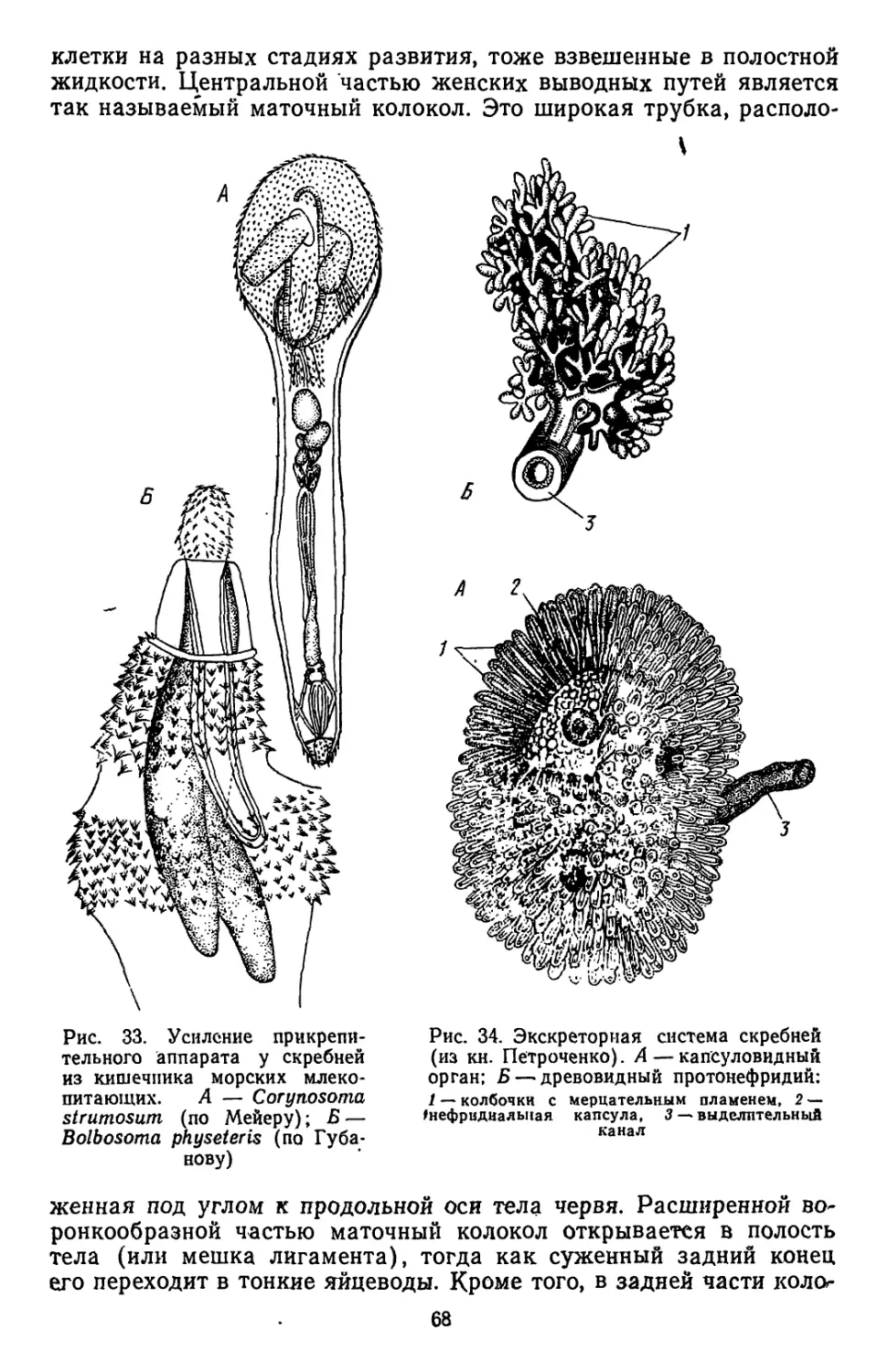
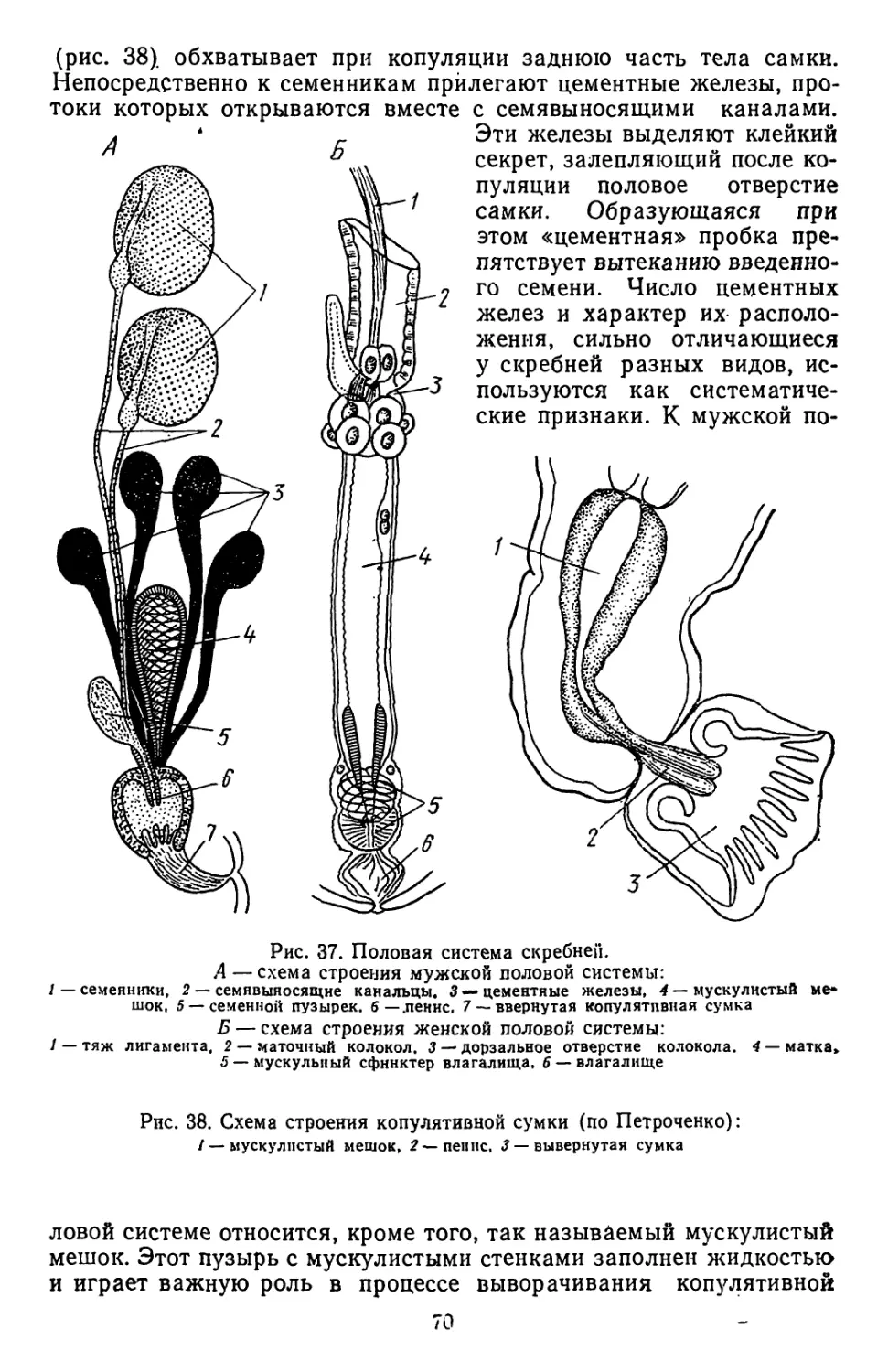
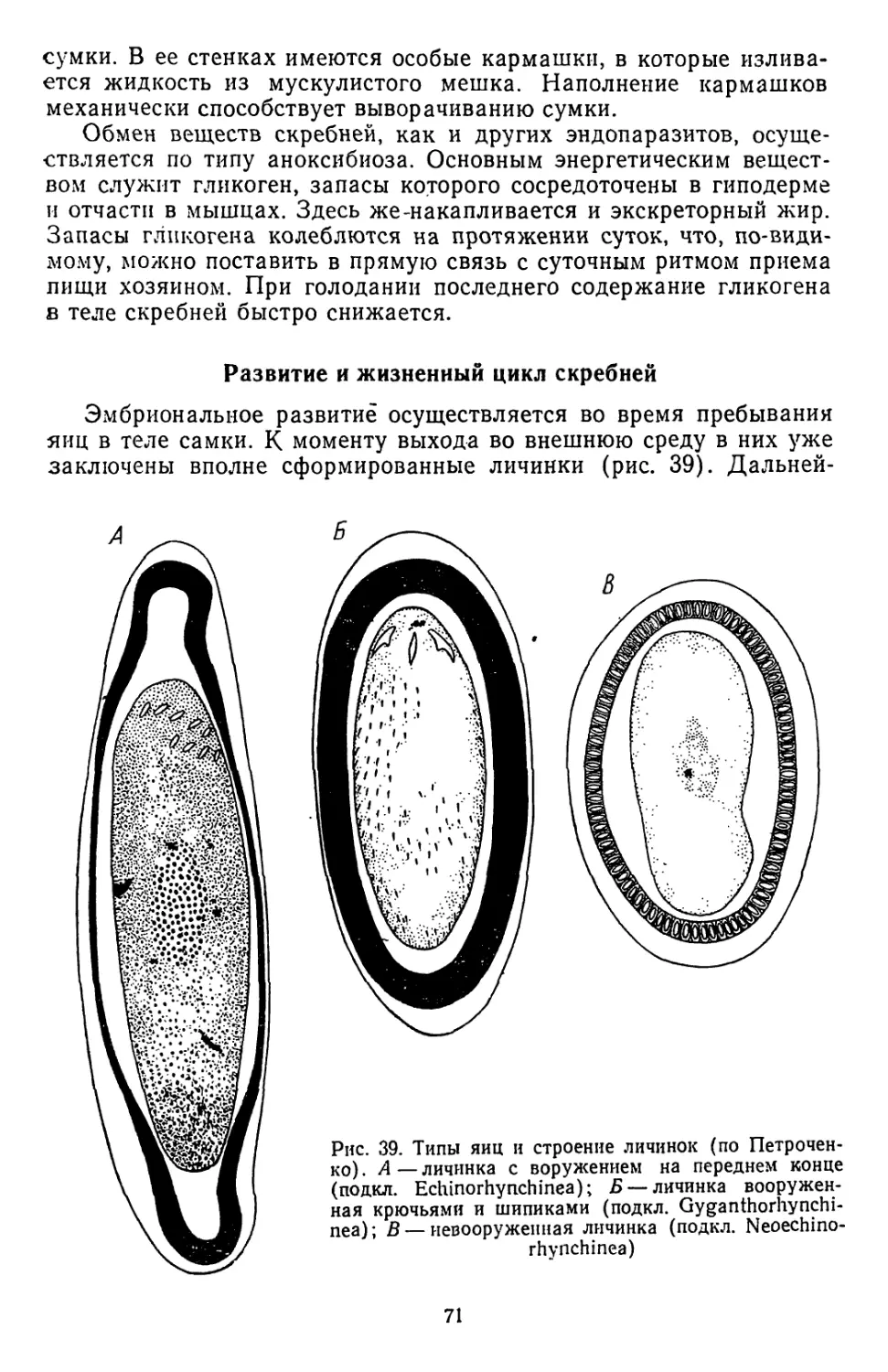
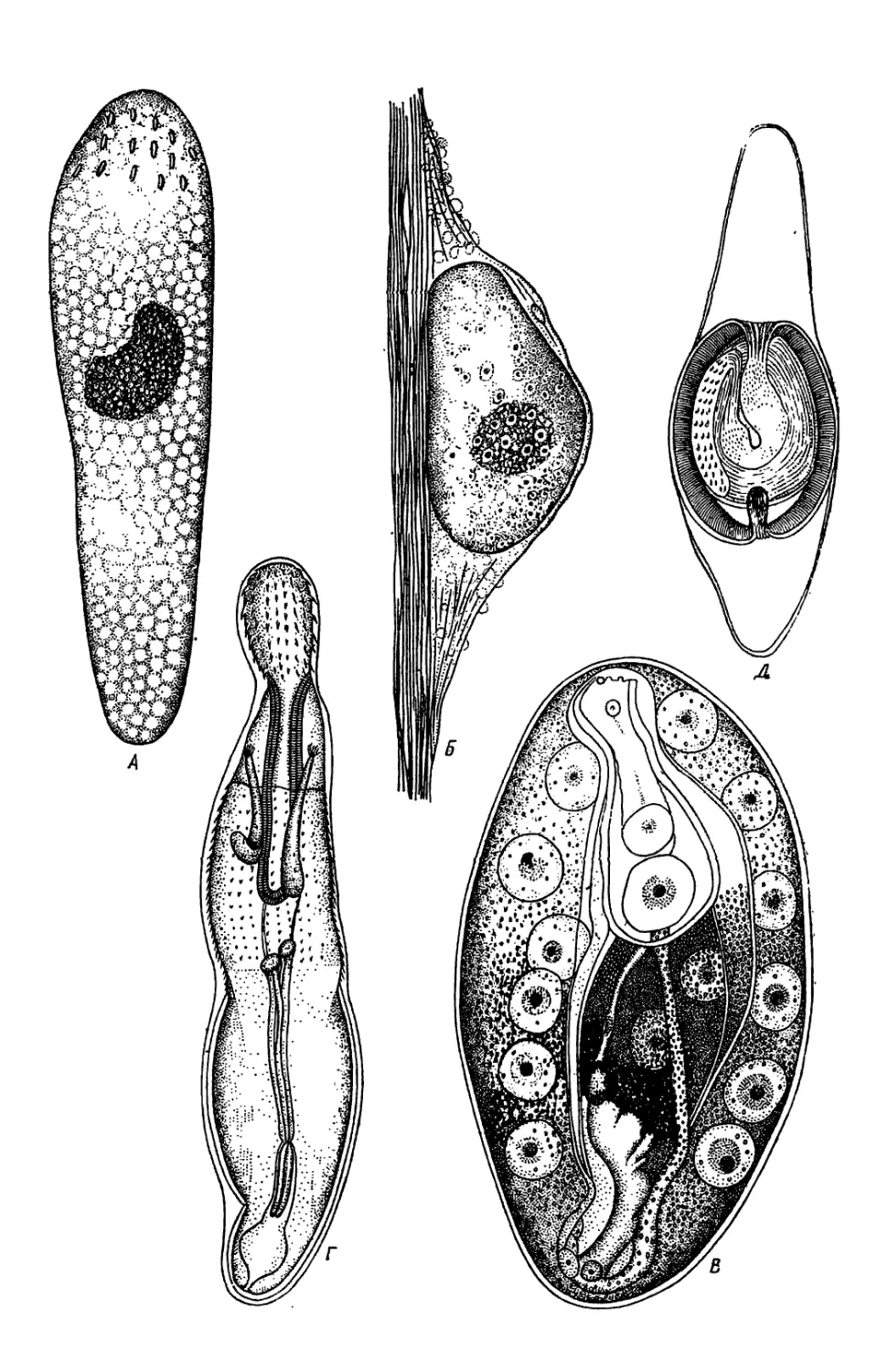
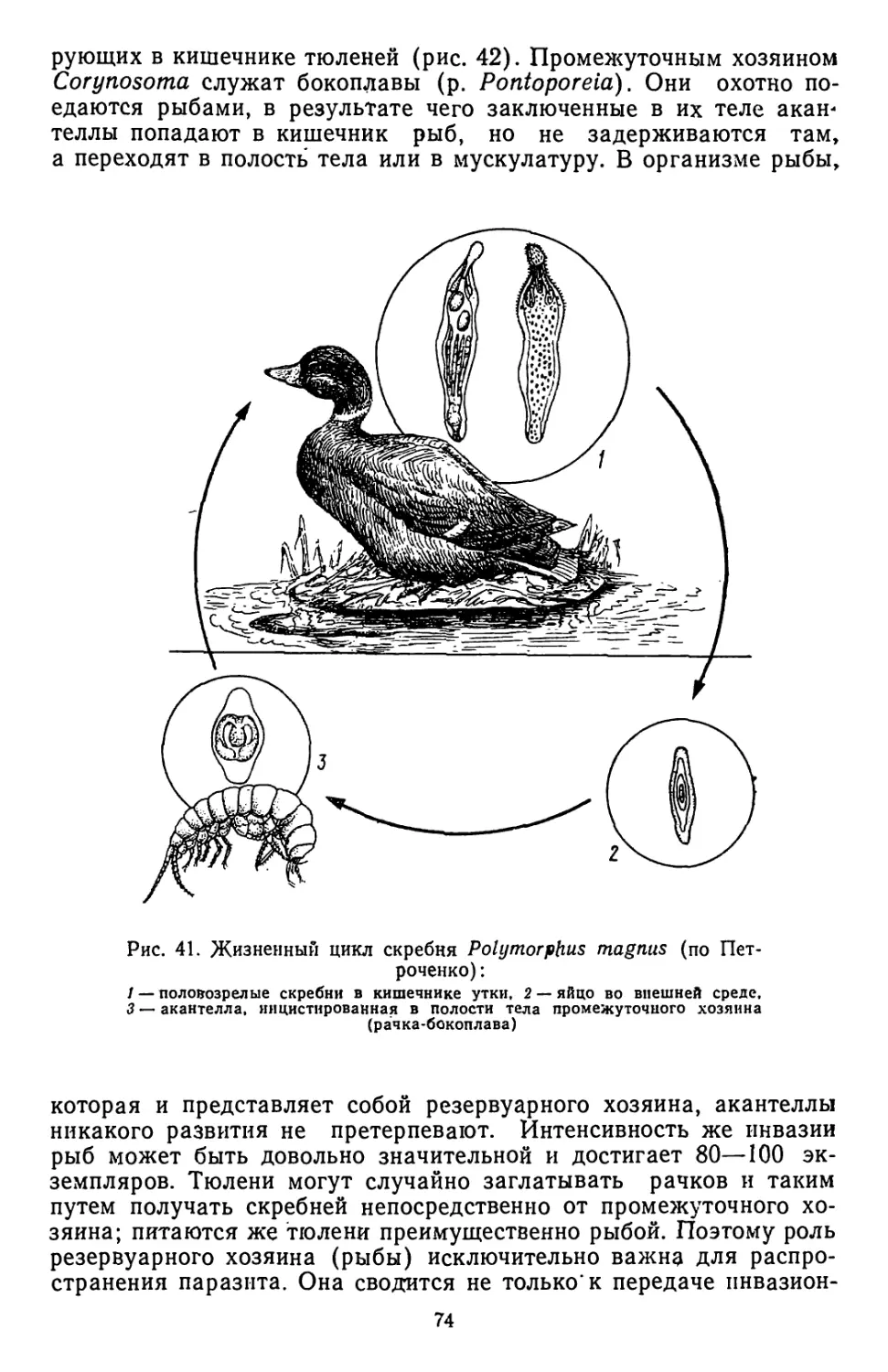
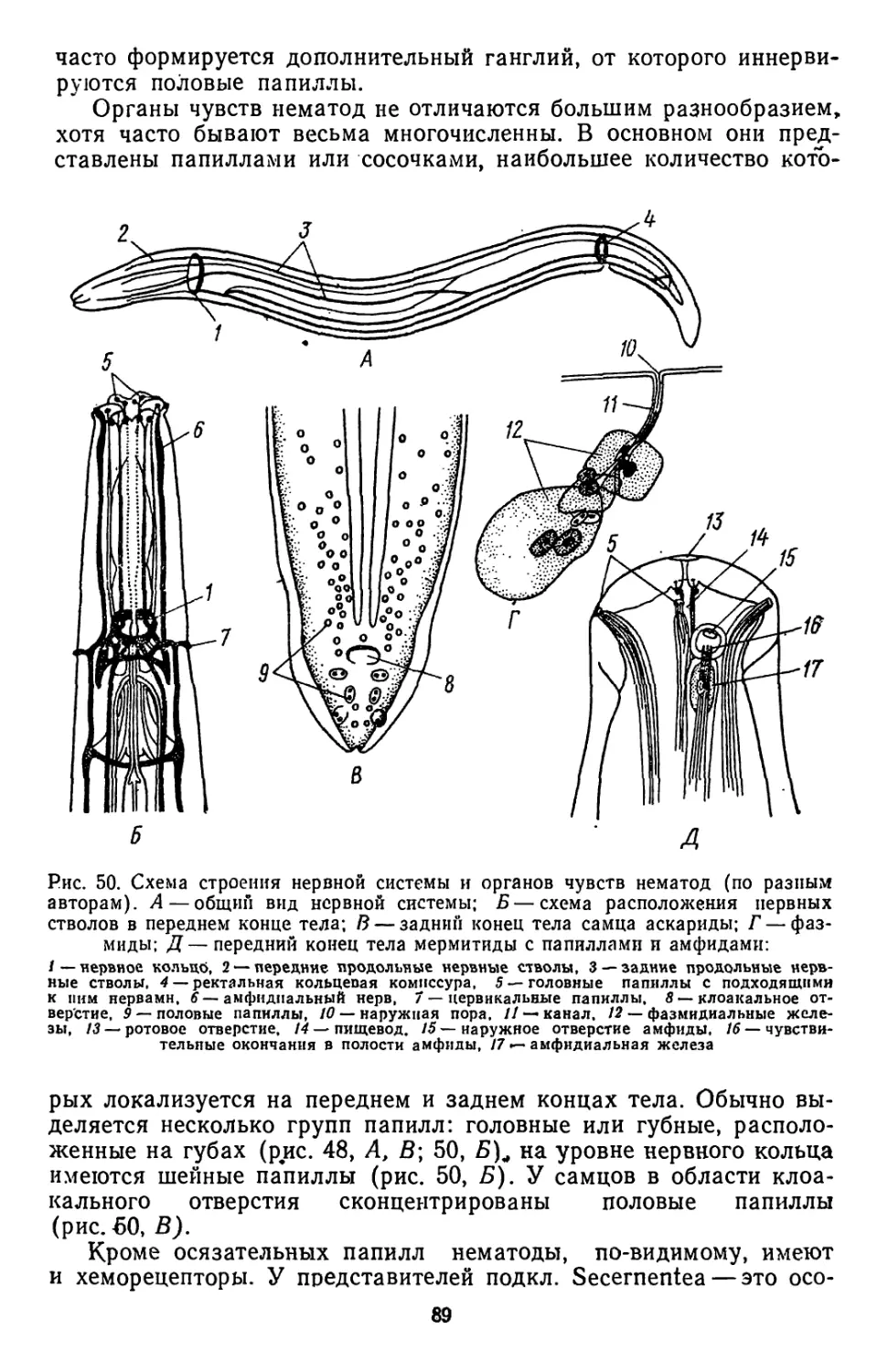
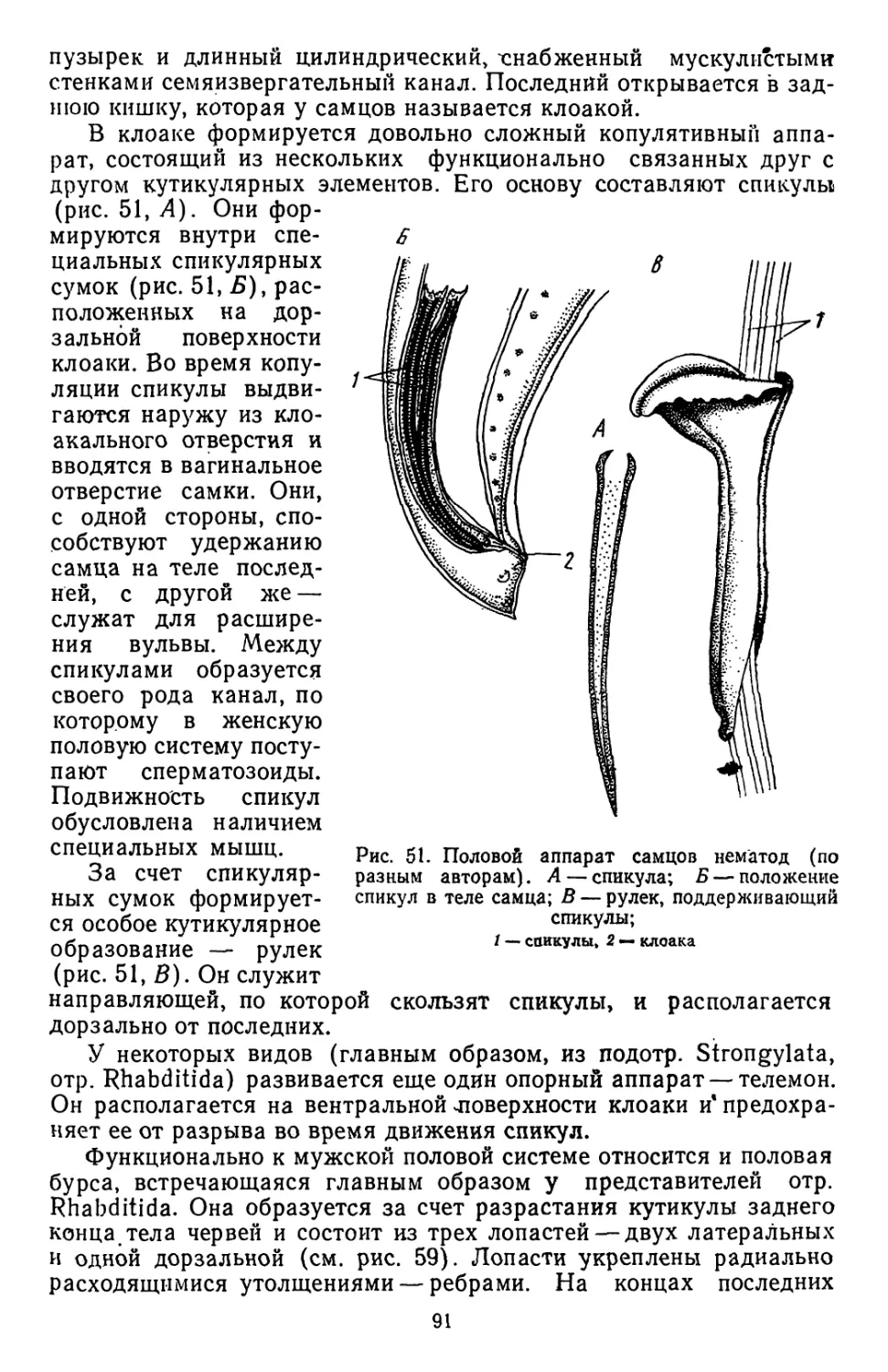
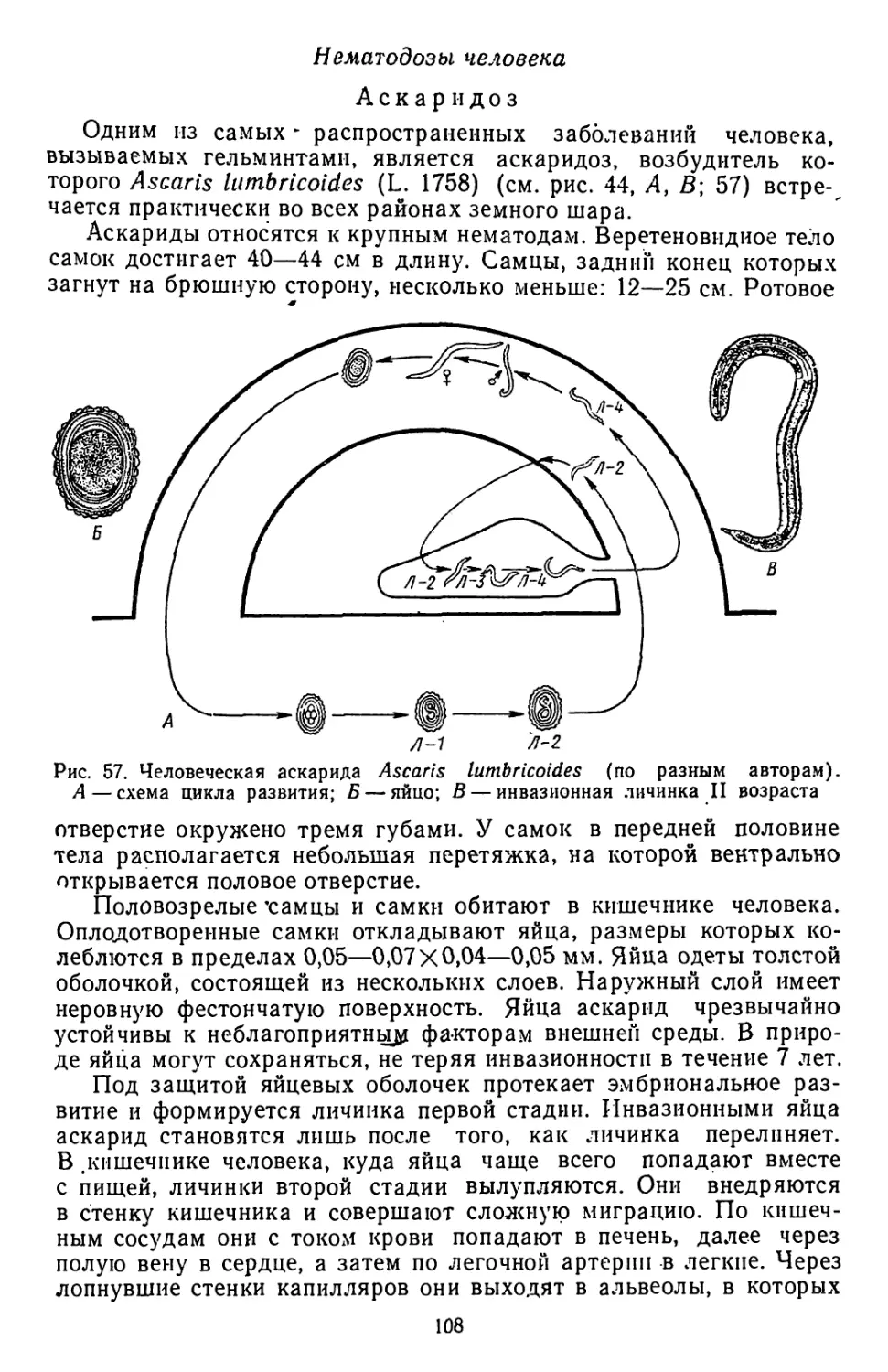
/




















































































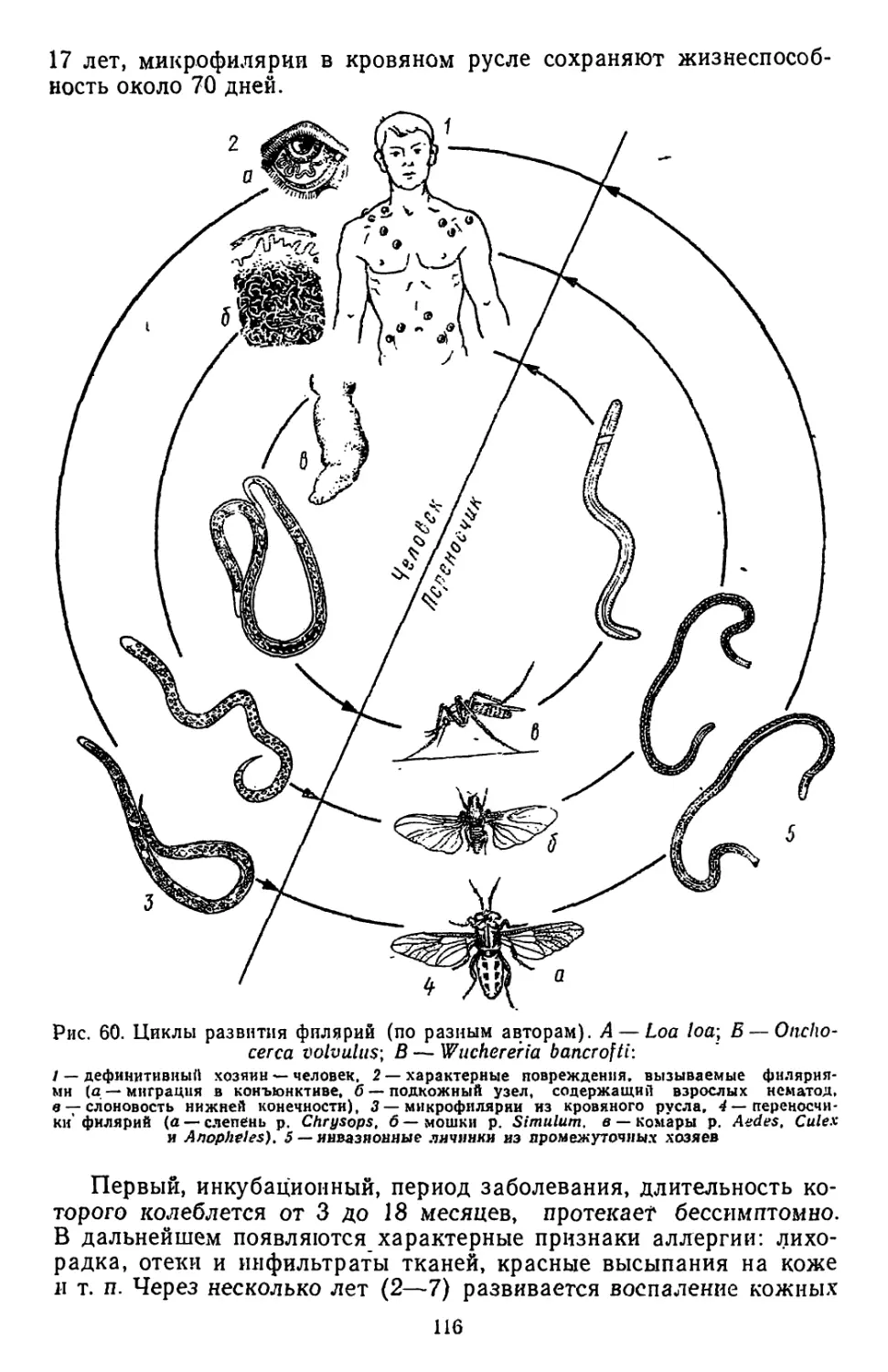



















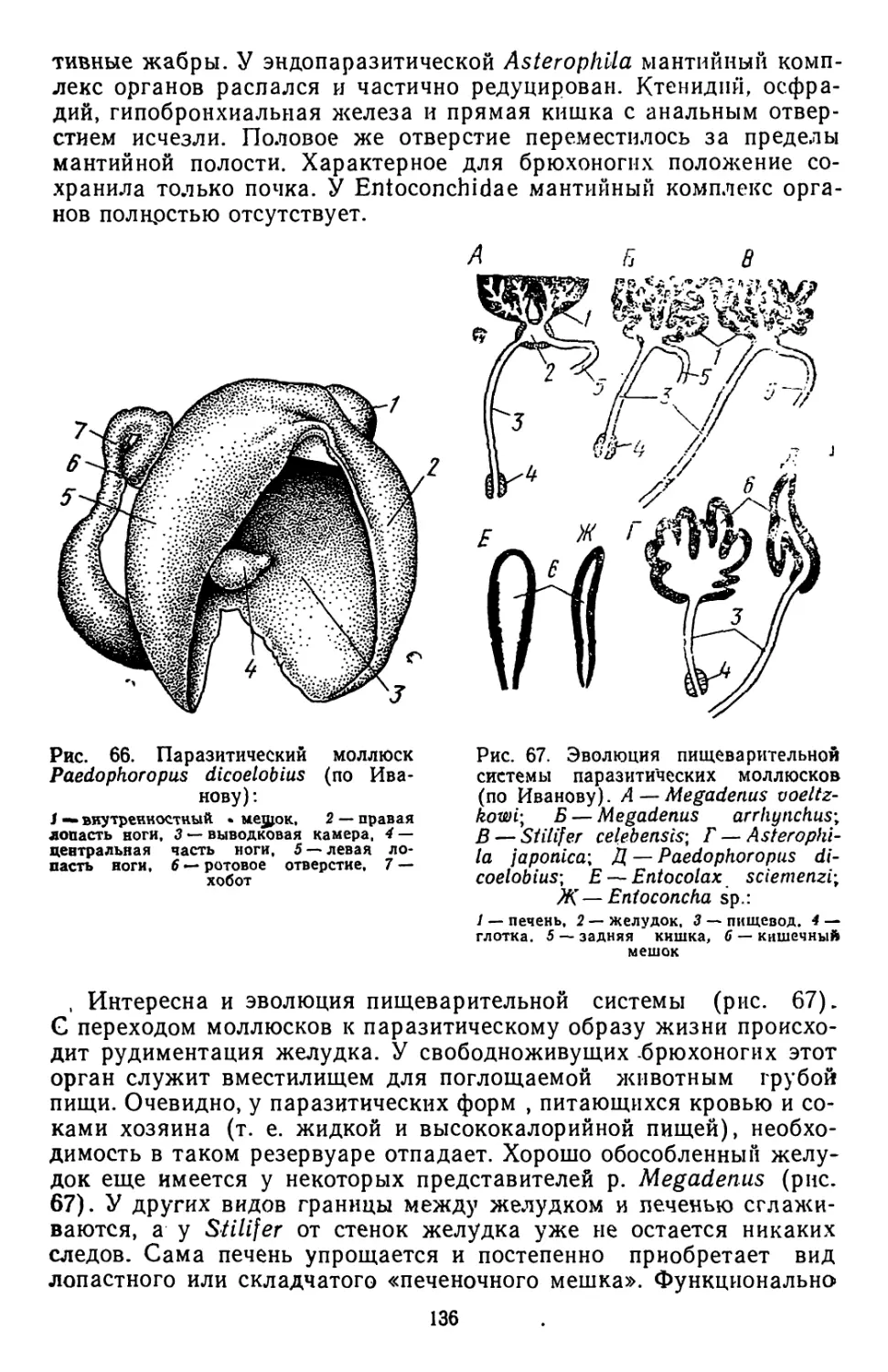







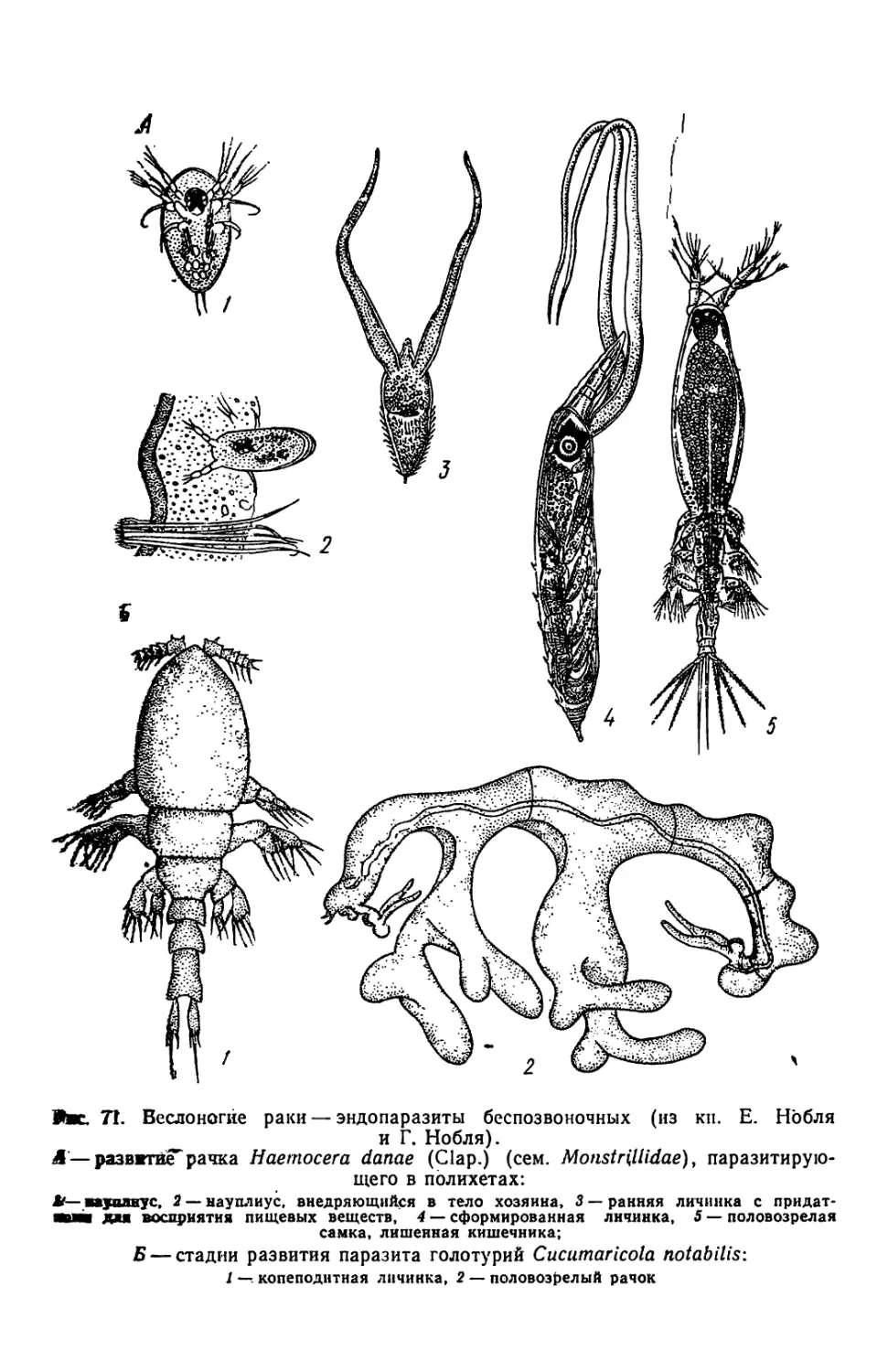

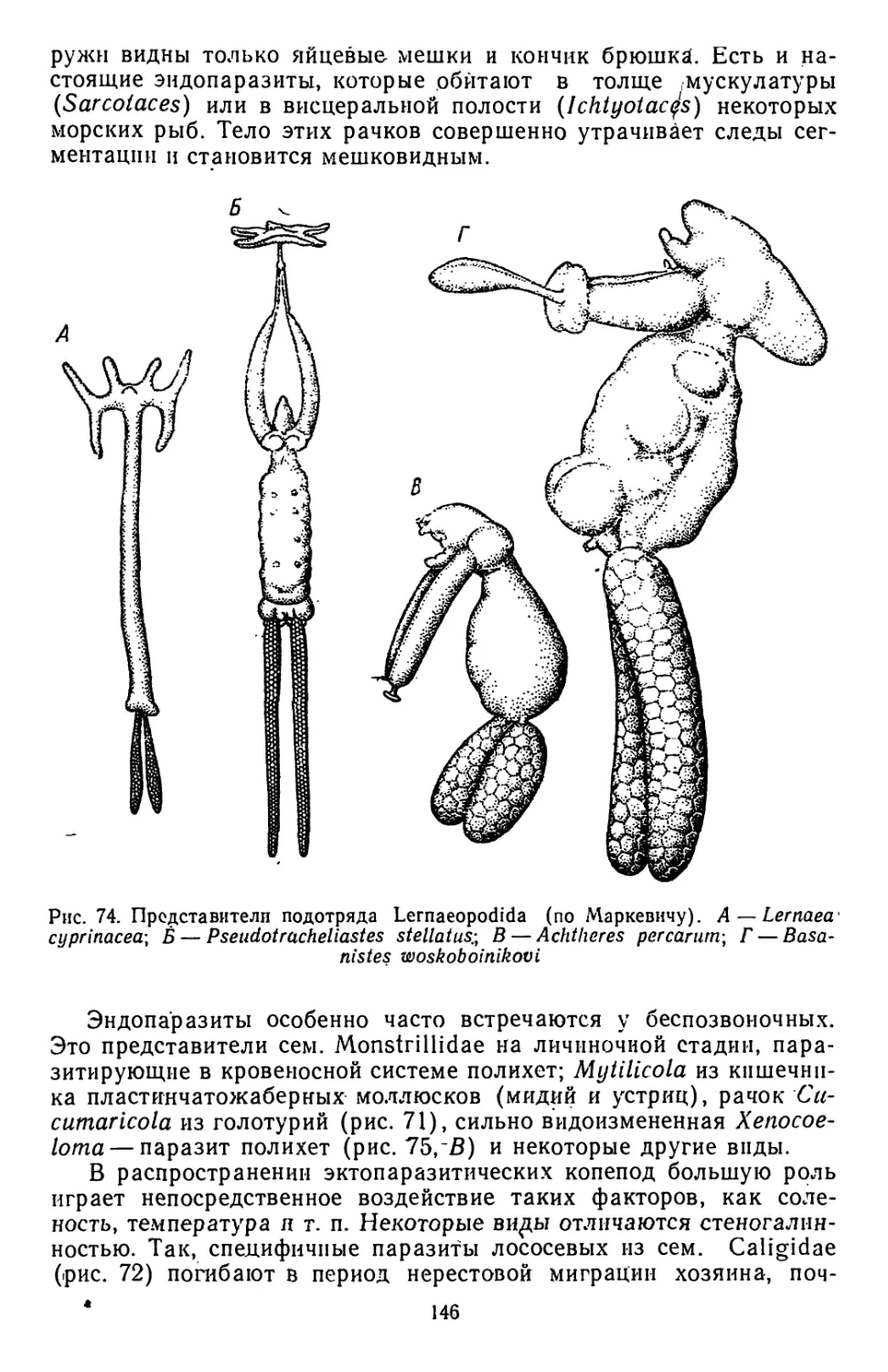







































































































































Similar




Text
ТА. ГИНИДИНСКАЯ
А.А.Д0ЬР0В0ЛЬСК1Т>!
ЧАСТНАЯ
ПАРАЗИТОЛОГИЯ
ТА. ГИНЕЦИНСКАЯ
А. А. ДОБРОВОЛЬСКИЙ
ЧАСТНАЯ
ПАРАЗИТОЛОГИЯ
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ,
моллюски И ЧЛЕНИСТОНОГИЕ
Под редакцией 10. И. Полянского
Допущено
Министерством высшего
и среднего специального
образования СССР
в качестве
учебного пособия
для студентов
биологических специальностей
высших учебных
заведений
ф
МОСКВА
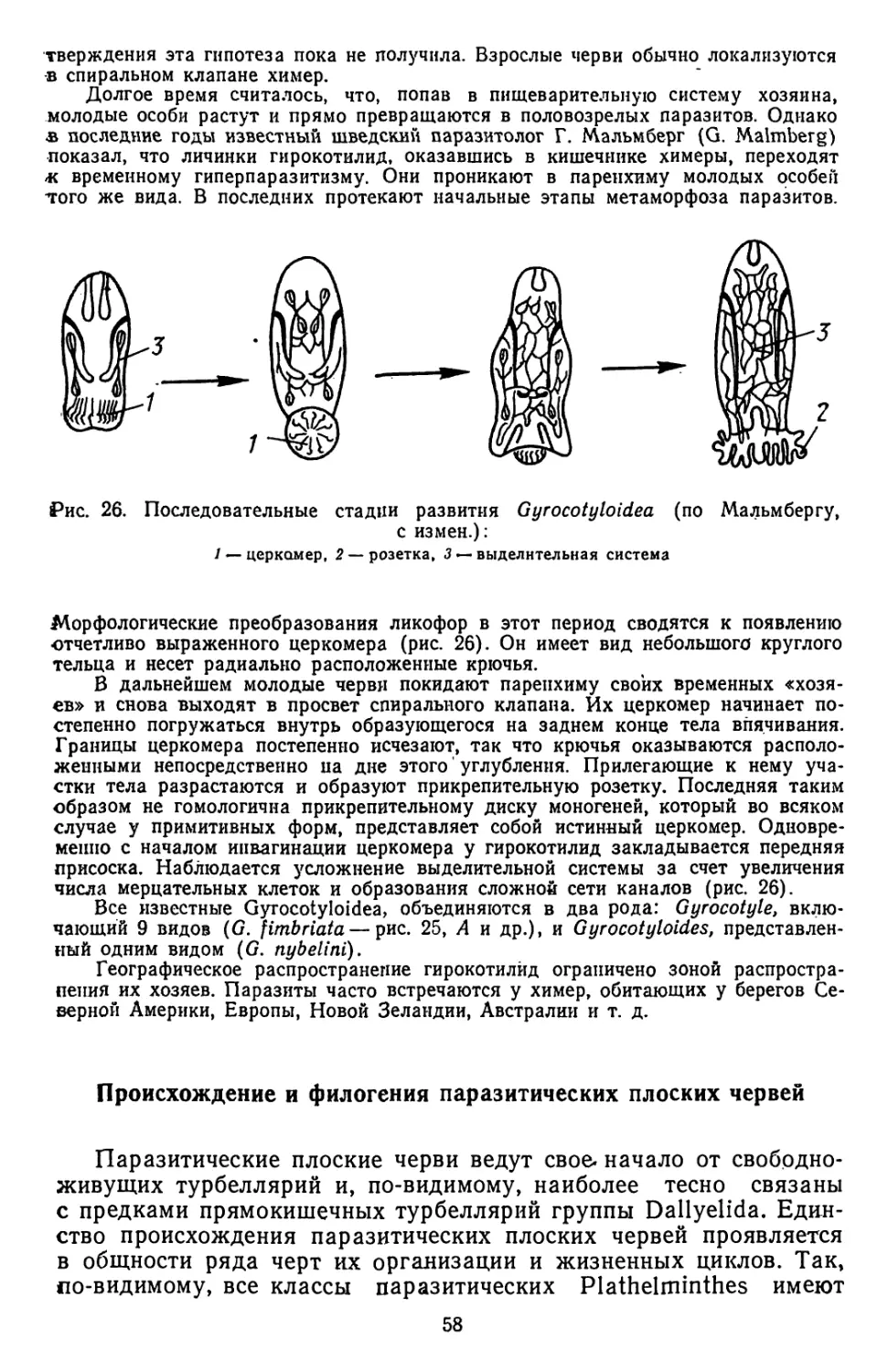
«ВЫСШАЯ ШКОЛА»
1978
ББК 28.083
Г49
Рецензенты учебного пособия:
кафедра зоологии Львовского университета
(зав. кафедрой проф. В. И. Здун);
проф, С. Л. Делямуре
(зав. кафедрой зоологии Симферопольского университета)
Гинецинская Т. А., Добровольский А. А.
Г 49 Частная паразитология = Паразитические черви, Мол-
люски и Членистоногие: Учеб, пособие для биолог, спец.
вузов/Под ред. Ю. И. Полянского.— М.: Высш, школа,
1978 — 292 с., ил.
В пер.: 1 р.
Книга содержит подробное описание паразитических червей: цестод, не-
матод и скребней. Приводятся морфофизиологическая характеристика, дан-
ные по биологии, жизненным циклам, эволюции и практическому значению.
Впервые в учебной литературе освещается вопрос о паразитических ра-
ках н моллюсках. В последнем разделе рассматриваются паразитические на-
секомые: вши, клопы, блохи и двукрылые. Описываются особенности их
строения, обусловленные паразитическим образом жизни, возможные пути
перехода к облигатному паразитизму. Рассматривается медицинское и вете-
ринарное значение насекомых, их роль в распространении инфекционных за-
болеваний.
40903-442
Г------------- оо—7о
001(01)—78
ББК 28.083
57.026.1
© Издательство «Высшая школа». 1978
z ПРЕДИСЛОВИЕ
Предлагаемая вторая книга по частной паразитологии включает
описание плоских и круглых паразитических червей, паразитических
моллюсков и членистоногих.
В главе, посвященной плоским червям, авторы придерживаются
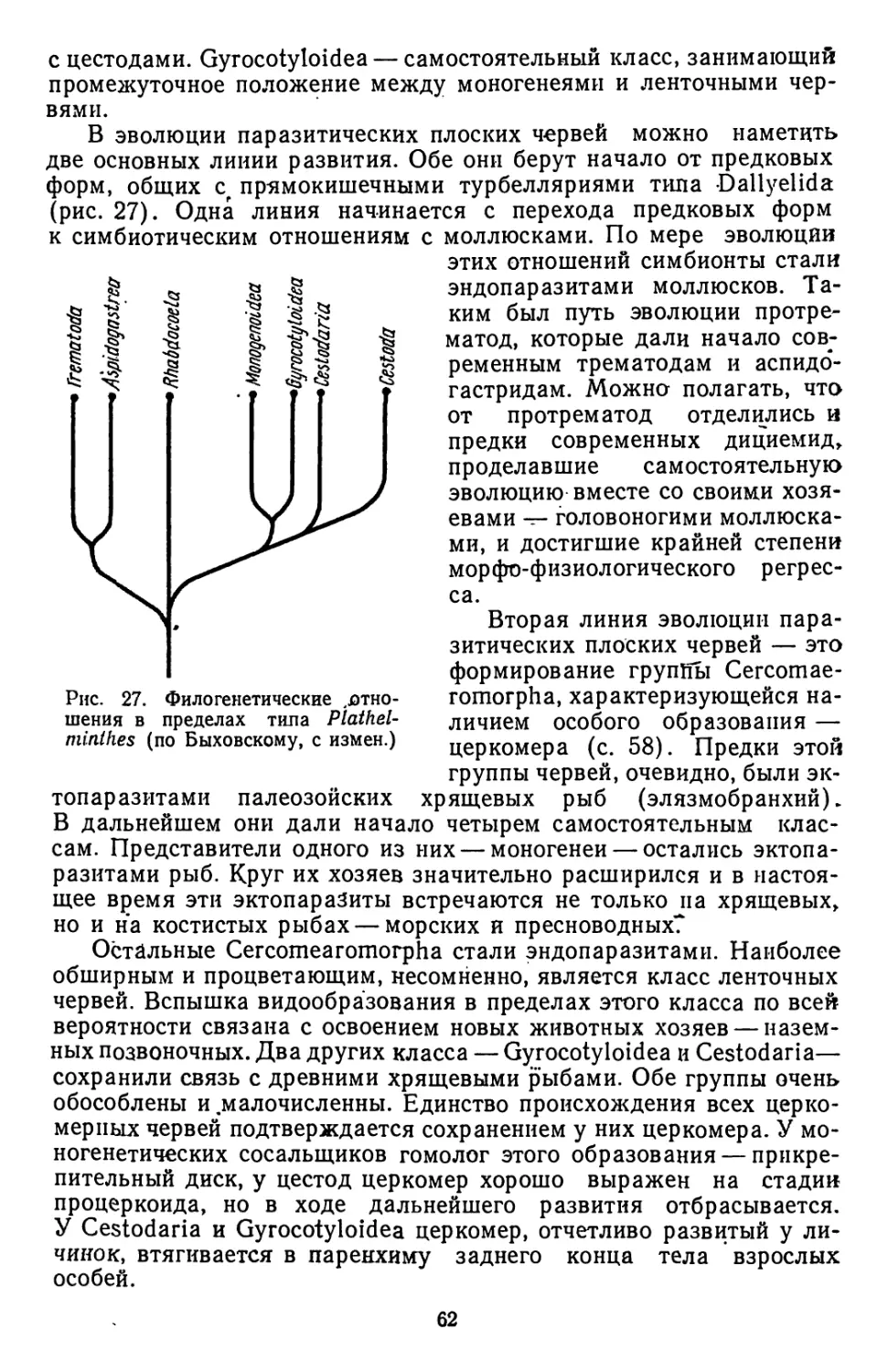
новой системы и рассматривают группы Cestodaria и Gyrococoty-
loidea как самостоятельные классы. При описании нематод особое
внимание уделяется вопросам их физиологии и эволюции. Впервые
в учебной литературе приводится довольно полная характеристика
паразитических моллюсков. Даются характеристики наиболее важ-
ных в практическом отношении групп членистоногих. Приводится
материал о паразитических ракообразных. В обширной группе кле-
щей особое внимание уделено иксодовым клещам, которые вследст-
вие их практической значимости изучаются сейчас наиболее интен-
сивно.
Материал по паразитическим- насекомые изложен довольно
сжато. Опущены паразитические перепончатокрылые (наездники).
В главах, посвященных паразитическим насекомым, приводятся
данные по их морфологии и анатомии, отмечаются особенности
строения, обусловленные паразитическим образом жизни и имеющие
адаптивное значение. Авторы исходили при этом из того, что сту-
денты получают достаточно полное представление об организации
насекомых в курсах зоологии беспозвоночных животных и энтомо-
логии, а также во время летних практик.
Список рекомендуемой литературы сознательно ограничен переч-
нем учебников, руководств и монографий по тем или иным парази-
3
тическим группам. Лишь в исключительных случаях в списки вклю-
чены статьи. Авторы искренне признательны профессору Ю. И. По-
лянскому, заведующему кафедрой зоологии беспозвоночных
Ленинградского государственного университета, за неизменное вни-
мание к их работе.
Ряд ценных советов авторы получили от доцента кафедры
А. А. Стрелкова и от доцента Брестского пединститута М. Я. Асса,
взявших на себя труд по просмотру отдельных глав рукописи. Ав-
торы благодарят также аспирантов и лаборантов кафедры зоологии
беспозвоночных Ленинградского университета — Б. И. Хари,
С. А. Соловьеву, О. Ю. Семенова и И. А. Тихомирова за помощь
при подготовке рукописи к печати.
Авторы
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ1
КЛАСС CESTODA RUDOLPH}, 1819 —ЛЕНТОЧНЫЕ ЧЕРВИ
Класс Cestoda объединяет около 3500 видов паразитических
плоских червей, обитающих главным образом в позвоночных живот-
ных. Характерной особенностью ленточных червей является отсут-
ствие у них пищеварительной системы. Свое название они получили
из-за характерной формы тела, которое уплощено и вытянуто
в-длину. В большинстве случаев оно еще разделено на многочислен-
ные членики — проглоттиды.
Цестоды, как правило, имеют сложный жизненный цикл с одной
или двумя личиночными стадиями. Паразитические фазы при этом
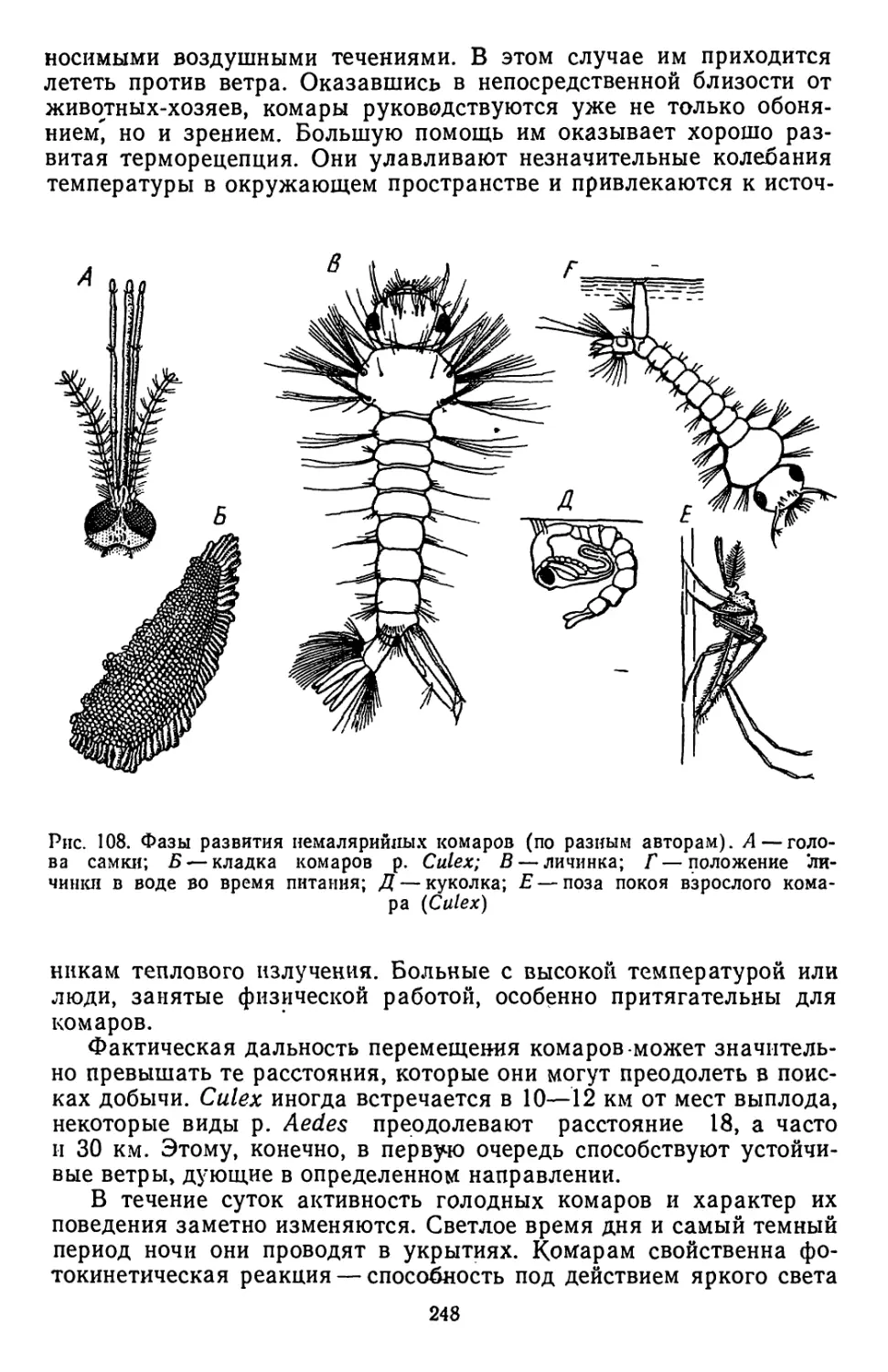
чередуются со свободно живущими. Широкое распространение полу-
чило и явление смены животных-хозяев. У некоторых видов( с. 28)
наблюдается вторичное усложнение развития за счет приобретения
их личиночными стадиями способности размножаться бесполым пу-
тем. Все ленточные черви во взрослой стадии обитают в пищевари-
тельной системе животных-хозяев. Исключение составляет так назы-
ваемый Archigetes— неотеническая форма, поселяющаяся в полости
тела пресноводных малощетинковых червей.
Цестоды были впервые описаны как самостоятельный класс
плоских червей еще Рудольфи (Rhudolphi, 1819). Большая заслуга
в ихдальнейшем изучении принадлежит Ван Бенедену (Van Вепе-
den), М. Брауну (М. Braun), И. Карусу (I. Carus), Р. Лейкарту
(R. Leuckart) и другим ученым XIX в., занимавшимся изучением
паразитических червей.
В XX в. особенно много было сделано швейцарским зоологом
О. Фурманом (О. Fuhrmann), автором известного руководства по
морфологии и биологии ленточных червей. Первая сводка фауны
?936)Д пРинадлежит Жуайе и Ж. Бэру (Ch. Joyeux et J. Baer,
В изучении фауны цестод России видное место занимают иссле-
дования В. О.. Клера и Н. А. Холодковского, работавших в начале
1 Продолжение, начало см. в кп.: Т. А. Гинецииокая, А. А. Добровольский,
частная паразитология, кн. I.
5
нашего столетия. Важную роль в развитии советской цестодологии
сыграл выход в свет многотомного издания «Основы цестодологии»,
выполненного под руководством акад. К. И. Скрябина, а также ра-
боты А. А. Спасского, М. Н. Дубининой, С. Л. Делямуре н др.
Построение естественной системы цестод еще не завершено.
Разными авторами выделяется от 4 до 11 отрядов. Столь же
различны взгляды исследователей и на филогению этой группы.
В настоящехМ учебном пособии принята за основу система, предло-
женная Р. С. Шульцем и Е. В. Гвоздевым в их книге «Основы общей
гельминтологии» (т. I, 1970). Однако Cestodaria рассматриваются
нами как самостоятельный класс. Таким образом, класс Cestoda со-
ответствует бывшему подкл. Eucestoda и включает 6 отрядов:
Caryophylla'eidea Beneden in Olsson — гвоздичники, Pseudophyllidea
Carus, 1863; Tetraphyllidea (Beneden, 1849)< Carus, 1863; Diphylli-
dea Beneden in Cams, 1863; Trypanorhyncha Diesing, 1863 и Cyclo-
phyllidea Beneden in Braun, 1900.
Морфология и анатомия цестод
Форма тела и размеры. Морфологическое разнообразие ленточ-
ных червей очень велико, хотя оно ни в коей мере не затрагивает
единого плана строения, характерного для всей группы в целом.
В большинстве случаев тело цестод подразделяется на три от-
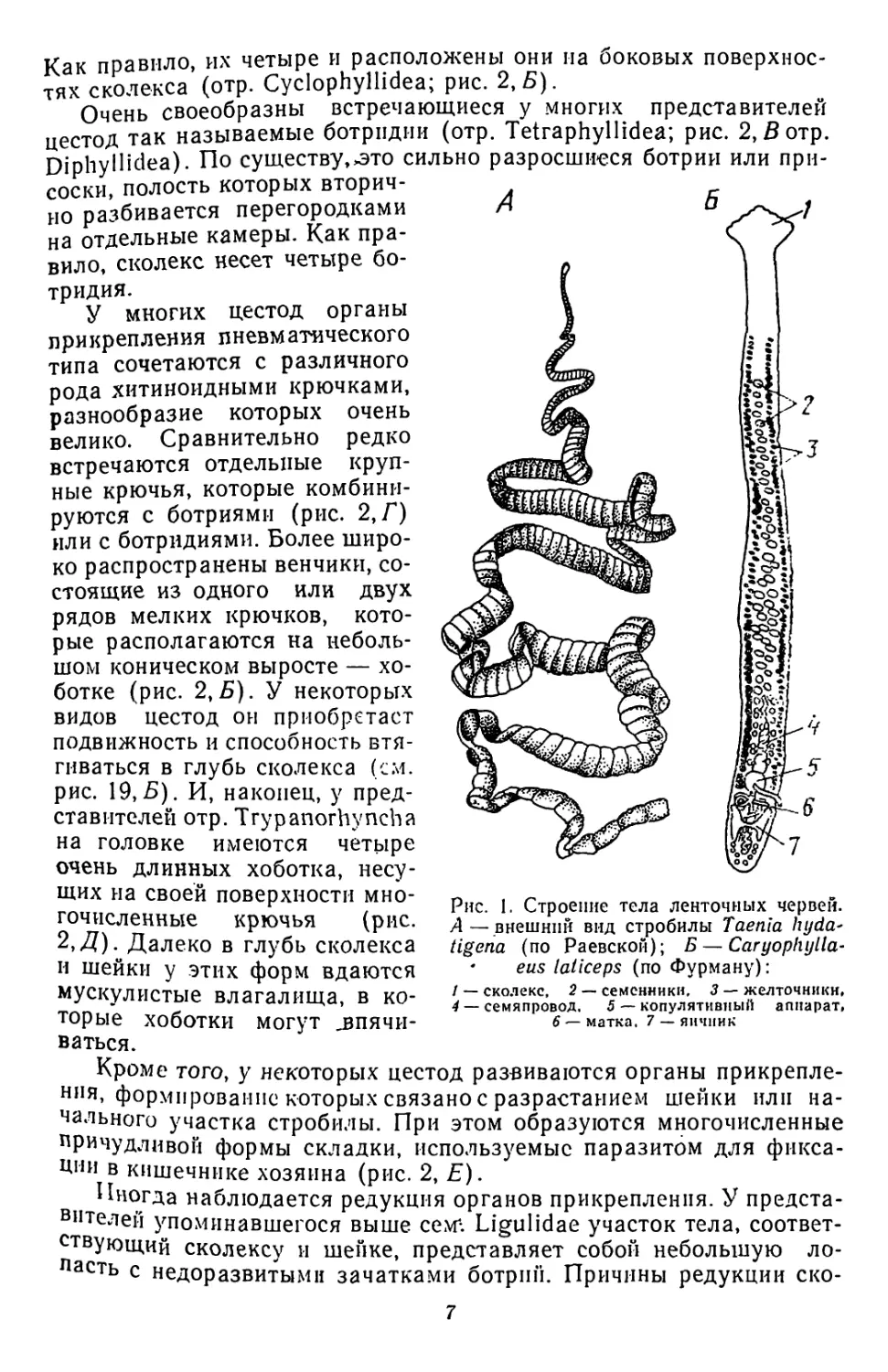
дела: головку — сколекс, шейку и членистую стробилу (рис. 1,А).
Формы, обладающие не расчлененным телом, немногочисленны.
Значительная их часть объединяется в отр. Caryophyllaeidea.—
гвоздичников. Это мелкие черви с одним половым комплексом
(рис. 1, 5; с. 32). Особый интерес представляет сем. Ligulidae —
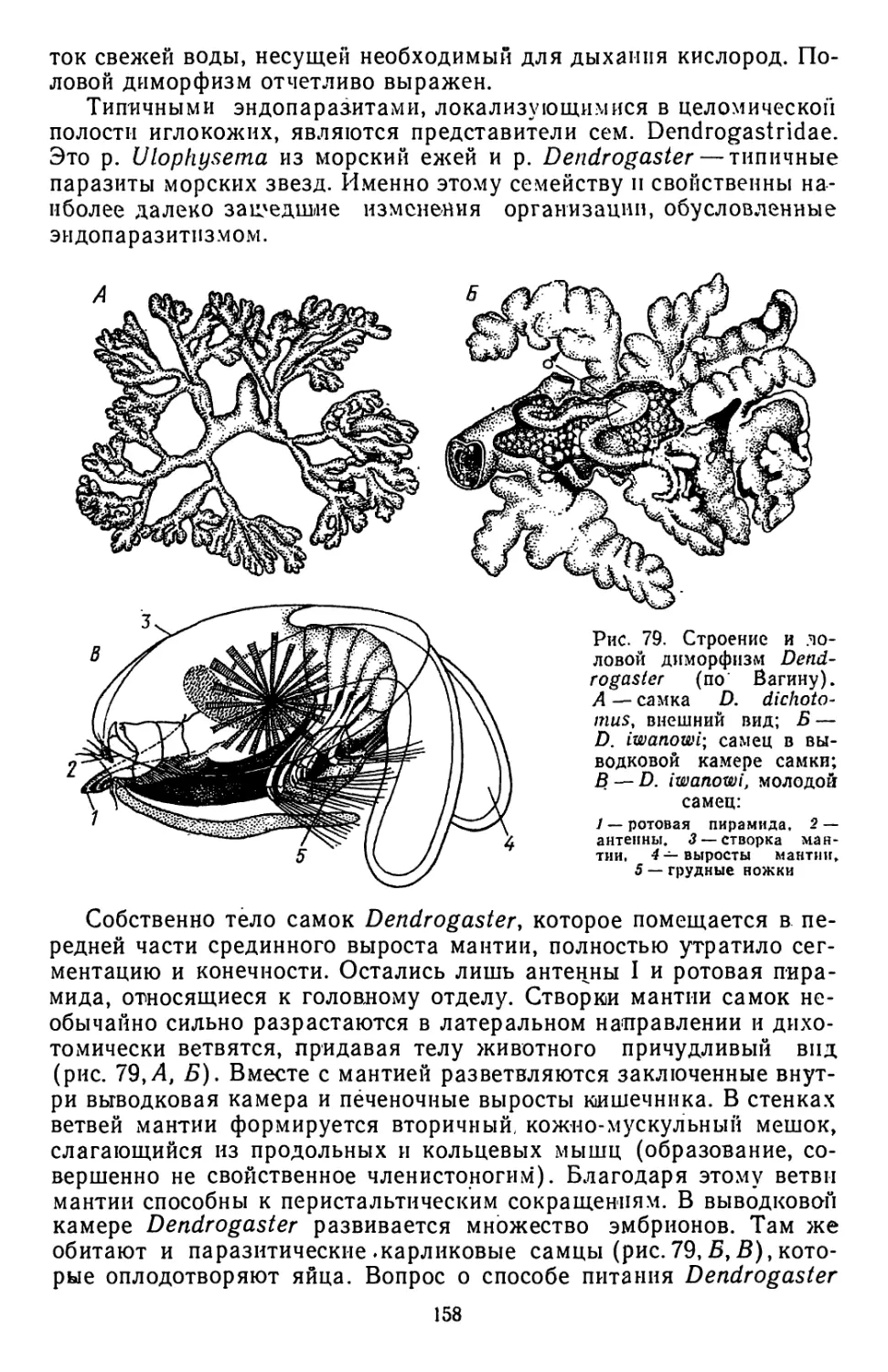
ремнецы (отр. Psedophyllidea), в которохМ наряду с формами, обла-
дающими настоящей стробилой (р. Schistocephalus), представлены
и нерасчлененные виды (р. Ligula, Digramma).
Линейные размеры цестод варьируют в очень широких преде-
лах. Среди них есть гиганты; средняя длина стробилы паразита че-
ловека Diphyllobothriurn latum составляет 6—10 м, однако известны
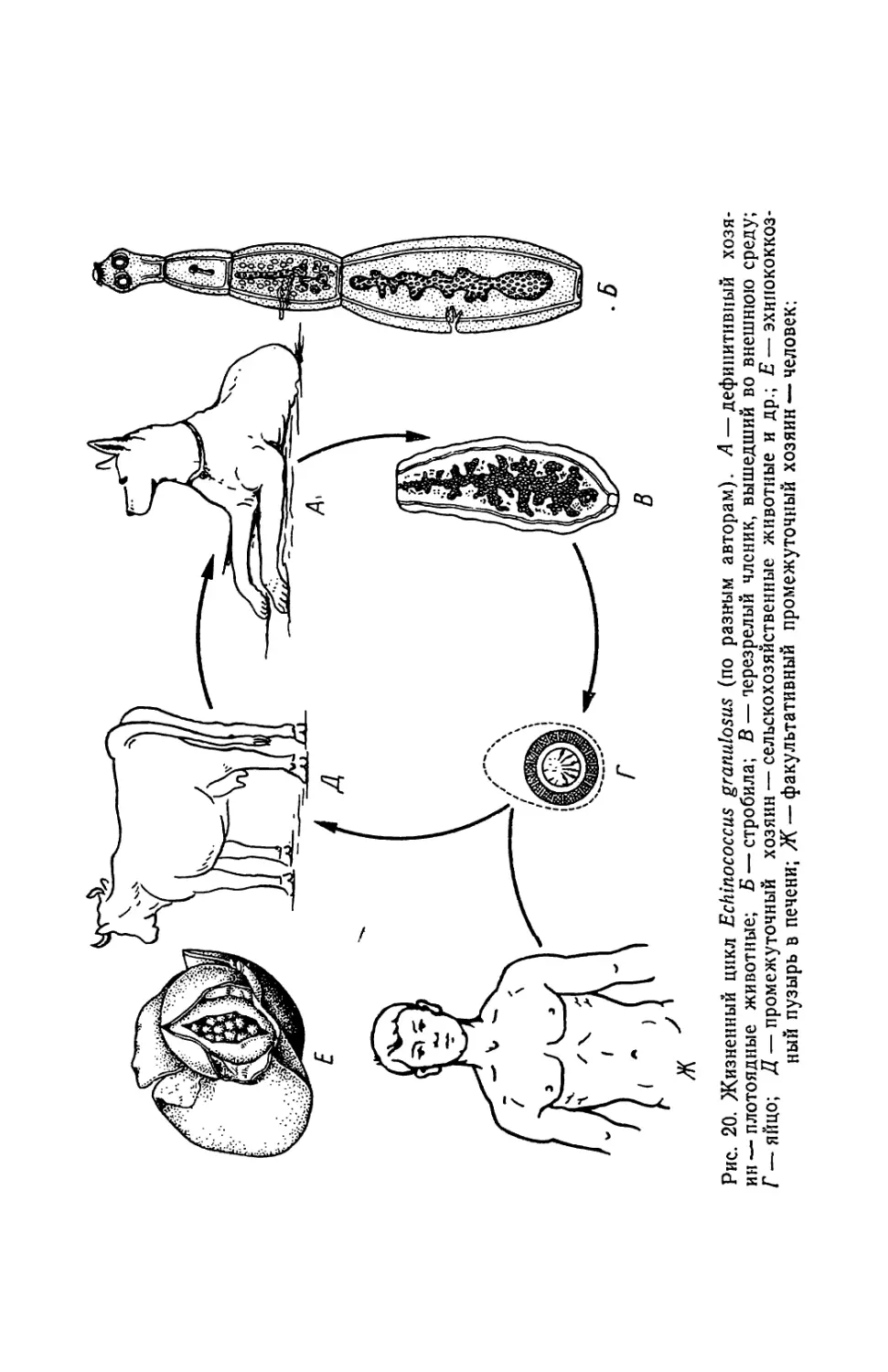
случаи, когда черви достигали 25 м. В то же время длина стробилы
Echinoccocus granulosus не превышает 8 мм (см. рис. 20,Б), а близ-
кого вйда Alveococcus multilocularis — 3,4 мм. Передний конец те-
ла цестод несет головку, или сколекс. На головке располагаются
иногда весьма совершенные органы прикрепления, позволяющие
паразиту надежно удерживаться в кишечнике хозяина. Прикрепи-
тельные образования ленточных червей по принципу действия под-
разделяются на две большие группы: присасывательные и заякори-
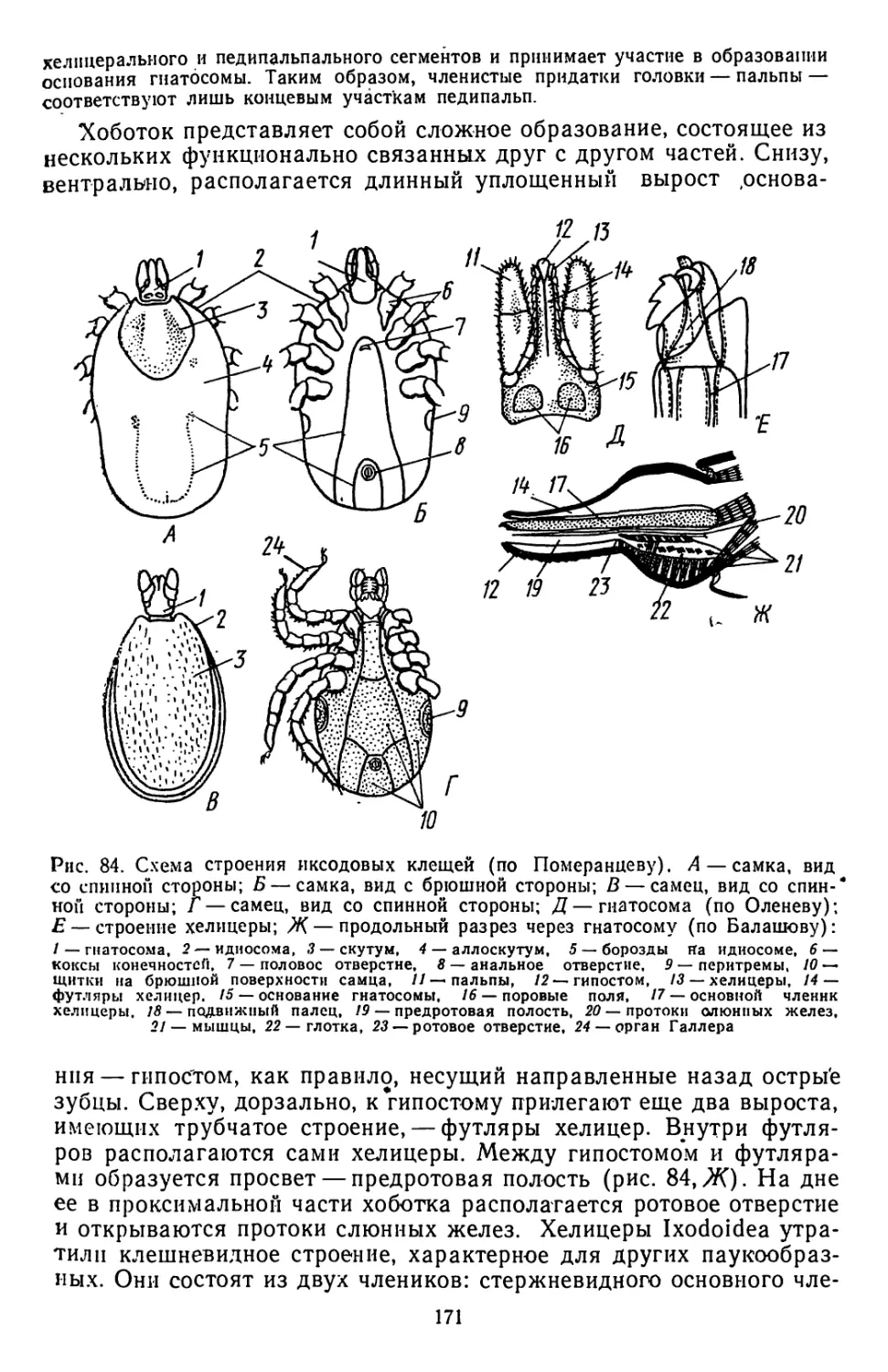
вающпе.
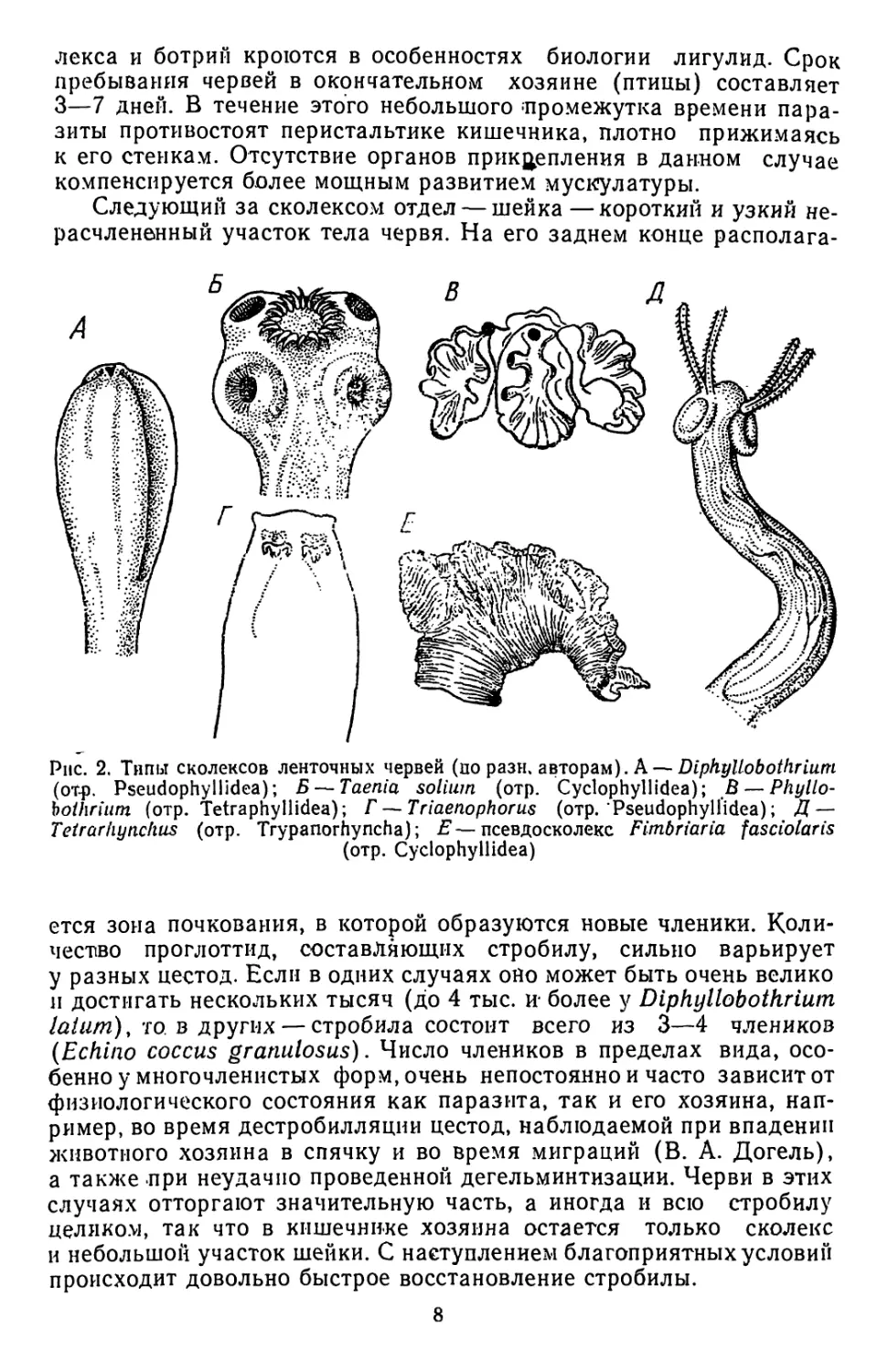
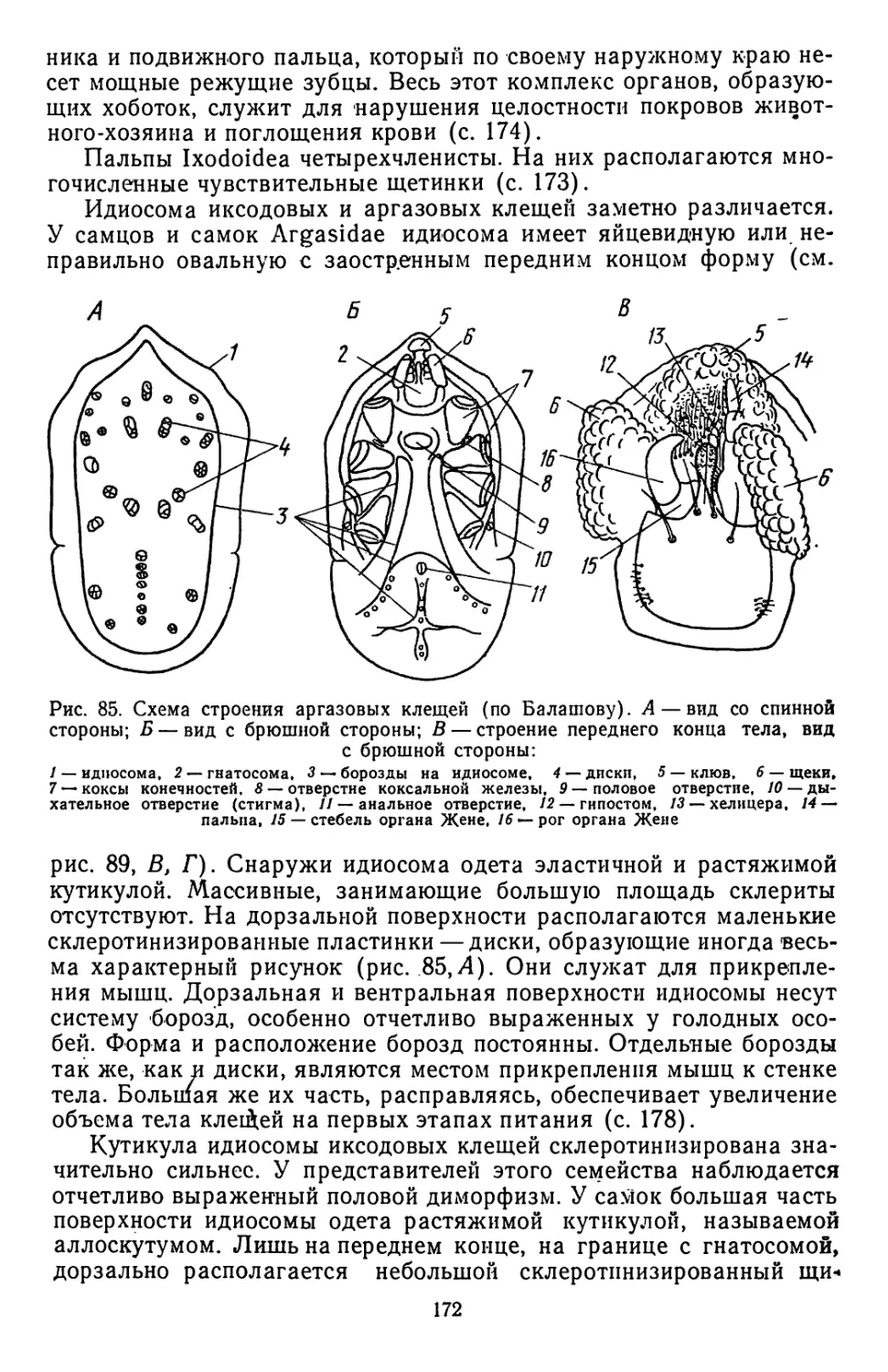
Примитивным пневматическим аппаратом прикрепления явля-
ются ботрии, встречающиеся главным образом у представителей отр.
Pseudophyllidea. Это просто устроенные присасывательные ямки, не
имеющие еще собственной мускулатуры. Чаще две ботрии в виде уз-
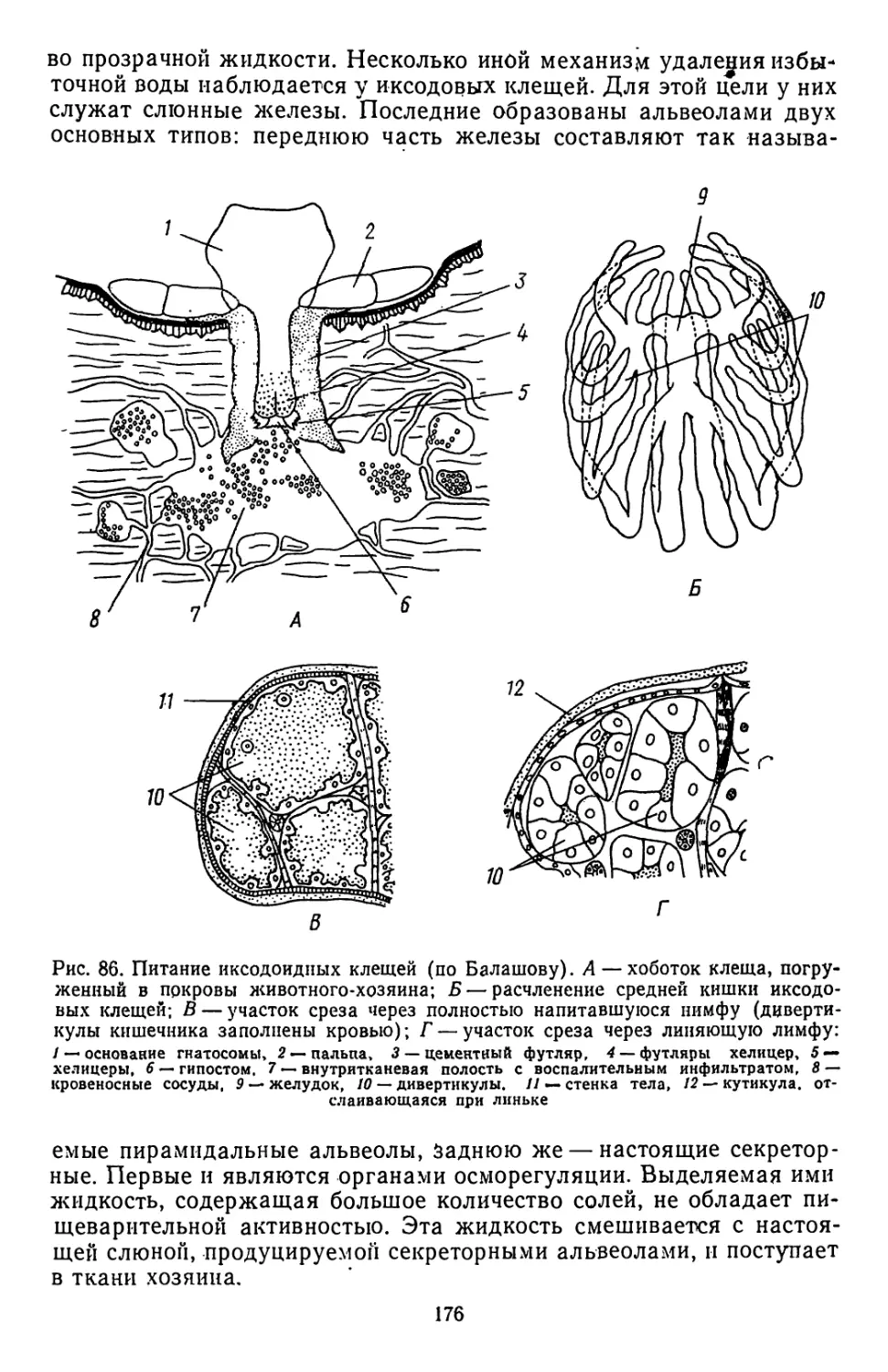
ких и глубоких щелей располагаются по бокам сколекса (рис. 2, А).
Значительно шире распространены у цестод настоящие присоски.
6
рме. 1. Строение тела ленточных червей.
А— внешний вид стробилы Taenia hyda^
tigena (по Раевской); Б — Caryophylla-
• eus laiiceps (по Фурману):
/ — сколекс, 2 — семенники, 3 — желточникн,
4 — семяпровод. 5 — копулятивный аппарат,
6 — матка. 7 — яичник
Как правило, их четыре и расположены они иа ооковых поверхнос-
тях сколекса’(отр. Cyclophyllidea; рис. 2,5).
Очень своеобразны встречающиеся у многих представителей
цестод так называемые ботрпдии (отр. Tetraphyllidea; рис. 2, В отр.
Diphyllidea). По существу,>это сильно разросшиеся ботрии или при-
соски, полость которых вторич-
но разбивается перегородками
на отдельные камеры. Как пра-
вило, сколекс несет четыре бо-
тридия.
У многих цестод органы
прикрепления пневматического
типа сочетаются с различного
рода хитиноидными крючками,
разнообразие которых очень
велико. Сравнительно редко
встречаются отдельные круп-
ные крючья, которые комбини-
руются с ботриями (рис. 2, Г)
или с ботридиями. Более широ-
ко распространены венчики, со-
стоящие из одного или двух
рядов мелких крючков, кото-
рые располагаются на неболь-
шом коническом выросте — хо-
ботке (рис. 2,Б). У некоторых
видов цестод он приобретает
подвижность и способность втя-
гиваться в глубь сколекса (см.
рис. 19,5). И, наконец, у пред-
ставителей отр. Trypanorhyncha
на головке имеются четыре
очень длинных хоботка, несу-
щих на своей поверхности мно-
гочисленные крючья (рис.
2,Д). Далеко в глубь сколекса
и шейки у этих форм вдаются
мускулистые влагалища, в ко-
торые хоботки могут впячи-
ваться.
Кроме того, у некоторых цестод развиваются органы прикрепле-
ния, формирование которых связано с разрастаниехм шейки или на-
чального участка стробилы. При этом образуются многочисленные
причудливой формы складки, используемые паразитом для фикса-
ции в кишечнике хозяина (рис. 2, Е).
Иногда наблюдается редукция органов прикрепления. У предста-
вителей упоминавшегося выше сем1. Ligulidae участок тела, соответ-
ствующий сколексу и шейке, представляет собой небольшую ло-
пасть с недоразвитыми зачатками ботрий. Причины редукции ско-
7
лекса и ботрий кроются в особенностях биологии лигулид. Срок
пребывания червей в окончательном хозяине (птицы) составляет
3—7 дней. В течение этого небольшого промежутка времени пара-
зиты противостоят перистальтике кишечника, плотно прижимаясь
к его стенкам. Отсутствие органов прикрепления в данном случае
компенсируется более мощным развитием мускулатуры.
Следующий за сколексом отдел — шейка —короткий и узкий не-
расчлененный участок тела червя. На его заднем конце располага-
Рпс. 2. Типы сколексов ленточных червей (по рази, авторам). А — Diphyllobothruun
(от-p. Pseudophyllidea); Б— Taenia solium (отр. Cyclophyllidea); В— Phyllo-
boihrium (отр. Tetraphyllidea); Г — Triaenophorus (отр. Pseudophyll’idea); Д —
Tetrarhynchus (отр. Trypanorhyncha); E—псевдосколекс Fimbriaria fasciolaris
(отр. Cyclophyllidea)
ется зона почкования, в которой образуются новые членики. Коли-
чество проглоттид, составляющих стробилу, сильно варьирует
у разных цестод. Если в одних случаях оно может быть очень велико
и достигать нескольких тысяч (до 4 тыс. и более у Diphyllobothrium
laium), чо в других — стробила состоит всего из 3—4 члеников
(Echino coccus granulosus). Число члеников в пределах вида, осо-
бенно у многочленистых форм, очень непостоянно и часто зависит от
физиологического состояния как паразита, так и его хозяина, нап-
ример, во время дестробилляции цестод, наблюдаемой при впадении
животного хозяина в спячку и во время миграций (В. А. Догель),
а также при неудачно проведенной дегельминтизации. Черви в этих
случаях отторгают значительную часть, а иногда и всю стробилу
целиком, так что в кишечнике хозяина остается только сколекс
и небольшой участок шейки. С наступлением благоприятных условий
происходит довольно быстрое восстановление стробилы.
8
Увеличение числа члеников и удлинение строоилы у оольшпнст-
ва видов осуществляется по принципу вставочного (интеркалярио-
го) роста. Новые проглоттиды образуются в зоне почкования
и сдвигают назад ранее сфомировавшиеся членики. Дальнейшая
судьба члеников различна у разных видов цестод. У одних форм пе-
пиод образования проглоттид ограничен определенным временным
промежутком, но зато все они
сохраняются вплоть до гибели
червя (сем. Ligulidae). У дру-
гих же (и таких большинство)
формирование члеников про-
должается в течение всей жиз-
ни паразита. Однако по дости-
жении какой-то определенной
и относительно постоянной для
данного вида величины даль-
нейшего увеличения числа чле-
ников все же не наблюдается.
Проглоттиды, составляющие
самый задний участок строби-
лы, постоянно отторгаются от
нее. Иногда они отрываются по
одной (Taeniarhynchus sagina-
tus), иногда же от стробилы от-
деляются целые куски по 5—6
члеников (Taenia solium). От-
Рис. 3. Схема строения тегумента цестод
(по Морзесу):
/ — микротрихии, 2 — наружный цитоплазма-
тический слбй тегумента. 3 —базальная мем-
брана, 4 — цитоплазматические отростки по-
груженных клеток тегумента, 5 — кольцевые
и продольные мускульные волокна, 6 — по-
груженная клетка тегумента
делившиеся членики часто спо-
собны к активному движению.
Более того, они могут длитель-
ное время самостоятельно су-
ществовать в кишечнике хозя-
ина (с. 29).
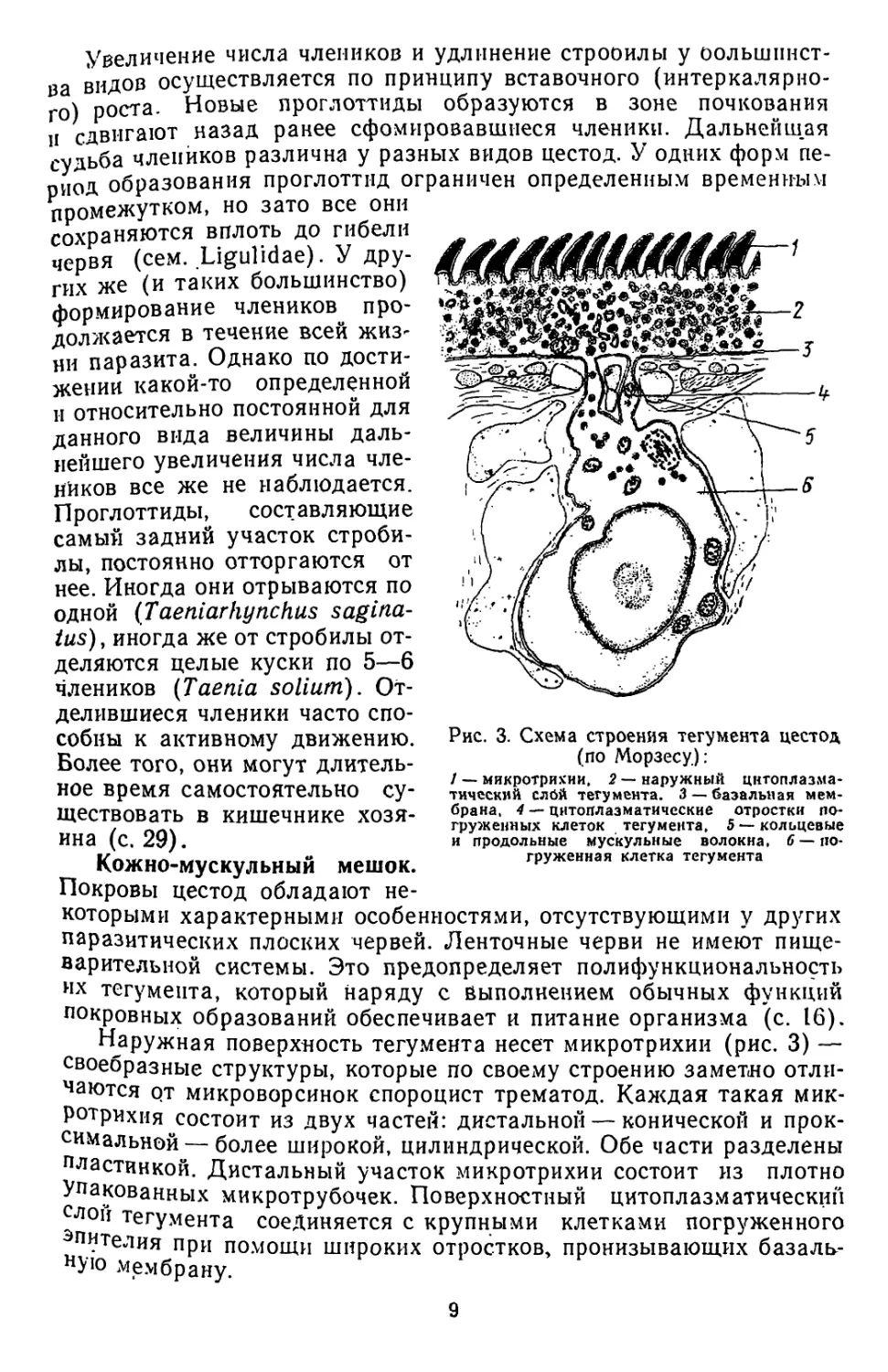
Кожно-мускульный мешок.
Покровы цестод обладают не-
которыми характерными особенностями, отсутствующими у других
паразитических плоских червей. Ленточные черви не имеют пище-
варительной системы. Это предопределяет полифункциональность
их тегумента, который наряду с выполнением обычных функций
покровных образований обеспечивает и питание организма (с. 16).
Наружная поверхность тегумента несет микротрихии (рис. 3) —
своебразные структуры, которые по своему строению заметно отли-
чаются рт микроворсинок спороцист трематод. Каждая такая мик-
ротрихия состоит из двух частей: дистальной — конической и прок-
симальной— более широкой, цилиндрической. Обе части разделены
пластинкой. Дистальный участок микротрихии состоит из плотно
Упакованных микротрубочек. Поверхностный цитоплазматический
слой тегумента соединяется с крупными клетками погруженного
пителия при помощи широких отростков, пронизывающих базаль-
ную мембрану.
9
Под базальной мембраной располагаются мускульные элемен-
ты. Субтегументальная мускулатура ленточных червей состоит из
двух слоев: наружного — кольцевого и более мощно развитого внут-
реннего— продольного. Кроме того, в паренхиме, ограничивая рас-
положение органов половой системы, располагается еще один —
внутренний — слой кольцевых мускульных волокон. У многих цес-
тод имеются еще довольно хорошо развитые дорзовентральные
мышцы. Мускулатура цестод, к$к и других паразитических плоских
червей, состоит из гладких мышечных волокон.
Непосредственно под кожно-мускульным мешком залегает па-
ренхима. Она построена из крупных неправильной формы клеток,
отростки которых переплетаются друг с другом. Как и у других
плоских червей паренхима цестод выполняет самые разно-
образные функции.
В< периферических слоях паренхимы наряду с погруженными
клетками тегумента располагаются так называемые известковые
клетки, в которых происходит образование «известковых» телец.
Последние'у многих цестод начинают формироваться на личиноч-
ных стадиях (процеркоид, плероцеркоид). При этом в цитоплазме
некоторых субтегументальных клеток появляются мелкие вакуоли,
которые, сливаясь, образуют крупную внутриклеточную полость. В
последней остается небольшое количество гранул, образующих центр
петрификации при формировании минерального тельца, рост кото-
рого идет за счет последовательного нарастания концентрических
слоев. Состав «известковых» телец отличается большой сложностью.
Они содержат белки, углеводы, липиды. Минеральные компоненты
состоят из карбоната кальция и магния. Кроме того, имеется не-
большое количество фосфора. х
«Известковые» тельца активно используются паразитами. По
мнению некоторых исследователей, они служат для забуферивания
кислой среды. Это особенно важно при попадании личинок (плеро-
церкоидов) в желудок животного-хозяина.
Кроме «известковых» телец в поверхностных слоях паренхимы
у многих видов ленточных червей располагаются одноклеточные
кожные железы. Особенно многочисленны они в сколексе и на гра-
ницах между двумя соседними члениками.
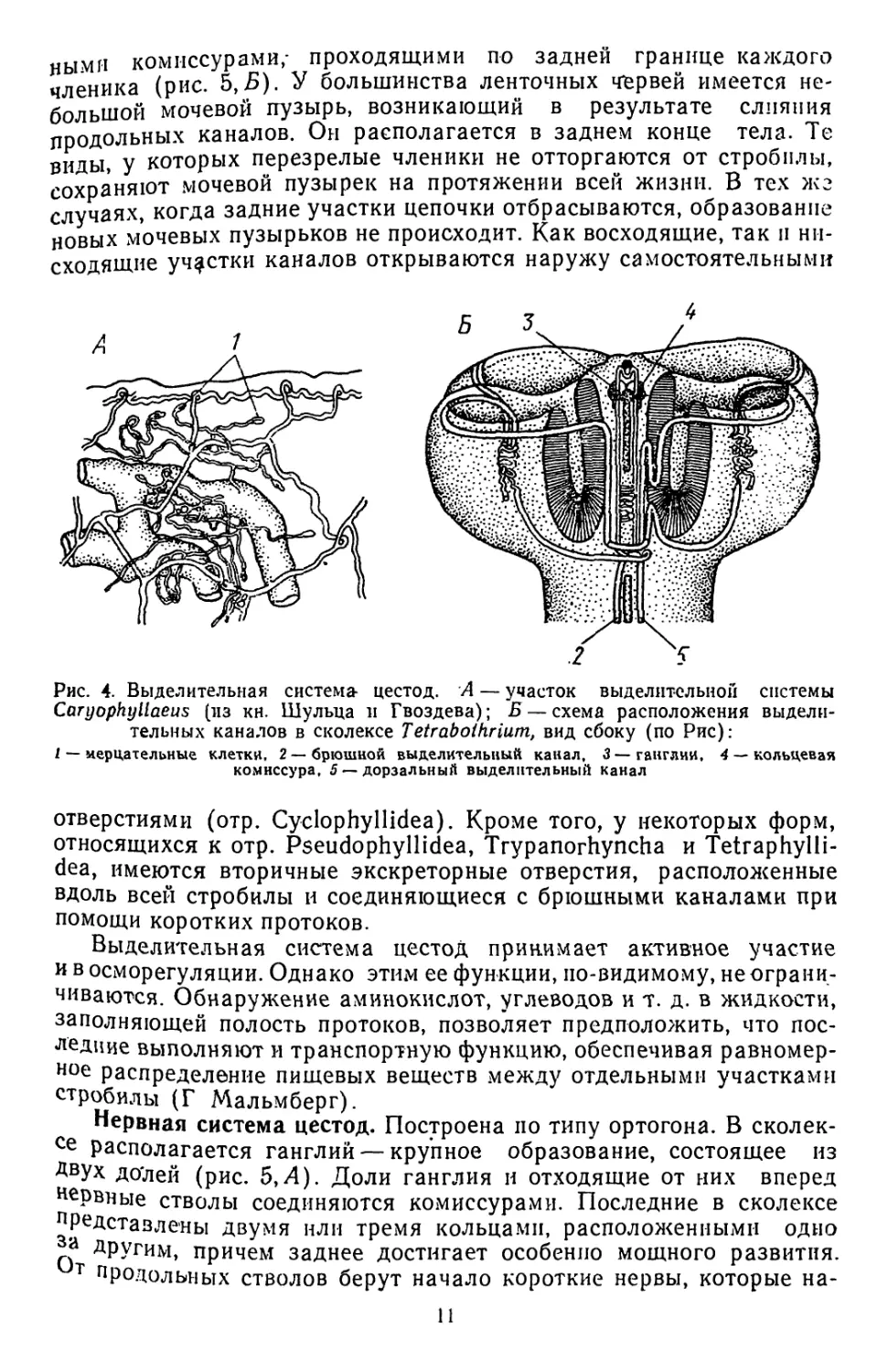
Выделительная система цестод. Протонефридиального типа; мер-
цательные клетки разбросаны по всей паренхиме червя. Отходящие
от них капилляры, сливаясь, дают начало более крупным протокам.
