 1959, vol. 78, p. 759.
Wood R„ Ferone R., Hitchings G. — Biochem. Pharmacol., 1961, vol. 6, p. 113.
Woodrojfe R., Wilkinson B. —J. Gen. Microbiol., 1966, vol. 44, p. 343.
Woods D. — Brit. J. Exper. Path., 1940, vol. 21, p. 74.
Woodward R., lacobucci G., Hochstein F. — J. Amer. Chem. Soc., 1959, vol. 81, p. 4434.
Woolf ]., Nixon J. — Biochemistry, 1981, vol. 20, p. 4263.
Woolley D. — Proc. Soc. Exper. Biol. Med., 1944, vol. 55, p. 179.
Woolley D. — J. Amer. Chem. Soc., 1950a, vol. 72, p. 5763.
Woolley D. — Proc. Soc. Exper. Biol. Med., 1950b, vol. 74, p. 747.
'Woolley D. A Study of Antimetabolites. — New York: Wiley, 1952.
Woolley D., Strong F., Madden R., Elvehjem C. — J. Biol. Chem., 1938, vol. 124, p. 715.
Woolley D„ White A. — Proc. Soc. Exper. Biol. Med., 1943, vol. 78, p. 489.
Woosley R., Draper D., Riedenberg M. et al. — New Engl. J. Med., 1978, vol. 298, p. 1157.
World Health Organization. Chemotherapy of Malaria and Resistance to Anti-malarials, Technical Report No. 529. — Geneva, 1973a.
415
World Health Organization. Pharmacogenetics, Technical Report No. 524. — Geneva, 1973b.
Worthing C. The Pesticide Manual: A World Compendium 7th edn. — Malvern (England): British Crop Protection Council; USA: State Mutual Books, 1983.
Wyss O., Rubin M„ Strandskov F.— Proc. Soc. Exper. Biol. Med., 1943, vol. 52, p. 155.
Wyss O., Strandskov F. — Ann. Biochem., 1945, vol. 6, p. 261.
Yaeger J., Munson S. — Science, 1945, vol. 102, p. 305.
Yagi K. — Adv. Enzymol., 1965, vol. 27, p. 1.
Yalkowsky S., Sinkula A., Valvoni S. (eds) Physical Chemical Properties of Drugs. — New York: Marcel Dekker, 1980.
Yamamura H., Enna S., Kuhar M. (eds) Neurotransmitter Receptor Binding.— New York: Raven Press, 1978.
Yamamura H„ Snyder S. — Proc. Natl. Acad. Sci., U.S.A., 1974, vol. 71, p. 1725.
Yendell A., Tupper R., Wills E.— Biochem. J., 1967, vol. 102, p. 23 P.
Youatt J. — Austral. J. Exper. Biol. Med. Sci., 1958, vol. 36, p. 223.
Youatt J. — Austral. J. Exper. Biol. Med. Sci., 1962, vol. 40, p. 201.
Yudkin M., Davis B.— J. Molec. Biol., 1965, vol. 12, p. 193.
Zanker V., Schnith H. — Chem. Ber., 1959, Bd 92, S. 2210.
Ziegler J., Hamm V., Berndt J. — Cancer Res., 1983, vol. 43, p. 5356.
Zimmerman A., Matschiner J. — Biochem. Pharmacol., 1974, vol. 23, p. 1033.
Zoncheddu A., Accomando R., Calendi E., Orunesu Л1. — Experientia, 1980, vol. 36, p. 1151.
Zribi A., Ben-Rachid M. — Bull. Soc. Path. Exot., 1973, vol. 66, p. 597.
