/


















































































































































































































Similar




Text
О. В.ЧЕКАНОВСКАЯ
ЧЕРВИ
ПОЧВО
ОБРАЗОВАНИЕ
А К АДЕ МИЛ НАУК
СССР
Дождевые черви на поверхности земли.
На переднем плане — Lumbricus terrestris, за ним справа —
Eisenia foe tide, сзади па траве — Allolobophora longa.
АКАДЕМИЯ НАУК СССР
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
НАУЧНО-ПОПУЛЯРНАЯ СИРИЯ
<». В. ЧЕКАНОВСКАЯ
ДОЖДЕВЫЕ ЧЕРВИ
И ПОЧВООБРАЗОВАНИЕ
ИвДАТ£ЛЬСТВО А К АДК МИИ И АУН СССР
Moettta tote Л «мин«раО
АННОТАЦИЯ
Книга знакомит читателя со строением, образом жизни,
систематикой и методами изучения дождевых червей. В ней
показана огромная положительная роль их в почвообразова-
нии и земледелии: жизнедеятельность этих червей является
существенным фактором в повышении плодородия почвы.
Обрисована роль дождевых червей в качестве переносчиков
паразитарных заболеваний домашних животных.
Книга рассчитана на широкий круг читателей, в част-
ности, агрономов, почвоведов и зоологов.
Главный редактор
академик Е. Н. ПАВЛОВСКИЙ
Редактор издания А. А, СТРЕЛКОВ
чПлуг принадлежит к числу древней-
ших и имеющих наибольшее значение изо-
бретений человека; но еще задолго до его
изобретения почва регулярно обрабатыва-
лась червями и всегда будет обрабаты-
ваться ими*.
Ч. Дарвин. «Образованно растительного
слон земли деятельностью дождевых червей. . . •
ПРЕДИСЛОВИЕ
Для успешной борьбы за повышение урожайности
должны быть полностью учтены и всемерно использо-
ваны все силы природы, имеющие отношение к жизни
растений. Одной из таких сил является деятельность
почвенных животных. Значение ее для почвообразования
до недавнего времени недооценивалось, несмотря на то,
что основные факты в этой области были уже давно уста-
новлены. Только в последнее десятилетие и у нас, п за
рубежом интерес к почвенной фауне оживился и стали
быстро накапливаться данные, показывающие с полной
несомненностью, что этот раздел зоологии и почвоведе-
ния заслуживает самого пристального внимания.
Среди почвенной фауны видное место занимают дожде-
вые черви; их большое значение в этом отношении с каж-
дой новой работой определяется все яснее, причем по
этому вопросу накопилась уже порядочная литература.
Изучались фауна, географическое распространение, раз-
витие и биологические особенности дождевых червей.
В последние годы в СССР появились две большие работы,
посвященные специальным вопросам почвообразователь-
ной деятельности дождевых червей (Соколов, 1956;
Зражевский, 1957). Переизданы отдельной книгой ра-
боты Н. А. Димо (1955), первого из русских почвоведов,
оценившего по достоинству почвообразовательную дея-
тельность животных, в частности дождевых червей.
Имеются и общедоступные статьи, и брошюры по до-
ждевым червям, например брошюра И. И. Малевича, из-
данная Зоологическим институтом Академии наук СССР
з
в 1950г., книга А. Л. Зеликмана и И. И. Малевича (1951)
и статья И. И. Малевича, опубликованная в издании
«Животный мир СССР» (1953). В первых двух работах при-
водится ценный определитель дождевых червей, и во всех
них содержится ряд новых сведении по биологии этих
животных и значению их в почвообразовании. Однако,
в силу очень небольшого объема этих работ и крайней
сжатости изложения, читатель, особенно не ориентирован-
ный в зоологии и почвоведении, не сможет получить
цельного представления об этой группе животных и их
почвообразовательной деятельности, не обращаясь к дру-
гой литературе.
Значительное внимание было уделено дождевым чер-
вям также на Первом Всесоюзном совещании по почвен-
ной фауне в декабре 1958 г.
В отличие от ряда сочинений, посвященных почво-
образовательной деятельности животных, в предлагаемой
книге отводится значительное место изложению анатомии
и физиологии дождевых червей. Это сделано нами на осно-
вании твердого убеждения в том, что только всестороннее
изучение этих животных может способствовать углублен-
ному пониманию их роли в почвообразовании. Кроме того,
приведение этих сведений казалось нам не лишним, по-
скольку они могут пригодиться всем, интересующимся
дождевыми червями по тому или иному поводу, т. е. уча-
щимся и учащим, работникам лабораторий, где поль-
зуются дождевыми червями для разных целей, агроно-
мам и другим лицам.
При окончательной редакции текста использованы цен-
ные указания М. С. Гилярова, И. И. Малевича и С. И. По-
номаревой, взявших на себя труд внимательного ознаком-
ления с рукописью.
ВВЕДЕНИЕ
Поверхность большей части суши покрыта почвами.
Свойства, почв (наряду с климатическими условиями)
определяют возможность заселения их той или иной ра-
стительностью, а последняя в свою очередь определяет
характер населения животных, обитающих в почве и на
ее поверхности в данном участке суши. Таким образом,
почва — это в полной мере «основа жизни»; недаром слово
«беспочвенный» означает «лишенный основания». А кому
же неизвестно, что растительный и животный мир нераз-
рывно связан с земледелием и скотоводством? Необхо-
димой предпосылкой для появления и развития этих
отраслей хозяйства является наличие плодородных почв.
Образование почв из горных пород и изменение свойств
уже существующих почв, т. е. почвообразование, — сово-
купность процессов, начавшихся, нужно полагать, одно-
временно с возникновением суши и идущих непрерывно
повсюду до настоящего времени. Первые шаги процесса
почвообразования совершаются силами неорганической
природы: солнечные лучи, вызывающие неравномерное
нагревание горных пород, атмосферный воздух (действие
ветра и кислорода воздуха) и вода (в виде осадков, ручьев,
рек) являются главными геологическими факторами
почвообразования. Параллельно с этим действуют и по-
степенно выходят на передний план биологические фак-
торы. В настоящее время общепризнано, что почва пред-
ставляет собой целостный комплекс минеральных и орга-
нических веществ с живыми организмами. Ее неживые
составные части, взятые отдельно от населяющих почву
я
организмов, уже не являются почвой, и равным образом
почвенное население без среды его обитания — только
отвлеченное понятие. Поэтому в известном смысле можно
говорить о жизни почвы. Если вещества почвы служат
источником существования населяющих ее организмов,
то свойства самой почвы определяются деятельностью
ее обитателей. Совокупность результатов жизнедеятель-
ности почвенных организмов и составляет комплекс био-
логических факторов почвообразования.
О том, что дождевые черви улучшают качество почвы,
было известно еще в древние времена. Однако роль до-
ждевых червей как животных-почвообразователей была
впервые освещена научно и понята во всем ее значении
знаменитым Чарльзом Дарвином. В результате своих
замечательных исследований, посвященных этому воп-
росу (Darwin, 1838, 1881), он пришел к заключению,
что «в ряд ли найдутся другие животные,
которые играли бы столь большую
роль в истории мира, как дождевые
черв и».1 Впоследствии сведения о дождевых червях и их
роли в почвообразовании были значительно углублены
и расширены трудами большого числа отечественных и
зарубежных ученых. Все усиливающийся поток этих
исследований приносит новые и новые доказательства
правильности мнения Дарвина о выдающейся роли до-
ждевых червей в экономике природы и истории мира.
Авторитетный почвовед, имеющий крупные заслуги в об-
ласти изучения деятельности почвенных животных,
Н. А. Димо (1938, стр. 520) характеризует значение до-
ждевых червей следующими словами: «Под поверхностью
почвы и в меньшей степени на ее поверхности совершается
громадная, пока еще, несмотря на целое столетие, про-
шедшее со времени первого сообщения Ч. Дарвина, не-
достаточно освещенная и оцененная по значимости для
почвообразования работа. Медленно, тихо и беззвучно
из года в год, из тысячелетия в тысячелетие накопляются
в почвах и грунтах черты сложения, структуры, химии,
физики, жизненных явлений и др., не воспроизводимые
никаким другим агентом природы*.
1 Имеются многочисленные издания полного собрания сочине-
ний Дарвина, куда вопли исследования над дождевыми червями.
Наиболее полное собрание трудов опубликовано в 1936 г.
б
Однако если вопрос о важном значении дождевых
червей для процесса почвообразования не вызывает сомне-
ний у специалистов, то эти сведения еще далеко не до-
статочно проникли в среду биологов и почвоведов, не
говоря о широких массах земледельцев и лиц, так или
иначе соприкасающихся с земледелием, садоводством и
лесоводством; они до сих пор обращают мало внимания
на неустанную деятельность огромных рабочих армий
червей, возделывающих землю бок о бок с ними. До сих
пор слово «червяк» служит для выражения чего-то низ-
менного и бесполезного. Фауст в одноименной трагедии
Гете, желая характеризовать ничтожество своего ученика
Вагнера, ученого-крохобора, не видящего за деревьями
леса, говорит про него:
«Он ищет клада жадною рукою,
Но рад, когда находит дождевых червей».
Это и многие другие аналогичные высказывания (у Дер-
жавина, Апухтина и др.), взятые изобиходных выражений
живой речи, имеют целью вызвать в воображении чи-
тателя образ дождевого червя. Иногда слово «червь»
служит даже символом вреда и зла (в этих случаях,
правда, по-видимому, имеются в виду паразитические
черви и личинки насекомых). Не поэтому ли довольно рас-
пространено мнение, что дождевые черви якобы могут
приносить какой-то вред растениям. По крайней мере
многие хозяева их старательно выбирают из огородных
гряд и уничтожают, не зная того, что их присутствие не
может принести растениям ничего, кроме пользы.
Некоторое практическое значение дождевые черви
имеют как корм для домашних птиц и как приманка для
рыб. Однако эффект деятельности дождевых червей в от-
ношении мелиорации почв столь велик, что гораздо бо-
лее перспективно, вместо того чтобы скармливать птицам
и рыбам этих «земледельцев», изыскивать способы для
усиления использования их в том направлении, в каком
их подготовила сама природа, т. е. как весьма совершен-
ные аппараты для переработки земли. В решении основ-
ной проблемы земледелия, т. е. интенсификации произ-
водства и повышения урожайности, дождевые черви как
наши союзники в обработке почв не должны быть забыты.
Эта книга написана зоологом. Она имеет целью
ознакомить читателя одновременно и с зоологическими
фактами, которые могут быть полезными для лиц, интере-
сующихся почвами, и с данными, взятыми из работ по
почвоведению, рассматривающих почвообразовательную
роль дождевых червей. Из сказанного выше ясно, что
почвообразование — это проблема, успехи в разработке
которой могут быть обеспечены только совместными
усилиями почвоведов и биологов. Это значит, что для
дальнейшей работы в этом направлении необходимо, чтобы
биологи (в частности, зоологи) всерьез заинтересовались
почвами, а почвоведы — животным населением почв.
Одной из глав почвенной зоологии и посвящена эта
книжка.
ГЛАВА J
СТРОЕНИЕ ТЕЛА
И ГЛАВНЫЕ ЖИЗНЕННЫЕ ОТПРАВЛЕНИЯ
ДОЖДЕВЫХ ЧЕРВЕЙ
I. ОБЩИЕ ЗАМЕЧАНИЯ. НАРУЖНЫЕ ПРИЗНАКИ
Начнем с ознакомления со строением тела дождевых
червей. Устройство тела — основа знаний о животных.
Хотим ли мы разобраться в разнообразии форм интере-
сующей нас почему-либо группы животных или ознако-
миться с образом их жизни, связью их со средой обитания
или подойти к решению тех или иных практических
вопросов, связанных с этими животными, и т. д. — во-
прос о строении тела является основной предпосылкой
для решения любых других. В частности, в отношении
дождевых червей, уже для того чтобы определить род
и вид какого-либо их представителя (а, как мы увидим
дальше, их существует немалое число), недостаточно знать
его наружные признаки, но необходимо путем вскрытия
установить ряд особенностей строения его внутренних
органов.
Параллельно мы ознакомимся с работой описывае-
мых органов и их значением в жизни червей.
В теле дождевого червя (рис. 1) можно отличить перед-
ний (или головной) конец тела, более толстый, с более
сильной мускулатурой и обычно темнее окрашенный,
и задний (или хвостовой), более тонкий и более бледный.
Задний конец червя часто бывает плоским. На головном
9
конце тела помещается рот, на хвостовом — заднепро-
ходное отверстие. Хорошо отличаются друг от друга
также спинная сторона, более выпуклая и обычно более
темная, и брюшная — более светлая и более плоская;
у червей, консервированных в спирту или в формалине,
Рис. 1. Lumbricus terrestris, вид
сбоку. (Ориг.).
/ — головная лопасть, 2 — поясок;
3 — мужское половое отверстие.
брюшная сторона может быть вогнутой местами или
по всей длине.
Все тело дождевого червя поделено поперечными пере-
тяжками на отдельные участки, которые называются
члениками, илп сегментами. Эта кольчатость, или сег-
ментация — ведущая черта их организации: каждый
из сегментов в принципе имеет одно и то же строение и
содержит в основном весь комплекс органов, свойствен-
ных этим животным. В передней части тела сегменты
более крупные, по направлению кзади их размер посте-.
10
4 б
Рис. 2. Формы головной лопасти.
(Из Фюллера).
А — эпилобическая; Б — танилобиче*
ская.
пенно убывает. Число сегментов у обычных видов варьи-
рует в пределах от 90 до 300; оно подвержено значитель-
ным колебаниям у разных экземпляров одного и того же
вида, но с возрастом в отличие от многих их водных
родичей не меняется. Только у некоторых тропических
видов число сегментов достигает 600. Внимательно при-
глядевшись к поверхности тела, можно увидеть, что
каждый сегмент подразделен на три части двумя неглу-
бокими бороздками. Это — так называемая вторичная
кольчатость, которая также отражает некоторые черты
внутренней организации
каждого сегмента. Сег-
менты тела нумеруются,
причем первым сегментом
считается головной.
Головной сегмент, кро-
ме ротового отверстия, об-
ладает еще одной харак-
терной особенностью: на
передней его части имеется
головная лопасть — под-
вижный, меняющий форму
придаток, нависающий над ртом. У дождевых чер-
вей головной сегмент может быть двоякого рода:
либо головная лопасть, вдаваясь на спинной стороне
в область первого сегмента, отделена от него поперечной
бороздкой, либо она доходит до борозды между 1-м и 2-м
сегментами. В первом случае головной сегмент называют
эпилобическим, во втором — танплобическпм. Эти раз-
личия в форме головной лопасти имеют важное значение
при определении видов червей (рис. 2).
Головная лопасть —• орган осязания и обоняния; ею
червь исследует встречающиеся на его пути предметы.
В передней части тела у взрослых особей имеется так
называемый поясок, X- утолщение, охватывающее
от 5 до 12 сегментов, обычно иначе окрашенное по срав-
нению с остальной частью тела (рис. 3). Кожные покровы
в области пояска содержат большое количество желез,
выделяющих питательное вещество для яиц при откладке
яйцевых коконов. Поэтому в период размножения поясок
выглядит сильно набухшим, а тогда, когда откладки
коконов нет, область пояска отличается от соседних
участков только цветом и иным характером поверхности
тела. Форма пояска может быть кольцевой, если он раз-
вит одинаково сильно со всех сторон, или седлообразной,
если с брюшной стороны он мало развит. По бокам брюш-
ной стороны пояска находятся удлиненные утолщения,
которые мы будем называть валиками зрелости (рис. 35).
Рис. 3. Передний
отдел тела дожде-
вого червя Luinb-
ricus terrestris со
спинной стороны.
(Из Фюллера).
1 — спинные поры;
2 — поясок.
У некоторых видов эти валики заме-
нены несколькими парами бугорков
зрелости. Форма, длина, цвет и рас-
положение пояска, валиков и бугор-
ков служат существенными видовыми
признаками дождевых червей.
По всей длине тела червя можно
заметить маленькие щетинки, которые
хорошо видны в лупу. Они находятся
на всех сегментах тела, кроме 1-го.
У дождевых червей фауны СССР ще-
тинки расположены по 8 на каждом
сегменте, попарно или поодиночке.
Щетинки образуют с каждой стороны
тела червя по 4 продольных ряда, ко-
торые принято обозначать буквами
латинского алфавита — a, b, с, d
(рис. 4). Их расположение имеет боль-
шое значение при определении червей.
Ряды щетинок а и 6, с и d обычно
сближены попарно. Степень их сбли-
жения у разных видов различна. При
определении червей обязательно учи-
тывается также отношение расстоянии
между рядами щетинок. Эти расстоя-
ния обозначаются буквами аа, ab, Ьс.
cd и dd (как принято обозначать от-
резки линий в геометрии). Имеет значение также отно-
шение расстояний между щетинками к величине наруж-
ного контура поперечного разреза через червя.
Щетинки — важные органы движения: червь может
зацепляться ими за частицы грунта или отталкиваться
от них при передвижении в почвенных норках и на по-
верхности земли. Можно также убедиться в их наличии,
проведя пальцем вдоль брюшной стороны тела от хвосто-
вого конца к головному. Если живого червя поместить
12
иа лист бумаги, то будет хорошо слышен характерный
шорох при его перемещении, обусловленный трением
твердых щетинок. На некоторых сегментах щетинки видо-
изменены в особые половые щетинки, имеющие значение
при спаривании червей.
На брюшной стороне тела, впереди от пояска, поме-
щаются половые отверстия. Сюда относится пара мужских
половых пор, обычно находящихся на возвышениях —
так называемых железистых подушках (рис. 34) и пара
женских половых пор, снаружи плохо различимых.
Рис. 4. Схема типов расположения щетинок на
поперечных разрезах через сегменты.
(По И. И. Малевичу).
А — щетинки не сближены попарно; Б — щетинки слабо
сближены попарно; В — щетинки сильно сближены попарно.
а — брюшные щетинки; Ъ — брюшно-боковые; с — спинно-бо-
ковые; d — спинные; аа — брюшной интервал; ab —брюшно-
боковой; he — боковой: cd — спинно-боковой; dd — спин-
ной.
Кроме того, у большинства видов имеется 2—3 пары пор
семеприемников. О значении всех этих отверстий будет
сказано далее.
На спинной стороне у консервированных червей’хо-
рошо видны в межсегментных бороздах спинные поры,
передняя граница расположения которых имеет значение
при определении видов червей.
Цвет тела дождевых червей зависит, с одной стороны,
от цвета их крови, с другой — от кожных пигментов.
Следует строго различать окраску тела червей, о которой
можно говорить только в отношении живых особей и ко-
торая зависит от комбинации кожного пигмента и цвета
крови, от пигментации кожи, которая обусловлена только
наличием пигментов. Черви, лишенные пигмента, имеют
при жизни розовую или красную окраску тела, а в консер-
вированном состоянии становятся белыми или Серова-
13
тыми, пигментированные же виды могут быть красных,
бурых, коричневых, желтых и синих тонов.
Длина тела дождевых червей СССР колеблется от 2
до 30 см при толщине от 2 до 12 мм. В тропических стра-
нах есть виды, достигающие в длину 3 м. Основная масса
червей, населяющих почвы во всем мире, представлена
видами, имеющими в длину 5—20 см.
Все дальнейшее изложение относится к дождевым
червям семейства люмбрицид (Lumbricidae). Черви дру-
гих семейств (если не считать ботанических садов, куда
черви иногда заносятся вместе с тропическими расте-
ниями) могут встретиться только в Уссурийском крае,
Средней Азии и в южной части Черноморского побережья
Кавказа.
2. ПОКРОВЫ ТЕЛА
Тело дождевых червей покрыто однослойным эпите-
лием. В его составе имеются поддерживающие, желе-
зистые и камбиальные клетки (рис. 5).
Рис.5. Кожный эпителий дождевого червя.
(По Стефенсону).
1 — белковая железа; 2 — кутикула; з — под-
держивающие клетки; 4 — слизистые клетки;
6 — камбиальные клетки.
Поддерживающими клетками осуществляется защит-
ная функция. Наружная часть этих клеток выделяет
вещество кутикулы — тонкой прозрачной пленки, по-
14
Рис. 6. Кутикула1 дождевого
червя с поверхности.
(Иа Штольте).
1 — волокна кутикулы; 2 — отвер-
стия желез.
крывающей эпителий. Кутикула состоит из двух систем
параллельных волокон, пересекающихся друг с другом
под прямым углом. В местах пересечения в кутикуле
могут быть отверстия. Направление волокон — диаго-
нальное по отношению к продольной оси тела (рис. 6),
что наилучшим образом обеспечивает прочность кутикулы
при растяжении изнутри (любопытно, что соединительно-
тканные волокна в коже млекопитающих имеют также
диагональное расположение по отношению к продольной
оси тела). Кутикула в тече-
ние жизни все время снаши-
вается и возобновляется дея-
тельностью эпителия. У кон-
сервированных экземпляров
кутикула может отставать,
и иногда ее можно снять це-
ликом, как чулок.
Кутикулой обусловлена
гладкость поверхности кожи,
которая облегчает скольже-
ние тела при движении по
твердым поверхностям. Опа
же обусловливает характер-
ную глянцевитость поверх-
ности тела.
Большое значение в жиз-
ни червей имеет деятельность
железистых клеток. Большая часть их выделяет слизистое
вещество, которым всегда смазана поверхность кути-
кулы; оно выходит на поверхность тела через отверстия
в ней (рис. 5 и 6). Это увеличивает легкость скольжения
по субстрату и предохраняет тело от высыхания. При лю-
бом сильном раздражении слизистые выделения выступают
на поверхность тела в огромных количествах: червь
мгновенно окутывается толстым слоем густой клейкой
слизи. Образование слизистого чехла на теле играет
важную роль при спаривании и образовании яйцевых
коконов. Кроме того, слизистые выделения покрывают
стенки ходов червей внутри почвы, что придает им зна-
чите л ьную прочность.
Помимо обычных слизистых клеток, в кожном эпите-
лии дождевых червей имеются на всей поверхности тела
15
так называемые белковые железистые клетки (рис. 5).
В области пояска (рис. 25), около щетинок половых от-
верстий и в других местах тела находятся кожные же
лезы, о значении которых будет сказано далее.
Рис. 7. Щетинковый мешок дож-
девого червя в продольном раз
резе. (Из Штольте).
1 — узелок; 2, з — мышцы; 4 — стенка
щетинкового мешка
Важным компонентом
кожного эпителия яв-
ляются маленькие клетки,
находящиеся в его глубо-
кой части, на границе с
иодл ежащей м уск ул ату
рой, и не соприкасаю-
щиеся с наружными частя
ми поддерживающих и же-
лезистых клеток (рпс. 5).
Это — камбиальные клет
ки, которые являются ре-
зервом; за счет них во-
зобновляются сношенные
функционирующие клетки
и происходит рост ткани
у молодых животных. Эти
клетки мобилизуются так-
же при заживлении ран
после ранений и иных по-
вреждений.
Из особых клеток кож-
ного эпителия образуются
также щетинки. На по-
верхности тела выступает
лишь наружная часть ще-
тинки. Своим внутренним
концом она глубоко по-
гружена в стенку тела
и может пронизывать ее на
сквозь, почти достигая полости тела. Щетинки помещаются
в щетпнковых мешках, представляющих собой врастания
внутрь тела кожного эпителия (рис. 7). Они состоят из ве-
щества, сходного с веществом кутикулы, непрочны и
быстро снашиваются. Поэтому в течение всей жизни в глу-
бине щетинковых мешков происходит образование новых
щетинок. Каждая щетинка образуется из одной клетки,
входящей в состав дна щртипкового мешка.
16
Щетинки дождевых червей неодинаковы но форме:
это палочки, иногда почти совсем прямые, иногда же с яв-
ственно загнутыми концами. На некотором расстоянии
от наружного конца щетинки находится небольшое утол-
щение — узелок, т. е. место, к которому прикрепляются
мышцы, втягивающие щетинку в глубь тела (мышцы
ретракторы; рис. 7). Кроме них, в щетинковых мешках
имеются мышцы протракторы, которые одним концом
прикрепляются к концу щетинки, а другим — к стенке
тела; их сокращением щетинка выталкивается наружу,
а также (при неодновременном их сокращении) может
совершать довольно разнообразные движения.
О половых щетинках см. ниже (стр. 54).
Говоря о покровах тела, упомянем об интересном
явлении свечения дождевых червей, которое с давних пор
привлекало внимание многих крупных натуралистов.
В частности, о светящихся дождевых червях писал из-
вестный исследователь жизни насекомых — Фабр. В раз-
ных странах описаны особые виды «фосфорических»
червей. Выяснилось однако. что свечение в темноте можно
наблюдать и у самых обыкновенных видов. Известный
чешский исследователь Вейдовский сообщил, что, раска-
пывая однажды ночью навозную кучу в поисках дожде-
вых червей, он увидел пятна мерцающего синевато-
белого света, который то появлялся, то исчезал в разных
точках. Оказалось, что свет исходил от обычных навоз-
ных полосатых червей, которых он собрал в большом
количестве. Он заметил, что его пальцы после того, как
он брал червей, начинали в темноте светиться. Таким
образом, светятся слизистые выделения червей, причем
только при наличии особых условий, так как свечение
наблюдается далеко не всегда. Имеются указания о све-
чении жидкости, выступающей из ротового и заднепро-
ходного отверстий.
Не может быть сомнения в том, что во всех этих слу-
чаях свечение вызвано бактериями, содержащимися
в выделениях червей. В процессе жизнедеятельности мно-
гих бактерий выделяется световая энергия, освобождаю-
щаяся при химических реакциях. Нужно сказать, что
почти всегда свечение животных обязано своим про-
исхождением бактериям, так или иначе связанным
с ним.
Некоторые исследователи полагают, что свечение по-
лезно для червей: одни думают, что вспышки света со-
действуют отыскиванию особями друг друга на поверх-
ности земли при спаривании (хотя у червей и нет глаз,
они все же способны воспринимать свет поверхностью
передней части тела); другие приписывают свечению
роль фактора, отпугивающего врагов; третьи думают,
что светящаяся слизь, оставляемая червями на их пути,
привлекает внимание врагов и делает их самих менее
заметными. Однако все это не более чем предположения,
не подкрепленные точными наблюдениями.
3. МУСКУЛАТУРА И ДВИЖЕНИЕ. ПОЛОСТЬ ТЕЛА
Основной частью двигательного аппарата дождевых
червей является мощно развитая мускулатура их стенки
тела (рис. 8). Она устроена по типу так называемого кожно-
Рис. 8. Схема строения сегмента дождевого червя
в поперечном разрезе и с поверхности. (Из Фюллера).
1 — эпидермис; 2 — кожно-мускульный мешок; з — брюш-
ная нервная цепочка; / — брюшной кровеносный сосуд;
£ — нефридий; в — кишечник; 7 — тифлоэол; 8 — спинной
кровеносный сосуд, а—d — щетинки.
мускульного мешка. Под кожным эпителием расположен
слой кольцевых мышц, сокращение которых уменьшает
диаметр червя. Слой кольцевой мускулатуры подстилается
18
слоем продольной мускулатуры (рис. 18), сокращение
которой уменьшает длину червя. На границе между этими
двумя слоями находится очень тонкий слой диагональных
мышечных волокон..
На большей части тела продольный слой мускулатуры
имеет значительно большую мощность, чем кольцевой,
но в передних 8—12 сегментах тела кольцевой слой мо-
жет достигать толщины продольного слоя. Эти сегменты
играют особенно важную роль
при вбуравливании червя в
землю (рис. 9).
Прежде думали, что ходы
червей в земле образуются по-
глощением ими земли, т. е. что
червь как бы въедается в землю.
Однако, как показал уже Дар-
вин, эти ходы — главным обра-
зом результат активной мышеч-
ной работы, благодаря которой
частицы даже очень твердых
грунтов могут быть раздви-
нуты. Заглатывание грунта во
время копания безусловно мо-
жет происходить, но оно имеет
второстепенное значение. Круп-
ным видам дождевых червей,
для того чтобы закопаться в
плотный грунт на всю длину
тела, достаточно 30—40 ми-
нут. Эта способность делать
Рис. 9. ДождевойЗ’чсрвь,
вбуравливающийся в зем-
лю. (По Фоке).
А — сокращением кольцевой
мускулатуры стенки тела червь
выбрасывает передний конец
тела вперед и погружает его в
почву; Б — сокращением про-
дольной мускулатуры стенки
тела ряд передних сегментов
втягивается в образовавшуюся
ямку.
ходы в грунте, позволяющая дождевым червям про
никать глубоко в землю, подчас на глубину 2 м и
более, в значительной мере и определяет космическую
роль дождевых червей как почвообразователей. Для
этого нужна большая мощность мускулатуры, каковой
онп и обладают. Мускулатура стенки тела составляет
38—44% от объема тела, а у наиболее сильных видов
цифра эта поднимается до 50%. В этом отношении черви
из беспозвоночных уступают только пиявкам, у которых
мускулатура тела может составлять до 65 % от объема тела.
На поверхности земли и внутри готовых подземных
ходов червь, так же как и при закапывании, двигается
путем закономерно чередующихся сокращений продоль-
ной и кольцевой мускулатуры, комбинируемых с движе-
нием щетинок (перистальтические движения). В спокой
ном состоянии черви передвигаются довольно медленно,
но при сильных раздражениях могут очень быстро со-
кращаться, совершая даже нечто вроде скачков, осо-
бенно когда приходится спасаться от преследования.
В этих движениях особую роль приобретает продольная
мускулатура, содействующая скорости поступательного
движения. Червп могут довольно быстро двигаться
вверх в вертикальных ходах, сделанных ими в грунте.
Опыты в стеклянных трубках с видами родов Lumbricus
и Allolobophora показали, что черви опираются спинной
поверхностью тела на твердую поверхность трубки.
Кроме того, движению червя помогает рот, действую-
щий как присоска (Japp, 1956).
Этим оправдывается не только несравненно бблыпая
толщина продольной мускулатуры по сравнению с кольце-
вым слоем, но и особенности ее строения. У многих видов
в продольной мускулатуре наблюдается своеобразная
упорядоченность расположения мышечных волокон. По-
следние укреплены на тяжах соединительной ткани парал-
лельными рядами, благодаря чему на поперечном разрезе
они кажутся составленными в «елочку». Такое располо-
жение волокон мускулатуры называется перистым. Оно
наблюдается не у всех видов; многим видам свойственно
обычное, пучковое расположение волокон продольной
мускулатуры.
Для эффективности работы мускулатуры имеет зна-
чение то обстоятельство, что под стенкой тела находится
полость, наполненная жидкостью. Эта полость сходна
по происхождению и характеру с брюшною полостью
высших позвоночных и человека, т. е., так же как у них,
в ней помещаются внутренности и она выстлана плоским
эпителием, носящим название «перитонеальный». У чер-
вей полость тела поделена соответственно сегментам тела
межсегментными перегородками. Кроме того, полость тела
делится на правую и левую стороны брыжейкой, соеди-
няющей брюшную сторону тела с кишечником. В общем,
тело червя представляет собой как бы две трубки, вложен-
ные одна в другую: стенка наружной трубки — стенка
тела, стенка внутренней — кишечник. Промежуток между
20
ними занят полостью тела, наполненной жидкостью.
Все жидкости, как известно, очень упруги и практически
несжимаемы при сколь угодно больших давлениях.
Поэтому полостная жидкость является антагонистом
действия мускулатуры и в известной степени заменяет
червю отсутствующий у него скелет. Прп сокращении
мускулатуры стенки тела давление на нее изнутри со сто-
роны полостной жидкости (тургор) возрастает, и, вслед-
ствие несжимаемости ее, поверхность червя приобретает
свойства упругого твердого тела. Это помогает ему при
движении, а особенно при рытье подземных ходов;
передним концом тела червь вбуравливается в грунт
наподобие твердого клина.
Упомянем еще раз, что при движениях дождевых чер-
вей очень большое значение имеет комбинированное
действие мускулатуры стенки тела и щетинок. Особое
значение работа щетинок (кроме вбуравливания в землю)
приобретает при лазапии по крутизне. Известно, что
многие виды червей могут влезать на деревья, что их на-
ходят в больших бочках, поставленных для собирания
дождевой воды, пли в зрелых кочнах капусты под наруж-
ными листьями, либо в середине кочна и т. д.
4. КИШЕЧНИК И ПИТАНИЕ
Рот, находящийся на переднем конце тела, ведет
в небольшую ротовую полость со складчатыми стенками,
за которой следует мускулистая глотка (рис. 10). Благо-
даря тому, что глотка соединена сложным переплетом
мышечных волокон со стенкой тела, она не только со-
вершает глотательные движения и прессует заглатывае-
мые вещества, но может также выворачиваться наружу
через широко открытый рот. Эти движения допускают
захватывание таких предметов, как листья, камешки
и пр., употребляемых для питания пли для других целей.
В толще глоточной стенки и за ее пределами находятся
многочисленные глоточные железы, протоки которых
открываются непосредственно в глотку или в особый
карман в спинной утолщенной части стенки ее. Глоточные
железы выделяют слизистую жидкость, обволакивающую
заглатываемые пищевые частицы. В этом отношении их
деятельность сходна с деятельностью слюнных желез
других животных. Но, кроме того, глоточные железы
вырабатывают вещество, переваривающее белки; оно ак-
тивно в щелочной среде и сходно по своему действию
с ферментом, поступающим в кишечник из поджелудочной
железы у позвоночных. Таким образом, химическая об-
работка белков начинается у дождевых червей уже в рото-
вой полости, что, вероятно, связано с необходимостью
возможно более полного извлечения белковых веществ
Рис. Ю. Схема строения переднего конца тела дождевого
черня. (По Фюллеру).
1 — надглоточный ганглий; 2 — рот; 3 — глотка; / — брюшная
нервная цепочка: 5 — пищевод с отверстиями известковых желеа
в 9-м—11-м и 13-м сегментах; 6 — зоб; 7 — мышечный желудок;
8 — средняя кишка: 9 — брюшной со?уд; 10 — спинной сосуд;
11 — кожный эпителий; 12 — мышцы стенки тела; 13 — места ответ-
влений кольцевых сосудов от спинного сосуда в 7-м—11-м сегментах;
14 — глоточный карман. Половая система не изображена.
из пищи, как правило, исключительно бедной этими ве-
ществами.
Глотка переходит в пищевод (рис. 10). Это догольно
узкая цилиндрическая трубка, стенки которой обладают
хорошо развитой мускулатурой. По бокам пищевода
имеются 1—3 пары боковых карманов (рис. 10) — так
называемые известковые железы. У некоторых видов они
расположены в толще стенки пищевода и потому снаружи
незаметны. Эти железы называются известковыми вслед-
ствие того, что в них под микроскопом находят кристаллы
углекислой извести. Что эти железы вырабатывают из-
весть — доказывается фактом значительного обогащения
ею пищевых масс при прохождении их через кишечник
(количество углекислой извести в содержимом кишечника
может возрастать от 0.8 до 1.3—1.8%). Предполагалось,
что роль этих желез состоит в нейтрализации кислот,
содержащихся в заглатываемом грунте. Это предполо-
жение стоит в хорошем соответствии с указанной выше
необходимостью наличия щелочной среды для деятель-
ности пищеварительных ферментов. Однако вряд ли
этим исчерпывается роль известковых желез. Относительно
их функции существует много других предположений,
притом самых разнообразных; уже это показывает, что
функцию известковых желез приходится считать не-
выясненной.
За- пищеводом находится объемистое расширение ки-
шечной трубки — так называемый зоб (рис. 10), занимаю-
щий 2—3 сегмента. В нем накопляется заглоченная пища,
которая оттуда порциями поступает в следующие отделы
кишечника. При отсутствии такого приспособления орга-
низм не успевал бы справиться с переработкой поступаю-
щего материала. Зоб имеет довольно тонкие эластичные
стенки, благодаря чему он хорошо растягивается.
Непосредственно сзади зоба находится другое расшире-
ние кишечной трубки — мышечный желудок. Внутри
он выстлан эпителием с толстой кутикулой, а стенка его
состоит из кольцевого и продольного слоев мускулатуры,
причем особенно сильно развит внутренний, кольцевой
слой, имеющий «перистую» структуру, наподобие продоль-
ного слоя мускулатуры стенки тела. Задача желудка —
растирание пищи; в этом процессе главную роль играет,
так же как у кур и других зерноядных птиц, трение друг
о друга минеральных частиц грунта, между которыми на-
ходятся органические пищевые вещества. Дарвин наблю-
дал, что песчинки и кусочки кирпича, прошедшие через
кишечник дождевых червей, принимают вместо угловатой
окатанную форму. Имеются новые наблюдения и опыты,
доказывающие значение минеральных частиц грунта для
перетирания пищи в кишечнике червей; при их отсутствии
(например, если поместить червей в торф) они голодают,
несмотря на обильный корм в виде листьев (Зражев-
ский, 1953).
За мышечным желудком следует средняя кишка, ко-
торая идет до заднего конца тела.
По всей длине средней кишки тянется глубокая спин-
ная складка, или тифлозол, благодаря которой на попе-
23
речных разрезах контур нолости кишечника принимает
подковообразные очертания (рис. 11). Физиологическое
аначение этой своеобразной черты организации кишечника
ясно: таким путем достигается увеличение всасывающей
Рис. И. Поперечный разрез через спинную часть кишечника
дождевого червя с тифлозол ом. (Из Штольте).
I - - хлорагогенная ткань; 2 — продольная мускулатура кишечника;
J ~ кольцевая мускулатура кишечника; 4 — эпителий кишечника;
з - кровеносный сосуд; 6 — мелкие кровеносные сосуды; 7 — спин-
ной кровеносный сосуд; 8 — кольцевые сосуды.
поверхности кишечника. Стенка кишечника содержит
большое количество железистых клеток, вырабатываю-
щих слизистый секрет и пищеварительные ферменты.
Среди последних, как и в глотке, имеются ферменты,
переваривающие белки, и, кроме того, ферменты, превра-
щающие крахмал в сахара (мальтозу и глюкозу); в ки-
24
шечнике происходит также превращение жиров в раство-
римое состояние. Таким образом, у червей, как и у поз-
воночных, питательные вещества в виде растворов вса-
сываются стенкой кишечника. Продвижение пищи со-
вершается действием мускулатуры кишечника, которая
состоит из внутреннего кольцевого и наружного продоль-
ного слоев мышц (следует обратить внимание на то, что
расположение слоев здесь обратное таковому в стенке
тела). У некоторых видов имеется несколько слоев мышц
в стенке кишечника.
На последних 10—15 сегментах тела кишечник лишен
спинной складки, а его эпителий приобретает реснички.
Эта часть называется задней кишкой. Всасывания в ней
уже не происходит, а только совершается процесс форми-
рования комочков испражнений, т. е. тех копролитов,
которые имеют столь важное значение для почвенной
структуры. На последнем сегменте тела кишечник откры-
вается наружу заднепроходным отверстием, имеющим
вид вертикальной щели.
Интересен спор двух знаменитых натуралистов прош-
лого века по вопросу о пище дождевых червей —Этьена
Клапареда (Франция), превосходного знатока беспозво-
ночных (в частности, кольчатых червей), и Чарльза Дар-
вина (Англия). Клапаред нашел в разных частях кишеч-
ника дождевых червей остатки измельченных листьев,
перемешанных с землей, и на этом основании полагал,
что землю черви заглатывают только с той целью, чтобы
заглоченные ими растительные остатки лучше растирались.
Дарвин, не отрицая того, что черви питаются опавшими
листьями и другими растительными остатками, в то же
время утверждал, что и заглатываемая земля исполь-
зуется ими для питания. Он паблюдал, что обильно за-
селены червями и те места, где они могут питаться только
землей, богатой органическими веществами (например,
аккуратно выметаемые дворы). Все дальнейшие исследо-
вания подтвердили правильность наблюдений Дарвина.
Вопрос о способности червей выбирать себе пищу мы
затронем в дальнейшем, когда будем говорить о функциях
их нервной системы и органов чувств.
Большое значение имеет количество земли, поглоща-
емой и перерабатываемой в кишечнике дождевых червей.
Оно оказалось огромным: взвешиванием копролитов уста-
25
новлено, что черви, населяющие культурные почвы,
пропускают через кишечник за 24 часа количество грунта,
равпое весу их тела.
Для того чтобы закончить рассмотрение кишечника,
упомяпем о характерной ткани, облекающей снаружи всю
среднюю кишку и спинной кровеносный сосуд и запол-
няющей спинпую складку кишечника. При вскрытии
живого или только что убитого дождевого червя привле-
кают внимание желтый цвет и рыхлая бархатистая поверх-
ность средней кишки, на которой контрастно выступают
красные кровеносные сосуды. Эта ткань называется хлора-
гогенной, или желтой. Связь ее с кишечником чисто
топографическая: она — измененная часть выстилки по-
лости тела (перитонеального эпителия), прилегающая
к кишечнику. Желтая ткань состоит из круппых клеток,
плазма которых заполнена каплями веществ, имеющих
желтоватую окраску. Происхождение и природа этого
вещества, а вместе с тем и функция самой ткани, не вполне
ясны. Одни исследователи считают эту ткань местом на-
копления запасных питательных материалов, наподобие
жировой ткани позвоночных. И действительно, вклю-
чения клеток желтой ткапп содержат жир, белок и ве-
щество, сходное с гликогеном (животным крахмалом).
В то же время известно, что эта ткань содержит большое
количество мочевой кислоты, что инородные вещества,
введенные в виде растворов в полость тела (краски),
скапливаются в клетках хлорагогенной ткани и что ко-
нечные азотистые продукты обмена веществ, подлежащие
выведению из организма, обычно имеют желтый или
бурый цвет. Все это заставляет думать о выделительной
функции этой ткани. Очень вероятно, что наряду с на-
капливанием запасных питательных веществ клетки жел-
той ткапи имеют способность извлекать из циркулирую-
щей в ней крови и жидкости, заполняющей полость тела,
отбросы, образующиеся в процессе обмена веществ. Попа-
дая внутрь клеток желтой ткани, эти вещества выклю-
чаются из тока крови и делаются безвредными. Постепенно
накапливаясь в клетках этой ткани, они могут длительно
там оставаться, но могут и выделяться из организма, так
как клетки желтой ткани часто отрываются и попадают
в полость тела, а оттуда выводятся наружу вместе с вы-
брызгиванием полостной жидкости через спинные поры.
26
а. КРОВЕНОСНАЯ СИСТЕМА. ФУНКЦИИ РАСПРЕДЕЛЕНИЯ
ПИТАТЕЛЬНЫХ ВЕЩЕСТВ И КИСЛОРОДА
Распределение пптательпых веществ, всосанных по-
верхностью кишечника, осуществляется у дождевых чер-
вей при помощи высокоразвитой кровеносной системы.
Схема расположения главных сосудов ее следующая
(рис. 8, 10 и 12). Вдоль всего тела проходят спинной (над
кишечником) и брюшной (под кишечником) сосуды.
Спинной сосуд снабжен мускулатурой, которая волно-
образными сокращениями гонит кровь от заднего конца
тела к переднему. В нескольких передних сегментах
(с 7-го по 11-й или, у других видов, с 7-го по 13-й) спинной
сосуд сообщается 5—7 парами поперечных сосудов с брюш-
ным сосудом. Эти сосуды снабжены особенно сильной
мускулатурой п называются сердцами. Они вполне оправ-
дывают это название, так как служат главным аппаратом,
обеспечивающим кровообращение. Кровь, поступающая
из сердец в брюшной сосуд, движется по направлению
к заднему концу тела. По пути она поступает в сосуды,
питающие стенку тела’, а также в сосуды, идущие к кишеч-
нику, к выделительным органам (рис. 13), а в соответ-
ственных сегментах и к половым органам.Во всех этих
частях тела сосуды распадаются на сеть микроскопических
капилляров. Из капилляров кровь поступает в попереч-
ные сосуды, которые собирают в конечном счете кровь
из всего тела в спинной сосуд.
Имеются и другие продольные и-поперечные сосуды,
которые можно видеть на рис. 8, 10, 12 и 13; на них мы
останавливаться не будем. Особое значение имеет густое
сплетение мелких сосудов вокруг кишечника (рис. 13).
Сюда поступают питательные вещества, всосанные ки-
шечником, и отсюда они разносятся по всему телу. За-
метим, что мускулатурой, хотя и не столь сильно развитой,
как в спинном сосуде и сердцах, обладают почти все со-
суды, чем предотвращается возможность застоя крови
в периферических частях кровеносной системы.
Кровь у дождевых червей, как уже было отмечено,
красная. Этот ее цвет обусловлен наличием вещества,
очень близкого к гемоглобину крови позвоночных.1
Однако у червей опо содержится не в тельцах крови,
1 Это вещество получило название «гемокруорпн».
27
Рис. 12. Дождевой червь, вскрытый со спиннои
стороны. (Из Н. А. Холодковского).
1 — надглоточный ганглий; 2 — глотка; 3 — сократимый коль-
цевой сосуд; / — пищевод; 6 — известковые железы; б — зоб;
7 — мускулистый желудок; 8 — нефридий; 9 — средняя кишка;
10 — спинной кровеносный сосуд; 11 — брюшная нервная
цепочка; 12 — яйцевая воронка; 13 — семепровод; 14 — семен-
ной мешок; 16 — яичник; 1в — семеприемники; 17 — семенник.
а растворено в жидкой части крови (кровяной плазме).
У дождевых червей имеются в крови только бесцветные
клетки нескольких типов, в общем тех же, что и типы
бесцветных клеток крови у позвоночных.
Как известно, гемоглобин у позвоночных обеспечи-
вает транспорт кислорода из органов дыхания ко всем
Рис. 13. Схема кровообращения в сег-
менте средней части тела дождевого
червя на поперечном разрезе.
(Из Штольте).
1 — спинной сосуд; У — спинно-кишечный
сосуд; з — кольцевой сосуд; / — сосуд тиф-
лозоля; 5 — кишечное сплетение; 6 — сосуды
нефридия; 7 — брюшной сосуд; 8 — сосуды
брюшной нервной цепочки. Черной линией
обозначено кишечное сплетение.
живым клеткам тела. Ту же роль играет вещество, сход-
ное с гемоглобином, и у дождевых червей. Однако спе-
циальных дыхательных органов у них нет: они дышат
всей поверхностью тела. Тонкая кутикула и нежность кож-
ных покровов дождевых червей, а также богатая сеть
кожных кровеносных сосудов, хорошо обеспечивают
возможность поглощения кислорода из окружающей
среды. Но отметим, что кутикула дождевых червей хорошо
смачивается водой и кислород, по-видимому, у них должен
29
сначала раствориться в воде, смачивающей кожные по-
кровы. Это влечет за собой необходимость сохранения
кожи во влажном состоянии. Уже это одно делает ясным,
насколько важными для жизни червей являются условия
влажности внешней среды. С подсыханием кожи дыхание
для них делается невозможным. Однако при попадании
в условия недостатка влаги в почве червь может долгое
время бороться с этим, используя запасы воды, имею-
щиеся внутри тела. В этих случаях ему на помощь при-
ходят кожные железы (см. стр. 15), а в случае острого
Рис. 14. Капилляры в кожном эпителии дождевого червя.
(Из Штольте).
/ - нпидермие; 2 — кольцевая мускулатура; 3 — продольная мускулатура.
недостатка влаги он утилизирует для этого полостную
жидкость, выбрызгивая ее из спинных пор.
Поглощение кислорода поверхностью тела облегчается
наличием очень богатой сети кровеносных капилляров,
проникающих даже в кожный эпителий (рис. 14) Отсюда
кровь через сосуды стенки тела и поперечные сосуды
попадает в главные стволы кровяного русла, чем дости-
гается снабжение кислородом всего тела. Красные тона
окраски большинства видов дождевых червей (не пигмен-
тация, см. стр. 15) определяются именно, наличием бога-
той сети кожных кровеносных сосудов.
Все это создает возможность жизни червей в условиях
очень пониженного содержания кислорода. В этом отно-
шении они приближаются к некоторым из своих отдален-
ных пресноводных родичей — червям-трубочникам (Ти-
bifex tubifex, Limnodrilus hoffmeisteri и др.), которые,
30
обитая в глубоким илу, могут выносить почти полное
отсутствие кислорода. Относительно дождевых червей
есть наблюдения, что они могут жить при содержании
кислорода в окружающем их воздушном пространстве,
равном 2.5% (как известно, обычно в воздухе его содер-
жится 21%). Даже при наличии 0.4% кислорода в воз-
духе черви могут поглотить половину количества кисло-
рода, необходимого им для поддержания жизни, и могут
находиться в этих условиях довольно долго. Кроме того,
попав в бескислородную среду, черви могут переключаться
на особый тип обмена веществ, в котором источником
энергии для жизненных процессов служат не реакции
окисления (для которых и нужен кислород), а распад
крахмалоподобного вещества — гликогена, который со-
вершается и в бескислородной среде. Однако запасы гли-
когена у червей не особенно велики, и, кроме того,
при этом способе обмена веществ выделяются кислоты,
вредно действующие на организм червей.
Находясь под водой, черви столь же хорошо могут
поглощать кислород, как и в воздушной среде. Известно,
что они могут жить в воде месяцами при наличии нужного
им минимума кислорода и других необходимых для них
условий. Этот факт имеет большое значение для понима-
ния многих явлений из жизни дождевых червей.
6. ОРГАНЫ ВЫДЕЛЕНИЯ. ПОГЛОЩЕНИЕ
И ВЫДЕЛЕНИЕ ВОДЫ
Функция выделения выполняется у дождевых червей
(как у всех кольчецов) трубчатыми органами, располо-
женными попарно в каждом сегменте, кроме самых перед-
них. Эти органы называются нефридиями, что означает
по-гречески «орган подобный почке». Нефридии располо-
жены в полости тела по бокам от кишечника (рис. 8 и 12).