Последние часто образуют сеть (рис. 4,А), напоминающую пара-
нефридиальный плексус трематод. Тонкие протоки впадают в
главные собирательные каналы, которые у многих видов начина-
ются на заднем конце тела и тянутся вперед по бокам стробилы
ближе к ее спинной поверхности.
В сколексе каналы образуют петли, иногда довольно сложной
конфигурации (рис. 4,5). Обе петли часто соединяются друг с дру-
гом- системой многочисленных тонких комиссур. Выйдя из сколек-
са, каналы направляются к заднему концу стробилы уже по ее вен-
тральной поверхности. У многих видов они соединяются попереч-
10
НЫ.МН комиссурами,- проходящими по задней границе каждого
членика (рис. 5,5). У большинства ленточных червей имеется не-
большой мочевой пузырь, возникающий в результате слияния
продольных каналов. Он располагается в заднем конце тела. Те
виды, у которых перезрелые членики не отторгаются от стробилы,
сохраняют мочевой пузырек на протяжении всей жизни. В тех же
случаях, когда задние участки цепочки отбрасываются, образование
новых мочевых пузырьков не происходит. Как восходящие, так п ни-
сходящие участки каналов открываются наружу самостоятельными
Рис. 4. Выделительная система цестод. А — участок выделительной системы
Caryophyllaeus (из кн. Шульца и Гвоздева); Б — схема расположения выдели-
тельных каналов в сколексе Tetraboihrium, вид сбоку (по Рис):
1 — мерцательные клетки, 2 —брюшной выделительный канал, 3—ганглии, 4 — кольцевая
комиссура, 5 —дорзальный выделительный канал
отверстиями (отр. Cyclophyllidea). Кроме того, у некоторых форм,
относящихся к отр. Pseudophyllidea, Trypanorhyncha и Tetraphylli-
dea, имеются вторичные экскреторные отверстия, расположенные
вдоль всей стробилы и соединяющиеся с брюшными каналами при
помощи коротких протоков.
Выделительная система цестод принимает активное участие
и в осморегуляции. Однако этим ее функции, по-видимому, не ограни-
чиваются. Обнаружение аминокислот, углеводов и т. д. в жидкости,
заполняющей полость протоков, позволяет предположить, что пос-
ледние выполняют и транспортную функцию, обеспечивая равномер-
ное распределение пищевых веществ между отдельными участками
стробилы (Г Мальмберг).
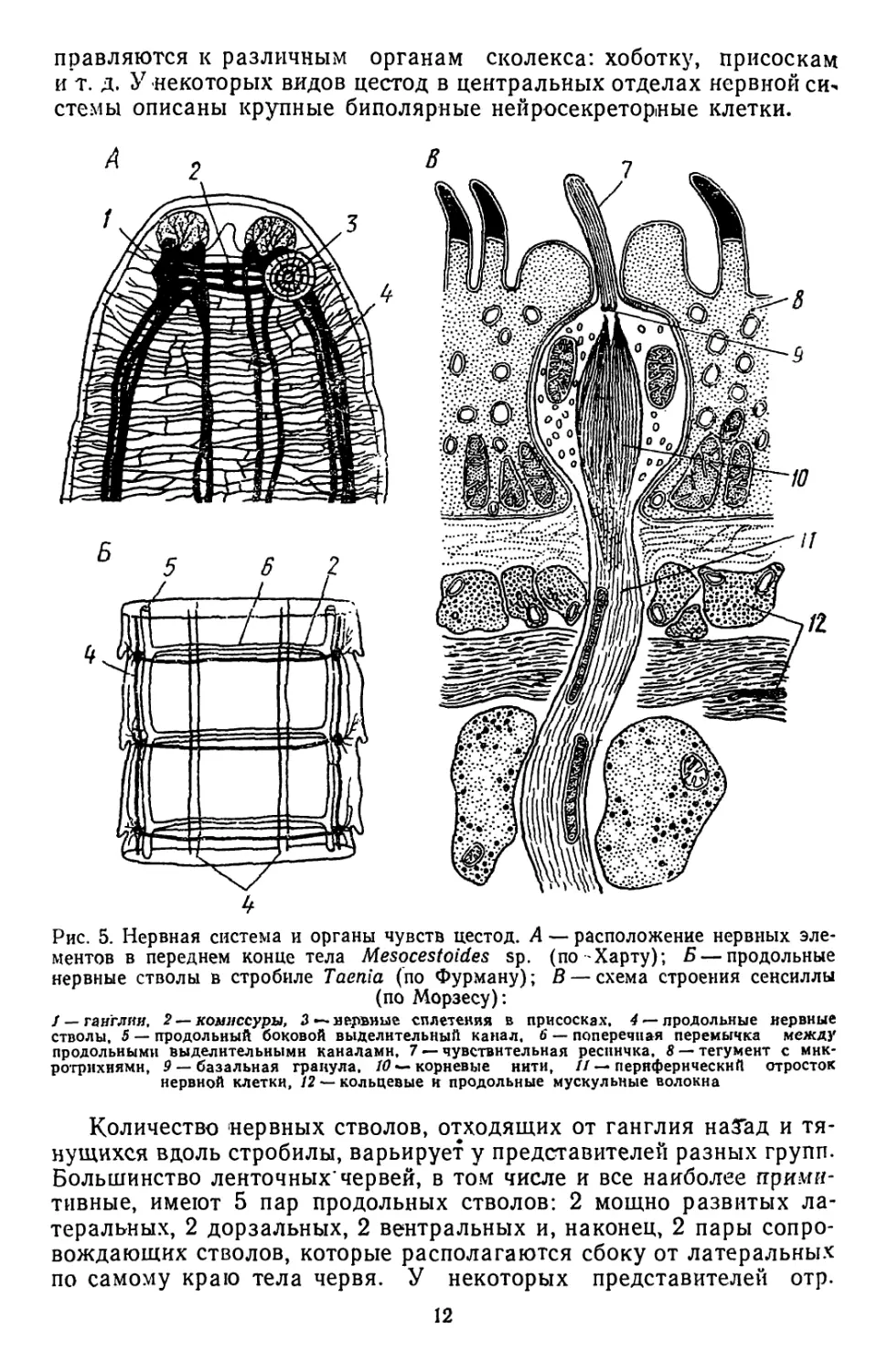
Нервная система цестод. Построена по типу ортогона. В сколек-
Се располагается ганглий — крупное образование, состоящее из
Двух до'лей (рис. 5,Л). Доли ганглия и отходящие от них вперед
нервные стволы соединяются комиссурами. Последние в сколексе
представлены двумя или тремя кольцами, расположенными одно
а Другим, причем заднее достигает особенно мощного развития.
т продольных стволов берут начало короткие нервы, которые на-
11
правляются к различным органам сколекса: хоботку, присоскам
и т. д. У некоторых вадов цестод в центральных отделах нервной си-
стемы описаны крупные биполярные нейросекреторные клетки.
Рис. 5. Нервная система и органы чувств цестод. А — расположение нервных эле-
ментов в переднем конце тела Mesocestoides sp. (по Харту); Б — продольные
нервные стволы в стробиле Taenia (по Фурману); В — схема строения сенсиллы
(по Морзесу):
/ — ганглия, 2 — комиссуры, 3 — нервные сплетения в присосках. 4 —продольные нервные
стволы. 5 — продольный боковой выделительный канал. 6 — поперечная перемычка между
продольными выделительными каналами. 7 — чувствительная ресничка. 8 — тегумент с мнк-
ротрихнямн, 9 — базальная гранула. 10 *- корневые нити, // — периферический отросток
нервной клетки, 12 — кольцевые н продольные мускульные волокна
Количество нервных стволов, отходящих от ганглия наТад и тя-
нущихся вдоль стробилы, варьирует у представителей разных групп.
Большинство ленточных* червей, в том числе и все наиболее прими-
тивные, имеют 5 пар продольных стволов: 2 мощно развитых ла-
теральных, 2 дорзальных, 2 вентральных и, наконец, 2 пары сопро-
вождающих стволов, которые располагаются сбоку от латеральных
по самому краю тела червя. У некоторых представителей отр.
12
Cvclophyllidea, к которому относятся 'наиболее прогрессивные
и специализированные виды, количество продольных стволов умень-
шается до 6 (рис. 5,5). В каждой проглоттиде продольные стволы
соединяются поперечной комиссурой, обычно расположенной в зад-
ней части членика. Кроме такой классической ортогональной нерв-
ной системы у некоторых представителей примитивных отрядов был
обнаружен поверхностно залегающий нервный плексус. Он состоит
из субтегументальных нервных волокон, анастомозирующих между
собой.
Какие-либо сложно устроенные специализированные органы
чувств у цестод отсутствуют. В большом количестве у них имеются
лишь сенсиллы, каждая из которых состоит из ригидной реснички,
воспринимающей раздражения и связанного с ней нервного оконча-
ния чувствительной клетки (рис. 5,5). Сенсиллы довольно редко
разбросаны по поверхности стробилы и только на сколексе и распо-
ложенных на нем органах прикрепления они образуют значитель-
ные скопления.
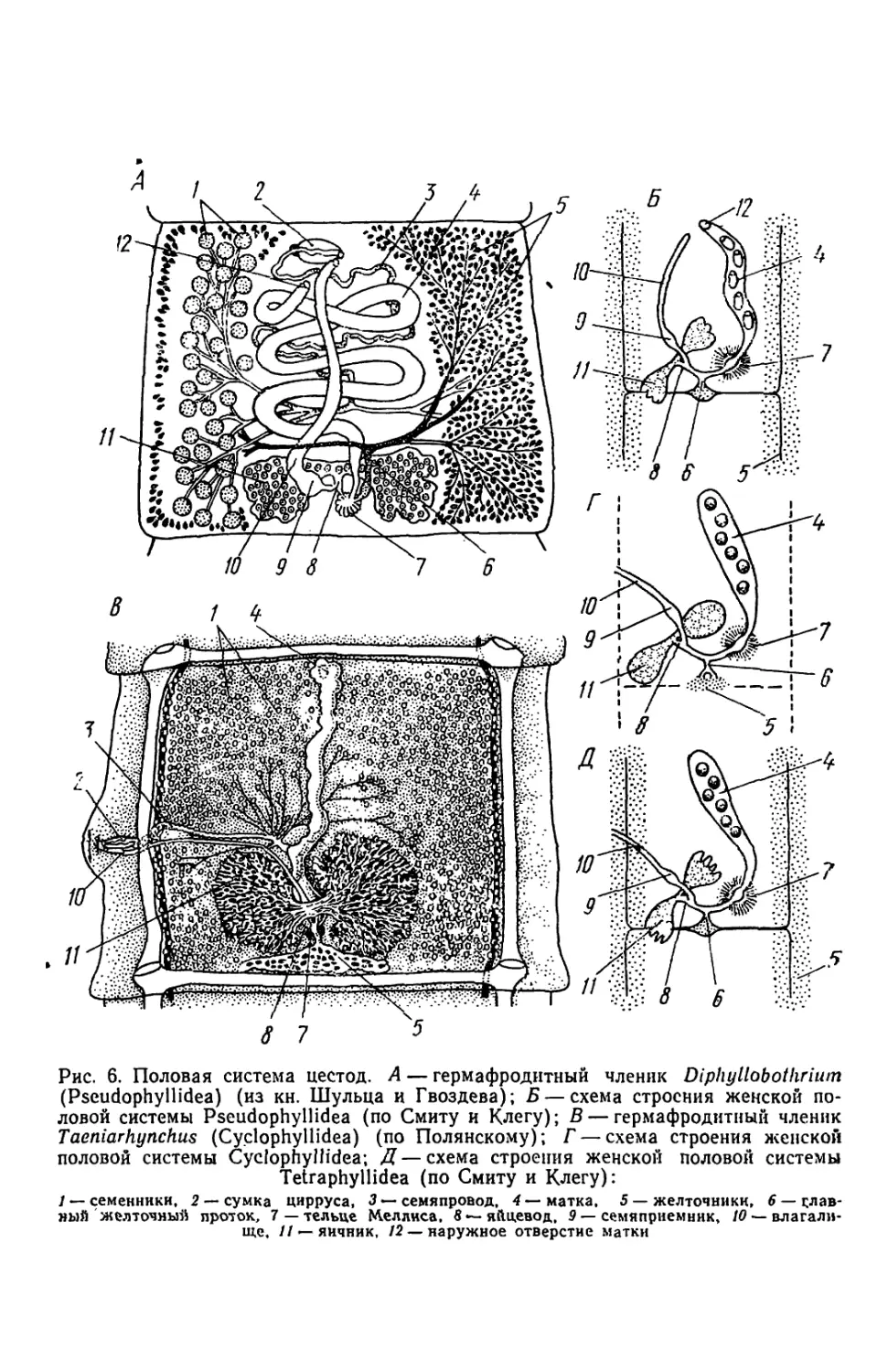
Половая система.- Цестоды, как и большинство плоских червей,
обладают сложно устроенной гермафродитной половой системой.
Лишь у представителей отр. Caryophyllaeidea имеется один «комп-
лект» половых органов (см. рис. 1,5). Для'всех остальных цестод
характерно наличие большого числа половых комплексов — по од-
ному (реже по два) в каждой проглоттиде. Степень развития поло-
вого аппарата, как правило, зависит от положения членикав стро-
биле. В молодых проглоттидах, составляющих начальный участок
тела паразита, происходит закладка и формирование органов поло-
вой системы. Среднюю часть стробилы составляют членики с хоро-
шо развитой функционирующей половой системой. Это так называ-
емые гермафродитные членики. И наконец, в заднем конце стробилы
у многих видов располагаются «перезрелые» членики, для которых
характерно сильное разрастание матки, заполненной яйцами, и ат-
рофия всех остальных органов половой системы. В большинстве
случаев именно эти проглоттиды отрываются от тела червя и выно-
сятся наружу.
Исходя из особенностей строения, половые аппараты цестод
можно условно разделить на две большие группы, которые в своей
наиболее характерной форме встречаются у представителей отр.
Pseudophyllidea и отр. Cyclophyllidea.
Половая система Pseudophyllidea (рис. 6, А, Б) обладает целым
рядом примитивных признаков. Мужская половая система состоит
из многочисленных семенников, расположенных в центральной зо-
не паренхимы. Они образуют два латеральных поля по бокам от ор-
ганов женской половой системы. От семенников отходят семенные
канальцы, которые, сливаясь, дают начало семяпроводу. Прокси-
мальный участок последнего обычно вздут и выполняет функцию се-
менного пузырька. Семяпровод в виде извитой трубки направляется
?Реред. Семяизвергательный канал пронизывает сумку цирруса.
Мужская половая система открывается наружу на дне полового ат-
Риума, который располагается медиально на вентральной поверх-
13
Рис. 6. Половая система цестод. Л — гермафродитный членик Diphyllobothrium
(Pseudophyllidea) (из кн. Шульца и Гвоздева); Б — схема строения женской по-
ловой системы Pseudophyllidea (по Смиту и Клегу); В — гермафродитный членик
Taeniarhynchus (Cyclophyllidea) (по Полянскому); Г — схема строения женской
половой системы Cyclophyllidea; Д—схема строения женской половой системы
Tetraphyllidea (по Смиту и Клегу):
/ — семенники. 2 — сумка цирруса, 3 — семяпровод, 4 — матка. 5 — желточники, 6 — слав-
ный желточный проток, 7—тельце Me л лиса, 8 — яйцевод. 9 — семяприемник, /0 —влагали-
ще, // — яичник, 12— наружное отверстие матки
пости в передней части членика. У некоторых видов (Eubolhrium)
он смещен на боковую поверхность теЛа.
Женская половая система имеет следующее строение. В задней
половине проглоттиды локализуется небольшой двулопастной яич-
ник. От перемычки, соединяющей лопасти, отходит яйцевод, в кото-
рый впадают влагалище с желточным протоком. Влагалище пред-
ставляет собой трубку, начинающуюся щелевидной порой на дне
полового атриума. Проксимальный конец влагалища вздут и функ-
ционирует как семяприемник. От желточных фолликулов, распола-
гающихся под тегументом, отходят тонкие желточные канальцы.
Сливаясь, они дают начало главному желточному протоку. Далее
по ходу яйцевода образуется небольшое расширение, в которое от-
крываются протоки многочисленных железок, — тельце Мелиса. На-
чинающаяся от тельца Мелиса матка образует несколько петель
и открывается наружу недалеко от полового атриума.
Половая система нерасчлененных цестод из отр. Caryophyllaei-
dea устроена сходным образом. В целом этот тип половой системы
почти полностью соответствует схеме строения органов размноже-
ния трематод. Если принять кажущуюся весьма убедительной гомо-
логизацию влагалища ленточных червей и Лаурерова канала со-
сальщиков (J. Smyth), то сходство это становится еще более полным.
Половая система Cyclophyllidea (рис. 6, В, Г) характеризуется
некоторыми особенностями. Количество семенников может сильно
варьировать — от 1—3 до 1000 и более. Семяпровод направляется
к половой клоаке, которая представляет собой углубление, располо-
женное на вершине сравнительно небольшого бугорка — полового
сосочка. Последний локализуется на боковой поверхности членика.
Отличия в строении женской половой системы гораздо заметнее.
Яичник, как правило, двухлопастной. Яйцевод на своем пути при-
нимает влагалищный и желточный протоки. Влагалище имеет вид
трубки с небольшим расширением — семяприемником на прокси-
мальном конце. Дистальный конец влагалища, открывается на дне
половой клоаки. Желточник Cyclophyllidea в отличие от псеудофил-
лид представлен более или менее компактной массой, расположен-
ной в задней части членика непосредственно за. яичником.
У цестод, так же,-как и у трематод, наблюдается зависимость
между степенью развития желточников и особенностями формиро-
вания личинок. У видов, имеющих многочисленные желточ'ные фол-
ликулы (Diphyllobothrium и др.), эмбриональное развитие личинок
осуществляется во внешней среде. В то же время у Cyclophyllidea,
которые обладают сравнительно небольшими желточниками, эмбри-
огенез и морфогенез личинки полностью завершаются еще во время
нахождения яйца в ма/ке.
Вслед за местом впадения желточного протока в яйцевод «скор-
луповые» железки образуют тельце Мелиса, от которого берет на-
чало матка. У всех Cyclophyllidea матка замкнута и не имеет само-
стоятельного отверстия. Форма матки не остается постоянной и ме-
няется в зависимости от «возраста» членика. В гермафродитных
проглоттидах матка представляет собой простое трубчатое образо-
15
ванне. По мере заполнения яйцами она сильно увеличивается в объ-
еме, чаще всего за счет появления боковых ветвей, количество кото-
рых— важный диагностический признак (с. 38). В некоторых слу-
чаях разросшаяся матка становится'мешковидной (сем. Hymenole-
pididae); иногда же ее полость разбивается на отдельные камеры,
превращающиеся в плотные фиброзные капсулы (р. Dipylidium,
р. Davaenia).
Половая система цестод, относящихся к отр. Trypanorhyncha
и Tetraphyllidea (рис. 6, Д), по особенностям строения занимает
промежуточное положение между двумя вышеописанными типами.
У них, как н у псеудофиллид, в женской половой системе сильно
развиты желточники. В то же время латеральное положение поло-
вой клоаки и, что самое главное, наличие слепо замкнутой матки
сближают их с циклофиллидами.
Очень своеобразное изменение в половой системе ленточных чер-
вей, связанное с усилением половой продуктивности, заключается
в увеличении (чаще всего удвоении) числа половых аппаратов
в каждом членике (см. рис. 19,В). Очень редко у цестод встреча-
ется раздельнополость, которая может возникать как в пределах
одной стробилы (Gynandrotaenia), так и касаться всей стробилы
в целом (Dioicocestus).
Особенности питания и обмена веществ цестод
Характерная особенность ленточных червей, как уже отмеча-
лось ранее, заключается в полной утрате этими паразитами пище-
варительной системы. Ни у личиночных стадий, ни тем более
у половозрелых особей не удается обнаружить никаких следов по-
следней. Таким образом, для поступления пищевых веществ в тело
червя остается один единственно возможный путь — через покровы.
По последним данным тегумент цестод содержит большое коли-
чество ферментов. Это говорит об активной роли тегу мента в раз-
личных метаболических процессах. По мнению Рида с соавт. (Read
et al.), микроворсинки слизистой и микротрихии тесно переплета-
ются друг с другом, что обусловливает тесный контакт тканей па-
разита и хозяина. Долгое время считалось, что цестоды способны
использовать только продукты пристеночного пищеварения живот-
ного-хозяина и не обладают собственными гидролитическими фер-
ментами. В последние годы было, однако, установлено, что ленточ-
ные черви далеко не полностью утратили ферментативные системы,
обеспечивающие переваривание белков и углеводов. Они не только
используют готовые продукты расщепления, но и сами в состоянии
активно их разлагать до аминокислот и сахаров.
Способность к самостоятельному пищеварению у разных видов
цестод выражена далеко не в одинаковой степени. Изучение про-
теиназ показало, что эти различия определяются главным образом
интенсивностью обменных процессов животного хозяина. НаиболЬ'
шей протеолитической активностью обладают цестоды, обитающие
в птицах, наименьшей — паразиты рыб. Значительные вариации ак-
16
тнвности гидролитических ферментов наблюдаются и на разных фа-
зах жизненного цикла ленточных червей. Протеиназы плероцерко-
идов Triaenophorus из печени промежуточного хозяина — налима,
значительно более активны, чем протеиназы взрослых особен * нз
кишечника щуки. Это, по-видимому, может служить примером тон-
кой физиологической адаптации паразитов к условиям обитания.
Половозрелые Triaenophorus не только сами переваривают белки,
но в большом количестве используют аминокислоты, освобождаю-
щиеся в результате ферментативной деятельности животного хозяи-
на. Личинки, поселяющиеся в тканях, могут использовать белки
только за счет активности собственных протеиназ.
Сорбция пищевых веществ у цестод осуществляется, по-видимо-
му, несколькими различными способами. Важная роль, по мнению
ряда исследователей, принадлежит пиноцитозу. Поступление неко-
торых сахаров и аминокислот в организм червей против градиента
концентрации позволяет предполагать существование механизмов
активного транспорта. Об этом же говорит и высокая ферментатив-
ная активность тегумента. Наконец, возможна и простая диффузия
некоторых химических соединений в тело паразита. Именно таким
способом цестоды получают пуриновые и пиримидиновые основания-
Спектр поглощаемых ленточными червями пищевых веществ
очень широк. Важная роль, несомненно, принадлежит углеводам,
к дефициту которых в окружающей среде паразиты особенно чув-
ствительны. Установлено, что уменьшение количества сахаров в ди-
ете хозяина приводит к резкому снижению веса обитающих в нем
паразитов. Значительная 'часть углеводов, по-видимому, немедленно
утилизируется червями для покрытия энергетических потребностей.
Обнаружение у многих цестод почти полного набора ферментов
пути Эмбден-Мейергофа говорит о том, что превращение глюкозы
у них осуществляется гликолитическим путем без участия кислоро-
да. Данные о наличии у цестод цикла Кребса не столь однозначны.
Если оксидативное расщепление глюкозы у -них и имеет место, то,
несомненно, оно играет подчиненную роль. Биологически это легко
объяснимо. Ленточные черви обитают в условиях острого дефицита
кислорода, хотя какое-то количество его и поступает в просвет ки-
шечника из слизистой за счет диффузии. Цестоды, вступающие
в тесный контакт с внутренней поверхностью кишечника, по-видимо-
му, могут использовать этот кислород. Более того, у ряда видов бы-
ли обнаружены и некоторые важнейшие ферменты цитохромной
системы дыхания, в частности цитохромоксидазы. Однако дыхатель-
ная функция цитохромов у ленточных червей многими подвергается
сомнению.
В энергетическом балансе ленточных червей, как и у некоторых
паразитических простейших, большое значение имеет и пентозо-фос-
фатный путь превращения глюкозы. Для Echinococcus, например,
было показано, что таким способом метаболизируется около 20%
Углеводов. Значительное количество глюкозы используется парази-
тами для синтеза гликогена — главного запасного энергетического
вещества, которое накапливается в клетках паренхимы. Наличие
17
больших энергетических резервов играет важную биологическую
роль. При малейшей нехватке экзогенных углеводов цестоды начи-
нают расщеплять собственные углеводы.
Большое значение в питании ленточных червей имеют белки,
частично, как уже отмечалось выше, перевариваемые хозяином,
.а частично самим паразитом. Кроме того, черви в состоянии исполь-
зовать аминокислоты, сецернируемые хозяином в просвет кишечни-
ка. Только за счет этого источника цестоды могут удовлетворять
свои потребности в азотистых продуктах. Во всяком случае исклю-
чение протеинов из пищевого рациона хозяина не сказывается на
паразитах столь катастрофически, как это имеет место в случае
с углеводами.
Значительная часть поступающих в организм паразитов амино-
кислот используется для построения белков. Синтезу последних, по-
видимому, часто предшествуют процессы трансаминирования. Од-
нако активность-трансамнназ у ленточных червей значительно ниже,
чем у животных хозяев. Объясняется это тем, что паразиты обита-
ют в среде, богатой самыми разнообразными аминокислотами,
и, следовательно, могут удовлетворять свои потребности без исполь-
зования сложных синтетических процессов.
Метаболизм липидов у цестод изучен еще недостаточно. Липиды,
обнаруживаемые в теле червей, имеют, по-видимому, различное
происхождение. Часть из них синтезируется паразитами de novo из
получаемых ими продуктов пищеварения. Другой источник липи-
дов— обменные процессы. Конечными продуктами превращения
глюкозы, так же как и у других гельминтов, обитающих в условиях
недостатка кислорода, являются жирные кислоты, которые затем
трансформируются в нейтральные жиры. Последние либо накапли-
ваются в организме человека, либо выводятся наружу через кана-
лы выделительной системы. Цестоды так ж;е, как и трематоды, по-
видимому, практически не могут использовать жиры в качестве
энергетического вещества.
У многих видов ленточных червей какая-то часть жиров все-таки
утилизируется, но не самими половозрелыми особями, а отрождае-
мыми ими личинками следующего поколения. Это происходит в тех
случаях, когда липиды в больших количествах накапливаются в
желточниках (Diphyllobothrium latum, Triaenophorus и др.). Вместе
с желточными клетками они входят в состав сложного яйца
и в дальнейшем служат источником энергии для-зародыша, разви-
тие которого протекает во внешней среде.
Размножение и жизненные циклы ленточных червей
Размножение и эмбриональное развитие
Копуляция и формирование сложного яйца. В большинстве слу-
чаев у ленточных червей наблюдается нормальная копуляция. При
этом возможны самые различные комбинации. Если в кишечнике
18
хозяина обитают две или более особей, имеет место перекрестное
оплодотворение. Однако часто совершается и самооплодотворение
как в пределах одной стробилы — между различными проглоттида-
ми, так и в одном членике, когда цнррус вводится во влагалище
той же самой проглоттиды.
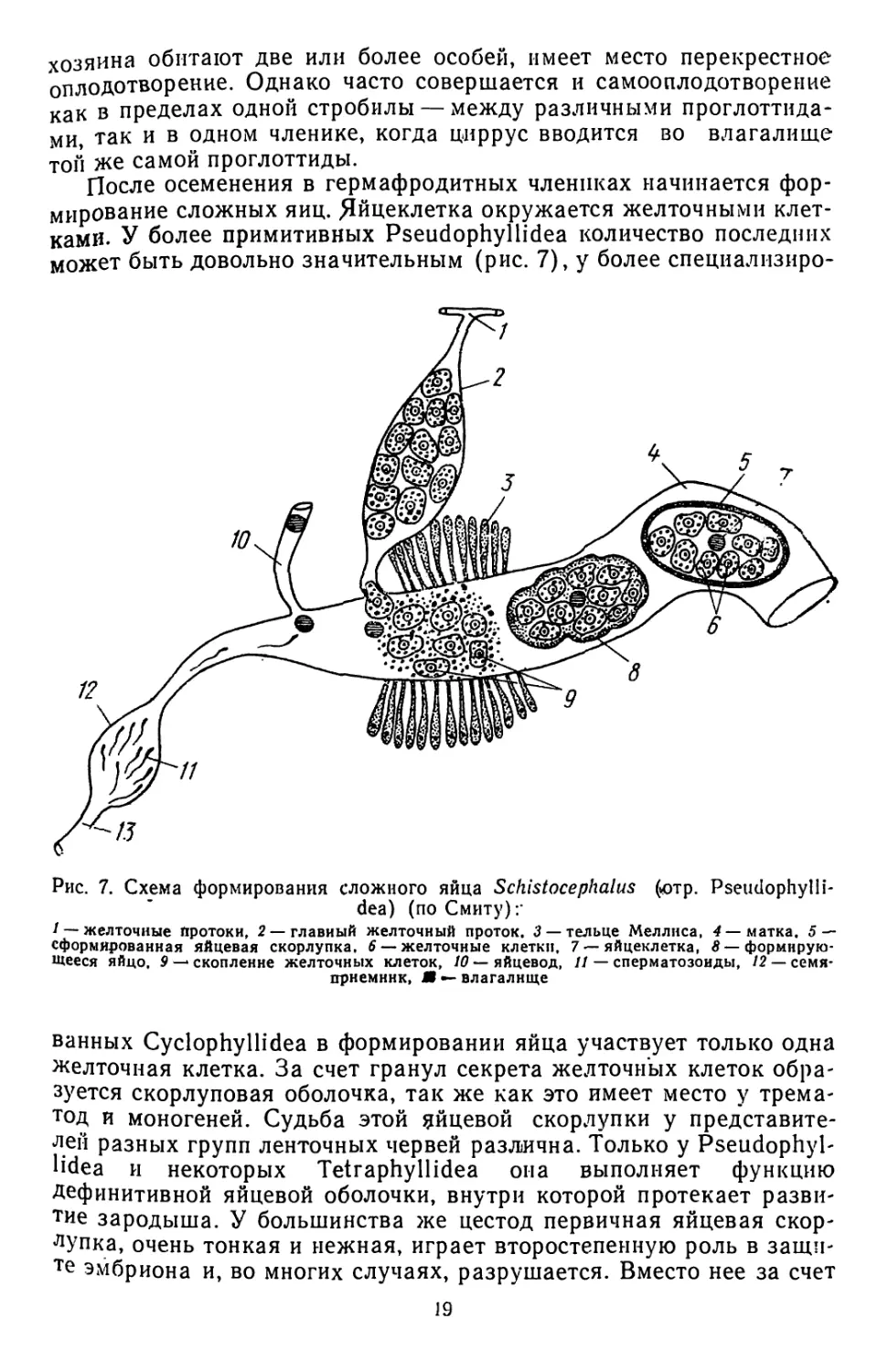
После осеменения в гермафродитных члениках начинается фор-
мирование сложных яиц. Яйцеклетка окружается желточными клет-
ками. У более примитивных Pseudophyllidea количество последних
может быть довольно значительным (рис. 7), у более специализиро-
Рис. 7. Схема формирования сложного яйца Schistocephalus (ютр. Pseudophylli-
dea) (по Смиту):'
/ — желточные протоки, 2 —главный желточный проток. 3 — тельце Меллиса, 4 — матка. 5 —
сформированная яйцевая скорлупка. 6 — желточные клетки, 7 — яйцеклетка, Л—формирую-
щееся яйцо, 9 — скопление желточных клеток, 10 — яйцевод, II — сперматозоиды, 12 — семя-
приемник, Я •- влагалище
ванных Cyclophyllidea в формировании яйца участвует только одна
желточная клетка. За счет гранул секрета желточных клеток обра-
зуется скорлуповая оболочка, так же как это имеет место у трема-
тод и моногеней. Судьба этой яйцевой скорлупки у представите-
лен разных групп ленточных червей различна. Только у Pseudophyl-
lidea и некоторых Tetraphyllidea она выполняет функцию
Дефинитивной яйцевой оболочки, внутри которой протекает разви-
тие зародыша. У большинства же цестод первичная яйцевая скор-
лупка, очень тонкая и нежная, играет второстепенную роль в защи-
те эмбриона и, во многих случаях, разрушается. Вместо нее за счет
19
клеток зародыша развиваются мощные эмбриональные оболочки
(с. 21).
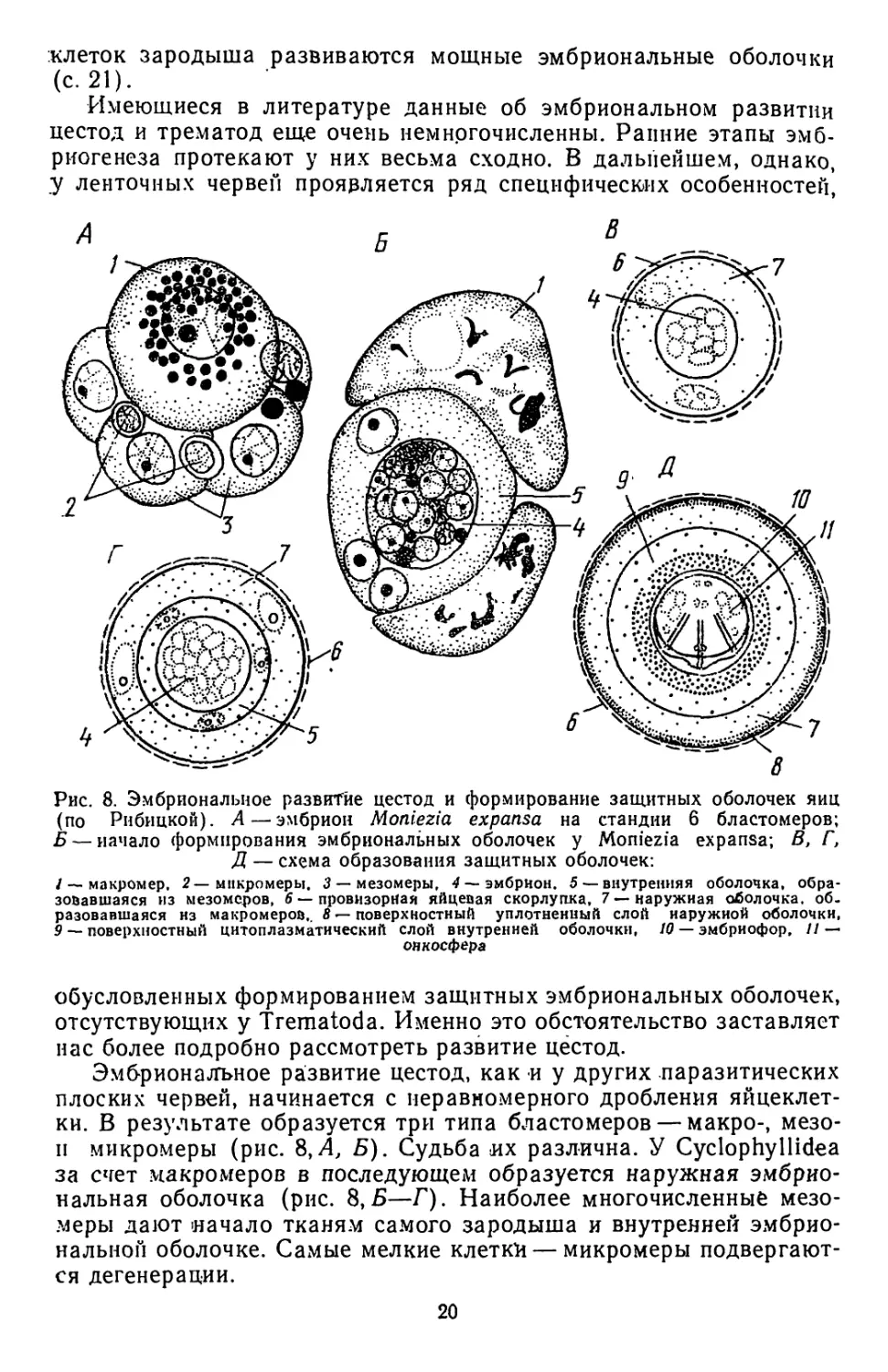
Имеющиеся в литературе данные об эмбриональном развитии
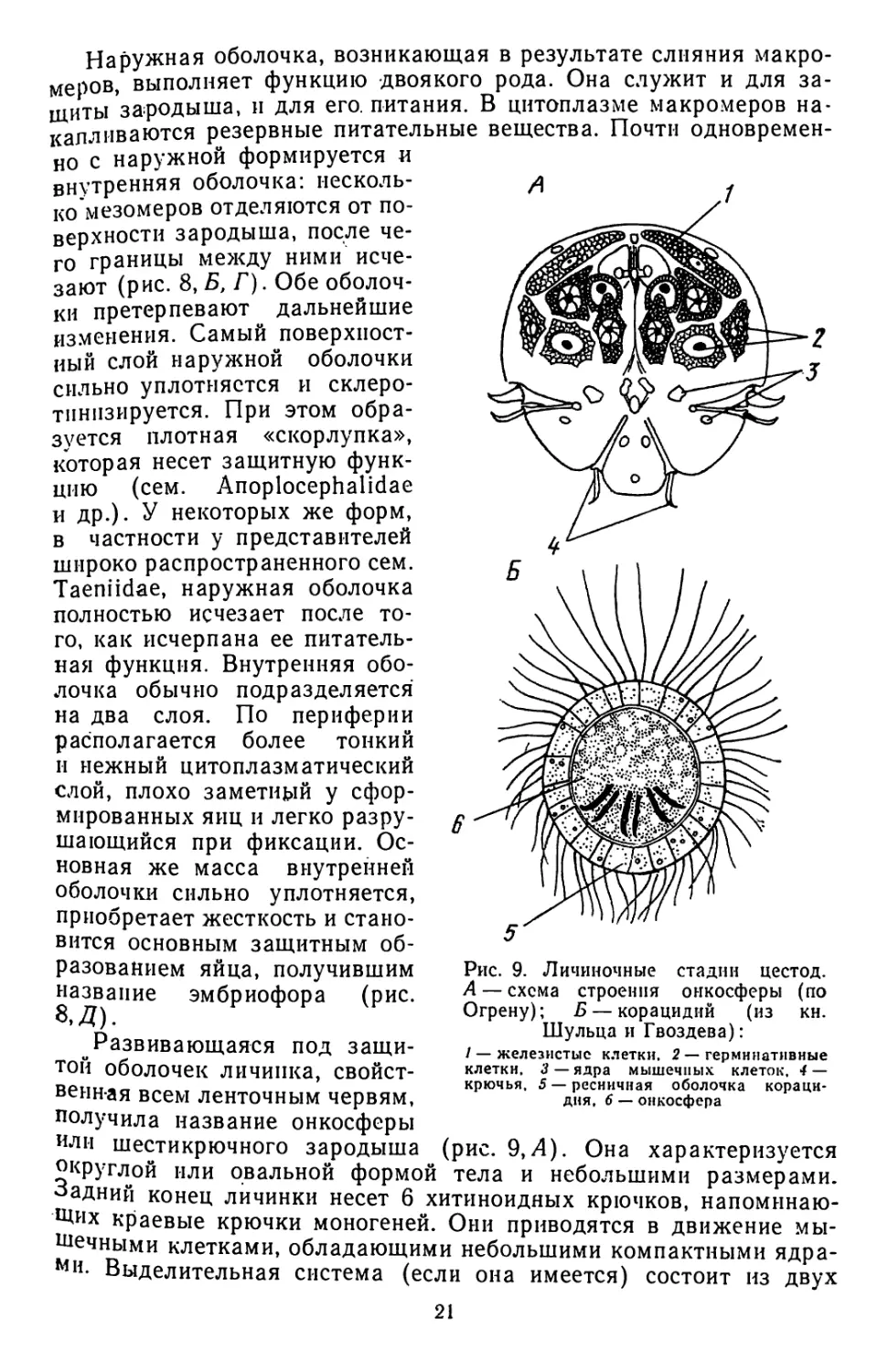
цестод и трематод еще очень немногочисленны. Ранние этапы эмб-
риогенеза протекают у них весьма сходно. В дальнейшем, однако,
у ленточных червей проявляется ряд специфических особенностей',
Рис. 8. Эмбриональное развитие цестод и формирование защитных оболочек яиц
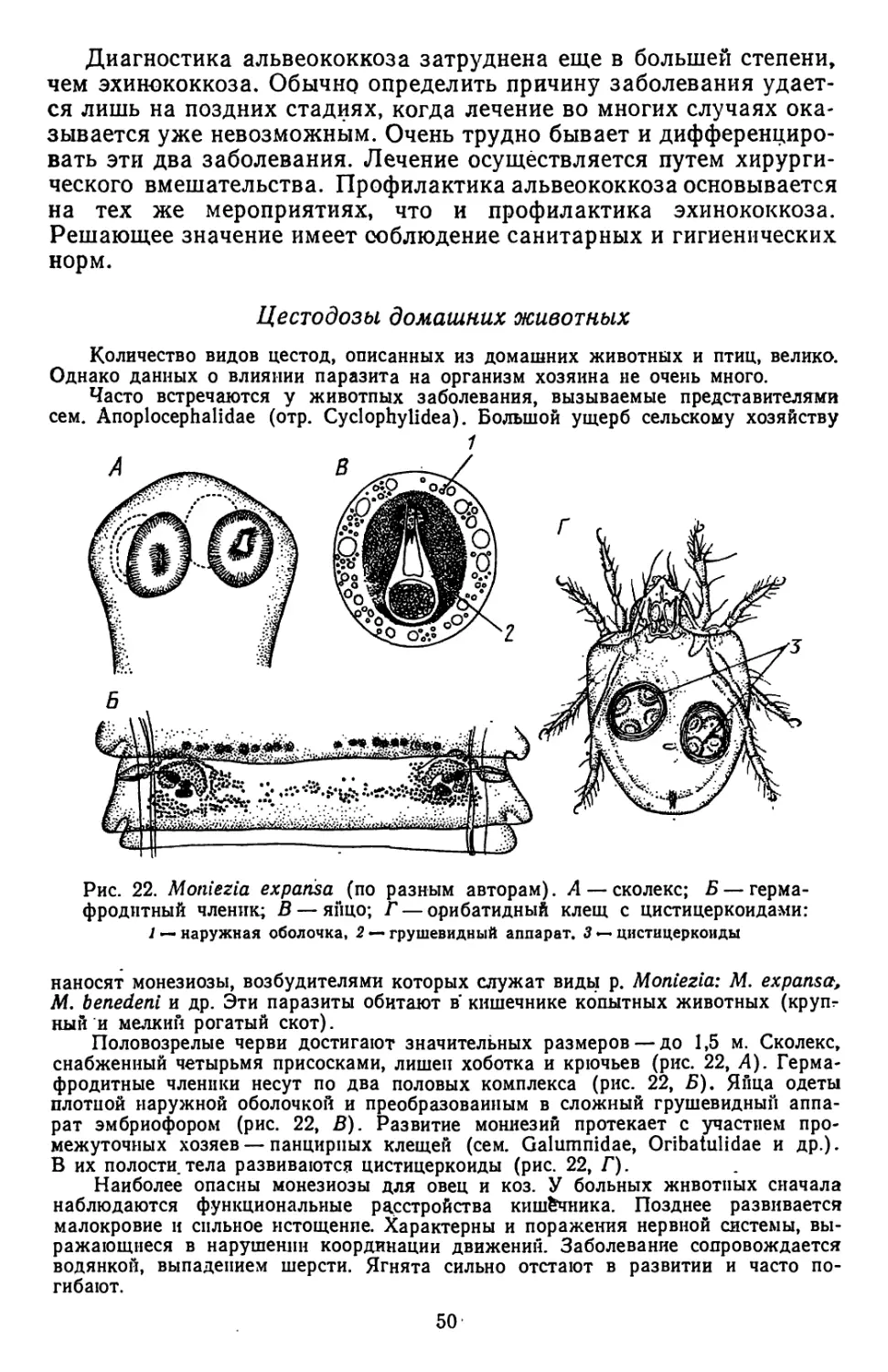
(по Рнбицкой). А—эмбрион Moniezia expansa на стандии 6 бластомеров;
Б — начало формирования эмбриональных оболочек у Moniezia expansa; В, Г,
Д — схема образования защитных оболочек:
/ — макромер. 2—микромеры. 3 — мезомеры, 4 — эмбрион. 5 —внутренняя оболочка, обра-
зовавшаяся из мезомеров, 6 — провизорная яйцевая скорлупка, 7 — наружная оболочка, об-
разовавшаяся нз макромеров,. 8 — поверхностный уплотненный слой наружной оболочки,
9 — поверхностный цитоплазматический слой внутренней оболочки, 10 — эмбриофор, // —
онкосфера
обусловленных формированием защитных эмбриональных оболочек,
отсутствующих у Trematoda. Именно это обстоятельство заставляет
нас более подробно рассмотреть развитие цёстод.
Эмбриональное развитие цестод, как и у других паразитических
плоских червей, начинается с неравномерного дробления яйцеклет-
ки. В результате образуется три типа бластомеров — макро-, мезо-
п микромеры (рис. 8,Л, Б). Судьба их различна. У Cyclophyllidea
за счет макромеров в последующем образуется наружная эмбрио-
нальная оболочка (рис. 8,Б—Г). Наиболее многочисленные мезо-
меры дают начало тканям самого зародыша и внутренней эмбрио-
нальной оболочке. Самые мелкие клетки — микромеры подвергают-
ся дегенерации.
20
Наружная оболочка, возникающая в результате слияния макро
меров, выполняет функцию двоякого рода. Она служит и для за
щиты зародыша, и для его. питания. В цитоплазме макромеров на
калливаются резервные питательные вещества. Почти одновремен
но с наружной формируется и
внутренняя оболочка: иесколь-
ко мезомеров отделяются от по-
верхности зародыша, после че-
го границы между ними исче-
зают (рис. 8, Б, Г). Обе оболоч-
ки претерпевают дальнейшие
изменения. Самый поверхност-
ный слой наружной оболочки
сильно уплотняется и склеро-
тпнпзируется. При этом обра-
зуется плотная «скорлупка»,
которая несет защитную функ-
цию (сем. Anoplocephalidae
и др.). У некоторых же форм,
в частности у представителей
широко распространенного сем.
Taeniidae, наружная оболочка
полностью исчезает после то-
го, как исчерпана ее питатель-
ная функция. Внутренняя обо-
лочка обычно подразделяется
на два слоя. По периферии
располагается более тонкий
и нежный цитоплазматический
слой, плохо заметный у сфор-
мированных яиц и легко разру-
шающийся при фиксации. Ос-
новная же масса внутренней
оболочки сильно уплотняется,
приобретает жесткость и стано-
вится основным защитным об-
Личиночные стадии
цестод.
Рис. 9. _ ______
А — схема строения онкосферы (по
Огрену); Б — корацидий (из кн.
Шульца и Гвоздева):
1 — железистые клетки, 2 — герминативные
клетки, 3 — ядра мышечных клеток, 4 —
крючья, 5 — ресничная оболочка кораци-
дня, 6 — онкосфера
разованием яйца, получившим
название эмбриофора (рис.
и Развивающаяся под защи-
той оболочек личинка, свойст-
венная всем ленточным червям,
получила название онкосферы
или шестикрючного зародыша (рис. 9,Л). Она характеризуется
округлой или овальной формой тела и небольшими размерами,
•задний конец личинки несет 6 хитиноидных крючков, напоминаю-
щих краевые крючки моногеней. Они приводятся в движение мы-
шечными клетками, обладающими небольшими компактными ядра-
ми. Выделительная система (если она имеется) состоит из двух
21
протонефридиев. Переднее полушарие онкосферы содержит не-
сколько пар так называемых «эмбриональных» клеток: за их счет
формируется тело последующих личиночных стадий, а потом и зре-
лого червя. Ближе к самому переднему концу тела онкосферы рас-
полагаются крупные железистые клетки. Их секрет используется
личинкой в процессе миграции по телу промежуточного хозяина.
Эмбриональное развитие Pseudophyllidea изучено значительно
слабее. Наиболее существенные отличия касаются судьбы эмбри-
ональных оболочек. Формирование зародыша Pseudophyllidea про-
текает во внешней среде под защитой настоящей яйцевой скорлуп-
кщ (с. 19). Обособляющиеся от эмбриона макромеры дадот начало
не наружной оболочке, а особой мембране, заключающей в себе
и желточные клетки и эмбрион. Внутренней же оболочке высших
цестод у Pseudophyllidea соответствует развивающийся за счет по-
верхностно расположенных мезомеров своеобразный ресничный
покров, иногда называемый ресничным эмбриофором (К. Рибицка).
Он выполняет нс защитную, а локомоторную функцию (см. ниже).
Личиночная стадия, состоящая из довольно типичной онкосферы,
заключенной внутри цитоплазматической оболочки, снабженной
ресничками, получила название корацидия (рис. 9, Б).
Жизненные циклы ленточных червей
Наиболее примитивные жизненные циклы, по-видимому, у пред-
ставителей отр. Pseudophyllidea. Их развитие осуществляется с уча-
стием трех животных-хозяев: двух промежуточных и одного дефи-
нитивного. Кроме того, они в большинстве случаев обладают сво-
бодноживущей расселительной личинкой и теснейшим образом
связаны с водной средой. Развитие яиц большинства Pseudpphyl-
lidea осуществляется во внешней среде. Под защитой скорлуповой
оболочки формируется корацидий, который, достигнув зрелости,
выходит в воду, откидывая крышечку яйца.
Ресничная оболочка корацидия имее! вид толстого цитоплазма-
тического слоя с большим количеством вакуолей и разнообразных
включений — главным образом каплями жира и гранулами гликоге-
на. Ее наружная поверхность несет многочисленные реснички, бие-
ние которых обеспечивает плавное и равномерное передвижение
личинки в толще воды. Внутренняя поверхность этой оболочки об-
разует глубокие щелевидные впячивания. В них заходят соответст-
вующие им по форме выросты мембраны, одевающей онкосферу
(рис. 10. Д). Последняя при этом может слегка поворачиваться из
стороны в сторону, сокращаться, расправляться. Эти движения ни-
как не отражаются на форме и характере поведения корацидия
в целом. Сроки жизни личинки ограничены и определяются количе-
ством запасных питательных веществ, полученных ею в процессе
формирования.
Для дальнейшего развития корацидий должен попасть в орга-
низм первого промежуточного хозяина. Таковыми в большинстве
случаев являются ракообразные — главным образом мелкие планк-
?2
тонные формы (р. Cyclops, р. Diaptomus и др.). Рачки, привлеченные
движениями личинок, охотно проглатывают их. В пищеварительной
системе рачка ресничная оболочка корацидия разрушается и пере-
варивается. Освободив-
шаяся онкосфера быстро
пробуравливает стенку
кишечника и мигрирует в
полость тела хозяина.
Здесь онкосфера превра-
щается в следующую ли-
чиночную стадию — про-
церкоид (рис. 11,4).
Личинка в хозяине ра-
стет, теряет более или ме-
не^ правильную сфериче-
скую форму, вытягивает-
ся’в длину. Ее задний по-
люс, несущий крючья, от-
деляется от остального
тела узкой перетяжкой.
Этот участок получил наз-
вание церкомера. Слож-
ной перестройке подвер-
гаются покровы процерко-
ида. В самом начале на
них образуются многочис-
ленные выросты, пред-
ставляющие собой настоя-
щие микроворсинки (рис.
10, Г, Д). Позднее, однако,
они замещаются микро-
трихпями. Через несколь-
ко дней после перехода к
паразитическому образу
жизни личинка приобре-
тает дефинитивные покро-
вы, сохраняющиеся .уже
на протяжении всей жиз-
ни червя. К моменту за-
вершения перестройки по-
кровов в поверхностных
слоях паренхимы появля-
ются многочисленные из-
Рис. 10. Схема формирования покровов
Triaenophorus в процессе превращения онко-
сферы в процеркоид (по Куперману). Д — уча-
сток корацидия; Б — поверхность онкосферы
в кишечнике рачка; В— поверхность процер-
коида в первые сутки развития; Г, Д — фор-
мирование микроворсинок; Е — закладка мик-
ротрихий; Ж — участок поверхности тела
сформированного процеркоида:
/ — ресничный покров корацидия, 2 — плотная мем-
брана. одевающая онкосферу, 3 — микроворсинкн,
4 — микротрихии
вестковые тельца. Увеличивается количество протонефридиев.
Между собирательными каналами образуются анастомозы, в ре-
зультате чего возникает густая сеть (рис. 11, Д). На переднем кон-
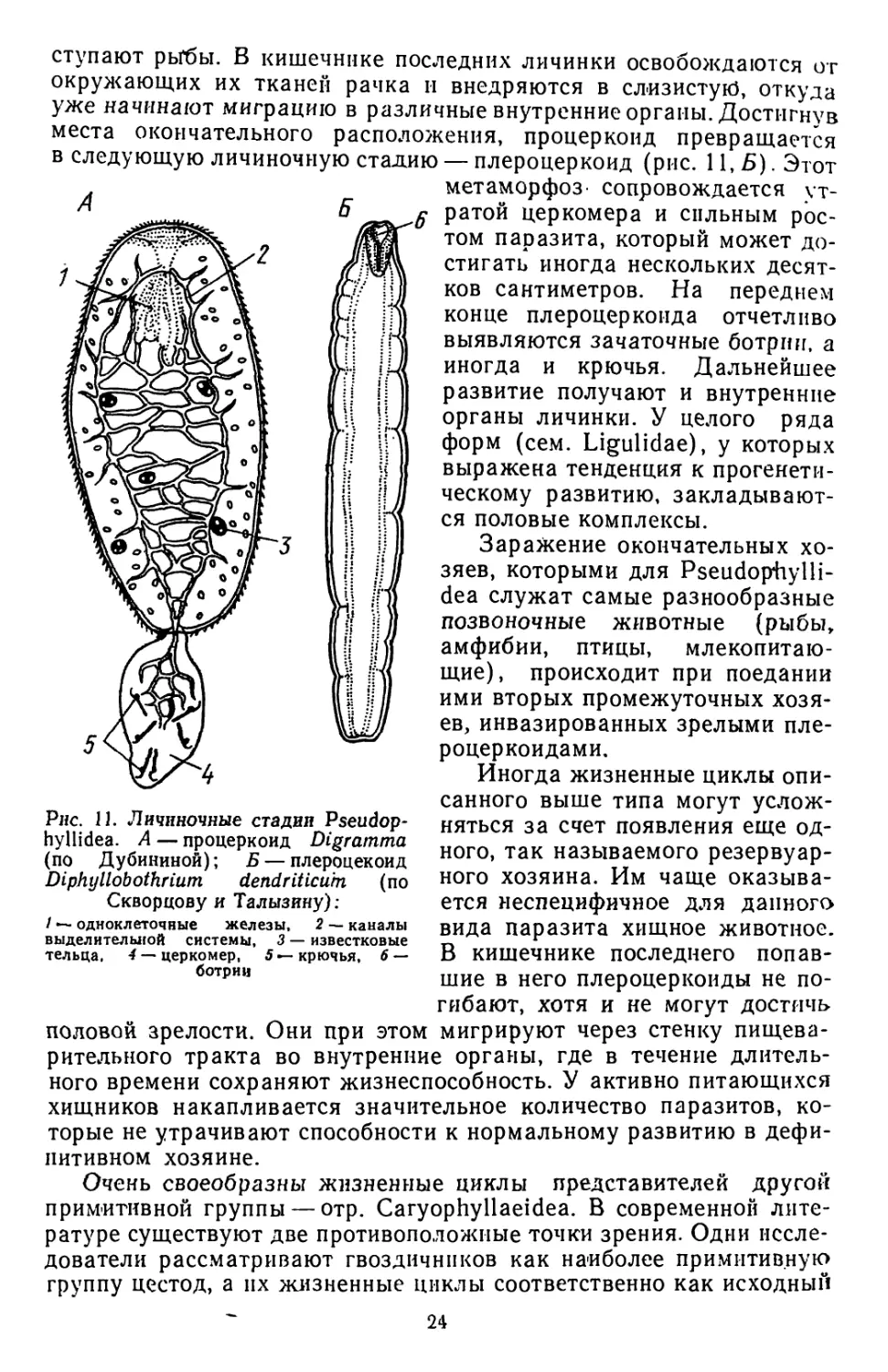
це тела личинки развиваются одноклеточные железы.
Дальнейшее развитие паразитов становится возможным только
во втором промежуточном хозяине, в роли которого чаще всего вы-
23
ступают рьГбы. В кишечнике последних личинки освобождаются от
окружающих их тканей рачка н внедряются в слизистую, откуда
уже начинают миграцию в различные внутренние органы. Достигнув
места окончательного расположения, процеркоид превращается
в следующую личиночную стадию — плероцеркоид (рис. 11,5). Этот
Рнс. 11. Личиночные стадии Pseudop-
hyllidea. А — процеркоид Digramma
(по Дубининой); Б — плероцекоид
Diphyllobothrium dendriticu'tn (по
Скворцову и Талызину):
/ — одноклеточные железы, 2 — каналы
выделительной системы, 3 — известковые
тельца, 4 — церкомер, 5 — крючья, 6 —
ботрии
метаморфоз- сопровождается ут-
ратой церкомера и сильным рос-
том паразита, который может до-
стигать иногда нескольких десят-
ков сантиметров. На переднем
конце плероцеркоида отчетливо
выявляются зачаточные ботрии, а
иногда и крючья. Дальнейшее
развитие получают и внутренние
органы личинки. У целого ряда
форм (сем. Ligulidae), у которых
выражена тенденция к прогенети-
ческому развитию, закладывают-
ся половые комплексы.
Заражение окончательных хо-
зяев, которыми для Pseudophylli-
dea служат самые разнообразные
позвоночные животные (рыбы,
амфибии, птицы, млекопитаю-
щие), происходит при поедании
ими вторых промежуточных хозя-
ев, инвазированных зрелыми пле-
роцеркоидами.
Иногда жизненные циклы опи-
санного выше типа могут услож-
няться за счет появления еще од-
ного, так называемого резервуар-
ного хозяина. Им чаще оказыва-
ется неспецифичное для данного
вида паразита хищное животное.
В кишечнике последнего попав-
шие в него плероцеркоиды не по-
гибают, хотя и не могут достичь
половой зрелости. Они при этом мигрируют через стенку пищева-
рительного тракта во внутренние органы, где в течение длитель-
ного времени сохраняют жизнеспособность. У активно питающихся
хищников накапливается значительное количество паразитов, ко-
торые не утрачивают способности к нормальному развитию в дефи-
нитивном хозяине.
Очень своеобразны жизненные циклы представителей другой
примитивной группы — отр. Caryophyllaeidea. В современной лите-
ратуре существуют две противоположные точки зрения. Одни иссле-
дователи рассматривают гвоздичников как наиболее примитивную
группу цестод, а их жизненные циклы соответственно как исходный
24
тип развития этих паразитов. Другие же считают отр. Caryophyllaei-
dea весьма специализированной группой, многие особенности кото-
рой носят явно вторичный характер.
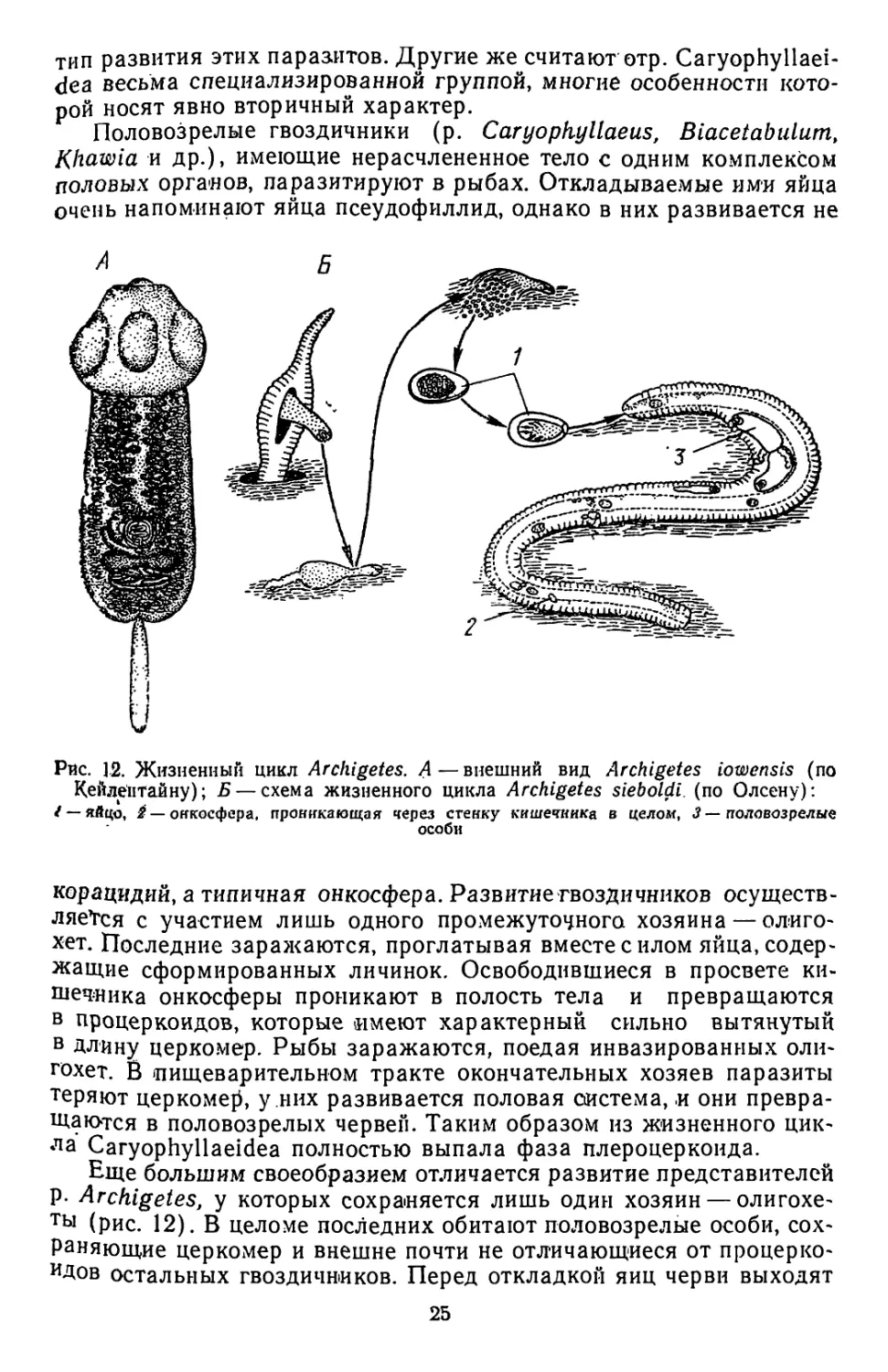
Половозрелые гвоздичники (р. Caryopfiyllaeus, Biacetabulum,
Khawia и др.), имеющие нерасчлененное тело с одним комплексом
половых органов, паразитируют в рыбах. Откладываемые ими яйца
очень напоминают яйца псеудофиллид, однако в них развивается не
Рис. 12. Жизненный цикл Archigetes. А — внешний вид Archigetes iowensis (по
Кейлептайну); Б — схема жизненного цикла Archigetes sieboldi. (по Олсену):
/ — яйцо, ^ — онкосфера, проникающая через стенку кишечника в целом, 3 — половозрелые
особи
корацидий, а типичная онкосфера. Развитие гвоздичников осуществ-
ляе*гся с участием лишь одного промежуточного хозяина — олиго-
хет. Последние заражаются, проглатывая вместе с илом яйца, содер-
жащие сформированных личинок. Освободившиеся в просвете ки-
шечника онкосферы проникают в полость тела и превращаются
в процеркоидов, которые имеют характерный сильно вытянутый
в длину церкомер. Рыбы заражаются, поедая инвазированных оли-
гохет. В пищеварительном тракте окончательных хозяев паразиты
теряют церкомер, у них развивается половая система, и они превра-
щаются в половозрелых червей. Таким образом из жизненного цик-
ла Caryophyllaeidea полностью выпала фаза плероцеркоида.
Еще большим своеобразием отличается развитие представителей
Р- Archigetes, у которых сохраняется лишь один хозяин — олигохе-
ты (рис. 12). В целоме последних обитают половозрелые особи, сох-
раняющие церкомер и внешне почти не отличающиеся от процерко-
идов остальных гвоздичников. Перед откладкой яиц черви выходят
25
во внешнюю среду через разрыв стенки тела хозяина. Завершив
кладку, они погибают.
Некоторые современные исследователи (Н. И. Демшин, Р. С. Шульц ц
Е. В. Гвоздев и др.) склонны видеть в архигетесах примитивные формы, сохра-
нившие исходный для всех цестод тип жизненного цикла. Олигохеты же рассмат-
риваются ими как филогенетически первичные хозяева всех ленточных червей.
Согласно другой точке зрения, кажущейся нам более обоснованной, Archigetes —
это классический пример иеотенического развития, когда личиночная стадия
(в данном случае процеркоид) приобрела способность к половому размножению
(R. Leuckart, W. Wisniewsky, J. Baer, H. Stunkard и др.). Вторая точка зрения
получила и прямое экспериментальное подтверждение. При скармливании некото-
рых архигетесов рыбам в последних удалось вырастить червей, относящихся к
р. Biacetabulum и Elaridacris. По-видимому, в отдельных случаях преждевремен-
ное развитие личинок еще не препятствует нормальному завершению жизненного
цикла.
Развитие цестод, относящихся к отр. Tetraphyllidea и Trypanor-
hyncha, несколько напоминает жизненные циклы Pseudophyllidea.
Чаще оно протекает с участием двух промежуточных хозяев. У не-
которых вадов сохраняется свободноживущая личинка типа корацп-
дия.
Наиболее специализированные паразиты несомненно объединя-
ются в отр. Cyclophyllidea. Это находит свое отражение и в особен-
ностях их развития.
Формирование личинки — онкосферы, как правило, протекает
еще в матке червя. Зародыш одевается эмбриональными оболоч-
ками (с. 20). Выпадение свободноплавающей личинки и развитие
мощного защитного образования — эмбриофора — позволило лен-
точным червям утратить зависимость от водоемов. Многие Cyclo-
phyllidea становятся настоящими сухопутными формами. Во внеш-
ней среде яйца длительное время сохраняют инвазионность и жпз-
неспособность^в фекальных массах, влажной почве, подстилке и т. д.
Дальнейшее развитие циклофилид осуществляется в организ-
ме промежуточного хозяина. У разных видов это самые различные
животные: и беспозвоночные (моллюскщ кольчецы, ракообразные,
насекомые), и представители всех классов позвоночных. В проме-
жуточном хозяине онкосфера освобождается от эмбриофора, про-
. никает через стенку кишечника в кровяное русло, а затем оседает
в каком-нибудь внутреннем органе. Здесь она дает начало новой ‘
личиночной стадии. Эти паразитические личинки Cyclophyllidea
называемые ларвоцистами, очень разнообразны и отличаются друг
от друга рядом признаков, позволяющих выделить несколько мор-
фологических типов.
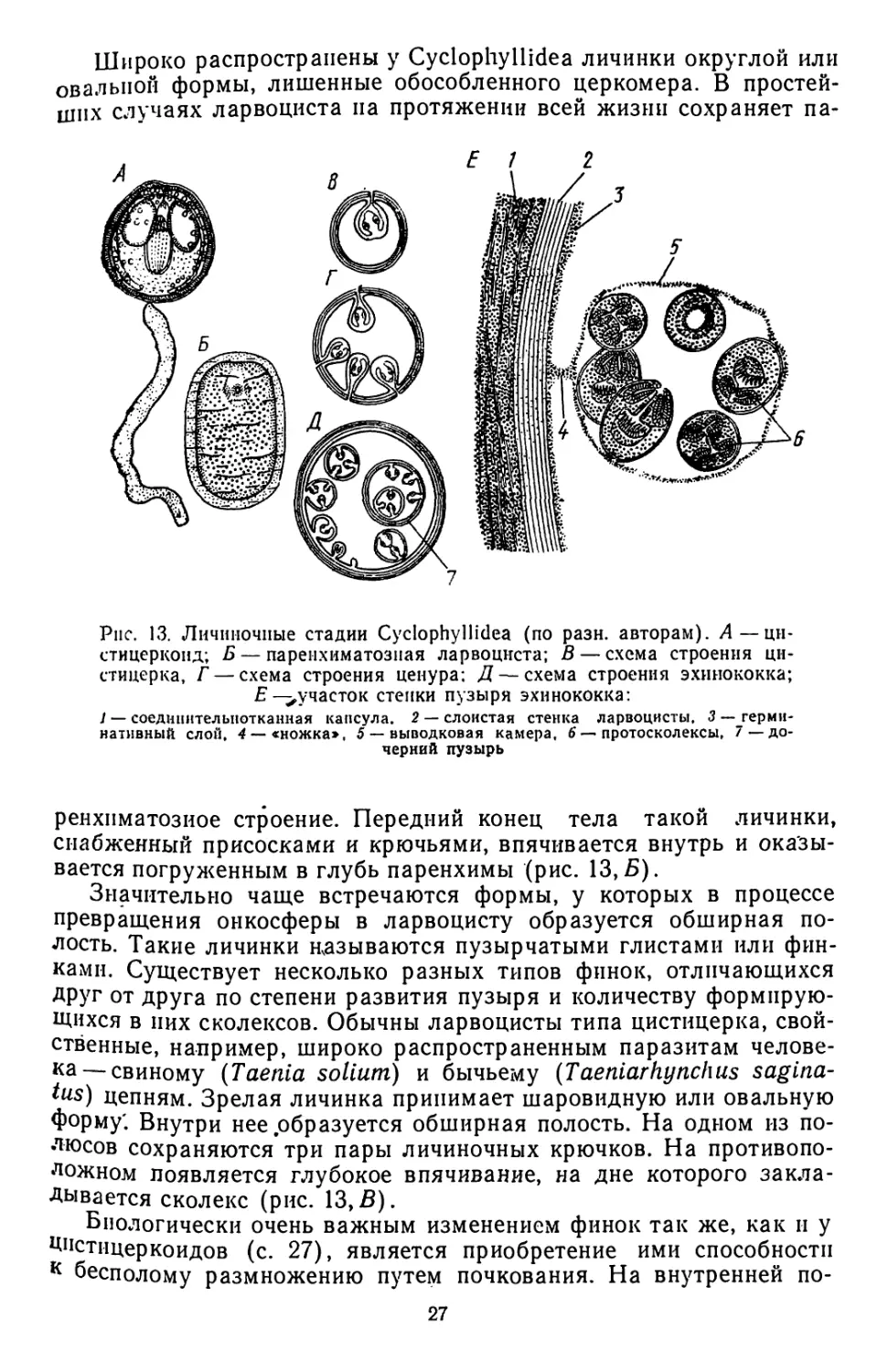
Наиболее примитивными личинками, по-видимому, являются
цистицеркоиды (рис. 13,Л), свойственные ряду семейств. Цистпцер-
коид состоит как бы из двух частей — длинного вытянутого хвосто-
вого придатка, снабженного тремя парами эмбриональных крючьев
(церкомера), и вздутой пузыревидной передней части. Самый пе-
редний конец, несущий зачаточный сколекс, ввернут в это расши-
рение. У некоторых представителей сем. Hymenolepididae от общего
церкомера отпочковываются несколько цистицеркоидов.
26
Широко распространены у Cyclophyllidea личинки округлой или
овальной формы, лишенные обособленного церкомера. В простей-
ших случаях ларвоциста па протяжении всей жизни сохраняет па-
Рис. 13. Личиночные стадии Cyclophyllidea (по рази, авторам). А—ци-
стицеркоид; Б — паренхиматозная ларвоциста; В — схема строения ци-
стицерка, Г — схема строения ценура; Д — схема строения эхинококка;
Е —^участок степки пузыря эхинококка:
1 — соединительнотканная капсула. 2 — слоистая стенка ларвоцисты, 3 — герми-
нативный слой, 4 — «ножка», 5 — выводковая камера, 6—протосколексы, 7—до-
черний пузырь
ренхпматозиое строение. Передний конец тела такой личинки,
снабженный присосками и крючьями, впячивается внутрь и оказы-
вается погруженным в глубь паренхимы (рис. 13, Б).
Значительно чаще встречаются формы, у которых в процессе
превращения онкосферы в ларвоцисту образуется обширная по-
лость. Такие личинки называются пузырчатыми глистами или фин-
ками. Существует несколько разных типов финок, отличающихся
друг от друга по степени развития пузыря и количеству формирую-
щихся в них сколексов. Обычны ларвоцисты типа цистицерка, свой-
ственные, например, широко распространенным паразитам челове-
ка— свиному (Taenia solium) и бычьему (Taeniarhynchus sagina-
tus) цепням. Зрелая личинка принимает шаровидную или овальную
форму'. Внутри нее .образуется обширная полость. На одном из по-
люсов сохраняются три пары личиночных крючков. На противопо-
ложном появляется глубокое впячивание, на дне которого закла-
дывается сколекс (рис. 13,В).
Биологически очень важным изменением финок так же, как и у
Цнстицеркоидов (с. 27), является приобретение ими способности
к бесполому размножению путем почкования. На внутренней по-
27
верхности пузыря закладывается не один, а множество сколексов,
каждый из которых в дальнейшем дает начало отдельной стробиле’
Таким образом, из одной онкосферы развивается большое число
паразитов. В наиболее простом случае это явление встречается
у Multiceps multiceps, ларвоцнсты которого получили название це-
нуров (рис. 13,Г).
Еще активнее почкование протекает у самых сложно устроенных
личинок—эхинококков (р. Echinococcus). Снаружи эти достигаю-
щие иногда крупных размеров ларвоцнсты (рис. 13, Д) одеты сое-
динительнотканной капсулой, образующейся за счет тканей хозяи-
на. Собственно стенки пузыря построены из нескольких отличаю-
щихся по своей структуре и функциям слоев. Самый внутренний
из них называется герминативным. Разрастаясь, он дает начало
небольшим замкнутым полостям — выводковым капсулам, в кото-
рых закладываются сколексы (их иногда называют протосколек-
сами). От герминативного слоя отпочковываются и бо-лее крупные
образования — дочерние пузыри. В них также образуются вывод-
ковые капсулы с протосколексамп. Таким образом, внутри эхинокок-
ка развивается огромное количество головок, каждая из которых
в дефинитивном хозяине даст начало отдельной стробиле.
Личинки тийа альвеококка также продуцируют большое число
дочерних пузырей. Последние, однако, отпочковываются не от внут-
ренней, а от наружной поверхности материнской особи. В резуль-
тате возникает довольно крупное скопление плотно прилегающих
друг к другу пузырей, в которых развиваются протосколексы.
В кишечнике дефинитивного хозяина под действием пищевари-
тельных ферментов сколекс выворачивается наружу. В зоне роста
начинается закладка члеников. У цестод не все тело личинки при-
нимает участие в образовании организма взрослой особи. Как пра-
вило, отбрасывается церкомер (у цистицеркоидов) или же участки,
в состав которых церкомер входит, в частности пузырь ларвоцист,
несущий на заднем полюсе личиночные крючья.
Гипотезы полизоичности и монозоичности цестод
Говоря о развитии цестод, необходимо помнить, что решение
вопроса о природе их жизненного цикла зависит от выбора той пли
иной гипотезы, объясняющей возникновение стробилы. В настоя-
щее время в литературе распространены две точки зрения. Соглас-
но одной из них членистое тело цестод возникло в результате пе-
доведенного до конца бесполого размножения и соответственно
представляет собой своеобразную колонию. Эта гипотеза получила
название гипотезы полизоичности. Сторонники другой гипотезы —
монозоичности, считают, что расчлененное тело ленточных червей не
колония, а целостный организм. «Членистость» же возникает вто-
рично как реакция на поселение в кишечнике хозяина (В. А. До-
гель). Увеличение количества половых комплексов рассматривает-
ся как проявление действия «закона большого числа яиц». В каче-
стве аргументов, подтверждающих правильность такого взгляда,
28
приводятся данные о строении нервной и выделительной систем.
Последние у цестод морфологически и функционально всегда еди-
ны. Однако подобные соображения на наш взгляд не имеют реша-
ющего значения, о чем будет сказано несколько ниже (с. 31).Вслед
за рядом исследователей (Б. Н. Беклемишев, Е. Н. Павловский,
д. В. Наумов и др.) мы принимаем гипотезу полизоичности и рас-
сматриваем процесс образования новых члеников в стробиле как
педоведенное до конца почкование.
Бесполое размножение в впде поперечного почкования встреча-
ется у некоторых турбеллярнй. Таким образом, нет ничего невоз-
можного в том, что цестоды сохранили свойство, присущее их дале-
ким предкам. Проглоттиды, пока они находятся в составе строби-
лы, действительно являются частью единого функционального,
целого. Очень часто, однако, картина резко меняется, когда члени-
ки отделяются от тела червя. У многих видов при этом отчетливо
проявляется «автономность» проглоттид.
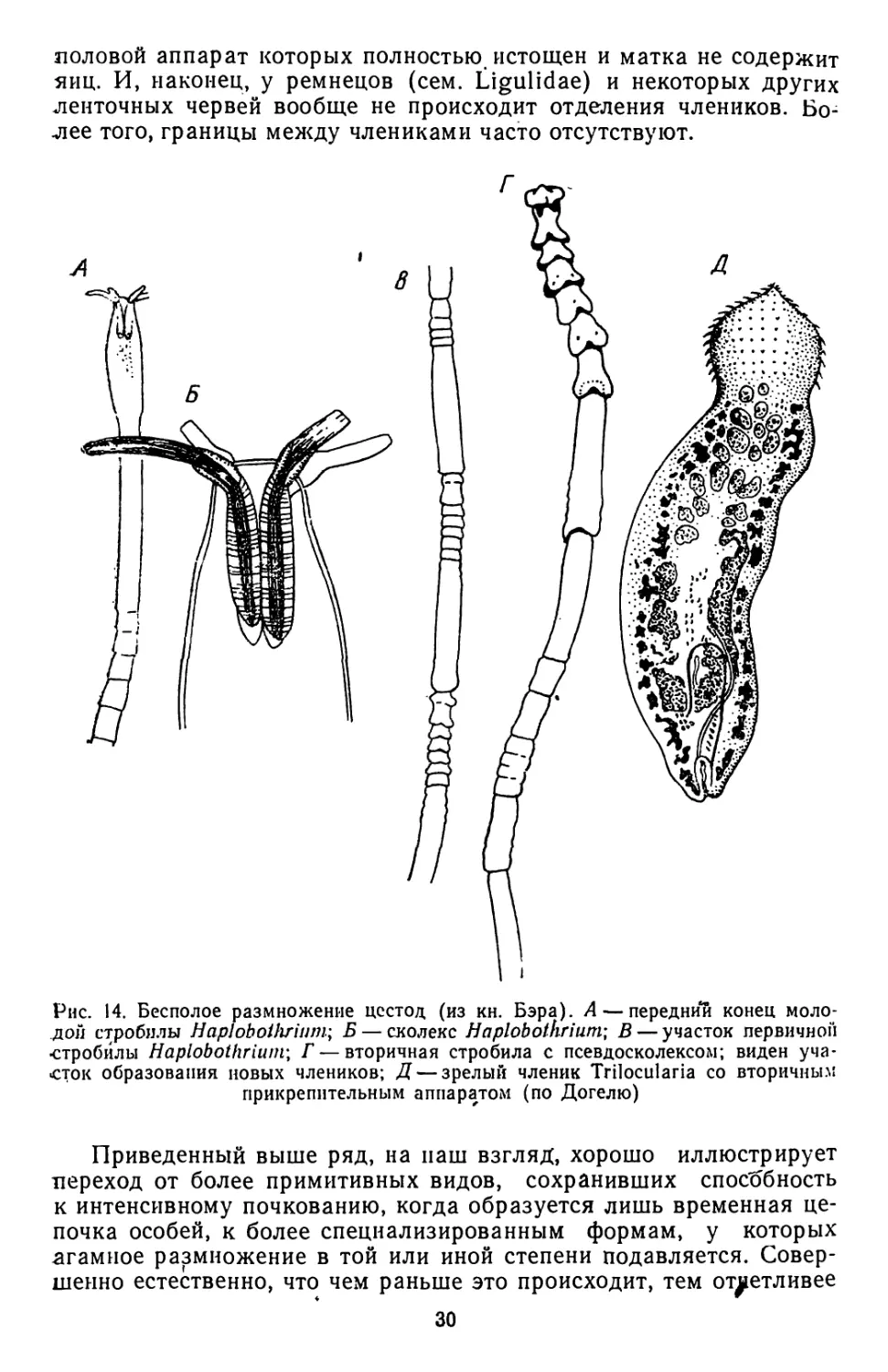
Наиболее ярким примером может служить р. Haplobothrium,
относящийся к отр. Pseudophyllidea1 (рис. 14). Haplobothrium па-
разитирует в очень древних и примитивных костных рыбах — Antia
calva. Образование члеников у этих цестод протекает по типу пара-
томпи. Их стробила очень напоминает цепочки особей турбеллярнй
р. Microstomum, размножающихся бесполым путем. Каждый чле-
ник еще до отделения от тела червя начинает образовывать вторич-
ные проглоттиды (рис. 14,В, Г). В дальнейшем вся стробила рас-
падается на серию более коротких «вторичных» стробил. На перед-
нем конце этих дочерних особей развивается новый сколекс-
Последний имеет иное строение, чем сколекс материнского организ-
ма. Он несет не хоботки (рис. 14, Л, Б), а ботрии (рис. 14,Г) и очень-
напоминает типичные сколексы псеудофиллид. Таким образом,
у Haplobothrium имеет место настоящее бесполое размножение.
Для многих видов характерно очень раннее отделение члеников
от стробилы, происходящее, задолго до того, как завершится созре-
вание их половой системы и начнется продукция яиц. Каждый чле-
ник при этом фактически превращается в самостоятельную особь,
у которой развиваются новые органы прикрепления, позволяющие
ей удерживаться в кишечнике хозяина {Echinobothrium, Trilocula-
ria и др.— рнс. 14, Д).
У целого’ряда Cyclophyllidea лишь- перезрелые, наполненные
яйцами членики, развитие которых завершилось в составе строби-
лы, отделяются от последней и сохраняют способность к движе-
нию. Они могут активно выходить через анальное отверстие хозяи-
на во внешнюю среду и ползать некоторое время по субстрату, рас-
сеивая при этом яйца {Echinococcus, Taeniarhynchus, Dipylidiurn
и др.). У свиного цепня {Taenia solium) от стробилы также отрыва-
ются только перезрелые членики, но уже совершенно неподвижные-
Многие Pseudophyllidea отторгают лишь отмирающие проглоттиды.
1 В настоящее время некоторые паразитологи склонны выделять самостоя-
тельно отр: Haplobothriadea.
29
половой аппарат которых полностью истощен и матка не содержит
яиц. И, наконец, у ремнецов (сем. Ligulidae) и некоторых других
ленточных червей вообще не происходит отделения члеников. Бо-
лее того, границы между члениками часто отсутствуют.
Рис. 14. Бесполое размножение цестод (из кн. Бэра). А — передний конец моло-
дой стробилы Haploboihrium; Б — сколекс Haplobothriufn; В — участок первичной
•стробилы Haplobothriuin; Г — вторичная стробила с псевдосколексом; виден уча-
сток образования новых члеников; Д — зрелыГ! членик Trilocularia со вторичным
прикрепительным аппаратом (по Догелю)
Приведенный выше ряд, на наш взгляд, хорошо иллюстрирует
переход от более примитивных видов, сохранивших способность
к интенсивному почкованию, когда образуется лишь временная це-
почка особей, к более специализированным формам, у которых
агамное размножение в той или иной степени подавляется. Совер-
шенно естественно, что чем раньше это происходит, тем отчетливее
30
выражена высокая степень интеграции возникающего «сложного»
организма, проявляющаяся в целостности выделительной и нерв-
ной систем.
Более серьезные возражения против теории полизоичНости мо-
гут быть связаны с широким распространением видов, обладаю-
плохо выраженной расчлененностью пли вообще иерасчленен-
ных, в отр. Pseudophyllidea, сохранившем множество примитивных
признаков. В то же время у наиболее специализированного отр.
Cyclophyllidea подобное явление никогда не встречается, хотя, ка-
залось бы, должно быть наоборот.
Объяснение этому кажущемуся противоречию, по-вндпмомуг
нужно искать в особенностях строения половой системы и связан-
ных с ними различиями в способах выведения яиц во внешнюю сре-
ду. Представители отр. Pseudophyllidea обладают трубчатой мат-
кой, открывающейся наружу самостоятельным отверстием. Проду-
цируемые паразитами яйца все время выделяются в просвет
кишечника и оттуда попадают во внешнюю среду. Таким образом,
биологическая необходимость в постоянном отторжении члеников
от стробилы у псеудофиллид отсутствует. Весьма вероятно, что
именно это обстоятельство позволило Pseudophyllidea встать на
путь повышения уровня целостности стробилы. Подобная тенден-
ция отчетливо проявляется у большинства колониальных организ-
мов и ведет к подавлению отдельных особей, которые зачастую
теряют свою индивидуальность, становясь частью интегрированной
сложной системы. Не представляют собой исключения из общего
правила и ленточные черви. Нерасчлененные Ligula и др. — это все
те же колонии, в которых границы между отдельными, входящими
в их состав особями (проглоттидами) полностью исчезли. У таких
наиболее специализированных видов вообще уже трудно, говорить,
о «почковании». Половые органы у них обособляются очень рано
и практически одновременно еще на стадии плероцеркоида, что дол-
жно рассматриваться как выражение склонности к прогенетическо-
му развитию, свойственной многим паразитам.
От некогда существовавшего бесполого размножения у нерас-
члененных ремнецов (р. Ligula, р. Digramma) остается лишь мно-
жественная закладка половых комплексов. Сохранение этого по-
следнего проявления очень древней особенности ленточных червей
обусловлено, по-видимому, действием уже неоднократно упоминав-
шегося «закона большого числа яиц». У'паразитических организ-
мов естественный отбор поддерживает существование видов с по-
вышенной плодовитостью. Таким образом, на определенном этапе
филогенетического развития Pseudophyllidea сталкиваются две эво-
люционные тенденции: одна из них направлена на полное подавле-
ние индивидуальности особей, составляющих стробилу, другая же
способствует проявлению отдельных признаков ранее отчетливо,
выраженной колониальности.
Совершенно иная картина наблюдается у Cyclophyllidea, по-
давляющее большинство которых обладает вторично измененной
маткой, лишенной самостоятельного отверстия. Развивающиеся
31
яйца накапливаются в ней на протяжении всего периода функцио-
нирования половой системы членика. Только отрыв последнего от
•стробилы и выход наружу обеспечивает их рассеивание во внешней
среде. Уже одно это исключает возможность полного стирания гра-
ниц между отдельными проглоттидами. Приведенные выше разли-
чия между Pseudophyllidea и Cyclophyllidea легко объясняются их
биологическими особенностями, и ни в коей мере не противоречат
гипотезе полизоичности.
Выбор той или иной гипотезы определяет, как уже говорилось
выше, и ответ на вопрос: что же представляет собой жизненный
цикл цестод? Сторонники монозоичности ленточных червей рассмат-
ривают их развитие как сложный метаморфоз. Если же принимать
гипотезу полизоичности, то проблема эта не может быть решена
однозначно. У большинства видов ленточных червей жизненный
цикл представляет собой метагенез: половому размножению гер-
мафродитных особей (.проглоттид) обязательно предшествует бес-
полое размножение путем почкования молодых кишечных форм. Это
.первичный метагенез,- исходно свойственный всем цестодам. У не-
которых видов (отр. Cyclophyllidea — MuLticeps-, Echinococcus, ряд
.представителей сем. Hymenolepididae) жизненный цикл усложня-
ется за счет появления еще одного этапа бесполого размножения
на личиночной стадии (см. 28). Специализация паразитов идет и в
другом направлении. Утрата членистости, одновременная закладка
половых органов на личиночных стадиях и, наконец, возникновение
видов, имеющих всего один набор половых органов, приводит ко
вторичному упрощению цикла развития, который действительно
можно рассматривать как обычный, хотя и довольно сложный ме-
таморфоз одного поколения.
Классификация цестод
Система класса Cesfoda в настоящее время еще не окончательно установи-
лась. Вслед за целым рядом специалистов мы принимаем существование 6 отря-
дов, для обоснования которых используются как морфологические признаки, так
и особенности жизненных циклов.
Среди примитивных цестод выделяется отр. Pseudophyllidea, один из самых
крупных по числу входящих в его состав рндов. Прикрепительные образования
представлены ботриями, к которым иногда добавляются крупные крючья. Боль-
шинство видов имеют расчлененную стробилу. У некоторых расчленение частично
или полностью исчезает (сем. Ligulidae, Triaenophoridae).
Развитие в большинстве случаев осуществляется с участием двух промежу-
точных хозяев. Личиночные стадии представлены корацидием, процеркоидом
и плероцеркоидом. Обитают в очень широком круге позвоночных животных: от
рыб до человека включительно.
Очень близок отр. Caryophillaeidea — гвоздичники, небольшая группа, пред-
ставленная почти исключительно паразитами пресноводных рыб. Передний конец
тела у них слегка расширен и несет несколько фестончатых складок. Тело нерас-
членено н несет один половой аппарат, сходный по своему строению с органами
размножения Pseudophyllidea. Развитие осуществляется с участием одного про-
межуточного хозяина (олигохеты). Отсутствие свободноплавающего корацпдня
говорит о вторичной специализации Caryophyllaeidea. Для некоторых гвоздични-
ков характерно неотеннческое развитие (с. 25).
Отр. Tetraphyllidea объединяет цестод, сколексы которых несут ботрндии-
Последние иногда замещаются присосками. Часто имеются и крючья. Развитие
32
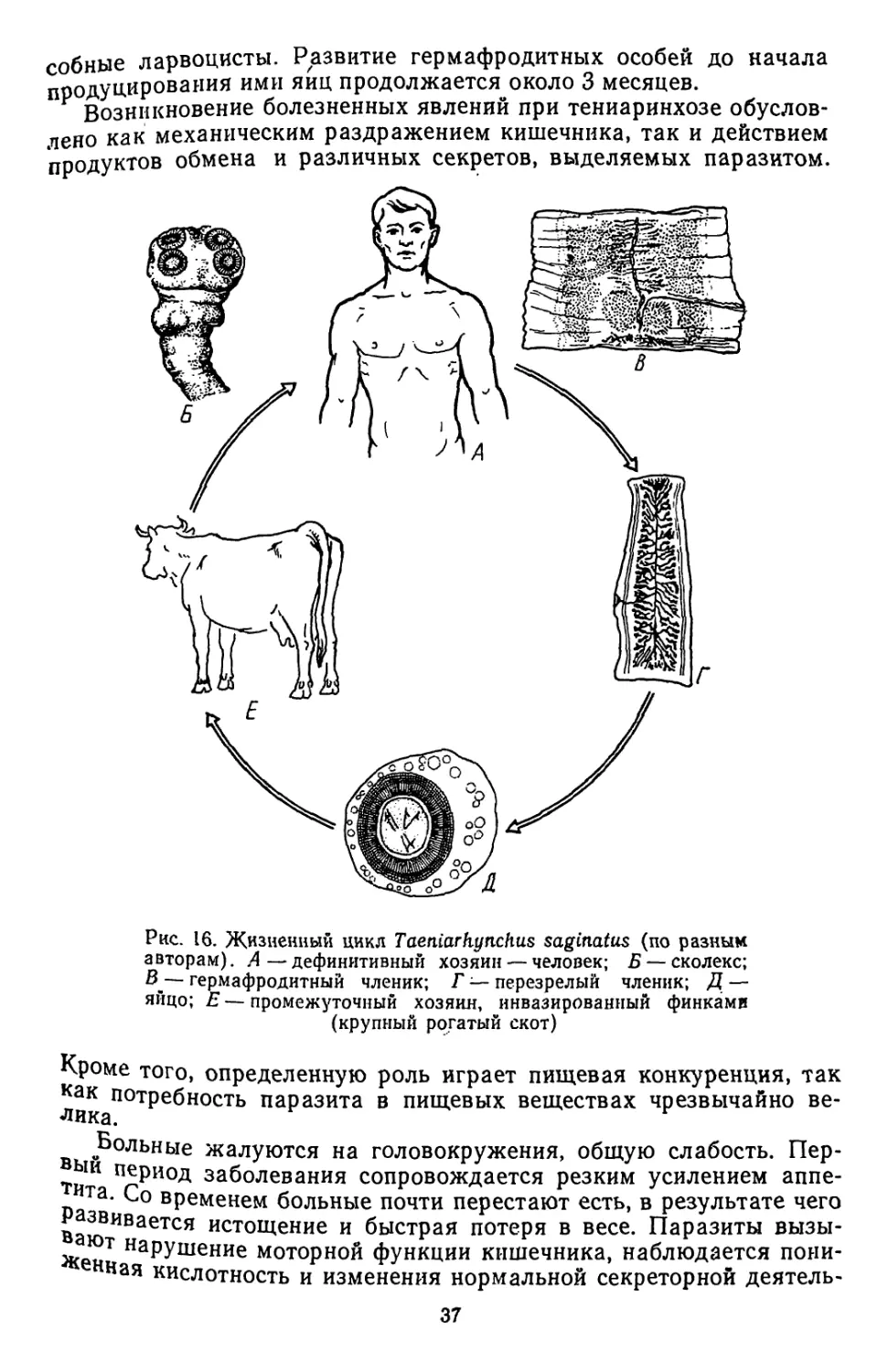
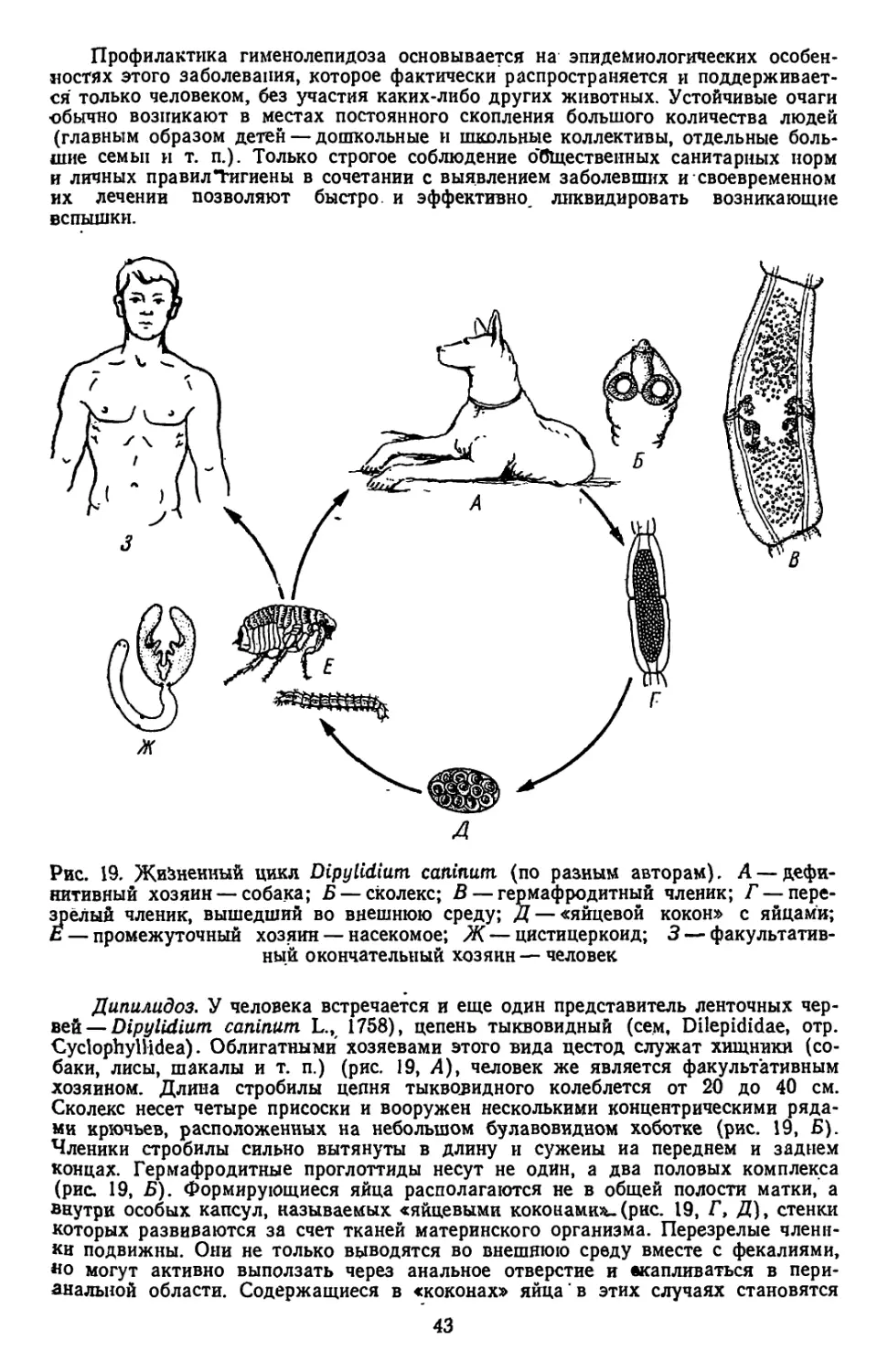
,цгесТвляется чаще всего с участием двух хозяев и включает смену нескольких
ичиночных стадий: онкосферы, процеркоида и плероцеркоида. Между двумя
лоследними у ряда видов вклинивается еще фаза цистнцеркоида. Tetraphyllidea
Ндпазитирует в рыбах, реже они встречаются в амфибиях, рептилиях, птицах
и млекопитающих.
Небольшой отр. Тгурапогпупспа представлен паразитами акул и скатов.
гКолекс несет 4 подвижных хоботка, вооруженных крючьями. Сведения об их
жизненных циклах немногочисленны и неполны. По-видимому, промежуточными
хозяевами служат морские костистые рыбы, ракообразные и головоногие мол-
люски, в которых обнаруживаются инкапсулированные личинки паразитов.
В* акулах и скатах встречаются представители небольшого отр. Diphyllidea.
Сколекс нх несет два мощно развитых ботридия и снабжен крючьями. Личинки
этих ленточных червей обнарухсены у моллюсков и ракообразных. Полные иссле-
дования по жизненным циклам отсутствуют.
Самый многочисленный отряд цестод, объединяющий наиболее специализи-
рованных представителей класса —отр. Cyclophylidea, приурочен главным обра-
зом к наземным позвоночным — рептилиям, птицам и млекопитающим. Сколекс
этих ленточных червей несет 4 хорошо развитые мускулистые присоски и часто
снабжен венчиками крючьев. Развитие Cyclophyllidea в подавляющем большин-
стве случаев осуществляется с участием одного промежуточного хозяина. Личи-
ночные стадии представлены онкосферой и различными типами ларвоцист. Отряд
включает большое количество паразитов человека и домашних животных.
Заболевания, вызываемые цестодами
Многие ленточные черви служат причиной тяжелых, а иногда
и просто опасных для жизни своих хозяев заболеваний, называе-
мых цестодозами. В одних случаях" наиболее патогенные полово-
зрелые черви, паразитирующие в кишечнике, в других же — личи-
ночные стадии.
Цестодозы человека
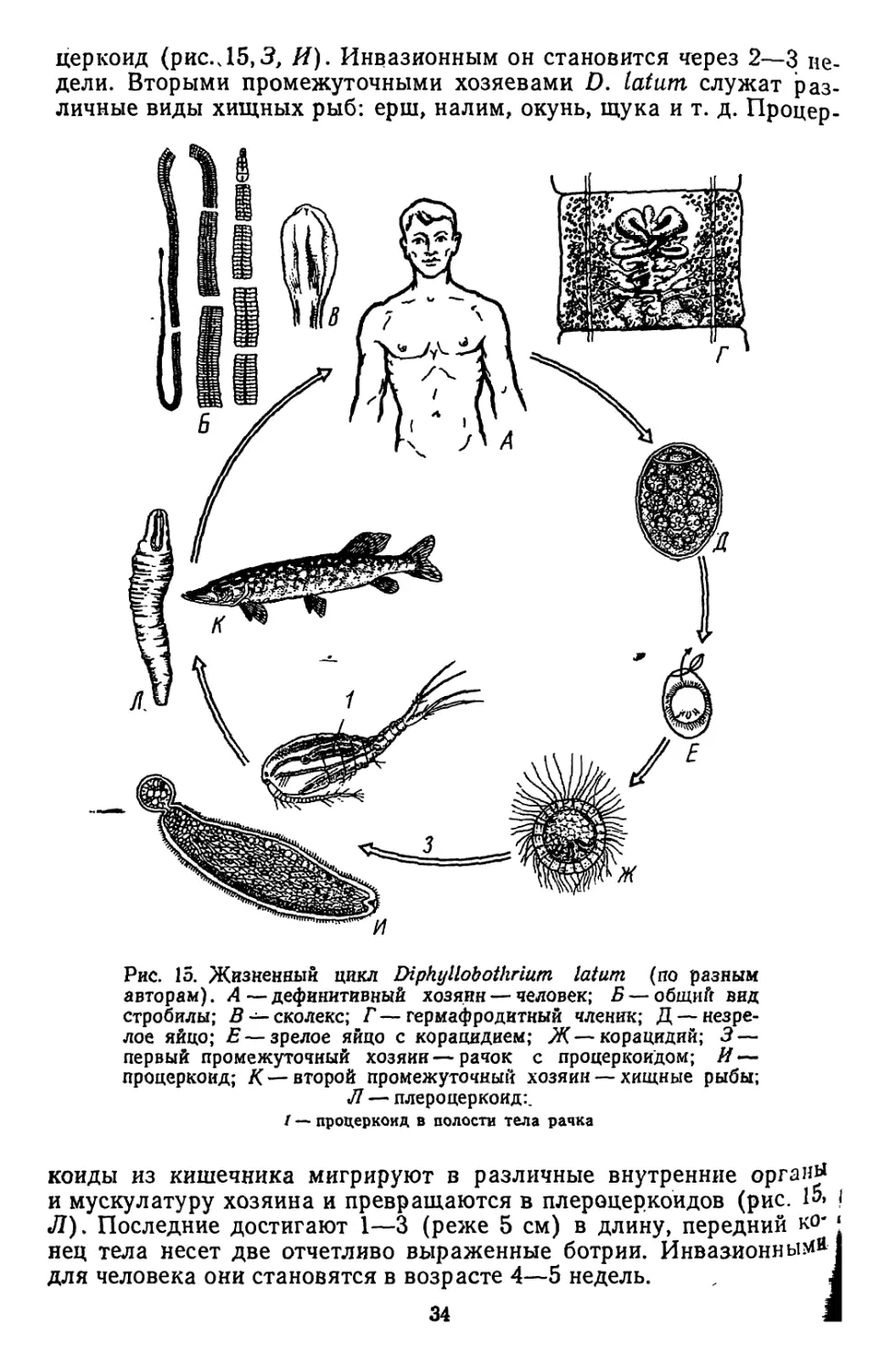
Дифиллоботриозы. Эти заболевания обусловлены паразитиро-
ванием в пищеварительной системе человека цестод, относящихся
к р. Diphyllobothrium (сем. DiphylLobothriidae, отр. Pseudophylli-
dea). Наибольшее медицинское значение имеет весьма распростра-
ненный вид Diphyllobothrium latum L., 1748, — лентец широкий
(рис. 15). Длина стробилы в среднем составляет около 10 м. Ско-
лекс вытянут в длину и несет две щелевидные ботрии (рис. 15, В).
Половые органы полностью сформированы, начиная с 60—70-го
членика. Сильно извитая и заполненная яйцами матка образует
характерную розетковидную фигуру (рис. 15, Г). Срок жизни этих
паразитов может быть весьма значительным — 20 и более лет.
Обычно в кишечнике человека обитает одна или в крайнем случае
несколько особей.
Продуцируемые паразитами яйца имеют овальную форму и на
Переднем конце несут крышечку. Развитие корацидия осуществля-
йся в воде. При оптимальной температуре формирование зароды-
ша продолжается 2—3 недели. Возможно перезимовывание яиц
ь водоеме. Вылупившийся из яйца корацидий должен быть прогло-
Чен промежуточным хозяином (рачки из отр. Copepoda: р. Eudiapto-
Acanthodiaptomus, Diaptomus, Cyclops и т. д.). В полости тела
"ачка из онкосферы, заключенной в корацидии, развивается про-
зз
церкоид (рис.Л5,3, И). Инвазионным он становится через 2—3 не-
дели. Вторыми промежуточными хозяевами D. latum служат раз-
личные виды хищных рыб: ерш, налим, окунь, щука и т. д. Процер-
Рис. 15. Жизненный цикл Diphyllobothrium latum (по разным
авторам). А — дефинитивный хозяин — человек; Б — общиГг вид
стробилы; В — сколекс; Г — гермафродитный членик; Д — незре-
лое яйцо; Е—зрелое яйцо с корацидием; Ж—корацидий; 3—
первый промежуточный хозяин—рачок с процеркоидом; И —
процеркоид; К— второй промежуточный хозяин — хищные рыбы;
Л — плероцеркоид:.
I — процеркоид в полости тела рачка
коиды из кишечника мигрируют в различные внутренние органу
и мускулатуру хозяина и превращаются в плероцеркоидов (рис. 1»> !
Л). Последние достигают 1—3 (реже 5 см) в длину, передний ко' ‘
нец тела несет две отчетливо выраженные ботрии. Инвазионными !
для человека они становятся в возрасте 4—5 недель. , J
34 Я
Жизненный цикл D. latum может усложняться за счет появле-
резервуарного хозяина, которыми также становятся хищные
оыбьг крупные окуни, щуки, налимы и т. п. Они заражаются, пое-
лая мелких, уже инвазированных плероцеркоидами хищников. Ли-
чинки при этом мигрируют в мускулатуру и внутренние органы
нового для них хозяина. Паразиты длительное время сохраняют
жизнеспособность и инвазионность. Это обстоятельство и обуслов-
ливает накопление большого числа плероцеркоидов в тканях и ор-
ганах активно питающихся крупных хищных рыб. Окончательными
хозяевами кроме человека служат й дикие животные, питающиеся
рыбой (песцы, некоторые морские млекопитающие и т. п.). Таким
образом, могут существовать и природные очаги, играющие извест-
ную роль в распространении дифиллоботриозов.
Клинические проявления заболевания, вызываемого широким
лентецом, очень разнообразны. Иногда внешние симптомы вообще
отсутствуют. Часто паразиты вызывают лишь слабые кишечные
расстройства. Однако во многих случаях наблюдается и очень тя-
желое течение болезни. Оно вызывается механическим воздействи-
ем цестод на слизистую кишечника и сенсибилизацией организма
зараженного человека продуктами их обмена. В основе развиваю-:
щегося авитаминоза лежит усиленное потребление паразитами
витаминов (В12), содержащихся в пище. Кроме того, продукты
обмена лентецов вызывают изменения микрофлоры кишечника
хозяина, что приводит к почти полному прекращению биосинтеза
фолиевой кислоты, осуществляемого кишечными бактериями,
и уменьшению количества витамина С. Развивается очень тяжелая
анемия. У больного появляются слабость, сильные головокружения,
сонливость. Очень характерны функциональные нарушения кишеч-
ника, понижение или, наоборот, резкое повышение кислотности,
бледность покровов, отечность. Часто развиваются патологические
изменения слизистой ротовой полости и языка. Наблюдаются уве-
личение печени и селезенки.