Zysk J., Bushway A„ Whistler R„ Carlton W. — J. Reprod. Fertil., 1975, vol. 45, p. 69.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Аватек, связь атома кислорода с катионами 324
Азасерин 51
Аденозинмонофосфат циклический 227
Адреналин 220
Адсорбция на поверхности 84
— неспецифическая амфифильных веществ 84
— специфическая гидрофильных веществ 85
Азот, свойства анестезирующие 342
Акридин (ы), зависимость бактериостатического действия производных от ионизации 9.1
----------механизм связывания с нуклеиновыми кислотами 104
— и их аналоги, биохимические действия 103
—, конкуренция между ионами 95
—, контур молекулы, равноудаленный от атомов углерода и азота 101
—, применение в современной медицине 98
— производные, зависимость бактериостатического действия от степени катионной ионизации 94
----чувствительность 22 видов бак-
терий к их действию 93
—, фунгицидное действие 98
Акрихин как противомалярийное средство 115
— , свойства боковой цепи 117
Активность биологическая, не связанная со структурой 335
Аламетицин, анализ рентгеноструктурный 321
Д-алаиин 13
4-Аминогекс-5-иноевая кислота 69
Алифатические заместители, константы Тафта 377
Алкилфенолы, расщепление стенки
бактериальных клеток 326
Алкилирующие агенты, этерификация анионов фосфорных и карбоновых кислот 295
Аллопуринол для снижения содержания мочевой кислоты 53
Аллостерический эффект 15
Альбумин сывороточный, связывание лекарственных веществ 314
Альдегиды как ингибиторы пептидазы 621
Алюминий 137
Амеса тест на канцерогенность 306
Амидная связь, гидролиз 60
Амин(ы) ароматические канцерогенные 304
Аминакрин 90
Аминоакридины, действие на клетки млекопитающих 108
пара-Аминобензойная кислота, структура 26
Аминоглутетимид 48
Аминозин, действие нейролептическое 261
Аминокислоты, аналоги 51
— как связывающие катионы тяжелых металлов 144
— установление последовательности в молекуле фермента 11
"f-аминомасляная кислота, (ГАМК) в качестве тормозного нейромедиатора 250
Амины алифатические, лизис клеточной мембраны ,109
Амитриптилин 262
Амитрол, действие на рост дрожжей и водорослей Prototheca 58
Амоксициллин 276
Ампициллин 276
Ампролиум для лечения кокцидиоза 46
27—734
417
Амринон, применение при сердечной недостаточности 192
Амсакрин, воздействие на костный мозг 110
— для лечения лейкоза 110
Амфифильные вещества на границе раздела масло — вода 311
Амфотерицин В 331
Анализ регрессионный множественный для нахождения биологически активных лекарственных веществ 352 Аналоги гормонов и нейромедиаторов 47
Анальгетики, независимость аналгети-ческого действия от конформации молекулы 254
Анаприлин 225
Анестетики, зависимость активности от температуры и давления 344 — местные, активность 125
Антагонизм на полипептидном уровне 21
Антагонисты (метаболитов), Ван-дер-ваальсовы радиусы заместителей 20 — и агонисты, взаимосвязь 70 —, природа химическая 17
—, создание для большой молекулы 18
— фолиевой кислоты 25
— Н2 — рецепторов 56
Антергап 55
Антибактериальные свойства, зависимость от плоской структуры молекулы 100
— средства катионные 99
и анионные 109
-----, создание введением в молекулы заместителей для увеличения плоской поверхности 101
Антибиотики ^-лактамные 281
Антигельминтные препараты 191
Антидепрессанты 262
—, зависимость действия от топологии трициклического скелета 263
—, повышение уровня норадреналина и 5-гидрокситриптамина 264
Антидоты, содержащие SH-группы 269
— фосфорорганических инсектицидов 292
418
Антиметаболиты 16
— адренергические 49
— , история изучения 24
— , определение 8
— пептидов 51
—, повышение терапевтической эффективности 63
Апофермент, определение 9
Апрессип 192
Ароматический цикл в молекуле лекарственного вещества замена 198
Аскорбиновая кислота, самоокисление 85
свойство изомеров 83
Ауринтрикарбоновая кислота для лечения отравлений бериллием 171
Ацетилирования реакции 72