Каждый из них представляет собой извитую трубку, на-
чинающуюся внутри тела отверстием в полость тела,
расположенным на головчатом расширении, клетки ко-
торого снабжены ресничками. Это расширение называют
по аналогии со сходными образованиями у более прими-
тивных кольчецов воропкой (рис. 15). Почти непосред-
ственно за воропкой нефридий прободает межсегментную
перегородку и проникает в следующий сегмент тела.
31
Гам он сначала образует сильно извитую тонкую трубку,
которая переходит в более широкую среднюю часть нефри-
дия, снабженную ресничками. Затем нефридий, делая
несколько петель, переходит в выводную часть, которая
кончается на брюшной стороне тела наружным отверстием
(рис. 15), или нефридиальной порой. Снаружи ее найти
очень трудно, так как края ее всегда плотно сомкнуты.
Недалеко от нефридиальной поры имеется расширение
Рис. 15. Строение нефридия дождевого червя. (Из Штольте).
i — воронка; 2 — диссепимент: 3 — извитая часть трубки нефридия; 4 — рес-
ничная часть трубки нефридия; 5 — «мочевой пузырь»: в — пора нефридия;
7 — наружная петля.
нефридиальной трубки, представляющее собой нечто
вроде мочевого пузыря. Нефридии снабжены очень бо-
гатой сетью кровеносных сосудов. Кровь, выходящая из
нефридия, поступает в поперечный, а из него в спинной
сосуд (рис. 16).
Следует заметить, что у одного из дождевых червей
(Allolobophora antipae) нефридиальные трубки не откры-
ваются независимыми друг от друга порами, а их наруж-
ные части впадают в продольные выделительные каналы,
которые проходят справа и слева вдоль всего тела и на
заднем его конце впадают в кишечник недалеко от аналь-
ного отверстия. Таким образом, здесь намечается соеди-
нение выделительного аппарата в одно анатомическое
32
целое и устанавливается связь с задним отделом кишеч-
ника.
Клетки тонкой части нефрпдиальной трубки улавли-
вают из крови, циркулирующей вне сети нефридиальных
капилляров, продукты азотистого обмена, подлежащие
выведению из организма. Эти вещества попадают в по-
лость нефридпальпой трубки и здесь смешиваются с по-
лостной жидкостью, поступающей через воронку на внут-
Рис. 16. Кровеносные сосуды нефридия дождевого червя.
(Из Штольте).
I — воронка; В — брюшной кровеносный сосуд; 8 — дисссппмент; 4 — брюш-
ная нервная цепочка; л — поднервный сосуд; 6 — трубка нефридия с крове-
носными сосудами.
рением конце нефридия. Полостная жидкость также со-
держит продукты выделения, отмершие клетки, туда
попадают сношенные щетинки и т. д. Жидкость нефридиаль-
ной трубки биением ресничек гонится по направлению
к выводному концу, откуда периодически выбрасывается
через наружную пору сокращением мускулатуры стенки
тела (Roots, 1955).
Имеются данные о том, что опорожнение концевого
пузыря нефридия происходит один раз в три дня. Другие
наблюдения говорят о том, что за сутки червь весом
в 1. —1.8 г выделяет 0.82 см3 экскрета. Такие количества
его должны выделяться из тела несколько раз в сутки.
3 О. В. ЧеканоЕСкая
33
В экскрете содержатся в общем те же вещества, что и
у млекопитающих, а именно: мочевина, аммиак, креати-
нин, соли и пр., но в значительно более низких концентра-
циях. Однако нормальный экскрет червей содержит
0.3% белка, тогда как у высших животных белок в про-
дуктах выделения отсутствует.
Клетки средней части нефридиальной трубки обла-
дают способностью к фагоцитозу, т. е. к активному за-
глатыванию из полости тела нерастворимых в воде ве-
ществ (отмершие клетки, свернувшийся белок, бактерии
и пр.). Эти вещества накапливаются там неопределенно
долгое время. Санитарную службу этого рода несут и дру-
гие клетки внутри организма: амебоидные клетки крови,
клетки полости тела и вышеупомянутые клетки хлораго-
гепной, или желтой ткани (см. стр. 26). Особенно много
амебоидных клеток, наглотавшихся посторонних телец,
встречается в полости тела. Сюда они попадают, активно
выползая из сосудов, протискиваясь между клетками со-
судистой стенки. Из полости тела эти клетки удаляются
разными путями. Во-первых, они проползают через стенку
кишечника и, попадая в его полость, выводятся наружу
вместе с испражнениями (это наблюдалось много раз);
во-вторых, как уже упомянуто, они могут с полостной
жидкостью выходить через нефридии и, в-третьих, могут
выходить вместе с полостной жидкостью, выбрызгиваемой
через спинные поры. Вообще можно думать, что полостная
жидкость довольно быстро сменяется. Поэтому она п
приобретает у червей такое большое значение в процессе
выделения. Роль ее в жизни червей станет яснее после
ознакомления с водным режимом их тела.
Мы уже указывали на значение воды в теле червей,
когда говорили о роли полостной жидкости (98.8 % ее
состава — вода) при мышечной работе и о необходимости
увлажнения кожи для дыхания (стр. 30). Вода в тело
червей непрерывно поступает и выводится обратно в на-
ружную среду указанными выше путями. Таким образом,
тело червя и, в частности, полость тела все время пропо-
ласкиваются водой. Поэтому для нормального выполне-
ния указанных физиологических функций червям необ-
ходимы условия среды, которые обеспечивали бы поступ-
ление в их тело воды в значительно большем количестве,
чем у большинства наземных животных.
34
Каким образом вода поступает в тело червей?
Заметим прежде всего, что черви никогда не пьют.
Они поглощают воду всей поверхностью тела; вода про-
ходит сквозь покровы и мускулатуру, накапливаясь
в полости тела. При этом черви могут использовать воду
только в жидком состоянии. Червь в среде, содержащей
пары воды, может погибнуть от высыхания, если не имеет
иного источника влаги.
При нормальных условиях тело червей содержит около
84% воды. Несмотря па столь значительный запас воды,
он оказывается далеко не предельным. Если червю предо-
ставить возможность еще увеличить запас воды в своем
теле, он это тотчас же сделает. В этом легко убедиться,
если положить дождевых червей в поду. Через несколько
часов вес их возрастет на 10—12?о за счет воды, всосан-
ной поверхностью тела. После вынимания из воды червь
возвращается к исходному весу, причем это происходит
за очень короткий срок (1—2 часа). Удаление избыточной
воды из организма происходит очень своеобразным путем:
она всасывается клетками кишечника, из них поступает
в полость последнего п удаляется главным образом через
заднепроходное отверстие, а отчасти и через рото-
вое.
При нормальных условиях жизни в почве функция
удаления избытка воды лежит на нефридиях. Наличие
постоянного тока воды через организм путем всасывания
ее поверхностью тела и выделения ее в избытке почками —
явление, весьма обычное среди водных животных. Оно
несомненно унаследовано дождевыми червями от их вод-
ных предков.
Обмен веществ водных животных совершается при уси-
ленной циркуляции воды через их тело; им не может
угрожать недостаток воды, тогда как в почвенных усло-
виях при таком типе водного обмена достаточное коли-
чество влаги делается основным фактором, обеспечиваю-
щим возможность существования. Поэтому условия влаж-
ности почв имеют главное значение в вопросе о засе-
лении их теми пли иными видами дождевых чер-
вей.
О способности дождевых червей терять воду в периоды
засухи и зимовки, что связано с переходом их в состояние
скрытой жизни, см. далее (стр. 105).
з
35
7. НЕРВНАЯ СИСТЕМА И ОРГАНЫ ЧУВСТВ. РЕФЛЕКСЫ
Вдоль средней линии брюшной стороны тела у дожде-
вых червей под мускулатурой тянется нервный ствол,
называемый брюшной нервной цепочкой. В каждом сег-
Рис. 17. Нервная система передних сегментов дож-
девого червя. Вид со спинной стороны. (Из Штольте).
1 — надглоточный ганглий; 2 — нервы от окологлоточной ко-
миссуры; з — окологлоточные комиссуры; 4 — сегментальные
нервы; 3 — брюшная нервная цепочка; 6 — подглоточный ган-
глий; 7 — нервы головной лопасти; I—VI — 1-й—6-й сегменты.
менте тела имеется нервный узел; или ганглпй, представ-
ляющий собой скопление нервных клеток и отдающий
от себя 3 пары нервов. Ганглии соединены друг с другом
перемычками, коннективами, которые, кроме нервных
36
волокон, также содержат нервные клетки. В переднем
конце тела, в 3-м сегменте, брюшная нервная цепочка
разделяется на правую и левую глоточные коннективы,
образующие окологлоточное нервное кольцо, соединяю-
щееся с надглоточным, или головным ганглием (рис. 17).
Этот ганглий — парный, состоит из правой и левой по-
ловин, плотно соединенных друг с другом. Но в цтличие
от всех остальных нервных узлов, лежащих на брюшной
стороне тела под’жиптечпнком.'’этот ганглий находится
Рис. 18. Брюшная часть поперечного среза через дождевого червя.
(По Ретциусу).
1 — опорные волокна внутри брюшной нервной цепочки; 2 — нервные клетки
в ганглии брюшной нервной цепочки; 3 — нерв; 4 — продольная мускула-
тура; 5 — кольцевая мускулатура; в — кожный эпителий с чувствительными
нервными клетками; 7 — щетинка; 8 — брюшной кровеносный сосуд.
на спинной стороне тела и лежит над кишечником. Ганг-
лий этот можно противопоставить всем прочим и вслед-
ствие того, что морфологически он сравним с головным
мозгом вышестоящих форм (членистоногих). От него идут
вперед многочисленные нервные стволы, обильно разветв-
ляющиеся и образующие густые нервные сплетения в пер-
вых трех сегментах. Под глоткой, в месте расхождения
глоточных коннектив, лежит подглоточный ганглий, пред-
ставляющий собой результат слияния нескольких ганг-
лиев брюшной нервной цепочки.
Как видно на поперечных разрезах, нервные клетки
лежат по периферии ганглия, а средняя его часть занята
сплетением отростков нервных клеток (рис. 18). В брюш-
ной нервной цепочке обращают на себя внимание три
87
очень толстых волокна, пробегающих по всей длине тела
червя под соединительнотканной капсулой нервной це-
почки на ее спинной стороне. Это так называемые нейро-
хорды, которые еще совсем недавно принимали за гигант-
ские нервные волокна. Однако теперь окончательно вы-
яснено, что они являются своеобразными опорными обра-
зованиями (Невмывака, 19476). Эти образования и по
строению, и по функции, и по положению между нервной
системой и кишечником сходны с хордой позвоноч-
ных.
Нервы, отходящие от ганглиев брюшной цепочки,
содержат двигательные волокна, заканчивающиеся в мус-
кулатуре, и чувствительные, по которым раздражения
поступают в нервную систему с периферии. Тела чувстви-
тельных нервных клеток находятся на периферии, в том
числе в наружном эпителии (рис. 18). Нервные клетки
здесь стоят в ряду эпителиальных клеток. Этот чрезвы-
чайно древний тип соотношений элементов нервной си-
стемы сохранился у дождевых червей от их далеких пред-
ков, примитивных многоклеточных животных. Очень инте-
ресно, что чувствительными нервными клетками здесь ста-
новятся не только клетки наружного эпителия, но, как это
недавно установлено, также и клетки кишечника, проис-
ходящие из внутреннего зародышевого листка (Невмыва-
ка, 1947а).
Чувствительные нервные клетки и их окончания
имеются и в других местах тела. Ими богато снабжены
также нефридии, щетинковые мешки и другие органы. Та-
ким образом, у дождевых червей, как и у высших живот-
ных, работа внутренних органов протекает под контролем
регулирующей и централизующей роли нервной системы.
Из рефлексов дождевых червей лучше всего известны
наблюдаемые в акте ползания. При движении червя
по всей длине тела, от переднего конца до заднего, пробе-
гают перистальтические волны комбинированных сокра-
щений мускулатуры. Они следуют друг за другом, и каждая
следующая волна может возникнуть еще задолго до того,
как первая дойдет до заднего конца тела. Казалось бы
очевидным, по аналогии с высшими животными, что при-
чиной этих воли сокращений служит последовательная
передача раздражений вдоль по брюшной нервной цепочке.
Однако, к удивлению исследователей, выяснилось, что
38
перерезание брюшного нервного ствола и даже выреза-
ние из нее нескольких нервных узлов не останавливают бе-
гущих волн сокращений мускулатуры: волна сокращения
проходит через место повреждения так же, как это было
и у нормального червя. С тем же результатом можно
вдобавок к разрыву брюшной нервной цепочки удалить
мускулатуру нескольких сегментов или повредить ее
кислотой.
Анализ этих и аналогичных
опытов показал, что поступатель-
ное движение червя представляет
собой длинную цепь рефлектор-
ных актов, в которой каждый
сегмент является в значительной
мере автономной физиологической
единицей. Раздражения с перифе-
рии приводят к сокращению му-
скулатуры данного сегмента.
Вследствие этого сокращения раз-
дражаются периферические аппа-
раты в соседнем сегменте, которые
вызывают и в нем сокращения
и т. д. Таким образом, комбиниро-
ванные сокращения мышц в каж-
дом сегменте могут представлять
собой самостоятельный рефлекс,
начинающийся с возбуждения чув-
ствительных клеток на перифе-
рии и кончающийся эффектом
сокращения мускулатуры этого
сегмента. Это наиболее прими-
тивный тип реакции на внешние
Рис. 19. Схема бисег-
ментального рефлекса
у дождевого червя.
(Из Штольте).
1 — брюшная нервная це-
почка; 2 — мускулатура
стенки тела; з — чувстви-
тельная нервная клетка,
воспринимающая импульс;
4 — оконч ние двигательной
нервной клетки в мышцах;
5 — диссепимент; 6 — нерв-
ная клетка, передающая
раздражение в соседний
сегмент.
воздействия. Некото-
рым его усложнением является передача полученного
раздражения по нервной цепочке в соседний задний
сегмент тела, в ответ на которое мускулатура этого сег-
мента сокращается. На рис. 19 изображена схема рефлекса
при дугообразном сгибании червя, когда волна сокращений
мускулатуры идет по одной стороне тела. Этот рефлекс
и является основным при осуществлении поступательного
движения червя вперед. Этот способ передачи раздражений
по телу, как указано выше, говорит о слабой централи-
зации его нервной системы.
39
О том же говорят и опыты с удалением надглоточного
ганглия. Выше отмечено, что морфологически надглоточ-
ный ганглий можно сопоставить с головным мозгом выше-
стоящих форм (членистоногих). У многих морских коль-
чецов надглоточный ганглий имеет довольно сложное
устройство. Однако у наземных червей надглоточный ганг-
лий подвергся упрощению и физиологическая роль его
очень невелика. После удаления надглоточного ганглия
удается отметить только некоторое общее расслабление
мускулатуры переднего отдела тела, изменения в восприя-
тии света; возможно, он играет роль и при размножении.
Но в движениях червя существенных изменений после
заживления раны подметить не удается: червь также
зарывается в грунт, также избегает опасности и осуще
ствляет все те довольно сложные рефлекторные реак-
ции, с которыми мы ознакомимся далее. Особенно удиви-
тельно, что способность к «научению», т. е., по современ-
ной терминологии, к условным рефлексам, у червей, ли-
шенных надглоточного ганглия, не пропадает.
Несколько большее значение имеет подглоточный
ганглий, так как после его удаления червь лишается
многих присущих ему способностей: очень страдают его
вкусовые способности (стр. 45).
Было бы ошибочно думать, что слабая централизация
нервной системы и относительная автономия отдельных
сегментов, обнаруживаемые при поступательном движе-
нии червя, означают отсутствие реакций организма как
целого. Заранее можно сказать, что таких реакций не
может не быть, и, действительно, их очень легко обнару-
жить. При слабом раздражении заднего конца тела (при
легком прикосновении) червь ползет вперед, при раз-
дражении спереди он быстро весь сокращается и ползет
в другую сторону, при сильном раздражении в любом
месте червь начинает сильно сокращаться дугообразно,
в разных направлениях (так называемые гимнастические
движения); черви обнаруживают быстрые реакции на
свет, на запахи и т. д. Таким образом, указанные выше
несовершенство нервной системы и ее слабая централи-
зация обнаруживаются только при внимательном наблю-
дении и в специально поставленных экспериментах.
Итак, нам известно, что червь располагает довольно
богатым арсеналом возможностей для осуществления тех
Рис. 20. Светочувствитель-
ная клетка из кожного эпи-
телия дождевого червя.
(Из Стефенсона).
1 — ядро; 2 — линза;
з - нерв; 4 — сеть нейрофи-
брилл.
или иных реакции на изменения, происходящие в окру-
жающей их среде.
Рассмотрим теперь, каким путем он может распозна-
вать эти изменения. Средствами для этого являются ор-
ганы чувств.
Как уже сказано, вся поверхность тела червя покрыта
огромным количеством чувствительных нервных клеток.
Эти клетки служат органами
осязания, развитого у червей
очень сильно. Известно, что
достаточно, осторожно прибли-
зившись, слабо дунуть на
червя, чтобы он ответил резким
сокращением продольной му-
скулатуры; с помощью такого
движения он прячется в норку.
Вдобавок к чувствительным
нервным клеткам в наружном
эпителии между клетками
имеется очень большое количе-
ство свободных нервных окон-
чаний, которые, скорее всего,
также обеспечивают функцию
осязания.
Как известно уже более ста
лет назад, дождевые черви, не-
смотря на отсутствие глаз, хо-
рошо воспринимают свет. Во-
сприятие света производится
особыми светочувствительными
клетками, которые по большей части располагаются
поодиночке между клетками наружного эпителия (рис. 20).
Внутри этих клеток, кроме ядра и густой сети тончайших
волоконец — нейрофибрилл, имеется прозрачное свето-
преломляющее тельце бобовидной или удлиненной формы;
его называют линзой по аналогии с хрусталиком глаза
более высоко организованных животных. От тела клетки
отходит нервный отросток, вступающий в подкожное
нервное сплетение и связывающий ее с центральной
нервной системой. Такая клетка несомненно представляет
собой простейший глаз, как бы изолированную и авто-
номную клетку сетчатки глаза высших животных. Свето-
41
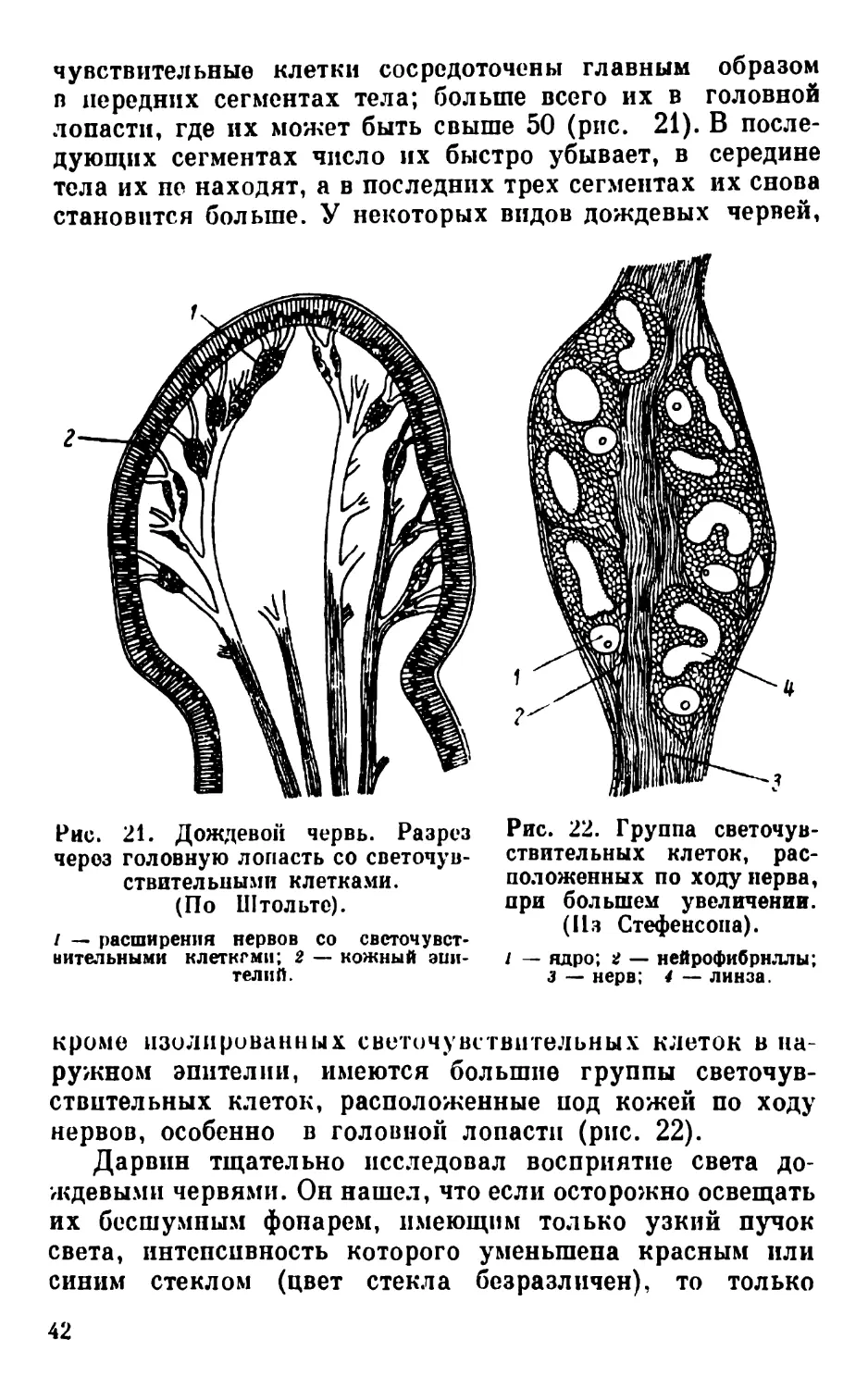
чувствительные клетки сосредоточены главным образом
в передних сегментах тела; больше всего их в головной
лопасти, где их может быть свыше 50 (рис. 21). В после-
дующих сегментах число их быстро убывает, в середине
тела их не находят, а в последних трех сегментах их снова
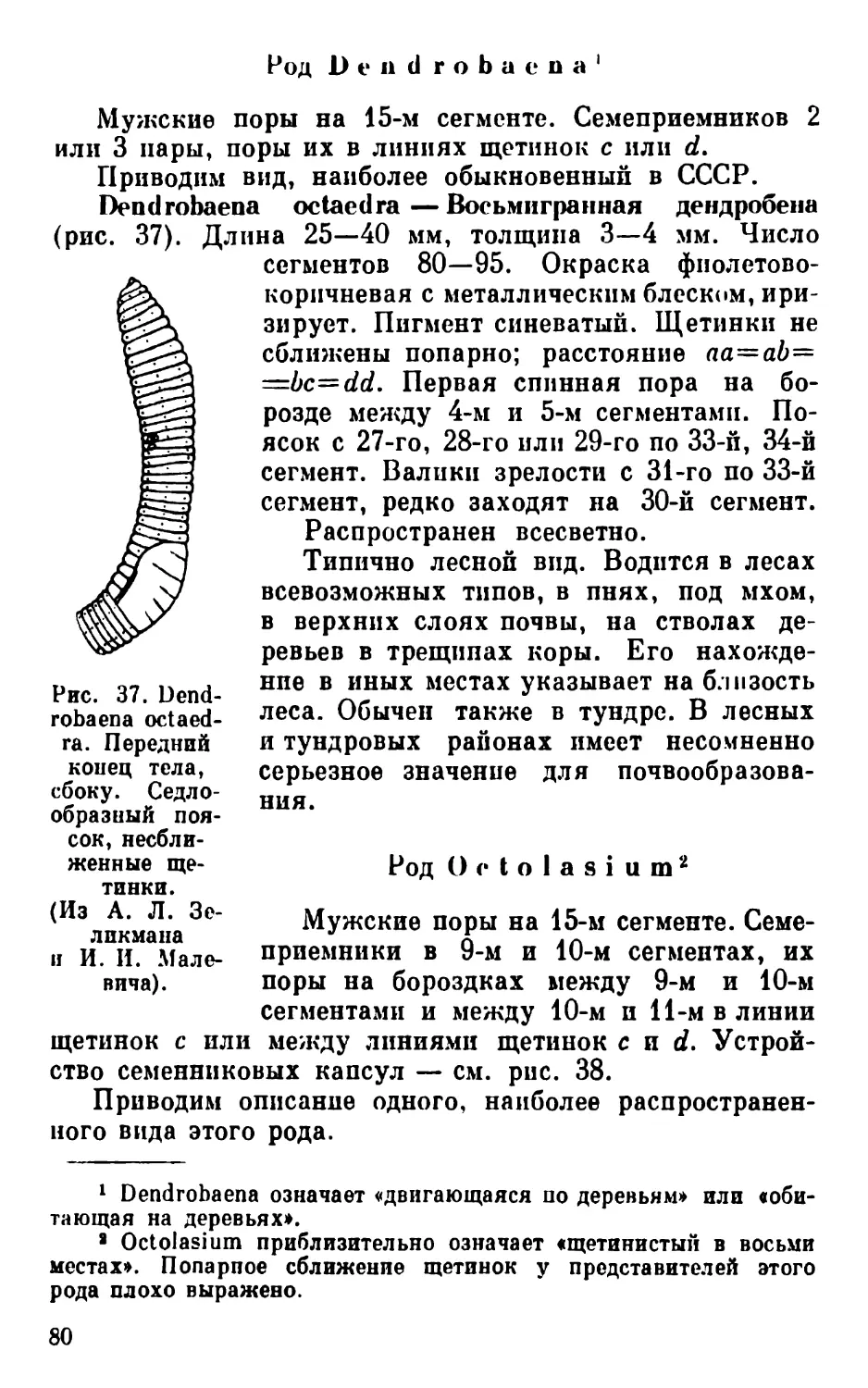
становится больше. У некоторых видов дождевых червей,
Рис. 22. Группа светочув
ствительных клеток, рас
положенных по ходу нерва,
при большем увеличении.
(Из Стефенсона).
i — ядро; 2 — нейрофибрнллы;
з — нерв; 4 — линза.
Рис. 21. Дождевой червь. Разрез
через головную лопасть со светочув-
ствительными клетками.
(По Штольте).
/ — расширения нервов со светочувст-
вительными клеткгми; 2 — кожный эпи-
телий.
кроме изолированных светочувствительных клеток в на-
ружном эпителии, имеются большие группы светочув-
ствительных клеток, расположенные под кожей по ходу
нервов, особенно в головной лопасти (рис. 22).
Дарвин тщательно исследовал восприятие света до-
ждевыми червями. Он нашел, что если осторожно освещать
их бесшумным фонарем, имеющим только узкий пучок
света, интенсивность которого уменьшена красным или
синим стеклом (цвет стекла безразличен), то только
42
очень немногие черви реагируют, а именно: уходят в свои
норы.1 Дарвин вел наблюдения над теми видами, которые
ночью выходят из нор в поисках пищи или для спари-
вания; это большой красный червь (Lumbricus terrestris),
длинный червь (Allolobophora longa) и некоторые другие.
Задний конец у них обычно остается в норке. При более
сильном освещении (особенно точные результаты полу-
чались при концентрации световых лучей с помощью
увеличительного стекла) черви, быстро сокращая про-
дольную мускулатуру, скрываются в норках, «как кро-
лики», замечает Дарвин, приводя выражение одной»
из своих приятелей, наблюдавшего за его опытами. При
этом Дарвин доказал, что реагируют черви именно на свет,
а не на лучистую теплоту, исходящую от источника света.
Опыты с приближением к червям накаленных кусков
железа показали, что они мало чувствительны к лучистой
теплоте. Однако, когда черви чем-нибудь «заняты», т. е.
когда они втаскивают листья в своп поры, едят и пр.,
то они не замечают света, даже когда свет концентрировали
на них при помощи большого зажигательного стекла.
Не реагируют на свет онц и во время спаривания. В даль-
нейшем было доказано, что очень слабый свет может
привлекать червей, так как они двигаются по направле-
нию его источника.
Способность ощущать свет играет очень большую роль
в жизни червей, так как солнечный свет на них действует
губительно (черви очень чувствительны к ультрафиоле-
товой части солнечного спектра). Реакция ухода в тем-
ноту спасает им жизнь (Smith, 1902).
Специальных органов слуха у червей нет. На очень
сильные звуки, передаваемые по воздуху, черви не реа-
гируют в том случае, если не вибрирует твердый субстрат,
с которым они имеют соприкосновение. Но дрожание твер-
дых тел, с которыми они связаны, вызываемое звуками,
они воспринимают очень тонко. Например, по наблюде-
ниям Дарвина, «когда горшок, заключавший в себе пару
червей, оказавшихся совершенно бесчувственными к зву-
кам рояля, был поставлен на самый инструмент, то при
взятии ноты до в басовом ключе оба моментально спрята-
1 Однако имеются наблюдения, что черпи более чувствительны
к синему свету, чем к красному.
43
лись в норы. Через некоторое время они снова показались
на поверхности ее, но, когда была взята нота соль в скри-
пичном ключе, они снова удалились». Эти вибрации крышки
рояля ^воспринимались,
Рис. 23. Поперечный разрез
через дождевого червя. Чув-
ствительная почка в кожном
эпителии. (По Гессе).
1 — чувствительные волоски;
2 — чувствительные клетки;
з — спинно-брюшное мышечное во-
локно; 4 — продольные мышцы;
5 — чувствительный нерв; 6 — ка-
пилляр; 7 — кольцевые мышцы;
8 — эпителиальные клетки; 9 — ку-
тикула.
очевидно, органами осязания
червей.
На высокоразвитом ося-
зании основан метод сбора
червей, практикуемый во
Флориде: доска или палка
втыкается в грунт, обильно
населенный червями, и по
ее верхнему краю водят дру-
гой палкой, как смычком
по скрипке (этот способ и
называется там «скрипич-
ным»), Пишут, что черви при
этом покидают норки и во
множестве выходят на по-
верхность.
В научной литературе из-
давна от времени до времени
появлялись сообщения о зву-
ках, издаваемых дождевыми
червями. Действительно, при
трении тела и щетинок о
грунт, при поршневых дви-
жениях во влажных норках,
при растирании пищи в глот-
ке, при затаскивании листьев
и камешков и т. д. могут воз-
никать звуки. Они тем лучше
ощутимы, чем червей больше
и чем они крупнее. Но очень
сомнительно, чтобы эти звуки
имели какое-нибудь биологи-
ческое значение.
Кроме чувствительных
нервных клеток, нервных
окончаний и светочувствительных клеток в наружном
эпителии рассеяно большое количество органов, пред-
ставленных комплексами клеток. Их называют иногда
чувствительными почками. Несколько десятков чувстви-
44
тельных клеток образуют цилиндрический или яйцевидный
комплекс (рис. 23). Это — чувствительные нервные клетки
и длинные нервные отростки, которые идут в брюшную
нервную цепочку. Поверхность кутикулы в области чув-
ствительной Ьочки несколько приподнята, а каждая клетка
снабжена чувствительным волоском. Эти микроскопиче-
ские органы распространены в большом количестве по
всему телу, но особенно многочисленны они в 1-м сегменте
и в его головной лопасти, где у крупных видов их насчи-
тывается около 1800. Функция их точно не установлена.
Как полагают некоторые исследователи, часть их может
нести осязательную функцию. Но вряд ли можно сомне-
ваться в том, что ими же осуществляются и функции обо-
няния и вкуса. В пользу такого заключения свидетель-
ствует тот факт, что эти органы имеются в большом
количестве в ротовой полости.
Чувство обоняния, т. е. способность к распознаванию
различных веществ в газообразном состоянии (что и яв-
ляется способностью воспринимать запахи), развито
у червей сравнительно ,слабо. В опытах Дарвина черви
не реагировали на запах табачного сока, духов, уксусной
кислоты, но куски лука (который они очень любят) и листья
капусты они находили по запаху. На эфир, поднесенный
на близком расстоянии к переднему концу тела, черви
реагировали отрицательно и немедленно устранялись
от него. Н л
Чувство вкуса, т. е."способность распознавать химиче-
ские различия веществ при контакте с ними, развито
у червей очень тонко и наряду с чувством осязания служит
для них главным источником восприятия событий внеш-
него мира. Опытами Дарвина, развитыми в последнее
время рядом исследователей, способность червей выби-
рать себе пищу доказана совершенно бесспорно и возра-
жения, высказанные некоторыми авторами по этому по-
воду (например, Тарнани, 1928), несомненно основаны
на ошибках.
Очень точная постановка опытов определения вкусо-
вых способностей червей, разработанная Мангольдом
(Mangold, 1924,1951), заключается в следующем. Свернутые
в трубку листья вишни или пучок из нескольких сосновых
игл перевязывается в нескольких местах нитками и ки-
пятится. Все вкусовые вещества, таким образом, из них
45
удаляются. Затем одна половина такого «испытателя
вкуса» окунается в чистый 20 %-й желатин, другая по-
ловина — в такой же желатин, в который добавлялось
испытуемое вещество — измельченные листья разных де-
ревьев и трав, кислоты, хинин и т. д. Такие испытатели
вкуса кладутся па ночь на поверхность земли цветочных
горшков, в которых культивируются черви. На утро
подсчитывают, сколько испытателей черви затащили
в норки и при этом отмечают, за какой из концов испытателя
ухватился червь. Нужно сказать, что черви, собирая корм,
попадающийся им на поверхности земли, никогда его
глубоко не вносили в норки, а оставляли его недалеко
от наружного отверстия или же только пододвигали к нему.
Поэтому вышеуказанный подсчет сделать нетрудно. Если
червь не различает концов испытателя вкуса, то при доста-
точно большом числе повторений опыта должно оказаться,
что как за тот, так и за другой его конец червь берется
одинаково часто. Если он предпочитает испытуемое ве-
щество чистому желатину, то пропитанный им конец
должен оказываться чаще впереди при затаскивании.
Наоборот, если вещество оказывается на вкус хуже, чем
чистый желатин, то за него червь должен ухватываться
реже. Этот опыт видоизменяется подкидыванием червям
испытателей вкуса, пропитанных разными веществами,
с последующим определением числа втянутых в норки
тех и других. Результаты обрабатывались статистически.
Проведенные опыты показали, что черви предпочитают
гниющие листья только что опавшим осенним; еще менее
нравятся им свежие зеленые листья и в еще меньшей
степени — высушенные зеленые. Чистый желатин их
привлекает более, чем высушенные листья. Гниющие
листья разных растений могут быть выстроены в такой ряд
в порядке убывающей склонности к ним червей: ива,
сладкий лупин, грецкий орех, белая акация, тополь, дуб,
горький лупин, липа, бук, клен, конский каштан. Све-
жие листья располагаются совсем в другой последователь-
ный ряд. От желатина с примесью хинина черви отказы-
ваются, причем чувствуют это вещество уже в концентра-
ции 0.07%. От минеральных кислот они отказываются
в любых концентрациях, но прибавка к желатину 1—2%
лимонной и фосфорной кислот им нравится. К сахарам
они относятся безразлично, но от очень крепких растворов
46
Сахара отказываются вовсе. К сахарину обнаруживается
отрицательная реакция уже начиная с ничтожных кон-
центраций.
Способность определять форму тел у червей, по-ви-
димому, отсутствует. Предпочтительное затаскивание ими
в норки листьев за передний конец, а сосновой хвои за
основание (факт, установленный Дарвином), дальнейшими
исследованиями подтвержден. Однако опытами Мангольда
установлено, что черви при этом руководствуются только
чувством вкуса, позволяющим им отличать верхушку
листа от черешка.
Говоря о рефлекторной деятельности дождевых чер-
вей, необходимо отметить, что у них давно доказана спо-
собность к научению и изменению поведения в связи
с пережитыми ранее ощущениями, т. е. условные рефлексы.
Не вдаваясь в подробности довольно сложных опытов,
установивших этот факт, упомянем, что черви могут
«запомнить» дорогу, на которой им не грозит удар элек-
трическим током, а если электрический удар сопрово-
ждать прикосновением наждачной бумаги, то черви на-
чинают избегать и наждачную бумагу без электрического
тока, хотя она сама по себе и не вызывает изменений
в направлении движения червей. В опытах с определением
вкусовых способностей червей выяснилось также, что
реакция на предлагаемое вещество меняется в связи с пред-
шествующими испытаниями. Черви обычно сначала от-
казываются от незнакомой им пищи, но потом часто при-
выкают и берут ее при наличии другой, им знакомой пищи.
Как уже отмечено (стр. 39), аппарат, обеспечивающий
наличие условных рефлексов, может быть локализован
и в частях нервной системы, не соответствующих голов-
ному мозгу вышестоящих форм. Определить, где именно
локализована эта функция у дождевых червей, — дело
будущих исследований.
Чтобы закончить рассмотрение рефлекторных реакций
у дождевых червей, коснемся еще вопроса о болевых ощу-
щениях у них.
Могут ли эти животные испытывать боль?
Замечательный русский зоолог В. Фаусек рассматри-
вал болевые ощущения как полезные приспособления,
роль которых состоит в сигнализации наличия повре-
ждений на теле. Оп постарался проследить возникновение
47
этого признака в эволюции мира животных и приводит
дождевого червя как пример животного, для которого
чувство боли еще недоступно. Если мы, укалывая до-
ждевого червя, замечаем его быстрые хлыстообразные
движения, то напрашивается аналогия с существом,
корчащимся от боли. Насколько необоснована, однако,
такая аналогия показывает следующий простой опыт:
если червя, спокойно ползущего вперед, разрезать брит-
вой пополам, то задняя половина будет хлыстовидно
сокращаться, имитируя болевые ощущения высших жи-
вотных, а передняя будет спокойно продолжать ползти
вперед, «не замечая» нанесенного повреждения. Припи-
сать чувство боли задней половине червя и отказать в нем
передней — явно нелепо. Но это значит, что и приписы-
вать чувство боли сокращающемуся цельному дождевому
червю мы не имеем права.
8. ОРГАНЫ ВНУТРЕННЕЙ СЕКРЕЦИИ
Упомянем о наличии у дождевых червей веществ, вы-
рабатываемых в определенных местах тела и служащих
химическими возбудителями разных проявлений жизне-
деятельности * организма. Такие вещества называются
гормонами (греческое слово, означающее «возбуждающий»),
а процесс их образования — внутренней секрецией.
У позвоночных выработка гормонов происходит частично
в специальных железах внутренней секреции (например,
надпочечники, щитовидная железа, гипофиз), а также
в органах, выполняющих одновременно и другую функцию
(например, половые железы, поджелудочная железа,
клетки мозга).
У дождевых червей специальных желез внутренней
секреции нет, но гормоны вырабатываются в разных ча-
стях нервной системы. Уже давно было известно, что
в ганглиях брюшной нервной цепочки червей имеются
так называемые хромаффинные клетки, которые выделяют
адреналин, т. е. вещество, вырабатываемое центральной
частью надпочечников высших позвоночных. Это вещество,
как известно, является специфическим возбудителем
нервных аппаратов, приводящих в движение мускулатуру
стенок кровеносных сосудов и служащих важным сред-
ством для регулирования ширины просветов сосудов кро-
48
веносной системы, а тем самым и кровяного давления.
У дождевых червей это вещество играет такую же роль.
В недавнее время обнаружено, что значительная часть
нервных клеток надглоточного ганглия также несет вну-
трисекреторную функцию (Herlant-Meewis, 1956). Разли-
чают два типа таких нервносекреторных клеток: одни из
них — с однородной протоплазмой, другие — с зерни-
стой. Первые, как полагают, служат регуляторами дея-
тельности половых желез, причем вещество, вырабаты-
ваемое ими, по-видимому, тормозит деятельность половых
желез: они начинают функционировать в те месяцы, когда
заканчивается размножение червей, и исчезают в периоды
размножения. Зернистые клетки имеют значение при за-
живлении ран и восстановлении утраченных частей тела
(регенерации): во время этих процессов секреция в них
особенно усиливается.
Деятельность пояска дождевых червей, которая заклю-
чается в выработке оболочек и питательного содержимого
яйцевых коконов, также несомненно регулируется гор-
монами. Одно время считалось, что гормоны, возбуждаю-
щие деятельность железистых клеток пояска, вырабаты-
ваются мужскими половыми клетками, созревающими
в семенных мешках. Однако это оказалось неверным. Но
деятельность пояска несомненно регулируется какими-то
гормонами: если пересадить кусок пояска от червя с неак-
тивным пояском к находящемуся в разгаре половой ак-
тивности, то пересаженный кусок быстро приобретает
свойства пояска последнего. Место выработки гормонов,
регулирующих деятельность желез пояска, пока неиз-
вестно.
9. ОРГАНЫ РАЗМНОЖЕНИЯ
Дождевые черви размножаются только путем откладки
яиц, заключенных в особые яйцевые коконы.
Рассмотрим, как устроены у пих органы, обеспечиваю-
щие образование яиц, их оплодотворение и откладку.
Совокупность этих органов образует половой аппарат
Мужские и женские половые органы находятся у дож-
девых червей в одной и той же особи; таким образом, среди
них нет мужских и женских особей, но все они — обоепо-
лые существа, или, как их принято называть, гермафро-
диты.
4. О. В. Чекановскан
4Р
Яйца образуются и паре очень маленьких женских
половых желез — яичниках, которые укреплены на пере-
городке между 12-м п 13-м сегментами с брюшной стороны1
(рис. 24). Яичники устроены очень просто. Они представ-
ляют собой комплексы развивающихся яиц; наиболее
ранние стадии развития находятся в части, прилегающей
L
Рис. 24. Схема строения половой системы дожде-
вого червя. (Из Стефенсона).
1 — нервная система; 2 — семенники; з — семеприемники;
4 — передняя и задняя семенные воронки; 5 — яичник;
в — яйцевая воронка и яйцевод; 7 — семепровод, 8 — се-
менные мешки; IX—XIV — 9-й—!4-й сегменты.
к межсегментной перегородке, где яичник состоит из
мелких клеток. Наиболее крупные клетки находятся на
свободном заднем конце яичника, обращенном в полость
тела. Здесь яйцевые клетки достигают своих окончатель-
ных размеров (около 0.1 мм в поперечнике) и падают
в полость тела. Яйца дождевых червей сферические или
слегка вытянутые. Они почти прозрачные, так как в их
1 Обращаем еще раз внимание на то, что описание анатомии
червей относится только к представителям семейства люмбрицид.
50
протоплазме имеется лишь очень небольшое количество
зерен питательного материала — желтка. Отсутствие до-
статочного количества питательного материала для раз-
вивающегося зародыша внутри яйца вызывает необходи-
мость снабжения его питанием извне при помощи белка
яйцевого кокона.
Яйца заканчивают созревание в так называемых яйце-
вых мешках. Это — слепые мешковидные выступы меж-
сегментных перегородок, в которые попадают яйца, ото-
рвавшиеся от задней части яичника.
Яйца выводятся наружу через короткие яйцеводы,
которые начинаются яйцевыми воронками в 13-м сегменте,
затем прободают перегородку между 13-м и 14-м сегмен-
тами и открываются на брюшной стороне 14-го сегмента
(рис. 24). Яйцевые воронки снабжены ресничками, рабо-
той которых яйца улавливаются из яйцевых мешков и
в нужный момент (во время образования яйцевого кокона)
выводятся по яйцеводу наружу.
Мужские половые железы — семенники — также очень
малы. В числе двух пар они помещаются на перегородках
между 9-м и 10-м сегментами и между 10-м и 11-м
(рис. 24). Мужские половые клетки — сперматозоиды —
только начинают развитие в этих крошечных маленьких
тельцах. Комплексы будущих сперматозоидов в виде
микроскопических комочков округлых клеток падают
в полость тела и оттуда попадают в семенные мешки,
представляющие собой объемистые разрастания межсег-
ментных перегородок. Количество, форма, расположение
и относительные размеры семенных мешков варьируют
и служат важным признаком при определении червей.
У некоторых видов дождевых червей (в родах Octola-
sium и Lumbricus) брюшная часть полости тела возле
семенников отгорожена особой стенкой от основной поло-
сти сегмента; получаются так называемые семенниковые
капсулы. Благодаря их наличию формирующиеся комочки
сперматозоидов не могут распространяться по всей полости
сегмента и им создается более прямой путь в семенные
мешки (рис. 24).
Для выведения сперматозоидов наружу служат семен-
ные воронки и семепроводы (рис. 24). Семенные воронки
обычно крупные; они хорошо заметны при вскрытии чер-
вей. Семепроводы, в которые поступают сперматозоиды
из семенных воронок, представляют собой очень тонкие
цилиндрические трубки, направляющиеся по брюшной
стенке тела кзади. Семепроводы от воронок 10-го и 11-го
сегментов в 12-м сегменте сливаются друг с другом, и
общая трубка семепровода с каждой стороны тела тянется
обычно до 15-го сегмента, где переходит через толщу стенки
тела и кончается мужским половым отверстием (порой),
обычно имеющим вид вертикальной щели.
Мужские половые отверстия сидят на более или менее
сильно развитых железистых подушках. Эти подушки,
кроме железистых клеток, содержат большое количество
сосудов, которые во время спаривания наполняются
кровью.
Оригинальной особенностью полового аппарата мало-
щетинковых кольчецов, к которым принадлежат дождевые
черви, являются семеприемники (рис. 24) — небольшие
сферические полые мешочки, плотно прижатые к стенке
полости тела. Протоки семеприемников проходят сквозь
толщу стенки тела и открываются наружными порами,
располагающимися в межсегментных бороздах. Стенки
семеприемников содержат мускулатуру, действием кото-
рой семенная жидкость может всасываться в семеприем-
ник и, наоборот, выбрызгиваться из него наружу. Эта му-
скулатура действует наподобие резинового колпачка
пипетки. Семеприемников бывает 2 или 3 пары; они могут
быть расположены сбоку, на брюшной стороне, или могут
быть (как в роде Eisenia) сдвинуты на спинную сторону,
вплоть до средней линии. Но нужно иметь в виду, что у не-
которых видов дождевых червей семеприемники отсут-
ствуют.
К органам, обеспечивающим размножение, следует
отнести поясок дождевых червей. У червей, достигших
половой зрелости, поясок всегда заметен, но его вид за-
висит от сезона и состояния упитанности. В периоды
размножения поясок сильно набухает. Его функция —
формирование яйцевых коконов.
Поясок представляет собой видоизменение наружного
эпителия. В области пояска наружный эпителий очень
сильно утолщен. Все клетки приобретают железистый
характер; среди них можно наметить три типа: 1) относи-
тельно мелкие клетки, не содержащие зерен, — слизи-
стые клетки; 2) клетки средней величины, содержащие
52
крупные зернышки, образующие оболочку яйцевого ко-
кона; 3) огромные мелкозернистые клетки, вырабатываю-
щие белковое вещество, составляющее содержимое яйце-
вого кокона и служащее питанием для развивающихся
Рис. 26. Форма щети-
нок. (Из Уде).
А — двигательная; Б — по-
ловая; В — она же в попе-
речном разрезе; Г — дру-
гая форма половой щетинки.
Рис. 25. Кожный эпителий в
области пояска. (Из Стефен-
сона).
L — кутикула; 2 — слизистые
клетки; з — кровеносные сосуды;
4 — крупнозернистые железы;
5 — мелкозернистые железы;
в — соединительнотканная перего-
родка; 7 — ядра соединительно-
тканных клеток; s кольцевая
мускулатура.
эмбрионов (рис. 25). Помимо железистых клеток, в пояске
можно видеть большое количество кровеносных сосудов
и нервных окончаний.
В функции размножения участвует также ряд других
желез на брюшной стороне тела между пояском и перед-
53
ним концом тела. Особенно заметны железы на 10-м и 11-м
сегментах, которые придают этой части поверхности тела
у половозрелых червей беловатый оттенок. Кроме того,
около брюшных щетинок в указанной части тела на неко-
торых сегментах, иногда только с одной стороны тела, бы-
вают развиты железы, заметные в виде небольших припух-
лостей. Часто сами щетинки изменены, превращены в так
называемые половые щетинки, функционирующие при
спаривании для удерживания партнера и для раздвигания
пор семеприемников. Иногда половые щетинки отли-
чаются от обычных только большими размерами, но у не-
которых видов они очень сильно отличаются и по форме
(рис. 26). Образуются, с одной стороны, острые стилеты,
очевидно вкалываемые при спаривании в кожные покровы
партнера, а с другой — щетинки, вводимые в поры семе-
приемников.
ГЛ AHA //
РАЗМНОЖЕНИЕ И РАЗВИТИЕ ДОЖДЕВЫХ ЧЕРВЕЙ
Дождевые черви размножаются обычно в течение всего
теплого времени года, в условиях умеренного климата —
с весны до осени. Размножение прекращается только во
время зимовки и засухи, когда черви перестают питаться
и уходят в глубокие слои почвы.
Несмотря на наличие мужских и женских половых
органов в одной особи, оплодотворение у червей обычно
перекрестное. Имеются некоторые данные о возможности
у них самооплодотворения (Evans a. Guild, 1947а). Однако
скорее всего факты, относимые к самооплодотворению,
следует толковать как случаи так называемого девствен-
ного размножения (см. стр. 59). Откладке яйцевых коко-
нов обычно предшествует спаривание. Процесс спарива-
ния прослежен у большого красного червя и навозного
червя. Имеются некоторые наблюдения по спариванию
длинного червя.
Рассмотрим процесс спаривания у навозного червя,
где он хотя и протекает с некоторыми усложнениями
вследствие расположения семеприемников на спинной
стороне тела, но зато более полно известен и лучше изо-
бражен.
Спаривание может происходить на поверхности земли,
но у навозного червя (а также у длинного червя) оно не-
сомненно совершается и внутри грунта. Два червя сбли-
жаются ротовыми концами и, двигаясь навстречу друг
другу, плотно соприкасаются брюшными сторонами в об-
55
ласти передних 30—40 сегментов. Долгое время (до 1 часа)
у обоих партнеров наблюдаются скольжение вперед в
назад и обильное слизеотделение. Вокруг каждого червя
образуется замкнутая трубка из слизи. Затем черви
плотно слипаются брюшными сторонами таким образом,
что сегменты 9-й—11-й, в которых расположены семе-
прпемники одного партнера, располагаются против пояска
другого партнера. Тогда’железы, расположенные на брюш-
ной стороне 9-го—11-го сег-
Рис. 27. Передние концы двух
дождевых червей, находя-
щихся в копуляции.