Главным источником распространения инвазионного начала во
внешней среде является человек. Определенное значение, вероят-
но, имеют и природные очаги, которые очень часто становятся ис-
точником возникновения новых синантропных очагов. Заражение
промежуточных хозяев становится возможным при попадании не-
обезвреженных сточных-вод в водоемы. Распространению заболе-
вания способствует, по-видимому, и гидростроительство. Замедлен
ние стока рек и возникновение больших мелководных участков по
берегам водохранилищ создают благоприятные условия для раз-
множения веслоногих рачков и концентрации молоди рыб. Наибо-
ее устойчивые очаги дифиллоботриоза существуют в районах, где
селение часто употребляет в пищу полусырую, а иногда и сырую
Р юу; строганину, приготавливаемую из свежезамороженной рыбы,
Дуегка п°Дс°ленную рыбу, малосольную щучью икру, рыбные про-
ЛЫ’жНе подвеРгшиеся- достаточной термической обработке, и т. д,
н ^Ифиллоботриоз пРиУРочен главным образом к северным стра-
и странам с умеренным климатом. Он отмечается в Северной
2*
35
Америке (центральные и северо-западные районы), Ирландии, Ис-
ландии, многих странах Европы (Финляндия, Швеция, Дания,
Польша и т. д.). Зарегистрировано заболевание и в некоторых рай-
онах Южной Америки и на востоке Австралии. В СССР дифилло-
ботриоз встречается в северных районах европейской части, в Си-
бири и на Дальнем Востоке. В последние годы появились новые
очаги в бассейне Волги и в центральных районах.
Диагноз на дифиллоботриоз ставится, как правило, на основе
копрологического анализа.
Борьба с дифиллоботриозом должна включать целый ряд меро-
приятий. Прежде всего необходимо создание условий, препятст-
вующих заражению водоемов необезвреженными сточными вода-
ми из населенных пунктов и курсирующих по рекам и озерам су-
дов. Индивидуальная профилактика заключается в употреблении
в пищу только хорошо приготовленной рыбы (просоленной, прожа-
ренной или проваренной). Не следует также кормить домашних
животных (кошек, собак, свиней) сырой рыбой. В неблагополуч-
ных районах необходима периодическая массовая проверка насе-
ления, позволяющая выявить носителей инвазии и их последующее
лечение.
Тениидозы. К распространенным цестодозам человека относятся
тениидозы — тениаринхоз, вызываемый Taeniarhynchus saginatus
(сем. Taeniidae, отр. Cyclophyllidea), и тениоз — возбудитель Tae-
nia solium (сем. Taeniidae, отр. Cyclophyllidea).
Тениаринхоз. Taeniarhynchus saginatus Goeze, 1782 (невоору-
женный, или бычий, цепень) — крупные черви, длина которых ко-
леблется в пределах 4—10 м. Сколекс несет только 4 присоски, во-
оружение отсутствует (рис. 16,Б). Гермафродитные членики имеют
почти квадратную форму (рис. 16,В). Яичник двухлопастный. Пе-
резрелые членики, слегка суженные на переднем и заднем концах,
внешне напоминают семена тыквы. Их матка с каждой стороны
несет 17—35 узких боковых ветвей (рис. 16, Г).
Перезрелые членики Т. saginatus отрываются от стробилы по-
одиночке. Проглоттиды некоторое время сохраняют подвижность.
Будучи выведенными во внешнюю среду, они некоторое время «пол-
зают» по субстрату, рассеивая при этом яйца (рис. 16, Д). Послед-
ние содержат уже инвазионные онкосферы. Промежуточными хо-
зяевами для невооруженного цепня служит крупный рогатый скот
(рис. 16, Е). В кишечнике животных онкосферы освобождаются от
яйцевых оболочек, проникают в кровеносную систему и током кро-
ви разносятся по организму хозяина. Личинки обычно оседают
в мускулатуре (межмышечная соединительная ткань скелетных
мышц, сердечная мышца, жевательные мышцы, мышцы языка
и т. д.). Превращение онкосферы в цистицерк занимает около 4—-
5 месяцев. Зрелая финка достигает 4—10 мм в поперечнике. В воз-
расте 8—9 месяцев личинки дегенерируют и погибают. Человек за-
ражается при употреблении в пищу мяса инвазированных живот-
ных, в котором по тем или иным причинам сохраняются жизнеспо-
36
собные ларвоцисты. Развитие гермафродитных особей до начала
продуцирования ими яиц продолжается около 3 месяцев.
F Возникновение болезненных явлений при тениаринхозе обуслов-
лено как механическим раздражением кишечника, так и действием
продуктов обмена и различных секретов, выделяемых паразитом.
Ркс. 16. Жизненный цикл Taeniarhynchus saginatus (по разным
авторам). А—дефинитивный хозяин — человек; Б — сколекс;
— гермафродитный членик; Г — перезрелый членик; Д —
яйцо; Е — промежуточный хозяин, инвазированиый финками
(крупный рогатый скот)
Кроме того, определенную роль играет пищевая конкуренция, так
ликаП°ТРебН°СТЬ паРазита в пищевых веществах чрезвычайно ве-
Больные жалуются на головокружения, общую слабость. Пер-
ми период заболевания сопровождается резким усилением аппе-
рата‘ С° временем больные почти перестают есть, в результате чего
ваюИВаеТСЯ истои*ение и быстрая потеря в весе. Паразиты вызы-
эк т НаРУшение моторной функции кишечника, наблюдается пони-
ная кислотность и изменения нормальной секреторной деятель-
37
ности желудка. Довольно серьезные патологические отклонения
отмечаются и со стороны печени. Прохождение отделившихся чле-
ников из тонкого кишечника в толстый часто сопровождается силь-
ными болевыми ощущениями, симулирующими приступы аппенди-
цита. Большинство болезненных явлений быстро исчезает после ус-
пешно проведенной дегельминтизации.
Единственным источником распространения Т. saginatus служит
человек. Выделяемые во внешнюю среду яйца устойчивы к небла-
гоприятным факторам н легко перезимовывают под снегом, сохра-
няя инвазионность. Разносимые со сточными, паводковыми или та-
лыми водами, яйца легко вместе с кормом (сеном, травой и т~ п.)
попадают к скоту. Очень велика опасность рассеивания яиц, зара-
женных людьми на животноводческих фермах и в индивидуальных
хозяйствах. Заражение человека происходит при употреблении в пи-
щу термически плохо обработанного мяса. Важное значение в рас-
пространении заболевания имеют некоторые профессиональные
навыки и национальные обычаи (распространенная привычка про-
бовать мясной фарш, употребление в пищу сырого рубленого мяса
и т. п.).
Тениаринхоз встречается практически во всех странах мира.
Особенно часто от тениаринхоза страдает население Африки, Ав-
стралии и некоторых районов Азии. В СССР очаги заболевания
имеются на Кавказе, в республиках Средней Азии, ряде областей Си-
бири и т. д. Благодаря активным лечебно-профилактическим меро-
приятиям, заболеваемость тениаринхозом в нашей стране постоян-
но снижается.
Для диагносцирования тениаринхоза используются методы ко-
прологического анализа. Однако значительно более надежен спо-
соб исследования перианально-ректальных соскобов. Лишь по фор-
ме матки перезрелых члеников можно точно установить видовую
принадлежность обитающего в человеке паразита. Лечение тениа-
ринхоза обычно сводится к дегельминтизации хозяина.
Борьба с тениаринхозом представляет собой комплекс меропри-
ятий, направленных на оздоровление местности (уничтожение яиц
на поверхности почвы и растительности, обеззараживание сточных
вод, строгое соблюдение санитарных норм и правил); периодиче-
ские массовые осмотры населения в очагах заболевания и лечение
выявленных больных, строжайший ветеринарный контроль за ус-
ловиями содержания крупного рогатого скота и за выполнением
правил по убою животных и обезвреживанию финозного мяса.
Тениоз. Возбудителем этого заболевания служит Taenia
solium L., 1758 (вооруженный, или свиной, цепень). От бычьего цеп-
ня Т. solium отличается несколько меньшими размерами стробилы,
длина которой в среднем составляет 2—3 м. Сколекс несет 4 при-
соски и короткий хоботок, вооруженный двумя рядами. крючьев
(рис. 17,Б). Гермафродитные членики (рис. 17,В) в средней час-
ти'стробилы имеют почти правильную квадратную форму, перезре-
лые же заметно вытянуты в длину. Яичник Т. solium в отличие от
Taeniarhynchus saginatus не двухлопастный, а трехлопастный. В пе-
38
резрелых члениках матка образует 7—12 пар толстых боковых вет-
вей (рис. 17, Г).
Выведение яиц во внешнюю .среду осуществляется при отделе-
нии от стробилы перезрелых члеников. Г------------;-------—
Последние отрываются не
Рис. 17. Жизненный цикл Taenia solium (по разным авторам).
Л — дефинитивный хозяин — человек; Б — сколекс; В — герма-
фродитный членик; Г—группа перезрелых члеников; Д—яйцо;
В — промежуточный хозяин, инвазированный финками; Ж—цн-
стицерк в мясе; 3 — цистицеркоз мозга при аутоинвазии челове-
ка; И — человек в роли факультативного промежуточного хозяина
п°одиночке, а группами. Подвижностью они не обладают. Яйца
одеты толстой защитной оболочкой (рис. 17, Д) и весьма устойчи-
ы к действию факторов внешней среды. Главными промежуточны-
и хозяевами вооруженного цепня служат свиньи (рис. 17, Е), зна-
чительно реже ими могут быть и другие животные (собаки, кошки
т- Д-). Возможно заражение личиночными стадиями и человека
39
(рис. 17, Л, И). Онкосферы в просвете кишечника быстро освобож-
даются от эмбриофора, внедряются в слизистую и с током крови
разносятся по телу хозяина. Паразиты локализуются практически
во всех тканях и органах, предпочитая, однако, межмышечную сое-
динительную ткань. У человека они часто располагаются в глазах
и центральной нервной системе. Формирование цистицерка закай*
чивается через 60—70 дней после заражения. В плотной мышеч-
ной ткани личинки, как правило, остаются сравнительно неболь-
шими (5—8 мм), тогда как в мягких паренхиматозных органах они
сильно разрастаются (до 1,5 см), не встречая сопротивления ок-
ружающих тканей. В кишечнике дефинитивного хозяина — челове-
ка— ларвоцисты дают начало гермафродитным стробилам.
Внешние клинические проявления заболевания выражены яр-
че, чем при тениаринхозе. У больного наблюдаются расстройства
аппетита, тошноты, нарушения стула, боли в ^ивоте, головокруже-
ния и головные боли, нарушения сна. Однако главная опасность
тениоза заключается не в этом. Выше уже отмечалось, что человек
может выступать не только в роли дефинитивного, но и промежу-
точного хозяина для Т. solium. Заражение человека финками, по-
лучившее название цистицеркоза, может осуществляться двумя спо-
собами. Во-первых, путем проглатывания рассеиваемых во внеш-
ней среде яиц. Интенсивность инвазии при этом обычно не бывает
особенно высокой. Во-вторых, очень часто .имеет место аутоипва-
зия, последствия которой могут быть чрезвычайно тяжелыми. От-
торгаемые от стробилы членики содержат огромное число вполне
инвазионных яиц. В большинстве случаев свободные проглоттиды
или кусочки, состоящие из нескольких проглоттид, выводятся во
внешнюю среду вместе с фекалиями. При рвотах, однако, которые
весьма обычны при тениозе, в результате интенсивной антипери-
стальтики кишечника такие участки стробилы могут попасть в же-
лудок. Здесь под действием желудочного сока происходит массо-
вое вылупление онкосфер, которые в дальнейшем разносятся по
всему телу, вызывая различные поражения. Развитие личинок
в подкожной клетчатке и скелетной мускулатуре, как правило, про-
ходит бессимптомно или же-сопровождается неприятными болевы-
ми ощущениями, которые часто неправильно диагносцируются как
ревматизм. Опасно попадание личинок в головной (рис. 17,3)
и спинной мозг и глаза, но именно в этих органах они локализуют-
ся чаще всего. Поражения центральных отделов нервной системы
сопровождаются сильными головными болями, судорожными при-
падками, парезами, параличами. Повышается внутричерепное дав-
ление. Иногда отмечается затемнение сознания, легкие психические
нарушения. Заболевание часто имеет летальный исход. Внутриглаз-
ной цистицеркоз, вызывающий ухудшение, а иногда и полную поте-
рю зрения, при отсутствии лечения приводит к атрофии поражен-
ного глаза.
Распространение инвазионного начала во внешней среде осу-
ществляется только зараженным человеком. Заражение человека
личиночными стадиями, как уже отмечалось выше, может проис-
40
ходить не только путем аутоинвазии, но и с пищевыми продуктами,
загрязненными яйцами паразитов. Наиболее часто это наблюдает-
ся в условиях сельской местности при несоблюдении санитарно-
гигиенических норм. Кишечными формами человек заражается при
употреблении в пищу свиного мяса, в котором при недостаточной
термической или иной обработке сохраняются жизнеспособные
финки.
Тениоз встречается практически во всех странах, в которых рас-
пространено свиноводство. В СССР отмечаются единичные случаи.
Данные о распространении заболевания бывают не всегда досто-
верны, если диагноз ставится не по отходящим проглоттидам, а по
яйцам с онкосферами, которые практически неразличимы у Т. solium
и Т. saginatus-, регистрируют тениаринхоз без последующей провер-
ки определения.
Точная же диагностика, которая проводится только по перезре-
лым членикам, играет важную роль. При любом, даже самом лег-
ком течении тениоза возможна аутоинвазия финками и развитие
тяжелого цистицеркоза. Последний распознается с известным тру-
дом. Обычно в практике используют неврологические показатели,
анализы спинномозговой жидкости, рентгеноскопию, реакцию свя-
зывания комплемента с антигеном из цистицерков, офтальмоско-
пию, биопсию подкожной клетчатки. Определенную роль для пра-
вильной постановки диагноза может сыграть и тщательно собран-
ный анамнез.
Лечение тениоза также сопряжено с рядом трудностей. Препа-
раты, с успехом применяемые при тениаринхозе, в этом случае не
употребляются. Не исключено, что вызываемое ими разрушение
члеников может явиться причиной цистицеркоза. Особое внимание
при лечении необходимо обращать на предупреждение рвоты, ко-
торая сильно повышает опасность проникновения онкосфер в же-
лудок.
Эффективные и надежные методы лечения цистицеркоза пока от-
сутствуют. Хирургическое вмешательство дает хорошие результаты
лишь при цистицеркозе глаза. Оперативное лечение при множест-
венном цистицеркозе других органов 'часто оказывается невозмож-
ным. В этих случаях используют рентгенотерапию в сочетании
с большими дозами антибиотиков.
Профилактика и меры борьбы с тениозом аналогичны таковым
при борьбе с тениаринхозом.
Гименолепидоз. Очень широко распространен, особенно у детей; гйменоле-
пидоз — заболевание, вызываемое Hymenolepis папа Siebold, 1852 — карликовым
цепнем (сем. Hymenolepididae, отр. Cyclophyllidea). Это сравнительно мелкие
цестоды (рис. 18, Д), длина стробилы которых едва превышает 4—5 см. Сколекс
несет 4 присоски и подвижный хоботок, вооруженный венчиком крючьев. Количе-
ство члеников в стробиле колеблется от 100 до 200. Матка неразветвленная, меш-
ковидной формы. ‘
Жизненный цикл карликового цепня протекает в одном человеке, который
таким образом одновременно выполняет функции и промежуточного и дефини-
тивного хозяина (рис. 18). Выходящие во внешнюю среду яйца выживают всего
лишь в течение нескольких часов. Человек заражается, проглатывая инвазионные
япца. В тонком кишечнике онкосферы (рис. 18, В) освобождаются от защитных
41
оболочек и внедряются в ворсинки. В последних они превращаются в цистицер-
коидов (рис. 18, Г), которые через некоторое время выходят из слизистой обрат-
но в просвет кишечника. Здесь личинки претерпевает последний этап метамор-
фоза и дают начало взрослым червям. Таким образом, у Н. папа наблюдается
заметное упрощение путей циркуляции паразита. У них исчезает феномен смены
животных-хозяев.
По данным некоторых авторов, Н. папа не утратил способности развиваться
и с участием промежуточных хозяев. При скармливании яиц с онкосферами жу-
кам из р. Ten^brio в последних развиваются типичные инвазионные цистицерко-
иды (рис. 18, Е).
Рис. 18. Жизненный цикл Hymenolepis папа (по разным авторам). Д—де-
финитивный хозяин — человек; Б — яйцо с онкосферой; В — онкосфера
в просвете кишечника; Г — цистицеркоид в ворсинке кишечника; Д—по-
ловозрелый червь; Е —цистицеркоид в гемоцеле факультативного проме-
жуточного хозяина — насекомого
Гименолипедоз как заболевание хотя в большинстве случаев и не представ-
ляет серьезной опасности, тем“не менее может протекать довольно остро. Он со-
провождается поносами, которые сменяются тяжелыми запорами, сильными бо-
лями в области живота, истощением. Во многих случаях к этому прибавляются
функциональные нарушения нервной системы: бессоница, конвульсии, припадки,
напоминающие эпилепсию, и т. п. В основе механизмов патогенеза лежит разру-
шение ворсинок кишечника развивающимися цистицеркоидами, механическое
раздражение слизистой и сенсибилизация организма продуктами обмена парази-
тов. От гимеполепидоза преимущественно страдают дети.
Карликовый цепень распространен практически пбвсеместно, ио чаще встре-
чается в южных районах.
Диагноз обычно ставится на основе данных копрологического анализа. Курс
лечения необходимо повторять 2—3 раза, так как после изгнания фишечных форм
в ворсинках,могут оставаться неповрежденные личинки. Последние через некото-
рое время дадут начало рецидиву заболевания.
42
Профилактика гименолепидоза основывается на эпидемиологических особен-
ностях этого заболевания, которое фактически распространяется и поддерживает-
ся только человеком, без участия каких-либо других животных. Устойчивые очаги
обычно возникают в местах постоянного скопления большого количества людей
(главным образом детей — дошкольные и школьные коллективы, отдельные боль-
шие семьи и т. п.). Только строгое соблюдение оЧйцествепных санитарных норм
и личных правил Тигпены в сочетании с выявлением заболевших и своевременном
их лечении позволяют быстро, и эффективно ликвидировать возникающие
их лечении позволяют быстро, и эффективно ликвидировать
вспышки.
Рис. 19. Жизненный цикл Dipylidium caninum (по разным авторам). А — дефи-
нитивный хозяин — собака; Б — сколекс; В — гермафродитный членик; Г — пере-
зрелый членик, вышедший во внешнюю среду; д— «яйцевой кокон» с яйцами;
Е— промежуточный хозяин — насекомое; Ж— цистицеркоид; 3 — факультатив-
ный окончательный хозяин — человек
Дипилидоз. У человека встречается и еще один представитель ленточных чер-
вей— Dipylidium caninum L., 1758), цепень тыквовидный (сем, Dilepididae, отр.
Cyclophyllidea). Облигатными хозяевами этого вида цестод служат хищники (со-
баки, лисы, шакалы и т. п.) (рис. 19, 4), человек же является факультативным
хозяином. Длина стробилы цепня тыквозидного колеблется от 20 до 40 см.
Сколекс несет четыре присоски и вооружен несколькими концентрическими ряда-
ми крючьев, расположенных на небольшом булавовидном хоботке (рис. 19, Б).
Членики стробилы сильно вытянуты в длину и сужены иа переднем и заднем
концах. Гермафродитные проглоттиды несут не один, а два половых комплекса
(рис. 19, Б). Формирующиеся яйца располагаются не в общей полости матки, а
внутри особых капсул, называемых «яйцевыми коконами»-(рнс. 19, Г, Д), стенки
которых развиваются за счет тканей материнского организма. Перезрелые члени-
ки подвижны. Они не только выводятся во внешнюю среду вместе с фекалиями,
«о могут активно выползать через анальное отверстие и вкапливаться в пери-
аналыюй области. Содержащиеся в «коконах» яйца ’ в этих случаях становятся
43
легкой добычен власоедов (насекомые из р. Trichodectes, с. 215), которые и служат
промежуточными хозяевами. Кроме того, паразиты могут развиваться и в ли-
чинках блох, поедающих яйца, рассеиваемые во внешней среде. В полости тела
промежуточного хозяина онкосферы превращаются в цистоцеркоидов (рис. 19, Ж).
У власоедов этот процесс совершается довольно быстро. У блох же он растяги-
вается на длительный промежуток времени. В личинке блохи онкосфера не пре-
терпевает никаких изменений; у куколки начинается формирование цистицеркоида,
и лишь у имаго эта личиночная стадия D. caninutn становится инвазионной. Та-
ким образом, наблюдается тесная зависимость развития паразита от развития
хозяина. Биологически это вполне оправдано, так как только взрослые блохи
вступают в тесный контакт с дефинитивным хозяином и могут быть прогло-
чены им.
Заражение человека D. canituun, как правило, происходит случайно. Чаще
от него страдают дети, вступающие в тесный контакт с домашними животными.
Паразитирование тыквовидного цепня в специфических хозяевах почти никогда
не сопровождается какими-либо болезненными симптомами. У человека же он
вызывает довольно серьезные функциональные расстройства пищеварительной
и нервной систем, сходные с аналогичными явлениями при гименолепидозах
(с. 42). После изгнания червей (для этого используются экстракт мужского па-
поротника, филиксан, феносал и т. д.) все признаки заболевания полностью
исчезают.
Эхинококкоз и альвеококкоз. Очень опасными цестодозами че-
ловека являются заболевания, вызываемые не стробиллярными,
а личиночными .стадиями (финками). Среди них по широте распро-
странения и исключительной патогенности выделяются эхинококкоз
и альвеококкоз.
Echinococcus granulosus 6atsh, 1786 — специфический паразит
собаки и диких представителей сем. Canidae — один из самых мел-
ких ленточных червей. Стробила, длина которой не превышает 8 мм,
состоит из 3—4 члеников (рис. 20, Б). Сколекс снабжен 4 присос-
ками и хоботком, несущим 2 венчика крючьев. За относительно
короткой шейкой располагается первый — незрелый — членик. Вто-
рой и третий членики обычно содержат функционирующую половую
систему; четвертый членик — перезрелый. В нем располагается
матка, образующая неправильной формы ответвления и заполнен-
ная сформированными яйцами.
Перезрелые членики (рис. 20,В), выходящие во внешнюю среду
вместе с фекалиями, подвижны. Они способны передвигаться на
расстояние до 25 см, выделяя во внешнюю среду содержащиеся
в них яйца через разрывы стенок.
Круг промежуточных хозяев Е. granulosus очень широк. Это
практически все домашние и дикие копытные животные, сумчатые
и некоторые грызуны. Кроме того, паразиты могут развиваться и в
человеке.
В пищеварительном тракте промежуточного хозяина онкосферы
освобождаются от яйцевых оболочек и внедряются в слизистую.
Чаще всего они попадают в печень, где и начинается разви-
тие ларвоцнсты, называемой также гидатидой. Паразиты могут ло-
кализоваться и в других органах, но это наблюдается значительно
реже. Второе место по частоте поражений после печени занимают
легкие. Ларвоцнсты бывают очень крупными, до 15 см в попереч-
нике. Материнский пузырь содержит большое количество дочерних
44-
Рис. 20. Жизненный цикл Echinococcus granulosus (по разным авторам). А—дефинитивный хозя-
ин— плотоядные животные; Б — стробила; В—перезрелый членик, вышедший во внешнюю среду;
Г—яйцо; Д — промежуточный хозяин — сельскохозяйственные животные и др.; Е — эхнпококкоз-
ный пузырь в печени; Ж — факультативный промежуточный хозяин — человек:
пузырей, выводковых капсул и отдельных протосколексов. Послед-
ние освобождаются при разрушении дочерних пузырей и выводко-
вых капсул. Протосколексы, количество которых в полостной жид-
кости пузыря может быть очень велико, образуют так называемый
«гидатидный песок». Иногда наблюдается множественный эхино-
коккоз. В одних случаях он обусловлен первичным заражением хо-
зяина не одной, а значительным числом онкосфер. В других же он
связан с явлением «обсеменения». При нарушении целостности ма-
теринского пузыря поступающие в полость тела хозяина протоско-
лексы и обрывки герминативного слоя могут дать начало новым
пузырям. Заражение дефинитивных хозяев происходит при поеда-
нии мяса инвазированных животных. Так как при этом количество
проглоченных протосколексов бывает очень велико, то и интенсив-
ность инвазии соответственно оказывается чрезвычайно высокой.
Человек в жизненном цикле эхинококка служит факультатив*-
ным промежуточным хозяином. На ранних стадиях развития пара-
зита заболевание протекает бессимптомно. Появление первых
клинических признаков, характер которых во многом определяет-
ся локализацией пузыря, часто отделено от момента заражения
периодом в несколько месяцев, а иногда и лет.
Патогенность эхинококка обусловливается сенсибилизацией
организма хозяина и механическим воздействием паразита на ок-
ружающие ткани. Аллергическая реакция в легких случаях прояв-
ляется в виде кожного зуда, крапивницы. При осложнениях, свя-
занных с разрывом пузыря, когда большая масса гидатидной жид-
кости поступает в организм хозяина, возникает шок. Особенно
опасно разрушение погибших и нагноившихся ларвоцист. Обычно
при этом развивается гибельный для больного гнойный перитонит.
При любом нарушении целостности материнского пузыря всегда
существует самая серьезная опасность возникновения большого ко-
личества новых пузырей.
При поражении печени наблюдаются постепенно усиливающие-
ся боли, увеличивается объем пораженного органа, появляются
признаки истощения, анемии, желтушное окрашивание покровов,
асцит. Развитие ларвоцисты в легких сопровождается кашлем
с мокротой, небольшими подъемами температуры, сильной одыш-
кой. По мере усиления симптомов клиническая картина все больше
и больше напоминает туберкулез. Разрыв легочного пузыря может
иметь тяжелые последствия. Если гидатидная жидкость поступает
в плевральную полость, то у больного развивается шок, сопровож-
даемый тяжелыми анафилактическими явлениями. При этом часто
наступает смерть.
В распространении эхинококкоза решающая роль принадлежит
дефинитивным хозяевам — собакам и, несколько реже, волкам, ша-
калам п другим хищникам и промежуточным хозяевам — крупному
и мелкому рогатому скоту, свиньям. Рассеивание яиц во внешней
среде осуществляется подвижными члениками. Последние, выпол-
зая из анального отверстия собаки, загрязняют яйцами ее шерсть.
При тесном контакте человека с инвазированным животным веро-
46
ятность заражения становится очень большой. Яйца, оставляемые
члениками на поверхности почвы и травы, легко попадают на
шерсть скота. При стрижке овец, доении они остаются на руках
и могут быть проглочены человеком. Источником заражения служат
также немытые овощи и фрукты. Устойчивости очагов эхинококко-
за способствует устойчивость яицгкоторые сохраняют жизнеспо-
собность в течение длительного времени.
Эхинококкоз чаще встречается в странах с развитым пастбищ-
ным животноводством (Австралия, Новая Зеландия, некоторые
районы Северной-Африки, целый ряд стран Азии, $Эжной Европы
и т. д.). В СССР он приурочен главным образом к южных райо-
нам— Северному Кавказу и Закавказью, республикам Средней
Азии и т. д.
Диагносцнрование эхинококкоза, особенно на ранних стадиях
заболевания, представляет большие трудности. В последние годы
широко применяется внутрикожная реакция: приготовленный из
ларвоцист антиген вводят в кожу обследуемого человека. У зара-
женных людей развивается положительная реакция. Хорошие ре-
зультаты дает и реакция преципитации протосколексов эхинококка
в сыворотке. Большое значение имеет рентгеноскопия.
Единственным по настоящему эффективным способом лечения
эхинококкоза в наше время остается хирургическое вмешательст-
во. Правда, в запущенных случаях и оно становится иногда невоз-
можным. Предлагаемые химиотерапевтические средства, как по-
казали результаты их применения, еще очень несовершенны.
Мероприятия по предупреждению заражения людей эхинокок-
козом основываются на строгом соблюдении правил личной и об-
щественной гигиены.. В эндёмичных очагах заболевания необходим
строгий ветеринарный контроль за собаками. При забое скота важ-
ное значение имеет-уничтожение или обезвреживание органов, по-
раженных ларвоцистами. Это же относится и к трупам павших
животных, которые ни в коем случае не должны поедаться со-
баками.
Alveococcus multilocularis (рис. 21,А) также паразитирует
у различных плотоядных животных. Чаще он встречается у лис
и песцов. Стробиллярные формы этого вида морфологически очень
близки к Echinococcus granulosus и отличаются от последнего
главным образом заметно меньшими размерами (с. 6), а также
строением матки перезрелого членика, которая не образует боковых
выростов. Промежуточными хозяевами альвеококка служат раз-
личные грызуны: полевки, лемминги, ондатры и др. Факультатив-
ным промежуточным хозяином этого паразита является и человек.
Ларвоцнсты A. multilocularis, как уже отмечалось ранее, ха-
рактеризуются наружным почкованием, в результате чего возни-
кает большое скопление мелких (3—5 мм в диаметре) пузырьков,
которые заполнены студенистой массой и содержат единичные
сколексы. Рост всего этого образования происходит по периферии,
тогда как в центральных ее участках возникают некротические оча-
ги. Пузыри альвеококка содержат большое количество гиалурони-
47
A
Рис. 21. Alveococcus multilocularis (по разным авторам с измен.).
А — стробила; Б — пути циркуляции A. multilocularis в природе
дазы, что обеспечивает, по-видимому, инфильтрирующий рост па-
разита и образование многочисленных метастазов.
Заражение человека происходит при случайном проглатывании
яиц, ^содержащих инвазионные онкосферы. Личинки совершают
миграцию, аналогичную миграции личинок эхинококка. Паразиты
первоначально, как правило, локализуются в печени. Вокруг раз-
вивающегося скопления пузырей возникает воспалительный очаг.
Между пузырьками появляются разрастание грануляционной, а по-
том и плотной фиброзной ткани. По плотности альвеококкозный
узел, часто прорастающий в соседние с печенью органы, напомина-
ет хрящ. Первичный очаг дает начало многочисленным метастазам,
которые обнаруживаются как в самой печени, так и в других орга-
нах: лимфатических узлах, легких, почках, селезенке, головном
мозгу и т. д. У больных наблюдается атрофия печеночной ткани,
застой желчи, абсцессы печени. В случае прорыва нагноивше-
гося альвеококкозного узла в брюшную полость развивается пери-
тонит.
Клинические признаки заболевания появляются, как правило,
очень поздно, иногда через несколько лет после заражения. Пер-
вым отчетливо выраженным симптомом является увеличение пе-
чени, позднее возникает ощущение тяжести и тупая ноющая боль.
В дальнейшем развивается желтуха, истощение. Больные могут
жить с альвеококкозом до 20 лет (в среднем, однако, этот срок зна-
чительно короче и составляет всего 8—9 лет) и погибают от желту-
хи или, что несколько реже, от метастазов в мозгу.
Эпидемиология альвеококкоза характеризуется наличием устой-
чивых природных очагов, существование которых поддерживается
плотоядными и различными грызунами. Возможно возникновение
и синантропных очагов, в которых роль главного дефинитивного
хозяина играют собаки, а промежуточного — синантропные грызу-
ны. Сельскохозяйственные животные, хотя в экспериментальных
условиях и заражаются альвеококкозом, по-видимому, не имеют
сколько-нибудь серьезного эпидемиологического значения.
Заражение человека, как правило, происходит в природных оча-
гах (рис. 21,5) и чаще всего встречается у охотников. Нередко на-
блюдается инвазия и у лиц, связанных с обработкой шкурок живот-
ных. Источником заболевания людей могут быть ездовые, охот-
ничьи и пастушечьи собаки. Серьезную опасность представляет
употребление воды из небольших непроточных лесных водоемов,
лесных ягод и съедобных растений, собранных в районах обитания
диких хищников (лисиц, волков и т. п.).
Альвеококкоз в отличие от эхинококкоза-довольно четко приу-
рочен к определенным географическим областям. Чаще он встре-
чается в северных районах (Северная Канада, Аляска, северные
районы Европы, о-ва Берингова моря и т. д.). Отдельные очаги за-
болевания зарегистрированы в странах центральной' и южной Ев-
ропы: Швейцария, Франция, ФРГ. В СССР альвеококкоз отмечен
в Якутской АССР, Западной Сибири, на Камчатке, республиках
Средней Азии, Башкирской АССР.
49
Диагностика альвеококкоза затруднена еще в большей степени,
чем эхинококкоза. Обычно определить причину заболевания удает-
ся лишь на поздних стадиях, когда лечение во многих случаях ока-
зывается уже невозможным. Очень трудно бывает и дифференциро-
вать эти два заболевания. Лечение осуществляется путем хирурги-
ческого вмешательства. Профилактика альвеококкоза основывается
на тех же мероприятиях, что и профилактика эхинококкоза.
Решающее значение имеет соблюдение санитарных и гигиенических
норм.
Цестодозы домашних животных
Количество видов цестод, описанных из домашних животных и птиц, велико.
Однако данных о влиянии паразита на организм хозяина не очень много.
Часто встречаются у животпых заболевания, вызываемые представителями
сем. Anoplocephalidae (отр. Cyclophylidea). Большой ущерб сельскому хозяйству
1
Рис. 22. Moniezia expansa (по разным авторам). А — сколекс; Б — герма-
фродитный членик; В — яйцо; Г— орибатидный клещ с цистицеркоидами:
1 — наружная оболочка, 2 — грушевидный аппарат. 3 ~ цистицеркоиды
наносят монезиозы, возбудителями которых служат виды р. Moniezia: М. expansa,
М. bene deni и др. Эти паразиты обитают в’ кишечнике копытных животных (крупт
ный и мелкий рогатый скот).
Половозрелые черви достигают значительных размеров — до 1,5 м. Сколекс,
снабженный четырьмя присосками, лишен хоботка и крючьев (рис. 22, А). Герма-
фродитные членики несут по два половых комплекса (рис. 22, Б), Япца одеты
плотной наружной оболочкой и преобразованным в сложный грушевидный аппа-
рат эмбриофором (рис. 22, В). Развитие моннезий протекает с участием про-
межуточных хозяев — панцирных клещей (сем. Galumnidae, Oribatulidae и др.).
В их полости тела развиваются цистицеркоиды (рис. 22, Г).
Наиболее опасны монезиозы для овец и коз. У больных животных сначала
наблюдаются функциональные расстройства кишечника. Позднее развивается
малокровие и сильное истощение. Характерны и поражения нервной системы, вы-
ражающиеся в нарушении координации движении. Заболевание сопровождается
водянкой, выпадением шерсти. Ягнята сильно отстают в развитии и часто по-
гибают.
50
Борьба с мопезиозами в основном строится на профилактических мероприя-
тиях и дегельминтизации заболевших животных.
Важное экономическое значение имеет Multiceps multiceps Leske, 1780, — це-
пень мозговик (сем. Taeniidae, отр. Cyclophyllidea). Дефинитивным, хозяином
этого вида служат плотоядные (собаки, волки и т. д.). Длина стробилы полово-
зрелых паразитов обычно не превышает 10 см. Выводимые во внешнюю среду
перезрелые членики обладают подвижностью. Рассеиваемые ими яйца остаются
на поверхности почвы и травы. Облигатные промежуточные хозяева — овцы зара-
жаются, проглатывая инвазионные яйца вместе с кормом или водой. Онкосферы
мигрируют из кишечника в кровяное русло и разносятся по всему организму
животного. Развиваются, однако, лишь те личинки, которые достигают головного
или спинного мозга. Формирующаяся ларвоцнста — ценур (cm.jjhc. 13, Г) содер-
жит большое количество протосколексов. Личинки становятся инвазионными на
третий месяц после заражения.
Вызываемое М. multiceps заболевание овец, получившее название церебраль-
ного ценуроза, или вертячки, наносит серьезный урон овцеводству. Первый этап
€олезни связан с воспалительными процессами, развивающимися в мозговой
ткани. Он совпадает с периодом миграции личинок в мозге. Животные становят-
ся легко возбудимыми или, наоборот, впадают в глубокую депрессию. Некоторые
яз них погибают. Следующий этап сопровождается видимым улучшением состо-
яния. Он соответствует периоду медленного развития ценура. Третий и послед-
ний этап сопровождается резким ухудшением состояния больного животного.
У овец появляются судороги, они начинают совершать беспорядочные движения.
Очень часто животные двигаются по кругу (откуда и пошло название болезни —
вертячка). В конце концов наступает смерть.
Распространение ценуроза приурочено к районам интенсивного овцеводства.
Важное эпидемиологическое значение имеют пастушеские собаки, которые явля-
ются главными источниками инвазии. Борьба с ценурозом основывается главным
образом на. профилактических мероприятиях. С одной стороны, это строгий ве-
теринарный контроль за пастушескими собаками и периодическая их дегельмин-
тизация, с другой же — уничтожение и обезвреживание трупов забитых и павших
животных, которые могут стать источником новых инвазий.
Серьезный ущер.б животноводству может приносить и описанный выше
Echinococcus granulosus (с. 44). Эхинококкоз столь же опасен для животных,
как и для человека.
Обращает на себя внимание ярко выраженная патогенность личиночных ста-
дий ряда цестод: Echinococcus granulosus, Alveococcus multilocularis,
Multiceps multiceps и некоторых других близких к ним видов. Все они во взрос-
лом состоянии являются паразитами хищных животных (главным образом пред-
ставителен сем. Canidae). Весьма вероятно, что в этом, случае мы имеем дело
со своеобразной биологической адаптацией. Ослабленные, истощенные животные,
часто страдающие и нарушениями двигательных функций, становятся легкой
добычей хищников. Это обеспечивает бесперебойную циркуляцию паразит# в си-
стеме хищник — жертва.
Цестодозы рыб
В рыбах паразитирует бгромиое количество цестод, представляющих все
отряды этого класса, причем одни используют рыб только как промежуточных
хозяев, другие как окончательных, у третьих же с рыбами связано все развитие.
Наибольшее практическое значение имеют представители отр. Pseudophyllidea.
Их патогенность проявляется главным образом на фазе плероцеркоида. В этом
отношении наиболее" показательны ремнецы — сем. Ligulidae. В половозрелом
состоянии паразиты поселяются в рыбоядных птицах (с. 8). Первыми промежу-
точными хозяевами, как и у большинства остальных Pseudophyllidea, служат
пресноводные рачки (отр. Copepoda). В них формируются процеркоиды, которые
для своего дальнейшего развития должны попасть в организм второго промежу-
точного хозяина. Для видов, относящихся к р. Ligula, это преимущественно раз-
личные карповые рыбы — лещ, густера, плотва, язь, карась и т. д. Плероцеркоиды
локализуются не в тканях, а непосредственно в брюшной полости хозяина (рис. 23,
А, Б). Паразиты растут и постепенно достигают размеров взрослых червей. От
51
Рис. 23. Цестоды — паразиты рыб (по
Дубининой). А — плероцеркоид Ligula;
Б — рыба, инвазированная плероцеркои-
дами Ligula; В — сколекс Bothriocepha-
lus gowkongensis; Г — участок стробилы
Boihriocephalus gowkongensis
своих специфических хозяев. Они обычно
последних они по сути дела отличаются только тем, что не продуцируют яйца.
Крупные плероцеркоиды Ligula и других ремнецов обладают отчетливо выражен-
ной патогенностью. Их присутствие задерживает рост хозяина, снижает его упи-
танность, вызывает атрофию печени, селезенки, половых желез. Зараженные ры-
бы держатся около поверхности водоема, часто плавают на боку или даже кверху
брюшком. Распространение лигулеза снижает уловы в естественных водоемах,
Кроме того, ухудшаются вкусовые и питательные качества рыбы. Иногда парази-
ты становятся причиной массовых эпизоотий. В условиях прудовых хозяйств
лигулез, как правило, особой роли
не играет. Большая часть прудо-
вых рыб (карп, сазан, белый амур,
толстолобик) не восприимчивы к
этим паразитам.
Серьезный экономический
ущерб могут наносить и представи-
тели сем. Triaenophdridae (р. Triae-
nophorus). Эти средних размеров
черви паразитируют в кишечнике
хищных рыб (щука, окунь, форель,
сиг, судак и» т. д.). Их сколекс
несет две крупные ботрии и 4
крючка, имеющих форму трезубца
(см. рис. 2, Г). Тело практически
не расчленено и содержит много-
численные половые комплексы. Раз-
витие Thiaenophorus осуществляет-
ся с участием двух промежуточ-
ных хозяев — веслоногих рачков и
рыб. В последних, чаще всего в пе-
чени, развиваются плероцеркоиды.
В природных водоемах личинки
обычно встречаются в молоди тех
же видов хозяев, в которых оби-
тают и половозрелые черви. Нор-
мальная циркуляция паразитов
обеспечивается широко •распро-
страненным у хищных рыб явле-
нием каннибализма.
В природных условиях, при не-
высокой интенсивности инвазии па-
разиты не очень патогенны для
инкапсулируются, а компенсаторные
механизм обеспечивают нормальное функционирование пораженной печени. Кар-
тина резко меняется в прудовых хозяйствах, где выращивается форель. Эти
обычно обитающие в водоемах с быстрым течением рыбы в естественных условиях
почти никогда не заражаются плероцеркоидами Triaenophorus. Соответственно
и иммунитетом к новым для нее видам паразитов форель не обладает. В прудо-
вых же хозяйствах, где проточность невелика, иногда возникает массовое зараже-
ние молодых рыбок плероцеркоидами. Заболевание протекает исключительно
остро. Рыбы быстро теряют в весе, их развитие задерживается, заметно увеличи-
вается отход.
В последние годы довольно широкое распространение получил ботриоцефалоз
рыб. Boihriocephalus gowkongensis (сем. Bothriocephalidae) (рис. 23, В, Г), спе-
цифический паразит дальневосточных карповых рыб: белого амура и толстоло-
бика. Жизненный цикл осуществляется с участием одного промежуточного хозяи-
на — веслоногих рачков, в которых обитают процеркоиды. Плероцеркоиды, а позд-
нее и половозрелые черви, развиваются в кишечпике рыбы. Акклиматизация
в прудовых хозяйствах амура и толстолобика (без должного ветеринарного кон-
троля) послужила причиной распространения паразитов сначала в прудовых хо-
зяйствах, а потом и в естественных водоемах. Особенно опасны паразиты
в условиях рыбопитомников, где в первую очередь заражаются мальки и сего-
52
летки. Иногда наблюдается массовая гибель молодых рыбок. Чаще всего страда-
ют карповые хозяйства.
Борьба с цестодозами рыб в основном строится на широком применении
профилактических мероприятий, направленных па разрыв путей циркуляции па-
разитов в уже существующих очагах, и препятствующих возникновению новых.
Активное лечение используется лишь в случаях заражения рыб кишечными поло-
возрелыми формами (Bothriocephalus и др.).
КЛАСС CESTQDARIA (MONTICELLI) — ЦЕСТОДООБРАЗНЫЕ
Класс Cestodaria, или цестодообразные,— очень небольшая группа паразити-
ческих плоских червей, насчитывающая всего 8 видов, которые объединяются
в один отр. Amphilinidea.
Долгое время Amphilinidea вместе с Gyrocotylidea (с. 55), в качестве подкл.
Cestodaria входили в состав кл. Cestoda. В 1952 г. Р. Уордл и Д. МакЛеод
(R. Wardle and J. McLeod) предложили рассматривать Cestodaria как отдельный
Класс. Эта точка зрения была поддержана и известными французскими парази-
тологами Ч. Жуане и Ж- Бэром (Ch. Joyeux et G. Bayer). Академик Б. E. Быхов-
ский показал, однако, что гирокотилнды представляют собой явно обособленную
группу, стоящую ближе к моногенеям, чем к цестодам. Его взгляды получили
широкое распространение среди отечественных паразитологов. В то же время
Amphilinidea в отечественной литературе продолжали рассматриваться как абер-
рантные ленточные черви. Большую роль в этом сыграл авторитет К. Япицкого,
1{рторый считал амфилин иеотеническнми плероцеркоидами цестод, некогда пара-
зитировавших в хищных мезозойских рептилиях. Вымирание дефинитивного хозя-
на повлекло за собой и элиминацию половозрелых стадий. В последние годы
некоторыми исследователями справедливость гипотезы К. Яницкого была по-
ставлена под сомнение. М. Н. Дубинина убедительно показала отсутствие непо-
средственной филогенетической связи между Amphilinidea и Cestoda. Полностью
принимая эту точку зрения, мы выделяем группу Amphilinidea в самостоятельный
класс, сохраняя за последним название Cestodaria.
ОТРЯД AMPHILINIDEA РОСНЕ
Морфология
Тело взрослых Amphilinidea сильно уплощено и в большинстве
случаев имеет листовидную форму (рис. 24, А). На переднем конце
Ъ небольшом углублении располагается подвижный хоботок, к ко-
торому подходят протоки многочисленных железистых клеток. Зад-
ний конец тела несет узкое впячивание, которое в ряде старых работ
трактовалось как мочевой пузырь. К выделительной системе оно,
однако, не имеет отношения. На дне впячивания даже у половозре-
лых особей сохраняются хитиноидные личиночные крючки — оста-
ток церкомера.
Покровы амфилинидей образованы тегументом, поверхность ко-
торого лишена, однако, и типичных микротрихий (с. 9) и типичных
микроворсинок.
.Пищеварительная система отсутствует.
Выделительная система состоит из многочисленных мерцатель-
ных клеток, разбросанных в паренхиме. Собирательные каналы
образуют густую сеть (рис. 24,Б). Это характерно, по-видимому,
Для наиболее специализированных видов (р. Amphilina).^/ р. Ge-
phyrolina выделительная система, как и у Trematoda, Monogenoidea
и др., сохраняет билатерально-симметричное строение, причем от-
четливо выделяются главные продольные собирательные каналы.
Нервная система ортогонального типа. Впереди располагается
небольшой, парный ганглий, широко
соединяются кольцевой комиссурой,
стволы, впадающие в заднее нервное
//
Рис. 24. Amphilina foliacea (по разным ав-
торам) : А — строение половозрелой особи;
Б — выделительная система; В — задний
конец тела ликофоры с крючьями:
/ — наружное отверстие матки. 2 — протоки одно-
клеточных' желез, 3 — матка, 4 — желточникя, 5 —
-семенники, 6 — яичник, 7 — влагалище, 8 — семя-
извергательный канал, 9 — пропульсаторный ор-
тан, 10 — семяпровод, 11 — сеть каналов выдели-
тельной системы
расставленные доли которого
В теле проходят продольные
кольцо.
Гермафродитная половая
система устроена по общему
для большинства паразити-
ческих’плоских червей типу.
Массивный неправильной
формы яичник расположен
в задней части тела. От него
отходит короткий яйцевод,
в который открываются вла-
галище и главный желточ-
ный проток. Наружное от-
верстие влагалища располо-
жено на боковой поверхно-
сти тела недалеко от его
заднего конца. В своей про-
ксимальной части влагали-
ще несколько расширено и
образует небольшой семя-
приемник. Главный желточ-
ный проток возникает в ре-
зультате слияния правого и
левого желточных протоков,
направляющихся к желточ-
ным фолликулам. Послед-
ние в виде двух латераль-
ных рядов тянутся вдоль
всего тела. Яйцевод впадает
в оотип, окруженный много-
численными одноклеточны-
ми железками (тельце Мели-
са), от которого берет нача-
ло длинная извитая матка. Последняя открывается наружу субтер-
минально на переднем конце тела.
Мужская половая система представлена многочисленными мел-
кими семенниками, образующими два широких продольных поля,
которые расположены ко внутри от желточников. Отходящие от
семенников семявыносящие канальцы, сливаясь, дают начало изви-
тому семяпроводу, который направляется к заднему концу тела.
Настоящий копулятивный орган (пенис, или сумка цирруса)
у Amphilinidea отсутствует. Семяпровод переходит в трубчатый се-
мяизвергательный канал. На границе между ними в.виде мускули-
стого сфинктера располагается пропульсаторный аппарат, сокраще-
нием которого семенная жидкость и выбрасывается из мужского
полового отверстия.
Жизненный цикл
Как уже отмечалось ранее, амфнлинидеи обитают в полости те-
ла своих хозяев. В процессе эволюции у них возникли специальные-
адаптации для выведения яиц во внешнюю среду. Виды, паразитиру-
ющие у осетровых рыб (Amphilina), для этой цели используют
брюшные поры хозяина. Черви вводят в пору передний конец свое-
го тела, на котором расположено отверстие матки, и выбрасывают
яйца в воду. Виды, обитающие в рыбах, лишенных брюшных пор,,
и в черепахах, передним концом прободают стенку тела хозяина..
Развитие яиц завершается в матке червя. В воду попадают яй-
ца, которые содержат сформированную личинку — ликофору. Пос-
ледняя, как правило, несет ресничный покров. В теле личинки рас-
полагаются крупные железистые клетки, протоки которых откры-
ваются на переднем конце. На заднем полюсе имеется 10 крючьев-
(рис. 24,В). Две пары крючьев, расположенных терминально, за-
метно отличаются от остальных по форме и размерам.
Ликофора в воду не выходит, и заражение промежуточного хо-
зяина происходит при поедании им яиц, содержащих инвазионных:
личинок. Для вида Amphilina foliacea, цикл развития которой изве-
стен, промежуточными хозяевами являются различные гаммарусы>
(р. Gammarus, Dikerogammarus и др.).
Личинка из кишечника мигрирует в полость тела хозяина, где
дает начало следующей личййочной стадии, соответствующей про-
церкоиду цестод. В отличие от ленточных червей и гирокотилид
(с. 57) у личинок Amphilina foliacea не происходит образования обо-
собленного церкомера. Распространенные в литературе данные
о наличии на этой стадии развития хвостового придатка, как пока-
зали исследования последних лет, ошибочны. Хитиноидные крючка
располагаются непосредственно на заднем конце тела личинки.
Дальнейшее развитие A. foliacea осуществляется в дефинитив-
*ном хозяине. Проглоченная вместе с рачком личинка пробуравли-
вает стенку кишечника осетра, мигрирует и локализуется в его-
брюшной полости и через некоторое время достигает половой зре-
лости.
Данные о географическом распространении амфилин не очень
многочисленны, хотя они и обнаружены на всех материках, кроме-
Антарктиды. Основное количество видов приурочено к тропической
и субтропической зонам. В СССР встречаются лишь представители
р. Amphilina: в осетровых европейской части — A. foliacea и на
Дальнем Востоке — A. japonica.
КЛАСС GYROCOTYLOIDEA BYCHOWSKY— ГИРОКОТИЛИДЬГ
Класс Gyrocotyloidea— небольшая группа плоских червей, паразитирующих
в'химерах (отр. Holocephala — цельпоголовые рыбы).
Как уже отмечалось ранее (с. 53), Gyrocotyloidea долгое время включались
либо в состав кл. Cestoda, либо в состав кл. Cestodaria. Лишь Б. Е. Быховскнй
выделил, гирокотилид в самостоятельный класс, показав при этом их филогене-
тическую близость с моиогенеями. „
55
Морфология
Гирокотплиды имеют уплощенное листовидное тело, края которого часто
несут многочисленные складчатые фестоны (рис. 25, А). На переднем конце рас-
полагается округлой или грушевидной формы присоска. Задний же конец тела
снабжен своеобразным прикрепительным аппаратом, имеющим вид сильно склад-
чатой розетки. В передней ее части на границе с туловищем дорзально распола-
гается небольшое впячпвание. иногда называемое воронкой. На дне этого углуб-
ления у взрослых червей сохраняются личиночные крючья.
Покровы гироктилид построены по типу тсгумента (рис.'25, Б). Поверхность
его несет микроворсинки, концы которых вытягиваются в тонкий бичевидный
придаток. В субтегумептальной зоне розетки располага!отся железистые клетки,
продуцирующие PAS-положительный материал (по-видимому, мукополисахари-
ды), который выделяется наружу и, вероятно, способствует более прочному при-
креплению паразита. У некоторых видов тегумеит переднего конца тела несет
мощные кутикулярные шипы (рис. 25, А). Непосредственно под тегументом рас-
полагаются базальная мембрана и два слоя мышц — кольцевые и продольные.
Пищеварительная система у гнрокотилид отсутствует. Выделительная система
представлена протонефридиями. В паренхиме разбросаны многочисленные мерца-
тельные клетки, протоки которых впадают в систему каналов, образующих гу-
стую сеть (см. рис. 26, А). Эта сеть в свою очередь соединяется с продольными
главными собирательными каналами, которые направляются вперед. Экскретор-
ные поры так же, как и у моногеней, располагаются на боковых поверхностях
в передней трети тела.
Нервная система ортогонального типа. От ганглиев, локализующихся по бокам
или позади передней присоски и соединенных поперечной комиссурой, берут на-
чало два боковых продольных ствола. В заднем конце тела они соединяются хо-
рошо развитым кольцом, от которого иннервируется прикрепительная розетка.
Органы чувств у гнрокотилид развиты слабо. Они представлены одиночными сен-
силлами. Чувствительные нервные клетки дают отростки, заходящие в толщу те-
гумента. Здесь они соединяются с основанием неподвижной реснички (рис. 25, Б).
Gyrocotyloidea гермафродиты. Женская половая система представлена не-
большим яичником, локализующимся в задней половине тела. В. короткий яйце-
вод открываются проток семяприемника, многочисленные скорлуповые железы и
влагалище, которое в виде простой трубки тянется вперед и открывается нару-
жу на дорзальной поверхности тела. От яйцевода берет начало извитая матка.
В дистальной части она образует расширение — маточный мешок. Отверстие
матки располагается вентрально в передней половине тела.
Мужская половая система состоит из многочисленных мелких семенников,
распространение которых ограничено передней половиной тела, семявыносящих
путей и слабо развитого копулятивного органа, расположенного вблизи наруж-
ного отверстия матки.
Жизненный цикл
Развитие эмбриона у разных видов гнрокотилид осуществляется либо во
внешней среде, либо в период прохождения яиц по петлям матки. Зрелая личин-
ка — ликофора покидает яйцевую скорлупку и переходит к *активному плаванию.
Ее тело слегка вытянуто в длину и одето ресничным покровом (рис. 25, В). На
переднем конце открываются протоки одноклеточных желез. Выделительная си-
стема представлена небольшим числом мерцательных клеток и отчетливо била-
терально-симметрична. Сеть анастомозирующих каналов еще отсутствует
(рис. 25, Г). На заднем конце тела личинки располагаются 10 одинаковых по
размеру и строению крючьев.
Каким путем ликофоры попадают в организм хозяина, еще не до конца ясно.
Американский исследователь Г. Мантер (Н. Manter) не исключает возможности
активного проникновения ликофор через жаберный эпителий в кровеносную си-
стему рыбы. В дальнейшем личинки мигрируют в сосуды слизистой кишечника
и уже оттуда в просвет последнего. Однако никакого экспериментального под-
56
Рис. 25. Организация гирокотилид (по разным авторам). А—строение Gyrocotyle
fimbriata; Б — схема строения тегумента; В — ликофора Gyrocotyle urna-t Г —
схема -строения выделительной системы лнкофоры G. игпа:
/ ~-присоска, 2 —ганглий, 3 — влагалище, 4 — семенники, 5—маточный мешок, 6 — матка,
7 — продольный нервный ствол, 8 — семяприемник. 9 — яичник. 10 — воронка, 11 — кольце-
вая комиссура, 12 — розетка, 13 — семяизвергательный канал, 14 — наружный слой тегумен-
та, 15 — микроворсинкн, 16 — базальная мембрана, /7 — мускульные слон, 18 — нервное
окончание, 19 — ресничка
тверждения эта гипотеза пока не получила. Взрослые черви обычно локализуются
в спиральном клапане химер.
Долгое время считалось, что, попав в пищеварительную систему хозяина,
молодые особи растут и прямо превращаются в половозрелых паразитов. Однако
в последние годы известный шведский паразитолог Г. Мальмберг (G. Malmberg)
показал, что личинки гирокотилид, оказавшись в кишечнике химеры, переходят
/К временному гиперпаразитизму. Они проникают в паренхиму молодых особен
того же вида. В последних протекают начальные этапы метаморфоза паразитов.
Рис. 26. Последовательные стадии развития Gyrocotyloidea (по Мальмбергу,
с измен.):
1 — церкомер, 2 — розетка, 3 — выделительная система
Морфологические преобразования ликофор в этот период сводятся к появлению
отчетливо выраженного церкомера (рис. 26). Он имеет вид небольшого круглого
тельца и несет радиально расположенные крючья.
В дальнейшем молодые черви покидают паренхиму своих временных «хозя-
ев» и снова выходят в просвет спирального клапана. Их церкомер начинает по-
степенно погружаться внутрь образующегося на заднем конце тела впячивания.
Границы церкомера постепенно исчезают, так что крючья оказываются располо-
женными непосредственно на дне этого' углубления. Прилегающие к нему уча-
стки тела разрастаются и образуют прикрепительную розетку. Последняя таким
образом не гомологична прикрепительному диску моногеней, который во всяком
случае у примитивных форм, представляет собой истинный церкомер. Одновре-
менно с началом инвагинации церкомера у гирокотилид закладывается передняя
присоска. Наблюдается усложнение выделительной системы за счет увеличения
числа мерцательных клеток и образования сложной сети каналов (рис. 26).
Все известные Gyrocotyloidea, объединяются в два рода: Gyrocotyle, вклю-
чающий 9 видов (G. fitnbriaia — рис. 25, А и др.), и Gyrocotyloides, представлен-
ный одним видом (6. nybelini).
Географическое распространение гирокотилид ограничено зоной распростра-
нения их хозяев. Паразиты часто встречаются у химер, обитающих у берегов Се-
верной Америки, Европы, Новой Зеландии, Австралии и т. д.
Происхождение и филогения паразитических плоских червей
Паразитические плоские черви ведут свое< начало от свободно-
живущих турбеллярий и, по-видимому, наиболее тесно связаны
с предками прямокишечных турбеллярий группы Dallyelida. Един-
ство происхождения паразитических плоских червей проявляется
в общности ряда черт их организации и жизненных циклов. Так,
по-видимому, все классы паразитических Plathelminthes имеют
58
единый план строения покровов, соответствующий таковому неко-
торых турбеллярнй. Единство исходных структур обусловливает
характер их изменений, вызванных паразитизмом. Именно этим
можно объяснить тот факт, что у цестод и у спороцист трематод,
утративших пищеварительную систему, возникли сходные адапта-
ции, выразившиеся в появлении на тегументе микроворсинок и ми-
кротрихий, рассматривающихся как структурной основа пристейоч-
ного пищеварения. Паразитические Plathelminthes сохраняют уна-
следованные от турбеллярний сенсиллы. Для всех классов
характерен общий план строения нервной, половой и экскреторной
систем и т. д.
Однако, несмотря на общность их происхождения, разные груп-
пы паразитических червей прошли совершенно различные пути эво-
люции и самый переход к паразитизму осуществлялся ими в разные
геологические эпохи. О хронологии паразитических Plathelminthes,
так же, как о хронологии других паразитов,'не оставляющих следа
в палеонтологической летописи, можно судить только по косвенным
данным. Эволюция паразитов и их хозяев является сопряженным
процессом. Исходя из этого, можно сделать некоторые заключения
о филогении паразитов, основываясь на филогении их хозяев. Тре-
матоды и близкий к ним класс аспидогастрид, по-видимому ведут
свое начало от тех рабдоцелиообразных предков, которые стали
сперва симбионтами, а потом паразитами моллюсков.
По современным представлениям две основные ветви развития
моллюсков — подтип боконервных (Amphineura) и раковинных
(Conchifera) —обособились очень рано. Отсутствие у боконервных
моллюсков партеногенетических поколений трематод позволяет
сделать вывод о том, что установление первых экологических свя-
зей между протрематодами и моллюсками началось уже'после то--
го, как от общего ствола развития моллцрсков отделились формы,
давшие начало современным Amphineura. Партеногенетические
поколения трематод никогда не- встречаются и у головоногих мол-
люсков. Однако последние являются хозяевами своеобразных па-
разитических организмов — дициемид, о систематическом положе-
нии которых до сих пор нет единого мнения.
Дициемиды (подкл. Dicyemide, относящийся к кл. Mesozoa) —
своеобразная группа вторично упростившихся плоских червей (Бек-
лёмишев, 1964), паразитирующих в почках головоногих моллюсков.
Их жизненный цикл представляет собой гетерогонию. Партеноге-
нетических самок (нематогены) можно сравнить с партенитами
трематод. Развивающееся из их зародышевых клеток гермафродит-
ное поколение подавлено и представлено очень кратковременной
стадией (инфузориген), которая, размножаясь, дает начало рассе-
лительной ресничной личинке (инфузориформ).
Большое сходство жизненных циклов дициемид и трематод на-
водит на мысль о существовании филогенетической близости меж-
ду обеими группами. Основываясь на этом, можно полагать, что
переход протрематод к паразитизму должен был начаться еще до
Того, как примитивные Conchifera разделились на Cephalopoda
59
и Prorhipidoglossa (гипотетические предки всех современных Соп-
chifera). Очевидно, протрематоды, паразитировавшие на предках
головоногих, проделали вместе с ними особый путь эволюции и да-
ли начало дициемидам. Предками же сосальщиков были, по-види-
мому, те протрематоды, эволюция которых шла параллельно эво-
люции Prorhipidoglossa. Доказательством этого предположения
можно считать тот факт, что партеногенетические поколения сов-
ременных трематод паразитируют только у потомков Prorhipidog-
lossa, т. е. у брюхоногих, пластинчатожаберных и лопатоногих мол-
люсков. Обособление головоногих как самостоятельной ветви раз-
вития моллюсков относят к кембрию. Очевидно, в этот же период
произошла и дивергенция протрематод, приведшая к образованию
дициемид, с одной стороны, и трематод — с другой.
Важным этапом эволюции жизненного цикла протрематод сле-
дует считать момент перехода их свободноЖивующего поколения
к паразитированию в организме позвоночных. Представители при-
митивных групп современных трематод (Paramphistomata) не встре-
чаются у элязмобранхий, но довольно широко распространены
у морских костистых рыб (Teleostei). Это дает основание предпо-
лагать, что не элязмобранхии, а костистые рыбы явились первыми
хозяевами трематод из числа позвоночных. Установление паразито-
хозяинных отношений между трематодами и позвоночными мож-
но, таким образом, отнести ко времени появления Teleostei, т. е.
к мезозойской эре (на границе триаса и юры).
Жизнь древнего палеозоя была сосредоточена в море. Несом-
ненно, что именно там и проходила совместная.эволюция протре-
матод и моллюсков на протяжении многих миллионов лет. Трудно
сказать, в какой период произошел выход протрематод в пресные
воды. Ясно лишь одно — они могли попасть туда только вместе со
своими хозяевами моллюсками. Первые пресноводные моллюски
(Prosobranchia) отмечены уже в карбоне. Однако широкое распро-
странение они получили лишь в мезозойскую эру. Для протрематод
выход в пресные воды стал переломным моментом эволюции, преж-
де всего потому, что при этом значительно расширился круг их по-
тенциальных дефинитивных хозяев. Особенно важным оказалось
приспособление к паразитированию в теплокровных животных.
Постоянство температуры их тела наряду с постоянством внутрен-
ней среды и изобилием пищи создало для существования паразитов
благоприятные условия.
Особого внимания заслуживает вопрос о филогенетических вза-
имоотношениях между трематодами и аспидогастридами. Послед-
ние ограничены в своем распространении пойкилотермными позво-
ночными и беспозвоночными, главным образом моллюсками. Объе-
динение их в один класс с трематодами, по-видимому, совершенно
искусственно: те общие черты строения, на основании которых они
объединяются, в равной мере свойственны турбелляриям. Органы
же прикрепления аспидогастрид, представленные своеобразным
диском, поделенным на множество присоскообразных ячеек, не
имеют гомологов у трематод. Особенно существенным следует счи-
60
тать характер жизненного цикла аспидогастрид, который в отли-
чие от трематод осуществляется без гетерогонии: даже в тех слу-
чаях, когда имеется смена.хозяев, чередования поколений нет. Воп-
рос о происхождении и систематическом положении аспидогастрид
является очень дискуссионным. Мы присоединяемся к тем авторам,
которые полагают, что аспидогастриды в своем становлении и эво-
люции связаны с моллюсками и являются боковой ветвью филоге-
нетического развития протрематод.
Совсем иным путем осуществлялся переход к паразитизму пред-
ков мопогенетических сосальщиков. Как полагает Б. Е. Быховский
(1957), данные по морфологии, биологии и развитию моногеней
позволяют судить о том, что их первыми хозяевами были ископае-
мые хрящевые рыбы — химерообразные, или Elasmobranchia. Пред-
ки моногеней поселялись на поверхности их тела как эктопарази-
ты. Возникновение этой группы плоских червей должно быть
приурочено к силуру или, скорее, к девону. Таким образом, Мопо-
genoidea, по-видимому, стали паразитами значительно позднее,
чем партеногенетические поколения трематод, и много раньше, чем
их гермафродитное поколение. Жизненный цикл промоногенетиче-
ских сосальщиков был примитивным и проходил без чередования
поколений. Система промежуточных хозяев в ходе эволюции моно-
геней не выработалась, так как для расселения эктопаразитов она
не дает особых преимуществ: заражение новых особей хозяина
обеспечивается специальными адаптациями в строении яиц и био-
логии личинок. Моногеней — эктопаразиты, весьма тесно связанные
с внешней средой, не пошли по пути морфо-физиологического рег-
ресса. Главным направлением их эволюции стало усложнение
и усовершенствование прикрепительного аппарата, обеспечивающе-
го надежность связи с хозяином.
Местом первичного поселения мопогенетических сосальщиков
служили покровы и жаберная полость рыб. Постепенное перемеще-
ние червей в ротовую полость, а затем в глотку и пищевод рыбы
можно рассматривать как переход к эндопаразитизму. Наличие
этой тенденции у моногеней объясняет возможность развития от
общего с ними предка таких типичных эндопаразитов, как ленточ-
ные черви. Возникновение цестод и моногеней было, по-видимому,
приурочено к одной .геологической эпохе. Таким образом, цестоды
должны были приспособиться к паразитированию в позвоночных
намного раньше, чем гермафродитное поколение трематод, видимо,
долгое время остававшееся свободным. Очевидно, это п послужи-
ло причиной далеко зашедшего морфо-физиологического регресса
цестод. Жизненный цикл этих последних проходит, к^к и у моноге-
ней, без гетерогонии, но с участием промежуточных хозяев, выпол-
няющих роль передаточного звена, необходимого для заражения
дефинитивного хозяина. Промежуточные и дефинитивные хозяева
цестод экологически объединены наличием пищевых связей.
Особое место в системе плоских червей занимает своеобразная
группа Gyrocotyloidea — эндопаразиты древних химеровых. Одни
черты организации роднят этих червей с моногенеями, другие —
61
с цестодами. Gyrocotyloidea — самостоятельный класс, занимающий
промежуточное положение между моногенеями и ленточными чер-
вями.
В эволюции паразитических плоских червей можно наметить
две основных линии развития. Обе они берут начало от предковых
форм, общих сг прямокишечными турбелляриями типа Dallyelida
(рис. 27). Одна линия начинается с перехода предковых форм
к симбиотическим отношениям с моллюсками. По мере эволюции
этих отношений симбионты стали
Рис. 27. Филогенетические .отно-
шения в пределах типа Plaihel-
minthes (по Быховскому, с измен.)
эндопаразитами моллюсков. Та-
ким был путь эволюции протре-
матод, которые дали начало сов-
ременным трематодам и аспидб-
гастридам. Можно полагать, что
от протрематод отделились и
предки современных дициемид,
проделавшие самостоятельную
эволюцию вместе со своими хозя-
евами — головоногими моллюска-
ми, и достигшие крайней степени
морфо-физиологического регрес-
са.
Вторая линия эволюции пара-
зитических плоских червей — это
формирование группы Cercomae-
romorpha, характеризующейся на-
личием особого образования —
церкомера (с. 58). Предки этой
группы червей, очевидно, были эк-
топаразитами палеозойских хрящевых рыб (элязмобранхий).
В дальнейшем они дали начало четырем самостоятельным клас-
сам. Представители одного из них — моногенеи — остались эктопа-
разитами рыб. Круг их хозяев значительно расширился и в настоя-
щее время эти эктопаразиты встречаются не только па хрящевых,
но и на костистых рыбах — морских и пресноводных?
Остальные Cercomearomorpha стали эндопаразитами. Наиболее
обширным и процветающим, несомненно, является класс ленточных
червей. Вспышка видообразования в пределах этого класса по всей
вероятности связана с освоением новых животных хозяев — назем-
ных позвоночных. Два других класса — Gyrocotyloidea и Cestodaria—
сохранили связь с древними хрящевыми рыбами. Обе группы очень
обособлены и .малочисленны. Единство происхождения всех церко-
мерпых червей подтверждается сохранением у них церкомера. У мо-
ногенетических сосальщиков гомолог этого образования — прикре-
пительный диск, у цестод церкомер хорошо выражен на стадии
процеркоида, но в ходе дальнейшего развития отбрасывается.
У Cestodaria и Gyrocotyloidea церкомер, отчетливо развитый у ли-
чинок, втягивается в паренхиму заднего конца тела взрослых
особей.
62
ТИП ACANTHOCEPHALES
(RUD., 1808) SKR., S C H„ 1931 —
СКРЕБНИ, ИЛИ КОЛЮЧЕГОЛОВЫЕ
Небольшая, хорошо обособленная группа паразитических чер-
вей, насчитывающая около 500 видов. Тело подразделяется на
туловищную часть и вооруженный крючьями прикрепительный хо-
боток, вворачивающийся в специальное влагалище._ Кожно-мус-
кульный мешок хорошо развит, полость тела — первичная. Выде-
лительная система устроена по типу протонефридиев, нервная —
характеризуется наличием дифференцированного мозгового
ганглия, пищеварительная —отсутствует. Скребни раздельнополы.
Их жизненный цикл проходит с метаморфозом и со сменой живот-
ных хозяев. К типу Acanthocephala относится только один класс
с тем же названием.
КЛАСС ACANTHOCEPHALA (RUD.) 1808
Первое упоминание о скребнях принадлежит Реди, который в 1684 г. обнару-
жил этих червей в кишечнике угря. Но самостоятельный отр. Acanthocephala,
насчитывающий около 50 видов, был обоснован Рудольфи только в начале
XIX в. Первое описание морфодогии, а отчасти и физиологии скребней было дано
А. Веструмбом (A. Westrumb); позднее Р. Лейкарт (R. Leuckart) отметил харак-
терное для их жизненного цикла явление смены хозяев. Основные принципы си-
стемы скребней были разработаны М. Люэ (М. Luhe).
В паше время наиболее важные исследования по морфологии, развитию и си-
стематике скребней принадлежат X. И. Ван-Кливу (Н. Van-Cleave) и А. Мейеру,
автору большой сводки по скребням (А. Меуег, 1932—1933). Среди русских ис-
следователей необходимо назвать Н. Н. Костылева и В. 'И. Петроченко.-
Морфологическая характеристика кл. Acanthocephala соответствует таковой
типа. Половозрелые черви паразитируют в кишечнике позвоночных. Выведенные
наружу яйца заглатываются водными или наземными членистоногими, в кишеч-
нике которых из яйца выходят личинки, проникающие через стенку кишечника
в полость тела хозяина. Там личинки претерпевают сложный метаморфоз и инцисти-
руются. Дефинитивный хозяин получает скребней, пожирая зараженных члени-
стоногих. Этот тип жизненного цикла может вторично усложняться за счет вклю-
чения еще одного хозяина — резервуарного (с. 73).
Класс скребней подразделяется на три подкласса, различающихся между
собоГг по строению ранних личиночных-стадий (см. рис. 39). Для подкл. Neoechi-
norhynchinea характерны невооруженные личинки. Подкл. Echinorhynchinea имеет
личинок, вооружение которых представлено крючьями, сгруппированными на пе-
реднем конце тела, и, наконец, личинки скребней, относящихся к подкл. Gi-
ganthorhynchinea, кроме крючьев несут многочисленные шипики по всей поверх-
ности тела.
Морфология и физиология скребней
Скребни (рис. 28) характеризуются вытянутым, слегка упло-
щенным телом-, на переднем конце которого находится хоботок,
вооруженный рядами кутикулярных крючьев. Размеры тела варьи-
руют от 1,5 (Pandasentis iracundus) до 650 мм (Macracanthorhyfi-
chus hirudinaceus). Окраска тела беловатая,’иногда ярко-оранже-
вая или коричневатая.
Кожно-мускульный мешок скребней (рис. 29) отличается очень
сложным строением. Он слагается из кутикулы, субкутикулярных
63
слоев и гиподермы. Снаружи тело одето тонкой пленкой мукополи-
сахаридов, играющей, по-видимому, защитную роль. Вся кути-
кула пронизана множеством микропор, ведущих в тончайшие
Рис. 28. Строение и половой димор-
физм скребня Polyinorphus tnagnus
(по Петроченко). А— самец; Б —
самка:
1 — хоботок, 2 — влагалище хоботка, 3 —
лемниски, 4 — яйца. 5 — семенники, б — це-
ментные железы, 7 — копулятивная сумка
канальцы, которые расположены
в субкутикулярном слое перпен-
дикулярно к поверхности тела.
В световом микроскопе эти ка-
нальцы видны как тонкая исчер-
ченность, и поэтому первый суб-
кутикулярный слой получил наи-
менование полосатого (или па-
раллельного). Далее следует вто-
рой субкутнкулярный слой, назы-
ваемый волокнистым и пронизан-
ный множеством переплетающих-
ся опорных фибрилл. Это слой
живой цитоплазмы, содержащий
митохондрии. Основную часть
толщи стенок тела скребней со-
Рис. 29. Поперечг/ый разрез стенки
тела скребня (по Петроченко):
/ — кутикула, 2 — параллельный слой суб-
кутикулы, 3 — войлочный слой, 4 — лаку-
на в гиподерме, 5 — гиподерма, б — слой
кольцевых мышц, 7 — ядра гиподермы
ставляет расположенная ниже гиподерма. Она тоже пронизана
фибриллярными структурами, играющими опорную роль, и несет
систему лакун, лишенных собственных стенок. Лакуны заполнены
жидкостью и, возможно, представляют собой транспортную систе-
му, осуществляющую перенос питательных веществ к разным участ-
64
кам тела животного. В лакунах находятся ядра. В толще гиподер-
мы тоже имеются крупные, иногда разветвленные ядра, называе-
мые гигантскими. Число их невелико и строго постоянно в преде-
лах вида. За гиподермой следуют слои продольных и кольцевых
мускульных волокон, отделенных от нее тонкой базальной мем-
браной.
Покровы скребней вооружены большим числом кутикулярных
крючьев, которые, по-видимому, являются производными волокни-
Рис. 30. Хоботки скребней и их вооружение (из кн. Петрочен-
ко). А — цилиндрический хоботок Echinorhynchus gadi\ Б —
шаровидный хоботок, Parafillicollis altmani-, В — хоботок
Rha'dinorhynchus horridus
стого слоя субкутикулы. Крючья локализуются не только на хо-
ботке, но и в передней части тела скребней.
Органом прикрепления служит хоботок, который имеет цилинд-
рическую, конусовидную или, реже, шаровидную форму (рис. 30J.
По всей его длине продольными рядами или по очень пологой спи-
рали расположены кутикулярные крючья. Последние имеют харак-
терную форму и состоят из корня, глубоко догруженного в толщу
гиподермы хоботка, и из острия, свободно выдающегося наружу
(рис. 31). Прикрепление осуществляется по принципу заякорива-
ния. Крючья, острия которых направлены назад, легко вводятся
в толщу тканей кишечника хозяина, глубоко вонзаясь при этом
в слизистую. Однако извлечь их оттуда можно лишь с трудом.
3—58
65
Число рядов крючьев и их форма строго постоянны в пределах
вида, и эти особенности используются как важный таксономический
признак.
Основание хоботка непосредственно переходит в мускулистый
мешок — хоботковое влагалище. Оно пронизано по всей длине мус-
кульными лентами (ретракторами хоботка), дистальные участки
которых прикреплены к стенке тела скребня. При сокращении рет-
ракторов хоботок вворачивается внутрь влагалища, как палец
Рис. 31. Строение крючьев хобот-
ка (по кн. Петроченко)":
/ — острие крючка, 2 —корень, погру-
женный в ткани
Рис. 32. Характер прикрепления сам-
ки (/) и самца (2) скребней р. Filicollis
к стенке кишечника хозяина (схема по
Оденингу)
перчатки. Между задней границей хоботка и собственно телом
скребня располагается небольшой участок, именуемый «шейкой»^
Отсюда отходят очень своеобразные и характерные для скребней
органы — лемниски. Это в большинстве случаев парные, широкие,
лентовидные образования, которые располагаются по обе стороны
хоботкового влагалища (см. рис. 28). По-видимому, они представ-
ляют собой выросты гиподермы, с ядрами и лакунами, в которые
заходит жидкость. Функция лемнисков не вполне ясна. Считается,
что они играют известную роль в процессе выворачивания хоботка,
так как при наполнении лакун жидкостью объем лемнисков увели-
чивается, и они сдавливают хоботковое влагалище.
Строение хоботка у скребней разного пола бывает различным.
Так, например, у вида FilicolUs anatis (паразиты водоплавающих
птиц) самки имеют мощно развитый пгаровидный хоботок, глубоко
66
внедряющийся в стенку кишки хозяина. Хоботок же самцов, напро-
тив, мал и слабо вооружен (рис. 32), вследствие чего самцы легко
могут быть выброшены наружу перистальтическими сокращениями
кишечника хозяина. Однако для жизни вида это не представляет
опасности. Гораздо важнее надежное закрепление самки, которая
после однократного оплодотворения живет долго и откладывает
множество яиц. Степень развития прикрепительного аппарата
скребней может быть связана и-с особенностями биологии хозяина.
Так, скребни, паразитирующие в морских млекопитающих, имеют
более мощный прикрепительный аппарат (рис. 33), чем паразиты
наземных позвоночных. Это, по-видимому, обусловлено повышенной
перистальтикой кишечника морских млекопитающих, заглатываю-
щих много воды с пищей. Для их паразитов опасность быть выбро-
шенными из кишечника особенно велика.
Пищеварительная система у скребней отсутствует. Пищевые
вещества воспринимаются всей поверхностью тела, в наружном
слое которого удается выявить активность щелочной фосфатазы.
По-видимому, важную роль в этом процессе играют микропоры
кутикулы (с. 64). Источником пищевых веществ, как показали
опыты с изотопом 14С, является содержимое кишечника хозяина.
Экскреторная система еще недостаточно изучена, а у некоторых
групп скребней и вообще неизвестна. В большинстве случаев она
устроена по типу протонефридиев и представляет собой либо дре-
вовидно ветвящийся орган, либо капсуловидное образование со
множеством колбообразных клеток мерцательного пламени
(рис. 34). Выводной капал протонефридия, как правило, соединен
с выводными протоками половой системы, открываясь наружу
общим отверстием.
Нервная система представлена шаровидным или овальным
ганглием (рис. 35), который заключен внутри хоботкового влага-
лища. Нервные стволы отходят от ганглия вперед к хоботку, где,
разветвляясь, иннервируют мускулы ретракторы, к боковым'стен-
кам и к задней части тела. У самцов в области полового отверстия
Имеется нервное кольцо, соединяющее продольные нервные стволы
и парное утолщение — половой ганглий. В строении нервной систе-
мы скребней имеются явственные следы ортогона.
Половая система находится в тесной связи с лигаментом. Это
узкий тонкостенный мешок, занимающий почти всю полость тела
червя. У некоторых видов лигамент представлен не одним, а двумя
мешками (вентральным и дорзальным), которые располагаются
по обе стороны проходящего по центру плотного осевого-тяжа
(рис. 36).
Половая система самки (рис. 37, Б) состоит из яичников и свое-
образно устроенного комплекса выводных путей. Яичники закла-
дываются в связи с тяжом лигамента, однако вскоре отрываются
от него и распадаются на многочисленные «яйцевые комки», т. е.
группы яйцеклеток, свободно плавающих в полости мешков лига-
мента или (если последние разрываются) непосредственно в поло-
сти тела червя. От этих своеобразных яичников отделяются яйце-
3* 67
клетки на разных стадиях развития, тоже взвешенные в полостной
жидкости. Центральной частью женских выводных путей является
так называемый маточный колокол. Это широкая трубка, располо-
Рис. 33. Усиление прикрепи-
тельного аппарата у скребней
из кишечника морских млеко-
питающих. А — Corynosoma
strumosum (по Мейеру); Б —
Bolbosoma physeteris (по Губа-
нову)
Рис. 34. Экскреторная система скребней
(из кн. Петроченко). А — капсуловидный
орган; Б — древовидный протонефридий:
1 — колбочки с мерцательным пламенем, 2 —
^нефрндиальная капсула, 3 выделительный
канал
женная под углом к продольной оси тела червя. Расширенной во-
ронкообразной частью маточный колокол открывается в полость
тела (или мешка лигамента), тогда как суженный задний конец
его переходит в тонкие яйцеводы. Кроме того, в задней части колот
68
кола имеется еще одно отверстие, сообщающееся с полостью тела.
Яйца через переднее отверстие поступают в маточный .колокол,
в котором происходит своеобразная сортировка. Зрелые яйца,
имеющие характерную веретеновидную форму, проталкиваются
в узкие яйцеводы, а оттуда в матку. Последняя ведет во влагалище,
которое открывается наружу половым отверстием. Только зрелые
Рис. 35. Центральная
часть нервной системы
скребней (из кн. Петро-
ченко) :
1 — стенки хоботкового вла-
галища, 2 — мозговой ганг-
лий, 3 — нервные стволы
Рис. 36. Схема организации
самки скребня (по Кестнеру,):
1 — хоботок с крючьями, 2— шейка,
3 — полость тела. 4 — вентральный
мешок лнгамента, 5 —тяж лнга-
мента, 6 — ничннк, 7— кожно-мус-
кульный мешок, 8 — матка, 9 —
экскреторный канал, 10 — протоне-
фридий. 11—дорзальный мешок
лнгамента, /2 —лемниски
яйца .могут пройти по этим очень узким протокам. Яйцевые комки
и незрелые яйца, имеющие шаровидную форму, выводятся из ма-
точного колокола через его заднее отверстие обратно в полость
тела. Там происходит их дозревание.
Половая система самца (рис. 37, 4) представлена двумя округ-
лыми семенниками, которые обычно расположены один за другим
и плотно облекаются лигаментом. Отходящие от них семяпроводы
соединяются в единый семявыносящий канал, который впадает
в совокупительный орган — пенис. Концевой участок семявынося-
щего канала расширяется, образуя семенной пузырек. Пенис от-
крывается в копулятивную сумку (бурсу), которая в спокойном
состоянии втянута в тело червя (рис. 37). Вывернутая бурса
69
(рис. 38). обхватывает при копуляции заднюю часть тела самки.
Непосредственно к семенникам прилегают цементные железы, про-
с семявыносящими каналами.
Эти железы выделяют клейкий
секрет, залепляющий после ко-
пуляции половое отверстие
самки. Образующаяся при
этом «цементная» пробка пре-
пятствует вытеканию введенно-
го семени. Число цементных
желез и характер их располо-
жения, сильно отличающиеся
у скребней разных видов, ис-
3 пользуются как систематиче-
ские признаки. К мужской по-
токи которых открываются вместе
А ‘ 5
Рис. 37. Половая система скребней.
А — схема строения мужской половой системы:
/ — семенники, 2 — семявыносящие канальцы. 3 — цементные железы, 4—мускулистый ме*
шок, 5 — семенной пузырек. 6—.пенис, 7 —ввернутая копулятивная сумка
Б — схема строения женской половой системы:
/ — тяж лигамента, 2 — маточный колокол. 3 — дорзальное отверстие колокола. 4 — матка»
5 — мускульный сфинктер влагалища, 6 — влагалище
Рис. 38. Схема строения копулятивной сумки (по Петроченко):
/—мускулистый мешок, 2 — пение, 3 —вывернутая сумка
ловой системе относится, кроме того, так называемый мускулистый
мешок. Этот пузырь с мускулистыми стенками заполнен жидкостью
и играет важную роль в процессе выворачивания копулятивной
70
сумки. В ее стенках имеются особые кармашки, в которые излива-
ется жидкость из мускулистого мешка. Наполнение кармашков
механически способствует выворачиванию сумки.
Обмен веществ скребней, как и других эндопаразитов, осуще-
ствляется по типу аноксибиоза. Основным энергетическим вещест-
вом служит гликоген, запасы которого сосредоточены в гиподерме
и отчасти в мышцах. Здесь же-накапливается и экскреторный жир.
Запасы гликогена колеблются на протяжении суток, что, по-види-
мому, можно поставить в прямую связь с суточным ритмом приема
пищи хозяином. При голодании последнего содержание гликогена
в теле скребней быстро снижается.
Развитие и жизненный цикл скребней
Эмбриональное развитие осуществляется во время пребывания
яиц в теле самки. К моменту выхода во внешнюю среду в них уже
заключены вполне сформированные личинки (рис. 39). Дальней-
Рис. 39. Типы яиц и строение личинок (по Петрочен-
ко). А — личинка с воружением на переднем конце
(подкл. Echinorhynchinea); Б — личинка вооружен-
ная крючьями и шипиками (подкл. Gyganthorhynchi-
пеа); В — невооруженная личинка (подкл. Neoechino-
rhynchinea)
71
г
шее развитие их становится возможным только после попадания
в организм промежуточного хозяина. Как теперь установлено, это
могут быть моллюски, ракообразные или наземные насекомые,
заглатывающие яйца скребней. В кишечнике хозяина личинка, ко-
торая на этой стадии развития называется акантором, выходит из
яйца и внедряется в толщу кишечной стенки (рис. 40, А, Б). Здесь
она держится в течение нескольких дней (иногда до двух недель),
после чего прорывает стенку кишечника и попадает в полость тела
хозяина. С этого момента начинается сложный метаморфоз. Ли-
чинка теряет крючочки и шипики, тело ее округляется, сильно уве-
личивается в размерах. Начинается закладка внутренних органов:
прежде всего хоботкового влагалища, затем хоботка, лигамента,
нервной системы. Последними появляются зачатки половой систе-
мы. Эта личиночная стадия, для которой характерен процесс ин-
тенсивного органогенеза, получила название преакантеллы
(рис. 40, В). После завершения органогенеза, занимающего 20—
30 дней и более, личинка переходит в следующую стадию разви-
тия— акантеллу. Этот переход сопровождается инцистированием.
Передний и задний концы впячиваются в полость тела личинки
(рис. 40, Г, Д), и оно принимает округлую или яйцевидную форму.
Снаружи развивается плотная оболочка, и таким образом личинка
переходит в покоящуюся стадию, которая сохраняет жизнеспособ-
ность в течение длительного времени.
Заражение окончательного хозяина происходит при поедании
промежуточных хозяев с акантеллами. В кишечнике окончательно-
го хозяина акантелла освобождается из цисты, хоботок ее вывора-
чивается, и она прикрепляется к стенке кишки. Сразу же начина-
ется интенсивный рост паразита и развитие его половой системы.
Сроки достижения половозрелости у разных видов скребней раз-
личны. Так, у утиного скребня (Polymorphus magnus) выделение
яиц начинается через 25—30 дней после попадания акантеллы
в кишечник хозяина. Напротив, у паразита свиней — гигантского
скребня (Macracanthorhynchus hirudinaceus) этот срок затягивает-
ся до 80 дней и более.
Таким образом, жизненный цикл скребней обычно проходит
с участием двух хозяев: промежуточного (различные беспозвоноч-
ные) н окончательного (позвоночные). Типичным примером явля-
ется жизненный цикл скребней рода Polymorphus (рис. 41), для
которых промежуточным хозяином служат рачки бокоплавы, а
окончательным — утиные птицы. Однако у многих скребней жиз-
ненный цикл может вторично усложняться за счет включения еще
одного хозяина, который не обязателен и называется резервуар-
ным. Таков жизненный цикл скребней рода Corynosoma, паразити-
Рис. 40. Стадии развития скребня Polymorphus magnus (по Петроченко).
А — акаптор; Б — акантор, проникший в стенку кишечника .рачка-хозяина; В —
преакантелла; Г — сформированная акантелла; Д — ннцнстировавшаяся акан-
телла
73
рующих в кишечнике тюленей (рис. 42). Промежуточным хозяином
Corynosoma служат бокоплавы (р. Pontoporeia). Они охотно по-
едаются рыбами, в результате чего заключенные в их теле акан-
теллы попадают в кишечник рыб, но не задерживаются там,
а переходят в полость тела или в мускулатуру. В организме рыбы,
Рис. 41. Жизненный цикл скребня Polymorptius magnus (по Пет-
роченко) :
/ — половозрелые скребни в кишечнике утки. 2 — яйцо во внешней среде.
3 — акантелла, иицистнрованная в полости тела промежуточного хозяина
(рачка-бокоплава)
которая и представляет собой резервуарного хозяина, акантеллы
никакого развития не претерпевают. Интенсивность же инвазии
рыб может быть довольно значительной и достигает 80—100 эк-
земпляров. Тюлени могут случайно заглатывать рачков и таким
путем получать скребней непосредственно от промежуточного хо-
зяина; питаются же тюлени преимущественно рыбой. Поэтому роль
резервуарного хозяина (рыбы) исключительно важна для распро-
странения паразита. Она сводится не только’к передаче пнвазион-
74
ных стадий дефинитивному хозяину, но и к увеличению интенсив-
ности его заражения, поскольку акантеллы могут накапливаться
в рыбах в большом количестве.
Рис. 42. Жизненный цикл скребня Corynosoma
strutnosurn (по Догелю):
/ — половозрелый скребень в кишечнике тюленя, 2 —
яйца во внешней среде, 3 — промежуточный хозяин
Pontoporeia affinls, 4 — резервуарный хозяин — рыба,
5 — и пикетированная акантелла
Патогенное значение скребней
Скребни с помощью своего вооруженного крючьями хоботка
могут внедряться глубоко в толщу стенки кишечника хозяина, до-
ходя при этом почти до наружной границы мускульного слоя.
Вокруг внедрившегося хоботка начинается некроз тканей и обра-
зуется плотная соединительнотканная капсула, которая отчетливо
выдается с наружной стороны кишечника (рис. 43). Вызываемые
скребнями повреждения носят в основном локальный характер
и выражаются в воспалительных процессах, возникающих вокруг
места внедрения хоботка. В отдельных случаях возможны разрывы
стенки кишечника и как следствие этого перитонит. При сильных
инвазиях начинаются расстройства пищеварения, животные теряют
аппетит, слабеют. Среди скребней, паразитирующих в рыбах, па-
тогенное значение имеют представители родов Pomphorhynchus
и Echinorhy.nch.us, в особенности Е. salmonis — частый паразит ло-
сосевых. Интенсивность инвазии бывает очень значительной: 200—
300 и более экземпляров. Паразитирование скребней вызывает
изъязвление стенок кишечника и приводит к снижению упитанности
рыб. Птицы, в особенности утиные, часто страдают от паразитиро-
вания скребней. Наиболее опасны представители родов Polymor-
phus, FilicolUs и др. Эти паразиты, поселяющиеся иногда в очень
больших количествах, вызывают изъязвление стенок кишки, появ-
75
ление некротических очагов, а в отдельных случаях нарушение
иннервации кишечника, его моторной и секреторной функций.
Сильные заражения приводят к гибели птиц, особенно домашних,
при условии нерационального ведения птицеводческих хозяйств.
Известен случай массовой гибели молодняка уток, вызванной па-
разитированием скребней Polymorhus magtius. Небольшое озеро
в течение нескольких лет использовалось для выращивания тысяч-
Рнс. 43. Кишечник кряквы, пораженный скребнями Filicollis
anatis (по Павловскому)
ных стад уток. С их фекалиями в воду попадало неисчислимое ко-
личество яиц скребней, в результате чего заражение бокоплавов
(Gatnmarus lacustris) промежуточных хозяев Р. magnus достигло
99%. Понятно, что когда на это озеро было выпущено еще одно
стадо утят, охотно питающихся рачками, началась эпизоотия, вы-
звавшая гибель нескольких тысяч птиц.
Из паразитов млекопитающих особого упоминания заслуживает
скребень-великан (Macracanthorhynchus hirudinaceus), паразитиру-
ющий в кишечнике свиней. Это очень крупный червь, до 650 мм
длиной. Промежуточным хозяином его служат личинки майского
жука (Melolontha melolontha) и других хрущей. В некоторых райо-
нах Белоруссии отмечалось до 60% зараженности личинок старших
возрастов при интенсивности инвазии до 130 акантелл на одну
личинку жука. Акантёллы не погибают во время их метаморфоза.'
так что не только личинки, но и куколки и взрослые майские жуки,
могут быть источником инвазии свиней. Последние заражаются,
выкапывая и пожирая личинок и другие стадии развития хрущей.
Скребни достигают половозрелости через 70—110 дней; продолжи-
тельность их жизни составляет от 10 до 23 месяцев. Поселение
скребней в кишечнике свиньи вызывает резкие кишечные расстрой-
ства, повышение температуры, исхудание. Особенно опасны случаи
абсцессов и разрыва стенки кишки, обычно вызывающие перитонит
76
и гибель животного. Макраканторинхоз свиней может приносить
свиноводству значительный ущерб. Это заболевание распростра-
нено в разных странах (в особенности в Югославии, Италии и др.).
В СССР отмечено на Украине, в Белоруссии, на Дальнем Востоке.
Лечение свиней от макраканторинхоза мало эффективно; применя-
ются отвары и настои багульника, четыреххлористый углерод
и другие антгельминтики, не дающие, однако, особенно обнадежи-
вающих результатов. Профилактические мероприятия основывают-
ся на необходимости «разрыва» жизненного цикла скребня. С этой
целью проводится термическая обработка завоза, при#которой уби-
ваются яйца скребней. Уничтожаются майские жуки, вводится ка-
рантин, предотвращающий возможность перевозки свиней из зара-
женных хозяйств в другие районы, и т. д.
Паразитирование скребней у человека отмечено в исключитель-
но редких случаях. Тем не мепее, потенциально человек может быть
хозяином этих' гельминтов. Экспериментально это было доказано
опытами по самозаражению, проведенными в конце прошлого сто-
летия. Итальянский ученый Каландруччио проглотил около 50 ли-
чинок скребня Moniliformis moniliformis (паразиты грызунов), из-
влеченных им из тела жуков чернотелок. Спустя 19 дней исследо-
ватель почувствовал признаки заболевания: начались сильные боли
в животе, кишечное расстройство, слабость, головокружение, звон
в ушах. Все эти симптомы исчезли после приема внутрь экстракта
мужского папоротника и последовавшего за тем изгнания паразитов,
«
Происхождение и эволюция
Вопрос о филогенетических связях скребней и их эволюции до
сих пор нельзя считать решенным. Широко распространенное мне-
ние о близости скребней к типу Nemathelminthes, к которому их
нередко и относят, основано на ряде черт внешнего сходства между
обеими группами. Это постоянство клеточного состава, наличие
первичной полости тела и раздельнополость. Однако постоянство
клеточного состава описано пока лишь для одного вида скребней,
а возникновение первичной полости могло осуществляться и неза-
висимо от типа круглых червей. Раздельнополость же сама по се-
бе ничего не доказывает, тем более, что в строении полового аппа-
рата скребней и нематод нет ничего общего.
Некоторый интерес представляют рассуждения о близости
скребней к классу приапулид (Priapulida). У обеих групп живот-
ных имеется вворачивающийся хоботок, полость тела — первичная.
Особое значение придается слиянию выводных путей выделитель-
ной и половой систем в единый урогенитальный канал. Однако та-
кой признак, как наличие вворачивающихся хоботков, вряд ли
может считаться серьезным основанием для утверждения филоге-
нетических связей. Известно, что вворачивающийся хоботок имеет-
ся и у некоторых нематод (р. Hystrichis), и у ряда плоских червей
(финны цестод, церкарии трематод и др.). Вопрос о первичном
характере полости тела у приапулид еще спорный. Серьезного вни-
77
мания заслуживает лишь урогенитальный канал, однако генезис
этого образования еще недостаточно изучен.
На данном этапе наших знаний наиболее логичными представ-
ляются взгляды, развйваемые В. И. Петроченко (1956), который
рассматривает скребней как самостоятельный тип, ведущий свое
начало от турбеллярий. Большое филогенетическое значение он
придает строению нервной системы скребней, несущей следы орто-
гона и органам выделения, устроенным по типу протонефридиев.
Возникновение первичной полости тела скребней легко может
быть объяснено развитием схизоцеля (щелевндных лакунарных
пространств в паренхиме турбеллярий). Известно, что и у некото-
рых турбеллярий и у партенит трематод за счет схизоцеля разви-
вается первичная полость тела.
В' организации скребней имеется целый ряд черт, указываю-
щих на далеко зашедшие адаптации этой группы животных к пара-
зитическому образу жизни. Они проявляются в полном исчезнове-
нии у скребней пищеварительной системы и в появлении специали-
зированных покровов, приспособленных к поглощению частично
переваренных пищевых веществ из кишечника хозяина; в сильном
развитии органов прикрепления, в высокой половой продуктивности
скребней и т. п. Характерно также, что эмбриональное развитие
завершается в теле самки и наружу выводятся яйца со сформиро-
ванными личинками внутри. Подвижные расселительные личинки
отсутствуют. Все это указывает на очень давний переход предков
скребней к паразитизму и на длительную эволюцию их, направлен-
ную в сторону узкой специализации к этому образу жизни.
ТИП NEMATHELMINTHESSCHNEIDER—
ПЕРВИЧНОПОЛОСТНЫЕ
Тип первнчнополостные включает животных с хорошо развитой
первичной полостью тела — схизоцелем. Покровы первичнополост-
ных образованы либо ресничным эпидермисом, либо кутикулой,
которая продуцируется нижележащим тканевым слоем — гиподер-
мой. Пищеварительная система в большинстве случаев состоит из
трех отделов: передней, средней и задней кишок. Последняя от-
крывается наружу анальным отверстием. Выделительная система
представлена протонефридиями или специальными видоизменен-
ными кожными железами. Нервная система ортогонального типа.
Первнчнополостные раздельнополы. Развитие прямое или с мета-
морфозом. У некоторых представителей'наблюдается явление ге-
терогонии.
В состав типа Nemathelminthes большинство исследователей
в настоящее время включают 5 классов: Gastrotricha, Kinorhyncha,
Rotatoria, Ncmatomorpha и Nematoda. Кроме того, очень часто
к пЪрвнчнополостным в качестве дополнения относят и класс Pria-
pulida.
Среди Nemathelminthes паразитизм получил широкое распрост-
ранение, правда не во всех группах. Два небольших класса'Gastro-
78
tricha n Kinorhyncha представлены только свободноживущими
формами. Из 1500 известных видов коловраток (кл. Rotatoria)
к паразитическому образу жизни перешло лишь около 20. Большая
часть их — комменсалы, поселяющиеся на поверхности тела различ-
ных водных животных (чаще на ракообразных).
Очень широко становление паразитизма происходило в самом
обширном среди Nemathelminthes классе Nematoda. Круглые чер-
ви, однако, сохранили и большое количество свободноживущих
видов. И, наконец, последний — сравнительно небольшой класс
Nematomorpha — волосатики — представлен исключительно пара-
зитическими и притом крайне специализированными видами.
КЛАСС NEMATODA RUDOLPHI, 1808 —КРУГЛЫЕ ЧЕРВИ
Круглые черви по числу представителей занимают второе место
после насекомых. По мнению А. А. Парамонова, этот класс насчи-
тывает свыше 500 тыс. видов. Очень многие нематоды обитают
в море, пресных водоемах, и в почве. Однако большая часть круг-
лых червей перешла к паразитическому образу жизни. Животные,
в которых нематоды паразитируют, очень разнообразны: от беспоз-
воночных до высших позвоночных. Многие виды нематод стали
парезитами растений.
Организация нематод характеризуется удивительной однотип-
ностью. Они полностью утратили ресничный эпителий и его про-
изводные. Развитие с метаморфозом: появлению половозрелых
особей предшествуют четыре сменяющие друг друга личиночные
стадии. Переход от одной стадии к другой осуществляется в про-
цессе линьки. Очень многие паразитические круглые черви — ти-
пичные геогельмйнты. Широко распространены и биогельминты,
в цикл развития которых включается один или даже два промежу-
точных хозяина. Наиболее специализированные виды полностью
утратили какие-либо свободноживущие стадии.
Важная-роль в изучении круглых червей — паразитов живот-
ных— принадлежит советским ученым: К. И. Скрябину, Н. П. Ши-
хобаловой, К. М. Рыжикову, М. Д. Сонину и многим другим. Осо-
бое место занимают исследования крупнейшего советского зоолога
А. А. Парамонова. В его работах, посвященных главным образом
фитогельмиитам, нашли свое решение многие теоретические вопро-
сы, связанные с происхождением и эволюцией всего класса.
Среди зарубежных исследований видное место занимают рабо-
ты Б. Г. Читвуд и М. Б. Читвуд (В. G. Chitwood, М. В. Chitwood),
Т. Камерона (Т. Cameron), Р. Гольдшмидта (R. Goldtschmidt),
В. Г. Инглиса (W. G. Inglis), Д. Ли (D. Lee) и др.
В соответствии с системой, принятой в многотомном капиталь-
ном издании «Основы нематодологии», кл. Nematoda подразделя-
ется на 2 подкласса: Adenophorea (syn. Aphasmidia) —аденофореи
и Secernentea (syn. Phasmidia) —сецерненты. Первый из них вклю-
чает большинство древних и примитивных форм (средн них много
79
свободноживущих видов), объединяемых в 5 отрядов: Chromadori-
da Chitwood, 1933, Enoplida Chitwood, 1933, Dorylaimida Pearce,
1942, Trichoccphalida Spassky, 1954, и Dioctophymida Schulz et
Gvosdev, 1970. Лишь три последних содержат паразитов животных.
Второй подкласс — сецерненты — подразделяется на 4 отряда:
Rhabditida Chitwood, 1933, Tylenchida Thorne, 1949, Ascaridida
Skrjabin et Schulz, 1940, и Spirurida Chitwood, 1933. Большинство
видов Secernentea — паразиты животных и растений, но есть среди
них и свободноживущие.
Морфология и анатомия нематод
Форма тела и размеры. Большинство нематод имеют круглое
(в поперечном сечении) удлиненное веретеновидное тело, передний
и задний концы которого заострены (рис. 44, А, Б). Отношение
Рис. 44. Представители класса Nematoda (из кн. Шульца и Гвоздева).
А — самка Ascaris lumbricoides; Б — самец Ascaris lumbricoides; В —
самка Loa loa; Г—самка Tetrameres sp.; Д— самец Telrameres sp.;
Е—самка и самец Syngamus trachea; Ж— самка Trichocephalus sp.
80
длины к ширине может сильно варьировать. Одни виды имеют
почти шаровидное (рис. 44, Г), другие — сильно вытянутое ните-
видное тело (рис. 44, В).
У большинства круглых червей отчетливо выражен половой ди-
морфизм. Самцы, как правило, меньше самок. Иногда они приоб-
ретают все признаки карликовых самцов, теряя/при этом способ-
ность к самостоятельному существованию. У широкораспространен-
ного паразита дыхательных путей птиц Syngamus trachea самцы,
размеры которых в два-три раза уступают размерам самок, прира-
стают к телу последних в области полового отверстия (рис.'44, £).
Микроскопические самцы Trichosomoides crassicauda становятся
гиперпаразитами: они поселяются в половых путях самок.
Столь же отчетливо выражены различия и в форме тела
(рис. 44, А, Б. Г, Д). У очень многих видов круглых червей на зад-
нем_конце тела самцов располагаются так называемые бурсальные
крылья (см. рис. 58), отсутствующие у самок и функционирующие
в процессе копуляции.
Размеры круглых червей варьируют в очень широких пределах.
Нематоды, как правило, микроскопически малы. Среди свободно-
живущих есть виды, не превышающие в длину 0,3 мм. Фитонема-
тоды несколько крупнее — до 8—10 мм. Самые же крупные черви
встречаются среди паразитов позвоночных животных. Человеческая
аскарида имеет в длину около 30—40 см, а обитающая в плаценте
кашалота Placentonetna gigantissima достигает 6—8 м.
Ца переднем конце тела терминально располагается ротовое
отверстие (рис. 45). Анальное отверстие (а оно имеется не всегда)
локализуется субтерминально на вентральной поверхности. Жен-
ское половое отверстие чаще всего лежит в передней половине те-
ла, также вентрально. Самостоятельное мужское половое отверстие
у всех видов’круглых червей отсутствует.
Кожно-мускульный мешок. Кожно-мускульный мешок нематод
представляет собой единое функциональное целое. Это сложно
устроенное образование, состоящее из кутикулы, гиподермы и му-
скульных клеток. Кутикула круглых червей является продуктом
жиднедеятельности подлежащей покровной ткани и по своей орга-
низации и происхождению принципиально отличается от тегумента
паразитических плоских червей.
Тонкое строение кутикулы, которая всегда состоит из несколь-
ких слоев, различно у разных видов нематод (рис. 46). В наиболее
полном случае их может быть 9—10. Наружные слои, количество
которых варьирует от 1 до 4, пронизаны системой тончайших ка-
нальцев. Последние заходят в верхнюю’ часть центрального слоя,
называемого гомогенным или матричным. Он служит основой всей
кутикулы, связывая все остальные элементы в одно целое. Под
матричным слоем лежит еще несколько слоев (до 5), подстилае-
мых тонкой базальной мембраной.
Количество слоев, образующих кутикулу того или иного вида
нематод, определяется его экологическими особенностями и в пер-
вую очередь характером локализации паразита в организме живот-
81
ного-хозяина. Кишечные виды обычно обладают 8—10 слоями
(кутикула представителей р. Ascaris состоит из 10 слоев). У легоч-
ных гельминтов, обитателей глазничных впадин, кровеносной си-
стемы и т. п., строение кутикулы упрощается: она образована всего
5—7 слоями.
Рис. 45. Схема строения круглых червей (по Ли). А— самка; Б — самец; В — по-
• перечный срез:
/ — головные папиллы, 2 — стома, 3 — прокорпус, 4 — метакорпальнЫй бульбус, 5 — пере-
шеек, 6 — нервное кольцо. 7 — кардиальный бульбус, 3 —средняя кишка, 9 — яйцевод. 10—*
яичник, // — ректальные железы, /2—задняя кишка. /3—матка с яйцами, 14 — влагалище.
/5 — фагоцитарная клетка, 16 — экскреторная железа (ренетта), П — семенник, 18 — семя-
провод, /£—семяизвергательный канал, 20 — спикулы, 21 — половая бурса, 22 — кутикула,
23 — гиподерма, 24 — боковые гиподермальные валики, 25— спинной гиподермальный валик.
26 — спинной нервный ствол, 27 — брюшной гиподермальный валик, 28 — брюшной нервный
ствол, 29 — канал выделительной системы
Химический состав кутикулы довольно сложен и включает ряд
соединений различной природы: альбумины, коллаген, кератин,
глюкопротеиды, липиды и т. д. Наличие кератина и коллагена, осо-
бенно в наружных слоях, обусловливает механическую прочность
кутикулы. Липиды, скорее всего' обеспечивают ее химическую
устойчивость. У некоторых видов в матричном слое, содержащем
большое количество влаги, обнаружены гемоглобин и эстеразы. Все
это заставляет изменить представление о кутикуле круглых червей
как о мертвом, индифферентном образовании.
Роль кутикулы в процессах жизнедеятельности паразитов очень
велика. С одной стороны, она выполняет барьерные функции, пре-
82
яятствуя проникновению химических агентов внутрь тела червя
« поддерживая тем самым постоянство внутренней среды организ-
ма. С другой же стороны, имеются данные об участии кутикулы
в поглощении пищевых веществ (с. 93). Наличие гемоглобина в ку-
тикуле у ряда видов рассматривается некоторыми исследователями
как признак участия кутикулы в дыхании (с. 97). Наконец, очень
важную роль кутикула играет как опорное скелетное образование.
К ней прочно прикрепляются мускульные клетки (с. 84), а наличие
полостного тургора (с. 85) придает упругость и эластичность всему
телу паразита.
Непосредственно под кутикулой залегает покровная ткань нема-
тод — гиподерма. Она образует сравнительно тонкий слой, приле-
гающий к внутренней поверхности кутикулы. Вдоль всего тела чер-
вя тянутся несколько утолщений гиподермы, имеющих вид сравни-
тельно узких валиков. У подавляющего числа видов их обычно 4:
дорзальный, вентральный и два латеральных (рис. 45, В).
У большинства видов гиподерма представлена сикпластом, в ко-
тором не прослеживаются границы между составляющими его
клетками. Гиподерма наиболее примитивных нематод сохраняет
клеточное строение.