Ацетилхолин, головка катионная 237
—, изменения катионной головки 238
Ацетилхолинэстераза 229
рАх для классификации лекарственных веществ 71
Барбитал 347
Барбитураты 347
—, повышение коэффициентов распределения 347
Бария хлорид и натрия сульфат, применение совместное 70
Бацитрацин 283
Бемегрид 348
Бензидин, канцерогенность 302
Бензилпенициллин 276
4-бензилфенол, действие бактерицидное в водных растворах лаурата калия различной концентрации 313
Бензодиазепины 252
Бензойная кислота, поглощение дрожжами 122, 124
Бензол, канцерогенность 302
Биаламикол 191, 192
Бикорбонат-иои, структура 58
Биологическая активность, зависимость от площади планарной части молекулы 197
Биоптерин 34
Биотин, структура 45
Блеомицин 190
Блокаторы кальциевых каналов 325
Блокирование последовательное 63 -----при лечении болезней домашней птицы 64
Борные кислоты, действие ингибирующее на пептидазы 61
Бриггса—Холдейна уравнение 23
Буринамид 57
Буферные растворы для проверки фармакологических исследований 129
Валиномицин 320
Ванадий для питания позвоночных 137
Ван-дер-ваальсовы силы притяжения молекул 310
Ванкомицин 283
Варфарин 47
Верапамил 325
Вещества, менее активные в ионизированном состоянии 120
Вода, понижение температуры замерзания 75
Водород, выделение из бактерий Bacillus bellus 96
-----, конкуренция с катионами акридинов 97
Газы инертные, действие анестезирующее 342
----, увеличение наркотического дей-
ствия 342
Галоксон для защиты крупного рогатого скота от нематод 292
ГАМК, действие физиологическое, обусловленное тремя рецепторами 251
Гаммета константы 374
— уравнение 373
— уравнения, константа р 378
Гексокиназа дрожжевая, состав 10
Гемоглобин 134
Гидразины с липофильными заместителями 50
6-Гидроксидофамин, применение в экспериментальной физиологии 52
5-Гидрокситриптамин (серотонин) 229
D-гистидин, ингибирование фермента гистидазы 18
Гистидин, основа для создания мо
дельного соединения небелковой породы 13
Гликозиды сердечные 316
— — безопасность применения 318
-----•, источники природные 316
----, наличие в молекуле лактонного>
кольца 318
-----, транспорт в бактериальные клетки 329
Гормоны половые млекопитающих 18
----- полиеновые водорослей 18
— тиреотропный, ацетилирование, образование аналога 21
Грамицидин, свойства разрушения мембран 328
Гриб Endomyces, устойчивость к действию пиритиамина 46
Гуанин, деградация 34
Декарбазин 299
Депрессанты, активность термодинамическая 339
—, влияние на митоз 350
—, гипотеза биологического действия 335
—, концентрации минимальные при подавлении подвижности головастиков 341
—, применение в медицине 336
Дефероксамин(ы) 148
— для лечения отравлений железом 171
Диаверидин 44
Диазинон, применение для защиты овец от падальных мух 289
Диаминобензил 29
Диаминобутилтриазанафталин, структура 42
2,4-диаминопиримидииы как противомалярийные средства для позвоночных 42
— повышение антибактериальной активности 43
Диафенилсульфон 29
Дигидрофолатредуктаза (ДГФР) анализ реитгеноструктурный 38
— как рецептор лекарственных веществ 10
—, катализ восстановления фолатов 38
27*
419
— L. casei, стереодиаграмма птери-динсвязывающего участка 39
—, применение при кокцидиозе 43
Дигидрофолиевая кислота 30
Дикумарол, антагонист фактора свертывания крови 46
— для профилактики тромбозов коронарных артерий и инсультов 47
—, структура 45
Димедрол, действие седативное 55
Димеркапрол для лечения отравления металлами 169
Диметилглиоксим 191
Диметоат 291
Дискенезия при длительном применении нейролептиков 262
Диуретики, основа действия 315
Дифосфотиамин 8
Дифторметилорнитии 68
Дихлон, структура 45
2,2-дихлорпропионат натрия, применение для уничтожения однодольных сорняков 46
Дихлофос для уничтожения домашних мух 289
—, как антигельминтный препарат действия для свиней, лошадей и собак 292
Диэтилдитиокарбамат натрия для лечения отравлений таллием 172