(По Каули).
Путь от мужских отверстий (/) до
семеприемников (?) обозначен
стрелками.
ментов, обопх червей начи-
нают усиленно выделять осо-
бое густое клейкое белое ве-
щество, облекающее со всех
сторон сегменты семеприем-
ников одного партнера и
сегменты пояска другого
партнера (рис. 27). Сегменты
пояска плотно присасы-
ваются к телу партнера ва-
ликами зрелости, и между
последними образуется жело-
бок, в который погружается
область тела 9-го—11-го сег-
ментов другого червя. В этих
местах тело обоих червей
перехвачено как бы двумя
повязками.
Железистые подушки 15-го
сегмента, на котором на-
ходятся мужские половые отверстия, сильно набухают,
наполняясь кровью, а на пространстве от 15-го сегмента
до конца пояска появляются с каждой стороны тела глу-
бокие складки — семенные бороздки. Образование семен-
ных бороздок происходит действием особой мускулатуры,
расположенной в стенке тела. В эти бороздки из мужских
половых пор изливается семенная жидкость, которая
движется назад по телу червя, к пояску. Достигнув
уровня отверстий семеприемников другого партнера, се-
менная жидкость изливается туда.
Процесс спаривания может длиться несколько часов.
Потревоженные черви (например, найденные в состоянии
спаривания в грунте) расходятся не сразу, и за ними не-
56
трудно наблюдать, если предохранить их от сильного
света и высыхания. После того как семеприемнпки обоих
партнеров оказываются наполненными семенной жидко-
стью, черви расходятся и выползают из ,своих слизистых
трубок. Такие популяционные трубки бывают находимы
в грунте. В них обычно остается избыточная часть семен-
ной жидкости.
Некоторые виды дождевых червей лишены семеприем-
ников. Как происходит спаривание у этих видов — пока
не выяснено. Но известно, что отсутствие семеприемников
заменяются у этих червей образованием сперматофоров.
Так называются капли семенной жидкости, заключенные
в засохшие слизистые выделения околощетпнковых же-
лез. Их находят прикрепленными в разных местах тела
к кутикуле червей. По-видимому, это прикрепление со-
вершается во время спаривания, причем сперматофоры,
образованные каждым из червей, приклеиваются к кути-
куле партнера при копуляции.
Япца дождевых червей откладываются, как уже упо-
мянуто, в яйцевых коконах. Раньше считалось, что коконы
образуются во время спаривания. В недавнее время выяс-
нено, что образование коконов — акт, не зависимый от
спаривания.
Процесс откладки кокона также начинается с образо-
вания слизистой трубки на участке тела примерно от
8-го до 35-го сегмента. Эта трубка захватывает все половые
отверстия и поясок (если откладка кокона начинается
сразу после спаривания, то, по-видимому, может исполь-
зоваться копуляционная трубка). Железы пояска начи-
нают усиленно функционировать, причем наибольшее
значение имеет деятельность зернистых клеток. Крупно-
зернистые клетки выделяют вещество оболочки будущего
кокона. Сначала оболочка кокона представляет собой
не что иное, как совершенно прозрачную коллоидную
пленку, расположенную под слизистой трубкой и охваты-
вающую всю поверхность пояска. Под нею быстро накап-
ливается густая опалесцирующая жидкость (выделение мел-
козернистых клеток пояска) — содержимое кокона, пред-
ставляющее собой питательный материал для яиц, отклады-
ваемых в кокон, так называемый белок яйцевых коконов.
Процесс выделения жидкостей, образующих кокон,
совершается поразительно быстро — в течение I —2 минут.
57
Очевидно, это рефлекс, возможность которого обеспечи-
вается наличпем большого количества нервных окончаний
в области пояска. Далее можно наблюдать, как по телу
червя начинают пробегать характерные волны сокращения
мускулатуры
Рис. 28. Только
что отложен-
ный яйцевой
кокон навоз-
ного дождевого
червя в слизи-
стом чехле.
(Из Фюллера).
от пояска к ротовому концу, причем червь
остается на месте. Образующийся кокон
в виде белого толстого кольца, некото-
рое время находится у переднего края
пояска, а затем начинает довольно бы-
стро катиться к переднему концу тела.
Мы употребляем слово «катиться» потому,
что есть указания на «винтовой» харак-
тер этого движения образующегося ко-
кона. При прохождении кокона через 12-й
сегмент в его белок поступают из яйце-
водов зрелые яйца, а при прохождении
кокона через сегменты с порами семе-
приемников туда впрыскивается семенная
жидкость, полученная при спаривании от
другой особи. Разрезами через белок ко-
конов установлено, что в момент образо-
вания около пояска белок не содержит
сперматозоидов, последние появляются в
нем только после прохождения формирую-
щегося кокона через сегмент с семепри-
емниками (Grove, 1925; Grove a. Cowley,
1926, 1927). Затем кокон сбрасывается
через голову. Весь процесс образования
кокона занимает 3—4 минуты.
Только что отложенный кокон одет
длинной слизистой трубкой, имеющей
следы наружной сегментации червя
(рис. 28). Оболочка кокона некоторое время остается бес-
цветной. При наблюдении за откладыванием кокона
кажется, что с головы сбрасывается бесформенная масса
мутной жидкости в слизистой трубке. На воздухе оболочка
кокоца быстро темнеет и твердеет; кокон приобретает вид
овальной капсулы с двумя концами: передним, более
широким и зубчатым, и задним, более тонким и обычно
более длинным. Широкий конец трубки представляет
собой канал, запечатанный, пробкой из ссохшегося белка.
Слизистая трубка сохраняется на коконах очень недолго;
58
ее наличие может служить указанием на то. что кокон
имеет самое большее суточный возраст.
Форма коконов у разных видов варьирует: она то более
геометрически правильная, то угловатая; характер конце-
вых трубочек также может быть различным. Окраска
коконов бывает зелено-желтой, желтой, коричневой и бу-
рой. Цвет характеризует не только вид, но и возраст
кокона: с течением времени его поверхность несколько
светлеет. Размеры коконов в длину колеблются от 2 до
6 мм.
Вопрос, как часто откладываются коконы одним червем,
изучен недостаточно. Имеются сведения, что пашенный
червь (Allolobophora caliginosa) откладывает коконы один
раз в 7—10 дней. У навозного червя откладка коконов
происходит несомненно чаще, но точных сведений об
этом пока нет. %
У большинства видов внутри кокона содержится не
более 2—3 яиц, чаще всего — только одно. У навозного
червя в коконе 6—20 яиц, но обычно развиваются далеко
не все.
Оплодотворение яиц совершается в коконе уже после
его откладки. Если вскрыть свежеотложенный кокон и
выпустить его содержимое на предметное стекло, то под
небольшим увеличением микроскопа в белке можно найти
яйцо, окруженное сперматозоидами. Один из сперматозои-
дов проникает в яйцо. После этого у некоторых видов
оболочка яйца (желточная оболочка) отстает от его по-
верхности. У других видов желточная оболочка бывает
не видна.
Прежде чем говорить о развитии яиц, отметим, что, как
установлено в последнее время, яйца ряда видов дожде-
вых червей могут развиваться без оплодотворения (Mul-
dal, 1950). Этот вариант полового размножения, называе-
мый обычно девственным размножением, широко распро-
странен среди беспозвоночных. Некоторые виды (напри-
мер, большой красный червь — Lumbricus terrestris,
длинный червь — Allolobophora longa) хотя и могут от-
кладывать яйцевые коконы без предварительного опло-
дотворения, но отложенные в них яйца не развиваются.
У других (например, у пашенного червя — Allolobophora
caliginosa) одни особи способны к девственному размноже-
нию, другие — неспособны. Однако, как правило, раз-
59
витие без оплодотворения у этих видов в природе не про-
исходит. Наконец, у водного дождевого червя (Eiseniella
tetraedra), розового червя (Eisenia rosea) и у некоторых
других видов, помимо особей, размножающихся с опло-
дотворением, есть особи, способные только к девственному
размножению. Свойство яиц развиваться без оплодотво-
рения облегчает червям заселение незанятых ими террито-
рий, так как случайное попадание в новое место всего
4 5 д
Рис. 29. Дробление яйца дож-
девого червя. (По П. Г. Свет-
лову).
А — яйцо до дробления; Б — ста-
дия 2 клеток; В — стадия 4 кле-
ток; Г — стадия 12 клеток; Д — бо-
лее поздняя стадия дробления
яйца. Пунктированы клетки, обра-
зующие стенку кишечника.
одного экземпляра может
привести к возникновению
впоследствии огромного чи-
сла особей.
При содержании в ком-
нате развитие червей внутри
коконов продолжается 2—3
недели. Но в естественных
условиях оно обычно растя-
гивается на значительно бо-
лее длительные сроки, при-
чем очень различные в зави-
симости от температурных и
иных условий. Так, на Ротам-
стэдской сельскохозяйствен-
ной станции в Англии раз-
витие зародышей пашенного
червя продолжалось (в зави-
симости от сезона) от 10 до
40 недель (Evans a. Guild,
1948).
Развитие начинается с того, что оплодотворенная яй-
цевая клетка многократно делится (рис. 29). В результате
этого процесса дробления яйца возникает двуслойная
пластинка, состоящая из клеток (рис. 30). Клетки буду-
щего кишечника уже на этой стадии развития начинают
поглощать окружающую их белковую массу, содержа-
щуюся в коконе. Затем образуется своеобразная сфериче-
ская личинка, приспособленная для жизни только внутри
яйцевого кокона. У нее имеется кишечник с ротовым и
глоточным аппаратом, при помощи которого белковая
жидкость накачивается в кишечник и, растягивая его, бы-
стро увеличивает объем личинки во много раз (рис. 30).
Нагнетание белковой жидкости в кишечник осущест-
60
вляется действием ресничек, а обратный выход ее делается
невозможным из-за особого клапана на границе между
глоткой и кишечником. Заднепроходного отверстия у ли-
чинки нет, да оно и не нужно, так как белковая жидкость
всасывается клетками кишечника без остатка.
Сферическая личинка вытягивается и мало-помалу
приобретает червеобразную форму. На ней становятся
видимыми границы между сегментами, размеры которых
убывают спереди назад, так как образование новых сег-
Рис. 30. Зародыш и личинка дождевого червя.
(По П. Г. Светлову).
А — плоский зародыш; нижний слой пластинки, состоящий из круп-
ных клеток, образует впоследствии стенку кишечника; Б — личинка,
питающаяся белком кокона: слева — ротовое отверстие, внутри ли-
чинки — полость кишечника.
ментов происходит на заднем конце личинки. Спинная
сторона формирующегося червя, развивается медленнее,
чем брюшная. Поэтому вытянутые зародыши скоро при-
обретают характерную форму запятых, так как тело их
сильно согнуто на спинную сторону, а толщина быстро
убывает сзади наперед (рис. 31). По мере роста червячки
постепенно приобретают цилиндрическую форму и у них
формируются все органы, кроме полового аппарата, кото-
рый появляется только у выросшего червя. Полное число
сегментов, характерное для данного вида (от 80 до 250),
образуется еще в коконе.
Издавна известно, что зародыши дождевых червей
на стадиях дробления могут делиться пополам и давать
начало однояйцевым близнецам. Имеются данные о том,
61
что у Allolobophora caliginosa f. Lrapezoides в кокон от-
кладывается всегда одно яйцо, но из него всегда выходят
два червя, которые являются однояйцевыми близнецами
(Omodeo, 1948; Omodeo a. Magalde, 1951). Таким образом,
можно говорить о наличии у дождевых червей полиэмбрио-
нии — особого способа размножения, свойственного не-
которым насекомым (наездники) и некоторым низшим
млекопитающим (армадилы).
Способность к активному движению черви получают
очень рано. Сначала они двигаются внутри кокона при
Рис. 31. Формирующийся червь
из яйцевого кокона. (По Фюллеру).
Слева — передний конец тела с рото-
вым отверстием и головной лопастью;
справа — задний конец тела с форми-
рующимися новыми сегментами.
помощи ресничек. Затем возникают конвульсивные сокра-
щения мускулатуры. Во вторую половину периода раз-
вития внутри кокона появляются типичные для них «чер-
веобразные» перистальтические движения.
Белок яйцевого кокона по мере развития зародышей
разжижается. В начале развития он представляет собой
густую клейкую массу, а у некоторых видов имеет кон-
систенцию густого желатина, так что его можно резать
почти на куски. Еще’’ на стадии сферической личинки
можно заметить, что вокруг этой массы находится капля
более жидкого белка. Это можно видеть в лупу через обо-
лочку неповрежденного кокона. Количество жидкости
в белке постепенно возрастает, и к концу развития чер-
вячки плавают в прозрачной, легко подвижной жидкости.
Вряд ли можно сомневаться в том, что белок кокона по-
62
степенно переходит в раствор под действием ферментов,
выделяемых телом зародыша. Таким образом, в кишечник
попадает уже подготовленная к усвоению пища.
Оболочка коконов играет роль барьера, охраняющего
содержимое от вторжения микробов и вредных веществ
из почвы. Но опа хорошо проницаема для воды и солей,
что имеет большое значение для жизни зародышей (Свет-
лов, 1928).
Вылупление червячков из кокона происходит при
помощи их активных движений; они выползают передним
концом вперед через более широкую трубочку кокона.
Очень вероятно, что пробочка из засохшего белка, заку-
поривающая выход из этой трубочки, предварительно
растворяется теми же ферментами, которые растворяют
белок кокона во время развития. Выползание молодых
червячков из кокона легко наблюдать, если положить
в воду кокон, содержащий червячков, готовых к выходу.
Такие коконы встречаются в земле нередко; их можно уз-
нать по красному цвету крови в кровеносных сосудах
молодых червячков, просвечивающей через прозрачную
оболочку кокона. При погружении в воду червячки полу-
чают, очевидно, сильное раздражение, начинают беспо-
койно двигаться и вскоре один за другим выползают
из кокона.
Вылупившиеся червячки имеют в длину 10—15 мм.
Они обычно еще лишены пигмента. Почти сразу после
выхода из кокона они начинают заглатывать замлю и оты-
скивать пищу в виде разложившихся листочков, стебель-
ков и пр.
О темпах роста червей после выхода их из яйцевого
кокона наука пока располагает очень скудными сведе-
ниями. В естественных условиях большинство видов дож-
девых червей достигает половой зрелости примерно
через год после выхода из кокона. У навозного червя этот
срок, по-видимому, значительно меньше. В росте дожде-
вых червей различают три периода: постнатальпый,
период быстрого роста до образования пояска и, наконец,
период медленного роста после образования пояска.
При старении вес дождевых червей падает (Michon,
1957). В общем, темпы развития очень сильно варьируют
в зависимости от внешних условий. У трех видов рода
Allolobophora и у розовой эйзении обнаружено,что в зим-
63
нее время происходит полная остановка развития молодых
червей (Evans a. Guild, 1948). У пяти видов Allolobophora
такого рода остановка в развитии, так называемая диа-
Рис. 32. Реге-
нерацияу дож-
девого червя.
Из середины
тела было вы-
резано 13 сег-
ментов. Сзади
восстановилась
хвостовая
часть. Спереди
образовались
два хвостовых
конца.
(По Кор-
шельту).
пауза, может происходить и в теплое
время года при понижении влажности
почвы. Диапауза может длиться меся-
цами; при наступлении более благоприят-
ных условий развитие продолжается
(Michon, 1954, 1957).
Сведения о продолжительности жизни
дождевых червей также довольно скудны.
Известно, что в неволе навозные черви
(Eisenia foetida) жили от 3% до 4J4 лет,
большой красный червь (Lumbricus ter-
restris) — 5—6 лет, длинный червь
(Allolobophora longa) — от 5% до 10%
лет. Есть менее надежные сведения, что
какой-то, ближе не определенный, дож-
девой червь прожил в неволе 15 лет.
Эти факты говорят об относительно боль-
шом сроке их индивидуальной жизни,
особенно если принять во внимание, что
никто из содержавших червей длительное
время в неволе не указывает на ухудше-
ние их состояния или на какие-либо при-
знаки старения. Таким образом, можно
думать, что максимальная продолжитель-
ность жизни дождевых червей значительно
превосходит приведенные выше цифры.
Дождевым червям в довольно значи-
тельном масштабе свойственна способность
к восстановлению утраченных частей тела,
т. е. к регенерации. Головной конец вос-
станавливается при утрате спереди не бо-
лее 15 сегментов, причем, сколько бы сег-
ментог не было отрезано, восстанавли-
вается только 4. Передняя отрезанная
часть не восстанавливает задней и поги-
бает; в редких случаях изолированные 5—6
головных сегментов восстанавливают на поверхности раз-
реза вторую голову, т. е. получается уродливый нежизне-
способный организм, который также погибает. После от-
резания заднего конца тела регенерация идет гораздо
лучше. На задней раневой поверхности разрезанного
червя всегда восстанавливается хвостовый конец, даже
если оставлено только 30 передних сегментов (в редких
случаях и при меньшем числе их). После заживления
раны, недели через 154—2, появляется тоненький вырост
(регенерат), размерами и цветом напоминающий червячка,
только что вышедшего из яйцевого кокона. Он тем длин-
нее, чем большее число сегментов было отрезано. Регене-
рат растет в длину и в толщину, и месяца через 154—2
червь делается неотличимым от нормального.
При разрезании червя пополам передняя половина вос-
станавливает хвостовой конец; головной конец на задней
половине не восстанавливается. Однако и здесь возможна
патологическая регенерация: па переднем конце задней
половины в некоторых случаях, после длительной паузы,
появляется регенерат, но оказывается, что это не регене-
рат головы, а второй хвост. Такие уродливые формы гиб-
нут (рис. 32).
Отметим, что хотя способность к регенерации у дожде-
вых червей гораздо лучше выражена, чем у позвоночных,
так как они могут восстанавливать довольно крупные
отделы тела, но ее не следует переоценивать. По сравне-
нию с низшими формами малощетинковых червей люмбри-
циды восстанавливают утраченные части в значительно
меньших масштабах и гораздо медленнее.
Г JI AB A III
ГЛАВНЫЕ ПРЕДСТАВИТЕЛИ ДОЖДЕВЫХ ЧЕРВЕЙ
СССР. СПОСОБЫ ИХ СБОРА И ОПРЕДЕЛЕННА
1. ПОЛОЖЕНИЕ В СИСТЕМЕ ЖИВОТНЫХ И
ПРОИСХОЖДЕНИЕ
Нам несколько раз приходилось упоминать, что дожде-
вые черви относятся к кольчатым червям, или кольчецам.
Кольчецы — группа высших червей; она делится на классы
многощетинковых (Polychaeta), малощетинковых (Oli-
gochaeta) и пиявок (Hirudinea), не считая небольших
групп, имеющих меньшее значение в экономике природы.
Предков малощетинковых кольчецов нужно искать среди
представителей обширного класса многощетинковых коль-
чатых червей, живущих в море. Указать более точно ис-
точник происхождения класса малощетинковых кольче-
цов вряд ли возможно. Ни одно из современных семейств
морских кольчецов мы не можем считать прямыми пред-
ками малощетинковых червей (Oligochaeta), так как все
они в течение эволюции успели получить те или иные
приспособительные черты, чуждые интересующей нас
группе. Очевидно, родоначальниками класса малощетин-
ковых червей явились более примитивные, не специали-
зированные представители морских многощетинковых
кольчецов, вероятно ныне вымершие.
В какую из геологических эпох произошло возникно-
вение малощетинковых кольчецов — сказать трудно.
Окаменелые остатки червей очень редки, так как все части
их мягкого тела, не исключая щетинок, после смерти жи-
вотного обычно очень скоро распадаются, превращаясь
ьв.
в бесформенную массу. Но остатки, похожие на малоще-
тинковых кольчецов, находят в очень древних геологи
ческих отложениях (каменноугольной и, может быть,
силурийской эпохи). Таким образом, происхождение
класса следует признать очень древним.
Не легче вопрос и о родственных отношениях внутри
класса малощетинковых червей. Во всяком случае черви
из моря не сразу вышли па сушу. Скорее всего предки сов-
ременных малощетинковых кольчецов сначала приспосо-
бились к жизни в устьях рек, заселили пресные воды, а
уже потом у них выработались приспособления к назем-
ному образу жизни. Об усиленной потребности в воде
наземных форм, их способности жить под водой и других
свойствах, указывающих на сравнительно недавний их
выход на сушу говорились на стр. 35. Сравнение орга-
низации и развития наземных форм малощетинковых чер-
вей с водными показывает несомненно большую прими-
тивность последних.
Выход на сушу водных малощетинковых червей со-
вершался неоднократно. В результате возникло несколько
семейств почвенных малощетинковых червей. Сюда отно-
сятся энхитреиды (Enchytraeidae) — обширное семейство,
представители которого, мелкие белые червячки, обильно
населяют верхние слои почв, но встречаются также в прес-
ных водах и в морях.1 Другими путями возникли дожде-
вые черви, среди которых зоологи различают также не-
сколько семейств. По сравнению с водными олигохетами
это гиганты. Но среди них, как было указано, в тропи-
ках встречаются сверхгиганты — черви до 3 м длиной.
Семейство люмбрицпд (Lumbricidae), к которому при-
надлежат почти все дождевые черви СССР, представляют
собой по ряду признаков молодую пластичную группу,
в которой и поныне продолжается интенсивный процесс
видообразования.
2. ОПРЕДЕЛЕНИЕ РОДА И ВИДА ЧЕРВЕЙ
Неспециалистам часто кажется удивительным, что
среди дождевых червей можно различать разные виды.
Им кажется, что все они одинаковы. Однако зоологи на-
1 В последнее время энхитреиды получили известность в рыбе
водстве как хороший корм для мальков.
5*
67
считывают не менее 18U видов семейства люмбрицид,
причем почти каждый год до сих пор еще обнаруживаются
и описываются виды, ранее неизвестные. В СССР водится
не более половины известных видов, а из них подавляющее
большинство приурочено к горным районам Кавказа и
Средней Азии. На большей же части территории нашей
страны насчитывается несколько деЛтков видов люмбри-
цид, из которых массовыми почти во всех областях явля-
ются не более 15—16 видов. Рассмотрением их мы и огра-
ничимся в дальнейшем, так как главная роль в почвообра-
зовании несомненно принадлежит представителям именно
этих видов.
Вопрос о видовых различиях дождевых червей может
показаться неинтересным и ненужным. Можно подумать,
что занятие ими самое большее должно составлять удел
немногих ученых специалистов и что систематика дожде-
вых червей не имеет отношения к вопросам, связанным с их
жизнедеятельностью, и, в частности, с вопросом об их
роли в почвообразовании. Ничего ошибочнее такого мне-
ния нельзя себе представить. Виды дождевых червей ха-
рактеризуются не только признаками формы и строения
их тела, но также местом обитания, образом жизни и ря-
дом особенностей, имеющих непосредственное отношение
к их почвообразовательной деятельности. Поэтому каж-
дый интересующийся почему-либо дождевыми червями
должен ознакомиться прежде всего с их видовым составом.
Любые наблюдения за жизнью и деятельностью дождевых
червей, сделанные без учета вида червя, к которому отно-
сятся эти наблюдения, в настоящее время следует признать
ненаучными. На этом основании мы помещаем ниже
таблицу для определения наиболее обыкновенных пред-
ставителей фауны дождевых червей СССР и их описа-
ние.
Все виды семейства люмбрицид сгруппированы в 8
родов. Согласно правила^м международной номенклатуры,
каждый вид имеет латинское название, состоящее из двух
слов: первое — обозначает род, к которому принадлежит
данный вид, а второе — собственное имя вида. Так,
в роде Lumbricus имеются виды Lumbricus terrestris
(большой красный червь), Lumbricus rubellus (малый
красный червь) и Lumbricus castaueus (каштановый
червь). По совокупности признаков они ближе друг
68
к другу, чем к другим видам, и поэтому объединяются
в один род.
Приводим таблицу для определения родов и видов люм-
брицид (Lumbricidae), имеющих широкое распространение
в СССР.
Принцип пользования определительными таблицами
мы считаем известным.
Заметим, что определение родов и видов по таблице
часто приводит к ошибкам. Поэтому, кроме таблицы (или
ключа, как ее иногда называют), мы приводим сжатые
описания родов и видов (диагнозы). Дойдя до того или
иного рода пли вида по таблице, необходимо проверить,
сходятся ли признаки исследуемого объекта с данными
диагноза. При этом ошибки, допущенные в ходе опреде-
ления, сразу обнаруживаются.
При составлении определительных таблиц и данных
по распространению видов люмбрицпд на территории
СССР нами широко использованы данные работ И. И. Ма-
левича (1950а, 19506, 1951, 1954).
СЕМЕЙСТВО LUMBRICIDAE
(Наиболее обыкновенные и распространенные повсюду виды)
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ СЕМЕЙСТВА
LUMBRICIDAE
1 (4). Семеприемники расположены на спинной стороне
тела около средней спинной линии или несколько
отступя от нее. Поры семеприемников одного сег-
мента либо почти соприкасаются друг с другом
на спинной стороне, либо несколько раздвинуты,
по в таком случае находятся между линией щетинок
d и спинной средней линией.
2 (3). Мышечный желудок занимает только один,
17-й сегмент. Поясок простирается кзади, самое
большее до 27-го сегмента включительно.............
..................................Род Eiseniella.
3 (2). Мышечный желудок занимает 2 или более сегмен-
тов. Поясок достигает по меньшей мере 32-го сег-
мента .................................Род Eisenia.
4 (1). Семеприемники и их поры расположены по линии
щетинок d либо сдвинуты от нее к брюшной стороне,
либо вовсе отсутствуют.
69
5 (8). 2 пары семенных мешков, располо/кенных в 11-м и
12-м сегментах.
6 (7). Поясок простирается кзади не дальше 32-го сег
мента, т. е. бороздки между 32-м и 33-м сегментами.
Щетинки слабо сближены; расстояппе между щетин-
ками одной пары немного меньше расстояния между
парами. Семепрпемпики обычно отсутствуют; если
они есть, то зачаточны и развиты несимметрично . .
......................................Род Bimastus.
7 (6). Поясок обычно простирается кзади значительно
дальше бороздки между 32-м и 33-м сегментами.
Щетинки обычно сближены попарно. Семеприем-
ники имеются, реже отсутствуют . . Род Eophila.
8 (5). 3 или 4 пары семенных мешков.
9 (12). Семенных мешков 4 пары, в 9-м, 11-м и 12-м
сегментах. Черви без пурпурной пигментации (ро
зовые, коричневые, бурые, синеватые).
10 (11). Семенники и семенные воронки находятся в се-
менниковых капсулах. Их имеется по паре в 10-м
и 11-м сегментах. Семенные мешки являются вы-
ростами этих капсул. Щетинки в передней части
тела обычно не сильно сближены попарно, в сред-
ней и задней частях почти совсем не сближены . . .
.......................................Род Octolasium.
11 (10). Семенниковых капсул нет: семенники и семенные
воронки лежат свободно в полостях 10-го и 11-го
сегментов. Щетинки всегда более или менее сильно
сближены попарно.....................Род Allolobophora.
12 (9). Семенных мешков, как правило, 3 пары, распо-
ложены они в 9-м, 1£-м и 12-м сегментах; если (редко)
имеется четвертая пара мешков в 10-м сегменте, то
они гораздо меньше всех остальных. Черви в боль
шинстве случаев различных оттенков красного цвета
благодаря наличию пурпурной пигментации.
13 (14). 3 пары семенКых мешков отходят от двух непар-
ных (срединных) семенниковых капсул, заключаю-
щих в себе семенники и воронки семепроводов:
передняя семенниковая капсула, расположенная
в 10-м сегменте, дает начало первой и второй парам
семенных мешков, расположенных в 12-м сегменте
(рис. 24). Щетинки всегда более или менее сильно
сближены попарно: пигментация темно-красная или
70
красно-коричневая, во всяком случае в передней
части тела; головной сегмент танилобический . . . .
........................................Род Lumbricus.
14 (13). Семенниковых капсул нет; семенники и воронки
семепроводов лежат свободно в полостях 10-го и
11-го сегментов. Щетинки или
попарно, или совсем не сближены.
Головной сегмент эпилобический
..........................................Род Dendrobaena.
слабо сближены
Род Eiseniella 1
Головной сегмент эпплобпческий.
Щетинки тесно сближены парами.
4 пары семенных мешков, располо-
женных в 9-м—12-м сегментах.
Приводим описание единственного
широко распространенного вида.
Eiseniella tetraedra — Четырехгран-
ная эйзениелла (рис. 33). Длина 30—
50 см, толщина 2—4 мм. Число сег-
ментов около 90. Пигментация корич-
нево-желтая до красно-бурой. Средняя
и задняя части тела четырехгранные.
Расстояние Ьс=аа и только немного
менее, чем dd. Первая спинная пора
на борозде между 4-м и 5-м сегментами.
Поясок занимает сегменты с 22-го по
26-й или с 23-го по 27-й, валики зре-
лости — с 23-го (с половины 23-го)
по 25-й, 26-й сегмент. Мужские поло-
вые поры на железистых подушках, у
типичной формы на 13-м сегменте.
Распространен повсеместно, но встре-
чается в сравнительно небольших ко-
личествах.
По берегам рек и озер, около уреза воды или в грунте
прибрежной зоны, по краям болот, в колодцах и т. п.
На юге и юго-западе европейской части СССР, кроме ти-
6
Рис. 33. Eiseniella
tetraedra.
(Из А. Я. Зелик-
мана и И. И. Ма-
левича).
А — передняя часть
тела; Б — задний ко-
нец тела. J — муж-
ское половое отвер-
стие; 9 — женское по-
ловое отверстие.
1 Названия Eiseniella и Eisenia происходят от фамилии швед-
ского зоолога Эйзена.
71
пичноп, встречается герцинская форма Eiseniella tetraedra
f. hercynia, отличающаяся от типичной только тем, что
мужские половые поры помещаются у нее, как и у всех
остальных люмбрицид, на 15-м сегменте. Она названа
«герцинской» по наименованию Герцинского леса (так
римляне называли юго-запад Германии).
Род Eisenia
Головной сегмент эпплобический. Щетинки сближены
попарно или широко расставлены. Семенных мешков
4 пары в — 9-м—12-м сегментах, или 3 пары — в 9-м,
11-м и 12-м сегментах. С 9-го по 11-й сегмент граница
пигментации размыта и сдвинута к спинной стороне.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА EISENIA
1 (6). Щетинки тесно сближены попарно.
2 (3). Межсегментные бороздки не пигментированы, бла-
годаря этому червь выглядит полосатым, особенно
в заднем конце тела. Поясок с 24-го, 25-го, 26-го
по 32-й сегмент (на 7—9 сегментах). Валики зре-
лости с 28-го по 30-й или 31-й сегмент...........
.............Eisenia foetida— Навозный червь.
3 (2). Пигментация сплошная, или ее'нет вовсе. Червь
не выглядит полосатым.
4 (5). Поясок с 27-го по 33-й сегмент занимает 7 сегментов.
Валики зрелости с 29-го по 31-й сегмент (очень
стойкие признаки, уклонения редки). Пигмента-
ция изменчива, но всегда есть, по крайней мере
на спинной стороне в передней части тела...........
. . Eisenia nordenskioldi— Эйзения Норденшельда.
5 (4). Поясок с 24-го, 25-го или 26-го по 31-й, 32-й или
33-й сегмент. Валики зрелости с 28-го по 30-й, 31-й
сегмент. Пигмента нет..............................
...............Eisenia rosea — Розовая эйзения.
6 (1). Щетинки не сближены попарно, или расстояния
ab и cd только в 2—3 раза меньше расстояния Ьс . .
Eisenia veneta — Венецианская эйзения.
Eisenia foetida — Навозный червь (см. цветной рисунок
в начале книги). Длина 60—130 мм, наибольшая толщина
0.75 мм. Число сегментов 80—110. Пигмент красный,
72
оранжево-красный, яркий. У молодых экземпляров пиг-
ментация более темная, и светлые межсегментные полосы
могут быть незаметны. Расстояние aa — bc, dd=V2 длины
окружности. Первая спинная пора на борозде между 4-м
и 5-м сегментами. Мужские поры в линии щетинок с на
больших подушках, которые, однако, не переходят гра-
ниц 15-го сегмента. 4 пары семенных мешков — в 9-м и
12-м сегментах, они меныпих размеров, чем в 10-м и 11-м.
Семеприемники возле спинной линии.
Распространение всесветное.
Чаще всего в навозе, в парниках, на свалках и т. п.,
где можно встретить до 1000 и более экземпляров на 1 м2.
Навозный червь — широко употребительное народное
название. В русском народе этот вид осознается как
нечто обособленное от других люмбрицпд, которых назы-
вают земляными, пли дождевыми червями, а некоторым
даются специальные наименования. Англичане также
дали этому виду совершенно особое обозначение: E.foetida
называется у них brandling, а все остальные виды дожде-
вых червей они именуют earthworms.
Латинское видовое название этого вида — foetida —
означает «вонючая». Червь действительно обладает свое-
образным, довольно неприятным запахом. При раздраже-
нии червь выдавливает из спинпых пор ярко-желтую
жидкость. Ее цвет обусловлен обильным содержанием
клеток желтой ткани.
Каково почвообразовательное значение этого вида
в природных условиях — пока установить трудно. Но
этот вид с практической стороны интересен тем, что обла-
дает очень высокой плодовитостью, очень г<шослив и
вполне удовлетворительно приспособляется к условиям
существования в культурных почвах.
Eisenia nordenskioldi — Эйзения Норден шельда. Раз-
меры сильно варьируют: в европейской части СССР это
небольшие черви длиной 25—80 мм при толщине 2.5—*
3.5 мм (редко до 120 мм). В Сибири достигают 300 мм
в длину, 6—7 мм в толщину. Число сегментов 80—125.
Пигментация варьирует; она может быть буро-красной,
красно-коричневой, красно-фиолетовой. Ее интенсивность
также может изменяться — от очень темных до очень
светлых тонов. Пигментирована только спинная половина
тела, притом равномерно, но задний конец более бледный.
Рис. 34. Eisenia
rosea. Передний
конец тела с брюш-
ной стороны; спе-
реди от мужских
(8) и женских (§)
половых отвер-
стий пятна кож-
ных желез.
(ПоМихаэльсену).
Семенных мешков 4 пары, с 9-го по 12-й сегмент. Мужские
поры на небольших железистых подушках.
Вид этот назван в честь шведского исследователя по-
лярных стран — Норденшельда, впервые прошедшего
на корабле «Вега» по великому северному пути от Атланти-
ческого до Тихого океана. В богатых
сборах*животных, сделанных им на се-
верном побережье Сибири, этот вид
оказался в больших количествах.
Распространен по всей Сибири, где
преобладает над всеми прочими видами
как в природных, так и в окультурен-
ных почвах, на Допу и на востоке Ук-
раины.
Водится в почвах разного типа.
Очень обычен на пойменных лугах, но
водится также в лесах, в пахотной
земле, огородах и других местах.
В восточном Алтае найден на больших
высотах. Роль этого вида в почвообра-
зовательных процессах несомненно зна-
чительна повсюду, где он указан, а в
Сибири она исключительно велика, так
как там это единственный крупный вид
дождевых червей (представители рода
Allolobophora в Сибири редки, а виды
Lumbricus вовсе отсутствуют).
Eisenia rosea — Розовая
эйзения
(рис. 34). Длина 25—80 мм, толщина
3—4 мм. Число сегментов 120—150.
Окраска розовая, фиксированные экзем-
пляры сероватые. Поясок при жизни
часто ярко окрашен — красный, розо-
вый, желтый. Боковые края его с валиками зрелости
сильно выдаются в стороны. Железистые подушки около
мужских пор хорошо развиты, но не переходят границ
15-го сегмента. Щетинки 9-го и 10-го или 12-го и 13-го
сегментов, все или только часть их, на каждом из назван-
ных сегментов на железистых сосочках, вытянутых в попе-
речном направлении, 0.8 мм длиной и 0.02 мм толщиной.
4 пары семенных мешков — с 9-го по 12-й сегмент.
Распространен всесветно.
74
Часто обильно заселяет садовую, огородную, нолевую,
лесную и луговую почвы. В Ивановской области в почвах
некоторых полей на 1 м2 приходится около 67 червей,
а в смешанных лесах с густым травостоем — около 63 на
1 м2. Глубина его ходов в почве достигает 1 м. Отсюда
ясно серьезное значение этого вида в почвообразователь-
ных процессах.
Eisenia vrneta—Венецианская эйзения.1
Длина 50 — 100 мм, толщина 5—6 мм. Число сегментов
120—155. Голова эпилобическая, но может быть почти тани-
лобической. Пигмент расположен кольцами в средней части
сегментов с бесцветными межсегментными бороздками.
Кольчатый характер окраски у типичной формы этого
вида выражен еще более резко, чем у навозного червя.
Цвет червей этого вида более темный, пурпуровый. Поясок
с 24-го, 25-го, 26-го или 27-го по 32-й или 33-й сегмент.
Валики зрелости заменены отдельными бугорками зре-
лости на 30-м и 31-м сегментах. Щетинки у типичной формы
в средней и задней частях вовсе не сближены попарно.
4 пары семенных мешков — с 9-го по 12-й сегмент. Муж-
ские половые поры на хорошо выраженных железистых
подушках, которые не переходят границ 15-го сег-
мента.
Типичная форма — самый обычным вид дождевых
червей на Кавказе и в Крыму. Встречается там всюду
вблизи домов, в огородах, садах, виноградниках, лесах;
довольно высоко идет в горы.
Имеется форма этого вида, названная садовой (Е. ve-
neta var. hortensis). Отличается от типичной формы более
узкими межсегментными светлыми полосками, некоторой
сближенностью щетинок попарно (ab : be : cd=5 : 9 : 5)
и наличием лишь 3 семенных мешков в 9-м, 11-м и 12-м
сегментах.
Распространение ее почти неизвестно; кроме Кавказа,
был найден в Ленинграде.
1 Название «венецианская* объясняется только тем, что впер-
вые этот вид был описан итальянским зоологом, который нашел
ее в Венеции. Впоследствии оказалось, что вид этот имеет широкую
область распространения и характерен для Венеции нр более, чем
для всего юга Европы и для Передней Азии.
75
Рис. 34. Eisenia
rosea. Передний
конец тела с брюш-
ной стороны; спе-
реди от мужских
(8) и женских (§)
половых отвер-
стий пятна кож-
ных желез.
(ПоМихаэльсену).
Семенных мешков 4 пары, с 9-го по 12-й сегмент. Мужские
поры на небольших железистых подушках.
Вид этот назван в честь шведского исследователя по-
лярных стран — Норденшельда, впервые прошедшего
на корабле «Вега» по великому северному пути от Атланти-
ческого до Тихого океана. В богатых
сборахЯкивотных, сделанных им на се-
верном побережье Сибири, этот вид
оказался в больших количествах.
Распространен по всей Сибири, где
преобладает над всеми прочими видами
как в природных, так и в окультурен-
ных почвах, на Дону и на востоке Ук-
раины.
Водится в почвах разного типа.
Очень обычен на пойменных лугах, но
водится также в лесах, в пахотной
земле, огородах и других местах.
В восточном Алтае найден на больших
высотах. Роль этого вида в почвообра-
зовательных процессах несомненно зна-
чительна повсюду, где он указан, а в
Сибири она исключительно велика, так
как там это единственный крупный вид
дождевых червей (представители рода
Allolobophora в Сибири редки, а виды
Lumbncus вовсе отсутствуют).
Eisenia rosea — Розовая эйзения
(рис. 34). Длина 25—80 мм, толщина
3—4 мм. Число сегментов 120—150.
Окраска розовая, фиксированные экзем-
пляры сероватые. Поясок при жизни
часто ярко окрашен — красный, розо-
вый, желтый. Боковые края его с валиками зрелости
сильно выдаются в стороны. Железистые подушки около
мужских пор хорошо развиты, но не переходят границ
15-го сегмента. Щетинки 9-го и 10-го или 12-го и 13-го
сегментов, все или только часть их, на каждом из назван-
ных сегментов на железистых сосочках, вытянутых в попе-
речном направлении, 0.8 мм длиной и 0.02 мм толщиной.
4 пары семенных мешков — с 9-го по 12-й сегмент.
Распространен всесветно.
74
Часто обильно заселяет садовую, огородную, полевую,
лесную и луговую почвы. В Ивановской области в почвах
некоторых полей на* 1 м2 приходится около 67 червей,
а в смешанных лесах с густым травостоем — около G3 на
1 м2. Глубина его ходов в почве достигает 1 м. Отсюда
ясно серьезное значение этого вида в почвообразователь-
ных процессах.
Eisenia venela — Венецианская эйзения.1
Длина 50 — 100 мм, толщина 5—6 мм. Число сегментов
120—155. Голова эпилобическая, но может быть почти тани-
лобической. Пигмент расположен кольцами в средней части
сегментов с бесцветными межсегментными бороздками.
Кольчатый характер окраски у типичной формы этого
вида выражен еще более резко, чем у навозного червя.
Цвет червей этого вида более темный, пурпуровый. Поясок
с 24-го, 25-го, 26-го пли 27-го по 32-й или 33-й сегмент.
Валики зрелости заменены отдельными бугорками зре-
лости на 30-м и 31-м сегментах. Щетинки у типичной формы
в средней и задней частях вовсе не сближены попарно.
4 пары семенных мешков — с 9-го по 12-й сегмент. Муж-
ские половые поры на хорошо выраженных железистых
подушках, которые не переходят границ 15-го сег-
мента.
Типичная форма — самый обычный вид дождевых
червей на Кавказе и в Крыму. Встречается там всюду
вблизи домов, в огородах, садах, виноградниках, лесах;
довольно высоко идет в горы.
Имеется форма этого вида, названная садовой (Е. ve-
ueta var. hortensis). Отличается от типичной формы более
узкими межсегментными светлыми полосками, некоторой
сближенностью щетинок попарно (ab : be : cd=5 : 9 : 5)
и наличием лишь 3 семенных мешков в 9-м, 11-м и 12-м
сегментах.
Распространение ее почти неизвестно; кроме Кавказа,
был найден в Ленинграде.
1 Название «венецианская* объясняется только тем, что впер-
вые этот вид был описан итальянским зоологом, который нашел
ее в Венеции. Впоследствии оказалось, что вид этот имеет широкую
область распространения и характерен для Венеции не более, чем
для всего юга Европы и для Передней Азии.
75
Род Allolobophora 1
Головной сегмент эпилобическпй.* Мужские норы на
15-м сегменте. Семеприемнпков 2 или 3 пары, по бокам
тела в липни щетинок d. Щетинки сближены попарно.
Из широко распространенных в СССР к этому роду от-
носятся два крупных вида, населяющих открытые места
с почвами, богатыми перегноем.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА ALLOLOBOPHORA
I (2). Поясковые валики с 32-го по 34-й сегмент. Окраска,
особенно па переднем конце тела, темная. Пигмен-
тация темпо-серая, иногда с синеватым оттенком,
иризирует .........................................
. . . Allolobophora longa — Длинная аллолобофора.
2(1). Поясковые валики с 31-го по 33-й сегмент. Окраска
светлая, серая с розовыми и желтыми оттенками,
пигментация незначительная, придающая фикси-
рованному червю светло-серую окраску..............
. . Allolobophora caligonosa — Пашенный червь.
Allolobophora longa — Длинная аллолобофора (см. цвет-
ной рисунок в начале книги). Длина 120—200 мм при
толщине 6—8 мм (в среднем крупнее, чем пашенный червь).
Число сегментов 160—200. Головной сегмент эпилобиче-
ский; часто от заднего края лопасти идут косые бороздки
кзади. Расстояния между щетинками: в передней части
тела йс=2/3 аа, в средней части /?с= % аа. Первая спинная
пора на борозде между 12-м и 13-м сегментами. Поясок
с 27-го или 28-го по 35-й сегмент. Мужские поры на 15-м
сегменте, их железистые подушки развиты. Половые ще-
тинки с 9-го по 11-й и па 31-м, 33-м, 34-м сегментах, по
форме и положению сходны с таковыми у пашенного червя.
Семенные мешки в 9-м и 10-м сегментах меныпих размеров,
чем в 11 -м и 12-м.
Распространен по всей Европе.
Преимущественно в огородах, дворах, парках, на
плотно укатанных площадках, где встречаются массами,
1 Название Allolobophora означает «несущая необычную ло-
пасть*. Название подчеркивает наличие эпилобической головы в от-
личие от более известного в старину рода Lumbricus, имеющего
танилобическую голову.
76
обычно вместе с пашенным червем. Ходы в почве очень
глубокие. Ночью выходит на поверхность почвы.
Allolobophora calliginosa — Пашенным червь (рис. 35).
Длина тела 60—170 мм, толщина 4—5 мм. Чи-
сло сегментов 100—240. Головной сегмент эпилобический.
Спинные щетинки теснее сближены, чем брюшные, аа
больше 6с, dd= У2 длины окружности. Первая спинная
пора на борозде между 9-м и 10-м
сегментами, реже между 8-м и 9-м
сегментами. Поясок длинный, сед-
лообразный, с 27-го, 28-го или 29-го
но 34-п или 35-й сегмепт занимает
7 или 8 сегментов. Мужские поры
на сильно развитых железистых по-
душках, захватывающих с 14-го по
16-й сегмент. Брюшные щетинки
(а, 6) на сегментах 9-м, 10-м и 11-м
представляют собой половые щетинки
на железистых сосочках; они длин-
нее и тоньше обычных щетинок,
концы их загнуты, а сбоку имеется
желобок.
Распространен всесветно.
Различают типичную и трапецоид-
ную формы этого вида. Отличия меж-
ду ними небольшие, но очень стой-
кие; к тому же каждая форма имеет
собственную область*, распростране-
ния. Поэтому лучше считать их под-
видами, различая A. caliginosa ty-
pica и A. caliginosa trapezoides.
Трапецоидный подвид отличается от
у него, во-первых, вместо валиков зрелости с 31-го по 33-й
сегмент на пояске имеются две пары бугорков зрелости
на 31-м и 33-м сегментах и, во-вторых, более темная окраска
тела. Трапециодной эта форма названа потому, что в по-
перечном разрезе контуры тела образуют фигуру, напоми-
нающую трапецию. Однако этот признак свойствен мно-
гим видам люмбрицнд. A. caliginosa trapezoides преобла-
дает на юге европейской части СССР.
Вид этот заслуживает особого внимания со стороны
почвоведов и агрономов. Хотя он встречается в неболь-
Рис. 35. Поясок и
примыкающие к нему
сегменты. Вид сбоку.
(По Граффу).
А — Allolobophora cali-
ginosa typica; Б — A. ca-
liginosa trapezoides.
J — мужские половые
отверстия с железистыми
подушками около них;
1 — валики зрелости;
2 — бугорки зрелости.
типичного тем, что
77
тих количествах в довольно различных почвах, в том
числе в лесных влажных почвах, но главным образом
он селится в богатых перегноем, обрабатываемых землях,
где разводится массами. Особенно богаты червями этого
вида глинистые почвы. Огромные площади пахотной
земли европейской части СССР заселены преимуще
ственно этим видом; все прочие виды дождевых червей
встречаются на пашнях в несравненно меньшем коли-
честве. Поэтому этот главный наш червь-земледелец
вполне заслуживает название «пашенный червь». Обла-
дая порядочными размерами и мощной мускулатурой,
он делает глубокие ходы, перемешивает землю, вы-
носит на поверхность значительные количества перерабо-
танной почвы в виде копролитов и вызывает другие изме-
нения в почве, благоприятные для ее урожайности.
Об эффективности почвообразовательной деятельности
червей этого вида можно судить по обилию их в полевых
почвах. Так, для полевых почв Курской и Орловской
областей отмечено от 20 до 200 экземпляров на 1 м2 (Ку-
лагин, 1889), для полей Ивановской области — от 16
до 114 на 1 м2 (Балуев, 1950), в Великом Анадоле для
огородной почвы — 200—300 экземпляров на 1 м2 (Мале-
вич, 1954). Ходы этих червей углубляются на 1.5—2 м.
но обычно черви держатся на глубине 6—15 см. Размно-
жение происходит с мая по сентябрь, а в неволе — и
значительную часть зимы.
Дальнейшие наблюдения над численностью, биоло-
гией и развитием червей этого вида чрезвычайно жела-
тельны ввиду его большой практической важности.
Род Е о р h i 1 а 1
Головной сегмент эпилобический. Мужские поры
на 15-м сегменте. Семенники и семенные воронки лежат
свободно в полости тела (семенниковых капсул нет).
В СССР широко распространен 1 вид.
1 Eophila означает «любящая утреннюю зарю*. Название
объясняется находками этих червей рано утром на поверхности
земли.
78
Eophiia оси lata — Глазчатая эофила (рис. 36). Длина
35—80 мм, толщина 1.3—2 мм. Число сегментов 95—150.
Окраска красная, обусловлена цветом крови. Пигмента
нет, фиксированные экземпляры бесцветны. Первая
снинная пора между 4-м и 5-м сегментами. Мужские поры
на железистых подушках, захватывающих с 14-го по 16-й
сегмент. Поясок длинный — с 21-го, 22-го по 32-й сегмент;
занимает 11-12 сегментов.
В Западной Европе встречается по-
всюду. У пас констатирован на Украине
и Северном Кавказе.
В сырых местах, встречается иногда
в грунте водоемов.
Рис. 36.
Еophila оси-
lata. Передний
конец тела с
брюшной сто-
роны.
(По Михаэль-
сену).
Род В i m a s t u s 1
Голова эпи- или танилобическая. Бу-
горки зрелости неявственны. Мелкие
черви с пигментом красного цвета.
В СССР известны 3 вида.
Bimastus tenuis — Тонкий бимаст.
Длина 20—80 мм, толщина 3 мм. Число
сегментов 90—105. Окраска в передней
части тела коричнево-красная, в средней
и задней—красная. Голова эпилобиче-
ская. Первая спинная пора на борозде
между 5-м и 6-м сегментами. Поясок с
25-го, 26-го по 31-й, половину 32-го сег-
мента. Валиков зрелости может не быть;
если они и есть, то на 29-м и 30-м сег-
ментах. Расстояние щетинок ab : be : cd =
= 5 : 10 :7.
Распространение всесветное.
Очень обыкновенен повсюду. В лесах, на поймах около
гниющей древесины, в строительном мусоре и пр. Дер-
жится близко от поверхности.