Цитоплазма гиподермы вакуолизирована и содержит большое
количество тонких опорных фибрилл. Количество ядер, обнаружи-
ваемых в гиподерме, как правило, постоянно для каждого отдель-
ного вида.
Функции гиподермы многообразны. Главная и наиболее важная
из них — формирование кутикулы. Детали этого процесса еще не
до конца ясны. Одни авторы считают, что кутикула возникает в ре-
зультате уплотнения самых наружных слоев цитоплазмы гиподер-
мы, которые претерпевают своего рода соединительнотканное
перерождение. Другие же рассматривают кутикулу как продукт
секреторной деятельности гиподермы. Очень возможно, что имеют
место оба этих процесса, и формирование различных элементов
кутикулы происходит за счет действия разных механизмов. Во вся-
ком случае построение кутикулы сопровождается усилением био-
синтетических процессов в гиподерме, о чем говорит наличие боль-
шого количества РНК в ее цитоплазме.
Вторая, очень важная, функция гиподермы — барьерная. Наря-
ду с кутикулой она препятствует проникновению в тело червя ряда
химических веществ. И, наконец, гиподерма служит своеобразным
депо, в котором накапливаются запасные питательные вещества:
жиры и гликоген.
Под гиподермой, примыкая к ней, располагается один слой мус-
кульных клеток. У нематод имеется только продольная мускулату-
ра. Количество клеток и их строение отличаются у разных предста-
вителей класса. У подавляющего большинства видов мускульные
клетки образуют не сплошной слой, а четыре ширркие продольные
ленты, разделенные гиподермальными валиками (рис. 45, В). Об-
щее количество клеток варьирует у разных представителей, однако
в пределах вида оно довольно постоянно.
83
В наиболее типичном случае мускульные клетки состоят из ве-
ретена, цитоплазматической сумки (иногда ее называют саркоплаз-
матическим мешком) и отростков (рис. 47, Д, Б), По периферии
веретена располагаются сократимые миофибриллы. Образуемый
ими слой изогнут в виде продольного’желобка (рис. 47, Б). Цент-
ральная часть веретена и сумка построены из цитоплазмы, прони-
занной многочисленными тонкими опорными фибриллами. Отходя-
Рис. 46. Схема строения слоистой кутикулы нематод
(по Ли):
/ — наружные слои. 2 — матричный или гомогенный слой. 3 —
внутренние слон, 4 — базальная мембрана, о — каналы, прони-
зывающие наружные слои
щие от сумок отростки направляются к гиподермальным валикам
(рис. 45. В). Отростки мускульных клеток, образующих вентраль-
цые ленты, — к вентральному валику; отростки дорзальных кле-
ток— соответственно к дорзальному. Свободные концы отростков
плотно прилегают к вершинной части валиков. Ядро мускульной
клетки обычно располагается на границе сумки и веретена.
Долгое время’ничего не было известно о механизме фиксации
мускульных клеток на внутренней поверхности стенки тела. Лишь
электронно-микроскопические исследования позволили решить этот
вопрос. От базальной части клеток отходят многочисленные фиб-
риллярные отростки (рис. 47, В). Они пронизывают гиподерму,
базальную мембрану и глубоко внедряются во внутренние слои
кутикулы.
С покровами у нематод связаны разнообразные железистые
клетки — так называемые кожные железы, протоки которых откры-
ваются на поверхности тела. Часть кожных желез функционально
относится к самым различным системам органов: пищеваритель-
ной— пищеводные железы (с. 86), выделительной — ренетты, или
шейные железы (с. 87), органам чувств — фазмиды (с. 90) и т. д.
Схизоцель. Как уже говорилось выше, внутреннее пространство
нематод представлено обширной первичной полостью тела — схизо-
84
целем, который заполнен водянистой жидкостью, имеющей слож-
ный химический состав (с. 95). Функциональное значение схизоцеля
очень велико. А. А. Парамонов рассматривает его появление в про-
цессе эволюции как крупное преобразование типа ароморфоза^
определившее все своеобразие организации нематод. В полости те-
ла заполняющая ее жидкость находится под известным внутренним
давлением, что в совокупности с наличием прочной и эластичной
кутикулы создает довольно значительный полостной тургор, при-
дающий телу червя большую упругость. Это позволяет говорить об
опорной функции схизоцеля, своеобразном гидроскелете у круглых
червей.
Полостная жидкость, омывающая кишечник, приобретает важ-
ное значение и как транспортная система. Через нее осуществля-
ется передача пищевых веществ от органов пищеварения к мус-
кульным клеткам и другим системам органов. Наконец, наряду
с трофической схизоцель берет на себя и экскреторную функцию.
В полостную жидкость поступают многие продукты обмена круг-
лых червей. Схизоцель выстлан тонкими мембранами (рис. 47, Б).
В работах последних лет (Ю. К. Богоявленский) показано, что это
весьма активное В1 физиологическом отношении образование. В мем-
бранах, например, обнаружено большое количество РНК. Это не-
сомненно говорит о том, что они имеют клеточное происхождение.
Ядра же клеток, за счет которых они возникают, подверглись в
процессе гистогенеза полной дегенерации.
Пищеварительная система. У нематод, как правило, пищевари-
тельная система хорошо развита. Она состоит из трех отделов:
передней, средней и задней кишок. Отличия в деталях строения от-
дельных участков пищеварительного тракта, наблюдаемые у раз-
ных представителей, определяются, с одной стороны, систематиче-
ской принадлежностью того или иного вида, а с другой — чисто
биологическими особенностями: локализацией в организме хозяи-
на, характером пищи, способом питания и т. п.
Ротовое отверстие обычно окружено специальными придатка-
ми— губами. У большинства примитивных видов их бывает &
(рис. 48, А), у более специализированных это число сокращается
до 3 (рис. 48, Б). Иногда губы редуцируются и замещаются так
называемыми головными буграми. У форм, лишенных губ или го-
ловных бугров, на переднем конце тела часто имеется кутикуляр-
ное вздутие — головная везикула. В одних случаях она целиком
окружает ротовое отверстие, в других же от нее остаются только
латерально расположенные гребни (рис. 48, Я).
Ротовое отверстие ведет в переднюю кишку, которая в специ-
альной литературе обозначается .как фарннкс (глотка). Фаринкс,
будучи эктодермальным образованием, выстлан тонкой кутикулой
и отчетливо подразделяется на два отдела: передний — стому и зад-
ний— более длинный пищевод. Строение стомы у разных предста-
вителей класса разнообразно. У многих видов она имеет относи-
тельно простую цилиндрическую форму. Иногда она принимает
вид сильно вздутой ротовой капсулы (рис. 59, Б, Г). Это характер-
85
но для видов, питающихся кровью и тканями животного хозяина
(р. Ancylostoma, Necator и др.). Примером крайне специализиро-
ванной стомы могут служить ротовые части некоторых фитонема-
тод. Имеющееся у них копье, служащее для прокладывания плотных
растительных тканей, представляет собой полностью преобразован-
ную стому.
е Следующий за стомой отдел фаринкса — пищевод выполняет
функции глоточного насоса. Его форма сильно варьирует. По-види-
мому, исходным для многих нематод (в частности, для подкл. Se-
Рис. 47. Мускульные клетки нематод. А—общий вид мускульной клет-
ки аскариды (по Дейнеке); Б — поперечный срез через мускульную
клетку Hetarakis sp. (по Богоявленскому); В — электронно-микроскопи-
ческая схема строения мускульной клетки Nippostrongylus sp. (по Ли):
1 *- сократимое веретено, 2 — саркоплазматическая сумка с ядром, 3 — отросток,
4 — опорные фибриллы, 5 — система пограничных мембран, 6 — гранулы глико-
гена, 7 — капли жира, 8 — митохондрии. 9 — опорные структуры. 10 — фибрилляр-
ные отростки базальной части мускульной клетки, // — гиподерма, /2—базальная
мембрана, 13 — внутренние слои кутикулы
•cemeritea) является пищевод, встречающийся у представителей
отр. Rhabditida (рис. 45, Д) и называемый рабдитовидным. Он со-
стоит из корпуса, подразделенного на узкую цилиндрическую часть
(прокорпус) и расширенную (метакорпальныйбульбус), перешейка
(истмуса) и заднего пли кардиального бульбуса. У многих видов
пищевод вторично упрощается и принимает трубчатую форму.
С пищеводом у нематод связаны особые пищеводные железы
(с. 84). У сецернентов их немного и они залегают в толще стенки
(рис. 48, Г), у аденофоре?! они могут быть весьма многочисленны-
ми. Крупные клетки, плотно примыкая одна к другой, располага-
ются вдоль узкого пищевода в виде сплошного клеточного столбика
(рис. 48, Д).
Непосредственно за пищеводом начинается эктодермальная
средняя кишка. Она в большинстве случаев имеет вид простой
трубки, тянущейся вдоль тела червя. Граница между средней и
86
эктодермальной задней кишкой (ректумом) определяется наличи-
ем сфинктера. Задняя кишка несет кутикулярную выстилку. У мно-
гих представителей подкл. Secernentea в ректум открывается от 3
до 6 ректальных одноклеточных желез. Функции этих образований
не известны. У всех Adenophorea они отсутствуют.
У самцов нематод в заднюю кишку открываются протоки муж-
ской половой системы. Здесь же образуются сложные кутикуляр-
ные образования (с. 91) копулятивного аппарата.
Рис. 48. Строение передних отделов пищеварительной системы нематод
(по разным авторам). А—схема расположения губ RJiabditis terricola\
Б — схема расположения губ у Ascaris luinbricoides; В — головная ве-
зикула Enterobius vermicular is; Г — пищеводные железы Heierakis
gallinarum.} Д— пищеводные железы Trichinella spiralis
В ряде случаев у нематод наблюдается в той или иной степени
выраженная тенденция к редукции пищеварительной системы. При
этом иногда отдельные части пищеварительной системы выполняют
не свойственные им функции. У представителей сем. Mermithidae
(подкл. Adenophorea) рудимент средней кишки превращается в
своеобразное «жировое тело», в котором накапливаются запасные-
питательные вещества. Поступление пищи в тело паразита у таких
видов осуществляется непосредственно через кутикулу (с. 94).
Выделительная система. Представлена у нематод образования-
ми двух типов. Главная роль принадлежит уже упоминавшимся ра-
нее (с. 84) шейным железам или ренеттам*. Их может быть две-
87
или одна. От железистых клеток, расположенных в передней поло-
вине тела, отходят длинные внутриклеточные каналы,-залегающие
в толще боковых гнподермальных валиков (рис. 45, В\ 49). В обла-
Рис. 49. Выделительная система нема-
тод. Д — схема строения выделительной
•системы стронгилят; Б — схема строения
выделительной системы Anisakis (по
Б. Г. Читвуд и М. Б. Читвуд); В — фа-
гоцита рнар клетка (из кн. Шульца
и Гвоздева):
./ — внутриклеточные каналы, 2—наружный
выводной проток, 3 — ренетты
сти расположения ренетт кана-
лы правой и левой половины
тела сливаются и дают начало
непарному короткому протоку,
открывающемуся наружу на
вентральной поверхности тела.
Предполагается, что эта систе-
ма кроме экскреторной выпол-
няет еще и осморегуляторную
функцию.
В удалении продуктов ме-
таболизма из тела червя уча-
ствуют и особые фагоцитарные
клетки. Последние располага-
ются в схизоцеле чаще всего по
ходу латеральных валиков ги-
подермы. Количество фагоци-
тарных клеток варьирует от
2 до 6. На поверхности клеток
образуются многочисленные
псевдоподии, при помощи ко-
торых они захватывают твер-
дые частицы из полостной
жидкости.
Нервная система и органы
чувств. Нервная система круг-
лых червей сохраняет все при-
знаки ортогона (рис. 50). В
передней части тела, в области
пищевода располагается нерв-
ное кольцо, связывающее два
боковых ганглия. От кольца
отходят продольные нервные
стволы. К переднему концу те-
ла вдоль пищевода обычно на-
правляются 6 стволов, иннер-
вирующих губные папиллы.
Кроме того, от боковых гангли-
ев берут начало так называе-
мые амфидальные нервы (с.’ 90). К заднему концу тела от кольца
•отходят от 8 до 12 нервных стволов: дорзальный, 2 пары субмеди-
альных, 1—3 пары латеральных и вентральный. Дорзальный и вент-
ральный стволы лежат в соответствующих гнподермальных вали-
ках, а остальные в толще гиподермы. В задней половине тела
залегают ректальная комиссура и анальный ганглий. У самцов
88
часто формируется дополнительный ганглий, от которого иннерви-
руются половые папиллы.
Органы чувств нематод не отличаются большим разнообразием»
хотя часто бывают весьма многочисленны. В основном они пред-
ставлены папиллами или сосочками, наибольшее количество кото-
Рис. 50. Схема строения нервной системы и органов чувств нематод (по разным
авторам). А — общий вид нервной системы; Б — схема расположения нервных
стволов в переднем конце тела; В— задний конец тела самца аскариды; Г — фаз-
миды; Д— передний конец тела мермитиды с папиллами и амфидами:
/ — нервное кольцо, 2 — передние продольные нервные стволы, 3 — задние продольные нерв-
ные стволы, 4 — ректальная кольцевая комиссура, 5 —головные папиллы с подходящими
к ним нервами, 6 — амфидпальный нерв, /—цервикальные папиллы, 8 — клоакальное от-
верстие, Р —половые папиллы, 10 — наружная пора, // — канал, 12 — фазмидиальные желе-
зы, /3 —ротовое отверстие, 14 — пищевод. /5—наружное отверстие амфиды. /б —чувстви-
тельные окончания в полости амфиды, 17 — амфндиальная железа
рых локализуется на переднем и заднем концах тела. Обычно вы-
деляется несколько групп папилл: головные или губные, располо-
женные на губах (рис. 48, А, В; 50, на уровне нервного кольца
имеются шейные папиллы (рис. 50, Б), У самцов в области клоа-
кального отверстия сконцентрированы половые папиллы
(рис. 50, В).
Кроме осязательных папилл нематоды, по-видимому, имеют
и хеморецепторы. У представителей подкл. Secernentea — это осо-
89
<5ые преобразованные кожные >келезы — фазмнды, расположенные
в хвостовой части тела (рис. 50, Г). У наиболее примитивных видов
фазмнды сохраняют вид небольших одноклеточных железок н об-
ладают чисто секреторной функцией. У специализированных видов
они становятся сенсорными образованиями. К ним подходят особые
фазмидальпые нервы, берущие начало от латеральных нервных
стволов. У всех Adenophorea фазмнды отсутствуют.
Второй тип хеморецепторов — амфиды распространен значи-
тельно шире и встречается у представителей обоих подклассов.
Амфиды располагаются на переднем конце тела, иннервируются
специальным амфидальным нервом (с. 88). В, наиболее типичном
случае они представляют собой узкое впячивание покровов — ам-
фидальный кармашек (рис. 50, Д), на дне которого располагается
чувствительное нервное окончание.
Половая система. У круглых червей половая система, сохраняя
общий план строения, характерный для всего класса, в деталях
сильно варьирует у разных представителей. Женская половая си-
стема в подавляющем большинстве случаев парная (рис. 45, А).
Она начинается двумя трубчатыми яичниками. Последние часто
имеют вид нитевидных, сильно извитых придатков (р. Ascaris
и др.). В дистальном слепо замкнутом конце яичников происходит
размножение половых клеток. Последние довольно правильно рас-
полагаются вокруг тонкого тяжа — рахиса, проходящего вдоль
яичника по его оси (см. рис. 45, В). Рахис формируется за счет
соединения проксимальных концов развивающихся яйцеклеток.
Считается, что он служит для передачи пищевых веществ половым
клеткам.
Яичник непосредственно переходит в трубчатый яйцевод, диа-
метр которого несколько увеличивается. Поступившие в яйцевод
яйцеклетки округляются и постепенно продвигаются в сторону мат-
ки. Последняя также имеет трубчатое строение, но характеризует-
ся еще большим диаметром. Граница между ней и яйцеводом, как
правило, выражена совершенно отчетливо. Стенки матки образова-
ны мускульными и железистыми клетками. В матке происходит
оплодотворение яйцеклетки и формирование защитных оболочек
яйца.
Две матки сливаются дистальными концами и дают начало
сравнительно короткому непарному протоку — влагалищу. Послед-
нее открывается наружу на вентральной поверхности тела червя.
Отклонения от приведенной выше схемы встречаются не столь
уже часто и главным образом касаются числа тех или иных поло-
вых органов. Многие Adenophorea (отр. Trichocephalida и др.)
обладают непарной женской половой системой. Иногда же количе-
ство половых органов (за исключением влагалища) может сильно
увеличиться (у р. Placentonema до 32).
Мужская половая система в отличие от женской почти всегда
одиночна (см. рис. 45, Б). Она начинается трубчатым семенником,
в котором происходит формирование сперматозоидов. Семенник
переходит в семяпровод. Далее следуют слегка вздутый семенной
90
Рис. 61. Половой аппарат самцов нематод (по
разным авторам). А — спикула; Б — положение
спикул в теле самца; В — рулек, поддерживающий
спикулы;
1 — спикулы» 2 — клоака
пузырек и длинный цилиндрический, снабженный мускулистыми
стенками семяизвергательный канал. Последний открывается в зад-
нюю кишку, которая у самцов называется клоакой.
В клоаке формируется довольно сложный копулятивный аппа-
рат, состоящий из нескольких функционально связанных друг с
другом кутикулярных элементов. Его основу составляют спикулы
(рис. 51, Л). Они фор-
мируются внутри спе-
циальных спикулярных
сумок (рис. 51, 5), рас-
положенных на дор-
зальной поверхности
клоаки. Во время копу-
ляции спикулы выдви-
гаются наружу из кло-
акального отверстия и
вводятся в вагинальное
отверстие самки. Они,
с одной стороны, спо-
собствуют удержанию
самца на теле послед-
ней, с другой же —
служат для расшире-
ния вульвы. Между
спикулами образуется
своего рода канал, по
которому в женскую
половую систему посту-
пают сперматозоиды.
Подвижность спикул
обусловлен а н аличием
специальных мышц.
За счет спикуляр-
ных сумок формирует-
ся особое кутикулярное
образование — рулек
(рис. 51, В). Он служит
направляющей, по которой скользят спикулы, и располагается
дорзально от последних.
У некоторых видов (главным образом, из подотр. Strongylata,
отр. Rhabditida) развивается еще один опорный аппарат — телемон.
Он располагается на вентральной поверхности клоаки и’предохра-
няет ее от разрыва во время движения спикул.
Функционально к мужской половой системе относится и половая
бурса, встречающаяся главным образом у представителей отр.
Rhabditida. Она образуется за счет разрастания кутикулы заднего
конца тела червей и состоит из трех лопастей — двух латеральных
и одной дорзальной (см. рис. 59). Лопасти укреплены радиально
расходящимися утолщениями — ребрами. На концах последних
91
располагаются папиллы. При копуляции лопасти плотно охваты-
вают тело самки в области полового отверстия.
В ряде случаев у нематод наблюдается становление гермафро-
дитизма (с. 98).
Некоторые особенности питания и обмена веществ нематод
Питание. Изучение особенностей питания и обмена круглых
червей представляет особый интерес. Среди представителей этой
группы имеются й свободноживущие формы, и факультативные
паразиты, обитающие как во внешней среде, так и в организме
животного-хозяина, и облигатные паразиты, морфологическая
и физиологическая специализация которых выражена в различной
степени.
Целый ряд паразитических нематод, поселяющихся в разных
отделах кишечного тракта позвоночных животных, сохраняет до-
вольно примитивный способ питания, свойственный их свободножи-
вущим предкам. Особенно отчетливо это бывает выражено у ви-
дов, локализующихся в таких участках пищеварительной системы,
в которых отсутствуют активные процессы переваривания и всасы-
вания пищевых веществ. В задней кишке растительноядных чере-
пах встречаются нематоды из подотр. Oxyurata (отр. Ascaridida).
Пищей им служат непереваренные хозяином растительные остат-
ки. Эти нематоды по характеру питания — настоящие комменсалы.
Многие круглые черви (ряд представителей сем. Trichostrongy-
lidae и др), поселяющиеся в желудке и тонком кишечнике копыт-
ных животных, также поглощают непереваренные частицы пищи,
бактерий и даже кишечных простейших. Не исключено, однако,
что наряду с этим они получают и какое-то количество пищевого
материала, уже частично переваренного животным-хозяином; Сле-
дующий шаг в этом направлении делают аскариды, питающиеся
преимущественно содержимым тонкого кишечника. Их пищей ста-
новится кишечный химус.
На этих этапах пищевой специализации у круглых червей еще-
не наблюдается серьезных морфологических и физиологических
изменений. Они касаются главным образом передних отделов пи-
щеварительной» системы: головной везикулы, стомы и пищевода.
У питающихся растительными' частицами оксиурат в сложно диф-
ференцированном пищеводе развивается кутикулярный аппарат,
служащий для измельчения пищи. Аскариды, поглощающие полу-
жидкую массу, утратили подвижность губ, а их сравнительно
просто устроенный пищевод выполняет функции глоточного на-
соса.
Заглатываемая паразитами пища переваривается в их кишеч-
нике, причем этот процесс, по-видимому, протекает по общей для
большинства многоклеточных животных схеме. Эпителиальные
клетки кишечника, снабженные многочисленными микроворсинка-
ми, образуют настоящую щеточную кайму. Рядом работ показано
я наличие у нематод многочисленных гидролитических ферментов.
92
Все это позволяет думать, что у них имеет место и полостное, и при-
стеночное пищеварение.
Роль покровов в поступлении пищевых веществ в тело паразита
на этом уровне специализации еще ничтожна. Проницаемость ку-
тикулы очень невелика. Однако аскариды, например, способны уже
к активному транспорту некоторых изомеров аминокислот (арги-
нина, триптофана и др.) через стенку тела.
Возникновение тесных связей с тканями хозяина, а в дальней-
шем и переход к тканевому паразитизму, неизбежно привели
к более узкой пищевой специализации нематод, а соответственно
и к появлению целого ряда иногда весьма своеобразных, морфоло-
гических и физиологических адаптаций. Все большее и большее
значение приобретало внекишечное пищеварение за счет выделе-
ния пищеварительных ферментов через стому в окружающие пара-
зита ткани. _
В наиболее простых случаях нематоды переходят к питанию
слизистой. Виды р. Nippostrongylus (подотр. Strongylata, отр. Rha-
bditidae) буквально срезают ворсинки ребрами кутикулы. Наруше-
ние целостности эпителия кишечника' приводит к тому, что он лег-
ко подвергается воздействию пищеварительных ферментов как
паразита, так и самого хозяина. Образующийся клеточный детрит
и служит пищей нематодам.
Несколько иной механизм питания наблюдается у власоглавов
(р. Trichocephalus, подкл. Adenophorea). Эти черви погружают
в глубь слизистой свой тонкий, почти нитевидный передний конец
тела, вокруг которого скапливается жидкий экссудат. Последний
и рлужит пищей нематодам. Власоглавы могут поглощать и кровь
хозяина, однако гёматофагия у них еще не стала обязательной.
Становление облигатной гематофагии — эволюционная тенден-
ция, характерная для многих групп паразитических организмов.
Отчетливо выражена она и у нематод. Питание тканевым клеточ-
ным детритом явилось хорошей предпосылкой для перехода к пи-
танию кровью. Примеры такого рода особенно многочисленны
у круглых червей из подотр. Strongylata (отр. Rhabditida). Послед-
ние часто имеют хорошо развитую стому, имеющую вид объемистой
ротовой капсулы с мощным кутикулярным вооружением — зубами,
крючьями н т. п. Червь втягивает в ротовую капсулу участки сли-
зистой и выделяет в просвет стомы пищеварительные ферменты,
разрушающие ткапи хозяина. Грубое нарушение целостности сте-
нок кишечника неизбежно сопровождается и повреждением зале-
гающих в ней кровеносных сосудов. По-видимому, паразиты внача-
ле лишь случайно поглощали небольшие порции крови в очагах
вызываемых ими местных кровоизлияний. Однако со временем этот
высококалорийный пищевой материал начал играть значительно
большую роль в их пйщевом рационе. Переход же к питанию толь-
ко кровью превратил нематод в настоящих облигатных гемато-
фагов. Именно таковы многие представители сем. Ancylpstomatidae:
р. Ancylostoma, Necator, Uncinaria и др. Погружая головной конец
глубоко в стенку кишечника, они разрушают кровеносные сосуды
93
и в больших количествах поглощают чистую кровь. Как и многие
другие гематофаги, нематоды приобретают способность продуци-
ровать антикоагулянты — вещества, препятствующие свертыванию
крови. У Ancylostoma, например, для этой цели служит пара ги-
пертрофированных гигантских железистых клеток, протоки которых
открываются в ротовую капсулу.
Увеличение роли внекишечного пищеварения сопровождается
целым рядом морфологических преобразований. Особенно отчетли-
во эти процессы выражены у фитогельминтов. В большинстве слу-
чаев у них (паразиты растений из подкл. Secernentea) стома преоб-
разуется в специальный колюще-сосущий аппарат, имеющий вил
выдвижного стилета. В последнем содержится узкий проток, по ко-
торому сначала в ткани растений поступает сложная смесь пище-
варительных ферментов (преимущественно различные гидролазы),
продуцируемых пищеводными железами, а затем уже в просвет
пищевода засасывается жидкая пищевая масса.
Многие паразитические нематоды в этой же связи обнаружива-
ют тенденцию к частичной, а иногда и почти полной рудимента-
ции пищеварительной системы. В первую очередь это характерно
для тканевых паразитов и форм, обитающих в полости тела, или
замкнутых системах органов: кровеносной, лимфатической и т. п.
Очень далеко атрофия пищеварительной системы заходит
у представителей сем. Mermithidae — крайне специализированных
паразитов членистоногих (с. 106). Полная утрата органов пищева-
рения наблюдается у Buddenbrockia-, паразитирующей в полости
тела пресноводных мшанок.
Редукция пищеварительной системы неизбежно должна сопро-
вождаться повышением проницаемости покровов тела. Реальность
существования этого пути поступления пищевых веществ в орга-
низм паразитов показана многочисленными наблюдениями. Кути-
кула многих видов нематод проницаема не только для воды и солей,
но и для целого ряда низкомолекулярных органических соединений
(аминокислот, глюкозы и т. и.).
Обмен веществ. Потребности нематод в пищевых веществах
очень разнообразны. Об этом позволяет судить наличие у нематод
широкого спектра гидролаз, способных расщеплять-белки, жиры
и углеводы.
Переваривание белков осуществляется с помощью протеиназ.
Не говоря уже о видах, питающихся непереваренными пищевыми
частицами, способностью синтезировать протеолитические фермен-
ты обладают и паразиты, поглощающие кишечный химус (аскари-
ды и др.). Очень высокая активность протеиназ характерна для
форм, перешедших к питанию кровью и тканями хозяина. Напро-
тив, фитогельминты, основу пищевого рациона которых составляют
углеводы (крахмал, клетчатка и т. п.), обладают довольно низкой
активностью ферментов, расщепляющих белки.
Конечным продуктом белкового обмена нематод, как и у многих
других низших беспозвоночных животных, является преимущест-
венно аммиак. В обычных условиях он выводится из организма
94
паразита через стенку тела и через кишечник за счет диффузии
в окружающую жидкую среду. Таким путем удаляется основная
масса экскретируемого азота. Превращение аммиака в мочевину,
как правило, ограничено. Картина резко меняется, когда выделе-
ние токсичного аммиака из тела паразита по каким-либо причинам
оказывается невозможным. Можно было бы ожидать, что он будет
накапливаться в тканях и органах червей. Этого, однако, не про-
исходит. Экспериментальные работы с аскаридами показали, что
у них начинается активный синтез значительно менее токсичной
мочевины, количество которой резко увеличивается. Образование
же аммиака снижается.
Нематоды, обитающие в условия» дефицита кислорода, так же,
как и другие гельминты, нуждаются в больших запасах углеводов.
В качестве основного энергетического вещества они используют
гликоген. Последний, как правило, откладывается в гиподерме
и в саркоплазме мускульных клеток. Количество запасаемого гли-
когена, как и у многих других гельминтов, находится в прямой за-
висимости от условий снабжения паразитов кислородом. Крупные
кишечные нематоды содержат много полисахаридов, тканевые
паразиты и формы, поселяющиеся в кровяном русле, обладают
небольшим количеством гликогена, а у паразитов крови — очень
мелких личинок Wuchereria bancrofti (микрофилярий) —он вооб-
ще не был обнаружен. Как правило, очень немного гликогена име-
ется и у свободноживущих личинок паразитических нематод. Кроме
глигокена в тканях и полостной жидкости круглых червей содер-
жатся и низкомолекулярные сахара. Некоторые из них характерны
только для нематод (например, аскарилоза) и не встречаются
у других животных.
Основным источником углеводов, в которых нуждаются круг-
лые черви, служит поглощаемая ими пища. Гликоген, крахмал
и другие полимерные соединения предварительно расщепляются
при помощи гидролитических ферментов. У нематод, в частности,
обнаружена весьма активная амилаза. В первую очередь это отно-
сится к «растительноядным» и фитонематодам.
Данные о характере превращений углеводов в теле нематод
очень противоречивы. Важную роль, по-видимому, играет гликолиз,
о чем свидетельствует обнаружение в тканях паразитов соответст-
вующих ферментов. ОднакЪ анаэробное превращение глюкозы
у круглых червей несколько отличается от аналогичных процессов,
протекающих у свободноживущих организмов.
Лишь у немногих круглых червей (главным образом это филя-
рии) конечным продуктом гликолиза является молочная кислота.
У большинства же нематод образуется смесь преимущественно ле-
тучих жирных кислот, среди которых очень часто (например,
у аскарид) доминирует валериановая кислота. Последняя, в част-
ности, в большом количестве накапливается в полостной жидкости
паразитов.
Многие нематоды, по-видимому, обладают нормальным циклом
Кребса. В литературе, однако, высказывается предположение
95
о существовании и какого-то другого пути расщепления углеводов,
обеспечивающего процессы окислительного фосфорилирования.
Очень различно отношение нематод к свободному молекуляр-
ному кислороду. Большинство из них потребляет содержащийся
в окружающей их среде кислород, как бы мало его ни было. В то
же время "для ряда видов, у которых преобладают анаэробные ме-
таболические процессы (аскариды и др.), высокое парциальное
давление кислорода оказывается губительным.
У многих круглых червей в тканях обнаружены ферменты цито-
хромной системы дыхания и флавины. Однако далеко »не всегда
эти системы активно функционируют. Так, у крупных кишечных
форм, живущих в условиях ревко выраженного недостатка кислоро-
да (например, аскариды), цитохромы хотя и имеются, непосредст-
венного участия в дыхании паразитов не принимают.
Характер обмена того или иного вида нематод часто подверга-
ется серьезным перестройкам в процессе индивидуального разви-
тия паразитов. Рядом исследователей было показано, что яйца
аскарид, находящиеся на самых разных стадиях дробления, а так-
же содержащие личинок, и личинки, совершающие миграцию по
телу животного-хозяина (кровеносная система —легкие, с. 102),
обладают отчетливо выраженным аэробным метаболизмом, тогда
как личинки, перешедшие к постоянному обитанию в просвете ки-
шечника, и взрослые черви становятся настоящими анаэробами.
Как уже отмечалось выше, их цитохромная система дыхания при
этом перестает функционировать.
Таким образом, у круглых червей, как и у многих других
гельминтов, наблюдается значительное разнообразие типов угле-
водного обмена. Большинство свободноживущих форм и фитонема-
тоды— аэробные организмы, сохранившие оксидативное расщеп-
ление глюкозы. У паразитов животных наблюдается Переход от
аэробиоза к анаоксибиозу, причем во многих случаях явно преоб-
ладают анаэробные процессы. Об этом говорит и утрата ими зна-
чительной части цикла Кребса, и отмечаемое многими авторами
сильное изменение системы терминальнрго окисления.
Поступление кислорода в организм паразита осуществляется
различными путями. Тканевые формы и паразиты, постоянно на-
ходящиеся в тесном контакте с внутренней поверхностью кишечни-
ка, используют кислород, диффундирующий из окружающих тка-
ней. Источником кислорода для гематофагов служит кровь живот-
ного-хозяина. Попадающий в пищеварительную систему паразита
оксигемоглобин прежде всего восстанавливается в обычный гемо-
глобин и лишь после этого подвергается действию протеолитиче-
ских ферментов. Освобожденный кислород диффундирует через
стенку кишечника.
Очень важной биологической особенностью многих паразитиче-
ских нематод является наличие у них дыхательных пигментов —
гемоглобинов. Последние свойственны, однако, только зоогельмнн-
там: ни у свободноживущих форм, ни у паразитов растений гемо-
глобины не обнаружены.
96
У нематод, как и у ряда паразитических плоских червей, описа-
но несколько видов гемоглобинов, которые локализуются в кутику-
ле н гиподерме паразитов, жидкости схизоцеля, нервной системе,
стенках кишечника. По мнению американских-паразитологов Д. Ли
и М. Смита (D. Lee and М. Smith), адаптивное значение имеет
большое содержание гемоглобина в нервной системе, которая наи-
более чувствительна к дефициту кислорода. Пигменты, обнаружен-
ные в кутикуле и гиподерме (с. 83), по-видимому, играют важную
роль в сорбции кислорода из окружающей паразита среды.
Следует, однако, отметить, что далеко не всегда удается легко
доказать дыхательную функцию гемоглобинов нематод. Некоторые
гемоглобины, как, например, содержащиеся в полостной жидкости
аскарид и ряда других круглых червей, практически ни при каких
условиях не отдают связанный с ними кислород. Соответственно,
участие таких пигментов в процессах дыхания паразитов становит-
ся весьма сомнительным.
Имеющиеся в литературе сведения о липидном обмене круглых
червей довольно ограничены. Жиры, как и углеводы, откладывают-
ся практически во всех тканях и органах паразитов, но особенно
много их в гиподерме, саркоплазме мускульных клеток и органах
системы размножения. Больше всего жиров обнаруживается
у форм, обитающих в условиях острого дефицита кислорода.
По-видимому, существует несколько источников накопления
жиров в теле паразита. Какую-то часть липидов круглые черви
получают вместе с пищей. У аскарид и целого ряда других видов
в кишечнике обнаружены липазы — ферменты, осуществляющие
гидролиз жиров. Большая часть жира нематод имеет, однако, дру-
гое происхождение. Многие исследователи указывают на их экскре-
торную природу и считают, что они возникают как продукты
углеводного обмена.
Взрослые черви использовать жиры в качестве источника энер-
гии в большинстве случаев не могут. Как правило, иначе обстоит
дело с ранними стадиями развития, которое у многих видов про-
текает во внешней среде. По мнению ряда исследователей, у сво-
бодноживущих личинок нематод именно липиды играю роль основ-
ного резервного вещества.
Размножение и жизненные циклы нематод
Размножение. Обычно в организме животного-хозяина поселя-
ются и самцы, и самки нематод, которые по достижении половой
зрелости копулируют друг с другом.
Мужские половые клетки — спермин — при копуляции поступа-
ют в органы женской половой системы и в течение определенного
срока (характерного для каждого конкретного вида) сохраняются
в просвете маток. Особенностью сперматозоидов круглых червей
является отсутствие жгутиков. Передвижение мужских половых
клеток по половым путям самки осуществляется с помощью псев-
доподий.
4—58
97
Б
Рис. 52. Схема строе-
ния личинок нематод
(из кн. Е. Нобля и
Г. Нобля). А — раб-
дйтрвидиая личинка;
Б — фйляриевпдная
личинка
Сразу же после проникновения спермин в женскую половую
клетку последняя формирует на своей поверхности первую защит-
ную оболочку. Несколько позже выделяется еще одна оболочка,
а на поверхности яйца за счет секреторной деятельности стенок
матки образуется самый наружный, так называемый белковый слой.
Таким образом, зрелое яйцо нематод коренным образом отличается
от. сложных яиц паразитических плоских червей (с. 19).
Отсутствие желточных клеток предопре-
деляет важную особенность половых клеток
круглых червей. У всех паразитических
Plathelminthes яйцеклетки не содержат за-
пасных питательных веществ, которые в
наиболее типичных случаях привносятся в
яйцо вместе с желточными клетками. У
большинства же нематод сама яйцеклетка
несет в себе питательный материал, необ-
ходимый для развития будущего заро-
дыша.
Хорошо прослеживается у нематод отме-
ченная еще В. А. Догелем тенденция к воз-
никновению гермафродитизма у паразитов,
берущих свое начало от раздельнополых
свободноживущих предков. Многие круглые
черви становятся физиологическими герма-
фродитами, фенотипически оставаясь при
этом самками. У одних видов они способны
производить мужские половые клетки лишь
в течение сравнительно короткого проме-
жутка времени в самом начале периода сво-
ей половой активности (протерандрический
гермафродитизм). У других же формирова-
ние сперматозоидов возобновляется не-
сколько раз на протяжении жизни паразита.
Накбпец, в очень редких случаях нематоды
переводят к партеногенетическому размно-
жению без оплодотворения.
Постэмбриональное развитие нематод представляет собой мета-
морфоз. Обычно имеется четыре личиночные стадии. Переход от
едной стадии к другой сопровождается линьками. Последователь-
но сменяющих друг друга личинок принято обозначать так: Л-1,
Л-2, Л-3 и Л-4.
По своему внешнему облику и внутреннему-строению личинки
во многом напоминают взрослых червей (рис. 52). Тем не менее
имеется ряд серьезных отличий, которые касаются как морфологи-
ческих, так и биологических особенностей. Первая и главная черта
личинок нематод — это отсутствие сформированной половой систе-
мы. Последняя представлена недифференцированным половым за-
чатком, расположенным в задней половине тела. Как правило,
недоразвита и пищеварительная система. Задняя кишка часто име-
98
ет вид простого клеточного тяжа, иногда отсутствует просвет
и в средней кишке. Исследования кутикулы личинок нематод пока-
зали, что она содержит меньшее количество слоев п отличается от
кутикулы взрослых червей по своим свойствам (проницаемости,
механической прочности и т. д.). Часто очень несхожи и особенно-
сти обменных процессов п физиологии половозрелых червей п ли-
чинок.
Личинок нематод по особенностям их строения иногда разделя-
ют на две морфологические группы «рабдитовидных» и «филярие-
Рис. 53. Схема цикла развития различных трихострон-
гилид (по Шульцу и Дикову). А—жизненный цикл
Haemonchus\ Б — жизненный цикл Marschallagia-, В —
жизненный цикл Nemaiodirus'.
1 — яйцо, выведенное во внешнюю среду, 2 — эмбриональное
развитие во внешней среде
видных» (рис. 52). К первой относятся личинки с коротким и срав-
нительно толстым телом. Их фаринкс снабжен бульбусом и по сво-
ему строению напоминает фаринкс нематод из отр. Rhabditida.
Вторые же обладают более узким и стройным телом. Длинный
и тонкий фаринкс филяриевидных личинок очень похож на тако-
вой многих филярий (отр. Spirurida).
У ряда видов нематод на протяжении их индивидуального раз-
вития наблюдается смена типов личинок.
Отношения личинок нематод с внешней средой очень различны.
Как и в других группах паразитических организмов, у круглых
червей проявляется тенденция к сокращению сроков пребывания
активных стадий во внешней среде, вплоть до полного их выпаде-
ния. Очень часто конкретные примеры этого положения могут быть
обнаружены даже в пределах сравнительно небольших таксономи-
ческих групп: семейств и надсемейств. Хорошей иллюстрацией
служит сем. Trichostrongylidae (подотр. Strongylata) (рис. 53)-
4*
99
В наиболее примитивных случаях (р. Haemonchus и др.) из попав-
шего во внешнюю среду яйца выходит личинка первой стадии. Она
претерпевает две линьки и лишь образовавшиеся в результате это-
го личинки-3 (Л-3) становятся инвазионными для позвоночного
животного. Очень часто первая линька протекает под защитой
яйцевой оболочки, так что во внешнюю среду попадают только ли-
чинки второй стадии (Л-2) (р. Marschallagia). Здесь они претерпе-
вают линьку и дают начало инвазионным личинкам-3. И, наконец,
обе линьки могут осуществляться в яйце, так что во внешнюю сре-
ду сразу же выходят личинки-3 (р. N'ematodirus).
Многие нематоды вообще утратили свободноживущие активные
стадии. Ивазионными для животного-хозяина являются рассеивае-
мые во внешней среде яйца, содержащие сформированных личинок.
У некоторых видов исчезают и они. Последнее становится возмож-
ным при появлении в жизненном цикле паразита промежуточных
хозяев, которые и осуществляют передачу инвазионного начала от
одного дефинитивного хозяина к другому (с. 106). Во многих слу-
чаях это сопровождается появлением яйцеживорождения
(р. Wuchereria, Trichinella и др.).
Личинки паразитических, нематод могут сохраняться во внеп!-
пей среде длительное время. Описаны случаи, когда в естественных
условиях они оставались жизнеспособными в течение двух и более
лет. Жизнедеятельность личинок обеспечивается запасами пита-
тельных веществ, полученных ими при развитии. В старых личин-
ках количество жира и гликогена по сравнению с молодыми замет-
но уменьшается.
Поведение личинок во внешней среде. Свободноживущие личин-
ки круглых червей обладают рядом биологических адаптаций, на-
правленных на заражение животных-хозяев. В первую очередь это
способность к миграциям. Как показывают многочисленные наблю-
дения, личинки нематод во внешней среде довольно активно
перемещаются в вертикальном направлении. Главными факторами,
вызывающими подобную активность, по-видимому, являются свет
и влажность. Яркое освещение и подсыхание верхних слоев почвы
вызывают уход личинок на довольно значительную глубину (до
1 м). Повышение влажности снова заставляет их мигрировать
к поверхности почвы. У многих видов личинки способны поднимать-
ся по траве и мелким кустарникам на высоту до 60 см, что и обус-
ловливает массовое заражение животных-хозяев. Движение личи-
нок по субстрату оказывается возможным только при наличии
тонкого слоя влаги. Они прижимаются к твердым предметам плен-
кой поверхностного натяжения и под ее защитой перемещаются,
изгибаясь в вертикальной плоскости. На активность личинок боль-
шое влияние оказывает температура. Даже в условиях подходящей
влажности при очень низких или, наоборот, слишком высоких тем-
пературах они остаются неподвижными.
Характер поведения личинок во внешней среде, вероятно, во
многом определяется присущими им таксисами. Однако имеющие-
ся литературные данные по этому вопросу очень противоречивы.
100
Скорее всего у разных видов личинок в различной степени выра-
жены фото-, гео-,, рсо- и хемотаксисы.
Попадание нематод в окончательных хозяев осуществляется
несколькими различными способами. В тех случаях, когда инвази-
онным началом служат яйца, содержащие зрелых личинок, зара-
жение животных возможно только пероральным путем. В кишеч-
нике хозяина личинки освобождаются от скорлуповых оболочек.
Механизм вылупления паразитов из яиц еще не до конца ясен.
По-видимому, как и у других гельминтов, он включает в себя не-
сколько этапов. Яйцо, попавшее в кишечник, подвергается дейст-
вию целого ряда факторов, среди которых, по мнению В. Роджер-
са (W. Rogers), главную роль играет высокая концентрация СОг
или углекислоты. Более поздние исследования показали, что име-
ют значение еще и pH среды, температура и наличие определенных
солей. Эти факторы стимулируют секреторную деятельность личи-
нок. Продуцируемые ими вещества, среди которых у аскариды, на-
пример, обнаружены такие ферменты, как хитиназа и эстераза,
выделяются в полость яйца и, по-видимому, воздействуют на его
скорлупку. В результате личинки освобождаются от одевающих
их оболочек и переходят к паразитическому образу жизни.
Заражаться животные могут и активными совободноживущимн
личинками. У одних видов этот процесс осуществляется также пе-
рорально. Хозяева проглатывают личинок вместе с травой, на кото-
рой последние скапливаются иногда в довольно большом количест-
ве (с. 104). Личинки многих видов проникают в животных через
кожу — перкутанный способ заражения. В этом случае они, как
показывают наблюдения над Ancylostoma,- Strongyloid.es и рядом
других видов, проникают в-толщу-кожи через волосяные фоллику-
лы. Иногда для этой цели ими используются участки, на которых
произошло слущивание поверхностного сл.оя ороговевших клеток.
Наконец, заражение окончательных хозяев может осуществлять-
ся при посредстве промежуточных и резервуарных хозяев. Чаще
всего передача паразитов от одного животного к другому происхо-
дит по пищевым цепям в системе хищник — жертва. Однако возмо-
жен и другой путь, когда промежуточными, хозяевами становятся
кровососущие насекомые (комары, москиты и т. п.). Заражение
в этом случае осуществляется контаминативным способом. Личин-
ки выходят из хоботка насекомого в тот момент, когда оно питает-
ся кровью на позвоночном животном и активно проникают через
кожу последнего в его кровяное русло.
Жизненные циклы паразитических нематод. Циклы развития
паразитических круглых червей характеризуются исключительным
многообразием. Последнее в значительной мере обусловлено тем,
что отдельные группы нематод переходили к паразитизму в разное
время и приспосабливались к существованию в самых различных
животных. Поэтому наряду с крайне специализированными форма-
ми имеется много примитивных, которые по сути дела должны рас-
сматриваться как факультативные паразиты. Их циклы развития,
как правило, характеризуются известной «неустойчивостью» и мо-
101
гут в зависимости от условий осуществляться различными спосо-
бами. <
На этой эволюционной стадии сейчас находятся многие предста-
вители подотр. Rhabditata (отр. Rhabditida), который включает
целый ряд свободноживущих видов, ставших факультативными
паразитами, и, наконец, формы, перешедшие к облигатному пара-
зитизму, но сохранившие и свободноживущие поколения.
Рис. 54. Цикл развития Strongyloides stercoralis (по разным авторам). Л—пара-
зитическая самка; Б — паразитический самец; В — рабдитовпдная личинка;
Г—филяриевидная личинка; Д —схема жизненного цикла
К числу факультативных паразитов можно отнести широко рас-
пространенный вид Strongyloides stercoralis (рис. 54). При благо-
приятных внешних условиях (температура, влажность и т. д.)
в почве обитают половозрелые самцы и самки. Они копулируют
между собой. Из оплодотворенных яиц, отложенных самками, вы-
ходят рабдитовидные личинки (Л-1), значительное большинство
которых линяет четыре раза и дает начало новому свободноживу-
щему поколению. Меньшая часть, однако, после второй линьки
превращается не в рабдитовидных, а в филяриевидных личинок
(Л-3), которые уже не могут нормально развиваться во внешней
среде. Для завершения цикла развития им необходимо проникнуть
в организм человека. Заражение последнего осуществляется глав-
ным образом перкутанно. Личинки обязательно мигрируют в кро-
вяное русло, по которому они сначала попадают в малый круг кро-
вообращения и оттуда в легкие. В легких личинки линяют еще два
раза и дают начало молодым самцам и самкам. Последние по воз-
духоносным путям поднимаются в гортань, из которой уже и
102
проникают в пищеварительную систему. Здесь они копулируют, и
самки откладывают япца. Возможен и пероральный путь зараже-
ния человека. Проглатываемые филяриевидные личинки, однако,
все равно проникают в слизистую носоглотки и совершают мигра-
цию по кровесноп системе и легким.
Из яиц, отложенных самкой прямо в кишечнике хозяина, вы-
лупляются рабдитовидные личинки первой стадии, которые вскоре
линяют. Судьба личинок второй стадии может быть различной.
Часть выводится во внешнюю среду, где они могут дать начало
свободноживущему поколению или же после линьки превращаться
в филярйевидных личинок, инвазионных для человека. Часть же
претерпевает вторую линьку прямо в просвете пищеварительной
системы. В этом случае становится возможным аутоинвазия хозя-
ина. Личинки внедряются либо непосредственно в слизистую ки-
шечника, либо в кожу перианальной области. И в том, и в другом
случае они снова проделывают сложную миграцию прежде, чем
достигнут средней кишки.
У относящегося к тому же подотряду паразита легких амфибий
Rhabdias bufonis наблюдается известная стабилизация цикла. Это
уже облигатные паразиты, которые сохраняют, однако, и свободно-
живущее поколение. В легких лягушек обитают гермафродитные
черви. Откладываемые ими яйца сначала попадают в глотку, а за-
тем через кишечник вместе с непереваренными остатками пищи
выводятся во внешнюю среду. Вылупившиеся из яиц личинки после
соответствующего числа линек дают начало особям свободноживу-
щего раздельнополого поколения, которые почти вдвое меньше
гермафродитных паразитических особей. Половозрелые черви ко-
пулируют, самки откладывают яйца, а вышедшие из них личинки
для дальнейшего развития снова должны попасть в позвоночное
животное. Заражение амфибий осуществляется перкутанно. Личин-
ки мигрируют по кровеносной системе в легкие, где, проделав
линьки, достигают половой зрелости.
У Rhabdias bufonis, таким образом, развитие приобретает ха-
рактер сложного жизненного цикла, протекающего по типу гетеро-
гонии. В нем закономерно чередуются свободноживущие раздель-
нополые и паразитические гермафродитные поколения.
Все представители подотр. Strongylata (отр. Rhabditida) пол-
ностью утратили способность достигать половой зрелости во внеш-
ней среде. Это облигатные паразиты, у которых свободный образ
жизни сохраняют в лучшем случае личинки хозяина. Виды, отно-
сящиеся к сем. Ancylostomatidae (см. рис. 59), обладают всего
двумя свободноживущими личиночными стадиями. Их самки, па-
разитирующие в кишечнике человека и ряда других животных,
откладывают яйца, из которых во внешней среде вылупляются раб-
дитовидные личинки первой стадии. Они проделывают две линьки
и дают начало фпляриевидным личинкам (Л-3). Последние чаще
всего проникают через кожу животного-хозяина и далее проделы-
вают почти тот же путь, что и инвазионные личинки Strongyloides
siercoralis. В отличие от последних, однако, личинки анкилостомид
103
в легких не линяют. Они сразу же совершают миграцию в кишеч-
ник, где, проделав две линьки, достигают половой зрелости.
У ряда видов ход подобного цикла развития усложняется за
счет вклинивания промежуточных хозяев. Примером- может слу-
жить р. Protostrongylus (сем. Protostrongylidae, подотр. Slrongy-
lata рис. 55). Яйца, откладываемые самками, паразитирующими
в легких овец, не сразу выводятся во внешнюю среду, а дозревают
в бронхах. Из них вылупляются личинки, которые проходят по пи-
щеварительному тракту и
выводятся во внешнюю сре-
ду с фекалиями. Дальней-
шее развитие паразитов, од-
нако, возможно только в
промежуточном хозяине —
наземных моллюсках. В
промежуточном хозяине ли-
чинки дважды линяют и на
стадии Л-3 снова выходят
во внешнюю среду. Они кон-
центрируются на траве, вме-
сте с которой и попадают в
пищеварительную систему
Рис. 55. Цикл развития Protostrongylus овцы. Из кишечника они
(по Шульцу и Дикову) мигрируют в лимфатическую
систему, далее в кровенос-
ную и через малый круг кровообращения в легкие. В легких личин-
ки линяют еще два раза и превращаются в половозрелых особей.
Как уже отмечалось выше, возможен и другой путь сокращения
сроков пребывания активных расселительных стадий во внешней
среде. Это смещение части линек на период нахождения личинки
в яйце (см. рис. 53), что характерно для ряда стронгилят (с. 125),
и, наконец, полное выпадение из цикла развития свободноживущих
личинок. Последнее свойственно очень многим специализированным
паразитам. Инвазионным началом становятся яйца, содержащие
личинок. Эмбриональное развитие и первая личиночная лииька
у многих видов осуществляются только при доступе кислорода во
время нахождения яйца во внешней среде. Именно на этом этапе
приспособления к паразитическому образу жизни мы застает мно-
гих представителей подотр. Ascaridata (отр. Ascaridida). Несмотря
на то что заражение яйцами осуществляется, главным образом,
перорально и вылупившиеся личинки сразу же попадают в.пище-
варительную систему, они в ряде случаев (р. Ascaris и др.) сохра-
няют характерную для многих более примитивных форм сложную
миграцию по организму хозяина (см. рис. 57). Личинки по крове-
носной системе мигрируют из кишечника в легкие, где претерпева-
ют две линьки. Образующиеся личинки четвертой стадии (Л-4)
поднимаются по дыхательным путям в носоглотку, а оттуда снова
попадают в кишечник, где линяют еще раз и дают начало самцам
и самкам.
104
Подобные миграции кишечных паразитов В. А. Догель называл
«реакцией бегства». По его мнению, которое разделяют многие
видные современные паразитологи, миграция осуществляется «не
в поисках каких-нибудь лучших условий жизни, а для избежания
гибельного действия содержимого кишечника, ставящего под угро-
зу жизнь очень мелких и нежных личиночных стадий на первых
фазах их эндопаразитического периода существования». Личинкам,
по-видимому, необходим известный период адаптации к существо-
ванию в животном-хозяине. Миграция личинок сопорвождается не
только их морфологическим развитием, но и серьезными физиоло-
гическими перестройками (с. 96), позволяющими им поселяться
в условиях острого дефицита кислорода и высокой химической
активности кишечной среды.
Последующая эволюция циклов развития круглых червей шла
по нескольким направлениям. С одной сторонй, личинки приобре-
тали все более ярко выраженную способность непосредственно по-
селяться в пищеварительном тракте хозяина. Первый шаг в этом
направлении наблюдается у упоминавшихся ранее представителей
сем. Ancylostomatidae (с. 112), личинки которых в легких не претер-
певают никакого развития. Последнее целиком осуществляется
в просвете кишечника (см. рис. 59). В этом случае при перкутан-
ном заражении животного-хозяина миграция, по-видимому, обес-
печивает попадание паразита к месту окончательного поселения
и поэтому сохраняется в процессе эволюции. У многих же специа-
лизированных форм, заражение которыми происходит перорально,
такая чисто «транспортная» миграция.утрачивает свое биологиче-
ское значение и со временем выпадает из цикла. Это характерно
для представителей самых разных групп нематод: ряда стронгилят
(см. рис. 53), аскаридат (рис. 56) и др.
Другое направление эволюционных изменений связано с сокра-
щением периода пребывания яиц во внешней среде, необходимого
для достижения ими инвазионного состояния. Если у тех же аска-
ридат способны заражать хозяина лишь яйца, содержащие личин-
ку. второй стадии (Л-2), то у Тrichocephalus trichiurus, по-видимому,
очень древних паразитов из подкл. Adenophorea, инвазионными
становятся яйца, содержащие личинок первой стадии, не прошед-
ших ни одной линьки.
Ряд нематод полностью утратил свободноживущие рассели-
тельпые стадии. Ни на одной фазе цикла развития паразйты не вы-
ходят во внешнюю среду. Передача инвазионного начала от одного
дефинитивного хозяина к другому осуществляется с помощью пе-
реносчиков или промежуточных хозяев. Очень далеко в этом на-
правлении зашла специализация филярий (подотр. Filariata, отр.
Spinirida). Многие представители этой группы (сем. Filariidae)
поселяются в органах и тканях, непосредственно не связанных
с внешней средой,— подкожная клетчатка, нервная, кровеносная
и лимфатическая системы и т. д. Самки примитивных видов откла-
дывают яйца, из которых сразу же вылупляются личинки, в то вре-
мя как у большинства форм развивается настоящее яйцеживорож-
105
дение. В этом случае самки отрождают большое количество мелких
личинок, называемых микрофиляриями. '
Микрофилярии чаще всего мигрируют в кровеносную систему,
в которой сохраняются длительное время. Кровососущие насекомые
поглощают личинок во время питания на зараженном животном
или человеке. Пребывание паразитов во внешней среде, таким об-
разом, полностью исключается (см. рис. 60).
Все приведенные выше приме-
ры иллюстрируют возможный
путь возникновения и последую-
щей эволюции имагинального па-
разитизма нематод. Однако у них
возможен и настоящий ларваль-
ный паразитизм, когда свободно-
живущими становятся не личи-
ночные стадии, а половозрелые
особи.
У нематод, относящихся к сем.
Mermithidae (подкл. Adenopho-
геа), временное пребывание ли-
чинок в других организмах стано-
вится обязательным условием за-
вершения всего цикла развития.
Самцы и самки мермитид обита-
ют в водоемах или во влажной
Рис. 56. Цикл развития Ascaridia почве. Они не питаются и выпол-
galU (по Шульцу и Дикову) няют в основном генеративную и
расселительную функции. Из от-
кладываемых самками яиц выводятся личинки. Заражение живот-
ных-хозяев, которыми могут быть различные водные и наземные
беспозвоночные (насекомые, моллюски, ракообразные), осущест-
вляется либо перкутанно, либо перорально. Во втором случае пара-
зиты мигрируют'в полость тела, пробуравливая стенку кишечника.
В организме хозяина личинки быстро развиваются. Морфогенети-
ческие процессы сопровождаются накоплением большого количе-
ства запасных питательных веществ. Созревшие паразиты покида-
ют хозяина, который при этом гибнет. После линьки они дают на-
чало самцам и самкам. Как и у многих других паразитов, цикл
развития мермитид теснейшим образом приурочен к особенностям
биологии и развитию животных-хозяев. Эти особенности мермитид
привлекают к ним большое внимание специалистов по биологиче-
ским методам борьбы с вредными насекомыми. В ряде стран сей-
час делаются попытки использовать мермитид для уменьшения
численности мошек и других кровососущих Diptera.
Классификация нематод
Разработка естественной системы класса Nematoda еще очень далека от сво-
его завершения. Особенно много противоречий вызывают вопросы, связанные
с установлением таксономического ранга и объема тех или иных групп круглых
106
червей. Очень различно трактуются паразитологами и филогенетические связи
в пределах класса.
Как уже отмечалось ранее, нематоды разделяются на два подкласса: Ade-
nophorea и Secernentea.
Многими исследователями признается, что аденофореи филогенетически бо-
лее древняя группа. Об этом говорит большое число свободноживущих видов,
поселяющихся в почве, пресноводных водоемах и морях. Наряду с этим имеется
и значительное количество паразитов животных (как позвоночных, так и безпо-
звоночных) и растений.
Морфологически Adenophorea характеризуются отсутствием фазмид, сильным
развитием кожных желез, разнообразными по своему строению и, как правило,
довольно крупными амфидами. В настоящее время в пределах подкласса выде-
ляется 5 отрядов, из которых лишь в трех паразитизмЯЮлучил широкое распро-
странение.
Отр. Trichocephalida включает виды, представители которых имеют более или
менее утонченный или вытянутый в нитевидный придаток передний конец. Пара-
зитируют в позвоночных животных. Цикл развития осуществляется как с участи-
ем промежуточных хозяев, так и без них. В состав отряда входят широкорас-
пространенные паразиты человека и животных, относящиеся к р. Trichocephalus
(сем. Trichocephalidae, с. 120), р. Trichinella (сем. Trichinellidae, с. 121).
Представители отр. Dioclophymida, как правило, довольно крупные нематоды,
паразитирующие в кишечнике, желудке и почках млекопитающих и птиц. Отр.
Dofylaimida объединяет как свободпоживущие морские, пресноводные и почвен-
ные’ виды, среди которых имеются настоящие хищники, так и паразитические
формы. Последние обитают в беспозвоночных животных (сем. Mermithidae,
с. 106) и в растениях.
Нематоды, вклтрчаемые в состав подкл. Secernentea, характеризуются нали-
чием фазмид. Типичные кожные железы отсутствуют. Амфидии, как правило,
очень мелкие, поровидные. Подавляющее большинство Secernentea — паразиты
животных. Виды, поселяющиеся в растениях, не столь многочисленны. Количество
свободноживущих форм и факультативных паразитов еще меньше. Подкл. Se-
cernentea включает 4 отряда. Наиболее примитивным, несомненно, является отр.
Rhabditida, в состав которого наряду с облигатными входят факультативные па-
разиты и свободпоживущие виды. Последние преимущественно представлены са-
пробионтами. К этому отряду относятся такие широкораспространенные паразиты
человека и домашних животных, как Ancylostoma (с. 112)$ Strong у loides (с. 102),
Protostrongylus (с. 104) и др. Очень своеобразен отр. Tylenchida. Он представлен
свободножнвущими хищниками и паразитами растений й‘беспозвоночных живот-
ных. Сохранив целый ряд примитивных черт строения, тиленхиды приобрели
в то же время признаки, говорящие о их глубокой специализации: их стома, на-
пример, превращается в подвижный колюще-сосущий орган — стилет (с. 86).
К отр. Spirurida относятся исключительно паразитические виды, развитие
которых осуществляется с участием промежуточных хозяев. В состав отряда
входят многие опасные паразиты человека и домашних животных: Wuchereria
bancrofti, Loa loa, Onchocerca volvulus и др., относящиеся к подотр. Filariata
(с. 115), Dracunculus medinensis из подотр. Camallanata (с. 107) и др.
Отр. Ascaridida также включает только облигатных паразитов, обитающих
в позвоночных и беспозвоночных животных (членистоногих). К аскаридидам от-
носится большое число патогенных видов, вызывающих заболевания человека
и домашних животных: р. Ascaris (с. 126), р. Ascaridia (с. 126) и др. из подотр.
Ascaridata; Enterobius vermicularis (с. 110), Heterakis gallinarum (с. 107) и др.,
входящие в состав подотр. Oxyurata.
Нематодозы
Паразитические круглые черви — возбудители многих опасных
болезней (нематодозов) человека и домашних животных. Экономи-
ческий ущерб, причиняемый нематодами в сельском- хозяйстве,
очень велик. Серьезную угрозу они представляют и для здоровья
людей.
107
Нематодозы. человека
Аскаридоз
Одним из самых - распространенных заболеваний человека,
вызываемых гельминтами, является аскаридоз, возбудитель ко-
торого Ascaris lumbricoides (L. 1758) (см. рис. 44, А, В\ 57) встре-
чается практически во всех районах земного шара.
Аскариды относятся к крупным нематодам. Веретеновндиое тело
самок достигает 40—44 см в длину. Самцы, задний конец которых
загнут на брюшную сторону, несколько меньше: 12—25 см. Ротовое
Л
Рис. 57. Человеческая аскарида Ascaris lumbricoides (по разным авторам).
А — схема цикла развития; Б — яйцо; В — инвазионная личинка II возраста
отверстие окружено тремя губами. У самок в передней половине
тела располагается небольшая перетяжка, на которой вентрально
открывается половое отверстие.
Половозрелые ’самцы и самки обитают в кишечнике человека.
Оплодотворенные самки откладывают яйца, размеры которых ко-
леблются в пределах 0,05—0,07x0,04—0,05 мм. Яйца одеты толстой
оболочкой, состоящей из нескольких слоев. Наружный слой имеет
неровную фестончатую поверхность. Яйца аскарид чрезвычайно
устойчивы к неблагоприятны^ факторам внешней среды. В приро-
де яйца могут сохраняться, не теряя инвазионностп в течение 7 лет.
Под защитой яйцевых оболочек протекает эмбриональное раз-
витие и формируется личинка первой стадии. Инвазионными яйца
аскарид становятся лишь после того, как личинка перелиняет.
В .кишечнике человека, куда яйца чаще всего попадают вместе
с пищей, личинки второй стадии вылупляются. Они внедряются
в стенку кишечника и совершают сложную миграцию. По кишеч-
ным сосудам они с током крови попадают в печень, далее через
полую вену в сердце, а затем по легочной артерии в легкие. Через
лопнувшие стенки капилляров они выходят в альвеолы, в которых
108
дважды линяют. Формирующиеся лЫинки четвертой стадии по
трахеям мигрируют через гортань в Пищеварительную систему.
Здесь они линяют еще раз п достигают половой зрелости. Все раз-
витие аскарид от момента инвазии до начала продуцирования сам-
ками яиц продолжается около 10—11 недель. Общий срок жизни
червей обычно .не превышает 11 —12 месяцев.
В последние годы появились сообщения о существовании у ас-
карид и другого пути развития. Было установлено, что в определен-
ных условиях (наличие в почве большого количества перегноя,
высокие температуры и т. д.) инвазйонные личинки могут в массе
выходить из яиц во внешнюю среду. Такие активно подвижные ли-
чинки легко внедрялись в кожу экспериментальных животных.
В течение аскаридоза отчетливо- выявляются два этапа: первый
связан с миграцией личинок, второй — с паразитированием полово-
зрелых особей в кишечнике. -Мигрирующие личинки травмируют
стенки кишечника, печень и особенно легкие. Важную роль играет
сенсибилизация организма чужеродными белками, источником ко-
торых являются продукты обмена личинок и сами погибающие ли-
чинки. При слабых инвазиях симптомы заболевания могут отсут-
ствовать, в случае сильного заражения у больного отчетливо про-
являются признаки аллергии: крапивница, зуд, насморк, кашель
и т. п. Иногда наблюдаются острые пневмонии, эксудатнвный плев-
рит и явления бронхиальной астмы.
Обитание взрослых червей в кишечнике хозяина сопровождает-
ся, как правило, общей интоксикацией организма и аллергически-
ми явлениями, связанными с усвоением экскретируемых паразита-
ми продуктов обмена. Во многих случаях болезнь протекает бес-
симптомно. Однако иногда аскаридоз принимает исключительно
тяжелые формы и может оказаться летальным. Клиническая карти-
на второго, этапа аскаридоза слагается из желудочно-кишечных
и нервных симптомов. Снижается аппетит, наблюдаются функцио-
нальные расстройства кишечника и острые боли в животе. Заболев-
ший становится раздражительным, у него снижается трудоспособ-
ность и нарушается сон. Нередко возникают менингиальные явле-
ния. Развивается острая анемия.
Тяжелые осложнения связаны с нарушением проходимости ки-
шечника и извращением локализации взрослых червей. Последние
могут мигрировать в протоки печени, поджелудочной железы, ды-
хательные пути, лобные пазухи. Очень опасен- аскаридоз печени,
часто сопровождаемый закупоркой желчных протоков, механиче-
ским разрушением тканей органа и острым перитонитом.
Источником распространения аскаридоза являются больные
люди, выделяющие вместе с фекалиями огромное число яиц. Устой-
чивость последних к действию вредных-факторов внешней среды
(с. 98) обусловливает широкое распространение инвазионного на-
чала в природе. Наиболее массовые заражения наблюдаются в теп-
лые периоды года. Особенно часто аскаридами заражаются дети
п люди, связанные с работой в сельском хозяйстве. Инвазионные
яйца попадают в организм человека с водой, немытыми овощами
109
л фруктами. По-видимому, важным агентом в распространении
аскаридоза являются мух#. Часто инвазионные яйца сохраняются
в консервированных и соленых овощах.
Как уже отмечалось выше, вероятно, существует и другой путь
заражения аскаридами. Способность личинок выходить из яиц во
внешней среде и внедряться в кожу животных (см. 109) позволяет
иредполагать и возможность перкутанной инвазии людей. Эпиде-
миологическое значение этого явления пока еще недостаточно изу-
чено.
Распространение аскаридоза чрезвычайно широко. Он домини-
рует среди прочих гельминтозов, встречающихся на территории
СССР. По данным 1965 г., 75% зарегистрированных случаев зара-
жения гельминтами приходилось на аскарид.
Диагноз устанавливается при обнаружении в фекалиях яиц ас-
карид. Для лечения аскаридоза в настоящее время применяется
широкий набор лечебных препаратов. Значительное распростране-
ние в последние годы получил метод оксигенотерапии, основанной
на том, что для половозрелых червей кислород токсичен.
Важное значение в борьбе с аскаридозом имеют профилактиче-
ские мероприятия: санитарно-гигиеническая пропаганда, массовое
обследование и дегельминтизация населения, обезвреживание фе-
. калиев и т. п.
Энтеробиоз
Возбудитель энтеробиоза Enterobius vermicularis' (L. 1758)
^отр. Ascaridida, подотр. Oxyurata), или острица (рис. 58, А), рас-
пространен не менее широко, чем аскариды. Взрослые черви дости-
1?ис. 58. Паразитические нематоды человека и домашних животных (по разным
авторам). А—острица Enterobius vermicularis, самка с маткой, переполненной
яйцами; Б — общий вид самца Haetnonchus
рают небольших размеров: самки 9—12 мм, самцы 3—5 мм. Перед-
ний конец остриц несет небольшие кутикулярные крылья головной
везикулы. Задний конец тела самца крючковидно изогнут и снаб-
жен двумя латеральными крыловидными пластинками.
Откладываемые самками яйца созревают в течение 4—6 ч. Во
внешней среде они остаются жизнеспособными около трех недель.
Человек заражается, проглатывая инвазионные яйца. В тонком
110
ППШСЧППЯС ирииьлидш виду 11J1CHHC днчииик, которые MHipnp^lVi
в начальные отделы толстой кишки. Оченц часто острицы обнару-
живаются в слепом отросл». Половой зрелости черви достигают
через 12—14 дней. Самцы после копуляции обычно погибают. Сам-
ки начинают продуцировать яйца, которые, однако, не выводятся
из' половых протоков, а накапливаются в матке. При этом паразиты
постепенно спускаются в прямую кишку. Откладка яиц происходит
в складках перианальной области, куда черви выходят через аналь-
ное отверстие. Одна самка выделяет от 5 тыс. до 17 тыс. яиц, после
чего она гибнет. Общая продолжительность жизни червей в кишеч-
нике не превышает одного месяца.
Острицы не относятся к числу самых опасных паразитов чело-
века, однако их патогенность не вызывает сомнений. Наиболее
часто встречающийся симптом энтеробиоза — острый зуд и жжение
в области анального отверстия и промежности, что обусловливает-
ся раздражением, вызываемым паразитами во время их выхода из
прямой кишки для откладки яиц. Обычно это происходит в ночное
время. У больных нарушается сон, развивается слабость и снижа-
ется трудоспособность. Дети, у которых энтеробиоз встречается
особенно часто, теряют аппетит и сильно худеют. У женщин и де-
вочек миграция остриц часто осложняется вульвовагинитами, воз-
никающими в результате попадания червей во влагалище и даже
в матку.
Для больного не проходит бесследно и пребывание паразитов
в кишечнике. Иногда черви способны глубоко внедряться в ткани,
проникая в брюшную полость и внутренние органы. По-видимому,
острицы часто способствуют развитию аппендицита. Кишечная фаза
энтеробиоза сопровождается острыми болями в животе, поносами
с обильным выделением слизи, а иногда и крови.
Источником распространения инвазионного начала во внешней
среде служит зараженный человек. Яйца паразитов в большом ко-
личестве скапливаются на постельном белье и одежде. Они легко
рассеиваются вместе с пылью и оседают на поверхности самых раз-
личных предметов. При расчесывании зудящих мест множество
яиц остается на. руках и под ногтями. Это обусловливает широкое
распространение при эптеробиозе явления аутоинвазии, в резуль-
тате чего болезнь иногда затягивается на длительное_время. Осо-
бенно широко энтеробиоз распространен у детей.
Диагноз ставится по результатам обследования соскобов, сде-
ланных в перианальной области. Решающая роль в борьбе с энте-
робиозом принадлежит профилактическим мероприятиям, основан-
ным на строгом соблюдении санитарно-гигиенических правил.
В этих случаях удается ликвидировать заболевание даже без при-
менения лекарственных средств. t
А п к и л о с т о м и д о з ы
Под этим названием объединяются два очень сходных заболе-
вания: анкилостомоз и некатороз. Возбудителем первого является
111
Ancylostoma duodenale (Dtibini, 1843) (рис. 59, Б, В), второго —
Necator americanus (рир. 59, Г, Д) (отр. Rhabditida, подотр. Stron-
gylata). Это сравнительно небольшие черви: размеры самок анки-
лостом не превышают 10—14 мм, а самцов — 8—11 мм. У Necator
americanus'—соответственно 9—42 и 5—9 мм. Передний конец те-
ла изогнут в виде крючка, чем н обусловлено русское название
этих паразитов — кривоголовки. Живые черви окрашены в бледно-
розовый цвет. Обширная ротовая капсула снабжена острыми ре-
Рис. 59. Развитие представителей-сем. Ancylostomidae. А— цикл
развития Necator americanus (по разным авторам); Б — головной
конец тела Ancylostoma duodenale-, В — хвостовой конец тела самца
Ancylostoma duodenale-, Г — головной конец тела Necator americanus-
Д — хвостовой конец тела самца Necator americanus (из ки. Е. Ноб-
ля и Г. Нобля)
жущи.ми зубами (рис. 59, Б, Г). Задний конец тела'самдов снабжен
бурсальными крыльями. Половозрелые черви поселяются в двенад-
цатиперстной кишке человека и в верхнем отделе тонкого кишеч-
ника. Оплодотворенные самки за сутки 'откладывают до 10 тыс. яиц,
которые выводятся во внешнюю среду вместе с фекалиями. При
благоприятных условиях через 1—2 суток из яиц вылупляются раб-
дитовпдпые личинки.
Первая линька обычно происходит через 2—3 дня. Еще через
1—2 дня личинки снова линяют и становятся филяриевидными. Эта
личиночная стадия характеризуется наличием чехлика, который
представляет собой несбрасываемый при линьке экзувий второй
личиночной стадии. Филяриевидные личинки в чехлике инвазион-
ны для человека. Они сохраняются в почве до 18 месяцев. Как уже
отмечалось выше, личинки совершают вертикальные миграции.
При похолодании зимой они опускаются на глубину до 1 м, где мо-
112
гут перезимовывать. При повышении температуры поднимаются
в верхние слон почвы и даже заползают на растения.
Заражение людей осуществляется двояким способом: перкутан-
но или перорально. В первом случае личинки активно проникают
в кожу при контакте человека с почвой. В момент внедрения пара-
зит сбрасывает одевающий его тело чехлик. Далее личинки совер-
шают сложную мнграцию.по кровеносной системе и легким (рис. 102),
прежде чем попадают в кишечник. При пероральном заражении
наблюдаются существенные различия между анкилостомой и не-
катором. Проглоченные инвазионные личинки- Ancylostoma duode-
nale, попав в кишечник, остаются там и развиваются без миграции.
У Necator americanus личинки внедряются в слизистую ротовой
полости и мигрируют обычным для них путем. Сравнительно недав-
ние наблюдения японских паразитологов показали, что заражение
анкилостомами осуществляется преимущественно путем заглаты-
вания личинок, тогда как для некатора характерен в.основном пер-
кутанный способ заражения.
Попавшие тем или иным путем в кишечник личинки линяют еще
два раза и дают начало взрослым червям. Последние глубоко
внедряются своим передним концом в слизистую и переходят к пи-
танию кровью. Половой зрелости черви достигают через 5—6 не-
дель после попадания в организм человека. Анкилостомы, по-види-
мому, живут около 5 лет, тогда как продолжительность жизни
некатора составляет 10—15 лет.
Возбудители апкилостомидозов очень патогенны. Проникнове-
ние личинок через кожу сопровождается появлением аллергиче-
ской сыпи. Кроме того, паразиты могут внести с собой бактериаль-
ную флору, что приводит к появлению воспалительных очагов
и экзематозных поражений кожи. Весьма серьезны и патологиче-
ские изменения в легких: инфильтраты, кровоизлияния, пневмони-
ческие очаги. Половозрелые черви в кишечнике вызывают наруше-
ние целостности слизистой, понижение кислотности, тяжелые
доудениты, а иногда и язвенный процесс. Часто наблюдается нару-
шение сердечного ритма и падение артериального давления. Од-
ним из самых тяжелых последствии анкилостомидозов несомненно
является очень острая анемия. При сильной инвазии часто наблю-
дается извращение вкуса: появляется стремление есть бумагу,
землю, уголь и т. д.
Источником распространения анкилостом и некаторов в приро-
де является зараженный человек. Выделяемые с фекалиями яйца
рассеиваются во внешней среде. Поддержание устойчивых очагов
заболевания обусловливается способностью личинок перезимовы-
вать глубоко в почве. Распространение анкилостомидозов обычно
ограничено районами с теплым и влажным климатом, в которых
годовой уровень осадков составляет 1000 мм и более.
Подобные условия могут искусственно возникать в результате
деятельности человека. Примером могут служить так называемые
подземные очаги заболевания, которые формируются в глубоких
1В
шахтах. Постоянная температура и высокая влажность создают-
необходимые предпосылки для развития червей.
Анкилостомидозы приурочены в основном к тропическому
и субтропическим поясам. Очаги заболевания были обнаружены
на Кавказе, где явно доминирует Necator americanus (Stiles, 1902),
и в Средней Азии, где встречается исключительно Ancylostoma
duodenale. Подземные очаги анкилостомидозов к 1960 г. в нашей
стране полностью ликвидированы.
Диагносцирование анкилостомидозов в период миграции личи-
нок затруднено. Присутствие же половозрелых червей в кишечнике
легко определяется по нахождению яиц в фекалиях. Лечение ведет-
ся по двум направлениям: изгнание червей обязательно должно со-
провождаться ликвидацией последствий анемии.
Как и при других гельмиптозах, борьба с анкплостомпдозамп
невозможна без выполнения целого ряда санитарно-профилактиче-
ских мероприятий. С этой целью проводят массовое обследование
и дегельминтизацию населения. Необходимо создание условий, ис-
ключающих загрязнение почвы фекалиями. В очагах распростране-
ния анкилостомидозов большое значение имеет личная профилак-
тика, которая заключается в предохранении открытых участков
тела от контактов с почвой.
Стронгилоидоз
Возбудителем этого заболевания является довольно распространенный вид
Sirongyloides stercoralis (Bavay, 1877) (отр. Rhabditida, подотр. Rhabdita). Жиз-
ненный цикл этих нематод отличается большой вариабельностью, (с. 102, рис. 54).
Он может целиком осуществляться во внешней среде или же частично или пол-
ностью протекать в организме хозяина. Половозрелые паразитические самки име-
ют нитевидное тело, длина которого составляет 2,0—2,2 мм. Свободпоживущие
самки несколько-меньше: их размеры не превышают 1 мм. И паразитические,
и свободноживущие самцы очень сходны по своему строению, их размеры не пре-
вышают 0,7 мм.
Клинические проявления стронгилоидоза обусловлены особенностями биоло-
гии червей. На первом этапе заболевания, как и при анкилостомидозах, мигриру-
ющие личинки травмируют ткани органов, в которых они находятся. Кроме того,
наблюдается сильная сенсибилизация организма. Развиваются крапивница, зуд,
инфильтраты в легких. В пищеварительном тракте паразиты, как правило, лока-
лизуются в верхних отделах тонкого кишечника. Присутствие паразитов вызывает
сильные поносы, чередующиеся с запорами, колиты, снижение кислотности, бо-
лезненность живота.- При сильных инвазиях черви обнаруживаются в желчных
протоках. Это сопровождается дуоденитами, раздражением печени и желчных
протоков. Очень часто при стронгилоидозах наблюдается нервно-аллергическая
реакция организма, проявляющаяся в функциональных расстройствах нервной си-
стемы. В особо тяжелых случаях истощающий понос приводит к обезвоживанию
организма, развивается тяжелая анемия, язвенные поражения кишечника и пе-
ритонит. Заболевание может быть очень длительным. Описаны случаи, когда
стронгилоидоз продолжался свыше 25 лег. Это обусловлено возможностью ауто-
инвазии (с. 102).
В природе основным источником распространения червей служит человек.
Кроме того, как считают некоторые авторы, определенную эпидемиологическую
роль играют собаки, в которых паразиты также могут развиваться. Заражение
осуществляется главным образом через кожу при контакте с почвой. Возможен
и пероральный путь попадания паразитов в организм человека при употреблении
в пищу немытых овощей и фруктов.
114
Как и анкилостомидозы, строигилоидоз приурочен в основном к тропическойг
и субтропической зонам. Очень широко он распространен в странах Латинской
Америки, Юго-Восточной Азии, южных штатах США. В европейских странах
и южных районах СССР имеются отдельные очаги заболевания. Широко рас-
пространены подземные очаги в шахтах.
Правильное н своевременное диагносцироваиие стропгилоидоза представляет
несомненные трудности. Этому способствует большая вариабельность симптомов
заболевания. Единственно надежным признаком является обнаружение живых
личинок паразитов в фекалиях, рвотных массах, дуоденальном содержимом или
мокроте. Борьба со строигилоидозом включает в себя те же профилактические
и санитарно-гигиенические мероприятия, что и при анкйлостомидозах.
Филярпатозы
Большая группа заболеваний, вызываемых нематодами, входя-
щими в состав подотр. Filariata (отр. Spirurida). Жизненные циклы
этих паразитов, называемых нитчатками, осуществляются с учас-
тием- дефинитивного и промежуточного хозяев (рис. 60). Полово-
зрелые черви обитают в тканях или замкнутых системах органов.
Личинки (микрофилярии) скапливаются либо в кровеносной систе-
ме, либо в коже хозяина. Переносчиками, в которых паразиты пре-
терпевают определенное развитие, служат различные кровососу-
щие насекомые.
Классическим примером фнляриатоза служит вухерериоз — за-
болевание, вызываемое видом Wuchererla bancrofti (Cobb., 1877)
(рис. 60, В). Это узкие нитевидные нематоды, самки которых дости-.
гают 10 см в длину при диаметре 0,24—0,30 мм, а самцы 4 см. Поло-
возрелые черви обитают в лимфатических протоках и узлах. Обыч-
но самцы и самки локализуются вместе, тесно переплетаясь друг
с другом. Половозрелые самки отрождают личинок второй стадии,
сохранивших экзувий —чехли к. Они мигрируют в кровяное русло.
Поведение микрофилярий в организме человека характеризуется
известной цикличностью. В подавляющем большинстве случаев они
в дневное время (период активности человека) находятся в сосу-
дах внутренних органов (чаще всего легких, сердца и т. п.),
а в ночные часы мигрируют в периферические сосуды. Здесь они
становятся легкой добычей комаров (р. Anopheles, Aedes и Culex),
нападающих на спящих людей для питания. В пищеварительном
тракте промежуточного хозяина, куда микрофилярии попадают
вместе с кровью, они сбрасывают чехлик и проникают через стенку
желудка в полость тела. Локализуются личинки обычно в мускула-
туре, насекомого. Здесь они дважды линяют и дают начала инвази-
онным личинкам четвертой стадии. Последние проникают в ротовой
аппарат комара и скапливаются в дистальном конце нижней губы.
Во время питания переносчика личинки активно выходят из хобот-
ка на поверхность кожи человека. Отсюда они через трещинки
в коже или ранку, образованную уколом комара, проникают в ор-
ганизм человека. Паразиты совершают миграцию и в конце концов
локализуются в лимфатической системе. Проделав еще одну линь-
ку, они достигают половой зрелости и начинают отрождать новые
поколения микрофилярий. Взрослые черви в человеке живут до
115
17 лет, микрофилярии в кровяном русле сохраняют жизнеспособ
ность около 70 дней.
Рис. 60. Циклы развития филярий (по разным авторам). А — Loa loa; В — Oncho-
cerca volvulus; В — Wuchereria bancrofti:
/ — дефинитивный хозяин — человек, 2 — характерные повреждения, вызываемые филярия-
ми (а —миграция в конъюнктиве, б —подкожный узел, содержащий взрослых нематод,
е —слоновость нижней конечности), 3 — микрофилярии из кровяного русла, 4 — переносчи-
ки’ филярий (а — слепень р. Chrysops, б — мошки р. Simulum, в — комары р. Aedes, Culex
и Anopheles). 5 — инвазионные лячянки из промежуточных хозяев
Первый, инкубационный, период заболевания, длительность ко-
торого колеблется от 3 до 18 месяцев, протекает бессимптомно.
В дальнейшем появляются характерные признаки аллергии: лихо-
радка, отеки и инфильтраты тканей, красные высыпания на коже
и т. п. Через несколько лет (2—7) развивается воспаление кожных
116
и глубоких лимфатических сосудов. Характерный признак послед-
него этапа заболевания — слоновость, представляющая собой па-
тологическое увеличение размеров конечностей, половых органов
и грудных желез. Взрослые черви вызывают закупорку лимфатиче-
ских сосудов, что приводит к замедлению лимфотока и застою лим-
фы. На коже пораженных участков тела появляются разрастания’
экземы, трофические язвы. Наблюдается атрофия мышц. Длитель-
ные лимфадениты и служат причиной развития слоновости.
Единственным источником распространения инвазии являются
зараженные люди. Животные не заражаются вухерериозом ни
в природных условиях, ни в эксперименте. Чаще всего, в качестве
промежуточных-хозяев Wuchereria. bancrofti в городских условиях
служат подвиды вида Culex pipiens. В сельской местности в Афри-
ке, Южной Америке и ряде стран Азии главная роль принадлежит
видам р. Anopheles, а на о-вах Тихого океана р. Aedes.
Вухерериоз широко распространен в ряде стран Африки, Азин,
Южной Америки и на о-вах Тихого и Индийского океанов. По дан-
ным Всемирной организации здравоохранения (ВОЗ) за 1963 г.,
наблюдается рост пораженности населения этим паразитом. Уве-
личению заболеваемости способствуют, по-видимому, главным
образом социальные факторы: рост крупных городов в странах
Азии и Африки и большая скученность населения в них, недостаточ-
ность или отсутствие санитарного контроля, тяжелые экономиче-
ские условия (плохое питание, изнурительная работа и т. д.), сни-
жающие сопротивляемость организма человека. В Советском Сою-
зе зарегистрировано всего два случая слоновости — в Таджикистане
и Грузии. Но так как оба раза микрофилярии в крови больных об-
наружены не были, истинные причины заболевания нельзя считать
достоверно установленными.
Диагностика вухерериоза осуществляется путем обнаружения
микрофилярий в крови. Иммунологические реакции с использова-
нием антигенов, к сожалению, не обладают достаточной эффектив-
ностью и специфичностью.
Комплексное лечение вухерериоза включает в себя дегельмин-
тизацию, ‘ подавление бактериальных инфекций, которые часто
сопровождают заболевание "и ослабление аллергической реакции.
Часто приходится прибегать и к оперативному вмешательству.
Особенности эпидемиологии вухерериоза определяют и харак-
тер мероприятий по борьбе с этим заболеванием. В первую очередь
проводится оздоровление населения как главного источника пива-
зпи: массовое обследование и дегельминтизация людей в очагах
распространения болезни. Не менее важную роль играет борьба
с переносчиками этой болезни (с. 266).
В странах Западной Африки встречается еще один филяриатоз, вызываемый
нематодой Loa loa (Cobb., 1864) (рис. 60, А). Это заболевание известно под на-
званием лоаоз. Взрослые паразиты обитают в подкожной клетчатке и серозных
полостях тела. Иногда они проникают в глаз и локализуются под конъюнктивой.
Отрождаемые ими микрофилярии сначала скапливаются в капиллярах легких
и лишь через лесколько педель начинают поступать в периферическую кровь. Ли-
чинки Loa loa покидают легкие в дневные часы. Эта особенность представляет
117
собой важную биологическую адаптацию. Переносчиками лоаоза служат слепни
из р: Chrysops (с. 252). Последние в отличие от комаров характеризуются днев-
ной активностью. Заболевание сопровождается сильной аллергической реакцией:
лихорадочным повышением температуры, зудом, крапивницей и т. д. Миграции
паразитов в подкожной клетчатке вызывают отечность и сильный зуд. Часто раз-
виваются абсцессы в мышцах и лимфатических узлах. Обитание паразитов в гла-
зах приводит к конъюнктивиту и ослаблению зрения.
Лечение и борьба с лоаозом осуществляются теми же методами, что и при
вухерериозе.
Важное медицинское значение имеет онхоцеркоз, вызываемый широкораспро-
страненным в Западной и Экваториальной Африке и Латинской Америке видом
Onchocerca volvulus (рис. 60, Б). Половозрелые нитевидные черви, достигающие
в длину 4—5 см, чаще всего локализуются под кожей человека, в области таза,
суставов конечностей и на голове. Вокруг паразитов образуются плотные соеди-
нительнотканные узлы. Микрофилярии постоянно обитают в толще кожи. Пере-
носчиками служат мошки из р. Simulium, в которых личинки достигают инвази-
онной стадии на 6—7-й день после заражения.
Клиническим проявлениям заболевания предшествует очень длительный (до
одного года) инкубационный период. Развивающиеся соединительнотканные узлы
очень болезненны. Кожа вокруг них сморщивается и затвердевает. Часто обра-
зуются медленно рубцующиеся язвы. Заболевание сопровождается общим недо-
моганием, лихорадкой и сильными головными болями. В качестве осложнений
Наблюдаются абсцессы, а иногда и слоновость Нижних конечностей и мошонки.
Попадание микрофилярий в глаза вызывает конъюигсгивиты, помутнение роговой
оболочки, атрофию зрительного нерва и потерю зрения. Лечение онхоцеркоза
затруднено из-за отсутствия эффективных препаратов. В настоящее время обыч-
но применяют диэтилкарбамазип в сочетании с антриполом. Исключительно важ-
ное значение имеет борьба с переносчиками (с. 275).
Дракункулез, или ришта
Возбудитель дракункулеза Dracunculus medinensis (L. 1758) —
медицинский струнец (отр. Spirurida, подотр. Camallanata)
(рис. 61) характеризуется отчетливо выраженным половым димор-
физмом. Самки достигают 120 см в длину при диаметре теда 0,5—
1,7 мм. Самцы значительно меньших размеров: длина тела 12—
30 см, диаметр 0,2—0,4 мм. Половозрелые самки локализуются
в подкожной клетчатке человека и некоторых плотоядных живот-
ных (собаки, кошки и т. п.). Вокруг паразита (который также как
заболевание, называется риштой) образуется пузырь, который со
временем разрывается. При соприкосновении с водой стенка тела
червя лопается в области, вульвы. В образовавшееся отверстие
выпячивается матка, из которой в воду выходит множество мелких
личинок, находящихся на второй стадии развития (рис. 61, Г).
Промежуточными хозяевами служат пресноводные рачки, относя-
щиеся к р. Cyclops, Eucyclops, Mesocyclops и др. Рачки проглаты-
вают личинок. Последние прободают стенку кишечника и локали-
зуются в гемоцеле хозяина. В дальнейшем личинки претерпевают
две линьки, и лишь после этого на 12-й день становятся инвазион-
ными. Дефинитивный хозяин заражается при питье сырой воды,
содержащей инвазированных циклопов. Освободившись от тканей
рачка, .личинки проникают в стенку кишечника человека или жи-
вотного и уже на 21-й день обнаруживаются в подкожной клетчатке.
Достижение половой зрелости, по;видимому, наступает лишь через
несколько месяцев. Самцы погибают вскоре после оплодотворения
самок.
118
Начальный период заболевания в большинстве случаев проте-
кает бессимптомно. Миграция паразитов сопровождается крапив-
ницей, одышкой, иногда развивается бронхиальная астма и поносы.
Примерно через год после заражения вокруг головного конца чер-
вя появляется пузырь, о котором уже говорилось выше. В некото-
Рпс. 61. Цикл развития Dracunculus medinensis (по разным
авторам). А — окончательный хозяин — человек; Б — сам-
ка; В — локализация паразита в ноге человека (рисунок с
рентгенограммы); Г — инвазионная личинка, отрождаемая
самкой; Д — циклоп, зараженный личинкамп:
1 — личинки в полости тела циклопа
рых случаях формируется настоящая киста, которая сохраняется
очень долго.
Источником распространения дракункулеза является человек.
Подобные синатропные очаги, по-видимому, могут поддерживать-
ся и собаками. В диких, не освоенных человеком районах встреча-
ются настоящие природные очаги, представляющие серьезную
опасность. Распространение дракункулеза ограничено преимуще-
ственно странами с теплым климатом (государства Африканского
континента, Южной Азии и Латинской Америки).
119
На- территории Советского Союза очаг дракункулеза существо-
вал в районе Старой Бухары. Городские пруды (хаузы) долгое
время служили фактически единственным источником питьевой
воды для населения. Но в этих же прудах люди купались и мыли
ноги. Собаки также имели свободный доступ к хаузам. Заражен-
ные животные и люди рассеивали в этих сравнительно небольших
водоемах огромное количество инвазионных личинок. А загрязне-
ние воды -большим количеством органических веществ создавало
благоприятные условия для массового размножения циклонов.
Вода же, используемая для питья и продаваемая разносчиками
в городе, обычно не подвергалась ни кипячению, ни очистке
(фильтрации). Так складывались условия, обеспечивающие посто-
янную циркуляцию паразитов и заражение все новых и новых хо-
зяев.
Большая работа, проводившаяся в конце 20-х годов под руко-
водством Л. М. Исаева (санитарное просвещение населения, запре-
щение мытья и купания в хаузах, отлов и уничтожение бродячих
собак, упорядочение торговли водой в разнос, очистка водоемов
и т. д.), позволила уже в начале 30-х годов полностью ликвидиро-
вать бухарский очаг дракункулеза. Начиная с 1932 г., это’заболе-
вание у населения в районе Старой Бухары больше не регистриру-
ется. Заметные успехи в борьбе с дракункулезом достигнуты и
в других странах, в частности в Бразилии.
Ранние стадии дракункулеза практически не диагносцируются.
После формирования пузыря определение заболевания и его при-
чин уже не вызывает затруднений. Лечение комплексное: ослабле-
ние аллергических явлений' и уничтожение молодых паразитов
с помощью химиопрепаратов. Зрелых червей удаляют следующим
образом: паразит захватывается за выступающий из ранки конец
и медленно наматывается на марлевый валик. Эта операция продол-
жается иногда до 20 дней. Быстрое извлечение паразита чревато
осложнениями. При случайном разрыве тела нематоды и излияния
полостной жидкости в ранку развивается тяжелая флегмона.
Профилактические мероприятия, разработанные еще Л. М. Иса-
евым, заключаются в своевременном выявлении и лечении больных,
периодическом обследовании- и очистки водоемов. Для уничтоже-
ния зараженных циклопов за рубежом широко применяются спе-
циальные химические препараты. Рачки к ним очень чувствитель-
ны, для человека же и домашних животных они совершенно без-
вредны.
Трихоцефалез
* ч*
Шлрокораспространениый пематодоз, вызываемый видом Trichocephalus
triduurus (L. 1771) (отр. Trichocephalida) (см. рис. 44, Ж). Передний конец тела
червей вытянут в длинный волосовидный придаток, что и послужило основанием
для русского названия власоглав, или хлыстовик. Длина паразитов может дости-
гать 4—5 см. У самок задний расширенный отдел тела изогнут в виде дуги, у
самцов же он закручен в спираль. Власоглавы локализуются в слепой кишке,
а при сильных инвазиях и в прилегающих отделах пищеварительного тракта.
Отложенные половозрелыми самками яйца одеты многослойной оболочкой и име-
120
ют характерную вытянутую форму. На полюсах скорлупки располагаются про-
зрачные «пробочки». Развитие яиц осуществляется во внешней среде. Хотя яйца
власоглава хуже переносят охлаждение, чем яйца аскарид, в умеренном климате
они могут перезимовывать под толстым слоем снега. Жизнеспособность и пива-
знойность яйца сохраняют 'до 1,5 лет.
Заражение человека осуществляется при проглатывании инвазионных яиц.
Вылупившиеся личинки обладают стилетом. Они проникают в ворсинки слизистом
где и находятся до 10 суток. После этого личинки снова выходят в просвет ки-
шечника и достигают слепой кишки. Передним концом они прикрепляются к сли-
зистой. В процессе развития личинки Т. trichiurus линяют 4 раза. Половой зрело-
сти они достигают примерно через месяц после заражения. В кишечнике человека
они, по-видимому, обитают около 5 лет.
При невысокой интенсивности инвазии заболевание протекает бессимптомно.
Сильное заражение часто влечет за собой серьезные нарушения работы пищева-
рительного тракта, сопровождающиеся тошнотой, болями, понижением кислотно-
сти. Нарушение целостности слизистой открывает ворота для вторичных бактери-
альных инфекций. Часто при трихоцефалезе наблюдается и реакция со стороны
нервной системы: головные боли, головокружения, а иногда и припадки.
Широкораспрострапенное мнение, что власоглавы могут быть причиной ап-
пендицита, в последние годы не подтвердилось. Черви в слепом отростке обнару-
живаются лишь в исключительно редких, действительно единичных случаях.
В эпидемиологии трихоцефалеза решающую роль играет зараженный чело-
век, который представляет собой единственный источник распространения инвази-
онного начала во внешней среде. Рассеянные в почве яйца власоглава легко по-
падают на овощи и фрукты. Возможно заражение и при помощи загрязненных
рук. Трихоцефалез встречается практически по всему земному шару, исключение
составляют лишь районы Крайнего Севера и засушливые районы. Особенно обы-
чен он в зонах с влажным умеренным или теплым климатом.
Для лечения широко применяются химиопрепараты. В последнее время, как
и при аскаридозах, используется метод лечения кислородом.
Трихинеллез
Заболевание, вызываемое видом Trichinella spiralis (Owen,
1835) (отр. Trichocephalida) (рис. 62), относится к числу тяжелых.
Половозрелые черви, обитающие в кишечнике человека, обладают
небольшими размерами. Длина неоплодотворенной самки состав-
ляет 1,5—1,8 мм, оплодотворенные же особи заметно увеличивают-
ся (до 4,4 мм). Самцы обычно не превышают 2 мм.
Цикл развития трихин отличается очень своеобразными особен-
ностями: главная заключается в том, что в качестве дефинитивного
и промежуточного хозяина выступает одна особь (рис. 62, А). Па-
разиты сначала достигают половой зрелости в кишечнике животно-
го, а затем их личинки инкапсулируются в его же мышцах. Таким
образом, ни одна из стадий развития паразита не выводится во
внешнюю среду.
Заражение хозяина осуществляется при поедании им мяса, со-
держащего инкапсулированные личинки трихинелл. Последние
в просвете кишечника освобождаются от окружающих их тканей
и в течение трех суток достигают половой зрелости. Оплодотворен-
ные самки прикрепляются головным концом тела к слизистой. Для
трихинелл характерно яйцеживорождение: из продуцируемых сам-
кой яиц еще в матке вылупляются личинки, которые и выходят
в просвет кишечника. Обычно за свою жизнь (около 50 дней) самка
отрождает до 2 тыс. личинок. Долгое время считалось, что самцы
121
погибают вскоре после копуляции. Сейчас, однако, установлено,
что иногда cftin паразитируют в кишечнике дольше самок.
Личинки через стенку кишечника проникают в лимфатические
сосуды, а позднее и в кровеносную систему. Они разносятся по все-
Рис. 62. Цикл развития Trichinella spiralis (по разным авторам). А—схема
цикла развития; Б— половозрелая самка; В — половозрелый самец; Г —
проникновение личинки в мускульное волокно; Д — инвазионная инкапсу-
лированная личинка:
/ — первичное заражение тканей личинками, 2 — вторичное заражение тканей личин-
ками, возникающее в результате аутоинвазни, 3 — поступление инвазионного начала
в организм животного-хозяина, 4 — половозрелые самцы и самки, 5 — инвазионные
инкапсулированные личинки
му организму и, спустя некоторое время, могут быть обнаружены
в самых различных органах. Окончательная локализация трихи-
нелл обычно приурочена к поперечно-полосатой мускулатуре. Ли-
чинки, выделяя гиалуронидазу, проникают в сарколемму мышечно-
го волокна. Последнее при этом разрушается. На 17—18-й день
личинки сворачиваются в спираль. Вокруг них постепенно в тече-
ние 2—3 месяцев формируется соединительнотканная капсула
122
(рис. 62, Д). Это сложное образование, выполняющее функции
двоякого рода: с одной ‘стороны, оно служит для защиты организ-
ма хозяина от паразита, с другой же — способствует нормальному
существованию последнего. Стенки капсулы прорастают кровенос-
ными сосудами и нервными окончаниями. Вместе с кровью паразит
получает необходимые ему пищевые вещества и кислород, с кровью
же удаляются и продукты обмена личинок. Начиная с шестого ме-
сяца, в большинстве случаев наблюдается обызвествление капсул.
Личинки при этом часто сохраняют жизнеспособность.
Локализуются паразиты, как правило, в поперечно-полосатых
мышцах. В сердечной мышце и гладкой мускулатуре личинки не
встречаются. Обычно они предпочитают мускулатуру, хорошо снаб-
женную сетью кровеносных сосудов: мышцы языка, предплечья,
диафрагмы, дельтовидные, межреберные, икроножные мышцы.
Очень часто они оседают в мышцах глазного яблока, мочеиспуска-
тельного канала, пищевода и т. д.
Заражение нового животного-хозяина (в том числе и человека)
происходит при поедании инвазированных трихинеллами тканей
«предыдущего» хозяина.
Приведенная выше классическая схема цикла развития трихи-
нелл, как показали исследования последних лет, может услож-
няться. При резком снижении сопротивляемости организма в силу
тех или иных причин (с. 125) продуцируемые самками личинки,
внедряясь в слизистую, чаще всего в ворсинки, быстро развивают-
ся в них и через сравнительно короткий промежуток времени воз-
вращаются обратно в кишечник, где и достигают половой зрелости.
Такая своеобразная аутоинвазия приводит к резкому увеличению
числа особей, паразитирующих в кишечнике. Зрелые черви в свою
©чередь отрождают новое поколение личинок. Все это обусловли-
вает, с одной стороны, удлинение сроков пребывания половозрелых
червей в кишечнике, а с другой стороны, соответственно и усиление
интенсивности инвазии мышц. Медицинское значение этого фено-
мена исключительно велико (с. 115).
Клиническому проявлению трихинеллеза человека обычно пред-
шествует инкубационный период, продолжительность которого
варьирует от нескольких дней (5—8) до 3—4 недель. В основе ме-
ханизма патогенеза трихинелл лежит аллергическая реакция орга-
низма, вызванная сенсибилизацией продуктами обмена и распада
гибнущих паразитов.
Один из наиболее характерных симптомов заболевания — отеч-
ность. Первыми появляются отеки век, которые при тяжелом тече-
нии болезни распространяются чпо всему лицу, на шею, туловище
и даже конечности. Одновременно развивается лихорадка, которая
достигает максимума через 2—3 дня. Не менее постоянным приз-
наком трихинеллеза являются мышечные боли. Они возникают,
как правило, в глазных, жевательных и шейных мышцах. Довольно
часто болезненность ощущается и в плечевых, поясничных и икро-
ножных мышцах. Очень сильные боли при движении могут приво-
дить к контрактуре мышц.
123
Весьма обычна при трихинеллезе и высокая эозинофилия. Од-
нако очень тяжелые случаи трихинеллеза сопровождаются резким
падением числа эозинофилов.
Иногда трихинеллез сопровождается расстройствами кишечно-
желудочного тракта: поносами, сильными болями в животе, рво-
тамп. Нередко наблюдаются бессонница, головные боли, депрес-
сия или, наоборот, возбуждение, галлюцинации.
Если заболевание не сопровождается осложнениями, то обычно
через 2—3 недели состояние больного заметно улучшается. Ослож-
нения, часто возникающие при высокой интенсивности заражения,
обычно связаны с поражениями миокарда (миокардиты, сердечная
недостаточность), развитием пневмонии, имеющей аллергическую
природу, и менинго-энцефалита. Прогноз при осложнениях очень
часто бывает неблагоприятен.
Трихинеллез относится к заболеваниям с природной очагово-
стью. В настоящее время выделяют два типа очагов: природные
и синантропные. Первые поддерживаются дикими животными:
волками, медведями, лисицами, енотовидными собаками, барсука-
ми и т. д. Реже встречается заражение рысей, мелких хищников,
мышевидных грызунов, насекомоядных, ластоногих. Вторые суще-
ствуют среди домашних животных и некоторых грызунов. Циркуля-
ция паразитов в очагах этих двух типов осуществляется в какой-то
мере независимо и самостоятельно. Синантропные очаги, однако,
постоянно пополняются за счет притока инвазионного начала из
природных Ьчагов. Важную роль в распространении трихинелл,
ло-видимому, играют хищные птицы. Поедая мелких зараженных
животных или склевывая мясо с трупов, они рассеивают в природе
инкапсулированных и освободившихся от капсул личинок, которые
еще некоторое время сохраняют инвазионность и способны зара-
жать новых хозяев. Личинки трихинелл, оказавшиеся в почве, мо-
гут быть проглочены не только позвоночными животными, но и не-
которыми беспозвоночными: личинками хищных мух, личинками
жуков, взрослыми хищными жуками и т. п. В пищеварительной
системе этих факультативных хозяев паразиты переживают около
недели. Таким образом, насекомые, играющие большую роль в пи-
щевых цепях природных биоценозов, становятся важным эпидемио-
логическим звеном на пути распространения паразитов.
Трихинеллез встречается практически повсеместно. Единствен-
ное исключение составляет Австралия, где за всю историю зареги-
стрированы единичные случаи заболевания. В большей степени он
приурочен к северному полушарию. До сих пор очаги заболевания
сохраняются в США и некоторых европейских странах (ФРГ, ГДР,
Польша). В последние годы, благодаря профилактическим меро-
приятиям, наблюдается резкое уменьшение числа регистрируемых
случаев трихинеллеза и исчезновение многих синантропных очагов.
В СССР отмечаются отдельные спорадические случаи трихинелле-
за, появление которых обусловлено несоблюдением санитарных
правил. Чаще всего это связано с употреблением в пищу мяса ди-
ких животных (кабанов, медведей и т. п.), не прошедшего ветери-
124
парного контроля. Последнее обстоятельство объясняет «микроочя-
говый» характер вспышек, приуроченных обычно к одной или ни-
скольким связанным друг с другом семьям и небольшим замкнутым
коллективам (экспедиционные группы, изыскательские партии
и т. д.).
Лечение трихинеллеза связано с целым рядом больших трудностей. Долгое
время вообще не существовало сколько-нибудь эффективных специфических пре-
паратов. С начала пятидесятых годов в медицинской практике стали широко
применяться стероидные гормоны. Первые результаты были весьма обнадежива-
ющими: гормонотерапия, обладая аитиаллергическнм действием, снимает острее
симптомы начального периода трихинеллеза, улучшает показатели крови и т. Д.
В практических рекомендациях содержалось указание на необходимость -как мота-
но более раннего, назначения больным высоких доз гормонов. В последние годы,
однако, появился ряд данных и об отрицательных последствиях гормонотерапии.
Будучи сильными иммунодепрессантами, кортикостероиды и другие аналогичное
по действию препараты вызывают ослабление защитных свойств организма. Это
приводит к удлинению сроков существ оз а ння кишечных форм паразитов, сокра-
щению сроков развития личинок, аутоинвазии, нарушению нормального хода ин-
капсулирования трихинелл в мышцах и т. д. Заболевание растягивается на дли-
тельный промежуток времени и соответственно часто обозначается в литературе
как «хронический трихинеллез». По мнению многих специалистов, назначение
гормонотерапии в ранние сроки заболевания крайне нежелательно. Применять ее
можно лишь после завершения кишечной н миграционной фаз развития пара-
зитов.
Решающая роль в борьбе с трихинеллезом принадлежит профи-
лактическим мероприятиям. К ним относятся: обязательное микро-
скопическое обследование мяса домашних животных и диких жи-
вотных, употребляемых в пищу; выбраковка и уничтожение или
техническая утилизация зараженных туш; создание санитарных
условий, исключающих возможность заражения скота и других до-
машних животных; уничтожение крыс и мышей, представляющих
собой важный компонент синантропных очагов трихинеллеза.
Нематодозы домашних животных
Еще более многочисленны, чем нематодозы человека, заболевания, вызывае-
мые круглыми червями у животных. Среди них встречаются формы, наносящие
серьезный ущерб сельскому хозяйству.
Очень широко распространены болезни, вызываемые представителями сеМ-
Ascarididae: Ascaris suum— у свиней, Parascaris equorum — у лошадей и т. Д.
Цикл развития этих паразитов и их биология очень сходны с циклом развития
A. lumbricoides — возбудителем, аскаридоза человека (с. 108). По-видимому, много
общего имеется и в характере этих заболеваний. У животных наблюдаются сен-
сибилизация организма продуктами обмена червей, интоксикация, непроходи-
мость кишечника, осложнения, связанные с нарушением функций печени, повреж-
дение легких мигрирующими личинками. Молодняк, который обычно страдает
больше всего, отстает в росте, животные слабеют, становятся вялыми. У лошадей
течение заболевания часто сопровождается функциональными нарушениями пи-
щеварительной и нервной систем.
Большое количество заболеваний скота вызывают представители подотр.
Strongylata (отр. Rhabditida). Виды р. Oesophagostomum, относящегося к сеМ-
Strongylidae, паразитируют в свиньях и жвачных. Заражение осуществляется
путем проглатывания хозяином инвазионных личинок. Последние внедряются
в слизистую толстой кишки. Через несколько дней они снова выходят в просвет
кишечника достигают половой зрелости Пребывание в пищеварительном тракте
и личинок и взрослых паразитов сопровождается серьезными нарушениями це-
125
лостиости слизистой, которые могут дать начало язвенному процессу. Животные
страдают тяжелыми колитами, отказываются от еды, худеют, часто развивается
малокровие.