Диэтилстильбэстрол, применение 212
ДНК, интеркаляция 106
— нормальная и с включением интер-калированных молекул профлавина 105
4-Додецилпиридин 83
• —, способность разрушать мембраны раковых клеток 331
Додин и глиодин, действие на мембраны грибов 329
Дофамин 227
Жгутиковые 112
Железо в непорфириновых ферментах 134
-----порфириновых ферментах 134
----- почве 135
Ивермектин 253
Изомерия геометрическая 208, 210
420
— оптическая 199
Изомеры аксиальные 215
— конформационные, образуемые двумя заместителями 213
— экваториальные 215
Изониазид, образование хелатных соединений с медью 187
— , открытие 186
—, применение в сочетании с рифампицином или этамбутолом 189
Имидазол для ускорения гидролиза сложных эфиров 13
— структура 14
Имизии при лечении депрессивных состояний 263
Ингибирование ДНК- и РНК-полиме-разы 95
— РНК-полимеразы Е. coli профлавином 98
— синтеза дигидрофолиевой кислоты сульфаниламидными препаратами 30
Ингибиторы, влияние на конформационные изменения фермента 60
— «переходного состояния» 59
Индекс ингибирования 22
Интал, противоаллергические свойства 333
Интеркаляция ДНК 106
—, теория 105
Инфекция протозойные, лекарственные вещества для лечения 112
Ионизация, значения констант при изучении биологических действий веществ 129
— слабых оснований 77
Ионизации степень, см. Степень ионизации 79
Ионы, прохождение через мембрану 86
Ионофоры анионные 323
— белки 324
— подвижные 319
— стационарные 319
Иприты азотистые 295
4-йодацетамидсалициловая кислота 66
Калпайн 139
Кальмодулин 139
Кальций, обеспечение твердости тканей 137
Кальций, концентрация 137
Кальций, накопление в митохондриях 138
Кальций, роль в сопряжении химического стимула с секрецией 138
Кальция этилендиаминтетраацетат для лечения отравления свинцом 170
Канцерогенез, двустадийность 301
Канцерогенность соединений для человека 302
Канцерогенность веществ, исследования 302
Канцерогенность, получение данных на крысах 306
Канцерогенные химические соединения 302
Канцерогены 300
Каптоприл для снижения повышенного давления крови 52
Карбаматы (уретаны), ацилирование АХЭ 293
Карбаматы, гидрофильность 294
Карбарил 294
Карбахол 294
Карбенициллин 278
Карбоангидраза, расположение активного центра 11
Карбоксипептидаза А, стереодиаграмма активного участка 12
Карбоксипептидаза А, фермент поджелудочной железы 11
Карцинофилин А, действие противораковое 112
Каталаза, снижение энергии активации разложения перекиси водорода 13
Катехоламины, активность биологическая 218
Катехоламины, изучение рецепторы 219
Катехоламины, синтез 16
Катионы, активность антибактериальная 89
Квантово-химические методы 358
Кислоты и основания, значения Ка и рКа 79
Кислород, введение атома в пролекарство 70
Кислород, положение атома в боковой цепи 241
Кислоты и основания 75
Кислоты слабые, действие на биологические объекты 121
Клетка, активность химическая 8
Кобальт, антагонизм к токсическому действию комплекса оксина с железом 176
Кобальт как необходимый кофактор ферментов 131
Кобальт, недостаток, вызывающий анемию у овец 131
Ковалентные связи в молекулах биологически активных веществ 266
Конканавалин А 77
Константа ионизаций, ее отрицательный логарифм рКа 77
Константа ионизации (Ка) 76
Конфигурация абсолютная аминокислот белка 203
Конфигурация абсолютная, определение 201
Коферменты органические 8
Коэффициент распределения газа 370
Коэффициенты распределения лекарственных веществ между собой и октанолом 373
Коэффициент распределения твердых и жидких веществ 370
Кремний 143
рКа, установление значений 78
Ксенон, действие депрессантное 343
Кумарины, действие антикоагулянтное 47
Кумафос 292
Лактатдегидрогеназа, апофермент 9
Леворфанол 255
Лейшманиозы, лечение диамидинами 115
Лекарственные вещества антигиста-миновые 54
-----, блокирующие действия дигидрофолатредуктазы 36
----- действие на поверхности 312