1 Bimastus означает «дважды жующий». Очевидно, название
указывает на наличие мышечного желудка и зоба. Название не-
удачное, так как двойным расширением кишечника обладают все
люмбрициды.
79
Род Dead г о b а о n а 1
Мужские поры на 15-м сегменте. Семеприемников 2
или 3 пары, поры их в линиях щетинок с или d.
Приводим вид, наиболее обыкновенный в СССР.
Dendrobaena octaedra —Восьмигранная дендробена
(рис. 37). Длина 25—40 мм, толщина 3—4 мм. Число
сегментов 80—95. Окраска фполетово-
коричневая с металлическим блеском, ири-
зирует. Пигмент синеватый. Щетинки не
сближены попарно; расстояние ла=ай=
—bc=dd. Первая спинная пора на бо-
VrS розде между 4-м и 5-м сегментами. По-
ясок с 27-го, 28-го или 29-го по 33-й, 34-й
сегмент. Валики зрелости с 31-го по 33-й
сегмент, редко заходят на 30-й сегмент.
Распространен всесветно.
//xj Типично лесной вид. Водится в лесах
Лг /у всевозможных типов, в пнях, под мхом,
в верхних слоях почвы, на стволах де-
ревьев в трещинах коры. Его нахожде-
Рис. 37. Dend-
robaena octaed-
ra. Передний
конец тела,
сбоку. Седло-
образный поя-
сок, несбли-
нпе в иных местах указывает на близость
леса. Обычен также в тундре. В лесных
и тундровых районах имеет несомненно
серьезное значение для почвообразова-
ния.
Род О с t о 1 a s i и ш 2
женные ще-
тинки.
Мужские поры на 15-м сегменте. Семе-
и И. И. Мале- приемники в 9-м и 10-м сегментах, их
вича). поры на бороздках между 9-м и 10-м
сегментами и между 10-м и 11-м в линии
щетинок с или между линиями щетинок с и d. Устрой-
ство семенниковых капсул — см. рис. 38.
Приводим описание одного, наиболее распространен-
ного вида этого рода.
1 Dendrobaena означает «двигающаяся по деревьям* или «оби-
тающая на деревьях».
8 OctoIasium приблизительно означает «щетинистый в восьми
местах». Попарное сближение щетинок у представителей этого
рода плохо выражено.
80
Ос to Iasi uni lacleum — Молочный октолазий. Длина
40—100 мм, толщина 3—5 мм. Число сегментов 100—165.
Пигмента почти нет, поэтому фиксированные черви мо-
лочно-белые, чаще светло-серые с синеватым оттенком.
Головной сегмент эпилобический, иногда танилобический.
Несколько сближены попарно на протяжении всего тела
только щетинки с п d\ щетинки а и 6 очень немного сбли-
,4
Рис. 38. Oclolasium lacteuin.
А — передний отдел тела с брюшной сто-
роны; Б — сх£ а строения половых органов
со спинной стороны. (А — из А. Л. Зелик-
мана и И. И. Малевича. Б — из Уде).
1 — семеприемники; 2 — семенники; 3 — яич-
ник; 4 — яйцевые воронки; 6 — семепро*
воды; в — семенные мешки; 7 — семенни-
ковые капсулы.
жены только в передней части тела; расстояние Ьс по се-
редине тела равно или меньше ab. Первая спинная пора
на бороздке между 8-м и 9-м, 9-м и 10-м или 10-м и 11-м
сегментами. Поясок с 30-го по 35-й сегмент, занимает
6 сегментов. Мужские поры на крупных железистых по-
душках, занимающих с 14-го по 16-й сегмент.
Распространен всесветно.
Встречается в сырых местах в почве, богатой перегноем,
на луговых поймах (в кустарниках, под камнями около
дорог и пр.). Водится повсюду, но в небольших количе-
ствах.
*’ О Н. .Чекановекин
Род Lumbricus1
Мужские поры на 15-м сегменте между линиями ще-
тинок b и с. Поясок седлообразный. Семеприемники в 9-м
и 10-м сегментах, их поры на бороздках между 9-м и 10-м
и между 10-м и 11-м сегментами в линии щетинок cd.
В некоторых местностях виды этого рода называются
выползками.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА LUMBRICUS
I (2). Поясок с 26-го, 27-го по 32-й сегмент (занимает
6 или 7 сегментов) .................................
. . . Lumbricus rubellus — Малый красный червь.
2 (1). Поясок доходит по крайней мере до 34-го сегмента.
3 (4). Поясок с 31-го, 32-го по 37-й сегмепт (занимает
6 пли 7 сегментов). Черви крупные...............
. . Lumbricus terrestris — Большой красный червь.
4 (3). Поясок с 28-го по 33-й сегмент (занимает 6 сегмен-
тов). Черви мелкие, не длиннее 50 мм................
.......Lumbricus castaneus — Каштановый червь.
Lumbricus rubellus — Малый красный червь. Длина
70—150 мм, толщина 4—6 мм. Число сегментов 95—150.
Пигментация на спинной стороне темно-красная с корич-
невым плп фиолетовым оттенками, особенно сильно пиг-
ментирован передний конец тела. Расстояния между ще-
тинками аа : ab : be : cd : dd=b : 1 : 5 : 6/e : 19. Первая
спппная пора на бороздке между 7-м и 8-м сегментами.
Валики зрелости с 28-го по 31-й сегмент (иногда на по-
ловину 31-го или 32-го). Мужские поры почти незаметны,
железистые подушки около них не развиты.
Распространен очень широко, но в Сибири, по-види-
мому, отсутствует.
В сырой земле, богатой перегноем. В огородах, в сы-
рых лесах, под гнилыми листьями, в парках. Для черно-
земных полей Курской и Орловской областей указы-
вается в очень больших количествах (до 170 экземпля-
ров на 1 м2, по Кулагину, 1889). Поэтому в процессе
почвообразования этому виду, по крайней мере в неко-
1 Слово Lumbricus у римлян означало «дождевой червь* (впро-
чем, также и «глист»).
82
горых местностях, принадлежит весьма значительная
роль. Держится обычно недалеко от поверхности. Глу-
бина ходов неизвестна.
Lumbricus terrestris — Большой красный червь (см.
цветной рисунок в начале книги и рис. 3). Длина 90—300 мм,
толщина 6—9 мм. Число сегментов 110—180. Пигмента-
ция спинной стороны темно-красная, кзади светлее, по
средней линии спинной стороны темная полоса. Задний
конец широкий, плоский. В средней части тела щетинки
тесно сближены попарно, на переднем и заднем концах
щетинки увеличены и менее тесно сближены; в переднем
конце тела расстояния между щетинками аа : ab : be : cd :
dd=4 : 1 : 31/2 : 3/4 : 16. Первая спинная пора на бо-
роздке между 7-м и 8-м сегментами. Мужские поры в виде
поперечных щелей на крупных железистых подушках,
простирающихся с 14-го по 16-й сегмент. Щетинки а и b
на 25-м и 26-м сегментах превращены в половые щетинки
длиной 1.6 мм п толщиной 0.045 мм, расположенные на
железистых сосочках.
Широко распространенный вид. В Сибири отсутствует.
Самый крупный представитель среди люмбрицид,
известных для СССР. Очень ценится рыбаками, особенно
профессионалами, для крупных крючков переметов. Ими
этот вид и назван «красным червем». Мы называем его
большим красным черь.м для отличия от Lumbricus
rubellus, которого рыбаки смешивают с L. terrestris.
Последний приведен нами выше под именем «малый крас-
ный червь». Излюбленный объект для анатомических и
физиологических исследований, а также для наблюдений
за различными проявлениями жизнедеятельности червей.
В земле, богатой перегноем, преимущественно в поч-
вах, усиленно удобряемых, т. е. в парниках, огородах,
старых свалках и т. п.
Lumbricus castaneus — Каштановый червь. Длина 30—
50 мм, толщина 4 мм. Число сегментов 90 — 120. Пигмен-
тация со спинной стороны темно-каштановая с коричнево-
фиолетовым оттенком, кзади светлее. Расстояние между
щетинками аа : ab : be : cd : dd=4 : 1 : 4 : 4/6 : 16. Пер-
вая спинная пора на бороздке между 6-м и 7-м сег-
ментами. Мужские поры почти незаметны.
В европейской части СССР встречен во многих ме-
стах — от Ленинграда до побережья Азовского моря и
S3
от Латвии до Ивановской области. Есть одно указание
для Сибири, но несколько сомнительное.
По-видимому, приурочен к лесным местностям. До-
вольно обыкновенен, но встречается в небольших коли-
чествах. Держится в верхних слоях почвы.
3. МЕТОДЫ ВСКРЫТИЯ
Для того чтобы определить вид, нужно детально озна-
комиться с наружными признаками червя и произвести
его вскрытие. Исследований простым глазом для этого
недостаточно. Нужно иметь штативную лупу или лучше
препаровальный бинокуляр и хороший источник света.
Современные советские бинокуляры МБС-1 и МБС-2
снабжены хорошими осветителями. В случае отсутствия
готовых осветителей их легко сделать, для чего можно
использовать любой источник света и линзу на штативе.
Для вскрытия червей нужна ванночка, в которую
наливается смесь воска с парафином, лучше подкрашенная
сажей; с прибавкой сала или вазелина. Ванночка должна
быть таких размеров, чтобы можно было удобно устано-
вить ее на предметный столик бинокуляра или лупы и
вести вскрытие при увеличении в 5—10 раз.
Из инструментов необходим обычный комплект для
вскрытия беспозвоночных, т. е. маленькие ножницы, пин-
цеты с тонкими концами (глазные), препаровальные иглы,
маленькие скальпели; полезны ножницы, употребляемые
для глазных операций, и лезвия для безопасной бритвы.
Для прикалывания червей ко дну ванночки нужны бу-
лавки. Очень удобны для этой цели энтомологические
булавки. И. И. Малевич рекомендует загнуть концы на
двух энтомологических булавках и пользоваться ими как
хирургическими крючками, т. е. растягивать ими в обе
стороны различные тонкие пленки в теле червя. Разрез
стенки тела необходимо делать строго по средней линии
спинной стороны червя. Достаточно вскрыть полость
тела от пояска до 1-го сегмента, так как в этой области
расположены все органы, признаки которых имеют зна-
чение для определения червей. Рекомендуемый в неко-
торых руководствах разрез стенки тела по боковой ли-
нии для ознакомления с видовыми особенностями внутрен-
ней организации люмбрицид мало пригоден. Разрез
84
следует делать с большой острожностыо, чтобы не задеть
кишечника и других внутренних органов. Удобнее всего
делать разрез стенки тела на воздухе, держа червя в ру-
ках. Производить разрез можно тонкими ножницами или
лезвием бритвы. После того как сделан разрез, червя
следует приколоть булавками за второй и предпояско-
вый сегменты ко дну ванночки и слегка растянуть его.
Затем в ванночку наливается вода и все дальнейшее
вскрытие ведется под водой.
При разведении краев разреза следует попутно осто-
рожно разрывать иглами межсегментные перегородки.
Когда стенка тела справа и слева будет свободной, ее
нужно приколоть булавками. Главное внимание при опре-
делении червей должно обратить на семенные мешки,
семеприемпики и семенниковые капсулы. Семенные во-
ронки обнаруживаются при раздвигании семенных меш-
ков. Расположение пор семеприемников относительно
линий щетинок (а, 6, с, d) очень легко определить, найдя
семеприемники, проколов в этом месте стенку тела тон-
кой булавкой и посмотрев на место выхода булавки с на-
ружной стороны стенки тела.
• К микроскопу приходится прибегать при определении
видовых особенностей тольгэ для изучения щетинок.
Представляют интерес также особенности строения мус-
кулатуры. Но для этого необходимо приготовлять тон-
чайшие срезы через тело червя, пользуясь сложным обо-
рудованием зоологической лаборатории.
4. СПОСОБЫ СБОРА И КОНСЕРВИРОВАНИЯ
Дождевые черви населяют почвы лугов, лесов, степей,
горных местностей, речных наносов, селятся и в грунтах
всевозможных пресноводных водоемов. Они избегают
лишь засоленных почв, засушливых местностей и кислых
болотных почв. Особенно обильно заселены дождевыми
червями культурные земли: поля, сады, огороды, дворы
около жилья, кучи навоза, гниющих листьев и т. д.,
причем в разных местностях эти «культурные» черви мо-
гут быть различными. Поэтому при ознакомлении с фау-
ной дождевых червей той или иной местности целесо-
образно прежде всего произвести сбор червей на куль-
турных землях и около домов. Но если ограничиться только
83
этим, то большая часть видов, населяющих данную мест-
ность, останется неизвестной. Для того чтобы ознако-
миться с ними, требуется произвести возможно большее
число сборов в самых разнообразных местах. Червей
нужно искать во всех влажных местах; так, летом в сухих
районах необходимо прежде всего обследовать почвы
возле воды, низины около канав, мох, пни, трещины коры
деревьев. Черви в активном состоянии находятся обычно
днем недалеко от поверхности почвы и не уходят глубже
чем на 10—15 см. Поэтому, переворачивая камни и дру-
гие предметы, пролежавшие на земле достаточно долго,
снимая кору с гнилого дерева и т. д., обычно можно об-
наружить червей. Этот способ вполне можно рекомендо-
вать как наименее трудоемкий.
Но как для массовых сборов с практическими целями,
так и для более углубленного ознакомления с фауной
необходимо использовать другие способы. Прежде всего
нужно копать землю, поднимать дерн, разламывать в лесу
гнилые пни. Хорошие результаты дает работа крепким
ботаническим совком, который на дальних экскурсиях
удобен своей портативностью. Однако более продуктивна
работа обычной лопатой; она может быть заменена лопат-
кой военного образца — с короткой ручкой. Лучше всего
выкапывать большие комья земли, переворачивать их
и разбирать руками, выбирая при этом из земли и яйце-
вые коконы.
Ходы червей простираются на значительную глубину.
Но копать глубже чем на 25—30 см приходится лишь
в тех случаях, когда преследуются специальные цели,
например точный количественный учет населения чер-
вей, исследование червей во время засухи, зимовки, или
в тех случаях, когда черви перед сбросом были чем-либо
потревожены. Например, вспахивание почвы заставляет
их уходить на значительную глубину.
При фаунистических сборах следует иметь в виду,
что определить можно только взрослых червей, имеющих
поясок (хотя бы следы его); лишь такие черви имеют глав-
ную ценность. Однако если у сборщика нет достаточного
навыка, то лучше собирать червей подряд, так как есть
виды, достигающие предельной величины и половой зре-
лости при размерах в 1.5—2.0 см, и при плохо развитом
пояске их можно принять за молодь крупных видов.
86
Для количественных сборов нужно на почве отмерить
четырехугольную площадку в 1 м2 (100x100 см), 0.5 м2
(50x100 см) или 0.25 м2 (50x50 см). Чем больше червей
в почве, тем меньших размеров можно брать площадку.
На отмеренной площадке земля снимается и складывается
на клеенку или иной материал, разостланный рядом, и
из каждого слоя в 10 см толщиной отдельно выбираются
и подсчитываются все черви. Все площадки в исследуемом
районе нужно выкапывать на одну и ту же глубину, же-
лательно не менее 40—50 см.
Существуют и другие способы сбора дождевых червей.
Для крупных червей, выходящих ночью на поверхность,
широко распространен старый способ ночного сбора с фо-
нарем. Фонарь должен быть не яркий, с узким пучком
света, а двигаться при сборе следует с величайшей осто-
рожностью, учитывая описанную выше чуткость червей
к сотрясению почвы. Напомним об описанном (см. стр. 44)
старинном «скрипичном» способе, изобретенном удиль-
щиками в Америке. В Англии поливают землю в местах,
где в изобилии водятся дождевые черви, раствором мар-
ганцевокислого калия в концентрации 1.5 г на 1 л
воды или 0.5°/о раствором формалина (Raw, 1959).
Для того чтобы выгонять дождевых червей на поверх-
ность земли, в Америке употребляют переносные электри-
ческие индукционные машинки (очевидно, наподобие
применяемых при подрывных работах). Электроды ин-
дуктора втыкаются в землю, и вращением ручки прибора
создаются индукционные удары электрическим током, ко-
торые заставляют червей выползать из своих ходов на-
ружу. А. Л. Бродский пользовался током осветительной
сети в ИОв и брал его со столба воздушной проводки,
регулируя силу тока ламповым реостатом. Практич-
ность электрического способа сомнительна.
Червей из каждого пункта сбора можно складывать
в отдельные стеклянные банки, крупные пробирки с проб-
ками или жестянки с крышками, снабдив каждую из них
временной этикеткой.
Чтобы сохранить червей для коллекции, определе-
ния или для пересылки, их необходимо фиксировать.
Для этой цели обычно употребляется 70° спирт и 4%-й
раствор формалина (продажный формалин, имеющий
крепость 40%, разводится в 10 раз). Формалин неприя-
Я7
тен своим запахом, а главное содержанием кислоты;
червей, фиксированных формалином, прежде чем брать
в руки, необходимо основательно отмыть в воде, так как
в противном случае кожа на пальцах пострадает. Выпрям-
лять, а особенно вскрывать червей, фиксированных фор-
малином, труднее, чем фиксированных спиртом. Но фор-
малин незаменим в экспедиционных условиях ввиду
своей портативности и дешевизны (в экспедицию берется
крепкий раствор). Следует иметь в виду, что для хорошей
фиксации нужно, чтобы объем фиксирующей жидкости
во много раз превосходил объем консервируемого объекта;
идеальным в микроскопической технике считается
отношение объемов 1:100. Соблюсти это отноше-
ние при фиксации червей на практике трудно. По-
этому приходится сменять фиксирующую жидкость не-
сколько раз, о необходимости чего следует твердо пом-
нить.
Сосуд, в котором хранятся фиксированные черви, не
должен быть заполнен ими более чем наполовину, при этом
фиксирующая жидкость должна быть сменена по крайней
мере два раза. Вообще собранный материал следует про-
сматривать и, если при встряхивании сосуда появляется
муть, сменять фиксирующую жидкость.
* г При недостатке времени можно, ополоснув собранных
червей в воде, сразу поместить их в фиксирующую жид-
кость. Но при этом онц очень сильно сокращаются, что
затрудняет их определение, монтаж для коллекции и пр.
Очень желательно поэтому сначала усыпить червей, для
чего можно использовать хлороформную воду, т. е. взяв
несколько капель хлороформа на бутылку воды, сильно
встряхнуть ее и влить в сосуд с червями. Черви в такой воде
сначала интенсивно сокращаются, а потом мускулатура
их совершенно расслабляется и они вытягиваются, впа-
дая в состояние наркоза. Вместо хлороформа можно
пользоваться эфиром или 10° спиртом. Во время возбужде-
ния перед наркотическим сном черви выделяют огромные
количества слизи, выбрызгивают полостную жидкость
из спинных пор и землю из обоих концов кишечника.
Поэтому перед перенесением наркотизированных чер-
вей в фиксирующую жидкость их необходимо быстро
сполоснуть в воде и осторожно снять с них слизь фильтро-
вальной бумагой.
bh
И. И. Малевич рекомендует для хранения червей после
фиксации их слабым формалином следующую смесь:
продажного формалина . 5 частей
глицерина 1 часть
воды . 94 части
Прибавка глицерина, по-видимому, делает фиксиро-
ванных червей менее хрупкими.
Очень полезно перед фиксацией записать характер
окраски <червя. Все прочие признаки червей удобнее
изучать на фиксированном материале.
Хранить червей следует в хорошо закупоренных бан-
ках или пробирках. Пробирки можно затыкать ватными
пробками и помещать в большие банки. Можно также упо-
треблять химические пробирки, закрывая их пробками.
Пробки полезно заливать расплавленным парафином или
стеарином, для чего можно использовать свечку. Каждая
проба должна быть помещена в отдельную пробирку и
снабжена этикеткой. На этикетке должно быть обозна-
чено: 1) дата сбора, 2) точное название географиче-
ского пункта с указанием области, к которой он отно-
сится, 3) характер места (огород луг, лес и т. п.), 4) тип
почвы, 5) фамилия сборщика. Если есть возможность
вести журнал экскурсий, то на этикетке необходимо ука-
зать номер пробы, а в журнале отметить руководящие
формы растений (деревья, кустарник, травы) и их назва-
ния, степень влажности почвы, наличи ерастительных
остатков на поверхности грунта и, кроме того, все моменты,
связанные со взятием пробы, например о нахождении дру-
гих почвенных животных (моллюски, многоножки, насе-
комые); интересны указания на присутствие кротовин
и т. д.
ГЛАВА IV
ОБРАЗ ЖИЗНИ И СРЕДА ОБИТАНИЯ ДОЖДЕВЫХ
ЧЕРВЕЙ
В предыдущих главах мы уже много раз останавлива-
лись на различных особенностях жизни дождевых чер-
вей. Нами были рассмотрены вопросы питания, размно-
жения, выкапывания подземных ходов, связи отдельных
видов червей с теми или иными условиями существования
и т. д. В этой главе мы коснемся значения дождевых чер-
вей для человека с тем, чтобы разобрать этот вопрос бо-
лее подробно в последующих главах.
1. НЕКОТОРЫЕ ОБЩИЕ СВЕДЕНИЯ О ПОЧВАХ И ИХ
НАСЕЛЕНИИ
Прежде всего остановимся на вопросе о том, что пред-
ставляет собой та среда, которую населяют черви, а
именно — почва.
В. В. Докучаев (1899) под почвой понимал естественно-
историческое тело, образовавшееся на поверхности зем-
ной коры под влиянием породы, климата, растительного
и животного мира, рельефа и геологического возраста.
В. Р. Вильямс, кроме того, считал обязательным для ха-
рактеристики почвы учитывать ее способность произво-
дить урожай растений. Здесь мы займемся вопросом о клас-
сификации почв и о некоторых общих их свойствах.
При классификации почв принимаются во внимание
самые разнообразные их свойства, т. е. их физические,
90
химические и структурные признаки, определяемые ус-
ловиями климата и местности, в которых образуется почва.
Большое значение в классификации почв имеет размер
частиц, их составляющих. Гравий и хрящ с частицами
размером 2—10 мм стоят на границе почв и рыхлых гор-
ных пород с еще более крупными частицами. Им про-
тивопоставляются истинные почвы, причем, если в них
преобладают частицы менее 2 мм, их называют мелкозе-
мом. По механическому составу в большей части почв
размер составляющих пх частиц колеблется от довольно
крупных, хорошо видимых простым глазом (песчинки),
до невидимых даже в микроскоп, размером 0.0002 мм и
даже меньше (тонкий ил и коллоидные частицы).
Очень важным признаком, характеризующим почву
в отношении условий существования в ней раличных орга-
низмов и произрастания на ней растений, т. е. ее плодо-
родия, является структура. Если взять осторожно кусо-
чек садовой почвы и посмотреть на него в сильную лупу,
то можно убедиться в том, что почва состоит из отдельных
комочков (или агрегатов), между которыми находятся
полости. Характер отдельностей (агрегатов) и взаимное
их расположение определяют почвенную структуру. По
величине отдельностей различаю? труктуры: зернистую —
с отдельностями размером 0.5—5.0 мм, ореховатую —
с плотными отдельностями различной формы, с попереч-
ником 5—20 мм, комковатую — с легко распадающимися
отдельностями размером 5—50 мм, и некоторые другие.
Для почв характерно наличие в них полостей, напол-
ненных воздухом, так называемая порозность (или скваж-
ность) почв. Поры могут составлять значительную часть
объема почвы. Так, в культурных землях объем полостей
составляет до 30—40%, а в верхних слоях до 60% объема
почвы. Чем больше порозность, тем условия для жизни
в почве благоприятнее. Крупные поры, размером около
0.3 мм, могут содержать воду; в то же время они обеспе-
чивают проникновение в • почву атмосферного воздуха,
т. е. вентиляцию и дыхание для обитателей почвы. Поры
меньших размеров (0.03—0.003 мм) играют, кроме того,
другую роль: они составляют очень важную систему ка-
пилляров в почве, по которым грунтовая вода подтяги-
вается снизу в верхние слои почв. Система узких щелей
в почве играет роль водопровода, снабжающего верхние
91
слои почвы водой за счет подпочвенных вод, подчас рас-
положенных на порядочной глубине. В засушливых рай-
онах это имеет особенно большое значение для обитате-
лей почв. Однако в условиях степных местностей подня-
тие грунтовых вод капиллярными силами может иметь
и отрицательные последствия: таким путем происходит
обогащение верхних слоев почвы солями, что приводит
к образованию засоленных почв и солончаков. Мелкие
поры, особенно минимальных размеров (менее 0.003 мм),
очень важны также и потому, что в них очень медленно
происходит испарение воды. Поэтому они могут служить
для мелких почвенных организмов местами хранения ре-
зервов воды, что особенно важно во время засухи. По-
лости в почве, как мы увидим далее, являются местом
обитания большей части микроскопической флоры и
фауны почв. Почвы, имеющие малую порозность, как
например болотные почвы, бедны животным населением.
Таким образом, система щелей и каналов в почве за-
нята отчасти водой, отчасти же воздухом, необходимым
для дыхания почвенных животных. Состав воздуха почв
отличается от атмосферного меньшим количеством кисло-
рода и главным образом значительно большим количе-
ством углекислоты. Это происходит вследствие поглоще-
ния кислорода недоокисленными составными частями
почвы, дыхания почвенных организмов, а также выделе-
ния углекислоты из углекислых солей почв под действием
почвенных кислот. Количество кислорода и углекислоты
зависит от типа почвы и от глубины почвенного слоя.
Количество углекислоты возрастает с глубиной и пони-
жением порозности. Поэтому жизнь в почвах для всех
организмов, дышащих воздухом (т. е. для всех животных
и растений, кроме анаэробных бактерий), должна по
преимуществу сосредоточиваться в верхних слоях почвы.
Во всех почвах это в действительности и наблюдается.
Большую роль в этом вертикальном распределении жизни
в почвах играет не столько уменьшение количества ки-
слорода в глубоких слоях почвы, сколько ядовитое дей-
ствие углекислоты, которое естественно увеличивается
с ее концентрацией.
Количество кислорода и углекислоты в почве изме-
няется также по сезонам. В верхних слоях почвы коли-
чество кислорода довольно постоянно в течение всего
92
года, но в глубоких ее слоях оно зимой значительно па-
дает, а с мая месяца довольно медленно возрастает, до-
стигая максимума только к августу. Количество угле-
кислоты также несколько убывает зимой.
Для того чтобы получить представление об условиях
жизни в почвах, следует ознакомиться с общими свой-
ствами почвенного климата. Он характеризуется преиму-
щественно водным и температурным режимами почвы.
Почва днем нагревается и ночью охлаждается. Охлажде-
ние почв происходит тем бы
стрее, чем больше она содержит
влаги (рис. 39). Те же соотно-
шения наблюдаются и в изме-
нениях температуры почв по
сезонам. Зимой температура
на поверхности почвы падает,
вследствие чего в умеренных
широтах верхний слои ее про-
мерзает и жизнь в ней на не-
который период прерывается.
Прерываются также все хими-
Зремя суток (6 часах)
ческпе процессы в почве и пере-
движение в ней воды. Но глу<
бокие слои почвы охлаждаются
гораздо меньше, они не про-
мерзают, и температура в них
держится круглый год на по-
стоянном уровне. Чем дальше
Рис. 39. Суточный ход тем-
пературы в почве летом.
(Из Н. П. Ремезова).
I — на поверхности; 2 — на
глубине 5 см; 3 — на глубине
10 см; 4 — на глубине 15 см;
6 — на глубине 20 см.
к северу, тем короче период, в который возможна актив-
ная жизнь в почве, а следовательно и процесс почвообра-
зования. На крайнем севере в течение короткого поляр-
ного лета земля едва успевает оттаять и почвообразо-
вание почти отсутствует.
Температура почвы зависит от растительности и сне-
гового покрова. Земля, покрытая травой, а особенно
древесной растительностью, прогревается и охлаждается
в поверхностных слоях значительно меньше, т. е. расти-
тельный полог является фактором, умеряющим почвен-
ный климат как в отношении суточных, так и годовых тем-
пературных колебаний. Большую роль в предохранении
от глубокого промерзания почвы зимой играет также,
как известно, снеговой покров.
93
Из изложенного видно, что условия жизни в почве,
по сравнению с наземными, хотя и более суровы в отно-
шении снабжения кислородом, но зато более постоянны.
Поэтому зимой почва и служит убежищем для очень мно-
гих животных.
Мы не упомянули еще об очень существенной части
почвы, именно — о перегное, или гумусе. Перегной —
это совокупность органических веществ почвы, материа-
лом для образования которых служат отмирающие части
растений, выделения животных и их трупы. Это было
известно еще Ломоносову, который в своем сочинении
«О слоях земных» (1763) писал: «Нет сомнения, что черно-
зем не первозданная материя, но происходит от согни-
тия животных и растущих тел» (растущие тела — это,
конечно, растения).
В настоящее время известно, что в образовании гу-
муса большую роль играют почвенные бактерии, грибы
и множество беспозвоночных животных. Образование
гумуса — очень сложный химический процесс, состав-
ными частями которого являются не только реакции
распада органических молекул, но и их синтез из более
простых соединений. Как известно, для корней растений
органические вещества сами по себе почти недоступны и
они поглощают только растворы минеральных солей.
Тем не менее, именно наличие перегноя определяет прежде
всего плодородие почв. Это объясняется тем, что органи-
ческие вещества почвы являются субстратом для жизни,
источником питания для бесчисленных растительных и
животных организмов. Используя гумус почвы для пи-
тания, почвенные организмы продолжают разрушение
органического вещества, входившего когда-то в состав
тела других живых существ. Конечными продуктами этого
распада являются неорганические соединения. Таким
образом, в процессе питания и обмена веществ почвенных
организмов просходит так называемая минерализация
органических соединений. Особенное значение имеет ми-
нерализация соединений азота, фосфора, калия и других
элементов, необходимых для высших растений. Главную
роль в конечных звеньях этого процесса играют почвенные
бактерии, по во всем процессе превращений органических
веществ в почве немалая роль принадлежит и живот-
ным. '
94
Если мы вспомним, что корни растений могут погло-
щать азот, фосфор, калий п ряд других элементов, необ-
ходимых для построения их тела, только в виде растворов
минеральных солей, то станет ясной созидательная роль
почвенных организмов в великом круговороте веществ,
непрерывно происходящем на поверхности земной коры.
При этом почва в конечном счете не обедняется органиче-
скими веществами, так как чем лучше развит на ее поверх-
ности растительный покров, тем больше растительных
остатков вновь и вновь поступает в почву. Наоборот,
если процесс минерализации гумуса задерживается, то
избыток его приводит к понижению плодородия почвы,
в особенности когда она при избытке влаги заболачи-
вается и превращается в торф.
Мощность почвенного горизонта и его морфологиче-
ские признаки в разных почвах очень различны. Для на-
глядности можно дать следующую схему вертикального
разреза через почву (рис. 40). Наверху с атмосферой
граничит растительный покров; у его основания лежит
слой отмерших листьев и стеблей на поверхности почвы.
Под ним находятся дерновина и слой перегноя (гумусный
слой горизонта Л). Это горизонт, наиболее богатый
почвенными организмами. Д?лее следует горизонт Б,
в котором количество перегноя быстро падает с глубиной.
Жизнь здесь сосредоточивается по преимуществу в тре-
щинах, в трубках, оставшихся от отмерших частей расте-
ний, и в ходах дождевых червей. Этот слой постепенно
переходит в горную породу (горизонт В), подстилающую
почву.
Бросим беглый взгляд на разнообразие населения почв,
чтобы уточнить место и удельный вес, занимаемые в нем
дождевыми червями.
Прежде всего сюда относится огромное многообразие
бактерий и грибов, которые населяют все промежутки
между почвенными отдельностями, вплоть до самых мел-
ких. Бактерии и грибы — постоянный и во всех отноше-
ниях весьма важный компонент почвенной фауны, пред-
ставленный в каждом кубическом миллиметре почвы
очень большим количеством особей. В полостях, содержа-
щих воздух, они в большом количестве находятся на их
стенках, покрытых пленками воды. В этих пленках оби-
тают также простейшие, т. е. микроскопические однокле-
95
Точные животные. Они представлены почвенными аме-
бами, корненожками, инфузориями и некоторыми жгу-
тиконосцами. Кроме простейших, жителями почвенных
вод и пленок жидкости, облекающих почвенные отдель-
Глубина (8 м)
Рис. 40. Схема разреза лесной почвы с пнями. (По Вилке).
Черные линии — ходы дождевых червей. Ао — слой гниющей листвы; А, —
почва, богатая перегноем; Б> — подпочва без камней; Б, — подпочва с кам-
нями; В — горная порода.
ности, являются некоторые низшие черви (коловратки,
нематоды) и другие группы беспозвоночных. В верхних
слоях почвы и гнилой листве эти водяные пленки населены
многочисленными нематодами, там же встречаются и рес-
ничные черви.
96
Обитателями воздушных пространств внутри почвы
являются моллюски, заползающие в трещины почвы, и
различные членистоногие: мокрицы (из ракообразных),
ложноскорпионы, много видов клещей (из паукообразных),
многоножки и насекомые. Из последних особенно много-
численны низшие бескрылые насекомые, обычные размеры
тела которых не превышают 1—2 мм, и многие виды выс-
ших насекомых; из них преобладают муравьи, личинки
жуков и мух, гусеницы бабочек. Наконец, многие насе-
комые зимуют в почве. По вычислениям энтомологов,
около 95% всех насекомых имеет то или иное отношение
к почве.
Особую группу обитателей почв составляют копающие
животные. Сюда, кроме дождевых червей, относятся черви,
причисляемые к тому же классу, — энхптрепды, очень
многочисленные во всех почвах. Это мелкие червяки
белого цвета, длиной редко более 1.5 см, обычно же ме-
нее. Сюда же из насекомых относятся медведки, делающие
длинные и, порой, глубокие ходы в почве, личинки жуков
и ряд других насекомых, а также некоторые пауки и мо-
крицы. Из позвоночных наиболее обычными роющими
животными являются кроты. Кроме того, многочисленные
млекопитающие, делающие норы в почве, особенно гры-
зуны (суслики, байбаки, хомяки, тушканчики и др.),
хотя и проводят в почве только часть жизни, все же имеют
серьезное значение в преобразовании почв.
Составить некоторое представление об относительной
численности разных групп животных, обитателей почвы,
можно из приводимых количеств особей на кубический
дециметр возделанной почвы в Средней Европе (Fran/,
1950).
Простейшие, бактерии . . 1 000 000 000 эка.
Нематоды . . 30 000
Клещи 2000 »
Низшие насекомые 1000
Прочие членистоногие . 100 »
Коловратки 500 *
Энхитрепды . 50 »
Люмбрициды 2 *
Таким образом, дождевые черви — группа беспозво-
ночных почвы, представленная наименьшим числом осо-
бей, населяющих единицу поверхности. Это стоит в связи
17
О В. Чекмнонскан
с их размерами: дождевые черви — самые крупные оби-
татели почв из беспозвоночных. Легко понять, что чем
размеры животного больше, тем максимальная плотность
заселения им той или иной среды меньше (Гиляров, 1944).
Плотность населения почвенных позвоночных (кроты,
землеройки и т. д.), естественно, еще гораздо меньше.
Примем во внимание, что вес дождевых червей в единице
объема почвы едва ли не превысит веса остальных живот-
ных, в ней обитающих, т. о. составит более половины
всей биомассы почвы. Само по себе это еще мало что го-
ворит об их значении в почвообразовании. Заметим, что
разные представители почвенной фации различным обра-
зом влияют на окружающую их среду. Этим вопросом
мы займемся подробнее в следующей главе. Пока отметим
также, что разные группы организмов, населяющих почвы,
в жизни отнюдь не являются независимыми друг от друга
компонентами. Наоборот, внимательное изучение пока-
зывает, что между ними существуют многочисленные и
довольно сложные связи, благодаря которым все почвен-
ное население данного участка почвы представляет со-
бой всегда, в известном смысле, нечто целое.
2. РАСПРЕДЕЛЕНИЕ ДОЖДЕВЫХ ЧЕРВЕЙ В РАЗНЫХ
МЕСТНОСТЯХ И ПОЧВАХ
Из рассмотренных нами условий жизни в почвах наибо-
лее общее значение для дождевых червей имеют некото-
рый минимум влажности и наличие перегноя в почве.
Большая часть поверхности суши в той пли иной степени
заселена червями. Они живут во всех странах света, ими
заселены и тундровые почвы, и луга всевозможных типов,
и степи, п полупустынные местности. Отсутствуют они
только в пустынях (если они не орошаются искусственно),
в сильно засоленных почвах и в торфяных болотах.
Самые северные пункты нахождения дождевых чер-
вей — п-ов Таймыр, о-ва Новая Земля и о-в Колгуев.
Имеются сведения и о нахождении их на островах около
Антарктического материка. Естественная область рас-
пространения люмбрицид, как уже указано, ограничена
на юге умеренными широтами северного полушария.
Субтропическая и тропическая зоны, а также все южное
полушарие, обильно заселены дождевыми червями дру-
гих семейств. Однако некоторые виды люмбрицид раз-
везены европейцами по всему миру, и теперь пег ни одной
страны, где бы они пе встречались. Хорошо известно,
что ни в Индии, ни в Южной Америке, ни в Австралии
дождевых червей семейства люмбрицид до колонизации
этих стран европейцами не было. В настоящее время почвы
всех этих стран густо заселены люмбрицидами. Они
попадали туда с землей на привезенных культурных
растениях, корнеплодах и иными пассивными, случай-
ными способами. Благодаря легкой приспособляемости
и выживаемости люмбрицид этого было достаточно, чтобы
в течение столетия огромные территории были заселены
ими. Особенно легко расселяются те из люмбрицид,
яйца которых не нуждаются в оплодотворении. Это как раз
и характерно для в? еветно распространенных видов,
именно для навозного червя, розовой эйзении, пашенного
червя и др. Для них достаточно попадания на новую тер-
риторию только одного яйцевого кокона, чтобы возможность
дальнейшего расселения их была обеспечена.
Кроме отмеченной выше способности к развитию яиц
без оплодотворения, эти виды обладают рядом особен-
ностей, облегчающих им завоевание новых мест обитания.
Нужно иметь в виду, что сожительство с человеком,
представляя богатые возможности для использования
гниющих субстратов, ставит в то же время перед приспо-
собляющимися к этому организмами нелегкие задачи.
Еще Дарвпн удивлялся способности дождевых червей
жить в почве дворов, мощеных плитами известняка.
Их копролиты он находил в щелях между плитами. Для
СССР имеются наблюдения, что пашенный червь и длин-
ная аллолобофора охотно селятся под булыжной мостовой
и под плитняковыми тротуарами. Нет сомнения, что уело-'
вия жизни в таких местах очень своеобразны и освоить
их могут лишь виды, обладающие большой выносливостью
и приспособляемостью.
Люмбрициды неприхотливы к температурным усло-
виям. Лабораторные опыты П. Ю. Шмидта (1920) с навоз-
ными червями (Eisenia foetida) показали, что они выносят
охлаждение до —1.2° в течение 2—3 часов, впадая в оце-
пенение, из которого возвращаются к жизни после ото-
гревания. Но при —1.6° черви погибают даже в подсу-
шенном состоянии, в котором все животные гораздо легче
7*
99
переносят низкие температуры. Однако наблюдения в при-
роде показывают, что другие виды дождевых червей,
по крайней мере в естественных условиях, выносят гораздо
более низкие температуры. Червей разных видов много
раз находили зимой на снегу или даже вмерзшими в лед,
причем при перенесении в теплое помещение они ожи-
вали. Особенно интересны в этом отношении дождевые
черви тундры и севера Сибири: эйзения Норденшельда
во мху и в верхних слоях почвы и восьмигранная дендро-
бепа встречаются здесь летом повсюду. Зимой эти черви
несомненно должны переносить очень сильное охлажде-
ние, так как в почве на глубине 40 см температура сни-
жается до —24° и даже до —27°, а на глубине 80 см уже
залегает слой вечной мерзлоты (Тихомиров, 1937). Усло-
вия зимовки червей в тундре пока неизвестны, но трудно
допустить, чтобы зимой здесь были горизонты не промерз-
шей почвы.
Заметим, что температура тела червей очень быстро
следует за изменениями температуры среды. Например,
после снижения температуры почвы на 10° температура
тела червя уже через несколько минут выравнивается
с температурой среды и отличается от нее не более чем
на 0.05°. То же можно сказать и об их способности при-
способляться к высоким температурам. Например, наш
обыкновенный пашенный червь (Allolobophora caiiginosa)
весьма обычен также в Пенджабе (Индия), где температура
воздуха в тени достигает летом 46—49°.
На Кавказе, в Средней Азии, в Альпах черви встре-
чаются на больших высотах и доходят до самых ледни-
ков. В Швейцарии в зоне выше 2600 м (в зоне снегов)
обитает 18% состава фауны дождевых червей Альп. При
этом фактором, ограничивающим их распространение,
является не суровый и изменчивый температурный режим
горных местностей, а недостаточная глубина почвенного
покрова. В скалистых местностях с повышением над уров-
нем моря прежде всего исчезают такие виды, как боль-
шой красный червь и длинная аллолобофора, делающие
в почве глубокие ходы. Но крупные виды люмбрицид
все же избегают холодного воздуха. Об этом говорит
одно замечательное явление в поведении червей, откры-
тое Дарвином, а впоследствии наблюдавшееся неодно-
кратно и другими. Оказывается, что черви после своего
100
ночного выхода на поверхность затыкают отверстие норки
листьями, палочками, камешками или другими подобными
предметами, находящимися поблизости. Иногда черви,
по наблюдению Дарвина, нарочно выкапывают камешки
из плотно утрамбованного грунта дорожки и кладут
их над отверстием норки. Это им удается делать при по-
мощи глотки, которой они могут присасываться к твер-
дым телам и переносить их с места на место. По-видимому,
черви прибегают к закрыванию отверстия норки посторон-
ними предметами в тех случаях, когда им не удается за-
крыть его кучкой своих копролитов. Последнее делается
чаще всего, но собирание кучек камешков и прутиков
иногда наблюдается в очень больших масштабах. Обсуж-
дая возможное биологическое значение этого инстинкта,
Дарвин пришел к выводу, что отчасти затыкание отвер-
стий норок может служить для сохранения влаги в под-
земных ходах, но главное значение их он видит в предот-
вращении попадания холодного воздуха в норки. С этим
трудно не согласиться, особенно приняв во внимание
возможность утренних заморозков. Наблюдениями уста-
новлено, что есть виды, для которых очень небольшие от-
рицательные температуры оказываются губительными.
Фактором, ограничивающим распространение дожде-
вых червей, является также состав почвенного раствора.
Там, где создаются условия для накопления солей в почве,
так называемого засоления их, фауна червей резко сокра-
щается. Концентрированных растворов любых солей,
например морской воды, дождевые черви совсем пе пере-
носят. Интересны в этом отношении воспоминания Ч. Дар-
вина, который пишет, что, будучи мальчиком, он очень
любил удить рыбу, но ему был так неприятен вид живой
приманки, извивающейся на крючке, что он для ловли
рыбы на удочку в море употреблял дождевых червей;
они моментально умирали при погружении в морскую
воду, и он мог спокойно следить за поплавком, не терзая
себя мыслью о мучениях приманки (о результатах таких
ловов он не пишет, но вряд ли они были особенно успешны,
так как приманкой при ужении в море служат обычно
морские черви, крабы, рыбки и т. д.). В степных солон-
чаках черви отсутствуют, но в слабо засоленных почвах
степей Западной Сибири встречается эйзения Норден-
шельда (Четыркипа, 1930).
101
Большое внимание исследователей привлекал вопрос
о значении кислотности почв для распространения в них
дождевых червей. Экспериментальные исследования по-
казали, что для дождевых червей характерен довольно
широкий диапазон кислотности растворов, так как они хо-
рошо выносят почвенные растворы с pH от 4.5 до 8.5.1
По новым данным, у дождевых Зервей снижается актив-
ность жизнедеятельности только в сильно кислых средах
(pH ниже 4) при прочих неизменных условиях (Зражев-
ский, 1949, 1954а). Однако в природных условиях кислот-
ность почв очень тесно связана с другими их свойствами,
поэтому показатель кислотности оказывается очень су-
щественным для характеристики почвы вообще, а тем са-
мым и условий существования люмбрицид. В почвах тор-
фяных лугов с низким значением pH черви также немно-
гочисленны. Но есть виды, лучше всего размножающиеся
в кислых почвах; такова американская эйзения Лен-
берга (Eisenia lohnbergi), предпочитающая почвы со зна-
чением pH 4.5—5.0 всем другим. Установлено, что наш
пашенный червь лучше всего размножается в почвах
со значением pH 7.0—7.8 (т. е. нейтральных и слабоще-
лочных). На большом материале показано, что в самом
Воронеже и Хоперском заповеднике эйзения Норден-
шельда чаще обнаруживается в несколько укисленных
почвах, чем розовая эйзения и белый октолазий (Петров,
1946). Вопрос этот осложняется тем, что не только та
или иная степень кислотности влияет на распространение
люмбрицид, но и, наоборот, наличие червей в почве из-
меняет кислотность, так как они способны нейтрализовать
кислоты почв.
Итак, дождевые черви населяют самые разнообразные
почвы всех стран света, причем разные виды приспособ-
ляются к различным условиям существования. При этом,
несмотря на очень большую приспособленность и пластич-
ность видов этой группы животных, разные страны насе-
лены представителями, относящимися к различным семейст-
вам, родам и видам. Происхождение этих географиче-
ских различий в фауне червей связано с медленностью
1 Как известно, величина pH характеризует степень кислот-
ности или щелочности растворов. При pH 7.0 раствор нейтрален.
Значения pH ниже этой цифры указывают на кислую реакцию,
выше — на щелочную.
102
их распространения и трудностями, с которыми сопря-
жено для них заселение новых территорий. Не только
моря и пустыни, но и горы служат непреодолимыми
препятствиями для их распространения. Последнее обсто-
ятельство накладывает характерный отпечаток на фауну
люмбрицид горных местностей. Альпы, Пиренеи, Лппенины,
а особенно горы Средней Азии и Кавказа, содержат
очень большое число видов, не встречающихся нигде
в других местах. Это связано с тем, что виды, образовав-
шиеся в процессе эволюции в горных местностях или
сохранившиеся там от предыдущих геологических эпох,
оказались изолированными горными цепями, преградив-
шими им путь к дальнейшему расселению.
Там, где нет указанных непреодолимых преград,
расселение червей происходит разными путями. К актив-
ным горизонтальным странствиям некоторые виды, по-вп-
видимому, совершенно неспособны. Большой красный
червь, длинная аллолобофора и другие крупные виды,
делающие глубокие подземные ходы, вероятно, редко
покидают свои норки; по ночам они высовываются пе-
редним концом тела из норки, а щетинками заднего конца
тела крепко зацепляются за стенки норки, чтобы при пер-
вом сигнале опасности спрятаться в нее. Если эти черви
ползают на поверхности земли, то это служит показате-
лем их плохого состояния: это либо болезнь, либо отсут-
ствие кислорода в почве, либо что-нибудь подобное.
Виды, не делающие особенно глубоких ходов и живу-
щие в более рыхлых грунтах, несомненно могут совер-
шать горизонтальные странствия. Так, навозных червей
(Eisenia foetida) можно найти под испражнениями скота
в таких местах, где заведомо их поблизости не было.
Особенно доказательны находки их в горах, на скалистых
грунтах.
Восьмигранная дендробена выходит из лесов на при-
легающие участки поймы пли пашни, по особенно далеко
от своих основных лесных местонахождений не удаляется.
Малый красный червь способен совершать странствия.
Имеются наблюдения, что особи этого вида с целины
переходят на опытные участки возделанных полей, по-
крытые соломой и навозом.
Есть немало указаний на то, что качественный состав
фауны дождевых червей меняется по сезонам и годам.
10В
Факты эти можно рассматривать как указание на возмож-
ность их активных перемещений. Но в общем можно ска-
зать, что способность к активным перемещениям у дож-
девых червей все-таки развита очень слабо, что для всех
видов она (особенно па свету!) может служить показа-
телем какого-то их серьезного неблагополучия.
Более существенны пассивные способы распространения
червей по земной поверхности. Можно думать, что пере-
несение червей и особенно их яйцевых коконов на копы-
тах животных должно играть роль в их расселении.
Вероятно, некоторое значение в качестве переносчиков
яйцевых коконов имеют роющие млекопитающие, к но-
гам которых могут приставать комки земли с яйцевыми
коконами люмбрицид и их молодью. Но главным образом
они переносятся водой рек и дождевых потоков. Длитель-
ное пребывание под водой, если она снабжена кислоро-
дом, для червей совершенно безвредно. Поэтому черви,
вымытые из почвы дождями, могут переноситься на очень
далекие расстояния, особенно если они попадают в по-
ток вместе с мхом, корой, кусками дерева, с корнями,
дерновинами и т. д. Большую роль в расселении люмбри-
цид должны играть весенние половодья рек, когда во-
дой затопляются огромные пространства речных поим.
Тогда черви, выползающие на поверхность, могут легко
подхватываться током воды. По этому вопросу имеются
наблюдения И. А. Четыркиной (1926).
Интересные наблюдения по пассивному расселению
червей при помощи токов воды сделаны в Средней Азии.
Как хорошо доказано, почвы неорошенной пустыни сов-
сем лишены червей. Их безусловно не было в Голодной
Степи до ее орошения. Грунтовые воды там залегали
на уровне 10—15 м, а поверхность почвы была пересохшей.
В 1899 г. из р. Сыр-Дарьи по системе каналов в глубину
Голодной Степп была отведена вода и орошенные участки
распаханы под хлопчатник. В 1917 г. Н. А. Димо, иссле-
дуя фауну этих участков, нашел ее густо заселенной
червями (1 600 000 экземпляров на 1 га!). Аналогичные
наблюдения сделал в 1926 г. А. Л. Бродский на других
участках пустыни, орошенной, как это точно известно,
в 1914—1916 гг., и также нашел там богатую фауну чер-
вей (Бродский, 1927). Таким образом, для заселения чер
вями новых мест в этих случаях оказался достаточным
104
промежуток времени в 10—12 лет. Е. С. Кирьянова (1936)
на участках целины, расположенных приблизительно
в 36 км от Ташкента в сторону Голодной Степи, находила
на глубине 40—50 см дождевых червей в количестве
до 22 200 000 экземпляров на 1 га.