Очень широко распространены у жвачных трихостронгилидозы (с. 100) возбу-
дителями которых служат представители р. Trichostrongylus, Ostertagia, Haemoti-
chus (см. рис. 58, Б), Nematodirus и др. (сем. Trichostrongylidac). Заражение жи-
вотных осуществляется при проглатывании инвазионных личинок. Последние
проделывают в тонком"- кишечнике хозяина две линьки и достигают половой зре-
лости. Степень патогенности разных видов трихостронгилнд различна. Наиболее
тяжелые поражения вызывают облигатные гематофаги, виды р. Haernonchus
и остертагии, локализующиеся в толще слизистой. Животные страдают гастро-
энтеритом, поносами с кровью, становятся вялыми, их упитанность снижается,
начинает выпадать шерсть. У молодняка наблюдается сильная задержка в росте.
Наиболее тяжелые формы трихостронгилидозов. встречаются у овец.
Другим часто встречающимся заболеванием мелкого и крупного рогатого
скота является диктиокаулез, вызываемый видами р. Dictyocaulus (сем. Dictyoca-
ulidae). Эти паразиты обитают в легких животного-хозяина. Пребывание парази-
тов в дыхательных путях вызывает развитие мощных воспалительных процессов,
часто охватывающих целые доли легких. Заболевание сопровождается кашлем,
который со временем постепенно усиливается. Иногда во время приступа из лег-
ких выбрасываются комки слизи с большим количеством червей. У животных бы-
стро наступает истощение, развивается анемия. При отсутствии лечения животные
часто погибают, причем смертность иногда может достигать 75%.
Крупный рогатый скот часто страдает от телязиоза — заболевания, вызыва-
емого р. Thelazia (сем. ,Thelaziidae, отр. Spirurida). Взрослые нематоды парази-
тируют в слезном канале, конъюнктивальной полости и под третьим веком.
Телязиоз сопровождается слезотечением, воспалением век, помутнением и
изъявлением роговицы. Часто наблюдается обильное отделение гноя. Животные
становятся беспокойными, плохо, едят.
Нематодозы птиц
Как и млекопитающие, птицы часто подвергаются заражению нематодами.
Многие из них представляют серьезную опасность для своих хозяев.
Очень широко распространен аскаридиоз кур — возбудитель Ascaridia galli
(Schr.) Freeb., 1923 (сем. Ascaridiidae, подотр. Ascaridata). В отличие от аскарид
виды р. Ascaridia развиваются без миграции (см. рис. 56). Аскаридиозом зара-
жаются преимущественно цыплята. Они теряют аппетит, перья обычно взъероше-
ны, крылья опущены. Очень часто наблюдается большой отход поголовья.
У гусей (домашних и диких) довольно обычен амидостоматоз. Возбудители
этого заболевания — нематоды Ainidostomum anseris (сем. Amidostomidae, подотр.
Strongylata) локализуются под кутикулой мышечного желудка птиц. В месте по-
селения паразита кутикула отслаивается и развивается острый воспалительный
процесс, сопровождающийся изъязвлениями стенки желудка. У зараженных птиц
наблюдается сильное истощение, потеря аппетита5, одышка. При высокой интен-
сивности инвазии болезнь заканчивается смертью хозяина.
Часто встречается у птиц сингамоз, вызываемый Syngamus trachea (Mont.)
Chap., 1925 (сем. Syngamidae, подотр. Strongylata). Эти паразиты обладают ши-
роким кругом хозяев: куры, индейки, фазаны, гуси и многие дикие птицы (осо-
бенно часто врановые). Последние поддерживают существование природных
очагов заболевания. Половозрелые нематоды, объединенные в парочки (см.
рис. 44, Е), локализуются в трахеях. Заражение хозяина осуществляется при
проглатывании яиц, содержащих инвазионных личинок. Паразиты могут исполь-
зовать в своем развитии и резервуарных хозяев — дождевых червей, в которых
они инцнстируются и сохраняют жизнеспособность до четырех лет, наземных
моллюсков, некоторых членистоногих и т. д. Из кишечника птицы, куда они по-
падают вместе с кормом, паразиты мигрируют в легкие по кровяному руслу.
В дальнейшем они переходят сначала из альвеол в бронхи, а затем и в трахеи.
У больных птиц наблюдаются затруднение дыхания, вялость, зевота, характерные
подергивания головой и сильное истощение. Массовое развитие паразитов иногда
является причиной падежа в птицеводческих хозяйствах.
126
Успешная борьба с нематодозами домашних животных является важнейшей
задачей современной ветеринарии. Как и борьба с заболеваниями человека, она
ведется по двум направлениям. Первое представляет собой лечение заболевших
животных — их дегельминтизацию. С этой целью в настоящее время успешно
используется, большое число лекарственных препаратов. Весьма перспективны, по-
видимому, поиски путей повышения иммунобиологической активности организма,
что приводит к нарушению нормальной жизнедеятельности паразитов и резкому
снижению их половой продуктивности.
Второе направление связано с разработкой и строгим соблюдением саиитарно-
профилактических мероприятий. Основная их цель — создание условий, при кото-
рых осуществление нормального цикла развития паразита становится невозмож-
ным. Это и изоляция животных от инвазионного начала (яиц, личинок, заражен-
ных промежуточных хозяев), ликвидация природных очагов, своевременное вы-
явление возникающих вспышек заболеваний и их подавление, оздоровление мест
содержания и выпаса животных и т. д.
КЛАСС NEMATOMORPHA VEJDOVSKY — ВОЛОСАТИКИ
Класс Nematomorpha (syn. Gordiacea) — сравнительно небольшая группа
очень специализированных паразитических первичнополостных червей, насчитыва-
ющая около 300 видов.
Для всех Nematomorpha характерен личиночный паразитизм. Свободный об-
раз жизни ведут лишь только что вылупившиеся из яиц личинки и взрослые черви.
Обитают волосатики в море и самых разнообразных пресных водоемах. В качест-
ве хозяев они используют насекомых, а морские виды — преимущественно рако-
образных.
В настоящее время не существует единого мнения о систематическом поло-
жении волосатиков. Одни авторы: А. А. Парамонов, Р. С. Шульц и Е. В. Гвоздев
и др.— включают их в состав типа Nemathelminthes bzкачестве самостоятельного
класса. При этом подчеркивается сходство с нематодами. Кроме того, личинки
волосатиков во многом напоминают представителей свободноживущего класса
Priapulida. Несмотря на это, многие исследователи рассматривают Nematomorpha
как самостоятельный тип. Нам, однако, кажется, что эта точка зрения малооп-
равдана. Принципиальной разницы - в организации Nematomorpha и остальных
первичнополостиых нет, а имеющиеся отличия легко объясняются древностью и
далеко зашедшей специализацией этой группы.
В составе класса Nematomorpha в настоящее время выделяют 5 отрядов.
Морфология и анатомия
Длинное нитевидное тело волосатиков (рис. 63, А) может достигать значи-
тельных размеров—1,5 м в длину. Многие виды, однако, не превышают 40—
50 см. Диаметр тела колеблется от 1 до 3 мм. Благодаря своей характерной
форме и плотным темным покровам, у многих народов Nematomorpha называ-
ются «живым волосом» или чаще «конским волосом».
Передний конец тела волосатиков закруглен и у некоторых видов несет не-
функциоиирующее ротовое отверстие. Па заднем копце, в первую очередь у сам-
цов, располагаются две лопасти (рис. 63, Б), в основании которых лежит отвер-
стие клоаки.
Покровы волосатиков образованы плотной многослойной' кутикулой, прони-
цаемость которой заметно меняется па протяжении жизни паразита (с. 128). Не-
посредственно под кутикулой залегает тонкий слой гиподермы, которая в.отличие
от нематод образует лишь одно продольное утолщение, соответствующее брюшно-
му валику (рис. 63, В). Так же, как и нематоды, волосатики обладают только
продольными мускульными клетками. Последние располагаются сплошным слоем.
Единая обширная полость тела у волосатиков отсутствует. Опа заполнена парен-
химой, состоящей из крупных полигональных клеток и соединительнотканных
прослоек и тяжей. От схизоцеля у Nematomorpha остаются лишь продольные ка-
налы — синусы, среди которых выделяют спинной, околокишечнып и два оваря-
127 ♦
альных. Лишь у морских форм, относящихся к р. Nectoneruuy слнзоцель выражен
хорошо.
Пищеварительная система волосатиков сильно редуцирована. Чаще всего ру-
диментарной бывает передняя кишка, которая лишена просвета и не соединяется
со средней кишкой. Иногда полностью недоразвитой остается задняя кишка. Ни
на одной из стадий развития пищеварительная система не принимает участил
в поглощении пищевых веществ. Молодые личинки, еще не перешедшие к парази-
тизму, существуют за счет энергетических запасов, полученных ими при развитии.
Паразитические лнчиночпые стадии питаются, но поступление пищи осуществил-
Рис. 63. Организация волосатиков (по разным авторам). А — внешний вид
волосатика Gocdius aquaticus-, Б — задний конец тела самца G. aquaticus;
В — поперечный разрез через тело волосатика; Г — личинки G. .aquaticus
в йоге водной личинки подеики; Д — личинка волосатика:
1 — кутикула, 2 — гиподерма, 3 — слой продольных мышечных клеток, 4 — паренхи-
ма, 5 —спинной синус шизоцеля, 6 — яичник. 7 — овариальный синус, 8 — средняя
кишка, 9 — кишечный синус, 10— брюшной нервный ствол в брюшном утолщении
гиподермы, // — хоботок, 12 — кутикулярные крючья
ется через кутикулу, проницаемость которой в этот период жизни червей оказы-
вается достаточно высокой. Волосатики, покинувшие хозяина, во внешней среде
не питаются. Проницаемость их кутикулы'заметно снижается. Для поддержания
своего существования, а главное, весьма интенсивного размножения они исполь-
зуют энергетические резервы, накопленные ими в течение паразитического перио-
да развития.
Органы выделения у волосатиков отсутствуют.
Нервная система представлена окологлоточным кольцом, от которого отходит
продольный нервный ствол, залегающий в брюшном валикообразном утолщении
гиподермы. Органы чувств развиты очень слабо. У самцов выделяется группа
половых сосочков. У некоторых видов на переднем конце тела имеются более или
менее четко ограниченные скопления темного пигмента, напоминающие глаза.
Как уже отмечалось выше, все Nematomorpha раздельнополы. Половой ди-
морфизм проявляется в размерах тела (самцы, как правило, заметно меньше
самок и в строении заднего конца тела (с. 127). Женская половая система со-
стоит из двух яичников, имеющих вытянутую мешковидную форму. От яичников
отходит множество боковых выпячиваний — дивертикулов. Задние концы яични-
ков переходят в короткие яйцеводы, которые впадают в непарную матку. У во-
128
лосатиков в отличие от нематод самостоятельное женское половое отверстие
отсутствует. Матка открывается в заднюю кишку, которая, таким образом, пре-
вращается в клоаку. С маткой соединен небольшой трубчатый семяприемник.
Мужская половая система состоит из двух колбасовидных семенников, тяжу-
щихся от переднего конца тела назад, где они переходят в семяпроводы. Послед-
ние открываются в клоаку. Самцы обладают совокупительным органом.
Цикл развития волосатиков
Зрелые волосатики копулируют во внешней среде. Оплодотворение у* ши.
внутреннее. Самка откладывает до 1 млп. яиц и более. Кладка обычно ммеег
вид длинных и тонких слизистых шнуров. Из яиц вылупляются личинки, рею
отличающиеся по своему виду от взрослых червей (рис. 63, Д). Поверхность як
тела несет отчетливо выраженную поперечную кольчатость, на переднем конце
располагаются два венчика крючков и терминально расположенный хоботом,
вооруженный тремя стилетами. Развитие волосатиков осуществляется с участяев
двух или реже одного хозяина. Проникновение личинок в насекомых осутрспяъ-
ется_ различными способами. В одних, случаях паразиты активно внедряются
в тело хозяина при помощи хоботка и крючьев. В других же — личинки выделяют
слизистое вещество, которое образует тонкую оболочку и в виде таких своеобраз-
ных цист проглатываются насекомыми. В кишечнике последних цисты разруша-
ются, а личинки мигрируют в гемоцель.
Развитие волосатиков чаще всего осуществляется с участием двух хозяев.
Первым хозяином обычно бывают различные мелкие водные насекомые (лячвнвв
поденок, комаров и т. и.). Паразиты проникают в их ткани (ряс. 63, Г) и яшде-
стнруются. Активное развитие начинается лишь во втором хозяине—крупный
хищных насекомых, в кишечнике которых личинки волосатиков освобождаются
от тканей первого хозяина, проникают в гемоцель и начинают быстро растя.
По своему внешнему облику они напоминают взрослых червей, ио никогда не
достигают половой зрелости. Последнее становится возможным только после вы-
хода червя в воду. Насекомое-хозяин при этом обычно погибает.
Смена хозяев характерна не для всех волосатиков. Развитие некоторых вцдев
целиком осуществляется в одном насекомом. Nematomorpha паразитируют не
только в водных, но и в некоторых наземных насекомых и многоножках, которые
держатся недалеко от водоемов. Иногда виды, обитающие в наземных хозяевах,
долгое Ьрёмя, уже будучи вполне к этому готовыми, не выходят наружу. Лашь
попадание хозяина в. воду стимулирует выход паразита. Массовое появление
волосатиков удается иногДа наблюдать во время сильных дождей.
Географическое распространение волосатиков очень широко. Они встречаются
и в тропиках, и в районах с умеренным климатом. Морские же виды могут прод-
вигаться далеко ла север, достигая широты Шпицбергена.
ПАРАЗИТИЧЕСКИЕ МОЛЛЮСКИ
И ЧЛЕНИСТОНОГИЕ
ТИП MOLLUSCA С U V1 Е R — М О Л Л Ю С К И
Тело моллюсков, как правило, подразделяется на три отдела:
голову, туловище (внутренностный мешок) и ногу. На голове име-
ются две пары щупалец и глаза. На нижней стороне головы нахо-
дится ротовое отверстие, которое у некоторых брюхоногих моллюс-
ков расположено на конце длинного хоботка. Нога представляет
собой мускулистый вырост тела, служащий для движения. Основа-
ние туловища окружено кожной складкой — мантией. Между ман-
тией и телом есть небольшая полость, называемая мантийной. В ней
лежат жабры (ктенидии) и орган химического чувства (осфрадий);
здесь же открываются анальное, выделительное и половое отвер-
стия. Все эти образования относятся к мантийному комплексу ор-
Еанов. Эпителий мантии выделяет известковую раковину, в кото-
рой и заключен внутренностный мешок. Большая часть моллюс-
ков— билатерально-симметричные животные, но для класса брю-
хоногих характерна асимметричность строения. Развитие моллюс-
ков обычно связано с метаморфозом. Из яйца выходит планктон-
ная личинка — парусник или велигер.
Тип моллюсков подразделяется на два подтипа, которые объе-
диняют 7 классов. Паразитические моллюски немногочисленны
я встречаются лишь среди пластинчатожаберных и брюхоногих. Бо-
лее существенна роль моллюсков как промежуточных хозяев раз-
личных паразитических червей, главным образом трематод.
В моллюсках развиваются также личиночные стадии некоторых
ленточных червей (сем. Hymenolepididae) и нематод (с. 104).
Для отдельных представителей класса пластинчатожаберных
(Lamellibranchia, или Bivalvia) известен личиночный (ларваль-
ный) паразитизм. Явление это характерно для пресноводных пер-
ловиц и беззубок (сем. Unionidae). Яйца этих моллюсков отклады-
ваются во внутрижаберную полость, образующуюся между наруж-
ным и внутренним листками, составляющими мешковидную жабру.
Здесь и происходит развитие яиц до стадии личинки — глохидия.
130
Личинка эта имеет очень тонкую двустворчатую раковину. Каждая
створка несет на свободном конце острый, усаженный шипами зу-
бец. Большая часть органов, жабры и нога личинки еще недораз-
виты; имеется биссусовая железа, выделяющая длинную и клейкую
нить биссуса. Сформированные личинки выходят в мантийную
полость моллюска. Оттуда они выталкиваются во внешнюю среду
через сифон вместе с водой, которая постоянно циркулирует в ман-
тийной полости, омывая жабры (рис, 64).
Рис. 64. Личиночный паразитизм пластинчатожаберных моллюсков
(из кн. Е. Нобля и Г. Нобля). А — вскрытая перловица; Б —глохи-
дий; В — обрастание глохидпев жаберным эпителием рыбы; Г — схе-
ма цикла, развития:
/ — наружная жабра с развивающимися внутри глохидиями. 2 — зубец рако-
винки глохидия, 3 — биссусовая нить, 4—моллюск, выбрасывающий чеоез
сифон глохидиев, 5 — заражение . рыбы, 6 — молодые моллюски на дне во-
доема
Дальнейшее развитие глохидиев возможно лишь в том случае,
если они попадут в организм хозяина, роль которого выполняют
рыбы. С помощью липкой нити биссуса и шиповатых зубцов ство-
рок личинки зацепляются за плавники и жабры рыб. Под влияни-
ем раздражения ткань жаберных лепестков начинает разрастаться
и вокруг глохидиев образуются опухоли. Паразиты, замурованные
внутри такой опухоли, интенсивно растут, питаясь за счет хозяи-
на. По истечении нескольких недель глохидий превращается в ма-
ленького, но вполне сформированного моллюска. Опухоль к этому
времени изъязвляется, ракушки вываливаются в воду и, попав на
дно, начинают вести свободный образ жизни. Временное паразити-
рование глохидиев на рыбе, сопровождается иногда довольно зна-
чительным поражением жаберного эпителия. При очень сильной
интенсивности заражения возможна даже гибель рыб.
Лишь среди брюхоногих моллюсков (Gastropoda) имеются об-
лигатные паразиты, связанные с хозяином на протяжении всей
жизни. Это чрезвычайно своеобразные организмы, паразитирую-
5* 131
цие на морских беспозвоночных (главным образом на иглокожих)
н выработавшие, в связи с паразитическим образом жизни, ряд мор-
фобиологическнх адаптаций.
Все паразитические брюхоногие моллюски относятся к подкл. Prosobranchia
(отр. Mesogastropoda, подотр. Aglossa) I Группа эта полифилетичиа и, по-видн-
мому, связана в своем происхождении с несколькими семействами свободноживу-
щих моллюсков, переходивших к паразитическому образу жизни разное время
ж независимо друг от друга. Очень древними паразитами являются* по всей веро-
ятности моллюски сем. Capulidae, характеризующиеся наличием небольшой рако-
вины в виде колпачка. Известны ископаемые представители этого семейства,
паразитировавшие на стебельчатых морских лилиях в период от кембрия до
триаса. По-видимому, совсем особняком стоит небольшое сем. Pyramidellidae,
представители которого являются эктопаразитами аннелид и пластинчатожабер-
ных моллюсков.
Наибольший интерес представляют сем. Melanellidae н Entoconchidae, парази-
тирующие на иглокожих. По Е. Н. Грузову (1965), к сем. Melanellidae относятся
ж считавшиеся ранее самостоятельными сем. Stiliferidae, Asterophilidae и Paedo-
^horopodidae.
Паразитические моллюски известны с середины прошлого века, когда И. Мюл-
лером (I. Muller, 1852) был впервые обнаружен н описан р. Entoconcha. Иссле-
дователь сначала принял паразита, плотно прикрепившегося к кишечнику хозяи-
на, за участок гонады голотурий. Обнаружив в теле паразита множество личинок
со спиральной раковинкой, Мюллер предположил, что имеет дело с необычайным
случаем чередования поколений. Лишь несколько лет спустя удалось разобрать-
ся в возникшей ошибке. Интерес к этим своеобразным животным возобновился
а начале нашего столетия, когда появилось несколько серьезных исследований,
посвященных описанию новых форм и изучению их адаптаций к паразитическому
существованию. В серии этих работ следует назвать исследования Бонневье
(К. Bonnevie, 1902), К. Ваней (С. Vaney, 1913) и А. Джонкер (A. Jonker, 1916),
внесших принципиально новые данные в изучение группы. Отечественная зоология
сведениями по паразитическим моллюскам, их строению и эволюции обязана
ученым Ленинградского университета. В 1917 г. Б. Н. Шванвич опубликовал ра-
боту по морфолргии р. Entocolax, эндопаразита голотурий. В дальнейшем пара-
зитические Prosobranchia обстоятельно изучались проф. А. В. Ивановым и его
учениками.
Особенности морфологии паразитических Prosobranchia
и их распределение по хозяевам
Большинство паразитических брюхоногих приурочено к разным
классам иглокожих. Мы ограничимся характеристикой лишь двух
близких семейств: Melanellidae и Entoconchidae.
К сем. Melanellidae относятся не только паразитические, но и
евободноживущие виды. Интересны в этом плане р. Mucronalia и
Metanella. последний включает в себя хищных моллюсков, коменса-
лов и, наконец, эктопаразитов. Среди них около 10 видов встре-
чается на морских лилиях, 9 видов — на голотуриях, 5—на морских
звездах и столько же на морских ежах. Есть среди них и эктопа-
разиты офиур. Все эти виды еще оченф мало изменены паразитиз-
мом. Раковина их хорошо развита (рис. 65,Л), на голове имеется
пара щупалец и длинный, втяжной хобот. Нога снабжена типичной
для Prosobranchia крышечкой. Хобот моллюска обычно глубоко
________i _
1 Некоторые авторы рассматривают паразитических брюхоногих как само-
стоятельный отр. Homoeostropha Golikov et Starobogatow, 1968.
132
погружен в ткани хозяина и служит паразиту как для прикрепле-
ния, так и для питания. На хоботе образуется небольшая кожная
складка — зачаток ложной мантии или псевдопаллнума. Эта склад-
ка имеется у всех представителей семейства и, по мере углубления
Рис. 65. Паразитические брюхоногие моллюски сем. Melanellidae. А—Mucronalia
palmipedis; Б — Megadenus arrhyfichus; В — Stilifer celebensis, раковина покрыта
ложной мантией; Г — тот же моллюск, ложная мантия удалена (Д—Г по Ивано-
ву); Д — Asterophila japonica, ложная мантия частично удалена (по Грузову):
/ — раковина, 2—щупальца, 3 —ложная мантия, 4 — хобот, 5 — крышечка, б — яйцевой
кокон, 7 — яичник, 8 — печень, 9 — ложно-мантийная полость
их адаптации к паразитизму, приобретает все большее развитие
и значение. Внутренняя организация представителей р. Melanelta
ничем не отличается от основных особенностей строения других
брюхоногих моллюсков.
Переход моллюсков от паразитирования на поверхности тела
иглокожих к эндопаразитическому образу жизни привел к глубо-
133
ким изменениям организации, связанным и с общей деградацией,
и с вторичным развитием адаптивных признаков.
Разные виды паразитических брюхоногих находятся в различ-
ных. взаимоотношениях с организмом животного-хозяина. Эктопа-
разиты типа Melanella удерживаются на его поверхности, погружая
в ткани свой втяжной хобот. В результате паразитирования мол-
люсков могут возникать изменения в строении покровов хозяина:
образуются более или менее глубокие впячивания или опухоли; из-
меняется структура покровных тканей и скелета. В открытых чаше-,
видных углублениях на теле морской звезды помещаются, напри-
мер, моллюски р. Megadenus (рис. 65,Б). Известковые пластинки
скелета’ морской звезды, непосредственно прилегающие к телу па-
разита, утолщены и деформированы. По краю углубления образу-
ются окаймляющие валики. Снаружи видны 3—4 оборота еще хо-
рошо развитой раковины моллюска. У Megadenus уже имеется
ложная мантия. Она выстилает дно чаши, в которой сидит моллюск,
и заворачивается иа наружную сторону его раковины.
Для некоторых представителей р. Stilifer (например, St. cele-
bensis) характерно еще большее развитие ложной мантии, которая
полностью обрастает раковину, оставляя свободными лишь один-
два верхушечных оборота (рис. 65,5).. Моллюски этого вида почти
целиком погружены в ткани хозяина, которые в этом месте утол-
щены и образуют подобие опухоли или галла. В центре имеется
маленькое отверстие, из которого высовывается наружу лишь вер-
хушка раковины. Очень длинный и утративший способность втяги-
ваться хобот моллюска (рис. 65, В, Г) вдается в полость тела мор-
ской звезды, откуда и черпает необходимые пищевые вещества.
Таким образом представители р. Megadenus и, в особенности,
Stilifer хотя и являются эктопаразитами, но уже находятся на гра-
ни перехода к эндопаразитизму. Настоящими эндопаразитами мож-
но считать моллюсков р. Gasterosiphon, обитающих в полости тела
голотурий, и Asterophila — из морских звезд. Для обоих родов ха-
рактерно полное исчезновение раковины. Ложная мантия обраста-
ет все тело животного. У Gasterosiphon внутренностный мешок еще
сохраняет спиральную закрученность, столь характерную для боль-
шей части брюхоногих. У Aste.rophila эта особенность полностью
утрачена. У Gasterosiphon имеется длинный, хорошо развитый хо-
бот, который прикрепляется к кровеносному сосуду голотурии: оче-
видно, моллюск питается кровью. Ложная мантия, окружающая
все его тело, на заднем конце суживается, образуя довольно длин-
ную трубку — сифон, прободающий стенку тела хозяина. С по-
мощью этого сифона паразит и сообщается с внешней средой. Мол-
люски р. Asterophila полностью изолированы от внешней среды.
Хобот у них отсутствует, нога рудиментарна, тело, одетое ложной
мантией, имеет почти шаровидную форму (рис. 65,Д). Еще более
изменен внешний облик моллюсков, относящихся к другому, близ-
кому сем. Entoconchidae (см. рис. 70). Все они эндопаразиты, ло-
кализующиеся в целомической полости голотурий. Тело этих мол-
люсков червеобразно вытянуто. Для них характерно очень сильное
134
разрастание ложной мантии, за счет которой образуется большая,
вместительная выводковая камера. Передним концом паразит сво-
бодно свешивается в целомическую полость хозяина, прикрепляясь
с помощью сифона к стенке его тела или к кишечнику.
Таким образом, представители рассмотренных семейств могут
быть расположены в определенный ряд, в котором прослеживают-
ся все переходы от свободноживущих видов до облигатных эндо-
паразитов. совершенно утративших не только связь с внешней сре-
дой, но и самый облик моллюска. По мере усиления связи паразита
с телом хозяина исчезает раковина и развивается новое образова-
ние— ложная мантия, представляющая собой разросшуюся кож-
ную складку головного конца или ноги.
Роль ложной мантии разнообразна. У Megadenus она способст-
вует закреплению паразита в чашевидном углублении, которое об-
разуется на месте его поселения. У Stilifer, возможно, имеет защит-
ную функцию. У Gasterosiphon мантия не только одевает все тело
моллюска, но и образует сифон, который прободает стенку тела
хозяина и осуществляет связь паразита с внешней средой. Наконец,
у Asterophila и у всех Entoconchidae за счет разрастания ложной
мантии формируется выводковая камера, которая по объему иног-
да значительно превышает размеры собственно тела моллюска.
Интересно проследить судьбу такого характерного для моллюс-
ков органа, как нога. Свободноживущие Melanella и относящиеся
к этому роду эктопаразиты имеют хорошо развитую ногу, которая
несет крышечку (признак многих переднежаберных). У других пара-
зитических брюхоногих нога претерпевает постепенную редукцию
и у некоторых видов полностью исчезает. В отдельных случаях
нога, хотя и сохраняется, но начинает выполнять совершенно осо-
бые функции. Так, например, у Megadenus часть ноги приобретает
вид желобка, несущего ресничный эпителий. По этому желобу во-
да, необходимая для дыхания, поступает в мантийную полость мол-
люска. У одного йз видов Stilifer хорошо развитая мускулистая
нога работает как поршень, нагнетая воду в ложно-мантийную по-
лость. Совсем особую роль выполняет нога у самок моллюсков
р. Paedophoropus. Чрезвычайно разрастаясь, боковые лопасти ноги
образуют обширную выводковую камеру (рис. 66), в которой про-
исходит развитие яиц.
Сильно изменяется головной отдел паразитических брюхоногих.
Прежде всего исчезают щупальца, а затем и глаза. У эктопарази-
тических видов щупальца, хотя и имеются, но носят рудиментар-
ный характер. Хобот, превращающийся у эктопаразитов в орган
прикрепления к хозяину, утрачивает способность втягиваться, а у
некоторых видов исчезает. У эндопаразитических Entoconchidae го-
лова, по сути дела, отсутствует.
Переход от эктопаразитизма к обитанию в полости тела хозяи-
на сопровождается редукцией мантийного комплекса и самой ман-
тии. У эктопаразитов мантийный комплекс еще имеет типичное
строение. Но у таких специализированных форм, как Gasterosiphon,
исчезает ктенидий, функционально его замещают вторичные адап-
135
тивные жабры. У эндопаразитической Asterophila мантийный комп-
лекс органов распался и частично редуцирован. Ктенидий, осфра-
дий, гипобронхиальная железа и прямая кишка с анальным отвер-
стием исчезли. Половое же отверстие переместилось за пределы
мантийной полости. Характерное для брюхоногих положение со-
хранила только почка. У Entoconchidae мантийный комплекс орга-
нов полностью отсутствует.
Рис. 66. Паразитический моллюск
Paedophoropus dicoelobius (по Ива-
нову) :
1 — внутренностный • медрк, 2 — правая
лопасть ноги, 3 — выводковая камера, 4 —
центральная часть ноги, 5 — левая ло-
пасть ноги, 6 — ротовое отверстие, 7 —
хобот
Рис. 67. Эволюция пищеварительной
системы паразитических моллюсков
(по Иванову). А— Megadenus voeltz-
kowi; Б — Megadenus arrhynchus;
В — Stilifer celebensis; Г — Asterophi-
la japonica; Д — Paedophoropus di-
coelobius; E — Entocolax sciemenzi;
Ж—Entoconcha sp.:
1 — печень, 2 — желудок. 3 — пищевод. 4 —
глотка. 5 — задняя кишка, б — кишечный
мешок
, Интересна и эволюция пищеварительной системы (рис. 67).
G переходом моллюсков к паразитическому образу жизни происхо-
дит рудиментация желудка. У свободноживущих брюхоногих этот
орган служит вместилищем для поглощаемой животным грубой
пищи. Очевидно, у паразитических форм , питающихся кровью и со-
ками хозяина (т. е. жидкой и высококалорийной пищей), необхо-
димость в таком резервуаре отпадает. Хорошо обособленный желу-
док еще имеется у некоторых представителей р. Megadenus (рис.
67). У других видов границы между желудком и печенью сглажи-
ваются, а у Stilifer от стенок желудка уже не остается никаких
следов. Сама печень упрощается и постепенно приобретает вид
лопастного или складчатого «печеночного мешка». Функционально
136
этот орган в основном соответствует печени свободноживущих мол-
люсков. У Entocolax этот мешок еще сохраняет несколько эпители-
альных складок, которые можно рассматривать как остатки слож-
ного строения печени. У р. Entoconcha исчезают и складки (рис.
67,Ж). Одновременно с изменениями центральной части пищева-
рительной системы происходит редукция глотки и пищевода (все
Entoconchidae); редуцируется задняя кишка и исчезает анальное
отверстие, так что пищеварительная система становится слепо зам-
кнутой. Моллюски р. Enteroxenos и Parenteroxenos вообще лишены
кишечника.
Нервная система характеризуется концентрацией ганглиев;
А. В. Иванов рассматривает эту особенность как следствие парази-
тического образа жизни. Исчезают ганглии; иннервирующие осфра-
дии и ногу, что, несомненно, связано с редукцией соответствующих
органов. У некоторых эндопаразитов, например Parenteroxenos,
нервная система редуцирована.
Интересны адаптации, связанные с размножением паразитичес-
ких моллюсков. У эктопаразитических Melanella половой димор-
физм слабо выражен и самцы по внешнему виду почти ничем не
отличаются от самок. По мере усиления связи с хозяином степень
полового диморфизма моллюсков заметно углубляется, что в конце
концов приводит к появлению карликовых самцов. Одновременно
развиваются приспособления, обеспечивающие перекрестное опло-
дотворение, затрудненное при паразитическом образе жизни. Кар-
ликовые (около 1,5 мм) самцы моллюсков р. Megadenus всегда
прикрепляются к телу хозяина рядом с самками (рис. 68, Б). Тем
не менее процесс копуляции может иметь место лишь благодаря
особому устройству совокупительного органа, отличающегося боль-
шой длиной (свыше 1,8 мм). В спокойном состоянии penis, превы-
шающий по длине размеры тела карликового самца, петлеобразно
сложен в его мантийной полости. Карликовые самцы Asterophila
прикрепляются непосредственно к телу самки, располагаясь вокруг
отверстия ложиомантийной полости (рис. 68,А).
Крайнюю степень редукции претерпевают карликовые самцы
р. Entocolax. Самки этих моллюсков довольно крупны; размер тела
колеблется у разных видов от 15 до 90 мм. В то же время почти
шаровидное тело карликовых самцов (рис. 68, Г) не превышает
0,6 мм в диаметре. Стенка тела самца очень тонка; под базальной
мембраной прослеживаются редкие мускульные волокна. В полости
тела лежит относительно крупный семенник с отходящим от него
семяпроводом. Все остальные системы органов отсутствуют. Руди-
ментарные карликовые самцы Entocolax способны совершать очень
слабые движения. Они паразитируют в выводковой камере самки,
локализуясь в заднем отделе камеры, вблизи сифона. Наконец,
моллюски р. Enteroxenos и Parenteroxenos — гермафродиты. В со-
ставе других семейств паразитических моллюсков тоже встреча-
ются отдельные виды, вторично приобретшие гермафродитную по-
ловую систему. -
137
Рис. 68. Половой димор(}?изм у паразитических брюхоногих.
Л — самка Asterophila japonica с прикрепившимися к пей самца-
ми (по Грузову); Б — самец и самка Megadenus arrhynchtis
в чашевидном углублении па тело хозяина (по Иванову); В—
карликовый самец Л. juponica (по Грузову); Г.— рудиментарный
карликовый самец Eniocolax rimsky-korsakowi (по Иванову):
/ — penis, 2 — внутренностный мешок, 3 — ложная мантия, 4 — семяпро-
вод, 5 — семенник
1
Рис. 69. Развитие паразитического
моллюска р. Entocolax (из кн. Е. Ноб-
ля и" Г. Нобля):
/—/—последовательные стадии внедрения
личинки в покровы голотурий. 5, б рост
•и развитие моллюска
Паразитические брюхоногие дают красивую иллюстрацию «за-
кона большого числа яиц», сформулированного В. А. Догелем, при-
менительно ко всем паразитическим организмам. Особенно показа-
телен в этом отношении вид Parenteroxenos dogieli. Огромная вы-
водковая камера этого моллюска переполнена коконами, в каждом
из которых содержится до 1000 яиц. Сами коконы формируются
Рис. 70; Паразитические моллюски сем. Entoconchidae (по Иванову с измен.).
4 — Parenteroxenos dogieli\ Б — Entocolax riinsky-korsakoui-.
2 — передний конец. 2—коконы с эмбрионами, 3 — сифон, 4— яичник, 5— семяприемник,
6 — яйцевод. 7 — кишечный мешок. 8— пищевод, 9 — ротовое отверстие, 10 — матка, It —
карликовые самцы в выводковой камере самки
еще в матке моллюска; их оболочка состоит из уплотнившегося
слизистого секрета, выделяемого стенками матки. В выводковой
камере крупных особей помещается до 4 тыс. коконов; таким обра-
зом, количество яиц в ней составляет 3,5—4 млн. Все яйца разви-
ваются до стадии лпчпнки, находясь в пределах выводковой каме-
ры. Таким образом, у Parenteroxenos и других представителей сем,
Entoconchidae имеется «забота о потомстве».
Несмотря на необычайное изменение морфологии взрослых па-
разитических брюхоногих, их личинки сохраняют черты строения
139
велигера. Они имеют, как правило; хорошо развитый парус и
головные щупальцы. Нога хорошо развита и снабжена крышечкой.
Тело личинки заключено в жесткую прозрачную раковину. Личин-
ки оседают на поверхности тела хозяина, например голотурии,
н, отбрасывая раковину, постепенно погружаются в толщу его тка-
ней, а затем и в полость тела (рис. 69). Положение паразита внут-
ри целома хозяина определяется особенностями биологии послед-
него. У роющихся голотурий, выставляющих из грунта лишь перед-
ний конец тела, личинки паразитов попадают вместе с пищей в ки-
шечник хозяина и оттуда в полость тела. В таких случаях паразит
прикреплен сифоном, к стенке кишечника. Если же хозяин свободно
лежит на грунте, личинки проникают в его целомическую полость
через покровы и прикрепляются сифоном к стенке тела.
Следует отметить, что наиболее изменен паразитическим обра-
зом жизни Par enter oxenos dogieli, описанный в 1945 г. проф. А. В.
Ивановым. Этот вид, относящийся к сем. Entoconchidae, совершен-
но утратил внешнее сходство с брюхоногими моллюсками. Его чер-
веобразное тело (рис. 70) достигает в длину более метра. Крупные
половозрелые экземпляры свободно лежат в целоме хозяина (го-
лотурия Cucumaria japonica), причем тело паразита снаружи одето
перитонеальным эпителием хозяина. В этом отношении Р ar enter о-
xenos напоминает своеобразного паразитического рачка — Хепосое-
loma (с. 147). Внутренняя организация Parenteroxenos предельно
упрощена. У этого моллюска нет дыхательной, кровеносной, выде-
лительной и нервной систем. Отсутствуют и органы чувств. Интен-
сивного развития достигает лишь половая система, которая ста-
новится гермафродитной. Сильно разросшаяся ложная мантия оде-
вает весь внутренностный мешок и позади него образует громад-
ную, заполненную коконами выводковую камеру. Последняя откры-
вается наружу сифональным отверстием. Внутренностный мешок
составляет не более !/з длины тела.
ТИП ARTHROPODA—ЧЛЕНИСТОНОГИЕ
Членистоногие — самый многочисленный по числу входящих
в него представителей тип животных. Он объединяет свыше
1500000 видов, обладающих сегментированным телом и членисты-
ми конечностями.
Сегментарный состав тела членистоногих очень разнообразен,
но в пределах различных филогенетических ветвей этой группы от-
четливо наблюдается тенденция к стабилизации числа сегментов
и слиянию сегментов в более крупные отделы—тагмы. У наиболее
специализированных форм наружная -расчлененность тела может
вторично исчезать.
Тело членистоногих одето хитиновой кутикулой, отдельные
участки которой уплотняются и приобретают вид твердых пласти-
нок— склеритов. Все членистоногие обладают хорошо выраженной
смешанной полостью тела — миксоцелем, заполненной полостной
жидкостью — гемолимфой. Пищеварительная система состоит из
140
трех отделов^ передней, средней и задней кишок. Строение .рото-
вого аппарата и отдельных участков пищеварительного тракта час-
то имеет адаптивный характер и во многом определяется особенно-
стями питания. Кровеносная система незамкнутого типа. У ряда
специализированных видов н некоторых мелких форм она вторичн©
исчезает. Дыхание осуществляется либо через всю поверхность те-
ла (виды, обладающие небольшими размерами, п многие парази-
ты), либо с помощью специальных органов (жаберные ножки, жаб-
ры, легочные мешки, трахеи). Выделительная система примитивные:
видов представлена видоизмененными целомодуктами (коксальиые
железы, антеннальные и максиллярные железы), у более прогрес-
сивных— развиваются мальпигиевы сосуды.
Нервная система построена по типу брюшной нервной цепочки,
которая в процессе эволюции подвергается сильной концентрации.
Органы чувств членистоногих часто имеют сложное строение и весь-
ма совершенны. Однако под влиянием образа жизни (и в первую
очередь у паразитов) наблюдается вторичная редукция сенсорных
аппаратов. Членистоногие раздельнополы. Им часто свойствен от-
четливо выраженный половой диморфизм. Развитие в большинства
случаев осуществляется с метаморфозом.
Тип Arthropoda включает 4 подтипа: Trilobitomorpha, Втаткйнн-
ta, Chelicerata и Tracheata. Первый из них объединяет исключи-
тельно вымершие формы. Остальные же широко представлены
в современной фауне. Биологические возможности членистоногих
оказались чрезвычайно широкими. Эти беспозвоночные смогли при-
способиться к существованию в самых различных условиях. Среди
них есть как морские, так и пресноводные виды. Огромное чиста
представителей превратилось в настоящих сухопутных животных.
Наиболее совершенные формы (кл. Insecta) приобрели способ-
ность к полету и освоили воздушную среду. Подобная пластичность,
несомненно, обусловила и возможность их перехода к паразитизму.
В разных филогенетических ветвях членистоногих этот переход осу-
ществлялся многократно в разное время и совершенно независим©
друг от друга. Паразитизм получил широкое распространение
и среди ракообразных, и среди хелицеровых, и среди насекомых.
В силу этих обстоятельств паразитические членистоногие чрезвы-
чайно многообразны. Наряду с крайне специализированными вада-
ми, принадлежность которых к типу Arthropoda устанавливается
лишь с помощью детальных исследований, широко встречаются
виды, на организацию которых паразитический образ жизни еще
не наложил сколько-нибудь заметного отпечатка. В одних случаях
паразитами становятся личинки, в других — взрослые организмы,
в третьих же весь цикл развития осуществляется на теле животно-
го-хозяина. В разных группах Arthropoda отчетливо прослеживают-
ся многочисленные переходы от факультативного паразитизма к об-
лигатному, от временного нападения па животных-хозяев для пи-
тания (большинство гематофагов) к постоянному эктопаразитиз-
му. Многие виды становятся настоящими эндопаразитами. Крайне
141
широк и круг хозяев, используемых паразитическим^ членистоно-
гими: от беспозвоночных до высших позвоночных животных.
Практическое значение паразитических членистоногих велико.
Ущерб, наносимый ими сельскому хозяйству и в частности живот-
новодству и рыбоводству, может быть очень значительным. Неко-
торые виды представляют серьезную опасность для здоровья че-
ловека. Но, пожалуй, главная роль в этом отношении принадле-
жит кровососущим Arthropoda, многие из которых становятся
переносчиками серьезнейших заболеваний человека и домашних
животных. .Однако не только этим паразитические членистоногие
привлекают к себе пристальное внимание исследователей. В послед-
ние годы все шире и шире применяются биологические методы
борьбы с разнообразными вредителями. В основе этих методов
лежит использование естественных врагов вредных организмов и в
частности их паразитов (главным образом, насекомых).
ПОДТИП BRANCHIATA —ЖАРОДЫШАЩИЕ
КЛАСС CRUSTACEA LAM. — РАКООБРАЗНЫЕ
Ракообразные относятся к подтипу жабродышащих. Это преи-
мущественно водные животные,'тело которых, покрытое хитиновой
кутикулой, подразделяется на три отдела — голову, грудь и брюш-
ко. Однако сегменты, входящие в состав этих отделов (тагм), часто
сливаются между собой, что искажает исходный характер расчле-
нения. В отдельных случаях (с. 148) внешняя сегментация почти
полностью исчезает. Для ракообразных характерно наличие двух
пар антенн, трех пар челюстных ножек и трех пар ногоч ел гостей—
грудных конечностей, принимающих участие в захватывании и удер-
жании пищи. Число грудных л брюшных ножек, осуществляющих
функции передвижения и дыхания, очень варьирует в пределах
разных отрядов. Для многих примитивных ракообразных харак-
терно отсутствие конечностей на сегментах брюшка. Анальная ло-
пасть (тельсон) часто песет вилообразно расщепленное образова-
ние— фурку. Все • конечности ракообразных исходно двуветвисты,
но в процессе эволюции претерпевают значительные изменения.
Двуветвистый характер конечностей сохраняется, как правило,
у примитивных форм.
Ракообразные обладают пищеварительной, выделительной
и кровеносной системами. Их нервная система представлена моз-
говым ганглием и брюшной нервной цепочкой. Органами дыхания
служат жабры, но некоторые мелкие виды способны воспринимать
.кислород путем диффузии через тонкие покровы тела. Как и все
членистоногие, ракообразные, за отдельными исключениями (с. 150),
раздельнополы. Значительное число ракообразных приспособилось
к паразитическому образу жизни. Это привело к глубоким преоб-
разованиям их организции, подчас совершенно изменяющим внеш-
ний облик животных (см. рис. 79).
142
ПОДКЛАСС MAX1LLOPODA
ОТРЯД С<^ PEPODA M.-EDWARDS, 1834, — В E С Л О H О Г И E
I
Веслоногие рачки — преимущественно мелкие свободноживу-
щие п паразитические виды, обитающие в пресных водах и в море.
Тело свободножпвущнх веслоногих подразделяется на сложную
голову (или головогрудь), грудь и брюшко. В состав.первой тагмы
входит акрон (головная лопасть), четыре головных сегмента и один
грудной. Все эти сегменты слиты между собой и одеты сплошным
хитиновым покровом. Грудь состоит из пяти, обычно свободных,
сегментов, четыре из которых несут плавательные ножки. Брюшко
четырехчлеииковое. На первом брюшном сегменте — половом или
генитальном, располагаются половые отверстия, а у самок, кроме
того, и характерные для Copepoda яйцевые мешки. Последние пред-
ставляют собой .пакеты яиц, склееных между собой особым секре-
том. Конечностей на брюшке нет. Анальная лопасть (тельсон) не-
сет фурку. У паразитических веслоногих характер расчленения тела
подвергается значительным изменениям.
Отряд веслоногих раков распадается на три подотряда: Cyclo-
poid, Galigoida и Lernaeoida. В первом подотряде, представлен-
ном в основном свободноживущими формами, имеются и парази-
тические семейства (сем. Ergasilidae, Monstrillidae). Второй и тре-
тий подотряды целиком паразитические.
Распространение, морфология и жизненный цикл паразитических
Copepoda
Паразитические копеподы — довольно обширная группа, насчи-
тывающая около 1000 видов. Большая часть их — эктопаразиты,
и лишь немногие виды поселяются во внутренних органах хозяина.
Изучение паразитических копепод началось еще в середине прошлого столе-
тия. Среди русских исследователей, по-видимому, первым обратил па них внима-
ние К. Ф. Кесслер, опйсавший в 1868 г. несколько видов паразитических рачков
с рыб Онежского озера. В дальнейшем эти исследования развивались зоологами
Петербургского университета — В. М. Шимкёвичем, Ф. Яржинским и Д. Д. Пе-
дашенко. изучавшими паразитических копепод на рыбах Белого моря.
Начиная с 30-х годов нашего столетия, когда во всех районах страны под
руководством В. А. Догеля широко развернулись исследования паразптофауны
рыб, паразитическим копеподам стало уделяться большое внимание. Результаты
обработки данных, полученных в многочисленных экспедициях, легли в основу
известной монографии акад. А. П. Маркевича, посвященной паразитическим вес-
лоногим рыб СССР. Эта монография, опубликованная в 1956 г., в отечественной
литературе остается пока единственной сводкой, в которой дана характеристика
морфологии, биологии и практического значения Copepoda.
Говоря о зарубежных работах, необходимо прежде всего назвать серию ис-
следований по морфологии и систематике паразитических копепод, проводивших-
ся в начале века К. Б. Вильсоном (С. В. Wilson). Эти исследования сыграли
очень большую роль в изучении рассматриваемой группы. В недавнее время
опубликована книга 3. Кабата (Z. Kabata, 1971), которая представляет собой
сводку данных по морфологии и физиологии паразитических копепод и тому
болезнетворному воздействию, которое они оказывают па рыб.
143
Жвс, 71. Веслоногие раки — эндопаразиты беспозвоночных (из кп. Е. Нобля
и Г. Нобля).
А — развитие"рачка Haernocera danae (Clap.) (сем. Monstrillidae), паразитирую-
щего в полихетах:
If— вауалнус. 2 — науплиус, внедряющийся в тело хозяина, 3 — ранняя личинка с придат-
ашвв ди восприятия пищевых веществ, 4 — сформированная личинка, 5 — половозрелая
самка, лишенная кишечника;
Б — стадии развития паразита голотурий Cucutnaricola notabilis:
1 —. копеподнтная личинка, 2 — половозрелый рачок
^Особенности биологии и распространения
Паразитические копеподы широко распространены в пресновод-
ных и морский водоемах. Многие виды поселяются на морских бес-
позвоночных (кишечнополостных, кольчатых червях, моллюсках, иг-
локожих) и низших хордовых. Встречаются на китообразных и, как
Рис. 72. Веслоногие раки подотряда
Caligoida Lepeophtheirus chatoni (по Гу-
севу) . А — самка; Б — самец
Рис. 73. Строение пило-
видных мандибул Cali-
gus curtus (из кн. Ка-
бата)
исключение, на земноводных. Однако основными хозяевами пара-
зитических веслоногих несомненно следует считать морских и прес-
новодных рыб.
Большинство паразитических Copepoda локализуется на жаб-
рах, коже, чешуе, плавниках рыб и в их ротовой полости. Но неко-
торые виды способны проникать глубоко в толщу тканей хозяина.
Таковы Penella, паразиты китообразных; рачок Lemaeocera bran-
chialis (см. рис. 76), поселяющийся на жабрах трески, но внедряю-
щийся своим передним концом в сердце рыбы, и некоторые другие
виды. В этих случаях нерасчлененное тело рачка свободно выда-
ется наружу. У р. Perodertna в полость тела хозяина втягивается не
только передний конец, но и весь туловищный отдел, так что сна-
145
ружн видны только яйцевые- мешки и кончик брюшка. Есть и на-
стоящие эндопаразиты, которые обитают в толще мускулатуры
(Sarcoiaces) или в висцеральной полости (Ichtyoiacqs) некоторых
морских рыб. Тело этих рачков совершенно утрачивает следы сег-
ментации и становится мешковидным.
Рис. 74. Представители подотряда Lernaeopodida (по Маркевичу). A— Lernaea'
cyprinacea\ Б — Pseudotracheliastes stellatus;, В — Achtheres регсагигщ Г — Basa-
nistes woskoboinikovi
Эндопаразиты особенно часто встречаются у беспозвоночных.
Это представители сем. Monstrillidae на личиночной стадии, пара-
зитирующие в кровеносной системе полихет; Mytilicola из кишечни-
ка пластинчатожаберных моллюсков (мидий и устриц), рачок Си-
cumaricola из голотурий (рис. 71), сильно видоизмененная Хепосое-
loma — паразит полихет (рис. 75,’В) и некоторые другие виды.
В распространении эктопаразитических копепод большую роль
играет непосредственное воздействие таких факторов, как соле-
ность, температура л т. п. Некоторые вцды отличаются стеногалин-
ностью. Так, специфичные паразиты лососевых из сем. Caligidae
(рис. 72) погибают в период нерестовой миграции хозяина, поч-
146
ju сразу же Ьосле захода рыб из моря в пресные воды. Примером
эвригалинный копепод может служить паразит осетровых Pseudo-
tracheliastes stellatus (рис. 74, Б), проделывающий вместе с хозяи-
ном весь путь миграции из моря в реку и обратно.
Or образа жизни рачков в значительной мере зависит степень
их специфичности. Так, временный паразит Ergasilus sieboldi
(рис. 75, Л) встречается на нескольких десятках видов рыб. То Же
Рис. 75. Упрощение организации веслоногих рачков под влиянием паразитиче-
ского образа жизни. А — Ergasilus sieboldi, мало измененный паразитизмом (по
Маркевичу); Б — Sphyrion lumpi (из кн. Кабата); В — Xenocoeloma brumpti
(по Коллери и Мениль):
/—антенны, преобразованные в органы прикрепления, 2 — расчлененное тело и плаватель-
ные ножки. 3 —яйцевые мешки, 4—выросты головного конца, внедряющиеся в ткани ры-
бы. 5 — нерасчлененное тело. 6— кишечник хозяина, 7 —эпителий, одевающий тело хозяина
и паразита, 8 — участок целомической полости хозяина, врастающий в тело паразита.
9 — семенник, 10 — яичник с яйцеводами
относится и ко многим -представителям Caligidae (Caligus lacustris
п др.). У постоянных же паразитов специфичность, как правило,
выражена. Некоторые виды рачков строго приурочены к опреде-
ленному семейству рыб. Так, Achteres percarutn (рис. 74, В) встре-
чается на окуневых, Pseudptraehellastes stellatus (рис. 74, Б) —на
осетровых и т. д. Есть и очень узко специфичные паразиты, парази-
тирующие только на одном виде хозяина, например Basanisies
huchonis — на дунайском лососе.
147
Строение и физиология
Переход к паразитическому образу жизни вызвал глубокие из-
менения в организации и, особенно, во внешнем облике копепод
(рис. 74, 75). Изменения эти выражены тем сильнее, чем теснее свя-
зан паразит с организмом хозяина. В большинстве случаев пара-
зитические Copepoda значительно крупнее своих свободноживущих
родичей — циклопов. Размеры их тела варьируют от 0,5 до 1 см,
а некоторые виды, например рачок Penella, паразитирующий на ки-
тообразных, достигает 32 см в длину.
Изменения внешнего облика рачков, обусловленные утратой
сегментации тела, появлением разного рода выростов и преобра-
зованием конечностей, носят часто адаптивный характер. Сохране-
ние более или менее отчетливо выраженной сегментации характер-
но только для паразитических представителей подотр. Cyclopoida,
например для различных видов р. Ergasllus. Это в сущности почти
неизмененные паразитизмом рачки, внешним обликом схожие с
циклопами. Большую часть своей жизни они проводят в воде, как
планктонные организмы. В подотр. Caligoida наблюдается отчет-
ливая тенденция к слиянию сегментов. У этих рачков обычно ос-
тается лишь два свободных сегмента груди (остальные членики
сливаются с головой или с брюшком). Брюшко состоит всего из
трех-четырех члеников, нередко тоже сливающихся. Тело рачков
уплощено, головогрудный отдел покрыт широким щитком (см.
рис. 72). Особенно отчетливо выражена тенденция к исчезновению
внешней сегментации у представителей подотр. L^rnaeoida, у кото-
рых иногда можно только наметить границу между слитным туло-
вищным отделом и брюшком. В ряде случаев брюшко отсутствует.
Некоторые виды, например паразитирующие на кольчатых червях
Xenocoelotna brumpti, совершенно утрачивают облик рачка (рис.
75, В).
Сильно видоизменяются антенны и конечности. У некоторых
представителей р. Caligus антенны I преобразуются в полулунные
присоски. У многих копепод они рудиментарны или полностью ис-
чезают. Антенны II большей частью превращаются в органы при-
крепления, имеющие вид острых саблевидных крючьев (р. Ergasi-
lus) или коротких когтевидных образований (р. Caligus). У Ler-
naeoida они, как правило, недоразвиты.
Судьба ротовых конечностей различна и зависят от способа пи-
тания. Так, у Caligoida развивается специализированный ротовой
аппарат, с помощью которого рачки разрезают кожу хозяина
(рис. 73), в других случаях ротовые части могут быть недоразвиты
или полностью отсутствуют (Xenocoelotna). Особое значение име-
ют органы прикрепления, с помощью которых эктопаразнтнческне
рачки удерживаются на теле рыбы. В ряде случаев эта роль вы-
полняется видоизмененными антеннами. У рачка Latnproglena силь-
ный и крепкий коготь развивается за счёт второй пары максилл.
Чрезвычайно своеобразно видоизменены максиллы многих Lerna-
eoida, превращающиеся в длинные выросты, соединенные своими
148
дистальными концами и по виду напоминающие сомкнутые над го-
ловой руки (рис. 74). В месте соединения этих рукообразных вы-
ростов находится конусовидное, звездообразное или пробковидное
образование — «булла», которое представляет собой застывший
секрет личиночных желез рачка. Снаружи булла обрастает эпите-
лием хозяина и иногда очень глубоко погружается в его ткани.
Прикрепление паразита может осуществляться также за счет вы-
А
Рис. 76. Половой диморфизм у паразитических веслоногих. А— Lernaeocera
branchialis (из кн. Догеля); Б — Chondr acanthus pinguis (из кн. Кабата):
/ — самка. 2 — самец (сильно увелнч.), 3 — карликовый самец, прикрепленный возле'поло-
вого отверстия самки
v
ростов переднего конца тела, бульбовидно вздутых у Sphyrion
(рис. 75, Б) или причудливо’разветвленных, похожих на оленьи
рога у Lernaeocera (рис. 76). Грудные плавательные ножки имеют-
ся и функционируют только у временных паразитов, способных от-
рываться от хозяина (Ergasilus, Caligus и др.). Переход к посто-
янному паразитизму приводит к редукции плавательных ножек.
Если у некоторых видов (Latnproglena, Lernaea) они еще сохраня-
ются в виде крошечных рудиментов, то во многих случаях (р. Ach-
teres, Lernaeocera, Xenocoeloma и др.) они утрачиваются полностью
(рис. 75,*Б, В; рис. 76).
Под влиянием паразитизма изменяется не только внешний об-
лик, но и внутренняя организация Copepoda. Покровы их становят-
ся очень тонкими; туловищная мускулатура претерпевает значи-
тельное упрощение, хотя и сохраняет некоторые черты метамерного
строения. Исчезает мускулатура, приводящая в движение плава-
тельные ножки, зато те мышцы, которые обеспечивают работу
149
органов прикрепления, претерпевают прогрессивное развитие. За-
метно редуцирована нервная система и органы чувств, в особенно-
сти у тех видов рачков, которые утратили способность к передви-
жению. У многих представителей сем. Lernaeidae п Lernaeopodidae
(см. рис. 74) во взрослом состоянии исчезают глаза; хорошо разви-
ты лишь органы осязания (а может быть, и химического чувства),
представленные чувствительными щетинками и сосочками.
Пищеварительная система паразитических копепод обычно раз-
вита хорошо, хотя известны-виды, у которых она подвергается час-
тичной или полной редукции. Питаются копеподы исключительно
за счет хозяина. Представители сем. -Lernaeopodidae поглощают
клетки покровного эпителия и слизь. У представителей р. Ergasllus,
ротовые части которых не приспособлены к питанию плотными тка-
нями, появляется внекишечное пищеварение. В месте‘прикрепления
рачка к телу хозяина всегда “есть участок разрушенной, полупере-
варепной ткани, которая и поглощается паразитом. Тканями
и кровью питаются представители сем. Caligidae. В их подвижном
ротовом хоботке лежат длинные вытянутые мандибулы, концевой
членик которых преобразован в пилу с острыми зубьями (рис. 73),
с помощью которых паразит разрезает кожу рыбы. Наконец, встре-
чаются и типичные гематофаги — распространенный паразит трески
Lernaeocera branchialls (plic. 76), паразит карповых и других прес-
новодных рыб Lernaea (рис. 74, А) и т. п. Некоторые рачки извле-
кают кровь из жаберных сосудов, другие (например, Spliyrion),
глубоко внедряясь в ткани, сосут кровь из спинной артерии. Нако-
нец, Lernaeocera branchialis, прободая ткани хозяина, добывает
кровь из бульбуса аорты или непосредственно из сердца рыбы.
Некоторые рачки, паразитирующие на беспозвоночных, лишены
пищеварительной системы. У них вырабатываются особые адапта-
ции к питанию. Так, Xenocoelotna brutnpti получает переваренные
вещества непосредственно из кишечника хозяина. Рачки же из сем.
Monstrillidae питаются лишь на стадии личинки; паразитируя
в крови полихет, они воспринимают пищевые вещества всей поверх-
ностью тела. Этому процессу, видимо, способствует- образование
длинных выростов, увеличивающих всасывающую поверхность
(рис. 71).
Половая система развита хорошо. Паразитические копеподы,
за исключением гермафродитной' Xenocoelotna brumpti, раздельно-
полы и для них характерен отчетливо выраженный половой димор-
физм. У представителей отр. Cyclopoida и Caligoida самцы отлича-
ются от самок меньшими размерами (см. рис. 72). Но особенно
резко выражены различия между полами в подотр. Lernaeoida.
В сущности все описанные выше особенности строения этих рако-
образных (увеличение размеров, исчезновение сегментации, руди-
ментация конечностей и т. п.) характерны только для самок. Сам-
цы же обычно сохраняют внешний облик рачка. Они обладают рас-
члененным телом, на грудных сегментах располагаются хорошо раз-
витые плавательные ножки (рис. 76,Б). Самцы в 10 и более раз
меньше самок. У некоторых видов имеются типичные карликовые
150
самцы. С помощью максилл вторых п ногочслюстей они прикреп-
ляются к телу самки и постоянно сидят возле ее полового отверстия
(рис. 76,В). Самцы живут недолго и обычно погибают вскоре пос-
ле оплодотворения. По-видимому, они не питаются. Их недоразви-
тый кишечник имеет вид слепо замкнутого мешка.
Жизненный цикл протекает следующим образом. На гениталь-
ном сегменте оплодотворенных самок находятся два яйцевых меш-
ка, в каждом из которых может быть заключено до 2000 яиц, скле-
енных между собой секретом специальных желез. Яйцевые мешки
прикрепляются возле полового отверстия и самка носит их на себе
вплоть до вылупления личинок.
Личинка, вышедшая из яйца, обычно представляет собой ти-
пичного науплиуса с хорошо развитым глазком и тремя парами
конечностей. После ряда линек, сопровождаемых ростом личинки,
опа переходит в так называемую копеподитную или циклопоидную
стадию, внешне напоминающую свободно живущих циклопов. Груд-
ные сегменты личинки обособлены, плавательные ножки хорошо
развиты, имеется расчлененное брюшко, заканчивающееся фур-
коп (см. рис. 71, Ж). Копеподитные личинки оседают на тело хо-
зяина, прикрепляются к нему и вскоре достигают половой зрелости.
Мужские и женские особи копулируют, после чего самцы вскоре
отмирают. Самки же, претерпевая метаморфоз, принимают харак-
терный облик паразитического рачка и начинают откладывать
большие количества яиц.
Интересно отметить, что личинка свободноживущих копепод
в процессе развития от иауплиуса до взрослого рачка претерпева-
ет 12 линек. У паразитических же видов весь ход развития сильно
сокращен. У многих Lernaeoida личинка на стадии науплиуса еще
заключена внутри яйцевых оболочек и вылупляется сразу метана-
уплиус, а иногда и копеподитная личинка. Такое сокращение числа
свободных личиночных стадий и длительности их жизни —черта,
вообще очень характерная для паразитических животных (Догель,
1947).
Патогенное воздействие на хозяина
Патогенность паразитических копепод проявляется как в ло-
кальных повреждениях пораженных органов, так и в общем токси-
ческом воздействии на организм хозяина. Мало что известно об их
болезнетворном влиянии на беспозвоночных. Приведем лишь све-
дения о том, что паразитирующие в кишечнике мидий рачки
Mytilicola вызывают задержку роста хМоллюсков, а при сильном за-
ражении могут стать причиной их гибели. Есть данные о значитель-
ном сокращении численности мидий, пораженных Mytilicola.
Более обстоятельно изучены вызываемые копеподами болезни
рыб. Особенно большое значение имеет причиняемое рачками по-
вреждение жабр.. Наиболее опасны в этом отношении представите-
ли сем. Ergasilidae. Рачки, прикрепляющиеся к жаберным лепест-
кам, сдавливают и сминают их, нарушая тем самым нормальную
151
циркуляцию крови, ь результате такого механического раздраже-
ния нередко возникают местные утолщения жаберного эпителия,
затрудняющие газообмен. Наконец, рачки, вонзая в них свои саб-
левидные антенны, разрушают жабры и частично переваривают их
изливаемыми наружу ферментами. Довольно часто это оказывает-
ся не только вредным, но и пагубным для хозяина, особенно если
принять во внимание тот факт, что на жабрах одной рыбы иногда
паразитирует до 3 тыс. рачков. Эргазилез может стать причиной
серьезных эпизоотий.
Поражения жабр и кожи наблюдаются и при паразитировании
представителей сем. Lernaeopodidae. Прикрепляясь с помощью
буллы (см. рис. 74), они вызывают патологическое разрастание
эпителия, приводящее к возникновению опухолей. Следствием па-
разитирования копепод бывают разнообразные повреждения кожи
и мышц хозяина. Предствители сем. Caligidae разрушают кожу
скоблящими движениями своих пиловидных мандибул (см. рис. 73),
в результате чего возникают кровоточащие ранки. Местные воспа-
ления и опухоли образуются в местах внедрения разветвленного
переднего конца тела рачков Lernaea и Sphyrion (см: рис. 74, Д,
75,5). После их гибели остаются открытые ранки и язвы, на кото-
рых поселяются губительные для рыб плесневые грибки Saproleg-
nia. Особенно серьезны поражения, вызываемые рачками р. Ler-
naea. Эти паразиты поселяются на карповых и лососевых рыбах,
культивируемых в рыбоводных хозяйствах, и могут стать причиной
гибели молоди. Поражаются и органы чувств. Целый ряд пресно-
водных рачков из р. Parergasilus локализуются в обонятельных
ямках рыб. Многие копеподы поселяются на поверхности глаз или
даже внутри глазного яблока хозяина — преимущественно морских
рыб (сельдевых и скумбрии). Относительно редко повреждаются
копеподами скелет, мускулатура и внутренние органы.
За последние годы получены некоторые данные и о влиянии ко-
пепод на общее состояние хозяина. Выяснилось, что при заражении
трески рачками Lernaeocera branchialis вес печени рыбы и содер-
жание в ней жира падает почти в два раза. У линей, зараженных
рачками Ergasilus, вес печени составляет всего 1,2% от общего
веса против 4—5% у незаражениых рыб и т. д. Отмечаются также
нарушения обмена, изменения крови (гемоглобин у зараженных
рыб падает почти в 2 раза), задержка в развитии гонад.
Лечение заболевании, вызванных копеподами, мало эффективно. Против
Lernaea cyprinacea применяются солевые, формалиновые и марганцевые ванны.
Все они действуют главным образом на личинок и молодых рачков, еще не за-
крепившихся в толще тканей рыбы. Марганцевые ванны способствуют гибели
и взрослых паразитов.
ОТРЯД BRANCHIURA BURM.
— КАРПОЁДЫ, ИЛИ КАРПОВЫЕ ВШИ
Карпоеды распространены довольно широко. Различные пред-
ставители их паразитируют не только на пресноводных, но и на
морских рыбах. Известен один вид, приспособившийся к парази-
152
Рис. 77. Карповая вошь — Argulus
foliaceus (из кн. Догеля):
1 — крючковидные антеннулы, 2 — руди-
ментарные антенны, 3 — колющий стилет.
4 — максиллы вторые, преобразованные в
присоски, 5 — ногочелюсти, б—кишечник,
7 — плавательные ножки
гированню на амфибиях. В состав Branchilira входит около 130 ви-
job, основная по численности часть которых (100 видов) относится
к р. Argulus Milller, 1785 (рис. 77). Это довольно крупные рачки
(от 3 до 15 мм), тело которых уплощено в дорзо-веитральном на-
правлении (адаптация к эктопаразитизму) и со спинной стороны
покрыто слегка вогнутым головогрудным щитом карапаксом. На
его поверхности расположена па-
ра крупных фасетированных глаз,
между которыми лежат три прос-
тых глазка. Карапакс, прикрыва-
ющий почти все тело, принадле-
жит головогрудному отделу рачка
(состоящему из слившихся го-
ловных и первого грудного сег-
ментов). Далее следуют 4—6 сво-
бодных сегментов груди, несущих
плавательные ножки и слитное
брюшко, образующее вместе с
тельсоном две лопасти, иногда
называемые хвостовым плавни-
ком. Наличие четырех пар дву-
ветвистых плавательных ножек
обеспечивает подвижность рач-
ков. Они могут не только пере-
мещаться по поверхности тела
хозяина, но и, отрываясь от него,
некоторое время плавать в воде.
Argulus нередко обнаруживается
в планктонных пробах, но вместе
с тем это облигатный паразит,
который не может жить без хо-
зяина.
Адаптации карпоедов к пара-
зитическому образу жизни нахо-
дят свое выражение не только в
приобретении уплощенной формы
и конечностей. Главным органом прикрепления этих рачков служат
максиллы (по мнению некоторых авторов, ногочелюсти), превра-
щенные в сильные бокаловидные присоски. С их помощью паразит
прочно закрепляется на теле рыбы. Роль дополнительных органов
прикрепления играют антенны I, преобразованные в короткие, но
сильные крючья. Они помогают рачку наиболее удобно ориентиро-
вать передний конец тела при питании.
Ротовой аппарат колюще-сосущего типа представлен мускулис-
той трубкой, имеющей вид хоботка, в котором, как в футляре, за-
ключены видоизмененные максиллы, мандибулы и острый стилет
(не гомологичный конечностям), с помощью которого рачок проре-
зает кожу хозяина. Процесс питания наиболее полно изучен на
примере широко распространенного паразита пресноводных рыб —
тела, но и в изменении антенн
153
Argulus foliaceus. Рачок разрезает кожу рыбы, несколько раз под-
ряд вводя в толщу покровов стилет, пронизанный тонким кана-
лом, полсоторому в ткани хозяина изливается секрет слюнных же-
лез. Канал этот очень узок (8 мкм в диаметре), поэтому рачок не
в состоянии поглощать эритроциты, размер которых крупнее от-
верстия, ведущего в канал стилета. Всасывается только плазма
крови хозяина. Не исключено, что в отдельных случаях имеет место
внекпшечное переваривание эритроцитов.
Кишечник карпоедов, как и у других гематофагов, имеет пару
сильно разветвленных выростов, служащих резервуарами, в кото-
рые поступает насосанная паразитом кровь рыбы. Это позволяет
рачкам длительное время (до трех недель) жить вне тела хозяина,
не питаясь п постепенно переваривая заглоченную пищу. Argulus
не имеет специальных органов дыхания, воспринимая кислород че-
рез тонкие покровы путем диффузии. Биение плавательных ножек
рачка, прикрепившегося к телу хозяина, обеспечивает непрерывный
ток свежей воды, богатой кислородом.
В период размножения самки Argulus покидают хозяина и от-
кладывают яйца на камнях, погруженных в воду ветках и других
подводных предметах. Кладки имеют вид слизистых комков или
шпуров, в которых заключено от 4 до 250 яиц. Молодые рачки
вылупляются через несколько недель и нападают на рыбу, дости-
гая половозрелости в течение месяца. При поисках хозяина важную
роль, по-видимому, играют хеморецепторы. Рачки обладают также
положительным реотаксисом, благодаря которому реагируют на
движение воды, вызываемое плывущей рыбой.
Патогенность карпоедов заключается прежде всего в том, что
они нарушают.'Целостность покровов хозяина. В местах прикрепле-
ния присосок остаются отчетливые следы. Эпителий, сдавливаемый
присосками, уплощается и даже может полностью разрушаться.
Argulus могут иногда поселяться на поверхности глаза рыбы, вы-
зывая раздражение роговицы.
Особенно значительные повреждения отмечаются в точке внед-
рения стилета, который, проникая в толщу тканей, приводит к об-
разованию ранок до 3 мм в диаметре. Большое значение имеет
при этом секрет слюнных желез, вызывающий острую воспалитель-
ную реакцию, гистолиз подкожной клетчатки и другие поврежде-
ния. Степень токсического воздействия слюны карпоедов зависит
от размера рыбы и от интенсивности ее заражения. Мальки и мел-
кие аквариумные рыбки гибнут в течение нескольких дней, даже
в случае заражения одним-двумя карпоедами. При массивном за-
ражении возможна и гибель крупных карпов.
В прудовых хозяйствах, в которых отмечено сильное заражение рыб карпо-
едамп, рекомендуется спуск п высушивание прудов, дезинфекция дна негашеной
известью. Иногда в прудах устанавливают специальные щитки, на которые рачки
откладывают яйца. Щитки периодически вынимаются и очищаются от кладок.
Этим способом удается значительно сократить численность карпоедов.
154
ОТРЯД ASCOTHORACIDA WAGIN, 1937, —
МЕШКОГРУДЫЕ РАКИ
Чрезвычайно своеобразная группа паразитических раков, при-
способившихся к обитанию в полости тела и гонадах различных
морских беспозвоночных, преимущественно иглокожих. Паразити-
ческий образ жизни вызвал глубокие изменения во внутренней
организации и внешнем облике аскоторацид.
Первые сведения об этих животных относятся к 1866 г., когда известный
французский зоолог Ж. Лаказ-Дютье (G. Lacaze-Duthiers) обнаружил рачка
(Laura gerardiae), паразитировавшего на кораллах. Семнадцать лет спустя этот
исследователь дал подробное описание морфологии найденного им вида, который
отнес к усоногим ракам (отр. Cirripedia). Почти одновременно профессор Петер-
бургского университета Н. П. Вагнер,' исследуя фауну беспозвоночных Белого
моря, нашел в полости тела морской звезды другого представителя этой группы
(впоследствии получившего название Dendrogaster}, по не смог определить его
систематическую принадлежность, столь необычен был внешний вид животного
(см. рис. 79). Не сделал этого и В. М. Шимкевнч, который писал о найденном
им паразите: «Это настолько загадочная форма, что почти ничего невозможно
сказать относительно ее природы». Загадка паразитов морских звезд была рас-
крыта Н/М. Книповичем, который в 1892 г. подробно описал морфологию и раз-
витие этих паразитов и присвоил им родовое название — Dendrogaster. Книпович
отпес их к отр. Cirripedia, в пределах которого выделил особый подотр. Ascotho-
racida, объединивший Dendrogaster и рачков, ранее описанных Ж. Лаказ-Дютье.
В дальнейшем изучении обоснованной Книповичем группы ракообразных большую
роль сыграли исследования О. Леруа (О. Le Rqi, 1907), японского зоолога
Н. Иосии (N. Jpsii, 1931) и в особенности В. Л. Вагина, который не только опи-
сал новые роды и виды, но и дал подробную сравнительно анатомическую
и эмбриологическую характеристику всей группы. На основании проведенных им
исследований В. Л. Вагин'выделил этих своеобразных паразитических рачков
в самостоятельный отр. Ascothoracida, связанный с Cirripedia только общностью
происхождения.
Морфология и развитие
К отр. Ascothoracida относится около 40 видов как экто-, так
и эндопаразитов. Характерные особенности их организации удобно
рассмотреть на примере типичного представителя — Ascoihorax
ophioctennis, паразитирующего в половых бурсах офиур. В одной
бурсе обычно можно найти самку и несколько самцов. Тело самки,
длиной 2—Змм, почти целиком покрыто свешивающимися с боков
двумя створками мантии (рис. 78, А), которая представляет собой-
видоизмененный и разросшийся грудной шит личинки (науплиуса).
Внутрь прилегающей к телу рачка утолщенной части мантии захо-
дят ответвления внутренних органов (гоанды и кишечника), тогда
как в ее заднем тонкостенном отделе образуются мешкообразные
вздутия — выводковые камеры (рис. 78,5). В последних заключе-
ны яйца и эмбрионы на разных стадиях развития. Собственно тело
самки S-образно изогнуто, так что брюшко подогнуто под грудь рач-
ка. Головной отдел состоит из акрона и четырех слившихся сегмен-
тов. Он несет короткие клешневидные антенны I и три пары ротовых
конечностей, окруженных, как футляром, верхней губой. Этот комп-
лекс, представляющий собой ротовой аппарат колюще-сосущего ти-
па, называется ротовой пирамидой (рис. 78).
155
Грудной отдел состоит из шести сегментов, каждый из которых
несет по варе рудиментарных листовидных ножек. Брюшко из пяти
сегментов и тельсона заканчивается хвостовой вилочкой — фуркой.
Рис. 78. Строение и половой диморфизм Ascothorax (по Вагину). А — самка, вид
с вентральной стороны; Б — сагиттальный разрез через тело самки; В — карли-
ковый самец; Г — личинка аскоторапидной стадии:
/ — створки мантии, 2 —тело рачка, 3 — ротовая пирамида, 4 — антенны. 5— мозговой
ганглий, 6 — выросты кишечника, 7 — выводковая камера с яйцами, 8 —кишечник, 9 —
брюшко, 10 — фурка, // — грудные ножки
Ротовое отверстие ведет в хорошо развитый кишечник, от которого
отходят .сильно разветвленные печеночные выросты, оплетающие
ветви яичника. Кишка заканчивается анальным отверстием. От не-
парного, сильно разветвленного яичника отходят два яйцевода;
имеется семяприемник. Яйца по мере созревания поступают в вы-
156
водковые камеры; предварительно они оплодотворяются поступаю-
щими из семяприемника спермиями. Имеются типично устроенные
максиллярные железы. Нервная система представлена мозговым
ганглием и слитной брюшной ганглиозной массой. Органы чувств
развиты очень слабо: имеются лишь чувствительные волоски (скорее
всего осязательные) , которые сконцентрированы на антеннах и вет-
вях фурки. Глаз нет. Самцы значительно меньше самок. Тело их
лиш*ь частично прикрыто боковыми складками мантии (рис. 78,В).
Грудные ножки хорошо развиты и служат для передвижения.
Жизненный цикл Ascothorax связан с метаморфозом. Из яиц вы-
ходит типичный науплиус, несущий три пары конечностей (одновет-
вистые антенны I, двуветвистые антенны II и мандибулы), но ли-
шенный глазка. После нескольких линек науплиус переходит в ста-
дию метанауплиуса, характеризующегося сильным развитием спин-
ного щитка — карапакса. Следующая стадия получила название
аскоторацидной личинки. В процессе ее формирования редуцируют-
ся антенны II, карапакс перегибается на спинной стороне и превра-
щается в две свешивающиеся с боков створки (мантия). На члени-
ках груди развиваются двуветвистые плавательные ножки. Внешне
аскоторацидная личинка (рис. 78, Г) напоминает половозрелых сам-
цов Ascothorax. Это подвижная расселительная стадия, свойствен-
ная всем представителям отряда. Личинка активно выходит из вы-
водковой камеры, а потом через щели половой бурсы хозяина — во
внешнюю среду.
Анализируя особенности строения и биологии известных к насто-
ящему времени представителей Ascothoracida, можно наметить ос-
новные направления эволюции этого отряда. Наиболее примитив-
ным, по-видимому, следует считать р. Synagoga. Рачки S. mira
Norm. — временные паразиты коралловых полипов — легко меняют
место своего прикрепления на хозяине, значительную часть жизни
проводят как свободно плавающие организмы. Тело их лишь отча-
сти прикрыто створками мантии, грудные сегменты несут хорошо
развитые двуветвистые плавательные ножки. Половой диморфизм
выражен слабо; самцы и самки почти не отличаются друг от друга,
но в створках мантии самок имеются небольшие углубления для вы-
нашивания молоди.
Другой вид того же рода — S. metacrinicola Okada — настоящий
эктопаразит, поселяющийся на внутренней поверхности диска мор-
ской лилии (Metacrinus rotundas). Однако и эти рачки обладают хо-
рошо развитыми конечностями и, по-видимому, еще способны пере-
мещаться. Питаются они соками хозяина, которые добывают с по-
мощью колюще-сосущего ротового аппарата. Половой диморфизм
у этого вида выражен сильнее. Самцы отличаются от самок мень-
шими размерами. Кроме того, все тело самки покрыто створками
мантии, в толщине которых формируются выводковые камеры.
Третьим представителем того же семейства (Synagogidae) являет-
ся уже знакомый нам Ascothorax. Рудиментарные грудные ножки
*самок этого вида уже не могут служить для плавания, но находятся
.в непрерывном движении, обеспечивая тем самым постоянный при-
157
ток свежей воды, несущей необходимый для дыхания кислород. По-
ловой диморфизм отчетливо выражен.
Типичными эндопаразитами, локализующимися в целомической
полости иглокожих, являются представители сем. Dendrogastridae.
Это р. Ulophysema из морский ежей и р. Dendrogaster— типичные
паразиты морских звезд. Именно этому семейству п свойственны на-
иболее далеко зашедшие изменения организации, обусловленные
эндопаразитнзмом.
Рис. 79. Строение и .по-
ловой диморфизм Dend-
rogaster (по Вагину).
А — самка D. dichoto-
musy внешний вид; Б —
D. iwanowi; самец в вы-
водковой камере самки;
В — D. iwanowi, молодой
самец:
/ — ротовая пирамида, 2 —
антенны. 3 — створка ман-
тии, 4 — выросты мантии,
5 — грудные ножки
Собственно тело самок Dendrogaster. которое помещается в пе-
редней части срединного выроста мантии, полностью утратило сег-
ментацию и конечности. Остались лишь антенны I и ротовая пира-
мида, относящиеся к головному отделу. Створки мантии самок не-
обычайно сильно разрастаются в латеральном направлении и дихо-
томически ветвятся, придавая телу животного причудливый вид
(рис. 79,Л, 5). Вместе с мантией разветвляются заключенные внут-
ри выводковая камера и печеночные выросты кишечника. В стенках
ветвей мантии формируется вторичный, кожно-мускульный мешок,
слагающийся из продольных и кольцевых мышц (образование, со-
вершенно не свойственное членистоногим). Благодаря этому ветви
мантии способны к перистальтическим сокращениям. В выводковой
камере Dendrogaster развивается множество эмбрионов. Там же
обитают и паразитические .карликовые самцы (рис. 79, Б, В), кото-
рые оплодотворяют яйца. Вопрос о способе питания Dendrogaster
158
не до конца ясен. В. Л. Вагин полагает, что самки заглатывают
целомические клетки хозяина, в изобилии оседающие на поверх-
ности тела паразита. Кишечник слепо замкнут, задняя кишка и
анальное отверстие отсутствуют. Выделительная система включа-
ет кроме макенллярной железы множество амебоидных клеток —
фагоцитов, — поглощающих твердые экскреты. Обильное ветвле-
ние мантии, возможно, играет некоторую роль в дыхании парази-
та, извлекающего кпелород из целомической жидкости хозяина.
Увеличение поверхности тела, вероятно, облегчает процесс газо-
обмена.
Карликовые самцы Dendrogaster очень напоминают аскотора-
цидную личинку'. На заднем крае створок их мантии образуются
мешковидные выросты (нередко очень длинные), в которые захо-
дят семенники, семяпроводы и печеночные выросты кишечника.
Жизненный цикл дендрогастрид часто бывает укорочен по сравне-
нию с другими Ascothoracida. Из яиц могут выходить личинки, на-
ходящиеся на разных стадиях развития — науплиуса, метанауплиу-
са и даже аскоторацидной личинки. Свободно плавающие личинки
оседают на поверхности тела морских звезд, проникают в гонаду хо-
зяина, а оттуда в целом, где и претерпевают регрессивный метамор-
фоз, выражающийся в слиянии грудного и брюшного тела и руди-
ментации конечностей.
Эволюция Ascothoracida
Ascothoracida ведут свое начало от предков, общих с усоногими
раками. Вероятно, это были свободноподвижные хищники, эволю-
ция которых частично пошла по линии приспособлений к прикреп-
ленному образу жизни (будущие Cirripedia), частично же по пути
адаптации к паразитизму. В этой второй группе, давшей начало
мешкогрудым ракам, можно проследить все переходы от временно^
го паразитизма (Synagoga mira) к постоянному эктопаразитизму
(S. metacrinicola) и, наконец, к эндопаразитизму. Главными тенден-
циями их эволюции (нашедшими свое крайнее выражение в р.
Dendrogaster) следует считать утрату сегментации и конечностей и
разрастание створок мантии, которая превращается в гигантскую
(по сравнению с размерами собственно тела рачка) разветвленную
выводковую камеру. Наблюдается также тенденция к редукции пи-
щеварительной системы. В ходе эволюции Ascothoracida очень уг-
лубляется половой диморфизм. От почти одинаковых по своей орга-
низации самцов и самок Synagoga mira через ряд промежуточных
состояний возникают настоящие карликовые самцы Dendrogaster.
Одновременно эволюция идет по пути роста половой продуктивности
самок. Временные паразиты Synagoga mira откладывают 30—40
яиц, Ascothorax— около 300, а в выводковой камере самки Dendro-
gaster одновременно развивается до 20 000 яиц.
Личинки Synagoga mira вылупляются из яйца на стадии науп-
лиуса, который тотчас же выходит во внешнюю среду. У Ascothorax
вылупляется метанауплиус, покидающий выводковую камеру обыч-
но на стадии аскоторацидной личинки. Наконец, у Dendrogaster
159
на этой стадии происходит ие только выход из выводковой камеры,
но нередко и вылупление личинки. Таким образом в эволюции отр.
Ascothoracida имеет место сокращение личиночных стадий, процесс
эмбриопизации.
В пределах отряда эволюция шла, по-видимому, в двух направ-
лениях. Часть семейств — Lauridae и Petrarcidae — приспособились
к паразитированию на кораллах, поселяясь под ценосарком и внут-
ри гастральной полости. Эндопаразитизм привел к глубоким измене-
ниям морфологии этих рачков, в частности их половой системы —
для р. Petrarca характерен гермафродитизм. Второе направление
эволюции оказалось связанным с переходом к паразитированию на
иглокожих. Наиболее древние и примитивные виды (например,
S. metacrinicola) поселялись на стебельчатых морских лилиях.
В дальнейшем, возможно, в связи с вымиранием этой группы Echi-
nodermata, начался переход мешкогрудых раков к паразитированию
на офиурах, а затегл на других иглокожих. Одновременно менялись
локализация и характер взаимоотношений с хозяином. От типично-
го эктопаразитизма рачки, перешли к поселению внутри опухолей,
образующихся вокруг паразита (Parascothorax) в полости органов,
сообщающихся с внешней средой (Ascothorax), и, наконец, к пара-
зитированию в целомической полости хозяина (Dendrogas'ter), т. е.
к типичному эндопаразитизму.
ОТРЯД С 1RRI PEDI А — У С ОН О Г И Е
ПОДОТРЯД RHIZOCEPHALA — КОРНЕГОЛОВЫЕ
К подотряду корнеголовые относятся ракообразные, пожалуй,
•наиболее сильно измененные паразитизмом. Типичным представите-
лем отряда можно считать саккулину (р. Sacculina), паразитиру-
ющую на различных крабах. Взрослая саккулина имеет вид мешка,
лишенного следов сегментации и конечностей и расположенного на
брюшке краба с нижней стороны. Мешок представляет собой лишь
часть тела паразита и сидит на разветвленном столоне, который
буквально прорастает все органы краба (рис. 80,Ж). С помощью
столона паразит извлекает необходимые для его существования ве-
щества из гемолимфы хозяина. Наружный мешок (иногда называ-
емый экзосаккулиной) несет слабо выраженные элементы нервной
системы и половые железы (как женские, так и мужские). В полос-
ти мешка содержатся массы развивающихся яиц, из которых выхо-
дят личинки типа науплиуса (рис. 80, А) с тремя парами конечно-
стей и простым глазком, но лишенные кишечника. Проплавав не-
сколько дней, науплиус линяет и превращается в метанауплиус, ко-
торый в свою очередь превращается в следующую стадию развития,
известную под названием циприсовидной (рис. 80,Б). Тело этой ли-
чинки одето двустворчатым панцирем, что делает ее похожей на
рачков р. Cypris (отр. Ostracoda). Личинка снабжена парой корот-
ких, но сильных антенн, грудной отдел ее'несет хорошо развитые
160
плавательные ножки, расчлененное брюшко заканичвается рудимен-
тарной фуркой.
Циприсовидная личинка плавает 3—4 дня, оседает на поверхно-
сти тела своего будущего хозяина — краба, прикрепляясь с по-
Рис. 80. Стадии развития Sacculina (по Деляжу). А — науплиус; Б —
циприсовидная личинка; В, Г — стадии метаморфоза диприсовидной
личинки; Д, Е — внедрение кентрогона в покровы хозяина—краба;
Ж — краб, несущий на нижней стороне брюшка половозрелую экзо-
саккулину (по Коллери):
/ — антенны, 2 —карапакс, 3—фурка, 4 —грудные плавательные ножки, 5 —
участок покровов краба-хозяина, 6 — тело саккулины, 7 — разветвления при-
крепительного стебелька в теле краба
мощью антенн. Затем линяет, отбрасывая при этом все грудные
сегменты и брюшко (рис. 80,В). Оставшийся головной отдел личин-
ки через некоторое время линяет вторично, освобождаясь от кара-
пакса (рис. 80,Г), и приобретает вид небольшого мешковидного
тельца, заполненного недифференцированными клетками. Эта ста-
дия развития саккулины называется кентрогоном. На переднем кон-
це его образуется небольшой, пронизанный каналом, вырост, с по-
6—58 161
мощью которого кентрогон внедряется под покровы краба. Через
канал этого выроста вся масса недифференцированных клеток пере-
мещается в тело краба и током гемолимфы заносится к его кишеч-
нику. Здесь на поверхности кишки хозяина начинается рост парази-
та, приводящий в конце концов к образованию упоминавшегося вы-
ше разветвленного столона. Через 7—8 месяцев на столоне образу-
ется мешковидное тело, постепенно опускающееся в брюшко краба.
Рис. 81. Почкующийся Peltogaster socialis (1) на
брюшке рака отшельника (ориг. рисунок М. Ос-
повата)
Достигнув 2 мм в диаметре, паразит прорывает тонкий хитиновый
покров между сегментами брюшка и оказывается таким образом
снаружи. Это и есть «экзосаккулина», которая растет, достигая
12—15 мм в диаметре. Приблизительно через 9 месяцев после осе-
дания циприсовидной личинки на теле хозяина паразит достигает
половозрелости и приступает к размножению.
Сходным образом идет развитие и другого представителя кор-
неголовых раков — Peltogaster, который паразитирует нс на крабах,
а на раках отшельниках. Некоторые представители р. Peltogaster
(Р. socialis) и паразитирующие на креветках Thompsonia приобре-
ли совершенно не свойственную членистоногим способность к веге-
тативному размножению. Столон этих видов отпочковывает мно-
жество гермафродитных мешковидных тел (рис. 81), усиливая тем
самым половую продуктивность паразита.
Обычно считается, что при линьке хозяина экзосаккулина сбра-
сывается вместе с экзувием, однако это не подтверждается новей-
162
щими исследованиями. По последним данным зараженные крабы
не линяют, так как саккулина выделяет вещества, которые подавля-
ют активность желез внутренней секреции краба, продуцирующих
линочные гормоны.
Паразитирование саккулины оказывает на организм хозяина
и другие воздействия, касающиеся главным образом состояния его
половой системы и вторичных половых при-
знаков. У зараженных самок краба нередко
наблюдается паразитарная кастрация;
иногда редуцируются конечности брюшка.
Однако гораздо более существенные изме-
нения происходят в организме зараженного
саккулиной самца. Прежде всего кардиналь-
но изменяемся состояние его гонады, кото-
рая перестает продуцировать сперматозои-
ды и через некоторое время превращается
в яичник. Брюшко становится широким, как
у самки, на нем развиваются ножки, к ко-
торым у самок обычно приклеиваются вы-
нашиваемые яйца. Нередко исчезает муж-
ское половое отверстие. Все это настолько
меняет внешний облик зараженного сакку-
линой самца, что его с трудом можно отли-
чить от самки. Причины всех этих измене-
ний заключаются, по-видимому, в гормо-
нальном воздействии паразита, который
подавляет активность андрогенных желез
краба. Обычно у зараженных особей эти
железы в значительной степени дегенериру-
ют, чем и обусловлена описанная реверсия
пола. Это не объясняет, однако, почему за-
раженные крабы-самцы приобретают неко-
торые особенности поведения, свойственные
самкам, которые вынашивают яйца. Созда-
Рис. 82. Усоногий рачок
Rhizolepas, прикрепив-
шийся передним концом
к телу хозяина, полихе-
ты (из кн. Е. Нобля
и Г. Нобля):
1 — тело рачка. 2 — раз-
ветвленный прикрепитель-
ный стебелек
ется впечатление, что паразит оказывает
на организм хозяина такое же воздействие, как зрелый, функцио-
нирующий яичник.
Корнеголовые раки, несомненно,— очень древняя группа, связан-
ная в своем происхождении с усоногими. Близость корнеголовых
раков к Cirripedia подтверждается рядом особенностей морфоло-
гии и биологии, свойственных обеим группам. К их числу относится,
например, наличие циприсовидной личинки и способ ее прикрепле-
ния к субстрату. Циприсовидная личинка всегда садится на субстрат
головным концом и прикрепляется с помощью антенн I. Но личинка
Cirripedia закрепляется с помощью выделений цементных желез,
тогда кйк головной отдел личинки Rhizocephala формирует стебе-
лек, проникающий в ткани хозяина. В этой связи представляет ин-
терес один из видов, относящихся к Cirripedia, — рачок Rhizolepas
(рис. 82). Это эктопаразит полихет. Рачок садится на тело хозяина
6*
163
головным концом, от которого отходит стебелек, внедряющийся в
ткани хозяина и разветвляющийся там. Rhizolepas представляется
нам неким прообразом саккулины — «переходной формог!», позво-
ляющей понять пути возникновения столь своеобразной группы, как
Rhizocephala.
ПОДКЛАСС MALACOSTRACA LATR.
ОТРЯД ISOPODA LATR. — РАВНОНОГИЕ
Равноногие — большая группа ракообразных, включающая и не-
которые паразитические формы. Тело обычно уплощено в дорзо-
вентральном направлении. Голова, в состав которой входит 1—2
сегмента груди, несет две пары антенн и сидячие фасетированные
глаза. Ротовой аппарат жующего или колюще-сосущего типа. Груд-
ные сегменты несут одноветвистые конечности. Одна или две пары
их преобразованы в ногочелюсти, остальные — ходильные ножки.,У
половозрелых самок в основании этих ножек развиваются листовид-
ные пластинки, которые черепицеобразно налегая друг на друга, об-
разуют выводковую сумку. В ней происходит развитие янц. Брюш-
ко несет листовидные двуветвистые ножки, служащие для дыхания.
Среди Isopoda имеется довольно большое число паразитических
видов, приспособившихся к обитанию на рыбах и морских беспоз-
воночных. Паразиты рыб (около 430 видов) внешке относительно
мало изменены по сравнению со свободноживущими родичами.
Большая часть их относится к подотр. Flabellifera, в рамках кото-
рого можно найти все переходы от подвижных активных хищников
до настоящих паразитов, всю жизнь проводящих на хозяине.
Тело Isopoda, паразитирующих на рыбах, как правило, отчетли-
во сегментировано. Грудные конечности их несут мощные коготки,
служащие органами прикрепления. Большое число видов — времен-
ные паразиты, многие из них гематофаги. Например, Aega — круп-
ные ракообразные, обладающие колюще-сосущим ротовым аппара-
том. Их кишечник адаптирован к кровососанию и имеет два боль-
ших карманообразных выпячивания, наполняемых кровью хозяина.-
Многим паразитическим Isopoda свойствен протерандрический
гермафродитизм. Такие рачки (например, Cymothoa) обычно при-
крепляются к хозяину на стадии самца. Через некоторое время, од-
нако, самец превращается в самку, которая начинает продуциро-
вать яйца; на брюшной стороне тела этого рачка формируется ха-
рактерная для самок выводковая сумка. Если самец прикрепляется
к рыбе, на которой уже паразитирует половозрелая самка, превра-
щение его в самку задерживается или вовсе не происходит. Пара-
зитические рачки локализуются главным образом на жабрах, коже
п плавниках рыб, часто поселяются в их ротовой полости. Своеоб-
разный рачок Livoneca amurensis Achm. (рис. 83, Д) вызывает обра-
зование тонкостенных впячиваний, глубоко вдающихся в полость
тела. Внутри такого впячивания, сообщающегося с внешней средой
164
узким отверстием, помещается крупная самка (до 3 см длиной), ко-
торая не может покинуть места своего обитания. Там же находится
и маленький самец.
Среди равноногих раков, паразитирующих на рыбах, следует особо отметить
своеобразный подотр. Gnathiidea, для представителей которого характерен лар-
вальный паразитизм. Личинки Gnathiidea (рис. 83, Л) — всегда временные пара-
зиты. Разрывая крючковидными конечностями ткани рыбы, они погружают
в ранку колюще-сосущий хоботок и сосут кровь. Взрослые рачки ведут свободный
образ жизни и не питаются. Их ротовой аппарат рудиментарен, кишечник атро-
фируется.
Рис. 83. Паразитические равноногие раки. А — Gnathia sp.; Б — Telotha siluri
(сем. Cymothoidae) (из кн. Кабата); В, Г — рачок Portunion, паразитирующий
в жаберной полости десятиногих раков (В — самец, Г — самка) (по Лангу
и Коллери); Д— Livoneca atnurensis, сидящая внутри «кармана» на теле амур-
ского язя (по Ахмерову):
J—• пластинки, прикрывающие выводковую камеру, 2—челюстные ножки. 3 ~ антенны,
4 — головной отдел, 5 —брюшко, 6 —тело рачка, 7 — отверстие «кармана>
Равноногие, паразитирующие на беспозвоночных, относятся
к подотр. Epicaridea и характеризуются далеко зашедшими из-
менениями организации. Самки р. Bopyrus и некоторых других ви-
дов, паразитирующих в жаберной полости креветок и крабов, еще
сохраняют в той или иной степени сегментацию тела. Но они обыч-
но асимметричны и их ротовые конечности превращены в колюще-
сосущий хоботок. В очень крупной выводковой сумке содержится
огромное количество яиц. Здесь же в сумке поселяются и карлико-
вые самцы, которые сохраняют типичный облик свободноживущих
Isopoda. В месте паразитирования Bopyrus образуется выпячивание
165
покровов хозяина — карман, в котором и локализуется паразит. Не-
которые представители подотряда становятся эндопаразитами и по-
селяются в полости тела крабов: Они сильно видоизменены, совер-
шенно утрачивают сегментацию и самый облик ракообразного (рис.
83,В, Г,). Самцы же и здесь сохраняют внешний вид равноногого
рачка.
Развитие всех Epicaridea связано с довольно сложным метамор-
фозом. В их жизненном цикле имеется несколько морфологически
различающихся личиночных стадий. Интересно, что определение по-
ла рачков приурочено к метаморфозу последней личиночной стадии.
В тех случаях, когда личинка садится непосредственно на тело хо-
зяина, она превращается в самку. Если же она оседает на самку
рачка соответствующего вида, то становится самцом. Механизм
влияния самки на определение пола личинки пока не ясен.
У ряда видов имеет место протерандрический гермафродитизм.
Интересен в этом отношении необычный жизненный цикл рачка
Danalia curvata (сем. Cryptoniscidae). Из яйца выходит расчленен-
ная личинка. Она прикрепляется к телу различных веслоногих раков
и, питаясь за счет хозяина, превращается в самца. Последний через
некоторое время отрывается, свободно плавая, отыскивает самку,
которую оплодотворяет, и после копуляции вновь переходит к па-
разитическому образу жизни, но уже на совсем другом хозяине.
Самец Danalia становится гиперпаразитом, так как начинает пи-
таться за счет саккулины, в свою очередь паразитирующей на крабе
Inachus. Поселившись в жаберной полости краба, самец Danalia
претерпевает регрессивный метаморфоз. Он линяет, отбрасывает
конечности, утрачивает все следы сегментации и, наконец, превра-
щается в бесформенную мешковидную самку. Последняя проникает
своим хоботком внутрь тела краба и внедряется в разветвленный
столон саккулины (с. 160), за счет которой и питается. По мере соз-
ревания яичника самка Danalia растет и достигает размера 12—
13 мм, становясь таким образом более чем в 10 раз крупнее, чем
она была в то время, когда выполняла функции мужской особи.
Длина тела самца Danalia составляет всего 1 мм.
Основные направления эволюции паразитических ракообразных
Среди паразитических Crustacea отчетливо прослеживается пе-
реход от временного эктопаразитизма к постоянному и, наконец,
к эндопаразитизму. В процессе этого перехода осуществляются ос-
новные тенденции эволюции паразитов, выражающиеся как в ха-
рактере морфологических изменений, так и в особенностях биологий.
Временные эктопаразиты по своему внешнему виду мало чем от-
личаются от свободноживущих ракообразных. Их адаптации к пара-
зитизму выражаются прежде всего в развитии органов прикрепле-
ния, которые позволяют рачку удерживаться па хозяине во время
поглощения пищи (крови, тканей). Это мощные присоски Argulus
(Branchiura), саблевидные антенны Ergasilus (Copepoda), крючксн
166
видные конечности Cymothoa, Aega и других равноногих раков
и т. п. Одновременно изменяется и строение ротового аппарата, ко-
торый превращается, как правило, в колюще-сосущий. Такой рото-
вой аппарат, имеющий вид хоботка, приспособленного к добыванию
крови п соков хозяина, характерен для представителен разных отря-
дов и встречается среди веслоногих (у Caligus), карпоедов (Argu-
lus), мешкогрудых (Synagoga) и равноногих раков (Aega и др.).
Если же указанное преобразование ротового аппарата не происхо-
дит, у паразитов развиваются адаптации к внекишечному перевал
риванню тканей хозяина (например, у Ergasilus).
Для постоянных эктопаразитов характерен резко выраженный
половой диморфизм. Более подвижные и часто очень мелкие сам-
цы, как правило, сохраняют внешний облик ракообразного. Зато
самки, проводящие всю жизнь на теле хозяина, видоизменяются
очень сильно. У них интенсивно развиваются разнообразные орга-
ны прикрепления (см. рис. 74). Рачки становятся неподвижными,
что ведет в свою очередь к рудиментации плавательных ножек, ко-
торые нередко полностью исчезают (например, у многих представи-
телей подотр. Lernaeoida). Одновременно утрачивается расчлене-
ние тела. Это происходит как путем слияния, так и за счет редук-
ции отдельных сегментов и даже тагм. Наибольшие морфологичес-
кие изменения претерпевают эндопаразитические ракообразные.
Они утрачивают сегментацию и конечности, что нередко (у Asco-
thoracida и. Rhizocephala) связано с отбрасыванием целых отделов
тела (с. 161). Наблюдается тенденция к редукции пищеварительной
системы: кишечник приобретает характер слепо замкнутого мешка
(Dendrogaster) или совсем отсутствует Овсе Rhizocephala, Хепосое-
loma и Monstrillidae из веслоногих и др.).
Особый интерес представляет изменение половой системы и со-
отношение полов у паразитических ракообразных. Временным эк-
топаразитам по большей части свойственны раздельнополость
и половой диморфизм. Исключение составляют некоторые Isopoda
(с. 164). У постоянных эктопаразитов появляются карликовые сам-
цы (Bopyrus из равноногих, Lernaeocera, Ascothorax и др.). У эндо-
паразитов карликовые самцы становятся паразитами самок (Den-
drogaster) и в ряде случаев развивается вторичный гермафродитизм
(Petrarca из аскоторацид, все Rhizocephala).
По мере усиления связи с организмом хозяина неизменно воз-
растает половая продуктивность рачков (с. 159). Характерна также
тенденция к эмбрионизации и сокращению числа свободно плава-
ющих личиночных стадий (с. 159).
Анализ морфологии и биологических адаптаций паразитических
ракообразных позволяет прийти к выводу о том, что на протяжении
эволюции Crustacea переход к паразитизму осуществлялся много-
кратно и >в разные геологические эпохи. Это положение, по-видимо-
му, справедливо не только для кл. Crustacea в целом, но и для
отдельных его отрядов. Так, разные представители веслоногих
рачков, очевидно, стали паразитами беспозвоночных гораздо
раньше, чем паразитами рыб, на которых перешли много
позднее и независимо. Об этом можно судить, сравнивая степень
адаптированности к паразитизму двух семейств подотр. Cyclopoida.
Несомненно, очень древними паразитами являются рачки сем.
Monstrillidae (см. рис. 71), паразитирующие в крови полихет. Это
эндопаразиты, утратившие кишечник и приобретшие специальные
адаптации к питанию через поверхность тела. В то же время сем.
Ergasilidae — филогенетически молодые паразиты. Они связаны
с хозяином — рыбой только в период питания, незначительно изме-
нены паразитизмом, очень патогенны для хозяина, и характеризу-
ются довольно широкой специфичностью по отношению к нему.
В пределах подотр. Lernaeoida филогенетически наиболее древними
паразитами, несомненно, следует считать р. Xenocoeloma (см. рис.
75,В). Эти рачки, утратившие какие бы то ни было следы сегмента-
ции и конечностей, паразитируют на полихетах. Они лишены орга-
нов чувств, нервная система у них не обнаружена, половая стала
гермафродитной. Перитонеальный эпителий хозяина обрастает це-
ликом мешковидное тело паразитка и, по некоторым данным, даже
замещает его собственные покровы.
К числу наиболее древних следует отнесли таких паразитов бес-
позвоночных, как мешкогрудые и корнеголовые раки, полностью ут-
ратившие морфологическое сходство с ракообразными (рис. 78—
80). Многие из них стали гермафродитами, а некоторые (Thompso-
nid) вторично приобрели совершенно не свойственную членистоно-
гим способность к вегетативному размножению путем почкования.
По-видимому, полифилетично происхождение паразитических рав-
ноногих раков. Так, паразитирующие на беспозвоночных представи-
тели родов Portunion, Danalia и других по уровню деградации и ха-
рактеру обусловленных паразитизмом изменений морфологии на
первый взгляд кажутся более похожими на мешкогрудых и корне-
головых, чем на других равноногих раков. Это, несомненно, очень
древние паразиты. В то же время Isopoda — временные паразиты ко-
стистых рыб (Cymothoa, Aega и др.) еще очень слабо изменены
паразитизмом и должны рассматриваться как филогенетически
молодые. Время перехода обеих групп, равноногих к паразитизму,
несомненно, различно и, возможно, разделено геологическими эпо-
хами. По мнению акад. А. П. Маркевича (1956), различия в строе-
нии самцов и самок паразитических Copepoda тоже объясняются
разновременностью их перехода к паразитизму. Самцы, сохраняю-
щие анцестральные признаки, по-видимому, стали паразитами по-
зднее, чем самки.
ПОДТИП CHELICERATA —ХЕЛИЦЕРОВЫЕ
Хелицеровые включают около 40 000 видов преимущественно на-
земных членистоногих. Тело их обычно подразделяется на голово-
грудь и брюшко. Головогрудь несет 6 пар конечностей: хелицеры,
педнпальпы и 4 пары ходильных ног. Брюшные конечности, как пра-
вилд, либо совсем отсутствуют, либо рудиментарны и преобразова-
168
ны в самые различные по своему функциональному назначению
органы: легочные мешки, половые крышечки, органы чувств и т. д.
Развитие прямое пли с метаморфозом.
Подтип Chelicerata объединяет 3 класса, из которых паразиты
встречаются лишь в кл. Arachnida.
КЛАСС ARACHNIDA LATR., 1832, — ПАУКООБРАЗНЫЕ
Класс Arachnida — самый многочисленный из всех хелицеровых.
Представлен почти исключительно наземными видами. Морфоло-
гическое разнообразие Arachnida очень велико. Вопрос о количе-
стве отрядов, в которых .они объединяются, еще окончательно не ре-
шен. В современной литературе эта цифра колеблется от 9 др 13.
Причем наибольшее количество противоречивых мнений высказы-
вается относительно «группы» клещей, к которым и принадлежат
все паразитические хелицеровые. Согласно взглядам одних авторов,
клещи подразделяются на три самостоятельных отряда (А. А. Зах-
ваткин, А. Б. Ланге и др.). Многие же исследователи рассматрива-
ют их'как вполне целостную группу — отр. Acari. Эта точка зрения
принимается и нами.
ОТРЯД ACARI LEACH, 1817,— КЛЕЩИ
В настоящее время известно около 10 000 видов клещей, однако
на самом деле их, вероятно, намного больше. Наиболее примитив-
ные представители группы обладают богато расчлененным телом,
тогда как специализированные виды практически полностью утрати-
ли сегментацию (с. 170). Сильным изменениям в процессе эволюции
подвергается и ротовой аппарат клещей —в одних случаях сохра-
няются типичные клешневидные хелицеры, предназначенные для
пережевывания и измельчения пищи, в других же образуется слож-
но устроенный колющий хоботок. Развитие клещей осуществляется
с метаморфозом. Из яйца вылупляется личинка, обладающая всего
тремя парами ходильных ног. За ней следует одна или несколько
фаз (чаще всего их 2), объединяемых под общим названием «ним-
фы». Нимфы несут уже 4 пары конечностей, но не имеют еще функ-
ционирующей половой системы. Последняя развивается лишь
у взрослых особей (имаго). Большинство клещей — свободноживу-
щие организмы, среди которых многие виды стали паразитами как
животных, так и растений.
Изучению клещей уделялось и уделяется большое внимание.