— —, ингибирование ферментов 16
-----, индексы ингибирования 24
421
----- и рецептор, адаптация взаимная 216
-----, литературные данные 385
----- пути поиска 351
-----, связывания сывороточным альбумином 314
Летальный синтез 307
Лиганды (металлосвязывающие вещества) 130
— константа устойчивости 155—159
—, логарифмы констант устойчивости 150
—, образование комплексов 1 : 1 152
-------- 2:1 152
—, распределение катионов металлов в нейтральных растворах 160
Лизоцим, действие на его субстрат 10
Липофильность, увеличение введением углеводородного радикала 371 Литературные данные о лекарственных веществах 385
Литий, лечение хронических психозов 140
— соли для контроля маниакальной фазы МДП 262
Магний 139
Малоновая кислота, индекс ингибирования 23 -----, структура 19 41-(имидазол-4-ил) масляная кислота, структура 14
Мафенид 29
Медь, влияние на ингибирование роста aspergillus niger диметилдитиокарбаматом натрия 183
— в церулоплазмине 133
—, действие избирательное иа беспозвоночных животных, грибы и водоросли 132, 133
— как составная часть фенолазы 132
—, накопление в нервных тканях 133
Мезатон 221
Мембраны плазматические 315
—, повреждение биологически активными агентами 326
2-меркаптопиридин-Ь1-оксид 181
Метаболиты, использование в качестве специфических блокаторов 21
—, положение метильной группы 17
—, превращение в аналог 17
Металл (ы) в живой клетке 131
-----качестве кофактора ферментов 145
—, нарушение баланса 140, 141
—, образование комплексов их катионов с компонентами 193
—, поддержание определенной концентрации 143
—, потенциал окислительно-восстановительный 164
—, радиусы катионов кристаллические 161
—, реакция организма при их получении 141
— тяжелые, необходимость для всех форм жизни 130
—, уменьшение токсического действия в результате хелатообразования 169, 173
Метараминол для выведения из шокового состояния 49
Метацнллины 276
Метенилтетрагидрофолиевая кислота 33
Метиамид, побочные действия 57
Метилдофа 223
Метиленовый синий 103
Метилентетрагидрофолиевая кислота 33
Метопролол 225
Метотрексат, амидиновая группа 40
—, для лечения лимфоцитарного лейкоза 36
------- хориокарциномы 36
— , индекс ингибирования 23
— , механизм действия 37
— , основность молекулы 40
— , связывание с ферментом, полученным из L. casei 38
— , токсичность 40
— , упрощение молекулы 40, 41
Метильная группа, замена хлором 20
Мефлохин, лечение малярии, вызванной falciparum и vivax 118
422
Миелосан, для лечения хронического миелолейкоза 296
Микроэлементы неметаллические 143
Миоглобин 134
Митомицин, токсичность 298
Михаэлиса константа 23
— акцептор, образование 69
Михаэля конденсация как тип реакции между ферментом и ФАНИ 67
Мицеллы ферментов, избирательность к субстратам 15
Мицеллярная концентрация 312
Мицеллярные равновесия 312
Мицеллярный катализ, эффективность 15
Молекулы (биологически-активных веществ) константы фрагментарные 372
—, конфигурация 361
— на границе раздела жидкость — жидкость, силы притяжения 311
— связь структура — активность 265
Молекулярная конфигурация, анализ Hopfinger 366
— рефракция, расчет величин 365
— связность, расчеты 359
Молекулярные модели лекарственных веществ 199
Молибден для фиксации азота 136
Молибдоктерин 34, 35
Молочная кислота 66
Монензин 324
Морфин, активность анальгетическая 255
—, алкалоид опия 253
Мышьяк, действия антибактериальные 270
— оксиды 268
— при лечении сифилиса, механизм действия 269
•—, применение в медицине 271
—, три уровня окисления 266
Надолол 225
Налидиксовая кислота 189
Налоксок 255
Наратиои 287
Натрий 140
Натрия додецилсульфат, как катализатор 15
Нейролептики, побочные действия 262
Необратимые ингибиторы активного центра ферментов (НИАЦФ) 65
Никель, обнаружение 137
Никотин, свойства холиномиметические в трех структурах у позвоночных 244
Никотинамидадениндинуклеотид 8