Вряд ли можно сомневаться в том, что во вновь оро-
шенные участки черви попали из долины Сыр-Дарьи,
где они всюду, из-за жаркого климата жмутся к арыкам,
легко в них попадают и уносятся водой на далекие рас-
стояния.
3. ЗИМОВКА И ПЕРЕНЕСЕНИЕ ЗАСУХИ
не одинакова в
разные сезоны.
Деятельность червей
При наступлении неблагоприятных погодных условий они
опускаются глубоко в почву и впадают в пассивное со-
стояние: перестают питаться,
все их жизненные функции при-
останавливаются. Слизистые вы-
деления их кожных желез об-
разуют толстую пленку; если
почва высыхает, то эта пленка
превращается в твердую обо-
лочку и червь оказывается за-
ключенным в капсулу (рис. 41).
Свертывание в клубок и обра-
зование капсулы способствуют
сохранению необходимого ми-
нимума влаги и уменьшают
теплоотдачу. Расширения под-
земных ходов червей, в кото-
рых они переживают стадию
покоя, называются камерами,
причем можно различать зим-
ние и летние камеры.1 Нередки
в одной камере двух
в один общий клубок. В таком состоянии, сходном с со-
стоянием зимней спячки, характерной для очень многих
свертываются в клубок,
11. Дождевой червь
Рис.
но время спячки. (По Фюл-
леру).
и более
случаи нахождения
червей, сплетшихся
1 Некоторые почвоведы называют эти камеры коконами, что
крайне нежелательно ввиду возможного сметливания их с яйцевыки
коконами.
105
животных, черви могут оставаться в течение многих ме-
сяцев.
Это чрезвычайно важное приспособление; благодаря
ему люмбрициды получили возможность заселить огром-
ные пространства во всех частях суши.
Как ясно из сказанного ранее, в полярных странах
черви вынуждены оставаться бблыпую часть года в ука-
занном покоящемся состоянии и «оживают» только на пе-
риод короткого полярного лета. В засушливых степных
районах период активной деятельности червей также
довольно короток, так как она возможна только весной
и осенью, во влажные и не морозные месяцы. Считается,
что в умеренных широтах период активной деятельности
червей продолжается б1/^—7 месяцев.
Черви используют все возможности для активной
деятельности, предоставляемые нм климатом. Они не ухо-
дят в глубокие слои почвы на спячку, пока почва не про-
мерзнет на 5—G см и не появится спеговоп покров в 8—10 см,
т. е. пока зима не установится окончательно. Кроме того,
достаточно оттепели, чтобы черви перешли в активное
состояние, причем они могут выползать даже на снег.
И. И. Малевич сообщил (на съезде энтомологов в феврале
1954 г.), что он под Москвой находил свежие яйцевые
коконы дождевых червей в декабре. Весной черви возобно-
вляют деятельность за 10—15 дней до псчезповения мерз-
лого слоя почвы, т. е. просыпаются сразу, как только
вешние воды и теплый воздух, проникнув к ним через
почвенные поры в глубокие слои почвы, сигнализируют
им возможность перехода к активному существова-
нию.
Отметим, что переход в покоящееся состояние про-
исходит не только при ухудшении условий существования.
Это наблюдалось для длинной аллолобофоры в Западной
Европе и для эофилы в Средней Азии (Гиляров, 1949),
а также и в других местах; последняя весной в изобилии
встречается в поверхностных слоях почвы, но в конце
весны и в начале лета, до наступления засухи, спускается
на значительную глубину (на 1 м и более), где впадает
в спячку, свертываясь в клубок и окружаясь капсулой.
Через некоторое время черви этого вида вновь появляются
в поверхностных слоях почвы. Причины описанного явлегг
ния неясны.
106
Переход в покоящееся состояние сопровождается по-
терей большей части воды, находящейся в теле живот-
ного, что характерно для зимнего сна насекомых и мно-
гих других холоднокровных животных. Это понятно,
так как вода в каждой клетке тела служит необходимой
внутренней средой, в которой совершаются все процессы
обмена веществ, являющиеся, как известно, основой
жизни. Уменьшить в клетке количество воды — значит
понизить уровень ее обмена вещества, а тем самым по-
низить ее восприимчивость к любым воздействиям со сто-
роны внешней среды. Обезвоживание организма наряду
с понижением жизненной активности означает как бы его
консервирование, изоляцию от воздействий среды, в том
числе от повреждающих воздействий.
Опыты показали, что дождевые черви наделены исклю-
чительно высоко развитой способностью терять воду без
вреда для организма. Черви, помещенные в стеклянную
трубку с землей, лежали в комнатном воздухе, без увлаж-
нения, с мая по ноябрь. Почва вокруг червей была сухая,
черви были свернуты в клубок, окружены обычной кап-
сулой и выглядели как высохшие мумии. Однако в воде
они ожили. В последующих опытах было установлено,
что навозные черви, помещенные в пробирках в закрытый
сосуд (эксиккатор), куда была поставлена баночка с по-
глотителем паров воды (хлористый кальций), в воздуш-
ной среде могут терять в весе до 78.5 % содержащейся
в них воды, что составляет 69.2% от веса червей. Подсу-
шивание производится в течение 1—3 суток. Черви сильно
уменьшаются в объеме, становятся жесткими, но, положен-
ные на мокрую фильтровальную бумагу, через 20—30 ми-
нут начинают двигаться, а через несколько часов стано-
вятся вполне активными. Чем медленнее ведется подсу-
шивание, тем больше воды может потерять червь без вред-
ных для него последствий (опыты П. Ю. Шмидта п его
сотрудников 1943 г.).
Если мы примем во внимание, что у млекопитающих
и у человека потеря воды более 10% от ее содержания
в организме уже опасна для жизни, то оценим по
достоинству удивительную способность люмбрицид
терять воду в столь больших количествах, несом-
ненно являющуюся полезным приспособительным свой-
ством.
107
Отметим, что не у всех видов люмбрицид способность
переносить засуху развита в одинаковой степени.
Интересны данные В. К. Балуева, который продол-
жал эксперименты II. Ю. Шмидта с разными видами люм-
брицид. Он вел опыты высушивания червей в почве,
сравнивая разные виды. Это давало ему возможность
наблюдать реакцию их на высушивание в виде сворачи-
вания и образования камер в почве. Оказалось, что
только пашенный червь и длинная аллолобофора реаги-
руют таким образом: 60% особей первого вида и 40%
особей второго вида на третьи сутки подсушивания обра-
зовали камеры. Кроме них, лишь у особей восьмигранной
дендробены в 10% случаев наблюдались камеры. Осталь-
ные виды при подсушивании гибли, но камер не образо-
вывали. Таким образом, наибольшая приспособляемость
оказалась у двух наиболее обычных жителей культурных
земель. Если сравнить среднюю продолжительность вы-
живания, обнаруженную в этих опытах у особей разных
видов, то получается следующий ряд:
Пашенный червь . 16 суток
Длинная аллолобофора . 12.2 ♦
Малый красный червь . . 8.0 *
Восьмигранная дендробена 6.2
Розовая эйзения . . . 5.3 ♦
Большой красный червь . 4.0 *
Белый октолазий 2.4 л
Навозный червь . 2.2 •>
Таким образом,’ и по этому признаку упомянутые два
вида далеко превосходят все остальные в отношении вы-
носливости, тогда как навозный червь оказывается
на последнем месте. Отсюда можно сделать, между про-
чим, заключение, что избрание П. Ю. Шмидтом навоз-
ного червя в качестве образца животного, способного
терять влагу и выносить отрицательные температуры,
было крайне неудачным. Этот пример показывает, как
важно при любых начинаниях, предпринимаемых с жи-
вотными, — будь то эксперименты или практические меро-
приятия — иметь точные знания о видовых особенно-
стях.
На какой же именно глубине происходят зимовка
и перенесение засухи дождевыми червями?
108
Общего ответа на этот вопрос дать невозможно. Мы уже
говорили, что черви уходят на разную глубину в зави-
симости от степени засухи и промораживания почвы;
кроме того, разные виды обладают различной способностью
к пробуравливанию глубоких слоев почвы. В местностях
с очень влажным климатом, например в Колхидской низ-
менности (Грузия), дождевые черви, по-видимому,
никогда не спускаются глубже 10—20 см. В умеренных ши-
ротах много ходов червей и их камер наблюдается на глу-
бине 1.5—2.0 м. Сюда они спускаются, не только при не-
благоприятных условиях; па этих глубинах встречаются
в порядочном количестве их копрол’ 'гы. В засушливых
степных районах черви спускаются на зпачительно боль-
шую глубину. Крупный червь мариупольская дендро-
бена углубляется на 8 м, причем на стенках ямы в 1 м2
Г. Н. Высоцкий насчитал па этой глубине 110 ходов.
Таким образом, в общем дождевыми червями может быть
освоена мощная толща почвы до подстилающей горной
породы или до грунтовых вод.
Вертикальные перемещения дождевых червей отнюдь
не обязательно связаны с полным прекращением их актив-
ной деятельности. На значительной глубине черви могут
продолжать заглатывать почву, содержащую органи-
ческие вещества, и делать ходы, но деятельность их при
этом становится менее интенсивной. Следовательно, между
полноценной деятельностью и состоянием покоя имеются
все возможные переходы.
Характер деятельности червей, в частности интенсив-
ность питания (что имеет особенно большое значение для
процесса почвообразования), может испытывать сезон-
ные изменения и независимо от их вертикальных переме
щений.
Чем выше температура почвы при достаточном
минимуме влажности, тем интенсивнее обмен веществ
у червей. Это объясняется тем, что с повышением темпе-
ратуры среды возрастает температура тела червей, а от-
сюда и темп всех физических и химических процессов
в их клетках. Отмечено, что одно и то же количество чер-
вей при более высокой температуре почвы (однако не выше
известного предела), но при прочих равных условиях
выбрасывает большее количество копролитов (т. е. пи-
тается более интенсивно).
юн
4. ЗАТОПЛЕНИЕ ПОЧВ. ПОЧЕМУ ЧЕРВИ НАЗЫВАЮТСЯ
ДОЖДЕВЫМИ?
Рассмотрим явление, обратное засыханию почвы, —
ее затопление дождями, водой от таяния снега, половодьями
рек.
Само по себе пребывание под водой для червей вполне
безопасно. Если в воде нет вредных для них веществ,
она пе перегрета и содержит некоторое количество ки-
слорода, черви могут в ней жить неопределенно долгое
время. Например, оказалось, что большой красный червь
может жить под водой около года (Roots, 1956а). Рядом
исследователей ставились опыты для определения спо-
собности червей жить под водой. Оказалось, что при
комнатной температуре они могут жить в аквариумах
месяцами, причем для этого не нужно особенно заботиться
о снабжении воды кислородом. Наоборот, энергичное
продувание воды воздухом действует на червей вредно,
так как сотрясение воды, производимое непрерывным
током воздуха, слишком сильно их беспокоит, как живот-
ных, чрезвычайно чутких к осязательным раздражениям.
В связи с этим на теле у них образуются глубокие пере-
тяжки, а затем посредством активного местного сокраще-
ния мускулатуры по середине тела или на заднем его
конце происходит отбрасывание кусков тела. Это так на-
зываемая автотомия — явление, наступающее у люмбри-
цид при весьма различных неблагоприятных воздействиях.
Она может быть полезной при повреждениях задней по-
ловины тела, когда червю приходится волочить за со-
бой отмирающие сегменты, ставшие лишними и задержи-
вающими движение.
Эти сегменты отбрасываются, рана заживает, а потом
происходит восстановление недостающей части тела пу-
тем регенерации (см. стр. 64). Возможно, что автотомия
может спасти червю жизнь при нападении врага (крота,
птицы, жабы).
Что касается кислорода, то дождевые черви, как и мно-
гие из водных их сородичей, могут довольствоваться
крайне небольшим его содержанием в воде. Доказано,
что они могут жить под водой при содержании растворен-
ного в ней кислорода всего в 2.5% (Dolk а. V. d. Paauw,
1929). Это объясняется, с одной стороны, наличием в их
110
крови гемоглобина (см. стр. 29), а с другой — крайне
замедленным по сравнению с другими беспозвоночными
почвы обменом веществ. В то время как нематоды (очень
мелкие нитевидные червячки) потребляют 890—1440 мм8
кислорода на 1 г веса в час, а энхитреиды — 50 мм8,
дождевые черви поглощают всего только 23.3—3G.6 мм3
кислорода па 1 г веса в час (Kiihnell, 1950, стр. 205).
Необходимым условием для выживания червей под
водой является защита их от света. Их беспокоит даже
рассеянный свет, а если держать червей в комнате на свету,
то у них начинается автотомия. Прямой же солнечный свет,
содержащий ультрафиолетовые лучи, для них губите-
лен: достаточно оставить дождевых червей на солнце
на короткое время, чтобы они погибли (Мегкег, 1926).
О возможности жить под водой свидетельствуют и
наблюдения за дождевыми червями в природе. Не говоря
уже о видах, типическим местом жизни которых являются
прибрежные группы водоемов (восьмигранная эйзениелла,
эйзения Ленберга), представители многих наземных
люмбрицид далеко не редко встречаются в грунтах водо-
емов, притом далеко от берега. Особенно часты такие
находки летом в засушливых районах, где все население
дождевых червей поневоле жмется к воде. При исследо-
вании донной фауны прудов одного из рыбных хозяйств
Краснодарской области оказалось, что чуть ли не глав-
ной составной частью этой фауны являются типично
наземные виды люмбрицид.
Выше мы уже упоминали о роли водных потоков
в расселении червей. Эти факты также свидетельствуют
в пользу широких возможностей для подводного сущест-
вования последних. Огромные площади лугов речной до-
лины находятся ежегодно под водой в течение месяца и
более. Затопление речных пойм начинается обычно только
в мае, т. е. спустя значительное время после того, как
черви очнулись от зимнего сна и перешли к активной дея-
тельности и откладке коконов. Пробы, взятые в разгар
половодья, показывают, что черви в этот период нахо-
дятся в общем на тех же местах, что и в другие времена
года. Они подвижны, в хорошем состоянии и держатся
во мху или в дерновом слое. На залитой пойме наблюдается
значительная скорость течения, вода холодная и содер-
жит много кислорода. В этих условиях черви прекрасно
111
переносят жизнь иод водой и население их в пойме после
спада воды не становится малочисленное (Беклемишев
и Четыркина, 1935).
Все вышесказанное свидетельствует как будто о том.
что .затопление почв водой не может иметь для червей
особого значения и что эти животные по своему образу
жизни могут быть охарактеризованы как земноводные.
Однако это совсем не так. С затоплением почв водой свя-
зано периодически вспыхивающее катастрофическое яв-
ление в жизни населения червей, а именно — их мас-
совая гибель. Широко известно, что после сильных
дождей на поверхности земли появляется огромное коли-
чество червей, частью еще живых, частью умирающих,
частью мертвых. В холмистых и горных местностях, там.
где дождевые потоки образуют временные ручейки, можно
видеть скопления трупов червей, которых можно насчи-
тать сотни и тысячи. Но п на ровном месте, часто после
дождей буквально нельзя ступить шагу, чтобы не уви
деть на поверхности почвы одного или нескольких
червей, находящихся в довольно жалком состоянии.
Это явление было хорошо известно Дарвину, который
писал: «После сильного дождя, следуемого за продолжи-
тельной засухой, замечается часто на поверхности гро-
мадное количество мертвых червей. М-р Гальтон сообщил
мне, что в один из таких случаев (1 марта 1881 г.) на одной
из аллей Гайд-Парка приходилось по одному мертвому
червю на расстояние в два с половиной шага. В одном
месте на протяжении шестнадцати шагов он насчитал
не менее 45 мертвых червей».1
Массовое выползание червей на поверхность после
дождей — очень обычное и широко известное явление.
Самое наименование «дождевые черви», очевидно, воз-
никло в связи с этим явлением у нас или в Германии,
где эти животные носят аналогичное название (Regen-
wurm; в других языках их название не связано с дождем).
По-видимому, когда-то существовало поверье, что черви
падают с дождем с неба. Такое происхождение животных
еще в недавнее время приписывалось неграмотным насе-
1 Приведенная цитата из книги Дарвина показывает, что это
явление наблюдается но только после «продолжительной засухи»:
в Лондоне не могло быть засухи в феврале!
112
лением также и лягушкам, которые во множестве появ-
ляются после дождя.
Естественно, ни один исследователь люмбрицид не
проходил мимо описанного явления, благодаря чему
о нем накопилась порядочная литература. Это внимание
безусловно заслужено, так как к массовой гибели этих
полезных животных нельзя отнестись равнодушно.
Однако до конца понять его причины пока трудно и еще
труднее поэтому сказать что-нибудь о возможностях борьбы
с этим явлением. Последний вопрос еще и не ставился.
Обычно считается, что выползание червей на поверх-
ность земли — следствие недостатка кислорода в почве,
наступающего там после дождей. Доказано, что почва,
богатая перегноем (особенно сухая), поглощает кислород
и поэтому до червей доходит вода, лишенная кислорода,
несмотря на то что пм насыщены дождевые капли. Воздух
же, имевшийся ранее в порах почвы, вытесняется водой.
В результате черви выползают на поверхность земли,
но раньше, чем они достигну! атмосферного воздуха, поги-
бают от недостатка кислорода.
Выше было сказано, что черви очень хорошо переносят
жизнь под водой. Однако этот факт сам по себе еще не
противоречит приведенному объяснению гибели червей
после дождей. Во-первых, черви только в холодной
воде могут жить долгое время, а во-вторых, — что самое
существенное, — вода и мокрая почва — далеко не одно
и то же. В опытах с личинками жуков (проволочниками)
установлено, что эти животные сравнительно хорошо пере-
носят жизнь под водой, но очень быстро гибнут в почвах,
обильно пропитанных водой. М. С. Гиляров (1949) по-
лагает, что это различие обусловлено тем, что во втором
случае смена воды возле животного совершается гораздо
медленнее и, кроме того, большая часть поверхности его
кожных покровов непроницаема для растворенного в воде
кислорода ввиду плотного соприкосновения ее с твердыми
частицами грунта. Очень вероятно, что эти соображения
применимы и к дождевым червям.
Гипотезы о том, что причиной выползания червей на
поверхность почвы служат массовые паразитарные или
инфекционные заболевания червей, отравления и т. д.,
высказывавшиеся в литературе, совершенно невероятны
и легко опровергаются фактами.
8 О. В. Чекановскан ИЗ
Совершенно ясно, что выползание и гибель червей
связаны с пропитыванием почвы водой и недостатком
кислорода. Поэтому отрицать нацело приведенное объясне-
ние, как делают некоторые исследователи (например,
Focke, 1930), невозможно. Однако удовлетвориться им
также нельзя. Прежде всего, очевидно, что это бедствие
наступает у червей только при комбинации каких-то
условии, ближе неизвестных. Сам по себе дождь, любой
силы и продолжительности, гибели червей не вызывает.
Выше уже было сказано, что комбинация выпавшего
дождя с предшествующей засухой — необязательна. Мас-
совая гибель червей наблюдается и после дождей, выпав-
ших на влажную почву, когда поглощение почвой кисло-
рода не может быть столь значительным, чтобы вызвать
внезапное массовое удушье червей. В частности, выпол-
занпе их очень часто наблюдается весной, когда почва
влажная. Предполагалось, что описанное явление про-
исходит, когда после сильного дождя наступает резкое
утреннее похолодание, но и эта гипотеза оказалась несо-
стоятельной. Оно может происходить в период времени
от ранней весны до поздней осени, при очень разнообраз-
ных метеорологических условиях. Следовательно, в чем
состоит та комбинация условий, при которых проис-
ходит внезапная массовая гибель червей, остается неиз-
вестным. Тем самым ни предсказать, ни предупредить это
явление пока невозможно.
Непонятно также, почему черви погибают от удушья
в почве, залитой водой, тогда как в бескислородной среде,
как показывают экспериментальные данные, они живут
по крайней мере часами, а при очень небольших коли-
чествах кислорода в среде неопределенно долго. Во время
дождя падение количества кислорода не может произойти
мгновенно, а много ли червям нужно времени, чтобы вы-
браться из норки на поверхность, принимая во внимание,
что они в нормальных условиях сосредоточены всегда
в верхних слоях почвы?
Интересными опытами Фокке (Focke, 1930) показано,
что вопросы о причинах выхода червей на поверхность
и о причинах их гибели должны быть рассмотрены от-
дельно. Если взять стеклянный аквариум, наполнить его
до половины землей, поместить в нее червей, а затем за-
лить его водой так, чтобы она стояла слоем над поверх-
114
ностью почвы, то черви сразу начинают бурно двигаться и
очень скоро все оказываются на поверхности. При этом
если создать такие условия, что заливание водой нс будет
сопровождаться понижением количества кислорода
в почве, то результат окажется тот же самый: после за-
ливания почвы водой все черви через короткий промежуток
времени выходят на поверхность. Если вводить воду
в почву не сверху, а снизу, все равно черви оказываются
на поверхности. Поэтому никак нельзя сказать, что их
влечет наверх кислород воздуха. Далее, если в закрытый
сосуд с землей вводить сверху, через пробку, азот и вы-
пускать газ через отверстие в дне сосуда, затянутое сет-
кой, то почвенный воздух вытесняется азотом и черви
оказываются в бескислородной среде. Если держать чер-
вей в атмосфере азота более 5 часов, то онп все поги-
бают, но ни один не выходит на поверхность. При этом
сколь медленно ни производилась бы замена воздуха азо-
том, черви на поверхность не выползают. Но если продер-
жать их в атмосфере азота 2% часа, а потом через трубку,
сквозь которую пропускался азот, облить землю водой,
то полузадушенные черви оказываются на поверхности,
а вынутые из сосуда быстро оправляются.
Все рассмотренные факты не позволяют ответить на
вопрос: чем же именно вызывается массовая гибель чер-
вей, происходящая время от времени после больших
дождей? Можно сказать только, что, как обнаружило
исследование, вопрос этот далеко не так прост, как еще
недавно казалось. Он относится к числу тех вопросов,
к решению которых должно быть привлечено внимание
как биологов, так и почвоведов.
5. ВРАГИ И ПАРАЗИТЫ
Кроме резкого нарушения нормальных условий суще-
ствования наводнениями, засухой, морозами и другими
аналогичными причинами, значительное влияние на со-
кращение численности червей имеют хищники и пара-
зиты.
Опасность быть съеденными подстерегает буквально
на каждом шагу этих животных, довольно беспомощных
в отношении активной борьбы с многочисленными вра-
гами, которыми они окружены. К врагам относятся кроты,
8*
115
Рис. 42. Головные концы
Lumbricus terrestris, по-
врежденные кротом. Пер-
вые пять сегментов были
откушены или очень сильно
повреждены, на их месте
образовались аномальные
регенераты. (По Эвансу).
VI — 6-й сегмент.
которые поедают дождевых червей в больших количе-
ствах. Известно, что кроты делают запасы пищи на зиму
в своих ходах. Оказалось, что эти пищевые запасы со-
стоят преимущественно из дождевых червей, причем из-
любленным видом является большой красный червь
(Evans, 1948). Интересно при этом, что кроты откусы-
вают или сильно повреждают головные концы у червей,
собранных ими впрок (рис. 42). В таком состоянии
черви не могут ни уползти, ни закопаться в землю
раньше чем у них закончится
процесс регенерации головного
конца, на что нужно несколько
месяцев. Таким образом, этим
приемом кроты обеспечивают
себе свежее продовольствие на
длительный срок. Кроты осо-
бенно опасны для населения
червей своей многочисленно-
стью.
Вероятно, нападают на чер-
вей и другие насекомоядные
млекопитающие, но об этом
пока мало что известно. За чер-
вями охотятся также прожор-
ливые жабы; они подстерегают
их по ночам у норок и хва-
тают червей, как только те
появляются на поверхности земли. Нередко черви ста-
новятся жертвой и лягушек. Из мелких почвенных жи-
вотных на червей нападают многоножки.1
Очень существенный ущерб населению червей наносят
птицы. Острое зрение позволяет им видеть концы тела
червей, располагающихся неподалеку от наружных от-
верстий норок. Дарвин пишет: «Привычка червей распо-
лагаться неподалеку от поверхности ведет к их гибели
в огромных количествах. Каждое утро в течение значи-
тельной части года разные виды дроздов и, в частности,
черные дрозды на каждой лужайке по всей стране выта-
скивают червей из их норок: этого не могло бы быть,
1 Это наблюдение одного из старейших исследователей дожде-
вых червей — Гофмейстера (1845).
116
если бы они не держались так близко от поверхности».
Современные специалисты подтверждают, что главной
пищей разных видов дроздов являются дождевые черви.
Систематически уничтожают дождевых червей и другие
птицы (Black, 1954). По данным американского Бюро
биологических исследований, не менее чем у 45 видов
птиц в желудках можно найти проглоченных червей.
При этом птицы охотятся не только за взрослыми червями,
но и собирают из земли их яйцевые коконы.
К врагам дождевых червей относятся также конская
пиявка (Haemopis sanguisuga). Она водится в водоемах
всевозможных типов — от озер и рек до пересыхающих
луж. Эту крупную пиявку обыкновенно смешивают с ме-
дицинской пиявкой (Hirudo medicinalis), и вид ее часто
внушает ужас купающимся. Однако это животное со-
вершенно безвредно для человека и крупных животных*
ее тупые челюсти не могут прокусить кожу человека»
да она п не пытается сделать это, так как является ти-
пичным хищником. Она питается не только мелкими вод-
ными животными, но и дождевыми червями. При вскры-
тии этих пиявок в кишечнике их обычно находят прогло-
ченных дождевых червей^ тли их переваренные остатки,
распознать которые можно по щетинкам.
Естественно возникает вопрос: как водное животное
может попасть в число врагов дождевых червей?
Это объясняется тем, что конская пиявка легко пере-
носит длительное пребывание на воздухе. Она может вы-
ходить на сушу, где и охотится на дождевых червей иногда
довольно далеко от берега. Известный английский иссле-
дователь кольчатых червей — Фрэнд (Friend, 1924) описы-
вает и изображает случай единоборства пиявки с дожде-
вым червем, который ему удалось наблюдать. Борьба эта
закончилась проглатыванием червя пиявкой.
Тело дождевых червей почти всегда заселено паразити-
ческими организмами. Излюбленным местом паразитов
являются семенные мешки. В них находили бактерий и
спирохет, однако заразных болезней, возбудителями ко-
торых являются сходные организмы у высших животных
и человека, у люмбрицид неизвестно.
Очень обычным паразитом семенных мешков являются
простейшие животные из класса споровиков — грега-
рины. Рассматривая под лупой семенные мешки любых
117
видов люмбрицид, обычно можно видеть в них белые
пятна, более или менее многочисленные. Это цисты г ре-
гарин. Содержимое семенных мешков является для них,
как и для многих других паразитов, превосходным пи-
тательным материалом. Существенного вреда червям гре-
гарины, по-видимому, не приносят и даже не приводят
к бесплодию, так как сами половые железы ими не по-
вреждаются, а уничтожается лишь некоторая часть
спермы, вырабатываемой, как всегда, в избытке. Однако
со стороны хозяина этих паразитов проявляется реакция
защиты: цисты грегарин окружаются блуждающими клет-
ками полости тела, способными к фагоцитозу, которые,
соединяясь друг с другом, образуют вокруг них капсулы.
Своеобразен способ выведения таких цист, содержащих бес-
численное количество спор грегарин, из организма червя.
Они освобождаются из семенного мешка через разрыв его
стенки п попадают в полость тела. Перистальтическими
сокращениями мускулатуры стенки тела цисты передви-
гаются к заднему концу тела, где могут накапливаться
в больших количествах (от 45 до^ЭО шт.). Из одного сег-
мента в другой цисты попадают через отверстия, имени
щиеся в межсегментных перегородках. Когда задние сег-
менты червя, переполненные цистами, вздуваются, то
это создает достаточные условия для автотомии (см стр. 29).
Задний конец тела отбрасывается и потом восстанавли-
вается путем регенерации. Этот процесс замечателен тем,
что, с одной стороны, при помощи его червь освобождается
от избытка паразитов (скорее всего, именно благодаря
этому грегарины относительно безвредны для червей),
а с другой он обеспечивает расселение паразитов и зара-
жение ими других червей, которые проглатывают с зем-
лей споры грегарин, выпавшие из цист после разложения
отброшенного куска червя. Таким образом, процесс этот
оказывается полезным приспособлением одновременно и
для паразита, и для его хозяина.
Почти столь же обычными паразитами дождевых чер-
вей являются нематоды — мелкие червячки, чаще всего
длиной около 1 мм и очень тонкие. Среди них имеются
полупаразиты, которые часть жизненного цикла проводят
в почве (Pelodera pellio); другие проводят всю жизнь
в теле червей, и только их яйца и личинки в цистах на-
ходятся в земле. Большинство нематод паразитирует также
11«
в семенных мешках, но иногда ими переполнена вся по-
лость тела. Находят их также в сосудах кровеносной
системы. Личинки нематод проникают в тело червей через
поры нефридиев и, быть может, через спинные поры.
Из организма червя нематоды выводятся тем же путем,
что и грегарины, т. е. путем скопления в заднем конце
тела с последующей автотомией.
Наиболее вредными паразитами червей являются ли-
чинки мух поллении (Pollenia rudis) и онезии (Onesia
sepulcralis), из которых лучше изучена первая. Эта муха
по величине и общему виду очень сходна с комнатной.
От последней ее отличает наличие желтого пушка на теле,
а также ряд некоторых признаков, которые указывают на
ее принадлежность к другому семейству. Полления встре-
чается повсюду в больших количествах и может залетать
в дома. Самки откладывают яйца в землю. Вылупляющиеся
из них крошечные червеобразные безногие личинки оты-
скивают дождевых червей и заползают в мужские половые
отверстия. Из видов люмбрицид, зарегистрированных
для СССР, в качестве ее хозяина известна розовая эйзе-
ния. Двигаясь по семепроводам, личинки достигают по-
лости тела половых сег^нтов, где остаются или проби-
раются оттуда в семеннйе мешки. Здесь они кормятся,
растут и проделывают линьку. В одном черве может быть
несколько личинок, но закончить развитие может только
одна из них. Остальные, как это бывает и с нематодами,
окружаются фагоцитами и погибают. Развивается далее
та личинка, которая опередила других и раньше других
закончила линьку. Эта личинка двигается к переднему
концу тела й пробирается в головную лопасть червя
(рис. 43). Здесь она прогрызает стенку глотки и устраи-
вается в переднем отделе кишечника задним концом к ро-
товому отверстию. Размеры личинки увеличиваются, и
вскоре ее задний конец становится видным снаружи: он
выступает из ротового отверстия, которое растянуто и
закрыто телом личинки. В связи с этим червь перестает
питаться и его кишечник постепенно освобождается от
земли, находящейся в нем. Личинка же не довольствуется
больше заглатыванием полостной жидкости с ее содержи-
мым, а попросту съедает всего червя, начиная с переднего
конца. Поперечник ее тела скоро сравнивается с таковым
червя. Личинка еще раз линяет и продолжает съедать
119
сегмент за сегментом черня, двигаясь таким образом к его
заднему * концу. Ткани червя долго остаются живыми,
поэтому растущая личинка вполне обеспечена свежим и
питательным кормом. Впрочем, если червь умирает, ли-
чинка поллении продолжает его съедать, так как она, но-
рме. 43. Личинка поллении в
теле дождевого червя. (По Кей-
лину).
А — задний конец личинки высо-
вывается из ротового отверстия
червя; Б — задняя половина вырос-
шей личинки выступает из тела
пожираемого ею дождевого червя.
1 — личинка; 2 — червь.
добно своим ближайшим ро-
дичам — мясным мухам, не
брезгует гнилым мясом.
Обычно полления съедает
червя до конца, после чего
превращается в куколку
(Keilin, 1915, 1925).
Онезия ведет такой же об-
раз жизни. Она паразити-
рует на навозных и пашен-
ных червях (Eisenia foetida
и Allolobophora caliginosa),
а также, вероятно, и на дру-
гих видах люмбрицид фауны
СССР. Она отличается от
поллении только строением
челюстного аппарата и неко-
торыми другими морфологи-
ческими признаками. Онезия
менее разборчива в отноше-
нии хозяев. Предполагают,
что она паразитирует не толь-
ко на дождевых червях, но и
на птенцах гнездовых птиц.
Личинку одной из мясных
мух (Sarcophaga haemorrhoi-
dalis) также находили в по-
лости тела дождевых червей.
6. ПОЛЬЗА И ВРЕД
Естественно, что столь распространенные и столь
многочисленные животные, как дождевые черви, не могут
не иметь отношения к человеческой культуре.
Рассмотрим сначала, какой вред для человека могут
приносить эти животные.
Небольшой реальный вред дождевые черви приносят
только как переносчики и распространители некоторых
120
паразитов домашних животных. Так, люмбрициды яв-
ляются промежуточными хозяевами опасных паразитов
свиней, червей из класса нематод — так называемых ме-
тастронгилид (три вида рода Metastrongylus). Во взрослом
состоянии эти черви паразитируют в легких свиней, где
часто насчитываются тысячи этих мелких червячков.
В таких случаях животные заболевают, так как у них
расстраивается функция дыхания и, кроме того, червями
выделяются ядовитые вещества, отравляющие организм
хозяев. Борьба с этим заболеванием свиней (метастронгиле-
зом) очень трудна. Особенно опасны эти паразиты для по-
росят, уже начавших самостоятельно питаться. Для них за-
ражение метастронгилидами часто оказывается гибельным.
Личинки метастронгилид развиваются в дождевых чер-
вях, которые проглатывают вместе с почвой яйца и ли-
чинок этих червей, попавших туда с мокротой и испраж-
нениями зараженных свиней. В пищеводе червей крошеч-
ные личинки метастронгилид (длина их 0.2—-0.3 мм) за-
держиваются и, прободая его стенку, попадают в сосуды
кровеносной системы червя, где очень скоро вырастают
до 0.60—0.65 мм. Однако половой зрелости они могут
достигнуть только в легк! : свиней. В кровеносных сосудах
червей личинки могут жить годами. Свиньи и поросята
заражаются метастронгилидами, поедая дождевых червей.
Таким образом, черви содействуют расселению этих вред-
ных животных.
Зараженность червей зависит от количества свиней,
больных метастронгилезом. В очагах заболевания свиней
от 20 до 90°/0 червей могут содержать личинок метастрон-
гилид. По-видимому, все распространенные виды люм-
брицид фауны СССР могут быть промежуточными хозяе-
вами метастронгилид, но наиболее легко подвергаются за-
ражению ими виды рода Lumbricus и навозный червь.
Дождевые черви являются также промежуточными
хозяевами практически важных паразитов птиц — син-
гамид (род Syngamus с шестью видами, в пределах СССР —
преимущественно Syngamus trachea), также относящихся
к классу нематод.1 Сингамиды паразитируют в разных
птицах, в том числе и в домашних (курица, утка, индюшка).
1 Птицы могут заражаться сингамидами и без посредства
дождевых червей, заглатывая яйца этих паразитов. На этом осно-
вании К. М. Рыжиков (1949) называет червей не промежуточными,
121
Цикл развития их очень сходен с таковым метастронги-
лид. Они также живут в легких и в дыхательных путях
своих хозяев. Яйца их с мокротой и пометом попадают
в почву и проходят дальнейшее развитие в теле дождевых
червей (Eisenia foetida, Lumbricus terrestris). Из яиц
сингампд, проглоченных дождевыми червями, вылупля-
ются подвижные личинки, которые прободают стенку
кишечника, попадают в полость тела, живут там некоторое
время, а потом проникают в мускулатуру и там переходят
в покоющееся состояние, окружаясь капсулами. В таком
виде они могут жить в теле червя несколько лет. Поло-
возрелости же сингампды достигают только в случае
заглатывания их птицами. Из кишечника они проби-
раются в кровяное русло, с током крови разносятся по
телу и, попав в легкие, задерживаются там. Паразиты
эти вызывают у кур болезнь, называемую сингамозом.
Особенно опасна она для цыплят, которые погибают при
симптомах удушья, при этом они часто раскрывают рот,
как бы зевая. Поэтому англичане и называют сингамоз
«зевательной болезнью». Очень ,страдают от этих пара-
зитов индюшата.
Главную роль в заражении дождевых червей сингами-
дами играют скворцы, легкие которых очень часто по-
ражаются ими. Как известно, эти птицы прилетают ран-
ней весной, и чаще всего приходится их видеть бродящими
по земле в поисках пищи. Как уже отмечено, их излюблен-
ной пищей являются дождевые черви. Но они не только
уничтожают червей, но наносят им и другой вред: расха-
живая по земле и роясь в ней клювом, они разбрасывают
яйца сингамид, которые проглатываются червями. А не-
сколько позже, к тому времени, когда куры-наседки вы-
ведут потомство и цыплята начинают рыться клювами
в земле, в теле червей их ждут уже готовые инкапсули-
рованные личинки сингамид. Глотая таких червей, цып-
лята подвергаются смертельной опасности заражения
этими паразитами.
а резервуарными хозяевами сингамид. Как увидим из дальнейшего
изложения, дождевые черви являются все-такп особого рода «резер-
вуаром* сингамид: последние проходят там строго определенную
часть своего развития и, кроме того, резервуар активно реагирует
на присутствие личинок сингампд, причем эта реакция неотли
чима от типичной реакции хозяина па присутствие паразита.
122
Другим обыкновенным паразитом домашних и охот-
ничьих промысловых птиц, которым они заражаются,
поедая дождевых червей, является нематода Capillaria.
Образ жизни ее, равно как п поражения, наносимые ею
птицам (капилляриоз) в общем сходны с таковыми синга-
мид (Гагарин, 1953).
Рис. 44. Личинки иорроцека в спинном
кровеносном сосуде дождевого червя.
(По А. Л. Мозговому).
Болезнь домашних уток — порроцекоз — вызывается
нематодой порроцеком (Porrocaecum crassum), паразити-
рующей в тонких кишках различных видов уток (Мозго-
вой, 1952). Массовое заражение этими паразитами может
повести к непроходимости кишечника уток. Промежуточ-
ными хозяевами порроцека служат дождевые черви (Lum-
bricus terrestris). Личинки, вылупляющиеся из яиц, про-
никают в кровеносные сосуды дождевых червей, где встре-
чаются иногда массами (рис. 44).
123
Дождевые черви служат промежуточными хозяевами
для некоторых ленточных червей. Из видов люмбрицид,
известных для СССР, в навозном черве (Eisenia foetida)
нередки финны (стадия развития ленточной глисты в виде
пузырька) цепеня Amoebotaenia sphenoides, паразити-
рующего в половозрелом состоянии в кишечнике домашних
кур, не принося им особенного вреда, а большой красный
червь (Lumbricua terrestris) и длинная аллолобофора
(Allolobophora longa) служат промежуточными хозяевами
для ленточного червя Dilepis undula из кишечника раз-
ных птиц (вороны, грачи, скворцы, дрозды и др.).
Отметим, что хотя некоторый вред, приносимый чело-
веку дождевыми червями как промежуточными хозяевами
паразитов свиней и птиц и несомненен, но все меры борьбы
с указанными заболеваниями домашних животных должны
быть направлены на уничтожение паразитов внутри жи-
вотных и на охрану пастбищ от заражения паразитами.
Отметим также, что борьба с метастронгилидами, сингами-
дами и другими перечисленными выше паразитами имеет
значение и как освобождение от'них населения дождевых
червей, что не может в конечном счете не сказаться благо-
приятно на растительности. Охрана пастбищ от сингамид
имеет значение и как защита страдающих от них ди-
ких птиц, так как червей едят преимущественно полезные
птицы, уничтожающие вредных насекомых. В итоге, как
ни полезна почвообразовательная деятельность дождевых
червей, все же пожертвовать некоторым их количеством
для поддержки полезных птиц, особенно скворцов, не
жалко. Наоборот, хотелось бы спасти этих птиц от син-
гамоза, предоставив им для корма здоровых дождевых
червей.
Не исключена возможность, что дождевые черви могут
приносить вред очень молодым растениям, корневая си-
стема которых может страдать от прокапывания червями
подземных ходов возле поверхности. Например, некоторые
кустики только что пикированной цветочной пли огород-
ной рассады, а также отдельные всходы после посева и
посадки семян могут "быть повреждены или уничтожены
проползающими люмбрицидами. Но в общем вред этого
рода ничтожен.
Что же касается вреда, якобы приносимого червями
хорошо укоренившимся растениям, о чем нередко при-
124
ходится слышать и читать, то нужно сказать, что хорошо
установленных научных данных по этому вопросу не
имеется (Heuschen, 1956). Указания А. О. Лаврентьева
(1958) на вред, приносимый дождевыми червями огород-
ным культурам и садам, нуждается в проверке. Так,
иногда думают, что трава под деревьями не растет потому,
что там водится много дождевых червей. Однако она плохо
растет там совсем по другим причинам. Вымышленным
является также мнение о вреде, приносимом люмбрици-
дами грядковым культурам. Изобретено даже несколько
способов для уничтожения червей в садах. Дарвин пишет
об «уничтожении дождевых червей садовниками» как
о регулярном бытовом явлении: «Когда садовники намере-
ваются уничтожать дождевых червей, то прежде всего
они сгребают с поверхности земли упомянутые куски
извержений для того, чтобы раствор негашеной извести
получил возможность свободно проникнуть в ходы чер-
вей». Дарвин ссылается при этом на изданные инструкции
по уничтожению дождевых червей. Эти инструкции из-
давались за границей еще говеем недавно. »««
Департамент сельского Хозяйства США опубликовал
популярную брошюру в серии «Бюллетень фермера» под
названием «Дождевые черви как наш бич и прочие их
свойства» (Walton, 1928), в которой описываются способы
борьбы с этими «вредителями» при помощи поливки земли
раствором извести, табачным настоем и даже сулемой.
Правда, там в качестве вреда, приносимого дождевыми
червями, на первом месте приводятся неровности, воз-
никающие на площадках для игры в гольф, на местах
кучек извержений червей, а далее следуют сомнительные
данные о повреждении цветов на клумбах. И тем более
странно, что Министерство сельского хозяйства в Америке
нашло возможным пропагандировать среди фермеров
уничтожение дождевых червей, т. е. проведение борьбы
с их лучшими союзниками в обработке почв!
Методы уничтожения дождевых червей можно найти
также в брошюрах по культуре комнатных растений
(см., например, Шипчпнский, 1949). В горшечных куль-
турах крупные черви могут приносить некоторый вред
растениям; однако в общем, как показали многочислен-
ные опыты, черви оказывали положительное влияние на
рост растений и в этих условиях.
125
Пастер указывал на возможность разнесения спор
бактерий сибирской язвы дождевыми червями с трупов
животных, павших от этой болезни и зарытых в землю
(см. также: Шимкевич, 1884). Такое мнение — образец
ошибок, в которые иногда впадают даже великие люди.
Пастер думал, что люмбрициды питаются трупами! Ра-
зумеется, возможность попадания спор бактерий сибир-
ской язвы в кишечник червя после того, как трупы окон-
чательно разложатся и превратятся в гумус почвы, не
исключена, но трудно назвать животных, которые имели
бы меньшее отношение к разделке трупов, попавших
в почву, нежели дождевые черви. Этой работой занято
множество совсем иных животных. Но и им в цивилизо-
ванных странах не удается разносить бактерий сибирской
язвы, так как повсюду закапывание в землю трупов жи-
вотных, павших от этой болезни, производится только
после дезинфекции.
Более серьезны данные о значении дождевых червей
в распространении эпидемий вирусного гриппа. После
эпидемии этого заболевания в 1918 г. в США возникла
гриппозная эпидемия у свиней. Полагают, что свиньи
получают вирус от дождевых червей, в теле которых дей-
ствительно он был обнаружен в межэпидемические пе-
риоды (Гражуль, 1957). Однако вопрос этот нуждается
в дальнейшем исследовании.
Переходя к положительному значению дождевых
червей для человека, прежде всего заметим, что уже из-
давна производилось использование дождевых червей
для разного рода практических надобностей. В Новой
Зеландии туземцы раньше употребляли их в пищу. Ис-
пользовались черви и как лекарственное средство в на-
родной медицине в разных странах, однако рациональных
оснований для применения их с целью лечения не
имеется.
Общеизвестно использование дождевых червей в ка-
честве приманки для рыбной ловли. По-видимому,
ужение рыбы на червя представляет один из древнейших
способов рыбной ловли. В XV в. в Англии появилось уже
руководство по рыбной ловле на удочку. В настоящее
время ловля рыбы на крючки с приманкой имеет не
только спортивное, но и серьезное промысловое значение.
На Средней Волге, например, ловля переметом занимает
126
важное место в промысловой добыче рыбы. При этом лучшей
приманкой считается большой красный червь (Lumbri-
cus terrestris).
Все народные названия дождевые черви получили от
рыбаков. Особенно многочисленны они в Англии, где очень
развит спорт ловли рыбы на удочку. Английский исследо-
ватель Фрэнд (Friend, 1924) приводит 53 народных назва-
ния дождевых червей! Это не означает, однако, что анг-
лийские рыбаки различают 53 вида люмбрицид. В на-
родной номенклатуре один и тот же вид может иметь
много разных наименований и, наоборот, разные виды
носят одно и то же наименование. Некоторые из названий
очень любопытны, например: «беличий хвост» (Lumbri-
cus terrestris), «лососевый червь» (Lumbricus rubellus)
и др.
Черви служат также для корма аквариальных рыб
и комнатных птиц. Поэтому в Западной Европе и в США
дождевые черви — обычный товар на рынке. Появились
промышленники, занимающиеся сбором и разведением
дождевых червей. Город Ноттингам (Англия) издавна
известен как центр промышленности и оптовой торговли
дождевыми червями. В последнее время, когда уже вполне
осознано значение дождевых червей в почвообразовании,
вопрос о разведении дождевых червей стал привлекать
еще большее внимание.
В СССР с недавнего времени дождевыми червями на-
чали интересоваться птицеводы. На многих птицефермах
стали применять кормление птиц дождевыми червями и
разводить их с этой целью. По-видимому, это начинание
имеет серьезные хозяйственные перспективы. Следует
только иметь в виду, что прежде чем давать червей в корм
птицам, необходимо проверять их на наличие в них
сингампд.
Наконец, в недавнее время получены данные о воз-
можной роли дождевых червей в самоочистке почв от
загрязнения радиоактивными изотопами. Как известно,
такое загрязнение может происходить не только от взрыва
атомных бомб, но и в результате неосторожного обращения
с радиоактивными веществами при мирном использовании
их. Растения, выросшие на такой почве, становятся опас-
ными для человека и животных, так как питание ими
может иметь грозное последствие в виде лучевой болезни;
127
хозяйственное же использование такой почвы делается
невозможным. Искусственные приемы очистки почв не-
известны, но автоматическая их очистка происходит пу-
тем промывания дождевой водой, путем эрозии и, главное,
путем накопления радиоактивных веществ в теле растений,
произрастающих на загрязненных почвах. Опыты пока-
зали, что поглощение радиоактивных веществ растениями
совершается несравненно интенсивнее в почвах с дожде-
выми червями, нежели в почвах, лишенных червей
(Передельский, 1958; Передельский и др., 1958).
Все приведенные выше примеры практического зна-
чения дождевых червей говорят о весьма небольшой цен-
ности его по сравнению с участием их в почвообразова-
нии. Эту роль мы имели в виду на протяжении всего
предыдущего изложения. Теперь же пришло время поды-
тожить все сказанное и, пополнив его новыми фактами,
формулировать окончательные выводы.
ГЛ А В А V
ДОЖДЕВЫЕ ЧЕРВИ И ПОЧВООБРАЗОВАНИЕ
I. ОБЩИЕ ЗАМЕЧАНИЯ
Проблема соотношения организма и среды имеет две
стороны: среда не только воздействует на организмы,
изменяя их, вызывая у ник разнообразные приспособле-
ния, обусловливая их закономерное распределение в раз-
ных местностях и различных условиях существования, но
и сама изменяется под влиянием населяющих ее организ-
мов. Поэтому недаром говорят о взаимоотношениях орга-
низма и среды.
В предыдущей главе был рассмотрен вопрос о раз-
ного рода влияниях со стороны среды на жизнь дождевых
червей. Теперь нам нужно рассмотреть, как изменяется
под влиянием дождевых червей та среда, в которой они
живут, т. е. почва. Вопрос этот, помимо значения его для
почвоведения и агрономии, имеет общебиологпческий ин-
терес, так как здесь перед нами очень демонстративный
пример быстрого и легко обнаруживаемого воздействия
на среду со стороны небольшой и четко ограниченной
группы животных.
Для того чтобы понять как следует процесс почво-
образования и правильно оценить роль дождевых червей
в этом процессе, нужно было бы учесть деятельпость дру-
гих животных и растительных организмов, связанных
с почвой, а также рассмотреть и другие факторы почво-
образования. Но сделать это мы не в состоянии, так как
9 О. В. Ченановсная 129
проблема почвообразования составляет основное содержа-
ние всего современного почвоведения — обширной науки,
которая при решении этой проблемы пользуется данными
смежных паук: геологии, бактериологии, ботаники и поч-
венной зоологии. Ограничивая свою задачу рассмотрением
деятельности дождевых червей в почвообразовании, мы
не хотели бы этим дать повод думать, что недооценка всех
прочих факторов почвообразования, которых мы по не-
обходимости касаемся лишь слегка, хоть сколько-нибудь
допустима. Для всестороннего ознакомления с проблемой
почвообразования существует ряд работ, ссылку на кото-
рые интересующийся данным вопросом читатель найдет
в литературном указателе, помещенном в конце книги.
С глубокой древности известно, что в образовании тон-
кого слоя почвы, покрывающей большую часть суши и
обеспечивающей возможность существования всех оби-
тающих на ней живых существ, принимали участие до-
ждевые черви. Аристотель называл червей «кишками
земли». Этим он хотел выразить мысль, что, подобно тому
как в кишечнике животных любые пищевые вещества
превращаются в тончайшую, легко всасываемую пищевую
кашицу, грубая земля, проходя через кишечник червя,
делается мягкой и нежной, благодаря чему растения по-
лучают возможность извлекать из нее нужные нм вещества.