Видное место в современной акарологии принадлежит советским
ученым. Под руководством акад. Е. Н. Павловского проводились
выдающиеся по своему значению исследования по биологии клещей
и их роли в передаче трансмиссивных заболеваний. Большой вклад
в изучение сложных и запутанных вопросов происхождения эволю-
ции и систематики клещей внесли А. А. Захваткин, В. Б. Дубинин,
Б. И. Померанцев, М. В. Поспелова-Штром, Э. Г. Беккер,
В. И. Волгин, Ю. С. Балашов, А. Б. Ланге и многие другие.
169
В настоящее время в пределах отр. Acari выделяется несколько
подотрядов, из которых ниже будут рассмотрены лишь некоторые,
имеющие наиболее важное практическое значение.
ПОДОТРЯД PARASITIFORMES — ПАРАЗИТИФОРМНЫЕ КЛЕЩИ
Паразитиформные клещи объединяют наиболее специализиро-
ванных представителей отряда, практически полностью лишенных
внешних проявлений сегментации. По ряду морфологических и биот
логических особенностей подотр. Parasitiformis разделяется на не-
сколько надсемейств. Среди них по практической значимости выде-
ляются два: Ixodoidea, включающее исключительно паразитические
формы, к которым относятся наиболее важные переносчики возбу-
дителей ряда заболеваний (с. 185), и Gamasoidea — свободножи-
вущие многоядные и хищные виды, а также паразиты.
Надсемейство Ixodoidea — иксодоидные клещи
Надсем. Ixodoidea представлено облигатными гематофагами,
паразитирующими на позвоночных животных: рептилиях, птицах
и млекопитающих. Они объединяются в два семейства: сем. Arga-
sidae Can., 1890, — аргазовые клещи и сем. Ixodidae — иксодовые
клещи/
Морфология
Иксодоидные клещи — наиболее крупные представители отр.
Acari. Размеры взрослых особей в зависимости от степени насыще-
ния могут варьировать от нескольких миллиметров до 2—3 см.
Голодные клещи имеют, как правило, сильно уплощенное овальное
тело, сытые же резко увеличиваются в объеме и приобретают
яйцевидную или каплевидную форму.
Тело разделено на два хорошо обособленных отдела: небольшую
головку — гнатосому и собственно туловище — массивную идиосому
(рис. 84, 85). Это расчленение возникло вторично и не соответству-
ет границам между сегментами. В состав гнатосомы входят лишь
часть акрона и первые два сегмента, несущие хелицеры и педипаль-
пы. Идиосома образована остальными сегментами тела и тем участ-
ком акрона, на котором развиваются глаза.
Иксодовые и аргазовые клещи заметно отличаются друг от дру-
га по характеру соединения гнатосомы и идиосомы. У первых —
головка прикреплена к туловищу терминально и направлена впе-
ред, у вторых — она располагается субтерминально и направлена
вниз. Спереди и с боков ее окружают три массивных выроста: клюв
и две щеки (рис. 85,В).
Гиатосома Ixodoidea состоит из двух частей — расширенного ос-
нования и вытянутого хоботка. По бокам от последнего располага-
ются членистые пальпы.
Пальпы Ixodoidea не полностью отвечают педипальпам остальных паукооб-
разных. Основной коксальпый членик пальп клещей срастается со склеритами
170
хелнцерального и педипальпального сегментов и принимает участие в образовании
основания гнатосомы. Таким образом, членистые придатки головки — пальпы —
соответствуют лишь концевым участкам педипальп.
Хоботок представляет собой сложное образование, состоящее из
нескольких функционально связанных друг с другом частей. Снизу,
вентрально, располагается длинный уплощенный вырост ,основа-
Рнс. 84. Схема строения иксодовых клещей (по Померанцеву). Л —самка, вид
со спинной стороны; Б — самка, вид с брюшной стороны; В— самец, вид со спин-*
нон стороны; Г — самец, вид со спинной стороны; Д—гиатосома (по Оленеву);
Е— строение хелицеры; Ж— продольный разрез через гнатосому (по Балашову):
/ — гиатосома, 2 —идиосома, 3 — скутум, 4 — аллоскутум, 5 — борозды На идиосоме, 6 —
коксы конечностей. 7 — половое отверстие, 8 — анальное отверстие, 9 — перитремы, 10 —
щитки на брюшной поверхности самца, // — пальпы, /2—гипостом, 13 — хелицеры, 14 —
футляры хелицер. 15 — основание гнатосомы, 16 — поровые поля, /7 — основной членик
хелицеры. 18— подвижный палец, 19 — предротовая полость, 20— протоки слюнных желез,
21 — мышцы, 22— глотка, 23 — ротовое отверстие, 24 — орган Галлера
нпя — гипостом, как правило, несущий направленные назад остры'е
зубцы. Сверху, дорзально, к гипостому прилегают еще два выроста,
имеющих трубчатое строение, — футляры хелицер. Внутри футля-
ров располагаются сами хелицеры. Между гипостомом и футляра-
ми образуется просвет — предротовая полость (рис. 84,Ж). На дне
ее в проксимальной части хоботка располагается ротовое отверстие
и открываются протоки слюнных желез. Хелицеры Ixodoidea утра-
тили клешневидное строение, характерное для других паукообраз-
ных. Они состоят из двух члеников: стержневидного основного чле-
171
ника и подвижного пальца, который по своему наружному краю не-
сет мощные режущие зубцы. Весь этот комплекс органов, образую-
щих хоботок, служит для нарушения целостности покровов живот-
ного-хозяина и поглощения крови (с. 174).
Пальпы Ixodoidea четырехчленисты. На них располагаются мно-
гочисленные чувствительные щетинки (с. 173).
Идиосома иксодовых и аргазовых клещей заметно различается.
У самцов и самок Argasidae идиосома имеет яйцевидную или не-
правильно овальную с заостр.енным передним концом форму (см.
Рис. 85. Схема строения аргазовых клещей (по Балашову). А — вид со спинной
стороны; Б — вид с брюшной стороны; В— строение переднего конца тела, вид
с брюшной стороны:
1 — идиосома, 2 — гнатосома, 3 — борозды на идиосоме, 4 — диски, 5 — клюв, 6 — щеки,
7 — коксы конечностей, 8 — отверстие коксальной железы, 9 — половое отверстие, 10 — ды-
хательное отверстие (стигма), // — анальное отверстие, 12 — гипостом, 13 — хелицера, 14 —
пальпа, 15 — стебель органа Жене, 16 — рог органа Жене
рис. 89, В, Г). Снаружи идиосома одета эластичной и растяжимой
кутикулой. Массивные, занимающие большую площадь склериты
отсутствуют. На дорзальной поверхности располагаются маленькие
склеротинизированные пластинки — диски, образующие иногда весь-
ма характерный рисунок (рис. 85,А). Они служат для прикрепле-
ния мышц. Дорзальная и вентральная поверхности идиосомы несут
систему борозд, особенно отчетливо выраженных у голодных осо-
бей. Форма и расположение борозд постоянны. Отдельные борозды
так же, как и диски, являются местом прикрепления мышц к стенке
тела. Большая же их часть, расправляясь, обеспечивает увеличение
объема тела клешей на первых этапах питания (с. 178).
Кутикула идиосомы иксодовых клещей склеротинизирована зна-
чительно сильнее. У представителей этого семейства наблюдается
отчетливо выраженный половой диморфизм. У самок большая часть
поверхности идиосомы одета растяжимой кутикулой, называемой
аллоскутумом. Лишь на переднем конце, на границе с гнатосомой,
дорзально располагается небольшой склеротпнизированный щи-
172
ток — скутум (см. рис. 84, Л). Кутикула идиосомы самцов, которые
значительно меньше самок, несет крупные склериты. Один такой
щиток покрывает практически всю дорзальную поверхность, остав-
ляя лишь узкую свободную полоску мягкой кутикулы по краям
тела. Вентрально располагается целая система щитков (см. рис.
84, В, Г).
Идиосома клещей несет 4 пары ходильных ног. Каждая конеч-
ность состоит из 6 члеников. Основной членик — кокса обычно при-
растает к идиосоме, но сохраняет при этом ограниченную подвиж-
ность. Лапка несет два коготка и нежную перепончатую присосоч-
ку.
На вентральной поверхности идиосомы на уровне основания вто-
рой пары конечностей локализуется половое отверстие, имеющее
вид поперечной щели. У самцов оно прикрыто небольшой кутику-
лярной пластинкой. Ближе к заднему концу лежит анальное отвер-
стие. Оно всегда снабжено системой склеритов, состоящей из плот-
ного кутикулярного кольца и двух подвижных полукруглых створок.
Во время дефекации створки раздвигаются; из образовавшейся ще-
ли выделяется капелька жидких экскрементов, которые на воздухе
почти моментально затвердевают. Створки усилием специальных
мышц сдвигаются обратно и своими острыми краями срезают обра-
зовавшийся шарик. Позади основания последней пары конечностей
по бокам.тела на особых кутикулярных пластинках — перитремах—
лежат отверстия трахейной системы — стигмы.
У представителей сем. Argasidae на вентральной поверхности
между основаниями первой и второй пар конечностей открываются
протоки коксальных желез. Последние выполняют функцию орга-
нов осморегуляции. Выделительная же функция полностью перешла
к мальпигиевым сосудам. У Ixodidae коксальные железы отсутству-
ют.
На поверхности тела клещей располагаются многочисленные ор-
ганы чувств. По строению и функциональным особенностям они
весьма разнообразны. Наиболее многочисленны осязательные волос-
ки, или хеты, подвижно сочлененные с кутикулой. К их основаниям
подходят отростки чувствительных нервных клеток. Среди обычных
довольно длинных волосков на теле клещей встречаются и более ко-
роткие. Им приписывается терморецепторная функция. Разнообраз-
ны у клещей хеморецепторы. Контактная хеморецепция — восприя-
тие вкусовых раздражителей — осуществляется, по-видимому, спе-
циальными тонкостенными хетами, расположенными на дистальном
членике пальп, а также на лапках первой пары ног. Органами обо-
няния (дистантная хеморецепция) служат органы Галлера, локали-
зующиеся на дорзальной поверхности лапок передней пары ног. Ор-
ган Галлера представляет собой глубокое впячивание кутикулы,
дно которого усажено специальными щетинками.
Органы зрения клещей устроены по типу простых глазков. Их
всего два. У Ixodidae они располагаются по краям скутума на иди-
осоме. У ряда видов (р. Ixodes и др.), а также у большинства
Argasidae глаза отсутствуют.
173
Иксодовым клещам свойственны еще и так называемые желе-
зисто-чувствительные органы, в состав которых, кроме чувствитель-
ных, входят и железистые клетки. Они располагаются поодиночке
пли же небольшими группами по две-три сенсиллы в каждой, но на
отдельных участках тела они образуют большие скопления. Таковы,
например, поровые поля на дорзальной поверхности основания гна-
тосомы (см. рис. 84, Д) и так называемые «фовеа» или дорзальные
ямки на аллоскутуме многих иксоднд. Единого мнения о функцио-
нальном назначении этих сенсилл в литературе нет.
Питание клещей и адаптации к кровососанию
Все иксодоидные клещи, как уже говорилось выше, — облигат-
ные гематофаги. Питание кровью — необходимое условие, без кото-
рого невозможно нормальное развитие клещей и их размножение.
У Ixodidae, как и у многих других кровососов, отчетливо выражены
гонотрофический цикл и гонотрофическая гармония (с. 179).
В большинстве случаев иксодоидные клещи на каждой фазе сво-
его развития питаются всего один раз (исключение составляют
взрослые клещи сем. Argasidae, с. 181), поглощая при этом значи-
тельную порцию крови. Перерывы между двумя приемами пищи
могут быть очень значительными (от нескольких недель до несколь-
ких лет). Все это невозможно без существования специальных очень
совершенных морфологических и биологических адаптации. Пита-
ние нксодоидных клещей — сложный процесс, сопровождающийся
глубокими морфологическими и физиологическими изменениями,
затрагивающими не только органы пищеварительной системы, но
фактически и весь организм паразита.
Поведение клещей во время питания и сами механизмы погло-
щения пищи несколько отличаются у представителей сем. Argasidae
и Ixodidae. Попав на тело хозяина*, аргазовые клещи очень быстро
закрепляются лапками на поверхности кожи и выдвигают вперед
хелицеры, подвижные пальцы которых совершают режущие движе-
ния? В образующийся надрез постепенно вводится весь хоботок.
Фиксация паразита на теле хозяина обеспечивается направленными
назад зубцами, расположенными по краю гипостома, и пальцами
хелицер, которые в глубине ранки клещ разводит в стороны, напо-
добие якорных лап. За счет повреждения капилляров, а иногда и бо-
лее крупных сосудов кожи в месте укуса возникает небольшая ге-
матома, из которой клещи и высасывают кровь. Главную роль в
этом процессе, по-видимому, играет мощная мускулистая глотка,
выполняющая функции глоточного насоса. Поглощение крови пе-
ремежается короткими перерывами, во время которых в ранку
стекает слюна, продуцируемая крупными сложно устроенными
слюнными железами (с. 176). Смешивание крови со слюной осу-
ществляется также непосредственно в предротовой полости.
Продолжительность сроков питания аргазовых клещей сравни-
тельно невелика. От нападения голодной.особи на хозяина до пол-
ного насыщения проходит обычно от нескольких минут до 2—3 ч.
174
Совещпенно иначе ведут себя иксодовые клещи. Как правило,
для них характерно многодневное питание, растягивающееся ино-
гда на 15—20 дней. В связи с этим меняется способ прикрепления
клещей- к телу хозяина, а главное, характер поглощаемой ими пи-
щи. Отличия проявляются уже в процессе внедрения хоботка в
кожу. После появления первого разреза, сделанного пальцами хе-
лицер, из хоботка начинает выделяться особая слюна, не обладаю-
щая пищеварительной активностью. Она быстро застывает вокруг
хоботка в виде плотного чехла. В результате разведенные в сторо-
ны пальцы хелицер, футляры хелицер и гипостом по всей своей
длине оказываются впаянными в этот «цементный футляр» (рис.
86, Л). Таким образом, достигается надежное прикрепление пара-
зита.
В месте внедрения хоботка Ixodidae так же, как и у аргазовых
клещей, наблюдается разрушение мелких кровеносных сосудов.
Однако этим дело не ограничивается. Вокруг конца хоботка доволь-
но быстро формируется значительный по размерам некротический
очаг. Его возникновение, по-видимому, обусловлено выделением в
ранку новых порций слюны. Последняя по своим химическим и фи-
зическим свойствам резко отличается от того клейкого секрета,
который использовался клещом для создания «цементного футля-
ра». Это настоящая активная пищеварительная слюна, обладающая
гистолитическими и гемолитическими свойствами и содержащая в
большом количестве антикоагулянты — вещества, препятствующие
свертыванию крови. В зоне некроза образуется внутритканевая по-
лость, заполненная экссудатом, состоящим из лимфы, большего
числа лейкоцитов, клеток лизированных тканей и крови. В первые
дин ее, как правило, не очень-много. В дальнейшем, однако, по мере
разрушения все новых и новых сосудов поступление крови в очаг
некроза резко усиливается, „и она начинает доминировать в пище-
вой массе, поглощаемой клещом.
Количество всасываемой клещами крови очень велико. Вес на-
питавшихся клещей более чем^в десять раз может превышать вес
голодных особей. Однако эта разница не отражает истинного коли-
чества проглоченной крови. Часть ее усваивается клещом еще во
время питания, часть же выделяется при дефекации. Кроме того,
значительное количество воды и солей, содержащихся в крови,
очень быстро удаляется обморегуляторными органами (с. 176).
Основным вместилищем поглощаемой пищи у клещей служит
передний отдел эктодермальной средней кишки, подразделяющийся
на центральный участок — желудок и отходящие от него боковые
выросты — дивертикулы (рис. 86,5). С поступлением крови в прос-
вет средней кишки последняя сильно растягивается, так что-ее стен-
ки приобретают вид тонкой клеточной мембраны. У представителей
обоих семейств клещей предварительные процессы начинаются
с удаления избыточной воды и солей, содержащихся в плазме кро-
ви. У аргазид для этого используются коксальные железы. Еще
в процессе питания и сразу же после его окончания из отверстий
коксальных желез (с. 172, рис. 85, 5) выделяется большое количест-
175
во прозрачной жидкости. Несколько иной механизм удаления избы-
точной воды наблюдается у иксодовых клещей. Для этой цели у них
служат слюнные железы. Последние образованы альвеолами двух
основных типов: переднюю часть железы составляют так называ-
Рис. 86. Питание иксодоидпых клещей (по Балашову). А — хоботок клеща, погру-
женный в покровы животного-хозяина; Б — расчленение средней кишки иксодо-
вых клещей; В— участок среза через полностью напитавшуюся нимфу (диверти-
кулы кишечника заполнены кровью); Г — участок среза через линяющую лимфу:
/ — основание гнатосомы, 2 —пальца, 3 — цементный футляр, 4 — футляры хелицер, 5 —
хелицеры, 6 — гипостом. 7 — внутритканевая полость с воспалительным инфильтратом, 8 —
кровеносные сосуды, 9 — желудок, 10 — дивертикулы. 11 — стенка тела, 12 — кутикула, от-
слаивающаяся при линьке
емые пирамидальные альвеолы, заднюю же — настоящие секретор-
ные. Первые и являются органами осморегуляции. Выделяемая ими
жидкость, содержащая большое количество солей, не обладает пи-
щеварительной активностью. Эта жидкость смешивается с настоя-
щей слюной, продуцируемой секреторными альвеолами, и поступает
в ткани хозяина.
176
Последующие этапы пищеварения также различны у аргазовых
п иксодовых клещей. Это во многом предопределяется характером
поглощаемой ими пищи. У Argasidae, быстро засасывающих почти
чистую цельную кровь, в просвете кишечника эритроциты подвер-
гаются гемолизу, в результате чего количество их постепенно умень-
шается. Клетки, еще не захваченные этим процессом, сохраняют все
свои свойства. Отдельные эритроциты можно обнаружить в содер-
жимом кишечника и через 20—25 дней после завершения питания.
У иксодовых клещей гемолиз эритроцитов осуществляется еще
в воспалительном очаге под действием слюны. Лишь на последних
этапах питания, когда количество поглощаемой крови возрастает
в содержимом кишечника клеща, можно обнаружить в течение 1—2
дней остатки эритроцитов.
Основу пищи клещей составляет поступающий с кровью гемо-
глобин. Его окончательное переваривание протекает преимуществен-
но внутриклеточно. Эти процессы сопровождаются морфологически-
ми и функциональными преобразованиями эпителиальных кишеч-
ных клеток. У голодных особей стенки желудка и дивертикулов
образованы преимущественно так называемыми резервными — не-
дифференцированными — клетками. Почти одновременно с началом
сосания клещом крови эти клетки начинают интенсивно размно-
жаться. Часть их превращается в крупные секреторные клетки, сек-
рет которых выделяется в просвет кишечника и, по-видимому, обес-
печивает цитолиз и начальные этапы переваривания. Другая же
часть дифференцируется в пищеварительные клетки. Последние не-
сут небольшое количество микроворсинок, не образующих настоя-
щей щеточной каймы. Это, по-видимому, связано с отсутствием у
клещей пристеночного пищеварения.
Пищеварительные клетки аргазовых клещей до завершения ге-
молиза способны заглатывать путем фагоцитоза целые эритроци-
ты. Однако главная роль, несомненно, принадлежит процессам пи-
ноцитоза, которые обеспечивают поглощение гемоглобина. При этом
образуются характерные трубчатые впячивания, по которым гемог-
лобин поступает в пиноцитозные вакуоли. Сама средняя кишка
представляет собой резервуар, в котором гемолизированная кровь
сохраняется очень долго. Обычно интенсивное усвоение основной
массы гемоглобина продолжается от нескольких недель до нес-
кольких месяцев. Остатки же полупереваренной крови использу-
ются голодными особями весьма экономно, что позволяет им выжи-
вать в течение длительного промежутка времени.
У иксодовых клещей часть пищеварительных клеток, как и у
арагазид, способна и к пиноцитозу, и к фагоцитозу. Они обеспе-
чивают усвоение пищевых веществ в начальный период питания.
Остальные же клетки в это время уже только путем ппйоцитоза ин-
тенсивно поглощают гемоглобин. В результате за короткий про-
межуток времени они настолько сильно увеличиваются в размерах,
что в средней кишке почти полностью исчезает просвет, в котором
к этому времени практически не остается никакого содержимого
(рис. 86, В, Г). Как и у многих других питающихся кровью орга-
177
низмовл гемоглобин усваивается не полностью. Используется лишь
глобиновая часть молекулы, гематин перевариванию нс подверга-
ется.
С процессом питания у клещей теснейшим образом связаны про-
цессы преобразования покровов, носящие отчетливо выраженный
адаптивный характер. Первое, что бросается в глаза, это слабая
склеротинизация кутикулы иксодоидных клещей. Исключение сос-
Рис. 87. Изменения кутикулы иксодовых клещей в процессе питания
(по Балашову). А — кутикула голодной самки; Б — кутикула самки
в середине питания; В — кутикула самки, окончившей питание:
/ — эпикутикула, образующая узкие складки, 2 — внутренние слои кутикулы,
3 — гиподерма, 4 — кожные железы
тавляют лишь самцы сем. Ixod'idae, многие из которых вообще не
питаются или же являются факультативными кровососами (виды
р. Ixodes).
Увеличение размеров тела во время питания клещей прежде все-
го достигается за счет расправления борозд на поверхности иди-
осомы. Дальнейший рост объема связан с изменениями, протекаю-
щими в самой кутикуле. Последняя состоит из нескольких слоев,
отличающихся по своим механическим свойствам. Самый наруж-
ный очень тонкий слой — эпикутикула характеризуется большой
гибкостью, но почти не способен к растяжению. Обычно у голодных
особей он образует той или иной формы гребни и складки
(рис/87,Л). Подстилающие его внутренние слои кутикулы, образу-
ющие основную массу покровов, наоборот, обладают хорошо вы-
раженной растяжимостью. У большинства аргазовых клещей, ко-
торым свойственно сравнительно быстрое насасывание крови, склад-
ки эпикутикулы расправляются, в результате чего поверхность тела
становится почти гладкой. Остальные же слои при этом сильно
растягиваются.
У иксодовых клещей и личинок некоторых видов арагазид эти
процессы усложняются за счет межлиночного роста кутикулы —
178
явления, исключительно редко встречающегося у других членистоно-
гих. В процессе питания эпикутикулярпые складки у них расправля-
ются так же, как и у взрослых аргазовых клещей. Внутренние же
слои кутикулы при этом растягиваются, однако вместо ожидаемого
истончения покровов наблюдается быстрое увеличение их толщины
(рис. 87,5). Это обусловливается нарастанием кутикулы за счет
интенсификации секреторной деятельности гиподермы. Лишь в са-
мый последний период питания рост кутикулы замедляется. На
этом этапе увеличение объема тела паразита обеспечивается -уже
главным образом за счет растяжения покровов, толщина которых
заметно уменьшается (рис. 87, В). Возникновение в процессе эво-
люции подобных механизмов и превратило иксодовых клещей в од-
них из самых совершенных и специализированных гематофагов.
Размножение и цикл развития клещей
Размножение клещей находится в тесной зависимости от пита-
ния кровью. Как и для многих других гематофагов, для IxodoWea
характерно наличие гонотрофического цикла и гонотрофической
гармонии. Последнее означает полную согласованность и зависи-
мость процесса созревания яиц от процессов переваривания и ус-
воения крови. Поглощение самками определенной порции крови
обеспечивает формирование, развитие и откладку одной порции
яиц. Новые яйцекладки возможны только после' повторного пита-
ния. Период, протекающий от одного акта кровососания до следую-
щего, между которыми и осуществляется размножение клещей, по-
лучил название гонотрофического цикла. Самки сем. Argasidae за
свою жизнь проходят несколько гонотрофических циклов (до 4—6).
Самки иксодовых клещей обладают только одним гонотрофическим
циклом и после первой и единственной откладки яиц погибают.
У самцов подобная зависимость созревания половых продуктов-от
питания выражена далеко не всегда (с. 178).
Аргазовые клещи, как правило, копулируют во внешней среде;
у иксодовых же этот процесс в большинстве случаев протекает па
животном-хозяине. Во время копуляции самец прикрепляется к
вентральной поверхности самки (рис. 88, А). Затем он вводит хо-
боток в половое отверстие последней и медленными движениями
постепенно расширяет его. Покончив с этим, самец подгибает гна-
тосому на вентральную поверхность и хоботком подхватывает выс-
тупающий из своего полового отверстия сперматофор. Сделав на
нем надрез, самец вводит сперматофор в половое отверстие самки.
Осемененные и насытившиеся самки, у которых произошло пол-
ное созревание половых продуктов, приступают к откладке яиц.
Для этого они выбирают затененные и хорошо защищенные места
с достаточно высокой влажностью: лесная подстилка, трещины в
почве, норы грызунов и т. п. Важную роль в процессе яйцекладки
играет так называемый орган Жене (рис. 88, В). Это очень харак-
терное образование, представляющее глубокое впячивание покро-
вов, расположенное дорзально на границе между гнато- и идиосо-
179
мой. Обычно орган Жене состоит из уплощенного мешковидного ство-
ла, па конце которого располагаются отростки — «рога». Во время
откладки яиц, благодаря повышению внутреннего давления гемолим-
фы, орган Жене выворачивается наружу и нависает над гнатосомой,
которая подгибается па брюшную сторону. Рога, располагаясь по
бокам хоботка, приближаются к половому отверстию и подхваты-
вают выступающее из него яйцо. Последнее покрывается защитным
слоем секрета специальных желез, расположенных па концах «ро-
гов».
8
Рис. 88. Цикл развития клещей. А—копуляция клещей Ixodes ricinus (по Бала-
шову); Б — личинка Dermacenior nuttail i; В — нимфа Dennacentor nuttalli
(по Павловскому)
Плодовитость иксодовых и аргазовых клещей очень различна.
Первые, как правило, откладывают от нескольких тысяч (р. Ixo-
des— 1500—2500 яиц; р. Rhipicephalus — 3000—4000) до 17 тыс.
(р. Hyalomma Koch, 1844). Самки же аргазовых клещей единовре-
менно откладывают от 50 до 200 яиц, но так как они проходят не-
сколько гонотрофических циклов, то за свою жизнь они продуци-
руют 1000—1500 яиц. Плодовитость самок определяется количест-
вом выпитой ими крови. Полностью насытившиеся самки отклады-
вают максимальное число яиц; чем меньше была порция, тем мень-
ше развивается яйцеклеток в яичнике.
Как уже отмечалось выше, стадии развития иксодоидиых кле-
щей представлены личинками и нимфами. Вылупляющиеся из яиц
личинки (рис. 88, Б) имеют не четыре, а только три пары ног. На-
ружное половое отверстие у них отсутствует. Личинки многих ви-
дов лишены трахейной дыхательной системы. Борозды на поверх-
ности идиосомы выражены у личинок не столь отчетливо, как у по-
ловозрелых клещей. Более просто устроен и рецепторный аппарат.
Личинки аргазовых клещей! отличаются от имаго еще и тем, что
гнатосома у них занимает терминальное, а не субтерминальное по-
ложение на идиосоме.
Следующая стадия развития — нимфа (рис. 88, В). После пер-
вой линьки молодые клещи приобретают уже 4 пары ног. Борозды
ца идиосоме выражены отчетливее. Развивается трахейная система
и появляются перитремы. У нимф Argasidae гнатосома, за редкими
180
исключениями, смещается на вентральную поверхность. Хотя нимфы
и крупнее личинок, они по своим размерам сильно уступают взрос-
лым особям. Кроме того, от последних они отличаются еще и отсут-
ствием полового отверстия.
Циклы развития иксодовых н аргазовых клещей различаются по
числу составляющих их фаз. у Argasidae число ннмфальных стадий
увеличивается. Их количество, однако, непостоянно даже в пределах,
одного вида. Развитие видов р. Argas осуществляется обычно с 2—4
нимфальными фазами, Ornithodorus — с 3—6 и т. д. Личинке и каж-
дой нимфальной стадии требуется для нормального развития и
линьки одноразовое питание. Личинки и нимфы многих видов спо-
собны к линьке даже при поглощении неполной порции крови. Од-
нако это приводит к тому, что последующие стадии развития
уменьшаются в размерах, а число их возрастает. Некоторыми ис-
следователями подобная пластичность развития аргазовых клещей
рассматривается как адаптация, позволяющая избежать появления
в популяции карликовых половозрелых особей.
Развитие Ixodidae значительно более стабилизировано. У них
произошло сокращение нимфальных фаз до одной. Из нимфы, сме-
нившей личинку, всегда сразу же развивается имаго.
Питание кровью личинок, нимф и имаго на позвоночных
животных обусловливает чередование в цикле развития клещей пе-
риодов свободного существования во внешней среде и периодов па-
разитирования на хозяине. По числу таких «переходов» от одного,
образа жизни к другому иксодоидные клещи заметно различаются.
Для большинства Argasidae характерен многохозяинный тип цикла
развития. Вылупившаяся из яйца личинка нападает на позвоночное
животное и питается кровью.' Насосавшись, она покидает его, и
последующее развитие, в том числе и линька, протекают во внеш-
ней среде. Сходным образом ведет себя первая и все последующие
нимфы. Соответственно число животных, на которых питаются кле-
щи по ходу своего цикла развития, может быть довольно значитель-
ным. Это усугубляется еше и способностью самок Argasidae про-
ходить несколько гонотрофических циклов. Их питание каждый раз.
осуществляется на новом животном-хозяине.
У иксодовых клещей цикл развития может протекать сходным
образом, однако максимальное число используемых хозяев не пре-
вышает трех. Такие циклы называются треххозяинными ’.«и харак-
терны они для целого ряда родов: Ixodes, Dermacentor Koch, 1844
и др. Как правило, личинки и нимфы нападают на мелких позво-
ночных— ящериц, грызунов, насекомоядных, птиц; половозрелые
же клещи питаются на более крупных животных — домашние и ди-
кие копытные, собаки, лисы, зайцы.
Сокращение числа хозяев приводит к возникновению двуххозя-
инных циклов (р. Rhipicephalus). В этом случае напитавшаяся ли-
чинка не покидает хозяина, на котором проходит все необходимое
1 Схемы типа циклов клещей приводятся в книге «Частная паразитология»,
т. I (рис. 91).
181
развитие паразита и линька. Освободившаяся из личиночного эк-
зувия нимфа сразу же приступает к кровососанию. Однако после
завершения питания она уходит во внешнюю среду, где и линяет
на имаго. Половозрелые клещи питаются уже на новых особях хо-
зяев. При двуххозяинном развитии и неполовозрелые фазы, и имаго
обычно паразитируют на одном или близких видах позвоночных
животных. Дальнейшая эволюция в этом- направлении привела к
возникновению клещей с однохозяинным типом развития (р. Воор-
hilus). Все три фазы — личинка, нимфа и имаго паразитируют на
одном животном-хозяине. Разделяющие их линьки осуществляются
здесь же. Во внешней среде протекает лишь откладка яиц напитав-
шимися самками, развитие яиц и начальный период существования
голодных личинок.
Переход от многохозяинных циклов через ряд последовательных
этапов к однохозяинным, по-видимому, совершался в процессе эво-
люции в связи с изменениями биологии клещей. Аргазовые клеши
в основном относятся к экологической группе так называемых гнез-
дово-норовых паразитов. Чаще всего они обитают в заселенных но-
рах и гнездах позвоночных животных (грызунов, птиц и т. д.) и
практически не покидают их. В ограниченном пространстве норы
возможность встречи паразита и хозяина достаточно высока. В та-
ких условиях отпадает необходимость в длительном пребывании
клещей на животных-прокормителях. В силу этого у Argas'idae от-
сутствуют механизмы, обеспечивающие надежное прикрепление к
телу хозяина (образование цементного футляра вокруг хоботкд
(с. 176) и позволяющие поглощать во время питания очень боль-
шие порции крови (межличиночный рост кутикулы, с 178).
Иксодовые клещи в процессе своей эволюции превратились в
пастбищно-подстерегаюшнх паразитов. Они, как правило, поселяют-
ся в подстилке и на поверхности почвы, что резко снижает вероят-
ность их встречи с потенциальными хозяевами. Сохранение много-
хезяинного цикла с многократным питанием на животных становит-
ся биологически невыгодным. Ixodidae утрачивают «лишние»
нимфальные фазы. Их развитие становится треххозяинным. Это,
по-видимому, было сопряжено с появлением адаптаций, обеспечива-
ющих поглощение очень большого количества пищи на каждой фазе
развития и увеличением сроков пребывания, на хозяевах (с. 176).
Однако и треххозяинный цикл мог сохраниться только в таких ус-
ловиях, когда возможности нападения на позвоночных животных
оставались достаточно большими: паразитирование на хозяевах,
обитающих в подстилке, — различные мелкие грызуны, вторичное
возвращение к гнездово-норовому типу паразитизма и т. д. Освое-
ние в качестве хозяев крупных подвижных животных, перемещаю-
щихся на большие расстояния, и особенно копытных, обитающих
на открытых пространствах (степи, полупустыни и т. д.), неизбеж-
но привело к появлению сначала двух, а потом и однохозяинных ви-
дов клещей.
Обнаружение животного-хозяина и нападение на него обеспечи-
вается наличием у клещей целого ряда поведенческих адаптаций.
182
Гнездово-норовые паразиты^ по-видимому, обладают наиболее про-
стым поведением. Перемещаясь в небольшом пространстве поры
или гнезда, они легко находят хозяина*? Совершать длительные
миграции они не могут. Лишь в отдельных случаях в ночное время-
они покидают нору и нападают на людей и животных, находящихся
поблизости. Расселение вида осуществляется обычно с помощью
животных, несущих на себе питающихся клещей.
Подавляющее большинство иксодовых клещей пассивно подсте-1
регает своих хозяев. Клеши, однако* концентрируются в таких точ-
ках пространства, где вероятность встречи оказывается наиболее вы-
сокой. Так, специальные наблюдения показали, что, не будучи в-
состоянии совершать большие миграции, они тем не менее в лесных
массивах собираются вдоль троп, по которым передвигаются живот-
ные. В ожидании хозяина клещи располагаются не на почве, а па
растительности, поднимаясь иногда на высоту до 1 м. При этом они
сидят на концах стеблей и ветвей. При приближении хозяина клещ,
принимает позу активного ожидания. Он вытягивает вперед перед-
нюю пару конечностей, богато снабженную органами чувств. У не-
которых обитателей открытых пространств—пустыни и полупус-
тыни (Нyalomma asiaticum) — пассивное ожидание животного-про-
кормителя сопровождается его активным преследованием. Клеши
обычно неподвижно сидя'Гна поверхности почвы. Иногда они совер-
шают поисковые движения в разных направлениях. При обнаруже-
нии хозяина они быстро устремляются в его сторону и могут актив-
но преследовать на расстоянии в несколько десятков метров.
Для ориентации во внешней среде клещи используют органы
чувств (с. 173). Большую роль, очевидно, играют специфические за-
пахи. животных-хозяев. Экспериментальные наблюдения показали,
что клещи реагируют на изменения концентрации углекислоты.
Источники СО2, а такими в природе чаше всего бывают живые ор-
ганизмы, привлекают клещей. По-видимому, они улавливают и»
изменения температуры, колебания субстрата, движение воздуха
и т. д. Некоторые виды обладают зрением.
На активность клещей большое влияние оказывают факторы
внешней среды. Они совершают вертикальные миграции на расти-
тельность только при достаточной влажности воздуха. Уменьшение
последней заставляет клещей концентрироваться в лесной подстил-
ке. Аналогичное действие оказывает температура. Так, широко-
распространенный вид Ixodes persulcatus проявляет наибольшую
активность при температуре 5—15° С и влажности воздуха 55—
80%. Отклонение этих показателей от оптимальных величин в ту
или иную сторону вызывает уход клещей в подстилку.
Внешние факторы, от которых во многом зависит активность
клещей, тем самым влияют и.на возможность их нападения на жи-
вотных-хозяев. А от такого условия, как нормальное питание, в
свою очередь зависит и продолжительность цикла развития парази-
тов. Выше уже отмечалась способность клещей к длительному голо-
данию. Это своеобразная адаптация, которая особенно важна для
видов емного- и треххозяинным типом развития, позволяет и личин-
183
кам, и нимфам, и имаго долгое время сохранять жизнеспособность
в ожидании животного-хозяина. По данным Е. Н. Павловского и
А. Н. Скрынник, голодные особи Ornithodorus papillipes могут выжи-
вать в течение 11 лет. Соответственно и цикл развития клещей (от
яйца до яйца) может растягиваться на очень большой юрок. В ус-
ловиях Московской обл. продолжительность каждой фазы цикла со-
бачьего клеща Ixodes ricinus составляет около года, так что весь
цикл занимает не менее 3 лет (немногим более 1000 дней). Мак-
симальные же сроки развития, известные для этого вида, составля-
ют свыше 3 тыс. дней. С другой стороны, в эксперименте при опти-
мальных условиях весь цикл завершается в течение 5 месяцев
(154 дня).
Клещи — эктопаразиты позвоночных животных
В большинстве случаев клещи надсем. Ixodoidea не проявляют
-строгой специфичности к определенным животным-хозяевам. Обыч-
но один и тот же вид клещей способен паразитировать на целой груп-
пе близких видов позвоночных животных. Упоминавшиеся же выше
Ixodes, Dermacentor и некоторые другие нападают на десятки са-
мых разнообразных организмов, включая и холоднокровных. Более
того, у некоторых видов аргазовых и иксодовых клещей довольно ши-
рокое распространение получило явление каннибализма. Голодные
особи могут питаться на сытых клешах того же самого или другого
вида. Они прокалывают хоботком. стенку тела и поглощают ге-
молимфу и содержимое кишечника последних. Гибели клещей-до-
норов при этом не наблюдается.
Способность нападать на разных животных-хозяев не означает,
•фщако, полного отсутствия пищевой специализации. Питание на
неспецифичных хозяевах часто приводит к серьезным нарушениям
в процессе последующего развития паразитов, в частности к сниже-
нию жизнеспособности потомства, уменьшению его размеров и т. д.
Влияние клещей на организм хозяина определяется действием
нескольких факторов. Выделяемая в ранку слюна вызывает мест-
ную кожную реакцию. Массовые нападения клещей и соответствен-
но попадание большого количества слюны в организм хозяина мо-
гут быть причиной лихорадочных явлений и нервных расстройств.
Аргазовыс клещи Alveonasus lahorertsis вызывают клещевые пара-
личи, а иногда и гибель молодых овец. Сходным действием на до-
машнюю птицу обладают укусы Argas persicus. Высокая интенсив-
ность питания клещей приводит иногда к значительным потерям
крови позвоночными животными. По подсчетам Ю. С. Балашова,
100 самок Dermacentor pectus поглощают о^оло 400 мл крови хо-
зяина. В литературе отмечались случаи анемии животных при мас-
совых нападениях иксодовых клещей. Это не может не сказываться
на продуктивность скота его упитанности, величине надоев и т. д.
На пребывание клещей организм хозяина, как правило, отвечает
выработкой защитных реакций. Установлено, что слюна клещей об-
ладает антигенными свойствами. Животные приобретают известную
184
устойчивость к новым нападениям паразитов. На иммунизирован-
ных животных клещи обычно уже не могут нормально питаться.
Иногда наблюдается даже инкапсуляция ротовых частей паразита,
что полностью перекрывает поступление в хоботок крови и лимфы.
Клещи — переносчики заболеваний
Наибольшее практическое значение клещи надсем. Ixodoidea
имеют как переносчики ряда тяжелых заболеваний человека и до-
машних животных. Большинство этих заболеваний, возбудители ко-
торых относятся к самым разным группам организмов: вирусы, бак-
терии, спирохеты, риккетсип, простейшие, представляют собой зо-
онозы и антропозоонозы с природной очаговостью.
Вирусные заболевания
В настоящее время установлена спонтанная зараженность иксо-
довых клещей большим количеством (более 20) различных вирусов.
Некоторые из них служат причиной опасных болезней человека.
Страдают от вирусов и домашние животные.
Наиболее широко распространен клещевой весенне-летний энце-
фалит. Это заболевание вызывается комплексом близкородственных
вирусов, встречающихся в Европе и Азии п несколько реже в Аме-
рике. Выдающаяся роль в изучении энцефалита принадлежит со-
ветским ученым. Первое описание течения и клинических проявле-
ний болезни было сделано в 1935 г. А. Г. Пановым. Начиная с
1937 г., в очагах энцефалита на Дальнем Востоке работали ком-
плексные экспедиции Наркомздрава СССР, в организации и прове-
дении которых огромная заслуга принадлежит акад. Е. Н. Павлов-
скому. Выдающемуся советскому микробиологу Л. А. Зильберу и
его ученикам в 1937 г. удалось выделить 29 штаммов вируса, а
несколько позднее Н. В. Рыжов и А. Н.. Скрынник доказали природ-
ную зараженность вирусом нескольких видов иксодовых клещей.
Изучение биологии клещей и путей циркуляции вируса в природных
очагах проводилось Е. Н.Павловскими его. сотрудниками. Резуль-
таты этих исследований в значительной мере послужили основани-
ем для создания учения о природной очаговости трансмиссивных
заболеваний. *
На территории СССР эндемичные очаги клещевого весенне-лет-
него энцефалита приурочены к таежной и лесным зонам. Основным
переносчиком вируса служат клещи р. Ixodes: в лиственных и сме-
шанных лесах европейской части СССР — I. ricinus (рис. 89,А),
в таежных лесах и по всей азиатской части страны — I.persulcatus.
Распространение энцефалита привело к его проникновению и в
лесостепную зону, где в роли переносчиков выступают представи-
тели р. Dermacentor (рис. 89, В) и Hyalotntna. Клещи выполняют
функции не только переносчиков, но и природных резервуаров ви-
руса. Возбудитель поражает клетки самых различных органов кле-
щей, в том числе слюнных желез и гонад. Это делает возможным
инокулятивный способ заражения позвоночных животных при напа-
185
дении клещей для питания и трансовариальную передачу вируса от
одного поколения переносчиков к другому. Это обеспечивает сохра-
нение вируса в природных очагах даже при отсутствии других под-
ходящих резервуарных хозяев (животные, сохраняющих вирус в
Рис. 89. Представители иксодоидпых клещей. А — голодная самка Ixodes riciftus-,
Б — самка Dermacentor pictus (по Померанцеву); В — самка Ornithodorus
papillipes] Г — самка Argas persicus (по Филипповой)
природе) круг которых, очень широк. Это различные мелкие живот-
ные, на которых питаются личинки и нимфьГ клещей: мыши, полевки,
землеройки, ежи, бурундуки. Спонтанная зараженность вирусами
отмечена и у ряда птиц: рябчиков, скворцов, поползней, береговых
ласточек и т. д. Эпидемиологическое значение птиц не ограничи-
вается, однако, только тем, что они выполняют функции резервуар-
ных хозяев. Совершая миграции и перелеты, птицы разносят на се-
бе питающихся клещей и тем самым способствуют возникновению
186
новых природных очагов. Роль крупных диких животных (лоси, оле-
ни, волки, медведи и т. д.) как резервуарных хозяев менее значи-
тельна. Восприимчивы к вирусу энцефалита и некоторые сельско-
хозяйственные животные, средн которых наиболее чувствительными
оказались козы.
Столь широкий круг резервуарных хозяев обеспечивает беспе-
ребойную циркуляцию вируса в природе. При этом, однако, необ-
ходимо помнить, что главная роль принадлежит все-таки клещам,
в которых вирус сохраняется значительно дольше, чем в позвоноч-
ных животных. Последние либо сравнительно быстро погибают от
заболевания, либо, что наблюдается значительно чаще, приобрета-
ют стойкий иммунитет.
Заражение человека происходит при посещении им природных
очагов. Какое-то время могут, по-видимому, существовать и синан-
тропные очаги, поддерживаемые домашними животными ( в част-
ности козами) и пастбищными видами иксодовых клещей. Чаще
всего передача вирусов человеку осуществляется клещами, однако
возможен и алиментарный путь заражения при употреблении не-
прокипяченного молока коз.
Клещевой энцефалит относится к одним из самых тяжелых за-
болеваний человека. Вирус поражает центральную нервную систе-
му. В начале заболевания наблюдаются лихорадочное состояние,
головные боли, тошноты. Позднее развиваются парезы и параличи,
появляются припадки. Очень характерны нарушения двигательных
функций и психические расстройства. Смертность при клещевом
энцефалите* может достигать 30%. °
К вирусным заболеваниям человека, распространяемым клещами п обладаю-
щим природной очаговостью, относятся и геморрагические лихорадки, сопровож-
дающиеся кровоизлияшргми, например Омская геморрагическая лихорадка.
Иксодовые клещи распространяют и вирусы, опасные для животных. При-
мером может служить шотландский энцефалит, или вертячка овец. Больные жи-
вотные, вследствие поражения мозжечка, страдают нарушениями двигательных
функций. Позднее у них развиваются параличи и парезы. В литературе описаны
эпизоотии, охватывающие до 50% поголовья овец.
Вирус шотландского энцефалита может поражать и человека.
Бактериальные заболевания
Среди бактериальных заболеваний, передаваемых иксодоидиы-
ми клещами, наиболее важное место, несомненно, принадлежит
туляремии. Его возбудитель Francisella tularensis 1 довольно широко
распространен в странах северного полушария. Пути циркуляции
микробов в природе очень разнообразны. В настоящее время F, tu-
larensis выявлена у 50 с лишним видов диких позвоночных живот-
ных и 10 видов домашних животных, относящихся к разным отря-
дам. Переносчиками туляремии могут быть и различные кровосо-
сущие членистоногие: блохи, вши, двукрылые и т. д. Специфический
перенос, однако, характерен лишь для иксодовых и аргазовых кле-
1 Многие авторы относят этот вид к р. Pasteurella, к которому относятся
и возбудители чумы (с. 237).
187
щей. Наиболее эффективно перенос туляремии осуществляется кле-
щами р. Dermacentor. Туляремийный микроб размножается в клет-
ках стенки средней кишки, проникает в гемолимфу, а оттуда в ган-
глий, мальпигиевы сосуды и слюнные железы. Предположения о су-
ществовании трансовариальной передачи Francisella tularensis
у клещей не подтвердились.
Заражение позвоночных животных может осуществляться как
инокулятивно, так и контаминативно — большое количество микро-
бов выводится во внешнюю среду с фекалиями и жидкостью кок-
сальных желез (у аргазид). Причиной возникновения заболевания
иногда служит и поедание животными инфицированных клещей.
Человек черпает инвазионное начало из природных очагов, при-
чем заражение может осуществляться не только клещами, но и не-
специфическими переносчиками. Среди последних важную роль
•играют слепни (с. 255) и мухи-жигалки (с. 255). Кроме того, воз*
будитель может передаваться с водой, при разделке тушек зара-
женных животных и респираторным путем, т. е. нетрансмиссивны-
ми способами без участия переносчиков.
Туляремия по своим клиническим проявлениям напоминает
легкие нелетальные формы чумы (с. 235). Подобно последней, она
может протекать в бубонной, кожной, кишечной и легочной фор-
мах. Наиболее широко распространена первая. Микробы, поселя-
ясь в лимфатических узлах, вызывают их увеличение7. На коже в
районе расположения такого узла отчетливо вырисовывается плот-
ная опухоль — бубон. Койсная форма туляремии сопровождается
образованием в местей проникновения возбудителя открытых язв.
Легочная форма туляремии сопровождается сильным кашлем
и обильным отделением мокроты с кровью. Попадание микробов
в кровяное русло и их размножение там приводит к общей инток-
сикации организма, последствия которой часто проявляются в виде'
функциональных нарушений сердечно-сосудистой системы. Туляре-
мия иногда принимает хроническое течение и способна давать ре-
цидивы, повторяющиеся даже через 15—20 лет после первичного
заражения.
Другим микробным -заболеванием, в эпидемиологии которого иксодоидные
клещи могут играть определенную роль, является чума. Во всяком случае в при-
родных условиях выявлена спонтанная зараженность клещей ее возбудителем —
Ipasteurella pestis. Специфическими переносчиками чумы служат блохи, резервуар-
ными хозяевами — грызуны (с. 238). Хотя в медицинской литературе не зареги-
стрировано случаев заражения человека чумой в результате укусов клещей, тем
не менее некоторые авторы (В. Н. Беклемишев) считают, что клещи являются
важным звеном, поддерживающим существования природных очагов этого за-
болевания.
Клещевые спирохетозы
Спирохетозами называются заболевания, вызываемые спирохе-
тами— своеобразными микроорганизмами, которые многими авто-
рами рассматриваются как сильно видоизмененная группа бакте-
рий.
188
Наибольшее практическое значение имеет клещевой возвратный
тиф, вызываемый спирохетами р. Borrelia. Лихорадки, вызванные
укусами клещей, были известны очень давно. Однако лишь в 1904 г.
Д. Россу и Милну (D. Ross, Milne) удалось обнаружить спирохет
в крови человека, страдающего лихорадкой. А еще через год Дат-
тон, Тодд и Р. Кох экспериментально доказали возможность пере-
дачи спирохет аргазовыми клещами. В последующие годы важное
значение имели работы русских исследователей: Е. П. Джунков-
ского, Е. И. Марциновского и др., изучавших спирохетозы человека
в Средней Азии и Иране. Начиная с 1927 г., эти исследования при-
обрели систематический и планомерный характер. Большая заслу-
га в этом принадлежала акад. Е. Н. Павловскому.
В настоящее время известно свыше 20 близких видов спирохет,
относящихся к р. Borrelia. Каждый из этих видов довольно строго
приурочен к определенному виду хозяина-клеща и соответственно
ограничен в своем распространении. В Азии Borrelia sogdiana пе-
редается видом Ornithodorus papillipes (рис. 89, В), Borrelia latis-
hevi—Ornithodorus tartakovskyi и т. д. В Европе Ornithodorus
verrucosa переносит В. caucasica, a Ornithodorus erraticas — В. his-
panica. В Африке широко распространена В. duttoni, переносчиком
для которой служит Ornithodorus moubata и т. д.
Заражение клещей спирохетами происходит при сосании ими
крови зараженных животных. Спирохеты интенсивно размножают-
ся в кишечнике хозяина, проникают в гемолимфу, а оттуда в боль-
шинство внутренних органов. Трансовариальная передача-позволя-
ет спирохетам сохраняться в популяции переносчиков неограничен-
но долгое время.
Спирохетозами в природе заражаются самые различные позво-
ночные животные — лисы, шакалы, крысы, мыши, песчанки, ежи,
летучие мыши, рептилии и др. Передача инвазионного начала мо-
жет происходить как инокулятивно, так и контаминативным спо-
собом. Выделяемая аргазидами во время и после питания коксаль-
ная жидкость в изобилии содержит спирохет, способных процикать
через кожу и слизистые.
Очаги заболевания отчетливо разделяются на две группы: при-
родные и синантропные — поселковые. Основным резервуаром
спирохет и в тех, и в других служат клещи. Позвоночные же живот-
ные, в том числе и человек,— лишь временные, носители возбуди-
телей и обеспечивают передачу последних от одной линии клещей
к другой.
Заражение человека спирохетами может происходить в очагах
любого типа. В природных биотопах люди, как правило, подверга-
ются нападению клещей при ночевках и во время выполнения раз-
личного рода работ — раскопка нор грызунов, земляные работы
и т. д. Формирование поселковых очагов обычно связано с заносом
инвазионного начала из природы. Они оказываются весьма устой-
чивыми в связи с тем, что некоторые виды аргазовых клещей, в том
числе п Ornithodorus papillipes, наряду с дикими представлены и
синантропными расами, поселяющимися в населенных пунктах.
189
Циркуляция спирохет в поселковых очагах осуществляется через
собак, кошек, различных грызунов. Источником распространения
инвазионного начала могут быть и уже заболевшие люди.- Скот
непосредственного участия в циркуляции возбудителей не прини-
мает. Однако, привлекая к себе клещей для питания, домашние
животные способствуют возникновению очагов с очень высокой
концентрацией переносчиков и тем самым повышают вероятность
заражения людей.
Заболевания, вызываемые разными видами р. Borrelia, очень
сходны между собой по всем клиническим проявлениям. В течение
инкубационного периода, который продолжается в среднем 6—8
дней, спир'охеты интенсивно размножаются в клетках ретикуло-эн-
дотелия. К концу этого срока они поступают в кровяное русло и пн-
Базируют различные органы. Течение болезни характеризуется
повторяющимися лихорадочными приступами с высокими подъема-
ми температуры. Часто наблюдаются патологические изменения
в легких: бронхиты, воспаление, а иногда и отек легких. Пораже-
ние центральной нервной системы проявляется в возбужденном
состоянии больных во время приступов, галлюцинациях, бреде
и т. д.
Заболевание обычно заканчивается выздоровлением без каких-
либо серьезных последствий через 2,5—4 месяца. Лишь в Восточной
Африку местный клещевой возвратный тиф сопровождается иногда
серьезными осложнениями: менингитами, параличами, желтухой
и т. д.
Среди спирохет р. Borrelia есть и специфические паразиты крупного рогатого
скота. В частности, довольно широко распространена Borrelia iheileri, передавае-
мая иксодовыми клещами (р. Boophilus, Rhipicephalus). Важное экономическое
значение имеет Borrelia anserinum — возбудитель спирохетоза птиц, передавае,мая
клещами р. Argas (рис. 89, Г).. В некоторых районах ущерб, причиняемый птице-
водству этим заболеванием, может быть очень значительным.
Риккетсиозы
Риккетсиозами называются заболевания, вызываемые своеоб-
разными микроорганизмами бактериальной природы — риккетсия-
ми. Однако от обычных бактерий риккетсий отличает ряд харак-
терных признаков: облигатное внутриклеточное паразитирование
и явная приуроченность к членистоногим животным. Большая часть
известных риккетсий, обитающих только в беспозвоночных хозяе-
вах, совершенно непатогенна для последних. Лишь незначительное
число видов, ставших паразитами кровососущих клещей и насеко-
мых, приспособилось к существованию и в позвоночных животных.
Именно эти формы и являются возбудителями ряда тяжелых забо-
леваний.
Группа клещевых пятнистых лихорадок
Под этим названием объединяются несколько риккетсиозов че-
ловека, течение которых сопровождается появлением обильной сы-
190
пи на теле больного. Среди них наибольшую известность получила
пятнистая лихорадка Скалистых гор.
История изучения этого заболевания по сути дела является началом истории
изучения самих риккетсий. Выдающаяся роль в этих исследованиях принадлежит
Г Т. Рпккетсу (Н. Т. Ricketts), который в 1906 г. установил, что острое лихора-
дочное заболевание, распространенное в некоторых районах США, передается
иксодовыми клещами. Три года спустя он обнаружил самого возбудителя в маз-
ках крови больных людей и в тканях переносчиков. В настоящее время установ-
лено, что пятнистая лихорадка Скалистых гор встречается не только на террито-
рии США, но п по всему Американскому континенту.
Возбудитель лихорадки Скалистых гор Rickettsia rickettsi посе-
ляется в цитоплазме пораженных клеток и в их ядрах. Спонтанное
заражение риккетсиями обнаружено у 13 видов иксодовых клещей,
относящихся к р. Dermacentor, Amblyomma, Haemophysalis и др.,
и у нескольких видов Argasidae. Клещи в природе служат не толь-
ко переносчиками, но и основными резервуарными хозяевами, чему
способствует трансовариальная передача возбудителя. В организме
клеща риккетсии в первую очередь проникают в клетки стенки ки-
шечника, а позднее и в гемолимфу. В дальнейшем они поражают
мышцы хозяина, его нервную систему, гонады, слюнные железы,
мальпигиевы сосуды.
Передача инвазионного начала позвоночным животным, в том
числе и человеку, осуществляется главным образом инокулятивным'
путем во время питания клещей. Выделение большого количества
риккетсий во внешнюю среду в процессе дефекации не представ-
ляет сколько-нибудь серьезной опасности: возбудитель очень чув-
ствителен к действию неблагоприятных факторов и в сам,ые корот-
кие сроки полностью теряет инразионность. Позвоночные животные
играют важную роль в эпидемиологии заболевания. К Rickettsia
rickettsi восприимчивы собаки, овцы, крупный рогатый скот, раз-
личные грызуны и т. д.
В организме человека риккетсии поражают стенки кровеносных
сосудов, что приводит к образованию некротических очагов. Про-
должительность инкубационного периода колеблется от 3 до 14
дней. Затем развивается перемежающаяся лихорадка, иногда с
очень высокой температурой. На 3—4-й день появляется характер-
ная сыпь, которая в наиболее тяжелых случаях принимает харак-
тер многочисленных кожных кровоизлияний. Иногда это приводит
к гангренозному поражению покровов. При тяжелом течении болез-
ни развиваются пневмония, внутренние кровоизлияния, нефрит,
а иногда и кома. Летальность может быть очень высокой. В неко-
торых районах она достигала 80%.
Значительно более спокойно протекает относящаяся к этой же группе риккет-
еиозов марсельская лихорадка, вызываемая Rickettsia conori. Это заболевание
довольно широко распространено в бассейнах Средиземного, Черного и Каспий-
ского морей. Кроме того, отдельные очаги обнаружены в Экваториальной и Юж-
ной Африке. Переносчиками R. conori служат иксодовые клещи, относящиеся к
р. Rhipicephalus, Haemophysalis и Amblyomma. Наличие траисоварнальцоп пере-
дачи превращает клещей и в резервуарных хозяев риккетсий в природе. Из по-
звоночных животных долгое время эти функции приписывались собакам. Однако
экспериментальные исследования показали, что собаки почти ие восприимчивы
191
к риккетсиям этого вида. Таким образом, вопрос о резервуарных хозяевах — по-
звоночных животных в очагах заболевания остается открытым.
Заражение людей марсельской лихорадкой, по-видимому, осуществляется
непосредственно в населенных пунктах. Целый ряд клещей, в частности, очень
широко распространенный Rhipicephalus sanguineus в изобилии встречается в не-
больших поселках. Переносчики держатся преимущественно во дворах и на при-
усадебных участках, поселяясь в поверхностных слоях почвы, трещинах фунда-
ментов и стен. Особенно многочисленны они там, где есть собаки. Риккетсии
попадают в организм человека чаще всего при укусе клещей инокулятивным
путем. Возможно проникновение возбудителей ч^рез кожу и слизистые (чаще
всего глаза) при случайном раздавливании инвазированных переносчиков.
У люден заболевание продолжается 2—3 недели. После короткого инкуба-
ционного периода начинается острая лихорадка с подъемом температуры до
39,5—40°. Через несколько дней после начала болезни появляется сыпь, которая
быстро распространяется по всему телу. Тяжелые случаи заболевания со смер-
тельным исходом наблюдаются редко.
На огромных территориях Средней, Северной и. Центральной Азии широкое
распространение получил сыпной тиф Северной Азии, острое риккетсиозное забо-
левание, сопровождающееся лихорадкой и обильной сыпью на коже. Болезнь
впервые была описана в 1936 г. на Дальнем Востоке. Принадлежность возбуди-
теля к риккетсиям удалось установить лишь три года спустя. Он получил назва-
ние Rickettsia sibericus. Сыпной тиф Северной Азии — типичное природно-очаговое
заболевание. Переносчиками (а в силу трансовариальной передачи и резерву-
арными хозяевами), являются клещи из р. Dermacentor D. silvarum, D. pictus
и др.). Среди позвоночных резервуарными хозяевами служат грызуны (суслики,
хомяки, полевки, бурундуки и т. д.).
Крупный рогатый скот в циркуляции риккетсий обычно большой роли не иг-
рает. Однако, привлекая взрослых клещей, домашние «животные способствуют
возникновению неблагоприятной эпидемиологической обстановки. Заражение че-
ловека осуществляется только при укусах инвазированными переносчиками.
Клиническая картина заболевания, вызываемого Rickettsia sibericus, довольно
близка остальным клещевым риккетсиозам из группы пятнистых лихорадок.
Сходные заболевания, вызываемые другими видами риккетсий, описаны в насто-
ящее время из Южной Африки, Индии и Австралии.
Группа пневмотропных риккетсиозов (группа Q-лихорадки)
Риккетсиозы этой группы характеризуются отсутствием сыпи.
Наиболее широко известна так называемая Q-лихорадка, впервые
описанная в 1937 г. в Австралии.
Природа этой болезни вначале оставалась неясной, что и нашло свое отра-
жение в названии (Q — первая буква английского слова «querry» — неясный,
неопределенный). Выделенный позднее возбудитель был отнесен к самостоятель-
ному роду риккетсий Coxiella и стал обозначаться как С. burneti. Первоначаль-
но предполагалось, что Q-лихорадка эндемична для Австралии. Позднее было
установлено, что это очень широко распространенное заболевание, встречающееся
практически во всех странах мира, в том числе и в Советском Союзе.
Возбудитель Q-лихорадки отличается рядом характерных осо-
бенностей', определяющих своеобразие ее очагов. В первую очередь
бросается в глаза чрезвычайно .широкая специфичность Coxiella
burneti. Она способна паразитировать в самых различных перенос-
чиках. Только на территории СССР среди иксодовых и аргазовых
клещей зарегистрировано свыше 30 .видов, в которых развиваются
эти риккетсии. Кроме того, спонтанная зараженность возбудителем
обнаружена в гамазовых (с. 195) и краснотелковых (с. 205) кле*
щах и у постельного клопа (с. 210). Це менее широк и круг позво-
192
ночных хозяев С. burneli. На территории СССР этот риккетсиоз
обнаружен более чем у 60 видов диких млекопитающих. Заражают-
ся и дикие птицы.
Вторая особенность Coxiella burneli заключается в удивитель-
ной устойчивости к действию факторов внешней среды. Риккетсии,
выведенные из организма хозяина, длительное время сохраняют ин-
вазионную способность. Все это вместе взятое и обусловливает ши-
рочайшее распространение Q-лихорадки и многообразие путей цир-
куляции возбудителя.
Очаги заболевания отчетливо подразделяются на природные и
синантропные. Состав позвоночных животных, служащих резерву-
арными хозяевами в разных районах земного шара, различен.
В Европе это крысы, зайцы, косули, лисы, воробьи, жаворонки
и др.; в Америке — дикие кролики, дикобразы, суслики; в Австра-
лии к списку резервуарных хозяев прибавляются сумчатые живот-
ные. Переносчиками в таких очагах служат разнообразные крово-
сосущие клещи, связанные с определенными биотопами, в которых
обитают позвоночные животные.
Первичные природные очаги Q-лихорадки дают начало вторич-
ным синантропным очагам, в которых главная роль принадлежит
домашним животным и в первую очередь крупному рогатому скоту
и собакам. Очень часто встречается заражение риккетсиями и у
птиц, причем не только у домашних, но и сопутствующих челове-
ку— ворон, воробьев, голубей и т. д. Переносчиками в данном слу-
чае выступают синантропные виды и расы клещей.
В организме клещей риккетсии интенсивно размножаются
в клетках стенки средней кишки. Позднее они проникают в гемо-
лимфу, а оттуда в различные внутренние органы. Однако наиболее
интенсивно возбудители размножаются лишь в рыхлой ткани жи-
рового тела и в яичниках, обеспечивая тем самым трансовариаль-
ную передачу. В то же время они полностью отсутствуют в цито-
плазме клеток и полости альвеол слюнных желез. Такая локализа-
ция риккетсий предопределяет и путь их. проникновения в
позвоночных животных, заражение которых осуществляется иск-
лючительно контаминативным способом.
Устойчивость С. burneti к действию факторов внешней среды
превращают Q-лихорадку из трансмиссивного, заболевания в фа-
культативно-трансмиссивное. Заражение позвоночных животных
действительно происходит в период питания переносчиков. Риккет-
сии, выделяемые с фекалиями и коксальной жидкостью (у Argasi-
dae), легко проникают в организм хозяина. В этих случаях можно
говорить о непосредственной передаче клещами инвазионного нача-
ла. Однако фекалии переносчиков опасны не только для животно-
го-прокормителя. Высыхая, они вместе с пылью рассеиваются во
внешней среде и могут стать источником заражения других орга-
низмов.
Источником рассеивания риккетсий во внешней среде служат не
'только клещи. В большом количестве возбудитель выделяется
с мочой зараженных животных, при отеле с околоплодной жидко-
7—58 193
стыо и плацентой и т. д. Очень важным в эпидемиологическом от-
ношении является обнаружение риккетсий в молоке домашних жи-
вотных (коров, овец, коз и т. д.).
Клинические проявления Q-лихорадки очень разнообразны,
Продолжительность инкубационного периода колеблется от 10 дс
26 дней. Лихорадка (часто с очень высокими подъемами темпера-
туры) держится около 3 недель. Очень характерны сильные голов-
ные боли и боли в мышцах и суставах. При респираторном способе
заражения развивается пневмония, сопровождающаяся сухим каш-
лем и отделением мокроты. Последняя содержит большое количе-
ство риккетсий, которые могут стать источником новых заражений.
Заболевание заканчивается полным выздоровлением, однако в ред-
ких случаях при остром течении возможен и смертельный исход.
Клещи передают также возбудителей целого ряда других рнк-
кетсиозов, медицинское значение которых, однако, не столь велико.
Особого внимания заслуживает лишь обнаружение спонтанной за-
раженности иксодовых клещей Rickettsia prowazeki — возбудителя
эпидемического сыпного тифа (с. 227). Роль клещей в эпидемиоло-
гии этого заболевания пока остается неясной.
Заболевания, вызываемые простейшими
Иксодовые клети служат переносчиками ряда простейших — кровепаразптов
позвоночных животных. Эти формы объединяют в отр. Piroplasmida и включа-
ются в состав кл. Sporozoa. Биология, особенности организации, пути циркуляции
в природе и хозяйственное значение пнроплазмид были подробно рассмотрены
ранее (Частная паразитология. Паразитические простейшие и плоские черви).
Борьба с клещами
Значение клещей как облигатных гематофагов и как переносчи-
ков и резервуарных хозяев возбудителей большого числа заболе-
ваний человека и домашних животных очень велико. Это обусло-
вило необходимость разработки эффективных методов борьбы
с клещами, что подразумевает не только уничтожение клещей, но
и широкую систему профилактических мероприятий, исключающих
возможность контакта с кровососами.
Широкое распространение получило использование акарицидных препаратов:
ДДТ, гексахлорана, хлорофоса, полнхлорпинена и некоторых других. Применение
акарицидов для обработки больших площадей (распыление с самолетов над паст-
бищами и лесными массивами), хотя и приносит желаемый эффект, как правило,
влечет за собой ряд вредных последствии. Большинство этих препаратов токсично
не только для клещей, но и для многих других организмов (как беспозвоноч-
ных, так и позвоночных). Кроме того, многие из них разрушаются очень медленно
(например, ДДТ). В результате они постепенно накапливаются в тканях живот-
ных и растений, передаются по пищевым цепям и в конечном счете попадают
в организм человека.
Несомненно, более рационально использование акарицидных препаратов для
обработки помещений, в которых живут люди или размещаются животные. Такие
помещения легко поддаются последующей очистке и уборке. Для уничтожения
клещей, уже напавших на животных, применяется купание последних в специ-
альных ваннах или опрыскивание растворами ядохимикатов. Все эти операции
необходимо производить, строго соблюдая меры предосторожности, чтобы исклю-
чить возможность отравления животных и обслуживающего персонала, а также
194
попадания используемых веществ в корм скоту п па хозяйственный инвентарь.
Эффективность подобного типа обработок увеличивается при периодическом их
повторении.
Значительно более безопасными для человека и окружающей его природы
являются мероприятия, основанные па уничтожении биотопов, в которых поселя-
ются клещи: перепахивание диких малопродуктивных пастбищ и превращение их
в культурные, уничтожение подстилки и сухой травы в загонах для скота, со-
блюдение санитарных норм на скотных дворах — своевременный ремонт и тща-
тельная очистка помещений, заделка щелей и т. д. Последнее в полной мере
относится к приусадебным участкам и жилым помещениям людей.
При поселении людей в диких неосвоенных районах (поселки лесозаготови-
телей, поисковых геологических групп и т. д.) следует проводить те же меропри-
ятия: па месте расположения временного поселка или палаточного городка вы-
жигаются сухая трава и подстилка, тропинки, по которым наиболее часто прохо-
дят люди, расчищаются и расширяются. Вдоль них вырубается мелкий кустарник.
Важное значение имеет борьба с животными — прокормнтелями клещей (грызу-
нами, насекомоядными и т. д.). Последние не только привлекают клещей, по и ча-
сто служат природными резервуарами возбудителен различных заболеваний
(с. 238). Особенно важна раскопка нор и гнезд животных, ибо именно там чаще
всего скапливается большое количество кровососов.
Наибольшей эффективностью, однако, обладают индивидуальные методы за-
щиты от нападения клещей, основанные на личной профилактике. Для этого ис-
пользуют специальные защитные костюмы (комбинезоны), исключающие возмож-
ность заползания клещей па поверхность тела, накидки, прототипом которых
послужили предложенные еще в 30-х годах сетки Павловского. Первые изготав-
ливаются из хлопчатобумажного тюля, вторые из рыболовной сети с ячеей около
2 см. Сетки закрепляются на головном уборе, так что они прикрывают шею
и плечи, оставляя свободным только лицо. Пропитывание костюма и накидки
репеллентами — веществами, отпугивающими клещей, делает такую защиту на-
дежной и эффективной. В настоящее время с успехом используется целый ряд
репеллентов, обладающих продолжительным действием и малотоксичпых для
человека. Комбинезоны, обработанные препаратом ДЭТА (днэтилтолуамид),
предохраняют от нападения клещей в течение 9—20 дней. Еще большей устой-
чивостью обладает булефнд (гексаметнлепбутапсульфамнд).
Даже самые надежные репелленты и защитная одежда не исключают воз-
можности попадания отдельных клещей на тело человека. Поэтому после работы
в лесу или других местах возможного обитания клещей необходимо проводить
тщательные осмотры (лучше всего взаимные). Присосавшихся "клещей следует
немедленно удалять.
В последние годы Для предупреждения заражения людей различными заболе-
ваниями, переносимыми клещами, широко применяют предохранительные при-
вивки соответствующих вакцин. Такие вакцины разработаны против энцефалита,
пятнистой лихорадки Скалистых гор, клещевого сыпного тифа Северной
Азин и т. д.
Надсемейство Gamasoidea — гамазовые клещи
Гамазовые клещи, вместе с иксодовыми и аргазовыми, входят в группу Ра-
rasitiformes. В противоположность Ixodoidea— это очень мелкие клещи, размер
тела которых колеблется от 0,2 до 2,5 мм. Тело их слабо хитинизироваио, несет
множество щетинок,/ окраска желтоватая или буроватая.
Надсем. Gamasoidea объединяет около 30 семейств, в которых насчитывается
несколько сотен видов, как свободноживущих, так и паразитических. Среди по-
следних встречаются временные и постоянные эктопаразиты позвоночных (реже
беспозвоночных) животных и немногочисленные^эндопаразиты (с. 197).
Тело гамазовых клещей подразделяется на гнатосому и идиосому. Хелицеры
по большей части клешневидные, лишь у наиболее специализированных парази-
тических видов сем. Dermanyssidae они видоизменены и превращаются в тонкие
стилеты, с помощью которых паразит прокалывает кожу хозяина. Детали стро-
ения хелицер в связи с различием выполняемых ими функций разнообразны
и широко используются систематиками как важный определительный признак.
7* 195
Собственно тело (идиосома) имеет овальную, реже удлиненную форму. Тонкая,
морщинистая хитиновая кутикула легко растягивается, и тело клещей, насосав-
шихся крови, может значительно увеличиваться в размерах. Сильно хитинизиро-
ванные участки—щитки имеются как на спинной, так и иа брюшной стороне.
Форма и характер расположения этих щитков используется в систематике.
Тело гамазовых клещей несет многочисленные и разнообразные по форме
щетинки, преимущественно выполняющие роль органов осязания. Расположение
щетинок (хетотаксия) имеет определительное значение. Ноги хорошо развиты
и у многих эктопаразитов снабжены специальными коготками и присосками,
Рис. 90. Представители гамазовых клещей (из кн. Брегетовой). А — Laelaps (со
спинной стороны); Б — Spinturnix (по Догелю); В, Г —самка и самец Dertnanys-
sus gallinae (с брюшной стороны) :
/ — хелицеры. 2—пальпы, 3 —дорзальный щит. 4— щетинки. 5 — брюшные щитки
обеспечивающими прикрепление к телу хозяина. Половой диморфизм выражен
отчетливо, самцы часто отличаются меньшими размерами (рис. 90). В матке
самки обычно созревает только одно яйцо. Откладываемые поодиночке или куч-
ками яйца приклеиваются к субстрату или непосредственно на теле хозяина.
Из яйца выходит шестиногая личинка. Ее покровы слабо хитннизировапы и бес-
цветны. Личинка линяет, превращаясь в нимфу первой, а потом и второй стадии.
Последняя отличается от взрослых клещей лишь отсутствием половых органов.
У многих паразитических гамазовых клещей наблюдаются отклонения от описан-
ного хода жизненного цикла, суть которых сводится к характерной для многих
паразитов эмбрионпзации. У многих видов развитие яйца идет в матке самки
до стадии личинки. У некоторых представителей р. Laelaps (рис. 90) материнская
особь отрождает нимфу первой стадии.
Паразитические гамазовые клещи питаются кровью хозяина. У самок наблю-
дается отчетливая связь между процессами переваривания поглощенной крови
п созреванием яиц (гонотрофический цикл). Временные паразиты питаются лишь
в определенные периоды суток. Так, например, куриный клещ Dermanyssus
196
(рис. 90) днем прячется в различных укрытиях и нападает на хозяина только
ночью. Постоянные паразиты (Laelaps и др.) сосут кровь часто и небольшими
порциями, независимо от времени суток. Их кишечник никогда не переполняется
кровью и тело не раздувается, как у временных паразитов. Интересно, что гама-
зовые клещи питаются не на всех стадиях своего жизненного цикла. Способность
сосать кровь хозяина часто приобретается лишь на стадии нимфы.
Свободноживущие гамазовые клещи в большинстве своем хищники или де-
тритофаги. Многие из них поселяются в норах грызунов и в гнездах птиц. Имен-
но от этих обитателей гнездовой подстилки и берут свое начало современные па-
разитические гамазиды. Возможные пути становления их паразитизма можно
проследить на примере обширного сем. Laelaptidae. Часть видов, относящихся
к этому семейству,— типичные обитатели гнездовой подстилки, но многие пере-
ходят к жизни на самих грызунах. Для этих видов характерны короткие и ши-
роко расставленные ноги, снабженные крепкими коготками. На теле имеются
многочисленные щетцнки, помогающие удержаться в шерсти. Вырабатывается
специфичность по отношению к виду животного хозяина, изменяется и способ
питания клещей. Постоянно находясь' на теле хозяина, клещи получают возмож-
ность слизывать капельки крови, выступающие при случайных повреждениях
кожи. Некоторые виды клещей приобретают способность активно повреждать
кожу грызунов; питание кровью становится для них обязательным. Однако
в строении ротового аппарата клещей р. Laelaps еще нет специальных приспособ-
лений к гематофагии. Их ротовые части ничем не отличаются от таковых у сво-
бодноживущих клещей, и паразиты просто прогрызают кожу млекопитающих
своими клешневидными хелицерами. Только у представителей сем. Haemogama-
sidae, Dermanyssidae и др., ставших облигатными гематофагами, хелицеры преоб-
разуются в колющие стилеты.
Среди гамазовых клещей имеются виды, приспособившиеся к эндопаразитиз-
му. Это представители сем. Entonyscidae, которые живут в воздушных мешках
змей, Rhynonyssidae из носовой полости птиц и, наконец, Halarachnidae — пара-
зиты трахеи и легких млекопитающих. Все эндопаразитические Gamasoidea отли-
чаются слабой хитинизацией покровов; их хелицеры преобразованы в колющие
щетинки. Взрослые клещи и нимфы — гематофаги.
Патогенность гамазовых клещей и их роль
в распространении инфекционных болезней
Важное практическое значение гамазовых клещей определяет интерес к этой
группе. Среди советских исследователей следует назвать имя Н. А. Брегетовой —
автора монографии по гамазовым клещам.
Многие виды гамазовых клещей могут вызывать сильные раздражения ко-
жи— дерматиты. Особенно неприятен в этом отношении куриный клещ — Derma-
nyssus gallinae (рис. 90). Паразитируя на домашних птицах (главным образом —
курах), он тормозит нормальный рост цыплят, может стать причиной снижения
яйценоскости, а при сильном заражении вызывает даже гибель птицы.
Многие виды гамазовых клещей имеют значение как переносчики различных
вирусных и бактериальных заболеваний человека и животных. В их числе следует
назвать эндемический, или крысиный, сыпной тиф. Заболевание это относится
к числу антропозоонозов и передается человеку крысиным клещом Ornitonyssus
bacoti. Клещи из сем. Laelaptidae (Laelaps и Haemolaelaps), паразитирующие на
водяных крысах, переносят туляремию; куриный клещ Dermanyssus gallinae —
переносчик Q-лихорадки, вызываемой риккетсиями (с. 192), и т. д.
ПОДОТРЯД SARCOPTIFORMES REUTER, 1909
Надсемейство Analgesoidea— перьевые клещи
Перьевые клещи — крайне специализированная группа эктопа-
разитов, приспособившихся к обитанию на перьевом покрове птиц.
7**-58 197
К надсем. Analgesoidea относится несколько семейств (Analges'idae,
Pterolichidae, Syringobiidae п др.). Все перьевые* клещи — очень
мелкие животные, около 1 мм длиной, поселяющиеся главным обра-
зом на первостепенных и второстепенных маховых, а также п на
рулевых перьях. Небольшая часть видов (сем. Dermaglyphidae,
Syringobiidae) приспособилась к жизни внутри очина пера. Есть
и гиперпаразиты. Это представители сем. Myalgesidae, паразити-
рующие на мухах-кровососках и пухоедах (с. 215).
Несмотря на мелкие размеры перьевых клещей, едва различимых невоору-
женным глазом, они известны очень давно. Первое изображение было сделано
итальянцем Реди еще в 1668 г. Сто лет спустя де Геер (de Geer, 1778) дал науч-
ное описание известных к тому времени видов и объединил их в самостоятельную
группу. Позднее известный естествоиспытатель Ш. Нищи (Nitzsch, 1818) присво-
ил этой группе родовое наименование Analges. В XIX в. появился ряд серьезных
исследований, среди которых- нужно назвать работы Е. Труассара (Trouessert),
А. Берлезе (Berlese) и Г. Канестрини (Canestrini). В 20—30-х годах нашего сто-
летия изучением этой своеобразной группы занимались А. Удеманс (Oudemans)
и Г. Фитцтум (Vitztum). В 50-х годах в серии «Фауна СССР» появился капи-
тальный трёхтомнын труд В. Б. Дубинина, посвященный морфологии, системати-
ке, распространению, экологии перьевых клещей и их адаптациям к паразитиче-
скому образу жизни. Этот труд и положен в основу настоящей главы.
Морфология
Тело перьёвых клещей отчетливо подразделяется на гнатосому
(комплекс ротовых органов) и идиосому, несущую две пары конеч-
Рис. 91. Половой диморфизм перье-
вых клещей. А — копулирующие
перьевые клещи Bonnetella (по Дуби-
нину); Б, В — самка и самец Piero-
lichus: *
/ — самец, 2—поздняя женская нимфа,
3 — щитки, 4 — готовое к откладке яйцо
в матке самки
198
ностеп. Гнатосома подвижна и может слегка втягиваться внутрь те-»
ла. Хелицеры имеют клешневидную форму. Гипостом представля-
ет собой короткую трапециевидную или вытянутую пластинку с ко-»
роткими щупиками (пальпами). Тело одето тонкой хитиновой кути*
кулой. На спинной стороне она образует утолщенные щитки
(рис. 91), а на брюшной — довольно сильно склеротинпзирован-
Рис. 92. Строение и локализация перьевых клещей (па.Дубинииу).
А — клещ р. Michaelichus с асимметричными конечностями, служащи-
ми для прикрепления к бородкам пера; Б — расположение перьевых
клещей между лучами пера и способы их прикрепления: при помощи
увеличенной левой ноги — функция «якоря», (наверху), при помощи
видоизмениой щетинки — заклинивание (внизу); В —отроение лапки;
Г — задний конец самца Pterolichus-.
/ — присоска на лапке» 2—половые присоски, 3 — мембрановидиые щетинки
ные валики. Последние несколько вдаются внутрь тела и служат
для прикрепления мышц, приводящих в движение конечности. Вали-
ки окаймляют места прикрепления тазиков ног и называются эпи-
мерами (если расположены кпереди от них) или эпимеритами (если
находятся сзади). Все утолщенные части покровов окрашены в ры-
жеватый или темно-бурый цвет. У некоторых видов клещей разви.
ваются тонкие и прозрачные хитиновые мембраны (см. рис. 93, Л),
которые располагаются либо латерально по краям тела, либо на
его заднем конце и в последнем случае приобретают вид листовид-
ных или мечевидных придатков.
На теле перьевых клещей имеются многочисленные щетинки,
иногда тоже приобретающие вид мембран (рис. 92,Г). Характер
расположения щетинок отличается большим постоянством в преде-
лах отдельных групп и потому широко используется в систематике.
Щетинки в основном несут осязательную функцию. В то же время
они наряду с мембранами играют известную роль в увеличении
площади соприкосновения тела клеща с пером. Последнее очень
199
важно для закрепления клещей, которые должны противостоять дей-
ствию токов воздуха, возникающих во время полета птицы. Ноги
перьевых клещей состоят- из 6 члеников. Тазики неподвижно сра-
стаются с покровами тела. На конце лапки располагается не кого-
ток, как свойственно большей части клещей, а округлая перепон-
чатая пластинка (рис. 92, В). Мягкие и эластичные, эти пластинки
действуют наподобие присосок, позволяющих клещам прочно удер-
живаться на гладкой поверхности некоторых участков пера.
Всем Analgesoidea свойствен хорошо выраженный половой ди-
морфизм (рис. 91). У самцов, как правило, имеются анальные ко-
пулятивные присоски (рис. 92, Г), представляющие собой видоизме-
ненные щетинки. Они служат для удержания самки во время ко-
пуляции. Ту же функцию выполняют и задние ноги самца, у некото-
рых видов клещей чрезвычайно увеличенные (рис. 91, А). У самцов
обычно сильнее, чем у самок, развиты располагающиеся на ногах
зубцы, шипы и щетинки; передние конечности нередко изогнуты,
иногда асимметричны (Michaelichus). Все это — дополнительные
адаптации к прикреплению, необходимость которых обусловлена
особенностями биологии самцов. Дело в том, что в период копуля-
ции самец должен не только удержаться на пере, но и удержать
самку, с которой он остается спаренным (рис. 91, А) в течение
продолжительного времени. Очень часто самцы копулируют не с
половозрелыми самками, а с нимфами последней стадии развития.
Размножение и жизненный цикл
В матке оплодотворенных самок обычно содержится лишь одно,
довольно крупное яйцо. Зрелые яйца откладываются на оперение
птицы и приклеиваются к бородкам опахала с помощью особого
секрета. У некоторых видов (например, у Freyana) развитие яйца
осуществляется в матке родительской особи, так что наружу выво-
дятся яйца, уже содержащие вполне сформированную личинку.
Жизненный цикл Analgesoidea проходит с метаморфозом. Из яйца
выходит шестиногая личинка, имеющая очень тонкие, слабо хити-
низированные покровы. После линьки личинка переходит в фазу
нимфы. Последние претерпеваютнесколько линек, благодаря кото-
рым можно различить нимф первой, второй и третьей стадии. Пос-
ледняя отличается от взрослой особи лишь отсутствием наружного
полового аппарата. На всех стадиях жизненного цикла перьевые
клещи неразрывно связаны с телом хозяина — птицы.
Экология и биология перьевых клещей
Питание и специфичность
Перьевые клещи относятся к числу узко специфичных парази-
тов, что в значительной мере определяется характером их жизни.
Они питаются в основном жироподобным секретом копчиковой же-
лезы, состав которого у разных видов птиц строго постоянен. Имен-
200
но с этим и связана, по данным В. Б. Дубинина, приуроченность оп-
ределенных видов перьевых клещей к определенным видам хозяев.
Перья здоровой птицы всегда смазаны секретом копчиковой желе-
зы. Возможно, что это н является причиной малой подвижности
паразитов, постоянно имеющих пищу в достаточном количестве.
Клещи поедают, кроме того, отмершие частички кожи и перьев, а
также споры грибов (паразиты водоплавающих птиц — и однокле-
точные водоросли), пристающие к оперению птицы.
Локализация на теле хозяина
Разные виды перьевых клещей строго приурочены к различным
участкам перьевого покрова птицы. Одни виды поселяются только
на первостепенных, другие — на второстепенных маховых, третьи —
Рис. 93. Перьевые клещи уток (Freyana anatina) и их поведение
во время линьки птиц (по Гинецинской). А — самец Freyana-, Б —
локализация клещей в опахале пера; В — вскрытая перьевая сум-
ка линяющей утки и положение клещей на растущем пере;
Г — то же, увеличено
локализуются в кроющих перьях крыла и т. д. Более того, даже на
самом опахале пера клещи занимают вполне определенное место.
Закономерности, обусловливающие такой характер распределения
клещей на хозяине, В. Б. Дубинин связывает с особенностями мик-
роструктуры пера и с различием условий, создающихся во время
полета птицы на тех или иных участках оперения. Зона поселения
клещей на опахале приходится на серединную часть пера и располо-
жена вдоль его стержня .(рис. 93, Б), т. е. имеет вид узкого, вытя-
нутого овала. Именно эта зона, как правило, характеризуется ти-
пичным устройством лучей и бородок пера, тогда как за ее преде-
лами находятся так или иначе измененные структуры.
Во время полета птиц возникают аэродинамические силы, состав-
ляющие физическую основу полета. Под воздействием этих сил из-
меняются и условия жизни перьевых клещей как на крыле в целом,
так и на отдельных участках оперения. Изменения эти зависят от
201
характера полета птицы. Разный угол поворота тех или иных ма-
ховых перьев при полете связан с различным распределением воз-
душных токов на поверхности пера. Так, маховые перья, располо-
женные ближе к вершине крыла, имеют больший угол поворота, чем
остальные перья. Вследствие этого опн сильнее «обдуваются» тока-
ми воздуха и лучше аэрируются. Именно на этих перьях и скапли-
вается основная масса клещей. У тех птиц, вершинные первостепен-
ные маховые перья которых не совершают вращательного движения
(такой полет свойствен большинству воробьиных), клещи распреде-
лены по всем маховым перьям более или менее равномерно.
Клещи, поселяющиеся на разных по структуре перьях, характе-
ризуются различными морфологическими адаптациями. Так, виды,
обитающие на жестких (первостепенных маховых) перьях — Fre-
yatia, Michaelichus и др., — имеют более крупные, по сравнению
с остальными перьевыми клещами, размеры. Их покровы сильнее
хитинизированы, скелет хорошо развит и может служить опорой
для сильных мышц, приводящих в движение ноги. По бокам тела
развиваются прозрачные хитиновые мембраны, появляются упло-
щенные и расширенные щетинки. Это увеличивает поверхность тела
паразита, соприкасающуюся с пером, а может быть, и придает те-
лу более «обтекаемую» форму. В то же время тонкие и длинные ще-
тинки, создающие большую «парусность» у этой группы клещей,
отсутствуют. Все это адаптации, обеспечивающие лучшее прикреп-
ление клещей. Они особенно важны в связи с тем, что именно пер-
востепенные маховые перья при полете наиболее сильно подверга-
ются воздействию токов воздуха, которые легко могли бы сдуть кле-
щей с поверхности пера.
На более мягких второстепенных маховых и кроющих перьях,
не подвергающихся такому сильному «обдуванию», поселяются дру-
гие виды перьевых клещей (Megninia, Analges и др.), морфологичес-
ки сильно отличающиеся от клещей первой группы. Они обладают
меньшими размерами. Покровы их слабо хитинизированы, на дор-
зальной стороне тела имеются длинные щетинки. Специальные ор-
ганы прикрепления к перу развиты слабее. Наконец у видов, посе-
ляющихся внутри очинов пера (сем. Dermaglyphidae), адаптации к
прикреплению, ютоль характерные для обитателей первостепенных
маховых перьев, практически отсутствуют. Клещи этой группы име-
ют узкое, вытянутое тела Их бесцветные, слабо хитинизирован-
ные покровы становятся мягкими. У многих видов совсем исчезают
щитки. На теле имеются многочисленные длинные и топкие ще-
тинки.
Зависимость биологии перьевых клещей
от особенностей биологии хозяина
и от климатических условий
Строгая приуроченность разных видов клещей к определенным
маховым перьям обычно нарушается в период, предшествующий
линьке хозяина. По-видимому, это связано ю изменением условий
202
питания. Незадолго да начала линьки птица перестает смазывать
оперение секретом копчиковой железы и клещи лишаются своей ос-
новной пищи; это и заставляет их покидать место своего обитания
и расползаться по всему телу птицы. Несмазываемые перья подсы-
хают, становятся ломкими и выпадают. Таким образом переселе-
ние клещей спасает часть их популяции от гибели, хотя значитель-
ное число паразитов все же остается на старом пере и отбрасывается
вместе с ним. Во время линьки клещи обычно переползают с махо-
вых и рулевых перьев на кроющие крыла и контурные перья спины,
а иногда и йепосредствешю на кожу птицы.
У утиных, у которых линька происходит бурно и связана с одно-
временным выпадением всех маховых перьев, наблюдаются особые
адаптации. Специфичные для уток перьевые.клещи р. Freyana пере-
ползают с опахала пера на его стержень, а потом в перьевую сумку.
Там они сидят на пеньках растущих новых перьев у самого края
сумки, но не выходя за ее пределы (рис. 93, В, Г). Когда прорыва-
ется чехьтик растущего пера и становится заметным развивающееся
опахало, клещи покидают перьевую сумку, переползают к основанию
опахала и сосредотачиваются на сосочке пера. Они сидят под прик-
рытием лоскутов разорванного рогового чехлика, образуя там жи-
вой копошащийся комок. Только после окончательного развития
опахала клещи занимают свою обычную стацию. Значительная
часть уток во время линьки все же освобождается от заражения
перьевыми клещами.
В период линьки хозяина наблюдаются очень существенные из-
менения возрастного состава популяции перьевых клещей. Еще до
наступления линьки погибают в большинстве старые, половозрелые
особи. Отчасти за счет этого, отчасти же за счет роста личинок к
началу линьки популяция клещей становится очень однородной. Она
почти целиком состоит из поздних нимф, уже готовых к метамор-
фозу. '
Весь период линьки сохранившаяся часть популяции проводит
в неблагоприятных условиях. Нарушается нормальное питание кле-
щей. Паразиты утиных — клещи р. Freyana, по-видимому, совсем не
питаются. Косвенным указанием на это служит характер их распо-
ложения на сосочке растущего пера, где они сидят сплошной мас-
сой друг на друге. Находясь внутри перьевой сумки, клещи испыты-
вают недостаток в кислороде; именно с этим, вероятно, и связана
их характерная локализация возле наружного отверстия сумки.
Неблагоприятные условия, видимо, задерживают последнюю линь-
ку, и только после того как клещи вновь занимают свое местооби-
тание на опахале, начинается массовое превращение их в половозре-
лую стадию. Одновременное появление большого числа самцов и
самок приводит к вспышке размножения, которое способствует пос-
тепенному восстановлению численности популяции перьевых кле-
щей, пострадавшей во время линьки хозяина.
Сезонные миграции тоже представляют собой очень важный мо-
мент в жизни паразитов перьевого покрова. Их приспособления к
перелетам хозяина зависят от биологических особенностей послед-
203
него. Так, у птиц, совершающих небольшие по расстоянию мигра-
ции, в поведении перьевых клещей и в состоянии их популяций не
наблюдается особых изменений. Клещи только переходят в глубже
лежащие слои оперения и скапливаются в его пуховых частях.
Перьевые клещи птиц, совершающих дальние миграции, могут ока-
заться в очень неблагоприятных условиях. В связи с этим в биоло-
гии и поведении клещей наблюдаются определенные адаптивные
изменения. Изменяется возрастной состав популяции паразитов. Не-
задолго до начала перелета резко возрастает количество половозре-
лых особей. Попадается много копулирующих пар, самок с готовыми
к откладке яйцами. Иными словами, идет интенсивное размножение
и к началу перелета на маховых и кроющих перьях крыла можно
видеть множество яиц, прикрепленных к бородкам. В фазе яйца
перьевые клещи переносят неблагоприятное для них время переле-
та, по окончании которого начинается вылупление личинок. У дру-
гих видов, например у представителей р. Ardeacarus, наблюдаются
иные адаптации. Жизненный цикл этих клещей приспособлен к био-
логии хозяина таким образом, что к началу миграции почти вся
популяция состоит из нимф второго возраста. Последние внедряются
под кожу птицы и в таком состоянии переносят перелет. Немного-
численные половозрелые особи клещей, оставшиеся во время пере-
лета на пере, как правило, погибают.
Перьевые клещи чувствительны к изменениям температуры и
влажности окружающей среды. Это теплолюбивые животные, опти-
мальной является для них температура 40° С. На понижение темпе-
ратуры или на резкое повышение влажности (дождь, туман) клещи
реагируют изменением своей локализации на хозяине. Они пере-
ползают с опахала пера к его основанию, на кроющие перья или да-
же на кожу птицы. При значительном повышении температуры па-
разиты расползаются по всей поверхности опахала, максимально
собираясь в его средней части. В зависимости от погоды такие из-
менения локализации перьевых клещей проявляются в течение дли-
тельного времени и носят характер сезонных адаптаций.
В зимнее время активность клещей резко понижается. Популя-
ция их большей частью состоит лишь из нимф, которые находятся
в состоянии диапаузы. Метаморфоз их обычно задерживается до
весны.
Надсемейство Sarcoptoidaea
Семейство Sarcoptidae Trouessart, 1892
Sarcoptes scabiei L. — чесоточный зудень
Чесоточные клещи — небольшая группа микроскопически мел-
ких паразитов, поселяющихся в толще эпидермиса млекопитающих.
Насчитывают около пяти самостоятельных родов чесоточных кле-
щей, встречающихся на разных хозяевах. Однако клещи эти не об-
204
ладают узкой специфичностью и легко переходят с одного хозяина
на другого. Клещи, паразитирующие на домашних и диких живот-
ных (лошади, козы, овцы, верблюды, кошки, собаки, волки, лисы
и ДР-), способны перейти на человека, вызывая характерные пора-
жения кожи.
Рис. 94. Чесоточный зудень (А — по Павловскому, Б, В — по Дубинину).
А — самка клеща; Б — самка и отложенное ею яйцо в ходе, прогрызенном в
толще эпидермиса; В — схема расположения ходов, проделанных нимфой (сле-
ва), самкой (в середине) и самцом (справа):
/ — входное, 2 — вентиляционные отверстия
Специфичный паразит человека — чесоточный зудень (рис.
94, Д) имеет 0,4 мм в длину. Тело его одето очень тонкой, почти бес-
цветной кутикулой, несущей характерную исчерченность. На спин-
ной стороне располагаются многочисленные шипы и щетинки. Ма-
ленькая округлая гнатосома несет трехчленистые пальпы п клешне-
видные хелицеры. Ноги очень короткие и толстые, с присосками на
концах лапок.
Зудни прогрызают ходы в толще эпидермиса, поселяются на гра-
нице рогового и мальпигиева слоев и питаются клетками последне-
го. Самцы чесоточных зудней копулируют не с половозрелыми сам-
ками, а с нимфами. Последние после оплодотворения обычно выхо-
дят на поверхность кожи и пробуравливают новые ходы, где
я линяют, превращаясь в самок*. Расселение клещей обычно бывает
приурочено к ночному времени, когда поверхность тела согревается
и несколько увлажняется за счет деятельности потовых и сальных
желез. При пониженной температуре клещи теряют активность.
Приступая к проделыванию хода, клещ отгибает хелицеры и сна:
чала прогрызает небольшую’ямку, в которую вводит лапки первой
пары ног. Затем, углубляя ямку работой хелицер, он вводит в нее
ноги второй пары и постепенно погружается в отверстие хода. Весь
процесс внедрения совершается быстро и занимает от 2,5 до 10 мин.
В течение суток самка прогрызает ход длиной в 1—2 мм; по мере
продвижения она проделывает вентиляционные отверстия — корот-
кие вертикальные ходы, сообщающие основную галерею с внешней
средой. Обычно возле этих отверстий и откладываются яйца. Вс:его
за 1,5—2 месяца жизни самка откладывает от 20 до 50 яиц. Эмбри-
ональное развитие продолжается 2—3 дня. Вышедшая из яйца ли-
чинка не способна прогрызать эпидермис'и питается клеточным дет-
ритом, который скапливается в ходах в результате жизнедеятель-
ности самки. После линьки личинка превращается в нимфу первой,
в затем и второй стадии. В среднем жизненный цикл чесоточного
зудня занимает около 15 суток.
Патогенность чесоточных зудней
Чесоточные зудни могут поселяться в любом месте кожи, но чаще
они используют для внедрения наиболее нежные ее участки. У жи-
вотных клещи поселяются на морде; у человека — в складках кожи
между пальцами, на сгибах рук и т. д. Пробуравливая эпидермис и
перемещаясь в его толще, клещи раздражают нервные окончания,
вызывая ощущение нестерпимого зуда, который особенно чувству-
ется ночью, очевидно, в связи с повышением активности клещей в
тепле. При расчесывании пораженных мест неизбежно вскрываются
ходы и клещи заносятся руками на соседние, еще не пораженные
участки кожи. Таким путем осуществляется дополнительное рассе-
ление паразитов.
Чесотка — мучительное заболевание, иногда принимающее эпи-
демический характер. Тяжесть болезни нередко усугубляется за-
несением инфекции в расчесываемые места. В запущенных случаях
на коже образуются толстые корки, иногда даже препятствующие
разгибанию суставов (норвежская чесотка). Заражение человека
происходит при непосредственном контакте с больными людьми
или животными. Можно заразиться и при пользовании одеждой
•больных, так как чесоточные зудни некоторое время (до двух не-
дель) могут оставаться живыми вне тела хозяина.
Лечение чесотки производится втиранием в кожу специальных
мазей, мыла <с серой, раствора гипосульфита с последующей обра-
боткой кожи слабой (6%) соляной кислотой и т. п. Для лечения до-
машних животных, тоже сильно страдающих от чесотки, применяют-
ся очищающие ванны, окуривание, кожи серой (в специально обору-
дованных камерах) и обработка другими препаратами.
206
ПОДОТРЯД TROMBID1FORMIS REUTER, 1909
Довольно разнородная группа, насчитывающая в своем составе около 70 се-
мейств и очень большое число видов. Сюда входят как свободноживущие, так
н паразитические клещи, в том числе и клещи, паразитирующие на растениях.
Сем. Eriophyidae — это клещи-галлообразователи, со своеобразной биологией
и сложным ходом жизненного цикла. Рассмотрение их не входит в задачу на-
стоящей книги. Паразитические представители Trom bidi form is характеризуются
расположением дыхалец (стигм) на переднем конце тела, в основании хелицер.
Последние в большинстве случаев преобразованы в колющие щетинки.
Особого внимания заслуживает сем. Trombiculidae Ew-ing, 1944, представите-
лям которого свойствен личиночный паразитизм. В половозрелой фазе эти клещи
обитают в почве. В большинстве случаев они окрашены в яркий киноварно-крас-
Рис. 95. Клещи подотряда Trombidiformis (из кн. Павловского). А, Б — красно-
телка Trombidiuin automnalis (А — личинка, Б — взрослый клещ); В — железница
угревая Demodex folliculorutn '
ный цвет. Тонкий хитиновый покров клещей песет бесчисленные волоски, придаю-
щие телу бархатистый вид. Личинки Trombiculidae (рпс. 95), так называемые
«краспотелки», паразитируют на наземных позвоночных, в том числе и на че-
ловеке.
Питаясь кровью хозяина, личинки вводят в место укуса токсический секрет
своих слюнных желез, который вызывает покраснение кожи, сопровождающееся
образованием волдырей (папулл) и ощущением зуда. Болезнь известна под на-
званием «осенней эритемы». Опа носит сезонный характер и обычно приурочена
ко второй половине лета и к началу осени, когда в массе появляются вышедшие
из яиц личинки клещей. При одновременном нападении большого числа личинок
зуд может быть очень сильным, особенно ночью. В результате расчесывания па-
пулл, образующихся в местах укуса, возможно внесение различных инфекций,
осложняющих течение болезни. Заражение происходит непосредственно в приро-
де — в лесу или при садово-огородных и полевых работах и не зависит от кон-
такта с больными людьми или животными. Некоторые японские красиотелки
р. Trontbicula распространяют тяжелое трансмиссивное заболевание, вызываемое
риккетсиями (с. 190), известное под названием цуцугамуши или «японская реч-
ная лихорадка». Личинки клещей близкого сем. Trombidiidae Leach, 1815,— пара-
зитируют на членистоногих, в том числе и на вредителях (например, на саранче).
Некоторые исследователи ставят вопрос о возможности использования этих кле-
щей для биологической борьбы с вредными насекомыми.
207
Довольно близка к названным семействам группа водных клещей — Hydra-
carina, объединяющая несколько самостоятельных семейств. Эти клещи, часто
окрашенные в яркий киноварный цвет,— в половозрелой фазе активные хищкккщ
Они питаются различными водными членистоногими (насекомыми и планктонны-
ми рачками); нападая па добычу, клещи прокалывают покровы жертвы хелпце-
рамп, вводят в ее тело пищеварительные ферменты и потом высасывают полупе-
реваренную кашицу. Тело гидракарин обычно одето эластичной мягкой кутикулой.
На длинных ногах плавательные волоски. Весной самки откладывают яйца
на различные подводные предметы (камни, ветки и т. п.). Кладки, как и сами
клещи, имеют ярко-красный цвет. Вышедшие из яиц личинки всплывают на по-
верхность и благодаря широко расставленным тонким ногам, поддерживающим
крошечное тело личинки, легко бегают по поверхностной пленке воды. Как только
вблизи появляется какое-либо насекомое (жук-плавунец, стрекоза, водный клоп
и др.), личинки быстро прикрепляются к поверхности его тела и, вонзив свой
хоботок в тонкие участки сочленовного хитина, начинают питаться за счет хо-
зяина. Паразит довольно быстро растет и, достигнув второй нимфальной стадии,
отрывается от хозяина и переходит к самостоятельной жизни.
Наконец, к Trombidiformis относится сем. Demodicidae; один из представителей
которого — Demodex folliculorum (Simon)—паразитирует в коже человека. Тело
клеща размером 0,38X0,44 мм червеобразно вытянуто (рис. 95), одето тонкой
прозрачной кутикулой. Гнатосома несет короткие пальпы и щетинковидные хе-
лицеры. Ноги очень короткие, заканчиваются парой коготков. Клещи поселяются
в сальных железах (откуда и происходит их русское наименование — железницы)
или в волосяных сумках. Они могут паразитировать в совершенно здоровой коже.
Однако в случае закупорки волосяной сумки или воспаления железы создаются
условия, по-видимому, наиболее благоприятные для клещей и они скапливаются
здесь в большом количестве. В таких случаях развиваются гнойные прыщики
(угри). Заражение железницами (или угрицами) осуществляется при непосредст-
венном контакте с больным человеком.
ПОДТИП TRACHEATA — ТРАХЕЙНОДЫШАЩИЕ
Подтип трахейнодышащие объединяет наземных членистоногих,
дыхание которых осуществляется с помощью хорошо развитой тра-
хейной системы. Для Них характерна редукция конечностей перво-
го сегмента и потому расположенные на голове усики соответст-
вуют антеннам I (антеннулам). Подтип Tracheata включает 2 клас-
са: Myriapoda — многоножки и Insecta — насекомые. Лишь во вто-
ром паразитизм получил широкое распространение.
КЛАСС INSECTA —НАСЕКОМЫЕ
Насекомые — самый обширный класс среди беспозвоночных жи-
вотных. Тело Insecta отчетливо подразделяется на три тагмы: голо-
ву, грудь и брюшко. Голова несет усики (сяжки), глаза и сложно
устроенный ротовой аппарат. Особенности организации последнего
в первую очередь определяются характером употребляемой пищи и
способом ее поглощения. У видов, перешедших к питанию соками
растительных тканей или кровью позвоночных животных, форми-
руются разного типа сосущие и колющие хоботки (с. 222,256). Сле-
дующий за головой отдел — грудь — у всех насекомых состоит из
трех сегментов, каждый из которых несет по паре ходильных ног.
У абсолютного большинства высших насекомых два последних груд-
ных сегмента снабжены крыльями. Лишь некоторые формы, пере-
шедшие к постоянному эктопаразитизму (вши, блохи, ряд двукры-
208
дых и т. д.), утратили их. Последний отдел тела — брюшко, как пра-
вило, лишен конечностей. Иногда сохраняются гомологичные им
придатки (церки и др.), выполняющие самые разнообразные функ-
ции.
Развитие большинства насекомых осуществляется с неполным
(Hemimetabola) или полным (Holometabola) метаморфозом.
Высокая биологическая пластичность насекомых позволила им приспосо-
биться к существованию в самых различных условиях и распространиться прак-
тически по всему земному шару. Эта же особенность обусловила и широкий
переход насекомых к паразитизму. Одни из них стали облигатными паразитами
растений (отр. Homoptera — тли, червецы, листоблошкн, ряд бабочек, двукрылых,
перепончатокрылых и т. д.), вызывающими специфические повреждения расти-
тельных тканей. Другие же приспособились к паразитированию на животных.
Превращение взрослых насекомых в паразитов животных в большинстве случаев
было связано с возникновением гематофагии. В разных отрядах отчетливо rfpo-
слеживаются основные этапы этого процесса: на смену факультативному пита-
нию кровью растительноядных или хищных форм приходит обязательное крово-
сосание, без которого оказывается невозможным нормальное размножение вида.
Временный паразитизм сменяется постоянным эктопаразитизмом. Последнее не-
избежно влечет за собой морфологическую и биологическую специализацию
(утрата крыльев, изменение формы тела и конечностей, появление специальных
адаптаций, направленных на расселение вида, и т. д.).
Не менее широко у насекомых распространен паразитизм и на личиночных
фазах развития, причем последние часто становятся облигатными эндопаразита-
ми. В некоторых случаях (отр. Hymenoptera) мы сталкиваемся даже с явлением
яйцевого и эмбрионального паразитизма. Классические примеры подобного рода
можно обнаружить в обширной группе наездников (Parasitica), входящей в со-
став подотр. Apocrita. Личинки более примитивных наездников ведут себя как
эктопаразиты. Развитие яиц осуществляется либо во внешней среде, и тогда по-
явившиеся на свет особи активно отыскивают своего хозяина, либо самки сразу
же откладывают яйца на поверхность тела насекомых соответствующих видов.
Й в том и в другом случае паразиты не проникают в глубь тела хозяина, а оста-
ются на его поверхности и питаются высасываемой гемолимфой.
Самки более специализированных Parasitica откладывают яйца непосредст-
венно в ткани своих хозяев, которыми для одних видов служат взрослые насе-
комые, для других — их личинки и куколки, для третьих же — яйца. Эндопара-
зитизм наложил глубокий отпечаток па весь ход развития наездников. Прежде
всего у них наблюдается уменьшение размеров яиц. Это связано с уменьшением
и даже полным исчезновением откладывающегося в яйцах желтка. Отсутствие
последнего приводит к замене поверхностного дробления, свойственного большин-
ству членистоногих, полным.
У одних видов оболочка яйца не претерпевает сколько-нибудь серьезных из-
менений. В этом случае уменьшение запасов желтка обусловливает необходи-
мость более раннего вылупления личинки, которая выходит из яйца, будучи еще
морфологически недоразвитой, но вполне способной к самостоятельному питанию
за счет организма хозяина. Происходит упрощение и самого процесса эмбрио-
нального развития, в частности полностью исчезают зародышевые оболочки.
У других же видов под влиянием паразитизма в первую очередь изменяется обо-
лочка яйца: она становится проницаемой для пищевых веществ, которые и ис-
пользуются развивающимися эмбрионами. Трофическую функцию берет на себя
сохраняющаяся у таких видов зародышевая оболочка — сероза.
В результате паразитического образа жизни у развирающихся личинок наезд-
ников нередко возникает типичная полиэмбриония — распадение массы эмбрио-
нальных клеток на многочисленные группы, каждая из которых дает начало са-
мостоятельному зародышу. Полиэмбриония приводит к значительному увеличе-
нию численности потомства.