Никотиновая кислоста, синтез аналогов в 1938 году 25
Никотиновые антагонисты 246
— лекарственные вещества, не обладающие ацетилхолиноподобной активностью 246
-------, обладающие высокой ацетилхолиноподобной активностью 247
Никотиновые молекулы, конформация 245
— холинорецепторы 242
Никотиновая кислота 250
Нистатин 331
Нитракрин для послеоперационного лечения рака толстой кишки 111
Нитрозепам 252
Нитрозомочевины 298
Нифедипин 325
Нонактин 320
Окрашивание живых систем 95
Оксидаза D-амииокислоты 13
Оксин в дистиллированной воде 175 — действие антибактериальное 173 — зависимость бактерицидного действия от концентрации 174
Олово 137
Онкогены 301
Опиатные рецепторы, классификация 257
Опиаты, содержащиеся в пище 258
—, термин 253
ПАБ, ингибирование конкурентное и неконкурентное 31
Пантотеновая кислота 47
Парафуксин 87
—, интеркаляция в ДНК 108
423
Пеницилламин для лечения болезни Вильсона 170
Пенициллины, выделенные из среды штамма Penicillum notatum 273 — как антиметаболит П-аланил-П-аланииа 275
—, механизм действия на бактерии 272
— модифицированные 276
D-пеницилламин 190
Пентагастрин 52
Пентобарбитал 347
Пептидная связь, замена для улучшения всасываемости лекарственных веществ 48
Пептнлдипептндаза (ПДП) 52
Пентостатии, действие ингибирующее по отношению аденозиндезамина-зе 61
2-[пирид-2-ил (этиламин)] структура 71
Пиридоксин, структура 19
Пириметамин для профилактики малярии 42
—, структура 42
Пиритиамин, структура 19
Пластоцианин, участие в фотосинтезе растений 133
Повреждение мембран, изучение на модельных системах 328
Полимиксин 327
Полнпептидиые гормоны, усиление действия катионами двухвалентных металлов 192
Полиэлектролиты 80
Полиэфиры циклические, («краун-эфиры») способность переносить катионы щелочные и щглочно-земель-иые металлы 322
Порин, выделенный из Е. coll 324
«Правило последовательности» для определения номенклатуры оптически активных молекул 203
— «пяти атомов в боковой цепи» 240
Пропоксур 294
Простациклин 18
Противовирусные лекарственные средства 189
Противогрибковые препараты полиеновые 329
424
Противовоспалительные средства, степень ионизации 125
Противораковые препараты ПО, 190
Противотуберкулезное действие гидразидов 184
Противотуберкулезные препараты хелатообразующие 188
Противоэлептические препараты 348
Протонный магнитный резонанс (ПМР) 379
--------, растворители для съемки спектров 380
Протозойные инфекции 191
Профлавин 90
—, механизм связывания с ДНК 104
Псевдооснования 87
—, образования при усложнении гетероциклического ядра 88
—, химизм образования 87
Психотерапевтические агенты 260
Птеридины как катализаторы синтеза пуринов и пиримидинов 36
—, как рецептор электронов 35
—, растворимость в воде 35
Птеридиновое кольцо, связи химические 35
Птеридиновый цикл нормального субстрата 39
Пурины и пиримидины, их аналоги 53
pH внутри клетки, определение 127 — для растворения и ионизации 75
Равновесие между четвертичными катионами и соответствующими псевдоосноваииями 88
Ранитидин, способность понижать секрецию кислоты в желудке 57
Растворители, применяемые для съемки спектром ПМР 380
Релаксационные методы для получения данных о стереохимии ключевого атома 382
Релаксация спин-спиновая 384
— спин-решеточная, определение времени Ti 382
fi-Рецепторы 224
— а1,а2-постсинаптические 221
— ГАМК и бензодиазепины 250
— внутриклеточные 127
— клеточные наружные 126
— мускариновые, их антагонисты 242
— симпатомиметические, природа 226
Рибоза 32
Рибонуклеаза 9
— получение из циклодекстрнна 14
—, размер молекулы 10
Рибофлавин, биосинтез 34
Рингер Сидней, обнаружение сбалансированных физиологических растворов 24
Ртуть, препараты 272
Салициловая кислота 66, 189
----зависимость ингибирующих
свойств от pH 120, 121
L-Сарколнзин, активность 296
Секвенирование 11
Селен 143
Снотворные средства и анестетики, коэффициенты распределения 337 ----корреляция минимальной дозы 338 ----место действия 343