Это представление взято Аристотелем скорее всего от
земледельцев древнего мира, для которых значение до-
ждевых червей в почвообразовании казалось очевидным.
Современный энтузиаст в деле пропаганды разведения до-
ждевых червей в целях улучшения качества почвы, аме-
риканский сельский врач-фермер Баррет рассказывает
в своей книге «Червь-земледелец» (Barret, 1947), что
в 1918 г. он встретил во Франции пожилого крестьянина,
который, сидя на корточках, собирал что-то с земли в мешок.
Когда Баррет, заинтересовавшись спросил, что крестья-
нин делает, тот ответил, что собирает кучки извержений
дождевых червей, чтобы положить их под лучшие из
цветов в своем саду, потому что это ни с чем несравни-
мое по качеству удобрение. Из дальнейшего разговора
выяснилось, что единственным источником сведений о по-
лезной деятельности дождевых червей у этого крестьянина
были устная молва среди односельчан и собственные
наблюдения.
130
В научной литературе мысль о положительной роли
дождевых червей в почвообразовании была впервые вы-
сказана английским натуралистом Гильбертом Уайтом
в его книге, опубликованной в 1789 г., где он пишет, что
земля без дождевых червей была бы «холодной и непита-
тельной».1 Однако основными исследованиями этого воп-
роса до сих пор являются работы Дарвина (Darwin,
1881), который заинтересовался дождевыми червями еще
в молодые годы. В 1837 г., когда Дарвину было 28 лет,
он сделал в Лондонском геологическом обществе доклад
на тему: «Об образовании почвенного слоя». Дарвин из-
ложил в ней теорию, согласно которой частицы почвы
все время выносятся дождевыми червями из глубипы на
поверхность, благодаря чему предметы, лежащие на земле,
оказываются по прошествии немногих лет на глубине
6—10 см под дерном. Таким образом, весь почвенный слой
оказывается прошедшим через кишечник червей. В даль-
нейшей своей жизни Дарвин не утратил интереса к этому
вопросу и продолжал делать наблюдения и собирать ма-
териалы по деятельности дц чдевых червей в почвообразо-
вании. В 1881 г., т. е. через 43 года после первого сообще-
ния и всего за год до смерти, Дарвин опубликовал новое,
более подробное сочинение под названием «Образование ра-
стительного слоя земли деятельностью дождевых червей и
наблюдения над их образом жизни». Эта работа, так же
как п первая, вызвала несколько откликов, но затем на
очень долгое время вопрос этот был забыт и ни почвоведы,
ни биологи к нему не обращались. Открытие почвообразо-
вательной деятельности бактерий отвлекло внимание уче-
ных от вопроса об участии других организмов в этом про-
цессе, и работы Дарвина если и упоминались в курсах
почвоведения, то лишь мимоходом и только затем, чтобы
перейти к более значительным факторам почвообразования.
В работах отца русского почвоведения В. В. Доку-
чаева проявился значительный интерес к почвенным жи-
вотным, а его ученик Г. Н. Высоцкий в конце прошлого
века (1898, 1899) посвятил почвообразовательной деятель-
ности дождевых червей уже специальные исследования и
таким образом явился непосредственным продолжателем
1 Уайт уже в 1770 г. в письме к Баррингтону высказал своп
мысли о значении дождевых червей в почвообразовании.
9*
131
работ Дарвина. Интерес к дождевым червям как почво-
образователям оживился после превосходной работы
Н. А. Димо (1938), а в последнее десятилетие и у нас,
и за границей ряд исследователей (и почвоведов, и зооло-
гов) стал усиленно заниматься разработкой этого вопроса.
С данными этих исследований нам и предстоит ознако-
миться.
Для того чтобы определить возможный масштаб воз-
действия дождевых червей на среду, в которой они оби-
тают, рассмотрим прежде всего с вопросом о численности
их в различных почвах.
2. СВЕДЕНИЯ О ЧИСЛЕННОСТИ ДОЖДЕВЫХ ЧЕРВЕЙ
В ПОЧВАХ
Из сказанного в главах I и IV о питании и образе жпзни
дождевых червей ясно, что деятельность их тесно связана
с поверхностью почвы. Поэтому плотность населения
дождевых червей в природных условиях целесообразно
определять на единицу площади поверхности почвы,
удобнее всего на 1 м2.
В табл. 1 сопоставлены некоторые цифровые данные
о плотности населения червей в разных местностях и
в разных географических пунктах.
Из данных табл. 1 можно видеть, что количество червей
в разных пунктах очень различно, но везде, где были
найдены черви и был произведен их учет, получились
величины в десятки и сотни червей на 1 м2. Большая
часть этих данных может характеризовать плотность на-
селения червей на значительных площадях только с при-
ближенной точностью. Но там, где условия однородны и
плотность населения червей значительна, установленные
цифры могут приблизительно указать число червей, при-
ходящихся на 1 га (умножив цифры таблицы на 10000,
получим плотность населения червей от 200 000 до
5 100 000 экз.). Величины этого же порядка получены
для Северной Америки и долины Большого Нила. Дарвин,
воспользовавшись данными Гензена для Германии, при-
водит в своем труде меньшие цифры, а именно — от
60 000 до 133 000 на 1 га. Однако более новые данные
говорят о том, что и в Западной Европе число червей на
1 га нужно считать сотнями тысяч и миллионами особей.
132
Таблица 1
Количество дождевых червей в разных условиях
и в разных районах СССР
Условия местности Коли- чество червей на 1 м» Район Автор
Луга । 1
Заливные луга, возвышенные места То же, пониже- ния 150 390 Камская пойма у г. Перми. Там же. 1 Четыркина (1926).
То же, пониже- ния Заболоченный луг Луг с мелкими осоками . . . 463 119 80 Ивановская обл. Белоруссия. С Ярославская обл. Балуев (1950). Малевич (19536). Работнов (1938).
Степные почвы
Пустынно-степ- ной глинистый грунт Солончак луго- вой Чернозем степ- ной 61 34 20 Узбекистан (окр. Ташкента). Урал (Троицкая степь). Приазовская степь. Димо (1938). Четыркина (1930). Высоцкий (1898).
Леса
Сосновый лес . . Сосново-дубовый лес Смешанный лес с подлеском и хорошо выра- женным траво- стоем То же, со слабо выраженным травостоем . . 16 96 122 21 Брянская обл. * » Ивановская обл. Шиперович (1937). Балуев (1950).
133
Таблица 1 (продолжение)
Условии местности Колн- ЧССТ о чер|ей на 1 м* Район Айтор
Горные леса . . Ольшанник . . Еловый лес . . Лесные насаждения Насаждения бука То же, с обиль- ной подстил- кой и затене- нием Поля и огороды Поле под трава- ми 2 го года пользования . Опытный участок Черный пар на суглинке . . . То же, на под- золистой почве Поля с почвами, богатыми пере- гноем Культурно-по- ливные степ- ные почвы . . Грунт дорожки около дома . . Парник ;о 350 144 61 ’ 64 192 179 180 64 5 463 512 620 700 Северо-западный Алтай Окр. Киева (Гло- сеево). Московская обл. >> » » »> >> » Зоосад в Ташкен- те. Ивановская обл. Московская обл. Средняя Азия. Сталинская обл. (Великий Ана- дол). Там же. Соколов (1953). Зражевский(1952>. Пономарева (1953). Малевич (1954). Пономарева (1953). Бродский (1937). , Балуев (1950). Гиляров (1942). Димо (1938). Малевич'(1954).
Эти цифры при выяснении численности червей на больших
территориях районов и областей должны быть снижены,
так как в ряде мест (болота, солончаки, пустыни, еловые
леса и др.) червей нет вовсе или они крайне малочисленны.
Кроме того, нужно принять во внимание, что большая
134
часть цифр табл. 1 основана на количественных сборах,
сделанных в сезоны, благоприятные для жизни червей;
в холодные и сухие сезоны их меньше, и не только потому,
что они уходят вглубь п поэтому их труднее находить,
но и вследствие их массовой гибели. Но, принимая во
внимание все эти оговорки, все же можно сделать безоши-
бочное заключение о том, что дождевыми червями более
или менее обильно заселены огромные территории суши.
Для определения значения воздействия на среду тех
или иных организмов необходимо знать не только число
особей, приходящихся на единицу ее объема среды, но
и их размеры. Чем животные крупнее, тем значительнее
следы их деятельности. Поэтому для разбираемого нами
вопроса большое значение имеют вес червей в единице
объема обрабатываемой ими почвы и отношение его
к весу всех находящихся в ней организмов, т. е. к био-
массе в данной почве. Как уже сказано, количество до-
ждевых червей в том или ином объеме почвы в конечном
счете определяется числом' гсобей, приходящихся на еди-
ницу поверхности почвы. Получить хотя бы приблизи-
тельный вес червей, исходя только из числа особей,
путем каких-либо вычислений невозможно, так как вес
червей зависит не только от видового и возрастного со-
става их населения, но и от условий обитания в каждом
отдельном пункте. Не говоря о влажности, температуре,
количестве пищи и т. д., имеются, например, сведения,
что представители одного и того же вида могут иметь раз-
ные размеры в зависимости от обитания их на той или
другой высоте над уровнем моря.
Вес отдельных червей может колебаться от сотых
грамма (мелкая молодь) до 5—6 г (крупные экземпляры
большого красного червя). Вес же червей на 1 м2 приво-
дится исследователями довольно редко. Для зауральских
степей при средней численности червей па луговом со-
лончаке в 34.4 особи на 1 м2 вес их при пересчете на 1 га
оказывается равным 49 кг, т. е. средний вес одного червя
составляет 0.11 г; очевидно, черви там очень мелкие.
В Ветлужских лесах Горьковской области (Шпперович,
1937), в бору с подстилкой лиственных мхов (кукушкин
лен — Polytrichum), где черви немногочисленны (6 экз. на
1 м2), их вес на 1 га равен И кг (средний вес одного червя
0.18 г). Гораздо более высокие цифры веса червей при-
135
водятся для лесов северо-западного Алтая (Соколов, 1956),
их там может быть до 3 т. на 1 га. Для луговых и возделанных
почв иногда указывались очень большие величины —
от 1300 до 6500 г на 1м2, т. е. от 13 до 65 т червей на 1 га
(France, 1921). Для полей штата Огайо (Северная Америка)
определено 2248 кг (2.25 т) червей на 1 га при 247 особях
на 1 м2 (средний вес одного червя 0.91 г).
Видовой состав дождевых червей Северной Америки
почти тот же, что и в большей части СССР. Поэтому на
наших полях и лугах, судя по данным, приведенным
в табл. 1, можно ожидать таких же или еще больших ве-
личин живой массы червей. По расчетам американских
исследователей, вес всех червей в США превосходит в 10 раз
вес людей, населяющих эту страну (цит. по Barret, 1947).
Вес червей составляет в лесах от 50 до 72% всей био-
массы почвы. Для Западной Европы имеется указание,
что вся биомасса в 1м2 почвы при наличии в ней 200 до-
ждевых червей равна 500 г (Franz, 1950). Если принять
во внимание, что количество» дождевых червей на 1 м2
и их видовой состав примерно те же, что и в штате Огайо,
то, произведя вычисление, можно прийти к выводу, что
и в исследованных местах Западной Европы дождевые
черви составляют примерно 50% биомассы почвы.
Как уже неоднократно упоминалось в предыдущих
главах, в любом вопросе, связанном с деятельностью
дождевых червей, очень большую роль играет точный
учет видового состава населения червей той или иной
территории. Отсюда ясно, что приведенные нами только
что суммарные данные о численности люмбрицид и их
весе могут служить только для самой общей ориентировки
в этом вопросе.
Мы не можем за недостатком места углубляться в рас-
смотрение численности разных видов в различных грунтах,
комплексов видов, сопутствующих друг другу, и т. д.1
Приведем только один пример, показывающий, как сильно
изменяются численность и видовой состав люмбрицид в за-
висимости от внешних условий и как мало говорит одна
суммарная величина численности червей на том или ином
участке почвы о земледельческой работе червей.
1 Нужно сказать, что эти вопросы и в специальной литературе
еще очень мало разработаны.
136
В Белоруссии был произведен точный количественный
учет червей в ряде пунктов на склоне, занятом картофель-
ным полем и переходящем в болотистый луг (Малевич,
1953а). Результаты учета изображены на рис. 45. Из
обнаруженных в этом участке четырех видов люмбрицид
пашенный червь (Allolobophora caliginosa) лучшие усло-
вия существования нашел в нижней трети склона; он идет
Рис. 45. Состав фауны дождевых червей и их
численность в разных частях склона и луга.
(По И. И. Малевичу).
1 — Allolobophora caliginosa; 2 — Lumbricus rubellite;
з — Dendrobaena octaedra; 4 — Octolaaium lacteum.
довольно далеко в верхние, более сухие и выщелоченные его
части, но лишь в ничтожных количествах заходит на сырой
луг внизу склона (см. кривую!). Дальше мы еще встре-
тимся с тем, что работа червей в верхних частях* склонов
имеет особенно важное значение. Малый красный червь
(Lumbricus rubellus) более влаголюбив: он почти отсут-
ствует в верхних частях склона, но зато далеко идет
на луг (см. кривую 2). Молочный октолазий (Octoiasium
lacteum) и восьмигранная дендробена (Dendrobaena octa-
edra) явно избегают склона с картофельным полем и со-
137
средоточены на сыром лугу (кривые 4 и 3). Лучший «земле-
делец» из этих видов несомненно — пашенный червь.
Значение в почвообразовании восьмигранной дендро-
бены ничтожно, так как это маленький червячок, обитаю-
щий преимущественно во мху и в самых поверхностных
слоях почвы. Малый красный червь хотя и полезен тем,
что поглощает неразложпвшиёся растительные остатки,
но глубоких ходов также не делает, и этим его значение
в почвообразовании ограничено. Что касается молочного
октолазия, то хотя о его образе жизни мало известно, но,
поскольку обычным местом обитания особей этого вида
являются почвы с избыточной влажностью и плохо
дренированные, можно думать, что и его значение как
почвообразователя не так велико. Таким образом, зона
с максимальным количеством червей в самом низу склона
(204 экз. на 1 м2) отнюдь не является зоной наиболее ин-
тенсивной деятельности дождевых червей в почвообразо-
вании.
Рассмотренные выше данные о численности дождевых
червей обосновывают лишь самую возможность постановки
вопроса о деятельности дождевых червей в почвообразо-
вании и в самой общей форме характеризуют ее масштабы.
Вспомним, что червь в день .пропускает через свой кишеч-
ник количество земли, равное весу его тела. Приняв
средний вес червя в 0.5 г, а количество их на 1 м2 в 50 осо-
бей (500 000 экз. на 1 га — цифра невысокая, как видно
из просмотра данных табл. 1), получим, что за сутки
через кишечник червей на площади в 1 га проходит
250 000 г, т. е. 0.25 т земли. Если принять далее, что
активная деятельность червей продолжается 200 дней
в году, то количество земли, прошедшей через кишечник
червей, выразится в 0.25x200=50 т на 1 га. Сделав соответ-
ственное вычисление для почв, содержащих 400—600 чер-
вей на 1 м2, получим колоссальные количества земли,
переработанной за год на 1 га, — 400—600 т.
Недоверие к столь большему значению деятельности
дождевых червей, как было указано еще Дарвином, пред-
ставляет собой «пример неспособности людей оценивать
результаты, получающиеся от постоянно действующих
причин». Дарвин настойчиво указывал также, что нельзя
не придавать значения совокупной деятельности мелких
факторов и что выражение «о ничтожном закон не бес-
138
покоится» к науке пе применимо. Суточная работа одного
червя, проглатывающего и извергающего свои 0.1—1.0 г
земли, безусловно ничтожна. Но ведь червей — бил-
лионы, причем они глотают и буравят землю не один
день, а проделывают эту работу неутомимо и непрерывно
в течение большей части года. С разными сторонами этой
грандиозной работы и с ее результатами мы сейчас озна-
комимся.
3. ВЛИЯНИЕ ДОЖДЕВЫХ ЧЕРВЕЙ НА ПРОВЕТРИВАНИЕ
И ДРЕНАЖ ПОЧВ
Уже самое наличие ходов дождевых червей в почве
изменяет ее свойства. Ранее мы отметили значение скваж-
ности почвы. Совершенно ясно, что чем больше дождевые
червп проделают ходов в почве, тем более благоприятные
условия будут созданы для проникания в нее воздуха
и воды. И то, и другое обязательно для ряда химических
процессов в почве, а, главное, воздух и вода составляют
непременные условия для жизни почвенных организмов,
в первую очередь бактерий и грибков, деятельность
которых играет выдающуюся роль в снабжении корневых
систем высших растений необходимыми для них веще-
ствами. Вещества перегиб почвы превращаются микро-
организмами в растворимые химические соединения, и
с помощью корней растения имеют необходимые для них
азот, фосфор, калий и другие элементы. Воздух в почве
имеет значение еще и как источник получения азотистых
соединений, что совершается особыми почвенными бакте-
риями. Таким образом, дождевые черви содействуют осу-
ществлению и этого замечательного звена в процессе
круговорота азота, облегчая циркуляцию воздуха в почве
и проникновение его в глубокие почвенные слои.
Мы уже видели также, что полости в почве, различного
происхождения и всевозможных размеров, представляют
собой главные места обитания разных групп и мелких поч-
венных животных, в свою очередь принимающих участие
как в изготовлении перегноя, так и в его дальнейшей
обработке.
Почвы, населенные дождевыми червями, бывают очень
обильно пронизаны их ходами (рис. 46). Это происходит
благодаря тому, что один червь может прорыть целую
139
систему ходов, сообщающихся друг с другом и выходящих
в нескольких местах на поверхность. Например, наблю-
дения за работой в почве одного пашенного червя в спе-
циально сделанном для этой цели террариуме со стек-
лянными стенками показали, что червь, сначала углубив-
шись в землю, вышел из нее обратно, проделав другое
Рис. 46. Ходы червей в почве и корни
растений. (По Вилке).
наружное отверстие (рис. 47). На другой день червь
вернулся в глубину, сделал там горизонтальный ход и
вышел опять на поверхность. Дальше он то пользовался
старыми ходами, то прорывал новые, и через 21 день
образовалась сложная система частью заброшенных и
засыпавшихся, частью действующих ходов.
Чтобы хорошо были видны отверстия ходов, нужно рас-
чистить их от мелких комков земли. Эту работу удобнее
140
всего произвести с помощью ручных мехов или каучукового
баллона с наконечником. Для подсчета отверстий в них
удобно вставлять соломинки. Ходы червей — довольно
прочные образования и сохраняются даже после очень
сильных дождей. Это объясняется тем, что стенки ходов
покрыты слоем слизи и испражнений червя (рис. 48),
которые лишь с большим трудом поддаются разрушению
водой.
Подробные исследования количества ходов червей
в почвах Средней Азии проведены Н. А. Димо. Он нашел
Phcs 47. Ходы, Сделанные пашенным чер-
вем (Allolobophora caliginosa) в узком тер-
рариуме со стеклянной стенкой с 9 по оО
декабря. Заштрихованы ходы, наполнен-
ные экскрементами. (По Эвансу).
весной на целинной почве (пустынно-степной тяжелый
глинистый светлозем) в среднем 176 ходов на 1м2 со сред-
ним диаметром в 0.57 см. В сумме площадь этих ходов
на 1 м2 составляла 50 см2, т. е. 0.5% от всего объема почвы
было занято ходами червей. На поливных участках куль-
турных земель Средней Азии число ходов оказалось
значительно большим, оно достигало 1692 на 1 м2. Под-
счет показал, что при этом больше 1% объема почвы
было занято ходами червей. На поливных землях в мяг-
ких, неутоптанпых местах количество ходов под поверх-
ностью почвы настолько велико, что поверхностный слой
представляет отделившуюся корку, разбитую на участки
в 2—3 см, под которой располагается взрыхленный и на-
141
Рис. 48. Участок хода
дождевого червя.
Степка его цементи-
рована засохшими
слизистыми выделе-
ниями червя и густо
оплетена тонкими
разветвлениями кор-
ней растений.
(По Вилке).
Полненный извержениями слом в 5—6 см (Димо* 1938).
В почвах северо-западного Алтая, где водятся очень круп*
ные черви, площадь, запятая их ходами, составляет
1.5—2.0 от всего объема почвы (Соколов, 1953).
Г. Н. Высоцким (1858) в При-
азовской степи произведены подсчеты
числа ходов червей на разной глу-
бине (табл. 2).
Из табл. 2 видно, что с глубиной
число ходов падает, но еще на глу-
бине 8 м на 1 м2 приходится значи-
тельное число ходов.
Не все виды червей способны
делать столь глубокие ходы; это
свойство мариупольской дендробены
(см. стр. 109). Таким образом, почва
пронизана густой и глубокой сетью
дренажных каналов, общее протя-
жение которых под поверхностью
почвы в 1 м2 составляет больше 8 км.
Почвы средней полосы европей-
ской части СССР дренированы хо-
дами червей не на столь большую
глубину, по так же очень значи-
тельно, причем на полях сеть их
ходов значительно гуще, чем в лесах
(табл. 3).
Ходы червей в земле идут в раз-
ных направлениях и под любым
углом к горизонту. Стенки их про-
питаны высохшими слизистыми вы-
делениями кожи, что придает им
большую прочность по сравнению
со случайными трещинами в почве,
разным причинам. Дренаж почвы,
дождевых червей, приобретает
Аэрация и дренаж —
по
возникающими
образованный ходами
особенно серьезное значение,
важные факторы плодородия почвы. Невентилируемые
и недренируемые почвы, которые сельские жители
называют мертвыми, лишены дождевых червей и аг-
рономически — очень низкого качества. Разумеется,
не отсутствие червей делает их такими. Наоборот
142
червей там нет в силу химических особенностей почвы.
Но если качества почвы таковы, что она является при-
годной для жизни червей, то ценность ее несомненно
повышается пробуравливанием червями.
Укажем еще на одно важное следствие наличия ходов
червей в почве. В степных засушливых районах расте-
ниям приходится добывать влагу с больших глубин и
корням, чтобы достигнуть воды, находящейся на глубине
нескольких метров, необходимо пробиться через толщу
совершенно высохшего грунта. Как установлено Г. Н. Вы-
Таблица 2
Количество и характер ходов дождевых
червей в Приазовской степи.
(Из Г. Н. Высоцкого, 1898)
Количество ходов
Глубина
(в м) общее на 1 м’ с корнями растений полых
1 525 80 100
2 400 90 150
3 350 /' 75 170
4 320 50 150
5 240 35 110
6 160 15 60
7 130 5 30
8 110 1 15
Таблица 3
Количество ходов червей в поле и в лесу.
(Из М. Е. Ткаченко, 1908)
Глубина (в м) Паровое поле Лес
дубовый еловый
0.1 232 152 12
0.2 228 152 8
0.3 272 88 52
0.4 208 64 20
0.5 96 24 20
143
соцким, корни дорастают до водоносных слоев, пользуясь
ходами червей. На глубине 2 м нет ни одного корня, кото-
рый пробил бы себе дорогу самостоятельно; они идут внутри
ходов дождевых червей (рис. 46). Часто корни спаиваются
вместе внутри хода, так как они оказываются сдавленными
окружающим их сухим грунтом. Вероятно, аналогичную
роль играют, кроме ходов червей, пустоты, образующиеся
на месте сгнивающих корней.
'По ходам червей проникают в глубокие слои почвы не
только воздух, вода и корни растений: стенки ходов и
граничащие с ними участки почвы заселяются микробами
и другими почвенными организмами; ходы червей служат
проводниками жизни в глубь почвы.
4. ВЛИЯНИЕ ДОЖДЕВЫХ ЧЕРВЕЙ НА ПЕРЕМЕШИВАНИЕ
ЗЕМЛИ И ВЫНОС ЕЕ НА ПОВЕРХНОСТЬ
Переходим к результатам заглатывания почвы дожде-
выми червями.
После прохождения через кишечник червей комочки
почвы выбрасываются в виде копролмтов. Это название
происходит от греческих слов kopros — что означает «на-
воз», «испражнения», и lithos — «камень», т. е. в целом —
«навозные камешки». Копролпты представляют собой
сферические или удлиненные комочки земли размером
1—5 мм. У свежевыброшенных копролитов — гладкая
поверхность; они могут склеиваться друг с другом в тельца
(агрегаты) размером до 20 мм и более. Копролпты выбра-
сываются червями в виде кучек высотой 0.3—1.5 см,
закрывающих обычно наружное отверстие хода червя»
Эти кучки называют выбросами червей; их можно наблю-
дать повсюду (рис. 49). Особенно хорошо они видны на
дорожках в парках, на немощеных открытых площадках
или под деревьями.
Как мы знаем, вся деятельность червей совершается
главным образом ночью. Большая часть выбросов обра-
зуется также ночью. Поэтому они особенно обращают
на себя внимание по утрам в хорошую погоду (в холод-
ную и дождливую погоду не активны). В фауне СССР
выбросы, сходные с таковыми дождевых червей, образуют
только мокрицы пустынь Средней Азии (Hemilepistes).
144
До известной степени сходны с ними кучки земли,
выносимые на поверхность почвы некоторыми видами
муравьев. Но эти кучки неправильной формы состоят из
мельчайших частиц почвы, и чего-либо похожего на копро-
литы в них не наблюдается.
Не следует думать, что все копролпты выносятся на
поверхность земли. Значительная часть их откладывается
в подземных ходах, которые оказываются заполненными
ими на значительном протяжении. Сначала рассмотрим,
однако, выбрасывание копролитов на поверхность; это
явление привлекло в свое время внимание Дарвина и
послужило объектом его многолетних наблюдений.
Рпс. 49. Кучка копролитов. (Ориг.).
Дарвин заметил, что предметы, лежащие на поверх-
ности земли, оказываются через известное время под
нею тем глубже, чем больший срок времени прошел с того
момента, когда они были наверху.
«Английским фермерам, — пишет он, — хорошо из-
вестен факт, что предметы, оставленные на поверхности
пастбища, исчезают по прошествии времени или, как они
выражаются, „закапываются44. Очевидно, они не зада-
вали себе вопроса, каким образом могут известковая
мука, шлаки и тяжелые камни проникнуть, и притом еще
с одинаковой скоростью, через покрытую дерном поверх-
ность». Интересно, что самая мысль о вынесении на по-
верхность земли извержений червей как о причине ка-
жущегося погружения тел, находящихся на поверхности
почвы, была подсказана Дарвину, как он пишет в преди-
словии к своей классической работе 1881 г., его дядей
Д. Веджвудом. Эта мысль настолько поразила Дарвина,
10 О. В. Чекановскан 145
что послужила толчком к его исследованиям данного
вопроса, которые он вскоре предпринял и не оставлял
в течение всей жизни. В одном из своих писем, написан-
ных уже в пожилом возрасте, он пишет: «Моя душа все
еще целиком поглощена червями». В этих словах ярко
отразилась его способность, свойственная гениям, —
видеть великое в малом.
Приведем одно из наблюдений Дарвина. «Участок
болотистой пустоши был обнесен оградою, дренирован
и покрыт в 1822 г. толстым слоем жженого мергеля и шла-
ков. Он был затем засеян семенами трав и представляет
собою в настоящее время хорошее пастбище. В 1837 г.,
т. е. через 15 лет, на нем были вырыты ямы, и на прила-
гаемом здесь рисунке [рис. 50] видно, что участок покрыт
дерном в 1.25 см толщины, а под ним слой земли, богатый
перегноем, в 6.25 см. Этот слой не содержал никаких
обломков, но под ним лежал слой почвы толщиной около
4 см, переполненный кусками жженого мергеля, бросаю-
щимися в глаза благодаря 'красному цвету. Этот слой
содержал, кроме того, куски каменноугольных шлаков
и белые кварцевые камешки. Под этим слоем на глубине
11.5 см залегал черный торфяной песок, содержавший
немного кварцевых камешков.1 Следовательно, здесь куски
жженого мергеля и каменноугольных шлаков были по-
крыты в течение 15 лет слоем почвы толщиной в 6.25 см,
не считая дерна. Шесть с половиной лет спустя это поле
было вторично исследовано, и почвенный слой из камней
был найден на глубине от 10 до 12.5 см. Таким образом,
в течение 6% лет поверхностный слой почвы стал толще
на 4 см. В среднем для всего периода времени почвенный
слой ежегодно утолщался приблизительно на % см».
Со скрупулезной точностью Дарвин приводит один
за другим аналогичные примеры. Крупные камни, упав-
шие на землю, также постепенно погружаются под слой
дерна медленно, но верно идущим процессом выбрасы-
1 Жженый красный мергель, — очевидно, отходы от приго-
товления негашеной извести из мергеля (рухляка). Очевидно, они
были выброшены вместе с каменноугольным шлаком на болотистый
лужок отчасти с целью его осушения, а отчасти имелось в виду
его использовать, как место свалки. В черной торфяной почве
бывает довольно много червей. Интересная выдержка из Дарвина
несколько сокращена, а толщина слоев, выраженная у Дарвина
в дюймах, приводится нами в сантиметрах. Прим, автора.
146
наиия червями на поверхность кучек копролитов с по-
следующим разравниванием их силами ветра и дожде-
вой воды (рис. 51). То же происходит со щебнем на до-
рожках, с кирпичами и другими предметами. При этом
скорость их погружения под дерн не зависит от их удель-
Рис. 50. Разрез почвы, сделанный 1 через
15 лет после того, как ее поверхность
была засыпана слоем мергеля и шлака.
В результате деятельности дождевых чср
вей мергель и шлак оказались зарытыми
в почву. (По Дарвпну)
а — дерновина; б — почва, богатая перегноем;
в — слой мергеля и шлака, бывший ранее на
поверхности; г — слой почвы, бывший некогда
на поверхности.
кого веса: шлак, известняк, кварц, кирпич закапываются
с одной и той же скоростью.
Установив эти факты, Дарвин естественно заинтере-
совался древними постройками и развалинами, которые
в случае справедливости созданного им представления
о погружении под землю всех предметов, находящихся
на ее поверхности, должны были дать показательные об-
10*
147
разцы этого процесса. В Англии очень много остатков
древних построек. Дарвин сам принял участие в раскоп-
ках римской виллы, разрушенной 1400—1500 лет назад,
чтобы сделать нужные наблюдения. Кроме того, сыновья
привозили ему из более отдаленных мест Англии точные
описания и чертежи, демонстрирующие большие толщи
земли, накопившиеся за ряд столетий над обломками
фундаментов, стен, полов и других остатков римских вилл,
монастыря, разрушенного Генрихом VIII, и т. д. Везде
Дарвин находил доказательства правильности своего
взгляда на значение деятельности дождевых червей в по-
гружении предметов в глубь почвы.
Рис. 51. Камень, лежавший на поверхности,
постепенно погружается в землю. (По Дар-
вину).
А А — исходный уровень почвы.
Самый факт накопления земли над лежащими на ней
предметами, как уже сказано, общеизвестен, и каждый
несомненно с этим явлением сталкивался: крупные
камни в поле (валуны) обычно лишь небольшой своей
частью выступают над поверхностью почвы, старые мо-
стовые оказываются глубоко под землей, места с остатками
всевозможных древностей археологи всегда добывают
путем раскопок. Что же касается причин этого явления,
то Дарвин хорошо сознавал, что здесь кроме деятельно-
сти дождевых червей, нужно принять во внимание и дру-
гие факторы, т. е. других животных (особенно кротов
и муравьев), а также ветер и потоки дождевой воды.
В местностях с сухим климатом и обилием незадернован-
ных поверхностей почвы главную роль в засыпании пред-
метов, лежащих на земле, играет ветер, поднимающий
и переносящий огромные количества пыли. Равным об-
разом в некоторых местах наибольшее значение в этом
148
явлении имеют потоки дождевой воды. Но в условиях
влажного климата и на ровных местах главная роль
в нем несомненно принадлежит роющим животным,
в основном дождевым червям.
Были поставлены многочисленные^опыты с целью
воспроизвести в искусственных условиях явления, ко-
торые наблюдал Дарвин; На (рис. 52) изображен резуль-
тат опыта, состоявшего в том, что в одну половину стек-
Рис. 52. Погружение белых фаян
совых пластинок в почву деист
вием роющихся червей. (Из Мп-
ха эльсена).
лянного сосуда с плоскими
стенками была насыпана
земля, в другую — круп-
ный гравий, не проходи-
мый для дождевых чер-
вей. Сверху почва и гра-
вий были покрыты прямо-
угольными белыми пла-
стинками из фаянса. В зем-
ле содержались дождевые
черви, которым давалась
пища, и в ней поддержи-
валась нужная влажность. ‘
По прошествии некото-
рого времени, как можно
видеть в левой части
рис. 52, керамические пла-
стинки оказались глубоко
под поверхностью почвы,
а в части сосуда, не до-
ступной для червей, они
остались в прежнем поло-
жении, на поверхности.
Ознакомимся с некоторыми данными о количестве
извержений, выбрасываемых дождевыми червями на по-
верхность почвы. Вес одной кучки выбросов может коле-
баться в очень широких пределах. Н. А. Димо приводит
для свежих кучек величины в 0.6—3.7 г при среднем
весе в 2.4 г, А. Л. Бродский для высушенных кучек из-
вержений — 0.2—0.27 г. В еще более сильной степени
варьирует количество кучек выбросов, приходящихся на
единицу площади. Безусловно, оно прежде всего нахо-
дится в зависимости от плотности населения червей (по
не только от нее).
149
Таблица 4
Количество выбросов дождевых червей на различных
участках дерново-подзолистой почвы в Московской области
(Из Пономаревой, 1953)1
Участок Выбросы, со- бранные на по- верхности (в т на 1 га) Выбросы в почве, в воне корневых систем Общее количест- во выбросов до- ждевых червей Число дождевых червей на 1 га
в т на 1 га 1 в % к весу всей почвы
Посевные травы 1-го года поль- I
зования .... 7.8 4.0 9.7 11.8 1520000
То же, 2-го года
пользования 26.2 25.8 48.1 52.0 1790000
Озимая пшеница 11.2 2.3 20.2 13.5 880000
Пар, занятый кар-
тофелем .... 3.2 Не опреде- лено Не опреде лено — 800000
Рожь с подсевом
трав 2.7 6.2 13.6 8.9 320000
Лес дубовый . . j 16.0 Не опреде- лено Не опреде- лено 2940000
Как можно видеть из табл. 4, количество выбросов
в одном и том же географическом пункте и на почвах
одного типа колеблется в чрезвычайно широких преде-
лах. Приводятся и другие данные, говорящие о возмож-
ности как значительно ббльших, так и меньших количеств
извержений люмбрицид на поверхности почвы, приходя-
щихся на единицу площади. Так, в Германии было со-
брано в сосновом лесу 5684 г копролитов на 1 м2: а на
задернованной площадке для игры в гольф 8125 г, что
при пересчете на 1 га дает 56.8 и 81.25 т (Stockli, 1929).
В то же время аналогичный подсчет количества изверже-
ний дождевых червей на луговой почве в долине Большого
Нила в Судане дал значительно меньшие цифры, а именно—
2.0—7.5 т на 1 га при численности червей 1 330 000 осо-
бей на 1 га (Боже, цит. по Димо, 1938). Пока заметим,
1 Подсчеты выбросов делались па основании однократных
сборов на площадках 0.25 м2 с последующим пересчетом на 1 га.
150
что эти цифры характеризуют количество земли, выне-
сенной на поверхность за какой-то длительный, но ближе
неопределенный промежуток времени. Но они дают пред-
ставление о том, что на поверхность земли червями вы-
брасываются очень значительные количества почвы, вы-
ражающиеся тоннами и даже десятками тонн на гектар.
Точный учет скорости процесса накопления выбросов
дождевых червей на определенной площадке был произ-
веден Дарвином. Для этого на старой насыпи, поросшей
редкой травой, им были выбраны две квадратные площадки
размером в 1 квадратный ярд (0.85 м2) и с них были
тщательно удалены все уже бывшие там извержения чер-
вей. Условия жизни для червей были не особенно благо-
приятными, так как на поверхности песка и шлаков на-
ходился слой темной почвы толщиной не более 10 см.
С одной из выбранных площадок регулярно собирались
извержения дождевых червей в течение года (с 9 X 1870
по 14 X 1871). Все извержения тщательно высушивались
и взвешивались. Оказалось, что за год они составили
в сумме 1600 г, т. е. при пересчете на 1 м2 дали 1867 г,
или 18.7 т на 1 га. С другой очищенной площадки сбор
был произведен однократно по истечении года. На ней
оказалось'при пересчете на 1 га 39.8 т извержений. Если
взять среднее из этих цифр, т. е. около 30 т на 1 га, то
общий объем выброшенных на поверхность извержений,
как показывает расчет, составит приблизительно величину,
необходимую для того, чтобы покрыть площадь в 1 га
ровным слоем в 0.24 см.1
Значительная часть выбросов смывается водой, уно-
сится ветром и т. д. Поэтому толщина слоя почвы, созда-
ваемого за год выбросами дождевых червей, в итоге зна-
чительно меньше. Например, Эванс (Evans, 1948b) вы-
числил, что на старом пастбище Ротамстэдской сельско-
1 Определение толщины слоя, образующегося при равномер
ном распределении 30 т почвы на площадь в 1 га, т. е. на 10 000 м2.
сделано следующим образом. Объем 1 т почвы принят равным 0.8 м9.
Значит, объем 30 т почвы равен приблизительно 24 м9. Так как
объем тела, определяемый произведением трех его измерений
(длины а, ширины Ь и высоты с) в данном случае равен 24 м3, т. е.
а • Ъ-с= 24, а произведение а • b (площадь в 1 га) задано равным 1000*),
24 24
то искомая толщина слоя (С) будет С = = уд ддд = 0.24 см
151
хозяйственной опытной станции (Англия) за 300 лет вы-
бросами червей образован слой почвы толщиной в 10.5 см.
Это показывает, что на участках с отнюдь не максималь-
ной плотностью населения дождевых червей интенсивность
их деятельности достаточна, чтобы обусловить измери-
мую величину ежегодного прироста толщины почвы.
Большое значение имеет вопрос о влиянии внешних
условий на интенсивность образования выбросов дожде-
вых червей на поверхность почвы. Н. А. Димо заметил,
Рис. 53. Количественные показа-
тели деятельности дождевых чер-
вей и их численность по месяцам
на площадке в 1 м2.
(По А. Л. Бродскому).
1 — количество кучек; 2 — вес копро-
литов (в г); з — количество червей.
тидневку опять возросло и <
до дождей.
что на одной и той же
площадке в 1 м2 количе-
ство образовавшихся кучек
извержении бывает очень
различным; оно испыты-
вает изменения в зависи-
мости от сезона и погоды.
В холодную и дождливую
погоду число кучек из-
вержений резко сокра-
щается.
Например, после про-
шедших дождей количе-
ство кучек извержений на
одной из площадок сокра-
тилось с 139 до 21 за 5
дней, а в следующую пя-
ало еще больше, чем было
Изменение количества выбросов дождевых червей
по сезонам прослежено А. Л. Бродским. По примеру
Дарвина он на выбранных метровых площадках, предва-
рительно очищенных от извержений червей, собирал
каждые 2—3 дня вновь образовавшиеся кучки, произ-
водил их подсчет, определял сухой вес и параллельно
на сходных по характеру площадках в 1 м2 выбирал и со-
считывал количество червей. Сопоставление этих данных
изображено на рис. 53. Как можно видеть, количество
червей меняется от 100 до 280 на 1 м2, вес кучек изверже-
ний меняется еще более значительно. Это относительное
постоянство плотности населения червей объясняется тем,
что исследование производилось на поливных участках
в Узбекистане и условия влажности поэтому были до-
152
вольно постоянными. Все же и при этих условиях кривая
веса выбросов на поверхность дает крутой подъем с ав-
густа по октябрь. Таким образом, интенсивность почво-
образовательной деятельности дождевых червей зависит
не только от количества особей, но и от других причин,
в частности от физиологического состояния червей, ко-
торое определяется рядом условий, в том числе сезонными
и погодными.
При учете скорости накопления извержений червей
на поверхности почвы нужно учесть еще и многое другое.
Прежде всего имеет значение видовой состав люмбрицид,
что видно из следующего опыта. В глиняные горшки с поч-
вой емкостью около 8—10 л помещались разные виды
дождевых червей , по 20 экземпляров и опавшие листья
различных древесных пород по 30 г. Через 4 месяца было
произведено взвешивание копролитов, образовавшихся
при разных условиях; результаты этого опыта приведены
в табл. 5.
Таблица 5
С
Вес извержений, выброшенных на поверхность почвы
дождевыми червями в лабораторном опыте, в зависимости
от вида и от характера пигцп (в граммах на 1 червя
за 4 месяца). (Из А. И. Зражевского, 1954а)
Виды Листья Контроль (без листьев)
дуба клена полевого ольхи черной
Пашенный червь (Allolo- bophora caliginosa . . . 13.8 28.1 27.8 8.1
Роговая эйзения (Eisenia rosea) 12.7 21.6 24.7 8.6
Малый красный червь (Lumbricus rubellus) . . 11.6 10.5 11.0 0.6 1
Как можно видеть (табл. 5), на первом месте при пи-
тании разными листьями оказывается пашенный, а на
последнем — малый красный червь. Отсюда ясно, на-
сколько различной должна быть почвообразовательная
деятельность на участках, заселенных разными видами
червей.
153
Не меньшее значение должен иметь возрастной состав
населения люмбрицид, но данных по этому вопросу пока
не имеется.
В чем же заключается почвообразовательное значение
выноса на поверхность извержений червей?
Оставляя пока в стороне важные качественные изме-
нения, претерпеваемые почвой при прохождении ее через
кишечник червей и превращении в копролиты, обратим
внимание на то, что благодаря этому процессу происхо-
дит перемешивание слоев почвы. Черви могут заглаты-
вать почву в любом ее слое, и вынесение ее на поверх-
ность представляет собой непрерывно идущий процесс
образования нового поверхностного слоя, в котором ча-
стицы почвы с разных глубин оказываются тщательно
перемешанными друг с другом.
Перемешивание слоев почвы идет не только благодаря
вынесению частиц почвы с глубины на поверхность; не
меньшее (если не большее!) значение имеет перемещение
частиц из поверхностных сдоев в глубину. Этот процесс
может происходить путем стекания вместе с водой размо-
ченного дождем гумусного слоя по ходам дождевых чер-
вей. Как уже указано, извержения люмбрицид отклады-
ваются не только на поверхности, но и в подземных ходах,
причем подчас на большой глубине. Заметим, что перед
свертыванием в клубок при впадении в спячку в зимних
и летних камерах обязательно опоражнивается кишечник,
причем обычно это происходит неподалеку от камер, т. е.
на значительной глубине.
Очень большое количество извержений червей нахо-
дится также в дерновом слое почвы. Об этом свидетель-
ствуют данные табл. 4. В некоторых случаях в зоне кор-
ней растений извержений червей оказывается не меньше,
чем на поверхности, а иногда даже больше (на ржаном
поле). Таким образом, если учесть не только дерновый,
но и более глубокие слои почвы, то, вероятно, окажется,
что бблыпая часть извержений кишечника червей не вы-
носится наружу, а остается внутри почвы (рис. 46 и 48).
Возможно, что наверх поступает только количество из-
вержений, необходимое для защиты норки от струи ут-
реннего холодного воздуха.
Очень показательны многочисленные опыты, ставив-
шиеся с целью изучения процесса перемешивания почвы
154
дождевыми червями. Вот иример такого опыта (Понома-
рева, 1953). В сосуд с плоской стеклянной стенкой был
насыпан слой очень мелкого светлого грунта (350 г), так
называемый лёсс.1 Сверху на него был насыпан такой же
объем просеянной через сито темной почвы (400 г), со-
держащей значительные количества перегноя (рис. 54).
В сосуд было пущено шесть дождевых червей (Lumbri-
cus terrestris и Allolobophora caliginosa); для питания им
клались листья салата и клевера; поддерживалась также
равномерная влажность почвы. Через 17 дней слой свет-
лого лёсса оказался пронизанным до дна ходами червей,
причем значительная часть их была заполнена копро-
литами темного цвета, а через 110 дней вся почва в сосуде
была полностью перемешана (рис. 54, /?), т. е. к этому
времени вся почва в сосуде прошла через кишечник чер-
вей и, может быть, не один раз. Исследование показало
полную однородность всего объема почвы в сосуде в от-
ношении структуры и химического состава. Общий
объем почвы увеличился. Это объясняется увеличением ее
скважности вследствие возникновения ходов червей и про-
межутков между образовав^ тмися структурными еди-
ницами.
Отметим, что в природе этот процесс идет с гораздо
меньшей скоростью, так как соотношение количества чер-
вей и массы почвы, взятое в опыте, превосходит обычное
соотношение в десятки раз. Расчет показывает, что 6 чер-
ней на взятые в опыт 750 г почвы соответствуют плотно-
сти населения, гораздо большей по сравнению с наблю-
даемыми в действительности; даже огородная и парнико-
вая земля содержит в 4—6 раз меньше червей в единице
объема. Но все же этот опыт, который является повторе-
нием классического опыта Гензена (1877). убедительно
свидетельствует о том, что если перемешивание почвы
дождевыми червями в природе идет даже в десятки раз
медленнее, то и тогда оно должно иметь колоссальное
значение в эволюции почв. Кроме того, описанный опыт
показывает, как велики возможности повышения интен-
1 Лёсс — мелкозернистый грунт, образующийся за счет вы-
ветривания горных пород или осаждения тончайшей мути в водое-
мах; термин «лёсс» происходит от французского слова laissp, что
означает «осадок».
155
сивности процесса перемешивания почвы при увеличении
плотности населения дождевых червей.
Изложенные данные
Рис. 54. Перемешивание дожде-
выми червями слоев почвы: буро-
зема (вверху) и лёсса (внизу).
(По С. И. Пономаревой).
А — до опыта; В — через 17 дней
после внесения дождевых червей;
В — через НО дней.
размера, принадлежащих к тому или иному виду.
Через 101 день (опыт длился с 11 XI по 19 II) об-
объясняют, почему гра-
ница между темным слоем
почвы, содержащим гу-
мус, и подлежащими, бо-
лее светлыми слоями в
природе никогда не бы-
вает резкой, а всегда очень
неотчетлива и размыта.
Заметим, что органические
вещества почвы прони-
кают в более глубокие слои
не только вследствие ак-
тивного переноса их туда
червями в кишечниках,
но и вследствие стекания
размягченной дождями гу-
мусной почвы в ходы чер-
вей.
Оказалось, что разные
виды дождевых червей
обладают неодинаковой
способностью к перемеши-
ванию слоев почвы (Franz,
1950). Опыты, аналогич-
ные только что описан-
ному, ставились следую-
щим образом. В литровые
стеклянные банки насы-
пался слой прокаленного
мелкого песка, а поверх
него слой просеянной са-
довой земли. Во всех бан-
ках было одинаковое коли-
чество почвы, и в каждую
из них было пущено оди-
наковое количество червей
приблизительно одного
156
наружилось, что в разных банках полнота перемеши-
вания была резко различной. Лучше всего земля
была перемешана в банках с молочным октолазнем
(Octolasium lacleum); в этих банках чистого белого песка
почти не осталось. Наоборот, в банках с малым красным
червем (Lumbricus rubellus) песок был смешан с пере-
гноем в очень незначительном количестве. Этот вид —-
житель верхних слоев почвы, питается крупными кусками
отмерших растений и, по-видимому, поглощает не так
много почвы, как другие виды. Промежуточное положе-
ние в отношении скорости перемешивания слоев почвы
заняли розовая эйзения (Eisenia rosea) и пашенный червь
(Allolobophora caliginosa). Таким образом, в отношении
работы по перемешиванию почвы наземные виды люмбри-
цид располагаются в следующий ряд: молочный октолазий,
розовая эйзения, пашенный червь, малый красный червь.
До сих пор очень мало оса.нана та большая роль,
которую играют дождевые черви в земледелии, переме-
шивая удобрения с почвой. Последнее относится как
к минеральным, так и к органическим удобрениям: из-
вестковые, калийные, фосфорные и азотистые удобрения,
насыпанные на поверхность, оказываются внутри почвы
и распределяются там равномерно действием люмбрицид.
Эту важную сторону работы иллюстрирует другой опыт
Франца (Franz, 1950). В литровых банках полевая почва
переслаивалась с навозом: слой земли, слой навоза,
сверху опять слой земли. В банки было пущено по 9 эк-
земпляров розовой эйзенип. Через 215 дней вся земля
в банках оказалась совершенно однородной, без всяких
признаков разделения на слои. Франц полагает, что
навоз в почвах, лишенных дождевых червей, может ле-
жать годами, не перегнивая и не смешиваясь с почвой;
он приобретает торфянистый характер и нисколько не
улучшает почвы, а скорее ухудшает ее. Это мнение можно
считать справедливым, если сделать оговорку, что в рас-
пределении навоза в почве играют роль не только дожде-
вые черви, но и почвенные насекомые, хотя роль послед-
них в данном случае несомненно второстепенная. Кроме
того, отсутствие червей ведет к образованию торфа лишь
при определенных климатических и флористических ус-
ловиях. В сухих степных или горных районах об этом
не может быть речи.
157
Известный почвовед и агроном Э. Россель (1933).
бывший директором знаменитой Ротамстэдской опытной
станции, предложил особый прием удобрения огородных
гряд, основанный на способности дождевых червей пере-
мешивать почву. Если приходится устраивать огород
там, где почва представляет собой грубую смесь глины,
песка и мелких камней, то особенно эффективными, по
мнению этого исследователя-практика, являются зака-
пывание в почву (на глубину 20 см) скошенной травы
п других веществ растительного происхождения и много-
кратное поверхностное удобрение травой и измельченным
известняком или мелом. Это агротехническое мероприя-
тие производится в расчете на перемешивающую дея-
тельность червей в почве: «Дождевые черви, переползая
из верхних слоев почвы в нижние, основательно пере-
мешивают и проветривают ее». В согласии с приведен-
ным выше мнением Франца, Рэссель пишет: «При актив-
ности дождевых червей в почве органическое вещество
распределяется по всей толще, в которой совершается
их работа. Когда же дождевых червей мало или их сов-
сем нет, перемешивание почвы задерживается; на поверх-
ности накопляется мертвое растительное вещество, пре-
вращающееся в частично разложившуюся кислую тор-
фянистую массу, в которой нормальные процессы распада
почвенных органических соединений не доходят до конца.