Окукливание личинок наездников происходит либо во внешней среде, либо
в теле хозяина. Взрослые насекомые ведут себя как типичные свободноживущие
организмы. Личиночный паразитизм распространен и среди отряда двукрылых,
209
паразитирующих па гусеницах различных бабочек или в тканях позвоночных жи-
вотных (с. 268).
Количество отрядов, выделяемых в настоящее время в кл. Insecta, превышает
30. Три из них: Hemiptera— Kjjonbi, Hymenoptera— перепончатокрылые н Dipte-
га — двукрылые — включают как свободноживущие. так и паразитические виды.
Отр. Mallophaga — пухоеды, Anoplura — вши и Siphonaptera — блохи целиком
представлены паразитами.
Обьем настоящей книги не позволяет с достаточной подробностью остано-
виться на всех группах паразитических насекомых. Ниже будут рассмотрены
лишь некоторые из них, имеющие наиболее важное медицинское значение.
ОТРЯД HEMIPTERA ( = HETEROPTERA LATR.,
1817)—ПОЛУЖЕСТКОКРЫЛ ЫЕ, ИЛИ КЛОЦЫ
Клопы составляют обширный отряд, насчитывающий 30—40 тыс.
видов в своем огромном большинстве—свободноживущих. Разви-
тие клопов идет с неполным превращением. Число паразитических
видов очень ограничено.
Рис. 96. Постельный клоп (из кн. Павловского). А — взрослое насекомое; Б —
ротовой аппарат клопа; В — яйцо
/ — крышечка яйца, 2 — клеевой секрет, 3—верхняя губа, -/—мандибулы, 5—мансиллы;
6 — нижняя губа
Отличительными признаками отряда могут служить особенности
строения крыльев и ротового аппарата. Передняя пара крыльев в
основной части сильно хитинизирована, тогда как вершинная часть
крыла, называемая перепоночкой, тонкая, прозрачная с хорошо за-
метными жилками. Крылья плоско складываются на спине. Ротовой
аппарат колюще-сосущего типа (рис. 96, Б). Максиллы и мандибул
лы, преобразованные в колющие щетинки, вложены, как в футляр,
в желобок видоизмененной и сильно вытянутой нижней губы, со-
стоящей из трех или четырех члеников. Максиллы, несущие по
внутреннему краю два желобка, на поперечном разрезе имеют вид
фигурной скобки. Складываясь вместе, они образуют два канала.
Один из них (верхний) служит для всасывания крови пли другой
жидкой пищи. Через второй ч (нижний) канал изливается секрет
слюнных желез. Мандибулы лежат кнаружи от максилл; обе пары
210
щетинок подвижны по отношению друг к другу. В покое колющий
хоботок всегда подогнут на брюшную сторону тела насекомого.
Характерной особенностью клопов является наличие пахучих же-
лез, выделяющих секрет с резким неприятным запахом (за исключе-
нием одного тропического вида, пахнущего гиацинтом). У взрослых
насекомых протоки желез открываются на заднегруди, возле тази-
ков последней пары ног. У личинок функционируют другие железыг
выделяющие такой же секрет, но открывающиеся на спинной сторо-
не тела, на границах между тергитами III—V члеников брюшка.
Отверстия пахучих желез всегда открыты.
Многие свободпоживущие клопы — хищники. Из наземных —
это представители сем. Reduviidae (хищнецы), из водных — гла-
дыш, водяной скорпион, водомерка и многие другие. Всем им свой-
ственно внекишечнос пищеварение. Нападая на мелких насекомых
и других членистоногих, клопы парализуют или убивают жертву
секретом своих слюнных желез, а затем, вводя в ее тело пищева-
рительные ферменты, переваривают и высасывают ее.
Эволюция способа питания хищных клопов могла идти в двух
направлениях. Прокалывая ткани растений, насекомые, по-видимо-
му, приспособились к питанию их соками. Эта линия эволюции
получила большое развитие. Огромное большинство клопов — рас-
тительноядны. С другой стороны, наличие сильных и жестких ко-
лющих щетинок, легко проникающих даже через покровы позво-
ночных, дало клопам возможность перейти к гематофагии. Такой
переход можно видеть в пределах сем. Reduviidae, многие предста-
вители которого стали временными паразитами ящериц, птиц и
млекопитающих. Некоторые хищнецы — гематофаги поселяются
при этом в норах грызунов (сусликов, песчанок и др.). Есть и си-
нантропные виды, обитающие в хозяйственных постройках и жилых
домах, где они питаются различными насекомыми. Появление та-
ких видов, вероятно, должно было предшествовать переходу кло-
пов к временному паразитированию на человеке. Вероятно, в поль*
зу этого предположения говорит способность некоторых хищнецов—
гематофагов наряду с питанием кровью пожирать постельных кло-
пов и других синантропных насекомых. По-видимому, паразитизм
хищнецов можно считать филогенетически относительно молодым.
Типичными эктопаразитами являются представители неболь-
шого семейства Cimicidae, насчитывающего около 20 видов. Внеш-
ний облик этих насекомых довольно сильно изменен под влиянием'
паразитического образа жизни. Тело широкое и сплющенное в дор-
зо-вентральМом направлении, имеет округлую или яйцевидно-оваль-
ную форму. Ноги широко расставлены. Крылья редуцированы
(рис. 96, X). Тело одето эластичным и растяжимым хитиновым пок-
ровом, несущим многочисленные волоски и щетинки. Все Cimici-
dae— облигатные гематофаги, нападающие на хозяина для соса-
ния крови. Они питаются на человеке, многих млекопитающих и
птицах, но активны только в ночное время. Днем клопы прячутся в
норах грызунов, в гнездовой подстилке, в различных укромных мес-
тах жилища человека. Насосавшись крови, клопы-могут длительное
211
время находиться вне тела хозяина, переваривая заключенную в
кишечнике пищу.
Имеется и еще одно семейство паразитических клопов, близко
родственное Cimicidae. Это очень мелкие насекомые — Polyc-
tenidae, живущие на теле тропических летучих мышей. Polycte-
nidae — постоянные эктопаразиты, всю жизнь проводящие на теле
хозяина. Они сидят, погрузив хоботок в кожу летучей мыши, и со-
сут кровь почти непрерывно. Подобно многим другим паразитичес-
ким организмам, эти клопы отрождают живых личинок.
Клопы — паразиты человека
Клопы — временные паразиты человека — относятся к двум се-
мействам. Среди хищнецов (сем. Reduviidae) насчитывается около
100 таких видов. Это представители р. Triatoma (например, Т. san-
guisuga — «поцелуйный» клоп) и некоторые другие клопы, распро-
страненные главным образом в странах Южной Америки. По боль-
шей части это крупные насекомые с хорошо развитыми крыльями,
нередко ярко окрашенные. Они ведут ночной образ жизни, днем пря-
чутся во всевозможных укрытиях и, будучи потревожены, очень
быстро убегают от света. Яйца откладывают кучками.. Одна опло-
дотворенная самка может отложить до 300 яиц. Эмбриональное
развитие продолжается 2—3 недели. Личинки проходят пять линек,
причем каждой линьке обязательно должно предшествовать полно-
ценное питание кровью. Весь цикл развития от яйца до имаго про-
должается год, а у некоторых видов — даже два года.
Клопы-хищнецы нападают ночью на спящих людей, вводя свой
хоботок в кожу открытых частей тела (лица, шеи, рук) и сосут
кровь долго, иногда на протяжении получаса. Момент введения хо-
ботка безболезнен, и неприятные ощущения, связанные с попада-
нием в кровь слюны- насекомого, возникают обычно позднее. У не-
которых людей укусы этих клопов могут вызвать тяжелую аллерги-
ческую реакцию. Наиболее опасными для человека видами считают-
ся: Pahtostrongylus megistus (крупный черный клоп с ярко-красны-
ми пятнами), распространенный в Бразилии, Triaioma infestans
(Аргентина), космополитный вид Т. rubrofasclata и др. Триатомо-
вые клопы имеют большое эпидемиологическое значение как пере-
носчики Trypanosoma cruzi—возбудителя болезни Чагаса. Зареги-
стрировано около 37 видов клопов, передающих это тяжелое забо-
левание.
Ко второму семейству (Cimicidae) относятся клопы р. Cimex.
Это широко распространенный постельный клоп — Cimex lectulari-
us L. и близкий к нему топический вид—С. hemipterus. Подобно
остальным паразитическим клопам, они временные облигатные па-
разиты, нападающие на хозяина для сосания крови, и активны в ноч-
ное время. Лишь очень голодные насекомые нападают на человека
днем или при искусственном освещении. Обычно постельный клоп
в светлое время суток прячется в щелях, под обоями, занавесками, в
старой мебели, книгах и т. д. Здесь же самки откладывают яйца;
довольно крупные (до 1 мм) и снабженные крышечкой (рис. 96,В),
212
они приклеиваются к субстрату секретом придаточных желез сам-
ки. В среднем за сутки откладывается около 12 яиц; общая же по-
ловая продуктивность самки (за весь срок ее жизни) составляет
несколько сот яиц. Скорость эмбриогенеза зависит от температуры
окружающей среды. При 35—37° С личинки выходят из яиц уже че-
рез 4—6 дней, тогда как при 14—18° С их развитие затягивается
почти до месяца, а ниже 10° С вообще приостанавливается. Превра-
щение личинки в имаго связано с линьками и при благоприятных
условиях (оптимум — 35° С) занимает около месяца, при комнат^
пой температуре — полтора-два месяца. Для осуществления мета-
морфоза личинки должны регулярно питаться кровью, однако они
способны выносить длительное голодание, оставаясь живыми до по-
лутора лет.
Взрослые клопы живут около 14 месяцев. Они тоже способны
долго (более полугода) голодать, особенно в холоде. Повышение
температуры и влажности неблагоприятно сказывается на жизне-
способности клопов. В поисках хозяина клопы ориентируются с по-
мощью обоняния. Насекомые нередко переползают из квартиры в
квартиру по стенам домов, преодолевая при этом значительные рас-
стояния. Двигаются они очень быстро. За минуту клоп может про-
ползти более метра.
Клопы хорошо адаптированы к гематофагии. Они извлекают
кровь непосредственно из сосудов, не разрушая их и не вызывая об-
разования гематомы. Их колюще-сосущий хоботок работает при
этом по принципу шприца. Приступая к сосанию крови, насекомое
иногда несколько раз подряд вводит свой хоботок в кожу хозяина,
пока не найдет подходящий кровеносный сосуд. Момент укола без-
болезнен, но введение слюны вызывает болевые ощущения и зуд, на
коже образуются довольно быстро проходящие красные пятна и
волдыри. Степень чувствительности к токсическому воздействию
слюны клопов очень индивидуальна. Некоторые люди почти не ощу-
щают его. В слюне клопов, так же как и у других гематофагов, со-
держатся антикоагулянты. Сосание продолжается от 7 до 15 мин,
за это время самка клопа поглощает около 7 мг крови. Постельный
клоп может питаться также кровью мелких млекопитающих и'птиц
(например, кур и голубей). По-видимому, это обстоятельство игра-
ет известную роль в том, что клопы иногда длительное время сохра-
няется в оставленных жилищах. С другой стороны, на человека
могут нападать клопы, паразитирующие на разных млекопитающих
(летучих мышах и др.) и птицах. Известно, что паразит голубей —
Cjmex columbarius нередко переползает в квартиры из гнезд, рас-
положенных под крышами домов.
Значение постельного клопа
В неопрятно содержащихся жилищах клопы могут размножаться
в несметных количествах. Нападая ночью на спящих, они нарушают
сон, лишают человека нормального отдыха. В этом главное значение
клопов как паразитов человека. Роль клопов в переносе инфекцион-
213
ных болезней, хотя и подозревалась неоднократно, но до сих пор
не доказана. Известно лишь, что в организме клопов могут длитель-
ное время оставаться живыми возбудители висцерального лейшма-
ниоза (кала-азар) и других трансмиссивных болезней. Так, напри-
мер, риккетсии Провачека (с. 228) и возбудители Q-лихорадки спо-
собны размножаться в желудке постельного клопа. Чумные палочки
остаются в его кишечнике живыми на протяжении двух, а спирохе-
ты — возбудители возвратного тифа, даже пяти месяцев. Длитель-
ное время (свыше полугода) сохраняются и возбудители туляре-
мии. Однако в распространении этих болезней клопы, по-видимому,
роли не играют. Возможно, что это связано с их биологией. Дефе-
кация клопов никогда не происходит на коже хозяина, и поэтому
контаминнтивный путь заражения затруднен или даже исключен.
В экспериментальных условиях постельный клоп может инокуля-
тивным путем передавать возбудителей крысиного сыпного тифа
(с. 239), гемолитический стрептококк и. вирус куриной оспы. Тем не
менее эпидемиологических данных, которые подтверждали бы
участие постельных клопов в* переносе этих и других инфекционных
болезней, пока нет.
Наличие в жилище человека клопов всегда рассматривается как
показатель санитарного неблагополучия. -Для истребления этих
насекомых в настоящее время применяются различные, контактные
яды длительного действия (хлорофос, метафос и др.), которыми об-
ливаются стены, мебель, книги и т. и. Клопы гибнут, соприкасаясь
с обработанной ядами поверхностью. Применение этих средств тре-
бует осторожности, так как все они, в той или иной мере ядовиты и
для человека.
ОТРЯД MALLOPHAGA NITZSCH, 1818, — П У X О Е Д Ы
Цухоеды представляют собой относительно небольшую группу
паразитических насекомых, насчитывающую немногим более 2500
видов. Они обитают в перьях птиц (пухоеды, пероеды или перовики)
и в волосяном покрове млекопитающих (власоеды). Развитие их
идет с неполным превращением. По своему внешнему виду пухоеды
напоминают вшей, с которыми их раньше путали. Основные приз-
наки различия между этими близкими отрядами сводятся к особен-
ностям строения ротового аппарата (грызущего у пухоедов и ко-
люще-сосущего у вшей) и некоторым деталям эмбрионального раз-
вития.
Относительно систематического положения пухоедов пет единого мнения.
В отечественной энтомологической литературе они фигурируют как самостоятель-
ный отряд. Однако зарубежные авторы нередко объединяют их вместе со вшами
в единый отр. Phiraptera. Такой же точки зрения придерживался в начале века
Н. А. Холодковский, объединявший обе группы в отряд • ложиохоботковых
(Pseudorhynchota).
Пухоеды известны очень давно. Первое упоминание о них можно найти
у Ф. Реди (1668), однако начало серьезному изучению этой группы было поло-
жено трудами Ш. Ницша (Ch. Nitzsch, 1818), который не только описал ряд ви.-
дов, но и дал первую классификацию, в основных чертах сохранившую свое
значение до наших дней. В середине прошлого века пухоедам были посвящены
214
капитальные исследования Г. Денни (Н. Denny, 1892), Г. Бурмейстера (Н. Bur-
meister, 1838), Г. Гибели (G. Giebel, 1874) и Е. Пьяже (Е. Piajet, 1880, 1885).
Монография, созданная последним автором, снабжена великолепным атласом,
включающим изображения всех известных к тому времени пухоедов. В дальней-
шем в изучении этой группы было многое внесено трудами американского ис-
следователя В. Келлога (V Kellog, 1896—1914). Из современных зарубежных
работ необходимо назвать монографию В. Энхлера (W. Eichler, 1963), в которой
не только дается подробный анализ морфологии пухоедов, но разбираются также
вопросы их эволюции, распространения и практического значения. В пашен стра-
не изучением пухоедов специально занимался Д. И. Благовещенский, кроме того,
отдельные сведения по биологии этих паразитов имеются в трудах В. А. Догеля
и его учеников (В. Б. Дубинина и др ).
Морфология пухоедов
Пухоеды — довольно мелкие насекомые, от 1 до 100 мм длиной.
Тело их дорзо-вентрально уплощено, имеет узкую и вытянутую или
овально-яйцевидную (рис. 97) форму. Голова чаще широкая и плос-
кая, иногда с хорошо выраженными височными выступами. Лоб
и наличник сильно развиты и образуют выступ, с помощью которо-
го пухоеды, ползая, раздвигают густой волосяной или пуховой
покров хозяина. Антенны укорочены и основания их смещены кза-
ди, что тоже облегчает движение паразитов в толще пухового по-
крова. Этому же способствуют и многочисленные щетинки, направ-
ленные свободными концами назад. Ротовые части грызущего типа.
Они смещены на^нижнюю поверхность головы, что позволяет пухое-
дам захватывать пищу с поверхности пера или кожи хозяина. Глаза
сильно редуцированы. Они представлены одним или двумя омма-
тидиями и, по-видимому, позволяют лишь отличать свет от темноты.
У некоторых видов редукция идет еще дальше и сохраняется только
хрусталик омматидия, тогда как рабдом и пигментные клетки
исчезают. Редукция глаз, несомненно, связана с паразитическим
образом жизни пухоедов, скрывающихся от света в пуху или шерсти
хозяина. I
Грудь довольно явственно расчленена. Каждый сегмент несет
по паре укороченных (рис. 97), слегка уплощенных, сильных ног.
В зависимости от образа жизни отдельных групп пухоедов это мо-
гут быть ноги бегательного типа (подотряд АтЫусега) или конеч-
ности, приспособленные к закреплению на пере или волосе (подотр.
Ischnocera). В последнем случае функцию прикрепления часто вы-
полняют сильно укороченные передние ноги. У многих пухоедов
на голенях развиваются разного рода выросты и шипы. Цепляясь
за бородки пера подвижным коготком лапки, паразит прижимает
их к этим шипам, защемляя таким способом участки пера и при-
крепляясь к опахалу. Сходным образом удерживаются и ползают
на волосах хозяина власоеды (сем. Trichodectidae). Характерной
чертой всех Mallophaga является отсутствие крыльев. Бескрылость
эта, несомненно, вторична и связана с паразитическим' образом
жизни.
Тело пухоедов одето тонкой, но очень прочной и эластичной хити-
новой кутикулой. Эластичность покровов рассматривается как адап-
215
тивная особенность строения пухоедов. Это позволяет им избегнуть
опасности быть раздавленными клювом птицы. На поверхности
тела располагается множество щетинок. Часть их связана с нервны-
ми окончаниями и, по-видимому, играет роль органов осязания.
Длинные щетинки, расположенные вокруг глаз, считаются защит-
ными.
Рис. 97. Представители отряда пухоедов (паразиты перьевого покро-
ва куликов). А—A ctor nithophilus (подотр. Amblycera); Б — Esthiop-
terum\ В — Docophoms (подотр. Ischnocera):
1 — антенны, 2 — глаза, 3 — копулятивный аппарат, 4 — «лобпыйэ вырост,
5 — мандибулы
Пухоеды обычно окрашены в светло-желтый, бурый и темно-бу-
рый цвета, причем интенсивность тона окраски зависит от степени
склеротинизации отдельных участков кутикулы. В некоторых слу-
чаях можно говорить о защитном характере окраски пухоедов.
Так, например, виды, поселяющиеся в черном оперении каравайкн
или лысухи, окрашены в темно-бурый, почти черный цвет. В то же
время пухоеды белой цапли совсем не пигментированы.
Классификация
Отряд Mallophaga подразделяется на два подотряда, довольно значительно
отличающихся друг от друга. Для подотряда Amblycera характерно наличие ниж-
нечелюстных щупиков. Антенны короткие (4—5 члеников), булавовидные н мо-
гут вкладываться в специальные ямки или бороздки, по-внднмому, играющие
216
защитную роль. АгпЫусега — очень подвижные паразиты; они держатся в пухо-
вой части покрова или непосредственно на коже птиц. Второй подотряд — Ischno-
сега характеризуется отсутствием нижнечелюстных щупиков. Антенны их ните-
видны и свободно выдаются по бокам головы. Паразиты мало подвижны, обладают
хорошо развитыми адаптациями к прикреплению. Держатся преимущественно
на жестких кроющих перьях спины и крыла. К этому же подотряду относятся
и паразиты волосяного покрова млекопитающих, представленные 'Небольшой,
по числу видов, группой Trichodectoidea.
Биология пухоедов
Данные, приведенные в этом разделе, относятся в основном
к паразитам перьевого покрова птиц, изученным более полно, чем
власоеды.
Пухоеды относятся к числу постоянных паразитов, тесно связан-
ных с хозяином на протяжении всей своей жизни. Они характери-
зуются чрезвычайно узкой специфичностью, и каждый вид пухое-
дов строго приурочен к определенному виду хозяина. Жизненный
цикл пухоедов проходит с неполным превращением. Самки откла-
дывают яйца, приклеивая их застывающим на воздухе секретом,
непосредственно к бородкам перьев или (власоеды) к волосам.
Из япца выходит личинка, рост и развитие которой продолжается
18—20 дней.
Питаются пухоеды главным образом частицами пера и воло-
сом. При этом они могут, иногда очень сильно, повреждать как
перья, так и шерсть хозяина. В связи с характером пищи наблюда-
ются адаптивные изменения в строении пищеварительной системы
пухоедов. Кератин пера относится к разряду очень стойких про-
теинов, с трудом поддающихся перевариванию; По-видимому, имен-
но с этим связано появление у пухоедов зоба, отделенного от дру-
гих разделов пищеварительной системы специальными хитиновыми
образованиями. Это исключает и выход заглоченной пищи обратно
в пищевод, и прохождение необработанных пищевых частиц в сред-
нюю кишку. Последнее, по-видимому, особенно важно, так как
предотвращает возможность повреждения пищеварительного трак-
та. В полости зоба происходит предварительная механическая
и химическая обработка пищи.
Многие виды пухоедов (преимущественно из отр. АтЫусега)
способны, кроме того, питаться кровью. Они «слизывают» капельки
крови, выступающие из случайных ранок и царапин на коже хозяи-
на. Некоторые виды могут активно повреждать кожу мандибула-
ми. Это характерно для представителей сем. Menoponidae и в осо-
бенности для паразита кур Eomenacanthus straumineas, который
предпочтительно держится не на пере, а непосредственно на коже
птицы. Настоящим облигатным гематофагом становится паразит
пеликана Piagetiella (-Tetraphthalmus) titan, поселяющийся в под-
клювном мешке хозяина. Это настоящие эндопаразиты, сохраняю-
щие связь с перьевым покровом птицы лишь во время размножения.
По наблюдениям В. Б. Дубинина, в этот период они выползают
из подклювного мешка и откладывают яйца на перьях головы
пеликана.
8—58
217
В отдельных редких случаях пухоеды локализуются в полости
очинов пера. Таковы Actornithophilus patellatus, паразиты куликов.
Эти пухоеды прогрызают отверстия в стержне маховых перьев
и проникают внутрь очинов, где питаются роговыми частицами
и откладывают яйца.
Пухоеды, обитающие непосредственно на перьях, приурочены
к различным участкам опе-
рения птицы, представляю-
щим собой своеобразные
мнкростации (рис. 98). Эта
приуроченность часто со-
провождается адаптивными
особенностями строенияхпа-
разитов. Так, например,
пухоеды, поселяющиеся на
кроющих перьях спины и на
маховых перьях (представи-
тели р. Lipeurus, р. Anatico-
1а и др.), характеризуются
узким,' вытянутым телом,
сильно уплощенным в дор-
зовентральном направлении.
Такая форма тела позволя-
ет этим паразитам забирать-
ся в толщу опахала, распо-
лагаясь между лучами пера.
Короткне передние ноги
этих видов (рис. 97, Б) дей-
ствуют как зажимы. Все это
обеспечивает паразитам воз-
можность противостоять то-
кам воздуха, возникающим
во время полета птицы.
Рис. 98. Распределение различных видов удержаться при этом на пе-
пухоедов на опереиии каравайки (по Ду- ре. Напротив, пухоеды типа
бинину) Docophorus (рис. 97, В),
обладающие коротким, ши-
роким телом и крупной головой с сильно развитыми челюстями,
хорошо приспособлены к жизни среди коротких, мелких перьев го-
ловы и шеи.
Зависимость биологии пухоедов от биологии их хозяев
Расселение пухоедов. Переход на новые особи хозяина обеспе-
чивается переползанием паразитов с одной птицы на другую. Это.
возможно только при тесном контакте птиц между собой и осуще-
ствляется,- главным образом, в периоды спариванья и гнездованья..
Зараженность птиц пухоедами, как правило, очень велика, од-
нако процент зараженных особей в значительной степени зависит
218
от образа жизни хозяина. Показательны в этом отношении данные,
полученные учениками и сотрудниками В. А. Догеля при изучении
паразитов птиц Волжской дельты. Наиболее сильно оказались за-
ражены так называемые колониальные птицы — цапли, бакланы,
каравайки и др., образующие при гнездовании тесные поселения —
колонии. Скученность гнезд в колониях и контакт птиц друг с дру-
гом в этих условиях так велики, что создаются возможности легкого
перехода пухоедов не только с одной птицы на другую, но и из гнез-
дй в гнездо. Все колониальные птицы заражены на 100%. Совсем
иная картина характерна для птиц, которые держатся поодиночке
или парами (пастушки, погоноши поганки и др.), зараженность
которых в дельте Волги редко превышает 35—50%. Известную роль
в расселении паразитов перьевого покрова играют мухи кровосос*
ки, часто нападающие на птиц. Пухоеды прикрепляется к брюшку
этих мух и переносятся ими на значительные расстояния. По сви-
детельству Г. С. Маркова, изучавшего паразитофауну скворцов,
около 25% найденных им мух кровососок (О г nit homy а) несли на се-
бе пухоедов.
В начале периода гнездованья обычно наблюдается вспышка
размножения пухоедов. В результате к моменту вылупления птен-
цов интенсивность заражения взрослых птиц бывает особенно высо*
кой. Это имеет важное значение для распространения паразитов,
так как молодые птицы заражаются от родителей. Сначала на птен-
цов переселяются пухоеды подотряда Amblycera. Это, как правило,
мелкие насекомые, приспособленные к жизни в пуху или даже
на поверхности кожи хозяина-, очень подвижные и быстрые. Птенцы
выводковых птиц заражаются через несколько часов после вылуп-
ления. У птенцовых срок заражения значительно отодвигается. Пу»
хоеды же подотр. Ischnocera, малоподвижные и поселяющиеся толь-
ко на пере, переходят на птенцов лишь после того, как оперение
уже почти сформировано. Птенцы кукушки, не имеющие контакта
с птицами своего вида, заражаются специфичными для них пухоеда-
ми только через год, во время спаривания (Догель, 1947).
Влияние линьки и миграций хозяина. Пухоеды, как и перьевые
клещи (с. 203), зависят от состояния оперения, на котором они жи->
вут и питаются. Во время линьки условия жизни паразитов в зна-
чительной мере меняются. Перья, которые перестают смазываться
секретом копчиковой железы, становятся сухими и ломкими. В ре-
зультате обламывания и выпадения таких старых перьев весь по-
кров становится более редким. Появляется множество растущих,
молодых перьев с кровенаполненными сосочками, что увеличивает
влажность оперенйя. Все это сказывается на пухоедах, которые, ос-
тавляя свои микростацин, беспорядочно расползаются по всему
телу птицы. Как и в случае с перьевыми клещами, это в какой-то
мере спасает популяцию паразитов от гибели. Тем не менее боль-
шое количество пухоедов выпадает вместе со старым пером. В ре*
зультате птицы в значительной мере освобождаются от пухоедов
и. процент их заражения снижается в два-три раза, Резко падает
и интенсивность инвазии. У колониальных птиц, зараженных осо-
8* 219
бенно сильно, столь заметного очищения от паразитов обычно
не наблюдается.
Вследствие ухудшения условий существования пухоедов, обус-
ловленного началом линьки, насекомые обычно прекращают отклад-
ку яиц. Зато с восстановлением нормальных условий жизни начина-
ется вспышка размножения паразитов.
Пухоеды, паразитирующие иа перелетных птицах, относятся
(как и перьевые клещи) к категории убиквистов 1. Перелет, свя-
занный с длительным пребыванием птицы в воздухе, создает
для паразитов перьевого покрова опасность быть сдутыми ветром.
Очевидно, с этим связана интенсивная откладка яиц пухоедами
незадолго до начала миграции. Птицы уносят на себе не только
самих пухоедов, но и большое количество их яиц, надежно при-
клеенных к оперению.
Патогенность пухоедов и борьба с ними
Пухоеды, иногда поселяющиеся на хозяевах в очень большом
количестве, вызывают раздражение кожи и зуд, а иногда и прямое
повреждение эпидермиса, открывающее «ворота инфекции». У птиц
нарушается целостность перьев, у зараженных власоедами млеко-
питающих клочьями выпадает шерсть. Вследствие этого в овцевод-
ческих районах власоеды могут наносить значительный материаль-
ный ущерб. Среди пухоедов домашней птицы наибольшее значение
имеет Eomenacanthus strautnineus, паразитирующий на курах.
При сильном заражении птицы теряют в весе, яйценоскость снижа-
ется, нарушается нормальный рост цыплят. Зараженные куры за-
метно выделяются по внешнему виду: оперенье их всегда взъеро-
шено, птицы беспокойны, непрерывно чистятся клювом, пытаясь
выловить паразитов. Не исключено, что пухоеды могут играть из-
вестную роль в передаче инфекционных болезней (риккетсиозов
и вирусных заболеваний) домашних животных.
Борьба с пухоедами относительно проста, так как весь жизненный цикл па-
разитов проходит на теле хозяина. Она сводится к применению различных инсек-
тицидов. Однако яйца паразитов устойчивы по -отношению к ним, так это обра-
ботка должна быть повторной. В борьбе с власоедами используется короткая
стрижка шерсти.
ОТРЯД ANOPLURA ( = S I Р Н U N С U L А Т А) - В Ш И
Вши — близкие к пухоедам кровососущие насекомые, паразити-
рующие только на млекопитающих. Это немногочисленный отряд,
насчитывающий около 300 видов.
1 Убиквистами В. А. Догель (1947) называет паразитов перелетных птиц, по-
стоянно присутствующих на теле хозяина. Их можно обнаружить на птицах как
в местах гнездования и зимовки, так и во время перелета.
220
Морфология вшей
Тело вшей уплощено в дорзальном направлении’. Маленькая го-
лова значительно уже груди, сегменты которой обычно слиты меж-
ду собой. У некоторых видов (например, у Phtirus pubis — площи-
цы) грудь почти не отграничена от брюшка (рис. 99), На голове
Рпс. 99. Вши, паразитирующие на человеке {А, В, Г — по Павлов-
скому; Б, Д— из ки. Ноблей). А — платяная вошь; Б — яйцо платя-
ной вши; В — головная вошь; Г, Д — лобковая вошь и ее яйцо;
Е—концевой членик лапки (пр Чендлеру):
/— крышечка, 2—секрет клеевых желез
располагается пара коротких антенн, основание которых (как
и у пухоедов) несколько смещено назад. Глаза рудиментарны или
совсем отсутствуют. Ротовой аппарат колюще-сосущего типа.
В процессе эволюции ротовые части вшей претерпели столь глубо«
кие изменения, что установить их гомологию с ротовыми конеч-
ностями других насекомых очень трудно. От хоботков остальных
кровососущих насекомых ротовой аппарат вшей тоже отличается
и не только малым числом колющих щетинок, но и тем, что в покое
он всегда втянут и целиком помещается внутри головы, лишь час-
тично выдвигаясь при сосании (рис. 100).
221
У человеческой вши (Pediculus hutnanus L.) ротовое отверстие
окружено особым воротничком, или ротовым конусом, который пе-
сет венчик коротких, но сильных крючочков, по пять с каждой сто-
роны. Когда паразит приступает к сосанию крови, воротничок вы-
ворачивается, крючочки выдвигаются наружу, их острия поверты-
ваются в стороны и служат для прикрепления головного конца на-
секомого к поверхности кожи хозяина. С нижней стороны головы
(под ротовым конусом) находится пара режущих склеритов, приво-
димых в движение специальными мышцами. С помощью этих скле-
ритов соскабливается поверхностный роговой слой кожи, что облег-
чает введение в нее тонких колющих щетинок.
Рис. 100..Схема строения ротового аппарата вши (из кп. Кестнера). А— в со-
стоянии покоя; Б — в момент сосания:
I —ротовое отверстие, 2 —втянутый колющий хоботок, 3 — влагалище колющих щетинок,
4 — проток слюнных желез, 5 — полость глотки, 6 — ротовая полость, 7 — ротовой конус,
В —дорзальные щетинки, 9 — вентральная щетинка, 10 — ретракторы хоботка, П —крючоч-
ки ротовора конуса
Ротовое отверстие ведет в ротовую полость, в которую открыва-
ется глотка, и расположенный под ней длинный мешок — влагали-
ще колющих щетинок (рис. 100, Д). Последних всего трц. Две дор-
зальные щетинки, по-видимому; гомологичны максиллам, тогда как
вентральная соответствует нижней губе. Она-то и служит для про-
калывания кожи хозяина, тогда как две желобообразные максиллы,
прикладываясь друг к другу, образуют трубку, по внутреннему
каналу которой и всасывается кровь Между максиллами и ниж-
ней губой лежит приросший к ней очень тонкий гипофаринкс;
он пронизан каналом, по которому проходит секрет слюнных желез
паразита, содержащий антикоагулянты. Комплекс колющих щети-
нок вводится насекомым непосредственно в просвет кровеносного
сосуда. Отсюда кровь сокращением мышц глоточного насоса пере-
качивается в пищеварительную систему- паразита. Благодаря про-
зрачности покровов вшей и красному цвету всасываемой крови,
этот процесс можно легко наблюдать на живом насекомом. Мышцы
глоточного насоса работают очень интенсивно, сокращаясь несколь-
ко раз в секунду.
1 Гомология колющих щетинок (стилетов) дается здесь по Б. Н. Шванвнчу
(1949), ио существуют и другие трактовки.
222
С гематофагией вшей связана слабая хитинизация их покровов,
лишь по бокам тела укрепленных склеротинпзированными участ-
ками. Когда кишечник паразита наполняется кровью, такая струк-
тура покровов обеспечшзаеТПзозможность их растяжения.
Вши совершенно лишены крыльев, что является следствием па-
разитического образа жизни. Сильные, слегка укороченные конеч-
ности вшей обычно широко расставлены (рис. 99). На концевом
членике лапки имеются крупные, подвижные коготки. Прижимая
коготок к специальному выросту голени (рис. 99, £), насекомое
может защемлять волосы хозяина и тем самым прочно удержива-
ется на нем.
Биология вшей и их приуроченность к хозяевам
Вши — типичные паразиты млекопитающих; на других живот-
ных никогда не встречаются. Характерной особенностью вшей явля-
ется узкая специфичность по отношению к виду животного хозяина.
Локализуются они обычно на поверхности кожи (у человека — в во-
лосистых частях тела). Многие виды вшей характеризуются значи-
тельной патогенностью. Очень важна их роль как переносчиков
инфекционных болезней (с. 227).
Вши довольно широко распространены на домашних животных. Паразитируя
на свиньях, рогатом скоте, верблюдах, собаках и др., они беспокоят животное
своими укусами, вызывающими сильный зуд. Места укусов часто становятся во-
ротами вторичной инфекции, что приводит к повреждению кожи.
Как правило, вши поселяются на наземных млекопитающих, однако есть
в виды, паразитирующие на водных животных. У таких видов в связи с перехо-
дом к жизни в водной среде появляются специальные адаптации к дыханию.
Развиваются многочисленные хитиновые волоски и щетинки, густо покрывающие
тело насекомого. В момент погружения хозяина в воду вокруг щетинок обра-
зуется воздушный пузырек, сквозь стенки которого происходит обмен газов
с окружающей средой. Кислород, растворенный в воде, диффундирует внутрь
пузырька через его стенку. Через нее же выходит наружу и выдыхаемая насеко-
мыми углекислота. Таким образом пузырек, облекающий тело вши,— это пе запас
воздуха, а своеобразное «физическое легкое». (Такой тип дыхания известен для
зимующих подо льдом жуков плавунцов и легочных моллюсков.) Это приспо-
собление наряду с большим диаметром трахей позволяет вшам ластоногих по-
селяться по всему телу хозяина, а не только на голове, как обычно указывается
(М. Я. Асс)‘.
Своеобразными особенностями биологии отличаются Lepidophtlrus macrorhi-
ni — паразиты морских слонов. В отличие от всех остальных вшей эти насеко-
мые локализуются не на коже, а внутри ходов, проделываемых ими глубоко
в толще кожного покрова хозяина. Такая локализация обусловлена способом
линьки морского слона, который сбрасывает не только волосы, но и ороговевший
поверхностный слой эпидермиса. Способность пробуравливать ходы избавляет
вшей от угрозы гибели в период линьки хозяина.
Вши — паразиты человека
Известно два вида вшей, являющихся специфичными паразита-
ми человека (рис. 99). Один из них — лобковая вошь, или площица
{Phtirus pubis Leach), локализуется непосредственно на теле чело-
века (в волосах лобка и подмышек, в бороде, бровях и даже рес-
ницах). Другой вид — человеческая вошь (Pediculus hutnanus L.)
223
представлен двумя сильно различающимися подвидами *. Один
из них—Pediculus humanus vestimenti (de Geer), или платяная
вошь, поселяется в одежде человека, тогда как другой — Pediculus
humanus capitis (de Geer), или головная вошь, паразитирует в во-
лосах головы. Оба подвида отличаются друг от друга не только
морфологически, но и рядом особенностей своей биологии и жиз-
ненного цикла. Тем не менее их рассматривают как один вид, так
как они способны скрещиваться, давая плодовитое потомство.
Размножение вшей
Вши обладают хорошо выраженным половым диморфизмом.
Самцы обычно меньше самок, задний конец их брюшка закруглен,
тогда как задний конец брюшка самки раздвоен. Плодовитость
вшей довольно велика. Платяная вошь откладывает в течение жиз-
ни около 300 яиц, по 5—14 яиц ежедневно. Площица (более тесно
связанная с телом хозяина) —всего 25—50 яиц в течение всей жиз-
ни. Оплодотворенные яйца вшей (гниды) одеты плотной блестя-
щей оболочкой и снабжены сложно устроенной крышечкой (рис.
99, Д), которая обеспечивает доступ воздуха внутрь скорлупки
яйца. Через имеющиеся в крышечке отверстия проникают спермато-
зоиды. Процесс откладки яйца занимает всего несколько секунд.
Самка захватывает волос или участок одежды специальными хити-
новыми придатками на конце брюшка (гоноподами) и выпускает
из полового отверстия каплю прозрачной жидкости. Это секрет
клеевых придаточных желез, сильно развитых у вшей. Капля
секрета выталкивается наружу готовым к откладке яйцом, во вре-
мя его выхода наружу. На волос, предварительно смоченный клей-
ким секретом, и попадает откладываемое яйцо. Уже через несколько
минут оно оказывается очень прочно приклеенным к субстрату, так
как секрет быстро застывает на воздухе.
Продолжительность эмбрионального развития вшей сильно зави-
сит от температуры; оптимальной является температура поверх-
ности тела человека, т. е. 30° С, при которой эмбриогенез продол-
жается 6—8 дней. Понижение температуры тормозит этот процесс.
Известно, что в складках одежды, периодически снимаемой и вновь
надеваемой, развитие яиц может затягиваться до шести недель.
Развитие вшей идет с неполным метаморфозом, поэтому вышед-
шая из яйца личинка отличается от имаго лишь малыми размерами
и отсутствием полового аппарата. Проделав в течение недели три
линьки, личинка превращается во взрослое насекомое, а через 1—2
дня после метаморфоза самка уже приступает к откладке яиц.
Таким образом, весь жизненный цикл платяной вши занимает
16 дней. Лобковая вошь развивается медленнее, и ее цикл осу-
1 До недавнего времени эти подвиды рассматривались как самостоятельные
виды. Некоторые авторы и сейчас придерживаются такой точки зрения (см. кни-
гу «Паразитология человека», 1975).
224
ществляется за 22—27 дней. Наибольшей продолжительностью жиз-
ни обладает платяная вошь (полтора-два месяца), тогда как голов-
.ная вошь живет около четырех, а лобковая — трех недель.
Питание вшей и другие особенности их биологии
Вши относятся к числу постоянных эктопаразитов, весь жизнен-
ный цикл которых проходит на теле хозяина. Поэтому в отличие
от других кровососущих членистоногих вши питаются по нескольку
раз в день, заглатывая кровь небольшими порциями. Особенно это
характерно для площиц, которые подолгу сидят на одном месте,
погрузив свой хоботок в кожу хозяина, и сосут кровь почти непре-
рывно, с частыми, но короткими промежутками. Кишечник платя-
ной вши вмешает до 1 мг крови. Для его наполнения насекомому
требуется от трех до десяти минут. Такое количество крови (при
температуре 30° С) переваривается в течение 10 ч. С понижением
температуры пищеварение замедляется, поэтому вши, оказавшиеся
вне тела хозяина, в течение некоторого времени обходятся без пи-
щи. В зависимости от условий температуры и влажности, насекомые
могут голодать от двух до десяти дней.
Вши чувствительны к температуре. Оптимальна для них тем-
пература поверхности кожи здорового человека (около 30°С). С по-
вышением температуры (например, у сильно лихорадящего больно-
го) вши переползают на одежду или постельное белье. После смерти
хозяина насекомые быстро покидают труп. Оказавшись во внешней
среде, они ползают в поисках нового хозяина, ориентируясь с по-
мощью антенн, на которых расположены органы обоняния. Зрение
не играет при этом особой роли.
При температуре 25—37° С вши могут быть очень активными.
Отмечено, что они способны вползти по вертикальной поверхности
на высоту до 1 м. На плоскости они за одцу минуту преодолевают
расстояние в 20—25 см. При тесном физическом контакте людей
между собой вши легко переползают с одного хозяина на другого.
Патогенность вшей и их роль в распространении болезней
Вши — паразиты человека и борьба с ними
Паразитирование вшей на теле человека сопровождается тягост-
ным зудом, вызываемым их частыми укусами. Неприятные, раздра-
жающие ощущения возникают и при ползании паразитов по поверх-
ности кожи. При укусе вошь вводит в ранку секрет слюнных желез.
Их два типа — подкововидные и бобовидные. Токсичными свойст-
вами обладает только секрет бобовидных желез. При введении
в кожу он вызывает ощущение зуда и жжения. У некоторых людей
как реакция на укусы вшей и введение их слюны повышается тем-
пература. В месте инъекции появляются плотные на ощупь возвы-
шения — папулы, характерного синеватого цвета. Они возникают
225
из-за поверхностных кровоизлияний и нарушения целостности маль-
пигиева слоя. Воспалительное расширение сосудов облегчает вшам
кровососаппе. При расчесывании мест укуса с папулы легко сди-
рается пленка эпидермиса. Таким путем открываются ворота вто-
ричной инфекции. В образующиеся ссадины часто попадают различ-
ные бактерии, в результате чего возникают поражения кожи. Осо-
бенно тяжелым и трудно излечиваемым являехся поражение воло-
систой части головы (колтун), при котором волосы склеиваются
в один сплошной ком, прикрывающий гноящуюся поверхность кожи.
В прошедшие века заражение вшами нередко представляло серь-
езную опасность, так как при сильной инвазии наступает и общее
ослабление организма. В гноящиеся ранки мухи могут откладывать
яйца, из которых выходят личинки, питающиеся живыми тканями.
Е. Н. Павловский (1948) приводит сведения о гибели некбторых
исторических лиц — испанского короля Филиппа II, римского дик-
татора Суллы и некоторых других, «заживо съеденных» вшами.
Паразитирование площиц вызывает особую форму заболева-
ния— фтириоз. Благодаря широкому, плоскому телу (рис. 99, Г)
ллощица очень плотно прикрепляется к поверхности кожи человека.
Погрузив свой колющий хоботок в толщу кожи, паразиты подолгу
неподвижно сидят на одном месте. Вызываемый ими зуд ощущается
непрерывно и днем и ночью. Площицы, сидящие на ресницах, неред-
ко становятся причиной воспаления век и тканей, окружающих глаз.
В местах их сосания долго (до 12 суток) сохраняются синеватые
пятна, вызванные воздействием секрета слюнных желез насекомого.
Эти пятна не возвышаются над уровнем кожи, не вызывают непри-
ятных ощущений. При массовых медицинских осмотрах (например,
во время призыва на военную службу) наличие пятен помогает
выявить носителей площиц, так как сами насекомые плохо замет-
ны. Заражение человека происходит при непосредственном контакте
с носителями паразитов, реже через белье. Вне тела хозяина пло-
щицы, плохо переносящие голодание, выживают всего 1—2 дня.
Площица не передает никаких инфекционных заболеваний.
В условиях скученности людей й невозможности выполнения пра-
вил личной гигиены платяная и головная вши могут размножаться
в несметном количестве. Борьба с ними сводится к тщательному
мытью тела горячей водой с мылом. Полезно использовать при этом
ядовитые для вшей препараты. Необходимо сбрить волосы на голо-
ве и других частях тела, так как это единственный способ изба-
виться от гнид. Секрет, с помощью которого яйца приклеены к во-
лосам, не поддается разрушению. Для уничтожения площиц при-
меняется втирание ртутной мази.
Е. Н. Павловский считает, что широко распространенное в дре-
вности натирание тела различными маслами и благовониями имело
•своей конечной целью избавление от вшей. Попадание на тело на-
секомых маслянистых веществ, залепляющих дыхательные отвер-
стия (стигмы), должно привести к их гибели. Это происходит, одна-
ко, не сразу. Известно, что вши способны жить без воздуха в тече-
ние одного-трех дней.
226
Вши — переносчики инфекционных болезней
Среди вшей, паразитирующих на диких и домашних животных,
есть виды, имеющие эпидемиологическое значение. Так, например,
специфичный паразит водяных крыс (р. Haplopleura) переносит
возбудителей туляремии (с. 187) от больных животных к здоровым.
В результате поддерживается постоянное существование природ-
ных очагов этого тяжелого заболевания. Однако наиболее важное
эпидемиологическое значение имеют вши человека — платяная
и головная, являющиеся переносчиками спирохет (Borrelia гесси-
rentis)— возбудителей возвратного тифа, риккетсий (Rickettsia
aDolhynica)—возбудителей волынской лихорадки, и риккетсий
(Rickettsia prowazeki), вызывающих эпидемический сыпной тиф.
Сыпной и возвратный тифы и история их изучения
Тифы — возвратный и в особенности сыпной, отнрсятся к числу
особо опасных инфекции и представляют собой тяжелейшие заболе-
вания человека. Для возвратного тифа характерны повторяющиеся
приступы изнурительной лихорадки с повышением температуры
до 40—41° При сыпном-тифе такая же лихорадка держится без ре-
миссий на протяжении 9—11 дней. Типично появление сыпи на ко-
же груди, спины и живота. Сыпной тиф вызывает тяжелые рас-
стройства кровообращения, головные боли и головокружения, нару-
шения сознания. Процент смертности колеблется в широких пре-
делах и зависит от условий и общего состояния больных. Причиной
смерти является специфическая интоксикация, а в конце болезни —
•осложнения, чаще всего пневмония.
История Европы изобилует сведениями об опустошительных эпидемиях сып-
ного тифа, как правило, связанных с различными социальными потрясениями пли
стихийными бедствиями. Только в течение XIX в. и первой половины XX в. эпи-
демии сыпного тифа унесли многие миллионы человеческих жизней. Огромную
роль в распространении этой страшной болезни сыграли войны. Перемещения
большого числа людей, создание концентрационных лагерей для беженцев и во-
еннопленных, антисанитарные условия жизни солдат в окопах — все это способ-
ствует распространению вшивости, а с нею и болезни.
Наполеоновские войны разнесли сыпной тиф по всей Европе. Известно, что
армия Наполеона за время наступления на Москву потеряла около трети своего
состава <?т эпидемий, в первую очередь от сыпного тифа. От тифа же погибло
и около трех пятых состава русской армии. Не менее впечатляющую картину дает
Крымская кампания. При осаде Севастополя французы потеряли от сыпного
тифа столько же солдат, сколько они потеряли уоитыми и ранеными. Интересно,
что в английской армии, где поддерживалась чистота и соблюдались правила
личной гигиены, потери, от тифа были неизмеримо меньшими. Во время русско-
турецкой войны тиф вывел из строя 200 тыс. солдат русской армищ причем число
умерших превышало число погибших от ран. Колоссальная вспышка сыпного тифа
сопровождала и первую мировую (империалистическую) войну. В период .же
гражданской войны и интервенции (1919—1920 гг.) эта страшная болезнь рас-
пространилась по всей территории России с небывалым по высоте показателем
заболеваемости (389 больных на 10 тыс. населения) >. По данным некоторых ав-
торов, за этот период переболело около 10 млн. человек. Сыпной тиф оказался
1 Эти и другие приведенные выше цифры заимствованы из книги Е. Н. Пав-
ловского «Руководство по паразитологии человека» (1948).
227
настоящим бедствием и для армии, и для гражданского населения молодой Со-
ветской Республики, и для борьбы с болезнью были мобилизованы все силы.
В период Великой Отечественной войны благодаря четкой организации меро-
приятий по борьбе со вшивостью (с. 230) советским органам здравоохранения
удалось избежать возникновения серьезных эпидемий. Успешное решение этой
проблемы стало возможным только в результате работы множества научных кол-
лективов, всесторонне изучавших риккетсий — возбудителей сыпного тифа и био-
логию вшей — их переносчиков. Разрабатывались и научно обоснованные методы
борьбы со вшивостью.
История изучения сыпного и возвратного тифов — это история самоотвержен-
ного, георического труда, сопряженного со смертельной опасностью. Немногим
более 100 лет назад (в 1873 г.) в крови человека, больного возвратным тифом,
были обнаружены спирохеты (Borrelia reccurentis) . Это открытие, сделанное Обер-
мейером, можно считать первым звеном в серии исследований, раскрывших при-
чины и пути распространения эпидемий тифа.
Вскоре после открытия Обермейера профессор Киевского университета
Г. Н. Мипх привил себе содержащую спирохеты кровь больного. В результате он
заболел тяжелой формой возвратного тифа и едва не погиб. Героический экспе-
римент Минха повторил врач О. О. Мочутковский, который привил себе кровь
сыпнотифозного больного и через 18 дней тяжело заболел, доказав тем самым,
что возбудитель сыпного тифа тоже локализуется в крови.
Таким образом была установлена заразность крови больных людей, и на этом
основании Г. Н. Мипх высказал предположение, что тнфы передаются от. челове-
ка к человеку при укусах кровососущих насекомых Одиако эта мысль оказалась
настолько впереди общепринятых зоологических и медицинских представлений
того времени, что не была оценена современниками и не получила дальнейшего
развития. Лишь, много лет спустя (в 1909—1914 гг.) идея Минха получила под-
тверждение в экспериментальных исследованиях французских ученых, работавших
в Тунисе (Nicoll с соавт.) и Алжире (Е. Sergent с соавт.). Минх ошибся лишь
в одном. Заражение человека как возвратным, так и сыпным тифом происходит
не инокулятивным, а контаминативным путем (см. ниже).
Для выяснения природы возбудителя сыпного тифа многое было сделано
американским ученым Г. Т. Риккеттсом/В 1909 г., изучая пятнистую лихорадку
скалистых гор, передаваемую клещами р. Dennacentor (с. 191), Г. Риккетс
(G. Ricketts) обнаружил возбудителя заболевания в крови больных и в теле кле-
ща переносчика. Это были очень мелкие, овальные и палочковидные тельца, непо-
хожие па известных до сих пор бактерий. Год спустя в 1910 г., Риккеттс нашел
сходные тельца в крови людей, больных сыпным тифом, и в кишечнике питав-
шихся на них вшей. Однако ученому не удалось довести свое исследование до
конца. Заразившись сыпным тифом, он тяжело заболел и скончался.
Между тем изучение возбудителей сыпного тифа широко развернулось в ла-
бораториях разных стран. Описанные'Риккеттсом микроорганизмы были найдены
и в Алжире (Sergent и др.), и в Европе. Многое было сделано известным чеш-
ским ученым С. Провачеком, который, работая с сыпнотифозными вшами, зара-
зился и погиб в 1915 г. Год спустя исследование, начатое Провачеком вместе
с Роха-Лима, было успешно завершено этим последним. Описанных им возбуди-
телей сыпного тифа Роха-Лима назвал в честь обоих погибших ученых — риккет-
сиями Провачека (Rickettsia prowazeki). Данные, полученные Провачеком и Роха-
Лима, составляют основу наших современных представлений об эпидемиологии
сыпного тифа.
Вши и эпидемический сыпной тиф
Вши, питающиеся на сыпнотифозном больном, всасывают рик-
кетсий вместе с кровью. Попадая в желудок насекомого, риккетсии
проникают в эпителий его стенок и интенсивно размножаются там.
1 Статья Минха «О высоком вероятии переноса возвратного и сыпного тифов
с помощью насекомых» опубликована в 1877 г. в журнале «Хирургическая ле-
топись». у
228
Пораженные эпителиальные клетки разрушаются, и массы риккет-
сий, высвобождаясь, оказываются в просвете желудка, а затем и ки-
шечника насекомого. Оттуда они выделяются наружу с фекалиями.
Для вшей заражение риккетсиями Провачека смертельно. В зави-
симости от интенсивности инфекции зараженные насекомые гибнут
в разные сроки, чаще всего на 15—18-й день, по-видимому, вслед-
ствие разрушения эпителиальных клеток стенки желудка.
Токсичная слюна вшей вызывает зуд и потребность расчесы-
вать кожу в местах укуса. В образующиеся при этом царапины лег-
ко могут быть занесены фекалии зараженных вшей, содержащие ри-
ккетсий, а в случае раздавливания насекомых — содержимое их
кишечника. Таким образом риккетсии проникают в организм че-
ловека через поврежденную кожу и заражение происходит контами-
нативным путем !.
Для борьбы с сыпным тифом необходимо знать пути распростра-
нения инфекции. Поскольку переносчиками служат только вши,
изучение их биологии приобретает особо важное значение. Уже от-
мечалось, что вши, в том числе и зараженные, легко переползают с
одного человека на другого. Они уходят с тела сыпнотифозного
больного при сильном повышении температуры, быстро покидают
остывающий труп. Оказавшись во внешней среде, насекомые ин-
тенсивно ползают в поисках нового хозяина. Все это способствует
распространению болезни, приводит к возникновению эпидемии.
Для оценки эпидемиологической роли вшей существенно знать
также, сколько времени остаются живыми вши и их яйца вне тела
человека во внешней среде. Этому вопросу посвящено множество
исследований. Кратко суммируя экспериментальные данные, можно
сказать, что максимальный срок выживания взрослых вшей состав-
ляет 10 дней. Оптимальна для них температура 10—25° О; при бо-
лее низкой и более высокой температуре вши погибают значитель-
но быстрее. Существенное значение имеет влажность. Во влажном
воздухе насекомые выживают дольше, чем в условиях значительной
сухости. Только что вылупившиеся личинки вшей способны жить
во внешней среде, пе питаясь, от 12 ч до 2—4 дней. Яйца вшей (гни-
ды) довольно стойки к неблагоприятным воздействиям; хотя опти-
мальной для их развития является температура 32—35 ° С, они вы-
живают при температуре +1—3 ° С в течение недели, а при повыше-
нии температуры до + 54 ° С остаются живыми более получаса.
Только +98° С убивает их в течение нескольких секунд. Без вреда
яйца выдерживают 10-минутное погружение в керосин, бензин, кар-
боловую кислоту и другие химические вещества.
Установлено, что риккетсии сохраняются долгое время в теле
мертвых высохших вшей. Эти сведения, а в особенности данные
о заразности фекалий вшей выявили некоторые новые аспекты эпи-
демиологии сыпного тифа. Оказалось, что риккетсии могут очень
1 Николль и другие ученые, изучавшие сыпной тиф, первоначально предпола-
гали наличие инокулятивного способа заражения, однако отсутствие риккетсий
в слюнных железах вшей полностью исключает эту возможность.
229
долго оставаться живыми в высохших фекалиях, хотя сроки эти
сильно зависят от температуры и влажности воздуха. Так, например,
при комнатной температуре в фекалиях платяной вши риккетсии
сохраняют инфекционность (для морских свинок) в течение многих
месяцев. В леднике они остаются живыми до года. Но при смачива-
нии фекалий сроки жизни риккетсий сильно сокращаются. Губи-
тельными для них являются воздействия различных дезинфицирую-
щих средств. На основании этих данных стало понятно, что зара-
зиться сыпным тифом можно и при отсутствии живых вшей.
Например — через одежду больного, в складках которой могут сох-
раниться сухие фекалии паразитов или мертвые насекомые. При
случайном попадании этого инфекционного начала в ссадины и ца-
рапины кожи или при занесении его загрязненными пальцами в
конъюнктиву глаза или на слизистые развивается заболевание
сыпным'тифом.
Борьба с эпидемическим сыпным тифом
Борьба с сыпным тифом — это борьба со вшивостью (педикулезом). Она про-
водится по трем основным направлениям: уничтожение вшей на теле человека,
уничтожение их в одежде и, наконец, во внешней среде. Первое строится на ос-
нове выполнения правил личной гигиены Вся одежда обрабатывается горя-
чим паром в специальных дезинсекционных камерах («санобработка»); для обез-
зараживания внешней среды необходимо тщательное мытье и очистка помещений.
Неукоснительное выполнение всех этих мероприятий приобретает особую важ-
ность при любом нарушении нормальных условий жизни людей (война, различ-
ные стихийные бедствия, связанные с массовыми переселениями, и т. п.). В ме-
стах возникновения сыпнотифозных очагов проводится вакцинация населения.
ОТРЯД S1PHONAPTERA (APHANIPTERA L A R.) — БЛОХИ
Блохи составляют отряд паразитических насекомых насчитыва-
ющий около полутора тысяч видов. Все они облигатные гематофаги,
паразитирующие только па теплокровных (преимущественно на
млекопитающих), и имеют важное эпидемиологическое значение
как дереносчики тяжелых инфекционных болезней человека (с. 235).
Развитие блох осуществляется с полным метаморфозом.
Морфология и жизненный цикл
Блохи — мелкие Насекомые от 1 до 5 мм длиной. Тело их сплю-
щено с боков и одето твердой, блестящей хитиновой кутикулой
коричневого или черного цвета. Края тергитов соседних сегментов
черепицеобразно налегают друг на друга, что сглаживает углова-
тость контуров и придает телу блохи обтекаемую форму. В сочета-
нии-с наличием большого числа щетинок, направленных остриями
назад, это облегчает блохам возможность быстрого передвижения в
густой шерсти млекопитающих или в перьевом покрове птиц.
У многих блох часть щетинок преобразована в довольно широкие,
сильно хитинизированные зубцы, образующие гребни (ктенидии),
которые располагаются на нижней части головы (рис. 101) или по
230 . - "
заднему краю переднеспипки. Наличие этих гребней помогает бло-
хе удержать в шерсти хозяина в момент введения в его кожу колю-
щего хоботка. Расположение щетинок на. теле п голове блох (хето-
таксия) и число зубцов, входящих в состав ктенидиев, имеют зна-
чение определительных признаков.
На голове блох находятся рудиментарные глаза, по-видимому,
гомологичные фасетированным глазам других насекомых. Лежащие
непосредственно позади глаз ко-
роткие булавовидные антенны, в
покое укладываются назад в спе-
циальные ямки.
Ротовые конечности сильно
видоизменены и преобразованы
в колюще-сосущий аппарат (рис.
101). По современным данным, у
блох полностью отсутствуют ман-
дибулы. Парные колющие щетин-
ки представляют собой максил-
лы1, от основания которых отхо-
дят листовидные выросты и
короткие нижечелюстные щупики.
Третья, непарная, колющая ще-
тинка является дериватом изме-
ненной верхней губы. Нижняя губа
редуцирована, но нижнегубные
щупики хорошо развиты. Они
имеют желобовидную форму и,
плотно прилегая друг к другу,
образуют футляр, в который за-
ключены колющие щетинки. Со-
Рис. 101. Голова и ротовой аппарат
блохи Ctenocephalides fells (по Ченд-
леру) :
/ — антенна в антеннальной ямке (2);
3 — зубцы ктенидия, 4 — листовидные вы-
росты максплл, 5 — нижнегубные щупики,
6 — макспллы, преобразованные в колю-
щие щетинки, 7 — верхняя губа, в —ниж-
нечелюстные щупики, 9 — глаз
санию крови предшествует ощупывание поверхности кожи хозяи-
на нижнечелюстными щупиками, после чего хоботок принимает
вертикальное положение и вводится в кожу на всю свою длину.
Обе пары щупиков и листовидные выросты максилл остаются при
этом на поверхности.
Блохи полностью лишены крыльев. Членики груди несут только
конечности. Последние хорошо развиты, в особенности третья пара,
представленная сильными прыгательными ногами. Брюшко состоит
из 10 члеников. На IX сегменте располагается своеобразный орган
чувств — пигидий. Это утолщенная хитиновая пластинка, на кото-
рой сосредоточено множество разнообразных сенсилл. У блох име-
ется явственно выраженный половой диморфизм. Самцы несколько
меньше -самок, задний конец их брюшка загнут кверху и несет слож-
но устроенный копулятивный аппарат. На заднем конце брюшка
самки имеется колбовидный хитиновый резервуар — семяприемник
(или сперматека). Форма и строение копулятивного аппарата, так
1 Колющие щетинки блох рассматривались раньше как мандибулы, а максил-
лами считались лишь их листовидные выросты.
231
же как и форма сперматеки, используются в систематикё блох как
важные определительные признаки.
Самки блох откладывают яйца в таких местах, где в достаточном
количестве имеются органические остатки, составляющие основ-
ную пищу личинок. Чаще всего — в норах млекопитающих, в гнез-
дах птиц, в различных хозяйственных постройках (сараи, курятники
и т. п.) и в жилище человека. Самка откладывает около 400—
Рис. 102. Блохи и стадии их развития (из кн. Е. Нобля и Г. Нобля).
А — личинка: Б — куколка в коконе, покрытом частицами пыли; В — чело-
веческая блоха Pulex irritans; Г — песчаная блоха Tunga penetrans
500 яиц, ио за один прием обычно не более 6—10. Период кладки,
обязательно связанный с питанием кровью, может растягиваться на
несколько месяцев. Блохи, некоторых видов размножаются в тече-
ние круглого года. Сроки развития яиц сильно варьируют как в за-
висимости от вида блох, так н под воздействием факторов внешней
среды, главным образом температуры п влажности. В среднем эм-
бриогенез продолжается от трех до пятнадцати дней.
Вышедшие из яиц безногие личинки, непигментированные и ли-
шенные глаз (рис. 102, Л), внешне напоминают личинок длинноусых
двукрылых. Они имеют хорошо развитую головную капсулу и рото-
вой аппарат грызущего типа. Личинки питаются детритом и высох-
шими фекалиями взрослых блох, всегда'содержащими остатки по-
лупереваренной крови. Период личиночного развития продолжается
232
в среднем около двух недель, но при неблагоприятных условиях (по-
ниженная температура, голодание) может затягиваться до трех ме-
сяцев. Личинки дважды линяют и перёд окуклением плетут легкий
паутиновый кокон, к которому приклеивают песчинки, частички дет-
рита. Замаскированный таким образом кокон становится совершен-
но незаметным. После третьей линьки личинка превращается в ку-
колку свободного типа. Вылупление имаго может произойти уже
через 8—14 дней, но при неблагоприятных условиях, главным обра-
зом при понижении температуры, задерживается на много месяцев.
Так, например, у человеческой блохи (Pulex irritans) стадия кукол-
ки иногда продолжается от 7 до 239 дней, у крысиной (Xenopsylla
cheopis) — от 1 до 182 дней и т. д.
Интересно, что в отсутствие хозяина, покинувшего нору или гнез-
до, вылупление блох не происходит в течение длительного времени.
В таких случаях внутри кокона заключена уже не куколка, а впол-
не сформированная, но совершенно неподвижная взрослая блоха.
Стимулом к вылуплению может служить малейшее сотрясение поч-
вы. Это своеобразная адаптация к заражению хозяина, неожидан-
ное появление которого в брошенной норе вызывает мгновенный
выход блох из всех находящихся поблизости коконов. Такую же
картину можно наблюдать и в заброшенных строениях (старых из-
бах, сараях, землянках и т. п.). Зашедшего туда человека букваль-
но облепляют вылупившиеся от сотрясения половиц блохи. Сведе-
ния о длительности жизни взрослых блох довольно разноречивы.
Считается, что в нормальных условиях блохи живут в среднем 3—
4 месяца. При понижении температуры длительность их жизни уве-
личивается, а в условиях эксперимента (по данным И. Г. Поффа)
блохи могут жить даже до пяти лет.
Некоторые особенности биологии блох
Блохи не могут долго существовать без хозяина, потому что
единственным источником пищи для них служит кровь. Введя свой
хоботок в ткани животного, паразиты остаются в таком положении
от нескольких минут до нескольких часов и даже месяцев (с. 234).
Интенсивность питания в значительной мере определяется темпера-
турой окружающей среды. Так, например, собачья блоха (Ctenocep-
halides canis) может сосать кровь лишь при температуре не ниже
13° С. Нормально блохи должны питаться на хозяине не реже од-
ного раза в сутки. Но есть данные, указывающие на способность
блох переносить длительное голодание в течение 3—4 месяцев,- а
при низкой температуре — даже до года и более. Питаясь, блохи вы-
сасывают больше крови, чем может поместиться в их кишечнике.
Поэтому капли непереваренной крови все время выступают из
анального отверстия насекомого, засыхая, они падают в гнездовую
подстилку и служат пищей находящимся там личинкам.
Большая часть блох (95% всех известных видов) паразитирует
на млекопитающих, преимущественно на грызунах. Блохи подвиж-
ны, но по поверхности почвы перемещаются на небольшие расстоя*
9—58 233
ния (не далее 1 м). Благодаря способности к прыжкам, они легко
переходят с одной особи хозяина на другую. Некоторые блохи, осо-
бенно те, которые паразитируют на крупных млекопитающих, со-
вершают прыжки до 20—30 см в высоту, преодолевая при этом рас-
стояние до 50 см. Человеческая блоха — Pulex irritans (рис. 102, В)
прыгает на 33 см. По-видимому, это необходимо паразитам, чтобы
попасть на хозяина. Блохи, поселяющиеся на мелких животных
(например, на грызунах), могут прыгать лишь на небольшие расстоя-
ния (до 15 см), а иногда и просто переползают с одного хозяина на
другого. Блохи, паразитирующие на птицах и летучих мышах, сов-
сем не прыгают. Многие виды блох характеризуются очень широкой
специфичностью. По данным И. Г. Иоффа, из 71 вида изучавшихся
им ‘блох птиц и млекопитающих 36 видов оказались способны на-
падать также и на человека и питаться его кровью. Это обстоятель-
ство имеет важное эпидемиологическое значение (с. 238).
По характеру связи с животным-хозяином блохи могут быть раз-
биты на временных (большинство) и постоянных (около 100 видов)
эктопаразитов. Пожалуй, наименее выражена связь с хозяином у
блох птиц. Это, по существу, обитатели гнезда, только в период гнез-
дованья нападающие на хозяина для сосания крови. Блохи многих
видов грызунов тоже проводят большую часть своей жизни в норе,
локализуясь в гнездовой подстилке. Они переходят иа хозяина в оп-
ределенные периоды года. К этой же экологической группе следует,
вероятно,'отнести и человеческую блоху, которая, хотя и питается
на человеке, в остальное время держится на постельном белье, в
щелях пола или в норах синантропных грызунов. Гораздо более тес-
но связаны с хозяином блохи кошек, собак и других хищников, по-
долгу сосущие кровь.
Промежуточное положение между временными стационарными
паразитами занимают своеобразные блохи р. Vermipsylla. Это экто-
паразиты копытных (овец, лошадей, крупного рогатого скота и др.),
которые, прикрепившись к телу животного, сосут кровь непрерывно
на протяжении нескольких месяцев. Тело паразитов, в котором на-
капливается за это время огромное количество яиц, сильно увеличи-
вается в размерах (до 1,6 см), принимая червеобразную форму. Это
становится возможным за счет растяжения тонких участков хитино-
вой кутикулы между тергитами. По окончании питания блохи по-
кидают хозяина и приступают к откладке яиц. Интересно, что в
этот период насекомые могут передвигаться лишь с помощью чер-
веобразных сокращений огромного раздувшегося брюшка.
К числу стационарных эктопаразитов следует отнести группу
тропических блох Echidnophaginae. Подобно вшам-площицам эти
насекомые всю жизнь проводят на теле хозяина, погрузив в его тка-
ни хоботок. Вероятно, именно в связи с этим нижнегубные щупики
таких блох слабо развиты, а максиллы зазубрены и служат не толь-
ко для сосания крови, но и для прикрепления.
Для тропических песчаных блох — Tunga (-Dermatophilus) - pe-
netrans (рис. 102, Г) характерен переход к эндопаразитизму. Это па-
разиты человека и млекопитающих. Нападают обычно оплодотво-
234
ренные самки, которые в ожидании подходящего хозяина держатся
на поверхности почвы. Локализуются они чаще на ногах (у человека
на подошвах ног, между пальцами или под ногтями) и довольно
быстро проникают в подкожную соединительную ткань. Они связа-
ны с внешней средой при помощи узкого отверстия, остающегося в
месте внедрения блохи. Питание кровью и лимфой хозяина приво-
дит к интенсивному развитию яичников. В теле блохи формируются
тысячи яиц, периодически с силой выбрасываемые наружу через
ранку в коже хозяина. Тело блохи в этот период очень сильно раз-
дувается за счет растяжения мягкой сочленовной кутикулы. Пара-
зит принимает вид шарика размером с горошину, вокруг которого
наблюдается опухолевидное разрастание тканей.
Патогенность блох
Питаясь кровью человека или животного, блоха вводит в ранку
декрет своих слюнных желез, который вызывает болевые ощущения,
Чувство жжения и зуда. Частое нападение блох нарушает нормаль-
ный отдых и сои ночью. Паразитирующие на овцах и других копыт-
ных блохи р. Vertnipsylla высасывают большое количество крови,
вызывая иногда сильное истощение хозяина. Куриные блохи Echid-
nophaga gallinacea (распространены в США) прикрепляются обыч-
но на коже гребня и непрерывно в течение двух месяцев сосут кровь.
Отмечено замедленное развитие зараженных этими паразитами
цыплят. Блохи р. Echidnophaga могут нападать также на грызунов
или домашних млекопитающих, у которых локализуются в ушных
раковинах. Иногда 'нападают на человека.
Особенно неприятны последствия паразитирования песчаной
блохи. Ранки, образующиеся в результате ее внедрения в кожу, ча-
сто воспалены и очень болезненны. Воспаление усиливается, если
случайно раздавить сидящую в коже блоху. Нередко развиваются
вторичные инфекции, которые могут привести к потере пораженного
пальца или даже всей конечности. В Южной Америке зарегистри-
рованы случаи смерти от гангрены и столбняка, явившихся следст-
вием паразитирования песчаной блохи. Родиной этого паразита яв-
ляется Южная Америка, но в 70-х годах прошлого века песчаная
блоха была завезена в тропическую Африку и теперь широко рас-
пространилась там.
Эпидемиологическое значение блох
Блохи и чума
Чума — тяжелейшая эпидемическая болезнь человека, в основ-
ном распространяемая блохами, хотя возможна и непосредствен-
ная передача инфекции от человека к человеку.
Заболевание начинается внезапно, резким подъемом'температу-
ры, необычайной слабостью и общей разбитостью, сразу же доводя-
щей больного до состояния полной прострации. Характерные симп-
9* 235
томы чумы проявляются на вторые сутки. Наиболее распространена
бубонная чума, связанная с появлением бубона, т. е. плотной, впос-
ледствии нагнаивающейся опухоли в лимфатических узлах, в кото-
рых происходит интенсивное размножение чумных микробов. Чаще
бубоны локализуются в паховых и подмышечных, реже в шейных
лимфатических узлах. Смертность от бубонной чумы велика, но еще
большую опасность представляет легочная форма чумы. Развиваю-
щееся при этом специфическое воспаление легких сопровождается
множественными кровоизлияниями и некрозом легочной ткани. При
отсутствии лечения это приводит к смерти почти в 100% случаев.
Наконец, кожная форма чумы характеризуется появлением гной-
ничков, местными омертвениями кожи и многочисленными крово-
излияниями. На последних этапах болезни кровоизлияния слива-
ются друг с другом, так что вся кожа покрывается крупными тем-
но-синими пятнами. Именно этот симптом и дал повод называть
чуму «черной смертью». Кожные кровоизлияния обычно считаются
показателем общего сепсиса, наступающего при интенсивном раз-
множении микробов в крови.
Кроме описанных местных явлений болезнь сопровождается тя-
желым бредОхМ, нарушением речи и другими расстройствами цент-
ральной нервной системы. Тяжелые нарушения, иногда становящие-
ся непосредственной причиной смерти, наблюдаются и в сердечно-
сосудистой системе. Болезнь развивается бурно и, как правило, не-
продолжительна. Больной погибает через 3—4 дня, а иногда даже
в первые сутки. Если же он остается жив, то заболевание обычно
заканчивается через 7—8 дней. Смертность при чуме необычайно
велика. Так, например, во время эпидемии 1894 г. в Гон-Конге она
составляла 60—97%.
Пожалуй, трудно назвать болезнь, которая принесла бы человечеству больше
страданий и бедствий, чем чума. По-видимому, первые достоверные сведения
о ней относятся к концу второго, началу третьего века нашей эры. Однако поми-
мо этих и множества последующих более или менее значительных эпидемий чумы
в истории запечатлены сведения о трех пандемиях, носивших характер всеобщего
бедствия и охватывающих целые страны и континенты.
Первая такая пандемия известна под названием «чумы Юстиниана». Она
вспыхнула в 542 г. в Египте, охватила Сирию, Малую Азию и Константинополь,
где в течение четырех месяцев ежедневно умирало от 5 до 10 тыс. человек.
В последующие века вспышки чумы, иногда довольно значительные, регистриро-
вались в разных странах Европы, но самая страшная пандемия, вошедшая в ис-
торию под названием «черная смерть», разразилась в XIV в. В Европу болезнь
проникла из Азии, где в 1334 г., по свидетельству китайских историков, от чумы
умерло 5 млн. человек. Отсюда, с караванами, чума двинулась через Индию
и Малую Азию в Константинополь, а потом через Аравию в северную Африку
п па Средиземное море. В 1348 г. почти полностью вымерло от чумы население
Кипра. В течение ближайших двух-трех лет болезнь распространилась по всей
Европе, вплоть до Норвегии, неся с собой страшные опустошения. Смертность
во время этой эпидемии превзошла все, что было известно до и после нее. Евро-
па потеряла, в среднем, четвертую часть всего населения (около 25 млн.). В от-
дельных же странах бедствие выглядело еще более ужасным. В Италии погибла
половина всех жителей, некоторые небольшие города и поселения вымерли пого-
ловно. В. 1351 г. «черная смерть» проникла в Россию п распространилась в Пско-
ве, Новгороде, Смоленске и других городах. Судя по летописям, в Смоленске
уцелело всего 10 человек.
236
После ужасающей пандемии XIV в. чума периодически повторялась, вспыхи-
вая то здесь, то там, но эпидемии не принимали таких грандиозных размеров.
3 России тоже отмечались вспышки чумы, иногда очень сильные. В их числе
нужно назвать эпидемию 1603 г. в Москве, 1684 г.— на больших пространствах
от Казани до Чернигова и от Астрахани до Новгорода. В XVIII В. от чумы силь-
но пострадала армия Петра I под Ригой, в XIX в. болезнь неоднократно вспыхи-
вала в Одессе, в Астрахани и т. д. Последняя, третья, пандемия, начавшаяся
в 1894 г. в Гон-Конге, охватила в основном Индию и другие Азиатские страны.
Именно в этот период были сделаны важнейшие открытия, положившие начало
научно обоснованной борьбе с чумой.
В 1893—1894 гг. француз Иерсен и японец Китасато одновременно и незави-
симо друг от друга описали возбудителя чумы — чумную палочку, которая впо-
следствии получила название Pasteurella pestis. Непосредственно вслед за этим
(в 1896 г.) работавший в Индии русский врач В. А. Хавкин создал первую про-
тивочумную вакцину. Предварительно попробовав ее на себе, Хавкин начал ши-
рокую вакцинацию населения в районах, где свирепствовала эпидемия, и достиг
резкого снижения заболеваемости, а среди забЬлевших — снижения смертности.
В 1897 г. М. Огата, а год спустя Зимонд получили экспериментальные доказа-
тельства участия блох в переносе чумы. Наконец, и это тоже очень важно,
в 1897 г. русский, врач Д. К. Заболотный впервые высказал мысль о том, что ис-
точником распространения чумы могут служить дикие животные.
В настоящее время эпидемиология чумы рисуется следующим
образом. Главным распространителем болезни являются блохи. Пи-
таясь на больных людях или животных, блохи всасывают вместе
с кровью и возбудителей чумы. В желудке и кишечнике насекомого
чумная палочка начинает интенсивно размножаться. В результате
бактерий становится так много, что они полностью блокируют же-
лудок блохи, образуя так называемые «чумные пробки», препятст-
вующие нормальному питанию насекомого. Зараженная блоха пы-
тается сосать кровь, но пища в ее желудок не проходит. Всасыва-
емая кровь наталкивается на чумную пробку, частично разрушает
ее и уносит с собой некоторое количество бактерий. При повторных
попытках кровососания эта порция крови нередко попадает обрат-
но в тело хозяина через ранку, сделанную колющими щетинками
блохиг-Этого уже достаточно для заражения. Голодная блоха мно-
гократно повторяет свои попытки сосать кровь на-одном или не-
скольких хозяевах, каждый раз внося инфекцию* Гибнут такие бло-
хи довольно быстро (через 7—14 дней), хотя иногда, вследствие ча-
стичного разрушения блока, проходимость их кишечника может
временно восстанавливаться. Столь интенсивное размножение Pas-
ieurella pestis в кишечнике имёет место далеко не у всех видов
блох. Поэтому не все они играют одинаковую роль в распростране-
нии чумы. Главным переносчиком служит крысиная блоха Хепор-
sylla cheopis Rothshild (рис. 103).
Блохи могут передавать чуму и при отсутствии «чумного блока»
в кишечнике. Доказано, что чумные бациллы всегда имеются в фе-
калиях блох, дефекация которых часто происходит во время крово-
срсанпя на хозяине: Случайно втирание выделенных таким путем
бактерий в ранки и царапины на коже, занесение их с загрязненны-
ми пальцами на слизистую глаз и носа приводит к заражению'чу-
мой. Таким образом блохи jdpryT передавать эту инфекцию и кон-
таминативным путем. Возможен также механический перенос чум-
237
ных микробов на ротовых частях блох, загрязненных содержащей
возбудителей кровью животного.
Теперь уже давно твердо установлено, что основной источник
чумы — зараженные грызуны. Различают антропургические (свя-
занные с человеком) и природные очаги чумы. В первом случае ис-
точником инфекции служат домовые крысы {Rattus norvegicus,
Rattus rattus). Вспышки чумы среди крыс могут возникать в любой
точке земного шара, но в тропической зоне они поддерживаются
Рис. 103. Крысиная блоха Xenopsylla
cheopis, главный переносчик чумы
(из Чендлера)
постоянно. Причины этого кро-
ются в сочетании температуры и
влажности, благоприятном для
размножения крысиных блох
Xenopsylla cheopis, играющих
первостепенную роль в переносе
возбудителей чумы. В умеренных
и северных широтах эти блохи не
бывают многочисленными, а дру-
гие виды, хотя и переносят чуму,
но далеко не так активно. Оче-
видно поэтому эпидемии чумы за
пределами тропиков редко быва-
ют продолжительными.
Важную роль в распростране-
нии болезни всегда играли торго-
вые связи. Болезнь разносилась
многочисленными судами, отправ-
лявшимися из тропической зоны
во все страны мира. Крысы, в
изобилии водившиеся в корабельных трюмах и в складских пост-
ройках любого торгового порта, играли при этом решающую роль.
Не случайно, что именно в портовых городах чаще вспыхивали
эпидемии. В России наиболее неблагополучна была Одесса, где
вспышки чумы были отмечены даже в начале XX в. (1910 г.).В на-
стоящее время, благодаря хорошо налаженной борьбе с грызунами
и с блохами, этот путь распространения чумы практически пре-
сечен.
Природные очаги чумы существуют и сейчас. Они поддержива-
ются сусликами, сурками, песчанками, тарбаганами и другими
грызунами. Основными видами блох, участвующими в переносе ин-
фекции, являются паразиты сусликов Ceratophyttus tesquorum,
Neopsylla setosa и др. При вспышках эпизоотий среди грызунов ес-
тественная зараженность блох может достигать 60%. Температура
окружающей среды действует на продолжительность жизни чумных
палочек в кишечнике блох. При 22—25° С бактерии выживают около
трех недель, а при более низкой температуре (7—8° С) —до 50 дней.
По-видимому, зараженные блохи могут перезимовывать в норах
сусликов. Возможность переноса чумных бактерий с больных грызу-
нов на человека, попавшего в зону природного очага, обеспечивает-
ся некоторыми особенностями биологии блох: в первую очередь
238
их широкой специфичностью и той легкостью, с которой они перехо-
дят с одного хозяина на другого. Большое значение для распрост-
ранения инфекции имеют также суточные миграции блох, к вечеру
выходящих из норы на поверхность почвы. И, наконец, то обстоя-
тельство, что блохи оставляют труп погибшего от чумы зверька
и быстро расселяются на новых хозяев.
Природные очаги чумы имеются во многих местах земного ша-
ра. В нашей стране — это прикаспийские степи, низовья р. Урала,
равнинно-предгорная часть Восточного Закавказья, пустыни и полу-
пустыни Средней Азии. Распространение очагов ограничено опре-
деленными географическими зонами и зависит от сочетания условий
климата, рельефа местности, типа почв, видового состава грызунов
и характера растительных сообществ. Это зоны сухих степей, паяу-
пустынь и пустынь.
В настоящее время вспышки чумы среди населения земного шара
довольно ограничены. Европа полностью свободна от чумы; в США
за 50 лет отмечено всего несколько десятков случаев, но в ряде
тропических районов Азии, Африки и Южной Америки эта страш-
ная болезнь все еще продолжает напоминать о себе. Так, в период
с 1952 по 1962 г. там ежегодно регистрировалось от 700 до 14 тыс.
случаев чумы. Эти факты заставляют органы здравоохранения все-
»го мира относиться со всей серьезностью к необходимости поддер-
живать постоянный и действенный контроль над чумой.
Роль блох в передаче других инфекций
Питаясь на других животных, блохи могут вместе с кровью по-
лучать возбудителей различных инфекционных болезней (энцефа-
литы, ложный сап, туляремия и др.), однако роль блох в их распро-
странении не доказана. Туляремийные микробы в кишечнике блох
не размножаются, поэтому и передача инфекции может носить толь-
ко случайный характер. Важную роль играют блохи в передаче че-
ловеку эндемического крысиного сыпного тифа. Это острое лихора-
дочное заболевание, впервые описанное лишь в 20-х годах нашего
столетия, относится к числу риккетсиозов (возбудитель — Rickettsia
mooseri). Источником и резервуаром инфекции служат домовые
крысы. Вспышки болезни часто приурочены к крупным портовым го-
родам, преимущественно южным, в которых крысы бывают особен-
но многочисленны. Переносчиком инфекции служат блохи — Хепор-
syllus cheopis, Pulex irritans и др. Передача риккетсий осуществля-
емся контаминнтивным путем. В сухих фекалиях блох риккетсии ос-
таются живыми ,и вирулентными длительный срок (до 4, 5 лет).
Борьба с блохами
В жилых помещениях борьба с блохами сводится главным образом к тому,
чтобы не допускать возникновения условий, благоприятных для развития личинок
блох. С этой целью необходимо тщательное заделывание щелей, регулярное мытье
полов и содержание помещений в чистоте. Необходимо также изолировать жи-
лые помещения от возможности попадания в них грызунов. Мероприятия/по унич-
тожению блох (различными инсектицидами) должны быть связаны с упичтоже-
239
иисм их основных хозяев — грызунов — как в антронургических, так и в при-
родных очагах. Очень важно, чтобы эти мероприятия проводились одновременно,
так как уничтожение только грызунов неизбежно приводит к расселению блох
(нередко зараженных),- что может вызвать опасные вспышки болезней среди
населения.
ОТРЯД DIPTERA LINN., 1767,—Д В У К Р Ы Л Ы Е
Двукрылые — один из самых крупных (свыше 80 тыс. видов)
и наиболее высокоорганизованных отрядов насекомых, представи-
тели которого имеют одну пару крыльев. Задние крылья превраще-
ны в небольшие булавовидные придатки — жужжальца. Ротовые
аппараты двукрылых имеют вид хоботка, предназначенного для вы-
сасывания жидкой пиши из растительных и животных тканей или
для ее слизывания.
Отряд Diptera разделяется на три подотряда. Подотр. Nemato-
сега — длинноусые — объединяет виды, обладающие многочленико-
выми сяжками (больше трех). У личинок длинноусых голова всегда
хорошо развита и обособлена от остального туловища. Куколки
покрытого типа и часто способны к активному движению за счет
сокращения мускулатуры брюшка. В ряде семейств, входящих
в состав Nematocera, встречаются кровососущие виды (мошки,
мокрецы, москиты, настоящие комары).
Подотр. Brachycera— Orthorrhapha— короткоусые прямошов-
ные, включает виды, у которых сяжки всегда состоят из трех чле-
ников. Голова у личинок сильно редуцирована. Куколки же, как и
у длинноусых, покрытого типа. Во время выхода имаго из куколоч-
ного экзувия на дорзальной поверхности последнего образуется
прямая щель. Из кровососущих форм к короткоусым прямошовным
двукрылым относится обширное сем. Tabanidae — слепни.
Наиболее специализированной группой, несомненно, являемся
подотр. Brachycera — Cyclorrhapha — короткоусые круглошовные.
Они, как и предыдущий подотряд, всегда имеют трехчленистые
сяжки. Дальнейшее морфологическое упрощение претерпевают ли-
чинки, головная капсула которых полностью редуцируется. В про-
цессе окукливания экзувий последней личиночной стадии не сбра-
сывается, уплотняется, темнеет и превращается в. ложный кокон-
пупарий. Внутри последнего располагается куколка свободного
типа. Имаго, покидая пупарий, отбрасывает его переднюю стенку
в виде круглой крышечки. Переход к паразитизму у представите-
лей этого подотряда осуществляется по нескольким направлениям.
С одной стороны, взрослые мухи приспосабливались к питанию
кровью (р. Glossina, Stomoxys и др.), а в дальнейшем и к постоян-
ному эктопаразитизму (сем. Hippoboscidae — кровососки и т. д.).
С другой же — паразитами (в том числе и эндопаразитами) стано-
вятся личинки (группа оводов и т. д.). Важное медицинское и ве-
теринарное значение имеют и- взрослые свободноживущие мухи,
способствующие распространению возбудителей ряда опасных за-
болеваний (с. 271). Все это послужило причиной для более широ-
кого изучения двукрылых. Работы, посвященные их систематике,
240
морфологии, патогенному значению, проводятся во всех странах
мира. Из русских исследователей особо следует отметить А. А. Шта-
кельберга, В. Н. Беклемишева, А. С. Мончадского и Н. Г. Олсуфь-
ева, а также А. И. Гуцевича, К. Я. Грунина, К. А. Бреева и
А. С. Лутта — авторов известных монографий и определителей по
кровососущим, паразитическим и синантропным двукрылым.
Кровососущие двукрылые
Наиболее широкое распространение гематофагия получила
в подотр. Neniatocera. Четыре семейства, входящих в его состав
(сем. Simuliidae— мошки, сем. Phlebotomidae— москиты, сем. Се-
ratopogonidae—мокрецы и сем. Culicidae — настоящие комары),
включают виды, превратившиеся в облигатных кровососов. Среди
Brachycera — Orthorfhapha обязательное питание кровью свойствен-
но только слепням (сем. Tabanidae). Все эти формы, объединяемые
под общим названием «гнус», обладают рядом сходных признаков:
они — временные паразиты, кровью питаются только самки, в раз-
витии половых продуктов отчетливо выражена гонотрофическая
гармония. Возникновение гематофагии у Brachycera — Cyclorrha-
pha не сопровождалось появлением гонотрофических циклов. Кро-
вью питаются и самцы, и самки.
Циклы развития и биология кровососущих двукрылых
ПОДОТРЯД NEMATOCERA MEIP, 1818
Семейство Simuliidae — мошки
Мошки — мелкие насекомые с плотным и сравнительно корот-
ким телом (рис. 104, А), длина которого колеблется от 2 до 6 мм.
Подогнутая вниз голова снабжена многочленистыми усиками и не-
сет короткие, но мощные ротовые части, приспособленные для со-
сания и слизывания жидкой пищи (с. 256). Сильно выпуклая грудь
снабжена широкими прозрачными, лишенными рисунка крыльями.
Конечности по сравнению с другими Nematocera толстые и не очень
длинные. Брюшко состоит из 9 видимых снаружи сегментов, слабо
склеротинйзировано и может сильно растягиваться при приеме
пищи.
Вылупившиеся из куколок мошки обычно вскоре после вылета
копулируют, что у некоторых видов сопровождается настоящим
роением. После этого они разлетаются в разные стороны в поисках
пищи. Самостоятельно мошки способны преодолевать лишь неболь-
шие расстояния, однако воздушные течения и ветер могут уносить
их на 200 км и более.
В пределах сем. Simuliidae наблюдается переход от фитофагии
к облигатной гематофагии. Самцы и самки ряда видов питаются
241
только соками растений. Некоторые виды в северных районах ведут
себя как докучливые кровососы, в то время как на юге они вообще
не нападают на людей и животных. Широко распространены фор-
мы, у которых первый цикл размножения осуществляется без пи-
тания кровью, тогда как последующие возможны только после
Рис. 104. Фазы развития мошек (по разным
авторам). Л—имаго (самка); Б — личин-
ка; В — куколка; Г — куколка в коконе
кровососания. И, наконец,
такие виды, как Gnus cho-
lodkovskii, Odastnia ornata
и др., способны откладывать
яйца лишь после поглоще-
ния порции крови и завер-
шения гонотрофического
цикла. Самцы всех видов
Simuliidae питаются только
соками растений.
На животных-хозяев кро-
вососущие мошки нападают
преимущественно в светлое
время суток. В дождливую
погоду, ночью и в сумерках
они не активны. Виды, пи-
тающиеся на крупных жи-
вотных, обычно разыскива-
ют и преследуют свою добы-
чу, виды же, которые пьют
кровь мелких животных,
большей частью сидят в ук-
рытых местах, подстерегая
своих хозяев.
От момента принятия пи-
щи до яйцекладки обычно
протекает около 10 дней.
Число продуцируемых сам-
кой яиц в первую очередь
определяется порцией выпи-
той крови. У отдельных ви-
дов оно может достигать
тысячи. Подавляющее боль-
шинство самок погибает
после первой откладки яиц. Лишь в редких случаях самки способ-
ны пройти несколько (до 5) гонотрофических циклов.
Развитие преимагинальных фаз целиком осуществляется в воде.
Большинство мошек для откладки яиц использует ручьи и реки
с быстрым течением. Самки по водным растениям и другим пред-
метам опускаются на дно водоема и приклеивают яйца к субстрату.
Чаще всего в одном месте скапливается столько яиц, что выходя-
щие из них личинки образуют огромные «колонии», насчитывающие
миллионы особей. Развитие яиц в зависимости от температуры про-
должается от 4 до 15 дней.
242
Личинки имеют червеобразное тело (рис. 104, Б). Голова хоро-
шо обособлена и снабжена ротовыми частями, близкими к ротовым
аппаратам грызущего типа. Наибольшим изменениям подвергает-
ся верхняя губа, на боковых частях которой развиваются подвиж-
ные пучки перистых щетинок — веера. Последние все время совер-
шают взмахивающие движения. На поверхности щетинок при этом
оседают находящиеся в воде многочисленные частицы органическо-
го детрита, обрывки водорослей, микроскопические организмы
и т. Д. — все это и составляет пищу личинок. Нерасчлененная грудь
па вентральной поверхности несет непарный конический вырост —
ложную ножку, на конце которой располагается венчик крючьев.
По бокам груди через тонкие покровы отчетливо просвечивают
зачатки дыхательных нитей куколки (см. ниже). На заднем конце
явственщэ расчлененного брюшка еще один венчик крючьев огра-
ничивает мощную присоску, при помощи которой личинки плотно
прикрепляются к субстрату и противостоят иногда довольно силь-
ному течению. Анальное отверстие располагается дорзальной неда-
леко от задней присоски. По бокам от него находятся два втяги-
вающихся тонкостенных придатка (анальные жабры), играющих
важную роль в процессах осморегуляции.
Большую часть времени личинки сидят на одном месте, прикре-
пившись к подводным предметам (водная растительность, камни,
коряги и т. д.). Передвигаются они шагающим способом наподобие
гусениц бабочек пядениц. Иногда они совершают дальние мигра-
ции на десятки километров. В этих случаях личинки выпускают
длинную паутинную нить, отделяются от субстрата и парят в тол-
ще несущего их водяного потока.
В своем развитии, которое продолжается 15—20 дней, личинки
проходят несколько стадий. Перед окукливанием личинки выбира-
ют участок с тихим течением и плетут кокон. Последний чаще име-
ет вид конического чехлика, открытого со стороны основания. Из
этого отверстия выступает конец тела куколки (рис. 104, В, Г),
несущий два пучка длинных дыхательных нитей. Обычно окукли-
вание так же, как и последующий выход имаго, происходит доволь-
но синхронно. Развитие куколки продолжается около недели. Вы-
ходящая из экзувия мошка окружена пузырьком воздуха, в кото-
ром она и всплывает на поверхность воды. В течение года у мошек
сменяется от одного до трех и более поколений. Зимуют они обыч-
но в фазе яйца или личинки.
Семейство Ceratopogonidae (syn. Heleidae —
мокрецы
Мокрецы — самые мелкие из кровососущих двукрылых
(рис. 105, А). Их размеры редко превышают 3—4 мм. Некоторые
виды имеют длину всего лишь 1 мм. Голова мокрецов несет длин-
ные многочленистые усики и вытянутый в хоботок ротовой аппарат.
Виды, относящиеся к наиболее широко распространенному р. Си-
243
licoides, имеют пятнистые крылья. У представителей р. Leptoconops
и р. Forcipomyia крылья прозрачные.
Далеко не все представители семейства питаются кровью. Толь-
ко р. Leptoconops представлен облигатными гематофагами. Еще
в четырех родах (из 20, входящих в
А
Рис. 105. Фазы развития мо-
крецов Culicoides (из кн. Гор-
ностаевой) . А — самка; Б — ли-
чинка; В — куколка
состав семейства) имеются наряду с
кровососущими и растительноядные
виды. Нападают на животных-хозяев
только самки. У некоторых видов на-
блюдается известная пищевая специа-
лизация— они предпочитают питаться
кровью определенных животных. Наи-
более активны мокрецы (в частности,
р. Culicoides) в утренние и вечерние
часы при температуре от 8 до 18° С.
Особенно сильными нападения мок-
рецов бывают во время слабого
дождя.
Процессы переваривания крови и
созревания яиц, подчиняющиеся пра-
вилу гонотрофической гармонии, обыч-
' но занимают около недели. Яйцеклад-
ки небольшие (до 150—170 яиц). Число
продуцируемых самкой яиц зависит от
количества выпитой крови.
Самки откладывают яйца в самых
различных биотопах. Одни виды пред-
почитают мелководные участки озер,
прудов, болот и т. п. Другие использу-
ют небольшие временные водоемы: лу-
жи, воду, скапливающуюся в дуплах,
и т. д. Третьи оставляют яйца просто
на сильно увлажненной почве, лесной
подстилке, илистых берегах, торфяни-
ках. Из яиц в благоприятных условиях
через 3—6 дней выходят узкие черве-
образные личинки с хорошо обособлен-
ной головой и, как правило, лишенные
каких-либо придатков (рис. 105, Б).
Они очень подвижны и все время ак-
тивно питаются. Пищей им служат,
главным образом, бактерии и органи-
ческий детрит. В своем развитии личинки мокрецов проходят четы-
ре стадии. Общая продолжительность личиночной фазы составляет
2—3 недели. Сильно вытянутые в длину куколки мокрецов почти
неподвижны (рис. 105, В). Их развитие продолжается 3—7 дней.
В течение сезона может сменяться до пяти генераций мокрецов.
Зимуют они в фазе яйца или личинки.
244
Семейство Ph 1 ebotomidae — москиты
Москиты — небольшое семейство; на территории СССР встре-
чается всего два рода — Phlebotofnus и Sergentomyia. Это очень
мелкие кровососущие насекомые (рис. 106, 4), размеры которых
колеблются от 1,3 до 3,5 мм. Все тело москитов сильно опушено.
Небольшая голова несет 16-члениковые длинные сяжки и хорошо
развитый колюще-сосущий хоботок (с. 257). Мощная грудь, обра-
зующая характерный «горб», снабжена парой прозрачных остро-
Рис. 106. Фазы развития москитов Phlebotomus (из кн. Дергачевой). А — самка;
Б — копулятивный аппарат самца; В — личинка; Г — куколка с личиночным
экзувием
конечных крыльев, которые, как и туловище, покрыты многочис-
ленными короткими волосками. Два последних сегмента брюшка
превращаются в наружные части полового аппарата. Последний
особенно сложно устроен у самцов (рис. 106, Б).
Москиты — облигатныСу.кровососы, однако некоторые виды спо-
собны производить откладку первой порции яиц без питания кро-
вью, только за счет энергетических запасов, накопленных еще ли-
чинкой. В поисках добычи в пустынной местности москиты совер-
шают перелеты до 1,5 км. В местах скопления животных и
в поселках они обычно отлетают от мест выплода не более чем на
несколько десятков метрОв. Днем москиты обычно прячутся в убе-
жищах, которыми чаще всего служат норы грызунов. Однако они
охотно используют и постройки человека. Активизируются насеко-
мые незадолго до захода солнца. Именно в этот период они напа-
дают на позвоночных животных для питания. Кровь поглощают
только самки москитов. Наряду с этим они, как и самцы, могут
сосать соки растений.
Кровь, проглоченная самкой, обычно не превышает ее собствен-
ного веса (0,4—0,5 мг). Этого количество крови хватает на разви-
тие 50—70 яиц. После первой кладки значительная часть самок
245
погибает, однако многие особи способны проходить несколько го-
нотрофических циклов.
Для откладки яиц москиты используют те же убежища, в ко-
торых проводят дневное время и периоды неблагоприятных усло-
вий. В природе это могут быть норы грызунов, дупла деревьев,
гнезда птиц, пещеры и т. д. Яйца москитов, обитающих в непосред-
ственном соседстве с человеком, можно обнаружить в любых за-
тененных и влажных местах, богатых органическими веществами,—
на скотных дворах, под полом жилых комнат, в погребах, на свал-
ках мусора. Яйца, а затем и личинки (особенно младших возра-
стов) хорошо развиваются только в условиях достаточно высокой
влажности. Излишняя сухость и яркий солнечный свет губительны
для москитов. При оптимальных условиях личинки вылупляются
из яиц примерно через неделю после откладки последних.
Червеобразное тело личинки (рис. 106, В) покрыто многочис-
ленными волосками, к которым обычно прилипают разнообразные
мелкие частички (пыль, песчинки и т. п.). На заднем конце тела
располагается пучок из нескольких особенно длинных хвостовых
волосков. Круглая голова личинок несет хорошо развитый ротовой
аппарат грызущего типа. Пищей им служит тот же органический
субстрат, на котором они обитают. В процессе своего развития,
продолжающегося при обычных условиях около двух месяцев, ли-
чинки проходят четыре стадии. Личинки IV возраста линяют на ку-
колку (рис. 106, Г). На концевых члениках ее брюшка часто сохра-
няется личиночный экзувий, при помощи которого куколка прикреп-
ляется к субстрату. Фаза куколки продолжается 10—12 дней.
На территории нашей страны москиты обычно дают две генера-
ции. В конце весны или в начале лета вылетают самки, развившие-
ся из перезимовавших личинок. Второй массовый вылет приходит-
ся на конец июля — август. Личинки, вылупившиеся из яиц, отло-
женных этим поколением, и уходят на зимовку.
Семейство Culicidae Steph., 1 82 9,—н а ст о я щ и е
комары |
Семейство настоящих комаров по современным данным насчи-
тывает свыше 2 тыс. видов, многие из которых являются облигат-
ными гематофагами. На территории Советского Союза наиболее
широко распространены представители р. Anopheles (подсем. Апо-
phelinae) и р. Culex, Aedes, Mansonia и Culiseta (syn. Theobatdia),
относящихся к подсем. Culicinae. Первые часто называются маля-
рийными комарами, вторые же — немалярийными.
Взрослые комары имеют стройное вытянутое тело (рис. 107, А)
со сравнительно маленькой головой и длинными тонкими ногами.
Длинные сяжки состоят из 15 члеников, у самок они покрыты ко-
роткими волосками, у самцов же густо опушены. На нижней по-
верхности головы прикрепляется совершенный колюще-сосущий
хоботок (с. 257). У немалярийных-комаров нижнегубные щупики
короткие и едва достигают одной трети длины хоботка, у малярий-
246
ных же — они равны длине хоботка (рис. 107, Б; 108, А). Мощно
развитая грудь несет удлиненно-овальные крылья. На поверхности
крыльев располагаются мельчайшие чешуйки, скопления которых
образуют характерный рисунок, свойственный некоторым видам
комаров.
Рис. 107. Фазы развития малярийного комара Anopheles (по разным авторам).
А — самка (внешний вид); Б— голова самки; В— яйцо; Г — личинка; Д—по-
ложение личинки в воде во время питания; Е — куколка; Ж—поза покоя взрос-
лого комара
Вылупившиеся из куколок комары проходят период физиологи-
ческого дозревания, который у самок р. Anopheles продолжается
около четырех дней. Развитие яичников на этом этапе обеспечива-
ется наличием запасов питательных веществ, унаследованных еще
от личинки, и питанием сахаристыми соками растений. Достигнув
определенной стадии зрелости, комары копулируют. В тихие без-
ветренные вечера самцы образуют многочисленные рои. Самки
влетают в такой рой на короткий промежуток времени и увлекают
за собой отдельных самцов.
Дальнейшее развитие гонад у осемененных самок возможно
только после поглощения определенной порции крови. Это застав-
ляет их искать животных, на которых они могут питаться. На
большие расстояния (до 3 км) самки привлекаются запахами, при-
247
носимыми воздушными течениями. В этом случае им приходится
лететь против ветра. Оказавшись в непосредственной близости от
животных-хозяев, комары руководствуются уже не только обоня-
нием^ но и зрением. Большую помощь им оказывает хорошо раз-
витая терморецепция. Они улавливают незначительные колебания
температуры в окружающем пространстве и привлекаются к источ-
Рис. 108. Фазы развития немалярийлых комаров (по разным авторам). А — голо-
ва самки; Б — кладка комаров р. Culex; В — личинка; Г—положение ли-
чинки в воде во время питания; Д — куколка; Е — поза покоя взрослого кома-
ра (Culex)
никам теплового излучения. Больные с высокой температурой или
люди, занятые физической работой, особенно притягательны для
комаров.
Фактическая дальность перемещения комаров-может значитель-
но превышать те расстояния, которые они могут преодолеть в поис-
ках добычи. Culex иногда встречается в 10—12 км от мест выплода,
некоторые виды р. Aedes преодолевают расстояние 18, а часто
и 30 км. Этому, конечно, в первую очередь способствуют устойчи-
вые ветры, дующие в определенном направлении.
В течение суток активность голодных комаров и характер их
поведения заметно изменяются. Светлое время дня и самый темный
период ночи они проводят в укрытиях. Комарам свойственна фо-
токинетическая реакция — способность под действием яркого света
248
активизироваться. В освещенных участках комары часто взлетают
п, отлетев на некоторое расстояние, снова садятся. Таким образом,
они постепенно концентрируются в затененных участках, где уже
ничто не стимулирует их активности. В то же время слабый свет
привлекает комаров: он заставляет их вылетать на открытый воз-
дух в вечерние сумеречные часы с мест дневок, а ночью влетать
с улицы в освещенные помещения. Наиболее интенсивное нападе-
ние голодных самок на позвоночных животных отмечается в суме-
речные утренние и вечерние часы, когда этому способствуют соот-
ветствующая температура и относительно высокая влажность воз-
духа.
В укрытиях нуждаются не только голодные, но и сытые самки,
которым необходим период покоя для нормального течения процес-
сов переваривания крови и созревания яиц. Однако и в это время
самкам часто приходится вылетать в поисках воды. В сухих и жар-
ких помещениях запаса воды в организме насекомого не хватает
до окончания пищеварения. Комары обычно испаряют большое ко-
личество влаги. Это заставляет их регулярно пить воду или соки
растений.
Выбор насекомыми тех или иных укрытий определяется микро-
климатическими условиями, которые в них создаются, и биологи-
ческими особенностями разных видов комаров. Как правило, это
хорошо затененные места с достаточно высокой влажностью воз-
духа и лишенные сквозняков. В природе разные виды комаров ис-
пользуют норы животных, гнезда птиц, дупла, пещеры и расщели-
ны в земле и камнях. Очень часто они в массе скапливаются в гу-
стой растительности, в зарослях осоки, камыша и кустарников по
берегам водоемов, в нижних ярусах подлеска в лесных массивах
п т. д. В населенных пунктах дневки комаров в изобилии встреча-
ются в скотных дворах, подвалах и чердаках различных построек,
в жилых помещениях.
Поза, принимаемая комаром, сидящим в укрытии, весьма харак-
терна. По этому признаку малярийные комары легко отличаются
от немалярийных. У первых брюшко всегда направлено вверх под
острым углом к субстрату, у вторых же оно расположено парал-
лельно субстрату или смотрит концом вниз (рис. 107, Ж\ 108, £).
По окончании созревания яиц самки вылетают из убежищ на
поиски пригодного для яйцекладки водоема. Комары р. Anopheles
п Culex предпочитают постоянные или длительно существующие,
временные водоемы (мелководные хорошо прогреваемые участки
озер, прудов, тихие заводи рек, глубокие лужи, заполненные дож-
девой водой, и т. д.).„Важными условиями является наличие спо-
койной поверхности воды, высокое содержание кислорода, развитие
водной растительности. В очень загрязненных водоемах с загни-
вающей водой, содержащей азотистые продукты распада органи-
ческих веществ, комары не развиваются.
Сигналом для откладки яиц служит смачивание кончика хобот-
ка, лапок или церок (у комаров р. Anopheles) — органов, на кото-
249
рых расположены хеморецепторы. Яйца Anopheles плавают по по-
верхности воды поодиночке или сцепившись небольшими группами.
Они имеют овальную форму и снабжены двумя боковыми поплав-
ковыми камерами (рис. 107, В). Кладки комаров р. Culex имеют
вид плотиков, составленных из большого числа вёртикально рас-
положенных яиц (рис. 108, Б). Они также плавают по поверхност-
ной пленке воды. Самки р. Aedes откладывают яйца не на воду,
а на сильно увлажненные участки субстрата, периодически зали-
ваемые водой. Как правило, их развитие осуществляется в неболь-
ших относительно быстро пересыхающих водоемах (лужи, скопле-
ния воды в дуплах и т. д.).
Сроки развития яиц в первую очередь определяются темпера-
турой окружающей среды и могут.колебаться от 2 до 10 суток. Со-
зревшие личинки Anopheles и Culex сразу же докидают яйцевую
оболочку. У видов р. Aedes закончившие развитие личинки могут
длительное время (до одного года) сохраняться в яйце в покоя-
щемся состоянии. Их вылупление происходит только после залива-
ния яиц водой.
Длина личинок (рис. 107, Г; 108, В), только что вышедших из
Ъйца, около 1 мм. В своем развитии они проходят четыре возраста,
сильно увеличиваясь за это время в размерах. Голова всегда хоро-
шо обособлена от туловища. У видов р. Anopheles, добывающих
пищу с поверхностной пленки воды, она вытянута в длину. У личи-
нок, относящихся к подсем. Cullcidae, питающихся планктоном или
соскребывающих пищу с субстрата, она заметно короче, но зато
более широкая. Голова несет палочковидные антенны, простые
глазки (лишь начиная с III возраста, появляется еще пара сложных
глаз) и ротовой аппарат грызущего типа. По бокам верхней губы
развиваются пучки тонких и длинных волосков — щетки, способные
к быстрым ритмическим движениям.