Соли ионизирующие 75
Сонная болезнь, действие сура мина и пентамидина 113
Соответствие между лекарственным веществом и его рецептором на основе объемных соотношений, 3 метода установления 362
Спирогерманий при раке толстой кишки и яичников 194
Споровики 115
Среды с незначительным содержанием металла 142
Статистическая значимость планирования 354
Степень ионизации, влияние на избирательность действия 82
<---для кислот 370
---- наркотических средств и их действие иа червя Arenicola 123 — — расчет по заданным величи-
нам рКа и pH 80, 370
--------при близких значениях рКа и pH 84
Стереохимический подход к определению конфигурации молекулы 361 Стеримолекуляриые параметры L и
В 365
Стерические параметры, включения их в уравнения регрессиоииого анализа 364
— факторы 197
Стерическое перекрывание, расчет объема 366
Стрептоцид 27
Сульфазин и хлоридин применение совместное при лечении токсоплазмоза 64
Сульфаниламиды, быстро выводящиеся из организма 27
— влииние величины рКа на бактериостатическое действие 126
— длительно циркулирующие в кровотоке 27
—, природа аитиметаболитная действия на бактерии 21
Сульфаниламины, действие антибактериальные 26
Сульфапиридин 27
Сульфатиазол 27
Суперокснддисмутаза 132
Сурьма, препараты 271
Тамоксифен при лечении эстроген-зависимых форм рака молочной железы 48, 49
Тафта константы для алифатических заместителей 377
Тест на Hi-антагонистическое действие 54
— трансформации клеток Styles на канцерогенность 306
Д9-Тетрагидрокаинабинол (ТГК), липофильность, наркотическое действие 349
Тетраметиламмоиий-хлорид 244
Тетрациклины 184
—, образование липофильного комплекса с магнием 185
— структура, активность 184, 185
Тимин, синтез 32
Тимолол 225
Тиоацетазои 188
425
Тиоглюкоза 59
Тиосемикарбазоиы 190
Тиосемикарбазон - 2 - ацетилпири-дин 188
Толерантность и лекарственная зависимость 258
— к опиатам у животных 258
Трансаминаза, получение из цикло-декстрина 14
Триазанафталниы, производные метотрексата упрощенные 41
Триамперен для подавления выведения калия из организма 316
Триметоприм, зависимость степени избирательности от замещения в бензольном кольце 43
—, индекс ингибирования 43
— н сульфазин, синергизм действия 65
—, применение в комбинации со стрептоцидом 44
Трипаносомоз, действие нифуртимок-са 113
Т-Трифосфатная группа 34
Трихлорфон, для защиты скота от оводов 290
Трополон и его производные 190
Фансидар, лечение методом последовательного блокирования 64
Фармакогенетика 72
Фармакодинамические средства 119
Фенамии, стимуляция ЦНС 49
Фенильная группа в молекуле лекарственного вещества, замена 198
Фениидион 47
Фенинтротион 287
Феноксиуксусная кислота 189
Фенол, действие на кожу, ослабление введением в бензольное кольцо липофильных "заместителей 314
—, зависимость ингибирующих свойств от ph 122
—, повышение растворимости в воде применением мыла 313
Феноляты, способность связывать железо 147
Фентоламин 221
Фенформин, действие антидиабетическое 192
Фергюсона принцип 338
Фермент(ы), активность каталипти-ческая, принципы 13
—, зависимость действия от мицеля амфифильных молекул 15
— и лекарственные вещества 16
— , модели простые 13
— , очистка 9
— , получение 9
— , регуляция биологическая 15
—, содержание в клетке 9
—, типы специфичности 13
—, ускорение химических реакций 12
Ферментативно-аитивируемые необратимые ингибиторы (ФАНИ) 66
Ферредоксины 134
Ферритин 134
Физиостигмии для лечения миастении и глаукомы 294
Флавинадениидииуклеотид (ФАНИ)-ФАД 13
—, наличие характеристической группы нормального субстрата 67
Фолиевая кислота, антагонисты 25
--------, применение при беременности 44
—: —, биосинтез и функции коферментов 34
-----и другие птеридины 32
-----производные величины рКа 40
Формиатдегидрогеназа 136
Фосфономицин 283
Фосфорилирование фермента, инактивация и реактивация 286