Растительность на поверхности претерпевает глубокие
изменения, так как корни лишь немногих растений бы-
вают в состоянии проникнуть сквозь массу мертвого
вещества; по мере отмирания их остатки тоже остаются
на поверхности и при медленном разложении могут
накопиться в таком количестве, что образуется пласт
торфа».
Итак, перемешивание слоев почвы и вынос ее частиц
на поверхность — результат заглатывания дождевыми
червями почвы в одном месте и извержения ее в другом.
Одним из важнейших следствий из всех фактов, изложен-
ных нами в этом разделе, является вывод, что вся масса
почвы, заселенной дождевыми червями (а таковыми яв-
ляется, как мы видели в предыдущих главах, большая
часть почв земного шара), за время своего существования
прошла через кишечник дождевых червей и, может быть,
много раз! Этот смелый вывод со всей определенностью
158
был сделай Дарвином. Он многим казался иарадоксаль
ным и неоднократно подвергался критике. Но современ-
ная наука располагает рядом фактов, бесспорно доказы
вающих правильность вывода Дарвина.
Обратимся снова к табл. 4. В ней приведены цифры,
выражающие процентное отношение веса извержений
червей, найденных в зоне корней растений, к весу всей
почвы. Эти цифры вообще очень значительны, а для поля
с посевными травами второго года пользования подни-
маются до 48.1%, т. е. половина всей массы этого слоя
почвы состоит из копролитов. Примем во внимание также,
что хотя копролиты представляют собой довольно стой-
кую структуру, но прочность ее относительна. Поэтому
можно думать, что та часть почвы, в которой усмотреть
копролитную структуру уже не удается, некогда все же
прошла через кишечник дождевых червей. П. У. Бахтин
и М. Н. Польский (1950) нашли, что некоторые почвы
Калининского района Московской области на 90% со-
стояли из копролитов, а в целинных землях процент их
еще выше. То же констатировано и для целинных почв
северо-западного Алтая (Соколов, 1956).
Эти и ряд других данных привели современных ис-
следователей к уверенности в правильности вывода Дар-
вина о том, что весь почвенный слой, т. е. практически
почти весь поверхностный слой суши земного шара, —
продукт деятельности дождевых червей. Остается откры-
тым лишь вопрос, какое время нужно для того, чтобы почва
на том или ином участке прошла вся целиком через ки-
шечник червя. Но на основании всего того, что нам из-
вестно о дождевых червях, можно сказать, что общего
ответа на этот вопрос быть не может. В разных почвах,
в разных климатах, при разной растительности и разной
плотности населения дождевых червей скорость этого
процесса должна быть различной. Н. А. Димо для почв
Средней Азии определяет ее в 100 лет.
Если его расчет правилен, то и эту скорость нужно
признать значительной, так как 100 лет для истории
земли — срок очень небольшой. Но можно думать, что
на самом деле эта скорость значительно выше. Н. А. Димо
исходил в своих расчетах из того, что скважность почвы,
являющаяся следствием наличия в ней ходов червей,
составляла 1% от объема почвы. А так как эти ходы про-
159
делываются в один сезин, то 100 лет достаточно, чтобы
весь объем почвы прошел через кишечники червей. Од-
нако этот расчет преуменьшает действительную скорость
прохождения почвы через кишечник, так как ходы червей
в верхних слоях почвы — образования крайне нестойкие,
и одни ходы быстро заменяются другими. Кроме того,
скважность взята в слое почвы метровой глубины, тогда
как заглатывание почвы червями происходит преимуще-
ственно в верхних слоях почвы, где протекает большая
часть их жизни. Наконец, делать заключение о количе-
стве почвы, прошедшей через кишечник, исходя из общего
обилия ходов червей в почве, невозможно потому, что, как
мы знаем, ходы не только проедаются червями, но соз-
даются раздвиганием частиц почвы.
Для учета скорости прохождения почвы через «кишки
земли», т. е. кишечники дождевых червей, нужны специаль-
ные исследования, которые пока не ведутся. Некоторое
представление о порядке цифр, определяющих эту ско-
рость, можно получить, если исходить из того, что червь
за сутки пропускает через кишечник количество почвы,
равное его весу.1 Вспомним, что на стр. 138 приведены
данные о том, что черви на площади в 1 га ежегодно про-
пускают через кишечник в разных почвенно-климатиче-
ских условиях от 50 до 600 т земли в год. Так как 1 м3
почвы весит примерно 1.25 т (см. стр. 151), то вес слоя
почвы в 30 см, где совершается вся деятельность люм-
брпцид, весит 3750 т на 1 га. Следовательно, весь этот
слой почвы должен пройти через кишечники червей в срок
от 6 до 75 лет. А. А. Соколов (1956) считает, что метровая
толща в горных почвах северо-западного Алтая должна
перерабатываться червями за 60—190 лет.
Таким образом, перемешивание почвы и непрерывное
образование его верхнего слоя совершаются со скоростью,
ощутимой для жизни одного поколения людей. При этом
земля, проходя через кишечники дождевых червей, не
только перемещается с одного места на другое, но и ка-
чественно изменяется. К рассмотрению этих изменений
почвы мы и перейдем.
1 С. И. Пономарева (1953) считает, что каждым дождевым чер-
вем аа сутки перерабатывается 0.4—3.0 г почвы.
160
5. ВЛИЯНИЕ ДОЖДЕВЫХ ЧЕРВЕЙ НА СТРУКТУРУ И
ХИМИЧЕСКИЙ СОСТАВ ПОЧВ
Земля, заглоченная червем, перетирается в его же-
лудке с листьями и другими растительными остатками,
а также подвергается химической обработке при помощи
веществ, выделяемых железами разных отделов кишеч-
ника. В результате получается мелкая однородная пи-
щевая кашица, из которой некоторая часть растворенных
веществ всасывается клетками кишечника. Червь ути-
лизирует, конечно, какую-то очень небольшую часть за-
глоченных веществ. Известную аналогию в этом отноше-
нии можно найти у домашних животных. Так, из 100 кг
зерна, скармливаемых им, примерно только 10 кг рас-
ходуется на увеличение веса их тела, а остальное превра-
щается в экскременты, мочу и углекислый газ. Поскольку
в пище дождевых червей содержится еще меньше белков
и углеводов, эти животные должны пропускать через
кишечнпк огромные количества растительных остатков
и почвы. Можно думать, что количество извергнутых ко-
пролптов лишь немногим меньше веса заглатываемых ими
веществ.
В чем же состоят изменения, претерпеваемые почвой
при прохождении ею кишечника червей, и чем отличается
такая почва от нетронутой червями?
Рассмотрим прежде всего, как отражается прохожде-
ние почвы через кишечник на содержании в ней органи-
ческих веществ, пли гумуса. Из только что сказанного,
а также из изложенного в предыдущей главе о характере
минерализующей деятельности почвенных животных
(стр. 94) ясно, что в результате заглатывания почвы
червями количество гумуса в ней должно уменьшаться.
Но мы знаем также, что червп питаются не только гуму-
сом и что важной составной частью их пищи (особенно
крупных видов люмбрицид) являются отмершие части
растений, лежащие вне почвы, па ее поверхности. Черви
затаскивают пх внутрь почвы, перемалывают, подвергают
химической обработке и выбрасывают их остатки обратно
в почву. Там они подвергаются дальнейшей обработке
бактериями и активными химическими веществами, имею-
щимися в почве, в результате чего превращаются в гу-
мус. Таким образом, дождевые черви являются одно-
11 О. В. Чекановскяя
161
временно н потребителями, и производителями гумуса
почв.
Нужно сказать, что роль ряда видов дождевых червей
в продукции гумуса, вероятно, очень скромная. Есть
указания, что эйзения Норденшельда (Eisenia norden-
skioldi) питается только внутри почвы; значит особи
этого вида не вносят в почву органических веществ с ее
поверхности. Относительно ряда видов, и в частности
розовой эйзепии (Eisenia rosea) и пашенного червя (Allo-
lobophora caliginosa), имеющих наибольшее почвообра-
зовательное значение в Европейской части СССР, точных
данных по этому вопросу нет. Большой красный червь
(Lumbricus terrestris), длинная аллолобофора, очень круп-
ная аллолобофора, недавно открытая А. А. Соколовым
(1956) в северо-западном Алтае (Allolobophora magnifica),
и виды рода октолазий (Octolasium) несомненно собирают
растительные остатки с поверхности земли. Этим видам
принадлежит роль гумификатора в лесах, где они соби-
рают до 50% опавшей листвы (гигантская аллолобофора
Алтая собирает 100?о листопада, и ей еще не хватает его).
Особое значение в этом отношении имеет малый крас-
ный червь (Lumbricus rubellus). Являясь обитателем
самых поверхностных слоев почвы, он питается преиму-
щественно опавшими листьями и другими крупными ра-
стительными остатками. Деятельность особей этого очень
распространенного вида несомненно обогащает почву
гумусом.
Копролиты большого красного червя, как показал
анализ, содержат большие количества гумуса по срав-
нению с почвой, в которой они жили. Имеются указания
па большее содержание гумуса по сравнению с окружаю-
щей почвой в облицовке ходов некоторых люмбрицид.
Однако в общем нужно сказать, что деятельность люм-
брицид как образователен гумуса не имеет особого зна-
чения. Не в этом заключается их главная роль в преобра-
зовании почв. Участие других олигохет, энхитреид (см.
стр. 97), а также всех остальных почвенных животных
и микробов в образовании гумуса несравненно больше.
Прибавим к этому, что дождевые черви питаются остатками
растений, в которых уже сильно продвинуты бактериаль-
ные процессы гниения (зеленый корм они берут крайне
неохотно); поэтому их нельзя считать пионерами образо-
162
вания гумуса, они обрабатывают продукты, уже являю-
щиеся, так сказать, полуфабрикатами.
Более важны косвенные последствия внесения дожде-
выми червями в почву органических веществ. Одним из
таковых является накопление в извержениях дождевых
червей аммиака, нитратов, фосфорной кислоты, кальция
и магния, констатированное химическими анализами.
Превращения этих веществ в почвах — очень сложный
комплекс процессов. Опавшая листва и другие отмираю-
щие части растений подвергаются сначала обработке со
стороны микроорганизмов, а затем попадают в кишечники
червей и переносятся ими в почвенную толщу. То же про-
исходит с отмирающими корнями, которые служат од-
ним из главных источников образования гумуса.
Весьма существенно накопление в кишечнике червей
кальция в виде биогенного кальцита. Кальцит — мине-
рал, представляющий собой кристаллы углекислой из-
вести; биогенным же он называется в силу того, что его
происхождение связано с деятельностью живых существ.
Такие кристаллы находил еще Дарвин в железках на
пищеводе дождевых червей^., Однако его взгляды на меха-
низм их образования и физиологическое значение под-
верглись существенным изменениям. Приходится ска-
зать, что мы не знаем в точности способа образования
этих кристаллов; по-видимому, он заключается в том,
что кальций, поступающий с листьями и почвой в кишеч-
ник червей в виде тончайшим образом распыленной (кол-
лоидной ) взвеси окиси кальция, превращается в угле-
кислую соль, которая кристаллизуется в пищеводе. Эти
кристаллы при движении по кишечнику растут, а затем
соединяются друг с другом, образуя компактные камешки
(сростки, или друзы) размером до 1.5 мм и более
(рис. 55).
Как показывают анализы С. И. Пономаревой (1953),
проводившиеся как в лабораторных условиях, так и в поле,
количества биогенного кальцита, поступающего в почву
с копролитами дождевых червей, очень значительны.
Содержание его в копролитах, собранных в разных участ-
ках дерново-подзолистой почвы, колеблется от 0.82 (ози-
мая пшеница) до 34.12 кг (посевные травы второго года
пользования) на 1 га. Обогащение почвы биогенным каль-
цитом имеет очень большое значение. Его присутствие
И*
163
содействует нейтрализации кислот в почве и и вменяет
ре структурные свойства.
Известно, что при разложении отмерших частей расте
ний получается значительное количество кислот. Ранее
уже отмечено, что почвы при отсутствии в них дождевых
червей становятся кислыми вследствие избытка распадаю-
щихся растительных остатков. Как показали измерения
активной реакции среды в Киевской области (табл. 6),
Рис. 55. Кристаллы кальцита и их сро-
стки из выбросов дождевых черве!!.
(По С. И. Пономаревой).
опавшие листья разных деревьев имеют сильно кислую
реакцию. Несмотря на это, реакция копролитов дожде-
вых червей оказывается заметно щелочной. Почти та-
кая же щелочность характерна для самых верхних слоев
почвы, которые недавно созданы копролитами червей.
Более глубокие слои лесных почв могут оказываться в зна-
чительной степени кислыми, что, вероятно, стоит в связи
с уменьшением в этих слоях количества дождевых чер-
вей и их извержений.
Аналогичные данные получены в опытах с дерново-
подзолистой почвой Московской области; черви, помещен-
164
Таблица 6
Активная реакция (pH) опада листьев деревьев, извержений
дождевых червей и разных слоев почвы.
(Из А. И. Зражевского, 1У49)
Древесные породы
дуб граб бук ольха
Листья . . 5.22 4.85 5.75 4.89
Капролиты . . . Почва на глу- бине: — 7.52 7.23 7.21
0-2 см 7.09 7.44 7.32 7.02
5-10 » 5.69 5.67 6.20 6.88
20-30 » 6.66 5.84 1 6.37 7.35
ные в почвы с кислой, реакцией, образовали копролиты
со щелочной реакцией (Пь^омарева, 1953). При этом было
обнаружено, что чём больше биогенного кальцита в из-
вержениях червей, тем сильнее оказывается сдвиг реак-
ции среды в щелочную сторону. Однако подщелачивание
почвы дождевыми червями идет не только за счет обра-
зования кальцита. Может быть, еще большую роль играют
щелочные выделения желез стенки средней кишки дожде-
вых червей, которые обязательно нейтрализуют кислые
пищевые массы, поскольку их пищеварительные ферменты,
как это было известно еще Дарвину, переваривают пищу
только в щелочной среде. Таким образом, одно из важней-
ших следствий прохождения почвы через кишечник чер-
вей — ее подщелачивание. Не будь его, все свойства
почв, прежде всего их плодородие, были бы совсем иными.
Сопоставление только что сказанного с тем, что было
изложено на стр. 102 о зависимости расселения дождевых
червей от кислотности почв, как нельзя лучше характери-
зует не одностороннюю, а обоюдную зависимость организма
и среды. Возможность существования дождевых червей
лишь при определенных значениях pH и наличие оптималь-
ной кислотности почвы для каждого вида — вполне реаль-
ные факты, но не менее реальным является способность
165
дождевых червей изменять условия в почве в сторону
приближения пх к оптимальным.
Однако самое важное воздействие дождевых червей
на повышение качества почв состоит в придании ей зерни-
стой структуры. В предыдущей главе упоминалось, что
структура имеет весьма важное значение для характери-
стики почв. Отмечалось также, что следует различать
понятия «частицы» почвы и ее «структурные отдельности»
или «агрегаты». Изучение того и другого в почвах, обра-
ботанных дождевыми червями, показывает, что эти жи-
вотные уменьшают размеры частиц почвы п одновременно
создают почвенные структурные единицы, т. е. отдельности,
со многими свойствами которых мы уже не знакомы.
В мышечном желудке происходят обкатывание и из-
мельчение частиц почвы (особенно мягких). Механи-
ческий анализ копролитов показывает, что по сравнению
с исходной почвой в них содержится большее количество
мелких, пылеватых частиц. Однако обнаружить их можно
только после искусственного разрушенпя структурных
отдельностей, в которые они слипаются в задних отделах
кишечника червей. Отдельные комочки извержений чер-
вей, как уже отмечалось, могут сливаться друг с другом.
Неоднократно ставились опыты с целью проследить, как
из мелкой почвы постепенно образуются довольно круп-
ные отдельности неправильной формы (рис. 56). В одном
из таких опытов в деревянные ящики с почвой, просеян-
ной через сито с ячейками в 1 мм, помещалось по семь
экземпляров большого красного и пашенного червей
(Lumbricus terrestris и Allolobophora caliginosa) на 1 кг
почвы. По прошествии 47 дней в почве оказалось 32.4%
отдельностей размером больше 15 мм, тогда как в контроль-
ных ящиках с почвой в тождественных условиях, но без
дождевых червей их было только 3.5%. Как следует из
табл. 7, количество частиц размером более 7 мм в почве
с дождевыми червями значительно выше, а количество
частиц размером менее 5 мм ниже, чем в контроле. Таким
образом, структура почвы из очень мелкозернистой ста-
новится крупнозернистой и ореховатой. В дальнейшем
размеры отдельностей делаются еще несколько больше.
Отдельности в почве образуются также под влиянием
деятельности других почвенных организмов, главным
образом грибков и микробов; но они отличаются от обра-
166
зовавшпхся копролитов дождевых червей более округлой
формой и более рыхлой консистенцией.
Но главная особенность копролитов заключается в том
их свойстве, которое почвоведы называют водопрочно-
стью. Это название означает способность структурной от*
дельности почвы противостоять размыванию ее водой.
Водопрочность копролитов может измеряться различными
Рис. 56.Образование почвенных агрегатов деятельностью дождевых
червей. (По С. И. Пономаревой).
А — образец почвы, взятой до опыта; Б — то же, через 47 дней после опыте.
методами, например количеством воды, падающей на
них каплями с определенной высоты, которое необходимо
для размывания их на отдельные почвенные частицы.
В описанном опыте С. И. Пономаревой оказалось, что для
размывания определенного размера выбросов дождевых
червей требовалось от 3.24 до 21.15 л воды, тогда как
для размывания такого же размера структурных отдель-
ностей, образованных деятельностью микроорганизмов, —
.от 0.005 до 1.53 л воды.
Эти данные подтверждены рядом других исследователей
(Бахтин и Польский, 1950; Мамытов, 1953; Соколов,
167
Таблица 7
Содержание отдельностей разных размеров (в %) в почве,
измененной деятельностью дождевых червей, и в почве
без червей. (Из С. И. Пономаревой, 1953)
Поперечник отдельностей (в мм)
боль- ше 15 10-10 г— 1 о 7 । ю ео 1 сч 1 1 со 7 менее 1 ' всего 1
Почвы, измененная дождевыми червя- ми 32.4 8.8 6.3 1.9 4.0 4.1 42.5 100
Почва без дождевых червей (контроль) 3.5 5.9 5.9 4.4 9.6 2.7 2.7 2.4 62.9 100
1956; Зражевский, 1957). Все они приводят очень высокие
цифры водопрочности почвенных агрегатов, образован-
ных разными видами люмбрицид в разных грунтах и при
разных условиях питания. Оказалось, что из трех видов,
обычных в лесных почвах, наиболее водопрочные струк-
турные единицы образует малый красный червь (Lumbri-
cus rubellus), за ним идет розовая эйзения (Eisenia rosea)
и на последнем месте стоит пашенный червь (Allolobo-
phora caliginosa).
Следя за процессом размывания структурных отдель-
ностей, образованных дождевыми червями, можно видеть,
что в нем находятся в большом количестве неразложив-
шиеся остатки растений (рис. 57). Очень вероятно, что
чрезвычайно высокая водопрочность копролитов и их
сростков объясняется наличием каркаса из структурных
частей растений, связывающих их частицы. Под микро-
скопом видно, что отдельные части почвенной отдель-
ности связаны спиральными волокнами (рис. 58). Эти спи-
ральные волокна — растительного происхождения.
В живых стеблях трав они выстилают внутреннюю поверх-
ность сосудов, проводящих воду и минеральные вещества
от корней к листьям; они не дают спадаться их стенкам
и как самые прочные части растений остаются целыми
дольше других частей; после разложения стеблей они
видны в изолированном состоянии. Но наличие этого орга-
нического каркаса внутри копролитов само по себе не
обеспечивает еще их водопрочности; этому содействует
присутствие в них соединений кальция, о которых ска-
зано выше. Влажные копролиты пропитаны растворами
окиси кальция и его углекислой соли, которые при под-
сыхании кристаллизуются и цементируют мелко растер-
тые почвенные зернышки. А если принять во внимание,
что*копролит пронизан мелкими соломинками и микро-
скопическими волоконцами, то его исключительная *одо-
прочность делается понятной.
6
Рис. 57. Агрегат, образованный деятельностью дождевых
червей. (По С. И. Пономаревой).
А — до рашывания; Б — в процессе равмывания проточной водой.
В чем же заключается значение водопрочности струк-
турных отдельностей почвы?
Ответить на этот вопрос нетрудно. Вспомним, как важна
крупнозернистая структура почвы для проникания в нее
воздуха и воды, для развития в ней микробов и мелких
животных. Учтем далее, что копролиты содержат в концен-
трированном виде ряд важных для растений веществ, как
органических, так и неорганических. Водопрочность
копролитов создает возможность более полного их исполь-
зования растениями, так как в противном случае эти ве-
щества быстро вымывались бы из почвы дождями. Водо-
прочностью обеспечивается дозированное поступление
в раствор минеральных составных частей копролитов,
что также выгодно для растений.
Отметим, что водопрочность не следует смешивать
с водонепроницаемостью (Бахтин и Польский, 1950).
Бели бы структурные отдельности, созданные дождевыми
J69
червями, были непроницаемы для воды, наподобие вера*
створимых минералов, то наличие такой структуры почвы
влияло бы на плодородие почвы только отрицательно.
Исследованиями обнаружено, что удельный вес копро-
литов меньше удельного веса других фракций почвы. Это
Рис. 58. Микроскопический препарат агрегата,
образованного дождевыми червями. Видны спираль-
ные волокна растительного происхождения, скреп-
ляющие мелкие частицы почвы. (По С. И. Поно-
маревой).
доказывает, что копролпты обладают большой скважно-
стью. Вода может проникать внутрь копролита и цирку-
лировать в нем, как по системе каналов, медленно под-
тачивая его и непрерывно вынося наружу небольшие пор-
ции веществ, утилизируемых микроорганизмами и выс-
шими растениями.
Закончим обзор качественных изменений почвы, про-
изводимых деятельностью дождевых червей, упомина-
170
нием о важном следствии создания ими структурных
отдельностей, обогащенных органическими веществами:
на них поселяются почвенные грибки и всевозможные
микроорганизмы в больших количествах, чем на других
фракциях почвы (табл. 8).
Таблица 8
Количество бактерий (в тысячах экземпляров)
на 1 г дерново-подзолистой почвы.
(Из С. И. Пономаревой, 1953)
Копролиты Почва без копролитов
Посевные травы 2-го года пользования . . Рожь с подсевом трав Дубовый лес . 3940 3430 740 2000 2530 450
На копролитах бактерий оказывается гораздо больше.
Но этим дело не ограничивается. Здесь же селятся
грибки, а затем появляется ряд животных, питающихся
грибками и бактериями. Это прежде всего—низшие бес-
крылые насекомые (Collembola), клещи и нематоды (см.
стр. 97). Здесь перед на^и один из примеров связей, сое-
диняющих организмы почвы в одно целое. Клещи и низ-
шие насекомые, питаясь микроорганизмами, оставляют
в почве свои извержения и являются строителями почвы,
обогащая ее органическим веществом — гумусом, ко-
торый в свою очередь утилизируется другими организ-
мами, в частности теми же дождевыми червями. Упомина-
ние о связях почвенных организмов друг с другом при-
водит нас к необходимости бросить взгляд на процесс
почвообразования в целом.
6. ЗАКЛЮЧЕНИЕ
Итак, дождевые черви являются очень важным факто-
ром почвообразования. Без них не могло бы быть на поверх-
ности суши почв в том виде, в каком мы их наблюдаем,
и во всяком случае не было бы того, что земледельцы на-
зывают «хорошей землей». Однако это не дает права перео-
171
цеиивать их роль в почвообразовании, так как если бы не
существовало большого количества других факторов, дей-
ствующих в том же направлении, то не только «хорошей»,
но и вообще никакой почвы на земле не могло бы быть.
Употребляя фигуральное сравнение, можно сказать,
что черви представляют собой как бы один цех того огром-
ного производства, каким является почвообразование.
Прежде всего деятельность дождевых червей становится
возможной только после того, как какая-то почва уже
имеется. Образовалась ли она на месте скалы в процессе
выветривания, или материалом для нее послужила пыль,
приносимая ветром, или разные наносы, или она появилась
в результате усыхания озер, отступания моря и т. д.,
эти субстраты остаются длительное время вовсе недо-
ступными для дождевых червей. Прежде чем последние
получат возможность заселить почву, должно достаточно
далеко пойти накопление в ней органических веществ,
возникающих за счет отмирания организмов, преимуще-
ственно растительных. Накопляющееся таким путем тем-
ное вещество называют, как мы уже говорили, перегноем,
или гумусом, а процесс его образования — гумификацией.
Этот процесс очень сложен; он состоит из множества
отдельных этапов, сменяющих друг друга, а отчасти из
идущих параллельно и независимо. Разные этапы гуми-
фикации совершаются различными организмами. Первыми
поселенцами на образующейся почво являются лишай-
ники, водоросли, бактерии. В более влажных местах к ним
присоединяются грибы и некоторые* микроскопические
низшие животные, которые и являются первыми гуми-
фикаторами. Отмирая, они дают возможность для посе-
ления различным мхам, под подушками которых находят
приют, кроме микроорганизмов, клещи и низшие насеко-
мые. Обрабатывая отмирающие части мхов, эти орга-
низмы создают более значительные количества гумуса,
что открывает возможность для поселения в таких ме-
стах высших растений. Их появление на поверхности
почвы дает мощный толчок дальнейшему развитию про-
цессов гумификации: их отмирающие корни и наземные
части служат обильным источником перегноя. Гумифи-
кация осуществляется деятельностью не только огром-
ного числа бактерий разных видов, но и множества мелких
животных, среди которых, кроме клещей и низших насе-
172
комых, важное значение приобретают мелкие червячки
(нематоды и энхитреиды), а также членистоногие (рако-
образные, паукообразные, многоножки, разные отряды
насекомых; особое место занимают личинки некоторых
мух). Чем бы они ни питались, органические вещества,
проходя через кишечник этих животных, обогащают почву
гумусом.
Дождевые черви включаются в процесс почвообразо-
вания, когда гумификация почвы уже в полном разгаре.
Мы знаем уже, что почвы, бедные гумусом, бедны и дожде-
выми червями; и, наоброт, чем его больше, тем обычно
больше и дождевых червей. Мы видели также, что до-
ждевые черви хотя и участвуют в продукции гумуса, но
значение их в этом отношении не так велико. В числе мно-
гих других гумификаторов они только вкладывают свою
долю в продуцирование гумуса.
В известном смысле всех наземных животных можно
считать гумифпкаторами, поскольку они обогащают почву
продуктами своей жизнедеятельности, а тела их после
смерти подвергаются разложению. Но их значение вы-
ступает па первый план на том этапе почвообразования,
когда гумус уже создан и возникают задачи о распреде-
лении его по разным слоям почвы, о разрыхлении ее,
о снабжении всей армии гумификаторов воздухом и водой,
о предохранении гумуса от быстрого вымывания из почвы,
об удалении избытков растительных остатков с поверх-
ности почвы, о нейтрализации кислот, возникающих
при образовании гумуса, о консервации питательных ма-
териалов для растений в водопрочных копролитах. Во
всех этих процессах принимают участие и другие живот-
ные, создающие структуру и тем самым обрабатывающие
почву, но главная роль в них принадлежит несомненно
дождевым червям как массовым животным, мощным земле-
роям и поглотителям почв.
Следует обратить особое внимание на то, что деятель-
ность дождевых червей в почвообразовании органически
связана с деятельностью других физических, химических
и биологических агентов, участвующих в этом про-
цессе. Вспоминаются слова В. В. Докучаева о том, что
«почвы и грунты есть зеркало, яркое и вполне правдивое
отражение, так сказать, непосредственный результат со-
вокупного. весьма тесного, векового взаимодействия между
173
водой, воздухом, землей, с одной стороны, животными и
растительными организмами и возрастом страны, с другой,
этими извечными и поныне действующими почвообразова-
телями». Это «вековое взаимодействие», в котором дожде-
вые черви играли и продолжают играть далеко не послед-
нюю роль, создало гумусные почвы земного шара, ко-
торые в свою очередь явились необходимой предпосыл-
кой для возникновения пышной растительности, а тем
самым и необходимой пищевой базой для всех наземных
животных и человека.
ГЛАВА VI
ДОЖДЕВЫЕ ЧЕРВИ И ЗЕМЛЕДЕЛИЕ
1. ВЛИЯНИЕ ДОЖДЕВЫХ ЧЕРВЕЙ НА ПЛОДОРОДИЕ
ПОЧВ
По выражению В. ₽. Вильямса, «понятие о почве и ее
урожайности — неразделимы». Урожайность — признак,
характеризующий самое существо попятил «почва»;
почва, по В. Р. Вильямсу, — «это рыхлый поверхностный
горизонт суши, способный производить урожай растений».
Мы рассмотрели в предыдущей главе, какие глубокие изме-
нения свойств почвы могут 611ть произведены дождевыми
червями. Ясно, что эти изменения должны как-то сказаться
на плодородии почв. Какие же данные имеются по этому
вопросу?
Как уже сказано раньше, многим земледельцам-
практикам с давних времен было известно положитель-
ное влияние дождевых червей на урожай. Г. Н. Высоц-
кий (1898) заметил, что в южнорусских степях раститель-
ность лучше там, где почва содержит большое количество
дождевых червей. Первые экспериментальные данные по
этому вопросу были сообщены в конце прошлого века
(Djemil, 1896; Wollny, 1897). В сосудах с почвой, удо-
бренной костяной мукой и каинитом,1 выращивались раз-
ные культурные растения, причем в одних сосудах в почве
жили черви, в других их не было. Оказалось, что в пер-
* 1 Минерал, содержащий калий и магний; употребляется как
минеральное удобрение.
175
вых при прочих равных условиях урожай гороха повы-
сился на 82%, рапса — на 168%, а льна — на 858%.
Опыты, произведенные М. П. Архангельским (1929),
Кравковым и Голубевой (Кравков, 1931), В. II. Лады-
киным (1941), В. К. Балуевым (1950) с различными расте-
ниями и в разных условиях, подтвердили и углубили
эти данные.
Особенно показательны опыты В. К. Балуева, которые
были поставлены с определенным видом люмбрицид
в естественной почве и в условиях, очень близких к по-
левым. Ведение опытов по урожайности в лаборатории
и в небольших сосудах дает менее ясные результаты, так
как, помимо влажности, температуры, освещения и т. д.,
экспериментальные условия резко отличаются от полевых
совсем иной флорой микроорганизмов, развивающейся
в сосудах. Опыт Балуева был осуществлен следующим
образом. В глиняные неглазированные горшки емко-
стью 5 л насыпалось по 3.5 кг просеянной почвы, либо
огородной, либо подзолистой супесчаной, взятой с поля,
занятого пропашными культурами (опыт велся в этих
двух вариантах). В каждом варианте было по четыре со-
суда, один из которых служи и контролем (без червей),
а в остальных трех на весь объем сосуда было 4, 8 и 16
червей, что соответствовало 880 000—3 540000 особей на
1 га. Все сосуды были закопаны в землю на поле вровень
с поверхностью почвы. 15-го мая во все сосуды были по-
сеяны зерна яровой пшеницы сорта Лютесценс 0.62.
Через 15 дней после посева в каждом из сосудов было
оставлено по 9 проростков пшеницы, а остальные выдер-
нуты. Осенью был собран урожай и учтены общий вес
растений, средний вес колоса, средний вес зерна и число
зерен. Каждый вариант опыта был повторен 5 раз. Как
можно видеть из табл. 9, все показатели урожайности
были выше в сосудах с червями, причем выяснилось, что
чем больше червей, тем урожай выше, т. е. эффект при-
сутствия в почве червей даже при очень больших плот-
ностях их населения, оказывается положительным.
Интересен опыт Андерсона (1940), проведенный в Ка-
лифорнии. В трех ящиках одновременно были посеяны
семена газонной травы мятлика (Роа trivialis): в одном
ящике червей вовсе не было, в другом была та же почва
с червями, в третьем грунт состоял только из копролитов,
176
Таблица 9
Влияние деятельности дождевых червей на урожай
пшеницы. (Из В. К. Балуева, 1950)
Количество черней в сосуде Количество черней в пе- ресчете на 1 га Общий вес растений в сосуде (н г) Вес собран- ных зерен (нг) Коли- чество собранных зерен
Без червей 0 110.6 27.6 873
i 880000 116.5 31.1 882
8 1790000 175.5 50.7 1313
16 3540000 183.0 55.6 1422
собранных в той же почве. Вес надземных частей растений
через одинаковый срок оказался, — если принять урожай
в первом ящике за 100%, — во втором 271 ?ь, в третьем
463%.
Очень важно изучение деятельности дождевых червей
для роста древесных пород, так как повышение плодоро-
дия лесных почв при помощи обычных агротехнических
приемов (перепахивание, удобрение) невозможно. В этом
отношении интересны опыты А. И. Зражевского (1957),
который выращивал разные породы деревьев в почве
с пашенными червями (Allolobophora caliginosa) и без
них. Молодые деревца в вазонах с почвой, содержащей
червей, росли заметно лучше, чем в почве без червей при
прочих равных условиях (рис. 59). При этом оказалось,
что влияние червей очень сильно зависит от того, какш*
листья даются им для корма. Опавшая листва разных
деревьев по-разному действует на рост сеянцев дуба
(табл. 10). Гниющие листья сами по себе, без червей, уси-
ливают рост сеянцев, удобряя почву при содействии
микроорганизмов (табл. 10). Но при наличии пашенных
червей влияние удобрения значительно повышается, при-
чем в весьма различной степени в зависимости от харак-
тера листьев. Можно думать, что это объясняется неоди-
наковой степенью пригодности листьев разных деревьев
для питания дождевых червей. Выше было сказано, что
черви обладают тонко развитой способностью восприни-
мать вкусовые ощущения и очень разборчивы в пище.
Кроме того, результат деятельности червей зависит от ха-
12 О. В. Чекаnonehiп 177
рактера микробиологических процессов в почве. Положи-
тельное влияние дождевых червей на рост сеянцев раз-
ных деревьев отчетливо выразилось уже в первый год,
Рис. 59. Вазоны с саженцами зеленого ясеня:
левый — с червями, правый — без червей, при
равных прочих условиях. (По А. И. Зражев-
скому).
а через два года (табл. 10) действие их сказалось еще силь-
нее.
Влияние дождевых червей на произрастание деревьев
и трав различно также в разных грунтах. Так, в опытах
с саженцами сосны положительное влияние червей на рост
саженцев проявилось па торфе, смешанном со сланце-
178
Таблица 10
Влияние дождевых червей на рост сеянцев дуба
в зависимости от корма для червей в виде опавших
листьев разных деревьев. Опыт велся в течение 2 лет.
(Из А. И. Зражевского, 1957)
Листья, подкладываемые на поверхность почвы в сосудах с сеянцами дуба Средняя высота сеянцев (в см) Средняя толпшна ствола у корненой шейки (в мм)
с червями [без червей с червями без червей
Контроль (без листьев) . . 1 37.5 1 31.2 | ' I 9.4 8.2
Дуб 39.8 29.9 9.0 1 8.6
Дуб + черная бузина . . 4°.7 j 31.6 9.9 8.9
Дуб + граб j i 49-* 42.1 9.2 8.7
Дуб + клен остролистный 1 51.3 । 39.9 11.1 8.9
вой породой, значительно сильнее, чем на песке и чистом
торфе. Наоборот, в некоторых опытах, например при под-
кладывании листьев желтой акации и граба на почву
в сосудах с сеянцами, черви или не оказывали благоприят-
ного действия на рост сеянцев дуба, или даже несколько
тормозили рост. Этот исход опыта очень поучителен
так как он свидетельствует, насколько важно всесторон-
нее знание вопроса для управления ростом растений при
проведении разного рода мелиоративных мероприятий.
Отрицательный результат в этих случаях объясняется,
как сказано выше, слабой активностью дождевых червей
при определенных видах корма.
Возможно, что значение дождевых червей для жизни
растений не исчерпывается их деятельностью в преобразо-
вании свойств почвы. В последнее время появились дан-
ные о том, что копролпты содержат вещества, повышаю-
щие скорость прорастания семян древесных и огород-
ных растений. Действие копролитов на прорастающие
семена настолько сильно, что избыточная прибавка
их к земле, в которую семена посеяны, может повлечь
за собой гибель последних; умеренное же добавление ко-
пролитов к земле с семенами значительно повышает их
всхожесть (Зражевский, 1957). Таким образом, следует
учитывать действие червей не только на почву, но и на
растительные организмы.
12*
17ч
В общем нужно сказать, что имеющиеся строго про-
веренные данные по влиянию деятельности дождевых
червей па плодородие почв пока крайне недостаточны.
Самый факт положительного значения дождевых червей
для жизни растений не вызывает никаких сомнений. Но он
порождает огромное количество вопросов, решение кото-
рых необходимо для разумного использования этих жи-
вотных в целях растениеводства. Особенности разных ви-
дов дождевых червей в отношении их потребностей, ха-
рактера работы в разных почвах и т. д. известны пока
очень мало. Приведенные данные показывают только
с полной ясностью необходимость и перспективность
дальнейших исследований в этом направлении.
2. МЕРОПРИЯТИЯ ПО РАЗВЕДЕНИЮ И ОХРАНЕ
ДОЖДЕВЫХ ЧЕРВЕЙ
Данные современной науки с несомненностью свиде-
тельствуют в пользу того, что деятельность дождевых
червей — существенный фактор плодородия почв. Однако
если изучение действия этого фактора, как мы видели,
продвинуто еще не особенно далеко, то разумное исполь-
зование его для сельского хозяйства и лесоводства на-
ходится в самом зачаточном состоянии.
Работа, осуществляемая миллиардами дождевых чер-
вей в почве, огромна. Если учесть только мышечную
работу, производимую червями по рытью норок и выбра-
сыванию земли на поверхность, то это одно составит
огромный энергетический фонд, использование которого
с нужной интенсивностью, в нужное время и в нужных
местах для возделывания земли — задача принципиально
разрешимая и сулящая значительные выгоды для всех
видов растениеводства. Но, как мы видели, деятельность
дождевых червей не ограничивается мышечной работой —
прокапыванием земли и переносом ее в кишечнике с од-
ного места на другое. При прохождении через кишечник
почва изменяется химически и, — что особенно важно, —
структурно. По словам В. Р. Вильямса, «наша задача —-
повсеместно превратить почвы Советского Союза в струк-
турные, т. е. самые плодородные почвы мира». Не может
быть сомнения, что овладение способами максимального
использования деятельности дождевых червей как агента,
180
«ничем не воспроизводимого» (Н. А. Димо), должно ока-
зать существенную помощь в решении этой грандиозной
задачи.
• В каком же направлении следует искать пути для все-
мерного использования дождевых червей в агротехни-
ческих целях?
Прежде всего приходит в голову мысль об искусствен-
ном разведении червей. Эта мысль усиленно пропаганди-
руется в США и отчасти в Англии. Одним из главных
защитников этого приема в США является упомянутый
уже нами Баррет, врач по профессии и собственник зе-
мельной фермы. Он начал заниматься разведением дожде-
вых червей в 1936 г. и в своей книге «Червь-земледелец»
(Barret, 1947) сообщает, что им достигнуты крупные ре-
зультаты в деле поднятия доходности своего хозяйства
при помощи «удобрения» земли дождевыми червями.
Эта книга, несмотря на рекламный характер, заслуживает
внимания.
Идея Баррета заключается в том, что необходимо сде-
лать дождевых червей домашними животными. Тело
червя — это «фабрика, изготовляющая быстро и в большом
количестве вещество, необходимое для человека» (имеется
в виду гумус). Увеличение народонаселения при ограни-
ченном пространстве суши, которым располагает1 челове-
чество, неминуемо должно привести к недостатку пита-
ния, если последнему не противопоставить интенсифика-
цию сельского хозяйства. Одним из неиспользованных
резервов в этом отношении, по мнению Баррета, является
усиление деятельности дождевых червей при помощи их
искусственного разведения и контроля за их биологией.
Это одомашнение, или доместикацию дождевых червей
он считает фактором мирового значения и пропаганду
этой идеи называет «новым фронтом» в борьбе за благо-
получие человечества. В такой подаче этой проблемы,
разумеется, много наивного. Помимо недоучета мощи со-
циального фактора, здесь очевидна недооценка других
агротехнических мероприятий.
Основным положением Баррета является необходи-
мость полнейшей утилизации всех органических веществ,
получающихся в хозяйстве, т. е. отходов кухни, сада,
огорода, а также опавшей листвы и т. д., для того чтобы
кормить этим дождевых червей. Земля может прокормить
181
5—6 миллионов червей на 1 га. На самом деле их в поле-
вых почвах гораздо меньше — 0.1—1.5 миллиона. Эту
«недогрузку» полей можно, по мнению Баррета, воспол-
нить, если разводить червей в особых культурах, где
в 1 кубическом футе земли может содержаться до 3000
червей, что соответствует плотности населения в 345 мил-
лионов особей на 1 га. Червями, выращенными в культу-
рах, «удобряется» садовая и полевая почвы, где они и
производят свою полезную деятельность.
Прежде чем дать оценку взглядам Баррета, ознако-
мимся с существующими приемами разведения дождевых
червей. Отметим способ, разработанный М. М. Исаковой-
Кео.
Выращивание дождевых червейдля
корма цыплят (способ М. М. И с а к о в о й - К е о).
Для разведения червей в земле выкапываются ямы дли-
ной 3.5—4 м и шириной 1.5—2 м. Если почва глинистая,
то вполне достаточна глубина в 1.2—1.3 м. Если же почва
песчаная, то глубина ям должна быть 1.5—2.0 м. В по-
следнем случае на дно ямы помещают слой глины, толщи-
ной 30—35 см; глиной покрываются и боковые стенки
ямы (слой глины 15—20 см). Далее в яму вносят слой
свежего конского навоза в 30—35 см, который покрывают
слоем земли в 20—35 см. На этот слой накладывают опять
навоз слоем в 20—25 см и мягкую траву (мокрица, лебеда
и др.). Затем навоз и траву покрывают еще одним слоем
земли в 20—25 см. Корм для червей (смоченные отруби,
картофельные очистки и другие кухонные отбросы, гни-
лой картофель, а также опавшие листья и трава) насыпают
в сделанные в верхнем слое земли ямки диаметром 10—
15 см и такой же глубины, на расстояние 15—20 см друг
от друга. Собранных червей раскладывают возле ямок
с кормом, распределяя их по 100—150 шт. на 1 м2. Ямы
для червей лучше закладывать с осени, что имеет следую-
щие преимущества: 1) еще с осени черви приползут в яму
из окружающей ее почвы; 2) если ямы заложены в поло-
вине августа или в сентябре, то наблюдается откладка
большого количества коконов; 3) до холодов успевают
вылупиться из коконов молодые черви, а очень рано вес-
ной снова начинается откладка коконов. Одновременно
закладывают ряд ям, что позволяет правильно их эксплуа-
тировать.
182
Землю время от времени поливают водой с таким рас-
четом, чтобы влажность ее верхнего слоя была не более
25%. Сверху ямы накрываются деревянными или соло-
менными щитами. На зиму для предохранения от промер-
зания все ямы покрывают конским навозом, причем
покрывают не только поверхность ямы, но и участок земли
вокруг нее в 30—40 см шириной. Весной, когда кончатся
заморозки, весь навоз с ям сгребают, также выгребают
несгнившие листья. Навоз и листья могут быть исполь-
зованы для закладки новых ям. Слегка перемешивают
верхний слой земли и червей подкармливают, как ука-
зано выше. Когда появится трава, можно червей подкарм-
ливать и ею. Скошенная трава вымачивается 5—7 дней
в бочке с водой, затем закапывается пучками по 500—
600 г в разных местах ямы в землю и поливается водой.
Гниющая трава сменяется несколько раз в лето.1
Методика культивирования дождевых
червей по ^арр£ту Баррет разработал два варианта
культивирования дождевых червей: ящичный и траншей-
ный. Для ящичных культур употребляются ящики разме-
ром 36x40x12 см. Они делаются из топких досок и
устанавливаются в стеллажи по 4—10 шт. Стеллажи
устраиваются таким образом, чтобы между ящиками, нахо-
дящимися друг над другом, оставались промежутки для воз-
духа. В дне ящиков делается 6—8 отверстий размером
2—3 см. Ящики наполняются компостом, состоящим из ло-
шадиного или кроличьего навоза (1 часть), просеянной земли
(1 часть) и торфяного мха (1 часть). Для приготовления
компоста рекомендуется устроить особый ящик размером
150x90x25 см; можно производить смешивание компоста
и в яме. Компост смешивается в почти сухом состоянии и
смачивается лишь перед укладкой в ящики. На компост кла-
дутся черви из расчета 500 шт. на ящик. Ящики можно за-
ряжать и яйцевыми кокопами червей: 200—300 коконов
рассыпаются в ящик на компост и посыпаются сверху
слоем земли в 2—3 см. Сверху ящики прикрываются меш-
ками или рогожами. Стеллажи с ящиками должны быть
защищены от прямых лучей солнца, п температура почвы
1 Излагаемая методика разведения дождевых червей с одер-
жиТся в статье А. Б. Рабиновича (1953), который не указывает
фамилии разработавшего ее автора,
1*3
в них не должна быть выше 15—17°. Умеренной полив-
кой поддерживается постоянная влажность компоста.
Сбор червей п коконов, накопившихся в ящиках,
производится следующим образом. Ящик вынимается
из стеллажа и опрокидывается над столом, величина ко-
торого должна быть достаточной для того, чтобы высы-
панное па него содержимое ящика при разборке не па-
дало па землю (т. е. ширина его должна быть около 75 см).
Все черви при этом сразу уходят в глубину и собираются
на поверхности стола под компостом. Сначала просма-
тривается компост слой за слоем, и из него выбираются
коконы. Затем собираются черви, оказавшиеся па дне
кучи. Крупные черви помещаются обратно в ящик как
производители, а собранная молодь кладется под нужные
растения. Коконы же используются для зарядки других
ящиков или для продажи. Эта операция производится
раз в месяц.
По-видимому, ящичные культуры червей используются
Барретом главным образом для плодовых деревьев. Он
считает, что однократный ящичный сбор достаточен для
обеспечения червями двух-трех деревьев на месяц. Из че-
тырех ящиков в год можно собрать 55 000 коконов.
Ящичные культуры дождевых червей успели приоб-
рести большую популярность в США. Это видно из того,
что Барретом приводятся рыночные цепы переносных
ящичных культур и коконов. За заряженный червями
ящик культуры фермеры платят 15 долларов, а коконы
ценятся по 1 центу за штуку. Таким образом, производ-
ство червей там делается особой отраслью промышленности.
Траншейный способ, употребляемый в Америке в не-
скольких вариантах, мало чем отличается от приведен-
ного выше способа М. М. Исаковой-Кео. Глубину траншеи
в перегнойной почве Баррет считает достаточной в 60 см,
так как большая глубина затрудняет проветривание почвы.
На дно траншеи кладутся дюймовые доски, стенки также
обкладываются досками, закрепляемыми брусками в 5х
10 см в поперечнике. Вся конструкция делается без гво-
здей. На дно кладется дренажный материал, сверху
насыпается смешанный компост того же состава, что и
для ящиков. Траншея размером 2.5x1.25x0.6. м может
прокормить 50000 особей одомашненных дождевых червей.
Каждый из приведенных способов культивирования
червей имеет свои преимущества. Нам представляется,
что в методе М. М. Исаковой-Кео очень важным является
указание на скошенную траву как хороший корм для чер-
вей. Это указание совпадает с приведенной выше методи-
кой Росселя (стр. 158) для использования дождевых червей
в целях повышения урожайности огородных культур.
В методике Баррета главный интерес представляют ящич-
ные культуры. Возможность переносить и перевозить
как угодно далеко такие культуры и, кроме того, удоб-
ство при сборах «урожая» червей, при которых черви
не ранятся лопатой, и выбирание их, как и их коконов,,
которое может быть проведено более полно, — являются
достоинствами этого способа.
Главный недостаток обоих методов заключается в том,
что в них совершенно не учитывается видовой состав куль-
тивируемых червей. Говорится о червях вообще как о не-
которой, якобы однородной совокупности. Но мы знаем,
что, даже главные представители дождевых червей в СССР
составляют семейство со многими родами и видами, ха-
рактеризующимися весьма различным образом жизни и
значением в почвообразовании.
, Разумный подход к делу требует продуманного выбора
из имеющегося состава фауны видов, наиболее подходящих
для культуры. Одни из них, наверное, окажутся малопри-
годными вообще, другие будут пригодны в разной степени.
При этом скорее всего в разных почвенных условиях и
для разных целей нужны будут культуры разных видов
люмбрицид (Grant, 1955).
Далее, остается неизвестной экономическая эффек-
тивность этих методов. Не произведен учет стоимости
работ по культивированию червей, и затраты на него
не сопоставлены с получаемыми выгодами. Однако рас-
пространение и успех культивирования червей в Аме-
рике н отчасти в Англии говорят в пользу того, что ка-
кая-то практическая польза от него имеется.
Что же касается теоретических положений Баррета,
то в той их части, в которой он настойчиво говорит о не-
обходимости всемерного использования дождевых червей
в целях интенсификации сельского хозяйства, считая их
энергетическим ресурсом, не использованным еще как
следует человеком, эти положения нужно считать про-
185
грессивными. В сущности они представляют собой не что
иное, как указание на необходимость сделать практиче-
ские выводы из соображений Дарвина о мировой роли
дождевых червей. В этом отношении взгляды Баррета
полностью совпадают со взглядами наших выдающихся
почвоведов и агрономов — Г. Н. Высоцкого. Н. А. Дпмо,
С. П. Кравкова и других исследователей.
Но нужно задать вопрос, является ли искусственное
разведение червей в «червятниках», организуемых в усадь-
бах, единственным приемом для разумного использования
этой силы природы?
Имеются все основания полагать, что это не только,
не единственный, но и отнюдь не главный прием.