Туловище личинки покрыто многочисленными волосками. Груд-
ной отдел, границы между сегментами которого не видны, сильно
вздут. Особенно отчетливо это проявляется у личинок IV возраста.
Членистое брюшко состоит из 9 члеников. Предпоследний сегмент
несет на себе дыхательные отверстия — стигмы, расположенные
дорзально на специальной стигмальной пластинке. У личинок ма-
лярийных комаров стигмальная пластинка лежит непосредственно
на поверхности сегмента, у немалярийных — она локализуется на
конце довольно длинного трубчатого сифона (рис. 108, В). Послед-
ний, IX, сегмент брюшка снабжен двумя хорошо заметными пуч-
ками щетинок. Дорзальные щетинки с крючковидно изогнутыми
концами выполняют якорную функцию; с их помощью личинки
удерживаются на поверхности подводных предметов. Вентральные
или рулевые щетинки, образуют плоский веерообразный пучок,
служащий для увеличения гребной поверхности при движении ли-
чинок, которые осуществляются за счет энергичных сокращений
мускулатуры брюшка. Терминально на конце последнего сегмента
располагается анальное отверстие, окруженное четырьмя анальны-
250
ми жабрами. Последние, как и у мошек, играют важную роль в про-
цессах осморегуляции.
Большую часть времени личинки проводят, прикрепившись к по-
верхностной пленке воды. У немалярийных комаров для этого слу-
жит только несмачивающаяся стигмальная пластинка, расположен-
ная на конце сифона. Сама личинка при этом свешивается вниз
под углом к поверхности воды (рис. 108, Г). Личинки малярийных
комаров располагаются горизонтально (рис. 107, Д). Для прикреп-
ления к поверхностной пленке используется не только стигмальная
пластинка, но и так называемые плечевые лопасти — мягкие выро-
сты спинной поверхности груди, и особые пальмовидные щетинки,
локализующиеся дорзально на брюшных сегментах.
Пищей для личинок Anopheles служат самые разнообразные
мелкие частицы и организмы (бактерии, простейшие, пыльца рас-
тений, органический детрит и т. п.), оказавшиеся на поверхностной
пленке и в прилегающих к ней слоях воды. Движением щеток со-
здаются токи воды, направленные к ротовому отверстию. Личинки
старших возрастов привлекают к себе пищу с площади, равной
8 см2. В связи с таким способом питания у личинок Anopheles раз-
вивается очень подвижное сочленение головы с грудью. Питающие-
ся личинки поворачивают голову на 180°, так что ротовые части
и щетки оказываются обращенными наверх. Механизм питания ли-
чинок Culex и Aed.es в основном такой же, с той лишь разницей,
что пищу они добывают себе из толщи воды, и голова у них сохра-
няет нормальное положение. Личинки ряда видов способны, кроме
того, соскребывать бактериальную пленку и органический детрит
с поверхности подводных растений.
Продолжительность развития личинок находится в прямой за-
висимости от условий питания и температуры окружающей среды.
В среднем оно колеблется от 10 дней до месяца.
У куколки комаров четкая граница между головой и грудью
отсутствует. Сквозь прозрачные покровы просвечивают зачатки
ротового аппарата, конечностей, крыльев (рис. 107, Е; 108, Д). Пе-
редний отдел тела, часто обозначаемый как «головогрудь», на
спинной поверхности несет два дыхательных сифона, соединяю-
щихся с трахейной системой. Брюшко, заканчивающееся анальны-
ми лопастями, обычно подогнуто под «головогрудь». Большую
часть времени куколки Неподвижно висят, прикрепившись к по-
верхностной пленке воды. Однако при малейшем раздражении они
способны опускаться на дно водоема за счет энергичных сокраще-
ний брюшка. Линька на имаго осуществляется на поверхностной
пленке. Молодрй комар, выбравшись из экзувия, некоторое время
обсыхает, сидя на поверхности воды. Благополучное завершение
линьки возможно только при тихой погоде.
Количество генераций комаров в течение одного года во многом
определяется конкретными местными условиями. В северных райо-
нах обычно в течение теплого сезона развивается 1—2 поколения,
на юге число генераций заметно возрастает (в условиях Закавка-
зья у Anopheles происходит смена 7—10 поколений).
251
ПОДОТРЯД BRACHYCERA (ZETT., 1892) — ORTHORRHAPHA
Семейстцо Tabanidae Leach, 1819-слепни
Сем. Tabanidae насчитывает свыше 3 500 представителей, боль-
шинство из которых является облигатными кровососами. Виды*
Рис. 109. Слепни (по/разным авторам). А — самка Tabanus aufamnalis; Б — сам-
ка Haematopota\ В — самка Chrysops; Г — кладка слепней; Д — личинка;
Е — куколка
встречающиеся на территории СССР, относятся к 12 родам. Наи-
более обычны Tabanus — собственно слепни (рис. 109, Д), Chry-
252
sops — пестряки (рис. 109, В) и Haematopota — дождевки’
(рис. 109, Б).
Слепни — наиболее крупные среди кровососущих двукрылых.
Их тело может достигать 3 см в длину. Усики состоят из трех чле-
ников. Концевой членик на поверхности несет вторичную кольча-
тость. Ротовые части соединяют в себе черты колюще-сосущего
хоботка Nematocera и лижущего аппарата высших мух (с. 257).
Грудь несет пару крупных крыльев. Последние либо прозрачны,
либо с рисунком в виде темных перевязей (р. Chrysops), серых
«мраморных» пятен (р. Haematopota) и т. д. Брюшко сплющено
в дорзовентральном направлении. Все тело слепней, как правило,
несет многочисленные тонкие волоски.
Появление взрослых слепней в природе связано с наступлением
теплой погоды и обычно наблюдается в мае. Исчезают слепни
осенью. Вылет отдельных видов приурочен к разным периодам. Как
правило, слепни хорошо и быстро летают, покрывая большие рас-
стояния в поисках животных-хозяев. Исключение составляют дож-
девки, полет которых обычно довольно непродолжителен. -
Первые дни после вылупления и самки и самцы питаются толь-
ко соками растений и сахаристыми выделениями тлей. Быстрое
испарение воды из организма заставляет их много пить. Иногда
насекомые слету'ударяются о поверхность воды и уносят на хобот-
ке несколько капель влаги; иногда же они просто опускаются к
водоему и некоторое время спокойно сосут воду при помощи хобот-
ка. Жажда крови появляется у самок только после копуляции,
однако и в этот период опи продолжают поглощать соки растений.
Голодные самки очень агрессивны: они нападают на самых разно-
образных позвоночных животных. Главным образом это крупные
млекопитающие, хотя слепни могут питаться на мелких грызунах,
птицах и. даже рептилиях. Часто источником пищи для слепней
служат трупы павших животных,
В большинстве случаев слепни теплолюбивые и светолюбивые
насекомые. Наиболее активны они в жаркие дневные часы, и лишь
отдельные виды нападают в сумерки. Дождевки в отличие от дру-
гих представителей семейства активны в пасмурную погоду и во
время не очень сильных дождей.
Созревание яиц завершается у самок в течение 3—4 суток после
поглощения крови. Разные виды обладают различной плодовито-
стью. Число откладываемых яиц колеблется от 200 до 1 тыс. Самки
слепней способны проделать несколько гонотрофических циклов,
однако после каждого очередного цикла значительная часть особей
данного возраста погибает. Яйца откладываются на нижней по-
верхности ветвей и листьев растений, нависающих над водой. Клад-
ки слепней (рис. 109, Г) очень характерны: яички плотно приклеи-
ваются друг к другу и располагаются в несколько слоев. Эмбрио-
1 Дождевки долгое время фигурировали под названием Chrysozona, которое
в настоящее время рассматривается как синоним названия Haematopota.
253
нальное развитие обычно занимает 4—8 дней. При неблагоприят-
ных условиях оно может растягиваться на 3—4 недели.
Вылупившиеся из яиц личинки падают в воду и обычно либо
по дну водоема, либо по поверхностной пленке передвигаются к бе-
регу. Они поселяются во влажной илистой почве у линии уреза во-
ды. Личинки многих видов слепней — активные хищники, питаю-
щиеся червями, мелкими членистоногими, моллюсками и себе по-
добными. В то же время личинки некоторых златоглазиков
(р. Chrysops) поглощают только органический детрит.
Зрелые личинки могут достигать 3—4 см в длину (рис. 109, Д).
Их тело сигаровидно и заострено с переднего и заднего концов.
Оно поделено на 12 сегментов, на границах между которыми рас-
полагаются валики, используемые в качестве ложноножек. Голов-
ная капсула очень маленькая и вытянута в длину. Из ротовых ча-
стей наибольшего развития достигают крючкообразная верхняя
губа и острые изогнутые верхние челюсти. На заднем конце тела
личинки лежат стигмы. Фаза личинки у слепней представлена не-
сколькими стадиями, количество которых варьирует у разных ви-
дов. Личинки последнего возраста перемещаются в более сухие
участки почвы выше уровня воды и окукливаются. Куколка вытя-
нута в длину и имеет характерную цилиндрическую форму
(рис. 109, Е). Продолжительность развития куколки колеблется от
5—7 дней до 2—3 недель. Перед вылетом имаго куколка поднима-
ется к поверхности почвы и наполовину высовывается наружу.
В районах с умеренным климатом слепни имеют одногодичный
цикл развития. Однако по направлению на север сроки формирова-
ния отдельных фаз увеличиваются, и весь цикл может занимать два
года.
Кровососущие представители подотряда
Brachycera — Cyclorrhapha
Среди короткоусых круглошовных двукрылых гематофагия не
получила очень широкого распространения. Тем не менее в разных
семействах этого подотряда наблюдается переход к питанию кро-
вью.
В сем. Muscidae — настоящие мухи, встречаются виды, для ко-
торых гематофагия еще не стала обязательной. Такова, например,
широкораспространенная на юге нашей страны базарная муха —
Musca sorbens. Взрослые мухи, имеющие лижущий ротовой аппа-
рат, как и многие другие представители этого и ряда близкородст-
венных семейств, питаются разлагающимся мясом, соками фруктов,
молочными продуктами и т. д. Одновременно они охотно нападают
на человека, слизывая пот и выделения слизистых оболочек. 'При
наличии открытых раневых поверхностей они пьют вытекающую
кровь и лимфу.
В сем. Muscidae встречаются, однако, и облигатные кровососы,
такие, как Stomoxys calcitrans L. — жигалка и представители
р. Glossina — мухи це-це.
254
Stomoxys catcitrans — осенняя жигалка, ‘отличается от осталь-
ных представителей сем. Muscidae в первую очередь наличием свое-
образного ротового аппарата, построенного по типу сверлящего
хоботка (с. 261). Основной пищей для мух служит кровь крупных
позвоночных животных, в том числе и человека. Питаются кровью
и самцы, и самки, причем у последних отсутствует гонотрофическая
гармония. Обычно мухи нападают на животных-хозяев раз в 1—2
дня. В течение жизни самки откладывают яйца несколько раз. Суб-
стратом для развивающихся личинок служит навоз, перемешанный
с соломой, а также гниющие растительные остатки. Окукливаются
личинки в этой же массе, обычно выбирая более сухие участки.
В умеренных широтах массовый вылет жигалок наблюдается осе-
нью, в странах с тропическим климатом они могут размножаться
круглый год.
Муха це-це (р. Glossind) широко распространенная в эквато-
риальной Африке, так же, как и жигалки, питается кровью
позвоночных животных. Соответственно они имеют и сходно уст-
роенный ротовой аппарат. Для глоссин характерно живорождение.
Яйца по-одному поступают в половые протоки самки, где и проте-
кает эмбриональное развитие. Сформированная личинка остается
в матке. Она располагается таким образом, что ее ротовое отвер-
стие оказывается рядом с отверстиями протоков особых придаточ-
ных желез, секрет которых и служит для личинки пищей. Вырос-
шая личинка выходит во внешнюю среду, зарывается в почву и сра-
зу же окукливается. В это время в теле самки начинает развивать-
ся следующая личинка.
Наиболее специализированными кровососами среди Brachyce-
ra — Cyclorrhapha, несомненно, являются представители мух-крово-
сосок, объединяемых в три семейства: Hippoboscidae, Nycteribiidae
и Streblidae.
Эти семейства насчитывают немногим более 300 видов облигат-
ных гематофагов, приспособленных к длительному пребыванию на
животном-хозяине. Взрослые мухи обладают сильно уплощенным
телом. Широко расставленные ноги и плотно прижатые к туловищу
крылья позволяют им надежно удерживаться на коже различных
видов птиц и млекопитающих. Ротовой аппарат кровососок по свое-
му строению близок к таковому жигалок и мух це-це.
Некоторые виды (лошадиная кровососка — Hippobosca equina,
рис. НО, А) сохраняют крылья на протяжении всей жизни, более
специализированная Lipopiena cervi — оленья кровососка (рис. 110,
Б) “утрачивает крылья сразу же после нападения на хозяина. Нако-
нец, имеются и полностью бескрылые виды, как, например, овечий
рунец — Melophagus ovinus (рис. ПО, В) и паразитирующие на ле-
тучих мышах представители р. Nycteribia. Морфологические преоб-
разования кровососок сопровождались и биологическими измене-
ниями. У всех представителей этой группы развитие личинок проте-
кает так же, как и у видов р. Glossina, внутри материнского
организма. У более примитивных видов отрождаемые поочередно
личинки окукливаются в почве, а вышедшие из них насекомые са-
255
состоятельно разыскивают животных-хозяев. Личинки же Meloptia-
gus ovinus не покидают хозяина. Они приклеиваются к шерстинкам
и сразу же превращаются в пупарий. Распространение паразитов
и переход их с одного животного на другое осуществляется лишь
при тесном контакте последних.
Рис. ПО. Кровососущие высшие мухи (по разным авторам). А—лошадиная
кровососка Hippobosca equina-, Б — оленья кровососка Lipoptena cervi; В — ове-
чий рунец Melophagus ovinus; Г — молодая самка Ascodipteron speiserianum;
Д — самка Ascodipteron speiserianum в толще кожи хозяина
Адаптация двукрылых к питанию кровью
и переход к постоянному паразитизму
Становление гематофагии у двукрылых неизбежно сопровожда-
лось появлением целого ряда специальных адаптаций. В первую
очередь морфологические преобразования должны были затронуть
ротовые части. Основные этапы этого процесса, приведшего к фор-
мированию совершенного колюще-сосущего хоботка настоящих
комаров, хорошо видны на примере современных Nematocera.
Наиболее примитивным ротовым аппаратом обладают мошки
(сем. Simuliidae). Во многом он сохраняет черты, свойственные ро-
товым конечностям грызущего типа. Все составляющие его части
очень коротки (рис. 111, А) и приспособлены не столько для прока-
лывания, сколько для прорезания кожи животного-хозяина. В этом,
по-видимому, участвуют широкие заостренные на концах мандибу-
лы, соединенные наподобие ветвей ножниц, и треугольные максил-
лы. Переднюю стенку хоботка образует широкая желобовидная
верхняя губа. Просвет между нею и мандибулами служит для вса-
сывания крови. Слюна же, вводимая в ранку, стекает по желобку,
256
идущему по передней поверхности гипофаринкса. Широкая мяси-
стая нижняя губа, прикрывающая хоботок сзади, заканчивается
двумя мягкими придатками — лабеллумами, которые представляют
собой видоизмененные нижнегубные щупики. При сосаний крови
нижняя губа отгибается и в ранку не входит.
Ротовые части мокрецов устроены сходным образом (рис. 111,
Б). Отличаются они лишь значительно большей длиной, что позво-
ляет вводить хоботок в кожу хозяина более глубоко. В прокалыва-
нии кожи участвует весь комплекс ротовых частей, включая верх-
нюю губу и гипофаринкс. Слюнной и пищевой каналы у мокрецоа
так же, как и у мошек, разделяются мандибулами.
Прямые стилетовидные придатки москитов (сем. Phlebotomidae}
образуют уже настоящий колющий хоботок (рис. 111, В), который
почти целиком погружен в желоб длинной и эластичной нижней
губы. Пищевой канал, правда, все еще образуется верхней губой
и мандибулами, но зато слюнной канал полностью изолируется за
счет замыкания краев желобка гипофаринкса.
Наиболее совершенным хоботком (рис. 111, Г) обладают пред-
ставители сем. Culicidae. Их ротовые конечности достигают макси-
мальной среди Nematocera длины. Боковые края верхней губы
смыкаются, в результате чего формируется замкнутый герметич-
ный пищевой канал, по которому кровь поступает в пищеваритель-
ную систему. Все колющие части в спокойном состоянии надежно
укрыты в глубоком желобе нижней губы. При нападении на добы-
чу комар сначала ощупывает поверхность кожи лабеллумами. Най-
дя подходящее место, он вонзает в него весь колющий комплекс.
Пиловидно зазубренные на концах максиллы при этом движутся
взад и вперед, расширяя ранку. Мандибулы же своими ланцето-
видно расширенными концами закрывают отверстие на косо сре-
занном конце верхней губы. Они отходят в стороны и открывают
доступ в пищевой канал, лишь после того как хоботок полностью
погружен в кожу й попадает в кровеносный сосуд. Нижняя губа
в ранку не вводится и изгибается в виде дуги. Кровь, засасываемая
комаром, предварительно смешивается со слюной, поступающей по
каналу гипофаринкса.
В приведенном выше ряду отчетливо прослеживаются основные
направления, по которым шло преобразование хоботков Nematoce-
ra: замена режущего аппарата прокалывающим, удлинение рото-
вых частей и, наконец, обособление и герметизация пищевого и
слюнного каналов.
Ротовой аппарат слепней (рис. 112, Д) во многих отношениях
более примитивен, чем хоботок Culicidae. Его широкие ножеобраз-
ные мандибулы и максиллы скорее всего не прокалывают, а про-
резают кожу. Кроме того, мандибулы вместе с верхней губой фор-
мируют пищевой канал. Главное отличие, однако, заключается в
совершенно ином строении нижней губы. Лабеллумы последней,,
как и у высших мух, формируют настоящий оральный диск, снаб-
женный псевдотрахеями и приспособленный для фильтрации и сли-
зывания воды и жидкой пищи (с. 260).
257
Все кровососущие Nematocera и приближающиеся к ним по це-
лому ряду признаков слепни поглощают пищевые вещества двояко-
го рода. Прежде всего это богатые углеводами соки растений и са-
харистые выделения тлей, слизываемые слепнями при помощи
лопастей нижней губы. Обычно во время питания эта пищевая мас-
Рис. 111. Ротовые аппараты Nematocera (из кн. Шванвича). А — ро-
товой ’ аппарат мошек (ротовые конечности и общий вид); Б — рото-
вой аппарат мокрецов (общий вид и поперечный разрез); В — рото-
вой аппарат москита (общий вид); Г — ротовой аппарат комаров
(общий вид и поперечный разрез):
/ — верхняя губа, 2— мандибула, 3 —максилла (нижняя челюсть), 4—нижне-
челюстные щупикн, 5 — гипофаринкс, 6 — нижняя губа, 7 — лабеллумы, 8 —
пищевой канал, 9 — слюнной канал, 10 — положение хоботка во время соса-
ния крови, П — сяжки (или их основания)
са поступает в особый тонкостенный придаток пищевода — зоб. Сю-
да же нагнетается и выпитая насекомыми вода. По мере надобно-
сти жидкое содержимое зоба небольшими порциями перекачивает-
ся через пищевод в желудок, где быстро переваривается и
усваивается. Расщепление глюкозы и других сахаров используется
насекомыми для покрытия больших энергетических затрат, связан-
ных с активным полетом в поисках животных-хозяев, мест дневок,
подходящих биотопов для откладки яиц. Вода, всасываемая стен-
ками желудка, поддерживает постоянство осмотического давления
гемолимфы, которое в связи с интенсивным испарением влаги через
258
покровы может заметно меняться. Подобные изменения весьма не-*
благоприятно сказываются на состоянии насекомых.
Наличие зоба как вместилища воды л сахаристой пищи позво-
ляет двукрылым быть постоянно-готовыми к нападению на живот-
ных-хозяев. Сколько-нибудь значительное заполнение желудка
препятствовало бы поглощению того количества крови, которое не-
обходимо для нормального созревания максимального числа яиц.
Рис. 112. Ротовые аппараты Brachycera (по разным авторам). А — ротовой ап-
парат слепня; Б — внешний вид хоботка мухи; В — положение сосательных ло-
пастей в покое; Г—положение сосательных лопастей при питании жидкой пи-
щей; Д — положение сосательных лопастей при питании твердой пищей; Е — уча-
сток поверхности сосательной лопасти с псевдотрахеями:
/ — верхняя губа, 2 — мандибула, 3— максилла (нижняя челюсть), 4 — гипофаринкс, 5 —
нижняя губа, 6 — сосательные лопасти, 7 — нижнечелюстные щупики, S — ротовое отверстие,
9 — псевдотрахея, 10 — «зубы»
В отличие от иксодоидных клещей двукрылые не в состоянии выса-
сывать очень много крови. Выпиваемая ими порция обычно не силь-
но превышает собственный вес их тела. Это ограничение, несомнен-
но, вызвано необходимостью для самок сохранять способность к
полету после кровососания.
Проглатываемая кровь еще в ранке смешивается со слюной,
которая содержит целый ряд ферментов: антикоагулянты гемоли-
зины, агглютинин и т. д. По пищеводу кровь поступает непосред-
ственно в среднюю кишку — желудок, где и происходит ее перева-
ривание. Этот процесс завершается сравнительно быстро и занима-
ет в среднем при оптимальных условиях не более 4—5 дней. Внача-
ле происходят агглютинация и гемолиз эритроцитов, сопровождаю-
щиеся всасыванием воды, которая содержится в плазме крови.
Содержимое кишечника уплотняется, начинается процесс .расщеп-
ления гемоглобина. Лишь после того как все белки переварены
и усвоены, неиспользованный гематин поступает из желудка в зад-
нюю кишку, а оттуда выводится наружу.
259
Как уже отмечалось выше, всем кровососущим Nematocera, а
также слепням свойственна гонотрофическая гармония.
Гематофагия Brachycera — Cyclorrhapha возникла вторично и
независимо от Nematocera и Tabanidae. Предпосылкой для перехо-
да к питанию кровью у высших мух явилась отчетливо выражен-
ная у многих видов склонность к всеядности. Они поглощают са-
мые различные пищевые вещества, начиная от растительных соков
и кончая фекалиями и разлагающимися животными тканями. На-
падения на позвоночных животных для слизывания пота и выде-
лений гноящихся ран явились первым шагом на пути превращения
мух в облигатных кровососов. В пищевом рационе упоминавшейся
выше Musca sorbens кровь играет явно второстепенную роль. Бо-
лее того, неповрежденные покровы животного-хозяина являются
непреодолимым барьером для базарной мухи. У ряда других фа-
культативных гематофагов тяга к поглощению крови проявляется
более отчетливо. Представители р. Hydrotaea (зубоножки) способ-
ны уже разрушать коросту на подсыхающих ранках и таким обра-
зом обеспечивать себе доступ к лимфе и крови. Для этой цели они
используют конечности. Замыкают этот ряд виды (жигалки, мухи
це-це, кровососки), ставшие облигатными гематофагами.
Переход к обязательному кровососанию у Brachycera — Cyclor-
rhapha так же, как и у Nematocera, сопровождался специализацией
их ротового аппарата. Последний, однако, имеет принципиально
иное строение, чем колюще-сосущие хоботки длинноусых. Он берет
начало от лижущего ротового аппарата (рис. 112, Б—Е), характер-
ного для большинства мух, в том -числе и факультативных гемато-
фагов. Основу их хоботка составляет массивная нижняя губа, об-
разующая глубокий желоб, который прикрыт верхней губой. Воз-
никающий таким образом пищевой канал открывается на конце
хоботка ротовым отверстием. По бокам от последнего располагаются
два крупных лабеллума, преобразованных в подвижные сосатель-
ные лопасти. Внутренняя поверхность лопастей, которые в спокой-
ном состоянии прижаты друг к другу, несет систему тонких каналь-
цев— псевдотрахей, сообщающихся с внешней средой по всей
длине при помощи узкой щели и небольших пор. Псевдотрахен
сходятся к ротовому отверстию, края которого вооружены заост-
ренными хитиновыми пластинками — «зубами». Во время питания
муха расправляет лопасти, прикладывает их к поверхности пище-
вого субстрата й начинает сосать. Жидкая пища при этом фильт-
руется через систему псевдотрахей. Поглощение твердых пищевых
веществ осуществляется иначе. Муха отгибает сосательные лопа-
сти кверху, так что «зубы», окружающие ротовое отверстие, оказы-
ваются снаружи. Поливая пищу слюной, которая стекает по каналу
гипофарнкса, муха одновременно скоблит ее «зубами». Под дей-
ствием активных пищеварительных ферментов, содержащихся в
слюне, образуется жидкая масса, которая и засасывается через
хоботок.
Изменения ротового аппарата облигатных гематофагов заклю-
чаются, главным образом, в уменьшении размеров лабеллумов и
260
более сильном развитии «зубов» (рис. 113). Последние на внутрен-
ней поверхности лопастей образуют продольные ряды, напоминаю-
щие зубчатый край пилы. Прикладывая кончик хоботка к покровам
животного-хозяина, кровососы быстрыми движениями попеременно
выворачивают лопасти наружу и втягивают их обратно. При этом
зубные ряды, как напильники, прорезают в коже отверстие.
Рис. 113. Ротовой аппарат Glossina (из кн.
Шванвича). А — внешний вид хоботка;
Б> В — последовательные стадии работы
лабеллумов; Г — схема поперечного разре-
за через хоботок:
/ — нижнечелюстные щупики, 2 — нижняя губа,
3 — лабеллумы, 4 — зубы, 5 — верхняя губа. 6
гипофаринкс, 7 — пищевой капая, 8 — слюнной
канал
Постоянное питание кровью наложило отпечаток и на особен-
ности развития и биологии наиболее специализированных предста-
вителей высших мух. Прежде всего это переход к живорождению
и последующему вынашиванию развивающихся личинок в половых
путях самок (с. 269). Высокая калорийность крови как пищевого
вещества позволяет облигатным гематофагам не только полностью
удовлетворять собственные энергетические потребности, но и обе-
спечивает возможность развития особей следующего поколения под
защитой материнского организма. Такое изменение биологии раз-
вития кровососущих насекомых, несомненно, должно рассматри-
ваться как прогрессивное по сравнению с обычной откладкой яиц
во внешнюю среду.
Возрастающая потребность в питании кровью обусловила необ-
ходимость увеличения продолжительности контактов с животным-
261
хозяином. Это по сути дела означало переход от временного пара-
зита к стационарному — сначала периодическому; а потом и по-
стоянному. Осенняя жигалка (Stomoxys calcitrans) и представители
р. Glossina нападают на животных только для сосания крови. Ма-
лая коровья жигалка (Liperosia irritans) проводит на хозяине
почти все время и покидает его только для откладки яиц. Крово-
соски, отрождающие готовых к окукливанию личинок, превраща-
ются в стационарных паразитов. Примитивные виды, у которых
только имаго поселяется на хозяине, а куколки развиваются во
внешней среде, можно рассматривать как периодических парази-
тов. Овечий рунец и ряд видов, обитающих на летучих мышах, ста-
новятся постоянными эктопаразитами. Все их развитие протекает
на животном-хозяине, а расселение осуществляется путем перепол-
зания насекомых с одного животного на другое при тесном контак-
те последних.
Переход к постоянному паразитизму неизбежно отразился и на
особенностях строения кровососущих мух. Сильное уплощение ту-
ловища, широко расставленные ноги с цепкими коготками, утрата
крыльев (см. рис. 110)—все это совершенные адаптации к экто-
паразитическому образу жизни.
Еще более глубокие изменения внешнего облика наблюдаются
у тех видов, которые по сути дела становятся эндопаразитами
и поселяются в толще кожи позвоночных животных. Самки Asco-
dipteron speiserianum — паразита летучих мышей, относящегося
к группе кровососок (сем. Streblidae), после копуляции садятся на
хозяина и глубоко внедряются в его покровы, так что наружу тор-
чит лишь самый задний конец тела. При этом они теряют крылья,
отбрасывают конечности, а голова и грудь глубоко впячиваются
в сильно увеличивающееся по размерам брюшко (см. рис. ПО,
Л Д).
Медицинское и ветеринарное значение кровососущих двукрылых
Практическое значение кровососущих двукрылых очень велико.
Количество насекомых, нападающих на человека и животных, мо-
жет быть значительным. Так, в течение часа на одном крупном
животном обычно питается от нескольких сотен до нескольких ты-
сяч слепней. Еще активнее ведут себя мелкие представители гнуса.
На юге Красноярского края, например, за 5 мин на человека напа-
дает до 8 тыс. мокрецов. Не менее агрессивны комары, мошки
и москиты. Массовые нападения кровососов часто влекут за собой
достаточно серьезные последствия. В местах укусов возникают зу-
дящие папулы, расчесывание которых приводит к повреждению
покровов и открывает ворота вторичным инфекциям. Введение в
кровяное русло относительно больших количеств токсичной слюны
вызывает со стороны организма сильную аллергическую реакцию.
Последняя в тяжелых случаях сопровождается лихорадочным по-
вышением температуры, потерей сна, истощением нервной системы,
сдвигом формулы крови и т, п. Большой ущерб кровососущие дву-
262
крылые, особенно слепни и жигалки, наносят сельскому хозяйству.
Нападение одних только слепней может вызвать снижение удоев
молока на 10—20%. Серьезную опасность Для овцеводства пред-
ставляет собой и овечий рунец. Мухи, развивающиеся иногда в
большом количестве, вызывают воспаление кожи и выпадение
шерсти у зараженных животных. Кроме того, большие потери кро-
ви последними приводят к сильному истощению.
Переход к питанию кровью позвоночных животных обусловил
превращение многих двукрылых в переносчиков возбудителей
большого числа тяжелых заболеваний животных и человека.
Заболевания, вызываемые вирусами
К опасным вирусным заболеваниям человека относится распро-
страненная в тропическом и субтропическом поясах желтая лихо-
радка. Это типичное трансмиссивное природно-очаговое заболева-
ние, передаваемое комарами из р. Aed.es. В природных очагах ре-
зервуарными хозяевами служат разнообразные дикие животные:
опоссумы, муравьеды, ежи, броненосцы, обезьяны и т. д. В синан-
тропных очагах возбудитель передается переносчиками непосред-
ственно от человека к человеку.
Заболевание протекает, как правило, очень остро с высокой
температурой. Часто сопровождается сильной желтухой, местными
кровоизлияниями, тяжелыми поражениями почек. Больной теряет
сознание, бредит. Нарушения со стороны пищеварительного трак-
та выражаются в частых и мучительных рвотах. Возможно доб-
рокачественное течение болезни, которая в этом случае, начиная
с 8—10 дня, идет на убыль. Однако часто наблюдается и летальный
исход. В периоды особо тяжелых эпидемий смертность может до-
стигать 20%.
В тропических и субтропических странах распространено еще
одно острое вирусное трансмиссивное заболевание — лихорадка
денге (костоломная, или суставная, лихорадка). Носителями виру-
са в природных очагах служат обезьяны. Передача возбудителя от
зараженных животных к человеку осуществляется комарами
р. Aedes. Трансовариальная передача вирусов отсутствует. В райо-
нах природных очагов у местных жителей лихорадка денге чаще
всего регистрируется в виде отдельных случаев или небольших
вспышек среди детей и приезжих. Более серьезное медицинское
значение имеют эпидемические вспышки, вызванные переездом
больных людей или завозом инфицированных комаров в местности,
где обычно это заболевание не встречается. Отсутствие иммуните-
та у коренного населения таких областей способствует быстрому
распространению болезни. '
Начало болезни сопровождается резким подъемом темпера-
туры до 39—40° С, которая держится 3—4 дня. После небольшого
перерыва возможен второй лихорадочный приступ. На коже появ-
ляется сыпь, ощущаются сильные боли в мускулатуре, сухожилиях,
суставах. Больной впадает в депрессивное состояние. Возможны
263
осложнения в виде менингоэнцефалитов, кровотечений и т. д., при-
водящие к летальному исходу.
Значительно шире распространена москитная лихорадка, или
лихорадка папатачи, очаги которой встречались и в Советском
Союзе: в Молдавии, Крыму, на Кавказе, в республиках Средней
Азии. Единственным хранителем вируса в природе служат моски-
ты (главным образом Phlebotomus papatasii), у которых существу-
ет трансовариальная передача возбудителя. Передача инвазионно-
го начала осуществляется инокулятивным путем.
Заболеваемость населения в очагах москитной лихорадки силь-
но колеблется в разные сезоны года и тесно связана с периодом
активности взрослых москитов. Местное население в силу приобре-
тенного устойчивого иммунитета страдает значительно меньше, чем
приезжие. Именно за счет последних заболевание иногда может
принимать характер эпидемических вспышек. Такие вспышки неод-
нократно наблюдались и у нас в стране: в рядах Красной Армии
в 1922 г. в Крыму, при освоении некоторых районов в Средней
Азии и т. д.
Заболевание обычно начинается внезапно с резкого подъема
температуры, которая держится в течение 3—4 дней. Очень харак-
терным признаком является покраснение лица и шеи, острый конъ-
юнктивит. После исчезновения основных симптомов долгое время
сохраняется изнуряющая слабость и угнетенное состояние психики.
Благодаря успешному проведению санитарно-профилактическпх
мероприятий в ряде районов нашей страны очаги москитной лихо-
радки успешно ликвидированы. Так, уже в течение многих лет за-
болевание не регистрируется в Крыму, Краснодарском крае, Ашха-
баде и т. д.
Среди заболеваний, переносимых комарами, видное место зани-
мают энцефалиты. В Японии, Китае, Индии, в Приморском крае
СССР и'ряде других стран встречается японский энцефалит. Очень
часто он принимает характер эпидемических вспышек, приурочен-
ных преимущественно к сельской местности. Иногда, однако, эпи-
демии возникают и в крупных городах (Токио, Осака и т. д.). В на-
шей стране первая вспышка японского энцефалита была зарегист-
рирована в 1938 .г. Природными резервуарными хозяевами служат
многочисленные виды диких птиц и грызуны. Существование очагов
в населенных пунктах также поддерживается птицами, в частности
воробьями. К ним присоединяются восприимчивые к вирусу домаш-
ние животные (лошади, свиньи, козы и т. д.). Комары (р. Culex
и Aedes) служат лишь переносчиками: трансовариальная передача
отсутствует. Заражение позвоночных животных осуществляется
инокулятивным путем.
Течение болезни сопровождается нарастанием мышечного тону-
са, ослаблением и утратой сухожильных рефлексов, судорогами.
Весьма обычны параличи и парезы. Больные часто теряют созна-
ние, а иногда впадают и в коматозное состояние. В мозгу развива-
ются очаговые кровоизлияния. Заболевание сопровождается ослож-
нениями в виде пневмоний, пролежней, тромбофлебита и т. д.
264
Летальность при японском энцефалите может быть очень высокой
и иногда достигает 70—80%. К японскому энцефалиту близок аме-
риканский энцефалит Сан-Луи, встречающийся преимущественно
на территории США.
Заболевания, вызываемые бактериями
Кровососущие двукрылые служат переносчиками бактериаль-
ных заболеваний, среди которых встречаются весьма опасные. К по-
следним относится сибирская язва, или антракс, вызываемая Ba-
cillus anthracis. Возбудитель поражает позвоночных животных,
в том числе и человека. В организм хозяина бактерии проникают
различными путями: с пищей и загрязненной водой, контактно че-
рез кожу, при вдыхании пыли, содержащей споры сибиреаязвенной
палочки, и т. д. В распространении заболевания активно участвуют
и кровососущие насекомые, среди которых главная роль принад-
лежит мухам-кровососкам, мошкам и слепням. Особенно опасны
последние, так как в природе они могут питаться на трупах жи-
вотных, павших от сибирской язвы, и пить воду из зараженных
водоемов. Перенос осуществляется чисто механически: никакого
развития в насекомых бактерии не претерпевают.
Сибирская язва протекает в нескольких формах: кожной, легоч-
ной, кишечной, менингоэнцефалитической. Последние три .особенно
тяжелы. Болезнь сопровождается высоким подъемом температуры,
ознобами, головными болями, рвотами. Часто развиваются пневмо-
ния, колики, кровавый понос. При поражении нервной системы
обычны явления энцефалита и менингита. Летальность при сибир-
ской язве исключительно высока. Смерть наступает через 4—5
дней, а в особо тяжелых случаях через 24—48 ч.
Важную роль кровососущие двукрылые играют в эпидемиологии
туляремии. Механический перенос Francisella tularensis осуществ-
ляется слепнями, которые сохраняют возбудителя в течение 2—3 су-
ток; комарами; мошками; мокрецами. Пб-видимому, определенное
значение мошки имеют и как распространители чумы и проказы.
Заболевания, вызываемые простейшими 1
Значение кровососущих двукрылых как промежуточных хозяев
и переносчиков ряда паразитических простейших очень велико. Ко-
мары р. Anopheles служат промежуточными хозяевами для кровя-
ных споровиков (р. Plasmodium)—возбудителей малярии челове-
ка. Развитие Leishmania tropica и L. donovani, вызывающих лейш-
маниозы человека (пендинскую язву и кала-азар), осуществляется
в москитах. Мухи це-це (р. Glossina) служат специфическими пе-
реносчиками сонной болезни. И, наконец, слепни в качестве меха-
нических переносчиков распространяют возбудителей сурры до-
машних животных.
1 Подробная характеристика заболеваний и биологии их возбудителей приве-
дены в книге «Частная паразитология», т. I.
10—58 265
Заболевания, вызываемые нематодами
Двукрылые принимают активное участие в циркуляции парази-
тических нематод. Они служат промежуточными хозяевами филя-
рий, вызывающих заболевания человека и домашних животных.
Микрофилярии возбудителя слоновости человека (элефантиазиса)
развиваются в комарах р. Anopheles и Aedes (с. 115). Лоаоз, рас-
пространенный в Африке, переносится слепнями (с. 117). В эква-
ториальной Африке и Латинской Америке мошки служат промежу-
точными хозяевами для личинок Onchocerca volvulus — возбудите-
ля онхоцеркоза (с. 117). Кроме того, в двукрылых развиваются
личинки и других видов филярий, вызывающих заболевания чело-
века и домашних животных.
Синантропные мухи
Среди высших мух, относящихся к подотр. Brachycera — Сус-
lorrhapha, ряд видов в той или иной степени связан с человеком.
Рнс. 114. Фазы развития комнатной мухи
Musca domestica (из кп. Дербеневой-
Уховой). А — взрослая муха, Б — яйца;
В — личинка; Г — пупарий
Их выделяют в особую группу
синантропных мух. В состав
этой группы входят представи-
тели разных семейств: Musci-
dae— настоящие мухи, Sarco-
phagidae — серые мясные му-
хи, С а 1 lip horid а е — падальные
мухи, группа оводов, объединя-
ющая три семейства — Gastro-
philidae, Hypodermatidae,
Oestriidae п др.
Характер связей, которые
возникают между синантроп-
ными мухами разных видов
и человеком, очень различен.
Многие виды обитают и размно-
жаются как в природных усло-
виях, так и в населенных
пунктах. К ним относятся па-
дальные мухи (р. Lucilia, Cal-
liphora и др.), серые мясные
мухи (р. Sarcophaga, Ravinia
и др.). Целый ряд представите-
лей сем. Muscidae довольно
постоянно сопутствует челове-
ку (Af usca domestica — ком-
натная муха, рис. 114; Fannia
canicularis — малая комнатная
муха и т. д.) и в естественных биотопах практически не встречает-
ся. Наконец, с некоторыми видами человек связан через домашних
животных. Это упоминавшиеся выше жигалки и группа пастбпщ-
266
ных мух, развитие которых осуществляется в навозе в местах вы-
паса скота, а имаго часто залетают в поселки.
Циклы развития и биология большинства синантропных мух,
хотя и отличаются некоторыми особенностями, характерными для
каждого отдельного вида, в то же время обладают рядом общих
черт. Имаго в большинстве случаев снабжены лижущим ротовым
аппаратом.
Пищей для мух служат самые разнообразные пищевые вещест-
ва, однако у многих видов проявляется, хотя и не очень строгая, но
достаточно^отчетливо выраженная пищевая специализация. Копро-
фаги (малая комнатная муха — Fannia canicularis, домовая му-
ха— Muscina stabulans, ряд серых мясных мух—сем. Sarcophagidae
и др.) главным образом поглощают экскременты человека и.до-
машних животных. В то же время они могут питаться мясом, мо-
лочными продуктами, ягодами, рыбой и т. д. Зеленые мясные мухи
(р. Lucilia), относящиеся ж группе некрофагов, чаще всего питают--
ся мясом и трупами животных. Дополнительными же источниками
пищи для них служат фекалии, кухонные отбросы и т. п. Входящая
в состав сем. Sarcophagidae вольфартова муха—Wohlfahriia mag-
nified кормится главным образом соками растений, хотя ее можно
обнаружить и на помете животных. Упоминавшаяся выше базар-
ная муха (Musca sorbens, с. 260) относится к факультативным
гематофагам. И, наконец, некоторым видам (Musca domestica
и др.) свойственна настоящая полифагия.
Размножение мух связано с проявлением своеобразной заботы
о потомстве. Самки откладывают яйца лишь на такой субстрат,
который в дальнейшем будет служить пищей для развивающихся
личинок. Яйца обычно одеты плотной защитной оболочкой и имеют
удлиненно-овальную форму (рис. 114, Б). У некоторых видов на-
блюдается живорождение (с. 269).
Выходящие из яиц личинки у большинства видов имеют черве-
образное тело (рис. 114, В), заостренное спереди и расширяющее-
ся к заднему концу. Голова личинок редуцирована. Ротовое отвер-
стие располагается субтерминально на вентральной поверхности.
Из него наружу могут выдвигаться серповидно изогнутые ротовые
крючки, которыми личинки скоблят пищевой субстрат. Задний ко-
нец тела обычно закруглен или тупо срезан. Он несет дыхальца,
которые у взрослых личинок располагаются на особой стигмальной
пластинке, отсутствующей у особей младших возрастов. У личинок
разных видов мух наблюдается известная приуроченность к опре-
деленным субстратам. Личинки одних видов (Musca domestica,
Fannia canicularis, некоторые Sarcophagidae) поселяются в пище-
вых отходах и фекалиях. В связи с этим местами их выплода часто
служат выгребные ямы и отхожие места, легко доступные для взрос-
лых мух. Падальные мухи развиваются в мясных отходах и на тру-
пах животных. Некоторые виды используют свежий, еще не под-
сохший навоз домашних животных в местах выпаса скота. И, нако-
нец, в ряде случаев личинки мух становятся факультативными или
облигатными паразитами человека и домашних животных (с. 268).
10*
267
В процессе питания личинок важную роль играет внекишечное
пищеварение. Из ротового отверстия они выделяют слюну, содер-
жащую активные пищеварительные ферменты. Последние разлага-
ют органические соединения и превращают их в жидкую каши-
цу. Этому способствует и работа ротовых крючков, которые способ-
ны измельчать плотные пищевые частицы. Полупереваренная пище-
вая кашица всасывается личинкой через ротовое отверстие. В этом
отношении дикие несинантропные виды играют определенную по-
ложительную роль, так как их личинки наряду с другими организ-
мами уничтожают растительные остатки и трупы животных.
Развитие личинок мух осуществляется в большинстве случаев
довольно быстро (у комнатной мухи.оно протекает за 3—4 дня).
Этому способствует высокая температура, которая часто возникает
внутри больших скоплений навоза, кучах мусора и пищевых отхо-
дов. Личинки последнего возраста часто называются предкуколкой.
Активно перемещаясь, предкуколки отыскивают участки с меньшей
температурой и влажностью. Окукливание происходит с образова-
нием пупария (рис. 114,Г), внутри которого располагается куколка
свободного типа. При оптимальных условиях эта фаза продолжает-
ся около 5 дней.
Вылупление молодой мухи из пупария осуществляется с по-
мощью так называемого лобного пузыря. У только что сформиро-
вавшегося насекомого передняя часть головы образована тонкой и
растяжимой кутикулой, которая под давлением нагнетаемой гемо-
лимфы может выдаваться вперед в виде широкого и массивного вы-
роста. Лобный пузырь упирается в переднюю стенку пупария и от-
брасывает ее. Через образовавшееся круглое отверстие муха выби-
рается наружу. Если вылуяление произошло в слое почвы или
в подсохшем субстрате, в котором развивалась личинка, муха под-
нимается на поверхность, прокладывая себе дорогу все тем же лоб-
ным пузырем и конечностями. Молодым насекомым требуется еще
от одного до двух часов для того, чтобы у них расправились крылья
и затвердела кутикула.
Факультативный и облигатный паразитизм личинок мух
Способность личинок существовать в разлагающихся органиче-
ских веществах явилась предпосылкой для возникновения у мух
ларвального паразитизма. Многие виды, которым в большей или
меньшей степени свойственна некрофагия, в качестве субстрата для
откладки яиц могут использовать не только мясо и трупы, но и гно-
ящиеся раны на теле животных и человека. В большинстве случаев
личинки не выходят за пределы раны и не повреждают здоровые
ткани животного-хозяина. Они поглощают гнойный экссудат и от-
мирающие ткани и, по сути дела, еще ничем не отличаются от сво-
их свободноживущих родичей.
Переход к питанию здоровыми тканями означал переход к фа-
культативному паразитизму. Личинки Musca domestica внедряются
в ткани, окружающие рану лишь в очень редких случаях. Личинки
268
же падальных (р. Lucilia, Calliphora) и серых мясных мух (ссм.
Sarcophagidae) делают это значительно чаще. Дальнейшее разви-
тие подобной тенденции привело к появлению облигатных парази-
тов, причем процесс этот совершался независимо в разных семейст-
Рис. 115. Короткоусые двукрылые, паразитирующие на личиночной фазе
(по разным авторам). А — самка wohlfahrtia magnified; Б — личинка
Wohlfahrtia magnified; В — самка Dermatobia hominis, откладывающая
яйца на кровососущую муху; Г — самка желудочного овода Gastrophilus
intestindlis; Д — личинка полостного овода Oestrus ovis; Е — личинка же-
лудочного овода Gdstrophilus intestindlis
Е
вах и сопровождался возникновением целого ряда адаптаций, спо-
собствующих успешному заражению животных. Упоминавшаяся
выше вольфартова муха—Wohlfahrtia magnifica (сем. Sarcopha-
gidae) (рис. 115, А, Б) размножается, отрождая одновременно
160—190 живых личинок. Сформированные, готовые к выходу ли-
чинки скапливаются в половых путях самки. С этого момента на-
секомые начинают назойливо кружиться около крупных животных.
269
При первой возможности они с силой выбрасывают личинок на по-
верхность небольших ссадин, ранок, а часто п просто на слизистые
глаз, ноздрей т. д. Даже небольшое промедление губительно для
самок: в этом случае личинки мигрируют по ее телу, что приводит
к гибели насекомого. Попав на тело позвоночного животного, ли-
чинки активно проникают через поврежденные участки кожи и сли-
зистые в подкожную клетчатку, а оттуда уже могут достигать гай-
моровой полости, лобных пазух и т. д. Миграции личинок сопро-
вождаются тяжелыми разрушениями встречающихся на их пути
тканей. На стадии предкуколки личинки покидают хозяина и Окук-
ливаются в почве.
Еще более совершенные приспособления к заражению человека
можно обнаружить у представителей сем. Calliphorida£. Dermato-
bia hominis для распространения потомства использует своеобраз-
ных переносчиков. Самки откладывают яйца, содержащие сформи-
рованных личинок^ на поверхность тела комаров или мух жигалок
(рис. 115,В). Когда эти кровососы нападают на человека или до-
машних животных, из яиц немедленно вылупляются личинки. Сиг-
налом к этому служит повышение температуры, связанное с пре-
быванием на поверхности тела теплокровного хозяина. Несомненно,
прогрессивный характер носит и способность личинок проникать
в подкожную клетчатку, где они обычно локализуются, через непо-
врежденные участки покровов.
Очень своеобразна группа оводов (рис. 115, Б, Г, Д'). Относящие-
ся к ним виды, хотя и отличаются друг от друга рядом морфологи-
ческих особенностей, характеризуются двумя общими признаками,
во-первых, на личиночной стадии все они являются облигатными
паразитами позвоночных животных, во-вторых, во взрослом состоя-
нии эти насекомые не питаются, в связи с чем у них происходит ре-
дукция ротового аппарата. Имаго таким образом сохраняют за со-
бой лишь генеративную и связанную с ней расселительную функ-
ции. Трофическая функция — питание, которое обеспечивает как
существование особей данной генерации, так и начальные этапы
развития личинок следующего поколения, — целиком ложится
на личиночную фазу.
Личинки желудочных оводов (сем. Gastrophilidae) паразитиру-
ют в желудке и других отделах пищеварительного тракта. Самки
обычно приклеивают яйца к волоскам на теле хозяина в строго оп-
ределенных местах. У одних видов это участки покрове®, непосред-
ственно примыкающие к ротовому отверстию: губы, щеки и т. д.
В этом случае вылупившиеся из яиц личинки мигрируют в ротовую
полость, а оттуда по пищеводу в желудок. У других же видов яйца
откладываются на такие части тела, которые животные могут выли-
зывать или вычесывать зубами. В частности, это характерно для
широко распространенного паразита лошадей — Gastrophilus intes-
tinalis. Сигналом для вылупления личинок из яиц обычно служит
прикосновение к последним влажной поверхности губ или языка жи-
вотного. Часть личинок при этом слизывается хозяином и попадает
в ротовую полость. Те же особи, которые случайно остались на по-
270
верхности тела, внедряются в кожу и, проделывая в ней ходы, вы-
зывают сильный зуд. Лошадь, расчесывая пораженное место зу-
бами, поглощает всех остальных личинок. В ротовой полости все
личинки сначала внедряются в язык, где и проходят первые этапы
их развития. Через месяц, после линьки, они покидают ткани, миг-
рируют »в желудок и прикрепляются к его стенкам при помощи ро-
товых крючков (рис. 115, Е). Паразиты, скапливающиеся иногда
в большом количестве, могут вызывать тяжелые изъязвления сли-
зистой желудка. Зрелые, завершившие свое развитие, личинки вы-
ходят из желудка; вместе с экскрементами выводятся во внешнюю
среду и окукливаются.
Развитие личинок представителей сем. Hypodermatidae — под-
кожных оводов осуществляется в подкожной клетчатке. Яйца отк-
ладываются самками на волосяной покров в самых различных уча-
стках тела. Вылупившиеся из них личинки обычно сразу же внед-
ряются в кожу. Паразиты, использующие в качестве хозяев круп-
ных позвоночных животных, обычно локализуются на их спинной
поверхности. Для этого личинкам приходится совершать длитель-
ную (до 9 месяцев) н сложную миграцию. В одних случаях они
не покидают клетчатки и прокладывают свои ходы только в ней.
В других же личинки передвигаются по мускулатуре и внутренним
органам. Обычно какое-то время они проводят в стенках пищевода,
вызывая нарушение целостности последнего, иногда попадают
в просвет спинно-мозгового канала, но в конце концов все-таки до-
стигают дорзальной поверхности тела животного. Здесь в подкож-
ной клетчатке личинки питаются и линяют. Вокруг них образуется
пузырь, на вершине которого через некоторое время открывается
свищ. Таким образом, личинки старших возрастов получают доступ
к наружному воздуху, необходимому им для дыхания. Полностью
сформированные личинки выходят через свищ во внешнюю среду
и окукливаются в почве.
Оводы, относящиеся к сем. Oestridae (носоглоточные, или поло-
стные, оводы), живородящи. Зрелые самки в своих половых путях
несут сформированных личинок, которых они вместе со струей жид-
кости с силой выбрасывают в нос, в рот или на поверхность глаз жи-
вотного-хозяина, которым может быть и человек. У нас наиболее
широко распространены русский (Rhiuoestrus purpureas) и овечий
(Oestrus ovis) оводы. Попавшие на поверхность слизистых личин-
ки (рис. 115, Д) мигрируют в носовую полость, где и осуществля-
ется их развитие. Очень часто, однако, они, разрушая ткани, прони-
кают в лобные пазухи, а оттуда иногда и в полость черепа. В силу
этого патогенность личинок очень велика. Перед окукливанием они
покидают хозяина и выходят во внешнюю среду через ноздри.
/
Медицинское и ветеринарное значение синантропных мух
Практическое значение высших синантропных мух, причем как
имаго, так и личинок, очень велико. Взрослые насекомые в силу
ряда биологических особенностей — развитие в разлагающихся ор-
271
ганических веществах (фекалии, трупы животных, хозяйственные
пищевые отходы и т. д.) и питание на аналогичных субстратах —
служат переносчиками возбудителей самых'разнообразных заболе-
ваний. Особенно большую роль играют мухи в распространении ки-
шечных инфекций, среди которых имеются и очень опасные: брюш-
ной тиф, холера, дизентерия и т. д. Свободный доступ мух к пище-
вым продуктам человека и может явиться причиной вспышки того
или иного заболевания. Очень велика роль мух, особенно тех ви-
дов, которые нападают на человека для питания потом и выделени-
ями слизистых, как переносчиков возбудителей инфекционных конъ-
юнктивитов и трахом. Инвазионное начало мухи в этом случае чер-
пают из гнойных выделении глаз больного человека. По-вйдимому,
мухи могут служить важным передаточным звеном в эпидемиологии
таких опасных заболеваний, как полиомиелит, туберкулез, сибир-
ская язва и т. д. Кроме возбудителей бактериальных заболеваний
синантропные виды мух активно распространяют цисты кишечных
простейших и яйца ряда гельминтов (аскарид, власоглавов и т. д.).
Важное медицинское и хозяйственное значение имеют личинки
мух — факультативные и облигатные паразиты человека и домаш-
них животных. Причем в одних случаях нх присутствие может ока-
заться весьма полезным для хозяина, в других же — чревато самы-
ми тяжелыми последствиями.
Еще во время наполеоновских войн было отмечено, что поселе-
ние личинок-некрофагов в ранах заметно ускоряет заживление
последних. Спустя сто лет, во время первой мировой войны, эти на-
блюдения были подтверждены. Американский хирург Бейер впер-
вые успешно применил личинок синей падальной мухи для лечения
неоперабельных случаев остеомиелита. Однако использование ли-
чинок из природных популяций всегда было сопряжено с опас-
ностью занёсения вторичных инфекций. Лишь с разработкой мето-
дики выращивания стерильных личинок Musca domestica, Lucilia,
Calliphora и некоторых других видов этот метод получил доволь-
но широкое распространение. Благотворное воздействие личинок
мух на состояние раны обусловлено причинами двоякого рода. Во-
первых, слюна личинок многих видов содержит эффективные бак-
терицидные вещества, которые вызывают стерилизацию раны. Во-
вторых, выедая отмирающие ткани, личинки очищают поверхность
раны, а выделяемые ими секреты стимулируют регенерационные
процессы. С появлением антибиотиков, однако, интерес к «хирур-
гическим» личинкам, как их называли долгое время, снизился,
и исследования в этой области почти полностью прекратились.
Однако, как уже отмечалось выше, далеко не всегда некрофаги
ведут себя как полезные и даже просто безобидные обитатели ран.
В некоторых случаях они могут внедряться,и в здоровые ткани хо-
зяина. Вызываемые ими при этом повреждения получили названия
миазов. Причиной факультативных тканевых* миазов человека мо-
гут быть личинки Musca domestica, М. stabulans, Calliphora, Luci-
lia, Sarcophaga и некоторых других видов. Чаще всего они не
причиняют особого вреда. Правда, в некоторых случаях могут воз-
272
никать осложнения гнойных заболеваний уха, носа, глаз (конъюнк-
тивиты) и т. д. Значительно большую опасность личинки этих
видов представляют собой как источники вторичных инфекций,
связано с загрязнением тела паразитов патогенными микроорга-
низмами, вызывающими гангрену, общее заражение крови и т. д.
Облигатные миазы вызываются личинками, для которых пара-
зитизм стал единственно возможной формой существования. Осо-
бенно патогенны личинки, которые быстро развиваются и интенсив-
но растут. Их миграции сопровождаются возникновением обширных
некротических очагов, обильных нагноений, гангренозных процес-
сов. Описаны случаи, когда личинки Wohlfahrtia magnified пол*
ностыо уничтожали глаза и покровы головы. Запущенное заболева-
ние может иметь летальный исход. Не менее опасны и личинки
Callitroga hominivorax (сем. Calliphoridae), распространенной
в ряде стран Америки.
Вред, наносимый личинками, развитие которых осуществляется
медленно, не столь велик. Вызываемые ими повреждения объеди-
няются в группу доброкачественных миазов. Примером может слу-
жить упоминавшаяся выше Dennatobia hominis (с. 270). Личинки
этого вида, проникнув под кожу человека, вызывают образование
небольшого желвака со свищом. Никаких миграций по телу хозяи-
на они не совершают. Развитие паразита продолжается около
70 дней. Взрослые личинки могут вызывать неприятные болевые
ощущения.
Человек может заражаться и личинками оводов. Личинки сем.
Gastrophilidae обычно могут проникать только в толщу эпидер-
миса и, передвигаясь, оставляют за собой тонкий и извитой след.
Это заболевание получило название ползучей болезни или «воло-
сатика». Личинки подкожных оводов мигрируют в толще подкож-
ной клетчатки. Это продолжается в течение нескольких месяцев
и обычно проходит незамеченным. Лишь после того как личинка
остановилась, вокруг нее образуется небольшая опухоль. Личинки
полостных оводов у человека чаще всего локализуются в глазах.
Наиболее опасны случаи, когда паразиты проникают внутрь глаз-
ного яблока. Это может привести к полной потере зрения.
Диагностика миазов обычно не представляет больших затруд-
нений. Лечение осуществляется, главным образом, путем вскрытия
ходов, прокладываемых личинками, и удаления последних.
Одной из форм факультативных миазов являются повреждения
кишечника, вызываемые личинками, проглоченными с водой или
пищей. У людей с пониженной кислотностью они не только могут пе-
реживать некоторое время в пищеварительном тракте, но и разви-
ваться. Присутствие паразитов вызывает раздражение слизистых
оболочек и воспалительныё процессы. Больные обычно жалуются
на острые боли в животе. Чаще всего кишечные миазы вызываются
личинками Musca domestica, Muscina stabulans, Fannia canicularis
и некоторых других видов.
Значительный ущерб паразитические личинки мух наносят
сельскому хозяйству. Личинки Wohlfahrtia magnified часто вызы-
273
яают массовые заболевания овец. Иногда наблюдается и значи-
тельный падеж, причем не только среди молодых, но и среди взрос-
лых животных. Очень опасны для овец и полостные оводы. Их ли-
чинки, проникая в лобные пазухи в полость черепа, служат
причиной тяжелого заболевания, известного под названием лож-
ной вертячки овец. Животные начинают кружиться на одном месте,
теряют координацию движений, перестают питаться и через не-
сколько дней погибают. Паразитирование личинок кожных оводов
в крупных домашних животных приводит к сильному истощению
последних, а кроме того, безнадежно испорченными оказываются
шкуры, так что использовать их в кожевенной промышленности
уже невозможно. Причиной сильного истощения и ослабления жи-
вотных часто бывают и желудочные оводы.
Борьба с гнусом и синантропными мухами
Большое практическое значение кровососущих двукрылых и си-
нантропных видов высших мух предопределило необходимость
разработки надежных и эффективных методов борьбы с ними.
Большие успехи в свое время были достигнуты с помощью широко-
го применения ДДТ. Распыление этого инсекцитида на огромных
площадях суши и по поверхности водоемов позволило в короткий
срок добиться больших успехов в борьбе с малярийными комарами
и практически полностью ликвидировать малярию в целом ряде
стран мира. Столь же эффективным ДДТ оказался и в борьбе про-
тив других представителей гнуса и синантропных мух. Химические
меры борьбы со взрослыми насекомыми и их личинками не утрати-
ли своего значения и по сей день. Однако высокая токсичность
большинства препаратов и появление форм, резистентных к ним,
привели к необходимости применения иных методов борьбы. Хоро-
ший эффект дает использование паразитов и врагов насекомых.
В частности, оздоровлению очагов малярии на Кавказе способст-
вовало широкое разведение в местных водоемах рыбки гамбузии,
которая питается личинками комаров. Обнадеживающие резуль-
таты были получены в опытах с заражением личинок комаров ми-
кроспоридиями. Последние, если и не вызывают массовой гибели
насекомых, повышают их чувствительность к инсектицидам, что
позволяет резко снижать дозы ядов, рассеиваемых в природе.
В настоящее время проводятся успешные опыты по использованию
личинок нематод (сем. Mermithidae, с. 106) для борьбы с мош-
ками.
Очень перспективным и эффективным оказался метод использо-
вания стерильных самцов, оправдавший*себя в борьбе с мухами из
р. Callitroga, которые наносили огромный ущерб животноводству
в США. Выращенные в лаборатории и стерилизованные радиоак-
тивным методом самцы в огромных количествах выпускались
в районах массового размножения мух. Самки, спариваясь с таки-
ми самцами, откладывают неоплодотворенные яйца, дальнейшее
развитие которых невозможно. За сравнительно короткий срок по-
274
пуляция этих опасных мух в США была полностью уничтожена.
Аналогичные способы борьбы начинают применяться и по отноше-
нию к другим вредным двукрылым, в частности комарам.
В борьбе с синантропными видами (главным образом мухами)
особенно важное значение приобретает строжайшее соблюдение
санитарных норм. Необходимо поддерживать чистоту и удалять на-
воз из животноводческих помещений, постоянно очищать мусоро-
прпемники от бытовых отходов, в неканализированных уборных
должны создаваться условия, исключающие возможность контакта
мух с фекалиями. Обезвреживание экскрементов, навоза, пищевых
отходов должно производиться в специальных компостерных рвах
или на полях запахивания. Ограничиваться простой засыпкой ор-
ганических веществ землей нельзя, так как вылупляющиеся из ку-
колок молодые насекомые легко прокладывают себе путь в рыхлой
почве: Необходимо хорошее утрамбовывание поверхностного слоя.
Если очаги выплода мух все-таки возникают, то хорошие результа-
ты дает применение трихлофоса.
Важное значение имеют методы индивидуальной защиты людей
от нападения двукрылых. В местах большого скопления гнуса сле-
дует -пользоваться специальными защитными костюмами и нако-
марниками, пропитанными репеллентами (с. 195). Необходимо
предупреждать появление насекомых в жилых помещениях с по-
мощью защитных сеток на окнах и дверях. При ночевке на откры-
том воздухе или в помещениях, в которые имеется доступ крово-
сосущим двукрылым, следует пользоваться марлевыми пологами.
Для предупреждения распространения кишечных и других инфек-
ционных болезней необходимо строго следить за тем, чтобы мухи
не имели доступа к пищевым продуктам. Это особенно важно не
только в домашних условиях, но и в организациях общественного
питания, в магазинах, на базарах и т. д.
Успешное проведение комплексных мероприятий и строгий са-
нитарный контроль позволили в -нашей стране ликвидировать, а в
некоторых случаях значительно уменьшить встречаемость целого
ряда опасных заболеваний, в распространении которых большую
роль играли двукрылые.
Основные направления эволюции паразитических клещей
и насекомых
Несмотря на глубокие различия между клещами и насекомыми
в эволюции их приспособлений к паразитическому образу жизни
можно найти много общего. Самый переход к паразитированию на
позвоночных был, по-видимому, связан с изменением «пищевого
поведения» членистоногих, обитавших в норах и гнездах птиц, гры-
зунов и других животных. От питания органическим детритом мел-
кие клещи и насекомые постепенно переходили в питанию орого-
вевшими частицами эпидермиса, пером или волосом животных,
а потом и к поглощению секрета кожных желез или живой ткани
(перьевые клещи, чесоточные зудни и др.). «Слизывание» капель
275
крови, выступающих из ранок и ссадин на коже позвоночных, или
случайное активное повреждение кожи привели к появлению ге-
матофагии, сначала факультативной, а затем и облигатной. Ве-
роятно, именно так возникла гематофагия различных паразити-
формиых клещей (с. 197) и многих насекомых (вшей, некоторых
короткоусых двукрылых и др.). В отдельных случаях (например,
у клопов) способность к кровососанию могла развиться из хищни-
чества (с. 211), а у некоторых временных паразитов (например,
у длинноусых двукрылых) даже из адаптаций к питанию соками
растений.
Переход к питанию кровью неизбежно должен был сопровож-
даться изменениями в строении ротового аппарата членистоногих.
Возникла необходимость в появлении таких органов, которые по-
зволили бы паразиту нарушать целостность кожных покровов поз-
воночного животного и извлекать кровь.
У членистоногих, еще слабо адаптированных в кровососанию,
обычно сохраняется тот тип ротового аппарата, который свойст-
вен их ближайшим родичам. Так, у многих гамазовых клещей (сем.
Lealaptidae) еще имеются клешневидные хелицеры; у питающихся
кровью пухоедов (Piagetielld)—типичные грызущие ротовые час-
ти и т. д. Однако в большинстве случаев у членистоногих — гема-
тофагов— развивается специализированный режущий или колю-
ще-сосущий ротовой аппарат. Ротовые конечности преобразуются
при этом в подвижные режущие пластинки (хелицеры иксодоидпых
клещей, максиллы и мандибулы слепней и т. д.) или в тонкие ко-
лющие стилеты (у некоторых гамазовых клещей и у большинства
кровососущих^насекомых). Взаиморасположение ротовых частей
обеспечивает образование каналов, по одному из которых течет
слюна, вводимая в ранку на теле хозяина, а по другому — высасы-
ваемая паразитом кровь. Процесс поглощения этой жидкой пищи
обеспечивается сокращением мускулатуры глотки, которая преоб-
разуется в интенсивно работающий насос. В состав секрета слюн-
ных желез всех облигатйых гематофагов обязательно входят ве-
щества, препятствующие свертыванию крови, — различные по .ха-
рактеру своего действия антикоагулянты.
Строение ротового аппарата кровососущих членистоногих пре-
терпевает значительную эволюцию, связанную с изменением спосо-
ба поглощения пищи. Можно проследить все переходы от грубого
разрывания кровеносных сосудов с последующим всасыванием
крови из образующейся гематомы (иксодоидные клещи, слепни
и др.) до появления совершенных колющих хоботков, вводимых
в просвет кровеносного сосуда и действующих по принципу насасы-
вающего кровь шприца (комары, вши, клопы, блохи и др.).
Многие гематофаги являются временными или периодическими
паразитами, в связи с этим их пищеварительный тракт бывает
снабжен карманообразными выпячиваниями и выростами (см.
рис. 86), которые наполняются кровью, всосанной за время питания.
Возможность единовременного наполнения кишечника обеспечива-
ется особенностями строения покровов членистоногих-гематофагов.
276
Всем йм свойственно наличие эластичной, растяжимой кутикулы,
благодаря чему тело может сильно раздуваться. У некоторых кле-
щей этому способствует и межлиночный рост кутикулы.
Питание кровью обусловливает многие особенности размноже-
ния паразитических членистоногих. Кровь — высоко калорийная
пища, обеспечивающая быстрое созревание яичников (гонотрофи-
ческая гармония, гонотрофический цикл, с. 179). Вероятно, именно
с питанием кровью связана и отчетливо выраженная у паразитиче-
ских членистоногих тенденция к яйцеживорождению. Развитие
зародыша в яйце, еще находящемся в матке родительской особи,
и даже отрождение живых личинок свойственно многим гамазовым
клещам (Laelaptidae и др.), перьевым клещам, клопам сем Poly-
ctenidae, многим кровососущим двукрылым (с. 279).
На примере членистоногих отчетливо прослеживается усиление
связи паразитов с хозяином, переход от временного паразитизма
к стационарному и, в конце концов, к эндопаразитизму. Интересны
в этом отношении иксодоидные клещи. Так, если аргазовые — ти-
пичные временные паразиты, многократно нападающие на различ-
ных хозяев и питающиеся на них очень кратковременно (от не-
скольких минут до нескольких часов)? то клещи р. Boophilus,
например, связаны лишь с одним хозяином и проводят на нем
свыше 20 дней. Среди гамазовых клещей можно проследить пере-
ход от типичных временных паразитов (сем. Laelaptidae) к ста-
ционарным, всю жизнь проводящим на теле хозяина,— примером
может служить паразит летучих мышей Spinturnix (см. рис. 90, Б).
Постоянный стационарный паразитизм свойствен п всем перье-
вым клещам.
Ряд переходов от временного паразитизма к стационарному
(периодическому и постоянному) имеется и среди насекомых. Осо-
бенно интересен в этом отношении отряд двукрылых, в пределах
которого прослеживаются все переходы от типичного временного
паразитизма (подотр. Nematocera) к стационарному (личиночному
или имагинальному) паразитизму многих представителей подотр.
Brachycera. Интересно, что эта тенденция конвергентно возникает
во многих семействах. Среди таких временных паразитов, как кло-
пы и блохи, тоже намечается переход к стационарному (даже к по-
стоянному) паразитизму. Постоянными паразитами являются пу-
хоеды и вши. Связь их с организмом хозяина иногда становится
очень тесной. Некоторые виды неподвижно сидят на теле животно-
го-хозяина, погрузив свой колюще-сосущий хоботок в его ткани.
Они сосут почти непрерывно с небольшими промежутками. Инте-
ресно, что такие паразиты конвергентно появляются в разных
отрядах. Среди вшей — это площица (Phtirus pubis), среди кло-
пов сем. Polyctcnidae, среди блох — сем. Echidnophagidae, среди
двукрылых — овечий рунец (Melophagus) и др.
Такне постоянные, малоподвижные паразиты характеризуются
рядом морфологических особенностей. Они обладают уплощенным
широким телом, широко раставленными, тоже слегка уплощенными,
конечностями (см. рис. 90, 5; 99, Г). Последние служат не столько
277
для движения, сколько для прикрепления к шерсти, перьям или во-
лосам хозяина. Поэтому они нередко снабжены сильными коготка-
ми и выростами, играющими роль зажимов (см. рис. 99, £), или
несут на концевых члениках лапки нежные эластичные присоски
(Analgesoidaea).
В то же время многие временные и даже стационарные парази-
ты (например, пухоеды) сохраняют подвижность и могут, иногда
с большой быстротой, передвигаться в шерсти или пуху, одевающем
тело хозяина. Этому способствует приобретение паразитами «обте-
каемой» формы тела, снижающей сопротивление густого волосяно-
го или пухового покрова, которое приходится преодолевать пара-
зитам при движении. У блох тело сплющено с боков, у клопов,
перешедших к облигатному временному паразитизму (Cimicidae),
сохраняются лишь небольшие рудименты крыльев. Полностью ис-
чезли крылья в отрядах вшей, пухоедов, блох. Переход наземных
членистоногих от эктопаразитизма к эндопаразитизму относитель-
но редок. Так, ряд гамазовых клещей (сем. Rhynonossidae и др.)
перешел к паразитированию в носовых полостях птиц. Эндопара-
зитами стали и некоторые другие клещи. Средн тромбидиформ-
ных — это Demodex, паразит волосяной сумки млекопитающих;
среди саркоптид—чесоточные зудни, проделывающие ходы в тол-
ще кожи млекопитающих и человека; некоторые перьевые клещи
(р. Ardeacarus и sp.) и др. Эндопаразиты встречаются и среди на-
секомых. Это песчаная блоха Tunga penetrans (с. 235) и своеобраз-
ный представитель двукрылых — Ascodipteron, паразитирующий
в коже летучих мышей (с. 262).
ЛИТЕРАТУРА
Руководства и учебники
Беклемишев В. Н. Бноценологические основы сравнительной паразитс^огии.
М., 1970.
Бэкер Э. и Уартон-Г. Введение в акарологию. М., 1955.
Деляму ре С. Л. Гельмпптофаупа морских млекопитающих в свете их эколо-
'гии и филогении. М., 1955.
Догель В. А. Курс общей паразитологии. Л., 1947.
Догель В. А. Общая паразитология, Л., 1962.
Догель В. А. Зоология беспозвоночных. М., 1976.
Дубинина М. Н. Ремнецы фаунььСССР. М.— Л., 1966.
Жизнь животных. Под ред. Л. А. Зенкевича, т. II. М.» 1968; т. III, 1969.
Крашкевич К- В., Тарасов В. В. Медицинская паразитология. М., 1969.
Основные проблемы паразитологии рыб. Под ред. В. А. Догеля, Г. К. Петру-
шевского и Ю. И. Полянского. М., 1958.
Павловский Е. Н. Руководство по паразитологии человека, т. I, М.— Л., 1946;
т. 1Г, 1948.
Парамонов А. А. Основы фитогельминтологии, т. I. М., 1962.
Паразитология человека. Под ред. Г. С. Первомайского п В. Я. Подоляпа.
Л., 1974. »
Подъяпольская В. П. и Капустин В. Ф. Глистные заболевания человека.
М., 1950.
Руководство по медицинской паразитологии. Под ред. В. П. Дербеневой-
Уховой. М., 1974.
Руководство по микробиологии, клинике и эпидемиологии инфекционных бо-
лезней. Под ред. П. Г. Сергиева, т. IX. М., 1968.
Скрябин К. И. и Шульц Р. С. Тельминтозы крупного рогатого скота и его
молодняка. М., 1937.
Шульц Р. С. и Гвоздев Е. В. Основы общей гельминтологии, т. I. М., 1970;
т. II, 1972; т. III, 1976.
Brand Th. Parasitenphysiologe. VEB. Gustav Fischer Verlag. Jena, 1972.
279
Chandler A. C. Introduction to parasitology. 9tn Ed. New-York — London, 1955.
Kaestner A. Lchrbuch der spezicllen Zoologie. В. 1, Wirbellose, 3 Tei|, G. Fischer
Verl. Jena, 1973.
Noble E. R. and G. A. Noble. Parasitology. The Biology of anfrnal parasites.
Philadelphia, 1964.
Smyth, J. D. Introduction to animal parasitology. The english University press,
London, 1962.
Монографии и статьи
Балашов Ю. С. Кровососущие клещи — переносчики болезней человека и жи-
вотных. Л., 1967.
Благовещенский Д. И. Определитель пухоедов (Mallophaga) домашних жи-
вотных. Фауна СССР, и. сер. 27. М.— Л., 1940.
Богоявленский Ю. К. Структура и функции покровных тканей паразитических
нематод. М., 1973.
Брегетова Н. Г. Гамазовые клещи (Gamasoidea). Краткий определитель.
М.—Л., 1956.
Вагин В. Л. Ascothorax ophioctcnnis Djakonov п положение отряда Ascothora-
cida Vagin в системе Entomostraca. Ученые записки ЛГУ, № 101, сер. биол.,
вып. 19, с. 12—70. Л., 1949.
Вагин В. Л. О новых паразитических ракообразных из сем. Dendrogasteridae
(отряд Ascothoracida). Труды Ленинградского общества естествоиспытателей,
т. LXX, вып.-4, с. 3—89. Л., 1950.
Вагин В. Л. Мешкогрудые раки. Казань, 1976.
Грузов Е. Н. Эндопаразитический моллюск Astcrophila japonica Randall, et
Heath (Prosobranchia, Melanellidae) и его связь с паразитическими брюхоногими.
Melacologia, 3 (1), 1965.
Грунин К. Я. Личинки оводов домашних животных СССР. Определители
по фауне СССР. М.— Л., 1953. «
Дубинин В. Б. Перьевые клещи (Analgesoidea), ч. I. Введение в их изучение.
Фауна СССР, т. VI, вып. 5. Паукообразные. М. — Л., 1951.
Дубинин В. Б. Чесоточные клещи. М-» 1954.
Здродовский П. Ф. и Голиневич Е. М. Учение о риккетсиях и риккетсиозах.
М., 1972.
Иванов А. В. Строение и развитие эндопаразитического брюхоногого моллюс-
ка Parenteroxenos dogieli A. Iwanow (семейство Entoconchidae), Изв. АН СССР,
сер. биол., № 1. 1947, с. 3—28.
Иванов А. В. Строение эктопаразитических брюхоногих Stiliferidae как ре-
зультат их образа жизни. Труды Ленинградского общества естествоиспытателей,
т. LXXI, вып. 4, 1952, с. 86—140.
280
Иванова-Казас О, М. Очерки по сравнительной эмбриологии паразитических
перепончатокрылых. Л., 1961.
Лутта А. С. Слепни Карелии. Л., 1970.
Маркевич А. П. Паразитические веслоногие рыб СССР. Киев, 1956.
Основы нематодологии. Под ред. К. И. Скрябина, т. 1—XXI. М. — Л., 1949—
1969.
Основы цестодологии. Под ред. К. И. Скрябина, т. I—VII. М., 195Г—1969.
Петроченко В. И. Акантоцефалы (скребни) домашних и диких животных,
т. I. М.» 1956; т. II, 1958.
Померанцев Б. И. Иксодовые клеши (Ixodidae). Фауна СССР. Паукообраз-
ные, т. IV, вып. 2. М,—Л., 1950.
Рубцов Я. А. Водные мермитиды, ч. I. Л., 1972; ч. II, 1974.
Штакельберг А. А. Синантропные двукрылые фа^ны СССР. Определители по
фауне СССР. М.— Л., 1956.
Филипова Н. Я. Аргасовые клещи (Argasidae). Фауна СССР, т. 4, вып. 3,
М.—Л., 1956.
Eichler Wd. Phthiraptera. 1. Mallophaga. In: Bronns Klassen und Ordnungen
des Tierreichs. B. 5, Abt. Ill, Buch 7. Leipzig, Akad. Verlag, Geest und Portig. 1963.
Kabata Z. Crustacea as enemies of fishes. Fish. Res. Board of Canada, 1970*.
Lee D. L. The physiology of nematodes. Edinburgh — London. 1965.
Lee D. L. The structure and composition of the helminth cutic ola. Adv. Para-
sitol., vol. 4, 1966.
Smyth J. D The physiology of Cestodes. Edinburgh, Oliver Boyd, 1969.
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ ЖИВОТНЫХ И ТЕРМИНОВ
Акаптелла 73
Акантор 73 -•
Аллоскутум 172
Альвеококкоз 44
Амидостоматоз 126
Амфиды— 90, 107
Анкилостомндозы 111
Анкилостомоз 111
Анокснбиоз 71
Антикоагулянты 94, 276
Антракс 265
А'нтропозооноз 197
Аскаридиоз 126
Аскаридоз 108
Аутоннвазия 41, 103, 111, 114, 123, 125
Блоха крысиная 233
— куриная 235
— песчаная 234, 235
— человеческая 233, 234
БлохЬ 187, 230, 276
Ботриоцефалез 52
Булла 149, 152
Бурса копулятивная 69
Везикула головная 85
Велигер 140
Вертячка овец 51, 187
----«ложная» 274
Власоеды 215
«Волосатик» 273
Волосатики 127
Вошь головная 223
— лобковая 223, 224
— платяная 223, 224
Вухерериоз 1.15
Вши 187, 214, 220, 276
Галлера орган 173
Гвоздичники 32
Гематофагия 93, 154, 164, 174, 179,
200, 213, 217, 222, 230, 241, 244, 245
246, 254, 256, 260, 267, 276
Гермафродитизм 98, 160
— протераидрический 98, 164, 166
Гетерогония 59, 6L, 103
Гиалуронидаза 47, 122
Гидатида 45
Гидатидная жидкость 46
Гидатидный песок 46
Гидракарины /208
Гименолепидоз 41
Гиперпаразит 1*66, 198
Глохидий 130
Гиатосома 170, 195, 198
Гнус 262, 274
Гонотрофическая гармония 174, 179,
244, 255, 260
Гонотрофический цикл .174, 179, 196,
241, 253
Двукрылые 240
Диктиокаулез 126
Дилипидоз 43
Дифпллоботриозы 33
Дициемиды 59
Длинноусые 240
Дракункулез 118
Жене орган 179, 180
Железница 208
Животные прокормители 182
Жигалка осенняя 255, 262
Жигалки 255, 263
Заболевания бактериальные 187, 265
— вирусные 1-85
— вызываемые простейшими 194, 265
Заражение ипокулятцвное 188, 191,
192, 228
— коптаминативное 188, 193, 228, 299,
237, 239
— перкутанное 102,'ПО, 113
— пероральное 105
Зудень чесоточный 204, 275
" Идиосома 170, 178, 195, 199
Инфузориген 59
Инфузориформ 59
Истмус 86
Кала-азар 265
Камера выводковая 135, 139, 155, 157
Капсула выводковая 28, 46
Карповая вошь 152, 167
Карпоеды 152
Кастрация паразитарная 163
Кентрогон 161
282
Клещевые пятнистые лихорадки 190
— спирохйозы 188
Клещи аргазовые 170, 174, 177, 187
— гамазовые 195, 276
— иксодовые 170, 174, 177, 183, 18Л
276
— иксодоидные 170
— паразитиформные 170
— перьевые 197, 275
Клопы 210, 276
Комары 257, 262, 276
— малярийные 274
— настоящие 246
Копрофаги 267
Корациднй 22, 33
Короткоусые круглошовные 240
— прямошовные 240
Краснотелкн 205
Кровососка лошадиная 255
— оленья 255
Кутикула 81
Лабеллум 257
Ларвоциста 26, 44, 47
Лейшманиозы 265
Лентец широкий 33
Ликофора 55, 57
Личинка аскоторацидная 157, 159
— копеподитная 151
— рабдитовидная 99, 112
— филяриевидная 99, 112
— циприсовидная 160
Личинки «хирургические» 232, 272
Лихорадка волынская 227 •
— денге 263
— желтая 263
— марсельская 192
— папатачи 264
— скалистых гор 191
— цуцугамуши 207
— японская’ речная 207
— Q 192, 197-
Лоаоз 117, 266
Малярия 265
Маития 155, 157
— ложная 133
Метанауплиус 159
Метакорпальный бульбус 86
Миазы 273
Микроворсинки 23, 59, 177
Микротрихии 9, 53, 59
Микрофилярии 106, 115, 1117, 118, 266
Мокрецы 243, 257, 262
Мониезиоз 57
Москиты 245, 257, 262
Мошки 241, 256, 262, 274
Муха базарная 267
— вольфартова 267, 269
— домовая 267
— малая комнатная 266
— комнатная 266
— падальная 266, 269
— серая мясная 266, 269
Мухи кровососки 255
— синантропные 266, 274
— це-це 255, 265
Насекомые 208
Науплиус 151, 155, 159, 160
Некатороз 111
Некрофаги 267, 272
Нематоген 59
Нематодозы 107
— домашних животных— 125
— птиц 126
— человека 108
Нимфа 169, 180, 200, 203, 204, 205, 208
Овод желудочный 270
Овода 266, 270, 271
Онкосфера 21, 44
Онхоцеркоз 118, 266
Ортогон 11, 88
Очаг антропургический 238
— природный 47, 124, 185, 189, 193,
238
— синантропный 49, 119, 124, 187, 189,
193
Папиллы — 89
Паразитизм временный 234, 262
— нмагннальный 106
— ларвальный (личиночный) 209, 268,
277
— облигатный 106, 268
— постоянный 262, 277
— стационарный 234, 262, 277
— факультативный 106, 268
— эмбриональный 209
Паразиты гнездово-норовые 182
— пастбищно-подстерегающие 182
— растений 94, 207, 209
Паукообразные 169
Партеногенез 98
Педикулез 230
Первнчнополостные 78
Переносчики 185, 187, 189, 192, 193,
194, 197, 223, 237
Перитремы 171, 180
Пиноцитоз 17, 177
Пищеварение внекишечное 268
—пристеночное 177
Пищеводные железы 86
Плероцеркоид 24
Площнца 223, 225, 226, 277
Полиэмбриония 209
Полужесткокрылые 210
Поровые поля 174
Прокорпус 86
Протосколекс 28, 46
Протрематоды 59, 60
283
Процеркоид 23
Псевдолаллиум 133
Пухоеды 214, 277, 278
Раки веслоногие 143, 167
— корнеголовые 160, 168
— мешкогрудые 155, 159, 167
— равноногие 164, 167
— усоногие 166, 168
Ракообразные 142
«Реакция бегства» 105
Ремнецы 6, 51i, 30
Ренетты 87
Репелленты 275
Риккетсии 190, 191, 193, 197, 227, 228,
229
Риккетсиозы 190, 239
Ришта 118
Рост иитеркалярный 9
— межлиночный 178
Ротовая пирамида 155
Рунец овечий 255, 277
Саккулина 160, 166
Самцы карликовые 137, 150, 159, 165,
167
Сингамоз 126
Сифон 134, 251
Скребень великан 176
Скребни 63
Скутум 172
Слепни 252, 257, 263, 265
Слоновость 117, 266
Сонная болезнь 265
Специфичность 147, 200, 223, 238
Спикулы 91
Стилет 94, 153
Столон 160, 161
Стома 85
Стронгилоидоз 114
Сумка выводковая 165
— копулятивная 69
— перьевая 203
Сурра 265
Схизоцель 84, 85
Телязиоз 126
Тениарннхоз 36
Тениидозы 36
Тениоз 38
Тиф возвратный 227
----клещевой 189
— сыпной крысиный 197, 239
— сыпной эпидемический 194, 227
— эндемический 197
Трансмиссивные болезни 165, 214, 263
Трансовариальная передача 186, 188,
189
Трахома 272
Трихинеллез 121
Трихостронгйлидоз 126
Трихоцефалез 118
Туляремия 187, 197, 227, 2Й9, 265
Убиквисты 220
Угрица 208
Фагоцитоз 177
Фазмиды 90, 107
Филяриатоз 115
Финка 27
Фйтогельминты 94
Фовеа 174
Фтироз 226
Фурка 156
Хелицеровые'168
Хетотаксия 196
Хобот 134
Хозяин резервуарный 24, 35, 73, 186,
192
Ценур 28
Ценуроз 51
Цепень бычий 36
— вооруженный 38
— карликовый 41
— мозговик 51
— невооруженный 36
— свиной 30—38
Цепень тыквовидный 43
Церкомер 23, 55, 58, 62
Цестоды 5, 61
Цестодозы 33
— домашних животных 50
— рыб 51
— человека 33
Цистицерк 27, 36
Цистицеркоз 40
Цистицеркоид 26, 42, 44
Чесотка 206
Чума 235
Чумная пробка 237
Шейные-железы 87
Экзосаккулина 160, 162
Эктопаразиты временные 166, 177, 234
— постоянные 167, 177, 178, 234
Элефантиазис 266
Эмбриофор 21/40
— ресничный 22
Энтеробиоз 110
Энцефалит весенне-летний 185
— клещевой 187
— шотландский 187
— японский 264
Эпикутикула 178
Эхинококк 28, 46
Эхинококкоз 44
Язва пендинская 165
— сибирская 265, 272
284
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
Acanthocephala 63
Acanthocephales 63
Acanthodiaptomus 33
Achtheres 149
— percarum 146 *, 147
Actornithophilus 216*
— p'atcllatus 218
Adenophorea 79, 87, 89, 93, 105, 106,
107
Aedes 115, 116*, 117, 246, 248, 250,
251, 263, 266
Aega 164, 166, 167, 168
Aglossa 132
Alveococcus multilocularis 6, 47, 48*,
51
Amblycera 216, 216*, 216, 217, 219
Amia calva 29
Amidostomidae 126
Amidostomum anseris 126
Amphilina 53, 55
— foliacea 54*, 55 •
— japonica 55
Amphilinidea 53, 54
Amphineura 59
Analges 202
Analgesidae 198
Analgesoidea 198, 200, 278
Anaticola 218
Ancylostoma 86, 93, 94, 101, 107
— duodenale 112*, 113, 114
Ancylostoma tidae 93, 103, 105
Anisakis 88*
Anopheles 115, 116*, 117, 246, 247*,
247, 249, 250, 251, 265, 266
Anophelinae 246
Anoplocephalidae 21, 50
Anoplura 210
Aphasmidia 79
Apocrita 209
Archigetes 5, 25, 25*, 26
— iowensis 25*
— sieboldim 25*
Ardeacarus 204, 278
Argasida’e 193
Argulus 153, 154, 166, 167
—Toliaceus 153*, 154
Arthropoda 141
Ascaridata 104, 108
Ascaridia 107, 126
— galli 106*, 126
Ascaridida 80, 92, 104, 107, 110
Ascarididae 125
Ascaris 89, 104, 107
— lumbricoides 80*, 87*, 108*, 125
— suum 125
Ascodipteron 278
— speiserianum 256*, 262
Ascothorasida 155, 159, 160, 167
Ascothorax 156*, 157, 159, 160, 167
— ophioctennis 155
Asterophila 134, 135, 136, 137
— japonica 133*, 136*, 138*
Asterophilidae 132
Bacillus anthracis 265
Basanistes huchonis 147
— woskoboinikovi 146*
Biacetabulum 25, 26
Bivalvia 130
Bolbosoma .physeteris 68*
Bonn etc Ila, 19’8*
Boophilus 277
Bopyrus 165, 167
Borrelia reccurentis 227, 228
Bothriocephalus 53
— gowkongensis 52*, 52
Brachycera 240, 259*
Brachycera — Cyclorrhapha 241, 255.
260, 266
Brachycera — Orthorrhapha 241
Branchiata 141
Branchiura 166
Buddenbrockia $4
Caligidae 146, 147, 150, 152
Caligoida 143, 145*, 148
Caligus 148, 149, 167
285
— curtus 145*
— lacustris 147
Calliphora 266, 269, 272
Calliphoridae 266, 270, 273
Callitroga 274
— hominivorax 273
Camallanata 107, 118
Canidae 44, 51
Copulidae 132
Caryophyllaeidea 6, 13, 15, 24^ 25, 32
Caryophyllaeus 11*, 25
— laticeps 7*
Cephalopoda 59
Ceratopogonidae 241
Ceratophyllus tesquorum 238
Cercomaeromorpha 62
Cestoda 5, 6, 26, 53, 56
Cestodaria 3, 6, 53, 56, 62
Chelicerata 141
Chondracanthus pinguis 149
Chromadorida 80
Chrysops 116*, 118, 252*, 252, 253, 254
Chrysozona 253
Cimex 212
— columbarius 213
— hemipterus 212
— lectularius 212
Cimicidae 211, 212, 278
Cirripedia 155, 159, 163
Conch if era 59, 60
Copepoda 33, 51, 145, 149, 166, 168
Corynosoma 73, 74
— strumosum 68*, 75*
Coxiella burneti 193
Crustacea 166, 167
Ctenocephalides canis 233
— felis 231*
Cucumaria japonica 140
Cucumaridola 146
— notabilis 144*
Culex 115; 116*, 246, 248*, 248, 249,
250, 251
— pipiens 117
Culicidae 241, 250, 257
Culicinae 246
Culicoides 243, 244, 244*
Culiseta 246
Cyclophyllidea 6, 7, 8*, 11, 13, 14*, 15,
20, 26, 27, 27*, 29, 31, 32, 33, 36,
41, 43, 50, 41
Cyclopoida 143, 168
Cyclops 23, 33, 118
Cymothoa 164, 166, 168
Cymothoidae 165*
Cypris 160
•
Dallyelida 58, 62
Danalia 166, 168
—curvata 166
Davaenia 16
Demodex 278
folliculorum 207*, 208
Dendrogastcr 155. 158, 168*, 160, 167
— dichotomus 158*
— iwanowi 158*
Dendrogastridae 158
Dermacentor 228
Dermaglyphidae 198, 202
Dermanyssidac 195, 197
Dermanyssus 196
— gallinae 196*, 197
Dermatobia hominis 269*, 270
Diaptomus 23, 33
Dictyocaulidae 126
Dictyocaulus 126
Dicyemida 59
Digramma 6, 31
Dikerogammarus 55
Dilepididae 43
Dioctophymida 80, 107
Dioicocestus 16
Diphyllidea 6, 7, 33
Diphyllobothriidae 33
Diphyllobothrium 8*, 14*, 15, 33
— latum 6, 8, 18, 33, 34*, 34, 35
Diptera 210, 240
Dipylidium 16, 29
— caninum 43*, 43, 44
Docophorus 216*, 218
Dorylaimida 80, 107
Dracunculus medinensis 107, 118, 119*
Echidnophaga 235
— gallinacea 235
Echidnophagidae 277
Echidnophaginae 234
Echinobothrium 29
Echinococcus 17, 28, 29, 32
— granulosus 6, 8, 44, 45*, 47, 51
Echinodermata 160
Echinorhynchinea 63, 71*
Echinorhynchus 75
— gadi 65*
— salmonis 75
Elasmobranchia 61
Enoplidae 80
Enterobius vermicularis 87*, 107, 110*
Enteroxenos 137
Entocolax 132, 137, 1,38*
— rimsky-korsacowi 138*, 139*
— sciemenzi 136
Entoconcha 132; 137
Entoconchidae 132, 134, 135, 139*, 139
Entonyscidae 197
Eomenacanthus straumineus 217, 220
Eoicaridea 165, 166
Ergasilidae 143, 151, 168
Ergasilus 148, 149, 150, 166, 167
— sieboldi 147, 147*
Eriophydae 207
Eucestoda 6
Eucyclops 118
286
Eudiaptomus 33
Fannia canicularis 266, 267
Filariata 105, 107, 115
Filariidae 105
Filicollis 66*, 75
— anatis 66, 76*
Fimbriaria fasciolaris 9
Forcipomyia 244
Francisella tularensis 265
Freyana 200. 201*, 202, 203
— anatina 201*
Galumnidae 50
Gamasordea 195, 197
Gammarus 55
— lacustris 76
Gaslerosiphon 134, 135
Gastrophilidae 266’, 270, 273
Gastrophilus intestinalis 269*, 270
Gastropoda 131
Gastrotricha 78
Gfephyrolina 53
Gigainthorhynchinea 63, 71*
Glossina 240, 254, 255, 261*, 262, 265
Gnathia sp. 165*
Gnathiidea 165
Gnus cholodkovskii 242
Gordiacea 127
Gordius aquaticus 128*
Gynandrotaenia 16 Z
Gyrocotyle 58
— fimbriata 57*, 58
— nybelini 58
— urna 57*
Gyrocotylidea 53
Gyrocotyloidea 3, 55, 56, 58*, 58, 61, 62
Gyrocotyloides 58
Haematopota 252*, 253
Haemocera danae 144*
Haemogamasidae 197
Haemolaelaps 197
Haemonchus 99*, 100*, 126
Halarachnidae 197
Haplobothrioidea 29
Haplobothrium 29, 30*
Haplopleura 227
Hemimetabola 209
Hemiptera 210
Heterakis gallinarum 87*, 107
Heterakis sp. 86*
Hippobosca equina 255, 256*
Hippoboscidae 255
Holocephala 55
Holometabola 209
Homoptera 209
Hydracarina 208
Hydrotace 260
Hymenolepididae 16, 27, 32, 41, 130
Hymenolepis nana 41, 42*, 42
Humenoptera 209, 210
Hypodermatidac 266, 271
Histrichis 77
Ichthyotaces 146
Insecta 141, 208, 210
Ischnocera 215, 216*, 217
Isopoda 164, 167, 168
Ixodoidea 195
Ixodidae 195
Kinorhyncha 78, 79
Khawia 25
Laelaps 196, 196*, 197
Laelaptidae 197, 277
Lamellibranchia 130
Lamproglena 148
Laura gerardiae 155
Lauridae 160
Leishmania donovani 265
— tropica 265
Lepeophtheirus chatoni 145*
Lepidophtirus macrorhini 223
Leptoconops 244
Lernaea 150, 152
— cyprinacea 146* 152
Lernaeidac 150
Lernaeocera 149, 167
— branchialis 145, 149*, 150, 152
Lernaeoida 143, 148, 150, 167, 168
Lernaeopodtda 146*, 150, 152
Ligula 6, 31, 51, 52, 52*
Ligulidae 6, 7, 9, 24, 30, 32 51
Liperosia irritans 262
Lipeurus 218
Lipoptena cervi 255, 256*
Livoneca amurensis 164, 165*
Loa loa 107, 116*, 117
Lucilia 266, 267, 269, 272
Macracanthorhynchus hirudinaceus 63,
73, 76
Mallophaga 210, 215, 216
Mansonia 246
Marschallagia 99*, 100*
Megadenus 134, 135, 136, 137
— arrhynchus 133*, 136*, 138*
— voeltzkowi 136*
Megninia 202
Melane.lla 132; 133, 134, 135, 137
Melanellidae 132, 133*
Melolontha meeolontha 76
Melophagus 277
— ovinus 255, 256, 256*
Menoponidae 217
Mermithidae 87, 94, 106, 107, 274
Mesocestoides 12*
Mesocyclops 118
Mesogastropoda 132
Mesozoa 59
287
Metacrinus rotundus 157
Michaelichus 199*, 200, 202
Microstomum 29
Moniezia 50
—benedeni 50
— expansa 20*, 50*, 50
Moniliformis moniliformis 77
Monogenoidea 54
Monstrillidae 143, 144*, 146, 167
Mucronalia 132
— palmipedis 133*
Multiceps 32
—’multiceps 28, 51
Musca domestica 266, 266*, 267, 268,
272, 273
— sorbens 254, 260, 267
Muscidae 254, 266
Muscina stabulans 267, 272, 273
Myriapoda 208
Mytilicola 146, 151
Necator 86, 93
— americanus 112\ 113, 114
Nectonema 128
Namathelminthes 77, 78, 79, 127
Nematocera 240, 241, 253, 256, 257, 258,
258*, 260, 277
Nematoda 78, 79, 80*, 106
Nematodirus 99*, 100, 126
Nematomorpha 78, 127, 128, 129
Neoechinorhynchinea 63, 71*
.Neopsylla setosa 238
Nippostrongylus 93
Nippostrongylus sp. 86*
Nycteribia 255
Nycteribiidae 255
Odagmia ornata 242
Oesophagostomum 125
Oestriidae 266, 271
Oestrus ovis 269*, 271
Onchtrus ovis 269*, 271
Onchocerca volvulus 107, 116*, 118, 266
Oribatulidae 50
Ornithomya 219
Ostertagia 126
Ostracoda 160
Oxyurata 92, 107, 110
Paedophoropodidae 132
Paedophoropus 135
— dicoclobius 136*
Pandasentis iracundus 63
Pantostrongylus megistus 212
Parafillicollis altmani 65*
Parascaris equorum 125
Parascothorax 160
Parasitica 209
Parasitiformes 195
Parenteroxenos 137, 139, 140
— dogieli 139, 139*, 140
Parergasihts 152
Pasteurella pestis 237
Pediculus humanus 222, 223
— humanus capitis 224
— himanus vestimenti 224
Peltogaster 162
— socialis 162, 162*
Penella 145, 148
Peroderma 145
• Petrarca 160, 167
Petrarcidae 160
Phasmidia 79
Phlebotomidae 241, 257
Phlebotomus 245, 245*
— papatasii 264
Phtiraptera 214
Phtirus pubis 221, 223, 277
Phyllobothrium 8*
Piagetiella 276
— titan 217
Piroplasmida 194
Placentonema 190
— gigantissima 81
Plasmodium 265
Plathelminthes 58, 59, 62*’, 98
Polyctenidae 212, 277
Polymorphus 73, 75
— magnus 64*, 73, 74*,.76
Pomphorhynchus 75
Pontoporeia 74
Pdrtunion 165*, 168
Priapulida 77, 127
Prorhipidogldssa 60
Prosobranchia 60, 132
Protostrongylidac 1*04
Protostrongylus 104, 107
Pseudophyllidea 6, 8*, 11, 13, 14*, 16»
22, 24, 26, 29, 30, 31, 32, 33, 51
Pseudorliynchota 214
Pseudotracheliastes stellatus 146 *, 147
Pterolichus 198*, 199*
Pulex irritans 232*, 233, 234, 239
Pyramidellidae Г32
Rattus rattus 238
— norwegicus 238
Ravinia 266
ReduViidae 211
Rhabdias bufonis 103
Rhabditata 102, 114
Rhabditida 80, 86, 91, 83, 99, 102, 103,
107, 112, 114, 125
Rhabditis terricola 87*
^Rhadinorhymchus horridus 65*
Rhinoestrus purpureus 271
Rhizoccphala 163, 164, 167
Rhizolepas 163, 163*,’164
Rhynonyssidae 197. 278
Rickettsia mooseri 239
— prowazeki 194, 227, 228
— wolhynica 227
288
Rotatoria 78, 79
Sacculina 160, 161*
Saprolegnia 152
Sarcotaces 146
Sarcophaga 266, 272
Sarcophagidae 266, 267, 269
Schistocephalus 6, 19*
Secernentea 79, 80, 86, 87, 89, 94, 107
Sergentomyia 245
Simuliidae 241, 242, 256
Simulium 116*, 118
Siphonaptera 210
Sphyrion 149, 150, 152
— lumpi 147*
Spinturnix 196*, 277
Spirurida 80, 99, 105, 107, 115, 118, 126
Sporozoa 194
Stilifer- 134, 135, 136
— celebensis 133*, 134; 136*
Stiliferidae 132
Stomoxys 240
— calcitrans 254, 255, 262
Streblidae 255, 262
Strongylata 91, 93, 99, 103, 104, 107,
125, 126
Strongylidae 125
Strongyloides 101, 107
— Stercoralis 102*, 104, 114
Syngamidae 126
Syngainus trachea 80*, 81, 126
Synagoga 157, 167
— metacrinicola, 157, 159, 160
— mira 157, 159
Synagogidae 157
Syringobiidae 198
Tabanidae 240, 241, 252, 260
Tabanus 252
— a utomn al is 252*
Taenia 12*
— hydatigena 7*
— saginatus 41
— solium 8*, 9, 27, 30, 36, 38, 39*,
40, 41
Taeniarhynchus 14*, 29
— saginatus 9, 27,. 36, 37*, 38
Taeniidae 21, 36, 51
Tele os к i 60
Telotha siluri 165*
Tenebrio 42
Tetrameres sp. 80*
Tetraphthalmus titan 217
Tetraphyllidea 6, 7, 8*, 11, 14*, 16, 19,
26, 32, 33
Tetrarhynchus 8*
Thelazia 126
Thelaziidae 126
Theobaldia 246
Thompsonia 106, 168
Tracheata 141, 208
Trematoda 54
Trombidiforniis 207, 207*, 208
Triaenophoridae 32. 52
Triaenophorus 8*, 17, 18, 23*, 52
Triatoma 212
— inlestans 212
— rubrofacsiata 212
— sanguisuga 212
Trichinella 100, 107
— spiralis 87*. 121, 122*
Trichinellidae 107
Trichocephalidae 80, 90, 107, 120, 121
Trichocephalus 93, 107
Trichocephalus sp. 80*
— trichiurus 105, 120, 121*
Trichodectes 44
Trichodectidae 215
Trjchodcctoidea 217
Trichosonioides crassicauda 81
Trichostrongylidae 92, 99,. 126*
Trichostrongylus 126
Trilobitoniorpha 141
Trilocularia 29, 30*
Trypanosoma cruzi 212
Trombicula 207
Trombiculidae 207
Trombidiidae 207
Trombidium automnalis 207*
Trypanorhyncha 6, 7, 8*, 16, 26, 33
Tunga penetrans 232 *, 234, 278
Tylenchida 80, 107
(Jncinaria 93
Unionidae 130
Ulophysema 158
Vermipsylla 234, 235
Wohlfahrtia magnifica 267, 269*, 269,
273
Wuchfereria 100
— bancrofti 95, 107, 115, 116*, 117
Xenocoeloma 140, 146, 148, 149, 167,
168
— brumpti 147*, 148, 150
Xenopsylla .cheopis 233, 237, 238*, 238,
239
содержание
Ст
Предисловие ........... ’.......................3
Паразитические черви
Класс Cestoda...........................................&
Морфология и анатомия цестод.......................&
Особенности питания и обмена веществ цестод . . . К>
Размножение и жизненные циклы ленточных червей
, Гипотезы полизончиости и монозоичности цестод . . 28-
Классификация цестод..............................32
Заболевания, вызываемые цестодами.................33
Класс Cestodaria.......................................53
Отряд Amphilinidea ..................................53
Класс Gyrocotyloidea...................................55
Происхождение и филогения паразитических плоских
червей . .........................................58
Тип Асапthocepha les
Класс Acanthocephala...................................63
Морфология и физиология скребней..................63
Развитие и жизненный цикл скребней................71
Патогенное значение скребней . ...................75
Происхождение и эволюция..........................77
Тип N em a t h е Imi n t h e s
Класс Nernafoda......................................79
Морфология и анатомия нематод...................80
Некоторые особенности питания и обмена веществ не-
матод ..........................................92
Размножение и жизненные циклы нематод .... 97
Классификация нематод..........................106
Нематодозы.....................................107
Класс Nematomorpha ......... ,.......................127
290
Стр..
Паразитические моллюски и членистоногие
Тип М о 11 u s с а
Особенности морфологии паразитических Prosobran-
chia и их распределение по хозяевам............132
Тип Arthropoda
Подтип Branchiata . . 142
Класс Crustacea..........................................142
Подкласс Maxillopoda.................................... 143
Отряд Copepoda........................................ 143
Распространение, морфология и жизненный цикл пара-
зитических Copepoda............................... 143
Патогенное воздействие на хозяина.................. 151
Отряд Branchiura . . •............................... 152
Отряд Ascothoracida................................... 155
Отряд Cirripedia.......................................160
Подотряд Rhizocephala................................. 160
Подкласс iMalacostraca.................................. 164
Отряд Isopoda..........................................164
Основ1гые направления эволюции паразитических рако-
образных .......................................... 166
Подтип Chelicerata...................................... 168
Класс Arachnida......................................... 169
Отряд Acari .......................................... 169
Подотряд Parasitiformis .............................. 170
Надсемейство Ixodoidea.............................. 170
Морфология иксодоидных клещей...................... 170
Питание клещей и адаптации к кровососанию . . .174
Размножение н цикл развития клещей................. 179
Клещи — эктопаразиты позвоночных животных . . 184
Клещи — переносчики заболеваний.................... 185
Надсемейство Gamasoidea............................. 195
Подотряд Sarcoptiformes............................... 197
Надсемейство Analgesoidea.......................... 197
Надсемейство Sarcoptoidaea.......................... 204
Сел/. Sareoptidae, Sarcoptes scabiei.................204
Подотряд Trombidiformis............................... 207
Подтип Tracheata...................................... 208
Класс Insect а........................................ 208
Отряд Hemiptera........................................210
Клопы — паразиты человека.......................... 212
Отряд Mallophaga . ............................ . 214
Отряд Anoplura..................................... 220
Морфология вшей.................................... 221
Биология вшей и их приуроченность к хозяевам . . 223
Патогенность вшей и их роль в распространении бо-
лезней ....................................... .... 225
Отряд Siphonaptera..................................- 230
Морфология и жизненный цикл........................ 230
Патогенность блох.................................. 235
Эпидемиологическое значение блох................... 235
Отряд Diptera......................................... 240
Кровососущие двукрылые............................. 241
Подотряд Nematocera................................... 241
Подотряд Brachycera — Orthorrhapha................... 252
291
Адаптация двукрылых к питанию кровью и переход
к постоянному паразитизму 256
Медицинское и ветеринарное значение кровососущих
двукрылых........................................ 262
Синантропные мухи . . . ..........................266
Медицинское и ветеринарное значение синантропных
мух...............................................271
Основные направления эволюции паразитических клещей и на-
секомых .................................................275
Литература...............................................
Указатель русских названий животных и терминов 282
Указатель латинских названий животных.............285
ТАТЬЯНА АЛЕКСАНДРОВНА ГИНЕЦИНСКАЯ
АНДРЕИ АЛЕКСАНДРОВИЧ ДОБРОВОЛЬСКИЙ
ЧАСТНАЯ ПАРАЗИТОЛОГИЯ
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ, МОЛЛЮСКИ И ЧЛЕНИСТОНОГИЕ
Редактор В. С. Капышева. Художник А. Е. Коленков.
Художественный редактор Т. А. Колен ков а. Технический редактор
Т. А. Новикова. Корректор С К- Марченко.
ИБ № 1143*
Изд. № Е—328 Сдано в набор 10.01.78. Поди, в печать 01.08.78.
Формат 60X90’/ie Бум. тип. № 3. Гарнитура литературная.
Печать высокая. Объем 18,5 усл. печ. л. 21,28 уч.-изд. л. Тираж 15 000 экз.
Зак. № 58. Цена 1 руб.
Издательство «Высшая школа»,
Москва, К-51, Неглинная ул., д. 29/14
Московская типография № 8 Союзполиграфпрома
при Государственном комитете Совета Министров СССР
по делам издательств, полиграфии- и книжной торговли.
Хохловский пер., 7.