Фосфорорганические антигельинтные-средства 291
— инсектициды системные 291
-----, токсичность для млекопитающих и насекомых 288
-------- сравнительная 290
Фосфорорганические соединения действие на примере параоксона 285> — избирательность 286
-----, способность ацилирования 285
-----, способность ацилирования
эстеразы 284
426
-------связывать металлы в клетках 146
-----, химизм действия 284
Фри — Фильсона метод аддитивный 358
3-фтор-аланин 51
D-3-фторалании, ингибирование ала-ниирацемазы 68
Фторуксусная кислота, метаболизм, термины «летальный синтез» 299
-----содержание в Dichapetalum 300
Фунгицидные препараты 190
Фуросемид для лечения отеков 316
Хелатирующие агенты антидоты 169 — — в качестве антидотов для удаления токсических веществ 166 ----- для восполнения недостаточного количества металла 166
— —, «кооперативный эффект» 167
—: —, механизмы биологического
действия 166
-----, проникновение через клеточные мембраны 168
-----, создание новых 195
Хелатные комплексы, конфигурация 130, 161
— соединения с курареподобиыми свойствами 193
Хелатообразование, химизм 151 а-Химотрипсин, структура 10 Химотрипсин, содержание аминокислот в активных центрах 12
Хингамин 115
Хинин, лечение малярии, устойчивой к действию хингамина 118
Хиральность 205
Хлозенид 252
Хлорбутин для лечения лимфоцитарного лейкоза 296
Хлоргексидииа глюконат для дезинфекции кожи и слизистых НО
Хлорфенамин 55
Холинорецепторы 231
Холинорецептор (ы) мускариновый 232
— третьего типа 250
Hepfinger метод анализа молекулярной конфигурации 366, 367, 368
Хром, усиление действия инсулина 136, 137
Цветтерионы 80
Цефалоспорин С, строение, активность биологическая 279
Цефалоридин 280
Цефалотин 279
Цефамандол 280
Цефокситин 281
Циклогексан, три конформации 214
Циклодекстрины, структура 14
Циклосерин 51
Циклофан и эмбихии, ингибирование
ДНК 297
Циклофосфан, действие противоопухолевое 297
Циметидин 56
Циик 136
Цис-дихлордиаминплатина для подавления опухолей саркомы 194
Цис- и трансизомеры 208
Цисилатин, активность биологическая при замещении атомов CI гидроксигруппами 194
Цитоплазма животных и растительных клеток, значение pH 128
ЦНС, стабилизация мембран 334
Шизофрения 260
Шизофрения, лечение нейролептиками 260
Электролиты 74
Электронные эффекты 373
Электропроводность воды, действие на нее веществ 74
Эллиптицин для лечения лейкозов
111
Эмбихии 295
Эналаприл 53
^-Эндорфин, содержание аминокислотной последовательности 256
Энкефалины, активность 256
427
Энтерохелин 147
Эстрогены как контрацептивное средство 48
Этаноламнн для получения антигистаминных препаратов 54
Этиловый спирт 17
Этиловый эфир карболин-3-карбоно-вой кислоты 252
Эфедрин, действие прн поллипозах н аллергических заболеваниях, побочные действия 49
Эффект прямого полярного сопряжения, влияние на величину ор 376 Эхиномицин, ннтерколяцпя в ДНК
108
Ядерный магннтаный резонанс (ЯМР) 381
--------в биологической н медицинской химии 383
Янтарная кислота, структура 19
Руководство
А. АЛЬБЕРТ
Избирательная токсичность. Физико-химические основы терапии
Том 2
Зав. редакцией В. С. Залевский Редактор Л. Б. Пиотровский Редактор издательства Г. Ф. Аншакова Переплет художника А. Е. Григорьева Художественный редактор В. Л. Фисенко Технический редактор Н. В. Сорокина
Корректор А. К. Баскакова
ЦБ № 5686
Сдано в набор 01.03.89. Подписано к печати 12.06.89. Формат бумаги бОХЭО'Лв- Бумага тип. № 1. Гарнитура Литерат. Печать высокая. Усл. печ. л. 27,0. Уел. кр.-отт. 27,0. Уч.-изд. л. 31,04. Тираж 7000 экз. Заказ 734.
Цена 2 р. 50 к.
Ордена Трудового Красного Знамени издательство «Медицина»
101000, Москва, Петроверигский пер., 6/8
Московская типография № 11 Союзполи-графпрома прн Государственном комитете СССР по делам издательств, полиграфии и книжной торговли.
113105, Москва, Нагатинская ул., д. 1.
ИЗБИРАТЕЛЬНАЯ ТОКСИЧНОСТЬ
А. АЛЬБЕРТ
ТОМ 2