Заметим, что расчеты Баррета, свидетельствующие,
по его мнению, о «недогрузке» полевых и садовых почв
в отношении возможной численности дождевых червей, —
неубедительны (см. стр. 182). Из того факта, что в полевых
почвах обычно плотность населения червей значительно
ниже максимально возможных количеств их в природных
условиях (0.1 —1.5 миллиона особей вместо 5—6 миллио-
нов на 1 га), никоим образом нельзя сделать вывод, что
искусственное добавление червей непременно окажет
благоприятное влияние на почву. Это произойдет только
в том случае, если черви найдут в данном участке подхо-
дящие для них условия существования. Если же черви,
искусственно внесенные в почву, там вскоре погибнут,
то никакой пользы от этого не будет. Погибнуть они могут
прежде всего от недостатка пищи. Ответить на вопрос,
сколько червей может прокормить данная почва при дан-
ных условиях — дело не такое простое. Поэтому есть ли
в том или ином случае действительная возможность
для увеличения плотности населения червей, решить не
так легко.
Следует отметить, что обилие особей разных групп
организмов в любой среде и в любом ее участке, равно
как отношения между количествами разных организмов, —
отнюдь не дело случая. Количественное соотношение
разных организмов в почве п плотность населения ка-
ждого из них — результат сложнейшего комплекса про-
цессов. Поэтому в общем случае количество • дождевых
червей, обнаруживаемое в той или иной почве, таково,
каким оно только и может быть при данных условиях,
186
Отсюда ясно, что если возникает вопрос о желательности
увеличения количества дождевых червей в каком-нибудь
участке почвы, то прежде всего нужно думать не о внесе-
нии их туда извне, а об изменении условий их существо-
вания в лучшую сторону.
Без сомнения, в случае падежа червей, происшедшего
вследствие болезней, паразитов, наводнений и прочих
бедствий, внесение их в почву извне может оказаться
очень полезным для нее. Далее, весьма вероятно, что
внесение в почву дождевых червей параллельно с вне-
сением в нее органических удобрений явится очень по-
лезным агротехническим приемом. Поэтому сказанное
не снимает вопроса об искусственном разведении червей,
а только намечает для пего надлежащее место. К тому же
повторим, что при внесении червей в почву необходимо
решить, какой именно вид люмбрицид наиболее подходит
для данной почвы, данной культуры растений и ожидае-
мой от него в данном случае деятельности (гумификация,
подщелачивание, созданпе структуры почвы и т. д.).
Принимая во внимание все указанные ограничения и
трудности, можно думать, что на огромных площадях
в экстенсивных полевых хозяйствах искусственное «очерв-
ление» вряд ли имеет большие перспективы. Но можно счи-
тать, что в интенсивных хозяйствах, т. е. в огородах, пло-
довых и ягодных садах, виноградниках и т. д., искусствен-
ному разведению червей предстоит сыграть видную роль.
Но все-таки основным в использовании дождевых чер-
вей как почвообразователей является не их искусствен-
ное разведение, а совсем другие мероприятия. В преды-
дущей главе было показано, в какой тесной зависимости
находится количество дождевых червей в почве от ее ха-
рактера и от растительности на ней. В частности, на полях,
истощенных культурами однолетних злаков, количество
червей очень невелико (см. табл. 1). Наоборот, число их
макспмальное в почвах, очень богатых органическими
удобрениями, на целине и в лиственных лесах с траво-
стоем. Уже из этих фактов становится ясным, какое зна-
чение для фауны люмбрицид на культурных землях имеет
тот или иной тип полевого хозяйства. Использовать до-
ждевых червей как союзников в земледелии — это значит
прежде всего заботиться о том, чтобы, во-первых, без
нужды не приносить им вреда и, во-вторых, по возмож-
187
ности охранять их от всевозможных неблагоприятных
воздействий. Рассмотрим, в общих чертах, в чем именно
то и другое может выражаться.
В зарубежной литературе высказывалось мнение о том,
что для дождевых червей якобы вредно внесение минераль-
ных удобрений в почву. Если бы это было действительно
так, то интересы полевого хозяйства стали бы в неприми-
римое противоречие с вопросом об охране населения до-
ждевых червей, так как в агрономии прочно установлено,
что наиболее высокие урожаи получаются при комбинации
минерального удобрения с органическим (В. Р. Вильямс).
С целью выяснить этот вопрос на Ротамстэдской станции
были проведены тщательные исследования па луговых
участках с разными типами минеральных удобрений.
Как видно из табл. И, минеральные удобрения угне-
тают деятельность червей только в тех случаях, когда
одновременно повышается кислотность почв (т. е. сни-
жается значение pH). В комбинации с известью, нейтра-
лизующей кислоты, они не оказывают вредного влияния
на червей, а калифосфатное удобрение с известью скорее
даже стимулирует деятельность червей в почве. Органи-
ческое же удобрение, как и можно было ожидать, имеет
положительное значение для дождевых, червей.
Из агрономических мероприятий, оказавшихся по-
лезными для дождевых червей, отметим действие укрытия
почвы соломой на зиму, особенно на период осенних
холодов до выпадения снега. М. П. Архангельский (1928)
сообщает, что на черноземной почве в Горьковской об-
ласти на паровом поле, укрытом соломой, он находил
80 червей на 1 м2, тогда как на непокрытых участках
поля их было 42 па 1 м2. Урожаи пшеницы на покрытых
участках были выше, что автор связывает с действием
ряда факторов, в том числе с деятельностью дождевых
червей. Г. Работнов (1938) наблюдал, что прикрытие соло-
мой и картофельной ботвой суходольных лугов в Ярослав-
ской области благоприятно действует на урожаи трав;
он также связывает это с предохранением дождевых
червей от холодов. Американские агрономы указывают,
что черви в ежегодно вспахиваемых землях умирают
массами при первых заморозках осенью, когда земля
не покрыта снегом, и советуют покрывать поля соломой
(Hopp a. Linder, 1947).
188
Таблица 11
Зависимость деятельности дождевых червей от удобрений,
вносимых в почву. (Из Франца (Franz, 1950) по работам
Ротамстэдской сельскохозяйственной опытной станции)
Удобрение pH (кислотность) Число кучек вы- бросов на акр 1
Вез удобрений 5.6 245
♦ > 4- известь.... 7.3 276
Органическое удобрение 4- из- 6.6 337
весть
КалифосЛатное удобрение . . . 5.1 136
На дифосфатное удобрение 4 из-
весть 6.7 252
Сернокислый аммоний 4-- кали- фосфатное удобрение 4.0 0
Сернокислый аммоний 4~ кали- фосфагное удобрение 4- из- 127 I I
весть 5.0
Следует обратить внимание на борьбу с врагами до-
ждевых червей. В главе IV отмечено, что одним из опасней-
ших врагов дождевых червей являются кроты, пожираю-
щие их в больших количествах. Очень вероятно, что унич-
тожением кротов следует заняться не только потому,
что они затрудняют применение машин при уборке трав
(что общеизвестно), но и вследствие вреда, наносимого
ими дождевым червям. Однако главные враги дождевых
червей, как мы знаем, не кроты, а птицы (стр. 116). Конечно,
никто не станет и думать об уничтожении насекомоядных
птиц, так как они очень полезны в сельском хозяйстве
и земледельцы обязаны им уничтожением огромных коли-
честв вредных насекомых. Но возможно применение меро-
приятий, защищающих червей от птиц. Как справедливо
заметил А. А. Соколов (1953), черви особенно страдают
от птиц во время перепахивания полей. Поэтому системы
земледелия, допускающие оставление земли в некоторые
годы без перепахивания, уже одним этим должны содей-
ствовать увеличению численности червей в почвах. Кроме
того, при вспахивании полей червей разрезает плуг, их
1 Акр — площадь, равная 4050 м2.
189
давит трактор, они гибнут от высыхания, действия сол-
нечных лучей, резкого изменения температурных усло-
вий и т. д.
Здесь мы подходим к вопросу о положительном зна-
чении для жизни дождевых червей многопольного сево-
оборота. Как известно, эта система земледелия со-
стоит в том, что земля в хозяйстве делится на известное
число участков (существуют 7-, 9-, 12-польные и другие
системы) и на каждом из них происходит установленное
чередование культур разных растений: зерновых злаков
(озимых, яровых), так называемых пропашных культур
(картофель, горох, корнеплоды и пр.) и многолетних
трав (клевер, тимофеевка, люцерна и др.). Благодаря
соответственному подбору сменяющих друг друга расти-
тельных культур почва не истощается, так как одни ра-
стения вносят в нее вещества, используемые впоследствии
другими, и таким образом осуществляется автоматиче-
ское восстановление почвы. Особенно важным для вос-
становления почвы является посев многолетних трав.
Исследования С. И. Пономаревой (1953, 1958), подтвер-
жденные А. Мамытовым (1953), А. А. Соколовым (1953,
1956), М. М. Алейниковой и В. В. Изосимовым (1958)
и другими исследователями, показали, что культуры много-
летних трав создают максимально благоприятные условия
для жизни, размножения и деятельности дождевых чер-
вей. Как было показано (стр. 134 и табл. 1 и 4), уже в пер-
вый год пользования посевными травами количество
червей и копролитов сильно возрастает, а на второй год*
достигает максимальных размеров, приближающихся
к наблюдаемым на целине. Это объясняется не только
отсутствием перепашки. Здесь большую роль играет
образующийся дерновый покров, создающий условия
большего постоянства температуры и влажности в верх-
них слоях почвы, обилие пищи (отмирающие корни и пр.)
и появление многочисленной микрофлоры и микрофауны
в почве.
Мы уже знаем, какова значимость дождевых червей
в образовании структуры почвы, как велика их роль именно
в создании прочных структур. Вряд ли можно сомне-
ваться в том, что очень важным посредствующим звеном
между травосеянием и восстановлением плодородия почвы
является деятельность дождевых червей.
190
Заметим, что и в лесах, как единогласно утверждают
исследователи (Шиперович, 1947; Соколов, 1956; Зражев-
ский, 1957, и др.), появление многолетних трав вызывает
увеличение плотности населения дождевых червей и уси-
ление их жизнедеятельности. Таким образом, у лесоводов
имеются широкие возможности для управления почво-
образовательными процессами. Регулируя густоту и со-
став лесных насаждений, они могут влиять на произра-
стание трав под пологом леса, а тем самым создавать благо-
приятные условия для жизни люмбрицид.
По словам Г. Н. Высоцкого (1930), вести хозяйство
следует, проявляя заботу о дождевых червях, т. е. «так,
чтобы способствовать размножению этих важных помощ-
ников в деле обработки почв, чтобы помочь их деятель-
ности и не уничтожать их». Как мы сейчас видели, это
облегчается тем, что важные агротехнические мероприя-
тия, направленные на улучшение жизни культурных ра-
стении, являются в то же время действенными факторами,
положительно влияющими на население дождевых червей
в почвах. Учет влияния па жизнь червей упомянутых
приемов земледелия несомненно должен иметь серьезное
значение для сельского хозяйства и лесоводства. Но за-
дача всестороннего разумного использования этих живот-
ных еще не может считаться решенной. Успешное'ее ре-
шение, т. е. выработка специальных мероприятий для этой
цели, требует совместных усилий всех работников, свя-
занных с земледелием и лесоводством.
,1 И Т Е Р А Т У Р А
А л е й н и к о в а М. М. и В. В. Изосимов. 1958. хМатериалы
по фауне и экологии дождевых червей в Татарской АССР.
Всесоюзн. совещ. по почв. зоол. АН СССР» Тез. докл.,
М. : 6-7.
А л и м д ж а и о в Р. А. 1946. Плотность почвенной фауны полив-
ных люцерников на севере Узбекистана. Тр. Сект. зоол.
АН УзССг, Ташкент : 41—51.
Архангельский М. II. 1928. К вопросу о влиянии мерт-
вого (соломистого) покрова на урожай озимой пшеницы.
Изв. Нижегор. гос. унив., 2 : 165—179.
Архангельский М. П. 1929. К вопросу о влиянии работы
дождевых червей на урожай овса и ячменя в связи с внесе-
нием в почву некоторых удобрительных веществ. Научно-
агрон. журн., 6, 12 : 849—862.
Балуев В. К. 1950. Дождевые черви основных почвенных раз-
ностей Ивановской области. Почвоведение» 4 : 219—227.
Б я л у е в В. К. 1958. Динамика беспозвоночных в почвах полей,
обрабатываемых по Мальцеву в условиях Ивановской обл.
Всесоюзн. совещ. по почв. зоол. АН СССР, Тез. докл.,
М. : 12.
Б а х т и п П. У. и М. И. Польский. 1950. О роли дождевых
червей в оструктурировании дерново-подзолистых почв.
Почвоведение, 3 : 487—491.
Беклемишев В. Н. и И. А. Четыркина. 1935. К биоло-
гии половодья. О судьбе дождевых червей во время весен-
него половодья. Вопр. экол. и биоценол., Сб. статей, 2,
Л. : 120-136.
Бродский А. Л. 1927. Материалы к изучению биоценозов
ирригационной сети в Голодной Степи. Вести, ирригации,
6 : 21—29.
Бродский А. Л. 1937. Исследования по фауне почв. Таш-
кент : 1—68.
Вильямс В. Р. 1946. Структура почвы и урожай. Сов. агро-
номия, 10:8—11.
192.
Виноградов М. П. 1927. Анабиоз при высушивании. Опыты
над дождевыми червями. Зап. Ленингр. с.-х. инет.,
4 : 689-702.
Высоцкий Г. Н. 1898. Растительность Велпкоанадольского
участка. Тр. Эксп. лесн. департ., 2 : 135—268.
Высоцкий Г. Н. 1899. Гидрологические и гео-биологические
наблюдения в Великом Анадоле. Почвоведение, 1, 2 : 36—
44.
Высоцкий Г. Н. 1900. Дождевой червь. Полная энциклопедия
русск. сельск. хоз., 2 : 12—39.
Высоцкий Г. Н. 1930. Агенти структуре грунпв. Тр. Науково-
дослщно! катедри грунтознавства, 1 : 141—143.
Высоцкий Г. Н. 1933. Общие положения об агрономическом
значении почвенной структуры. Тр. Сов. секц. Междунар.
ассоц. почвоведов, 1 : 115—117.
Гаврилов К. А. 1950. Влияние состава лесонасаждений на ми-
крофлору и фауну лесных почв. Почвоведение, 3 : 129—141.
Гаврилов К. А. иТ. С. Перель. 1958. Дождевые черви
и другие беспозвоночные в почвах лесов Вологодской обл.
Почвоведение, 8 : 133—140.
Гагарин В. Т. 1953. К изучению цикла развития Capillaria
caudiinflata и терапии капилляриоза у кур. Тр. Всесоюзн.
£ инет. Гельминтол. им. акад. К. И. Скрябина, 5 : 160—162.
Г а р к у ш а И. Ф. 1954. Почвоведение. Сельхозгиз, М.—
Л. : 1-424.
Г^и^Гя р о в М. С. 1939. Почвенная фауна и жизнь почвы. Поч-
S воведение, 6 : 3—15.
Гиляров М. С. 1942. Сравнительная заселенность почвенными
животными темноцветной и подзолистой почв. Почвоведение,
9-10 : 3-15.
Гиляров М. С. 1944. Количество почвенных животных на еди-
ницу площади почвы. Докл. Акад, наук СССР, 43,
№ 6 : 283-285.
Гиляров М. С. 1947. Распределение гумуса корневых систем
и почвенных беспозвоночных. Докл. Акад, наук СССР, 55,
№ 1 : 53-56.
Гиляров М. С. 1949. Особенности почвы как среды обитания
и ее значение в эволюции насекомых. Изд. АН СССР,
М.-Л. : 17-197.
Г и л я р о в М. С. 1951. Роль почвенных животных в формирова-
нии гумусового слоя почвы. Успехи совр. биол., 31,
2 : 161-169.
Гиляров М. С. 1953. Почвенная фауна и задачи почвенной зоо-
логии. Вести. Акад, наук СССР, 8 : 12—21.
Глинка К. Д. 1924. Русское почвоведение. Зап. Ленингр. с.-х.
инет., 1 : 1—24.
Глинка К. Д. 1935. Почвоведение. Сельхозгиз, М. : 1—631.
Горизонтова М. Н., Л. А. Красная, Т. С. Перель.
1957. Наблюдения над распределением и численностью
дождевых червей в почве в течение года. Уч. зап. Моск. гор.
педаг. инет. им. В. П. Потемкина, 65 : 161—178.
ГражульВ. 1957. Пандемия гриппа. Газ. «Медицинский работ-
ник», 27 : 3.
13 о. В. Чекановская
193
Гриб А. В. 1950. Дождевые черви Lumbricidae. ЖивотнЛмир
СССР, 3, Изд. АН СССР, М.-Л., 3 : 471-477. Ml ,
Гурьянова О. 3. 1940. Влияние дождевых червей и органи-
ческих удобрений на структурообразование черноземных
почв. Почвоведение, 4 : 99—107. м
Дадыкин В. П. 1941. О деятельности дождевых червей в почвах
крайнего севера. Почвоведение, 4 : 42—47.
Д и м о Н. А. 1938. Земляные черви в почвах Средней Азии. Почво-
ведение, 4 : 494—526. «м
ДимоН. А. 1941. Деятельность животных в почвах Алазанской
долины. Почвоведение, 6 : 12—20.
Д и м о Н. А. 1955. Наблюдения и исследования по фаупе почв.
Изд. 2-е. ГИЗ Молдавии, Кишинев : 1 —140.
Докучаев В. В. 1899. К учению о зонах природы. СПб. : 1—
28. См. также: Собр. соч., 6, Изд. АН СССР, М,—Л.,
1951 : 398-414.
3 е л и к м а н А. Л. и И. И. На л евич. 1951. Эксперименталь-
ные работы по курсу зоологии беспозвоночных, ч. 1. Учпед-
гиз, М. : 1—136?
(3 р а ж е в с к и й А. И.) Зражевський А. I. 1949. Нейтра-
лизуюча д!яльн1сть дощовикчв Lumbricidae у л!совому
грунтк Пращ Ihct. niciBii., 1, Изд. АН УРСР : 144—151.
Зражевский А. И. 1950. Характер распространения дожде-
вых червей в лесной почве. Сб. «Вторая экологическая кон-
ференция по проблеме: Массовые размножения животных
и их прогнозы», Тез. докл., 1, Киев : 80—83.
Зражевский А. И. 1952. Роль дождевых червей в поднятии
плодородия лесных почв. Массивное лесоразведение и выра-
щивание посадочного материала. Изд. АН УССР,
Киев : 53—67.
(Зражевский А. И.) Зражевський А. 1.1953. Про вплив до-
niQBHKiB на схожкть нас1ння. Украинск. Бот. журн., 10,
№ 2 : 66—72.
Зражевский А. И. 1954а. О значении фауны беспозвоночных
в повышении плодородия лесных почв. Тр. Инет, леса АН
СССР, 23 : 237-265.
Зражевский А. И. 19546. Фауна почвенных беспозвоночных
как составное звено леса и фактор плодородия почв. Третья
эколог, конфер., Тез. докл., 4, Киев : 138—142.
Зражевский А. И. 1954в. Роль дождевых червей в формиро-
вании профиля почв на каменистых россыпях горных скло-
нов Карпат. Третья эколог, конфер., Тез. докл., 4,
Киев : 142-145.
Зражевский А. И. 1957. Дождевые черви как фактор плодо-
родия лесных почв. Изд. АН УССР, Киев : 1—271.
Качинский Н. А. 1925. Корневая система растений в почвах
подзолистого типа. Изд. Моск. обл. с.-х. Оп. ст., М. ; 1—89.
Кашкаров Д. Н. 1933. Среда и сообщество. Госмедиздат,
М. : 1-242.
Кирьянова Е. С. 1936. К вопросу о вертикальном и горизон-
тальном распределении фауны беспозвоночных в почвах
окрестностей Ташкента. Тр. Зоолог, инет. АН СССР,
3 : 309-352.
194
КобахидзеД. Н. 1941. Количественное распределение дожде-
вых червей в почвах центральной части Колхидской низмен-
ности. Сообщ. Акад, наук ГрузССР, И, 10:657—662.
Кононова М. М. 1943. Применение микроскопического метода
К
К
Л
л
м
м
м
м
м
м
м
м
м
м
м
м
при изучении вопроса о происхождении гумусовых веществ.
Почвоведение, 6 : 27—41.
р а в к о в С. П. 1931. Курс общего земледелия, 1. Гос. изд. с.-х.
и» колхозн.-кооп. лит., М.—Л. : 1—332.
улагин Н. М. 1889. Материалы по естественной истории
дождевых червей (сем. Lumbricidae). Изв. Общ. люоит.
естеств., антропол. и этногр., 58, 2 : 1—65.
аврентьев А. И. 1958. Дождевые черви как вредители и
паразиты. Первая зоолог, конфер. Белорусской ССР, Тез.
докл., Минск : 138—141.
омоносов М. 13. 1763. Первые основания металлургии или
рудных дел. О слоях земных. В Санкт-Петербурге печатаны
при имп. Акад, паук : 237—416. См. также: Сочинения, 7.
Изд. АН СССР, Л., 1934 : 168-271.
а л е в и ч И. И. 1937. К фауне Oligochaeta Белорусской ССР.
Тру Гос. зоолог, музея при Моск. гос. унив., 4 : 127—
алевич И. И. 1950. Дождевые черви как промежуточные
хозяева метастронгилид. IS юл л. Моск. общ. испыт. прир.,
Отд. биол., 49, 1 : 33—36.
алевич И. И. 1941. Материалы к познанию фауны дождевых
червей (Lumbricidae) СССР. Сб. тр. Гос. зоолог, музея при
Моск. гос. унив., 6 : 273—276.
алевич И. И. 1949. Материалы к познанию дождевых чер-
вей орехово-плодовых лесов Южной Киргизии. Докл. Акад,
наук СССР, 67, 2 : 397—400.
алевич И. И. 1950а. Новые и малоизвестные виды дождевых
червей в фауне европейской части СССР. Докл. Акад, наук
СССР, 70, № 6 : 1083—1086.
алевич II. И. 19506. Собирание и изучение дождевых червей-
почв ообра зова тел ей. Изд. АН СССР, М.—Л. : 1—37.
а л е в и ч И. И. 1951. Дождевые черви окрестностей Галичской
биостанции. Уч. зап. Моск. гор. педаг. инет. им. В. П. По-
темкина, 18, Каф. зоол., 1 : 85—114.
алевич И. И. 1952. Дощовики (Lumbricidae) лкових насад-
жепь Украшп. Пращ 1нст. JiiciBii., 3 Изд. АН УРСР : ISO-
164.
алевич И. И. 1953а. Дождевые черви. Животн. мир СССР,
4 Изд. АН СССР : 577-589.
алевич И. И. 19536. Материалы по фауне и экологии дожде-
вых червей Белоруссии. Бюлл. Моск. общ. испыт. прир.,
Отд. биол., 58, 5:39—49.
а л е в и ч И. И. 1954. Некоторые особенности распространения
дождевых червей в районах полезащитного лесоразведения.
Уч. зап. Моск. гор. педаг. инет. им. В. П. Потемкина, 28,
Каф. зоол., 2 : 5—32.
а л е в и ч И. И., А. А. К а ч а н о в а и С. П. С а п р ы к и н а.
1954. Сравнительный анализ фауны и распределения дожде-
вых червей в Голоссевском и Велико-А на дольском лесниче-
13*
195
ствах Украины. Уч. зап. Моск. гор. педаг. инет. им. В. П. По-
темкина, 28, Каф. зоол., 2 : 41—54.
Малевич И. И. и Т. С. II е р о л ь. 1958. Дождевые черви
Теллермановс кого пргцичества и их распределение в нагор-
ном дубраве и лесах поймы. Уч. зап. Моск. гор. педаг.
инет. им. В. II. Потемкина, 84, Каф. зоол., 7 : 257—268.
М а м ы т о в А. 1953. Влияние дождевых червей на водопрочность
горно-долинных сероземных почв. Почвоведение, 8 : 58—60.
Мозговой А. А. 1952. Биология Porrocaecum crassum — нема-
тоды водоплавающей птицы. Тр. Гельминтолог, лабор.,
6 : 114-125.
Мюллер Э. П. 1907. Влияние дождевых червей на произраста-
ние растений с корневищами, главным образом, в буковых
лесах. Леси, журн., 10 : 1446—1469.
Невмывака Г. А. 1947а. Иннервация кишки у дождевого
червя. Докл. Акад, наук СССР, 56, 5 : 533—536.
Невмывака Г. А. 19476. Брюшной мозг дождевого червя.
Докл. Акад, наук СССР, 58, 7 : 1483—1486.
Передельский А. А. 1958. Актуальность изучения роли
почвенных беспозвопочных в судьбе загрязнений почвы
радиоактивными изотопами. Всесоюзн. совещ. по почв. зоол.
АН СССР, Тез. докл., М. : 74-75.
Передельский А. А., Н. А. II о р я д к о в а, Л. 3. Р о-
дионоваиС. В. Тарчевская. 1957. Роль дождевых
червей в очистке почвы от загрязнений радиоактивными
изотопами. Докл. Акад, наук СССР, 115, 4 : 809—812.
Перель Т. С. 1958. Зависимость численности и видового со-
става дождевых червей от породного состава лесонасаждений.
Зоолог, журн., 37, 9 : 1307—1315.
Петров В. С. 1946. Активная реакция почвы (pH) как фактор
распространения дождевых червей. Зоолог, журн., 25,
2 : 107-110.
Петров В. С. 1947. Материалы по фауне и экологии сем. Lum-
bricidae (Oligochaeta) Воронежской области. Бюлл. Моск,
общ. испыт. прир., Отд. биол., 52, 1 : 51—53.
Пономарева С. И. 1949а. Влияние деятельности дождевых
червей на создание устойчивой в эрозионном отношении
структуры почв. Тр. Юбил. сессии, иосвящ. 100-летию со
дня рожд. В. В. Докучаева, М. : 475—483.
Пономарева С. И. 19496. Скорость образования в почвах
кальцита дождевыми червями. Докл. Акад, паук СССР, 61,
3 : 505—507.
Пономарева С. И. 1950. Роль дождевых червей в создаппи
прочной структуры в травопольных севооборотах. Почво-
ведение, 8 : 476—486.
Пономарева С. И. 1953. Влияние жизнедеятельности дожде-
вых червей на создание устойчивой структуры дерново-
подзолистой почвы. Тр. Почв. инет. им. В. В. Докучаева,
41 : 304-378.
ГГо номарева С. И. 1958. Роль жизнедеятельности дождевых
червей в повышении плодородия дерново-подзолистых почв.
Всесоюзн. совещ. по почв. зоол. АН СССР, Тез. докл.,
М : 80—81.
196
Потемкина В. А. 1953. Основные гол ьми птозы домашних
птиц. Сельхозгиз, М. : 1 — 168.
Рабинович А. Б. 1953. Разведение дождевых червей. Птице-
водство, 6 : 17—18.
РаботновГ. 1938. Дождевые черви в луговых почвах. Природа,
9 : 73—75.
Ремезов Н. П. 1952. Почвы, их свойства и распространение.
Учпедгиз, М. : 1—268.
Рыжиков К. М. 1949. Сингамидоз домашних и диких живот-
ных. Основы нематодологии, 1. Изд. АН СССР, М.—Л. : 1—
164.
РэссельЭ. Д. 1933. Почвенные условия и рост растений. Гос.
изд. колх. и совх. лит., М. : 1—388.
Светлов П. Г. 1928. Исследования над развитием дождевых
червей. Тр. Особ, зоолог, лабор. и Севаст. ст. АН СССР,
сер. 2, № 11-13 : 95—329.
Соколова. А. 1953. Роль дождевых червей в образовании почв
Сев.-Зап. Алтая. Тр. Почв. инет. КазССР, 2 : 5—83.
Соколов А. А. 1956. Значение дождевых червей в почвообразо-
вании. Изд. АН КазССР, Алма-Ата : 1—263.
Тарнани И. К. 1928. О деятельности дождевых червей. Тр.
3-го Всеросс. съезда зоол., анат., гистол., Л. : 406.
Тихомиров В. А. 1937. Об условиях обитания дождевых чер-
вей в тундровых условиях. Природа, 5 : 52—58.
Ткаченко М. Е. 1908. О роли леса в почвообразовании. Изв.
Лесн. инет., 18 : 85—198.
Цыганов М. О. 1947. К вопросу о роли дождевых червей
в жизни почвы и образовании почвенной структуры. Реф.
н.-исслед. раб. за 1945 г. Отд. биолог, наук АН СССР,
М.—Л. : 125-126.
Четыркина И. А. 1926. Почвенно-зоологический профиль
поймы левого берега р. Камы. Изв. Биолог, н.-исслед. инет,
и Биолог, ст. при Пермск. гос. унив., 5, 2 : 61—92.
Четыркина И. А. 1930. Распределение Lumbricidae по поч-
вам Троицкого округа Уральской области. Тр. Биолог,
инет, при Пермск. гос. унив., 2, 4, ч. 1 : 433—468.
Шимкевич В. М. 1884. Польза и вред, приносимые дождевыми
червями. Изв. Общ. любит, естеств., антропол., этногр.,
44, 2 : 24—27.
Шиперович В. Я. 1936. Лесная зоология. Гослестехиздат,
Л. : 1-200.
Шиперович В. Я. 1937. Почвенная фауна в разных типах
леса. Зоолог, журн., 16, 2 : 301—309.
Шиперович В. Я. 1947. Деятельность дождевых червей. Есте-
ствознание в школе, 2 : 36—45.
Шипчинский Н. В. 1949. Комнатные растения. Лениздат^
Л. : 1-84.
Шмидт П. Ю. 1920. Анабиоз дождевых червей. Тр. Петрогр.
общ. естествоиспыт., 50, 1 : 1—19.
Шмидт П. Ю. 1955. Анабиоз. Изд. АН СССР, М.-Л. : 1-436.
Ш м и д т П. Ю. и Т. В. Щ е п к и н а. 1917. К вопросу об анабиозе.
Опыты над дождевыми червями. Изд. Петрогр. биолог,
лабор., 14 : 1—6.
197
Ш у л ь ц Р. С. 1938. Дождевые черви как промежуточные хозяева
метастронгилид. Тр. Башкирок, гельминтолог, экспед.,
Уфа : 187—223. *
Anderson W. Л. 1940. Earthworms in role of great benefacion
of human race. South Pasadem Revew (Calif.).
В a г r e t T. J. 1947. Harnessing the earthworm. Boston: 1—166.
В e a u g u e Ch. 1912. Les vers de terre et la fertilitt) du sol. Journ.
agric. prat., Paris, 23 : 506—507.
В 1 a c k J. D. 1954. Biological conservation with particular empha-
sis on wildlife. New York : 1—328.
Bretsch er K. 1900. Oligochaetenfauna der Schweiz. Rev. Suisse
Zool., 8 : 1—44 435-465.
CernosvitovL. and A. С. E v a n s. 1947. Lumbricidae. Linn.
Soc. London, Syn. Brit. Fauna, 6 : 136.
С 1 a p h a m P. A. 1935. On the experimental transmission of Syn-
gamus trachaea from starlings to chickens. Journ. Helmin-
tology, 13 : 1—12.
Darwin Ch. 1838. On the formation of mould. Geol. Soc. Proc.,
2 : 574-576.
Darwin Ch. 1881. The formation of vegetable mould through the
action of worms, with observations on their habits. London :
1-326.
Dawson R. 1947. Earthworm microbiology and formation of
waterstable soil aggregates. Soil Sci. Soc. Amer. Proc.,
12 : 512-516.
D jem i 1 M. 1896. Untersuchungen uber den Einfluss der Regen-
wiirmer auf die Entwicklung der Pflanzen. Halle a/S.
Dolk H. E. and F. V. d. P a a u w. 1929. Die Leistungen des
Hemoglobins beim Regen wurm. Ztschr. vergl. Physiol.,
10 : 324-343.
(Evans А. С.) А. К. Эванс. 1948a. О влиянии земляных чер-
вей на почву. Британский союзник, 12/293 : 9.
Е v a n s А. С. 1948b. The identity of earthworms stored by moles.
Proc. Zool. Soc. London, 118, 2 : 356—359.
E v a n s A. C. and W. J. Guild. 1947a. Some notes on reproduc-
tion in British earthworms. Ann. Magaz. Natur. Hist. 11,
ser. 14, 117:654-659.
E v a n s A. C. and W. J. Guild. 1947b. Studies on the relation-
ships between earthworms and soil fertility. Biological stu-
dies in the field. Ann. Appl. Biol. Cambridge, 34, 3 : 307—
330.
E v a n s A. C. and W. J. G u i 1 d. 1948. Studies on the relationships
between earthworms and soil fertility. On the life cycles of
some British Lumbricidae. Ann. Appl. Biol. Cambridge, 35,
4 : 471-484.
F о c k e F. 1930. Experiments und Beobachtungen uber die Biolo-
gie des Regenwurms, unter besonderer Beriicksichtigung der
Frage nach der Raumorientirung. Ztschr. wiss. Zool.,
136 : 376—421.
Franz H. 1950. Bodenzoologie als Grundlage der Bodenpflege.
Berlin : 1—316.
Friend H. 1924. The story of British Annelids (Oligochaeta).
London ; 1—278.
198
Fuller H. 1954. Die Regenwurmer. Neue Brehm-Bucherei, 140,
Wittenberg : 1—56.
Graff O. 1953. Regenwurmer Deutschlands. Hannover : 1—81.
Grant W. 0. 1955. Earthworm Breeding. Farms. Sci., 121,
3134 : 107-108.
Grove A. J. 1925. On reproductive processes of the earthworm
L. terrestris. Quart. Journ. Mier. Sci., 69; 245—290.
G г о v e A. J. and L. F. С о w 1 e y. 1926. On the reproductive pro-
cesses of the brandling worm Eisenia foetida. Quart. Journ.
Mier. Sci., 70 : 559—581.
G г о v e A. J. and L. F. С о w 1 e y. 1927. The relation of the glan-
dulare elements of the clitellum of brandling worm to the
secretion of the cocoon. Quart. Journ. Mier. Sci., 71 : 31—45.
Heim burger H. V. 1924. Reactions of earthworm to tempera-
ture and athmospheric humidity. Ecology, 5 : 276—282.
Hensen V. 1877. Die Tatigkeit des Regenwurms fur die Frucht-
barkeit des Erdboden. Ztschr. wiss. Zool., 28 : 354—364.
H e г 1 a n t - M e e w i s H. 1956. Phenomenes neurosecretoires et
ponte chez Eisenia foetida. C. R. Acad, sci., 243, 11 : 823—825.
Hess H. 1925. Nervous system of the earthworm. Journ. Morph.,
40 : 235-259.
Heuschen J. 1956. Regenwurm und Bodenfruchtbarkeit. Mitt.
Dtsch. Landwirtsch. Ges., 71, 48 : 1243—1244.
Hopp H. 1947. The ecology of earthworms in cropland. Soil Sci.
Soc. Amer. Proc., 12 : 503—507.
H о p p H. and P. J. Linder. 1947. A principle for maintaining
earthworms in farm soils. Science, 105, Lancaster : 663—664.
Hopp H. and G. S. Slater. 1948. Influence of earthworms on
soil productivity. Soil Sci. Baltimore, 66, 6 : 421—428.
J a p p W. B. 1956. Locomotion of worms. Nature, 177, 4509 : 614—
615.
К e i 1 i n D. 1915. Recbercbes sur les larves de dipteres Cyclora-
phes. Bull. Sci., France-Belg., 49 : 15—118.
К e i 1 i n 1). 1915. Parasitic autotomy of the host as a made of libe-
ration of coelomic parasites from the body of the earthworm.
Parasitology, 17 : 170—172.
Kellmansperger E. 1934. Die Oligochaeten der Bellinchen-
gebietes, eine okologische, ethologische und tiergeographische
Untersuchung. Diss. Berlin.
Korsch elt E. 1925. Ueber Ruhezustande der RegenwQrmer.
Zool. Anz., 64 : 53—55.
К u b i e n a W. 1943. Die mikroskopische Humusuntersuchung.
Ztschr. Weltforstwirtsch., 10 : 387—410.
К u h n e 1 t W. 1950. Bodenbiologie. Wien : 1—369.
Lunt H. A. and H. G. J а с о b s о n. 1944. The chemical composi-
tion of earthworm casts. Soil Sci., 58, 5 : 367—375.
Mangold O. 1924. Beobachtungen und Experimente zur Biologie
des Regenwurms. Ztschr. angew. Physiol., 2 : 57—81.
Mangold O. 1951. Experimente zur Analyse des chemischen
Sinns des Regenswmrms. Zool. Jahrb., Abt. allg. Zool., Phys.,
62, 4 : 441-512.
Merk er E. J. 1926. Die Empfindlichkeit feuchthautiger Tiere
im Lichte. II. Warum kommen Regenwurmer im Wasserlachen
199
um und warum verlassen si© bei Regen ihre Wohnrohre? Zool.
Jahrb., Abt. allg. Zool., 42. 4 : 487—555.
Meyer L. 1943. Experimented Beitrag zur makrobiologischen
Wirkung auf Humus uns Bodenbildung. Ztschr. Bodenkunde
u. Bodenerniihrung., 29 : 119—140.
M i c b a e 1 s e n W. 1928. Oligochaeta. W. Kiikenthal — T. Krum-
bach's Handbuch d. ZooTogie, Eine Naturg. der Stamme der
Tierreichs., 2, Lief. 2 : 1 — 112.
M i c b о n J. 1954. L’influence de la temperature sur les phases du
developpment postembryonaire Lumbricidae amphodynames.
C. R. Acad. Sci. Paris, 238 : 2358-2360.
M i c h о n J. 1957. Contributions experimentales a 1'etude de la
biologie des Lumbricidae. Annee bioL, 3 sdr., 33, 7—
8 : 367-376.
M u 1 d a 1 S. 1950. Cytology, genetics and classification. Nature,
166 : 769-770.
Olson H. 1928. The earthworms of Ohio. Bull. Ohio Biolog., Sur-
vey, 17, 4, 2 : 1-90.
О m о d e о P. 1948. La poliembrionia e la anomalie di sviluppo
presso un commune lombrico Allolobophora caliginosa tra-
pezoides. Arch. zool. ItaL, 33 : 1—87.
О m о d e о F. and A. M a g a 1 d i. 1951. Variabilita nei gemelli
uniovulari di lombrico. Sci. Genet., 4 : 13—21.
Ramsay J. A. 1949. The osmotic relations of the earthworm.
Journ. Exper. BioL, 25 : 46—56.
R a w F. 1959. Estimating earthworm pupulation by using for-
malin. Nature, 184, 4699 : 1661—1663.
R о о t s B. J. 1955. The water relations of earthworms. I. The acti-
vity of the nephridiostome cilia of L. terrestris and A. chloro-
tica in relation to the concentration of the bathing medium.
Journ. Exper. BioL, 32, 4 : 765—774.
R о о t s B. J. 1956a. The water relation» of earthworms. II. Resis-
tance to dissication and immersion and behaviour when sub-
merged and when allowed a choice of environment. Journ.
Exper. BioL, 33, 1 : 29-44.
Roots B. J. 1956b. Famous animals. 7. The earthworm. New
BioL, 21 : 102-117.
R о u d e G. 1954. Vorkommen, Haufigkeit und Arten von Regen-
wiirmern in verschiedenen Waldboden und unter verschiedenen
Bestockungen. Forstwissensch. ZbL, 3/4 : 113—126.
Smith A. |C. 1902. The influence of temperature, odors, light and
contact on the movement of the earthworm. Amer. Journ.
Physiol., 6 : 459-486.
Spannagel G. 1954. Modellversuch mit Regenwurmern zur
Frage der Bodenbildung und Bodenfrucbtbarkeitssteigerung.
Ztschr. Pflanz., Diing. u. Bodenk. 64, 3 : 217—222.
Stephenson J. 1930. The Oligochaeta. Oxford : 1—
978.
S t о с к 1 i A. 1929. Studien uber den Einfluss des Regenwurmes auf
die Beschaffenheit des Bodens. Landwirtsch. Janrb. Schweiz.,
42 : 1-121.
Stolt e H. A. 1933—1955. Oligochaeta. Bronn’s Klassen und
Ordnungen des Tierreiches., 4, 3, 1 : 1—890.
200
Taylor E. L. 1928. Syngamus trachea from starling etc. Ann.
Trop. Med. Parasitol., 22 : 307—318.
W a 1 t о n W. R. 1928. Earthworms as pests and otherwise. U. S. De-
part. Agric., Farmers'bull., 1569 : 1—14.
White G. 1789, 1802. The worms in natural history. London»
1 : 1-392; 2 : 1-200.
W i 1 с к e D. 1951. Bemerkungen zu dem erdzeitlichen Alters der
Regenwurmer (Oligochaeta, Opistophora). Zool. Anz , 154,
7/6 : 149-156...
W i 1 с к e D. 1953. Uber die vertikale Verteilung der Lumbriciden
im Boden. Ztschr. Morph, u. Okol. Tiere, 41, 4 : 372—385.
Wittich W. 1943. Untersuchungen iiber die Streuzersetzung auf
einem Boden mit Mullzustand. Forstarchiv, 19 : 1—18.
W о 11 n у E. 1897. Die Zersetzung der organischen Stoffe und die
Humusbi Id ungen. Heidelberg.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Автотомия червей 110, 111
агрегат 91
активные перемещения червей
104
аммиак 163
аэрация почвы 142
Бактерии 97, 139
«беличий хвост» 127
белый октолазий 102, 108
беспозвоночные 97
биогенный кальцит 163
болевые ощущения 47
большой красный червь 59, 68,
82, 83, 100, 103, 108, 116,
127, 162
брюшная нервная цепочка 36,
37
брюшной сосуд 27
бугорки зрелости 12
Валики зрелости 12
венецианская эйзения 72, 75
вес кучек извержений 152
вкусовые способности 45, 177
водный дождевой червь 60
водонепроницаемость 169
водопрочность копролитов 167,
169
восьмигранная дендробена 80,
103, 108, 137
восьмигранная эйзепиелла 111
вторичная кольчатость И
выбрасывание копролитов 145
202
выбросы 144, 150, 151
выползки 82
Ганглий 36
гемоглобин 29, 111
гермафродит 49
глазчатая эофила 79
глотка 21
глоточные железы 21
головная лопасть И
горизонтальные странствия дож-
девых червей 103
гормоны 48
грибки 139
грунтовые воды 104
гумификатор 162, 173
гумификация почвы 172
гумус 161, 162, 171
гусеницы бабочек 97
Девственное размножение 69
диапауза 64
длинная аллолобофора 76, 100,
103, 106, 108
длинный червь 59
дренаж почвы 142
дробление яйца 60
друзы 163
Железистые клетки 15
желудок 23, 69
женские половые поры 13
Закладки ям для разведения
червей 183
заселение почвы червями 172
зевательная болезнь 122
зимние камеры 105
зоб 23
зона корней растений 154
Известковые железы 22
известняк 147
интенсивность почвообразова-
тельной деятельности червей
153
Кальций 163 •
камбиальные клетки 16
каинит 175
капилляриоз 123
капсула 105
каштановый червь 68, 82, 83
кварц 147
кирпич 147
кислотность почвы 102
классификация почв 90
клещи 97, 171
кожные железы 16
количественный учет червей 137
количество извержений 149
количество ходов червей 141
коловратки 97
кольчецы 66
компост 183
коннектива 36
коиролиты 25, 109, 144
корм для червей 182
костяная мука 175
кровь 27
круговорот веществ 95
кутикула 15
Летние камеры 105
лёсс 155
личинка 60
личинка жуков и мух 97
лососевый червь 127
Магний 163
малощетинковые черви 66
малый красный червь 68, 82,
103, 108, 137, 153, 162, 168
мариупольская дендробена 109
массовое выползание червей 112
межсегментные перегородки 20
мета стронгил иды 121, 123
механический анализ копроли-
тов 166
минерализация органических
соединений 94
минерализующая деятельность
червей 161
минеральные удобрения 188
многоножки 97
многопольный севооборот 190
многощетинковые черви 66
мокрицы 97, 144
моллюски 97
молочный октолазий 81, 137,
138, 157
мужские половые поры 13
мясная мука 120
Навозный червь 59, 63, 72,
73, 99, 103, 108, 120, 121
насекомые 97
нематоды 97, 118, 171
нефридий 31, 35
низшие насекомые 97
нитрат 163
Однояйцевые близнецы 61
окраска тола 13
онезия 119, 120
оплодотворение 55
опыт Гензена с перемешива-
нием почвы червями 155
опыты Балуева и Андерсона
по влиянию червей на уро-
жайность 176
опыт Зражевского по влиянию
червей на рост саженцев 178
органическое вещество 158, 161
откладка коконов 182
Надёж червей 187
пассивное расселение 104
пашенный червь 59, 76, 77,
100, 102, 108, 120, 137,
140, 141, 153, 157, 168
перегной 94, 139
перемешивание почвы червями
154
пигменты 13
пища дождевых червей 155,
161
пищевод 22
пиявки 66
203
плодородие почвы 94, 180
погружение под землю пред-
метов в результате деятель-
ности червей 147
поддерживающие клетки 14
полиэмбриония 62
полления 119, 120
половые щетинки 54
порозность почвы 91, 92
порроцекоз 123
почва 90, 91
почвенная кислота 92, 102
почвенная структура 91
почвенная фауна 95
почвообразование 171, 172
почвы торфяных лугов 102
поясок 11, 52, 70
прирост толщины почвы 152
проволочник 113
продолжительность жизни 64
продукция гумуса 162
простейшие 95, 97
Разведение червей 181, 182
реакция копролитов 164
регенерация 64, 118
рефлекс 39
розовая эйзения 72, 74, 102,
108, 153, 157, 162, 168
розовый червь 60
рост древесных пород 177
Самоочистка почвы 127
сбор червей 184
светочувствительные клетки 41
свечение 17
сегмент 10
семенники 51
семенниковые капсулы 51, 70
семенные мешки 70
семеприемники 52, 69
семепроводы 51
сипгамиды 121, 123
сингамоз 122
скважность почвы 139, 159, 170
снеговой покров 93
спаривание 55, 56
сперматозоиды 51
сперматофоры 57
спинной сосуд 27
спинные поры 13
спиральные волокна 168
старение червей 63
Температура тела червя 100
температура почвы 93
тифлозол 23
тонкий бимаст 79
тургор 21
Узелок 17
укрытие почвы 188
урожайность 175
условный рефлекс 47
Фосфорная кислота 163
Хлорагогенная ткань 26
ходы червей 86, 139
Целинные почвы 159
цисты грегарин 118
«Червятник» 186
четырехгранная эйзениелла 71
численность люмбрицид 136
чувствительные почки 44
чувство обоняния 45
Шлак 147
Щелочность 164
щетинки 12, 16
Эволюция почв 155
эйзения Ленберга 102, 111
эйзения Норденшельда 72, 73,
101,102, 162
энхитреиды 67, 97
эофила 106
эффективность траншейных ме-
тодов для разведения червей
185
Яйцеводы 51
яйцевые воронки 51
яйцевые клетки 50
яйцевые коконы 49, 57
яйцевые мешки 51
ящичные культуры цщщай 1-84
ОГЛАВЛЕНИЕ
Стр.
Предисловие 3
Введение................................................ 5
Глава I. Строение тела и главные жизненные отправления
дождевых червей............................ 9
1. Общие замечания. Наружные признаки . . 9
2. Покровы тела...............................ч 14
3. Мускулатура и движение. Полость тела ... 18
4. Кишечник и питание................... 21
5. Кровеносная система. Функции распределения пита-
тельных веществ и кислорода.............. 27
6. Органы выделения. Поглощение и выделение воды 31
7. Нервная система и органы чувств. Рефлексы ... 36
8. Органы внутренней секреции........... 48
9. Органы размножения................... 49
Глава II. Размножение и развитие дождевых червей . ... 55
Глава III. Главные представители дождевых червей СССР.
Способы их сбора и определения..................... 66
1. Положение в системе животных и происхождение 66
2. Определение рода и вида червей................. 67
3. Методы вскрытия................................ 84
4. Способы сбора и консервирования................ 85
Глава IV. Образ жизни и среда обитания дождевых червей 90
1. Некоторые общие сведения о почвах и их насе-
лении ............................................. 90
2. Распределение дождевых червей в разных мест-
ностях и почвах.................................... 98
3. Зимовка и перенесение засухи.................. 105
4. Затопление почв. Почему черви называются дожде-
выми? ....................................... . . 110
5. Враги и паразиты . 115
6. Польза и вред................................. 120
Глава V. Дождевые черви и почвообразование ... 129
1. Общие замечания................................. 129
2. Сведения о численности дождевых червей в почвах 132
205
3. Влияние дождевых червей на проветривание и дре-
наж почв............................................. 139
4. Влияние дождевых червей на перемешивание земли
и вынос ее на поверхность............................ 144
5. Влияние дождевых червей на структуру и химиче-
ской состав почв . . . ... 161
6. Заключение ...................................... 171
Глава VI. Дождевые черви и земледелие..................... 175
1. Влияние дождевых червей па плодородие почв ... 175
2. Мероприятия по разведению и охране дождевых
червей..................................... 180
Литература................................................ 192
Предметный указатель...................................... 202
Ольга Витольдовна Чекановская
ДОЖДЕВЫЕ ЧЕРВИ И ПОЧВООБРАЗОВАНИЕ
«
Утверждено к печати
Редколлегией научно-популярной литературы
Академии наук СССР
•
Редактор издательства А. А. Стрелков
Технический редактор В. Т. Бочевер
Корректоры Я. Я. Журавлева, А. И. Кац и И. А Кириллова
♦
Сдано в набор 10 XII 1959 г. Подписано
к печати В/ III 1960 г. РИСО АН СССР М 39-140В.
Формат бумаги 84Х1О8,/Л. Бум л. 3\. Печ. л.
6\j=l0,66 усл. печ. л. Уч.-ияд. л. 10,57+1 вкл.
(0,04 Уч-изд. л.). Изд. М 1011. Тип. ван. М4 456.
М-56547. Тираж 3500.
Цена 3 р. 30 к.
Ленинградское отделение Издательства Академии
паук СССР. Ленинград, В-164, Менделеев-
скан лин., д. 1
1-я тип. Издательства Академии наук СССР.
Ленинград, В-34, 9 линия, д. 12.
ИСПРАВЛЕНИЯ И ОПЕЧАТКИ
Страница Строка Напечатано Должно быть
66 Заглавие, 2 сверху он ределения определение
70 13 снизу 10 11
183 18 сверху б а р р о т у Баррету.
198 4 » benefacion benefaction
О. В. Чекановекак. Дождевые черви и почвообразование.